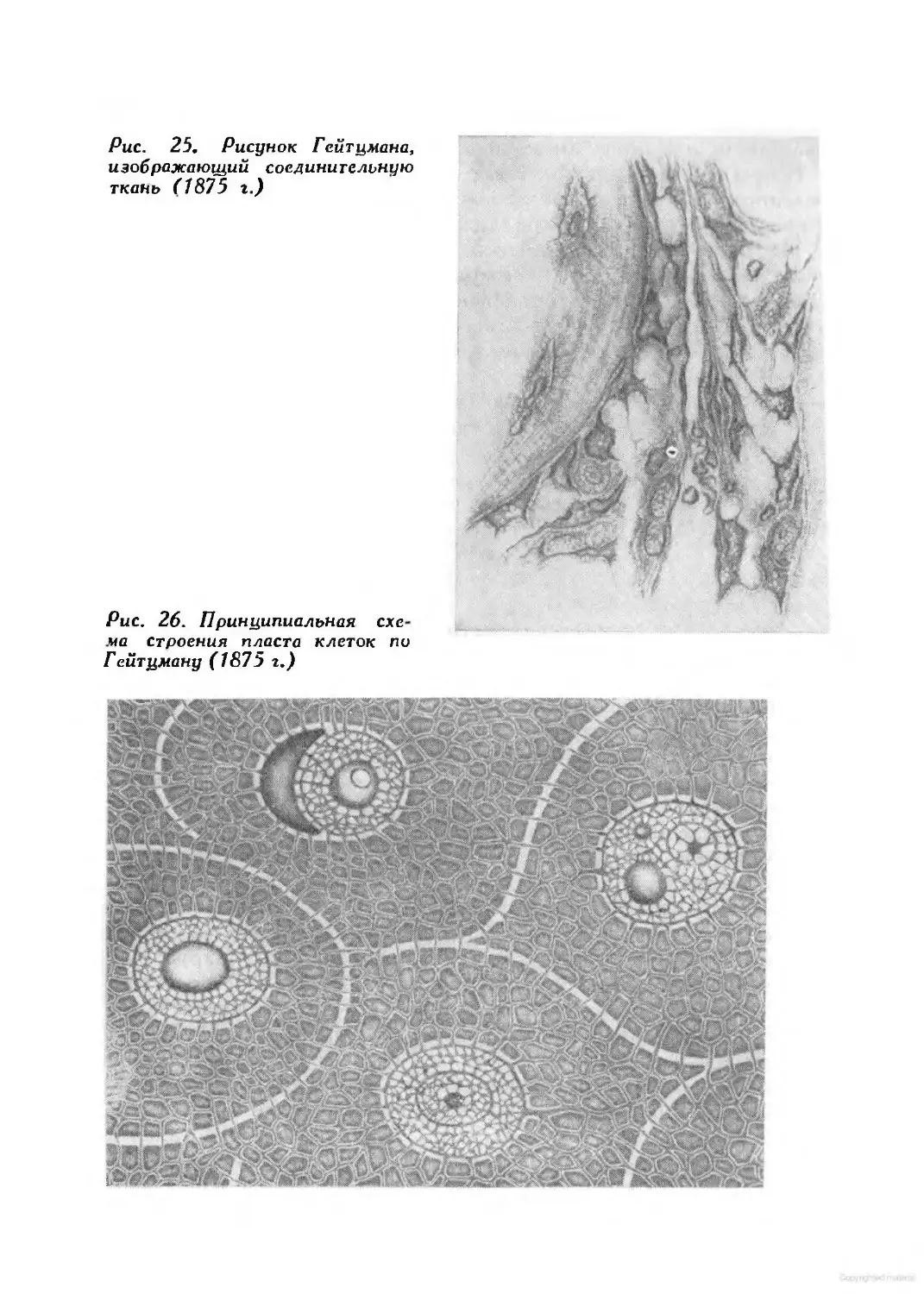
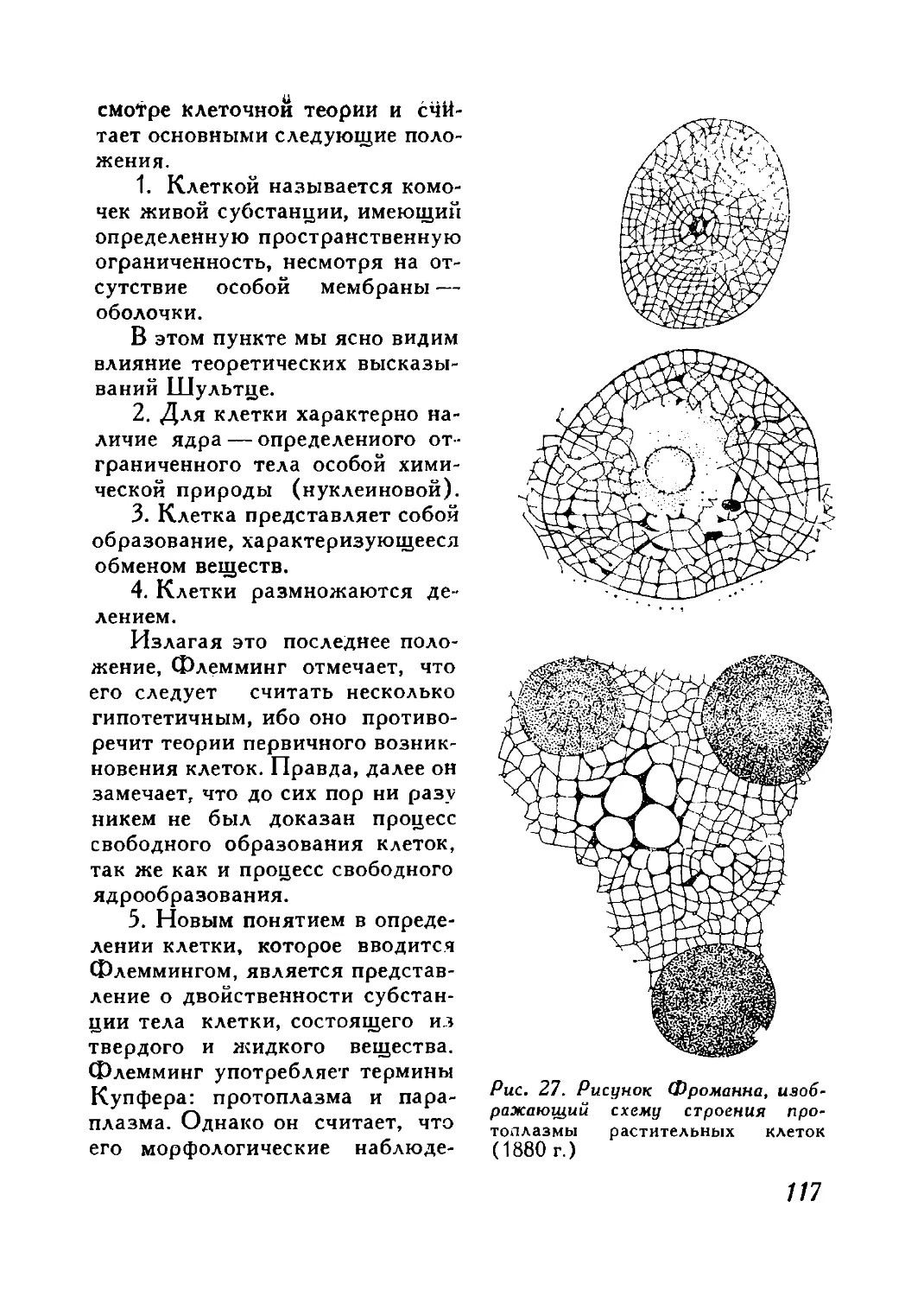
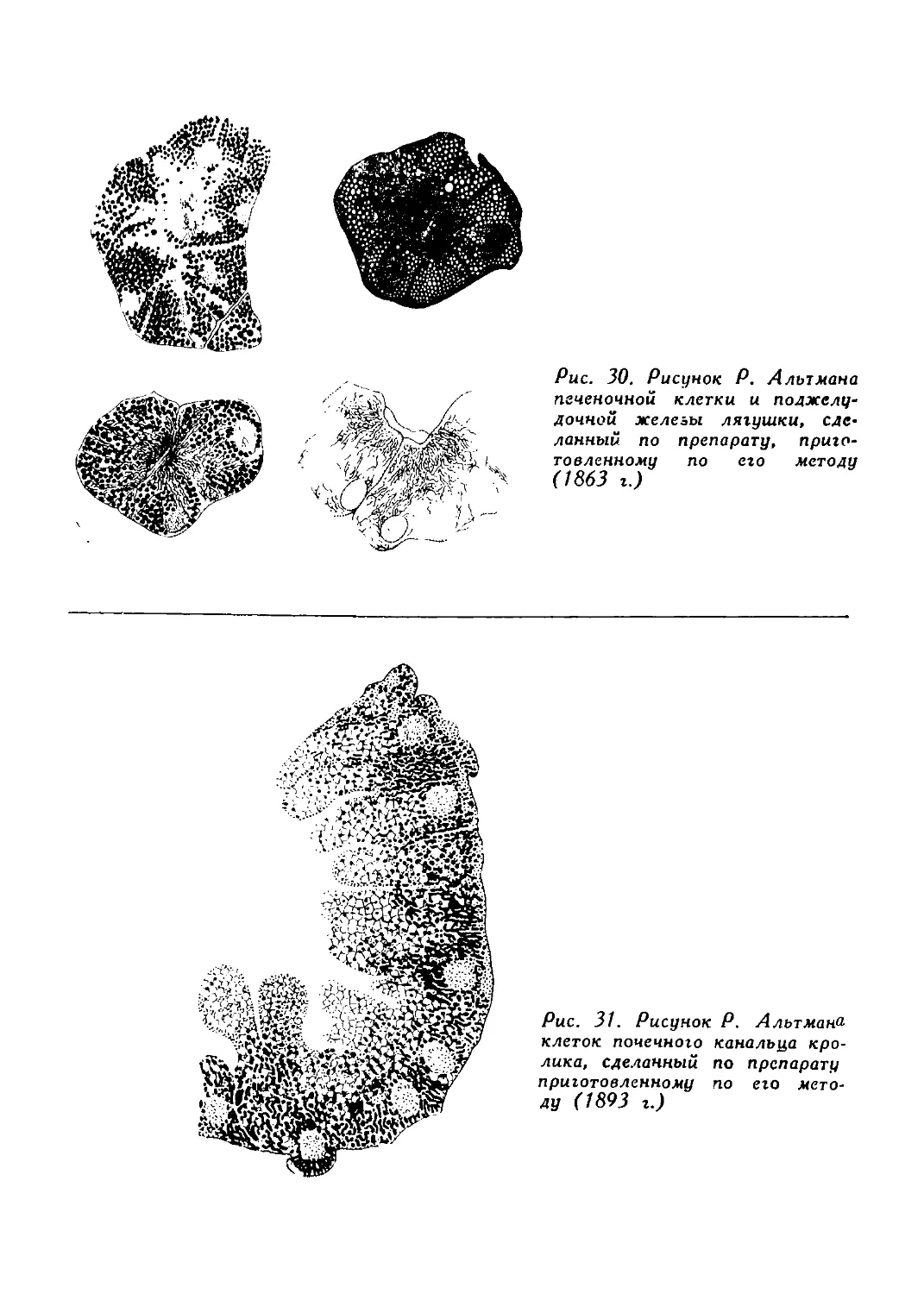
/







































































































































































































































































Текст
Г.МШЫЕЛЬ
ИСТОРИЯ
УЧЕПИЯ
ОКЛЕТКЕ
АКАДЕМИЯ НАУК СССР
ВСЕСОЮЗНЫЙ ИНСТИТУТ
НАУЧНОЙ И ТЕХНИЧЕСКОЙ ИНФОРМАЦИИ
«Я РАССКАЖУ ВАМ ОТКРОВЕННО. КАК БЫЛО ДЕЛО. НЕ ПРИБАВИВ НИЧЕГО ОТ СЕБЯ. ЧТО СТОИТ ИСТОРИКУ НЕ МАЛОГО ТРУДА».
Ф. М. ВОЛЬТЕР
Микромегас. 1752.
«КАКАЯ МЫСЛЬ. РАЗУМНАЯ ИЛЬ ГЛУПАЯ, НАЙ-
ДЕТСЯ, КОТОРУЮ Б НИКТО НЕ ВЕДАЛ ДО НЕГО».
И. В. ГЕТЕ Фауст. 1773—1775.
Е. М. Вермель
ИСТОРИЯ
УЧЕНИЯ
О КЛЕТКЕ
В
Издательство «Наука»
Москва 19 7 0
УДК (091) 576.3
Вермель Е. М. История учения о клетке. Изд-во «Наука», 1970 г.
В первой части книги излагается история изобретения микроскопа и первых научных открытий, сделанных с его помощью в XVII и XVIII вв. Освещаются взгляды на происхождение микроорганизмов и клеток растений и животных, развивавшиеся К. Вольфом и Л. Оке-ном. Подробно рассмотрены микроскопические исследования начала XIX в. и создание клеточной теории Т. Шванном на основе представления о единстве их происхождения путем цитогенеза, а также дальнейшее развитие этой теории в трудах Р. Вирхова.
Г лавам предшествует перечень открытий в хронологическом порядке, что составляет историю вопроса.
Вторая половина книги посвящена критике классической клеточной теории, начавшейся в начале XX в.
Книга рассчитана на широкий круг биологов.
Библ. 172 назв., рис. 74
2-10-2
490-69(1)
ПРЕДИСЛОВИЕ
Проблема клеточного строения организмов интересует ученых и широкие круги интеллигенции на протяжении более чем 100 лет. Это не удивительно, ибо данный раздел науки о жизни касается не частного биологического вопроса, а основного свойства живых существ. Выдающийся мыслитель, один из основоположников ма-териали|стической диалектики Фридрих Энгельс высоко ставил значение клеточной теории. Он прямо указывал, что в XIX веке решающее значение имели три великих открытия. Первым из них было доказательство превращения энергии, вторым — хотя .по времени и более ранним — открытием является открытие Шванном и Шлейденом органической клетки, как той единицы, нз размножения и дифференциации которой возникают и вырастают все организмы Третье великое открытие — теория развития, которая в систематическом виде впервые была разработана и основана Дарвином ’.
История отчетливо показывает, что как сам факт открытия клеточной организации живых существ, так и клеточная теория были поняты и признаны биологами далеко не сразу. Понадобилось почти 200 лет с того времени, как была впервые описана клетка (Гук, 1665), чтобы эту структуру признали основным элементом, из которых состоит и развивается организм. Клеточная теория дискутируется уже более 100 лет, но особенно интенсивными были споры в начале XX века, когда отчетливо выявилась ошибочность механистической трактовки этой теории. Характерной особенностью дискуссии явилось то, что в ней не участвовали защитники теории в ее классическом механистическом виде, а выступали преимущественно одни критики. Тем не менее, практически механистическое представление о клетке продолжало еще долгое время жить в умах очень многих биоло-
1 Энгельс Ф. 1964, Диалектика природы. М., Изд-во политической литературы,
5
гов, не занимавшихся специально этой проблемой. В результате клеточная теория во всех учебниках по биологическим дисциплинам (если не считать специальных пособий по цитологии и гистологии) часто излагалась в том же духе, как и в конце XIX века. Не следует полагать, что авторы были убежденными сторонниками механистической клеточной теории: они просто по традиции излагали вопрос упрощенно и впадали поэтому в вульгаризацию.
Вместе с тем следует указать, что некоторые ученые, признавая все историческое значение клеточной теории, выступали со столь резкой критикой, что по существу сводили ее значение на нет.
Ясно, что понимание такой большой и глубокой биологической проблемы, как организация живых существ, невозможно без знания истории вопроса. В. И. Ленин („Ленинский сборник", ХИ, 290) указал, что было бы чрезвычайно важно конкретно и подробно проследить общий ход человеческого познания на примере истории отдельных наук. Однако книг по истории учения о клетке мало как на русском, так и на иностранных языках. Автор пытается в какой-то мере заполнить этот пробел.
Первая попытка в этом отношении была автором сделана еще в 1940 г., когда была напечатана небольшая книга «Основные этапы в развитии учения о клетке» (Е. М. Вер<мель. Москва, Учпедгиз, 1940), изданная к столетию клеточной теории. Предлагаемая книга представляет собою переработанное и значительно дополненное издание первой книги.
Эта книга рассчитана на читателя, сравнительно мало подготовленного в специальных биологических вопросах, и поэтому мы ограничивались далеко не исчерпывающимся изложением проблемы и не могли останавливаться на вопросах, требующих специальных знаний по истории цитологии, гистологии и микроскопической анатомии. Автор поставил целью изложить не всю историю учения о клетке, а только лишь основные этапы этой истории.
Автор надеется, что его научно-популярный очерк истории учения о клетке будет полезен также в кружках по изучению диалектического материализма. По этой причине подробнее изложены такие вопросы, как системность в организации живых существ, организменная теория, теория биологического поля и др. Эти вопросы в общедоступной литературе отображены очень слабо. Кроме того, автор полагал, что изложение проблемы клеточного строения актуально в настоящее время еще и по
б
той причине, что в недавнем прошлом именно в этой области у нас в стране было сделано немало фактических ошибок и теоретических искажений.
И еще одно обращение к читателю. В своей книге автор излагает преимущественно историю клеточной теории. Что касается истории открытий деталей организации клетки, ее морфологии, физиологии и биохимии, то этих вопросов автор почти не касался, ибо для этого потребовалось бы написать еще одну книгу, может быть даже большую, чем эта. По этой причине совершенно не рассмотрены вопросы истории цитогенетики.
Конечно, излагая современное состояние и понимание вопросов, трудно оставаться на совершенно объективных позициях, и автор в какой-то степени невольно привносит свое понимание проблем, хотя он и стремился «ничего от себя не прибавлять» (Вольтер).
Автор с благодарностью примет все замечания и пожелания всех тех, кто возьмет на себя труд прочитать эту небольшую книгу.
Проф. Е. М. В е р м е л ь
Антон ван Левенгук (1632—1723)
ГЛАВА ПЕРВАЯ
ИСТОРИЯ МИКРОСКОПА
Основные даты по истории микроскопа
1608 Изобретение Галилео Галилеем в Италии подзорной трубы (телескопа).
1609—1610 Изобретение Г алилеем первого микроскопа.
1617—1619 Корнелием Дреббелем (Англия) сконструирован микроскоп по типу кеплеровской зрительной трубы.
1625 Иоганн Фабер (Германия) по аналогии с термином «телескоп» предложил называть прибор Галилея «микроскопом».
1665 Роберт Гук (Англия) усовершенствовал микроскоп путем введения третьей (собирательной) линзы.
1667 Евстахин Днвинн (Италия) предложил окуляры из двух линз, что создавало плоское поле зрения.
1715 X. Г. Гертель (Германия) применил осветительное зеркало для освещения микроскопических объектов.
1747 Леонард Эйлер (Санкт-Петербург) изложил идею конструирования системы линз хроматической аберрации.
1758—1760 Дж. Доллонд (Англия) применил для объективов два сорта стекла, чем добился дхроматнзацнн.
1827 Джамбаттиста Амичи (Италия) впервые применил плоскую фронтальную линзу для объектива.
1850 Дж. Амичи сконструировал первую водную иммерсию.
1873 Эрнст Аббе (Германия) создал систему осветительных линз к столику микроскопа.
1878 В. Стефенсон (Англия) сконструировал первые гомогенные масляные иммерсии.
1882 Фон Эбнер положил начало изучению анизотропных веществ (поляризационный микроскоп, темное поле).
1886 Э. Аббе сконструировал апохроматы, достигшие предела разрешающей способности световых микроскопов.
1908 А. Кёлер и Г. Зидентопф сконструировали люминесцентный микроскоп.
1928-1931 Е. Р у с к а, М. Кнолль и Б. Боррие предложили идею и сконструировали первый электронный микроскоп.
1934 Ф. Цернике разработал принцип фазового контраста.
9
§ /. Введение. Увеличительные стекла в качестве инструмента научных исследований начали применяться, как показывает история науки, только в конце XVII в., однако изобретены они были значительно раньше. Значение луп и микроскопов долгое время недооценивалось учеными. Микроскопические наблюдения как научный метод получили всеобщее признание только в XIX в. Основную роль в этом отношении сыграло создание клеточной теории. Именно теория определила интерес к микроскопическим наблюдениям и заставила ученых всего мира признать учение о клетке одним из кардинальных фактов, лежащих в основе общих биологических представлений.
Микроскоп — это, вероятно, самый широко распространенный инструмент биолога. Микроскоп внес в биологию очень много нового и специфического, с его помощью был создан особый раздел науки о жизни — микроскопическая анатомия животных и растений (цитология и гистология). Таким образом была открыта новая область видения: микроскоп, как нередко говорят, «сделал доступным наблюдению новый мир». Микроскопические наблюдения после открытия живой клетки заставили резко изменить взгляд на природу материи. Но понадобилось более двух веков, чтобы прийти к представлениям о живой клетке как элементарной структуре.
По этим причинам история открытия и развития микроскопа как физического прибора представляет для истории биологии выдающийся интерес.
§ 2. Изобретение оптических линз. В древности (это можно считать в настоящее время твердо установленным) оптические линзы не были известны. В то время геометрической оптики не существовало, и ученые не могли объяснить даже действия «зажигательных» стекол.
Оптическая линза была изобретена только в самом конце XIII или в начале XIV в. и была использована в качестве очков, сразу же получивших широкое практическое применение. В попытках установить личность изобретателя очков недостатка нет, но все они остались безуспешными. Единственно, что можно сказать определенно, это то, что изобретены очки были в Италии — в Венеции или Падуе — центрах стекольной промышленности того времени. Очки эти представляли длиннофокусные линзы, применявшиеся при дальнозоркости. Вогнутые же стекла для устранения близорукости появились только в XVI в.
10
Существует мнение, что Нерон (I в. н. э.) употреблял «очко» из кусочка зеленого смарагда. Однако эта версия по ряду причин несостоятельна. Нерон, обладая слабым зрением, очевидно, просто ослаблял силу света плоскопараллельными дымчатыми очками.
Вместе с тем интересно отметить, что хрусталик глаза был известен уже в I в. н. э., когда он был описан Клавдием Галеном (ок. 130 — ок. 200) и (особенно подробно) Руфусом Эфесским, которому принадлежит и сам термин «хрустали к». Однако о его функции в то время ничего не знали. Знаменитый арабский ученый Альхасен (965—1038) считал, что хрусталик превращает световое раздражение в зрительное ощущение. Это воззрение держалось до конца эпохи Возрождения, когда швейцарский врач Феликс Платтер (1583) показал, что хрусталик является телом, преломляющим лучи.
Спрос на очки особенно возрос в XV в., и это привело к тому, что появилось много профессионалов этого дела. Материалом для шлифовки очков было прежде всего стекло, но стекло того времени отличалось низкими качествами, и (в лучшем случае) оно было зеленоватого оттенка. Поэтому употреблялся также горный хрусталь и полудрагоценный прозрачный камень — берилл (отсюда и немецкое название очков — Brillen).
Первые научные исследования, в которых были использованы линзы (лупы), принадлежат энтомологам Томасу Моуффет и Георгу Гуфнагелю (1589—1590). Первый из них изучал мелких клещей (0,2—0,4 мм), в частности, он со всей отчетливостью установил различия между чесоточным и сырным клещами. В тот же период были описаны и такие труднонаблюдаемые процессы, как движение гемолимфы у вшей и др.
Лупами, или (как их позднее называли) «простыми микроскопами», пользовался и основоположник микроскопической биологии Антон ван Левенгук (1632—1723).
Интересно указать, что идея оптического инструмента, позволяющего отчетливо видеть отдельные, очень мелкие, объекты, была высказана еще в XIII в. ученым и фолософом Роджером Бэконом (ок. 1214—1294), а в XV—XVI вв. она уже многократно повторялась рядом выдающихся ученых и мыслителей: Леонардо да Винчи (1452—1519), Иеронимом Фракасторо (1478—1553), Диггсом, Борном и многими другими. В тот же период с полной отчетливостью была высказана мысль о существовании мира не видимых простым глазом существ — возбудителей заразных болезней (Фракасторо и др.). Мысль эта была,
11
следовательно, высказана за 125 лет до работ Левенгука, но обосновывалась она частично теоретически и была по существу фантастична.
Уже в XVI в. была высказана идея создания оптического прибора, состоящего из двух, а не из одной линзы. Среди математиков и физиков она была даже довольно широко распространена, хотя практически оставалась неосуществленной, видимо, из-за технических трудностей. Только в начале XVII в. знаменитый Галилей сначала сконструировал телескоп, а затем (в 1609 г.) и микроскоп.
§ 3. Изобретение сложною микроскопа. Честь создания первого микроскопа до последнего времени приписывали голландским шлифовальщикам очковых стекол Гансу и Захарию Янсенам из Миддельбурга и датировали это изобретение 1590 годом. Это вошло во многие учебники, но эти сведения, видимо, следует считать ошибочными. Документы, которыми обосновывается данное утверждение, как оказалось, доверия не заслуживают, и имеются основания полагать, что они даже намеренно искажают действительность
Твердо установленным можно считать, что первой была сконструирована «земная зрительная труба», которая получила название «голландской», или «галилеевой», трубы, а позднее — телескопа. «Земная (подзорная) труба», как считают, была изобретена независимо друг от друга шлифовальщиками оптических стекол Липерсгеем и Мециусом в Голландии (1608) и Галилеем (1608—1609) в Италии.
Первый микроскоп (он возник как видоизменение зрительной трубы, о чем имеются достоверные сведения) был сконструирован в 1609—1610 гг. самим Галилеем; линзы ему шлифовал мастер Бацци, осуществлявший все его замыслы. Сложный микроскоп состоял из объектива, дававшего действительное увеличенное И Обратное изображение мелкого объекта, и окуляра.
• 1 С о б о л ь С. Л. Очерки по истории микроскопа. Диссертация на соискание степени доктора биологических наук. Защищалась в 1944 г. в Академии наук СССР (Москва).
Соболь С. Л. 1949. История микроскопа и микроскопических исследований в России в XVIII в. М., Изд-во АН СССР.
Многие данные по истории микроскопа взяты нами из этих сочинений.
12
Первый микроскоп Галилея представлял собою длинную трубу и был очень неудобен в обращении. Для научной работы этот микроскоп никем не употреблялся и был утерян, но тем не менее получил известность. Вскоре в Голландии были построены аналогичные микроскопы.
Несколько позднее был сконструирован микроскоп физиком и астрологом (при дворе английского короля Якова I) Корнелиусом Дреббелем (1617—1619), голландцем по национальности. Трудно сказать, имел ли он в качестве образца голландский микроскоп или же Дреббель построил свой прибор сам в соответствии со сведениями, которые он получил о микроскопе Галилея (рис. 1). Последнее считается даже более вероятным. Дреббелевский микроскоп отличался от микроскопа Галилея тем, что был построен по типу кеплеровской трубы, т. е. он имел выпуклые линзы как объектива, так и окуляра. Этот первый микроскоп Дреббеля также не послужил для научной работы и был утерян, но именно микроскопы такого типа вскоре получили довольно широкое распространение.
Сложный микроскоп возник как видоизменение зрительной (подзорной) трубы, причем он был сконструирован уже через несколько лет после ее появления, однако дальнейшая их судьба очень различна. Значение подзорной трубы было тотчас же понято, и правящие круги Голландии, Бельгии и Италии пытались даже засекретить этот инструмент. Очевидное значение подзорной трубы в военном деле, мореплавании и в астрономии
Рис. 1. Реконструкция микроскопа Клиффорда Дреббеля, установленного на трех дельфинах- Выполнена по описанию В. Бореля. 1619 г- (из С- Л- Соболя)
73
привлекло большое к ней внимание; производство подзорных труб и телескопов немедленно приняло широкие масштабы. На микроскоп же очень длительное время смотрели как на занятную игрушку. Сам Дрсббель вместе со своим зятем Куффлером наладил производство подобных «сложных микроскопов»; они широко рекламировались и быстро распространились по всей Европе.
В 1624 г. Галилей переконструировал свой микроскоп, значительно его усовершенствовал, и им можно было уже пользоваться с практическими целями. Он имел значительно меньшие размеры, чем первая модель, и давал, по-видимому, максимальное увеличение в 35—40 раз.
§ 4. Первые микроскопические наблюдения. В Италии микроскопу было придано несколько большее значение, чем в Англии. Галилей подарил свой микроскоп в 1624 г. «Академии проницательных» в Риме2, членом которой он состоял. Уже в 1625 г. член Академии Стеллути опубликовал свои наблюдения о строении органов пчелы, сделанные с помощью микроскопа. В частности, он впервые увидел и описал фасеточное строение глаза насекомых. Ф. Чези (1628) использовал микроскоп Галилея, изучая спорангии папоротника, и описал с его помощью споры, считая их, однако, семенами.
Термин «микроскоп» впервые употребил Иоганн Фабер в 1625 г. (в письме к князю Ф. Чези), предложив так называть инструмент Галилея, предназначенный (в противоположность телескопу) для рассматривания мелких предметов. Термин этот удержался до настоящего времени и прочно вошел в обиход.
Однако число исследователей, пользовавшихся микроскопом в XVII в., было очень невелико, да и те прибегали к нему только от случая к случаю. В Германии микроскопы выпускались в большом количестве мастерскими игрушек (нюрнбергские мастера) и, будучи сделаны из картона и дерева, были дешевы. В Англии в начале XVIII в. они делались гораздо тщательнее и много лучше, в частности, они имели макрометрический винт и
" «Akademia dei lincei» была учреждена в 1603 г. князем Ф. Чези. Иногда название Академии переводится у нас как «Академия рысьиглазых», одиако перевод не может быть дословным: Lynx (лат.) действительно значит рысь, но в итальянском языке это слово применяется и в смысле зоркий или проницательный.
14
делались и& бронзы. Правда, эти микроскопы стоили очень дорого — до 40 фунтов стерлингов, по тем временам это были большие деньги. Оптика всех этих микроскопов оставалась весьма примитивной: объективы давали значительную хроматическую и сферическую аберрации, поэтому при более значительных увеличениях изображение было неотчетливым. Ко всем микроскопам обычно прилагались готовые коллекции препаратов, состоявшие из высушенных мелких насекомых (вши, блохи, головки мух и др.), а также различные дополнительные приборы, вроде микроаквариумов, зажимчиков для удержания мальков, чтобы можно было вщдеть движение крови в сосудах и т. д.
«Микроскопическое любительство» было в XVIII в. широко распространено. На многих гравюрах того времени с изображением аристократических салонов можно видеть, как кто-либо, сидя за столиком, что-то рассматривает в микроскоп.
В конце XVIII в. микроскопы делались уже исключительно мастерами-профессионалами, которые его значительно усовершенствовали. Центром микроскопической индустрии был Лондон (мастерская Джона Кеффа), откуда микроскопы расходились по всем странам. Объективная оценка микроскопов того времени показывает, что они давали уже удовлетворительное изображение и с их помощью различимы даже очень мелкие структуры (до 5 микронов). Аберрации во всяком случае в глаза не бросаются, что достигалось введением кольцевых диафрагм, срезавших краевые лучи.
Увеличения микроскопов XVIII в. были весьма разнообразны. Больше всего изготовлялись системы с увеличением в пределах от 10 до 150 раз, но встречались и значительно большие. Однако их разрешающая способность была невелика, так что большие увеличения являлись по существу иллюзорными. Интересно, что теоретические расчеты объективов и окуляров были в то время уже довольно хорошо разработаны (Байкер, Кеплер, Эйлер, Гюйгенс и многие другие). Однако техника значительно отставала, и, в частности, с большим трудом достигалось центрирование линз.
Микроскописты-любители сделали много интересных и важных открытий. К таковым по существу должен быть причислен и знаменитый Антон ван Левенгук — мануфактурный торговец по профессии. Его наблюдения были столь систематичны и многообразны, что должны считаться настоящим научным исследованием, положившим начало науке о микроскопических организмах. Левенгук вел свои наблюдения в продолжение более чем 50 лет
/5
Рис. 2. Фронтиспис к книге А. ван Левенгука «Тайны природы»
ARCANA
N A T U R /Е
Deteda
Ab
Antonio van Leeuwenhoek.
DELPHIS BATAVORUM, Apud HENRICUM a KROONEVEI.D. cio io c xcv.
Puc. 3. Титульный лист книги А. ван Левенгука «Тайны природы»
Рис. 4. «Микроскоп» А. ван Левенгука, в натуральную величину (слева схема, показывающая его устройство) (из С. Л. Соболя)
и сообщал их Лондонскому Королевскому Обществу в ряде писем, которые делались широко известными. Впоследствии они были объединены и напечатаны отдельной книгой, называвшейся «Тайны природы» (1696) 3 4, а сам Левенгук, еще в 1680 г., был избран членом Королевского Общества (соответствует нашей Академии наук) (рис. 2, 3).
Работы Левенгука привлекли серьезное внимание и явились ценнейшим вкладом в науку: они дали могучий толчок к даль-неишим исследованиям
Левенгук пользовался двояковыпуклыми линзами, которые он сам шлифовал и вставлял в пластинки. Эти увеличительные
3 Leewenhoek van. 1719. Arcana naturae ope et beneficic exquisitissimo-rum microscopicorum Lugd. Batavorum 1696. Epistole physiologicae super compluribus naturae arcanis etc. ad alios cl. vinos doctos. Delphis («Тайны природы, открытые с помощью совершеннейших микроскопов. Физиологические письма к другим ученым мужам о множестве тайн природы»).
4 О Левенгуке рекомендуется прочитать более подробно в замечательной книге Поля де К р ю и «Охотники за микробами», а также в небольшой книге Такжниа Н. В. «Левенгук, его жизнь и деятельность (по его письмам)». Л-, 1946.
18
стекла («простой микроскоп», или лупа) едва достигали величины булавочной головки, их употребление требовало немалого искусства и очень хорошего зрения. Левенгук добивался настолько больших увеличений, что мог видеть даже красные кровяные клетки (их диаметр у человека равен 7—8 микронам, т. е. 0,007 миллиметра) и многие другие объекты. Наибольшую славу ему доставило, однако, открытие мира микроскопических животных, которые он назвал инфузориями ( от слова Infusum, что по латыни значит «настой») (рис. 4).
Открытия Левенгука обратили на себя внимание не только ученых, но и всего культурного общества того времени. Он имел много подражателей, приобретавших увеличительные приборы и работавших с ними весьма упорно и настойчиво.
Многие любители настолько серьезно занимались микроскопическими наблюдениями, что сами сделали ряд важных открытий, вошедших в науку. Особую известность приобрели замечательно иллюстрированные книги юриста Мартина Ледермюллера (1719—1769). Одна из книг носит очень характерное для того времени название: «Микроскопические забавы для глаз и души» (1762). На рис. 5 и 6 приводится титульный лист книги, а на рис. 7 и 8 — таблица из нее с изображением ряда инфузорий, многие из которых были впервые описаны. Подобные исследователи-любители, не ставя себе никакой определенной цели, описывали самые разнообразные объекты. Они приносили большую пользу, обогащая науку сведениями о низших формах живых существ. Привлекая к себе большой интерес, эти наблюдения пропагандировали область микроскопических исследований.
§ 5. Отражение микроскопических открытий в художественной литературе. В настоящее время, когда о микроскопе мы слышим с детских лет, трудно себе даже представить, какое колоссальное впечатление производили в то время микроскопические исследования. Не надо забывать, что микроскоп открывал новый мир — мир микроскопических существ, о существовании которого ранее ничего не знали. Именно поэтому микроскопические животные всех так интересовали. Даже все хорошо известные предметы, будучи значительно увеличены, представляются нам совершенно иными.
Интерес широких кругов к микроскопическим исследованиям отразился в художественной литературе того времени. Приведем прежде всего одни наиболее яркий пример.
19
Рис. 5. Фронтиспис к книге М. Ледермюллера «Микроскопические забавы...» (1762 г.)
fi&drtin SrobtniuS $ebermu(Itr5, Shababs) 3«!®V!*•<$«, аИсаафМ &*>
W«H arikw hr tofcfb » »<t SMf*<a MdMaft и HMf аьщйм, 2Rifro(topi($e
unb
OTiiaen = <£r й&| к n g: SJeftebtnb t in Gilt $unbert паф bet Statue gejeitfintten unb mit Sarben trltufttctcn Jtupftrfaftln > 6<aat teten grtUrung.
3)»7ltgt von ШЬат ©offfgeng ®intrrfd)mibtt fspftrOncr it Ricabctn acbriirft non Cfcrlftlan de Lauooy.
। 7 i г.
Рис. 6. Титульный лист книги М. Ледермюллера «Микроскопические забавы...»
Рис. 7. Таблица XXXI из книги М. Ледермюллера «Микроскопические забавы...»
На рисунке стеклянный оплавленный шарик, его отшлифовывали на камне с обеих сторон для получения очень маленьких линз с большим увеличением; остальные рисунки — различные оправы, в которые вставляли линзы (простые «микроскопы»)
Рис. 8. Таблица LXXXVIII из книги М. Ледермюллера «Микроскопические забавы...» с изображением преимущественно инфузорий (под названием «полипов»), в большинстве впервые описанных автором
Имеются основания считать, что Джонатан Свифт (1667— 1745) написал свое бессмертное произведение «Путешествие в некоторые отдаленные страны света Лемюэля Гулливера» под влиянием открытий Левенгука. Сочинение это было опубликовано в 1726 г., в период, когда о микромире особенно много говорили. Известно также, что Свифт читал книги Левенгука и знал его лично5. В основу «Путешествия Гулливера» автором положена идея значительного увеличения илн уменьшения всей окружающей человека обстановки, при которой он как бы рассматривает все через уменьшающую илн увеличивающую систему линз.
В сочинении Дж. Свифта Гулливер сначала попадает в страну лилипутов: создается положение, при котором рассматривают человека в увеличительный прибор. Затем, когда Гулливер оказывается в стране великанов, происходит обратное явление, т. е. человек как бы рассматривает все окружающее в сильную лупу. В одном месте этого сочинения мы читаем, что король страны великанов приказал своим ученым дать заключение о Гулливере. Рассмотрев его в лупу, онн пришли к выводу, что он является вполне взрослым человеком. Далее Гулливер обращает внимание на то, что все предметы выглядят так, как будто нх рассматривают при сильном увеличении. Очень любопытно его замечание о коже девушки из страны великанов, которая ему очень понравилась. Оказалось, что вблизи кожа девушки «была отвратительна». Это навело Гулливера на размышления о том, что «нежная кожа английских дам кажется нам красивой только потому, что ее изъяны можно видеть не иначе, как в микроскоп, который ясно показывает, как груба, толста н скверно окрашена самая нежная и белая кожа» 6 (стр. 174).
Аналогичный прием избрал Ф. М. Вольтер в своем сатирическом сочинении «Мнкромегас» (1752). Путешествуя, великан Микромегас выловил в Балтийском море корабль, на котором «ватага ученых возвращалась нз полярной экспедиции». «Микроскоп, прн помощи которого он едва мог различить кита и корабль, не годился для наблюдения таких крошечных тварей, как люди» (стр. 119). Далее у Вольтера мы читаем, что Микромегас и сопровождавший его ученый секретарь Сатурнской Академии наук продолжали свои микроскопические наблюдения
5 Singer Ch. 1931. A short history of biology. Oxford (Краткая история биологии).
6 Свифт Джонатан. 1928. Путешествия Гулливера. Л., изд-во «Academia».
24
«Левенгук и Хартсекер, которые первые увидели или полагали, что увидели, семя, из коего мы образуемся, сделали открытие сравнительно далеко не столь удивительное» (стр. 120)7. «Обитатель Сатурна, перейдя от крайнего скептицизма к крайнему легковерию, подумал, что он наблюдает их (людей) размножение. «Ах,— сказал он,— я застиг природу на месте преступления». Но он ошибся, ибо судил только по внешности, что случается слишком часто, как при употреблении микроскопа, так и без него» (стр. 121) 8.
В целом ряде сочинений, написанных другими авторами, также фигурируют микроскоп и ученые-микроскописты. Укажем хотя бы еще на «Повелителя блох» Эрнста Теодора Амадея Гофмана (1776—1822). Идея этого произведения построена на увеличении микромира до размеров, обычных в нашем обиходе. Далее интересны «ученые» рассуждения барона Мюнгаузена. Содержание их, например, таково: «... я предавался различным размышлениям, так как мои довольно обширные познания способствуют тому, что самые разнообразные вещи приходят мне в голову одновременно. Я нашел, что инфузории, быт которых, между прочим, занимает меня в последнее время, представляют собою в сущности недоразвившихся карпов и обладают памятью» 9.
§ 6. Особенности микроскопических исследований. Уже в XVIII в. многие крупные ученые понимали, что область микроскопических исследований сложна именно своим своеобразием, специфичностью своих картин. Интересные мысли по этому вопросу мы находим у одного итальянского естествоиспытателя XVIII в.— Феликса Фонтана (1723—1805). Он писал, что «посмотреть в микроскоп может каждый, но лишь немногие могут судить о виденном» (1781 г.). Эти замечательные по своей глубине слова и в настоящее время со-
7Хартеекер (в современной русской транскрипции Гартсекер) — физик, претендовавший иа открытие сперматозоидов в 1678 г. В действительности сперматозоиды человека впервые были открыты студентом Гаммом в 1675 г., работавшим под руководством Левенгука. Всего этого Вольтер не знал. Подробнее см. в книге Некрасова А. Д. «Оплодотворение в животном царстве». М.— Л., 1930.
* Вольтер Ф. М. А. 1934. Микромегас. Философская история. Л., изд-во «Academia».
*Иммерман Карл. 1838—1839. Мюигаузен, история в арабесках.
Русское издание. Л., изд-во «Academia».
25
краняют свою актуальность. Фонтана специально указывает, что все наши суждения о различных предметах основаны на сравнениях и сопоставлениях, но поскольку микромир совершенно своеобразен, то весьма опасно пользоваться представлениями, которые получены нз наблюдений над макромиром, тем более, что «картина, открываемая микроскопом, может подходить одновременно к разнообразным предметам». Этим мыслям, которые мы здесь передаем, конечно, очень кратко, Фонтана посвятил в своей большой книге 10 специальную главу под названием «О микроскопических ошибках, которые происходят при микроскопических наблюдениях».
Фонтана оказался прав, и, может быть, нигде не было сделано прн установлении фактов стольких ошибок, сколько их было при микроскопическом изучении организмов. Даже в основном вопросе (о природе и строении клетки) мы встречаем чрезвычайно много противоречивых данных. Встает вопрос: не зависит ли это от несовершенства микроскопической техники и недостаточно быстрого ее прогресса. Такая мысль как будто напрашивается сама собою. Однако это не совсем так. Оказывается, что микроскоп— главный инструмент исследования — почти до самого последнего времени не ограничивал биологические наблюдения. В подавляющем большинстве случаев оптические приборы представляли большие возможности, чем те, которые были использованы. Укажем только на один пример: ядро в клетке было открыто только в 1833 г. Р. Броуном, а между тем уже микроскопы XVIII в. позволяли его прекрасно видеть.
Все это показывает, насколько важны в конкретных исследованиях теоретические представления, которых придерживается ученый, и какое значение имеет уровень его знаний. История биологических наук знает немало примеров, когда достаточно было одного указания авторитетного ученого, чтобы тотчас все обнаруживали факт, который проглядывали десятки лет. Но бывало и такое, что мнение авторитетного ученого, хотя и совершенно ошибочное, заставляло видеть то, чего в действительности не было, н подтверждать абсолютно неправильные представления. Примеры того и другого мы в дальнейшем увидим во множестве.
10 Fontana F. Traite sur le venin de la vipere etc. Florence. 1787 («Сочинение о яде гадюки, американских ядах, лавровишневом яде и некоторых других растительных ядах, а также наблюдения над простейшим строением организма, строением нервов и описание нового глазного канала»), Оригинал написан на итальянском языке в 1781 г.
26
Рис. 9. Микроскоп Роберта Гука (из С. Л. Соболя)
§ 7. Первые микроскопические исследования ученых. Число настоящих научных исследований, проводившихся с помощью микроскопа профессиональными учеными, было в XVII и XVIII вв. очень невелико.
Первое исследование принадлежит секретарю Лондонского Королевского Общества, разностороннему ученому (физику, астроному, геологу и биологу) Роберту Гуку (1635—1703). Он был в крупной ссоре с Исааком Ньютоном, оспаривая его корпускулярную теорию света. Известно также, что он был очень некрасив и не позволял себя рисовать, поэтому-то его портрета нигде нет.
27
Гук был в свое время крупным ученым, физиком по специальности. Он сам переконструировал микроскоп и технически значительно его усовершенствовал. Его микроскоп состоял из трех элементов: собирательной линзы (коллектор), линзы окулярной и линзы объективной (рис. 9). По этому типу и строились в основном микроскопы в XVIII в. Их дальнейшие усложнения заключались во введении в окуляр и объектив дополнительных линз. Работать с этими микроскопами все же было очень трудно, так как микроскопирование требует сильного освещения. Плоское зеркало с этой целью применил только в 1716 г. оптик X. Г. Гертель из г. Галле (Германия). «Зайчик» от зеркала значительно облегчил работу, и с тех пор зеркало становится обязательной составной частью микроскопов.
Свою известную монографию «Микрография или физиологическое описание мельчайших тел, исследованных при помощи микроскопа», Гук опубликовал впервые в 1665 г.'1 Таким образом, прошло болре 50 лет с того времени, как Галилеем был сконструирован первый микроскоп, пока, наконец, появилось первое фундаментальное исследование, сделанное с помощью этого прибора.
«Микрографией» назвал Гук описание микроскопического строения различных предметов. Этот термин держался довольно долго, и микроскопистов этого периода до снх пор называют микрографами. Сам Гук изучал очень многие предметы, при этом он не ставил никакой определенной цели или задачи. В этом легко убедиться, просмотрев названия глав его книги, которые называются: о кончике острия маленькой итлы, о тонком батисте, о песке в моче, о фигурах, образуемых морозом, о семенах мака, о муравье и т. д.
В числе других предметов (наблюдение № 18) Гук изучал тонкие срезы растений, и именно ему принадлежит честь открытия клеточного строения растений. К этому мы вернемся ниже и остановимся на этцх наблюдениях подробнее, поскольку они сыграли большую роль в развитии науки о тончайшем строении организмов.
Познание микроскопического строения организмов получило свое дальнейшее развитие в классических работах Марчелло Мальпиги (1628—1694) и Неемии Грю (1641—1712). Однако
11 Hooke R. 1665. Micrographia: or some physiological descriptions of minute bodies made by magnifying glasses with observations and inquiries thereupon. London.
28
это направление исследований в XVIII в. не было оценено, а работ по выяснению тонкой структуры тканей растений и особенно животных было очень мало. Как мы уже указывали выше, причина заключается не в технических несовершенствах микроскопов того времени. Стоит только взять в руки микроскопы XVIII в. или начала XIX в. и посмотреть с их помощью препараты, чтобы стало ясно, как многое в них можно видеть.
Отсутствие интереса к данной проблеме должно объясняться тем, что наука XVII и XVIII вв. еще недостаточно созрела для понимания значение микроскопической структуры организмов, того, что теперь называют «клеточным уровнем организации материи».
Значительно большую роль сыграло изобретение микроскопа в разрешении проблемы самозарождения организмов, особенно сильно волновавшей умы ученых того времени. Открытие Левенгуком «наливочных анималькулей» (инфузорий) поставило вопрос об их природе и возникновении. После знаменитых работ Ладзаро Спаланцани (1729—1799), когда стала очевидной невозможность самозарождения мух, клещей, червей, мышей н т. д., возник вопрос о самозарождении микроорганизмов, а поэтому большой интерес вызывали их строение и природа. Мы не станем останавливаться на данной проблеме, ибо это выходит за рамки нашей темы — истории учения о клетке, тем более, что вопрос этот хорошо освещен в литературе12 13. Укажем только, что окончательно он был разрешен лишь в XIX в. классическими исследованиями Луи Пастера (1822—1895), причем речь шла уже не о простейших, одноклеточных животных, а о бактериях, которые в XVIII в. вообще не были известны. Вопрос о самозарождении инфузорий отпал после того, как было показано их сложнейшее строение. Первое исследование, проведенное над «наливочными анималькулями» (инфузориями), принадлежит М. М. Терехов-скому (1740—1796). Оно представляет интерес еще и в силу того, что является первым экспериментальным исследованием русского ученого на биологическую тему |3. М. М. Тереховский путем ряда остроумных опытов показал, что инфузории являются настоящими сложно построенными организмами микроскопи-
12 Л у и к е в и ч В. В. 1940. От Гераклита до Дарвина. Очерки по истории биологии, т. II. М. Изд-во АН СССР.
13 Т е р е х о в с к и й Мартын. 1775. Зоолого-физиологическая диссертация о Chaos infusorium Лиинея. Перевод с латинского напечатай в книге С. Л. Соболя «История микроскопа». Изд во АН СССР, 1949.
29
Рис. 10. Микроскоп Евстахия Дивини (1672 г.)
ческой величины, поэтому вопрос об их возникновении должен решаться так же, как и в отношении всех остальных живых существ. Весьма существенно, что Тереховский резко возражал против учения Ж. Л. Л. Бюффона (1707—1788), имевшего в конце XVIII в. большую популярность. Согласно Бюффону, микроорганизмы представляют собою «органические молекулы», освобождающиеся при распаде (гниении) органической материи. Однако эта точка зрения защищалась отдельными исследователями еще и в XIX в., в частности, натурфилософами (Лоренц Окен и др.).
В XVIII в. микроскопы производились в большом количестве. «Простые микроскопы» (лупы) с небольшими увеличениями, правда, уже широко использовались как зоологами, так и ботаниками для изучения мелких объектов, но принципиально нового это в науку не вносило. «Сложные микроскопы» для анатомических работ были излишни, пользоваться ими было трудно, так как несовершенной была не только оптическая часть, но и меха
50
рис. 10, 11, 12
второй половине
начали выпускаться
ническая (см.
13). Во XVIII в. значительно более удобные для
работы микроскопы, наилучшие их типы были сконструированы в Англии Джоном Кеффом (1744—1770 гг.), позднее —
Г. Адамсом. Эти микроскопы (рис. 14 и 15) как по своей оптической конструкции, так и по механическому устройству были значительно рациональнее всех предшествовавших моделей
Таким образом, материально-техническая база (в виде достаточно совершенного инструмента— микроскопа) была создана в XVIII в. Теоретические работы физиков дали научно обоснованный расчет оптических систем, что тогда же привело к созданию ахроматических объективов (Доллонд, 1760; Эпинус, 1784; ван Дейль, 1807). Однако биологическая наука в теоретическом отношении не могла еще
Рис. 11. Микроскоп астронома Ге-велия гуковского типа (1673 г.), отличающийся более усовершенствованным передвижением тубуса
освоить и обобщить фактические
наблюдения, выделить из них
существенное и отбросить случайное и недостоверное. В то время не было достаточного понимания наблюдаемых явлений,
а поэтому отсутствовал стимул к систематическим микроскопи
ческим исследованиям.
14 Богатейшая коллекция старинных микроскопов собрана в «Кабинете по истории микроскопа» при Академии наук СССР (Москва).
31
Рис- 12. Микроскоп Джона Маршала, позволяющий рассматривать объекты в проходящем свете (1663 г.)
Рис. 13. Микроскоп Гертеля (1716 г.). Освещение объекта достигается зеркалами
2 Е. М. Вермель
Рис. 15. Большой микроскоп Адамса конца XVIII века
Рис 14. Первичная модель .микроскопа Джона Неффа; середина XVIII века
J 8. Первые микроскопы в России. Первые «зрительные трубы» были завезены в Россию еще во времена царя Алексея Михайловича, т. е. вскоре после их изобретения. Они предназначались для оснащения армии. Первые микроскопы были привезены только Петром I. В мае 1698 г. он посетил в Дельфте (Голландия) Левенгука, который демонстрировал ему, в частности, кровообращение в кровеносных капиллярах угря. Петр I настолько заинтересовался микроскопическими исследованиями, что не только впоследствии распорядился купить микроскоп, но и вывез из Голландии опытного мастера по шлифовке оптических стекол (Л. Шеппера). При Академии наук в Петербурге была организована специальная мастерская, конструирование увеличительных приборов велось уже самостоятельно на протяжении почти 100 лет. Как показало изучение архивных материалов |5, наши русские мастера И. Е. и И. И. Беляевы (отец и сын), Матвеев, Ремезов, Кулибин и др., начиная с 1726 г., сами сконструировали с большим мастерством н энтузиазмом собственные микроскопы в соответствии с лучшими образцами того времени. Однако судьба этих микроскопов была печальна: для научной работы «господа академики», как жаловались конструкторы, использовать их не хотели н, очевидно, не умели. До нас эти замечательные микроскопы не дошли: все они погибли при пожаре академических мастерских.
Высоко оценивал микроскопические исследования М. В. Ломоносов. Сам он микроскопом никогда не пользовался, но в «Письме о пользе стекла» (1752) он писал:
Хоть острым взором нас природа одарила, Но близок оного конец имеет сила.
Коль многих тварей он еще не досягает, Которых малый рост пред нами сокрывает! Но в нынешних веках нам микроскоп открыл, Что Бог в невидимых животных сотворил!
Коль много микроскоп иам тайностей открыл, Невидимых частиц и тонких в теле жил!
15 С о б о л ь С. Л. 1949. История микроскопа и микроскопических исследований в России в XVIII веке. М., Изд-во АН СССР.
2*
35
Ломоносов изучением биологических объектов не занимался, однако ему принадлежит неоспоримый приоритет в применении микроскопических исследований в области химии. Им впервые было использовано изучение явления кристаллизации с помощью микроскопа и описана форма микрокристаллов. Этот метод в настоящее время широко вошел в практику химических исследований. Ломоносов имел в своем распоряжении два микроскопа, изготовленных специально для него в мастерской Академии наук мастером И. И. Беляевым. Ломоносов сам проектировал технические усовершенствования в конструкции микроскопа и работал над улучшением качества сплавов стекла для шлифовки линз.
ГЛАВА ВТОРАЯ
ИСТОРИЯ ОТКРЫТИЯ КЛЕТКИ
Основные даты по истории открытия клетки
1665 Роберт Гук впервые описал клеточное строение пробки, стеблей и др. и ввел термин «клетка».
1671—1679 Марчелло Мальпиги дал первое систематическое описание микроструктуры органов растений, положившее начало анатомии растений.
1671—1682 Неемия Грю также очень подробно описал микроструктуры растений; ввел термин «ткаиь» для обозначения понятия совокупности «пузырьков», или «мешочков» («клеток»).
1676—1719 Антон ваи Левенгук открыл мир микроскопических животных, а также описал впервые красные кровяные клетки и сперматозоиды.
1759 Каспар Фридрих Вольф создал первую теорию клеткообразоваиия в растительных тканях.
1781 Феликс Фонтана первый увидел и нарисовал клетки животных с ядрами.
1801 Бриссо де Мирбель положил начало сравнительному изучению клеток растений, защищая, однако, старый взгляд на них как на пузырьки, разделенные общей стенкой.
1804 Г. Лиик и К. Рудольфи получили премию Геттингенской Академии наук за разрешение вопроса о природе клеток, которые, как они доказали, имеют самостоятельные оболочки.
1812 И. Мольденгауер окончательно доказал индивидуальность клеток путем их изоляции (мацерация).
1830 Франц Мейеи написал первую сводку по анатомии растений и сформулировал представление о клетке как о «пространстве, замкнутом вегетативной мембраной».
1830—1845 Школой Яна Пуркиия была значительно усовершенствована микроскопическая техника и правильно описаны клетки в многочисленных органах животных.
37
§ 9. Описание растительной клетки и ткани Р. Гуком, М. Мальпиги и Н. Г рю. Растительные клетки были впервые описаны Робертом Гуком. К своему труду «Микрография» (1665) он приложил большое количество таблиц с прекрасно выполненными рисунками. Никакого общего плана исследования у Гука не было; в дальнейшем к микроскопическим наблюдениям он больше не возвращался.
В истории науки сыграла огромную роль гл. XVII «Микрографии», озаглавленная «О схематизме или строении пробки и о клетках и порах в некоторых других рыхлых телах». Изучая тонкие срезы через пробку, Гук поставил себе прежде всего определенную узкую задачу: он стремился узнать, чем объясняется легкость пробочной ткани, ее эластичность, упругость и неспособность впитывать воду. На срезах пробки Гук под микроскопом обнаружил, что она имеет весьма своеобразное строение и состоит из вполне замкнутых пузырьков, или ячеек (cellula), которые и были переведены термином «клетка». На рис. 16 дано воспроизведение подлинного рисунка Мальпиги. Еще Гук сделал первую попытку подсчитать количество клеток в определенном объеме пробки. В одном кубическом дюйме (24 мм) их оказалось 125 712000. Гук писал: «это могло бы казаться невероятным, если бы в этом не убеждал нас микроскоп с очевидной наглядностью».
Следовательно, Гук представлял себе пробку, построенную из большого числа вполне изолированных, разделенных плотной (но очень тонкой) стенкой ячеек. А отсюда уже становилось понятным, почему пробка так мало весит и почему она легко поддается сжатию; этим же объясняется и ее водонепроницаемость.
Итак, задача была разрешена, но Гук пошел дальше. Он сам ставил вопрос о том, насколько широко распространено ячеистое строение, не является ли оно «схемой», распространяющейся на все органы растений.
С этой целью он изучал срезы стеблей различных живых растений и обнаружил в них аналогичные ячейки, разграниченные перегородками. Отличие этих ячеек-клеток от клеточных ячеек пробки состояло в том, что они не были пусты, а оказались заполненными, по его предположению, «питательным соком», который, как допускал Гук, перетекает из одной клетки в другую, хотя необходимых для этого пор в стенках ячеек он не видел.
Самым существенным в исследованиях Гука является то, что им сделаны также выводы общего характера. Гук сформулировал представление о клетке, как о ячейке, точнее как о пузырьке,
38
Рис. 16. Рисунки М. Мапъпиги срезов различных растительных тканей из книги «Анатомия растений» (1679 г,)
Марчелло Мальпши (1628—1694)
полностью замкнутом со всех сторон. Соседние пузырьки, как он считал, разделены-одной общей перегородкой. Затем Гук установил факт широкого распространения клеточного строения растительных тканей.
Именно эти два основных вывода и определили направление дальнейших исследований в этой области.
Уже в 1671 г. в Лондонское Королевское Общество поступили два доклада о микроскопическом строении растений. Это были исследования итальянца Марчелло Мальпиги и англичанина Неемия Грю. Оба эти исследователя (они работали совершенно независимо друг от друга) изучили большое количество объектов и дали изумительные по точности описания и рисунки. Любопытно, что их сочинения носят почти одинаковое заглавие, а именно: «Анатомия растений» *, хотя один писал по латыни,
' Malpighi М. 1675 (I), 1679 (П) Anatome plantarum idea; Grew N. The anatomy of plant's begun with general account of vegetation grounded thereupon. 1671. London, 2 ed. 1682.
40
а другой на английском языке. Под анатомией растений они понимали изучение ячеистого (клеточного) строения различных органов растений, описанием которого они систематически и занимались.
Прежде всего отметим, что оба эти исследователя пришли к одному и тому же выводу относительно всеобщности построения растительной ткани из п у з ы р ь к о в. Грю сравнивал их с пеной в кружке пива. Само название «ткан ь» введено впервые в употребление Г рю. Он считал, что растительная масса по своей структуре напоминает ткани одежды. Далее ими было обосновано положение, что все органы растительного организма всегда имеют определенное, типичное для них строение (см. рис. 10).
После исследований Гука, Мальпиги и Грю, которые были в свое время широко известны, сам факт существования клеток-ячеек в растительных тканях не вызывал сомнений. О клетках упоминали различные авторы, но должного значения им не придавалось, и они рассматривались как одна из структур, которая (наряду с трубочками и сосудами) обнаруживается при изучении растительных тканей под микроскопом.
Следует еще добавить, что несколько позднее (в XVIII в.) получил широкое распространение взгляд швейцарского физиолога и поэта Альбрехта Галлера (1708—1777), который защищал теорию волокнистого строения организмов. Сам он пришел к ней в основном умозрительным путем, но многие исследователи подтверждали ее прямыми наблюдениями. Тот же Феликс Фонтана, который высказал приводимые выше глубокие мысли относительно условной ценности микроскопических наблюдений, уверял, что он видел эти волоконца решительно везде. Некоторые исследователи описывали волоконца даже в камнях. В значительной степени все это оказалось фантазией, во всяком случае нитчатые структуры различных тел между собой не могут быть сравниваемы, а сходство, если оно даже иногда и имеет место, является чисто внешним.
§ 10. Теория возникновения клеток-мешочков К. Вольфа. Рассматривая и описывая клетки, исследователи начала XVIII в. не ставили вопроса об их возникновении.
Впервые попытка осмыслить факт постоянства клеточной структуры была сделана петербургским академиком Каспаром Фридрихом Вольфом (1733—1794) в его знаменитой дис-
41
И семи я Грю (1641—1712)
сертации «Теория происхождения»2. Вольф изучал эмбриональное развитие организмов и боролся против преформистской теории развития. Исследованием же клеточной структуры он занимался лишь попутно, хотя и придавал ей большое значение. В отличие от всех более ранних исследователей, Вольф говорит
2 W о 1 f С. F. 1759. Theoria generationis; Вольф К. Ф. 1950. Теория зарождения. М. См. также Гайсинович А. Е. «К. Ф Вольф и учение о развитии организмов», 1961.
42
о кЛетках в связи с явлениями роста или распределения веществ в организме. Он считал, что молодые органы растений (листочки, корешки и др.) состоят из гомогенной, вязкой или студневидной массы. Их рост осуществляется, по Вольфу, таким образом, что в них из более старых частей выпадают капли жидкого вещества, пограничный слой которого загустевает и капля превращается в ячейку-клетку (сам Вольф применял термин «utriculus», что значит «мешочек»). Если капля движется в основном вязком веществе медленно, то ее стенки успевают затвердевать, таким образом возникает трубочка-сосуд. По мере того как все новые и новые капли вдвигаются между уже возникшими, создается обычная пузыристая структура растительной ткани. Таким образом, Вольф представлял себе, что не клетки образуют сосуды, а, напротив, сосуды клетки.
Таково представление Вольфа о росте органа. В связи с этим относительно строения клеток он пишет, что, по его убеждению, клетки-пузырьки соединены друг с другом без особого порядка и соседние пузырьки разделены общей стенкой, а, следовательно, они не могут быть изолированы. Последнее относится также к сосудам растений, т. е. он полагал, что и сосуды не имеют самостоятельных стенок. Однако он знал, что в ряде случаев как клетки, так и сосуды (например, в зрелых плодах) могут быть сравнительно легко выделены. Тогда они, по его выражению, «представляют собой более или менее замкнутые истинные пузырьки (мешочки) различной величины и формы».
Тем не менее теория довлела над фактами, и Вольф рассматривал все, что не укладывалось в его представления, лишь как случайные исключения, не имеющие принципиального значения. Теория возникновения клеток-мешочков Вольфа в свое время пользовалась широким признанием.
Таким образом, именно представления Вольфа следует, пожалуй, считать первой (весьма, конечно, условной и своеобразной) «клеточной теорией», ибо она предлагала какую-то концепцию, какое-то толкование и объяснение широко наблюдаемого явления клеточно-ячеистой структуры растительных тканей.
§77. Первые описания животных клеток. Выше речь шла исключительно о структуре растительной ткани. Микроскопическое изучение органов животных значительно отставало. Это объясняется тем, что клетки животных увидеть в микроскоп значительно труднее, ибо, во-первых, в огромном большинстве случаев
43
Рис. 17. Рисунки Феликса Фонтана, изображающие слущив-шийся кусочек кожи угря (слева) и две клетки крови (справа), из книги 1787 г.
животные клетки много мельче, а во-вторых, они не имеют столь резко выраженных границ.
В силу этих причин их практически долгое время не видели.
Любопытно, что Вольф в самом конце своей «Теории происхождения» вновь возвращается к вопросу о клеточном строении и говорит на этот раз уже о клетках животных. Он пишет:
«§ 257. Что касается образования клеточной ткани у животных, то этот вопрос ... обойден молчанием, ибо он не представляет никаких трудностей. Эта ткань образуется так же, как и клеточная, пузыристая ткань растений. Кости не представляют ничего специфического, кроме их субстанции.
§ 258. Образование мышц, нервов и мозга требует специальных исследований и требует в качестве предпосылки глубокого знания мозга, что столь же важно, сколь и темно...»
Изображения клеток животных Вольф не дает, а из текста нельзя понять, что именно он видел. По существу он правильно определил свое отношение к данной проблеме: она оставалась для него «столь же важной, сколь и темной».
Если формально подойти к вопросу о приоритете описания клеток животных, то прежде всего следует назвать Левенгука, видевшего красные кровяные клетки (эритроциты) человека. Однако в истории развития учения о клетке эти наблюдения почти не сыграли никакой роли, как и более поздние наблюдения над клетками — от патера Делла Торре и до Феликса Фонтана. Оба исследователя рассматривали, в частности, кусочки слущивающегося эпидермиса кожи угря. На рис. 17 приводятся изображенные в сочинении Фонтана клетки. Нетрудно видеть, что даже ядра не ускользнули от его внимания.
Итак, несомненно, что в XVIII в. многие видели настоящие клетки животных, но никто не сделал и не мог сделать вывода
44
о всеобщности Клеточного строений, так как в то время не стоял еще вопрос о гомологичности (сопоставимости) клеток. Поэтому описанные и зарисованные клеточные структуры разных организмов оставались в представлении ученых лишь частной и несопоставимой структурой.
Сказалось здесь также отрицательное влияние теории волокнистого строения организмов, которую отстаивал Галлер (XVIII в.). Мы уже приводили заявление Фонтана, что он видел волоконца везде, даже в минералах. Это, конечно, была в большинстве случаев фантазия. Однако волоконца в животных тканях он мог видеть и, судя по его прекрасным рисункам, действительно видел их в соединительной ткани (коллагеновые волокна), в мышцах, нервах и т. д. По его мнению, именно эти волокна образуют основную массу животного организма, а все остальные структуры не имеют принципиального значения. Этот вывод, однако, был ошибочным, но он импонировал, так как находился в согласии с теорией Галлера, авторитет которого был очень высок.
§ 12. Новое представление о клетке в начале XIX века. Подытоживая результаты исследований XVII и XVIII вв. в области познания микроскопической структуры живых существ, можно сказать, что клеточная структура описывалась отдельными учеными неоднократно. Больше того, в отношении растительных тканей был накоплен уже настолько значительный фактический материал, что эта область знания выделилась в особую дисциплину— «Анатомию растений». Тем не менее, клеточному строению принципиального значения не придавали и ставили клетку в один ряд с другими структурами, например, волокнами и сосудами растений, волоконцами («цилиндрики») животных тканей. Клетка как элементарная живая единица еще никем не рассматривалась. Единственной попыткой понять возникновение этой столь широко распространенной структуры была теория Вольфа.
Сами клетки рассматриваются как простые пузырьки жидкости, ничего общего не имеющие с окружающей их оболочкой. Отсюда понятно происхождение и широкое применение самого термина «клетка», дословно передающего вкладываемый в него смысл. Полагали, что клетки разделены общей стенкой и, таким образом, не могут быть изолированы друг от друга.
В дальнейшем развитии науки большую роль сыграли исследования французского ботаника Бриссо де Мирбеля (1776—
45
Альбрехт фон Галлер (1708—1777)
1854), в которых впервые была высказана идея сравнительного изучения растительных тканей. Вместе с тем им защищался все тот же взгляд на природу клеток. Мирбель 3 продолжал считать клетки мешочками-пузырьками и полагал, что соприкасающиеся пузырьки разделены общей перегородкой — мембраной. Против
3 Mirbel Br de 1801. Trane d'anatomie el de physiologie vegetable («Трактат по анатомии и фичиологии растений»).
46
этой точки зрения выступили многие немецкие in < i < mi и вопрос возник в такой резкой форме и привлек к с <•(>< пш шм, ко большое внимание, что Геттингенская академия объяви mi д<-нежную премию за ее разрешение (1804).
Премия была поделена между ботаниками Г. Линком и К. Ру дольфи. Оба эти исследователя пришли к заключению об обособ ленности клеток, о наличии у них собственных мембран, окру жающих их со всех сторон. Тот же вывод был сделан затем и Л. X. Тревиранусом (1779—1864). Следует отметить, что исследования последнего «О внутреннем строении растений и о движении в них соков» (1806) были особенно точными и обстоятельными4. Всякие сомнения о том, что клетки могут быть выделены и изолированы, были окончательно оставлены после работы ботаника И. Молденгауера (1766—1827), из которой следовало, что клетка является самостоятельной структурой («коробоч-к о й»), покрытой непрерывной оболочкой 5.
Особенно убедительно было доказательство, полученное с помощью метода мацерации. Линк добился полного выделения клеток из тканей длительным кипячением растительной ткани в воде. Позднее этого же достигали много быстрее и проще, воздействуя на ткань разведенной азотной кислотой.
Итак, было создано новое представление о клетке. Наиболее четко оно было сформулировано несколько позже Ф. Мейеном (1804—1840), написавшим первую сводку по анатомии растений. Его определение клетки таково: «Клетка растительного организма представляет собой пространство, вполне замкнутое вегетативной мембраной» 6.
Мейен ясно сформулировал этот взгляд, который в начале XIX в. сменил первоначальное воззрение на клетку как на пору. Но конкретное содержание анатомии растений по-прежнему принципиально мало чем отличалось от старого. По-прежнему продолжали систематически описывать строение различных органов растений, а также детализировать и уточнять изученное. Этот период справедливо считается периодом собирания материала, периодом накопления многочисленных сведений о тончайшей структуре растений.
4 Treviranus L. С. 1806. Von invendigen Bau der Gewachse und von der Saftbewegung in denselben.
5 Moldenhiwer J. P. 1812. Beilrage zur Anatomie der Pflanzen. Kiel («Сообшения по анатомии растений»).
6 M e у e n F. 1830. Phytotomie. Berlin («Фитотомия»).
47
Итак, сам факт наличия клеток в различных тканях сомнения, не вызывал. Но исчерпывается ли все разнообразие структур) растительных организмов только клетками, оставалось еще неясным. Более того, большинство ученых отвечало на этот вопрос отрицательно, указывая наряду с клетками на сосуды и волокна растений как на вполне своеобразные структуры. Обобщая все знания в этой области, Мейен прямо писал, что растительные ткани состоят из клеток, волокон и сосудов. Возникновение этих последних из метаморфизирующихся клеток было окончательно установлено лишь несколько позднее — в 30-х годах прошлого века.
§ 13. Клеточная структура животных тканей. Изучение клеточной структуры тканей и органов животных оказалось делом гораздо более трудным и сложным. Выше мы упоминали, что отдельные животные клетки видели и описали уже Левенгук (красные кровяные клетки) и Фонтана (клетки кожи угря и эритроциты). Видели животные клетки и многие другие ученые, но эти данные оказались еще совершенно недостаточны, чтобы можно было делать вывод о всеобщности клеточной структуры. Изучать животные ткани трудно, техника того времени не позволяла получать тонкие срезы через мягкие ткани животных, а метод фиксации и уплотнения органов известен еще не был. Животные клетки (как не имеющие клеточного сока) относительно очень мелки, их границы часто весьма неотчетливы, ибо их оболочка представляет собою только тонкую «кожицу» — уплотненную протоплазму, которая и современными методами обнаруживается нередко с большим трудом. Кроме того, в тканях (особенно в тканях позвоночных животных) имеется большое количество межуточного вещества, характеризующегося студневидной консистенцией и пронизанного волокнами. Генез межуточного вещества и до сих пор не может считаться изученным до конца. Клеток в нем, как правило, сравнительно мало, и именно межуточное вещество явилось, можно сказать, камнем преткновения для многих гистологов в период пересмотра клеточной теории уже в начале XX в.
Не удивительно, что животные клетки были изучены и вообще обнаружены далеко не сразу, хотя многие ученые и подходили к исследованию уже с предвзятым мнением, будучи наперед убеждены, что клетки в животных тканях должны быть. Правда, эта же предубежденность заставляла искать животные клетки,
48
Анри Милън-Эдвардс (1800—1885)
сходные по своему строению с растительными, что было ошибочным.
Даже такой крупнейший ученый своего времени, как французский зоолог Анри Мильн-Эдвардс (1800—1885) 7 достиг небольшого успеха в своих микроскопических изысканиях. Он имел в своем распоряжении хороший микроскоп, но готовил препараты, раздавливая ткани между двумя стеклами, и в силу этого наряду с настоящими клетками он на своих рисунках большей частью изображал капельки жира, отдельные ядра, комочки зернышек и т. д., принимая и их за клетки. Ясности эти работы не внесли.
Значительно большее значение имели работы другого французского ученого — Анри Дютроше (1776—1847), который опи-
7 М i 1 n е-Е d w aids. 1823. Memoire sur la structure elementaire des principally tissus organiques des animaux. Arch, gener. de medecine 3 («Об элементарной структуре основных тканей животных»).
49
Рис. 18. Рисунки Я. Пуркиня, изображающие «зернышки» (клетки), из которых состоят ткани органов животных
сал ряд клеток из животных тканей, но и его работа в основном была посвящена растениям 8.
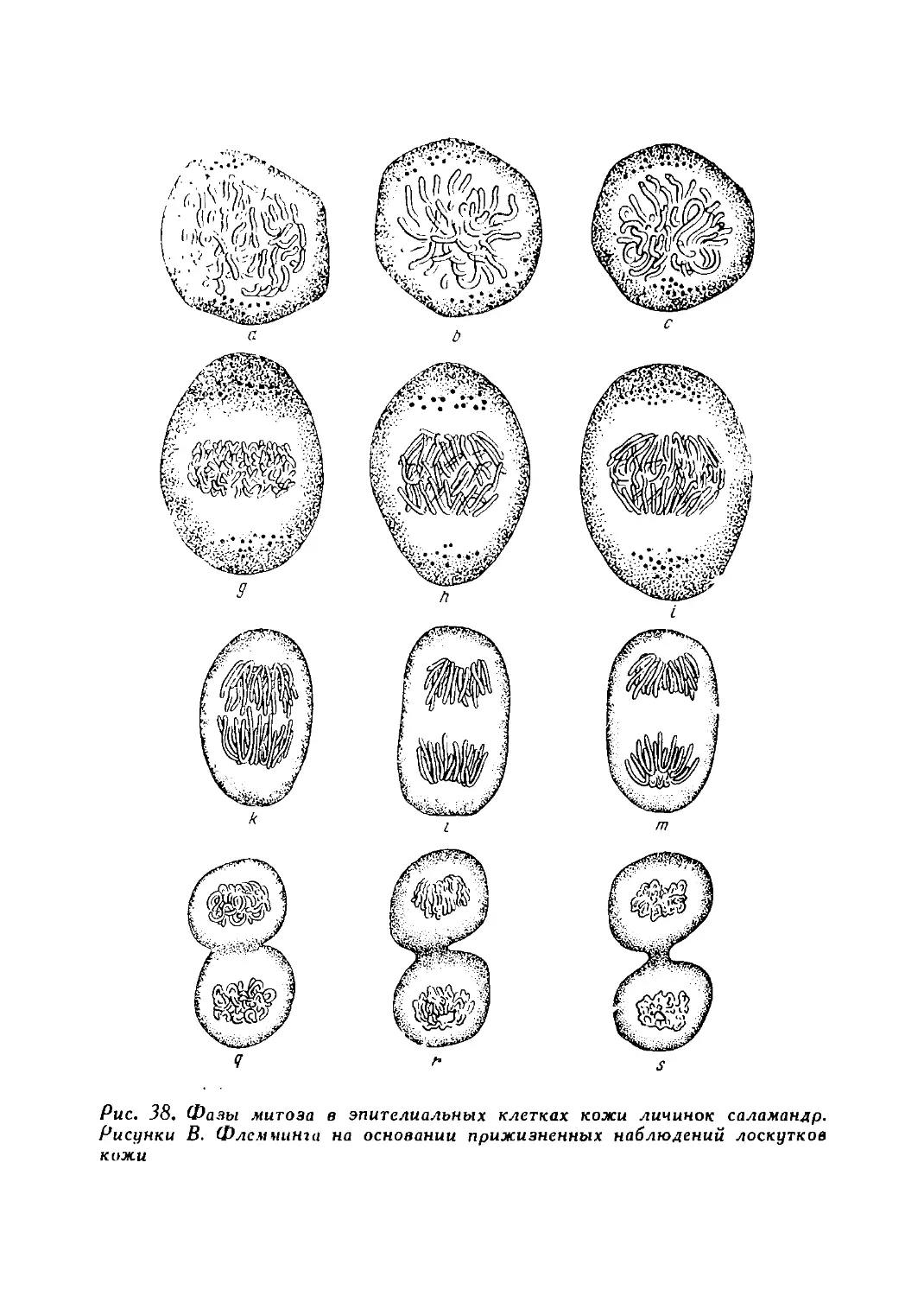
По существу животные клетки были впервые описаны только в 30-х годах XIX в. замечательным чешским ученым Яном Пуркиня (1787—1869) и его многочисленными учениками (Валентин, Рашков, Розенталь, Бернгард, Меккауер, Дейтч, Вендт и др.). Описания клеток, сделанные этими учеными в ряде статей, отличались большой точностью. Во всех тканях они обнаружили клетки, называя их, однако, зернами или шариками. Ими был открыт, в частности, реснитчатый эпителий, причем было описано даже движение ресничек. Они изучили также нервные клетки и дали очень неплохие их рисунки (рис. 18).
§ 14. Первые описания строения содержимого клетки. Заканчивая главу, посвященную истории открытия клетки, необходимо еще отметить, что в ряде исследований описывалась не только клеточная оболочка, которая признавалась главным анатомическим элементом клетки, но и ее содержимое.
Уже в конце XVIII в. итальянец Бонавентура Корти (1774) видел и описал активное движение жидкого содержимого в растительной клетке. Еще подробнее протоплазматические токи были изучены Л. Тревиранусом (1811). В клеточном содержимом
BDutrochet Н. 1824. Recherches anatomiques et physiologiques sur la structure inlime des animaux el des vegetaux, et leur mobilite. Paris («Ана-томические и физиологические исследования над тонкой структурой животных и растений»).
50
Роберт Броун (1773—1858)
было обнаружено наличие слизи, клееобразных веществ, сахара, были уже известны также хлорофилловые зерна, различные кристаллы, зерна крахмала и др. Зернам крахмала в свое время большое внимание было уделено Куртом Шпренгелем (1766— 1833), который полагал, что из них путем набухания возникают клетки.
Гипотеза эта, однако, успеха не имела и была вскоре полностью опровергнута и забыта.
Наконец, было обнаружено клеточное ядро. Впервые его описал, надо считать, сам Пуркиня (1830) под названием «зародышевого пузырька». Его объектом была куриная яйцеклетка (яйцо), т. е. клетка весьма специализированная9. Любо-
" Purkinje J. Е. 1830. Symbolae ad ovi avium bistoriam aule incubatio-nem Linsiae («Сообщение к истории птичьих яиц перед инкубацией»). Обычно фамилия Пуркиня пишется по-немецки: Johann Evangelist Purkinje, однако настоящая ее транскрипция на чешском языке иная: Jan Evangelista Purkyne; более точным на русский был бы перевод и транскрипция — Ян Пуркиия, которой мы и придерживаемся.
51
nbiTiio, что этот термин («зародышевый пузырек») употреблялся эмбриологами еще очень долгое время — вплоть до нашего времени.
Несколько позднее, в 1831 —1833 гг., шотландский путешественник и физик (открывший «броуновское движение») Роберт Броун (1773—1858) обнаружил ядро в растительных клетках10. Он дал ему название «Nucleus» или «Areola». Первый термин стал общепринятым и сохранился по настоящее время, второй же распространения не получил и забыт. Весьма важно, что Броун настаивал на постоянном наличии ядра во всех живых клетках. Роль и значение клеточного ядра не были в то время известны. Полагали, что оно представляет собой «конденсированную в комочек слизь, а возможно, и запасное питательное вещество» (Мейен, 1837).
'’Brown R. 1833. On the organs and mode of fecundation in Orchideae and Asclepiadeae. Transaction of the Linnean Society of London, 16 («Об op ганах и способах размножения орхндиых и ластовениых»).
ГЛАВА ТРЕТЬЯ
КЛЕТОЧНАЯ ТЕОРИЯ
Основные даты развития клеточной теории
1809 Л о реи ц Окен создал умозрительную теорию строения и развития организмов, в которой элементами являлись «инфузории» — клетки.
1824 1827 1829 Анри Дютроше, ФравсуаРаспайль, Пьер Тюрпеи. Эти французские ученые высказывали и защищали тезис, что клетки (мешочки, пузырьки) являются элементарными структурами всех растительных и животных тканей.
1834—1847 П. Ф. Горянинов сформулировал принцип, согласно которому клетка является универсальной моделью организации живых существ.
1837 Яи Пуркиия в докладе обществу естествоиспытателей (Прага) высказал теорию «ядросодержащих зернышек» (клеток).
1838 Иоганнес Мюллер на основании изучения ткани хорды высказал представление о соответствии в клеточном строении растений и животных.
1838 Матиас Шлейден создал теорию цитогенеза.
1839 Теодор Шванн обосновал, исходя из генетического принципа, клеточную теорию строения всех организмов.
1859 Рудольф Вирхов пересмотрел и развил клеточную теорию, заменив представление о цитогеиезе законом: «всякая клетка от клетки».
5?
Лоренц Окен (1779—1851)
§ 1 к Воззрения натурфилософа Л. Окена на клетку и ее возникновение на Земле. Микроскопические исследования начала XIX в. характеризуются не только тем, что клетки были изучены и описаны гораздо более подробно и на большом числе разнообразных объектов. Значительно важнее то обстоятельство, что клеткам стали приписывать значение простейших органических структурных элементов, в силу чего данные о микроскопическом строении организмов начинают играть все большую роль в построении общетеоретических воззрений того времени.
Уже давно ученые (и особенно философы) искали элементарную биологическую единицу (например, «монады» Лейбница, 1646—1716), но все эти искания были лишены конкретности. Впервые, вероятно, таковыми стал считать именно клетки естествоиспытатель и глава школы немецких натурфилософов Лоренц Окен (1779—1851). В своем известном «Учебнике натурфилософии» (1809) и в других сочинениях представление о клетке Окен поднял на большую принципиальную высоту, причем клетками он считал как ячейки и пузырьки растительных и животных тканей, так и микроскопические свободно живущие организмы —
54
«инфузории». Таким образом, Окен по существу высказал к>чиу зрения, которая стала господствующей значительно позднее. Надо специально отметить, что книги Окена были хорошо из вестны и широко изучались. Мы имеем все основания считать, что его сочинения оказали значительное влияние на развитие как теоретической мысли, так и на постановку конкретных исследований.
Коротко говоря, Окен считал, что сложные организмы являются суммой элементарных организмов, которые, войдя в его состав, живут общей жизнью целого, но в то же время продолжают оставаться в известной степени независимыми. Эти элементарные организмы, по его представлениям, являются теми пузырьками с плотной оболочкой и жидким содержимым, которые можно различать с помощью микроскопа в тканях организмов. Окен говорит, что они «в философском смысле могут быть названы инфузориями». Довольно сходное воззрение еще ранее высказал также известный французский натуралист Жорж Луи Леклерк Бюффон (1707—1788).
Основным недостатком всех сочинений Окена (в том числе и изложения его представлений о тончайшем строении организмов) является то обстоятельство, что он выступает прежде всего как натурфилософ. Он не изучает природу как исследователь, который критически разрешает конкретные вопросы, а стремится делать заключения о сущности явлений на основании одних общих априорных положений и умозаключений. Многие его обобщения имеют более чем странный характер. Так, Окен выводит «пузырчатое строение» элементарных единиц (клеток), которые он видел под микроскопом, как необходимое следствие шарообразной формы планет. К тому же он излагает свои мысли в виде ряда изречений, не допускающих ни критики, ни проверки. Обосновывать свои заключения и выводы конкретными наблюдениями Окен считал излишним. Однако имеются все основания думать, что Окен был хорошим наблюдателем и сам много и упорно изучал как растительные, так и животные ткани с помощью микроскопа. Он видел, вероятно, очень многое, иначе он не мог бы так хорошо и правильно понимать фактический материал. Не надо забывать, что это было первое десятилетие XIX в., когда еще только ставился вопрос о том, является ли клетка пузырьком в студневидной массе или же она представляет собой изолированную коробочку, имеющую собственные стенки. Из литературы Окен почти ничего не мог почерпнуть, ибо ее и не было.
55
Приведем для примера несколько параграфов («Учебник натурфилософии», 1809) ', в которых излагаются положения о микроскопическом строении организма:
«§ 901. Все органическое произошло из слизи и представляет собою слизь, принявшую различные формы.
§ 902. Первичная слизь, из которой было образовано все организованное, есть морская слизь.
§ 904. Морская слизь возникла вследствие развития планеты.
§ 908. Первично-органическое есть слизистая точка.
§ 934. Вследствие окисления воздуха в органической точке происходит противоположение составных частей: жидкого и твердого, и эти части взаимно обусловливают друг друга. Но жидкое и твердое не могут определять друг друга иначе, как таким образом, что твердое становится содержащим, а жидкое содержимым ... твердое не может находиться в другом месте, как на границе между воздухом и жидкостью. Поэтому оно с самого своего возникновения окружает жидкое. Физическим основанием этому процессу является, конечно, окисление слизи по ее поверхности. Шар, середина которого жидка, а поверхность тверда, называется пузырьком.
§ 935. Первые органические тела представляют собою пузырьки.
§ 936. Первичный пузырек слизи в философском смысле может быть назван инфузорией ...
§ 940. Если органическая масса состоит из инфузорий, то и весь органический мир должен возникать из инфузорий. Растения и животные могут быть только лишь метаморфозами инфузорий.
§ 941. Гниение есть не что иное, как распадение организмов на инфузории, приведение высшей жизни к жизни первичной.
§ 943. Организм представляет собою синтез инфузорий. Воспроизведение есть не что иное, как соединение воедино бесконечного множества слизистых точек — инфузорий».
Несмотря на все указанные выше недостатки, точка зрения Окена изложена вполне ясно. Если учесть, что цитированные выше строки писались на заре микроскопических исследований, нельзя не поражаться глубине его мыслей. Те фактические данные, которыми мог располагать Окен, были по существу чрезвычайно примитивны, а их количество ничтожно. И все же в 1809 г.
1 О k е n L. 1809. Lehrbuch der Naturphilosophie,
56
он предвидел многое из того, что позднее было развито па иной, гораздо более широкой, естественнонаучной основе - чсрс i 30 лет (в 1839 г.) Теодором Шванном (1810—1882) и через 50 с лишком лет (в 1866 г.) Эрнстом Геккелем (1834—1919). По существу ведь Окен, с одной стороны, формулировал принцип сведения строения сложных организмов (как теперь мы бы сказали — многоклеточных) к элементарным единицам (клеткам, по выражению Окена,— инфузориям), а с другой — во всей этой концепции совершенно отчетливо выражена эволюционная идея, хотя сам он, как и все натурфилософы, развития во времени (в нашем понимании) не признавал. Отдельные и даже многочисленные заблуждения Окена, вроде его представления о том, что гниение есть выделение «пузырьков» (клеток) из тканей и превращение их в «инфузорий», не имеют в этом отношении принципиального значения.
Воззрения Окена не могли, однако, стать общепризнанными, ибо фактическая основа естествознания того времени не располагала сколько-нибудь убедительными данными. Но теоретическая концепция Окена определенным образом направляла мысль ученых и создавала предпосылки к исследованию, подготавливая почву в этом направлении. Труды Окена были широко известны. В 1833 г. он выпустил «Общую естественную историю» — настольную книгу ученых того времени, где повторил все свои основные положения, высказанные им еще в 1809 г. Шванн, в частности, хорошо знал эту книгу2. Прогрессивное значение Окена часто недоучитывается. Надо прежде всего подчеркнуть, что, по Окену, натурфилософия должна была показать, каким образом возникли элементы и тела природы, как они развивались до высших форм. Окен писал, что натурфилософия является «историей развития мира и историей его творения».
Мы здесь не можем останавливаться на критике натурфилософии и на ее отрицательных сторонах, ибо это завело бы далеко от нашей темы. При изучении истории биологии, точнее истории учения о микроскопическом строении организмов, мы прежде всего руководствовались желанием выявить положительные, а не отрицательные моменты концепций различных исследователей, ибо, как мы убеждены, в истории науки представляет интерес прежде всего история прогресса, а не история заблуждений, хотя их и было во все времена много.
2 О k е n L. Allgemeinc Naturgeschichte fiir alle Slande («Общая естественная история для всех людей»).
57
§ 16. Идеи Горянинова о значении клеточной структуры. Дальнейшим развитием натурфилософии является концепция П. Ф. Горянинова (1796—1865), профессора Медико-хирургической Академии в Петербурге. В период с 1834 по 1847 г. им был написан ряд интереснейших книг, обобщавших и развивавших общебиологические представления. В свое время они не сыграли значительной роли и были вскоре забыты. Возможно, что решающее значение имело то, что книги эти были написаны автором по латыни и не получили сколько-нибудь широкого распространения, тем более, что и напечатаны они были малым тиражом на средства самого автора 3.
Горянинов по сравнению с Океном значительно выигрывает ясностью своих представлений, отношением к эмпирическому материалу и систематичностью. Будучи натуралистом-универсалом, он, однако, преимущественно работал по ботанике. Микроскопическим строением организмов сам он не занимался, но, будучи широко образованным ученым, хорошо зная литературу, следил за развитием микроанатомии, и в его концепции представление о клетке играет немаловажную роль.
Прежде всего надо отметить, что Горянинов четко делил мир живых существ на два царства, или круга (orbis). Первое царство-—-царство бесформенное, или молекулярное, второе — органическое, или клеточное (regnum cellulare). Так, в своей книге «Система природы», изданной в 1834 г., он писал: «Органические тела имеют клеточное строение и проявляются высшие ступени жизни от растений до человека. Органический мир есть прежде всего клеточное царство...»4
Затем в курсе зоологии, напечатанном в 1837 г., Горянинов дает такое определение: «Естественные тела, построенные из сочных пузырьков или трубочек и продолжающие жизнедеятельность до определенной степени и после момента зачатия, называются органическими». Еще более определенно говорится в книге 1843 г.: «Все органическое царство представлено телами кле-, точного строения, т. е. его тела состоят из сочных или зернистых пузырьков или трубочек».
В соответствии со своей концепцией на природу Горянинов развивал также представление о возникновении живых существ из
3 Козо-Поляиский Б. М. 1947. Натурфилософ П. Ф. Горянинов.— Тр. Воронежского Гос. ун-та, т. XIV, вып. 2.
4 Horaninow Р. (Горянинов П.). 1834. Primae lineae systematis naturae etc. Petropoli.
58
неорганического мира. Он считал, что зерна слизи, скученные (coacervata) вокруг первичного маленького пузырька, образуют ядро (nucleus), или цитобласт. Это ядро способно развиваться в клетку. Так возникают (как бы сами собою) наиболее просто организованные тела.
Таким образом, Горянинов связал проблему возникновения жизни с происхождением клетки и этим опередил свой век на 30 с лишним лет. К этой проблеме вновь вернулся только Геккель в 1866 г.
Взгляды Горянинова представляют собою оригинальную концепцию, в которой мы видим соединение элементов натурфилософии Окена (развитие клетки) и теории клеткообразования Шлейдена и Шванна (цитогенезис). Но Горяни|нов шел дальше и утверждал, что, раз возникнув, клетка далее воспроизводит себя уже сама. В работе 1847 г. он говорит, что существует два способа размножения клеток: один — путем образования перегородок, а другой — почкованием. Таким образом, он на 10—20 лет предвосхитил Р. Вирхова. Далее мы находим у Горянинова еще и следующую мысль. Утверждается, что «все органическое начинается пузырьком», а поскольку «все живое из яйца», то, следовательно, яйца должны рассматриваться как клетки. Таким образом, Горянинов в теоретическом отношении значительно опередил многих ученых (Бишоф, Келликер и др.), которые, конечно, сочинений Горянинова не знали.
Вероятно, все же Б. М. Козо-Полянский (1947) и Б. Е. Райков (1951) были правы, когда утверждали, что Горянинов несколько раньше немецких ученых формулировал некоторые положения клеточной теории. В самом деле, он четко изложил основной ее принцип, согласно которому клетка является универсальной моделью организации живых существ. Он утверждал, что все живое состоит из клеток и развивается из клеток.
В последние годы появился ряд исследований, согласно которым взгляды Горянинова следует считать ложной «клеточной» теорией, ибо он клеток не видел и будто бы даже принимал за таковые только пузырчатую межуточную соединительную ткань (Кацнельсон, 1956; Щербакова, 1957). Эти авторы, вероятно, правы в том отношении, что значение работ Горянинова многими авторами, боровшимися за приоритет русских ученых, было сильно преувеличено. Но все же утверждение, что он не отличал клетки от структур подкожной клетчатки, вряд ли правильно. Взгляды Горянинова следов в истории науки почти никаких не оставили.
59
В описанное время воззрения в стиле клеточной теории, что называется, носились в воздухе. В конечном итоге ведь и Т. Шванн трактовал клетки превратно, а Пуркиня, который знал их несравненно лучше, может быть, именно поэтому и не сделал убедительных обобщений.
§17. Оценка клеточной структуры в 20-х и 30-х годах XIX века. В 20-х годах XIX в. наиболее значительные работы в области микроскопического изучения тканей растений и животных принадлежали французским ученым. В первую очередь надо назвать А. Дютроше (1776—1847), П. Ж. Тюрпена (1775—1840) и Ф. Распайля (1794—1878) 5.
Эти исследователи доказывали, что именно клетки являются наиболее распространенной микроструктурой как растений, так и животных, и лишь из клеток возникают другие более сложные микроорганы (сосуды или волокна). Однако роль и значение этих ученых историками биологии также сильно преувеличивались, ибо за клетки они большей частью принимали любые пузырьки. Над ними довлело учение Г. Лейбница о монадах и взгляды натурфилософов. Тем не менее, у Дютроше мы находим много важных выводов и правильных, точных наблюдений. Так, например, в 1824 г. он писал: «Сосуды образуются путем исчезновения поперечных перемычек, разделяющих клетки друг от друга. Большие сосуды у различных двудольных образуются путем исчезновения перегородок между многими мелкими и смежными сосудами, соединяющимися таким образом в один общий». А далее он обобщает: «Все ткани, все органы животных являются не чем иным, как клеточной тканью».
Ведь по существу именно этот последний тезис является одним из основных положений клеточной теории, формулированной
5 Rasp ail F. V. 1827. Recherches chimiques et physiologiques destinees a expliquer non-seulement la structure et le development de la feuille, du tronc, ainsi que des organes qui n’en sont qu’une transformation, mais encore la structure et le development des tissus animaux. Memoire de la Soc. d’Histoi-re nat. 3, 17—88. Paris («Химические и физиологические исследования, имеющие целью выяснить не только строение и развитие листьев, как и органов, являющихся не чем иным, как их трансформацией, но также еще строение и развитие тканей животных»).
Dutrochet Н. 1824. Recherches anatomique et physiologique sur la structure intime des animaux et des vegetaux, et leur mobilite. Paris ^Анатомические и физиологические исследования над внутренним строением животных и растений»).
60
столь убедительно значительно позже Шванном. I ккот<>|....... ш
торики биологии (Райч, 1926) 6 даже полагают, что Шванн мин гое заимствовал у Дютроше, хотя нигде на него не ссылался. Однако эта точка зрения очень спорна и преувеличена. В самом деле, представление о клеточном строении организмов высказывалось в той или иной форме многими исследователями, но никем до Шванна оно не было логически доведено до конца и разработано с необходимой полнотой, чтобы стать общепризнанной.
Укажем, что еще в 1806 г. известный ботаник Людольф Тре-виранус в работе «О внутреннем строении растений и о движении в них соков»7 достаточно отчетливо говорил о развитии водоносных сосудов из клеток, сопровождая свои описания вполне убедительными, с нашей точки зрения, рисунками. Но это были лишь отдельные наблюдения, принципиальное значение которых никому, в том числе и самому автору, не было ясно. В результате эта работа прошла незамеченной и была забыта.
Необходимо еще упомянуть о сочинении ботаника Пьера Жана Тюрпена (1775—1840), которое носило следующее длиннейшее название: «Элементарная и сравнительная микроскопическая органография растений. Наблюдения над происхождением клеточной ткани, над пузырьками, составляющими эту ткань; из них каждый является отдельным индивидуумом, имеющим свой особый жизненный центр произрастания и перемещения, и предназначенный к тому, чтобы в соединении с другими, ему подобными, образовывать особи тех растений, организация массы которых состоит более чем из одного пузырька» 8.
Название это является, можно сказать, целым тезисом, из которого, казалось бы, ясна и принципиальная концепция автора. Однако наряду с совершенно правильными положениями в работе защищается взгляд на развитие клеток, исходящий по существу из теории вложения. Эта старинная, изживавшая себя точка зрения, длительное время заслоняла собою все новое. Однако было бы неверным сказать, что работа Тюрпена не сыграла своей положительной роли.
6 Rich A. R. 1926. The place of Dutrochet in the development of the celltheory. Bull, of the Hopkins Hospital. Vol. 39 («Роль Дютроше в развитии клеточной теории»).
7Treviranus L. С. 1806. Vom invendigen Bau der Gewachse und von der Saftbewegung in denselben. Gottingen («О внутреннем строении растений и о движении соков в них»),
8 Turpin Р. J. 1829. Organographie microscopique, elementaire et compare des vegetaux etc. Memoires du museum d'histoire natur, 18, 161—212.
61
Еще интереснее мысли, развивавшиеся Франсуа Распайлем (1794—1878) 9. Этот ученый, судя по его описаниям и рисункам, видел многое, но еще гораздо больше он домысливал, стараясь найти вместе с тем компромисс между теорией волокнистого строения (А. фон Галлер) и теорией «зернистого» (клеточного) строения животных тканей. Ф. Распайль пытался (как отчасти и Дютроше) провести сравнение между «растительными клетками» и «пузырьками животных тканей». Ои видит и находит это единство структуры, но мало заботится о том, чтобы дать строгие научные доказательства, развивая вместе с тем в достаточной мере фантастические натурфилософские идеи.
Все эти исследования, которые 3. С. Кацнельсон 10 по существу правильно называет «ложными клеточными» теориями, подготавливали, так сказать, почву для клеточной теории: они заставляли теоретическую мысль развиваться в определенном направлении, но сами по себе были недостаточны, чтобы заставить отказаться от старых представлений как теории волокнистого строения организмов А. Галлера, так и натурфилософских идей.
Большую трудность для формирования клеточной теории в тот период представляла, как уже в предыдущей главе отмечалось, неизученность микроскопической анатомии животных, чему в основном виною была примитивная и недостаточная техника исследования. Любопытно, что гистология животных в тот период уже существовала. Сам термин «гистология» был введен в употребление еще в 1819 г. К. Майером, учеником знаменитого французского анатома Мари Франсуа Ксавье Биша (1771—1802), который макроскопически прекрасно уже различал ткани и даже дал их классификацию и. Биша различал 21 ткань и учил, что органы животных образуются путем комбинации различных тканей, которым он, таким образом, приписывал значение элементарных структур.
9 Raspail F. V. 1827. Sur la structure intime des tissus de nature animal. Repert. general d’anatomie, 4 («О тонкой структуре тканей животных природы»),
19 Кацнельсон 3. С. 1963. Клеточная теория в ее историческом развитии. Л., Медгиз.
1! Bichat Xavie. 1801. Anatomie generate appliquee a la physiologic et a la medecine, I. Paris («Общая анатомия, ориентированная на физиологию и медицину»).
Mayer С. 1819. Lieber Histologie und cine Eintheilung der Gewebe des menschlichen Korpers. Bonn bei Ad. Marcus («Гистология и классификация тканей организма человека»).
62
Ксавье. Ьиша (1771—1802)
Гистология животных была разработана, как мы уже выше отметили, чешским ученым Яном Пуркиня и его многочисленными учениками. Современная микроскопическая техника находит свое начало в работах школы Пуркиня. Он первый применил в ряде случаев окраску (индиго и др.), он первый ввел просветляющие среды для препаратов, в том числе и канадский бальзам, используемый повсеместно и до настоящего времени. Его ученик Ошатц сконструировал первый микротом, однако этот термин принадлежит Пуркиня. Без преувеличения можно утверждать. что гистология животных создана Пуркиня и его школой. Кроме того, в 1837 г. и в 1839 г. Пуркиня выступил в научных обществах с обобщающими докладами. Они напечатаны в виде 19
коротких отчетов , в которых он изложил свою «теорию з е р-
*2 Purkinje J. Е. (Vortrage) 1838. Bendit iiber die Versammlung deutscher Naturforscher und Aerzte in Prag im September 1837. Prag.
Purkinje J. E. 1848. Lieber die Analogien in den Struktur-de-menten des thierischen und pflanzlichen Organismus. Uebersicht der Arbcilen und Veranderungen des schles. Ges f. vaterl. Kultur. im Jahre 1839. Bres-
63
Ян Пуркиня (1787—1869)
н ы ш е к». Зернышками Пуркиня называл клетки; он говорил об аналогии «клеток» растений и «зернышек» животных (рис. 12). Его представления о строении клеток не идут ни в какое сравнение с тем, о чем писали французские исследователи, принимавшие за клетки очень часто капельки жнра, обрывки волокон, различные артефакты и т. д. Фактические данные, на которые опирался Пуркиня, по своей точности не имели себе равных, тем более неудачен был сам термин «зернышки», что также, вероятно, сыграло свою роль. Данные даже Шванна не могут ставиться в один ряд с исследованиями школы Пуркиня.
Пуркиня, несомненно, был близок к формулировке клеточной теории, но он не рассматривал клеточную структуру в генетическом аспекте, поэтому клетки не были для него все же единственной структурой. Идея гомологии клеток растений и животных у Пуркиня отсу гствовала. Он слишком хорошо знал уже гистоло-
lau, Opera Selecta, I, 114—115 («Об аналогиях структурных элементов животных н растительных организмов»).
64
гию животных, поэтому было неясно, как сопоставить, например, волокна соединительной ткани и клетки (как, впрочем, и иски торым современным нам ученым). Основная его заслуга гогшит не в формулировке общебиологической теории, а в фактических данных, касающихся морфологии клеточных элементов животных тканей. Кроме того, Пуркиня выдвинул положение (являющееся основным тезисом клеточной теории) относительно построения тела животных именно из клеток, о чем (отметим еще раз) до работ Пуркиня существовали лишь туманные представления.
В 30-х годах XIX в. было опубликовано довольно много отдельных работ, в которых под разными названиями описывались клетки в различных тканях и органах животных. Но все они не шли дальше констатации отдельных фактов. Школа Пуркиия была на пороге формулировки клеточной теории, но ее опередила другая школа — школа И. Мюллера, много занимавшаяся микроскопической анатомией. Хотя И. Мюллер и был физиологическим идеалистом, за что был подвергнут справедливой критике В. И. Лениным в „Материализме и эмпириокритицизме" (Соч., 4 изд., т. 14, стр. 290), все же его многочисленные конкретные исследования имеют выдающееся значение. Поэтому о Иоганнесе Мюллере мы должны сказать несколько подробнее, тем более что о нем в нашей литературе почти ничего не говорится.
Иоганнес Мюллер (1801 —1858) был сыном сапожника из г. Кобленца; он очень нуждался, ему стоило больших трудов кончить медицинский факультет. Его блестящий талант проявился очень рано, и в 19 лет он получил премию Боннского университета за работу по экспериментальной эмбриологии. Сначала он работал на медицинском факультете в Бонне, а в 1833 г. получил освободившуюся кафедру анатомии в Берлинском университете. Получил он эту кафедру не совсем обычно. Мюллер написал министру народного просвещения свое, ставшее позднее знаменитым письмо, в котором настоятельно требовал предоставления заведования кафедрой именно ему. Письмо было полно собственного достоинства и говорило об огромной любви и преданности его автора науке. Оно произвело впечатление, и Мюллер получил кафедру в самом крупном университете Германии, которую и занимал 25 лет. Он умер 57 лет. После его смерти, по свидетельству нашего знаменитого хирурга Н. И. Пирогова13, ходили слухи, что он принял яд. Причину самоубийства приписывали
13 П и р о г о в Н. И. 1910. Вопросы жизни. Дневник старого врача, т. II.
Изд. Пироговского товарищества. Киев.
3 Е. М. Вермель
65
Иоганнес Петер Мюллер (1801—1858)
какому-то сделанному И. Мюллером открытию, поколебапик-мv его религиозные убеждения ревностного католика. Однако его биограф, друг и ученик, Э. Дюбуа-Реймон, отрицал это.
Влияние и авторитет Мюллера в области физиологии и анатомии были чрезвычайно великн. Н. И. Пирогов в своих воспоминаниях записал: «Это было время, когда И. Мюллер властвовал почти один в области физиологии». Это объяснялось тем, что он дал новое, сразу ставшее общепризнанным направление в биологических науках, базировавшееся иа «микроскопических исследованиях, истории развития, точного физиологического эксперимента и химического анализа...» (там же, стр. 497). Вполне естественно допустить, что И. Мюллер мог прийти к выводам об отсутствии божественного начала и это привело к крушению веры и жизни. Не случайно все же, что многие из его учеников в дальнейшем стали материалистами-механистами.
Н. И. Пирогов оставил замечательное описание Мюллера, которого он знал лично, у которого учился в Берлинском университете. Он писал: «Лицо Иоганнеса Мюллера поражало вас своим классическим профилем, высоким челом и двумя межбровными бороздами, придававшими его взгляду суровый вид и делавшими несколько суровым проницательный взгляд его выразительных глаз. Как на солнце, неловко было новичку смотреть прямо в лицо на Мюллера» (там же, стр. 501).
Мюллер оставил после себя огромную школу. Однако в обычном смысле этого слова ее нельзя назвать школой. Ум Мюллера отличался необычайной широтой взгляда по всем проблемам и совершенно необыкновенной проникновенностью в понимании степени важности разных проблем. Но он их только, как правило, начинал разрабатывать, а затем передавал своим ученикам. Мюллер с поразительным ясновидением, как писал Дюбуа-Реймон, присущим только гениальным людям, узнавал наиболее способных к научной работе молодых людей, приближал их к себе и умел направить их деятельность иа всю их жизнь. Дюбуа-Реймон писал про Мюллера: «Он действовал, как выражался Гете о красоте,— одним лишь своим присутствием». Но мы знаем, что он следил и направлял научную деятельность своих учеников, особенно пока они оставались при его лаборатории.
Мюллер привлекал к себе молодых ученых и своих сверстников не только силой своего таланта и знаниями, но и своей исключительной внешностью, напоминавшей, как говорили, римского воина. Он был постоянно окружен многочисленными учениками, а ими можно было гордиться. Вряд ли можно найти еще кого-
3*
67
либо, у кого было бы столько таких крупнейших ученых среди учеников. Это были — Я. Генле, Т. Шванн, Р. Ремак, К. Людвиг, А. Келликер, В. Вальдейер, Р. Вирхов, Э. Брюкке, Э. Дю-буа-Реймои, В. Гельмгольц (три последних были наиболее близкими и любимыми учениками) и др. Как мы уже отметили, слушал лекции и учился у Мюллера и Н. И. Пирогов, на которого производили, как он сам отметил, большое впечатление демонстрации на лекциях различных опытов на животных и микроскопических препаратах (там же, стр. 500).
Как мы отметили, Мюллер щедрой рукой раздавал своим сотрудникам не только сделанные им фактические наблюдения, но и прежде всего свои идеи, которые, попадая в руки талантливых людей, претворялись и|ми в жизнь. Так перечисленные выше ученики Мюллера стали основателями ряда новых наук или больших их разделов и вошли в историю науки. Отдать же должное И. Мюллеру часто забывают, ибо, очевидно, плохо знают историю.
Микроскопическим исследованиям, как отмечает в своих воспоминаниях Н. И. Пирогов, Иоганнес Мюллер придавал огромное значение. Н. И. Пирогов приводит случай острого спора между Шлеммом и Арнольдом о природе ушного узелка (ganglion oticum). За разрешением вопроса обратились к Мюллеру, и он «...уладил спор, доказав Шлемму микроскопом, что узелок был действительно нервный...» (там же, стр. 499).
Работая по сравнительной анатомии круглоротых, Мюллер нашел, что по своей микроструктуре хорда имеет много общего с клетками растительных тканей. Он, несомненно, видел клеточные структуры и других тканей. Ему были известны и работы Я. Пуркиня и его учеников. И. Мюллер понимал все важное значение этих исследований и поручил дальнейшую разработку микроскопической анатомии трем своим ученикам: Иоганну Мишеру (J. Miescher, 1811—1871), сделавшим диссертацию по строению кости, Фридриху Якобу Генле (F. J. Henle, 1809—1885)—он стал известным анатомом, ему, в частности, принадлежит первая систематическая работа по эпителию, и, наконец, Теодору Шваину (Th. Schwann, 1810—1882), который до этого работал по физиологии пищеварения и ему принадлежит честь открытия фермента желудочного сока, названного им пепсином. Интересно указать, что все свои работы, так широко его прославившие, Шванн сделал всего за пять лет пребывания в лаборатории Мюллера; больше он, по-видимому, научно не работал.
Остановимся' еще коротко на развитии патологической анатомии. Мало кто знает, что именно Мюллер в 1838 г. впервые опуб-68
ликовал исследование «О тонком строении и форме опухолей», в котором он описал с помощью микроскопа нераковыс опухоли молочной железы под общим названием цистосаркомы (кистозная мастопатия). Сам Мюллер более к патогистологии не возвращался, хотя и читал некоторое время этот курс в Берлинском университете. Он передал все эти направления работ своему замечательному ученику Вирхову, которого по праву и считают «отцом патогистологии» (целлюляриой патологии)) и патологической анатомии.
Основная заслуга Шванна, обеспечившая ему успех в работе и придавшая достоверность и убедительность клеточной теории, состоит в том, что он связал представление о клетке с вопросом ее происхождения. Именно тезис, что клетки как растений, так и животных принципиально сходны (гомологичны) между собой, ибо все они возникают единообразным путем, является тем совершенно новым, что внес Шванн. Исходил Шванн из работ Матиаса Шлейдена (1804—1881), с которым он познакомился в октябре 1837 г., а уже в 1838 г. Шванн опубликовал три небольших сообщения, в которых были сформулированы основы клеточной теории, подробно изложенной в книге, вышедшей в 1839 г. Сам Шванн писал, что именно его беседа со Шлейденом (работа последнего была в тот момент еще не опубликована) сыграла в формулировании его идеи огромную роль. Существует крылатая фраза: «Шванн стоял на плечах Шлейдена», она по существу правильно отражает дело. Шванн сам признавал, что в разработке клеточной теории он исходил из открытия Шлейдена. Именно общность происхождения клеток явилась тем основным фундаментом, который был положен в основу клеточной теории, оказавшей столь большое положительное влияние на развитие биологии. В г. Нейсе Теодору Шванну воздвигнут памятник; на его открытие (6 июня 1909 г.) съехалось много ученых. Приведем краткую выдержку из речи его друга, крупного ученого своего времени В. Вальдейера, произнесенной иа открытии памятника. Он сказал: «Не Шванн открыл те микроскопически малые структуры, которые мы обозначаем как «клетки». Но только Шванн научил нас понимать их значение..!» 14
Итак, мы видели, что Шванн имел много предшественников. Вопрос изучался в течение 2—3 десятилетий. Однако только
14 Her twig О., W aide yer W. 1909. Theodor Schwann zum Gediich-tnis. Arch. f. mikrosk. Anat., 74, 469 («Памяти Теодора Шванна»).
69
Маттиас Шлейден (1804—1881)
Шванн рассмотрел учение о микроструктурах с необходимой широтой и построил научно обоснованную теорию. Повторим в связи с этим известные слова французского математика Алексиса Клода Клеро (1713—1765), что существуют разные истины: есть истины, о которых только догадываются, есть истины, которые утверждают, и есть истины, которые обосновывают и доказывают.
История науки не раз показывала, что высказанная, но недостаточно логически обоснованная гипотеза или теория не могут
7Q
иметь серьезного значения для развития науки, однако отличип. истину от домыслов и фантазий не так просто. П р е д с т а н л <• ние о гомологичности клеток, о чем говорил Шванн, оказалось теоретически верным, а фактические данные о возникновении| клеток путем цитогенеза из зернышек, на которые он опирался, были ошибочными.
§ 18. Теория клеткообразования М. Шлейдена. М. Шлейдеи изучал возникновение клеток в процессе роста различных частей растений, причем эта проблема являлась для него самодовлеющей. Что же касается собственно клеточной теории в том смысле, как мы ее понимаем в настоящее время, то он ею не занимался. Основной заслугой Шлейдена является отчетливая постановка вопроса относительно возникновения клеток в организме. Эта проблема приобретала принципиальное значение, поскольку она толкала исследователей на путь изучения клеточной структуры под углом зрения процессов развития. Наиболее существенным является представление Шлейдена о природе клетки, которую он, по-видимому, впервые назвал организмом. Так он писал: «Нетрудно понять, что как для физиологии растений, так и для общей физиологии жизнедеятельность отдельных клеток является главнейшей и совершенно неизбежной основой, и поэтому прежде всего встает вопрос, как же собственно возникает этот маленький своеобразный организм — клетка».
Теория клеткообразования Шлейдена была им несколько позднее названа теорией цитогенезиса. Весьма существенным является то обстоятельство, что она впервые связала вопрос возникновения клетки с ее содержимым и (в первую очередь) с ядром; таким образом, внимание исследователей было перенесено с оболочки клетки на эти несравнимо более важные структуры.
Сам Шлейден считал, что он впервые ставит вопрос о возникновении клеток, хотя ботаники и до него описывали, правда, далеко не отчетливо, размножение клеток в форме клеточного деления, однако эти работы были ему, вероятно, неизвестны до 1838 г.
Возникновение клеток, согласно теории Шлейдена, в схеме протекает следующим образом. В слизи, которая составляет живую массу, возникает маленькое круглое тельце. Вокруг него конденсируется сферический сгусток, состоящий из гранул. По поверхности эта сфера покрывается мембраной — оболочкой. Так
71
Рис. 19. Схема процесса возникновения клеток по представ лениям М. Шлейдена (1838 г.)
Рис. 20. Бактерии и почкующиеся дрожжевые клетки, изображенные на рисунках М. Шлейдена (1824 г.)
возникает округлое тело, известное под названием клеточного ядра. Вокруг последнего в свою очередь собирается студенистая зернистая масса, которая также окружается новой оболочкой. Это будет уже оболочка клетки. На этом процесс развития клетки заканчивается (рис. 19).
Тело клеток, которое мы теперь называем протоплазмой, Шлейден (1845) обозначал словом цитобластема (термин принадлежит Шванну). «Цитос» по-гречески значит «клетка» (отсюда наука о клетке — цитология), а «бластео» — образовывать. Таким образом, на протоплазму (вернее, на клеточное тело) Шлейден смотрел как на клеткообразующую массу. По Шлей-дену, следовательно, новая клетка может образовываться исключительно в старых клетках, причем центром ее возникновения является конденсирующееся из зернышек ядро, или, по его терминологии, цитобласт.
Несколько позднее, описывая возникновение клеток в 1850 г., Шлейден отмечал также размножение клеток и путем их поперечного деления, ссылаясь при этом на наблюдения ботаника Гуго фон Моля (1805 —1872). Шлейден, не отрицая правильности тщательных наблюдений Моля, считал этот способ развития клеток мало распространенным.
Представления Шлейдена можно обобщить следующим образом: молодые клетки возникают в клетках старых путем конденсации слизистого вещества. Схематически Шлейден изобразил
73
это следующим образом. Он считал данный способ возникновения клетки из цитобластемы всеобщим принципом. Свои представления он довел, если можно так сказать, до абсурда, описывая, например, размножение дрожжевых клеток. На рис. 20 мы приводим изображение почкования дрожжевых грибков, описанное Шлейдеиом. При рассматривании этого рисунка для нас в настоящее время не остается никаких сомнений, что он видел типичное почкование дрожжевых клеток. Сам же Шлейдеи, вопреки очевидности, все же утверждал, что образование почек происходит лишь путем слияния в комочки зернышек около уже имеющихся дрожжевых клеток.
Возникновение дрожжевой клетки Шлейден представлял себе так. Он говорил, что в соке из ягод, если его оставить в комнате, уже через сутки можно заметить маленькие зернышки. Дальнейший процесс заключается в том, что эти взвешенные зернышки увеличиваются в числе и, слипаясь, образуют дрожжевые клетки. Новые дрожжевые клетки образуются из тех же зернышек, но преимущественно вокруг старых дрожжевых клеток (см. рис. 20). Шлейдеи склонен был объяснить появление инфузорий в гниющих жидкостях аналогичным образом. Его описания, как и приложенные к ним рисунки, ие оставляют никаких сомнений в том, что эти мельчайшие загадочные зернышки, из которых «образуются» дрожжи и инфузории, представляют собою не что иное, как размножившиеся в той же жидкости бактерии, не имеющие, конечно, непосредственного отношения к развитию дрожжей.
Теория цитобластемы была в дальнейшем признана с фактической стороны ошибочной, но вместе с тем она имела серьезное влияние иа дальнейшее развитие науки. Этих взглядов отдельные исследователи придерживались еще в течение ряда лет. Однако все они делали ту же ошибку, что и Шлейден, забывая, что, подбирая ряд отдельных микроскопических картин, мы никогда не можем быть вполне уверены в правильности вывода о направлении процесса. Мы уже цитировали (§ 6) слова Феликса Фонтана (1787) о том, что картина, открываемая микроскопом, может подходить одновременно к очень разнообразным явлениям. Эти слова сохраняют все свое значение и до сего времени.
§ 19. Клеточная теория Т. Шванна. Выше мы отмечали, что теория цитогенеза сыграла большую положительную роль, ибо, поставив во всей широте вопрос о возникновении клеток, она подвела таким образом базу под проблему их гомологичности.
74
Теодор Шванн (1810—1882)
Теодор Шванн считал, что в явлении цитогенеза впервые был найден общий принцип развития микроскопических структур всех организмов, позволяющий делать заключение о принципиальном сходстве клеток всех тканей и органов. Как мы уже видели выше, сама идея всеобщности клеточной структуры в той или иной форме высказывалась целым рядом исследователей. Но эта мысль нуждалась в четкой формулировке и (что особенно важно) в убедительных доказательствах. Это было блестяще
75
выполнено Шванном, напечатавшим в 1839 г. свой классический труд под названием «Микроскопическое исследование о соответствии в структуре и росте животных и растений» 15. Эта книга быстро получила широкую известность.
Первая часть книги носит чисто описательный характер Шванн старается доказать, что клетки животных морфологически принципиально сходны с клетками растительными. Основным объектом Шванна, из которого он исходил, была ткань хряща и ткань хорды. Действительно, нетрудно видеть (рис. 21), что клеточное строение этих тканей не оставляет никаких сомнений. Клетки повсюду хорошо отграничены оболочками как друг от друга, так и от окружающей среды. Важно отметить, что Шванн, как и все остальные ученые того времени, придерживался взгляда, что наиболее важной структурой клетки является ее оболочка, и поэтому, доказывая всеобщность клеточного строения тканей животных, он всюду стремился доказать наличие четкой и ясной оболочки. Немалые трудности поэтому доставили ему такие ткани, как мышечная, которая состоит из многоядерных волокон. Большие трудности для объяснения представляло также наличие большого количества межуточного вещества в соединительной ткани с типичными для нее волокнами (коллагеновыми). Для последних Шванн принимал, что они образуются путем превращения целых клеток.
Доказав всеобщность клеточного строения, Шванн устанавливает, что все клетки, сколь бы они различны ни были, возникают всегда принципиально сходным образом, а именно путем цитогенеза. Небольшое разногласие между представлением Шлейдена и Швайна заключается в том, что последний считал возможным возникновение новых клеток не только внутри старых за счет зернышек их содержимого, но также из живой бластемы, которая может находиться и между клетками. Принципиального значения эти разногласия, однако, не имеют. Установление самого факта — признание цитогенеза—важно потому, что таким образом выдвигается принцип, позволяющий говорить о сходстве развития клеток всех многоклеточных организмов — как растений, так и животных.
15 Schwann Th. 1839. Mikroskopische Untersuchungen uber die Ueber-einscimmungen in der Struktur und dem Wachstum der Thiere und Pflanzen. Berlin («Микроскопические исследования о соответствии в строении и росте животных и растений»). В русском переводе под редакцией проф. 3. С. Кацнельсона издана в 1939 г. Изд-вом АН СССР.
16
Рис. 21. Рисунок Т. Шванна, изображающий клетки в срезе через хрящ,
Отсюда уже следовал и вывод о принципиальной сравнимости всех клеток, поскольку признавалось наличие общего принципа развития для всех элементарных структурных частиц организма (клеток).
Работа Шванна богата мыслями, она впервые привела в систему различные факты. Для клеточной теории в целом на данном этапе было неважно, как конкретно надо трактовать процесс возникновения клеток, их генезис. Существенно было установить сам принцип, что все клетки возникают одним и тем же путем.
В своей книге Шванн не ограничился только перечислением ряда фактических данных. В ней мы находим главу «Теория клеток». Она начинается рассуждением о значении теории для конкретного исследования и о значении телеологической и физической (материалистической), как он их называет, точек зрения на природу вещей.
77
Основные положения клеточной теории, сформулированной Шванном, таковы: первое положение заключается в утверждении, что в с е ткани состоят из клеток. Второе говорит об общем принципе развития этих структур. Но здесь же Шванн ставит вопрос и о том, какова природа тех основных сил организма, которые обусловливают данный процесс. С телеологической точки зрения, как подчеркивает сам Шванн, силы, существующие в организме, формируют его развитие согласно предсуществующей идее, и основная сила организма (или «душа») развивается согласно определенной цели. Сам он относится отрицательно к признанию нематериального, наделенного сознанием принципа развития. Он прямо пишет, что силы организма совпадают с силами неорганического мира и действуют по законам необходимости без всякой связи с особой целью. В качестве аналогии ои приводит в пример силы, действующие в пределах нашей планетной системы. Эти силы действуют по законам необходимости, как и все физические явления, но тем не менее планетная система представляется нам целесообразно организованной. Причина этой целесообразности лежит, как говорит Шванн, «не в самих этих силах, а в Том, Кто так создал материю с ее силами, что они, следуя слепым законам, создают целесообразное целое».
Таким образом, Шванн четко отличал естественные закономерности, по которым протекают все процессы в природе, от «божественной причины». Так, процесс цитогенеза Шванн сравнивает с процессами кристаллизации и отводит им в свой книге довольно много места. Отсюда следует вывод, что поскольку в неорганическом мире (кристаллизация) и в органическом мире (цитогенез) мы наблюдаем сходные процессы, то, следовательно, надо заключить, что все эти процессы обусловливаются одними и теми же силами. Другими словами, процесс клеткообразования Шваии ставит наряду с физико-химическими процессами, отмечая лишь, что эти процессы более сложны и требуют определенных благоприятных условий для своего осуществления. Одиако Шванн специально указывал, что не следует ставить знака равенства между процессом кристаллизации и пластическими силами клеток.
Наконец третьим пунктом клеточной теории Шванна является представление о самостоятельной жизнедеятельности каждой отдельной клетки. Шванн представлял себе совокупную деятельность организма как сумму жизнедеятельности отдельных клеток.
78
Любопытно, однако, отметить, что Шванн предвосхитил и такое возражение, что отдельная клетка, будучи изолирована от организма, существовать не может. Он писал, что это возражение против теории биологической самостоятельности клетки аналогично тому, как если бы мы отрицали самостоятельную жизнь пчелы, которая, как известно, вне семьи также быстро гибнет. Этому сравнению нельзя отказать в остроумии, однако логика показывает, что аналогия не есть еще доказательство. Аналогий, при желании, можно подобрать сколько угодно, но они не двигают науку вперед, так как не выявляют сущности явлений.
Итак, третий пункт клеточной теории Шванна можно вкратце формулировать и так, что вопрос о свойствах организма сводится по существу к арифметической сумме свойств отдельных клеток.
§ 20. Клеточная теория в оценке современников. Значение исследований Шванна и сформулированной им клеточной теории для биологии заключается прежде всего в том, что им впервые была дана обоснованная обобщающая и ведущая идея трактовки строения организма. Она стала общепризнанной, поэтому Шванна нередко называют основателем новой гистологии. Историческое значение этой теории заключается еще и в том, что она дала колоссальный толчок к дальнейшему развитию этой области. Приведем небольшую цитату из обзора исследований по микроскопической анатомии за 1839—1840 гг., написанного ученым того времени Карлом Рейхертом (1811—1883). Он писал: «Микроскопическая анатомия пережила и переживает до сих пор удел наук, которые обогащены крупными открытиями. Интерес к ней стал всеобщим и разносторонним после того, как открытие клетки дало основание к планомерному развитию микроскопической анатомии. Тем не менее успехи микроскопической анатомии, развивающейся по пути, указанному Шванном, сравнительно еще не велики. Известные естествоиспытатели опубликовали наблюдения, которые привели к изменению и ограничению принципа клеточной теории как общего принципа развития разнообразнейших элементарных частей животных и растительных организмов. Некоторые шли настолько далеко, что ставили под сомнение правильность этого принципа — принципа, который теоретически был высказан еще Океном, Майером, особенно Распайлем и Дютроше. С другой стороны, многие другие исследователи привели ряд доказательств в пользу этой точки зрения, но не пронизы
79
вали последних общей идеей, которая столь мастерски как с теоретической, так и с фактической стороны была обоснована Шванном и Шлейденом» 16.
Следовательно, в 1841 г., когда писались эти строки, теория Шванна уже считалась, как это можно понять, общепризнанной и ведущей идеей.
Следует указать, что в обзоре, составленном Ш. Рейхертом и перечисляющим, следовательно, исследования, опубликованные только в течение двух лет, мы находим указание на десятки работ, касающихся почти всех систем органов. Это еще раз показывает, какое большое внимание было уделено этой области науки в результате обобщений, сделанных Шванном. До этого времени лишь очень немногие естествоиспытатели занимались микроскопическим строением тканей животных.
Признавая за Шванном огромные заслуги в формировании учения о микроскопическом строении организмов, мы все же должны отметить, что в его концепции имелся и ряд положений, отрицательно повлиявших на биологию. Особенно ясно они выявились в последующий период, задерживая развитие целого ряда дисциплин.
Прежде всего отметим, что в представлении Шванна клетка оставалась все тем же самым пузырьком, т. е. пространством, окруженным оболочкой, и именно этой оболочке придавалось исключительно важное значение в характеристике и определении клеточной структуры; в этом отношении Шванн остался на позиции таких своих предшественников, как Мейен, Дютроше и др. Отрицательное влияние теории цитогенеза Шлейдена сказалось также на понимании конкретного механизма процесса клеткообразования. Эти представления были позже признаны полностью ошибочными, а между тем они являлись одним из краеугольных камней всей концепции Шванна, и ей уделялось чрезвычайно большое внимание. Огромное большинство исследователей того времени пыталось конкретно подтвердить процесс цитогенеза и тратило много труда на ее обоснование. Эти исследователи настолько были убеждены в ее реальности, что
16 Reichert К. В. 1841. Bericht Gber die Fortschritte der mikroskopischen Anatomie in den Jahren 1839—1840. Arch. f. Anat., Physiol, u. wiss. Med. Jahrg («Обзор успехов микроскопической анатомии за 1839— 1840 гг.»).
80
Карл Рейхерт (1811—1883)
даже, не находя на своих препаратах подтверждающих ее фактов, все же безоговорочно ее принимали.
Наконец, ошибочной в принципе была также трактовка организма как своеобразного «государства» клеток. Это последнее представление держалось до начала XX в.
Положительное и отрицательное значение каждой теории или даже отдельных ее положений должно оцениваться в зависимости от того влияния, которое они оказывали на последующее развитие науки. Теория «клеточного государства» легла в основу руководящей точки зрения на организм, и на этом этапе развития биологии способствовала выделению гистологии и цитологии в самостоятельную науку. Но как только определенный этап формирования знаний оказывался более или менее завершенным, сразу же давали себя знать и те отрицательные, ошибочные положения, которые эти представления несли.
Прежде всего остановимся кратко на дальнейшей разработке проблемы клеткообразования и критике цитогенеза.
Из исследований того времени большой интерес представляет статья Габриэля Валентина (1810—1883), ближайшего и наибо-
81
Габриэль Валентин (1810—1883)
лее выдающегося ученика Пуркиня. Ему принадлежит обзор под названием «Ткани человеческого и животного тела»17, помещенный в «Физиологической энциклопедии» (1842), издававшейся под редакцией известного физиолога Рудольфа Вагнера (1805— 1864). Этот обзор интересен не столько фактическими данными, сколько общей концепцией, представляющей собою дальнейшее развитие представлений о возникновении клеток в связи с проблемой организации животных существ.
Согласно представлениям Валентина, отражающим наиболее распространенные взгляды того времени, элементарными частями сложного животного организма являются наряду с клетками различные кристаллы, зернышки, ядра, волокна и прозрачные мембраны, соединение которых и образует ткани животных. Но он все же признает, что основным морфологическим элементом живых существ, в первую очередь растений, являются образова
17 V а I е т t i n G. 1842. Gewebe des menschlichen und thierischen Korpeis. «Handworterbuch der Physiologie» von Rud. Wagner. Bd. I (-Ткани человеческого и животного организмов»).
82
ния, ограниченные тончайшими стенками, т. е. клетки. Но вместе с тем отсутствие четкой и ясной точки зрения на природу клетки приводит к тому, что Валентин сопоставляет совершенно несравнимые структуры, как, например, мелкие артерии животных со спиральными сосудами растений, перепонки и клетки (рис. 22).
Признав, что клетки являются основным типом элементарных структур как растений, так и животных, Валентин, однако, проводит идею относительности самого понятия клетки, поскольку история развития самой клетки не позволяет, с его точки зрения, провести принципиальное отличие последней от ядра. Детализируя процесс клеткообразова-ния, Валентин перечисляет четыре основных типа возникновения клетки. Из них три являются разными вариантами того же цитогенеза, и только последний заключается, по его мнению, в размножении клеток делением (приводятся наблюдения ботаников, в первую очередь Гуго фон Моля, над делением клеток различных водорослей).
Любопытно привести определение, которое дает Валентин ядру (1842). Согласно представлениям того времени, оно рассматривается как индивидуализированное тело, заключенное в клетке. При этом он отмечает, что все остальные отличня между ядром и клеткой могут настолько варьировать, что стирается всякая грань между ними.
Рис. 22. Микроскоп фабрики Росса (1831 г-), с которым работал Г. Валентин
81
В самом деле, согласно теории цитогенеза, как ядерная, так и протоплазматическая оболочки возникают сходным образом из сливающихся по сфере зернышек, одно поверх другого. Формально Валентин был прав, поскольку в определении клетки основное значение продолжали придавать ее оболочке. Развитие этой теории привело к положению, когда понятия как клетки, так и ядра оказывались по существу неразграниченными.
В приводимой выше статье Валентин, не удовлетворяясь простой систематизацией фактов, пытался также сформулировать и общие закономерности развития клеток, используя при этом различные варианты описывавшегося цитогенеза. Возникновение клетки он трактовал как различного рода «физико-химические комбинации», развитие которых находится под руководящим влиянием «организационной идеи». Такова общетеоретическая установка Валентина, которая конкретно сводится к следующим закономерностям.
1. Закон обособленного возникновения микроморфологических структур (как пример приводится возникновение в цитобластеме изолированных клеток).
2. Закон концентрации возникающих микроструктур вокруг уже существующих согласно предопределенному организационному плану.
3. Явление соединения клеток в более сложную структуру согласно тому же организационному плану.
Валентин считал, что развитием организма руководит «направляющая идея», распределяя его структурные компоненты по соответственным участкам тела и создавая тем самым гармонию целого. Сущность этого «руководящего принципа» будет, по мнению Валентина, навсегда скрыта для понимания естествоиспытателя, но, несмотря на это, он считал, что наука должна изучать эти процессы, чтобы приближаться к пониманию и к искусственному воспроизведению тех физико-химических явлений, которые природа применяет для создания органов. Очень интересно отметить, что Валентин цитирует работу Ашерсона, который, по-видимо-му, впервые пытался приготовить искусственные клетки. Последний изготовлял модели клеток, создавая эмульсии жира в белке и получал, таким образом, несливающиеся капельки. Сам Валентин также готовил модели клеток, энергично встряхивая ртуть в белке; при подсыхании подобной эмульсии возникали структуры, напоминавшие клетки. Он, однако, относился отрицательно к этому направлению исследования, считая подобные явления по самому своему существу отстоящими очень далеко от процессов
84
Возникновения клеток в организме. Валентин защищал нагл ид. согласно которому силы неорганической природы недостаточны для создания органических структур; поэтому он и утверждал, ч то развитие организма — и прежде всего осуществление его целостности— должно идти под влиянием организующего принципа («организующей идеи»).
Концепция Валентина виталистична. Но его заслуга состояла в том, что он очень ясно и отчетливо представлял себе проблему целостности развивающегося организма, хотя и трактовал ее чисто идеалистически. Тем не менее, как это будет показано ниже, именно исходя из подобной концепции, значительно позднее (в самом конце XIX в.) был нанесен сокрушительный удар по механистической теории «клеточного государства». Взгляды Валентина успеха не имели, и развитие клеточной теории в дальнейшем пошло по механическому пути, указанному в работе самого Шванна, а в дальнейшем еще более четко развитому Вирховым.
Мы остановились здесь несколько подробнее на высказываниях Валентина с целью показать, насколько значительно отличались представления о клетке в тот период от нашего времени. Природу клетки по существу не знали, и поэтому все конкретные данные о ее происхождении были мало доказательными, а теоретические трактовки натянутыми и произвольными.
§ 21. Цитогенез и эмбриология. Среди ряда исследований, якобы подтверждающих возникновение клеток из зернышек (цитогенез), можно найти довольно много публикаций, в которых разные авторы того времени писали, что им лично никогда не удавалось наблюдать первые фазы возникновения клеток из зернышек. Считалось поэтому общепризнанным, что увидеть этот процесс чрезвычайно трудно. Тем не менее, большинство исследователей полагали, что явление цитогенеза вполне доказано, хотя, как мы видим, главным доказательством оставался авторитет Шванна и Шлейдена.
Отрицательную роль эндогенная теория возникновения клеток сыграла в развитии эмбриологии. В самом деле, с точки зрения представлений о развитии отвергается самый принцип преемственности в развитии. В результате некоторые ученые стали допускать чуть ли не возникновение целых органов прямо из бесструктурной массы. В настоящее время трудно себе даже представить, как много труда понадобилось для того, чтобы опровергнуть теорию Шлейдена, не имевшую успеха у ботаников, но пользо
85
вавшуюся признанием зоологов благодаря пропаганде Шванна, авторитет которого стоял очень высоко.
Мы не можем на этом вопросе останавливаться сколько-нибудь подробно и отметим только, что в 40-х годах самый процесс дробления яйцеклеток был уже неоднократно описан на многочисленных объектах, однако спор о клеточной природе шаров дробления (бластомеров) продолжался еще очень долгое время. Даже такой известный ученый, как Теодор Людвиг Бишоф (1807—1882), оспаривал клеточную природу бластомера и утверждал, что сложные эмбриональные процессы могут быть объяснены только при допущении наличия своеобразных законов развития для каждого млекопитающего18. Много понадобилось труда для того, чтобы доказать, что «шары дробления» ранних стадий развития эмбриона являются не чем иным, как клетками.
Большая заслуга в выяснении клеточной природы ряда тканей и особенно в доказательстве процесса деления как единственного пути размножения клетки принадлежит Роберту Ремаку (1815— 1865) 19. Его исследования и|мели большое влияние как на современников, так и на дальнейшее развитие цитологии на протяжении нескольких десятилетий. Однако окончательный удар теории цитогенеза был нанесен только выдающимся патологом Рудольфом Вирховым (1821 —1902), который прямо и категорически писал, что там, где возникает новая клетка, должна предсуществовать клеточная ткань. Он базировался главным образом на исследованиях Ремака, но и сам привел многочисленные доказательства повсеместного распространения клеточного деления как в нормальных, так и в патологических тканях.
§ 22. Пересмотр клеточной теории Р. Вирховым. Рудольф Вирхов оказал очень большое влияние на дальнейшее развитие клеточной теории и вообще на учение о клетке. Его заслуга заключается прежде всего в том, что он свел воедино все отдельные, довольно многочисленные, но разрозненные факты и с большой убедительностью показал, что никем ни разу не было дано
18 Bischoff Th. 1852. Entwicklungsgeschichte des Meerschweinchens. Giessen («История развития морской свинки»).
19 R е m a k R. 1852. Uber extracellulare Entstehung thierischer Zellen und fiber Vermehrung derselben durch Theilung. Muller's Arch. Anat., Physiol, wiss. Med., 2, 47 («О внеклеточном образовании животных клеток и об их размножении путем деления»).
86
убедительных доказательств в пользу возникновении клетки de novo (заново) из неоформленной массы. Далее Вирхов показал в специальной области его исследования — в патологии — все щ»<•-имущество представления о размножении клетки путем деления. Основное сочинение Вирхова носит название «Целлюлярная патология, как учение, основанное на физиологической и патологической гистологии» (1855—1859) 20.
Первая глава этой книги — чисто теоретическая и посвящена рассмотрению клеточной теории. Вирхов доказывал, что клетки представляют собой самый низкий морфологический элемент и только из их совокупности слагаются все живые существа, что вне клетки мы не можем предполагать существования настоящей жизни.
Таким образом, Вирхов специально подчеркнул, что клетке присуща целостность и се жизнедеятельность своеобразна. Он писал: «Жизни приписывается нечто вполне своеобразное. Некоторым это может показаться чем-то вроде биологической мистики, поскольку жизнь таким образом ... не сводится вполне на химию и физику. Из дальнейшего изложения каждый убедится, что вряд ли кто может представить себе протекающие процессы более механистичными, чем это делаю я, когда речь идет о процессах жизнедеятельности в элементарных морфологических элементах».
По мнению Вирхова, любой процесс, протекающий в клетке, представляет собой процесс чисто физико-химический, но все же жизнедеятельность исходит от клетки, как от целого. Клетка является живой единицей, сохраняющейся только до тех пор, пока она остается единым целым. Этот взгляд представляет собой совершенно новую точку зрения.
Переходя к конкретному морфологическому описанию клетки, Вирхов не вносит чего-либо особенно нового. В общем он излагает данные о ее строении довольно трафаретно и по существу повторяет то, о чем уже много ранее писал Пуркиня. Он отмечает, что основное различие между растительной и животной клеткой заключается в отсутствии у последней целлюлозной оболочки. Он пишет, что «оболочка животной клетки соответствует так называемому первичному мешочку растительной клетки» (так назывался в то время загустевший поверхностный слой протоплазмы). Целлюлозная оболочка им справедливо рассматри
20 Virchow R. 1858. Die Cellularpathologie usw. Berlin. Имеется несколько русских переводов; первый вышел в 1859 г.
87
валась как нечто, возникшее в результате позднейшего развития из тела самой клетки.
Итак, важно отметить, что Вирхов рассматривал клетку как структуру «по-видимому, весьма своеобразную, хотя элементарно построенную и повторяющуюся с удивительным постоянством во всех живых организмах». Вирхов считал клетки постоянной структурой и утверждал, что они возникают только путем размножения. Это положение он формулировал так: «всякая клетка от клетки» (omnis cellula е celluiae).
Выступая против широко распространенной теории цитогенеза, Вирхов отмечал, что она обосновывается весьма недостоверными фактами. Поскольку теория цитогенеза придавала чрезвычайно важное значение в процессе размножения ядру клетки, то естественно было, что и Вирхов уделял ему много внимания. Но он пришел к другим выводам и доказывал, что как раз во время размножения клетки ядро в ней не может быть обнаружено. 'Тем не менее, он неоднократно подчеркивал, что клетки, полностью теряющие свои ядра, нежизнеспособны. Он прямо писал: «Из соединения ядра и оболочки возникает та морфологическая структура, которую всегда можно узнать во всякого рода живых как растительных, так животных формах, и которая является субстратом всех процессов жизнедеятельности».
Развенчав цитогенез и поставив на его место процесс деления клетки как единственный способ ее размножения, Вирхов считал, что он не только не опровергает клеточной теории, а наоборот, подводит под нее твердый фундамент. Он писал, что поскольку единство и характерные особенности жизни могут быть обнаружены лишь в постоянно повторяющейся клеточной организации, то отсюда следует делать вывод, что строение сколько-нибудь значительного по величине живого существа, так называемого индивидуума, следует характеризовать как своего рода «коллективную организаци ю», которая и придает организму своего рода «социальную» природу. Он подчеркивал, что все попытки отыскать какие-либо другие органические элементы, которые можно было бы поставить на место клетки, остаются тщетными, поэтому возникновение высших растений и животных следует рассматривать как процесс прогрессивного суммирования большего или меньшего числа клеток. Таким образом, организм представляет собою множество отдельных самостоятельных единиц, поставленных в их жизнедеятельности в тесную зависимость друг от друга. Однако такой единице (клетке) свойственна, по представлению Вирхова, самостоятельная жизненная активность,
88
Рудольф Вирхов (1821—1902)
и, хотя побуждения к этой деятельности она и получает от других частей, но все же все свои отправления она совершает самостоятельно.
Эта система представлений Вирхова, являющаяся дальнейшим развитием клеточной теории Шванна, получила название теории «клеточного общества», или теории «клеточного государств а». Нельзя, однако, не отметить, что формулировки этих представлений у позднейших исследователей звучат значительно механистичнее, нежели у самого Вирхова.
89
Франц фон Лейдиг (1821— 1908)
В свете изложенных представлений ясно, почему Вирхов придавал столь большое принципиальное значение закону «всякая клетка от клетки». Ведь таким образом устанавливается закон преемственности развития, и прежде всего приводится наиболее убедительное доказательство в пользу признания клетки в качестве элементарной живой структуры.
Нередко говорят о клеточной теории Шванна — Вирхова, и это совершенно правильно, поскольку именно Вирхов пересмотрел все общие положения этой теории в свете новых фактов. С другой стороны, он на материале патологической анатомии человека, одним из основателей которой он по праву считается, показал всю прогрессивность нового (целлюлярного) подхода к этим проблемам по сравнению с предыдущими умозрительными теориями. Вирхов не случайно назвал свою книгу «Целлюлярной патологией». Идеи, изложенные в ней, являлись тем знаменем, иод которым патология XIX в. добилась огромных успехов.
Вирхов был выдающимся реформатором теоретической и практической медицины; его целлюлярная патология пришла на
90
смену гуморальной патологии, наиболее известным iipt’Ai ннннг лем которой был австрийский патолог Карл Рокитапскии (1804—1878). В этом заключается большая заслуга Вирхова перед наукой и человечеством, в этом прогрессивное значение его клеточной теории в середине и конце прошлого века. Клеточная теория дала патологам (а также и биологам) определенную руководящую- точку зрения, в целом ряде отношений совершенно правильную, хотя во многом и не исчерпывающую, а в методологическом отношении и вовсе несовершенную.
Дальнейшее развитие биологических познаний потребовало кардинального пересмотра клеточной теории в целом и, прежде всего, установления новых представлений о природе самой клетки. Эта ревизия основ учения о клетке привела в конце концов к опровержению руководящей идеи того времени — теории «клеточного государства».
Нередко при критике клеточной теории в том виде, в каком ее формулировал Вирхов, указывается, что закон «всякая клетка от клетки» является как бы исключающим эволюционное представление об историческом возникновении клеток. С нашей точки зрения, такое толкование является произвольным. Закон преемственности возникновения клетки констатирует широко распространенное явление в органическом мире. Это есть прежде всего констатация факта, и уже современники Вирхова правильно отмечали, что его достоверность такая же, как и достоверность закона «в с е живоеот живог о». Крупнейший гистолог того времени Франц Лейдиг (1857) 2I, например, писал: «Откуда и как возникла первая клетка, столь же трудно решить путем исследования, как и вопрос, откуда произошел человек: мы убеждаемся, что клетки, подобно тому как и люди, возникают только путем размножения, т. е. происходят только друг от друга. Самозарождение клетки доказать не удается».
Следует отметить, что цитированные строчки писались еще в 1857 г., т. е. до появления в 1859 г. «Происхождения видов» Чарльза Дарвина (1809—1882). Вопрос об эволюционном происхождении клетки Вирховым не ставился вовсе, поэтому в данной плоскости его критиковать нельзя. Вопрос во всей широте был впервые поставлен лишь в 1866 г. Э. Геккелем (1834—1919), с> чем ниже будет сказано подробнее.
21 Leydig F. 1857. Lehrbuch der Histologie des Menschen und der Tliiere, Frankfurt («Учебник гистологии человека и животных»).
ГЛАВА ЧЕТВЕРТАЯ
КЛЕТКА — ЭЛЕМЕНТАРНЫЙ ОРГАНИЗМ
Основные даты развития представлений о клетке как об организме
1855 Роберт Ремак экспериментально доказал наличие оболочек у бластомеров дробящегося яйца.
1861 Макс Шультце дает новое определение клетки: «Клетка — комочек протоплазмы, внутри которого лежит ядро».
1861 Эрнст Брюкке рассматривает клетку как элементарный организм.
1866 Эрнст Геккель изучает развитие клеток с исторической (эволюционной) точки зрения.
1866 Эрнст Геккель предложил первую теорию происхождения многоклеточных организмов путем дифференциации клеток.
1866 Эрнст Геккель создал концепцию возиикиовеиия клеток нз монер (безъядерных кусочков первичной протоплазмы).
1872 Эрнст Геккель предложил термин «синцитий» — со-клетки.
1874 Жан Кариуа вводит понятие — «Биология клетки», чем кладет начало цитологии как науки о форме, структуре, функции и эволюции клеток.
92
§ 23. Определение понятия клетки М. Шультце. Пересмотрен ряд положений и дав новое обоснование клеточной теории, Рудольф Вирхов поставил в центре внимания биологических коп цепций клетку с гораздо большей определенностью, чем это имело место у предшествующих ученых. Клетка в его представлении является ие только морфологической (анатомической), но и физиологической единицей, имеющей преемственное развитие. В соответствии с этим понятно, что центр тяжести микроскопических исследований оказался направленным на изучение природы и свойств клетки, а это вызывало, естественно, как пересмотр самого определения клетки, так и вообще потребовало большего уточнения представлений о природе последней. Вместе с тем состояние биологических знаний того времени повлекло необходимость рассмотрения проблемы в эволюционном аспекте. Все это явилось основным содержанием гистологической науки в 60— 80-х гг. прошлого века.
Новое определение клетки было дано в 1861 г. протистологом и неврогистологом Максом Шультце (1825—1874), который по существу стремился обосновать всеобщность принципа клеточной организации для всех органов животных. Не менее яркое определение клетки в особенно убедительной форме было дано венским физиологом и гистологом Эрнстом Брюкке (1819—1892) в том же 1861 г. Именно исследования этих двух ученых и определили все дальнейшее развитие науки того периода.
Работа Шультце называлась «О мышечных тельцах и о том, что следует называть клеткой» '. Работа эта носила теоретический характер и почти не содержала нового фактического материала. В этом исследовании автор исходил из рассмотрения конкретного факта многоядерности мышечного волокна. Поскольку само явление многоядерности расходилось с представлениями того времени о природе клетки, он стремился дать новую, более общую трактовку сущности этого понятия.
Напомним, что при доказательстве единства в клеточной организации растений и животных много было потрачено труда для обнаружения клеточных оболочек животных клеток. Считалось, что именно оболочка является наиболее характерной структурной особенностью клетки. Эта проблема занимала даже таких крупных исследователей, как Рейхерт (1811—1885) и Ремак. Послед-
1 Schultze М. 1861. Ueber Muskelkorperchen und das, was man eino Zelle zu nennen habe. Arch. f. Anat. u. Physiol. Jahrg. 1—27 («О мышечных тельцах и о том, что следует называть клеткой»).
93
пий, например, доказывал наличие оболочки на бластомерах дробящихся яиц, которые он для этой цели обрабатывал смесью спирта, древесного уксуса и медного купороса, после чего ему удавалось легко сдирать оболочки, и таким образом, доказывать их наличие.
Объект Шультце — волокно поперечно-полосатой мышцы — уже давно привлекал внимание гистологов, ибо в нем классическую клеточную структуру обнаружить, конечно, нельзя. Надо сказать, что и до сих пор продолжается спор о том, как следует называть волокно поперечно-полосатой мышцы. Одни полагают, что их следует считать симпластом, или синцитием, т. е. соклетием, а другие рассматривают его как своеобразную высокоспециализированную многоядерную клетку. Для того, чтобы дать клеточную трактовку мышечному волокну, Шультце пришлось решительно пересмотреть господствовавшее представление о клетке, как о пузыревидном образовании с обязательным наличием оболочки. Поэтому свое исследование он начинает с постановки вопроса о том, что же является в клетке наиболее существенным.
Чтобы ответить на такой вопрос, Шультце изучает эмбриональные клетки и констатирует, что для них особенно характерно наличие ядра — почти гомогенного, округлого, сравнительно плотного тела, расположенного в центре клетки. Ядро окружено так называемой клеточной субстанцией — тягучей, полужидкой протоплазмой, содержащей много зернышек белковой и жировой природы. Сама протоплазма состоит из прозрачной основной массы, а гетерогенный ее характер является результатом накопления в ней разнообразных включений. Однако самый наружный слой протоплазмы состоит сплошь из гомогенной субстанции. В результате всего изложенного Шультце пришел к выводу, что протоплазма представляет собой шаровидный комочек и эта ее форма сохраняется благодаря свойственной ей консистенции. Исходя из этих данных, он дает свое известное определение клетки, вошедшее во все старые учебники: «клетка является комочком протоплазмы, внутри которого лежит ядро. Клетка характеризуется полноценностью жизненных отправлений, субстратом которых является преимущественно протоплазма. Однако также и ядро играет значительную, хотя все же до сих пор неизвестную роль».
Таким образом, мы видим, что из данного определения клетки исчезло упоминание об оболочке, и именно на это обстоятельство Шультце особенно обращает внимание, заявляя, что клетка
94
Макс Шультце (1825—1874)
представляет собой комочек протоплазмы с ядром, а наличие оболочки не обязательно.
Но кроме того, что Шультце четко формулировал новое определение клетки, он изложил его в очень убедительной для того времени форме. Далее будет показано все его преимущество перед старым определением и представлением о структуре клетки. Таким образом, весьма существенным моментом в обосновании представлений Шультце о строении клетки является доказательство отсутствия оболочки, но в этом отношении он, образно говоря, впадает в другую крайность. Некоторые его утверждения (например, об отсутствии оболочки у яйцеклеток животных) просто ошибочны.
Весьма интересно отметить, что Шультце уделял много внимания слиянию клеток. Он полагал, например, что соединительнотканные клетки сливаются друг с другом своим наружным гомогенным слоем, а затем этот гомогенный слой постепенно увеличивается в объеме. Позднее в этом бесструктурном веществе образуются каким-то образом фибриллы, и таким путем возникает межуточное вещество соединительной ткани, которое представля-
95
ет собой, следовательно, продукт разрастания эктоплазмы клетки. Однако клетки при этом не уничтожаются, поскольку вокруг каждого ядра остается небольшое количество прилежащей к нему зернистой протоплазмы (эндоплазма). Интересно отметить, что современные взгляды на развитие межуточного вещества и волокон в нем очень близки к представлениям Шультце. Этому ныне даны убедительные доказательства, особенно советскими гистологами А. А. Заварзиным, А. В. Румянцевым и др.
Отрицая обязательность наличия клеточной оболочки, Шультце писал, что протоплазма клетки является достаточно густой и вязкой, чтобы обеспечить свою морфологическую самостоятельность. Далее он специально останавливается иа значении анастомозов (межклеточных протоплазматических перемычек). Он либо отрицал их, либо категорически оспаривал мнение, что наличие связи между клетками наносит ущерб самостоятельности в их жизнедеятельности. Этот взгляд также неоднократно подтверждался в настоящее время. Однако все уже давно забыли, что это утверждал еще Шультце.
Физиологическое оправдание анастомозов Шультце видит в необходимости поддержания быстрого и активного обмена веществ, поскольку клетки (особенно клетки соединительной ткани) часто разделены большим количеством непроницаемого или плохо проницаемого межуточного вещества. Следует признать, что Шультце предвосхитил проблему транспорта веществ внутри тканей и что до сих пор еще никто лучше его не выразил этой мысли. Экспериментальные работы последних десятилетий как нельзя лучше подтвердили, что даже наличие широких связей между клетками нисколько не свидетельствует о потере ими их самостоятельности. Метод тканевых культур показал, например, что в целом ряде случаев клетки легко входят в связь с другими клетками, но так же легко могут и отделяться. Показано, что перетекания протоплазмы из клетки в клетку не происходит, и циркуляция протоплазматических токов идет лишь в очень ограниченных участках, не выходя за пределы протоплазматического тела данной клетки.
Все это говорит против правомерности применения термина «синцитий»2 для клеток, связанных в сети, если, конечно, не
2 Сам термин «синцитий» введен Э. Геккелем (1872) для обозначения «тканевой массы», возникшей путем слияния клеток эктодермы личиночных стадий развития известковых губок (Haeckel Е. 1872. Die Kalkschwamme. Berlin) («Известковые губки»).
96
придавать этому термину иного смысла, чем ;>г<> <><>tn<-ii|........inn,
Возвращаясь к вопросу о клеточной организации mi.iiiii-mii.h > волокна, Шультце дает последнему следующее толкование. К а а. дое ядро с прилежащей к нему протоплазмой он рассматривает как отдельную клетку, что находится в соответствии с его определением клетки как комочка протоплазмы с ядром. Фибриллярную структуру, занимающую основную массу волокна, он истолковывает (как и в соединительной ткани) в форме своего рода межуточного вещества, возникшего в периферическом слое протоплазмы. Далее он указывает, что все, даже самые крупные мышечные волокна возникают из отдельных хорошо ограниченных одноядерных миобластов (эмбриональных мышечных клеток). Отсутствие резких границ (мембран) между ядрами с окружающей их саркоплазмой (протоплазмой) в данном случае не представляется Шультце существенным.
Итак, представления Шультце о клетке еще более подчеркнули индивидуальность клетки. Они находились в полном соответствии со взглядом, который развивал Брюкке о клетках, как об элементарных организмах. Если Шультце обосновал морфологическое единство клетки, то Брюкке подкрепил эту идею с физиологической стороны.
§ 24. Клетка — элементарный организм в представлении Э. Брюкке. Брюкке был по своей специальности физиологом. Его известная работа «Элементарные организмы» (1861)3, как и работа Шультце, не носит характера экспериментального исследования, а является прежде всего теоретической статьей, обосновавшей физиологическую самостоятельность клеток — элементарных организмов, из которых, по его представлениям, построено тело животных. Он начинает свои рассуждения ссылкой на главу из книги Шванна (1839) «Клеточная теория», отмечая при этом, что считает необходимым назвать клетки элементарными организмами, ибо они являются неразложимыми организованными частицами, подобно тому, как неразложимы атомы химических элементов.
Слабым местом всей клеточной теории в целом оставалось само представление о клетке, ибо конкретных данных о ее строе-
3 Briicke Е. 1861. Die Elementarorganismen. Sitz-ber. d. Akad. d. Wis-senschaften. Wien. Math-Naturwiss. Cl., Bd. 44, 2, 381 («Элементарные организмы»).
4 E. M. Вермель
97
Эрнст фон Брюкке (1819—1892)
нии было все еще очень мало, и полагали, что она имеет очень простую структуру. Брюкке выступает с утверждением, что поскольку клетке присуща организованность, она должна быть сложно построенной единицей, ибо именно своеобразие ее организации и отличает живое от неживого. В этом заключается большая заслуга Брюкке. Эту мысль он оттенил в самом заглавии работы «Элементарные организмы». Однако сам Брюкке вовсе не настаивал на замене термина «к лет-к а» термином «элементарный организ м», считая, что первый из них настолько укоренился, что вряд ли имеет смысл делать какую-либо замену.
Первым и основным выводом, который Брюкке делал из изложенного выше положения относительно организованности клеточной структуры, является утверждение, что поскольку жизненные отправления очень сложны и охватывают весь комплекс явлений, характеризующих жизнь, клетка не может быть просто построенной. Он говорил: «утверждают, что протоплазма пред
98
ставляет собой гомогенную массу». Но по его мнению, ,»п> дюл но означать, что «протоплазма лишь п ре дета и л и <• i ся или кажется нам гомогенно й». В вопросе о струи туре клетки Брюкке не считал возможным удовлетворяться пред ставлением об элементарном молекулярном строении материи. Он подчеркивал, что простой раствор белка никогда не может иметь тех свойств, которые мы называем жизненными процессами, и поэтому мы вынуждены приписывать клетке сверхмолекулярную структуру, структуру более сложного порядка, которую он называл организацией живых существ. Таким образом, сложные молекулы органических соединений являются только материалом, который слагается не как однообразная масса, а имеет особую структуру, в результате чего создается живой организм. В живом существе, по Брюкке, наблюдается целый ряд сложнейших процессов. Особое внимание он уделял процессам раздражимости и движения протоплазмы.
Поскольку у сложных организмов различные функции выполняются различными органами, то и в отношении клетки Брюкке также полагал, что мы вряд ли имеем право предполагать принципиально иное положение вещей. Другими словами, он считал, что различные функции выполняются различными структурами клетки. Он подчеркивал также, что нельзя ожидать, чтобы органы и системы органов, которые мы находим в человеческом организме, могут или должны повториться в клетках в том же самом виде. Он считал, что с уменьшением размеров природа изменяет средства, с помощью которых она заставляет служить организму все силы неорганического мира.
Рассматривая клетку как маленькое живое существо, Брюкке указывал, что к ней надо подходить так же, как мы подходим к изучению мельчайших животных. В частности, он считал, что одноклеточные растения указывают на прямую связь между свободно живущим организмом и элементарными живыми единицами (т. е. клетками), которые могут поддерживать свое существование только как интегральные части большого единого целого.
Изложенные мысли мы находим уже во вводной части работы Брюкке. Важно подчеркнуть, что в ней, может быть, впервые с особой ясностью говорится о качественном различии между живым и неживым. Может быть, именно Брюкке впервые столь ясно проводил мысль об аналогии между клеткой и свободно живущим организмом. Но основной идеей, повторяем, у него было представление о сложности клетки и ее особой организации. Надо еще отметить, что в этом отношении представления Брюкке
99
4*
находятся на значительно большей высоте, чем взгляды более поздних исследователей, пытавшихся с помощью различных неорганических моделей имитировать сущность поотоплазмы, как например, известный зоолог механицист Отто Бючли, Румблер и многие другие, на работах которых мы остановимся несколько подробнее ниже. Он ссылается на описания гомогенности клеточных компонентов (протоплазмы, ядра), но в его представлении отсутствие видимых структур не увязывается с представлением о сложности строения клеток. Поэтому он и считал необходимым их отыскивать и специально подчеркивал, что неудача в этом отношении, возможно, объясняется недостаточной разрешающей способностью микроскопа.
В заключение остановимся на характеристике протоплазмы, данной в работе Брюкке. Прежде всего он ставит вопрос об ее агрегатном состоянии и отмечает, что его нельзя назвать ни жидким, ни твердым. Он пишет: «Вопрос о том, является ли живое тело твердым или жидким, по существу столь же абсурден, как если бы спросить, твердое или жидкое (в том смысле, как это употребляется в физике) тело медузы или улитки». А далее читаем еще: «Если мы будем протоплазму клетки характеризовать просто как слизистую или студенистую массу, то это будет нисколько не лучше, чем если кто-нибудь, кто ничего не знает о строении медузы, будет ее характеризовать как оживленный студень».
Эти строки, написанные Брюкке о протоплазме более 100 лет назад, сохраняют свою актуальность и правильность и поныне. В самом деле, что пишется о составе протоплазмы в большинстве учебников? Перечисляются поостейшие химические вещества, которые в ней открываются, затем добавляется несколько слов о коллоидном ее состоянии, в лучшем случае пишется о гипотетических мицеллах. Вопрос о качественном своеобразии живой субстанции в отличие от вещества «мертвого» по существу обычно не ставится. Сравнение студня с медузой в данном контексте нельзя не признать исключительно удачным. Понятие «студень» не может ничего дать для понимания медузы как организма, для которого прежде всего характерно определенное морфологическое строение, т. е. определенная структура. Предвидение Брюкке, требовавшее отыскания микроскопически видимой структуры протоплазмы, явилось направляющей идеей исследований последующих трех-четырех десятилетий. Можно сказать, что только в самое последнее время электронно-микооскопическая техника открывает новое в строении протоплазмы. Оно оказалось
100
Эрнст Геккель (1834—1919)
проще, чем предполагал Брюкке, но значительно сложнее, чем это считали до середины XX в. Об этом ниже (в гл. V) будет сказано подробнее.
§ 25. Значение учения о клетке для эволюционных построений в работах Э. Геккеля. Представление о клетке, как об элементарном живом организме, получило в 60—70-х годах всеобщее признание, поэтому вполне естественно было ожидать, что эта КОН
ГО/
цепция должна была быть как-то увязана с бурно развивавшейся эволюционной морфологией.
Эволюционная идея неудержимо проникала во все области биологической науки. В микроморфологии эта точка зрения впервые нашла яркое выражение в трудах виднейшего морфолога дарвиниста Эрнста Геккеля. В своей основной и, может быть, лучшей работе «Общая морфология организмов»4 он уделил очень много места проблеме клеточной теории с точки зрения интересовавшей его схемы эволюционного развития организмов («филогенетическое древо»). В филогенетических построениях Геккеля клетка играет весьма значительную роль, являясь основой в определении индивидуальности живых существ, а также как исходная форма развития жизни на Земле. Для Геккеля уже было ясно, что клетка сама по себе является сложной структурой. Так, он писал, что «элементарный организм по существу далеко не элементарен в своем строении». Поэтому Геккель искал также переходные ступени между сложным элементарным организмом и неорганической материей. Предложенная им схема развития организмов представляет теперь только исторический интерес, как первая попытка использовать данные о микроскопическом строении организмов для филогенетических построений.
Геккель разделял мир органических существ на три царства. Кроме животных и растений, он ввел третье царство — царство протистов, которое он в свою очередь разделил на две группы: на ц и т о д ы (или монеры) и на клетки. Цитоды им характеризовались как «участки протоплазмы без ядра». Он считал, что именно существа подобного строения и являются переходными ступенями между живым и неживым. Клетки являются в эволюционном отношении уже вышестоящими организмами, поскольку они представляют собой «протоплазматические комочки с ядром».
Интересно здесь отметить, что цитоды и клетки были объединены им под именем пластид (в дословном переводе это греческое слово означает «образователи»). Геккель их называет также индивидуумами первого порядка. По его представлениям, пластиды являются теми простейшими единицами, из которых составляются индивидуумы высшего порядка. Агрегаты пластид
4 Heckel Е. 1866. Generelle Morphologic Organismen. Bd. I. Allge-meine Anatomie der Organismen. Verl. von G. Reimer («Общая морфология организмов, т. 1. Общие проблемы анатомии организмов»).
102
создают ткани, органы и т. д. Вместе с тем Геккель псрный ,ы \ теорию возникновения многоклеточности. Он исходил из коло ниальных форм одноклеточных организмов, которые, по его пред ставлениям, при дальнейшем эволюционном развитии диффереп цировались таким образом, что составляющие их клетки становились качественно неравнозначными элементами. Так, колониальные организмы (группа сходных клеток) превращались в многоклеточные существа (Metazoa), состоящие из клеток, отличающихся различной структурой и функцией.
Геккель подробно останавливается также на вопросе о возникновении самой клетки. Его теории, конечно, чисто умозрительны, но заслуга Геккеля заключается прежде всего в том, что, использовав данные учения о клетке, он пытался показать в достаточно вероятной форме, как именно можно себе представить эволюционное происхождение живых существ. Он полагал, что первым полноценным организмом должна быть структура, аналогичная клетке, и пытался заполнить пробел между неживым миром и клеткой. Он выделил группу цитод, или монер (безъядерных комочков). Геккель по существу имел на это право, так как микроскопическая техника того времени не позволяла выявить клеточную природу этих «сверхпростых» организмов. В настоящее время, когда группа цитод давно уже развенчана, представление о родоначальных живых организмах сводится к клеткам. Конкретно никто не знает, чем заполнить пропасть между неорганизованной материей и клеткой, образовавшуюся после того, как существование монер было опровергнуто. Являются ли вирусы такой фазой развития, сказать очень трудно. Возможно даже, что вирусы вообще никакого отношения к возникновению жизни не имеют, а являются «веществом», а не «существом». В гипотезах, конечно, недостатка нет, но мы на них здесь останавливаться не можем5. Сам Геккель представлял себе процесс таким образом, что бесформенные комочки белковой субстанции — монеры — возникли в морях в очень отдаленные эпохи путем чисто химических и физических взаимодействий движущихся молекул или подобно тому, как возникает кристалл в маточном растворе. Он подчеркивал, что чисто химические и физические причины должны были обусловливать возникновение сложных углеродистых соединений. Эти вещества давали уже не растворы, а взвеси из маленьких комочков, которые оказывались
5 См., например, Опарин А. И. 1966 «Возникновение и начальное развитие жизни». М., изд-во «Медицина».
103
способными к накоплению разных субстанций, что допускало рост путем ассимиляции сходных соединений. Но такой рост оказывался возможным только до известных пределов, ибо аттракционный (притягивающий) центр, создававший такой индивидуум, расщеплялся на два или несколько центров. Таким образом возник процесс размножения делением. Лишь позднее в результате дифференцировки этих бесформенных кусочков и выделения самостоятельного ядра возникли, согласно представлениям Геккеля, истинные клетки.
Любопытно отметить, что Геккель продолжал приводить представления Шлейдена — Шванна о цитогенезе как конкретное обоснование его теории образования клеток. Он продолжал также приводить сравнения Шванна между клеткообразованием и процессом кристаллизации, но, повторяем, в его построении существенна не столько конкретная расшифровка данной проблемы, сколько важна ее постановка и попытка материалистического ее решения. От многих элементов этой теории, как, например, сравнения клеткообразования с кристаллизацией, уже давно отказались, но, с другой стороны, теория возникновения многокле-точности, так, как она понимается большинством исследователей и по настоящее время, принципиально ничем не отличается от теории, сформулированной Геккелем. Мы и до сих пор не без основания полагаем, что в филогенезе промежуточными формами между Protozoa и Metazoa могли быть колониальные организмы.
§ 26. Биология клетки как самостоятельная проблема. Точка зрения, приравнивавшая клетку к целому сложному организму, изменила общий подход и к ее изучению. Гистология становится наукой не только о своеобразной клеточной структуре органов и ткани живых существ. Ее содержание с этого времени определяется прежде всего как изучение формы и функций элементарных организмов (клеток), а также из взаимоотношений в тканях и органах. Именно такова была наиболее распространенная точка зрения, установившаяся в последнюю четверть XIX в. Она особенно ясно была сформулирована французским гистологом Жаном Батистом Карнуа (1836—1899), который ввел в соответствии со сказанным в употребление понятие биологии клетки. Его книга как раз и носит заглавие «Биология клетки» 6. В ней отведено много места обоснованию задач микроскопического ис-
6Carnoy J. В. 1884. «La biologie cellulaire» («Биология клетки»).
Рис. 23. Типичная клетка эпителия земляного червя из книги Карну а «Биология клетки» тс — оболочка клетки;
рс — протоплазма, в ней. имеются лучистый ретикулум и эн-хилемма, заключенная в ее петлях;
рп—плазма ядра, в нем также виден ретикулум;
Ьп — непрерывная ядерная кишка, собранная в центре ядра
следования. Он отмечал, что слово «биология», введенное в 1802 г. Готфридом Тревиранусом (1776—1837) И одновременно Жаном Батистом Ламарком (1774—1829)8, означает «изучение жи зн и», а поскольку клетка является живым элементарным организмом, то, следовательно, мы и должны заниматься ее биологическим изучением. В сущности зарождается цитология, как наука о клетке. Собрав большой фактический материал о строении клетки, который к тому времени уже накопился в научной литературе, Карнуа определяет три направления в изучении биологии клетки. Он говорит об общей биологии, имея в виду установление общих закономерностей, которые могут быть выявлены при изучении клеток и тканей; далее он говорит о сравнительной биологии, предполагающей сравнительное изучение клеток в пределах какой-либо группы организмов, и, наконец, он говорит о специальной биологии клеток, тканей и органов, относящейся к какому-либо частному объекту. Эти рассуждения о подходе к исследованию клетки являются общими для любой отрасли биологического исследования. Интересно отметить, что эти идеи развивались в теоретическом обзоре, посвященном исследо-
’Treviranus G. R. 1802—1816. «Biologie oder Philosophic der lebcn-den Nalur» («Биолсгия или философия живой природы. 6 томов»),
8 Lamark J. В. 1802. Hydrogeologie ...Paris.
105
ванию клетки. Таким образом, наука о клетке поднялась на такую же принципиальную высоту, как и другие крупнейшие разделы естествознания. Выходом в свет книги Карнуа подводился, собственно говоря, первый итог предшествовавшим разрозненным данным, в результате чего цитология оформилась как самостоятельная биологическая дисциплина со всеми тремя основными признаками, характеризующими любую науку, а именно: объектом, содержанием и методом исследования.
Высказывания Карнуа не отличаются большой оригинальностью. Мы на них останавливаемся потому, что они отражают взгляды большинства передовых ученых того времени и, следовательно, хорошо характеризуют тот период науки о клетке. Карнуа стоит на той точке зрения, что клетку следует называть организмом, или элементарной индивидуальной единицей сложных существ. Он подробно развивает идею об особой архитектонике живой массы протоплазмы (рис. 23). Называя клетку элементарным организмом, Карнуа поясняет, что под этим следует понимать биологическую автономность клетки прежде всего в смысле ее способности самостоятельно проявлять свою жизнедеятельность. Таким образом, клетка характеризуется истинной индивидуальностью, причем это справедливо как в отношении одноклеточных существ, так и в отношении клеток высших многоклеточных организмов, несмотря на то, что необходимые условия, при которых может проявляться их жизнедеятельность, большей частью не могут быть созданы без участия других клеток или соседних тканей, или даже целого организма. Правда, Карнуа указывает и приводит с большой подробностью ряд случаев, когда индивидуальность клетки может подавляться и даже исчезать. Конкретно приводятся примеры образования плазмодиев и многоядерных структур (миксомице-ты, млечные сосуды и спорангии многих растений), представляющих собой многоядерные протоплазматические массы.
Итак, руководящей идеей того периода было представление о клетке как о престейшем индивидууме. У Карнуа этот взгляд нашел свое завершение, поскольку его мысли в конечном итоге сводятся к тому, что клетка должна изучаться аналогично изучению многоклеточных организмов. Именно это и лежит в основе понятия «биология клетки», понятия, впервые четко сформулированного Карнуа,
ГЛАВА ПЯТАЯ
СТРОЕНИЕ ПРОТОПЛАЗМЫ
И КЛЕТОЧНАЯ ТЕОРИЯ
Основные даты по истории изучения протоплазмы
1830 Я, Пуркиня впервые употребил термин «протоплазма».
1835 Ф- Дюжардеи дал первое тщательное описание содержимого клетки простейших, называя его словом «саркода».
1846 Гуго фои Моль ввел в широкое употребление термин «протоплазма» для обозначения тела растительной клетки.
1850 Ф- Кон доказал сходство «саркоды» и «протоплазмы».
1852 0- Ремак применил термин «протоплазма» для обозначе-
ния тела клеток животных.
1861 Э. Брюкке развил теоретическое представление о необходимости признать структурную сложность протоплазмы.
1867 В. Гофмейстер дал подробное объективное описание строения и свойств протоплазмы живой растительной клетки.
1872 К. Гейтцмаи описал протоплазму после фиксации клетки и развил теорию сетчатого ее строения.
1875 К. фои Купфер впервые применил метод прижизненной окраски клетки.
1880 И. фон Г анштейн ввел термин «протопласт» для обозначения совокупности живой массы клетки.
1890 Р- Альтман развил теорию гранулярного строения протоплазмы, элементами которой являются «биобласты» — самостоятельные микроорганизмы.
1891 О. Б ю ч л и развил теорию пеиисто-ячеистого строения протоплазмы.
1890 В. Г а р д и доказал явление «артефактов» — структур, возникающих в протопласте под влиянием внешних воздействий.
1907 М. Гейдеигайн развил «теорию промеров».
1921 И. Шпек обосновал современную теорию субмикроииой структуры протоплазмы.
1955 Ф. Шестрэнд, Паллад и др. описали субмикронное строение протопласта, открыв систему мембран, гранул (рибосом) и других структур.
$ 27. Первые описания строения протоплазмы. Идея брюкке относительно сложной организации клетки отразилась, может быть, сильнее всего на учении о строении протоплазмы и, соответственно, па учении о строении ядра. Исходя из теоретического представления о том, что сложная функция предполагает и сложное строение, был сделан по существу априорный вывод, что морфология клеточной субстанции должна находиться в пределах разрешающей способности микроскопа, т. е. может и должна быть обнаружена морфологическими методами. Именно поэтому с такой легкостью описывались самые разнообразные микроструктуры и создавались столь многочисленные и разнообразные гипотезы строения протоплазмы. Предубежденность, вытекавшая из догматического следования за руководящей идеей, создавала некритическое отношение к тем фактам, которые должны были ее подтвердить.
Мы уже указывали, что первые исследования, описывавшие протоплазму, говорили о ее гомогенности. Отметим кстати, что самый термин «протоплазма» был введен в 1830 г. Пуркиня. Он называл этим термином неструктированную, как ему казалось, ткань эмбрионов животных. Но в его представлении протоплазма не обязательно связывалась с клеточной структурой. Во всеобщее употребление этот термин был введен ботаником Гуго фон Молем (1846), изучавшим ротационные (вращательные) движения в клетке и обозначавшим протоплазмой тело клетки. Несколько позднее в том же смысле — для обозначения тела клетки — этот термин стал применяться и зоологами (Ремак, 1852—1855).
Впервые подробное описание протоплазмы было дано Феликсом Дюжарденом (1801 —1860), изучавшим строение различных простейших организмов *. Для обозначения субстанции клетки он пользовался другим термином — «с а р к о д а». Его описания отличаются большой объективностью н находились на очень высоком уровне для того времени. Так, он охарактеризовал протоплазму как клейкую прозрачную гомогенную массу, преломляющую свет немного сильнее воды и слабее масел. Он описывал ее эластичность, сократимость, способность образовывать внутри себя вакуоли. Дюжарден отметил, что сар-кода нерастворима в воде и только лишь набухает в ней. Он от-
1 Dujardin F. 1835. Sur les pretendus eslomacs des animalcules infusoi-ries et sur une substance appele sarcode. Ann. de Soc. Natur, t. II («О так называемом желудке инфузорий и субстанции, называемой саркодой»),
108
МбтйЛ йалйчйе й ней ЗйачйтеЛьйоГо количества белка и Легкую ее коагулируемость азотной кислотой и спиртом. Дюжарден специально подчеркнул, что саркода не имеет никаких видимых структур или более тонкого сетчатого строения, ибо в ней наблюдаются сложные процессы, вроде передвижения гранул, ее растягивания, стягивания и т. д. Термин Дюжардена «саркода» для клеточного содержимого простейших употреблялся довольно долго, пока, наконец, в 1850 г. Фердинанд Кон (1828—1898) с большой убедительностью доказал, что понятие саркоды простейших совпадает с понятием протоплазмы растительных клеток. Кон, как и другие ботаники, стоял на точке зрения гомогенности протоплазмы.
Одно из лучших описаний протоплазмы принадлежит ботанику Вильгельму Гофмейстеру (1824—1877), опубликовавшему в 1867 г. сводку о строении растительной клетки2. Надо отметить, что наблюдения ботаников отличались большей точностью, чем наблюдения зоологов. Это объясняется тем, что прижизненное изучение растительных тканей значительно легче гистологического исследования тканей животных. И это, конечно, давало возможность ботаникам получать более точные данные, чем и отличались работы Гофмейстера. Этот исследователь дал наиболее правильное описание клеточной оболочки. Он указывал, что отделить плотный периферический слой протоплазмы, который только и следует считать истинной оболочкой, невозможно, ибо она представляет собой уплотнившийся слой эктоплазмы, постоянно образующийся на границе с внешней средой. Гофмейстер выступил против представления о возможности существования голой клетки, т. е. клетки, у которой нет оболочки. Он отмечал, что всюду, где находится протоплазма, она постоянно окружается эластической мембраной — оболочкой. Вместе с тем Гофмейстер доказывал, что целлюлозную оболочку ни в какой мере нельзя считать истинной протоплазматической оболочкой. Наконец, Гофмейстер дал многочисленные и тщательные описания агрегатного состояния и свойств протоплазмы. В частности, он указал на значение накопления воды в протоплазме и показал, что сама протоплазма имеет предел ее имбибирования (пропитывания, накопления) водой, а если этот предел переходит какую-то границу, то вода накапливается в протоплазме в виде больших или мень-
2 Hofmeister W. 1867. Die Lehre von der Pflanzenzelle. Bd. I. Handb. d. physiol. Bot. («Учение о растительной клетке»).
109
В. Гофмейстер (1824—1877)
ших капель, которые могут быть из клетки удалены хотя бы путем помещения ее в концентрированный раствор соли (опыт плазмолиза).
§ 28. Теории строения протоплазмы. Идея Брюкке о том, что элементарный организм (клетка) не может иметь простого строения, поскольку он является организмом, явилась ведущей для большинства цитологических исследований конца XIX в.
Брюкке (1861) считал, что гомогенная структура протоплазмы (а гомогенность рассматривалась им как признак простой структуры) не может обеспечить всей сложности жизненных отправлений. Но как было выяснено позднее, оптическая гомогенность вовсе не предполагает структурной простоты. Целым рядом новейших исследований показано, что протоплазма имеет исключительно сложное, не дифференцируемое даже с помощью лучших объективов микроскопа (мицеллярное и сверхмицеллярное) строение Другими словами, ее структура находится за пре
110
делами видимости светового микроскопа. Но эта точка зр<чi и и, подкрепленная многими фактическими данными, явилась резуль татом (исследований только последних десятилетий. Ученым же конца XIX в. идея Брюкке представлялась чрезвычайно убедительной. Считали, что структура протоплазмы должна быть видимой, поэтому и было описано очень большое количество разнообразных структур.
Весьма характерным является как раз то обстоятельство, что единства мнений в этом вопросе не удалось достигнуть. Каждый автор представлял себе строение протоплазмы по-своему и, прежде всего, как мы теперь видим, они определялись методикой, которой он пользовался.
Уже сам Брюкке в 1863 г. описывал ячеистое строение протоплазмы животных клеток, в особенности так называемых слюнных телец (лимфоцитов, обнаруживаемых в слюне).
Не упоминая более мелких работ из специальной литературы того времени, мы прежде всего остановимся несколько подробнее на исследованиях Юлиуса Гейтцмана (1847—1922) 3.
Гейтцман в своих построениях исходил из наблюдений над амебами и белыми кровяными тельцами человека. Он описывал в них сетчатое строение протоплазмы и высказал точку зрения, согласно которой протоплазма в основном состоит из двух субстанций: из субстанции плотной (в форме сеточки и зернышек) и из субстанции жидкой. Сеточке Гейтцман приписывал свойства живой сократимой материи, а содержащуюся в ячейках жидкость считал несократимой, неживой субстанцией. По Гейтцману, механизм сокращения протоплазмы заключается в укрупнении зернышек (мест пересечений нитей сеточки) и соответственном укорачивании находящихся между ними ниточек. При этом зернышки должны сближаться друг с другом. При растяжении протоплазмы (ее расслаблении) происходит как раз обратное: зернышки могут уменьшаться вплоть до своего полного исчезновения, одновременно при этом происходит растягивание нитей. Таким образом, была уже создана теория, объяснявшая механизм подвижности протоплазмы (рис. 24).
3Heitzmann С. 1873. Untersuchungen uber das Protoplasma. I. Bau des Proloplasmas. II. Das Verhaltniss zwischen Protoplasma und Grundsubs-tanz im Thierkorper. Sitz-ber. d. Wiener Acad. d. Wissenschaften Math.-natur. Cl. Bd. 67 («Исследования иад протоплазмой. I. Строение протоплазмы. II. О соотношении между протоплазмой и основной субстанцией в животном организме»).
111
Рис. 24. Схема строения протоплазмы, по Гейтиманд (1873 г.) А — протоплазма в состоянии покоя;
Б — в состоянии нормального сокращения;
В—«тетанус» протоплазмы;
Г—состояние расслабления
Весьма важно остановиться иа методике, которой пользовался этот исследователь. Знакомство с ней объясняет нам полностью, каким образом он пришел к изложенным выше взглядам. Дело в том, что Гейтцман изучал кровь, подогревая ее мазок на предметном стекле. Он отмечал, что структуру протоплазмы увидеть очень трудно и что она отчетливо выявляется только после нагревания объекта. Впоследствии было выяснено, что описанная Гейтцманом структура является результатом коагуляции белка под влиянием высокой температуры; изменяя свое коллоидное состояние и образуя хлопья, белок высаждается, создавая сетчатую структуру.
Гейтцман не остановился на приведенных выше теоретических представлениях; он пошел дальше и развил ряд обобщений, трактующих о построении организма в целом. Он различал, как уже указывалось, в клетке живую субстанцию (сетчато-гранулярную) и субстанцию неживую (жидкую). Отсюда Гейтцман заключил, что структура, которую называют клеткой, оказывается только частично построенной из живой субстанции, а затем следовал вывод, что клетка является не элементарным, а сложным
112
Рис. 25. Рисунок Гейтцмана, изображающий соединительную ткань (1875 г.)
Рис. 26. Принципиальная схема строения пласта клеток по Гейтцману (1875 г.)
организмом. «Элементарными живыми частицами являются, следовательно, описанные им зернышки на пересечении нитей.
Естественно возникли и такие вопросы: 1) является ли клетка последней организованной неделимой биологической единицей; 2) какова структурная природа тканей и органов?
На первый вопрос (изучая обособленные клетки — амебы, белые кровяные клетки) Гейтцман дает в своем исследовании отрицательный ответ. Второй вопрос он разрешает, изучая уже ткань (орган), которая фиксировалась им хромовой кислотой и обрабатывалась на срезах азотнокислым серебром или хлориым золотом. При этом он обнаруживал ячеистость строения тканевых клеток и связь с помощью нитей не только между зернышками одной клетки, но и непосредственную связь с помощью перемычек между всеми соседними клетками. А отсюда уже делался вывод, что клетки не существуют как изолированные индивидуумы и что их живая субстанция (плотные ниточки) непосредственно переходит из одной клетки в другую (рис. 25).
Общий вывод из всех этих наблюдений Гейтцман делает такой: животный организм представляет собой единую протоплазматическую массу. Подобно тому как амеба является протоплазматическим комочком, заключающим живую субстанцию в форме густой ячеистой сети, тело любого высокоорганизованного животного представляет собой единую массу, пронизанную живой ячеистой структурой с узловыми точками живой материи в форме протоплазматических комочков. На рис. 26 приводится схема ткани, данная Гейтцманом. На ней нетрудно увидеть полную связь всех ниточек ячеистой протоплазматической сети.
Схема клетки и схема тканевой структуры, предложенная Гейтцманом, в значительной степени, конечно, надуманна и исходит из предвзятой идеи. Однако известное основание для построения данной теории этот исследователь имел. Пользуясь методикой изучения мертвого, фиксированного препарата, он видел артефактную структуру. С другой стороны, ои обнаруживал настоящие анастомозы между соединительноткаными клетками, хотя это второе обстоятельство играло в его работах значительно меньшую роль. Следовательно, основная ошибка Гейтцмана заключается в том, что ему оставалась неизвестной суть явления артефактов — структур, возникающих в результате фиксации (гибели) клетки веществами, осаждающими белки (спирт, сулема и многие другие). Он слишком некритически относился к препарату, вернее — к применяемому методу исследования.
114
Карл фон Купфер (1829—1903)
Исследования Гейтцмана были далеко не одиноки. Почти одновременно с ним описание структуры протоплазмы было дано Карлом Купфером (1847—1922) 4. Этот автор пользовался очень оригинальным методом, по-видимому, впервые им и примененным. Купфер прижизненно вводил в кровь кроликам и лягушкам растворы разных красителей и затем изучал отложения их в различных клетках, в печени и почках в первую очередь. Идея Купфера, которую он развивает, такова: тело клетки состоит из гомогенной субстанции, пронизанной тончайшими канальцами, соединяющими кровеносные сосуды печени с желчными ходами, а в почке — с мочевыми канальцами. Инъецируя в кровь краситель, он стремился выявить с большей отчетливостью клеточные канальцы.
Конкретные наблюдения Купфера сводятся к описанию зернышек или окрашенных вакуолек в теле клетки, которые будто бы
* Kupffer С. 1875. Ueber Differenzierung des Protoplasma an den Zel-len thierischer Gewcbe. Schr. d. Naturwiss. Veieins f. Schleswig-Holstein. Bd 1. H. 3 («О дифференцировке протоплазмы клеток животных тканей»)
115
йс&гДа связаны между собой тончайшими канальцами. Он также различает две субстанции тела клетки: прозрачную гиалиновую субстанцию (он назвал ее параплазмой) и мелкозернистую нитчатую — протоплазму.
Но дело в том, что введенные в организм красители всегда откладываются в клетках только в виде гранул- и, таким образом, никакой нитчатой протоплазмы этот метод выявить не может. Тем не менее, в то время наблюдения Купфера казались очень убедительными в том смысле, что данный метод будто бы позволял четко разграничивать две различные субстанции тела клетки.
Нужно добавить, что и веществу ядра как Гейтцманом, так и Купфером в общем приписывалась аналогичная структура.
Целую серию работ по этому же вопросу опубликовал ботаник Фромман5 (1798—1880). Принципиально он придерживался аналогичной точки зрения и говорил о разделении протоплазмы на твердую и жидкую составные части, давая им, однако, иную конкретную трактовку. На рис. 27 приводится несколько фотографий с его рисунков, из которых ясно, что протоплазма изображается в виде грубоячеистой массы.
Принципиально того же взгляда придерживался и такой крупный цитолог, как Вальтер Флемминг (1843—1905), применивший значительно более совершенный метод исследования. Флемминг дал свою трактовку строения протоплазмы, известную под названием филярно й (нитчатой) теории. В свое время она пользовалась большим успехом и постоянно излагалась в учебниках вплоть до 20-х годов нашего века. Сущность теории Флемминга сводится к тому, что тело клетки состоит из гомогенной массы и большего или меньшего количества нитей (fila — нить). Мы не останавливаемся подробно на конкретных представлениях Флемминга, ибо они не представляют большого интереса в своей фактической части. Значительно важнее то, что Флемминг в гораздо более ясной и отчетливой форме, чем кто-либо другой, использовал новые данные о тонкой структуре протоплазмы для пересмотра общетеоретических представлений о клетке. Флемминг написал большую книгу, посвященную строению клетки6. В одной из первых глав он останавливается на пере-
Е Frommann С. 1880. Beobachtungen fiber Struktur und Bewegungserschei-nungen des Protoplasmas der Pflanzenzellen («Наблюдения над структурой и подвижностью протоплазмы растительных клеток»),
s Flemming W. 1882. Zellsubstanz, Kern und Zellteilung. Leipzig («Клеточная субстанция, ядро и клеточное деление»).
116
смоТре клеточной теории и считает основными следующие положения.
1. Клеткой называется комочек живой субстанции, имеющий определенную пространственную ограниченность, несмотря на отсутствие особой мембраны — оболочки.
В этом пункте мы ясно видим влияние теоретических высказываний Шультце.
2. Для клетки характерно наличие ядра — определенного отграниченного тела особой химической природы (нуклеиновой).
3. Клетка представляет собой образование, характеризующееся обменом веществ.
4. Клетки размножаются делением.
Излагая это последнее положение, Флемминг отмечает, что его следует считать несколько гипотетичным, ибо оно противоречит теории первичного возникновения клеток. Правда, далее он замечает, что до сих пор ни разу никем не был доказан процесс свободного образования клеток, так же как и процесс свободного ядрообразования.
5. Новым понятием в определении клетки, которое вводится Флеммингом, является представление о двойственности субстанции тела клетки, состоящего из твердого и жидкого вещества. Флемминг употребляет термины Купфера: протоплазма и параплазма. Однако он считает, что его морфологические наблюде-
Рис. 27. Рисунок Фроманна, изображающий схему строения протоплазмы растительных клеток (1880 г.)
117
йия, открывающие в клетке йаличйе отдельных ййтбчёк, дают основание пользоваться другой терминологией. Он говорит о ф и-лярной массе, обозначая этим всю массу нитей, и затем об и и т ерфилярно й, или межуточной, субстанции. Он употреблял также термины «мигом» и «интермитом», беря для словообразования не латинский, а греческий корень. Дело, конечно, не в том, какие термины вводил Флемминг; важно понять, что именно он наблюдал и насколько близки к действительности его описания. В настоящее время наше познание природы клетки шагнуло значительно вперед, и мы можем дать довольно точную оценку наблюдениям Флемминга. Если посмотреть на его рисунки с живых хрящевых и печеночных клеток (прижизненные наблюдения), то на них нетрудно видеть нитчатые образования (рис. 28). Они являются не чем иным, как хондриосомами (митохондриями), изученными и описанными подробно лишь 15—20 лет спустя. Однако эти структуры непосредственного отношения к строению самой протоплазмы (точнее — цитоплазмы), как это, видимо, считал и сам Флемминг, не имеют. На рис. 29 мы видим эти нити, нарисованные по фиксированному в хромовой кислоте препарату, поэтому они сильно изменены, а многие из них являются просто артефактами. Правда, в ядре часть нитей, как считают многие современные исследователи, можно отнести за счет остатков хромосомных скелетов, особенно типично выявляющихся в незрелых яйцах амфибий (так называемые «ламповые щетки»).
Принципиально сходные взгляды на строение тела клетки были в то время широко распространены и в какой-то степени разделялись большинством цитологов. Однако в конкретных описаниях структуры протоплазмы никакого единства мнений не было. Именно этот разнобой в конкретных наблюдениях и указывал, что описываются не реальные структуры, а структуры, возникающие вторично в результате воздействия на клетку различными агентами (так называемые артефакты). Поскольку единство методики в работах ряда авторов не сохранялось, естественно, что наблюдения различных исследований не совпадали.
Вопрос о структуре протоплазмы продолжал занимать умы исследователей еще и в 90-х годах, ибо именно в разрешении проблемы организации протоплазмы надеялись раскрыть сущность жизни. Эти чаяния хорошо выразил Отто Бючли. В своей речи на съезде зоологов в 1891 г. он говорил, что «подобно тому, как все дороги ведут в Рим, все биологические вопросы сводятся 118
Рис. 28, Рисунок хрящевой клетки, сделанный В. Флеммингом по живому препарату (*882 г.)
Рис. 29. Рисунок молодой яйцеклетки аксолотля, сделанный В. Флеммингом по фиксированному и окрашенному препарату (1882 г.)
в конце концов к этой полной тайн субстанции, которая составляет тело клетки и обозначается нами как протоплазма» 7.
Чем обширнее и обоснованнее будут познаны особенности и деятельность этой субстанции, тем глубже и яснее будут, как считалось, представляться нам все процессы, происходящие в сложных организмах. Говоря о представлениях гомогенного строения протоплазмы, Бючли категорически отрицал теорию гомогенного строения, считая ее отвергнутой. Однако Бючли полагал, что конкретное разрешение вопроса о структуре тела клетки остается крайне неудовлетворительным. Он подчеркивал, что организация клетки не представляет собой ничего мистического и непознаваемого и что в ней надо искать различные конструктивные и различно функционирующие образования, проводя, таким образом, аналогию с построением высших организмов. Бючли, как и другие исследователи, считал протоплазму резко гетерогенной структурой, но приписывал ей (в противоположность другим) пенисто-ячеистуюструктуру. Он исходил из своих наблюдений над протоплазмой простейших. Количество исследований очень значительно, и их число все больше и больше возрастало, поскольку Бючли создал большую школу учеников, работавших в этом же направлении. Его учениками являются такие крупные исследователи, как протозоологи В. Т. Шевяков, Шуберг и др.
Наблюдения Бючли и его школы принципиально не отличались от всего того, что было сделано ранее. Опровергая сетчатое и нитчатое представление о структуре протоплазмы, он заменил его пенисто-ячеистым. Так возникла еще одна теория строения протоплазмы. Но Бючли считал, что он сделал дальнейший шаг к познанию протоплазмы, который, как ему казалось, решал проблему ее организации. Он стал на путь создания моделей протоплазмы из различных субстанций. Он готовил тончайшие пеиы и достиг в этом большого искусства, добиваясь действительно огромного внешнего сходства комочков пены с клетками. Его пеиы, которые он изготовлял путем омыления различных масел, продолжали двигаться иногда по нескольку суток. Движения эти Бючли совершенно правильно объяснял изменением поверхностного натяжения в капле при разрушении ячеек пены и выхода при этом раствора мыла. Другими словами, он объяснял подвижность своих моделей элементарными, чисто физико-хими-
Buts chi i О. 1891. Uber die Struktur des Protoplasmas. Verh. d. deutsch. Zool. Ges. (<-O структуре протоплазмы»).
120
ческими закономерностями. Выводы, которые сделал из этих аналогий Бючли, принципиально ошибочны с методологической стороны и могут служить классическим примером механистического понимания жизни. Он был убежден в принципиальном сходстве между ротационным движением протоплазмы и наблюдавшимся им переливанием капель жиромыльной пены и даже считал, что ему удавалось изготовлять модели, ничем не отличающиеся от настоящей протоплазмы. Способ создания таких моделей, как считал Бючли, открывает перед исследователями путь к познанию возникновения протоплазматических структур. Он был вполне убежден также в том, что, идя этим путем, в скором времени можно будет изучить все свойства протоплазмы и ядра и, следовательно, окончательно отвергнуть мистические воззрения на явления жизнедеятельности, поскольку все процессы будут полностью сведены к физико-химическим реакциям.
На критике этих воззрений не стоит сколько-нибудь подробно останавливаться, ибо ясно, что свести процессы жизнедеятельности к простым физико-химическим явлениям невозможно. Идея эта порочна в корне. Сравнение комочка пены с протоплазмой является только лишь дидактической аналогией, ничего не говорящей по существу вопроса. Все явления, которые наблюдаются в комочке пены, обусловлены только одной-единственной реакцией (в данном случае омылением жира), следствием которой является диффузия мыла и изменение поверхностного натяжения капли, ведущее к изменению ее формы. Предлагая свою модель, Бючли не учел и не понял ее принципиального отличия от живой протоплазмы. Живое черпает энергию из лежащих в его основе процессов ассимиляции и диссимиляции, процессов построения и разрушения своей специфической субстанции. Живое обладает целым рядом признаков, которые в полной мере отсутствуют у такой модели. Например, модель не обладает явлением раздражимости. Любопытно отметить, что Бючли пытался раздражать свои движущиеся комочки пены электрическим током, но сам отмечал полную неудачу этих опытов. Тем не менее точка зрения Бючли в свое время пользовалась большим успехом.
Здесь уместно вспомнить, что еще Г. Валентин, говоря в 1842 г. о принципиально совершенно сходных опытах Ашерсона, дал в общем правильную критику этого сугубо механистического направления исследований и указал на всю его недостаточность при трактовке проблемы возникновения органических структур.
121
# 29. Протопласт И. Ганштейна. Несмотря на свой авторитет, Бючли пс смог убедить в своей правоте многих крупных ученых. Так, Вильгельм Вальдэйер (1830—1921), один из крупнейших ученых того времени, по вопросу о структуре протоплазмы выска' зывался иначе, хотя сам специально над этим вопросом не работал4. Он считал, что протоплазма построена однообразно и никакой структуры уловить в ней нельзя. Наблюдаемую ячеистость следует объяснить накоплением в протоплазме различных веществ (как жидких, так и более плотных).
С общетеоретической точки зрения мы, следовательно, должны сказать, что попытки отыскания структуры организованной единицы — «элементарного организма», т. е. клетки,— неизменно приводили к ее расчленению на более простые структуры, будь то зернышки, будь то ниточки, будь то ячейки пены и так далее. Распространение механистических воззрений в биологии еще более способствовало разрушению представлений о клетке как об индивидуальной организованной единице. За таковые признавались более элементарные структуры, из которых, как предполагали, построена клетка.
В этом отношении интересно отметить противоположный взгляд, который развивался ботаником Иоганнесом фон Ган-штейном (1822—1880) 9. Он предложил новый термин для обозначения клеточного тела (вернее, всей его живой мае-с ы) без расчленения на ядро и протоплазмы, термин, может быть, наиболее удачный из всех предлагавшихся за последние десятилетия XIX в. и сохраняющий значение и поныне. Ган-штейн называет индивидуализированную живую единицу (клетку) словом «протопласт». Этот термин не дублирует ни понятия протоплазмы (поскольку он говорит не о веществе, а лишь о том, что клетка является определенной биологической единицей), ни понятия клетки (ибо говорит только о живой его массе). Нам кажется, что словом «протопласт» и в настоящее время удобно пользоваться для обозначения всей живой массы клетки. Таким образом, в это понятие не будут включаться неживые структуры: оболочки, вакуоли, клеточный сок, а также неживые включения (жир, гликоген и др.).
8 Waldeyer W. 1895. Die neueren Ansichten iiber den Bau und das We-sen der Zelle. Deutsche med. Wichenschr. («Новые взгляды на строение и сущность клетки»),
9 На ns t ein Joh., von. 1880. Das Protoplasma als Trager der Pflanzli-chen und thierischen Lebensverrichtungen («Протоплазма как носитель жизнедеятельности растений и животных»),
122
1 анштеин совершенно не упоминает о различных теориях строения протоплазмы из твердой и жидкой фазы. Однако и его собственные представления можно характеризовать в известной их части как физико-химические. Он продолжает развивать взгляды Карла Нэгели (1817—1891) и Вильгельма Гофмейстера (1824—1877). Так, например, проницаемость протоплазмы он трактует как простую проходимость частичек в межмицеллярных и межмолекулярных пространствах, так что оболочка протоплазмы сравнивается им с ситом. Чисто физико-химически объясняются им также процессы набухания и роста протоплазмы.
Протопласт, согласно Ганштейну, представляет собой неделимый органический индивидуум, не сравнимый ни с молекулой, ни со структурой типа мицеллы. Ганштейн особенно подчеркивал целостность клетки. Эта целостность характерна для всего живого, для каждого живого индивидуума. В пользу этого положения он приводил известный факт, что отдельные клетки могут существовать как самостоятельные, законченные существа. В этих случаях протопласт сам регулирует все свои отправления. Однако гораздо чаще известное число клеток образует индивидуумы высшего порядка, в котором миллионы протопластов лишь представляются самостоятельными. С его точки зрения, они в своей жизнедеятельности зависят от общего плана, воли и потребности целого организма, как федерации клеток, для которой последние являются субстратом. «Чем выше организм,— писал Ганштейн,— тем яснее выявляется могущество нематериальных и психических сил на субстанцию с ее силами».
По существу концепция Ганштейна логически гораздо более продумана, чем теоретические представления материалистов-механистов. Исследуя клетку, как обособленную (Protozoa), так и заключенную в тканях (Metazoa), и обнаруживая при этом ряд процессов, которые могут быть объяснены чисто физическими закономерностями, Ганштейн ясно понимал, что основные движущие силы этих жизненных процессов настолько сложны, что они не могут быть сведены к элементарным физико-химическим закономерностям. Доводя эту мысль до логического конца и не находя, однако, никакого иного объяснения, он в силу ограниченности своей формальной методологии был вынужден прийти к витализму.
§ 30. Теория биобластов Р. Альтмана. Глава о строении протоплазмы будет неполной, если мы еще вкратце не остановимся на
123
Гранулярной теории Рихарда Альтмана (1852—1900), которая рассматривает клетку, состоящей из значительного числа зернышек-гранул. Альтман гомологизировал гранулы с микроорганизмами. Согласно этому представлению, клетка является своеобразной зооглеей бактериоподобных организмов, выделивших вокруг себя слизистое вещество. Если другие исследователи, как мы видели выше, в осторожной форме говорили о более элементарных, чем клетка, структурах, то именно это обстоятельство Альтман делает центральным пунктом всей своей концепции. Его взгляд, одиако, не является совершенно новым.
Альтман имел предшественников. Ряд французских исследователей начиная с 60-х годов прошлого века защищал взгляд на протоплазматические зернышки как на низшие организмы, причем высказывалась даже гипотеза, что именно они являются теми элементарными организмами, которые первыми возникли на Земле. Несколько позднее различные гранулы, встречаемые в клетке, уже прямо начали сравнивать с микрококками. Подобный взгляд также обосновывает представление Брюкке о сложности строения клетки и о необходимости априорно принять наличие в ней различных аппаратов, выполняющих различные функции.
Последнюю четверть XIX в. можно характеризовать как период изучения структуры клетки, при этом обнаруживалась все большая и большая ее сложность. В особенности значительную роль в этом отношении сыграло открытие кариокинеза. Познание процесса митотического деления с особенной ясностью показало как морфологическую, так и физиологическую сложность клетки. Вместе с тем господствовавшее в это время механистическое мышление требовало от аналитического исследования отыскания действительно элементарных структур, т. е. требовало сведения сложного к элементарному, притом в возможно более простой форме. Интересное в этом отношении высказывание мы находим, например, у русского исследователя Г. Г. Шлятера 10. Он писал, что уже чистая логика с необходимостью требует признания возможности и, более того, необходимости существования более элементарных биологических единиц, чем клетка, даже если бы мы в настоящее время их не смогли доказать. Сходную мысль мы находим и у такого выдающегося ученого, как Оскар Гертвиг (1849—1922). В первом издании своего учебника «Клетки и тка
10 Schlater G. 1899. Der gegenwartige Stand der Zellentheorie. Biolog
Zbl. Bd. 19 («Современное состояние клеточной теории»).
124
ни» 11 он развивал широко распространенную в то время мысль, что способность клеток к осуществлению сложных химических процессов заставляет признать ее построенной из многочисленных мельчайших и очень разнообразных компонентов. Другими словами, клетка не признавалась уже последней живой единицей, и последнюю старались во что бы то ни стало найти. Альтман с особой ясностью, на большом фактическом материале, с прекрасной для того времени техникой исследования доказывал, что все клетки состоят из гранул.
На рис. 30 и 31 приводятся некоторые иллюстрации из его книги «Элементарные организмы» 12. На одном из рисунков мы видим репродукцию клетки печени и поджелудочной железы, а на другом — почечный каналец кошки. Одного взгляда на эти рисунки достаточно, чтобы убедиться, что на них изображены прекрасно известные в настоящее время хондриосомы (митохондрии). Таким образом, Альтман открыл новые, до того времени неизвестные органоиды клетки, однако истолковал он их совершенно неправильно. Изучены детально они были лишь несколько позднее. Не обнаруживая в клетках гранул при обычной методике исследования, Альтман должен был разработать свои методы фиксации и окраски, которые ему действительно позволили открывать зернистые или нитчатые структуры во всех или (вернее) почти во всех клетках. Методика Альтмана применяется и до сих пор как дающая весьма совершенные результаты для обнаружения хондриосом. Особенно четко это было доказано в исследовании русского гистолога В. Самсонова 13.
Если внимательно прочитать исследования Альтмана, то нетрудно будет убедиться, что он занимался по существу лишь отысканием гранул во всех частях самых разнообразных клеток. Он сумел увидеть гранулы даже в ядре и разработал для этого специальные методы. Но Альтман совершенно не занимался изучением природы описываемых им зернышек, ибо был вполне убежден, что все открытые им гранулы являются не чем иным, как своего рода микрококками, элементарными живыми единицами, названными им в общей форме биобластами. Другими словами, все гранулы, по мнению Альтмана, между собою принципиально гомологичны.
11 Гертвиг О. 1911. Общая биология. Клетки и ткани. 3-е русское нзд., значительно дополненное. Изд-во К. Риккерта.
12 А1 t m a n n R. 1890. Die Elementarorganismen und ihre Beziehungen zu der Zelle («Элементарные организмы и их отношение к клеткам»).
l3Samsonoff W. 1910. Arch. f. mikrosk. Anat. Bd. 75.
725
Рис. 30. Рисунок Р. Альтмана печеночной клетки и поджелудочной железы лягушки, сделанный по препарату, приготовленному по его методу (1863 г.)
Рис. 31. Рисунок Р. Альтмана клеток почечного канальца кролика, сделанный по препарату приготовленному по его методу (1893 г.)
Итак, теория биобластов развенчивает клетку как последний неделимый живой элемент, и поэтому общей для всего живого мира элементарной единицей Альтман предлагает считать микрококк (биобласт).
Природу самого биобласта Альтман трактует как кристаллическую органическую структуру. Он отмечает, что было бы весьма удивительным, если бы биобласт не был кристаллом, ибо природа не имеет двух лиц, а имеется всегда общий закон, господствующий как над живым, так и над мертвым. Но разница заключается в том, что «организованный кристалл» (выражение самого Альтмана) не возникает путем «высажде-ния», а может получиться только путем размножения предшествующего индивидуума.
Альтман считал, что он возрождает старое учение об элементарных зернышках на принципиально новых началах. Отвлеченные неопределенные зернышки, или шарики Г. Валентина, а в 1823 г.— Анри Мильн-Эдварса (1800—1885), получают у него значение структур, гомологичных с биобластами.
Другими словами, в теории Альтмана мы видим концепцию, аналогичную клеточной теории, согласно которой сама клетка является элементом, построенным из еще гораздо более мелких живых структур.
§ 37. Современные представления о строении протоплазмы и клеточная теория. За последние 50—60 лет представления о строении протопласта резко изменились. Идея необходимости допущения большой сложности строения живой материи не была конечно, поколеблена, но было совершенно очевидно, что все ранее описанные структуры — ошибки наблюдения и метода исследования. Ясно, что они могут представлять интерес только для истории науки. Интересно отметить, что не было двух научных школ, которые сходились бы в своих представлениях о тонкой структуре протопласта, каждая школа отстаивала свои взгляды. Такая ситуация всегда указывает, что спор идет не о реальном, а о воображаемом. Когда делается существенное новое, но реальное открытие, то споры быстро кончаются, если же речь идет о гипотезах и толкованиях, то единства взглядов достичь невозможно, ибо личные предвзятые взгляды ученых всегда были источником несогласия. Поэтому, если долго нет единства во мнениях, то это значит, что надо искать новый подход к разрешению проблемы.
127
Рис. 32. Современный бинокулярный микроскоп фирмы К. Цейсса с микрофотокамерой; образец, 1968 г.
Было показано, что любая фиксация живой клетки не может не вести к образованию артефактов, а наблюдение и изучение живой протоплазмы — дело очень нелегкое. Так, например, инш -нилось, что сулема осаждает белки протоплазмы в виде грубых хлопьев, спирт и формалин — в форме округлых гранул, а ось-миевая кислота дает чрезвычайно мелкий зернистый осадок. Последняя в виде 1—2%-ных растворов остается и до сих пор лучшей из всех известных фиксирующих жидкостей, сохраняющих протопласт в состоянии, по-видимому наиболее близком к живому.
В 20—30-х годах XX в. было опубликовано много исследований над прижизненным строением протоплазмы. И. Шпек был, видимо, первым, показавшим еще в 1921 г., что в микроскопе • живая протоплазма (ее основа) должна выглядеть гомогенной, лишенной всяких структур, ибо свойственное ей строение неразличимо, поскольку оно субмикронно. По теории микроскопа Эрнста Аббе (1840—1905) (мы имеем в виду световой микроскоп), разрешающая способность микроскопа имеет предел и зависит от длины используемой световой волны. Таким образом, пользуясь лучами видимого человеческим глазом света, можно различать частички только порядка 0,2 |л. Линейное увеличение объектива микроскопа не может быть поэтому выше 120 X; увеличение же, даваемое окуляром, только растягивает его, но число видимых деталей при этом не возрастает (рис. 32). Если пользоваться оптикой, рассчитанной для более коротковолновых лучей (ультрафиолетовые лучи), можно несколько повысить разрешающую способность, но тогда нужно уже пользоваться фотопластинкой. Однако и она недостаточна для изучения субмикронных частиц. Еще более мелкие частицы можно обнаружить, применяя так называемое «темное поле», что нередко называют ультрамикроскопией. Но и с ее помощью субмикроиные частицы все же не видны.
Таким образом, стало ясно, что сложную структуру протопласта глазом в световой микроскоп увидеть нельзя. Только в самые последние годы с изобретением электронного микроскопа наши знания о строении протопласта значительно шагнули вперед, и каждый год приносит все новые и новые сведения.
Электронная микроскопия продолжает употреблять термин — «цитоплазматический матрикс»; именно его считают наиболее важной частью живой клетки. Ему приписывают гомогенную структуру, а в ней различают органоиды (митохондрии, цитоцентр и др.), рибосомы (рибонуклеиновые агрегаты), просто макромолекулы рибонуклеиновой кислоты, различные другие
5 Е. М. Вермель
129
Рис. 33. Первый электронный микроскоп 1931 г., сконструированный Е. Руска и М. Кноллем
крупные молекулы, а также систему внутриклеточных мембран разной сложности.
Надо полагать, что еще рано писать историю развития современных взглядов на структуру протопласта, сложившихся и а основе электронной микроскопии (рис. 33 и 34). Постоянно выясняются все новые важные детали, по-иному оцениваются обнаруженные структуры. Если еще 5—7 лет тому назад различали на электроннограммах, так называемую, эрга-стоплазму и ретикулум, то теперь на них смотрят как на в известной мере артефакты, и мы не можем быть уверены, что и дальше взгляды не изменятся. Коротко говоря, наши представления о строении протопласта еще далеко не «отстоялись», но важно, что ныне пределы разрешающей способности микроскопа резко повысились, и мы видим несравненно больше, чем ранее. Но живую протоплазму мы все же изучать не умеем и ныне. Электронный пучок ее убивает и не может не вызвать в ней артефактов. Приходится пользоваться фиксированным материалом, что, конечно, в первую очередь отражается на понимании тонкой структуры.
Кроме электронной микроскопии, позволившей изучать клетку при гораздо больших увеличениях, чем световая микроскопия, 130
Рис. 34. Новейший влектронный микроскоп фирмы ^Сименс* образца 1968 г.
много интересного внесла в понимание клеточных включении еще топографическая гистохимия и. особенно, дифференциальное центрифугирование. Принципиально этот метод состоит в том, что из тканей готовят кашицу, причем все клетки разрушают, а затем эту массу (гомогенат) центрифугируют при разных скоростях. Показано, что различные структуры клетки (ядро, ми-тохрондрии, разные гранулы) в вязком гомогенате расслаиваются и могут быть, следовательно, разделены. Это позволяет изучать их уже чисто биохимическими методами. Оказалось, что можно выделить слой мельчайших гранул — микрозом, получивших ныне название рибосом; они содержат рибонуклеиновые кислоты (РНК), но в них нет дезоксирибонуклеиновых кислот (ДНК), находящихся только в ядре клетки. Как показали исследования последнего времени, именно в рибосомах происходит биосинтез видоспецифических белков и в них «записан» информационный код наследственности.
Наряду с рибосомами из протоплазмы выделены также Другие ранее неизвестные структуры — лизосомы. Их величина составляет всего 0,4 |Л в диаметре, они содержат гидролитические ферменты, но их покрывает плотная оболочка, препятствующая поступлению в протоплазму. Если бы это случилось, эти активные ферменты разрушили бы и убили клетку. Поэтому лизосомы называют даже «мешочками самоубийства». Они были открыты лишь в 1929—1930 гг. К. де Дув 14, хотя их видели и раньше. Лизосомы окрашиваются на препаратах в клетках подобно митохондриям, поэтому при изучении клеток в световом микроскопе их трудно различать. Выходя из клетки или выпадая во внутриклеточные пищеварительные вакуоли, лизосомы теряют свою оболочку, освобождая ферменты и переваривая органическую пищу. Именно это явление уже описывалось ранее, но считали, что пищеварительные ферменты несут сами митохондрии|. В ряде протозоологических работ, например, при изучении цитологии амеб, подобные картины наблюдали, но большинство считало это маловероятным. По этой причине сами наблюдения подвергались сомнению и были забыты15. Правильно написал Гёте: «Какая мысль — разумная иль глупая — найдется, которую б никто не ведал до него» («Фауст»). История науки показывала уже много раз, что видеть — еще не значит правильно понять,
14 D u v е С. de. 1929. Subcellular particles («Субклеточные структуры»),
13 Вер мель. Е. М. 1925. Исследование иад Amoeba hydroxena. Русский арх. протистол., 4, Хе 1—2, 95—120.
132
а без понимания факты просматриваются, недооцениваются или толкуются произвольно.
Электронный микроскоп, фракционное центрифугирование при больших скоростях, рентгеноструктурный анализ и ряд других новых методов исследования создали возможность изучения уже молекулярной структуры клетки. Это направление в науке получило в наше время название молекулярной биологии. Она изучает форму, агрегацию и пространственное расположение молекул тех основных компонентов, которые образуют первую систему организации, которая получает новое свойство, называемое жизнью. В настоящее время мы все же еще далеки от понимания этого, несомненно, самого важного для биолога понятия, но наука о жизни к нему приближается уверенными, хотя и медленными, шагами. Как мы увидим ниже (гл. IX), представление о системности полностью опровергает виталистические взгляды и помогает найти ясный путь к пониманию возникновения нового качества за счет специфической организованности.
Современные тонкие методы исследования клетки и в первую очередь электронная микроскопия подтверждают, что свойство живого скрыто именно в его структуре, главная особенность которого состоит в том, что клетка, во-первых, сама себя питает и, во-вторых, сама себя воспроизводит, расходуя собственное вещество. Таким образом, именно клетка (разной степени сложности организации: от микрококка до оплодотворенной яйцеклетки высших многоклеточных) со своей качественно иной (чем в неорганическом мире) структурой вещества — протопласта является предельной, неделимой живой единицей, вне которой мы не знаем жизни. Очень важно, что и на современном методическом уровне -подтвердился единый принцип строения как животных, так и растений. Именно клетка есть общая принципиально сходная для организмов элементарная структура.
ГЛАВА ШЕСТАЯ
ЯДРО И ДЕЛЕНИЕ КЛЕТКИ
Основные даты по истории изучения ядра и деления клетки
1781 Ф. Фонтана впервые изобразил на рисунках ядра в эритроцитах и в клетках эпителия кожи угря.
1830 Я. Пуркння под названием «зародышевый пузырек» впервые описал ядро в яйцеклетке курицы.
1832 Б. Дюмортье описал размножение клеток нитчатых водорослей образованием перегородок.
1833 Р. Браун описал ядра как постоянную структуру в растительных клетках и предложил термин Nucleus.
1835 Г. фон Моль вновь описал размножение клеток делением.
1838 М. Шлейден связал процесс клеткообразования с вопросом возникновения ядра из «зернышек» цнтогенезом.
1839 Т. Шванн, развивая теорию цитогенеза М. Шлейдена, дал ядру название «ц и т о б л а с т».
1852 Р. Р е м а к описал последовательность деления клетки по этапам: разделение ядрышка, затем ядра и далее тела клетки.
1867 В. Гофмейстер установил закономерность процесса растворения ядра перед началом деления материнской клетки и появления новых ядер в дочерних клетках.
1873 А. Шнейдер впервые описал картины непрямого деления ядра в клетках дробящихся яиц равнореснитчатого червя.
1876 Е. ван Бенеден открыл клеточный центр.
1879 В. Шлейхер предложил термин «кариокинез».
1879—1882 В. Флемминг дал описание последовательности процесса непрямого деления; этим окончательно оформились представления о митозе. Предложил термины: «хроматин», «Митоз», «амитоз», — а также ввел в употребление термин «кариокинез».
1883 В. Вальдейер ввел термин «хромосомы».
134
1883
1884
1884
1885
1890
1894
1896
1898
1903
1905
В. Р у разработал теорию МИтоза, Дав биологическую трактовку процесса.
О. Гертвиг и Э. Страсбургер независимо друг о г друга высказали гипотезу о значении ядра как носителя наследственных свойств.
Э. Страсбургер предложил термины: «профаза», «метафаза», «анафаза», «гаплоидное и диплоидное число хромосом».
К. Р а б л ь формулировал закон постоянства числа хромосом.
Оскар Гертвиг высказал гипотезу индивидуальности хромосом.
М. Гейдеигайи ввел термин «телофаза».
Э. Вильсои написал сводку, оформившую цитогенетику как самодовлеющее направление в науке.
С. Г. Навашин открыл явление двойного оплодотворения у растений.
Рихард Гертвиг формулировал закон постоянства ядерно-плазмеиного отношения.
Дж. Фармер и Дж. Мур предложили термин «мейо-зис» (мейоз).
135
§ 32. Первые описания ядра и деления клетки. История развития представлений о размножении клеток может быть разделена на три периода.
1. До 1873—1874 гг. господствовал взгляд относительно свободного возникновения ядер, взгляд, идущий от воззрений М. Шлейдена (1838).
2. С 1873 по 1882 г.— период первых описаний ядерных превращений (митоз) в процессе деления клетки (кариокинез).
3. С 1882 г. и до наших дней — период дальнейшего детального описания кариокинеза (митоза) и мейозиса (делений созревания гамет), когда была дана также теоретическая трактовка добытых данных, позволивших, в частности, понять такое важнейшее биологическое явление, как оплодотворение.
В настоящее время процесс деления ядра изучен чрезвычайно подробно во всех разнообразных группах как животных, так и растений. Этот раздел учения о клетке может считаться одним из наибольших достижений биологической науки, представляющих не только выдающийся теоретический интерес, но имеющих и большое практическое значение. Можно не сомневаться, что современная генетика не могла бы существовать, если бы она в свое время не получила подтверждения своих выводов из цитологических данных. В настоящее время генетика и цитология в ряде вопросов настолько тесно слились, что уже не могут быть расчленены, и поэтому вполне обоснованным является выделение цитогенетики как особого направления науки.
Вместе с тем эти данные имеют большое значение и для клеточной теории в целом.
Как показывает история учения о клетке, представления о расчленении живой массы клетки на протоплазму и ядро, о разделении клетки поперечной перегородкой (т. е. размножении клетки делением), о роли ядра в этом процессе и т. д. возникли разновременно и были связаны в единое целое далеко не сразу. Напомним, что ядро впервые описал еще в 1830 г. Пуркиня. Броун же в 1833 г. показал, что эта структура, которой он дал наименование «Nucleus» (термин вошел во всеобщее употребление и в настоящее время не имеет синонимов), имеется во всех живых растительных и животных клетках. Значение и роль ядра в то время, конечно, известны не были. Так, Мейен (1837) полагал, что оно, видимо, является конденсированной слизью или представляет собою питательное запасное вещество.
Всеобщее внимание к ядру было привлечено в связи с теорией цитогенеза Шлейдена (1838), поскольку таким образом во-136
прос возникновения клетки связывался с процессом ядрообразо-вания (см. гл. III, § 18), но по существу до 70-х годов ядром как таковым интересовались очень мало.
Размножение клеток путем их поперечного деления представлялось никак не связанным с процессами, протекающими в ядре. Деление клеток путем образования поперечной перегородки было известно уже давно. Впервые этот процесс был описан, по-видимому, еще Дюмортье в 1832 г., наблюдавшим развитие нитчатых водорослей *. Несколько позднее, в 1835 г., подобные же наблюдения сделал известный ботаник Г. фон Моль, также занимавшийся изучением развития водорослей. Важно подчеркнуть, что Дюмортье противопоставлял свои наблюдения над разделением клеток перегородкой различным фантазиям о возникновении клеток из «зернышек» (Тюрпен, Распайль и многие другие). Надо сказать, что эти наблюдения над делением клеток, хотя и отличались большой точностью, никого не убедили и никакой роли в развитии науки о клетке не сыграли, по крайней мере на данном этапе. Высказывания авторитетных ученых того времени имели гораздо большее значение. Странным образом, даже сам Г. фон Моль поддерживал взгляды Шлейдена о свободном образовании ядер и клеток, он продолжал высказываться в этом же аспекте еще в 40-х годах. Подобные взгляды вошли в учебники, где разные варианты цитогенеза были тщательно классифицированы. Отметим, что Шванн также знал работы, описывавшие деление клеток с помощью перегородок, но он не придавал им значения, считая такой способ размножения клеток исключением, а не правилом.
Выше (гл. III) нами было уже показано, что в 40—50-х годах XIX в. постепенно накапливался большой фактический материал относительно широкого распространения явления размножения клеток путем их разделения на две дочерних клетки и подрастания последних до размеров материнской. В особенности большую роль сыграли в этом отношении работы К. Нэгели1 2. Это дало возможность Вирхову в 1855 г. формулировать свой закон: «Всякая клетка от клетки». Необходимо подчеркнуть, что от конста-
1 Dumortier В. С. 1832. Recherches sur la structure comparee et de developpement des animaux et des vegetaux. Med. Acad. Sci. Bruxelles. 16, 219—311 («Сравнительное изучение строения и развития животных и растений»),
2 Nagel i С. 1844. Zellkerne, Zellenbildung und Zellwachstum bci den Pflanzen. Zeitschr. f. wiss. Bot., N 1, 34—188 («Клеточные ядра, образование клетки и рост клеток в растениях»).
137
§ 32. Первые описания ядра и деления клетки. История развития представлений о размножении клеток может быть разделена на три периода.
1. До 1873—1874 гг. господствовал взгляд относительно свободного возникновения ядер, взгляд, идущий от воззрений М. Шлейдена (1838).
2. С 1873 по 1882 г.— период первых описаний ядерных превращений (митоз) в процессе деления клетки (кариокинез).
3. С 1882 г. и до наших дней — период дальнейшего детального описания кариокинеза (митоза) и мейозиса (делений созревания гамет), когда была дана также теоретическая трактовка добытых данных, позволивших, в частности, понять такое важнейшее биологическое явление, как оплодотворение.
В настоящее время процесс деления ядра изучен чрезвычайно подробно во всех разнообразных группах как животных, так и растений. Этот раздел учения о клетке может считаться одним из наибольших достижений биологической науки, представляющих не только выдающийся теоретический интерес, но имеющих и большое практическое значение. Можно не сомневаться, что современная генетика не могла бы существовать, если бы она в свое время не получила подтверждения своих выводов из цитологических данных. В настоящее время генетика и цитология в ряде вопросов настолько тесно слились, что уже не могут быть расчленены, и поэтому вполне обоснованным является выделение цитогенетики как особого направления науки.
Вместе с тем эти данные имеют большое значение и для клеточной теории в целом.
Как показывает история учения о клетке, представления о расчленении живой массы клетки на протоплазму и ядро, о разделении клетки поперечной перегородкой (т. е. размножении клетки делением), о роли ядра в этом процессе и т. д. возникли разновременно и были связаны в единое целое далеко не сразу. Напомним, что ядро впервые описал еще в 1830 г. Пуркиня. Броун же в 1833 г. показал, что эта структура, которой он дал наименование «Nucleus» (термин вошел во всеобщее употребление и в настоящее время не имеет синонимов), имеется во всех живых растительных и животных клетках. Значение и роль ядра в то время, конечно, известны не были. Так, Мейен (1837) полагал, что оно, видимо, является конденсированной слизью или представляет собою питательное запасное вещество.
Всеобщее внимание к ядру было привлечено в связи с теорией цитогенеза Шлейдена (1838), поскольку таким образом во-
136
прос возникновения клетки связывался с процессом яд|....... и>
вания (см. гл. III, § 18), но по существу до 70-х годов ядром к.и. таковым интересовались очень мало.
Размножение клеток путем их поперечного деления представ лялось никак не связанным с процессами, протекающими в ядре. Деление клеток путем образования поперечной перегородки было известно уже давно. Впервые этот процесс был описан, по-видимому, еще Дюмортье в 1832 г., наблюдавшим развитие нитчатых водорослей *, Несколько позднее, в 1835 г., подобные же наблюдения сделал известный ботаник Г. фон Моль, также занимавшийся изучением развития водорослей. Важно подчеркнуть, что Дюмортье противопоставлял свои наблюдения над разделением клеток перегородкой различным фантазиям о возникновении клеток из «зернышек» (Тюрпен, Распайль и многие другие). Надо сказать, что эти наблюдения над делением клеток, хотя и отличались большой точностью, никого не убедили и никакой роли в развитии науки о клетке не сыграли, по крайней мере на данном этапе. Высказывания авторитетных ученых того времени имели гораздо большее значение. Странным образом, даже сам Г. фон Моль поддерживал взгляды Шлейдена о свободном образовании ядер и клеток, он продолжал высказываться в этом же аспекте еще в 40-х годах. Подобные взгляды вошли в учебники, где разные варианты цитогенеза были тщательно классифицированы. Отметим, что Шванн также знал работы, описывавшие деление клеток с помощью перегородок, но он не придавал им значения, считая такой способ размножения клеток исключением, а не правилом.
Выше (гл. III) нами было уже показано, что в 40—50-х годах XIX в. постепенно накапливался большой фактический материал относительно широкого распространения явления размножения клеток путем их разделения на две дочерних клетки и подрастания последних до размеров материнской. В особенности большую роль сыграли в этом отношении работы К. Нэгели 1 2. Это дало возможность Вирхову в 1855 г. формулировать свой закон: «Всякая клетка от клетки». Необходимо подчеркнуть, что от конста-
1 Dumortier В. С. 1832. Recherches sur la structure comparee et de developpement des animaux et des vegetaux. Med. Acad. Sci. Bruxelles. 16, 219—311 («Сравнительное изучение строения и развития животных и растений»),
2 Nagel i С. 1844. Zellkerne, Zelltnbildung und Zellwachstum bei den Pflanzen. Zeitschr. f. wiss. Bot, N 1, 34—188 («Клеточные ядра, образование клетки н рост клеток в растениях»).
137
тации самого факта размножения клеток делением до познания процесса деления было еще очень далеко. В самом деле, закон Вирхова говорит только о пере ш н уро в ани и клетки, или ее перегораживании поперечной оболочкой, и, таким образом, речь идет по существу именно об ограничивающей клетку оболочке. Ядру Вирхов в тот период значе-ни|я не придавал, ибо в момент деления клетки ядро «исчезает».
Растворение ядра в момент деления отмечалось многими учеными, ио только В. Гофмейстер (1867) в своей известной книге «Учение о растительной клетке» 3 занялся этим вопросом детально и описал это явление как постоянную закономерность. Он писал: «Во всех молодых клетках растений, способных к образованию новых клеток, ... в протоплазме имеется совершенно постоянное округлое тело ... Это тело называется ядром (Nucleus) клетки... Полное исчезновение четких границ клеточного ядра, его растворение и образование им жидкости, заполняющей центральную часть клетки, может быть установлено с большой точностью при изучении истории развития пыльцы покрытосеменных и спор у сосудистых скрытосеменных. Возникновение ядра следует понимать как выделение из общей массы протоплазмы участка, богатого белком, при этом происходит соединение частичек этого вещества в сфероидные комочки илн капли ... Нигде с достоверностью нельзя установить, что ядро делится путем шнурования или распада... Отсюда следует, что способность к самостоятельному размножению клеточным ядрам вообще не свойственна».
Из приведенной цитаты и изложенного выше нетрудно видеть, что, с одной стороны, постоянное наличие ядра в клетке было общепризнанным, а с другой — устанавливалось, что ядро при делении клетки растворяется, чтобы затем возникнуть вновь путем конденсации массы в центральной части протоплазмы. Этим наблюдениям нельзя отказать в точности, но ясно, что сущность этого процесса ускользнула от внимания исследователей. Вряд ли простая ссылка на технику того времени будет правильной и исчерпывающей. Можно не сомневаться, что ученый нашего времени на тех же самых препаратах увидел бы гораздо больше, но он вооружен знанием результатов исследований своих предшественников и поэтому видит больше и оценивает иначе.
В тот же период эти наблюдения были общепризнанными и вполне удовлетворяли исследователей. Л. Ауэрбах в 1874 г. пред-
3 Hofmeister W. 1867. Die Lehre von der Pllanzenzelle. I Band.
Handbuch der physiol. Bot. Leipzig («Учение о растительной клетке»), 138
АоЖИл даже специальное название «к а р й о л И з й с» для обозначения процесса растворения ядра в момент деления клетки. Этот термин в настоящее время применяется для обозначения процесса растворения ядра, но при его дегенерации.
§ 33. Первые описания митоза в делящихся яйцеклетках. Клеточное деление хорошо было известно также эмбриологам. Дробление яйцеклеток описывалось начиная уже с 40-х годов. Однако ядерные процессы просматривались, и поэтому говорили даже о безъядерной фазе клеток иа разных стадиях развития организмов. Процессы, которые происходят в клеточных ядрах при дроблении (делении яйцеклеток), еще долгое время оставались неясными, хотя сам факт перешнуровки яйцеклеток был уже хорошо описан в 40—50-х гг. (Р. К. Келликер и многие другие).
Всякий, кто работал с крупными яйцеклетками (амфибии и др.), хорошо знает, что найти бластомеры с покоящимися ядрами довольно трудно, тем более, что среди желтка они мало заметны. Надо иметь значительный опыт, чтобы найти также и фазы митоза. Не удивительно поэтому, что процесс деления был обнаружен и описан далеко не сразу. Интересно еще указать, что представление о безъядерности яйцеклеток и бластомеров дало основание для филогенетических спекуляций в смысле биогенетического закона. Геккель в 1874 г. трактовал безъядерное состояние яйца и бластомеров как предковую стадию развития жизни, сходную с первичной безъядерной клеткой-моие-р о й. Он назвал эту фазу яйца монерулой4. С открытием митоза эта теория рухнула и была забыта; в настоящее время мало кто даже знает о ее существовании.
Первыми объектами, на которых было описано непрямое деление (митоз), были как раз дробящиеся яйца. И это не удивительно, ибо в них количество делящихся клеток очень велико и при тщательном изучении, если на это направлено внимание, очень легко обнаружить все стадии митоза.
Следует отметить, что, по-видимому, первое описание сложности процессов деления ядра яйцеклеток было дано в работе Н. А. Варнека, впоследствии профессора Московского университета. В 1850 г. им было опубликовано исследование относительно первых фаз развития прудовика (Lymnaea) и одного наземного слизня (Limax). Варнек видел, что «зародышевый
4 Haeckel Е. 1874. Anthropogenie («Антропогення»).
139
пузырек» (ядро) яйца после оплодотворения теряет свои ясные контуры, но полностью, однако, не исчезает, образуя светлую зону. Он видел также перетяжку этой зоны и образование двух светлых шаров и т. д. Таким образом, он открыл исчезновение ядерной оболочки в момент деления и превращения ядра в светлое поле, разделяющееся на две половины. Наблюдения Варнека отличаются большой точностью: из его дальнейших описаний следует, что он видел даже два пронуклеуса. Однако в то время он не мог правильно истолковать свои наблюдения, а несовершенная техника исследования не позволила ему обнаружить такие более мелкие структуры, как хромосомы5. Подобные описания были позднее опубликованы и другими авторами, но митоза никто еще не видел.
Работа Варнека появилась слишком рано, и она была или не замечена, или забыта. Ее оценили значительно позднее, когда она была, можно сказать, открыта французским эмбриологом А. Фо-лем. Он писал, что Варнек по существу видел все, что можно увидеть на живом объекте: процесс созревания яйцеклетки, оплодотворение и первые стадии дробления, но сущность явления ему понять не удалось.
Детальное описание процесса деления было дано еще только через 20—25 лет, и лишь с этого момента оно привлекло всеобщее внимание ученых, ибо довольно быстро была доказана всеобщность явления и постепенно была дана правильная теоретическая его оценка.
Описания отдельных стадий митоза можно найти в работах большого числа ученых того времени. Так, картинки с изображением метафазы и анафазы можно видеть в работах В. Гофмейстера, опубликованных в 18496 и в 1867 гг. Многое видел также талантливый ботаник И. Д. Чистяков в 1871 г.7, но и он не смог связать отдельные стадии деления, которые наблюдал, в единый процесс и многое трактовал совершенно ошибочно8. Более точные изображения и описания хромосом в процессе деле-
5 Варнек Н. А. 1850. Об образовании и развитии зародыша у брюхоногих.— Бюлл. об-ва испытат. природы, 23, 90—194. М.
’ Hofmeister W. 1849. Die Entstehung des Embryo der Phanerogamen. Leipzig.
7 Чистяков И. Д. 1871. История развития спорангиев и спор высших тайнобрачных, пыльников и пыльцы явнобрачных. — Изв. Об-ва любит, естествозн., антропол. и этногр., 9, 98.
“Бляхер Л. Я. 1960. Возникновение клеток в онтогенезе.— Тр. Ии-та истории естеств. и техники, 32, 3—57.
140
Рис. 35. Первое изображение митоза в дробящихся яйцах плоского червя. Рисунки А. Шнейдера (1873 г.)
ния даны в работе доцента Дерптского университета Э. Руссова, посвященной морфологии высших растений. Он настаивал, что образования, названные нм «палочковые пластинки» (это были, конечно, хромосомы), не могут быть артефактами, а представляют собою закономерные изменения клеточного ядра; он наблюдал их также прижизненно. Однако восстановление дочерних ядер из хромосом от его внимания ускользнуло. Все эти картины митоза не произвели на современников особого впечатления, как^ впрочем, и последующие работы, привлекшие к себе внимание только спустя 100 лет после их опубликования в исследованиях историков науки с целью установить приоритет открытия.
Первое описание последовательных картин митоза по существу. принадлежит зоологу А. Шнейдеру; оно было сделано в 1873 г. 9 Его работа посвящена дроблению яиц одного плоского червя (Mesosternum Ehrenbergii). Он отметил, что при делении бластомеров происходит полный «метаморфоз», которому подвергается ядро («зародышевый пузырек»). «Весьма вероятно,— пишет он далее,— что этот метаморфоз имеет место всегда, когда наблюдается кажущееся исчезновение ядра» (рис. 35).
Исследование Шнейдера не обратило на себя внимания биологов общего профиля. Оно было в основном посвящено чисто зоологическим вопросам и опубликовано в малораспространен
9 Schneider А. 1873. Untersuchungen fiber Plathelminthen. XIV. Beri-chte der oberhessischen Ges. f. Natur. u. Heilkunde. Giessen («Изучение плоских червей»).
141
ном журнале. Вряд ли оно было сколько-нибудь широко известно, хотя, например, Бючли упоминает о нем и ставит очень высоко.
Из описаний и особенно рисунков ясно следует, что Шнейдер правильно понял последовательность этапов непрямого деления. Он видел профазу, метафазу, образование веретена, видел, как «зернышки — нити» (так он называл хромосомы) «собираются в меридиональной плоскости» (в другом месте он применяет даже слово «экваториальная плоскость») и увеличиваются в числе, а затем расходятся к противоположным полюсам. Он заканчивает свое описание фразой: «Эти наблюдения дают давно желаемое объяснение клеточного деления и особенно процессов дробления. Они впервые ясно показывают, какие сложные превращения может при клеточном делении испытывать ядро (зародышевый пузырек)». Шнейдер видел митозы не только при дроблении, но и на более поздних стадиях развития. Для него гомология яйцеклетки и клеток более поздних стадий была несомненна.
Годом позже, в 1874 г., появилась работа Бючли (впоследствии ставшего одним из крупнейших зоологов Германии) о созревании яиц круглых червей и улиток. На его рисунках особенно хорошо видна анафаза. Отсюда, между прочим, вполне ясно, что микроскопическая техника того времени сама по себе не могла уже служить препятствием к изучению процесса кариокинеза в любых тканях. Но беда в том, что Бючли изучал процесс выделения полярных телец и вывод его исследования заключался в утверждении, что ядро полностью выталкивается из клетки (рцс. 36). Однако в этом он был, видимо, не вполне уверен, ибо в тексте его работы мы читаем: «Мои наблюдения привели меня к выводу, что зародышевый пузырек целиком элиминируется из яиц, тем не менее я не имею оснований не допускать, что в известных случаях только одна его часть подвергается этой участи, остающаяся же часть продолжает существовать в желтке в качестве ядра» 10.
Нельзя не признать, что деление созревания — мало удачный объект для первоначального изучения митоза. Тем не менее
1Э В й t s с h 1 i О. 1874. Vorlaufige Mitteilungen uber Untersuchungen betref-fend die ersten Entwicklungsvorgange im befruchteten Ei von Nematoden und Schnecken. Zeitschr. f. wiss. Zool. 25 («Предварительные сообщения об исследованиях, касающихся первых стадий развития в оплодотворенном яйце круглых червей и улиток»),
142
Бкочли видел многое; он по существу первый описал и по-настоя-щему нарисовал хромосомы. Несколько позднее Фоль предлагал их даже называть «бючлевские вздутия и нити». Но вместе с тем Бючли описывал выталкивание делящегося ядра и считал, что новообразование ядер происходит из протоплазматических вакуолек; это последнее, конечно, совершенно ошибочно.
Наиболее правильное понимание процесса деления ядра яйцеклетки в тот период было дано Оскаром Гертвигом (1849—1922), одним из самых крупных ученых конца XIX в.
В одной из его ставших классическими работ по развитию яиц морского ежа, которыми была разъяснена сущность явления оплодотворения (1876 г.), Гертвиг писал, что при делении яйцеклеток никакого кариолиза не наблюдается, а имеет место ядер-ный метаморфоз, состоящий в своеобразном изменении структуры ядерного вещества и.
§ 34. Описание фаз кариокинетического деления. Почти одновременно с первыми описаниями митоза в яйцевых клетках и в бластомерах процесс деления ядра продолжали изучать и в растительных клетках. Выше мы уже указывали на исследование И. Д. Чистякова, посвященное развитию спор у плауна и хвоща, оно было напечатано в 1874 г. в итальянском ботаническом журнале 11 12. Чистяков, несомненно, видел отдельные картины непрямого деления. Основное значение, однако, приобрела монография известного ботаника Э. Страсбургера «О клеткообразования и клеточном делении»13, опубликованная в 1875 г. Страсбургер доказывал, что представление об исчезновении ядра в момент деления клетки основано на недостаточно точных наблюдениях. В действительности же имеет место сложная перестройка ядра, приводящая в конце концов к разделению материнского ядра на два дочерних. Однако точной последовательности фаз кариокине-
11 Hertwig О. 1878. Beilragc zur Kenntniss der Bildung, Befruchlung und Theilung des tierischen Eies. HI. Abh. Morphol. Jahrb. Bd. 4 («Сообщения к позиаиию возникновения, оплодотворения и деления яйца животных»).
12Tschistjakoff J. D. 1874. Maleriaux pour servir a 1’histoire de la cellule vegetale. Nuovi Giornale botanico Italiano. 10. 1 («По русски напечатаны лишь выдержки в сборнике «Русские классики морфологии растений» в серии «Классики естествознания», т. XII, стр. 15—24, 1923).
l3Strasburger Ed. 1875. Ueber Zellbildung und Zelllhcilung. Jena H. Dabis. S. 156 («О возникновении и делении клеток»).
143
Рис. 36. Выделение «полярных телец» в яйцах круглого червя. Рисунки О. Бючли (1874 г.)
за не установил и Страсбургер, хотя основные этапы процесса были поняты им правильно.
В том же 1875 г. вышли в свет еще два сообщения Бючли, видевшего и изобразившего на рисунках все главные фазы митоза и выделение «полярного тельца» (рис. 36).
В 1875 г. появилась также работа ассистента кафедры гистологии Варшавского университета В. Майзеля, описывавшего процесс деления клеток эпителия. Майзель полнее и точнее, чем кто-либо до него, описал стадии непрямого деления 11 * * 14. Однако он оговаривался, что оставляет открытым вопрос, действительно ли процесс совершается в указанном им порядке. Кроме того, Майзель допускал и «свободное образование ядер», т. е. он недооценивал сущность наблюдавшегося им явления.
В последующие годы появилось уже много исследований, исходивших из наблюдений и толкований Страсбургера. Из них упомянем только статью В. Шлейхера15, изучавшего деление
11 Mayzel W. 1875. Uber eigenthiimliche Vorgange bei der Teilung der Kerne in Epithelzellen. Centralbl. med. Wiss., 50, 848—852 («О своеобразных явлениях при делении ядер эпителиальных клеток»).
15 Schleicher W. 1878. Uber die Teilungsprozesse der Knorpelzellen. Centralbl. med. Wiss., 23, 418—419 («О процессе деления хрящевых
клеток»),
Schleicher W. 1879. Die Knorpellzellteilung. Ein Beitrag zur
Lehre der Teilung von Gewebezellen. Arch. mikr. Anat., 16, 248—300 («Деление клеток хряща. К вопросу к. учению о делении клеток в тканях»),
144
хрящевых клеток амфибий. Им был предложен ныне широко применяющийся термин — кариокинез — с целью обозначения ядерных компонентов, образующих звездообразную экваториальную пластинку.
Тщательное описание процесса деления ядра в соматических тканях животных принадлежит также киевскому гистологу П. И. Перемежко, изучавшему деление в клетках эпителия кожи личинок тритона. Он прижизненно наблюдал почти все стадии митоза, но и ему не удалось установить точную последователь-и 1 fi ность превращении .
Подробно и очень убедительно изложил все стадии кариоки-нетического деления В. Флемминг в том же 1879 г. Интересно отметить, что статьи Перемежко и Флемминга были опубликованы в одном и том же томе (XVI) немецкого журнала «Архив микроскопической анатомии» — одна непосредственно за другой. Флемминг изучал кариокинез в клетках кожи саламандр, отличающихся крупными размерами и потому являющихся прекрасным объектом для изучения процесса непрямого деления клетки. Исследование Флемминга имеет преимущество перед всеми остальными работами, ибо в нем не только констатирован своеобразный метаморфоз ядра в период деления клетки, но и впервые были выяснены и точно установлены последовательность фаз превращения хроматина в нити (хромосомы), их продольное расщепление и дальнейшее поведение,7. Заметим, что сам термин хромосомы был предложен уже позднее Вальдейером (1883).
Заслуга Флемминга состоит не только в том, что он сделал определенные и очень точные наблюдения (рис. 37, 38). В 1882 г. он выпустил большую книгу «Клеточная субстанция, ядро и клеточное деление»18, в которой привел в систему все сведения о строении и непрямом делении ядра. Флеммингу принадлежит и большинство терминов: митоз, амитоз, хроматин и др., вошед-ши/х в общее употребление. Отметим только, что термином м и-т о з Флемминг называет совокупность только ядерных превращений в течение деления, а весь процесс
10 Peremeschko Р. 1879. Uber die Theilung der thierischen Zellen.
Arch. f. mikr. Anal. Bd. 16, 437—463 («О делении животных клеток»). 17 Flemming W. 1879. Beitrage zur Kenntniss der Zelle und ihrer Lebens-erscheinungen. Arch. f. mikr. Anat., 16, 302—436 («Сообщения к познанию клетки и ее жизненных проявлений»).
18 F 1 е m m i n g W. 1882. Zellsubstanz, Kern und Zelltheilung. Leipzig. F. C. W. Vogel. S. 424 («Вещество, составляющее клетку, ядро н клеточное деление»).
145
Вальтер Флемминг (1844—1905)
разделения клетки называет кариокинезом. Этот последний термин он заимствовал у Шлейхера (1879), употреблявшего его, как указывалось выше, в несколько ином смысле, но давшего Флеммингу свое согласие на приложение его для обозначения процесса деления клетки в целом. Итак термины митоз и кариокинез не синонимы, как это нередко считают.
Необходимо подчеркнуть, что полнота и убедительность изложения вопроса в книге Флемминга была такова, что все старые представления получили уже чцсто исторический интерес, а конкретные исследования — вполне определенную направленность.
§ 35. Дальнейшее изучение митоза. Интерес к детальному изучению структуры клетки, обусловленный теоретической предпосылкой понимания клетки как элементарного организма и берущий свое начало от работ Шультце и Брюкке, привел, как мы видели, к открытию, в частности, процесса кариокинеза. Монография Флемминга завершила период фактического обоснования
146
данного открытия. Этим процессом сразу начало заниматься очень большое число ученых, и количество работ по всем вопросам клеточного деления резко возросло. Не будет преувеличением сказать, что «ядро затмило протоплазму». Литература по вопросам деления клетки в настоящее время почти необозрима. Процесс деления ядра мы знаем гораздо детальнее, чем любые другие явления из области цитологии.
Работами целой плеяды первоклассных ученых (Рабль, Герман, Фоль, Костанецкий, Бонневи, Горожанкин, Навашин и др.) были выяснены многочисленные морфологические детали процесса деления не только соматических, но и половых клеток. Эти исследования позволили понять такие процессы, как явление созревания половых клеток, оплодотворение и др. Особо большое значение имели работы бельгийского ученого Эдуарда ван Бене-дена (1845—1910), который нашел исключительно удачный объект для изучения кариокинеза — яйца лошадиной аскариды (Ascaris megalocephala), отличающиеся крупной величиной и малым количеством хромосом (всего 4, а в некоторых расах даже 2).
Огромное внимание, которое было привлечено к процессу непрямого деления, вызвано, конечно, не только тем, что для цитологов открывалось широкое поле деятельности. Познание митоза позволило построить ряд важнейших теоретических обобщений принципиальной значимости.
Из всех явлений митотического процесса наибольший теоретический интерес на первых порах вызывал факт продольного расщепления хромосом и расхождения обеих половин к разным полюсам клетки, а также вопрос о постоянстве их числа. Надо сказать, что сам Флемминг не дал настоящей теоретической трактовки митотического процесса. Так, он писал, что смысл открытого им продольного расщепления хромосом продолжает для него оставаться неясным (1882). Сам этот факт был тотчас же подтвержден всеми исследователями того времени (Страсбургер, Ретциус и др.), но объяснение явления «повисло в воздухе» (выражение Рабля), и оно было дано в 1883 г.19 уже крупнейшим теоретиком того времени Вильгельмом Ру.
Он исходил в построении своей гипотезы из предположения, что хроматиновые нити (хромосомы) построены из зернышек, которые качественно между собою различны. В покоящемся ядре
19 Roux W. 1883. Uber die Bedeutung der Kernteilungsfiguren. Eine hupothetische Erorterung. Leipzig («О значении фигур деления ядра. Одна гипотетическая трактовка»).
147
Рис. 37. Фазы митоза в эпителиальных клетках кожи личинок саламандры. Рисунки В. Флемминга (1882 г.) по фиксированному и окрашенному препарату
А—от профазы до формирования метафазы (стадия «звезды»);
Рис. 31 (окончание)
Б — от метафазы до телофазы
с b
9
Рис. 38. Фазы митоза в эпителиальных клетках кожи личинок саламандр. Рисунки В. Флемчиню на основании прижизненных наблюдений лоскутков кожи
эти зернышки раскиданы в беспорядке; при митозе они объединяются в нить, состоящую, таким образом, из качественно различных гранул. Отсюда понятно, что только продольное расщепление может обеспечить правильное и равномерное разделение вещества ядра на две равные и идентичные половины. Именно в этом Ру и видел биологический смысл митоза: значение возникновения хромосом, их продольного расщепления, постоянного числа и др. В наше время, т. е. через 85 лет (I), трактовка кариокинеза осталась той же самой.
Таким образом, уже в 1883 г. Ру была дана теоретическая трактовка митотическому процессу20. Решающее слово в оценке достоверности этого представления оставалось за фактическими наблюдениями над продольным расщеплением хромосом. Мысль была дана, тема для исследования поставлена, и масса исследователей устремилась проверять это наблюдение Флемминга. Однако он считал это положение не совсем доказанным. Подтверждения начали поступать почти тотчас же. Первые данные были опубликованы Гюйаром уже в 1883 г., а Гейзером и Страсбурге-ром — в 1884 г. Наиболее убедительно это было доказано ван Бе-неденом в его работах над аскаридой (1884—1887 гг.).
Весьма серьезным испытанием для теоретической трактовки митоза Ру была проверка закона постоянства числа хромосом, который вытекал из этих гипотетических построений.
Надо отметить, что уже сам Флемминг обратил внимание на постоянство числа хромосом, но не решался постулировать это явление как закон, ибо, с одной стороны, он обнаружил, что в клетках кожи тритона и в других соматических тканях количество хромосом может несколько варьировать, а с другой — он знал, что в семенниковых клетках число хромосом значительно меньше. Более поздние исследователи не придавали значения факту известного колебания числа хромосом в соматических клетках (они действительно сравнительно невелики и обнаруживаются лишь у небольшого числа клеток).
Было показано, что клетки разных тканей одного и того же вида имеют одно и то же число хромосом. Вполне определенно и ясно это было высказано Раблем (1885), и поэтому следует счи- 21
21 Интересно отметить, что этот факт мало кому известен. Даже в такой полной и хорошей книге по истории учения о клетке, как книга 3. С. Кацнельсона «Клеточная теория» (1963, М., Медгиз), ни слова не сказано о работе Ру 1883 г. н о ее большом значении В развитии представлений о митозе.
75/
Вильгельм Ру (1850—1924)
тать, что именно он впервые формулировал закон постоянства числа хромосом21. Обоснование же закона индивидуальности хромосом (кариотип) принадлежит Т. Бовери, которым была опубликована целая серия экспериментальных работ (1888—1907 гг.).
Факт уменьшенного числа хромосом в зрелых половых клетках, столь смущавший Флемминга, был вскоре разъяснен ван Бе-иеденом. Было показано, что половые клетки (гаметы) получают половинное число хромосом (гаплоидный набор) и именно в этом состоит конечная сущность делений созревания (мейозис). Таким образом, было выяснено также явление «элиминации ядер» из яйцеклеток, которое впервые видел Бючли в 1873 г. Все эти факты позволили понять явление оплодотворения, открытое незадолго перед этим О. Гертвигом (1876) и состоящее, как он показал, в слиянии ядер. Эти ядра имеют половинное (п) число 21
21 Rabi К. 1885. Uber Zellteilung. Morph. Jahrb. 10, 214-230 («О клеточном делении»).
/52
хромосом, и их объединение в единое целое восстанавливает, таким образом, нормальное число хромосом в зиготе (2п).
Процесс клеточного деления привлекал всеобщее внимание биологов еще и в силу того, что уже в 1884 г. Оскаром Гертви-гом и Страсбургером (независимо друг от друга) была высказана гипотеза, что именно клеточное ядро является носителем наследственности. Особенно яркое развитие эта гипотеза получила в теоретических работах Августа Вейсмана (1834—1914), в его учении о зародышевой и соматической плазме. Дальнейшим развитием этой же концепции является вся современная теория гена Т. Г. Моргана. Фактический материал, трактующий данную проблему, в настоящее время настолько велик, что для того, чтобы в нем разобраться, ему нужно посвятить всю свою деятельность. Это есть содержание той области науки, которая получила наименование цитогенетики. Первая сводка данного материала, приведшая его в единое более или менее стройное целое, принадлежит известному американскому цитологу Э. Вильсону; она впервые появилась в 1896 г.22
§ 36. Значение открытия кариокинеза для учения о клетке. Рассматривая историю учения о клетке, важно прежде всего подчеркнуть, что детальный анализ вскрыл сложность клеточной организации с совсем неожиданной стороны. Ее искали в сложности структуры протопласта, а была показана сложность процесса размножения клетки, одного из основных жизненных явлений. Общебиологическое значение этого открытия понято было не сразу, оно осознавалось лишь постепенно.
Исследования по делению ядра в период с 1879 по 1890 г. подняли цитологию на новую ступень. Они были сделаны на столь высоком техническом уровне, что мало отличаются уже от современного, многие рисунки того времени до сих пор приводятся в учебниках. Мы имеем в виду прежде всего работы Флемминга, О. Гертвига, Страсбургера, ван Бенедена и др.
Правило постоянства числа хромосом и развитая (исходя из него) теория индивидуальности хромосом имеют целый ряд важнейших теоретических следствий. Прежде всего надо сделать
22 W i 1 s о n Е. 1896. The cell in development and heredity. Вильсон Э.
Клетка в развитии и наследственность, ч. I (1936), ч. П (1940). Био-медгиз.
153
Рихард Гертвиг (1850—1937)
вывод, что если действительно набор хромосом постоянен, то, следовательно, индивидуально и ядро, а соответственно— и клетка в целом. Уже давно отмечалось, что ядру определенной величины соответствует определенный размер протоплазматического тела. Это явление формулируется обычно как закон постоянства ядерно-плазменного отношения (закон, высказанный впервые Рихардом Гертвигом в 1903 г.). Он выражается обычно простой формулой
МК ~МР1
= const,
т. е. соотношение между массой (Л/) ядра (К) и массой протоплазмы (МРГ) является величиной постоянной. В общей форме это явление следует считать доказанным (фон Марк, 1942).
Итак, закон соответствия между величиной ядра и заключающей его протоплазмой дает законченное представление о клетке как единой живой единице.
154
Но вместе с тем нельзя не отметить, что устремление основного внимания цитологов на изучение ядра и его роли в делении привело в ряде случаев к переоценке значения ядерного компонента клетки. В настоящее время многие считают возможным говорить о ядре как о единственной, основной организующей структуре клетки. Однако правильнее говорить, что все элементы, не возникающие de novo, с принципиальной точки зрения одинаково важны для построения и существования клетки. Какая же из них является ведущей, в настоящее время вряд ли можно установить, ибо физиология клетки на молекулярном уровне остается по существу почти неизвестной. Достаточно сказать, что мы, например, не знаем, в чем конкретно заключается роль ядра в ассимиляционных процессах, а об его участии в них мы судим, исходя из факта, что в клетках, лишенных ядра (опыты на амебах), не происходит усвоения веществ. О роли хондриома, значении сетчатого аппарата Гольджи и т. д. мы также имеем лишь более или менее достоверные гипотезы. Наконец, новые данные все больше и больше свидетельствуют о важнейшем значении самой цитоплазмы в разнообразных целлюлярных процессах. В силу этого нет оснований говорить об абсолютном примате ядра, однако здесь, конечно, не место для дискуссии.
Изучение митотического деления ядра приводит к заключению относительно сложности построения клетки — этой элементарной живой единицы. Но к тому же заключению приводит и детальное изучение отдельных протоплазматических структур, на истории исследования которых подробнее мы не можем останавливаться, так как это завело бы нас слишком далеко в область специальных вопросов.
Необходимо еще отметить, что закон Вирхова («всякая клетка от клетки»), по мере того как сведения по цитологии углублялись, детализировался все больше и больше. Раньше всего эта формула была распространена на ядро («всякое ядро от ядра» — Э. Гертвиг), затем на хромосомы («хромосома от хромосомы» — Ру, Бовери), на хондриосомы (Мевес и др.), с большой вероятностью также на клеточный центр, в ряде случаев, возможно, на аппарат Гольджи и т. д.
Все эти факты говорят в пользу все того же общебйологиче-ского вывода (Брюкке, 1861), что клетка является сложнейшей структурой и одновременно элементарной живой единицей (элементарной в том смысле, что она уже не может быть разложена на части без нарушения жизни). Что касается сложности организации клетки, то ее не
755
следует понимать так упрощенно, как это в свое время делал Брюкке, предполагая сложность видимой морфологии. Сложность заключается не только в субмикронной и микронной сложности протопласта в каждый данный момент, а прежде всего в его потенциях к развитию. Сложность заключается в том, что клетки способны быстро создавать целый ряд сложнейших структур по мере возникновения в них надобности, причем очень часто эти процессы могут быть обратимы. Функции в клетках могут быстро сменять одна другую.
На современном этапе познания клетки должно быть акцен-трировано, что абстрактной клетки нет, а всякая схема схемой и остается, поскольку она является лишь суммарным описанием явления. Другими словами, перед нами всегда должны стоять конкретные вопросы о развитии клетки, об ее дифференцировке и детерминации, об ее специализации и т. д. Далее, должен возникать вопрос о превращении одних типов клеток в другие. Таким образом, появляются представления о клетках эмбрионального (камбиального) типа и клетках, использовавших все свои способности к развитию или утерявших их в связи со своей специализированной функцией. Но вместе с тем все вопросы развития клетки как элементарной живой единицы неразрывно связаны с развитием организма как единого целого. Так, собственно, как мы ниже увидим, и возникла наиболее, может быть, кардинальная проблема современной биологии организма — проблема «части и целого», с точки зрения которой клетка должна рассматриваться как часть, однако, с другой стороны, она должна расцениваться и как единое целое, но более элементарного порядка.
ГЛАВА СЕДЬМАЯ
УЧЕНИЕ О КЛЕТКЕ
НА РУБЕЖЕ ДВУХ СТОЛЕТИЙ
Основные даты в развитии методологии клеточной терии
на рубеже XIX и XX веков
1881 В. Ру переносит учение Ч. Дарвина о борьбе за существование на борьбу клеток и частей организма в процессе онтогенеза.
1892 И. И. Мечников создает сравнительную патологию воспаления.
1895 В. РУ создает механику развития (экспериментальную эмбриологию) животных.
1895 М. Ферворн выдвигает концепцию организма как «клеточного государства».
1895 М. Ферворн предлагает считать, что теоретическая физиология должна сводиться к сумме клеточных физиологий.
1895 Ю. Сакс выдвигает представление об энергиде как элементарной биологической единице.
1895 Э. Вильсон устанавливает синтез цитологии, оитогене-тики и генетики в книге «Клетка в развитии и наследственности».
1903 И. И. Мечников создает концепцию геронтологии на основе фагоцитарной нейрофагии
/57
§ 37. Энергиды Ю. Сакса и проблема специфичности живой материи. Как бы в противовес воззрениям на протоплазму как на конгломерат или сочетание каких-то элементарных частиц, что особенно резко и ясно выражалось в учении Альтмана, ботаник-физиолог Юлиус Сакс (1832—1897) в те же самые годы выдвинул новое представление о клетке как о неделимой единице — энергиде1. Конкретно под энергидон Сакс понимал отдельное ядро с участком протоплазмы, над которой, по его представлениям, оно господствует. Таким образом, он рассматривал ядро с окружающей его протоплазмой как нечто представляющее собой органическую единицу как в физиологическом, так и в морфологическом отношениях. В понятие энергида Сакс включал только живые микроморфологические структуры, обладающие энергией (т. е. жизнедеятельностью), что он многократно подчеркивал в ряде своих публикаций. Клеточные стенки, крахмальные и алейроновые зерна, а также кристаллы и прочие структурные образования клеток, по его представлениям, к энергидам не относятся. Они остаются тем, чем они были, если на них не воздействует обладающая жизнедеятельностью масса энергиды; в противоположность пассивным клеточным продуктам живая энергида подвержена постоянным изменениям. Наиболее важным в представлениях Сакса является утверждение, что специфические жизненные явления (и специально формообразование) зависят прежде всего от внутренней наследственной природы энергиды и от того, как она реагирует на внешние воздействия. В соответствии с его пониманием специфичности живой субстанции он энергично возражал против простого перенесения представлений о кристаллическом строении целлюлозной оболочки или крахмальных зерен на массу, составляющую энергиду. Таким образом, его взгляды резко отличались от представлений, например, ботаника Нэгели (1844—1867) и многих других, мысливших механистически. Сакс указывал, что раствор белка не может быть идентифицирован с протоплазмой. Они отличаются друг от друга как неорганизованное от организованного. И поэтому Сакс утверждал, что из простого раствора белка «сама по себе» протоплазма никогда образоваться не может. Увеличение числа протоплазматических частиц происходит только в организованной энергиде, между молекулами ее субстанции.
1 Sachs J. Physiologische Notizen. IX. Weitere Betrachtungen uber Energiden und Zellen. «Flora» (Allg. bot. Ztg.) Bd. 81 Erganzbd. («Физиологические заметки. Дальнейшее рассмотрение вопроса об энергидах и клетках»).
158
Сакс специально подчеркивал, что если бы даже химикам удалось создать белковые растворы, входящие в состав живого вещества, то это, тем не менее, не означало бы создания протоплазмы со всеми ее жизненными свойствами. Сакс постоянно указывал на различие, которое существует между живой субстанцией с ее особыми качествами и веществом, хотя бы и органическим. Как одно из важнейших свойств энергиды он, между прочим, отмечал постоянство ее величины. Он исходил из наблюдения, что никогда клетка не превышает определенной средней величины, и считал, что никакого определенного соотношения между величиной клетки и размерами целых растений не существует. Пытаясь дать этому явлению причинное объяснение, Сакс пришел к выводу, что оно сводится к тому, что сфера действия ядра имеет лишь небольшой радиус, а следовательно, одна и та же масса вещества выигрывает в энергии и силе жизнедеятельности, если она разделена на отдельные многочисленные участки — энергиды.
Представления о сфере действия ядра Сакса сводятся к тому, что в основном последнее, вероятно, заключается в поверхностных силах, действующих, как известно, в пределах только очень небольшого расстояния. Таким образом, Сакс пытался построить представление об энергиде как об определенной неразложимой единице, отличающейся специфическими свойствами. Он писал, что материальный субстрат жизни (т. е. энергиды) должен иметь определенные изначальные свойства в той же самой степени, как и каждое элементарное вещество, как каждая соль и каждый минерал. Он подчеркивал, что появление всех специфических особенностей можно объяснить, только исходя из изначальных свойств как в онтогенезе, так и в филогенезе растений и животных, т. е. лишь на основе изначальных (наследственных) свойств. Сакс писал, что в такой же степени необходимо привлекать для объяснения специфические свойства и для химических реакций, как химические элементы могут образовывать соединения с новыми свойствами. Сакс говорил далее о саморазвитии эиер-г и д ы, употребляя выражение «ауто-морфоз» энергиды и противопоставляя его «механоморфозам», возникающим как вторичное явление. Таким образом, он приходил к выводу, что каждая органическая форма (вид) возникает в результате взаимодействия аутоморфозов и механоморфозов.
Резюмируя сказанное, мы должны отметить, что наиболее существенным в высказываниях Сакса является утверждение на-
/59
Юлиус Сакс (1832—1897)
линия специфических свойств живого, характеризующегося саморазвитием. Вместе с тем он с особой ясностью впервые проводил взгляд на системность в архитектонике живых существ, взгляд, который по существу очень близок и к современным воззрениям, исторически, вероятно, от него и исходящим. Однако Сакс не дал никакой стройной концепции, ибо он не говорил ничего определенного о том, как надо себе представлять возникновение изначальных (вернее — имманентных) свойств самой клеточной структуры, а также о том, как возникает многоклеточность, в каких взаимоотношениях находятся энергиды друг с другом в организме, и т. д. Тем не менее, развивавшаяся им мысль о качественном своеобразии живой материи была мыслью очень глубокой, опередившей свой век и оставшейся поэтому у Сакса все же недостаточно конкретизированной и мало убеждавшей его современников.
Сакс по существу не касался проблемы взаимоотношений клеток. Поскольку его воззрения обосновывают новое представление только лишь об энергиде как об определенной биологической индивидуальности, они не затрагивают основных моментов целлю-
160
Лярной концепции в целом. Сакс вовсе не касался конкретных вопросов построения организма. Его точка зрения на природу клеток теоретически стоит на значительно более высоком уровне по сравнению со взглядами, сводившими ее жизнедеятельность к функциям суммы биобластов (Альтман и др.). Он решил кажущееся противоречие между видимой простотой строения протопласта и сложностью его отправлений таким образом, что признавал в нем наличие специфической системности, обусловливающей качественное своеобразие живой материи, но что это за силы, Сакс ничего не говорил.
§ 38. Теория «клеточного государства» в трактовке современников. В предыдущих параграфах рассматривались представления о природе клетки в конце XIX в. Мы видели, что этот вопрос решался различно. Одни считали клетку состоящей из целого ряда более элементарных частиц, обладающих самостоятельной жизнедеятельностью, другие признавали ее простейшей неразложимой единицей. Но все сходились на том, что клетка представляет собой определенную структурную единицу. Вторым положением клеточной теории является утверждение относительно гомологичности клеток всех живых существ. Наконец, третье ее положение трактует уже взаимоотношение клеток — этих элементарных структурных единиц в сложном многоклеточном организме. Именно эта последняя часть клеточной теории имеет наиболее важное, принципиальное значение, именно вокруг нее шла и идет до сих пор дискуссия, затухая в одни периоды и разгораясь с новой силой в другие.
Развитие клеточной теории в конце XIX в. являлось по существу лишь дальнейшей детализацией положений, сформулированных Шванном и Вирховым. Поскольку эти представления являются характерными для механистической концепции, их дальнейшее развитие и должно было привести к ряду ошибочных взглядов. Остановимся прежде всего на чрезвычайно типичной для того времени книге «Борьба частей в организме» одного из ведущих теоретиков того времени Вильгельма Ру 2.
В чисто умозрительных высказываниях Ру ведущей идеей было перенесение принципов дарвиновской борьбы за существование и принципов выживания наиболее приспособленных в область микроструктуры организма. Такая мысль возможна только
2 Roux W. 1881. Der Kampf der Theile im Organismus («Борьба частей в организме»).
б Е. М. Вермель
161
при том условии, если считать клетки самостоятельными и независимыми живыми существами. Исходя из позиций классической клеточной теории, рассматривающей организм как сумму равноправных биологических единиц (клеток), Ру построил теорию активной борьбы между всеми структурными единицами, входящими в состав организма. Он исходил из априорного положения известного неравенства всех органических единиц, постулировав отсюда борьбу между клетками, тканями и органами. Он писал, что более сильные клетки должны оставлять большее число потомков, чем клетки более слабые. Далее, он развивал идею, что те клетки будут иметь более существенное преимущество, которые при действии раздражения наименее изнашиваются и в то же время стимулируются к накоплению пищи и к регенерации. В качестве доказательства борьбы клеток он приводил патологические процессы, где клетки одной тканн прорастают в другую ткань, как это наблюдается, например, при деструкции мышечных волокон или кости под влиянием клеток злокачественных опухолей.
Три категории процессов, по мнению Ру, являются полем действия борьбы клетки: 1) их размножение, когда идет борьба за место; 2) накопление и усвоение питательных веществ; 3) освобождение клеток от ядовитых продуктов обмена веществ, накопление которых ведет к их ослаблению,
Принципиальная ошибка Ру заключается в прямом перенесении закономерностей и явлений одного порядка (порядка экологических взаимоотношений организмов друг с другом и со средой) на явления, лежащие совсем в иной плоскости — в плоскости организации живых существ. Регулятивные и коррелятивные процессы Ру были известны, но он им в то время не придавал принципиального значения. На современном этапе наших знаний О размножении клеток мы ясно себе представляем ошибочность этой концепции. Нам известно, что никакого хаоса в процессах размножения клеток нет и ни о каком соревновании за пространство между ними не может быть и речи. Точка зрения Ру является типичной ошибкой механистического воззрения.
Аналогичные взгляды мы находим и у других крупных ученых того времени). Укажем на представления, например, И. И. Мечникова (1903 г. и позже) о воспалении и старости. Он представлял себе воспаление как борьбу фагоцитов с раздражающим агентом или инфекционным началом. Фагоцитирующие клетки Мечников называл санитарами или активными борцами, захватывающими неприятеля. Эта, пусть даже фигуральная, антропо-
162
Макс Ферворн (1863—1921)
морфическая терминология была отражением неправомерного перенесения закономерностей одного порядка в область качественно отличных явлений другого порядка. В теории старости Мечникова мы находим рассуждение о «нападении фагоцитов на благородные клеточные элементы — нервные клетки», причем этот процесс трактуется как фактор старения. На самом же деле нейро-фагия есть лишь следствие гибели нервных клеток.
Подобная точка зрения разделялась почти всеми естествоиспытателями того времени. Но свое, быть может, наиболее законченное развитие клеточная теория получила в книге физиолога Макса Ферворна (1863—1921) «Общая физиология»3. Книга эта выдержала большое число изданий. По философским убеждениям Ферворн был кондициалист. В книге проводится точка
’Verworn М. 1895. Allgemeine Physiologie. Ein Grundriss der Lehre vom Leben.
Ферворн M. 1897. Общая физиология. Основы учения о жизни. Перевод М. А. Мензбира и Н. А. Иванцова.
Ферворн М. 1910, Общая физиология. Основы учения о жизни. Под ред. проф. И. Г. Кожевникова.
6*
163
зрения, согласно которой все отправления организма должны рассматриваться только как совокупность жизнедеятельности самостоятельных клеток, Ферворн развивал теоретическое обоснование этой концепции и пошел в этом отношении дальше своих предшественников. В заключительной главе он писал: «Поскольку мы знаем, что все крупные организмы являются «клеточными государствами», причем государствами, организованными очень рационально, то было бы весьма благодарной темой рассмотреть современную физиологию с точки зрения фактических форм устройства «клеточных государств».
Он даже полагал, что проекты социальных реформ имели бы совершенно иной успех, если бы учитывались данные биологии.
«Клеточное государство» растительных организмов Ферворн называл республиканским, организацию же тела животных он сравнивал с монархическим строем. Монархическим он его называл в связи с тем, что чем выше организовано животное, тем яснее обнаруживается влияние нервных клеток на все ткани тела, причем самостоятельная жизнедеятельность клеток этих тканей нередко понижается до минимума.
Вряд ли стоит подробно останавливаться на критике механистических представлений Ферворна. Ошибочность его концепции заключается прежде всего в том, что он не различает специфических особенностей сосуществования клеток в организме и индивидуумов в обществе. Между тем, поскольку мы имеем явления, качественно совершенно различные, не может быть никакой речи не только о выведении общих закономерностей для этих в корне отличных явлений, ио даже о простом проведении параллелизма между явлениями биологическими и явлениями социального порядка. В силу этого глубоко ошибочна и сама фразеология Ферворна, когда он говорит о «клеточном государстве человеческого тела» и т. п.
Нельзя отрицать некоторую положительную роль клеточной теории даже в ее наиболее механистической форме. Прежде всего, поскольку она была материалистична (хотя и механистична), она соответственно направляла конкретные исследования ученых. Постепенно накапливались многочисленные и точные наблюдения, а потому сама теория должна была быть рано илн поздно пересмотрена.
Критических работ накапливалось все больше и больше, и клеточная теория на рубеже XX в. вновь подверглась энергичной дискуссии и пересмотру. Но самого Ферворна критика того времени нисколько не убедила. В 5-м издании своей книги, 164
вышедшей уже в 1908 г., он держался все тех же положений, как и 10 лет до этого. Он писал: ... «поскольку изучение каждого отдельного отправления органа всегда указывает на клетку, то, следовательно, в клетке и заключается загадка деятельности органа в целом. В мускульной клетке заключается загадка сердечной деятельности, в эпителиальных клетках заключена проблема принятия пищи и всасывания, а в ганглиозных клетках покоится тайна духовных процессов».
Именно поэтому он и считал, что теоретическая физиология может быть только клеточной физиологией.
В такой предельно завершенной форме механистическая концепция учения о клетке вряд ли кем излагалась. Однако именно эта ее логическая завершенность неминуемо приводит к еще одной крупнейшей методологической ошибке. В самом деле, поскольку Ферворн сводил всю жизнедеятельность организма к сумме жизнедеятельности клеток, то он не только игнорировал целостность организма, но также и решение вопроса об индивидуальном развитии организмов оказалось в свете этой концепции по существу лишенным какого-либо материалистического содержания: онтогенез любого Metazoa заключается в том, что на разных его этапах возникают различно дифференцированные клетки. Приняв теорию «клеточного государства», причину дифференцировки можно толковать либо как результат воздействия нематериальных факторов (витализм), либо же рассматривать ее как следствие изменений условий существования клеток, приводящее к усилению одних родов ее жизнедеятельности и к подавлению других. Другими словами, причины (каузальность) заменяются условиями (кондиционализм), что также совершенно неприемлемо.
ГЛАВА ВОСЬМАЯ
КРИТИКА КЛЕТОЧНОЙ ТЕОРИИ
Основные даты в развитии критики клеточной теории
1888—1897 А. Се дж ви к, Ив Де ля ж и К. Уайтман предлагают плазматическую теорию строения организмов.
1891 Г. Д р и ш развивает представление о целостности организма на виталистических началах.
1904 А. Г. Гур внч развивает концепцию об иидивидуалнзн-рсванности и координированной системности организма.
1907 М. Гейд енганн выдвигает принцип функциональной дифференцировки организма взамен целлюлярного принципа.
1911 К. Добелл предлагает считать простейших неклеточными организмами.
1916 Э. Роде выдвигает концепцию плазмоднальной структуры организма.
1927 Ф К. Студннчка предлагает концепцию синцитиальной структуры организма.
1932—1934 В. Я. Рубашкин, 3. С. Кацнельсон н А. В. Немилое отрицают взгляд на клетку как общую для всех организмов элементарную биологическую структуру.
166
39. Целостность организма в трактовках Г. Дриша и A. Fypj вича. Биологи и медики последних трех десятилетий XIX в. в общем вполне удовлетворялись клеточной теорией Шванна — Вирхова — Шультце. Конкретные исследования были направлены на детальное изучение структуры .протоплазмы и ядра в связи с тем, что клетка признавалась элементарным организмом и к ее жизнедеятельности сводилась вся деятельность организма. Этот ход развития науки о клетке был закономерен. Для кардинального пересмотра общетеоретических положений было еще слишком мало фактических данных, и вполне естественно, что, пока клетка оставалась недостаточно изученной, она должна была оставаться в центре внимания исследователей. Одновременно совершенствовались как микроскопическая анатомия, так и эмбриология, которая должна была дать ключ к пониманию гистогенеза. Вместе с тем накапливался ряд фактов, из которых рождались представления о целостности организма, о его системности и системности его органов. Как описательный материал, так в особенности и экспериментальный все с большей и большей ясностью показывали, что системность не сводится к простой сумме клеточных построений и отправлений. Не только физиологическая сторона вопроса, но и анатомическая его сторона показывали, что существуют какие-то общие регулятивные процессы, которые представляют собой нечто более сложное и вполне специфическое. Для своего объяснения они требовали уже другой, новой, более глубокой теоретической трактовки. Примитивное представление о сложном организме как арифметической сумме низших, или элементарных, организмов не могло удовлетворить исследователей, а отсюда возникала необходимость создания и новых теоретических трактовок.
Остановимся прежде всего на понимании и трактовке клеточной теории школой известного эмбриолога и наиболее последовательного виталиста начала XX в. Ганса Дриша (1867—1941). Сам он специально по данному вопросу не выступал, но отрицательно относился к клеточной теории и высказывался по этому вопросу при рассмотрении общих проблем развития организма.
Именно это последнее вскрывает виталистическую сущность концепции Дриша поскольку из нее следует, что свойства организма как целого не обязательно находятся в связи с субстратом последнего. Положи|тельным в его критике является то, что
1 Driesch Н. 1928. Phylosophie des Organischen. 4 Aufl. («Философия органического мира»).
167
Дриш привел экспериментальные доказательства, опровергающий трактовку организма как агрегата клеток, при этом сам факт наличия клеток во всех органах сомнению им не подвергался.
Положительные элементы этой концепции в разбираемом вопросе изложены очень логично и последовательно Александром Гавриловичем Гурвичем (1874—1954) в его известной книге «Морфология и биология клетки»2. В определении организма Гурвич развивает представления об его индивидуализированно-сти и координированной системности. Системность, как он отмечает, имеет место и в мире неорганическом, но постоянное и закономерное взаимодействие структурных единиц в целом характерно только для живых существ. Гармоническое взаимодействие системных компонентов он называет координацией. Наличие последнего обусловливает устойчивость органических систем, стабильность которых преобразуется вместе с тем в самостоятельный фактор их существования.
Итак, основной заслугой школы Дриша, а также Гурвича является то, что для трактовки клеточного учения было привлечено понятие системы. К пониманию всего значения такой постановки проблемы подошли только в последнее время. Следует считать, что с принципиальной стороны этот путь является единственно правильным. Однако в свое время эта концепция не сыграла той роли, на которую она безусловно имела право. В этом отношении имело значение прежде всего то обстоятельство, что во всех представлениях Дриша решающим фактором выставлялась энтелехия (дословно — нечто несущее в себе цель), т. е. признание трансцендентных сил. Борьба ученых-1матери|али-стов с витализмом как принципиальной концепцией заслонила собой и то, что являлось положительным в представлениях Дриша об организме. Имело значение также и то обстоятельство, что его точка зрения в отношении клеточной теории не была изложена с необходимой полнотой и ясностью.
Концепция Гурвича может быть в немногих словах изложена следующим образом: жизнедеятельность многоклеточного организма не является простой функцией составляющих его клеточных элементов, а отправления отдельных клеток, как таковых, не предопределяют всю жизнедеятельность сложного организма. Таким образом, постулируется особый тип эндогенных процессов, являющихся свойством не отдельных
2Gurvitsch А. 1904. Morphologic und Biologie der Zelle («Морфология и биология клетки»).
168
элементов организма, т. е. клеток, а лишь органа или даже организма в целом. Основной фактический материал, который рассматривает Гурвич, касается преимущественно эмбриологических процессов, в основном из области развития первых фаз дробления. Эти фактические данные с необычайной ясностью показали, что целостность организма развивающегося эмбриона не может быть сведена к свойствам и особенностям отдельных клеток. Такая постановка вопроса, вне всякого сомнения, являлась шагом вперед. С несомненностью, в частности, было доказано, что в случае недетерминированного дробления каждый бластомер оказывает направляющее воздействие на развитие и судьбу всех остальных — прежде всего он ограничивает их потенции. Достаточно в качестве примера указать на опыты по разделению бластомеров на стадиях 2, 4 и даже 8 клеток при развитии морского ежа или лягушки. Как известно, из выделенного из общей связи бластомера развивается целый эмбрион, но только лишь меньших размеров. То же наблюдается и с отдельными зачатками: их расщепление приводит ж образованию двух, например,
169
голов вместо одной, двух лап на месте одной и т. д. Взаимодействие двух сближенных соприкасающихся клеток влияет не только на их развитие, но даже на их форму и форму их ядер — они принимают иногда бобовидную форму, прилегая выемками друг к другу. Подобных фактов известно очень много. Они легли в основу целой науки — экспериментальной эмбриологии, или механики развития организмов. Мы, конечно, не можем здесь на них останавливаться. Эти исследования впервые были развернуты Ру и Г. Шпеманном (1869—1941) в Германии, а у нас в СССР — Д. П. Филатовым (1876—1943) 3.
Фактические данные экспериментальной эмбриологии позволили Гурвичу сформулиоовать и обосновать понятие «биологического поля» (см. § 54) и сделать попытку установить определенные закономерности воздействия клеток в эмбрионе и в органе друг на друга (1944 г.). В таком подходе к биологическим явлениям, конечно, ничего виталистического уже нет. Более того, подобное направление в исследовании, на наш взгляд, прогрессивно, ибо оно ведет к научному изучению и прогнозированию процесса развития и жизнедеятельности организма. Надо только уметь объективно делать заключения и не спешить называть непознаваемым непознанное и особенно непонятое.
Следует указать, что именно Дришу принадлежит формулировка закона «Funktion der Lage» (функция положения), т. е. определяющее значение местоположения зачатка в эмбрионе на его дальнейшую судьбу. Экспериментальное перемещение лоскута кожи зародыша поверх зачатка глаза приводит к тому, что именно из этого лоскута возникает хрусталик (линза глаза); таким образом, можно утверждать, что нервная закладка глаза индуцирует образование линзы из кожи. Приведем еще один яркий пример: зачаток регенерирующего хвоста тритона, будучи пересажен в область конечности, превратится в ногу, а не в хвост. Таким образом, новая область тела, куда пересажена регенери-оующая ткань, определяет развитие зачатка именно в конечность. Подобных примеров известно очень много, и эта область знания в настоящее время энергично продолжает развиваться.
К сожалению, изложенная выше концепция была недооценена ни в то время, ни даже в должной мере и теперь. Ее сущность сводится к признанию того, что организм как целое имеет иные свойства, несводимые к свойствам
3 Филатов Д. П. 1939. Сравнительно-морфологическое напоавление
в механике развития, его объект, цели и пути. М-, Изд-во АН СССР, по
его частей. При этом представлены доказательства того, что организм как целое действительно оказывает сильное влияние па поведение частей развивающейся особи.
В вопросе о том, как нужно понимать это регулирующее влияние целого, у Гурвича в 1904 г. не было ясной позиции. С одной стороны, он говорил о координациях, обусловливающих собой новые качества, а с другой, разбирая конкретные биологические явления, считал, что биологический смысл многих процессов едва ли может быть понят, так как ряд этих явлений не находится в прямой связи с морфологической структурой. Дриш совершенно четко стоял на этих позициях, прямо утверждая невозможность материалистического объяснения ряда основных биологических явлений. Гурвич был более осторожен в высказываниях, но и у него непознаваемость, как можно понять, занимала, вероятно, определенное место в защищаемой им концепции.
Несмотря на все это, за школой Дриша нельзя не признать определенных серьезных заслуг. Стоя на целлюлярной точке зрения, эти исследователи убедительно показали ограниченное значение клетки как самостоятельной единицы в составе ткани многоклеточных организмов. Вместе с тем Гурвич выступил с критикой механистического принципа целлюлярной биологии и патологии. Он указывает, что эта точка зрения не объясняет целого ряда фактов и даже находится с ними в прямом противоречии. Опровергая классическую клеточную теорию «кирпичиков» и давая доказательство зависимости поведения клетки от целого организма, Гурвич в своей монографии 1904 г. не предлагает взамен никакой другой ясной концепции. При всем нашем отрицательном отношении к нематериалистическим теориям мы обязаны отметить, что положенные в их основу факты представляли собой новые, важные открытия. Механистами-материалистами эти факты были просмотрены, вернее, не поняты, а потому и оставлены без внимания.
§ 40. Представление М. Гейденгайна о целостности организма и его критика клеточной теории. Недостаточность клеточной теории Шванна — Вирхова с особой полнотой была показана выдающимся гистологом начала XX в. Мартином Гейденгайном (1864— 1949) в его широко известной книге «Плазма и клетка»4. Старая клеточная теория была названа им трудно переводимым на рус-
4Heidenhain М. 1907. Plasma und Zelle («Плазма и клетка»).
171
ский язык термином «Bausteintheorie», что в условном переводе означает «теория кирпичиков», сущность которой в изложении Гейденгайна сводится к следующим положениям.
1. Живая субстанция сконцентрирована в форме клеток, и вне клетки она существовать не может; все клетки являются однозначными структурами сходного морфологического порядка. Тканевые клетки как в морфологическом, так и в физиологическом отношении представляют собой индивидуумы, а тело животного является агрегатом этих клеток, представляющих собой как бы «кирпичики» («Bausteine») многоклеточного организма-здания.
2. В соответствии с принципом разделения функций, жизнедеятельность целого организма может быть разложена на разнообразные отправления, распределяющиеся между различными типами и видами клеток. Общая жизнедеятельность организма рассматривается как содружественная работа клеток, вернее, как арифметическая сумма их отдельных отправлений.
Заслуга Гейденгайна прежде всего заключается в том, что он придал клеточной теории четкую формулировку, хотя и не совсем объективную, а затем выступил с критикой ее основных положений. Гейденгайн с особой ясностью справедливо указал, что на развитие клеточной теории большое влияние оказали онтогенетическая и филогенетическая точки зрения, тогда как ги-стофизиологическая сторона вопроса осталась недооцененной. Он считал, что этим путем не может быть создано удовлетворительной теории строения организма, поскольку логически она не может быть выведена только из одного генеза. Помимо того, Гейденгайн дал несколько иную формулировку понятия клетки, указав, что определение Шультце является скорее только морфологическим ее описанием. Под клеткой Гейденгайн понимает ограниченный комочек живой субстанции, который как в морфологическом, так и в физиологическом отношении обладает характерными свойствами элементарного организма-индивидуума. Отсюда ясно, что наиболее важной проблемой является вопрос о том, что надо понимать под словом «индивидуум». Индивидуальность клетки, которую биологи уже давно, следуя за химиками, считали элементарной «живой органической молекулой», обосновывалась следующим образом. Проводилось сравнение между клетками и одноклеточными организмами. Далее, со времени Шванна и особенно Валентина указывалось, что выделенные из тканей клетки отмирают не сразу. Это явление хорошо видно на изолированных клетках реснитчатого эпителия, когда ясно удается уста-
172
Мартин Гейденгайн (1864—1949)
новить по движению ресничек, что их деятельность прекращается лишь постепенно, отдельными участками. В организме Metazoa имеются свободно живущие клетки — белые кровяные тельца (лейкоциты). Далее, как важнейшее доказательство их самостоятельности, вернее — индивидуальности, приводилось явление размножения клеток. Наконец, в настоящее время можно еще указать на действительно в значительной степени индивидуализированный характер в жизнедеятельности клеток злокачественных опухолей, доказанный, в частности, экспериментально мсто-
173
Дикой культивирования тканей вне организма (Фишер, 1925, и многие другие).
На развитие клеточной теории, несомненно, очень большое влияние оказала эволюционная теория, что особенно ясно было выражено Геккелем, использовавшим учение о клетке для построения филогенетической схемы развития организмов от простейших, одноклеточных, через колониальные формы, состоящие из равнозначных клеток, к истинным многоклеточным существам, построенным из разнообразных многообразно дифференцированных тканей.
Представление об организме как о «клеточном государстве» в конце XIX в. по существу было общепризнанным, и мы его можем найти в любом учебнике и популярной книжке того времени, касающихся проблемы строения организма. Успех этой теории объясняется еще и тем обстоятельством, что, как указано Гейнденгайном, получить ясное доказательство взаимозависимости и соподчинения клеток в большинстве случаев оказывается делом очень трудным и нужно быть образованным человеком, чтобы это понять. Вместе с тем «Bausteintheorie» очень удобна, ибо с этой точки зрения можно очень просто изложить принцип разделения труда, т. е. разделения функций между различными клетками, принцип, который был известен уже очень давно, но который кажется простым только по незнанию проблемы и фактических данных.
Отсюда родилось учение о «клеточном государстве» и сравнение его с человеческим обществом. Все это есть следствие, как правильно отметил Гейденгайн, переоценки цел-люлярного принципа или, вернее, слишком прямолинейной и формальной его трактовки.
Гейденгайн возражал против клеточной теории, которую он сформулировал очень механистично, указывая, что понятие живой субстанции имеет более широкую природу, чем понятие клетки. Исходил он при этом из двух положений. Он полагал, что нельзя считать доказанной клеточную организацию мельчайших микроорганизмов—бактерий (величиной до 0,5 р. в диаметре) и затем доказывал, что свойства межклеточного вещества не укладываются в рамки клеточной организации, хотя вместе с тем оно является живым. Эти два положения, особенно второе из них, нашли себе в гистологической литературе широкий отклик. В дальнейшем к ним добавилось еще и третье — синцитиальное строение ряда тканевых структур. Вторичность же строения этих структур во внимание не принимали.
174
Очень многие исследователи указывали на ряд наблюдений, из которых следовало, что межуточное вещество соединительной ткани обладает рядом признаков живой материи. Полагали, что, возникнув как производное клетки, оно существует и растет самостоятельно. Однако это последнее никогда никем доказано не было, и, несомненно, взгляд, изложенный в такой форме, ошибочен.
Значительно более приемлема в этом отношении позиция самого Гейденгайна, считавшего, что межуточное вещество находится в коррелятивной зависимости от регулирующих влияний окружающих клеточных структур, обусловливающих его рост и деятельность. Интересно указать, что природа регулирующего фактора была неясна для Гейденгайна. Сам он отмечал, что вопрос этот в данный момент остается нерешенным. Но отсюда еще не следует, что проблема, неразрешенная сегодня, если она правильно поставлена, не будет решена позднее.
Итак, основное возражение Гейденгайна против клеточной теории сводится к тому, что в организме констатируется наличие значительных живых масс неклеточного строения, генетически, однако, связанных с клетками. Ошибкой старой клеточной теории Гейденгайн считал также положение, согласно которому все клетки являются равнозначными, гомологичными единицами. Он полагал, что такой взгляд представляет собой абстракцию, настолько далеко отходящую от реального положения вещей, что ее научная ценность становится совершенно ничтожной, ибо с гистофизиологической точки зрения о равнозначности клеток говорить не приходится. Другими словами, Гейденгайн считал, что эта ошибка происходит в силу переоценки генетического, а в сущности филогенетического, подхода к изучаемому вопросу. Поскольку из клеток возникают структуры, которые уже перестают быть частью клетки, а представляют собой вполне своеобразное образование, то, следовательно, нельзя говорить о том, что организм может быть разложен на клетки, т. е. на одинаковые по своему значению структурные элементы. Непонятно, почему при подобного рода рассуждениях и построениях забывают, что межуточное вещество всегда имеет очень ограниченную продолжительность жизни и заменяется другим за сравнительно короткие сроки.
Более подробное изучение микроструктур организма, согласно представлениям Гейденгайна, показывает, что тело многоклеточных складывается не просто из суммы сходных биологических Индивидуумов-клеток, ибо сами клетки создают более высокие
775
биологические единицы (гистосистемы). Он развивал представление, согласно которому процесс развития зависит от размножения клеток, но, после того как образование формы и частей организма закончено, роль клетки, как таковой, при специальной физиологической деятельности, органа уже не имеет решающего значения.
Таким образом, представление о самостоятельной индивидуализированной деятельности клеток в целом организме следует считать ошибочным, что было обосновано Гейденгайном на большом и, казалось бы, убедительном фактическом материале. В этом и заключается его основная заслуга. Серьезным недостатком его концепции является утверждение, что и сама клетка разложима на еще более элементарные единицы, названные им протомерами. Он считал, что жизнь является определенным свойством самой живой субстанции, нисходящей в своей архитектонике до отдельных молекулярных соединений (протомеров).
Изложенная точка зрения Гейденгайна исходит, следовательно, из констатации факта разнообразия клеток и их производных. Так как он не видел возможности как-либо свести это разнообразие к единому клеточному прототипу, он поэтому считал, что должна быть найдена еще более примитивная, ниже стоящая, более общая структура. Все это и приводило к логической необходимости построения гипотезы протомеров. По существу эта гипотеза мало оригинальна, и сам ее автор указывает, что он исходил из представлений немецкого ботаника Ю. Визнера (1838— 1916) об элементарных живых частицах — элементарных органах, развитых им в книге «Элементарная структура и рост живого вещества» 5.
Таким образом, концепция Гейденгайна сводится к тому, что он считает необходимым трактовать наблюдаемые в организме структуры как производные каких-то элементов, организованных проще, чем клетки. По его представлениям, элементарными единицами являются гипотетические протомеры. Определенное их сочетание создает новую систему — клетку. Определенное количество клеток создает еще более сложную структурную единицу— гистомер (например, альвеола) (рис. 39). Последние в свою очередь, объединяясь определенным образом, создают более сложную гистосистему (например, первичную дольку железы и т. д.).
’Wiesner J. 1892. Die Elementarstruktur und das Wachstum der leben-den Substanz («Элементарные структуры и рост живого вещества»).
176
Рис. 39. Развитие поджелудочной железы по Гейденгайну. Показаны альвеолы (ацинусы) железистой части и их деление; гистосистемы 1-го порядка
Основным недостатком представлений самого Гейденгайна, не говоря уже о том, что гипотеза протомеров ни в какой степени не может считаться доказанной, является то обстоятельство, что она недостаточно учитывает проблему качественной специфичности системы любого порядка.
Укажем далее на отдельные любопытные моменты из критики клеточной теории Гейденгайном. Так, он считает, что принцип физиологического разделения отправлений между клетками гораздо правильнее обозначить как принцип функциональной дифференцировки, что ни в какой мере не является в его представлении результатом или свойством, присущим лишь клеточной структуре. Принцип функциональной дифференцировки имеет более общий характер, чем целлюлярный принцип. При этом Гейденгайн добавляет, что главное значение клетки, как таковой, выявляется в процессе развития организма, тогда как ее самостоятельная жизнедеятельность в функции сформированного органа отходит на второй план. Такая точка зрения является
/77
определенным шагом вперед в развитии клеточной теории, независимо от развиваемой Гейденгайном теории системности гистологических структур. Он совершенно прав, когда указывает, что представление о физиологическом разделении труда между различными клетками, функционирующими содружественно, представляющее собой один из основных принципов теории «клеточного государства», является по существу ясно выраженным антропоморфизмом. Гейденгайн много раз подчеркивает, что проводимый им принцип функционального разделения по своей природе значительно шире, чем принцип клеточный. И поэтому, если его сужать, как того требует теория «клеточного государства», и сводить все на разделение труда между клетками, не принимая во внимание функциональной дифференцировки частей как единых систем, что и привносит новые качества, то мы неизбежно будем приходить к ошибочным взглядам. С его точки зрения, л ю б а я функция есть функция органа в целом, ибо, будучи разложенной на клетки, она уже в действительности не существует. Другими словами, определенное отправление органа возможно только до тех пор, пока он представляет собой единое целое. Отсюда Гейденгайн делает совершенно правильный вывод, что соотношение единиц, составляющих животный организм, совершенно иное, чем в человеческом обществе и в государстве (или в любом биоценозе организмов).
Мы остановились на воззрениях Гейденгайна лишь в самой общей форме, не задерживаясь на конкретном изложении его представлений относительно организации тканей из гистомеров и гистосистем. Нас интересовал прежде всего его взгляд на клеточную теорию. Его критика была аргументирована в такой убедительной форме, что с этого момента можно было считать старую клеточную теорию по существу оставленной. Книга Гейденгайна «Plasma und Zelle» была широко читаема, и влияние ее на современников было очень велико. В заключение отметим, что почему-то наибольшим успехом пользовалась и пользуется до сих пор идея о недостаточности клеточного принципа, аргументируемая жизненными свойствами неклеточной, межуточной субстанции. Дискуссия шла (да и продолжается по настоящее время) по вопросу: действительно ли межуточное вещество является живым или не вполне живым? Ниже мы увидим, как эта проблема решается в самое последнее время (см. гл. IX, § 53). Вместе с тем идея целостности организма привлекала гораздо меньшее внимание, несмотря на то, что она глубже и принципиально важнее первой.
178
§ 41. Основатели плазматической теории строения организмов: А. Седжвик, Ив Деляж и К. Уайтман. Недостаточность целлю-лярного принципа для объяснения развития функций организма отмечалась многими учеными. Именно эта идея лежала в основе критики как Гурвича, так и Гейденгайна, указавших, что свойствами отдельных клеток ие исчерпываются жизнедеятельность и особенности многоклеточного организма в целом. Но вместе с тем оба указанных исследователя ни в какой степени не подвергали сомнению самый факт наличия клеток во всех без исключения тканях. Оба этих выдающихся ученых развивали идею системности, однако ни тот, ни другой не дали ее решения, которое удовлетворяло бы широкие круги биологов. Но часто постановка проблемы имеет не меньшее, если не большее, значение, чем ее конкретное решение. Неважно даже, если сам автор, поставивший проблему, решает ее неправильно. Мы видели, что концепция А. Гурвича (хотел этого сам автор или нет) по существу приводит к признанию нематериальных факторов, которые якобы и обусловливают специфический характер системы. Гейденгайн вопрос о специфичности системы вообще затрагивает мало. Но поскольку эти оба исследователя говорили о системах, они должны были признавать клетку конструктивной единицей.
Многие другие ученые пытались объяснить противоречия между признанием клеточной организации всего живого и несводи-мостью жизнедеятельности организма к сумме функций клеток совершенно другим путем. Они пытались опровергнуть реальность клетки как структурной единицы живых существ и считали, что по существу организм представляет собой анатомически единую структуру. Другими словами, отмеченное выше противоречие решается ими необычайно упрощенно и утверждается, что представление о клетке, как какой-то конструктивной единице, просто не соответствует действительности. Таким образом, сама проблема несводимости жизнедеятельности целого (организма) к свойствам его частей (клетки) лишается конкретного содержания, ибо организм трактуется как единое анатомическое целое. Впервые подобная точка зрения была выдвинута Адамом Седжвиком (1888—1892) 6. Фактиче-
s Sedgwick А. 1888. Monograph on the cape species of Pcripatus Quart. Jr. microsc. Sci. («Монография о капском виде перипатусл»). 1892. On the inadequancy of the cellular theory of development ets. Quarl. Jr. microsc. Sci. («О недостаточности клеточной теории развития»).
179
ское доказательство этой точки зрения А. Седжвик усматривал в том, что в мезенхиме, в нервной ткани, в дробящемся яйце некоторых члсиистоногирс и др., по его мнению, при объективном изучении не представляется возможным установить наличие клеток. Аналогичный взгляд высказывал также французский зоолог Ив Деляж (1896—1897) 7, утверждавший, что клетки являются вторичной структурой. Он полагал, что первичными организмами являются многоядерные простейшие; Metazoa же возникают таким образом, что тело Protozoa, имеющего несколько ядер, распадается на отдельные клетки, причем этот процесс не всегда идет до конца.
Наибольшее внимание в свое время привлекла, пожалуй, работа К. Уайтмана8, в которой автор очень энергично выступил с опровержением целлюлярного принципа, базируясь прежде всего на своих наблюдениях над строением нефридий у кольчатых червей. Он утверждал, что клеточные границы в этом органе крайне непостоянны; клеточное и плазматическое строение нефридия легко сменяют друг друга. Отсюда следовал вывод, что клетка не является определенной структурной единицей и поэтому она не может рассматриваться как какой-то константный элемент развития организма. По этой причине Уайтман ставил вопрос и о том, что любое простейшее, являясь организмом, не может считаться клеточной структурой. Он предложил клеточную теорию заменить «организменной точкой зрения», т. е. понятием морфологической целостности и признанием непрерывности структурных элементов организма. «Деление на клеточные территории имеет, в его понимании, второстепенное значение, а развитие, как и другие жизненные феномены, представляет собой функцию организма как целого» (Уайтман).
Однако сколько-нибудь стройного представления и цельной системы воззрений Уайтман не дал. Оставалось неясным, что он понимает под организационным принципом, о котором пишет. Вместе с тем он говорит об «элементарных живых элементах», для которых он даже предложил название «идиозом». Подобная точка зрения, как мы видели, не оригинальна и показывает лишь 7 Delage Y. et Labre. 1896. La conception polyzoique des etre. Rev. Sci. N 21 («Полизоидная концепция живых существ»). 1897. La diffe-renttiation des organismes. Rev. Sci. N 25. («Дифференцировка организмов»),
“Whitman С. O. 1893. The inadequancy of the cell-theory of development. Journ. of MorphoL, 8, 639—658. («Недостаточность клеточной теории развития»).
180
бессилие исследователя объяснить твердо установленные наукой факты, в силу чего он вынужден прибегать к признанию гипотетических биологических единиц.
Любопытно отметить, что этого не избежали даже крупнейшие биологи. Гипотетические «живые частички» разными исследователями всегда назывались новыми терминами. Одно их перечисление может занять довольно много места. Приведем некоторые из них: «идиоплазматические частички» К. Нэгели (по его представлениям, они в свою очередь состоят из мицелл), «физиологические единицы» Г. Спенсера, «пангены» де Фриза, «биофоры» А. Вейсмана, «геммулы» Ч. Дарвина, «биогены» М. Ферворна, «метаструктурные частички» В. Ру, «плазо-мы» Визнера, «идиобласты» О. Гертвига, «протомеры» М. Гейденгайна и др.
Итак, всякий раз, когда автор не знает, как ему привести в систему имеющийся фактический материал и построить теорию взаимоотношений клеток как определенных индивидуумов с организмом как единым целым, он обращается к какой-либо гипотезе, более или менее вероятной с его точки зрения. Логика вещей заставляет искать простейшую биологическую единицу, и если клетка почему-либо как элементарная структура отвергается, то волей-неволей мысль направляется в сторону невидимых субмикронных частиц. Другими словами, на место клетки ставится другая, но уже гипотетическая структура. Кстати сказать, современные данные электронной микроскопии никаких подтверждений существования субмикроиных биологических единиц, состоящих ниже клетки, не принесли 9.
§ 42. Одноклеточные как клетка и организм. Из развивавшихся Седжвиком и Уайтманом представлений наибольшее значение имело утверждение относительно принципиальной сравнимости протиста только с организмом Metazoa в целом, а также указание на синцитиальность строения целого ряда органов. Почти невыполнимой задачей было бы перечисление всех исследователей, которые продолжали развивать эти мысли. Укажем лишь на немногих авторов.
Одним из наиболее серьезных ученых, развивавших подобные мысли, был К. Добелл (1911). Он и его последователи доказыва-
9 См. книгу Робертнс Э. де., Новинский В., Case Ф. 1967. Биология клетки. М., изд-во «Мир».
181
Лй, что такое сопоставление является более содержательным также в том отношении, что к простейшим понятие клетки не приложимо ив силу этого их следует считать н е к л е-точными организмами. Действительно, на первый взгляд сравнение отдельных органелл простейших с целыми органами Metazoa имеет известный смысл. Однако еще никто не мог показать, что эти сравнения имеют большую ценность, чем простая аналогия. Вместе с тем при сравнении простейших с целым многоклеточным организмом возникает при детальном анализе целый ряд трудностей, которых не знает клеточная теория. Ограниченность такого сопоставления очевидна. Можно сравнивать лишь наиболее дифференцированных Protozoa вроде инфузорий; если же мы возьмем амеб или жгутиконосцев и поставим их рядом с наиболее низко организованными Metazoa (например, губки), то получим иную картину. Совершенно ясно, что сравнение органелл простейших с органами губок не имеет никакой цены и мы должны сравнивать эти организмы исходя только из клеточного принципа. Особенно это справедливо по отношению к воротиичковым клеткам (хоаноцитам) губок с их воротничком, жгутиком, сократимой вакуолью, с одной стороны, и хоанофля-геллятами (биченосцами, крайне сходными с этими клетками)— с другой. Всякое сравнение Protozoa с Metazoa искусственно, ибо оно может производиться лишь в самых общих чертах, не выходя притом из рамок ни к чему не обязывающих аналогий. Гораздо естественней и проще признать, что наблюдаемое сходство в структуре специализированных Protozoa с гистосистемами и органами Metazoa развивалось исторически независимо.
Признание простейшего действительно одноклеточным ни в какой мере не противоречит, как считает большинство ученых, признанию его одновременно целым организмом. Вряд ли стоит доказывать, что морфологический анализ и особенно изучение онтогенезов со всей отчетливостью показывают несравнимость органелл простейших с дифференцированными тканевыми структурами Metazoa. Однако подобные заблуждения повторялись нередко до самого последнего времени. Так, еще в 1934 г. А. В. Немилое в своей речи на 1-й гистологической конференции в Москве говорил: «В них (инфузориях) есть нечто от клетки, но они, если можно так выразиться, «переросли» клетку. «Ведущим» для них является не то, что их можно приравнивать к клетке, а высокая степень дифференцировки при отсутствии расчлененности. Недаром именно со стороны протистологов, которым специфика этих органов известна лучше, чем кому-нибудь друго
782
му, было предложено назвать их «бесклеточными». Но в самом слове «б е с к л е т о ч н ы е», «н е к л е т о ч н ы е» есть некоторый элемент пройденной стадии одноклеточности. Поэтому более удобно, мне представляется, название «надклеточные организм ы».
Однако все эти упражнения в составлении терминов лишены какого-нибудь серьезного смысла, ибо, сколь бы сложна ни была клетка, она все же остается клеткой, а поэтому нет реальных оснований для того, чтобы говорить о «перерастании» клетки в «надклеточные» организмы. Это прекрасно было понято в свое время еще Ф. Энгельсом. Придавая этому вопросу принципиальную значимость, он писал следующее 10 11: «инфузории это «одноклеточные» существа, которые «представляют собой, хотя и сравнительно низкие, но уже существенным образом дифференцированные организмы, ...снабженные довольно развитыми органами. Все они, по меньшей мере, одноклеточные».
Смысл приведенного означает, что инфузории надо рассматривать как дифференцированные клетки, совмещающие в себе все функции, и в этом сущность проблемы. Отсюда следует, что любое простейшее являетсяи клеткой, и организмом в целом, а еще проще — представляет собой свободноживущую клетку.
§ 43. Значение синцитиев и учение К. Студнички о единстве организма. Исходя из факта тесной анатомической связи клеток в ряде тканей, а в особенности исходя из наблюдений над много-ядерностью некоторых тканевых структур, в этой области было предложено много спекулятивных теорий. При ближайшем изучении выяснилось, что подобные образования весьма разнообразны как по своей морфологии, так и по их генезу. В связи с этим, в частности, возникла разнообразная и запутанная терминология. Возникли термины: «плазмодий», «синцитий», «синцелий», «синдесмий», «симплазма», «симпласт» и многие другие. Иногда они употребляются как синонимы; но нередко им пытаются дать определенное, ограниченное значение. Чаще других применяется термин «синцитий» п, введенный Геккелем (1872). Он определял-
10 Энгельс Ф. 1964. Диалектика природы, стр. 168. М., Изд-во политической литературы.
11 Приставки «сии» и «сим» следует переводить как слово «вместе»; отсюда — «соклетие» («сив» — вместе и «цитос» — клетка).
183
ся самим автором так: «Синцитием у известковых губок я называю тканевую массу, которая произошла путем слияния жгутиковых клеток... синцитий отличается от плазмодия наличием клеточных ядер...; плазмодий представляет собой колонию цитод» 12.
Обычно в гистологической литературе под словом «с и н ц и-т и й» понимают более или менее рыхло соединенные анастомозами (перемычками) клеточные сети. «С им пласт ом» в последнее время называют более или менее значительные протоплазматические массы с большим числом ядер. Ряд исследователей пытался внести ясность и провести определенную классификацию этих понятий. Однако эти попытки не получили сколько-нибудь широкого признания, ибо все подобные структуры по существу очень разнородны. Многоядерные плазматические массы могут возникнуть как вследствие слияния клеток, так и в результате запаздывания и вообще выпадения процесса разделения тела клетки при размножении ее ядер. Далее, в одних случаях — это полноценные потенциальные элементы, разделяющиеся позднее на клетки целиком или частично, в других — это необратимо дифференцированная ткань. А между тем в критике клеточной теории именно последняя категория многоядерных структур играла и продолжает подчас играть значительную роль и до сих пор.
Любопытно указать, что как противоречие закону Вирхова («всякая клетка от клетки») приводилось соображение, что клетки могут возникнуть путем раздробления симпласта, а следовательно, не из клетки. Подобные соображения не имеют, однако, принципиального значения, ибо всякий симпласт, если уж называть этим термином протоплазматическую многоядерную массу, сам является производным клеток. Таким образом, и эти «исключения» подтверждают по существу пра.вило и не заслуживают сколько-нибудь серьезного внимания.
Ряд исследователей пытался подтвердить представление о целостности организма, основываясь на анатомической связи некоторых групп клеток, но эта точка зрения переоценивает значение анастомозов (клеточных перемычек, мостиков), которым, по мнению ряда авторов, следует придавать принципиальное значение с точки зрения морфологического единства организма.
Остановимся коротко еще на учении об организме как е д и н с т в е («Einheitslehre»), сформулированном сравнительно
12 Haeckel Е. 1872. Die Kalkschwamme. Berlin («Известковые губки»). 184
Недавно Крупным чешским ученым К. Студничкой. Во главу угла он ставит представление о тесной морфологической связи всех живых структур. Студничка считает, что учение об организме как единстве отличается от клеточного учения (в смысле «Ваи-steintheorie») главным образом тем, что оно наряду с клетками различает также синцитии и симпласты, а также считает живой основную субстанцию (основное вещество) соединительной ткани 13. Таким образом, он полагает, что протоплазма — не един-< гвенная форма живой материи.
Студничка рассматривает многоклеточные организмы как единый комок «ж ивой масс ы», в которой различные плазмы дифференцированы по-разному. Он допускает, что в отдельных участках организма, а именно, в ядросодержащих участках («клетках») или только в клеточных ядрах должно сохраняться своего рода «осознание целого», которое при известных обстоятельствах (при размножении и регенерации) может себя проявить. Признание за клетками «осознания целого» является откровенно выраженной виталистической концепцией. Вместе с тем это представление о структуре организма, несмотря на то, что оно называется учением об организме как единстве, имеет в конечном итоге много общего с механистической теорией «клеточного государства».
В самом деле, по этой концепции получается, что «осознающие целое» структурные единицы (ядра, клетки либо симпласты— это безразлично) направляют свою жизнедеятельность так, чтобы создавался и функционировал сложный организм.
§ 44. Оценка плазмодиалъных структур в работах Э. Родэ. Антицеллюлярная концепция пользовалась одно время большим успехом. Причину ее успеха следует искать в том, что многие авторы не знали, как совместить неопровержимо доказанную целостность организма Metazoa с фактом расчленения их органов и тканей на клетки. Эти исследователи пошли по линии наименьшего сопротивления и «пожертвовали» клеткой. Другими словами, утверждалось, что организм потому целостен, что таково его анатомическое строение, а расчленение его на клетки принципиально несущественно и вообще является чуть ли не кажущимся. Нетруд
13 Studnizka F. К. 1929. Die Organisation der lebendigen Masse. Handb.
d. mikr. Anat. d. Menschcn. I, 421—568. («Организация живой массы»).
185
Но видеть всю примитивность подобных рассуждений, игнорирующих реальные факты.
Дальше всех в этом отношении пошел Э. Родэ, утверждавший, что тело многоклеточного организма представляет собой единую плазматическую массу, которая в целом соответствует телу одноклеточного существа14. Он утверждал, что дифференцировка органа не является продуктом развития клеток и что имеет место как раз обратное: тканевые клетки представляют собой следствие гистологической дифференцировки целого организма. Делая столь далеко идущий теоретический вывод, сводящий на нет взгляд иа клеточную организацию всех организмов, автор, казалось бы, должен был привести исчерпывающий фактический матерал и убедительные широкие теоретические обобщения развиваемых им положений. Между тем, читая его работу, можно только поражаться легкомыслию, с каким оперирует автор над фактическим материалом. Мы не можем поэтому не согласиться со справедливостью замечания Ю. Шакселя, который писал, что если бы Родэ дал полное, ясное и понятное изложение своей точке зрения, то ее можно было бы легко опровергнуть и этим была бы внесена ясность. Но в таком виде, как она изложена, она бессмысленна 15. Тем не менее работы Родэ импонировали многим гистологам.
Одним из основных примеров, на которых базируется Родэ, является поверхностное дробление яиц членистоногих. Действительно, дробление яиц большинства насекомых на первых фазах не сопровождается разделением на отчетливые клеточные территории (так называемое, поверхностное дробление). Но при этом Родэ упускает из виду, что этот тип дробления имеет лишь очень узкое распространение в животном мире и, вне всякого сомнения, является вторичным, характерным для высокоспециализированных организмов в связи с лециталь-ностью (богатством желтком). Алецитальные же (бедные желтком) яйца насекомых характеризуются полным дроблением. Еще гораздо менее убедительны другие примеры, часто мало прове-
14 Rohde Е. 1916. Histologische Differenzierung, Zellbildung und Entwicklung bei Protozoen bzw. Protophyten und Metazoen bzw. Metaphyten. Zeitsch. f. wiss. Zool., 115, 155—200. («Гистологическая дифференци-ровка, клеткообразование и развитие протозоа соотв. протофита и ме-тазоа соотв. метафита»). 1914. Zelle und Gewebe im neuen Licht. Vortr. u. Aufs. Entw.-Mech., H. 20. («Клетка и ткань в новом освещении»).
15 Schaxel J. 1915. Die Leistungen der Zeller, bei der Entwicklung der Metazoen, Jena («Роль клеток при развитии многоклеточных»).
186
репные и вообще вызывающие большое сомнение в их фактической точности.
Остановимся также на развиваемых Рода представлениях относительно функциональных структур и дифференцировок; они понадобились ему для того, чтобы объяснить существование и возникновение специализированных тканей, состоящих в огромном большинстве случаев из ясно выраженных клеток, реальность которых отрицать все же никак не удается. Рода писал, что определяющую роль при гистологической дифференцировке животных играют не клетки, а многоядерные плазмодии, и поэтому не образование и развитие клеток, а функциональная дифференцировка целостной живой массы является ведущей в развитии организмов. Другими словами, при развитии ткани клетки не играют никакой роли, а возникают они как следствие все той же функциональной дифференцировки целого.
С теоретической стороны это значит, что как и в концепции Ферворна, причины заменяются условиями (типичный кондици-онализм).
Фактический материал, долженствующий подтвердить эти положения, состоит в перечислении некоторых органов, которые имеют симпластическое строение. Необходимо отметить, что почти все случаи, в которых несомненно отсутствие клеточных границ, относятся не к ранним стадиям развития, а к структурам более поздних фаз развития. Мы считаем поэтому, что Шаксель (1915) был вполне прав, когда, критикуя Родэ, отмечал, что плазмодиальную связь, возникающую как клеточная связь, нет никакого основания рассматривать как нечто своеобразное или новое. Все структуры, происходящие путем слияния, являются, вне всякого сомнения, производными клеток, результатом их специальных дифференцировок и взаимоотношений. Совершенно ясно, что аналитическое изучение тканеобразования не дает никакого основания к построению теории гистогенеза, исходя из конечных стадий развития структур, развивающихся из клеточных детерминированных закладок. Именно с первыми клеточными закладками определяется строение комплекса компонентов, который принципиально в такой же мере, как клетки, можно было бы распространить на все образования.
В заключение этого раздела укажем, что бесспорные положения, свидетельствующие о связи формы и функции, еще ни в какой мере не позволяют заключать, что именно только функция определяет возникновение структур в эмбриональный период (гистогенез, в частности), а между тем представления Родэ
187
о функциональной дифференцировке возводят функцию в примат, игнорируя взаимосвязь функции и формы. На самом же деле ни о какой функции не может быть и речи, пока нет структуры.
§ 45. Некоторые современные представления об относительности понятия клетки. Еще более странные формы, чем у Э. Родэ, приняла критика клеточной теории у некоторых исследователей XX в. Если К. Добелл (1911), считал, что название «клетка» нельзя распространять только на простейших, то, например, В. Я. Рубашкин (1878—1932), А. В. Немилое (1879—1942), 3. С. Кацнельсон и др. пошли еще далее. Так, 3. С. Кацнельсон (1932) в свое время решительно отрицал взгляд на клетку как на определенную общую структуру организма и считал, что понятие «клетка» можно сохранить лишь для определенных структур, образующихся в форме относительно ограниченных, плазматических, ядросодержащих участков. Он считал невозможным называть клетками зрелые гаметы (яйца и сперматозоиды), а также зиготу (оплодотворенное яйцо) и бластомеры. Он полагал, что все они являются особыми структурами, которые не могут быть поставлены в ряд с тканевыми клетками. Правда, позднее Кацнельсон (1939, 1963) отказался от этих крайних взглядов, но все же он продолжает говорить о не вполне ясной - 16
«монолитной расчлененности» .
Вместе с тем необходимо отметить, что в своей критике большинство перечисленных исследователей обрушивается на клеточную теорию, идентифицируя ее с теорией «клеточного государства», которая уже давным-давно никем не защищается. Опровергнув теорию Шванна — Вирхова, они полагают, что ими вообще доказана невозможность существования какой бы то ни было клеточной теории. В своей критике они нередко исходят из правильного положения, что старая клеточная теория не учитывала (или почти не учитывала) целостности организма при всей его расчлененности. Но, как мы указывали, это было уже давно отмечено Дришем, который и ввел первым в употребление столь распространенное ныне выражение: «организм как целое». Таким образом, и в этом отношении «новаторы» последнего времени, хотя они и пытаются доказать материалистичность своих
16 Кацнельсон 3. С. 1939. К столетию клеточной теорни.— Под знаменем марксизма, 5, 116—129.
188
взглядов, не оригинальны, а идут вслед за виталистами, .....а
гораздо более низком теоретическом уровне.
Рубашкин и Кацнельсон обосновывали целостность оргапи i ма, в частности, наличием протоплазматических мостиков апа стомозов между клетками, в том числе и между бластомерами амфибий.
Однако это обстоятельство не имеет принципиального значения, ибо известно, как легко могут распадаться, например, бластомеры иглокожих в морской воде, лишенной солей кальция, а кроме того, также известно, что протоплазма из одной клетки в другую по анастомозам не перетекает.
Сущность всей проблемы, как это совершенно ясно, заключается в трактовке целостности развивающегося эмбриона или зрелого организма. И в этом отношении как Немилов, так и Кацнельсон по существу правильно устремляли свои помыслы. Последний вводит понятие «расчлененности и монолитности организма», но он считает, что оно не совпадает с понятиями цел-люлярного и симпластического строения. Кацнельсон развивал в 1932 г. представление, согласно которому расчлененность может выражаться не только в клеточном строении. Поэтому он считает нужным вовсе отказаться от какой-либо теории клеточного строения. Свою теорию он называл «теорией монолитно-расчлененной структуры организмов»17. Кацнельсон (1932) подчеркивал, что понятие «клетка» получает в его определении иной смысл, чем тот, который оно имеет в настоящее время. Он называл клеткой не общебиологическую единицу, а конкретную (специальную) микроструктуру, возникающую в организме согласно еще малоизученным закономерностям, и считал в связи с этим, что основной проблемой современной гистологии является изучение факторов образования этих структур (клеток).
Подобные взгляды, несомненно, глубоко ошибочны, и поэтому в своей полемической статье еще в 1933 г. мы писали (и продолжаем считать и поныне), что отрицание клетки как сравни-
17Katznelson Z. S. 1932. Histoligische Beobachtungen fiber die frfihen Entwicklungsstadien der Wirbeltiere. Zweiter Beitrag zur Kritik der Zellen-lenre. (Aus d. Histol. Labor, d. Univ. Petersburg (Leningrad). Leiter: Prof. Dr. A. W. Nemiloff): Zeitschr. f. mikr-ant. Forsch. 30, 407 («Гистологические наблюдения над ранними стадиями эмбрионального развития позвоночных». Второе сообщение к вопросу о критике учения о клетке. Из гистологической лаборатории ун-та Петербурга (Ленинград), руководитель проф. А. В. Немилов»),
189
мой общебиологической единицы недопустимо даже как рабочая гипотеза18.
Кацнельсон и Немилов держались по существу воззрений, сходных с представлениями Рубашкина, который даже в своем «Учебнике гистологии» (1931) писал, что на первое место следует выдвинуть ткань или весь организм как целое 1а. К сожалению, остается неясным, что же стоит на втором месте, что же является частью, если организм представляет собой только единое целое. Нельзя сказать, чтобы подобные концепции отличались ясностью и убедительностью, когда частью являются «специальные структуры, которые возникают в организме, согласно еще малоизученным закономерностям» (Кацнельсон).
Ныне Кацнельсон (1963) уже признает, что «клетка в органической природе является... несомненно главной формой структурной организации живого мира» (стр. 288), но все же считает ее не единственной и не придает ей как структурной единице принципиального значения.
Данная идея в работах перечисленных авторов является не только ведущей, но прежде всего и преобладающей. Отсюда становится понятным и самый подбор аргументирующих идею фактов, а также ряд выводов, делаемых автором:
1. Утверждается несравнимость простейших с клеткой, а организм Protozoa приравнивается к Metazoa в целом.
2. Проводится мысль относительной реальности клеток, которые переводятся в малоопределенную категорию «функциональных структур».
3. Чрезмерно переоценивается принципиальное значение клеточных протоплазматических перемычек.
4. Межуточным веществам приписывается не соответствующее действительности значение. Они рассматриваются как полноценное (а не коротко живущее) живое вещество.
5. Утверждается, что не в клетках сущность морфологического выявления жизненных процессов, а в протоплазме, и отсюда уже делается вывод, что всеобщим является «надмицеллярная организованность», т. е. понятие о морфологической структуре живых существ неправомочно сводится к субмикронной структуре их субстанции.
15 Вермель Е. М. 1933. К вопросу о критике учения о клетке.—Усп. совр. биол., т. II, вып. 6.
19 Р у б а ш к и и В. Я. 1931. Основы гистологии и гистогенеза человека, М., Медицинское издательство.
190
Ограниченность всего данного направления в критике к м1 точной теории заключается в том, что проблема ставится в виде дилеммы: либо механистическая клеточная теория «клеточного государства», либо признание абсолютной монолитности орга низма и отрицание клетки как общей элементарной живой единицы. По существу такое представление о монолитности является лишь новым изданием старой и давно оставленной гипотезы Пфлюгера (1845—1910), трактовавшей любой организм как единую гигантскую полимеризованную молекулу. Гипотеза монолитности, по сути дела, ничего не объясняет, а конкретные исследования она направляет по ложному пути. Формальное признание расчлененности ничего не меняет по существу, если отдельные структуры — клетки (ипритом не только они) — признаются лишь временными, преходящими образованиями. Поэтому вполне правильное утверждение Кацнельсона о том, что расчлененность и монолитность не являются несовместимыми и что их нужно, наоборот, рассматривать как диалектическое единство, в его истолковании лишается конкретного содержания.
Критикуя клеточную теорию, как мы уже отметили, ряд авторов, очевидно, для того, чтобы добиться легкой победы, изображали ее в таком виде, в каком в настоящее время никто ее не отстаивает. Например, Немилов считал основными положениями клеточной теории следующее20:
1. «Все живое состоит из клеток, и вне клеток не может быть и жизни».
Немиловым отсюда делается вывод, что как будто бы из этого положения должно постулироваться появление жизни на Земле сразу в виде клеточных организмов.
Далее Немилов пишет об этом в следующих образных выражениях: «Клетка не появилась сразу на нашей планете, как какая-нибудь Афина Паллада из головы Зевса, а сама является результатом длинного пути развития».
В более академических тонах совершенно то же самое написано в любом самом популярном современном учебнике биологии. Вместе с тем вряд ли можно отрицать, что о появлении живой материи на Земле и об ее эволюционных этапах мы, собственно, еще мало что знаем достоверно. Но поскольку мы стоим на ис-
20 Немилов А. В. 1935. Клеточная теория и ее современное состояние.— Тр. Первой гистологической конференции. М., Биомедгиз.
191
ТОрйЧескои точке зрения, МЫ ДОлЖнЫ йрйзйатЬ ЁозмоЖййсТЬ существования форм с более примитивным строением, чем современная нам клетка.
2. «В принципе все клетки равнозначны между собой, и поэтому клетка ни онтогенетически, ни Ф илогенетически не изменяется, и процесс развития сводится к простому перекомбинированию и количественному усложнению равнозначных и сравнимых между собой клеточных элементов».
Подобное мнение нельзя считать хоть сколько-нибудь широко распространенным. В огромном большинстве исследований постоянно затрагивается вопрос о качественном своеобразии клеток и тканей даже у близких видов. Но вместе с тем. представление о сравнимости всех клеток следует считать общепризнанным. Все это ничего не имеет общего с признанием их абсолютной равнозначности в понимании А. В. Немилова. В силу этого вывод, что из признания принципа сравнимости клеток должно следовать отрицание их изменяемости, просто-напросто представляется нелогичным и произвольным.
3. «Клетка является морфологической и физиологической единицей, и соответственно с этим жизнь многоклеточного организма представляет собой простую сумму составляющих его клеточных ж и з н е й».
Признание определенной биологической индивидуальности клетки, признание ее определенной организованной единицей есть действительно центральное положение любого варианта клеточной теории. Но делать отсюда вывод, что организм есть только простая сумма составляющих его клеток, логически неправомерно. Подобное утверждение основано либо на простом игнорировании фактов и известных теоретических положений, либо на непонимании их.
Отрицая клетку как первую н важнейшую структурную единицу, Немилов, Кацнельсон и другие не знают по существу, что же поставить на ее место. Немилов, например, утверждает, что «всеобщим для живой природы является не строение из клеток, а надмицеллярная организованность, живая микроскопическая структурность, созидающаяся через разрушение и только, и существующая на базе этих противоположностей— созидания и разрушения» (там же, стр. 27).
Итак, мы видим, что данная концепция сводит специфичность жизни к структуре вещества. Эта точка зрения ничем не отли
192
чается в принципе от протомерной гипотезы ГейденгаГпы, и<> справедливости считающейся механистической теорией.
Заметим в заключение, что основные положения, разиинап шиеся Кацнельсоном, столь резко критиковавшим клеточную теорию, исходили из ряда конкретных наблюдений над строением тканей, в которых он не мог обнаружить клеточных границ. Естественно, что поскольку все эти выводы имеют столь важное принципиальное значение, они не могли не вызвать серьезного внимания и попыток их проверить. В частности, это было проделано И. Зайнером21, учеником Студнички, который применил метод прижизненного изучения на тех же самых объектах, которые изучал Кацнельсон. Он пришел к выводам, что изучение живых объектов и эксплантатов не подтверждает выводы Кацнельсона. Зайнер в соответствии с данными других авторов устанавливает, что во всех изученных им случаях легко обнаруживаются клетки, но никогда не наблюдаются плазмодии.
§ 46. О трактовке понятия клетки. Невольно возникает вопрос: почему вместо того, чтобы привести клеточную теорию в соответствие со всеми новыми наблюдениями и фактами, некоторые гистологи столь легко и просто отказываются от величайшего достижения научной мысли — от представления о клетке, оказавшего столь плодотворное влияние на развитие разнообразных биологических дисциплин. Пытались даже уверить, что принцип клеточного строения организмов просто ошибочен, а сам термин «клетка» пытались объявить анахронизмом (Немилов, 1932).
Этим вопросом задавался еще в 1917 г. Шаксель 22, критикуя исследования Родэ. Его объяснения сводятся к тому, что обилие новых объектов изучения (и соответственно с этим обилие частных проблем) является для большинства исследователей препятствием к тому, чтобы заняться обобщающими идеями, которые должны стать ведущими. Т от факт, что ведущие принципы только расширяются вместо того, чтобы углубляться, является безусловно опасным моментом и часто приводит к тому, что идея оставляется раньше, чем она будет исчерпана во всем ее значении.
21 S a j n е г J. 1933. Beitrage zur Histogenese der Vertebraten. Publ. de la Fac. de med., Brno. T. XII. N 9 («К вопросу о процессах гистогенеза у позвоночных»).
22 S с h а х е I J. 1917. Zellcn und Plasmodien. Zool. Jhrb., 40, 341—382 («Клетки и плазмодии»).
1Д 7 Е. М. Вермель
193
Это может привести к тому, что сложившиеся концепции будут поставлены под сомнение преждевременно и поэтому будут потеряны для разрешения проблем, которые они могли бы выяснить. Именно такое критическое положение, как считал Шаксель, и имеет место в обсуждении клеточной теории.
Выше мы видели, что обоснованная Шванном клеточная теория, доказавшая общее, принципиальное значение клеточного учения, оказала положительное влияние на многие области науки о жизни. Она являлась ведущей идеей в патологии (Вирхов), дала очень много для познания эмбрионального развития (Ковалевский и Мечников), а также для филогенетических построений (Геккель), позволила ближе подойти к ряду проблем физиологии и т. д. Ошибки, к которым привело сугубо механистическое истолкование клеточной теории, ие являются еще доказательством порочности самой идеи. Отрицание клетки к а к по л но-ценнойисравнимойбиологическойединицыесть методологически пагубное направление в науке о жизни.
Эти теории, вроде учения Студнички об организме как единстве, которые в своем конкретном изложении нередко предлагают рассматривать живое существо как единый протоплазматический комок, представляют собой шаг назад в развитии науки. В теоретических построениях, когда синтициальные или симпла-стические структуры ставятся во главу угла при суждении о строении организма, делаются по существу три ошибки: 1) из исключений (которые, как известно, нередко подтверждают правило) делаются слишком широкие обобщения, вовсе из них не вытекающие; 2) конечные формы дифференцировки признаются за исходные и 3) как-то забывается, что все же клетки являются реально существующей действительностью, органической структурой, которая весьма отчетливо обнаруживается почти всюду. Считать синцитии новыми или своеобразными структурами, стоящими наравне с клетками, нет абсолютно никакого основания, ибо, как правило, они являются производными, дальнейшими формами развития специализированных в определенных направлениях групп клеток. Совершенно неправильно также противопоставлять клетки межуточному веществу, в которое они бывают нередко заключены. Эти неклеточные структуры являются производными клеток (см. § 54).
Все эти соображения элементарны и не раз высказывались, но все же в дискуссиях и нашего времени они не теряют своей актуальности.
194
Клетка несомненно является реально существующей элементарной биологической структурой. Но признавая ее -за конструктивную единицу, никто не имеет намерения воскрешать в какой-либо форме механистическую теорию арифметического суммирования клеток. Мы прежде всего констатируем факт, который говорит, что клеточные массы являются субстратом онтогенетического развития. Рассматривая процессы роста, развития и жизнедеятельности организма, мы должны вместе с тем ясно себе представлять, что эти явления гораздо сложнее, чем те, которые представляют собой поле деятельности отдельных клеток. Наиболее актуальная задача науки, заключается, по нашему мнению, в том, чтобы изучить и понять, в чем именно состоят явления, регулирующие клеточные процессы, на основе которых создается организм как единое целое. Вероятно, при таком подходе в качестве хотя бы рабочей гипотезы весьма плодотворной может оказаться разработка «теории биологического поля», предложенная впервые А. Г. Гурвичем. Эти теоретические представления должны, конечно, разрабатываться дальше, должны совершенствоваться. Надо искать новые, реальные параметры влияния клеток друг на друга, организма — на зачатки органов или их взаимодействий. Важно, что эта теория позволяет получать количественные показатели и конкретно выражать влияние частей организма друг на друга.
8 Е. М. Вермель
ГЛАВА ДЕВЯТАЯ
СОВРЕМЕННОЕ ПОЛОЖЕНИЕ
КЛЕТОЧНОЙ ТЕОРИИ
Основные даты 1-й половины XX века по истории учения о клетке и тканях.
1899 Г. Дриш формулирует закон постоянства клеточных размеров.
1904 Р. Гертвиг возрождает ошибочную концепцию о хроми-диях.
1909 О Гертвиг трактует организм, как «биологическое соединение» клеток.
1915 Ю. Ш а к с е л ь выдвигает представление об организме как системе клеток.
1921—1944 А. Г. Г у р в и ч выдвигает теорию биологического поля.
1921—1925 А. В. Румянцев доказывает ошибочность хромидиаль-ной концепции Р. Гертвига.
1925 В. Якоби и М. Гейденгайн открывают существование размерных клеточных классов.
1932 Л. фои Берталанфи развивает организменную теорию биологических систем.
1934 Е. М. Вермель вносит поправку в закон Г. Дриша, формулируя его как закон постоянства минимальных размеров клетки.
1934 А. А. Заварзин устанавливает закон параллельных рядов эволюции тканей.
1925—1941 А. А. Заварзин создает эволюционную гистологию.
196
§ 47. О методологической порочности виталистической и механистической точек зрения в оценке клеточной теории. Еще Шванн понимал, что объяснить деятельность целого организма суммой жизнедеятельности клеток в целом ряде случаев не удается. Не признавая особых свойств организма, он говорил о гармоническом целесообразном взаимодействии частей (ср. стр. 79). Но его учителе— знаменитый физиолог Иоганнес Мюллер (1801— 1858) — утверждал наличие специфической «жизненной силы», целесообразно действующей в организме и свойственной только ему в целом.
Итак, мы видим, что уже при зарождении клеточной теории предлагалось два пути объяснения целостности: позиция Шванна дала начало механистической трактовке, в то время как трактовка Мюллера представляла собой виталистическое понимание.
Можно сказать, что за последние 40—50 лет одной из основных теоретических проблем биологии было изучение деятельности организма как целого. Конкретно эту проблему можно формулировать так: является ли организм только агрегатом элементарных единиц, или же имеются факторы, не сводимые к простой сумме жизнедеятельности многочисленных клеток? Заслуга такой постановки, как мы уже указывали выше, принадлежит прежде всего Дришу и его школе (см. § 39). Но, доказав принципиальную правильность представления о целостности организма, исследователи! не сумели дать материалистического истолкования найденным ими закономерностям и пришли поэтому к выводу о существовании нематериального фактора. Другими словами, после ряда убедительных экспериментальных исследований (преимущественно эмбриологических) было доказано наличие факторов индивидуального развития, определяемых не просто отдельными клетками, а организмом в целом. Виталисты думали найти объяснение этому явлению введением понятия «энтелехии»— нематериального фактора, направляющего процесс к определенной цели (Дриш). Другие считали, что они разрешили противоречие, отрицая реальность клетки и провозглашая анатомическую целостность организма (Седжвик, Родэ, Немилов и др.). В последнем случае, собственно, никакой проблемы (проблемы соотношения части и целого) уже не оставалось. История науки, однако, убедительно уже не раз показывала, что упрощенчестве отнюдь не является решением вопроса.
Оба эти направления в науке ошибочны как с точки зрения оценки фактических данных, так и в теоретическом отношении, ибо понятие целостности не получает удовлетворительного тол
8*
197
кования. Единственно правильным путем преодоления противоречий традиционной клеточной теории является признание наличия системности как в организме, так и в самой клетке. Сущность этой точки зрения заключается в признании того, что любая так или иначе объединенная группа биологических единиц организма не может рассматриваться как простая их сумма, а является уже качественно новой ступенью организации. Каждая такая ступень системности представляет собой в определенной степени индивидуализированную единицу более сложной организации, чем предыдущие ступени; ей свойственны уже новые, специфические качества, не сводимые к особенностям составляющих ее элементов. Понятие системы требует признания возникновения нового качества за счет взаимного влияния составляющих ее единиц.
Одним из первых эту точку зрения высказывал в 1915 г. Шаксель. Доказывая ведущую роль клеток в детерминационных (определяющих) процессах онтогенеза, он развивал мысль, что многоклеточный организм является сложной саморазвивающейся системой, элементарными частицами которого являются клетки ’. Следовательно, механистическая трактовка организма как суммы клеток заменяется представлением об организме как системе клеток. Таким образом, введенное виталистами понятие организма как целого получает определенное материалистическое объяснение и содержание. При ближайшем рассмотрении вопроса мы видим, что механистическая и виталистическая точки зрения неизбежно терпят фиаско, ибо они не в состоянии дать полноценной, соответствующей фактам трактовки биологической специфичности сложного организма. Только философия диалектического материализма, полностью отражающая законы объективного развития материи, справляется со всеми трудностями, с которыми всегда сталкиваются в конечном итоге все механистические и виталистические воззрения.
Одним из основных положений диалектического материализма является признание принципа развития материи, которое рассматривается как результат свойственного ей самодвижения.
Правильность той или иной конкретной гипотезы, выдержанной в этом смысле, должна быть подтверждена критерием практики (наблюдениями и экспериментами). Клеточное строение само по себе является своеобразным качеством многоклеточного
1 Sch axel J. 1915. Die Leisl.ungen der Zellen bei der Entwicklung der Metazoen. Jena («Роль клеток в развитии»).
198
организма, которое, несомненно, играет выдающуюся роль в развитии. И естественно, что исторический процесс органической эволюции, приводящий к образованию многоклеточности, решительным образом должен отражаться на свойствах гистологических элементов, т. е. самих клеток и их производных (в том числе и межуточном веществе).
В связи со сказанным чрезвычайно интересно указать на следующие мысли Ф. Энгельса; он писал: «Главный факт, революционизировавший всю физиологию и сделавший возможной сравнительную физиологию, это — открытие клетки... Все есть клетка. Клетка есть гегелевское в-себе-бытие и в своем развитии проходит именно гегелевский процесс, пока из нее, наконец, не развивается «идея», данный завершенный организм...
Гегелевская история с качественным прыжком в количествен-
9 ном ряде прекрасно сюда подходит ...»
Чрезвычайно существенно, что Энгельс проводит резкую разницу между клеткой и «завершенным организмом». Несмотря на то, что «все есть клетка», последняя должна пройти процесс, который создает качественно иной индивидуум — многоклеточный организм, или систему клеток. Другими словами, качественное отличие создается самим явлением системности. Отсюда следует сделать уже и дальнейшие выводы. Мы считаем, что любая клетка, несмотря на то, что она остается определенной биологической единицей, является вместе с тем частью целого организма Metazoa. Ясно также, что всякие изменения свойств клетки (нормальные либо патологические) будут отражаться на организме в целом, как и обратно.
§ 48. 77редставление о «биологическом соединении» О. Гертвига. Одна из первых материалистических попыток истолковать специфичность биологических явлений принадлежит Оскару Герт-вигу2 3. Он исходил из представления, что никакие «кирпичики» не могут создать сложного органического единства. «Кирпичики» (клетки) могут построить только агрегат. Клетки, с его точки зрения, нельзя поэтому сравнивать с «кирпичиками», ибо они друг
2 К. Маркс и Ф. Энгельс. Соч., т. XXII. Изд. 1-е, стр. 345 546. Письмо Энгельса Марксу от 14 июля 1858 г.
3 Hertwig О. 1909. Der Kampf um die Kernfragen der Enlwicklung und Vererbungslehre. Jena. J. Fischer («Борьба вокруг вопроса о роли ядра в учении о развитии и наследственности»).
199
Оскар Гертвиг (1849—1922)
с другом создают сложное органическое единство. И так как создаваемый ими комплекс не является ни химическим, ни физиологическим, то совокупность клеток в организме он и предложил назвать «биологическим соединением». При этом отдельные клетки такого соединения в большей или меньшей степени утрачивают свою автономность, переходящую в целое, которое уже и определяет деятельность клеток. Другими словами, клетки, если так можно сказать, становятся интегрированными частями целого. Гертвиг считал, что нет ни
200
какого основания отказываться от целлюлярного принц,ина, но вместе с тем он признавал роль клеток довольно ограниченном, ибо их жизнедеятельность зависит от организма в целом. Соот ношение клетки и организма есть проблема части и целого. Для нас эти понятия являются столь же различными, сколь и связанными. Ни одно из них немыслимо без другого, и каждое из этих понятий требует существования другого. В силу этого, как считал и Гертвиг, выражения «растение образует клетки» и «клетка образует растение» не являются взаимно исключающими. Обе эти формулировки можно употреблять, если только правильно будут пониматься соотношения, в которых состоят друг к другу клетка как часть и растение как целое.
Гертвиг проводил еще и такое сравнение. Он отмечал, что между агрегатами клеток в смысле «Bausteintheorie» и их биологическим соединением имеет место столь же значительное различие, как между смесью двух объемов водорода с одним объемом кислорода и их химическим соединением в молекулах воды. Проводя подобное сравнение, Гертвиг приводил для иллюстрации пример е процессом светового раздражения сетчатки глаза; он отмечал, что конечный эффект нельзя рассматривать как результат суммарного действия клеток; он может быть правильно объяснен только исходя из общих закономерностей, основанных на всей организации животного, интегрированными частями которого являются составляющие его клетки.
Эти мысли Гертвига не раз повторялись и другими исследователями, но ни им самим, ни кем-либо другим не были развиты с необходимой полнотой. В частности, пример соединения атомов в молекулу был повторен Б. Фишером для проведения аналогии с процессом объединения белковых молекул в более высокую единицу— клетку, откуда следовал вывод, что Свойства клетки не могут выводиться из свойств составляющих е е элементов4.
Еще позднее ту же мысль развивал известный английский эмбриолог Нидхэм. Он в своей книге «Порядок и жизнь» указывает, что без понятия уровня организации и признания особых, более сложных и специфических свойств системы элементов не может быть понята современная биология с ее закономерностями, отличными от таковых в неорганическом мире. Другими
4 Fisher В. 1924. Vilalismus und Pathologic. Roux’s Vortr. u. Aufs. Bd. 34 («Витализм и патология». Есть русский перевод под ред. В. А. Обуха, Изд. Мосздравотдела, 1926).
201
словами, развивается все та же мысль, что биологическое целое всегда больше и выше, чем сумма частей 5.
К той же мысли возвращается современный цитолог де Ро-бертис6. Он приводит для ее иллюстрации пример, совпадающий с тем, который мы находим у О. Гертвига, хотя его работы он, по-видимому, не знает. Он пишет, что свойствами хлористого натрия по отдельности не обладают ни атом хлора, ни атом натрия, а только молекула хлористого натрия в целом. Нельзя также говорить о свойствах полисахарида (например, гликогена) на основании свойств его компонентов (сахаров), но это уже пример не соединений (уровня организации) атомов в молекулу, а молекул в полимер. Аналогично и система («соединение») полимеров дает нечто качественно новое. Может быть, это новое на каком-то этапе развития — уже клетка.
Мы считаем приведенные соображения О. Гертвига, Фишера, Нидхема и ряда других авторов не только полностью материалистичными, но и стихийно диалектичными, а поэтому не оставляющими никакого места витализму. Диалектический материализм как философское воззрение находится, по нашему убеждению, в полном соответствии с представлением об уровнях организации, создающих качественно новые системы со своеобразными закономерностями.
§ 49. Системность в строении организмов и представление о клетке. Механистические теории не объясняют специфических особенностей живых существ, основной особенностью которых является целостность. Витализм же, давая свое толкование органической целостности, сводит ее к нематериальным факторам, отрицая, таким образом, возможность их естественнонаучного понимания. Мы же исходим из положения, что истинная трактовка должна оставаться материалистической и вместе с тем диалектически объяснять явление целостности.
Ясно, что любая попытка свести без остатка жизненные феномены к процессам отдельных независимых слагаемых не может иметь никакого успеха, и с этой точки зрения нетрудно видеть, в чем заключается принципиальная ошибочность таких теорий, как гипотеза «клеточного государства», теория детерми
5 Needham J. 1936. Order and life («Порядок в организации жизни»).
6Робертис Э. де, Новинский В., Сазе Ф. 1967. Биология клетки, М., изд-во «Мир».
202
нантов (Вейсман), гипотеза сведения жизненных явлений к физико-химическим процессам, протомерная гипотеза Гейденгайна, надмицеллярность Немилова и т. д. Одним из основных положений современной биологии является утверждение, что жизненные процессы свойственны только лишь организмам, характерной особенностью которых является особая системная организация вещества, что и создает новое качество материи — жизнь. Понятие системы представляет собой особую проблему, и, следовательно, при изучении даже наиболее простых живых существ наряду с вопросом относительно причинности отдельных процессов должен ставиться также вопрос об органической целесообразности этих явлений.
О «загадках системы» говорили и писали многие крупные исследователи последнего времени. Эту идею мы встречаем, например, у крупного голландского физиолога Г. Ж. Иордаиа (1877—1943), считавшего, в частности, что биологическое исследование должно вестись «синтетическим методом»7. Однако в его конкретной работе все дело в конце концов все же сводится по существу к более сложному комплексному описанию явлений.
Число фактических наблюдений, обосновывающих представление системности организма, непрерывно возрастает. Живой организм следует рассматривать как индивидуализированную единицу, составленную из целого ряда более простых, соподчиненных систем; таким образом, надо говорить об иерархии систем. Например: клетка—>- гистосистема первого порядка (альвеола, ацинус, остеон и т. п.)—гистосистема второго порядка (долька железы, гаверсова система кости и т. п.)—>- гистосистема третьего порядка (железа, кость и т. п., т. е. орган)—>-организм. Конечно, на самом деле построение организма может быть гораздо сложнее, ибо системы составляются ие только из равноценных единиц, но в их состав входят и другие качественно различные структуры, между которыми создаются разнообразные взаимоотношения (например, нефрон). Все это создает условия, определяющие стабильность системы, но система живых элементов сохраняет свои особенности и свойства независимо от колебании внешних условий (конечно, в известных пределах), или же оиа гибнет.
Вышеприведенное понимание проблемы проводит резкую грань между живым существом, агрегатом и машиной, хотя по-
7 Jordan Н. 1929. Allgcmeine vergleichende Physiologie der Tierc («Общая сравнительная физиология животных»).
203
Людвиг фон Берталанфи
следняя тоже может пониматься как система, но система гетерономная, т. е. подчиняющаяся закономерностям, вытекающим не из ее существования. Любая машина работает только под влиянием внешних воздействий и производит только определенные процессы, не имеющие никакого отношения к поддержанию той системы, которую она представляет.
Представление о клетке как биологической системной единице является лишь одним из звеньев в общей цепи иерархических ступеней развития материи. Однако клеточная ступень организации имеет для биологии, может быть, наибольшее принципиальное значение. В силу этого с точки зрения истории развития (эволюции) вопрос о возникновении клетки особенно важен, ибо он совпадает с проблемой происхождения жизни на Земле. Труд-ность проблемы усугубляется тем, что представления о предкле-точных стадиях эволюции живой материи до сих пор остаются в области гипотез, которые подтвердить фактическими данными практически, по-видимому, пока невозможно.
204
§ 50. Организменная теория Л. Берталанфи и клеточная структура. Значительно ближе к развиваемой нами точке зрения стоит концепция, защищаемая в последнее время выдающимся биоло-гом-теорети^ком Людвигом фон Берталанфи (1928—1932) и известная под названием организменной, или организ-мической теории8. Ее сущность сводится прежде всего к тому, что особая специфичность физико-химических процессов, совершающихся в организме, определяет качественное своеобразие биологической системности (ниже на этом мы остановимся поподробнее).
Для иллюстрации сказанного Берталанфи приводит высказывания известного физика Г. Рейхенбаха, который считает, что в настоящее время никто не может утверждать, что известные нам физические законы уже достаточны для понимания жизненных процессов. Он отмечает, что при открытии любой новой физической области оказывалось необходимым установление новых физических закономерностей (например, для явления радиоактивности). Круг же жизненных процессов настолько оставался вне поля зрения физиков, что было бы большой и невероятной случайностью, если бы известные иам физические закономерности уже содержали все необходимое для Понимания жизни. По мнению Рейхенбаха, следовало бы считать, что должны существовать особые закономерности физического типа, являющиеся основой жизненных отправлений. Вместе с тем он отмечает, что нельзя предвидеть, не произойдут ли при изучении органической жизни даже изменения типа установленных теперь физических закономерностей 9. Но эта концепция говорит не о специфике живого, а только о недостаточности наших знаний в области физических закономерностей для объяснения сущности жизни.
Берталанфи идет дальше и указывает, что во все времена наиболее характерным моментом механицизма было утверждение, что кругом известных закономерностей неорганической природы исчерпываются вообще все явления, в том числе и процессы жизни. Однако современная ступень знания не подтверждает этого. Берталанфи совершенно прав, когда, указывая на существование целого ряда структурных ступеней (электроны и протоны, атомы,
sBerlalanffy L. von. 1932. Theoretische Biologic. Bd. I («Теоретическая биология»).
Reichenbach H. 1931. Ziele und Wege der heutigen Naturphilosophie. («Цели и пути современной натурфилософии»).
205
молекулы, мицеллы, коацерваты, свобод но-живу щие клетки, многоклеточные организмы), говорит, что уже a priori надо ожидать наличия специфических закономерностей, действующих в пределах только каждой из н и х. Другими словами, делается вывод, что если мы говорим об особой организации живых существ, то должны предполагать наличие в иих собственных характерных и специфических закономерностей. Такая постановка проблемы не несет в себе ничего виталистического, но вместе с тем и не является механистической. Признание же качественного своеобразия явлений сближает данную концепцию, с нашей точки зрения, с позициями материалистической диалектики.
Из такой постановки проблемы вытекает еще один вывод: поскольку организм следует рассматривать как своеобразную систему со специфическими закономерностями, то, следовательно, мы должны также выработать и специфические методы исследования, а также строить особые рабочие гипотезы, которые вовсе не должны базироваться на элементарных законах «неорганических» наук. Материалистическая расшифровка регистрируемых биологических процессов является конечной задачей исследования, которая вследствие недостаточности наших знаний в настоящее время не может быть еще во многих случаях решена окончательно. В качестве одного из таких существенных биологических представлений следует считать и клеточную теорию, а также теории наследственности, эволюции и др.
Берталанфи в своей книге делает одно остроумное замечание. Он пишет, что биологи в своем стремлении стать химиками и физиками перестают по существу заниматься своей специальностью и пытаются рассматривать живое существо не как организм, а как агрегат. Организм же является особой ступенью развития материи — организацией, стоящей над «уровнем» химии. Если это так, то совершенно ясно, что организм должен изучаться на всех уровнях и изучение одного из них ие может заменить изучения другого. Это остается справедливым независимо от того, что в будущем мы будем иметь возможность выражать свойство всех высших уровней в связях более низших ступеней.
Недостаточность элементарной механистической, физико-химической трактовки «органических» феноменов понимается и отмечается большинством ученых. И в силу этого с необходимостью вводятся чисто биологические понятия.
206
Если системные закономерности Организма не могут быть в настоящий момент сформулированы с физико-химической точки зрения, то тем более необходимо особое биологическое их понимание. Вопрос о возможном выражении в конечном итоге биологических (системных) закономерностей в иных более простых связях играет в конкретном исследовании, как нам кажется, в настоящее время второстепенную роль. Поскольку основой жизни является ее организация (порядок), то, следовательно, изучение отдельных процессов и превращения веществ в организме, даже при их самом глубоком элементарном физико-химическом анализе, не может рассматриваться как исчерпывающее разъяснение жизненных явлений. На этом пути изучения от нас ускользают как раз самые существенные особенности живого, а именно: упускается из виду тот факт, что процессы в организме направлены таким образом, что они обусловливают создание целостности. Признание направленности жизненных явлений также ни в коей мере не является виталистической гипотезой, а представляет собой прежде всего фактическое наблюдение, которое не может быть понято путем простого анализа отдельных явлений. Таким образом, понимание закономерностей организма как системного целого всегда должно ставиться во главу угла при объяснении отдельных явлений жизни. К этому вопросу мы еще вернемся в последних параграфах книги.
§ 51. О происхождении живой материи. Простейшей формой живой органической субстанции, которую мы знаем в настоящее время, является клетка. Мы уже отмечали, что подобный взгляд отнюдь не отрицает историчности клеточной формы существования жизни. Но этапы жизни, предшествующие ее клеточной ступени, пребывают до сих пор в области гипотез.
Сколь бы проста ни была клетка (или любая «предклетка»), ее организация неизмеримо сложнее любой неживой структуры. Признание наличия скачка между живым и неживым в настоящее время вряд ли может оспариваться. Эта мысль высказывалась целым рядом исследователей. Общепризнанным представляется взгляд, что носителем жизни является не какое-либо одно (хотя бы и очень сложное и лабильное) белковое соединение, а также и не смесь органических соединений. Жизнь представляет собой особую форму существования определенных органических веществ, из которых глав
207
нейшим являются белок и нуклеиновые кислоты. Эта точка зрения может быть легко обоснована рядом фактических данных и примеров.
Особенно ясно эта концепция была изложена Ф. Энгельсом в его широко известной формулировке: «Жизнь есть способ существования белковых тел, и этот способ существования состоит по своему существу в постоянном самообновлении химических составных частей этих тел.
Белковое тело понимается здесь в смысле современной химии, которая этим термином охватывает все тела, аналогичные по составу с обыкновенным белком и называемые также протеиновыми телами...
Повсюду, где мы встречаем жизнь, мы находим, что она связана с каким-либо белковым телом, и повсюду, где мы встречаем какое-либо белковое тело, не находящееся в процессе разложения, мы без исключения встречаем и явления жизни... В живом организме необходимо присутствие также и других химических соединений.
... Дефиниция жизни, разумеется, весьма недостаточна, поскольку она далека от того, чтобы охватить все явления жизни, а, напротив, ограничивается самыми общими и самыми простыми среди них» |0.
Эта формулировка Ф. Энгельса предусматривает качественное отличие белка «мертвого» от белка «живого» — отличие, которое мы теперь называем «порядком его организации». Современная химия показала, что классическими методами можно познать только одну сторону изучения белков. Эмиль Фишер (1852—1919) установил, что белки распадаются на аминокислоты, между которыми имеет место особая (пептидная) связь. Но даже чисто химическое исследование показало, что рассматривать белки как элементарное соединение аминокислот не представляется возможным. Белок значительно более сложен и обладает рядом специфических для него качеств (например, так называемые ассоциативные связи, основанные на особых добавочных валентностях). Эти особенности резко отличают белок от других органических соединений и обусловливают его гораздо большую стабильность и одновременно лабильность. Другими словами, в белке играют роль два типа связей: первая (низшая) —пептидная и вторая (высшая) —• ассоциативная, определяющая уже
Э и г е л ь с Ф. 1967. Анти-Дюринг, стр. 78—80. М., Изд-во политической литературы.
208
Фридрих Энгельс (1820—1895)
специфические особенности субстанции. Таким образом, уже на этой ступени организации материи устанавливается, если можно так сказать, «иерархический порядок связей». Несмотря на значительный прогресс наших сведений в области химии белка, вряд ли можно утверждать, что мы очень близки к разрешению проблемы его синтеза и пониманию его структуры. Тем более это справедливо для «живого» белка. Вывод отсюда вполне ясен: узкохимическое представление о белке недостаточно, ибо эта субстанция имеет качественно специфические особенности, которые обусловлены особой структурой, характеризующейся иными закономерностями.
Ф. Энгельс ясно формулировал признание качественной специфичности различных ступеней организации материи. Фактические исследования последних десятилетий все более и более подтверждают эту мысль. Мы видим, что даже в химических исследованиях приходят к выводу о качественно различных группах соединений. Тем более это справедливо для организации живых существ.
§ 52. Эволюция клетки и данные о степени самостоятельности клеток многоклеточных организмов. Сам факт клеточной организации всех живых существ находится вне всякого сомнения, и дискуссия в этом отношении является, бесспорно, анахронизмом.
Неясной, конечно, следует считать «клеточность» бактерий, спирохет, синезеленых водорослей и некоторых других. Мы также недостаточно знаем природу фильтрующихся вирусов и бактериофагов. Некоторые ученые продолжают рассматривать большинство вирусов даже не как «существ о», а как «веществ о». Но это совершенно особая проблема, относящаяся к области эволюции жизни, а не к клеточной теории. Однако мы не можем вовсе обойти вопрос об эволюции клетки.
Мы полагаем, что именно бактерии, спирохеты и синезеленые водоросли, хотя и являются клетками, но построены по иному типу, чем ядерные клетки простейших и многоклеточных. Но значит ли это, что все эти формы следует рассматривать как переходные фазы между истинной клеткой и предклеточными структурами? Ведь формула «можно думать» совсем еще не значит «необходимо считать». По нашему мнению, гораздо логичнее признать полифилию живых существ, а не монофилию. Последняя, как мы полагаем, только лишь маловероятная догадка, осно
210
ванная на весьма спорном положении, согласно которому сходство пропорционально родству. Если жизнь должна была возникнуть на Земле при определенных условиях, то она, конечно, возникала многократно в разных местах, причем условия не могли быть всюду идентичными. Утверждение же, что жизнь возникла только однажды, а лишь затем под влиянием внешних воздействий начала развиваться в разных направлениях, очень походит на «библейские рассуждения». Монофилитические представления, в частности, никак не объясняют «параллельные ряды» и конвергенции. Как, в самом деле, понимать удивительное сходство глаза некоторых моллюсков и позвоночных, сходство строения поперечнополосатых мышц у моллюсков (запирательная мышца пектена), насекомых и позвоночных, сходство в нервных связях в мозге (Заварзин, 1913, 1941, и мн. др.) и. Правда, в этих примерах речь идет о полифилии органа, но это же относится и к полифилии организмов в целом.
Нет сомнения, что развитие живых структур должно было пройти длинный путь до возникновения клетки. Протозоолог Е. А. Минчин в 1916 г. образно выразил это так: «...путь от неорганизованной материи до амебы должен был быть гораздо длиннее, чем от амебы до человека» |2. Длину этого пути мы, конечно не знаем, но где он длинен, там можно ожидать и тупиков развития. Именно таковыми следует, очевидно, считать синезеленые и сифонниковые водоросли и некоторые другие. Берталанфи по этому поводу писал, что природа неоднократно делала эксперименты создания крупных организмов без клеточной их дифференцировки, однако только принцип клеточной организации оказался прогрессивен.
Как известно, все одноклеточные организмы имеют микроскопические размеры. Очевидно, именно это свойство ставит пределы в их эволюции, т. е. оиа идет иначе, чем у многоклеточных, и своеобразно. Современные бактерии и спирохеты, оставаясь .микроскопическими, в конечном итоге приобрели все же очень сложное строение. У них нет истинного морфологически целостного ядра, но есть хроматин, им свойствен генно-мутационный процесс, в них совершаются очень сложные и многообразные биохимические процессы. Эти организмы оказываются весьма
11 Заварзин А. А. 1941. Очерки по эволюционной гистологии нервной системы. М., Медгиз
12 Minch in Е. А. 1916. The evolution if the cell. Русский перевод: M и и ч и н E. A. 1923. Эволюция клетки.
211
приспособленными к условиям существования на нашей планете. Они тоже совершенствуются, но по иному пути, не создавая систем дифференцированных функций. Все же нет никаких оснований микробам приписывать принципиально иную структуру, и поэтому мы убеждены, что столь часто употребляемое микробиологами выражение «бактериальная клетка», совершенно правильно и законно, хотя клетка и построена несколько иначе.
Однако только клеточная расчлененность, несущая с собою определенную индивидуализированность частей, открывает широкие перспективы к совершенствованию и эволюции. Именно клеточность легко позволяет замену изношенной в результате функции ткани (физиологическая регенерация); она же легко обеспечивает рост организма и дифференцировку его частей в самых разных направлениях. Следует указать, что, как правило, только необратимая дифференцировка ведет к образованию сим-пластических структур (мышечные волокна, клетки Лангханса), образованию межуточного вещества из эктоплазмы фибробластов (а также остеогенез и хондрогенез) или резко отклоняющихся по строению клеток (мегакариоциты, лейкоциты; клетки Пуркиня коры мозжечка, пирамидные нервные клетки коры мозга и др.). Называть такие проблемы неясными, как это делает, например, Кацнельсон 13, нет совершенно никаких оснований и не нужно дезориентировать в этом отношении биологов и других специалистов. Основная проблема биологии заключается не в этих вопросах клеточных преобразований при деятельности организма, а в познании законов создания системности организма на всех уровнях его организации.
Беря в основу положение о системном развитии материи, мы должны считать клетку той первой ступенью организации, которая обладает всем комплексом свойств, характеризующим явление полноценной жизни. Эволюция организации жизни может идти разными путями: с одной стороны, усложнением структурности самой клетки, а с другой — путем соединения клеток в комплексы большей или меньшей сложности. Вместе с тем, как показывают многочисленные конкретные исследования, клетки, как таковые, находясь в ткани или в органе любой сложности, постепенно обезличиваются в целом, т. е. они интегрируются 14.
13 Кацнельсон 3. С. 1963. Клеточная теория в ее историческом развитии. Л.
14 Интеграция, собственно, обозначает соединение в одно целое отдельных самостоятельных величин, причем происходит их дифференциация, т. е.
212
Во всяком случае, совершенно ясно, что понимание основ жп i-ненных явлений без познания клетки немыслимо. Но эта мысль, как мы видели, приводила нередко к тому, что значение клетки в общей жизнедеятельности организма переоценивалось. Ошибка исследователей заключалась в том, что они не понимали, что клеточная структурность Metazoa есть лишь одна (и притом первая в своем роде) из ступеней их организации. Но все это нисколько не снижает значения изучения клетки в свете общебиологических проблем. В настоящее время учение о клетке в основном идет по двум руслам: 1) познание самой клетки как элементарной живой системы и 2) изучение их взаимоотношений в многоклеточном организме.
Когда мы утверждаем, что клетка является «элементарной» живой единицей, то этим мы хотим сказать, что она представляет собой предельную биологическую индивидуальность. Ядро и протоплазма сами по себе, взятые в отдельности, не являются полноценными живыми единицами. Поэтому вопрос о том, что является в клетке «живым», а что «мертвым», неправилен по существу. Проблема эта должна ставиться так: что является в клетке ее основной, обязательной частью, а что — факультативной.
Надо сознаться, что клетку мы все еще знаем очень недостаточно, хотя ее изучала и изучает армия ученых. Сводку по любому (даже малозначительному) вопросу приходится делать по десяткам (в лучшем случае), а то и многим сотням работ. Приведем только один пример. Такой частный, казалось бы, вопрос, как величина клетки, специально обсуждался более чем в 750 специальных работах. Описательная сторона в большинстве случаев изучена достаточно хорошо, но этим, как известно, задачи гистологии как науки еще далеко не исчерпываются. Например, мы точно знаем, что огромное, подавляющее большинство клеток имеет ядро (или эквиваленты этих структур). Морфология и < ;аже химия клеточных ядер достаточно хорошо изучены, но об их роли в обмене веществ мы фактически почти ничего не знаем и судим о них лицпь косвенно. Так, известно, что амеба, лишенная ядра, пищу заглатывает, но ее не ассимилирует; безъядерная растительная клетка не строит целлюлозной оболочки; в некоторых энергично работающих клетках ядро изменяет свою форму
постепенное увеличение различия между первоначально однородными и самостоятельными единицами, которые при этом становятся все более и более зависимыми друг от друга.
9 Е. М. Вермел!
213
и структуру. Этим'почти исчерпываются наши сведения о конкретной роли ядра в метаболических процессах клетки.
Другими словами, мы сравнительно хорошо знаем морфологию клетки, но почти не знаем «собственно биохимии» клетки (прежде всего в смысле физиологического значения ее компонентов). Но, может быть, это и вполне закономерно, ибо знание физиологии клетки является ведь по чисто техническим причинам последним этапом в познании одной из важнейших ступеней жизни, а сопредельные с биологией дисциплины, как и сама биология, не располагают достаточным фактическим материалом и достаточными обобщениями.
Что касается изучения проблемы взаимоотношения (структурного и функционального) клеток, то в этом отношении сделано также довольно много. Однако все эти данные носят большей частью весьма несистематический характер и нигде в целом, будучи объединены общей точкой зрения, не сведены. Но даже самое беглое их перечисление показывает, насколько многообразны фактические сведения, свидетельствующие в пользу системности клеток. Интереснейшие данные в этом отношении мы находим в исследованиях из самых разнообразных областей гистологии.
Может быть, основным вопросом в этой области является степень самостоятельности клетки сложного многоклеточного организма. В этом отношении весьма ценные данные получены методом культивирования тканей вне организма. А. Фишер15 и М. Т. Берроуз (1925) показали, что отдельные изолированные клетки нормальных тканей, будучи поставлены даже в наилучшие условия, не размножаются. Это было несколько позднее подтверждено еще более совершенной методикой Э. Майером 16 и многими другими.
Отсюда можно сделать вывод, что в тканевой культуре мы имеем дело не с независимыми клетками и что последние создают общую тесно связанную систему. Таким образом, клетка многоклеточного организма несравнима в этом отношении с отдельной бактерией или клеткой протиста, обладающими полной самостоятельностью. Эту особенность клетки — невозможность ее
15 F i с s h е г А. 1925. Beitrag zur Biologic der Gewebezellen Roux. Arch.
Bd. 104 («Сообщение по биологии тканевых клеток»).
15 Mayer E. 1933. Formbildung und Wachstum von geziichteten Zellverban-Roux Arch. Bd. 130 («Формообразование и рост культивируемых кле-клеточных пластов»).
214
полноценной жизнедеятельности в изолированном состоянии ине тканевого комплекса — Фишер трактовал в духе представления о физиологической интеграции (Спенсер, 1820— 1903) 17. Однако тот же Фишер показал, что в потенции нормальные клетки все же сохраняют способность индивидуализироваться, и тогда они при определенных условиях растут и размножаются независимо от тканевого комплекса. Этот вывод сделан на основании сопоставлений с ростом в культурах клеток злокачественных опухолей, которые, как оказалось, будучи изолированными, могут продолжать развиваться и делиться.
Опыты культивирования нормальных тканей вне организма убедительно показали, что рост тканевой культуры является прежде всего ростом тканевого комплекса (своеобразной системы клеток). Это выражается прежде всего в том, что рост представляет собой периодическое явление и клеточные деления распределены в ткани| не случайно, а располагаются по периферии зоны роста. Опыты с регенерацией тканевых культур показали, что при поранении растущей культуры восстанавливаются все изъяны, причем скорость роста по раневой поверхности всегда идет быстрее, чем по неповрежденному краю. Следовательно, вырезание участка из зоны роста вызывает нарушение какого-то равновесия, которое затем вновь восстанавливается. Надо считать, что влияние целого (в данном случае всей культуры) на часть — регенерирующий участок ткани — имеет принципиально ту же природу, как и влияние целого организма на любую его часть 18.
Аналогичные ограничивающие и побуждающие влияние организма как целого на составляющие его клетки могут быть легко обнаружены специальными исследованиями. Примеры можно взять из различных областей; мы остановимся опять на явлениях роста.
Показано, что даже такой элементарный признак, как величина клетки, определяется в значительной мере организмом в целом. Это выражается в том, что каждый орган, каждая его система имеет клетки в норме вполне определенной для каждого вида животных и растений величины. Стоит только ослабить или
17 Spencer Н. 1876. Principles of biology («Основания биологии»). Есть несколько русских переводов.
1S Эти вопросы рассматриваются, например, в цитированной выше работе Mayer’а (1933) и в исследовании В. Ephrussi 1933. Arch, d’auat. microsc. T. 29 («Рост и регенерация в культурах тканей»).
9*
2/5
изменить влияние организма, например путем слабого отравления или перерезки нерва, как тотчас же клетки начинают укрупняться.
В этом случае начинается выявляться свойственная клеткам тенденция к росту. Приведем еще один показательный пример влияния целого на часть. Известно, что у болотной лягушки (Rana ridibunda) клетки кожи в 2 раза мельче, чем у травяной лягушки (Rana temporaria). У головастиков сравнительно легко удается пересаживать лоскуты кожи от одного вида к другому. Можно было надеяться, что, исходя из размеров клеток, можно будет легко следить за судьбой пересаженной ткани. Однако оказалось, что размеры клеток трансплантата в очень короткие сроки принимают размеры, характерные для кожи хозяина, т. е. клетки пересаженного лоскута кожи головастика болотной лягушки на теле головастика травяной лагушки становятся в 2 раза крупнее и, наоборот, при обратной пересадке от R. temporaria к R. ridibunda клетки становятся в 2 раза мельче. Таким образом, можно считать доказанным, что именно организм в целом контролирует величину клеточных элементов |9.
Еще более отчетливые результаты получены при изучении распределения делящихся клеток в различных органах. Например, В. Корнфельд (1922), одним из первых изучая клеточные деления в роговице глаза саламандр, установил определенные ритмы митотической деятельности20. Много писал об «эпид е-миях» митозов Гурвич; то же отмечали и другие. Экспериментально установлено, что митозы в роговице глаза проходят в определенном направлении (градиент) и что распределение митозов зависит, в частности, от давления века. В коже животных (мышей, крыс и многих других) обнаружен суточный ритм делений; так оказалось, что в 12 часов дня митозов в 3 раза больше, чем в 8 часов вечера. Интенсивность клеточных делений находится в определенной связи с половым созреванием животного, половым циклом и его возрастом.
14 Сводку по этому вопросу см. в сборнике «Рост животных», Биомедгиз, 1935, ст. Е М. Вер мел я «Величина, размножение и рост клеток», его же работу «Исследование о клеточных размерах».— Уч. зап. Моск, гос. пед. ин-та им. В. И. Ленина, т. 25, 1940 и книгу Я. Е. Хесина «Размеры ядер н функциональное состояние клеток». М., изд-во «Медицина», 1967.
2,1 Kornfeld W. 1932. Ueber den Zellteilungsrhythmus und seine Rege-iung. Arch. f. Entw. 50. («О ритмах клеточных делений и их регуляции»).
216
Число подобных примеров легко можно значительно увеличить. В заключение мы укажем только еще на одну группу фактов, касающихся накопления запасных веществ клетками печени грызунов. Установлено, что максимумы накопления гликогена, с одной стороны, и жиров и желчи — с другой, постоянны и закономерно чередуются в течение суток. Оказалось, что суточный ритм работы свойствен не только клеткам печени, но и ряду других органов. Так, показано, что интенсивность всасывания пищи (жцр) эпителиальными клетками кишечника также подчиняется дневному ритму. Поэтому, если скармливать мышам (а возможно, и человеку) пищу в неурочное время, то всасывание, у предварительно голодавшего животного, почти не происходит. Отсюда ясно, как важен правильный распорядок дня в питании.
Всеми перечисленными исследованиями устанавливается зависимость работы отдельных органов, тканей и клеток от общих координаций частей организма, в конечном итоге, как нередко говорят, от «организма как целого».
Краткие сведения, которые мы выше привели, свидетельствуют о различно выраженной интеграции клеточных элементов в органах Metazoa, о большем или меньшем «растворении» части (клетки) в целом (организме), т. е. примате высшей ступени системности (организации) над ступенью низшей, соподчиненной. Исходя именно из этого вывода, мы и получаем конкретную расшифровку положения что клеточный комплекс Metazoa представляет собой нечто большее, чем простая сумма клеток. Вместе с тем именно эти данные являются блестящим доказательством правильности закона перехода количественных изменений в качественные, объясняющего своеобразие организованной системы по сравнению со свойствами составляющих ее единиц.
§ 53. Проблема межуточного вещества. Остановимся еще на одном весьма существенном, хотя и более частном, вопросе, имеющем, однако, непосредственное отношение к клеточному учению, а именно — на проблеме межуточного вещества. Она сложна по целому ряду причин. Во-первых, надо отметить, что вещества эти разнообразны как по своему составу, так и по структуре; во-вторых, эта проблема должна решаться только в свете данных об их генезе, в отношении которою общепринятой точки зрения до сих пор нет, хотя и несомненно, что межуточное вещество продуцируется клеточными элементами.
217
Может быть, одним из самых важных вопросов, на котором нередко спекулировали отдельные критики клеточной теории, является решение уже давно стоящей дилеммы: «живо» или «мертво» основное вещество соединительной ткани?
Фактические данные приводились как в пользу той, так и другой точек зрения. Если исходить из отдельных признаков живого (рост, формообразование, раздражимость и т. д.), то ни до чего договориться невозможно, ибо эти явления в отдельности наблюдаются в той или иной форме также и в мире неорганическом, но только в живых индивидуумах мы наблюдаем синтез этих явлений. Поскольку мы этого в основной (межуточной) субстанции никогда не находим, то, следовательно, считать ее полноценной живой системой нельзя. Это положение подтверждается прежде всего тем фактом, что межуточное вещество не может длительное время существовать, будучи лишено клеток. Мы имеем все основания думать, что обмен межуточного вещества находится под очень значительным влиянием клеточных элементов, но и это надо понимать так, что ведущую роль играют не отдельные индивидуализированные клетки, а вся ими образуемая система. Исходя из этой позиции, собственно, снимается и проблема: «живо» или «мертво» основное вещество? Решение этой проблемы должно быть очень простым и может трактоваться в том смысле, что живой сколько-нибудь длительные сроки является вся система в целом, состоящая как из клеток, так и основного вещества. Но при этом вовсе не является необходимым, чтобы все ее составляющие элементы были полноценными и равноценными живыми компонентами.
В основе этих рассуждений лежит положение, что ничто живое нельзя рассматривать вне представлений о системности живых существ, а поэтому, когда мы утверждаем, что клетка является элементарной единицей, мы хотим сказать, что она представляет собой наименьшую элементарную самостоятельную живую систему.
В свете рассуждений о полноценности жизненных свойств межуточной субстанции важно указать на явление старения тканей. Известно, что жизнеспособность неклеточной межуточной субстанции очень ограничена и она в организме быстро и постоянно заменяется новой, начиная чуть ли не с эмбрионального состояния организма. Всем известно, что кость, например, постоянно обновляется путем перестройки и образует при этом так называемые Гаверсовы системы. С возрастом животного этот
218
процесс замедляется, поэтому накопление в органическом веществе (оссеин) неорганических солей возрастает в процентном отношении. Это и объясняет значительную хрупкость (ломкость) костей стариков. Таким образом, несомненно, межуточное вещество имеет какой-то свой собственный обмен веществ, все же контролируемый клетками, но он неполноценен и постепенно ослабляется, поэтому организм «вынужден» периодически заменять свое межуточное вещество. Клетки же, если они способны к размножению, как это показал опыт культивирования тканей вне организма, растут и делятся неограниченно долго. Есть культуры из сердца цыпленка, которые перевивают из одной капли питательного раствора в другую уже более 50 лет! Отсюда следует, что только клетка является полноценной биологической единицей, а старость — это свойство организма, но не составляющих его элементарных единиц, пока они не подверглись необратимой специализации (дифференцировке).
Основным соображением, которое было особенно сильно акцентировано Гейденгайном и которое произвело на большинство гистологов в свое время огромное впечатление (его до сих пор, в 1963 г., повторяет Кацнельсон 21 Hi ряд других исследователей), сформулировано было так: «Не все живое (в организме) сконцентрировано в клетке». Повторяем еще раз, против этого положения возражать нельзя, но жизнь межуточного вещества, как мы сказали, всегда кратковременна, а жизнь клетки, не утратившей способность к делению,— потенциально безгранична. Это великолепно выявляет метод тканевых культур, когда из пересаженного в питательную среду кусочка ткани мигрируют действительно жизнеспособные клеточные элементы и образуют зону роста (гистотипический рост), а все остальное быстро гибнет. Таким образом, жизненность клетки, сим-пластов и межуточного вещества неравноценна.
§ 54. Некоторые факты, подтверждающие целостность организма. В предыдущих параграфах приведен целый ряд фактов, подтверждающих идею целостности организма, которую мы себе представляем, следовательно, так: организм анатомически составлен из элементарных биологических единиц — клеток, которые слагаются в более или менее сложные комплексы (системы)— ткани, характеризующиеся уже новыми свойствами. Та
21 1. с., стр 212
2/9
ким образом, как развитие каждой клетки, так и ее поведение (функция) должны направляться и обусловливаться свойствами уже живых систем разного порядка, морфологически более сложных, чем клетка.
Для того чтобы этой концепции придать конкретность, остановимся несколько подробнее на примере истории изучения такого, казалось бы, второстепенного признака, как размер клетки и ее ядра (объемы протоплазматического тела и клеточного ядра — величины прямо пропорциональные).
Впервые вопрос о величине ядра был поставлен ботаниками Саксом, Страсбургером и Амелунгом. Еще в 1893 г. ими были проделаны большие и тщательные исследования, из которых явствовало, что величина растения зависит только от числа клеток, а размер их остается постоянным. Несколько позднее это было подтверждено и на тканях животных. Так, например, Рабль (1853—1917) измерял клетки хрусталика собак очень разных размеров и нашел, что их величины одинаковы. Бовери (1904) сопоставил клетки, слущивающиеся с языка человека, причем он взял гиганта ростом в 200 см и карлика, рост которого равнялся 90 см. Размеры клеток были совершенно сходными (1904).
Закономерность эта была сформулирована впервые Дришем » 1899 г. Формулируя закон постоянства клеточных размеров, он писал: «Величина клеток столь же постоянна, как форма кристаллов». В доказательство этого закона им был проделан следующий очень убедительный опыт: он взял дробившиеся яйца морского ежа и разделил бластомеры в одном случае на стадии двух клеток в другом — четырех клеток, далее на стадии восьми и шестнадцати клеток. Из изолированных клеток-бластомеров развивались вполне нормально сформированные личинки, но соответственно меньших размеров. Дриш сопоставил величину клеток соединительной ткани как нормальных, так и всех этих экспериментально полученных личинок и, мог убедиться, что их размеры во всех случаях оставались идентичными.
Однако когда подошли к данному вопросу путем вычисления средних размеров большого числа ядер и стали сопоставлять величины клеток (ядер) от животных разных возрастов, разных физиологических состояний и т. д., то довольно быстро установили, что величина клеток может отклоняться от типичной и иногда довольно значительно. В этом отношении большое внимание привлекли работы С. М. Лукьянова (1897), А. В. Арапова (1898) и особенно А. А. Березовского (1910, 1911).
220
Исследователей постоянно приводило в смущение также то обстоятельство, что клетки разных органов очень разнообразны по своей величине. Так, нервные клетки из коры мозжечка (клетки Пуркиня) в 1—2 тысячи раз крупнее лимфоцитов крови. Таким образом, закон постоянства клеточных размеров свидетельствовал о постоянстве величины клетки только при сравнении определенных тканей или органов, а не организма, как такового.
Этому последнему явлению было дано объяснение, исходя из способности клеток к делению. Оно принадлежит Дж. Леви, который в ряде многих исследований (1906—1925) разделил все клетки на 3 категории: клетки лабильные — легко размножающиеся в процессе роста, клетки стабильные — делящиеся с трудом и осуществляющие рост в значительной степени увеличением своих размеров, и клетки неизменные — утратившие способность к размножению и растущие только путем возрастания их величины (нервные клетки и др.).
Совершенно по-иному стали понимать эту проблему после работ В. Якоби (1925—1931), который, применив элементарную вариационно-статистическую методику, показал прежде всего, что печень, например, белой мыши состоит из четырех различных размерных групп клеток, названных им клеточными классами. Самым интересным, однако, оказалось то, что величины этих четырех клеточных Классов находились по своему объему в отношении 1:2:4:8, т. е. составляли ряд геометрической прогрессии. Далее было показано, что вообще все клетки одного вида животных находятся друг с другом в тех же объемных соотношениях. Для человека наименьший размерный клеточный класс равен 36 р3, а все остальные более крупные размеры клеток кратны этой величине и составляют ряд удвоения (геометрическая прогрессия с множителем 2). Эти данные были подтверждены очень многими исследователями на самых различных объектах, в том числе и на растениях
Данное открытие заставило пересмотреть правило пос т.о-янства клеточных размеров, и оно было формулировано уже как закон постоянства минимальных клеточных размеров (Вермель, 1934). Другими словами, это можно изложить так: для каждого вида организмов характерна совершенно определенная минимально возможная величина клетки (ядра); она может быть названа квотой, ибо рост клетки идет только путем ее последовательного удвоения. Таким образом, минимальный размер клетки мы рассматриваем как величину
227
постоянную. Однако клетка может расти, увеличивая свой объем, этот процесс обусловлен влиянием условий ее существования, в том числе и влияний клеточных систем или — в общей форме — координаций организма как целого. Все это уже обусловливает конкретные размеры клеток каждой ткани всех органов.
Само явление роста (увеличение массы протопласта) представляет собою элементарное свойство живой материи. Закон роста клеток путем удвоения их объема только лишь описывает процесс, но ничего не говорит о факторах. Можно поставить вопрос, в какой мере рост является свойством клетки как элементарной единицы и как он координируется с целостностью организма?
Мы уже отметили, что в каждом органе мы находим клетки типичных и постоянных для него размеров. Спрашивается, почему клетки не укрупняются? Потому ли, что нужны особые стимулирующие факторы, или же, наоборот, тенденция к росту есть элементарное свойство клетки, а целостная система организма оказывает на него в определенном пределе тормозящее влияние?
Уже простое рассмотрение явления изменчивости величины ядер самых разнообразных клеток позволило предположить, что, по-видимому, речь должна идти именно о сдерживающих рост факторах (Вермель, 1932). Эксперимент подтвердил представление о тенденции клеток к укрупнению: клетки, перенесенные в культуру вне организма (т. е. выделенные из организма), клетки в организме после слабых отравлений (отравление вызывает ослабление именно общих координаций), клетки злокачественных опухолей (клетки, слабо реагирующие на влияние организма), клетки денервированных органов (ослабление общих влияний как следствие перерезки нервов) — все они оказываются « 22 значительно укрупненными по сравнению с нормой .
Мы несколько подробнее остановились на рассмотрении данной проблемы для того, чтобы на конкретной области цитологии показать, как переплетаются свойства клетки и свойства организма. Из приведенного примера далее следует, что анализ явлений, связанных с клеточной организацией, должен вестись таким образом, чтобы выявлять и разграничивать свойства и закономерности части и целого.
22 Вермель Е. М. 1940. Исследование о клеточных размерах.— Уч. зап. Моск. гос. пед. ин-та им. В. И. Ленина, т. 25.
Хесии Я. Е 1967. Размеры ядер и функциональное состояние клеток. М., изд-во «Медицина».
222
Александр Гаврилович Гурвич
(1874—1954)
§ 55. Теория биологического поля А. Г. Гурвича. Изложенное в предыдущих параграфах ясно показывает, что элементарное представление о клетке как независимой биологической единице уже давно себя изжило. Фактически понятие целостности органа и организма приобрело общее признание, и в силу этого во всеобщее употребление вошли такие обороты речи, как «система клеток», или «система тканей». Правда, далеко не всегда в эти слова вкладывается один и тот же смысл. Однако для нас важно здесь прежде всего отметить, что на современном этапе развития ученця о клеточном строении организмов общепризнанным и ведущим является разработка представления о системности. Разные ученые подходят к нему с различных сторон и по-разному его интерпретируют.
Мы не можем взять на себя задачу изложить сколько-нибудь полно ни историю проблемы учения о целостности орга
223
низма, ни его современного состояния — это могло бы явиться темой специальной монографии. Тем не менее, рассматривая развитие представлений о клеточной организации живых существ, мы неизбежно должны были прийти к проблеме целостности, поскольку она возникла по существу как следствие разработки клеточной теории.
Мы уже довольно подробно останавливались на представлениях о «биологическом соединении» Гертвига, о гистосистемах Гейденгайна, об «уровнях организации» Берталанфи, но всякий раз, когда говорится о создании клеточного комплекса (или в более общей форме — системы клеток), всегда возникает вопрос не только о сущности явления, но и о факторах этого процесса. С этой точки зрения большой интерес представляет «теория биологического поля» Гурвича, впервые высказанная им в 1921 г. и заново пересмотренная в 1944 г.23
К теории эмбрионального и биологического поля Гурвич пришел, опираясь на ряд фактических данных из области изучения факторов, определяющих кариокинетический процесс. Эта теория пытается прежде всего развить понимание специфических закономерностей в развитии организма, т. е. таких понятий, которые распространялись бы только на живые системы и; были бы для них совершенно обязательными.
Надо отметить, что в настоящее время почти не созданы системы специфических для биологии универсальных теоретических представлений. С а м о осознание факта специфического уровня о р га н и заци и живых систем —• дело лишь последних десятилетий. Не удивительно поэтому, что подобные теории нередко, мы бы сказали, принципиально встречаются в штыки и плохо понимаются значительным большинством. Таково механистическое наследие конца XIX в., на котором воспитаны последние поколения ученых. Привычнее и «надежнее» кажется свести явление на.физические, физико-химические и химические факторы, чем попытаться рассмотреть его как следствие специфических закономерностей.
Гурвич ставил себе целью найти общую специфическую закономерность хода эмбрионального развития. Единственной независимой переменной величиной он принимает координаты участвующих в процессе элементов (т. е. клеток) и в силу этого считает, что искомая им функция является выражением поля
23 Г у р в и ч А. Г. 1944. Теория биологического поля. М., изд-во «Со-ветская наука».
224
в том смысле, как это понятие применяется в физике. Другими словами, эмбриогенез выражается путем установления векториального поля, в котором Протекают определенные — пространственные — процессы.
Исходным моментом в новой теории поля (теории биологического поля; ранее, в 1921 г., Гурвич говорил об эмбриональном поле, представляя его себе как надклеточный фактор, определяющий судьбу клеточных элементов) является представление о поле, создаваемом самой клеткой, и автор говорит, таким образом, о клеточном поле. Это понятие Гурвич рассматривает как принцип универсального значения.
Объектом клеточного поля являются органические вещества и комплексы их молекул. Именно поле создает и поддерживает определенную молекулярную упорядоченность, специфическую для живой системы. Таким образом, здесь идет речь о таком пространственном распределении молекул протопласта, которое не вытекает просто из их химической структуры или состояния равновесия (химические связи, силы Ван-дер-Ваальса и пр.). Поле есть фактор, создающий и поддерживающий «устойчивое неравновеси е», отличающее живое от неживого (Бауер) 24. Здесь речь идет, конечно, о субмикронных структурах молекулярного порядка, обеспечивающих устойчивость жизненного состояния. Эти структуры нельзя противопоставлять процессам, и правильнее было бы поэтому говорить о структурированных процессах. Именно молекулярная упорядоченность в пространственных взаимоотношениях (констелляции) объясняет два основных замечательных свойства живой материи: ее чрезвычайное многообразие и вместе с тем строгую консервативность. Но вместе с тем все возникающие в живых системах молекулярные комбинации непрерывно заменяются новыми, а для поддержания этого процесса необходима постоянная трата энергии и наличие фактора, направляющего процесс совершенно определенными путями.
Изложенное выше в очень краткой форме представление Гурвича о субмикронном строении протопласта само по себе имеет большое принципиальное значение. Надо специально подчеркнуть, что эта концепция основана на эмпирических данных, которые и были положены в основу уже дальнейшего умозрительного развития учения о поле. Гурвич открыл в 1937 г. общий для
24 Б а у э р Э. 1935. Теоретическая биология. Изд. ВИЭМ.
225
всех живых систем феномен «деградационного излучения»*’. Им показано, что в момент резкого торможения метаболизма или при механическом нарушении пространственного распределения субмикронных частиц протопласта имеет место сильная вспышка митотического излучения. Совершенно независимо В. В. Лепешкин показал, что в аналогичных условиях нарушения структуры клеток происходит освобождение тепловой энергии.
Гурвич объясняет это явление таким образом, что распад молекулярных констелляций ведет к освобождению потенциальной энергии, отдаваемой в виде «деградационного излучения».
Дальнейшее логическое построение таково: неравновесная констелляция не может создаваться путем простого взаимодействия между молекулами, ибо это привело бы к равновесному состоянию системы и потере ею свойств живого. Поддержание постоянной равновесности может быть, следовательно, обусловлено лишь воздействием внешнего по отношению к ней фактора, который и отождествляется Гурвичем с клеточным полем. Этому понятию он придавал значение общебиологического принципа.
Клеточному полю А. Г. Гурвич приписывает далее следующие свойства: оно анизотропно, и именно характер анизотропии определяет его специфичность, а значит и особенности клеток данного вида. Поле непрерывно и преемственно, т. е. нет состояния живой системы без поля, при делении клетки делится и ее поле.
Биологическое поле в понимании Гурвича является инвариан-той клетки и представляет собою чисто динамический фактор, но по совокупности целого ряда цитологических данных предполагается, что оно связано как-то с. хроматином. За центр поля Гурвич принимает приблизительно геометрический центр клеточного ядра. Поле использует энергию экзотермических реакций в живых системах для сообщения их молекулам упорядоченного, направленного движения. Направление этих векторов определяется на основании уже анализа эмпирических данных, в частности процессов раннего формообразования при эмбриональном развитии.
Поскольку само понятие поля заимствовано из физики, то отсюда следует также, что клетка создает поле и вокруг себя, т. е. его действенность распространяется и за пределами клетки. Таким образом, клетки должны оказывать влияние друг на друга своими полями. Следовательно, собственное поле клетки прояв-
15 Гурвич А. и Гурвич Л. 1937. Деградациоииое митотическое излучение.— Бюлл. эксп. биол. и медицины.
226
ляет свои эффек! на протопласт в чистом виде только в случае полной его изоляции. В организме же любая клетка подпадает под влияние синтезированного, или, как Гурвич его называет, актуального или общего поля, которое будет изменяться при каждом пространственном перемещении клеток. Другими слонами, общее поле будет зависеть как от величины тканевого комплекса, так и от его конфигурации.
Мы здесь могли лишь коротко остановиться на общих положениях теории биологического поля, но должны отметить, что они, неся в себе, конечно, много умозрительных моментов, обосновываются и весьма значительным фактическим материалом.
Большой интерес, в частности, представляет работа А. В. Аникина— ученика А. Г. Гурвича, объясняющая изменение формы клеточного ядра соединительнотканных клеток в конечности тритона и многое другое. Самое важное заключается в том, что, приняв гипотезу поля, как показали Гурвич и его сотрудники, можно построить ту или иную схему общего поля и выявить определенную закономерность в развитии тканевого комплекса или органа. Основное положение экспериментальной эмбриологии (доказанное бесконечное число раз), согласно которому судьба клетки в эмбриональном процессе определяется ее местоположением, получает свое конкретное объяснение. Мало того, исходя из той или иной формы зачатка, мы имеем возможность построить поле и даже предсказать следующую стадию развития.
Значит ли это, что процессы эмбрионального развития можно считать уже «понятными» или «ясными» в обычном смысле слова? Сам Гурвич отмечает, что, конечно, нет. Однако вряд ли понятие поля по существу труднее для понимания, чем идея зародышевой плазмы, гена, процессов ассимиляции и т. п.
Мы не сомневаемся, что само представление о клеточном поле и его суммации в общее, актуальное поле в дальнейшем еще проделает значительную эволюцию. Здесь мы сравнительно подробно остановились на этих теоретических построениях, придавая им большое принципиальное значение, поскольку ставится задача конкретизации понятия системности клеточного комплекса, исходя из специфических биологических закономерностей.
Наличие явления, которое названо Гурвичем биологическим полем, является, как нам представляется, фактом, с которым уже нельзя не считаться. Несомненно, что все теоретические выводы, которые должны быть отсюда сделаны, еще очень трудны для понимания, и многих исследователей это ставит в тяжелое положение, поскольку приходится пересматривать старые и, казалось 221
бы, установившиеся положения. Самый «простой» выход — это отрицание самого факта и объявление нового подхода тавтологией. Но история науки уже не раз показывала бесплодность такого отношения к новым открытиям и взглядам.
Вряд ли можно спорить о том, что само понятие взаимовлияния клеток в тканевом комплексе, явление клеточной интеграции, представление о градиентах и ритмах, детерминационные процессы, стойкое сохранение клетками гистомерами и гистосистемами определенных размеров и параметров и ряд других биологических явлений требуют (и притом очень настоятельно) новой теоретической трактовки.
Необходимость исходить при этом из специфических биологических представлений чисто теоретически также как будто никем из передовых биологов не отрицается.
Ни идеалистические схемы развития современных западноевропейских ученых (А. Мейер, Л. Вудреф и др.), ни попытки воскрешения механистических концепций не смогут двинуть науку вперед. Нельзя считать большим прогрессом также многочисленные исследования, ставящие перед собою цель уточнить или расширить наши конкретные познания в той или иной специальной области (например, экспериментальной эмбриологии, называемой нередко механикой развития организмов).
В настоящее время — во второй половине XX в. — особенно чувствуется необходимость новой концепции в понимании комплексности и системности организма с точки зрения специфических биологических закономерностей.
Конкретной попыткой именно в этом направлении, как нам представляется, и следует считать теорию биологического поля Гурвича в том виде, как она им сформулирована в 1944 г. Теорию эту, с нашей точки зрения, следует считать вполне материалистической, хотя о природе самого динамического фактора мы пока еще ничего не знаем. Непознанность нередко путают с непознаваемостью и в силу этого предпочитают игнорировать факты, что всегда тормозило развитие науки. Такое отношение показывает недостаточность абстрактного мышления у некоторых биологов. Физики, имеющие хорошую математическую подготовку, в этом отношении прогрессирнее. Ведь если подходить с такой установкой, что нельзя заниматься явлением, поскольку неизвестна его природа, то невозможно было бы развитие, например, оптики, ибо неизвестна природа света, нельзя было бы изучать магнитные свойства, ибо 'Неизвестна природа магнетизма, и т. д.
228
§ 56. Философские основы представления об уровнях организации материи. В связи со всем изложенным следует в заключение остановиться на современных взглядах в отношении уровней биосистем, развиваемых за последнее время философами в СССР.
Долгие годы данной проблеме не уделялось почти никакого внимания. Однако за последние 10 лет в этом отношении произошел значительный поворот. В 1967 г. секцией философских проблем Академии наук СССР была созвана специальная конференция по проблеме «Структурные уровни биосистем» 26. Подчеркивалось, что идея структурных уровней в биологии должна ставиться в ряду теоретических обобщений наравне с клеточной теорией и теорией эволюции.
В докладе М. Ф. Веденева, В. И. Кремянского и А. Т. Шаталова было указано, что еще древнегреческий философ Платон развивал положение, согласно которому «целое больше суммы своих частей». Как предшественник представлений о системной организации авторами назван также философ Ф. В. Шеллинг, создавший философскую систему так называемого объективного идеализма. Именно Шеллинг писал об «иерархии организованностей, показывающей лишь различные моменты в эволюции мироздания».
Несомненно, Шеллинг не видел путей объяснения сложнейших явлений жизни элементарными законами материального мира; следует иметь в виду, что он был современником Иоганнеса Мюллера, говорившего о «жизненной силе» и, очевидно, оказавшего на него влияние.
В философской литературе за рубежом представления об уровнях организации материи (от космогонических явлений до развития общества) начали энергично развиваться с начала XX в. Рассмотрение этих проблем, однако, не входит в наши задачи, и мы ограничимся лишь вопросами биоструктуры живого существа и роли клетки как его основной анатомической единицы.
Структурные уровни уже и ранее рассматривались философами как ступени развития материи (холизм Я. Смэтса и А. фон Мейер-Абиха). Они пытались разобраться в категориях разви-
23 Очень показательны были сами заглавия докладов иа этой конференции; вот некоторые из иих: «Становление и конкретизация идеи структурных уровней в биологии», «Главные структурные уровни биологических систем», «О философском аспекте концепции уровней», «Проблема взаимосвязей структурных уровней биологических систем» и др. в сб. «Структурные уровни биосистем» (материалы к конференции). М., изд-во «Наука», 1967.
10 Е. М. Вермель
229
тия материи и внести ясность в применяемую терминологию и в создаваемые понятия.
Основное положение этих представлений состоит в том, что качественная специфика структурного уровня не сводится к сумме свойств ее компонентов, взятых порознь или вне данной системы, а создается при ее становлении. Своеобразие организованности ведет к возникновению новых закономерностей и появлению новых свойств и сил. Особенности каждого уровня организации определяются не столько даже составляющими его элементами, сколько особенностями их взаимодействия между собой и с общим комплексом. Отсюда следует представление о значении организации и структуры материи, о закономерностях соотношения части и целого, элементов и системы, специфичных для качественно различных ее ступеней. По мере усложнения каждой высшей ступени возникают качественно новые свойства. Новые уровни создаются посредством соединения и интеграции сходных или подобных единиц в общую систему.
На многих проблемах в предыдущих параграфах мы уже останавливались, рассматривая конкретные вопросы клеточной структуры организмов, в частности на представлении об иерархическом их соподчинении. В истолковании этой проблемы мы, естественно, следовали индуктивному методу, исходя из реальных наблюдений и экспериментов. К аналогичным взглядам пришли и философы, излагая эту проблему дедуктивно, в абстрагированном виде, и пытаясь дать развернутую логическую классификацию этих явлений.
Так, например, в докладе на упомянутой конференции Н. Т. Абрамова предложила для живых систем следующий иерархический ряд звеньев: молекулярное—>-органоидное—>-клеточное —>- органо-тканевое —* система органа ->- организменное —* -> популяционное.
В общей форме это правильно, но кое с чем биолог согласиться вряд лц может, необходимо сделать ряд уточнений. Прежде всего встает вопрос, в какой степени сопоставимы указанные уровни между собой. Сравним ли скачок между молекулярным звеном и клеткой, с одной стороны, и скачком клетка -> органотканевое звено, с другой, и далее со звеном организм —популяция (или «сверхорганизмом»—стадом, семьей, колонией)? Мы, как биологи, недостаточно понимаем и знаем хщмию, чтобы говорить о наличии 'звеньев в молекулярных системах, но ряд от клетки до многоклеточного организма, несомненно, существенно и принципиально отличается от перехода молекулы к клетке и
230
от организма к популяции. Другими словами, надо уточнить нс только терминологию, но и сами понятия степеней различия иерархических этапов усложнения органических (и, вероятно, и других) систем.
Возвращаясь к предложенному Н. Т. Абрамовой ряду этапов живых систем, надо сказать, что молекулярное звено выделено правильно. В настоящее время, когда наблюдается огромный интерес к молекулярной биологии, рассмотрение этого вопроса особенно актуально. На молекулярном уровне в клетке продолжают действовать законы «элементарных» частиц (законы химии), хотя ведущими становятся уже законы биологического целого, создающие высокоорганизованную самоуправляющуюся систему. Именно по этой причине вирусы не следует рассматривать как существа, а лишь как вещества, поскольку их свойства проявляются только после проникновения и включения вирусов в клетку.
Следующее восходящее звено названо Н. Т. Абрамовой органоидным. Реально такого особого уровня жизни не существует. Для биолога «органоидный» уровень равнозначен цитологическому, ибо только клетка действительно представляет собой принципиально (качественно) новую живую систему по сравнению с биомолекулярным уровнем. Дело в том, что ни ядро, а тем более ядрышко илн хромосомы, ни центросома, ни митохондрии, ни лизосомы, ни аппарат Гольджи, ни любые другие так называемые «ооганоиды» вне клетки не существуют и существовать не могут. И если, например, митохондрии вне клетки (будучи выделены центрифугированием) еще некоторое время «работают», то только до тех пор, пока не израсходован их запас ферментов или других активных веществ. Органоиды — это сложные молекулярные биоагрегаты особой архитектоники, сохраняющиеся только в составе клетки и создающие в своей специфической совокупности индивидуализированную, морфологически неделимую элементарную единицу — клетку. Таким образом, клетку надо характеризовать как систему гетерогенных биокомпонентов, находящихся в состоянии устойчивой неравновесности.
Сочетание клеток образует звено, удачно названное Н. Т. Абрамовой органо-тканевым. Его следует расценивать как гистосистему гомогенного строения, т. е. состоящую из сходных элементов, образующих морфологическое и функциональное единство. Эти гистосистемы (гистомеры) могут быть разного порядка, и в этом также состоит их специфичность, пока они не создадут органы, являющиеся уже гетерогенными элементами для о р-
10* 231
ганизма как целого. Таким образом, каждое звено в цепи, создающей организм многоклеточного существа, имеет свои характерные особенности в смысле генеза и структуры. Но сами эти отличия иного «уровня», чем различия между неживым (веществом) и живым (клеткой, организмом).
Мы хотим здесь еще раз указать, что не только большинство биологов, но и теоретики — философы, защищающие позиции диалектического материализма, пришли ныне к убеждению, что элементарной биологической системой могла быть только клетка, ибо только клетка обладает свойством самоуправления, самосохранения и самовоспроизводства. Правда, в многоклетчном организме процессы жизнедеятельности клетки становятся ограниченными, зависимыми от системы целого; ее регуляция подчиняется более высоким уровням структуры организма (В. Г. Афанасьев, А. А. Заморский и др.). С этой точки зрения одноклеточность (Protozoa) и многоклеточность организма (Metazoa)—это лишь разные формы эволюции «первоклетки», если будет позволено применить такой термин. Не случайно И. П. Павлов писал, что только организм (а он может быть как одноклеточным, так и многоклеточным) представляет собой саморегулирующуюся, поддерживающую себя и даже совершенствующуюся систему.
Из сказанного следует, что методологическое значение принципа структурности особенно возрастает в свете теоретической оценки клеточной теории (М. Ф. Веденев, В. И. Кремян-ский и А. Т. Шаталов).
Клетка, несомненно, главный структурный уровень организма, основной и ведущий элемент его строения и развития (А. А. Заморский).
В заключение необходимо еще раз подчеркнуть, что понятие целостности как следствия системной организации нельзя рассматривать вне материалистической концепции жизни. Теория уровней организации является неотъемлемой частью материалистической диалектики о формах движения материи от космологических процессов до социальных явлений. Конечно, все проблемы возникновения и развития жизни, форм ее существования должны решаться в свете этой концепции о высших и низших ступенях ее организации— иерархии звеньев и интеграции компонентов.
Эти идеи начали развиваться давно, их высказывал еще Г. Гегель, а затем и Ф. Энгельс в своих представлениях о формах движения материи. В то же время многие весьма серьезные ученые-естествоиспытатели (начиная с И. Мюллера и до наших дней)
232
видели и понимали, что с совершенствованием организации возникает упорядоченность живой материи, 'которая необъяснима законами элементарной химии и физики. Однако сущность явления осталась для них загадочной, и потому они привлекали к объяснению жизненных явлений нематериальный фактор (жизненная сила, энтелехия, душа и т. д.). Все это показывает, как важна в конкретном Исследовании философская концепция. До конца качественная сторона уровня систем может быть объяснена материалистически и понята только марксистской философией — диалектическим материализмом.
§ 57. Эволюционная гистология А. А. Заварзина. Наиболее неразработанным вопросом в учении о клетке до последнего времени остается проблема эволюции тканевых структур и возникновение клетки.
Сравнительная гистология понималась обычно как сопоставление гистологического строения гомологичных и аналогичных тканевых структур. Предпринимались подобные исследования преимущественно в рамках эволюционной морфологии (сравнительной анатомии), но они практически ничего существенного в построение филогенетических рядов не внесли. Больше того: гистологические данные в значительном числе случаев оказались стоящими в противоречии с эволюционной морфологией, ибо они обнаруживали сходство в строении между такими организмами, которые ни палеонтологически, ни сравнительно-анатомически и ни эмбриологически ничего общего не имеют. На это явление специально указывал в свое время В. фон Эбнер (1911) и делал отсюда вывод, что при современном положении наших знаний приложение эволюционной теории к рассмотрению гистологических знаний невозможно27. Практически гистологические факты зоологами-эволюционистами игнорировались.
Заслуга создания нового направления в гистологии — эволюционной гистологии — принадлежит А. А. Заварзину (1886— 1945).
Отправным моментом в теоретических построениях для решения проблемы эволюционной гистологии у А. А. Заварзина было как раз то, перед чем, образно выражаясь, большинство ученых бессильно опустило руки. Он исходил как раз из тех противоре-
2Г Е b n е г V. von. 1911. Gewebeentwicklung und Phylogenese. Anat. Anz.,
Erg.-heft zum Bd. 38 («Развитие тканей и филогенез»).
233
Алексей Алексеевич Заварзин (1886—1945)
чий, в которые вступают гистологические факты с филогенетическими схемами эволюционной морфологии. Классическим примером такого противоречия является сопоставление, например, гистологического строения мышечной ткани членистоногих и позвоночных (а также некоторых мышц моллюсков). Представители этих двух типов животных эволюционно ничего общего не имеют, между тем структура их мышечных волокон сходна до ряда мелких деталей, а наблюдаемые различия принципиального значения не имеют. Таким образом, мы встречаемся с фактом чрезвычайно большого сходства гистологического
234
строения ткани и ее физиологических свойств у форм, не имеющих никаких сколько-нибудь близких родственных в эволюционном смысле отношений.
Таких фактов констатировано очень и очень много. Они ско рее правило, чем исключение. Отсюда Заварзин сделал заключение, что именно разрешение этого противоречия и должно быть ключом к эволюционному объяснению гистологического фактического материала.
За 30 лет работы в этом направлении Заварзиным и его сотрудниками было проделано очень большое число исследований. Он исходил из общеизвестного факта, что вообще ткани всех типов животных можно свести к четырем основным типам: эпителию, соединительной ткани с кровью, мышцам и нервной ткани. С другой стороны, он принимал, что вся эволюция организмов носила приспособительный характер к условиям существования. Таким образом, одниитеже факторы среды привел и, с одной стороны, к многообразию видов животных, а с другой стороны — обусловили однообразие.строе-ния их тканей.
Заварзин специально подчеркивал, что тканевые системы нельзя рассматривать вне факторов, определяющих взаимодействие организма со средой. Он развивает эту свою мысль следующим образом: организм любой степени сложности имеет ткань, отграничивающую его тело от внешней среды, он их называет пограничными тканями. Именно пограничность является ее основным и наиболее общим свойством, определяемым как развитием, так и условиями существования. Пограничная функция ткани в соответствии с усложнением организации животного дифференцируется: обособляются поверхности, несущие различные функции,— дыхательную, пищеварительную, защитную и выделительную. Как бы различны животные ни были, эти их поверхности должны развиваться в указанных совершенно определенных направлениях. В силу этого сам тканевой материал и должен приобретать сходство. Наибольшее разнообразие в соответствии с разнообразием среды обитания животных должна, естественно, иметь защитная поверхность.
Внутри организма находится ткань, непосредственно с внешней средой не соприкасающаяся. Ее Заварзин назвал тканью внутреннейсреды, она осуществляет распределение по организму питательных веществ и кислорода, она поддерживает стабильность физико-химических условий, отличных от внешней среды, и т. д. Отсюда ясно, что свойства внутренней среды долях-
235
ны быть у всех организмов Относительно стойкими и вместе с тем в какой-то степени сходными. Естественно, что эволюция тканей внутренней среды должна идти по пути усложнения вместе с эволюцией организмов, но эволюция ее пойдет у самых разнообразных животных в общих чертах сходно. Это обусловлено, следовательно, относительным однообразием среды и сходством процессов внутреннего обмена даже у очень разных животных.
Ткани мышечная и нервная, по представлениям Заварзина, появились в эволюции в системе пограничных тканей, но затем они погрузились во внутреннюю среду, где они развивались уже во все более и более стабилизующихся условиях. Именно это способствовало все возраставшей специализации этих структур, но вместе с тем определяло и их сходство, ибо оно шло в близких
** 9Я
условиях независимо от видовой принадлежности организмов .
По существу перечисленными выше четырьмя тканями — пограничной (эпителием), тканью внутренней среды (соединительная ткань), мышечной и нервной — исчерпывается состав тела всех животных организмов. Заварзин показал на огромном фактическом материале, что ткани как своим происхождением, так и своим развитием обязаны взаимодействию организма с внешней средой. Явление сходства в строении тканей у филогенетически очень далеко отстоящих животных (например, клеточных элементов сетчатки глаза головоногих моллюсков и позвоночных и многих других) Заварзин еще в 1934 г. назвал законом параллельных рядов тканевой эволюции.
Таким образом, гистология, или наука о тканях животных организмов, впервые приобретает свое эволюционное содержание. Совершенно очевидно, что основной ее задачей является установление путей тканевой эволюции, а также выявление ее частных закономерностей, которые представляют собою закономерности филогенетической дифференцировки живого вещества. Другими словами, эволюционная гистология, как определил ее задачи сам Заварзин, изучает элементарные закономерности органической эволюции, лежащие в основе развития частей, но которые вместе с тем обусловлены закономерностями эволюции целых организмов.
Заварзин и его ученики дали ряд блестящих примеров анализа тканей различных животных; в первую очередь следует ука-
28 Заварзин А. А. 1941. Очерки по эволюционной гистологии нервной системы. М., Медгиз, 1945. Очерки эволюционной гистологии крови и соединительной ткани. М., Медгиз.
236
зать на их работы по нервной системе, крови и соединительной ткани насекомых и др.
Наиболее слабым моментом в концепциях Заварзина, по нашему представлению, является разработка понятия «тканевой системы»—термин, который он применяет очень часто. Термин «система», по А. А. Заварзину, следует понимать прежде всего как качественное определение разнообразия дифференцировок клеточных элементов. По существу, как нам кажется, здесь кроется даже некоторое логическое противоречие: само слово «ткань» не может указывать на нечто иное и|ли большее, чем на качество, а под понятием «система» должна подразумеваться известная пространственная характеристика. Система в конкретном понимании биолога есть прежде всего структура определенных свойств и параметров. Заварзин как-то никогда не придавал большого значения «микроскопической анатомии», полагая, что изучение микроструктур даже не входит, собственно, в задачу гистологии как науки, являясь предметом анатомии. Таким образом, понятие «тканевой системы» у него обозначает, собственно говоря, только разнообразие клеточных элементов. Это видно и из формулированного им определения ткани: «ткань есть филогенетически обусловленная элементарная система, объединенная общей структурой, функцией и камбиальностью или развитием» (1. с. 1941, стр. 56).
Другими словами, ткани представляют собою известную совокупность разнообразных клеточных форм, а между тем что такое клеточная форма или клеточный вид (выражения, очень широко применяемые всеми гистологами), остается неясным. Впрочем, определения клеточной форме вообще никем не дано, и мы даже не знаем серьезных попыток поставить этот вопрос.
Вместе с тем, если мы хотим говорить об эволюции тканевых структур (а необходимость этого ясна) и если мы признаем за клеткой значение индивидуализированной элементарной единицы, то мы обязаны ставить вопрос и о таксономических понятиях. Такова, как нам кажется, назревшая задача в учении о клетке, и ее разрешение находиться на очереди.
Несмотря на отсутствие определения понятия клеточного вида (клеточной формы, сорта или др.), практически мы им вынуждены пользоваться, как нередко для обозначения определенного типа клеток пользуются словом ткань.
В свете проблем эволюционной гистологии во всей широте встала задача внести ясность в явление многообразия клеточных форм и тканей. Исторический подход (как в свете филогенети
237
ческих построений, так и онтогенеза организма) не позволяет сводить задачу к составлению определительной таблицы разнообразных клеточных видов, основное требование — создание генетической классификации клеточных элементов и построение схемы наподобие «эволюционного древа».
В этом отношении мы должны указать на работу другого замечательного русского гистолога Н. Г. Хлопина (1897—1961), разрабатывавшего естественную классификацию тканей на эволюционной основе. Опираясь на огромный изученный им и его учениками фактический материал, Хлопин убедительно показывает, что определенные ткани стойко сохраняют присущие им свойства. Он считает, что единственно рациональным и научно обоснованным принципом классификации тканей является генетический принцип. Н. Г. Хлопиным29 впервые разработана подробная схема взаимоотношения тканей, которой можно придавать не только онтогенетический смысл, но и известное филогенетическое значение. Основным ее недостатком, лишающим ее должной четкости, надо считать, является неразработанность и неясность низших и высших таксономических понятий. Без них никакая классификация биологических элементов невозможна.
Практически столь часто применяемое слово «ткан ь», очень может быть удобно, но это еще не термин и не определенное понятие, а привычный оборот речи. Совершенно необходимо ясно и четко разграничивать понятия тектологические (качественное, видовое своеобразие) и понятия архитектоники (структуры в узком смысле этого слова). Не делая дифференциацию этик понятий, представления о микроскопических структурах лишаются определенной конкретности.
Огромной заслугой Заварзина, работы которого, несомненно, создали новый этап в развитии учения о клетках, является то. что он впервые показал, как надо понимать сравнительный метод в гистологии и как надо подходить к пониманию филогенеза гистологических структур. Эти представления Заварзина вытекают из его многочисленных исследований по микроскопическому изучению беспозвоночных и позвоночных животных.
Именно Заварзиным создана эволюционная гистология.
23 Результаты своих исследований и исследований своих учеников Н. Г. Хлопни обобщил в книге «Общебиологические и экспериментальные основы гистологии». Изд-во АН СССР, 1946.
238
§ 58. Еще о заблуждениях а изучении клетки и клеточной теории. Заблуждения в науке бывают двоякого рода: одни исходят Из ошибочной методологии исследования, а также неправильной методики изучения, хотя и правильных наблюдений, другие основаны на неверной теоретической предпосылке, что ведет к недооценке или переоценке фактических Данных, бот в этом последнем случае оправдывается печально знаменитое утверждение: «если факты расходятся с теорией, тем хуже для фактов». Действительно, вера в определенную концепцию нередко заставляет видеть то, чего нет, либо толковать увиденное в желаемом свете, искренне считая излишним проверять свои данные.
К категории методических ошибок, относится, в частности, все учение о так называемом хромидии, развивавшемся известным зоологом Рихардом Гертвигом (1850—1957) в начале XX в. Он исходил из уже давно известного факта наличия в протоплазме многих простейших структур, окрашивающихся, так же как и ядро, «ядерными» красителями (в то время пользовались преимущественно борным кармином). Часто эти образования лежат около клеточного ядра, что и послужило основанием создать теорию выхода веществ ядра (хроматина) в протоплазму в виде зернышек (хромидии(). Особенно подробно это явление было описано Гертвигом в 1904 г. при изучении корненожки арцеллы и солнечника Actinosphaerium30. В течение последовавших 15 лет этот взгляд получил очень широкое распространение и был подтвержден большим числом ученых, в том числе выдающимся протозоологом Ф. Шаудиным (1871—1906), открывшим возбудителя сифилиса — бледную спирохету. Авторитет Р. Гертвига и Ф. Шаудина стоял очень высоко, и их последователи стали видеть гораздо больше, чем сами авторы этой теории. «Хромидии» были найдены решительно у всех простейших. Их начали описывать уже в клетках многоклеточных (Гольдшмидт). По данным некоторых авторов, «хромидии», выйдя из ядра в протоплазму, даже копулировали и во всяком случае давали начало новым ядрам. Этими наблюдениями ставились под сомнения многочисленные и безусловно очень точные данные о митотическом процессе, законах уменьшения числа хромосом в мейозе и т. д. Другими словами, ставился вопрос о необходимости пересмотра представлений о клетке!
30 Her twig R. 1904. Die physiologische Degeneration bei Actinosphaerium Eichhornii. Feslschr. f. R. Haeckel («О физиологической дегенерации у солнечника»).
239
По счйстЫо, почти тотчас (уже в 1912—1914 гг.) хромидий в клетках Metazoa были полностью развенчаны, было показано, что за таковые принимали остатки плохо фиксированных структур (митохондрии и др.), которые никакого отношения к ядру пэ своему геиезу не имеют. Аналогичные работы начали появляться И в отношении простейших. Именно в это время среди цитологов стала особенно модной фраза: «одно около другого не значит одно из другого» (Neben einander ist nicht auseinander). В самом деле, метод микроскописта состоит в том, что ткань или орган фиксируют (консервируют в спирте, формалине, сулеме и т. д.), далее готовят тонкие срезы, которые обрабатывают растворами красителей и затем изучают под микроскопом. Как известно, вещество ядра имеет кислую реакцию, и поэтому оно закрашивается основными красителями значительно сильнее, чем протоплазма. Нередко оказывалось, однако, что ядро и различные зернышки в протоплазме окрашивались с одинаковой интенсивностью. В случаях, когда эти зернышки прилежали к оболочке ядра, то казалось. что вещество ядра выступает из пего. По фиксированному, т. е. мертвому, объекту установить направление даже реально существующего процесса часто очень трудно. Это ведь как кинолента, которую можно просматривать как спереди назад, так и наоборот! Подбирая последовательность стадий входа или выхода зернышек неизвестной природы из ядра, можно легко все перепутать. В случае описания выхода хромидий из ядра простейших оказалось, что это просто ошибка, основанная на порочности самой методики, ибо даже очень различные вещества могут окрашиваться сходно, тем более что гистологическое окрашивание— не простая химическая реакция, а процесс очень сложный.
Большая заслуга в опровержении хромидиальной гипотезы принадлежит советским исследователям, особенно А. В. Румянцеву (1899-—1947). Он и его ученики применили для изучения цитологии Protozoa простые методы гистохимического анализа. Оказалось, что никакого «хромидия» в понимании Гертвига не существует: в одних случаях это были жиро-белковые включения (липопротеиды), в других — углеводно-белковые (гликопротеиды), а в третьих—-структуры sui generis вообще отсутствовали Румянцев, в частности, изучал тот же объект, что и Гертвиг31.
si Румянцев А. В. 1925. Исследование строения протоплазмы Rlii-zopoda.— Тр. н.-и. ин-та Зоологии МГУ, т. I. Rumjanzew А. и. Е. Wermel. 1925. Protoplasmabau von Actinosphaerium Eichhornii. Arch, f. Protistenk, 52, 217—264 («Строение протоплазмы солнечника»).
240
Алексей Всеволодович Румянцев (1899—1947)
Следует подчеркнуть, что реакция ученого мира на «разоблачение» хромидий была удивительно правильной и объективной. К ним сразу утратили всякий интерес, и ни в учебниках, ни в большинстве специальных работ о них больше совсем не упоминали. О хромидиях забыли. Положительным результатом всех этих исследований можно считать то, что цитология простейших оказалась хорошо изученной. Было показано, что принципиально клетка-организм Protozoa построена так же, как клетки Metazoa.
Другим примером серьезных заблуждений, на котором надо остановиться, является представление о «живом веществе», лишь недавно (с 1932 г.) энергично развивавшееся в ряде работ О. Б. Лепешинской32.
Как следует из всего изложенного в этой книге, узловой проблемой клеточной теории является представление о клеткообразо-вании. Шлейден и Шванн в 30—40-х годах прошлого века пред
32 Лепешинская О. Б. 1950. Происхождение клеток из живого вещества и роль живого вещества в организме. М.
241
ставляли себе возникновение путем слияния «зернышек» по сферам. Это было совершенно ошибочно, но это можно было объяснять несовершенной оптикой и главным образом отсутствием надежных методов гистологического исследования. Несостоятельность этих наблюдений была вскоре достоверно показана. Плеяда крупных ученых установила, что клетки возникают только путем деления. Далее более 100 лет тщательно изучали этот процесс, при этом все варианты «свободного» образования ядер и клеток быстро отвергались.
Однако идея возникновения клеток из зернышек или из бесструктурной массы оказалась живучей. В 1899 г. М. Д. Лавдов-ский и Н. П. Тишуткин вновь к ней вернулись33. Они изучали развитие куриного эмбриона — объект этот очень сложен и технически необычайно труден. Только этим и можно объяснить ошибочность наблюдений и выводов этих авторов, которые скорее (Путем догадок пришли к заключению о возникновении, клеток из. так называемых «желточных шаров». Эти последние являются запасным питательным веществом (липоиды и белки), за счет которых живет и развивается зародыш, а поэтому эти «шары», конечно, ие могут стать живой клеткой.
В 30-х годах XX в., т. е. через 100 лет после работ Шлейдена и Шванна (!), О. Б. Лепешинская вновь пришла к выводу о в и е-клеточном возникновении клеток. Она изучала развитие некоторых рыб и куриного зародыша и утверждала, что клетки образуются заново из желтка («желточных шаров»).
Удивительно, что нашлось много гистологов, в частности А. Г. Киорре (1954), которые поддерживали Лепешинскую своими конкретными исследованиями и тоже писали, что они будто бы видели образование клеток из «желточных шаров» 34. Однако тщательное изучение этого вопроса (С. И. Щелкунов исотр.) решительно опровергло эти данные, и тот же Кнорре в 1955 г. отказался от подобных представлений, хотя и продолжал считать бластомеры за какие-то «монероподобные» (?) состояния35.
33Lavdovsky М. u. Tischutkin N. 1899. Von den Beziehungen der Dotterelementen zu den Keimblattercellen. Biol. Zentralbl., 19 («О соотношении между элементами желтка и клетками зародышевых листков»).
34 Киорре А. Г. 1954. К вопросу о процессах развития яйцевых клеток у птиц и амфибий.— Тр. коиф. «Новые данные по проблеме развития клеточных и неклеточиых форм живого вещества». М.
33 Кнорре А. Г. 1955. Морфологические особенности элементов желтка куриного яйца.— Докл. АН СССР, 103, № 1.
242
Наряду со своим главным объектом изучения — яйцеклетками, богатыми желтком,— Лепешинская же приводила доказательства в пользу существования «доклеточного» вещества, способного превращаться в клетки, из области патологии. Она считала, что некроз и распад клеток приводит к образованию «живого вещества», из которого вновь возникают клетки. Эти фантастические утверждения имели успех. Например, такой крупный ученый, как И. В. Давыдовский, в 1951 г. писал: «Возможно, что так называемый кариорексис есть процесс размножения клеток, который начинается распылением ядерного вещества и заканчивается образованием новых ядерно-плазменных комплексов» 36. Правда, Давыдовский— патологоанатом, и он был далек от чисто биологических проблем, хотя нередко апеллировал к иим. Удивительно, что некоторые биологи — гистологи и эмбриологи, как А. Н. Студитский, защищали подобные взгляды. Кроме того, полагали, что регенерационная бластема (совокупность недифференцированных клеток, из которых образуется регенерирующий орган) в опытах на амфибиях возникает из разрушенной ткани 37. Отсюда следует делать вывод, что при травмах хирург должен не убирать разрушенные ткани, а, наоборот, еще больше их травмировать, чтобы создавать «живое вещество», из которого будут возникать клетки регенерата38 39. Все это совершенно ошибочно и ничего общего с действительностью не Имеет.
Лепешинская, вопреки очевидности, чрезвычайно воинственно выступала против положения «всякая клетка от клетки» (Вирхов), считая своей главной задачей развенчание этого закона. Другими словами, восстанавливалась идея внеклеточного образования клеток, а поэтому иное понимание получала и клеточная теория. Основным ее положением была новая формулировка, а именно: «Каждая клетка развивается из живого вещества». Однако и «живое» вещество, по ее представлениям, могло просто создаваться из «неживых» субстанций. Представление об эволюции жизни, таким образом, упрощалось до примитива.
36 Давыдовский И. В. 1951. Совещание по проблеме живого вещества и развития клеток. Изд-во АН СССР.
’’Полежаев Л. В. 1956. Изменение регенераторной способности у жи-
вотных.— Изв. АН СССР, сер. биол.
39 Студитский А. Н. 1954. Основы биологической теории регенерации. Тр. Ин-та морфологии животных им. А. Н. Северцева, вып. 2. М., Изд-во АН СССР.
£43
Само определение «живого вещества», даваемое Лепешинской, весьма неотчетливо. Она пишет: «Под живым веществом мы понимаем не только массу вещества, не имеющую форму клеток, но даже вещество на разных стадиях его развития, начиная от живой молекулы...» (1 с., стр. 207). Позднее в статье «Живое вещество» (БСЭ, 1952) она расширяет это понятие, и в ней мы читаем, что под живым веществом следует понимать «сложное, содержащее белок образование, основным и необходимым условием существования которого является постоянное самообновление...»
Вряд ли стоит подробно рассматривать и вдумываться в это определение, как и во все выводы и «факты», приводимые О. Б. Лепешинской.
В настоящее время даже непонятно, как об этом можно было серьезно говорить. Между тем гистолог А. Н. Студитский в 1952 г. писал в специальных журналах, что открытие О. Б. Лепешинской происхождения клеток из живого вещества является исключительным достижением науки и служит «основой для разработки новой передовой теории строения и развития животных и растительных организмов» 39. Академик Т. Д. Лысенко в предисловии к монографии О. Б. Лепешинской «Происхождение клеток из живого вещества» (1950) заявлял: «Клетка и все ее содержимое могут происходить не только из себе подобных, т. е. из клетки же..., а и и|з соответствующих веществ, не имеющих структуры, присущей клетке. Это принципиально новое положение в биологической науке блестяще показано О. Б. Лепешинском в ее тонких экспериментах. В этом и заключается ее большая научная заслуга...» И тем не менее все это было опровергнуто.
Как уже отмечалось, Студитский был одним из самых энергичных прозелитов «учения» Лепешинской о живом веществе, диких представлений Бошьяна о возникновении микроорганизмов из неживых веществ, а также псевдомичуринских представлений Лысенко о скачкообразном возникновении видов из неклеточного «живого вещества». Вместе с тем Студитский яростно выступал против научной, классической генетики, называя ее буржуазным учением — вейсманизмом, менделизмом и морганизмом, что звучало тогда почти как ругательство. Он напечатал до 39
39 Студитский А. Н. 1952. За творческую разработку учения о живом веществе.— Арх. анат., гистол. и эмбриол. 29, № 4.
244
двух десятков статей по этим вопросам, в том числе в широком прессе, пропагандируя подобные ненаучные взгляды (укажем лишь некоторые) 40.
Не менее восторженно выступил на «Совещании по проблеме живого вещества» в 1950 г. патологоанатом И. В. Давыдовский. Он оказал, что Лепешинская находится на верном пути. По его мнению, «клеточная патология и идея целлюляризма не встанут на ноги после нанесенных им ударов». Однако и это уже давно забыто, а большинство патологоанатомов никогда и не разделяло мнения И. В. Давыдовского по этим вопросам.
Сама идея, упрощавшая до предела представление о жизни и ее эволюции, несомненно, подкупала многих, а демагогическое орудование философской терминологией «убеждало» невзыскательных ученых. Поэтому вновь начинали искать «живое некле-точиое вещество» и ... «находили» его! Не будем приводить фамилии многих наших современников, поддерживавших эту искажающую биологию концепцию.
Между тем «открытия» продолжались. Нет смысла перечислять здесь все эти измышления. Однако выступления сторонников Лепешинской имеют известное поучительное значение (методические, методологические и философские ошибки). Поэтому укажем для примера на немногие, но едва ли не на самые невероятные. Так, например, доказывалось образование клеток даже в белке куриного яйца, представляющем собой лишь секрет железистых клеток стенки яйцеводов. За великое открытие выдавалось наблюдение, будто бы клетки образуются в соке растений при прибавлении к нему нуклеиновой кислоты. А профессор В. Г. Шипачев из Иркутска утверждал даже, что при пересадке проросших семян пшеницы в брюшную полость кролика происхо-дит превращение растите\ьных клеток в клетки животные! . И это Лепешинская энергично поддерживала. Она сама ставила под сомнение опровержение гениальным Луи Пастером самозарождения микроорганизмов или, например, писала, что «...партеногенез как таковой вообще не существует, а процесс оплодотворения происходит в доклеточном периоде в живом вещест-
49 Студитский А. Н. 1953. За творческую разработку проблемы видообразования.— Усп. совр. биол., 35. 1950. Новое в науке о жизни (по поводу книги Г. М. Бошьяиа «О природе вирусов и микробов»). «Огонек», № 22, и мн. др.
41 Шипачев В. Г. 1954. Об исторически сложившемся эволюционном пути развития животной клетки в свете новой диалектико-материалистической теории. Иркутск.
24г)
ве...»42. Никаких фактических доказательств в последнее время ею уже не приводилось; так поступали 150 лет назад натурфилософы 43.
В заключение следует добавить, что воззрения Лепешинской по существу откровенно механистичны, поскольку игнорируется исторически сложившаяся системность клеточной организации и эволюционный принцип. Вся эта печальная история, не принесшая с собой ничего положительного, правильно и подробно изложена в хорошо и убедительно написанной книге С. И. Щелку-нова 44.
Не нужно думать, что взгляды Лепешинской не вызывали и в свое время серьезных возражений. Особенно в ранний период критика была даже ожесточенной. Всю несостоятельность ее работ одним из первых показал Б. П. Токин45, затем А. А. Заварзин, Д. Н. Насонов и Н. Г. Хлопин 46 и многие другие. Но несмотря на то, что это были 'крупнейшие гистологи того периода, к ним не прислушались.
$ 59. Основные положения учения о клеточном строении организмов. Попытаемся подвести некоторые итоги учению о клеточном строении организмов. Основными моментами в представлении о клетке, как надо полагать, являются следующие положения.
1. Клетку следует считать важнейшим этапом в развитии материи от неживого к живому, ибо она является элементарной живой единицей или, точнее, элементарной полноценной живой системой.
42 Лепешинская О. Б. 1952. Клеточная теория на новом этапе.
43 Пропагандистами представлений О. Б. Лепешинской были также профессор Л. И. Фалин и особенно член-корр. АН СССР проф. Г. К. Хрущев, а также академик Т. Д. Лысенко. См. Фалин Д. И. 1952. Новое учение о клетке. М. 1954. О роли живого вещества и генезе тетраплоидных опухолей. М.
Хрущев Г. К. 1952. Проблема развития неклеточных форм живого вещества. М., 1953. Новые данные в развитии учения о живом веществе. М., и др.
Лысенко Т. Д. 1952. Работы О. Б. Лепешинской и превращение видов. В ки. «Неклеточиые формы жизни». М.
44 Щелкунов С. И. 1958. Клеточная теория и учение о тканях. Л., Медгиз.
45 Т о к и и Б. П. 1936. Клетка и организм.— Под знаменем марксизма, № 8.
43 Заварзин А. А., Насонов Д. Н. и Хлопин Н. Г. 1937. Об одном «направлении» в цитологии-— Арх. биол. наук, 56.
246
Несколько особая организация бактерий и сииезеленых водорослей не представляется существенной, ибо последние следует рассматривать либо как результаты регрессивных изменений, либо как особую, очень древнюю, специализированную ветвь эволюции. Нельзя пока что сказать ничего определенного также о природе фильтрующихся вирусов.
2. Клетка как с морфологической, так и с физиологической точек зрения является структурной единицей как наиболее простых, так и наиболее сложных Metazoa. Такова принципиальная сторона вопроса, и поэтому все клетки гомологичны.
3. Как в процессе филогенеза, так и онтогенеза возникновение многоклеточности сопровождается постепенным ограничением и даже потерей клетками их физиологической активности и генетической потенции. Другими словами, можно сказать, что, оставаясь гомологичными структурами, клетки перестают быть друг другу аналогичными. Образование тела многоклеточного организма обязательно должно сопровождаться детерминацией, дифференцировкой и специализацией составляющих его элементарных живых клеток, что ие может не отражаться, конечно, и на ик свойствах.
4. Понятие «клетка как таковая» есть лишь абстракция, отвлеченная схема. Любая клетка имеет свои характерные особенности и способна лишь к определенному кругу функций. Развитие (дифференциация) клеток в онтогенезе организма представляет собою характернейшую особенность эволюции Metazoa. Изучая этот процесс с точки зрения выявления свойств клеток, мы должны отметить возникновение большого их разнообразия («видообразование» клеток). Процесс этот, как правило, необратим, и идет он от клеток высокопотенциальных к клеткам более специализированным, параллельно с этим теряющим способность к дальнейшим превращениям. Но самое важное заключается в том, что специализация рано или поздно ведет к ограничению жизненной полноценности. Пути развития клеточных элементов в процессе их специализации необычайно разнообразны: иногда в результате этих процессов возникают вполне изолированные клетки (лейкоциты), а иногда, развиваясь, они дают многоядерные массы (путем слияния или нерасхождения плазматических тел после деления ядер). В ряде случаев тело клетки так или иначе растрачивается на образование межуточных или кутикулярных веществ, ограниченной жизнедеятельности.
Все специализированные структуры имеют ограниченную жизненность, т. е. им присущ не весь комплекс свойств, харак
247
Терный для полноценной живой системы. Однако известное количество свойств живого в них постоянно присутствует, каковые мы и констатируем даже в неклеточном межуточном веществе.
5. Онтогенез следует рассматривать как процесс последовательно возникающих специализаций и дифференцировок, а также нарастающего числа клеточных элементов, образующих гистосистемы. Этот процесс является следствием взаимовлияния как клеток, так и закладок органов зародыша единого целого.
6. Ни одна клетка не может рассматриваться вне той среды, в которой она существует. А таковой прежде всего является постепенно создающаяся система клеток определенной архитектоники (структуры). Именно это положение является одним из важнейших в теории клеточного строения организмов. На современном этапе развития науки можно считать установленным, что комплексы клеток представляют собой своеобразные, в какой-то определенной степени индивидуализированные системы (гистосистемы). Эта системность отражается на свойствах самих составляющих ее клеточных элементов таким образом, что они как в морфологическом, так и особенно в физиологическом отношении в значительной степени теряют свою индивидуальность, т. е. клетки теряют способность к независимой жизнедеятельности. Именно это явление и называют клеточной интеграцией.
Образуя в определенном числе гистосистему, клетки ие просто суммируют свои свойства, а образуют нечто целое, характеризующееся новыми качествами. Взаимоотношения клеток элементарной гистосистемы (например, альвеолы) можно называть «биологическим соединением», поскольку при этом возникают новые качества, свойственные системе (порядку организации) как целому.
7. Без понятия системности не может быть создано представление об организме. Каждый этап возникновения системности (системы низшего и высшего порядка, например, клетка, альвеола, дольки, орган — железа) с необходимостью привносит новые качественные моменты. В этом заключается особенность организации тела многоклеточного организма.
Без понятия системности в структуре организма не может быть создано правильное представление и оклетке, представляющей собой единое самостоятельное целое — элементарную живую единицу, но вместе с тем она является и частью целого организма. Следовательно, вся жизнедеятельность клетки определяется как ее собственными особенностями, так и свойствами тех более сложных, вышестоящих систем, частью которых она является. Далее следует сказать, что, оста
248
ваясЬ определенными морфологическими и физиологическими единицами, клетки оказывается в теле Metazoa в результате длительной исторической эволюции интегрированными частями целого.
Отсюда ясна ошибочность взгляда на организм как на сумму клеток, отсюда ясно также, почему организм, который мы, следовательно, представляем себе построенным из ряда систем различных порядков, находящихся друг с другом в иерархическом подчинении, имеет ряд особенностей, не сводимых просто на отдельные свойства клеток. Итак, первая элементарная биологическая система — это сама клетка, а последняя высшая система — это организм многоклеточного в целом.
Как мы выше уже отмечали, многократно деклалировалась необходимость коренного пересмотра клеточной теории, или, как нередко писали, «вирховианства». По этому вопросу, в частности, патологоанатом Л. Ашофф в своей статье, посвященной 100-летию клеточной теории (1939), правильно написал, что «требование во что бы то ни стало заменять клеточную теорию каким-то другим представлением бессмысленно».
Надо, как он отметил, лишь понять своеобразиесуще-ствования клеток в комплексе. В общетеоретических статьях многих исследователей можно найти изложение взгляда, согласно которому организм нечто большее, чем сумма составляющих его частей. Наиболее отчетливо и обоснованно, как мы полагаем, эти мысли изложил и продолжает развивать и поныне Бер-талаифи. Именно он подчеркивает, что в целом организме проявляются свойства, отсутствующие в изолированных частях, а поэтому невозможно «растворить явления жизни в элементарных единицах», каковыми являются клетки. Проблема жизни — это проблема организации, и она характеризуется системами различной сложности. Берталанфи предлагает даже закон «каждая клетка от клетки» дополнить еще другим: «каждая организация от организации» 47. Однако это вряд ли нужно.
Изложенная нами выше концепция, как нам представляется, исключает как механистическое, так и виталистическое понимание клеточной теории строения организмов. Вместе с тем эта точка зрения (поскольку она во главу угла ставит материалистическую трактовку явления качественного своеобразия живых систем организма) оказывается в соответствии с передовой философией современности — философией диалектического материализма.
17Berlalanffy L. von. 1952. Problemes of life («Проблемы жизни»),
24l)
§ 60. Этапы в развитии представлений о строении клетки. Подведем краткие итоги истории развития взглядов на природу клетки, которые, как мы видим, неоднократно менялись.
Первой должна быть названа схема, предложенная Гуком (1665) и Мальпиги (1675—1679), разделявшаяся К. Вольфом (1759) и защищавшаяся еще Мирбелем (1801—1808). Эта ^трактовка чрезвычайно примитивна: к л е т к и рассматриваются Как ячейки, сравнимые с «пеной на кружке Пива» (Грю, 1682), стенки, которых затвердевают. Ясно, что при таком понимании клетки общепринятым было и представление об общности стенок двух соседних ячеек. Единственное представление о развитии клеток, которое связано с этим этапом,— гипотеза Вольфа о возникновении ячеистости в результате образования жидких капель в студенистой массе живого вещества с последующим затвердением пограничного слоя.
Второй этап развития представлений о микроструктуре организмов связан с именами Линка и Рудольфи (1804), Треви-рануса (1807—1821), Мейена (1830) и др. Этими исследователями, во-первых, было показано, что каждая ячейка-клетка покрыта своей особой оболочкой и потому клетка отделима от других ей подобных; во-вторых, было констатировано наличие клеточного содержимого, обладающего самостоятельной подвижностью. Никакой особой теории с этим этапом развития в науке не связано, если не считать того, что в то время считалась уже бесспорной чрезвычайно широкая распространенность клеточного строения. Наряду с клетками как элементарные структуры рассматривали также волоконца и сосуды растений; их клеточное строение клеточный генез оставались неизвестны.
Третий период клеточного учения является переломным, Он связан с именами Шлейдена (1838) и Шваниа (1839). Прежде всего как обязательный элемент рассматриваются ядро и ядрышко, возникающие путем агломерации элементарных зернышек.
Вместе с тем впервые формулируется клеточная теория, основным моментом которой является принцип общностиклеткообразования во всех частях живой материи. Способ клеткообразования из сливающихся зернышек признается всеобщим, а отсюда постулируется уже сравнимость всех клеток как растений, так и животных. Основными элементами всех клеток признаются: оболочка, протоплазма и ядро с ядрышком.
250
Итак, все живые существа, согласно этим воззрениям, состоят из клеток или их производных, но сами клетки возникают из зернышек первично-бесструктурной массы.
Четвертый период характеризуется прежде всего накоплением обильного нового фактического материала, который очень скоро встал в противоречие с рядом теоретических положений Шлейдена, Шванна и др. Доказывалась лишь относительная ценность оболочки клетки как неотъемлемой части клетки; при этом была окончательно выяснена разница между растительной и животной клетками. Затем (и это основное) была установлена ошибочность теории цитогенезиса, а вместо этого доказывалась преемствен ностьклеточной структуры, что особенно четко было сформулировано Вирховым (1857) в виде закона «omnis cellulae cellulae». В результате ряда замечательных для своего времени работ как ботаников (Кон и др.), так и зоологов (Ремак и др.). Шультце было сформулировано новое определение клетки как «комочка протоплазмы с ядром». Вместе с тем клеточная теория иа новой фактической основе получила свое дальнейшее углубление, в особенности с введением представления о клетке как об «элементарном организме» (Брюкке, 1861). Представление об организме как о сумме клеток стало неопровержимым и привело в дальнейшем к теории «клеточного государства» (Вирхов, Ферворн). Этот период может быть характеризован как господство механистических взглядов в учении о клеточном строении организмов.
Пятым по счету этапом в истории учения о клетке может быть названо направление, пытавшееся разложить клетку на еще более простые жив ы е элементы, сопоставлявшиеся и даже гомологизировавшиеся с бактериями. Этот взгляд развивался рядом французских ученых (Бешан, 1860—1883 гг. и др.) и затем (казалось, в особенно убедительной форме) Альтманом (1890—1894). Достаточных доказательств эта точка зрения, однако, не получила. Тем не менее сходные взгляды в той или иной форме всплывают и до самого последнего времени (Иван Валлин, 1926).
Шестым этапом в изучении клетки условно можно назвать с х ем у, предложенную Вильсоном (1896—1925) в его известной сводке. Она по существу стремится лишь свести все наши сведения о морфо чогических структурах клетки, не внося никакой принципиально новой точки| зрения.
Наконец, последним, седьмым этапом представлений а клетке следует считать схему, которая может быть выведена 25/
исходя из современных исследований о природе протоплазмы, ядра и различных включений клетки. Основная сущность этой схемы заключается в том, что структуру клетки мы себе представляем исключительно сложной, несмотря на то, что при применении даже наилучших световых микроскопических объективов субстанция протоплазмы (цитоплазма и ядро) представляется нам гомогенной. Современная методика морфологи|ческого исследования клетки, в связи с освоением электронной микроскопии, находится на большой высоте; хуже разработаны гистохимические методы.
Итак, современная схема строения клетки в и з-вестной мере сходна с представлениями Дюжардена (1835), Гофмейстера (1867) и др., которые также писали о гомогенности протоплазмы. Говоря о схеме строения клетки, мы и теперь должны по существу ограничиваться лишь очень немногим; мы указываем, что клетка состоит из протоплазмы, одетой тонкой мембраной. В протоплазме мы постоянно обнаруживаем хонд-риосомы, а также постоянное присутствие ядра. Электронная микроскопия обнаружила в протоплазме еще так называемый ретикулум, рибосомы и некоторые другие мембранные структуры. О более тонком строении ядра мы еще ничего не узнали.
Однако современное понимание клетки резко отличается от всех более ранних представлений тем, что мы пришли к «про с-той» схеме строения, убедившись в такой ее чрез-fl ычайнойс ложное г ни своеобразии, которая современными методами не может быть достаточно полно анализирована. Правда, мы знаем многочисленные включения и органоиды клеток, которые изучены довольно тщательно.
Другими словами, клетка лишь внешне представляется нам просто организованной системой, тогда как ее внутренняя, интимная структура лежит за пределами видимости и нашего понимания. К этому убеждению современная цитология пришла в результате исследований целой плеяды ученых на протяжении последних 100 лет. Исследования, проведенные при использовании электронного микроскопа, ныне это подтвердили и внесли много нового, но клетка осталась клеткой, и никаких более простых и мелких полноценных живых структур не найдено.
В заключение необходимо еще раз подчеркнуть, что меньше всего мы знаем о возникновении клеток. Эта проблема практически совпадает с вопросом о возникновении жизни из неживой материи. Впервые данный вопрос был поставлен Геккелем в 1866 г., но он пока остается целиком в области гипотез.
АВТОРСКИЙ УКАЗАТЕЛЬ
Аббе Э. 9, 129
Абрамова Н. Т. 230—232
Адамс Г. 31, 34
Альтманн Р. 107, 123—127, 161, 252
Альхасец 10
Амелунг Е. 220
Амичн Д. 9
Аникин А. В. 227
Арапов А. В. 220
Ауэрбах Л. 138
Ашерсон 84, 121
Ашоф Л. 249
Афанасьев В. Г. 232
Бауэр Э. 225
Беляев И. И. 35, 36
Беиеден Е. фон 134, 147, 151, 152
Березовский А. А. 220
Берроуз М. Т. 214
Берталанфи Л. фон 196, 204—207,
211, 224, 249
Бешан 252
Биша Кс. 62, 63
Бишоф Т. 59, 86
Бляхер Л. Я. 140
Бовери Т. 152, 155, 220
Боррие Б. фон 9
Броун Р. 26, 52, 134
Бошьяи 244
Брюкке Э. 68, 92, 97—101, 107, 108, 110, 111, 124, 251
Бэкон Роджер 11
Бюффон Ж. Л. Л. 30, 55
Бючлн О. 100, 107, 118, 120, 121, 142—144, 152
Вагнер Р. 82
Вальдейер В. 68, 69, 122, 134
Валентин Г. 50, 81—85, 121, 127
Валлин И. 252
Вариек Н. А. 139, 140
Веденев М. Ф. 229, 232
Вейсман А. 153, 181
Вермель Е. М. 6, 132, 191, 196, 221, 222, 241
Визнер ГО. 171, 181
Вильсон Э. 135, 153, 157, 252
Винчи Леонардо да 11
Вирхов Р. 53, 59, 68, 69, 85—91, 155, 161, 251
Вольтер (Аруэ) Ф. М. де 2, 7, 24
Вольф К. Ф. 37, 41—44, 250
Гайсннович А. Е. 42
Галей К. 11
Галлилей Г. 9, 12—14
Галлер А. фон 41, 45, 46, 62
Гансштейн И. фон 107, 122, 123
Гарди В. 107
Гевелий 31
Гегель Г. 232
Гейденгайн М. 107, 135, 166, 171 — 178, 181, 196, 203
Гейцмаи К. 107, 111—113
Геккель Э. 57, 59, 91, 92, 96, 101—
104, 183, 253
Генле Ф. Я. 68
Гельмгольц В. 68
Гертвиг Оскар 124, 135, 143, 152, 153, 155, 181, 196, 199—203
Гертвиг Рнхард 135, 154, 196, 239
Гертель X. Г. 9, 28, 33
Гольджи К. 155, 156
Гольдшмидт Р. 239
Горянинов П. Ф 53, 58, 59
Гофман Т. А. 25
Гофмейстер В. 107, 109, 110, 123, 134, 140
Грю Н. 28, 37, 40—42, 250
Гук Р. 5, 9, 27, 28, 37, 38, 250 Гурвич А. Г. 166—170, 179, 195, 196, 216, 223—228
Гуфнагель Г. 11
Гюйар 151
Гюйгенс X. 14
253
Давыдовский И. В. 243, 245
Дарвин Ч. 5, 91, 181
Дейль ван 31
Дсляж Ив. 166, 178, 179
Дивини Е. 9, 30
Добелл К. 166, 181, 188
Доллонд Дж. 9, 31
Дреббель К. 9, 13, 14
Дриш Г. 166—170, 196, 197, 220
Дув К. де 132
Дюбуа-Реймон Э. 67, 68
Дюжарден Ф. 107—109, 252
Дюмортье Б. 134, 136
Дютроше А. 49, 53, 60, 79
Заварзин А. А. 96, 196, 211, 233— 238, 239, 246
Зайнер И. 194
Заморский А. А. 7, 232
Зидентопф Г. 9
Зингер Ч. 24
Иммермаи К. 25
Иордан Г. 203
Карнуа Ж. Б. 92, 104—106
Кацнельсон 3. С. 59, 62, 151, 166, 188—191, 193, 212
Кёлер А. 9
Келликер Р. фон 59, 68, 139
Кеплер И. 14
Кёфф Дж. 15, 31, 33
Клеро А. К. 70
Киолль М. 9, 130
Кнорре А. Г. 242, 243
Козо-Полянский Б. М. 58, 59
Кон Ф. 107, 251
Корнфельд В. 216
Корти Б. 50
Крюн П. де 18
Кремянский В. И. 229, 232
Купфер К. фон 107, 115, 116
Куффлер 14
Лавдовский М. Д. 242
Ламарк Ж. Б. 105
Левенгук А. ван 11, 12, 15, 18, 19,
ЭЗ, 37, 44, 48
Леви Дж. 221
Ледермюллер М. 19—21
Лейбниц Г. 54, 60
Лейднг Ф. фон 90, 91
Ленин В. И. 6, 65
Лепешинская О. Б. 241—246
Лепешкин В. В. 226
Линк Г. 37, 47, 250
Липперсгейм И. 12
Ломоносов М. В. 35
Лукьянов С. М. 220
Лункевнч В. В. 29
Людвиг К. 68
Лысенко Т. Д. 244, 245
Майер Е. 214
Майер К. 62
Майзиль В. 144
Мальпиги М. 28, 37, 39, 40, 250
Марк А. фои 154
Мевес Ф. 155
Мейен Ф. 37, 47, 52, 80, 136, 250
Мециус Я. 12
Мечников И. И. 157, 162
Мильн-Эдварс А. 49, 127
Мннчни Е. А. 211
Мнрбель Б. де 37, 45, 46, 250
Мишер И. 68
Молденгауер 37, 47
Моль Г. фон 73, 83, 107, 108, 134,136
Морган Т. Г. 153
Мур Дж. 135
Моуффет Т. 11
Мюллер Иоганн 53, 65—69, 197,232
Навашин С. Г. 135, 147
Насонов Д. Н. 246
Немилов А. В. 166, 182, 188—193
Нидхэм И. 201, 202
Ньютон И. 27
Нэгели К. 123, 136, 158, 181
Окен Л. 30, 53—57, 79
Опарин А. И. 103
Ошатц 63
Павлов И. П. 232
Паллад 107
Пастер Луи 29, 245
Перемежко П. И. 145
Платон 230
Платтер Ф. 11
Полежаев Л. В. 243
Пирогов Н. И. 65, 67
Пфлюгер Е. 191
Пуркиня Ян 37, 50, 51, 53, 60, 63—
65, 87. 107, 108, 134, 211
254
Рабль К. 135, 147, 151, 152, 220
Райков Б. Е. 59
Райч А. 61
Распанль Ф. 53, 60, 62, 79
Рейхенбах Г. 205
Рейхерт К. 79, 93
Ремак Р. 68, 86, 92, 93, 107, 108,
134, 251
Ретциус 147
Робертис Э. де 182, 202
Рокитанский К. 91
Рода э. 166, 185—188
Ру В. 134, 147, 151, 152, 157, 162,
170, 181
Рубашкин В. Я. 166, 188, 190
Рудольфи К. 37, 47, 250
Румблер 100
Румянцев А. В. 96, 196, 240
Руска Е. 9. 130
Руссов Э. 141
Руфус Эфесский 11
Сакс ГО. 157—162, 220
Самсонов В. 125
Свифт Дж. 24
Седжвнк А. 166, 179
Соболь С. Л. 12, 33
Спаланцани Л. 29
Спенсер Г. 181, 215
Стелутти Ф. 14
Стефенсон В. 9
Страсбургер Э. 135, 143, 220
Студитскнй А. Н. 243—245
Студничка Ф. К. 166, 183, 185
Тереховский М. М. 29, 30
Тишуткин Н. П. 242
Токии Б. П. 246
Торре делла 44
Тревираиус Г. 105, 250
Тревиранус Л. X. 47, 50, 61
Тюрпен П. 53, 61
Уайтмен К. 166, 179—181
Фабер И. 9, 14
Фармер Дж. 135
Ферворн М. 157, 163—165, 181, 251
Филатов Д. П. 170
Фишер Альберт 174, 214, 215
Фишер Б. 201
Фишер Эмиль 208
Флемминг В. 116—119, 134, 145
152
Фриз Г. де 181
Фоль А. 140, 143
Фонтана Ф. 25, 26, 37, 41, 44, 45, 48, 76, 134
Фракасторо Дж, 11
Фроманн К. 116, 117
Хесин Я. Е. 216, 223
Хлопин Н. Г. 238, 246
Чезн Ф. 14
Чистяков И. Д. 140, 143
Церникс Ф. 9
Шаксель Ю. 186, 193, 194, 196, 198
Шаталов А. Т. 229, 232
Шаудинн Ф. 239
Шванн Т. 5. 53, 57, 60—62, 68, 74—80, 134, 161, 194, 197, 242, 251
Шевяков В. Т. 120
Шеллинг Ф. 229
Шестранд Ф. 107
Шлейден М. 5, 53. 69-74, 134, 242, 251
Шлейхер В. 134, 144, 146
Шлятер Г. Г. 124
Шнейдер А. 134. 141, 142
Шпек И. 107, 129
Шпеман Г. 170
Шпренгель К 5 1
Шульце М. 92—97, 251
Щелкунов С. И. 242, 246
Щипачев В. Г. 245
Эбнер В. фон 234
Эйлер Л. 9, 14
Энгельс Ф. 5, 184, 199, 208—210, 232
Эпинус Ф. 31
Эфруссн Б. 215
Якоби В. 196, 221
Янсен Г. 12
Янсеи 3. 12
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
Амитоз (В. Флемминг, 1880) 134, 145, 146
Анатомия растении (М. Мальпиги, 1679) 40, 41, 45
Areola (Р. Браун, 1833) 52
Биобласт (Р. Альтман, 1890) 125
Биология (Г. Тревнранус, 1802;
Ж. Б. Ламарк, 1802) 105
Гистология (К. Майер, 1819) 62, 79
Зародышевый пузырек (Я. Пуркиня, 1830) 51, 139, 141, 142
Инфузории (А. ван Левенгук, 1696) 19, 56
Кариокинез (В. Шлейхер, 1879) 134, 145, 146
Кариолизнс (Ауэрбах, 1874) 139
Клетка, определение (Р. Гук, 1665)
38, 45, 47, 50, 59, 63, 64, 71, 173
Клетка, деление 88
Клеточная теория (Т. Шванн, 1839) 43, 172—178
Лизосомы (К. де Дув, 1929) 132
Мейозис (Дж. Фармер и Дж. Мур, 1905) 135, 152
Metazoa 103, 104
Микрография (Р. Гук, 1665) 28
Микроскоп (И. Фабер, 1625) 7, 13, 128—131
Микротом (Ошатц, 1825—1838) 63
Митоз (В. Флемминг, 1880) 139, 141, 145
Митохондрии (Ф. Мевес, 1900) 118, 125
Монеры (Э. Геккель, 1866) 102, 103, 139
Nucleus (Р. Браун, 1833) 52, 59, 138
Пластиды (Э. Геккель, 1866) 102
Полярные тельца (О. Бючлн, 1874) 142
256
Протисты 102
Протомер (М. Гейденгайн, 1904) 177
Протоплазма (Я.Пуркиня, 1830) 108
Происхождение клетки 103, 104, 204, 207,210—212
Протопласт (Г. фон Гансштейн, 1880) 122
Пузырек первичный (Я. Пуркиня, 1831) 56
Рибосомы 132
Саркода (Ф. Дюжарден, 1835) 108
Симпласт 94, 184
Синцитий (Э. Геккель) 94, 96, 174, 183
Системность 160, 161, 168, 178, 212, 217
Ткань (Н. Грю, 1671) 41
Utriculus (К. Вольф, 1759) 43
Хондриосомы (К. Дюсберг, 1907) 118, 125
Хроматин (В. Флемминг, 1879) 134, 239
Хромидни (Р. Гертвнг, 1904) 239
Хромосома (В. Вальдейер, 1883) 134
Cellula (Р. Гук, 1665) 38, 69
Целлюлярная патология (Р. Вирхов, 1857) 69
Цитобласт (М. Шлейден, 1838) 59
Циотобластема (М. Шлейден, 1838) 73, 74
Цитогеиезис (М. Шлейден, 1838) 71, 75, 78, 84, 104
Цитогенетика 136, 153
Цнтоды (Э. Геккель, 1886) 102, 103, 184
Энергнда (Ю. Сакс, 1895) 158—161
Ядро (Р. Браун, 1833) 52, 59, 136
ОГЛАВЛЕНИЕ
Предисловие 5
Глава первая ИСТОРИЯ МИКРОСКОПА Основные даты по истории микроскопа............................. 9
§ 1. Введение ................................................. 10
§ 2. Изобретение оптических линз............................... 10
§ 3. Изобретение сложного микроскопа........................... 12
§ 4. Первые микроскопические наблюдения........................ 14
§ 5. Отражение микроскопических открытий в художественной литературе ........................................................ 19
§ 6. Особенности микроскопических исследований................. 25
§ 7. Первые микроскопические исследования ученых............... 27
§ 8. Первые микроскопы в России................................ 35
Глава вторая
ИСТОРИЯ ОТКРЫТИЯ КЛЕТКИ
Основные даты по истории открытия клетки.................... 37
§ 9. Описание растительной клетки и ткани Р. Гуком, М. Мальпиги
и Н. Грю............................................... 38
§ 10. Теория возникновения клеток-мешочков К. Вольфа .... 41
§11. Первые описания животных клеток....................... 43
§12. Новое представление о клетке в начале XIX века....... 45
§13. Клеточная структура животных тканей *................. 48
§ 14. Первые описания строения содержимого клетки........... 50
Глава третья
КЛЕТОЧНАЯ ТЕОРИЯ
Основные даты развития клеточной теории..................... 53
§15. Воззрения натурфилософа Л. Окена на клетку и ее возникно-
вение на Земле......................................... 54
§ 16. Идеи Горянинова о значении клеточной структуры .... 58 § 17. Оценка клеточной структуры в 20-х и 30-х годах XIX века . . 6() § 18. Теория клеткообразования М. Шлейдена.................. 71
251
§ 19. Клеточная теория Т. Шванна.............................. 74
§ 20. Клеточная теория в оценке современников................. 79
§ 21. Цитогенез н эмбриология................................. 85
§ 22. Пересмотр клеточной теории Р. Вирховым.................. 86
Глава четвертая
КЛЕТКА — ЭЛЕМЕНТАРНЫЙ ОРГАНИЗМ
Основные даты развития представлений о клетке как об организме .
§ 23. Определение понятия клетки М. Шультце................
§ 24. Клетка — элементарный организм в представлении Э. Брюкке .
§ 25. Значение учения о клетке для эволюционных построений в работах Э. Геккеля ................................
§ 26. Биология клетки как самостоятельная проблема ......
Глава пятая
СТРОЕНИЕ ПРОТОПЛАЗМЫ И КЛЕТОЧНАЯ ТЕОРИЯ
Основные даты по истории изучения протоплазмы..........
§ 27. Первые описания строения протоплазмы.............
§ 28. Теории строения протоплазмы......................
§ 29. Протопласт И. Ганштейна..........................
§ 30. Теория биобластов Р. Альтмана....................
§ 31. Современные представления о строении протоплазмы и клеточная теория..........................................
Глава шестая
ЯДРО И ДЕЛЕНИЕ КЛЕТКИ
Основные даты по истории изучения ядра и деления клетки .... 134
§ 32. Первые описания ядра и деления клетки..................... 136
§ 33. Первые описания митоза в делящихся яйцеклетках .... 139
§ 34. Описание фаз кариокинетического деления....................143
§ 35. Дальнейшее изучение митоза................................ 146
§ 36. Значение открытия кариокинеза для учения о клетке . . . 153
Глава седьмая
УЧЕНИЕ О КЛЕТКЕ НА РУБЕЖЕ ДВУХ СТОЛЕТИИ
Основные даты в развитии методологии клеточной теории на рубеже
XIX и XX веков................................... '. . . 157
§ 37. Энергиды Ю. Сакса и проблема специфичности живой материи 158
§ 38. Теория «клеточного государства» в трактовке современников . 161
Глава восьмая
КРИТИКА КЛЕТОЧНОЙ ТЕОРИИ
Основные даты в развитии критики клеточной теории ............ 166
§ 39. Целостность организма в трактовках Г. Дриша н А. Гурвича 167 § 40. Представление М. Гейденгайиа о целостности организма и его
критика клеточной теории ................................ 171
§ 41. Основатели плазматической теории строения организмов:
А. Седжвнк, Ив Деляж и К. Уайтман........................179
§ 42. Одноклеточные как клетка и организм......................181
§ 43. Значение синцитиев и учение К. Студннчки о единстве орга-
низма ................................................... 183
§ 44. Оценка плазмодиальных структур в работах Э. Родэ . . . 185
§ 45. Некоторые современные представления об относительности по-
нятия клетки.............................................188
§ 46. О трактовке понятия клетки ............................. 193
Г лавадевятая
СОВРЕМЕННОЕ ПОЛОЖЕНИЕ КЛЕТОЧНОЙ ТЕОРИИ
Основные даты 1-й половины XX века по истории учения о клетке и тканях . . . . ................................................196
§ 47. О методологической порочности виталистической и механи-
стической точек зрения в оценке клеточной теории .... 197
§ 48. Представление о «биологическом соединении» О. Гертвига . 199
§ 49. Системность в строении организмов и представление о клетке . 202
§ 50. Организменная теория Л. Берталанфи и клеточная структура . 205
§ 51. О происхождении живой материи............................207
§ 52. Эволюция клетки и данные о степени самостоятельности кле-
ток многоклеточных организмов........................... 210
§ 53. Проблема межуточного вещества........................... 217
§ 54. Некоторые факты, подтверждающие целостность организма . 219
§ 55. Теория биологического поля А. Г. Гурвича.................223
§ 56. Философские основы представления об уровнях организации
материн . ................................................229
§ 57. Эволюционная гистология А. А. Заварзина..................233
§ 58. Еще о ваблуждениях в изучении клетки и клеточной теории . 239
§ 59. Основные положения учения о клеточном строении организмов 246 § 60. Этапы в развитии представлений о строении клетки .... 250
Авторский указатель . . 253
Предметный указатель . .................................. . 256
Евгений Матвеевич Вермель
ИСТОРИЯ УЧЕНИЯ О КЛЕТКЕ
Утверждено к печати
Всесоюзным институтом научной и технической информации
Редактор
В. Ф. Мирек
Редактор издательства
Э. А. Фролова Технический редактор Э. Л. Кунина
Сдано в набор 29/IX 1969 г.
Подписано к печати 13/V 1970 г.
Формат 60X84‘/j6. Бумага № 1
Усл. печ. л. 14,46. Уч.-изд. л. 13,9
Тираж 9.600. Т-07934
Тип. эак. 3043. Цена 1 р. 07 к.
Издательство «Наука». Москва, К-62, Подсосенский пер., д. 21
2-я типография издательства «Наука». Москва, Г-99, Шубииский пер., 10