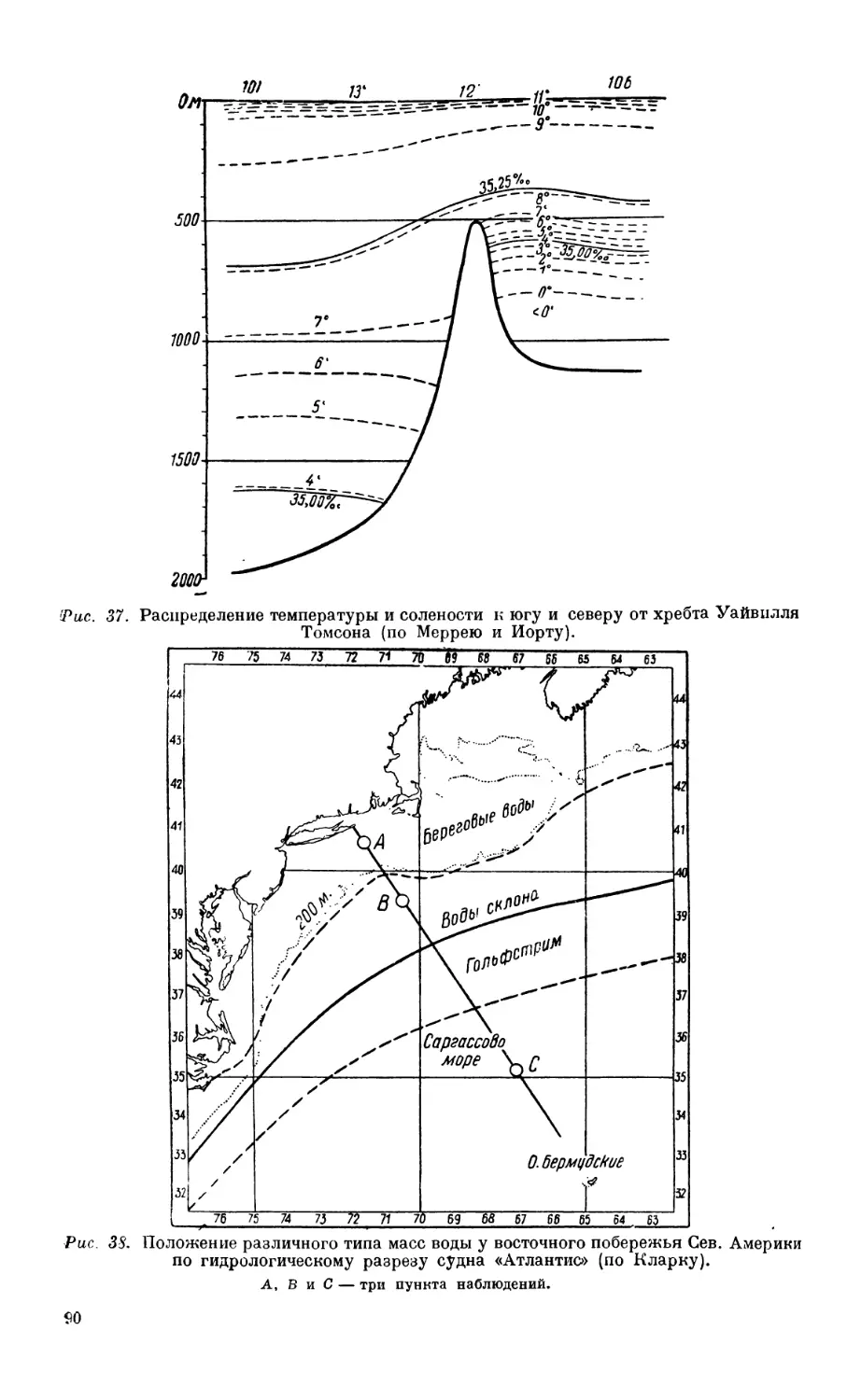
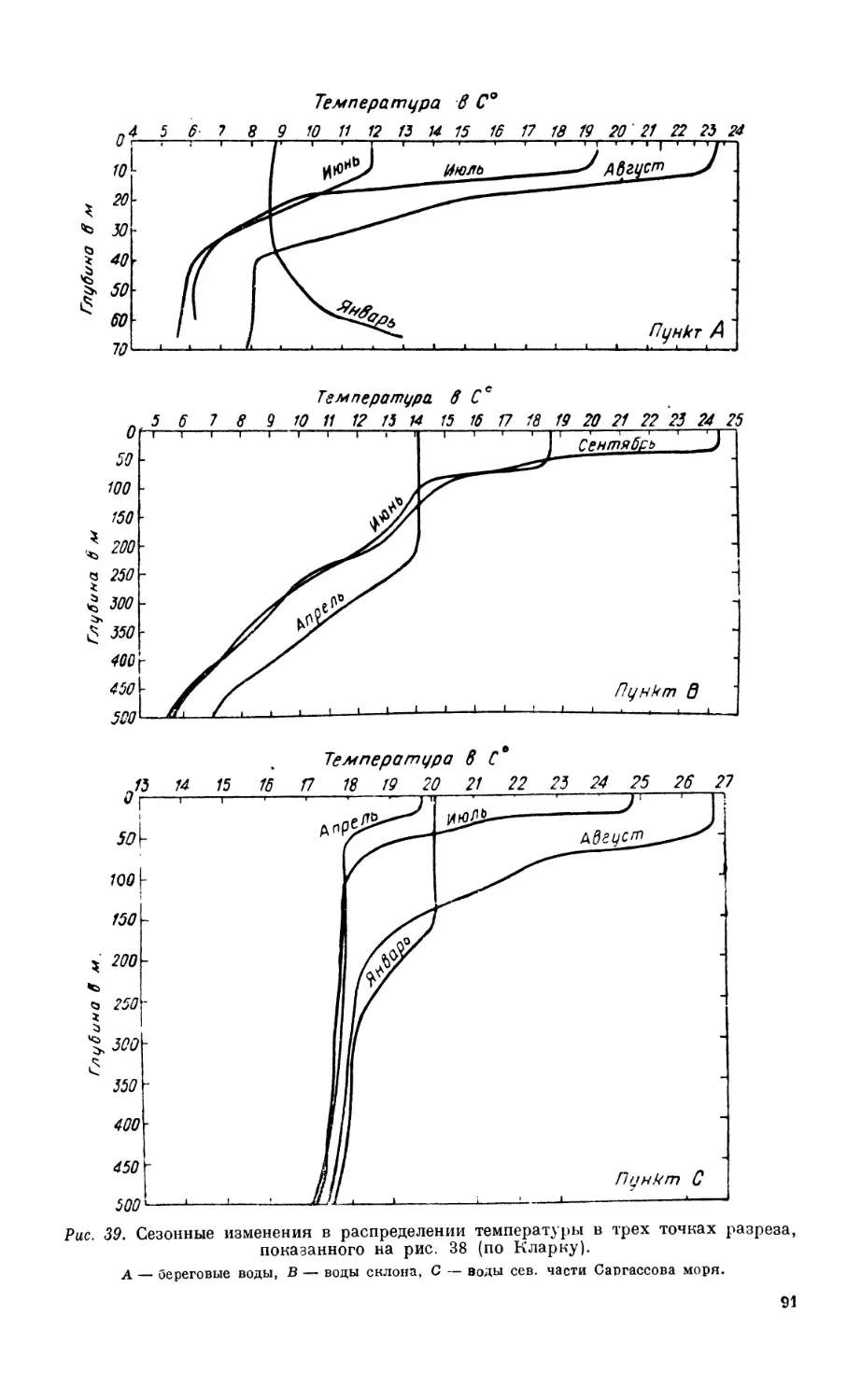
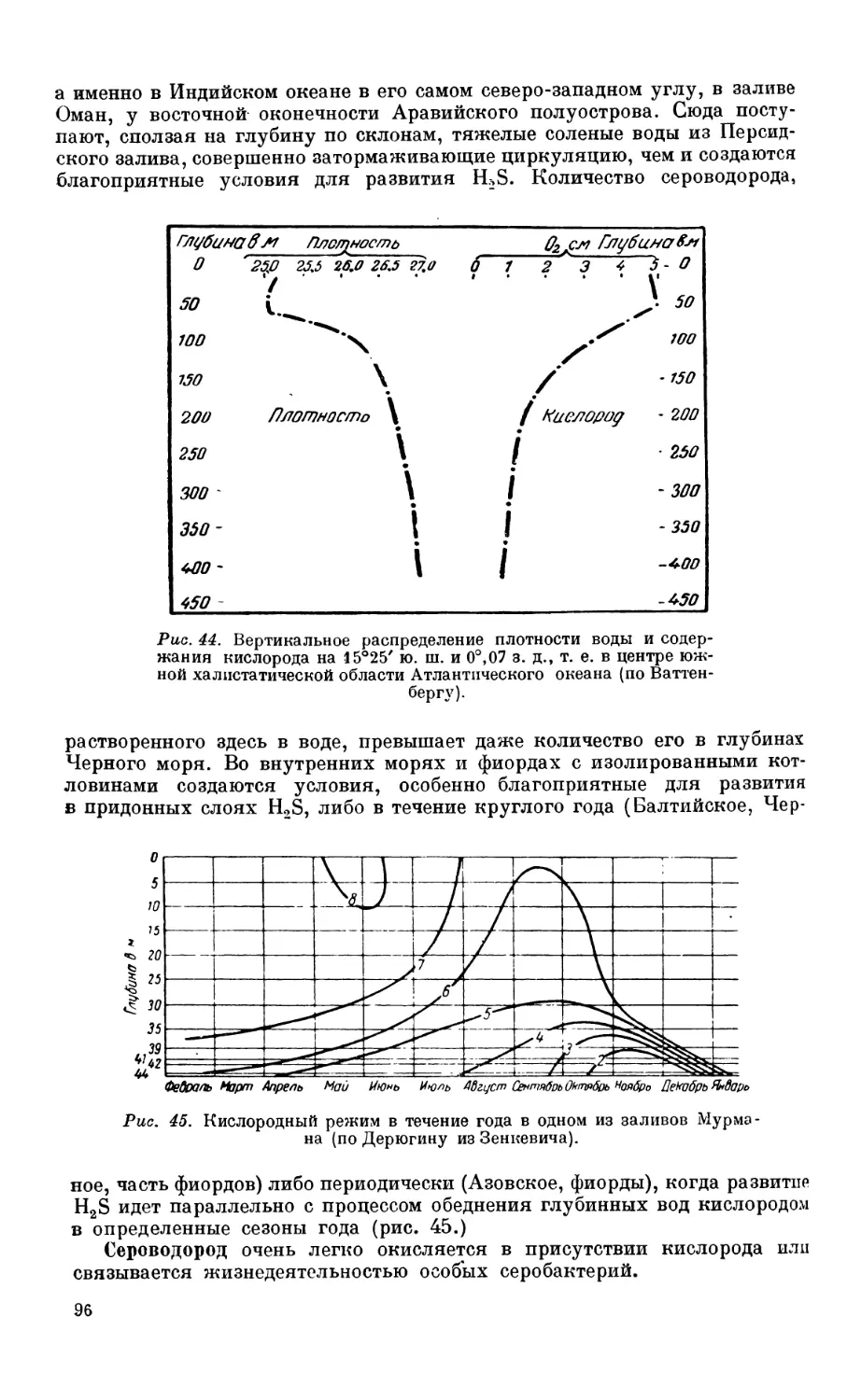
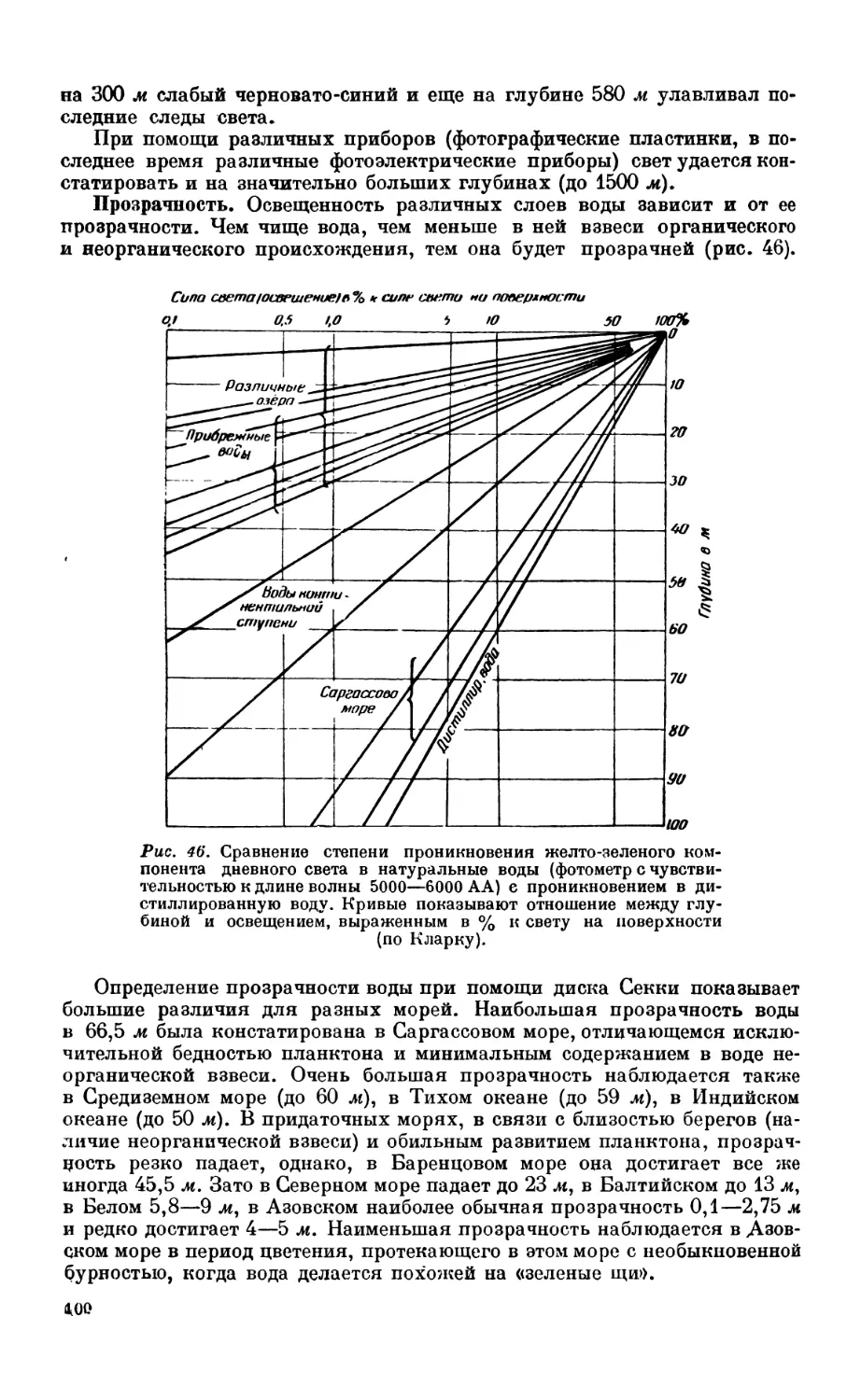
/










































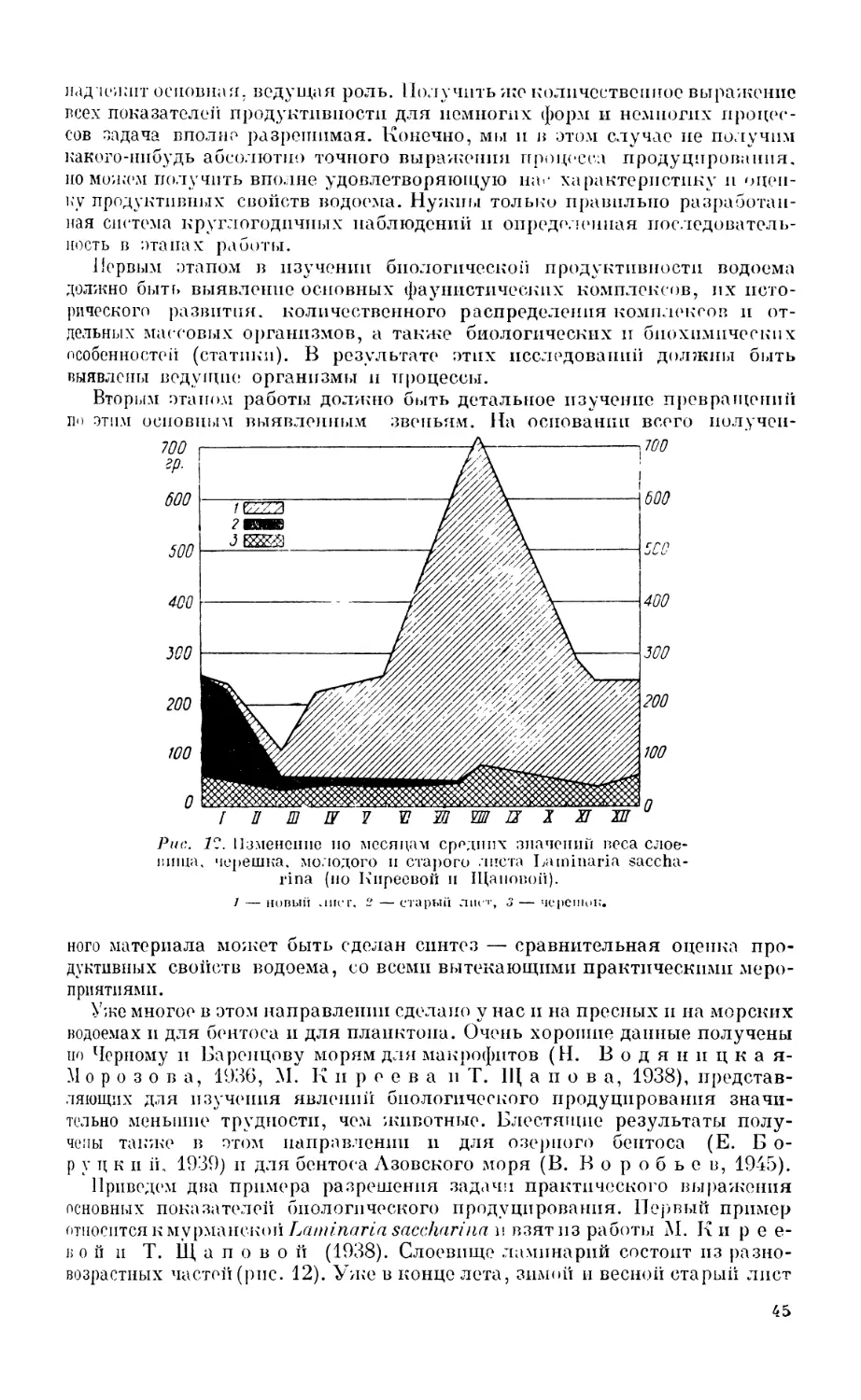








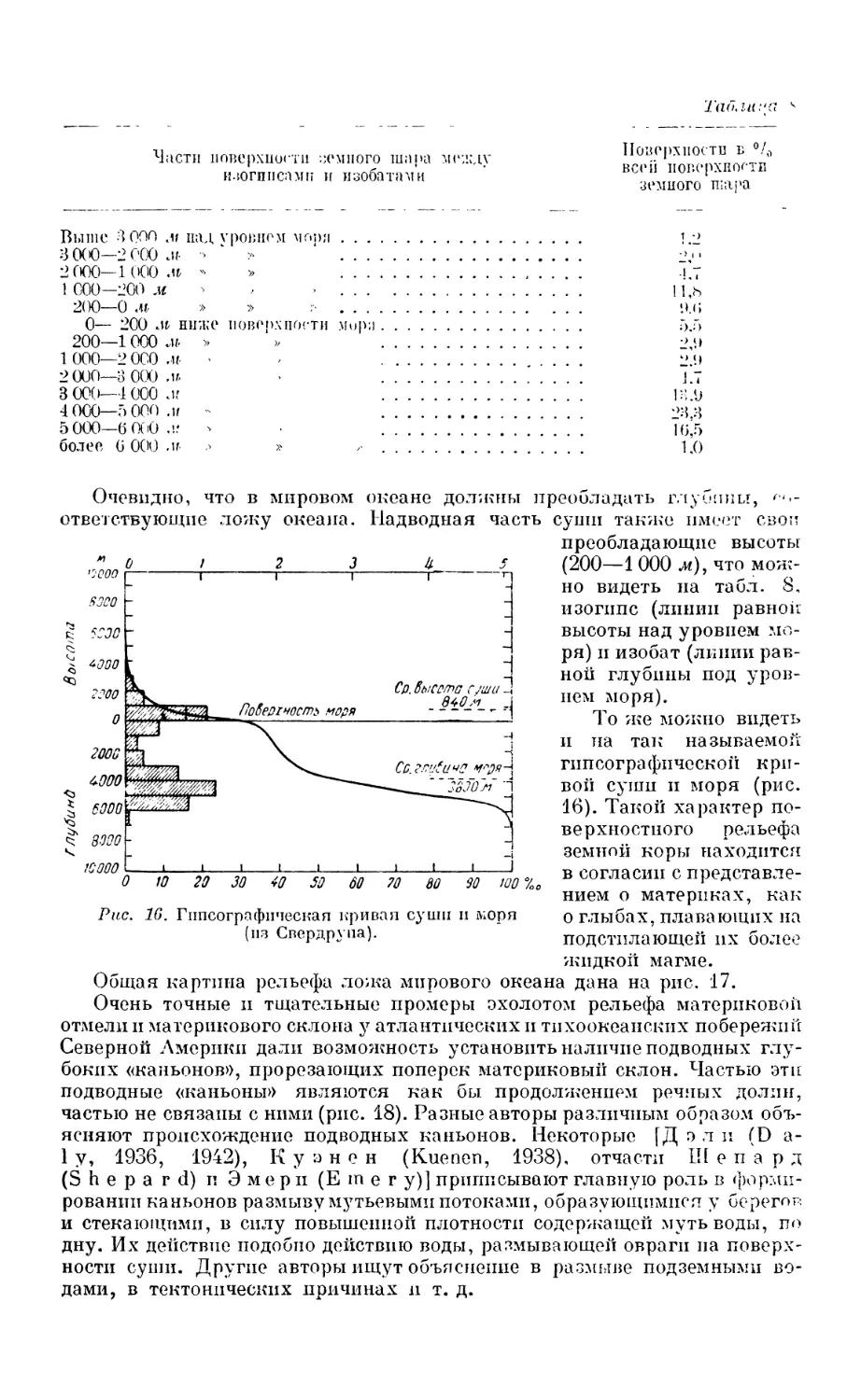






































































































































































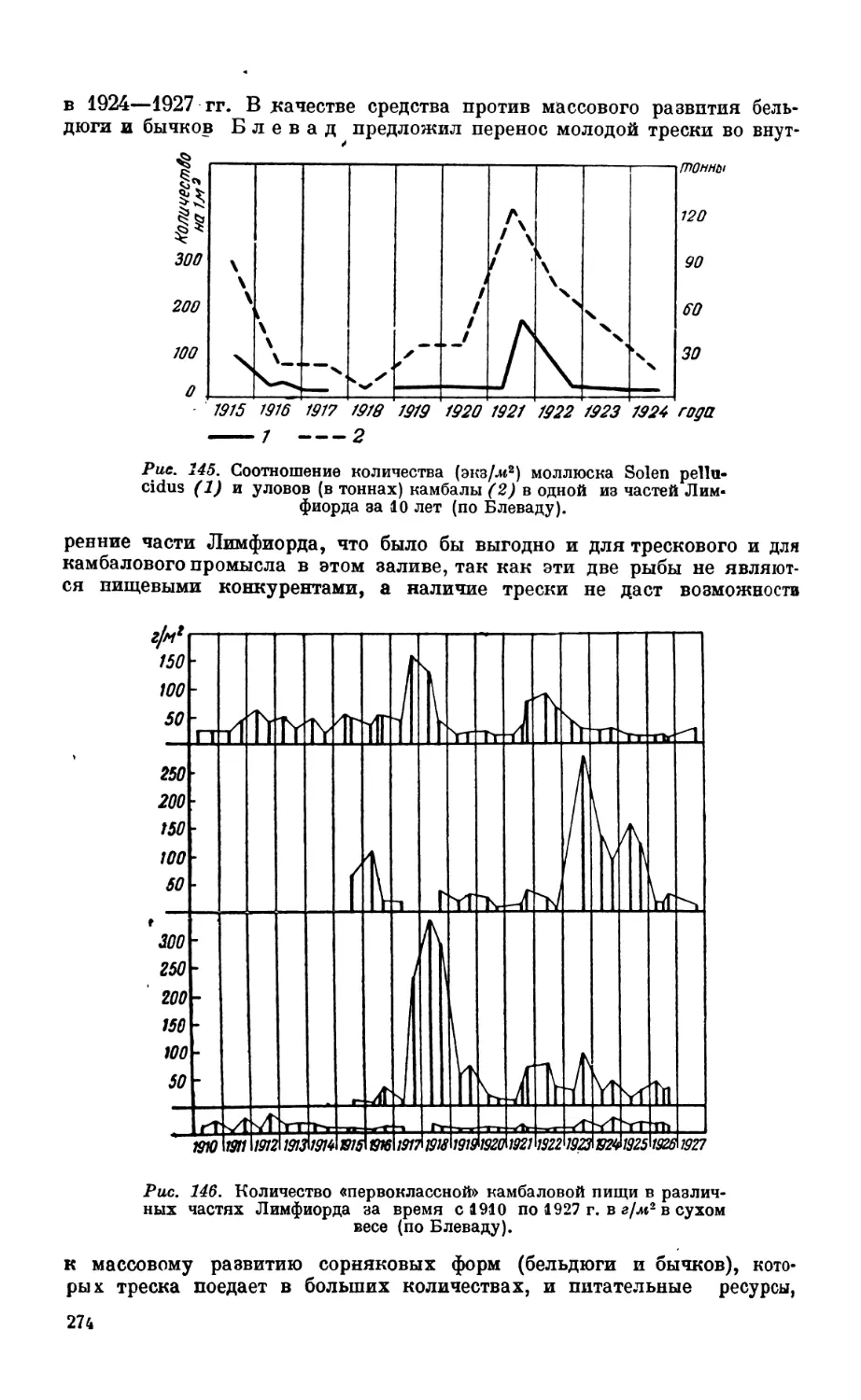
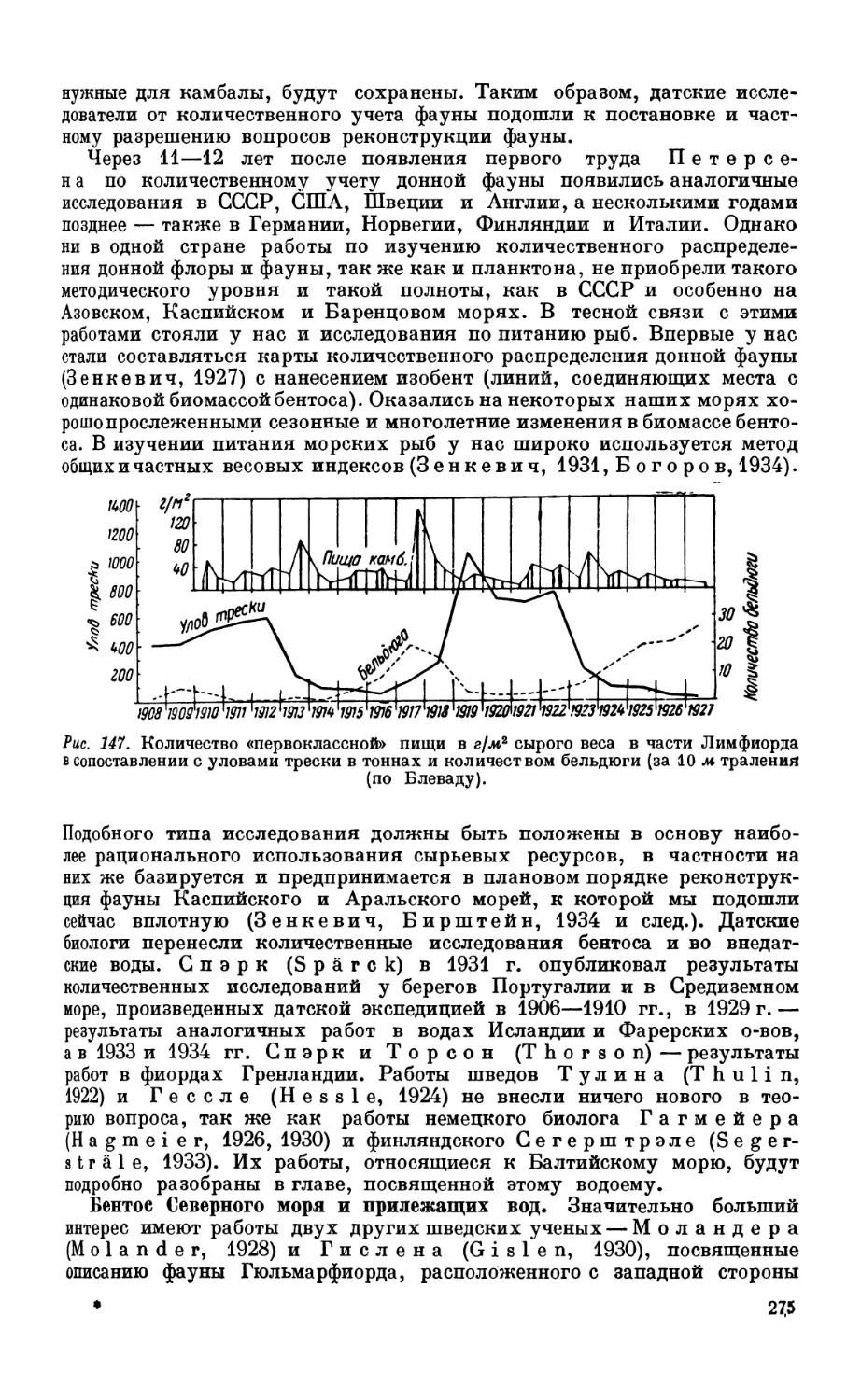




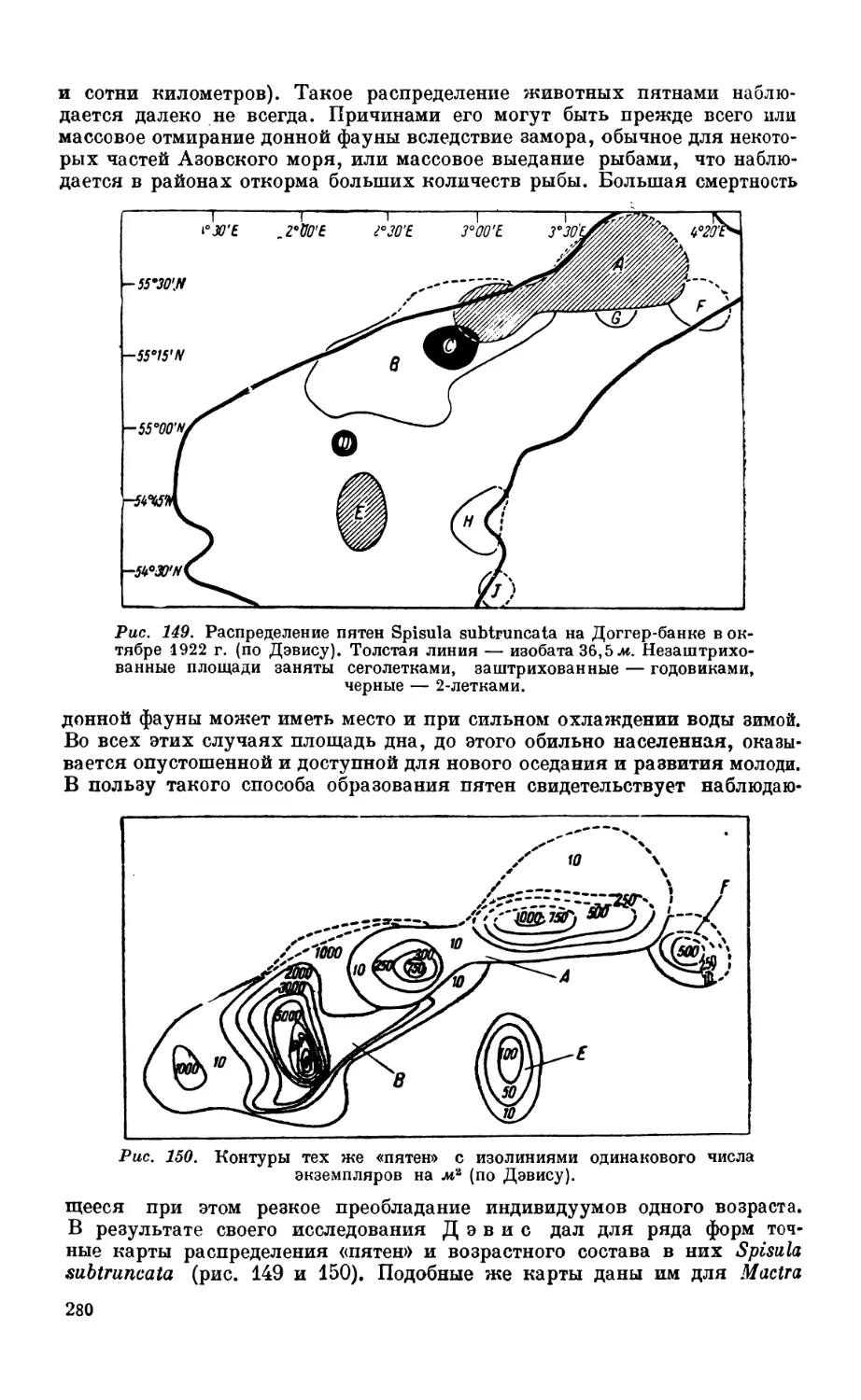












































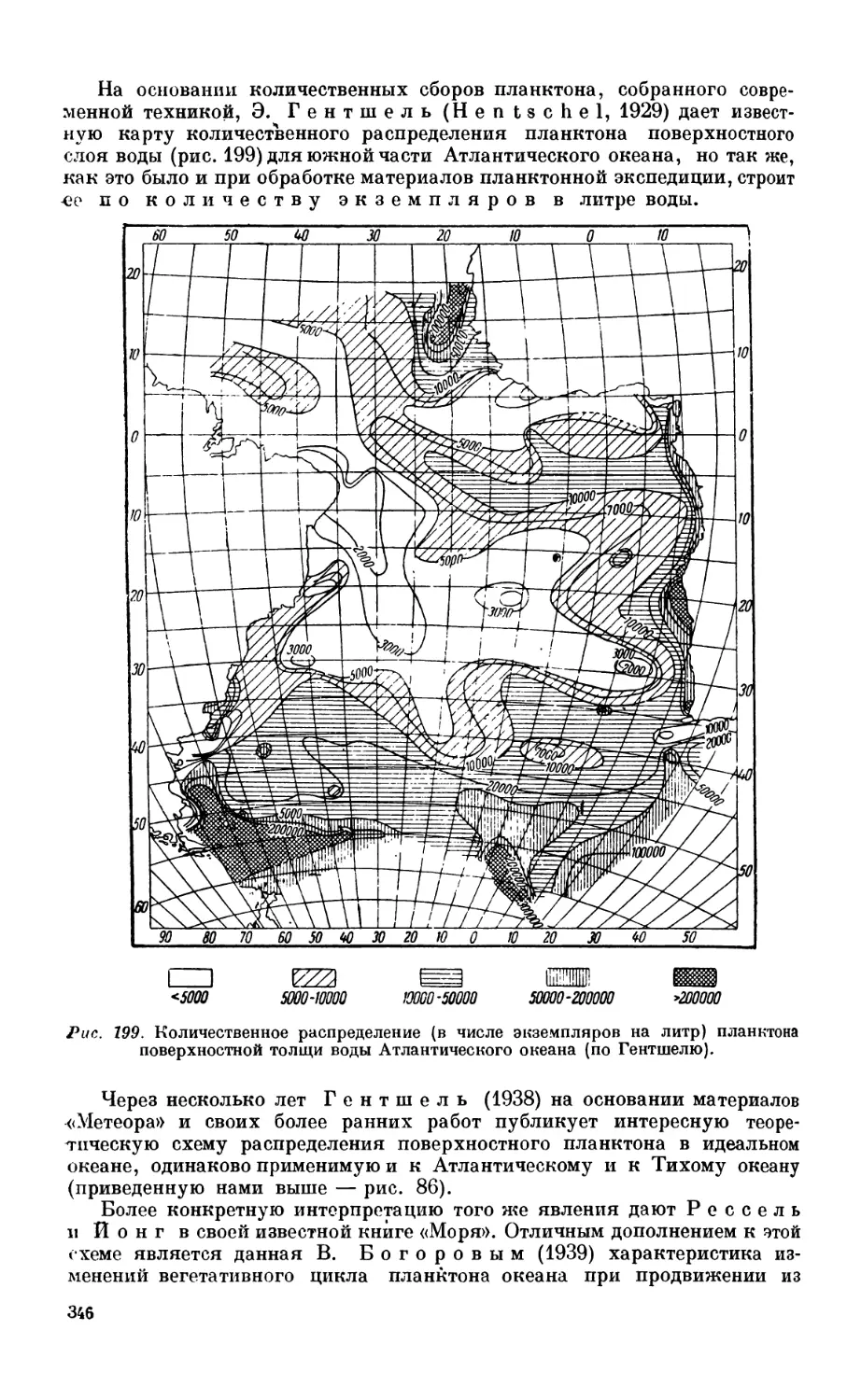




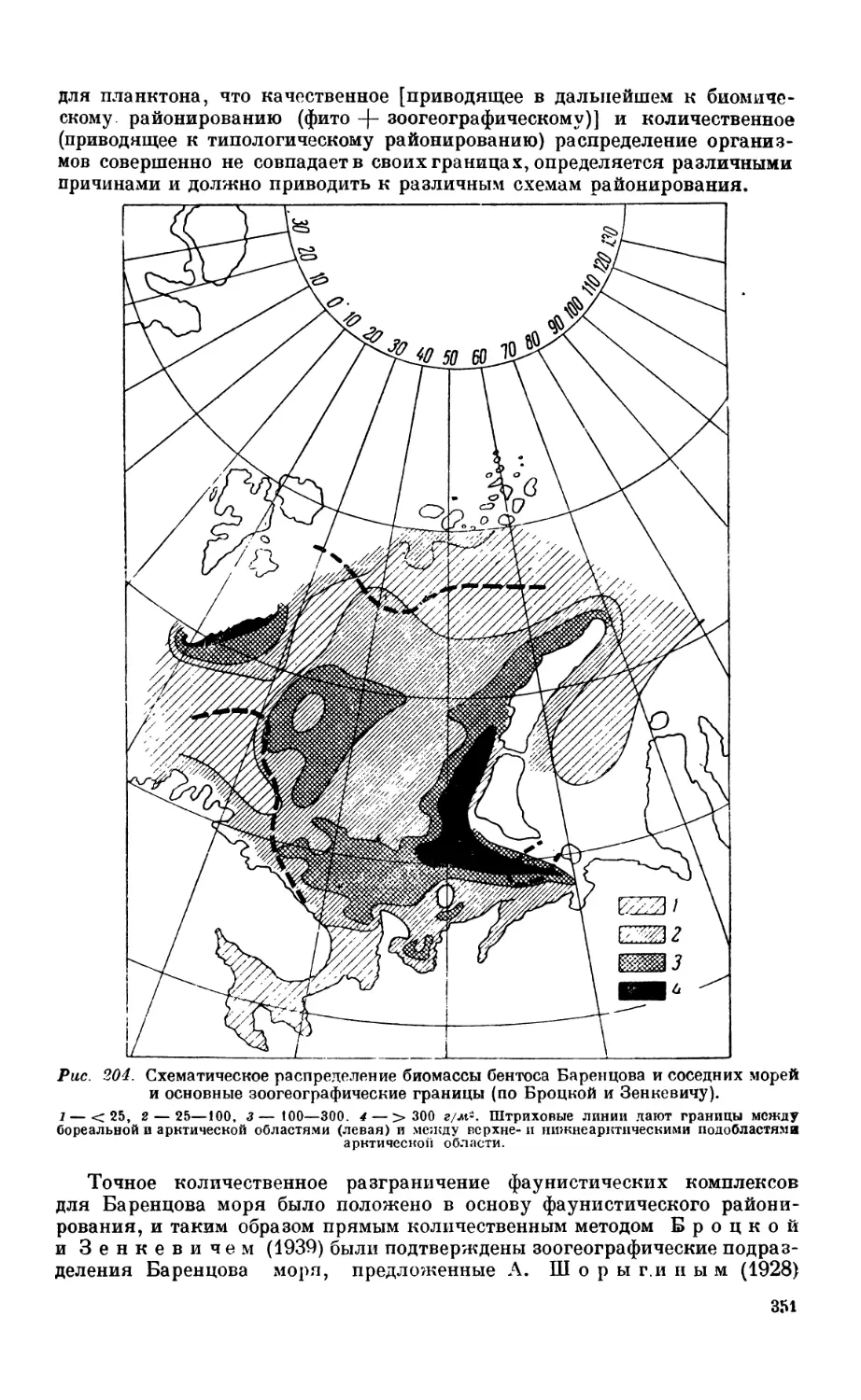




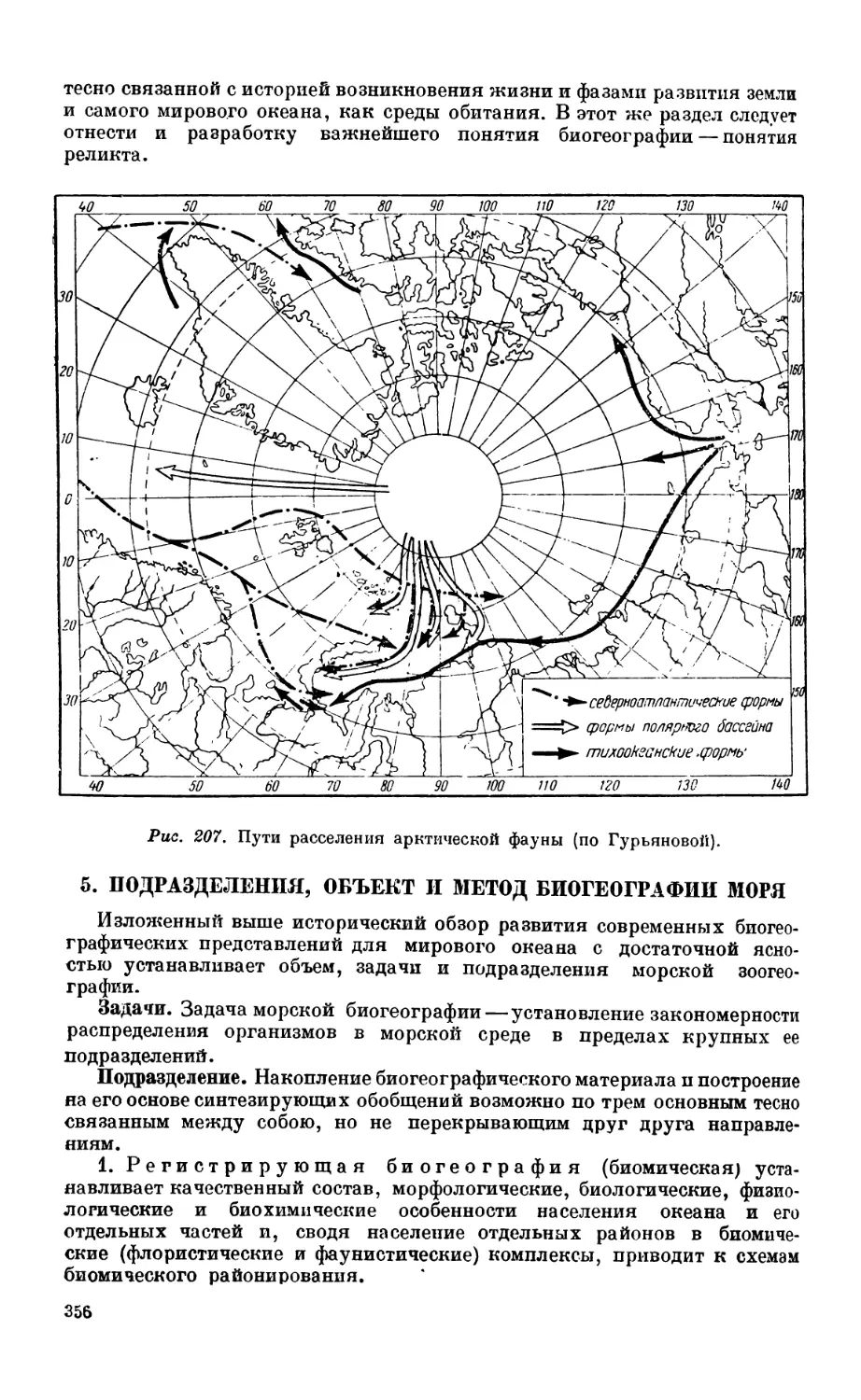







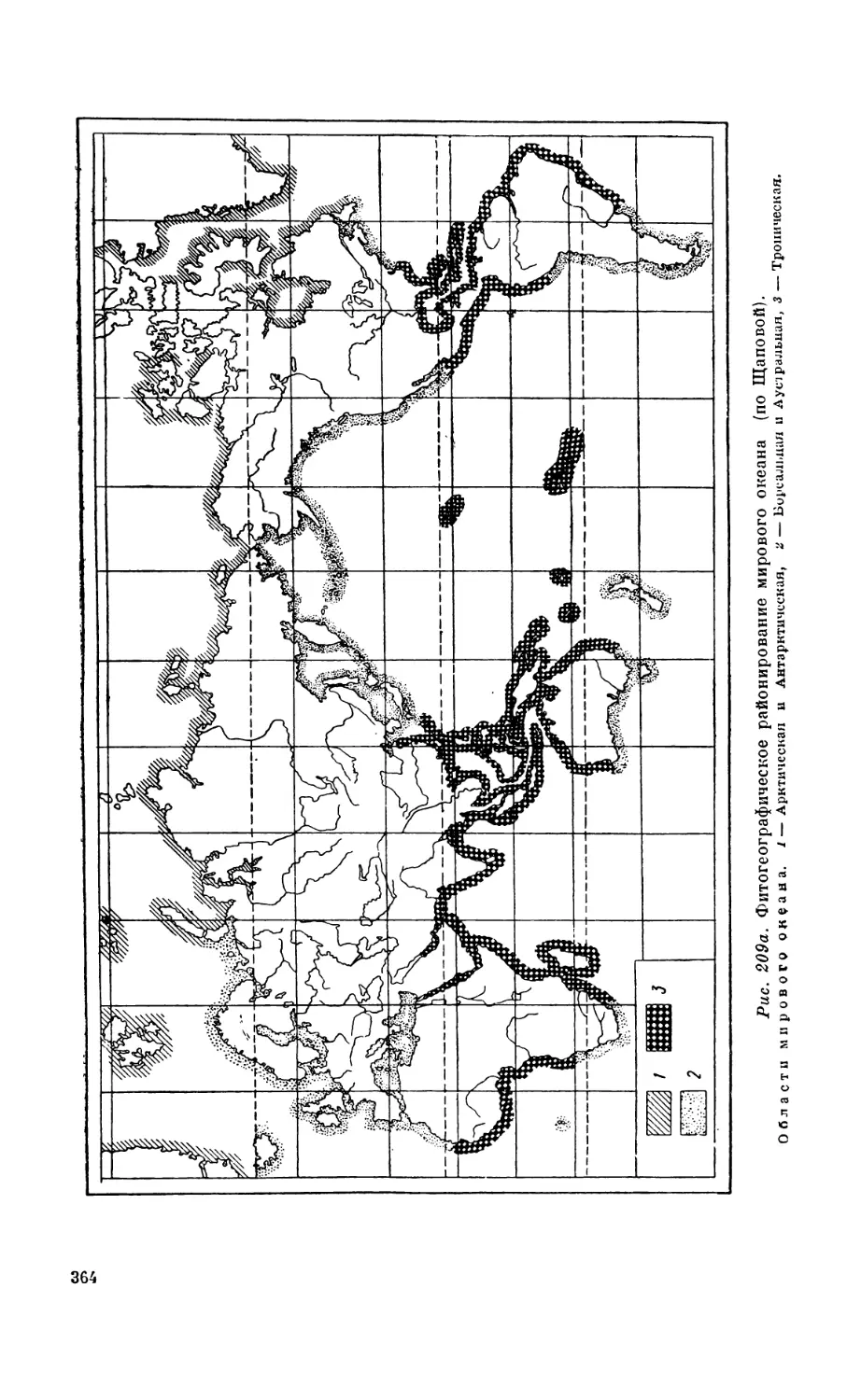























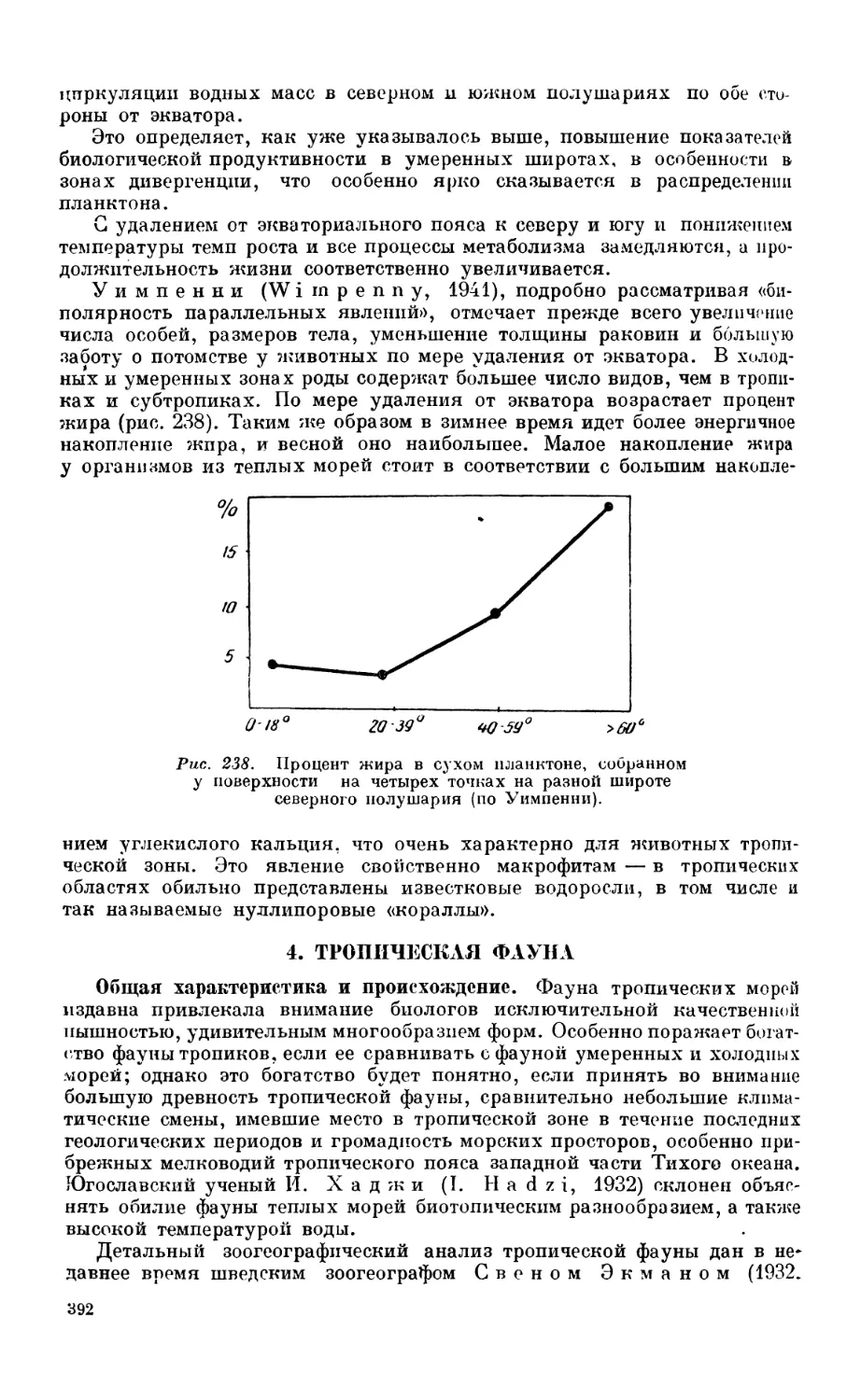

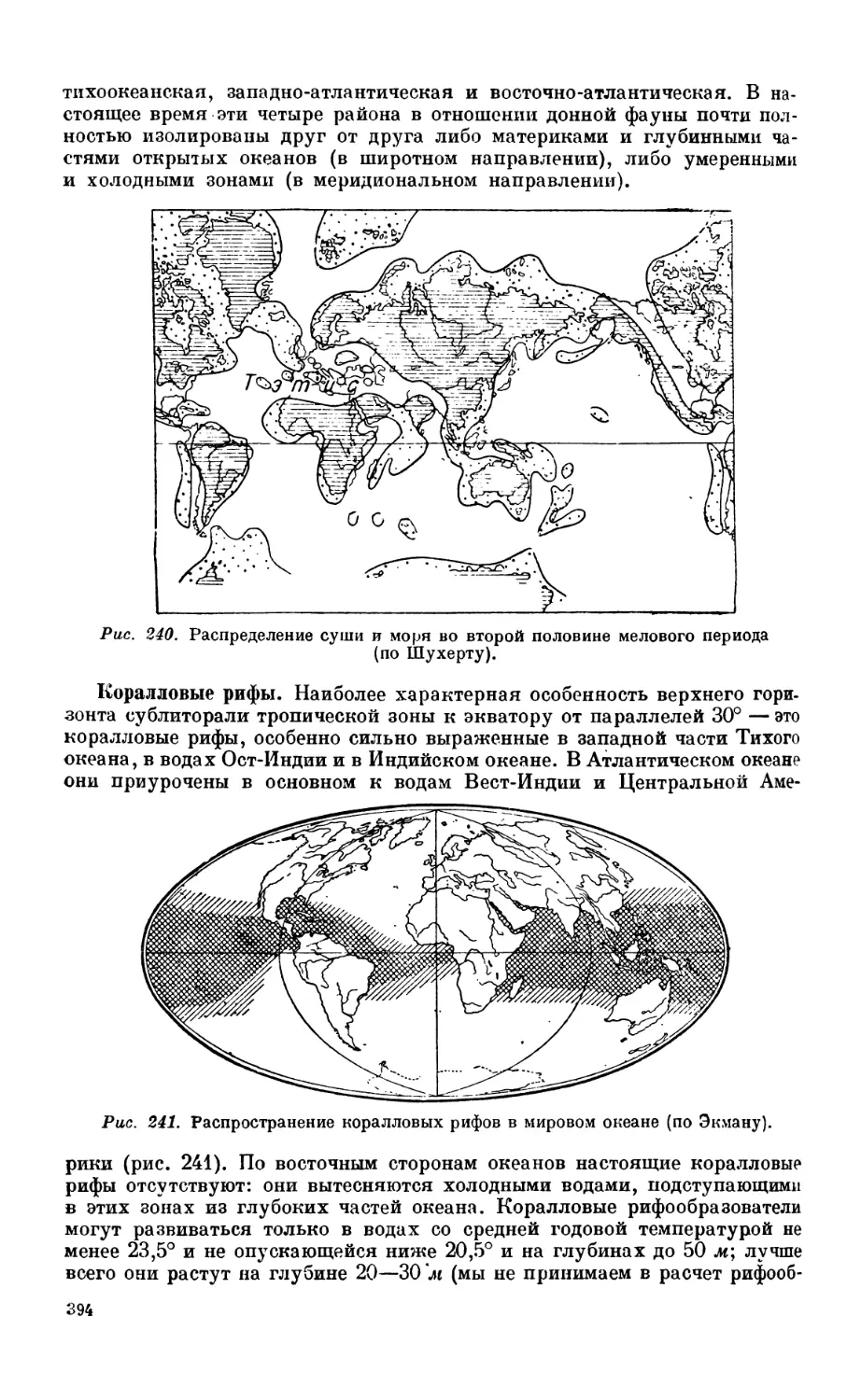










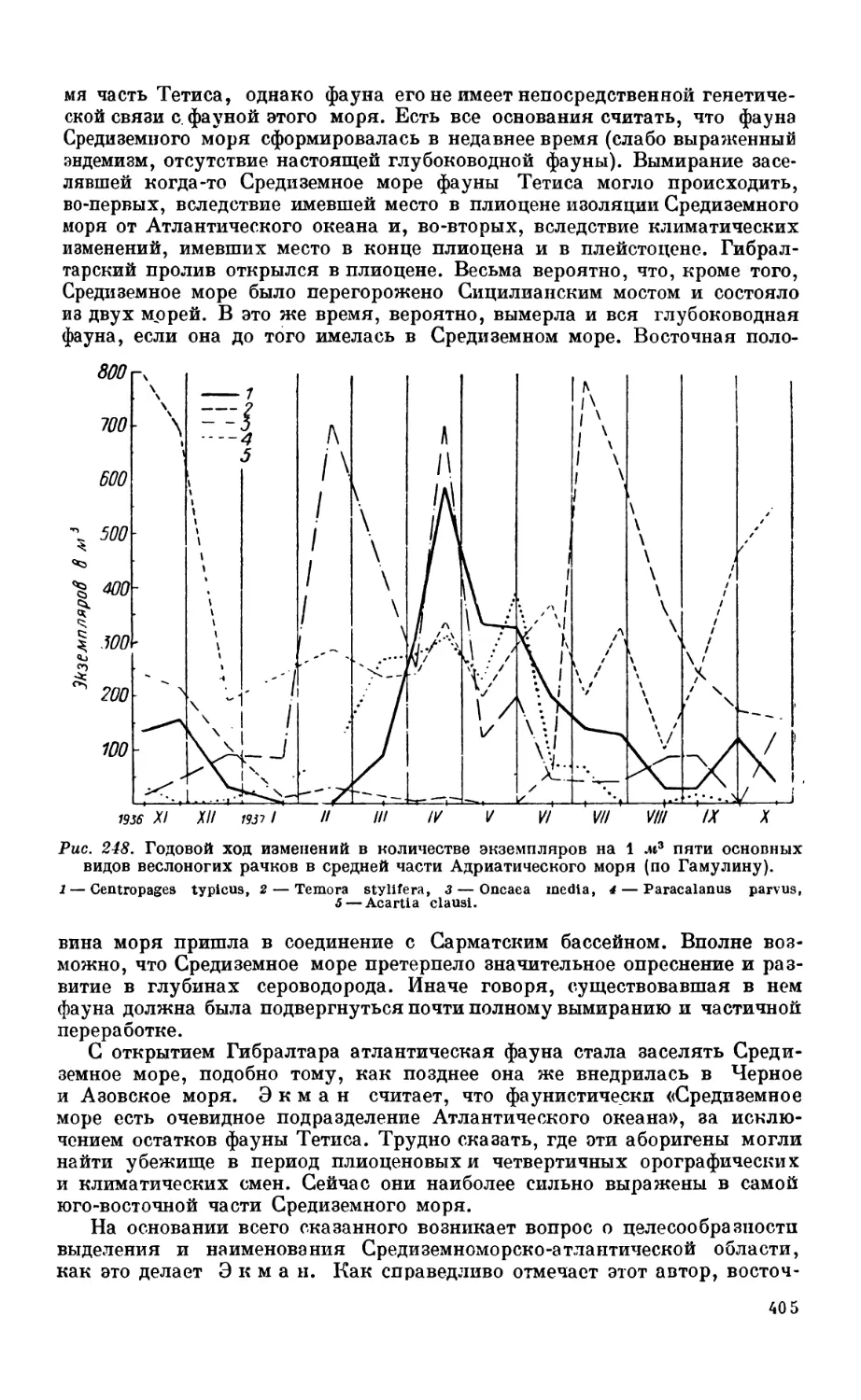



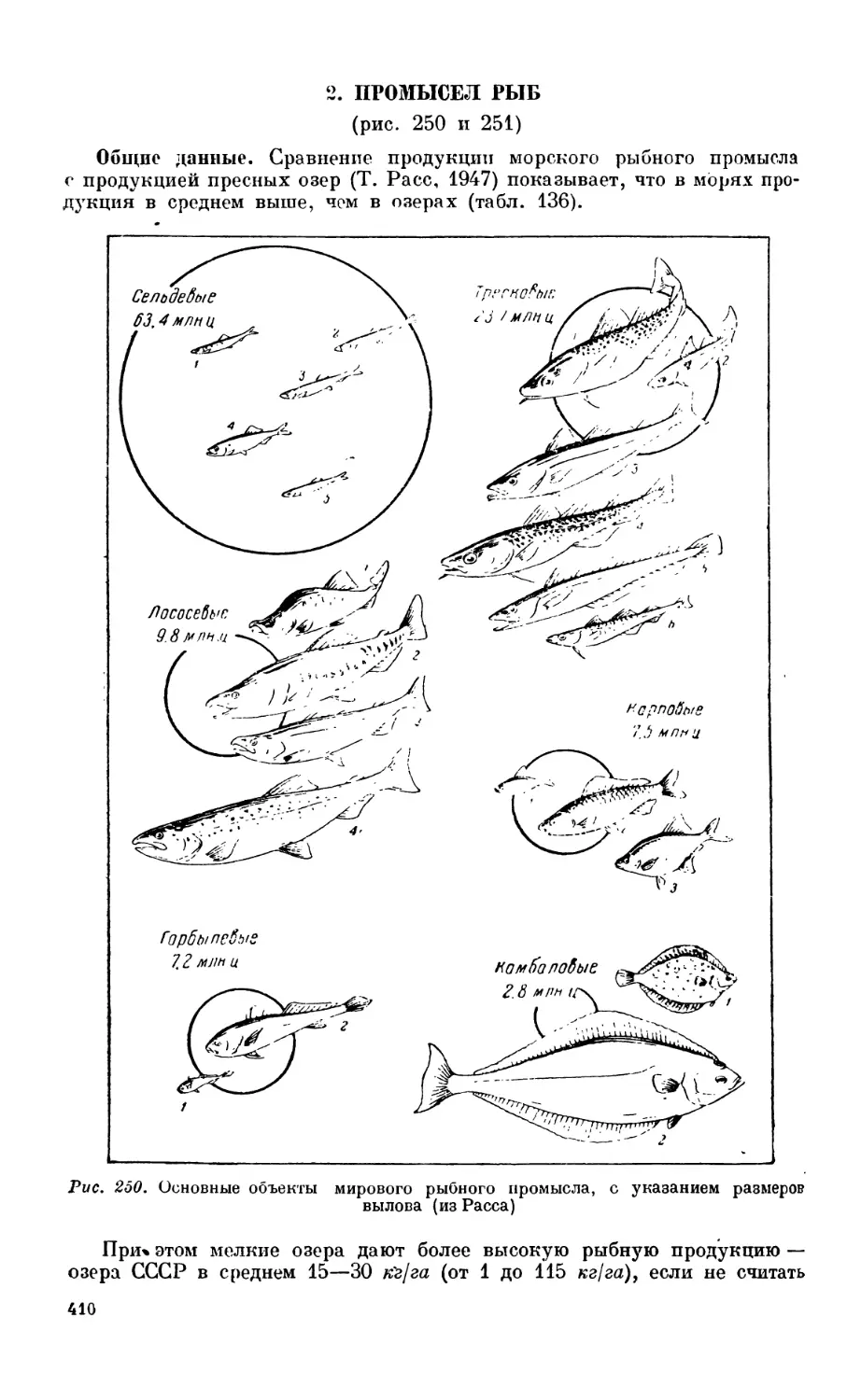































































































Текст
Л.А.ЗЕНКЕВЙЧ
МИРОВОЙ
ОКЕАН
том
Памяти учителя и друга
ИВАНА ИЛЛАРИОНОВИЧА МЕСЯЦЕВ А
автор посвящает этот труд
ПРЕДИСЛОВИЕ
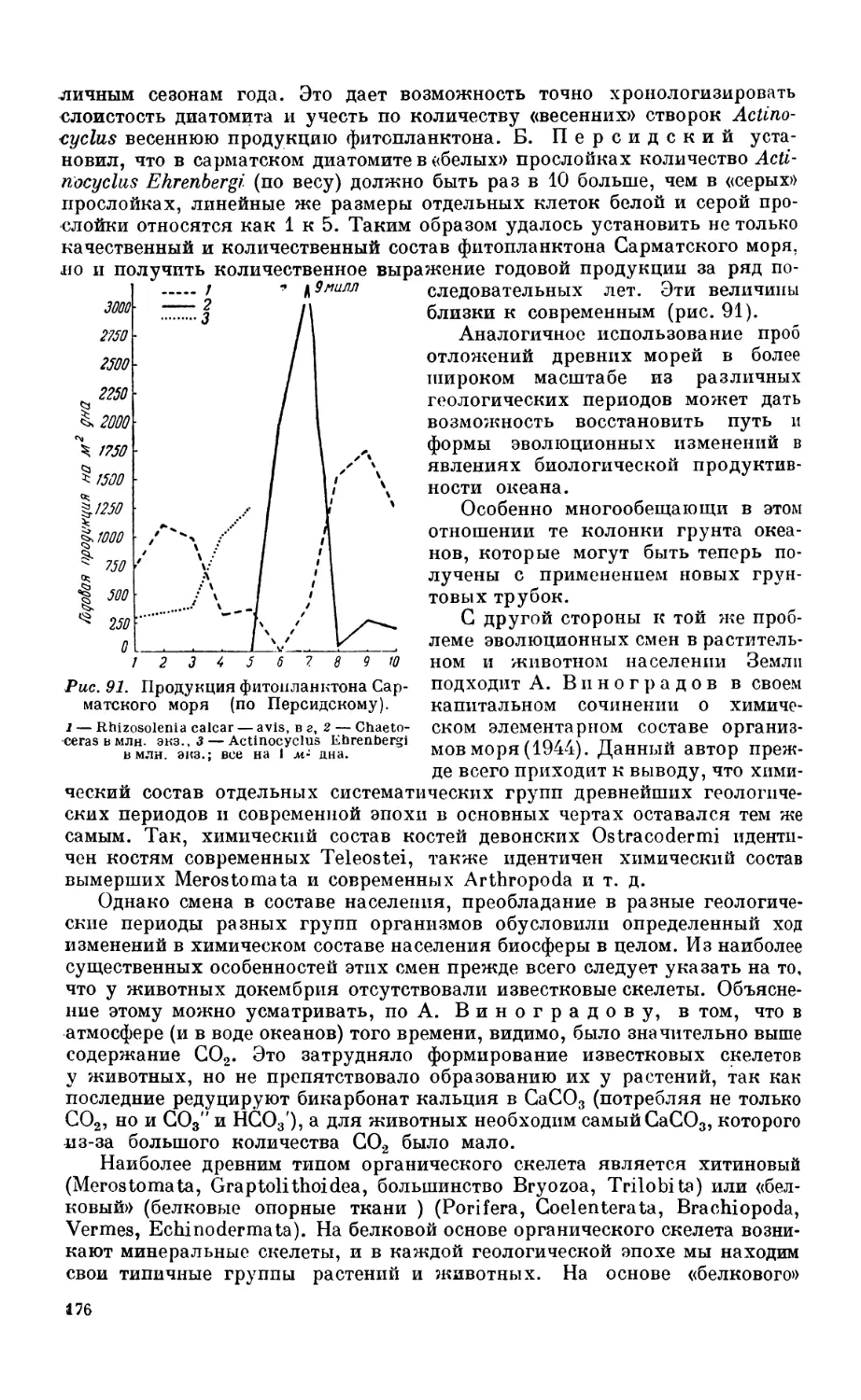
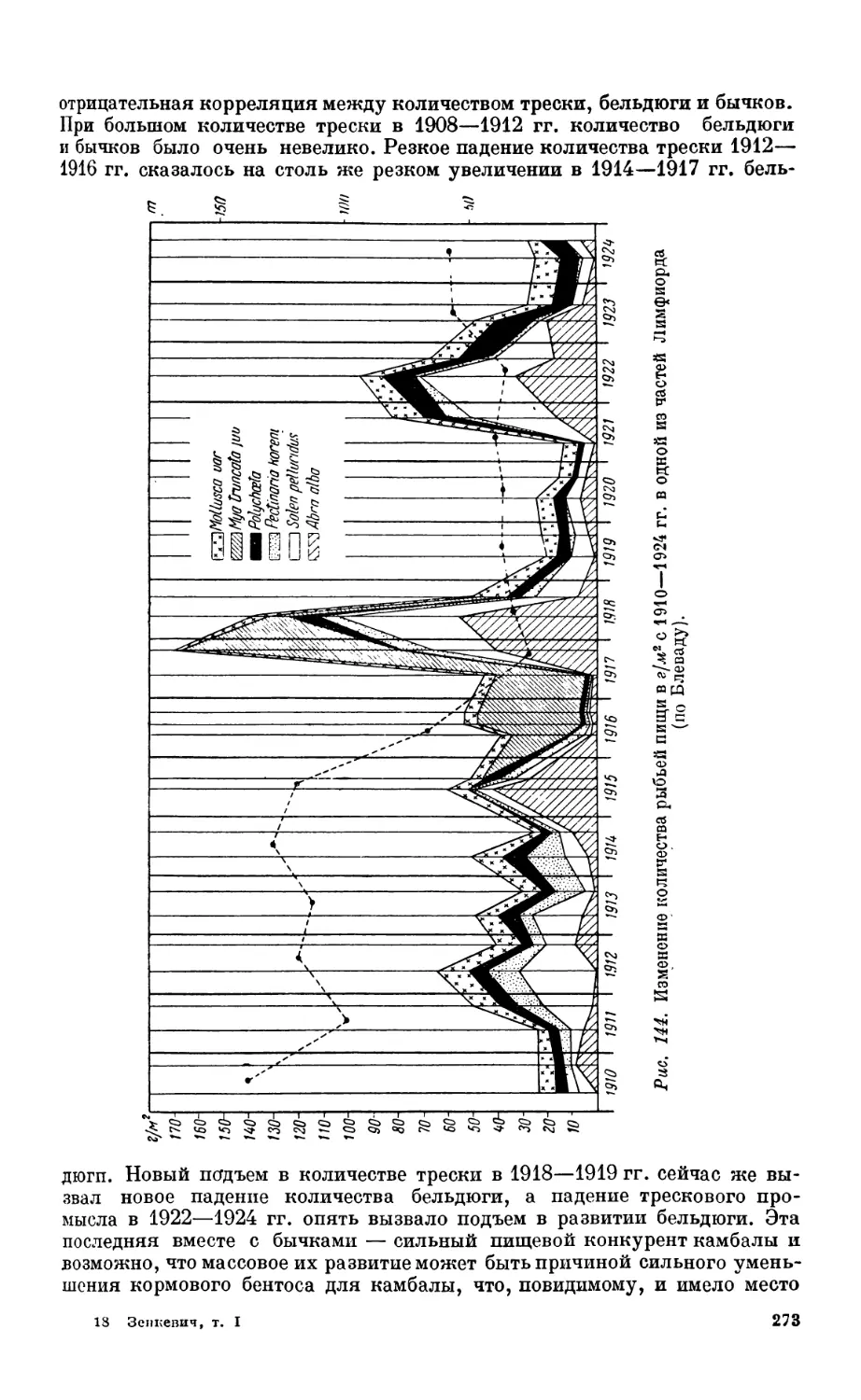
Настоящая работа подводит итоги накопившимся знаниям о составе,
распределении и хозяйственном значении организмов, населяющих моря.
Автор надеется, что эта книга будет полезна для развития следую-
щего этапа данной науки, посвященного созданию планомерного повы-
шения продуктивных свойств моря, наиболее рациональных способов
использования сырьевых ресурсов моря и переделки населения океанов
и морей в интересах социалистического строительства нашей великой
Родины.
Три обстоятельства побудили автора к изданию настоящего обшир-
ного труда, хотя автор хорошо сознает многие его недостатки.
Прежде всего к этому побуждали автора двадцатипятилетние соб-
ственные исследования по фауне наших морей и глубокий интерес ко всем
явлениям, совершающимся в море. Всему этому автор хотел найти отра-
жение в своей книге.
С другой стороны, оправданием автору в предпринятом труде было от-
сутствие сводных работ подобного типа по фауне наших морей при нали-
чии громадной отечественной литературы, в значительной части еще
ждущей опубликования. Особенно нуждаются в такой сводке много-
численные кадры молодых специалистов в различных областях океано-
логии.
И, наконец, можно указать также и на чрезвычайно недостаточное
знакомство зарубежных ученых с русской литературой, примером чего
могут быть и вышедшая 15 лет тому назад «Зоогеография моря» швед-
ского зоолога Экмана (1935) и совсем уже недавно опубликованная свод-
ная работа американских океанологов Свердрупа, Флеминга
и Джонсона «Океаны» (1942).
Ни одна страна мира не обладает таким обилием и разнообразием
морских водоемов, как наше государство. Только небольшая часть наших
побережий омывается непосредственно открытым океаном, большинство
берегов опоясано придаточными морями трех океанов — Северного
Ледовитого, Атлантического и Тихого.
Особенно резко выступает это своеобразие евразиатского материка,
ости его сравнить с другими материками, обычно почти лишенными
придаточных морей.
Двенадцать наших морей сохранили более пли менее широкую связь
с открытыми океанами, два величайших озера-моря — Каспийское и
Аральское в настоящее время от них отделены.
Все эти 14 морей, по площади составляющие около 5% поверхности
мирового океана, поражают исследователя разнообразием физико-геогра-
фических условий, богатством и разнообразием фауны и сложностью
своего геологического прошлого, наложившего неизгладимую печать
на состав, биологические особенности и распределение фауны.
Фауна наших морей представляет собой богатейший объект для ис-
следования п поистине неисчерпаемый ресурс животного сырья.
С большим удовлетворением мы можем отметить два факта. Первое, —
что имеющимися знаниями о фауне наших морей мы в преобладающей
части, а для большинства из них почтя полностью обязаны работам рус-
ских ученых.
Второе — это то, что в результате упорных многолетних исследований
некоторые наши моря можно считать для настоящего времени наилучше
изученными водоемами мира (Каспийское, Азовское, Баренцово).
Совершенно исключительны, даже на общем фоне отмеченной высокой
степени изученности, по масштабам и полученным результатам предпри-
нятые нашей страной в течение двух последних десятилетий исследо-
вания Арктического бассейна и особенно наименее доступных его централь-
ных частей. Эти исследования составят в истории географической науки
одну из наиболее блестящих страниц.
Автор имеет все основания считать, что только помощь многочислен-
ных товарищей и учеников дала ему возможность завершить этот нелег-
кий труд.
Книга была закончена и сдана в печать еще в 1940 г. Война задержала
ее печатание. В 1947 г. пришлось коренным образом переделать весь
текст и внести много дополнений.
Глава о зоопланктоне написана для этой книги В. Г. Богоровым
еще в 1939 г. и лишь в малой степени изменена и дополнена автором.
Глава о промысловом значении морских животных написана Я. А. Бир-
штейном совместно с автором. Автор выражает свою глубокую благо-
да рность обоим товарищам. Особенно автор обязан большой работе,
проделанной над рукописью, своим редакторам В. Броцкой и Я. Бир-
штейну.
Зенкевич
СОДЕРЖАНИЕ
Стр»
Предпсловпе.; . ;............................................................ 3
Введение................................................................... И
1. Морская фауна как объект изучения............................_........... 13
Целостность океанологии как науки —13. Основные проблемы морской био-
логии — 13.
2. История изучения моря ....................................... . . 15
История морских экспедиций — 15. Первые плавания — 15. Дочелл’щд-
жеровский период — 15. Экспедиция «Челленджера» — 17. Период конца XIX и
начала XX столетия— 17. Современный период — 19.
3. Научные учреждения, еедугцие изучение фауны моря..........; . . . ;..... 22
Организация первых морских лабораторий — 22. Русские морские паучные учреж-
дения— 23. Иностранные морские научные учреждения — 28. Основные издания,
посвященные вопросам океанологии—28. Русские издания — 28. Иностранные
издания — 30. Основные этапы изучения морской фауны — 31.
4. Биогеография, экология и гидробиология и их роль в изучении фауны моря... 33
Биогеография — 33. Экология — 33. Гидробиология — 34. Сравнение задач, объекта
и метода биогеографии, экологии и гидробиологии.—35. Процесс биофильтра, как
пример комплексного явления в водоеме — 35.
5. Основные понятия учения о биологической продуктивности . . . . .......... 36
Возникновение учения о биологической продуктивности водоемов—36. Понятие
продуктивности, биомассы и продукции — 38. Проблема биологической продук-
тивности в трактовке различных авторов — 38. Терминология, применяемая в
связи с проблемой биологической продуктивности — 42. Методы количественного
определения продуктивных свойств водоема — 42.
ЧАСТЬ ПЕРВАЯ
РАСЧЛЕНЕНИЕ МИРОВОГО ОКЕАНА И РЕЛЬЕФ ЕГО ЛОЖА
Г л а в а 1. РАСПРЕДЕЛЕНИЕ МОРЯ II СУШИ НА ПОВЕРХНОСТИ ЗЕМЛИ И ОБЩИН БАЛАНС
ГИДРОСФЕРЫ
Объем мирового океана — 53. Баланс гидросферы — 53.
Глава II. РАСЧЛЕНЕНИЕ МИРОВОГО океана и рельеф его ложа
Океаны — 54. Придаточные моря — 54. Рельеф дна океана — 54.
ЧАСТЬ ВТОРАЯ
ФАКТОРЫ АБИОТИЧЕСКОЙ СРЕДЫ, ОПРЕДЕЛЯЮЩИЕ
РАСПРЕДЕЛЕНИЕ ОРГАНИЗМОВ В МОРЕ
Глава III. ФИЗИКО-ХИМИЧЕСКПП РЕЖИМ мирового океана
1. Основные факторы внешней среды ............................. 61
Питательные вещества — 61. Газы — 61. Исторические причины — 62.
Стр-
2. Со твой состав морской воды................................................. 62
Элементарный состав — 62. Постоянство солевого состава—62. Солевой состав
морской и речной воды — 62. Общее количество солей в океане — 63. Общая со-
леность— 64. Происхождение солености мирового океана — 65.
3. Химический состав морских организмов........................................ 67
Влияние организмов на химизм морской воды — 67. Элементарный состав мор-
ских организмов—67. Способность некоторых морских организмов аккумулиро-
вать отдельные элементы— 69. Известковая функция морских организмов — 70.
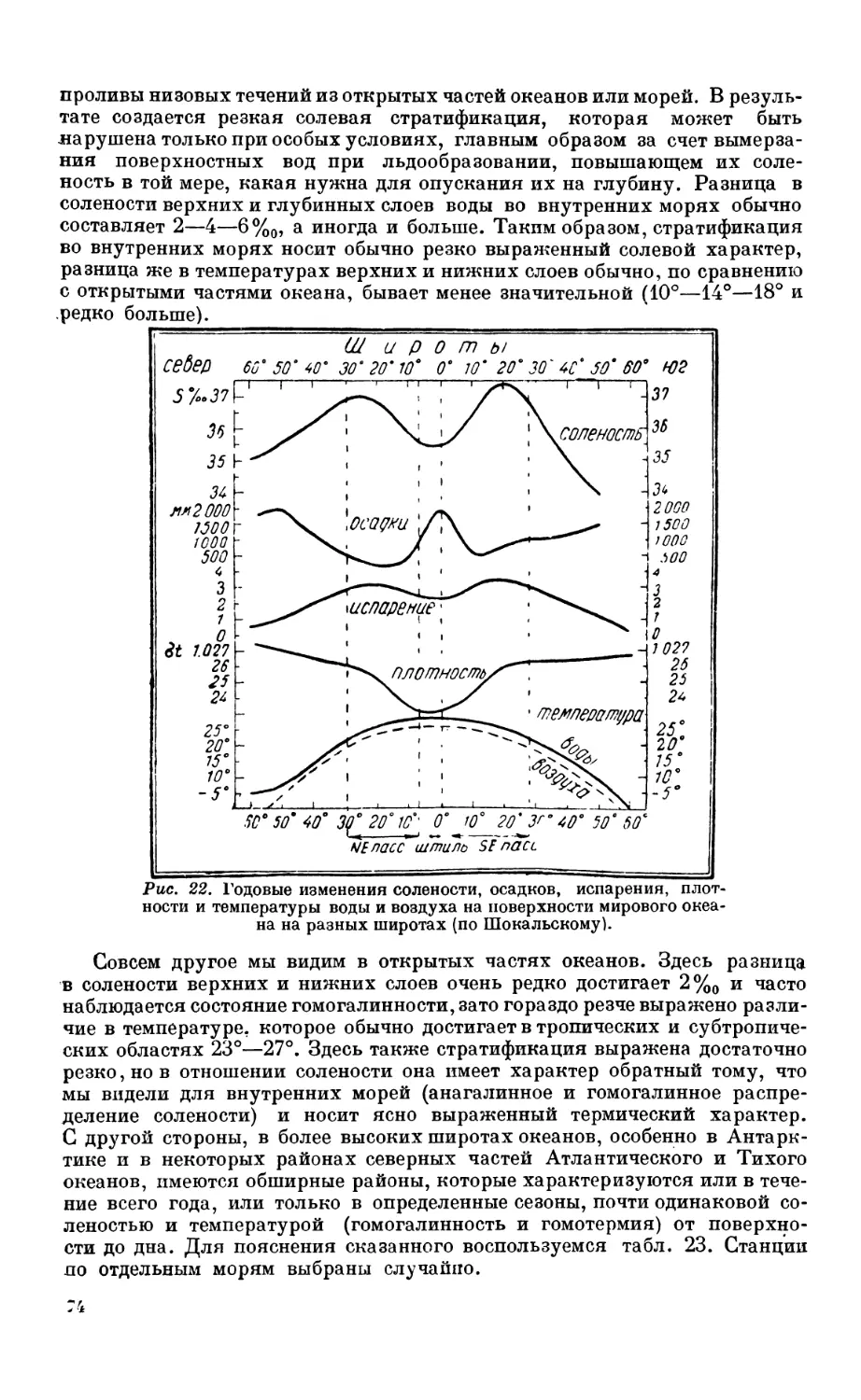
4. Распределение солености в мировом океане.................................... 71
Основные факторы, воздействующие на соленость океанических вод — 71. Зави-
симость распределения солености от течений — 73. Вертикальное распределение
солености—73. Соленость внутренних морей умеренных широт — 79.
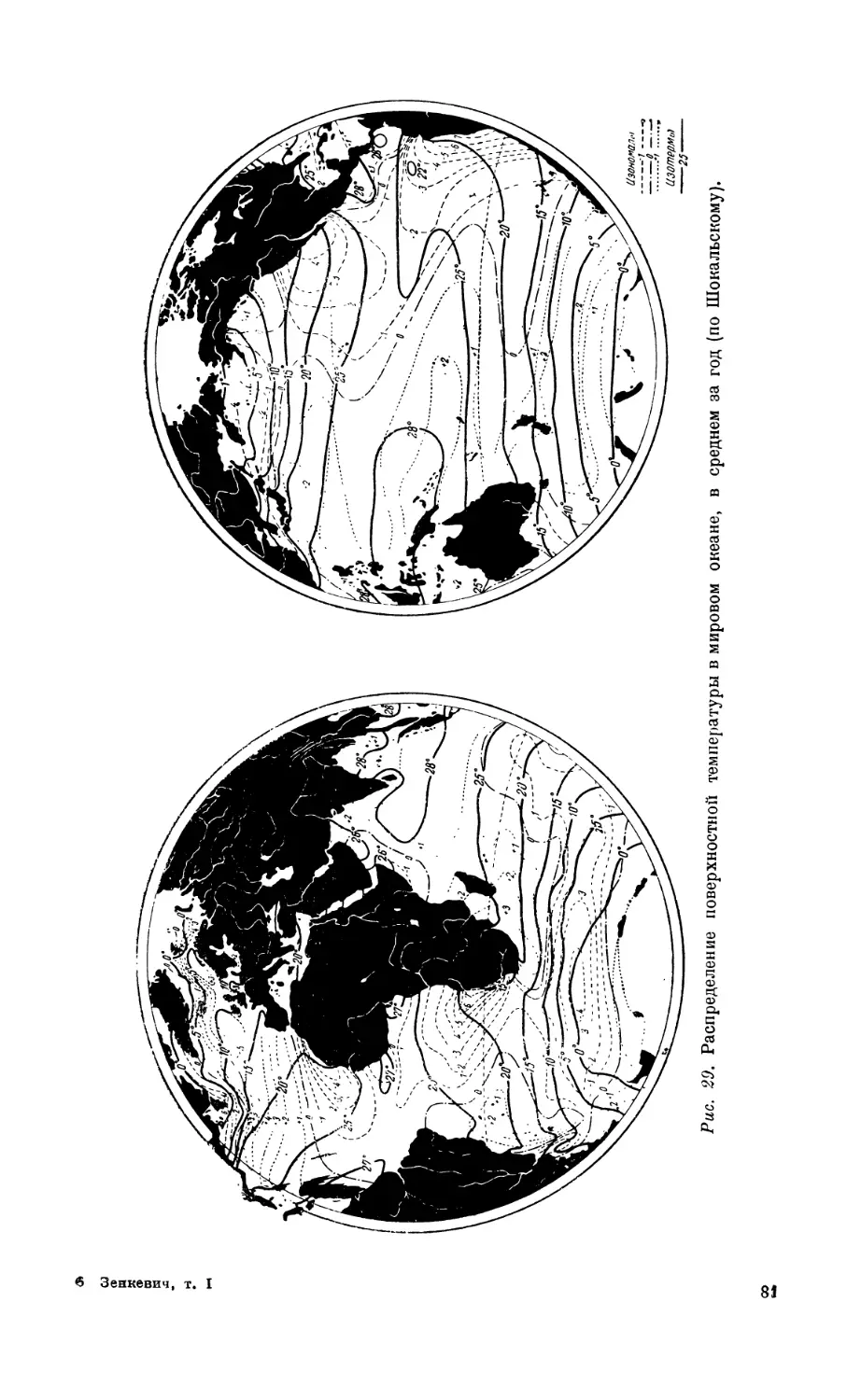
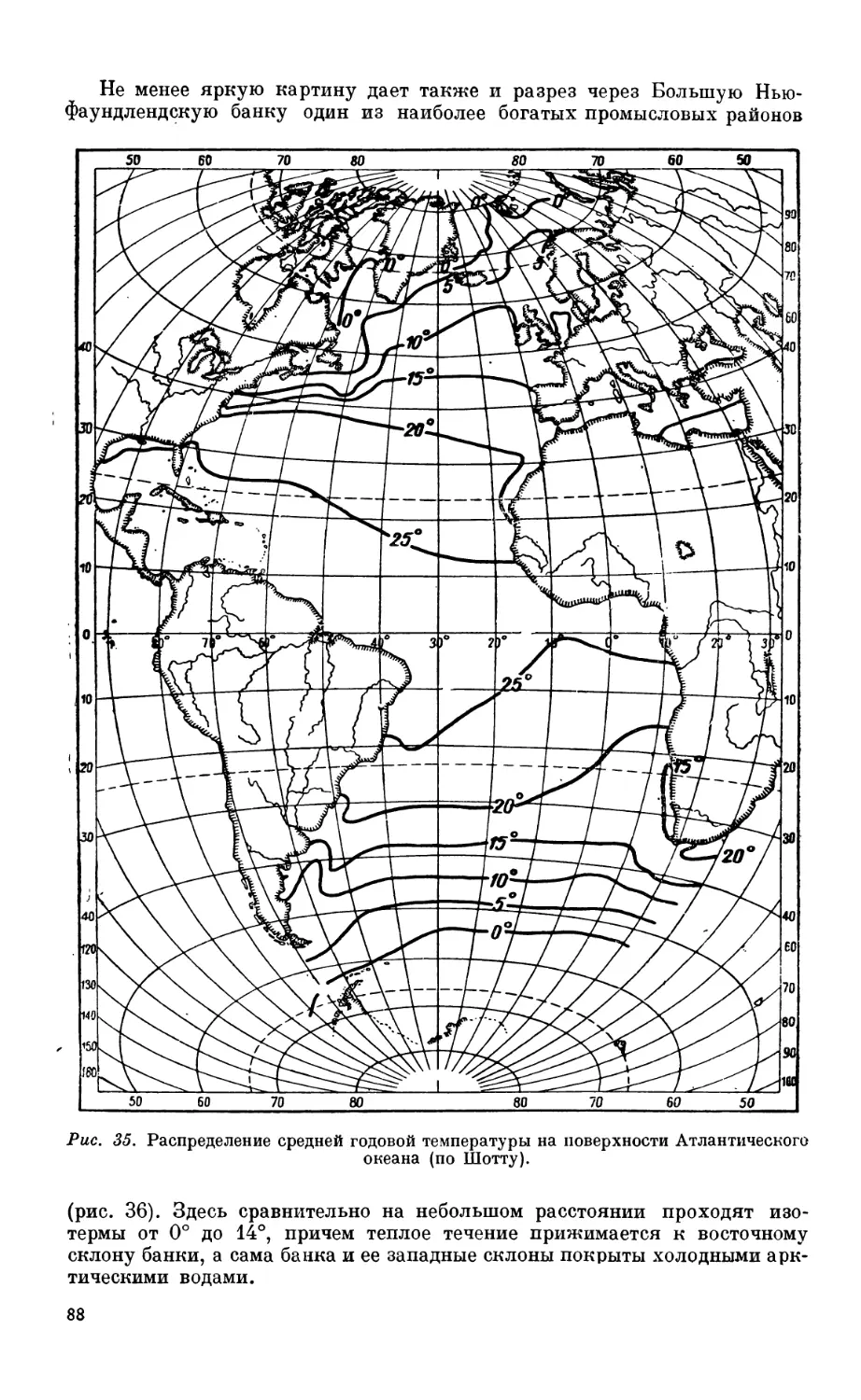
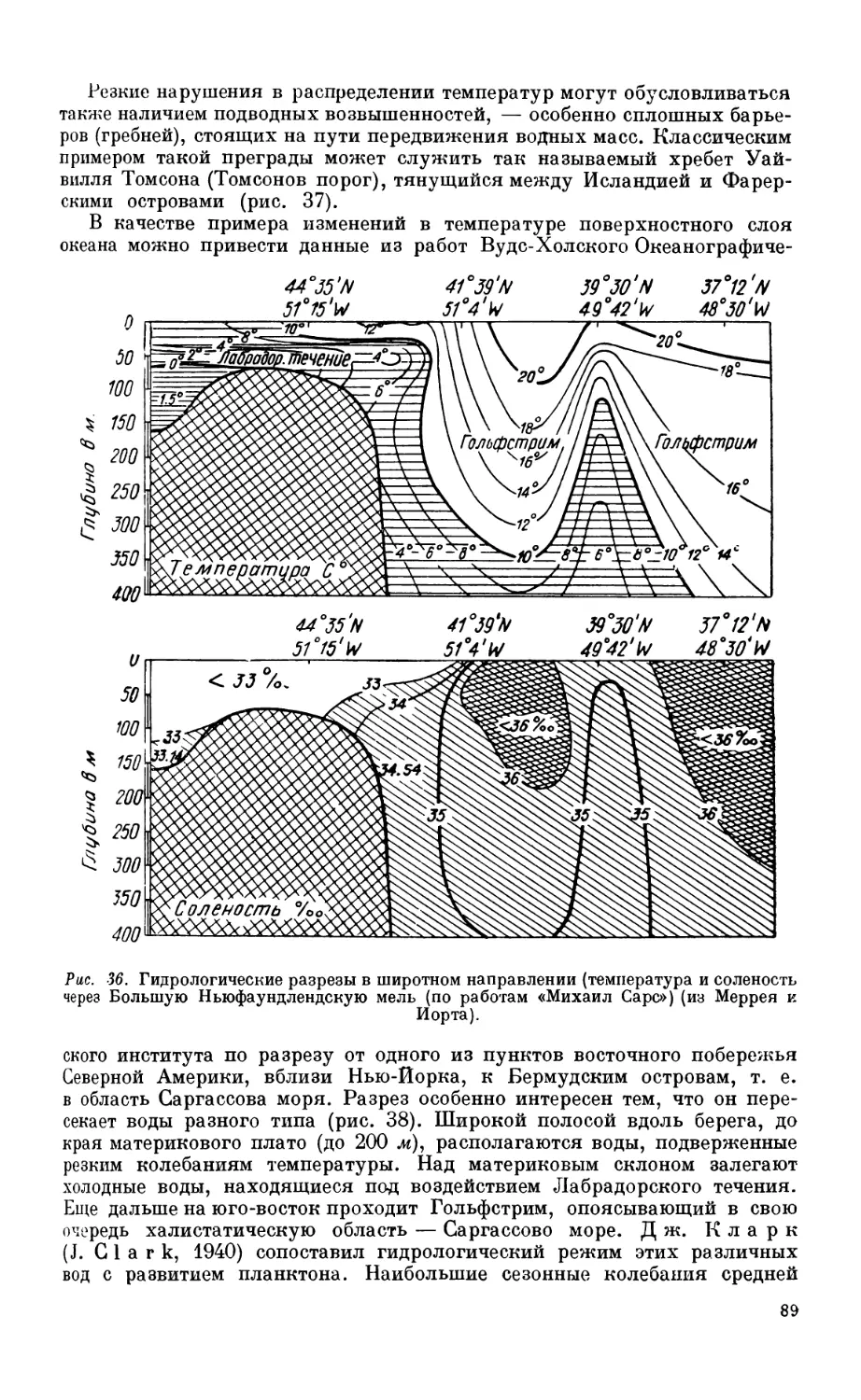
5. Распределение температуры в мировом океане.................................. 80
Распределение температуры па поверхности мирового океана — 80. Годовые ампли-
туды поверхностной температуры — 80. Вертикальное распределение темпера-
туры — 83. Особенности температурного режима Средиземного и Красного
морей — 84. Влияние температуры па распределение организмов — 87.
6. Газовый режим мирового океана..........................................; . . 92
Соотношение основных газов в воздухе и в воде — 92. Кислород — 93. Углекис-
лота— 95. Сероводород — 96. Аммиак—97. Метан — 97. Газовый режим придон-
ного слоя — 97. Вертикальная зональность в океане — 97.
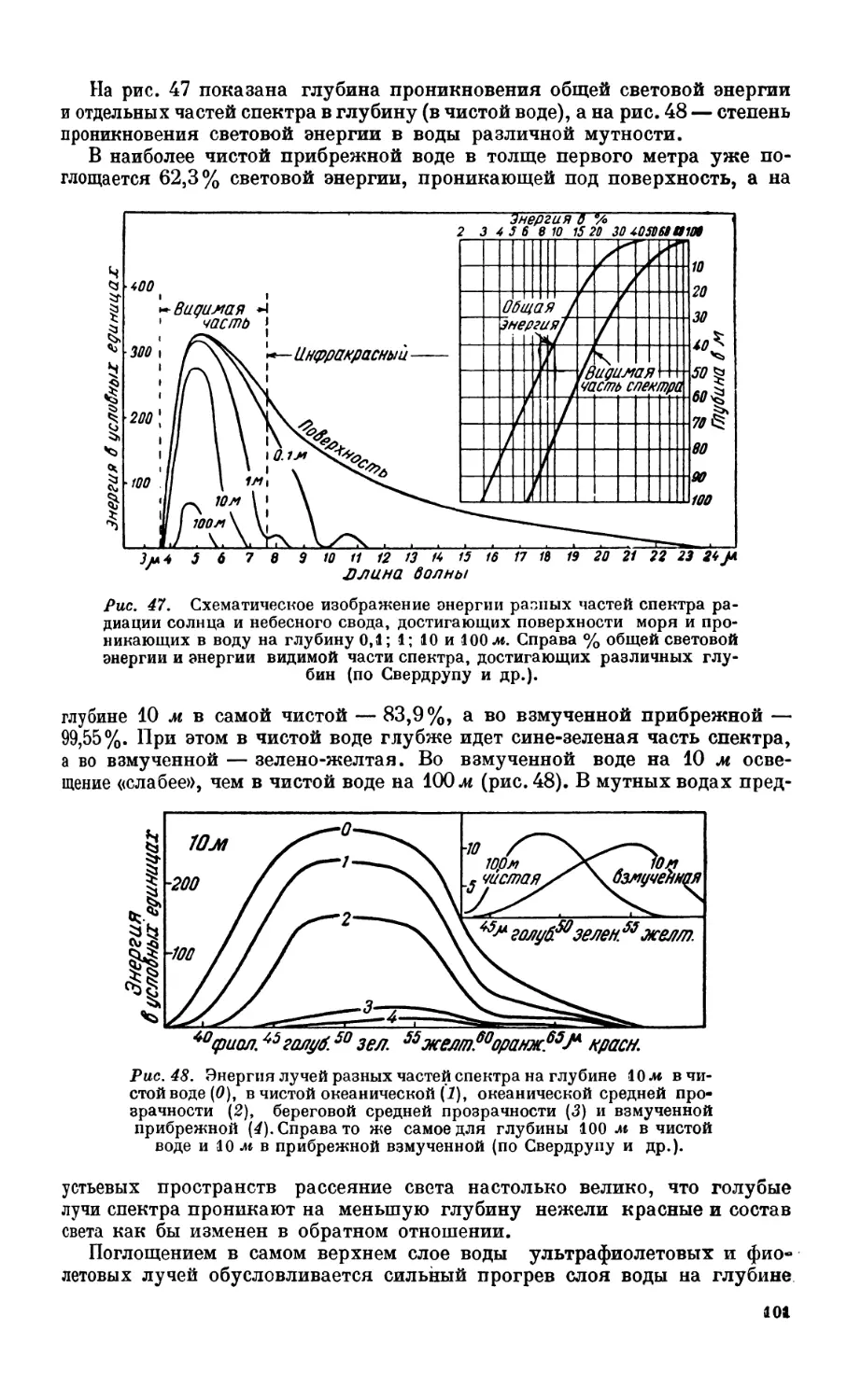
7. Свет....................................................; -................. 98
Проникновение света в толщу морской воды — 98. Проникновение в глубину раз-
личных частей спектра— 99. Прозрачность—100. Влияние интенсивности освеще-
ния на распределение растительных организмов —103. Окраска организмов на раз-
ных глубинах —104.
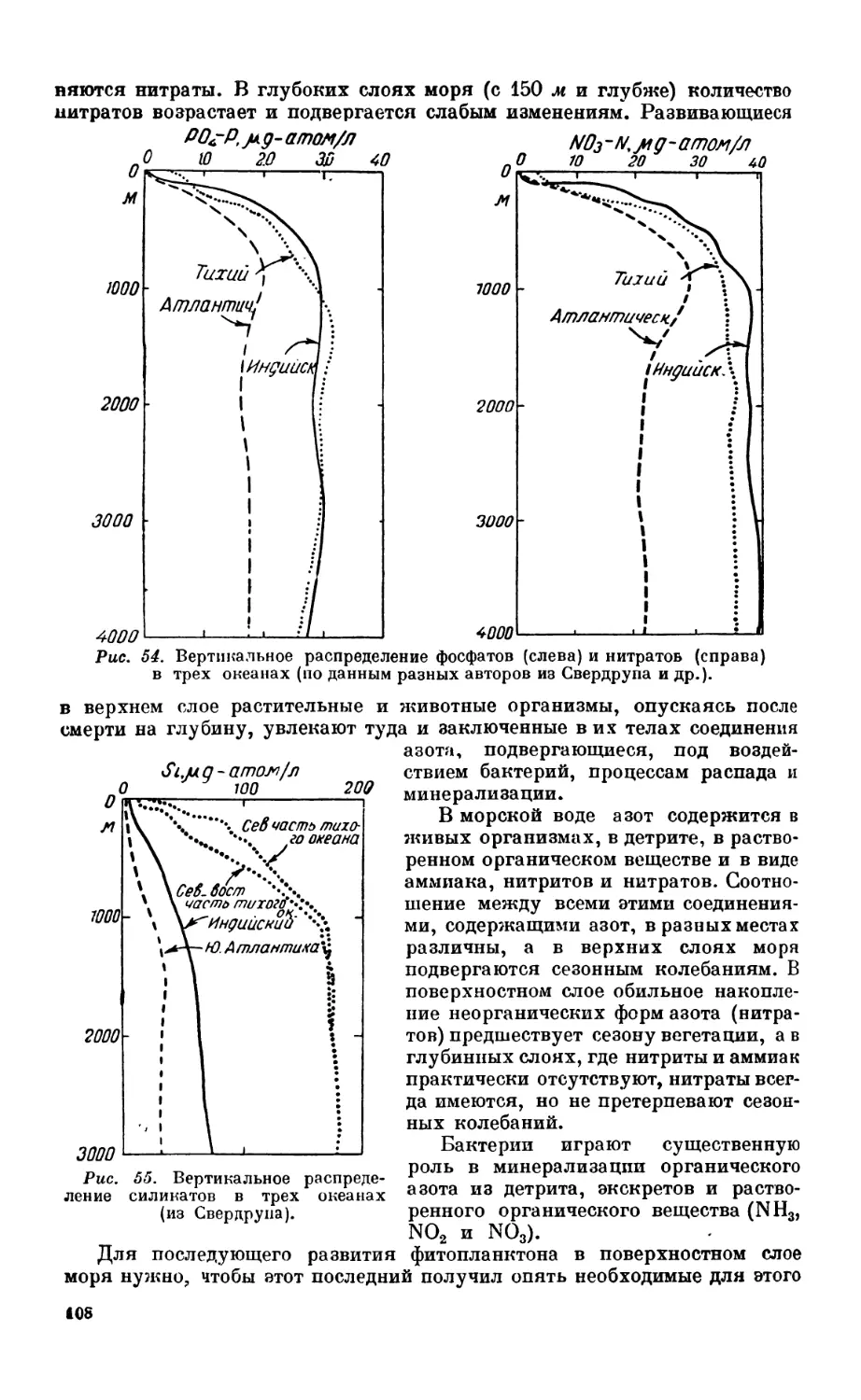
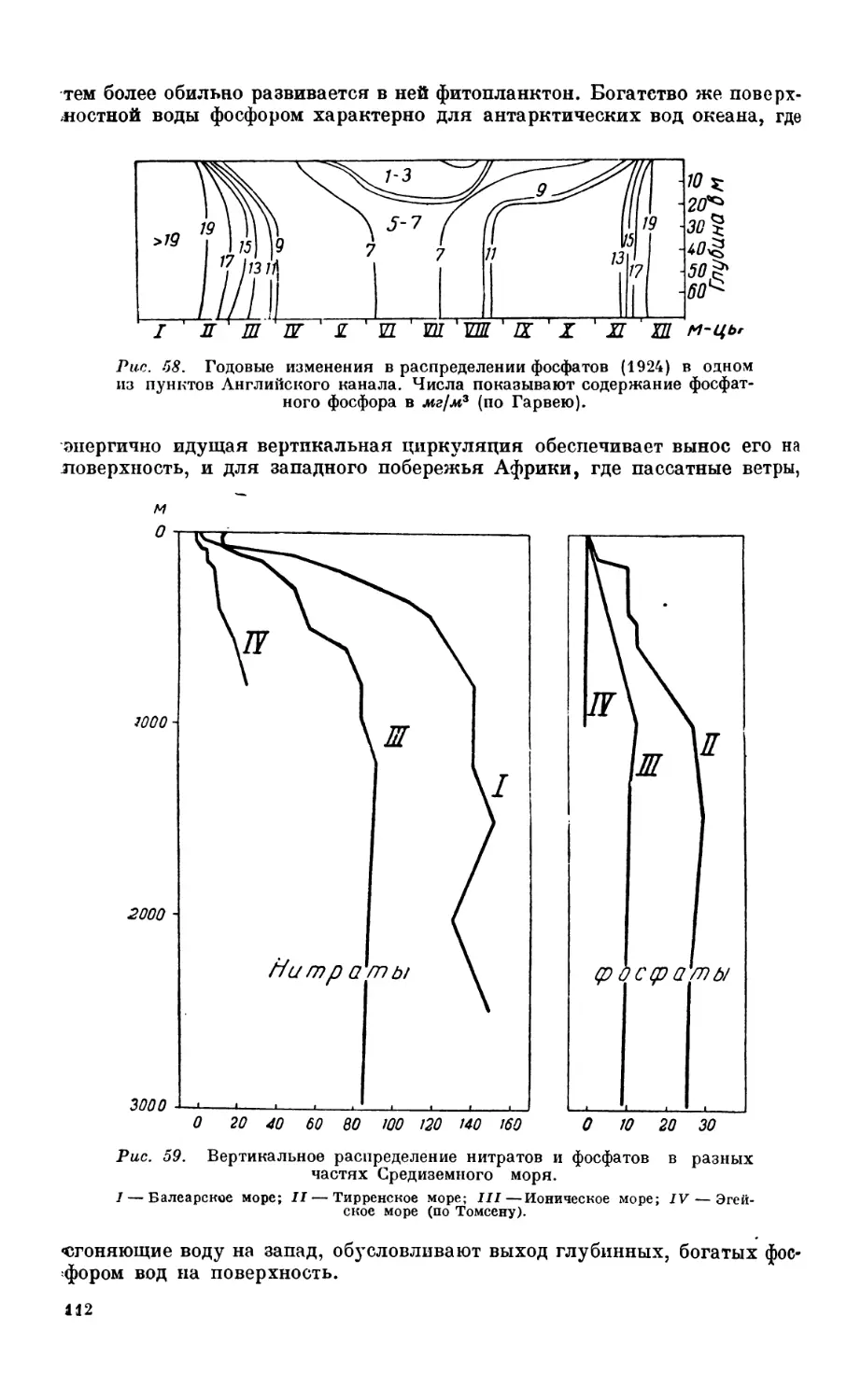
8. Биогенные элементы и растворенное органическое вещество.................... 105
Минимум соли — 105. Азот и его соединения—105. Фосфор — 109. Кремпеки-
слота — 113. Особенности распределения биогенных элементов в некоторых водо-
емах—113. Цикл соединений фосфора и азота —114. Растворенное органическое
вещество—114
Глава IV. ДИНАМИКА ВОД МИРОВОГО океана
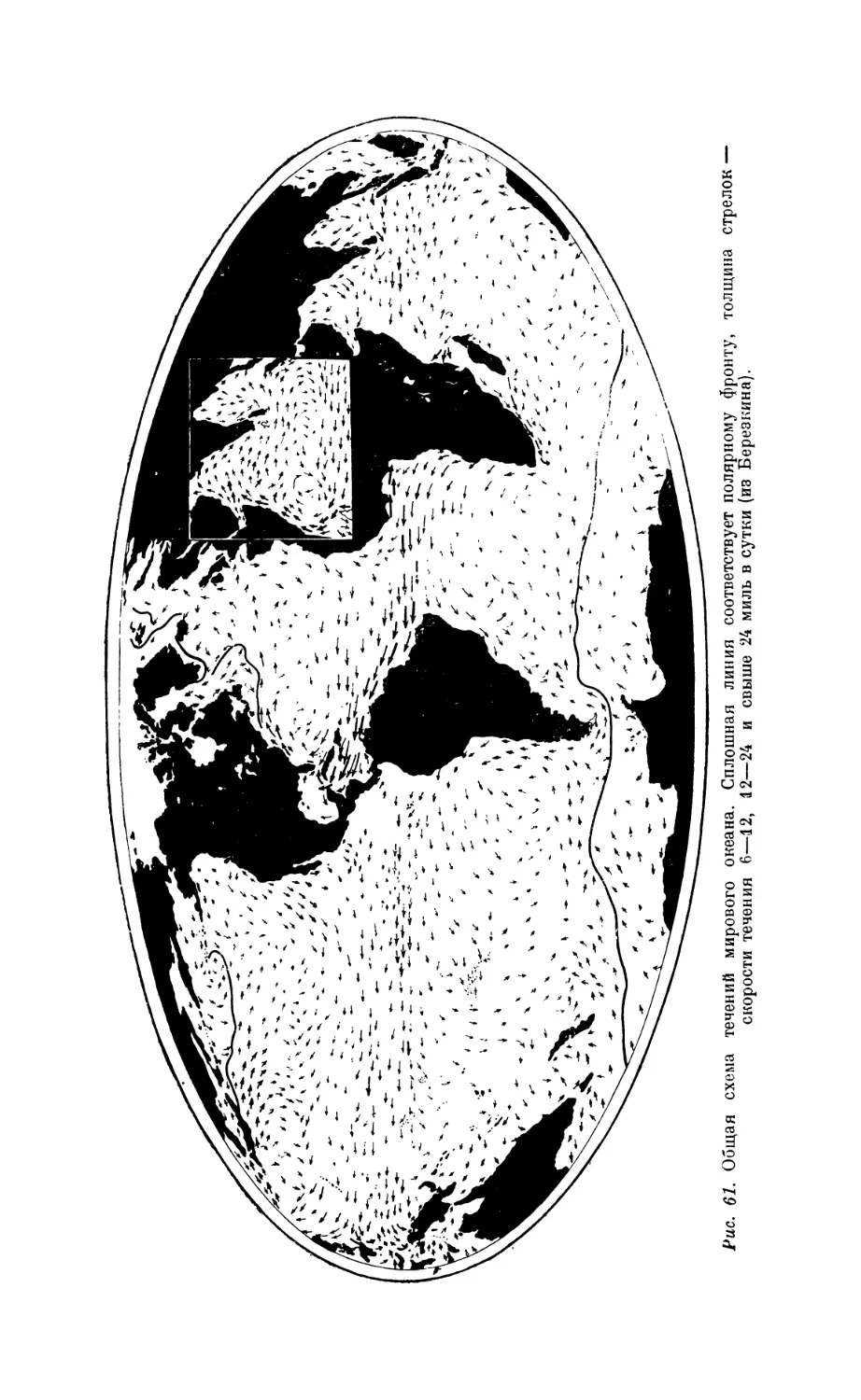
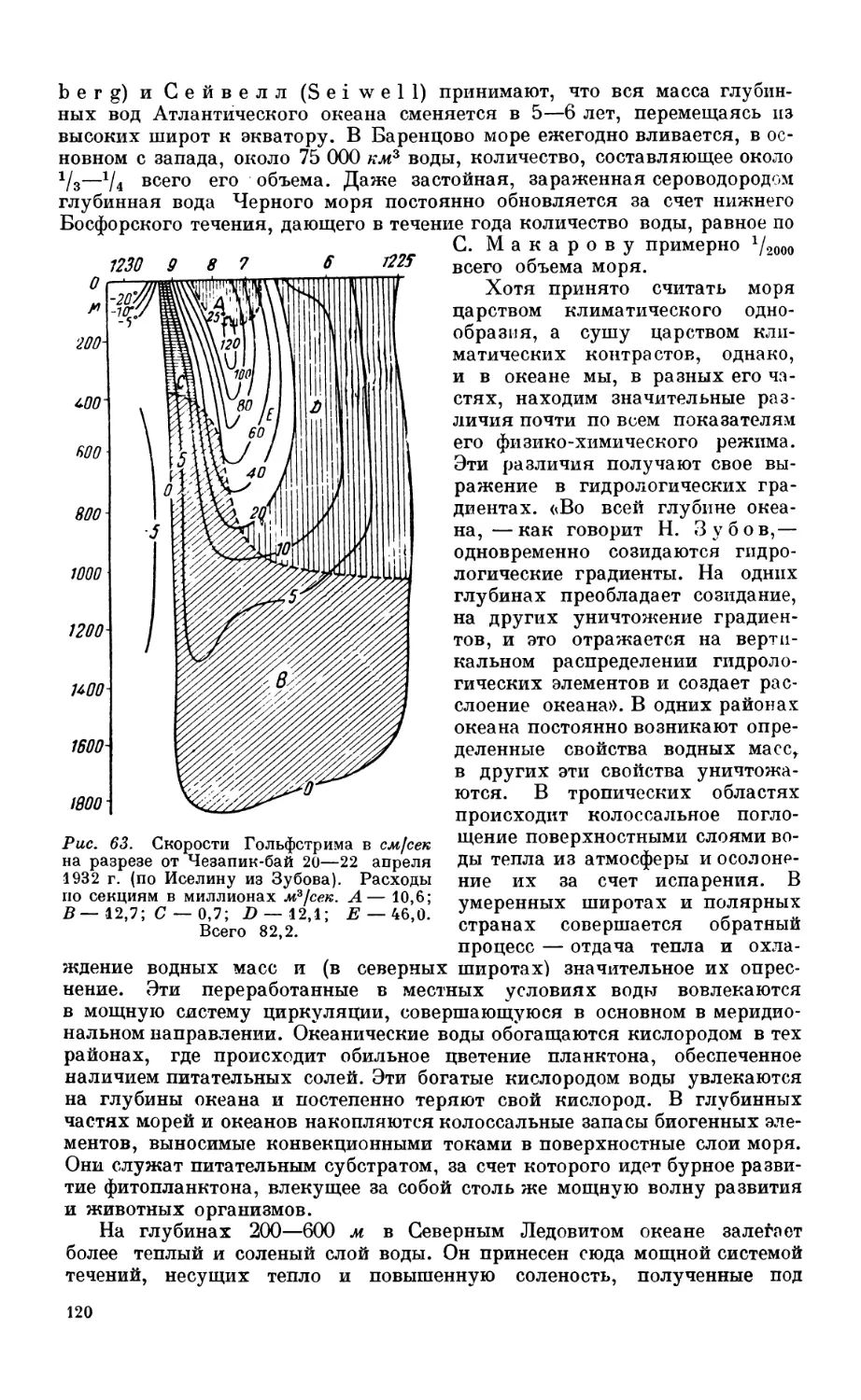
1. Течения.................................................................... 117
Значение перемешивания водных масс для развития жизни —117. Стратификация
как показатель степени перемешиваемости вод — 117. Типы морских течений и при-
чины, их вызывающие—117. Основная система горизонтальных океанических
течений —121.
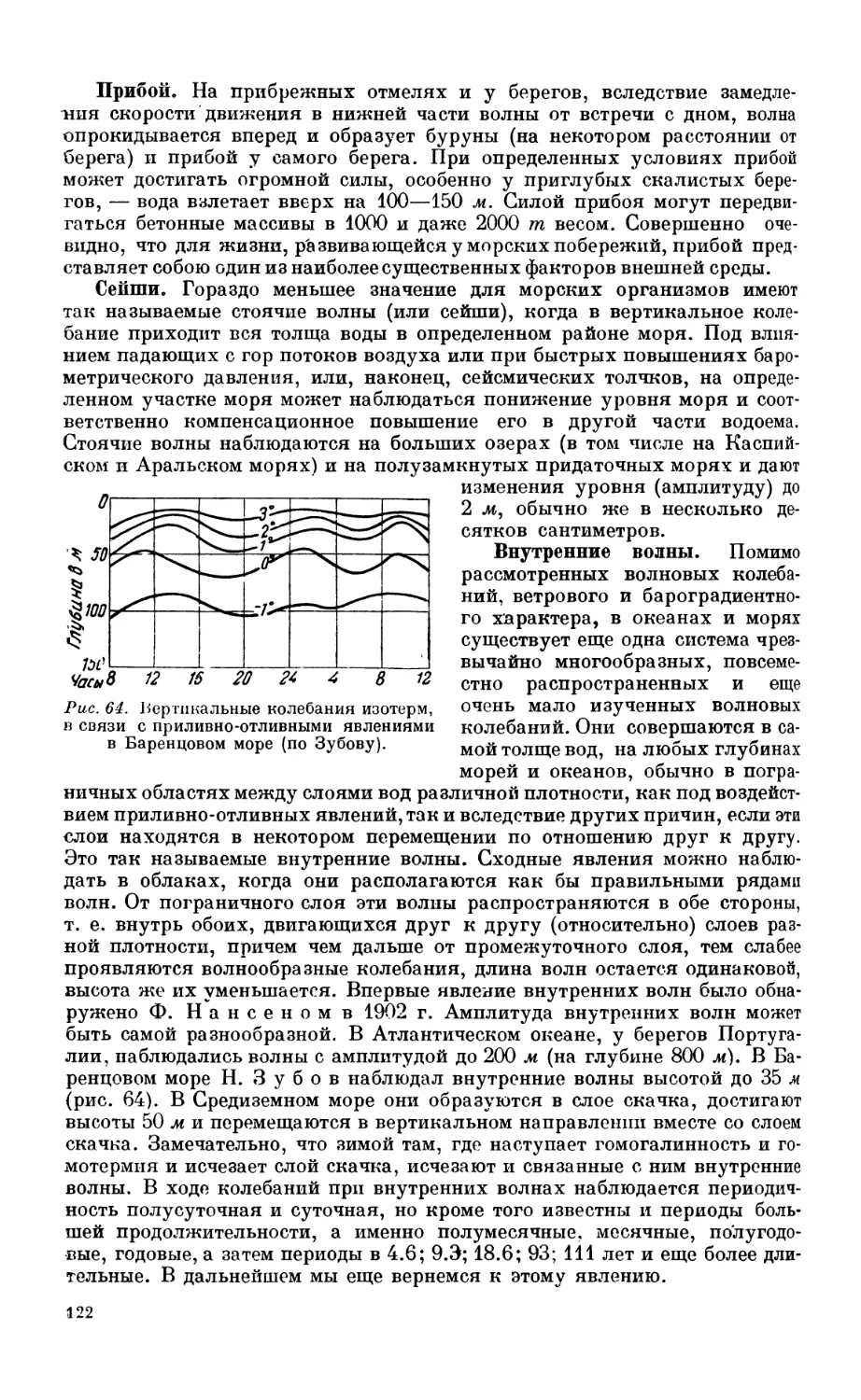
2. Волнение и другие виды перемешивания воды в океане......................... 121
Волнение — 121. Прибой — 122. Сейши — 122. Внутренние волны — 122. Значение
внутренних волн для морских организмов —123.
3. Вертикальная циркуляция и полярный фронт................................... 123
Явления полярного фронта и его значение для развития жизни — 123. Верти-
кальная циркуляция —123.
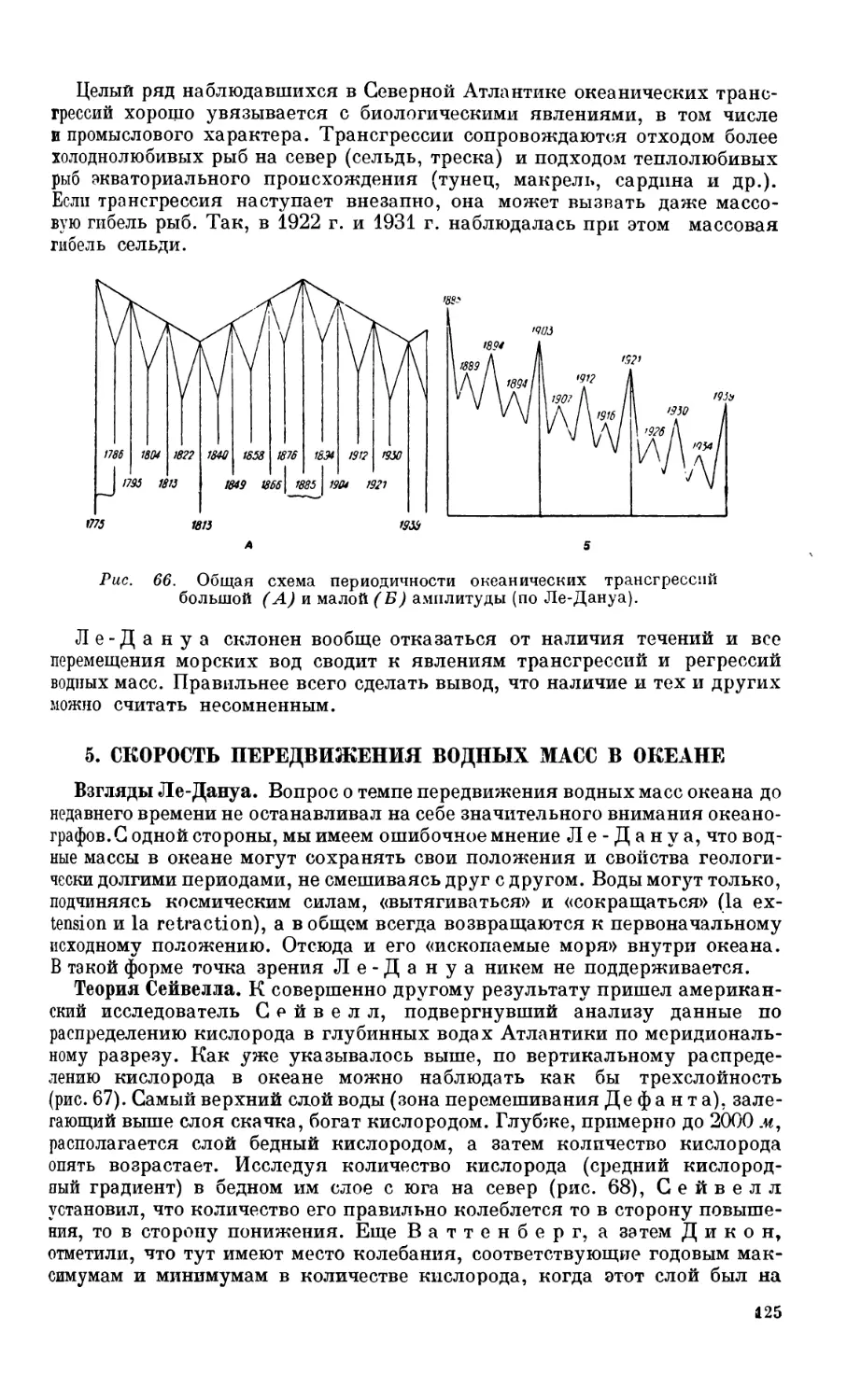
4. Океанические трансгрессии.................................................. 124
Понятие океанических трансгрессий — 124.
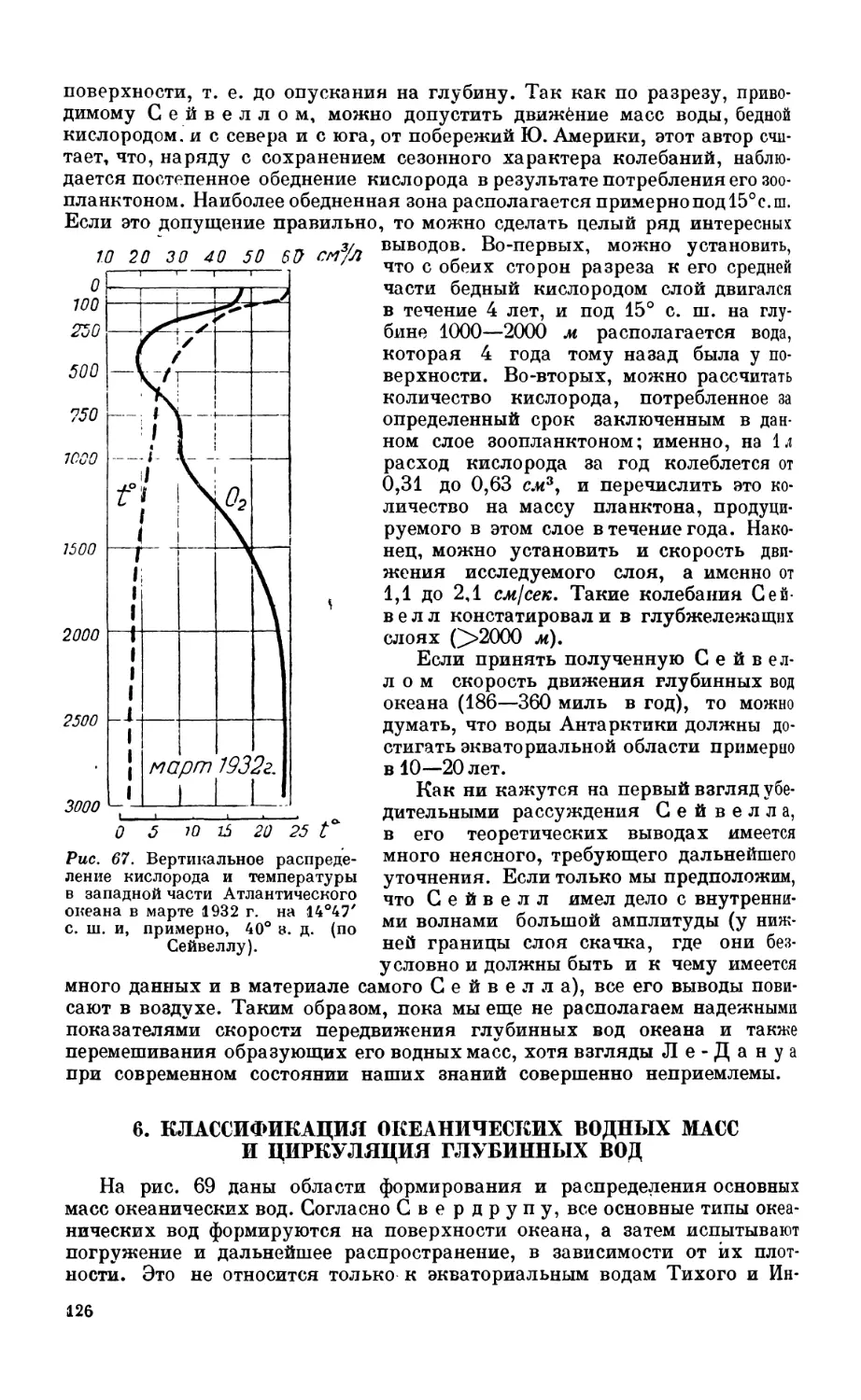
5. Скорость передвижения водных масс в океане................................. 125
Взгляды Ле-Дапуа—125. Теория Сейвелла—125.
6. Классификация океанических водных масс и циркуляция глубинных вод.......... 126
Формирование «центральных» водных масс—127. Формирование и распростра-
нение «промежуточных» вод — 129. «Глубинные» и «придонные» воды — 130. Цир-
куляция водных масс—130. Классификация водных масс — 131.
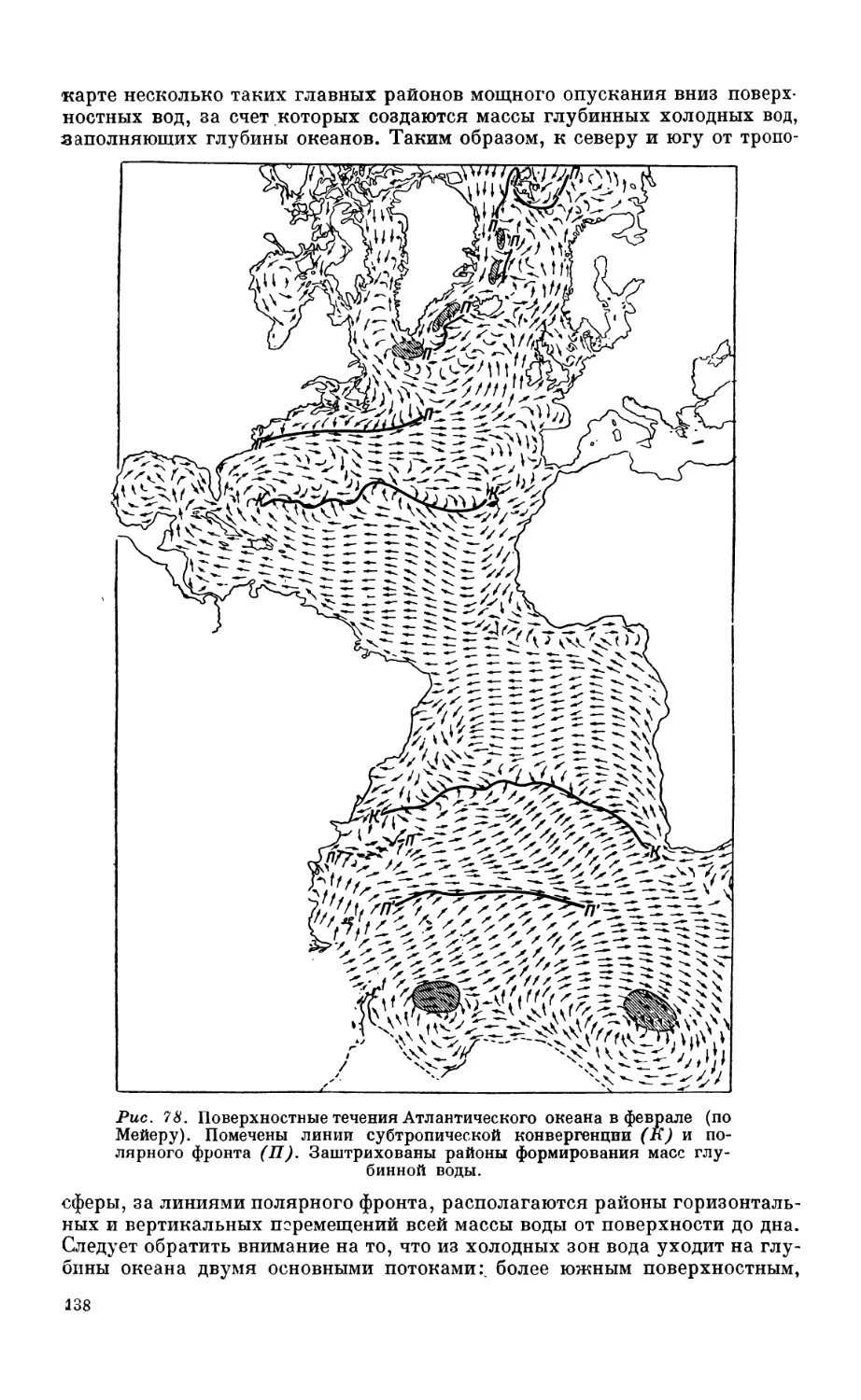
7. Общая схема циркуляции вод Атлантического океана........................... 132
Деление вод Атлантики на четыре райопа по типу стратификации—132. Общий
характер циркуляции — 136. Атлантическое течение — 139.
Глава V. грунты мирового океана
1. Механический состав грунтов................................................ 144
Связь донпых отложений с населением моря — 144. Механический состав —144.
Происхождение морских отложений —145.
2. Распространение различных отложений на дне океана.......................... 145
Глубоководные отложения литоригепного характера—146. Известковые грун-
ты— 146. Кремпеземовые грунты — 147.
3. Зависимость животных от механического состава грунта....................... 147
Жизнь на твердых грунтах —147. Жизнь па мягких грунтах—149. Груптоеды —149.
Стр.
4. Химические процессы, протекающие в грунтах.............................. 149
Общий характер химических процессов —149. Кислород —149. Роль марганца —150.
Органическое вещество—150. Соединения фосфора — 151.
3. Темп отложения морских осадков.......................................... 151
Мощность и возраст различных отложений —151. Слои, характерные для леднико-
вого и межледникового времени — 152.
ЧАСТЬ ТРЕТЬЯ
РАСПРЕДЕЛЕНИЕ ЖИЗНИ В МИРОВОМ ОКЕАНЕ
Глава VI. БИОСФЕРА И РАСПРЕДЕЛЕНИЕ организмов в море
1, Понятие о биосфере...................................................... 155
Состав —156. Геохимическая энергия организмов — 156. Распределение ауто-
трофных организмов на суше и в море —156. Особенности условий существования
на суше и в море — 157.
J. Основные закономерности распределения организмов в мировом океане....... 159
Неоднородность водной массы океана —159. Симметричное распределение явле-
ний —159. Биполярное и амфиокеаническое распространение организмов —160.
Характер распределения организмов по зонам —161. Миграции —162.
3. Состав морской фауны.................................................... 162
Общее количество видов — 162. Руководящие группы и формы — 1С5.
4. Океан — колыбель жизни.................................................. 167
Жизнь зародилась в море—167. Свойства воды, благоприятствующие развитию
жизни — 167. Правило гомотопности—167. Происхождение пресноводной и назем-
ной фаун —169.
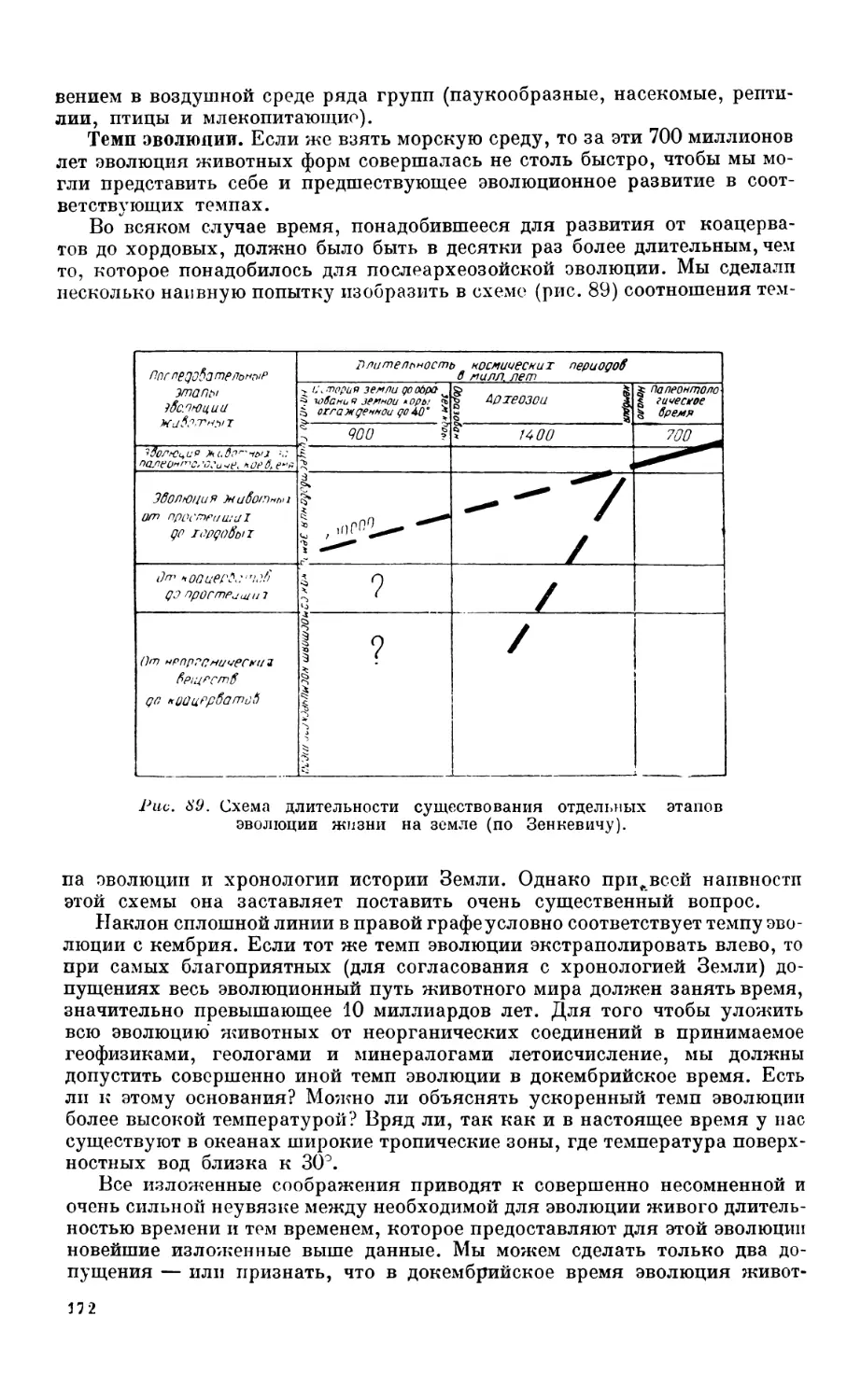
5. Вопрос о происхождении и развитии жизни на земле........................ 170
Постановка вопроса —170. Возраст Земли—170. Время зарождения жизни на
Земле—171. Темп эволюции—172. Эволюция живого населения биосферы — 173.
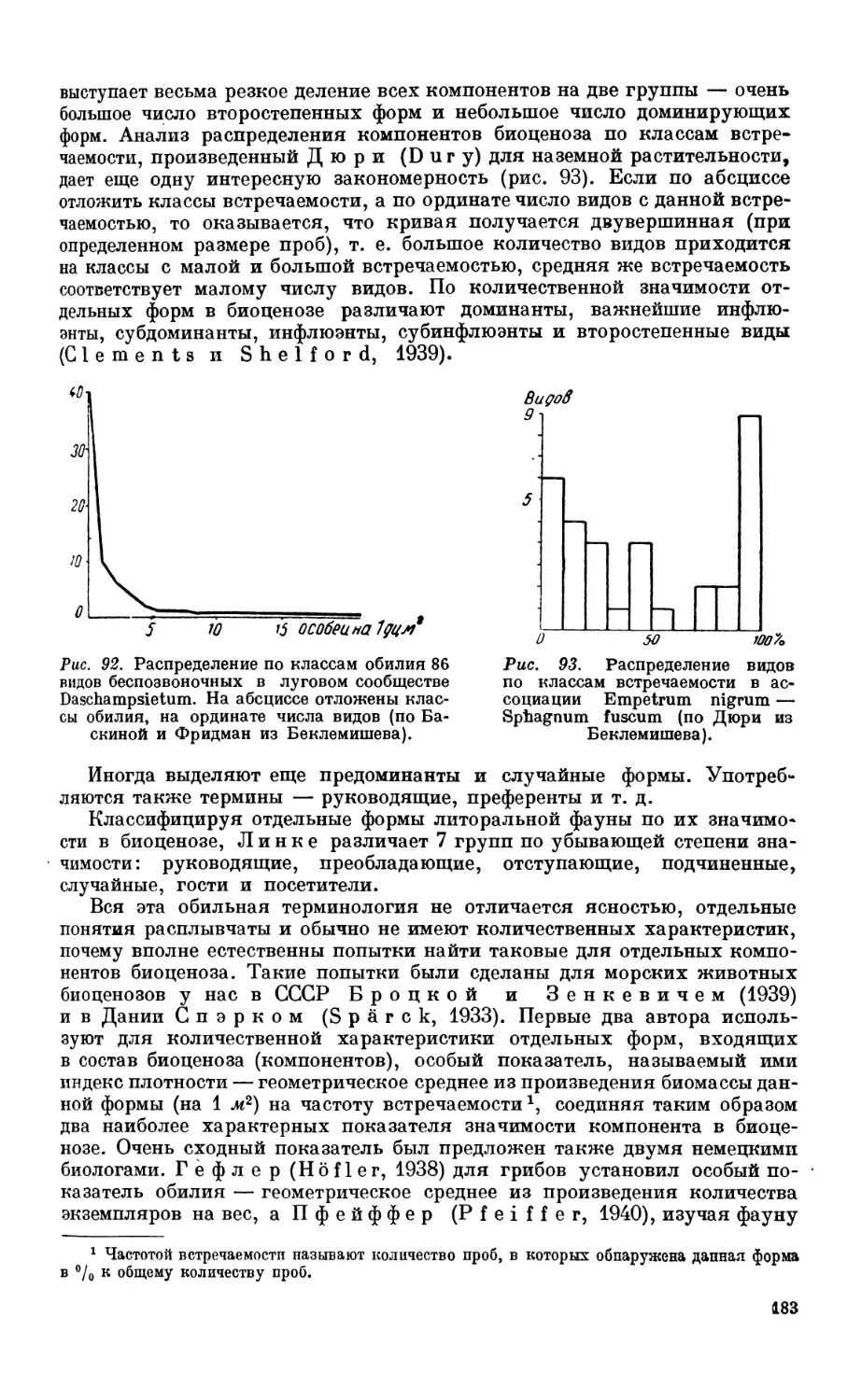
6. Понятие биоценоза и основные методы его анализа......................... 177
Основные проблемы биоценологии—177. Биоценоз и биотоп—177. Состав био-
ценоза—181. Методы изучения биоценоза—182.
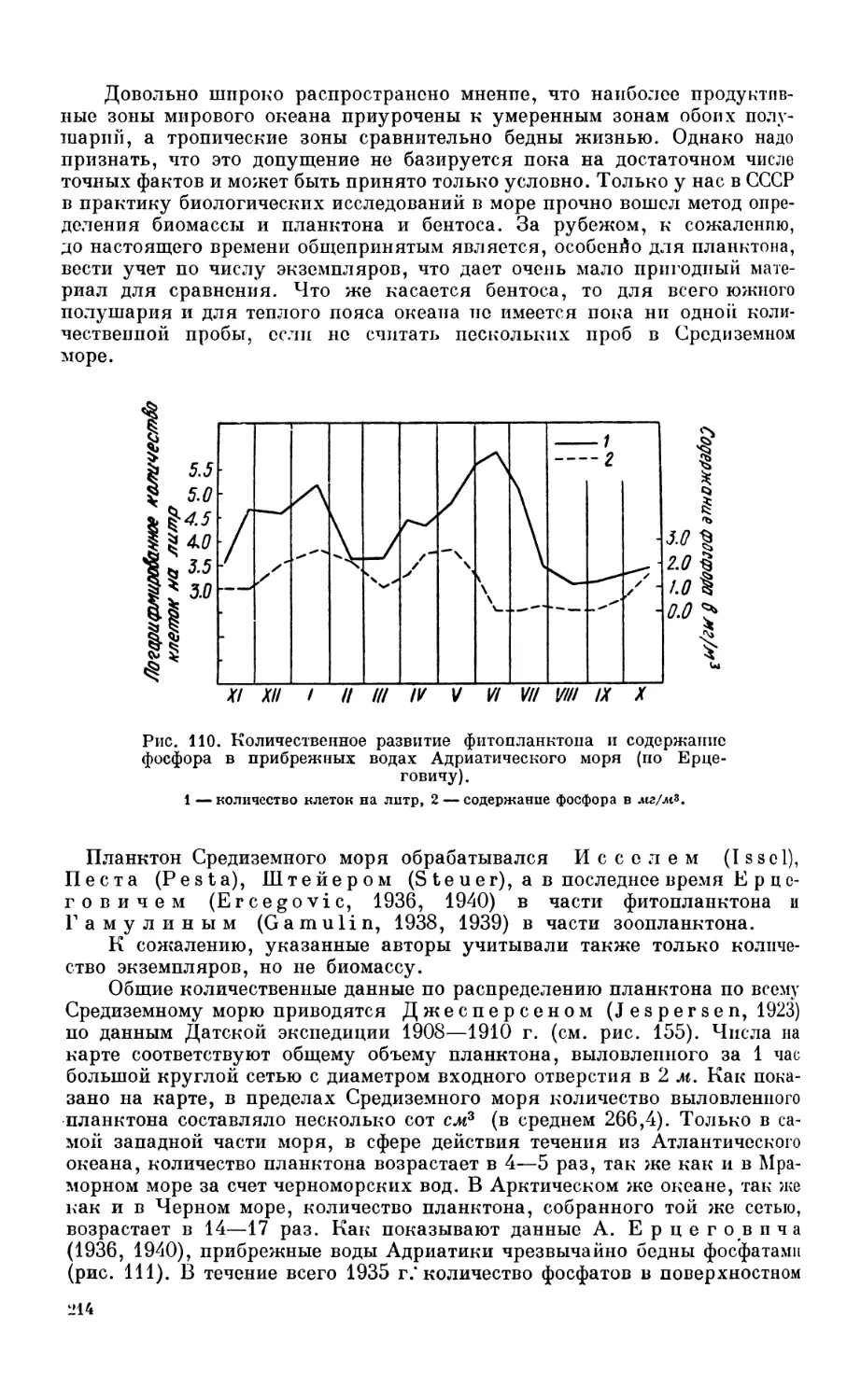
7. Деление морских организмов на основные биологические группы............. 188
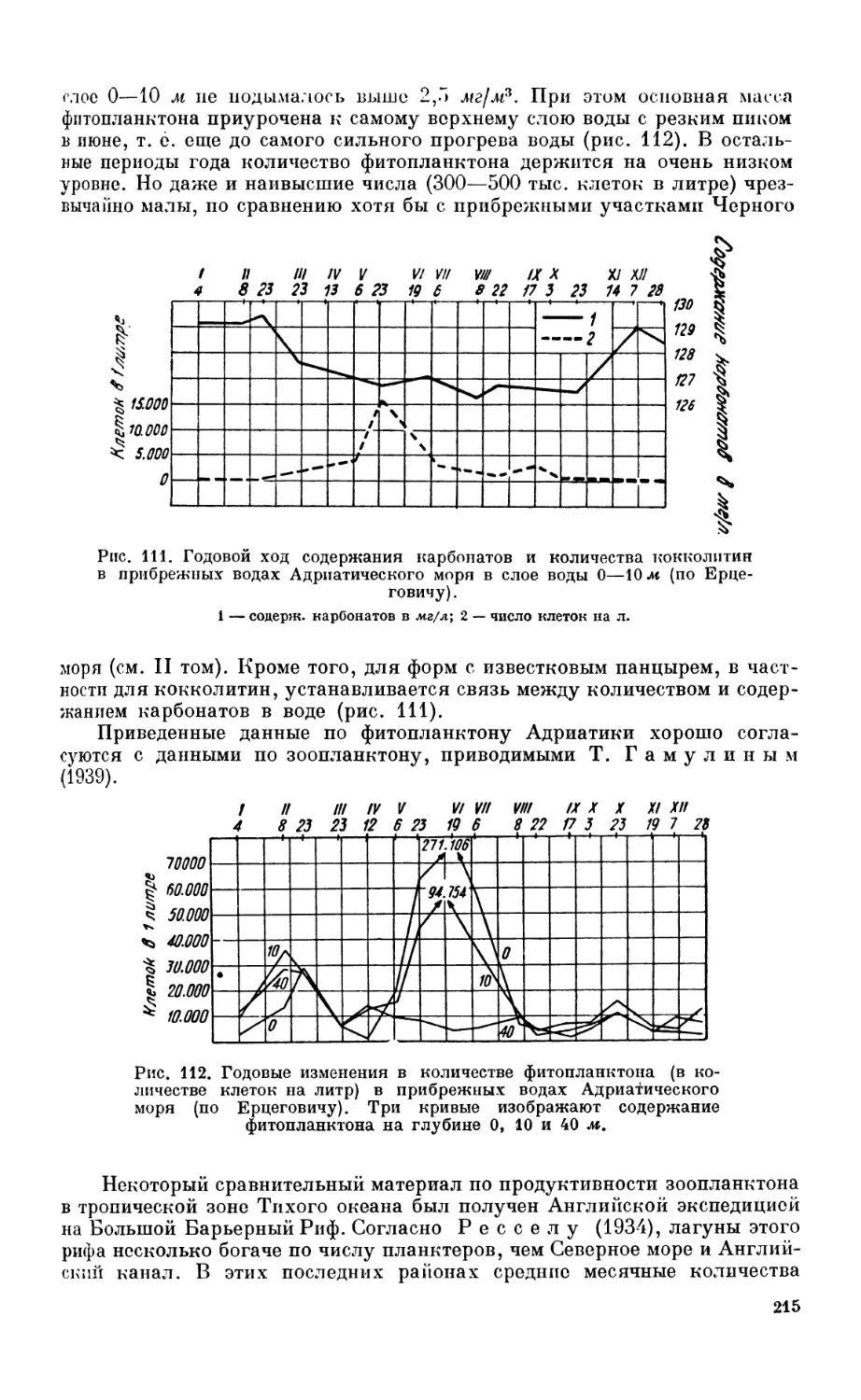
Планктон и бентос —188. Деление по размерам тела —188. Деление по харак-
теру обитания — 190.
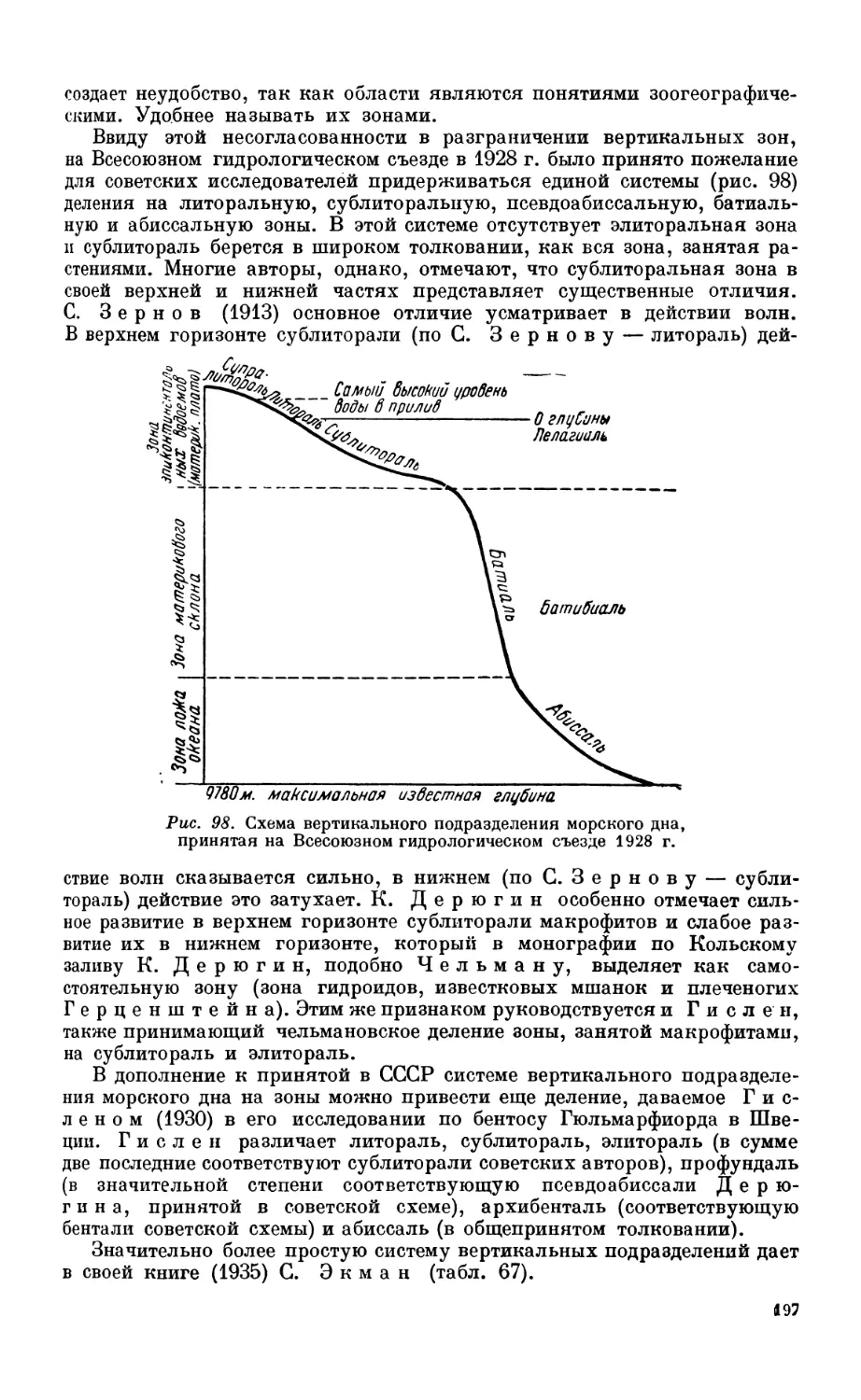
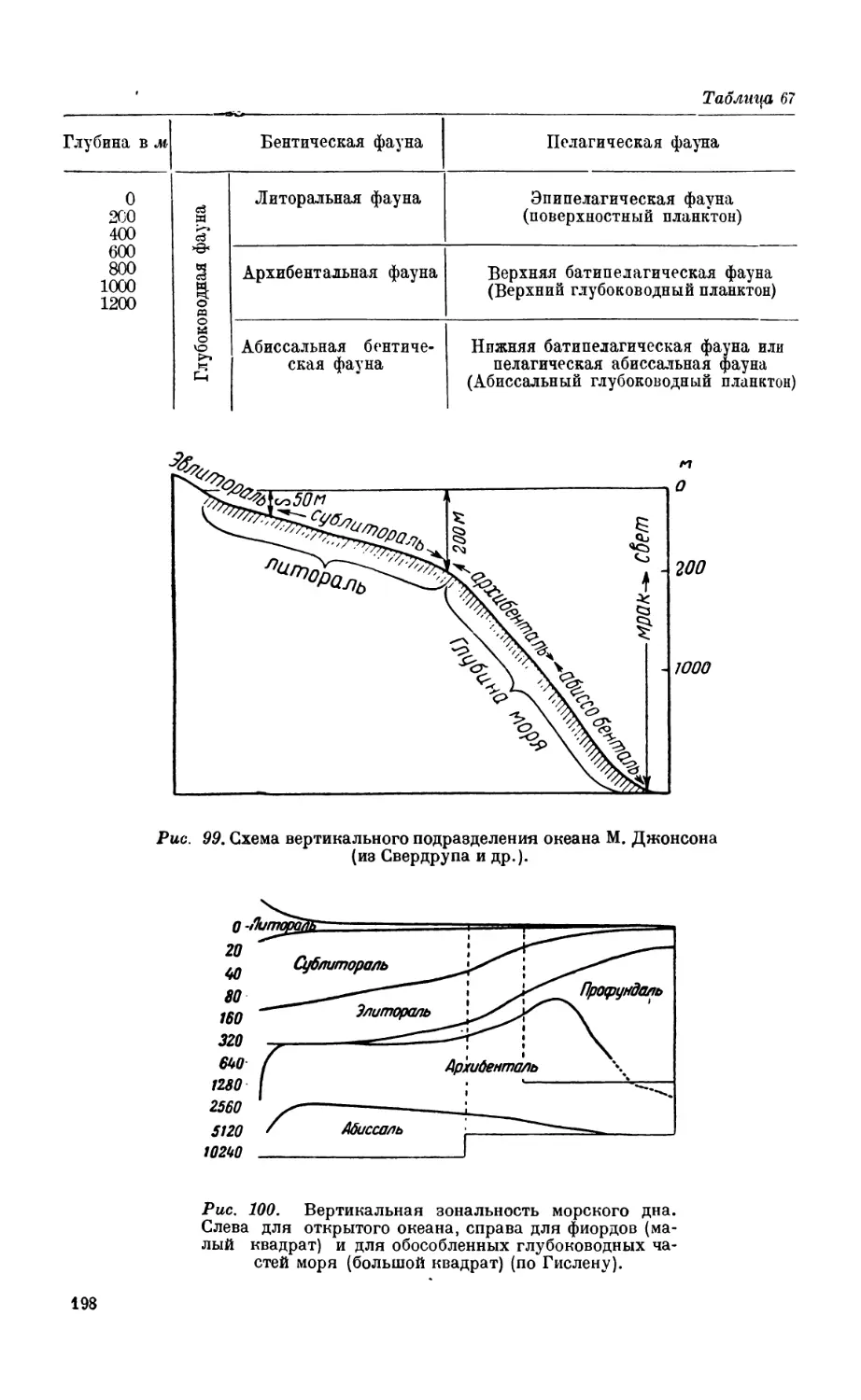
< Подразделение морской фауны по вертикальным зонам........................ 190
Исследования XIX века —190. Работы XX столетия — 193. Системы вертикаль-
ных подразделений бентоса —196.
Глава VII. планктон
1. История и методика изучения............................................. 200
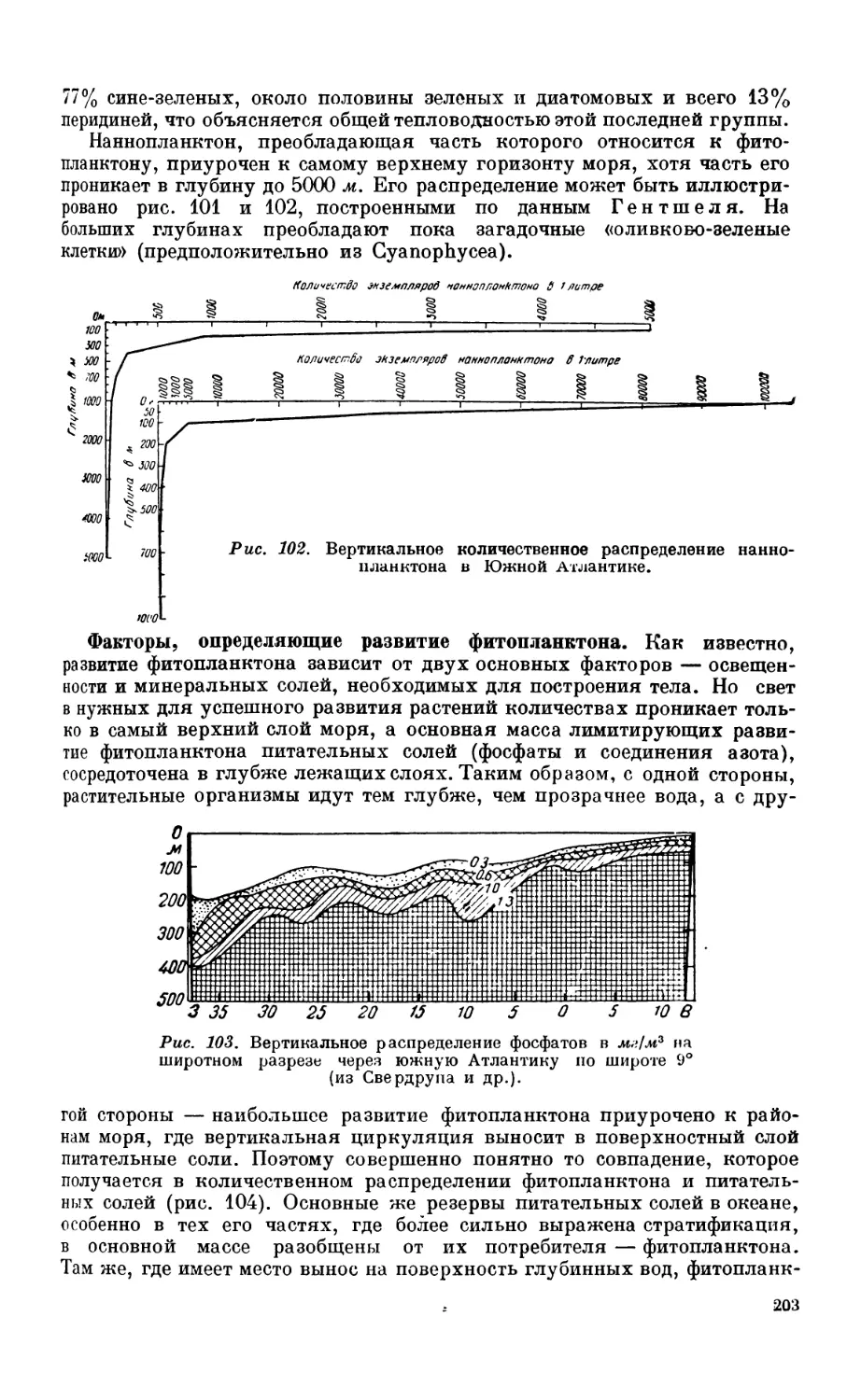
2. Фитопланктон............................................................ 201
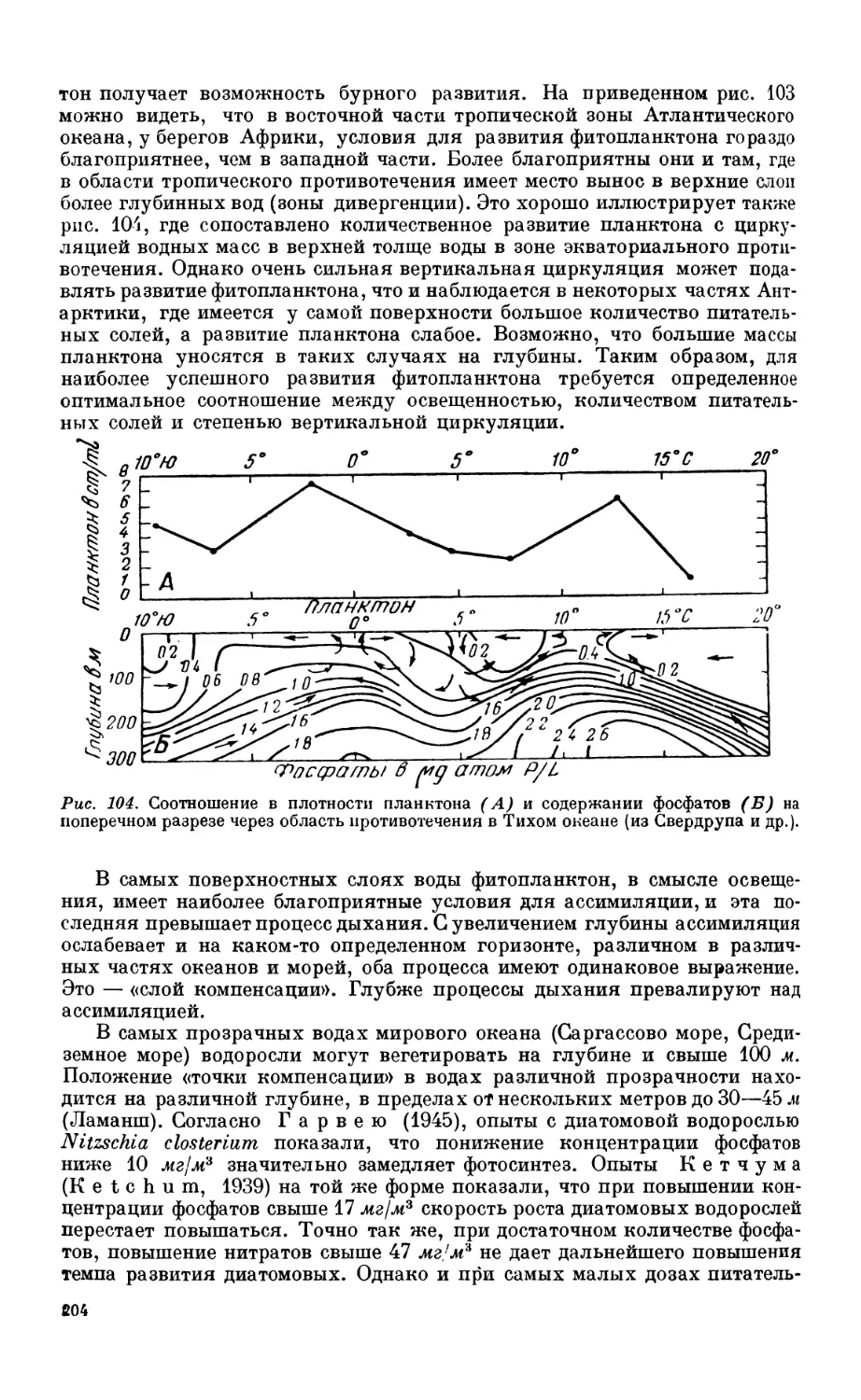
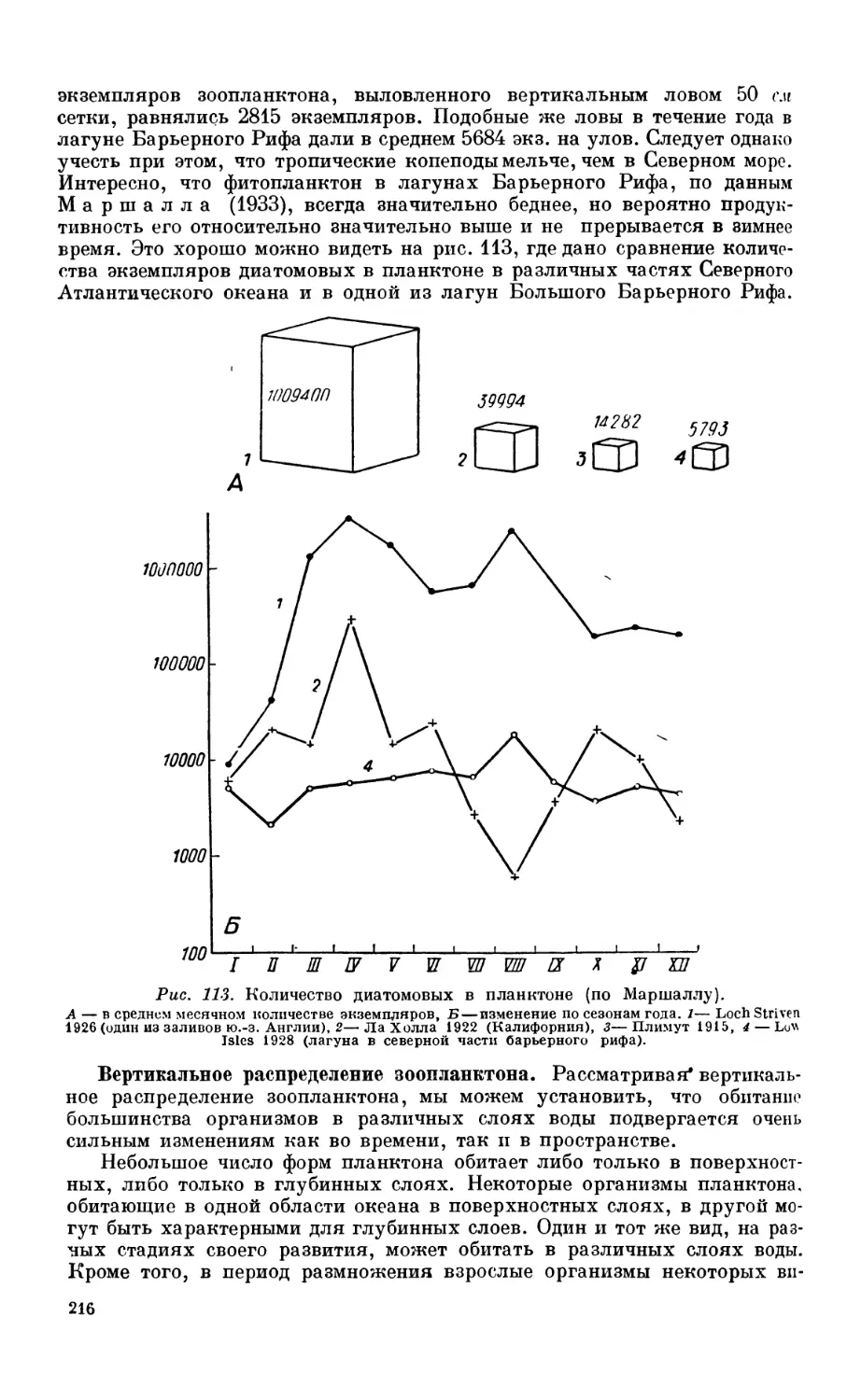
Состав фитопланктона —201. Факторы, определяющие развитие фитопланктона—203.
Соотношение фито- и зоопланктона—205. Продукция фитопланктона — 206.
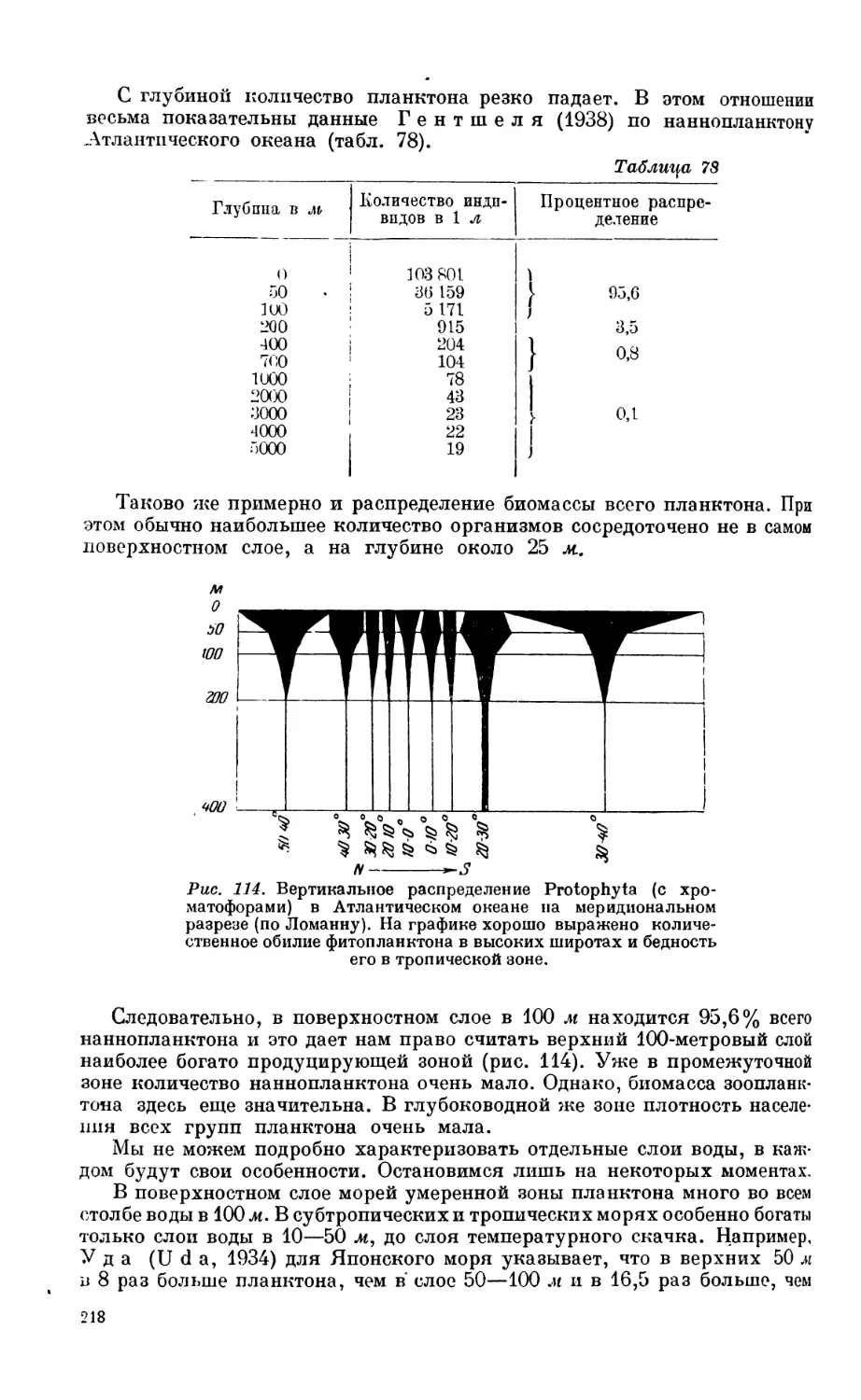
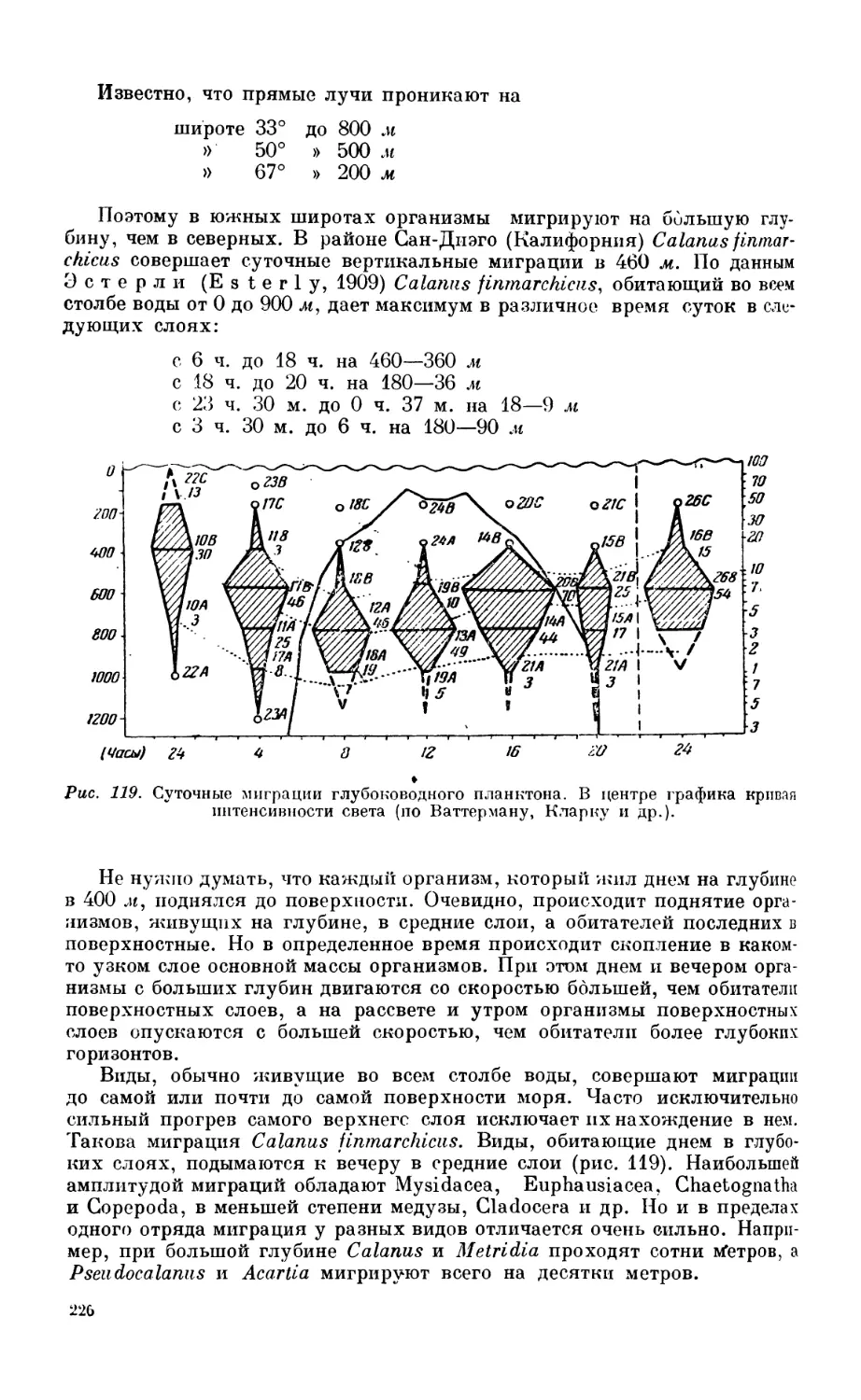
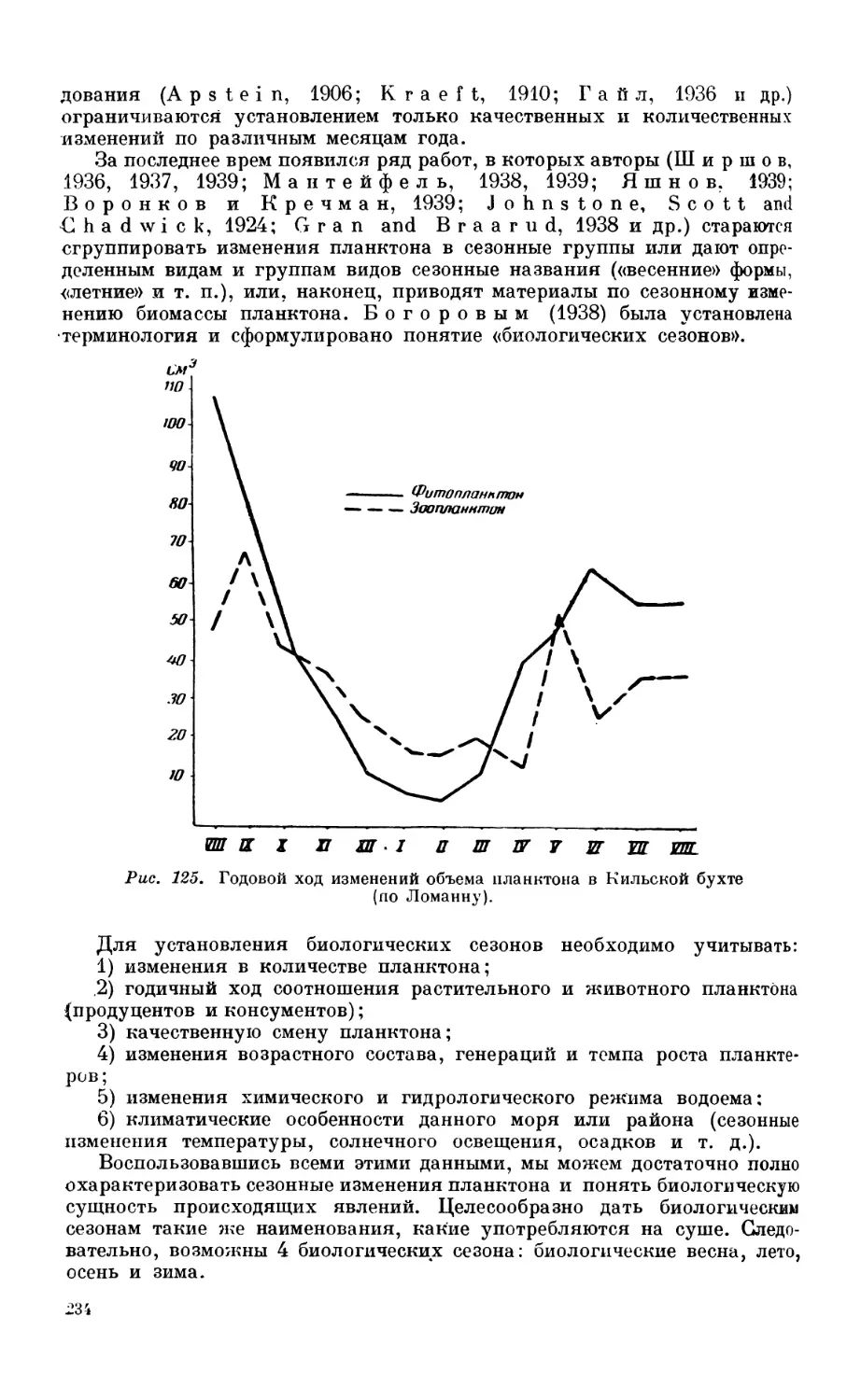
3. Зоопланктон............................................................. 208
Состав зоопланктона—208. Горизонтальное распределение зоопланктона — 210. Вер-
тикальное распределение зоопланктона — 216. Вертикальные миграции зоопланк-
тона — 224. Биологические сезоны — 233. Сезонные изменения планктона в раз-
ных морях — 237.
Глава VIII. бептос
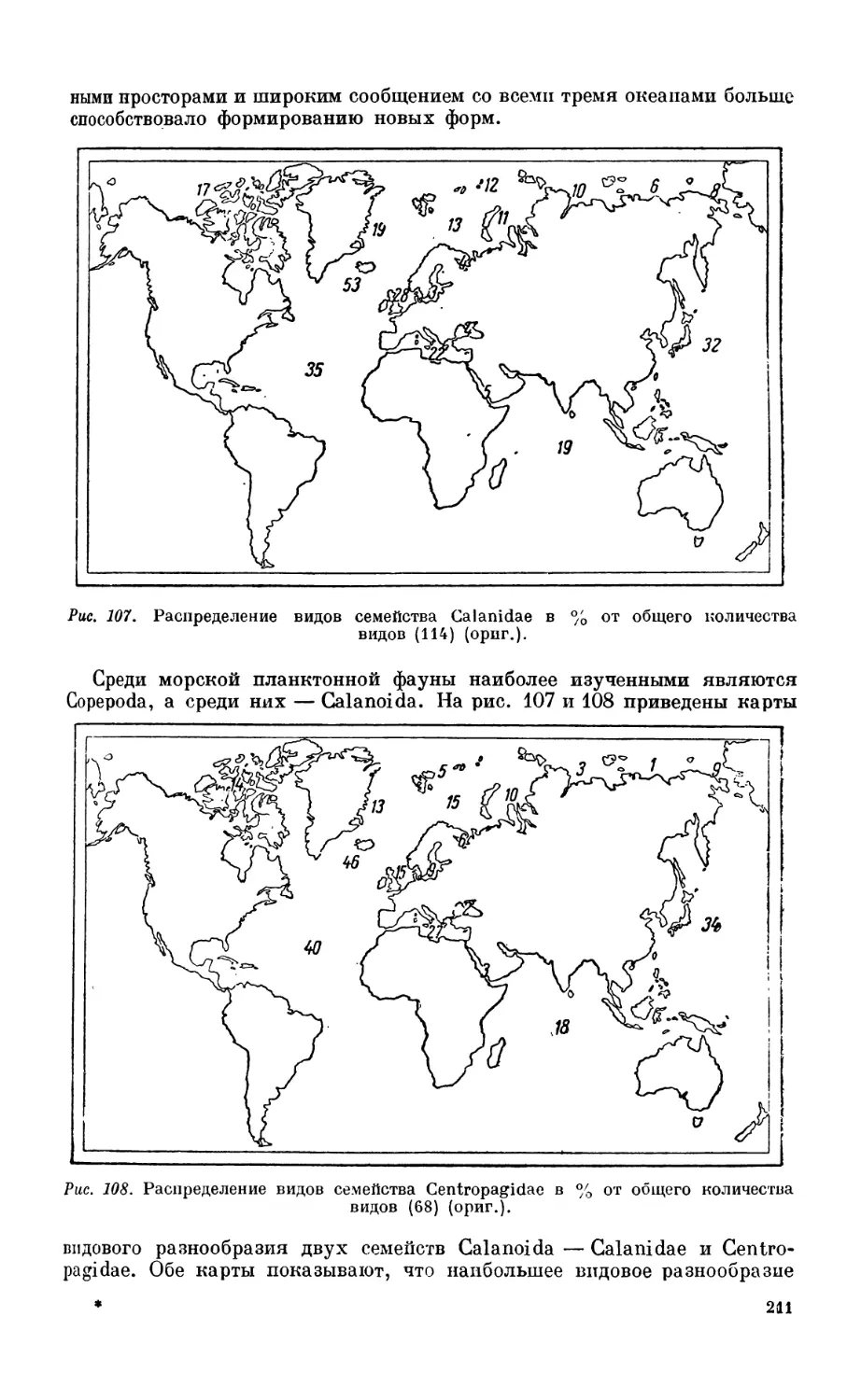
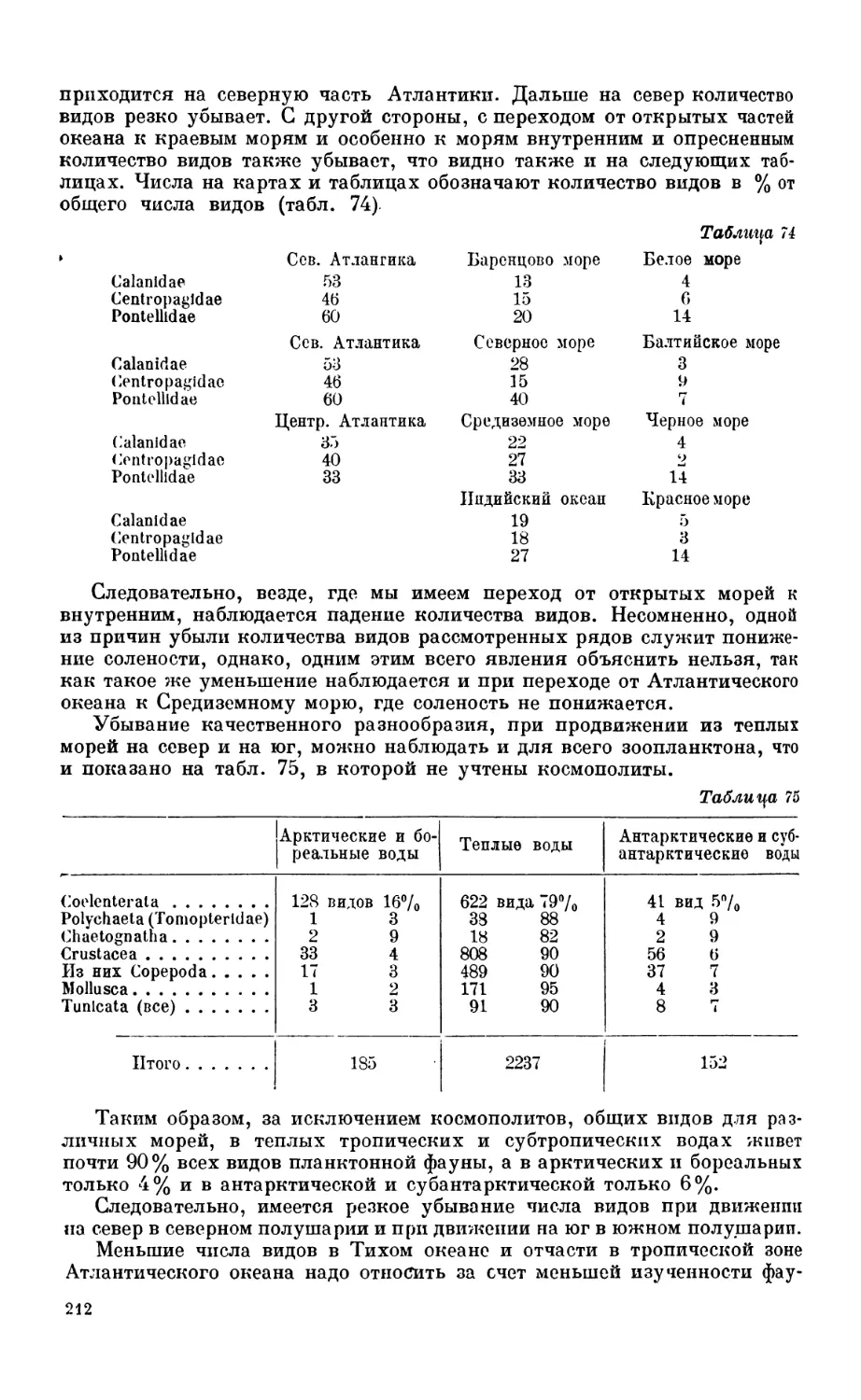
1. История и методика изучения.....................................;........241
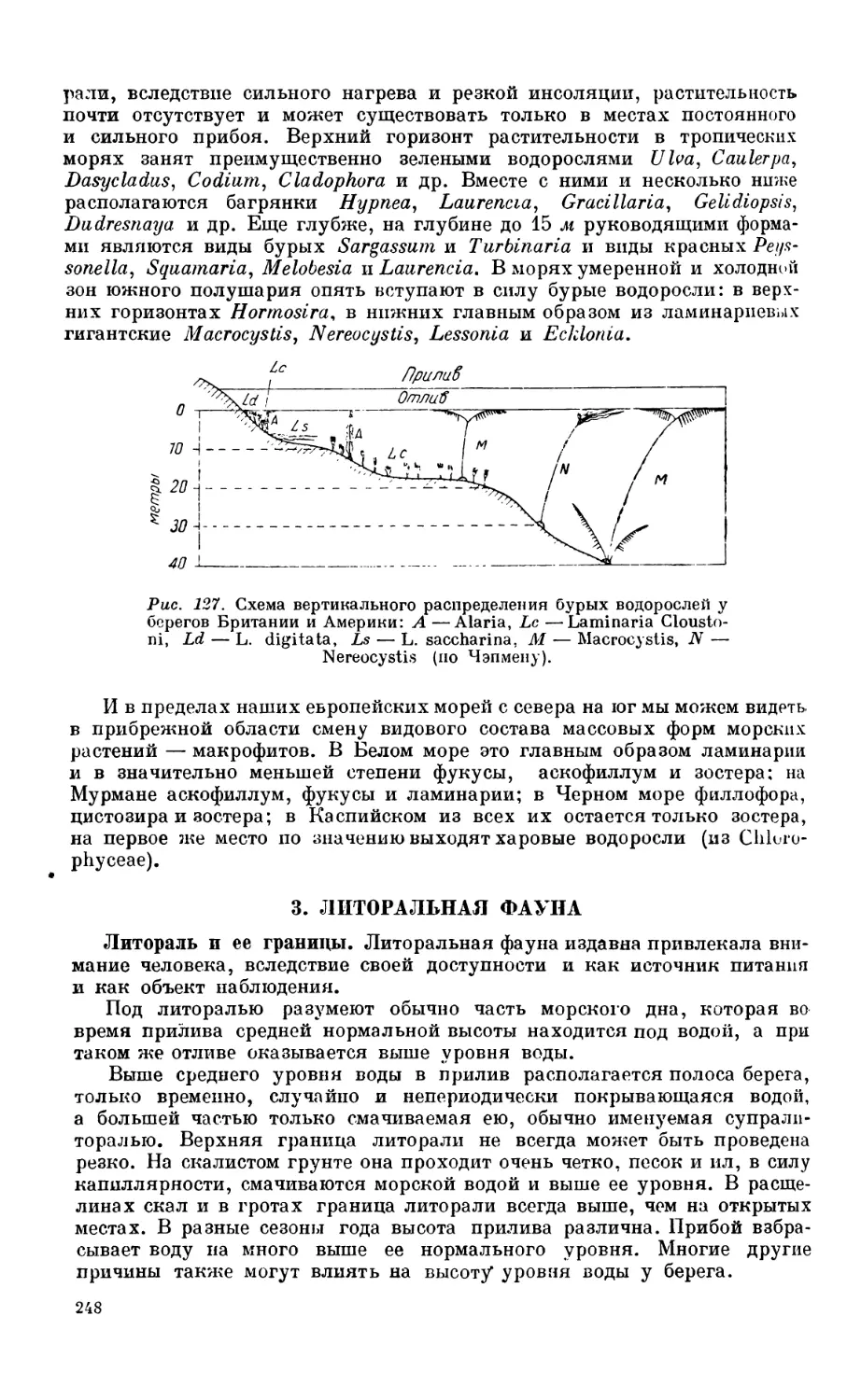
2. Фитобентос.............................................................. 246
Состав фитобентоса — 246. Вертикальная зональность — 246. Горизонтальное рас-
пределение — 247.
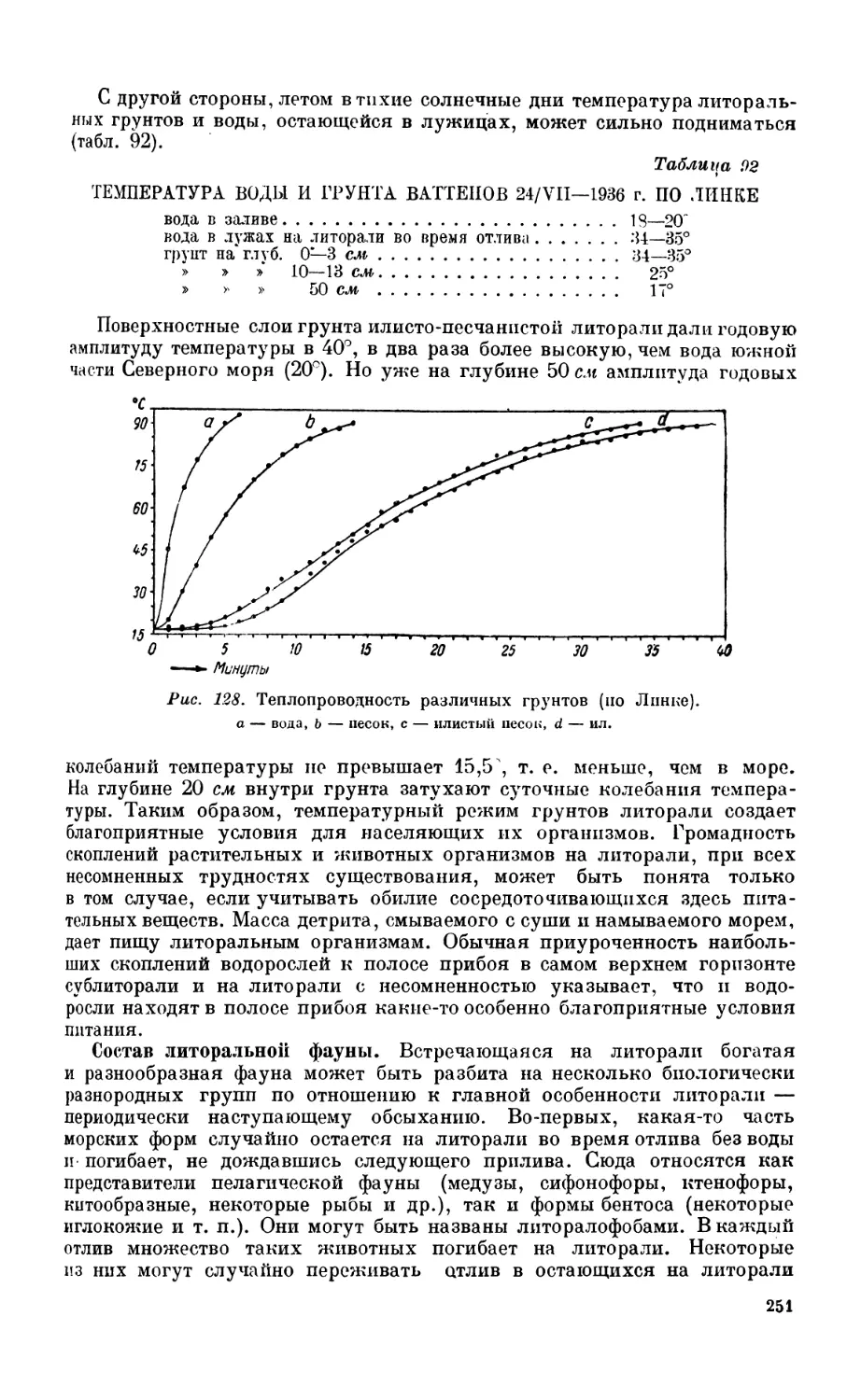
3. Литоральная фауна....................................................... 248
Литораль и ее границы—248. Супралитораль—249. Условия существования на
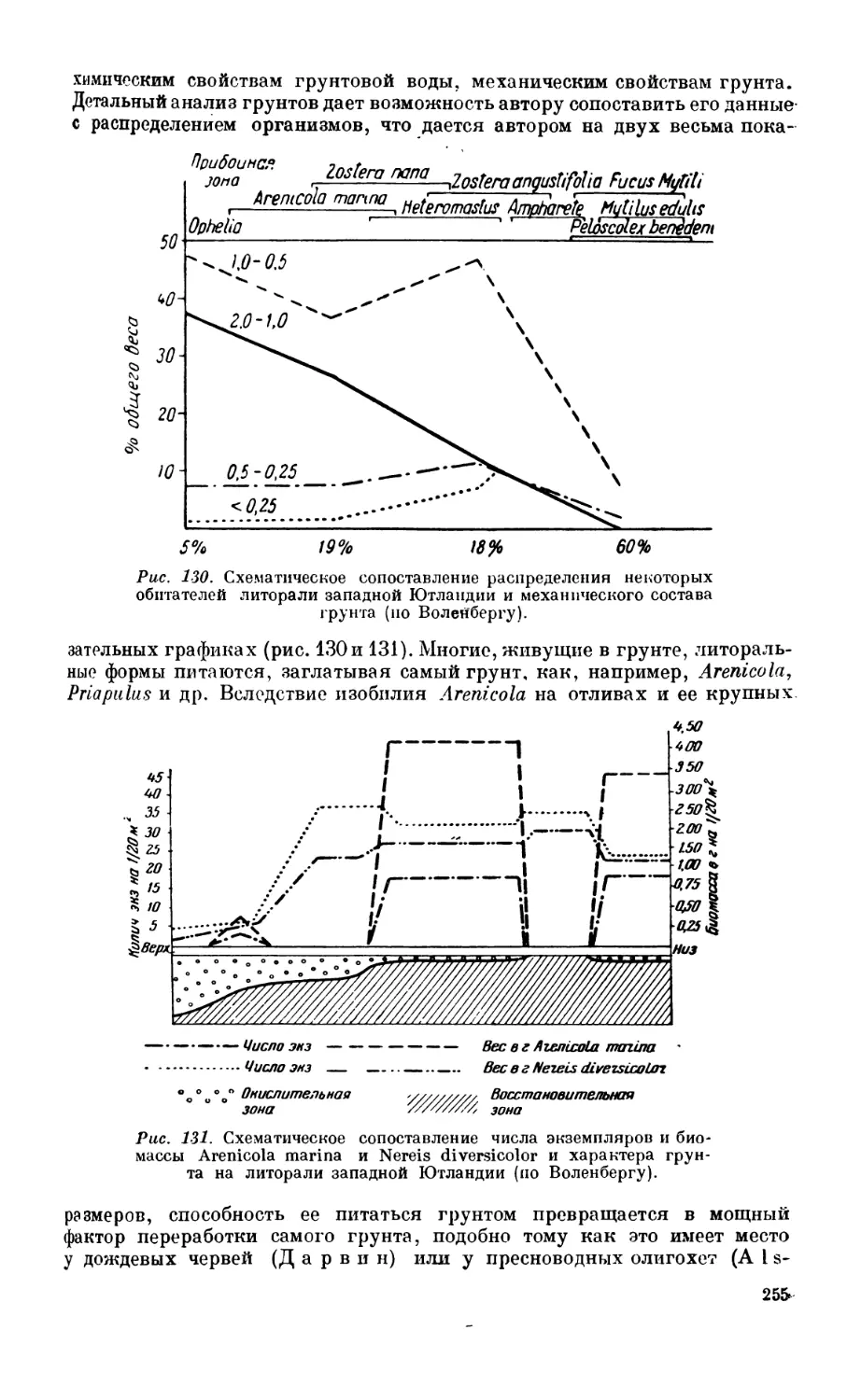
литорали — 250. Состав литоральной фауны — 251. Собственно литораль — 252.
Географические изменения состава литоральной фауны—256.
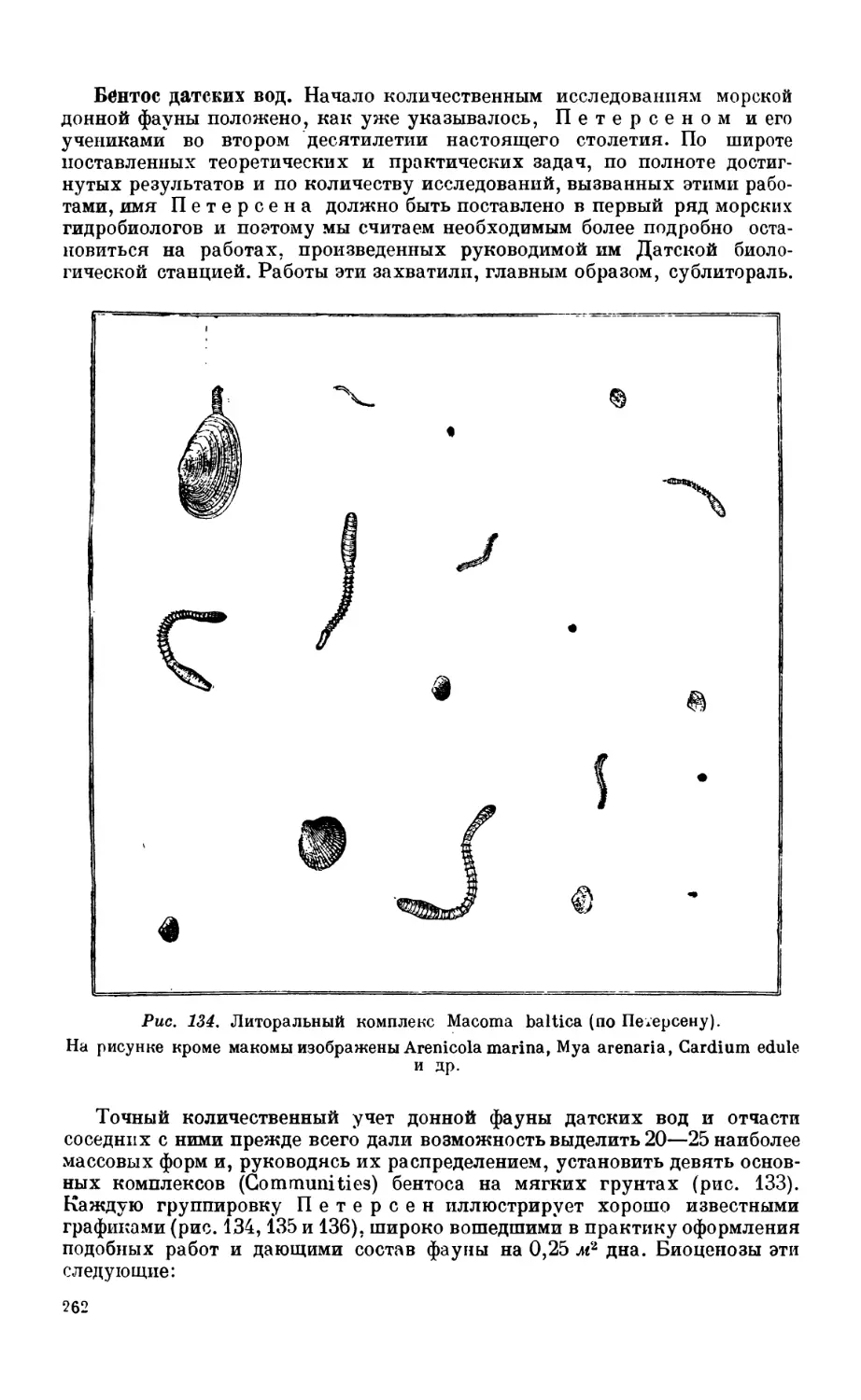
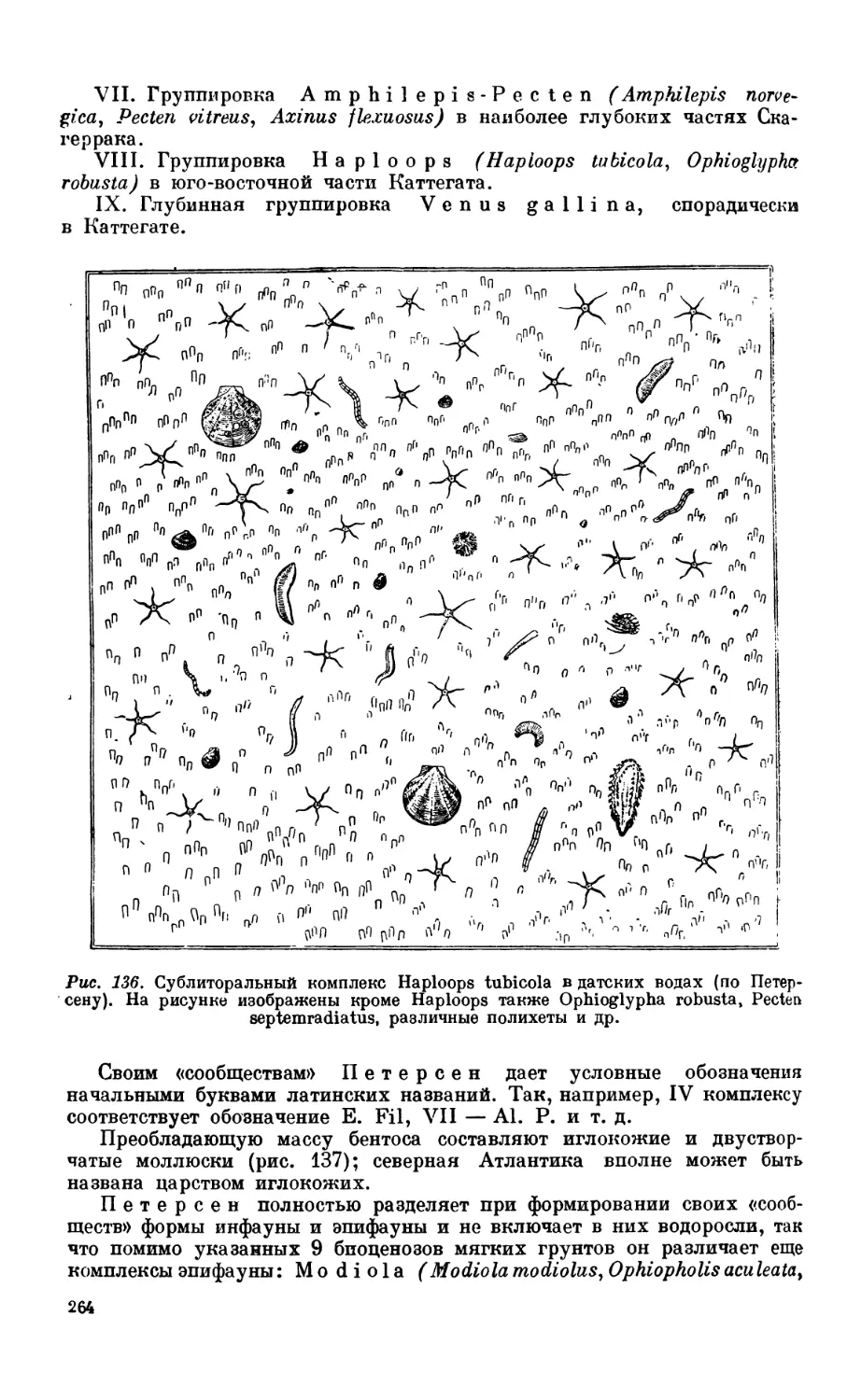
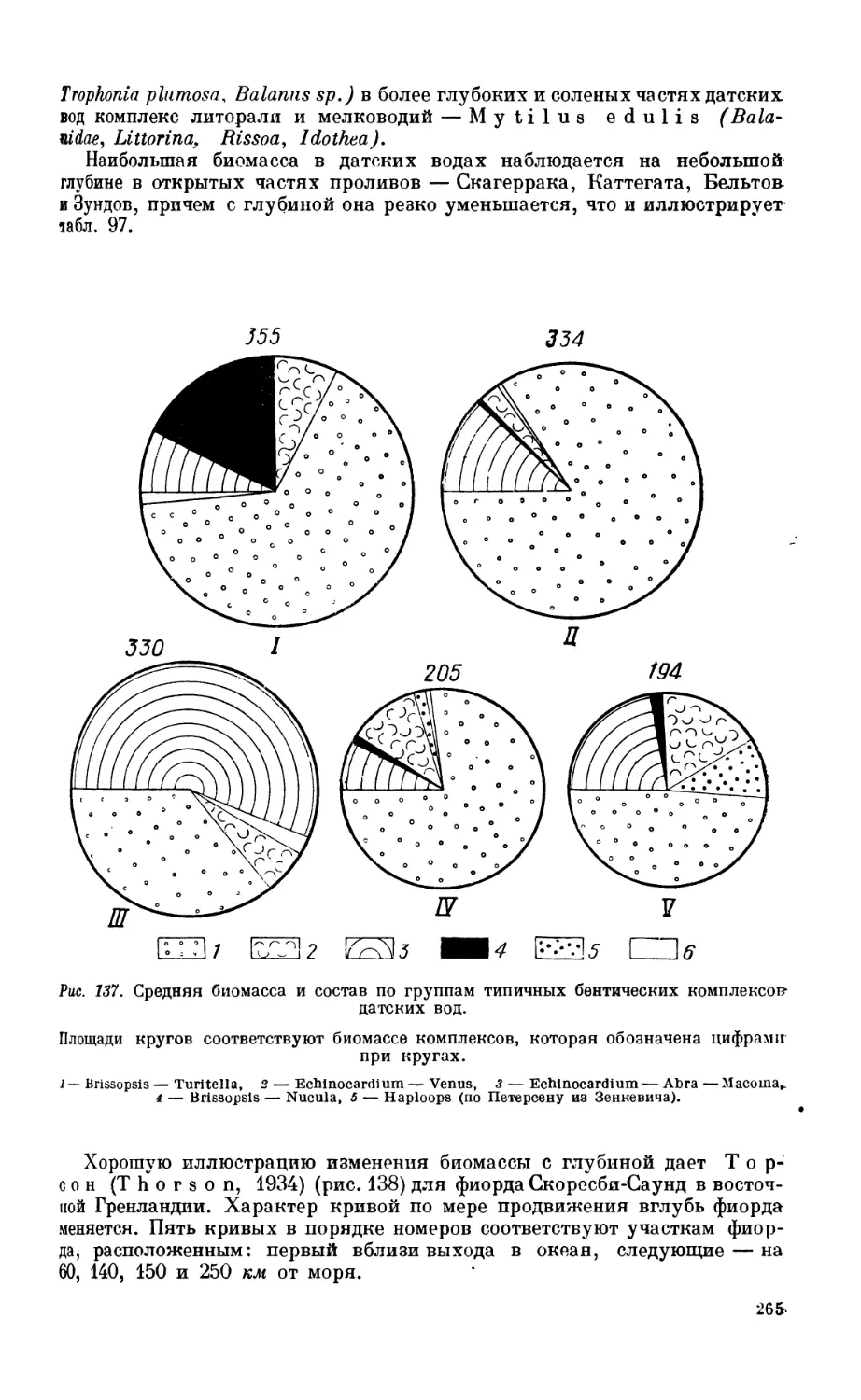
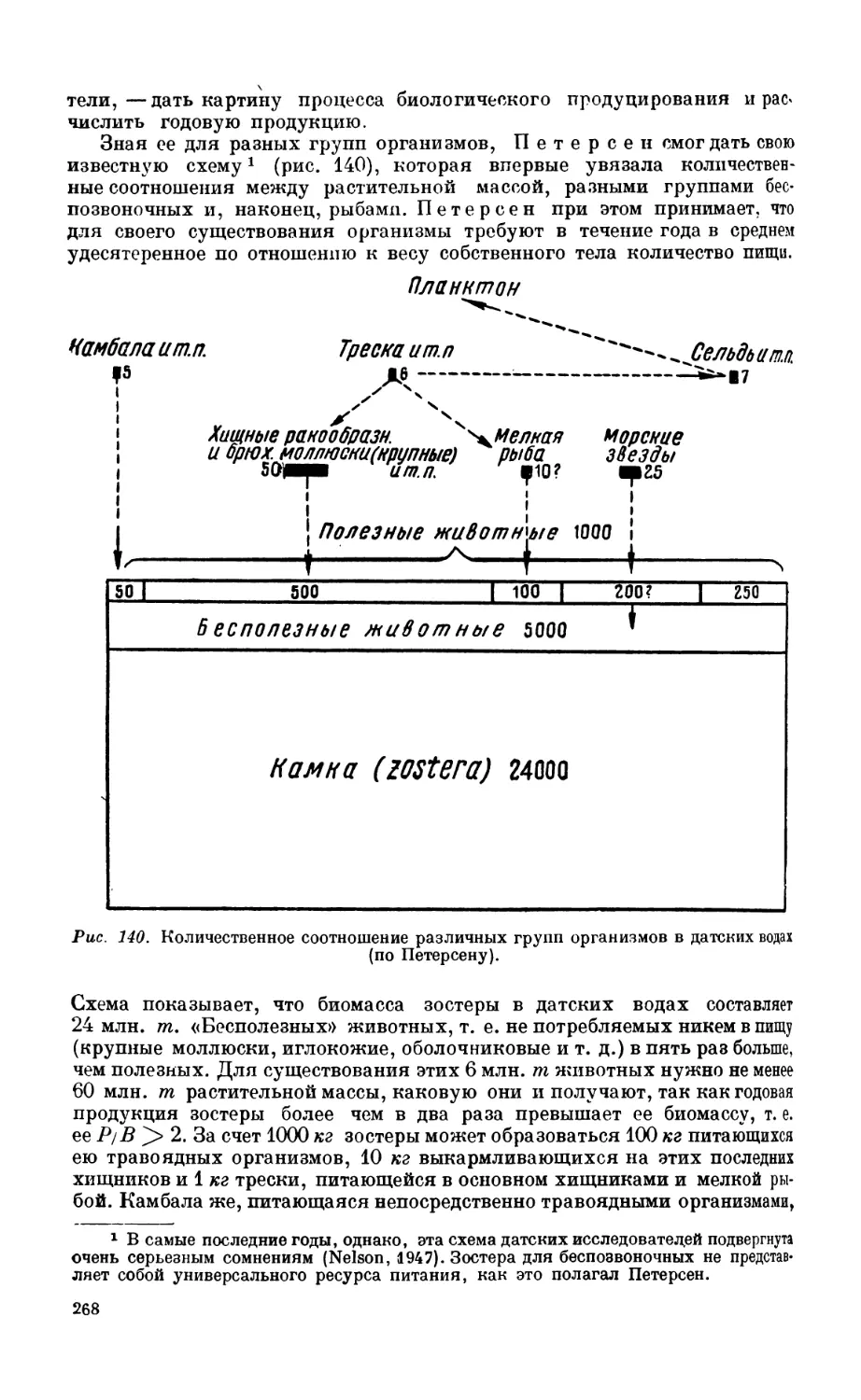
1. Сублиторальная фауна.................................................... 260
Сублиторальная зона—260. Бентос датских вод — 262. Бентос Северного моря
и прилежащих вод—275. Бептос побережья Исландии — 281. Бентос восточно-
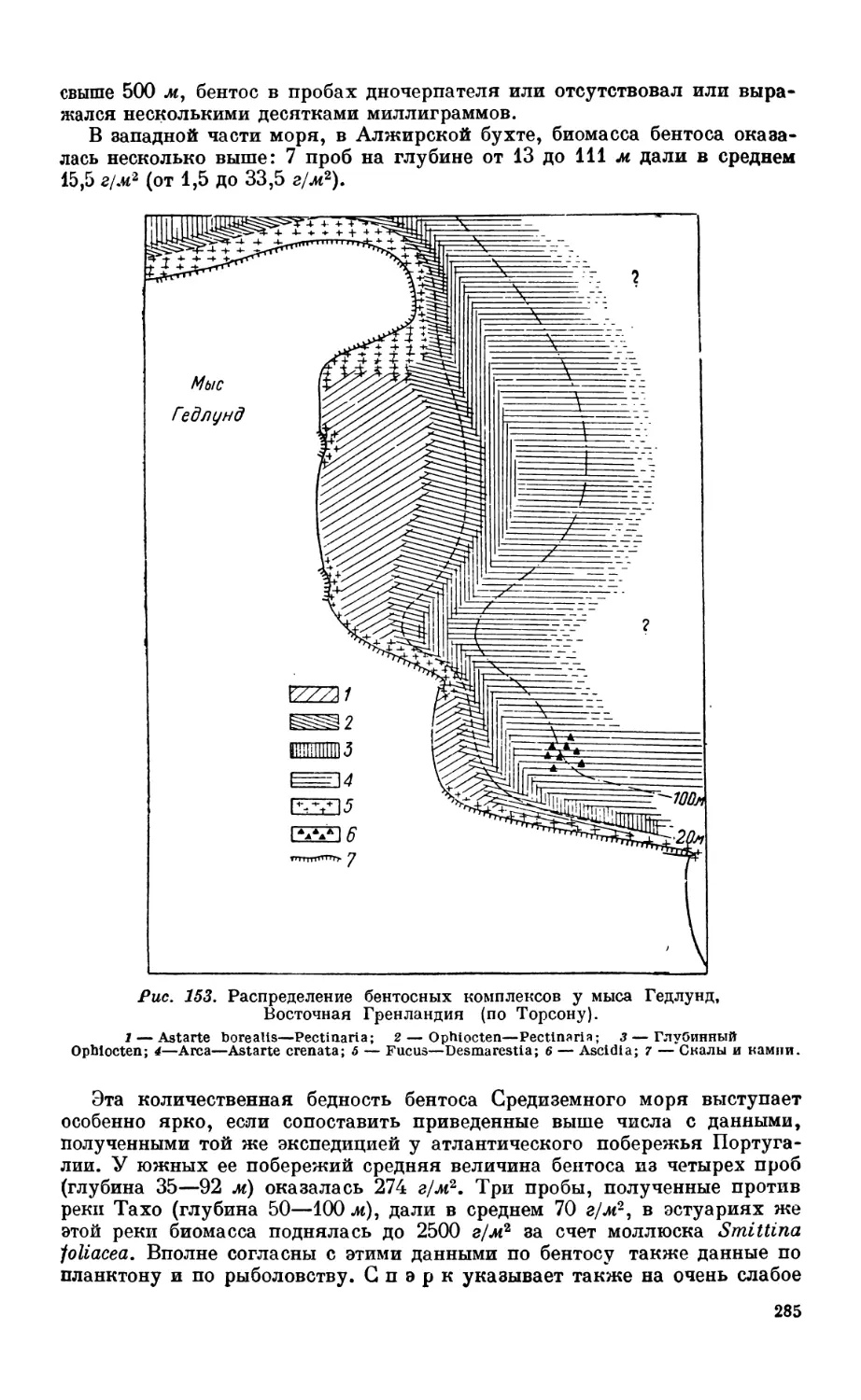
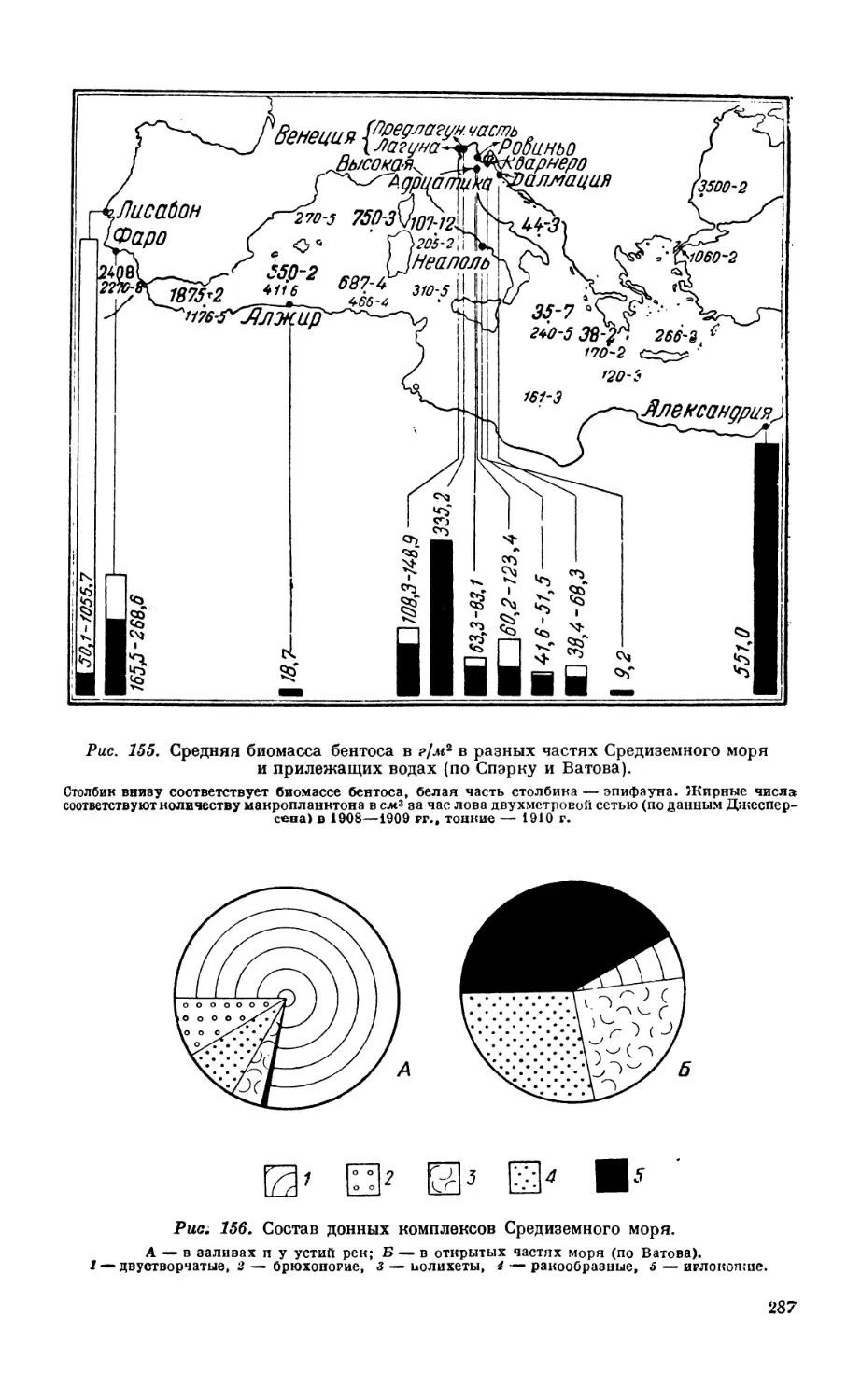
гренландского побережья—283. Бентос Средиземного моря — 284. Бентос неко-
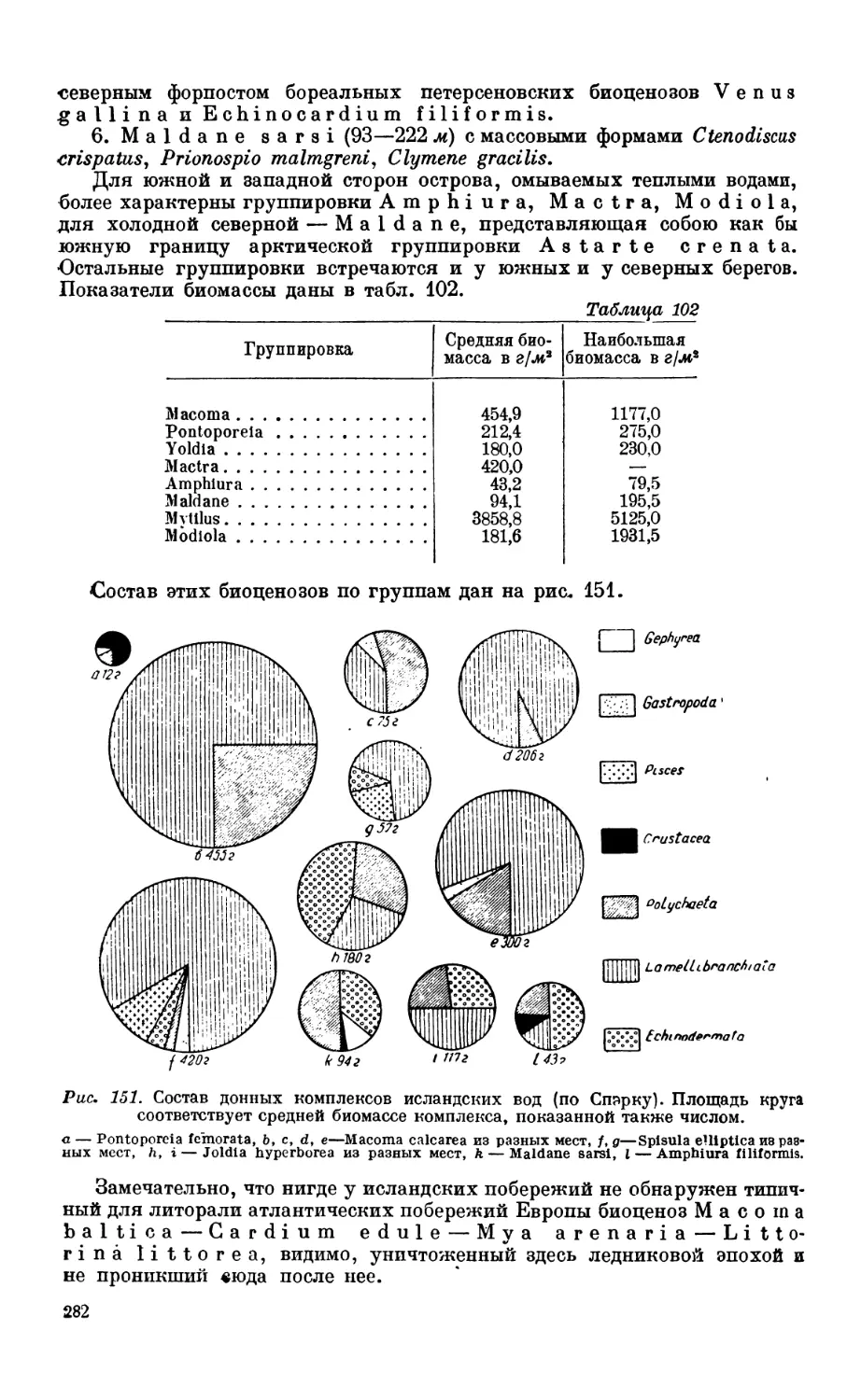
торых районов Тихого океана—288. Максимальная биомасса разных форм — 292.
7
Стрл
5. Фауна псевдоабиссали ...................................................... 294
6. Фауна батиали.............................................................. 294
Батиальная зона — 294. Состав фауны батиали — 295.
7. Фауна абиссали............................................................. 296
Зона абиссали—296. Состав и происхождение фауны абиссали—296. Пути эволю-
ции абиссальной фауны — 298.
Глава IX. бактерии
Значение бактерии в круговороте веществ в море — 301. Количество бактерий — 3<)2.
Химические процессы, протекающие при участии бактерий —304.
Глава X. океанические птицы............................................ ЗОС
Глава XI. СОЛОНОВАТЫЕ водоемы и их фаун а
Обеднение морской й пресноводной фаун в солоноватых водах — 309. Условия су-
ществования в солоноватых водах—312. Рост животных в солоноватых водах—3.
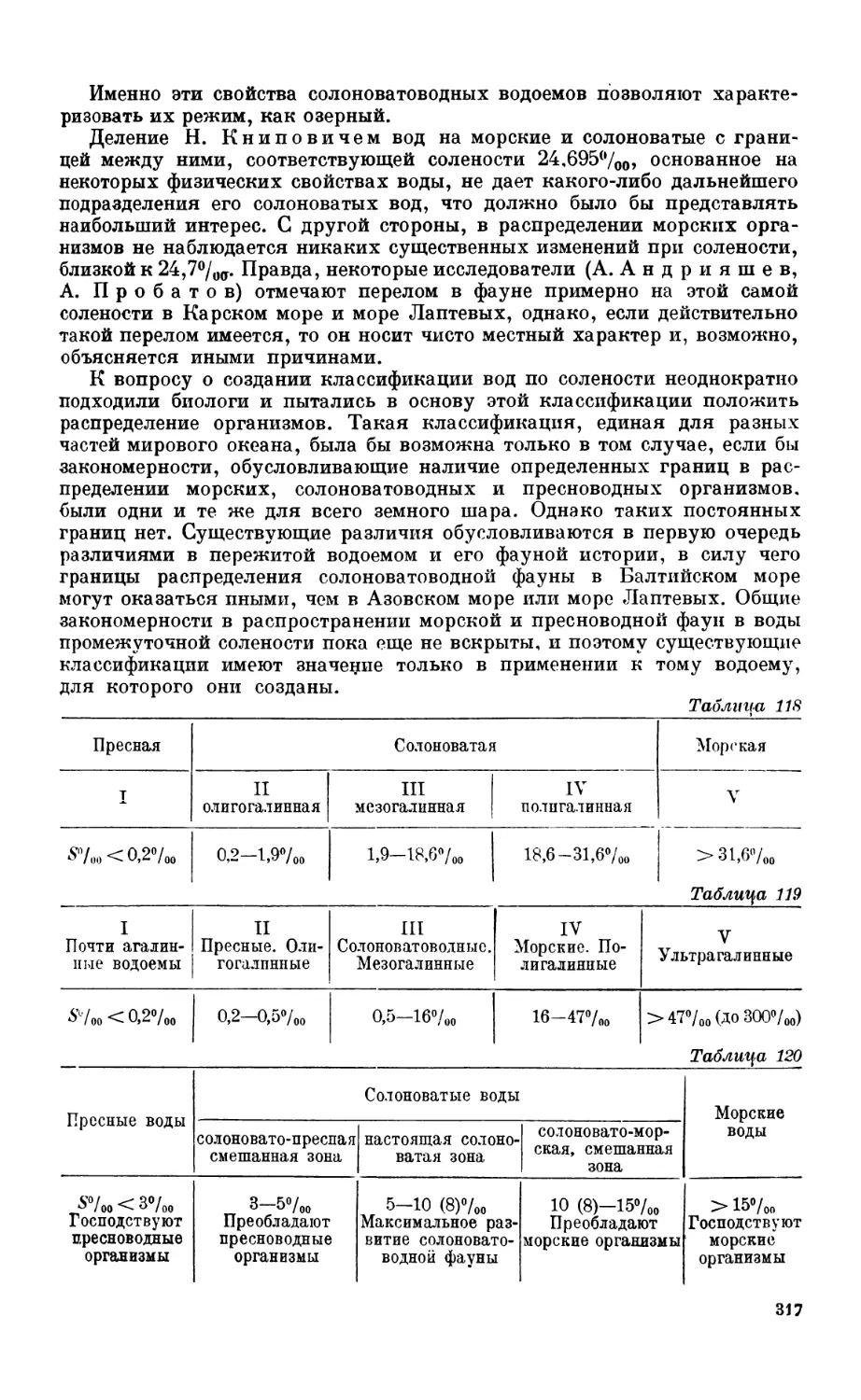
Специфическая солоповатоводная фауна—314. Классификация солоноватых вод—313.
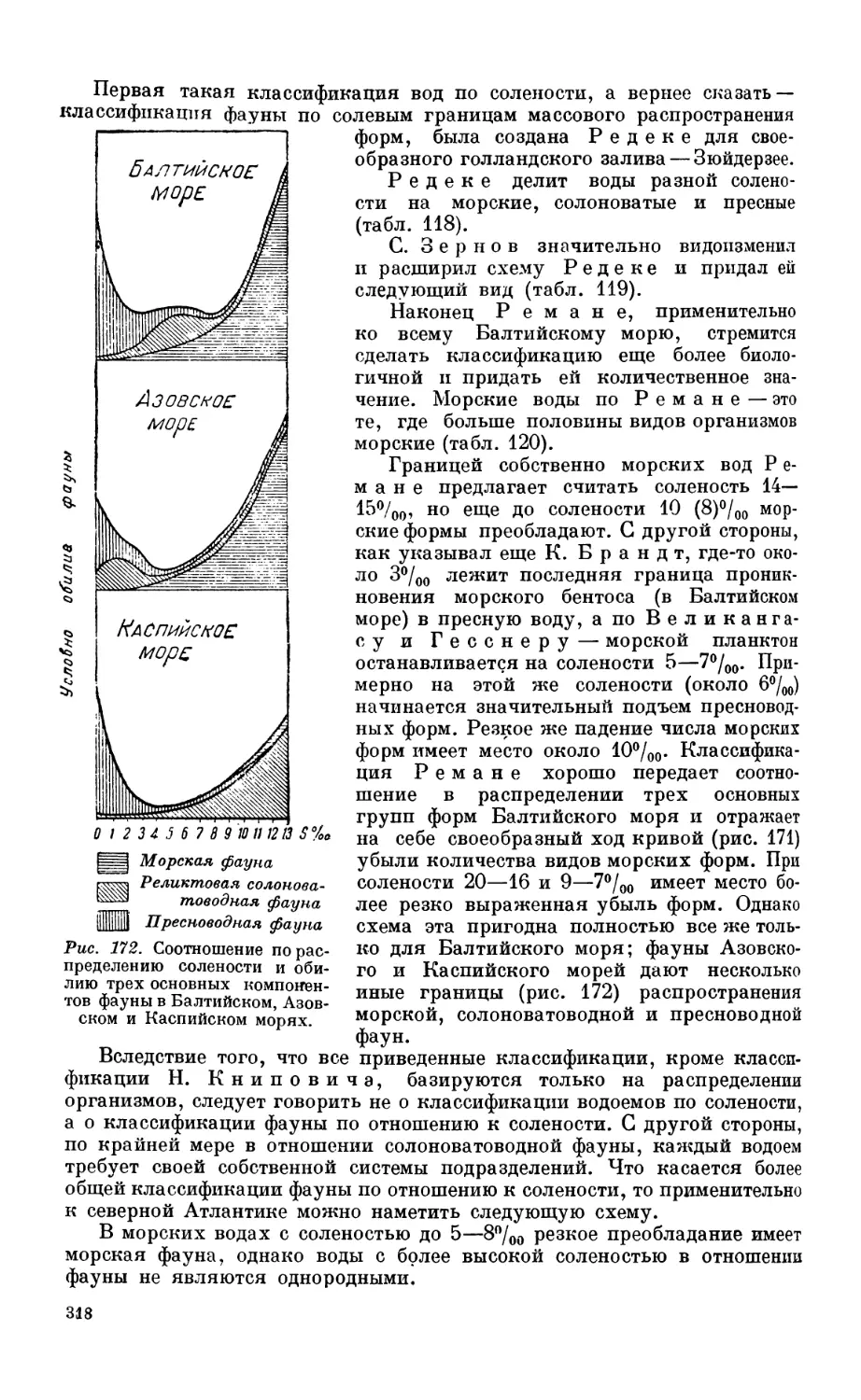
Глава XII. ПИТАНИЕ МОРСКИХ животных, преимущественно рыб
I. История и методика исследовании.......................................... 2С
Методы изучения питания беспозвоночных—320. Методы изучения питания рыб— /2'
Советские исследования питания рыб — 322.
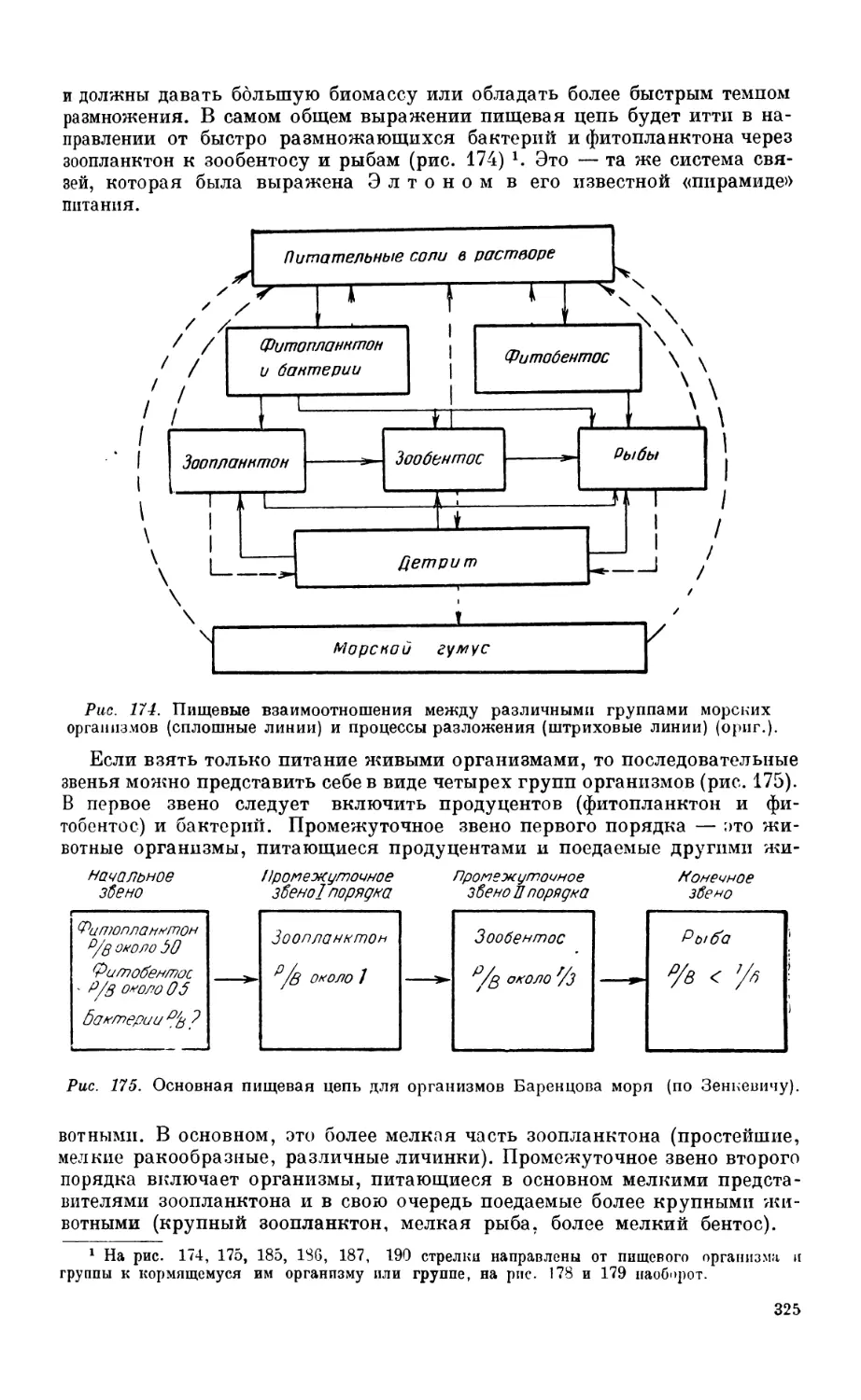
2. Общие закономерности пищевых взаимоотношении в морс....................... 32
Постановка вопроса—324. Пищевые отношения и P/В коэффициент — 324.
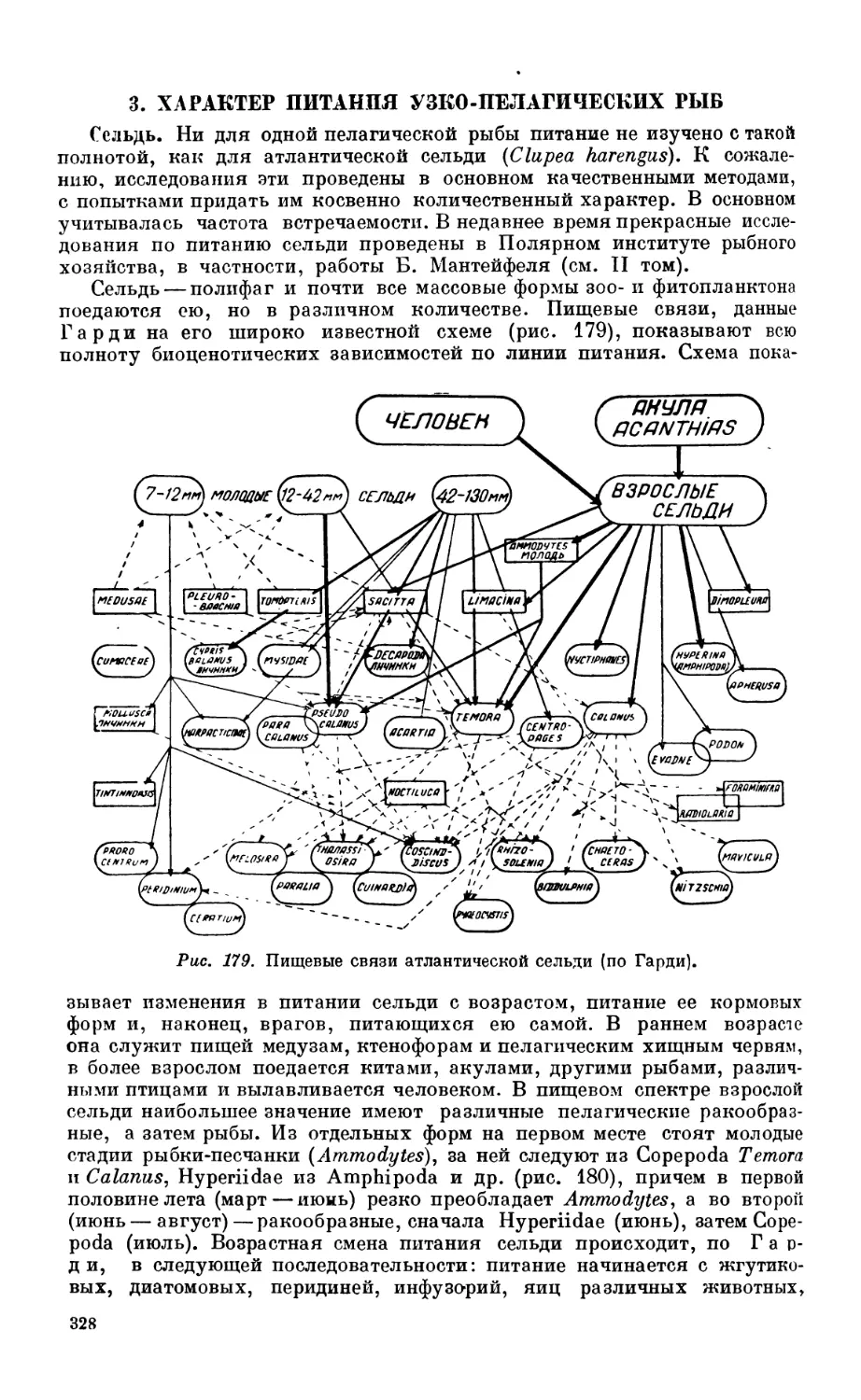
3. Характер питания узко-пелагических рыб................................... 32.
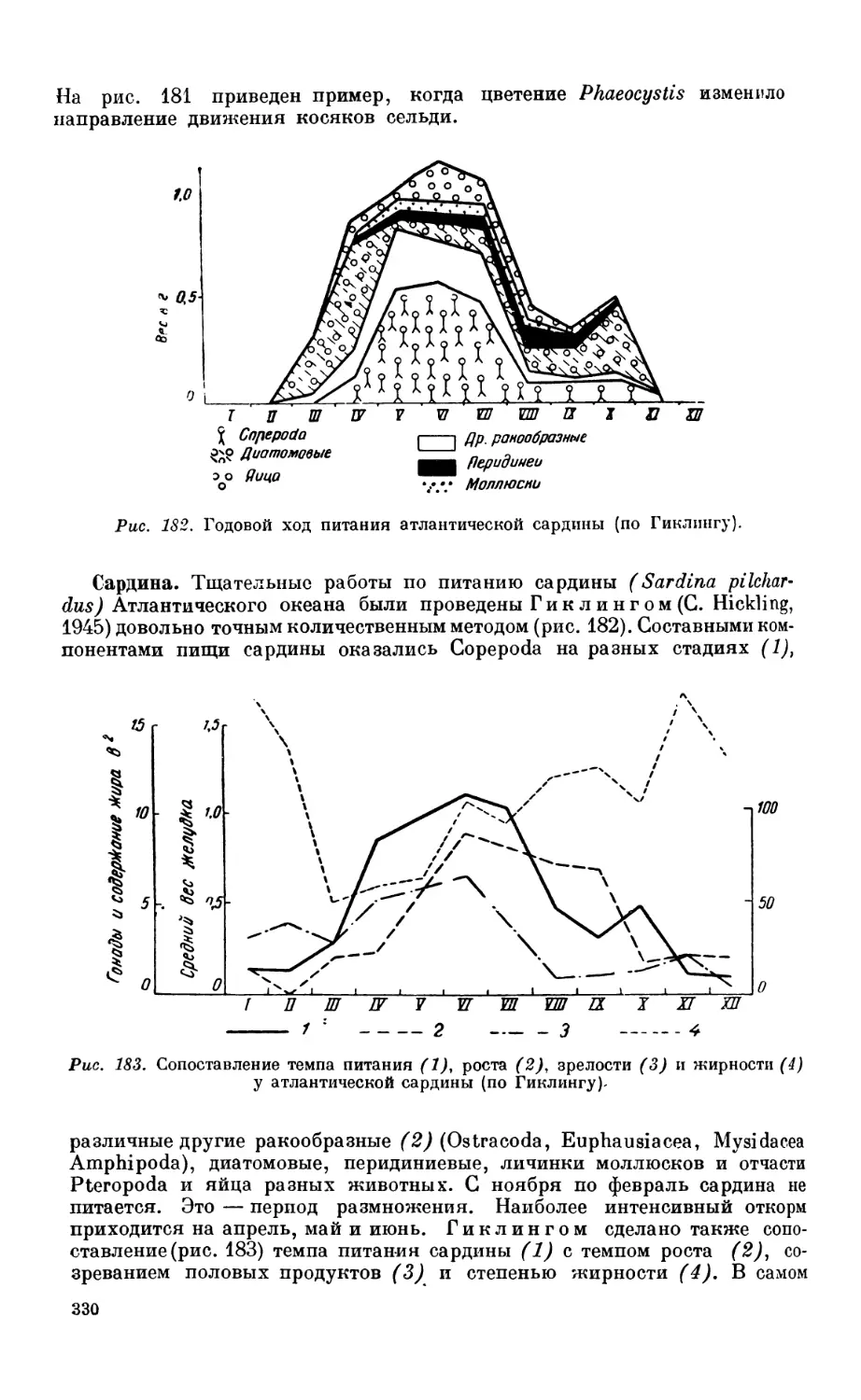
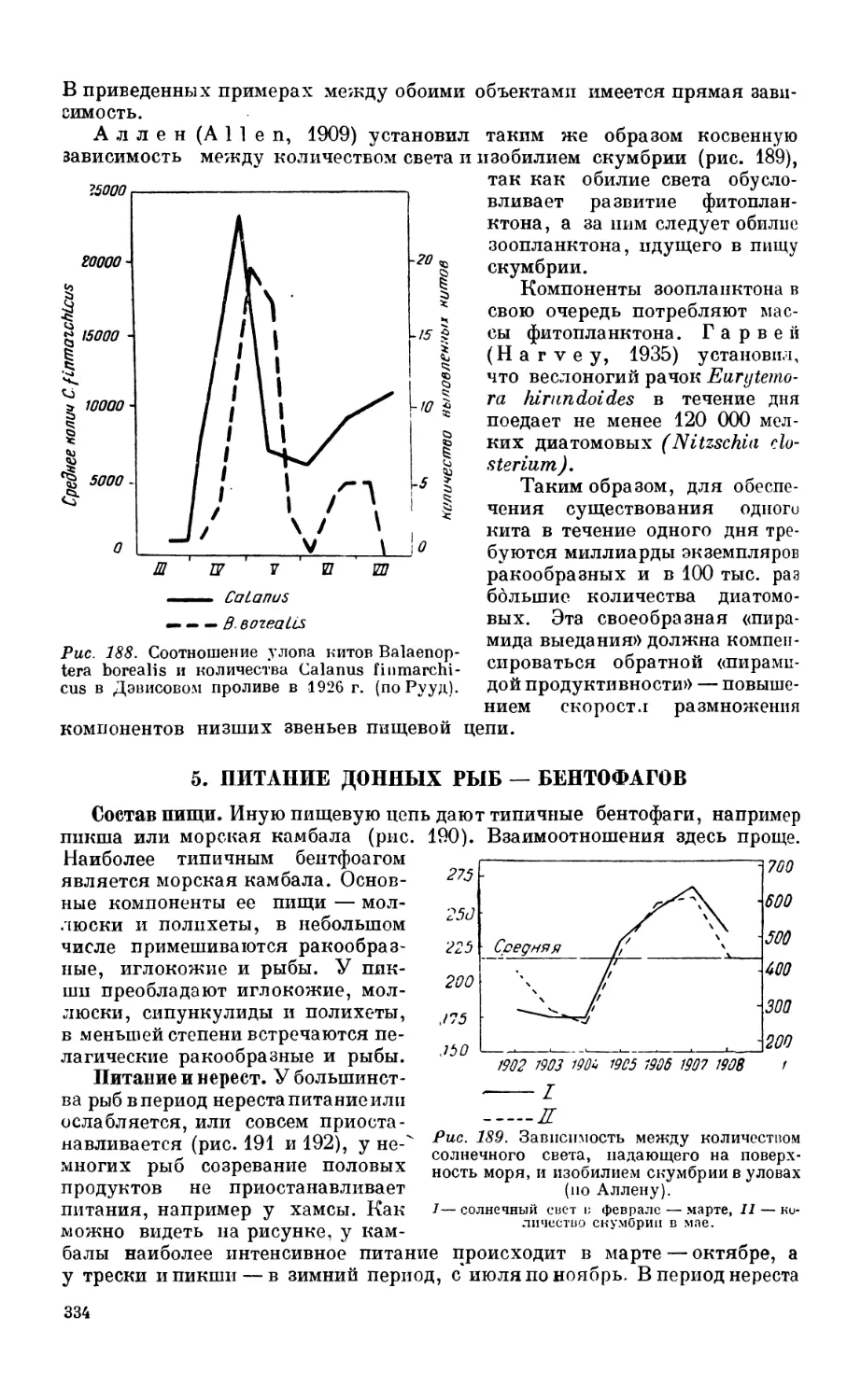
Сельдь — 327. Сардина — 330.
4. Характер питания гаироко-пелагических рыб................................ 331
Треска—331. Neoplatyccphalus—332. Пищевая цепь пелагиали—332.
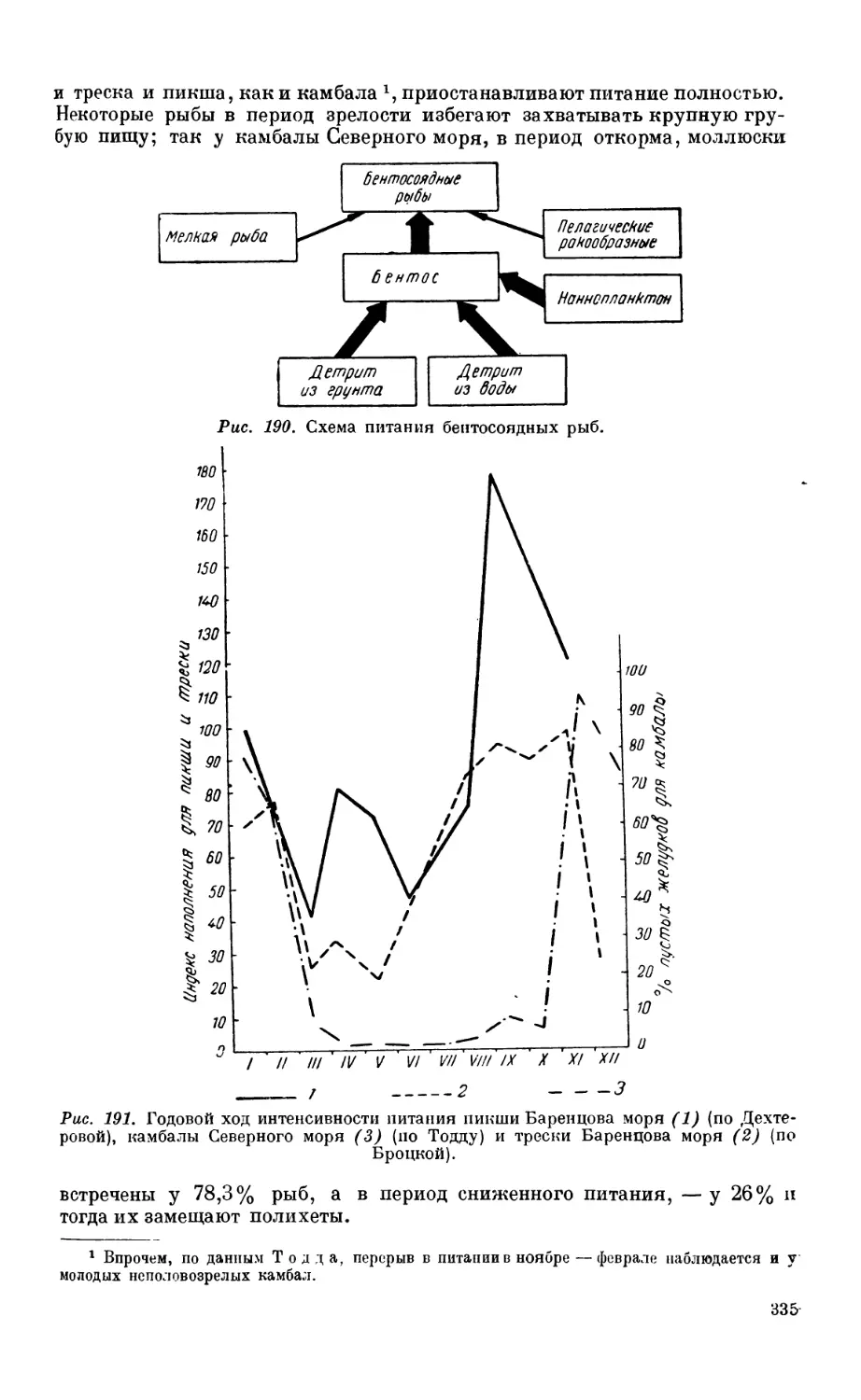
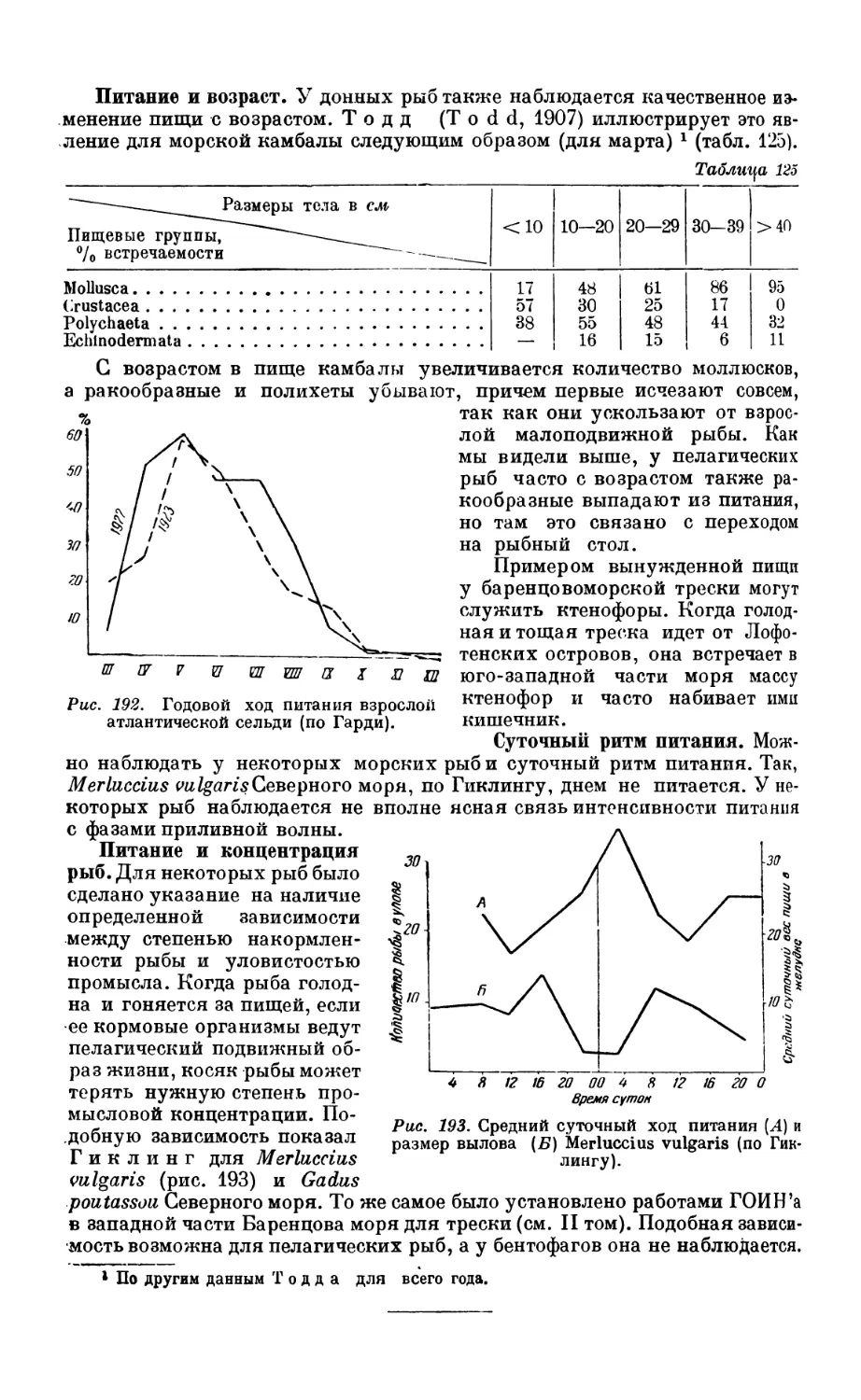
5. Питание донных рыб — бентофагов ........................................ 334
Состав пищи—334. Питание и перест—334. Питание и возраст — 336. Суточный
ритм питания—336. Питание и концентрация рыб — 336.
ЧАСТЬ Ч Е Т В Е Р Т А Я
БИОГЕОГРАФИЯ МИРОВОГО ОКЕАНА
Глава XIII. история, предмет п метод биогеографии моги
1. Введение.......................................................................33°
2. История изучения.............................................................. 340
Первые исследования—340. Работы Дана—340. Историческая школа второй по-
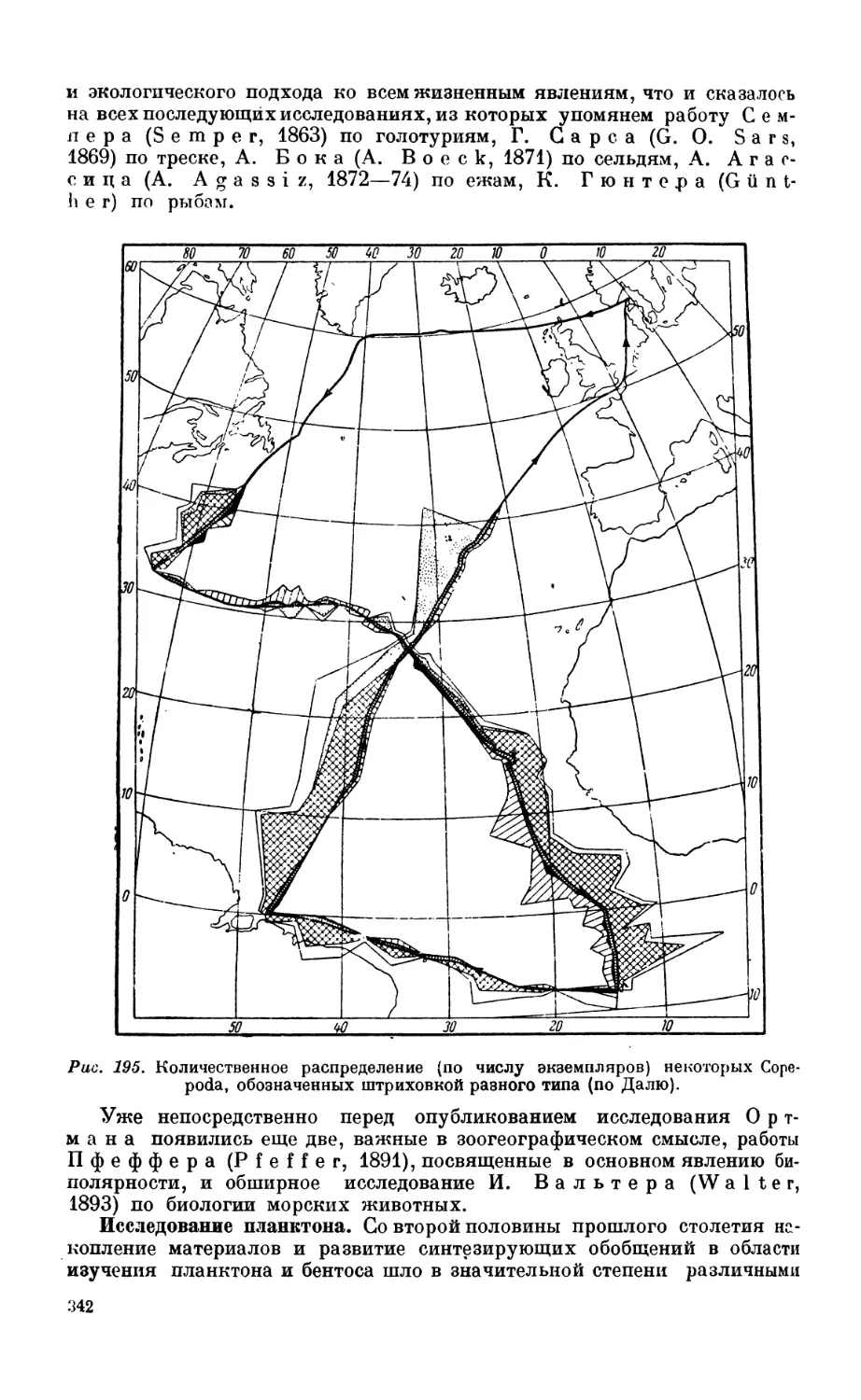
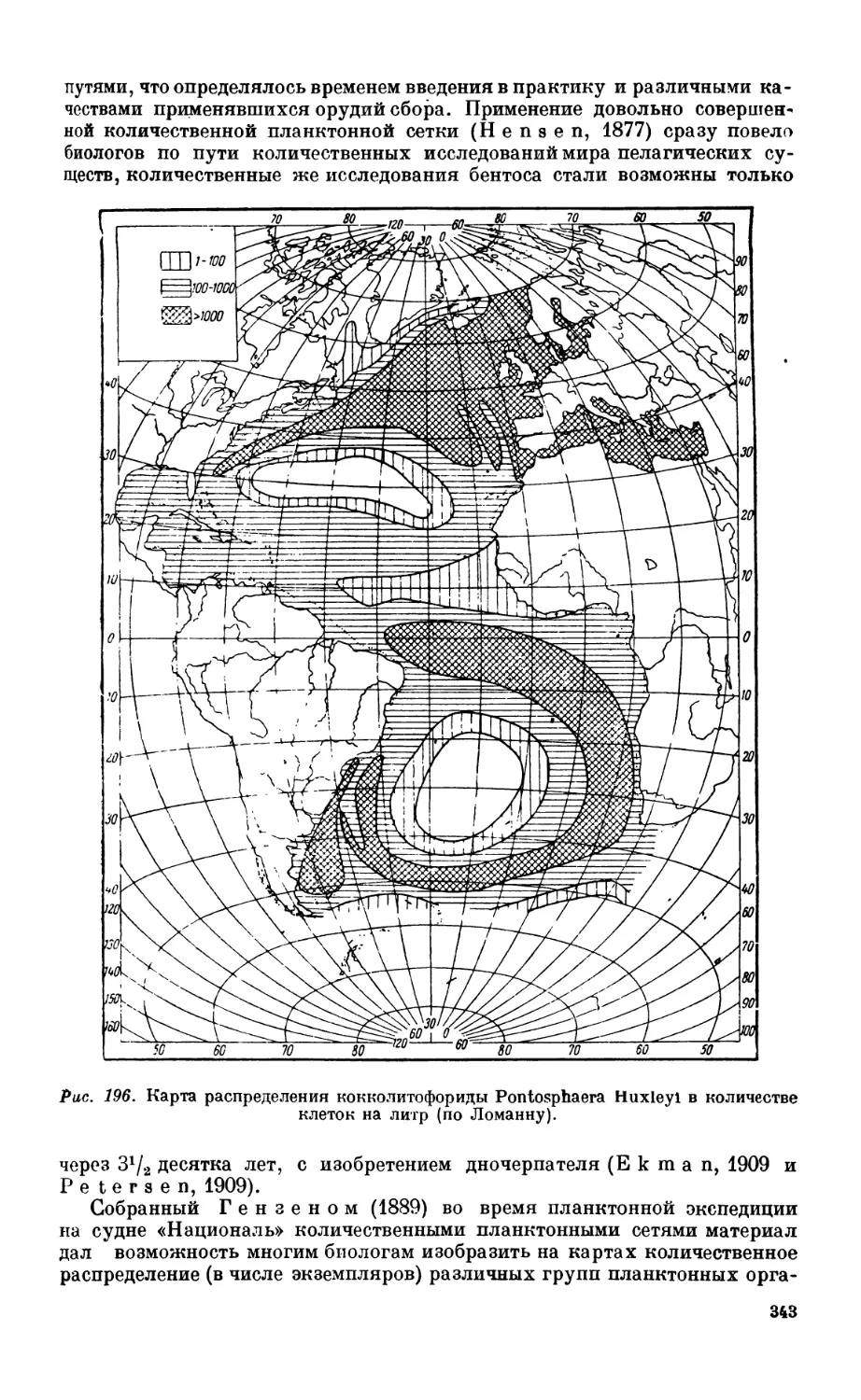
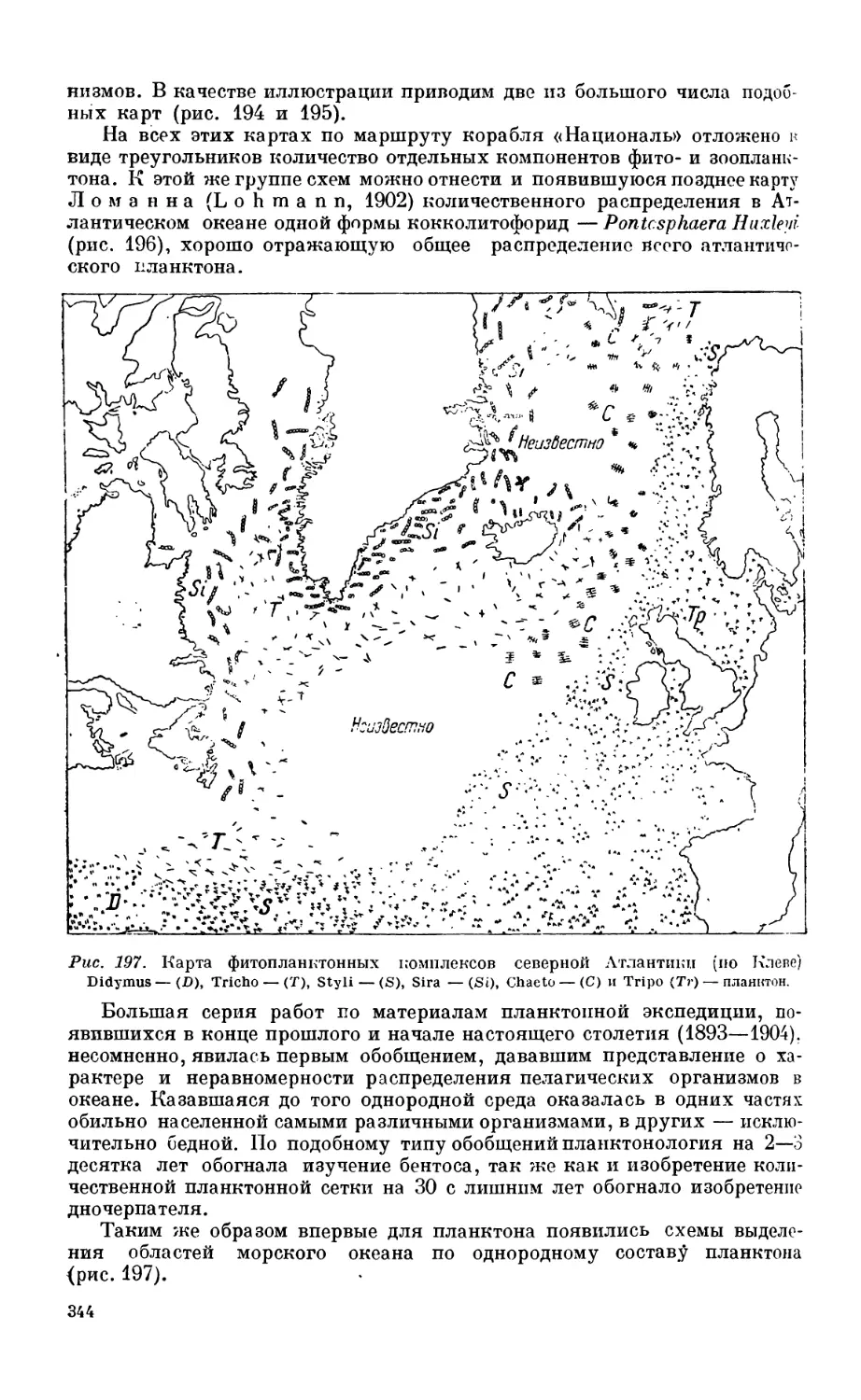
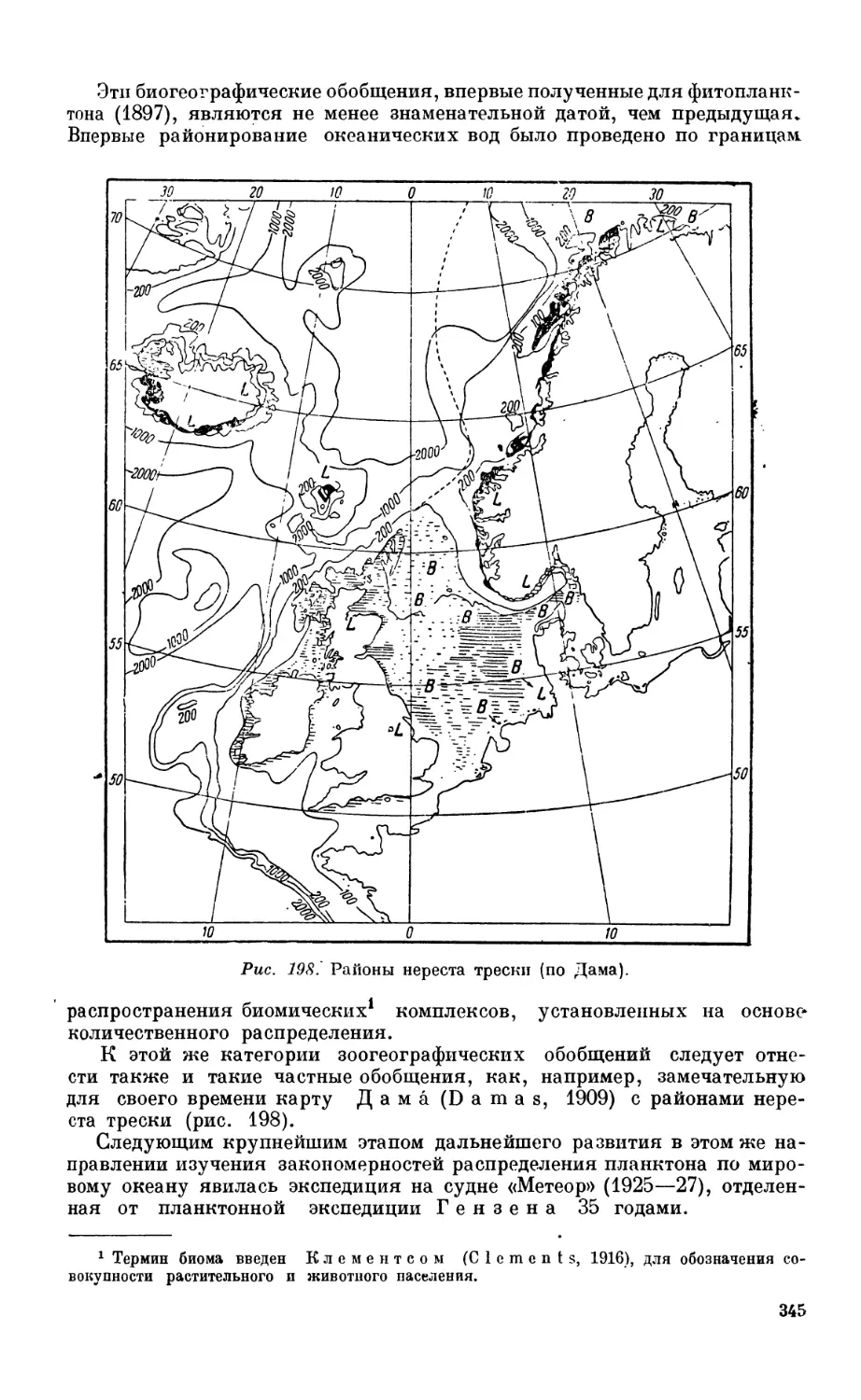
ловины XIX века —341. Исследование планктона — 342. Исследование бентоса — 347.
3. Биогеографическая зональность мирового океана................................. 352
Приполярные области — 352. Умеренные зоны—353. Экваториальная зона—353.
Биологическая асимметрия —353.
4. Основные проблемы биогеографии моря........................................... 353
Роль количественного метода —353. Проблемы регистрирующей зоогеографии — 354.
Проблема реконструкции фауны— 354. Проблемы исторической зоогеографии —355.
5. Подразделения, объект и метод биогеографии моря............................... 356
Задачи — 356. Подразделения — 356. Объект — 357. Методы — 358.
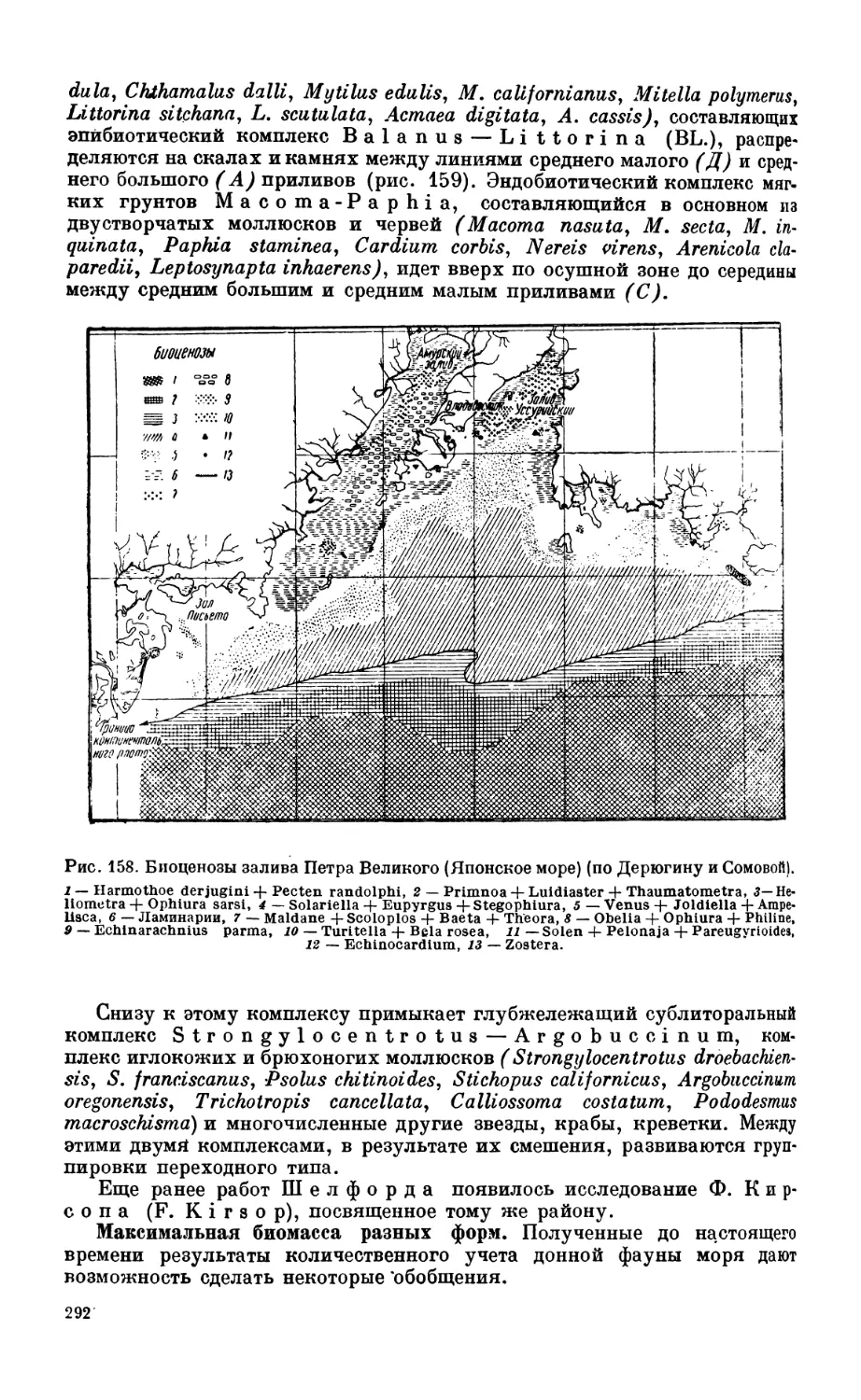
Глава XIV. биогеографическое районирование
1. Деление оксана на зоогеографические области ................................ 359
Схема Ортманна —359. Несовпадение зоогеографических границ для литорали, пе-
лагиалп и абиссали —359. Схема И. Шмидта — 361.
2. Современное положение районирования мирового океана......................... 361
Дополнение к схеме Ортманна — 361. Биогеографическое районирование пела-
гиалп—362. Схема Экмана—363. Районы резких смен фауны—363. Биогеогра-
фические критерии — 366.
3. Основные факторы, определяющие распространение организмов в океане.......... 367
Общие положения —367. Температура —368. Течения — 368.
4. Зоогеографическая принадлежность морей СССР................................. 369
Северные моря—369. Балтийское море — 370. Черное море — 370. Дальневосточ-
ные моря — 370.
8
Стр.
Глава XV. ареалы морских организмов
1. Тиш ареалов ............................................................. 372
Космополиты — 372. Формы с ограниченным ареалом—373. Эндемики—374. Осо-
бенности биогеографической опенки планктона — 375.
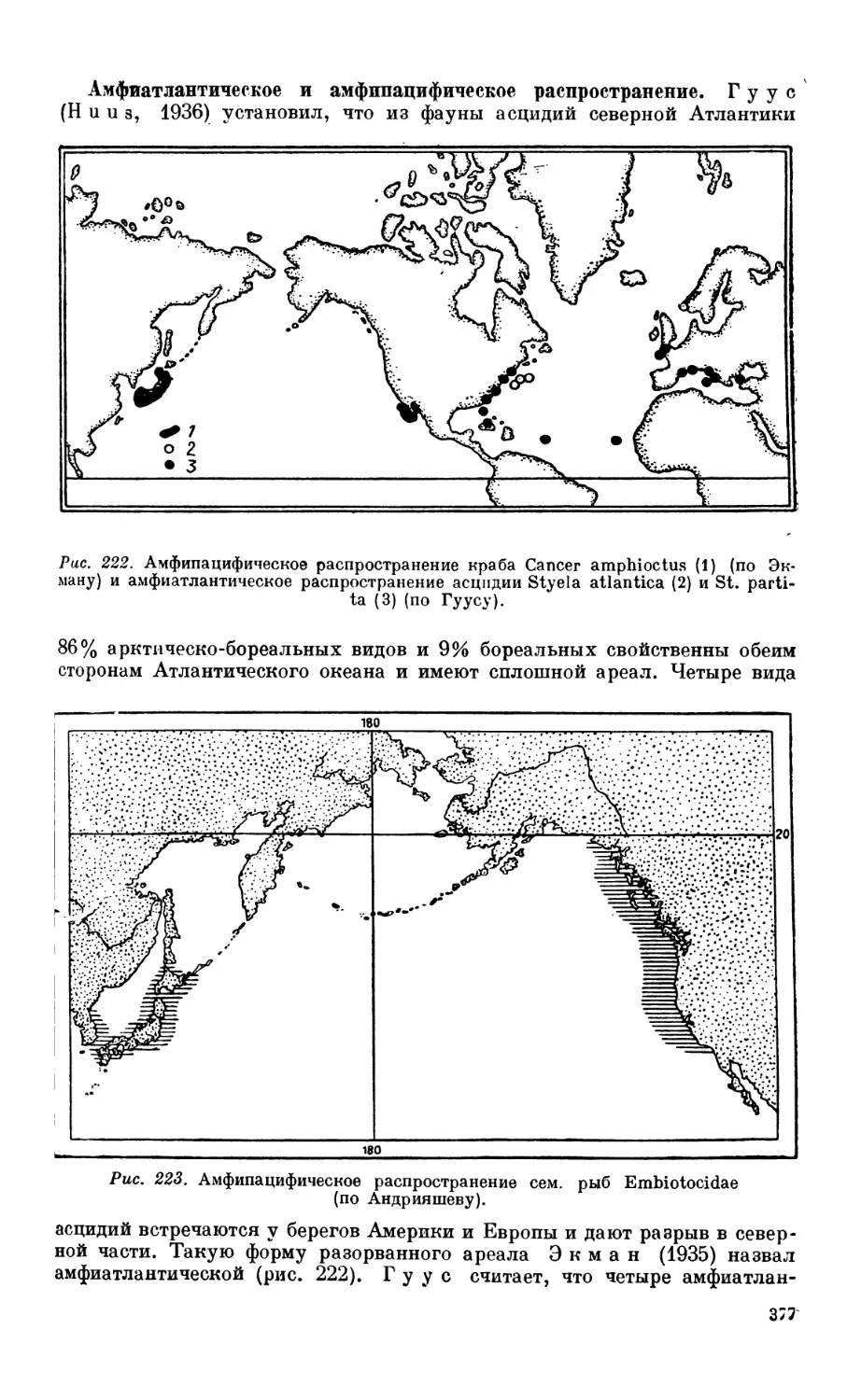
2. Широтные разрывы ареалов в северном полушарии.............................375
Сходство фауны Атлантического и Тихого оксанов — 375. Амфиатлаитпческое п
амфипацифпческое распространение — 377. Причины разрывов ареалов — 378.
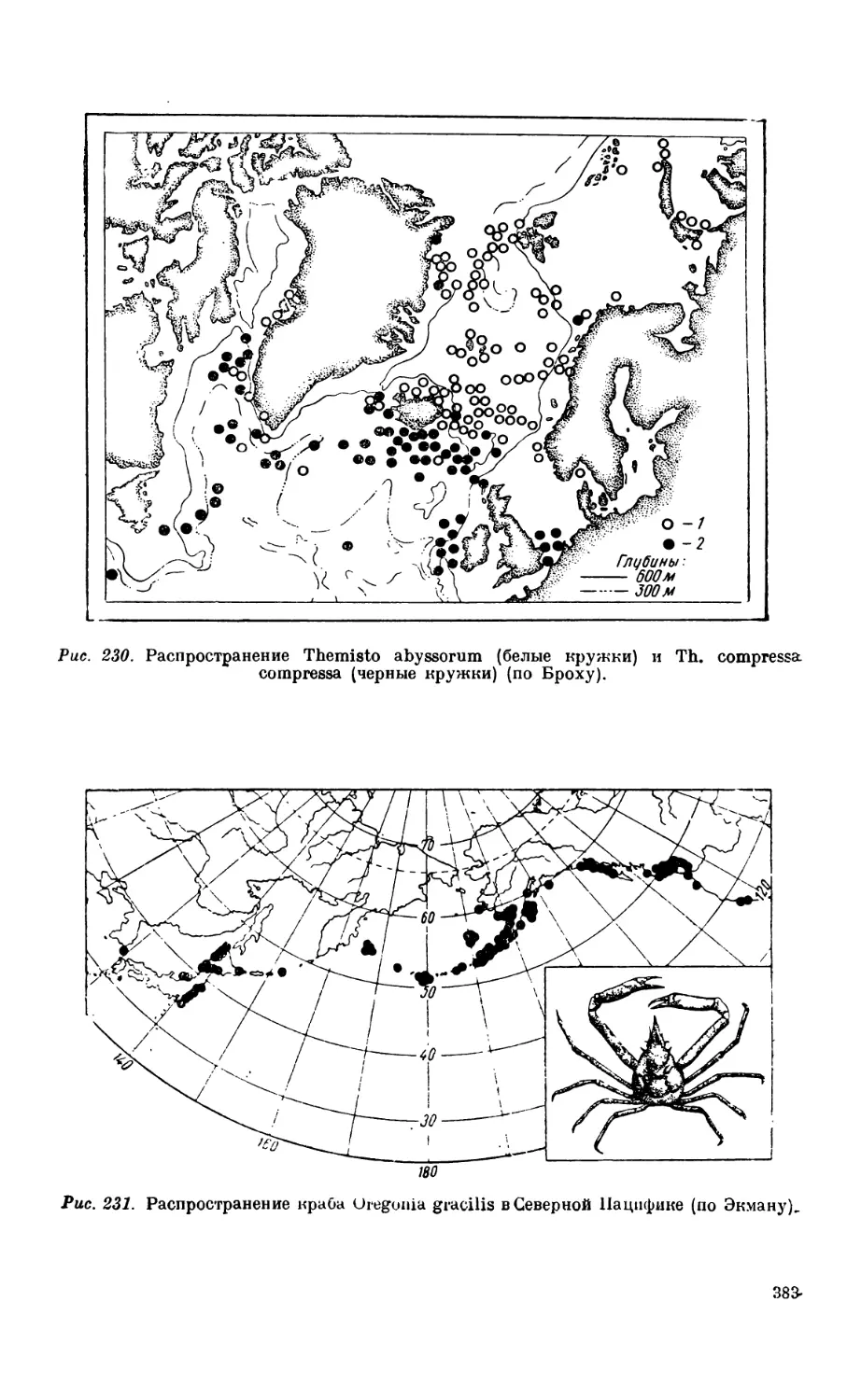
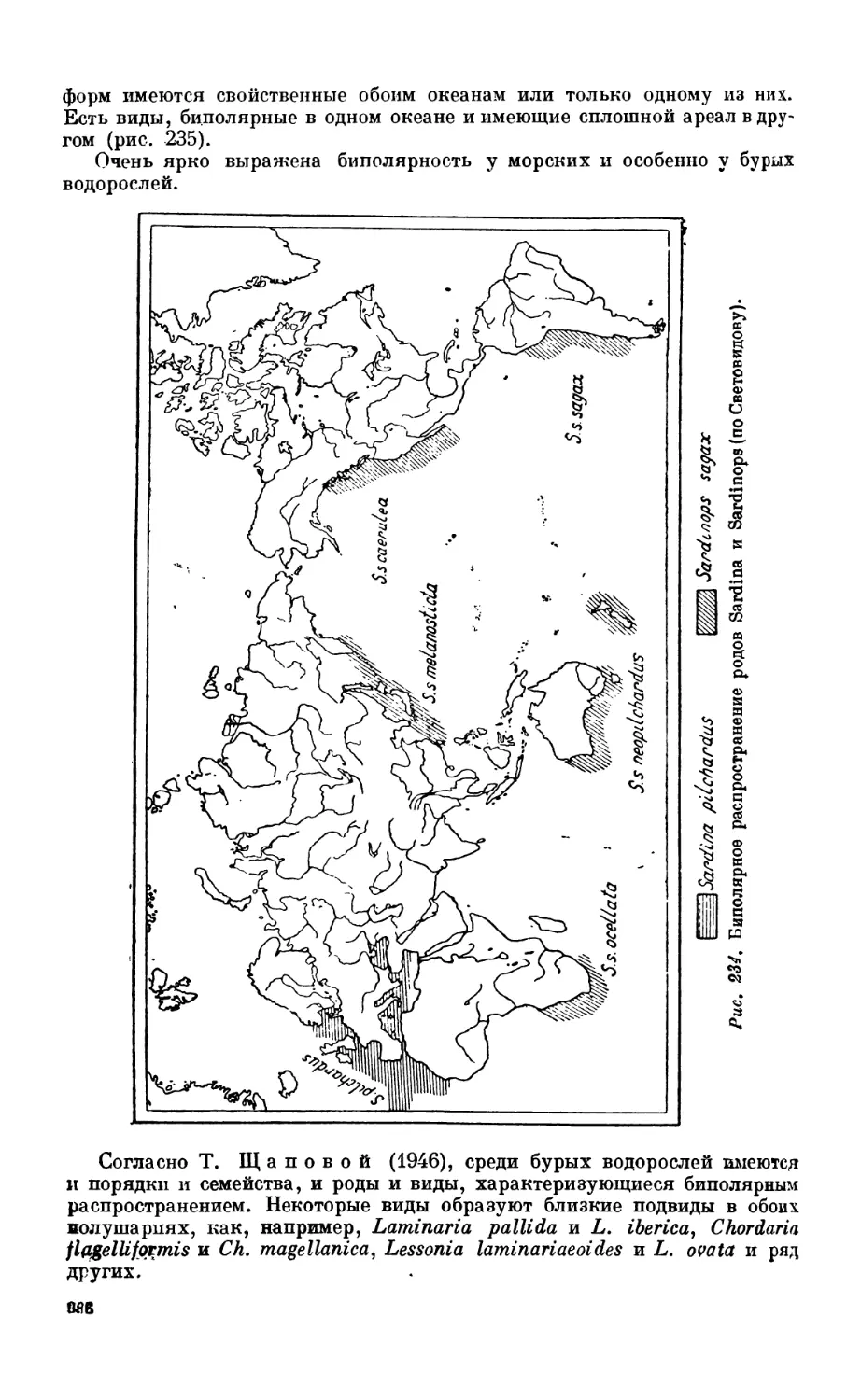
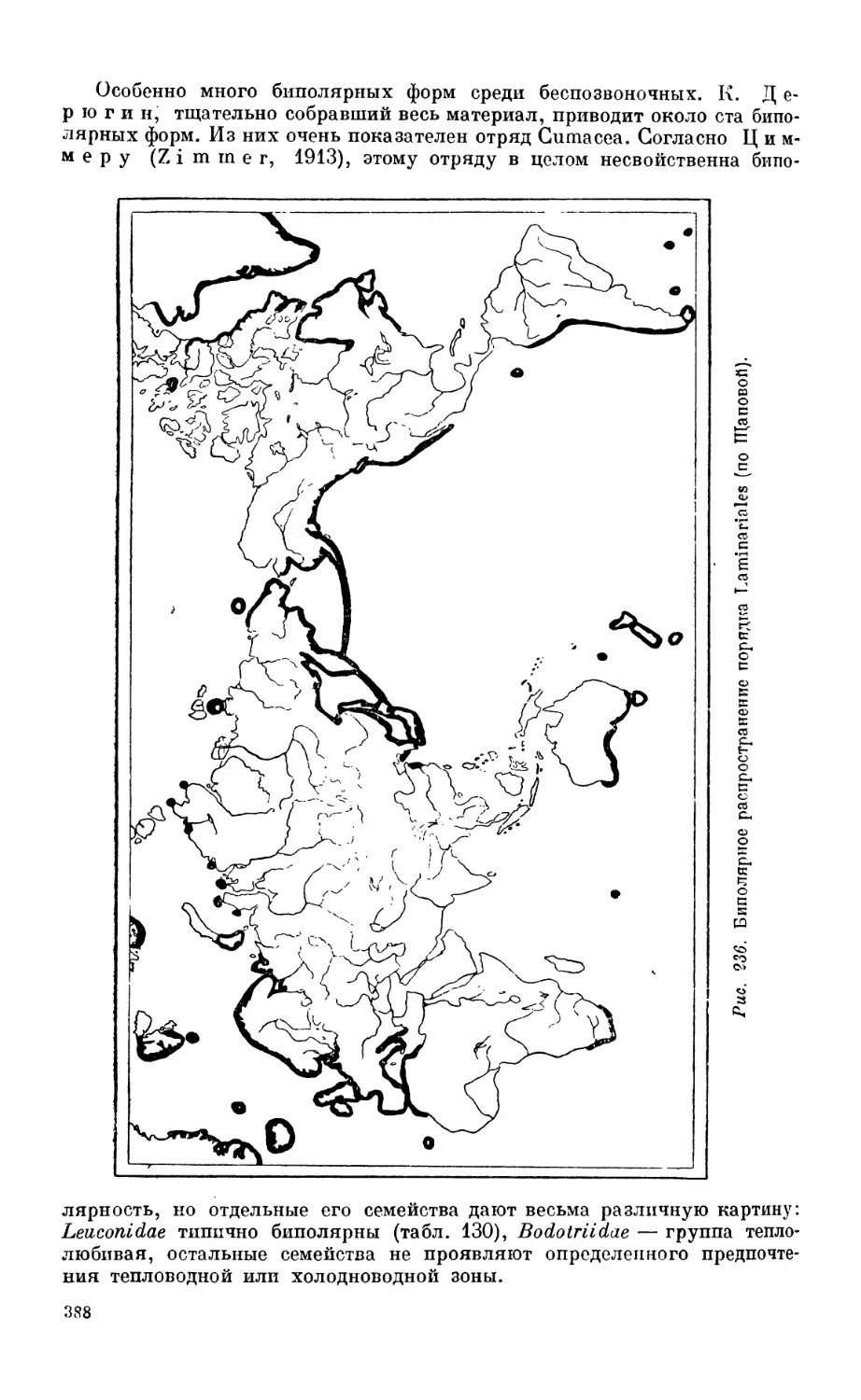
3. Биполярность............................................................. Зб5
Биполярное распространение организмов — 385.Происхождение биполярности —383.
Биологическая биполярность — 391.
4. Тропическая фауна ....................................................... 392
Общая характеристика и происхождение—292. Коралловые рифы—391. Мангро-
вые заросли—395. Фауна Ппдо-вестпацг.фики — 393. Фауна тропической Аме-
рики— 397. Атлантическая тропическая фауна—400. Саргассово море—нЮ.
’). Фауна Средиземного моря................................................. 402
Зоогеографическое положение—402. Тропические элементы — 402. Бореальные
эчемеиты—402. Эндемики — 403. Термическая неоднородность фаупы — 4(>3. Исто-
рия фауны—404.
3. Другие области оксана.................................................... *06
Ч А С Т Ь II :1 Т \ Я
ПРАКТИЧЕСКОЕ ЗНАЧЕНИЕ МОГСКПХ ЖИВОТНЫХ II РАСТЕНИИ
Глава XVI. НРОМЫСЕ I МОРСКИХ организмов
1. Введение................................................................. 409
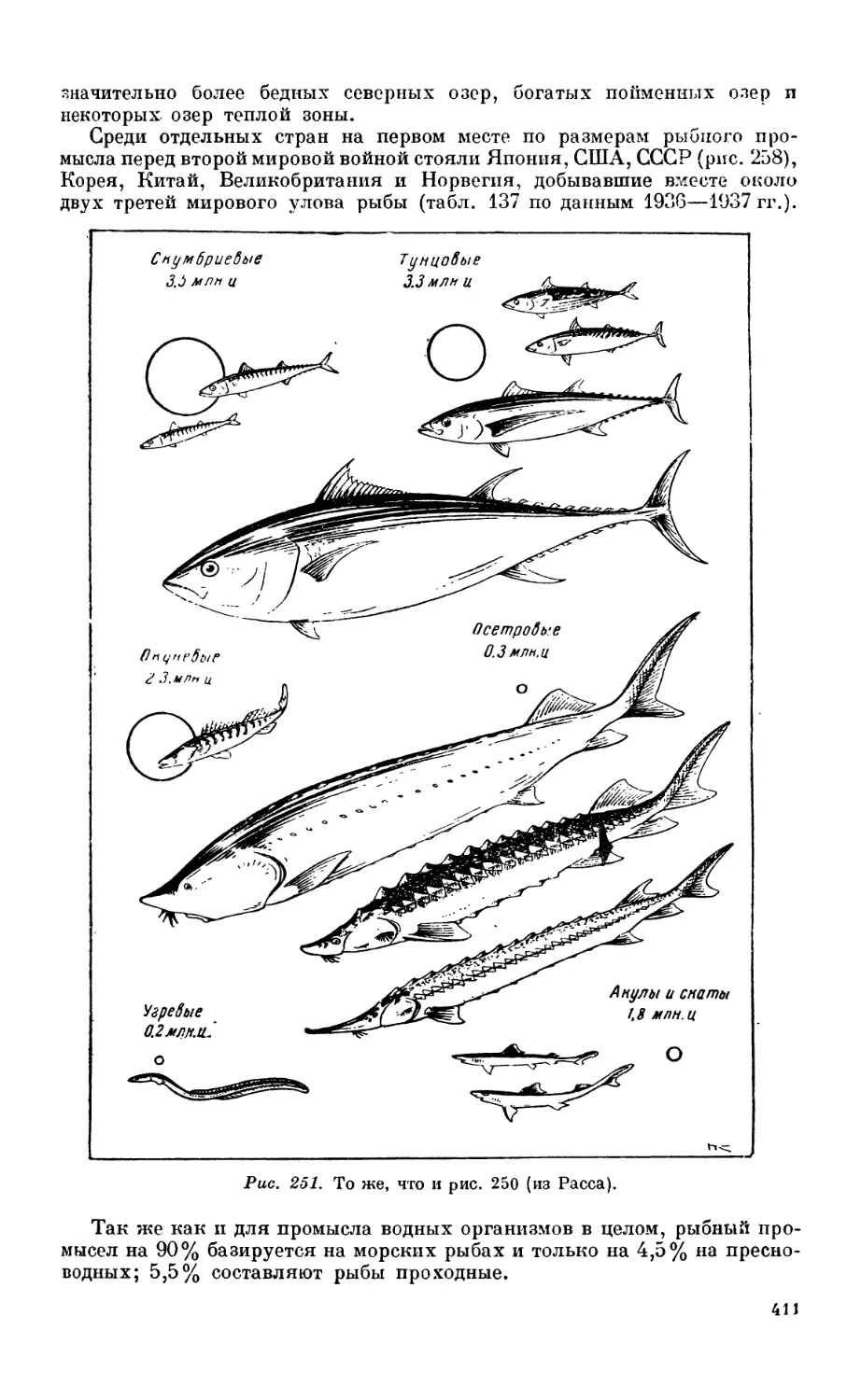
2. Промысел рыб............................................................. 410
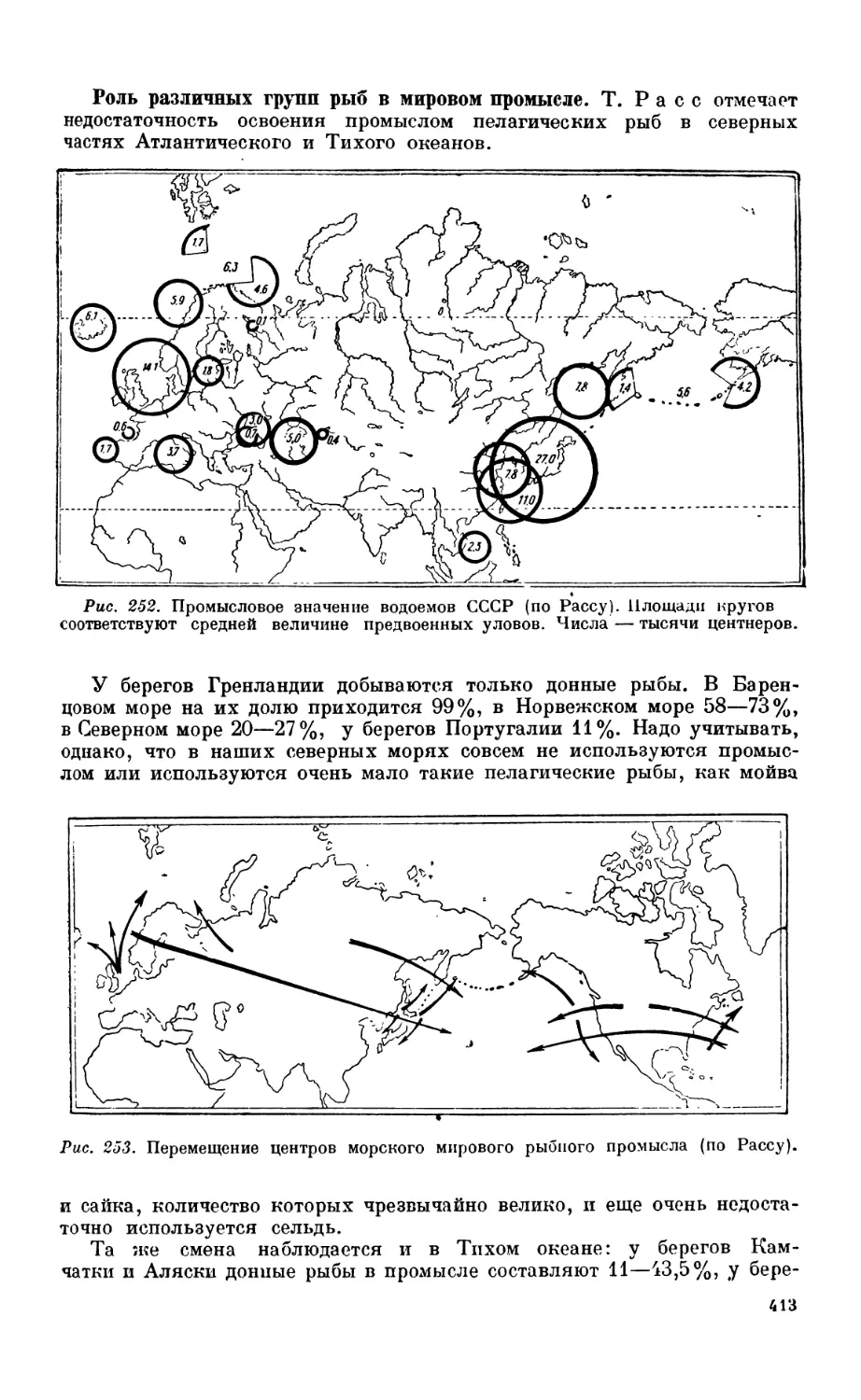
Общие данные — 410. Роль различных i рунп рыб в моровом промысле—413. Дина-
мика мирового рыбного промысла — 414.
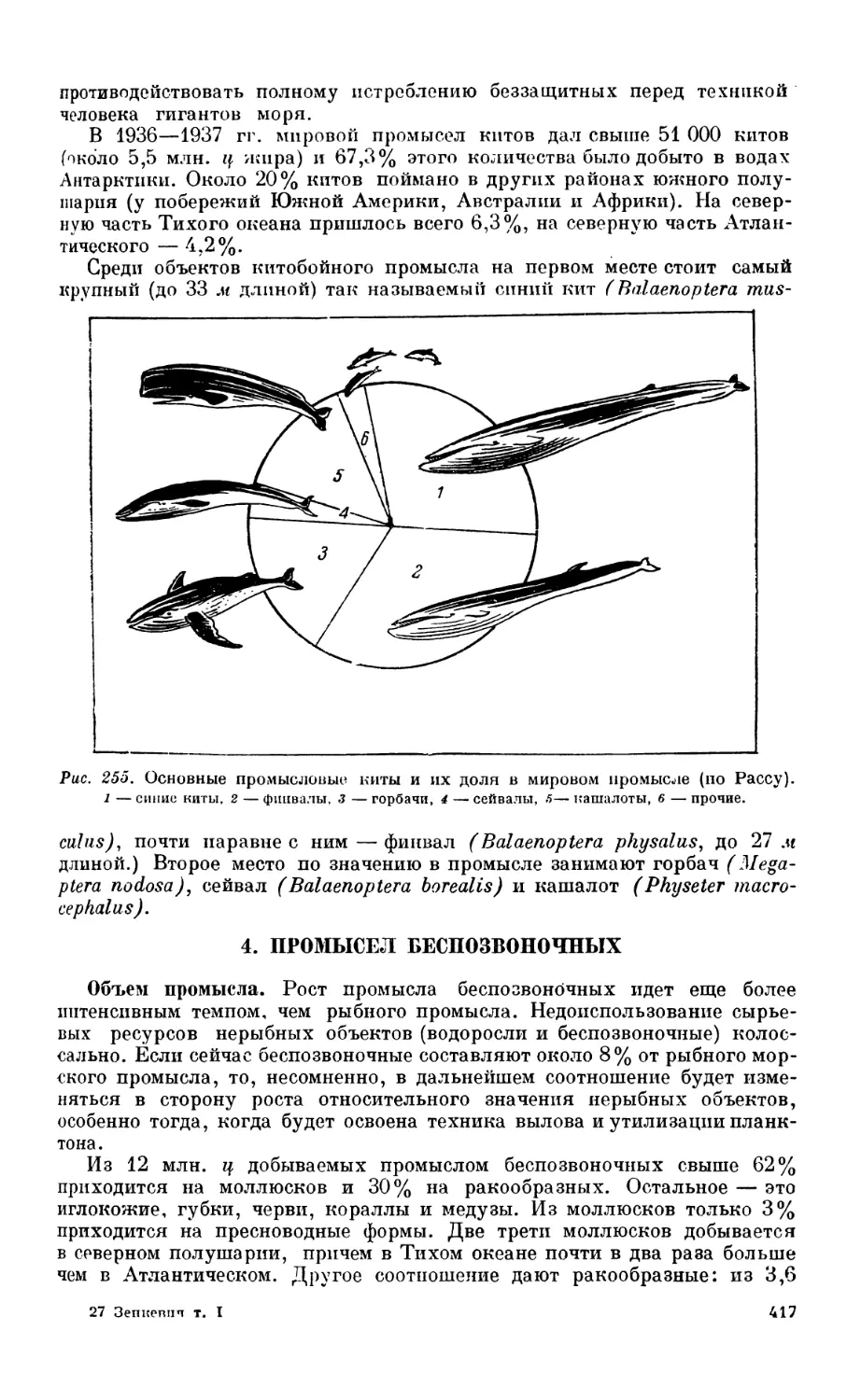
3. Промысел китов.......................................................... 116
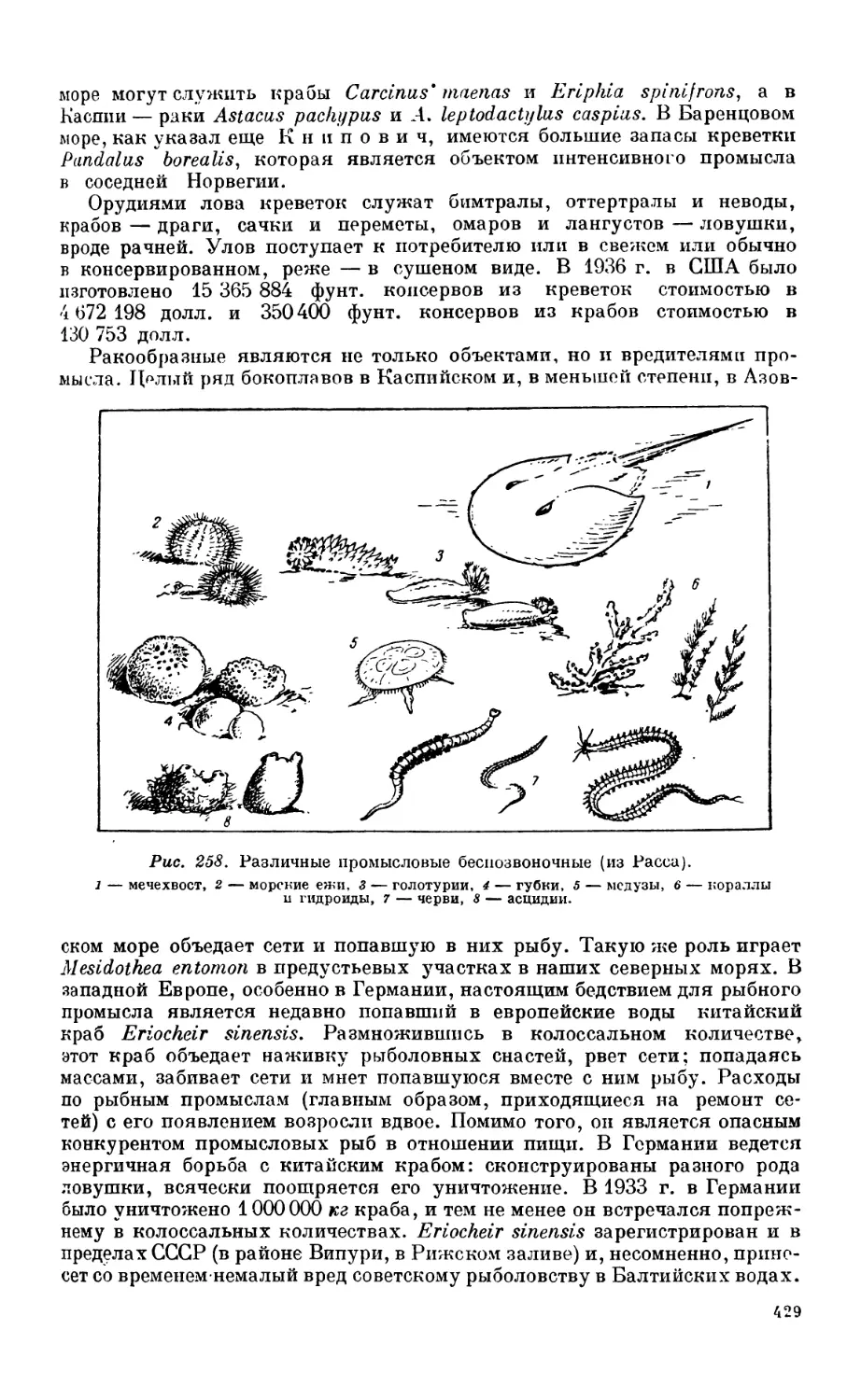
4. Промысел беспозвоночных.................................................. 417
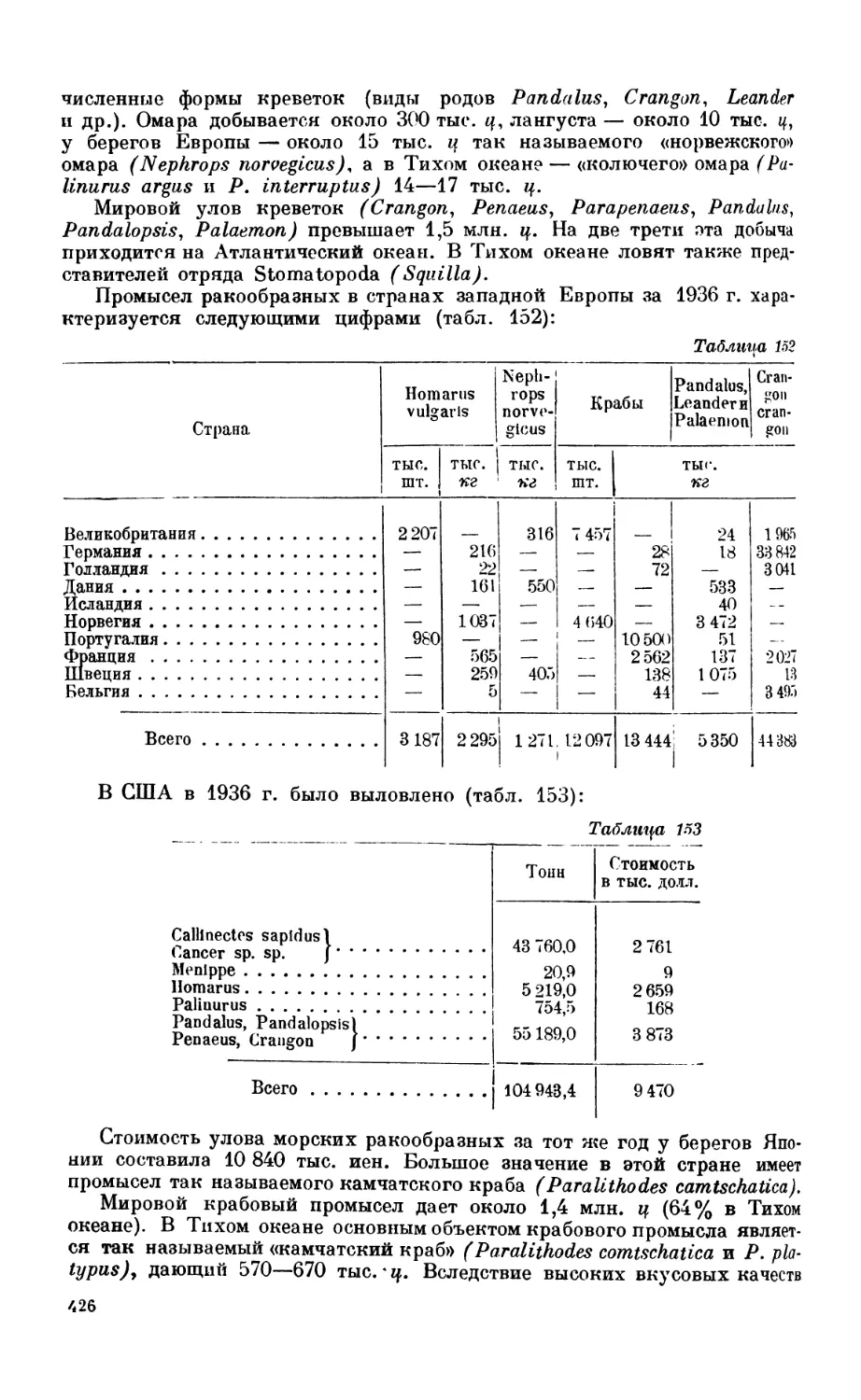
Объем промысла—4 17. Моллюски—120. Ракообразные—123. Иглокожие— !•?).
Губки, кишечнополостные и др. —130. Планктон—130.
5. Промысел морских растении ............................................... 431
Глава XVII. значение морских организмов в судоходстве
Обрастания —133. Древоточцы —433. Ледовые прогнозы, роль свечения и цветения
моря — 434.
Глава XVIII. проблема акклиматизации в море............................ 136
Литература.................................................................. 442
Предметный указатель......................................'..................461
Указатель латинских названий................................................ 488
Указатель авторов русский................................................... 500
Указатель авторов иностранный............................................... 504
ВВЕДЕНИЕ
1. МОРСКАЯ ФАУНА КАК ОБЪЕКТ ИЗУЧЕНИЯ
Целостность океанологии как науки
пология моря должна развиваться в самом тесном кон-
такте со всеми другими отраслями океанологии и, наобо-
рот, все отрасли этой науки в отрыве от биологии будут
неполноценны. Успешное развитие океанологии возмож-
но только при теснейшей связи составляющих ее дис-
циплин, в том числе п биологии. Это объясняется тем
положением, что развитие жизни в морях и океанах в большей или
меньшей степени зависит от всех факторов среды, а эти последние в свою
очередь в большей или меньшей степени зависят от населяющих водоем
организмов.
Таким образом, целостность океанологии как науки в той ;ке мере,
в какой целостен океан или отдельный морской водоем, как естественно-
исторический и народнохозяйственный объект, должна быть основным
положением современной океанологии.
Основные проблемы морской биологии
Однако мы при этом не должны забывать, что, помимо всего выше-
указанного, животные и растения, населяющие моря, в силу своего исклю-
чительного разнообразия, дают богатейший материал для изучения
многим биологическим дисциплинам — систематике, морфологии, фило-
генетике, физиологии, биогеохимии и др. Без приобщения морской фауны
п флоры, как объекта самых разносторонних биологических исследова-
ний, развитие биологии, как полноценной науки, было бы невозможно.
Из этих дисциплин особенное значение для океанологии имеет систе-
матика. Систематическая обработка (инвентаризация) морской фауны
и флоры — важнейшая проблема океанологии и одна из фундаменталь-
ных основ всего последующего изучения жизни моря и еще долгое время
систематика будет оставаться важным разделом морской биологии.
Вторая проблема — биогеографическая, она опирается па данные
по качественному и количественному распределению населения океана,
приводит к биогеографическому районированию и включает в свои за-
дачи выяснение истории фауны и флоры.
Изучение количественного распределения жизни в гидросфере и
основных показателей биологического продуцирования позволяет раз-
решить проблему типологического (регионального) районирования ми-
рового океана и отдельных морских водоемов.
Количественное выражение жизненных явлений в море дает нам
возможность выявить наиболее массовые и характерные формы и про-
13
цессы, которые должны быть подвергнуты широкому и углубленному
биоэкологическому и гидробиологическому изучению. Идя по этому пути,
мы можем свести все многообразие развертывающихся в океанах п морях
биологических явлений к более простым закономерностям и получить ту
характеристику отдельных частей галосферы, которая составляет основу
рационального народнохозяйственного освоения морских недр.
Большое внимание исследователи моря должны уделять вопросам
взаимосвязи его населения с окружающей средой и организмов друг с
другом. В первую очередь имеет значение в этом разрезе химический
состав морской воды и газовый режим. Между водой и населяющими
ее организмами имеется постоянная и напряженная взаимосвязь, для
которой характерны как процессы циклического характера (в системе
продуценты —консументы — редуценты или декомпозиторы), так и некото-
рые постоянные показатели, подвергающиеся лишь многолетним или
вековым колебаниям (растворенное органическое вещество, основной
солевой состав воды).
Не следует придавать жизненным явлениям в море чрезмерного зна-
чения, но не следует и умалять их роли. Например, переработка воды
животными-фильтраторами, несомненно, входит существенным компо-
нентом в динамику физических и химических свойств воды.
В круге этих проблем существенное значение имеет вопрос о формах-
индикаторах. Биологические методы в оценке гидробиологического ре-
жима, в различении водных масс, в океанологических прогнозах быстрым
темпом завоевывают все большее и большее место в самых различных
разделах океанологии.
Требуют уточнения характер и масштаб участия организмов в фор-
мировании осадочных пород (учение о биолитах, в частности генезис
горючих ископаемых, участие организмов в процессах денудации и акку-
муляции донных отложений и в процессе их переработки).
Важнейшей задачей в биологии моря было и всегда будет изучение
объектов промысла, их распределения и запасов, отыскание новых объек-
тов хозяйственного использования и обоснование рациональной си-
стемы эксплоатации растительного и животного сырья.
Большой раздел в изучении моря составляет проблема «биология
моря и флот» — выявление всех тех биологических явлений, которые
имеют положительное или отрицательное значение для торгового и воен-
ного мореплавания (древоточцы, обрастания, свечение и цветение моря,
береговые заросли и т. п.).
Накопление громадного фактического материала по всем разделам
энологии моря привело также и к проблеме реконструкции морской флоры
и фауны, в целях повышения возможностей хозяйственного их исполь-
зования.
Если исходить из представления о населении моря, как о некоторой
системе соподчиненных звеньев пищевой цепи, в основной части харак-
теризующейся циклическими процессами, то определенный хозяйствен-
ный эффект может быть получен в результате воздействий, направляемых
на различные звенья этой цепи, начиная от продуцентов и кончая ко-
нечным звеном — рыбой.
Наиболее применимый в практике прием — воздействие на конечное
звено -- рыборазведение и акклиматизация самих объектов промысла.
Однако в последние годы уже приступлено на морях к реконструкции
и предпоследнего звена пищевой цепи — к акклиматизации организмов,
служащих кормом для рыб. Мысльисследователей направляется также
па способы воздействия на самые начальные звенья пищевой цепи, на
удобрение морских водоемов, путем обогащения их соединениями азота
и фосфора. С другой стороны, все шире входят в практику и акклиматиза-
ционные мероприятия. Проблема реконструкции морской фауны путем
акклиматизации основывается преимущественно на зоогеографических
и палеогеографических представлениях в широких масштабах всего ми-
рового океана, путем использования тех поистине гигантских возмож-
ностей, которые предоставляют родственные пли сходные, но ныне разоб-
щенные фауны по линии использования их потенциальных ареалов,
исходя из их биологической и хозяйственной неравноценности.
Все вышесказанное подтверждает выставленное выше положение,
что все особенности природного явления, именуемого нами — океан,
находятся в теснейшем и непрерывном взаимодействии и наиболее успешно
могут изучаться только в таком разрезе. Этой основной идее автор стре-
мился подчинить весь тот фактический материал, который составляет
содержание настоящего труда.
2. ИСТОРИЯ ИЗУЧЕНИЯ МОРЯ
ИСТОРИЯ МОРСКИХ ЭКСПЕДИЦИЙ
Первые плавания. История изучения жизни моря неразрывно свя-
зана с большим количеством экспедиций, причем в XVI и XVII веках
морские экспедиции носили чисто географический характер и были вы-
званы стремлением к овладению морскими путями в связи с колониаль-
ной политикой морских держав. Однако уже в XVIII веке экспедиции
начинают проявлять интерес и к изучению самого океана, а в XIX и
XX вв. это становится их основной задачей. Первой в этом отношении
следует считать детище Петра Великого, замечательную почти 16-летнюю
русскую экспедицию Витуса Беринга (1725—1741), по справед-
ливости считающуюся началом научного изучения Тихого океана, в том
числе и его фауны. Имена биологов Стеллера п Краше-
н и н н и к о в а, участников экспедиции Беринга, одни из наиболее
славных имен биологов того времени.
Весьма плодотворны биологическими наблюдениями были англий-
ская экспедиция Д ж е м с а Кука, искавшего северо-западный
проход (1772—1773), и русская экспедиция Билингса и Сары-
чева по Тихому океану (1785—1793), в которой принимал участие
натуралист Мерк.
Дочелленджеровский период. XIX столетие — это настоящий век мор-
ских экспедиций, начавшийся замечательными русскими кругосветными
плаваниями Крузенштерна и Лися некого (1803—1806)
и Коцебу (1815), которые с полным правом можно считать началом
науки океанологии. В этих экспедициях были произведены первые точ-
ные измерения температуры на больших глубинах океана (до 1200 м).
Почти одновременно проходят знаменитые плавания англичанина Д же м-
< а Росса (1818) и русские экспедиции Беллинсгаузена
и Лазарева (1819—1821), открывших неизвестный до того Антарк-
тический материк, и не менее знаменитое кругосветное плавание на-
шего замечательного географа Литке (1826—1829). В этих экспедициях,
как правило, принимали участие опытные биологи.
В 1831—36 гг. совершает свое замечательное кругосветное плавание
па «Бигле» Чарльз Дарвин, произведший множество биологи-
ческих наблюдений.
В 1837 г. петербургский академик К. Бэр совершает поездку на
Новую Землю и привозит обширные коллекции по фауне Баренцева и
15
Ъетот морей, а в 1839—1813 гг. английская экспедиция Джемса
Р о с с а ведет исследования в Антарктике. В этой экспедиции
участвовал известный биолог Г укор (Нооке г) и были впер-
вые произведены глубоководные драгпровкп на нескольких тысячах
метров.
В 1853—56 гг. протекает первая обширная и по поставленным за-
дачам и по полученным результатам научно-промысловая экспедиция
нашего академика Бэра и Данилевей ого на Каспийское море.
Из экспедиций дочелленджеровского периода необходимо указать
также английскую экспедицию на корабле «Поркьюпайн» («Porcupine»),
Академик К. Бэр.
проделавшую в 1869 г. три рейса при участии Жеффрейза (Jef-
freys), Карпентера (Carpenter) и У а й в и л я Том-
сона (W yville Thomson) п обогатившую биологию многими
новыми и важными открытиями.
В течение четырех десятков лет перед плаванием «Челленджера»
целый ряд знаменитых русских, английских, французских, норвежских,
шведских, немецких, американских исследователей ведет изучение морской
фауны, однако, преимущественно «с берега». Среди этих славных имен
петербургские академики Бэр, М и д д е н д о р ф и Ш р е н к и зоологи
Вознесенский и Гримм и из иностранных ученых Эдуард
16
Рис. 1. Корвет < Витязю.
Форбс (Edward Forbes), О Д у э и и М и л ь и-Э д в а р д с
(А и d о u i п и Milne Edwards), Михаэль Gape (М i с h а е I
Sars), Л о в е н (L о v е п), Г. О. Cape (G. О. Sa г s), П у р т а л е
(Р о u г t а 1 ё s), А г а с с и ц (Aga ssi z) и многие другие.
Экспедиция «Челленджера». К началу 70-х годов зоологи располага-
ли уже довольно полными знаниями о составе и распределении морской
прибрежной фауны, открытые же части океанов были очень мало затро-
нуты исследованиями, и на долю английской экспедиции на корабле
«Челленджер» выпала большая честь снять еще оставшееся таинственное
покрывало с морских глубин. Три с половиной года работы этой
экспедиции — знаменательная дата в истории изучения фауны океа-
нических просторов. 50 больших томов трудов этой экспедиции — не-
отъемлемая часть каждой библиотеки
учреждений, ведущих изучение морской
фауны.
Период конца XIX и начала XX сто-
летия. После плавания «Челленджера»
количество морских экспедиций чрезвы-
чайно возросло — началось своего рода
соревнование между различными стра-
нами в организации этих экспедиций,
бороздивших все океаны и моря.
Наибольшее значение из них для
развития биологии имели Норвежско-
русская экспедиция на корабле «Форин-
геп» (1876—78) в северную часть Атлан-
тики, американские экспедиции на ко-
рабле «Блек» в Караибское море и Мек-
сиканский залив (1877—80) и на корабле
«Альбатрос» в западную часть северной
половины Атлантического океана, заме-
чательная шведская экспедиция II о р-
д е н ш е л ь д а на корабле «Вега», про-
шедшая северо-восточным проходом через
Северный Ледовитый океан (1878—79),
принца Монакского по Средиземному
морю (начиная с 1885 г.) и кругосветное
плавание русского корвета «Витязь» (рис. 1)
крупнейшего океанолога адмирала С. М
Заслуги этой экспедиции перед мировой наукой очень велики, что
нашло свое отражение и в том, что на фронтоне Океанографиче-
ского института в Монако среди десяти наиболее знаменитых в исто-
рии развития океанологии кораблей значится и Макаровский «Ви-
тязь». В начале 90-х годов работали широко известные по своим резуль-
татам экспедиции Андрусова и Л е б е д и и ц е в а на Черном
море, открывшие в нем явление зараженности сероводородом глубин-
ных вод.
В 1889 г. в северной Атлантике работала, при участии В. Гензена,
немецкая экспедиция на судне «Националь», собиравшая материалы
по распределению планктона.
Почти к этому же периоду относится и замечательный дрейф пансенов-
ского «Фрама» (1893—1896) через Северный Ледовитый океан.
Большой биологический материал собрала в северной Атлантике
датская экспедиция на корабле «Ингольф» (1895—1896) и в самом конце
прошлого столетия (1898—1899) немецкая глубоководная экспедиция
под командованием нашего
а к а о о в а
2 Зенкевич, т. I
17
на судне «Вальдивия» по Атлантическому и Индийскому океанам, под
начальством К. X у н a (Carl Chun).
Особенно следует выделить 10-летние работы на Баренцевом море
«Экспедиции для научно-промысловых исследований у берегов Мурмана»,
организованной нашим знаменитым океанографом-биологом академиком
Н. Книповичем и начавшей работы в 1898 г. Замечательно то, что
построенный Книповичем для экспедиции корабль «Андрей Первозван-
ный» (рис. 2) был в то же время и тральщиком. В результате К н и-
п о в и ч у удалось не только произвести обширные общеокеанографиче-
ские исследования Варенцова моря, но и открыть в нем колоссальные
промысловые скопления рыб — трески, пикши, морского окуня и сельди.
Подобно экспедиции Бэра на Каспийское море, экспедиция К н и п о-
Рис. 2. Экспедиционное судно «Андрей Первозванный».
вич а по поставленным научно-промысловым задачам выделяется среди
других экспедиций того времени.
В последующие годы Книпович проводил столь же углубленные
и ценные по результатам океанологические и научно-промысловые ис-
следования на Каспийском, Азовском и Черном морях.
На грани двух столетий (1899—1900) работала американская экспе-
диция под начальством зоолога А. Агассицана судне «Альбатрос»
и одновременно голландская экспедиция под начальством М. Вебера
на корабле «Зибога». К началу нашего века относятся русские экспеди-
ции То л ля на «Заре» вдоль северных берегов Азии (1900—1901)
и Корейско-Сахалинская экспедиция П. Шмидта (1900—1901),
которая так же, как и экспедиции Бэра и Книповича, ставила
своей основной задачей изучение промысловых богатств. К этому
же периоду (1901—1903) относятся также две большие антарктиче-
ские экспедиции, давшие новые обширные фаунистические мате-
риалы— немецкая южнополярная экспедиция Д риг а л ьс кого на
судне «Гаусс» и шведская антарктическая экспедиция О. Норден-
шельда.
Из последующих экспедиций, до первой мировой воины, наибольшее
значение для изучения морской фауны имели работы норвежского
18
судна «Михаэль Саре» в 1900—1910 гг. в северной Атлантике, датская
океанографическая экспедиция на судне «Дана» (1908—1910) и ряд
других.
Современный период. Громадный расцвет экспедиционной дея-
тельности в целях изучения всего мирового океана, столь характер-
ный для второй половины прошлого столетия и начала нашего (пер-
вое десятилетие), на время затухает; исследователи почти всех разделов
океанологии изменяют характер работы и переносят основное внимание
на систематическое и углубленное изучение сезонных изменений в био-
логических явлениях в отдельных морях и частях открытого океана.
Рис. 3. Шхуна «Россинантэ».
Из экспедиций периода между двумя мировыми войнами следует отме-
тить замечательные и многочисленные советские экспедиции в Северный
Ледовитый океан и по изучению омывающих наши берега морей — Барен-
цева, Белого, Черного, Азовского, Каспийского, Чукотского, Берингова,
Охотского и Японского. Можно с уверенностью сказать, что такого углуб-
ленного и разностороннего океанологического и научно-промыслового
изучения отдельных морей, какое проводилось и проводится в СССР, ни
одна другая страна не знает.
Блестящий расцвет советской океанологии связан прежде всего
с многолетней деятельностью крупных русских ученых Ю. Шокальского,
Н. Книповича и К. Дерюгина. Первый из них работал в области
физической океанологии и под влиянием его прекрасных исследований
и обобщений сформировались многочисленные современные советские
исследователи-гидрологи. Научно-промысловое направление работ
П. Книповича, сочетавшего глубокое теоретическое изучение морских
водоемов с разработкой конкретных путей народно-хозяйственного
использования их природных ресурсов, нашло свое яркое завершение
в ого посмертно изданной в 1938 г. книге «Гидрология морей и соло-
новатых вод».
К. Дерюгин широко известен своими блестящими исследованиями
Варенцова и Белого морей и их фауны, а также развернувшимся впо-
2*
19
следствии изучении наших дальневосточных морей, проводившихся в
основном на шхуне «Россинантэ» (рис. 3). Н. Книпович и К. Дерюгин
воспитали каждый по большой школе учеников, продолжающих сейчас,
после их смерти, начатое ими дело. Велика также заслуга и И. Месяцева,
основателя Государственного океанографического института и строителя
«Персея» (рис. 4 и 6). И. Месяцев, его сотрудники и ученики много
сделали для развития океанологии на новом ее советском этапе.
Из иностранных экспедиций наибольшее значение имели норвежские
п английские экспедиции в Антарктику, связанные, главным образом,
Рис. 4. Станции советского исследовательского судна «Персеи» в Баренцевом
и Белом морях. 1923—1934 гг.
с китобойным промыслом, немецкая экспедиция в южную половину
Атлантического океана на судне «Метеор» (1923—1927), американская
кругосветная экспедиция на судне «Карнеги» (1933),м!1оголетние и си-
стематические работы норвежского экспедиционного бота «Армауор Ган-
зен» в северо-восточной Атлантике (с 1913 г.), работы голландской
экспедиции на судне «В. Снеллиус» (1929—1930), английская экспедиция
на большой барьерный риф (1928—1929), работы экспедиционных кораб-
лей американских океанографических институтов в Вудс-Холе (на э/с.
«Атлантис») (с 1931) и в Калифорнии па «Е. W. Scripps» (с 1938 г.),
датского экспедиционного корабля «Dana II» (с 1921 г.), английского —
«Discovery II» (с 1930).
20
Рис. б. Институты и станции, ведущие изучение биологии моря (перед 2-й мировой войной).
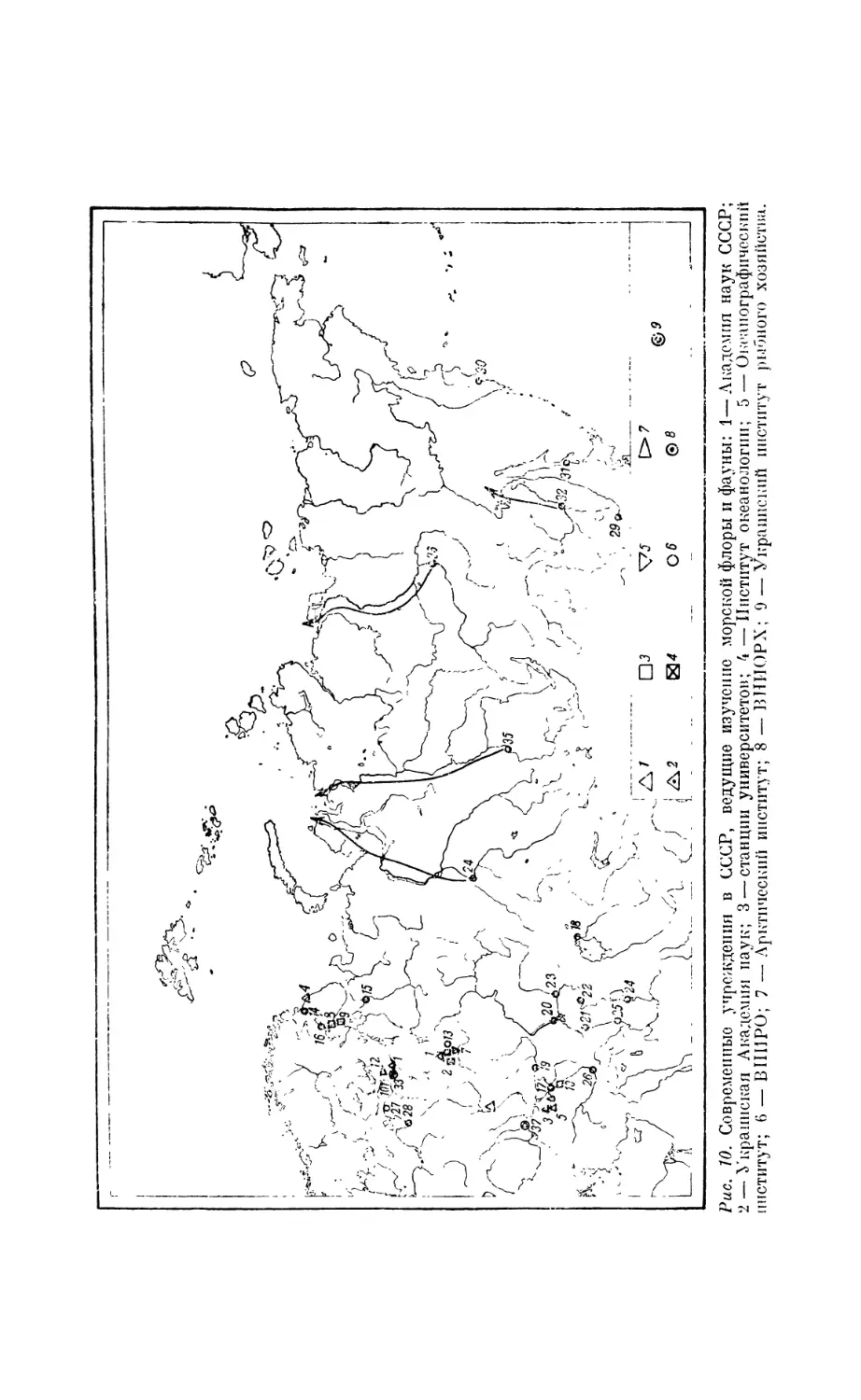
3. НАУЧНЫЕ УЧРЕЖДЕНИЯ. ВЕДУЩИЕ ИЗУЧЕНИЕ ФАУНЫ МОРЯ
Изучение морей и океанов и, в частности, их флоры и фауны требует
затраты больших материальных средств, так как для этой работы необхо-
димы расположенные на побережье лаборатории с аквариумами и спе-
циально оборудованные экспедиционные суда с большим персоналом,
обладающим многолетним опытом. Отдельные экскурсии ученых на моря
давали некоторое знакомство с морской фауной, но они ограничивались
только районом побережий.
В настоящее время на земном шаре существует около 250 постоянных
учреждений, занятых изучением моря и возникших в течение последних
Академик Л. Ковалевский.
100 лет. Часть из них перед второй мировой войной представляла собой
организации международного характера, как, например, Постоянный
Международный совет по изучению моря (с 1899 г.), Международная
комиссия по изучению Средиземного моря (с 1919 г.), Международный
Тихоокеанский комитет (с 1923 г.). Из всех этих многочисленных
учреждений 180 ведут в большей или меньшей степени биологиче-
ские исследования, а 95 являются в основном биологическими (рис. 5).
Организация первых морских лабораторий. Первая морская лабора-
тория была организована французским зоологом Марионом (Ma-
rion) в 1834 г. в Марселе, а следующая — зоологом Костом (Coste)
в 1859 г. в Конкарно. В 1863 г. возникла биологическая станция в Арка-
шоне. Эти три французские биологические станции были и сейчас оста-
ются маленькими университетскими лабораториями местного значения.
С 1872 г. начался быстрый рост морских станций и организация многих
22
Рис. 6. Экспедиционное судно «Персей» во льдах Варенцова моря.
в uxr, плимутская в
Рис. 7. Севастопольская био-
логическая станция, 1914 г.
ю зоологическую станцию.
из них связана с именами крупнейших зоологов. Некоторые из них
представляют собой целые обширные институты, как например, Сева-
стопольская (рис. 7) и Мурманская (рис. 8)
глии, Неаполитанская в Италии, Вудсхолл-
ская в США. В 1872 г. А. Ковалев-
с к п м по инициативе нашего замечатель-
ного путешественника Н. М и к л у х а-
М а к л а я и согласно постановлению 1-го
съезда естествоиспытателей и врачей в 1868 г.
была организована Севастопольская биоло-
гическая станция и в том же году фран-
цузским зоологом ЛаказДютье (L а-
с a z e-D u t h i е г) — станция в Роскове.
Двумя годами позднее немецкий зоолог
А. Дорн (A. Dorn) па свои средства
создал широко известную Неаполитанску
В 1881 г. начала свои работы Соловецкая биологическая станция,
организованная Петербургским обществом естествоиспытателей по ини-
циативе Н. В а г н е р а, перенесенная в 1899 г. в г. Александровск на
Мурмане и получившая название Мурманской биологической станции.
В 1897 г. возникла первая в нашей стране морская рыбохозяйствен-
ная станция в Астрахани, проводившая в течение 50 лет изучение
Каспийского моря и его рыбных богатств.
Русские морские научные учреждения. В России до революции было
6 морских станций, из которых две (Астраханская и Бакинская) имели
рыбохозяйственное значение, одна принадлежала Академии наук (Сева-
23
стопольская), одна — Петербургскому обществу естествоиспытателей
(Мурманская), Ковдеиская на Белом море — Юрьевскому университету
Рис. 8. Мурманская биологическая станция Академии наук СССР, 19'i0 г.
и шестая — университетская биологическая станция в Одессе. В настоящие
время мы располагаем 43 морскими научными институтами, лаборатори-
Рис. 9. Русская зоологическая станция в Виллафрапке.
ими и станциями, ведущими исследования в области биологии моря
и промыслового дела (рис. 10 и табл. 1).
24
Рис. 10. Современные учреждения в СССР, ведущие изучение морской флоры и фауны: 1—Академия наук СССР;
2 — Украинская Академия наук; 3 — станции университетов; 4— Институт океанологии; 5 — Океанографический
институт; 6 — DIII1PO; 7 — Арктический институт; 8 — ВНИОРХ; 9 — Украинский институт рыбного хозяйства.
Таблица 1
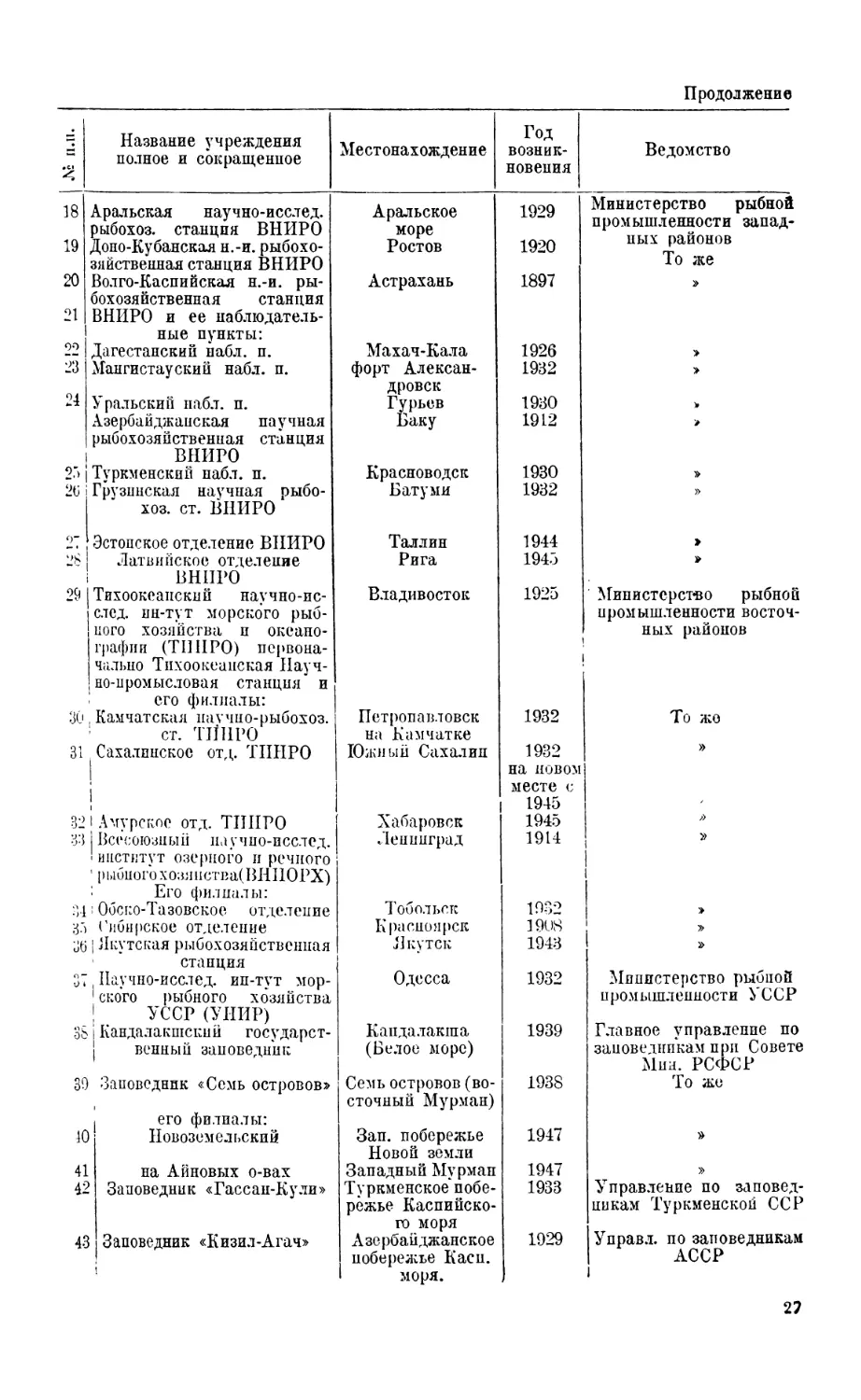
СОВЕТСКИЕ УЧРЕЖДЕНИЯ, ВЕДУЩИЕ ИССЛЕДОВАНИЯ МОРСКОЙ ФАУНЫ
И ФЛОРЫ
№ п.п. Название учреждения полное и сокращенное Местонахождение Год возник- новения Ведомство
1 Академия наук СССР Москва и Ленинград 1725 Академия наук СССР.
2 Институт океанологии (ИОАН) Москва 1941 Отд. геолого-географи- ческих наук
3 Севастопольская биологиче- ская станция Севастополь 1872 Академия паук СССР. Отд. биологических наук
4 Мурманская биологическая станция (II) 1 Мурман, губа Даль- няя Зеленецкая 1936 То же
5 Черноморская гидрофизиче- ская станция Крым, Кацивели 1929 Академия паук СССР. Отд. физико-математи- ческих наук
6 Карадагская биологическая станция Крым, Отузы 1914 Украинская Академия наук
7 Государственный океаногра- фический институт (ГОИН II) 1 2 Москва 1942 Центр, у правя, единой гидромет. службы (ЦУЕГМС)
8 Беломорская биологическая станция Кандалакшский залив, Великая Салма 1938 Моск. гос. университет
9 Гридинская биологическая станция Кандалакшский залив, Гридипо Новороссийск 1945 Петрозаводский гос. университет
10 Новороссийская биологиче- ская станция им. В. М. Ар- но льди 1921 ( Ростовский гос. университет
11 Петергофский научно-иссле- довательски й инети тут Ленинград 1920 I I Ленинградский гос. университет
12 Всесоюзный арктический институт (ВАП) Ленинград 1919 ! ! Главное управление Сев. : морского пути (ГУСМИ)
13 Всесоюзный * научно-исслед. институт морского рыбного хозяйства и океанографии (ВНПРО) Его филиалы: Москва i 1921 Министерство рыбной промышленности запад- ных районов
14 Полярный и.-и. ин-т мор- ского рыбного хозяйства и океанографии (И ПН ГО) при нем: Мурманск 1933 3 о же 1
15 ! Научная Беломорская рыбо- ! хозяйственная станция Архангельск 1930 I ।
16 j Кандалакшский наблюда- | тельный пункт Кандалакша 1931 »
17 . Азовско-Черноморский н.-и. ! ин-т морского рыбного хо- 1 зяйства и океанографии (АзЧерППРО) первоначально Керченская ихтиологическая лаборатория Керчь 1921 »
1 Мурманская биологическая станция (I), организованная в 1899 г. в г, Алексан-
дровске, в 1930 г., была присоединена к Государственному океанографическому инсти-
туту и в дальнейшем получила название Полярного отделения ГОИН-а, а с 1933 г. По-
лярного Института Рыбного хозяйства и океанографии (ПИНРО). Указываемая в таб-
лице Мурманская станция в губе Зеленецкой возникла наново без непосредственной
связи со старой станцией (рис. ‘8).
2 ГОИН-1 — Государственный океанографический институт был организован в 1921 г.
в Москве. В 1933 г. ГОИН был соединен с Институтом рыбного хозяйства и преобра-
зован во Всесоюзный институт рыбного хозяйства и океанографии (ВНПРО). Указан-
ный в таблице ГОИН (II) возник в 1942 г. без связи с ГОИН (1).
26
Продолжение
ЛЬ п.п. Название учреждения полное и сокращенное Местонахождение Год возник- новения Ведомство
18 19 20 21 22 23 24 25 26 9" 28 29 30 31 32 33 34 ;<) 36 31 38 39 1С 41 42 43 Аральская научно-исслед. рыбохоз. станция ВНИРО Доно-Кубанская н.-и. рыбохо- зяйственная станция ВНИРО Волго-Каспийская н.-и. ры- бохозяйственная станция ВНИРО и ее наблюдатель- ные пункты: Дагестанский набл. п. Мангистауский набл. п. Уральский набл. п. Азербайджанская научная рыбохозяйственная станция ВНИРО Туркменский пабл. п. Грузинская научная рыбо- хоз. ст. ВНИРО Эстонское отделение ВНИРО Латвийское отделение ВНИРО Тихоокеанский научно-ис- след. ин-тут морского рыб- ного хозяйства и океано- графии (Т1ШРО) первона- чально Тихоокеанская Науч- но-промысловая станция и : его филиалы: , Камчатская научно-рыбохоз. ст. ТИНРО । Сахалинское отд. ТИНРО 1 Амурское отд. ТИНРО I Всесоюзный научно-исслед. !институт озерного и речного ' рыбногохозяиства(ВНПОРХ) : Его филиалы: । Обско-Тазовское отделение Сибирское отделение | Якутская рыбохозяйственная станция , Научно-исслед. ип-тут мор- 1 ского рыбного хозяйства ; УССР (УИИР) 1 Кандалакшский государст- । венный заповедник Заповедник «Семь островов» его филиалы: | Новозем ел некий на Айновых о-вах ! Заповедник «Гассан-Кули» ( Заповедник «Кизил-Агач» Аральское море Ростов Астрахань Махач-Кала форт Алексан- дровск Гурьев Баку Красноводск Батуми Таллин Рига Владивосток Петропавловск на Камчатке Южный Сахалин Хабаровск Ленинград Тобольск Красноярск Якутск Одесса Кандалакша (Белое море) Семь островов (во- сточный Мурман) Зап. побережье Новой земли Западный Мурман Туркменское побе- режье Каспийско- го моря Азербайджанское побережье Касп. моря. 1929 1920 1897 1926 1932 1930 1912 1930 1932 1944 1945 1925 । 1932 1932 на новом месте с 1945 1945 1914 1932 1908 1943 1932 1939 1938 1947 1947 1933 1929 Министерство рыбной промышленности запад- ных районов То же > > Министерство рыбной промышленности восточ- ных районов То жо » Л> » Министерство рыбной промышленности УССР Главное управление по заповедникам при Совете Мни. РСФСР То же » Управление по заповед- никам Туркменской ССР Управл. по заповедникам | АССР 1
27
Иностранные морские научные учреждения. В 1882 г. возникла первая
морская ихтиологическая лаборатория в Англии, в Абердине,
а в 1885 г. английским биологом и океанографом Джоном М е р р е е м
(J . М и г г а у) была организована биологическая станция в Милль-
порте (Шотландия). В 1888 г. английские зоологи Гексли Huxley)
и Р э й-Л а и к е с т е р (Ray La nkes tor) организовали Плимут-
скую биологическую станцию, а в 1889 г. датчанин Петерсен —
Датскую биологическую станцию. Еще двумя годами позднее возникла
также широко известная Бергенская рыбохозяйственная станция. В 1886 г.
Киевский профессор А. К о р о т и е в создал на своп средства русскую
зоологическую станцию на Средиземном море в Villefranche sur Мег,
ныне находящуюся в ведении Парижского университета (рис. 9).
В США в настоящее время имеется 28 учреждений, ведущих изучение
моря, в Великобритании с доминионами—30, во Франции с колониями —
21, в Японии —21, в Италии — 11, в Германии — 5. в СССР — 43. Послед-
ние десятилетия и у нас и за рубежом наблюдается тенденция к возник-
новению морских лабораторий рыбохозяйственного характера, резко
преобладающих в настоящее время над другими, и центральных комп-
лексных океанографических институтов.
Основные издания, посвященные вопросам океанологии. В течение
последних десятилетий большую роль в развитии биологических раз-
делов океанологии, помимо трудов перечисленных выше экспедиций,
сыграли специальные журналы н серии и отдельные монографии,
посвященные вопросам океанологии.
Русские издания Ч Из наиболее важных работ на русском языке сле-
дует назвать в первую очередь монографии В. С о в и н с к о г о «Вве-
дение в изучение фауны Понто-Касиийско-Аральского морского бассейна»
(1904), С. Зернова «К вопросу об изучении жизни Черного моря»
(1912) и две монографии К. Дерюгина «Фауна Кольского залива
и условия ее существования» (1915) и «Фауна Белого моря и условия
ее существования» (1928), а также издания различных научных институ-
тов, станций и труды важнейших экспедиций: «Труды Арало-Каспийской
экспедиции» (1875—1905); «Труды экспедиции для научно-промысловых
исследований у берегов Мурмана» (1906—1915); «Труды Каспийской экспе-
диции» (1907—1914, 1921); «Труды Азовско-Черноморской научно-промыс-
ловой экспедиции» (с 1926 г.); «Бюллетень Всекасппйской научной рыбо-
хозяйственной экспедиции» (с 1932 г.); «Труды Каспийской комиссии
Академии паук СССР» (с 1937 г.); «Записки по гидрографии» (с 1900 г.);
«Труды Севастопольской биологической станции» (с 1915 г.); «Работы
Мурманской биологической станции» (1925—1929); «Труды научной
рыбохозяйственной и биологической станции Грузии» (с 1934 г.); «Труды
Ленинградского научно-исследовательского ихтиологического инсти-
тута» (с 1929 г.). Наибольшее значение имели издания нескольких круп-
нейших институтов, возникших в первые годы после Октябрьской рево-
люции:
Арктического научно-исследовательского ин-
ститута (первоначально Северная научно-промысловая экспедиция,
позднее Институт по изучению Севера):
1. «Труды Северной научно-промысловой экспедиции» (1920—1925).
2. «Труды Института по изучению Севера» (1925—1931).
1 Приведенный ниже перечень изданий отнюдь не претендует на исчерпывающую полноту.
В нем приведены лишь основные издания, сыгравшие ведущую роль в развитии биологического
раздела океанологии в нашей стране.
28
3. «Труды Арктического научно-исследовательского института» (с
1931 г.).
4. «Материалы по изучению Арктики» (1931—1935).
5. «Arctica» (1933—1937).
6. «Бюллетень Арктического института» (1931—1936).
7. «Проблемы Арктики» (с 1937 г.).
Государственного гидрологического инсти-
тута (до 1926 г. —Российский гидрологический институт):
1. «Известия Государственного гидрологического института» (с 1921 г.).
2. «Бюллетень Российского гидрологического института» (1920—1922).
3) . «Исследования морей СССР» (с 1925 г.).
4. «Записки Государственного гидрологического института» (с 1926 г.).
Всесоюзного научи о-и с с л е д о в а т е л ь с к о г о ин-
ститута морского рыбного х о з я й с т в а и о к е а н о-
г р а ф и и и его филиалов (возник в 1933 году путем слияния Государ-
ственного океанографического института и Центрального научного
института рыбного хозяйства):
1. «Труды Плову чего морского научного института» (1923—1926).
2. «Труды Морского научного института» (1926—1930).
3. «Труды Государственного океанографического института» (1931 —
1934).
4. «Бюллетень Государственного океанографического института»
(1931—1934).
5. «Труды Научного института рыбного хозяйства» (1924—1932).
6. «Труды Всесоюзного научно-исследовательского института мор-
ского рыбного хозяйства и океанографии» (с 1935 г.).
7. «Труды Азовско-Черноморского научно-исследовательского инсти-
тута рыбного хозяйства и океанографии (первоначально «Труды Керчен-
ской научной рыбохозяйственной станции», впоследствии «Труды Азов-
ско-Черноморской научной рыбохозяйственной станции» (с 1926 г.).
8. «Труды Волго-Каспийской научно-исследовательской рыбохо-
зяйственной станции» (первоначально «Труды Астраханской ихтиологи-
ческой лаборатории», впоследствии «Труды Астраханской научной рыбохо-
зяйственной станции») (с 1909 г.).
9. «Труды Полярного научно-исследовательского института морского
рыбного хозяйства и океанографии» (с 1938 г.).
10. «Труды Аральской научной рыбохозяйственной станции» (с 1933 г.).
И. «Труды Азербайджанской научной рыбохозяйственной станции»
(с 1932 г.) (ранее «Известия Бакинской ихтиологической лаборатории» —
< 1922 г.).
12. «Бюллетень Туркменской научно-рыбохозяйственной станции»
(1931).
13. «Известия Тихоокеанского научно-исследовательского института
рыбного хозяйства» (ранее Тихоокеанская научно-промысловая станция)
(с 1928 г.).
В 1941 году Академия наук начала издавать серию «Исследования
Дальневосточных морей», пока вышел в свет только I том.
С 1944 года началась регулярная публикация «Докладов Государ-
ственного океанографического института», основанного в Москве в
1942 году, а с 1947 г. подобная же серия «Докладов Всесоюзного ин-
ститута морского рыбного хозяйства и океанографии» (ВНИРО).
Помимо вышеуказанных изданий, целиком посвященных морю и
морской фауне, необходимо указать ряд журналов и серий, в которых,
наряду с другими, были опубликованы некоторые руководящие статьи
и монографии по морской биологии.
23
Это прежде всего «Бюллетень Московского общества испытателей
природы» (издается с 1829 года), «Ежегодник Зоологического музея Ака-
демии наук» (с 1896 года), «Труды Ленинградского общества есте-
ствоиспытателей» (с 1870 года) (ранее «Труды С.-Петербургского
общества естествоиспытателей»), «Русский гидробиологический журнал»
(с 1921 по 1930 год), «Зоологический журнал» (с 1916 года) (до 1930 года
«Русский зоологический журнал»), «Ученые записки Московского госу-
дарственного университета» и ряд других изданий.
Очень большим событием в развитии русской зоологии явилось изда-
ние обширной серии монографий по самым различным группам живот-
ных: «Фауна России и сопредельных стран», начало которой положил
выход в свет в 1911 году монографии Л и н к о «Гидроиды», второй том
этой монографии был выпущен уже после смерти автора Куделиным
в 1914 году. Впоследствии в этой серии были опубликованы следующие
монографии по морским группам: Редикорцев «Оболочники»
(1916), М и л а ш е в и ч «Моллюски Черного и Азовского морей»
(1916), Дьяконов «Иглокожие (Echinoidea)» (1923).
С 1929 года эта серия получила название «Фауна СССР и сопредель-
ных стран». В этой новой серии были изданы: Ш и м к е в и ч «Рап-
topoda» (1929—1930); Гурьянова «Равноногие дальневосточных
морей» (1936); Макаров «Апотига» (1936); Светов и дов «Рыбы»
(Triglidae) (1936).
Кроме того, Зоологическим институтом Академии наук СССР издаются
«Определители по фауне СССР», где были опубликованы: «Морские арк-
тические равноногие раки» Г. Гурьяновой (1932) и «Иглокожие
Северных морей» А. Дьяконова (1933).
Иностранные издания. Из иностранных изданий следует указать
в Англии Журнал Плимутской станции («Journal of the Marine Biological
Association» c 1895 г.) и серию «Fishery Investigations», издания Между-
народного совета по изучению морей: «Journal du Conseil» (с 1926 г.)т
«Rapports et Proces Verbaux des Reunions и Publications des circonstan-
ces». Первые отчеты Международного совета вышли в 1902 г.
В США издается журнал Вудс-Хольского института («Biological Bul-
letin» с 1898 г.), издания Калифорнийского океанографического института
и серия «Bulletins U. S. Bureau of Fisheries», в Германии журнал «Inter-
national Revue der gesamten Hydrobiologie und Hydrographie» (c 1908 r.),
обширная серия «Wissenschaftliche Meeresuntersuchungen» (c 1894 r.),
перешедшая c 1936 г. в две серии: Кильскую («Kieler Meeresforschun-
gen») и Гельголандскую («Helgolander Wissenschaftliche Meeresuntersu-
chungen») и столь же обширные серии—«Tierwelt der Nord- und Ostsee».
начавшая выходить с 1925 г., «Handbuch der Seefischerei Nordeuropas»
(с 1928 г.), «Fauna Arctica» (1900—1933), «Nordisches Plankton» (c 1901 r.),
в Италии большая серия Неаполитанской зоологической станции «Fauna
und Flora des Golfes von Neapel» (c 1880 г.), во Франции «Revue des
Travaux des Peches maritimes» (c 1886 г.) и издание биологической
станции в Роскове («Travaux de la Station biologique de Roscoff»
(c 1922 года), в Монако «Bulletin de 1’Institut Ocoanographique de Monaco»
(c 1905 г.), в Дании «Reports of the Danish Biological Station» (c 1895) и
«Meddelelser fra Kommission for Danmarks. Fiskeri og Havundersogelser»
(c 1904 г.), аналогичное шведское издание — «Svenska Hydrografisk-bio-
logiska Kommissionens skrifter» (c 1902 г.) и многие другие более мелкие
серии и журналы.
Нельзя не упомянуть также тех отдельных книг, которые в тече-
ние 70 лет знаменуют собой все глазные этапы развития биологии моря.
Начать это перечисление нескольких основных сочинений следует, по-
30
жалуй, с книги немецкого исследователя К. Мебиуса (К. Mobius)
«Die Auster und die Austerwirtschaft» (1877), в которой впервые было
сформулировано содержание понятия биоценоз. Через полтора десятка
лет (1893—94) вышли труды И. Вальтера (J. Walther) «Bionomie
des Meeres» а в 1896 г. появилось сочинение, давшее основы морской
зоогеографии, «Grundzuge der marine Tiergeographie» немецкого зоолога
А. Ортманна (A. Ortmann, 1896), до сих пор сохраняющее
научное значение.
В 1910 г. появилось обширное сочинение другого немецкого ученого
А. Штейера (A. Steuer) «Planktonkunde», посвященное специально
морскому планктону.
В 1912 г. английский ученый Дж. Меррей (Jonu Murray) и норвеж-
ский ученый Порт (Johan Hjort) в известной книге «The Derptus of
Ocean» на базе обработки обширных материалов норвежского ко-
рабля «Michael Sars» развернули сложившиеся к тому времени оке-
анологические представления, в частности в отношении биологии
океана.
В 1923 г. вышла в свет полезная сводка американца Трес-
слера (D. Tressler) «Marine Products of Commerce», а в 1924 г.
появилось сочинение немецкого зоолога Р. Гессе (Richard Hesse)
«Tiergeographie auf okologischer Grundlage», посвящающее большой раз-
дел зоогеографии моря.
Сводная работа по биохимии моря английского биолога Гарвея
(Н. Harvey) «Biological chemistry and physics of Sea Water», переве-
денная и на русский язык, появилась в 1928 г.
В 1931г. вышла книжка американского биолога Бидже л оу
(Н. В. Bigelow) «Oceanography its Scope, Problems, and Economic
Importance», в 1935 г. вышла обширная, оригинальная и глубоко
содержательная книга шведского зоолога и зоогеографа С. Экмана
(Sv. Ekman) «Tiergeographie des Meeres», а в 1937 г. чрезвычайно
полезная сводка американца Вогана (Т. W. Vaughan) «Inter-
national Aspects of Oceanography».
В последние годы был опубликован обширный труд американских
исследователей Свердрупа, Джонсона и Флеминга (Sverd-
rup, Johnson, Fleming) «The Oceans» (1942), подводящий итоги
больших экспедиций периода между двумя мировыми войнами, в
котором, к сожалению, быть может сознательно игнорируется обширная
русская литература. Сводкой специально по вопросам химии и био-
логии служит небольшая книжка упомянутого выше Гарвея «Recent
Advances in the Chemistry of Sea Water» (1945). Этот перечень можно
закончить упоминанием книги Цо-Белла (Zo-Bell) «Marine Microbio-
logy», вышедшей в 1946 г.
В каждой из крупных стран издаются специальные периодические
издания со статистикой морского промысла, а в некоторых при-
водятся данные по мировому промыслу. Такими являются статистиче-
ские ежегодники Лиги Наций («Annuaire statistique de la Societe de
Nations»), Международного Совета по изучению морей («Bulletin
Statistique») и издание Сев.-Американского Статистического бюро
(«Fishing Gazette»).
ОСНОВНЫЕ ЭТАПЫ ИЗУЧЕНИЯ МОРСКОЙ ФАУНЫ
Систематико-фаунистический этап. Некоторые представления об орга-
низмах, населяющих море (сначала применительно к позвоночным и оби-
тающим у берегов беспозвоночным), имелись у человека с чрезвычайно
31
древних времен, однако возникновение систематических знаний о фауне
моря связано с развитием практических и научных интересов к мор-
ской фауне п с применением для исследовательских целей специаль-
ных орудий лова — планктонных сетей и драг. Впервые драга была при-
менена для этой цели итальянскими зоологами Марс и л ь и и Д о-
натп (Marsigli и Donati) в середине XVIII столетия (1750)-
Постепенио сведения о фауне моря стали расширяться; и французские
исследователи Оду эн и М и л ь н - Э д в а р д с (Au do u in и Milne
Edwards) в 1832 г. уже опубликовали первую работу
зоогеографического •характера по морской фауне. Почти одновре-
менно начал опубликовывать свои работы норвежский исследователь
М. С а р с (М i ch а е 1 S ar s) (с 1835). Английский биолог Э. Форбс
(Е. Forbes) изучал фауну морского дна уже до глубины 300 са-
жен и в 1844 г. начал опубликовывать своп блестящие исследова-
ния. Первые иолвека чисто фаунистических исследований дали
С а р с у возможность построения некоторых зоогеографических обобще-
ний (1850).
Систематическое изучение пелагических организмов началось несколь-
ко позднее исследований по донной фауне, что, естественно, совпало
с первым применением в исследовательской практике планктонной сетки
зоологом 11. Мюллером (Johann Muller) (1845). Постепенно
биологи стали завоевывать глубины океанов, и некоторым образом завер-
шающей фазой этого первого систематико-фаунистического эта-
па в изучении морских организмов явились исследования английской экс-
педиции на судне «Челленджер» («Challenger»), проводившиеся в течение
трех с половиной лет (с декабря 1872 по май 1876 г.). Исследования
«Челленджера» и ряда других глубоководных экспедиций заложили
прочные основы наших представлений о морской флоре и фауне. В эти
же годы возникло несколько крупнейших морских биологических стан-
ций (Севастопольская, Неаполитанская и др.), продолжавших те же ис-
следования. Все эти фаунистические исследования дали громадный ма-
териал для развития наших современных представлений о распростра-
нении морских растений и животных.
Экологический этап. Постепенно экологические исследования все
сильнее и сильнее стали внедряться в биологию и знаменуют собою
второй этап в изучении морской фауны — этап экологический.
Наиболее яркое выражение этот этап получает в исследованиях Лоренца
(Lorenz, 1863), Мебиуса (Moebius, 1877), Вальтера
(Walther, 1893—94) и Прюво (Pruvot, 1896—98).
В русской литературе наиболее характерными для этого этапа
исследованиями являются классические работы С. А. Зернова
по Севастопольской бухте Черного моря (1912) и К. М. Дерюгина
по Кольскому заливу Варенцова моря (1915).
Количественно-топологический этап. На то же десятилетие падает
и другое крупное событие, знаменующее собою то новое направление, кото-
рому суждено было в дальнейшем стать характернейшей чертой треть-
его этапа в изучении фауны моря, начинающего четко оформляться
только в течение последних 15—20 лет. Немецкий исследователь В. Г е н-
зен (Victor Hensen) в 1877 г. впервые применил в Кильской
бухте специально приспособленную им планктонную сеть для количе-
ственного изучения икры некоторых рыб. В 1877 г. появилось в свет
известное сочинение того же исследователя но количественному изучению
планктона.
Значение количественного метода в изучении морской флоры и фауны,
введенного для планктона В. Гензеном в 1877 г., а для бентоса
директором Датской биологической станции в Ныоборге Петерсе-
32
н о м (С. G. J )ii. Ре terse и) в 1908 г. было исключительно велико.
Количественный метод ио всем разделам изучения жизни моря создал
тот фундамент, на котором базируются наши современные представления
и совокупности биологических процессов, совершающихся в водоемах,
п о путях их хозяйственного использования.
Именно в нашей стране, в советское время, на всех морях были
развернуты в очень широком масштабе количественные исследования
фауны и флоры. Этот громадный количественный материал дал возмож-
ность учесть все основные качественные и количественные показатели
биолого-продуктивных свойств водоема, что в свою очередь ложится
в основу рационального промыслового использования сырьевых ре-
сурсов морских недр.
Этот третий этап в изучении (фауны моря мы могли бы назвать коли-
чественно-типологическим.
1. БИОГЕОГРАФИЯ. ЭКОЛОГИЯ И ГИДРОБИОЛОГИЯ
И ИХ РОЛЬ В ИЗУЧЕНИИ ФАУНЫ МОРЯ
Изучением распределения жизни в морях и обусловливающих его зако-
номерностей занимаются три науки, близко соприкасающиеся и частично
перекрывающие друг друга — биогеография, экология и гидробиология.
Однако каждая из этих наук имеет свой самостоятельный объект и свои
методы и каждая подходит к морской фауне со своих особых позиций.
Биогеография в основном освещает вопросы состава фауны и ее геогра-
фического распределения, возникновения и истории развития различных
фаун и, наконец, имеет конечной целью биогеографическое районирование,
биогеография изучает закономерности распространения организмов и
различных свойственных им особенностей в рамках больших ареалов
морей, островов, частей материков, климатических поясов, растительных
зон и т. п.
В новейшее время, обычно, различают несколько основных направ-
лений в зоогеографии:
Регистрирующая (описательная, фаунисти-
ческая) зоогеография или фаунистика дает опи-
сание фауны того или иного района по систематическому составу и сравни-
вает фауны отдельных районов друг с другом (сравнительная фауни-
стика). Долгое время зоогеография в основном имела фаунистическое
содержание.
Каузальная (аналитическая) зоогеография
выясняет причины, обусловливающие современное распределение жи-
вотных в биосфере. Анализ может итти либо в направлении выяснения
исторических причин современного распределения, либо причин эколо-
гических (историческая и экологическая зоогеография). По линии этих
последних особенное внимание обращается па условия существования
и условия распространения.
Синтетическая (таксономическая, региональ-
ная, б и о и е и о т и ч е с к а я) зоогеография на основе
всего предыдущего материала дает зоогеографическое районирование*
биосферы.
Экология1 изучает взаимоотношения организма со средой его обитания,
приспособление к ней, и противоречия между особенностями вида и абиоти-
ческими и биотическими элементами этой среды, именуемыми факторами;
1 Термин введен Геккелем в 18(36 г.
3 Зенкевич, т. J
33
задачей экологического исследования является познание условий суще-
ствования вида, т. е. тех факторов среды, которые являются необходи-
мыми для существования вида (К а ш к а р о в, 1938). Это наиболее
широкое толкова ине задач экологии включает в нее также и изучение
взаимоотношения организма с другими организмами, называемое син-
экологией, в противопоставлении к аутэкологии (экологии отдельного
организма) и воздействие организмов на среду. Задачей экологии яв-
ляется также описание и объяснение распределения организмов во внеш-
ней среде, но, в противоположность биогеографии, главным образом
в рамках топографических подразделений мелкого порядка.
Разделение аутэкологии и синэкологии или биоценологии в разные
науки, как это делал Геккель (1866), совершенно неправильно. Аут-
экологию он считал основным содержанием экологии, а синэкологию
переносил в биогеографию (хорологию но Геккелю). Очень многие аутэко-
логические особенности отдельных организмов могут быть нами поняты
только в системе бноцепотических связей и наоборот — эти последние
без аутэкологического анализа останутся в большинстве непонятны.
Однако другой крайностью было бы утверждение, что все взаимоотно-
шения отдельного организма с абиотической внешней средой могут
быть изучаемы и поняты только в системе биоценоза и вне такового
теряют свое реальное значение.
Гидробиология представляет собою наиболее молодую из трех рас-
сматриваемых наук, с наиболее спорным объемом и содержанием.
Еще совсем недавно многие исследователи склонны были чрезмерно
расширять рамки гидробиологии и относить к ней любое исследование
систематического, анатомического, экологического или физиологического
характера, если объектом его является водный организм. Гидробиология
представляется в таком случае некоторой всеобъемлющей наукой, исполь-
зующей для своих целей! целый ряд смежных наук. В последнее время
часто ее рассматривают только как часть экологии, а именно, как эко-
логию водных организмов (С. Зерно в, Д. К а ш к а р о в) и иногда
противополагают ее аэробиологии (С. 3 е р ков) — экология наземных
животных.
В связи с этим С. 3 е р н о в в своей «Общей гидробиологии» придер-
живается такой структуры гидробиологии: гидробиология имеет три основ-
ные понятия: водный организм, биоценоз и тип водного бассейна и
рассматривает их как три последовательные ступени — идиографп-
ческую, ценографическую и океанологическую (соответственно лимно-
логическую).
Можно, однако, рассматривать гидробиологию как самостоятельную
от экологии науку. Основной ее задачей является изучение совокупности
массовых жизненных явлений, протекающих в водоемах, в рамках ком-
плексного их (водоемов) изучения, как целого. Гидробиология должна
дать оценку всей совокупности явлений прихода, превращений, нако-
пления и расхода органического вещества в водоемах с точки зрения вы-
явления и количественного учета ведущих массовых процессов, скорости,
напряжения и динамики их и степени накопления органического веще-
ства живого и неживого (Л. Зенкевич и В. Б р о цк а я), иначе
говоря, изучение явлений биологической продуктивности водоемов.
Гидробиология должна создать теоретическую базу для регулирования
человеком и направления в желательную сторону процессов биологи-
ческого продуцирования водоемов и наметить основные пути этого воздей-
ствия. Как правильно говорит G. М у р а в е й с к и й, «только ^а теория
биологической продуктивности, которая исходит из принципа возмож-
ности применения человеческого труда к повышению (или вообще к пзме-
34
пению) биологической продуктивности в водое?1ах, явится действительной
подлинной теорией».
Итак, объект гидробиологии это водоем со всей совокупностью жиз-
ненных явлений, протекающих в нем как в целом, в их взаимосвязи и
суммарном выражении; основная задача ее — изучение основных массовых
биологических явлений, протекающих в водоеме, и в первую очередь
явлений биологической продуктивности и изыскание способов управления
ими в хозяйственных целях; основной метод — количественный учет»
В связи с этим мы можем рассматривать три последовательные ступени
гидробиологии С. Зернова как последовательные исторические
этапы ее развития.
С. Муравей с к и й (1936) считает возможным долить совре-
менную гидробиологию на две самостоятельные науки: гидробиологию
собственно — науку о закономерностях между водной средой и населяю-
щими ее организмами, причем на первом месте должен быть всегда орга-
нпзм, и биогидрологию (как часть гидрологии) —науку, изучающую водо-
емы как целое с биологических позиций (по аналогии с биохимией,
биофизикой и т. п.), с последующим делением этой последней на биоокеапо-
логпю. бполимнологпю, бпопотамологпю. Нам же кажется, что то, что
М у р а в е й с к и й разумеет как гидробиологию, представляет собою
Ч.ТТ11 других наук п в первую очередь экологии, физиологии, морфологии
п зоогеографии водных организмов.
Сравнение задач, объекта и метода биогеографии, экологии и гидро-
биологии. Различия между тремя рассматриваемыми науками можно
характеризовать различием основных задач, объекта и метода, что сум-
мировано на основании вышесказанного в следующей таблице (табл. 2).
Таблица 2
Основная проблема
Объект
Метод
Биогео-
графия
Зоогеографическое райониро-
вание на основе закономерно-
стей современного распреде-
ления, основных особенностей
и генезиса фауны
Биогеографически и
ареал
Качественно-коли-
чественный анализ
флоры и фауны
Па \ ка
Экология
II з\ ченге взаимоотношений
организмов н внешней среды
друг к другу
Элементы связей | Полевые наблюдения
' и эксперимент
Гидро- I Изучение биологической про-
биология дуктивности водоемов (баланс
органического вещества)
Массовые процессы
в водоемах, связан-
ные с жизнедеятель-
ностью организмов
Количественный
учет
Процесс биофильтра, как пример комплексного явления в водоеме.
В качестве хорошего примера, поясняющего высказанные положения,
можно взять хотя бы явление биофильтра в море.
Многие морские животные — губки, двустворчатые моллюски, усоно-
гие и веслоногие ракообразные, оболочниковые, пропускают через осо-
бые полости тела (парагастральная, мантийная, жаберная) или вдоль тела
большие количества воды, отфильтровывая при этом из нее органическую
и неорганическую взвесь, производя, таким образом, осветление воды.
з*
35
Оказалось, что количество воды, пропускаемое через мантийную по-
лость мидии, образующих мощные барьеры у побережий, соизмеримо но
объему с массами воды неритической зоны (К. В о с к р е с е и с к и й, 19\8).
Если в какой-то степени справедливо допущение (К. u е н е н. 1938,
Daly, 1942) о роли мутьевых потоков в формировании подводных кань-
онов, то несомненно будет также и значение барьеров бпофильтраторов,
способных осветлять мутьевые потоки п тем изменять их плотность,
направление движения и силу специфического действия.
С другой стороны, моллюски биофильтраторы, наряду с фекалиями,
могут выделять в еще больших количествах псевдофекалии — агглю-
тинированные частицы взвеси, склеенные слизью. Многометровые толщи
илистых отложений у морских побережий, под грядами мидий, а также
в открытых частях океанов могут быть нацело сложены из их фекалий
и псевдофекалий.
Наконец, большое значение может приобретать при атом также и
воздействие на планктон и в частности на личинок других животных,
вовлечение их в биофильтрациопный поток.
При помощи данного примера мы хотим оттенить три основных
момента.
Во-первых, комплексность явления биофильтра в море. Способность
животных пропускать через свое тело громадные массы воды и, осаждая
взвеси, изменять ее плотность оказывает влияние на направление токов
воды и меняет конфигурацию сил, воздействующих на формирование
рельефа дна; одновременно ото ведет к образованию богатых органиче-
ским веществом илистых отложений и к повышению прозрачности воды.
Также значителен должен быть и биоценотический аспект этого явления.
С другой стороны, все это явление приобретает для нас интерес только
в силу его массовости. Если бы явление биофильтра имело весьма ограни-
ченное распространение и было связано с немногими формами, не образую-
щими плотных поселений, вся проблема осталась бы в области морфоло-
гии и экологии и для гидробиологии не представляла бы особенного
интереса.
Наконец, в третьих, все это явление приобретает полноценное значение
только в рамках какого-нибудь конкретного водоема, имеющего определен-
ные очертания берегов, рельеф дна и объем водной массы.
Массовость явления и его комплексная связь со многими разделами
океанологии, в рамках конкретного морского бассейна — три основных
критерия для включения его в рассмотрение морской гидробиологии.
Таким образом, морфолога и физиолога в рассмотренном явлении
будет интересовать строение и функции фильтрационного аппарата.
Эколог основное внимание обратит на распределение организмов био-
фпльтраторов в зависимости от условий среды обитания и воздействия
бпофильтраторов на поселение планктона фильтруемой воды. Гидробиолог
будет анализировать значение биофпльтрационного аппарата в цикли-
ческих процессах, совершающихся в водоеме. Распределение же бпофиль-
трационных систем в мировом океане явится частью биогеографии.
5. ОСНОВНЫЕ ПОНЯТИЯ УЧЕНИЯ О БИОЛОГИЧЕСКОЙ
ПРОДУКТИВНОСТИ
Возникновение учения о биологической продуктивности водоемов.
Только начинающее развиваться учение о биологической продуктивности
водоемов имеют едва тридцати летнюю историю и основным своим объек-
том имеет воду водоемов, как особое природное тело и его харак-
тернейшее свойство— плодородие. 40 гедами ранее эти же основные
36
понятия возникли применительно к почве в почвоведении, науке, со-
зданной в последние десятилетия прошлого столетия В. Докучаевым,
II. Костычевым и Н. Сибирцевым, а в наше время блестяще развитой
В. Вильямсом. Только эти два природные тела — вода и почва — обла-
дают свойством плодородия. И в том и в другом случае в основе лежит
минеральное тело, переработанное длительным процессом в особые при-
родные тела, именуемые В. Вернадским биокосными. «Накопленные
отличия, говорит Вильямс, слагаются в повое существенное свойство —
плодородие, которое диаметрально противоположно по своему значению
свойству первоначальной горной породы — бесплодию».
Б о й с е и - И е н с с и (J. В о j s е п - J е n s е и, 1919) первый дал
основные понятия биологической продуктивности и правильно расчле-
нил их (применительно к морской фауне). Эти обобщения явились резуль-
татом предшествующих десятилетних количественных исследований дон-
ной фауны, начатых в 1909 г. Петерсоном и его учениками
на Датской биологической станции. Б о й с е н - И е н с е и различает,
во-первых, «запас» (Stock). Это понятие близко к биомассе Д е м о л л я
и выражается в граммах сырого веса на 1 м2 или в количестве экземпля-
ров на 1 м2. Остаточной биомассой (rest stock) Б о й с е н - И е н с е и
называет то, что остается от исходной биомассы через год (от весны до
весны). Выедание (consumption), прирост исходной биомассы (growth
increment), учитывая при этом и смертность, и новое поколение, или
0-группу (Upgrowth) дополняют два первые понятия.
Исходную и конечную биомассу, а также новое поколение Б о ii с е ri-
ll е н с е н определял непосредственным учетом. Выедание определялось
U , V
по формуле (я—а1) , где а и aY — исходное и конечное число эк-
земпляров, а Б и У3, — исходная и конечная биомасса. Прирост исходной
биомассы определялся добавлением к выеденной биомассе исходной и от-
нятием конечной. Однако Б о й с е н - И е и с е и за годовую продукцию
принимает сумму из прироста исходной биомассы и нового поколения.
На примере моллюска Solen Б о й сен- И е н с е н иллюстрирует свой
метод (рис. 11). Подобным образом начатые С. Экманом одновре-
менно с Петерсеном (1909) количественные исследования дон-
ной фауны на озерах также привели, но только несколько позднее, к пер-
вым теоретическим обобщениям явлений биологической продуктивности
для пресных водоемов (Т hi one ma п п, 1926 и 1931, Naumann,
L u n d he с к, 1926, D е т о 1 1, 1927 и М ii n s t е г - S I г б т,
Запасй!
а--число жз.
V-- Сырой бес
Выедание
Продукция
Прирост
исходной
биомассы
о-группа
Остаточные
за 'io 7.
у-
Рис. И. Динамика популяции моллюска Solen в датских водах
(по Бопсен-Пенсену).
1928 к 1932). И на море и на пресных водах начало количественных
исследований донной фауны совпадает с введением в практику количе-
ственных орудий лова — дночерпателей, независимо друг от друга при-
мененных Э к м а н о м на шведских озерах, а Петерсеном в дат-
ских морских водах. В дальнейшехМ проблема биологической продук-
гивности водоемов стала содержанием соответственного раздела со-
ветской науки и в пашей стране поднята на новый высокий уровень.
Понятие продуктивности, биомассы и продукции. При рассмотрении
вопросов биологической продуктивности водоемов, применительно к
живым организмам, пользуются тремя основными понятиями — про-
дуктивность, биомасса и продукция. Под продуктив-
ностью (плодородием) разумеют все явление в целом, во всем его разнооб-
разии, как свойство водоемов обусловливать тот или иной характер разви-
тия вещества в живых организмах и темп воспроизведения этого вещества.
Когда же мы разрешаем проблему применительно к определенному
пространству и к определенному отрезку времени, то мы оперируем с
понятиями биомасса (Д е м о л ь, 1927) и продукция (Б о й с е и - И е н-
иеп, 1919). Под биомассой разумеют количество вещества в живых орга-
низмах на определенную площадь, выраженное в весовых единицах. Био-
масса может быть разовая, средняя, максимальная, минимальная и
постоянно претерпевает более или менее значительные изменения во вре-
мени. Продукция (урожай) есть выражение того прироста, который дает
биомасса за определенный промежуток времени, и который складывается
из: 1) естественного увеличения биомассы за счет роста организмов,
2) отрождения новых индивидуумов и 3) убыли биомассы, обусловливае-
мой естественным отмиранием особей, отходом в результате биологиче-
ских превращений, миграций, вылова человеком и гибели от врагов и от
наступления неблагоприятных для жизни условий. Позднее часть понятий,
сформулированных Б о й с е н - И е н с е н о м (1919), анализировал
Т и н е м а н н (1931) и детальному теоретическому рассмотрению под-
верг Мюнстер-Стрем (1932). В СССР, в связи с многочисленными
количественными исследованиями фауны и флоры водоемов, за послед-
ние годы эти понятия также неоднократно подвергались обсуждению
как с точки зрения общих принципиальных установок, так и в отдель-
ных деталях (Е. Б о р у ц к и й 1938, В. Б р о ц к а я и Л. 3 е и к е-
в и ч 1936, К. Б роде к и й 1934, Г. В и н б е р г 1936, Г. К а р-
з и н к и н 1936, С. М у р а в е и с к и и 1936, Л. Р о с с о л и м о
1934, В. Жадин 1910).
Основными показателями продуктивных свойств водоема должны
быть: 1) количественное распределение живых организмов, органиче-
ского детрита, растворенных органических веществ и питательных солей,
как по поверхности дна, так и в толще воды водоема (статика); 2) ка-
чественные и количественные показатели динамики всего органического
вещества; биологические особенности и в первую очередь показатели
темпа роста для основных составляющих население водоема групп и форм:
3) хозяйственный выход промысловых объектов. Система возможных
мероприятий, направляемых человеком к повышению, изменению и регу-
лировке продуктивных свойств водоема, всегда должна быть в центре
внимания биолога.
Проблема биологической продуктивности в трактовке различных авто-
ров. Несколько более расширительное толкование проблеме биологи-
ческой продуктивности водоемов дает Л. Р о с с о л и м о (1934): «для
полного понимания явления необходимо изучение всех элементов его,
всех фаз и состояний, проходимых органическим веществом и заключен-
ной в нем энергией, всех физикохимических и биологических процессов,
связанных с этими превращениями и перемещениями, без отрыва от ка-
чественной специфичности процессов накопления и превращения энергии»,
иначе говоря, баланс их вод, в том числе и органический.
Почти столь же широкие рамки теории биологической продуктив-
ности, только в несколько ином смысле, ставит и С. М у р а в е й с к и й
38
(1936J. Она должна разрешить, по его мнению, вопросы рационального
использования организмов, населяющих водоем и биологических процес-
сов, в нем протекающих. Г. К а р зи н к и н склонен рассматривать
теорию биологической продуктивности более ограничительно и приме-
нительно к «четкому и ясному представлению о продукте», а продуктив-
ность определяет как способность воспроизводить про-
дукт. «Любой процесс, — говорит он, — связан с объектами, в которых
он протекает. Каждая последующая фаза процесса есть продукт преды-
дущих фаз. Следовательно, понятие продукта есть понятие относитель-
ное, относительная фаза данного процесса. Продукт становится продуктом
лишь по отношению предыдущих фаз своего развития, так, например,
.мезосапробная вода есть продукт процессов, разыгравшихся в полпсапроб-
ной воде». Таким образом, Г. К а р з и и к и и продуктом считает не только
тот конечный продукт, который непосредственно представляет хозяйствен-
н ли интерес, а «любую фазу процесса». Тем самым стираются существен-
ные отличия такой трактовки от более широких определений других авто-
ров. При этом первой фазой разрешения всей проблемы в целом Г. К а р-
з и н к п н считает вопрос кормноети водоемов.
В качестве примера отвлеченной трактовки понятий апологической
продуктивности водоемов можно привести взгляды М ю и с т е р - С т р ё м а.
Этот автор различает понятия продуктивность и продую
ц и я и прежде всего конструирует свои понятия на некоторых идеаль-
ных водоемах, которые отличаются отсутствием всякой смены воды и не-
растворимым ложем. Установив основные понятия па этих «идеальных»
водоемах. Стрём переносит их на реальные водоемы. Вторая харак-
терная особенность построений Стрёма заключается в том, что он опе-
рирует только с питательными солями в зоне фотосинтеза и с продуцен-
тами. т. е. растительными организмами.
Стрём оперирует понятием п о т е н ц п а л ь н о й 1 и ф а к-
т и ч е с к о й про д у к т и в н о с т и и под первой разумеет коли-
чество органической субстанции, соответствующей всей сумме минпмум-
солей воды п организмов в слое фотосинтеза, а под второй — то же са-
мое, что обычно именуется биомассой. В данном случае Стрём имеет
в виду биомассу растительных организмов в слое фотосинтеза. Что ка-
сается продукции, то под валовой продукцией Стрём ра-
зумеет сумму всех особей, отродившихся за определенный период времени,
а под чист о й п р о д у к ц и е й — величину, слагающуюся из ор-
ганических отложений в грунте, не возвращающихся в питательный цикл,
и организмов, покидающих водоем тем пли иным путем, т. е. это — сумма
органических веществ, выходящих за пределы циклических процессов
водоема. Таким образом, по Стр ё м у, если организмы родились, вы-
росли и живут в водоеме, включать их в сумму продукции нельзя.
Коренных недочетов определений Стрёма два — во-первых, он
из всего громадного разнообразия факторов, слагающих физиономию во-
доема. берет только один —материальную основу для построения тела
продуцентов в зоне фотосинтеза. Во-вторых, он пытается дать харак-
теристику процессов биологического продуцирования только по двум фа-
зрм — «прпвходу» и «выходу» и оставляет в стороне самое существенное —
динамику процессов, которая именно все и определяет п только исходя из
которой для нас представляют интерес эти две фазы, теряющие иначе свой
основной смысл. При одних п тех же количествах «минимум» солей и ор-
1 Блоцкая п Зенкевич (1936) понятию потопциальпой продукции придают чисто практиче-
ское хозяйственное значение, это —возможная степень повышения продуктивных свойств водо-
ема в результате мелиоративных мероприятии, допускаемых совокупностью наличных факторов
39
ганических отложений в грунте, в водоеме могут разыгрываться весьма
различные процессы продуцирования. Водоемы с пониженной продук-
цией могут отлагать на дне обильные массы органических веществ и на-
оборот. В зависимости, хотя бы от температуры, при одном и том же коли-
честве солей, процессы будут развертываться с совершенно различным
темпом и эффектом. Но меньшее значение имеют различия в составе фауны
и флоры со всеми свойственными им особенностями. Нам кажется, что для
биолога основной интерес именно и представляет, как и в какой форме раз-
вертываются процессы продуцирования, т. е. что представляет собой
основная масса органического вещества во всех его фазах, каковы ка-
чественные и количественные показатели динамики этой массы в посе-
зонной смене, при более длительных изменениях в палеогеографическом
разрезе и при изменениях, связанных с хозяйственной деятельностью
человека. Также не удовлетворяет нас и стремовское понятие потенциаль-
ной продукции, так как в основу определения ее ложится то максималь-
ное выражение, которое мы получим, если всю сумму мшшмум-солей в
зоне фотосинтеза перечислим на органическое вещество. Эта величина
представляется нам чистой абстракцией, не выражающей основного мо-
мента того колоссального конкретного разнообразия, которое мы имеем
в природе. Взять хотя бы тот факт, что количество минимум-солей в з-ше
фотосинтеза —величина, тесно связанная с запасами солей в глубинных
слоях. Если в водоеме лимитирующие соли нацело перешли в живое
органическое вещество, то «потенциальная» продуктивность Стрёма
будет выражаться разовой биомассой на данный момент и не будет от-
ражать напряжения циклических процессов и характера отдельных их
фаз.
Принимая стремовскпе понятия потенциальной и фактической про-
дуктивности, М у р а в е й с к и й (1936) в отношении первой к послед-
ней считает возможным видеть меру напряжения биологических процессов
в водоеме, определяющих биологическую продуктивность. Основным
явлением в «биогидрологическом» аспекте М у р а в о й с к и й считает
динамику биогенных элементов, а фауну водоема во всех его проявлениях,
циклические процессы и отдельные их фазы рассматривает как произ-
водное динамики биогенных элементов (веществ).
По указанным выше соображениям, такая трактовка вопроса неиз-
бежно должна оставить без учета не только состав фауны данного водоема,
но и всю совокупность свойств этой фауны в комбинации со всеми усло-
виями внешней среды (температура, свет и т. д.). Все это может считаться
производным динамики биогенных элементов только условно. Приведем
хотя бы только два примера. Рыболовство Северного моря знает случаи
массового цветения воды одноклеточной водорослью Phaeocyslis, захваты-
вающего обширные части водоема. Английские рыбаки называют это
«вонючей водой». Рыбы не используют Phacocystis в пищу, а громадные
косяки сельди, встретив зоны цветения Phaeocystis. совершенно изменяют
направление своих миграционных путей. В 1934—37 гг. в Каспийском
море в колоссальных массах развилась случайно попавшая сюда из Чер-
ного моря диатомовая водоросль Rlrizosolenia calcar aw я. мало употреб-
ляемая в пищу рыбами и частично вытеснившая другие, более полезные
для рыб компоненты планктона.
В обоих приведенных случаях мы имеем очень высокие показатели фо-
тосинтеза, но для целого ряда последующих явлений развитие РНане-
су stis и Rhizosolenia весьма различны. Различна и дальнейшая судьба
тех органических веществ, которые накоплены обеими водорослями.
В Каспийском море большая масса их, опустившись на глубины, выйдет
надолго из круговорота, в Северном море в большей своей части посту-
40
пит уже в последующие годы в новые циклические процессы. Как можно
видеть, явленна, сходные по напряжению фотосинтетической деятельности,
влекут за собою весьма различные последующие фазы биологического
продуцирования. Мощные начальные фотосинтетические звенья пищевых
рядов не всегда обеспечивают и мощные конечные звенья — «выход» по-
лезной человеку продукции.
В своей «Проблеме стока» Мураве й с к и й (1936) дает очень инте-
ресную интерпретацию явления биологической продуктивности водое-
мов: «Сток,— говорит он, — это сложный процесс, определяющий взаи-
моотношение между океаном и водами суши, процесс обмена водных масс
океана и суши, процесс исторический, в котором факторы геологические,
географические, физические, химические, биологические не только опре-
деляют самы!’! обмен и его характер, но и обусловливают круговорот
веществ во всем процессе стока и в каждом водоеме (водном объекте),
в частности... каждый водоем со своей качественной специфичностью
представляет собой одну из фаз (частей) этого процесса стока, следо-
вательно, и круговорот веществ в данном водоеме является в основ-
ном функцией круговорота веществ в результате процесса стока в це-
лом». Эта формулировка представляет собой ценную попытку найти
некоторое синтетическое единство всех процессов, совершающихся в гид-
росфере.
В самое последнее время В. Ж а д п и ы м (1910) сделана попытка
построить единую теорию биологической продуктивности для донной
фауны водоемов всех типов, пресноводных и морских. Конечной задачей
своей теории автор считает выработку способов управления процессом
биологического продуцирования. Теория В. Ж а д и и а в основном за-
ключается в следующем: биологическая продуктивность есть функция
аккумуляции органического вещества и бпоэкологической обеспеченности.
Это последнее понятие, к сожалению, совсем неразработанное автором.
В. Ж а д и н выражает количественным богатством видов. Слабому на-
коплению органических веществ в водоеме, по Ж а д и н у, соответствует
качественная бедность фауны, при возрастании аккумуляции возрастают
качественное разнообразие фауны и биомасса, но только до определенного
предела. При дальнейшем возрастании аккумуляции органического ве-
щества видовое разнообразие резко падает, а биомасса продолжает уве-
личиваться, пока не наступает,наконец, п падение биомассы. При явлении
гппераккумуляцпп органического вещества оба показателя биологиче-
ской продуктивности падают до пуля.
Нам кажется, что эта теория прежде всего представляет собою несколько
своеобразную формулировку известной типологической классификации
озер Наумана — Т и н е м а н н а, построенную на ряде олпготро-
фня — эгтрофпя — дистрофия, с распространением ее на все водоемы зем-
ного шара. К морским водоемам, во всяком случае, теория В. Жадин а
мало при .пенима.
Что касается морских водоемов, то значительное качественное обед-
нение, имеющее место при переходе от тропических областей океанов
к полярным, так же как и при переходе от открытых морей к внутренним
(например, Белое или Азовское), ни в какой связи с аккумуляцией орга-
нического вещества не находится, так же как и усиление процессов биологи-
ческого продуцирования в областях так называемого полярного фронта.
Отсутствие жизни в глубинах Черного моря и почти полное отсутствие
<‘с в глубинах Каспийского моря не являются результатом гппераккуму-
л:щшт. а являются вместе с этой последней следствием характера дина-
мики водных масс в этих водоемах. За исключением только стагнирующих
котловин некоторых морских водоемов мы нигде не знаем случаев, чтобы
61
гнпераккумуляцпя органических веществ ооусловила качественное ооед-
нение и падение биомассы. Вряд ли вообще правильна попытка строить
единую теорию биологической продуктивности для всех водоемов: чем
более коротка н проста по виду формулировка такой теории, тем неизбежно
меньше ее теоретическая и практическая значимость, ио зато неуклонно
возрастает ее абстрактность, не говоря уже о том, что трудно построить
единую теорию биологической продуктивности без учета планктона и дон-
ной растительности.
В построениях В. Ж а дина очень ценно то. что он еще раз резко
подчеркивает значение для явлений биологического продуцирования на-
личия и возможности использования питательных веществ и различает
два состояния органического вещества в водоеме — состояние оседания
и состояние переноса. Нужно при этом оговориться, что, невидимому,
В. Жадин под органическим веществом разумеет не растворенное ор-
ганическое вещество, а детрит.
Терминология, применяемая в связи с проблемой биологической про-
дуктивности. На приводимой ниже таблице (табл. 3) мы делаем попытку
дать сводку терминологии, применяемой в связи с проблемой биологической
продуктивности, сейчас же отметим только некоторые наиболее существен-
ные моменты. Некоторые наши авторы (В о р о б ь е в, 1943, Ш о р ы г и и.
1939) вводят понятие остаточной п р о д у к ц и и, разумея под
этим то, что Б о й сен- И е н с е п именовал остаточным запасом (био-
масса по Д е м о л л ю), т. е. разницу между исходной и конечной биомас-
сой за определенный промежуток времени. Значительное неудобство
этого понятия заключается в том, что изменение биомассы за определен-
ный срок именуется п р о д у к ц и е й, понятием, выражающим про-
цесс. Лишь в очень редких случаях разница в биомассе будет действи-
тельно соответствовать продукции, в большинстве же случаев это буд^т
только разница в биомассе, и ничего больше. Эта разница ни в какой мере
не может характеризовать тех процессов продуцирования, которые имели
место за данный отрезок времени. Это неудобно еще и потому, что мы мо-
/КОхМ получать отрицательное выражение «остаточной продукции», тогда
как отрицательная и р о д у к ц и я сама по себе немыслима
и при отрицательном изменении биомассы на самом деле может иметь по-
ложительное значение.
К этой же группе неудачных терминов следует также отнести и
proof-production Г и с л е н a (Gislen. 1930), точно соответст-
вующий тому, что все другие разумеют под биомассой.
В. Я ш н о в и его ученики [В. Богоров (разные работы),
А. Ок ул (1939) и др.] пользуются термином титр, заимствованным у
Э. Наумана и измененным Я ш н о в ы м, Под титром В. Я ш н о в
разумеет вес организмов па единицу площади или объема, а иод биомассой
вес организмов на весь водоем или его часть. Такое деление имеет неудоб-
ство в искусственности разграничения понятий титр и биомасса. Нам ка-
жется более целесообразным ограничиться одним понятием биомассы,
каковое может одинаково применяться и ко всему водоему и к отдельным
ого частям, и к тем или иным единпцахМ площади или объема.
Методы количественного определения продуктивных свойств водоема.
Совершенно очевидно, что практическое разрешение всех вопросов био-
логической продуктивности и особенно наиболее сложного из них —
динамики всего органического вещества —может встретить применительно
к тому или иному водоему очень большие трудности и в первую очередь
в виду очень большого числа видов растений и животных, населяющих не-
которые водоемы. Определение для каждого из них указанных показателей
продуктивности было бы задачей едва ли разрешимой. Особенно возрастает
Таблица 3
СОДЕРЖАНИЕ ОСНОВНЫХ ПОНЯТИЙ БИОЛОГИЧЕСКОЙ ПРОДУКТИВНОСТИ
__________________________ВОДОЕМОВ___________________________
Явлсине "Гермин I । Автор
Вес. организмов на единицу площади или объема, или на весь водоем или его часть Запас (Slock) Биомасса Фактическая ироду к- ( тивиость । Proof-production Bojsen-Jensen, 1919 (также и количество эк- земпляров) Dcmoll, 1927 Большинство авторов Munster-Strom, 1932 (в отношении фитопланк- тона) Gislen, 1930
Вес организмов на весь водоем или часть его Биомасса Яшнов. Богоров, Усачев и некоторые другие
Вос организмов на 1 квадратный или куби- ческий метр Титр Яшнов. Богоров, Усачев и некоторые другие
Разница в биомассе в на- чале ив конце определен- ного отрезка времени Изменение биомассы во времени Остаточная продукция (?) Реальная продукция Броцкая и Зенкевич Воробьев, Шорыгин, Яшнов
Отношение разницы в био- массе за определенный про- межуток времени к исход- ной биомассе Остаточна я удельная продукция (?) Воробьев 1 i 1
Максим 1льпая сумма мипи- мум-солей в воде и в орга- низмах и солей, долженст- вующих поступить в водоем за данный отрезок времени, в гн,с фотосинтеза, пере- численная на органическое вещество 11 (>тен ц и ал ьпа я п роду к- тявность (?) Валовая продукция Miinsler-Strom, 1932 Карзипкип, 1936
Явление военроизведения органического вещества в живых организмах в во- доемах (Биологическая про- дуктивность) Большинство исследова- телей (в то)! или иной форме)
Прирост, даваемый naceie- н.юм водоема за год и сла- гающийся из прироста ор- ганизмов, родившихся в пре- дыдущие годы и от рожде- ния этого года (О-группа) Годовая продукция 1 । Bojsen-Jensen, 1919
Весь прирост, даваемый биомассой за год Годовая продукция i Валовая продукция 1 Бронкс и Зенкевич, 1936 Боруцкий, 1939 Воробьев, 1939 iMunsler-Strom, 1932
43
Пиление
Термин
Продолжение
Автор
Сумма органических отло-
жений в грунте, не возвра-
щающихся обратно в пита-
тельный цикл, и организмов,
покидающих водоем том или
иным путем (унос реками,
вылет насекомых, рыболов-
ство)
Чистая прод\ кция
Munster-Strom, 1932
Возможная степень повыше-
ния продуктивных свойств
водоема, в результате мел ио-
рати вп ых меройри яти й
Потенциальная продук-
тивность
Броцкая и Зенкевич, 1936
Скрытые в водоеме продук-
тивные способности, не мо-
гущие непосредственно быть
переветонными в продукт
С и особность, за к. iк»чинная
в ресурсах водоема, могущих
при данных условиях среды,
непосредственно превра-
щаться в продукт
Отношение годовой продук-
ции к средней годовой био-
массе. или продукции за
определенный отрезок вре-
мени к исходной биомассе
Потенциальна и прод\ к-
тявность
Актуальная иродх ктив-
НО'-ТЬ
Карзипкип, 1936
Карзипкип, 1936
I
I
P/В коэффициент । Demoll, 1927
1 Вроцкая и Зенкевич, 1936
। и другие
трудность при желании дать определение продукции для целых биоцено-
зов, для всего бентоса или планктона, или, наконец, для всего водоема
в целом. Однако на выручку исследователю приходит сама природа.
Как хорошо известно среди большого числа организмов, населяю-
щих водоемы, лишь очень немногие имеют массовое значение. Многие био-
ценозы дна Азовского моря состоят на 85—95%, в отдельных случаях на
100%, из одной единственной (Card turn, eclule. Sy tides шуа ovata и др.)
формы \ Планктон Варенцова моря па 85% состоит из Calunus Unniar-
chicus. В любом водоеме, где в состав фауны и флоры входят тысячи раз-
личных форм, лишь несколько десятков из них играют основную роль.
То же самое можно сказать и об основных биохимических процессах,
совершающихся в водоеме. При в г ем качественном многообразии совер-
шающихся в водоеме химических процессов лишь немногим из них при-
1 Такие биоценозы мы назвали олпгомнкстнымн, т. е. состоящими из малого числа ком-
понентов (Л. Зенкевич. 1931).
44
надлежит основная. ведущая роль. Получить же количественное выражение
всех показателей продуктивности для немногих форм и немногих процес-
сов задача вполне разрешимая. Конечно, мы и в этом случае не получим
какого-нибудь абсолютно точного выражения процесса продуцирования,
но можем получить вполне удовлетворяющую нас характеристику и оцен-
ку продуктивных свойств водоема. Нужны только правильно разработан-
ная система круглогодичных наблюдений и определенная последователь-
ность в этапах работы.
Первым этапом в изучении биологической продуктивности водоема
должно быть выявление основных фаунистических комплексов, их исто-
рического развития, количественного распределения комплексов и от-
дельных массовых организмов, а также биологических и биохимических
особенностей (статики). В результате этих исследований должны быть
выявлены ведущие организмы и процессы.
Вторым этапом работы должно быть детальное изучение превращений
по этим основным выявленным звеньям. На основании всего иолучен-
Рис. I?. Изменение по месяцам средних значений веса слое-
вища, черешка, молодого и старого листа Laminaria saccha-
rina (по Киреевой и Щаповой).
1 — новый лис г. 2 — старый лист, з — черешок.
него материала может быть сделан синтез — сравнительная оценка про-
дуктивных свойств водоема, со всеми вытекающими практическими меро-
приятиями.
Уже многое в этом направлении сделано у нас и на пресных и на морских
водоемах и для бентоса и для планктона. Очень хорошие данные получены
по Черному и Варенцову морям для макрофитов (Н. В о д я н и ц к а я-
Морозова, 1936, М. К и р е е в а и Т. Щапова, 1938), представ-
ляющих для изучения явлений биологического продуцирования значи-
тельно менынне трудности, чем животные. Блестящие результаты полу-
чены также в этом направлении и для озерного бентоса (Е. Б о-
р у ц к п й, 1939) и для бентоса Азовского моря (В. Воробьев, 1945).
Приведем два примера разрешения задачи практического выражения
основных показателей биологического продуцирования. Первый пример
относится к мурманской Laminaria saccharina и взят из работы М. К и р е е-
в о й п Т. Щ а п о в о й (1938). Слоевище ламинарий состоит из разно-
возрастных частей (рис. 12). Уже в конце лета, зимой и весной старый лист
45
разрушается и постепенно сбрасывается, а вместо него весной и летом нара-
стает новый лист, являющийся продукцией одного года. Наоборот, сте-
бель ламинарии представляет собою многолетнее образование. На рисунке
показан годовой ход изменений в весе нового и старого листа и стебля.
Имея, кроме того, данные по количественному распределению ламинарий
на дне того или иного района, мы можем без труда определить годовой
прирост продукции на всю площадь.
Гораздо более сложен процесс определения годовой продукции д m
животных. Однако вполне удовлетворительное решение задачи уже дано
рядом авторов. Наиболее точное практическое решение получено Е. В о-
р у ц к и м на озерах применительно к населению дна профундали. Е. В о-
р у ц к и й прежде всего дает динамику биомассы водоема в виде рабочей
схемы (рис. 13), на которой учтены биомасса живых организмов определен-
ного вида в данной водной и воздушной среде (кружки), биомасса есте-
ственно отмерших особей (черные треугольники), биомасса организмов,
Рис. 13. Схема динамики биомассы
бен юса озера (по Боруцкому).
Рис. 1/. Динамика биомассы профунда-
ли Белого озера (в Косине), выраженная
в кг сухого вещества (по Боруцкомч).
съеденных другими организмами (белые треугольники), биомасса орга-
низмов, перешедших в другую биологическую фазу, в данном случае окук-
ливание и вылет (миграции), вылупление из яиц (отрождение) (черные квад-
раты) и, наконец, увеличение биомассы за счет роста.
По этой схеме Е. Б о р у ц к и й определяет количественное выра-
жение отдельных моментов для всех основных компонентов бентоса, а
затем дает сводную диаграмму для всего бентоса (рис. 14) профундали
Белого озера в Косине, в килограммах сухого вещества. Мы можем счи-
тать, что на настоящий момент работа Е. Б о р у ц к о г о дает наилуч-
шее разрешение вопроса и совершенно достаточную количественную ха-
рактеристику продуктивных свойств глубинного бентоса озер. Само собой
понятно, что впервые полученное определение годовой продукции важней-
ших составных частей рыбьего корма имеет большое практическое зна-
чение.
Для сравнения продуктивных свойств разных организмов между со-
бою и, главным образом, для сравнения друг с другом различных водое-
мов может бытье успехом использован так называемый Р В коэффициент
46
(D е ш о 1 1, 1927) — отношение прироста к биомассе (Production — Bio-
masse). P/В коэффициент может иметь различное выражение. Ото может
быть: отношение годовой продукции к средней годовой биомассе или
отношение годовой продукции к исходной биомассе. Наконец, продукция
может определяться не только на год, но и для любого промежутка вре-
мени. хотя бы для одних суток 1.
Не раз делались попытки найти количественное выражение процессов
продуцирования путем учета совершающихся в водоеме биологических про-
цессов, однако все они учитывают только процессы, связанные с жизнедея-
тельностью фитопланктона. Планктон, бентос и рыбные богатства водоема
отнюдь не находятся в постоянном одинаковом количественном соотноше-
нии и поэтому продукция фитопланктона еще не характеризует продукции
других групп организмов. Одна из таких попыток принадлежит Е. К р о п-
с у и Н. В е р ж б и и с к о й (1930) в отношении Варенцова моря.
Ими установлено общее количество фосфатов, которое было использовано
на развитие фитопланктона, по разнице между весенним и летним коли-
чеством данной миннмум-соли. Перечисляя это количество на живое ве-
щество, Е. К р е и с и И. Вер ж описка я. таким образом, устанав-
ливают, что годовая продукция фитопланктона в Баренцевом море дол-
ита составлять 300—500 тонн на квадратный километр. Одл iko расчет,
сделанный данными авторами как бы предполагает, что на перешедшем из
виды в организмы фосфоре развилась только одна генерация организмов.
Все попытки суммарного определения продукции планктона, начатые еще
Аткинсом, имеют еще один дефолт, а именно, невозможность устано-
вить роль фотосинтезирующих форм планктона и роль других планктон-
ных организмов, потребляющих эти же биогенные элементы, и в порву к
очередь бактерий.
Гораздо более реальное значение для определения продукции фито-
планктона имеет метод, использующий суточные колебания количества
растворенного в воде кислорода. Зная количество кислорода, выделенного
в толще воды за сутки, можно вычислить, какое соответственно создано
количество вещества планктонными растениями, так как между этими ве-
личинами существует строго пропорциональная зависимость. При этом
должно быть также учтено количество кислорода, поглощаемого планк-
тонными организмами, в результате дыхания и окисления.
Впервые точный метод такого учета был разработан Пюттером
(А. Р ii tter, 1924), выдерживавшим в склянках на свету и в темноте
пробы морской воды, взятые из моря. В светлой банке продолжались
процессы фотосинтеза, а в темной шли только процессы потребления кисло-
рода. Складывая обе величины, Пюттер получил суммарное количе-
ство кислорода, выработанное фитопланктоном за сутки. С различными из-
менениями методикой П ю т т е р а пользовался Гран (G. Gran, 1927
и 1929). У нас этот весьма ценный и точный метод был применен с
изменениями Г. В и и б е р г о м на озерах (1934, 1935, 1939, 1946),
П. Ширшов ы м (1934) и С. Бруевичем (1939) — на море. Про-
изводимые достаточно часто в течение года подобные определения су-
точной продукции фитопланктона дают весьма точные показатели годовой
его продукции.
Хотя числа, получаемые таким способом, характеризуют только продук-
цию фитопланктона и в большинстве не показательны для бентоса и мало
показательны для нектона, они представляют интерес, так как уточняют
одну из последовательных фаз биологического продуцирования.
1 С. Бруевич заменяет очень удачный P/В коэффициент Де чо л л я мало удачным терми-
ном «оборачиваемость».
47
Новые пути дальнейшего изучения явления биологической продук-
тивности открывают последние работы И в л е в а, Готчи нсо и а
(Hu L с 11 i n s о п), Л п н д см а н a (Lin d е in а и), Г а е в с к о й и др.
В. Л и и д е м а н (1942, 1945) считает, что изучение системы и ди-
намики трофических связей в биоценозе количественными методами «по-
новому освещает динамику смены биоценозов» и в атом отношении является
одной из основных частей количественного изучения биологической про-
дуктивности. Действительно трофология с этой точки зрения является
промежуточной областью исследования между экологией и гидробиологией.
Л и н д е м а и делает попытку дать общетеоретическое обосно-
вание данной проблеме. Следуя Г е т ч и н с о н у (Н и t с h i n s о n.
1942), Л и и дема и обозначает продуцентов планктона как первый
уровень использования солнечной энергии (AJ. Питающийся фито-
планктоном зоопланктон есть второй уровень использования (А2).
Те компоненты планктона (хищники), которые питаются сами зоо-
планктоном, представляют третье звено (А;{) и т. д. до конечных
звеньев-уровней (Azl). Энергия поступает в каждое последующее звено
и покидает его, переходя в новое звено, при этом количество энергии,
включенной в живые организмы, уменьшается. Скорость изменения за-
паса энергии Г е т ч и и с о и определяет формулой с^а =1п %-,
где — положительная величина, представляющая скорость поступления
энергии от Ап-1 к Ал, а — отрицательная величина, представляющая
собой сумму скорости энергии, убывающей с уровня Ал и скорости запаса
энергии, передаваемого следующему уровню Azz;4. Таким образом, % опре-
деляет истинную продуктивность уровня п, причем следует учитывать
потери на рассеяние энергии, на дыхание, хищничество и посмертное раз-
ложение. Используя также данные В. И в л е в а (1939) по подсчету энер-
гии ассимиляции, роста и дыхания у малощетинковых червей Tubifex и
годовиков карпа, Л и н д е м а н дает для первых дыхание в процентах
роста 62,3%, а для второго объекта — 140%. т. е. гораздо более высокий
коэффициент. Если считать максимальную эффективность роста в 60—70%,
то 140% можно считать за среднюю величину этого коэффициента для ды-
хания водных хищников. Что же касается продуцентов, то для них со-
ответственный коэффициент Л и и д е м а н принимает в 33%, таким об-
разом получается ряд — 33 : 62 : 140.
Так как никакой вид питания не дает полного энергетического исполь-
зования пищевого материала, то следует еще учитывать недопереваривае-
мость пищи, которая, видимо, меньше на более высоких пищевых уровнях.
В результате Линдеман дает расчет продуктивности разных
пищевых групп для Каспийского моря (поданным С. Бруевича, 1940)
и двух американских озер (табл. 4).
Группа организмов по уровню питании
Фитопланктон Ат.............................
Фитобентос Ат...............................
Зоопланктон А2 .............................
бентосные травоядные А2.....................
бентосные хищники А3........................
Планктонные хищники А.».....................
Промысловые рыбы А3 (л.А-,?)................
Карп А;. (+А.,?)............................
Рыбы-охотники А, <+ А.,?)...................
Тюлени А5...................................
Таблица /
Каспий 1 Оз. Мендота Оз. Цедар- Бог
59,5 j 299 25.8
0,3 i 22 ' 14.6
20,0 I | 22 ! 6.1
: 1.8 О.8
20,6 0.9‘ , < ).2
1 1 I о.ь
0 6 1 1 - - 1 0,3
0.1) ; 0,2 | 0,0
(>.6 0.1 ! 0.0
Од) Г о?» ; 0,0
являются травоядными.
* Па основании допунщпня,
что 2/3 донной фауны
I
;8
Хотя эти данные имеют пока только предкарительяып характер,
весьма схематизируют явление и не учитывают индивидуальных особен-
(<<)'rreii различных видов, однако они передают порядок величин и
показывают и перспективный путь для исследований в области изучения
проблемы продуктивности.
Согласно Г е т ч и н с о н у эффективность продуктивности любого
уровня (Ал) относительно продуктивности любого предыдущего уровня (А/л)
можно вычислить по формуле А-и пользоваться коэффициентом• 100.
Это дает возможность определять для разных звеньев пищевой цепи «про-
грессивную эффективность». Эта последняя, как величина относительная,
отличается от продуктивности, представляющей собой выражение процесса
:'табл. 5). Однако и здесь следует отметить чрезвычайную схематич-
ность построения и оторванность от тех конкретных явлений, которые
имеют место в водоемах.
Все приведенные данные из работы Линдемана уточняют смысл
тех соотношений разных групп в биоценозе, которые Элтон (Elton,
1927) выразил своей «пирамидой»: по мере перехода от первичных звеньев
лшцевой цепи к последующим количество экземпляров (соответственно
и биомасса и продуктивность) уменьшается, а размеры увеличиваются.
Таблица б
ПРОДУКТИВНОСТИ В е-?шл/сж2/годах И ПРОГРЕССИВНЫЕ ЭФФЕКТИВНОСТИ
ПИЩЕВЫХ ЦЕПЕЙ ДВУХ АМЕРИКАНСКИХ ОЗЕР
(По Линдеману)
Звенья пищевой цепп
Радиация..........................
Продуценты At....................
Первичные консументы А2...........
Вторичные *» А,...........
Третичные » А4...........
Оз. Мендота
II роду к- । Эффектив-
тнвпость । ность в °/0
___Оз. Цедар-Бог
И роду к- ! Эффектпв-
тивность | ность в %
118.872 I -
1П.З ! 0,1
14,8 । 13,3
3,1 । 22,3
i
118.872
48(С
41.7
ПЗ
(»40
87
5.5
13,0
Конечно, «элтоновская пирамида» требует некоторых оговорок. Био-
масса первичных пищевых звеньев может быть меньше, чем у последующих.
Так в мелководных морях бентос количественно всегда богаче планктона,
а зоопланктон большую часть года по биомассе больше фитопланктона.
Эта неувязка вполне перекрывается гораздо более высоким темпом раз-
множения у фитопланктона. С другой стороны, макрофиты моря нередко
очень велики по размерам.
’ Повидпмому, слишком высока.
** Нов удимому, слишком низка.
4 Зенкевич, т. I
МИРОВОГО
ГЛ ABA L
РАСПРЕДЕЛЕНИЕ МОРЯ И СУШИ НА ПОВЕРХНОСТИ
ЗЕМЛИ И ОБЩИЙ БАЛАНС ГИДРОСФЕРЫ
идросфера, водная часть земной оболочки, состоя-
щая примерно на 99,5% нз вод соленых — галосферьг
и всего на 0,5% из вод пресных—лпмносферы, имеет
поверхность около 363 млн. /;д% причем поверхность
мирового океана равна 361 млн. «.п2, а поверхность ма-
териковых вод от 1,7 до 2,5 млн. км2 (включая Ка-
спийское п Аральское моря). Большое количество
поды находится на поверхности земли в твердом состоянии, в виде
льда. Около 16 млн. км2 представляют площадь современного оледенения,
т. е. 3% всей поверхности земного шара. Кроме того, громадную массу
составляют пловучие льды Арктиче-
ского бассейна и Антарктики с их об-
щей площадью свыше 15 млн. км2.
Объем мирового океана. Объем во-
ды мирового океана составляет около
1 370 млн. км? (3,5% по весу згой
воды составляют растворенные в ней
соли). Объем материковых льдов в об-
ластях современного оледенения оп-
ределяется величиной 20,9 млн. км2.
Если растопить весь этот лед, уровень
мирового океана стал бы выше на
50 м. Объем льдов последнего ледни-
кового периода определяют в 34,3
млн. ттл?3; за счет этого льда, если бы
он растаял, уровень океана поднялся
бы па 85 м.
Баланс гидросферы. Кроме того,
около 334 тысяч км? воды ежегодно
океана в атмосферу путем испарения, 297 000 км2 возвращается обратно
в океан в виде осадков, а 37 000 км2, вместе с 62 000 км2, испаряемыми
с поверхности суши и материковых вод, дают осадки на сушу (99 000 км
Общее количество воды, испаряющейся в течение года с поверхности
.'.емкого шара и поступающей в атмосферу, определяют величиной в
428 тыс. км2 [по Бюсту (1936) 379,2 тыс. яти3]. Речной сток в iu-
ченпе года дает в оксан около 30 000 км2 воды (рис. 15).
Рас. 15. Циркуляция воды в ирир« де
(по ^1отьн).
поступает с поверхности мирового
ГЛАВА 11
РАСЧЛЕНЕНИЕ МИРОВОГО ОКЕАНА
И РЕЛЬЕФ ЕГО ЛОЖА
Океаны. Поверхность океана представляет непрерывное целое, однако
материковые массы подразделяют ее на более или менее обособленные
части. До недавнего времени географы различали 5 самостоятельных океа-
нов (Северный Ледовитый, Атлантический, Индийский, Тихий и Антарк-
тический). В настоящее время водам, омывающим северные полярные стра-
ны и антарктический материк, иногда не приписывают характера самостоя
тельного океана; однако общепринято океанами считать Атлантический,
Тихий, Индийский и Северный Ледовитый.
Придаточные моря. Краевые части океанов, прилежащие к материкам,
могут быть более или менее значительно отчленены от океана, вдаваться
в сушу и образовывать придаточные моря. Последние в свою очередь под-
разделяются на моря средиземного типа, если они вдаются глубоко между
материками и соединены с океаном более пли менее узкими проливами.
Если придаточное море не имеет резкого отчлененпя от океана, его обычно
называют краевым или окраинным. По своим основным показателям
(поверхность, объем, средняя и максимальная глубина, характер склона)
моря резко различаются между собой. Наряду с такими громадными водое-
мами средиземноморского типа, как Северный Ледовитый океан, с его по-
верхностью в 14 млн. км-. объемом в 17 млн. средней глубиной около
1200 м и наибольшей свыше 5000 лг, существуют такие моря-пигмеи, как,
например, Азовское, имеющее поверхность в 33 000 км- и наибольшую глу-
бину 13,5 м. На табл. 6 даны основные гидрографические показатели
для океанов п морей по Э. К о с с и н а (К о s s i n a, 1921) (с дополне-
ниями).
Рельеф дна океана может быть разделен на три основные части:
континентальная отмель, склон материка и
л о ж е океана. Континентальная пли материковая отмель (сту-
пень, шельф) с очень слабым наклоном дна ограничена со стороны моря
континентальным гребнем и обычно достигает глубины 200 м (иногда
значительно больше). Ширина континентальной отмели весьма различна,
иногда несколько километров, а иногда много сотен. Это как бы затоплен-
ная морем часть поверхности материка. На континентальной отмели
располагаются иногда целые обширные водоемы (так называемые
зппконтииентальные), как все наши краевые моря, омывающие с севера
Евразию, начиная от Варенцова до северной половины Берингова моря.
54
Таблица 6
Оксаны и моря Поверхность в тысячах tt.W2 Объем в тыся- чах ялг' н- ! А. 1 В%к поверхн. 1 миров, океана
Паибол biiraj глубина в . i Средняя г.г бина в л-
\тладтический океан . . ) Без при- 82 441 323 613 8526(7370) 3 926 26
Индийский океан > даточных 73 443 I 291 030 7 450 3 963 23
Гихий океан ,1 морен 165 246 707 555 10 800 4 282 51
Океаны без придаточных морен 321 130 | 1 322 198 10800 4 117 100
Л лантический океан . . 1 106 463 354 679 8 526 3 332 29
Индийский океан > n q 74917 291 945 7 450 3 897 21
ТихпП океан .1 морями 119 679 723 699 10 800 4 028 50
Мировой океан 361 059 1370323 10 800 3 795 । 100
11 р и д а т о ч п ы е мо р я Г'евериый Ледовитый океан । , 14 690 ; [ i , 16 980 5 440 I 1 205
Европейское Средиземное с Черным и Азовским 1 2 966 ' 4 238 1 4 Ю4 : 1429
Америк. Средиземное море 4 319 i 9 573 6 269 ; 2 216
Японское море 1 008 ! 1361 3 743 1 1350
Охотское море : 1 528 . 1279 I 3 500 838
Берингово море ! 2 268 3 259 4 273 ! 1437
Баренцево мире 1360.0 271 548 ! 199,3
белое море 89,6 5,4 340 1 60
Балтийское море 385,0 22 427 । 56
Азовское море 37.6 0,256 13,25 6,8
Черное море 411,54 492,5 2 243 1 1197
Каспийское морс 436,34 77,0 1000 1 184
Лралыкое море 64,49 1,0 68 j 15,8—16,2
Вее придаточные моря . . . 39 928 S 48125 | 6 269 1 205
Нередко эпиконтинентальные моря имеют глубины, превышающие
200 м. Эпиконтинентальные моря представляют собой водоемы с наиболее
благоприятными условиями для развития растительных и животных
организмов. За краем континентальной отмели (континентальным гребнем)
начинается крутой склон (батиаль) к большим глубинам океана. Это
как бы крап (бок) материка. Материковый склон имеет уже гораздо более
крутое падение (4—7°, иногда 13—14° и больше).
На глубине 3—4 тысяч м материковый склон переходит в ложе миро-
вого океана (абиссальные г л у б и н ы), в отдельных местах имею-
щее более глубокие впадины (табл. 7).
Площадь, занимаемая этими частями морского дна (в%), показана (по
Ко с си на, 1921) на табл. 7 (включая придаточные моря)
Таблииа 7
>
Части дна океана Атлант и-1 ческий океан Индий- 1 | ский 1 океан | Тихий океан Мировой океан
Чггериковая отмель (0—200 ло I 13.3 4,2 1 5,7 7,6
Материковый склон (200—3 000. и) [ 21.2 13,9 , 12.2 15,3
Абиссальные глубины (> 3 000 .и) 1 ! 65,л 81,9 -ч2,1 17,1
55
Таблица
Части поверхности семного шара ?4сжду
изогипсами и изобатами
Поверхности в °/<
всей поверхности
земного шара
Выше 3 090 л/ над уровнем моря...........................
3 000-2 000 аг > > ........................ 2 < •
2 000—1 000 * » ................... -1,7
1000—200 л/ > , > ................................ 11, ь
200—О М » » ; ........................ *>Ji
О— 200 м ниже поверхности мира.......................... 5.5
200—1000 м » >/ 2,9
1000—2 000 м > > 2.9
2 000—3 000.1а > 1.7
3 000—1000 .и .................... 1:’.9
4 000—5 000. и .................... 23.3
5 000—6 000 .и > • .................... 1б/>
более 6 000 .п > » , ........................... 1,0
Очевидно, что в мировом
ответствующпе ложу океана.
океане должны преобладать глубины,
Надводная часть суши также имеет свои
Рис. 16. Гипсографическая кривая суши п моря
(из Свердрупа).
преобладающие высоты
(200—1 000 м), что мож-
но видеть на табл. 8,
изогипс (линии равной
высоты над уровнем мо-
ря) и изобат (линии рав-
ной глубины под уров-
нем моря).
То же можно видеть
и на так называемой
гипсографической кри-
вой суши и моря (рис.
16). Такой характер по-
верхностного рельефа
земной коры находится
в согласии с представле-
нием о материках, как
о глыбах, плавающих на
подстилающей их более
жидкой магме.
Общая картина рельефа ложа мирового океана дана на рис. 17.
Очень точные и тщательные промеры эхолотом рельефа материковой
отмели и материкового склона у атлантических и тихоокеанских побережий
Северной Америки дали возможность установить наличие подводных глу-
боких «каньонов», прорезающих поперек материковый склон. Частью эти
подводные «каньоны» являются как бы. продолжением речных долин,
частью не связаны с ними (рис. 18). Разные авторы различным образом объ-
ясняют происхождение подводных каньонов. Некоторые [Дэли (D а-
1 у, 1936, 1942), К у эне н (Kuenen, 1938), отчасти Ш е п а р д
(Shepard) и Эмери (Emery)] приписывают главную роль в форми-
ровании каньонов размыву мутьевыми потоками, образующимися у берегов
и стекающими, в силу повышенной плотности содержащей муть воды, по
дну. Их действие подобно действию воды, размывающей овраги па поверх-
ности суши. Другие авторы ищут объяснение в размыве подземными во-
дами, в тектонических причинах и т. д.
‘(О ЮЧ'НГ'»'!!''»] ]] f И ' ViIV'1'ТО ГИОП'-'iniV РЧ'^Т'’ фрпт^(у ‘
Рис. 18. Топография шельфа и материкового склона восточ-
ного побережья США с подводными каньонами (из Сверд-
рупа). Глубины в метрах.
А — ход изобат. Б — на разрезе.
ГЛАВА TH
ФИЗИКО-ХИМИЧЕСКИП РЕЖИМ МИРОВОГО ОКЕАНА
1. ОСНОВНЫЕ ФАКТОРЫ ВНЕШНЕЙ СРЕДЫ
з основных факторов внешней среды, обусловливающих
в море распределение организмов и направление про-
цесса биологического продуцирования, могут быть вы-
делены некоторые, имеющие особенно важное значе-
ние. Это — питательные вещества (свет для растений),
температура и растворенные в воде газы.
Питательные вещества. Без определенного коли-
чества питательных веществ никакие организмы су-
ществовать не могут; фотосинтезирующие организ-
мы не могут развиваться при отсутствии света. В морях имеются
районы, где жизнь ничтожно мала вследствие почти полного отсутствия
питательных ресурсов; таковы поверхностные слои воды в тропической зоне
центральных частей океана к северу и к югу от экватора. В общем распро-
странение плотности населения в море находится в зависимости от рас-
пределения пищи. Пища, нужная животным, присутствует либо в виде
организмов, либо в виде органического детрита.
Если для существования животных необходимы в качестве пищи го-
товые органические вещества, то растения требуют света и растворенных
минеральных веществ.
Часто соединения азота, фосфаты и кремнекислота имеются в недо-
статочных количествах и лимитируют развитие фитопланктона. Это яв-
ление выражено особенно резко в тропических областях океанов.
Газы. Громадное значение фактора, способствующего или тормозя-
щего развитие жизни, имеют некоторые растворенные в воде газы (кис-
лород, углекислота, сероводород). Хорошая вентиляция способствует
богатому развитию жизни. Слабое поступление кислорода ее тормозит,
например на глубинах Каспийского моря. В глубинах Черного моря и
в норвежских фиордах отсутствие кислорода и наличие сероводорода со-
вершенно устраняют возможность развития животной жизни.
Обычно резкое уменьшение количества кислорода сопровождается
очень значительным накоплением углекислоты (глубины Черного моря)
и иногда значительных количеств сероводорода (впадины фиордов, глу-
бины Черного моря). Углекислота, оказывающая ядовитое действие на
большинство животных, однако, необходима для развития растений.
61
Кроме указанных факторов, развитие жизни в море определяется так-
же другими химическими и различными физическими свойствами воды
(солевой состав, концентрация водородных ионов и др.) и грунта (механи-
ческий состав, химические свойства), системой циркуляций, перемеши-
вающих слои воды, освещенностью моря, покрывающими поверхность
моря льдами и рядом других моментов.
Исторические причины. Наконец, те климатические и орографические
изменения, которые имеют место в морских бассейнах и совершаются в
масштабе геологических периодов, могут также определять собою харак-
терные особенности явлений биологической продуктивности в морях (и с-
торический фактор). Это воздействие складывается из отбора
тех или иных форм (состав фауны и флоры), выработки их индивидуальных
особенностей и воздействия на формирование биоценозов. Таким образом,
все рассмотренные факторы и обусловливаемый ими эффект должны всегда
рассматриваться в исторической смене, в зависимости от палеогеографиче-
ских изменений. Это особенно ярко сказывается на наших южных морях
и на Балтийском море, в силу той богатой событиями истории, которую
они пережили в третичном и четвертичном периодах и которая наложила
неизгладимый след на все детали биологических явлений, развертываю-
щихся в них.
2. СОЛЕВОЙ СОСТАВ МОРСКОЙ ВОДЫ
Элементарный состав. В воде океанов и морей найдены 34 химических
элемента: О, Н, Cl, Na, Mg, S, Са, К, Вг, С, Sr, В, F, Si, Rb, N, Zn, Ba,
Li, I, Fe, P, As, Al, Gu, Mn, Cs, V, Ag, Au, Ra, Pb, Go, Ni.
Количественное значение отдельных элементов весьма различно, что
можно видеть на прилагаемой таблице, где дан (по Вернадскому
и Виноградову) элементарный состав морской воды в весовых про-
центах (табл. 9).
Таблица 9
Кислород 85,80 Кальций 0,045
Водород 10,67 Калий 0,038
Хлор 2,00 Бром 0,0065
Натрий 0,07 Углерод 0,0035
Магний 0,14 Стронций 0,0010
Сера 0,09 Бор 0,0004э
Другие элементы растворены в морской воде в зничтожном количестве,
например, кремний 2.10~5, иод, фосфор и железо по5.10-6, медь — 2.10'6,
золото — 2.10“9, радий — 2.10-12.
Постоянство солевого состава. Океанические воды обладают замеча-
тельным постоянством солевого состава — соотношения между главными
ионами, определяющими свойства морской воды, остаются постоянными.
Общая же концентрация солей в океанической воде составляет примерно
3,5% (или как обычно обозначают 35°/00) и колеблется в пределах от 0 до
4%. Эта особенность солевого раствора морской воды привела датского
ученого М. К н ю д с е н а (К п й d s е п) к выведению формулы опре-
деления общей солености морской воды по содержанию только одного
хлора: S%0 = 0,03 1,805 G1.
Среднее весовое содержание главных ионов (по Виноградову)
показано на табл. 10.
Солевой состав лоре кой и речной воды. Речная вода содержит солей
немного меньше 0,05% и при этом в другом количественном соотношении,
чем в море, что показано на табл. 11.
62
Таблииа 10
Катионы °/о I i Анионы 0/ /0
Натрин Магний Кальции Калий 30.59 3,72 1,20 1,00 Хлор .... Сульфат-ион (SO4). . Карбонатный ион (СО-) Бром i 55,29 7,69 0.21 0.19
Состав солеи
Хлорпды.........................
Сульфаты........................
Карбонаты.......................
Соединения азота, фосфора, кремния
и органическое вещество..........
Таблииа 11
I
Морская I Речная
вода I вода
в % । в °/о
88,64 i ;>,2
10.80 9,9
0,34 | 60,1
0,22 [ 24.8
100,0 *1 ; и о,о
Если же взять только хлориды, сульфаты и карбонаты, то соотноше-
ние будет следующее (табл. 12).
Таблииа 12
>
Морская Речная
вода вода
В % в °/о
88.8
10.8
0,4
6,9
7(k9
100,0
Состав солей
Хлориды .......................
Сульфаты.......................
Карбонаты......................
100,0 i
Там, где имеется значительный приток речных вод, особенно в полу-
замкнутых придаточных морях, соотношение отдельных солей в морской
воде может претерпевать некоторый сдвиг в сторону речной воды, что мы
и имеем в Черном, Азовском и некоторых других морях. Еще большие раз-
личия наступают, если водоем совсем теряет связь с мировым океаном,
например, в Каспийском и Аральском морях.
Однако соли, растворенные в мировом океане, постоянно приносятся^
речной водой и выпадают из атмосферы, состав же их различен в море и
в реках (табл. 13).
Общее количество солей в океане. Общий объем солей, растворенных
в морях (5ХЮ16 тп) можно представить себе из следующего сопоставления:
если эту соль распределить на всю поверхность земного шара, полу-
чится слой в 45 м толщиной, а если только на сушу —153 ле.
Таким образом, соли, приносимые в течение года в океан реками, со-
ставляют миллионные доли от количества солей, заключающихся в воде
океанов.
63
Таблииа 13
>
Составные части морских солей1 Содержится в океане (в 1012 тонн) Приносится еже- годно реками (в 10е ТОНН)
со3 96 961
S04 3 553 332
Cl 25 538 155 -
Br 86 —
N03 — 25
Na 14 130 258
К 511 58
Ca 553 558
Mg 1721 93
(Al, Ee)O3 1 75
Si(J2 — 319
46188
(у Свердрупа
5хЮ1в)
А. Виноградов (1944) считает, что солевая масса современного
океана на 50% составлена из веществ, образовавшихся за счет разрушения
магматических и других пород, и на 50% за счетпривноса из атмосферы.
Очевидно, что в океане количество отдельных солей не сохраняется
в том соотношении, как в речной воде. На дне морей и океанов идет мощный
процесс седиментации, в который отдельные растворенные в воде соли во-
влекаются (преимущественно со скелетами организмов и их телами) не
в том соотношении, в котором они растворены в воде.
Общая соленость. Мы утке указывали, что общая соленость мирового
океана близка к 35°/00, со слабыми колебаниями « 4°/00). Однако вблизи
устьев рек и особенно в придаточных морях (тем больше, чем сильнее они
отчленены от океана) соленость может значительно понижаться. С дру-
гой стороны, более или менее изолированные участки моря, подвергаю-
щиеся сильному испарению и лишенные речного стока, могут претерпе-
вать очень сильное осолонение, вплоть до предела насыщения, например
в Сиваше и Кара-Богаз-голе.
С. Зернов (1934) в своей известной книге подразделяет по соле-
ности все воды на 5 групп (табл. 14).
Таблица 14
I II III IV V
Почти несоленые (Почти агалинные) <0,2<700 Пресные (Олигогалинные) 0,2—О,5°/оо Солоновато- водные (Мезогалинные) 0,5—16°/оо Морские (Полигалинные) 16-47%0 Пересолоненные (Ул ьт рагал инн ы е) 47—300°/О(< и выше
Солоноватоводным бассейнам посвящена одна из следующих глав.
Ультрагалинных водоемов мы касаемся в главах об Азовском и Каспий-
ском морях (том II). Вся же основная масса океанических вод попадает
в рубрику морских полигалинных.
1 Из Н. Зубова (1938).
64
Происхождение солености хМирового океана. Большое значение в реше-
нии ряда вопросов, связанных с историей развития морской фауны, возник-
новением наземной и пресноводной фаун и основными особенностями всех
трех фаун, имеет изучение тех изменений в общей концентрации и соотно-
шении солей, растворенных в морской воде, которые имели место за период
существования в море организмов. Вопрос этот неоднократно вставал пе-
ред учеными, но и сейчас еще нет никакого общепринятого его решения.
В. Вернадский (1926) считал, что данный вопрос «еще лежит вне
поля современной науки». Проще всего представить себе, что соли, рас-
творенные в морской воде, вынесены реками и что первичный океан был
пресным, а затем постепенно соленость возрастала, пока не достигла
современной концентрации. Одновременно шла перестройка в соотноше-
нии солей от «речного» типа к «морскому».
Последнее время в нашей литературе этот вопрос подвергли очень
серьезному пересмотру (с двух разных точек зрения) Л. Блинов
(1946) и Я. Бирштейн (1947).
В решении данного вопроса следует исходить из следующих основных
положений: в настоящее время главная масса солей, поступающих в море,
приносится туда реками, главная масса солей, теряемых морской водой,
возвращается из воды в литосферу в виде донных отложений. Таким об-
разом они из литосферы поверхности материков переносятся в литосферу
морского дна. Этот процесс Б. Л и ч к о в (1946) иллюстрирует схемой
(рис. 19) В. Вернадского (1926), несколько ее видоизменяя.
Горные породу — — ।
ПочЬы----► Пресные боды--Морские боды----Морские илы
Герно/е породы Горные породы
Рис. 19. Схема переноса водой твердых частиц (по Личкову).
Однако, как мы видели выше, в солевом составе материкового стока
п морской воды имеется существенное различие. В речной воде относи-
тельно преобладают кальций, карбонаты и сульфаты, а в морской — нат-
рий и хлориды.
Для того чтобы объяснить изменение солевого состава воды в море,
необходимо допустить извлечение из воды в осадки громадных масс карбо-
ната кальция. Л. Блинов полагает, что данные солевого состава от-
ложений палеозойского Цехштейнового моря указывают как бы на не-
которое промежуточное положение солености его воды.
G другой стороны, А. Виноградов (1944) высказывает мнение,
что «мощность осадочных пород докембрия во много раз превышает мощ-
ность осадков всех последующих периодов, вместе взятых. Нужно счи-
тать, что основной процесс разрушения магматических пород протек в
докембрии, а в последующие периоды осадочные толщи докембрия по-
вторно и, может быть, многократно разрушались, перемывались и от-
лагались вновь. Поэтому мы считаем, что уже в кембрии существовало
установившееся, огромное по масштабам, солевое равновесие между
морем и сушей. С этого времени и до наших дней мы не можем ни в харак-
тере осадков, нив изменении химического состава организмов моря, ни в
составе атмосферы найти следов каких-либо крупных значительных
изменений состава солевой массы окейна».
5 Зенкевич, т. I
65
С иной точки зрения к решению того же вопроса подошел Я. Бир-
штейн (1947). Данный автор исходит из положения, что пресноводная
фауна (первичноводная) сохраняет гораздо больше древних и реликтовых
(консервативных) групп, нежели морская, и выражает его следующим об-
разом: пресноводные группы преимущественно мезозойские, а морские —
кайнозойские (табл. 15). На приводимой таблице дано для разных групп
(пресноводных и морских) количество родов, известных в ископаемом со-
стоянии, причем это количество принято за 100%.
Таблииа 15
Группа и число родов,
известных в ископае-
мом состоянии
3
я
Периоды
Пресноводные
»=5
О
Sc
о
Я
S g
л
о <D
к з
0 я
Морские
со
я
я
я
я
В
со
Силур. . .
Девон . . .
Карбон . .
Пермь. . .
Триас . . .
Юра ....
Мел ....
Третичный
16
16
84
31
30
50
20
67
17
16
50
50
50
50
93
3
13
35
49
4
14
15
16
44
32
49
3
16
51
22
12
20
63
11
22
67
к
и
я
Я
3
Я
я
я
и
я -S
я
1
3
я
2
3
2
6
я
я
м
о 9
8
5
Для определения сходства качественного состава современных пресно-
водной и морской фаун Я. Бирштейн пользуется отношением разно-
водных (т. е. содержащих и морские и пресноводные роды или
виды) семейств и родов ко всем семействам и родам, зарегистрированным
в пресной воде (табл. 16). Анализируя данную таблицу, автор делает вы-
вод, что сходство между пресноводной и морской фаунами имеет тенден-
цию возрастать вверх по системе. Для примитивных групп обе фауны пол-
ностью разобщены, у высоко организованных наблюдается большее
сходство.
Таблица 16
Группа
% разноводных
семейств родов
Губки .............
Кишечнополостные. .
Коловратки.........
Олигохеты.........
Мшанки.............
Пластинчатожаберные
Брюхоногие.........
Веслоногие ........
Бокоплавы..........
Десятиногие .......
0
0
21
33
12
28
58
73
83
55
О
0
4
9
7
6
4
33
53
17
Из этих данных делается вывод, что переход многих групп пресноводной
фауны из моря в материковые воды совершался в отдаленные времена, когда
66
разница в солености была еще очень невелика. Морская фауна в дальнейшем
эволюционировала и развивалась в постепенно повышавшем соленость
океане, а пресноводная сохранила свои древние черты, и в результате раз-
рыв между обеими фаунами становился все резче и резче. Таким образом,
Я. Бирштейн считает, что только малая соленость древнего океана
дает возможность объяснить формирование пресноводной фауны и совре-
менный разрыв между нею и морской. Несмотря на высказанные сообра-
жения, мы все же еще не располагаем достаточными доказательствами очень
малой солености палеозойского океана. Если в то время море и материко-
вые воды мало различались по солености, то почему же из моря в пресную
воду перешло так мало групп, тем более, что, как это показывает Я. Бир-
штейн, материковые водоемы больше способствовали сохранению древ-
них форм.
С другой стороны, в морфологических изменениях древнепалеозой-
ских морских форм в последующие эпохи мы пока не можем указать
таких, которые можно было бы объяснить повышением солености, и, на-
конец, среди морской фауны имеется очень много групп, совершенно не
переносящих опреснения, что не очень понятно, если принять, что предки
этих форм перенесли, правда, очень длительный эксперимент перехода
к существованию из пресной воды в морскую. Таким образом, правильнее
присоединиться к взглядам тех ученых, которые считают, что за время,
протекшее с кембрия, океан не претерпел в своей солености существенных
изменений.
3. ХИМИЧЕСКИЙ СОСТАВ МОРСКИХ ОРГАНИЗМОВ
Влияние организмов на химизм морской воды. Организмы, населяющие
моря и океаны, оказывают весьма сильное воздействие на химизм морской
воды и сами в значительной степени от него зависят.
Они выбирают из воды для построения своего тела определенные ве-
щества и после смерти возвращают их обратно в окружающую среду,
либо в воду, либо в донные отложения. Кроме того, они в течение жизни
обогащают воду своими выделениями.
Элементарный состав морских организмов. Капитальная сводка по хи-
мическому составу морских организмов дана учеником Вернадского —
А. Виноградовым (1935, 1937,1938,1944). В телах морских орга-
низмов установлено наличие 60 элементов. 39 из них более обычны и только
18 постоянно входят в состав организмов; их относительное количество
в % к весу тела показано на табл. 17.
Таблица 17
Первичные (1-60%) Вторичные (0,05- 1®/о) Микроконетитуенты «0,05%)
Водород Натрий Бор
Углерод Магний Фтор
Азот Сера Кремнезем
Кислород Хлор Марганец
Фосфор Калий Медь
Кальций Железо Иод
Если расположить приведенные в табл. 17 элементы по их количествен-
ному содержанию в морской воде, то они соответственно займут (исключая
газы) 8, 17, 2, 3, 4, 1, 6, 5, 21, 10, 12, 11, 22, 23 и 19 места. Как известно,
при этом содержание отдельных элементов, входящих в состав тел орга-
низмов, различно в скелетах, живых частях тела и в полостной жидкости
(табл. 18, 19 и 20).
б* 167
Таблица 18
СРЕДНИЙ СОСТАВ ОРГАНИЧЕСКИХ ВЕЩЕСТВ
(из Свердрупа по Роджерсу, 1938)
Процентный состав Относительный вес, если за 100 принято содержание углерода
Элементы Углеводы Жиры Белки Элементы Морская вода Жиры Белки
О 49,38 17,90 22,4 С 100 100 100
С 44,44 69,05 51,3 P 0,05 3,1 1,4
11 6,18 10,00 6,9 N 0,5 0,88 34,7
р 2,13 0,7 S 3150 0,45 1,6
N 0,61 17,8 Fe 0,07 0,2
Таблица 19
ПРОЦЕНТНЫЙ СОСТАВ СКЕЛЕТНЫХ МАТЕРИАЛОВ (пз Свердрупа)
Вещество Группа организмов
Foraminifera (Orbttolites margin alls) Коралл Ocullna diffusa Известковая водоросль (Lithophvl- lum antilla- rum) Омар (Homarus s₽.) Плеченогое (Discinisca lamellosa) с фосфор- ным скеле- том Кремневая губка (Euplectella speclosa)
Са 34,90 38,50 31,00 16,80 26,18 0,16
Ng 2,97 0,11 4,36 1,08 1,45 0,00
со3 59,70 58,00 62,50 22,40 7,31 0,24
so4 — — 0,68 0,52 4,43 —
РО4 следы следы следы 5,45 34,55 —
SiO2 0,03 0,07 0,04 1 0,30 0,64 88,56
(Al, Fe)2O3 Органиче- 0,13 0,05 0,10 J 0,44 0,32
ское веще- ство и др. 2,27 3,27 1,32 53,45 25,00 10,72
Таблица 20
ОТНОСИТЕЛЬНЫЙ СОСТАВ полостной жидкости
(если количество натрия принять за 100)
(из Свердрупа по Робертсону)
Элементы Морская вода Морской еж Echinus esculentus Омар (Homarus vulgaris) Краб (Cancer pagurus)
Cl 180 182 156 156
Na 100 100 100 100
Ng 12,1 12,0 1,5 5,7
SbSO4 8,4 8,5 2,2 6,7
Ca 3,8 3,9 50 4,8
К 3,6 3,7 4,7 4,0
Как можно видеть из приведенных таблиц, полостная жидкость мор-
ских животных в общем сохраняет соотношения, близкие к морской воде,
что обусловливается постоянно идущим осмотическим обменом между
водой и полостной жидкостью животных.
Скелеты же животных по составу наиболее резко отклоняются* от мор-
ской воды и дают наибольшие различия между отдельными группами.
68
Способность некоторых морских организмов аккумулировать отдель-
ные элементы. Избирательная способность организмов в отношении раз-
личных компонентов морской воды очень велика и многие вещества,
растворенные в морской воде в ничтожно малых дозах (10~5%), концентри-
руются в теле животных и растений в относительно гораздо большем
количестве. Из таких веществ можно назвать хотя бы иод, ванадий, алю-
миний, стронций, кремнезем, цинк и мн. др. Иногда имеет место (как ука-
зывает А. Виноградов, 1944) равномерное и недиференцированное
накопление во всех частях тела того или иного элемента, например, цинка
у кишечнополостных, галоидов у всех примитивных беспозвоночных (Pro-
tozoa, Porifera, Coelenterata) и водорослей. У более высоко организованных
групп (Vermes, Mollusca, Brachiopoda, Arthropoda) в определенных тка-
нях и органах могут концентрироваться тяжелые элементы.
В этом процессе выборочного накопления некоторых веществ, раство-
ренных в морской воде, может наблюдаться некоторая регио-
нальная неоднородность. Так было установлено, что некоторые моллюски,
иглокожие и рыбы норвежских прибрежных вод богаче иодом, чем те же
организмы по другую сторону Атлантического океана, что связано, види-
мо, с большей продуктивностью планктона в северо-восточной части
Атлантического океана.
Способность морских организмов к выборочной концентрации отдель-
ных элементов, растворенных в морской воде, становится особенно нагляд-
ной из сопоставления, по Виноградову, состава морской воды
с составом организмов (табл. 21).
Таблица 21
Натрий принят за 100 Натрий принят за 1
Эле- менты Balan us по Виногра- дову (1938) Рыба (в среднем) по Jowett и Davies (1938) и Clements и Hutchinson (1939) Голожа- бернпк Archidoris britannica по Мс. Сапсе и Masters (1937-38) Морская вода Фактор концентрации
Cope- pod а Рыба Голожа- берные моллюски
С1 194 180 180 1,1 1,0
Na 100 100 100 100 1,0 1,0 1,0
Mg 5.6 36 156 12,1 0,46 3,0 12,9
S 25,9 259 7,1 8,4 3,1 31 0,85
Са 7,4 52 262 3,8 1,9 13,7 69
К 53,7 383 20 3,6 15 109 5,5
Вг 1,7 — — 0,6 — — 3
С 1113 ок. 4100 ок. 480 0,26 4300 15 800 1850
Sr — — 11 0,12 — — 92
Si 1,3 — — 0,001 * 13 000 — —
F — — 69 0,01 — — 6 900
N 280 1276 107 0,001 * 28 0000 1 276 000 10 7000
P 24,1 256 6 0,0001 * 24 1000 2 560000 60000
J 0,04 — — 0.0005 80 — —
Fe 1,3 1,3 0,23 0,0002 6000 6000 1000
Nn — 0,0008 — 0,0001 — 8 —
Cu — 0,008 0,43 0,0001 — 80 4 300
Из приведенной таблицы видно, как это уже указывалось и выше,
что морские организмы весьма различны в отношении концентрации от-
дельных веществ в своем теле (табл. 22).
* Зимой в Английском канале.
69
Таблица 22
Относительный состав планктонных организмов
Углерод принят за 1С0 Углерод принят за 1
Эле- менты Морская вода Диато- меи Пери- динеи Соре- poda Диато- меи Перидинеи Copepoda
С 100 100 100 100 1 1 1
N 0,5* 18,2 13,8 25 36 28 50
Р 0,05* 2,7 1,7 2,2 54 34 44
Fe 0,07* 9,6 3,4 0,13 137 49 2
С а 1420 12,5 2,7 0,66 0,01 0,002 0,005
Si 0,4* 93,0 6,6 0,13 232 16 0,1
Приведенная таблица показывает, что азот и фосфор концентрируются
во всех трех группах в сходных количествах, остальные элементы в весьма
различных.
Известно значение для развития моллюсков содержания меди в морской
воде. У североамериканских побережий Атлантического океана установ-
лена зависимость в распределении и биологических особенностях некоторых
моллюсков (устрица) от распределения в воде Fe, Си, Мп, в связи с выно-
сом речными водами этих веществ в море.
Известковая функция морских организмов. Как показывает А. Вино-
градов (1944), особенное значение для распределения организмов
в гидросфере имеет содержание СаСО3. В тропических областях, где океани-
ческие воды находятся в состоянии насыщения (или даже перенасыщения)
углекислым кальцием (при высокой температуре и уменьшении парци-
ального давления СО2), имеются особенно благоприятные условия для су-
ществования организмов-концентраторов углекислого кальция — рифо-
образующих кораллов с целой группой других организмов, участвующих
в создании коралловых рифов (известковые водоросли, фораминиферы,
губки, мшанки, иглокожие и др.). Здесь быстро и легко растет массивный
известковый скелет.
В холодных областях океана, где имеет место значительное недона-
сыщение воды СаСО3, известковая функция организмов выражена неиз-
меримо слабее. Кокколитофоридовый и фораминиферовый планктон сме-
няется диатомовым и рачковым, выпадает большое число групп бентоса,
известковые скелеты становятся легче.
Такое же уменьшение форм карбонатонакопителей наблюдается и при
переходе от морских зон в солоноватоводные.
Так же хорошо известна потеря известкового скелета корненожками,
гидрозоями, губками, мшанками и другими организмами, перешедшими
к обитанию в пресной воде. Все они заменяют известковый скелет хити-
новым или кремневым.
В значительной степени тот же процесс испытывают и пресноводные
моллюски. Количество СаСО3 в значительной степени заменяется в их
скелете конхиолином и уменьшается. У пресноводных моллюсков скелет
не кальцитовый, а арагонитовый.
Та же закономерность наблюдается и в океане с переходом от его
поверхности в область больших глубин. Понижение температуры и уве-
личение давления увеличивают растворимость СаСО3 и обусловливают
недонасыщение воды карбонатами. В результате с увеличением глубин
* Для Английского капала зимой.
70
не только изменяется 1 характер населения, в смысле исчезновения форм
с известковым скелетом — фораминифер, кораллов, известковых губок
и других беспозвоночных и замещения их организмами с органическим
скелетом (голотурии, губки) или скелетом из SiO2—кремневые Tri ахо-
nidae, Radiolaria, но происходит изменение состава скелета у видов с из-
вестковым скелетом. У тех же Foraminifera — глубоководных форм —
скелет становится агглютинирующим.
Глубоководные Xenophyophora имеют в скелете не СаСО3, а нераствори-
мый BaSO4. У Brachiopoda раковины истончаются и теряют известь. У Seal-
pellum степень обизвествления раковин с глубиной уменьшается. У Мо1-
lusca с глубиной, как наблюдалось для Buccinum, Pecten и др. в Японском
море, раковины становятся менее обизвествленными и т. п. Наконец, у
некоторых глубоководных рыб из Teleostei не происходит полного око-
стенения скелета. Мопсообразные головы этих рыб и мягкость костей
позволили R о u 1 е назвать это явление «абиссальным рахитом».
4. РАСПРЕДЕЛЕНИЕ СОЛЕНОСТИ В МИРОВОМ ОКЕАНЕ
Основные факторы, воздействующие на соленость океанических вод,
связаны с их поверхностными слоями. Этих факторов пять — испарение,
образование льдов, осадки, таяние льдов и приток пресных вод с суши.
В связи с этим могут быть поняты те различия, которые наблюдаются в
распределении солености на поверхности мирового океана (рис. 20).
Насколько мощен фактор испарения, можно судить не только по резко
обособленным заливам, расположенным в засушливых областях (Сиваш,
Карабогаз-Гол и др.), но и по более открытым частям океана. В ю с т
(W ii s t) определяет среднюю мощность слоя, испаряемого с поверхности
мирового океана за год, в 85 сл, что соответствует массе воды в 304 тыс. км3
(объем Варенцова моря). Наиболее значительное осолонение поверхност-
ной воды открытых океанов, за счет испарения, наблюдается в областях
развития пассатных ветров до 36,9 и 36,5°/ОО в Тихом и Индийском океа-
нах и до 37,9 %0 в Атлантическом океане. В Тихом и Атлантическом океа-
нах имеется по две таких осолоненных области к северу и югу от эквато-
риальной зоны, в Индийском океане осолоненная зона только одна —
южная. Все эти зоны ограничены на севере и юге параллелями 40° и об-
разуют пять громадных халистатических областей мирового океана. Мень-
шая соленость Тихого океана объясняется, возможно, большим количе-
ством осадков. Наибольшей соленостью из всех морей обладают Среди-
земное и Красное, и северо-западная часть Индийского океана, тогда
как северная и северовосточная его части, наоборот, под влиянием
тропических ливней испытывают в определенные сезоны года сильное
опреснение.
Вся восточная половина Средиземного моря имеет соленость выше
38 %0, а у берегов Сирии выше 39 %0. Еще выше соленость в узком, сжатом
с обеих сторон пустынями, Красном море (до 43%0). Более обычные со-
лености в Красном море 40—40,5 %0, а в Суэцком канале в зимнее время
до 52%О (рис. 21). Повышенная соленость северо-западной части Индий-
ского океана — результат влияния сильно осолоненных вод Красного
моря и Персидского залива.
Значительное осолонение поверхностных вод океана, в результате об-
разования льда, захватывает в зимнее время большие пространства в при-
полярных областях и совместно с сильным охлаждением этих вод обусло-
1 А. Виноградов, 1944.
71
вливает, как мы увидим дальше, опускание на глубины громадных масс
воды.
Наибольшее значение, в смысле опреснения поверхностных вод океана,
имеют осадки, дающие в течение года массу воды в 267 тыс. «л3, что со-
ответствует слою воды мощностью в 74,2 см. Опресняющему действию
осадков обязаны своим происхождением воды района экваториальных про-
Г~~1/ CZZJ2 E2SP ЕШШ>
Рис. 20. Распределение поверхностной солености Атлантического
океана (по Шотту). 1— < 33°/ОО; 2—33—34°/ooU—34—35°/ОО; 4—
35—36°/ОО; 5—36—37°/ОО; 6— 37°/оо.
тивотечений, тянущихся в обоих океанах несколько севернее экватора.
В этих поясах тропических дождей соленость опускается до 34 %0 и ниже.
То же имеет место и в северо-восточных частях Индийского океана.
Значительное опреснение поверхностных вод в приполярных областях
может обусловливаться летним таянием пловучих льдов, частично выно-
симых течением за их пределы. Восточно-Гренландское течение выносит
далеко на юг массы льда, понижающие своим таянием соленость воды
до 31—32 %0.
72
водоемах полузам-
в
о
s с
о '
= к
1-0 Q.
5 ©
S- s
в Тихом океане,
зонах открытых ча-
Приток пресных вод с суши составляет в общем балансе вод океана не-
большую величину в 30 тыс. км3 за год и значительное опресняющее дей-
ствие оказывает только в Арктическом бассейне и особенно в краевых его
морях, прилежащих к побережьям Сибири, а также
кнутого характера, как Балтий-
ское, Белое, Черное и Азов-
ское моря.
Зависимость распределения
солености от течений. Соотно-
шение солености, осадков, ис-
парения, плотности воды и тем-
пературы воды и воздуха на
поверхности океана на разных
широтах может быть показано
на рис. 22. Правильное широт-
ное распределение соленостей в
океанах нарушается течениями.
Особенно сильно это нарушение
в северной части Атлантики,
широко сообщающейся с Аркти-
ческим бассейном. Теплые ат-
лантические воды, подчиняясь
рельефу дна и берегов, уносятся
далеко на север и даже в глубь
самого Арктического бассейна,
в результате чего изогалины (и
изотермы) идут здесь в северо-
восточном и частью даже в се-
верном направлении. В обрат-
ном направлении движется Во-
сточно-гренландское течение,
прижатое к берегам Гренлан-
дии, и течение из Дэвисова про-
лива, выносящие, по расчетам
Шокальского, ежегодно
около одной трети всего поляр-
ного пака, объемом около 12 700
км3 восточнее Гренландии и
5 000 км3 западнее Гренландии,
через Дэвисов пролив. Эти мас-
сы льда и холодной и опреснен-
ной воды идут далеко на юг,
вдоль берегов Северной Амери-
ки, и достигают 40° сев. ши-
роты. Аналогичная комбинация
холодного течения, идущего с
северо-востока, и теплого, дви-
жущегося на северо-восток,
имеет место и у восточных побережий Евразии,
Вертикальное распределение солености в теплых
стей мирового океана и в придаточных морях дает черты резкого
различия. В краевых придаточных морях мы имеем, как правило, увели-
чение солености с глубиной (катагалинное распределение солености).
Объясняется это обычно опреснением поверхностных слоев за счет при-
тока вод с суши и осолонением глубинных вод за счет поступающих через
73
проливы низовых течений из открытых частей океанов или морей. В резуль-
тате создается резкая солевая стратификация, которая может быть
нарушена только при особых условиях, главным образом за счет вымерза-
ния поверхностных вод при льдообразовании, повышающем их соле-
ность в той мере, какая нужна для опускания их на глубину. Разница в
солености верхних и глубинных слоев воды во внутренних морях обычно
составляет 2—4—6%0, а иногда и больше. Таким образом, стратификация
во внутренних морях носит обычно резко выраженный солевой характер,
разница же в температурах верхних и нижних слоев обычно, по сравнению
с открытыми частями океана, бывает менее значительной (10°—14°—18° и
редко больше).
Рис. 22. Годовые изменения солености, осадков, испарения, плот-
ности и температуры воды и воздуха на поверхности мирового океа-
на на разных широтах (по Шокальскому).
Совсем другое мы видим в открытых частях океанов. Здесь разница
в солености верхних и нижних слоев очень редко достигает 2%0 и часто
наблюдается состояние гомогалинности, зато гораздо резче выражено разли-
чие в температуре, которое обычно достигает в тропических и субтропиче-
ских областях 23°—27°. Здесь также стратификация выражена достаточно
резко, но в отношении солености она имеет характер обратный тому, что
мы видели для внутренних морей (анагалинное и гомогалинное распре-
деление солености) и носит ясно выраженный термический характер.
С другой стороны, в более высоких широтах океанов, особенно в Антарк-
тике и в некоторых районах северных частей Атлантического и Тихого
океанов, имеются обширные районы, которые характеризуются или в тече-
ние всего года, или только в определенные сезоны, почти одинаковой со-
леностью и температурой (гомогалинность и гомотермия) от поверхно-
сти до дна. Для пояснения сказанного воспользуемся табл. 23. Станции
по отдельным морям выбраны случайно.
74
Глубина в м Зап. часть Атлантиче- ского океана 20° ю. ш. Тихий океан у Филип- пинских о-вов Антарктика, Атлантич. океан, 67°47' ю. пт. Северный Ле- довитый океан к северу от Се- верной Земли
V- 1930 6/1-1912 20/V1I1-1935
t° S°/oo t° S7oo t° S°/oo t° s°/ ° /00
0 25,00 37,00 28,80 34,44 — 1,64 34,06 — 1,70 31,60
50 22,00 36,5 28,20 34,31 — 1,77 34,40 — 1,74 33,98
100 36,4 25,90 34,71 — 1,69 34,44 — 0,37 34,33
150 — —- — — — — 1,91 34,74
200 19,00 36,00 Г),15 34,60 — 0,19 34,66
250 2,12 34,83
300 15,00 35,00 — — 0,37 34,67
1000 4,00 34,50 4,45 34,55 0,22 34,69 -0,16 34,85
2 000 3,30 34,90 2,25 34,61 — 0,19 34,68 — 0,67 34,85
3000 2,80 34,90 1,64 34,66
4000 1,80 34 83 1,60 34,67
5000 0,70 34,70 1,72 34,67
6000 1,87 34,67
Колеба- 24,30 2,30 27,20 0,40 2,14 0,63 3,86 3,15
ния 1
Таблица 23
Баренцево море Балтийское море Белое море Черное море вблизи Босфора
30/VIII-1928 15/VII-1922 6/VIII-1922 10/VIII-1910
1° S7oo S°/oo S7oo t° S°/oo
8,60 34,33 12,40 7,00 14,65 23,80 24,10 17,59
5,40 34,72 3,50 8,68 0,95 28,37 8,22 18,30
5,20 34,72 4,30 9,85 -1,10 29,36 7,61 19,65
4,00 34,94 — 1,40 30,10 8,74 21,24
— 1,40 30,34 8,54 21,29
3,С0 34,90 8,68 21,77
8,85 22,27
5,00 0,57 8,10 2,85 15,75 i 6,54 i 16,49 4,66
Таким образом, между параллелями 40° северной и южной широты по-
верхностные воды океана хотя и имеют вследствие усиленного испарения
несколько более высокую соленость (на 2,0—2,3%0), однако, они не могут
опуститься вниз, так как настолько нагреты, что их плотность меньше,
чем плотность нижележащих слоев, хотя и менее соленых. Но достаточно
охладить эти воды (что и происходит в более высоких широтах, куда эти
воды переносятся течениями), чтобы они начали безудержно опускаться
вниз на дно океана. В более высоких широтах, в Антарктике почти вплоть
до льдов, в северном полушарии вплоть до Гренландско-Шпицбергенского
порога (порог Нансена) господствуют условия, приближающиеся к со-
Рис. 23. Распределение солености на меридиональных разрезах.
А — в западной половине Атлантического океана; Б — в западной половине Тихого океана
(по Вюсту).
стоянию гомогалинности и гомотермии. Дальше на север, в Арктическом
бассейне и придаточных морях океанов господствуют катагалинность и
анатермия, причвхМ для Арктического бассейна характернейшим признаком
является мезотермия — самые теплые слои находятся на некоторой глу-
бине, а сверху и снизу располагаются более холодные слои воды. Соб-
ственно говоря, это распределение температуры можно рассматривать как
анатермию, нарушенную сильным опреснением речными водами самого
поверхностного слоя. В качестве иллюстрации ко всему сказанному может
служить рис. 23, на котором дано распределение солености в западных по-
ловинах Атлантического и Тихого океанов на вертикальных разрезах.
На приведенном продольном разрезе через западную половину Атлан-
тического океана можно видеть слой осолоненной поверхностной воды тро-
76
пической области между 40° ю. ш. и 45° с. ш., с двумя субтропическими
халистатическими областями слабой циркуляции и с перерывом в области
экваториального противотечения, со свойственными ему гораздо бо-
лее низкими соленостями (34,0—34,5%0). Менее соленые воды этого
противотечения залегают на подстилающих их более соленых водах суб-
тропического нагрева, соприкасающихся с поверхностью океана на ши-
ротах 10—30° ю. ш. и 20—40° с. ш. Таким образом, и здесь мы видим
сдвинутость этих границ на север.
Рис. 24. Распределение солености в Атлантическом океане на продольном разрезе
через восточный желоб (по Дефанту).
Подробнее детали аналогичного разреза через восточный желоб Ат-
лантического океана между параллелями 20° сев. и южн. широты в по-
верхностном слое в 300 м можно видеть на рис. 24. В южном полушарии
теплые воды халистатической области залегают на слое холодных вод
малой солености (34,0—34,5%0), притекающих сюда из Антарктики
сначала поверхностным течением (до 42—43° ю. ш.), а затем идущим на
Рис. 25. Глубины северной части Атлантического океана (по Свердрупу и др.).
глубине 600—1300 м. Хотя эти воды легки, однако они обладают низкой
температурой и тяжелее сильно нагретых тропических вод, почему и
уходят вниз. Глубже 2000 м соленость медленно падает с глубиной, из-
меняясь в пределах едва 0,3 %0. С юга на север, на глубинах Атланти-
ческого океана, соленость равномерно возрастает, но также в очень слабых
пределах (до 0,3 %0). Котловина океана заполняется холодными и менее
77
солеными водами, все время поступающими из Антарктики и из областей,
расположенных к югу от Гренландии, не отгороженных никакими барье-
рами от глубин Атлантического океана. Таким образом, холодные воды
Антарктики чрезвычайно активно поступают в Атлантический океан двумя
течениями — менее соленым, более поверхностным и более соленым глу-
бинным. Арктические воды могут поступать подобным же образом на глу-
бины Атлантического океана в гораздо более ограниченной степени, так
как глубины Северного Ледовитого океана, Гренландского и Норвежского
морей и Баффинова залива отгорожены от глубин Атлантического океана
более или менее высокими барьерами — Шпицбергенским, Исландским,
Нью-фаундлендским и Дэвисовым (рис. 25).
Распределение соленостей в толще Индийского океана весьма сходное та-
ковым для южной части Атлантического океана: холодные глубинные воды
Рис. 26. Понижение солености от Северного моря к Балтийскому и убывание коли-
чества видов морской фауны (по Зенкевичу).
поступают только из Антарктики. Иную картину (рис. 23) распределения
солености мы находим в Тихом океане. Для глубин свыше 2000 м харак-
терна замечательная однородность глубинных вод: все колебания со-
лености укладываются между 34,6О°/оо и 34,67°/00. Так же мало, как мы уви-
дим, меняется здесь и температура. Глубинные воды поступают в Тихий
океан, в противоположность двум другим океанам, и с севера и с юга, чго
хорошо можно видеть на разрезе. Отсутствие высоких порогов между глу-
бинами Тихого океана и Берингова и Охотского морей обеспечивает про-
никновение холодных вод с севера и вся картина распределения солено-
стей более покойная.
Как можно видеть по приведенным выше фактам, соленость открытых
частей мирового океана меняется сравнительно очень мало, во всяком
случае в тех пределах, в которых это не может ограничивать распроотра-
78
нения в море организмов. Только в местах впадения больших рек и с
переходом к придаточным морям мы встречаемся с более резкими измене-
ниями солености, которые ограничивают распространение растений и
животных. Большие области Северного Ледовитого океана, его эпиконти-
нентальные моря, особенно прилежащие к сибирскому побережью, имеют
значительно пониженные солености, начиная уже с юго-восточной части
Варенцова моря, и значительные ее колебания.
Значительное опреснение поверхностных вод океана имеет также место
в некоторых частях его, в районах обильных тропических ливней, напри-
- ’i
-so ъ
6аренце8о море
Рис. 27. Схематическое изображение изменения солености от Варенцова моря к Бе-
лому вдоль Воронки и Горла.
мер, в северных частях Индийского океана (Бенгальский залив) и может
иметь сезонный характер.
Соленость внутренних морей умеренных широт. Наибольшее опресне-
ние характерно для вод внутренних морей умеренных широт, имеющих
сравнительно ограниченное сообщение с мировым океаном и принимающих
в себя значительные массы речных вод. Такими морями являются в первую
очередь Балтийское, Черное с Азовским и Белое. Области проливов,
соединяющих эти моря с открытым океаном, имеют высокие солевые гра-
диенты, и морская фауна и флора обнаруживают резкое обеднение, с значи-
тельным понижением солености. В качестве, пожалуй, наплучшего
примера можно привести умень-
шение солености в проливах,
ведущих в Балтийское море
(рис. 26), сопоставив его с ко-
личественным обеднением фау-
ны. Столь же резкое обеднение
морской фауны имеет место в
Черном и Азовском морях, по
мере продвижения от Эгейского
моря до Азовского.
В менее резкой форме выра-
жено понижение солености от
Рис. 28. Продольный разрез вдоль Гибрал-
тарского пролива (из Свердрупа и др.).
Варенцова моря к Белому
(рис. 27), однако и здесь оно
обусловливает значительное
обеднение фауны. Как указывает К. Дерюгин (1928), около 140
форм, обычных в соседних районах Варенцова моря, отсутствуют в Бе-
лом море, причем именно понижающаяся соленость кладет предел рас-
пространению этих форм.
Наибольшие градиенты солености могут наблюдаться также в обла-
стях, прилежащих к проливам, соединяющим моря с резко различной
степенью солености. В проливах тогда устанавливается система течений —
поверхностного в сторону более соленого водоема и глубинного в обратном
направлении.
В подавляющем большинстве случаев соленость внутренних морей бы-
вает меньше, чем в открытом океане и поэтому в проливе, соединяющем их,
глубинное, более соленое течение движется в сторону внутреннего моря,
а поверхностное менее соленое в сторону океана. В редких случаях
система течений в проливах имеет обратный характер, как, например, в
Гибралтарском проливе. Так как в Средиземном море соленость выше,
чем в Атлантическом океане, то массы средиземноморской воды выносятся
глубинным течением в Атлантический океан и передвигаются далеко на
север вдоль берегов Европы. Это, пожалуй, наиболее крупное явление
подобного типа (рис. 28).
5. РАСПРЕДЕЛЕНИЕ ТЕМПЕРАТУРЫ В МИРОВОМ ОКЕАНЕ
Распределение температуры на поверхности мирового океана показано
на рис. 29. В общем температура убывает от экватора к полюсам и поверх-
ностные изотермы имеют широтное направление, нарушаемое течениями.
Наиболее сильные нарушения изотерм наблюдаются в северной части
Атлантики, под воздействием двигающегося на северо-восток теплого те-
чения. Таким же образом, под воздействием расходящихся веером течений,
раздвинуты с востока на запад изотермы 25° в тропической полосе Атлан-
тического океана. В Индийском и Тихом океанах имеет место несколько
больший прогрев поверхностных вод, нежели в Атлантическом. Весьма
характерным моментом является также значительное понижение темпе-
ратуры в южной части западного побережья Африки, где в результате
постоянных пассатных ветров, отгоняющих верхние слои воды на запад,
происходит в очень большом масштабе подъем на поверхность более
глубинных холодных вод. В меньшей степени это явление имеет место и
у западного побережья Южной Америки. Западные части Атлантического
и Тихого океанов прогреты лучше, чем восточные, вследствие сгона на
запад ветрами теплых поверхностных вод.
Вычисление средних поверхностных температур для трех океанов
показывает (табл. 24), что наиболее высокой температурой обладает Ти-
хий Океан, а наиболее низкой — Атлантический, что следует объяснить
особенностями циркуляции водных масс.
При этом в северном полушарии более прогреты поверхностные воды
Атлантического океана, а в южном — Тихого.
Таблица 24
Северное полу- шарие с. ш. Средняя температура Южное полу- шарие с. ш. Средняя температура
Атлантиче- ский океан Индий- ский океан Тихий океан Атланти- ческий океан Индий- ский океан Тихий океан
70°—60° 5,60 70°—60° 1,30 1,50 1,30
60—50 8,66 — 5,74 60—50 1,76 1,63 5,00
50—40 13,16 — 9,99 50—40 8,68 8,67 11,16
40—30 20,40 — 18,62 40—30 16,90 17,00 16,98
30—20 24,16 26,14 23,38 30—20 21,20 22,53 21,53
20—10 25,81 27.23 26,42 20—10 23,16 25,85 25,11
10—0 26,66 27,88 27,20 10—0 25,18 27,41 26,01
Средняя для всей по- верхности обоих полушарий Атл. ок. Инд. ок. Тих. ок.
16,9 17,0 19,1
Средняя температура для мирового океана —17,4
Годовые амплитуды поверхностной температуры. Не меньший интерес
представляет распределение годовых амплитуд колебаний температуры
80
Зенкевич
Рис. 29. Распределение поверхностной температуры в мировом океане, в среднем за год (по Шокальскому).
GO
Рис. 30. Годовые амплитуды колебаний поверхностной температуры мирового океана (по Шокальскому),
на поверхности океана (рис. 30, 31, 32). Как можно видеть, в тропических
областях всех океанов имеются районы, где годовая амплитуда колебаний
температуры не выходит за пределы 1°. Основные области океанов имеют
годовые колебания температуры воды с амплитудой 2—6° и только в огра-
ниченных районах в северной части Атлантического и Тихого океанов
Рис. 31. Средние годовые колебания поверхностной температуры в различных океанах
и разных широтах и соответственные изменения солнечной радиации (гонкие линии)
(по Свердрупу и др.).
*2
' 7
0
- 7
2
-3
Рис. 32. Годовые колебания температуры
воды на различных глубинах поверхностно-
го слоя (0,25,50 и 100 м) в Бискайском за-
ливе (47°N и 12°W) (по Свердрупу и др.).
амплитуда колебаний температуры воды составляет 7—15° и выше. Весьма
замечательно, что в южном полушарии амплитуда только в редких слу-
чаях превышает 7°, что стоит в связи со значительно меньшим количеством
материковых масс. Области наибольших температурных амплитуд (сред-
них годовых) приходятся на 30°—50° с. ш. и на 30°—40° ю. ш. (табл. 25).
Дальше на юг и север ампли-
туда опять падает и в полярных
водах не превышает 2°.
У восточных берегов Северной
Америки и Азии наблюдаются
амплитуды в 20° и даже 30°, что
создается резкими температурны-
ми различиями сходящихся здесь
теплых и холодных течений.
Колебания не средних, а .абсо-
лютных температур на поверхно-
сти океана констатированы от —2°
до 4-36°, т. е. полная амплиту-
да укладывается в 38°.
Вертикальное распределение
температуры. С глубиной температура в океане быстро падает и уже на
глубине 100 м в тропиках опускается до 4—5°.
Общий характер распределения температуры в толще мирового океана
может быть показан на рис. 33 и на табл. 26 (в том и в другом случаях по
меридиональному разрезу вдоль Атлантического океана).
Придонные слои имеют температуру близкую к нулю. На этих разрезах
хорошо заметно, что глубинные воды Атлантического океана обновляются
83
главным образом со стороны Антарктики, тогда как в Тихом океане п
с севера и с юга.
Таблииа 25
Северное полушарие Южное полушарие
Широта 50° 40° £0° 20° 10° , 0° 10° 20° 30° 40° 50°
Годовая амплитуда температуры воды. 8,4 10,2 6,7 3,6 2,2 2,3 2,6 3,6 5,1 4,8 2,9
Таблица 23
Глубина в м Г е о г р а ф п ч е с к а я ш и р о т а
80° с. ш. 60° , с. ш. 1 40° с. ш. 20° 1 С. Ш. | 0° 20° ю. ш. 40° ю. ш. 60° ю. ш.
0 2° 9° 16° 20° 27° 17° 15° <0°
200 2 6 12 15 15 11 10 <1
400 1 9 12 12 9 9 10 <1
800 0 8 11 8 5 5 4 <1
1000 <0 4 9 6 5 4 3 <1
2000 — 1 3 4 4 3,3 3 2 <0
Рис. 33. Распределение температуры в глубине океана.
А — мера тональный разрез вдоль восточного желоба Атлантического океана;
Б — то же для западной стороны Тихого океана (по Бюсту).
Особенности температурного режима Средиземного и Красного морей.
Весьма своеобразный гидрологический облик имеет Средиземное море.
Малый сток и сильное испарение обусловливают повышенную соле-
ность. В холодное время года в водоеме устанавливается гомогалинность
и гомотермия и полное перемешивание всей толщи воды, что можно видеть
на табл. 27.
84
Таблица 27
30.1.1910 г. i
Глубина в м ° 1 50 100 200 300 400 500 600 800 1000 1500 2000
Температура. . . . 12,40 12,49 12,53 12,67 13,08 13,01 12,97 19,94 12,92 12,91 12,94 13,01
Соленость Условная плот- 38,24 38,24 38,28 38,33 38,48 38,42 38,42 38,39 38,39 38,39 38,35 38,35
ность 1 29,04 29,02 29,04 29,06 29,08 29,06 29,07 29,05 29,05 29,05 29,02 29,00
Интересно сравнить также распределение температуры с глубиной
в Средиземном море и в Атлантическом океане (табл. 28).
Таблица 28
Глубина в м Средняя годовая температура
В Атлантическом океане к югу от Азорских о-вов В Средиземном море между Сицилией и Палестиной
0 19—20 19—20
100 15—18 15—17,5
200 14—16 14—14,7
400 12—14 14—14,5
600 10,5—11 13,5—14,0
800 8,5 14
1000 8,0 13,8
2000 3,8 13,5—13,7
3000 2,7 13,6—13,7
4000 2,5 13,5
И соленость и температура в Средиземном море как на поверхности,
таки на глубинах различна в разных частях моря, а на поверхности ис-
пытывает также и сезонные колебания. В общем поверхностная темпера-
тура возрастает в направлении с запада на восток, исключая зимний пе-
риод, когда наиболее низкая t° поверхностной воды наблюдается у се-
верных побережий моря, где годовые колебания температуры достигают
13°—-14°, а в остальном море —9°—10°. Различия в вертикальном распре-
делении температуры и солености в Тирренском и Ионическом морях по-
казаны на рис.. 34.
Общее нарастание температуры и солености с запада на восток было кон-
статировано в Средиземном море различными экспедициями (табл. 30).
Таблица 30
Район моря Средние показатели на глубине 2000 м
Температура Соленость
Алжир-Провансачьский 13,00 38,39
Тирренский 13,10 38,44
Ионический 13,57 38,65
Левантский 13,62 38,66
1 Вместо плотности 1,02904 пишут условно 29,04, что и обозначается как условная плот-
ность.
85
Очень сходен с описанным режим Красного моря; та же особенность
выражена в нем в еще более резкой форме. Постоянная температура глу-
Рис. 34.
А — вертикальное распределение температуры (Т°) и солености (S) в
Средиземном море по данным экспедиции «Дана» в Тирренском и
Ионическом морях (из Свердрупа и др.). Б — колебания температуры
на разных глубинах Средиземного моря в течение года (февраль,
август и ноябрь) (из Фольца).
1—Тирренское море; 2—Ионическое море.
бинной части, соответствующая степени охлаждения поверхностных вод
в самое холодное время года, еще выше, чем в Средиземном море (табл. 29).
Эти два водоема по температурному режиму глубинных слоев стоят
совершенно особняком среди всех морских водоемов Земли.
86
Таблица 29
Температура
Глубина в м
В Красном море
(по Люкшу)
О
200
400
600
800
1000
1500
2000
25—29
21-25
21-22
21,4—21,8
21,5
21,5
21,5
21,5
В Индийском океане
близ Маледивских
о-вов (по Шотту)
28—29
13—15
10,2—11
9
7,5—8
6—7
4
2,5—3
Подобного же типа вертикальное распределение температуры свой-
ственно и нашим Черному и Каспийскому морям — глубинные воды
у них нагреты более чем во всех открытых морях кроме Средиземного
и Красного и их температура близка к средней температуре воды
поверхностного слоя в зимнее время.
Влияние температуры на распределение организмов. Температура
имеет гораздо большее значение для распределения фауны и флоры
в океане, чем соленость. Как на поверхности, так и в глубинах океана
наблюдаются резкие смены температуры, кроме того, в умеренных широ-
тах северного полушария температура поверхностных вод значительно
меняется и по сезонам. Большие градиенты температуры, в пространстве
и во времени, существенным образом сказываются на распределении жизни.
В районах близкого схождения масс теплой и холодной воды, где имеются
высокие температурные градиенты, создаются как бы непроходимые барье-
ры для распространения организмов. Наконец, очень большое значение
для развития жизни имеет та общая сумма тепла, которую получает орга-
низм из окружающей среды. Рифообразующие кораллы не могут суще-
ствовать при температуре ниже 20,5°, с другой стороны, в наших поляр-
ных морях многие животные существуют при температуре, никогда не
подымающейся выше 0°.
Еще более удивительно то, что определенная температурная огра-
ниченность, иногда очень узкая, может, видимо, сохраняться не изме-
няясь в течение геологически длительных периодов.
Особенно резкие температурные барьеры имеют место там, где про-
исходит встреча холодных и теплых течений. Два района мирового океана
особенно выделяются в этом отношении — это береговые воды у Атлан-
тического побережья Северной Америки, севернее мыса Гаттераса, и
примерно на тех же широтаху Тихоокеанского побережья Азии. У берегов
Америки встречаются холодное Лабрадорское течение и Гольфстрим,
в Тихом океане — холодное Оиа-сиво и теплое Куро-сиво. На сравни-
тельно коротком расстоянии от Флориды до Лабрадора близко сходятся
средние годовые изотермы: 25°, 20°, 15°, 10°, 5° и 0° (рис. 35).
В восточной половине Северной Атлантики эти изотермы расходятся
веером. Особенно близко сходятся изотермы 5°, 10°, 15° и 20°. Именно
в этом месте происходит резкая смена фауны и близко сходятся и частично
смешиваются тепловодные и холодноводные формы. Как мы увидим да-
лее, в подобных местах стыка теплых и холодных вод фауна поражает
не только качественным, но и количественным богатством.
87
Не менее яркую картину дает также и разрез через Большую Нью-
фаундлендскую банку один из наиболее богатых промысловых районов
Рис. 35. Распределение средней годовой температуры на поверхности Атлантического
океана (по Шотту).
(рис. 36). Здесь сравнительно на небольшом расстоянии проходят изо-
термы от 0° до 14°, причем теплое течение прижимается к восточному
склону банки, а сама банка и ее западные склоны покрыты холодными арк-
тическими водами.
88
Резкие нарушения в распределении температур могут обусловливаться
также наличием подводных возвышенностей, — особенно сплошных барье-
ров (гребней), стоящих на пути передвижения водных масс. Классическим
примером такой преграды может служить так называемый хребет Уай-
вилля Томсона (Томсонов порог), тянущийся между Исландией и Фарер-
скими островами (рис. 37).
В качестве примера изменений в температуре поверхностного слоя
океана можно привести данные из работ Вудс-Холского Океанографиче-
44°35'N 41*39*» 39*30'» 37°12^
*
Рис. 36. Гидрологические разрезы в широтном направлении (температура и соленость
через Большую Ньюфаундлендскую мель (по работам «Михаил Саре») (из Меррея к
Иорта).
ского института по разрезу от одного из пунктов восточного побережья
Северной Америки, вблизи Нью-Йорка, к Бермудским островам, т. е.
в область Саргассова моря. Разрез особенно интересен тем, что он пере-
секает воды разного типа (рис. 38). Широкой полосой вдоль берега, до
края материкового плато (до 200 м), располагаются воды, подверженные
резким колебаниям температуры. Над материковым склоном залегают
холодные воды, находящиеся под воздействием Лабрадорского течения.
Еще дальше на юго-восток проходит Гольфстрим, опоясывающий в свою
очередь халистатическую область — Саргассово море. Дж. Кларк
(J. Clark, 1940) сопоставил гидрологический режим этих различных
вод с развитием планктона. Наибольшие сезонные колебания средней
89
Рис. 37. Распределение температуры и солености к югу и северу от хребта Уайвплля
Томсона (по Меррею и Иорту).
Рис. 38. Положение различного типа масс воды у восточного побережья Сев. Америки
по гидрологическому разрезу судна «Атлантис» (по Кларку).
А, В и С — три пункта наблюдений.
$0
Глубина б м, Глубина б м Глубина
Рис. 39. Сезонные изменения в распределении температуры в трех точках разреза,
показанного на рис. 38 (по Кларку).
А — береговые воды, В — воды склона, С — воды сев. части Саргассова моря.
91
месячной температуры свойственны береговым водам в поверхностном
слое в 50 м (рис. 39). В береговых водах амплитуда годовых колебаний
составляет 17,5°, в водах материкового склона — 10,3° и в водах Саргас-
сова моря всего 7°. При этом наивысшая температура в береговых водах
23,4°, на склоне 24,6° и в Саргассовом море 26,7°.
Количество планктона в теплое время года в береговых водах оказа-
лось в 20—40 раз больше, чем зимой, в водах склона — только в 10 раз
Рис. 40. Колебания в количестве Calanus finmar-
chicus (все стадии) по разрезу, показанному на
рис. 38 (по Кларку), 1—8—номера станций.
больше, а в водах Саргассова
моря колебания в количестве
планктона обнаружены не
были. Кроме того, средний
объем планктона в береговых
водах был в 4 раза больше,
чем в водах склона, а в этих
последних в 4 раза больше,
чем в водах Саргассова моря.
Различия в обилии планк-
тона в береговых водах и в
водах материкового склона
по тому же разрезу могут
быть показаны на примере
одной из массовых форм —
Calanus jinmarchicus (рис. 40).
Конечно, эти различия в
количестве планктона только
в какой-то части могут быть
отнесены за счет температуры
и зависят также от целого
ряда других моментов.
6. ГАЗОВЫЙ режим
МИРОВОГО ОКЕАНА
В воде пресных и мор-
ских водоемов находятся в
растворенном состоянии раз-
личные газы (NO2, СО2, H2S,
NH3, Н, СН4 и др.).
Соотношение основных
газов в воздухе и в воде,
при условии равновесия с
атмосферой, различно, что
можно видеть на табл. 31.
Заключенные в воде газы либо поступают в нее с поверхности
(из атмосферы, с речнымЦ водами, осадками), либо образуются в самой
Таблица 31
Газы при 0° °/о В атмо- сфере В пресной воде В морской воде
N(+Ar) 79,0 63,0 63,0
О. 21,0 35,0 35,0
сб2 0,03 1,7 1,9
толще воды, либо в покрывающих дно грунтах. Газовый режим в водое-
мах в значительной степени обусловливается организмами и в свою оче-
92
редь коренным образом воздействует на распределение и развитие жизни.
Только азот, так же как и в воздухе, нейтрален для организмов.
Кислород представляет собою один из наиболее важных факторов
среды для развития жизни. Основная масса кислорода в море создается
растительными организмами.
Содержание кислорода в морской воде обычно выражается в см3 на
литр, либо в % к состоянию насыщения (равновесия с атмосферой) при
данной температуре.
Нередко, при очень сильном развитии растительных организмов или
прп длительном действии солнечного света (полярные страны), может
93
наблюдаться пересыщение кислородом. В отдельных случаях оно может
достигать 200, 300 и даже 400%. Так, в поверхностных слоях Азовского
моря, в период особо бурного цветения фитопланктона, количество кисло-
рода может превышать 300%. В мелководных озерах Новой Земли с дном,
сплошь покрытым густыми зарослями растений, при условиях полярного
летнего дня, наблюдалось содержание кислорода в 322,2%. Обычно же в
поверхностных водах количество кислорода близко к 100% насыщения.
В океанических водах пересыщение также может наблюдаться, но
лишь в ничтожной степени, причем оно может быть результатом бурной
деятельности растений, а может вызываться и нагревом водных масс.
Поверхностные воды оксанов и морей, за очень немногими исключе-
ниями, имеют количество кислорода, близкое к состоянию насыщения.
На глубине 10—20—40 ж, в зоне наибольшего развития растительного
планктона очень обычно небольшое пересыщение. В кислородном реяшме
более глубоких слоев между придаточными морями и открытыми океанами
наблюдается довольно резкая разница. В придаточных морях обычно,
с глубиной, количество кислорода уменьшается, иногда в слабой степени,
иногда в очень сильной, вплоть до полного его исчезновения. В тропи-
ческих зонах океанов (рис. 41) с их своеобразной динамикой водных масс,
с глубиной (50—300 м) сначала количество кислорода падает, иногда очень
значительно (меньше 1 см{л), а затем на глубине примерно в 1500 м снова
достигает довольно высоких показателей (5,0—5,5 см3/л). Обедненная кисло-
родом толща воды залегает в тропических областях (главным образом между
*25° ю. ш. и 35° с. ш.) в виде громадной двойной линзы. Как мы увидим
дальше, эти обедненные кислородом массы воды соответствуют халиста-
тическим районам океана и сохраняются в силу слабой вертикальной
и горизонтальной циркуляции и потребления кислорода планктоном
(рис. 43). Глубинные, богатые кислородом, воды приходят из высоких ши-
рот и сохраняют принесенный с собой кислород, вследствие очень слабого
его потребления организмами. С переходом в высокие широты, за парал-
лели 50°, обедненный кислородом слой сходит на нет.
В обоих океанах обедненные кислородом промежуточные слои при-
платы к восточным сторонам океанов (рис. 42), что можно рассмат-
ривать в первую очередь как результат наличия мощных поверхностных
течений, движущихся в обратном направлении с востока на запад. Откло-
нение масс этой воды в восточной части Тихого океана может рассматри-
ваться как явление компенсационного характера по отношению к явле-
ниям сгонного характера на поверхности океана.
Насколько резка изоляция слоев с малым количеством кислорода от
вышележащих можно видеть на графике (рис. 44), сопоставляющем распре-
деление с глубиной плотности воды и количества растворенного кислорода
в центре южной ха листа тической области Атлантического океана. Слой
скачка, располагающийся на50л«, служит верхней границей изолирован-
ных слоев. На 300 м глубины насыщение воды кислородом падает до 2%,
а на 400—500 м были обнаружены слои воды с количеством кислорода,
почти падающим до нуля (1,1—0,5 см3)л). Значительное понижение (до
0,2—0,3 см3/л) количества кислорода на глубинах нескольких сот метров
констатировано также в северных частях Тихого океана, к северу и югу
от Алеутских островов.
В открытых частях мирового океана мы не знаем больше значительных
районов с малым содержанием кислорода. Это явление получает сильное
развитие только в придаточных морях (Черное, Балтийское), в частях океа-
на, глубоко вдающихся в сушу (фиорды), если их котловины отделены от
моря подводными барьерами, где могут развиваться явления застойности
глубинных вод, и в северо-западной части Индийского океана под воз-
54
действием соленых вод большой плотности, поступающих из Персидского
залива.
Рис. 42. Распределение кислорода в см6/л в глубинах Атлан-
тического океана.
Из других газов, которые могут находиться в морской воде в раство-
ренном состоянии, следует упомянуть СО2,
Углекислота является продуктом
жизнедеятельности организмов. Поступая
в воду из атмосферы и из организмов,
углекислота может находиться в воде в
свободном состоянии и в соединении с дру-
гими веществами, главным образом, в со-
стояний карбонатов и бикарбонатов (свя-
занная углекислота, или щелочной резерв).
Имеющаяся в воде свободная углекислота
потребляется растениями, в какой то части
она может выделяться в атмосферу и мо-
жет переходить в связанное состояние.
Сероводород содержится в океаниче-
ских водах в ничтожных количествах и
только в грунтах присутствует постоянно.
В воду он может поступать в результате
жизнедеятельности особых бактерий (груп-
пы Microspira), производящих восстанов-
ление сульфатов (см. II том, глава Черное
море) из разлагающихся белковых веществ и, наконец, из донных
отложений, где он создается в результате тех же двух процессов.
Только в одном месте мирового океана в открытых его частях конста-
тировано значительное содержание сероводорода в глубинных водах,
H2S, NH3 и СН4.
о + s
6СМ3/л.
^кислорода
$
100
200-
300
35*07'N
V-66'25'W
I
I
Рис. 43. Вертикальное распреде-
ление кислорода в халистатиче-
ской зоне Северной части Атлан-
тического океана.
95-
а именно в Индийском океане в его самом северо-западном углу, в заливе
Оман, у восточной оконечности Аравийского полуострова. Сюда посту-
пают, сползая на глубину по склонам, тяжелые соленые воды из Персид-
ского залива, совершенно затормаживающие циркуляцию, чем и создаются
благоприятные условия для развития H2S. Количество сероводорода,
Глдбинав М Плолрос/пь
25.0 25.5 26.0 26.5 27,0
150 \
200 Ллотносто \
250 \
300 - \
350- |
400- |
Глубина&н
/ Кислород - 200
I 250
I -ЗОО
I -350
I -400
450 -
-450
Рис. 44. Вертикальное распределение плотности воды и содер-
жания кислорода на 45°25' ю. ш. и 0°,07 з. д., т. е. в центре юж-
ной халистатической области Атлантического океана (по Баттен-
бергу).
растворенного здесь в воде, превышает даже количество его в глубинах
Черного моря. Во внутренних морях и фиордах с изолированными кот-
ловинами создаются условия, особенно благоприятные для развития
в придонных слоях H2S, либо в течение круглого года (Балтийское, Чер-
Рис. 45. Кислородный режим в течение года в одном из заливов Мурма-
на (по Дерюгину из Зенкевича).
ное, часть фиордов) либо периодически (Азовское, фиорды), когда развитие
H2S идет параллельно с процессом обеднения глубинных вод кислородОхМ
в определенные сезоны года (рис. 45.)
Сероводород очень легко окисляется в присутствии кислорода или
связывается жизнедеятельностью особых серобактерий.
96
Аммиак, образующийся в океанических водах за счет разложения
белковых веществ и при восстановлении нитратов денитрифицирующими
бактериями, обычно содержится в ничтожно малых количествах. Только
в некоторых придаточных морях аммиак может присутствовать в воде
в очень больших количествах либо постоянно (Черное море), либо в опре-
деленные сезоны (Азовское море) (см. II том).
Метан. В некоторых придаточных морях констатировано образование
метана (СН4) и водорода в результате жизнедеятельности некоторых
групп бактерий, разлагающих клетчатку. Процессы эти идут и в воде
и в грунте. В некоторых районах Каспийского моря процесс выделения
болотного газа из грунта иногда принимает столь бурный характер, что
вода как бы кипит.
Газовый режим придонного слоя. Рассмотренные нами условия газо-
вого режима в мировом океане создают достаточно определенное впечат-
ление о толще воды, но не могут вполне характеризовать газовый режим
самого придонного слоя, тех 5—10—15 см прилежащей ко дну воды,
в которой собственно и обитает донное население. Незнание нами газо-
вого режима этого слоя объясняется только непреодоленными пока труд-
ностями чисто технического характера. Океанография не располагает
в своей аппаратуре батометрами той конструкции, которые позволяют
брать пробы тонкого придонного слоя воды, не нарушая его стратификации.
Несомненно, что в придонном слое происходит особенно сильное по-
требление кислорода, как под воздействием жизнедеятельности орга-
низмов, часто очень плотно заселяющих дно, так и в результате разложе-
ния накопляющихся на дне органических осадков. Наконец, прилежащий
к дну слой воды вступает с самим дном в химическое взаимодействие,
в результате которого также идет потребление кислорода. Укажем хотя
бы на тот своеобразный круговорот в придонном слое, который связан
с марганцем и железом. В самом грунте они находятся в восстановленном
состоянии (гидраты закиси), а стало быть в растворимом виде. Поступая
в виде раствора в придонную воду, гидраты закиси окисляются и выпадают
в виде осадка (гидраты окиси). В грунте они опять восстанавливаются
и могут поступать в воду в виде раствора. Таким образом, они играют
здесь роль, по образному сравнению Я. Самойлова, гемоглобина
в крови животных, постоянно мигрируя из грунта в придонный слой воды
и перенося в грунт связываемый ими кислород, с тем, чтобы затем опять
поступить в воду в восстановленном виде.
Донные отложения должны отдавать в воду также и значительное
количество углекислоты, повышая тем концентрацию водородных ионов
и способность воды растворять целый ряд веществ. При сильном обедне-
нии придонного слоя кислородом в воде может оказаться и H2S, выделен-
ный грунтом. Газовая стратификация придонного слоя воды может суще-
ствовать длительно, а может носить только сезонный или даже очень быстро
протекающий характер в период ослабления циркуляции. Толщина при-
донного слоя, с резко измененным газовым режимом, может быть весьма
различна.
Вертикальная зональность в океане. Основные моменты газового со-
стояния толщи океанических вод, рассмотренные выше, дают основа-
ние наметить общую схему вертикального распределения слоев воды
(табл. 32).
Верхняя граница восстановленной зоны, обычно расположенной
внутри самого верхнего слоя грунта (под поверхностной пленкой грунта),
может либо постоянно, либо в определенные сезоны года (Азовское, Кас-
пийское, Черное моря, норвежские фиорды, северо-восточная часть Оман-
ского залива) подыматься более или менее высоко над грунтом.
7
Зенкевич, т. I
97
Таблица 32
Золы Подзоны Характеристика Глубина в м
Окислительная Верхняя Обогащение воды кислородом пу- тем фотосинтеза и обмена с атмо- сферой 0—100
Нижняя Слабое потребление кислорода зоо- планктоном >100
Окислительно-вос- становительная Значительное потребленпе кисло- рода животными, в процессах гни- ения и за счет обмена с грунтом Обычно придон- ный слой и самая верхняя пленка грунта
Восстановитель- ная Обычно верхний слой лонных отложений
7. СВЕТ
Двумя основными моментами, определяющими развитие фитопланктона
и береговой растительности в море, являются свет и питательные соли.
Проникновение света в толщу морской воды. Солнечный свет и свет
небесного свода, достигающий поверхности моря, частично отражается,
а в определенной части проникает в толщу воды и освещает ее на большую
или меньшую глубину. Освещенность поверхностных слоев моря зависит
от многих причин: от географической широты места, от сезона года, от
облачности, от высоты солнца над горизонтом, от количества и состояния
влаги, находящейся в воздухе.
Зависимость количества света, отражаемого горизонтальной водной
поверхностью, от высоты солнца над горизонтом показана на следующей
таблице (табл. 33):
Таблица 33
Высота солнца над горизонтом (°) Свет от солнца п небесного свода, отраженный поверхностью моря в ясный день (в °/о от общего количества)
5 10
10 25
20 12
30 6
40 4
50 3
60 3
70 3
80 3
90 3
Таким образом, количество света, проникающего под поверхность
моря в северных широтах относительно всегда меньше, чем в южных.
Это в значительной степени компенсируется в полярных морях длитель-
ностью дня, особенно в период непрерывного полярного дня, когда солнце
98
не заходит за горизонт. Как показывают данные, приводимые К и м б е л-
лом (Kimball, 1928), в июне поверхность полярного моря получает
в общем то же количество света, что и под экватором (табл. 34).
Таблица 34
Местоположение Месяцы
Широта- | Долгота XII III VI IX
60 N 56 W 0 .126 .292 .147
52 » 10 » .041 .148 .267 .160
42 » 70 > .086 .212 .329 .230
30 » 77 > .142 .238 .310 .239
10 » 69 » .239 .299 .276 .287
0 » 12 Е .235 .244 .196 .220
Зато в зимнее время, в период непрерывной полярной ночи, количество
света, проникающее под поверхность полярных морей, или равно нулю,
или весьма ничтожно.
Чем ближе к экватору, тем глубже солнечный свет проникает под по-
верхность моря. Примерно одинаково освещены слои воды на глубине:
800 м под 33°, 500 м под 50° и 200 м под 67°.
Потеря света, при прохождении через толщу воды, выражается (по
Кларку, 1939) уравнением:
где Jo — исходная интенсивность света, J — конечная интенсивность,
к — коэффициент затухания (поглощения), L — толщина слоя воды в
метрах, а е — 2,7 (рис. 46).
Проникновение в глубину различных частей спектра. Хотя в открытых
морях свет и проникает на очень большую глубину, однако уже в толще
первого метра он теряет половину своей силы, причем красные и ультра-
фиолетовые лучи поглощаются быстрее всего, а голубые и зеленые прони-
кают всего глубже.
Таблица 35
ПРОНИКНОВЕНИЕ ЛУЧЕЙ ТРЕХ ЧАСТЕЙ СПЕКТРА
ВГЛУБЬ МОРСКОЙ ВОДЫ ВБЛИЗИ ЮЖНОЙ АЛЯСКИ
ПО АТ ТЕР БЕКУ (UTTER В А СК, 1936) в °/0
Глубина в м Части спектра и длина волны
Желтый бооо А Зеленый 5300 А Голубой 4800 А
0 100 100 100
5 18 35 26
10 1,8 16 7,8
15 0,53 7,6 3,9
20 0,27 5,7 2,3
30 0,012 0,12 0,082
В дистиллированной воде на 100 м глубины доходят только зеленые,
голубые и фиолетовые лучи. Даже человеческий глаз способен улавливать
свет еще на очень большой глубине. Американец Биб (В е е Ь), опустив-
шийся в изобретенной им батисфере на глубину 900 м в районе Бер-
мудских островов, в верхних 50 м наблюдал зеленую окраску воды,
на 60 м зеленовато-синюю или сине-зеленую, на 180 м ясный синий цвет,
*
99
на 300 м слабый черновато-синий и еще на глубине 580 м улавливал по-
следние следы света.
При помощи различных приборов (фотографические пластинки, в по-
следнее время различные фотоэлектрические приборы) свет удается кон-
статировать и на значительно больших глубинах (до 1500 зс).
Прозрачность. Освещенность различных слоев воды зависит и от ее
прозрачности. Чем чище вода, чем меньше в ней взвеси органического
и неорганического происхождения, тем она будет прозрачней (рис. 46).
Сипа света /освещение! в % к сипе света ни поверхности
Рис. 46. Сравнение степени проникновения желто-зеленого ком-
понента дневного света в натуральные воды (фотометр с чувстви-
тельностью к длине волны 5000—6000 АА) е проникновением в ди-
стиллированную воду. Кривые показывают отношение между глу-
биной и освещением, выраженным в % к свету на поверхности
(по Кларку).
Определение прозрачности воды при помощи диска Секки показывает
большие различия для разных морей. Наибольшая прозрачность воды
в 66,5 м была констатирована в Саргассовом море, отличающемся исклю-
чительной бедностью планктона и минимальным содержанием в воде не-
органической взвеси. Очень большая прозрачность наблюдается также
в Средиземном море (до 60 зг), в Тихом океане (до 59 м)9 в Индийском
океане (до 50 м). В придаточных морях, в связи с близостью берегов (на-
личие неорганической взвеси) и обильным развитием планктона, прозрач-
ность резко падает, однако, в Баренцевом море она достигает все же
иногда 45,5 м. Зато в Северном море падает до 23 зс, в Балтийском до 13 зс,
в Белом 5,8—9 з«, в Азовском наиболее обычная прозрачность 0,1—2,75 м
и редко достигает 4—5 м. Наименьшая прозрачность наблюдается в Азов-
ском море в период цветения, протекающего в этом море с необыкновенной
бурностью, когда вода делается похожей на «зеленые щи».
0(00
На рис. 47 показана глубина проникновения общей световой энергии
и отдельных частей спектра в глубину (в чистой воде), а на рис. 48 — степень
проникновения световой энергии в воды различной мутности.
В наиболее чистой прибрежной воде в толще первого метра уже по-
глощается 62,3% световой энергии, проникающей под поверхность, а на
Энергия б
2 3 4 5 6 8 10 15 20 30 4050680100
3/л4 5 6 7 8 9 Ю 11 12 13 14 15 16 17 18 19 20 21 22 23 24JA
длина волны
Рис. 47. Схематическое изображение энергии разных частей спектра ра-
диации солнца и небесного свода, достигающих поверхности моря и про-
никающих в воду на глубину 0,1; 1; 10 и 100 м. Справа % общей световой
энергии и энергии видимой части спектра, достигающих различных глу-
бин (по Свердрупу и др.).
глубине 10 м в самой чистой — 83,9%, а во взмученной прибрежной —
99,55%. При этом в чистой воде глубже идет сине-зеленая часть спектра,
а во взмученной — зелено-желтая. Во взмученной воде на 10 м осве-
щение «слабее», чем в чистой воде на 100 м (рис. 48). В мутных водах пред-
40(риал.45 галуй.50 зел. 55желт.60оранж63Н краем.
Рис. 48. Энергия лучей разных частей спектра на глубине 10 м в чи-
стой воде (0), в чистой океанической (1), океанической средней про-
зрачности (2), береговой средней прозрачности (3) и взмученной
прибрежной (4). Справа то же самое для глубины 100 м в чистой
воде и 10 м в прибрежной взмученной (по Свердрупу и др.).
устьевых пространств рассеяние света настолько велико, что голубые
лучи спектра проникают на меньшую глубину нежели красные и состав
света как бы изменен в обратном отношении.
Поглощением в самом верхнем слое воды ультрафиолетовых и фио-
летовых лучей обусловливается сильный прогрев слоя воды на глубине
101
1—2м, показанный Гаардером и Сперком (Gaarder and
S р а г с к, 1931, 1932) на примере мелководных норвежских устричных
парков (рис. 49). В развитии мо-
f8" 20° 22" 24° 26°С лоди устриц этот сильный про-
^9
/30 июня / 15июля
1927г / 1927г.
грев воды имеет очень важное зна-
чение (эффект парника).
Исследования по проникнове-
нию света внутрь вод океана
вскрыли одно чрезвычайно суще-
ственное явление в распределе-
нии планктона, которое остава-
лось до того неизвестным. Если
Рис. 49. Распределение температуры в
верхнем слое воды в норвежских устрич-
ных бассейнах (по Гаардеру и Сперку).
изучать метр за метром прозрач-
ность последовательных слоев во-
ды от поверхности в глубь моря,
то оказывается, что слои более
прозрачные чередуются с
было отмечено Кларком
вопрос разрабатывал
А. Трофимов (рис. 50).
Как можно видеть по
приведенным графикам,
прозрачность воды в об-
щем возрастает с глуби-
ной, причем в верхних
10 м возрастание идет
очень быстро, а в гори-
зонте от 10 до 50 м
(для Гюльмар-фиорда)
или 10—30 (для Каспия)
возрастание прозрачно-
сти очень сильно замед-
ляется. С глубины 80 м
и больше для Каспий-
ского моря степень про-
зрачности как бы оста-
ется на одном уровне.
Помимо этой общей за-
кономерности в ходе
кривой, применимой,
вероятно (только с из-
менением высоты от-
дельных горизонтов), к
любому водоему, мож-
но видеть отдельные
нарушения плавного хо-
да кривой. Кроме того,
на графике Петтер с-
с о н а, кривая про-
зрачности дает в общем
обратный ход кривой
рассеяния, а кривая
плотности воды дает
менее прозрачными. Впервые это явление
на Вудс-Холлской станции. У нас этот
Глубина
Глубина
Рис. 50. Изменение прозрачности слоев воды;
Вверху — в Гюльмар-фиорде, а) прозрачность, б) рассеяние, то-
чечная кривая — плотность воды (по Петтерссону из Трофи-
мова). Внизу — профиль прозрачности моря (а) и прозрачность
для дневного света в Каспийском море (б) (по Трофимову).
некоторое совпадение переломов с уменьшением прозрачности. А. Тро-
фимов приходит к выводу, что ход кривой прозрачности опре-
102
деляется распределением микро- и наннопланктона и всякой другой взве-
си (био- и а биосестон). >
Таким образом, эти данные позволяют заключить, что, во всяком слу,-
чае, мелкие компоненты планктона и абиосестона накапливаются в опре-
деленных слоях и тем уменьшают прозрачность и увеличивают рассея-
ние света этих слоев. Весьма вероятно, что слои концентрации планктона
приходятся на горизонты изменения плотности, а возможно и других по-
казателей состояния среды. Планктон постоянно совершает некоторые
вертикальные перемещения и может задерживаться в горизонтах более
значительных градиентов по некоторым физическим и химическим фак-
торам и в первую очередь по плотности.
Эти явления в распределении планктона должны учитываться при
выработке методики сбора планктона, когда пользуются батометром и
результаты отдельной пробы пересчитываются на весь столб воды.
Влияние интенсивности освещения на распределение растительных ор-
ганизмов. Очень интересен устанавливаемый Гисленом (Gislen,
1930) факт, что растения, в общем, распределены в море гораздо более
равномерно, нежели на суше. Это находится в полном согласии с равно-
мерным распределением в воде света и растворенных питательных солей,
нужных растениям. Пища, нужная животным, распределена гораздо
менее равномерно, и в поисках за ней равномерность распределения жи-
вотных нарушается.
Хотя в процессе фотосинтеза наибольшее значение имеют красные,
оранжевые и желтые лучи, быстро поглощаемые в самом верхнем слое
воды, наибольшее скопление планктонных водорослей в морях имеет место
обычно не в самом верхнем горизонте, а на глубине 10—50 глубже
количество фитопланктона резко убывает.
Нижняя граница распространения донной флоры в Балтийском море
20 з«, у берегов Исландии 50 в Средиземном море 130—160 м. Те же
различия наблюдаются и в распределении планктона. У побережий се-
веро-западной Европы наибольшее скопление фитопланктона имеет место
на глубине 10—30 л«, у берегов Калифорнии 25—55 лс, в Средиземном
море у Сиракуз на 50 м. Обычно наблюдаемое слабое развитие планктона
в поверхностных 10 м может быть следствием неблагоприятного воздей-
ствия прямых солнечных лучей, иногда сильного прогрева, иногда зна-
чительного опреснения и свойственных поверхностному слою сильных
волнений. Во всяком случае массовая гибель планктона после штормов,
а также в районах сильных токов воды — явление обычное. С другой
стороны, накопление фитопланктона на глубине 20—50 м может быть ре-
зультатом неуклонно идущего опускания и отмирания.
Тж „ фотосинтез л
Интенсивность освещения, при которой --------= 1, называется
’ r г дыхание ’
интенсивностью точки компенсации. По новым данным (Кларк, 1939),
она равна 350—500 люксов (или 0,3% солнечного освещения в полдень).
В разных частях моря, в зависимости от прозрачности воды, точка
компенсации находится на разной глубине (табл. 36).
В озерах из падающего на поверхность воды света растениями утили-
зируется не более 15 %, вся же остальная часть поглощается водой и дет-
ритом.
Очень существенное значение для обилия и развития в морях расти-
тельной жизни имеет интенсивность прямого солнечного освещения, опре-
деляющего собою фотосинтетическую деятельность, о чем уже говорилось
выше. Как указывает Л. Иванов, суточный максимум интенсивности
света, выраженный в грамм-калориях, под 68° с. ш. приблизительно равен
таковому в тропиках, т. е. калорический эффект тропического солнца не
103
больше, чем у нас на дальнем севере. Л. И в а н о в, учитывая значительную
облачность на севере, полагает, что в среднем интенсивность рассеянной ра-
диации, получаемой от небесного свода, в высоких широтах не меньше, чем
Таблица 36
Район
Нижнее положение точки
компенсации в м
Саргассово море............
Английский канал...........
Майнский залив.......... . .
Западное побережье Канады .
Район Вудс-Холла...........
Ниже 100
45
24—30
10—19
7
в низких. Что же касается количества радиации, получаемой от прямых
солнечных лучей, то в северном полушарии оно убывает с севера на юг.
Если учесть, что температурный фактор в море сам по себе не определяет
интенсивности процесса продуцирования растительной массы, а непрерыв-
ный полярный день обеспечивает получение растениями такого количе-
ства солнечного излучения, то эффект продуцирования в полярных стра-
нах может вполне стать в ряд с тем же процессом в более южных
широтах. К а л и т и н приводит следующие данные, для характеристики
Таблица 37
Месяц Апрель .... Май Июнь Июль Количество малых калорий, получаемых одним см9 горизонтальной поверхности
Земля Франца Иосифа Ленинград Феодосия
7860 14870 16080 9720 8130 10940 12260 12690 10700 15340 15240 16350
Всего . 48530 44020 1 57630
этого явления (табл. 37). Если это справедливо в отношении наземной ра-
стительности, то еще более это применимо к водной растительности. Это
может объяснить нам то, казалось бы удивительное явление, что про-
дукция прибрежной растительности (макрофитов) Варенцова, Белого,
Каспийского и Черного морей дает удивительно сходные величины, не-
смотря на различие в широтном положении в 25°.
Окраска организмов на разных глубинах. С неравномерным проникно-
вением в толщу воды лучей разной длины волны связано явление верти-
кальной зональности в преобладающей окраске растении и животных.
Последние очень часто бывают окрашены в дополнительный цвет к той
части спектра, которая проникает на данную глубину, в результате чего
приобретают защитную окраску — кажутся черными. На больших глу-
бинах, лишенных света, животные бывают большей частью или окра-
шены в черный цвет, или лишены окраски. Водоросли также дают смену
окраски с глубиной, давая от поверхности вглубь смену зеленых на бурые
п дальше бурых на красные. Эта окраска растений играет не защитную
роль, а способствует использованию соответственных лучей спектра.
404
БИОГЕННЫЕ ЭЛЕМЕНТЫ И РАСТВОРЕННОЕ ОРГАНИЧЕСКОЕ
ВЕЩЕСТВО
Многие из веществ, растворенных в морской воде, необходимы оби-
тающим в ней организмам для построения своего тела. Животные полу-
чают эти вещества из тела растений или других животных, которыми они
питаются, и лишь в очень малой степени (простейшие, губки) могут исполь-
зовать для питания органическое вещество, растворенное в воде, хотя
и это подвергается сомнению. Растения берут их непосредственно из
воды.
Минимум-соли. Среди веществ, растворенных в воде и необходимых
растениям для построения тела, большинство всегда присутствует в мор-
ской воде в достаточных количествах. Другие находятся в растворе в очень
небольшом количестве и по использовании их развивающимся фитопланк-
тоном развитие последнего приостанавливается. Таким образом, ограни-
чивающее (лимитирующее) действие на развитие растений в море оказы-
вает недостача только в некоторых солях, называемых в силу этого мини-
мум-солями. К ним обычно относят соединения фосфора, азота и для части
растений — кремния.
По очень удачному выражению Гарвея(Наггеу, 1942) соленость
морской воды является показателем ее физического прошлого, а со-
держание питательных солей является показателем ее биологического
прошлого.
На водоемы вообще и морские в частности вполне могут быть перене-
сены те обобщения, которые были сформулированы Либихом и Мит-
че р л и х о м для наземных растений. Либих установил так называе-
мый закон минимума, по которому развитие растения ограничивается тем
из нужных ему веществ, которое находится в настоящий момент в мини-
муме. В дальнейшем это правило было изменено Митчерлихом,
установившим правило совокупного действия факторов, заключающееся
в том, что рост растения ограничивается не только веществом, находя-
щимся в минимуме, но и всеми другими веществами, нужными растению
и имеющимися в недостаточном количестве. В настоящее время закон со-
вокупного действия факторов понимают в более расширенном толковании,
учитывая не только химические вещества, но и все другие факторы роста—
свет, температуру и т. п. В. Вильямс формулирует это так: «растения
для своей жизни требуют одновременного и совместного наличия... всех
без исключения условий или факторов своей жизни». Кильский гид-
робиолог К. Брандт (К. В г a n d t) в самом начале настоящего сто-
летия впервые перенес на развитие морского фитопланктона законо-
мерности соотношения количества питательных солей и темпа роста,
установленные для развития наземных растений.
Азот и его соединения. Количество азота, растворенного в воде, относи-
тельно меньше, чем в воздухе. В атмосфере количества кислорода и азота
относятся как 1 : 4, а в воде как 1 : 2. Однако этот азот, так же как и на
суше, непосредственно растениями усваиваться не может. Накопление
элементарного азота в воде происходит путем растворения воздуха из
атмосферы и в процессе денитрификации. Количество элементарного азота
в морских водах, в противоположность кислороду, весьма постоянно,
так как растительные организмы не способны его усваивать. Процесс
образования свободного азота, в результате восстановления нитратов и
нитритов сероводородом, имеет весьма ограниченное значение (например,
в Черном море). Содержание свободного азота в морских водах выражается
величиной 10,8—14,6 см3 на литр. Так же, как и в наземных почвах,
в морской воде может иметь место процесс связывания азота микроорга-
105
Рис. 51. Схема круговорота азота в
море. Площадями квадратов пока-
зано среднее количественное соот-
ношение нитратного, нитритного и
аммиачного азота (ориг.).
низмами (бактерии типа Azotobacter^ Clostridium и некоторые дрожжевые
грибки).
Большие количества азота в связанном состоянии содержатся в рас-
творенном в воде органическом веществе (аминокислоты). Это так называе-
мый альбуминоидный азот. Количество альбуминоидного азота в морской
воде выражается числами порядка десятков, до сотни, а иногда нескольких
сот мг/мг.
Наибольшее значение для развития в море фитопланктона имеет азот
нитратов, нитритов и аммиака.
Общее количество этих трех типов соединений азота очень невелико
и исчисляется обычно сотнями миллиграммов в кубическом метре, т. е.
дает концентрацию в 50 000 раз более
слабую, чем концентрация основного
солевого состава. Однако организмы
обладают способностью извлекать из
окружающей их воды вещества, нахо-
дящиеся в ничтожно малых концентра-
циях, и даже в еще гораздо более сла-
бых, чем соединения азота. Возможно,
что в море, как и в почвах, одновремен-
но идут процессы денитрификации и
нитрификации. Белковые вещества в
процессе минерализации, под действи-
ем микроорганизмов дают аммиак. Про-
цессы нитрификации переводят аммиак
через нитриты до нитратов. Процесс
денитрификации идет в обратном на-
правлении. Однако, присутствие в мор-
ской воде нитрифицирующих бактерий
с точностью но установлено п некото-
рые микробиологи, например, В. Бут-
кевич, считают более вероятным, по
крайней мере в поверхностных водах,
образование нитритов за счет восста-
новления нитратов. Эти процессы мож-
но пояснить прилагаемой схемой
(рис. 51).
Количество аммиака в морской во-
де очень незначительно, от нуля до
0,02—0,13 мг/л. Только в отдельных
случаях, в придаточных морях, коли-
чество аммиака может достигать очень
больших величин, в результате энер-
гично идущего процесса распада органического вещества. На глуби-
нах Черного моря содержание NH3 достигает 1,46 мг)л. В южной части
Азовского моря также констатировано накопление аммиака до 0,8—0,9л«г/л,
но только как периодически наступающее явление. Повышенное количество
аммиака констатировано также на глубинах в Средиземном и Балтийском
морях.
В качестве примера вертикального распределения аммиака в океане
можно привести данные меридионального разреза через Атлантический
океан по Брандту (рис. 52) и данные Баттенберга по Ирмин-
герову морю (рис. 53), расположенному между Исландией и Гренландией.
Нитриты встречаются в морской воде в очень малых концентрациях
(редко больше 10 мг нитритного азота на л«3): очень часто вода не содержит
45 мг
106
их совсем или только в виде следов* Большинство исследователей рассмат-
ривают нитриты как промежуточную стадию между аммиаком и нитратами
в процессе окисления. Во всяком случае, между количеством нитратов и
нитритов существует определенная обратная зависимость и кроме того не-
которые наблюдатели констатировали массовое появление нитритов,
север
40° 30°
Ум « 25 40
54 40
5ООм-
по
59
1000м
20“ 10° ' 0°
во 60 2026 70 40
26
в1 90
нгг
10° 20° 30°
69 51 41178320 QQ
50
19 37 ^°30
70
34 15 26 ,п
40
1500м-
50 20
50 20
ои 60
90
60
Рис. 52. Вертикальное распределение аммиака [мг/л) в Атлантическом
океане с севера на юг (но Брандту из Гарвея).
как стадии, предшествовавшей появлению нитратов (Ж у к о в, Азовское
море).
Аммиачный азот потребляется растениями в очень малой степени и
основным непосредственным азотным фондом для развития водорослей в
море служат нитраты, количество которых значительно превосходит
нитриты и аммиак.
Рис. 53. Вертикальное распределение фосфатов, нитратов и
аммиака в Ирмингеровом море (по Баттенбергу).
В поверхностной толще вод мирового океана, в слое фотосинтеза (выше
50—150 л«) количество нитратов обычно невелико и подвергается очень боль-
шим колебаниям. Сильное развитие планктона приводит к резкому уменьше-
нию и очень часто к полному использованию нитратов. Только в немногих ме-
стах мирового океана в течение круглого года в поверхностных водах сохра-
107
няются нитраты. В глубоких слоях моря (с 150 м и глубже) количество
нитратов возрастает и подвергается слабым изменениям. Развивающиеся
Рис, 54. Вертикальное распределение фосфатов (слева) и нитратов (справа)
в трех океанах (по данным разных авторов из Свердрупа и др.).
в верхнем слое растительные и животные организмы, опускаясь после
смерти на глубину, увлекают туда и заключенные в их телах соединения
азота, подвергающиеся, под воздей-
Рис. 55. Вертикальное распреде-
ление силикатов в трех океанах
(из Свердрупа).
Для последующего развития
ствием бактерий, процессам распада и
минерализации.
В морской воде азот содержится в
живых организмах, в детрите, в раство-
ренном органическом веществе и в виде
аммиака, нитритов и нитратов. Соотно-
шение между всеми этими соединения-
ми, содержащими азот, в разных местах
различны, а в верхних слоях моря
подвергаются сезонным колебаниям. В
поверхностном слое обильное накопле-
ние неорганических форм азота (нитра-
тов) предшествует сезону вегетации, а в
глубинных слоях, где нитриты и аммиак
практически отсутствуют, нитраты всег-
да имеются, но не претерпевают сезон-
ных колебаний.
Бактерии играют существенную
роль в минерализации органического
азота из детрита, экскретов и раство-
ренного органического вещества (NH3,
NO2 и NO3).
фитопланктона в поверхностном слое
моря нужно, чтобы этот последний получил опять необходимые для этого
108
количества нитратов. В открытом океане это может происходить либо
в результате подъема снизу из богатых нитратами слоев, либо приноса
со стороны. В береговых же водах большое значение имеет и принос с суши
речными водами.
Фосфор растения берут из фосфатов — соединений фосфорной кис-
лоты (Р2Об). Не весь фосфор в море заключается в фосфатах (неорганиче-
ский фосфор), он содержится также в телах организмов живых и мертвых
и в продуктах, ими выделяемых. Органического фосфора обычно бывает
несколько меньше, чем неорганического.
Характер вертикального распределения фосфатов, нитратов и кремпе-
кпслоты показан на табл. 38 и 39 и па рис. 54 и 55.
Таблица 38
РАСПРЕДЕЛЕНИЕ АЗОТА НИТРАТОВ И НИТРИТОВ И ФОСФОРА ФОСФАТОВ
В жДи3 В ВОДЕ АТЛАНТИЧЕСКОГО ОКЕАНА НА РАЗРЕЗЕ ПО МЕРИДИАНУ
30° 3. Д. В АПРЕЛЕ —МАЕ 1931 г. ПО ДАННЫМ ЭКСПЕДИЦИИ <ДИСКОВЕРИ>
Глубина в м 57°36' S 46°43' S 38° 10' S 2ГТЗ' S 9°00' S
N Р N 1 р N | Р N Р N 1 р
0 510 62 210 47 60 5 2 0 2 0
40 490 63 210 47 42 7 0 0 2 0
80 510 76 240 52 42 7 0 3 27 14
100 530 77 250 55 89 15 1 4 47 33
200 530 85 380 65 131 . 18 7 6 210 45
400 530 83 450 67 260 45 100 31 220 52
800 530 83 480 81 310 58 240 64 250 80
1500 510 80 450 79 360 86 170 54 210 53
2500 510 83 420 73 330 78 180 42 210 45
3200 500 85 — — — 82 * — 41 — —
4500 — — 440 81 — — 55 210 48
Таблица 39
РАСПРЕДЕЛЕНИЕ НИТРАТОВ И ФОСФАТОВ В ВОДЕ ТИХОГО II ИНДИЙСКОГО
ОКЕАНОВ в мг/м*
Глу- бина в м Индийский океан Тихий океан
12°05' ю. ш. 96°45' в. д. 10/Х-1929 3°40' ю. ш. 137°40' з. д. । 12/VII-1929 1 i 0°18' ю. ш. 1 99°07' з. д. 18/IX-1928 7°34' ю. ш. 167°03' в. д. 31/Х-1928
t°C n2 РА t°C N2 РА 1°С N, РА t°C n2 РА
0 25,20 30 0 28,40 0 0 19,10 150 50 28,46 ! 5 15
25 25,24 8 0 28,58 0 10 18,02 175 70 28,46 5 15
75 24,91 15 5 27,76 45 30 16,35 175 80 28,42 5 15
100 24,70 30 15 26,73 85 35 14,09 175 90 27,58 35 20
200 17,14 225 80 19,18 150 65 12,46 350 125 23,18 80 30
400 9,53 475 120 8,5 525 145 9,4 500 160 9,03 450 90
800 6,42 600 150 5,94 600 150 5,68 500 160 5,76 500 110
1000 5,46 600 150 4,68 600 175 4,50 500 160 | 4,74 550 125
2000 2,74 600 145 2,40 600 165 2,32 500 175 2,26 550 150
3000 1,68 600 130 1,65 600 165 1,77 500 175 1 1,681 । 550 150 .
3500 1,44 600 130 1,60 600 165 — — i 1,56: 550 150
.4100 1 • 1 _ 1 1,4-1 400 125
109
Приведенные таблицы показывают, что в верхних слоях количество
питательных солей невелико и возрастание их начинается с глубины 75—
150 м. Иногда, однако, у поверхности количество солей весьма значительно.
Обогащение фосфатами и нитратами поверхностных слоев моря имеет место
в районах облегченной вертикальной циркуляции и при понижении тем-
пературы поверхностного слоя. Левый столбец табл. 38 может служить
примером равномерного распределения нитратов в антарктических водах
в период усиленной циркуляции.
Рис. 56. Среднее содержание фосфора в мг!м* в слое от 0 до 50 м в южной части
Атлантического океана (по Баттенбергу).
Средний столбец для Тихого океана (табл. 39) показывает обогащение
поверхностного слоя воды питательными солями в зоне тропической ди-
вергенции. Такое же обогащение наблюдается в тепловодной зоне у бере-
гов Южной Америки и Африки, где имеет место подток более холодных
вод с юга и подъем глубинных вод к поверхности. Так, у берегов Африки
поверхностные воды содержат 13—26 мг фосфора в кубическом метре.
Отношение нитратного азота к фосфатному фосфору в морской воде
в среднем составляет около 6—7, хотя и может претерпевать довольно зна-
чительные колебания. При малом общем содержании питательных солей
110
данное отношение может падать до 1—2—3, при большом содержании
солей подыматься до 8.
В водах Антарктики отношение N/Р выше (5,9—8,2), по направлению
к экватору понижается (3—5,5), а дальше на север опять повышается,
у Бермудских островов в среднем до 5,8, в Ирмингеровом море до 6,6.
В Баренцовом море вариирует от 6,5 до 14. Средиземноморские воды также
дают высокое отношение N/P — около 10 в западной части моря и около
16 в восточной.
Рис. 57. Среднее содержание наннопланктона в тысячах особей на мл в южной
части Атлантического океана (по Гентшелю).
Соотношения С : N : Р в морской воде и в планктоне довольно близки
между собой. Среднее для планктона по Флемингу (Fleming,
1942) — 41 : 7,2:1, в морской воде по Куперу (1938) отношение азата
к фосфору 6,8 : 1. В донных же отложениях отношение углерода к азоту
выше чем в организмах, что указывает на то, что детрит, отлагающийся
в грунте, теряет больше азота, чем углерода.
Горизонтальное распределение фосфора в верхнем слое воды (0—50 м)
южной части Атлантического океана было дано Баттенбергом
(W a 11 е n b е г g, 1929) по работам экспедиции на судне «Метеор» и со-
поставлено Гентше л ем (Hentschel, 1936) с содержанием планк-
тона (рис. 56 и 57). Чем богаче вода фосфатами (до определенного предела),
ш
тем более обильно развивается в ней фитопланктон. Богатство же поверх-
ностной воды фосфором характерно для антарктических вод океана, где
Рис. >58. Годовые изменения в распределении фосфатов (1924) в одном
из пунктов Английского канала. Числа показывают содержание фосфат-
ного фосфора в мг/м3 (по Гарвею).
энергично идущая вертикальная циркуляция обеспечивает вынос его на
поверхность, и для западного побережья Африки, где пассатные ветры,
Рис. 59. Вертикальное распределение нитратов и фосфатов в разных
частях Средиземного моря.
I — Балеарское море; II — Тирренское море; III—Ионическое море; IV — Эгей-
ское море (по Томсену).
«сгоняющие воду на запад, обусловливают выход глубинных, богатых фос-
фором вод на поверхность.
112
Прибрежные воды и мелководные моря обычно значительно беднее
питательными солями, чем глубинные воды океанов, где происходит их
накопление, но богаче, чем поверхностные воды всей тепловодной зоны
океана. В зимнее время в мелководных зонах устанавливается обычно
равномерное распределение питательных солей от дна до поверхности,
в летнее — стратификация, при общем обеднении всего столба воды
питательными солями (рис. 58).
Кремнекислота. Многие планктонные водоросли (например диатомовые
пли кремнежгутиковые) и простейшие (например радиолярии), а также
донные животные (губки) нуждаются для создания своих скелетов в крем-
некислоте. Хотя она обычно не исчезает в той полной мере, как фосфаты
и нитраты, однако в периоды бурного цветения диатомовых может сильно
уменьшаться в количестве, что сказывается на интенсивности развития
фитопланктона. В верхних слоях океанов кремнекислота обычно содер-
жится в количестве нескольких сот миллиграммов на 1 м3 воды, а в глу-
бинных до 1000 и больше.
Особенности распределения биогенных элементов в некоторых водое-
мах. Весьма своеобразным водоемом, с точки зрения интенсивности раз-
вития в нем жизни и существующих для этого условий является Среди-
земное море. Хорошо известна высокая прозрачность воды Средизем-
ного моря, обусловливаемая малым развитием планктона. Все это нахо-
дится в связи с общими условиями питания и в первую очередь с недо-
статочным количеством в воде фосфатов и нитратов от поверхности до дна.
На табл. 40 дано распределение фосфатов и нитратов в толще средизем-
номорской воды. Количество их резко убывает с запада на восток, так как
через Гибралтар постоянно привносятся с поверхностным течением бога-
тые питательными солями воды.
Таблица 40
ИЗМЕНЕНИЕ В РАСПРЕДЕЛЕНИИ НИТРАТОВ
И ФОСФАТОВ В ТОЛЩЕ ВОДЫ СРЕДИЗЕМНОГО МС)РЯ
С ЗАПАДА НА ВОСТОК ПО ДАННЫМ ДАТСКОЙ
ЭКСПЕДИЦИИ в мг/м3
Глубина в м Гибралтарский пролив и Альборанское море Ионическое море
N, р205 N, р2о3
0 3 0 2 0
10 7 0 1 0
25 18 0 0 0
50 72 8 4 0
100 75 13 17 0
150 85 13 32 0
200 93 17 38 1
300 110 20 49 2
400 125 18 54 3
500 118 23 58 4
600 157 25 76 6
800 — 25 85 8
1000 — — 86 12
1200 — — 92 10
1500 — — 92 11
2000 — — 89 9
2500 — < — 87 8
3000 — .— 85 10
3500 .— — 90 8
4000 — — 85 8
8 Зенкевич, т. I
113
То же самое показано и на графике (рис. 59). Очевидно сильное зимнее
перемешивание всей толщи воды в Средиземном море препятствует накоп-
лению значительного количества питательных солей на глубинах. Принос
пх речными водами в Средиземное море очень незначителен, с другой
стороны, беспрерывно происходит определенный выход фосфора и азота
из круговорота и при этом в немалой степени и с вылавливаемыми чело-
веком промысловыми животными. Расход питательных солей видимо пере-
крывает принос их реками и основным источником остается атлантическая
вода, вливающаяся через Гибралтар.
Еще меньше количество питательных солей в придаточных морях
Средиземного моря. Так Ерцегович в своих обстоятельных работах
указывает, что в средних частях Адриатического моря (у Сплита) содер-
жание фосфора в поверхностном слое в течение года колеблется от 0,2
до 3 мг/м3, а на глубине 90 м от 1 до 3,2 мг)м3.
Изучение годового цикла содержания нитратов и нитритов в поверх-
ностных прибрежных водах (гавань Фрайди в заливе Сен-Жуан, Тихоокеан-
ское побережье США), произ-
веденное Фифер иТомпсо-
н о м (1937), показало (рис. 60),
что накопление нитритов прихо-
дится на период особенно энер-
гичного потребления фитопланк-
тоном нитратов, т. е. весной
и летом.
Накопление аммиака в общем
соответствует накоплению нитри-
тов и приходится на время, когда
нитратов меньше всего. Это ука-
зывает на то, что в процессе ре-
генерации нитратов из органиче-
ского вещества азот проходит че-
рез эти стадии.
Цикл соединений фосфора и азота. Скорость, с которой фосфаты и со-
Рис. 60. Годовой цикл содержания нит-
ратов и нитритов в поверхностной воде бух-
ты Фрайди, Вашингтон (по Фиферу и
Томпсону из Свердрупа и др.).
единения азота используются растениями, возвращаются в морскую воду
и снова используются, и количество этих соединений могут служить пря-
мым мерилом первичной продуктивности.
Гарвей (Harvey, 1945) различает прямую и непрямую регене-
рацию фосфора. Под первой он понимает те значительные количества
фосфатов, которые прямо выделяются в воду травоядным планктоном
вместе с экскретами (что установлено экспериментальным путем Гар-
динером, 1937). Непрямая регенерация фосфора это минерализация
фосфатов из отмирающих и разлагающихся тел растений и животных,
совершающаяся при помощи бактерий. Прямая регенерация идет быстро,
непрямая — более или менее медленно.
Гораздо сложнее протекает в морской воде цикл соединений азота.
Фитопланктон способен утилизировать различные соединения азота,
диатомеи могут усваивать нитраты, нитриты, аммиак, а также мочевину
и мочевую кислоту, причем различные группы бактерий и выделяемые
ими энзимы способны переводить одни из указанных соединений в дру-
гие.
Растворенное органическое вещество. Мы уже указывали на то, что
в морской воде растворено также и органическое вещество, поступающее
в раствор от распада тел животных и растений. Известный датский физи-
олог А. К р о г (A. Krogh) дал по этому вопросу следующие вы-
воды (табл. 41):
114
Таблица 11
СОДЕРЖАНИЕ АЗОТА В ОРГАНИЧЕСКОЙ СВЯЗИ (АЛЬБУМИНОИДНОГО)
В ТОЛЩЕ ОКЕАНА НО К РОГУ
Глубина в м 0 1000 2000 3250 4250 4750
Количество азота в рг-ато- мах на л 0,258 0,248 0,240 0,229 0,250 0,238
Первое, что установил К р о г, это постоянство количества азота
в органической связи на разных глубинах океана. Среднее его коли-
чество 0,244+0,010 ре-атомов на литр.
Параллельно производилось также определение и углерода и полу-
чился результат, приведенный на табл. 42.
Таблица 42
СОДЕРЖАНИЕ УГЛЕРОДА В ОРГАНИЧЕСКОЙ СВЯЗИ В ТОЛЩЕ ОКЕАНА
ПО КРОГУ
Глубина в м 25 2000 3250 3750 4250 4750 Среднее
Количество углерода в рг-атомах 2,40 2,35 2,32 2,36 2,17 4 38 2,35 ±0,9
Таким образом, количество углерода (25—30 мг)л) также имеет весьма
константный характер. Произведя соответственный пересчет А. К р о г
определяет количество органического азота в 1,3 яг и органического угле-
рода в 12,7 кг под 1 м2 поверхности океана, принимая среднюю его глу-
бину в 5,4 км. Как указывает К р о г, это количество «по крайней мере
в 300 раз, а вероятно более чем в 1000 раз превышает количество органи-
ческого вещества, заключающегося во всех живых морских организмах
вместе взятых».
Данные А. К р о г а относятся к глубинным водам открытого океана.
Бонд (1933), произведя аналогичные анализы в прибрежных богатых
планктоном водах, установил, что количество растворенной фракции орга-
нического вещества примерно то же, что и у Крога, но соотношение
органического вещества в растворе и в организмах (-^детрит) иное (табл. 43).
Таблица 43
Вещество планктона Углерод в цг-атомах/л
минимум максимум среднее
Сетяной планктон 0,0С8 0,06 0,03
Наннопланктон 0,005 0,11 0,03
Растворенное вещество 0,13 0,25 0,20
В сумме 0,143 0,42 0,26
Таким образом, в богатых планктоном прибрежных водах растворен-
ное вещество превышает вещество, заключенное в организмах (-|- детрит)
только в 3—10 раз.
*
115
Принимая, что наибольшая величина углерода в живых организмах
констатирована для вод к югу от Гренландии, и определяя его величиной
600 мг на 1 м‘л и расчисляя его опять-таки на весь столб воды, К р о г
получает величину 7,3 рг-атомов углерода на 1 л/3, т. е. 1/300 органического
углерода в растворенном состоянии. К р о г полагает, что количество рас-
творенного в воде океанов органического вещества изменяется чрезвы-
чайно медленно и представляет собою результат весьма длительного про-
цесса. Вещества, о которых идет речь, представляют собою смесь амино-
кислот и углеводов, количество первых 1,5 рз-атомов на литр, вторых —
3,9 рл?-атомов. В связи с полученными данными А. К р о г ставит очень
существенный вопрос: в какой степени растворенное органическое ве-
щество может быть использовано животными? Считая точку зрения Пют-
тера о большой доступности для животных (от простейших до рыб и ам-
фибий) в качестве пищи растворенного в воде органического вещества
неверной, К р о г допускает это только для некоторых животных, и то
в весьма ограниченной степени. Что касается бактерий, то К р о г пола-
гает, что они связаны в своей жизнедеятельности в основном с фитопланк-
тонными организмами и существенной роли в переработке органического
вещества, растворенного в воде, играть не могут. В конце концов К р о г
ставит вопрос об источнике питания глубоководного бентоса, сомневаясь,
чтобы ему могло хватить тех органических веществ, которые падают сверху
в виде детрита и так называемого «дождя трупов», и считает возможным
поставить вопрос о существовании на дне океанов и морей особых орга-
низмов, еще неизвестных науке, которые, возможно, обладают способ-
ностью питаться растворенными в воде органическими веществами и уже
сами являются пищей для глубоководного бентоса.
С этой точки зрения могут привлекать внимание, пока еще очень мало
изученные, «оливково-зеленые клетки», регулярно попадавшиеся в уловах
планктонных сетей экспедиции на «Метеоре» в глубинных водах южной
Атлантики (Hentschel, 1936). Максимальное количество их было
приурочено к слоям ниже эвфотической зоны. Гентшель полагает,
что это гетеротрофные организмы. Возможно, что они могут усваивать
растворенное органическое вещество и в свою очередь служить пищей
животным фильтраторам обитателям средних глубин.
В морских водоемах известны случаи массовой гибели рыб и донной
фауны при наступлении неблагоприятных условий существования. Иногда
эта массовая гибель зависит от низкой температуры. Довольно обычна
в Азовском море гибель хамсы, не успевшей уйти через Керченский про-
лив в Черное море и застигнутой зимним охлаждением воды.
Данневиг описал случаи массовой гибели трески у берегов Нор-
вегии в те редкие зимы, когда температура воды опускалась ниже 6°.
Известны также случаи массовых заморов фауны при наступлении
неблагоприятного газового режима. В Азовском море обычно явление
летних заморов донной фауны, а частично и рыбы иногда на площади в ты-
сячи квадратных километров в штилевую погоду, когда в придонном слое
воды быстро исчерпывается кислород п появляется сероводород.
Хорошо известны случаи массовой гибели сельди в заливах Мурман-
ского побережья при быстром наступлении явлений кислородного де-
фицита. Недавно описайы явления массовой гибели рыбы, морских чере-
пах и беспозвоночных (моллюсков и ракообразных) в заливах побережий
Флориды при наступлении неблагоприятного газового режима. Количе-
ство погибающей при этом рыбы исчисляется десятками миллионов штук.
ГЛАВА IV
ДИНАМИКА ВОД МИРОВОГО ОКЕАНА
1. ТЕЧЕНИЯ
Значение перемешивания водных масс для развития жизни. Если бы
воды мирового океана находились в неподвижном состоянии, жизнь в толще
его вод была бы невозможна, за исключением только самой тонкой поверх-
ностной пленки и пространств, прилегающих к устьям рек. В течение
короткого срока кислород был бы использован, а биогенные элементы ока-
зались бы унесенными вместе с трупами организмов на глубины и навсегда
там погребенными. Процесс диффузии дает очень слабый эффект и не мог
бы обеспечить распределение нужных для развития жизни элементов
в водных массах.
Стратификация как показатель степени перемешиваемости вод. На
самом же деле мы имеем в океане очень сложную систему перемешивания
вод, в разной степени и форме выраженную в различных его частях,
обусловливаемую самыми разнообразными причинами и нарушающую
состояние покоя. Однако, всякое такое нарушение, действующее более
или менее длительный срок, дающее толчок к перемешиванию вод, в ко-
нечном счете затухает, если обусловливающая его сила не действует не-
прерывно. Земное тяготение действует в обратном направлении к силам
нарушения и, затормаживая перемешивание, способствует установлению
различного типа стратификации. Из всех факторов среды только проник-
новение света вглубь воды относительно мало обусловливается системой
перемешивания. Распределение всех других факторов — температуры,
солености, кислорода и других газов, питательных солей и т. д., при ослаб-
ленном перемешивании, быстро переходит в стратификацию, все более
и более охватывающую все стороны жизни водоема. Если воды Антарк-
тики мы могли бы взять как крайний пример области океана с сильным
перемешиванием и весьма равномерным распределением всех факторов,
то противоположным примером крайней степени стратификации может
служить Черное море, в котором почти 7/8 его объема представляют
собой мертвую, зараженную сероводородом, глубинную часть. Таким
образом, степень и характер стратификации может служить самым луч-
шим показателем перемешиваемости вод.
Типы морских течений и причины, их вызывающие. Перемешивание
водных масс достигается путем горизонтальных, вертикальных (называе-
мых конвекционными) и смешанных течений и сопутствующего им турбу-
лентного перемешивания.
117
Рис. 61. Общая схема течений мирового океана. Сплошная линия соответствует полярному фронту, толщина стрелок —
скорости течения 6—12, 12—24 и свыше 24 миль в сутки (из Березкина).
Горизонтальные течения возникают от разных причин. Они создаются
ветрами (дрейфовые), притяжением водных масс луной и солнцем (приливно-
отливные), неравномерным и постоянно меняющимся распределением
атмосферного давления (бароградиентные), впадением с суши речных
потоков, и, наконец, различиями в плотности масс воды, обусловливае-
мыми различиями в солености и степени нагретости.
Конвекционные перемещения (иначе говоря вертикальные течения)
возникают или при увеличении плотности поверхностного слоя воды, полу-
чающемся, например, при охлаждении, а иногда и вымерзании или как
компенсационные восходящие и нисходящие течения, при явлениях сгона
пли нагона у побережий. Все перечисленные виды течений, перемещающие
в определенном направлении
водные массы, вызывают систе-
му разнообразных компенса-
ционных течений.
Первоначально полученное
направление во всех этих ви-
дах течений, под влиянием от-
клоняющей силы вращения зем-
ли (сила кориолиса), силы тре-
ния, рельефа дна и очертания
берегов, претерпевает сложную
систему отклонений и деформа-
ций. Различают течения по-
стоянные, периодические, слу-
чайные; теплые и холодные;
поверхностные, глубинные и
прибрежные; прямолинейные,
циклонические и антициклони-
ческие и т. д. В дальнейшем
нам придется иметь дело с при-
мерами самых разнообразных
течений. На рис. 61 дана общая
схема течений мирового океана.
Громадность масс воды, увле-
каемых океаническими тече-
ниями, может быть показа-
Рис. 62. Схема, показывающая источники
(прерывистые линии) и пути (сплошные ли-
нии) течений системы Гольфстрима. В запад-
ной Атлантике каждая линия течения соот-
ветствует расходу приблизительно 43,2 км3/час
(По Иселину из Зубова).
на на примере Гольфстрима. Проходя Флоридский пролив, это те-
чение имеет скорости, достигающие 257 см в секунду (9,2 км в час). Глу-
бина его около 700 м и ширина свыше 30 км. В один час через Флоридский
пролив проносится масса воды в 91 км3, или около 800 000 км3 в год, что
соответствует почти двум объемам Черного моря. При этом, после выхода
из Флоридского пролива, Гольфстрим принимает в себя еще более мощные
массы вод Антильского течения (рис. 62) и здесь несет уже, вдоль побере-
жий Северной Америки массы воды, превышающие 2у2 млн. км3. Громад-
ные скорости движения водной массы, подобные скорости Гольфстрима
во Флоридском проливе, представляют собою один из примеров наиболь-
ших скоростей морских течений. Обычно скорости значительно меньше
даже и в периферических частях Гольфстрима (рис. 63). Обычная скорость
движения воды в океане определяется несколькими сантиметрами в се-
кунду.
В результате горизонтальных и вертикальных перемещений в толще
океанических вод и вследствие обмена водами придаточных морей через
проливы с океаном, достигается, ускользающая на первый взгляд, боль-
шая сменность вод океанов и морей. Так, Баттенберг (W a 11 е п-
119
berg) и Сейвелл (Sei well) принимают, что вся масса глубин-
ных вод Атлантического океана сменяется в 5—6 лет, перемещаясь из
высоких широт к экватору. В Баренцово море ежегодно вливается, в ос-
новном с запада, около 75 000 км3 воды, количество, составляющее около
г/з—V4 всего его объема. Даже застойная, зараженная сероводородом
глубинная вода Черного моря постоянно обновляется за счет нижнего
Босфорского течения, дающего в тес
1230 9 8 7 8 722Т
Рис. 63. Скорости Гольфстрима в см/сек
на разрезе от Чезапик-бай 20—22 апреля
1932 г. (по Иселину из Зубова). Расходы
по секциям в миллионах мР/сек. А— 10,6;
В— 12,7; С — 0,7; D — 12,1; — 46,0.
Всего 82,2.
ie года количество воды, равное по
С. Макарову примерно 1/2Ооо
всего объема моря.
Хотя принято считать моря
царством климатического одно-
образия, а сушу царством кли-
матических контрастов, однако,
и в океане мы, в разных его ча-
стях, находим значительные раз-
личия почти по всем показателям
его физико-химического режима.
Эти различия получают свое вы-
ражение в гидрологических гра-
диентах. <<Во всей глубине океа-
на,— как говорит Н. Зубов,—
одновременно созидаются гидро-
логические градиенты. На одних
глубинах преобладает созидание,
на других уничтожение градиен-
тов, и это отражается на верти-
кальном распределении гидроло-
гических элементов и создает рас-
слоение океана». В одних районах
океана постоянно возникают опре-
деленные свойства водных массг
в других эти свойства уничтожа-
ются. В тропических областях
происходит колоссальное погло-
щение поверхностными слоями во-
ды тепла из атмосферы и осолонр-
ние их за счет испарения. В
умеренных широтах и полярных
странах совершается обратный
процесс — отдача тепла и охла-
ждение водных масс и (в северных широтах) значительное их опрес-
нение. Эти переработанные в местных условиях воды вовлекаются
в мощную систему циркуляции, совершающуюся в основном в меридио-
нальном направлении. Океанические воды обогащаются кислородом в тех
районах, где происходит обильное цветение планктона, обеспеченное
наличием питательных солей. Эти богатые кислородом воды увлекаются
на глубины океана и постепенно теряют свой кислород. В глубинных
частях морей и океанов накопляются колоссальные запасы биогенных эле-
ментов, выносимые конвекционными токами в поверхностные слои моря.
Они служат питательным субстратом, за счет которого идет бурное разви-
тие фитопланктона, влекущее за собой столь же мощную волну развития
и животных организмов.
На глубинах 200—600 м в Северным Ледовитом океане залегает
более теплый и соленый слой воды. Он принесен сюда мощной системой
течений, несущих тепло и повышенную соленость, полученные под
120
тропиками и постепенно утрачиваемые в Северном Ледовитом океане.
У западных побережий южной Африки и Южной Америки имеет место
пышное развитие фитопланктона, использующего при этом нитраты и
фосфаты, подымающиеся вместе с водными массами на поверхность
с глубин.
Таким образом в физико-химических циклах в океане можно разли-
чать согласно Н. 3 у б о в у четыре основных фазы — 1) созидание гидро-
логических градиентов, 2) созидание водных масс путем перемешивания,
3) перераспределение водных масс при помощи течений и, наконец, 4) уни-
чтожение гидрологических градиентов.
С областями высоких гидрологических градиентов совпадают и наи-
более резко выраженные границы в распространении морских организмов.
Основная система горизонтальных океанических течений сводится
к двум пассатным экваториальным течениям, идущим с востока на запад
и к движущимся между ними в обратном направлении экваториальным
противотечениям, к двум циклоническим и двум антициклоническим вра-
щениям вод. В северном полушарии сначала идет система антициклони-
ческого круговорота, а дальше к северу циклонического, в южном полу-
шарии они расположены в обратном порядке. В центральных частях этих
круговоротов располагаются так называемые халистатические области
со слабыми показателями перемещения вод. Халистатические области,
окруженные, как правило, кольцевыми течениями, встречаются во всех
морях и обладают своими характерными особенностями физико-химиче-
ского и биологического режима. Так, в Атлантическом океане, к северу
от экватора, между параллелями 20° и 40° и меридианами 30° и 60° распо-
лагается одна из наиболее известных халистатических областей мирового
океана, так называемое Саргассово море.
2. ВОЛНЕНИЕ И ДРУГИЕ ВИДЫ ПЕРЕМЕШИВАНИЯ ВОДЫ
В ОКЕАНЕ
Волнение. Помимо течений, при которых происходит перенос масс
воды в определенном направлении на более или менее значительное рас-
стояние, существует еще система колебаний частиц воды, при которых они
описывают только круговые колебания (орбита волны с определенным
радиусом), возвращаясь затем в исходное положение. Это — совокуп-
ность колебаний, именуемых волновыми. Наиболее обычный вид волновых
колебаний, захватывающих поверхностные слои моря, это — ветровые
волны, достигающие иногда громадных размеров. Предельные наблю-
давшиеся элементы волн, указываемые Шоттом,следующие: высота —15 ж,
длина 824 л«, скорость — 38,5 м в секунду и период — 23 секунды.
Обычно же волновые колебания значительно меньше. Для развития жизни
в верхних слоях моря сильные волнения, так же как п сильные течения,
имеют очень существенное значение и при определенных условиях могут
вызывать массовую гибель планктонных организмов, особенно если при
этом имеют место резкие градиенты солености и температуры, как это
наблюдается, например, в Горле Белого моря. Не меньшее значение для
организмов имеет и глубина, на которую распространяются волновые
колебания. С глубиной ветровые волны затухают: при возрастании глу-
бины в арифметической прогрессии, радиусы кругового движения частиц
воды уменьшаются в геометрической прогрессии. На глубине нескольких
сотен метров затухает самое сильное океаническое волнение. На мелко-
водьях, во внутренних морях, волнения не достигают таких размеров,
как в океанах.
121
Прибой. На прибрежных отмелях и у берегов, вследствие замедле-
ния скорости движения в нижней части волны от встречи с дном, волна
опрокидывается вперед и образует буруны (на некотором расстоянии от
берега) и прибой у самого берега. При определенных условиях прибой
может достигать огромной силы, особенно у приглубых скалистых бере-
гов, — вода взлетает вверх на 100—150 м. Силой прибоя могут передви-
гаться бетонные массивы в 1000 и даже 2000 т весом. Совершенно оче-
видно, что для жизни, развивающейся у морских побережий, прибой пред-
ставляет собою один из наиболее существенных факторов внешней среды.
Сейши. Гораздо меньшее значение для морских организмов имеют
так называемые стоячие волны (или сейши), когда в вертикальное коле-
бание приходит вся толща воды в определенном районе моря. Под влия-
нием падающих с гор потоков воздуха или при быстрых повышениях баро-
метрического давления, или, наконец, сейсмических толчков, на опреде-
ленном участке моря может наблюдаться понижение уровня моря и соот-
ветственно компенсационное повышение его в другой части водоема.
Стоячие волны наблюдаются на больших озерах (в том числе на Каспий-
ском и Аральском морях) и на полузамкнутых придаточных морях и дают
Рис. 64. Вертикальные колебания изотерм,
в связи с приливно-отливными явлениями
в Баренцевом море (по Зубову).
изменения уровня (амплитуду) до
2 л, обычно же в несколько де-
сятков сантиметров.
Внутренние волны. Помимо
рассмотренных волновых колеба-
ний, ветрового и бароградиентно-
го характера, в океанах и морях
существует еще одна система чрез-
вычайно многообразных, повсеме-
стно распространенных и еще
очень мало изученных волновых
колебаний. Они совершаются в са-
мой толще вод, на любых глубинах
морей и океанов, обычно в погра-
ничных областях между слоями вод различной плотности, как под воздейст-
вием приливно-отливных явлений, так и вследствие других причин, если эти
слои находятся в некотором перемещении по отношению друг к другу.
Это так называемые внутренние волны. Сходные явления можно наблю-
дать в облаках, когда они располагаются как бы правильными рядами
волн. От пограничного слоя эти волны распространяются в обе стороны,
т. е. внутрь обоих, двигающихся друг к другу (относительно) слоев раз-
ной плотности, причем чем дальше от промежуточного слоя, тем слабее
проявляются волнообразные колебания, длина волн остается одинаковой,
высота же их уменьшается. Впервые явление внутренних волн было обна-
ружено Ф. Нансеном в 1902 г. Амплитуда внутренних волн может
быть самой разнообразной. В Атлантическом океане, у берегов Португа-
лии, наблюдались волны с амплитудой до 200 м (на глубине 800 м}. В Ба-
ренцевом море Н. Зубов наблюдал внутренние волны высотой до 35 м
(рис. 64). В Средиземном море они образуются в слое скачка, достигают
высоты 50 м и перемещаются в вертикальном направлении вместе со слоем
скачка. Замечательно, что зимой там, где наступает гомогалинность и го-
мотермия и исчезает слой скачка, исчезают и связанные с ним внутренние
волны. В ходе колебаний при внутренних волнах наблюдается периодич-
ность полусуточная и суточная, но кроме того известны и периоды боль-
шей продолжительности, а именно полумесячные, месячные, полугодо-
вые, годовые, а затем периоды в 4.6; 9.3; 18.6; 93; 111 лет и еще более дли-
тельные. В дальнейшем мы еще вернемся к этому явлению.
122
Значение внутренних волн для морских организмов. Для морских
организмов роль внутренних волн очень велика. Как мы увидим далее,
на границах между слоями вод разной плотности происходит значитель-
ное накопление планктона. Волнообразные колебания этого слоя будут
то приподнимать, то опускать вместе с водой на десятки метров и планк-
тонные организмы, создавая как бы явление миграций, однако это чисто
пассивные перемещения, не являющиеся активными миграциями. Если
эти волнообразные колебания идут со стороны моря по направлению
к суше, то периодически будет происходить то подход, то отступание
низовых более соленых вод. Таким образом, через мелководные пороги
фиордов и проливов, ведущих во внутренние моря, в котловины и тех
и других могут попадать массы свежей соленой богатой кислородбм мор-
ской воды и приносить с собой свойственную им фауну и флору. На таких
периодических поступлениях внутрь Балтийского моря свежих масс
глубинной океанической воды нам придется остановиться в главе, посвя-
щенной этому морю (т. II).
3. ВЕРТИКАЛЬНАЯ ЦИРКУЛЯЦИЯ И ПОЛЯРНЫЙ ФРОНТ
Явление полярного фронта и его значение для развития жизни. Очень
важным для биолога понятием является полярный фронт. Вследствие
очень хорошего перемешивания водных масс за пределами полярного
фронта и вообще в местах соприкосновения более теплых и соленых вод,
идущих с юга, с местными более холодными (и в северном полушарии
менее солеными), обеспечивается обильное поступление снизу питатель-
ных солей и хорошее проникновение сверху кислорода. В результате
этого перемешивания создаются исключительно благоприятные условия
для развития фитопланктона, за счет которого развивается обильный
зоопланктон, богатого бентоса и, наконец, для скоплений промысловых
рыб. Именно поэтому наиболее известные районы скопления жизни и,
в частности, промысловые районы Северной Атлантики располагаются
вблизи линии полярного фронта, а именно у Ньюфаундленда, у юго-
восточной Гренландии, у Исландии, на Медвежинском и Центральном
мелководьях Варенцова моря и в юго-восточной части Варенцова моря,
там, где имеется хорошая вентиляция всей толщи моря. Хорошо аэрируе-
мым районом является также и Северное море, где местные более холодные
воды встречаются с более теплыми, атлантическими, проникающими сюда
и через Английский канал и с севера вокруг Шотландии.
Вертикальная циркуляция. В районах хорошей вертикальной цирку-
ляции, как, например, на Ньюфаундлендской банке, или в районе о. Мед-
вежьего, в Баренцовом море, вода перемешивается до самого дна, созда-
вая тем благоприятные условия для развития планктона и бентоса.
В некоторых случаях режим сильного перемешивания устанавливается
в проливах, соединяющих внутренние моря с открытыми частями океана,
особенно, если, помимо двух течений, идущих в обратных направлениях,
добавляется еще и сила приливно-отливных колебаний. Проливы с таким
режимом, как, например, Воронка и Горло Белого Моря, могут являться
очень серьезным препятствием в распространении организмов (К. Д е-
р ю г и н), создавая в крайней степени неблагоприятные физические усло-
вия существования для нежных планктонных форм и личинок донных
животных и рыб.
Иногда вертикальная циркуляция почти не захватывает придонного
слоя, например в Черном море или в отгороженных от моря порогом фиор-
дах Норвегии (вернее сказать, она идет с чрезвычайной медленностью).
В других районах она затруднена или имеет место только в определенные
123
сезоны года. Взгляды Дефантаи многих других современных океано-
графов можно пояснить схемой (рис. 65) распределения водных масс Атлан-
тического океана. Несомненно бросается в глаза большое преобладание
в океане масс холодной воды над теплой. Средняя температура воды океа-
нов должна быть значительно ниже средней температуры нижних слоев
атмосферы. Это результат своеобразия общей циркуляции с малоподвиж-
ным нагретым верхним слоем тропического пояса, играющим роль изоля-
тора, и легким опусканием на глубины холодных полярных вод, запол-
няющих все глубинные части океанов.
Рис. 65. Схематическое изображение динамики водных масс Атлантического океана
на меридиональном разрезе. 1 — теплые тропические воды и атлантическое течение;
2 — холодные воды (по Дефанту).
L ОКЕАНИЧЕСКИЕ ТРАНСГРЕССИИ
За последние 2—3 десятка лет среди некоторых океанологов стало скла-
дываться новое представление о характере океанических течений. Осно-
ванием к этому явилось открытие еще в начале этого столетия Ф. Нан-
сеном внутренних волн, детальнее изученных в дальнейшем им самим,
Г е л л а н д-Г а н з е н о м и О. Петтерссеном*. О внутренних
волнах мы говорили уже выше. Крайнее выражение этому взгляду дал
французский океанограф Эд. Ле-Дануа.
Понятие океанических трансгрессии. Наряду с этим Ле-Дануа
разработал, вместе с другими океанографами, вопрос об океанических
трансгрессиях северной части Атлантического океана.
Трансгрессии, как полагает Ле-Дануа, являются основой океани-
ческой циркуляции и вызываются космическими причинами — притя-
жением луны и солнца и солнечными пятнами. Эти приливно-отливные
колебания перемещают отдельные водные массы то в ту, то в другую сто-
рону и создают явление, которое может быть смешано с течениями иного
рода.
Океанические трансгрессии известны также для Тихого и Индий-
ского океанов и представляют собою явления временные и периодические.
Амплитуды периодических колебаний трансгрессивного характера весьма
различны — 1 год, 4 г. 6 м; 9 л. 3 м., 18 л. 6 м., 93 года, 111 лет (рис. 66).
В подобного же типа колебания некоторые (Миланкович, 1920) склонны
укладывать и климатические колебания геологического порядка.
124
Целый ряд наблюдавшихся в Северной Атлантике океанических транс-
грессий хорошо увязывается с биологическими явлениями, в том числе
в промыслового характера. Трансгрессии сопровождаются отходом более
холоднолюбивых рыб на север (сельдь, треска) и подходохм теплолюбивых
рыб экваториального происхождения (тунец, макрель, сардина и др.).
Если трансгрессия наступает внезапно, она может вызвать даже массо-
вую гибель рыб. Так, в 1922 г. и 1931 г. наблюдалась при этом массовая
гибель сельди.
Рис. 66. Общая схема периодичности океанических трансгрессий
большой (А) и малой (Б) амплитуды (по Ле-Дануа).
Ле-Дануа склонен вообще отказаться от наличия течений и все
перемещения морских вод сводит к явлениям трансгрессий и регрессий
водных масс. Правильнее всего сделать вывод, что наличие и тех и других
можно считать несомненным.
5. СКОРОСТЬ ПЕРЕДВИЖЕНИЯ ВОДНЫХ МАСС В ОКЕАНЕ
Взгляды Ле-Дануа. Вопрос о темпе передвижения водных масс океана до
недавнего времени не останавливал на себе значительного внимания океано-
графов. G одной стороны, мы имеем ошибочное мнение Ле-Дануа, что вод-
ные массы в океане могут сохранять свои положения и свойства геологи-
чески долгими периодами, не смешиваясь друг с другом. Воды могут только,
подчиняясь космическим силам, «вытягиваться» и «сокращаться» (la ex-
tension и la retraction), а в общем всегда возвращаются к первоначальному
исходному положению. Отсюда и его «ископаемые моря» внутри океана.
В такой форме точка зрения Ле-Дануа никем не поддерживается.
Теория Сейвелла. К совершенно другому результату пришел американ-
ский исследователь С е й в е л л, подвергнувший анализу данные по
распределению кислорода в глубинных водах Атлантики по меридиональ-
ному разрезу. Как уже указывалось выше, по вертикальному распреде-
лению кислорода в океане можно наблюдать как бы трехслойность
(рис. 67). Самый верхний слой воды (зона перемешивания Д е ф а н т а), зале-
гающий выше слоя скачка, богат кислородом. Глубже, примерно до 2000 .к,
располагается слой бедный кислородом, а затем количество кислорода
опять возрастает. Исследуя количество кислорода (средний кислород-
ный градиент) в бедном им слое с юга на север (рис. 68), С е й в е л л
установил, что количество его правильно колеблется то в сторону повыше-
ния, то в сторону понижения. Еще Баттенберг, а затем Дикон,
отметили, что тут имеют место колебания, соответствующие годовым мак-
симумам и минимумам в количестве кислорода, когда этот слой был на
125
Рис. 67. Вертикальное распреде-
ление кислорода и температуры
в западной части Атлантического
океана в марте 1932 г. на 14°47z
с. ш. и, примерно, 40° з. д. (по
Сейвеллу).
поверхности, т. е. до опускания на глубину. Так как по разрезу, приво-
димому Сейвеллом, можно допустить движёние масс воды, бедной
кислородом, и с севера и с юга, от побережий Ю. Америки, этот автор счи-
тает, что, наряду с сохранением сезонного характера колебаний, наблю-
дается постепенное обеднение кислорода в результате потребления его зоо-
планктоном. Наиболее обедненная зона располагается примерно под 15°с.ш.
Если это допущение правильно, то можно сделать целый ряд интересных
выводов. Во-первых, можно установить,
что с обеих сторон разреза к его средней
части бедный кислородом слой двигался
в течение 4 лет, и под 15° с. ш. на глу-
бине 1000—2000 м располагается вода,
которая 4 года тому назад была у по-
верхности. Во-вторых, можно рассчитать
количество кислорода, потребленное за
определенный срок заключенным в дан-
ном слое зоопланктоном; именно, на 1л
расход кислорода за год колеблется от
0,31 до 0,63 с.и3, и перечислить это ко-
личество на массу планктона, продуци-
руемого в этом слое в течение года. Нако-
нец, можно установить и скорость дви-
жения исследуемого слоя, а именно от
1,1 до 2,1 см)сек. Такие колебания Сей
в е л л констатировал и в глубжележащих
слоях 02000 л<).
Если принять полученную Сейвел-
лом скорость движения глубинных вод
океана (186—360 миль в год), то можно
думать, что воды Антарктики должны до-
стигать экваториальной области примерно
в 10—20 лет.
Как ни кажутся на первый взгляд убе-
дительными рассуждения Сейвелла,
в его теоретических выводах имеется
много неясного, требующего дальнейшего
уточнения. Если только мы предположим,
что С е й в е л л имел дело с внутренни-
ми волнами большой амплитуды (у ниж-
ней границы слоя скачка, где они без-
условно и должны быть и к чему имеется
много данных и в материале самого Сейвелла), все его выводы пови-
сают в воздухе. Таким образом, пока мы еще не располагаем надежными
показателями скорости передвижения глубинных вод океана и также
перемешивания образующих его водных масс, хотя взгляды Ле-Дануа
при современном состоянии наших знаний совершенно неприемлемы.
6. КЛАССИФИКАЦИЯ ОКЕАНИЧЕСКИХ ВОДНЫХ МАСС
И ЦИРКУЛЯЦИЯ ГЛУБИННЫХ ВОД
На рис. 69 даны области формирования и распределения основных
масс океанических вод. Согласно Свердрупу, все основные типы океа-
нических вод формируются на поверхности океана, а затем испытывают
погружение и дальнейшее распространение, в зависимости от их плот-
ности. Это не относится только к экваториальным водам Тихого и Ин-
126
дпйского океанов, которые формируются в результате перемешивания
в подповерхностных слоях (рис. 70).
Формирование «центральных» водных масс. Во всех океанах «централь-
ные» водные массы формируются в областях субтропической конверген-
ции (между 35° и 40° ю. ш. и 35° и 40° с. ш.), о чем говорят сходные в тех
и других соотношения t° и S°/00. Созидание их обусловливается атмосфер-
ной циркуляцией и процессами нагрева и охлаждения поверхностных вод.
Соленость «центральных» водных масс северной Атлантики выше, чем
северной Пацифики, мощность их обычно не превышает в северной части
Тихого океана 200—300 м.
Рис. 6Я Средний кислородный градиент в бедном кислородом слое, на ме-
ридиональном разрезе от Северного побережья Южной Америки на се-
вер, примерно по меридиану 40° до 35° с. ш. (по Сейвеллу).
В Атлантическом океане северная и южная центральные водные массы
разделены узкой переходной зоной, тогда как в Тихом и Индийском океа-
нах Свердруп выделяет широкую зону «экваториальных» вод.
В области Саргассова моря мощность этого слоя достигает 900 м.
«Центральные» водные массы Свердрупа, с включением его «эквато-
риальных» вод, соответствуют «тропосфере» Дефанта (1928) (рис. 71).
«Субантарктические» воды располагаются между центральными вод-
ными массами и зоной антарктической конвергенции и опоясывают земной
шар. Они имеют низкую соленость и, возможно, образуются путем сме-
шивания и вертикальной циркуляции в области между зонами субтропи-
ческой и антарктической конвергенции. В северной Атлантике «субаркти-
ческие» воды имеют ограниченное распространение и относительно высо-
кую соленость, а в Северной Пацифике образуют большую зону и имеют
пониженную соленость.
Соответственно и зона арктической конвергенции, в отличие от ант-
арктической, имеет ограниченное протяжение и приурочена только к за-
падной части океана; в восточной «субарктические воды» не имеют ясной
127
гэ
00
Рис. 69. Распределение основных поверхностных водных масс мирового океана (по Свердрупу и др.). Квадратиками
показаны районы формирования центральных водных масс, крестиками—линии, вдоль которых погружаются антаркти-
ческие и арктические промежуточные воды. Черным обозначены районы массового опускания вниз и формирования глу-
бинных и природных вод.
северной границы. «Субарктические» воды очень сходны с «арктическими
промежуточными» водами, а в Антарктике они резко различны.
Рис. 70. Схематическое изображение вертикальной цирку-
ляции в экваториальной области Атлантики. W и Е обо-
значают области течений, двигающихся на запад и на
восток (по Свердрупу).
Формирование и распространение «промежуточных» вод. Во всех океа-
нах под «центральными» водными массами залегают «промежуточные»
Таблица 44
РАСПРЕДЕЛЕНИЕ t°, S%o и 02 (в мг на литр) НА ГОРИЗОНТАХ 2000, ЗСОО
и 4000 м ПО РАЗЛИЧНЫМ ЭКСПЕДИЦИЯМ 1925—1938 гг. (из Свердрупа)
Широта Атлантический океан Индийский океан Тихий океан
X ! S“/oo 0, 1° s»/oo o2 t° S%o I o2
60° 50° 40° 30° 20° 10° 5СР27, 3,32 2,97 2,38 33°1 3,62 2,95 2,45 19°1( 3,54 2,76 2,39 5' N 40°J 34,92 34,93 34,95 9' N 68°1Е 34,97 34,96 34,92 У N 27°21 34,98 34,94 34,89 1,5' W 6,30 6,17 6,34 Г W 6,08 5,99 6,06 V W 5,07 5,27 5,42 1 50°3( 1,88 1,61 28°0: 2,13 1,83 3°18 2,24 )' N, 175c 34,58 34,64 2' N, 129C 34,6*1" 34,64 ' x\, 129°0 | 34,67 16' w I 1,64 j 2,58 08' W 1 1,89 j 2,72 2'E 2,56
по i
V
10° 20° 80° 40° 50° 60° S 32°4 2,97 2,86 0,77 58°; —0,26 —0,42 —0,55 9' S 40°01 34,77 34,92 34,69 1 >3' S, 4°5' 34,67 34,66 34,64 L' W 4,71 5,53 4,88 1' Е 3,37 3,59 ’ 3,79 1°4с 2,68 1,84 1,71 36°41 2,70 1,93 1,25 604 0,90 0,34 —0,09 j' N, 71с0‘ 34,79 34,79 34,74 L' S, 114Д 34,60 34,72 34,74 0' S, 63c5 34,72 34,68 34,67 41 ui I i 1 й и -14- III O-l| -:i OO ь—1 00 1 1,65 1,57 ЗГ2Г 2,42 1,89 1,22 59°0; 1,70 1,09 0,87 34,69 34,70 )' S 176'2 34,60 34,68 34,72 VS, 163°4 34,73 34,72 34,70 3,15 3,27 5' W 3,32 3,75 4,52 6' W 4,27 4,37 4,06
9 Зенкевпч, т. I
129
воды, широко распространенные в Антарктике и образующиеся в зоне
антарктической конвергенции. Они также в Антарктике опоясывают
земной шар, а в северной части Атлантического океана приурочены только
к западной половине. После погружения «промежуточные» воды распро-
страняются в сторону экватора. В Индийском и Тихом океане антаркти-
5вМЗ,5 35,0 Св0 5 10 15
~-lJ-.l-.LJ !_L_1—J L -1- '-LLLX l_l XU. J
~ Сцбтропичес_______'кие &рды_
Субполярные проме
воды fарктическое и анг/арктическое
течение)
к Воды глубин
\ (Глубинное
течение)
1
20$
500
Ю00
1500
2000
2500
3000
3500
4000________
S%o34,5
Тропосфере
те плодная.
циркуляр.
Страто-
сфера
I Полярные пр идо
~ А ^Полярное прйдо
н.чые воды
иное течение}
Холодно-
водная
циркуля-
ция
-1000
-1500
-2000
-2500
-3000
-3500
-4000
10 15
35,0 С°0
Рис. 71.
А —схематическое строение водных масс Тихого океана и
циркуляция их (по Вюсту 1929, из Агенорова) в сопоста-
влении с вертикальным изменением температуры (Б).
ческие промежуточные
воды простираются к се-
веру до 10° ю. ш., а в
Атлантическом до 20°
с. ш.
В северной Атлан-
тике соответственный
«промежуточный слой»
образуется к востоку от
Большой Ньюфаунд-
лендской банки, но в от-
носительно небольших
количествах, вследст-
вие ограниченности рай-
она формирования.
В северной части Ти-
хого океана арктиче-
ские «промежуточные»
воды расположены меж-
ду 20° и 43° с. ш., ис-
ключая западное побе-
режье Америки, где
«субарктические» воды
продвигаются на юг до
20° с. ш. Формирова-
ние «промежуточных»
вод, вероятно, имеет
место к северо-востоку
от Японии.
Значительно мень-
шие массы «промежу-
точных» вод создаются
за счет вод Красного и
Средиземного морей.
Таким образом
Свердруп рисует кар-
тину распределения
«промежуточных» океанических вод и районы их формирования.
«Глубинные» и «придонные» воды. Что касается «глубинных» и «при-
донных» вод океанов, залегающих на глубинах свыше 2000 м. то в основ-
ном им свойственна температура от 3,5 до—0,5° и соленость от 35 %0 до
34,6°/00 (табл. 45). Приобретя большую плотность во время соприкосно-
вения с атмосферой в субарктической и антарктической областях, эти воды
погрузились под воды меньшей плотности и заполнили все глубины
океана.
Циркуляция водных масс. Северно-атлантические глубинные и придон-
ные воды распространяются на юг. Существенную роль при этом играют
присоединяющиеся к ним средиземноморские воды большой плотности,
выходящие низовым течением через Гибралтар. Годовое их количество со-
ставляет величину порядка 50 000 км3. В момент формирования «глубин-
130
ные» и «придонные» океанические воды обильно снабжены кислородом,
а затем постепенно его теряют (табл. 44).
Антарктические глубинные воды, движутся в противоположном напра-
влении, с юга на север, и могут быть прослежены до 35° с. ш. [В ю ст
(Wiis t), 1935].
Глубинные воды субарктического генезиса, проникая в антарктику,
распространяются поверх более холодных антарктических придонных
вод и достигают 45° ю. ш. и в свою очередь перекрываются антарктическими
«промежуточными» водами.
В южной части Атлантического океана большое количество вод «при-
донных» и «промежуточных» возвращается обратно в Антарктику и пере-
мешивается с движущимися на юг «глубинными» водами.
На основании соответственных вычислений Свердруп (1942)
определяет баланс вод, проносимых через экватор и 9 млн. лг3/сек.
(табл. 45).
Таблица 45
Количество вод, проносимых с юга на север и обратно в Антлантиче-
ском океане через экватор и через сечение ио 30° ю. ш.
(по Свердрупу)
Шпрота
Водные массы
В миллионах .м3/сек.
30° ю. ш. Поверхностные Промежуточные Глубинные Придонные по направлению к северу 23 9 3 по направлению к югу 17 18
0°
Поверхностные...............
Промежуточные...............
Глубинные ..................
Придонные ..................
6
2
1
9
При этом также около 9 млн. л//3сек. антарктических вод возвращается
обратно в Антарктику. В экваториальной зоне соленость североатланти-
ческих глубинных вод в среднем 34,91°/00 (на глубине 2000—3500 м),
а промежуточных и придонных антарктических вод 34,7°/00; при смеше-
нии их в равных количествах должна получаться средняя соленость 34,81°/00,
что и имеет место в глубинных водах южной части Атлантического океана
по сечению 40° параллели ю. ш.
Сходную картину циркуляции дает и Тихий океан — «промежуточ-
ные» и «придонные» воды движутся на север, а глубинные воды на юг.
Кроме того в Тихом океане имеется общее движение вод с запада на восток.
Классификация водных масс. В заключение мы приводим классифика-
цию водных масс мирового океана, взяв ее из работы Н. Зубова
(табл. 46).
От общей площади Атлантического океана отдельные классы вод со-
ставляют :
Экваториальные и тропические 4%
Собственно атлантические 11%
Глубинные атлантические 45%
Субантарктические 7%
Антарктические 23%
131
Таблица 46
КЛАССИФИКАЦИЯ ВОД АТЛАНТИЧЕСКОГО ОКЕАНА
(по Зубсву 1938)
Класс Соленость Температура Содержание кислорода в см* на литр Глубина В .11-
1. Экваториальные 34,0—35,3 20—32 4,0—5,<) 0—100
2. Тропические 3. Атлантические 35,5—37,0 15—32 1,0—4,0 0—400
а) собственно 35,0—35,5 5—15 4,0—5,0 0—2500
б) глубинные 34,8—35,0 2—5 5,0—6,^) 0—6000
в) европейские 35,0—35,4 2—10 5,0—7,5 0—500
г) арктические 35,0—35,2 0—3 7,5—8,0 100—700
< Субантарктические 5. Антарктические 34,0—34,5 2—5 4,0—7,5 0—1500
а) поверхностные 32,0—34,0 —1,8—3,0 >7,0 0—200
б) глубинные 6. Арктические 34,5—34,7 <0 4,0—6,0 0—6800
а) поверхностные 31,0—34,0 —1,8—3,0 >8,0 0-200
б) глубинные 34,9—35,0 <0 >8,2 200—3500
7. Средиземноморские 36,3—37,3 12—14 4,0—5,3 200—1000
8. Прибрежные 32,0 — 0—200
7. ОБЩАЯ СХЕМА ЦИРКУЛЯЦИИ ВОД АТЛАНТИЧЕСКОГО
ОКЕАНА
Общую схему циркуляции вод мирового океана и некоторые ее детали
удобнее всего рассмотреть на примере Атлантического океана, для которого
это явление лучше изучено.
Деление вод Атлантики на четыре района по типу стратификации.
Мы уже указывали, что при продвижении из Антарктических морей
в Арктический бассейн можно установить четыре основных района, ха-
рактерных особыми показателями стратификации.
I. Область антарктических холодных вод с более или менее равномер-
ным распределением гидрологических и гидрохимических факторов, что
обусловливается постоянно идущим вертикальным перемешиванием
и опусканием на глубины поверхностных вод в результате низкой их тем-
пературы и отсутствия опреснения.
Область эта, расположенная южнее 40° ю. ш., неоднородна (рис. 72
и 73). Ее можно подразделить на две части. Наиболее южная (южнее
60 ю. ш.), с поверхностной температурой близкой к 0°, это зона сильней-
шей вертикальной циркуляции, с наиболее полно выраженными гомотер-
мией и гомогалинностью от поверхности до дна. Севернее этой зоны рас-
полагается вокруг всего Антарктического материка широкий пояс (между
60° и 40° ю. ш.) западных ветров и кольцевого дрейфового течения с запада
на восток. Примерно на широте 60—55° ю. ш. и дальше к северу получает
начало весьма слабо развитая температурная стратификация, дающая
на широте 42—44э ю. ш. разницу между поверхностными и придонными
температурами до 10—12°. На широте 60—50° ю. ш., также кольцом
охватывая Антарктику, располагается зона антарктической конверген-
ции, где поверхностные воды испытывают опускание. Вторая аналогичная
зона конвергенции (субтропическая конвергенция) следует параллели
40эю. ш., где также поверхностные воды испытывают частичное опускание.
Между двумя зонами конвергенции располагается субантарктическая
область (Свердруп, 1942). Изложенное выше может быть иллюстри-
а 32
Рис 72. Схематическое изображение течений и водных масс антарктической области
и распределение температуры (по Свердрупу и др.).
60*
Рис. 73. Положение зон антарктической и субтропический конвергенций. Заштри-
хованы районы с глубинами меньше 3000 м (по Дикону из Свердрупа).
133
ровано также гидрологическими разрезами от Австралии к Антарктике
(рис. 74).
Антарктические циркумполярные воды имеют соленость около 34,7°/00
и температуру между 2° и 0э. Антарктические воды с этими показателями
t° и S°/oo в придонных (придонная вода) и глубинных (глубинная вода)
слоях продвигаются на север и подстилают более теплые поверхностные
воды — антарктические промежуточные с t от 3—7° и субантарктиче-
ские с t° до 8—9°.
Рис, 74. Распределение температуры (Л) и солености (Б) на раз-
резе от юго-западного угла Австралии к Антарктике (по Дикону из
Свердрупа и др.). СТК — субтропическая конвергенция; АК — ант-
арктическая конвергенция.
II. Между 40° и 30° ю. ш. мы вступаем в другую область, имеющую
в меридиональном направлении протяжение в 70—80 градусов и полу-
чающую наиболее полное свое выражение в обеих халистатических обла-
стях. Это область тепловой стратификации. Поверхностный слой воды
значительно осолонен, но он при этом так нагрет, что опускаться в глубину
не может. Тонкий нагретый слой залегает на массах глубинных холодных
вод. Между ними находится переходный слой (слой скачка). Изображение
этих трех слоев тропических областей океанов дают в виде известной
графической схемы (рис. 71). Вся глубинная вода, с однообразным распре-
134
делением температуры, примерно начиная с 800 м, названа им (по аналогии
со строением атмосферы) стратосферой. Располагающаяся выше вода с
высокими температурой и соленостью названа им тропосферой и подраз-
делена на тонкий 200-метровый поверхностный слой, зону перемешивания
и глубже лежащий 600-метровый слой.
соленость %о
о2м/л
Р а том/л
Рис. 75. Распределение температуры (А), солености (Б), кислорода (В) и фос-
фатов (Г) на разрезе поперек экваториальной зоны ст 10° ю. ш. до 20° с. ш. в
Тихом океане (по данным экспедиции Карнеги из Свердрупа и др.).
На юге, в антарктических холодных морях, тропосфера выклинивается
и на поверхность выходит стратосфера. Для тропосферы характерно пре-
обладание течений, идущих в широтном направлении, для залегающей
под ней стратосферы — перемещение водных масс в меридиональном
направлении. Самый верхний слой тропосферы в экваториальной зоне
в области экваториальных течений и противотечений имеет весьма
характерную вертикальную циркуляцию. Согласно Д е ф а н т у (1929)
экваториальный слой воды на 25—30э к северу и югу и мощностью
50—200 м представляет собой (рис. 70) как бы четыре вращающихся
«стержня» с горизонтальными осями, имеющими широтное направление.
У южной границы самого южного «стержня» находится зона тропической
конвергенции, а у северной воды испытывают подъем у самого экватора
135
(линия дивергенции). На границе этого второго «стержня» и следующего,
представляющего собой тропическое противотечение, имеется опять линяя
конвергенции, а у ее северной границы опять зона дивергенции и, нако-
нец, у северной границы рассматриваемой области опять находится
линия тропической конвергенции, т. е. три линии конвергенции и две —
дивергенции. Дефантовская схема циркуляции находит хорошее подтвер-
ждение в распределении основных гидрологических факторов, на мериди-
ональных разрезах (рис. 75) через верхний слой Тихого океана по дан-
ным «Карнеги» (Свердруп, 1942). Зоны дивергенции (0° и 10э
с. ш.) обнаруживают подъем к поверхности более глубинных вод, содер-
жащих питательные соли, и для этих зон характерно более обильное
развитие фитопланктона. Вертикальное распределение температуры и
солености в области экваториальных течений показаны на рис. 76.
Рис. 76. Распределение течений (А) в тропической зоне Тихого океана и вер/и-
кальное распределение t° и S°/oo на шести станциях (В), показанных на рис. Л
точками (по данным экспедиций «Дана» и «Карнеги» из Свердрупа и др.).
III. Севернее 40° с. ш. тропосфера опять выклинивается, и страто-
сфера достигает поверхности океана. Это опять область сильного охлажде-
ния поверхностных слоев и вертикального перемешивания всей толщи
воды (рис. 77). Так же как и в первой области (в антарктических водах),
здесь преобладают в поверхностном слое течения, идущие в меридиональ-
ном направлении, и мощные перемещения вод в вертикальном напра-
влении.
IV. Еще дальше на север, за 80° с. ш., т. е. уже в центральных частях
Северного Ледовитого океана, залегает опять область стратифицирован-
ных вод, но здесь стратификация носит характер не тепловой, а солевой,
так как на поверхности залегает сильно опресненный 200-метровый слой,
препятствующий развитию вертикальной циркуляции, и часто на глуби-
нах находится более теплая вода, чем на поверхности. В этой области опять
преобладают горизонтальные перемещения водных масс, в основном кру-
гового характера. •
Общий характер циркуляции. Границы между этими областями Атлан-
тического океана даны на рис. 78, а в виде идеальной схемы на меридио-
нальном разрезе на рис. 65. Теплые тропические воды, попадая в более
высокие широты, еще в области тропосферы, несколько охлаждаясь и
встречаясь с более легкими, идущими с севера, холодными водами, пере-
136
врываются ими и уходят на сравнительно небольшую глубину (несколько^
сот метров), описывают полукруг и основной массой возвращаются к эк-
ватору, создавая замкнутое поверхностное вращение вод тропосферы.
Районы опускания этих вод названы Дефантом зонами субтропи-
ческой конвергенции. Меньшая часть опускающейся воды направляется
в стороны от экватора, доходит до краев тропосферы и обусловливает до-
полнительные зоны вертикального вращения (на рис. 78 между К и 11).
По краям тропосферы, где к поверхности моря выходит стратосфера,
тянется линия полярного фрота (П.-П.), за которым располагаются
области вертикального перемешивания. Мейер помечает на своей
137
карте несколько таких главных районов мощного опускания вниз поверх-
ностных вод, за счет которых создаются массы глубинных холодных вод,
заполняющих глубины океанов. Таким образом, к северу и югу от тропо-
Рис. 78. Поверхностные течения Атлантического океана в феврале (по
Мейеру). Помечены линии субтропической конвергенции (к) и по-
лярного фронта (П). Заштрихованы районы формирования масс глу-
бинной воды.
сферы, за линиями полярного фронта, располагаются районы горизонталь-
ных и вертикальных перемещений всей массы воды от поверхности до дна.
Следует обратить внимание на то, что из холодных зон вода уходит на глу-
бины океана двумя основными потоками: более южным поверхностным,
138
уходящим под тропосферу, и более северным, сразу опускающимся на
большие глубины. Д е ф а н т полагает, что первый поток в экваториаль-
ной области, встречаясь с идущими ему навстречу, погружается на глу-
бину и уже в слое 1000—3000 м движется обратно в высокие широты.
Это те воды, которые приносятся на юг постоянными холодными течениями,
как, например, Восточно-Гренландское и Лабрадорское.
Другой поток, более мощный, формируется из тех вод, которые при-
носятся из теплых областей в северные широты, здесь охлаждаются в зим-
нее время и, обладая сравнительно высокой соленостью, получают боль-
шую плотность и неудержимым потоком устремляются вниз. Предполо-
жительные места формирования их показаны на карте Мейера (рис.
78). Прекрасную иллюстрацию к этому дал Ф. Нансен для Гренланд-
ского моря, по материалам своей экспедиции 1912 г. в Шпицбергенские
воды на судне «Веслемё» (рис. 77). В левой части графика более теп-
лые атлантические воды, в правой части типичная для Арктического бас-
Рис. 79. Течения в северной части Атлантического океана. Толщина линий соот-
ветствует силе течений, а числа показывают проносимые объемы воды в миллионах
№ в секунду. Штриховыми линиями показаны холодные течения. Заштрихованы
районы с положительными температурными аномалиями (по Свердрупу и др.).
сейна стратификация с опресненным поверхностным слоем и глубже
лежащим слоем более теплой воды, приносимой сюда Атлантическим (соб-
ственно уже Норвежским) течением.
Центральная часть графика соответствует месту охлаждения и опу-
скания в глубину атлантических вод. Интересно отметить, что это те же
атлантические воды, что и заполняющие теплый промежуточный слой
Полярного бассейна. Разница между ними только в одном: промежуточные
теплые воды безостановочно двигались на север и донесли до Арктического
бассейна относительно высокую температуру и поэтому и не опустились
на большие глубины. Первые же были вовлечены в круговые течения
в зоне стыка с идущими с севера холодными водами, потеряли поступа-
тельный темп на север и подверглись нужной степени охлаждения, чтобы
опуститься на глубины. Весьма существенной деталью схемы Д е ф а н т а
является отсутствие подъема глубинных холодных вод на поверхность
в экваториальной зоне, как это обычно принимали ранее.
Атлантическое течение. Сложность явления, при встрече двух мощных
течений, двигающихся в разных направлениях и обладающих различными
139
показателями гидрологического режима, может быть показана на примере
Атлантического течения (рис. 79). Как мы уже указывали, мощный поток
сильно нагретой воды, выйдя из Флоридского пролива и соединившись с
еще более мощным Антильским течением, движется на северо-восток.
Это Гольфстрим. Примерно на широте 40° левым своим краем Гольфстрим
соприкасается с наступающими с севера от Ньюфаундленда массами хо-
лодных вод соединенных течений Лабрадорского и Восточно-Гренланл-
Рис. 80. Морские течения в Норвежском и Гренландском морях.
На карту нанесены также поверхностные изотермы (по Мейнар-
ДУСУ).
7 —главная ветвь Атлантического течения, II — Западно-гренландское те-
чение, III — течение Английского канала, IV — Норвежское течение, V —
Полярное течение, VI — Восточно-исландское течение, VII — Восточно-
гренландское течение (из Шотта по Мейнардусу).
ского. Под влиянием этого «удара во фланг» Гольфстрим раскалывается.
Часть уходит дальше на север и северо-восток, огибая холодные воды,
нажимающие с севера, и идя навстречу им и бок о бок с ними. Это так
называемое Атлантическое течение. Другая часть гольфстримных вод,
оторвавшись от Атлантического течения, частью замыкает круговым те-
чением халистатическую область Саргассова моря, частью же уходит
дальше на восток, погружается под верхние слои воды и движется обратно
140
в экваториальные области к берегам Африки, на глубине нескольких сот
метров.
Однако и Гольфстрим (как и Куро-Сиво) не движется вдоль самых
побережий, занятых прибрежными водами. Береговые воды континенталь-
ного плато и воды материкового склона образуют свои циркуляционные
системы, находящиеся отчасти под воздействием идущих с севера вдоль
берега более холодных вод. (
Двигающееся к берегам Европы Атлантическое течение подвергается
сложному расчленению и примерно на широте 50 и долготе 30° отделяет
сначала ветвь в Английский канал, а затем к Исландии (рис. 80).
Идя вдоль берегов Норвегии, Атлантическое течение, называемое
здесь уже Норвежским, отделяет на широте Тронтгеймфиорда большую
ветвь на север и северо-запад, к Гренландии, а у северо-западного побе-
режья отделяет на северо-восток так называемую Нордкапскую ветвь,
огибающую Скандинавию и вливающуюся в Баренцово море. Проходя
между Фарерскими островами и Шотландией, течение приобретает боль-
шую скорость (40—50 сл*/сек), на других же частях пути движение его
гораздо слабее (4—8 сле/сек). Расстояние от Америки до Европы течение
проходит примерно в 2 года. От Ньюфаундленда до середины океана тече-
ние доходит в 8—9 месяцев, а отсюда до берегов Европы нужен еще год.
Двигаясь дальше к Шпицбергену, Норвежское течение проходит по его
западной стороне и, перейдя через порог Нансена, уходит под поверхност-
ный слой опресненной воды и на глубине 200—600 м распространяется
по Полярному бассейну, двигаясь на восток севернее Земли Франца-
Посифа, Северной Земли и Новосибирских островов, уже как глубинное
течение.
Норвежское течение вступает с идущим вдоль Гренландии из Поляр-
ного бассейна холодным течением в сложную систему взаимодействия,
образуя в пограничных областях системы круговых течений в основном
антициклонического характера.
В качестве примера можно взять район Датского пролива, между
Исландией и Гренландией, где на узком пространстве сталкиваются воды
Восточно-Гренландской и Западно-Исландской ветви Атлантического те-
чения с различными показателями температуры, солености и плотности
(рис. 81). Взаимодействие двух течений сказывается на высоких градиен-
тах, сложной системе завихрений, вызывающих, конечно, еще более
сложную систему турбулентного перемешивания. Подобного типа районы
не являются в силу этого благоприятными для развития планктонной
ЖИЗНИс
Таблгща 47
Глубина в м 121 ст. «Метеора» 9/III-1935 56°37' с. ш. 44°54,5' з. д. 79 ст. «Метеора» 30/Ш-1933 59“38' c. in. 40°42,5' з. д.
t° s»/oo ct O2 (»/o) t° S°/oo °t o2 (°/o)
0 2,82 34,87 27,80 96 4,07 34,96 27,76 95
50 3,01 34,90 27,81 97 4,07 34,97 27,77 96
100 3,09 35,92 27,82 97 4,06 34,96 27,77 94
200 3,17 34,93 27,82 90 3,98 34,97 27,77 94
800 3,26 34,96 27,83 94 3,76 34,95 27,77 90
1000 3,20 34,95 27,83 93 3,33 34,89 27,77 89
1500 3,17 34,93 27,83 94 3,28 34,94 27,82 86
2000 3,23 34,93 27,82 i — 2,84 34,96 27,88 l 87
141
Рис. 81. Распределение поверхностных течений (А) и температуры (Б)
в Датском проливе (по Гентшелю и Вагтенбергу).
l'*2
Рис. 81. (В). Распределение солености в Датском проливе (по Генште-
лю и Баттенбергу).
Несколько южнее Датского пролива в так называемом Ирмингеровом
море влияние Гольфстрима почти не сказывается. Этот район оказывается
в стороне от его согревающего действия (табл. 47) и сохраняет весьма
устойчивый режим. Для всей толщи воды характерны ярко выраженная
гомогалинность и гомотермия. жЭто дает возможность массового опуска-
ния их на глубины при зимнем охлаждении поверхностных вод. На осно-
вании изложенных особенностей северная часть Атлантического океана
может быть подразделена на ряд естественных районов, характеризую-
щихся водными массами, обладающими определенными свойствами, в том
числе и специфической фауной.
ГЛАВА V
ГРУНТЫ МИРОВОГО ОКЕАНА
1. МЕХАНИЧЕСКИЙ СОСТАВ ГРУНТОВ
Связь донных отложений с населением моря. Грунты океанов и морей
представляют для нас интерес в нескольких отношениях. Грунты являются
тем механическим субстратом, на котором и в котором обитают донные
организмы. Для части донных организмов грунт при этом является пище-
вым веществом.
Мягкие илистые грунты часто как в прибрежных зонах (мидиевые
илы), так и в открытых частях океана (диатомовые илы) могут слагаться
нацело из фекалий и псевдофекалий донных и пелагических организмов.
Еще в 1853 г. М. Gape (Michael Sars) указал, что донные жи-
вотные в своем распределении часто зависят от свойств грунта больше,
чем от глубины, температуры или солености.
Кроме того, как это будет показано ниже, донные отложения — это
сложная лаборатория мощных химических процессов.
Механический состав. Под механическим составом разумеют соотно-
шение частиц различной крупности, составляющих грунт. Как общее
правило, по мере продвижения от берега к центральным частям морей и
океанов, средний размер частиц, составляющих грунт, мельчает.
В настоящее время в СССР пользуются следующей классификацией
донных отложений по механическому составу (классификация зерен по
Таблица 49
Таблгица 48
Название грунта Количество ча- стиц диаметром <0,01 мм в %
Песок < 5
Илистый песок .... 5—10
Песчанистый пл . . . 10—30
Пл : 30—50
Глинистый ил > 50
Название грунта
Размер зерна в мм
Глыбы ................
Валуны ...............
Галечник (щебень) ....
» крупный ....
» средний ....
» мелкий ......
Гравий крупный........
» средний .........
» мелкий ..........
Песок крупный.........
» средний.........,
» мелкий ..........
Ил (сильт) крупный . . .
» мелкий ....
Глина (пелит) крупная .
» мелкая . /........
>1000,0
>100,0
100,0—50,0
50,0—25,0
25,0—10,0
10,0—5,0
5,0—2,5
2,5—1,0
1,0-0,5
0,5—0,25
0,25—0,10
0,10—0,05
0,05—0,01
0,01^-0,001
<0,001
«44
размерности), разработанной (М. Кленова, 1929) в Государственном
Океанографическом институте и принятой также Гидрографическим управ-
лением. Для крупнозернистых грунтов название дается по преобладающей
фракции, для мелкозернистых по % содержания фракции с диаметром
зерен менее 0,01 мм (табл. 48 и 49).
Происхождение морских отложений. Все отложения дна мирового
океана можно подразделить на две основные группы — терриген-
ные (или литоригенные), представляющие собою в основном продукты
разрушения береговых пород и выноса реками, и пелагические,
образованные в основном из остатков пелагических организмов, прово-
дящих жизнь в толще воды, от крошечных силикофлагеллят, диатомей,
радиолярий и фораминифер до рыб и морских млекопитающих. Грунты
переходного типа залегают между отложениями двух основных типов. На
дне мирового океана преобладают грунты пелагического происхождения,
они покрывают 75,5% всей площади дна, прибрежные отложения — 24,5%.
В отложении океанических грунтов всегда принимают участие также и ча-
стицы вулканического или даже космического происхождения, попадающие
на поверхность океана из атмосферы. В самых глубоких частях океаниче-
ского ложа этот тип отложений может получать преобладающее значение.
2. РАСПРОСТРАНЕНИЕ РАЗЛИЧНЫХ ОТЛОЖЕНИЙ НА ДНЕ
ОКЕАНА
Соотношение в % площади дна мирового океана, занятой различными
типами грунтов, и глубина их залегания показаны на табл. 50, 51
и52изМерреяиРенара (с некоторыми изменениями).
Таблица 50
Вертикальные зоны Характер грунта Границы глубин в м Вещества органи- ческого происхож- дения в % Про-1 цент к| общей площ.! Генезис
Вме- сте —— __ _ А* **
из- вест- ковые Крем- незем даа ।
на |
1. Приливно- отливная зона (литораль) Гравий, песок, ил Меньше 0 глубины > 9,1 1 Прибреж- ные отло- жения (литори- генные) 24,5%
2. Мелководье до глубины 2'30 м. Матери- ковая отмель (сублитораль) Гравий, песок, ил 0—200
3. Глубоковод- ная. Материко- вый склон. Ло- же океана и глубоководные ложбины Коралловый ил . Вулканический ил Зеленый ил... . Красный ил ... Синий ил 200-3300 500—5100 200—2300 200—2200 200—5100 86 20 25 32 13 1 2 14 1 3 87 22 39 33 16 1 1 } 15,4 1 Пелаги- ческие . отложе- ния 75.50/0
Птероподовый ил Глобигерпновый » Диатомовый » Радиоляриевый » Красная глина 700—2800 700—5400 1100—3600 4300-8200 4100—7200 79 64 23 4 7 3 2 41 54 2 82 66 64 58 9 0,4 29,2 6,4 3,4 36,1
10 Зепкевич, т. I
145
Таблица 51
Характер отложений в 0 /о Индийский океан Тихий океан Атлантиче- ский океан
Известковые илы 54,3 36,2 67,5
Кремнеземовые илы . . . 20,4 14,7 6,7
Красная глина 25,3 49,1 25,8
100,0 100,0 100,0
Таблица 52
Характер отложений Глубина залегания в м
Наимень- шая Наиболь- шая Средняя
Глобигериновый ил... . 777 6006 3612
Птероподовый ил 713 3519 2072
Диатомовый ил 1097 5733 3900
Радиоляриевый ил ... . 4298 8184 5292
Красная глина 4060 8282 5407
Глубоководные отложения литоригенного характера в основной своей
части (зеленый, красный, синий и вулканический илы) характеризуются
преобладанием мелких частиц минерального характера, представляющих
собою мельчайшую муть в воде, выносимую реками и морскими тече-
Рис. 82. Вертикальное распределение СаСО3 в грунтах океана (по Меррею).
ниями далеко за пределы берегов, но не достигающих, однако, центральных
частей океана. Среди них имеется, однако, коралловый ил, продукт раз-
рушения коралловых рифов, на 87% состоящий из остатков органиче-
ского происхождения, в том числе и остатков известковых водорослей
(Lithot hamnion, Corallina и др.).
Известковые грунты. В пелагических отложениях резко преобладают
углекислый кальций (до 80%) или кремнезем (до 54,4%). Эти грунты об-
разованы в значительной части скелетами планктонных организмов,
причем в распределении этих скелетов наблюдается определенная зональ-
ность — большим глубинам (1100—8200 м) свойственны кремнеземовые
отложения, меньшим (700—5400) — известковые. Этот характер распре-
деления в отношении СаСО3 показан на рис. 82 и на табл. 52. И угле-
кислый кальций и кремнезем растворяются в морской воде и первый
146
быстрее второго. Поэтому известковые скелеты не могут достигать боль-
ших глубин, они растворяются, не достигнув дна.
Наибольшее распространение (29,2%) имеет глобигериновый ил,
образованный скелетиками пелагических фораминифер, в основном мно-
гочисленных видов рода Globigerina. Важное участие в формировании
кальциевых отложений дна принимает также и другая группа пелаги-
ческих одноклеточных, именно группа Coccolithophozidae (известковые
жгутиконосцы), известковые коккосферы и рабдосферы, раковинки ко-
торых, несмотря на их незначительные размеры (несколько микронов),
могут образовывать донные отложения. Как показал Пратье (Рга-
t j е, 1939), различные глубоководные донные отложения океана на 30—
45% могут состоять из раковинок кокколитофорид и на 20—25% из ра-
ковинок фораминифер.
Существенное участие в образовании морских отложений известкового
характера принимают также и раковины пелагических брюхоногих мол-
люсков групп Pteropoda (Limacina, Clio, Cavolinia и др.) и Heteropoda
(Carinaria, Atlanta и др.), составляющих, однако, не свыше 4% грунта
даже в тех илах, которые носят название птероподовых.
Кремнеземовые грунты. В образовании кремнеземовых грунтов наи-
большую роль играют диатомовые водоросли и радиолярии. К ним следует
еще добавить группу Silicoflagellata. На рис. 83 дано общее распределе-
ние разных типов грунтов по ложу океанов. Основную массу отложений
в Атлантическом и Индийском океанах составляют глобигериновые илы,
а в Тихом — красная глина. Радиоляриевые илы отсутствуют в Атланти-
ческом океане и хорошо выражены в экваториальной зоне Индийского
и Тихого океанов. В высоких широтах севера и юга широкой полосой
тянутся диатомовые илы, причем в Атлантике они имеются только в Антарк-
тике. Наконец, небольшими пятнами, главным образом в Атлантическом
океане, располагаются птероподовые илы.
Само собою разумеется, что не только пелагические животные, но
и многие донные участвуют своими телами в образовании грунтов в основ-
ном мелководной зоны. В кремнеземных отложениях участвуют кремне-
вые губки, образуя в иных местах большие скопления своих спикул.
В образовании известковых грунтов, наряду с упомянутыми пелагическими
организмами, принимают участие также и аннелиды, ракообразные, игло-
кожие, плеченогие, мшанки и моллюски. Скелеты и тела рыб и морских
млекопитающих также участвуют в создании донных отложений. Весьма
обычны в глубоководных илах зубы акул и слуховые косточки китов.
3. ЗАВИСИМОСТЬ ЖИВОТНЫХ ОТ МЕХАНИЧЕСКОГО СОСТАВА
ГРУНТА
Для живущих на грунте и в грунте животных механический состав
грунтов имеет очень существенное значение тем более, что в мелководных
районах механический состав грунта есть результат горизонтального дви-
жения вод.
Жизнь на твердых грунтах. На скалах и крупных камнях, если они
не обросли макрофитами, могут жить только плотно присасывающиеся
к камням или прирастающие к ним организмы. В россыпях гальки, гра-
вия и крупнозернистого песка животные или отсутствуют, или представ-
лены некоторыми особенно выносливыми формами. На мелкозернистых
песках количество животных возрастает. Наиболее благоприятные условия
существования животные находят на песчанистоилистых и илистопесча-
нистых смешанных грунтах; на мягких илах жизнь опять слабо выражена.
147
На примере фауны одного из районов Варенцова моря это можно пояс-
нить следующим образом (рис. 84). Количество экземпляров населяющих
грунт организмов падает от ила к песку и гравию, общий вес населения
также падает, но эпифауна дает увеличение биомассы на гравии, так как
имеет здесь нужный для прикрепления субстрат. Что же касается среднего
148
веса одного экземпляра, то он больше всего на гравии для эпифауны, и
меньше всего для инфауны.
Жизнь на мягких грунтах. Мягкие полужидкие илы и вязкая глина
или, наоборот, очень плотная глина могут представлять собою также небла-
гоприятный субстрат для развития жизни. Известно, что обитатели мяг-
ких илистых грунтов обладают различными приспособлениями, чтобы
Рис. 84. Зависимость количества экземпляров, биомассы и среднего веса одного
организма бентоса на разных грунтах (по Зенкевичу).
Слева — средний вес одного экземпляра в jwa; посредине — средняя биомасса в г/лс2; справа —
количество экземпляров на 1 лс2.
не тонуть в грунте (например, голотурии), или чтобы возвышаться над
грунтом (восьмилучевые кораллы, морские лилии и др.). Во всяком слу-
чае многие группы животных не встречаются на очень мягких грунтах
и биомасса здесь обычно очень низка.
Грунтоеды. Многие формы, обитающие над грунтом (рыбы), на грунте
и в грунте, питаются самим грунтом. Грунтоеды встречаются среди мно-
гих наиболее типичных, морских групп животных — червей, моллюсков,
иглокожих и рыб. При этом способе питания используется органическое
вещество грунта.
4. ХИМИЧЕСКИЕ ПРОЦЕССЫ, ПРОТЕКАЮЩИЕ В ГРУНТАХ
Общий характер химических процессов. Химические процессы в грун-
тах характеризуются обычно преобладанием явлений аккумуляции (на-
копления). Верхние слои грунтов, заселенные жизнью, находятся в по-
стоянном химическом взаимодействии с придонным слоем воды; глубже
лежат слои, где идут процессы диагенезиса — перехода морских грунтов
в осадочные породы, выходящие, как правило, из химических процессов
циклического характера между водой и грунтом. Иногда в морских грун-
тах резко преобладают процессы накопления, либо в результате большой
глубины водоема и отсутствия значительных движений масс воды, как
в глубоководных впадинах мирового океана, либо в результате специфи-
ческого явления стагнации, с развитием сероводорода и исчезновением
кислорода, как в некоторых фиордах Норвегии и особенно в Черном море.
Кислород. Сложные химические процессы, совершающиеся в толще
грунта, обычно идут в восстановительной среде и только самая поверх-
ностная, часто очень тонкая пленка грунта окислена. Иногда кислород
149
в придонном слое может исчезать и восстановительная зона может выхо-
дить из грунта и захватывать придонный слой воды. Иногда эти явления
носят периодический характер и тогда происходит отмирание всей донной
фауны (явления замора), за исключением только некоторых наиболее
эвриоксибионтных и специально приспособленных к тому форм. Так,
двустворчатый моллюск Syndesmya, при исчезновении кислорода в при-
донном слое, может вытягивать вверх, на высоту 20—30 см вводной си-
фон и брать воду, содержащую кислород. Наконец, ряд форм может без
вреда для себя находиться несколько дней в бескислородной среде. Заме-
чательно развитие на некоторых участках дна Каспийского моря бакте-
риальной пленки тионовых, метановых и водородных бактерий, препят-
ствующей поступлению в воду из грунта вредных для животных газов
(H2S, СН2, Н2).
Роль марганца. Особенно большое значение в круговороте кислорода
в придонном слое придается марганцу. Я. Самойлов (1929) высказал
мнение, что марганец верхнего слоя грунта находится все время в подвиж-
ном состоянии. Выпадая в грунт в виде нерастворимых окисных соеди-
нений, марганец в восстановительной среде грунта переходит в раство-
римые закисные соединения и поступает в раствор в самом придонном слое.
Снова окисляясь, марганец вновь выпадает в грунт, где опять восстанав-
ливается. По этой роли в газовом обмене Я. Самойлов образно срав-
нивает марганец придонного слоя с гемоглобином крови.
Теоретическое предположение о роли марганца в химическом режиме
придонного слоя, высказанное Я. Самойловым, нашло подтвержде-
ние в работах Перселла (Purcell, 1939) в Америке и А. Фран-
цева (1941) у нас на водохранилищах в первые годы их обводнения.
Марганец в закисном состоянии переходит в воду при отсутствии свобод-
ного кислорода и низком показателе pH. Достигая слоев с наличием кисло-
рода, марганец окисляется и выпадает из раствора на дно, после чего,
восстанавливаясь, может снова поступать в циклические процессы транс-
портирования кислорода в грунты.
Органическое вещество. В морских грунтах постоянно идет процесс
накопления органического вещества, хотя количество его обычно бывает
очень невелико (0,5—1,0—1,5%) и только в отдельных районах достигает
значительных величин (4—7% и больше). В замкнутых и полузамкнутых
бассейнах в грунтах всегда накапливается более значительное количество
органического вещества, что иногда даже связывают с образованием нефти.
В открытых морях, в более мелководных районах, на более мелкозерни-
стых грунтах, органического вещества обычно бывает больше.
Органическое вещество грунтов формируется в основном за счет планк-
тона в более открытых частях и за счет береговой растительности и жи-
вотных организмов бентоса в закрытых участках. Наиболее характерными
показателями, при определении общего количества органического ве-
щества, считают С, N и отношение . При образовании органического
вещества грунтов за счет планктона, это отношение будет близким к 7—
9, а за счет береговой растительности (Zostera) — 13,4. Таким образом
соотношение отчасти дает возможность установить (как показал Б о й-
сен- Йенсен) и генезис органического вещества в грунтах. Как видно
на карте, составленной Траском (Trask, 1932) по содержанию
азота в океанических грунтах и даваемой нами в переделке М. Клено-
вой (рис. 85), значительные количества органического вещества , при-
урочены в общем к береговым областям и особенно к западным побе-
режьям обеих Америк и Африки, т. е. к областям постоянного подъема
150
к поверхности глубинных вод, вследствие сгонного влияния пассатных
ветров. Суммарное количество органического вещества в 14 раз превы-
шает количество азота (Траск считает наиболее типичным показателем,
при определении количества органического вещества, именно азот).
Соединения фосфора. Интересно отметить, что с этими же районами
подъема глубинных вод связано и накопление в грунтах соединений фос-
фора. А. Казаковым высказано предположение, что генезис фосфорных
месторождений связан с аналогичными зонами аккумуляции их в морях.
Казаков считает, что тут имеет место чисто химический процесс, иду-
щий без участия организмов, на шельфах, путем химического осаждения
избытка фосфатов из глубинной воды в районах ее поднятия.
Рис. 85. Количество органического азота в грунтах мирового океана
(по Траску в переделке Кленовой) в % сухого веса.
1 — >20; 2 — 10—20; 3 — 5—10; 4 — <5.
В общем к тому же выводу о химических процессах выпадения фос-
фатов в отложениях без участия микроорганизмов пришли и американские
авторы (R. S. D i е t z, К. О. Е ш е г у and F. Р. S h е р а г d, 1942). Они
считают этот процесс неизбежным, так как количество фосфатов в глубин?
ных водах океана значительно превышает предел насыщения при малом
давлении.
5. ТЕМП ОТЛОЖЕНИЯ МОРСКИХ ОСАДКОВ
Мощность и возраст различных отложений. Большой интерес представ-
ляет вопрос о темпе отложения морских осадков. Чем ближе к берегу, тем
в общем быстрее идет накопление грунтов. У устьев рек, при определенных
комбинациях течений, могут итти процессы быстрого образования мощных
наносов в одних местах и размывов в других. В общем же темп этого про-
цесса нам известен очень мало. Еще меньше известно о темпе отложения
глубоководных осадков. Шотт (Schott, 1935) принимает, что в сред-
нем слой в 1 см отлагается за 1000 лет. Американским геологам при
сборе проб грунта, взятых особыми трубками на глубине 1250—4750 м
в основном на глобигериновом илу северных частей Атлантики, удалось
получить колонки в 3 м длиной, соответствующие нескольким леднико-
вым и межледниковым периодам. Послеледниковые отложения имеют мощ-
кость в 30 сж, что соответствует возрасту примерно в 20000 лет.
151
Если для открытых частей океана темп седиментации замедлен, то
ближе к берегам и во внутренних морях он идет гораздо быстрее. По
расчетам С. Бруевича (1948) в открытых частях Черного и Каспийского
морей слой грунта в 1 см отлагается в среднем в 20—40 лет, а в бере-
говой зоне всего в 5—8—23 года.
Если принять (п о Т у л е) (Т h о u 1 е t) скорость оседания мертвых
фораминифер (при размере 0,4 мм) в 2,11 см/сек, то в несколько суток они
должны достигать самых больших глубин и относ в сторону от места оби-
тания должен быть незначительным. Таким образом, районы накопления
в грунте скелетов организмов в общем соответствуют и районам их мас-
сового развития в океанах.
Слои, характерные для ледникового и межледникового времени. Изу-
чение длинных колонок грунта из северной части Атлантического океана
(Пеггот, Peggott, 1935) показало, что в отложениях обнаружи-
вается чередование трех слоев. Верхний слой, богатый корненожками,
относится к современным осадкам, под ним лежит слой бедный корненож-
ками, образование которого авторы относят к последнему ледниковому
периоду. Еще глубже залегают слои, опять богатые корненожками, видимо
соответствующие межледниковой эпохе. Оказывается, что ледниковая
эпоха сказалась в донных отложениях и экваториальной зоны океана,
обедненных остатками фораминифер (Schott, 1935).
Та^ое же чередование слоев богатых и бедные раковинками форами-
нифер обнаружено (Горшкова, 1942) и в грунтах центральных частей
Сев. Ледовитого океана по материалам дрейфующей Станции «Северный
Полюс» 1937—38 г., с тем отличием, что в Полярном бассейне самый
верхний слой также обеднен остатками фораминифер, но под ним залегает
слой богатый ими; Т. Горшкова делает предположение, что этот бо-
гатый корненожками слой соответствует теплой фазе послеледникового
времени, а современной фазе свойственно обеднение полярных вод форами-
ниферами.
Шведские исследователи на судне «S k a g е г а к», во время подго-
товки к предстоящей кругосветной глубоководной экспедиции и испытания
новых приборов (Pettersson, 1946) при помощи трубки новой кон-
струкции получили в шведских территориальных водах колонки грунта
до 20 м, соответствующие по времени отложения нескольким миллионам
лет. Оригинальность конструкции новой трубки заключается в том, что
из плотно закрытой камеры выкачан воздух. Когда трубка вонзается
в грунт, она силой давления загоняется в грунт на большую глубину.
Кроме того, применением особого типа эхолота шведские исследова-
тели получили возможность определения не только поверхности дна, но
и глубины залегания всей толщи донных отложений до коренных пород.
Петерсон опубликовал несколько данных, полученных судном «Ска-
герак» в Средиземном море (табл. 53).
Таблица 53
Глубина места в м Длина полученной колонки грунта (в м) Мощность донных отложений (в м)
870 0,0 0,0
1030 0,9 0,9
2810 10,5 470,0
3620 10,0 2730,0
2450 15,0 —
ГЛАВА VI
БИОСФЕРА И РАСПРЕДЕЛЕНИЕ ОРГАНИЗМОВ В МОРЕ
1. ПОНЯТИЕ О БИОСФЕРЕ
отя организмы (бактерии) проникают в глубину земной
коры на несколько километров, однако, в основной массе
на границе литосферы и атмосферы, организмы распре-
делены тонкой пленкой — соответствующей слою био-
сферы мощностью в несколько метров. При определен-
ных условиях микроскопические организмы и их споры и
цисты могут подниматься в высокие слои атмосферы, однако
организмов, полностью приуроченных к воздушному образу жизни (аэро-
планктона) и не нуждающихся в твердой поверхности земли, повидимому,
не существует. Только в гидросфере толщина слоя, достаточно плотно засе-
ленного жизнью, достигает относительно большой мощности — порядка не-
скольких сот метров (глубже и в гидросфере жизнь существует в весьма раз-
реженном состоянии). Это обусловливается свойственной водной среде спо-
собностью к перемешиванию, ее прозрачностью, ее механически легкой про-
ницаемостью, однообразием в распределении факторов среды, плотностью
воды, дающей организмам возможность парения, и всей совокупностью
свойств воды, как чрезвычайно благоприятного субстрата для развития
жизни. Э. 3 ю с с (1877), разработавший представление о земных оболоч-
ках, ввел также в науку и понятие о биосфере, разработка же этого
понятия приходится уже на XX век и связана главным образом с именем
нашего замечательного ученого академика В. В. Вернадского. Био-
сфера, как среда жизни, захватывает все три оболочка, «живое вещество
биосферы — единственной области планеты, закономерно связанной с кос-
мическими просторами, есть совокупность ее живых организмов... ее жи-
вого вещества, как планетного явления» (Вернадский, 1939).
«Все живое вещество в целом можно представлять себе, как единую,
особую область накопления... химической энергии в биосфере, превра-
щения в нее световых излучений солнца» (Вернадский, 1926).
«Биосфера представляет собой определенную геологическую оболочку,
резко отличную от всех других геологических оболочек нашей планеты.
И это не только потому, что она заселена живым веществом, которое яв-
ляется огромного значения геологической силой, ее совершенно перера-
батывающей, меняющей ее физические, химические и механические свой-
ства. Но это единственная оболочка планеты, куда проникает заметным
образом космическая энергия, ее еще более живого вещества изменяю-
щая. Главным источником этой энергии является солнце. Его энергия —
тепловая, световая и химическая — наряду с энергией химических эле-
ментов есть первоисточник создания живого вещества. Живое вещество
проникает всю биосферу и ее в значительной степени создает. Оно акку-
155
мулирует энергию биосферы, главным образом ее тепловую и химиче-
скую энергию солнечных лучей и химическую энергию земных атомов»
(Вернадский, 1939).
Состав. В биосфере В. Вернадский различает: 1) главную массу,
не входящую в живое вещество, ее косное вещество; 2) совокупность жи-
вых организмов планеты — ее живое вещество и 3) тела биокосные, со-
стоящие из косных и живых тел одновременно (например, почва, озерная
вода, морская вода и т. п.). Эта совершенно правильная мысль находит
себе не мало подтверждений. Та или иная часть биосферы, находящаяся под
постоянным воздействием живых организмов, насыщаемая продуктами
их жизнедеятельности и распада, резко отличается от косных, изолиро-
ванных от организмов, частей земных оболочек и представляет собой «осо-
бые естественные тела».
Геохимическая энергия организмов. В. Вернадский ввел и раз-
работал понятие о геохимической энергии организмов. Поскольку размно-
жение всех организмов выражается геометрической прогрессией, оно
может быть выражено формулой:
2пДу 1Чп,
где п — число дней с начала размножения, Д — показатель прогрес-
сии, Nп — число неделимых, существующих благодаря размножению
через п дней. В зависимости от показателя Д «темп растекания» живого
вещества в процессе размножения для разных организмов будет различен,
и за предельное расстояние для всех организмов, по которому может рас-
пространяться передача жизни, В. Вернадский принимает земной
экватор, т. е. 40075 721 м. Для каждого вида может быть определено макси-
мальное количество неделимых при достижении этого предела. Это число
(Nmx) В. Вернадский называет стационарным числом однород-
ного живого вещества. Скорость достижения этого предела (7) также будет
характеризовать каждую живую форму. Эта скорость связана со ста-
ционарным числом следующей формулой:
_ 13963 • ЗхД
lgNmx •
Для шаровой бактерии объемом в 10-12 м скорость — и определяется
В. Вернадским в 33 100 см/сек. Для наиболее крупных млекопитаю-
щих скорость передачи жизни в сотни тысяч раз меньше, чем для бактерий,
например, для индийского слона она определяется величиной 0,09 см/сек.
Помимо скорости передачи жизни, определяемой величиной р, необходимо
получить еще значение кинетической геохимической энергии живого
вещества, выражаемое формулой у- , где р — средний вес организма.
Таким образом может быть получено количество (вес) органического ве-
щества, которое может быть создано размножением и ростом организмов
на единицу поверхности. Однако этот критерий геохимической энергии
не был никем больше использован, так как имеет чисто абстрактный харак-
тер. а
Распределение аутотрофных организмов на суше и в море. Раститель-
ные организмы обладают гораздо большей геохимической энергией, не-
жели животные. В. Вернадский допускает, что по биомассе живое
вещество биосферы делится на две примерно равные части — на зеленое
аутотрофное и на питающееся уже готовым органическим веществом —
гетеротрофное. В. Вернадский также совершенно правильно ука-
зывает на характерные отличия между строением и особенностями размно-
жения «зеленого трансформатора солнечной энергии» на суше и в море.
156
На суше водоросли и особенно одноклеточные составляют ничтожную
часть растительной массы, в океане главную массу составляют именно
одноклеточные водоросли, гораздо слабее представлены водоросли —
макрофиты (зеленые, бурые и красные) и почти отсутствуют высшие расте-
ния.
Главную же часть растительной массы на суше составляют травяни-
стая, кустарниковая и древесная растительность, с сильно развитой кор-
невой системой и характерной для нее скоростью размножения (коэффи-
циент о Вернадского), вряд ли превышающий сантиметры в се-
кунду, тогда как для зеленых протистов она достигает тысяч сантиметров.
Хотя на суше отношение массы растений к массе животных выше, чем
в море (на суше значительно преобладают растения, а в море животные
дают несколько большую биомассу), зато первые обладают гораздо более
высокой способностью к размножению и вследствие этого они обеспечи-
вают гетеротрофньш организмам быстро восстанавливающиеся запасы
пищи.
Особенности условий существования на суше и в море. Эти характер-
ные различия между растениями суши и to по-
нятны, если отметить некоторые особенности среды существования.
Воздушные почвы, ни в какой степени не пропускающие в себя сол-
нечного света, могут предоставить растениям только тонкую поверх-
ность, обращенную к свету, и вот на смену водорослям являются
травы, кустарники и деревья, с их сложной мозаикой листьев. В водную
среду свет проникает на сотни метров, обеспечивая развитие взвешенным
в воде одноклеточным растениям, также прекрасно приспособленным
К своеобразию условий водной среды, как травы, кустарники и деревья
приспособлены к условиям существования на суше.
Поверхность суши как бы покрыта зеленой краской хлорофилла,
В море эта краска находится в мельчайшей взвеси в двухсотметровом слое.
Весьма возможно, что суммарная поверхность всех особей фитопланктона
в период цветения окажется близкой к суммарной ассимилирующей зе-
леной поверхности растительного покрова суши в весеннее время.
Выход растительных организмов из моря на сушу обусловил даль-
нейшую их морфологическую (и физиологическую) эволюцию. В морях
обитают только простейшие и наиболее примитивные формы растений
(Thallophyta). Все вышестоящие, от мхов до покрытосемянных, возникли
в воздушной среде. Представители этих последних групп, обитающие
в морях, представляют собой вторичных выселенцев с суши в море (вто-
ричноводных). Для животных мы имеем совсем другую картину — только
4 класса типа хордовых и четыре класса членистоногих не обитают
в морях или являются в них вторичноводными. Таким образом чуждых
морской воде по своему генезису среди животных имеется только 8 клас-
сов, или 13%, а среди растений (из общего количества 33 классов)1,18 клас-
сов, или 55%.
33 класса ныне живущих растений распределяются следующим обра-
30хМ (табл. 54):
Таблица 54
только в море 5 классов (15%)
и в море и в пресной воде 10 классов (30%)
только в пресной воде 3 масса (9%)
наземных 15 классов (46%)
Всего 33 класса 100%
1 Эти числа даны по Л. К у р с а н о в у.
157
Объяснение этому следует искать в глубоком различии между расте-
ниями и животными в основном способе питания. Для растений это в пер-
вую очередь и главным образом свет и минеральные вещества. Свет нахо-
дится в воде в сильно рассеянном состоянии, а минеральные вещества рас-
творены в окружающей растение среде. Иными словами, растение в море
получает и то и другое всеми сторонами своего тела. В воздушной среде
оба источника питания как бы резко поляризованы — свет сверху (при
значительно более слабом рассеянии), минеральное питание снизу (из
почвы). Эта гетерополярность, видимо, и была основным фактором вытя-
нутой формы тела растений, со всей последующей системой морфо-физио-
логической дифференциации, приведшей к формированию многих новых
классов. Вряд ли будет ошибкой утверждение, что именно смена водной
среды обитания на воздушную вызвала в истории развития растений бур-
ный процесс дивергентного и прогрессивного развития.
Пока растения оставались в море, указанные выше причины способ-
ствовали сохранению у них тела в форме клеточки, нити, клеточной пла-
стинки, комочка или клеточного тела. Удлиненная форма тела у прибреж-
ных морских растений (зеленые, харовые, бурые и красные водоросли),
с чисто внешним конвергентным сходством в расчленении тела на корень,
стебель и лист, возникает в результате необходимости получить опору
в грунте и увеличить размеры тела в прибрежной полосе прибоя. Но при-
брежная растительность в общей массе растений моря составляет незна-
чительную часть.
По-иному протекала эволюция животных. Гетеротрофное питание, ма-
лая зависимость от света и подвижное существование способствовали в море
дивергентной прогрессивной эволюции и образованию многих классов
многоклеточных с высокой организацией. Однако и у животных выход
в воздушную среду (резкая смена среды обитания) повел к энергичному
процессу дивергентного формообразования и возникновению высоко
специализированных классов среди обеих ветвей животного мира — пер-
вичноротых и вторичноротых.
Мы пока не располагаем сколько-нибудь достаточными данными, чтобы
дать общую схему количественного распределения растительных и жи-
вотных организмов в биосфере и определить количество вещества, сосре-
доточенного в них. Согласно В. Вернадскому масса зеленого
вещества биосферы составляет в каждый отдельный момент 1020—1021 г
(иначе говоря 105—106 ил3), годовая же продукция его не должна пре-
вышать 5\1018 г (5\103 ял3). Соответственные же величины для животного
вещества В. Вернадский считает близкими к 1016 г и 5ХЮ15—7/
X 1015 (10 км3 и 5—7 км3).
Что касается животных, то приведенные в отношении их числа В. В е р-
надского, возможно, близки к действительности, растительная же
биомасса должна быть несколько менее указываемой В. Вернадским
и вряд ли превышает 1019 г, что же касается годовой продукции растений,
она вряд ли меньше ее биомассы. Животная биомасса в литосфере в сред-
них величинах только в редких случаях достигает килограмма на 1 л2,
иногда выражается величинами 200—400 г (за счет Oligochaeta), часто
составляет немногие граммы (за счет членистоногих), большею же частью
не превышает нескольких десятков или немногих сотен миллиграмм на
1 м2 (также в основном за счет насекомых и паукообразных). В гидросфере
плотность животного населения обычно превышает животную биомассу
y Zoa g*
литосферы и таким образом отношение по биомассе в море значи-
тельно выше. Объясняется это исключительно благоприятными свойствами
воды, как среды обитания и прежде всего чрезвычайно высокой способ-
158
юстью самых разнообразных веществ к растворению в воде и подвиж-
ностью воды, обусловливающей перемешиваемость. Таким образом, про-
дуцируемое в воде органическое вещество, равномерно в ней распреде-
ляясь и длительно удерживаясь в растворенном виде, обеспечивает наи-
более полное ее использование растительными организмами. Поэтому
и в морских грунтах количество органического вещества значительно
меньше, чем в почвах. В стоячих пресных водоемах, по сравнению с морем,
отношение также понижается и обычно идет накопление значительных
масс неиспользованных растительных остатков.
2. ОСНОВНЫЕ ЗАКОНОМЕРНОСТИ РАСПРЕДЕЛЕНИЯ ОРГАНИЗМОВ
В МИРОВОМ ОКЕАНЕ
Неоднородность водной массы океана. Водная масса морей и океанов
(галосфера) по своим абиотическим и биотическим показателям неодно-
родна в разных своих частях. Морскую среду принято называть «царством
однообразия» в противоположность наземной среде — «царству контра-
стов». Однако это справедливо только в некоторых отношениях и совсем
неприменимо в других. По резкости различий между некоторыми био-
топами (поверхностные и глубинные слои, пелагиаль и бенталь, открытые
просторы моря и литоральная зона, область полярных льдов и тропиче-
ская зона) морская среда не только не уступает суше, но в некоторых от-
ношениях и превосходит ее. Это особенно следует учитывать еще и в том
отношении, что воздух, облекающий организм наземного животного сам
представляет собой прекрасный изолятор для целого ряда физических
и химических явлений, тогда как в водной среде организм становится
как бы игрушкой свойств этой среды, гораздо легче воздействующих на
организм, чем в воздушной среде.
Современная океанология, как указывалось выше, базирует свои
общие представления на подразделении галосферы на отдельные водные
массы, характеризующиеся определенными свойствами и своим биологи-
ческим обликом.
Симметричное распределение явлений. Конфигурация океанов и кли-
матическая зональность создают некоторые явления симметричного рас-
пределения в меридиональном и широтном направлении. Особенно сильно
выражено симметричное распределение явлений в обе стороны от эквато-
риальной зоны вплоть до приполярных областей, представляя собой в не-
котором приближении как бы зеркальное изображение явлений, разви-
вающихся в обоих полушариях. Сходны в обоих полушариях в обе сто-
роны от экватора, как мы видели, распределение и циркуляция водных
масс, распределение солености, температуры, кислорода, питательных
солей, прозрачности и цветности воды, донных отложений, качественное
и количественное распределение биологических явлений и в частности
миграционных.
Наличие указанной выше симметрии, а также форма и рельеф ложа
обоих великих океанов (а отчасти и Индийского), вытянутых в меридио-
нальном направлении и ограниченных с обеих сторон материками, обусло*
вливают симметрический характер явлений и по обеим сторонам океанов —
восточной и западной. Эта широтная симметрия связана главным образом
с наличием в каждом океане двух неритических зон, разделенных океани-
ческими просторами. Многие из указанных выше факторов претерпевают
сходный тип изменений от центральных частей в обе стороны к побережьям
материков в широтном направлении.
159'
Наряду с наличием этой двойной симметрии галосфера дает в целом
ряде моментов резко выраженную асимметрию, прежде всего в верти-
кальном направлении — от поверхности ко дну.
В широтном направлении наибольшее выражение асимметрии нахо-
дит в схеме горизонтальной циркуляции водных масс. Однако смена
океанического на неритический режим водных масс все же находит и
в широтном направлении свое достаточное выражение, чтобы с достаточной
четкостью сказывались симметрические явления.
Биполярное и амфиокеаническое распространение организмов. Под
таким общим углом зрения рассмотрим некоторые биологические явления,
^свойственные водным массам мирового океана.
Рис. 87. Схема распределения неритической
флоры и фауны океана (по Зенкевичу).
1 — тропическая фауна, 2—арктическая фауна,
3 — фауна умеренных широт.
Рис. 86. Схема распределения
плотности планктона в океане
(по Гентшелю с некоторыми
изменениями).
Э. Гентшель (Е. Hentschel) впервые дал схему распреде-
ления плотности планктона, одинаково применимую и к Атлантическому
и к Тихому океанам (рис. 86). В разделе, посвященном планктону, эта
схема иллюстрируется конкретными примерами. Широтная зональность
в качественном распределении планктона находит себе хорошее отражение
в донных отложениях, особенно в Тихом океане (см. рис. 83). Для тропи-
ческой зоны характерны радиоляриевые илы. С продвижением на север
и юг большое развитие получают глобигериновые илы и красная глина, и
еще дальше в умеренные и холодные зоны в широтном направлении тя-
нутся диатомовые илы.
Мы в настоящее время еще не можем дать для распределения донного
населения океана схему, подобную Гентшелевской, но для населения не-
ритической области может быть дана иная схема, также отображающая
на себе особенности симметрического распределения организмов, приме-
нимая в равной мере к планктону и бентосу, к растениям и животным
(рис. 87). Население неритической зоны океана дает две в значительной
степени биполярные арктические и частично бореальные группы, а также
две тропические, две бореальные и две потальные группировки, имею-
щие амфиокеаническое (амфибореальное, амфиатлантическое и амфипаци-
фическое) распространение.
160
Характер распределения организмов по зонам. По мере продвижения
от экваториального пояса в обе стороны, можно наблюдать резкую убыль
в количестве видов, населяющих океан (рис. 88).
Если принять общее количество видов морских животных равным
150 000, то оказывается, что в море Лаптевых обитает из них менее 0,3%,
а в водах Малайского архипелага не менее 27%, т. е. почти в 100 раз больше.
По мере продвижения из Арктики на юг меняется и состав фауны —
одни группы резко возрастают (ктенофоры, сифонофоры, головоногие мол-
люски, крабы, морские лилии и др.), другие возрастают слабее. Так, в море
11 Зенкевич, т. I
161
Лаптевых обитает 0,06% десятиногих ракообразных, 0,2% двустворчатых
моллюсков, 1,0% полихет. 0,7% иглокожих, 1,3% гидроидов, 0,05%
антозоев, 2,1% амфипод и 0,07% брюхоногих моллюсков, 0,18% рыб.
Есть основания думать, что для бентоса, так же как для планктона,
экваториальные зоны относительно беднее по количеству организмов,
чем зоны умеренных широт, то есть по биомассе изменения в меридио-^
нальном направлении идут в направлении обратном изменениям в видо-
вом обилии.
Миграции. Подавляющее большинство населяющих моря животных
способно к значительным перемещениям, пассивным или активным, перио-
дическим или непериодическим.
Большинство донных животных отрождает пелагическую икру и ли-
чинок, уносимых течениями часто на большие пространства, осуществляя
таким образом заселение новых пространств. Колоссальные массы зоо-
планктона каждые сутки совершают периодические вертикальные переме-
щения в несколько десятков, иногда сотен метров.
Очень многие организмы (и особенно десятиногие ракообразные, иногда
некоторые иглокожие и моллюски) совершают значительные сезонные пере-
мещения, большей частью к берегу и от берега.
Наиболее показательны нерестово-кормовые перемещения, совершаемые
лучшими пловцами моря — китами, рыбами, ластоногими и, возможно,
кальмарами. Эти миграции охватывают часто расстояния в тысячи ки-
лометров, обычно в направлении с севера на юг (сардина, сельдь, треска),
или по системам водоемов (пеламида, скумбрия, хамса в Средиземном,
Черном и Азовском морях), или в пределах отдельного моря (каспийские
сельди).
Все отмеченные явления в целом показывают присущую подавляю-
щему большинству хорошо подвижных морских животных способность
использовать в целях питания, размножения и расселения преимущества,
предоставляемые разными участками биосферы.
Кормовые миграции всегда направлены в те части моря, где обильнее
корм, нерестовые же туда, где более обеспечено наиболее успешное раз-
витие икры и мальков.
Имеется много данных, указывающих на то, что при низкой температуре
с большей эффективностью идет накопление жира в теле животного.
Таких животных, которые проводили бы всю свою жизнь в ограничен-
ном ареале галосферы, очень немного.
3. СОСТАВ МОРСКОЙ ФАУНЫ
Общее количество видов морской фауны, известных науке, можно
принять примерно в 140—150 тысяч. Среди морских групп животных
имеются и весьма обильные видами, как моллюски, ракообразные, кишечно-
полостные, но имеются и такие, в состав которых входит очень малое число
видов — Xiphosura, Phoronida, Echiurida, Priapulida и др. (табл. 55).
Интересное сопоставление численности разных групп фауны в разных
районах моря приводит Саутерн (Southern, 1915). К таблице Сау-
терна (табл. 56) мы добавили данные по Северному, Варенцову и морю
Лаптевых (цифры в скобках — дополнительные данные по другим авто-
рам).
Конечно, эти списки очень неполны, однако понятие о порядке чисел
и о соотношении отдельных групп они дают. При этом бросается в глаза:
1) большее разнообразие фауны по западную сторону Великобритании и
Ирландии, где подходят теплые атлантические воды и 2) обедненность фау-
ны у холодных побережий Канады.
<62
Таблица 55
КОЛИЧЕСТВО ВИДОВ МОРСКИХ ЖИВОТНЫХ
Группа Количе- ство видов Группа | Количестве* видов
Foraminifera 1200 Mysidacea >300
Radiol aria 4400 Comacea >400
Clliata 700 Euphausiacea 85
Porifera 4000 Amphipod a 3000
Hydrozoa 2700 Isopoda 3000
Scyphozoa 200 Stomatopoda 200
Anthozoa 6100 Decapoda 8000
Ctenopbora 80 Pantopoda 400
Nemerhni 1500 Xlpbosara 5
Nematoda 1500 Amphineura 630
Bryozoa 3000 Scaphopoda 200
Brachiopoda 120 Gastropoda 49000
Phoronlda 12 Pteropoda 90
Chaetognatha 30 Heteropoda 90
Pogonophora 1 Lamellibranchlata 10000
Polychaeta 4000 Cephalopoda Holothurioldea 400
Echlurida 20 650
Prlapullda 3 Asteroidea 1100
Sipuocullda 250 Ophiuroldea 1600
Ostracoda (вместе с пресновод- Echlnoldea 600
ными) 2000 Crlnoidea 800
Copepod а (вместе с пресновод- Tunlcata 1000
ными) 6000 Pisces около 16000
Clrrlpedia 500 Pinnipedia 32
Cetacea 58
Если принять, что действительное число видов на 50% превышает чи-
сла, приведенные в таблице, то можно определить видовое богатство для
морской фауны различных участков Северной Атлантики в 1500—3000 ви-
дов. Можно думать, что соответственно для Тихого океана эти величины
должны быть в 2—3 раза больше.
Значительно возрастает видовое богатство также и с продвижением
на юг, в тропические области. Так, например, у Канарских о-вов обитает
около 790 видов моллюсков (вместе с наземными). Из них морских двуствор-
чатых 147 видов, а брюхоногих 363 вида. Для Средиземного моря известно
около 500 видов рыб, из которых 144 сублиторальных, а в южной Японии
сублиторальных рыб 334 вида, в Австралии 344 вида. По обе стороны тро-
пической Америки живет около 600 видов крабов — около 250 видов
с тихоокеанской стороны и 350 — с атлантической. Для Индо-вест-паци-
фики известно около 5000—6000 видов моллюсков, около 400 видов
лилий, 86 видов ежей только из группы Cydaroidea и т. д. Интересно от-
метить, что некоторые группы находятся всегда примерно в одинаковом
числовом соотношении. Так, количество видов двустворчатых моллюсков
примерно в два раза меньше, чем брюхоногих.
Аналогичное явление можно наблюдать и на макрофитах по подсче-
там Т. Щаповой (1947) (табл. 57).
Всего для Тихого океана известно 3794 вида зеленых, бурых и красных
водорослей, причем тропическая зона в данном случае не дает такого рез-
кого видового преобладания, как для животных. В качественном отноше-
нии растения гораздо равномернее заселяют океан. Флора макрофитов
Малайского архипелага составляет всего 23% от всей флоры Тихого оке-
ана, а у Командорских о-вов обитает 5% макрофитов Тихого океана, у
Ю. Австралии — около 9%.
163
Таблица 561
Группа Район оз. Клэр, зап. побережье Ирландии Ирландское море Плимут
286 239 109
69 (+15) 54 (+1) 6 58 18
129 (+1) 6 121
8
оСурнолиа 24 24 34
45 27 62 (+2)
31 24 (+2) 35
76 136 ЮЗ (+1)
Toil ЛплНагш я f я ..«•••••••• 41 35 36
ИС111ИОО VI11* ала 253 88 (+2) 119 (+1) 3
10 1(+D
UCpiiyiva 65 57 (+1) 195 6
163 25
клОририид, 6 (4-1) 37 (+1) 95 10 10
24 30
129 52
14 17 6
Qnhi7nnnrla 22 16 24
65 61 71
"PnnfonoHa > 8 12 8
Lamelllbranchiala A mnlilnPllTA 1 90 7 108 (+3) 10 86 6
f’ я ct rnnod a ! 1S8 207 (+3) 164
Cephalopoda ! 5 ' 31 (+7) 45 (+14) 11 36
98 134 136
140 (+5) 17 16
• *Rpot4> 1929 1816 (+27) 1365 (+4)
1 Настоящая таблица имеет, конечно, только ориентировочное значение.
2 Koramlnifi ra.
Северное море 1 Баренцево море Море i Лаптевых | Вудсхолл 1 Канада, Атлантиче- ское побе- режье Средизем- ное морс Черное море
р 1902 46 2 99 (+5) 64 9 р.
40 135 8 12 (+7) 36 (+12) >110 42
96 87 36 132 (+8) 66
9 2 1 5(+D 5 1 208 } 44
15 20 3 14 (+3) 44
9 27 9 40 (+1) 9 J 93 79
9 19 (+3) 9 25 (+1) 20 (И) 65 18
70 220 io 76 (+5) 115 306 12
70 62 33 24 (+1) 71 101 4
271 143 (+70) 36 136 (+10) Ю5 (+1) 516 126
15 И 9 3 5(+D 29 0
? ? Р 26 29 (+9) 125 25
70 38 10 83 (+3) р 304 77
8 6 0 15 (+2) 10 43 4
80 41 8 25 (+3) 26 159 32
330 76 63 71 (+6) 70 (+4) 223 70
р 9 р 8 (+2) 11 22 12
р 21 2 5 7 40 22
90 25 5 51 (+4) 34 251 35
р 24 7 5(+D 11 47 5
170 64 23 70 (+6) 100 358 49
р 8 0 2 8 22 2
>150 150 32 129 (+10) 160 965 74
9 9 1 4 13 72 0
Р 50 24 22 (+10) 27 (+1) 2i'0 16
> ПО 114 28 247 (+6) 9 549 180
133 (+24) 14 383 185
1594 1542 (+-73) 376 1462 (+119) 1051(+29) 5191 1113
Части Тихого океана
Таблица 57
Количество
видов
макрофитов
Берингов пролив и о. Лаврентия............
Командорские о-ва.........................
Японское море (побережья СССР)............
Восточное побережье Японии................
Малайский архипелаг.......................
Южн. Австралия............................
Антарктика ...............................
54
174
269
445
860
340
81
Руководящие группы и формы. Среди тех 53 классов животных, кото-
рые обитают в море (табл. 58), лишь немногие, благодаря массовости
своего развития, играют существенную роль в динамике органического
вещества. К таковым следует отнести всего 20 классов (табл. 59).
Главная, ведущая роль в биологических процессах в море из этих 20
классов принадлежит только восьми. С другой стороны, из большого числа
видов, входящих в состав каждого из указанных классов, также только
немногие выделяются своим большим значением.Так, например, в зоопланк-
тоне северных частей Атлантического океана и Арктического бассейна
исключительную роль играет рачок Calanus finniarchicus (Gopepoda).
Несколько видов глобигерин (Globigerina ballot des, G. inflata и др.) и
широко распространенные кокколитофориды: Pontosphaera huxleyi, Сое-
colithophora jragilis, Calyptosphaera oblonga — главные образователи мощ-
ных отложений глобигеринового ила. Колоссальные саргассовые скопле-
ния макрофитов образованы всего несколькими видами, а столь же колос-
сальное скопление красных водорослей в Черном море — всего одним ви-
дом — Phyllophora nervosa и т. д.
Одним словом, если мы подходим к массовым проявлениям жизни в оке-
ане, мы всегда сталкиваемся всего лишь с несколькими формами, которым
и принадлежит главная роль в этих явлениях. Весьма вероятно, что 90%
всей совокупности биологических процессов, совершающихся в миро-
вом океане, обязаны всего нескольким сотням видов растений и животных,
вряд ли больше 1000. Для отдельного моря или части океана это количество
определяется несколькими десятками и едва ли достигает сотни. Это об-
легчает путь исследователя в изучении явлений биологической продуктив-
ности.
Таким образом, первый этап этих исследований — выявление руко-
водящих и характерных форм, достигаемое количественным учетом, вто-
рой этап — углубленное изучение этих форм. Хорошим примером тому
может быть всестороннее изучение рачка Calanus jinmarchicus, предпринятое
в целом ряде морских лабораторий. Изучается его питание, обмен, раз-
множение, темп роста, возраст и т. п.
Часть перечисленных выше групп морских животных в морях всех
широт распределена более или менее равномерно, т. е. примерно в одном
и том же соотношении ко всей фауне. К таким группам можно отнести
Foraminifera, Porifera, Hydrozoa, Nematoda, Nemertini, Polychaeta, Bryo-
zoa, Gastropoda, Lamellibranchiata, некоторые группы ракообразных
(Copepoda, Amphipoda, Isopoda, Cumacea), некоторые классы иглокожих
(Ophiuroidea, Asteroidea, Holothurioidea), некоторые группы рыб.
Другие группы, наоборот, очень слабо представлены или совсем от-
сутствуют в одних районах и обильно населяют другие, например свой-
ственные теплым морям крабы, все группы кишечнополостных, за исклю-
чением Hydrozoa, радиолярии, головоногие моллюски, усоногие, многие
165
Из 62 классов животных Встречаются в море i В основном живут в море Живут только в море Встречаются в пресной воде В основном живут в прес- ной воде Живут в прес- ной воде и, как правило, не встречают- ся в море •
5 7 30 6 1 __ 2
1 i Oligochae- te Insecta Arachnol- dea Reptilia Mammalia Spongla Hydro2oa Nemertinl Polychaeta Bryozoa Gastropoda Lamelllbran- chiata Dlcyemlda Orthonectlda Scyphozoa Antnozoa Ctenophora Kinorhyncha Echlurida Prlapulida Sipuncullda Brachiopoda Phoronoldea Entoprocta Chaetognata । Po^onophora Palaeostraca Solenogastres Amphlneura Scaphopoda Cephalopoda Asteroiaea j Echinoldea | Ophluroldea . Holothurioldea Crlnoldea i Enteropneusta I Appenalcularia | Ascidia ( Salpa Pterobranchla Acranla Spongla Hydrozoa Nemertinl Polychaeta Bryozoa Arachnoldea 1 i 1 1 1 Rotatoria Insecta Amphibia
1 Совершенно очевидно, что отдельные деления носят условный характер.
Таблыца 581
Живут и в море и в прес- ной воде Встречаются в воздушной среде В основном живут в воз- душной среде Живут только в воздушной среде Живут в прес- ' ной воде и в воздушной среде Живут в море, в пресной воде и в воздушной । среде
14 3 3 3 4 3
Flagellata Hlrudlnea Arachnol- Protra- Gastropo- Nematoda
Rhlzopoda Crustacea dea cbeata da Oligo-
Clllata Turbella- Reptilia Mvriapo- Insecta chaete
Suctorla ria Mammalia *da Amphibia Gastropo-
Turbellarla Gordlacea Lamelllbran- chlata G astro trIcha Hlrudlnea Rotatoria Crustacea Cyclostomata Pisces Tardigrada । ! 1 I i Aves Ollgochae- ta i j 1 i i i i i i da
Таблица 59 1
Группы животных, играющие существенную роль в динамике органического вещества в море Число видов Группы животных, играющие существенную роль в динамике органического вещества в море Число видов
Flagellata ? 6000 ! 700 4000 2700 , 200 6100 1500 1500 Poly chaeta 4000 3000 49000 10000 400 400 20000 5000 1000 16000 90
Rhizopoda Bryozoa ...............
Clllata ! P&rifera Hydrozoa Scyphozoa Gastropoda Lamellibranchiata Cephalopoda Pantopoda
Antnozoa Crustacea
Nematoda Echinodermata
Nemertini Tunicata ..............
Pisces Mammalia
группы иглокожих (например лилии), большинство оболочниковых. Наряду
с этим киты и ластоногие свойственны, в основном, высоким широтам обоих
полушарий и только кашалот обычен и в тропической полосе.
4. ОКЕАН — КОЛЫБЕЛЬ ЖИЗНИ
Жизнь зародилась в море. Распространено мнение, что океан яв-
ляется прародиной для пресноводной и наземной фаун. Мы уже ука-
зывали выше на большее разнообразие форм животных, обитающих в море,
по сравнению с сушей, включая пресные водоемы (табл. 58). В пресной
воде живет, не встречаясь в море, около 3% от всего количества клас-
сов животных (причем насекомые являются вторичноводными), а толь-
ко в воздушной среде — всего 8%, тогда как только в море—50% всех
классов.
Если же прибавить те классы животных, которые в основном живут
в море и в пресной воде встречаются немногими представителями (немер-
тины, полихеты, губки и пр.), то мы получим 61 %, а добавив и те, которые
обитают и в море и в пресной воде, будем иметь 87 % всех классов.
Свойства воды, благоприятствующие развитию жизни. Эта особенность
объясняется не только тем, что море, повидимому, представляет собою
колыбель жизни, но теми чрезвычайно благоприятными для развития
жизни свойствами, которыми обладает вода, по сравнению с воздушной
средой. Вода обладает очень высокой теплоемкостью, вследствие чего тем-
пературные колебания в природных водах относительно невелики (равно-
мерность распределения факторов среды). Это свойство воды имеет и гро-
мадное климатическое значение. Поверхностные воды тропических частей
океана поглощают массы тепла и, унося его затем в высокие широты, от-
дают воздуху, производя этим очень большой термический эффект, так
как теплоемкость воздуха по сравнению с водой очень мала.
Вода обладает исключительной способностью к растворению других
веществ, сохраняя при этом химическую инертность.
Правило гомотопности. Заслуживает большого внимания также и тот
факт, что отдельные группы приурочены к какой-нибудь одной определен-
ной среде. Если мы возьмем более мелкие таксономические единицы, чем
класс (отряды, семейства), процент групп, заселяющих разные среды,
1 Набраны жирным шрифтом наиболее значимые группы, курсивом—средние по значимо-
сти.
167
резко упадет. Совершенно особенное место в этом отношении занимают
только классы рыб, ракообразных и нематод, захвативших, благодаря
исключительно прочной кутикуле и некоторым другим особенностям, все
среды обитания, но и у них в более дробных группах правило гомотопности
находит свое подтверждение. При этом следует отметить, что наиболее
значимые в филогенетическом отношении группы, как ктенофоры, поли-
хеты, первично- и личиночнохордовые, бесчерепные и др. являются оби-
тателями моря.
В качестве другого доказательства происхождения воздушной и прес-
новодной фаун от морской указывают на солевой состав крови, предста-
вляющий собою как бы наследие морского обитания предков.
В этом отношении особенный интерес представляет развиваемый в
последнее время некоторыми исследователями (Ромер и Г р о в,
Ватсон, Л. Берг) взгляд, что вся группа рыб имеет пресно-
водный генезис, и все морские рыбы ведут свое начало от пресноводных.
Ромер и Гров приводят замечательную табличку подсчета соотно-
шения морских и пресноводных рыб в различные геологические эпохи
(табл. 60).
Таблица 60
। Пресновод-
• ные в %
Морские
в 7о
Силур ..........................
Нижний девон....................
Средний » ......................
Верхний » ......................
100
13
29
0
23
87
71
Как указывает Л. Б е р г, основной переход рыб из пресных вод в море
можно приурочивать к середине девона. Необходимо, однако, выяснить
один очень существенный момент: мы пока не знаем в пресной воде ни
одного организма ни современного, ни ископаемого, от которого могли бы
развиться рыбы. Все без исключения известные нам низшие хордовые и.
родственные им группы представляют собой морских животных, чрезвы-
чайно болезненно относящихся к понижению солености морской воды.
Какой-то предок рыб все равно должен был переселиться из моря в прес-
ную воду, но об этом мы пока ничего не знаем, никаких следов от этой груп-
пы не сохранилось.
Если взять основные группы морской фауны — губок, кишечнополост-
ных, немертин, полихет, ракообразных, моллюсков, мшанок, иглокожих,
оболочниковых и рыб, то только ракообразные и рыбы образуют группу,
обильно представленную и в пресной воде часто близкими родами, даже
видами. Очевидно, для первых плотный хитиновый панцырь, для вторых
чешуя, многослойный эпидермис и покрывающая тело слизь сыграли
благоприятствующую роль при переходе в пресную воду. В противопо-
ложность морю, главную массу животного живого вещества суши и прес-
ных вод составляют насекомые, клещи и пауки из членистоногих и олиго-
хеты из червей. Значительно уступают им рыбы, птицы и млекопитающие.
Такое несходство в распределении животных в море и в остальной биосфере
объясняется тем, что лишь немногие обитатели моря смогли захватить
пресную воду и воздушную среду; те же группы, которые сумели это
сделать, благодаря тем или иным сложным изменениям, в дальнейшем
захватили воздушную и пресноводную среду и дали в них большое видо-
вое разнообразие. Следует также отметить, что основные по массе группы
пресноводного бентоса — личинки насекомых, большинство брюхоногих
168
моллюсков и, возможно, малощетинковые черви и пиявки, некоторые тур-
беллярпи являются вторичноводными, т. е. все они попали в прес-
ную воду, перейдя в нее из воздушной среды. Вторичноводные орга-
низмы в море, если говорить о группах, имеющих массовое значение,
представлены лишь млекопитающими, и только в морях с пониженной
соленостью приобретают некоторое значение и другие группы.
Происхождение пресноводной и наземной фаун. Вопрос о том, какими
путями шло выселение организмов из моря и захват других биотопов,
пока выяснен еще очень недостаточно. Морские беспозвоночные в подав-
ляющем большинстве пойкилосмотичны, т. е. их внутренняя соленость
и солевой состав крови очень близки к морской воде. Наземные и пресно-
водные организмы, в отличие от морских, гомойосмотичны, т. е. сохраняют
постоянную солевую концентрацию, вследствие наличия у них особых
осморегуляторных способностей. Выйти из родной морской стихии могут
только те организмы, которые выработали в себе эти способности. Во вся-
ком случае путь непосредственно через солоноватые воды из моря в прес-
ные воды отнюдь не является единственным и даже, возможно, главным.
Переход в пресную воду, так сказать, водным путем, для подавляющего
числа групп животных невозможен, очевидно, в первую очередь из-за
тех нарушений в солевом составе полостных и тканевых жидкостей, кото-
рые происходят, при изменении наружной солености у животных, тело
которых не имеет на поверхности плотных защитных оболочек. Это заклю-
чение можно сделать потому, что те группы, которые несомненно и много
раз переходили из одной водной среды в другую — рыбы и ракообразные —
обладают хорошо защищающими их покровами, непроницаемыми для воды
и солей. Нематоды, широко распространенные и в море и в пресной воде,
также имеют непроницаемые покровы.
Однако даже для хорошо защищенных покровами организмов переход
из моря в пресную воду чрезвычайно труден, так как к этому имеются и
другие препятствия. Н и д г эм (Needham, 1930) полагает, что для раз-
вития в воде яиц и личинок морских организмов необходимо определен-
ное количество минеральных солей (например, фосфора), которые в прес-
ной воде отсутствуют.
С другой стороны, переход в пресную воду связан с приспособлением
к водоемам малого размера и, как правило, сопровождается выпадением
личиночных стадий и переходом к криптометаболии. Наконец, в пресной
воде гораздо труднее идет процесс дыхания, нежели в морской.
Переход из моря в пресную воду водным путем только частично мог
совершаться через устьевые зоны рек. Для большинства перешедших та-
ким образом животных можно установить, что этому способствовало дли-
тельное (геологически) существование солоноватоводных бассейнов, раз-
вивавшихся из морских и претерпевавших значительное или даже полное
опреснение. Вот в таких обширных водоемах, по ряду моментов сходных
с морскими, в результате чрезвычайно медленно совершающегося опрес-
нения, происходила переработка части фауны в солоноватоводную и прес-
новодную. Такие примеры дают нам наши южные водоемы, Арктический
бассейн и Балтийское море.
Гораздо большее значение в заселении животными воздушной и прес-
новодной среды имела литоральная фауна, вырабатывавшая в течение мно-
гих миллионов лет приспособления к более или менее длительному пре-
быванию в воздушной среде. Таким путем должны были формироваться
предки многих современных наземных животных (брюхоногих моллюсков,
паукообразных, насекомых). Часть из них в дальнейшем вторично проникла
в пресную воду (Insecta, Pulmonata), а некоторые даже в морскую (млеко-
питающие, рептилии). Некоторые организмы могли совершать выход из
169
моря и иным путем. Фауна грунтов литорали, являясь сама производным
фауны морских подводных грунтов (галоэдафона), могла в свою очередь
давать начало фауне воздушных почв (аэроэдафону), а в дальнейшем и
фауне грунтов озер (лимноэдафону). Таким путем, вероятно, возникли
гастротрихи, тубеллярии, олигохеты и, может быть, некоторые ракообраз-
ные, первичнотрахейные, многоножки и пресноводные двустворчатые мол-
люски. Наконец, имеется еще один своеобразный биотоп, где легче про-
исходил переход из морской воды в пресную, — это пещеры.
Весьма вероятно, что для некоторых форм через ту же морскую лито-
раль легче мог происходить и переход из морской воды в пресную, через
амфибиальное существование. Организм при этом вырабатывает способ-
ность стабилизировать внутреннюю соленость в воздушной среде и тогда
для него легче становится переход в пресную воду. Возможно, что именно
таким путем совершался переход в пресную воду жабернодышащих брю-
хоногих моллюсков (например, сем. Hydrobiidae) и некоторых высших
ракообразных (Amphipoda, Isopoda, крабы).
5. ВОПРОС О ПРОИСХОЖДЕНИИ И РАЗВИТИИ ЖИЗНИ НА ЗЕМЛЕ
Постановка вопроса. В наших представлениях об истории развития
жизни на земле имеется много существенных пробелов. На вопросы о том,
как и когда возникла жизнь на земле, мы не имеем еще вполне научно обос-
нованной и общепринятой концепции, которая могла бы разъяснить основ-
ные трудности и неувязки. Если мы в общем представляем себе эволюцион-
ный путь развития растительных и животных форм, то мы почти ничего не
знаем о том, как развивались в целом явления жизни в биосфере в преж-
ние геологические эпохи, и какие формы они принимали. Мы совсем ничего
не можем сказать о том, как изменялось общее количество организмов в био-
сфере, как усложнялась система взаимоотношений между основными биоло-
гическими группами населения биосферы — продуцентами, консументами
и редуцентами — и как создавалась та система циклических процессов
в биосфере, которую мы знаем ныне.
Мы не можем здесь останавливаться подробно на вопросе о возникно-
вении жизни на земле и хотим отметить только, что эта проблема до тех пор
не будет достаточно разъяснена, пока нам не будет ясно, что собою пред-
ставляют и как возникли основные функции живого — обмен, рост, размно-
жение, ощущение и движение. Пока что о природе этих функций мы знаем
очень немного. Даже столь блестящие работы, как исследования Э н-
гельгардта по миозину, пока не разъяснили нам самой сущ-
ности процесса сокращения. Трудно объяснить происхождение того, сущ-
ности чего мы пока еще не понимаем.
Возраст земли. Очень мало может удовлетворить биологов и современ-
ное решение вопроса о том, когда возникла жизнь на земле. А. Ферс-
ман (1946) считает, что «физики и химики... наконец, нашли точные часы»
времени, которые дают возможность измерить длительность существования
земли, как самостоятельного космического тела. Эти «точные часы пла-
нетного времени» заключаются в так называемом «свинцовом методе»,
наиболее совершенном из всех ныне существующих. Металлы уран и
торий еще с того времени, когда земля находилась в огненно-жидком со-
стоянии, неизменно и с одним и тем же темпом разрушаются и из них обра-
зуются гелий и свинец. Оказывается, что за 100 миллионов лет 1000 г
урана должны дать 13 г свинца и 2 г гелия. Г. Г а в е ш и (1935) говорит,
что «с начала образования уранового свинца в огненно-жидкой земной
массе... должно было пройти... около 3000 миллионов лет... Одновре-
менно мы можем также считать 3000 миллиона лет за верхний предел
170
возможного возраста твердой земли. Однако возможно, что фактический
возраст лежит ближе к нижнему пределу — 1600 миллионов лет, — чем
к вышеупомянутому числу». Нижний предел — это тем же способом рас-
численный возраст циркона из Мозамбика, соответствующий по возникно-
вению нижнему прекембрию.
Многие геологи (напр., К. Ш у х е р т, 1931), минералоги (В. Вер-
надский, 1935) и геофизики сходятся на возрасте земли с начала па-
леозоя в 500—700 миллионов лет.
Также и А. Ферсман считает, что «около 3 миллиардов лет отде-
ляет нас от того момента космической истории, когда зародилась наша
солнечная система, т. е. когда от солнца отделились планеты и среди них
наша Земля, в виде раскаленной массы паров и газов... не менее 1 * или
2 миллиардов лет прошло от зарождения первой жизни на земле». Боль-
шинство геологов считает, что со времени начала кембрия прошло 700 мил-
лионов лет. Итак, по общему мнению, найдено точное исчисление возраста
Земли и основных стадий ее истории (табл. 61).
Таблица 61
Стадия истории Земли
Возраст в
миллионах лет
Палеонтологическое время (с начала кем-
брия) .................................
Археозой (с момента формирования земной
коры)..................................
Доархеозойское время...................
Общий возраст Земли как само-
стоятельного космического тела . . .
700
1400
900
3000
Время зарождения жизни на Земле. Теперь сделаем попытку уложить
в эту хронологию то, что нам известно об эволюционном развитии организ-
мов на Земле.
Если определять, как это делают сейчас геологи, время «от зарождения
первой жизни на Земле» в 1——2 миллиарда лет, то на период, предше-
ствующий кембрию, приходится примерно 700—1300 миллионов лет.
В эти 700—1300 миллионов лет нужно уложить всю предшествующую
кембрию историю развития живого от формирования «коацерватов» (О п а-
р и н, 1936). Развитие животных на Земле можно условно представлять
в виде 4 основных последовательных фаз (рис. 89):
1) от неорганических веществ до коацерватов;
2) от коацерватов до простейших;
3) от простейших до хордовых;
4) от первых хордовых до нашего времени.
В начале кембрия на Земле уже существовали все 11 типов. Весьма
вероятно, что уже существовали и примитивные позвоночные, так как
в силуре уже существовали панцырные рыбы, а во второй половине силура
уже появились амфибии. С кембрия известны уже фораминиферы, радио-
лярии, губки, кишечнополостные, брюхоногие и головоногие моллюски,
кольчецы, трилобиты, палеостраки, низшие и высшие ракообразные,
мшанки, плеченогие и иглокожие. Совершенно очевидно, что еще в архео-
зое выработалось все основное многообразие животных обитателей моря.
Основная эволюция животных (да и растений тоже) в послекембрийскоег
время не выходит за пределы уже имевшихся типов и связана свозникно-
171
вением в воздушной среде ряда групп (паукообразные, насекомые, репти-
лии, птицы и млекопитающие).
Темп эволюции. Если же взять морскую среду, то за эти 700 миллионов
лет эволюция животных форм совершалась не столь быстро, чтобы мы мо-
гли представить себе и предшествующее эволюционное развитие в соот-
ветствующих темпах.
Во всяком случае время, понадобившееся для развития от коацерва-
тов до хордовых, должно было быть в десятки раз более длительным, чем
то, которое понадобилось для послеархеозойской эволюции. Мы сделали
несколько наивную попытку изобразить в схеме (рис. 89) соотношения тем-
Рис. 89. Схема длительности существования отдельных этапов
эволюции жизни на земле (по Зенкевичу).
па эволюции и хронологии истории Земли. Однако при^всей наивности
этой схемы она заставляет поставить очень существенный вопрос.
Наклон сплошной линии в правой графеусловно соответствует темпу эво-
люции с кембрия. Если тот же темп эволюции экстраполировать влево, то
при самых благоприятных (для согласования с хронологией Земли) до-
пущениях весь эволюционный путь животного мира должен занять время,
значительно превышающее 10 миллиардов лет. Для того чтобы уложить
всю эволюцию животных от неорганических соединений в принимаемое
геофизиками, геологами и минералогами летоисчисление, мы должны
допустить совершенно иной темп эволюции в докембрийское время. Есть
ли к этому основания? Можно ли объяснять ускоренный темп эволюции
более высокой температурой? Вряд ли, так как и в настоящее время у нас
существуют в океанах широкие тропические зоны, где температура поверх-
ностных вод близка к 30°.
Все изложенные соображения приводят к совершенно несомненной и
очень сильной неувязке между необходимой для эволюции живого длитель-
ностью времени и тем временем, которое предоставляют для этой эволюции
новейшие изложенные выше данные. Мы можем сделать только два до-
пущения — или признать, что в докембрийское время эволюция живот-
172
ных совершалась в 10—20 раз более быстрым темпом, к чему у нас нет ни-
каких данных, либо признать неправильность современного летоисчисле-
ния истории Земли. Необходимо обратить самое серьезное внимание на
это пока непримиримое противоречие.
Существенное удлинение срока существования жизни на Земле дает
новая теория происхождения Земли П. Шмидта (1946), допускающая
существование твердой земной коры в течение 7 миллиардов лет. Однако
и такой срок недостаточен для того, чтобы уложить в него всю историю
жизни. Нельзя не согласиться с Л. Давиташвили (1943), что «орга-
нический мир... автотрофных растений, способных к фотосинтезу (под-
черкнуто мною Л. 3.) формировался в течение колоссального промежутка
времени», если даже не рассматривать первых этапов развития живых
существ на Земле.
Эволюция живого населения биосферы. Не меньшее значение имеет
также и вопрос вступления на жизненную арену новых групп организмов
и связанные с этим изменения в биосфере.
Так, Г и р м е р в своем Руководстве палеоботаники (1927) указывает,
что в то время, как почти все высокоорганизованные животные, видимо,
существовали в археозое, развитие растительных форм шло значительно
позднее.
Наиболее древними группами морских растений являются сине-зеле-
ные, известные с докембрия. Зеленые и красные водоросли возникли в
кембрии (о р д о в и ц и й), бурые — с силура, но из них Chordacea
и Fucacea известны только с триаса, диатомовые и перпдиниевые — с юры,
Laminariacea с палеогена, a Desmarestiacea с неогена.
Как мц должны представлять себе, если верны эти данные, круговорот
веществ в море без основных продуцентов современного океана — диато-
мовых, перидиней, ламинариевых и фукусовых? Кто в палеозойском море
выполнял их роль?
За последние годы в советской литературе возникли две попытки пред-
ставить себе эволюцию живого населения биосферы, развитие его массы,
взаимосвязей между частями и характера основных циклических про-
цессов. Мы имеем в виду работы Б. Л и ч к о в а (1945) и Л. Давита-
швили (1943).
Б. Л и ч к о в не усматривает в эволюции живого населения поверх-
ности Земли единого и целостного эволюционного процесса развития от
менее сложного и дифференцированного к более сложному и дифференци-
рованному. Б. Л и ч к о в приходит к выводу, что «на протяжении истори-
ческой части бытия нашей планеты сменилось шесть геологических циклов,
из которых... каждый начинался расцветом органической жизни, а закан-
чивался фазой ее кризиса. Цикл можно разделить на три фазы: леднико-
вую, фазу с максимально контрастным рельефом и максимальной денудаци-
ей, умеренную, когда рельеф падал несколько в своей интенсивности и
падала денудация, приобретая средние размеры, и ксеротермическую, когда
интенсивность рельефа была минимальной и минимальной была водная
денудация. Засушливые ксеротермические фазы и были фазами вььмира-
ний животного мира и сокращения растительности... Вымирание в ксеро-
термических фазах охватывало не все население планеты, а касалось менее
пластичных форм живых существ; формы жизни более пластичные в это
именно время выдвигались вперед и с этой точки зрения фазы вымирания
являлись фазами усиленного творчества новых порядков и классов в жи-
вотном мире; это были вместе с тем фазы творчества нового. Для леднико-
вой эпохи, сменявшей ксеротермическую, характерно уже не столько
вымирание, сколько переселения живых существ. Наконец, в среднюю
фазу осуществлялось нормальное видообразование». Эта своеобразная
173
«теория катастроф» Б. Л и ч к о в а, однако, рассматривает процесс раз-
вития населения биосферы не как единый процесс, так сказать, по
спирали, а разбивает его на ряд циклов.
Очень интересную попытку дать целостную интерпретацию эволюцион-
ного развития живого населения поверхности Земли с позиций дарвинизма
делает Л. Давиташвили в своей работе «Дарвинизм и проблема
накопления горючих ископаемых» (1943). Если даже многое в концепции
данного автора пока еще гипотетично, то и общие положения, из которых
исходит автор, и оформление проблемы, и отдельные вопросы, на которые
автор разбивает проблему, представляют для нас громадный интерес. Это
именно раздел палеобиологии, который разъясняет, как исторически сло-
жилась современная система биологических явлений в биосфере; частью
его является и палеобиоокеанология. Л. Давиташвили ставит перед
геологией следующие вопросы: место и формы возникновения растительных
организмов, пути расселения и преобразования растительного покрова
океана и воздушной среды, в частности, время и формы выселения растений
в воздушную среду, соотношение аутотрофной и гетеротрофной групп
организмов (продуцентов и консументов) и количество той и другой в раз-
личных зонах мирового океана, в более или менее обособленных морских и
лагунных бассейнах, а также в различных областях материков.
В результате соответственного анализа имеющихся пока еще весьма
недостаточных фактов Л. Давиташвили приходит к выводу, что
«древнейшие гетеротрофные организмы, разрушавшие накоплявшееся
на дне морских бассейнов органическое вещество, действовали в этом на-
правлении в общем несравненно медленнее и слабее, чем организмы со-
ответствующих групп, живущие в настоящее время». В отношении про-
цесса накопления на дне древних морей органического вещества данный
автор полагает, что при малом количестве гетеротрофных организмов
донное накопление могло совершаться при обыкновенном режиме
в мелководной полосе. В современном океане это возможно только
в условиях особого газового режима, при котором гетеротрофные
организмы не могут существовать (напр. H2S). В морях же и океанах
с нормальным режимом, в результате чрезвычайно длительного про-
цесса эволюции и некоторых особенностей морской среды, использо-
вание органического вещества гетеротрофными организмами достигло
чрезвычайно высокой степени.
На суше сравнительно молодая гетеротрофная часть населения, воз-
можно, еще не выработала достаточно напряженной системы использо-
вания растительной массы.
Если принять, что до конца докембрия первичным биотопом, обильно
заселенным жизнью, была мелководная хорошо прогреваемая зона теплых
морей, то отсюда жизнь распространилась и в другие зоны океана и на сушу.
В докембрии поверхность материков еще не была заселена растительностью.
Первая из известных наземных флор — псилофитовая «характерная для
нижнего и среднего девона, но появившаяся, по всем признакам, не позже
верхнего силура, была флорой «земноводной», связанной в своем распро-
странении с полосой низменной суши, непосредственно прилегавшей к во-
доемам, и образовывавшей массу в десятки тысяч раз, а может быть, в сотни
тысяч раз меньше биомассы нынешней наземной флоры. Растительность
каменноугольного периода заселяла также небольшую часть суши в низ-
менных болотистых местностях, чаще всего примыкавших к морям. Ге-
теротрофное население суши в то время было еще биологически слабо и
проходило только первые фазы своего развития. В дальнейшем наземная
флора распространялась все более в глубь материков, а гетеротрофное1 на-
селение приобретало все большее значение в круговороте органического
174
вещества, хотя еще и в настоящее время далеко не достигло возможного
предела». .
Эта стройная концепция, которую можно принять как плодотворную
рабочую гипотезу в изучении прошлого живого населения биосферы,
исходит из того основного положения, что современное распространение
основных групп населения биосферы и количественное соотношение в уча-
стии их в основных био-
логических процессах,
совершающихся в био-
сфере, есть только по-
следний этап сложного
эволюционного процес-
са, от простых мало диф -
ференцированных си-
стем соотношений к
сложным и весьма диф-
ференцированным.
Интересную попыт-
ку сравнить продук-
цию современного Кас-
пийского моря с Сар-
матским бассейном сде-
лал Б. П е р сидски й1.
Б. Персидский под-
верг анализу створки
диатомовой одноклеточ-
ной водоросли Actinocy-
clus Ehrenbergi в слоях
диатомита из окрестно-
стей Новороссийска,
соответствующих Са р-
матскому бассейну (рис.
90). Как известно, в
весеннее время створки
диатомей имеют наи-
большие размеры. В
летнее время, при веге-
тативном размножении,
делящиеся клетки мель-
чают, так как новая
половинка створки всег-
да меньше старой. Од
нако такие деления с
уменьшением размеров
клеток прекращаются в
Рис. 90. Сверху — чередование мелких и крупных ство-
рок диатомеи Actinocyclus Ehrenbergi в слоях диатомита
сарматских отложений окрестностей Новороссийска (по
Персидскому). Снизу—слоистые диатомиты из Ахал-
цыха (Грузия) (фото Б. Персидского).
размеры. Таким образом, в слоистых диато-
осеннее время, когда на-
ступает иной процесс
размножения с образо-
ванием ауксоспор - кле-
ток, восстанавливаю-
щих первоначальные
митах можно установить наличие чередующихся слоев с крупными
и мелкими створками Actinocyclus, очевидно соответствующих раз-
1 Сообщено Б. Персидским устно.
175
Рис. 91. Продукция фитопланктона Сар-
матского моря (по Персидскому).
1 — Rhizosolenia calcar — avis, в г, 2 — Chaeto-
ceras в млн. экз., 3 — Actinocyclus Ehrenbergi
в млн. экз.; все на i ju- дна.
личным сезонам года. Это дает возможность точно хронологизировать
слоистость диатомита и учесть по количеству «весенних» створок Actino-
cyclus весеннюю продукцию фитопланктона. Б. Персидский уста-
новил, что в сарматском диатомите в «белых» прослойках количество Acti-
nocyclus Ehrenbergi (по весу) должно быть раз в 10 больше, чем в «серых»
прослойках, линейные же размеры отдельных клеток белой и серой про-
слойки относятся как 1 к 5. Таким образом удалось установить не только
качественный и количественный состав фитопланктона Сарматского моря,
но и получить количественное выражение годовой продукции за ряд по-
следовательных лет. Эти величины
близки к современным (рис. 91).
Аналогичное использование проб
отложений древних морей в более
широком масштабе из различных
геологических периодов может дать
возможность восстановить путь и
формы эволюционных изменений в
явлениях биологической продуктив-
ности океана.
Особенно многообещающи в этом
отношении те колонки грунта океа-
нов, которые могут быть теперь по-
лучены с применением новых грун-
товых трубок.
С другой стороны к той же проб-
леме эволюционных смен в раститель-
ном и животном населении Земли
подходит А. Виноградов в своем
капитальном сочинении о химиче-
ском элементарном составе организ-
мов моря (1944). Данный автор преж-
де всего приходит к выводу, что хими-
ческий состав отдельных систематических групп древнейших геологиче-
ских периодов и современной эпохи в основных чертах оставался тем же
самым. Так, химический состав костей девонских Ostracodermi иденти-
чен костям современных Teleostei, также идентичен химический состав
вымерших Meros to ma ta и современных Arthropoda и т. д.
Однако смена в составе населения, преобладание в разные геологиче-
ские периоды разных групп организмов обусловили определенный ход
изменений в химическом составе населения биосферы в целом. Из наиболее
существенных особенностей этих смен прежде всего следует указать на то,
что у животных докембрия отсутствовали известковые скелеты. Объясне-
ние этому можно усматривать, по А. Виноградову, в том, что в
атмосфере (и в воде океанов) того времени, видимо, было значительно выше
содержание СО2. Это затрудняло формирование известковых скелетов
у животных, но не препятствовало образованию их у растений, так как
последние редуцируют бикарбонат кальция в СаСО3 (потребляя не только
СО2, но и СО3" и НСО3), а для животных необходим самый СаСО3, которого
из-за большого количества СО2 было мало.
Наиболее древним типом органического скелета является хитиновый
(Merostomata, Graptolithoidea, большинство Bryozoa, Trilobita) или «бел-
ковый» (белковые опорные ткани ) (Porifera, Coelenterata, Brachiopoda,
Vermes, Echinodermata). На белковой основе органического скелета возни-
кают минеральные скелеты, и в каждой геологической эпохе мы находим
свои типичные группы растений и животных. На основе «белкового»
176
скелета развился и белково-апатитовый скелет (Vertebrata). Для современ-
ной эпохи наиболее характерны хитиновые опорные ткани членистоногих
и белково-апатитовые скелеты позвоночных.
G. ПОНЯТИЕ БИОЦЕНОЗА И ОСНОВНЫЕ МЕТОДЫ ЕГО АНАЛИЗА
Основные проблемы биоценологии. Углубленное изучение сложной
системы закономерностей, определяющих все стороны совместного существо-
вания совокупности живых организмов, населяющих определенный уча-
сток биосферы, как целого, представляет собою важнейшую научную
проблему и центральную задачу синэкологии (биоценологии).
Потребность в таком изучении диктуется все более и более проявляю-
щимся воздействием хозяйственной деятельности человека на окружаю-
щую природу, все более растущим использованием человеком животного и
растительного сырья.
Знания эти нужны для того, чтобы научиться управлять органической
природой, чтобы привести существующую систему использования природ-
ных ресурсов, через организованную систему реконструктивных меро-
приятий, на путь наиболее рационального хозяйственного их использо-
вания.
Основные объекты этого изучения следующие:
1. Закономерности структуры. Качественное и количественное изуче-
ние состава биоценоза. Зависимость его от окружающих условий.
2. Система взаимоотношений между отдельными компонентами био-
ценоза и составляющими его биологическими группами.
3. Характерные особенности биоценоза, как целого.
4. Формирование биоценозов и характер изменений, происходящих
в них во времени под влиянием внутренних и внешних факторов.
5. Характер изменений в биоценозе, происходящих под влиянием воз-
действия хозяйственной деятельности человека.
6. Система возможных, со стороны человека, мероприятий реконструк-
тивного характера.
Биоценоз и биотоп. Два основных понятия экологии, с которыми при-
ходится сталкиваться при изучении фауны водоемов,это биоценози би-
отоп. Фридерикс (Friederichs, 1927) основной единицей
биоценологии считает голоцен. «Голоцен — фактор единства — комбинация
местных факторов, объединенная взаимодействием их друг с другом»...
«можно также говорить... о жизненном сообществе, как о биоцене и, в про-
тивоположность ему, о всей неживой части голоцена, как об абиоцене».
То же самое Элтон (Elton) именует экосистемой, Гек (Heck) —
ценом. Биоцен Фридерикса соответствует биоме или био-экологиче-
ской формации Шелфорда (Shelford, 1940).
Гагмейер и Линке (A. Hagmeier, 1930, О. Linke,
1939) дают следующие биоценотические подразделения для морского бен-
тоса. Они различают, во-первых, эпибиоз (фауна над грунтом) и эндобиоз
(фауна в грунте). Такое подразделение бентоса на две самостоятельных
системы биоценозов принимают и некоторые другие авторы (С. J. Р е-
t е г s е п 1911, 1914 и его ученики, G i s 1 е п 1930 и др.).
В качестве самой мелкой биоценотической единицы принимают иногда
фаунулу (Friederichs, 1930). Это несамостоятельная часть био-
ценоза, подвергающаяся различным изменениям, без того, чтобы суще-
ственно изменялся весь биоценоз. Однако фаунула может быть подразде-
лена на еще более мелкие деления — «поселения», обычно одновидовые.
Их можно было бы еще назвать популяциями. Несколько поселений
могут быть объединены еще в биоценозы низшего порядка, отличающиеся
12 Зенкевич, т. I
177
главным образом тем, что та или иная массовая форма становится господ-
ствующей. Таким образом, в нисходящем порядке получается следующий
ряд: биоценоз I порядка-> фаунула-> биоценозы II порядка—► поселе-
ние. В содержании, вкладываемом в приведенные понятия биоценоза и
биотопа разными исследователями, имеется много условного, противоре-
чивого и ошибочного. Проистекает это от того, что, как правильно ука-
зывает Д. Кашкаров (1938), «в науке еще слишком мало разработано
это понятие (биоценоз), слишком мало вскрыто его реальное, конкрет-
ное содержание». Как известно, термин биоценоз был впервые сформули-
рован в 1877 г. немецким биологом Мебиусом на примере устричной
банки, если не считать более ранних, еще недостаточно выработанных по-
нятий, дававшихся применительно к морской фауне Ф о р б с о м (Е. For-
bes, 1844) иЛоренцом (Lorenz, 1865).
По Мебиусу, биоценоз — это сообщество живых существ, «кото-
рые соответствуют решительно всем данным внешним условиям, которые
взаимно обусловливают друг друга и путем размножения поддерживают
свое существование в определенной области». В настоящее время нас
уже не может удовлетворить подобное определение.
Мы здесь не можем входить в детальное рассмотрение дальнейшей исто-
рии развития понятий биоценоза и биотопа и ограничимся только некото-
рыми указаниями. С очень большой тщательностью и подробностью весь
вопрос изложен в «Основах экологии» Д. Кашкарова (1938).
Иногда два эти понятия определяются взаимно одно другим: био-
ценоз есть население биотопа, биотоп есть часть пространства, заселенная
определенным биоценозом. Такие определения мало что дают. Если же
исходить из одного из этих двух понятий, то необходимо создать класси-
фикацию исходного понятия. В большинстве исходят от биотопа и пытаются
создать иерархическую систему для него. Биотопом высшего порядка будет
вся биосфера, как среда жизни нашей планеты. Ему соответствует все
население биосферы, биоценоз высшего порядка, или, как его иногда назы-
вают, геомерида (Беклемишев). Затем следуют категории низшего
порядка — биоциклы, биохоры, биотопы, констации и, наконец, стации,
как элементарные единицы среды, так сказать, линнеоны этой системы.
Д. Кашкаров, давая этому оценку, говорит: «Получается путани-
ца, в которой невозможно разобраться». Трудность проистекает оттого,
что границы между этими категориями совершенно произвольны и что
может быть добавлено любое число и промежуточных и более низших
категорий. С другой стороны, совершенно невозможно сравнивать одно-
именные категории в разных системах биоценозов. Возникающие при этом
трудности усугубляются тем, что большинство экологов в классификации
биоценозов исходит как раз от кажущейся им более определенной системы
биотопов. Но эта определенность только кажущаяся. В действительности,
в природе мы не находим иерархической системы биотопов, исходя из кото-
рой можно было бы создать иерархическую систему и для биоценозов.
Также неправильно было бы пытаться построить систему биоценозов без
учета факторов среды.
Правильнее всего рассматривать совокупность организмов и окружаю-
щих условий в той тесной взаимосвязи, которая фактически и существует,
не отрывая одного от другого. Именно это К. Дерюгин первоначально
называл фацией, разумея под ней (применительно к бентосу) комбинацию
определенного типа грунта с определенным комплексом животных (оче-
видно, и растительных. Л. 3.) форм, т. е. принимал термин фация в смысле
Вальтера (1893—1894).
Очевидно, исследуя тот или иной участок биосферы и установив от-
носительное единообразие в населении и условиях среды, мы имеем осно-
178
вание различать разные типы и формы биоценозов и биотопов и подрази
делять население на более мелкие группировки. Мы можем их объединять
в более крупные, в зависимости от того, с какой детальностью мы будем
изучать данный участок биосферы и его население, исходя из конкретного
состава населения, взаимоотношений каждой формы с условиями среды
и с другими формами, населяющими данный биотоп, во всем многообразии
этих связей, не стремясь искать единых систем и подразделений для раз-
ных частей биосферы.
Очень часто, особенно в старой литературе, склонны были трактовать
биоценоз, как сообщество, что нашло, однако, уже соответствующую кри-
тику в нашей советской литературе. Очень многие наши исследователи
(зоологи) склонны не усматривать различий в понятиях биоценоз и
население. К. Дерюгин (1915) прямо указывает, что употребляя
термин биоценоз, имеет в виду «не сообщество, а лишь комплекс форм или
население, соответствующее той или иной фации». Очень близок к такому
пониманию биоценоза и С. Зернов (1912), считающий его крайне под-
ходящим и удобным для обозначения тех комплексов животных и расти-
тельных форм, «которые мы постоянно встречаем в данном море в самых
разных его пунктах, при наличности одинаковых условий существования».
Нередко зоологи склонны совсем отказываться от термина биоценоз,
заменяя его чисто формальными терминами комплекс или группировка
населения.
С другой стороны, делались необоснованные попытки разделить поня-
тия населения и биоценоза: все формы, обитающие на данном участке, со-
ставляют население, а биоценоз это совокупность форм, связанных пря-
мыми или косвенными отношениями друг с другом. Население — не обя-
зательно биоценоз; могут быть участки без биоценозов, но с населением,
если между формами этого комплекса нет прямых или косвенных отноше-
ний. На одном и том же участке может присутствовать любое количество
биоценозов, а отдельные биоценозы могут быть открытые, замкнутые,
простые, сложные, чистые и смешанные и т. д. и т. п. Все эти рассуждения
грешат полным отрывом от действительности. Это — неудачная попытка
решить большую и сложную проблему путем чисто умозрительных рассу-
ждений.
Другой момент, вызывающий много противоречий и неправильных
толкований в понимании биоценоза, возникает из попыток рассматривать
биоценоз как некоторый организм, способный к самосохранению и саморегу-
ляции. Нет сомнений в том. что комбинации животных и растений, свой-
ственные тому или иному участку биосферы, обладают определенной устой-
чивостью состава и повторяемостью цикла, однако им свойственна при
этом и весьма широкая пластичность. Изменение внешних условий, вклю-
чение в биоценоз новых компонентов, выпадение прежде в нем существо-
вавших (в рамках обычных явлений) не вызывает «умирания» биоценоза
п на этом месте не «рождается» новый биоценоз. Повторяемость определен-
ных форм соотношений и циклов в биоценозе далеко еще не вскрыта в своей
причинной зависимости. Явления саморегуляции и самосохранения, свой-
ственные телу отдельного организма и обеспечиваемые, как их теперь на-
зывают, регулирующими силами, свойственными целому, приурочены
к определенным частям этого целого и на отдельные части целого воздей-
ствуют через определенные передаточные системы. Вот эта-то характерная
особенность целостного организма отсутствует в биоценозе.
Понимание степени устойчивости и пластичности биоценоза должно
явиться в результате тщательного и конкретного изучения его состава,
соотношений отдельных компонентов с окружающей средой и друг с дру-
гом п сезонных п многолетних изменений. Несомненно, что биоценоз не
*
179
является только суммой составляющих его слагаемых, что он представляет
собою особое качество, но качество совершенно отличное от того, которое
мы связываем с представлением об организме. Для выявления этого осо-
бого качества, еще не вскрытого, предстоит большая работа, в которой
в равной мере должны найти свое применение и метод полевых наблюдений
и метод эксперимента.
Хороший анализ биоценозов методом полевых наблюдений на примере
донной фауны Азовского моря дан В. Воробьевым (1945).
Когда пытаются установить границы того или иного зооценоза, обычно
только констатируют совместное существование тех или иных форм в опре-
деленном биотопе и оставляют в стороне всю сложную систему связей,
которыми каждый организм связан с окружающим биотопом и насе-
ляющими его организмами в целях питания, роста и размножения. Тем
более такое игнорирование невозможно, если пытаться искать в биоце-
нозе качеств «организованного целого». Поясним это некоторыми приме-
рами. Определенная часть зоопланктона, обитающего в дневные часы на
глубине нескольких сот метров, в ночное время поднимается в самые верх-
ние слои моря, получая здесь обильную пищу. В этих же верхних слоях
нередко обитают молодые стадии, взрослые же появляются здесь только
ночью, а днем опускаются на глубины. Когда они находятся наверху, их
во множестве поедают рыбы, а на глубине могут использовать в пищу более
крупные компоненты планктона. Рыбы, откормившиеся ими, могут затем
переместиться совсем в другие части моря. Очевидно, что мигрирующий
ночью в верхние слои зоопланктон гораздо более связан с питающимися
им рыбами и со своим собственным кормом, чем с теми компонентами планк-
тона, вместе с которыми он держится днем на глубине 200—300 м. Если
мы возьмем в качестве примера не планктон, а бентос, отношения останутся
те же. В большинстве случаев питание донных организмов зависит от тех
животных и растительных остатков, которые падают сверху, из богатых
жизнью слоев моря. Отрождаемая донными животными молодь в большин-
стве случаев подымается в верхние слои моря и переносится течениями
часто на большие пространства. Мигрирующая на тысячи километров рыба
может, откармливаясь в данном районе, выесть большую часть донного
населения и затем передвинуться дальше на сотни километров.
Следовательно, существование отдельных компонентов того, что на-
зывают биоценозом, очень часто обеспечивается организмами, сосуще-
ствующими не вместе с ними, а обитающими далеко за пределами данного
биоценоза. Таким образом, биоценоз следует рассматривать
как систему совместного использования определенного
пространства обитания и заключенных в нем пищевых
ресурсов совокупностью организмов, составляющих на-
селение данного пространства в целях питания, роста
и размножения.
Учитывая все вышесказанное, биоценоз 1 можно определить как исто-
рически сложившуюся группировку организмов, населяющих тот или иной
участок биосферы (биотоп), в совокупности всех их свойств, взаимоотно-
шений друг с другом и с окружающей средой, способную к устойчивости
при относительной стабильности внешней среды и весьма способную к раз-
нообразным сменам в результате изменений во внешней среде и в составе
1 При рассмотрении конкретного материала по отдельным морям, в специальной части этой
книги, автор часто воздерживается от употребления термина биоценоз, ввиду недостаточной
его конкретной разработанности, и пользуется более формальным термином — комплекс,
разумея под ним ту или иную группировку населения со всеми входящими в его состав формами,
независимо от тех взаимосвязей, которые существуют между ними.
180
самого биоценоза, причем ведущая роль в биоценозе принадлежит немно-
гим руководящим формам.
Поэтому биоценоз не может рассматриваться как сумма слагаемых.
Биоценозы представляют собой особую категорию явлений и обладают
особыми, свойственными им качествами. Биоценоз не может быть ограни-
чен во времени, так как он претерпевает постоянные и глубокие изменения.
В биоценозе господствует только система взаимоограничений, а не само-
регуляции. Биоценоз только в редких случаях может быть ограничен в про-
странстве.
Состав биоценоза. В состав биоценоза (населения определенной части
биосферы) входят организмы, находящиеся в самой различной степени
связи и взаимозависимости. Весьма значительную часть биоценоза (не-
редко основную) могут составлять формы, наименее зависимые друг от
друга, в наибольшей степени нейтральные друг к другу, наименее мешаю-
щие друг другу, в наибольшей степени способные существовать рядом и
наилучшим образом использовать совместно заселяемое пространство
и источники питания. Каждая из таких форм может вводить в биоценоз
«свиту» других форм, связанных с ней связями комменсализма, симби-
оза, паразитизма или отношениями хищник — жертва.
Очень существенным моментом, при количественно-биоценотическом
изучении определенной части биосферы, является установление классифи-
кации отдельных входящих в состав населения форм по их значимости
в биоценозе. На данном уровне наших знаний о соотношениях между от-
дельными компонентами биоценоза мы можем пользоваться пока только
такими формальными показателями как биомасса, количество экземпляров
(или обилие), количество видов, встречаемость, равномерность распределе-
ния и коэффициент общности х.
Количество различных видов животных, входящих в состав биоценоза,
будет различно в зависимости от размеров того биотопа, которым мы огра-
ничиваем данный биоценоз, и от его свойств. Все население земного шара
также можно рассматривать как некоторый целостный биоценоз. Если же
взять сравнительно ограниченные биотопы с более или менее однообраз-
ными условиями существования, состав населения которых мы определяем
методом учета отдельных площадок, то количество входящих в него видов
(исключая микрофауну) по Элтону обычно колеблется в пределах
от 30 до 173 и редко превышает 200 (в наземных биоценозах). Для макро-
бентоса Варенцова моря мы получили для разных биоценозов мягких грун-
тов от 82 до 171 вида. При всей условности этих величин, они дают некото-
рое представление о составе биоценозов. Однако нередко, при наличии от-
носительно неблагоприятных условий (пониженная или повышенная со-
леность, неблагоприятный газовый режим, особые свойства грунта и т. п.),
биоценозы могут иметь значительно обедненный состав. Таким образом
можно различать биоценозы полимикстные и олигомикстные. Биоценозы
с большим числом видов (которые удобно называть полимикстными) ха-
рактерны для нормальных биотопов, т. е. различных частей открытых
морей (экологически оптимальных), однако отдельные виды не выделяются
особым преобладанием над другими. Биотопы же, резко отличные в каком-
нибудь отношении от нормальных, населены биоценозами с небольшим
набором видов (олигомикстные), но при этом часто наблюдается резкое
преобладание в биоценозе отдельных видов. В качестве примера олиго-
микстных биоценозов можно привести донные биоценозы Азовского моря
1 Детальный анализ понятий биоценологии можно найти в работах В. Беклемишева
(1931) и Г. Г а у з е (1936).
181
или сильно осолоненных култуков и сивашей наших южных морей,
состоящие иногда всего из нескольких видов.
Отдельные виды, входящие в состав биоценоза (компоненты или кон-
ституенты), дают весьма различные количества экземпляров и биомассы
на единицу площади или объема. Нередко бывает, что один какой-нибудь
вид составляет по весу свыше 90% всего биоценоза (например, Cardium
edule в некоторых биоценозах Азовского моря или Mytilus edulis, Bala-
nus balanoides- в обрастаниях скал на Мурмане). Другие виды, наоборот,
участвуют в биоценозе с очень малыми показателями числа экземпляров
и биомассы. Одни виды присутствуют в биоценозе примерно в одина-
ковых количествах в течение всего года, другие дают иногда громадное
преобладание только в определенное время года или время суток и схо-
дят на нет в другое время года или суток (например, цветение планктона,
миграции рыб, вертикальные миграции планктона). И те и другие могут
при этом или равномерно распределяться по биоценозу, или встречаться
в виде отдельных скоплений или пятен (степень дисперсности).
Методы изучения биоценоза. Закономерности распределения компо-
нентов биоценоза нам в основном неясны и иногда исследователи ста-
новятся на путь чисто статистических методов, которые констатируют
факты, но не разъясняют причинных связей. Так, в 1922 г. Сведберг
(S v е d b е г g, 1922) предложил метод учета равномерности распределе-
ния в биотопе отдельных форм и ввел понятия нормального рассеяния,
сверхрассеяния, недорассеяния и коэффициента дисперсии. Нормальное
рассеяние в наибольшей степени соответствует такому распределению,
которое определяется случайностью и может быть вычислено по законам
теории вероятности. В определенной группе случаев, наблюдаемой в
естественных условиях, характер распределения очень близок к вычи-
сленному. В. Беклемишев приводит пример такого распределения
для паука Oxyptila trux (табл. 62).
Таблица 62
Сколько Наблюдаемое Вычисленное -
экземпляров найдено на площадке число площадок число площадок Уклонение
0 66 66 0
1 13 12,4 +0,6
2 1 1,2 —0,2
В других случаях мы имеем отклонения от такого распределения, либо
в сторону неравномерности, когда животные собраны пятнами, либо в сто-
рону резко выраженной равномерности. В первом случае число отклонений
от среднего значительно возрастает (в обе стороны), коэффициент диспер-
сии больше единицы, почему такое распределение и называется сверх-
рассеянием. Во втором случае резко вырастет совпадение с некоторой сред-
ней и коэффициент дисперсии будет меньше 1. Это будет недорассеяние.
Сведберт дает математический путь расчета этих трех состояний
дисперсности.
Таким образом, по количественному участию отдельной формы в био-
ценозе могут быть созданы три системы классификации — по постоянству
встречаемости, по количественной значимости и по дисперсности. В. Б а с-
к и н а и Г. Фридман дали анализ распределения по классам оби-
лия составляющих биоценоз компонентов (на примере лугового сообще-
ства Deschainpsietuin) и выразили это особыми кривыми (рис. 92). При этом
182
выступает весьма резкое деление всех компонентов на две группы — очень
большое число второстепенных форм и небольшое число доминирующих
форм. Анализ распределения компонентов биоценоза по классам встре-
чаемости, произведенный Д ю р и (D иг у) для наземной растительности,
дает еще одну интересную закономерность (рис. 93). Если по абсциссе
отложить классы встречаемости, а по ординате число видов с данной встре-
чаемостью, то оказывается, что кривая получается двувершинная (при
определенном размере проб), т. е. большое количество видов приходится
на классы с малой и большой встречаемостью, средняя же встречаемость
соответствует малому числу видов. По количественной значимости от-
дельных форм в биоценозе различают доминанты, важнейшие инфлю-
энты, субдоминанты, инфлюэнты, субинфлюэнты и второстепенные виды
(Clements и Shelford, 1939).
Рис. 92. Распределение по классам обилия 86
видов беспозвоночных в луговом сообществе
Daschampsietum. На абсциссе отложены клас-
сы обилия, на ординате числа видов (по Ба-
скиной и Фридман из Беклемишева).
по классам встречаемости в ас-
социации Empetrum nigrum —
Sphagnum fuscum (по Дюри из
Беклемишева).
Иногда выделяют еще предоминанты и случайные формы. Употреб-
ляются также термины — руководящие, преференты и т. д.
Классифицируя отдельные формы литоральной фауны по их значимо*
сти в биоценозе, Линке различает 7 групп по убывающей степени зна-
чимости: руководящие, преобладающие, отступающие, подчиненные,
случайные, гости и посетители.
Вся эта обильная терминология не отличается ясностью, отдельные
понятия расплывчаты и обычно не имеют количественных характеристик,
почему вполне естественны попытки найти таковые для отдельных компо-
нентов биоценоза. Такие попытки были сделаны для морских животных
биоценозов у нас в СССР Броцкой и Зенкевичем (1939)
и в Дании Спэрком (Sparc к, 1933). Первые два автора исполь-
зуют для количественной характеристики отдельных форм, входящих
в состав биоценоза (компонентов), особый показатель, называемый ими
индекс плотности — геометрическое среднее из произведения биомассы дан-
ной формы (на 1 м2) на частоту встречаемости1, соединяя таким образом
два наиболее характерных показателя значимости компонента в биоце-
нозе. Очень сходный показатель был предложен также двумя немецкими
биологами. Гёфлер(Нб£1ег, 1938) для грибов установил особый по-
казатель обилия — геометрическое среднее из произведения количества
экземпляров на вес, аПфейффер (Pfeiffer, 1940), изучая фауну
1 Частотой встречаемости называют количество проб, в которых обнаружена данная форма
в °/0 к общему количеству проб.
183
эпифитных микроаквариумов на Dipsacus lacinatus, применил показатель
п, е, где п —количество нахождений, а е — средняя длина формы. Д. Мор-
духай-Болтовской предложил некоторое изменение индекса
плотности Броцкой и Зенкевича, называемое им индекс до-
минирования, а именно V рУЬ (р — встречаемость, Ь — биомасса).
В. Воробьев добавляет еще один показатель — геометрическое сред-
нее из произведения числа экземпляров на встречаемость (У ар).
Используя вычисленные для всех компонентов биоценоза индексы
плотности, Броцкая и Зенкевич вычертили для всех устано-
вленных ими донных биоценозов Варенцова моря особые кривые, которые
дают им основание подразделять компоненты биоценоза на три основные
группы, которые они называют руководящими, характерными или второ-
Рис. 94. Кривые индексов плотности биоценозов Варенцова моря
(по Броцкой и Зенкевичу).
степенными формами. По вертикальной осп откладывается значение ин-
декса плотности, по горизонтальной — все виды, входящие в состав биоце-
ноза со своими индексами в убывающем порядке. Рассмотрение некоторых
из этих графиков (рис. 94) действительно показывает, что кривые имеют
два довольно ясные перелома, отделяющие формы с большими показате-
лями от форм со средними показателями и эти последние от форм с малыми
показателями (руководящие, характерные и второстепенные формы).
Кроме того, формы характерные и второстепенные могут быть еще подраз-
делены на формы первого и второго порядка. Особенно легко удается
провести такое деление на полимикстных биоценозах, олигомикстные дают
часто более простую картину. Таким образом, разные группы форм полу-
чают количественную оценку не абсолютного характера, а относительно
к значимости всех других компонентов. Совершенно очевидно, что коли-
чество форм, входящих в состав каждой из вышеуказанных групп, будет
обратно значимости группы: наибольшее количество видов будет входить
в состав групп второстепенных форм и наименьшее — в состав руководя-
щих.
184
Эта методика установления значимости форм в биоценозе была принята
также рядом других советских исследователей (Морозова-Во-
д я н и ц к а я, Воробьев, Мордухай-Болтовской, Фи-
латова и др.).
Несколько иначе подошел к этому вопросу G п э р к (1936). Во-первых,
С п э р к выделяет характерные формы (соответствуют нашим руководя-
щим), которые должны удовлетворять нескольким требованиям. Они дол-
жны занимать достаточно обширный ареал (батиметрический, географиче-
ский или по типу грунта), быть константами в смысле ботаников-экологов-
(т. е. должны обладать встречаемостью не менее 50%), и вес данного вида
должен составлять не менее 15% общего веса всех видов в исследуемой
Рис. 95. Зависимость встречаемости от размеров пробных
площадок для наземной улитки Vallonia pulchella в сооб-
ществе Filipenduletum (по Баскиной и Фридман).
Доминантными формами G п э р к называет такие, биомасса которых
в общей биомассе составляет не менее 5%, инфлюэнтами — с биомассой 2—
5% и резидентами — с биомассой менее 2%. Между прочим, Спэрк
отмечает неудобство классификации компонентов биоценоза по значимо-
сти только по биомассе, без учета частоты встречаемости и выбирает
биомассу, как наиболее характерный показатель.
Понятие встречаемости совпадает с коэффициентом постоянства Б р о к-
м а н-Е р о ш а (1936), при этом можно считать константами виды, встре-
чающиеся более чем на 50% станций, второстепенными — от 25 до 50%
и случайными, встречающиеся меньше чем на 25% станций.
Если какая-нибудь форма дает равномерно-случайный характер рас-
пределения внутри биоценоза, то показатель встречаемости может очень
хорошо характеризовать и обилие (количество экземпляров).
Зависимость между частотой встречаемости и размером пробной пло-
щадки была очень хорошо показана В. Б а с к и н о й и Г. Фридман
на примере наземной улитки Vallonia pulchella (рис. 95): при определенном
размере площадки встречаемость устанавливается на постоянном уровне.
Весьма существенным моментом при количественном учете фауны (так
же как и флоры) является размер пробных площадок и число их, необхо-
димое для правильной оценки количественной структуры биоценоза.
Единых величин ни для того ни для другого не может быть не только для
разных биоценозов, но и для различных компонентов, входящих в один и
тот же биоценоз. Чем мельче форма, тем меньше, как правило, должна быть
п пробная площадка, однако размер площадки зависит также и от ха рак-
185
тера распределения (от степени дисперсии). Особенные трудности в этом
отношении представляет водная фауна, так как мы пока лишены возмож-
ности проверить наши данные непосредственным наблюдением и не можем,
кроме того, проконтролировать само качество взятия пробы прибором.
Попытку ответить на этот вопрос сделал шведский исследователь Гис-
лен (Gislen, 1930) применительно к морскому бентосу. Г и с л е н при
своих подводных исследованиях в водолазном аппарате пользовался рам-
ками в 0,1 и 0,25 м2 и установил, что для растений площадка 0,1 м2 доста-
точна, а для животных мала, так как многие из них имеют более обшир-
ный минимальный ареал. Этот вывод Г и с л е н иллюстрирует очень пока-
зательной таблицей (табл. 63).
Таблица 63
Биоценозы Количество проб Среднее количество форм в сериях проб
0,1 м2 0,25 м2 0,1 м2 0,25 м2
раст. жив. всего раст. жив. всего
15 различных биоценозов суб- лнторали Гюльмар-фиорда. . 33 107 78,1 138,6 216,7 74,96 204,11 279,1
Из этой таблицы можно видеть: число растительных форм (видов) с уве-
личением размеров площадки от 0,1 к 0,25 м2 не увеличивается, а животных
увеличивается на 48,4 %. В последние годы морские биологи стали работать
с дночерпателями 0,20 и 0,25 м2. К сожалению, пользование дночерпате-
лями большего размера по ряду соображений (вес прибора, объем матери-
ала, свойства грунта) не представляется возможным.
Что касается числа проб, то, как правильно говорит В. Беклеми-
шев, число необходимых площадок или вообще уловов (так же как и раз-
мер или объем их — Л. 3.) определяется изменчивостью уловов и нужной
для целей исследования точностью средних величин. Мерой точности яв-
ляется в данном случае средняя ошибка. Увеличивая число уловов, мы
можем уменьшить эту ошибку до желаемого предела. Предел этот опре-
деляется чаще всего степенью различия между сравниваемыми обилиями:
чем эта разница больше, тем большая необходима точность при вычисле-
нии обилия.
Применительно к морскому бентосу указывают также иногда на то, что
если брать подряд несколько проб дночерпателя, то разные пробы дают
значительные расхождения для руководящих форм. Так, Б р о х
(Н. В г о с h, 1927) произвел подсчет числа экземпляров пяти массовых
Таблица 64
Формы
9 10
Scoloplos armlger .
Scalibregma infla-
tum................
Pectinaria koreni .
Thyasira sarsf . . .
Macorn a calc are a .1
0
370
30
10
490
30
20
60
10
70
20
80
30
40
30
50
10
30
222
40
28
50
20 0 20
290
0
0
70
440
30
60
10
140
50
70 380
0 20
50 30
80( 50
264
20
30
46
12
243
30
29
48
1 2
3 4
7 8
186
форм в десяти последовательных пробах дночерпателя 0,1 м2, взятых не-
посредственно одна за другой у Линдеснесс (Южная Норвегия) (табл. 64).
Если пользоваться дночерпателем 0,25 м2 и брать на каждой станции
не 1, а 2 пробы, ряды чисел получались бы гораздо ровнее.
Мы получили бы таким образом из каждого пункта сбор в 0,5 л«2, равно-
ценный 5 пробам Б р о х а, т. е. как если бы мы соединили 10 при-
веденных проб по пяти. Результат показан в двух правых графах со
средними из каждых пяти проб и может считаться вполне удовлетво-
рительным.
Рис. 96. Экологические ареалы некоторых донных животных Варенцова моря
(по Зенкевичу и Броцкой).
1 — Pelonaja corrugata, 2—Spiochaetopterus typicus, 3 — Macoma calcarea, 4— Schizaster fragilis,
5 — Cardium groenlandicum, 6 — Portlandia lentlcula.
Совершенно ясно, что размер площадки или размер улова и число взя-
тых проб будут совершенно различны для бактерий, для крупных моллю-
сков или медуз и для рыб, для сидячих и для свободно подвижных форм.
Так как в состав биоценоза входят весьма различные по биологии и по раз-
мерам организмы, нужно руководствоваться показателями средней ошибки
для массовых форм (руководящих), играющих в биоценозе наиболее важ-
ную роль (таковых обычно немного), пренебрегая всей массой второстепен-
ных и частью характерных форм.
При количественном анализе состава населения может иметь также
применение метод вычерчивания так называемых экологических ареалов,
предложенный Л. Зенкевичем и В. Броцкой (1937) (рис. 96).
Метод этот заключается в следующем. Если мы располагаем достаточно
обширным материалом для большого района с значительными колебани-
ями в каких-нибудь факторах (t°, S°/oo5 глубина, грунт), мы можем вычер-
тить график изменений по двум факторам в пределах нашего водоема,
187
например по глубине с колебаниями для Варенцова моря от 0 до 500 м
и по температуре с колебаниями от —2° до —8°. В такую решетку у нас
уместится все разнообразие для этих двух факторов в Баренцовом море.
Затем мы распределяем по полученным квадратам решетки все данные по
биомассе, соответствующие данной комбинации температуры и глубины, и
для каждого квадрата, из всех попавших в него данных, вычисляем сред-
нее арифметическое. По полученным таким образом числам можно прове-
сти изолинии, соответствующие той или иной биомассе.
Вычерчивание подобных графиков по двум факторам производилось и
другими авторами. Еще в 1916 г. Пирс (W. Pearse) иллюстрировал сход-
ным образом отношение разных биологических фаз долгоносика к опре-
деленным комбинациям температуры и влажности, а позднее Паркер
(J. Parker, 1930)—отношение между этими двумя факторами и вылу-
плением прямокрылых из яйца. По тому же типу построены климограммы
Стеггерда (Steggard, 1929) и Джонсон (Johnson,
1924).
Сходный вид имеют и «гидрограммы» для некоторых рыб Черного моря,
приводимые С. Малятским (1938).
Применяемый ботаниками коэффициент общности показывает сходство
в видовом составе на разных участках биосферы и выражается процентом
общих видов в разных биоценозах.
7. ДЕЛЕНИЕ МОРСКИХ ОРГАНИЗМОВ НА ОСНОВНЫЕ
БИОЛОГИЧЕСКИЕ ГРУППЫ
Планктон и бентос. По характеру распределения и передвижения в во-
доеме различают прежде всего пелагические организмы (население тол-
щи воды, пелагиали) и бентические (население дна, бентали) или бентос.
Обычно население пелагиали подразделяют, кроме того, на планктон
и нектон. В таком случае под планктоном разумеют совокупность мелких
пелагических организмов, не обладающих достаточно активными органахми
передвижения и в силу этого носящихся вместе с массами воды. Нектон,
объединяющий в себе в основном рыб и млекопитающих, включает формы
с сильными активными органами движения, обеспечивающими перемеще-
ние в водной среде на значительное пространство. Правда, такое деление
весьма условно. Казалось бы, такой постоянно планктонный рачок, как
калянус (Calanus finmarchicus), не может быть причислен к хорошим
пловцам, однако он за сутки, во время суточной вертикальной миграции,
проплывает несколько сот метров вверх и вниз. Если отнести путь, который
проделывает этот рачок, к размерам его тела, то мы получим удельную
скорость настолько большую, что обычное определение планктона, как
пассивных обитателей, нужно считать весьма условным. С другой сто-
роны, некоторые рыбы обладают весьма несовершенными органами пере-
движения.
Поэтому в настоящее время деление это должно быть признано искус-
ственным и лучше в понятии планктон объединять все организмы, насе-
ляющие пелагиаль (рис. 97). И планктон и бентос могут быть разбиты на
растительную и животную часть (фито- и зоо-), а кроме того, можно
различать и для планктона и для бентоса несколько групп.
Деление по размерам тела. Планктон различают главным образом по
размерам тела, бентос — по характеру обитания, «на» или «в» грунте.
Наиболее мелкие организмы планктона (бактерии, многие одноклеточ-
ные водоросли и жгутиковые) с длиной тела, не превышающей 50—60 у,
называются наннопланктоном. Микропланктон объединяет формы с раз-
188
мерами тела от 50 р до 1 мм (большинство одноклеточных водорослей, про-
стейшие, коловратки, наиболее мелкие ракообразные, многие личиночные
формы).
К мезопланктону относят формы с размерами 1—10 мм (главная масса
взрослых Copepoda и Gladocera, многие личиночные формы, многие гидро-
медузы). Формы макропланктона имеют тело, измеряемое сантиметрами
I класс - БентОС
Рис. 97. Основные группы населения водоема (по Зернову).
(сцифоидные медузы, моллюски, сальпы, щетинкочелюстные). Наконец
мегалопланктон объединяет в себе наиболее крупные формы планктона
(многие сцифоидные медузы, некоторые ктенофоры, головоногие моллюски,
сифонофоры). Рыбы, млекопитающие и наиболее крупные головоногие,
особенно кальмары, могут быть выделены как нектопланктон или некти-
ческий планктон.
Бентос в свою очередь может быть подразделен на микро- и макро-
бентос, причем общепринятой размерной границы нет. Примерно формы
мельче 1 мм могут быть отнесены к микробентосу. В море в состав микро-
189
бентоса входят в основном, не считая бактерий и диатомей, форамини-
феры, часть нематод, некоторые другие черви, Ostracoda, Harpacticoida,
Turbellaria, Gastrotricha и молодые формы различных представителей
бентоса.
Деление по характеру обитания. По характеру обитания в грунте можно
различать фауну, обитающую над грунтом и на грунте (английское —
onfauna, немецкое — epibiose), и фауну, обитающую в самом грунте (англий-
ское — infauna, немецкое — endobiose). Животные, обитающие над грун-
том, или онфауна, в свою очередь могут быть подразделены на планкто-
бентос или формы, плавающие у дна, как, например, крупные ракообраз-
ные (в параллель можно говорить о нектобентосе или придонных рыбах,
как камбалы, скаты, бычки и мн. др.), формы, ползающие по дну, или ва-
гильный бентос (крабы, брюхоногие моллюски, многие иглокожие, многие
черви и т. д.), свободнолежащие на дне (некоторые моллюски), воткнутые
в грунт (многие черви в трубках, Pennatularia и др.), и, наконец, формы,
прикрепленные к грунту, или так называемая эпифауна (губки, гидроиды,
мшанки, некоторые черви, кораллы, усоногие, некоторые моллюски,
некоторые иглокожие и т. д.). Одни представители эпифауны прикреплены
совершенно неподвижно, другие сохраняют некоторую возможность пере-
движения (подвижное прикрепление).
Инфауна, составленная в основном из червей и моллюсков, может
быть подразделена на формы, живущие в трубках, и на свободно роющиеся
в грунте. Некоторые из этих форм приобрели способность делать ходы не
только в мягком грунте, но и в более твердых породах: некоторые моллю-
ски и ракообразные сверлят дерево (древоточцы), другие моллюски, губки,
черви, иглокожие сверлят скалы (камнеточцы). Большое число форм (мол-
люски, мшанки, гидроиды, усоногие, асцидии) могут образовывать сплош-
ное обрастание подводных предметов, достигающее иногда 30—40 и даже
90 к?1м2.
Планктон подразделяют обычно на планктон береговой (нерити-
ческий) и планктон открытого моря (океанический), находящийся уже
вне воздействия берега. По вертикали различают планктон поверх-
ностного слоя (до 200 л«), так называемый эпипланктон, и планктон
глубже лежащих слоев — батипланктон, а этот последний, в свою оче-
редь, делят на мезопелагический планктон (200—800 м) и абиссопелаги-
ческий 0800 л«).
8. ПОДРАЗДЕЛЕНИЕ МОРСКОЙ ФАУНЫ ПО ВЕРТИКАЛЬНЫМ
ЗОНАМ
Псследовапия XIX века. Впервые вертикальная зональность для мор-
ской фауны была установлена французскими исследователями О д у э-
ном и Миль н-Э двардсом (Audouin и Milne-Ed-
wards, 1832) для побережья Франции, где они различали 5 следующих
зон: баланусов, фукусов, кораллипы, ламинарий и устриц. Сходные деле-
ния были установлены М. С а р с о м (М. S а г s, 1835) для побережья
Норвегии и Фогтом (F о g t, 1848) для Сен - Мало. Э. Форбс
(Edward Forbes, 1840) разработал вертикальное распределение
бентоса для побережий Англии, причем различал 4 зоны —литоральную,
ламинариевую, кораллиновую и зону глубоководных кораллов. В 1844 го-
ду Э. Ф о р б с опубликовал также данные по вертикальному распределе-
нию бентоса в Эгейском море.
Подобные форбсовским деления установил также Ловен/Loven,
1845) для берегов Норвегии. В 1846 г. Ф о р б с подразделил свою корал-
линовую зону на верхнюю и нижнюю подзоны, а в 1853 г. вместе с Г е н-
190 ---
леем (Hanley) дал детальную схему вертикального распределения
бентоса у побережий Англии. Этими авторами различаются:
I. Зона литоральная (приливно-отливная).
1. Верхняя подзона (Littorina rudis и Littorina neritoides), соответ-
ствующая зоне баланусов других авторов.
2. Средняя подзона (Mytilus edulis, Littorina rudis).
3. Промежуточная подзона (Littorina littorea и Purpura lapiUus).
к. Нижняя подзона (Littorina littorea, Rissoa sp. sp.).
II. Зона циркумлиторальная или ламинариевая (до 27 л).
III. Зона средняя или кораллиновая (27—91 м).
IV. Зона нижне-средняя, или глубинных кораллов (глубже 91 м\
V. Абиссальная зона.
Эту схему, с небольшими изменениями, принял в дальнейшем и Г е й л ь-
прин (Heilprin, 1887—1894). В 1863 г. появилось обстоятельное
сочинение Лоренца по распределению фауны Кварнерского залива
(Адриатическое море) в зависимости от различных факторов среды (со-
леность, прибой, приливы, свет и т. д.). Им же введены понятия сублито-
раль и супралитораль. Очень много для дальнейшей разработки вертикаль-
ной зональности в распределении европейской морской фауны дали фран-
цузские и скандинавские авторы. В 1888 и 1891 гг. В э й а н (V a i 11 a n t)
установил деление на четыре зоны: литораль, береговую область (0—
300м), абиссальную область и пелагическую область. Особенную же роль
сыграли работы Мариона (Marion, 1883, 1886) и П р ю в о (Р г и-
v о t, 1894—1898), давших впервые описание с очень большой детализацией
топографического распределения фауны различных частей побережья
Франции.
В русских морях аналогичные исследования качественно-биоценоти-
ческого характера в данном периоде производились Герценште й-
н о м (1885), К. Хворостанским (1892) и Н. Книповичем
(1892) на Мурмане и Белом море, В. Ульяниным (1868—1886) и
А. Остроумовым (1892—1897) на Черном море, Н. Зенгером
(1869) и Г. Кожевниковым (1890) на Балтийском море.
Герценштейн для Мурманского побережья различал 4 зоны:
супралиторальную, литоральную, ламинариевую и брахиоподовую, а
Хворостанский иКнипович для Соловков — литоральную,
ламинариевую и гидроидную. Для Черного моря Остроумовым
были намечены 6 вертикальных зон (по распределению моллю-
сков).
Ж. П р ю в о (1894, 1895, 1898) установил для побережий Бретани
6 поясов растительности: 1. Pelvetia (Р. canaliculata). 2. Фукусов (Fucus
platy carpus, F. oesiculosus, F. serratus, Ascophyllum nodosum). 3. Himantha-
lia (H. lorea, Bifurcaria tuberculata, Chorda jilum). Цистозиры (Cysto-
seira fibrosa, C. granulata, C. ericoides, Halidrys siliquosa). 5. Ламинарий
(Laminaria saccharina, L. digitata, L. bulbosa). 6. Морской травы (Zostera
marina).
Затем П p ю в о дает и общую* схему вертикального подразделения
жизни дна (табл. 65, см. стр. 192).
Очень существенно в работах П р ю в о то, что он дает для побережий
Бретани, с их очень большими приливами, и для Лионского залива, где
приливы отсутствуют, единую схему вертикального подразделения. Зер-
нов позднее (1912) объединил в единую литоральную зону все зоны и го-
ризонты П р ю в о до глубины в 40—80 м.
Из скандинавских исследований конца прошлого века следует упо-
мянуть обширные флористические работы Челльмана (К jellman,
1877—1878), введшего понятие элиторали для обозначения зоны, где водо-
191
Таблица 65
Области
Зоны
Горизонты
Литоральная (до глубины
40—80 м)
Субтеррестровая
(супралиторальпая)
Литоральная
Верхний
(литораль в современном
смысле, выше 0 глубины)
Средний
Нижний
Прибрежная
(от нижней границы лито-
рали до 250 м)
Песчанистая
Илистая
Глубинная (>250 м)
Зона глубинных кораллов.
Зона ‘глубинных илов
росли сходят на нет, и лежащей ниже сублиторали, а затем исследования
Аппелефа (Appellof, 1892—1896), Ay ривиллиуса (Aurivil-
1 i u s , 1895) иЛеннберга (Lonnberg, 1893, 1903).
Плавание Норденшельда на «Веге» через Арктический бас-
сейн расширило сведения о распределении морской фауны далеко на се-
вер и восток [работы Стуксберга (A. S t u х b е г g), 1880 и 1882].
В Северной Америке первым солидным трудом по зональному биоце-
нотическому распределению морской фауны, в зависимости от условий оби-
тания, является обширное исследование В е р и л л я (А. V е г г i 11 е,
1873), проведенное у побережья Веньярд (близ мыса Код). Затем необхо-
димо отметить работу Девенпора (Davenport, 1903), давшего
тщательно детализированную экологическую картину распределения
фауны песчанистого побережья Атлантического океана. Он различает
три зоны: 1) верхний пояс приливно-отливной зоны (прибойная зона), от
средней высоты прилива до верхнего края прибоя (upper beach); 2) нижний
пояс приливно-отливной зоны, между нулем глубины и средней высотой
прилива (lower beach) и 3) подводную зону (submarine zone).
Что касается тропической области, то здесь внимание исследователей
сосредоточивалось главным образом на изучении коралловых рифов.
Еще Эренберг (Ehrenberg, 1834) дал описание коралловых
рифов Красного моря. Хорошо известно также блестящее исследование
Дарвина (1842), явившееся результатом его кругосветного плавания
на «Бигле», впервые давшее теорию происхождения и развития разных
типов коралловых рифов и повлекшее за собою большое число работ не-
мецких и английских исследователей (Dana, Gardiner, Kii ken-
t h а 1 и др.), получивших свое завершение в работах большой английской
экспедиции на Австралийский барьерный риф.
Из работ по фауне антарктических морей следует отметить исследо-
вания Штудера (Th. Studer, 1879—1889), давшего описание
фауны побережия Кергелена и распределение ее по зонам: 1) приливно-
отливной, 2) родофицей (до 10 м) и 3) илистых грунтов (10—40 м).
Таким образом, к началу настоящего века выработалось, вертикальное
подразделение населения морского дна на основные зоны — супралито-
раль, литораль, сублитораль, элитораль и абиссаль. В пределах каждой
зоны разные авторы различали то или иное количество, группировок ор-
ганизмов, в основном в зависимости от распределения грунтов. Наилучше
«92
изученным районом открытых океанских побережий была северная
часть Атлантического побережья и особенно побережья Франции, Англии
и Скандинавских стран.
Работы XX столетия. В XX столетии в различных странах сильно воз-
рос интерес к морским исследованиям. G одной стороны, продолжалось
развитие работы в том же направлении экологического изучения зональ-
ного качественно-биоценотического распределения донной фауны, с дру-
гой стороны, со второго десятилетия этого века начало развиваться новое
направление, количественно-биоценотическое, ставшее на путь изучения
явлений биологической продуктивности. Наряду с этим, усиленным тем-
пом стало развиваться изучение отдельных форм водорослей, моллюсков
и ракообразных, имеющих промысловое значение.
Качественно-биоценотпческое направление особенно успешно развер-
нулось в России, во Франции, Скандинавских странах, Англии, Германии
и Северной Америке. Из большого ряда исследований мы остановимся
только на наиболее крупных.
За первые десятилетия настоящего столетия в нашей стране было
проведено несколько блестящих исследований по морской фауне в от-
ношении ее зонального качественно-биоценотического распределения.
Подробно их результаты будут рассмотрены в главах, посвященных
отдельным нашим морям, здесь же отметим только некоторые основные
работы. Первая из них по времени опубликования—обширная моногра-
фия С. Зернова (1913) — посвящена фауне Черного моря (в основ-
ном бентосу). Через три года появилось не менее замечательное иссле-
дование К. Дерюгина (1915) по фауне Кольского залива. Обе эти
работы сыграли исключительно большую роль в развитии нашей морской
биологии, на них воспитывались и воспитываются многие поколения
морских фаунистов. Оба автора дали чрезвычайно полную картину ка-
чественно-биоценотического распределения бентоса, от уреза воды до
наибольших глубин его распространения. С. Зернов подразделяет
склон дна Черного моря, заселенный животными, на литоральную и суб-
литоральную зоны, с границей на глубине 15—30 сажен. Самый верхний
горизонт литоральной зоны, прибойная зона, не выделяется С. Зерно-
вым в самостоятельную зону. Главнейшие биоценозы литоральной зо-
ны: биоценозы скал, песка, ракушечника, зарослей зостеры, илистых
берегов, мертвой зостеры и водорослей и филлофорного поля; сублито-
ральной: мидиевского ила, теребеллидного ила и фазеолинового ила —
всего 10 основных биоценозов. Названия биоценозам даны по основному
субстрату. От понятия биоценозов С. Зернов резко отделяет понятие
фации, субстрата, характеризующегося определенными физическими
свойствами и мертвыми остатками организмов. Все биоценозы Черного
моря очень легко сопоставляются с биоценозами Средиземного моря л
Ламанша (по работам Прюво). С. Зерновым установлено инте-
ресное явление подъема границ биоценозов, а частично и полного их
выклинивания при переходе от открытого моря в заливы и бухты. Гра-
ница между литоралью и сублиторалью в заливах проходит по С. Зе р-
н о в у всего на глубине 4—9 сажен.
К. Дерюгин для Мурманского побережья принимает вертикаль-
ное деление берега на 4 зоны — литоральную (область прилива и отлива),
сублиторальную (до 30—35 саж.), элиторальную (до 100—125 саж.)
и псевдоабиссальную. В пределах каждой зоны К. Дерюгин разли-
чает фации и подфации, разумея под ними комбинацию определенного типа
грунта с определенным комплексом животных форм. Дерюгин раз-
личает фации скал и камней, песка, ила, раку ши и ветвистого литотамния.
Каждой отдельной фации или подфации может соответствовать то или
13 Зенкевич, т. I
193
иное количество биоценозов, которые именуются Дерюгиным по
руководящим формам.
В течение 1925—1930 гг. опубликовали результаты своих многолет-
них работ по литорали Мурманского побережья, а также Японского и
Охотского морей Е. Гурьянова, И. Закс и П. Ушаков. Все эти
многочисленные работы сделаны в том же стиле чрезвычайно детального и
тщательного описания зонального качественно-биоценотического распре-
деления фауны. К сожалению, эта серия работ содержит только каче-
ственное описание фауны1. Подобно К. Дерюгину они подразделяют
литораль на фации скал, камней, илистого песка, глинистого ила, а затем
в пределах каждой фации различают то или иное количество биоцено-
зов. Кроме того, в вертикальном направлении эти авторы подразделяют
литораль на три горизонта и 7 этажей.
Наконец, в 1928 г. вышло второе капитальное сочинение К. Дерю-
гина, посвященное фауне Белого моря и условиям ее существования
и дающее полное монографическое описание водоема. В этой работе
К. Дерюгин принимает подразделение на литоральную (до 0 глубины),
сублиторальную (до нижней границы распространения водорослей) и псев-
доабиссальную зоны, с последующим делением их на горизонты и этажи.
В самое последнее время К. Дерюгин дал по той же схеме де-
тальную картину зонального распределения биоценозов залива Петра
Великого в Японском море.
Во Франции следует отметить серию исследований Дебошана
(Р. de Beauchamp), начная с 1913 г. и его учеников — Лами
(R. L а m i) и И. Закса.
В первой из серий его работ, посвященных литорали, с исключитель-
ной подробностью описываются 11 различных типов биоценозов ила,
песка, гравия и зарослей зостеры. Второе его большое исследование,
посвященное литорали у Роскова, и последующее, совместное с
Лами, по литорали северных побережий Франции, могут считаться наи-
лучшими в серии подобных работ французской школы. Д е б о ш а н исполь-
зовал понятие фации для обозначений свойств грунта.
Целый ряд других французских авторов (Dollfus, Labbe,
Sauvageau и др.) дополнили и расширили работы de Beauchamp.
По фауне английских побережий подобный анализ зонального распре-
деления фауны от супралиторали до глубины 600—700 м, были даны
Джонстоном (Johnston, 1908), Кингом и Ресселем
(King и Russel, 1909), Саутерном (Southern, 1915),
Ферраном (Farran, 1915), Флаттели и Уолтоном
(Flattely и Walton, 1922).
Много нового и оригинального дала работа Кинг и Рессела (К i n g и
Russel), отличающаяся тщательностью экологического анализа мор-
ских биоценозов литорали. Они показали, что его следует вести, учи-
тывая смену дня и ночи, дождливой и сухой погоды, температуры, баро-
метрического давления, направления и силы ветра, степени опреснения,
влажности, затененности, явлений эпифитизма и т. д.
В дальнейшем интересные наблюдения над воздействием на фауну
мелководий лунного света были произведены Элмхирстом (R. Elm-
hirst, 1932), показавшим, что в лунную и темную ночь, в при-
лив, на литорали можно наблюдать резкие различия в составе фауны
высших ракообразных. Впрочем явления «лунатизма» у морских организ-
1 Некоторые количественные данные содержатся только в работе И. Закса по фауне
литорали Шантарского моря.
194
мов были констатированы и ранее на ряде морских организмов (например,
появление палоло, сроки размножения у разных форм и др.).
Для датских вод исследования принадлежат Варминг (Е. War-
ming) и Везенберг-Лунду(С. Wesenberg-Lund, 1904)
и Мортенсену (Mortensen, 1922). Обширный зоологический
материал по зональному распределению фауны у Норвежского побережья
был собран шведом Аппелёфом (Appellof) в его статье в книге
Murray и Hjort (1912). А п п е л ё ф делит морское дно на лито-
ральную зону (до 40 л«), сублиторальную (до 100—150 лс) и континенталь-
ный склон (до 1200 л«). Шведские исследователи Однер (N. Odhner,
1915) и Гофстен (N. Н о f s t е n, 1916) дали первые сведения по
зональному распределению фауны в Шпицбергенских водах. Данные эти
были затем пополнены исследованиями англичан Саммерхейса
(W. Summerhayes) и Элтона (С. Elton, 1923 и 1928),
указавшими на слабое развитие в Шпицбергенских водах литорали под
воздействием пловучих льдов и на развитие ее там, где берег не под-
вергается воздействию льдов.
Уже в самые последние годы, почти одновременно, появляются три
обстоятельных исследования, посвященные описанию фауны, так назы-
ваемых ваттенов — обширных осушных пространств по южному побе-
режью Северного моря и по западной стороне Ютландского полуострова
с мягкими песчанистыми и илистыми грунтами. Эта обширная песчани-
стая и илистая литораль занимает свыше 10 000 км1 и достигает в ширину
20 км. В датских водах эти исследования велись Темдрупом
(Н. Т h a m d г и р, 1935), в немецких — Воленбе ргом (Е. Wo h-
1 е n b е г g, 1937) и Линке (О. Linke, 1939).
Для Северной Америки хорошее описание распределения донных био-
ценозов района биологической станции в Пюджет-Саунд (острова Сан-
Хуан, Тихий океан, близ границы США и Канады) дали Ш е л ф о р д
и Таулер (Shelford и Towle г, 1925). Воган (Vaughan,
1916), изучая кораллы мелководий окрестностей Флориды и Багамских
островов, показал, что их можно разделить на формы прибойной полосы
и спокойной воды, а эти последние — на формы, существующие на или-
стых грунтах и благоденствующие только в наиболее чистых водах. Сле-
дует отметить также работы Ньюкомба (С. Newcombe, 1935)
по песчанистой и Хьювотта (W. Н е w a 1 1, 1937) по каменистой
литорали, первая в штате Нью-Бренсуик, вторая в заливе Монтери (Ка-
лифорния). Вне европейских и американских вод продолжалось изуче-
ние коралловых рифообразователей. Финк (Finck h, 1904) дал опи-
сание кораллового атолла Фунафути (о-ва Эллиса). Он показал, что с
наружной стороны атолла доминируют представители Lithothamnion и
Madrepora, а с внутренней Helipora coerizlea и Fungidae. На самой плат-
форме встречаются представители рода Porites. Имеются различия в со-
ставе фауны также на наветренной и подветренной сторонах. Финк
различает 4 зоны: эрозии, коррозии, морской травы и литотамния. Глав-
ные образователи рифов — литотамний, Halimeda, кораллы и форамини-
феры. Изучая распределение Madreporaria у Гавайских и Лейзанских
о-вов, V a g h а п (1907) сделал интересное открытие, что имеется две
коралловых фауны — мелководная (мельче 12 м) и глубинная (глубже
30 м).
Для австралийских вод имеется описание флоры и фауны литорали
у Сиднея, данное Хедлеем (Hedley, 1915), а для Квинсленда —
Джонстоном (Johnston, 1917). Зональное подразделение жи-
вотных литорали и сублиторали о-вов Кермадек (к северо-востоку от
Новой Зеландии) дано в работе Оливер (Oliver, 1915). В следую-
*
195
щей своей работе (1923) этот автор дал описание фауны и флоры литорали
Новой Зеландии.
Системы вертикальных подразделении бентоса. Что касается системы
вертикальных подразделений для донного населения, то, несмотря на
большое число работ, общепринятой системы еще не имеется (табл. 66).
Таблица 66
ВЕРТИКАЛЬНЫЕ ПОДРАЗДЕЛЕНИЯ МОРСКОГО ДНА И ЕГО ФАУНЫ
ПО РАЗЛИЧНЫМ АВТОРАМ
Глубина в м Чельман 1877 Герцен- штейн 1885 Аппел- лёф 1912 Зернов 1913 Дерюгин 1915 2-й Все- союзн. ги- дрология, съезд 1928 Гарт- мейер 1911
Выше среднего уровня прилива От 0 глубины до среднего уровня прилива 0 10 20 30 40 50 СО 70 60 90 100 125 150 200 250 300 400 500 1000 2000 3000 4000 5000 Элитораль Сублито- Литораль До нижней границы раль водорослей Глубоководный ил Зона гидроидов, Ламинарис- Литоральная зона известков. мша- во-литотам- нок и плече- ниевая зона ногих § и Континентальн. Сублитораль Литораль g g4 оз 3 и глубоководная ^9 § ? зона ₽э • Сублитораль Литораль Псевдо- Элитораль- Сублиторальная Литоральная • абиссальная ная д г- > t= я § > о - о 3 »=£ , § g К Ja Батиаль . Абис- саль Абиссаль Копти- Литораль неиталь
Иногда литоралью называют весь склон берега до нижней границы рас-
пространения водорослей (200 м), тогда приливно-отливную зону назы-
вают супралиторалью или просто рассматривают ее как верхний горизонт
литорали. Обычно крупные подразделения в вертикальном распределении
бентоса (литораль, сублитораль и т. д.) именуют областями, что, однако,
196
создает неудобство, так как области являются понятиями зоогеографиче-
скими. Удобнее называть их зонами.
Ввиду этой несогласованности в разграничении вертикальных зон,
на Всесоюзном гидрологическом съезде в 1928 г. было принято пожелание
для советских исследователей придерживаться единой системы (рис. 98)
деления на литоральную, сублиторальную, псевдоабиссальную, батиаль-
ную и абиссальную зоны. В этой системе отсутствует элиторальная зона
и сублитораль берется в широком толковании, как вся зона, занятая ра-
стениями. Многие авторы, однако, отмечают, что сублиторальная зона в
своей верхней и нижней частях представляет существенные отличия.
С. Зернов (1913) основное отличие усматривает в действии волн.
В верхнем горизонте сублиторали (по С. Зернову — литораль) дей-
Рис. 98. Схема вертикального подразделения морского дна,
принятая на Всесоюзном гидрологическом съезде 1928 г.
ствие волн сказывается сильно, в нижнем (по С. Зернову — субли-
тораль) действие это затухает. К. Дерюгин особенно отмечает силь-
ное развитие в верхнем горизонте сублиторали макрофитов и слабое раз-
витие их в нижнем горизонте, который в монографии по Кольскому
заливу К. Дерюгин, подобно Чельману, выделяет как само-
стоятельную зону (зона гидроидов, известковых мшанок и плеченогих
Герценштейна). Этим же признаком руководствуется и Г и с л е н,
также принимающий чельмановское деление зоны, занятой макрофитами,
на сублитораль и элитораль.
В дополнение к принятой в СССР системе вертикального подразделе-
ния морского дна на зоны можно привести еще деление, даваемое Гис-
леном (1930) в его исследовании по бентосу Гюльмарфиорда в Шве-
ции. Г и с л е н различает литораль, сублитораль, элитораль (в сумме
две последние соответствуют сублиторали советских авторов), профундаль
(в значительной степени соответствующую псевдоабиссали Дерю-
гина, принятой в советской схеме), архибенталь (соответствующую
бентали советской схемы) и абиссаль (в общепринятом толковании).
Значительно более простую систему вертикальных подразделений дает
в своей книге (1935) С. Экман (табл. 67).
197
Таблица 67
Глубина в м Бентическая фауна Пелагическая фауна
0 260 400 600 800 1000 1200 Глубоководная фауна Литоральная фауна Эпипелагическая фауна (поверхностный планктон)
Архибентальная фауна Верхняя батипелагическая фауна (Верхний глубоководный планктон)
Абиссальная бентиче- ская фауна Нижняя батипелагическая фауна или пелагическая абиссальная фауна (Абиссальный глубоководный планктон)
Рис. 99. Схема вертикального подразделения океана М. Джонсона
(из Свердрупа и др.).
Рис. 100. Вертикальная зональность морского дна.
Слева для открытого океана, справа для фиордов (ма-
лый квадрат) и для обособленных глубоководных ча-
стей моря (большой квадрат) (по Гислену).
198
От всех предыдущих схем вертикальной зональности отличается также
и схема Джонсона (1942), приводимая в океанографии Сверд-
рупа и др. (рис. 99).
Джонсон выделяемую им литоральную зону (до глубины 200 л«)
подразделяет на эвлиторальную зону (до 50 м) и сублиторальную (50—
200 м).
Во внутренних морях и в обособленных частях морей границы между
отдельными вертикальными биоценотическими зонами обычно лежат выше,
чем в открытых морях. Г и с л е н дает хорошую иллюстрацию этого
(рис. 100) и поясняет ее следующим образом (табл. 68).
Таблица 68
В наибольшем выражении В наименьшем выражении (фиорды и
(открытые моря) обособленные части морей)
1. Литораль. Приливы выражены сильно. Длительное обсыхание. Чистая вода. Сильная инсоляция. Приливы выражены слабо. Малая про- зрачность воды. Сильная инсоляция.
2. Суб литораль. Слабое действие прилива, незначительное обсыхание, чистая вода, сильная инсоляция (высокое солнце- стояние). Сильное действие прилива (значи- тельное обсыхание), мутная вода, слабая инсоляция.
3. Элитораль. Чистая вода. Значи-
тельная инсоляция.
Мутная вода, слабая инсоляция.
4. Профундаль. Сильное осаждение
растительного детрита (отсутствие свето-
вого фактора).
Полоса оседания мертвых растительных
остатков выражена слабо.
5. Ар х ибенталь. Питание растительностью сходит на нет. Грунты слагаются из
неорганических терригеновых осадков. Монотонность физических факторов среды.
6. Абиссаль. Только животная пища. Скудное питание. Мягкие грунты пелаги-
ческого генезиса. Однообразие физических факторов среды. Различия обусловливаются
только различием грунтов. .
К. Дерюгиным на примере Варенцова и Белого морей, М о-
ландером на примере Гюльмар-фиорда также показано явление
вертикального смещения и целых комплексов и отдельных форм не только
при переходе из южных широт в северные, но и от открытых частей моря
в глубь заливов, причем этот процесс идет в обоих направлениях: одни
организмы подымаются в более высокие горизонты, другие опускаются
вниз. Опускание литоральной фауны в сублитораль принимает характер
массового явления, при переходе от открытых морей к внутренним без-
ливным. Замечательный пример — уход на глубины в Балтийском море
целого литорального комплекса в составе Mytilus edulis, Масота baltica,
Cardium edule, Pygospio elegans, Halicryptus spinulosus, Priapulus caudatus
и многих других. В Средиземном, Черном и Азовском морях Mytilus и
Cardium также уходят на глубины.
ГЛАВА VII
ПЛАНКТОН
1. ИСТОРИЯ И МЕТОДИКА ИЗУЧЕНИЯ
Начало изучения планктона положено применением в качестве ору-
дия лова планктонной сетки в восьмидесятых годах прошлого столетия
(В. Гензен). Так как планктонная сетка давала возможность и коли-
чественного учета планктона, то в те же годы появилось и специальное
исследование о количественном распределении планктона (В. Гензен,
Hensen, 1887).
Сразу же был выдвинут ряд важных проблем биологической продук-
тивности — условия массового развития фитопланктона, питание зоопланк-
тона и в частности рыб, возможность количественного учета рыб по выме-
танной икре ит. д. Ломанном (Lomann) был открыт мир мель-
чайшего планктона — наннопланктона, проходящего через планктонную
сетку, и установлены закономерности сезонной смены планктона. К тому
же времени Брандтом (К. Brandt) было выяснено значение план-
ктона в круговороте органического вещества в море.
В 1910 г. появилась обширная сводка Ш т е й е р a (A. S t е и е г) по
планктону и планктонология стала специальным разделом гидробиологии.
Существенным дефектом подавляющего большинства зарубежных иссле-
дований по морскому планктону является отсутствие точных количествен-
ных данных — весовых или объемных. Обычно приводятся только дан-
ные по количеству экземпляров отдельных компонентов планктона, что
устраняет возможность сравнения материала, полученного из разных
мест. Советские исследования в этом отношении стоят на более высоком
уровне, так как базируются на точном количественном методе. Кроме того
в советскую практику исследований планктона В. Богоровым введен
метод использования «стандартных» весов для массовых форм планктона
на разных возрастных стадиях. Этот метод даст в дальнейшем возможность
перечислить нужные материалы прежних работ с числа экземпляров
на весовые показатели и таким образом использовать их для сравнения
с данными советских экспедиций.
До самого последнего времени все планктонологи продолжают широко
пользоваться планктонными сетками, хотя они далеко не представляют
собою безупречных количественных орудий сбора планктона. Количест-
венные планктонные сетки сконструированы с таким расчетом, чтобы
при сравнительно небольшом входном отверстии и большой боковой
фильтрующей поверхности вся вода, попадающая в горловину, про-
ходила сквозь сетку. Тогда, зная размер входного отверстия, скорость
хода судна и продолжительность лова, можно определить, в каком коли-
честве воды были распределены организмы, оставшиеся в сетке.
200
Хорошо известно, однако, что количественная планктонная сеть не
является универсальным прибором сбора морских организмов и имеет
ряд крупных недочетов. Основные недочеты планктонной сетки — про-
ходимость материала, из которого она сшита, для более мелких организмов
(наннопланктон), входящих в состав планктона и часто составляющих
основную его массу, и практическая невозможность взять пробу со строго
определенного горизонта. С другой стороны, сеть из очень мелкоячеистого
материала быстро забивается и перестает с нужной скоростью пропускать
воду. В результате у горловины сетки образуются завихрения воды и
планктон может вымываться из сетки. Наконец, более крупные и актив-
ные компоненты уходят от горловины сетки и попадают в нее в ничтожнохМ
количестве. Поэтому в дальнейшем, при количественных исследованиях
планктона стали пользоваться взятием целостных образцов воды с помощью
гидрологических батометров, с последующим отстаиванием пробы и иссле-
дованием полученного осадка (отстойный или осадочный планктон).
В последние годы для этих целей употребляют специальные батометры
большой емкости (8—10 л), так называемые планктонные батометры. Для
получения проб, больших по объему, с точно фиксированного горизонта
стали конструировать сетяные планктонные батометры большей емкости
(0,5 м3) (В. Б о г о р о в, В. Я ш н о в).
Этот метод имеет свои недочеты. Планктон не дает однообразно-сплош-
ного распределения и часто сосредотачивается в тонком слое воды, осо-
бенно в зонах температурного или солевого скачка. Отдельная проба
батометра в силу этого может дать неправильное представление о распре-
делении планктона во всем столбе воды.
Однако сборы планктонными батометрами (Богорова) дают в 5—6 раз
больше материала, чем соответственные ловы планктонной сетью.
2. ФИТОПЛАНКТОН
Состав фитопланктона. По подсчетам, произведенным П. Усаче-
вым, в северном полушарии в состав фитопланктона входит 1668 видов
и главнейших вариететов, относящихся к 189 родам (табл. 69).
Таблица 69
Группа Количество, родов Количество видов
общее число В % общее число В °/о
Flagellata 5 3 9 0,5
Silicoflagellata 7 4 87 5
Coccoll thine ае 21 11 106 6
Perldlneae 82 43 1090 66,5
Chlorophyceae + Heterocontae 12 7 30 2
Diatomeae 50 26 * 329 19
Cyanophyceae 12 6 17 1
Всего 189 100 1668 100
В фитопланктоне Средиземного моря резко выделяются кокколитины,
перидинеи и диатомовые. Так, для Адриатического моря А. В а т о в а
(А. V a t о v а, 1928) и А. Ерцегович (1936) дают следующий ка-
чественный состав фитопланктона (табл. 70).
Таким образом, первые три группы, приведенные в списке, составляют
около 90%. С другой стороны, довольно значительно и количество сине-
201
зеленых. При этом диатомеи преобладают в холодное время года, а пери-
динеи и кокколитины в теплое. Данные по количественному составу сре-
диземноморского планктона невелики.
Таблица 70
Количество Количество
Группа видов Группа видов
Кокколитины 40 Сине-зеленые 10
Перидинеи 129 Кремнежгутиковые 5
Диатомовые 74 Жгутиковые 1
Вольвоксовые и протококковые.... 12 Разножгутиковые 1
Всего 272
В морях СССР зарегистрировано по данным П. И. Усачева 375
видов и главных вариететов, входящих в 96 родов (табл. 71).
___________________________________________________________Таблица 71
Группа Количество родов Количество видов
общее число в 7« общее число В 7.
Flagellata 5 5 9 3
Slllcoflagellata 4 4 7 2
Coccohthineae 5 5 14 4
Peridlneae 22 23 140 37
Chlorophyceae 4- Heterocontae 10 11 15 4
Dfatomeae 42 43 177 47
Cyanophyceae 8 9 13 3
Всего 96 100 375 100
о
10
20
30
20
50
50
Ю
во
10
юо
Зона_оптимольного развития_фитоплан^т6на_
Среднее положение топки компенсации
для фитопланктона
Зона опускающегося
отмирающего фитопланктона
450
А
Рис. 101. Распределение наннопланктона в толще моря в северной Атлантике
А — вертикальные зоны развития планктона. Б — мощность слоев моря по освещенности
Изтабл. 71 следует, что из всех известных морских форм фитопланктона
-северного полушария в наших морях найдены почти все жгутиковые,
202
77% сине-зеленых, около половины зеленых и диатомовых и всего 13%
перидиней, что объясняется общей тепловодностью этой последней группы.
Наннопланктон, преобладающая часть которого относится к фито-
планктону, приурочен к самому верхнему горизонту моря, хотя часть его
проникает в глубину до 5000 м. Его распределение может быть иллюстри-
ровано рис. 101 и 102, построенными по данным Гентшеля. На
больших глубинах преобладают пока загадочные «оливково-зеленые
клетки» (предположительно из Gyanophycea).
развитие фитопланктона зависит от двух основных факторов — освещен-
ности и минеральных солей, необходимых для построения тела. Но свет
в нужных для успешного развития растений количествах проникает толь-
ко в самый верхний слой моря, а основная масса лимитирующих разви-
тие фитопланктона питательных солей (фосфаты и соединения азота),
сосредоточена в глубже лежащих слоях. Таким образом, с одной стороны,
растительные организмы идут тем глубже, чем прозрачнее вода, а с дру-
Рис. 103. Вертикальное распределение фосфатов в мг!м3 на
широтном разрезе через южную Атлантику по широте 9°
(из Свердрупа и др.).
гой стороны — наибольшее развитие фитопланктона приурочено к райо-
нам моря, где вертикальная циркуляция выносит в поверхностный слой
питательные соли. Поэтому совершенно понятно то совпадение, которое
получается в количественном распределении фитопланктона и питатель-
ных солей (рис. 104). Основные же резервы питательных солей в океане,
особенно в тех его частях, где более сильно выражена стратификация,
в основной массе разобщены от их потребителя — фитопланктона.
Там же, где имеет место вынос на поверхность глубинных вод, фитопланк-
203
тон получает возможность бурного развития. На приведенном рис. 103
можно видеть, что в восточной части тропической зоны Атлантического
океана, у берегов Африки, условия для развития фитопланктона гораздо
благоприятнее, чем в западной части. Более благоприятны они и там, где
в области тропического противотечения имеет место вынос в верхние слои
более глубинных вод (зоны дивергенции). Это хорошо иллюстрирует также
рис. 101, где сопоставлено количественное развитие планктона с цирку-
ляцией водных масс в верхней толще воды в зоне экваториального проти-
вотечения. Однако очень сильная вертикальная циркуляция может пода-
влять развитие фитопланктона, что и наблюдается в некоторых частях Ант-
арктики, где имеется у самой поверхности большое количество питатель-
ных солей, а развитие планктона слабое. Возможно, что большие массы
планктона уносятся в таких случаях на глубины. Таким образом, для
наиболее успешного развития фитопланктона требуется определенное
оптимальное соотношение между освещенностью, количеством питатель-
ных солей и степенью вертикальной циркуляции.
Рис. 104. Соотношение в плотности планктона (А) и содержании фосфатов (Б) на
поперечном разрезе через область противотечения в Тихом океане (из Свердрупа и др.).
В самых поверхностных слоях воды фитопланктон, в смысле освеще-
ния, имеет наиболее благоприятные условия для ассимиляции, и эта по-
следняя превышаетпроцесс дыхания. С увеличением глубины ассимиляция
ослабевает и на каком-то определенном горизонте, различном в различ-
ных частях океанов и морей, оба процесса имеют одинаковое выражение.
Это — «слой компенсации». Глубже процессы дыхания превалируют над
ассимиляцией.
В самых прозрачных водах мирового океана (Саргассово море, Среди-
земное море) водоросли могут вегетировать на глубине и свыше 100 м.
Положение «точки компенсации» в водах различной прозрачности нахо-
дится на различной глубине, в пределах от нескольких метров до 30—45 м
(Ламанш). Согласно Гарвею (1945), опыты с диатомовой водорослью
Nitzschia closterium показали, что понижение концентрации фосфатов
ниже 10 мг}м6 значительно замедляет фотосинтез. Опыты Кетчума
(Ketchum, 1939) на той же форме показали, что при повышении кон-
центрации фосфатов свыше 17 мг]м3 скорость роста диатомовых водорослей
перестает повышаться. Точно так же, при достаточном количестве фосфа-
тов, повышение нитратов свыше 47 мг,'м3 не дает дальнейшего повышения
темпа развития диатомовых. Однако и при самых малых дозах питатель-
ео4
ных солей развитие фитопланктона не приостанавливается полностью.
Как говорит Гарвей (1942), «низкая концентрация растворенных в
воде солей замедляет рост растений во многих участках моря, но нет
никаких оснований предполагать, что он когда-либо совсем приостанав-
ливается, так как фосфор и усвояемые азотистые соединения непрерывно
восстанавливаются».
Соотношение фито- и зоопланктона. Главный потребитель диато-
мовых — взрослые Copepoda. Следующее место в их питании занимают
перидинеи, а затем кокколитофориды. Молодь веслоногих питается
в основном бактериями, жгутиковыми и пр.
Copepoda отличаются чрезвычайной прожорливостью и в условиях
опыта быстро очищают воду от диатомовых, даже при очень большом их
количестве. Как указывает Гарвей (1942), кишечники ракообразных
интенсивно выделяют фекалии, часто наблюдаемые в воде в очень
большом количестве и способные к быстрому опусканию на дно.
Герт (Hart, 1934) установил, что диатомовый ил Южной Атлантики
видимо слагается из фекалий и содержащихся в них масс панцырей диа-
томовых. Таким образом, ракообразные планктона в целом создают еще
более мощный биофильтрационный аппарат, чем мидии и другие моллюски
прибрежной зоны и так же, как они, седиментируют взвесь в своих фека-
лиях и формируют иловые отложения.
Само собой разумеется, что пока фекалии падают на дно, они могут
быть использованы для питания другими организмами и отдать в воду
часть заключенных в них веществ.
Весьма существенен вопрос скорости опускания отмирающих компо-
нентов планктона и выделяемых ими фекалий. Диатомовые Chaetoceras
в спокойной воде опускаются со скоростью около 1 м в 33/4 часа. Планк-
тонные животные, как сальпы, опускаются со скоростью 4000 м в 2 дня
7 часов. Опускание копепод происходит со скоростью нескольких тысяч
метров в течение одного дня.
Как мы уже указывали, Гертом установлено, что диатомовые илы
южных частей Атлантического океана сплошь состоят из фекалий копе-
под и эвфаузиид. Таким образом: 1) створки диатомовых опускаются на
дно с гораздо большей скоростью, чем если бы они опускались самостоя-
тельно и 2) находясь внутри комочка фекалий, диатомовые изолированы
от растворяющего действия морской воды. Таким образом, и темп оседа-
ния и процесс растворения могут претерпевать существенные изменения
в связи с попаданием планктона в фекалии животных.
Большой интерес представляет количественное соотношение расти-
тельной и животной части планктона. Известны случаи одновременного
обпльного развития обеих частей планктона (Nielsen, 1937), однако,
обычно наблюдается обратное соотношение в количестве диатомей и ра-
кообразных, что может в отдельных случаях зависеть от усиленного вы-
едания зоопланктоном фитопланктона. Гарди и Гунтер (Н а г-
d у and Gunther, 1935) указывают, что места с особенно густыми
скоплениями диатомовых лишены обильного зоопланктона и что, якобы,
в ночное время Copepoda не поднимаются в верхние слои моря, если в
них много диатомовых. Согласно этим авторам, фито- и зоопланктон в
основном распределяются разобщенными скоплениями: совместные
скопления встречаются гораздо реже. Гарвей (1942, 1944) дает этому
следующее объяснение. Массовое развитие диатомовых и ракообразных
разделены во времени — размножение диатомовых определяется усло-
виями, существующими в данный момент, а ракообразных — пред-
шествующими, почему обычно и не наблюдается их одновременного
массового развития.
205
Продукция фитопланктона. Первые попытки вычислить суммарную
продукцию фитопланктона были сделаны Аткинсом (Atkins,
1922 и 1923) в Ламанше, а затем продолжены рядом исследователей (С о о-
рег, R i 1 е у и др. в 1932—41 гг.), а у нас на Баренцовом море Креп-
сом и Вержбинской (1932). Эти исследователи вычисляли про-
дукцию фитопланктона по балансу СО2 и питательных солей, а затем и
кислорода, главным образом весной и в начале лета, т. е. в период макси-
мального развития фитопланктона. Обычно при таких расчетах принимали,
что за счет использованных веществ развивается одно поколение водо-
рослей и не учитывали процессы регенерации и повторного использо-
вания питательных веществ, а также деятельность бактерий и других
организмов. В дальнейшем расчету продукции фитопланктона весьма
помогло определение соотношения С : N: Р в диатомовом планктоне, ока-
завшееся равным в среднем 100 : 16 : 1,67 (Р е д ф и л ь д, 1934, В а к с-
м а н, 1937).
В результате всех этих расчетов для северо-восточной Атлантики были
получены числа порядка в среднем 60 г углерода на столб воды под 1 лА
Для западной части Варенцова моря Крепе и Вержбипская
определили продукцию фитопланктона в 92 г углерода под 1 м2. Однако
эти величины могут быть приняты только как минимальные.
Сейвелл (Seiwell, 1935) дал иной метод расчета годовой продук-
ции фитопланктона для тропической части северной Атлантики, где под
слоем фотосинтеза находятся слои воды, чрезвычайно обедненной кисло-
родом. Принимая, что этот дефицит кислорода—следствие полного окисле-
ния накопившихся В’ зоне фотосинтеза за год органических веществ, и
учтя количество кислорода, которое должно было поступить в обеднен-
ную кислородом зону из соседних слоев в результате вертикального пере-
мешивания, Сейвелл определил годовую продукцию фитопланк-
тона под каждым м2 в 278 г углерода.
В дальнейшем был широко использован метод темной и светлой склян-
ки, введенный Граном (Gran, 1927), а затем Маршаллом
и Орром (Marshall and О г г, 1930) и Рэлеем (Riley,
1938—41), а у нас на Каспийском море С. Бруевичем (см. II том) и
широко разработанный на пресных водоемах Г. Винбергом (1946).
Этот значительно более точный метод заключается в том, что пробы
планктона, помещенные в темную и светлую склянку (иногда подвешен-
ные в том же слое, из которого взята проба), через определенный промежу-
ток времени естественно давали разницу в содержании кислорода, так как
в светлой склянке идет и процесс ассимиляции и процесс дыхания, а в
Таблица 72
Место наблюдения Метод Продукция С в г/Зи2/г°Д
Пролив Лонг-Айленд По выделению кислорода 400—700
По потреблению фосфора 440—875
Зап. Атлантика По потреблению соединений азота 140—365
(23—41° С. Ш.)
Залив Мэйн:
январь По выделению кислорода 50
март 190
апрель » » » 950
май > » » 540
июнь » » » 630
сентябрь > » » I 140
206
темной только последний. Если сложить величины, показывающие про-
исшедшие изменения в содержании кислорода в обеих банках, можно
получить общую продукцию кислорода, а путем пересчета (за вычетом
кислорода, пошедшего на дыхание) общее количество углерода, образо-
вавшегося в растительных клетках. В результате подобных наблюдений
Рэлей получил (для пролива Long Island Sound) продукцию в 1000 г
углерода под 1 л«2.
Рэлей (1941) приводит ряд данных, характеризующих годовую^
продукцию фитопланктона в граммах углерода под 1 м2 (табл. 72).
Если в тропических водах биомасса планктона, возможно, мала, то
в некоторой степени это компенсируется более значительной глубиной слоя,
фотосинтеза и круглогодичной вегетацией фитопланктона.
Компенсационно* — ,, Положение точни
точна оптимального роста
Рис. 105. Положение компенсационной точки в разных частях океана.
Рэлей (1941) определил биомассу фитопланктона в заливе Мэйнг
в период максимального развития, в 2 г углерода под 1 м2. Гарвей
(1942) указывает, что у побережий Южной Георгии, в зоне океана, харак-
теризующейся чрезвычайной продуктивностью, в верхнем слое в 50 м,
содержание углерода в фитопланктоне равнялось 7,2 г под 1 м2.
Если в Средиземном море макрофиты, благодаря большой прозрачно-
сти воды, могут существовать на глубине, несколько превышающей 100 лц
то также глубоко лежит граница вегетационной зоны и в Саргассовом
море. Эксперименты, произведенные Кларком (Clark, 1936) в
Саргассовом море с диатомеями, показали, что интенсивный процесс
фотосинтеза совершается и глубже 100 м. Но других таких мест в мировом
океане нет. При приближении к побережьям и во внутренних частях, вдаю-
щихся в сушу морей, нижняя граница растительной жизни повышается
вплоть до нескольких метров (рис. 105). Согласно опытам Гардена
и Грапа (1927), в Осло-фиорде в марте «компенсационная точка» для
диатомовых находилась на глубине 10 м\ глубже дыхание превалировало
над ассимиляцией. В заливе Loch Striven, по данным Маршалла
и 0 р р а (1928), «компенсационная точка» (для диатомеи Coscinosira poly-
chorda) находилась летом на глубине 20—30 м, а зимой близко к поверхно-
207
сти. В Английском канале «точка компенсации» летом опускается до 45 м
(Jenkin, 1937), а зимой подымается к самой поверхности.
Совершенно очевидно, что количество питательных солей в верхнем
слое океанических вод обусловливает степень развития фитопланктона.
Также совершенно очевидно, что развитие зоопланктона стоит в прямой
зависимости от развития фитопланктона. Поэтому очень большой интерес
представляет сделанное Ресселем (Russel, 1935) сопоставление
количественного соотношения фосфатов и личинок рыб за ряд лет (1924—
1937) в одной и той же точке Английского канала (рис. 106).
Рис. 106. Соотношение в отклонении от среднего
зимнего максимума (горизонтальная линия) содер-
жания фосфатов (заштрихованные столбики) и сред-
него количества личинок нерестящихся летом рыб (бе-
лые столбики) (по Ресселу).
Иногда и в открытых частях океана могут наблюдаться весьма большие
•скопления фитопланктона. Так, Аллен (Allen, 1936) наблюдал
в северной части Тихого океана рекордное количество — 220 000 диато-
мовых в одном литре воды. Однако в береговой зоне фитопланктон, как
правило, богаче, как считает Ломани, в 5 раз. Л о м а п н также высчи-
тал, что хотя растительная биомасса в Кильской бухте может опускаться
ниже биомассы животных, однако продукция фитопланктона в летнее
время обычно превышает потребление животными и этот излишек может
достигать 29 мм6/100 л воды. В течение же зимы продукция фитопланк-
тона может даже не перекрывать потребность животных в питании.
3. ЗООПЛАНКТОН
Состав зоопланктона. В составе животного планктона можно различать
две основные группы: временные компоненты планктона (меропланк-
тон) — яйца и личинки различных рыб и донных животных, и постоян-
ные компоненты (голопланктон) — жгутиковые, фораминиферы, радио-
лярии, инфузории, копеподы, эвфаузииды, амфиподы, полихеты, крыло-
ногие, оболочниковые, некоторые головоногие и разные другие группы
беспозвоночных.
1) Жгутиковые. Эта наименее изученная группа морских ор-
ганизмов имеет, очевидно, большое значение в жизни моря. Их количество
бывает огромно. Так Phaeocystis pouchetii обусловливает цветение обшир-
ных пространств моря. В Черном море жгутиковое — Nocliluca miliar is,
развиваясь в массе, вызывает свечение моря.
2) Фараминиферы. Из них особенно сильно распространены
несколько видов рода Globigerina, встречающихся в таком количестве,
что скопления их скелетов составляют обширнейшие зоны морских от-
ложений.
3) Радиолярии. Также весьма обширная группа планктона. Бес-
численные скелеты радиолярий входят в состав распространенного в
океане радиоляриевого ила.
208
4) Инфузории. Среди инфузорий значительного развития в планк-
тоне достигает только группа Tintinnoidea. Количество их бывает так
велико, что в 1 м* воды можно встретить более 100 000 экземпляров
различных Tintinnoidea.
5) Кишечнополостные. Наиболее распространенными яв-
ляются различные медузы, сифонофоры и гребневики, последние часто в
громадном количестве.
б) Черви. Типично планктонных (голопланктонных) червей мало.
Можно указать полностью планктонных щетинкочелюстных (Ghaetogna-
tha), некоторых немертин (Pelagonemertes), некоторые группы полихет.
Большей частью черви появляются в планктоне только в определенный
период года, или в стадии личинок, или, наоборот, взрослых форм, поды-
мающцхся в верхние слои моря для размножения.
Иногда весьма значительную часть планктона могут составлять сагит-
ты (Ghaetognatha).
7) Ракообразные. Это наиболее разнообразная и многочислен-
ная часть животного планктона. Особенно сильное развитие имеет группа
Copepoda. Буквально нельзя взять ни одной пробы планктона в любом
море, чтобы там не было веслоногих рачков на всех стадиях развития.
В настоящее время известно не менее 750 видов морских Copepoda. Наи-
более широко распространено в открытом море семейство Calanidae.
Рачок Calanus jinmarchicus распространен от тропиков до Северного полю-
са и обитает на самых различных глубинах. Кроме Copepoda в планктоне
встречаются Cladocera, Ostracoda, Cirripedia (личинки), Euphausiacea,
Mysidacea, Amphipoda, Isopoda и Decapoda. Часть из них ведет полно-
стью планктонный образ жизни (голопланктонные), другие встречаются
в планктоне только на ранних стадиях развития (меропланктонные).
8) Моллюски. Этот класс представлен в планктоне преимущест
венно группами Pteropoda, Heteropoda и Cephalopoda. Все другие группы
донных моллюсков встречаются в планктоне в личиночном состоянии.
9) Оболочники. Среди оболочников имеются настоящие планктон-
ные организмы. Таковы Appendicularia и Salpae.
10) Рыбы. Помимо многочисленных пелагических рыб, в планктоне
постоянно присутствуют икра и мальки различных донных рыб.
Кроме того, почти все обитатели дна дают в планктон молодые стадии
развития, что имеет двоякое значение: в верхних горизонтах моря ли-
чинки находят для себя обильное питание, с другой стороны, они перено-
сятся течениями в новые районы и таким образом осуществляют расселе-
ние. Чем ближе к берегам, тем больше в планктоне личиночных стадий
донных организмов. Видовое разнообразие зоопланктона, так же как и
количество входящих в его состав классов, сравнительно очень невелико.
В качестве настоящих планктонных обитателей фигурирует, если не счи-
тать паразитические формы, всего 14 классов. Это может быть показано
на планктоне тропических и субтропических частей мирового океана.
Основные группы, составляющие зоопланктон, это — кишечнополо-
стные (свыше 400 видов), ракообразные (около 1200 видов), причем глав-
ную роль играют веслоногие (около 750 видов), амфиподы (свыше 300 ви-
дов) и Euphausiacea (свыше 80 видов), крылоногие моллюски и Heteropo-
da (около 180 видов) и оболочниковые (свыше 100 видов). Количественно
иногда значительную роль в планктоне играют щетинкочелюстные, отли-
чающиеся небольшим видовым разнообразием (около 30 видов). Общее же
число видов зоопланктона, если не считать простейших, пелагических рыб,
головоногих, икру и личиночные стадии рыб и донных беспозвоночных,
составляет свыше 2000 видов, иначе говоря, видовое разнообразие планк-
тонных беспозвоночных раз в 70 меньше видового разнообразия бентоса,
14 Зенкевич, т. I
209
причем многие основные группы бентоса либо совсем (кроме личиночных
стадий) не образуют пелагических форм (губки, двустворчатые моллюски,
мшанки, плеченогие, иглокожие), либо дают их в очень небольшом коли-
честве (полихеты, десятиногие и равноногие ракообразные и др.).
Горизонтальное распределение зоопланктона. С переходом из высоких
широт в тропические области можно наблюдать весьма значительное уве-
личение качественного многообразия планктона, что можно отлично
иллюстрировать сводной таблицей Р е с с е л а (1935) (табл. 73).
Таблица 73
ЧИСЛО ВИДОВ ГОЛОПЛАНКТИЧЕСКИХ ЖИВОТНЫХ В РАЗЛИЧНЫХ ЧАСТЯХ
Группы
Трахи- и наркомедузы . 4
Calico phorae..........
Physophorae ...........
Ктенофоры .............
Немертины .............
Tomopterldae (Polychaeta) I
Chaetognatha...........i
Cladocera..............;
Copepoda ..............
Hyperiidae.............
Gammarldae.............
Euphausiacea...........
Thecosomata .
Gymnosomata
Heteropoda . .
Appendicularia
Doliolldae . . .
Salpidae....
Pyrosomidae .
ОКЕАНА (ИЗ РЕССЕЛА)
2
90 21
64
29
69(?)
38 '
18
3
489
250
20
46
! 44
37
90
48
11
21
8
134
72
34
80(?)
52
44
30
7
754
292
44
85
51
41
90
61
12
25
8
Всего.........
1378 338 1916
Приведенная таблица показывает, что тепловодные формы составляют
72% от общего количества известных голопланктических форм. Глубо-
ководные составляют 19%, космополиты — несколько меньше 1% (0,8%),
столько же дают и биполярные формы (0,9%). Кроме того, из приведен-
ной таблицы можно сделать еще один любопытный вывод: антаркти-
ческая (-f- субантарктическая) планктическая фауна значительно богаче
арктической (Ц- арктическо-бореальной и бореальной). Первая даст
4,7%, а вторая несколько меньше 3% от общей массы голопланкти-
ческих форм, при этом в первую группу не входят потальные формы, а
во вторую входят бореальные. Если бы не это, разница была бы еще резче.
Причины такого различия ясны: южное полушарие с его громадными вод-
210
ними просторами и широким сообщением со всеми тремя океанами больше
способствовало формированию новых форм.
Рис. 107. Распределение видов семейства Calanidae в % от общего количества
видов (114) (ориг.).
Среди морской планктонной фауны наиболее изученными являются
Copepoda, а среди них — Calanoida. На рис. 107 и 108 приведены карты
Рис. 108. Распределение видов семейства Centropagidae в % от общего количества
видов (68) (ориг.).
видового разнообразия двух семейств Calanoida — Calanidae и Centro-
pagidae. Обе карты показывают, что наибольшее видовое разнообразие
* 211
приходится на северную часть Атлантики. Дальше на север количество
видов резко убывает. С другой стороны, с переходом от открытых частей
океана к краевым морям и особенно к морям внутренним и опресненным
количество видов также убывает, что видно также и на следующих таб-
лицах. Числа на картах и таблицах обозначают количество видов в % от
общего числа видов (табл. 74)
Таблица 74
Calanldae Centropagldae Pontellldae Сев. Атлангика 53 46 60 Баренцево море 13 15 20 Белое море 4 G 14
Calanidae Centropagidae Pontellidae Сев. Атлантика 53 46 60 Северное море 28 15 40 Балтийское море 3 9 7
Calanidae Centropagidae Pontellidae Центр. Атлантика 35 40 33 Средиземное море 22 27 33 Черное море 4 2 14
Calanldae Centropagidae Pontellidae Индийский океан 19 18 27 Красное море 5 3 14
Следовательно, везде, где мы имеем переход от открытых морей к
внутренним, наблюдается падение количества видов. Несомненно, одной
из причин убыли количества видов рассмотренных рядов служит пониже-
ние солености, однако, одним этим всего явления объяснить нельзя, так
как такое же уменьшение наблюдается и при переходе от Атлантического
океана к Средиземному морю, где соленость не понижается.
Убывание качественного разнообразия, при продвижении из теплых
морей на север и на юг, можно наблюдать и для всего зоопланктона, что
и показано на табл. 75, в которой не учтены космополиты.
Таблица 75
Арктические и бо- реальные воды Теплые воды Антарктические и суб- антарктические воды
Coelenterata Polychaeta (Tomopteridae) Chaetognatha Crustacea Из них Copepoda Mollusca Tunlcata (все) 128 видов 16% 1 3 2 9 33 4 17 3 1 2 3 3 622 вида 79% 38 88 18 82 808 90 489 90 171 95 91 90 41 вид 5% 4 9 2 9 56 6 37 7 4 3 8 7
Итого 185 2237 152
Таким образом, за исключением космополитов, общих видов для раз-
личных морей, в теплых тропических и субтропических водах живет
почти 90% всех видов планктонной фауны, а в арктических и бореальных
только 4% и в антарктической и субантарктической только 6%.
Следовательно, имеется резкое убывание числа видов при движении
па север в северном полушарии и при движении на юг в южном полушарии.
Меньшие числа видов в Тихом океане и отчасти в тропической зоне
Атлантического океана надо относить за счет меньшей изученности фау-
212
5°сш 3°сш
гз°зо г8°зл
о
50
ЮО
150
§гоо
\250
300
30°зд 32°лд 3<Лп
Рис. 109. Количественное распределение планк-
тонной водоросли Coccolithophora fragilis
на разрезе поперек южноэкваториального
течения Атлантического океана (по Ломанну).
В количестве экз. на литр.
ны, так как для подавляющего числа организмов Тихий океан дает зна-
чительно большее видовое разнообразие. Если небольшие полузамкнутые
и замкнутые водоемы в определенном смысле могут представлять благо-
приятные условия для формирования новых видов, то открытые просторы
океанов и морей всегда должны характеризоваться большим видовым раз-
нообразием флоры и фауны, нежели ограниченные краевые моря.
Неритический и океанический планктон резко различаются по своему
составу. Из Copepoda для океанического планктона Атлантического океа-
на характерны Centropages typicus, Oithona atlantica, из медуз Aglantha
digitalis. Сифонофоры и летучие рыбы также типичные обитатели океани-
ческих просторов, а коловратки и бесчисленные личиночные стадии дон-
ных животных столь Яле типичны для неритического планктона. На рис.
109 дано распространение плотности океанической планктонной водо-
росли Coccolithophora fragi lis,
характерной для южноэквато-
риального течения Атлантиче-
ского океана.
Горизонтальное количест-
венное распределение планк-
тона в мировом океане весьма
неравномерно, что было впер-
вые установлено В. Гензе-
ном по материалам экспеди-
ции на судне «Holsatia» (1885)
и главным образом планктон-
ной экспедиции (1889 г.). В об-
щем, как данные двух этих
экспедиций, так и все после-
дующие, установили полную за-
висимость распределения плот-
ности планктона от близости
к побережьям и от гидрологи-
ческих условий, в основном от
степени вертикального пере-
мешивания. Это совершенно
понятно, так как развитие
тона, а этот последний требует для развития достаточное коли-
чество питательных солей. Чем ближе к берегам, чем сильней вертикаль-
ная циркуляция, тем обильнее может развиваться планктон. Совершенно
очевидно такя^е, что при всех прочих равных условиях продуктивность
планктона будет тем выше, чем более благоприятны для его развития тем-
пературные условия. Таким образом, наилучшие условия для развития
планктона будут иметь место при обилии питательных солей и высокой тем-
пературе (20—30°).
Низкая температура не останавливает развития планктона, но только
понижает темп его продуцирования. Отсутствие питательных солей
останавливает развитие планктона. Помимо этих основных закономерно-
стей, степень развития планктона зависит и от всех других факторов сре-
ды — освещения, газового режима, солености, скорости течения, выеда-
ния другими организмами и т. д.
Основной характер распределения плотности планктона в Атланти-
ческом океане был дан на рисунках, приведенных выше.
Олиготрофные районы — это районы со слабой горизонтальной и осо-
бенно вертикальной цикруляцией. Политрофные районы характери-
зуются хорошей вертикальной и горизонтальной циркуляцией.
зоопланктона зависит от фитопланк-
213
Довольно широко распространено мнение, что наиболее продуктив-
ные зоны мирового океана приурочены к умеренным зонам обоих полу-
шарий, а тропические зоны сравнительно бедны жизнью. Однако надо
признать, что это допущение не базируется пока на достаточном числе
точных фактов и может быть принято только условно. Только у нас в СССР
в практику биологических исследований в море прочно вошел метод опре-
деления биомассы и планктона и бентоса. За рубежом, к сожалению,
до настоящего времени общепринятым является, особенйо для планктона,
вести учет по числу экземпляров, что дает очень мало пригодный мате-
риал для сравнения. Что же касается бентоса, то для всего южного
полушария и для теплого пояса океана по имеется пока ни одной коли-
чественной пробы, если но считать нескольких проб в Средиземном
море.
Рис. 110. Количественное развитие фитопланктона и содержание
фосфора в прибрежных водах Адриатического моря (по Ерце-
говичу).
1 — количество клеток на литр, 2 — содержание фосфора в лсгЛиЗ.
Планктон Средиземного моря обрабатывался И с с с л е м (I ssel),
Песта (Pest а), Штейером (Steuer), ав последнее время Е р ц с-
говичем (Ercegovic, 1936, 1940) в части фитопланктона и
Г а м у л и н ы м (G а m uli п, 1938, 1939) в части зоопланктона.
К сожалению, указанные авторы учитывали также только количе-
ство экземпляров, но не биомассу.
Общие количественные данные по распределению планктона по всему
Средиземному морю приводятся Джесперсеном (Jespersen, 1923)
по данным Датской экспедиции 1908—1910 г. (см. рис. 155). Числа на
карте соответствуют общему объему планктона, выловленного за 1 час
большой круглой сетью с диаметром входного отверстия в 2 м. Как пока-
зано на карте, в пределах Средиземного моря количество выловленного
планктона составляло несколько сот см3 (в среднем 266,4). Только в са-
мой западной части моря, в сфере действия течения из Атлантического
океана, количество планктона возрастает в 4—5 раз, так же как и в Мра-
морном море за счет черноморских вод. В Арктическом же океане, так же
как и в Черном море, количество планктона, собранного той же сетью,
возрастает в 14—17 раз. Как показывают данные А. Е р ц е г о в и ч а
(1936, 1940), прибрежные воды Адриатики чрезвычайно бедны фосфатами
(рис. 111). В течение всего 1935 г.’ количество фосфатов в поверхностном
214
слое 0—10 м не подымалось выше 2,5 мг]м2. При этом основная масса
фитопланктона приурочена к самому верхнему слою воды с резким пиком
в июне, т. е. еще до самого сильного прогрева воды (рис. 112). В осталь-
ные периоды года количество фитопланктона держится на очень низком
уровне. Но даже и наивысшие числа (300—500 тыс. клеток в литре) чрез-
вычайно малы, по сравнению хотя бы с прибрежными участками Черного
Рис. 111. Годовой ход содержания карбонатов и количества кокколитин
в прибрежных водах Адриатического моря в слое воды 0—10 м (по Ерце-
говичу).
1 — содерж. карбонатов в м.г/л\ 2 — число клеток на л.
моря (см. II том). Кроме того, для форм с известковым панцырем, в част-
ности для кокколитин, устанавливается связь между количеством и содер-
жанием карбонатов в воде (рис. 111).
Приведенные данные по фитопланктону Адриатики хорошо согла-
суются с данными по зоопланктону, приводимыми Т. Г а м у л и н ы м
(1939).
Рис. 112. Годовые изменения в количестве фитопланктона (в ко-
личестве клеток на литр) в прибрежных водах Адриатического
моря (по Ерцеговичу). Три кривые изображают содержание
фитопланктона на глубине 0, 10 и 40 м.
Некоторый сравнительный материал по продуктивности зоопланктона
в тропической зоне Тихого океана был получен Английской экспедицией
на Большой Барьерный Риф. Согласно Р е с с е л у (1934), лагуны этого
рифа несколько богаче по числу планктеров, чем Северное море и Англий-
ский канал. В этих последних районах средние месячные количества
215
экземпляров зоопланктона, выловленного вертикальным ловом 50 см
сетки, равнялись 2815 экземпляров. Подобные же ловы в течение года в
лагуне Барьерного Рифа дали в среднем 5684 экз. на улов. Следует однако
учесть при этом, что тропические копеподы мельче, чем в Северном море.
Интересно, что фитопланктон в лагунах Барьерного Рифа, по данным
Маршалла (1933), всегда значительно беднее, но вероятно продук-
тивность его относительно значительно выше и не прерывается в зимнее
время. Это хорошо можно видеть на рис. ИЗ, где дано сравнение количе-
ства экземпляров диатомовых в планктоне в различных частях Северного
Атлантического океана и в одной из лагун Большого Барьерного Рифа.
Рис. 113. Количество диатомовых в планктоне (по Маршаллу).
Л — в среднем месячном количестве экземпляров, Б—изменение по сезонам года. 1— Loch Striven
1926 (один из заливов ю.-з. Англии), 2— Ла Холла 1922 (Калифорния), 3— Плимут 1915, 4 —
Isles 1928 (лагуна в северной части барьерного рифа).
Вертикальное распределение зоопланктона. Рассматривая* вертикаль-
ное распределение зоопланктона, мы можем установить, что обитание
большинства организмов в различных слоях воды подвергается очень
сильным изменениям как во времени, так п в пространстве.
Небольшое число форм планктона обитает либо только в поверхност-
ных, либо только в глубинных слоях. Некоторые организмы планктона,
обитающие в одной области океана в поверхностных слоях, в другой мо-
гут быть характерными для глубинных слоев. Один и тот же вид, на раз-
ных стадиях своего развития, может обитать в различных слоях воды.
Кроме того, в период размножения взрослые организмы некоторых вп-
216
дов меняют свое местообитание. Многие компоненты планктона в различ-
ное время суток и в различные сезоны года обитают в различных слоях
воды.
Уже во время экспедиции «Челленджера» было установлено, что даже
в ловах с глубины свыше 3500 м было еще довольно много (относительно)
живых организмов. Трудами экспедиции на «Вальдивии» планктонная
жизнь была обнаружена на всех исследованных глубинах.
По характеру планктона всю водную толщу океанов и морей различ-
ные исследователи разделяют по-разному. Наибольшей известностью
пользуется деление, предложенное Далем (Dahl, 1894) и Фо у-
лером (Fowler, 1898) (табл. 76).
Таблииа 76
_______________________'__________________________________._____J___
Глд бины 0—200 200—1000 > 1000 Дио
Даль Поверхностная область Срединная область Глубинная область
Фоулер Epiplankton Mesoplankton 80 м над дном Hypoplankton
Учитывая громадное значение для организмов освещенности разных
слоев моря и распределение фитопланктона, можно дать следующую
схему вертикального подразделения зоопланктона (табл. 77).
Таблица 77
Глубина в .и Название Световые условия (по Зернову) Характер планктона Назва- ние планк- тона
зоны слои значе- ние фито- планктон зоопланктон
0 10 50 100 200 500 1000 Поверхностная верхний освещенный i Продуцирующая область Очень много Интенсивн. фотосинтез. Очень много. Особенно яиц и личинок. Обычно большое разнообразие видов. Имеются специ- альные поверхностные виды. Поверхностный
средний
порядочно
нижний
Переходная верхний полуосве- щенный Потребляющая область мало Много. Преимущест- венно «средние» и стар- шие возрасты. Специ- альных видов почти нет. Среднеглу- бинпый
нижний Живых растений пет
Глубоководная верхний 1 малоосвсщен- п ый Мало и очень мало. Прп глубинах более 1000 м. появляются специаль- ные глубоководные виды. Глубоцоводпый
нижний неосвещенный
50 м над дном Наддонная Виды, живущие в самых придонных слоях. |п ; Наддон- 1 ный
217
С глубиной количество планктона резко падает. В этом отношении
весьма показательны данные Гентшеля (1938) по наннопланктону
Атлантического океана (табл. 78).
Таблица 78
Глубина в м Количество инди- видов в 1 л Процентное распре- деление
! О 1 103 801 1
50 • ; 36 159 95,6
100 ! 5 171
200 915 3,5
400 | 204
700 ' 104 и,о
1000 ; 78
2000 i 43
3000 ; 23 0,1
4000 . 22
5000 19
Таково же примерно и распределение биомассы всего планктона. При
этом обычно наибольшее количество организмов сосредоточено не в самом
поверхностном слое, а на глубине около 25 м.
Рис. 114. Вертикальное распределение Protophyta (с хро-
матофорами) в Атлантическом океане на меридиональном
разрезе (по Ломанну). На графике хорошо выражено количе-
ственное обилие фитопланктона в высоких широтах и бедность
его в тропической зоне.
Следовательно, в поверхностном слое в 100 м находится 95,6% всего
наннопланктона и это дает нам право считать верхний 100-метровый слой
наиболее богато продуцирующей зоной (рис. 114). Уже в промежуточной
зоне количество наннопланктона очень мало. Однако, биомасса зоопланк-
тона здесь еще значительна. В глубоководной же зоне плотность населе-
ния всех групп планктона очень мала.
Мы не можем подробно характеризовать отдельные слои воды, в каж-
дом будут свои особенности. Остановимся лишь на некоторых моментах,
В поверхностном слое морей умеренной зоны планктона много во всем
столбе воды в 100 м. В субтропических и тропических морях особенно богаты
только слои воды в 10—50 л, до слоя температурного скачка. Например.
Уда (U d а, 1934) для Японского моря указывает, что в верхних 50 м
в 8 раз больше планктона, чем в слое 50—100 м и в 16,5 раз больше, чем
218
в слое 100—200 м. Бродский (1937) также подтверждает это, по в
меньших масштабах — верхние слои до 50—75 м в 5 раз богаче нижних.
Необходимость выделения верхнего слоя в поверхностной зоне (0—
10 м) объясняется тем, что в полярных морях этот слой является особен-
но богатым в отношении фитопланктона, а в южных морях, наоборот,
сильное освещение солнцем препятствует массовому развитию растений
и скоплению животных в поверхностном слое. Кроме того, сильный про-
грев поверхностного слоя также служит препятствием для массового раз-
вития многих видов планктона.
Средний слой (10—50 м) поверхностной зоны — это главный проду-
цирующий слой: здесь особенно много фитопланктона и зоопланктона.
Помимо водорослей в нем в массе можно встретить яйца, личинок и моло-
дые стадии различных живущих обычно глубже видов. Много здесь про-
стейших — жгутиковых, инфузорий и др.
Переходная зона (100—500 м) не имеет своего специфического состава
планктона. Среди фитопланктона в ее верхнем слое (100—200 л<) обычно
присутствуют опустившиеся из поверхностного слоя клетки водорослей.
Обилие животного планктона сильно варьирует в течение суток; зоопланк-
тона много только днем.
Различные слои глубоководной зоны весьма отличны по обилию планк-
тона и по специфичности фауны. Чем глубже, тем меньше количество планк-
тона, зато разнообразие не всегда уменьшается. В ЛедовитОхМ океане коли-
чество видов с глубиной обычно возрастает. Так Дамаси Кефэд,
(Damas and К о е f о е d, 1909) для Гренландского моря указывают
следующее количество видов (табл. 79):
Таблица 79
Глубина....... 0—20 — 100 — 200 — 500 — 1000 — > 100J
Количество видов . 6 11 13 15 17 21
То же явление отмечает В. А. Я ш н о в для центральной части Се-
верного Ледовитого океана (см. II том).
В обоих приведенных случаях увеличение качественного разнообра-
зия связано с находящимися на глубине атлантическими водами, пере-
крытыми арктическими.
При этом количество организмов закономерно падает с глубиною (это
характерно вообще для всех морей!). В той же работе Дамас и Ке-
фэд указывают на изменение численности С alarms hyperboreus
в связи с глубиной (табл-. 80).
Таблица 80
Глубина............. 0—10 20—100 210—400 м
Количество организмов . . . 1000 2 4
В некоторых случаях, как показал Б. Л и в и т, количество мезо- и
макропланктона на больших глубинах может несколько возрастать. Сборы
производились у западного побережья Северной Америки (в районе
Вудс-Холла). Оказалось, что крупный планктон в главной своей массе
(до 94%) сосредоточен в верхнем слое в 800 м. Однако, после минимума
(на глубине 300—700—1000—1200 м) имеет место опять некоторое по-
вышение за счет типичных глубоководных обитателей.
Разница между верхним и нижним слоем глубоководной зоны весьма
условна и в очень большой мере зависит от общей глубины моря и его
происхождения. Так, например, в Японском море, в связи с его происхо-
ждением, отсутствует специфическая глубоководная фауна, а рядом, в
Тихом океане, на соответственных глубинах она хорошо представлена.
На глубинах свыше 3000 м в Атлантическом океане, как указывает Б. Л и-
в и т (1935), обитает еще довольно обильный планктон из Copepoda (Met-
219
ridia), Acanthephyridae и др. Decapoda, пелагических Ostracoda, Euphau-
siacea, си фонофор и рыб. Большинство из этих форм типичные глубо-
ководные обитатели.
Вообще же, в составе глубоководной фауны имеются представители
различных групп животных. Многие из них своеобразно окрашены
и фосфоресцируют.
Наддонный планктон, состоящий из обитателей небольшого слоя воды
у самого дна, очень сильно отличается в зависимости от общей глубины
места. Наддонная фауна в глубоких морях совершенно не изучена. Для
глубин до 100—200 м и особенно для более мелких мест характерны мас-
сы мизид, кумацей и других высших раков, веслоногих рачков — Награ-
cticoida, червей и других животных, которые то находятся над дном,
то ползают по дну. Ночью многие представители гипопланктона поды-
маются в более верхние слои, а в мелководных районах даже в самые по-
верхностные слои, как, например, в Каспийском море.
Явления суточной и сезонной вертикальной миграции нарушают
четкое разграничение фауны по вертикали.
Несмотря на отмеченные трудности в определении вертикальной при-
уроченности различных видов, мы можем указать ряд представителей,
которые преимущественно обитают в том или другом слое воды. Для
примера возьмем северную Атлантику. К видам, предпочитающим поверх-
ностную зону, относятся: Oithona similis, Paracalanus parvus. Centropages
hamatus и C. typicus, Evadne nordmanni, Podon leukarti, молодь Calanus
finmarchicus, Pseudocalanus elongatus, Oikopleura и Fritillaria. К ви-
дам, предпочитающим переходную зону, относятся: Calanus jinmarchicus,
Metridia longa, M. lucens, Pseudocalanus elongatus, Aglantha digitalis,
Sagitta elegans. В нижнем слое переходной зоны часто можно найти:
Euchaeta norvegica, Microcalanus pusillus, Undinella oblonga, Gaidius sp.
sp., Chiridius sp. sp., Krohnia hamata и др. В глубоководной зоне в верх-
нем слое обычны виды родов Gaetanus и Scolecithrix, а в нижних слоях ви-
ды родов Heterorhabdus, Augaptilus, Copilia и др.
Довольно точно распределяются различные виды радиолярий по раз-
личным глубинам (Н а е с к е г, 1908). Поверхностной зоне особенно
свойственны виды Collidae. Переходной зоне свойственны виды Challen-
geria. Верхние слои занимают виды рода Tuscarora, а глубже Aulographis
pandora. Еще глубже главное значение переходит к видам рода Aulospa-
this. Наконец, нижние слои глубинной зоны населены преимущественно
видами рода Pharyngella (Ph. gastula), а также видами родов Porcupinia,
Porospathis, Couchopis и др.
Таблица 81
Глубина в м
0-50 50—100 100—200 200—500 500—1000
Calanus finmarchlcus...............
Calanus hyperboreus................
Oithona simllls....................
Oithona plumifera..................
Euchaeta norvegica.................
Chiridius armatus..................
» obtusifrons..................
Oncaea conifera....................
Anomalophora niagna................
Microcalanus pusillus..............
Euchaeta glaclalis.................
220 z
Все приведенные примеры являются в значительной мере условными.
Так, например, в относительно неглубоком море, как Баренцово, мы мо-
жем на глубинах в 200—300 м встретить большинство видов, которые в
Северной Атлантике живут на глубине в 500 м. В Английском канале,
при общей глубине всего около 50 .и, можно встретить почти всех предста-
вителей переходной зоны. Очень часто можно встретить поверхностные
виды и на больших глубинах. Так, Дамас и Кефэд для Норвеж-
ского моря указывают следующую встречаемость различных видов
(табл. 81).
Следовательно, в глубоководной зоне (верхнем ее слое), наряду с не-
которыми батипланктонными видами, живут и представители поверхност-
ной и среднеглубинной фауны. Понятно, эти виды находятся здесь в
меньшем количестве, чем в вышележащих слоях.
Так же трудно провести различия между видами, обитающими в про-
межуточной зоне и в глубоководной. Если мы возьмем различных пред-
ставителей макропланктона, которые обитают в большом количестве в
промежуточной зоне, то оказывается, что они же живут и в глубоковод-
ной зоне и иной раз в таких же количествах, как и в промежуточной зоне.
Огромное большинство обитателей поверхностной и переходной зоны
являются эврибатными видами. В различных морях они могут обитать на
различных глубинах, в каждом же отдельном водоеме имеется обычно
хорошо выраженная дифференцировка в вертикальном распределении,
даже при относительно малых глубинах. Так, например, Р е с се л
(Russell, 1926) приводит очень наглядное изображение миграций
планктона (рис. 115) и различий в вертикальном распределении некоторых
видов, характерных для определенного времени года в районе Плимута
в ясную и пасмурную погоду (рис. 116).
Таким образом, вертикальная дифференциация компонентов планктона
существует как в отношении многокилометровых толщ моря, так и в
отношении сравнительно небольших глубин.
Несмотря на то, что видов, специально приуроченных к определенным
глубинам (стенобатных), за исключением настоящих эпи- и батипланктон-
ных, очень мало, мы можем все же в отношении ряда организмов, обита-
телей поверхностной и промежуточной зоны и верхнего слоя глубинной
зоны, установить их тенденцию в избирании определенного слоя. Так,
например, в районе Фарерского канала (по Fowler) частота встре-
чаемости различных видов на разной глубине весьма различна
(табл. 82 и 83).
Таблица 82
0—100 м | > 100 м
Acartia clausi 70% 30%
Calanus finmarchtcus 47 53
Eucalanus attenuatus 28 72
Metrldia longa 26 74
Euchaeta norvegica 24 86
Pleuromamma abdominaHs 9 91
Таким образом у различных видов имеется определенно выраженная
тенденция к предпочтению какого-либо одного слоя. Одни виды, как,
например, Acartia clausi предпочитают поверхностную зону, другие рас-
пространены равномерно в различных слоях, как Calanus finmarchiciis,
третьи главным образом сосредоточены в слое глубже 100 м.
221
222
Таблица S3
КОЛИЧЕСТВО ЭКЗЕМПЛЯРОВ РАЗЛИЧНЫХ ВИДОВ EL’PIIAUSIACEA НА РАЗЛИЧНЫХ ГЛУБИНАХ АТЛАНТИЧЕСКОГО ОКЕАНА,
У СЕВЕРО-АМЕРИКАНСКОГО ПОБЕРЕЖЬЯ (ПО ЛИВИТУ)
Глубина в м 200 400 603 800 1000 1200 1400 1600 1800 2000 2200 2400 2600 2800 3000
Количество организмов в лове: 7 8 8 7 6 6 6 7 5 6 6 4 3 3 2
Thysanopoda orient alts 8 11 1 1 I 1 1 1 1 1 1
Thysanopoda aequalls 8 1 61 i I 1
Euphausia tcncra 1 92 110 97 11 16
» krolinll 3,064 49 4 4 3 3 1
» bcmigfbba 4 753 243 24 12 4 10
» pseudogibba 17 69 12 14
Thysanocssa gregarla 2,439 77 230 1 16 4 5 4
> parva 2 262 1,911 466 699 926 2,804 91 88 1,540 480 1 1 4
» longlcaudata.... 3 1,504 222 201 328 94
Nematoscelis megalops 11,670 29,694 2,043 7 1 466 8
» microps 82 231 92 62 24 307 23 2 7 2
» tenella 8 6 14 12 1
Slylochcrlon longlcorne 555 347 14 5 2 1 1 8
» maximum 2 5 8 i
Bcnthbuphausia amblyops .... 1 1 । > । 4 1 5 7 i 3 3 l i 2 2 1 I 1 1
Необходимо всегда помнить, что вертикальное расположение различ-
ных видов характерно лишь для определенного места и времени (года,
п суток).
и
нал масса планктонной фауны является эв-
в разное время предпочитает самые различ-
оатную и эврибатную. Ochi
рибатной и в разных морях
ные слои
воды.
<Я7
Рис. 116. Вертикальное распределение (в %) различных планктонных организ-
мов в районе Плимута летом, в ясную (А) и пасмурную (Б) погоду (по Ресселу).
В распределении организмов
видеть несколько основных типов,
морях в поверхностных слоях, а в
второй группы (эврибатных) можно
Обычны виды, живущие в холодных
умеренных и теплых районах препму-
щественно в промежуточной и даже глубоководной зоне (например, Cala-
nus finmarchlcus). Таково же распределение и Eukrohnia hamata в Атлан-
тическом океане. В арктической и антарктической областях этот рачок
живет в поверхностной зоне, в субтропической и тропической — в глубин-
ной зоне и может служить примером зависимости распределения от темпе-
ратуры. Часто на вертикальное распределение влияют течения. Например,
в Гренландском море Calanus hyperboreus и другие высокоарктические
виды в восточной части моря живут преимущественно в промежуточ-
ной и глубинной зонах, а в западной части они же являются типичными
представителями планктона поверхностной зоны.
Замечательный пример влияния течений на вертикальное распреде-
ление планктона мы увидим в дальнейшем на примере Северного Ледови-
того океана.
Вертикальные миграции зоопланктона. Многие формы планктона на
различных стадиях развития обитают на различной глубине. У большин-
ства видов, предпочитающих во взрослом состоянии глубинные слои,
яйца и ранние личиночные стадии держатся в поверхностных слоях.
В связи с этим в период размножения в поверхностные слои воды поднима-
ются виды, обычно живущие на глубине нескольких сот метров. Поэтому
в поверхностной зоне главную биомассу зоопланктона составляют яйца
и различные личинки как планктонных, так и донных животных.
Но бывает и наоборот. У сифонофоры Velella spirans, обитательницы
самого поверхностного слоя моря, молодые стадии развития живут на
большей глубине. Иногда разные полы одного животного (Metridia longa)
живут на разных глубинах (Б о г о р о в, 1933).
Изменяется вертикальное распределение различных видов также и в
связи с периодом размножения. Наиболее типичным является поднятие
глубоководных животных в поверхностную зону для размножения. Часто
в этот период даже придонные животные (например, палоло) подымаются
к самой поверхности моря, где и размножаются.
Сезонные изменения в вертикальном распределении планктонных ор-
ганизмов находятся обычно в связи с размножением и фазами развития.
Но существуют также сезонные изменения вертикального распределения,
зависящие от изменений гидрологических условий. Летне-осенний прогрев
поверхностных слоев моря заставляет всю холоднолюбивую фауну жить
в этот период на большей глубине, чем зимой и весной. Так, Calanus jinmar-
chicus (helgolandicus) в Черном море живет летом на 50 м глубже, чем
зимою (Никитин, 1929).
Нужно отметить, что сезонные изменения в вертикальном рас-
пределении различных гидрологических условий оказывают огромное
влияние на вертикальное распределение планктона. Например, весною
массы пресной воды, вносимые в море реками, занимают не только боль-
шую площадь, но и имеют большую мощность. Соответственно на большей
глубине можно встретить солоноватоводных животных, а ниже будут оби-
тать морские формы.
Большое влияние на вертикальное распределение планктонных форм
могут оказывать и метеорологические факторы, особенно сгонно-нагонные
ветры. Под их воздействием в прибрежной области сильно меняется гори-
зонт нахождения поверхностных и глубинных животных.
Особенно сильно изменяется вертикальное распределение планктона
в различное время суток. Большинство видов зоопланктона совершает
регулярные суточные вертикальные миграции. Размах (изменения гори-
зонта обитания) этих миграций у различных видов меняется очень сильно,
от нескольких метров до сотен метров. Это явление исключительно сильно
меняет наше представление о приуроченности того или другого вида к
224
определенному слою. Ночью мигрирующие организмы скапливаются в
поверхностной зоне, часто даже в верхнем слое, утром эти же виды могут
оказаться на глубине в 50 jw, а днем на глубинах более 100 м. Большинство
животных, обитающих на глубине до 500 активно участвуют в суточных
Рис. 117. Схема вертикальной миграции планктонных организмов.
А — населяющих весь столб воды (например Calanus finmarchicus) и Б — при-
уроченных к узкому слою (например, Cosmetira pilosella) (по Ресселу).
Минимум
Количество
экземпляров
вертикальных миграциях. Часто мигрируют даже обитатели наддонной
фауны. В мелководных районах, например, в Северном Каспии ночью
к поверхности поднимаются кумацеи, мизиды и другие рачки, днем за-
капывающиеся в ил. В глубоких морях наддонная фауна поднимается
до срединных слоев, а фауна срединных слоев в поверхностные. Среди
видов, обитающих в по-
верхностных слоях, есть и
такие, которые обычно не
мигрируют (Oithona simi-
lis, Paracalanus parous,
Microsetella atlantica и др.).
В настоящее время
установлено, что суточные
вертикальные миграции
зоопланктона имеются во
всех морях (рис. 117 и
118). Амплитуда миграций
сильно меняется в зависи-
мости от общей глубины
места, от географического
положения места, от того,
какие виды и какого возра-
ста участвуют в миграции,
от времени года, гидроло-
гических и метеорологи-
ческих особенностей и
от других условий.
Так как суточная миграция зависит главным образом от света, то,
в связи с изменениями в проникновении света в глубь моря, сильно ме-
няется амплитуда миграций на различных широтах.
Рис. 118. Связь между амплитудой миграций планк-
тона и проникновением света в воду (по Ресселу).
А — среднее распространение света вводу, Б — вертикаль-
ное распределение эврибатного организма, В — то же для
стенобатного организма.
15 Зенкевич, т. I
225
Известно, что прямые лучи проникают на
широте 33°
» 50°
» 67°
до 800 м
» 500 м
» 200 м
Поэтому в южных широтах организмы мигрируют на билыпую глу-
бину, чем в северных. В районе Сан-Диэго (Калифорния) Calanus jinmar-
chicus совершает суточные вертикальные миграции в 460 м. По данным
□стерли (Е s t е г 1 у, 1909) Calanus finmarchicus, обитающий во всем
столбе воды от 0 до 900 м, дает максимум в различное время суток в сле-
дующих слоях:
с 6 ч. до 18 ч. на 460—360 м
с 18 ч. до 20 ч. на 180—36 м
с 23 ч. 30 м. до 0 ч. 37 м. на 18—9 м
с 3 ч. 30 м. до 6 ч. на 180—90 м
Рис. 119. Суточные миграции глубоководного планктона. В центре графика кривая
интенсивности света (по Ваттерману, Кларку и др.).
Не нужно думать, что каждый организм, который жил днем на глубине
в 400 ,и, поднялся до поверхности. Очевидно, происходит поднятие орга-
низмов, живущих на глубине, в средние слои, а обитателей последних в
поверхностные. Но в определенное время происходит скопление в каком-
то узком слое основной массы организмов. При этом днем и вечером орга-
низмы с больших глубин двигаются со скоростью бдльшей, чем обитатели
поверхностных слоев, а на рассвете и утром организмы поверхностных
слоев опускаются с большей скоростью, чем обитатели более глубоких
горизонтов.
Виды, обычно живущие во всем столбе воды, совершают миграции
до самой или почти до самой поверхности моря. Часто исключительно
сильный прогрев самого верхнего слоя исключает их нахождение в нем.
Такова миграция Calanus finmarchicus. Виды, обитающие днем в глубо-
ких слоях, подымаются к вечеру в средние слои (рис. 119). Наибольшей
амплитудой миграций обладают Mysidacea, Euphausiacea, Chaetognatha
и Gopepoda, в меньшей степени медузы, Cladocera и др. Но и в пределах
одного отряда миграция у разных видов отличается очень сильно. Напри-
мер, при большой глубине Calanus и Metridia проходят сотни Петров, а
Pseudocalanus и Acartia мигрируют всего на десятки метров.
226
Далю разные половые стадии и стадии развития одного вида мигрируют
по-разному. Николь с (Nicholls, 1933) указывает, что чем
старше стадия Calanus finmarchicus^ тем больший размах миграций ей
свойственен (рис. 120 и 121). В Белом море у Metridia longa первые три
копеподитные стадии вообще не мигрируют, а начинается миграция только
со стадии IV, причем с дальнейшим ростом амплитуда меняется мало
(Богоров, 1941). Известно также, что одни и те же виды мигрируют
в разные периоды года с разной силой.
Рис. 120. Суточные миграции Calanus finmarchicus в июле в Атлантическом океане
(по Никольсу).
А — копеподигцые стадии I, II и III; Б — взрослые самцы; В — взрослые самки.
Таблица 84.
ГЛУБИНА В м НАХОЖДЕНИЯ МАКСИМУМА CALANUS FINMARCHICUS
В РАЗЛИЧНЫХ СЛОЯХ ВАРЕНЦОВА МОРЯ
Время взятия проб 1 17.50 18.10 | 20.55 0.00 2.57 | 6.00 I 9.20 15.00
Осенью (X) . . 100-50 50—25 10—0 25—10 22—23 1 125—10 150—100 i 150—100 150—100
Время взятия пробы .... 14—15 1 4-6 13-14
Летом 1160—100 i 1 1 1 160—100 160-100 160—100
227
Очень большие отличия в характере миграции наблюдаются в поляр-
ных морях. Летом при круглосуточном дне организмы держатся в преде-
лах одного слоя и совершают миграции для удержания себя в этом слое
(Б о г о р о в, 1938) (рис. 122). Осенью во время смены дня и ночи они
совершают регулярные миграции обычного типа (рис. 123). Таким
образом, положение максимума количества у того же Calanus finmarchicus
летом и осенью меняются следующим образом (табл. 84).
Стадия
I
м
0-Ю
Ю~25
25-50
50-100
100160
Стадия
С <
/ тодия
Ш
Стадия
W -
Стадия
V ’
Стадия
И 1
Самки
О Ю
Ю 15
25-50
50100
ЮО160
О 10
10-25
25-50
50 100
100'160
0-10
10 25
2550
50100
ЮО-ЮО
0-10
10-25
25 50
50100
100160
0-10
1025
25-50
50-100
100-160
10 О 020304050%
Рис. 121. Вертикальное распределение разных стадий Calanus finmarchicus в течение
суток летом в Баренцовом море (по Богорову).
Соответственно изменению амплитуды миграции у различных видов
и в различных условиях меняется и скорость плавания мигрирующих
животных. Вообще же она очень велика, если принять во внимание вели-
чину мигрирующих животных. Так, Calanus finmarchicus в районе Сан-
Диего (Тихий океан) плавает со скоростью не менее 20—30 см в минуту.
В районе Плимута С. finmarchicus мигрирует со скоростью почти до 10 см
в минуту. Длина тела Calanus finmarchicus, как известно, не превышает
в этих районах размера в 2 мм. следовательно удельная скорость плава-
ния у Calanus очень велика. В Каспийском море Eurytemora grimmi (рис.
124) плавает с еще большей скоростью — 15 см в минуту (Б о г о р о в,
1939). Ко всему этому надо добавить, что плавание из глубипы к поверх-
ности и обратно идет активно. За это говорит то, что скорости поднятия
и опускания животных почти равны между собою. Опыты Гардинера
228
(Gardiner, 1933) показали, что пассивное опускание не может про-
исходить с такой большой скоростью
<Ъ • **•—** —
дно
ДНО
Рис. 122. Вертикальное распределение Calanus finmarchicus в Варенцовой море
в светлое и темное время года (по Богорову).
Что же происходит в море, в результате суточной вертикальной миг-
рации планктона? Очевидно, что миллионы тонн живого вещества,
о зо 40 м ао юо
Рис. 123. Вертикальное распределение взрослого Calanus finmarchicus в течение
суток осенью в Баренцовом море (по Богорову).
подымающиеся регулярно (ежедневно) круглый год к поверхности вече-
ром и ночью и опускающиеся на глубину днем, влияют на жизнь планк-
ЧасьЛ
HOU7S] 18 — _
/ААДЛ A i'^l
Z0
23-24 2
85
/7
/4
8
°/о
30
го
5
18200 г 3 811 /4 18200 29811 14 1820О 2 5 8 111918 200 2 5 811 /4
Рис. 124. Суточная вертикальная миграция Eurytemora grimmi в Каспийском море
(по Богорову).
А — вертикальное распределение Е. grimmi в разные часы суток, Б — изменение биомассы Е. grimmi
родном слое в разные часы суток. Цифры у кривых обозначают % от всей биомассы в данном слое.
тона, на газовый режим водоема, на поведение и жизнь рыб. К сожалению,
по этому вопросу мы имеем ничтожно малое количество исследований.
Только в отношении рыб имеется ряд данных.
229
В жизни планктона суточная вертикальная миграция имеет, очевидно,
огромное значение, учитывая повсеместное распространение этого явле-
ния. В результате миграций в течение суток происходит очень резкое
скопление животных ночью в поверхностном слое и совершенно обратное
днем. Затратив большие усилия для подъема вверх, животные начинают
здесь интенсивно питаться. Очевидно, наннопланктон, бактерии и фито-
планктон (D a k i п, 1908; Le bо иг, 1922; Marshall, 1935; Clarke,
1933) терпят значительный урон от этой массы животных, скопляю-
щихся в поверхностном слое вечером и ночью. Организмы, опустившиеся
в глубину, попадают уже не в ту самую воду, из которой они вышли, плы-
вя наверх, так как течения на разных глубинах могут иметь разную ско-
рость и даже различное направление. Таким образом, благодаря суточ-
ной вертикальной миграции животные расселяются с течениями раз-
личных слоев в различном направлении (Hardy и Gunther,
1935).
Следовательно, для всей жизни планктона суточная вертикальная миг-
рация имеет первенствующее значение.
Немалое значение это явление имеет и для газового режима водоема.
В результате огромных изменений в массе живых организмов, населяю-
щих в разное время суток различные слои воды, происходит резкая убыль
кислорода и, наоборот, увеличение углекислоты в этих слоях. Так, по
данным Богорова (1939), если принять, что все потребление кисло-
рода рачком Eurytemora grimmi во всем столбе воды равнялось 100%, то
ночью и днем в различных слоях воды потребление кислорода выразится
в следующих показателях (табл. 85):
Таблица 85
Часы суток Глубины в м 18 23—24 1 5 1 11
10—0 15% 32»/0 4‘7о 0,2%
25—10 39» 44 » 8 » 0,2 »
50—25 46 » 24» 80 » 0,6 »
83—50 — 2 » 8 » 99 »
Аналогично этому обогащение углекислотой в результате
Eurytemora grimmi происходит в следующем объеме (табл. 86):
Таблица 86
дыхания
Часы суток Глубины В 18 23—24 5 11
10—0 14% 32»/, 27о 0,2»/»
25—10 40» 44 » 8 » 0,2 »
50—25 46 » 22 » 80» 0,6 »
83—50 2 » 8» 99 »
Исключительно велика роль суточной вертикальной миграции планк-
тона для жизни рыб и водных млекопитающих.
230
Некоторые важнейшие для питания рыб организмы (Euphausiidae,
Mysidacea, Calanus и др.) днем полностью покидают поверхностные слои.
Наоборот, исключительно богатыми пищей становятся днем глубинные
слои. В связи с миграцией изменяется величина биомассы планктона всех
слоев воды, через которые проходят эти животные. Таким образом, кор-
мовое значение планктона для рыб в различных слоях меняется все время
в течение суток и будет больше в тех слоях воды, в которых дольше со-
храняются большие количества мигрирующих организмов. Глубинные слои
часто тоже довольно сильно обогащаются за счет опускающихся днем
организмов и подымающейся вечером наддонной фауны. Таково типичное
кормовое значение различных слоев воды в случае, если глубина района
не превышает 300 м.
Планктоноядные рыбы поверхностного слоя моря интенсивно питаются
обычно в вечерние и зоревые часы. В подповерхностных слоях (10—25 м)
особенно много корма вечером от подымающихся к поверхности организ-
мов и на заре от опускающихся вниз. Очевидно, в этих слоях и происхо-
дит откорм рыбы. На вертикальную миграцию рыб действуют и другие
причины — общие и для планктона и для рыб, как, например, освещение.
Каковы же причины этого удивительного явления? По этому вопросу
было проведено много исследований как в лабораторных условиях, так
и наблюдений в природе.
Тот факт, что суточная вертикальная миграция планктона имеет вполне
закономерный характер, в результате которого организмы днем опус-
каются вниз и вечером плывут вверх, побудил уже первых исследователей
искать объяснения этого явления в отрицательном отношении животных
к свету. Это простое естественное объяснение в результате эксперимен-
тальных исследований подверглось сомнению (Franz, 1910, 1912;
Е s t е г 1 у, 1912, 1914, 1919), хотя такие крупнейшие экспериментаторы
как Леб (Loeb, 1910) настаивали на нем. Действительно, при экспе-
риментальных работах было установлено, что свет действует отрицательно
лишь при определенных условиях (Rose, 1925). Например, организмы
при высокой температуре из отрицательно фототропичных делаются
положительно фототропичными. Изменение солености, газового режима
и других условий также могут менять знак фототропизма. Несмотря на
это, первенствующая роль света очевидна. Проведенные за последнее время
наблюдения над вертикальной миграцией и изменением интенсивности
света на различных глубинах (Russell, 1933; Clarke, 1934) полностью
подтвердили роль света в этом явлении. На это указывают и исследования
в полярных морях.
Полярные условия представляют собою своеобразную лабораторию
в природе, в которой можно изучать явления суточной вертикальной миг-
рации планктона, при естественном круглосуточном освещении летом,
при смене дня и ночи осенью и при полной круглосуточной темноте зи-
мою. В результате этих исследований было установлено, что летом, в усло-
виях круглосуточного освещения, организмы находятся все время в одном
слое, несмотря на то, что приливо-отливные явления перемещают частицы
воды в течение суток вверх и вниз. Таким образом, этим организмам
(Calanus finmarchicus, Metridia longa, Sagitta eleganswRp.) дважды в течение
суток приходится плыть вверх при отливных явлениях и дважды вниз
при приливных. Сохраняя определенный уровень обитания, эти организмы
претерпевают в течение суток значительные изменения в гидрологиче-
ских условиях (особенно термине), происходящие на том же горизонте
в результате вертикальных перемещений водных масс под воздействием
приливо-отливных явлений. Следовательно для того, чтобы сохранить
определенную степень освещения организмы совершают полярным летом
231
постоянные путешествия вверх и вниз. Но чтобы это было еще более
очевидным, были проведены исследования во время смены дня и
ночи. Осенью в том же Баренцовом море те же самые виды, которые в усло-
виях круглосуточного освещения предпочитают находиться неизменно
в течение суток в одном слое, совершают суточные миграции обычного типа,
т. е. вечером они скапливаются в верхних горизонтах, а днем в нижних.
Таким образом, исследования в «природной лаборатории» полностью под-
твердили первенствующую роль света во всем этом поразительном явле-
нии (Б о г о р о в, 1938).
Интересно отметить то, что миграции вниз начинаются до восхода солн-
ца. Явление предрассветного опускания вниз наблюдали Э сте р л и,
Р е с с е л и др. Первый в связи с этим построил сложное предположе-
ние о том, что это опускание вызывается геотропизмом. Более прав, од-
нако, Р е с с е л, показавший, что ряд организмов начинает опускание
еще среди ночи и, следовательно, здесь имеет значение отсутствие рассеян-
ного света, являвшегося возбудителем поднятия вверх.
Значение света в явлении суточной вертикальной миграции не подле-
жит сомнениям. Свет является отрицательным фактором, препятствую-
щим поднятию многих организмов в верхние слои днем и, кроме того,
отрицательно действующим на нормальное для мигрирующих видов
течение всех жизненных процессов.
Что же может привлекать различные организмы мигрировать в поверх-
ностный слой ночью? Главное значение в данном случае имеет улучше-
ние условий питания в верхних слоях. Действительно, известно, что по-
верхностные слои (0—25 м) более плотно населены бактериями, нанно-
планктоном, фитопланктоном, личинками различных животных, чем
более глубокие. Таким образом, поднятие вверх стоит возможно в связи
с потребностью улучшения питания, что подтверждается данными ряда
авторов (Dakin, 1908: Marschall. 1924 и др.).
Гарди и Патон (Hardy and Neil Paton, 1947) путем остро-
умных опытов показали, что планктонные организмы сохраняют биоло-
гический миграционный ритм в определенные часы суток даже в том
случае, если они изолированы от воздействия света.
То, что совершающие вертикальные миграции организмы не остаются
круглые сутки в поверхностных слоях, а опускаются днем в более глубо-
кие слои, можно объяснить как приспособление, выработавшееся в ре-
зультате борьбы за существование. Если в темное время суток мигрирую-
щие снизу организмы в верхних слоях находят более благоприятные для
питания условия, то в дневное время в поверхностном слое они сами могут
подвергаться выеданию другими организмами. Известно, что в юго-за-
падной части Варенцова моря в летнее время Calanus finmarchicus под-
вергается массовому выеданию различными животными и, в первую оче-
редь, сельдью и ктенофорами. Уходя в глубину, вытесняемые туда борьбой
за существование планктонные организмы в этой первоначально чуж-
дой им среде вырабатывают в процессе эволюции ряд характерных осо-
бенностей. Прежде всего они приспособляются к существованию в темноте,
утрачивая те морфологические и физиологические особенности, которые
давали бы им возможность жить на свету. Кроме того, у них вырабаты-
вается приспособление к существованию при низкой и мало изменяю-
щейся температуре.
Если такое объяснение можно считать удовлетворительным для
суточных миграций, то оно явно недостаточно, если иметь в виду
сезонные миграции. Правда Р е с с е л (1926), описывая сезонные вер-
тикальные миграции Calanus finmarchicus у Плимута, указывает, что
опускание в глубину весной и поднятие к поверхности осенью главной
232
массы этого рачка хорошо согласуется с изменениями в интенсивности
освещения. Однако на примере черноморского планктона В. Ник и-
тиным (1929) наглядно показана зависимость посезонных вертикаль-
ных перемещений ряда планктонных форм от большей или меньшей на-
гретости поверхностного слоя. Очевидно там, где сезонные колебания тем-
пературы поверхностного слоя очень велики (в Черном море 18—19°),
сезонные вертикальные перемещения планктонных организмов будут в
значительной степени испытывать на себе влияние этих колебаний.
Если эти колебания незначительны (у Рессела — 6—7°), на первое
место выступает фактор света. Как Показал Э с т е р л и (1919) Calanus
finmarchicus при низкой температуре (меньше 10°) становится положительно
фототропичным, а при высокой (больше 14°) отрицательно фототропичным.
Все это будет совершенно понятно, если мы учтем, что планктон более
глубоких слоев адаптирован к низкой температуре и отсутствию света.
Некоторыми исследованиями показано, что на распределение морских
организмов воздействует не только солнечный, но и лунный свет.
Так, Эльмгирст (Elmhirst, 1932) показал, что в лунную
и темную ночь, в прилив, на литорали можно наблюдать резкие разли-
чия в составе фауны высших ракообразных.
Биологические сезоны. Сезонные явления в биологии моря распро-
странены повсеместно. Проявляясь по-разному, они существуют на всех
широтах от тропиков до полюсов.
Сезонные явления в биологии моря самым тесным образом связаны с
гидрологическими изменениями (гидрологическими сезонами). Эти по-
следние находятся в такой же тесной зависимости от сезонных изменений
климата (климатических сезонов). Получается последовательная цепь
зависимости в сезонных явлениях. Таким образом, определенные сезон-
ные изменения в биологии моря, которые Б о г о р о в (1938) называет
биологическими сезонами, зависят от гидрологических сезонов, а эти —
от климатических. Биологические сезоны в планктоне Богоров
рассматривает на основе учения Т. Лысенко о стадийном развитии.
Биологические сезоны проявляются в жизни всего населения моря,
начиная с бактерий и кончая китами. Наиболее изучены сезонные явле-
ния в жизни планктона.
Первые, же исследования показали, что и видовой и возрастной состав
планктона сильно меняется в течение года. В определенные периоды года
в планктоне появляются личинки донных животных и рыб и происходит
размножение и развитие молоди всех планктонных и планктоноядных
организмов. Далее оказалось, что виды, которые живут длительный пе-
риод или даже круглый год, имеют максимум своего количественного раз-
вития в определенный период.
Наиболее полно стали изучать сезонные изменения планктона с появ-
лением работы Гарстанга (Garstang, 1894) «Calender of the
floating fauna».
Замечательно название этой работы. Это название побудило целый ряд
исследователей начать составлять планктонные календари (см. S t е и е г,
1910; Зернов, 1914 и др.). Для этого пришлось давать все более и более
подробные сведения об изменениях в планктоне. Первоначально такие
работы ограничивались наблюдениями за изменениями качественного
состава планктона. В 1908 г. вышла работа Ломанна. Ломанн не
ограничился описанием качественных изменений. Он приводит также по-
дробные данные о количественных изменениях планктона (рис. 125) и впер-
вые указывает на изменение объема планктона в течение года, т. е. под-
ходит к вопросу изменения биомассы. К сожалению, биологических сезонов
в планктоне Кильской бухты он не устанавливает. Дальнейшие иссле-
233
дования (А р s t е i n, 1906; К га е f t, 1910; Гай л, 1936 и др.)
ограничиваются установлением только качественных и количественных
изменений по различным месяцам года.
За последнее врем появился ряд работ, в которых авторы (Ш и р ш о в,
1936, 1937, 1939; Мантейфель, 1938, 1939; Я ш н о в, 1939;
Воронков и Кречман, 1939; Johnstone, Scott and
Chadwick, 1924; Gran and В r a a r u d, 1938 и др.) стараются
сгруппировать изменения планктона в сезонные группы или дают опре-
деленным видам и группам видов сезонные названия («весенние» формы,
«летние» и т. п.), или, наконец, приводят материалы по сезонному изме-
нению биомассы планктона. Богоровым (1938) была установлена
терминология и сформулировано понятие «биологических сезонов».
Рис. 125. Годовой ход изменений объема планктона в Кильской бухте
(по Ломанну).
Для установления биологических сезонов необходимо учитывать:
1) изменения в количестве планктона;
.2) годичный ход соотношения растительного и животного планктона
(продуцентов и консументов);
3) качественную смену планктона;
4) изменения возрастного состава, генераций и темпа роста планкте-
ров;
5) изменения химического и гидрологического режима водоема;
6) климатические особенности данного моря или района (сезонные
изменения температуры, солнечного освещения, осадков и т. д.).
Воспользовавшись всеми этими данными, мы можем достаточно полно
охарактеризовать сезонные изменения планктона и понять биологическую
сущность происходящих явлений. Целесообразно дать биологическим
сезонам такие же наименования, какие употребляются на суше. Следо-
вательно, возможны 4 биологических сезона: биологические весна, лето,
осень и зима.
234
Биологическая весна характеризуется 1 обилием всего планктона. Осо-
бенно много фитопланктона. Количество его обычно так велико, что мож-
но говорить о «цветении моря». Биомасса зоопланктона меньше биомассы
фитопланктона. Часто биомасса фитопланктона в десятки раз превосхо-
дит биомассу зоопланктона. Сезонный показатель- равен 10—100. Каче-
ственный состав в различных морях, естественно, отличается очень сильно.
Общим является для всех морей то, что: 1) среди зоопланктона в большом
количестве находятся яйца и личинки донных животных и сами донные
черви и ракообразные; большинство видов зоопланктона приступает к
интенсивному размножению; 2) среди фитопланктона в большом количе-
стве находятся преимущественно виды, требующие для своего массового
развития больших концентраций питательных солей. Когда концентра-
ция питательных солей начинает уменьшаться, то главное значение пере-
ходит к видам рода Chaetoceras. Биологическая весна характеризуется
наибольшим количеством биогенных веществ в воде. Этому периоду часто
свойственно сильное опреснение (особенно в поверхностном слое) моря
и начало прогрева воды. В полярных морях появление больших разводьев
или начало освобождения моря от сплошных льдов. Из числа климати-
ческих особенностей нужно отметить увеличение продолжительности и
интенсивности освещения солнцем и потепление воздуха.
Биологическая весна обычно весьма кратковременна (около 1 месяца).
Биологическое лето характеризуется уменьшением общего количества
планктона. Биомасса зоопланктона почти равна или больше биомассы
фитопланктона (сезонный показатель близок к единице). Биомасса зоо-
планктона максимальна. Качественный состав зоопланктона характери-
зуется появлением летних форм. Из планктона исчезают весенние личинки
и появляются летние. Наблюдается общее «повзросление» копеподитных
стадии и других длительно развивающихся молодых стадий. В южных
морях копеподы дают обычно 2—3 и более генераций. Размер организмов
летних генераций обычно меньше, чем у весенних. Планктоноядные рыбы
и китообразные в течение биологической весны и лета совершают актив-
ные пищевые миграции. Среди фитопланктона руководящая роль пере-
ходит к видам, требующим для своего массового развития незначитель-
ных концентраций питательных солей. Такими среди диатомовых яв-
ляются различные виды рода Chaetoceras.
Повышается температура воды. Из числа климатических особенностей
отметим увеличение количества и интенсивности солнечного освещения
и температуры воздуха. Нужно особо отметить, что этот период характе-
ризуется наиболее сильным солнечным освещением. (
Длительность биологического лета в различных морях весьма различна
(от 1 до 7 месяцев).
Биологическая осень в различных морях весьма сильно отличается
по своему характеру. В полярных морях наступает резкое уменьшение
планктона, особенно фитопланктона. В морях умеренных и более южных
количество планктона, особенно фитопланктона, возрастает. Биомасса
его увеличивается настолько, что можно говорить о втором «цветении»
моря. Как правило, этот второй максимум фитопланктона значительно 1 2
1 Необходимо помпить, что однообразно охарактеризовать все моря посезонно очень трудно.
Обычно один и тот же фактор в различных морях (на различных широтах) меняется не одинаково.
Поэтому даваемая нами характеристика весьма обща. Каждое море (поширотно) надо характери-
зовать особо, так как видовой состав различных морей отличается так сильно, что оказалось
невозможным дать в этой общей характеристике посезонные изменения видового состава в
различных морях.
2 Сезонным показателем называют (Б о г о р о в, 1938) отношение количества фитопланктона
к зоопланктону, выраженное в весе или объеме.
235
меньше, чем первый. Сезонный показатель в полярных морях равен 1
или меньше, в умеренных и южных морях он больше 1 и часто при-
ближается к 10.
Качественный состав зоопланктона характеризуется дальнейшим «по-
взрослением» фауны. Обычно в полярных морях в этот период максималь-
ное количество копепод переходит в копеподитные стадии IV и V, т. е.
в зимующие стадии. В умеренных морях в этот период развивается послед-
няя генерация копепод и других организмов, которая перезимовывает и
дает новое поколение будущей весною.
Большого развития биологической осенью достигает и теплолюбивая
фауна, поскольку прогрев воды особенно сказывается к концу лета и на-
чалу осени.
Качественный состав фитопланктона резко изменяется. Главную роль
в планктоне играют наряду с диатомовыми перидиниевые водоросли.
Осенью, в отличие от летнего периода, сначала количество питатель-
ных солей увеличивается (за счет разложения весеннего и летнего планк-
тона), а затем резко уменьшается.
Гидрологический режим характеризуется максимальными годовыми
температурами в начале и начинающимся понижением температуры воды
в конце. Из числа климатических особенностей нужно отметить уменьше-
ние солнечного освещения и начало похолодания воздуха.
Продолжительность биологической осени не велика (около 1 меся-
ца).
Биологическая зима характеризуется общим минимумом планктона.
Биомасса зоопланктона больше биомассы фитопланктона. Сезонный по-
казатель меньше единицы. Особенно мало растительного планктона. Раз-
личные виды водорослей часто находятся в стадиях покоя или в зимую-
щих клетках. Среди животного планктона главное значение имеют взрос-
лые формы или зимующие стадии (например, копеподитная стадия V).
Новых генераций в планктоне не образуется.
Химический режим характеризуется постепенным увеличением био-
генных веществ. Количество их увеличивается все время от осени до весны
и в конце зимнего периода оно достигает максимальной величины. Обилие
биогенных веществ образуется за счет разложения планктона, поднятия
богатых органическими веществами глубинных вод в поверхностные слои
и в результате вертикальной циркуляции (Зубов, 1931; Зенке-
вич, 1932). Количество биогенных веществ все время увеличивается,
особенно потому, что нет потребления этих солей — фитопланктон почти
полностью отсутствует.
Гидрологический режим характеризуется все время нарастающим
похолоданием, во многих морях поверхность моря покрывается льдом.
Интенсивность солнечного освещения наименьшая, а в полярных морях
оно вовсе отсутствует.
Такова весьма краткая и общая характеристика биологических сезо-
нов в планктоне.
Совпадение биологического состояния (подготовленности) и соответ-
ствующих внешних условий определяет время наступления того или дру-
гого биологического сезона. Эти совпадения не случайны, а являются ре-
зультатом эволюционного процесса, приведшего к адаптации организма
к сезонным изменениям окружающей среды.
Из всего сказанного мы видим, что в течение года происходят все вре-
мя количественные изменения планктона. Этот годовой биологический
процесс совершается постепенно, но сумма этих изменений в определен-
ный период настолько отличает один период года от другого, что можно
говорить о переходе на новую ступень, в новую стадию развития годич-
.236
ных изменений планктона. Этими стадиями развития планктона в тече-
ние года и являются биологические сезоны.
Сезонные изменения планктона в разных морях. Попытаемся теперь
проследить, как протекают сезонные изменения планктона — биологи-
ческие сезоны — на различных широтах от северного полюса до экватора
(рис. 126).
1) Биологические сезоны в морях на различных широтах приходятся
на различные месяцы.
2) Продолжительность биологических сезонов (особенно биологиче-
ской зимы и лета) в различных широтах очень сильно колеблется.
3) В течение года в различных морях имеется то один, то два макси-
мума развития фитопланктона, а отсюда п зоопланктона.
4) Продолжительность времени между главными максимумами на раз-
личных широтах изменяется.
Месяцы Моря 1 а ш /г v пт ют ш к 1 и т
/ Полярные а Полярный бассейн б Высоноарктические УСибирские) в Нижнеарктические (Мурман) \ЛетЛ ''////У///Зйма7/////А^сна tyiemo\ Осень
П Умеренные а Северные /Норвежские воды) 6 Южные (Английские воды) Вест Д////7Лето7///Д Осень 'СДДЗимё'//// 'З'имаДА Весна У///////Ш7////^ W/МйЬ.
Ш Субтропические ',Эима& Весна W////7/^jfem66 Осень Мима
ИТ- Тропические BecnaV/////////^ Весна
Рис. 126. Продолжительность биологических сезонов планктона в разных
широтных зонах океана (по Богорову).
5) Вегетационный период сокращается с 12 месяцев в тропических
и умеренных морях до 1—2—3 месяцев в полярных.
6) С продвижением на север срок наступления биологической весны
запаздывает. В тропических морях планктонная весна падает на зимние
месяцы, а в полярном бассейне на август.
7) Продолжительность биологического лета укорачивается с 7 меся-
цев в тропиках до 1—2 месяцев в полярных морях.
8) С продвижением на север биологическая осень наступает кален-
дарно все ранее и ее продолжительность уменьшается. В высокоарктиче-
ских морях осень почти мимолетна.
9) Биологическая зима все время удлиняется. В тропиках она почти
не выражена. Зато в полярных морях она продолжается 10 месяцев.
10) Мы можем разделить моря на два основных типа: моря с одним
максимумом развития планктона — моноциклические и моря с двумя
максимумами — бициклические. Моноциклическими морями являются
высокоарктические и тропические моря, бициклическими — нижнеарк-
тические, умеренные и субтропические моря.
В связи с особенностями сезонных изменений планктона в разных мо-
рях находится число генераций различных компонентов планктона, что
можно иллюстрировать на примере Copepoda (табл. 87).
237
Таблица 87
Моря Количество генераций у Copepoda в разных морях
теплые умеренные холодные
Каспийское море английск. воды норвеж- ские воды нижпеарк- тич. Барен- цево высокоарк- тич. Сибир- ские
сл весна i 1 1 1 1
лето — 2 1 | 1 ! .
Л а i осень — 1 1 1 J
зима — — — , — —
Итого — 4 i 3 2 1
весна 1 1 1 1 1
о -о 5 о лето 5 3 2 1 ( 1
1 } 1 J
—и осень 1 1 1
.9
зима 1 1 — —
Итого 8 6 1 4 2 1
Следовательно: 1) чем теплее море, тем больше генераций у Copepoda
(вероятно и у всех других планктонных видов); 2) с продвижением на
север количество генераций всех Copepoda уменьшается; 3) количество
генераций наиболее изменчиво в течение биологического лета, причем,
чем длиннее лето, тем больше генераций дают Copepoda; 4) биологиче-
ской зимою Calanus finmarchicus вообще не дает новой генерации, а мел-
кие Copepoda дают ее только в южных морях.
Таблица 88
Моря Районы Средняя био- масса 1 Авторы
I. Полярные а) высокоарктические Чукотское Восточно-Сибирское . . Море Лаптевых 200 250 300 Богоров (средняя за период максимального развития)
б) нижнеарктические II. Умеренные а) северные б) южные Средн, для трех морей Баренцево море с 69° 30—74° 00 с. ш. . . . Охотское море Японское море Каспийское море .... 250 300 400 300 190 Яшнов (средняя с VI по VIII) Кусморская (средняя с V по VIII) Кусморская (средняя с IV до XI) ! Идельсон (средняя за 1 IV и VIII)
Цифры округлены. Средняя биомасса для поверхностного слоя моря.
238
У нас нет возможности оценить это явленно для тропических морей,
так как мы не располагаем соответственными данными. Очевидно, там мы
вправе ожидать 10 генераций. Само собою разумеется, что количество ге-
нераций у планктонных форм имеет очень большое значение для оценки
продуктивности моря. Известно, что биомасса планктона Черного и Ка-
спийского морей сравнительно с северными морями мала. Однако, огромное
количество планктоноядных рыб — хамсы, кильки, сельди говорит об
огромной продуктивности планктона и в первую очередь Copepoda. Мень-
шая биомасса Copepoda (например, Каспийского моря, по сравнению
с полярными морями) вовсе не означает меньшую продуктивность планк-
тона (табл. 88).
Следовательно, средняя биомасса зоопланктона Каспийского моря
значительно ниже средней биомассы его в Баренцовом, Охотском,
Японском и даже высокоарктических морях. Но продуктивность Кас-
пийского моря по мелким Copepoda в 8 раз больше полярных морей и,
хотя в полярных морях главную биомассу в открытых, удаленных от
берегов, районах составляет С alanus finmarchicus, мы все же вправе счи-
тать, что продуктивность зоопланктона Каспийского моря будет во много
раз больше, чем продуктивность полярных морей. Если мы примем про-
дуктивность высокоарктических морей равной пх средней биомассе за
период максимального развития (исходя из того, что в этих морях все
Copepoda — главнейшие компоненты зоопланктона — имеют одну гене-
рацию), то получим следующие «коэффициенты продуктивности по гене-
рациям», а отсюда и возможную продукцию основного компонента кор-
мового планктона — Copepoda (табл. 89):
Таблица 89
Коэф, продукт, по гене- рациям Полярные Умеренные
высоко- | арктич. । ни жне арк- тич. северные южные
1 i 2 3 ! 1200 ! 5 __ 1500
Продукция в среднем в ,иг‘на 1 at3 250 1 600
Следовательно, Каспийское море по продуктивности зоопланктона не
уступает таким исключительно богатым моряхм, как Охотское и Японское и
в два с половиной раза превосходит Баренцово. Это находится в соот-
ветствии с наличием в Каспии громадных масс планктоноядных рыб и,
в первую очередь, сельди и кильки.
Определение общей продуктивности планктона кажется нам задачей
в настоящее время непосильной. Пока мы даже не имеем вообще доста-
точно объективного и не вызывающего сомнений метода решения этой
задачи. Более того, решение этого вопроса в целом, в среднем для всего
планктона, кажется нами неверным, если мы будем определять продук-
цию планктона для целей рыбного прогноза или определения сырьевой ба-
зы. Важно не общее количество планктона, а его кормовая ценность.
Например, Мантейфель (1941) указывает, что в некоторые годы
общая биомасса планктона в Баренцовом море может быть достаточно
велика, но состоять в основном из гребневиков, выедающих всех Copepoda.
В таких случаях пришедшая на откорм в Баренцово море сельдь не находит
себе пищи, продолжает питаться и держаться в разреженном состоянии
всю осень и зиму. Следовательно, для рыбопромысловых целей важно
знать не продукцию планктона вообще, а продукцию кормового для рыб
23&
планктона. От продукции кормового планктона будут зависеть запасы
рыб, и, базируясь на кормовом планктоне, мы можем увеличить точность
прогноза.
Изучение биологических сезонов дает наиболее общую развернутую
картину жизни моря. В силу этого невозможно делать большинство
теоретических выводов без знания сезонных изменений биологии моря.
Учет существования биологических сезонов имеет очень большое методи-
ческое значение при сравнительном изучении планктона. Ведь различные
моря или даже отдельные районы моря в одно и то же календарное время
переживают различные биологические сезоны, причем различия планктона
между сезонами в одном и том же море могут быть даже большими (особенно
в отношении биомассы планктона), чем отличия между различными морями.
Отсюда, при суждении о богатстве или характере (составе) планктона,
нужно проводить сравнение между морями по одноименным биологи-
ческим сезонам. Это особенно важно при обработке больших экспедицион-
ных материалов. Часто даже на одном разрезе мы можем встретить раз-
личные биологические сезоны.
С сезонными изменениями в биологии моря связана и жизнь всех
промысловых объектов. Изучение сезонных изменений необходимо для
установления биологических основ морского рыбного промысла. Для
того, чтобы удачно ловить рыбу, надо знать, когда она собирается
в промысловые косяки или рассеивается по необозримым пространствам
моря. Все это связано с биологическими сезонами. По сути дела все био-
логические индикаторы промысла являются ничем иным, как показателями
сезонного состояния моря (особенно в отношении планктона и планктоно-
ядных организмов). Установленные рядом авторов биологические инди-
каторы для различных сельдей и китов (Мантейфель, 1939; Бол-
до в с к и й, 1939; Hardy, 1924, 1926; Hardy and Gunther,
1935, Savage, 1926,1931.1932; Savage and Hardy, 1935;
Savage and Wi mpenny, 1936 и многие другие) являются ярки-
ми примерами того, что знание сезонных изменений планктона (биоло-
гических сезонов) имеет большое значение для рационализации про-
мысла.
Биологические сезоны в планктоне полярных морей могут быть исполь-
зованы в качестве индикаторов ледового режима (Ш и р ш о в, 1936,
1937; Б о г о р о в, 1938, 1939). В последнем случае особенно важно то,
что сезонные изменения в планктоне являются своеобразным следом,
«отпечатком» ледового состояния данного моря. Значение сезонного
состояния планктона позволяет лучше оценить общее океанографическое
состояние моря в связи со льдами и дать, таким образом, полезный совет
при плавании во льдах.
Биологические сезоны в планктоне моря могут служить основой
для установления океанологической фенологии — особого раздела на-
уки о море, дающей возможность по степени развития планктона судить
о сезонном состоянии моря. Даваемые на основе океанологической
фенологии прогнозы для рыбной промышленности и плавания в море
(во льдах) послужат на пользу всему нашему народному хозяйству.
ГЛАВА VIII
БЕНТОС
1. ИСТОРИЯ П МЕТОДИКА ИЗУЧЕНИЯ
Только через 24 года после применения планктонной сетки для коли-
чественного изучения планктона был изобретен аналогичный прибор
для количественного изучения бентоса. Одновременно и независимо друг
от друга такие приборы, так называемые дночерпатели, были введены
в практику шведом Св. Экманом (Sv. Е k m а п) и датчанином
Петерсеном (С. G. 1 о h. Petersen). В 1911 году появилось
первое исследование Петерсена и его ученика Б о й с е н - И е п-
с е н а (Р. В о у s е п - I е n s е п) по количественному изучению живот-
ных морского дна и их питанию в датских водах, в котором авторы ука-
зывают, что подобно исследованиям Гензена и его продолжателей
по продуктивности планктона количественные исследования бентоса
должны войти составной частью в изучение метаболизма моря.
В последние годы различными исследователями (Kniidse n,E k m а п,
Гордеев и др.) предложено несколько новых моделей дно-
черпателей, но пока основным прибором при работах в море остается
модель Петерсена.
Несомненно наилучшей моделью из всех существующих остается
дночерпатель Гордеева (1945), дающий гораздо лучшие результаты,
нежели петерсеновский. Основные его преимущества заключаются в том,
что он даже на плотном песке глубоко входит в грунт и не захлопы-
вается при опускании, не достигнув дна.
Дночерпатель падает на дно прямо сверху и закрываясь вырезает
более пли менее толстый слой грунта (в 1—5 см на песке и в 10—20 см
на илистом грунте) со всеми животными, находящимися в нем и на нем.
Обычно морские дночерпатели имеют площадь покрытия 0,1 или 0,2
(0,25) м2. Сборы дночерпателя дают возможность пересчета полученных
показателей на ту или иную площадь морского дна.
Основные дефекты дночерпателя следующие: малая площадь захвата,
при невозможности значительно ее увеличить; в силу этого только бо-
лее пли менее равномерно распределенные по дну организмы удовлетво-
рительно облавливаются дночерпателем, т. е. в основном организмы
мягких грунтов. Мало пригоден дночерпатель для сбора организмов на
скалистых и каменистых грунтах. Недостаточно удовлетворительно
захватывается дночерпателем Петерсена чистый песок, в силу
16 Зспкевпч, т I
241
большой плотности залегания. Этот последний дефект устранен в
оригинальном дночерпателе М. Кнюдсена, дающем монолит песка,
глубиной в 30 см. Наконец, быстро опускающийся па дно дночерпатель
может распугивать подвижных животных, как, например, ракообразных.
Все эти недочеты, однако, кроме последнего, исчезают при сборах
дночерпателя на илисто-песчанистых или илистых грунтах.
Очень существенен вопрос о том, в каких весовых показателях сле-
дует давать биомассу организмов при обычных бонитировочных иссле-
дованиях. Сырой вес как будто представляет собой наиболее натуральный
показатель, однако неодинаковое у разных животных количество воды
и частей минерального скелета делают эти показатели не вполне надеж-
ными при оценке фауны, как кормовой базы. Совершенно справедливо
поэтому указание Петерсена, что предпочтение следует отдавать
«сухому» весу, когда вода удалена. Соотношение сырого и сухого веса у
разных животных различно, и Петерсен дает для некоторых
животных количество сухого вещества в процентах от сырого, причем
моллюски взвешивались без раковин.
Эти величины колеблются от 3,8% (мшанка Alcyonidium digitatum
декальцинированная) до 14,0% (моллюск Philine apertd). Для Lamel-
libranchiata, в среднем, сухой вес составляет 7,8% от сырого, для брюхо-
ногих 9,9 %, для полихет 14,8%, для десятиногих ракообразных (декальци-
нированных) 12—20%, для декальцинированных морских ежей 0,7—1,0%.
Очень большие трудности возникали также в связи со скелетными
частями организмов и понятно, что нередко исследователи задавались
вопросом, следует ли взвешивать червей с их трубками, моллюсков с их
раковинами и т. п. Обычно моллюсков взвешивают вместе с их ракови-
нами, а полихет с их трубками только в том случае, если эти трубки
органического происхождения, как, например, трубки Spiochaetopter<is
или Spirorbis. Еще более неясное положение создается с так называемой
мантийной жидкостью моллюсков и некоторых других животных, которая
может иногда составлять существенную часть веса тела. Кроме того для
некоторых животных (часть иглокожих, полихет и др.), особенно питаю-
щихся грунтом, вес тела может значительно меняться в зависимости от
степени наполнения кишечника. У некоторых голотурий и гефиреп
содержимое кишечника может составлять 40—50% веса тела, а в иных
случаях и больше. Все эти обстоятельства, даже при получении сухого
веса, не устраняют часто больших ошибок и поэтому нельзя не признать
совершенно справедливым указание Б л е в а д а, что в изучении про-
дуктивности надо перейти на определение калорийности, тем более, что
тогда мы будем иметь гораздо более ценный материал и для учета кормо-
вого значения организмов.
Таким образом, методы обработки количественного материала могут
быть расположены в следующий ряд по повышению качества результа-
тов: спиртовой вес — формалиновый вес — сырой вес — сухой вес —
калорийность. Вероятно в дальнейшем будут пользоваться определе-
нием сырого веса с последующим перечислением массовых форм на кало-
рийность по установленным для них стандартным показателям, подобно
тому как у нас планктонологи (по идее В. Богорова) пересчитывают
при обработке планктона численность на вес по весовым стандартам.
Это дает также возможность перечислить на калорийность и весь прежний
материал, для чего придется только установить показатели калорий-
ности для сотни форм.
В настоящее время в методике фиксации и обработки кроется наиболее
слабое место количественного метода учета явлений биологической про-
дуктивности;
242
Довольно детальный, но недостаточно обоснованный фактическим
материалом критический анализ работы с дночерпателем дал ученик
Экмана А. Линдрот (Lindroth, 1935) в работе, посвящен-
йой Гюльмар-фиорду. Свои критические замечания он начинает с указа-
ния на ненадежность проб дночерпателя для фаунистической характе-
ристики района. Резкую критику этой же стороны работы дночерпателя
дал и И. Месяцев (1930). Как было показано на том же самом мате-
риале (Б р о ц к а я и Зенкевич, 1939), критика М е с я ц е в а
нисколько не компрометирует дночерпателя, как орудия сбора по
некоторым группам бентоса даже и для фаунистических целей. В очень
наглядной форме это же показали Г. Г у р в и ч и И. Иванов
в своей работе по Белому морю. Как можно видеть на следующей, взятой
у них таблице, Amphipoda лучше облавливаются драгой, а полихеты
дночерпателем. Несомненно также, что двустворчатые моллюски тоже
облавливаются дночерпателем лучше, чем драгой или тралом (табл. 90).
Таблица 90
В уловах драги В уловах дночерпателя
количество количество количество количество
видов экземпляр. видов экземпляр.
Amphipoda 20 212 17 112
Polychaeta 24 85 43 1 706 1
Линдрот ненадежность дночерпателя Петерсена как орудия
сбора видит в «сдувании» верхнего слоя грунта, при быстром опускании
прибора, в захватывании только нижнего слоя грунта, в частичной
потере грунта, при работе прибора. Все эти технические недочеты
могут быть устранены комбинацией дночерпателей различных систем (П е-
терсена, Кнюдсена, Экмана, Болтовского, Гор-
деева и др.). С другой стороны, нет ни одного другого прибора для
сбора бентоса, которому эти свойства не были бы присущи в гораздо
более значительной степени.
В 1922 г. К и р с о п (F. К i г s о р) опубликовал результаты
весьма интересных и тщательных наблюдений по сравнению работы
трех орудий сбора бентоса — дночерпателя, обычной драги и особого
«кэмбриджского» трала, приспособленного для скалистых грунтов (rock-
trawl). Особенно интересно то, что К и р с о п одновременно с этим
в некоторых случаях эмпирическим путем устанавливал количество
обитателей на выбранном для опыта участке литорали. Результаты полу-
чились следующие: на песчанистом и илистом грунтах, на глубине 15—20 .и
rock-trawl дал лишь 1% того, что дал дночерпатель с соответствующей
площади дна. Особенно слабо в улове трала были представлены черви.
С другой стороны, для каждого из указанных трех орудий лова К и р с о п
устанавливает, применительно к скалистому грунту, общий коэффициент
уловистости, вычисленный эмпирически, и на соответственный коэффи-
циент умножает действительный улов прибора. Для «кэмбриджского»
трала коэфициент равен 43,8 (так как его улов составляет только 2,28%
наличной фауны), для обычной драги 33,5, для дночерпателя — 6. Данных
по сравнительной уловистости приборов на мягких грунтах К и р с о п,
к сожалению, не дает.
В одной из своих работ Петерсен (1913) также давал сравнение
работы дночерпателя и драги. Было взято 5 проб дночерпателем площадью
243
0,2 м~ и перечислено на 75 м-. Драгу тащили три минуты, что должно
соответствовать облову сю той же поверхности дна в 75 м*, результаты
двух опытов даны на табл. 91.
Таблица 91
Грунт Количество экз. животных на 75 .и2 Улов драги в % от улова дночерпателя
дночерпатель драга
Песок с детритом около 11175 311 ’ 3
Песчанистый пл около «550 340 4
Детальный сравнительный анализ работы дночерпателя и качествен-
ных орудий сбора (драга, трал) можно найти в работах Месяцева
(1930), Броцкой и Зенкевича (1939).
Указание Л и и д р о т а на недостаточные размеры дночерпателя
опровергается непосредственными наблюдениями Г и с л е н a (G i s 1 е п),
установившего полную пригодность площадок в 0,1 м2 для изучения расти-
тельности морского дна. Простым увеличением числа проб дночерпателя
на каждой станции можно свести до минимума этот недочет. Кроме того
Л и н д р о т считает необходимым резко различать методику фаунисти-
ческих исследований, которые приводят к описательно-фаунистическому
знакомству с населением дна и объяснительно-экологическому (okolo-
gisch-verklarend). Для первой цели дночерпатель вполне пригоден, для
второй его данные неудовлетворительны. Л и н д р о т считает, что
совместное проживание и конкуренция за место и пищу совершенно недо-
статочны для объяснения формирования биоценозов, для понимания ко-
торых в первую очередь необходим анализ воздействия внешних факторов.
Нельзя с фаунистической методикой притти к экологическим выводам
и т. д. Все эти рассуждения Линдрота не вполне понятны. Ведь
ботаник-биоценолог тоже работает методом площадок и однако при-
ходит к экологическим выводам.
Во всяком случае, даже при количественном изучении донной фауны
нельзя отказываться от сборов и качественными орудиями сбора (драги,
тралы), дающими чрезвычайно ценное дополнение к сборам дночерпа-
теля. Мы уже указывали выше, что с переходом к количественно биоцено-
тическим исследованиям водной фауны, ставшим возможными с момента
применения количественных орудий сбора, коренным образом изменилась
и методика исследований и объем проблем, ставших в центр внимания
гидробиолога. Наиболее существенным моментом являлась возможность
выделения из всей фауны небольшого числа массовых форм, которым
принадлежит ведущая роль в биологических процессах, совершающихся
в водоеме, иначе говоря, явилась возможность отделить главное от
второстепенного. Эта возможность разгрузила биолога от необходимости
определения всей массы пойманных животных, что часто представляет
непреодолимые трудности, тогда как массовые формы обычно хороню
всем известны. Сравнительно небольшой по объему материал, получаемый
дночерпателем в количестве нескольких сот проб, дает хорошую картину
количественного распределения донной фауны и вполне доступен для
быстрой обработки одному-двум работникам. Сами работы разгрузились
от бесконечных фаунистических списков, в которых трудно разобраться
даже специалисту, и загромождения материала перечислениями форм,
где редчайшие фигурируют рядом с массовыми.
244
Ио, конечно, не в этих методических моментах суть дела. Главное в том,
что первые же данные по количественному распределению организмов
как в пресных водах, так и в море подвели вплотную к основным проб-
лемам современной гидробиологии. Во-первых, исследователи получили
довольно точную картину горизонтального и вертикального количествен-
ного распределения организмов в водоеме, как в какой-нибудь опреде-
ленный момент, так и в сезонной смене. Во-вторых, явилась возможность
выделить из всей фауны ведущие массовые формы и затем подвергнуть
их уже более детальному количественному изучению, и основное внимание,
конечно, могло быть обращено на посредственно или непосредственно
полезные человеку организмы. Количественный учет флоры и фауны,
населяющей водоемы, по отдельным ее группам (фитопланктон, зоопланк-
тон, фитобентос, зообентос, рыбы и т. п.) дал возможность подойти
вплотную к проблеме биологической продуктивности, как в отношении
зависимости массового развития тех или иных форм от факторов среды,
так и со стороны обеспеченности развития определенных групп организ-
мов наличием питательных ресурсов, определяющих то или иное коли-
чественное соотношение отдельных групп организмов.
Вопросы распределения в пространстве, питания, роста, размножения,
смертности, акклиматизации и т. п. получили в свете проблемы биоло-
гической продуктивности свое новое содержание и целеустремленность.
Поэтому чрезвычайно велика в гидробиологии заслуга исследова-
телей, применивших впервые количественные орудия лова для изуче-
ния жизни в водоемах. Еще до Петерсена делались попытки
перейти к количественному изучению донных организмов, но до введе-
ния в практику дночерпателя эти попытки не давали нужного резуль-
тата. Таким образом, важнейший этап в гидробиологии определяется
казалось бы чисто техническим моментом. Впрочем на 150 лет ранее
первые этапы развития морской фаунистики также были связаны с тех-
ническим моментом, применением драги для сбора морских животных.
Еще в 1857 г. датчанин Г р о в (Grove) приводил данные по частоте
встречаемости некоторых морских животных на определенной площади
дна, Даль (F г. Dahl, 1893) был первым исследователем, который
произвел под влиянием работы Гензена учет донных животных
литорали устья Эльбы на определенной площади дна по числу экземпля-
ров и по объему. Англичанин Гердмаи (Herdman) в 1896 г.,
производя драгировки, подсчитывал и взвешивал отдельные уловы.
Подобным образом шведский исследователь Тиль (Т h е е 1, 1907)
подсчитывал число экземпляров отдельных форм в уловах драги в Гюль-
мар-фиорде. Наконец англичанин Джонстон (Johnstone, 1908)
приводил расчеты годовой продукции мидии и сердцевидки на 1 м2. Однако
все эти попытки не развернулись в обширные исследования, так как ору-
дия сбора не соответствовали поставленным задачам. Введение Петер-
сеном дночерпателя дало эту возможность. Большое число датских
исследователей, учеников и продолжателей Петерсена дали ряд
исследований, углубивших и расширивших работы Петерсена.
Среди них следует назвать Бойсен-Иен се на, Блевада,
Спэрка, Мортенсена, Торсона, Тамдрупа. Йен-
сена, П а у л ь с е п а, Иогансена, Ларсена и Лунда.
Особенно ценно в исследованиях школы Петерсена направ-
ленность всей работы на разрешение вопросов практического характера,
связанных с рыбным промыслом, так как Петерсен является,
помимо своих исследований бентоса, крупнейшим знатоком и во многом
основоположником современных рыбопромысловых исследований. Флора
п фауна рассматриваются им как кормовая база для рыб, поэтому с самого
245
начала изучение питания промысловых рыб вошло как составная часть
исследований его школы, поэтому бентос подразделяется ими на полезный
и бесполезный (как корм для промысловых рыб). Петерсен и Б о й-
с е н-И е н с е н подвергают тщательному исследованию зостеру в
датских водах, дающих в прибрежных датских водах, по их мнению,
основной пищевой резерв, в качестве растительного детрита.
2. ФИТОБЕНТОС
Состав фитобентоса. Чрезвычайно большое значение для развития
животной жизни в мелководных зонах морей имеет береговая раститель-
ность \ мощным бордюром опоясывающая побережья морей. В береговой
зоне макрофиты являются основными продуцентами органического
вещества. Но не только в этом значение макрофитов для животных.
Среди зарослей водорослей в подводных лесах и лугах многочисленные
животные находят для себя благоприятные условия существования.
Для многих из них стебли растений служат субстратом, к которому они,
а также водоросли, прикрепляются, а корневые ризоиды дают приют
массе животных. Плавающие скопления макрофитов, наиболее крупные
из которых известны в Саргассовом море, также дают приют богатой
и своеобразной фауне.
На первом месте по значению в морях стоят бурые водоросли (Phaeopliy-
сеае), а именно фукусовые (Fucus, Ascophyllum, Cystoseira, Sargassuni,
Turbinaria) и ламинариевые (Laminaria, Macrocystis, Nereocystis, Lcssonia,
Ecklogia, Hormosira). На втором месте должны быть поставлены зеленые
водоросли (Chlorophyceae) (Viva, Cladophora, Enteromorpha, Caulerpa,
Dasycladus, Codium и др.) и на третьем багрянки (Rhodophyceae) (Phyl-
lophora, Polysiphonia, Laurensia, Hypnea, Gracillaria, Gelidiopsis, Pey-
sonella, Squamaria, Melobesia, Udotea, Nemalion, Porphyra, Corallina,
Lithothamnion, Farcellaria, Chondras, Ptilota, Delesseria, Ceramium).
Наконец, очень большое, иногда преобладающее значение в некоторых
частях мирового океана и отдельных морях получают обитающие в соленой
воде цветковые растения и в первую очередь Zostera, Phyllospadix, Rappia
и др. Например, в датских водах именно Zostera, как продуцент, стоит
на первом месте. Только для Каттегата годовая продукция зостеры опре-
делена Петерсеном в 50 млн. т.
В течение последних 15 лет наблюдается массовая гибель морской
травы — Zostera marina. Впервые гибель зостеры была констатирована
в 1931 г. у берегов Виргинии (США), а в 1932 г. и у берегов Европы
и быстро распространилась по всему атлантическому побережью п
в Средиземном море. В 1938 г. гибла зостера и в Черном море и
в Каркинитском заливе. Возбудителем болезни оказался одноклеточный
организм из группы слизистых грибов, названный Labyrinthula. Общее
количество погибшей зостеры по атлантическим побережьям несомненно
превышает 200 млн. т. Интересно, что Zostera папа не подвержена
заболеванию.
Вертикальная зональность. Обычно наблюдается вертикальная посте-
пенная смена преобладающего значения макрофитов по группам. Выше
всего располагаются зеленые водоросли, за ними следуют бурые, глубже
красные. Таким образом, имеет место цветная вертикальная зональность
(зеленый, бурый, красный), сходная с тем, что отмечается и у животных.
1 Большинство данных по распределению водорослей взяты из неопубликованной работы
Т. Щаповой (1938) «Количественные исследования морских донных водорослей».
246
Как уже указывалось выше, вода в различной степени поглощает лучи
с различной длиной волны. На глубину 10 м доходит только 1—2% крас-
ных лучей и 45 % синих. Таким образом, на некоторой глубине преобладает
зеленое и синее освещение и наблюдается окрашенность в дополнительный
цвет. У животных такая окраска имеет, видимо, основной смысл в том,
чго, будучи окрашены в дополнительный цвет, например в красный при
синем освещении, они делаются мало заметны. Другое у растений. На
глубинах, в освещении разного цвета, фотосинтез будет итти тем лучше,
чем больше лучей будет поглощено хроматофорами.Таким образом, красные
водоросли сильнее ассимилируют при синем освещении, а зеленые — при
красном. Однако это только самая общая схема. Многие зеленые водоросли
опускаются на большие глубины (Cladophora в Черном море опускается
на 120 .и), багрянки (например, Porphyrd) часто играют существенную
роль на литорали и даже на супралиторали (например, Corallina), бурые
водоросли нередко опускаются на глубину в несколько десятков метров,
а иногда совсем отсутствуют, и красные водоросли граничат прямо с зеле-
ными. Так, например, в Тенсберг-фиорде пояс ламинарий отсутствует
и ниже 30HbiFtfcws serratus располагается пояс Furcellaria (Rhodophyceae).
Цветковые растения занимают самый верхний горизонт сублиторали,
несколько верхних метров, и нередко обнажаются во время отлива. Иногда
Zostera обитает на самой литорали, образуя здесь полянки карликовой
зостеры, в основном, на наиболее влажных пониженных ее участках.
Горизонтальное распределение. Интересна географическая, смена видо-
вого состава и распределения основных форм фитобентоса. В высокоаркти-
ческих широтах, в области постоянного действия пловучих льдов, литораль
лишена растительности и только на глубине нескольких (4—5) метров
встречаются отдельные кустики Fucus evanascens. Основная масса фито-
бентоса— это ламинарии с примесью красных (Ptilota, Odonthalia,
Delesseria, Lithothamnion).
Начиная от юго-западной части Варенцова моря (Мурманское побе-
режье) и Нью-Фаундлендской банки по американскому побережью для
всей Северной Атлантики, так же как и для северной Пацифики, харак-
терно наиболее мощное развитие береговой растительности, как литораль-
ной, так и сублиторальной. На литорали резко преобладают бурые водо-
росли, несколько видов фукусов и аскофпллум, образующие четко обо-
собленный и сверху и снизу пояс водорослей. В сублиторали их сменяют
ламинарии и ряд красных водорослей, как Furcellaria, Gelidium, Ahn-
jeltia, Corallina, Chondrus, Lithothamnion, Delesseria, Rhodymenia и др.
В Тихом океане, у берегов Камчатки, Аляски и затем на юг от Кали-
форнии в сублиторали растут гигантские ламинариевые Macrocystis,
Nereocystis и Lessonia, отсутствующие в Атлантике (рис. 127).
Далее на юг, в тропических морях, как общее правило, по сравнению
с морями умеренных и холодных климатических зон, растительная жизнь
на литорали и сублиторали очень бедна. В тропической полосе мы уже
не можем рассматривать сублитораль как зону доминирования водорослей.
Такое доминирование встречается только на отдельных участках; на
мягких грунтах в массах может получать развитие Caulerpa, на жестких
Halimeda на самих коралловых рифах литотамний. При переходе к тро-
пическим морям фукусы и ламинарии сходят на нет, их сменяют другие
бурые — Cystoseira, Sargassum и Turbinaria. Кроме того, в тропических
морях наблюдается массовое развитие на песчанистых и илистых грунтах
зеленых водорослей, опускающихся здесь иногда на глубину до 40 м. Udo-
tea, Laurencia и Peyssonella опускаются на глубину даже в 100 м. В высоких
широтах уже на глубине 30—40 м флора макрофитов практически сходит
на нет. В тропических морях на литорали и в верхнем горизонте сублито-
247
ради, вследствие сильного нагрева и резкой инсоляции, растительность
почти отсутствует и может существовать только в местах постоянного
и сильного прибоя. Верхний горизонт растительности в тропических
морях занят преимущественно зелеными водорослями Ulva, Caulerpa,
Dasycladus, С odium, Cladophora и др. Вместе с ними и несколько ниже
располагаются багрянки Hypnea, Laurencia, Gracillaria, Gelidiopsis,
Dudresnaya и др. Еще глубже, на глубине до 15 м руководящими форма-
ми являются виды бурых Sargassum и Turbinaria и виды красных Реуя-
sonella, Squamaria, Melobesia и Laurencia. В морях умеренной и холодной
зон южного полушария опять вступают в силу бурые водоросли: в верх-
них горизонтах Hormosira^ в нижних главным образом из ламинариевых
гигантские Macrocystis, Nereocystis, Lessonia и Ecklonia.
Рис. 127. Схема вертикального распределения бурых водорослей у
берегов Британии и Америки: А —Alaria, Lc —Laminaria Clousto-
ni, Ld — L. digitata, Ls — L. saccharina, M — Macrocystis, N —
Nereocystis (по Чэпмену).
И в пределах наших европейских морей с севера на юг мы можем видеть,
в прибрежной области смену видового состава массовых форм морских
растений — макрофитов. В Белом море это главным образом ламинарии
и в значительно меньшей степени фукусы, аскофиллум и зостера: на
Мурмане аскофиллум, фукусы и ламинарии; в Черном море филлофора,
цистозира и зостера; в Каспийском из всех их остается только зостера,
на первое же место по значению выходят харовые водоросли (из Chloro-
phyceae).
3. ЛИТОРАЛЬНАЯ ФАУНА
Литораль и ее границы. Литоральная фауна издавна привлекала вни-
мание человека, вследствие своей доступности и как источник питания
и как объект наблюдения.
Под литоралью разумеют обычно часть морского дна, которая во
время прилива средней нормальной высоты находится под водой, а при
таком же отливе оказывается выше уровня воды.
Выше среднего уровня воды в прилив располагается полоса берега,
только временно, случайно и непериодически покрывающаяся водой,
а большей частью только смачиваемая ею, обычно именуемая супрали-
торалыо. Верхняя граница литорали не всегда может быть проведена
резко. На скалистом грунте она проходит очень четко, песок и ил, в силу
капиллярности, смачиваются морской водой и выше ее уровня. В расще-
линах скал и в гротах граница литорали всегда выше, чем на открытых
местах. В разные сезоны года высота прилива различна. Прибой взбра-
сывает воду на много выше ее нормального уровня. Многие другие
причины также могут влиять на высоту уровня воды у берега.
248
Не во всех морях имеется осушная полоса, вернее сказать, в некоторых
морях она настолько мала, что не приобретает тех специфических особен-
ностей, которые имеет в открытых океанах. Средиземное, Черное, Азовское,
Каспийское, Балтийское моря—это безливные моря, приливно-отливные
колебания в них очень незначительны (несколько сантиметров). Еще С е р-
нандер (Serna n de г, 1917) указал, что в безливных морях лито-
ралью следует считать полосу берега, расположенную выше нормального
уровня воды, нерегулярно подвергающуюся покрытию водой, набегаю-
щими волнами, прибоем или при повышении уровня, вызываемом иными
причинами. Собственно говоря литораль безливных морей вполне соот-
ветствует супралиторали открытых морей l. С другой стороны, и в откры-
тых частях морей и океанов приливно-отливные явления могут быть
выражены совершенно по-разному. Амплитуда приливно-отливных коле-
баний, основной причиной которых является совместное притяжение
воды луной и солнцем, зависит от направления приливной волны по отно-
шению к берегу, от конфигурации берегов, от отклоняющей силы под вли-
янием вращения земли и от ряда других причин. Наибольшие приливно-
отливные колебания (16,2 м) известны для залива Фундп (Америка).
У южной оконечности Ю. Америки они достигают 12—14 м. У атланти-
ческого побережья Франции и на юге Великобритании амплитуда прилива
достигает 11—12 м. В других частях Атлантики приливно-отливные
колебания значительно слабее и обычно не превосходят 2 м. В Тихом оке-
ане наибольшие известные приливы достигают только 11 м (в Охотском
море). При слабом уклоне дна, даже при небольшой приливно-отливной
амплитуде, могут обсыхать обширные участки дна. Так, например, гро-
мадные пространства, обнажающиеся при отливе, прилежат с запада
к Ютландскому полуострову (так называемое Wattenmeer).
В открытых морях умеренной полосы литоральная фауна выражена
наиболее богато. Несколько сотен видов растений и животных входят
в ее состав. В безливных морях литоральная фауна частью приурочена
к так называемой зоне прибоя, частью погружается в более глубокие
горизонты, не обсыхающие во время прилива. По мере продвижения на
север литоральная фауна беднеет. В высоко-арктических широтах, где
льды большую часть года загромождают литораль, остающиеся предста-
вители литоральной фауны уходят в сублитораль. Наконец, слабо выра-
жена литоральная фауна на открытых побережьях в тропической полосе,
где очень сильный нагрев солнца мешает ее развитию и где также проис-
ходит опускание ряда литоральных форм в сублитораль. Только мангровые
заросли дают приют своеобразной тропической литоральной фауне.
Супралитораль. Верхний горизонт литорали, расположенный выше
линии среднего нормального прилива и покрываемый водой нерегулярно,
частью только в самые большие приливы, частью во время прибоя, а
частью только смачиваемый брызгами морской воды, многими авторами
выделяется как супралитораль, рассматриваемая иногда как самостоя-
тельная зона, иногда как верхний горизонт литорали. Свойственные ей
растительные и животные формы обладают способностью длительное время
оставаться без воды. Характерны для этой зоны в северном полушарии
лишайники (Verra с aria, Lithoicea), водоросли (Scytosiphon, Ceramium,
Enteromorpha, Corallina и др.), некоторые моллюски (Littorina, Patella),
некоторые ракообразные (Talitridae, Chthamalus. Balanus, Lygia, раз-
личные крабы). Представители семейства Talitridae (Amphipoda) могут
довольно далеко уходить от края воды, даже е самое жаркое и сухое время
года, совершенно обсыхая с поверхности и долго оставаясь без воды.
1 Л. А р н. о л ь д и предлагает называть ее псевдолиторалыо.
24£>
Удивительно видеть амфипод, с большой быстротой носящихся в пол-
день по раскаленному песку средиземноморского побережья. По данным
И. Закса (1929) на Дальнем Востоке Talitridae зимой уходят даже в
прибрежные леса, на чердаки домов и там зимуют под бревнами, пнями
и т. п. Мы видим здесь пример как бы наполовину совершившегося
перехода от водного образа жизни к наземному. В грунте супралиторали
и в выбросах водорослей в большом количестве обитают олигохеты и
насекомые, как в состоянии личинок (Staphylinidae, Diptera), так и
взрослые.
В верхних горизонтах литорали наблюдается любопытное смешение
морской инфауны (галоэдафон) с наземной фауной и фауной воздушных
почв (аэроэдафон), примеры чего мы указывали выше.
Очень красочное описание этого явления для эпибиозов верхнего
горизонта литорали, приводимое С. Зерновым, дает П р ю в о
для окрестностей Баньюльса (Средиземное море): «Это есть место сопри-
косновения морской и наземной фаун... Крысы бегают по скалам и соби-
рают остатки, выброшенные морем, и ловят иногда мелких крабов Pachij-
grdpsus marmoratus. На песке у Розаса цицинделлиды охотятся за тали-
тридами, которыми они питаются... видов там мало, но индивидуумов
масса. На скалах и на россыпях камней находятся: Pachygrapsus татто-
ratus, Lygia italica, Chthamalus stellatus из ракообразных, Littorina ne-
ritoides и Patella lusitanica представляют моллюсков».
В устьях рек такое же взаимное проникновение может происходить
между инфауной морской литорали (галоэдафон) и пресноводной фауной
прибрежной полосы (лимноэдафон).
Условия существования на литорали. Население литорали представ-
ляет собою весьма сложный и многообразный комплекс форм, наиболее
характерная общая особенность которых — способность оставаться на
определенные периоды суток без воды и переносить резкие колебания
внешних факторов и в первую очередь температуры и солености, причем
у разных форм, обитающих на литорали, эти способности выражены
в весьма различной степени. Таким образом, литоральная флора и фауна
отбираются среди наиболее выносливой (эвритопной) части морской флоры
и фауны. Вряд ли можно указать такие формы, которые встречались бы
исключительно и повсеместно только в литоральной зоне, но многие
литоральные формы именно здесь находят наилучшие условия для пышного
развития. Значительная часть литоральной фауны живет в самом грунте
и поэтому свойства грунта могут объяснить характер ее распределения.
Интересный анализ термического режима илисто-песчанистых грунтов дал
Л. Линке (1939) для вышеупомянутых ваттенов.
Грунты отличаются слабой теплопроводностью и тем большей, чем
мельче составляющие их частицы (рис. 128). Хорошо нагреваемые во время
отлива грунты осушных зон затем удерживают тепло и препятствуют
резким изменениям температуры. Кроме того некоторое повышение тем-
пературы внутри грунта может обусловливаться наличием гниющих
веществ. Во время зимних холодов поверхностные слои грунта на отливах
могут замерзать и понижать температуру до 5° ниже нуля, но с глубиной
температура повышается. Так, 10 февраля 1939 года Л и н к е наблюдал
на литорали следующие температуры в грунтах:
вода в заливе — 1.8°
грунт на глубине 0—3 см — 1.8°
» » » 21—23 см -|- 0.5°
» » » 39—42 см 4- 1,5°
250
С другой стороны, летом в тихие солнечные дни температура литораль-
ных грунтов и воды, остающейся в лужицах, может сильно подниматься
(табл. 92).
Таблииа 92
ТЕМПЕРАТУРА ВОДЫ И ГРУНТА БАТТЕНОВ 24/VII—1936 г. ПО ЛИНКЕ
вода в заливе.......................................18—20"
вода в лужах на литорали во время отлива............ 34—35°
грунт на'глуб. 0—3 см............................... 34—35°
» » » 10—13 см............................... 25°
» > » 50 см ............................. 17°
Поверхностные слои грунта илисто-песчанистой литорали дали годовую
амплитуду температуры в 40°, в два раза более высокую, чем вода южной
части Северного моря (20°). Но уже на глубине 50 см амплитуда годовых
Рис. 128. Теплопроводность различных грунтов (по Линке).
а — вода, Ь — песок, с — илистый песок, d — ил.
колебаний температуры не превышает 15,5т. е. меньше, чем в море.
На глубине 20 см внутри грунта затухают суточные колебания темпера-
туры. Таким образом, температурный режим грунтов литорали создает
благоприятные условия для населяющих их организмов. Громадность
скоплений растительных и животных организмов на литорали, при всех
несомненных трудностях существования, может быть понята только
в том случае, если учитывать обилие сосредоточивающихся здесь пита-
тельных веществ. Масса детрита, смываемого с суши и намываемого морем,
дает пищу литоральным организмам. Обычная приуроченность наиболь-
ших скоплений водорослей к полосе прибоя в самом верхнем горизонте
сублиторали и на литорали с несомненностью указывает, что и водо-
росли находят в полосе прибоя какие-то особенно благоприятные условия
питания.
Состав литоральной фауны. Встречающаяся на литорали богатая
и разнообразная фауна может быть разбита на несколько биологически
разнородных групп по отношению к главной особенности литорали —
периодически наступающему обсыханию. Во-первых, какая-то часть
морских форм случайно остается на литорали во время отлива без воды
и погибает, не дождавшись следующего прилива. Сюда относятся как
представители пелагической фауны (медузы, сифонофоры, ктенофоры,
китообразные, некоторые рыбы и др.), так и формы бентоса (некоторые
иглокожие и т. п.). Они могут быть названы литоралофобами. В каждый
отлив множество таких животных погибает на литорали. Некоторые
пз них могут случайно переживать отлив в остающихся на литорали
251
озерках, «ваннах» и т. п. Во вторую группу можно отнести те организмы,
которые часто остаются на отливе, обычно у его нижнего края и в углуб-
лениях между камнями, под камнями, в зарослях макрофитов, или просто
на влажном илистом грунте и, не погибая, пережидают время отлива. Эти
формы могут быть по аналогии названы литоралоксенами, так как лито-
ральная зона не является для них нормальной средой обитания. Третью
группу составляют организмы, которые могут быть названы литоралофп-
лами. Они обычно присутствуют на литорали во время отлива в более влаж-
ных ее частях и главным образом под камнями и среди зарослей макрофитов.
Литораль привлекает их своими богатыми кормовыми ресурсами. Так,
очень часто нижний горизонт литорали бывает усеян бесчисленным
множеством морских звезд, откармливающихся мидиями и брюхоногими
моллюсками. Весьма интересно, что помимо морской фауны, кормовые
богатства литорали привлекают также и наземных животных. В отлив
литораль собирает множество птиц, находящих здесь обильный корм.
Наконец четвертая группа, это литоралебионты, т. е. специфические
обитатели литорали, именно здесь встречающиеся в наибольших коли-
чествах и находящие здесь наилучшие условия для существования.
В состав этой группы в первую очередь входят двустворчатые и брюхо-
ногие моллюски, усоногие ракообразные, различные черви, главным обра-
зом полихеты и гефиреи, некоторые актинии, некоторые мшанки и гидро-
иды и др. Однако и эта группа наиболее типичных форм литорали биоло-
гически неоднородна. Большинство литоральных форм хотя и остается
на литорали во время отлива, но фактически находится в воде. В первую
очередь это грунтовая фауна (рис. 129). Хотя грунты литорали во время
отлива и дренируются, в них однако остается еще очень много воды и ходы
червей и моллюсков, трубки червей и некоторых ракообразных наполнены
водой, так что обсыхания поверхности тела животных не происходит.
Вероятно, именно в силу этого наилучшее выражение инфауна литорали
получает на песчанистых грунтах, обладающих свойством удерживать
наибольшее количество воды (Брюс, 1928). С другой стороны, плотно
полегшие кусты фукусов, часто совершенно обсыхающие с поверхности,
внутри сохраняют достаточно влаги, дающей приют червям, ракообраз-
ным, моллюскам, мшанкам, гидроидам, иглокожим и другим животным.
Наконец, в ямках под камнями, особенно в нижнем горизонте литорали,
всегда держится вода, дающая приют многим амфиподам и червям, а также
некоторым видам рыб. Сравнительно небольшое число видов остает-
ся на литорали во время отлива на открытом воздухе, — преимущественно
на теневых сторонах камней, или прямо на песчанистом или илистом грунте.
Большинство из них одето раковиной, способной плотно замыкаться
(усоногие ракообразные, брюхоногие и двустворчатые моллюски). Многие
присасываются к камням (боконервные и некоторые брюхоногие моллюски)
и защищаются спинными створками, некоторые сильно сжимаются и,
хотя не обладают плотными покровами, хорошо противостоят высыханию
(актинии, голожаберные моллюски и др.).
Во время прилива на литораль заходит большое число животных из
сублиторали и особый интерес представляет массовый заход рыбы, преи-
мущественно молоди, для откорма. На кормовое значение литоральной
фауны Северного моря для рыб указывали Тамдруп (Thamdrup,
1935) и Линке (1939). Более детальный количественный анализ этого
явления был дан для Мурманской литорали (см. Баренцево море).
Собственно литораль. На всем громадном протяжении, от Англий-
ского канала до Варенцова и Белого морей — литораль заселена сход-
ной по составу фауной. Вследствие указанных выше особенностей грунтов
бореальная фауна литорали подвигается далеко на север и восток.
252
см
Из растительных форм наиболее характерны (в последовательном
порядке от верхних горизонтов литорали) бурые водоросли Pelvetia сапа-
liculata, Fucus spiralis (в более южных широтах), Fucus vesiculosus, Asco-
phyllum nodosum, Fucus serratus, F. inflatus (в более северных широтах)
и в самом нижнем горизонте Chorda filum. Из зеленых водорослей, играю-
щих гораздо меньшую роль, чем бурые, можно указать в той же после-
довательности: Cladophora fracta, С. gracillis, Enteromorpha, Monostroma
juscuin, Dictyosiphon и др. Животное население литорали дает два основных,
везде повторяющихся комплекса — на скалах и камнях Cirripedia,
Gastropoda, Mytilus (онфау-
на), на мягких грунтах — La-
melli branchia ta, Polychaeta
(инфауна).
Для фации скал наиболее
типичны Balanus balanoides,
В. crenatus, Mytilus edulis,
множество гастропод, как по-
крытых раковиной, так и
голожаберных; из первых
следует указать Littorina
littorea, L. rudis, L. obtasa-
ta. L, palliata, Purpura lapil-
his, Acinaea testudinalis, Ske-
nea planorbis, Rissoa aculeus,
Hydrobia ulvae; в южных рай-
онах — Patella vulgata, из
вторых—представителей ро-
дов Aeolis, Doris, Dendro-
notus, Lamellidoris, Acantho-
doris, Limapontia и др. Вся
эта обильная фауна гастро-
под либо сидит прямо на
камнях, либо прячется сре-
ди зарослей фукусов и других
водорослей. Обычны на кам-
нях и некоторые предста-
вители актиний: в более се-
верных широтах Actinia
equina, в более южных—Metridium dianthus и Urticina crassicornis.
Во влажной среде на зарослях макрофитов обитает много видов гидро-
идов (Sertularia, Clava, Gonothyrea, Laomedea, Dynamena и др.), мшанок
(Flustrella, Alcyonidium, Bowerbankia, Membranipora и др.), ракообразных
(Gammarus marinus, G. locusta, laera marina, Orchestia littorea, Idothea
granulosa). Листья фукусов часто усеяны известковыми трубками
полпхет Spirorbis. Немало среди зарослей макрофитов на литорали немер-
тин, турбеллярий и полихет. В самом нижнем горизонте литорали в боль-
шие отливы появляются Hyas araneus, Pagurus pubescens, а в более
южных широтах—Pagurus bernhardus и Carcinus maenas.
Выползают также на литораль и некоторые иглокожие и иногда в очень
больших количествах, например Asterias rubens. Под камнями и под
дерновинами фукусов на литорали прячутся мелкие рыбки; более обычные
из них Pholis gunellus и живородящая бельдюга (Zoarces viviparus).
Песчанистые и илистые грунты литорали дают приют плотным популя-
циям двустворчатых моллюсков и червей. Наиболее массовые из моллю-
сков—промысловыеCardiumedule и Myqarenaria^ также Масота baltica и в
Рис. 129. Схема распределения в грунте основ-
ных компонентов фауны илисто-песчанистой ли-
торали Западной Ютландии (по Торсону).
1 — Hydrobia. 2 — ходы Pygospio, 3 — трубки Со-
rophium, 4 — ходы Arenlcola; 5 — Cardium; в—
Macoma; 7 — Scrobicularia; 8 — Mytilus; 9 — Littorina;
10 — Mya.
253
меньшей степени М. calcarea и Scrobicularia plana. Из червей наиболее
массовые: Arenicola marina. Nereis diversicolor (в более южных широтах),
Nephthys hombergi, N. ciliata, Scoloplos armiger, Pygospio elegans, Hetero-
mastus jilijormis, Polydora quadrilobata,Fabricia sabella-, в отдельных рай-
онах— Manayunkia, Ophelia, Traoisia, Nainereisnm немертин—Cerebratulus,
Amphiporus, Lineus. Часто в изрядных количествах обитают в мягких
грунтах литорали Priapulus caudatus и Halicryptus spinulosus. Из
ракообразных иногда в массовых количествах встречается амфипода
Corophium volutator.
Зональное распределение биоценозов на мягких грунтах литорали
Северного моря показано па рис. 132. Как показал Т а м д р у п, боль-
шинство форм, роющихся в грунте отливов, очень выносливо в отношении
кислорода и долгое время может находиться в анаэробных условиях,
не теряя жизнеспособности, и,что особенно интересно, часть их может совер-
шать дыхательный обмен в воздушной среде. Согласно с более ранними
данными Везенберг - Лунда (W esenberg-Lund, 1904) и
Мортенсена (Mortensen, 1922) Т а м д р у п различает
на литорали ваттенов лежащий выше пояс Corophium volutator и ниже —
пояс Arenicola marina. Для первого, в свою очередь, можно выделить
участки с Pygospio elegansn Hydrobia ulvae, а для второго — c Cardium edule.
Автор отмечает также весьма интересный факт — на литораль в прилив
заходят на откорм громадные количества мелкой камбалы в 2—3 см
длиной. В заключение Т а м д р у п приводит для массовых форм пока-
затели биомассы, приводимые на табл. 93, пополненной данными
Линке.
Таблгта 93
Название формы
Количество экземпляров
на 1 м2
Сырой вес в г
на 1 м2
обычно до I B 0TX
до
Ulus edulis . . . .
Cardium edule ... .
M acom a baltlta . . «
Llttorina littoroa. . <
Mya arenaria..........
Hydrobia ulvae . . .
Scrobicularia plana .
Arenicola marina . .
Nereis diversicolor .
Fabricia sabella . . .
Pygospio elegans . .
Nephthys hombergi .
Scoloplos armiger . .
Corophium volutator
Isotomurus palustris.
2000—12000 — 9500—32000 и до
75000
4675 • 11000 5809 и до 8250
300—500 3500 300—400
230—330 — 160—260
7—13
46300 271920 411
20—60 1082400 —
20—35 70 80—160 и до 172
130—150 320 25—34 и до 78
— >100000 —
20300 35000 —-
90 — 16
374—460 — 18—22
2060—7125 44251 7—22
— > 264000 —
Эти цифры показывают очень большое обилие фауны указанной лито-
рали, превышающее, видимо, биомассу бентоса в сублиторали. По основным
установкам и по применению количественного учета работа Тамдрупа
должна быть признана лучшей из всех работ по литорали Северного моря,
однако и в ней количественный учет не производится с нужной последова-
тельностью. Две другие работы по литорали Северного моря (Linke и
Wohlen berg) используют количественный учет в меньшей степени
и имеют общеэкологический интерес. Воленберг значительную
часть работы посвящает физико-географическому описанию ваттенов,
254
химическим свойствам грунтовой воды, механическим свойствам грунта.
Детальный анализ грунтов дает возможность автору сопоставить его данные-
с распределением организмов, что дается автором на двух весьма пока-
Прибойная j t
jona ,, A-Oslera попа Moslem angustifolia FucusMt/ttli
<—--- —'г0 ° manna , Heteromastuf Amaharete^ Mutilutedulis
Ophelia' Peldscolex berridem
5% 19% 18% 60%
Puc. 130. Схематическое сопоставление распределения некоторых
обитателей литорали западной Ютландии и механического состава
грунта (по Воленбергу).
зательных графиках (рис. 130и 131). Многие, живущие в грунте, литораль-
ные формы питаются, заглатывая самый грунт, как, например, Arenicola,
Priapulus и др. Вследствие изобилия Arenicola на отливах и ее крупных
------------Число экз Вес в г Arenicola marina
------------Число экз — Вес в г Nereis diversinoUn
VoV Окислительная ////////// Восстановительная
зона //////////, зона
Рис. 131. Схематическое сопоставление числа экземпляров и био-
массы Arenicola marina и Nereis di versicolor и характера грун-
та на литорали западной Ютландии (по Воленбергу).
размеров, способность ее питаться грунтом превращается в мощный
фактор переработки самого грунта, подобно тому как это имеет место
у дождевых червей (Дарвин) или у пресноводных олигохет (A I s-
25S
с е г b е г g). Как показал Линке, при плотности поселения 40 экз.
на 1 jh2, популяция Arenicola пропускает ежедневно через кишечники
1,5 кг на 1 м2, а в течение года 540 кг или ’/4 ле3, грунта, т. е. почти весь
тот слой, в котором обитает.
Воленберг так же, как Т а м д р у п, наблюдал биоценотическую
зональность (рис. 132). Характерно при этом, что некоторые неморские
организмы обильно заселяют верхнюю часть отливной зоны, в основном
супралитораль, и вплотную подходят к морской фауне литорали п часто
проникают друг в друга. Воленберг наблюдал в изобилии росшие в супра-
литорали солелюбивые ра-
стения и массами засе-
лявших ее жучков рода
Blpdius (стафилинпды) и
личинок мух, и среди них
можно было встретить так-
же нереид и офелий и не-
которых других полихет.
Наконец, последняя и на-
иболее обширная из работ,
посвященных фауне лито-
рали Северного моря, при-
надлежит Линке. Этот
автор также отмечает зна-
чительное количественное
обилие населения «ватте-
нов», по сравнению с суб-
литоралью, и большое кор-
..or, ~ мовое значение их фауны
Рис. 132. Схематическое изображение зональности J
в распределении литоральной фауны на «ваттенах» Для Рыо? заходящих на
Северного моря (по Воленбергу). них во время прилива.
1—зимняя полоса прибоя; 2—летняя полоса прибоя; 3—по- Большое Внимание
селение жуна Bledius spectabilis; 4 — Corophium; 5 — пес- яптпп пппятттяпт пя ттптяттт.
чаные росы с Bredius arenarius; 6 — поселения Volutator; автоР ООращает На ДСТаЛЬ-
7 — грядна с Arenicola; 8 — Pygaspio elegans. НОе изучение фаКТОрОВ Сре-
ды — физико - химических
свойств воды и грунта и приуроченность к тем или иным условиям
обитания определенных группировок населения. Особенно интересен
анализ плотности различных грунтов и сцепления частиц грунта. Это
последнее обусловливается жизнедеятельностью организмов, характером
содержания воды в грунтах, температурным режимом и т. д.
Л инке отмечает количественное обилие фауны ваттенов, привле-
кающее во время прилива на кормежку многих гостей из сублиторали,
в том числе промысловую креветку Crangon crangon и рыб — камбалу и
угря. Во время отлива обильный корм на литорали находят для
себя многочисленные птицы. Сравнительно маленький залив Jade Busen.
с дном, на 3/4 обнажающимся вовремя отлива, имеющий площадь 166,4 км\
из которой 121,7 км2 обсыхают во время отлива, дает в течение
года общий вылов креветок в 500—1000 т. Линке приводит сводную
таблицу биомассы фауны восьми биоценозов для ваттенов (табл. 94).
Географические изменения состава литоральной фауны. По западной
стороне Ирландии и Англии и в западной части Английского канала
в качественном составе литоральной фауны наступают существенные
изменения. У южных берегов Европы, в Средиземном и Черном морях
родовой и видовой состав литоральной фауны (Лузитанско-Средиземно-
морская подобласть) оказывается уже другим, хотя биологические соот-
ношения, биономические типы и ха'рактер зональности остаются в общем
256
Таблица 94
Комплексы Площадь в км- Средняя био- масса в г!м2 Общая био- масса в т
Scoloplos * 45,861 30 1375,83
Pygospio 33,2 300 9960
Scrobicularia 22,133 1200 26559,6
Corophium 18,592 40 743,68
X . “ i bcolecolepis 0,2656 5 1,328
Mytilus 1,6821 5000 8400,5
Карликовая зостера 5,312 2500 13280
Для всего залива 131,2077 555 78803,938
темп же, — в очень большой степени выступают явления биологического
впкарирования. В Черном море наиболее типичны для полосы прибоя —
Chthamalus stellatus, Littorina neritoides, Pachygrapsus marmoratus, Lygia
brandtii, Patella pontica, Mytilus gallorpovincialis, Alexia myosothis. Водо-
росли в этой полосе, расположенной выше уровня воды, также не те,
что в Северной Атлантике, а именно: Scytosiphon, Ceramium, Enteromor-
pha, Bangia, Raljsia и Corallina.
Особенно много сходства в распределении литоральной фауны дают
северные части Атлантического и Тихого океанов, несмотря на то, что
тихоокеанская фауна гораздо более богата и в подавляющем большинстве
представлена другими видами и родами, при обилии биологических вика-
риатов и сходных биономических типов.
Весьма тщательное описание литоральной фауны залива Петра Вели-
кого (Японское море) и побережий Шантарских островов (южная часть
Охотского моря) сделано И. 3 а к с о м (1927 и 1929) и позднее К. Д е-
рюги н ы м (1939). Основные группировки организмов на литорали,
а также сравнение тихоокеанской и баренцовоморской литоральной
фауны даны на 95 и 96 таблицах, взятых у И. Закса, с дополнениями по
К. Дерюгину ис некоторыми изменениями.
Литоральная фауна в районе залива Петра Великого, особенно на скалах
п камнях, вследствие перетирающего действия льдов зимой и вследствие
незначительной приливно-отливной амплитуды (не свыше 50 см в сизигий),
относительно бедна.
Литораль Шантарских о-вов дает ряд резких отличий от литорали
залива Петра Великого. Амплитуда приливно-отливных колебаний здесь
очень велика (до 5 л«). Исчезают саргассы, зостера, ульва и другие харак-
терные водоросли Японского моря. Исчезает свойственное Японскому морю
многообразие звезд и ежей. Мощный пояс фукусов на литорали и ламина-
рий на границе ее и сублиторали создает сходство с северно-европейским
побережьем. Рассматривая литоральную фауну американского побережья
Тихого океана, мы находим черты большого сходства (биологического)
и значительных отличий (главным образом видового состава) по сравне-
нию с противоположным побережьем Тихого океана и обоими берегами
Атлантического океана. Впрочем и здесь много родов общих: Balanus,
Mytilus, Chthamalus, Littorina, Acmaea, Purpura и др.
Преобладающее значение на скалисто-каменистой литорали в северной
части Тихого океана по американскому побережью вплоть до побережий
Канады имеют из Girripedia Balanus cariosus, В. glandula, Chthamalus
dalli, Mitella polymerus, из брюхоногих моллюсков — Littorina sitchana,
L. scutulata, Acmaea digitalis, A. cassis, из двустворчатых—Mytilus edulis,
M. californianus и актиния Cribrina xanthogrammica. Кроме того, весьма
257
17 Зенкевич, т. I
Таблица 95
Высота над уровнем моря в см Гори- зонты Японское море, залив Петра Великого Гори- зонты Охотское море, о-в Большой Шантар
фация скал и камней фация мягких грунтов фация скал фация мелко- зернистых грунтов
600 5С0 400 300 I Супралитораль Talitrldae
Мертвый | горизонт Мертвый при- брежный галеч- ный склон
200 100 Супра- лито- раль Rlvularia, Rhi- zoclonlum, Lit- torina squallda, L. sitchana. Li- gla clnerescens Валы выбро- шенных водо- рослей и зосте- ры с Orchestla, Oligochaeta, Lycastls, Forflculldae, Coelopa, Staphy- linldae II Balanus, Mytilus edulis, Fucus evanescens
I ! Glolopeltis, Bala- nus, Littorina squa- llda, L. sitcha- na, Patella, Tur- bo sangarensls, Chthamalus chal- lengerl, Brachl- notus Arenicola crl- stata, Polydora, My a arenaria III I Заросли Delesserla Mlddendor- fil и др. Биоценозы Oridla, Molgula, Chlrldota disco- lor, Macoma baltlca, Venus, Parasabella maculata, Hali- chondrla
II Corallina, Laurentla, Littorina squa- llda, L. sitchana, Thais, Actiniae, Patlrla, Aphela- sterlas 1
обильные и часто массовые компоненты этой литорали — гастроподы Thais,
Purpura, Amphissa и в нижнем горизонте литорали — крабик Hemigrapsus
rudis, звезда Pisaster ochraceus и рыбки Xiphister mucosus и Epigeichthys
atro-purpureus. Эта фауна образует несколько биоценотических сочетаний.
На мягких грунтах такое же преобладающее значение имеют двуствор-
чатые моллюски Масотъ, nasuta, М. secta, М. inguinata, Paphia staminea,
Cardium corbis, полихеты Nereis virens и Arenicola claparedii, голотурия
Leptosynapta inhaerus и др.
Литораль калифорнийских побережий, расположенных на границе
с тропической областью, дает другой набор массовых форм, чем у побе-
режий, расположенных дальше на север. Для залива Монтери Мас
G i п i t i е указывает для каменистого грунта следующие наиболее
обычные формы сообщества: Mytilus californianus, Cirolana harfordi,
Petrolisthes cinctipes, Physcosoma agassizi, Thais emarginata, Amphipoda.
Выше его располагается биоценоз Balanus glandula — Acmaea. Кроме
того, в биоценоз входят Balanus tintinnabulum californiciun, В. nobilis,
Hulosydna insignis, Saxicaoa arctica, Nereis.
258
Таблица 96
Высота вад уровнем моря в см Баренцево море Кольский залив
горизонт фация скал фация мелко- зернистых грунтов
Супралитораль
‘Oligochaeta
600
500
400
300
Мертвый горизонт
II
Balanus
Mytilus
edulis
Fucaceae
Wacom a
halt lea
Fabrlcia
sabella
200
100
III
F ucus
serratus
Pygospio
(Megans
Песчанистая литораль калифорнийских побережий имеет более огра-
ниченное число видов, но зато эти виды встречаются в громадном числе
экземпляров. В подавляющем большинстве это роющие формы. Очень
типичен здесь биоценоз крабика Emerita. Ему сопутствуют другой краб —
хищный Blepharipoda occidentalis и крошечная изопода Cirolana lingutfrons,
выполняющая роль падальщика. Также характерен для песчанистых лито-
ралей калифорнийского побережья крупный промысловый моллюск Pis то
clam (Tivela stultorum). Дальше на север, вдоль побережий штатов
Орегон и Вашингтон Emerita и Pismo clam заменяются другим промыс-
ловым очень крупным моллюском Razor clam (Siliqua patula). Для
самого нижнего горизонта песчанистой литорали всего западного
побережья Америки очень характерен неправильный ежик Dendraster
excentricus («Sand dollar»). Весьма интересно то, что структура биоцено-
зов литорали в различных частях океана и в более или менее сходных
географических зонах дает весьма сходную картину при видовом разли-
чии составляющих биоценозы видов растений и животных. Сходство
наблюдается не только в видовом составе, но и в количественных показа-
телях для отдельных биоценозов и для составляющих биологических
викариатов. Как мы видели выше, очень сходна структура биоценозов
литорали Мурманского побережья и Шантарских островов.
Согласно данным Брэди (Brady, 1943) весьма сходная картина
получена им для литоральной фауны Нортемберлендского побережья
Англии (восточное побережье). Видовой состав фауны почти тот же, что
ина Мурмане: Масота baltica, Arenicola marina, Cardium edule, Scoloplos
armiger, Nephthys hombergi, Nereis pelagica. Кроме них имеется ряд
новых форм. Судя по приводимым автором данным по количеству
экземпляров, должна быть примерно та же и биомасса. Гораздо больше
различия в составе дает литоральная флора и фауна залива Монтери
♦ 259
у побережий Калифорнии (Не wa tt, 1937). Основные обитатели литорали
здесь усоногие ракообразные — Balamis balanoides и Mitella polyinerus\
из моллюсков — Mytilus californianus; из брюхоногих — Littorina planaxis,
Lottia, Acanthina; из изопод — Cirolana; из амфипод — Allorchestes; вместо
наших звезд — PisasternLeptasterias. В супралиторали—множество Ligia
occidentalism Резко выраженный структурный и биологический парал-
лелизм биоценозов литорали обеих сторон Тихого и Атлантического
океанов в умеренной зоне и большое число общих родов и близнецовых
видов с несомненностью подтверждают общий генезис для всех четырех
ныне разобщенных литоральных фаун. В плиоцене это была единая фауна
северных частей обоих океанов. Похолодание климата и ледниковая
эпоха отодвинули ее на юг и разобщили на четыре дочерние фауны. Воз-
можно, что в теплые фазы послеледникового времени (литориновое время)
эти фауны вновь вступили в соединение.
Очень интересно исследование по фауне тропической литорали ман-
гровых зарослей, произведенное Гармсом (Harms, 1934). Автор
отмечает многочисленные особенности приспособления фауны мангровых
зарослей к амфибиотическим условиям существования, в частности, пере-
ход некоторых морских грунтовых форм (полихеты, сипункулиды) к суще-
ствованию в почвах уже вне пределов влияния морского прилива.
Описание населения тропической литорали (мангровые заросли) дано
также в разделе зоогеографии.
Литораль и ее население наилучше изучены среди всех вертикальных
зон, поэтому мы остановились на ней наиболее подробно.
4. СУБЛИТОРАЛЬНАЯ ФАУНА
Сублиторальная зона. Ниже литорали располагается сублптораль.
У северных и западных побережий Европы и в Охотском море граница
между поясом фукусов и ламинарий совпадает с наиболее низким уровнем
отлива, т. е. с границей литорали и сублиторали. Только в самые большие
отливы (сизигийные) может обнажаться пояс ламинарий.
Нижняя граница сублиторали проходит на разной глубине в разных
морях в зависимости от прозрачности воды и качественного состава макро-
фитов. Значительные скопления водорослей обычно приурочены только
к верхнему горизонту сублиторали. Громадные скопления филлофоры
(Rhodophyceae) в Черном море на глубине 30—60 м — редкое явление.
Часто уже на глубине 5—10 м макрофитов становится мало и масса живот-
ных начинает превалировать над массой растений.
Очень часто для отдельных морей и их частей можно указать преобла-
дающую группу животных. Так, например, восточная часть северной
Атлантики, северные части Варенцова и Карского морей — это царство
иглокожих: юго-восточная часть Варенцова моря, Белое, Балтийское и
Азовское моря — царство двустворчатых моллюсков. Преобладание дву-
створчатых моллюсков находится в связи с накоплением оседающего пита-
тельного органического вещества в грунте. Преобладание ракообразных
указывает на нахождение питательного детрита в состоянии взвеси в пере-
носимом состоянии. В Каспийском море это находится в соответствии
с громадным распространением песчанистых и ракушечных грунтов,
в Средиземном море — с общей недостаточностью питательного детрита,
который легче могут добывать подвижные ракообразные. Объяснить
преобладание полихет и иглокожих уже не так легко.
Более детальное описание фауны сублиторали мы приведем по данным
количественного учета, собранным за последние 30 лет.
260
Рис. 133. Основные
комплексы донного
населения датских вод
и прилежащих райо-
нов (по Петерсену).
1—Macoma baltica, 2 —
Abra alba, з — Venus,
4 — Echionocardium +
Amphiura, 5—Haploops,
6—Brissopsis4-Amphiura,
7— Brissopsis-|-Ophiogly-
pha, 8 — Amphilepis +
Pecten, м—Modiola mo-
diolus, a—Astarte bore-
alis, c—Macoma calcarea.
261
Бентос датских вод. Начало количественным исследованиям морской
донной фауны положено, как уже указывалось, Петерсеном и его
учениками во втором десятилетии настоящего столетия. По широте
поставленных теоретических и практических задач, по полноте достиг-
нутых результатов и по количеству исследований, вызванных этими рабо-
тами, имя Петерсена должно быть поставлено в первый ряд морских
гидробиологов и поэтому мы считаем необходимым более подробно оста-
новиться на работах, произведенных руководимой им Датской биоло-
гической станцией. Работы эти захватили, главным образом, сублитораль.
Рис. 134. Литоральный комплекс Macoma baltica (по Педерсену).
На рисунке кроме макомы изображены Arenicola marina, Муа arenaria, Gardium edule
и др.
Точный количественный учет донной фауны датских вод и отчасти
соседних с ними прежде всего дали возможность выделить 20—25 наиболее
массовых форм и, руководясь их распределением, установить девять основ-
ных комплексов (Communities) бентоса на мягких грунтах (рис. 133).
Каждую группировку Петерсен иллюстрирует хорошо известными
графиками (рис. 134,135 и 136), широко вошедшими в практику оформления
подобных работ и дающими состав фауны на 0,25 м2 дна. Биоценозы эти
следующие:
262
I. Литоральная группировка Macoma baltica ( M. baltica,
Mya arenaria, Cardium edule, Arenicola marina) в Балтийском море и
южном побережьй Дании.
II. Группировка Abra alba (A. alba, Macoma calcarea, Astarte
banksii, A. borealis, A. elliptica, Echinocardium cordatum), особенно в Бельте
и фиордах.
III. Группировка Venus gallina — Echinocardium
cordatum (V. gallina, Tellina fabula, Cyprina islandica, Echinocar-
Puc. 135. Сублиторальный комплекс Echinocardium cordatum — Amphiura filiformis
в датских водах (по Петерсену). На рисунке изображены также Cyprina islandica,
Turitella terebra, разные полихеты и др.
dium cordatum) на открытых песчанистых побережьях в Каттегате и
Северном море.
IV. Группировка Echinocardium-filiformis (Cyprina
islandica, Turitella terebra, Amphiura filiformis, Echinocardium cordatum)
на средних глубинах Каттегата.
V. Группировка Brissopsis-chiajei (Amphiura chiajei,
Brissopsis lyrifera) в самых глубоких частях Каттегата.
VI. Группировка Brissopsis-sarsi (Abra nitida, Axinus
flexuosus, Leda pernula, Nucula tenuis, Myriochele heeri, Ophioglypha sarsi,
Brissopsis lyrifera) в глубоких частях Скагеррака.
263
VII. Группировка A m р hi 1 е pi s-P е с t е n (Amphilepis norve~
gica, Pecten oitreus, Axinus flexuosus) в наиболее глубоких частях Ска-
геррака.
VIII. Группировка На ploops (Haploops tubicola, Ophioglyphct
robusta) в юго-восточной части Каттегата.
IX. Глубинная группировка Venus gallina, спорадически
в Каттегате.
Рис, 136. Сублиторальный комплекс Haploops tubicola в датских водах (по Петер-
сену). На рисунке изображены кроме Haploops также Ophioglypha robusta, Pecten
septemradiatus, различные полихеты и др.
Своим «сообществам» Петерсен дает условные обозначения
начальными буквами латинских названий. Так, например, IV комплексу
соответствует обозначение Е. Fil, VII — Al. Р. и т. д.
Преобладающую массу бентоса составляют иглокожие и двуствор-
чатые моллюски (рис. 137); северная Атлантика вполне может быть
названа царством иглокожих.
Петерсен полностью разделяет при формировании своих «сооб-
ществ» формы инфауны и эпифауны и не включает в них водоросли, так
что помимо указанных 9 биоценозов мягких грунтов он различает еще
комплексы эпифауны: Modiola ( Modiola modiolus, Ophiopholis acu leata,
264
Trophonia plumosa, Balanus sp.) в более глубоких и соленых частях датских
вод комплекс литорали и мелководий — Mytilus edulis (Bala-
nidae, Littorina, Rissoa, Idothea).
Наибольшая биомасса в датских водах наблюдается на небольшой
глубине в открытых частях проливов — Скагеррака, Каттегата, Бельтов-
и Зундов, причем с глубиной она резко уменьшается, что и иллюстрирует
аабл. 97.
Рис. 137. Средняя биомасса и состав по группам типичных бентических комплексов*
датских вод.
Площади кругов соответствуют биомассе комплексов, которая обозначена цифрами
при кругах.
1 — Brissopsis — Turitella, 2 — Echinocardium — Venus, 3 — Echinocardium — Abra —Macoma,.
4 — Brissopsis — Nucula, 5 — Haploops (по Петерсену из Зенкевича).
Хорошую иллюстрацию изменения биомассы с глубиной дает Т о р-
сон (Thorson, 1934) (рис. 138) для фиорда Скоресби-Саунд в восточ-
ной Гренландии. Характер кривой по мере продвижения вглубь фиорда
меняется. Пять кривых в порядке номеров соответствуют участкам фиор-
да, расположенным: первый вблизи выхода в океан, следующие — на
60, 140, 150 и 250 км от моря.
265-
Таблица 97
Комплексы
JMytilus еdulls.........
Modlola modiolus ....
Macoma baltica..........
Venus gallina...........
Echlnocardlum-Turitella
Abra alba...............
Echinocardium cordatum
Haploops tublcola ....
Brissopsis lyrifera ....
Amphiura chiajei........
Brlssopsis-sarsll.......
Amphllepls-Pecten ....
Глубина в м Биомасса в г/м2
осушпая по- лоса и до 2 м глубины 10—30 50—75 186 320
! 18784 1 193—275 11318 281—396 318 98 512 108 486 1 420 I 159 1 ч 295 20
6 417 1690 290 295 20
Среднее...............
Рис. 138. Изменение биомассы с глубиной в пяти различных пунктах в фиорде
Скоресби-Саунд, Восточная Гренландия (по Торсону).
Значительная биомасса бентоса (150—350 г/м2) характерна также и для
Лимфиорда. Это — своеобразная кормушка для рыбы. В глубоко вдающихся
в сушу мелководных (до 2 м) заливах Дании (например, Дибсофиорд),
так же как и в шведском Гюльмарфиорде, биомасса бентоса на мягких
грунтах невелика, в среднем 50—60г/м2. Таким образом от открытых частей
проливов вглубь заливов и фиордов биомасса бентоса обычно падает, что
обусловливается, вероятно, опреснением и сильным заилением дна. Здесь
резко преобладают выносливые формы литорали — Macoma baltica,
266
ffi/tilus edalis, Cardiuni edule, Hydrobia ulvae, Luttorina rudis, Nereis
diversicolor, составляющие свыше 85% всей фауны по весу. Уменьшение
биомассы вглубь заливов показано в табл. 98.
Таблица 08
Общая биомасса бентоса мяг- ких грунтов в г/лг2 Открытые ча- сти проливов Дании, Шве- ции н Норвегии Лимфиорд в । более про- дуктивных частях Гюльмар- фиорд Дибсофиорд
200—500 150—350 6С—SO 57
Посезонные и многолетние наблюдения над количественными изме-
нениями в бентосе дали возможность Петерсену и его сотрудникам
наблюдать большие изменения в количественном составе биоценозов
Рис. 139. Изменение количества экземпляров Abra alba (1) и So-
len pellucidus (2) на м2 в одном из расширений Лимфиорда в Да-
нии за 13 лет (по Бойсен-Иенсену).
(сукцессии), связанные прежде всего с массовым появлением молоди
у большинства видов в наиболее теплое время года. Изменения в коли-
честве того или иного представителя фауны бывают иногда очень значи-
тельными и от года к году. В качестве примера можно привести (рис.
139) изменение числа экземпляров двух моллюсков, Abra alba п Solen
pellucidus, в одном из расширений Лимфиорда в течение 13 лет. Оба
моллюска, особенно Solen, mrnt в корм некоторым рыбам. Как можно
видеть, за эти 13 лет количество Abra alba 7 раз падало практически
до нуля. В некоторые годы (1915, 1917) массовое отрождение молоди
резко поднимало количество моллюсков, и рыба не могла уничтожить
его целиком. Отрицательную роль в уменьшении количества полезных
моллюсков играют некоторые питающиеся ими хищные беспозвоночные,
как, например, Asterias rubens, некоторые виды офиур, крабы, брюхоногие
моллюски (Buccinum, Fusus, Nassa).
Количественный учет, проводимый через определенные промежутки
в течение всего года, дал возможность датским исследователям, следя за
популяцией того пли иного организма, весьма точно определить такие
важнейшие биологические показатели, как возраст, годовой прирост,
отрождение молоди, выедание рыбами, смертность, а, зная все эти показа-
267
тели,—дать картину процесса биологического продуцирования и рас*
числить годовую продукцию.
Зная ее для разных групп организмов, Петерсен смог дать свою
известную схему1 (рис. 140), которая впервые увязала количествен-
ные соотношения между растительной массой, разными группами бес-
позвоночных и, наконец, рыбами. Петерсен при этом принимает, что
для своего существования организмы требуют в течение года в среднем
удесятеренное по отношению к весу собственного тела количество пищи.
Планктон
Камбала и т.п. Треска и т.п ^'"""-^Сельдьит.п.
уз ---------:------------
'• Хищные ранообразн. ^'^мелная морение
и брюх, моллюски(нрипные) рыба звезды
। 5О'йув и т.п. фЮ? «р25
! । [ •
I ! Полезные жиВот н\ые woo I
i----------------1-----------Л------1--------1------------S
Рис. 140. Количественное соотношение различных групп организмов в датских водах
(по Петерсену).
Схема показывает, что биомасса зостеры в датских водах составляет
24 млн. т. «Бесполезных» животных, т. е. не потребляемых никем в пищу
(крупные моллюски, иглокожие, оболочниковые и т. д.) в пять раз больше,
чем полезных. Для существования этих 6 млн. т животных нужно не менее
60 млн. т растительной массы, каковую они и получают, так как годовая
продукция зостеры более чем в два раза превышает ее биомассу, т. е.
ее Р/В 2. За счет 1000 кг зостеры может образоваться 100 кг питающихся
ею травоядных организмов, 10 кг выкармливающихся на этих последних
хищников и 1 «г трески, питающейся в основном хищниками и мелкой ры-
бой. Камбала же, питающаяся непосредственно травоядными организмами,
1 В самые последние годы, однако, эта схема датских исследователей подвергнута
очень серьезным сомнениям (Nelson, 1947). Зостера для беспозвоночных не представ-
ляет собой универсального ресурса питания, как это полагал Петерсен.
268
в некотором смысле выгоднее трески, так как за счет 100 кг зостеры
может образоваться 1 кг камбалы. Для прокармливания в течение года
одного человека, питающегося треской, необходимо обеспечение в 10000 кг
зостеры на 1 кг его веса. Иначе говоря, чем длиннее и многочисленнее
пищевые цепи, тем больше требуется обеспечение их массой продуцен-
тов. Петерсен вполне четко поставил вопрос о необходимости при
Рис. 141. Географическое распределение донных комплексов сев.-
вост. Атлантики (по Петерсену).
I — комплекс Macoma baltica, II — комплекс Venus gallina, III — комп-
лекс Brissopsis lyrifera, IV — комплекс Br. lyrifera глубоководный, V —
северные сообщества Macoma calcarea—Astarte borealis, Va — c Portlan-
dia arctica, Vb — c Pecten frigidus, VI — комплексы лузитанской обла-
сти, VII — глубинная фауна Атлантики.
изучении явления биологической продуктивности подходить к фауне
дифференцированно и, после того как выделены массовые формы и основные
ряды пищевых взаимоотношений, вести изучение этих форм по отдельности,
так как они могут быть совершенно неравноценны по своей биологической
значимости, а иногда и прямо противоположны. Моллюски для рыб
могут быть ценнейшим кормом до определенного возраста, а затем,
перерастая определенные размеры, переходить в разряд «бесполезного»
бентоса. Мелкие иглокожие могут потребляться рыбами, а крупные ста-
269
новиться их злейшими конкурентами в истреблении двустворчатых
моллюсков.
Исследования Петерсена показали, между прочим, изобилие
кормовых ресурсов в Лимфиорде при недостаточности рыбы. G 1912 г.
была начата ежегодная пересадка молоди морской камбалы из моря в Лим-
фиорд в количестве около полумиллиона штук каждый год. При этом точно
учитывалось выедание камбалой донных животных и обеспеченность ее
развития их запасами и одновременно был поставлен и обоснован вопрос
о возможности проведения в Лимфиорде борьбы с вредными для рыбо-
ловства организмами, а именно — с крупным брюхоногим хищным
моллюском Buccinum, который обитает в Лимфиорде сотнями мил-
лионов и истребляет громадные количества двустворчатых моллю-
сков.
В дальнейшем Блевадом (Blegvad) была обоснована также
переброска молоди камбалы через Данию из Северного моря в Белы,
где количество камбаловой пищи оказалось значительно больше. В тече-
ние ряда лет, начиная с 1928г., производились эти переброски свыше 1 млн.
штук ежегодно и оказалось, что в Бельте камбала растет быстрее, чем в Се-
верном море. С другой стороны, детальному количественному изучению
подвергались и полезные формы, как промысловые (устрицы, сердце-
видка, мидия), так и кормовые объекты для рыб.
Количественное изучение донной фауны датских вод и прилежащих
районов дало возможность Петерсену впервые составить карту
распределения донных биоценозов в северо-западной Атлантике (рис. 141).
Сам Петерсен считал эту карту (вне датских вод) только рабочей
гипотезой и, как мы увидим дальше, карта подверглась в дальнейшем
значительным изменениям.
Биоценоз Ma со ma baltica ( М. baltica, Муа arenaria, Cardium
edule, Arenicola marina) приурочен в основном к приливно-отливной
полосе и только в Балтийском море в значительно измененном составе
распространяется на значительную глубину. На север он идет не только
до западного Мурмана, но заселяет также (с небольшими изменениями)
и литораль Белого моря.
Биоценоз Venus gallina дает две разности: а) мелководную
(Echinocardium cordatum, Tellina fabula, Mactra subtruncata, Cyprina
islandica) и б) глубоководную (Spatangus purpureus, Echinocardium flaves-
cens, Abra prismatica, Psammobia faeroensis, Mactra elliptica, Cyprina
islandica. В биоценозе Venus gallina существенную роль играют
также Modiola modiolus, Ophiopholis aculeata, Alcyonium digitatum и др.
в более мелководных районах и глубоководный коралл Lophohelia, Pecten
vitreus, Terebratulidae, Lima и др. в более глубоких районах.
Биоценоз Brissopsis lyrifera (Amphiura chiajei, Calocaris
marandreae, Nucula sulcata, Eumenia crassa) обитает на мягких глинистых
грунтах. Биоценоз располагается глубже предыдущего (Pecten vitreus,
Abra longicallis) также на мягких глинистых грунтах; это так
называемые «северные» сообщества Петерсена (Масота calcarea —
Astarte borealis). В глубинах Норвежского и Гренландского морей этот
биоценоз характеризуется массовым появлением Pecten jrigidus, а в наибо-
лее холодных и неглубоких районах — массовым появлением Portlandia
arctica. Биоценозы Лузитанской области, подходящие с юга к западным
побережьям Британских о-вов и к западной части Ламанша, отмечены в
своих северных пределах распространением моллюска Haliotis. Биоце-
нозы открыты» частей Атлантики распространяются на юг от биоце-
нозов Brissopsis, окаймляющих с юга Шетландско-фаррерско-
исландский гребень.
270
Направленность исследований школы Петерсена обусловила
появление (с 1914 г.) ряда отличных исследований по питанию беспо-
звоночных и рыб. Этими работами, принадлежащими в основном ученику
Петерсена — Блеваду, было заложено новое направление (коли-
чественное, весовое) в изучении питания рыб, нашедшее в последнее десяти-
летие, главным образом у советских исследователей, свое дальнейшее
развитие. Исследования Б л е в а д а смогли дать так много только пото-
му, что автор располагал уже детальной картиной количественного распре-
деления кормовых организмов в датских водах. Он же первый применил
для характеристики количества съедобной пищи отношение веса содержи-
мого желудка к весу тела, —тот показатель, который в дальнейшем лег
в основу количественного изучения питания рыб у советских исследова-
телей (индекс наполнения). Блеваду принадлежит и первая схема
пищевых связей различных рыб (рис. 142), в которой отражена и коли-
чественная сторона явления. По этому типу строились в дальнейшем.
Рис. 142. Схема пищевых цепей некоторых промысловых рыб датских вод
(по Блеваду).
схемы пищевых цепей многими авторами. В течение первого десятилетия
этой плодотворнейшей деятельности Датской биологической станции она,,
однако, не нашла отражения в работах морских биологов в других стра-
нах. Что же касается Петерсена и его продолжателей, то в последую-
щие годы они тщательно разработали ряд важнейших деталей в количе-
ственном изучении донной фауны, как кормовой базы для промысловых
рыб. В 1920 г. появилась работа Бой сен-Иенсена (Boysen-
Jensen), подводившая итоги девятилетним исследованиям кормового
бентоса в Лимфиорде. В этой работе автор, впервые в гидробиологической
литературе, дает правильное расчленение понятия биологической продук-
тивности и анализ составных элементов того, что принято называть
годовой продукцией (см. введение). Бойсе н-И е н с е н дал коли-
чественное выражение годового прироста, отрождения молоди, выедания
рыбами, соотношения между кормовой базой и выловом рыбы и измене-
ний в биоценозах в течение ряда лет. Несколькими годами позднее
Б левад (Blegvad, 1926) установил для каждого биоценоза и для
отдельных частей датских вод ту часть его фауны, которая пригодна для
рыб в качестве пищи. В результате полной увязки результатов количе-
271
ственного учета содержимого кишечников камбалы, точного учета имею-
щихся в море кормовых ресурсов и вычисления их годовой продукции
Бле вад мог довольно точно определить, какое количество морской кам-
балы может выкормиться на данных пищевых ресурсах. Так, для Кат-
тегата годовая продукция «перво-
2/S12
Рис. 143. Пригодная (сверху) и непригод-
ная (внизу) в качестве пищи для рыб части
бентоса для разных комплексов датских
вод (по Блеваду).
классного» пищевого бентоса опре-
делена Блевадомв 230 620т,
а «второклассного»—в 377 000 т.
Применяя вычисленный коэффи-
циент для камбалы (18 кг пищи
дают 1 кг рыбьего мяса), Бле-
вад определяет количество кам-
балы, которая ежегодно может
выкормиться только за счет пищи
первого класса, в 12 812 т, что
превышало в 5 раз сумму факти-
ческого вылова (2600 т) в 1928 г.
Речь идет о площади в 8866,5 км-\
таким образом, фактический вылов
камбалы составляет 21 кг на гек-
тар.
Весьма интересно, что богатей-
шая фауна датских вод оказывает-
ся в значительной части «беспо-
лезной», т. е. не идет в корм рыбам.
Петер сен полагал, что «по-
лезные» составляют только одну
шестую всего бентоса. Б л е в а д
в дальнейшем подверг тщательно-
му количественному анализу ос-
новные биоценозы сточки зрения
их пригодности как корма рыбам
(рис. 143). Это тем более интерес-
но, что в некоторых водоемах,
как, например, в Азовском море,
за немногими исключениями бен-
тос целиком оказывается съедоб-
ным для рыб.
Б л е в а д установил также
многолетние колебания в коли-
честве и составе рыбной донной
пищи (рис. 144) и зависимость
уловов камбалы от концентрации
отдельных ее кормовых форм (рис.
145), а затем (1928) также и от
всей пищи (рис. 146).
Очень характерно выступает на этих графиках годовой ход биомассы
кормовых форм. Весной количество пищевых организмов обычно меньше,
чем осенью, так как зимой может происходить массовое отмирание бентоса.
Обычное возрастание биомассы к осени обусловливается сильным ростом
отродившейся в начале лета молоди, несмотря на выедание бентоса рыбами.
Первый перекрывает последнее. Не менее интересно сделанное Бле-
вадом (1928) сопоставление за 20 лет количества пищевых организмов
(в граммах сырого веса на 1 м2), трескового промысла и количества рыбки-
бельдюги (Zoarces viviparas) (рис. 147). Прежде всего бросается в глаза
272
отрицательная корреляция между количеством трески, бельдюги и бычков.
При большом количестве трески в 1908—1912 гг. количество бельдюги
и бычков было очень невелико. Резкое падение количества трески 1912—
1916 гг. сказалось на столь же резком увеличении в 1914—1917 гг. бель-
дюги. Новый подъем в количестве трески в 1918—1919 гг. сейчас же вы-
звал новое падение количества бельдюги, а падение трескового про-
мысла в 1922—1924 гг. опять вызвало подъем в развитии бельдюги. Эта
последняя вместе с бычками — сильный пищевой конкурент камбалы и
возможно, что массовое их развитие может быть причиной сильного умень-
шения кормового бентоса для камбалы, что, повидимому, и имело место
18 Зенкевич, т. I
273
в 1924—1927 гг. В качестве средства против массового развития бель-
дюги и бычков Б л е в а д предложил перенос молодой трески во внут-
Рис, 145. Соотношение количества (экзДи2) моллюска Solen pellu-
cidus (1) и уловов (в тоннах) камбалы (2) в одной из частей Лим«
фиорда за 10 лет (по Блеваду).
ренние части Лимфиорда, что было бы выгодно и для трескового и для
камбалового промысла в этом заливе, так как эти две рыбы не являют-
Рис. 146. Количество «первоклассной» камбаловой пищи в различ-
ных частях Лимфиорда за время с 1910 по 1927 г. в г/м2 в сухом
весе (по Блеваду).
к массовому развитию сорняковых форм (бельдюги и бычков), кото-
рых треска поедает в больших количествах, и питательные ресурсы,
274
нужные для камбалы, будут сохранены. Таким образом, датские иссле-
дователи от количественного учета фауны подошли к постановке и част-
ному разрешению вопросов реконструкции фауны.
Через 11—12 лет после появления первого труда Петерсе-
на по количественному учету донной фауны появились аналогичные
исследования в СССР, США, Швеции и Англии, а несколькими годами
позднее — также в Германии, Норвегии, Финляндии и Италии. Однако
ни в одной стране работы по изучению количественного распределе-
ния донной флоры и фауны, так же как и планктона, не приобрели такого
методического уровня и такой полноты, как в СССР и особенно на
Азовском, Каспийском и Баренцовом морях. В тесной связи с этими
работами стояли у нас и исследования по питанию рыб. Впервые у нас
стали составляться карты количественного распределения донной фауны
(Зенкевич, 1927) с нанесением изобент (линий, соединяющих места о
одинаковой биомассой бентоса). Оказались на некоторых наших морях хо-
рошо прослеженными сезонные и многолетние изменения в биомассе бенто-
са. В изучении питания морских рыб у нас широко используется метод
общих и частных весовых индексов (Зенкевич, 1931, Б о г о р о в, 1934).
Рис. 147. Количество «первоклассной» пищи в г/м2 сырого веса в части Лимфиорда
в сопоставлении с уловами трески в тоннах и количеством бельдюги (за 10 м траления
(по Блеваду).
Подобного типа исследования должны быть положены в основу наибо-
лее рационального использования сырьевых ресурсов, в частности на
них же базируется и предпринимается в плановом порядке реконструк-
ция фауны Каспийского и Аральского морей, к которой мы подошли
сейчас вплотную (Зенкевич, Бирштейн, 1934 и след.). Датские
биологи перенесли количественные исследования бентоса и во внедат-
ские воды. Спэрк (Sparc к) в 1931 г. опубликовал результаты
количественных исследований у берегов Португалии и в Средиземном
море, произведенных датской экспедицией в 1906—1910 гг., в 1929 г.—
результаты аналогичных работ в водах Исландии и Фарерских о-вов,
а в 1933 и 1934 гг. Спэрк и Торсон (Thorson) — результаты
работ в фиордах Гренландии. Работы шведов Тулина (Thulin,
1922) и Гессле (Hessle, 1924) не внесли ничего нового в тео-
рию вопроса, так же как работы немецкого биолога Гагмейера
(Н a g m е i е г, 1926, 1930) и финляндского Сегерштрэле (Sе ger-
str ale, 1933). Их работы, относящиеся к Балтийскому морю, будут
подробно разобраны в главе, посвященной этому водоему.
Бентос Северного моря и прилежащих вод. Значительно больший
интерес имеют работы двух других шведских ученых — М оландера
(М о 1 а n d е г, 1928) и Гис лена (Gislen, 1930), посвященные
описанию фауны Гюльмарфиорда, расположенного с западной стороны
2?5
южной Швеции и открывающегося в Скагеррак. Первая работа дает опи-
сание фауны мягких грунтов, вторая — биоценозов эпифауны (эпибиозов—
по терминологии автора). Моландер отмечает, что значительное
преобладание в биоценозе по весу каких-нибудь крупных, но встречаю-
щихся в 1—2 экз. на 1 л2 животных не должно побуждать нас исключать
из числа ведущих форм более мелкие организмы, дающие, может быть,
менее значительную общую биомассу, но встречающиеся постоянно и
в большом количестве экземпляров. Впротивоположность данным датских
исследователей, придававшим полихетам малое значение, Моландер
указывает на их массовое развитие в Гюльмарфиорде и выделяет неко-
торые из них в качестве руководящих форм. Он различает 9 следующих
фиордовых биоценозов (в скобках дана средняя биомасса в г/л2):
1. Melinna-tenuis (Melinna cristata, Maldane sarsi, Nucula
tennis, Syndesmya nitida, Axinas flexuosus,Lumbriconereis fragilis, Euchone
papillosa и Brissopsis lyrifera) (48—86).
2. Terebellides-flexuosus (Terebellides stroemi, Axinus
flexuosus, Melinna cristata) (23—31).
3. Eumenia-nucleus (Eumenia crassa, Nucula nucleus,
N. tenuis) (119,7).
4. Bri s s о psi s-c hi a j ei (Brissopsis lyrifera, Amphiura chiajei,
A. filiformis, Maldane sarsi, Turitella communis, Rhodine loveni) (69—
142).
5. Echinocardium-filiformis (Echinocardium cordatum,
Amphiura filiformis, Turitella communis, Pectinaria auricoma) (54—94).
6. Nephthys-nitida (Nephthys incisa, Nucula nitida, Turitella
communis) (34—353).
7. Scoloplos-cordatum (Scoloplos armiger, Echinocardium
cordatum, Pectinaria koreni) (26, 6).
8. А с г о ni d а-p г i s ma ti ca (Acronida branchiata, Syndesmya pris-
matica) (10).
9. Spatangus-montagui (Spatangus purpureus, Astarte mon-
tagui) (52).
Таблица 99
Глубина вл
16 20 30 40 50 60 TO 80 90 100 110 120
S. cor.
N. nit. -|-0
N. nit. -j- T
Sp. Mon.
eTIilVo
E.'ftL + P
A. prls
B. chT+T
E. fil '+ T
B. ch. + T.
B. chT+Mal. '
Eu. nucl.
Ter, flex. -J- L.
Ter, flex. + Mel.
M. ten. + Mai.
M. ten. + B.
276
Как мы видели, биомасса бентоса мягких грунтов в Гюльмарфиорд©
(в среднем 60—80 г/м2) значительно меньше, чем в открытых частях при-
лежащих проливов. Моландер дает схему вертикальной смены
биоценозов (табл. 99).
S. cor. = Scoloplos armiger—Echinocardium cordatum (21, 6)1
N. nit.—f— 0. = Nephthys incisa — Nucula nitida — Ophiura texturata (33,9).
N. nit. + T. = Nephthys incisa — Nucula nitida — Turitella commu-
nis (252, 6).
Sp. mont. = Spatangus purpureus — Astarte montagui (52, 3).
E. fil. 4- O.= Echinocardium cordatum — Amphiura filiformis — Ophiura
texturata (89, 1).
E. fil. 4- P. = Echinocardium cordatum — Amphiura filiformis (94, 1).
A. pris. = Acronida branchiata — Syndesmya prismatica (10, 1).
B. ch. 4- R. = Brissopsis lyrifera —Amphiura chiajei — Rhodine loveni
(69, 6).
E. fil. 4- T. = Echinocardium cordatum — Amphiura filiformis — Turi-
tella communis (54, 8).
B. ch. 4~ T. = Brissopsis lyrifera — Amphiura chiajei — Turitella
communis (142, 3).
B. ch. -J- Mai. = Brissopsis lyrifera —Amphiura chiajei —Maldane sarsi
(128, 6).
Eu. nucl. = Eumenia crassa — Nucula nucleus (119, 7).
Ter. flex. 4- L. =Terebellides stroemi — Axinus flexuosus — Leda per-
nula (30, 9).
M. ten. 4~ Mai. = Melinna cristata — Nucula tenuis — Maldane sarsi
(48, 7).
Ter. flex. 4- Mel. = Terebellides stroemi —Axinus flexuosus —Melinna
cristata (22, 7).
M. ten. 4- B. = Melinna cristata — Nucula tenuis — Brissopsis lyri-
fera (86, 5).
Работы Моландера не захватили глубины меньше 5—10 м.
В другой своей работе Моландер дал интересные результаты работы
с новым дночерпателем, сконструированным О. Петтерссоном
(0. Р е 11 е г s s о п).
Этот дночерпатель дает возможность вырезать монолит грунта и уста-
навливать в нем вертикальное послойное распределение организмов.
Оказалось, что наибольшая часть животных обитает в самом верхнем
слое грунта.
В год окончания работ Моландера в том же Гюльмарфиорде
начал свои чрезвычайно интересные и оригинальные исследования Гис-
лен. Самое замечательное в работах Г и с л е н а то, что большая часть
их проведена в водолазном аппарате самим автором. Кроме того, следует
отметить, что Г и с л е н одинаковое внимание уделил и растениям
и животным. В противоположность Моландеру Гислен подверг
детальному изучению эпибиозы и на твердых грунтах.
Спуск под воду давно привлекал внимание биологов. Еще М и л ь н-
Эдвардс (Miln е-Е d w а г d s) в 1845 г. опускался у берегов Сици-
лии на глубину 25 футов в особом водолазном костюме. В последнее время
американцы уделили особенно много внимания изучению жизни в теплых
морях путем ее непосредственного наблюдения в водолазных приборах
вплоть до ставших широко известными опусканий Биба в батисфере
до глубины свыше 900 м. Весьма интересные наблюдения в водолазном
аппарате над жизнью коралловых рифов были проделаны Кэри (Сагу,
В скобках биомасса в г/м2.
277,
1918) у о. Тортуга (близ о. Гаити), специально по росту и развитию горго-
ниевых кораллов, и Майор (Mayor, 1920, 1924) в заливе Паго-паго
(Марианские о-ва, Полинезия). Эти исследования дали прекрасную кар-
тину качественного и количественного распределения фауны коралловых
рифов до глубины 12 м. Что касается вод умеренной зоны, то можно отметить
только небольшие наблюдения Крампа (К ramp, 1925) у берегов
Дании. Необходимо также упомянуть о непосредственных наблюдениях
подводной жизни в водолазном аппарате молодого талантливого советского
исследователя А. Савилована Черном и Японском морях в 1938
и 1939 гг. Гислену же принадлежит честь выполнения, в основном
при помощи водолазного аппарата, в течение ряда лет очень широко
задуманного и тщательно проведенного изучения фауны и флоры, обра-
стающих жесткие грунты в Гюльмарфиорде.
Прежде всего Г и с л е н делит биоценозы на две группы по характеру
грунта: эпибиозы мягких грунтов и эпибиозы жестких грунтов. Г и с л е н
устанавливает десять, как он их называет, основных growth-forms, типов
формы тела и скелета у морских организмов. Общую форму тела и харак-
тер скелета Г и с л е н считает закономерно обусловленной основным
характером физических факторов среды, в основном—подвижности воды,
характера субстрата и распределения света.
Интересны приводимые Гисленом данные по отношению веса
растительной массы к массе животных на разной глубине и по количеству
видов животных и растений в эпибиозах на разных глубинах во внутренних
частях фиорда (табл. 100).
Таблица 100
Гюльмарфиорд Глубина в м
0,3 0,3—10 10—20 20—50 50-75
Отношение биомассы растений к био- массе животных в % (без Balanus и Mytilus) ................ 97,2 75,3 27,1 0,25 0,0
Количество видов животных и расте- ний (в скобках) в среднем в одном биоценозе 17(6) 57(23) 113 (27) 58 (0,3) 51 (0,0)
Средняя биомасса растений и живот- ных внутренних частей фиорда в г Ди2 10 8С0 5 000 3 015 2 500
То же, для биоценозов внешней части фиорда в г/м2 8 755 4621 13 807 3 080 —
Количество видов с глубиной уменьшается, что может быть иллюстри-
ровано графиком (рис. 148), составленным Соот-Р пеном (Soot-
Rye п) для моллюсков Рамфиорда (близ Тромсе). Наибольшее число
видов приходится на глубину 20 м.
Таблица 101
Биоценозы Наибольшая наблюдавшаяся биомасса на кг/м2 Биоценозы Наибольшая наблюдавшаяся биомасса на кг/м1*
Fucus vcsiculosus 12.9 . Chaetopterus 17,58
Ascophyllum nodosum . . 24,46 Mytilus edulls 49,50
Fucuc serratus 28,61 Halldrys siliquosa . . . 19,71
Chorda filum 10,35 Laminaria dlgitata. . . 11,42
Ascidia ob’dqua 17,58 Balanus balanus .... 28,71
278
Рис. 148. Количество
видов моллюсков на
разных глубинах в
Рамфиорде, у Тромсе
(по Соот-Риену).
1—общее число видов,
2—арктические виды,
3—бореальные виды, 4—
лузитанские виды. Глу-
бина в м.
Некоторые отдельные биоценозы дают особенно высокие максимальные
разовые показатели биомассы (табл. 101).
Норвежские исследователи добавили мало нового к сделанному ранее
по количественному анализу донной фауны. Соот-Риен (1924) обсле-
довал донную фауну Рамфиорда (близ Тромсе), а Б р о х (ВгосЬ)
работал на юге Норвегии, у Линдеснесс.
В английских водах первое исследование по количественному учету
бентоса было проведено Фордом (Е. Ford) в Плимутском районе.
Автор установил наличие пяти петерсеновских группировок бентоса
мягких грунтов, но, к сожалению, приводит данные только по количеству
экземпляров, но не по биомассе.
Больший интерес представляют две работы Ф. Д э в и с a (F. D a v i s,
1923, 1925), хотя и этот автор оперирует только с количеством экзем-
пляров и не приводит результатов взвешивания.
Районом исследований Дэвиса была так назы-
ваемая Доггер-банка — мелководье, расположенное
в центральной части южной половины Северного
моря, один из его наиболее важных промысловых
районов. В течение 1921—1924 гг. Дэвис сде-
лал 20 выездов на Доггерскую банку с общим ко-
личеством станций около 1400. Этот громадный мате-
риал, собранный со сравнительно небольшого района
площадью около 30 000 км2 в разные сезоны года,
дал возможность Дэвису с большой тщательно-
стью изучить распределение бентоса и установить
изменения в количественном составе в течение ряда
последующих лет.
Доггер-банка имеет небольшие глубины (18—
19 лс, более обычно — 25—35 м) и покрыта одно-
образным мелким песком с примесью битой раку-
ши. По краям банки с переходом на большие глуби-
ны песок переходит в песчанистый ил (с южной и во-
сточной стороны) и ил (с западной и северной сторо-
ны). На песчанистых грунтах банки фауна качест-
венно очень бедна. Резкое преобладание над всеми
другими формами имеет двустворчатый моллюск Spi-
sula subtruncata. На втором месте стоит другой пред-
ставитель Lamellibranchiata — Mactra stultorum,
количество которой в 30 раз (по числу экземпляров)
меньше чем S. subtruncata. Затем следует гастропода Natica alderi,
полихета Nephthys соеса и морской ёж Echinocardium. Остальные
организмы встречаются уже в значительно меньшем количестве, хотя
иногда попадаются их скопления. Дэвис отмечает, что одни виды
распределены чрезвычайно равномерно (например, Nephthys соеса),
другие дают пятнистое распределение (например, Spisula subtruncata);
количество их может то подниматься до нескольких тысяч экземпля-
ров на 1 м2, то падать до нескольких штук или даже до нуля. Для учета
этой второй группы Дэвис применил тот метод оконтуривания, которым
пользовались Бьюкенен-Уолластон (Buchana n-W о 11 a fl-
ton, 1923) для учета пелагической икры камбалы в Северном море.
Метод заключается в том, что через обследуемый район проводят кресто-
образно ряд станций, а затем проводят изолинии равных количеств орга-
низмов, пользуясь методом интер- п экстраполяции. Само собой разу-
меется, что этот метод пригоден только в том случае, если «пятна» иссле-
дуемой формы имеют достаточно большое протяжение (десятки, а иногда
279
и сотни километров). Такое распределение животных пятнами наблю-
дается далеко не всегда. Причинами его могут быть прежде всего или
массовое отмирание донной фауны вследствие замора, обычное для некото-
рых частей Азовского моря, или массовое выедание рыбами, что наблю-
дается в районах откорма больших количеств рыбы. Большая смертность
Рис. 149. Распределение пятен Spisula subtruncata на Доггер-банке в ок-
тябре 1922 г. (по Дэвису). Толстая линия — изобата 36,5ле. Незаштрихо-
ванные площади заняты сеголетками, заштрихованные — годовиками,
черные — 2-летками.
донной фауны может иметь место и при сильном охлаждении воды зимой.
Во всех этих случаях площадь дна, до этого обильно населенная, оказы-
вается опустошенной и доступной для нового оседания и развития молоди.
В пользу такого способа образования пятен свидетельствует наблюдаю-
Рис. 150. Контуры тех же «пятен» с изолиниями одинакового числа
экземпляров на м* (по Дэвису).
щееся при этом резкое преобладание индивидуумов одного возраста.
В результате своего исследования Дэвис дал для ряда форм точ-
ные карты распределения «пятен» и возрастного состава в них Spisula
subtruncata (рис. 149 и 150). Подобные же карты даны им для Mactra
280
stultorum и Nalica alderi— злейшего врага наиболее значимых в практи-
ческом отношении двустворчатых. Как указывает Дэвис, в этих пят-
нах имеет место и значительная смертность молодых моллюсков, видимо,
вызываемая перенаселением. Процент мертвых Spisula 0-группы достигал
в отдельные сезоны до 77%, I группы —до 47% и 2 группы до 76%.
Большая смертность наблюдалась в июне и в октябре. Смертность за год
Дэвис определяет в 90%. Spisula становится половозрелой уже в конце
первого года жизни (I группа), количество экземпляров этой группы
на дэвисовском пятне площадью около 600 кв. миль свыше 500 млрд.г
а 0-группа на пятне площадью 700 кв. миль имела свыше 4500 млрд,
штук.
Сухой вес Spisula в пятнах А и Е (I группа) равнялся (без раковин)
17 000 иг, что составляет 5—6% сырого веса.
Повсюду на банке Дэвис находил очень большое количество мертвых
Venus и Donax и мало живых и относит население центральных частей
банки к петерсеновскому комплексу Venus, а более глубокие — к ком-
плексу Echinocardium — filiformis.
Заселение площадей дна, освободившихся от обитавших на них попу-
ляций Spisula, Дэвис объясняет массовым оседанием переносимых
течениями личинок моллюска в период отрождения, если они попадают
на благоприятные для развития грунты (Soil-current Hypothesis).
Количественное изучение бентоса в шотландских водах Северного
моря было проведено Стефеном (Stephen, 1923), а в Карди-
ганском заливе — Лаури (Lawrie) и Уэткином (W a t k i п,
1922).
Три датских исследователя — Р. Спэрк (Sparc к, 1929, 1933,
1937), Г. Торсон (Thorson, 1933, 1934) и Г. М а д с е н (Mad-
sen, 1936) — провели ряд исследований по количественному учету
прибрежных вод Фарерских о-вов, Исландии и Гренландии. У Фарерских
о-вов фауна сохраняет еще бореальный характер. Спэрк различает
здесь три основные биоценоза: Ma со ma calcarea, Axinus
flexosus и Mactra elliptic а. Дальше на север, в исландских
водах, фауна приобретает переходный характер от бореальной к аркти-
ческой.
Бентос побережий Исландии. Р. Спэрк описал в мелководной
зоне (до 150 л«) у берегов Исландии 6 группировок инфауны и 2 эпифауны:
эти последние — уже известные нам комплексы Mytilus edulis.
(1—10 j£)nModiola modiolus (23—72 .u). Однако здесь комплекс
М о d i о 1 а дает биомассу, едва достигающую 2 кг!м2. Биоценозы мягких
грунтов следующие.
1. Macoma calcarea (2—50 л«) с наиболее массовыми фор-
мами: Nephthys ciliata, N. longisetosa, Lumbriconereis fragilis, Pectinaria
koreni, Yoldia hyperborea, Clymene gracilis, Thyasira sarsi, Prionospio malm-
greni, Euchone rubrocincta.
2. Pontoporeia femorata (1—4 м) в солоноватых зонах,
с массовыми формами: Gammarus locusta, Nephthys ciliata, Nereis di versi co-
lor, Halicryptus spinulosus.
3. Yoldia hyperborea (45—162 м) с массовыми формами:
Ctenodiscus crispatus, Cardium ciliatum, Nephthys longisetosa, Myriochele
heeri, Clymene gracilis, Prionospio malmgreni.
4. Mactra elliptica (31—103 м) с массовой формой Psammo-
bia tellinella.
5. Amphiura filiformis (90—120 ж) с массовыми формами:
Echinocardium flavescens, Mya truncata,Astarte sulcata, Aphrodite aculeata,
Melinna cristata. Последние группировки могут счиуаться крайним
281
северным форпостом бореальных петерсеновских биоценозов Venus
gallina и Echinocardium filiformis.
6. Maldane sarsi (93—222 м) с массовыми формами Ctenodiscus
crispatus, Prionospio malmgreni, Clymene gracilis.
Для южной и западной сторон острова, омываемых теплыми водами,
более характерны группировки Amphiura, Mactra, Modiola,
для холодной северной — Maldane, представляющая собою как бы
южную границу арктической группировки Astarte crenata.
Остальные группировки встречаются и у южных и у северных берегов.
Показатели биомассы даны в табл. 102.
Таблица 102
Группировка Средняя био- масса в г/м3 Наибольшая биомасса в г/м3
Мacornа 454,9 1177,0
Pontoporela 212,4 275,0
Yoldia 180,0 230,0
Mactra 420,0
Amphiura 43,2 79,5
Maldane 94,1 195,5
Mytilus 3858,8 5125,0
Modiola 181,6 1931,5
Состав этих биоценозов по группам дан на рис. 151.
Рис. 151. Состав донных комплексов исландских вод (по Спэрку). Площадь круга
соответствует средней биомассе комплекса, показанной также числом.
а — Pontoporcia femorata, b, с, d, е—Macoma calcarea из разных мест, /, д—Splsula elliptica из раз-
ных мест, h, i — J old! a hyperborea из разных мест, k — Maldane sarsi, I — Amphiura filiformis.
Замечательно, что нигде у исландских побережий не обнаружен типич-
ный для литорали атлантических побережий Европы биоценоз М а с о ш а
baltica — Cardium edule — Муа arenaria — Litto-
rina littorea, видимо, уничтоженный здесь ледниковой эпохой и
не проникший $юда после нее.
282
Бентос восточно-гренландского побережья. По восточно-гренланд-
скому, омываемому холодными полярными водами, побережью фауна
приобретает уже вполне арктический характер, во многом чрезвычайно
сходный с фауной Варенцова моря. По существу, это одна и та же фауна.
Тщательное количественно-биоценотическое ее описание дают G п э р к,
Торсон и Мадсен.
Как показали исследования Мадсена, литоральная фауна здесь
крайне скудна. Фукусы или совсем отсутствуют, или выражены очень
слабо. Нет здесь литорин, мидии, баланусов. На литорали обитают оли-
гохеты, гидракарины, турбеллярии, амфиподы, гарпактициды.
По западной стороне Гренландии литоральная фауна значительно
богаче. В сублиторали побережий Гренландии Торсон различает
следующие основные подразделения: комплексы и более дробные
подразделения — «зоны». Комплексы и зоны Торсона показаны в
табл. 103.
Таблица 103
№ по пор. Комплексы Зоны Глубина оби- тания в м
I Macoma calcarea 1. Halcompoides 2. Astarte borealis 4» Pectinaria granulata 3. Ophiocten sericeum + Pecti- naria granulata 4. Ophiocten sericeum 0—50
II Venus fluctuosa 7—30
III Portlandia arctica 10—60
IV V Area glacialls + Astarte crenata . Foramlnifera 50—550 50—550
Наиболее обычные формы, кроме перечисленных: из моллюсков— Astarte
borealis, A. elliptica, A. banksi, Муа truncata, Nucula tenuis, Axinus flexuo-
sus, Cardium ciliatum, Leda pernula, Saxicava arctica, Portlandia lenticula,
P. frigida, P. intermedia, Pecten groenlandicus, Lepeta coeca; из полихет —
Pectinaria granulata, Nereis zonata, Harmothoe imbricata, Nephthys ciUata'
из иглокожих— Ophiocten sericeum, Ophiopleura borealis, Ophiocantha biden-
4ata, Strongylocentrotas droebachiensis*, из гефирей—Priapulus caudatus.
На рис. 152 дан состав некоторых гренландских комплексов по груп-
пам, а на рис. 153 — их зональное распределение у побережий Грен-
ландии.
Таблица 104
Комплексы Максималь- ная биомас- са в г/м2 Средняя биомасса в г/м2 Примечание
Arca-Astarte Macoma calcarea + Halcampoides . Astarte + Pectinaria Ophiocten -f- Pectinaria Ophiocten Venus fluctuosa Portlandia arctica Foraminlfera 308,1 559 977,7 1172,5 121,5 744 109 138,1 117,5 368 154,6 236 280 372,6 33,8 54,0 Большая биомасса комп- лекса Macoma в значи- тельной степени прихо- дится на долю Saxlcava
283
Торсон дает также краткое описание гренландских комплексов
эпифауны. Общее их распределение можно видеть на рис. 154. Среди
зарослей Desmarestia множество Margarita groenlandica, М. helicina, Modi-
olaria laevigata и молоди Saxicava arctica, Mya truncata, а также немертин.
Фукусы дают приют, кроме перечисленных выше форм, также игло-
кожим: Stephanasterias albula,
Рис. 152. Состав некоторых грен-
ландских донных комплексов (по
Торсону).
1 — пластинчатожаберные, 2 — поли-
хеты, 3 — иглокожие, 4 — ракообраз-
ные , 5 — брюхоногие.
Strongylocentrotus droebachiensis и Ophiura
robusta. В зарослях красных водорослей
обитают наиболее глубоководные из пере-
численных и, кроме того, добавляется
много губок, мшанок, из иглокожих —
Solaster papposus. Antedon; из моллю-
сков— Buccinum hydrophanum, В. groen-
landicum, Pecten islandicus. В зарослях
ламинарий своя специфическая фауна не
обособляется. Saxicaoa arctica образует
громадные гроздья, подобно мидии и мо-
диоле в бореальных водах, викариатом
которых она как бы является в этих арк-
тических водах.
Показатели средней и максимальной
биомассы для восточно-гренландских дон-
ных комплексов приведены в табл. 104.
Бентос Средиземного моря. Исключи-
тельный интерес приобретают данные по
количественному учету бентоса Средизем-
ного моря при сравнении их с данными
для Атлантического океана и Черного
моря. В 1931 г. Спэрк опубликовал
соответственные материалы, полученные
Датской океанографической экспедицией
1908—1910 гг; позднее появился ряд
работ по количественному учету бентоса
Средиземного моря, принадлежащих Ва-
това (А. V a t о v а, 1931—1936).
Еще Фогт (С. V о g t) указывал на
большую количественную бедность сре-
диземноморской фауны по сравнению с
северо-западным побережьем Европы и
объяснял это неблагоприятными свойства-
ми грунтов. Только в небольших районах
побережий, главным образом вблизи устий
рек (устье Нила, Венецианская лагуна и
т. п.), биомасса достигает сотен граммов
на 1 м2. Совсем другое во всем остальном
море.
В своей работе К. Спэрк (1931) приводит данные по соседним
Салернскому и Неаполитанскому заливам, по Алжирскому заливу и для
сравнения по побережьям Португалии (у Faro) и у реки Тахо (рис. 155).
В очень редких случаях биомасса бентоса выражается десятками граммов
(10—50 а/лс2) на глубине менее 100 м. Обычно биомасса составляет не-
сколько граммов (1—10 г/м2) и часто меньше 1 г (0—1 а/л<2), преимуще-
ственно на глубине свыше 100 м. Средняя из 30 проб в Салернском и Неапо-
литанском заливах равняется 6 г/м2.
На станциях с глубиной свыше 100 м средняя биомасса (из 13 проб)
составляет менее 2 а/лс2, а в нескольких пробах, полученных с глубин
284
свыше 500 м, бентос в пробах дночерпателя или отсутствовал или выра-
жался несколькими десятками миллиграммов.
В западной части моря, в Алжирской бухте, биомасса бентоса оказа-
лась несколько выше: 7 проб на глубине от 13 до 111 м дали в среднем
15,5 з/.u2 (от 1,5 до 33,5 г/м2).
Рис. 153. Распределение бентосных комплексов у мыса Гедлунд,
Восточная Гренландия (по Торсону).
1 — Astarte borealis—Pectinaria; 2 — Ophiocten—Pectinaria; з— Глубинный
Ophlocten; 4—Area—Astarte crenata; 5 — Fucus—Desmarestia; 6 — Ascidia; 7 — Скалы и камни.
Эта количественная бедность бентоса Средиземного моря выступает
особенно ярко, если сопоставить приведенные выше числа с данными,
полученными той же экспедицией у атлантического побережья Португа-
лии. У южных ее побережий средняя величина бентоса из четырех проб
(глубина 35—92 м) оказалась 274 г/м2. Три пробы, полученные против
реки Тахо (глубина 50—100 лс), дали в среднем 70 г/м2, в эстуариях же
этой реки биомасса поднялась до 2500 г/м2 за счет моллюска Smittina
'foliacea. Вполне согласны с этими данными по бентосу также данные по
планктону и по рыболовству. Спэрк указывает также на очень слабое
285
развитие в Средиземном море берегового пояса растительности и малое
количество донных рыб. Очень скудное химическое питание фитопланк-
тона и донной растительности ограничивают развитие бентоса и рыб.
Интересно и то, что в Средиземном море только у устий рек и в заливах
преобладание в бентосе получают моллюски и аннелиды, т. е. грунтоеды
и детритоеды; в остальном море резко преобладают подвижные живот-
ные, активно разыскивающие пищу, особенно ракообразные и иглокожие
(рис. 156). Наиболее обычные формы бентоса, по С п э р к у, Amphiura
chiajei, A. jiliformis, Echinocardium cordatum, Astropecten jonstoni — из игло-
кожих, Venus gallina, Cultellus pellucidus, Syndesmya (Abra) nitida, Cor-
bula gibba, Pteroides griseus, Turitella terebra — из моллюсков.
Рис. 154. Схематическое изображение распределения макрофитов
у восточных берегов Гренландии (по Торсону).
Батова (1934—1936) путем детального изучения некоторых от-
дельных районов Средиземного моря (Адриатическое море и особенно Вене-
цианская лагуна и окрестности Александрии) добавил много ценного
к предварительным данным С п э р к а. Наибольшая биомасса бентоса
констатирована им у Александрии, против устья Нила. Здесь выносимый
рекой детрит дает возможность пышному развитию моллюсков, состав-
ляющих 99,6% всей биомассы. В основном это Tapes auratus (93%) и
Murex trunculus (5,8%). Другим районом с повышенной биомассой яв-
ляется Венецианская лагуна, где также обильный детрит благоприят-
ствует развитию двустворчатых моллюсков. Доминирующее значение
и здесь имеет Tapes auratus, а из других моллюсков — Nassa marnmil*
lata, Cyclonassa neritaea, Chlamis sulcata, Nucula nucleus, Modiola barbata,
286
Рис. 155. Средняя биомасса бентоса в ?/м2 в разных частях Средиземного моря
и прилежащих водах (по Спэрку и Батова).
Столбик внизу соответствует биомассе бентоса, белая часть столбика — эпифауна. Жирные числа:
соответствуют количеству макропланктона в см3 за час лова двухметровой сетью (по данным Джеспер-
сена) в 1908—1909 гг., тонкие — 1910 г.
Рис. 156. Состав донных комплексов Средиземного моря.
А — в заливах п у устий рек; Б — в открытых частях моря (по Ватова).
1 — двустворчатые, 2 — брюхоногие, з — полихеты, 4 — ракообразные, 5 — иглокожие.
287
Ensis siliqua и др.; из полихет—Sternaspis scutata и Branchiommd vesiculosumy
из ракообразных Carcinus maenas, Upogebia litoralis и Paguridae и иногда
в очень больших количествах асцидия Microcosmus. Кроме этого комплекса
Tapes auratus-j-MicroGOsmiis, Батова1 выделяет еще
комплексы Tapes с Nucula nucleus, Cyclonassa neritaea и несколько
других.
Изучение северной части Адриатического моря позволило В а т ов а1
дать карты донных комплексов и изобенты (рис. 157). Он различает шесть
основных комплексов: Schizaster canaliferus — Amphi-
ura chiajei (Sch. ch.), Turitella communis (T.), T e 1-
lina distorta (TL), Lima inflata — Ophiothrix f г a-
g i 1 i s (L. fr. i.), Ghione gallina (Ch. g.), Lima hians-
Ophiothrix fragilis (L. fr. h.), Amphioxus (Amph.)
и несколько других, имеющих менее широкое распространение. Кроме
перечисленных форм, здесь наибольшее значение имеют из моллюсков—
Syndesmya fragilis, S. alba, Nucula nucleus, Corbula gibba, Ensis ensis,
Tellina distorta, Cultellus adriaticus, Cyclonassa neritaea, Nassa mammil-
lata; из полихет — Hyalinaecia fauveli, Maldane globifex. Owenia fusiformis,
Notomastus latericeus, Marphysa bellii, Nephthys hombergi, Sternapsis scu-
tata; из иглокожих— Ophipsila aranea, Cucumaria elongata, Ophiura albida,
Echinocardium mediterraneum.
Более детальное описание тех же комплексов В а т о в а дает для
района Ровиньо, где биомасса бентоса опять повышается, поднимаясь
для комплекса Chione verrucosa до 505,5 г/м2, для большинства колеблясь
между 117—227 г/лг2, а для некоторых — между 9—60 г/м2.
Таким образом, Средиземное море в целом представляет собою мало-
продуктивный водоем с очень низкими показателями биомассы, резко
падающей от района Гибралтара, все время понижающейся с запада на
восток и повышающейся только в Эгейском море, у входа в Мраморное,
под влиянием черноморских вод. В Черном море биомасса опять сильно
поднимается. Особенно показательны в этом отношении изменения в био-
массе планктона (см. т. II). Замечательна еще одна особенность фауны
Средиземного моря. Многие авторы отмечали измельчание размеров тела,
особенно заметное также при продвижении с запада на восток, как для
беспозвоночных, так и для рыб. Это явление только подтверждает, что
основная причина всех этих явлений — недостаточность питания. Именно
бедность пищевых ресурсов обусловила развитие массовой миграции рыб
восточной части Средиземного моря на откорм в Черное море, куда массы
хамсы, скумбрии, ставриды, луфаря, пеламиды, тунца, шпрота, сардины
и некоторых других устремляются весной через Босфор.
Бентос некоторых районов Тихого океана. Нам остается еще
коснуться работ по количественному учету донной фауны в Тихом океане.
И. Закс (1929) приводит некоторые количественные данные для лито-
рали Шантарскпх островов. Различные группировки литоральной фауны
губы Якшиной на о. Б. Шантар дают биомассу от 126 до 2798 г/м2 в алко-
гольном весе, в большинстве свыше 300 г/м2, и пять группировок из две-
надцати имеют биомассу свыше 1000 г)м2. Наибольшую биомассу (1000—
2798 г/м2), дают группировки онфауны с Parasabella maculata и Venus
fluctuosa, наименьшую (126—450 г/м2) — группировки Chiridota dis-
х А. Ватова — первый после советских исследователей стал пользоваться изобентами. Гра-
фическим изображением состава комплексов, заимствованным также у советских исследовате-
лей, пользовались, кроме В а т о в ы, также Спэрк иТорсон. Итон другое введено у
нас Л. Зенкевичем. Кроме того, Ватова вводит термин изобии—линии, разграничиваю-
щие отдельные комплексы (биоценозы).
288
19 Зенкевич,
Рис. 157. Северная часть Адриатического моря с границами донных комплексов (А) и изобентами (Б) (по Батова).
А. 1 — комплекс Schlzaster — chiajei, 2 — Turitella. 3 — Telllna distort», 4 — Chione gallina, 4 — Lima tragllls, в — Amphloxus. Б, 1 — 5—30, 2 — 30—100, 3 — 100—
color. И. 3 а к с считает, что средняя биомасса литорали Шантар не может
быть менее 1000 г/л<2.
Обширные количественно-биоценотические исследования бентоса за-
лива Петра Великого (Японское море) проведены К. Дерюгиным и его
учениками в 1925—1933 гг. (Дерюгин 1939, Дерюгин и Сомова 1941).
Залив Петра Великого занимает площадь примерно в 10 000 км 2 и в своей
внешней части выходит за пределы континентального плато, охватывая
не только батиаль, но и абиссаль до глубины в 3 500 м (рис. 158). В своем
биоценотическом описании К. Дерюгин захватывает все зоны от супра-
литорали до абиссали.
В то время как литоральная жизнь в этом районе бедна, вследствие
действия льдов, сублиторальное население очень разнообразно и богато.
В Амурском и Уссурийском заливах, на глубинах 10—40 лг, на пес-
чанисто-илистом грунте громадное преобладание дают полихеты (биоценоз
Maldane sarsi), составляющие свыше 60, иногда 80% биомассы.
Среди них Maldane sarsi, Polydora соеса, Sosane gracilis, Ancistrosyllis
robusta, Asabellides sibirica и др. Из иглокожих особенно обычна Ophiura
sarsi vadicola из моллюсков Nucula, tenuis, Axinopsis orbiculata. Биомасса
этого биоценоза составляет 150—200 г]м2 и иногда за счет крупных мол-
люсков до 600 г/л£2.
В Уссурийском заливе на тех же глубинах на илистопесчаном грунте
преобладание получают моллюски (биоценоз Turitella), из которых
особенно характерны Turitella jortilirata, Joldia johanni, Axinopsis orbi-
culata и Масотпа calcarea, из полихет те же Maldane sarsi, Scoloplos armi-
ger, Magelone longicornis, Chone cincta, из иглокожих Ophiura sarsi vadi-
cola и Amphiodia craterodmeta. Биомасса данного комплекса около
100 г/ж2. Иногда резкое преобладание среди моллюсков получают Nucula
tenuis, Joldia johanni и Axinopsis orbiculata и их биомасса достигает
650 г]м2, причем на долю двустворчатых приходится более 90%.
При выходе из бухт и глубоких частей заливов с их биоценотической
пестротой и мозаичностью мы попадаем в район более спокойного зональ-
ного распределения комплексов.
Ближе всего к берегу широкой каймой тянутся заросли ламинарий
на глубинах 12—50 м (L. saccharina, L. bullata, L. japonica), с их очень
пестрой и качественно богатой фауной.
За пределами зоны ламинарий на глубине 50—80 м тянется сплошная
полоса очень своеобразного комплекса песчанистого грунта с амфипо-
дами Ampelisca macrocephala, Byblis gaimardi, Lembos arcticus, Haploops
tubicola, и большим числом кумацей и капреллид, асцидиями Pelonaja
corrugata и Pareugyrioides japonica и моллюсками Venus fluctuosa, Joldi-
ella derjugini, Macoma. calcarea и др.
При общей биомассе 130—200 г)м2 и в отдельных случаях до 700,
на долю ракообразных иногда приходится 70—95%.
Иглокожие в этом комплексе выражены очень слабо. Иногда в этом
комплексе резкое преобладание получает асцидия Pareugyrioides japo-
nica, дающая до 80% биомассы, достигающей в таких случаях 500 г/м-.
Описанный комплекс опоясывается комплексом Solariella — Eu-
pyrgus— Stegophiura, простирающимся на глубины 80—200 м и
занимающим плотные пески. По биомассе (в среднем 80—90 г/м2) черви,
иглокожие и моллюски представлены примерно поровну, а ракообраз-
ные выражены слабо.
Из моллюсков здесь следует указать Solariella oaricosa, Joldiella derju-
gini, Venus jluctuosa, из иглокожих Stegophiura brachiactis, St. nodosa,
Amphiodia craterodmeta, Eupyrgus pacijicus, Myriotrochus mitsukui,
а из полихет — Scoloplos armiger, Travisia jorbesi, Asychis punctata и др.
290
Глубже, уже за пределами континентального плато, на верхнем склоне
батиали, на глубинах 200—600 м и плотных песках, распространен до-
вольно узкой каймой комплекс иглокожих Heliometra glacialis и
Н. maxima и Ophiura s а г s i и двустворчатого моллюска Verticar-
dia nadina.
Средняя биомасса — 87 г/ж2, но иногда подымается до 180. Здесь
уже преобладание получают иглокожие и полихеты, из которых можно
указать Travisia farbesi, Amage anops, Polycirrus eous, Lumbriconereis
jragilis и др.
У нижнего края данного горизонта (на глубине 500—600 и до 1300 м)
грунты становятся илистыми и значительно изменяется фауна. Из игло-
кожих здесь обитают кроме уже упомянутых Т haumatometra tenuis и
Ctenodiscus crispatus, Luidiaster tuberculatus появляются горгониевые
кораллы Primnoa resedaeformis pacifica, одиночные мадрепоровые кораллы
Caryophyllin claous, плеченогое Terebratula coreanica, гидроид Lafoeina
maxima, полихеты Nephthys longisetosa, Harmothoe impar, Jasmineira
pacijica и др., несколько декапод и в том числе Nectocrangon dentata,
Spirontocaris linguis, Chionocoetes elongatus bathyalis и масса гефирей.
Моллюсков здесь уже ничтожное количество.
Еще глубже, уже в абиссали (1300—3500 м), на коричневых илах
с железпсто-марганцевыми конкрециями, жизнь бедна и качественно и
количественно.
Здесь прежде всего наблюдается чрезвычайное обилие корненожек
Hyperammina fragilis, Haplophragmoides canarieizsis, Stevelina, Rhab-
dammina и др., в чем большое сходство с населением глубинной зоны
Карского моря и северной части Варенцова. Из моллюсков следует ука-
зать Pecten randolfi и Dentalium, из иглокожих Ophiura leptoctenia и
Myriotrochus, из амфипод Tmetonyx cicada, из изопод Eurycope spinifrons
п Typhlotanais sp., из полихет — Harmothoe derjugini, Н. impar, Sca-
libregma inf latum, Jasmineira sp. и др.
К. Дерюгин отмечает, что фауна больших глубин Японского моря
«не абиссального происхождения, а батиального или частично даже
сублиторального. Все формы по существу свойственны более высоким гори-
зонтам и спустились в абиссаль вторично, приспособившись к жизни
на больших глубинах». Это объясняется, как мы указываем ниже, истори-
ческими причинами и мелкостью проливов, соединяющих Японское море
с океаном.
Дерюгин принимает среднюю биомассу бентоса континентального
плато Японского моря в 175 г/м2, т. е. величину, близкую к величине
биомассы южной половины Варенцова моря.
Резкий подъем биомассы бентоса по азиатскому побережью Тихого
океана наблюдается в самой северной его части — в прилежащей к Берин-
гову проливу части Берингова моря, где по данным В.Макарова (1937)
наблюдается биомасса до 890 г/м2, а в южной части Чукотского моря
даже свыше 1000 г/м2.
В северной же части Японского моря, как указывает К. Дерюгин,
по данным 3. Кобяковой биомасса бентоса составляет в среднем 100—
125 г/м2.
Количественно-биоценотические исследования на другой стороне Ти-
хого океана проведены в проливе Сан-Хуан (район биологической стан-
ции в Puget-Sound) под руководством В. Шел форда (Shel-
ford, 1935), и нужно только пожалеть, что авторы ведут учет коли-
чества экземпляров и не приводят никаких данных по биомассе. Это лишает
возможности проводить сравнение с другими частями мирового океана.
Несколько видов ракообразных и моллюсков (Balanus cariosus, В. glan-
291
dula, Chthamalus dalli, Mytilus edulis, M. californianus, Mitella polymerus,
Littorina sitchana, L. scutulata, Acmaea digitata, A. cassis), составляющих
эпйбиотический комплекс Balanus — Littorina (BL.), распре-
деляются на скалах и камнях между линиями среднего малого (Д) и сред-
него большого (А) приливов (рис. 159). Эндобиотический комплекс мяг-
ких грунтов Macoma-Paphia, составляющийся в основном из
двустворчатых моллюсков и червей (Масота nasuta, М. secta, М. in-
quinata, Paphia staminea, Cardium corbis, Nereis virens, Arenicola cla-
par edit, Leptosynapta inhaerens), идет вверх по осушной зоне до середины
между средним большим и средним малым приливами (С).
Рис. 158. Биоценозы залива Петра Великого (Японское море) (по Дерюгину и Сомовой).
1 — Harmothoe derjugini 4- Pecten randolphi, 2 — Primnoa + Luidiaster 4- Thaumatometra, 3— He-
liometra + Ophiura sarsi, 4 — Solariella 4- Eupyrgus 4-Stegophiura, 5 — Venus 4- Joldiella + Ampe-
lisca, 6 — Ламинарии, 7 — Maldane 4- Scoloplos 4- Baeta 4- Theora, 8 — Obelia 4- Ophiura 4- Philine,
9 — Echinarachnius parma, 10 — Turitella 4- Bela rosea, 11 — Solen 4- Pelonaja 4- Pareugyrioides,
12 — Echinocardium, 13 — Zostera.
Снизу к этому комплексу примыкает глубжележащий сублиторальный
комплекс Strongylocentrotus — Argobuc cinum, ком-
плекс иглокожих и брюхоногих моллюсков (Strongylocentrotus droebachien-
sis, S. franciscanus, Psolus chitinoides, Stichopus californicus, Argobuccinum
oregonensis, Trichotropis cancellata, Calliossoma costatum, Pododesmus
macroschisma) и многочисленные другие звезды, крабы, креветки. Между
этими двумя комплексами, в результате их смешения, развиваются груп-
пировки переходного типа.
Еще ранее работ Шелфорда появилось исследование Ф. К и р-
с о п a (F. К i г s о р), посвященное тому же району.
Максимальная биомасса разных форм. Полученные до настоящего
времени результаты количественного учета донной фауны моря дают
возможность сделать некоторые обобщения.
292
Наибольшая биомасса, констатированная до настоящего времени,
относится к щеткам мидии в Гюльмарфиорде и выражается в 49,5 кг/м\
Впрочем, В. Никитин (1947) указывает для обрастаний в Черном
море биомассу в 94,5 кг/м*. В других местах (датские воды, Мурманское
побережье) биомасса мидии достигает только 10—22 кг/м\ Биомасса
поселений Balanus balanoides достигает 28,7 кг/м\ Примерно этими же
величинами выражается предельная биомасса и некоторых зарослей
макрофитов (Ascophyllum, Fucus, Laminaria). Таким образом, предельную
биомассу животных и растений на литорали и в самом верхнем горизонте
Комплене Эктон Комплекс
Strongyloceritrotus - А рдоЪисйпит Macoma-Paphia
Рис. 159. Донные комплексы архипелага Сан-Хуан, близ биологической станции
в Пюджет-Саунд (по Шелфорду).
BL — комплекс Balanus—Littorina.
сублиторали можно определить в 20—30 кг/м2. Как правило, эта громад-
ная биомасса на 90% приходится на какого-нибудь одного представителя
эпифауны (Mytilus, Balanus, Chaetopterus, Ascidia) или на растительный
организм. Организмы инфауны никогда не дают такой высокой биомассы
даже на литорали. Чрезвычайно высокие показатели биомассы приводит
И. 3 а к с как для всей инфауны на литорали Охотского моря (до 2800 г/м2),
так и для отдельных форм (Venus fluctuosa до 1744 г/м2, Parasabella таси-
lata до 2150 г/м2). Обычно эти величины бывают меньше. Так, Масота
baltica на Мурманской литорали дает биомассу до 554 г/м2, в Охотском
море 275 г/м2, у берегов Исландии в среднем 455 г/м2, а в отдельных слу-
чаях— до 1177 г/м2. В сублиторали биомасса бентоса резко падает, и такие
случаи, как нахождение в губе Машигиной на Новой Земле биомассы
бентоса в 3600 г/м2 (Saxicava arctica, Муа truncata, С ar dium cilia turn),—
очень редкое явление. Обычно на глубинах до 200 м преобладает биомасса
в 100—200—300 г/м2 в среднем и редко больше. Отдельные пробы могут
давать и значительно больше за счет представителей эпифауны (Myti-
las, Modiola, Cucumaria, Saxicava и т. п.). Предельные величины биомассы
инфауны сублиторали, обнаруженные в открытых частях Варенцова
моря, не превышают 700 г/м2. В заливах, во внутренних морях с особо
благоприятными условиями питания разовая биомасса может значи-
293
тельно возрастать. Мы привели уже пример губы Машигиной; можно
указать также на Азовское море, где Cardium edule и Syndesmya ovata
дают биомассу свыше 1700 г/м2.
Наряду с этим отдельным видом в различных местах свойственна
примерно одинаковая биомасса и одинаковое количественное соотношение
с другими компонентами комплекса. Для примера сравним наибольшую
биомассу для ряда форм из Варенцова моря и восточных фиордов Грен-
ландии (табл. 105).
Таблица 105
Отдельные формы Наибольшая разовая биомасса в г/м2
Баренцово море Гренландия
Astarte borealis 457,0 540,0
Astarte montagui 80,0 75,0
Astarte elliptlca 174,0 307,0
Macoma calcarea 243,0 225,4
Axinus flexuosus 4,4 2,0
Ophiocten sericeum 17,0 9,7
Ophiopleura borealis 36,0 57,0
Мы приводили уже примеры изменения биомассы бентоса с глубиной.
Наибольшая биомасса приходится или на литораль или на верхний гори-
зонт сублиторали. Количественные данные с глубин свыше 1000 м от-
сутствуют, но можно думать, что биомасса бентоса в батиали и особенно
абиссали не превышает в среднем долей грамма на 1 м2.
5. ФАУНА ПСЕВДОАБИССАЛИ
Очень удачное понятие псевдоабиссали, введенное К. Дерюги-
н ы м (1916), охватывает нижние горизонты эпиконтинентальных водо-
емов, если край континентального склона опускается ниже 200 м, как,
например, в Баренцовом море, где он находится на глубине около 500 м.
В таком случае нижние горизонты континентальной ступени лежат далеко
за пределами зоны водорослей, т. е. сублиторали. Фауна приобретает
некоторые характерные признаки глубоководной. Более детально псевдо-
абиссальная фауна будет рассмотрена на примере Варенцова моря.
6. ФАУНА БАТИАЛИ
Батиальная зона. Область континентального склона, располагаю-
щаяся ниже края континентальной ступени, характеризуется прежде
всего резким падением глубин. Угол наклона дна здесь обычно очень
велик. Так, в Средиземном море он достигает 43°, у о. Кубы 35,5°, у за-
лива Петра Великого в Японском море 24°. Но это только крайние
примеры. Обычно угол падения меньше (4—8—12°). Средний же угол
падения для всего склона мирового океана между изобатами в 200
и 5000 м определяется в 2,3°. Батиали свойственна очень слабая седимен-
тация, это — нижняя граница терригеновых отложений. Поступление
растительного детрита с побережий очень мало и ограничивает развитие
фауны. Еле уловимы здесь колебания температуры и солености. Также
ничтожно малы становятся с нарастанием глубин и движения водных
294
масс. Волновые колебания сюда уже не достигают, а горизонтальные пере-
мещения воды редко превышают доли сантиметра в секунду.
Как указывает Аппеллеф (Appelloff), тралирование на боль-
ших глубинах по обеим сторонам гребня Уайвиль-Томсона в пределах
батиали дает весьма различные результаты. Если по северную сторону
гребня трал приносил более или менее обычные арктические формы, то
по южную сторону от него, на расстоянии 5 часов хода судна, уловы давали
обильную фауну глубоководных беспозвоночных и рыб, свойственных
и более южным широтам. Из 383 видов донных животных, собранных
рядом экспедиций по обе стороны гребня, только 48 видов (12,5%) оказа-
лись общими.
Состав фауны батиали. Для батиали северной Атлантики очень харак-
терны в первую очередь стеклянная губка Pheronema carpenteri, восьми-
лучевые кораллы (Anthoptillum, Umbellula, Acanthogorgia, Strophogorgia),
настоящие кораллы (Stephanotrochus, Amphiphelia, Flabellum, Oculina),
иглокожие — много видов ежей и в том числе замечательные мягкие
ежи семейства Echinothuriidae, звезды (например, Zoroaster fulgens). Много
различных десятиногих раков. Из рыб следует назвать придонных Масги-
ridae и очень большое число пелагических, часто светящихся, рыб. Что
касается фауны того же горизонта батиали к северу от Шетландско-
Исландского гребня, то наибольшее значение имеет из восьмилучевых
кораллов (Pennatulacea)—Umbellula encrinus; из звезд — Tylaster wil-
led Psilaster andromeda, Astrogonium pareli, Pteraster multipes; из ежей —
Pourtalesia jeffreysi, Spatangus raschi, Brizaster fragilis; из голотурий —
Elpidia glacialis, из офиур — Ophiocantha abyssicola, Ophiactis abyssicola,
Ophiocten sericeum, Gorgonocephalus lincki\wa лилий—Ilyocrinus carpenteri,
Rhizocrinus lofotensis, из изопод — Mesidothea megalura, Gnathia stygia\
из мизид — Boreomysis scyphops и В. nobilis9, из десятиногих раков —
Bythocaris leucopis и В. payeri, Pontophilus norvegicus, Pagurus pubescens,
Munida tenuimana* из моллюсков — Pecten frigidus*, из полихет — Aphro-
dite aculeata, Laetmonice filicornis, Lumbriconereis fragilis; из плеченогих —
Waldheimia septata* из рыб—Lycodes frigidus, Paraliparis bathybii и
Rhodichthys regina. Для глубоководной зоны Норвежского и Гренланд-
ского морей характерно отсутствие большинства наиболее типичных
элементов настоящей самобытной глубоководной фауны мирового океана
п очень большого числа форм, обитающих и в сублиторали и только
опускающихся на большие глубины. Как показала Е. Гурьянова
(1939) по материалам советских экспедиций, глубоководная фауна Аркти-
ческого бассейна представляет собой самостоятельно выработавшуюся
и сравнительно недавно заселившую глубины сублиторальную фауну,
имеющую мало общего с чрезвычайно древней единообразной и глубоко-
водной фауной мирового океана. К тому же выводу в отношении рыб
арктической абиссали ранее пришел Йенсен (Jensen, 1935). Шет-
ландско-Гренландский и Шпицбергенско-Гренландский гребни препят-
ствуют проникновению глубоководной фауны из Атлантического океана
в Полярный бассейн. Только немногие формы, достигающие горизонта
в 500—600 л«, получают возможность доступа через мелководные гребни
на север.
Многие формы из таких, которые могут подниматься и в более высокие
горизонты, представлены в «холодной» и «теплой» батиали по обе стороны
Шетландско-Исландского гребня близкими видами (или вариететами).
Таковы столь характерные для псевдоабиссали и батиали Норвежского
и Гренландского морей пикногон Colossendeis, восьмилучевой коралл
Umbellula, офиура Ophiocten, звезды Bathybiaster и Pontaster, неправиль-
ный ёж Pourtalesia, моллюск Pecten и др.
295
ФАУНА АБИССАЛИ
Зона абиссали. Ложе мирового океана занято абиссалью. Наибольшее
выражение здесь получают мягкие илистые и глинистые грунты, создаю-
щиеся только за счет скелетов пелагических организмов и вулканических
продуктов. Постоянство температуры и солености и почти полная неиз-
меняемость и других факторов среды характеризуют абиссаль. Чрезвы-
чайно слабо выражены также и колебания водных масс. Донная жизнь
количественно исключительно бедна.
Однообразие условий существования в глубоководных зонах мирового
океана является причиной значительного однообразия глубоководной
фауны, тем большего, чем больше температурное однообразие в этих слоях
моря. Если в верхних горизонтах батиали эти различия в температуре
и в фауне еще имеются, то в ее нижних горизонтах и особенно в абиссали
они исчезают.
Состав и происхождение фауны абиссали. Весьма характерны для
абиссальной фауны северной Атлантики прежде всего из иглокожих
обширная группа своеобразных голотурий, так называемых Elasipoda,
с длинными отростками ( Peniagone^ Deima^ Oneirophanta), оригинальная
звезда Freyella, ежи из сем. Echinothuriidae, маленький ёж с очень длин-
ными иглами Salenia hastigera, стебельчатые лилии (Rhizocrinus, Bathy-
crinus^ Pentacrinas и др.) и кремневые губки (Eaplectella, Hyaloneina,
Monorhaphis,, Disyringa). He менее характерно также для абиссали север-
ной Атлантики и абиссали Арктики
близнецов:
наличие некоторого числа видов
Атла нтические
В. robustus
Р. tenuispinus var.
О. sericeum var.
U. lindahli
Р. undatus (fragilis).
Арктические
Bathybiaster vexillifer
Pontaster tenuispinus
Ophiocten sericeum
Umbellula encrinus
Pecten frigidus
Три указанных вида иглокожих в арктической области обитают также
и в сублиторали. Как указывает С. Экман, трудно решить вопрос,
в какую сторону шел процесс расселения этих форм, является ли для них
обитание в арктической сублиторали конечной или начальной стадией
расселения. Взгляд на арктический генезис абиссальной фауны высказы-
вался еще первыми морскими зоогеографами (Thomson, 1877, О г t-
m a n п, 1896) и подтверждался последующими. Локар (L о с а г d,
1898) пришел к выводу, что среднеатлантическая абиссальная фауна
имеет скандинавский характер. Мартенс (Martens) и Тиле
(Thiele, 1903), анализируя моллюсков Индийской абиссали, пришли
к выводу, что они ближе к мелководной фауне холодных морей, нежели
к прибрежной фауне Индийского океана. Михаельсен (Micha-
el s е п) пришел к тому же заключению в отношении глубоководных
асцидий. То же можно сказать и о некоторых группах глубоководных
рыб (Cottidae, Zoarcidae, Liparidae) и ракообразных (Lithodidae). Однако,
наряду с этим, многие особенно типичные для океанской абиссали группы
или отсутствуют или очень слабо представлены в холодных морях. Часть
из них совсем не имеет родственных форм в мелководной фауне (Hexacti-
nellidae, Elasipoda и др.), другие близко связаны систематически с мел-
ководной фауной теплых морей, как, например, Gorgonaria, Реппаtu-
laria, Antipatharia, Madreporaria, крабы, Palinura, Astacura, многие
Anomura, стебельчатые лилии, многие ежи, большинство глубоководных
рыб и в первую очередь Macruridae. В результате подробного анализа
296
С. Э к м а н приходит к выводу, что «в большинстве случаев глубоковод-
ный животный мир теплых морей следует выводить от береговой фауны
теплых морей и в меньшей части от фауны холодных вод».
Итак, исходным материалом для создания глубоководной фауны,
как и для литоральной, должна была служить сублиторальная фауна.
В происхождении глубоководных батиальной и абиссальной фауны
наибольшее значение имеет ее древность. Абиссальная фауна Тихогог
Индийского и Атлантического океанов имеет наибольшую древность,
и здесь наиболее далеко зашел процесс формирования морфологиче-
ски и биологически своеобразной, обособленной систематически абиссаль-
ной фауны.
Абиссальная океаническая фауна почти отсутствует на глубинах
Арктического бассейна, но там формируется своя, более молодая глубоко-
водная фауна из сублиторальной. Тот же процесс мы находим в Среди-
земном и Японском морях.
Средиземное море, отделенное от Атлантического океана барьером
с глубиной 320 л, и Красное, отделенное от Индийского океана барьером
с глубиной всего 185 .к, имеют очень слабо выраженную глубоководную
фауну. Большие глубины этих морей заселены спускающимися из субли-
торали формами, т. е. здесь сформировалась своя собственная молодая,
еще очень бедная и нетипичная абиссальная фауна. Океаническая абис-
сальная фауна в этих морях почти отсутствует. Историческое прошлое
этих водоемов, возможно, уничтожило остававшуюся в нем глубоководную
фауну Тэтиса, новому же проникновению в эти моря океанической абис-
сальной фауны препятствуют мелководные барьеры проливов, а, может
быть, также высокая температура и соленость глубинных вод.
Отсутствует абиссальная фауна в Черном море, где она и не может
формироваться, вследствие заражения глубоких слоев моря сероводо-
родом. Едва намечающееся развитие глубоководной фауны имеет место
в Каспийском море. Замечательно отсутствие океанической абиссальной
фауны в Японском море, в то время как в соседнем, Охотском море, она
выражена очень ярко. Еще совсем недавно Японское море было внутрен-
ним, вероятно, опресненным водоемом, может быть, с развитием серо-
водорода; Сахалин и Японские о-ва составляли одно целое с материком.
После соединения Японского моря с океаном глубоководная фауна не
смогла проникнуть сюда из океана, так как все проливы для нее мелки.
Проливы же, ведущие в Охотское море, очень глубоки. В Японском море
уже формируется своя новая, молодая глубоководная фауна. Как указы-
вает К. Дерюгин, абиссальная фауна Японского моря «не абиссаль-
ного происхождения, а батиального, или частично даже сублиторального.
Все формы, по существу, свойственны более высоким горизонтам и спу-
стились в абиссаль вторично, приспособившись к жизни на больших
глубинах... Настоящие тихоокеанские элементы абиссальной фауны,
как, например, глубоководные губки, голотурии, декаподы, иглокожие,
моллюски и др., которые обнаружены в абиссали Охотского и Берингова
морей, здесь отсутствуют. Здесь нет глубоководных светящихся рыб
и настоящих глубоководных элементов планктона. Здесь обитает бедная
количественно и качественно однообразная фауна». Дерюгин выде-
ляет в абиссали Японского моря комплексы полихет — Harmothoe derju-
gini и Н. impar; моллюска — Pecten randolfi, Tmetonyx cicada (амфипода),
изоподы Euricope spinifrons и Typhlotanais (Tanaidacea).
Когда мы говорим о глубоководной фауне, следует помнить, что на
больших глубинах мы находим не только ту специфическую фауну, ко-
торая резко адаптирована к существованию в этих условиях, но и очень
много эврибатных животных из разных групп. Они способны оби-
297
тать от самой поверхности моря до глубин в 3-—5 тыс. лг, а вероятно,
и глубже. Процент этих форм в каждой группе весьма велик. По С. Э к-
м а н у (1935), среди морских лилий их не менее 20% (табл. 106), среди
морских звезд — около 30%, среди оболочниковых — около 25%.
Таблица 106
Группа животных Чисто литораль- ные1 виды Литораль- ные или ли- торальные и архибеп- тические виды Лито- ральные и архи- бентиче- ские От лито- рали до абиссали Архибен- тические Архибен- тические и абис- сальные
Морские лилии 31 10 11 35
Морские звезды 13 28 97
Оболочников ые 62 14 11 3 17
Падение видового разнообразия донного населения было хорошо
иллюстрировано для Атлантического океана Дж. Мёрреем (М и г-
г а у) еще в конце прошлого столетия (табл. 107).
Таблица 107
Вертикальные зоны в м Число станций Среднее количество видов на одну станцию Среднее число инди- видуумов на одну станцию
<180 70 62,8
180—900 40 51,2 150
900—1800 23 30,9 87
1800—2760 25 24,0 80
2700—3600 32 15,6 39
3600—4500 32 10,6 25,6
>4500 25 9,4 24
Уменьшается качественное разнообразие донной жизни и по мере
удаления от побережья, на что также указал М ё р р е й. На глубинах
от 1800 до 3650 м вблизи берега он собрал в одном улове 121 экз. живот-
ных, относящихся к 39 видам, а на расстоянии 500 км от берега — 21 экз.,
относящийся к 10 видам. М ё р р е й объясняет это уменьшением пищевых
ресурсов по мере удаления от берега.
Пути эволюции абиссальной фауны. Весьма интересно решение
вопроса о возникновении и формировании глубоководной фауны с эволю-
ционной точки зрения. При нашем современном знании физиологических
особенностей фауны, обитающей на больших глубинах, мы вряд ли можем
полностью ответить на этот вопрос, но попытаемся несколько осветить его.
Жизнь на больших глубинах моря связана с весьма специфическими
условиями существования. Прежде всего, это существование при низкой
температуре, не превышающей 4—5°, редко 6° (на глубине 1000 м). Дает ли
существование при низкой температуре какие-нибудь преимущества?
В последнее время накапливается все больше данных, указывающих на то,
накопление
что при низкой температуре в теле животных соотношение —расход—
получает более выгодное выражение. С этой точки зрения будет понятно
столь энергичное накопление жира у рыб, приходящих на летний период
1 В данном случае литораль—до 200 м глубины, архибенталь 200—1000 м и
абиссаль > 1С00 м ‘(см. рис. 98).
298
в Баренцево море. Весьма вероятно, что оно зависит не столько от обилия
пищи, сколько от температурного режима. Кроме того, имеется несколько
указаний на то (см. выше), что организмы, обитающие в высоких широтах
(планктон), обладают более высокими показателями жирности.
Если эти соображения верны, то глубоководная фауна, возможно,
существует при более благоприятном энергетическом режиме.
Второе, что имеет, возможно, еще большее значение, — это существо-
вание в условиях постоянной температуры. Даже там, где в поверхностных
слоях воды годовые колебания температуры наиболее велики, на глу-
бине 100 м они уже обычно не превышают Зэ, а часто 1°. В более глубоких
слоях моря практически температура остается в течение круглого года
неизменной. Дает ли это для организмов какие-нибудь преимущества?
Общепринятым является мнение, что гомойотермные животные (птицы
и млекопитающие) с физиологической точки зрения, сохраняя постоян-
ство температуры тела, находятся в особо благоприятных условиях для
всех процессов физиологического обмена. Не распространяются ли эти
благоприятные условия и на глубоководных животных, тело которых
также существует при постоянной температуре? С этой точки зрения мы
могли бы назвать их псевдогомон отермными.
Возможно, что в этом смысле несколько иначе мы должны взглянуть
на примеры особо резко выраженной стенотермии. Portlandia ar die а,
основная популяция которой существует при почти неизменяющейся
температуре—1,5, —1,8Э, или мадрепоровые кораллы, не выдерживающие
опускания температуры ниже 21,5°, может быть, также уклоняются в сто-
рону псевдогомойотермности, выгодной для них в каких-то отношениях.
Не следует ли думать, что потребность в сокращении амплитуды темпе-
ратурных колебаний — очень важный фактор, определяющий в какой-то
степени распределение организмов в биосфере и их морфофизиологиче-
скую эволюцию?
Третий момент неменьшего значения — это существование глубоко-
водной фауны в условиях темноты. Если в одних отношениях это момент
неблагоприятный, то в других, несомненно, благоприятный.
Вертикальные миграции планктона в море — это самое крупное по
масштабу явление перемещения массы организмов в биосфере — свя-
заны с отрицательным фототропизмом. Возможно, что в формировании
этого явления играет какую-то роль и защита от врагов, но вряд ли глав-
ную. Основная роль принадлежит свету, как фактору, определенным
образом воздействующему на поведение организмов. Физиология пока
что не дает нам ответа на вопрос о том, какой нагрузкой ложится на живот-
ный организм свет, а также и световые колебания. Мы знаем только о смер-
тельном вреде ультрафиолетовых лучей для мелких организмов и об анти-
биотическом действии прямого солнечного освещения. Таким образом,
уход морских животных на большие глубины разгружает их от термо-
и фоторегуляторного аппарата и дает какой-то эффект в смысле эконо-
мики энергетического баланса. В этом отношении между глубоководной
фауной имеется определенная принципиальная аналогия с фауной пещер
и подземных вод и частичная с паразитами \ Мы не знаем, перекрываются
ли получаемые при существовании на океанических глубинах выгоды
неблагоприятными моментами, как скудость и специфический характер
питания, сведенная до минимума способность «видеть», разреженное
существование и т. д. и т. п. Следует упомянуть еще об одном факте, ко-
1 Можно также напомнить, что и в наземной фауне существует большое число животных,
жизнедеятельных только в ночное время. Глаза у летучих мышей имеют, видимо, совсем ничтож-
ное значение.
299
торый имеет немаловажное значение для понимания рассматриваемого
явления. В полярных областях Арктики, там, где большую часть года
существуют пловучие льды, в северных частях морей Карского и Лаптевых,
фауна больших глубин Атлантического океана, Норвежского и Гренланд-
ского морей и центральной впадины Северного Ледовитого океана поды-
мается в совершенно несвойственные ей верхние горизонты и обильно
заселяет глубины в несколько сот метров. С полной уверенностью можно
сказать, что то же явление имеет место и в сублиторали Антарктического
материка.
Что же дает возможность этой фауне выйти на малые глубины? В рай-
онах, где это происходит, имеются условия, имитирующие условия боль-
ших глубин (за исключением давления) — постоянная и низкая темпера-
тура и слабый доступ света из-за малой прозрачности воды и ледяного
покрова.
Если все изложенное выше справедливо, то следует отказаться от
общераспространенного, не лишенного антропоморфических представле-
ний, взгляда на очень тяжелое существование глубоководной фауны и ее
реликтовый характер.
Это — высокоспециализированная фауна, прекрасно приспособленная
к условиям существования на больших глубинах, сформировавшаяся
из той части сублиторальной фауны, которая оказалась способной, в по-
рядке свойственного всем организмам стремления к расширению ареала,
выработать особенности, слагающие морфофизиологический облик
глу боков одно сти.
ГЛАВКА IX
БАКТЕРИИ
Значение бактерий в круговороте веществ в море. Бактерии в море
представляют собою одну из важнейших групп организмов, присут-
ствующих повсюду и выполняющих громадного масштаба биохимическую
роль во всех звеньях биоциклических процессов в море.
Изучение бактерий в море, особенно выполняемых ими массовых
процессов, началось совсем недавно и в силу методических моментов
находится в самой начальной стадии. Можно, однако, с полной уверен-
ностью утверждать, что без понимания роли бактерий во всех деталях
круговорота органических процессов в море едва ли могут быть поняты
и сами эти процессы.
Бактерии обладают громадной биохимической энергией, свойственные
им относительные показатели биологической продуктивности (Р/В-коэф-
фициент) во много раз выше, чем у растительных и тем более у животных
организмов. Бактериям свойственны и многие разнообразнейшие процессы
прототрофного и аутотрофного характера, и в этом отношении с ними
не может итти в сравнение ни одна другая группа организмов. Бактерии
чрезвычайно активны не только в живом состоянии, но многие выделяемые
ими вещества сохраняют громадную активность и тогда, когда произвед-
шие их бактериальные организмы уже погибли.
Бактерии присутствуют повсюду: в море, и во взвешенном состоянии
в морской воде, и на планктонных организмах, и около них, вступая
с ними в сложные биоценотические отношения, и в поверхностных слоях
грунтов, где они также тесно связаны в своей жизнедеятельности с живым
населением дна. Отмирающие и отмершие организмы и органический
детрит представляют собой субстрат для развития и деятельности разно-
образных бактерий. В сложной структуре бактериоценозов развиваются
и группы сильнейших антагонистов, ограничивающих деятельность друг
друга.
Как очень удачно говорит Ваксман (W a k s m а п, 1937) «когда
два организма способны утилизировать один и тот же пищевой ресурс,
но различно приспособлены к факторам среды, один из них найдет более
подходящие условия для быстрого развития и подавления другого орга-
низма». Цо Белл (Z о Bell, 1945) называет такие отношения между
противоположно «настроенными» группами микроорганизмов антибиозом.
Хотя все это в равной степени применимо и ко всем другим организмам,
населяющим морскую среду, однако у микроорганизмов п грибков явление
антибиоза находит наиболее яркую форму и значительное своеобразие.
301
Бактерии выделяют токсины, тормозящие развитие других организмов, как
защитные продукты метаболизма, «вырабатываемые микроорганизмами
Бактерии М2 х 100
в борьбе за существование» (Ваксман,
1937).
Рис. 160. Вертикальное распре-
деление бактерий (штрихо-
вая линия) и фитопланктона
(диатомеи) у побережья южной
Калифорнии (из Цо Белла).
Широкое распространение в морских
водах имеет и бактериофаг, констатирован-
ный повсеместно в прибрежных водах, но
обнаруженный в очень слабых дозах также
и в открытом океане.
В совокупности явлений, дающих воз-
можность выдвинуть рабочую гипотезу —
морская вода как коллоид, бактериям при-
надлежит ведущая роль; именно они созда-
ют основной фон этих явлений.
Количество бактерий. Количественная
связь мейщу фитопланктоном и бактериями
очень хорошо показана Цо Беллом
(1942) для побережий южной Калифорнии
(рис. 160).
При содержании проб воды с диатомо-
выми и бактериями в искусственных усло-
виях количество и тех и других возрастает,
и при последующем уменьшении количества
диатомей количество бактерий продолжает
увеличиваться и только в дальнейшем также
начинает уменьшаться (табл. 108).
G глубиной количество бактерий в мор-
ской воде резко убывает и только в верхнем
слое грунта имеет место колоссальное их
скопление (табл. 109).
Впрочем, еще Бертелем (Bertel,
1912) было показано для Средиземного мо-
ря, что самый верхний слой моря беден бактериями, погибающими от
действия солнечных лучей, но в глубь грунта, в пределах верхних 50 см,
количество их также резко убывает (табл. 110).
Таблица 10S
СРАВНИТЕЛЬНЫЕ КОЛИЧЕСТВА ДИАТОМОВЫХ И БАКТЕРИЙ НА 1 мл 1
МОРСКОЙ ВОДЫ В ТЕЧЕНИЕ ИНКУБАЦИИ ПРИ 25°С
(Ваксман, 1933)
Диатомеи Период инкубации
0 3 дня 7 дней 12 дней
177 346 1026 20600 178 22 800 I 165 I 13 900
Бактерии
Таблица 109
Глубина в морских саженях Количество бактерий в водах Вест- Индии по Дрью (Drew, 1912)
0—50 Очень много
200 160 на 1 мл
400 16 » 1 »
600 • 17 > 1 >
800 0—2 » 1 »
Грунт 160 000 0С0 на 1 мл
мл — миллилитр
302
Таблица 110
КОЛИЧЕСТВО БАКТЕРИЙ НА 1 г ИЛА В КОЛОНКАХ, ПО-
ЛУЧЕННЫХ ГРУНТОВЫМИ ТРУБКАМИ (Цо Белл, 1942)
32°26,4'N 117°41,3'W Глубина места 3120 фут._
Глубина в колонке грунта в дюймах Количество бактерий
0—1 38000000
1—2 940 000
4—5 88 000
9—10 36 000
14—15 2 400
19—20 400
29—30 180
39—40 330
59—60 250
79—80 130
99—100 290
Зависит количество бактерий в донных отложениях и от характера-
грунта (Цо Белл, 1938): чем мягче грунт и, тем самым, чем больше в нем
органических остатков, тем больше и бактерий (табл. 111).
Таблица 111
Характер грунта Средний диаметр частиц В Содержание азота в °/о Содержание воды в °/о Количество бактерий в 1 г
Песок 50—1000 0,09 33 22000
Пл 5—50 0,19 56 78 000
Глина 1—5 0,37 82 390 000
Коллоид <1 1,00 98 1510000
Мэр (Маг е, 1942) вычислила количество бактерий в грунте по
весу в отношении с другими группами организмов бентоса на 1 м2 (табл. 112).
Таблгща 112
Крупный макробентос (до 10 см)............... 75,00 г
Мелкий макробентос (до 5 см).................. 33,00 »
Мезобентос (до 0,5 см).......................... 1,15 »
Микробентос..................................... 0,02 »
Донные диатомеи................................. 0,05 »
Бактерии........................................ 0,36 »
Согласно этому автору, в 1 г морского ила (считая по сухому весу)^
содержится 0,3—2,0 мг живых бактерий. Количество бактерий в поверх-
ностном слое воды убывает по мере удаления от берега.
В чистых береговых водах бактерии содержатся в количестве несколь-
ких сот или более (в 1 см3) экземпляров, относящихся к 25—35 видам.
Таблица 113
СРЕДНЕЕ КОЛИЧЕСТВО БАКТЕРИЙ В 1 мл ВОДЫ ИЗ
ЗАЛИВА САН-ДИЕГО НА НИЗКУЮ ВОДУ ПО НАПРА-
ВЛЕНИЮ В ОТКРЫТОЕ МОРЕ
Местоположение
Количество бактерий
в 1 мл
Около мили в глубь залива .......
У выхода из залива...............
5 миль от входа в залив..........
10 миль от входа в залив ........
480000
67 000
430
510
303
В открытом океане в тОхМ же объеме воды содержится их меньше десятка
{табл. 113).
Естественно, что количество бактерий в прибрежных водах зависит
и от фаз прилива и отлива (табл. 114). В прилив количество бактерий
уменьшается, в отлив увеличивается.
Таблииа 114
КОЛИЧЕСТВО БАКТЕРИЙ В 1 мл У ВХОДА В ЗАЛИВ
МИССИОН (Цо Белл и Фельтем, 1942)
В прилив В отлив
Среднее для 34 дней 1472 294 800
Минимум 194 31000
Максимум 9 600 4400000
Химические процессы, протекающие при участии бактерий. Бакте-
рии, обитающие в морской воде, производят самые различные процессы
.{в море или в культурах). Гарвей (Harvey, 1945) дает следующий
их перечень.
Разложение хитина, углеводородов нефти, лигнина, мочевины.
Окисление аммиака до нитритов^
нитритов до нитратов,
сульфидов до сульфатов.
Редукция нитратов до нитритов,
нитратов до газообразного азота,
сульфатов до сульфидов (анаэробно),
сульфидов до серы.
Утилизация газообразного азота,
аммиака и мочевины,
нитратов и нитритов,
фосфатов.
Освобождение азота и фосфора (последнего в виде
фосфатов) после гибели бактерий.
Формирование железомарганцевых конкреций.
|Бактериальные клетки относительно богаче азотом и особенно фосфо-
ром, если их сравнивать с фитопланктоном (табл. 115).
Таблица 115
Бактерии..............
Планктонные диатомовые
Среднее (по различным авторам)
соотношение углерода, азота и
фосфора в теле бактерий и диато-
мовых водорослей
(из Гарвея, 1945)
С N Р
100 20 4,4
100 16 1,67
Бактериальные клетки, подобно всем прочим организмам, дышат,
потребляют кислород и выделяют углекислоту. Цо Белл (1946) ука-
зывает, что 1 г активно растущих морских бактерий потребляет 30 cmz
кислорода за час при 22° С, а морские животные за то же время на 1 г
живого вещества — только 0,002—1,0 еле3.
•304
Выделяемый животными и бактериями в процессе жизнедеятельности
аммиак или непосредственно утилизируется фитопланктоном, либо оки-
сляется при помощи бактерий до нитритов или нитратов. Растения могут
Рис. 161. Смены в соединениях азота в морской воде в сопоста-
влении с развитием планктона (по Brand и др. из Цо Белла).
в одинаковой степени потреблять разные соединения азота. В опытах
Бранда и др. (1937) формирование нитритов и нитратов из аммиака,
освобожденного из разлагающегося планктона, происходило только
в присутствии нитрифицирующих бактерий (рис. 161).
Весьма ценные данные по распространению бактерий в морях СССР и
по обусловленным ими процессам даны советскими исследователями
(Б. Исаченко, В. Буткевич). Результаты их работ излагаются
во II томе.
20 Зенкевич, т. I
ГЛАВА X
ОКЕАНИЧЕСКИЕ ПТИЦЫ
В нескольких местах мирового океана, там, где имеет место энергичное
перемешивание вод, обогащение верхних слоев питательными солями
Рис. 162» Холодное течение Гумбольдта, омывающее западное
побережье Южной Америки (из Мэрфи).
и обильное развитие планктона, там, где при этом поблизости находятся
удобные для гнездовий побережья, — иногда наблюдаются громадные
306
скопления птиц, жизнь которых неразрывно связана с океаном. Наиболее
известны в этом отношении западные побережья южной Америки, омы-
ваемые идущим с юга холодным течением Гумбольдта (рис. 162). Обуслов-
ленная им высокая продуктивность моря, совсем вблизи берегов,
способствовала формированию в течение многих тысяч лет колоссальных
скоплений птиц, откладывавших на местах своих гнездовий переработан-
Рис. 163. Разработка «гуано» в Перу (из Мэрфи).
ный планктон в виде мощных отложений помета («гуано») до 55 м толщи-
ной. Основными их кормовыми объектами являются мелкая рыба (особенно
Engraulis} и крупные ракообразные (главным образом Schizopoda).
Уже в течение 100 лет гуано американских побережий добывается
в промысловых целях и в первую очередь, как удобрение (рис. 163).
В 1933—1934 гг. ежегодный вывоз гуано из Перу превысил 160 000 пг.
Основные производители этих «горных хребтов» гуано—пеликаны,
буревестники, фаэтоны, фрегаты, чайки глупыши, пингвины и альбатросы.
Вообще в водах умеренных широт гораздо больше морских птиц,
чем в тропических, что находится в прямой зависимости от про-
307
дуктивности верхних слоев моря и от обилия в них мелких пелаги-
ческих рыб и крупных ракообразных. С п и с с (Spiess, 1926) иллю-
стрировал это очень наглядной схемой (рис. 164). Штот (Stott, 1936)
Рис. 164. Относительное обилие морских птиц в высоких
и низких широтах океана (по Списсу).
также описывает для западного Шпицбергена массовый выкорм птиц
с прибрежных птичьих базаров на воде за счет развивающихся здесь
в колоссальном множестве Thysanoessa inermis (Euphausiacea). (О пти-
чьих базарах Новой Земли и птицах Каспийского моря см. II том.)
ГЛАВА XI
СОЛОНОВАТЫЕ ВОДОЕМЫ И ИХ ФАУНА
Ни в одной стране мира вопрос о солоноватоводности не имеет такого
большого теоретического и практического значения, как в СССР. Лишь
немногие из тех 14 морей, которые омывают границы Союза, имеют пол-
ную морскую соленость. Большинство характеризуется той или иной
степенью опреснения, а соленость 5 морей — Балтийского, Черного,
Азовского, Каспийского и Аральского — составляет 1/i—нормальной
Рис. 165. Амплитуда солености наиболее опресненных морей СССР с указанием их
средней солености (ориг.).
морской (рис. 165). Кроме того, и в других морях имеются обширные
районы, подвергающиеся очень сильному опреснению, например, пред-
устьевые зоны по северному побережью Сибири. В этих морях и отдель-
ных их районах обитает своеобразная солоноватоводная фауна, не встре-
чающаяся ни в морях, ни в пресных водах. Нередко именно она оказы-
вается основной частью фауны и имеет иногда большое промысловое
значение.
Обеднение морской и пресноводной фаун в солоноватых водах.
Как мы уже указывали выше, с наступлением опреснения наступает
резкое качественное обеднение морской фауны; чем большему опресне-
нию подвергается тот или иной район моря, тем меньшее число морских
форм мы в нем находим. В наиболее опресненные участки морей прони-
кают только немногие наиболее эвригалинные морские формы, доходящие
иногда до пресной воды.
309
То же явление мы наблюдаем и на пресноводной фауне: с переходом
в осолоненные зоны также сохраняются
Рис. 166. Взаимное проникновение морской
и пресноводной фаун Балтийского моря на-
встречу друг другу (по Ремане).
люсков). Хорошая иллюстрация этого была дана Иогансеном
(J о b a ns е п, 1918) на примере моллюсков (рис. 168) в одном из зали-
вов Балтийского моря. В
Балтийском море в этой
зоне наименьшего видового
обилия, по Р е м а н е (R е-
m а п е, 1935), обитает 68 форм
беспозвоночных (без олиго-
хет и нематод), среди кото-
рых имеются и морские и
пресноводные в отношении
3 : 2 по числу видов. Полови-
на из этих 68 форм ракооб-
разные, четверть — коло-
вратки. Следует иметь в ви-
ду, что, несмотря на такое,
казалось бы, интенсивное
взаимное проникновение в
устьях рек морской и пре-
сноводной фаун, полного пе-
рехода к существованию в
чуждой среде не происходит,
или если и происходит, то
с чрезвычайной медленно-
стью, и во всяком случае
основные пути и способы
внедрения морской фауны
в пресную воду иные.
только наиболее эвригалинные
пресноводные формы. Таким
образом в слегка осолоненных
частях морей (2—5°/00) могут
смешиваться морская и пресно-
водная фауны (рис. 166).
Насколько беднеет при этом
качественно морская флора и
фауна можно видеть в табл.
116 на примере разных наших
морей по некоторым группам
фауны.
С большой детальностью
этот процесс обеднения можно
видеть на примере Балтийско-
го моря, наиболее опресненного
из всех наших морей (см. табл,
на стр. 226 т. II и рис. 167).
Весьма замечательны при этом
два момента: наименьшее число
видов приходится не на какую-
то среднюю соленость (близкую
к 17,5°/оо), а на гораздо более
низкую (для Балтийского моря
между 5 и 8°/00 для всей фауны
и между 3 и 5°/00 для мол-
Рис. 167. Убывание числа видов морских жи-
вотных, по мере продвижения из Северного мо-
ря через проливы вглубь Балтийского моря,
в сопоставлении с соленостью. Цифры на кар-
те — число видов.
310
Если солоноватоводная фауна в каждом водоеме имеет свои солевые гра-
ницы распространения, то качественное обеднение морской фауны,
Таблица 116
Количество видов в группах Средиземное море Черное море
Зеленые водоросли 63 54
Бурые » 93 64
Красные » 267 103
Всего 423 221
Кишечнополостные 208 42
Ракообразные 1174 283
Полихеты 516 126
Моллюски 1457 125
Иглокожие 101 4
Оболочниковые 200 16
Рыбы 549 180
Всего 4205 776
Кишечнополостные................
Полихеты........................
Двустворчатые моллюски..........
Иглокожие.......................
Рыбы............................
Всего ....................
Северное море Центральная часть Балтийского моря Финский и Бот- нический заливы
123 3 (1)
271 12 3
170 5 4
70 1 0
110 26 17
744 47 25
Баренцово море Белое море
Кишечнополостные 114 81
Двустворчатые моллюски 64 38
Иглокожие 62 22
Рыбы 114 50
Всего ....................
354 191
с переходом в менее соленые внутренние водоемы, подчиняется, видимо,
одним и тем же закономерностям. Для наших морей обеднение морской
фауны имеет следующий характер:
Соленость уменьшается на
Фауна беднеет на
Соленость уменьшается на
Фауна беднеет на
Соленость уменьшается на
Фауна беднеет на
7-8°/оо
50% (Белое море)
18°/оо
8О°/о (Черное море)
26—27%0
94—95% (Балтийское и Азовское моря)
311
Условия существования в солоноватых водах. Как указывалось
выше, основным тормозом в проникновении морской фауны в пресные
водоемы являются две основные группы факторов. Во-первых, сово-
Солрнпстъ в%о
Рис. 168. Число видов моллюсков в
Рандерс-фиорде, при различных соле-
ностях. Различная штриховка соответ-
ствует пресноводным (слева) и морским
(справа) формам (по Иоганнсену из Ре-
мане).
купность явлений, связанных с пе-
реходом от существования в усло-
виях изотонии и сходства в солевом
составе внешней и внутренней среды
к условиям гипертонии и необходи-
мости поддерживать строго опреде-
ленные степень и состав внутренней
солености. Большое значение могут
также иметь недостаток в опреде-
ленных солях во внешней среде и
изменение процессов дыхания и пи-
тания с переходом в пресноводную
среду. Вторая группа препятствую-
щих факторов — это резкая смена
физико-географических условий су-
ществования: малые размеры водое-
мов, незначительность их пелагиче-
ской части, резкие смены климати-
ческих и прочих условий, быстрые
токи воды и многое другое.
Гораздо труднее объяснить, почему пресноводная фауна не может
приспособиться к существованию в морской воде, или, во всяком случае,
почему процесс этот идет с чрезвычайной трудностью. Единственное пока
Колич
Рис. 169. Плотность планктона против устья Ла-Платы (по Тиманну).
I — морские диатомеи, II — пресноводные водоросли, III — все многоклеточные. А. — Cope-
poda, Б,— Nauplii, В.— Appendicularia. Внизу рельеф дна и соленость.
объяснение этого явления предложено Р е м а н е, который считает, что
при переходе в соленую воду, в условиях изотонии, должны переставать
работать органы выделения и должно происходить накопление в теле
продуктов распада.
31
Только рыбы и, вероятно, некоторые ветвистоусые ракообразные,
коловратки и инфузории выселились из пресной воды в море.
Таким образом устья рек и прилежащие к ним пространства моря,
иногда целые морские водоемы, более или менее отграниченные от моря
и принимающие в себя массы речной воды, характеризуются переход-
ными соленостями, одинаково неблагоприятными для существования
и морской и пресноводной фаун в их основной массе, доступными только
для наиболее эвригалинных форм. Тем более замечательно, что в этих
условиях переходной солености некоторые пресноводные и морские орга-
низмы находят особенно благоприятные условия для массового развития,
чему, видимо, способствует благоприятный режим биогенных элементов.
Так, например, в Азовском море колоссального развития достигают
моллюски Cardium edule^ Syndesmya ovata, Corbulomya maeotica, а в Бал-
тийском море — Macoma baltica. К солоноватым водам вполне приме-
нимо правило, что биотопы, уклоняющиеся от основного типа место-
обитания, населены немногочисленными видами, представленными, од-
нако, большим числом особей.
Некоторые морские и пресноводные планктонные формы также находят
наилучшие условия для развития в прилежащих к устьям рек зонам,
имеющим промежуточные солености. На это явление впервые обратил
внимание финский гидробиолог Великангас (Valikangas,
1935). Им установлено, что в Балтийском море многие пресноводные формы
речного планктона (коловратки, ракообразные) дают пышное количе-
ственное развитие уже вне реки в слегка соленой воде (3,5—5,4°/00). Ранее
Кольбе (Kolbe) указывал на стимулирующее действие слабо-
соленой воды на пресноводных диатомовых.
Сходное явление Н. Чугунов обнаружил перед дельтой Волги.
Недавно появилось обширное исследование Тиманна (К. Thie-
mann, 1934) по планктону предустьевых и устьевых зон некоторых
рек, впадающих в Атлантический океан (Эльба, Рио-Пара, Ла-Плата
и др.). Т и м а н н устанавливает значительное количественное увели-
чение некоторых групп морского планктона (диатомовые, аппендикуля-
рии, Copepoda) при подходе к устьям рек (рис. 169).
Рост животных в солоноватых водах. Давно уже известно, что ряд
моллюсков, населяющих Балтийское море, по мере продвижения из
Северного моря вглубь Балтийского мельчают. Данные для четырех
наиболее распространенных форм приведены в табл. 117 и на рис. 170.
Таблица 117
Формы В Кильской бухте В Финском заливе В Ботническом заливе
5°/oo = 157oo £°/00 = 5 2°/ 00 №/о.=5-27О(,
^лина в мм
Mytilus edulis 110 27 21
Муа arenarla 100 77—75 36,5
Cardium edule 44 22 18
Macoma baltica 22 17 15
Тот же процесс измельчания, при переходе в менее соленую воду,
наблюдается у брюхоногих, большинства рыб, иглокожих, Scyphozoa и
отчасти ракообразных. Это правило оказывается неприменимым для
большинства ракообразных (Amphipoda, Ostracoda, Copepoda), полихет,
313
оболочниковых, турбеллярий, нематод, коловраток и др. Часть форами-
нифер в Кильской бухте оказывается даже крупнее, чем в океане. Ре-
мане считает, что уменьшение размеров с понижением солености происхо-
дит у тех организмов, которым требуется известь для построения скелета
(Mollusca, Echinodermata, Pisces). Корненожки же с переходом в пресную
воду обычно заменяют известь в скелете песчинками. В океане песча-
нистые раковинки у Foraminifera составляют около 20%, а у пресно-
водных — около 61%. Раковины моллюсков с переходом в опресненные
части Балтики делаются иногда тонкими, как бумага. Р е м а н е по-
лагает, что уменьшение кальция обусловливает и замедленный рост
и уменьшение раковины (а следовательно, и тела). Параллельно в том же
направлении может действовать и возрастание содержания в более пресной
воде СО2.
Рис. 170. Изменение размеров различ-
ных морских и пресноводных моллюсков
в промежуточных соленостях Балтий-
ского моря (по Ремане).
Рис. 171. Схематическое изображение
соотношения пресноводных, солонова-
товодных и морских форм, при измене-
нии солености (по Ремане).
' По наблюдениям В. Воробьева, Cardium edule в лиманах Азовского
моря, более соленых, чем само море, значительно увеличивается в раз-
мерах при том же возрасте. Чрезвычайно интересно, что наряду с этим
пресноводные брюхоногие моллюски Bithynia и Theodoxus при проникно-
вении в солоноватоводные районы Балтики также мельчают. Замкнутые
водоемы морского генезиса могут, в зависимости от климатических
и геологических факторов, изменять соотношение растворенных в воде
солей. Таковы, например, наши Каспийское и Аральское моря. Нынешний
состав солей каспийской и аральской воды сказывается очень значи-
тельно на качественном и на количественном составе их фауны.
Специфическая солоноватоводная фауна. Уже давно было известно,
что солоноватые воды, не везде в равной мере, но почти всегда, кроме
эвригалинных морских и пресноводных форм, имеют еще и свое собствен-
ное, специфическое население, которое только здесь и обитает, т. е. не
встречается ни в морях, йи в пресных водах (рис. 171).
•314
Не менее яркую иллюстрацию обособленности морского и солоновато-
водного компонентов фауны дают черноморские мизиды (по В а с е s с о,
1940). И здесь мы имеем ту же двувершинную кривую. Морские (среди-
земноморские) мизиды дают максимум видов при солености 14—2О°/оо,
черноморско-каспийские автохтоны — при солености 1—2О°/оо.
Далеко не во всех частях мирового океана солоноватоводная фауна
выражена в равной степени. Богатством солоноватоводной фауны резко
выделяются моря так называемой южно-русской геосинклинали (Черное,
Азовское, Каспийское и Аральское), моря, примыкающие к Арктическому
бассейну (включая также и Балтийское море), и береговые воды юго-
восточного угла Азии и в меньшей степени устьевые зоны рек тропи-
ческой зоны.
В Балтийском море и морях Арктики солоноватоводная фауна состоит
в главной массе из лососевых, сиговых и корюшковых рыб и некоторых
отрядов высших ракообразных (Amphipoda, Isopoda и Mysidacea). В южно-
русских морях помимо преобладающих и здесь рыб и ракообразных, среди
настоящих обитателей солоноватых вод, мы находим двустворчатых мол-
люсков, губок, кишечнополостных, полихет, мшанок и ряд других групп.
Из рыб здесь выделяются сельдевые, осетровые, бычковые и др., а из
ракообразных — Amphipoda, Cumacea и Mysidacea.
Описанная из береговых лагун Индии (оз. Чилька) солоноватоводная
фауна замечательна тем, что здесь представлены группы, труднее всего
переносящие опреснение и совершенно отсутствующие в двух других
районах. К таким группам относятся Anthozoa (Virgtzlaria, некоторые
актинии, даже одна пресноводная), Polyclada, Echiuroidea (два вида
Talassema), мшанки из Gheilostomata.
Паниккар и др. (Р a n i к к а г, 1937) указывают, что река
Adyar у Мадраса, наполняющаяся обильными водами только в октябре —
декабре, уже в марте теряет связь с морем и устье ее отгораживается
от моря сплошным валом, представляя собой обширную лагуну с соле-
ностью, опускающейся до 16—17°/00. В этой солоноватоводной лагуне
констатировано 56 морских позвоночных и 92 вида беспозвоночных всех
морских групп, кроме иглокожих.
Своим происхождением солоноватоводная фауна в основной своей
части обязана историческому прошлому водоема. Если морские водоемы
или части их подвергаются, вследствие нарушения связи с мировым
океаном, медленно идущему процессу опреснения, исходно морская фауна
получает наиболее благоприятные условия для перехода к существова-
нию в слабо соленой или даже пресной воде. Водоем сохраняет свой как бы
морской облик, а вода становится все менее и менее соленой. Даже при
таком процессе опреснения, совершающемся с геологической медлен-
ностью и охватывающем часто весь водоем, многие морские группы вы-
мирают и лишь небольшая часть исходной фауны приспособляется к су-
ществованию в солоноватой и пресной воде. Именно такой процесс пере-
жили в своей третичной и четвертичной истории наши южные водоемы,
не раз подвергавшиеся почти полному опреснению. Как сами моря (Черное,
Азовское, Каспийское и Аральское), так и озера и реки их бассейнов
насыщены обильной реликтовой солоноватоводной и пресноводной фауной.
Менее разнообразная, но столь же резко выраженная солоноватоводная
реликтовая фауна свойственна наиболее опресненным частям Аркти-
ческого бассейна и Балтийскому морю и многочисленным рекам и озерам
их бассейнов. Эта фауна также есть след опреснений больших морских
пространств в четвертичное время.
Что касается юго-западной части Азиатского материка, то формиро-
вание здесь солоноватоводной фауны объясняют длительным существо-
315
ванием больших пространств моря, подвергающихся сильному опресне-
нию реками и тропическими дождями.
Согласно данным Саразинова и Михаельсена, палео-
географическая история остиндского архипелага, Индокитая и Индостана
протекала весьма бурно. Михаельсен полагает, что сама Индия
в свое время была разбита на отдельные острова. При формировании
современного рельефа суши могли образовываться различного типа
полузамкнутые и замкнутые водоемы, подвергавшиеся опреснению и пере-
рабатывавшие заключенную в них фауну.
Классификация солоноватых вод. Своеобразие солевого и гидро-
логического режима солоноватых вод уже давно ставило перед исследо-
вателями вопрос об их классификации, но только в последние годы на-
метилось правильное решение вопроса. Прежде всего следует различать
понятия солоноватые воды и солоноватоводная фауна; для того и для
другого могут существовать самостоятельные классификационные си-
стемы, не совпадающие друг с другом.
На такой точке зрения стоял Н. К н и п о в и ч, давший первую
попытку найти границу между «морской» и «солоноватой» водами безотно-
сительно населяющей их жизни. Н. Книпович обосновывает это
следующим образом.
Если пресная вода замерзает при 0° и наибольшую плотность имеет
при 4°, то с возрастанием количества растворенных в воде солей темпе-
ратура замерзания и температура наибольшей плотности понижаются,
однако, не равномерно — понижение температуры наибольшей плот-
ности идет быстрее и в некоторой точке, соответствующей 1,33°, обе кривые
пересекаются, т. е. вода солености 24,7°/00 (точнее — 24,695 °/00) замер-
зает и имеет наибольшую плотность при одной и той же температуре 1,33°.
Н. Книпович предлагает называть солоноватоводными водоемами
такие, «вода которых имеет температуру замерзания более низкую, чем
температура наибольшей плотности воды», т. е. существующие при усло-
виях, соответствующих левой половине нашего чертежа (рис. 165). Морские
водоемы в собственном смысле слова будут иметь температуру замерза-
ния воды более высокую, нежели температура наибольшей ее плотности.
«Явление вертикальной циркуляции протекает в солоноватой воде, —
говорит Н. Книпович, — более сложно, чем в воде морской, или
воде пресной. По отношению к водоемам с соленой водой учитывают
обыкновенно две основные формы этого процесса, которые обозначают
иногда терминами летняя и зимняя циркуляция. При летней вертикаль-
ной циркуляции содержание солей подымается в поверхностном слое,
вследствие испарения, и частицы воды опускаются в глубину, содей-
ствуя повышению в глубоких слоях и температуры, и содержания солей,
и содержания кислорода. При зимней вертикальной циркуляции опу-
скаются в глубину частицы, ставшие более тяжелыми, вследствие охлажде-
ния, причем уносят в глубокие слои сравнительно много кислорода
и оказывают более или менее сильное влияние на температуру глубоких
слоев. В воде солоноватых водоемов может иметь место, очевидно, еще
одна форма вертикальной циркуляции: весною при повышении температуры
до температуры наибольшей плотности возникает, как мы видели, вер-
тикальная циркуляция, вызываемая не понижением температуры и не
повышением солености, а повышением температуры без какого-либо по-
вышения солености, т. е., в сущности, то же самое явление, которое наблю-
дается в пресной воде, при изменении ее температуры от 0 до -|-40.
Отличие от вертикальной циркуляции в пресных водах заключается в том,
что в последних, конечно, не может иметь места циркуляция типа лет-
ней».
316
Именно эти свойства солоноватоводных водоемов позволяют характе-
ризовать их режим, как озерный.
Деление Н. Книповичем вод на морские и солоноватые с грани-
цей между ними, соответствующей солености 24,695°/00, основанное на
некоторых физических свойствах воды, не дает какого-либо дальнейшего
подразделения его солоноватых вод, что должно было бы представлять
наибольший интерес. С другой стороны, в распределении морских орга-
низмов не наблюдается никаких существенных изменений при солености,
близкой к 24,7°/0(Г. Правда, некоторые исследователи (А. А н д р и я ш е в,
А. Пробатов) отмечают перелом в фауне примерно на этой самой
солености в Карском море и море Лаптевых, однако, если действительно
такой перелом имеется, то он носит чисто местный характер и, возможно,
объясняется иными причинами.
К вопросу о создании классификации вод по солености неоднократно
подходили биологи и пытались в основу этой классификации положить
распределение организмов. Такая классификация, единая для разных
частей мирового океана, была бы возможна только в том случае, если бы
закономерности, обусловливающие наличие определенных границ в рас-
пределении морских, солоноватоводных и пресноводных организмов,
были одни и те же для всего земного шара. Однако таких постоянных
границ нет. Существующие различия обусловливаются в первую очередь
различиями в пережитой водоемом и его фауной истории, в силу чего
границы распределения солоноватоводной фауны в Балтийском море
могут оказаться иными, чем в Азовском море или море Лаптевых. Общие
закономерности в распространении морской и пресноводной фаун в воды
промежуточной солености пока еще не вскрыты, и поэтому существующие
классификации имеют значение только в применении к тому водоему,
для которого они созданы.
Таблица 118
Пресная Солоноватая Морская
т II 111 IV V
олигогалинная мезогалинная | полигалинная
£"/„„< О,2%о О,2-1,9<>/оо 1,9-18,б’/о. 18,6—31,6°/00 >31,6'700
Таблица 119
I Почти агалин- ные водоемы II Пресные. Оли- гогалпнные III Солоноватоволные. Мезогалинные IV Морские. По- лигалинные V Ультрагалинные
$700 < 0,2°/00 0,2—О,5°/оо 0,5- 16%о 16—47*/^ >470/оо(ДоЗОО°/01))
Таблица 120
Пресные воды Солоноватые воды Морские воды
солоновато-пресная смешанная зона настоящая солоно- ватая зона солоновато-мор- ская, смешанная зона
J7oo<3°/oo Господствуют пресноводные организмы 3-5°/оо Преобладают пресноводные организмы 5-10 (8)°/оо Максимальное раз- витие солоновато- водной фауны 10 (8)-15°/оо Преобладают морские организмы > 15°/оо Господствуют морские организмы
317
Первая такая классификация вод по солености, а вернее сказать —
классификация фауны по солевым границам массового распространения
О I 2 3 4 5 6 7 8 9 10111213 S%0
—-| Морская фауна
Реликтовая солонова-
товодная фауна
lillliliill Пресноводная фауна
форм, была создана Р е д е к е для свое-
образного голландского залива — Зюйдерзее.
Р е д е к е делит воды разной солено-
сти на морские, солоноватые и пресные
(табл. 118).
С. Зернов значительно видоизменил
и расширил схему Редеке и придал ей
следующий вид (табл. 119).
Наконец Р е м а н е, применительно
ко всему Балтийскому морю, стремится
сделать классификацию еще более биоло-
гичной и придать ей количественное зна-
чение. Морские воды по Р е м а н е — это
те, где больше половины видов организмов
морские (табл. 120).
Границей собственно морских вод Р е-
м а н е предлагает считать соленость 14—
15°/00, но еще до солености 10 (8)°/00 мор-
ские формы преобладают. С другой стороны,
как указывал еще К. Брандт, где-то око-
ло 3°/оо лежит последняя граница проник-
новения морского бентоса (в Балтийском
море) в пресную воду, а по Великанга-
с у и Гесснеру — морской планктон
останавливается на солености 5—7°/00. При-
мерно на этой же солености (около 6°/00)
начинается значительный подъем пресновод-
ных форм. Резкое же падение числа морских
форм имеет место около 1О°/оо. Классифика-
ция Р е м а н е хорошо передает соотно-
шение в распределении трех основных
групп форм Балтийского моря и отражает
на себе своеобразный ход кривой (рис. 171)
убыли количества видов морских форм. При
солености 20—16 и 9—7°/00 имеет место бо-
лее резко выраженная убыль форм. Однако
схема эта пригодна полностью все же толь-
ко для Балтийского моря; фауны Азовско-
го и Каспийского морей дают несколько
иные границы (рис. 172) распространения
морской, солоноватоводной и пресноводной
фаун.
Рис. 172. Соотношение по рас-
пределению солености и оби-
лию трех основных компонен-
тов фауны в Балтийском, Азов-
ском и Каспийском морях.
Вследствие того, что все приведенные классификации, кроме класси-
фикации Н. Книповича, базируются только на распределении
организмов, следует говорить не о классификации водоемов по солености,
а о классификации фауны по отношению к солености. С другой стороны,
по крайней мере в отношении солоноватоводной фауны, каждый водоем
требует своей собственной системы подразделений. Что касается более
общей классификации фауны по отношению к солености, то применительно
к северной Атлантике можно наметить следующую схему.
В морских водах с соленостью до 5—8°/Оо резкое преобладание имеет
морская фауна, однако воды с более высокой соленостью в отношении
фауны не являются однородными.
318
Уже при уменьшении солености от 35 до 27°/00 число видов уменьшается
примерно на 30% и возможно, что уже при солености 25—27°/00 ее мор-
ская фауна должна приобретать некоторую, хотя и очень слабую, гипер-
тонию внутренней среды по отношению к внешней.
Дальнейшее уменьшение солености до 17—18°/00 обусловливает еще
большее обеднение фауны (примерно на 70%), причем преобладание
получают формы, обладающие осморегуляторнымп способностями.
Рис. 173. Экологические ареалы основных групп водной фауны по отношению
к солености.
I — морские стеногалинные формы, II — морские эвригалинные с центром ареала в морских во-
дах, Ш — морские эвригалинные с центром ареала в солоноватых водах, IV — истинная солопо-
ватоводная фауна, V — пресноводные формы с центром ареала в солоноватых водах, VI —
эвригалинные пресноводные формы с центром ареала в пресной воде, VII — пресноводные стено-
галинные формы (ориг.).
В Черном море резкое обеднение фауны имеет место также при соле-
ности 14—12°/00 и, наконец, при солености 8—6°/00 исчезает большинство
морских форм, от них остается не более 3—4%, и вступает в силу, если
он имеется, солоноватоводный комплекс. В соответствии с этим водная
фауна может быть распределена в семь основных групп по характеру
экологического ареала в отношении к солености (рис. 173). У некоторых
морских (группа III) и пресноводных (группа V) центр экологического
ареала может находиться в зоне переходных соленостей.
ГЛАВА XII
ПИТАНИЕ МОРСКИХ ЖИВОТНЫХ, ПРЕИМУЩЕСТВЕННО
РЫБ
1. ИСТОРИЯ И МЕТОДИКА ИССЛЕДОВАНИЙ1
Данные по качественному анализу пищи различных морских живот-
ных, в первую очередь рыб, стали накапливаться очень давно.
Методы изучения питания беспозвоночных. Что касается беспозвоноч-
ных, то изучение их питания ограничивалось либо изучением механизма
захвата пищи, с чрезвычайной тщательностью разработанного для неко-
торых форм (например, работы И о н г а — Yong), либо физиологической
стороной процесса переваривания, либо указанием качественного состава
пищи. Этими исследованиями выяснено, что кишечнополостные, тур-
беллярии, немертины, щетинкочелюстные, большинство морских звезд
и офиур, некоторые брюхоногие моллюски, Septibranchiata из двуствор-
чатых, головоногие, часть полихет и Stomatopoda — хищники, губки,
мшанки, плеченогие, часть полихет, двустворчатые моллюски, часть
брюхоногих, низшие ракообразные, некоторые голотурии и оболочнп-
ковые питаются наннопланктоном (преимущественно фитопланктоном),
голотурии (большинство), некоторые ежи и звезды, офиуры, часть поли-
хет, часть сипункулид — детритом и грунтом. Некоторые беспозвоночные
всеядны, как, например, многие морские ежи, часть полихет, некоторые
брюхоногие и из ракообразных Isopoda, Amphipoda и Decapoda.
В разное время предлагались различные классификации морских
животных по характеру питания и по способу захвата пищи. Различают
плотоядных, растительноядных, грунтоедов, планктоно-(сестоно-)едов,
хищников и т. п., или щупалыциков, глоталыциков, фильтровальщиков,
седиментаторов, охотников, засадчиков и т. д.
Однако изучение питания морских беспозвоночных почти никогда не
переходило на методы количественного характера. Только для некоторых
форм (устрица, мидия) определялось количество пропускаемой через тело
воды. В противоположность этому для рыб уже с 30-х годов прошлого
столетия исследователи стали искать количественных показателей пи-
тания, и в дальнейшем мы будем говорить в основном о рыбах.
Методы изучения питания рыб. Еще во второй половине прошлого сто-
летия в работах американца Форбса (Forbes, 1878—1888), осно-
1 Для составления исторического обзора мы использовали превосходную сводку А. Шо-
ры ги на в его работе «Питание и пищевые взаимоотношения некоторых бентофагов Северного
Каспия».
320
воположника всего современного количественного направления в изуче-
нии питания рыб, появился метод учета встречаемости того или иного сорта
пищи в кишечнике животного (в скольких кишечниках от общего коли-
чества обследованных найден определенный вид), сначала просто по ко-
личеству кишечников, а затем (Scot t, 1888) в процентах от общего
количества (учет частоты встречаемости пищевых компонентов). Одновре-
менно с этим кормовые объекты, по их значимости в пищевом комке,
стали подразделять на несколько групп (очень мало, мало, средне, много,
очень много).
Сто лет назад (впервые К г б у е г, 1838—1853) начали производить
также и подсчет экземпляров кормовых животных, найденных в кишеч-
нике, а в дальнейшем определять и процентное соотношение различных
компонентов питания. Упомянутый выше Форбс устанавливал и от-
носительный объем содержимого кишечника. Несколько позднее стали
применять весовой анализ пищи (Marion, 1889). В самые последние
годы в методику количественного изучения питания рыб стали вводить
пересчет с количества экземпляров на вес по таблицам средних весов.
Для всех основных кормовых объектов вычисляется среднее соотношение
размеров тела и веса, а затем достаточно проводить только подсчет числа
экземпляров; перевод же на веса производится прямо по таблицам.
Этот метод, введенный В. Богоровым дает возможность вести точ-
ный весовой учет самых мелких и животных, и растительных компонентов
пищи, устанавливать общий вес съеденной пищи и устанавливать вес каж-
дой отдельной группы. Не менее важно, что, перечисляя вес пищевого комка
по указанным таблицам, мы избегаем той ошибки, которая неизбежна
в силу частичного переваривания содержимого кишечника. Наконец, эти
же таблицы используются при обработке планктона для перевода данных
по количеству экземпляров на весовые показатели. И в том и в другом слу-
чае метод пересчета с количества экземпляров на вес чрезвычайно плодо-
творен и в дальнейшем станет основным методом количественных исследо-
ваний планктона и питания рыб.
Также в середине прошлого столетия при изучении питания рыб стали
пользоваться глазомерными определениями наполненности кишечника
(пусто, слабое, среднее, сильное наполнение и т. д.), иногда в процентах
наполнения.
Большое значение для развития изучения питания рыб имели ра-
боты немецких исследователей во главе с Шименцем (Schiemenz)
и Вальтером (W al ter), относящиеся к началу настоящего сто-
летия. Работы эти, построенные на качественном анализе питания рыб и ко-
личественном анализе пищевой базы, были направлены, главным образом,
на выяснение «пищевых взаимоотношений рыб и соотношения между раз-
мерами водоема, его кормовой мощностью, с одной стороны, и размерами
рыбного стада и его приростом, с другой» (А. Ш о р ы г и н). Работы
Ш и м е н ц а (1900—1935) 1 и Вальтера (1895—1936) привели к
решению большого числа чисто практических вопросов в отношении хо-
зяйственного использования кормовой базы и особенностей отдельных про-
мысловых пород в прудовом и озерном рыбном хозяйстве.
Питание морских рыб в основном было изучено английскими и дат-
скими исследователями. При этом обращалось внимание, главным обра-
зом, на изменение питания в разные сезоны года, изменения возрастные
и локальные и на зависимость расцределения промысловых рыб от распре-
1 Шименцем была предложена также классификация видов пищи, в различной степени
предпочитаемой рыбами. В наиболее удобной формулировке ее дал в дальнейшем Б л е в а д, под-
разделивший пищу на основную, заменяющую, дополнительную, случайную и вынужденную.
21 Зенкевич, т. I
321
деления кормовых и некормовых организмов (Brook, Calder-
wood, Не rd man n, Riddel, Scott, Hicklin g, Hardy,
S a v a g e и др.).
। В результате этих работ Гарди был сконструирован особый прибор —
планктонный индикатор, а Севеджем объяснены замечательные
соотношения между сельдью и планктонной водорослью Phaeocystis.
В Дании большое число исследований по питанию донных рыб было
проведено Петерсеном и его учениками (1910—1940), в первую
очередь Блевадом и Бойсе н-И е н с е н о м. Эти исследования,
охватившие все стороны питания рыб, оказались особенно плодотворными,
так как базировались на прекрасной изученности кормовой базы и проводи-
лись путем хотя и неполного, но косвенного количественного метода.
Однако зарубежные исследования питания морских промысловых рыб
не подняли этого вопроса на должную высоту, так как не применили
точного и последовательного количественного метода. Это было осу-
ществлено у нас в большом количестве образцовых исследований.
Советские исследования питания рыб. Из работ советских авторов,
которые будут детально рассмотрены по отдельным морям (см. т. II), сле-
дует выделить работы А. Державина иН. Чугунова по пи-
танию севрюги, сельдей, воблы и других рыб Каспийского моря, проведен-
ные хотя и качественным методом, но выдающиеся по глубине и тщатель-
ности анализа, и работы А. ШорыгинаиВ. Воробьева, пред-
ставляющие собою лучшие образцы полевых исследований на основе
последовательного применения количественного метода. Исследование
А. Ш о р ы г и н а по питанию каспийских рыб может считаться высоким
образцом широко поставленного экологического исследования, охватив-
шего большой круг вопросов по использованию рыбами кормовой базы
и по анализу отношений конкуренции между отдельными рыбами.
Работа В. Воробьева дает, на основе анализа питания рыб Азов-
ского моря, прекрасно разработанную систему промысловых прогнозов.
Сейчас на всех морях Союза мы имеем целый ряд обстоятельных исследо-
ваний по питанию рыб. На Баренцовом море — это работы Л. Зенке-
вича и его сотрудников (1932) и В. Зацепина, на Азовском —
В. Воробьева (1936, 1938), на Каспийском — А. Ш о р ы г и н а
(1938, 1940), на Аральском — Г. Никольского (1940).
На основе метода учета частоты встречаемости тех или иных организ-
мов в пище рыбы было проведено несколько прекрасных исследований.
Образцом работ, проведенных по методу учета числа экземпляров отдель-
ных видов в пищевом комке, несомненно, являются работы Г з р д и по
питанию сельди. Как совершенно правильно замечает А. Ш о р ы г и н,
«основным недостатком метода учета числа экземпляров пищевых орга-
низмов, так же как и метода учета частоты их встречаемости, является
то, что он, хотя и слабее чем последний, искажает истинное пищевое
значение отдельных организмов. В его чистом виде он учитывает только
число организмов, совершенно не учитывая их хотя бы относительных
размеров. Это же неизбежно ведет к значительному преувеличению зна-
чения мелких организмов и к недоучету значения крупных. Так же как
и при учете частоты встречаемости, это искажение будет различным для
рыб с разным характером питания. Сравнение питания разных рыб, а,
следовательно, и разработка вопросов пищевой конкуренции будут за-
труднены. Затруднено будет и сравнение состава питания с составом
кормовой базы. Способ учета числа пищевых организмов для нас, как
таковой, следовательно, тоже непригоден».
Наиболее совершенным методом обработки материала следует считать
определение отношения веса пищи к весу тела как для всего содержимого
322
кишечника (общий индекс наполнения), так и для отдельных компонен-
тов пищи по группам или даже по отдельным видам (частные индексы
наполнения). Впервые отношение веса пищи к весу тела некоторых рыб
было приведено, так сказать между прочим, у Блевада (1916),
однако не было им использовано и положено в основу обработки материала.
В работах ГОИН (с 1930 г.), а затем и ряда других наших учреждений это
было сделано. За последние годы (с 1932 г.) этот метод стал применяться
и за границей.
Неоднократно разные авторы отмечали, что все рассмотренные выше
методы анализа питания рыб имеют один коренной недостаток: они опери-
руют со всеми видами пищи, как с равноценными по своему кормовому
значению. Однако это далеко не так. По содержанию скелетных неоргани-
ческих частей, по количеству жиров, белков и углеводов, наконец, по
усвояемости разными рыбами, кормовое значение различных организмов
весьма различно. Если бы мы могли получить для всех массовых кормовых
форм в водоеме (а их не так много) средние показатели калорийности, мы
могли бы перевести полевые методы анализа питания рыб на более высо-
кую ступень. В стремлении удовлетворить этой настоятельной потребно-
сти стали проводиться уже довольно многочисленные исследования по хи-
мическому составу различных массовых кормовых объектов (G е n g,
Mann, Raben, Brandt, Knauthe, Delff, Меуеги
ДР-)-
Для установления более точной оценки значимости в питании рыбы от-
дельных компонентов было введено понятие «индекса плотности» (за гра-
ницей Tester, 1932, у нас В. Б о г о р о в, 1934 и В. Воробьев,
1936, 1938), представляющего собою произведение частоты встречаемости
данного компонента в пищевом комке на относительное значение его (в %)
по объему (Tester) или весу (Б о г о р о в). Показатель этот близок
к тому, который был предложен Л. Зенкевичем и В. Броцкой
для выявления руководящих и характерных форм бентоса (корень квад-
ратный из произведения биомассы на частоту встречаемости).
Важный шаг вперед в методике полевого анализа питания рыб сделан
в результате работ А. Шорыгина (1939) по Каспийскому морю. Про-
изведенное им сопоставление сезонных карт количественного распределения
пищевых организмов с сезонными же картами интенсивности питания рыб
(по индексам наполнения), как общего, так и по отдельным пищевым
организмам, дало возможность подойти к числовому определению изби-
рательной способности и степени межвидовой конкуренции. Интенсивность
конкуренции определяется, исходя из количества пищи, потребной дан-
ной породе рыбы. Эти исследования, имеющие очень большое и теорети-
ческое и практическое значение, будут рассмотрены детально в главе, по-
священной Каспийскому морю. Совершенно очевидно, что рассмотренные
методы, представляя собой всего лишь полевые методы наблюдений, ос-
вещают еще далеко не все стороны сложного явления питания рыб, так как
дают нам только статику, почти не вскрывая динамики процесса. Не имея
показателей темпа переваривания пищи, чрезвычайно значительно из-
меняющегося в зависимости от температуры окружающей среды, мы не
можем ничего сказать о суточном рационе, который нам необходим для
учета количества пищи, потребной рыбе. Полевая методика может дать
на этот вопрос только косвенный и притом неточный ответ. Если полевая
методика вскрывает нам соотношения, существующие в водоеме, то по-
лученные применением ее результаты должны быть проверены, оценены
и уточнены экспериментальными работами. Поэтому многие авторы уже
давно стали ставить исследования в целях получения некоторых точных
показателей питания рыб, а именно: скорости переваривания, суточного
323
рациона и усвояемости пищи в разном возрасте, в разные периоды жиз-
ненного и сезонного цикла, при разной температуре и т. д. (К па u the,
1892—1907; Z и п t z, 1906—1911; Wohlgemut, 1916 и др.). У нас
эти физиологические работы были начаты Г. Карзинкиным на
пресноводных рыбах, а А. Карпевич и Е. Боковой — на мор-
ских. С другой стороны, некоторые исследователи, углубляя физиологи-
ческие анализы питания рыб, стали проводить различие между количе-
ством пищи, потребным на поддержание процессов, обеспечивающих жизнь
(поддерживающий корм), и на механическую работу, рост и образование
полезных продуктов (продуктивный корм). Эта установка, заимствован-
ная из животноводства, нашла наибольшее выражение в работе Бен-
Да¥Эса(Веп Dawes) по питанию морской камбалы. Наконец,
В. Й в л е в ы м (1938—1940) сделана весьма плодотворная попытка по-
дойти к питанию рыб с точки зрения энергетического баланса. Так. на-
пример, для карпа В. Ивлев приводит следующие данные по энерге-
тическому балансу (табл. 121).
Таблица 121
Количество поглощенной энергии...........1829 кал (100%)
Энергия прироста......................... 573 » (31,3%)
» выделений ....................... 454 » (24,8%)
» первичной теплоты.................G22 » (34,1%)
‘ > внутренней работы..................150 » <8,2°/0>
> внешней работы.................... 30 » (1,7%)
2. ОБЩИЕ ЗАКОНОМЕРНОСТИ ПИЩЕВЫХ ВЗАИМООТНОШЕНИЙ
В МОРЕ
Постановка вопроса. Питание — это одна из наиболее важных функ-
ций организма. Строение организма, его физиология и биология в силь-
нейшей степени приспособлены к захвату пищевых ресурсов. Стремление
к добыванию пищи создало сложнейшую систему взаимоотношений между
отдельными организмами как в отношении выработки средств к отыска-
нию и захвату пищи и способов защиты от хищника, так и многообразных
явлений конкуренции за однородную пищу.
Формирование этой системы взаимоотношений подчинялось некоторым
общим свойствам организмов и окружающей среды (размеры тела, темп
роста, химический состав организмов, характер кормовых веществ, окру-
жающая температура и т. д.).
Пищевые отношения и P/В коэффициент. Хорошо известно, что исход-
ной пищевой базой являются растительные организмы и что за счет их,
как продуцентов, существует вся масса животных организмов. Бактерии
и одноклеточные растения поедаются целиком, макрофиты — главным
образом в виде детрита. Животные, питающиеся другими животными,
обычно поедают более мелкие организмы, чем они сами. При этом обычно
чем крупнее организмы, тем медленнее они растут; однако это правило
может быть высказано лишь в самой общей форме, так как имеется очень
много случаев, не укладывающихся в него. Гиганты моря киты отли-
чаются изумительно быстрым ростом: они делаются половозрелыми уже
на втором году жизни. Их темп роста, учитывая их массу, неизмеримо
больше роста всякого другого многоклеточного. С другой стороны, неко-
торые моллюски и рыбы характеризуются очень медленным ростом.
С большей четкостью может быть сформулировано другое правило:
группы организмов, представляющие собою пищевую базу для других
организмов, всегда обладают, по сравнению с этими последними, бдлыпим
темпом размножения, иначе говоря — более высоким показателем Р/В,
324
и должны давать большую биомассу или обладать более быстрым темпом
размножения. В самом общем выражении пищевая цепь будет иттп в на-
правлении от быстро размножающихся бактерий и фитопланктона через
зоопланктон к зообентосу и рыбам (рис. 174) Ч Это — та же система свя-
зей, которая была выражена Элтоном в его известной «пирамиде»
питания.
Рис. 174. Пищевые взаимоотношения между различными группами морских
организмов (сплошные линии) и процессы разложения (штриховые линии) (ориг.).
Если взять только питание живыми организмами, то последовательные
звенья можно представить себе в виде четырех групп организмов (рис. 175).
В первое звено следует включить продуцентов (фитопланктон и фи-
тобентос) и бактерий. Промежуточное звено первого порядка — это жи-
вотные организмы, питающиеся продуцентами и поедаемые другими жи-
начальное
звено
Промежуточное
звено! порядка
Фитопланктон р/в около 50 Фитобентос ' Р/3 около 05 Бактерии ? Зоопланктон ?/в около /
Промежуточное
звено Л порядка
Конечное
звено
Зообентос Р/в около '/з — Рыба р/в < р
Рис. 175. Основная пищевая цепь для организмов Варенцова моря (по Зенкевичу).
вотными. В основном, это более мелкая часть зоопланктона (простейшие,
мелкие ракообразные, различные личинки). Промежуточное звено второго
порядка включает организмы, питающиеся в основном мелкими предста-
вителями зоопланктона и в свою очередь поедаемые более крупными жи-
вотными (крупный зоопланктон, мелкая рыба, более мелкий бентос).
1 На рис. 174, 175, 185, 186, 187, 190 стрелки направлены от пищевого организма и
группы к кормящемуся им организму пли группе, на рис. 178 и 179 наоборот.
325
Рис. 176. Соотношение биомассы и про-
дукции для разных групп населения Ва-
ренцова моря (по Зенкевичу).
1 — бактерии, 2 — фитопланктон, 3 — зооплан-
ктон, 4 — бентос, 5 — рыба.
Наконец, в четвертое, или ко-
нечное, звено входят те животные,
которые в живом виде обычно не
поедаются другими животными
(крупные моллюски, крупные иг-
локожие, губки, большинство ки-
шечнополостных, крупные рыбы,
большинство морских млекопита-
ющих). Если взять при этом для
каждого из четырех звеньев пока-
затель отношения продукции к
биомассе на примере Варенцова
моря, мы получим ряд чисел
(рис. 176 и 177) от 1000 (бакте-
рии) до Ve (рыбы), а для отдель-
ных пород рыб — до х/15 и даже до
1/30. Если мы будем переходить от холодного Варенцова моря к более
теплым морям, тот же показатель для четырех основных групп будет сдви*
питана? трупами
Рис. 177. Соотношение биомассы и продукции по пищевой цени
при разных типах питания (ориг.).
гаться влево, т. е. увеличиваться. Сравнение приближенных величин Р В
для Варенцова и Каспийского морей для основных групп организмов
Таблица 122
Группа организмов Приближенное значение P/В моря
Варенцова Каспийского
Фитопланктон 50 300
Зоопланктон 1 30
Зообентос 7з 0,5 0,16 4
Фитобейтос 1,0 0,5
Рыба
показывает значительное увеличение P/В коэффициента в последнем
(табл. 122).
Не лишено интереса, что эти соотношения (рис. 177) характерны
326
только при явлении свободного питания живыми организмами. Пи-
щевые соотношения конструируются так, что чем ниже по пищевой
лестнице стоят организмы, тем большей способностью к размножению
и пополнению запасов должны они обладать, чтобы обеспечить суще-
ствование питающихся ими организмов. Следует также отметить, что
в определенной степени отдельные компоненты третьего звена могут ба-
зироваться в своем питании и на первом звене, а четвертого — на втором
и частично даже на первом.
Совсем другие соотношения, как бы зеркально противоположные,
устанавливаются при паразитарном питании (рис. 177): крупные мед-
ленно растущие животные служат пищевой базой менее крупным и быстрее
растущим и, наконец, и те, и другие идут в пищу наиболее быстро размно-
жающимся микроорганизмам, причем и здесь эти последние могут, минуя
промежуточные звенья, питаться наиболее медленно растущими и круп-
ными организмами (бактерии — позвоночные). Обычно же связи более
короткие: цестоды паразитируют в позвоночных, грегарины — в беспо-
звоночных, ракообразные —на рыбах, низшие ракообразные —на высших
и т. д. При питании трупами
эта система соотношений рас-
страивается и в использова-
нии их равно участвуют все
группы.
Вероятно, во всех морях
продукция планктона превы-
шает продукцию бентоса во
много раз, и поэтому планк-
тон представляет собою боль-
шую по мощности пищевую
базу, нежели бентос. В пол-
ном согласии с этим нахо-
дится как бы «повернутость»
всей массы рыб в питании в
основном на пелагические
организмы (рис. 178). Весьма
знаменательно также и то, что
низшие группы многоклеточных животных почти не используются в
пищу другими животными. В основной своей массе губки, кишечно-
полостные и низшие черви, в смысле пищевых цепей, представляют
собою слепые ветви. Из простейших, если не считать жгутиковых, суще-
ственную роль в питании других организмов играют только инфузории.
Foraminifera и Radiolaria в основной массе—также слепые ветви. Бакте-
рии, одноклеточные водоросли и жгутиковые, составляющие в морях основ-
ную массу пищевых организмов, сохраняются от полного выедания, благо-
даря громадному темпу размножения. Foraminifera, Radiolaria, Porifera,
Coelenterata и низшие черви защищены от поедания либо массивным
скелетом, либо защитными ядовитыми выделениями. Основными кормо-
выми группами во всех морях являются ракообразные, двустворчатые
моллюски и полихеты, т. е. группы, относящиеся к наиболее высоко
организованным беспозвоночным.
Все вышесказанное объясняет те общие черты пищевых взаимоотноше-
ний в совершенно различных, казалось бы, частях мирового океана, ко-
торые можно наблюдать. Этот биологический параллелизм в пищевых ря-
дах обусловлен также и тем, что преобладающие в планктоне разных морей
группы (как, например, диатомовые или перидиниевые водоросли, илиCope-
poda) одни и те же.
Рыба
широко - п елаги чес -
кая
,. придонная
Рыба
поверхностны! слоев
______моря_________
Рыба
тесно связанная
_____с дном_____
Рис. 178. Кормовая зависимость разных групп рыбы
от пелагических организмов и от бентоса (ориг.)
бентос
и
донная
рыба
Планктон
и
пелагическая
оыба
327
3. ХАРАКТЕР ПИТАНИЯ УЗКО-ПЕЛАГИЧЕСКИХ РЫБ
Сельдь. Ни для одной пелагической рыбы питание не изучено с такой
полнотой, как для атлантической сельди (Clupea harengus). К сожале-
нию, исследования эти проведены в основном качественными методами,
с попытками придать им косвенно количественный характер. В основном
учитывалась частота встречаемости. В недавнее время прекрасные иссле-
дования по питанию сельди проведены в Полярном институте рыбного
хозяйства, в частности, работы Б. Мантейфеля (см. II том).
Сельдь — полифаг и почти все массовые формы зоо- и фитопланктона
поедаются ею, но в различном количестве. Пищевые связи, данные
Гарди на его широко известной схеме (рис. 179), показывают всю
полноту биоценотических зависимостей по линии питания. Схема пока-
Рис. 179. Пищевые связи атлантической сельди (по Гарди).
зывает изменения в питании сельди с возрастом, питание ее кормовых
форм и, наконец, врагов, питающихся ею самой. В раннем возрасте
она служит пищей медузам, ктенофорам и пелагическим хищным червям,
в более взрослом поедается китами, акулами, другими рыбами, различ-
ными птицами и вылавливается человеком. В пищевом спектре взрослой
сельди наибольшее значение имеют различные пелагические ракообраз-
ные, а затем рыбы. Из отдельных форм на первом месте стоят молодые
стадии рыбки-песчанки (Ammodytes), за ней следуют из Copepoda Тетога
и Calanus, Hyperiidae из Amphipoda и др. (рис. 180), причем в первой
половине лета (март — июнь) резко преобладает Ammodytes, а во второй
(июнь — август) — ракообразные, сначала Hyperiidae (июнь), затем Cope-
poda (июль). Возрастная смена питания сельди происходит, по Гар-
ди, в следующей последовательности: питание начинается с жгутико-
вых, диатомовых, перидиней, инфузорий, яиц различных животных,
328
науплиусов и молодых стадий Copepoda. Начиная с размеров 10—12 мм,
сельдь переходит на Copepoda (в основном Pseudocalanus elongates),
а с 30—32 мм — на Темога
longicornis. По окончании ме-
таморфоза (40 мм) основными
компонентами пищи сельди
становятся Copepoda (Centro-
pages, Eurytemora, Temora),
личинки Cirripedia и Deca-
poda, а несколько позднее
(50 — 70 мм) — Schizopoda.
При размерах около 80 мм
сельдь уже начинает захва-
тывать п мелкую рыбу.
Гарди полагает, что
сельдь обладает большой из-
бирательной способностью и
активностью в захвате пищи
и что распределение ее в зна-
чительной степени опреде-
ляется распределением планк-
тона, что было подтверждено
также нашими исследовате-
лями для Варенцова моря.
Эта связь дала возможность
разработать простую мето-
дику прогноза сельдяного промысла.
бедные планктоном, — неблагоприятный
Рис. 181. Районы цветения водоросли Phaeocystis
в Северном море и изменение путей хода косяков
сельди (показаны стрелками) (по Севеджу). Кре-
стиками показаны районы слабого развития фео-
цистис.
2, з, 4 — различная степень интенсивности цветения.
явление, имеющее для сельдяного промысла в Северном море
поистине громадное значение, было ‘подробно изучено С э в е д ж е м.
Рис. 180. Средний пищевой спектр взрослой
сельди (по Гарди).
Темнозеленая, и голубая вода,
признак для промысла; на-
оборот, серая, желтая, жел-
то-зеленая и красная во-
да, соответствующие скопле-
ниям кормового планктона,
благоприятны для про-
мысла. Особый сконструиро-
ванный Гарди прибор —
планктонный индикатор —
дает возможность, не умень-
шая хода судна, все вре-
мя облавливать планктон
и чисто визуально опре-
делять возможность кон-
центрации сельди в промы-
словых количествах. Рыба-
кам Северного моря хоро-
шо известно, что скопле-
ния сальп, диатомеи Rhi-
zosolenia sty li for mis и осо-
бенно одноклеточной водо-
росли Phaeocystis poucheti
(«вонючая вода») застав-
ляют сельдь менять ме-
сто и даже пути мигра-
ций. Это интереснейшее
329
На рис. 181 приведен пример, когда цветение Phaeocystis изменило
направление движения косяков сельди.
I Copepoda
Диатомовые
о о Яйца
о
।---1 Др . ранообразные
Рис. 182. Годовой ход питания атлантической сардины (по Гиклингу).
ХИ
Деридинеи
Моллюски
Сардина. Тщательные работы по питанию сардины (Sardina pilchar-
dus) Атлантического океана были проведены Гик ли нг ом (С. Hickling,
1945) довольно точным количественным методом (рис. 182). Составными ком-
понентами пищи сардины оказались Copepoda на разных стадиях (1),
Рис. 183. Сопоставление темпа питания (1), роста (2), зрелости (3) и жирности (4)
у атлантической сардины (по Гиклингу)-
различные другие ракообразные (2) (Ostracoda, Euphausiacea, Mysidacea
Amphipoda), диатомовые, перидиниевые, личинки моллюсков и отчасти
Pteropoda и яйца разных животных. С ноября по февраль сардина не
питается. Это — период размножения. Наиболее интенсивный откорм
приходится на апрель, май и июнь. Гиклингом сделано также сопо-
ставление (рис. 183) темпа питания сардины (1) с темпом роста (2), со-
зреванием половых продуктов (3) и степенью жирности (4). В самом
330
простом виде пищевые связи в океанической пелагиали могут быть изо-
бражены наглядной схемой (рис. 184).
Рис. 184. Упрощенная схема пищевых связей в океанической пелагиали.
I — диатомовые, 2 — перидиниевые 3 — бактерии, 4 — веслоногие, 5 — мелкая рыба, 6 — крупная
рыба, 7 — высшие ракообразные, 8 — киты, 9 — птицы, 10 — промысел.
4. ХАРАКТЕР ПИТАНИЯ ШИРОКО-ПЕЛАГИЧЕСКИХ РЫБ
Треска. В качестве примера широко-пелагической рыбы приведем,
во-первых, треску — основной промысловый объект Варенцова моря
Таблица 123
Размеры трески
Планктон 15 см 0 группа 15—30 см 1 группа 30—50 см 50 см
Основные: Cope- poda, Mysidacea Мальки и молодь рыб Сельдь Сельдь и треска
Бентос Позднее добавля- ются: Gammari- dae, Polychaeta Polychaeta, Gam- maridae, Decapoda Cumacea, Poly- chaeta. Gammart- dae, Рыбы Разные рыбы и крупные рако- образные
331
(рис. 185). Основная пища этой широко-пелагической рыбы прибрежных
частей океана — мелкая рыба, замещающая крупных пелагических ра-
кообразных. Если есть мелкая рыба (мойва и сельдь), треска будет пи-
таться ею. Дополнительная пища трески — бентос, главным образом,
придонные ракообразные и головоногие моллюски. Мелкая рыба кормится
пелагическими ракообразными (Copepoda и Mysidacea), донными ракооб-
разными (Gammaridae) и полихетами.
Как показал Поульсен (Poulsen, 1931), с возрастом харак-
тер питания трески в Северном море меняется — она как бы проходит с
возрастом от нижних частей схемы к верхним (табл. 123).
Рис. 185. Схема питания взрослой
трески Варенцова моря (по данным
Броцкой).
Рис. 186. Питание взрослого Neo-
platycephalus в австралийских водах
(по данным Дэкина).
Neoplatycephalus. Можно сравнить питание сходной по биологии с трес-
кой рыбы из других частей мирового океана. Для этого может быть ис-
пользован материал, приводимый Д э к и н ом (Dakin, 1931) для южно-
австралийских вод. Недавно там получил большое развитие траловый про-
мысел, причем основным объектом этого промысла является сравнительно
небольшая рыба (около 50 см) —Neoplatycephalus macrodon, относящаяся
к сем. Platycephalidae (близкому KCottidae). Эта весьма далекая от трески
в систематическом отношении рыба близка к ней по биологии и чрез-
вычайно сходна по характеру питания (рис. 186). Весьма замечательно при
этом то, что отдельные компоненты пищевой цепи в австралийских во-
дах другие, часто далеко стоящие в систематическом отношении от
североатлантических. Так, например, вместо наших сельди и мойвы там
обитают совершенно чуждые нашей фауне Prototroctes тагаепа, Сеп-
toscus scolop'ix, Apogonops anomalus и Anguilla australis. Иные также и
беспозвоночные, но общий характер питания тот же. Neoplatycephalas
претерпевает сходные с треской возрастные изменения питания (табл.
124).
Возрастные изменения в питании рыб подобного типа (рис. 187)
идут в направлении: одноклеточные растения и животные, личинки раз-
личных животных, мелкие ракообразные, мелкая рыба, причем обычно,
в среднем возрасте, рыба дополнительно питается за счет бентоса.
Пищевая цепь пелагиали. Прямой пищевой ряд, — диатомовые — рако-
образные — рыба — киты, — требует громадных масс пищевых резервов
каждого предыдущего звена для -каждого последующего. Кишечник
332
одной сельди может содержать более 60000 копепод, содержимое
желудка большого кита может весить несколько тонн и, если оно со-
стоит в основной массе из копепод, то в желудке кита находятся милли-
арды экземпляров Calanus finmarchicus. Обычно же беззубые киты пи-
таются в большей части Euphausiacea и Pteropoda.
Рис. 187. Пищевые цепи широкопелагических морских рыб и изменение их с воз-
растом (Gadus callarias, Merluccius vulgaris, Neoplatycephalus macrodon и др.)(ориг.).
Хорошей иллюстрацией к сказанному может служить отмеченное
Руудом (Ruud, 1929) соотношение в количестве копепод и до-
бычливости китобойного промысла в Дэвисовом проливе и отмеченное
Таблица 124
Размеры рыбы, см
0,5—2 25 -30 >35
Фитопланктон Инфузории Личинки ракообраз- ных и моллюсков Мелкая рыба (преимущест- венно пелагическая) Крупные пелагические ракообразные Головоногие моллюски Бентос Мелкая рыба (преимуще- ственно пелагическая) Головоногие моллюски
имиИортом (Ruud&Hjort, 1929) подобное же совпадение рай-
онов массового развития эвфаузиид и обильного вылова китов (рис. 188).
333
Си Lan us
— — — В. eozeaLLs
Рис. 188. Соотношение улова китов Balaenop-
tera borealis и количества Calanus finmarchi-
cus в Дэвисовом проливе в 1926 г. (по Рууд).
В приведенных примерах между обоими объектами имеется прямая зави-
симость.
Аллен (Allen, 1909) установил таким же образом косвенную
зависимость между количеством света и изобилием скумбрии (рис. 189),
так как обилие света обусло-
вливает развитие фитоплан-
ктона, а за ним следует обилие
зоопланктона, идущего в пищу
скумбрии.
Компоненты зоопланктона в
свою очередь потребляют мас-
сы фитопланктона. Гарвей
(Harvey, 1935) установил,
что веслоногий рачок Eurytemo-
ra hirundoides в течение дня
поедает не менее 120 000 мел-
ких диатомовых (Nitzschia do-
sterium).
Таким образом, для обеспе-
чения существования одного
кита в течение одного дня тре-
буются миллиарды экземпляров
ракообразных и в 100 тыс. раз
большие количества диатомо-
вых. Эта своеобразная «пира-
мида выедания» должна компен-
сироваться обратной «пирами-
дой продуктивности» — повыше-
нием скорости размножения
компонентов низших звеньев пищевой цепи.
5. ПИТАНИЕ ДОННЫХ РЫБ — БЕНТОФАГОВ
Состав пищи. Иную пищевую цепь дают типичные бентофаги, например
пикша или морская камбала (рис
Наиболее типичным бентфоагом
является морская камбала. Основ-
ные компоненты ее пищи — мол-
люски и полихеты, в небольшом
числе примешиваются ракообраз-
ные, иглокожие и рыбы. У пик-
ши преобладают иглокожие, мол-
люски, сипункулиды и полихеты,
в меныпей степени встречаются пе-
лагические ракообразные и рыбы.
Питание и нерест. У большинст-
ва рыб в период нереста питание или
ослабляется, или совсем приоста-
навливается (рис. 191 и 192), у не-'
многих рыб созревание половых
продуктов не приостанавливает
питания, например у хамсы. Как
можно видеть на рисунке, у кам-
190). Взаимоотношения здесь проще.
------Z
-----К
Рис. 189. Зависимость между количеством
солнечного света, падающего на поверх-
ность моря, и изобилием скумбрии в уловах
(по Аллену).
1— солнечный свет в феврале — марте, 11 — ко-
личество скумбрии в мае.
балы наиболее интенсивное питание происходит в марте — октябре, а
у трески и пикши — в зимний период, с июля по ноябрь. В период нереста
334
и треска и пикша, каки камбала \ приостанавливают питание полностью.
Некоторые рыбы в период зрелости избегают захватывать крупную гру-
бую пищу; так у камбалы Северного моря, в период откорма, моллюски
Рис. 191. Годовой ход интенсивности питания пикши Варенцова моря (1) (по Дехте-
ровой), камбалы Северного моря (3) (по Тодду) и трески Варенцова моря (2) (по
Броцкой).
встречены у 78,3% рыб, а в период сниженного питания, —у 26% и
тогда их замещают полихеты.
1 Впрочем, по данным Тодда, перерыв в питании в ноябре — феврале наблюдается и у
молодых неполовозрелых камбал.
335
Питание и возраст. У донных рыб также наблюдается качественное из-
менение пищи с возрастом. Тодд (Todd, 1907) иллюстрирует это яв-
ление для морской камбалы следующим образом (для марта) 1 (табл. 125).
Таблица 125
" ——______ Размеры тела в см <10 10—20 20—29 30-39 >40
Пищевые группы, ’—-— °/о встречаемости
Mollusca 17 48 61 86 95
Crustacea 57 30 25 17 0
Polychaeta 38 55 48 44 32
Echinoderm ata 16 15 6 11
G возрастом в пище камбалы увеличивается количество моллюсков,
а ракообразные и полихеты убывают, причем первые исчезают совсем,
Рис. 192. Годовой ход питания взрослой
атлантической сельди (по Гарди).
так как они ускользают от взрос-
лой малоподвижной рыбы. Как
мы видели выше, у пелагических
рыб часто с возрастом также ра-
кообразные выпадают из питания,
но там это связано с переходом
на рыбный стол.
Примером вынужденной пищи
у баренцовоморской трески могут
служить ктенофоры. Когда голод-
ная и тощая треска идет от Лофо-
тенских островов, она встречает в
юго-западной части моря массу
ктенофор и часто набивает ими
кишечник.
Суточный ритм питания. Мож-
но наблюдать у некоторых морских рыб и суточный ритм питания. Так,
Merluccius vulgarisQ^B^BOBO моря, по Гиклингу, днем не питается. У не-
которых рыб наблюдается не
с фазами приливной волны.
Питание и концентрация
рыб. Для некоторых рыб было
сделано указание на наличие
определенной зависимости
между степенью накормлен-
ности рыбы и уловистостью
промысла. Когда рыба голод-
на и гоняется за пищей, если
ее кормовые организмы ведут
пелагический подвижный об-
раз жизни, косяк рыбы может
терять нужную степень про-
мысловой концентрации. По-
добную зависимость показал
Г и к л и н г для Merluccius
вполне ясная связь интенсивности питания
Время сушам
Рис. 193. Средний суточный ход питания (Л) и
размер вылова (Б) Merluccius vulgaris (по Гик-
лингу).
vulgaris (рис. 193) и Gadus
poutassou Северного моря. То же самое было установлено работами ГОИН’а
в западной части Варенцова моря для трески (см. II том). Подобная зависи-
мость возможна для пелагических рыб, а у бентофагов она не наблюдается.
1 По другим данным Тодда для всего года.
22 Зенкевич, т. 1
ГЛАВА XIII
ИСТОРИЯ, ПРЕДМЕТ И МЕТОД БИОГЕОГРАФИИ МОРЯ
1. ВВЕДЕНИЕ
закономерностях исторического развития п современ-
ного распределения сухопутных и пресноводных мор-
ских растений и животных имеются весьма существен-
ные различия, так же как существенно различны и пу-
ти развития и достижения морской, пресноводной и
сухопутной зоогеографии.
Это определяется различиями в систематическом
составе преобладающих групп растений и животных в
трех основных средах обитания, орографическим харак-
тером контуров и рельефа материковых глыб и ложа океанической вод-
ной массы и физико-химическими условиями среды.
Биогеография оперирует с закономерностями крупного масштаба, охва-
тывающими либо всю биосферу, либо основные ее подразделения (макро-
хоры).
В этом отношении биогеография прежде всего вплотную примыкает к
экологии, изучающей закономерности распределения организмов в ос-
новном в пределах микрохоров и системы взаимоотношений между ор-
ганизмом и внешней средой (аутэкология) и между организмами друг с
другом (синэкология, биоценология). Биогеография, используя данные
экологии, основное внимание уделяет обобщающим закономерностям
распределения организмов в биосфере и основных ее подразделениях.
В учете массовых проявлений жизненных процессов в биосфере
биогеография также вплотную подходит к гидробиологии.
Все три науки тесно соприкасаются и частично перекрывают друг
друга в областях взаимных интересов.
В настоящее время нет оснований существенно изменять определение
биогеографии как науки, изучающей закономерности распределения
организмов в биосфере. Нет оснований также существенно изменять и
деление биогеографии на: 1) регистрирующую (флористика и фаунистика,
инвентаризация), 2) каузальную или аналитическую, изучающую—а) ис-
торию возникновения и развития и б) современные условия существо-
339
ванпя организмов и 3) синтетическую (биогеографическое районирование)
(Dahl, 1921,-1923; Hesse, 1924; Е k m a n, 1935).
В этом обзоре необходимо подчеркнуть, что по всем указанным
направлениям уже сделаны обобщения широко-географического харак-
тера, что эти направления характеризуются своими особенностями,
не перекрывают друг друга и каждое дает ценнейший научный материал.
Ни одно из них не является пройденным этапом и по каждому необходимо
продолжать сбор материала.
Без гармоничного развития всех этих направлений биогеография
не сможет занять то важное положение, которого она вполне заслужи-
вает не только как наука теоретических обобщений, но и как наука, со-
здающая обширный научный базис для построения системы рационального
использования растительного и животного сырья морей и океанов.
В отличие от прежних схем, в настоящее время целесообразнее реги-
ональную зоогеографию не противопоставлять остальным трем ее разделам,
а считать, что она входит в каждую из них. И регистрирующая, и
историческая, и экологическая зоогеографии в равной степени сводят
свои данные в региональные схемы.
Нельзя исчерпывать задачи зоогеографии восстановлением истории
фауны. Как ни важна эта сторона вопроса, как много ни дает выяснение
генезиса и путей формирования современной фауны, как ни тесно связано
с ними современное распространение морских организмов, однако это
последнее, совершенно естественно, имеет для нас большее значение.
Историческое развитие фауны для зоогеографии важно в меру воз-
можности объяснить им распространение и особенности современной
фауны.
Для того, чтобы яснее сформулировать современные задачи, объект п
метод зоогеографии, полезно взглянуть на историю развития морской
зоогеографии.
2. ИСТОРИЯ ИЗУЧЕНИЯ
Первые исследования. В середине XVIII столетия изобретение драги
(Marsigly et Donati, 1750) и планктонной сетки (О. Muller,
1777) привлекло внимание многих биологов к сбору и изучению морских
организмов, и в первой половине прошлого столетия стали появляться
капитальные работы (A u du in et Milne-Edwards, 1832: M. Sara
1835, E. Forbes, 1844, Schmarda, 1853), содержавшие уже много
существенных данных о закономерностях зонального (вертикального) п
географического (горизонтального) распределения морских организмов.
Так, еще в 1837 г. Мильн-Эдвардс впервые наметил региональное
деление жизни океана и различал три области европейской фауны —
Средиземноморскую, Кельтийскую и Скандинавскую, каждая, как
он полагал, со своим самостоятельным «центром творения». М. Саре
(1853) добавил Арктическую область. Ее южную границу он проводил
у Лофотенских о-вов. Этому исследователю не чуждо было уже и кау-
зально-аналитическое толкование современного распространения орга-
низмов, так же как его современнику Дана (James D. Dana).
Работы Дана. Дана, изучая распространение кораллов, показал,
что предел распространения коралловых полипов объясняется не столько
высотой средних или максимальных температур, сколько степенью по-
нижения температуры в холодное время года. Анализируя распределение
кишечнополостных и ракообразных, Дана установил следующие гео-
графические подразделения моря: •
340
зона коралловых рифов.........20°
умеренная зона................20 —1,7°
холодная зона.................^>1,7°
Каждую зону Дана подразделял на более мелкие единицы. Боль-
шое значение работ Дана для дальнейшего развития морской биогеогра-
50_____________ч0_____________30_____________20______________Ю
Рис. 194. Количественное распределение (по числу экземпляров) представителей
рода Trichodesmium (Schizophyceae) (по Вилле). Волнистая линия — северная граница
распространения. Кружки — встречаемость в очень малом количестве.
фии заключается в том, что он впервые выделил ведущие факторы в рас-
пределении организмов и последовательно проводил анализ их воздей-
ствия.
Историческая школа второй половины XIX века. Знаменитый труд
Ч. Дарвина (1859) сыграл крупную роль в развитии зоогеографии, так
как поставил в центр внимания зоогеографов необходимость исторического
341
и экологического подхода ко всем жизненным явлениям, что и сказалось
на всех последующих исследованиях, из которых упомянем работу Сем-
п е р a (Semper, 1863) по голотуриям, Г. С а р с a (G. О. S а г s,
1869) по треске, А. Бока (A. Boeck, 1871) по сельдям, А. Ага с-
с и ц a (A. Agassiz, 1872—74) по ежам, К. Г ю н т е р a (G u n t-
h е г) по рыбам.
Рис, 195. Количественное распределение (по числу экземпляров) некоторых Cope-
poda, обозначенных штриховкой разного типа (по Далю).
Уже непосредственно перед опубликованием исследования О р т-
м а н а появились еще две, важные в зоогеографическом смысле, работы
Пфеффера (Pfeffer, 1891), посвященные в основном явлению би-
полярности, и обширное исследование И. Вальтера (Walter,
1893) по биологии морских животных.
Исследование планктона. Со второй половины прошлого столетия на-
копление материалов и развитие синтезирующих обобщений в области
изучения планктона и бентоса шло в значительной степени различными
342
путями, что определялось временем введения в практику и различными ка-
чествами применявшихся орудий сбора. Применение довольно совершен-
ной количественной планктонной сетки (Hensen, 1877) сразу повело
биологов по пути количественных исследований мира пелагических су-
ществ, количественные же исследования бентоса стали возможны только
Рис. 196. Карта распределения кокколитофориды Pontosphaera Huxleyl в количестве
клеток на литр (по Ломанну).
через Зт/2 десятка лет, с изобретением дночерпателя (Е к та п, 1909 и
Ре tersen, 1909).
Собранный Гензеном (1889) во время планктонной экспедиции
на судне «Националь» количественными планктонными сетями материал
дал возможность многим биологам изобразить на картах количественное
распределение (в числе экземпляров) различных групп планктонных орга-
343
низмов. В качестве иллюстрации приводим две из большого числа подоб-
ных карт (рис. 194 и 195).
На всех этих картах по маршруту корабля «Националь» отложено к
виде треугольников количество отдельных компонентов фито- и зоопланк-
тона. К этой же группе схем можно отнести и появившуюся позднее карту
Ломан на (Lohmann, 1902) количественного распределения в Ат-
лантическом океане одной формы кокколитофорид — Pon tcsphaera Huxlem
(рис. 196), хорошо отражающую общее распределение всего атлантиче-
ского планктона.
Рис. 197. Карта фитопланктонных комплексов северной Атлантики (по Клеве)
Didymus — (О), Tricho — (Т), Styli — (S), Sira — (Si), Chaeto — (С) и Tripo (Tr) — планктон.
Большая серия работ по материалам планктонной экспедиции, по-
явившихся в конце прошлого и начале настоящего столетия (1893—1904).
несомненно, явилась первым обобщением, дававшим представление о ха-
рактере и неравномерности распределения пелагических организмов в
океане. Казавшаяся до того однородной среда оказалась в одних частях
обильно населенной самыми различными организмами, в других — исклю-
чительно бедной. По подобному типу обобщений планктонология на 2—3
десятка лет обогнала изучение бентоса, так же как и изобретение коли-
чественной планктонной сетки на 30 с лишним лет обогнало изобретение
дночерпателя.
Таким же образом впервые для планктона появились схемы выделе-
ния областей морского океана по однородному составу планктона
{рис. 197).
344
Эти биогеографические обобщения, впервые полученные для фитопланк-
тона (1897), являются не менее знаменательной датой, чем предыдущая.
Впервые районирование океанических вод было проведено по границам
Рис. 198. Районы нереста трески (по Дама).
распространения биомических* комплексов, установленных на основе*
количественного распределения.
К этой же категории зоогеографических обобщений следует отне-
сти также и такие частные обобщения, как, например, замечательную
для своего времени карту Дама (Damas, 1909) с районами нере-
ста трески (рис. 198).
Следующим крупнейшим этапом дальнейшего развития в этом же на-
правлении изучения закономерностей распределения планктона по миро-
вому океану явилась экспедиция на судне «Метеор» (1925—27), отделен-
ная от планктонной экспедиции Гензена 35 годами.
1 Термин биома введен
вокупности растительного и
Клементсом (Clements, 1916), для обозначения со-
животного населения.
345
На основании количественных сборов планктона, собранного совре-
менной техникой, Э. Гентшель (Hentschel, 1929) дает извест-
ную карту количественного распределения планктона поверхностного
слоя воды (рис. 199) для южной части Атлантического океана, но так же,
как это было и при обработке материалов планктонной экспедиции, строит
«ее по количеству экземпляров в литре воды.
<5000
5000-10000
10000-50000
50000-200000
>200000
Рис. 199. Количественное распределение (в числе экземпляров на литр) планктона
поверхностной толщи воды Атлантического океана (по Гентшелю).
Через несколько лет Гентшель (1938) на основании материалов
«Метеора» и своих более ранних работ публикует интересную теоре-
тическую схему распределения поверхностного планктона в идеальном
океане, одинаково применимую и к Атлантическому и к Тихому океану
(приведенную нами выше — рис. 86).
Более конкретную интерпретацию того же явления дают Россель
и И о н г в своей известной книге «Моря». Отличным дополнением к этой
схеме является данная В. Богоровым (1939) характеристика из-
менений вегетативного цикла планктона океана при продвижении из
346
высоких широт в экваториальную зо-
ну (рис. 200). Полярным и экватори-
альным зонам свойственен моноцик-
лический тип годового развития
планктона, умеренным зонам — дици-
клический. Одновременно с этими
широкими и весьма ценными для
дальнейшего развития биогеографии
обобщениями появляется и карта
районирования мирового океана
Штейера (Steuer, 1933) (рис.
201) по составу планктона, на 37 лет
позднее подобной же карты Орт-
манна, давшего карту зоогеогра-
фического районирования океана пре-
имущественно для бентоса. Зоогеогра-
фическая схема Штейера не явля-
ется большим шагом вперед. Она дает
мало нового по сравнению с картой
Ортманнаи, с другой стороны, по-
Рис. 200. Циклы количественного раз-
вития океанического планктона на раз-
ных широтах (по Богорову). Сплошная
линия — фитопланктон, пунктирная —
зоопланктон.
планктоном картину развития зоо-
I изучение морского бентоса; имею-
строена на основе чисто качественных
характеристик, без учета очень боль-
шого накопленного планктонологией
материала по количественному рас-
пределению планктонны х комплексов.
Исследование бентоса. Сходную с
географических представлений дает :
Рис. 201. Области распространения океанического планктона (по Штейеру).
щиеся различия ограничиваются только последовательностью развития
отдельных этапов.
А. Ортманн (Ortmann, 1896) в своей известной книге «Осно-
вы морской зоогеографии» привел карту биогеографического райониро-
347
вания мирового океана (рис. 208) на 37 лет раньше подобной же карты
А. Штейера. Карта Ортманна и по сей день сохраняет свое значение.
Все последующие схемы зоогеографического районирования мирового
океана, составленные на основе качественной оценки фауны, с большими
или меньшими изменениями повторяют карту Ортманна, на-
столько удачно она была составлена, хотя, как известно, основным
материалом автору послужили только десятиногие ракообразные. Орт-
манн придает особое значение двум моментам биологии, определяю-
щим географическое распространение: во-первых, топографической
изоляции в процессе видообразования, во-вторых, миграциям и спо-
собствующим и препятствующим им факторам. Оба эти момента выдвига-
ют вопрос о центрах формирования отдельных компонентов фауны. Пре-
пятствуют расселению факторы климатологические и топографические,
впервые четко выдвинутые в биологической литературе, посвященной мо-
рю. Схемы Ортманна (1896), П. Шмидта (1904), Э. Г е н т-
шелй (1926) показывают неустойчивость проводимых зоогеографиче-
ских границ даже применительно к наилучше изученному Атлантическо-
му океану.
В отличие от планктона, в изучении которого схемы количествен-
ного распределения на 40 лет обогнали качественную схему общеоке-
анического районирования (Ш т е й е р а), для бентоса первая схема
районирования дана именно на основе качественного распределения,
на 15 лет ранее первых обобщений, явившихся результатом коли-
чественного изучения бентоса.
Следующим очень существенным этапом в зоогеографическом райони-
ровании донного населения моря явились работы Петерсена и его
учеников (начиная с 1909 г.), составленные по данным количественного
учета донной фауны, собиравшейся при помощи дночерпателя.
Прежде всего Петерсен (1911) картировал распределение фау-
нистических комплексов бентоса, характеризируя их соответственными
количественными показателями веса и числа экземпляров. Эта карта
Петерсена вполне соответствует картам Клеве и Грана
для планктона, появившимся почти 20 годами ранее, только сделана с
применением более точного количественного метода.
В дальнейшем, используя имевшиеся сведения о распространении
фауны северной Атлантики, Петерсен экстраполировал донные
фаунистические комплексы далеко за пределы датских вод до Гренлан-
дии, Шпицбергена, Новой Земли и в Балтийское море. Конечно,
сейчас мы не можем уже считать эту карту вполне удовлетворительной,
но дело не в этом. Петерсен 35 лет назад для бентоса сделал то же, что
сделали Клеве иГран за 10—15 лет до этого для планктона. Они
дали впервые географическое районирование больших пространств моря
по биомическим (флористическими фаунистическим)комплексам,выделен-
ным на основании количественного метода.
Правильные идеи названных исследователей, однако, медленно входили
в практику сбора и обработки морского материала и в теоретические обоб-
щения зоогеографов. Только в нашей стране в течение последних 25 лет
количественные исследования морской флоры и фауны на всех наших мо-
рях приняли систематический характер и легли в основу изучения био-
логии моря. Именноу нас впервые были продолжены работы по методике
Петерсена сначала на Каспийском (1923) и Азовском (1926) морях
Н. Чугуновым, а затем многочисленными исследователями на Ба-
ренцовом, Белом, Каспийском, Азовском, Аральском и Японском морях.
Особенно высокого уровня достигли эти исследования на Азовском море
(Воробьев, 1945).
348
В дальнейшем исследования датской школы нашли своих продолжа-
телей во многих странах (Финляндии, Швеции, Норвегии, Германии,
Англии, Италии, США), но преимущественно в европейских. До настоя-
щего времени мы не имеем никаких количественных данных по распреде-
лению донной фауны у побережий Австралии, Африки (за исключением
нескольких проб в Средиземном море), Южной Америки и Антарктиче-
Рис. 202. Распространение арктических и бореальных фаунистических комплексов
бентоса северной Атлантики (по Спэрку).
Сплошные черные обозначения соответствуют арктическим комплексам, половинные — боресь
арктическим переходным, с точкой—внутри-бореальным и светлые—бореально-средиземноморским.
ского материка. За исключением датских вод и наших советских морей
нет больше достаточно обширных акваторий, по которым имелись бы более
или менее детальные данные по количественному распределению донной
фауны. Несколько лучше обстоит дело с планктоном, по до сего времени
за рубежохм в исследованиях морского планктона господствует явно по-
рочный метод количественного учета компонентов планктона только по
числу экземпляров.
В 1936 г., через 25 лет после Петерсена, Спэрк повторяет
попытку Петерсена экстраполировать фаунистические комплек-
сы датских вод далеко за их пределами, присоединяя к старым дан-
349
ным Датской биологической станции новые данные по Исландии, Грен-
ландии, Ян-Майену, Шпицбергену ~
II
Рис. 203. Соотношение бореальных,
арктических и арктическо-бореальных
видов в комплексах донной фауны юго-
западной части Варенцова моря (по
Филатовой).
1 — бореальные. 2 — арктические, 3 — арк-
тическо-бореальные.
Баренцовому морю (рис. 202).
Спэрк считает необходимым
положить в основу зоогеографиче-
ского районирования фаунистиче-
ские комплексы, установленные
количественными методами, и ра-
боту свою называет «Зоогеографи-
ческое значение фаунистических
комплексов Петерсена».
В 1938 г. 3. Филатова
чрезвычайно удачно показывает
возможность точного разграни-
чения отдельных фаунистических
комплексов, используя количе-
ственные данные (рис. 203).
Можно вспомнить также, что
в 1934 г. Ремане в своей ста-
тье о проблеме солоноватоводно-
сти границу между морской и
солоноватой зонами Балтийского
моря проводит по преобладанию
видов той или иной группы, так
что граница идет там, где и морские
и солоноватоводные формы дают
50% общего числа видов.
В наших отечественных иссле-
дованиях методика Петерсе-
н а не только была применена со
всеми деталями, но и расширена
и углублена.
Впервые именно для всего Ва-
ренцова моря (1928,1939) была да-
на карта распределения фаунисти-
ческих комплексов и биомассы
бентоса при помощи изобент
(рис. 204).
Количественное изучение дон-
ной фауны Варенцова, Белого и
Карского морей, во-первых, по-
зволило произвести подразделе-
ние на фаунистические компле-
ксы, соответствующие аналогич-
ным комплексам Петерсена
и С п э р к а. Распределение
этих комплексов (не приводимое
нами на рисунках) дало возмож-
ность провести точные границы
между бореальной и арктической
областями и подобластями послед-
ней (рис. 204). С другой
стороны, количественное рас-
пределение, впервые данное для
донной фауны обширной акватории океана (около 1.5 млн. «.и2), указывает
также, как это было показано одновременно на схемах Гентшеля
350
для планктона, что качественное [приводящее в дальнейшем к биомиче-
скому районированию (фито зоогеографическому)] и количественное
(приводящее к типологическому районированию) распределение организ-
мов совершенно не совпадаете своих границах, определяется различными
причинами и должно приводить к различным схемам районирования.
Рис. 204. Схематическое распределение биомассы бентоса Варенцова и соседних морей
и основные зоогеографические границы (по Броцкой и Зенкевичу).
1—< 25, 2— 25—100, 3 — ЮО—300. 4 — > 300 г/м'2. Штриховые линии лают границы между
бореальной и арктической областями (левая) и между всрхне- и нижнеарктическими подобластями
арктической области.
Точное количественное разграничение фаунистических комплексов
для Варенцова моря было положено в основу фаунистического райони-
рования, и таким образом прямым количественным методом Броцкой
и Зенкевичем (1939) были подтверждены зоогеографические подраз-
деления Варенцова моря, предложенные А. Шор ы г.и и ы м (1928)
351
на основании косвенного количественного учета, и внесено коренное по-
правление в прежде существовавшие схемы районирования. Нашими
работами на Баренцовом море было установлено, что районирование
по количественному распределению не совпадает с границами фаунисти-
ческих комплексов.
Таким образом, на основе количественного изучения фауны устанавли-
ваются два пути районирования: по фаунистическим комплексам и по
продуктивным свойствам. Материалы по количественному распределению
населения мирового океана выявили основной характер широтной зо-
нальности, тесно увязывающий продуктивные свойства морских организ-
мов с физико-химическими факторами океанической среды.
3. БИОГЕОГРАФИЧЕСКАЯ ЗОНАЛЬНОСТЬ МИРОВОГО ОКЕАНА
Приполярные области. Приполярные области Арктики и Антарктики1
одинаково бедны жизнью. Это обусловливается темнотой зимней ночи.
Рис. 205. Распределение полярного фронта в северной части Атлантики и районов
особо обильного скопления жизни и промысловых объектов (из Зенкевича).
постоянной низкой температурой, длительным существованием ледо-
вого покрова и, в большинстве сезонов года, наличием солевой стагнации,
препятствующей выносу на поверхность питательных солей. Это — об-
ласти очень короткого, часто очень сильного и остро выраженного веге-
тационного периода, особенно в зонах, приуроченных к кромке полярных
льдов.
1 Впрочем, относительно Антарктики мы еще не имеем точных данных о границах и даже
наличии этой олиготрофной зоны. Вероятно, она очень узка. В северном же полушарии эта при-
полярная область тянется в иных местах на 11/2—2 тыс. км к югу от полюса. В результате колос-
сальный по размерам бассейн в 10—12 млн. км2 выпадает из возможностей промыслового исполь-
зования. Мы не имеем права, однако, снимать с перспективы хозяйственных проблем будущего
человечества изменение режима арктического бассейна в целях его навигационного, промысло-
вого и всякого иного использования.
352
Умеренные зоны. К экватору от этих олиготрофных зон располагаются
в обоих полушариях пояса обильного развития жизни. Эти политрофные
зоны характеризуются значительными сезонными колебаниями темпера-
туры, обычно при отсутствии очень низких температур с хорошим летним
прогревом, бурно идущими процессами вертикального перемешивания,
выносящими на поверхность питательные соли, и длительным вегетатив-
ным периодом. Это — наиболее плодородные зоны мирового океана. К ним
приурочены и богатейшие мировые промысловые районы (рис. 205).
Экваториальная зона. От обеих политрофных зон к экватору распо-
лагаются беднейшие, в смысле биологической продуктивности, зоны ми-
рового океана. Эти океанические пустыни обязаны своим существованием
резко выраженной температурной стагнации вод, ничтожным количествам
питательных солей, возможно, постоянным очень высоким температурам
и резкому действию лучей солнца.
Почти в самой экваториальной зоне, в области экваториальных проти-
вотечений, существует весьма узкий пояс несколько повышенной продук-
тивности в зонах дивергенции тропических вод. Таким образом, можно раз-
личать 7 широтных зон мирового океана, резко различных по показателям
биологической продуктивности.
Биологическая асимметрия. Однако эти 7 зон не дают картины полной
симметрии — все соответственные зоны северного и южного полушария име-
ют разную ширину. Еще более резка асимметрия в широтном направлении.
Основная причина биологической асимметрии океана — асимметрия рас-
пределения материковых глыб и следствие ее — асимметрия океаниче-
ских течений, а также асимметрия в распределении климатических зон.
4. ОСНОВНЫЕ ПРОБЛЕМЫ БИОГЕОГРАФИИ МОРЯ
Роль количественного метода. Изучение распределения областей ко-
личественного обилия населения океана дает возможность выделить
зоны, особенно богатые жизнью, и установить, что многие участки этих
зон являются также и богатейшими районами рыбного промысла. Наме-
чаются и некоторые другие закономерности, связанные с количественным
распределением жизни в океане. Мы уже указывали на замечательное яв-
ление использования биологических выгод, предоставляемых разными рай-
онами океана в разные сезоны года наиболее подвижным компонентом
пела гнали — рыбами, китами, ластоногими и, возможно, некоторыми го-
ловоногими и ракообразными. В летний период массы этих животных
устремляются в политрофную зону на откорм, с наступлением зимы отхо-
дят обратно на размножение в более теплые, но менее продуктивные зоны.
Эта биологическая поляризованность морского нектона — одно из харак-
тернейших явлений в биогеографии океана.
К той же группе зоогеографических обобщений можно отнести и райо-
нирование комплексов объектов мирового морского промысла, удачно
произведенное в последнее время Т. Рассом (1947).
Фаунистическое районирование, первоначально проводившееся (О р т-
ма нн, Ш т е й е р, Экман и др.) на базе качественной оценки
фауны, и идущее ему на смену районирование на базе количественного
выделения фаунистических комплексов (Петерсен, Спэрк, Б р о ц-
кая и Зенкевич, Филатова и др.) не дают основания уде-
лять меньшее внимание вопросам регистрирующей биогеографии. Мате-
риал этот не только требует дальнейшего внимательного изучения, но и
дает возможность построения многих важных обобщений.
23 Зенкевич, т. I 353
Проблемы регистрирующей зоогеографии. Накопление материалов по
регистрирующей зоогеографии прежде всего должно привести к созданию
картины распределения в галосфере видового и группового состава
организмов.
Экваториальный пояс дает приют наиболее богатой качественно фауне,
высокие широты, наоборот, характеризуются качественно очень бедной
фауной. Размах колебаний очень велик — по крайней мере в 100 раз
(рис. 88).
Мы пока не можем дать объяснение этому явлению п не знаем, следует
ли искать причину в температурных различиях, в разнообразии биотопов
или в различной древности тепловодной и холодноводной фаун, связанной
с климатическими изменениями на поверхности земли.
Совершенно несомненна такая же неоднородность качественного раз-
нообразия (видового и группового) между двумя океанами, между восточ-
ными и западными частями обоих океанов, между отдельными морями и
частями морей. Известно, что арктический и антарктический планктон
богаче диатомовыми, тепловодный — перидинеями, кокколитофоридами,
фораминиферами, радиоляриями, сифонофорами, сальпами, ктенофорами,
и т. д. Так же велики различия в бентосе: известно громадное преобладание
в теплых морях крабов, морских лилий, мадрепоровых кораллов и т. д.
Темп роста, возраст, размеры тела претерпевают также в пределах гало-
сферы резкие изменения. Явления эти многообразны и разнородны. Мно-
гие морские организмы, по мере продвижения в высокие широты, увели-
чивают размеры тела и сроки жизни, особенно моллюски и ракообраз-
ные. Некоторые формы северных широт отличаются гигантизмом (коралл
умбеллула, камчатский краб, бореохитон Японского моря и многие дру-
гие). Но также хорошо известны и диаметрально противоположные при-
меры крупных размеров тела многих тепловодных форм (тридакна, каль-
мары, морские черепахи и т. д.).
Претерпевают изменения, по мере продвижения из высоких в низкие
широты, биологические циклы, темп роста и возраст, плодовитость и т. д.
и т. п.
Какие-то закономерные изменения претерпевают морские организмы
и в химизме скелетов, полостной жидкости и самого тела. Так, теплые моря
характеризуются резким повышением способности к накоплению карбо-
натов. Во всех этих отношениях океан не однороден и, вероятно, в его пре-
делах могут быть выделены различные химические провинции.
Для ответа на все подобные вопросы, представляющие собою одну из
увлекательнейших страниц биологии, потребуется громадная работа, в
результате которой возникнут многие синтезирующие обобщения райо-
нирования галосферы по качественному составу, морфологическим, био-
логическим и физиологическим особенностям и химическому составу мор-
ских организмов.
Проблема реконструкции фауны. Зоогеографическое районирование,
вытекающее из распределения фаунистических комплексов, приводит
нас, между прочим, к широкой обобщающей схеме акклиматизационных
путей в общеокеаническом масштабе. Это — как раз одно из тех зоогео-
графических обобщений, которое открывает поистине колоссальные воз-
можности рациональной реконструкции морской промысловой базы.
В основу указанной схемы можно положить существование в неритической
зоне обоих океанов 14 ныне разобщенных фаун (арктическая, антаркти-
ческая, по две амфиатлантических и амфипацифических бореальных, по
две таких же нотальных и, наконец, четыре тропических). За время раз-
дельного существования эти фауны дивергировали в различных направ-
лениях и, несомненно, дают богатейший фонд для акклиматизационных
354
переносов в сходные климатические зоны. Таких генеральных линий
в обоих направлениях можно насчитать 46.
Проблемы исторической зоогеографии. История развития морской
фауны потребует в дальнейшем не меньше кропотливой работы исследова-
теля, но уже и сейчас она дает ряд ценнейших обобщений и для открытых
частей мирового океана, и для отдельных морей, вдающихся в материки.
Экманом разработана схема путей развития четырех тропиче-
ских ныне разорванных фаун (рис. 206) из целостной тропической фауны
моря Тэтис, простиравшегося в третичное время в широтном направ-
лении от области Центральной Америки через Евразию до Тихого океана.
Несомненно, к этому же разделу зоогеографии следует отнести и раз-
работку таких явлений, как сходство фаун северных частей Атлантиче-
ского и Тихого океанов и анализ биполярного, амфибореального, амфипа-
Рис. 206. Главные области литоральной фауны по Экману.
Большие кружки обозначают тепловодную фауну (пустые — индовестпацифическую, большие
сплошные — тропическую американскую, большие полукруги — западно-африканскую). Малень-
кие—фауну умеренных и холодных зон (пустые—северную и южную умеренные зоны, сплошные—
арктическую и антарктическую фауны).
цифического, амфиатлантического, амфиарктического и иного типа гео-
графического распространения, на что в свое время обратил пристальное
внимание Л. Берг (1918—1934), который блестяще суммировал соответ-
ствующие факты и нашел им совершенно правильное объяснение в исто-
рическом прошлом.
Трудами нескольких наших морских зоогеографов (Горбунов, Гур ь-
янова, Дьяконов, Шмидт и др.) разработана история возникнове-
ния и путей формирования фауны Арктического бассейна (рис. 207).
Из отдельных морей особенно тщательно разработана история проис-
хождения и развития фауны Балтийского моря и морей южно-русской гео-
синклинали — Черного, Азовского, Каспийского и Аральского.
Большое внимание исследователей привлекает в настоящее время и
проблема возникновения и развития фаун воздушной, солоноватоводной,
пресноводной и пещерной. Если принимать, что все эти фауны генетически
связаны с морской, то очевидно, что весь вопрос в целом должен решаться
в рамках морской зоогеографии.
В связи с этим встает и другая весьма крупная проблема, пока еще
совершенно не разработанная, — истории самой океанической фауны, столь
*
355
тесно связанной с историей возникновения жизни и фазами развития земли
и самого мирового океана, как среды обитания. В этот же раздел следует
отнести и разработку важнейшего понятия биогеографии — понятия
реликта.
Рис. 207. Пути расселения арктической фауны (по Гурьяновой).
5. ПОДРАЗДЕЛЕНИЯ, ОБЪЕКТ И МЕТОД БИОГЕОГРАФИИ МОРЯ
Изложенный выше исторический обзор развития современных биогео-
графических представлений для мирового океана с достаточной ясно-
стью устанавливает объем, задачи и подразделения морской зоогео-
графии.
Задачи. Задача морской биогеографии—установление закономерности
распределения организмов в морской среде в пределах крупных ее
подразделений.
Подразделение. Накопление биогеографического материала п построение
на его основе синтезирующих обобщений возможно по трем основным тесно
связанным между собою, но не перекрывающим друг друга направле-
ниям.
1. Регистрирующая биогеография (биомическая) уста-
навливает качественный состав, морфологические, биологические, физио-
логические и биохимические особенности населения океана и его
отдельных частей и, сводя население отдельных районов в биониче-
ские (флористические и фаунистические) комплексы, приводит к схемам
биомического районирования.
356
2. Типологическая биогеография, анализируя ко-
личественное распределение всего населения океана и отдельных состав-
ных его групп, вскрывает общие закономерности и причины количе-
ственного накопления жизни в целом, отдельных групп и форм в водной
массе океана и его частей и приводит к схемам типологического
районирования.
3. Историческая биогеография, изучая генезис и пути фор-
мирования океанической флоры и фауны и их составных частей, дает
синтезирующие схемы генетической структуры.
Следующие вопросы, требующие внимательного анализа, — это до-
статочно четкое представление об основных объектах и методах зоогео-
графии.
Объект. Основным объектом биогеографии можно считать качественные
(биомические) и количественные (типологические) ареалы морских орга-
низмов, а также общих им морфологических, физиологических, био-
логических или химических особенностей для отдельного вида, более вы-
соких систематических категорий и, наконец, фаунистических комплек-
сов, т. е. пространственное распространение определенных биологических
характеристик.
Очень важной единицей биогеографического анализа является биоми-
ческий комплекс, представляющий собою качественно и количественно
охарактеризованную группировку, сохраняющую на достаточно обширном
ареале некоторое постоянство состава и структуры. Наилучшее выражение
фаунистические комплексы нашли в работах Петерсена и его уче-
ников и ряда советских исследователей.
Менее ясно пока понятие типологического ареала. Разъяснить его можно
следующим образом: каждый вид и более высокая систематическая ка-
тегория могут быть охарактеризованы не только качественным распре-
делением, но и количественно. Для отдельных видов это выполнено мною
и В. Броцкойв форме графиков, названных нами экологическими
ареалами. Ареалы такого же типа могут быть составлены и для более
высоких систематических категорий. Это осуществлено пока только нами
для полихет, моллюсков и иглокожих Варенцова моря, а затем В. В о-
робьевым — для Азовского, Куличенко — для Аральского и
Гордеевой — для восточной части Охотского моря.
Суммируя все эти частные экологические ареалы, йы получаем схемы
типологического районирования, показанные нами на примере гентше-
левских схем для планктона Атлантического океана и наших для бентоса
и рыб северной Атлантики и для Варенцова моря. Как мы уже указывали
выше, типологическое и биомическое районирование (биомические и эко-
логические ареалы) не совпадают.
В связи с развернутой концепцией мы совершенно не можем согласиться
с тем определением фаунистического комплекса, которое дает Г. Николь-
ский (1947): «группа видов, связанная общностью своего географиче-
ского происхождения».
Нет никаких оснований очень нужное и вошедшее в обиход научное
понятие наполнять новым содержанием: под фауной мы все разумеем
все животное население каждого биотопа полностью, а в состав фауны
могут входить — и почти всегда входят — генетически разнородные
компоненты. «Фаунистическим» комплексам Никольского должно
быть дано другое название, чтобы отразить в термине содержание поня-
тия и не вносить путаницу в существующие понятия.
Методы. Теперь нам остается ответить еще на вопрос, каковы методы
зоогеографии, если следовать и дальше развертываемой здесь концепции.
357
Из всего предыдущего должно быть совершенно ясно, что современная
морская биогеография оперирует двумя основными методами —качествен-
ным (регистрирующая и историческая биогеография) и количественным
(типологическая биогеография) описанием фауны. Может быть, не будет
ошибкой считать эти два метода одним качественно-количественным
методом, тем более, что в этой последней формулировке мы оттеняем
внутреннюю связь обеих оценок явления — качественной и количественной.
Я полагаю, что все изложенное выше является совершенно убедитель-
ным и решительным возражением против положения Г. Никольск о-
г о (1947), что «основная задача современной зоогеографии — это анализ
генетического состава фауны»..., а... «зоогеографическое районирова-
ние — в значительной степени пройденный этап в развитии зоогеографии,
ибо выделение крупных районов в основном закончено».
ГЛАВА XIV
БИОГЕОГРАФИЧЕСКОЕ РАЙОНИРОВАНИЕ .
1. ДЕЛЕНИЕ ОКЕАНА НА ЗООГЕОГРАФИЧЕСКИЕ ОБЛАСТИ
Схема Ортманна. Обзор современного понимания вопроса о морских
зоогеографических областях правильнее всего начать с делений, предло-
женных А. Ортманном в 1896 г. (рис. 208).
I. Литоральная зона жизни (литоральная здесь в самом широком
смысле).
1. Арктическая область.
а) Циркумполярно-арктическая подобласть.
в) Атлантическо-бореальная подобласть (с двумя локальными фау-
нами).
с) Тихоокеанско-бореальная подобласть (весьма однородная).
2. Индопацифическая область (весьма однородная).
3. Западноамериканская область (весьма однородная).
4. Восточноамериканская область (возможно, с локальными фаунами).
5. Западноафриканская область.
а) Средиземноморская подобласть.
в) Гвинейская подобласть.
6. Антарктическая область (с многими локальными фаунами).
II. Абиссальная зона жизни (без подразделения на области и подоб-
ласти).
III. Пелагическая зона жизни.
1. Арктическая область.
а) Циркумполярно-арктическая подобласть.
в) Атлантическо-бореальная подобласть.
с) Тихоокеанско-бореальная подобласть.
2. Индопацифическая. область.
3. Атлантическая область.
4. Антарктическая область.
а) Потально-циркумполярная подобласть.
в) Циркумполярно-антарктическая подобласть.
Несовпадение зоогеографических границ для литорали, пелагиали
и абиссали. Весьма важным моментом этой схемы является проведение
частично самостоятельных зоогеографических делений для фауны лито-
рали, пелагиали и абиссали. Иногда границы могут совпадать, но чаще
они не совпадают. Это явление не вызывает никаких затруднений в своем
объяснении.
359
Условия расселения организмов в абиссали и в сублиторали совсем иные,
иные и условия обитания, — и поэтому не могут быть общими и зоогео-
графические границы. Литоральная фауна северных берегов Франции и
Северного моря идет на север и восток до Белого моря, мало меняя
свой основной состав, но существенно меняется в южной части атланти-
ческого побережья Франции. Сублиторальная фауна Северного моря на се-
вер доходит только до северных частей Скандинавии, но на юг идет гораздо
далее в нижних слоях мало прогреваемой воды. Наконец, абиссальная
фауна всей центральной части Атлантического океана оказывается еди-
ной от Шетландско-Исландского гребня до Антарктики. Еще более яр-
кий пример дает Балтийское море: его глубинная часть заселена аркти-
ческими животными, а мелководная — бореальными; таким образом
360
оно в зоогеографическом смысле двуслойно. В Белом море литораль но-
сит бореальный характер, а животные, населяющие наибольшие глубины.,
имеют ясно выраженный высокоарктический характер. Таким образом,
в наиболее крайнем выражении литораль, верхний горизонт сублиторали,
ее самый нижний горизонт, и, наконец, псевдоабиссаль могут иметь раз-
личный зоогеографический характер.
Тем более, вряд ли возможно искать единых зоогеографических гра-
ниц для морской и наземной фаун. Для этого слишком различны условия
существования и расселения, а также исторические пути развития морской
и наземной фауны. Северное полушарие, более однообразное в отношении
наземной зоогеографии, дает наибольшее зоогеографическое разнообра-
зие морской фауны. Совершенно обратное соотношение имеет место в юж-
ном полушарии.
Схема П. Шмидта. Через 8 лет после выхода в свет работы Орт-
манна появилось обширное исследование русского ученого П. Шмид-
та (1904), посвященное рыбам восточных морей. Излагая взгляды Орт-
манна, Шмидт дает свою карту зоогеографического районирования ми-
рового океана и вносит в схему Ортманна существенные изменения.
Совершенно справедливо П. Шмидт считает необходимым выделить само-
стоятельную умеренную область, равноценную арктической и тропиче-
ской, т. е. возвести в ранг самостоятельной области бореальную подоб-
ласть арктической области Ортманна. Это и было принято в даль-
нейшем большинством морских биогеографов.
В целом ряде частностейП. Шмидт отступает от схемы Ортманна.
Так, большая часть Берингова и все Охотское море у Шмидта от-
носятся к Арктической области, а по Ортманну они должны быть вклю-
чены в бореальную область, с чем согласны и многие современные со-
ветские исследователи (Гурьянова, Ушаков, Щапова, Ви-
ноградов и др.). Согласно Ортманну даже восточная часть
Чукотского моря относится к бореальной подобласти, что в последнее
время признается также П. Ушаковым (1945). С другой стороны,
П. Шмидт южную половину Японского моря относит к субтропической
переходной области, что сейчас не считается правильным (Андрияшев,
1940; Щапова, 1948). Японское море и даже побережья Кореи и се-
верную часть Желтого моря сейчас относят к бореальной области. Менее
существенны расхождения биогеографических границ Ортманна и
П. Шмидта в Атлантическом океане. Ортманн отнес Средизем-
ное море к тропической области, П. Шмидт правильно изъял его
из тропической области, но склонен относить к субтропической пере-
ходной области. Однако сейчас, видимо, правильнее причислять Среди-
земное море к бореальной области.
2. СОВРЕМЕННОЕ ПОЛОЖЕНИЕ РАЙОНИРОВАНИЯ МИРОВОГО
ОКЕАНА
Дополнение к схеме Ортманна. За последнее десятилетие мысль зоогео-
графов идет различными путями. Большинство принимает с большими
или меньшими изменениями деления Ортманна. Наиболее существен-
ные дополнения к ортманновским делениям сводятся к следующему: арк-
тическую область стали подразделять на нижнеарктическую и высоко-
арктическую подобласти. По крайней мере в атлантической части Арктики
граница между этими подобластями очень резка. Баренцово море в своей
основной части принадлежит к нижнеарктической подобласти. К востоку,
за Вайгачоми Новой Землей и к северу за Шпицбергеном и Землей Франца-
361
Иосифа, простирается высокоарктическая подобласть со свойственными
ей подавляющую часть года или в течение всего года отрицательными
температурами. Многие авторы вставляют между арктической и бореаль-
ной областями еще переходную область, именуемую обычно или бореоарк-
тической, или субарктической \ В последнее время, однако, от этой пе-
реходной области отказываются. Затем оказалось, что абиссальная зона
не имеет того однообразия, какое допускал А. О р т м а н н, и ее под-
разделяют на арктическую, атлантическую, тихоокеанскую и антаркти-
ческую. Наконец, оказалось целесообразным также выделить тропиче-
скую область и к северу и к югу от нее — переходные южную и северную
субтропические. Между антарктической и южной субтропической встав-
ляют еще, по аналогии с северным полушарием, переходные: нотальную
область, или, как ее назвал Г. А у г е н е р, антибореальную и суб-
антарктическую.
Для северных частей океанов Н. Гофстен предложил очень удобную
схему деления организмов по их географическому распространению
(табл. 126):
Таблица 126
I
Арктически-
бореальные
виды
Нижнеарктиче-
ски-бореальные
виды
Бореальные
виды
Арктическая область Высокоарктическая подобласть! (зона но Гофстену) 1
Нижнеарктическая подобласть (зона по Гофстену)
Бореоарктическая (субарктическая) ' переходная область i
Высокоарктиче'
скис виды
Панарктические
виды
Нижнеарктичс-
ские виды
Бореоарктиче:
ские виды *
Бореальная область
Биогеографическое районирование пелагиали. Значительно проще зоо-
географические деления, принимаемые планктонологами. Так, Штейер
(1933) различает всего три области (рис. 201).
I. Циркумполярная арктическая область с субаркти-
ческой подобластью. В последнюю входят моря с умеренным климатом
(северная переходная область).
Субарктическая подобласть распадается на:
а) Атлантическую подобласть и
б) Тихоокеанскую.
II. Циркумэкваториальная тропическая область
распадается на:
а) Атлантическую область и
б) Индо-пацифическую область.
В состав этой области входит и субтропическая подобласть, которая
распадается на две части:
1. Северную субтропическую подобласть, состоящую из:
а) Атлантической (включая Средиземноморско-Понтическую про-
винцию) и
б) Тихоокеанской.
1 Субарктическая область К. Дерюгина полностью соответствует бореоарктической
области А п п е л л е ф а (1912) и Гофстена (1915). Бореоарктические формы, .по Г о ф-
стену, это те, которые в основном обитают в переходной бореоарктической области, а по
К. Дерюг ину—одинаково широко распространенные и в бореальной ив арктической областях.
Во избежание путаницы К. Дерюгин стал называть эти формы арктическо-бореальными.
362
2. Южную субтропическую подобласть, состоящую из:
а) Атлантической,
б) Индийской,
в) Тихоокеанской подобластей.
III. Циркумполярная антарктическая область,
в которую включается и циркумполярная субантарктическая подобласть
(южная переходная область).
Схема Экмана. С. Экман в своей зоогеографии моря дает очень
простую схему зоогеографического районирования (рис. 206), считая,
что существующая система зоогеографических подразделений и границы
между отдельными районами должны быть в корне пересмотрены. Поэтому
Экман говорит только об отдельных фаунах. Он различает для лито-
рали (= сублиторали — до 200 м) следующие фауны:
Арктическую, состоящую из панарктических (циркумполярных,
циркумарктических), высокоарктических и нижнеарктических видов.
Атлантическую бореальную с делением на европейскую, балтий-
скую и американскую.
Средиземноморско-атлантическую Са рматскую
и Северно-тихоокеанскую умеренные.
Тропическую с делением на индо-вест-пацифическую, восточно- и
западноамериканскую и западноафриканскую.
Умеренную южного полушария, южнее тропика, с
делением на южно- и юго-западноафриканскую, южноавстралийскую
и новозеландскую и перуанско-северночилпйскую.
Фауну холодных морей южного полушария с
делением на кергуэленскую, антибореальную южноамериканскую и ан-
тарктическую.
Придонную глубоководную фауну Экман, подобно Аппелёфу,
делит на атлантическую, индо-пацифическую, арктическую и антарктиче-
скую. Фауну эпипелагиали (до 200 л) Э к м а н делит на северный холод-
новодный планктон, тепловодный планктон и южный холодноводный
планктон.
Еще более проста рассмотренная выше схема зоогеографических подраз-
делений Гентшеля, сводящая их, по существу, к трем зонам.
Очень интересная схема фитогеографического районирования миро-
вого океана (по бурым водорослям) дана Т. Щаповой (рис. 209 А и Б).
Обращает на себя внимание тот факт, что границы отдельных областей
и подобластей для малоподвижных донных водорослей и наиболее под-
вижных организмов, как, например, рыбы, оказываются в общем одни
и те же.
Районы резких смен фауны. Хотя названия областей и меняются у раз-
ных авторов, но районы резкой смены фаун в большинстве одни и те же,
только их зоогеографическая трактовка иная. По побережью Северной
Америки такими пунктами являются южная оконечность Гренландии и
Нью-Фаундлендская мель, мыс Код и мыс Гаттерас. К востоку от Грен-
ландии это мелководный гребень, идущий через Исландию, Фарерские
и Шетландские о-ва. У берегов Европы и Африки эти границы не так резки,
но можно все же наметить северную оконечность Скандинавии, о-ва Ве-
ликобритании, северное побережье Африки. Такие же границы имеются
и в других частях океанов. Всегда это либо подводные гребни, либо рай-
оны высоких температурных градиентов, у которых задерживаются в своем
дальнейшем распространении как южные, так и северные формы. Граница
бывает тем резче, чем большее количество форм у нее задерживается.
В общей части мы приводили примеры районов с высокими температурными
и солевыми градиентами.
363
364
Рис. 209а. Фитогеографическое районирование мирового океана (по Щаповой).
Области мпровогр океана. 1 — Арктическая и Антарктическая, а — Бореальная и Аустральная, з — Тропическая.
365
Рис. 2096. Фитогеографическое районирование мирового океана (по Щаповой).
Подобласти Тихого океана. 1 — Северно-бореальная, 2 — Верхне-умеренпо-бореальная, 3 — Нижне-умеренно-бореальпая, 4 — Южно-
бореальная, 5 — Северно-аустральная, 6 — Умеренно-аустральная, 7 — Южно-аустральная.
По североамериканскому побережью Атлантического океана, вслед-
ствие наступления с юга теплых гольфстримных вод, а с севера — холод-
ного Лабрадорского течения, температурные градиенты высоки и имеют
место резкие смены фаун. Зоогеографические области здесь сходятся очень
близко. Сходное положение имеет место и по Тихоокеанскому побережью
Азиатского материка: на небольшом пространстве Японского моря сходятся
фауны разных зоогеографических областей. По восточным сторонам обоих
океанов границы между зоогеографическими областями расходятся по-
добно изотермам, веерообразно, и установление этих границ гораздо
труднее.
Хорошие примеры, иллюстрирующие ограниченность географических
ареалов у побережий северо-западной Европы, дают макрофиты. Наряду
Рис. 210. Распространение арктической во-
доросли Lithothamnion foecundum (по Нин-
бургу).
с настоящими космополитами,
как U Iva lactuca и Етеготот-
pha compressa из зеленых и Сега-
mium rubrum и Phyllitis fascia
идо красных, большинство
Рис. 211. Распространение арк-
тическо-бореальной водоросли
Elachista ficiola (по Нинбургу).
морских макрофитов имеет более или менее ограниченный ареал. В ка-
честве примера арктического элемента флоры можно взять известковую
водоросльLithothamnion foecundum (рис. 210). Южные побережья Исландии,
согреваемые теплыми атлантическими водами, для нее недоступны.
Некоторые арктические формы, однако, проникают далеко на юг до
Бискайского залива (рис. 211) и могут быть названы арктическо-бореаль-
ными. Умеренно северно-бореальная Polysiphonia (рис. 212) на север до-
ходит до Финмаркена, на юг — только до Бискайского залива. Типич-
ным широкобореальным видом является Bryopsis plumosa (рис. 213),
на север доходящий до северной Норвегии, а на юг — до Средиземного
моря и северо-западной Африки. Наконец, Scinaia furcelata (рис. 214)
южно-бореальная форма, доходящая на севере только до южной части Се-
верного моря.
Биогеографические критерии. Отсутствие общепринятой системы биогео-
графического районирования находит объяснение в субъективном подходе к
решению этой задачи у разных исследователей и в отсутствии каких-нибудь
точных биогеографических методов. Не разработан также вопрос о том, ка-
366
кие систематические группы должны преимущественно использоваться для
целей биогеографии — самые мелкие (подвиды, виды) или более крупные
Рис. 212. Распространение уме-
ренно-северно-бореальной крас-
ной водоросли Polysiphonia vio-
lacea (по Нинбургу).
Рис. 213. Распространение ши-
роко-бореальной водоросли Вгу-
opsis plumosa (по Нинбургу).
(роды, семейства). В последние годы уже не раз высказывалась мысль
(Спэрк, Ватова, Зенкевич) о необходимости положить в основу
биогеографического районирования точ-
ный количественный метод изу-
чения фауны. Применение этого метода к
задачам биогеографии не представляет
больших трудностей.
Если, например, фауна какого-нибудь
водоема или его части состоит по биомас-
се больше чем на 5О°/о из типично арк-
тических форм, то ни к какой другой об-
ласти, кроме как к арктической, нельзя
отнести обследуемый район.
С другой стороны, центр внимания зоо-
географы должны перенести на домини-
рующие массовые формы. «Четкие грани-
цы между зоогеографическими областями,
говорит Спэрк, легче установить,
если главное внимание обратить на фак-
тически, в природе, доминирующие фор-
мы». Иначе говоря, рассмотренные нами
во введении количественно-биоценотиче-
ские методы должны быть перенесены на
биогеографию.
Рис. 214. Распространение
южно-бореальной водоросли
Scinaia furcelata (по Нинбур-
гу).
3. ОСНОВНЫЕ ФАКТОРЫ, ОПРЕДЕЛЯЮЩИЕ РАСПРОСТРАНЕНИЕ
ОРГАНИЗМОВ В ОКЕАНЕ
Общие положения. Каждый организм стремится к расширению своего
ареала; однако топографические, физико-химические и биоценотические-
3G7
факторы ограничивают это стремление, но и те и другие с ходом времена
претерпевают изменения. В результате этих изменений создаются новые
условия и возможности для расселения. Очень существенна, например,
роль течений в распространении морской фауны, а течения — наиболее
подвижная и легко реагирующая на самые разнообразные климатические
и орографические изменения система в океане. Испытывая изменения
внешней среды, фауна также находится в постоянном изменении, она ме-
няется в своем составе и в распределении. Отдельные виды,
комплексы форм, целые фауны всегда по-разному и с разной быстротой
реагируют на изменение внешней среды. Одни виды реагируют на изме-
нение среды бурным процессом видообразования, другие — вымиранием,
третьи удивительным образом проходят как бы совершенно неизменны-
ми через ряд геологических эпох. Иногда вымирают целые фауны и остает-
ся только какая-то их часть, как было, например, с фауной Тетиса в
Сарматском бассейне или с фауной Арктического бассейна, а может быть
и всего Атлантического океана в ледниковую эпоху. Было бы, однако,
неправильным сводить всю зоогеографию к вопросу об истории фаун,
когда-то возникших, а затем только путешествующих с места на место.
Каждый водоем со всей совокупностью свойственных ему особенностей,
воздействуя на фауну даже разнородного генезиса, придает ей некоторый
единообразный облик, наиболее приспособленный к данной совокупно-
сти условий и тем более определенный, чем более длительный период
времени воздействуют на фауну эти условия.
Таким образом, каждый этап в развитии морской фауны во всем ми-
ровом океане или в отдельных его частях может рассматриваться и как
фаза развития, и как центр формирования.
Температура. Среди факторов внешней среды, обусловливающих рас-
пределение животных организмов, несомненно, наибольшее значение
имеет температура. Среди морских организмов имеются многие, ограничен-
ные весьма узкими границами температурных колебаний; практически
они существуют в неизменяющихся температурных условиях. Столь резко
выраженная стенотермия, свойственная морским животным, тем более
своеобразна, что сухопутные животные ее не знают, если не считать оби-
тателей пещер и подземных вод. Если в морях имеются организмы, при-
уроченные к высоким температурам тропической зоны, как, например, мад-
репоровые кораллы, неспособные размножаться при температуре ниже
22°, то имеются и такие, которые приурочены к температуре 1,5—1,8°
ниже нуля и не могут выйти за пределы этой температуры, как, например,
Portlandia arctica. Вся глубоководная фауна существует практически при
не меняющейся температуре 1—2°.
Не менее удивительно и то, что некоторые морские организмы, видимо,
способны сохранять свойственную им стенотермность в течение целых
геологических эпох. Незначительное изменение температурного режима
Варенцова моря в сторону потепления, измеряемое 1—2°, обусловливает
существенную перестройку его фауны — наступление тепловодной и от-
ступание холодноводной фаун.
Возможно, что организмы, существующие при неизменной температуре,
лишены физиологических механизмов, позволяющих приспосабливаться
к колебаниям температуры.
Течения. Немаловажное значение для распространения, главным об-
разом, планктонных и личиночных стадий донных организмов в океане
имеют течения. Значение течений следует учитывать в качестве фактора
не только современного распространения, но и как фактора, действовав-
шего и в прежние геологические эпохи. Согласно С. С ъ ю э л у
(S. S е w е 1, 1940), современное распространение кораллов следует объяс-
368
нять течениями моря Тетис и теми изменениями в них, которые произошли
в конце третичного времени. С ъ ю э л присоединяется к мнению К р о с-
с ленда (Grossland) о том, что бедность фауны кораллов в восточ-
ной Пацифике определяется направлением течений Тетиса с востока на
запад, препятствовавших, таким образом, заносу пелагических стадий
в Тетисе из западной части Тихого океана в восточную. Обеднение ко-
раллами Вест-Индии он также объясняет прекращением течений из Ти-
хого океана в Атлантический после гималайского и центральноаме-
риканского поднятий, положивших предел существованию Тетиса.
Еще более существенное значение имеют течения, по С ъ ю э л у,
для планктона.
Используя данные Ш т е й е р а для мирового планктона, G ъ ю э л
видит большое совпадение его распространения с существующей
системой течений не только поверхностных, но и глубинных.
Между прочим, данный автор отмечает для многих пелагических живот-
ных и донных в личиночном состоянии занос в Атлантику из Индийского
океана с юга, вокруг мыса Доброй Надежды вместе с течениями (губ-
ки, иглокожие, сальпы и др.).
Все основные потоки масс океанических вод — атлантическое течение,
восточногренландское, лабрадорское, Куро-сиво, тропические дрейфо-
вые и тропические противотечения, бразильское и все прочие — непре-
рывно несут вместе с собой свойственный им планктон. Глубинные те-
чения, двигающиеся во все три океана на север из Антарктики, также
уносят с собой обитающую в них фауну.
С ъ ю э л, кроме того, отмечает, что, благодаря суточным вертикальным
миграциям, планктеры могут попасть в иные водные массы двигающиеся
в другом, а иногда и в прямо противоположном направлении. Этим можно
объяснить известные случаи заноса в Индийский океан таких типичных
атлантических форм, как Calanus finmarchicus или Pontella atlantica.
Подобное же сопоставление течений и распределения водных масс Ат-
лантического океана с распределением шести видов батипелагических
немертин дает К у (N. R. Сое, 1946). Различные виды немертпн свой-
ственны арктическим и антарктическим промежуточным водам, средиземно-
морским в Атлантике и т. п.
Продвижение атлантического течения на северо-восток в Норвеж-
ское и Гренландское моря, а восточно-гренландского — на юго-запад,
значительно воздействует на распределение и бентоса и планктона. Осо-
бенно хорошо это может быть иллюстрировано некоторыми планктонными
формами. Гренландское холодное течение выносит далеко на юг многие
типичные арктические холодноводные формы, как Paraeuchaeta glai'ialis,
Undinella oblonga, Mertensia ovum и др. Наряду с этим многие южные
формы заносятся далеко на север к Шпицбергену и в Баренцово море.
4. ЗООГЕОГРАФИЧЕСКАЯ ПРИНАДЛЕЖНОСТЬ МОРЕЙ СССР
Существующие зоогеографические подразделения дают возможность
установить зоогеографическую принадлежность морей СССР.
Северные моря. Наши сибирские моря — Карское, Море Лаптевых
Восточно-Сибирское и Чукотское входят в состав высокоарктической под-
области Арктической области, а Баренцово море в основной своей части,
за исключением литоральной фауны, имеющей бореальный облик, —
к нижнеарктической подобласти той же области. Фауна Белого моря в ли-
торальной зоне имеет бореальный характер, в сублиторальной — нижне-
арктический, а в псевдоабиссальной — высокоарктический. В целом его
следует отнести к арктической области.
24 Зенкевич, т. I
369
Балтийское море в более мелководной части может быть отнесено
к бореальной области, а в глубинной — к арктической, в целом же —
к бореальной.
Черное море следует относить к Атлантическо-Средиземноморской под-
области бореальной области.
Дальневосточные моря. Вопрос о зоогеографической принадлежности
наших дальневосточных морей не вполне ясен. Ортманн проводил гра-
ницу между арктическо-циркумполярной и тихоокеанско-бореальной под-
областями несколько севернее Берингова пролива. В дальнейшем сравни-
тельно немногие авторы поступали таким же образом. Подавляющее
большинство, в том числе и Экман, относит Берингово и Охотское
моря и самую северную часть Японского к арктической области либо пол-
Рис. 215. Схема фаунистического районирования Берингова и Чукотского морей
(но Андрияшеву).
1—Чукотская (умеренно-арктическая) провинция; 2а— Северно-беринговский (субарктического
характера) район. Анадырский округ; 2Ь—то же, Нортонский округ; За—Восточно-намчатсная
(оэреальная) провинция, Авачинский (умеренно-бореальный) округ; ЗЪ — то же, Командорский
(умеренно-бореальный) округ; Зс — то же, Коряцкий (северно-бореальный) округ; 4 — Алеутская
провинция.
ностью, либо с некоторыми ограничениями. Г о ф с те н причислял
к нижнеарктической подобласти даже глубины Японского моря. В самое
последнее время советские исследователи (А. Ан д рияш ев, Е. Гурь-
янова, П. Ушаков) стали решительно высказываться за точку
зрения О р т м а н н а, т. е. за отнесение и Берингова и Охотского морей
к бореальной области и только частично (самая северная часть Берингова
моря) к субарктической.
Согласно этим авторам, северная часть Чукотского моря может быть
отнесена к нижнеарктической подобласти. Андрияшев (1939),
обосновывая свою точку зрения, указывает, что «для Атлантического
океана характерна его широкая непосредственная связь с арктическими
водами... обратная картина наблюдается в Тихом океане, который факти-
чески изолирован от влияния Северного Ледовитого океана: Тихий океан
был изолирован от Северного Ледовитого океана в течение большей части
его третичной и четвертичной истории... Дальневосточные моря имеют рай-
оны арктического характера (по физическим условиям), фауна которых раз-
вивалась в значительной степени изолированно от влияния северных мо-
37С
рей и имеет мало сходства с автохтонной арктической фауной, элементы
которой в северной части Тихого океана незначительны... По своему со-
ставу и характеру ихтиофауна Берингова моря является безусловно бо-
реальной (рис. 215) и лишь район северной части Берингова моря... имеет
смешанную фауну субарктического характера, наиболее близко стоящую
к западно-беринговской фауне». К такому же заключению о бореальном
характере северной тихоокеанской фауны, вплоть до Берингова пролива,
приходит иЕ. Гурьянова (1939) на примере Amphipoda и Isopoda.
Весьма интересно, что географическое районирование для макрофи-
тов (Щ а п о в а, 1947) оказывается в основном тем же, что и для живот-
ных. Для северной части Тихого океана Т. Щапова принимает те же
географические подразделения, что А. Андрияшев для рыб. Таким
образом, наиболее подвижные компоненты населения (рыбы) и наименее
подвижные (макрофиты) зависят в своем географическом распределении не
от способности к передвижению, а от распределения факторов среды,
ограничивающих их в стрехмлении к расселению.
ГЛАВА XV
АРЕАЛЫ МОРСКИХ ОРГАНИЗМОВ
1. ТИПЫ АРЕАЛОВ
Подобно растениям, подавляющее большинство животных форм, оби-
тающих в море, также заселяет только ограниченную часть мирового
океана.
Космополиты. Сравнительно небольшой процент может быть отнесен
к космополитам, т. е. расселенным по всему или почти по всему океану,
каковы, например, эвритермные космополиты — спфонофора Dimophyes
Рис. 216. Распространение сифонофоры Dimophyes arctica (по Мозеру из Экмана).
arctica (рис. 216), рачок Oithona similis, ктенофора Вегоё cucumis. Наряду
с ними имеются также космополиты, придерживающиеся определенных
температур. К их числу относится, например, Calanus finmarchicus, —
один из главнейших компонентов морского животного планктона. В холод-
372
вых водах северной Атлантики и полярных морей он является обычным
обитателем поверхностных вод, а в теплых морях живет преимущественно
на глубине. Так, частота встречаемости Calanus finmarchicus на различных
широтах следующая (табл. 127).
Таблица 127
Широта, °C Поверхностный слой о/ /0 Глубинный слой о/ /0
80—90 До 100 До 100
70—80 > 100 > 100
60—70 » 95 Нет данных
50—60 » 58
40—50 > 33,3 33,3
30—40 > 7,7 18,6
14—30 » 2,4 Не обнаружен пь
малочисленности
наблюдений
Кроме калануса, такое распространение имеют обычные в наших
северных морях: амфипода—Hyperia galba, naGhaetognatha—Eukrohnia
hainata, характерная для Тихого океана сифонофора Hippopodius serratus,
из веслоногих рачков — Rhinocalanus nasutus (рис. 217). Этот послед-
ний встречается в больших количествах в поверхностных слоях в север-
ных и антарктических водах и в глубинных — в умеренных и субтропи-
ческих областях. В районе экватора этого рачка нет. Его место заменяет
другой вид — Rh. cornu tus.
Сравнивая распространение холодноводных космополитов, мы часто
видим примеры формирования видов близнецов, особенно при наличии
разорванности общего ареала. Так, например, широко распространен-
ному в Антарктике Calanus acutus в северном полушарии как бы соот-
ветствует Calanus hyperboreus (биологические викариаты).
Формы с ограниченным ареалом. Обычно же ареалы бывают более
или менее ограниченными. Приведем в качестве примера две асцидии рода
Microcosmus — М. exasperatus и М. claudicans (рис. 218). Одна из них
занимает довольно ограниченный ареал в Средиземном море и у атланти-
ческого побережья Европы, другая широко расселена в тропических
водах всего земного шара. Очень многие формы приурочены только к од-
ному из океанов и являются эндемиками либо Тихого, либо Атлаптиче-
373
ского океана. Распространение других очень широкое, но имеет широт
ный характер. Одни из них обитают в северных или южных широтах —
панэрктические и арктическо-бореальные на севере, панантарктические
Рис. 218. Распространение двух асцидий рода Microcosmus (по Гуусу).
Черные кружки М. exasperatus, наполовину черные — М. claudicans/
и антарктическо-антибореальиые на юге, другие связаны с теплыми зо-
нами всех трех океанов (тепловодные космополиты).
Эндемики. Наконец, в фауне морей имеется более или менее значи-
тельное количество эндемиков — форм, приуроченных только к какому-
нибудь одному району или водоему. Каждый водоем имеет таких узких
Рис. 219. Распространение некоторых планктонных форм в Северном море (по Клеве).
— Triposplankton; Т — Trichoplankton; Si — Siraplankton; — Halosphaeroplankton;
Nc — Concinusplankton.
эндемиков. В нашем Белом море, с его молодой фауной, имеется, однако,
ряд эндемиков и среди них подвид трески, особая форма сельди,
своеобразная люцернария — Lucetnosa saint-hilairi и ряд других.
374
Особенности биогеографической оценки планктона. В противополож-
ность бентосу, для планктона характерна смена состава по сезонам,
с резким преобладанием в разные сезоны года разных форм. При этом
неритическому и океаническому планктону свойственны свои циклы
развития. Это обстоятельство настолько заметно, что на него обратили
внимание почти 50 лет
назад. Особенно широ-
ко это использовал в
своих работах извест-
ный фитопланктон о лог
Клеве (рис. 219),
который и предложил
называть типы планк-
тона по доминирующим
в данное время видам.
Так, им установле-
ны типы — Tri р os-plank-
ton, Halosphaerо-plank-
ton, (Jhae to-plank ton,
по внешней форме тела,
как S11/Н-plank ton, Si-
Рис. 220. Распределение типов планктона в южных ча-
стях Норвежского и Гренландского морей (по Грану).
I — район Tripos, II — район Asterionella.
ra-plankton и т. п.
Норвежский планк-
тонолог Гран для
летнего планктона Нор-
вежского моря устанавливает три района (рис. 220) с преобладанием в каж-
дом определенного типа планктона: Clio-region, Tripos-region, Asterionella-
region. Нужно только всегда иметь в виду, что эти районы весьма измен-
чивы во времени; кроме того, их границы в разные годы сильно изменяются.
2. ШИРОТНЫЕ РАЗРЫВЫ АРЕАЛОВ В СЕВЕРНОМ ПОЛУШАРИИ
Сходство фауны Атлантического и Тихого океанов. Фауны Атланти-
ческого и Тихого океанов проявляют определенную степень сходства,
которую можно объяснить только генетическим, родством. На это явле-
ние обращали внимание Ч. Д а р в и н и американский биолог Дана
еще в середине прошлого столетия. Несколько позднее А. Гюнтер
в деталях показал поразительную степень сходства фауны рыб Среди-
земного и Японского морей. В дальнейшем примеров сходства в фауне
обоих океанов накоплялось все более и более. При этом все с большей
ясностью стала выясняться относительная качественная бедность атлан-
тической фауны по сравнению с тихоокеанской и, что не менее замеча-
тельно, очень четко выступающая генетическая зависимость атлантической
фауны от тихоокеанской, отсутствие у первой из них в подавляющей части
населяющих групп самобытных черт. Очень немногие группы имеют боль-
шее видовое разнообразие в Атлантическом океане, и, видимо, в нем и
сформировались. К таким принадлежит, например, семейство тресковых
рыб, 46 видов которого свойственны Атлантике и Арктическому бассейну
и только 12 видов — Тихому океану. С. Фрейман считает, что группа
тюленей (Phocidae) имеет атлантическое, а не тихоокеанское происхо-
ждение. Тресковые и тюлени, конечно, не единственные группы, сфор-
мировавшиеся в Атлантическом океане, однако подавляющее количество
растительных и животных групп дают резко выраженное видовое раз-
нообразие в Тихом океане по сравнению с Атлантическим. Может быть,
один из самых замечательных примеров — это порядок Laminariales.
375
В Тихом океане обитает (по Т. Щаповой, 1947) 27 родов этого по-
рядка и 21 из них являются тихоокеанскими эндемиками, тогда как
Атлантическому океану свойственно только 8 родов и только один энде-
мичный род. Род Sargassum представлен в Тихом океане 174 видами,
а в Атлантическом — 20. И таких примеров можно было бы приводить
множество.
Возможно представить себе два пути обмена фаун обоих океанов —
путь южный и путь северный. Теория А. Вегенера могла бы хорошо
объяснить это явление так же, как она объясняет и ряд других зоогео-
графических загадок, но, к сожалению, она далеко не является обще-
признанной. С другой стороны, теория Вегенера могла бы дать
объяснение только сродству тепловодных, но- не холодноводных фаун.
Другое объяснение предлагает Экман. Большее богатство тихоокеан-
ской тропической фауны он объясняет тем, что в Индо-вестпацифике име-
лись наиболее благоприятные условия для пышного развития мелковод-
ной тропической фауны. С другой стороны, сравнительную бедность
Рис. 221. Распространение палтуса (Hippoglossus hippoglossns).
В Тихом океане обитает подвид Н. h. stenolepis (по Экману).
атлантической фауны Экман считает возможным рассматривать, как
результат частичного вымирания во время ледниковой эпохи. Зависимый
характер атлантической фауны от тихоокеанской П. Шмидт объяс-
няет разъединением Тихого и Атлантического океанов в области Цен-
тральной Америки только в третичном периоде (эоцен-миоцен). До этого
фауна обоих океанов должна была быть весьма схожей, а климат
океанов был равномерно тропический. С началом охлаждения у полю-
сов и дифференциацией климатических поясов в каждом океане из од-
ного и того же исходного материала появились сходные фауны умерен-
ных и холодных зон. Иначе говоря, для объяснения сходства арктической
и бореальной фаун обоих океанов П. Шмидт склонен прибегать к
допущению параллельного развития.
Подвергший это объяснение критике Л. Берг правильно указал,
что «климатические пояса дифференцировались гораздо раньше третич-
ного времени» и что объяснение П. Шмидта пригодно только для
объяснения сходства тропических фаун, но не бореальных; наибольшее
же сходство фаун обнаруживают как раз не тропические части обоих
океанов и не высокоарктические, а промежуточные северные или уме-
ренные (рис. 221). Этому явлению Л. Берг (1934) дал название амфи-
бореальности.
Подобно П. Шмидту, некоторые другие авторы также пытались
объяснять это сходство параллельным развитием.
376
Амфиатлантическое и амфипацифическое распространение. Г у у с
(Hung, 1936) установил, что из фауны асцидий северной Атлантики
Рис. 222. Амфипацифическое распространение краба Cancer amphioctus (1) (по Эк-
ману) и амфиатлантическое распространение асцидии Styela atlantica (2) и St. parti-
ta (3) (по Гуусу).
86% арктическо-бореальных видов и 9% бореальных свойственны обеим
сторонам Атлантического океана и имеют сплошной ареал. Четыре вида
Рис. 223. Амфипацифическое распространение сем. рыб Embiotocidae
(по Андрияшеву).
асцидий встречаются у берегов Америки и Европы и дают разрыв в север-
ной части. Такую форму разорванного ареала Экман (1935) назвал
амфиатлантической (рис. 222). Г у у с считает, что четыре амфиатлан-
377
тических вида асцидий обязаны своим происхождением политопии. Од-
нако подавляющее большинство исследователей находит явлениям амфи-
бореальности и амфиатлантичности вполне правильное объяснение
и не считает нужным прибегать к мало вероятному допущению парал-
лельного развития. Амфиатлантическому распространению вполне
соответствует в Тихом океане амфипацифическое распространение.
Явление это может относиться к отдельным видам (рис. 222) и к це-
лым семействам (рис. 223). Термин этот предложен А. А н д р и-
я ш е в ы м (1939), приведшим большое число примеров амфипацифиче-
ского распространения рыб и беспозвоночных.
Рис. 224. Распространение высокоарктической Portlandia arctica (по Мосевичу
из Экмана).
Причины разрывов ареалов. Наилучшее объяснение явлениям амфи-
бореальности — прерванного в северной части распространения морских
организмов, свойственных умеренным широтам Атлантического и Тихого
океанов,— сформулировано Л. Бергом (1934). Это объяснение может
быть использовано в равной мере и для объяснения явлений амфиатлан-
тического и амфипацифического распространения. Основная причина
разрыва — ледниковая эпоха. Прежде всего Л. Берг указывает, что
все формы с прерывистым распространением упомянутого типа «принад-
лежат к числу субарктических, бореально-арктических и бореальных,
а не субтропических, а тем более не тропических форм; поэтому распро-
странение вокруг южной окраины Азии для них невозможно... Обмен
378
фаунами мог происходить вдоль северного побережья Азии в те эпохи,
когда на месте Берингова пролива было море, и когда на севере
оно было теплее, чем сейчас. А это имело место: а) в плиоценовое
время, б) в теплое, послеледниковое (анциловое и литориновое время)».
Ряд новых палеонтологических данных действительно подтверждает, что
в доледниковое время (плиоцен) в районах Берингова пролива, Северной
Америки и северной Атлантики обитала гораздо более теплолюбивая и
более однообразная фауна, чем нынешняя, в дальнейшем частично вымер-
шая, частично продвинувшаяся на юг. «В плиоценовое время, — говорит
Л. Берг (1918), — средняя годовая температура воды в области Бе-
рингова пролива у поверхности была на 5—10° выше, чем теперь», тогда
как в послеледниковую фазу потепления в Скандинавии летом темпе-
Рис. 225. Распространение арктической Eunephthya rubiformis (по Броху).
ратура воды была всего на 2—2,5° выше, чем теперь. Подтверждается это
нахождением в ископаемом состоянии на Шпицбергене в послеледнико-
вых отложениях ныне вымерших здесь форм — Mytilus edulis, Littorina
littorea, Cyprina islandica, Modiola modiolus и др., а в бассейне Печоры
Cardium edule, ныне доходящего только до западного Мурмана. С другой
стороны, и современное распространение фауны в Арктическом бассейне,
дающее все последовательные переходы от сплошных ареалов к разор-
ванным самого различного типа, дает указание к толкованию явления
в целом.
Во-первых, можно выделить ряд форм, обитающих в высокоаркти-
ческой подобласти арктической области и почти не выходящих за ее пре-
делы (рис. 224). Одним из наиболее типичных представителей этой фауны
является моллюск Portlandia arctica. В Баренцовом и Белом морях он
встречается, как правило, в районах, лишенных какого бы то ни было про-
гревания, при температуре, держащейся в течение круглого года ниже —1°.
379
Рис. 227. Распространение морских тараканов рода Mesidothea
(из Экмана по Гурьяновой).
1 — М. entomon, 2 — М. slblrlca, 3 — М. sabini и 4 — М. megalura.
380
Масса подобных форм и составляет наиболее типичную часть высо-
коарктическбй фауны; другие формы, будучи столь же типичны для высо-
коарктических районов, заселяют также и нижнеарктическую зону, как,
например, кишечнополостные Eunephthya rubiformis (рис. 225). Весьма
обычно арктические формы дают разрыв в наиболее суровой по режиму
части Северного Ледовитого океана (моря Карское, Лаптевых, Восточно-
Сибирское). А. Дьяконов (1945) предлагает такие ареалы называть
амфиарктическими. Некоторые арктические формы могут при этом очень
далеко продвигаться на юг, как, например, офиура Ophiocantha biden-
tata, опускающаяся, правда, в южных широтах на большие глубины.
Она доходит на юг почти до 20° северной широты. Другой аналогич-
ный пример краб Hyas araneus (рис. 226).
Рис. 228. Распространение трех видов рыбки Artediellus в Арктике
(по Гофстену).
Кружочки —A. uncinnatus, крестики — A. scaber, штриховка — A. pacificus.
Иногда, как в предыдущих примерах, мы имеем дело с одним и тем*
же видом; большей же частью исходный вид распадается на ряд дочер-
них видов, соответственно различным климатическим зонам. Так. напри-
мер, род Mesidothea в своем составе (рис. 227) имеет высокоарктический
вид М. entomon, просто арктический М. sabini и глубоководный М. mega-
lura. Другим примером может быть рыбка Artediedlus. Ее исходный вид
распался ныне на три вида: высокоарктический A. scaber, нижнеаркти-
ческий атлантический A. uncinnatus и северотихоокеанский A. pacificus
(рис. 228). Идущий еще дальше процесс приводит к тому, что из централь-
ных частей Арктического бассейна форма исчезает и формируется амфи-
бореальный ареал, в иных случаях того же самого вида, в других — раз-
ных видов одного и того же рода (рис. 229).
На рис. 229 можно видеть, что какая-то исходная форма распалась
на несколько видов, причем один вид Chionocoetes opilio проник через
Арктический бассейн, вдоль берегов Северной Америки к ее Атланти-
ческому побережью, но не достиг, берегов Европы. Затем похолодание
вытеснило этого краба из центральных частей Арктического бассейна,
381
т. е. образовалось амфибореальное распространение; в северной же Пацп-
фике произошло меридиональное распадение на несколько видов — от наи-
более арктического Ch. opilio до наиболее тепловодного из них С/г. tanneri,
достигающего Калифорнии. Не менее показателен и пример, приведен-
ный на рис. 230 для северной части Атлантики. Типично арктическая
амфипода Themisto Ubellula (на рисунке показана южная граница ее
распространения) в северной части бореальной области сменяется Th.
compessa. Кроме того, вместе с Th. Ubellula в южной части ареала обитает
глубинный вид Th. abyssorum^ очень характерный для глубинной зоны
Норвежского и Гренландского морей.
Рис. 229. Распространение крабов рода Chionocoetes (из Экмана).
Черные кружки —Ch. opilio, белые — Ch. an^ulatus, кружки с крестиком — севернее 50е сев. шир.
Ch. bairdi — южнее Ch. tanner!.
Формы, оттиснутые похолоданием в северной части того или иного
океана, иногда имеют целостный ареал, захватывающий берега и Америки
и Евразии. В качестве такого примера можно привести краба Oregonia
gracilis (рис. 231), заселяющего все северное тихоокеанское побережье
от южных частей Японского моря до вод Калифорнии. Идущий еще дальше
процесс приводит к распадению первоначально единого вида на несколько
и к формированию явления амфиатлантического или амфипацифического
распространения как отдельных видов, так и более высоко стоящих систе-
матических групп.
В качестве примера бореальной формы, правда, далеко продвигаю-
щейся на север, можно привести краба Hyas araneus, доходящего на юге
до берегов Бретани (рис. 226).
Амфибореальная, амфипацифическая и амфиатлантическая фауны дают
прекрасный пример дивергирующих групп при наличии изоляции в двух
или нескольких частях разорванного, первоначально целостного, ареала.
Достаточно возникнуть такой изоляции, чтобы вслед за этим процесс
видообразования стал создавать систематическую обособленность, в основ-
ном тем более резкую, чем более длителен период изоляции. Еще Л о в е н
382
Рис. 230. Распространение Themisto abyssorum (белые кружки) и Th. compressa
cornpressa (черные кружки) (по Броху).
Рис. 231. Распространение краба Oregonia gracilis в Северной Нацпфике (по Экману)^
383-
'(Love n, 1846) обратил внимание на то, что бореальные европейские
виды в большинстве не обитают у американского побережья, а арктиче-
ские — свойственны обеим сторонам Атлантического океана. Из аркти-
ческо-бореальных моллюсков, известных в то время, 75% оказываются
общими для обеих побережий, тогда как из преимущественно бореальных
и бореальных у Америки живут только 8%. Это установленное Л о в е-
н о м правило в дальнейшем было подтверждено почти для всех других
литоральных и сублиторальных групп. Как можно видеть из табл. 128
(по Экману), эндемизм фауны резко возрастает с севера на юг, при-
чем родовой эндемизм выражен гораздо слабее видового. На этом примере
мы также видим степень эндемизма как результат разобщенности.
Рис. 232. Распространение видов Eisenia и Pelvetia (по Щаповой).
1 — Eisenia blcyclus. 2 — Е. arborea, 3 — Е. Cooker!, 4 — Е. Masonii и Е. desmarestioides,
5 — Pelvetia frisigiata, 6 — Р. Wright!!, 7 — Р. canaliculata, 8 — Е. galapagensis.
Таблгща 12$
Фаунистические группы Роды и виды европейской бореальной фауны, встречающиеся у берегов Америки, °/0'
Decapoda natantla Decapoda reptantla Асцидии Рыбы
виды виды виды виды роды
Арктическо-бореальные 70—100 ICO 88 70 93
Субарктическо-бореальные Бореальные к 90 16 12 80—90 100
Средиземноморско-бореальные Средиземноморско-лузитанско-южяо- 1 И 9 15 56
бореальные : 0 G 0 5 38
Подчиняясь тем же закономерностям, многочисленные случаи амфи-
бореальности, амфиатлантичности и амфипацифичности дают и морские
водоросли — макрофиты. В качестве примера можно взять род Eisenia.,
относящийся к бурым водорослям (рис. 232). В северных частях Тихого
-384
океана ареалы этого рода, по Т. Щ а п о в о й, разорваны. В северной
Атлантике (по Европейскому побережью) обитает один вид Pelvetia
(Р. canaliculate,), а в Тихом океане — остальные два.
3. БИПОЛЯРНОСТЬ
Биполярное распространение организмов. Уже около ста лет биоло-
гам известно явление биполярности. Под этим понятием разумеют такой
тип меридионального распространения, когда данный вид, род или группа
более высокого таксономического значения встречается в умеренных
широтах обоих полушарий и отсутствует в промежуточной теплой зоне.
Впервые на это явление обратил внимание Джемс Росс и за
ним Дана, Гукер, Форбс. Своеобразным сходством фауны
южного умеренного пояса с фауной берегов Европы были поражены участ-
* Рис. 233. Биполярное распространение бурой водоросли Macrocystis pyrifera
(по Щаповой).
ники Челленджеровской экспедиции. Так, Вайвилль Томсон
(VV. Т о m s о п), пишет: «Эти драгировки на мелководьи вокруг Тристан-
да-Кунья дали большое количество материала, причем фауна была
почти такого же характера, как и фауна несколько более мелких вод на
севере».
Биполярный тип разорванного географического ареала распространен
чрезвычайно широко и среди растений, и среди животных, и у наземных,
и у морских организмов. Чрезвычайно важным моментом является то,
что основная масса биполярных организмов относится не к высокоаркти-
ческим формам, а к субарктическим и бореальным и соответственно суб-
антарктическим и нотальным. Представление о биполярном типе распро-
странения могут дать рис. 233 и 234, первый для вида, второй — для
рода. В обзорных работах, посвященных этому вопросу (К. Дерюгин,
1915, 1927, Л. Берг, 1920, С. Экман, 1935), приводится большой
конкретный материал. Биполярные формы встречаются среди тюленей,
котиков, китов (не менее 12 видов); очень много их среди морских рыб.
Биполярны анчоусы, кильки, сардины, акулы и др. Среди биполярных
25 Зенкевич, т. I 385
форм имеются свойственные обоим океанам или только одному из них.
Есть виды, биполярные в одном океане и имеющие сплошной ареал вдру^
гом (рис. 235).
Очень ярко выражена биполярность у морских и особенно у бурых
водорослей.
Согласно Т. Щаповой (1946), среди бурых водорослей имеются
и порядки и семейства, и роды и виды, характеризующиеся биполярным
распространением. Некоторые виды образуют близкие подвиды в обоих
полушариях, как, например, Laminaria pallida и L. iberica, Chordaria
jlagelUfprmis и Ch. magellanica, Lessonia laminariaeoides и L. ovata и ряд
других.
MB
В известном примере с Priapulus мы также имеем два подвида: типич-
ный на севере Р. caudatas и его подвид Р. tuberculato-spinosus в Антарк-
тике.
Рис. 235. Распространение акулы Alopias vulpes.
В умеренных зонах обоих полушарий господствуют Fucaceae, а в тро-
пиках— Sargassum. Порядок Laminariales целиком биполярен (рис. 236).
Биполярность порядка ламинариевых может быть показана и в
табл. 129 (Щ а п о в а, 1947).
Таблица 129
Всего Северное полушарие Южное полушарие
северная часть Тихого океана северная часть Атлантического океана Арктика
родов видов родов ВИДОВ родов ВИДОВ родов видов родов ВИДОВ
30 130 27 84 6 27 6 25 1 6 22
100% 100% 90% 65% 20% 21% 20% 19% 1 20% п%
В том числе эндемиков
| 19 | 68 | 0 | 10 | 1 | 11 | 2 | 17
Таблица 130
МЕРИДИОНАЛЬНОЕ РАСПРОСТРАНЕНИЕ ВИДОВ РАЗНЫХ СЕМЕЙСТВ
ОТРЯДА CUMACEA ПО ЦИММЕРУ
Семейство Арктиче- ские + суб- арктические Бореаль- ные Тропиче- ские Ноталь- ные Антаркти- ческие + субантаркти- ческие
Bodotriidae Leuconidae Nannastacldae Lampropldae Dlastylidae 2 16 10 9 31 23 18 26 10 45 26 0 27 2 9 19 8 5 3 18 7 11 7 5 11
Все Cumacea 68 123 74 54 41
387
Особенно много биполярных форм среди беспозвоночных. К. Д е-
р ю г и н, тщательно собравший весь материал, приводит около ста бипо-
лярных форм. Из них очень показателен отряд Cumacea. Согласно Ц и м-
м е Р У (Zimmer, 1913), этому отряду в целом несвойственна бипо-
лярность, но отдельные его семейства дают весьма различную картину:
Leucontdae типично биполярны (табл. 130), Bodotriidae — группа тепло-
любивая, остальные семейства не проявляют определенного предпочте-
ния тепловодной или холодноводной зоны.
388
Биполярных видов сравнительно немного, но они имеются часто
и среди наиболее вульгарных форм, каковы, например, сипункулида
Phascolosoma margaritaceuin, приапулида Priapulus caudatus, полихеты:
Tomopteris septentrionalis, Terebellides stromi, Maldane sarsi, Fabricia
sabella, Pectinaria belgica, ракообразные: Microcalanus pygrneus, Metridia
longa, Podon leuckarti, Evadne nordmanni, Balanus porcatus, Nebalia, Thy-
sanoessa gregaria, моллюски: Mytilus edulis, Limacina helicina и многие
другие.
Происхождение биполярности. Явление биполярности в целом вряд
ли может подвергаться в настоящее время сомнениям. Однако некоторые
ученые его отрицают. Так, отрицал биполярное распространение Д о л л о
(Doll о, 1904), а затем в недавнее время С т и а с н и (S t i a s п у,
1934). Ошибка обоих исследователей, как отмечает Л. Берг в отно-
шении первого из них, заключается в том, что они сравнивали фауны высо-
коарктических областей, а не более умеренных.
Гипотезы, пытающиеся объяснить явление биполярности, можно раз-
делить на три группы. Во-первых, ряд авторов пытается объяснить его
политопным развитием сходных форм в сходных условиях приполярных
областей, из единого исходного материала. Дана (1854) связывал
это явление с самостоятельными божественными творческими актами.
Эттингаузен (Ettinhausen, 1894) и частично Экман
(1935) склонны искать объяснение биполярности в политопном развитии
видов, родов и т. д. из единого исходного материала.
Экман рассматривает явление биполярности, как один из видов
прерванного распространения, подобного амфибореальному или амфи-
атлантическому. Биполярность дает разрыв ареала в меридиональном
направлении, амфибореальность — в широтном. Экман считает, что
всякие случаи разорванного ареала необходимо рассматривать, учитывая
возможности политопного образования. Начальную фазу политопного
развития биполярных форм могут давать виды со сплошным рапростра-
нением и сходными вариететами в северной и южной холодных зонах.
Так, например, радиолярия Protocystis harstoni обитает в экваториальных
областях на глубинах 400—1500 м, а в холодных областях обоих полу-
шарий поднимается и в верхние слои, но только уже в виде «биполярного»
вида Р. sloggetti, который, видимо, произошел от материнской Р. hars-
toni, дав и на севере и на юге однотипную морфологическую разность,
именуемую как Р. sloggetti.
Таких примеров Экман приводит несколько, а затем говорит,
что «родство видов и вариететов, обозначаемых как биполярные, не пря-
мое, но устанавливается через общую исходную тропическую форму.
Когда же идущее дальше развитие отводит все больше и больше северные
п южные формы, но в одну сторону от тропической материнской формы,
создаются в конце концов хорошо обособленные виды, соответственно и ро-
ды, имеющие больше сходства между собой, нежели с исходной формой».
В этом процессе, по мнению Экмана, невозможно отделить биполяр-
ности параллельных биологических явлений от биполярности таксо-
номически близких групп. Таким образом, Экман приходит к широ-
кому признанию возможности политопного генезиса форм при разорван-
ном ареале.
Во вторую группу надо отнести гипотезы, дающие биполярности
реликтовое объяснение (Т е е л ь, П ф е ф ф е р, Мюррей, Д е р ю-
г и н). Согласно этим теориям, биполярность развилась из космополи-
тизма путем вымирания исходных форм в тропическом поясе и сохранения
их в виде реликтов в высоких широтах обоих полушарий. Одни из сторон-
ников реликтовой теории полагают, что прежняя космополитическая
389
фауна изменилась или вымерла в тропическом поясе, так сказать, еди-
новременно, при формировании климата. По мнению других, с охлажде-
нием приполярных областей вымерла населявшая их фауна и эти области
вновь заселились глубинной фауной океана. Наконец, К. Дерюгин
считает, что формирование биполярной фауны есть длительный процесс,
связанный вообще с постепенным исчезновением космополитизма, ранее
имевшего более универсальный характер.
Наконец, третью группу составляют миграционные гипотезы (Росс,
Ортманн, Л. Берг). Ортманн полагал, что обмен между
фаунами высоких широт постоянно происходит через глубины и вдоль
западных побережий Америки и Африки, где температура воды подвер-
гается некоторому охлаждению. В связи с этим Ортманн вообще
склонен был считать прерывистое распространение биполярных форм
нереальным.
Правильную критику всем изложенным выше взглядам и наилучшее
объяснение происхождению биполярности дал Л. Берг. «Биполяр-
ность. — говорит Л. Берг, — есть следствие ледниковых эпох. В лед-
Рис. 237. Биполярное распространение красной водоросли Iridaea cordata
(по Фрич).
никовое время охлаждением были затронуты не только арктические
и умеренные широты, но и тропики. Тогда же многие северные формы
нашли путь к экватору, а затем и в южное полушарие. Когда ледни-
ковое время прошло, в тропиках температура снова поднялась, и
проникшие сюда северные формы должны были здесь или вымереть, или
удалиться к северу и к югу. Таким путем создалось прерывистое рас-
пространение... с этой точки зрения понятно, почему явления биполярно-
сти обнаруживаются только среди форм умеренной, но не арктической зо-
ны: понижение температуры в тропиках было не настолько значитель-
ным, чтобы позволить обитателям полярных областей пересечь
экватор». Л. Берг отмечает, что сходную точку зрения вы-
сказывал еще Ч. Дарвин применительно к явлениям «би-
полярного» распространения у наземных растений. Подтвержде-
ние правильности своих взглядов Л. Берг видит в том, что
по побережьям Средиземного моря, на берегах Сенегала и в других местах
обнаружены морские четвертичные отложения с ископаемой фауной север-
ного характера, не свойственной этим местам в настоящее время. Очень
важное соображение в пользу своего толкования Л. Берг усматривает
в том, что, по его мнению, биполярные организмы имеют северное про-
исхождение, т. е. миграции морских животных в ледниковые эпохи шли
от северного полушария к южному.
390
Однако есть много примеров и обратного проникновения. Очень хоро-
шим примером формирования биполярного ареала формой южного полу-
шария является красная водоросль Iridaea cordata (рис. 237). Путем
весьма детального анализа распространения и морфологии биполярной
флоры макрофитов Т. Щапова (1946) приходит к заключению, что
для многих бурых водорослей, имеющих биполярное распространение,
родиной является Антарктика. К таким следует относить Macrocystis
pyrijera и Lessonia laminariaeoides, как и весь род Lessonia. Данный автор
склонен миграцию многих форм южного генезиса относить еще к третич-
ному времени (эоцен-олигоцен). К совершенно аналогичным выводам
пришел ранее А. О к с н е р (О х п е г, 1944) применительно к лишай-
никам. Он считает, что обусловившие биполярное распространение мигра-
ции северян к югу и южан к северу не совпадают во времени. С севера
на юг переселение происходило в четвертичное время, а в обратном на-
правлении — в третичное. При этом большая часть биполярных лишай-
ников — северяне, меньшая — южане.
Интересно отметить еще и следующее. По Скотсбергу (Scotts-
b е г g, 1907), бурые водоросли представлены в донной флоре Антарк-
тики и Субантарктики 44 родами. Число родов, общих с северным полу-
шарием, составляет 55%: из общих родов только один Lessonia представлен
большим числом видов в южном полушарии, 11 отличаются большим видо-
вым разнообразием в северном, остальные представлены примерно оди-
наковым числом видов в обоих полушариях. Согласно Т. Щаповой
(1946), эти данные служат доказательством того, что донная флора южного
полушария содержит гораздо более значительное число северных имми-
грантов, чем северная флора —южных.
Гидрологический режим океана и некоторые особенности современ-
ного распространения макрофитов позволяют думать, что основным путем
переселения были западные побережья обеих Америк и в меньшей степени
Африки и Европы.
Анализируя биполярную фауну, Л. Берг устанавливает, что для
некоторых рыб биполярность простирается не на виды или роды, а на
семейства и даже группы семейств (табл. 131).
Таблица 131
Северное полушарие Южное полушарие
Osmeridae
гр. Salmonoidei
Gadidae
Pelromyzonidae
Rctroplnnldae
Haplochitonidae
Galaxltdae
Muraenolepldae
Geortla, Mordacia
Л. Берг полагает, что здесь мы также' имеем явление биполяр-
ности, но гораздо более древнего происхождения (чем четвертичное оле-
денение). Такие оледенения были и в предшествующие геологические
эпохи.
С. Экман (1935) весьма справедливо замечает, что в конечном счете
миграционные теории биполярности являются в то же время и релик-
товыми.
Биологическая биполярность. Явление биполярности не ограничи-
вается только географическим распространением организмов, оно распро-
страняется на большой круг явлений, обусловливаемых как распреде-
лением температур, так и некоторой-симметрией в распределении системы
391
циркуляции водных масс в северном и южном полушариях по обе сти-
роны от экватора.
Это определяет, как уже указывалось выше, повышение показателей
биологической продуктивности в умеренных широтах, в особенности в
зонах дивергенции, что особенно ярко сказывается в распределении
планктона.
G удалением от экваториального пояса к северу и югу и понижением
температуры темп роста и все процессы метаболизма замедляются, а про-
должительность жизни соответственно увеличивается.
У и м п е н н и (W i m р е n п у, 1941), подробно рассматривая «би-
полярность параллельных явлений)), отмечает прежде всего увеличение
числа особей, размеров тела, уменьшение толщины раковин и большую
заботу о потомстве у животных по мере удаления от экватора. В холод-
ных и умеренных зонах роды содержат большее число видов, чем в тропи-
ках и субтропиках. По мере удаления от экватора возрастает процент
жира (рис. 238). Таким же образом в зимнее время идет более энергичное
накопление жира, и весной оно наибольшее. Малое накопление жира
у организмов из теплых морей стоит в соответствии с большим накопле-
Рис. 238. Процент жира в сухом планктоне, собранном
у поверхности на четырех точках на разной широте
северного полушария (по Уимпенни).
нием углекислого кальция, что очень характерно для животных тропи-
ческой зоны. Это явление свойственно макрофитам — в тропических
областях обильно представлены известковые водоросли, в том числе и
так называемые нуллипоровые «кораллы».
4. ТРОПИЧЕСКАЯ ФАУНА
Общая характеристика и происхождение. Фауна тропических морей
издавна привлекала внимание биологов исключительной качественной
пышностью, удивительным многообразием форм. Особенно поражает богат-
ство фауны тропиков, если ее сравнивать с фауной умеренных и холодных
морей; однако это богатство будет понятно, если принять во внимание
большую древность тропической фауны, сравнительно небольшие клима-
тические смены, имевшие место в тропической зоне в течение последних
геологических периодов и громадность морских просторов, особенно при-
брежных мелководий тропического пояса западной части Тихого океана.
Югославский ученый И. Хаджи (I. Hadzi, 1932) склонен объяс-
нять обилие фауны теплых морей биотопическим разнообразием, а также
высокой температурой воды.
Детальный зоогеографический анализ тропической фауны дан в не*
давнее время шведским зоогеографом Свеном Экманом (1932.
392
1934, 1935). Экман допускает, что все другие фауны мирового океана
в конечном счете ведут свое начало от тропической фауны. При всем том
при изучении тропической фауны ярко выступают ее генетическое един-
ство, гомогенность и вместе с этим ее циркумэкваториальность (циркум-
тропичность), т. е. сохранение того же единого фаунистического типа
во всех частях тропического пояса, хотя в настоящее время она разделена
материковыми массивами на четыре разобщенные фауны.
Очень большое количество родов, семейств и групп более высокого
систематического порядка либо целиком, либо почти полностью огра-
ничены в своем распространении тропиками. При этом особенно бросается
в глаза, что явление циркумтропичности ярко выражено не только у групп
более высокого систематического порядка, но и у отдельных видов. Все
это с несомненностью указывает на то, что тропическая фауна до недав-
Рис. 239. Циркумтропическое распространение Holothuria atra и ее
вариететов (по Паннингу из Экмана).
него времени существовала как единое фаунистическое целое, а то разъеди-
нение ее, котороемы имеем в настоящее время, образовалось уже в послед-
ние геологические периоды.
В качестве примера циркумтропического вида можно привести голо-
турию Holothuria atra с ее вариететами (рис. 239), замечательных
крабиков Planes minutus, Grapsus grapsus, Plagusia depressa, рыбу Mu-
gil cephalus^ черепах-каретт (Chelonia, Caretta) и многих других, часто
даже не образующих подвидов в различных частях тропической области.
Историю тропической фауны Экман связывает с историей моря
Тетис. В настоящее время материки имеют меридиональное протяжение,
так же как и океаны. Еще до середины третичного времени распределе-
ние моря и суши было иным. Океаны в основном имели широтное протя-
жение. В верхнемеловом периоде (рис. 240) море Тетис тянулось через
Центральную Америку, Атлантической океан, область Средиземноморья
и дальше на восток, а позднее находилось в широкой связи и с Тихим
океаном. Экман считает, что именно в этом море Тетис и создалась
та тропическая фауна, с которой мы сейчас имеем дело. Только в середине
и в конце третичного периода закончился процесс обособления тихоокеан-
ской и атлантической частей тропической морской области.
Экман различает четыре основных дочерних прибрежных фауны
от фауны Тетиса (рис. 206): основная индо-вестпацифическая, восточно-
393
тихоокеанская, западно-атлантическая и восточно-атлантическая. В на-
стоящее время эти четыре района в отношении донной фауны почти пол-
ностью изолированы друг от друга либо материками и глубинными ча-
стями открытых океанов (в широтном направлении), либо умеренными
и холодными зонами (в меридиональном направлении).
Рис. 240. Распределение суши и моря во второй половине мелового периода
(по Шухерту).
Коралловые рифы. Наиболее характерная особенность верхнего гори-
зонта сублиторали тропической зоны к экватору от параллелей 30° — это
коралловые рифы, особенно сильно выраженные в западной части Тихого
океана, в водах Ост-Индии и в Индийском океане. В Атлантическом океане
они приурочены в основном к водам Вест-Индии и Центральной Аме-
Рис. 241. Распространение коралловых рифов в мировом океане (по Экману).
рики (рис. 241). По восточным сторонам океанов настоящие коралловые
рифы отсутствуют: они вытесняются холодными водами, подступающими
в этих зонах из глубоких частей океана. Коралловые рифообразователи
могут развиваться только в водах со средней годовой температурой не
менее 23,5° и не опускающейся ниже 20,5° и на глубинах до 50 м; лучше
всего они растут на глубине 20—30 'м (мы не принимаем в расчет рифооб-
394
разные разрастания кораллового полипа Lophophelia, встречающегося
набольших глубинах в морях умеренной зоны, и тем более скопления
горгонарий).
Основные коралловые рифообразователи — это представители от-
ряда мадрепоровых (Anthozoa). Обычно к ним добавляются Foraminifera,
Milleporidae (Hydrozoa), Alcyonaria (особенно Tubiporidae) из восьмилу-
чевых кораллов и в особенно большом количестве отлагающие на поверх-
ности таллома углекислый кальций красные водоросли (Л'ullipures),
главным образом род Lithothamnion, а из зеленых водорослей — На li-
me da. И фораминиферы, и известковые водоросли по массе могут пре-
восходить сами каменистые кораллы.
Все эти формы образуют мощные пористые густоветвистые разрастания
п дают в образуемых ими пустотах и полостях приют бесчисленным чер-
вям (в том числе и палоло), крабам, губкам, иглокожим, моллюскам и
многим другим организмам, начиная от гигантских тридакн и кончая
микроскопическими фораминиферами.
Коралловые рифы — самый богатый по разнообразию видов биоценоз
моря. Замечательно широкое распространение у обитателей коралловых
рифов получили явления симбиоза с зооксантеллами не только у простей-
ших, но также у кишечнополостных, моллюсков и аспидий; у кораллов
зооксантеллы живут в клетках энтодермы. В углублениях между частями
коралловых рифов скопляется известковый ил и в нем также поселяется
богатая инфауна — черви и голотурии, в том числе и известный съедоб-
ный трепанг (Stichopus ), служащий объектом промысла. К этому добав-
ляется чрезвычайно своеобразная фауна красивых коралловых
рыб, составляющаяся из самых разнообразных систематических
групп.
Мангровые заросли. Если коралловые рифы — наиболее характерная
черта верхнего горизонта тропической сублиторали, то для литорали столь
же характерны мангровые заросли с их фауной.
В тропических морях на открытых побережьях литоральная фауна
выражена слабо. Сильнейшая солнечная инсоляция и высокая температура
создают неблагоприятные условия для развития растений и животных выше
уровня воды. И только среди мангровых зарослей, этих своеобразных
литоральных лесов почти всех тропических побережий, находит приют
бедная видами, но богатая количественно амфибиальная фауна. Состоят
мангровые заросли в основном из манглей (Rhizophora,Aoicennia), деревьев,
растущих на сложной системе воздушных корней, опускаемых в боло-
тистые или илистые грунты мелководных побережий и дающих для организ-
мов удобный субстрат и защиту от солнца. Это своеобразное литоральное
население состоит из смеси морских, пресноводных и наземных растений
и животных. Среди последних большое число приспособилось к рытью нор.
Мангровая литораль — приют маленькой актинии (Thelaceros rhizo-
phorae), бесчисленных десятиногих ракообразных (Uca, Sesarma, Car-
diosoma, Thalassina, Coenobita), одного из мечехвостов (Limulus mollu-
canas), моллюсков, часто целых щеток устриц, роющихся в грунте
двустворчатых (Psammobiidae, Solenidae), брюхоногих (Cerithium, Potami-
des, Littorina) и замечательных вылезающих на воздух рыбок (Periophthal-
mus и Boleophthalmus).
Фауна Индо-вестпацифики. Наиболее богатая и разнообразная тро-
пическая фауна, населяющая Индо-вестпапифическую подобласть, захва-
тывает ареал от восточных побережий Африки (от Красного моря до север-
ной границы Капской земли) до берегов Японии, островов Гавайских,
Маркизских и Паумоту и восточных побережий Австралии, с включением
всего Индийского океана до «35° ю. ш.
395
В качестве примера ареала индо-вестпацифических форм можно при-
вести одно из семейств горгониевых кораллов — Melitodidae (рис. 242).
Громадное богатство видов губок, кораллов, крабов, моллюсков, игло-
кожих и рыб приурочено к этой подобласти. Еще в 80-х годах прошлого
столетия считали, что 5000—6000 видов моллюсков свойственно фауне
Индо-вестпацифики. Cornacuspongida из губок имеют здесь основное место-
обитание. Все 9 семейств отряда Alcyonaria обитают здесь, причем 3 семей-
ства живут только в Индо-вестпацифике, как, например, так называемые
голубые (Helioporidae) и органчиковые (Tubiporidae) кораллы. Наиболь-
шее количество родов остальных 6 семейств приурочено также к Индо*
вестпацифике. Сходное распространение имеют горгонарии и пеннатуля-
рии. Особенно же выделяются каменистые кораллы. Специфическими
индо-вестпацифическими родами из них являются, например, грибовид-
ные кораллы (Fungia). Семейство Fungiidae, куда относятся еще 4 дру-
гие рода, включает 46 видов, из которых только один встречен вне Индо-
вестпацифики. Даже среди десятиногих раков, пелагические личинки
Рис. 242. Распространение индо-вестнацифического семейства горгониевых кораллов
Melitodidae (но Экману).
которых могут переноситься на большие расстояния, имеется около 100
родов, характерных для Индо-вестпацифики. Среди крабов—это подсе-
мейства Leucosiinae, Polydectiпае, Hexapodinae и Scopimerinae. Энде-
мичны для этой подобласти пелагический краб Portunus pelagicus и паль-
мовый вор Birgus latro. К ним можно добавить два рода мечехвостов
(Tachypleus и Сarcinoscorpius) с четырьмя видами. 40 родов моллюсков —
и в том числе единственный представитель четырехжаберных головоно-
гих Nautilus — встречаются исключительно в Индо-вестпацифике. Из
морских лилий группа Comatulida дает здесь 69 родов и 350 видов. Только
11 родов встречаются и в Атлантическом океане, а 14 имеют там близкие
формы. Около 400 видов морских лилий ограничены в своем распростра-
нении Индо-вестпацпфикой. Род морских звезд Ast го pecten дает здесь
67 видов, а в области центральной Америки их всего 14—15. Из 118 видов
морских ежей группы Cidaroidea 80 видов обитают только в Индо-вестпа-
цифике. Из 192 родов офиур для нее эндемичны 57. Вся группа морских
змей (15 родов и 60 видов) за исключением одной свойственны только
Индо-вестпацифике. Из млекопитающих только здесь обитают дюгони.
Экман подразделяет индо-вестпацифическую фауну на малайскую,
южно-тихоокеанскую, гавайскую, южно-японскую, австралийскую, северо-
и западно-тихоокеанскую. *
396
Рис. 243. Распространение эндемичного
амфиамериканского рода горгоний Muri-
сеа (из Экмана).
Фауна тропической Америки. Фауна тропической Америки значи-
тельно беднее индо-вестпацифической. До соединения обеих Америк
Панамским перешейком, в конце третичного периода, фауна прилежащих
частей — Пацифики и Атлантики — была одинаковой. За истекшее время,
после образования перешейка, в силу неизбежной дивергенции, общих
видов на запад и восток от перешейка осталось очень мало. Главная масса—
это виды-близнецы (викарирующие виды), иногда чрезвычайно близкие:
значительно меньше разных родов, и отсутствуют разные семейства. Это —
замечательный пример правильности дарвиновского принципа дивергенции.
Особенно ярко развитие видов близнецов в центральной Америке
сказалось на крабах, морских ежах и рыбах. Среди крабов их свыше 100
над (62% всех родов крабов центральной Америки дает близнецовые виды),
не менее 100 среди рыб. Роды с близкими видами по обе стороны Америки
Экман называет амфиамери-
канскими. Пример амфиамерикан-
ского распространения мы даем
на рис. 243 для рода горгониевых
кораллов — Maricea.
Крабы могут служить прекрас-
ным материалом для более деталь-
ного рассмотрения характера
сходства и различия тропических
фаун по обе стороны от Панам-
ского перешейка. Фауна крабов
этого района состоит из 157 ро-
дов и 582 видов. В изобилии кра-
бов наиболее резкое отличие тро-
пической фауны мелководных
районов от фауны умеренных и
высоких широт, где количество
их ничтожно мало. На рис. 244
дан подсчет родов и видов крабов
по обеим сторонам центральной
Америки. Общих для западной и
восточной стороны видов крабов
5%, а родов 40%.
Что же касается общих родов
и видов с другими районами тро-
пической области, то их очень немного, причем и здесь количество об-
щих родов значительно выше (25,6%), чем видов (4%). Количество цпр-
кумтропических форм еще меньше. По восточному побережью Америки
всего 2,4% циркумтропических видов и 6,3% циркумтропических родов.
Среди тихоокеанских крабов центральной Америки эндемичных видов
84%, а родов 12—13%.
Что касается западной, тихоокеанской, стороны тропической централь-
ной Америки, то для ее фауны очень характерны большая системати-
ческая близость к фауне восточной стороны Америки, относительно мень-
шее качественное богатство и довольно резкая обособленность от индо-
вестпацифической фауны. Глубинные части Тихого океана являются
непреодолимой преградой для донного населения мелководных районов
Индо-вестпацифики. Из 9 семейств индо-вестпацифических мелководных
морских лилий 4 представлены в Атлантическом океане и ни одного нет
у американских западных побережий. В семействе Antedonidae Индо-
вестпацпфики имеется 6 семейств, из которых 3 сублиторальных пред-
ставлены в Атлантике, но отсутствуют к западу от Америки, а 3 глубо-
397
Рис. 244. Распространение одной группы тропическо-американских крабов
(по Рэтбен из Экмана).
Числа в простых кружках показывают количество обитающих здесь родов или видов, не встречаю-
щихся на другой стороне. Количество общих для обеих сторон родов или видов дано в четыреуголь-
никах. Двуконтурные кружки и четыреугольники дают количество эндемичных родов или видов.
Стрелки показывают распространение во вне-тропическо-американских водах. В-Атл. — Восточная
Атлантика, Индо-ВП — Индовестпацифика. Границы тропической области даны штриховкой и тол-
стой линией; А — для родов, Б — для видов.
398
ководные подсемейства здесь имеются, так как глубины Тихого океана
не являются для них препятствием. Таких примеров можно было бы при*
Рис. 245. Распространение голотурий рода Pseudocucumis
(по Энгелю из Экмана).
вести немало. То же можно иллюстрировать и картами ареалов различных
форм. На рис. 245 изображено распространение голотурий Pseudocucumis,
, Рис. 246. Распространение рода Anguilla (угри) в береговой области
(по Шмидту).
а на рис. 246 — распространение угрей (Anguilla). И те и другие имеют
очень широкое циркумтропическое распространение, но отсутствуют у
западных берегов Америки.
39»
Атлантическая фауна тропической Америки богаче своего западного
соседа и после Индо-вестпацифики представляет наиболее богатый фауна-
стически район океана. Она тянется на север до Бермудских островов
и до мыса Гаттераса и американского побережья (35° с. ш.). Южная гра-
ница ее неясна (20—35° ю. ш.).
Атлантическая тропическая фауна. Атлантическая тропическая фауна
характеризуется высокой степенью видового и родового эндемизма, что
можно показать на примере некоторых групп беспозвоночных (табл. 132).
Таблица 132
Группы
Процент энде-
мичных видов
Процент энде-
мичных родов
Gorgonarla (по различным источникам)..............' 90
Brachyura (по Rathbun)................................... SO
Echinodern^ita верхней зоны сублиторали (по (Hark) SO
Crlnoldea (по Clark).................................... 100
Ophiuroidea (no Clark)............................
Euryale (no Doderlein)................................... 85
Holothurioldea (no Deichmann).....................; 80
55
25
80
23
75
Особенно велико здесь видовое богатство горгонарий (не менее 70
эндемичных видов в сублиторали) и антипатарий, а также крабов (свыше
300 видов). Замечательны некоторые отрицательные черты этой фауны,
как, например, отсутствие рода Madrepora, основного рифообразователя
Индийского и Тихого океанов. При всем том фауна тропической Атлан-
тики очень близка к индо-вестпацифической, если только учесть, что все
это видовое и родовое своеобразие есть результат длительного существо-
вания в изолированном положении дочерних фаун, происходящих от
одной и той же фауны третичных морей.
У западных берегов Африки тропическая зона наиболее сужена,
вследствие подступа и с севера и с юга более холодных вод, и занимает
только Гвинейский залив от Зеленого мыса на севере до 15—17° ю. ш.
на юге. Этот район имеет наиболее обедненную тропическую фауну,
со слабее выраженным эндемизмом. Здесь прежде всего отсутствуют
настоящие коралловые рифы, как думает Тиль, вследствие несколько
менее нагретой воды и большого развития песчанистых грунтов. Все же
моллюски дают здесь 63% эндемичных видов, асцидии—60%, игло-
кожие 50%, Decapoda — около 40%. Другие группы характеризуются
меньшим эндемизмом. Отрицательные черты выражены при этом очень
резко, особенно в отношении фауны отсутствующих здесь коралловых
рифов. Так, например, здесь почти полностью выпадает богатое видами
семейство Porcellanidae (Anomura), столь характерное для центральной
Америки. Другие группы животных имеют большое сходство с американ-
скими тропиками и несколько меньшее с индо-вестпацифической фауной,
но, как и в других частях тропического пояса, генетическое родство дочер-
них тропических фаун устанавливается по аналогии общих родов и еще
больше семейств, но не видов.
Саргассово море. Если для мелководной и литоральной зон тропи-
ческой области весьма характерны коралловые рифы и мангровые заросли,
то для океанической пелагиали Атлантического океана столь же харак-
терно образование, получившее название Саргассова моря. Саргассово
море располагается между 20 и 40° с. ш. и 30 и 60° з. д. и занимает область,
ограниченную антициклоническим круговым течением (халистаза), обра-
зованным на юге Антильским течением, на западе и севере — Гольфстри-
мом и Атлантическим, на востоке — Канарским. Шотт (Schott,
400
1936), определяет его площадь в 8 635 000 км2. Под Саргассовым морем
глубина в 4—6 тыс. м. Внутри Саргассова моря отсутствуют постоянные
течения и господствуют высокая температура воды (в среднем 20—25°;
зимой 17—23°, летом 23—27°) и довольно высокая соленость (не менее
36,5°/ОО, а в отдельных пунктах 37,О°/оо). Саргассово море характеризуется
исключительной качественной и количественной бедностью фауны, чрезвы-
чайной прозрачностью воды и постоянным присутствием массы плаваю-
щих бурых фукусовых водорослей из рода Sar^assum, дающих приют
очень своеобразному комплексу животных из нескольких десятков видов.
Как оказывается по последним исследованиям, саргассы не приносятся
сюда От берегов Вест-Индии, а постоянно пребывают в плову чем состоянии
и размножаются только вегетативным путем. Основные формы плаваю-
щих саргассов, видимо, другие, чем те, которые растут у берегов Вест-
Индии. Количество их не так велико, как это иногда думают. По подсче-
там Г е н т in е л я (1922—1923), количество кустов саргассов (между
Азорскими о-вамп и о. Гаити) составляло 2 000 на 1 км2, или один куст
на 83 м2, т. е. всего несколько граммов на 1 лг21. Резкий дефицит в пита-
тельных солях ограничивает как развитие фитопланктона, так п саргас-
сов. Несколько десятков видов (50—60) составляют население Саргассова
моря, причем кусты саргассов, так же как заросли любых других макро-
фитов, дают приют сидячим, ползающим и плавающим организмам.
В первую группу входят два вида морских уточек (Lepas), мелкие поли-
хеты в известковых трубочках (Spirorbis), известковая мшанка Membra-
nipora, сложная асцидия Diplosoma, брюхоногий моллюск Litiola. Все
это — очень мелкие животные. Так же мелки представители и второй
группы — ползающих по саргассам животных. Особенно известны среди
них крошечный крабик Planes minutus, несколько видов голых моллю-
сков, амфипод, бокоплавов, капреллид, пантопод и несколько видов мел-
ких рыбок. Наиболее многочисленна, но и наименее специфична третья
группа — плавающих форм. Среди них морские черепахи, летучие рыбы,
анчоусы, тунец и др., пелагический краб — Portunus.
Исключительно беден и планктон Саргассова моря (в 50—80 раз по
биомассе беднее планктона северноевропейских побережий). В большом
количестве в планктоне попадаются лептоцефалусы — личинки угрей,
нерестящихся тут же, на глубине нескольких сот метров. Зато чрезвы-
чайно богат качественно, а вероятно, и количественно (по сравнению
с другими районами абиссали) мир глубин Саргассова моря.
Опускающиеся на дно отмершие водоросли и трупы населяющих
саргассы животных дают здесь глубоководной фауне более обильное пита-
ние, чем в других районах тропической абиссали. Замечательно, что здесь
вместе с удивительным миром глубоководных рыб обитают и представи-
тели далеких и холодных морей северной Атлантики, находящие здесь
для себя благоприятные температурные условия, как, например, весло-
ногий рачок Calanus jinmarchicus и крылоногий моллюск CUone limacina.
Но особенно богата и разнообразна фауна удивительных глубоководных
рыб, изучаемых в последние годы американцем Бибом (Beebe) при
помощи батисферы у Бермудских островов.
Анализ фауны саргассовых скоплений показывает, что она ближе
к фауне Средиземного моря, чем к вестиндской.
Скопления плавающих водорослей в халпстатических областях из-
вестны и в других местах мирового океана и отдельных морей, но нигде
они не получают такого грохмадного развития.
1 Другие авторы определяют общую биомассу плавающих в Саргассовом море водорослей
в 20 млн. т, а среднюю на 1 м2 — примерно в 100 8.
26 Зенкевич i. I
401
5. ФАУНА СРЕДИЗЕМНОГО МОРЯ
Зоогеографическое положение. Зоогеографическое положение фауны
Средиземного моря, а вместе с тем и Черного, до сего времени остается
очень спорным. В большинстве случаев ее относят пли к бореальной или
к субтропической области, иногда же и к тропической, что объясняется
двойным ее генезисом и неясностью северных границ тропической и суб-
тропической областей.
В основной своей части это бореальная фауна, в меньшей части —
тепловодная тропическая, наследие того же Тетиса. G. Экман счи-
тает возможным выделять самостоятельную Средиземноморско-атланти-
ческую область, в которую он включает так называемую Лузитанекую
провинцию (от Гибралтара на север до входа в Английский канал), районы
Марроканский п Мавританский и о-ва Зеленого мыса, Канарские, Ма-
дейра и Азорские. Однако северная граница Средиземноморско-атлан-
тической области неясна. Как было показано еще С а р с о м (G. О. S а г s,
1878), 68% двустворчатых моллюсков и 61% брюхоногих, обитающих
у берегов Норвегии, встречаются и в Средиземном море. Примерно такие
же соотношения дают и другие группы. Даже средиземноморские крабы
вдут далеко на север. До западного устья Английского канала доходит
из них 27% родов и 28% видов. Из 123 видов гаммарид, обитающих
в Средиземном море, 76 видов (61,8%) встречаются и у побережий северо-
западной Европы. Моллюски, и тем более иглокожие, продвигаются
на север значительно дальше. С другой стороны, 45% средиземноморских
моллюсков доходит до Зеленого мыса. Такое фаунистическое разнооб-
разие, захватывающее 30° по меридиану, т. е. свыше 3000 км, в основ-
ном находит свое объяснение в однообразны температуры. У южной гра-
ницы этой области температура лишь на 3—8° (для разных сезонов года)
выше, чем у Английского капала.
Тропические элементы. Границей с тропической индийской фауной
является Суэцкий канал. Фауны Средиземного и Красного морей (эрит-
рейская) очень различны, несмотря на географическую близость. Ш т е й-
ниц (Steinitz, 1929) отмечает, что до прорытия Суэцкого канала
оба моря имели только 13 общих форм, в том числе 7 видов рыб и дель-
фина. С открытием канала (1869) через него идет вселение некоторых тро-
пических эритрейских форм из Красного моря в Средиземное. Так,
Бальсе (В а 1 s s, 1936), в водах, прилежащих к Александрии,
обнаружил 10 видов крабов (из 63) — вселенцев через Суэцкий канал.
Около 10 видов рыб (S t е i п i t z, 1927; G г u v е I, 1929) проникли
тем же путем.
Среди тропических элементов средиземноморской фауны мы находим
и циркумтропические формы, как, например, почти циркумтропический
род Microcosmus и тропические атлантические и, наконец, индо-вестпа-
цифические. Примером последних может служить голотурия рода Pseu-
docucumis (рис. 245).
Бореальные элементы. Гораздо многочисленнее в Средиземном море
формы, общие с побережьями западной и северо-западной Европы. Среди
них выделяется ряд видов, обычных у побережий Норвегии, Исландии
и в Баренцовом море, как, например, моллюск Saxicava arctica, ракооб-
разные М eganyctiphanes norvegica, Nephrops norvegicus, рыбы Molva molva,
Pleuronectes platessa, Gadus virens и др. Среди крабов побережья Египта
Бальсе различает 32 вида кельтическо-бореальных, 12 тепловодных,
5эндемикови несколько индо-пацифических форм—остатков фауныТетиса.
В эпоху похолодания по берегам Средиземного моря обитали и такие формы,
402
как Pecten islandicus, Cyprina islandica, Buccinum undatum, Panopea
norvegica, Mallotus villosus и другие, ныне вымершие.
Смена фауны у северных границ Лузитанско-средиземноморской про-
винции выявляется особенно резко на населении литорали. У северных
побережий Бретани еще в полной силе фукусы и ламинарии, весь набор
бореальных литорин, Purpura lapillas,Actinia equina, Gammarus locusta,
Mytilus edulis, Mya arenaria, Macoma baltica, Arenicola marina и др.,
однако уже добавляется много форм более южного характера. В западной
части Ламанша и дальше на юг фукусы и ламинарии сходят на нет, их
сменяют Cystoseira и Posidonia, исчезают Mya arenaria, Macoma baltica,
Arenicola marina, Mytilus edulis и др. и все больше и больше входит в силу
лузитанско-средиземноморскаяфауна: Chthamalus stellatus, Balanus per-
forates, Orchestia mediterranea, Pachygrapsus marmoratus, Lygia italica,
Talitrus locusta, Arenicola grubei, Littorina neritoides, Patella lusitanica
и др. При сравнении приводимых Прюво (Pruvot) списков
фауны Роскова (северная Бретань) и Баныольса на Средиземном море
(близ границы Франции и Испании) оказывается, что из указываемых
Прюво 865 форм для первого и 1264 для второго — 505 оказываются
общими, причем в их числе находится большинство массовых форм.
В более глубоких горизонтах граница теряет свою четкость. Комплекс
Venus gallind в мелководной части также имеет южную границу в районе
Ламанша, но на больших глубинах тянется дальше на юг. Расположенный
еще глубже комплекс Brissopsis lyrifera идет на юг уже до Средиземного
моря и западных берегов Африки. Фауна Средиземного моря качественно
несколько богаче фауны северных частей бореальной области, хотя отдель-
ные участки Средиземного моря, особенно его восточная часть, значительно
беднее ее. Исключительно богата здесь фауна рыб, включающая около
530 видов, из которых 144 прибрежных.
Эндемики. Эндемичных видов в Средиземном море немного, еще меньше
эндемичных родов. Так, для восточной части моря Бальсе указывает
только 5 эндемичных видов крабов из 92 видов, т. е. немногим больше
5%. Эндемизм фауны Средиземного моря, по Экману, — молодое
образование, объясняющееся изоляцией в третичное время. Это — пере-
работанные остатки фауны Тетис. Только некоторые группы фауны дают
большое число эндемиков, как, например, иглокожие (табл. 133).
Таблица 133
Классы иглокожих Эндемичные виды в Средиземном морс
общее количество видов из них эндемичных В 7о
Лилии 24 3 15
Звезды 24 3 12
Офиуры 21 5 23
t Ежи 21 8 38
Голотурии 30 12 40
Всего 120 1 31 1 26
Из 530 видов рыб эндемиков 30%, в том числе хамса (Engraulis епега-
sicholus).
Термическая неоднородность фауны. Еще 100 лет назад Форбс
(Forbes, 1844) заметил, что в Средиземном море с увеличением глу-
бины обитания животные приобретают все более и более «северный
403
характер». Мнение Форбса в дальнейшем нашло подтверждение во взгля-
дах других исследователей (S с h m а г d а, 1853, Lorenz, 1863,
Volz, 1940 и др.). П. Фольц (1940) в подтверждение мнения Форбса
приводит довольно показательные данные по вертикальному распреде-
лению иглокожих в районе, прилежащем к Монако (табл. 134).
Таблица 134
Горизонты Глубина в м Средиземно- морские эндемики «Атлантиче- ские» виды (не идут в североборе- альную об- ласть) «Северные» виды (идут в Субаркти- ку и арк- тику) Всего
Литоральные виды 0—50 15 18 0 33
Сублиторальные виды 50—250 6 15 8 29
Глубинные виды >250 5 6 20 31
Всего 26 39 28 1 93
Формы, приуроченные в Средиземном море к «литоральной» зоне,
размножаются в самое теплое время года и являются либо эндеми-
ками этого моря, либо тропическими формами.
Отмеченная вертикальная зональность животного населения
Рис. 247. Вертикальные изменения температуры в
Средиземном море в феврале, августе и ноябре (по
Окснеру и др. из Фольца).
хорошо согласуется с вертикальным распределением температуры
в Средиземном море
(рис. 247). Поверхност-
ный слой воды в 40—
50 м испытывает силь-
ное летнее прогревание,
а с глубины в 200—250м
температура близка к
13° и не меняется по
сезонам.
Термопатическая не-
однородность фауны
Средиземного моря и
особенно его северных
«заливов» может быть
хорошо иллюстрирова-
на приводимой Т. Г а-
мулиным (1938)
схемой сезонной смены массовых веслоногих ракообразных планктона
Адриатического моря (рис. 248). Зимняя форма Рагасаlaniisparvus сме-
няется весенней Опсаеа media. На смену ей приходят раннелетние Centro-
pages typicus и Acartia clausi и, наконец, в самое теплое время года
массовое развитие получает Temora stylifera.
Исключительно интересно указание Штамм е р a (Stammer, 1932) на
массовое нахождение в районе Тимаво (Северная Адриатика, близ Триеста)
в пресной воде Mysis oculata var. relicta. Вряд ли следует рассматривать
М. relicta в северной Адриатике, как остаток той морской фауны, кото-
рая в ледниковое время могла спуститься далеко на юг в Атлантическом
океане. Вероятнее, это, подобно близкой форме в Каспийском море,
иммигрант с севера, через речные системы из Балтийского моря. Но
почему М. relicta оказался единственным иммигрантом с севера?
История фауны. История Средиземного моря и его фауны не вполне
ясна. Хотя Средиземноморский бассейн и составлял в третичное вре-
404
мя часть Тетиса, однако фауна его не имеет непосредственной генетиче-
ской связи с фауной этого моря. Есть все основания считать, что фауна
Средиземного моря сформировалась в недавнее время (слабо выраженный
эндемизм, отсутствие настоящей глубоководной фауны). Вымирание засе-
лявшей когда-то Средиземное море фауны Тетиса могло происходить,
во-первых, вследствие имевшей место в плиоцене изоляции Средиземного
моря от Атлантического океана и, во-вторых, вследствие климатических
изменений, имевших место в конце плиоцена и в плейстоцене. Гибрал-
тарский пролив открылся в плиоцене. Весьма вероятно, что, кроме того,
Средиземное море было перегорожено Сицилианским мостом и состояло
из двух морей. В это же время, вероятно, вымерла и вся глубоководная
Рис. 248. Годовой ход изменений в количестве экземпляров на 1 м3 пяти основных
видов веслоногих рачков в средней части Адриатического моря (по Гамулину).
1 — Centropages typicus, 2— Temora stylifera, 3 — Oncaea media, 4 — Paracalanus parvus,
5 — Acartia clausi.
Средиземном море. Восточная поло-
вина моря пришла в соединение с Сарматским бассейном. Вполне воз-
можно, что Средиземное море претерпело значительное опреснение и раз-
витие в глубинах сероводорода. Иначе говоря, существовавшая в нем
фауна должна была подвергнуться почти полному вымиранию и частичной
переработке.
С открытием Гибралтара атлантическая фауна стала заселять Среди-
земное море, подобно тому, как позднее она же внедрилась в Черное
и Азовское моря. Экман считает, что фаунистически «Средиземное
море есть очевидное подразделение Атлантического океана», за исклю-
чением остатков фауны Тетиса. Трудно сказать, где эти аборигены могли
найти убежище в период плиоценовых и четвертичных орографических
и климатических смен. Сейчас они наиболее сильно выражены в самой
юго-восточной части Средиземного моря.
На основании всего сказанного возникает вопрос о целесообразности
выделения и наименования Средиземноморско-атлантической области,
как это делает Э к м а н. Как справедливо отмечает этот автор, восточ-
40 5
пая Атлантика представляет собою наиболее обширную и самостоятель-
ную во всем мире переходную субтропическо-умеренную зону. Пока мы
не располагаем достаточными данными, чтобы приравнивать ее к восточ-
ной половине атлантической северной субтропической области или счи-
тать южной восточно-атлантическо-средиземноморской провинцией (пли
подобластью) бореальной области. Нам кажется более правильным послед-
нее. Ее можно было бы называть также лузитанско-средиземноморской
провинцией (или подобластью) бореальной области.
6. ДРУГИЕ ОБЛАСТИ ОКЕАНА
Значительно слабее наши знания о морях южного полушария.
Мы не имеем возможности входить здесь в их рассмотрение и укажем
Рис. 249. Распространение морских ежей рода Abatus в водах Антарктики
(по Экману).
только, что и южная субтропическая область и расположенные южнее,
вплоть до Антарктики, части океана обладают своими характерными
фаунами и эндемичными формами. Приведем в качестве примера распро-
странение морских ежей рода Abatus в водах Антарктики и Субантарк-
тики (рис. 249).
ПРАКТИЧЕСКОЕ ЗНАЧЕНИЕ
МОРСКИХ ОРГАНИЗМОВ
ГЛАВА XVI
ПРОМЫСЕЛ МОРСКИХ ОРГАНИЗМОВ
1. ВВЕДЕНИЕ
о ре всегда имело громадное значение в жизни чело-
вечества и как неисчерпаемый источник пищевого и,
отчасти, медицинского и технического сырья и как
наиболее легкий и дешевый путь сообщения. По мере
развития человеческой культуры все шире и разносто-
роннее становилось использование моря, но и в на-
стоящее время еще далеко не все возможности в этом
отношении исчерпаны. Наиболее полно используются
сейчас запасы рыб и китов; наряду с этим колоссальные запасы морских
растений и беспозвоночных, особенно в СССР, затронуты промыслом еще
в очень слабой степени. С другой стороны, мало разработаны меры борьбы
с вредными для судоходства беспозвоночными и недостаточно учитывается
их громадное значение в этой отрасли хозяйства.
Новейшую сводку материалов по мировому промыслу водных орга-
низмов дал в своей книге Т. Расс (4948). По его данным, общий вылов
водных (морских и пресноводных) животных перед второй мировой вой-
ной превысил 180 млн. ц (не считая китов и дельфинов), причем свыше
12 млн. if, или 7%, приходилось на беспозвоночных. Из этого общего
количества 50% вылавливается в Азии, 31% — в Европе, 17% — в Аме-
рике и немного менее 2% в остальных странах. Бассейн Тихого
дает 53%, Атлантического — 42% и Индийского — всего 5% от общего
улова.
Из 182 млн. ц 170 млн. if, или 93%, приходится на моря и только-
12 млн. и, или 7%, на материковые воды.
Главная масса мирового промысла водных объектов (72%) приходится
на умеренную зону северного полушария (между Полярным кругом и
тропиком Рака), а все северное полушарие дает свыше 90% мирового
промысла (по Рассу). Промысловая продуктивность отдельных морей
колеблется в очень широких пределах (табл. 135, по данные 1936—
1937 гг.).
океана
409
2. ПРОМЫСЕЛ РЫБ
(рис. 250 и 251)
Общие данные. Сравнение продукции морского рыбного промысла
с продукцией пресных озер (Т. Расс, 1947) показывает, что в морях про-
дукция в среднем выше, чем в озерах (табл. 136).
Рис. 260. Основные объекты мирового рыбного промысла, с указанием размеров
вылова (из Расса)
При* этом мелкие озера дают более высокую рыбную продукцию —
озера СССР в среднем 15—30 кг) га (от 1 до 115 кг/га), если не считать
410
значительно более бедных северных озер, богатых пойменных озер п
некоторых озер теплой зоны.
Среди отдельных стран на первом месте по размерам рыбного про-
мысла перед второй мировой войной стояли Япония, США, СССР (рис. 258),
Корея, Китай, Великобритания и Норвегия, добывавшие вместе около
двух третей мирового улова рыбы (табл. 137 по данным 1936—1937 гг.).
Рис. 251. То же, что и рис. 250 (из Расса).
Так же как п для промысла водных организмов в целом, рыбный про-
мысел на 90% базируется на морских рыбах и только на 4,5% на пресно-
водных; 5,5% составляют рыбы проходные.
411
Таблица 135
Водоем Площадь в ты- сячах теле2 Общий улов в млн. ц Улов в кг 1га
Азовское море 38 3 80
Японское » 1008 27—30 26,8-29,8
Северное » 575 14,1 24,5
Каспийское» 424,5 5 11,8
Восточно-Китайское морс 735 10—12 13,6—16,3
Аральское море 63,8 0,36-0,38 5,6—5,9
Охотское » 1528 7,0—7,8 4,6—5,1
Баренцево » 1400 6,3 4,5
Берингово » 2268 5,6 2,5
Черное » 412 0,7 1,6
Средиземное » 2516 3,7 1,5
Белое » 95 0,1 1,2
Среднее 14
Таблица 136
Озеро Площадь в тысячах км9 Общий вылов в тыс. и ) Улов в кг 1га
Ильмень 1,1 35 31,3
Балхаш 17,3 147—175 8,5—10,1
Севан 1,4 10,2—12,1 7,3—8,7
Эри 25,7 189 7,3
Боденское 0,5 3,6—5,1 7,2—10,2
Охрид 0,3 2 6,7
Байкал 31,5 94 3,0
Ладожское 18,4 54,5 3,0
Мичиган 58,0 120 2,1
Иссык-Куль 6,2 10 1,6
Гурон 59,6 88 1,5
Онтарио 19,5 18 0,9
Среднее 7
Таблица 137
Страна (без колоний)
Улов в млн. ц
Япония.........................
США............................
Корея..........................
СССР...........................
Китай..........................
Великобритания ................
Норвегия.......................
Германия ......................
Канада.........................
Франция........................
Исландия.......................
33—37
20—22
17—21
16—16,3
14,4
11
10—11,8
6—6,7
4,9—5,0
3,2—3,6
2,6—3,3
412
Роль различных групп рыб в мировом промысле. Т. Расс отмечает
недостаточность освоения промыслом пелагических рыб в северных
частях Атлантического и Тихого океанов.
Рис. 252. Промысловое значение водоемов СССР (по Рассу). Площади кругов
соответствуют средней величине предвоенных уловов. Числа — тысячи центнеров.
У берегов Гренландии добываются только донные рыбы. В Барен-
цевом море на их долю приходится 99%, в Норвежском море 58—73%,
в Северном море 20—27%, у берегов Португалии 11%. Надо учитывать,
однако, что в наших северных морях совсем не используются промыс-
лом или используются очень мало такие пелагические рыбы, как мойва
Рис. 253. Перемещение центров морского мирового рыбного промысла (по Рассу).
и сайка, количество которых чрезвычайно велико, и еще очень недоста-
точно используется сельдь.
Та же смена наблюдается и в Тихом океане: у берегов Кам-
чатки и Аляски донные рыбы в промысле составляют 11—43,5%, у бере-
413
гов Японии и Канады 10—11%, у берегов Калифорнии—всего 4%.
В Тихом океане значение пелагических рыб в вылове вообще выше, чем
в Атлантическом.
Весьма интересно также, что основную массу рыб в море (во всяком
случае, если судить по промысловому выходу) составляют рыбы не осед-
лые, а мигрирующие, совершающие в течение жизни перемещения из одних
частей моря в другие, главным образом, в целях питания и размножения.
Для питания более пригодны одни части моря, для размножения — дру-
гие. Иногда эти миграции охватывают и материковые воды (лососевые,
сельдевые, угревые и др.), а миграционные пути растягиваются на тысячи
километров.
Таблица 13S
Семейство рыб В % от мирового 1 промысла В Тихом и Индий- ском океанах В Атлантиче- ским океане
Сельдевые 37,3 40,7 20,7
Тресковые 14,0 •>9 20,8
Лососевые 5,8 8,3 1,5
Наибольшее значение в мировом рыбном промысле имеют сельдевые,
тресковые и лососевые рыбы, составляющие вместе около 60% от общего
Рис. 254. Вылов пикши в Северном море на день
плавания английских тральщиков, в центнерах
(по Ресселу из Ионга).
улова. При этом между дву-
мя океанами наблюдается
существенное различие: в
Тихом океане преобладают
сельдевые и лососевые, а в
Атлантическом тресковые
(табл. 138).
В морском рыбном про-
мысле резко преобладает не-
сколько видов рыб: на пер-
вом месте стоит сардина (в се-
верной части Атлантического
океана Sardina pilchardus, в
Тихом — Sardinops sagax с
подвидами), дававшая (1936—
1939) от 30,6 до 33,6 млн. и
добычи. На втором месте сто-
ит сельдь Clupea harengus ha-
rengus в Атлантическом оке-
ане и С. h. pallasi — в Тихом,
дающая 21—30 млн. ц, на третьем — атлантическая треска Gadus тот-
hua morhua в Атлантическом океане и близкий тихоокеанский подвид
Ст. щ. macrocephalus, дающие до 15 млн. н, прячем на долю атлантиче-
ской трески приходится 13 млн. ц.
Современное размещение мирового морского рыбного промысла только
в малой степени соответствует распределению сырьевых запасов рыб.
В отдельных частях мирового океана сырьевая база рыбного промысла
хорошо изучена и интенсивно используется, в других ее освоение нахо-
дится в начальной фазе, в третьих оно еще не начато (рис. 252).
Динамика мирового рыбного промысла. Если во второй половине прош-
лого столетия и в нынешнем центр тяжести промысла перемещался из
материковых вод в береговые районы моря, то в настоящее время про-
мысел все более и более отходит от берегов в открытые части моря (рис. 253).
414
Наряду с этим, как указывает Т. Рас с, наблюдается рост рыбопромыс-
лового значения Тихого океана по сравнению с Атлантическим. В 1897 г.
рыбный промысел США был сосредоточен преимущественно по побережью
Атлантического океана (около 80%), а в 1936 г. на его долю приходилась
только одна треть (37,3%) морского промысла. Значение тихоокеанского
побережья в промысле США за тот же период поднялось с 12,5 до 59%.
Так же и в нашем промысле: в прошлом столетии на первом месте было
Каспийское море, затем на первое место вышло Баренцово море, а сейчас
громадный рост нашего промысла на Тихом океане должен обеспечить
ему в дальнейшем неоспоримое первое место. Наши уловы в Охотском
море с 1911 до 1936 г. возросли в 13 раз, у берегов Камчатки—в 2 раза,
у Сахалина — в 5 раз, в Приморье — в 9 раз.
Кроме того, наблюдается значительный рост рыбного промысла в юж-
ном полушарии, особенно в Тихом океане; в северном же полушарии
происходит перемещение центров промысла в более северные широты.
Если в Северном море за пятнадцатилетие (1922—1936 гг.) промысел
возрос на 50%, то в Баренцевом и Норвежском — на 120%. Причиной
этого является открытие новых богатых рыбой районов, как, например,
Шпицбергенско-Надеждинского мелководья в северо-западной части Ва-
ренцова моря или западно-гренландского района (табл. 139), где промы-
Таблииа 13!)
сел начался только в 1911 г.
Годы
Вылов в ш
1911—1916
1917—1925
1926—1929
1930
18—125
250—1000
2000-5500
8160
Наряду с отмеченными перемещениями центров мирового рыбного
промысла происходит и радикальная смена объектов промысла. Заме-
чательный пример этому дает развитие нашего промысла тихоокеанской
сардины (иваси) в Японском море за время с 1925 по 1940 г. (табл. 140):
Таблица 140
Годы Улов рыбы ' в тыс. и Годы । Улов рыбы В ТЫС. if
1925 4,4 1933 । 1 867,9
1926 13,0 1934 969,0
1927 128,0 1935 120(»,0
1928 354,5 1936 1054,9
1929 527,3 1937 1397,0
1930 777,2 1938 958,0
1931 898,9 1939 1095,5
1932 838,4 1940 1146.0
Однако в военные годы промысел иваси в Японском море совершенно
прекратился, так как иваси исчезла из Японского моря. Причины этого
замечательного явления неизвестны. В самые последние годы констати-
ровано вновь появление иваси в Японском море.
Громадное развитие рыбного промысла характерно также и для нашего
Берингова моря за последние несколько десятков лет. Если в 1913 г.
общий вылов давал здесь около 150 тыс. zz, то в 1937 г. он составлял около
3 млн. ц.
415
Причины всех этих изменений заключаются не только в открытии и
освоении новых районов и объектов промысла и усовершенствовании
техники лова, но иногда и в массовых изменениях географического рас-
пределения промыслового объекта. Как указывает Расс, «наименее
стойкими к воздействию промысла оказываются среди морских рыб виды,
более тесно связанные с жизнью на дне, — камбаловые и из тресковых
пикша. Значительно более устойчивы запасы трески. Наиболее же стой-
кими по отношению к промыслу являются, повидимому, запасы пелаги-
ческих рыб: сельдевых и скумбриевых. Колебания их урожайности и
изменчивость в подходах к побережьям пока, повидимому, перевешивают
влияние индустриализованного промысла».
В перемещении центров мирового рыбного промысла большую роль
может играть и подрыв сырьевой базы промысла в результате перелова
(Russell, 1945). Проблема перелова имеет две стороны: с одной сто-
роны может произойти уменьшение промыслового стада до той степени,
когда промысел становится нерентабельным, но объект промысла остается
во вполне жизненном состоянии и при запуске способен восстановить
первоначальное количество. При очень высокой технике лова и доступ-
ности объекта промысла может создаваться иное положение, при кото-
ром запасы будут подорваны до такой степени, что восстановление
стада окажется невозможным или потребует весьма длительного времени.
Может произойти и полное истребление промыслового объекта, как
это случилось со стеллеровой морской коровой и, безусловно, произо-
шло бы с ценными породами ластоногих, если бы не охранные меро-
приятия. Запасы китов в северной Атлантике, в шпицбергенских водах и
Баренцовом море чрезвычайно подорваны и, если в Антарктике не
будут проведены строгие правила охраны, киты будут уничтожены,
так как техника их вылова достигла очень высокой степени совершенства
(самолет и радары). Гораздо труднее определить состояние перелова
какого объекта, который пока недоступен для прямого наблюдения,
учета и очень эффективных способов вылова. Однако и в данном случае
возможно констатировать явление перелова. Так, например, 3. Р э с-
с е л л (1945) устанавливает состояние перелова пикши по статистике
промысла (рис. 254). Несмотря на неизменно идущее усовершенствование
техники тралового лова и изученность его объекта, вылов пикши не-
уклонно падает с 15,8 ц в 1919 г. до 2,6 ц в 1937 г. на день плавания
английского тральщика.
3. ПРОМЫСЕЛ КИТОВ 1
Киты — едва ли не самый доходный объект морского промысла (рис.
255). Китобойный промысел, начавшийся несколько сот лет назад у бере-
гов Европы и Японии, передвинулся затем в северные части обоих оке-
анов, а в настоящее время переместился в южное полушарие и сосредо-
точен, главным образом, в Антарктике. Техника китобойного промысла,
помимо давно уже применяемой знаменитой гарпунной пушки Фойна и
с недавних лет аэропланов, получила теперь еще мощное орудие—радар.
Только решительно проводимыми охранными мероприятиями можно
1 Мировой промысел ластоногих п дельфинов здесь не рассматривается ввиду отсутствия
сколько-нибудь полных статистических данных. Некоторые сведения по этому вопросу для
морей СССР приводятся во II томе.
’416
противодействовать полному истреблению беззащитных перед техникой
человека гигантов моря.
В 1936—1937 гг. мировой промысел китов дал свыше 51 000 китов
(около 5,5 млн. ц жира) и 67,3% этого количества было добыто в водах
Антарктики. Около 20% китов поймано в других районах южного полу-
шария (у побережий Южной Америки, Австралии и Африки). На север-
ную часть Тихого океана пришлось всего 6,3%, на северную часть Атлан-
тического — 4,2%.
Среди объектов китобойного промысла на первом месте стоит самый
крупный (до 33 м длиной) так называемый синий кит (Balaenoptera mus-
Рис. 255. Основные промысловые киты и их доля в мировом промысле (по Рассу).
1 — синие киты, 2 — финвалы, 3 — горбачи, 4 — сейвалы, 5— кашалоты, 6 — прочие.
cuius), почти наравне с ним — финвал (Balaenoptera physalus, до 27 м
длиной.) Второе место по значению в промысле занимают горбач ( Mega-
ptera nodosa), сейвал (Balaenoptera borealis) и кашалот (Physeter macro-
cephalus).
4. ПРОМЫСЕЛ БЕСПОЗВОНОЧНЫХ
Объем промысла. Рост промысла беспозвоночных идет еще более
интенсивным темпом, чем рыбного промысла. Недоиспользование сырье-
вых ресурсов нерыбных объектов (водоросли и беспозвоночные) колос-
сально. Если сейчас беспозвоночные составляют около 8% от рыбного мор-
ского промысла, то, несомненно, в дальнейшем соотношение будет изме-
няться в сторону роста относительного значения нерыбных объектов,
особенно тогда, когда будет освоена техника вылова и утилизации планк-
тона.
Из 12 млн. ц добываемых промыслом беспозвоночных свыше 62%
приходится на моллюсков и 30% на ракообразных. Остальное — это
иглокожие, губки, черви, кораллы и медузы. Из моллюсков только 3%
приходится на пресноводные формы. Две трети моллюсков добывается
в северном полушарии, причем в Тихом океане почти в два раза больше
чем в Атлантическом. Другое соотношение дают ракообразные: из 3,6
27 Зепкеппч т. I
417
млн. ц мирового улова в северном полушарии в Тихом океане вылавли-
вается 1,6 млн. zf. а в Атлантическом —1,9 млн. ц.
Рис. 256. Промысловые моллюски (из Расса).
1 — устрицы, 2 — мидии, 3 — петушок или тапес, 4 — сердцевидка, 5 — гребешок, 6 — сино-
ванула, 7 — морское ухо, 8— арка, 9— турбо, 10— хамагури, 11 — веиус. 12— песчаная ра-
кушка, 13 — литторина. 14— трубач, 15— морская бритва, 16— осьминог. 17 — кальмар, 18—
каракатица, 19 — пресноводные (перловица, беззубка и др.).
О размерах промыслов морских беспозвоночных в странах западной
Европы можно составить представление по данным за 1936 г. (в тыс. шил-
лингов), опубликованным в Статистическом бюллетене Международного
совета по изучению морей, издайном в 1938 г. (табл. 141).
418
Таблица 141
Страны Рыбы Морские беспозвоночные % от рыбного промысла
Бельгия 16 351 2 044 12,3
Великобритания 329 543 11744 5,5
Г ермания 172 128 5142 2,9
Голландия 45 853 4 985 10,9
Дания 35 488 1432 3,9
Норвегия 84 231 5 222 6,1
Португалия 36 409 1294 3,6
Франция 178 843 20 355 11,3
Швеция 28 271 1717 6,0
Гораздо большее значение имеют беспозвоночные в морском промысле
США. По данным Фидлера (Fiedler, 1938), в США и на Аляске
в 1936 г. было добыто 1 951 997 тп рыбы на сумму в 60 889 тыс. долларов
и 160 036 m беспозвоночных на сумму в 22 252 тыс. долларов. Таким обра-
зом, стоимость улова беспозвоночных в этой стране менее чем в три раза
уступает стоимости улова рыбы. Приблизительно такие же соотношения
получаются при сопоставлении данных по Японии, опубликованных
в Japan Jear Book за 1938—1939 г. Стоимость улова рыбы в Японии в
1936 г. по этим данным составляла 159 764 тыс. иен, стоимость улова бес-
позвоночных — 40 575 тыс. иен и стоимость водорослей —12 308 тыс. иен.
В СССР промысел морских беспозвоночных и водорослей проявляет
рост и сосредоточен почти исключительно на Дальнем Востоке, где,
по данным Т а р а н ц а (1939), в 1937 г. было добыто 246,5 тыс. ц
беспозвоночных и водорослей, из которых на долю камчатского краба
(Paralithodes camtschatica) приходится 186,1 тыс. ц. Добыча нерыб-
ных объектов составляла 9,5% по весу от добычи всего морского промысла.
Главный объект промысла среди беспозвоночных — моллюски и
ракообразные. Гораздо меньшее значение имеют губки, кораллы и игло-
кожие. Моллюски и ракообразные используются прежде всего как пи-
щевой продукт. Их питательность довольно значительна; по калорийности
они превосходят таких рыб, как сазан, треска и судак.
По Га в и н га (Having а, 1930), анализ мяса моллюсков л
ракообразных дает следующие результаты (в %) (табл. 142):
Таблица 142
Животные Вода Белки Жиры Углеводы 1 Зола 1 Б. кал. в 1 кг
Омар 81,84 14,49 1,84 0,12 1,71 842
Краб 78,81 15*83 1,32 2,42 1,62 939
Креветка свежая 79,29 14*88 0,80 2,19 2.84 847
Креветка сушеная 15,66 73*25 2,58 2,06 6,45 3692
Устрицы 80.52 9,04 2,04 6,44 1,96 863
Мидия сырая 86,74 8,66 1,31 2,16 1,43 607
Мидия вареная 77,30 16,80 2,36 1,45 2,09 1050
Сердцевидка 92,00 4,16 0,29 2 32 1,23 311
Морской гребешок 80,32 14,75 0,17 3,38 1,38 829
Пищевая ценность мяса моллюсков и ракообразных определяется
также высоким содержанием некоторых минеральных веществ, в част-
ности, иода (табл. 143).
419
Таблииа 143
СОДЕРЖАНИЕ ИОДА (частой пода на 1 млн. частей тела)
(но Ilavinga)
Консервиро- ванные Свежие
Омар 5 320 11 500
Краб 3 150
Креветка 1140 2 250
Устрица 4000 6000
Треска 1410 1000
Масло, мясо, хлебные злаки, фрукты. менее 250
Мясо устриц оказалось очень богатым витаминами А и В, несколько
меньше в нем витаминов С и D.
В мясе Pecten констатировано наличие витаминов А и D, причем
концентрация первого выше, чем в мясе млекопитающих и рыб.
Из раковин моллюсков, употребляемых в пищу, и из панцырей! рако-
образных изготовляется кормовая мука для птиц и удобрение.
Моллюски (рис. 256). Моллюски употреблялись в пищу человеком
еще в доисторические времена. Их раковины постоянно встречаются в так
называемых «кухонных кучах» доисторического человека, жившего у
берега моря. Так, например, при раскопках стоянок палеолитического
человека в Крыму было обнаружено большое количество раковин устриц,
мидий и гребешка, т. е. промышляемых и в настоящее время видов.
В качестве объектов мирового устричного промысла в разных районах
мирового океана имеют значение около 10 видов устриц (О. edulis, О. ап-
gulata, О. taurica, О. pUcata, О. cristata, О. virginica, О. lurida, О, gigas,
О. laperousi и О. chiloensis). Промысел дает около 1,6 млн. ц, которые
распределяются примерно поровну между Тихим и Атлантическим оке-
анами. Количество устриц, добываемых по западному и восточному побе-
режьям Атлантического океана, приблизительно одинаково; что же ка-
сается Тихого океана, то у его западных побережий добывается в 20 раз
больше устриц, чем у восточных (табл. 144).
Таблица 111
Страна
Улов
в тыс. и
Страна
Улов
{ в тыс. и
Япония ................
США..................
Франция .............
Китай................
716
(84°/0 путем
искусственного
разведения)
375
313
112
Новая Зеландия..........| 70
Австратя................! 30—45
Канада..................1 1
Великобритания..........1 J , ппо. о
Испания.................: ) 10-12
Таблица 146
Страна
Вылов
в тыс. и
Страна
Вылов
в тыс. и
Голландия.................I
Франция....................
Италия.....................
Великобритания ............
615
170
77
65
Испания
Германия
Дания . .
50
on
!
420
Второй по значению промысловый объект из двустворчатых моллю-
сков — мидия (М. edalis, М. cunaliculatas, М. dunckeri, М. caltforniantis,
М. magellanicus и М. ungulatus) — дает свыше 1 млн. ц вылова и свыше
90% из этого количества добывается в европейских морях (табл. 145).
На третьем месте стоит «петушок» (Tapes decussatus и Т. philippiiw-
тит), дающий так же, как и мидия, около 1 млн. причем свыше 90%
добывала Япония — на 50% за счет искусственного разведения. Осталь-
ная часть промысла приходится на Испанию и Португалию. Прочие
двустворчатые моллюски значительно уступают трем первььм (табл. 146).
Таблица 146
Моллюск
Мировой вылов ! Основные страны и их вылов
в тыс. ц i в тыс. ц
Cardium edule и С. corbis
Pecten maximus. Р. Irradians,
Р. aequisulcatus, Р. magella-
nicus и Р. jessoensls
Slnovacula constrlcta
Около 300
(9О°/о в Атлантике)
175
(26% в Атлантике)
136
Португалия (1171 Великобрита-
ния (ИЗ)
США (29), Великобритания (11),
Япония (117) и СССР (9)
Китай (136)
Кроме того, в Тихом океане существенными объектами промысла
являются «морское ушко» (Haliotis giganteus и другие виды), «ковчег»
(Area inflata), «турбо» (Turbo cornutus) и«хамагури»’(Meretrix meretrix),
добываемые главным образом в Китае и Японии и дававшие по 60—
80 тыс. ц. В гораздо меньших количествах промышляются многие другие
двустворчатые и брюхоногие моллюски.
Головоногие моллюски по своему промысловому значению конкурируют
с устрицей, мидией и «петушком». Мировой их промысел выражается
величиной 1,7 млн. причем свыше 90% этого количества приходится
на Тихий океан, главным образом, на его азиатское побережье (табл. 147).
Таблица 117
Вед Мировой npo- | мысел в тыс. it \ 1 Основные страны и их I вылов в тыс. ц
Осьминоги (Octopus vulgaris, О. tuberculatus, О. punctatus и О. hongkongensis) 260 Япония (230)
Кальмары (Lollgo vulgaris, L. media, L. opa- lescens, L. pealel, Todarodes sagittalus, Todaropsls veranyi, Ommatostrephes sloanei >300 Япония (230), Китай (20), США (30), Европа (20)
pacificus) Каракатицы (Seplella japonica, Sepia offici- nalis, Sepiola rondeleti, Rossia macrosoma) >1100 Китай (660), Япония (100), Европа (40—50)
О размере промысла моллюсков в европейских водах можно судить
по данным за 1936 г. (табл. 148, в тыс. шт., гектолитров и кг)>
Гораздо разнообразнее ассортимент промысловых моллюсков США 1
(табл. 149).
1 Суммированные данные по Новой Англии и Атлантическому побережью ва 1935 г., по
заливу Чезеиик, южно-атлантическому и тихоокеанскому побережьям и по Аляске за 1936 г.
421
Таблица 148
Страна Ostrea Mytilus edulls Саг- dlum edule Buccl- num unda- tum Pecten m axi- oms Litto- rina litto- rea
шт. 1 кг гкл | кг
Великобритания Германия Голландия Дания Норвегия Португалия Франция Швеция | 23297 1 565 I 400 I 6344 520945 259 7 1075 335 । 7 181 2 037 61506 2 074 464 11311 — 1 292 12 352 = 1766 — 1 374 1 1 1 i 1 1 1 £ 2196 1 1 - 1
Всего 551810 1084 335 173 862 23 955 2140 312 2196
Таблица 149
flallotis..........................................
Donax variables....................................
Tivela и Venus.....................................
Slllqua patula................................
Mya arenaria..................................
Mactra........................................
Pecten....................................
Mytilus........................................
Ostrea.........................................
Cardlum и Littorina............................
Strom bus и Busycon.........................
Loltgo......................................
Octopus.....................................
Всего ... .............
Улов в m Стоимость в тыс. долл.
| 299,6 93
; i’8 1
6 530,8 1853
1038,7 194
5295,9 710
380,4 37
2893,8 924
133,0 11
37 460,9 7 533
72,2 7
7,7 1
1956,7 575
73,5 8
56 145,0 11907
В СССР моллюски промышляются в настоящее время только на Даль-
нем Востоке, преимущественно в южном Приморье и в незначительной
степени на Черном море. По данным Разина (1934), добыча мол-
люсков на ДальнехМ Востоке выражается в следующих цифрах (табл. 150).
Таблица 150
ДОБЫЧА ПО ГОДАМ (в центнерах)
1927 1928 1 1929 I 1931 | 1932 1933
Pecten jessoensis 397 430 417 2 584 2194 6100
Mytilus dunkeri 88 397 1 862 3000
Spisula sachalinensls 85 246 3 420 3 102 2 350
Ostrea 47 36 О
Ommatostrephes i 867 31
Octopus 1 — 6
Прочие моллюски — 13,6 — —
Всего 617 1 725,6 | 417 (?), 6 401 J j 8025 [ It 487
(22
В последующие 5 лет, по данным Т а р а н ц а, в среднем за год
добывалось 9204 Pecten jessoensis и 1303 ц Mytilus dunkeri.
Как видно из этих данных^ улов моллюсков в СССР по сравнению
с заграницей ничтожен. Между тем возможности для развития моллюс-
кового промысла громадны, как показывают проведенные в советских
морях работы по учету сырьевой базы промысловых моллюсков.
По подсчетам Разина, в заливе Петра Великого (Японское море)
132—145 га площади дна занято промысловыми скоплениями устриц,
запасы которых составляют здесь 6—9 млн. экз. (из них 4—5,5 млн.
экз. промыслового размера). Pecten jessoensis в Южном Приморье встре-
чается на площади в 100 тыс. га; на площади 15 226 га сосредоточены
промысловые его скопления. Запасы этого моллюска определяются
Разиным в 60 млн. экз. Spisula sachalinensis занимает меньшую
площадь (1418 га), но запасы ее также довольно значительны (около
52 млн. экз.). Помимо перечисленных видов, в дальневосточных морях
весьма многочисленны другие моллюски, которые могут иметь немалое
промысловое значение (Macrocallista chishimana, Mercenaria stimpsoni^
Spisula grajana, Cardium calijorniense, Paphia philippinarum), но промы-
сел их еще не налажен и запасы не учтены.
Довольно значительные запасы промысловых моллюсков сосредо-
точены в Черном море. По данным Воробьева, в Черном море
запасы мидий составляют много миллионов тонн, из которых несколько
миллионов могут иметь промысловое значение. Никитин (1933),
занимавшийся черноморскими устрицами, описал устричные банки в
северо-западной части моря, в районе Севастополя, Керченского пролива
и на кавказском побережье у Гудаут. Общая площадь устричных банок
у черноморских берегов СССР составляет 33 000 га, населенных 75 млн.
устриц промыслового размера. Биомасса устриц равняется 26 000
а вес чистого их мяса —4000 ц. В 900-х годах, по данным Карпова,
добыча устриц по черноморскому побережью России достигала 9—
10 млн. шт. в год. Однако в результате хищнического лова устричные банки
угасали, устрицы мельчали, и во время империалистической и гра-
жданской войн промысел совершенно прекратился. По сведениям Ш п а р-
линского, в 1929 г. в районе Севастополя был восстановлен один
устричный завод и зимой 1929/30 г. на нем имелось около 1 млн. устриц,
но «история восстановления севастопольского устричного промысла не
блещет, к сожалению, какими-либо успехами» (Ш парлинский,
1932). Причины угасания черноморского устричного промысла, несо-
мненно, заключаются в примитивном способе ведения хозяйства. Никаких
мероприятий по разведению и выращиванию устриц не проводилось;
в лучшем случае взрослые устрицы выдерживались на так называемых
заводах для улучшения качества мяса и, главным образом, для регули-
рования рынка путем равномерного выпуска продукции. Тем не менее,
значительные запасы черноморских устриц позволяют и сейчас организо-
вать их промысел, который должен базироваться не только на вылове
устриц, но и на их разведении.
В Баренцовом и Белом морях можно без труда организовать промы-
сел мидий и литорин, в громадном количестве обитающих на литорали.
Лов промысловых моллюсков производится при помощи разного
рода драг, сачков, граблей и щипцов. В некоторых случаях (Haliotis
в США, Pecten) моллюски добываются водолазами. Обычно для лова
моллюсков применяют мелкие суда, но в США устриц промышляют с
больших пароходов, снабженных шестью большими драгами. Для ловли
головоногих моллюсков применяются ставные снасти, в которых крючки
заменены горшками, куда залезают головоногие.
423
Довольно сильно развито и разведение промысловых моллюсков.
Здесь прежде всего надо упомянуть об устричном хозяйстве, имеющем
Рис. 257. Промысловые морские ракообразные (из Расса).
1 — омар и лангуст. 2 — различные другие «морские раки»: а — «норвежский омар», б — «колю-
чий омар», .3 — креветки: а — Crandon, б — Penaeus, в — Pandalus, г — Leander. О — Palaemun,
е — Artemisia. 4— крабы: а—камчатский краб, б — крабы-плавунцы Charybdis, Callinectes,
в — Cancer, г— Menippc, Cheiragonus, Chionocoetes и др., 5 — речные раки (Astacus, Cambarus).
6—морская уточка Pollicipcs cornucopia.
многовековую историю. Еще до основания римской империи римляне раз-
водили устриц, которые доставлялись из Бретани. Китайцы начали зани-
маться устричной культурой еще раньше.
424
Устричная культура в настоящее время заключается в том, что моло-
дых устриц пересаживают на специально подготовленные отмели, где они
попадают в значительно лучшие условия, чем в своих естественных био-
топах. Другой, еще более совершенный способ — это широко применяю-
щееся в западной Европе, США и Японии погружение в море в районе
размножения устриц фашин, черепиц, пустых устричных створок и т. д.,
на которые оседают планктонные личинки устриц. Затем осевшие личинки
вместе с искусственным субстратом переносятся на отмели или, если
условия существования молоди в местах оседания личинок на искус-
ственный субстрат удовлетворяют предъявляемым требованиям, оста-
ются на месте. Устричные банки нуждаются в постоянном уходе. На них
периодически производится при помощи специальных швабр вылов
нападающих на устриц морских звезд, иногда для защиты от хищных
рыб дно огораживается частой изгородью. Устричное хозяйство дает,
как правило, больше продукции, чем лов «диких» устриц. В 1936 г. Япо-
ния получила от культурного хозяйства 59 028 750 кг устриц на сумму
1857 тыс. иен.
Кроме устриц, в довольно широких масштабах культивируются и
другие виды промысловых моллюсков: мидия, жемчужница, Tapes phi-
lippinarum. Особенно развито разведение моллюсков в Японии, где
в 1936 г. таким образом было получено:
раковин жемчужницы 139 751 тыс. шт. на сумму 905 тыс. иен
жемчуга 26 520 » » » » 983 » »
Tapes philippi пагит 42 675 » кг » » 890 » »
Методика искусственного получения жемчуга в раковинах разрабо-
тана японцем Микимото. Она заключается в том, что какой-нибудь
твердый предмет зашивается в кусок мантии и подкладывается под мантию
другого моллюска. Таким способом удается получать крупные и ценные
жемчужины, неотличимые от возникших естественным путем.
Промысловые моллюски поступают на рынок в свежем виде или в виде
консервов, реже —в сушеном и соленом виде. Особенно большое развитие
получило консервирование моллюсков в США, где в 1936 г. было изго-
товлено приводимое* ниже количество консервов (табл. 151):
Таблица 151
Устрицы . . . . 3 600,5 т 2180869 долл.
Суп из устриц .... 772,0 181201 »
Кальмар .... 175,8 30708 >
Прочие моллюски . . . . 8 681,9 2 976 297 »
Всего .... 13 230,2 т 5 369 075 долл.
Остающиеся при производстве консервов раковины стоили 5844094 долл.
Они были употреблены на производство пуговиц, кормовой муки,
удобрения и т. д.
Ракообразные. Промысел морских ракообразных (рис. 257) имеет
также большое значение. Крабы, омары, лангусты и креветки, как высо-
кокачественный пищевой продукт, находят себе многочисленных потре-
бителей. Использование отходов промысла в качестве технического сырья
еще не получило достаточного развития. Однако в США из панцырей рако-
образных изготовляется кормовая мука для птиц; в США и в Японии
делались существенные по своему значению и интересу попытки перера-
ботки хитина ракообразных в шелк (преимущественно для нужд военной
промышленности).
Наибольшее промысловое значение среди ракообразных имеют омар
(Homarus vulgaris и Н. americanus),‘лангуст ( Palinurus vulgaris) и много-
425
численные формы креветок (виды родов Pandalus, Crangon, Leander
и др.). Омара добывается около 300 тыс. лангуста — около 10 тыс. ц,
у берегов Европы — около 15 тыс. ц так называемого «норвежского»
омара (Nephrops norvegicus ) „ а в Тихом океане — «колючего» омара (Ра-
linurus argus и Р. interrupts) 14—17 тыс. ц.
Мировой улов креветок (Crangon, Penaeus, Parapenaeus, Pandalus,
Pandalopsis, Palaemon) превышает 1,5 млн. ц. На две трети эта добыча
приходится на Атлантический океан. В Тихом океане ловят также пред-
ставителей отряда Stomatopoda (Squilla).
Промысел ракообразных в странах западной Европы за 1936 г. хара-
ктеризуется следующими цифрами (табл. 152):
Таблица 152
Страна
Homarus vulgaris Neph- rops norve- gicus Крабы Pandalus, Leanderи Palaemon Cran- gon cran- gon
тыс. тыс. i I тыс. тыс. тыс.
шт. кг кг шт. кг
Великобритания..................
Германия .......................
Голландия ......................
Дания...........................
Исландия........................
Норвегия........................
Португалия .....................
Франция.........................
Швеция..........................
Бельгия ........................
Всего....................
2 207 — 316 i 457 — 24 1965
— 216 — — 28 18 33 842
—. 22 — —. 72 — 3041
— 161 550 — — 533 —
— — — — — 40 —
— 1037 — 4 640 — 3 472 —
980 — — — 10500 51 —-
— 565 — — 2 562 137 2 027
— 259 405 — 138 1 075 13
— 5 — — 44 — 3 495
3187 2 295 1271. 12097 13 444 ; 5350 44 383
Таблица 163
В США в 1936 г. было выловлено (табл. 153):
Тонн
Стоимость
в тыс. долл.
Calllnectes sapldus|
Cancer sp. sp. J......................
Menlppe...............................
Ilomarus..............................
Palinurus.............................
Pandalus, Pandalopsis)
Penaeus, Crangon J....................
43 760,0
20,9
5 219,0
754,5
55189,0
2 761
9
2 659
168
3 873
Всего.....................
104 943,4
9 470
Стоимость улова морских ракообразных за тот же год у берегов Япо-
нии составила 10 840 тыс. иен. Большое значение в этой стране имеет
промысел так называемого камчатского краба (Paralithodes camtschatica).
Мировой крабовый промысел дает около 1,4 млн. ц (64% в Тихом
океане). В Тихом океане основным объектом крабового промысла являет-
ся так называемый «камчатский краб» (Paralithodes comtschatica и Р. pla-
typus )9 дающий 570—670 тыс.4 ц. Вследствие высоких вкусовых качеств
426
издавна человек добывал из моря многие виды крабов (Charybdis,
Cluonocoetes, Erimacrus, Cheiragonas, Eriocheir, Cancer, Callinectes, Menippe,
Lithodes и др.). Кроме ракообразных, существует также промысел мече-
хвостов (Limulus), вылавливаемых в количестве 14—15 тыс. ц.
О размерах промысла в 1936 г. можно судить по следующим данным,
заимствованным из «Japan Year Book» за 1933—1939 гг. (табл. 154):
Таблица 151
Число судов.......................... 9
Их тоннаж......................... 36 737
Рыбаков............................ 3 243
Число выловленных крабов (в шту-
ках) ......................* . 13 948 000
Количество ящиков консервов. . . 184000
Стоимость продукции............ 9 490000 иен
Из этого количества в советских водах было поймано 6 565 000 кра-
бов, из которых изготовлено 50 000 ящиков консервов на 2,6 млн. иен.
Промысел камчатского краба в полной мере освоен и в СССР. Он воз-
ник сравнительно недавно (в 1928 г.) и сначала на советских краболовах
работали японские рабочие. По мере освоения техники промысла японская
рабочая сила постепенно сменялась русской. Параллельно с этим воз-
растала мощность советского краболовного флота; японские капиталисты
оказались не в состоянии конкурировать с этой молодой отраслью совет-
ского хозяйства и были вынуждены уменьшить численность своего фло-
та. Этот процесс хорошо иллюстрирует табл. 155, приведенная Зна-
менским (1931):
Таблица 155
Годы Число советских краболовов Число японских краболовов
с лионской рабо- 1 чей силой : с русской рабо- чей силой всего
1928 2 2 22
1929 4 3 7 19
1930 4 7 11 15
1931 2 8 10 9
В настоящее время весь советский промысел обслуживается исклю-
чительно русской рабочей силой. Число занятых им рабочих рук прибли-
зительно такое же, как во всем тресковом промысле наших северных
морей. Основным районом краболовного промысла является западное по-
бережье Камчатки. Насколько быстро растет советский лов крабов в этом
районе, видно из данных, приводимых Поповым (в штуках) (табл.
156).
Таблица 156
»
1923 696 630 1927 5121110
1924 964430 1928 6 759050
1925 2963100 1929 7 845 360
1926 4 454 090
Помимо крупных пловучих крабоконсервных заводов (краболовов),
краба ловят и заготовляют также береговые заводы. Динамику продук-
ции береговых заводов Приморья можно иллюстрировать следующими
данными, приводимыми Заксом (в ящиках с консервными банками
из расчета 55—60 крабов на ящик) (табл. 157)<
427
Таблица 157
Годы | Ящики | Годы Ящики
1928 2 746 1931 6 512
1929 8 405 1932 5 678
1930 6 500 1933 5 333
Общая добыча камчатского краба в СССР составляла в 1935 г. 128 тыс. ii
(по Виноградову), в 1937 г. — 186 тыс. ц (по Та р а н ц у),
Лов камчатского краба производится при помощи крупноячеистых
ставных сетей, которые ставятся нижней подборой с грузилами на дно
и наплавами на верхней подборе поддерживаются в вертикальном положе-
нии. Крабы запутываются в этих сетях и затем вместе с сетями выбираются
на борт судна.
Сети выставляются и выбираются мелкими моторными баржами —
кавасаки, для которых пароход-краболов (или береговой завод) служит
базой. Во время переходов краболова, ночью и в шторм кавасаки, в коли-
честве 8—10 шт., висят на шлюпбалках вдоль его бортов. Утром и днем
они привозят на краболов сети с запутавшимися крабами и уже на палу-
бе краболова производится выпутывание крабов из сетей, отрыванье ног,
которые только и годятся для консервирования, первая варка ног, их
рубка, вытряхивание из них мяса и, наконец, сортировка мяса из разных
члеников ног. В каждую банку мясо идет, согласно требованиям стан-
дарта, в строго определенном соотношении видов и кусков по точному
контрольному весу. В трюме краболова, где находится консервный завод,
на конвейере отбирается ассортимент для каждой банки и производится
укладка в них мяса, их предварительная закатка, вакуумизация, окон-
чательная закатка и стерилизация в автоклаве.
Помимо камчатского краба, на советском Дальнем Востоке, преиму-
щественно в южном Приморье, в очень небольшом количестве промы-
шляют креветок (главным образом Pandalas kessleri и Р. hypsinoUis},
О размерах этого промысла можно судить по следующим данным, заим-
ствованным у разных авторов (табл. 158):
Таблица 158
1927 4 246 кг
1928 10 600 »
1929 7 417 >
1935 38000 кг
1933—38
(в среднем
за год) 45 000 >
Промысел креветок на Дальнем Востоке может быть сильно увеличен
как за счет уже промышляемых видов, так и за счет освоения громадных
сырьевых ресурсов еще не затронутых промыслом видов, в первую оче-
редь крупного (до 30 см) Sclerocrangon salebrosa. Кроме того, вполне воз-
можно использование в качестве промысловых объектов некоторых мас-
совых дальневосточных крабов—Chionocoetes opilio, Charybdis japonica,
Thelmessus cheiragonas, Erimacras isenbecki, Eriocheir japonicus и др.
Сырьевые ресурсы промысловых ракообразных в других морях СССР
также довольно значительны, но они остаются до сих пор неиспользо-
ванными. По подсчетам Никитина, промысловая биомасса наиболее
многочисленной креветки Азовского и Черного морей Leander adsper-
sus составляет около 40 тыс. ц, т. е. может служить базой для весьма
интенсивного промысла. Попав недавно в Каспийское море вместе с дру-
гим видом — L. squilla, эта креветка и там встречается сейчас в промы-
словых количествах. Кроме L. adspersus, объектами промысла в Черном
428
море могут служить крабы Carcinus' maenas и Eriphia spinifrons, а в
Каспии — раки Astacus pachypus и Л. leptodactylus caspias. В Баренцевом
море, как указал еще Книпович, имеются большие запасы креветки
Pandalus borealis, которая является объектом интенсивного промысла
в соседней Норвегии.
Орудиями лова креветок служат бимтралы, оттертралы и неводы,
крабов — драги, сачки и переметы, омаров и лангустов — ловушки,
вроде рачней. Улов поступает к потребителю или в свежем или обычно
в консервированном, реже — в сушеном виде. В 1936 г. в США было
изготовлено 15 365 884 фунт, консервов из креветок стоимостью в
4 672 198 долл, и 350400 фунт, консервов из крабов стоимостью в
130 753 долл.
Ракообразные являются не только объектами, но и вредителями про-
мысла. Црлый ряд бокоплавов в Каспийском и, в меньшей степени, в Азов-
Рис. 258. Различные промысловые беспозвоночные (из Расса).
1 — мечехвост, 2 — морские ежи, 3 — голотурии, 4 — губки, 5 — медузы, 6 — кораллы
и гидроиды, 7 — черви, 8 — асцидии.
ском море объедает сети и попавшую в них рыбу. Такую же роль играет
Mesidothea entomon в предустьевых участках в наших северных морях. В
западной Европе, особенно в Германии, настоящим бедствием для рыбного
промысла является недавно попавший в европейские воды китайский
краб Eriocheir sinensis. Размножившись в колоссальном количестве,
этот краб объедает наживку рыболовных снастей, рвет сети; попадаясь
массами, забивает сети и мнет попавшуюся вместе с ним рыбу. Расходы
по рыбным промыслам (главным образом, приходящиеся на ремонт се-
тей) с его появлением возросли вдвое. Помимо того, он является опасным
конкурентом промысловых рыб в отношении пищи. В Германии ведется
энергичная борьба с китайским крабом: сконструированы разного рода
ловушки, всячески поощряется его уничтожение. В 1933 г. в Германии
было уничтожено 1000000 кг краба, и тем не менее он встречался попреж-
нему в колоссальных количествах. Eriocheir sinensis зарегистрирован и в
пределах СССР (в районе Випури, в Рижском заливе) и, несомненно, прине-
сет со временем немалый вред советскому рыболовству в Балтийских водах.
429
Иглокожие. Из других промысловых беспозвоночных (рис. 258) сле-
дует упомянуть об иглокожих. Из иглокожих промысловое значение
имеют прежде всего морские ежи, которых добывают, главным образом,
в Средиземном море (Echinus esculentus, Strongylocentrotus lividus, Loxe-
chinus albus) в количестве свыше 10 млн. шт. (Франция, Алжир, Тунис,
Испания), а также по тихоокеанскому побережью Америки.
Промысел голотурий — трепанга (Stichopus japonicus, Thyone bnu-
rcus, Cucumaria frondosa) существует в Китае, Японии, США и СССР
и дает 6—10 тыс. и. В США в последние годы стали использовать морских
звезд (Asterias) для изготовления кормовой муки для домашней птицы.
Вылавливание морских звезд производится вблизи устричных банок,
где морские звезды скопляются в колоссальных количествах.
В советских дальневосточных водах существует небольшой промысел
голотурий — трепанга (Stichopus japonicus) и морского огурца (Сиси-
maria japonica). Голотурии добываются водолазами, затем улов сушат
и экспортируют в Китай и Японию. За 1933—1938 гг. добывалось в среднем
5 663 ц трепанга ежегодно. Голотурий в большом количестве промы-
шляют в Японии и США. Во Франции и США объектом небольшого про-
мысла являются морские ежи, гонады которых употребляются в пищу.
Губки, кишечнополостные и др. Промысел губок сосредоточен, глав-
ным образом, в Средиземном и Адриатическом морях (3 тыс. ц), отчасти
в США, на островах Багамских и Кубе (2,3 тыс. и)- Губки добываются
или специальными драгами или при помощи водолазов, затем губки сушат
и в таком виде продают.
Объектами губочного промысла является несколько видов роговых
губок (Euspongia ojjicinalis, Hippospongia equina, H. canaliculata, Spon-
gia graminea).
Промысел благородного коралла (Corallium rubrum), так же как и
промысел туалетных губок, издавна процветал па Средиземном море,
а в последнее время был перенесен и в Японию. Кроме коралла, из ки-
шечнополостных в Китае промышляют медуз (Rhopilema esculenta),
которых употребляют в пищу в соленом виде, а в Германии — гидро-
идов (Sertularia и Hydrallmania). У островов Самоа употребляют в пищу
пелагические стадии донного червя Eunice viridis из полихет.
Большое количество беспозвоночных и мелких непромысловых рыб
вылавливается для употребления в качестве наживки. Среди полихет
известностью в этом отношении пользуются пескожил (Arenicola marina)
и некоторые другие (Nereis, Glycera).
Существует также небольшой промысел асцидий ( Microcosmus, Руига).
Суммарная промысловая продуктивность водных животных морей
Евразии показана на рис. 252.
Планктон. Большое будущее принадлежит использованию морского
планктона как для целей питания, так и для получения технического
сырья и медицинских препаратов (витамины, лечебный жир и т. д.).
В северных частях Атлантического и Тихого океанов, а также в Антарк-
тике, у нас в Баренцовом море и в морях Дальнего Востока в периоды
наибольшей биомассы планктона в 1 м* воды может содержаться 0,5—0,8
еле и более зоопланктона.
Если принять, что суточный пищевой рацион человека составляет в
среднем 3000 кал, то это соответствует примерно 750 г сухого планктона,
который можно получить из 6—8 тыс. л/3 воды. При наиболее благопри-
ятных условиях в Баренцовом море такое количество планктона может
быть получено планктонной сетью диаметром в 1 м в течение шестича-
сового лова. Этот расчет показывает, что едва ли может быть рентабельна
добыча планктона сетями, как бы велики они ни были.
430
Перед войной в «Известиях» (14/ХП 1940 г.) появилась информацион-
ная заметка о том, что профессор Вашингтонского университета К и н-
кейд разрабатывает метод получения китового жира из планктона.
В лабораторных условиях Кинкейдом это уже сделано. Сообщалось
также, что в Берингово море будет послан пароход с мощными компрес-
сорами, помпами, центрифугами и другим оборудованием для добычи
и переработки планктона.
5. ПРОМЫСЕЛ МОРСКИХ РАСТЕНИЙ
Помимо беспозвоночных, большое хозяйственное значение в некоторых
странах имеют морские водоросли и цветковые растения. Они исполь-
зуются как пищевой продукт (Laminaria, Viva, Porphyra), как сырье
для производства агар-агара (Ahnfeltia) и иода (Phyllophora, Laminaria)
и в мебельной промышленности для набивки матрацов, диванов и пр.
(Zostera, Phyllospadix). В Японии в 1936 г. было добыто 482 569 тыс. т
водорослей стоимостью в 12 308 тыс. иен. Кроме того, путем искусствен-
ного разведения было получено водорослей на 8 565 тыс. иен. Основными
странами, добывающими иод из водорослей, являются Чили (80%),
Ява (9%) и Япония (7%).
В СССР промысел морских водорослей существует на Дальнем Востоке,
на Белом и Черном морях и в меньшей степени на Каспийском море.
По данным Таранца, рост водорослевого промысла на советском Даль-
нем Востоке выражается в следующих цифрах (в центнерах) (табл. 159):
Таблица 159
— . Годы Виды водорослей -- 1926 1933 1934 1935 1936 1937
Laminaria Ahnfeltia Zostera и PInllospadix .... l 6 239 53 990 156 5 800 40 448 1 118 5 467 43 576 1882 1684 22013 6 647 4605 16 939 5 410 9 797
Всего | 6 239 59946 | 47 033 47 142 33265 32 146
Дальневосточный промысел Ahnfeltia позволил целиком обеспечить
страну агар-агаром и освободиться в этом отношении от импорта. В на-
стоящее время на наших северных морях добывается только Laminaria
(Белое море). По данным Ведр и некого, в год здесь промышляется
около 50 000 т ламинарий. Из ламинарий добывают альгины, применя-
емые в металлообрабатывающей промышленности. Ламинарии исполь-
зуются при изготовлении изоляционных плит, из них получают также
клеевое вещество и иод. Запасы ламинарий в Белом море определяются
величиной порядка 1,5 млн. т сырого веса (М е й е р) при средней биомассе
8—10 яг/.и2.
На Белом море1 имеется также значительная сырьевая база для про-
мысла Ahnfeltia, во всяком случае не менее 11 000 т. Количество лами-
нарий на Мурманском побережье — не менее 500—600 тыс. /п, а в Белом
море — до 800 тыс. т.
Из цветковых морских растений большое промысловое значение
имеют «морская трава» (Zostera marina и Z. папа в Черном и Азовском
морях, Z. папа в Каспийском море, Phyllospadix Scouleri в Японском море).
1 Данные по Белому морю любезно сообщены директором Водорослевого института в Архан-
гельске К. П. Г е м п.
431
Морская трава находит применение в мебельном производстве и как упа-
ковочный материал. Данные о разведанных сырьевых запасах морской
травы в морях СССР приведены в табл. 160 (в тыс. т сухого веса).
Таблица 160
Водоем | Pin llospadlx i Zostera । marina Z. nana 1 Всего
Белое море . | i 50 — 50
Черное море . . . — 25 .—.
Азовское (Генический район) . . . . . । — 8 17 25
Каспийское море . . — i 70 70
Дальневосточное побережье . . . . . .1 13 — 13
Всего ' 13 । 83 57 1 153
Промысловые участки зостеры и филлофоры на Черном море со-
средоточены, главным образом, в северо-западной части и в Каркинит-
ском заливе. Запасы Phyllophora rube.ns исчисляются здесь величиной
порядка 10—15 млн. т сырого веса. В Каспийском море зостера в зна-
чительном количестве встречается в Северном Каспии (о. Долгий, о.Ку-
лалы) и на западном побережье в районе Апшеронского п-ва и у о. Че-
чень. Ежегодно близ Баку добывается около 4000 т сухой травы; при-
мерно такое же количество морской травы добывается и в Азовском море
(Утлюкский лиман и Сиваш).
Нет никакого сомнения в том, что советский водорослевый промысел
в очень слабой степени использует имеющиеся сырьевые ресурсы. По
данным Г а й л а, в Приморье «ежегодно эксплоатируется лишь 20—25%
промысловой производительности района». Запасы дальневосточных водо-
рослей выяснены Г а й л о м (цитировано по Т а р а н ц у). Для
Laminaria получены следующие показатели (табл. 161):
Таблица 161
Районы Площадь в га Ежегодная промыс- ловая производи- тельность сухого вещества в 'тыс. ц
Приморье Сахалин Охотское побережье Камчатка 15 000 1400 37 500 50000 375,0 22,5 937,5 1250,0
Всего 103900 1 2 585,0
Phyllospadix занимает в Приморье площадь в 1696 га; Ahnfeltia, содер-
жащая 12—15% агар-агара, в заливе Петра Великого растет на площади
в 4000 га и запасы ее исчисляются в 12 тыс. ц. Кроме того, в дальне-
восточных морях имеются значительные запасы Ulva lactaca, богатой
витаминами и способной заменить салат, а также съедобной Porphyra.
В Белом море сосредоточены громадные запасы Laminaria, составляющие
по Мейеру 1,5 млн. т. Если учесть, что Laminaria saccharina содер-
жит 0,264% иода (на сухое вещество), a L. digitala — 0,347%, то можно
себе легко представить, какими неограниченными возможностями' рас-
полагает наша иодная промышленность.
ГЛАВ А XVII
ЗНАЧЕНИЕ МОРСКИХ ОРГАНИЗМОВ В СУДОХОДСТВЕ
Обрастание Ч Вследствие обрастания днищ судов морскими организ-
мами увеличивается трение о воду, значительно снижается скорость
судна и, следовательно, резко возрастают расходы по его эксплоатации.
Особенно резко сказывается на уменьшении скорости судов то обстоя-
тельство, что наиболее сильно обрастают участки днища около винта
н руля. По подсчетам, произведенным в США в 1929 г., убыток от обра-
станий по торговому флоту этой страны составляет не менее 100 млн.
долл, ежегодно. Суда несут потери в скорости до 50%, в горючем — до
40%, во времени — до 50%.
Обрастанию благоприятствуют следующие условия: 1) длительность
пребывания в гаванях; 2) гидрологически однородный режим плаванья;
3) высокая температура и постоянная соленость воды; 4) наличие в га-
ванях скал, молов и пр., на которых имеются обрастания; 5) редкое до-
кование; 6) темная окраска днища; 7) неровная поверхность днища.
Это делает понятным результаты анализа интенсивности обрастания
различного типа судов, который показал, что менее всего обрастают суда,
проводящие минимум времени в портах, т. е. пассажирские суда, затем —
грузовые суда, крейсеры, угольщики, линкоры и в наихудшем положении
оказываются мало подвижные плову чие маяки.
Об интенсивности обрастания можно судить по тому, что на Мурмане
вес обрастания в течение 3 месяцев достигает 710,6 г/м2, а в Черном море
в 15—25 раз превосходит эту величину. В тропических морях процесс
обрастания совершается еще более интенсивно. В Черном море В. Н и-
к и т и н ы м были констатированы многолетние обрастания мощностью
свыше 90 кг!м\ Основными компонентами обрастания являются
усоногие раки, мшанки, гидроиды, асцидии, моллюски и водоросли.
Меры борьбы с обрастанияхми до сих пор мало разработаны. Приме-
нение различных, подчас очень дорогих защитных красок не дает нужного
эффекта. Поэтому приходится периодически прибегать к докованию,
при котором обрастания механически удаляются. В умеренных широтах
суда каждые 6—8 месяцев ставят в сухой док; в тропиках это прихо-
дится делать чаще или в промежутках между докованиями производить
очистку днища при помощи водолазов.
Обрастание судов, вызывающее уменьшение их хода, имеет особенно
большое значение для военного флота. В военное время докование бы-
1 Этот вопрос разобран Н. И. Тарасовым, которому мы и следуем в дальнейшем
изложении.
28 Зенкевич, т. I 433
вает нередко чрезвычайно затруднено, а между тем предельная скорость
того или иного судна часто целиком решает успех военной операции.
Так, например, 5 мая 1917 г. быстроходный британский крейсер «Bristol»
не смог догнать менее быстроходное (по регистровым данным) австро-
венгерское судно, так как днище крейсера оказалось сильно обросшим.
Поэтому иногда военные эскадры предпочитают более длительный путь
в тех случаях, когда переход можно сделать по морям, где обрастание
судов незначительно. Таким образом, суда выводятся из строя часто
на довольно продолжительное время.
Древоточцы. Чрезвычайно большой вред приносят морскому транс-
порту различные организмы, разрушающие дерево и бетон. Морские древо-
точцы внушали ужас еще античным мореплавателям, называвшим их
«summa calamitas navium>> (наивысшее бедствие кораблей). Овидий
с поэтической образностью сравнивает разрушительную деятельность
древоточцев со своими тяжкими душевными муками.
Интересно, что во время осады Севастополя в 1854—1855 гг. древоточцы
нанесли англо-французскому флоту существенный вред. В настоящее
время, когда подавляющее большинство судов имеет железный корпус,
деятельность древоточцев распространяется, главным образом, на порто-
вые сооружения.
Основными вредителями дерева являются так называемые морские
древоточцы —моллюски и ракообразные. Главная роль принадлежит
корабельным червям — моллюскам из сем. Teredinidae, протачивающим
ходы в дереве, погруженном в море. Наибольший вред приносят в Чер-
ном море Teredo navalis и Т. utriculus, на Дальнем Востоке — Bankia
setacea, В. sibirica и Teredo sinensis. Баренцово и Белое моря практиче-
ски свободны от корабельного червя, вследствие низкой температуры
воды. В теплых морях сем. Teredinidae представлено сотней видов, на
север же проникают лишь немногие виды. У западных берегов Исландии
и Шпицбергена были найдены отдельные экземпляры Teredo megotara
и Т. norvegica; заходят они по берегам Норвегии и в самую юго-западную
часть Варенцова моря, куда постоянно могут проникать с плавниковым
деревом и завозиться судами, однако развития здесь не получают. В Бал-
тийское и Азовское моря корабельный червь не проникает, вследствие
низкой солености. Нет его и в Каспийском море. В Японском море два
вида Bankia, единственного рода, имеющего холодноводный облик,
доходят до Татарского пролива. В Охотском и Беринговом морях кора-
бельный червь может быть встречен отдельными экземплярами, но
практического значения иметь здесь не может. Деятельность корабельного
червя на Черном и Японском морях носит чрезвычайно интенсивный харак-
тер, и ничем не защищаемая свая может быть уничтожена в течение одного
сезона. Уже в апреле появляются в воде личинки корабельного червя
и исчезают только в октябре-ноябре. С наступлением похолодания воды
личинки оседают на дерево и вбуравливаются в него. Взрослый кора-
бельный червь дерева уже не покидает.
Гораздо меньший вред причиняется в морской воде ракообразными
Limnoria lignorum flsopoda) и Chelura terebrans (Amphipoda), разрушаю-
щими древесину только с поверхности.
Наиболее эффективная мера защиты дерева от корабельного червя —
пропитка его креозотом. Практикуются также различные способы меха-
нической защиты подводных частей судов и свай: покрывание их цинком,
железом, цементом, обжигание и осмолка. Деревянные суда обшиваются
в подводной части металлическими листами.
Ледовые прогнозы, роль свечения и цветения моря. Значение морских
организмов для водного транспорта не ограничивается вредом, прино-
434
симым древоточцами, обрастаниями и т. д. Как уже указывалось выше,
биологические данные можно использовать для составления ледовых
прогнозов (В. Б о г о р о в, П. Ш и р ш о в) и таким образом способ-
ствовать безаварийному плаванию судов в высоких широтах (см. II том).
Недостаточно изучено, но, несомненно, велико значение свечения
и цветения моря в боевых действиях морского флота. Свечение моря для
подводных лодок является демаскирующим фактором. 11 сентября 1917 г.
английское судно «British Trader» пошло по светящемуся следу, оставлен-
ному немецкой подводной лодкой И-49, и в конце концов настигло и про-
таранило ее. Известны случаи, когда погружавшиеся подводные лодки
окружались как бы светящейся оболочкой, видимой с парохода на зна-
чительной глубине, что позволяло бить по ним прямой наводкой. Цве-
тение моря, наоборот, маскирует подводные лодки, так как при цвете-
нии резко понижается прозрачность воды.
Известную роль при прибрежных морских операциях (атаки минных
катеров, десантные операции и пр.) могут играть водоросли, которые
часто наматываются на винт и служат причиной остановки судов.
ГЛАВА XVIII
ПРОБЛЕМА АККЛИМАТИЗАЦИИ В МОРЕ
(Биологическим метод реконструкции морской фауны)
В плановом народном хозяйстве, при все возрастающей степени ис-
пользования природных ресурсов, в том числе и морских, вопросы рекой-
струкции фауны путем акклиматизации должны будут встать на очередь
дня и занять весьма важное место в системе хозяйственных мероприятий,
направленных к наиболее рационально построенному морскому промыслу.
Акклиматизация новых организмов в морских водоемах должна пред-
ставлять собою не случайный набор отдельных более или менее проду-
манных и удачных перебросок, а целостную и плановую систему хозяйствен-
ных мероприятий, построенную на серьезной теоретической базе, обосно-
ванную в своей хозяйственной целесообразности и приводящую к опре-
деленному хозяйственному эффекту — к увеличению и улучшению сырье-
вой базы морского промысла.
Если применительно к акклиматизации наземных животных и ра-
стений человеком уже проделана громадная практическая работа, то
в отношении морской фауны она находится еще в самом начале.
Поскольку в настоящее время вопросы акклиматизации, в применении
к морским водоемам, возникают в связи с их рыбопромысловым значе-
нием, изучение каждого конкретного водоема должно ответить на следую-
щие вопросы:
1. Что представляет собою фауна и флора водоема как кормовая база
для промысловых объектов?
2. Как, в какой степени и какими промысловыми и непромысловыми
объектами используется эта кормовая база в сложной системе пищевых
взаимоотношений?
3. Каковы основные биологические и промысловые показатели глав-
ных кормовых и промысловых объектов (количественное распределение,
темп роста, плодовитость и т. д.)?
На основе всех указанных сведений могут быть сделаны выводы:
1) о недостаточном использовании промысловыми объектами кормовых ре-
сурсов водоема в целом, 2) о недостаточно высоких кормовых качествах
фауны, служащей пищей промысловым объектам, 3) о недостаточно вы-
соких качествах промысловых объектов в смысле использования ими
кормовых ресурсов данного водоема и, наконец, 4) о недостаточно вы-
соких качествах используемых промыслом объектов.
Другая, не менее важная, сторона проблемы — это выбор желатель-
ных кормовых или промысловых объектов акклиматизации. Вселяемые
формы должны обладать высокими, в смысле поставленных задач, ка-
чествами и либо должны заполнить пустующую нишу, либо вытеснить
менее ценную биологически викарируютую форму.
433
Установление вероятных акклиматизационных фондов для данного
водоема и выбор из них наиболее желательных объектов акклиматиза-
ции— большая и сложная эколого-зоогеографическая задача, правиль-
ное решение которой возможно только при наличии полных и деталь-
ных знаний о флоре и фауне мирового океана.
Существенное понятие, на котором следует прежде всего остановиться,
это потенциальный ареал (Ш лыков, 1936).
Под потенциальным ареалом следует разуметь те части биосферы,
в которых, по всей сумме факторов внешней среды, можно предполагать
существование данной формы и которые находятся за пределами есте-
ственного ее ареала. Фактические ареалы очень часто гораздо меньше
потенциальных, что является следствием наличия непереходимых, так
называемых импедитных, границ.
Обыкновенно импедитные границы возникают в тех случаях, когда
отдельные формы или целые фауны, связанные единым происхождением,
в силу возникшей изоляции были разобщены и, развиваясь в сходных
условиях, претерпели при этом неизбежный процесс дивергенции. Эти раз-
вившиеся дивергентно фауны и могут служить друг для друга основным
акклиматизационным фондом. Таким образом, установление акклима-
тизационных фондов для отдельных частей биосферы, в том числе и для
отдельных морей, проблема в первую очередь палео-биогеографическая.
Значение акклиматизационных фондов становится особенно большим
тогда, когда одна из двух изолированных частей невелика по сравнению
с другой, как, например, Каспийское море по отношению к открытым
морям, Аральское — по отношению к’ Каспийскому и т. п. «Хотя малые
ограниченные области в известном смысле и представлялись крайне бла-
гоприятными для образования новых видов, но, тем не менее, в обшир-
ных областях изменения в большинстве случаев совершались быстрее и,
что еще важнее, формы, вновь образовавшиеся на больших территориях
и уже победившие многих соперников, были более способны к широкому
расселению и... играли более выдающуюся роль в изменчивой истории
органического мира» (Дарвин).
Приведем несколько примеров.
Как известно, современная мелководная тропическая фауна разви-
лась из единой фауны моря Тетис, простиравшегося в шпротном направ-
лении. После установления меридионального расчленения моря Тетис
в третичное время из единой исходной фауны образовались четыре дочер-
ние фауны: западная и восточная атлантические, западная и восточная
тихоокеанские, разобщенные с одной стороны материковыми массами,
с другой — центральными глубоководными частями океанов, а с севера
и юга — низкими температурами высоких широт. Несмотря на то, что эти
четыре дочерних фауны существуют раздельно сравнительно недавнее
время и условия их существования в общем чрезвычайно сходны, каждая
из них характеризуется резко выраженным эндемизмом, причем чем
ниже систематическая группа по таксономическому положению, тем резче
эндемизм.
Вряд ли можно сомневаться в том, что различия в арктической и
антарктической фаунах изчезли бы, если бы существовала возможность
беспрепятственного обмена между обеими фаунами, если бы их не разде-
ляла широкая и непереходимая в настоящее время для большинства пред-
ставителей зона теплых морей. При всем том даже те биполярные формы,
которые проникли в Антарктику совсем недавно, а именно — в ледниковую
эпоху, успели образовать здесь викарирующие виды, подвиды и формы.
Совершенно прав Мичурин, когда он говорит, что «никакой сорт
иностранного происхождения, если он не имеет еще на родине способ-
437
нести выдерживать пониженную температуру, одинаковую с минимальной
температурой той местности, куда растение было перенесено, не может
акклиматизироваться... Не следует обманывать себя ложной надеждой».
Действительно, акклиматизационный материал должен быть уже на
своей родине подготовлен к основным особенностям климата (гидрологи-
ческий режим) того водоема, куда организм переносится.
Несомненно, что большинство животных высоких широт северного
полушария могло бы существовать в соответственных климатических
зонах южного полушария, если бы были сюда перенесены кем-либо,и наобо-
рот; так, например, треска могла бы отлично акклиматизироваться в
южноавстралийских водах. Имеются еще два больших морских района,
фауны которых, значительно различаясь по систематическому составу,
существуют при сходных условиях внешней среды в течение длительного
времени и почти не имели возможности обмена. Это—фауна северных
частей Тихого и Атлантического океанов, разобщенные сейчас неблаго-
приятными для расселения холодными арктическими и теплыми южными
морями. В этих двух фаунах человек имеет неиссякаемые источники
для взаимной акклиматизации. Подобные примеры уже известны.
Американцы производили успешные пересадки атлантической уст-
рицы на тихоокеанское побережье. Весьма интересно, что случайно при
этом была завезена и ракушка Mya arenaria. успешно расселившаяся по
американскому побережью Тихого океана. Этот случай особенно замеча-
телен тем, что Му a arenaria. в районе Калифорнии известна из отложении
плиоцена, но затем по каким-то причинам вымерла. Делались подобные опы-
ты и у нас: в Белое море завозились представители дальневосточных лосо-
сей, а в Баренцево, правда, только в опытном порядке, — камчатский краб.
Замечательный пример аутоакклиматизации того же типа дает ки-
тайский краб Eriocheir sinensis, который, проникнув 30 лет назад с кораб-
лями из китайских морей в северо-западную Европу, колоссально здесь
размножился, заселив солоноватые и пресные воды бассейнов Северного
и Балтийского морей.
Наконец, большие взаимные акклиматизационные фонды представ-
ляют фауны амфпбореальные, амфиатлантические и амфипацифические,
т. е. те фауны, которые в плиоцене, а частично и в теплые фазы после-
ледникового времени, находились в сообщении, а сейчас разобщены и успе-
ли образовать большое число самостоятельных видов, подвидов и форм.
Помимо такой наиболее широкой постановки вопроса об акклимати-
зационных фондах для морских водоемов, в рамках, так сказать, всего
мирового океана, применительно к каждому водоему в отдельности вопрос
может ставиться и более узко. Так, например, для Каспийского моря,
а частично и для Аральского, прекрасным акклиматизационным фондом
могут служить наиболее эвригалинные атлантические формы, в массах
заселяющие Азовское и Черное моря. Дополнительные акклиматиза-
ционные фонды для Каспийского моря могут дать фауны Балтийского
моря и опресненных районов Японского моря.
До настоящего времени различными способами в Каспийское море
были занесены из фауны и флоры Черного и Азовского морей 12 форм,
в том числе 4 рыбы, 7 представителей беспозвоночных и 1 планктонная
водоросль. Большинство из них успешно акклиматизировалось, некоторые
вымерли (см. т. II).
Для Варенцова и Белого морей очень ценный акклиматизационный
материал могут дать моря Дальнего Востока, а для Белого моря до-
полнительный фонд может дать и Балтийское море.
Новый вводимый в фауну водоема объект, если он получает более или
менее массовое развитие, вступает в сложную систему взаимоотношений
438
с местной фауной. Используя для своего существования определенное
количество питательных веществ, новая форма может сама использо-
ваться в пищу другими организмами или прямо промысловыми, или кор-
мовыми формами для них. Если вселенная форма входит, таким образом,
как звено в пищевую цепь промыслового объекта и при этом представляет
более высокие кормовые качества, чем существовавшие ранее, имея доста-
точно высокий темп роста и размножения, акклиматизация может счи-
таться удачной, так как она дает больший эффект в смысле перевода
первичных пищевых ресурсов в конечный промысловый продукт. Но может
получиться п обратный результат —новая форма может лишь в малой
степени использоваться в пищу другими формами, тогда она с рыбохо-
зяйственной точки зрения будет сорняком. Этот вопрос приобретает
Рис. 259. Акклиматизационные мероприятия в отношении тихоокеанских
лососей (род Oncorhynchus) (ориг.).
1 — область исходного распространения, 2 — места перевозок, 3 — удачная акклиматизация.
особо большое значение, если вновь вселенные объекты получают осо-
бенно массовое развитие, как это имело место в Каспийском море с водо-
рослью Rhizosolenia и моллюском Mytilaster, биомасса которых, возможно,
достигала в отдельные годы 80—85% всего населения моря.
Если акклиматизация прошла успешно, то развитие организма в но-
вом ареале должно итти сначала по типу S-образной кривой. После первой
фазы мало заметного развития (инкубационный период) обычно насту-
пает фаза бурного размножения, не ограничиваемого пока естественными
врагами (паразиты и хищники). Для некоторых форм, например, выше-
упомянутых Rhizosolenia, Mytilaster, Eriocheir темп этого первоначаль-
ного развития можно уподобить только как бы некоему биологическому
взрыву. Когда в результате массового развития во второй фазе форма
приблизилась к насыщению нового ареала, темп развития замедляется
(третья фаза), причем ограничение в развитии в первую очередь насту-
пает благодаря ограниченности пищевых ресурсов и пространства.
439
После этого некоторое время форма будет существовать при тех же
высоких показателях плотности (четвертая фаза). В пятой фазе должно
наступить, слабое вначале, уменьшение популяции (биомассы вида).
Вступают в силу явления конкуренции и появляются естественные враги,
обусловливающие в шестой фазе резкое снижение популяции. В седьмой
фазе темп снижения замедляется и, наконец, в восьмой фазе устанавли-
вается на некотором более или менее длительно существующем уровне.
Вряд ли можно сомневаться, что все эти фазы будут иметь место при всякой
удачной акклиматизации, но длительность их и высота по ординате могут
очень сильно колебаться. Первые семь фаз нашей гипотетической кривой
могут быть названы периодОхМ освоения, восьмая фаза — периодом стаби-
лизации.
Моллюск Mytilaster и креветка Leander в Каспийском море, повеем
данным, находятся сейчас в третьей фазе, а диатомея Rhizosolenia — в пя-
той или даже шестой фазе.
Наиболее существенны размах и достигнутые результаты акклимати-
зационных мероприятий для наземных растений и животных. Довольно
много сделано и в отношении пресноводных рыб (форели, сиги и др.),
но еще очень мало в отношении морских организмов.
Из рыб особенное внимание было уделено лососевым (проходные рыбы).
Распространение тихоокеанских лососей (род Oncorhynchas} ограни-
чено бореальными водами северной Пацифики. Отдельные местонахожде-
ния отмечены также и в восточной части советской Арктики. Как указы-
вает Давидсон и Гетчинсон (Davidson and Н u I-
с h i n s о n, 1938), с 1872 по 1930 гг. миллионы яиц и молоди тихоокеан-
ских лососей были перевезены с тихоокеанских побережий США на атлан-
тические. Перевозки были произведены также на Гавайские о-ва, Австра-
лию, Тасманию, Новую Зеландию, Чили, Аргентину, Англию, Голландию,
Ирландию, Францию, Германию, Финляндию и Италию. Таким образом,
была сделана попытка использовать потенциальные амфнбореальность и
биполярность. Эти попытки акклиматизации оказались удачными для
Новой Зеландии, Чили, штата Майн, провинций Нью-Брансвик и
Онтарио, где развились местные популяции лососей; таким образом,
лососей удалось превратить в биполярные и амфибореальные формы
(рис. 259).
Из беспозвоночных много акклиматизационных мероприятий было
проведено со съедобной устрицей. В Англию ввозились устрицы не только
из ближних районов Франции, но и из США (использование потенциаль-
ной амфиатлантичности). Из Франции перевозились не только О. edulis,
но и «португальская» устрица (О. angiilata), успешно колонизированная
в последние десятилетия по французскому атлантическому побережью.
Однако этот последний вид хорошо растет на английских устричных пар-
ках, но не становится половозрелым и, таким образом, не может здесь
акклиматизироваться. Местная устрица О. edulis начинает нереститься,
когда температура достигает 15°, а для О. angiilata необходимо не менее 20°.
То же сначала получилось и при перевозке «японской» устрицы (О. gigas)
на тихоокеанское побережье (о. Ванкувер) США (использование потен-
циальной амфипацифичности), которая была предпринята с целью заме-
нить мелкую местную форму (О. virginica). Температура здесь редко
бывает достаточно высока для естественного нереста. Однако оказалось
возможным получить нужный эффект при содержании устриц в верхнем
более теплом слое воды. В дальнейшем было использовано открытие
Т. Гальцова (Galtsoff, 1938), показавшего, что устрицы могут
нереститься при более низкой температуре, если они стимулируются
присутствием в воде их собственных половых продуктов. На о. Ванкувер
440
«японская» устрица регулярно побуждается к нересту обрызгиванием
эмульсией из зрелых яиц и спермы.
Таким образом, применением особых приемов можно расширить
потенциальные ареалы полезных нам форм при акклиматизации. Были
проделаны также попытки к переносу японской устрицы с западных по-
бережий Америки на восточные, в Атлантический океан (потенциальная
амфибореальность).
Уже указанные проводившиеся в пределах СССР попытки акклимати-
зировать некоторые тихоокеанские формы в бассейнах Белого и Варен-
цова морей (дальневосточные лососи и камчатский краб) представляют
собою использование потенциальной амфибореальности.
Из акклиматизационных мероприятий меньшего географического мас-
штаба надо упомянуть систематические пересадки молоди морской кам-
балы, проводимые в датских водах (см. выше), и предпринятые в нашей
стране успешные акклиматизационные мероприятия по переносу ряда
черноморско-азовских форм в Каспийское море.
ЛИТЕРАТУРА
(сокращение названий см. во II томе)
Агеноров В. К. 1944. Об основ-
ных водных массах и гидросфере. Тр.
научно-исслед. учреждений Гл. упр.
Гидрометеослужбы СССР. Серия V. Гид-
рология моря* 10.
Андрияшев А. II. 1939. Очерк
географии и происхождения фауны рыб
Берингова моря и сопредельных вод.
Изд. ЛГУ.
Архангельский А. Д. 1932.
Геологическое строение СССР.
БазикаловаА. Я. 1931. Морские
промысловые моллюски.
Баскина В. П. и Фридман Г. М.
1928. Статистическое исследован не живот-
ного населения двух сообществ Камской
поймы. Тр. Биол. н.-иссл. ин-та при
Пермском гос. ун-те, 1, 2—3.
Беклемишев В. Н. 1928. Орга-
низм и сообщество. Там же, 1, 2—3.
— 1931. Основные понятия биоцено-
логии в приложении к животным компо-
нентам наземных сообществ. Тр. защ.
раст., 1, 277.
Берг Л. С. 1918. О причинах сход-
ства фауны северных частей Атлантиче-
ского и Тихого океанов. Изв. АН.
— 1920. Биполярное распространение
организмов и ледниковая эпоха. Там же.
— 1932, 1933. Рыбы пресных вод СССР.
— 1934. Об амфибореальном распро-
странении морской фауны в северном
полушарии. Изв. Геогр. о-ва, 66, 1.
Березкин В. А. 1938. Динамика
моря.
Биб В. 1936. В глубинах океана.
Бируля А. А. 1906. О литораль-
ной области в береговой полосе Шпиц-
бергена. Еж. 3. М.
Бирштейн Я. А. 1947. Реликты
в пресных и солоноватых водах СССР
(диссертация, рукопись).
— 1947. Понятие «реликт» в биологии.
Зоол. журн. XXVI, 4.
— 1949. Некоторые проблемы проис-
хождения и эволюции пресноводной фау-
ны. Усп. совр. биол. XXVI.
Блинов Л. К. 1946. К вопросу
о происхождении солевого состава мор-
ской воды. Метеор, и гидрол. Информ,
сборн., 4.
Богоров В. Г. 1933. Изменение
биомассы с возрастом у Calanus finmar-
chicus. Бюлл. ГОИН, 8.
— 1938. Суточное вертикальное рас-
пределение планктона в полярных усло-
виях. Тр. ПИНРО, II.
— 1938. Биологические сезоны Поляр-
ного моря. ДАН, XIX, 8.
442
Богоров В. Г. 1939. Суточная вер-
тикальная миграция, Eurytemora grimmi
в Каспийском море. Сб., поев, научи,
деят. Н. М. Книповича.
— 1939. Особенности сезонных явле-
ний в планктоне полярных морей и их
значение для ледовых прогнозов. Зоол.
журн., XVIII, 5.
Болдовский Г. В. 1939. Пища
сельди. ПИНРО.
— 1939. Питание мурманской сельди.
ДАН, XXIV, 3.
Бору цк ий Е. В. 1939. Динамика
биомассы Chironomus plumosu^ профун-
дали Белого озера. Тр. лимнол. ст.
в Косине, 22.
Б р и с к и н а М. М. 1939. Инструк-
ция для сбора и первичной обработки
бентоса. В НИРО.
Бродский К. А. 1934. Проблема
продуктивности водоемов (бентос, кон-
спект). Вести. Д. В. АН, 10.
— 1937. Планктонные исследования в
северо-западной части Японского моря.
Изв. ТИНРО, 12.
Броцкая В. А. и Зенкевич
Л. А. 1936. Биологическая продуктив-
ность морских водоемов. Зоол. журн.,
XV, 1.
— 1939. Количественный учет донной
фауны Варенцова моря. Тр. ВНИРО, IV.
Бруевич С. В. 1937. Гидрохимии
Среднего и Южного Каспия. Тр. Комисе,
компл. изуч. Каспийского моря. АН, VI.
— 1939. Распределение и динамика
живого вещества в Каспийском море.
ДАН, XXV, 2.
Верещагин Г. Ю. 1932. Лимно-
логия и пути ее современного развития.
Иссл. озер СССР, 1.
Вернадский В. И. 1923. Живое
вещество в химии моря.
— 1926. Биосфера.
— 1932. Проблемы биогеохимии.
— 1932. Проблема времени в совре-
менной науке. Изв. АН.
— 1933—1936. История минералов зем-
ной коры т. II — История природных вод.
— 1935. Очерки геохимии.
— 1935. Радиоактивность и новые про-
блемы геологии. Основные идеи геохи-
мии, 3.
— 1939. Проблемы биогеохимии.
— 1940. Биогеохимические очерки.
— 1944. Несколько слов о ноосфере.
Усп. соврем, биол., XV111,,2.
Винберг Г. Г. 1936. Некоторые
общие вопросы продуктивности озер.
Зоол. журн., XV, 4.
Винберг Г. Г. 1946. Биотический
баланс вещества и энергии озер (дис-
сертация, рукопись).
Виноградов Л. Г. 4941. Камчат-
ский краб.
Виноградов Л., Разин А.,
Гайл Г. 1933. Экспортные морепро-
дукты: краб, моллюски, водоросли.
Виноградов А. П. 1935, 1938,
1944. Химический элементарный состав
организмов моря. Ч. I, ч. II, ч. III.
Тр. биогеохим. лаб. АН.
— 1938. Химический состав морского
планктона. Тр. ВНИРО, VII.
— 1940. Химический состав планктона.
Тр. биогеохим. лаб. АН, V.
Воробьев В. II. 1938. Мидии
Черного моря. Тр. Аз.-Чер. НИРО, 11.
— 1945. Бентос Азовского моря (дис-
сертация, рукопись).
Воронков П. II. и Кречман
Г. В. 1939. Сезонные изменения био-
массы и физико-химических условий сре-
ды северо-восточной части Кандалакш-
ского залива Белого моря. Тр. ГГИ,
8, раб. Беломорской ст.
Воскресенский К. А. 1946.
Биофильтры литорали и соображения
к построению биогидрологии прибрежья.
Докл. Гос. океаногр. инет., 58.
— 1946. О влиянии организмов на
циркуляцию вод. Там же, 105. >
— 1946. Об интенсивности реакции
населенного дна с водными массами.
Вести. МГУ, 1.
— 1948. Пояс фильтраторов, как био-
гидрологическая система моря. ТОНН,
6 (18).
Вульф Е. В. 1932. Введение в ис-
торическую географию растений.
Г а веши Г. 1935. Возраст элемен-
тов. Основные идеи геохимии, 2.
Гайл. 1936. Анфельция в водах залива
Петра Великого. Вести. ДВФ АН, 20.
— 1936. Распределение фитопланктона
в поверхностных слоях прибрежных вод
северо-западной части Японского моря.
Там же, 18.
— 1936. Ламинариевые водоросли
дальневосточных морей. Там же, 19.
Гарвей X. В. 1933. Биохимия и
физика моря.
Г а у з е Г. Ф. 1936. О некоторых
основных проблемах биоценологии. Зоол.
журн., XV, 3.
ГендерсонЛ. 1924, Среда жизни.
Гептнер В. Г. 1936. Общая зоо-
география.
Герасимов И. П. и Марков
К. К. 1939. Четвертичная геология.
Герцен штейн С. 1885. Фауна
Мурманского побережья и Белого моря.
Тр. СПБ ОЕ, XVI.
Горбунов Г. П. 1939. Донное
население морей Советской Арктики.
«Проблемы Арктики».
Горшкова Т. И. 1942. Осадки
Полярного бассейна по материалам дрей-
фующей станции «Северный Полюс», 1937,
1938 гг. (рукопись).
Гурьянова Е. Ф. 1939. К во-
просу о происхождении и истории раз-
вития фауны Полярного бассейна. Изв.
АН, сер. биол., 5.
— 1947. Фауна Полярного бассейна
и пути ее обмена с фаунами соседних
районов мирового океана. 2-й Всесоюзный
Географ, съезд 1947 г. Тезисы.
Гурьянова Е. Ф., Закс И.,
Ушаков II. 1925. Сравнительный
обзор литорали и т. д. Раб. Мурм.
биол. ст., 1.
Давиташвили Л. Ш. 1943. Дар-
винизм и проблема накопления горючих
ископаемых. Вестн. Гос. Муз. Грузии,
XII, А.
Дарвин Ч. 1939. Происхождение
видов путем естественного отбора.
— Путешествие вокруг света на корабле
«Бигль».
— Дерюгин К. М. 1915. Фауна
Кольского залива и условия ее суще-
ствования. Зап. АН, физ.-мат. отд., 34
— 1928. Фауна Белого моря и условия
ее существования. И сел. морей СССР,
7—8.
— 1939. Зоны и биоценозы залива
Петра Великого (Японское море). Сборн.,
поев, научи, деят. Н. М. Книповича.
Драгунов. 1940 Промышленное
использование мидий Черного моря. Рыбн.
хоз. СССР, 4.
Дьяконов А. М. 1945. Взаимо-
отношения Арктической и Тихоокеан-
ской морских фаун на примере зоогео-
графического анализа иглокожих. Жури,
общ. биол., VI, 2.
Жадин В. И. 1940, Фауна рек
и водохранилищ. Тр. ЗИН АН, V, 3—4.
Закс И. Г. 1927. Предварительные
данные о распределении фауны и флоры
в прибрежной полосе залива Петра Ве-
ликого в Японском море. Тр. конф,
изуч. произв. сил Дальнего Востока.
— 1929. К познанию донных сообществ
Шантарского моря. Изв. Тихоок. научно-
нромысл. ст., III, 2.
— 1936. Биология и промысел краба
в Приморье. Вестн. ДВФ АН, 18.
Зенкевич Л. А. 1930. Количест-
венный учет донной фауны Приканин-
ского района. Тр. МНИ, IV, 3.
— 1932. К вопросу о вентиляции при-
донного слоя при помощи вертикальной
циркуляции. Бюлл. ГОИН, 6.
— 1934. Некоторые моменты зоогео-
графии Северного Полярного бассейна
в связи с вопросом о его палеогеографи-
ческом прошлом. Зоол. журн., XII, 4.
—1935. Некоторые наблюдения по
обрастанию в Екатерини некой бухте
(Кольский валив). Бюлл. МОИ И, биол.,
XLIV, 3.
— 1937. Успехи изучения морской фау-
ны СССР за 20 лет. Зоол. журн., XVI, 5.
— 1947. Фауна и биологическая про-
дуктивность моря, т. II.
— 1947. О задачах, объекте и методе
морской биогеографии. Зоол. журнал,
XXVI, 3.
443
3 е н к е в и ч Jl. А. и Б р о ц к а я В. А.
И937. Материалы по экологии руково-
дящих форм бентоса Варенцова моря.
Уч. зап. МГУ, 3. 1937.
Зернов С. А. 1913. К вопросу об
изучении жизни Черного моря. Зап. АН.
— 1924. Научные исследования судна
«Персей» в юго-восточном углу Варен-
цова моря летом 1924. Бюлл. Рыбн.
хоз., 15—16.
— 1933. Изучение донной произво-
дительности морей СССР. Тр. Ноябрь-
ской юбил. сессии, посвящ. 15-й годов-
щине Октябрьской революции 12—19/XI
1932.
— 1934. Общая гидробиология.
Зубов. Н. Н. 1931. К вопросу
о вентиляции придонных вод моря. Иссл.
морей СССР, 12.
— 1935. О биологических свойствах
морского льда. Северный морской путь, II.
— 1938. Морские воды и льды.
— 1940. В центр Арктики.
— 1941. Океанографические таблицы.
Иванов Л. 1928. Солнечная радиа-
ция как экологический фактор. Тр.
прикл. бот., генет. и селекции, XVIII, 2.
Ивлев В. С. 1939. Энергетический
баланс карпов. Зоол. журн., XVIII, 2.
Инструкции (Сборник инструкций) для
стандартных работ во время срочных
рейсов судов системы В НИРО, 1935,
а также многочисленные инструкции по
разным разделам биологии, изданные
в более поздние годы.
Инструкции по биологическим исследо-
ваниям вод, ч. 1. Биология морей. Изда-
ние ГГИ, 1931.
Карзинкин Г. С. 1936. Теория
биологической продуктивности водоемов
как рабочая схема. Зоол. журн., III.
К а ш к а р о в Д. Н. 1945. Основы
экологии животных.
Келлер К. 1905. Жизнь моря.
Киреева М. С. и Щапова Т. Ф.
1938. Темп роста, возраст и спороноше-
ние Laminaria saccharina и т. д. Тр.
ВНИРО, VII.
------1939. Запасы и промысел морской
травы. Рыбн. хоз. СССР, 3, 1939.
Классен Ф. Е. 1912. Заметки
о новых методах морских исследований.
Вести, рыбопром. 4 и 7.
Кленова М. В. 1949. Геология
моря (в печати).
Книпович Н. М.в 1938. Гидроло-
гия морей и солоноватых вод.
Кожевников Г. А. 1890. О вер-
тикальном распределении беспозвоночных
у русских берегов Балтийского моря.
Йзв. об-ва люб. Е. А. и Э., 67.
Коштоянц X. С. 1940. Основы
сравнительной физиологии.
К р о г А. 1935. Жизнь в океаниче-
ских глубинах. Усп. совр. биол., IV, 6.
Лёб Ж. 1910. Динамика живого
вещества.
Линдеман Р. Л. 1943. Трофико-с
динамическое направление в экологи-
444
чрском исследовании. Усп. совр. бпол.,
XVI, 5.
Личков Б. 1945. Геологические
периоды и эволюция живого вещества.
Журн. общ. биол., VI, 3.
Малятский С. М. 1938. Мате-
риалы по экологии белуги Черного моря.
Зоол. журн., XVII, 4.
Мантейфель Б. П. 1938. Крат-
кая характеристика основных закономер-
ностей в изменениях планктона Барен-
цева моря. Тр. ПИНРО, 1.
— 1939. Зоопланктон прибрежных вод
Мурмана по материалам 1931—1932 гг.
Тр. ВНИРО, IV.
— 1941. Планктон и сельдь в Барен-
цовом море. Там же, VII.
Материалы по биологии, промыслу и
обработке камчатского краба. Изв.
ТИНРО, 19, 1945.
Миланкович М. 1939. Матема-
тическая климатология и астрономиче-
ская теория колебаний климата. ГОНТП.
Морозова-ВодяницкаяН.В;
1936. Опыт количественного учета донной
растительности в Черном море. Тр. Се-
васт. биол. ст. V.
Муравейский С. Д. 1936. Пути
построения теории биологической про-
дуктивности водоема. Зоол. журн.,
XV, 4.
Нансен Фр. 1939. Шпицберген.
Никитин В. Н. 1929. Вертикаль
ное распределение планктона в Черном
море, II. Тр. Севаст. биол. ст., 1.
— 1933. Устрицы, мидии и креветки,
как объекты промысла в Черном и
Азовском морях. Рыбн. хоз. СССР, 3.
— 1947. Обрастание судов в Черном
море, их биология и практическое зна-
чение (в печати).
Никольский Г. В. 1947. К позна-
нию биологической специфики фаунисти-
ческого комплекса и ее значение для
зоогеографического анализа фауны. 2-й
Всесоюзн. географический съезд 1947 г.
Тезисы.
— 1947. О биологической специфике
фаунистических комплексов и значение
ее анализа для зоогеографии. Зоол.
журн., XXVI, 3.
О к ул А. В. 1939. Питание и пища
планктоноядных рыб Азовского моря
(диссертация, рукопись).
Опарин А. 1936. Возникновение
жизни на земле.
О к с н е р А. 1944. О происхождении
ареала биполярных лишайников. Бот.
журн. СССР, IX, 6.
Попов А. 1930. Материалы по кра-
бовому промыслу в водах западной Кам-
чатки. Соц. реконстр. рыбн. хоз. Даль-
него Востока, 5—6.
— 1936. Производство крабовых кон-
сервов на Дальнем Востоке. Рыбн. хоз.
СССР, 5.
Разин А. 1934. Морские промыс-
ловые моллюски южного Приморья. Изв.
Тихоок. научн. ин-та рыб. х-ва, 8.
Расс Т. С. 194^. Мировой промысел
водных животных.
Р е с с е л ь Ф. С. и 11 о и г Ч. М.
1934. Жизнь моря.
Россолимо Л. Л. 1934. Задачи
и установки лимнологии как науки.
Тр. лимнол. ст. в Косине, 17.
Рох Ф. 1934. Teredinidae морей
СССР. Зоол. журн., XIII, 3.
Руководство по зоологии. Под ред.
В. Догеля, Л. Зенкевича и
Б. Матвеева, т. I, 1937, т. II, 1940.
Самойлов Я. В. 1929. Биолиты.
— и Титов А. Г. 1922. Железо-
марганцовые желваки со дна Черного,
Балтийского и Варенцова морей. Тр.
геол, и минер, муз. им. Петра Вел.
АН, III, 2.
С о в и н с к и й В. 1904. Введение
в изучение фауны Понто-каспийско-араль-
ского морского бассейна, Зап. Киевск.
об-ва естеств., XVIII.
Солдатов В. К. 1928. Рыбы и
рыбный промысел.
— 1934, 1938. Промысловая ихтиоло-
гия, I и И.
Съюорд /V Ч. 1936. Века и растения.
Т а н ф и л ь е в Г. И. 1931. Моря
Каспийское, Черное, Балтийское, Ледо-
витое, Сибирское и Восточный океан.
Таранец А. Я. 1938. Морские и
пресноводные богатства ДВК.
Тарасов Н. И. 1938. О колонках
донных отложений Сев. Атлантики. При-
рода, 7—8.
— 1939. Саргассово море. Там же, 5.
— 1943. Биология моря и флот. Воен-
мориздат.
Трофимов А. В. 1938. О верти-
кальном распределении прозрачности в
море. Метеорол. и гидрол., 6.
У и м пенни Р. С. 1945. Органи-
ческая полярность с экологической и
физиологической точек зрения. Усп. совр.
биол., XIX, 2.
Ушаков П. В. 1934. Некоторые
особенности фауны и гидрологического
режима Охотского моря. Природа, 11.
— К бентонической фауне Чукотского
моря. Научи, раб. эксп. ледок. «Красин»
1935 г., изд. ГУСМП.
— 1945. Донное население Чукотского
моря, как показатель течений. Докл.
ГОИН, 31 и 32.
— 1947. Чукотское море и его донная
фауна (в печати).
Ферсман А. Е. 1946. Атом и время.
Наука и жизнь, 1.
Филатова 3. А. 1938. Количе-
ственный учет донной фауны юго-за-
падной части Варенцова моря. Тр.
ПИНРО, 2.
Францев А. В. и Лебедева
С. К. 1941. Химизм воды Учинского
водохранилища. Тр. ЗИН АП, VII, 1.
Ф р и дер икс К. 1932. Экологи-
ческие основы прикладной зоологии и
энтомологии.
Ч у гунов Н. Л. 1923. Опыт коли-
чественного исследования продуктивности
донной фауны в Северном Каспии и ти-
пичных водоемах дельты Волги. Тр.
Астрах, ихт. лаб., V, 1.
— 1926. Предварительные результаты
исследования продуктивности Азовского
моря. Тр. Аз.-Черн. эксп., 1.
Ширшов П. П. 1936. Планктон,
как индикатор ледового режима. Сборн.
научн. раб. ледок. «Красин».
— 1937. Сезонные явления в жизни
фитопланктона полярных морей. Тр. Всес.
арктпч. ин-та, 82.
— 1939. Опыт определения продуктив-
ности фитопланктона полярных морей
по фотосинтезу. Там же.
Шмидт О. 10. 1946. Новая теория
происхождения земли. Природа, 7.
Шмидт II. Ю. 1904. Рыбы восточ-
ных морей Российской империи. Научн.
рез. Корейско-Сахалинской экспед.
ИРГО, 1900—1901.
— 1947. Миграции рыб.
— 1948. Рыбы Тихого океана.
Шокальский Ю. М. 1933. Физи-
ческая океанография.
Ш о р ы г и н А. А. 1928. Иглокожие
Варенцова моря. Тр. МНИ, III, 4.
— 1939. Питание и пищевые взаимо-
отношения некоторых бентофагов Север-
ного Каспия (диссертация).
Ш п а р л и н с к и й В. 1932. Новые
объекты промысла, моллюски и ракооб-
разные.
Штокман В. Б. 1947. Исследование
влияния ветра и рельефа дна на резуль-
тирующую циркуляцию и распределение
масс в неоднородном океане или море
(рукопись).
III у л е й к ин В. В. 1941. Физика
моря.
Щапова Т. Ф. 1937. Количествен-
ные исследования морских донных водо-
рослей (диссертация).
— 1946. Амфипацифическое распро-
странение некоторых видов бурых водо-
рослей. ДАН, LII, 2.
— 1946. Биполярное распространение
некоторых видов бурых водорослей. Там
же, L1I, 5.
— 1948. Географическое распростране-
ние представителей порядка Laminaria-
]es в северной части Тихого океана.
Тр. И-та океанол. /Ш, II.
— 1948. Географическое распростране-
ние и пути расселения порядков Lami-
nariales, Fucales и Desmarestimales в се-
верной части Тихого океана (диссерта-
ция).
Элтон Ч. 1934. Экология живот-
ных.
Энгельс Ф. 1933. Диалектика при-
роды.
Яковлев С. 1927. Трепанг.
Я ш н о в В. А. 1940. Планкти-
ческая продуктивность северных морей
СССР.
445
Adams, G h. C. 1913. Valuation
of the Sea. Amer. Nat., XLV11, 558.
Agassiz A. 1872—1874. Revision
of the Echini. Mem. Mus. Comp. Zool.
Allee W. G. 1923. Studies in marine
ecology. I. The distribution of common
littoral invertebrates of the Woods Hole
region. Biol. Bull,. 44.
— 1931. Animal aggregations.
Allen E. J. 1899. On the fauna and
bottom deposits near the 30-fath-line
from the Eddystone Grounds to Start
Point. Journ. Mar. Biol. Ass., 5.
— 1909. Mackerel and Sunshine. Ibid.,
8.
Allen W. E. 1936. Surface plank-
ton diatoms in the North Pacific Ocean
in 1934. Madrono, V.
d’Ancona U. 1931. Faune et flore
des eaux saumatres. I. Rapp. Proc. Verb.
Gomm. Intern. Exp. Sci. Mediterr. (N. S.),6.
A p p e 1 1 d f A. 1892. Om Bergens-
fjordens faunistiske praeg. Bergens Mu-
seum Aarbog, 1891, 2.
— 1895. Faunistiske undersogelser i
Herlofjorden. Ibid. 1894—1895, 11.
— 1896. Faunistiske undersogelser i
Osterfjorden. Ibid., 13.
— 1906. Die decapoden Crustaceen.
Meeresfauna von Bergen, Hf. 2.
— 1912. Invertebrate bottom fauna of
the Norwegian Sea and North Atlantic.
В книге Murray and Hjort, The
Depths of the Ocean.
Apstein A. 1904. Die Schiitzungs-
methode in der Planktonforschung. Wiss.
Meeresunters., Abst. Kiel, N. F., 8.
— 1905. Plankton in Nord- und Ostsee
auf den deutschen Terminfahrten, I Theil.
Ibid., 9.
A r 1 d t Th. 1919. Handbuch der Palao-
geographie.
Atkins W. R. S. 1922. Hydrogcn-
jon concentration of sea water in its
biological relation. Journ. Mar. Biol.
Assoc., XII.
— 1923. Phosphate content of waters
in relationship to growth of algal plank-
ton. Ibid., XIII.
— 1925, 1926. Seasonal changes in the
phosphate content of sea water in rela-
tion to the growth of the algal plankton
during 1923 and 1924. Part II. Ibid.,
XIII, XIV.
— 1926. A quantitative consideration
to some factors concerned in plant growth
in water. Part I — II. Some Chemical
Factors. Journ. Cons. 1, 1—2—3.
— 1930. Seasonal variations in the
Phosphate and Silicate Content of Sea
Water in Relation to the Phytoplankton.
J. Mar. Biol. Ass., XVI, 3.
Atlas fur Temperature, Salzgehalt
und Dichte der Nordsee und Ostsee.
Deutsche Seewarte, 1927*.
A u d о u i n A. et M i 1 n e - E d-
wards H. 1832—1834. Recherches pour
servir a 1’histoire naturelle du littoral
de la France, Paris.
446 1
Baldwin E. 1937. An Introduc-
tion to comparative biochemistry.
В a 1 s s H. 1936. Decapoda and Schizo-
poda. Fish. Res. Direct. Egypt, Notes
and Mem., 15.
Battle H. 1., Huntsman A. G.
and oth. 1936. Fatness digestion and food
of Passamaquoddy young hering. J. Biol.
Board Canada, II, 4.
De Beauchamp P. 1914. Les
greves de Roscoff. Paris.
— 1914. Apercu sur la repartition des
etres dans les zones des marees a Roscoff.
Bull. Soc. Zool. France, 39.
— 1914. Une tourbiere sous-marine com-
me milieu biologique. Ibid.
— 1923. Etude de bionomie interco-
tidiale. Les lies de Rd et d’Yeu. Arch.
Zool. Exp. Gen., 61.
— et La mi R. 1921. La bionomie
intercotidiale de File de Brehat. Bull,
biol. France Belg., 55.
— et Zachs J. 1913. Esquisse d’une
monographic bionomique de la plage de
Terrenes. Мёт. Soc. Zool. France, 26
Berg L. S. 1933. Die bipolare Ver-
breitung der Organismen und die Eiszeit.
Zoogeographica, I.
Bernard F. 1939. Etude sur les
variations de fertilite des eaux mediter-
raneennes. J. Cons., 14.
Bertel R. 1912. Sur la distribu-
tion quantitative des bacteries plancto-
nique des cotes de Monaco. Bull. Inst.
Ocean. Monaco, 224.
Bigelow H. B. 1926. Plankton
of the offshore waters of the Gulf of Maine
Bull. U. S. Bur. Fish., 40.
— 1931. Oceanography.
В 1 e g v a d H. 1915. Food and Con-
ditions of Nurishment among the com-
munities of invertebrate animals found
on or in the Sea bottom in Danish waters.
Rep. Dan. Biol. St., XXII.
— 1917. On the Food of fishes in the
Danish waters within the Skaw. Ibid.,
XXIV.
— 1922. Animal Communities in the
Southern North Sea. Proc. Zool. Soc.
London, April.
— 1926. Continued studies on the Quan-
tity of Fish-Food in the Sea-Bottom.
Rep. Dan. Biol. St., XXXI.
— 1927. Methoden der Untersuchung
der Bodenfauna des Meereswassers. Hand,
biol. Arbeitsmethoden. Abt. IX, Teil.
5, Hf. 2.
— 1928. Quantitative Investigations
of Bottom Invertebrates in the Limfjord
1910—1927. With special Reference to
the Plaice-Food. Rep. Dan. Biol. Stat.,
XXXIV.
— 1930. Quantitative Investigations
of Bottom Invertebrates in the Kattegat
with Special Reference to the Plaice
Food. Ibid., XXXVI.
Bogorov B. G. 1934. Seasonal
changes in Biomass of Calanus finmar-
chicus in the Plymouth Area in 1930.
J. Mar. Biol. Ass. Plymouth. N. S., XIX,
Bond R. M. 1933. A contribution
to the study of the natural food-cycle
in aquatic environments. Bingham Ocea-
nogr. Goll. Peabody Mus. Bull., 4, 4.
В 6 h n e с к e G., II e n t s c h e 1 E.
und Wattenberg H. 1930. Uber
die hydrographische, chemische und bio-
logische Verhaltnisse an der Meeres-
oberflache zwischen Island und Gronland.
Ann. Hydr. Mar. Meteorol., LVI1I Jahrg.,
VII.
— F о у n und Wattenberg H.
1932. Beitrage zur Ozeanographie des
Oberflachenwassers in der Danemarkstrasse
und Irminger See, II Teil., Ibid., LX.
Boysen-Jensen P. 1915. Studies
concerning the Organic Matter of the
Sea Bottom. Rep. Dan. Biol. St., XXII.
— 1920. Valuation of the Limfjord.
Ibid., XXVI.
Brady F. 1943. The distribution
of the Fauna of some intertidal sands
and muds on the Nortumberland coast.
Journ. Animal Ecol., 12, 1.
Brandt K. 1897. Die Fauna der
Ostsee, insbesondere die der Kieler Bucht.
Verh. D. Zool. Gesellsch., 19.
— 1899, 1902, 1916—1920, 1919. Uber
den Stoffwechsel im Meere. Wiss. Meeres-
unters. Kiel 1, 4, 6, 18, 19.
— 1905. Ueber die Produktion und
die Produktionsbedingungen im Meere.
Rapp. Proc. Verb. Cons. I nt., III.
— 1925. Die Produktion in den hei-
mischen Meeren und das Wirkungsgesetz
der Wachstumsfaktoren. Ber. deutsch.
wiss. Kommission Meeresforsch., N. F., I.
— 1927. Stickstoffverbindungen im
Meere. Wiss. Meeresunters., XX.
— 1929. Phosphate und Stickstoffver-
bindungen als Minimumstoffe fur die
Produktion im Meere. Rapp. Proces-
verbaux Rёun. LIII.
— und R a b e n E. 1921. Zur Kennt-
niss der chemischen Zusammensetzung des
Planktons und einiger Bodenorganismen.
Wiss. Meeresunt., N. F., 19, 4, Abt. Kiel.
В г о c h H j. 1910. Die Hydroiden
der arktischen Meere. Fauna Arctica, V, I.
— 1922. Riffkorallen im Nordmeer einst
und jetzt. Naturwissensch.
— 1927. Methoden der marinen Bio-
geographic. Abderhalden Handb. biol.
Arbeitsmeth., Abt. 9, T. 5.
— 1928. Untersuchungen uber die ma-
rine Bodenfauna bei Lindesness im Juni,
1926. Avhandl. Norske Vid-Akad. Mat.-
Naturvet. KI.
— 1933. Einige Probleme der biogeo-
graphischen Abgrenzung der arktischen
Region. Mitt. Zool. Mus. Berlin, 19.
Brockmann Chr. 1935. Diato-
meen und Schlick im Jade-Gebiet. Abh.
Senkenberg. Naturf. Ges., 430.
В г и g e J. R. 1928. Special factors on
the sandy beach. I. Tidal, climatic, eda-
phic; II—Chemical changes, carbon con-
mntration and sulphides. J. Mar. BioL
Assoc., 15.
Buch K. 1920. Ammoniakstudien an
Meer- und Hafenwasserproben. Meren-
futkimuslaitoksen Julkaisu Havsfors-
kningsinstitutets Skrift, 2.
— 1932. Untersuchungen liber geloste
Phosphate und Stickstoffverbindungen in
den Nordatlantischen Meeresgebieten. Ibid.,
86.
Buchanan-Wollaston H. J.
1923. The spawning of Plaice in the Sou-
Ihern Part of the North Sea in 1913—1914.
Fish. Invest., Ser. II, IV, 2.
— 1937. Estimation the Total or the
Mean Number of Organisms under a given
Area of Sea or on a given Area in Sea
Bottom. Journ. Cons. XII, 3.
Bulletin statistique des peches
maritimes des pays du nord et de 1’ouest
de ГЕигоре, XXIV, 1938.
Caiman W. T. 1936. Marine Bo-
ring Animals injurans to submerged Struc-
tures. British Museum.
Cary L. R. 1918. The Gorgonaceae
as a factor in the formation of a Coral
reef. Carnegie Inst. Publ. (Pap. Mar.
Biol., 9), 123.
Chapman R. N. 1931. Animal
Ecology.
Chapman V. J. 1944. Methods
of surveying Laminaria beds. J. Mar.
Biol. Ass., 26, I.
Chun C. 1888. Die pelagische Tierwelt
in grosseren Meerestiefen und ihre Beziehun-
gen zu der Oberflachenfauna. Bibl. Zool., I.
Chworostansky C. 1892. Uber
die Zonen des Kiistenstriches der Solo-
wetski Inseln. Zool. Jahrb., 15.
Clarke F. 1924. Date of Geoche-
mistry.
Clarke G. L. 1939. Diurnal migra-
tion of plankton in the Gulf of Maine
and its correlation with changes in sub-
marine irradiation. Biol. Bull., 65.
— 1936. Light penetration in the wes-
tern North Atlantic and its application
to biological problems. Rapp. Comm. int.
Mer. Medit., 101, 3.
— 1939. The Utilisation of solar energy
by aquatic organisms. Problems of Lake
biology. Public. Amer. Ass. Advancement
Science, 10.
— 1939. Plankton as a food source
for man. Science, 89.
— 1940. Comparative richness of zoo-
plankton in coastal and offshore Areas
of the Atlantic. Biol. Bull., 78, 2.
— and Ostler В. H. 1934. The
penetration of the blue and red compo-
nents of daylight into Atlantic coastal
waters and its relation to phytoplankton
metabolism. Ibid., 67.
— and Zinn G. J. 1937. Seasonal
production of zooplankton of Woods Hole
with special reference to Calanus finmar-
chicus. Ibid., 73.
Clements F. E. 1916. Plant succes-
sions. Carnegie Inst. Washington Publ., 242.
447
G 1 e m e n t s F. E. and S helford V. E.
1939. Bioecology.
Gleve P. T. 1897. A Treatise on
the Phytoplankton of the Atlantic and
its Tributaries and on the Periodical
Ghanges of the Plankton of Skagerrak.
Upsala.
— 1900. The Seasonal Distribution of
Atlantic Plankton Organisms.
Coewes A. J. 1938. Phosphate and
Silicate in the Southern Ocean. Disco-
very Rep., 19.
Goe W. R. 1932. Season of attach-
ment and Rate of Growth of sedentary
marine organisms at the pier of the Scripps
institution of oceanography, La-Jolla, Ca-
lifornia. Univ. Gal. Bull. Scripps Inst.
Oceanogr., Techn. Ser. Ill, 3.
— 1946. The means of dispersal of
bathypelagic animals in the North and
South Atlantic Oceans. Amer. Nat., 80,
793.
Colman J. 1933. The Nature of
the Intertidal Zonation of Plants and
Animals. Ibid., 18.
Colton H. S. 1917. On some varie-
ties of Thais lapillus in the Mount De-
sert region. Proc. Acad. Nat. Sci. Phila-
delphia, 68.
Cooper L. H. N. 1938. Redefini-
tion of the anomaly of the nitrate-phos-
phate ratio. J. Mar. Biol. Ass. V. 23.
Dahl F. 1894. Uber horizontale und
vertikale Verbreitung der Copepoden im
Ozean. Verb, deutsch. Zool. Ges. 1894.
— Untersuchungen liber die Tierwelt
der Unterelbe. Ber. Komm. Unters. deutsch.
Meere, 6.
— 1921—1923. Grundlagen einer okolo-
gischen Tiergeographie.
Dakin W. J. 1908. Notes on the
alimentary canal and food of Copepoda.
1RHH, I.
— 1931. Migrations and the producti-
vity in the Sea. Au^tr. Zool.
Daly R. A. 1936. Origin of subma-
rine «canyons». Amer. Journ. Sci., 31.
— 1942. The floor of the Ocean.
Damas D. 1909. Contribution a la
Biologic des Gadides. Rapp. Proces-ver-
baux, 18.
— and Koefoed E. 1909. Le plank-
ton de la Mer du Gronland. Grois. Ocea-
nogr. «Belgica» dans la Mer du Gronland.
1905.
Dana J. D. 1848. Zoophytes. U. S.
Explor. Exp.
— 1852. Crustacea. Ibid.
— 1890. Coral and Coral Islands.
Dannevig A. 1924. On Morta-
lity among Fish (Cod, Eel) and Lobster
in severe winters. Aarsberetn. vedk. Nor-
ges Fiskerier for 1924.
Darwin Ch. 1842. The structure
and distribution of Coral reefs.
Davenport С. B. 1903. The ani-
ma) ecology of the Cold Spring Sand
Spit etc. Decenn. Publ. Univ. Chicago,
Ser. I, 10.
•448
Davis F. M. 1923. Preliminary I in
vestigediion of the Dogger Bank. Fish.
Invest., Ser. II, VI, 2. 1923.
Davidson F. A. and Hutchin-
son S. J. 1938. The geographic distri-
bution and environmental limitation of
the pacific salmon. Bull. Bureau of Fish.,
48.
— 1925. Quantitative studies in the
Fauna of the Sea Bottom. No 2. Results
on the Investigations in the Southern
North Sea, 1921—24. Min. Agric. Fish.
Invest., II. 8. 34.
D e а к о n G. E. R. 1937. The hydro-
logy of the Southern ocean. Discovery
Reports, vol. 15 (CambridgeJUniv. Press ).
Defant A. 1928. Die systematised
Erforschung des Weltmeeres. Zs. Ges.
Erdk., Sonderband Hundertjahrfeier Ge-
sellschaft.
— 1936. Die Troposphare. Deutsche
Atlantische Exped. «Meteor» 1925—1927.
Wiss. Erg., VI, Th. 1, 3.
— 1929. Einfuhrung in die Geophysik.
III. Dynamische Ozeanographie.
D e m о 1 1 R. 1927. Betrachtungen liber
Produktions-Berechungen. Arch. Hydrob.,
XVIII, 3.
D e r j u g i n K. 1927. La distribu-
tion bipolaire des organismes marins.
Bull. Inst, oceanogr. Monaco, 495.
Dollfus A. 1914. Les zones subter-
restres et littorales a I’ile Tatihou et
dans la region de Saint-Vaast-la-Hogue
(Manche). Bull. Mus. Hist. Nat., 20.
Dietz R. S., E m e г у, К. O. and
Shepard, P. 1942. Phosphorite depo-
sits of the sea Floor of southern Califor-
nia. Bull. geol. Soc. America, 53 (Scripps
Inst. Oceanogr., Gontr. 160).
Drew, G. H. 1912. On the precipita-
tion of calcium carbonate in the sea by
marine bacteria etc. J. Mar. Biol. Ass., 9.
Eckman W. 1905. An Apparatus
for the Collection of Bottom Samples.
Cons, perman. int. explor. mer. Publ.
Circonst., 27, Copenhague.
Edmondson С. H. 1928. The eco-
logy of a Hawaiian Reef. Bishop Mus.
Bull., 45, Honolulu.
Ehrenberg H. 1834. Uber die
Natur und Bildung der Corallenbanke
des Roten Meeres. Abh. Akad. Wiss.
Berlin, 1832, Theil I.
E i c h e 1 b a u m E. 1910. Uber Nah-
rung und Ernahrungsorgane von Echi-
nodermen. Wiss. Meeresunters, Abt. Kiel,
N. F., XII.
E к m a n Sv. 1913. Studien liber
u. s. w. II — Die Variation der Kopf-
form bei Limnocalanus grimaldii und L.
macrurus IRHH, VI, 4—5.
— 1915. Vorschlage und Erorterungen
zur Reliktenfrage in der Hydrobiologie.
Ark. Zool., IX, 17.
— 1932. Prinzipielles liber die Wande-
rungen und die tiergeographische Stel-
lung des europaischen Aales Anguilla
anguilla (L.). Zoogeographica, L
Ekma n Sv. 1934. Indo-Wes tpazifikuncl
Ailanto?Ostpazifik, eine tiergeographische
Studie. Ibid., 2.
— 1935. Tiergeographie des Meeres.
— 1940. Begriindung einer statistischen
Methode in der regionalen Tiergeographie
Nova acta, reg. soc. sci. upsaliensis, Ser. IV,
12. 2.
Elmhirst R. 1932. Quantitative
studies between tide marks. Glasgow Na-
turalist.
Elton Ch. 1927. Animal Ecology.
— 1930. Animal Ecology and Evolu-
tion.
E r c e g о v i c A. 1934. Temperature,
sdinite, oxygene et phosphates dans 1’eau
rd Here de FAdriatique. Acta Adriatica
Inst. Oceanogr. Split, 5.
— 1936. Fizikokemiska i biologiska
ispitivanja u obalnum vodama istocnog
Jadrana tokom Godine 1934. Prirodos-
lovna istrazivanja Kraljevine Jugoslavije,
20.
— 1936. Etudes qualitative et quanti-
tative du phytoplancton dans les eaux
coheres de 1’Adriatique oriental moyen au
fours de Гаппёе 1934. Acta Adriatica.
Instit. Oceanogr. Split. (Jugoslavia), 9.
— 1940. Weitere Untersuchungen iiber
oinige hydrographische Verhaltnisse und
iiber die Phytoplanktonproduktion in den
Gewassern der osllichen Mitteladria. Ibid.,
II, 3.
E s t e r 1 у С. О. 1912. The occurence
and vertical distribution of the Copepoda
of the San Diego region etc. Univ. Cali-
fornia Publ. Zool., IX, 6.
— 1914. The vertical distribution and
movement of the Schizopoda of the San
Diego region. Ibid., XIII, 5.
— 1919. Reactions of various plankton
animals with reference to their diurnal
migrations. Ibid., XIX.
— 1909. Diurnal Migrations of Cala-
пи s finmarchicus in the San Diego Region.
IRHH, IV.
Far ran G. P. 1914, 1915. Results
of a biological Survey of Blacksod Bay
Сю Mayo. Dept. Agr. Techn. Inst. Ireland
Fish. Sci. Invest., 1914, 1915.
Ferronnieres G. 1901. Etudes
biologiques sur les zones supralittorales
de la Loire inferieure. Bull. Soc. Sc. nat.
Quest, XI.
Fiedler R. 1938. Fishery Indu-
stries of the United States, 1937. Bureau
Fish. Administr. Rep. U. S. Dep. Com-
merce. 32.
Field J. A. 1922. Biology and
economic value of the Sea-Mussel Myti-
lus edulis. Bull. U. S. Bur. Fish., 36.
Finckh A. 1904. Biology of the
reef formating organisms at Funafuti
Atoll. Rep. Coral Reef Comm. Roy. Soc.,
Sect. 6.
Fish C h. J. 1925. Seasonal distri-
bution of the plankton of the Woods
Hole region. Bull. U. S. Bur. Fish., 41,
91.
F i s h. C h. 1936. The biology of Calanus
finmarchicus in the Gulf of Maine and
Bay of Fundv. Biol. Bull, 70, 2.
Flattely F. W. and Walton
C. L. 1922. The biology of the Sea shore.
Flora en fauna der Zuiderzee.
Monographic van een brakwatergebied.
Dem Herder, 1922.
Food values о f S h e 11 f i s h.
Fishing Gasette, 57, 7, 1940.
Forbes E. 1840. On the associa-
tions of Molluscs on the British Coasts
etc. Edinburg, Academic Annual for 1840.
— 1844. Report of the Mollusca and
Radiata of the Aegean Sea etc. Report
13-th Meeting. Bri. Ass. Adv, Sci, 1843.
— 1844. Report on Aegaean Inver-
tcbrata. Brit. Ass. Adv. Sci. for 1843.
— 1844. On the light thrown on Geology
bv submarine researches. Ann. Mag. Nat.
Hist (ser. I), 13.
— 1851. Report on the investigation
of British Marine Zoology, Part I. R,ept.
Brit. Ass. Adv. Sci, 1850.
— and H a n 1 e y. 1853. History of
British Molluscs.
— and Godwin Austen, R. 1859.
The natural history of the European Seas.
Forbes S. A. 1888. On the food
relations of fresh-water fishes. State Lab.
Nat. Hist., 2.
— 1888. The food of the fishes of the
Mississipi Valley. Trans. Amer. Fish.
Soc., 7.
— 1884—1889. Studies of the food
of fresh-water fishes. Bull. Ill, State
lab. Nat. Hist., 2.
Ford E. 1923. Animal communities
of the level Sea-bottom in the waters
adjacent to Plymouth. J. Mar. Biol. Ass.,
N. S., 13, I.
Fowler G. H. 1898. The plankton
of the Faroe Channel. Proc. Zool. Soc.
Fraser J. H. 1932. Observations
on the Fauna and Constituents of an
Estuarine Mud in a Polluted Area.
J. Mar. Biol. Ass., N. S, 18.
Fritsch. 1945. The structure and rep-
roduction of the algae, vol. II, 1945.
Fuller J. L. and Clarke G. L.
1936. Further experiments on the feeding
of Calanus finmarchicus. Biol. Bulletin,
LXX, 2.
Gaarder T. and Gran, H. 1927.
Investigation of plankton in Oslo Fiord.
Rapp. Comm. I nt. Mer. Medit. 42, 3.
— and S p а г с к R. 1931. Bioche-
mical and Biological Investigations of
the Variations in the Productivity of the
West Norwegian Oyster Pools, Rapp,
et Proces-Verb., LXXV.
— 1932. Hydrographisch-biochemische
Untersuchungen in norvegischen A us tern -
Pollen. Bergens Museums Aarbok. Natur-
vidensk. rekke, I.
Galtsoff P. S. 1930. The role
of chemical stimulation in the spawning
reactions of Ostrea itrginica and Ostrea
gigas. Nat. Acad. Sci. Proc., 16.
29 Зенкевич, т. I
449
Galtsoff P. S. 1938. Physiology of
reproduction of Osirea virginica. Spawning
reaction of the female and male. Biol.
Bull. 47.
— and Loosanoff V. L. 1939.
Natural history and method of control-
ling the Starfisch (Asterias forbesi Desor)
U. S. Bur. Fisheries. Bull. 49.
Gamulin T. 1939. Kvalitativna
i kvantitativna istrazivanja planktonskih
kopepoda u istocnim obalinim vodama
srednjega Jadrana-tokom godine 1936—
1937. Prirodoslovna istrazivanja Kralje-
vine Jugoslavije, 22.
Gardiner A. C. 1933. Vertical-
distribution in Calanus finmarchicus. Jour.
Mar. Biol. Ass. 18.
— 1937. Phosphate production by plan-
ctonic animals. J. Cons., 12.
Gardiner Stanley J. 1898.
The coral reefs of Funifuti, Rotuma and
Fiji. Proc. Cambridge Phil. Soc., 9.
— 1903. The Maiadive and Laccadive
groups with notes on other Coral forma-
tions in the Indian Ocean. In Gardiner,
S. Fauna and Geogr. of the M. and L.
Archipelago, I.
G a r s t a n g. 1894. Calender of the
floating fauna.
G e n g. 1925. Der Futterwert der na-
tiirlichen Fischnahrung. Z. Fisch, 23, 2.
Giltay L. 1934. Notes sur 1’asso-
ciation de Balanus concavus pacificus
<*t Dendraster excentricus. Bull. Mus.
Hist. Nat. Beige, 18.
Gislen T. 1930. Epibioses of the
Gullmar Fjord Part II. Kristinebergs
Zoolog. Station, 4.
Goldschmidt V. M. 1933. Grund-
lagen der quantitativen Geochemie. Fort-
si hri tie der Mineral. Kristal und Petrogra-
phie, 17.
G о s s e P. H. H. 1853. A Naturalists
rambles on the Devonshire Coast, London.
— 1856. Tenby; a Sea-side holyday,
London.
Gourret P. 1897. Les etangs sau-
matre du Midi de la France. Ann. du
Mus. d’Hist. Nat. de Marseille, Zool.,5, I.
— 1907. Topographic zoologique des
etangs de Caronte etc. Ann. du Mus-
d’Hist. de la France, II.
Gran H. H. 1902. Das Plankton
des Norwegischen Nordmeeres. Rep. Norw.
Fish, and Marine-Invest., 2.
— 1915. The plankton production of
North European Waters in the Spring
of 1912. Bull. Planctonique pour Гаппёе
1912, publie par le Bureau du Conseil.
— 1927. The production of plankton
in Coastal waters off Bergen. Rep. Nor-
weg. Fish. Invest. 3.
— 1929. Quantit. Plankton Investi-
gations carried out during the Expedition
with the «Michael Sars» VII — IX, 1924.
Rapp, et Proc.-verb. LVI.
— 1931. Qn the Conditions for the
Production of Plankton in the Sea. Ibid.
LX XV.
450
Gran II. II. 1932. Phytoplankton,
methods and problems. Journ. Cons. Ill, 3.
— and В r a a r u d T. 1935. A quan-
titative Study of the Phytoplankton in
the Bay of Fundi and the Gulf of Maine
Journ. of the Biol. Board of Canada, I, 5.
Grave E. R. 1857, Naturen paa
Slesvigs Vest-Kyst. Tidskrift for populaere
Fremstilln, av. Naturvidenskaben, ufg.
Fogh, Liitken og Vai pell. 4.
Gruvel M. 1929. De 1’influcnec
du percement du Canal de Suez sur !<•
faune marine des Cotes de Syrie. C. 11.
Acad. Sci. Paris, 188.
G u e r i n-G a n i v e t J. 1906. Notos
preliminaires sur les gisements de mol-
lusques comestibles des cdtes de France —
Le golfe du Calvados. Bull. Mus. Ocean,
de Monaco, 67.
— 1909. Notes preliminaires sur le
gisement des mollusques comestibles des
cotes de France — La cote des Landes de
Gascogne, le bassin d’Arcachon. Bull,
de Flnstit. Осёап, 135.
Gunther A. 1880. An Introduction
to the study of Fishes.
— 1886. Handbuch der Ichthyologic.
Gunter G., Smith F. and W i 1-
1 i a m s R. 1947. Mass mortality of
marine animals on the lower west Coast
of Florida November 1946—January 1947.
Science, 105, 2723.
Hadzi J. 1932. Oktuda toliki broj
ribljih vrsta u Jadrany. Ribarski Kalen-
dar Jadranske Straze. Split.
Haeckel E. 1866. Generelle Morpholo-
gic.
H a e с к e г V. 1906. Challengerider.
Arch. f. Protistenkunde, 7.
Hagmeier A. 1930. Die Ziichtung
verschiedener wirbelloser Meerstiere. In
Abderhalden Handb. d. biol. Arbeitsme-
thoden, Abt. IX, 5.
— 1930. Die Besiedlung des Felsstran-
des und der Klippen von Helgoland. Wiss.
Meeresunters. Helgoland, 15.
— und Kandler R. 1927. Neue
Untersuchungen im Nordfriesischen Watten-
meer etc. Wiss. Meeresunters. N. F., 16, 6.
Hansen С. P. 1865. Das Schleswig-
sche Wattenmeer, Glogau.
— 1899. Die Cladoceren und Cirri -
pedien der Planktonexpedition. Ergeb-
nisse der Planktonexpedition, 2, G. d.
Hardy A. C. 1924. Report on the
Food and Feeding habits of the Herring.
Fish. Investig. Ser. IL VII, 3.
— 1926. The herring in Relation to its
animate Environment. Part II Ibid. VIII,7.
— 1936. Plankton ecology and the
hypothesis of animal exclusion. Proc.
Linn. Soc. 148—th Session.
— and Gunther, E. R., 1935.
The plankton of South Georgia whaling
grounds and adjacent waters. Discovery
Rep. II, 1.
Hartmey er R., 1910. Tunicata.
Bronn’s Klassen und Ord nungen des Tier-
reichs, Ill, Suppl.
Hartmpyer R. 1911. Die geographische
Verbreitungder Ascidien. Verh. d. Deutsch.
Zool. Gesell.
Harms. 1934. Wandlungen des Artge-
fiiges.
Hart J. J.. 1934. Phytoplankton of
the South-west Atlantic. Discovery Rep. 8.
Harvey H. W. 1926. Nitrate in
the Sea. Part. I. Journ. Mar. Biol. Assoc.,
14.
— 1928. Nitrate in the Sea. Part. II.
Ibid. 14.
— 1928. Biological chemistry and phy-
sics of Sea water.
— 1933. On the rate of Diatom Growth
Journ. Mar. Biol. Ass. 19.
— 1942. Production of life in the Sea.
Bio log. Rev. 17, 3.
— 1945. Recent advances in the che-
mistry and biology of Sea Water.
— Cooper L. H. N., Lebour
M. V. and. Russel F. S. 1935. Plank-
ton production and its control. Journ.
Mar. Biol. Ass. 20.
Havinga B. 1922. Marine Mollus-
ken, Fauna u. Flora der Zuiderzee.
— 1929. Krebse und Weichtiere. Hand-
buch d. Seefisch. Nordeuropas, III, 2.
Hedley C. 1915. An ecological
sketch of the Sydney beaches. Journ.
Roy. Soc. N. S? Wales, 49, Sydney.
Heilprin A ng. 1894. The Geo-
graphical and Geological Distribution of
Animals. The Intern. Scient. Series 58.
Helland-Hansen B. 1908. Die
Austernbassins in Norwegen. I. R. H. H.. I.
— 1909. The Norwegian Sea. Rep.
on Norw. Fish. Invest., II.
Hensen V. 1887. Uber die Bestim-
mung des Planktons u. s. w. V. Bericht
d. Kommission zur Wissensch. Unters. d.
Deutschen Meere in Kiel.
— 1911. Das Leben in Ozean nach
Zahlungen seiner Bewohner. Ubersicht
und Resultate der qhantitativen Unter-
suchungen. Ergebnisse d. Planktonexp., 5.
Hentschel E. 1916. Biologische
Untersuchungen uber den tierischen und
pflanzlichen Bewuchs im Hamburger Ha-
fen. Mitteil. Zool. Mus. zu Hamburg, 33.
— 1923. Der Bewuchs an Seeschiffen.
4. R. H. H., 11.
— 1923. Grundzuge der Hydrobiologie.
— 1926. Глава из книги Schott G.
Geographie des Atlantischen Ozean.
— 1928. Die Grundziige der Plankton-
verteilung im Siidatlantischen Ozean.
I. R. H. H. 21.
— 1938. Das biologische Werk der
«Meteor» — Expedition. «Der Biologe», 1.
— 1936. Allgemeine Biologic des siid-
atlantischen Ozeans. Part I. Das Pela-
gial der obersten Wasserschicht. Wiss.
Ergebn. d. Atlant. Exped. «Meteor», II.
— 1933. Untersuchungen fiber das Klein-
plankton an den Kiisten von Island.
Berichte d. Deutsch, wiss. Komm. fiir
Meeresforschung, VI.
— und W a t t e n b e r g II., 1930.
Plankton und Phosphat in der Oberfla-
chenschicht des Siidatlantischen Ozeans.
Ann. d. Hydrogr. u. Marit. Meteorol.
LVIII, 8.
Herdman W. A. 1896. The 9-th
Annual Report of the Liverpool Mar.
Biol. Committee.
— 1919. Spolia Runiana, IV. Notes on
the Abundance of some common Marine
Animals and a preliminary quantitative
Survey of their Occurence. Journ. Linn.
Soc. Zool., 34.
— 1920. Notes on the Abundance of
some Common Marine Animals and a pre-
liminary Quantitative survey of their
Occurence. Ibid. 34.
H e r 11 i n g H. 1926. Untersuchungen
des Darminhalte von Fischen. Ber. d.
Deutsch. Wiss. Komm. Meeresf. N. F.,
II, 4.
— 1928. Untersuchungen fiber die Er-
nahrung von Meeresfischen. I. Quanti-
tative Nahrungsuntersuchungen an Pleu-
ronectiden und einigen anderen Fischen
der Ostsee. Ibid. N. F., IV, 2.
H erub el M. 1907. L’unite biono-
mique primordiale.
Hesse R. 1924. Tiergeographie auf
okologischer Grundlage.
Hessle Chr. 1924. Bottenbonite-
ringar i ihre Ostersjon. Meddelanden
fran Kungl. Lantbrukstyrelsen, 250.
Hewatt W. G. 1935. Ecological
Succession in the Mytilus californtanus
habitat as observed in Monterey Bay,
California, Ecology, 16.
— 1937. Ecological studies on. selected
marine intertidal communities of Monterey
Bay, California. Amer. Midi. Nat., 18.
Hickling C. F. 1927. The Natu-
ral History of the Hake. Fish. Invest.,
Ser. II, X, 2.
H i r m e r. 1927. Handbuch der Palao-
botanik.
Hjort Joh. 1914. Fluctuations in
the Great Fisheries of Northern Europe.
Rapp, et proces-verbaux, 29, Cons. Perm.
Intern.
— and Ruud Joh. 1929. Whaling
and fishing in the North Atlantic. Ibid. 56.
Hoffer K. 1938. Pilzsociologie. Berl.
Deutsch, bot. Ges. 55 (1937).
H о f k e r J. 1930. Faunistische Beo-
bachtungen in der Zuidersee wahrend der
Trockenlegung. Zs. Morph. Oekol. d.
Tiere, 18.
H о f s t e n N. 1915. Die Echinoder-
men des Eisfjordes. Zool. Ergeb. der
schwed. Exped. nach Spitzbergcn 1908.
Teil II, Kungl. Svenska Vetensk. Akad.
Handl. 54.
— 1916. Die DecapodeYi Crustaceen des
Eisfjords. Ibid. 54, 7.
Hunt O. D. 1925. The Food of the
Bottom Fauna of the Plymouth Fishing
Grounds Journ. Mar. Biol. Ass., N. S.,
XIII, 3.
Hutchinson G. E. 1942. Recent
Advances in Limnology.
451
Iselin C. 1936. Л study of
the circulation of the western North Atlan-
tic. Papers in Physical Oceanography and
Meteorology Mass. Inst. Tech, and Woods
Hole Ocean. Inst. 4.
Ivlev V. S. 1939. Transformation of
energy by aquatic animals. I. R. H. H. 38.
Jacubowa L. und Malm E. 1931.
Die Beziehungen einiger Benthos For-
men des Schwarzen Meeres zum Medium.
Biol. Zentralbl., 51.
Jenkin P. M. 1937. Oxygen pro-
duction by the diatom Coscinodiscus excen-
tricus in relation to Submarine illumina-
tion. J. Mar. Biol. Ass. 22, 1937.
Jenkins J. T. 1921. A Textbook of
oceanography.
Jensen A. J. C. 1928. The Rela-
tion between the Size of the Plaice Stock
nd the Quantity of «First Class Plaice-
Food» in certain Parts of the Limfjord,
Rep. Dan. Biol. St. XXXIV.
Jespersen P. 1923. On the quan-
tity of Macroplankton in the Mediter-
ranean and the Atlantic. Rep. on the
Danish Ocean. Exp. 1908—10 to the
Medit. and Ad. Seas, 7.
— 1928. Investigations on the Food
of the Herring in Danisch Walers. Medd.
fra Kom. for Danm. Fisk, og Havund.
Ser. Plankton, II, 2.
— 1932. On the Food of the Herring in
Icelandic Waters. Ibid. Ser. Plankton, 11,3.
— 1935. Quantitative Investigations on
*he Distribution of Macroplankton in
different oceanic Rigions. «Dana» Rep. 7.
Japan year-book, 1938—39.
Johnstone J. 1908. Conditions
of life in the sea. Cambridge.
— Scott A. and Chadwick H. C.
4924. The marine plankton. Liverpool,
Univ. Press.
Johnston T. H. 1917. Ecological
notes on the littoral flora and fauna Ca-
loundra. Quensl. Naturalist.
Jordan D. S. 1905. A Guide to the
Study of Fishes.
J о u b i n L. 1906. La repartition
des animaux marins sur les cotes fran-
caises de la Mediterrannee. Bull, du Mu-
see Ocean, de Monaco, 74.
— 1906. Consideration sur la Faune
des Cdtes de France. Ibid. 71—72.
— Just Th. 1939. Plant and Animal
Communities.
К a n d 1 e г R. 1928. Verbreitung und
Wachstum der Austernbrut im Watten-
meer. Wiss. Meeresunters. Abt. Helgo-
lang, 17.
Ketchum В. H. 1939. The absorb-
tion of phosphate and nitrate by illu-
minated cultures of Nitschia closterium.
Amer. J. Bot. 26.
К i m b a 1 H. H. 1928. Amount of
Solar radiation etc. Monthly Wea, 56.
King L. A. and Russel E. S.
1909. A method for the study of the ani-
mal ecology of the shore. Proc, of the
Roy. Physiol. Soc., 17.
К i r s о p 1922. Preliminary study
of the Methods of Examining the Life
of the Sea-Bottom. Publications Puget
Sound Biol. St., Ill, 64—67.
К je 11 ma n R. 1877. Ueber die
Algenvegetation des Murmanschen Meeres.
Nova acta. Reg. Soc. Scient. Ups. Ser. HI.
Knipovitsch N. 1892. Etudes
sur la repartition verticale des animaux
le long du littoral des lies Solowetzki
etc. Congres Internal, de Zool. Sess.
2, Part 2.
Knudsen M. 1901. Hydrographical
Tables.
Kojevnikov G r. 1892. La faune
de la mer Baltique orientate et les pro-
blemes des explorations prochaines de
cette faune. Congres intern, de zoologie
a Moscou.
К о p p e n W. und Wegener A.
1924. Die Klimate der geologischen Vorzeit.
К о s s i n a E. 1921. Die Tiefen des
Weltmeeres. Institut fUr Meereskunde, 9.
Kraefft F. 1910. Uber das Plank-
ton in Ost- und Nordsee u. s. w. Wiss.
Meeresunters. Kiel. 20.
К r a m p P. L. 1925. Biologiske Dyk-
kerundersogelser for Vestkysthanvneanlaeg-
gene. Naturens Verden, Aarg. 9, Kopen-
hagen.
К r e p s E. and Verjbinskaja
N. 1930. Seasonal Changes in Phosphate
and Nitrate Content in the Barents Sea.
Journ. Cons. 5.
Krogh A. 1930. Uber die Bedeu-
tung von gelosten organischen Substan-
zen bei der Ernahrung von Wassertieren.
Zeitschr. f. vergl. Physiol. XII, 3—4.
— 1931. Dissolved substances as food
of aquatic organisms. Rapp. Proces-Ver-
baux, LXXV.
— 1934. Conditions of life in the ocean.
Ecolog. Monographs, 4.
— 1934. Conditions of life at great
depths in the ocean. Ibid.
Krummel O. 1907—1911. Hand-
buch der Ozeanographie.
К u e n e n P. H. 1938. Density cur-
rents in connection with the problem
of submarine canvons. Geolog. Magazine,
75, 888.
Kukenthal W. 1896. Ergebnisse
einer zoologischen Forschungsreise in den
Molukken und Borneo. i\bh. Senckenberg.
Naturforsch. Ges., 22.
— 1906. Die marine Tierwelt des arkti-
schen und antarktischen Gebietes in ihren
gegenseitigen Beziehungen. Veroffentl. d.
Inst, fiir Meereskunde u. d. geograph.
Inst. Univ. Berlin, II.
Labbe A. 1922. Les variations de la
concentration en ions hydrogene dans
les marais salants comme facteur biolo-
gique. C. R. des Sciences de 1’Acad. des
Sci, 175.
Larsen K. 1936. The Distribution
of the Invertebrates in the Dybso Fjord,
their Biology and their Importance as
Fish Food. Rep. Dan. Biol. Stat., XLI.
Laurie R. D. and W a t к i n E. E.
1922. Investigations into the fauna of
the sea-floor of Cardigan Bay. Aberyst-
wyth Series by the Memb. of the Univ.
Coll, of Wales 4.
Leavitt В. B. 1935. A quantita-
tive study of the vertical distribution
of the larger zooplankton in deep water.
Biol. Bull. LXVIII, I.
L e b о u г M. V. 1918. Feeding Ha-
bits of some Young Fish. Journ. Mar. Biol.
Ass. XII.
— 1919—1920. The Food of Young
Fish. Ibid. XII, 3. No 3, 1919, 1920.
— 1922. The food of plankton orga-
nisms. Ibid. XII.
— 1922—1923. The food of Plankton
organisms, I. Ibid. XII.
L e D a n о i s Ed. 1934. Les trans-
gressions oceaniques. Revue des Travaux
des Peches maritimes, VII, 4.
Lindeman R. L. 1942. The trophic-
dinamic aspect of Ecology. Ecology.
Lindroth A. 1935. Die Assozia-
tionen der marinen Weichboden, Zool.
Bidrag fran Uppsala, 15.
Linke O. 1939. Die Biota des Ja-
debusenwattes. Helgolander Wissensch.
Meeresunters. I, 3.
Lohmann H. 1896. Appendicularien.
Erg. d. Pl. Exped. 2, E. c.
— 1900. Uber das Fischen mit Netzen
a us Miillergaze N 20 zu dem Zwecke
quantitative!! Untersuchungen. Wissensch.
Meeresunt. Abt. Kiel. N. F. 7.
— 1902. Die Coccolithophoridae, eine
Monographic der Coccolithen bildenden
Flagella ten. Arch, fiir Protistenkunde, I.
— 1903. Neue Untersuchungen uber
der Reichtum des Meeres an Plankton und
fiber die Brauchbarkeit der verschiedenen
Fangmethoden. Wissensch. Meeresunters.
N. F. Abt. Kiel. 7.
— 1904. Eier und sog. Cysten. Erg. d.
Pl. Exped., 4.
— 1908. Untersuchungen zur Feststel-
lung des vollstandigen Gehaltes des Meeres
an Plankton. Wiss. Meeresunters., N. F.
Abt. Kiel, X.
— 1912. Die Probleme der modernen
Planktonforschung. Verhandl. Deutsch,
zoolog. Gesellsch. auf der 22 Jahresver-
samrnl. zu Halle.
— 1916. Neue Untersuchungen uber die
Verteilung des Planktons im Ocean. Sit-
zungsber. Gesellsch. naturforsch. Freunde,
Berlin.
— 1919. Die Bevolkorung der Ozeans
mit Plankton nach den «Ergebnissen der
Zentrifugenfange wahrend der Ausreise
der «Deutschland», 1911. Archiv f. Bion-
tologie, IV, 3.
Lonn be rg E. 1898. Undersoknin-
gar rorande Oresunds djurliv. Meddel. fr.
Kungl. Lantbruksstyr, I. Arg. Uppsa-
la.
— 1903. Undersokningar rorande Skel-
dervikens och angrinsande Kattega tomia -
des djurliv. Ibid., 2, 1902, Uppsala.
Lorenz I. R. 1863. Physikalische
Verbaltnisse und Verteilung der Organis-
men im Quarnerischen Golfe. Wien.
Lotka Alfred. 1925. Elements
of physical biology.
Maas O. 1893. Craspedote Medusen.
Ergcbn. Plankt. Exped., 2 K. c.
Mac Ginitie G. E. 1935. Ecolo-
gical aspects of a California marine estuary.
Amer. Midi. Nat., 16.
— 1938. Notes on the natural history
of some marine animals. Ibid. 19.
— 1939. Littoral Marine Communi-
ties. In Plant and Animal Communities,
ed. by Theodor Just.
Madsen H. 1936. Investigations
on the Shorefauna of East—Greenland
with a survey of the Shores of other Arctic
Regions. Medd. om. Gronland, 100. 8.
Mare M. F. 1942. A study of a marine
benthic community with special reference
to the microorganisms. Journ. Mar. Biol.
Ass. 25.
Marine Biological Asso-
ciation. Plymouth Marine Inverteb-
rate Fauna. Journ. Mar. Biol. Ass. N. S.
7, 1906.
Mario n. 1883. Esquisse d’une topo-
graphie zoologique du golfe de Marseille.
Ann. du Musee d’hist. nat. de Marseille, I.
— 1886. Les faunes des etangs saumat-
res des Bouches-du-Rhone. Mem. Acad,
de Marseille.
Marshall S. M. 1924. The food of
Calanus finmarchicus during 1923. Journ.
Mar. Biol. Ass. 13, 2.
— 1933. The production of MicroplaTi-
eton in the Great Barrier Reef Region.
Greet Barrier reef Exped. 1928—29, II, 5.
— and О г r A. P. 1927. The Relation
of the Plankton to some Chemical and
Physical Factors in the Clyde Sea Area.
Journ. Mar. Biol. Ass., 14.
-------- 1928. The photosynthesis of dia-
tom cultures in the sea. Journ. Ibid. 15.
-------- 1930. A study of spring diatom
increase in Loch Striven. Ibid. 16.
— Nicholls A. G. 1934. On the
biology of Calanus finmarchicus V Seasonal
distribution, size, weight and chemical
composition in Loch Skriven in 1933, and
their relation to the phytoplankton. Ibid.
XIX, 2.
-------- 1935. On the biology of Calanus
finmarchicus. Ibid. 20.
Mayor A. G. 1918. Ecology of the
Murray Island coral reef. Carnegie Inst.
Publ., 213 (Pap. Marine Biol. vol. 9),
Washington.
— 1920. The reefs of Tutuila. Samoa, in
their relation to coral reef theories. Ibid.,
59 (10).
— 1924. Structure and ecology of Sa-
moan reefs. Ibid., 19 (340) No 1.
Mayne C. 1918. Report of the Ed-
ward Forbes Exhibitions. Mar. Biol. Stat.
Port Erin. 32 nd Ann. Rept. Liverpool,
Mar. Biol. Comm.
Meek A. 1916. The Migrations of Fish.
453
Meseck G. 1933. Die tierischen
Vorrats- und Geratsschadlinge in der
Fischerei. Zeitschr. fur Fischerei, XXXI, 2.
Messjatzew J. I. 1930. Trawl
und Bodengreifer als Fangwerkzeuge der
Bodentiere. Arch, fiir Hydrobiologie, XXI.
Meyer J. A. 1914. Beitriige zur
Kenntnis der chemischen Zusammensct-
zung wirbelloser Tiere. Wiss. Meeresunter-
such. N. F. Abt. Kiel., 16.
Meyer P. F. 1936. Die Nordsee-
krabbe Crangon vulgaris Fabr. im Jaden
busen Zschr. f. Fischerei, 34.
Milankovitch. 1920. Theoria
mathematique des phenomenes thermiques
produits par ]a radition solaire.
Milne Edwards H. 1837. His-
toire naturelie des Crustacees, 3.
Mobius K. 1877. Die Auster und
Austerwirtschaft.
— 1893. Uber die Tiere dor Schleswig-
holsteinischen Austernbanke, ihre phy-
sikalischen und biologischen Lebensver-
haltnisse. Sitz. ber. Akad. der. Wiss., I.
Molander A. 1928. Animal com-
munities on soft bottom areas in the
Gullmar fjord Kristinebergs Zool. Stat.
1877—1927, 2.
— 1928. Investigations into the verti-
cal distribution of the fauna of the bottom
deposits in the Gullmar fjord. Svenska
Hydrogr.-Biolog. Komm. Skrifter N. S.
Ilydrografi, VI, 6.
— 1933. Investigations into the verti-
cal distribution of the fauna of the bottom
deposits in the Gullmar Fjord. A preli-
minary study carried out by means of
a novel bottomsampling apparatus desig-
ned by prof. O. Pettersson. Svenska Hyd-
rogr. Biol. Komm. Skrifter. N. S. Hyd-
rografi VI.
Monod Th. 1926. La region de la
dasse Seulle. fitudes bionomique. These
bour le doctorat, Paris.
Mortensen Th. 1921. Biologiske
studier over Sandstrandfaunen, saerling
ved de danske Kyster. Videnskabl. Meddel.
Dansk. Naturhist. Foren. Kobenhavn, 74.
Murphy R. C. 1936. Oceanic birds,
of South America.
Murray J. 1895. Summary of the
scientific results, obtained at the sounding,
dredging and trawling stations of «Chal-
lenger», Reports, Summary, pt. 2.
— et H j о r t J. 1912. The Depths of
the Ocean.
Nansen Fr. 1902. The oceanography
of the North Polar Basin. Norw. North
Polar Exp. 1893—96, Sci. Res., III.
— 1906. The Northern Waters.
— 1913. The waters of the North Eastern
North Atlantic. I. R. H. H.
— 1915. Spitzbergens Waters.
Nathansohn A. 1906. Verticale
Wasserbewegung und quantitative Vertei-
lung des Planktons im Meere. Annal. d.
Hydr. u. Marin. Meteor.
— 1908. Ueber die allgemeinen Produk-
tionsbedingungen im Meere. 1. R. H. H., 1.
454
Naumann E. 1924. Ueber einige
neue Begriffe des Sestonkunde. Sunds
Univ. Aarskr. N. F., 20, N 3.
Newcombe C. L. 1935. Certain
environmental factors of a sandbeach in
the St. Andrews region, New Brunswick,
with a preliminary designation of the
intertidal Communities. Journ. of Ecol. 23.
Nicholls A. C. 1933. On the
biology of Calanus /inmarchicus. Journ.
Mar. Biol. Ass., N. S., 19.
Nicol E. 1935. The Ecology of
a Salt-Marsh. Ibid. N. S. 20.
Nielsen S. E. 1937. On the rela-
tion between the quantities of phytoplank-
ton and zooplankton in the Sea. Journ.
Cons. 12.
Nordgard O. 1905. Hydrographi-
cal and biological investigations in Norwe-
gian fjords. Bergens Mus. Skr., Bergen.
Oceanographic Discussion. Proc. Linn.
Soc. London, 158, 2, 1947.
Odhner N. 1915. Die Mollusken-
fauna des Eisfjords. Kungl. Svenska Ve-
tensk. Akad. Handl., 54, 1.
Oliver W. R. B. 1915. The Mol-
luska of the Kermadec Islands. Trans.
etProc. New Zealand Inst., 47, Wellington.
— 1923. Marine littoral plant and ani-
mal communities in New Zealand. Ibid.
54.
О r t m a n ii A. 1892. Die Korall-
riffe von Dar-es-Salaam und Umgegend.
Zool. Jahrb. Abt. Syst, 6.
— 1893. Decapoden und Schizopoden.
Erg. d. Plankt. Exp. 2, G. b.
— 1896. Grundziige der marinen Tier-
geographie.
Orton J. H. 1914. Preliminary
Account of a Contribution to an Evalua-
tion of the Sea. Journ. Mar. Biol. Ass., 2.
— 1925. On the efficiency of the Peter-
sen Grab. Nature.
Ostroumov A. 1892. Distribu-
tion verticale des Mollusques dans la
Mer Noire. Congres Intern. Sess. 2, Part 2.
Panikkar, N. Kesawa and
A i у a r, R. G о p a 1 a. 1937. The
brackisch water fauna of Madrass. Proc.
Ind. Ac. Sc., 6, 5.
Parker J. R. 1930, Some effect
of temperature and moisture upon the
Melanopus mexicanus, Saussure, and Cam-
nula pellucida Scudder. Mont. Agr. Exp.
Sta. Bull., 223.
Parker S. H. 1924. The growth
of marine animals on submerged metals.
Biol. Bull., 47.
Pearse A. S. 1913—1914. Observa-
tions on the fauna of the rock beaches
at Nahant Mass. Bull. Wisconsin Nat.
Hist. Soc. Ser. 2, vol. IL
— 1926. Animal Ecology.
— 1936. The Migrations of Animab
from Sea to Land.
Pelseneer. 1906.. L’origine des
animaux d’eau douce.
Percival E. 1929. Report on the
Fauna of the Estuaries of the River Tamar
'nd the River Lynher. Journ. Mar
Biol. Assoc. (N. S.), 16, I.
Peters N., Panning A. und
Schnakenbeck W. 1933. Die chi-
nensische Wollhandkrabbe in Deutschland.
Zool. Anz. Erg., Bd. zu Bd., 104.
-------Thiel H., Schubert K.
und andere. 1938. Neue Untersuchungen
uber die chinensische Wollhandkrabbe in
Europa. Mitt. Hamburg. Zool. Mus. und
Inst., 47.
Petersen C. G. 1913. Valuation
of the Sea II. The Animal Communities
of the Sea Bottom and their importance
fur marine Zoogeography. Rep. Dan.
Biol. St. XXI.
— 1914. Notes to Charts I and II.
Appendix to Report XXI. Ibid. XXII.
— 1915. On the Animal Communities
of the Sea Bottom in the Skagerrack,
Christiania Fjord and the Danish Waters.
Ibid. XXIII.
— 1918. The Sea bottom and its Produc-
tion of fish-food. Ibid. XXV.
— 1924. The necessity for quantitative
methods in the investigations of the ani-
mal life on the Sea-bottom. Proc. Zool.
Soc. London, July 9.
— 1924. A brief survey of the Animal
Communities in Danish Waters based
upon quantitative samples, taken with
the bottom sampler. Amer. Journ. Sci, 7.
— 1926. On works concerning the Valua-
tion of the Southern North Sea. Journ.
Cons. I, 2.
— and Boysen -Jensen P. 1911.
Valuation of the Sea I. Animal Life of
the Sea-bottom, its food and quantity
(Quantitative Studies). Rep. Dan. Biol.
St., XX.
Pettersson H. 1946. Oceanogra-
phic work in the Mediterranean. The
geograph. Journ. 57, 3—4
Pfeffer G. 1891. Versuch iiber die
erdgeschichtliche Entwicklung der jetzigen
Verbreitungsverhiiltnisse unserer Tierwelt.
Phifer, LymanD. and Thomp-
son T. G., 1937—38. Seasonal varia-
tions in the surface waters of San-Juan
Channel during the five-years period.
Journ. Marine Research, I.
P h 1 e g e r S. B. and Hamil-
ton W. A. 1946. Foraminifera of two
submarine core from the North Atlantic
basin. Geolog. Soc. Amer. Bull, 57, 10.
Piggot C. S. 1938. Core samples of
the Ocean bottom and their significance.
Carn. Inst. Suppl. publ., 36.
Pierce W. 1916. A new interpreta-
tion of the relationships of temperature
and humidity to insect development.
Journ. agric. Res: 5.
P i e r r i e M. E., Bruce J. R.
Moore H. B. 1932. A Quantitative Study
«of the Fauna of the Sandy Beach of Port
Erin. Journ. Mar. Biol. Ass. N. S., 18.
Pierron R. P. and Hung U. C.
1925. Animal Succession on denuded
rocks. Publ. Puget Sound Biol. Stat., 5.
P о p о v s к у A. 1904. Acanthome-
tri den. Erg. d. Plankton-Exped., 3, Lief,
f. d.
Portier. 1938. Physiologie des Ani-
maux marins.
Poulsen E. M. 1931. Biological
investigations upon the Cod in Danish
Waters. Part IV. The Food of the Cod.
Medd. Komm. f. Danmarks Fisk, og
Havundersog., Fiskeri, IX.
P r a t j e O. 1939. Die Sedimente
des sudatlantischen Ozeans. Wiss. Ergebn.
deutsch. Atlant. Exp. «Meteor» 1925—27,
III, Th. 2, Lief. 2.
P r e n a n t M. 1927. Notes ethologi-
ques sur la faune marine sessile des envi-
rons de Roscoff IL Travaux Stat. biol.
de Roscoff, 6.
P r u v о t S. 1894. Essai sur la topo-
graphic et la constitution des fonds sous-
marines de la region de Banyuls Arch, de
Zool. Exper. et Gen. Ser. 3, II.
— 1895. Coup d’oeil sur la distribution
generale des invertebres dans la region
de Banyuls. Ibid. Ser. 3, III.
— 1897. Essai sur les fonds et la faune
de la Manche Occiden tale (cotes de Bre-
tagne) etc. Ibid. Ser., 3, V.
— 1896—1898. Conditions generales de
la vie dans les mers et principes de
distribution des organismes marins. L’an-
nee biologique, 2.
Purcell L. T. 1939. The Aging
of Reservoir Waters. Journ. of the Amer,
water work Ass. 31 (N 10).
Quatrefages, A. 1854. Souve-
nirs d’un Naturaliste. Paris.
R e d e к e. 1922. Zur Biologic der
niederliindischer Brackwassertypen. Bijdr.
Dierk. Amsterdam, 22.
Redfield A. C. 1934. On the pro-
portions of organic derivatives in sea
water and their relation to the composi-
tion of plankton. James Johnstone Memo-
rial Volume.
— Smith H. P. and К e t-
chum, B. 1937. The cycle of organic
phosphorus in the Gulf of Maine. Biol.
Bull. Woods Hole, 73.
R e g n a r d P. 1891. Physique biolo-
gique. La vie dans les eaux.
Reibisch J. 1895. Pelagische Phyllo-
dociden u. Typhloscoleciden. Ergebn.
Plankton-Exped., 2, H. c.
R e m a n e A. 1934. Die Brackwas-
serfauna. Zool. Anz. 7, Supplementband.
Richard J., Oxner M., S i-
went L. 1923. Observations metho-
diques de temperature et de salinite a di-
verses profondeurs au large de Monaco.
Bull. Inst, oceanogr. Monaco, 436.
Riley G. A. 1938, 1939. Plankton
studies I and II J. Mar. Res., 1, 2.
— 1941. Plankton studies III and IV.
Bull. Bingham Oceanogr. Coll., 7, 3—4.
— 1943. Physiological.aspect of spring
diatom flowerings. Ibid. 8, 4.
R i n к e H. 1937. Ueber die chemi-
sche Zusammensetzung einiger Bodentiere
455
der Nord- und Ostsee und ihre Heizwcrt-
bestimmung. Helg. Wiss. Meeresunt., 1, 2.
Rogers C. G. 1938. Textbook of
comparative physiology.
Rose M. 1925. Contribution a
1’etude de la Biologic du Plankion. Le
probleme des migrations verticales journa-
lieres. Arch, de Zool. exper. et gener., 64.
— 1928. La temperature ct la circula-
tion d’Aquiculture et de Peche de Castigli-
one. Alger.
Russel F. S. 1942. The Overfi-
shing Problem.
Russel F. S. 1925, 1926, 1928,
1931. The Vertical Distribution of Marine
Macroplankton. Observations on Diurnal
Changes. Journ. Mar. BioL Ass., 13,
14, 15, 17.
— 1927. The vertical distribution of
plankton in the sea. Biol. Reviews, II, 3.
— 1933. The seasonal distribution of
Macroplankton as shown by catches in
the 2-m stramin Ringtrawl in offshore
waters of Plymouth. Journ. Mar. Biol.
Ass., 19, I.
— 1934. The Zooplankton III. A Com-
parison of abundance of zooplankton in
the Barrier Reef Lagoon, e. c. Great
Barrier Reef Exp. 1928—29. Sci. Repts., 2.
— 1935. The seasonal abundance of
young fish in the offshore waters of the
Plymouth area. Journ. Mar. Biol. Ass., 20.
— 1935. A review of some aspects of
zooplankton research. Rapp, et Proc.
Verb., 95.
— and J о n g e С. M. 1928. The Seas.
Sars G. O. 1869. Indberetningar
til Departement for det Indre fra Cand.
G. O. Sars om de af ham i Aarene 1864—
1869 anstillende praktiskvidenskabelige
Undersogelser angaaende Torskefiskeriet
i Lofoten.
Sars M. 1835. Beskrivelser og lakt-
tagelser over nogle merkelige eller nye
i Havet ved den Bergenske К у st levende
Dyr.
— 1853. Bemaerkninger over det adria-
tiske Ha vs Fauna sammenlignet med
Nordhavets. Nyt. Magazin for Naturvi-
denskaberne 7.
Savage R. E. 1926. The plankton
of a Herring Ground. Fish. Invest. Ser.
II, 9, I.
— 1930. The Influence of Phaeocystis on
the Migrations of the Herring. Ibid., Ser.
II, XII, 2.
— 1931. The relation between the
Feeding of the Herring of the East Coast
of England e. c. Ibid. Ser. II, XII. 3.
— 1932. Phaeocystis and Herring Shoals.
Journ. of Ecology, XX, 2.
— and Hardy A. C. 1935. Phyto-
plankton and the Herring. Fish. Invest.,
Ser. II, XIV, 2.
— and Wim penny R. S. 1936.
Phytoplankton and the Herring. Ibid.
Ser II, XV, I.
S a v i 1 I e -K e n t W. 1893. The greajt
barrier reef of Australia, London.
Schiemenz P. 1905. Die Nahrung
unsercr gewohnlichen Wildfische. Allg.
Fischereizeit. N. F., XX.
S c h m a r d a L. K. 1853. Die geo-
graphische Verbreitung der Tiere. Wien.
Schmidt Johs. 1931. Oceano-
graphical expedition of the «Dana», 192b—
1930. Nature, March.
Schott G. 1926. Geographic des
Atlantischcn Ozeans.
— 1927. Das Meer, Grundzuge der phy-
sischen Erdkunde von A. Supan.
— 1935. Geographic des Indischen und
Stillen Ozeans.
Schott W. 1935. Die Fora mini fen n
in dem iiquatorialem Teil des Atlantischcn
Ozeans. Wiss. Ergebn. d. Deutsch. Atlant.
Exped. «Meteor», 1925—1927, 111, 13 B.
Sears M. and Clarke G. L. 19-io.
Annual fluctuation in the abundance of
marine Zooplankton Biol. Bull.
Sei we 11 H. R. 1931. Observations
on the Phosphate-Content and Hydrogen i-
on Concentration of the North Sea, the
Southern Entrance to the Norwegian Sea,
and the Water South of Iceland. Journ.
Cons. 6.
— 1931. A consideration of some exter-
nal factors governing the production of
plankton in the Sea. Journ. of Ecology,
XIX, I.
— 1934. The distribution of oxygen in
the western basin of the North Atlantic. ’
Papers in Phys. Ocean, and Meteorul.,
Ill, I.
— 1935. The annual organic production
and nutrient phosphorus requirement in
the tropical western North Atlantic. Journ.
Cons. X, 1.
— 1935. The cycle of phosphorus in the
western basin of the north Atlantic. 1.
Phosphate phosphorus. Ibid., Ill, 4.
S e r n a n d e r R. 1917. De nordeuro-
peiska havens vaxtregioner. Svensk. But.
Tidskr., IL
Sewel S. 1940. The extent to
which the distribution of marine organisms
can be explained by and is dependent un
the hydrographic conditions present in
the great oceans, with special reference
to the plankton. Proceed. Linn. buc.
152-nd Session.
Shelford V. E. 1930. Geographic
extent and Succession in Pacific North
American Intertidal (Balanus) Communi-
ties. Publ. Puget Sound Biol. St. 7.
— 1940. Bioecology.
— and T о w 1 e r E. D. 1925. Animal
communities of the San-Juan Channel and
adjacent Seas. Publ. Puget Sound Biul.
St., 5.
— Weese O. A., Rice L. A.,
Rasmussen D. J., Maclean A.
1935. Some marine biotic communities
of the Pacific coast of North America.
Ecol. Mon.
Shepard F. P. and Emery KO.
1941. Submarine topography off the
California coast. Geol. Soc. Amer., 31.
456
Simpson S. 19'j'i. Tempo and
Mode in Evolution.
Simroth II. 1895. Gastropoden.
Erg. Plankt-Exp. 2, F. d.
Skottsberg K. 1907. Wissensch.
Ergebn. Schwed. Siidpol. Exped. 1901 —
1903, 4, 6.
Sollaud E. 1925. Les associations
vegetates et animates des terrains sates
de I’embouchure de la Slack. Gian tires
Biologiques. Wimereux. Trav. Stat.
Biol., 9.
Soo t-R у e n, T. 1924. Faunistische
Untersuchungen im Ramfjorde. Tromso
Mus. Arshefter, 45, 6.
Southern R. 1915. Marine Eco-
logy. Proceed. Royal Irish Academy,
XXXI, 67.
S p i e s s F. 1927. Meteorfahrt.
S p а г с к R. 1926. On the food prob-
lem in relation to marine zoogeography.
Physiol.Papers ded. A.Krogh, Kopenhagen.
— 1929. Preliminary survey of the
results of quantitative bottom-investiga-
tions in Iceland and Faroe waters 1926—
1927. Gons. perm, internet, pour 1’explor.
de la mer.
— 1923—1931. Studies on the Biology
of the Oyster (Ostrea edulis). Rep. Dan.
Biol. St., XXX—XL.
— 1931. Some quantitative Investi-
gations on the bottom fauna at the west
coast of Italy in the Bay of Algiers, and
at the coast of Portugal. Rep. Dan. Ocea-
nogr. Exp. 1908—1910, III, 7.
— 1933. Contributions to the animal
ecology of the Franz-Joseph Fiord and
adjacent waters, 1—II. Medd. om Gronland,
100, 1.
— 1935. On the importance of quanti-
tative investigation of the bottom fauna
in marine biology. Journ. Cons., 10.
— 1936. Ueber die zoogeographische
Bedeutung der Petersenschen Tiergemein-
schaften. Zoogeographica, 3.
— 1936. Review of the Danish Inves-
tigations of the quantitative Composition
of the bottom fauna in Iceland and East
Greenland waters. Rapp. Proc. Verb., 99.
— 1937. The Ben tonic Animal Com-
munities of the Coastal Waters. The Zoology
of Iceland, I.
Stammer H. 1932. Die Fauna des
Timavo. Zool. Jahrb. Abt. f. Syst., 63.
S t e g g e r d a M. and Essex H.
1925. Rock bottom fauna of a restricted
area near Friday Harbor, Washington.
Publ. Puget Sound Biol. St., 5.
— and Morris 1929. The Relation
of Domestic Fowls to Climate. Ecology,
X, 3.
Steinitz W. 1929. Die Wande-
rung indo-pazifischer Arten ins Mittel-
meer seit Begin der Quartiirperiode,
1. R. H. H., 22.
Stephen A. C. 1923. Preliminary
survey of the Scottish Waters of the North.
Sea dy the Petersen Grab. Fisheries,
Scotland, Scient. Invest, 1922, 3.
Stephen A. C. 1928. Notes on the
Quantitative Distribution of Molluscs and
Polychaetes in Certain Intertidal Areas on
the Scottish Coast. Proc. Roy. Phys. Soc.,
V, 21. ,
— 1928. The Bottom Fauna of the
Moray Firth. Investig. with the Petersen
bottom sampler. Rapp. etProc. Verb., Lil.
— 1930. Studies on the Scottish Marine
Fauna. Additional Observations on the
Fauna of the sandy and muddy Areas of
the Tidal Zone. Trans. Roy. Soc. Edinb.,
56.
— 1933. Studies on the Scottish Ma-
rine Fauna. The Natural Faunistic Di-
visions of the North Sea as shown by
the Quantitative Distribution of the Mol-
luscs. Trans. Roy. Soc. Edinb., 57, part
HI (22).
— 1935. Notes on the intertidal Fauna.
The Scottish Naturalist.
S t e u e r A. 1910. Planktonkunde.
— 1933. Zur planmassigen Erforschung
der geographischen Verbreitung des Hali-
planktons, besonders der Copepoden. Zo-
ogeographica, I, 3.
— 1935. Preliminary report. Fish. Res.
Direct., Egypt. Notes and Mem., ;8.
S t i a s n у G. 1934. Das Bipolar!-
tatsproblem. Arch. Neerland. Zool., I,
1—2.
Stott F. C. 1936. The marine food
of birds in an inland fjord region in West
Spitzbergen. Part I. Plankton and inshore
benthos. Journ. Amer. Ecology, 5.
Strom Munster K. 1932. Ty-
rifjord. A limnological study. Skrift.
Norsk. Vidensk. Akad. i Oslo. I Ma tern.
Naturv. Klasse, 3.
Struve K. und К a i r i e s W.
1929—1930. Die chemische Zusammen-
setzung einiger als Fischnahrung wichti-
ger wirbelloser Bodentiere der Nordsee.
Wiss. Meeresunters. N. F., XVII, 6.
Studer Th. 1879. Die Fauna von
Kerguelensland. Frose he 1, Archiv. f. Na-
turg. 1.
— 1889. Die Forschungsreise S. M. S.
«Gazelle». 3 Theil. Zoologie und Geologie.
Stuxberg A. 1880. Evertebratfau-
nan i Sibiriens ishav. К. V A.; s Bihang,
5, 22, Stockholm.
— 1882. Evertebratfaunan i Sibirian
ishav. Forelopande Meddelande Vega Exp.
Vetenskapl. lakttagelser utg. Nordens-
kjold, 1.
Svedberg. 1922. Ett bidrag till
de statistiska metodernas anvanding inom
Vaxtbiologien. Svensk. Bot. Tidskrift, 16.
Sverdrup H. U. 1931. The origin
of the deep-water of the Pacific ocean as
indicated by the oceanographic Work
of the «Carnegie». Gerlands Beitr. zur
Geophysik, 29.
Sverdrup H. U., J о h n s о n M. W.
and F 1 e m i n g R. H. 1942. The Oce-
ans.
Summerhayes V. S. and E 1-
ton C. S. 1923. Contributions to the
Weber Max. 1922. Biologic der
Zee. De Zeeёn van Nederlandsh Oost
Indie. Leiden.
Wegener A. 1922. Die Entstehung
d<^r Kontinente und Ozeane. (Русский
перевод, 1925).
Wesenberg-Lund С. 1904. In
Eug. Warming. Bidrag til Vadernes,
Sandens og Marskens Naturhistorie. Kg.
Danske Vid. Sclsk. Skrifter, 7 Rk. Natur-
vidensk. og Mat. Afd. 2, I.
W i m p e n n у R. S. 1941. Organic
Polarity — some ecological and physio-
logical aspects. The Quart. Rev. of Biology,
76, 4. (Русский перевод 1945).
W i s m e r H. M. and S wa n-
s о n J. H. 1935. A study of the animal
communities of a restricted area of mud
bottom in San Juan Channel, Part II.
Ecol. Mon. 5.
Wohlenberg E. 1937. Die Wat-
tenmeer — Lebensgemeinschaften im Ko-
nigshafen von Sy It. Helg. Wiss. Meere-
sunters., I, H. 1.
Wiist G. 1928. Der Ursprung der
Atlantischen Tiefenwasser. Berlin. Zs. Ges.
Erdk. Sonderband zur Hundjahrfeier der
Gesellschaft.
— 1929. Schichtungen und Tiefenzir-
kulation des Pazifischen Ozean. Veroff.
Inst. Meeresk. A. Heft. 20.
— 1935. Die Stratosphere. Deutsche
Atlant. Exp. «Meteor», 1925—27. Wiss.
Ergebn., VI, I Teil, 2.
— 1936. Oberflachensalzgehalt, Verdiin-
stung und Niederschlag auf dem Welt-
meere. Festschrift Norbert Krebs. Stutt-
gart.
Yon go С. M. 1931. Digestive Pro-
cesses in Marine Invertebrates and Fishes.
Journ. Cons., VI, 2.
— 1947. The influence of man on marine
life. Endeavour, VI, 21.
Zenke witch L., Brotskv V.
and I de Ison M. 1928. Materials for
the study of the productivity of the Sea
bottom in the White, Barents and Kara-
Seas. Journ. Cons. Ill, 3.
Zimmer C. 1900. Die Arktische
Cumaceen. Fauna arctica. I, 3.
1904. Die arktischen Schizopoden. Ibid.,
Ill, 3.
— 1909. Schizopoden. Nordisches Plank-
ton, 12.
— 4 913. Die Cumaceen der Deutschen
Siidpolar-Expedition 1901—1903. Deutsche
Sudpolar.-Exped. (1901—1903), XIV, Zo-
ologie, VI, 3.
— 1940. Die Verbreitung der Cumaceen.
Arch. f. Naturg. N. F., 9, 2.
Z о Bell С. E. 1938. Studies on the
bacterial flora of marine bottom sediments.
Journ. Sediment. Petrology, 8.
— 1942. Changes produced by micro-
organisms in sediments after deposition.
Ibid., 12.
— 1942. Bacteria of the marine world.
Scient. Month., 55.
— 1946. Marine Microbiology.
— and F e 1 t h a m К. B. 1934. Pre-
liminary studies on the distribution and
characteristics of marine bacteria. Bull.
Scripps Inst. Oceanogr. Techn. Ser., 3.
-------- 1942. The bacterial flora of a
marine mud flat as an ecological factor.
Ecology, 23.
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
А
Абиосестон 103
абиоцен 177
абиссаль 192, 196, 197, 199, 290, 291,
294, 298, 359, 360
— арктическая 295, 296
— индийская 296
— Северной Атлантики 296
абиссальная область 191
Авачинский округ 370
Австралийский барьерный риф 192
Австралия 134, 163, 420, 439, см. также —
побережья
— Южная 163, 165
агар-агар 431, 432
Адриатика — см. море Адриатическое
Адриатическое море — см. море
Азия 18, 315, 366, 378,409
Азовское море — см. море
Азорские острова — см. острова
азот 14, 61, 70, 105—111, 114,115, 150, 151,
203, 303, 304
— альбуминоидный 106, 115
— аммиачный 107
— газообразный 304
— концентрация в морских организмах
70
— органический 115, 151
— свободный 105
— связанный 106
— элементарный 105
азотистые соединения 205
акклиматизационные пути 356
акклиматизационный фонд 435—437
акклиматизация 14, 15, 435—439
— тихоокеанского лосося 437—439
аккумуляция донных отложений 14
— органического вещества 41
— элементов морскими организмами 69
активная реакция 93
активин 252,253, 257, 315, 395
акулы 328, 385, 387, 411
Александрия 286, 402
Александровск на Мурмане 23
Алеутская провинция 370
Алеутские острова 94
Алжир 430
Алжир-Провансальский район 85
Алжирская бухта 285
Алжирский залив 284
Аляска 247, 413, 419, 421
Аляска южная 99
«Альбатрос» э. с. 17, 18
альбатросы 307
Альборанское море ИЗ
альгины 431
Америка 150, 382, 38'1. 409, см. также —
побережья
— Северная 56, 192, 193, 195, 219, 363,
379
— тропическая 163, 397—400
— Центральная 355, 376, 393, 394, 396,
397, 400
— Южная 110, 249
аминокис. юты 106, 116
аммиак 97, 106—108, 114, 304, 305
амплитуда миграций 225—228
— приливо-отливных колебаний 249,
257
— колебаний температуры 80—84
Амурский залив 290
амфиатлантичность 378, 384
— потенциальная 439
амфибиальное существование 170
амфибореальность 376, 378, 384
— потенциальная 439
амфипацифпчпость 384
— потенциальная 439
амфиподы 162, 208, 250, 252, 254, 260,
283, 290, 373, 382, 401, см. также боко-
плавы
анагалинное распределение солености 74
Анадырский округ 370
анатермия 76
Английский канал — см. капал
Англия 19, 193, 256, 275, 349, 439
«Андрей Первозванный» э. с. 18
Аннелиды 147, 286
аномалии температурные 139
Антарктика 15, 16.,*20, 53, 75—78, 84, 111,
117, 129—132, 134, 147, 165, 204,300,
352, 360, 369, 373, 387, 391, 406, 416,
417, 430, 436
Антарктическая область — см. область
антарктические моря — см. моря
антибиоз 301
антибореальная область 362
антипатарии 400
антозои 162
анчоусы 385, 401
аппендикулярии 313
Аише ронский полуостров 432
Аральское море — см. море
461
Аргентина 439
ареалы (морских организмов) 357, 372
и сл.
— амфиарктические 381
— амфиатлантические 377
— амфибореальные 381
— батиметрические 185
— биогеографические 35
— биомпческие 357
— географические 185, 366
— качественные 357
— количественные 357
— ограниченные 373
— потенциальные 15, 436, 439
— по типу грунта 185
— разорванные 377, 379, 385, 389
•-----биполярные 385
— сплошные 379
— типологические 355
— широтные разрывы их 375
— экологические 187, 319, 357
арка 418
Арконский район 78
Арктика 161,300,352,361,381, 387, 439
Арктическая область — см. область
Арктический бассейн — см. бассейн
— океан 214
«Армауер Ганзен» э. с. 20
ароморфозы 168
Археозой 171, 173
архибенталь 197, 199, 298
архипелаг Малайский 161,, 163, 165
«— Остиндскпй 316
— Сан Хуан 293
асимметрия климатических зон 353
— океана биологическая 353
— океанических течений 353
аскофиллум 247, 248
ассимиляция 204
асцидии 190, 288, 290, 373, 377, 378, 384,
395, 400, 429, 430, 433
— глубоководные 296
— сложные 401
Атлантика 20, 73, 125, 129. 152, 397, см.
также океан Атлантический
— восточная 405—406
— западная 119, 206
— северная 19, 87, 123, 125, 127, 163,
202, 212, 220, 221, 264, 269, 296, 318,
344, 348, 349, 352, 357, 377, 379, 382,
385, 401,416
— северо-восточная 206
— северо-западная 270
— тропическая 400
— центральная 212
— южная 116, 203, 205
«Атлантис» э. с. 20, 90
Атлантическая область — см. область
Атлантический океан — см. океан
атмосфера 155, 176
атолл коралловый Фунафути 195
ауксоспоры 175
Аустральная область 364
аутотрофные организмы 156, 173, 174
— процессы 301
аутэкология 34, 35, 339
Африка ПО, 141, 204, 366, см. также —
побережья
Ахалцых 175
аэробиология 34
аэропланктон 155
аэроэдафон 170, 250
Б
Багамские острова 195, 430
багрянки — см. водоросли красные
базары птичьи 308
Байкал оз. 4Г2
бактериальная пленка 150
бактерии 47, 95, 97, 106, 108, 114, 116,
150, 155, 156, 187, 188, 190, 205, 206,
230, 232, 233, 301—305, 324--327, 331
— водородные 150
— денитрифицирующие 97
— количество их в морской воде 302—
304
— метановые 150
— нитрифицирующие 106, 305
— потребление кислорода 304
— разлагающие клетчатку 97
— тионовые 150
бактериофаг 302
бактериоценозы 301
Баку 432
баланс вод 131
— гидросферы 53
— органического вещества 35
баланусы 190, 191, 283
Балеарское море 112
Балтийское море — см. море
Балхаш оз. 412
банка Большая Ньюфаундлендская 88,
130
— Доггерская 279, 280
— Ньюфаундлендская 123, 247
банки устричные 178, 430
Баныольс 250, 403
Баренцово море — см. море
барьер Дэвисов 78
— Исландский 78
— Ньюфаундлендский 78
— Шпицбергенский 78
барьеры для распространения орга-
низмов 87
— мелководные 297
— мидиевые 36
— подводные 89, 94
— температурные 87
бассейн Арктический 53, 73, 76, 132,
139, 165, 169, 192, 295, 297, 315, 353,
355, 368, 375, 379, 381
— Печоры 379
— Полярный 139, 141, 152, 295
— Сарматский 175, 368, 405
— Средиземноморский 404
бассейны лагунные 174
— солоноватоводные 169
— устричные 102
батиаль 55, 196, 290, 291, 294—296
батипланктон 190
батисфера 99, 277, 401
батометр 97, 103, 201
Баффинов залив 78
бедность средиземноморской фауны
284, 285
беззубка 418
белки 68
462
Белое море — см. море
— озеро 46
Бельгия 419, 426
бельдюга 253, 272—275
Бельт 263, 265, 270
Бенгальский залив 7 9
бенталь 159, 188, 197
бентос 46, 49, 123, 150, 162, 180, 188,
189, 241 и сл., 332, 333, 347—351,
354
— Азовского моря 45
— вагильпый 190
— вертикальные подразделения 196---
199
— глубоководный 116
— датских вод 262 и сл.
— кормовой 271—273
— методика изучения 241—24 5
— морской 177, 186
— озерный 45
— побережий Гренландии 283
---Исландии 281
— пресноводный 168
— Северного моря 275 и сл.
— Средиземного моря 284—28<8
— Тихого океана 288, 290—292
бентофаги 334, 336
берега — см. побережья
Берингов пролив — см. пролив
Берингово море — см. морс
Бермудские острова — см. острова
бесчерепные 168
«Бигль» э. с. 15, 192
бикарбонат кальция 176
бикарбонаты 95
бимтрал 429
биогеографические критерии 366
биогеография 33—36, 339 и сл.
— аналитическая 339
— биомическая 356
— историческая 357, 358
— каузальная 339
— регистрирующая 339, 353, 356, 358
— синтетическая 340
— типологическая 357
биогидрология 35
биокоспые тела 156
биолимпология 35
биолиты 14
биология моря 13, 14
биома 177, 345
биомасса 37—43, 148, 149,156, 158, 161,
181, 184, 185, 188,208, 214,254—257,
272, 278, 325, 326
— бентоса 46, 265—267, 275, 277, 281—
294, 350, 351,
---максимальная 292—294
— зостеры 268
— исходная 37, 42, 44, 47
— конечная 37, 42
— остаточная 37
— падение ее 42
— планктона 207, 218, 235, 236, 238,
239, 430
— промысловых креветок 428
— разовая 293, 294
— саргассовых водорослей 401
— средняя годовая 44, 47
— устриц 423
биомасса фитопланктона 207
бноокеанелогия 35
биопотамология 35
биосестон 103
биосфера 33, 155 и сл., 339
биотоп 159, 177—182
биофильтр 35, 36
биофильтраторы 3G
биофильтрациопный поток 36
биофильтрация 205
биохимия моря 31
биохоры 178
биоцен 177
биоценоз 31, 34, 47, 49, 62, 177 и сл.. 194-
— как организм 179
— методы изучения 182
— состав его 181
биоценозы Азовского моря 44, 181, 182
— архипелага Сан-Хуан 293
— Баренцева моря 181, 184
— вертикальная смена 277
— выклинивание 193
— Гюльмарфиорда 276
— донные 44, 262, 270, 293
— залива Петра Великого 292
— Ламанша 193
— литоральные 254, 258, 259
— мягких грунтов 262—264, 281, 282,
290
— наземные 181
— олигомикстпые 44. 181, 184
— побережий Исландии 281, 282
— подъем границ 193
— полимикстные 181, 184
— Средиземного моря 193
— сублиторали 186
— Черного моря 193
— эпифауны 276
биоценология 34, 177, 181, 339
биоциклы 178
биоэкологическая обеспеченность 41
— формация 177
биполярность 342, 385—392
— потенциальная 439
— происхождение ее 389—391
Бискайский залив 83, 366
«Блек» э. с. 17
Боденское озеро 412
бокоплавы 66, 401, 429, см. также амфи-
поды
Большой Барьерный риф 215, 216
Большой Шантар остров — см. остров
бореальная область — см. область
бореарктическая область 362
бореохитон 356
Борнгольмский район 78
борьба с обрастаниями 433
Босфор 75, 288
Ботнический залив — см. залив
Бретань 191, 382, 403
Британские острова 270, 363
буревестники 307
буруны 122
бухта Алжирская 285
— Кильская 32, 208, 234, 313, 314
— Севастопольская 32
— Фрайди 114
бычки 190. 273, 274
бычковые 315
463
в
Вайгач остров 361
валуны 144
«Вальдивия» э. с. 18
Ванкувер остров 439
ваттепы 195, 249—251, 254, 256
Вашингтон 114
«Вега» а. с. 17, 192
Великобритания 162, 249, 411, 412,
419—422, 426
Венецианская лагуна 284
венус 418
Веньярд побережье 192
вертикальная зональность — см. зо-
нальность
вес спиртовый 242
— сухой 242
— сырой 242
— формалиновый 242
« Be с л ем о ё» э. с. 139
весна биологическая 234, 235, 237
Вест-Индия 302, 369, 394, 401
ветры пассатные 80, 112, 151
— сгошю-пагонпые 224
вещества белковые 95, 97, 106
— биогенные 235, 236
— минеральные 61, 158
— органические 14,38—42,68,105.114—
116, 149—151, 156, 159, 174, 260
— органического происхождения 145
— питательные 61
вещество живое 155, 156
— зеленое биосферы 158
— косное 156
взаимопроникновение морской п прес-
новодной фаун 310
взвеси неорганические 100
— органические 100
взятие проб (методика) 186, 187
видообразование 173, 382
виды — см. формы
— близнецы 260, 296, 373, 397
викариат биологический 259, 284, 373
впкариировапие биологическое 257
Виргиния 246
витамины в промысловых беспозво-
ночных 420
«Витязь» э. с. 17
вобла 322
водные массы 117 и сл., 159, 1G0
--- Атлантического океана 369
---динамика 117 и сл.
— — классификация 126, 128, 131
---регрессии их 125
— — центральные 127—129
--- циркуляция их 130
водоемы агалипные 317
— материковые 67
— мезогалинные 64, 317
— морские 64, 317
— олигогалинные 64, 317
— пересоленные 64
— полигалинные 64, 317
— почти агалинные 64
— пресные 64, 159, 206, 317
— еолоноватоводные 64, 317
— солоноватые 309 и сл.
— ультрагалинные 64, 317
водоемы эпиконтинентальные 294
— южные СССР 169
водолазные приборы 277, 278
водолазы (добыча промысловых бес-
позвоночных) 423, 430
водород 97
водоросли 40, 104, 107, 157, 193, 206,
219, 236, 246, 249, 251, 257, 384, 417,
419, 433, 435
— арктические 366
- - арктическо-бореальные 366
— бурые 104, 157, 158, 163, 173, 246—
248, 253, 311, 363, 384, 386, 391, 401
— вольвоксовые 202
— диатомовые 40, 70, ИЗ, 1 14 , 145, 147,
173, 175, 190. 201—208, 216, 235, 236.
302—304, 312, 313, 327, 332, 334, 334,
438
— жгутиковые 202
— зеленые 104, 157, 158, 163, 173,
246—248, 253, 311, 366, 395
— известковые 68, 70. 146, 366, 392, 395
— красные 104, 157, 158, 163, 165, 173,
246—248, 284, 286, 311, 366, 390, 391,
395
— кремпежгутиковые 113, 202
— ламинариевые 387
— одноклеточные 157, 188, 189, 327, 329
— перидиневые 70, 173, 201—203, 205,
236, 327, 328, 330, 331,354
— планктонные 103, ИЗ, 322, 437
— пресноводные 312
— промысловые 431, 432
— протококковые 202
— разножгутиковые 202
— сине-зеленые 173, 201—203
— фукусовые 401
—• харовые 158, 248
воды австралийские 195
— адриатические 215
— английские 238, 279
— антарктические 110—112, 117, 126,
128, 130—132, 134, 212, 369, 373, 406,
407
— арктические 78, 88. 128, 129, 130,
132, 212, 219, 369, 370
— атлантические 73, 114, 123, 132,139,
162, 219, 366
— Атлантического океана (классифика-
ция) 132
— балтийские 429
— береговые 87, 90—92, 141, 303, 315
— бореальные 212, 284, 439
— Вест-Индии 302, 394
— глубинные 78, 83, 87, 110, ИЗ, 116,
128—132, 134, 138, 151, 203, 204, 236,
273. 373
— гольфстримпые 140, 366
— датские 37, 195, 246, 261 и сл., 293,
348, 349, 439
— европейские 132, 421
— исландские 275, 282
— Малайского архипелага 161
— материкового склона 90—92, 141
— морские 62—65, 317—319
— немецкие 195
— норвежские 238
— океанические 62, 64, 71, 72, 123
— Ост-Индии 39'i
464
воды поверхностные 71, 72, 7G,80, 94,
110, 113, 114, 127, 123, 131, 132, 134,
143, 373’
— подземные 56, 368
—• полярные 124, 283
— предустье вых пространств 101
— пресные 167—169, 309, 314, 317—
319
— прибрежные ИЗ, 114, 132, 141, 215,
302, 304
— придонные 128, 130, 131, 167
— промежуточные 128—131, 134, 139
— речные 62—65, 76, 79, 109, 114
- северные 373
- североатлантические 130, 131
— солоноватые 169, 309, 312—319
---классификация 316
— средиземноморские 111, 130, 132, 369
— субантарктические 127, 132, 134, 212,
406
— субарктические 127, 129, 130, 131
— субтропические 212
— теплые 77, 123, 136, 139, 162, 212,
366, 373
— тропические 124, 132, 136, 207
— холодные 77, 78,88, 89, 110,123,124,
134, 137,138, 141, 283, 373, 394,400
— циркумполярные (антарктические)
134
— черноморские 214, 288
— шведские 152
— шотландские 281
— шпицбергенские 139, 195, 416
— экваториальные 126, 127, 132
— южно-австралийские 332, 436
возраст земли 170
— отложений 151
волнение 121
волны 121, 122, 197
— ветровые 121
— внутренние 122, 124, 126
— стоячие 122
«вонючая вода» 329
Воронка Белого моря 123
Восточно-американская область 359
Восточно-Камчатская провинция 370
Восточно-Китайское море 412
Восточно-Сибирское море 238, 369, 381
впадины глубоководные 149
вращение вод 121, 137
вредители рыбного хозяйства 429
время анциловое 379
— доархеозойское 171
— докембрийское 172
— доледниковое 379
— ледниковое 152, 390, 404
— литториповое 260, 379
— межледниковое 152
— плиоценовое 379
— послекембрийское 171
— послеледниковое 152, 260, 379, 437
— третичное 355, 369, 383, 391,403, 404,
436
— четвертичное 315, 391
встречаемость 181, 183—185
вторичноводпые животные 157, 169
гторичноротые 158
Вудс-Холл 104, 164, 219
выбросы водорослей 250, 258
выедание 37
— бентоса 272
вылов камбалы 272
— китов 416
вымерзание поверхностных вод 74
вымирание фауны 173, 368, 37b, 389,
390, 405
выпадение личиночных стадий 169
Высокоарктическая область 389
«вытягивание» вод 123
Г
Гавайские острова — см. острова
гавань Фрайди 114
газ болотный 97
газы 61, 92, 117, 150
Гаити остров 278, 401
галечник 144
галоиды 69
галосфера 14, 53, 159, 356
галоэдафон 170, 250
галька 147
гаммариды 402
гарпактициды 283
гарпунная пушка 416
гастроподы — см. моллюски брюхоно-
гие
гастротрпхи 170
Гаттерас мыс. — см. мыс
«Гаусс» э. с. 18
Гвинейский залив 400
Гедлунд мыс 285
гелий 170
генезис абиссальной фауны 296
— литоральных фаун 260
географические подразделения моря 340,
341
геомерида 178
Георгия Южная 207
геосинклиналь южно-русская 315, 355
геотропизм 232
Германия 193, 275, 349, 412, 419, 420,
422, 426, 429, 430, 439
гетеротрофные организмы 116, 157, 174
гефиреи 242, 252, 291
гибель морских животных 116
— планктона 103, 121
Гибралтарский пролив — см. пролив
гигантизм 354
гидракарины 283
гидробиология 3,3—36, 48, 200, 339
гидрограммы 188
гидроиды 30, 162, 190, 252, 253, 291, 429,
430, 433
гидрологические разрезы 89, 90
гидрология 35
гидромедузы 189
гидросфера 53, 155, 158
гипераккумуляция органического веще-
ства 41, 42
гипертония внутренней среды организ-
мов 312, 319"
гипопланктон 217, 220
гипсографическая кривая суши и моря
56
глина 144, 303
— вязкая 149
30 Зенкевич, т. I
465
глина красная 145- 148» 160
— плотная 149
глобигерины 165
глоталыцики 320
глубины 55, 58
— абиссальные 55
— Атлантического океана 77
— Тихого океана 20
глубоководная фауна —см. фауна
глубоководные формы — см. формы
гнездовья птиц 306. 307
ГОНИ — см. Институт
Голландия 419, 420, 422, 426, 439
голопланктон 208—210
голотурии 71. 149. 242, 296, 297,342. 393.
395, 399, 403, 429, 430
— питание 320
голоцен 177
Гольфстрим 87, 89, 119, 120, 140—143.
400
гомогалинностк 74,84,122,132,143
гомойосмотич ность 169
гомойотермвость 299
гомотерм и я 74, 84, 122, 132, 143
гомотопность 167, 168
1т>рбылевые 410
горгонарии 395, 396, 400
горло Белого моря 121, 123
гравий 144, 145, 147, 148
градиент кислородный 125, 127
градиенты гидрологические 120, 121
— солевые 79
— температурные 87, 363
границы зоогеографические 359—361
гребень континентальный 54, 55
— Уайвиля Томсона 295
— Шотландско-Гренландский 295
— Шотландско-Исландский 295, 360
— Шотландско-Фаррерско-Исландский
270, 363
— Шпицбергенско-Гренландский 295
гребешок 418—420
гребневики 209, 239
гребни мелководные 295, 363
грегарипы 327
Гренландия 19, 73,78, 106, 116, 123, 141.
281, 283, 286, 294, 348, 350, 363, 413
• — восточная 265, 266, 285
Гренландское .море — см. море
грибки 301
— дрожжевые 106
грибы 183
— слизистые 246
Грузия 175
грунтоеды 149, 286, 320
грунты 97, 144 и сл., 190, 302, 303
— глинистые 296
— жесткие 276
— известковые 146
— илисто-песчанистые 147, 250
— илистые 144, 149, 286, 291
— кремпеземовые 147
— механический состав 144, 147
— мягкие 144, 149, 270, 276, 277, 279,
286, 291^ 292
— песчанистые 252, 279, 400
— песчано-илистые 147
— распределение в мировом океане Г«8
— твердые 147, 277
466
грунты химические процессы в них Г»9
группы населения водоема 189
— фаунистические 384
гуано 307
туба Машигина 293, 294
— Якшина 288
гхбки 35, 66, 70, 71, 105, ИЗ, 167, 168,
171, 190, 210, 284, 315, 326, 327, 369.
395, 396, 417, 419, 429
- - глубоководные 297
— известковые 71
кремневые 68, 147, 296
питание их 320
промысловые 430
- роговые 430
- стеклянные 295
— туалетные 430
Гудауты 423
Гурон оз. 412
Гюльмар-фиорд - - см. фиорд
д
Дальний Восток 250, 419, 422, 428, '<31,
434
«Дана» э. с. 86, 136
«Дана II» э. с. 20
Дания 183, 270, 322, 419, 420, 422, 426
Датский пролив 141, 143
девон 168, 174
декаподы — см. ракообразные десятино-
гие
де композиторы 14
дельфины 402, 409
денитрификация 105, 106
денудация 14, 173
детрит 38, 42, 61, 108, 111, 116, 251, 260,
286, 301, 324
детритоеды 286
дефицит кислорода 116, 206
— питательных солей 401
диагепезис 149
диатомеи — см. водоросли диатомовые
диатомит 175, 176
Дибсофиорд 266, 267
дивергенция фаун 436
динамика биогенных элементов 40
— биомассы водоема 46
— органического вещества 38, 42
— популяции 37
диск Секкп 160
«Дисковерп» э. с. 109
«Дисковери II» э. с. 20
дистрофия 41
диффузия 117
дночерпатель 37, 186, 187, 241, 243, 244,
343, 348
— Гордеева 241
— Киюдсепа 2'<2
— Петерсена 241
— Пёттерссопа 277
Доггер-банка — см. банка Доггерская
«дождь трупов» 116
докембрий 65, 173, 174, 176
Долгий остров 432
долгоносик 188
долины речные 56
доминанты 183
дочерино вилы 381
дочерние фауны 400
драга 32, 243, 244, 340
драгировки глубоководные 16
древность тропической фауны 392
— фаун 66
древоточцы 14, 190, 433, 434
дрейф «Фрама» 17
Дэвисов пролив — см. пролив
дюгонь 396
дыхание 48
— фитопланктона 20\
Е
Евразия 355, 382
Европа 141, 246, 409, 421, см. также —
побережья
— Западная 418, 425, 426, 429
— северо-западная 437
ежи морские 68, 242, 257, 279, 295, 296,
342, 396,397,403,406,429,430
--- мягкие 295
---неправильные 259, 295
— •— питание их 320
Ж
жгутиковые 188, 205, 208, 327, 328
жгутиконосцы известковые 14 7
железо 97
Желтое море 361
жемчуг 425
жемчужница 425
жизненное сообщество 177
жир китовый (получение из планктона)
431
— накопление его у организмов 392
жирность организмов 299
— планктона 392
— сардины 330
жиры (состав их) 68
жолоб восточный 77
--- Атлантического океана 84
жуки 256
3
закон минимума Либиха 105
— совокупною действия факторов 105
залив Алжирский 284
— Амурский 290
— Баффинов 78
— Бенгальский 79
— Бискайский 83, 366
— Ботнический 78, 311, 313
— Гвинейский 400
— Jade Busen 256
— Кара-Богаз-гол 64, 71
— Кардигепский 281
— Каркинитский 246, 432
— Кварперский 191
— Кольский 28, 32, 193, 197, 259
— Лионский 191
— Loch Striven 207
— Майнский 104
— Мексиканский 17
— Миссион 304
— Монтери 195, 258, 259
— Мэйн 206, 207
залив Неаполитанский 284
— Оманский 96, 97
- Персидский 71, 95, 96
— Петра Великого 194, 257, 258, 290.
292, 294, 423, 432
— Салернский 284
— Сан-Диего 226, 228, 303
— Сен-Жуан 114
— Уссурийский 290
— Финский 78, 311, 313
— Фу иди 249
— Чезепик 120, 421
заливы Дании 266
— Мурманского побережья 116
— побережий Флориды 116
заморы 116, 150, 280
Западно-американская область 359
Западно-африканская область 359
«запас» 37, 43
— остаточный 42
— энергии 48
запасы анфельцпи 432
— водорослей 432
— ламинарии 432
— морской травы 432
— промысловых моллюсков 423
— филлофоры 432
зарождение жизни на земле 171
заросли береговые 14
— водорослей 246
— зосте ры 194
— ламинарий 290
— макрофитов 252, 253. 293
— мангровые 249, 260. 395, 400
— фукусов 253
«Заря» э. с. 18
засадчики 320
заселение новых пространств 162
застойность глубинных вод 94
защита от древоточцев 434
защитная окраска 104
звезды морские 252, 257, 258, 260, 292,
295, 296, 298, 396, 403, 425, 430
---питание их 320
Зеленый мыс 400, 402
Земля Капская 395
— Северная 75, 141
— Франца Иосифа 104, 141, 361
«Зибога» э. с. 18
зима биологическая 234, 236—238
змеи морские 396
значимость форм в биоценозе 185
зона абиссали 191, 196, 197, 359, 362
— антарктической конвергенции 127,130,
132—134
— арктической конвергенции 127
— архибентальная 196
— ба л анусов 190, 191
— батиальная 197, 294
— береговая 208
— брахиоподовая 191
— вегетационная 207
— вертикального вращения вод 137
— вертикальной циркуляции вод 132
— восстановительная 97, 98
— гидроидная 191
— гидроидов, известковых мшанок и нле-
чено гих 196, 197
— глубинных илов 192
467
зона глубоководная 218—221, 224, 295,
296
— глубоководных кораллов 190—192
— дивергенции 204, 392
— илистых грунтов 192
— континентально-глубоководная 196
— кораллиповая 190
— коралловых рифов 341
— коррозии 195
— ламипариево-литотамнпевая 196
— ламинарии 190, 191, 290
— литоральная 159, 190—197, 199, ’59
— мелководная ИЗ, 174
— морской травы 195
— неритическая 36, 159, 354
— окислительная 98
— окислительно-восстановительная 98
— олиготрофная 352
— пелагическая 359
— перемешивания вол 125
— переходная (но глубине) 219—221,
224
— поверхностная 219—221, 224
— подводная 192
— политрофпая 352, 353
— полярная 347
— прибойная 192, 193, 249
— приливно-отливная 145, 191, 192,
196, 256
— продуцирующая 218
— промежуточная (по глубине) — см. з.
переходная
— псевдоабиссальная 193, 194, 196, 197
— родофиций 192
— солоноватая 281
— сублиторальная 193—197. 199, 358
— супралиторальпая 191, 192
— субтеррестровая 192
— субтропической конвергенции 132—
134, 137, 138
— субтропическо-умеренная 406
— тепловодная ПО, ИЗ, 436
— тропическая 61, 94,136,159,160,163,
172, 204, 212, 214, 215, 315, 368, 392
— тропической дивергенции 110,136, 353
---конвергенции 135, 136
— умеренная 162, 214, 341, 347, 352,
387, 392, 409
— устриц 190
— фотосинтеза 39, 40
— фукусов 190
— халиластатическая 95.
— холодная 341, 3V2
— циркумлиторальная 191
— эвфотичная 116
— экваториальная 131, 135, 139, 152,
159, 162, 347, 353
— элиторальная 193, 196, 197, 199
— эрозии 195
зональность биогеографическая 352
— биоценозов литорали 254, 256
— вертикальная в океане 97, 98
--- в окраске организмов 104
---макрофитов 246, 247
---населения Средиземного моря 404
---фауны 190
---фЛОрЫ 19о—199
зоогеографическое районирование миро-,
вого океана 361 и сл.
зоогеография 33, 340, 341, 358
— историческая 355
— каузальная 33
— региональная 340
— регистрирующая 33, 356
— синтетическая 33
зооксантеллы 395
зоопланктон 48, 49, 98, 123, 126, 162,
165, 180, 188, 200, 205, 208 и сл., 325,
326, 334, 347, 430
— вертикальное распределение 216 пел.
— горизонтальное распределение 210—
216
зооценоз 180
зостера 193, 194, 246, 248, 257, 268, 269,
432
— карликовая 257
зубы акул 147
Зунды 265
И
иваси 415, 416
иглокожие Зи, 69, 70, 147, 149, 162, 165,
167, 168, 171, 190, 210, 242, 251—253,
260, 264, 268, 269, 283—287, 290—292,
295—297, 311, 313, 326, 334, 357, 369,
395, 396, 400, 402—404, 417, 419
— промысловые 430
известковая функция организмов 70
издания, посвященные вопросам океано-
логии 28—31
изменения в составе биоценозов 267
— климатические 62
— орографические 62
— палеогеографические 62
— температуры сезонные 91
измерения температуры 15
изобаты 56, 58
изобенты 275, 288, 289, 350
изобии 288
изогалины 72, 73
изогипсы 56
изоляция топографическая 348
— фауны 370, 382
изоподы — ем. ракообразные равпонщие
изотермы 73, 80, 87, 88, 122, 140
изотопия 312
икра камбалы 279
— пелагическая 162
— рыб 32, 200, 209
ил 144, 148, 303
— вулканический 145, 146
— глинистый 144
— глобигериновый 145—148, 152, 160,
165
— глубоководный 196
— диатомовый 146—148, 160, 205
— зеленый 145, 146
— известковый 146, 395
— коралловый 145, 146
— коричневый 291
— красный 145, 146
— кремнеземовый 146, 148
— мидиевый 144
— мягкий 147, 14 9
— песчанистый 144
— полужидкий 14 9
— птеро подовый 145—148
468
ил радиоляриевый 145—148, 160, 208
— синий 145, 146
Ильмень оз. 412
иммигранты 391, 404
пмпедитные границы 436
инвентаризация фауны и флоры 13
индекс доминирования 184
— наполнения кишечника 271, 323
— плотности 183, 184, 323
индексы весовые 275
Индийский океан —см. океан
индикаторы биологические 240
— ледового режима 240
Индия 316
Пндо-вест-пацифика 163, 376, 396, 397
Индокитай 316
Индопацифическая область 359, 362
Индостан 316
инкубация 302
Институт всесоюзный паучно-исследова-
тельский рыбного хозяйства и океано-
графии (ВНИРО) 29
— Государственный гидрологический 29
----океанографический (ГОИН) 19, 26,
145, 323, 336
— научно-исследовательский Арктиче-
ский 28
— океанографический в Вудс-Холле 20,89
---- в Калифорнии 20
----Югославский в Сплите 214
интенсивность точки компенсации 103
— обрастаний 433
— освещения 103
— света 99, 226
инфауна 149, 190, 250, 252, 253, 264, 281,
293, 395
инфлюэнты 183, 185
инфузории 208, 209, 219, 313, 327, 328,
333
иод 69, 419, 420, 431, 432
Ионический район 85
Ионическое море — см. море
Ирландия 162, 164, 256, 439
Ирландское море 164
Ирмивгерово море — см. море
Исландия 89, 103, 106, 123, 141,275, 281,
293, 350, 363, 366, 402, 412, 426
Испания 403, 420, 421, 430
испарение 71, 73, 74, 76, 84
Иссык-Куль оз. 412
история фаун 355
истребление промысловых объектов 416,
417
Италия 275, 349, 420, 439
ихтиофауна Берингова моря 371
К
кавасаки 428
каланус 188, 373
Калифорния 103, 195, 216, 226, 247, 258—
260, 302, 382, 414, 437
калорийность 242
— промысловых беспозвоночных 419
кальмары 162, 189, 354, 418, 421, 425
кальции 314
— углекислый 65, 70, 146, 392, 395
камбала 190, 254, 256, 268—270, 272,
275, 279, 322, 335, 336
камбала морская 324, 334, 336, 439
— питание 324, 334—336
камбаловые 410/416
каменноугольный период 174
кампеточцы 190
камни 147
Камчатка 247, 413, 415, 427, 432
Камчатский краб — см. краб
Канада 104, 162, 164, 195, 257, 412, 414,
420
капал Английский 69, 104, 112, 123, 141,
208, 215, 221, 252, 256, 402
— Суэцкий 71, 73, 402
— Фарерский 221
Канарские острова 163, 402
каньоны подводные 36, 56, 58
капре л л иды 290, 401
Капская земля 395
Кара-Богаз-гол 64, 71
Караибское море 17
каракатица 418, 421
карбонат кальция — см. кальций угле-
кислый
карбонатонакопители 70
карбонаты 95, 354
Кардиганский залив 281
Каркинптский залив 246. 432
«Карнеги» э. с. 20, 135, 136
карп 48, 324
карповые, 410
Карское море — см. море
Каспий Северный 225
Каспийское море — см. море
ката талинное распределение солености 73
катагалинность 76
Каттегат 78, 246, 263—265, 272
кашалот 167, 417
Кварнерский залив 191
Квинсленд 195
Кельтийская область 340
кембрий 65, 171—173
Кергуэлен 192
Кермадек острова 195
Керченский пролив 116, 423
килька 239, 385
Кильская бухта — ем. бухта
кислород 47, 61, 93—98, 116, 117, 123.
125—127, 129, 131, 135, 141, 149, 150,
206, 207, 230
— дефицит его- 116, 206
— отсутствие его в глубинах 61
кислота мочевая 114
— фосфорная 109
кит горбач 417
— сейвал 417
— синий 417
— финвал 417
Китай 411, 412, 420, 421, 430
китообразные 235, 251
киты 147, 162, 167, 233, 240, 324, 331.
353, 385, 409, 416, 417
— беззубые 333
— питание 328, 332, 333
кишечнополостные 66, 69, 162, 165, 168,
171, 209, 311,315,326,327,340,381, 395
— питание 320
— промысловые 430
классификация вод по солености 64, 317,
318
;бо
классификация озер типологическая 41
— солоноватых вод 316
— фауны по отношению- к солености 318
классы встречаемости 183
— обилия 183
клещи 168
климатические пояса 376
климограммы 188
Клэр оз. 164
коацерваты 171, 172
«ковчег» 421
Код мыс 192, 363
кокколитипы 202
кокколитофориды 1'17, 165, 205. 313. 341,
354
коккосферы 147
колебания волновые 121, 122
— изотерм 122
— приливо-отливные 123, 121, 249. 257
— температуры воды годовые 83, 84
количественный учет — см. учет
количество солей в океане 63
коллоиды 302, 303
коловратки 66, 189, 213, 310, 313, 314
Кольский залив — см. залив
Командорские острова 163, 165
Командорский округ 370
Комиссия Международная по изучению
Средиземного моря 22
КохМитет Международный Тихоокеан-
ский 22
комменсализм 181
компенсационная точка 208
комплекс 179, 180
комплексы — см. также биоценозы
— абиссальные 297
- - бентические 261—266, 269, 272, 292,
348, 349
— Адриатического моря 288, 289
----Гренландии 283—285
— Средиземного моря 287
-----литоральные 257, 262
-----сублиторальные 262—266, 2G9.
282—294
- биомические 345, 348, 356, 357
- донные — см. комплексы бентические
— фаунистические 45, 348—357, 403
— фитопланктонные 344
— флористические 348, 356
— эпифауны 264
компоненты биоценоза 182
конкреции железисто-маргапцевые 291,
304
конкуренция межвидовая 323
— пищевая 273, 274
консервирование моллюсков 425
— камчатского краба 428
консервы из ракообразных 428, 429
коистации 178
конституенты 182
консументы 14, 49, 170, 174
Континенталь 196
конхиолин 70
концентраторы углекислого кальция 70
концентрацпя водородных ионов 97
— рыб 336
копеподы —см. ракообразные веслоногие
коралл благородный 430
кораллипа 190
коралловые полипы 395
•— рифы — см. рифы
кораллы 68, 71, 190, 195 340, 368, 3G9,
395, 396, 417, 419, 429, 430
— восьмилучевые 149, 295, 395
— глубоководные 190—192
— голубые 396
— горгониевые 278, 291, 396, 397
— грибовидные 396
— каменистые 395, 396
— мадрепоровые 291, 299, 354, 368, 395
— настоящие 295
— пуллипоровые 392
— органчиковые 396
— рифообразующие 70, 87
Корея 361, 411, 412
Кориолиса сила 119
корм поддерживающий 324
— продуктивный 324
кормпость водоемов 39
корхмовая база для рыб 271, 321, 322
- - мука для птиц 425, 430
кормовое значение литорали 256
---различных организмов 323
кормовые организмы 270, 271, 321, 323
корненожки 70, 152, 291, 314
корюшковые 315
Коряцкий округ 370
космические силы 124, 125
космополитизм 389, 390
космополиты 366, 372—374
котики 385
коэффициент дисперсии 182
— затухания 99
— общности 181, 188
— P/В 44, 46, 47, 268, 301, 324, 326
— поглощения света — см. к. затуха-
ния
— постоянства 185
— продуктивности 239
краб камчатский 354, 419, 424, 426—
428, 437, 439
— китайский 429, 437
краболовы 427, 428
крабы 68, 161, 163, 165, 170, 190, 249,
250, 258, 259, 267, 292, 296, 354, 377,
380—382, 393, 395—397, 400, 401, 403,
419, 420, 424—429
- - дальневосточные 428
— пелагические 396, 401
— плавунцы 424
— средиземноморские 402
— тропическо-ахмериканскпе 397, 398
Красное море — см. море
креветки 256, 292, 419, 420, 424—429,
438
кремнезем 14 5, 146
кремнекислота 61, ИЗ
кремний 105
криптометаболия 169
круговорот азота в море 106
— вод — см. вращение вод
ктенофоры 161, 168, 189, 210, 232, 251,
328, 336, 354, 372
— питание 328
Куба остров 430
Кулалы остров 432
кумацеи 220, 225, 290
Куро-Сиво — см. течение
470
куги пула нематод 168
«кухонные кучи» доисторического чело-
века 420
Л
Лаборатория ихтиологическая в Аберди-
не 28
— морская в Конпарно 22
---в Марселе 22
Лабрадор 87
лагуна Б. Барьерного рифа 215, 216
- - Венецианская 284, 286
лагуны береговые Индии 315
— солоноватоводные 315
Ладожское оз. 412
Ламанш 3 93, 204, 206. 270. 403
ламинариевые 173, 246, 248
ламинарии 45, 46, 190, 191, 24 7, 248,
257, 260, 290, 292, 403, 431
— промысловое использование 431
лангуст 424—426
Ла-Плата р. 312, 313
Лаптевых море — см. море
ластоногие 162, 167, 353, 416
Ла Холла 216
- (евантский район 85
.:едовые прогнозы 434
ледовый режим 240
Лейзенские острова 195
Ленинград 104
лептоцефалус 401
лето биологическое 234, 235, 237
летучие мыши 299
ливни тропические 71, 72, 79
лигнин 304
лилии морские 149, 161, 167, 295, 296,
298, 354, 396, 397, 403
лиман Утлюкский 432
лиманы Азовского моря 214
лимносфера 53
лимноэдафоп 170, 250
Лимфиорд — см. фиорд
Линдеснесс 187, 279
Лионский залив 191
литоралебионты 252
литоралоксены 252
титора лофилы 252
литоралофобы 251
литораль 145, 191—197, 199, 247, 248,
250 и сл., 293, 234, 298, 359, 361, 363
— безливпых морей 249
- - илисто-песчанистая 253
— каменистая 195
— кормовое значение ее 251, 252, 256
— мангровая 395
— мурманская 252, 293
• — мягких грунтов 253—256
— песчанистая 195, 259
- - Северного моря 254, 256
— скалисто-каменистая 253, 257
— тропическая 260
— устья Эльбы 24 5
литорины 283, 403, 418
литосфера 65, 155, 158
литотамний 195, 247
— ветвистый 193
личинки 162, 219. 224, 232, 332, 396
— декапод 223, 329
личинки донных животных 123. 208, 213,
233, 235, 368, 369
— корабельного червя 434
— моллюсков 209, 281, 330, 333, 425
— мух 256
— насекомых 168, 250, 256
— пелагические 332, 396
— планктонные 325, 332, 396
— ракообразных 209, 223, 312, 329, 333
— рыб 123, 208, 233, 401
— хгрей 401
— усоногих 209, 329
— устриц 425
— червей 209
л ичиночнохордовьте 168
личиночные стадии — см. личинки
лишайники 249, 391
лов камчатского краба 428
ложе океана 54, 145, 296
.ложбины глубоководные 145
.Лонг-Айленд пролив 206, 207
лососевые 315, 410, 414, 439
лосось дальневосточный (тихоокеанский)
437—439
Лофотенские острова 336, 340
Лузитанская область 269, 270
— провинция 402
Лузитанско-средиземпоморская провин-
ция 403, 406
«лунатизм» морских организмов 194, 233
луфарь 288
лучи ультрафиолетовые 101, 299
люцерпария 374
льдообразование 74
льды 73, 235, 236, 240, 249, 290
— материковые 53
— перетирающее действие 257
— пловучие 53, 72, 195, 247, 300
М
Мавританский район 402
магма 56
Мадейра остров 402
Мадрас 315
Майн (штат) 439
Майнский залив 104
макома 262
макрель 125
макробентос 181, 189, 303
макропланктон 189, 219, 221, 287
макрофиты 45, 49, 104, 147, 157, 163,
197, 207, 246—248, 260, 286, 324, 366,
371, 384, 391, 392
макрохоры 339
Малайский архипелаг — см. архипелаг
мальки рыб 209, 331
мангли 395
мангровые заросли — см. заросли
марганец 97, 150
Марианские острова 27 8
Маркизские острова 395
Мароканский район 402
материк Азиатский 366
— Антарктический 15, 132, 300
материки 54—56
Машигина губа — см. губа
мегапланктон 189
Медвежий остров 123
471
медузы 187, 209, 213, 226, 251, 'Л7,
'<29, 430
• — питание 328
— сцифоидные 189
медь 70
мезобентос 303
мезопланктон 189, 217, 219
мезосапробность 39
мезотермия 76
Мексиканский залив 17
мелководье Медвежипское 123
— Центральное Баренцева моря 123
— Шпицбергенско-Надеждпнское 415
мелководья 121
— прибрежные тропиков 392
меловой период 394
мель Большая Ньюфаундлендская 89
— Ньюфаундлендская 363
Мендота оз. 48, 49
меропланктон 208, 209
метан 97
«Метеор» э. с. 20, 111, 141, 345, 346
метод изучения питания рыб 321—324
— оконтуривания 279
— _учета поглощения кислорода планкто-
ном 47
— учета площадок 181
мечехвост 395, 396, 427, 429
миграции 162, 225—228, 348
— активные 162
— зоопланктона вертикальные 103, 182,
220, 221, 223—233, 299
-------сезонные 220, 232, 233
------- суточные 180, 188, 220, 224—
232, 369
— китообразных пищевые 235
— морских животных 390
— непериодические 162
— нерестово-кормовые 162
— пассивные 123, 162
— периодические 162 *
— рыб 180, 182, 288, 414
--- вертикальные 231
---пищевые 235
— флоры 391
мидии 205, 245, 252, 270, 283, 284, 293,
320, 418—421, 423, 425
мизиды 220, 225, 295, 315
микроаквариумы эпифитпые 184
микро бентос " 303
микроорганизмы 301, 302
— питание 327
микропланктон 103, 188
микрохоры 339
минерализация 106
— органического азота 108
— фосфатов 114
минимум — соли 105
миозин 170
миоцен 376
Мпссион залив 304
«Михаэль Саре» э. с. 19, 89
Мичиган оз. 412
млекопитающие морские 145, 147, 156,
168, 169, 172, 188, 189, 230, 299, 326
многоножки 170
модиола 284
Мозамбик 171
мойва 332, 413
472
моллюски 30, 36, 37, 69, 70, 116, 147, 149.
162, 163, 168, 171, 187, 189—191, 193,
205, 242, 249, 268, 278, 281, 286, 288.
290, 291, 295, 297, 310—313, 324, 326,
334—336, 354, 357, 389, 395, 396, 400,
402, 417, 419, 433, 438
— арктическо-бореальные 384
— боконервные 252
— брюхоногие 66, 162, 163, 168—171,
190, 242, 252, 253, 257, 258, 260, 267.
270 , 27 9 , 284 , 28 7 , 2 9 2 , 3 1 3 , 314 , 320.
395, 401, 402, 421
----- пелагические 147
— головоногие 161, 165, 171, 189, 208.
320, 332, 333, 353, 396, 421, 423
— голожаберные 69, 252, 253
— голые 401
— двустворчатые 35, 66, 150, 162, 163,
170, 210, 243, 252, 253, 257, 258, 260,
264, 270, 279, 284, 286, 287, 291, 292,
311, 315, 320, 327, 395, 401, 402, 421
— древоточцы 434
— использование раковин 314, 420, 425
— кормовая ценность 269
— крылоногие 208, 209, 401
— питание 320
— планктонные 209
— пластинчатожаберные — см. моллю-
ски двустворчатые
— пресноводные 70, 170, 418
— промысловые 253, 418, 420—425
— хищные 270
молодь донных животных 180
Монако 404
Монтери залив — см. залив
море Адриатическое 114, 191, 201, 215,
286, 288, 289, 404, 430
— Азовское 18, 19, 30, 41, 44, 45, 55.
63, 73, 79, 94, 96, 97, 100, 106, 116.
162, 180—182, 199, 249, 260, 272, 275,
280, 294, 309, 311, 313—318, 322, 348,
355, 357, 405, 412, 428—432, 434,
437
— Альборанское ИЗ
— Аральское 55, 63, 122, 275, 309, 314,
315, 322, 348, 355, 357, 412, 436, 437
— Балеарское 112
— Балтийское 55, 62, 73, 75, 78, 94, 96,
100, 103, 106, 123, 169, 191, 199, 212,
249, 260, 263, 270, 275, 309—318, 348,
350, 355, 360, 370, 404, 434, 437
— Баренцово 15, 18—20, 32, 44, 45, 47,
54, 55, 75, 79, 100, 104, 111, 120, 122,
123, 141, 148, 162, 164, 181, 184, 187,
188, 199, 206, 212, 221, 228, 229, 232,
239, 247, 252, 259, 260, 275, 283, 291,
293, 294, 299, 311, 322, 325, 326, 329,
332, 335, 336, 348, 350—352, 357, 361,
368, 369, 379, 402, 412—416, 423, 429,
430, 434, 437, 439
— Белое 16, 19, 24, 28, 41, 55, 73, 75,
79, 100, 104, 121, 123, 191,194, 199,
212, 227, 238, 243, 248, 252, 260, 270,
291, 311, 348, 350, 360, 361, 369, 374,
379, 412, 423, 431, 432, 434, 437, 439
— Берингово 54, 55, 78, 165, 227, 291,
297, 361, 370, 412, 416, 431, 434
— Восточно-Китайское 412
— Восточно-Сибирское 238, 369, 381
море Грепландское 78, 137, J39, 140, 219,
224, 270, 295, 300, 369, 375, 382
— Желтое 361
— Ионическое 85, 8G, 112, ИЗ
— Ирландское 164
— Ирмпнгерово 106, 107, 111, 143
— Караибское 17
— Карское 260, 291, 300, 317, 350, 369,
381
— Каспийское 16, 18, 19, 23, 40, 48, 55,
61, 63, 87, 97, 102, 104, 122, 150, 152,
175, 206, 220, 225, 228, 229, 238, 239,
248, 249, 260, 275, 297, 309, 314, 315,
318, 322, 323, 326, 348, 355, 404, 412.
415, 428, 429, 431, 432, 434, 436—439
— Красное 71, 73, 84, 86, 87, 130, 192,
212, 297, 395, 402
— Лаптевых 161, 162, 164,238,300,317,
369, 381
— Мраморное 214, 288
— Норвежское 78, 140, 221,270, 295, 300,
369, 375, 382, 413, 415
— Охотское 19, 55, 78, 194, 238, 239,
249, 257, 258, 260, 293, 297, 357, 361,
370, 412, 415, 434
— Палеозойское Цехштейновое 65
— Саргассово 89—92, 100, 104, 121, 127,
140, 204, 207, 246, 400, 401
— Сарматское 176
— Северное 40, 78, 100, 123, 162, 164,
•195, 212, 215, 216, 251, 252, 254, 256,
263, 270, 275, 279, 281, 310, 313, 329,
332, 335, 336, 360, 366, 370, 374, 412,
413, 415, 437
— Средиземное 17, 71, 80, 84—87, 100.
103, 106, 111—114, 122, 130, 152, 162—
164, 193, 199, 201, 204, 207, 212, 214,
246, 249, 250, 256, 260, 275, 284—288,
294, 297, 302, 311, 349, 361, 366, 373,
375, 390, 401—405, 412, 430
-----Американское 55
-----Европейское 55
— Тирренское 85, 86, 112
— Тэтис 297, 355, 368, 369, 393, 402,
404, 405, 436
— Черное 17—19, 28, 30, 32, 41, 45, 55,
61, 63, 73, 75, 79, 87, 94, 96, 97, 104—
106, 116, 117, 120, 123, 149, 152, 162,
164, 165, 188, 191, 193, 199, 208, 218,
214, 215, 224, 233, 239, 246—249, 256,
257, 260, 278, 284, 288, 293, 297, 309,
311, 315, 319, 355, 370, 402, 405, 412,
422, 423, 428, 431—434, 437
— Чукотское 19, 238, 291, 361, 369, 370
— Шаптарское 194
— Эгейское 79, 112, 190, 288
— Японское 19, 55, 71, 165, 194, 218,
219, 238, 239, 257, 258, 278, 290—292,
294, 297, 348, 354, 361, 366, 370, 375,
382, 412, 415, 416, 423, 431, 434, 437
морская биология — см. биология моря
— бритва 418
— корова Стеллера 416
— трава 191
— уточка 401, 424
морское ухо 418, 421
моря Антарктические 132, 135, 192
— Арктики 315, 317
— безливные 199, 249
моря бициклические 237
— внутренние 74, 79, 96, 121, 123, 152,.
199, 212, 293
— восточные 361
— высокоарктические 237—239
— дальневосточные 19, 370, 423, 430, 437
— древние 176
— европейские 248
— «ископаемые» 125
— китайские 437
— краевые 54, 73, 212, 213
— мелководные ИЗ
— моноциклические 237
— нижнеарктические 237—239
— окраинные 54
— открытые 150, 181, 199, 249
— палеозойские 173
— полярные 87, 99, 219, 228, 231, 235—
240, 373
— придаточные 54, 55, 73, 76, 79, 94.
97, 100, 106, 114, 119
— — полузамкнутые 63, 122
— северные 238, 239, 369, 373, 413, 429'
— сибирские 238, 369
— СССР 62, 182, 309, 315, 322, 348,
349, 369, 373, 413, 429, 432
— субтропические 218, 237
— теплые 238, 277, 296, 297, 354, 392,
434, 436, 437
— третичные 400
— тропические 218, 237, 239, 247, 248,
392, 395, 433
— умеренные 218, 235—239, 247. 392
— холодные 238, 247, 296, 392, 437
— эпиконтинентальные 55, 79
— южные 219, 235—239, 437
---СССР 62, 182, 315
мочевина 114, 304
мощность отложений 151
Мраморное море — см. море
Мурман 96, 182, 191, 248, 259, 433
— западный 270, 379
мутность воды 101
мутьевые потоки 36, 5G
мхи 157
мшанки 66, 70, 147, 168, 171, 190, 210,.
242, 252, 253, 284, 315, 401, 433
— питание 320
Мэйн залив 206, 207
мыс Гаттерас 87, 363, 400
— Гедлунд 285
— Доброй Надежды 369
— Зеленый 400
— Код 192, 363
Н
наннопланктон 103, 111. 115, 188, 200 —
203, 218, 230, 232, 320
наносы 151
наркомедузы 210
насекомые 158, 167—169, 172, 250
науплиусы 329
«Националь» э. с. 17, 343, 344
Неаполитанский залив 284
недорассеяние 182
нейстон 189
нектобентос 190
нектон 188
473
нектон ланкто! i 189
нематоды 168, 169, 190, 314
немертины 167, 168, 209, 210, 253, 284,
369
— питание 320
неоген 173
нереиды 256
нерест рыб 334, 335
— трески 345
— угрей 401
неритическая область 160
нефть 150
Нил р. 284, 28<‘>
нитраты 97, 105- 110, 112—114, 121,204,
304, 305
нитриты 105—109, 114, 304, 305
нитрификация 10(4
Новая Англия 421
Новая Зеландия 195, 196, 420, 439
Новая Земля 15, 293, 308, 348, 361
Новороссийск 175
Новосибирские острова 141
Норвегия 19, 141, 149, 190, 195, 275, 279,
316, 349, 402, 411, 412, 419, 422, 426,
429, 434
— Южная 187
Норвежское море —см. море
Нортонский округ 370
Потальная область 362
Нуллипоровые «кораллы» 392
Пью Бренсуик (штат) 193
-----(провинция) 439
Пью-Иорк 89
Ньюфаундленд 123, 140, 141
Ньюфаундлендская банка — см. банка
О
обеднение кислородом 94, 9G, 126
— флоры и фауны 78, 79, 309—311. 319
обилие 181, 185, 186
— видовое 162
области вертикального перемешивания
вод 136, 138
— зоогеографические 359, 363—366
— переходные 361—363
— приполярные 71, 72, 159, 300, 352.
389, 390
— субтропические 74, 77, 224, 361, 362,
373, 402, 406
— субтропической конвергенции 127
— тропические 70, 74, 77, 83, 94, 120.
124, 163, 192, 224, 361, 362, 364, 392,
393, 398, 402
— умеренные 361, 373, 389
— фаунистические 340
— халистатические 71, 77,89, 94,96,121,
134, 140, 401
— холодные 70, 389
— циркумполярные 362, 363
— экваториальные 126,129,139,141,389
область абиссальная 191
— антарктическая 130, 224, 359, 362—
364
— антарктических холодных вод 132
— аптибореальная 362
— Арктическая 224, 296, 340, 350, 351,
359, 361, 362, 364, 367, 369, 370, 379,
404
область Атлантическая 359, 362, 406
— Аустральная 364
— береговая 191
— бореоальная 350, 351, 361, 362, 364,
370, 382, 402, 406
— борсоарктическая 362
— Восточно-американская 359
— высокоарктическая 389
— глубинная 192
— Западно-американская 359
— Западно-африканская 359
— Индопацифическая 359, 362
— Кельтийская 340
— литоральная 192
— Лузитапская 269, 270
— неритическая 160
— нотальная 362
— пелагическая 191
— полярных льдов 159
— прибрежная 192
— северобореальпая 404
— Скандинавская 340
— солевой стратификации 13G
— Средиземноморская 340, 393
— Средиземноморско-атлантическая
402, 405
• — субантарктическая 132, 362
— субарктическая 130, 362, 370, 404
— тепловой стратификации 134
— экваториального противотечения 77
обмен фаунами 376, 378, 379
обогащение вод кислородом 120
оболочниковые 30, 35, 167, 168, 208, 209.
268, 298, 311, 314
— питание 320
образование грунтов 147
— льдов 71
обрастания 14, 182, 190, 293, 433
Общество естествоиспытателей Петербур!-
ское 23, 24
объекты кормовые 321
объем океанов и морей 55
озера 122, 410, 411
— американские 48, 49
— Новой Земли 94
— СССР 410, 412
— шведские 37
озеро Байкал 412
— Балхаш 412
— Белое 46
— Боденское 412
— Гурон 412
— Ильмень 412
— Иссык-Куль 412
— Клэр 164
— Ладожское 412
— Мендота 48, 4 9
— Мичиган 412
— Онтарио 412
— Охрид 412
— Севан 412
— Цедар-Бог 48, 49
— Чилька 315
— Эри 412
Ока-Сиво течение 87
океан Арктический 214
— Атлантический 17, 18, 54. 55,69, 71,
72, 74—78. 80. 83—85, «8, 93—96,
106—111, 120—131, 136, 139, 143, 146,
474
147, 152, 160, 1G5, 192, 204, 205,212—
214, 21G, 218, 219, 222, 224, 227, 247,
2'19, 257, 260, 284, 295, 297, 298, 300,
313, 330, 344, 346, 348, 355, 357, 360,
361, 368, 370, 373, 375—378, 384, 387,
393, 394, 396, 397, 404, 405, 409, 414 —
418, 420, 426, 430, 437, 439
— Индийский 18, 54, 55, 71, 72, 78—80,
83, 94, 96, 100, 109, 124, 126, 127, 129,
130, 146, 147, 212, 296, 297, 369, 394,
395, 409, 414
— палеозойский 67
- первичный 65
- Северный Ледовитый 17, 19, 54, 55,
75, 78—80,120,121, 136, 152, 219, 224,
300, 370, 381
— Тихий 15, 20, 54, 55, 71, 73—76, 78,
80, 83, 84, 87, 94, 100, 109, ПО, 124,
126, 127, 129—131, 135, 136, 146, 147,
160, 163, 165, 195, 204, 208, 212, 213,
215, 219, 228, 247, 249, 257, 260, 288,
291, 297, 346, 355, 365, 369, 370, 371,
273, 375, 376, 378, 384, 385, 387, 392—
394, 397, 399, 409, 413—421, 426, 430,
437
океанология 13—15, 19, 28, 159
окисление аммиака 30'*, 305
— - нитритов 304
сульфидов 304
окраска организмов 104
округ Авачинский 370
— Анадырский 370
— Командорский 370
— Коряцкпй 370
— Нортонский 370
окуневые 411
окунь морской 18
оледенение четвертичное 391
— современное 53
«оливково-зеленые клетки» 116, 203
олиготрофия 41
олиготрофные районы 213
олигохеты 66, 168, 170, 250, 283
— пресноводные 255
олиго цен 391
Оманский залив 96, 97
омар 68, 419, 420, 424—4 26
— «колючий» 424, 4 26
— «норвежский» 424, 426
Онтарио оз. 412
— провинция 439
енфаупа 190, 253, 288
определение солености по хлору 6*2
опреснение морей 309, 315, 316
— поверхностных вод 71, 72, 76, 103
опускание водных масс 72
— литоральной фауны 199
— поверхностных вод 73, 132
ордовиций 173
орудия лова 32, 37, 243, 244, 423, 429
осадки 71—74
— глубоководные 151
— известковые 148
— ледникового периода 152
— межледниковой эпохи 152
— морские 151
— современные 152
— терригенные 148
осветление воды 35, 36
освещение солнечное прямое 103, 10'*
освещенность 98, 100, 203
осень биологическая 234—237
осетровые 315, 411
Осло-фиорд 207
осморегуляторные способности 169, 319
осолонение глубинных вод 73
— поверхностных вод 71, 76
Ост-Индия 394
Остиндский архипелаг — см. архипелаг
остров Б. Шантар 258: 288, 290
— Вайгач 361
— Ванкувер 439
— Гаити 278, 401
- - Долгий 423
— Куба 430
К ул алы 432
- - Мадейра 402
- - Медвежий 123
- - Сахалин 415, 432
— Сицилия 85, 277
- Тортуга 278
- - Чечень 432
острова Азорские 85, 401, 402
— Алеутские 94
• — Багамские 195, 430
— Бермудские 89. 99, 111, 400, 401
- - Британские 270, 363
- Гавайские 195, 3,95, 439
• — Зеленого мыса 402
— Канарские 163, 402
— Керма дек 195
— Командорские 163, 165
— Лейзенские 195
— Лофотенские 336, 340
- Марианские 278
— Маркизские 395
- - Новосибирские 14 д
- - Пау моту 395
Самоа 430
- Сан-Хуан 195
Соловецкие 191
Фарерские 89, 141, 275. 28С 363
— Филиппинские 75
— Шаптарские 257, 239, 288
— Шотландские 363
Эллиса 195
— Японские 297
осьминог 418, 421
откорм рыбы 231
отлив 248, 252, 256, 304
отложения глубоководные 146
— донные 65, 95, 144
---классификация 144
— известковые 146
— илистые 36
— кремпеземовые 14 G
— литоригенные 145, 14G
— органические 39, 40
— пелагические 145, 146
— плиоценовые 437
— послеледниковые 152, 379
— прибрежные 145
— сарматские 175
— терригенные 145, 294
— четвертичные (морские) 390
отмели прибрежные 122
отмель континентальная 54, 55
— материковая 56, 145
475
отмирание донной фауны 150, 272, 280, 281
отношение C/N 150
отражение света 98
отрицательные черты фауны 400
отсутствие жизни в глубинах 41
оттертрал 429
офелия 256
офиуры 267, 320, 381, 396, 403
охотники 320
Охотское море — см. море
охрана промысловых объектов '• 1G
Охрид оз. 412
И
пагон 189
падение биомассы 42
пак полярный 73
палеобиология 174
палеобиоокеанология 174
палеоген 173
палеозой 171
палеозойский океан 67
палеостраки 171
Палестина 85
палоло 195, 224, 233, 395
палтус 376
пальмовый вор (краб) 396
Панамский перешеек 397
пантоподы 401
панцырь ракообразных 420, 425
паразитизм 181
паразиты 299, 327
параллелизм биоценозов 260
параллельное развитие фаун 376
парение 155
пауки 168, 182
паукообразные 158, 169, 172
Паумоту острова 395
Пацифика — см. океан Тихий
— восточная 369
— северная 127, 382, 393, 439
пелагиаль 159, 188, 331, 359, 400
— биогеографическое районирование 362
пелагическая область 191
пеламида 162, 288
пеликаны 307
пелит 144
пеннатулярии 396
первичпоротые 158
первичнотрахейные 170
первичнохордовые 168
перелов рыбы 416
перемешивание водных масс 117 и сл.,
123, 132
— турбулентное 117, 141
пересадка камбалы 270, 439
— трески 274
пересыщение кислородом 94
перешеек Панамский 397
период вегетационный 237
— верхнемеловой 393
— каменноугольный 174
— меловой\394
— третичный 62, 376, 397
— четвертичный 62
периоды геологические 392, 393
— ледниковые 152
— межледниковые 152
перловица 418
«Персей» э. с. 19, 20, 23
Персидский залив — см. залив
Перу 307
пескожил 430
песок 144, 145. 147, 148, 303
— илистый 144
— плотный 291
Петра Великого залив — см. залив
петушок (тапес) 418, 421
Печора р. 379
пещеры 170
пикногон 295
пикша 18, 334, 335, 416
пингвины 307
пирамида «выедания» 334
— «продуктивности» 334
— Элтона 49, 325
питание беспозвоночных 271, 320, 328
— глубоководного бентоса 116
— гетеротрофное 158
— донных рыб 334—336
— животных растворенным органическим
веществом 105, 116
— китов 328
— копепод 205, 334
— морских животных 320 и сл.
— паразитарное 327
— роющихся червей 225
— рыб 271, 275, 320 и сл., 332, 334—3:15
— сардины 330
— сельди 327—330
— трупами 327
— узко-пелагических рыб 327—331
— широко-пелагических рыб 331—334
пиша вынужденная 336
— рыб 321, 323
пищевая ценность промысловых беспо-
звоночных 419
пищевой спектр сельди 328, 329
пищевые взаимоотношения 324—327
— цепи — см. цепи
пиявки 169
планкто-бентос 190
планктон 31,48, 49,69, 92,100,102,103.
107, 111, 113, 115, 123, 150, 162, 180.
182, 188, 200 и сл., 307, 313,342—346,
349, 354, 369, 392, 401,417
— абиссопелагический 190
— Варенцова моря 44
— береговой 190
— вегетативной зоны 346, 347
— влияние температуры 213
— воздействие на него биофильтраторов
36
— глубоководный 198, 217, 297
— диатомовый 70, 206
— кокколитофоридовый 70
— кормовой 239, 240
— метод «стандартных» весов 200
— мезопелагический 190
— наддонный 217, 220
— нектический 189
— неритический 190, 213, 375
— океанический 190, 213, 375
— осадочный 201
— открытого моря 190
— отстойный 201
— плотность его 160, 213, 312
— поверхностный 217
476
л.laHKTOH промысловое значение 430, 431
— размеры 188, 189
— распределение 160
— рачковый 70
— Саргассова моря 401
— сезонные изменения 200, 233 и сл.,
404, 405
— среднеглубинный 217
— тепловодный 363
— типы его 375
— травоядный 114
— фораминиферовый 70
— холодноводный 363
планктонный индикатор 322, 329
планктоноеды 320
планктонология 200, 344, 347
пластичность живых существ 173
плато континентальное 290, 291
— материковое 89
плейстон 189
плеченогие 66, 68, 147, 171, 210, 291,
295, 320
Плимут 164, 216, 221. 223, 228, 232
Плимутский район 279
плиоцен 260, 379, 405, 437
плотность воды 73, 74, 76, 85, 96, 102,
103, 130, 316
— условная 85
плотоядные 320
площадь морей 412
— озер 412
побережье Австралии 349, 395, 417
— Азии 83, 87, 291, 366, 379, 421
— Аляски 413
— Америки атлантическое 247, 377, 384
---тихоокеанское 130, 150, 257, 259,
390, 391, 397, 399, 430, 437, 439
— Англии 190, 191, 194
---Нортумберлендское 259
— Антарктического материка 349
— Африки 141, 349, 363, 417
--- восточное 395
--- западное 112, 150, 390, 391, 400,
403
---южное 212
— Бретани 191, 382, 403
— Веньярд 192
— Гренландии 283, 413
— Дальневосточное СССР 165, 432
— Дании 263
— Евразии 73
— Европы 103, 141, 246, 256, 260, 282,
363, 366, 373, 377, 381, 385, 391, 402,
416, 426
— Египта 402
— Ирландии 164
— Исландии 281, 282, 366, 434
— Кавказское черноморское 423
— Калифорнии 258—260, 302, 414
— Камчатки 413, 427
— Канады 104,162,164, 257,414
— Кергуэлена 192
— Кореи 361
— Мурманское 191, 193, 194, 247, 259,
293, 431
— Норвежское 195, 402, 434
— Охотское 432
— Португалии 122, 284, 285, 413
— Северной Америки 56, 58, 73, 83, 87,
89, 90, 114, 119, 219, 222, 363, 366, 381,
415, 439
— Сибири 73, 79, 309
— Скандинавии 193
— Средиземноморское 250
— Франции 190, 191, 249, 360, 439
— штата Орегон 259
---Вашингтон 259
— Южной Америки 80, 121, 126, 127,
306, 307, 349, 417
— Японии 165, 414, 416
поверхность океанов и морей 55
поднятие Гималайское 369
— центрально-американское 369
подобласти Тихого океана 365
подобласть арктическо-циркумиолярная
370
— Атлантическая 362, 363
- - Атлантическо-бореальная 359
— Атлаптическо-Средиземноморская 370
— бореальная 361
— верхне-умеренно-бореалытая 365
— высокоарктическая 351, 361, 362, 369,
379
— Гвинейская 359
— Индийская 363
— Ипдо-вестпацифическая 395
— Лузитапско-Средиземпоморская 256
— Пижнеарктическая 351, 361, 362, 369,
370
— нижне-умеренно-бореальная 365
— нотально-циркумполярная 359
— северная субтропическая 362
— северо-аустральная 365
— северо-бореальная 365
— средиземноморская 359
— субтропическая 362
— Тихоокеанская 362, 363
— тихоокеанско-бореальная 359, 370
— умеренно-аустральная 365
— циркумполярно-антарктическая 359
--- арктическая 359
--- субантарктическая 363
— южно-аустральная 365
— южная субтропическая 362
нодфации 193
пойкилосмотичность 169
показатель п. е 184
— обилия 183
покрытосемянные 157
Полинезия 278
полисапробность 39
полигонное развитие сходных форм 389
политрофные районы 213
полифаги 327
полихеты 162, 167, 168, 208—210, 2'<2,
243, 252, 253, 256, 258, 260, 264, 276,
279, 283, 284, 287, 288, 290, 291, 295,
297, 311, 313, 315, 327, 332, 334—336,
359, 389, 401,403
— питание 320
ноловозрелость сардины 330
полостная жидкость (состав ее) 68
полуостров Апшеронский 432
— Ютландский 195, 249
поляризованпость биологическая 353
полярная ночь 99
полярные страны 104, 120
полярный бассейн — см. бассейн
477
полярный лень 98, 104
— кр)г 409
— фронт 41, 118, 123, 138, 352
популяции 177
«Поркьюпайн» э. с. 16
порог Гренландско-Шпицбергенски;! 76
— Нансена 76, 141
— Томсонов 89
пороги мелководные 123
породы магматические 65
— осадочные 65
Португалия 122, 275, 284, 285, 413, 419,
421, 422, 426
«поселения» 177, 178
послеледниковая фаза потепления 379
потоки мутьевые 36, 56
потребление кислорода 97, 98, 126, 304
почвы воздушные 157
пояс водорослей 247
— ламинарий 257
— тропический 124
— фукусов 260
— экваториальный 354
пояса растительности 191
правило гомотопности 167, 168
— совокупного действия факторов Мкт-
черлиха 105
предки наземных животных 169
предомипанты 183
нрекембрий 171
преференты 183
прпапулиды 389
прибой 122, 248, 251, 257
прилив 199, 248, 252, 304
приливо-отливные колебания 122, 231,
249
Приморье 415, 422, 427, 432
— Южное 423, 428
прирост биомассы 37
приток преспых вод 63, 71, 73
притяжение луны и солнца 124
пробные площадки 185—187
провинции океана химические 354
провинция Восточно-Камчатская 370
— Алеутская 370
— Лузитане кая 402
— Лузитанско-Средиземноморская 403,
406
— Чукотская 370
— южная восточно-атлантпческо-сре ди-
земноморская 406
прогнозы промысловые 322
— сельдяного промысла 329
прогрев воды 101—103
«прогрессивная эффективность» звеньев
пищевой цепи 49
продукт 39
продуктивность актуальная 4 4
— биологическая 34 и сл., 49, 167), 176,
193, 200, 245, 269, 271, 301, 392
— зоопланктона 215
— истинная 48
— первичная 114
— планктона 69, 213, 216, 239
— потенциальная 39, 40, 43, 44
— промысловая 409
— расчет ее для разных пищевых групп 48
— фактическая 39, 40, 43, 44
продуктивные свойства водоема 38, 42
478
продукция 38—40, 42, 44, 46, 47, :.2и,
327
— весовая 39, 43
— годовая 37, 43, 44, 46, 47, 159, 17G,
206, 207, 245, 246, 268, 271, 272
— зостеры 246, 268
— кислорода 207
— копепод 239
— макрофитов 104
— остаточная 42, 43
— отрицательная 42
— реальная 43
— планктона 47
— потенциальная 39, 40
— рыбная 410
— углерода 207
— фитопланктона 47, 176, 206—208
--- суточная 47
— чистая 39, 44
продуценты 14, 39, 48, 49, 170, 174, 269.
324, 325
продуцирование биологическое 18, 39,
42, 45, 47, 61, 104
прозрачность воды 36, 100—103, ИЗ, 435
происхождение жизни на земле 170 н с.!.
— морских отложений 145
--- рыб 168
— наземной фауны 168, 169
— пресноводной фауны 168, 169
пролив Вельт — см. Вельт
— Берингов 291, 370, 371, 379
— Босфор 75, 288
— Гибралтарский 79, 80, 113, 114, 13о,
288, 402, 405
— Датский 141, 143
— Дэвисов 73, 333, 334 *
— Каттегат — см. Каттегат
— Керченский 116, 423
— Ламанш — см. .Чамапш
— Лонг-Айленд 206, 207
— Сан-Хуан 291
— Скагеррак — см. Скагеррак
— Татарский 434
— Флоридский 119, 140
проливы Дании, Швеции и Норвегии 2G7
промысел беспозвоночных 402, 417 и сл.
— водорослей 419
— головоногих моллюсков 421
— губок 430
— иваси 415
— иглокожих 430
— камбалы 274
— камчатского краба 427, 428
— кардиум 421
— китобойный 333, 334. 416, 417
— кишечнополостных 430
— крабов 426—428
— креветок 426, 428
— мечехвостов 47
— мидий 420, 421
— мировой водных организмов 409
— моллюсков 417, 4 20—425
— морских растений 431, 432
— пектена 421
— петушка (тапес) 421
— ракообразных 417, 425—429
— рыбный 240, 353, 410—416, 419
— — вредители его 429
— сардины 414
промысел сельди 414
— синовакулы 421
— трепанга 430
— трески 272—274, 414
— устриц 420
промысловое значение водоемов СССР 413
проникновение морской фауны в пресные
воды 310, 312, 318
— пресноводной фауны в море 312, 313
— света в толщу воды 98, 99, 101
простейшие 105, ИЗ, 171, 189, 219, 325,
327, 395
противотечение тропическое 135, 136,
204, 363
— экваториальное 72, 77, 121, 204, 353
прототрофные процессы 301
профундаль 46, 197, 199
прямокрылые 188
псевдоабиссаль 197, 294, 295, 361
псевдогомойотермность 299
псевдолитораль 2'19
псевдофекалии 36, 144
птицы 168, 172, 252, 256, 299, 306 -308,
328, 331
пустыни океанические 353
пути расселения арктической фауны 356
Р
рабдосферы 147
равновесие солевое между морем и сушей
65
радар 416
радиация солнечная 83
— рассеянная 104
радиолярии ИЗ, 145, 147, 165, 171, 208,
220, 354, 389
разведение водорослей 431
— петушка (тапес) 421
— промысловых моллюсков 421, 423—
425
— устриц 424, 425
развитие жизни па земле 170 и с.г.
— морских организмов 169
разложение органических веществ 106,
304
размножение водорослей 401
— зоопланктона 235
— организмов 156
— устриц 439
размывы 151
разрезы гидрологические 89, 90
район Алжир-Провапсальский 85
— Арконский 78
— Борнгольмский 78
— Вудс-Хол ла 104
— Ионический 85
— Левантский 85
— Мавританский 402
— Мароканский 402
— Плимутский 279
— Северо-Беринговский 370
— Тирренский 85
районирование биогеографическое 13, 33,
340, 347—348. 350 и сл., 367
— биомическое 351, 356, 357
— зоогеографическое 33, 35, 348, 350
3 5 2 , 354 , 3 57 , 360
— мирового океана 345—348, 359 и гл.
районирование типологическое 351, 357
— фаунистическое 351, 353, 370
— фитогеографическое 364, 365
районы олиготрофные 213
— политрофные 213
— промысловые 123
— резких смен фауны 363
раковины моллюсков 314, 420, 425
— форамипифер 314
ракообразные 116, 147, 162, 165, 168—170,
189, 190, 193, 235, 249, 252—254, 260,.
284, 286—288, 290, 307—311, 313, 315,
325, 330, 334, 336, 340, 353, 35'*, 389,.
402, 417, 419, 420
— веслоногие 35, 66, 205, 208, 209, 216.
220, 231, 233, 235—238, 334, 3.73, 401,
404
— ветвистоусые 313
-- вредители рыбного хозяйства 4 29
-- высшие 170, 171, 194, 220, 315, 327,
331
— глубоководные 296
— десятиногие 66, 162, 210, 242, 29!,
295, 297, 348, 395, 396
— древоточцы 434
— использование панцырей 420, 425
— кумовые 220, 225, 290
— листоногие 66
— низшие 171, 320, 327
— пелагические 328, 332 -32.4
— питание 320
— планктонные 205, 209
— придонные 332
— промысловые 424—429
— равноногие 30,66, 210, 260, 295
— ракушковые 66
— усоногие 35, 165, 190, 252, 260, 433
— эвфаузпевые 205, 208, 333
ракушка песчаная 418
Рам фиорд 278, 279
Рапдерс-фиорд 312
распад органического вещества 106, 304
распределение бактерий вертикальное 302
— бентоса 349, 350, 369
— биогенных элементов ИЗ
— биомассы бентоса 350, 351
— животных в биосфере 166, 168
— жизни в морях 33
— организмов в море 87, 155 и сл.,
341—34 4
— планктона 17, 346, 369
— солености 73—79, 85, 86, 90
-----ана талинное 74
-----гомо га липкое 74
— — катал игаппое 73
— температуры 80 и сл., 102
— фптобентоса 247, 248
— фитопланктона 302
распространение организмов 121, 367—
369
-----амфиамериканское 397
-- — амфиарктическое 355
-----амфиатлантическое 3.55, 377, 378,
382, 389
— — амфибореалытое 355. 382, 389
— амфиокеаническое 16.0
— амфппацнфическое 355, 377, 378,
382
— — биполярное 160, 355, 385—292
479
распространение организмов циркумтро-
пическое 393, 399
рассеяние нормальное (организмов) 182
— света 101—103
расслоение океана 120
растения вторичноводпые 157
— одноклеточные 324
— цветковые промысловые 43J, 432
растительность береговая 150, 246, 247
— наземная 157, 183
растительноядные животные 320
«рахит абиссальный» 71
рацион суточный рыб 323, 324
реакция активная 93
регенерация нитратов 114
— фосфора 114
редукция органических веществ 304
редуценты 14, 170
резерв щелочной 95
резиденты 185
режим газовый 92 и сл.
— гидрологический 236
— кислородный 94
— температурный 84—87
река Ла-Плата 312, 313
— Нил 284, 286
— Печора 37 9
— Рио-Пара 313
— Тахо 284, 285
— Эльба 245, 313
реконструкция морской фауны 14, 275,
354, 435 и сл.
реликтовость 300
реликты 66, 356, 389
рельеф 173
— дна (ложа) мирового океана 36, 54,
56, 57
— земной коры 56
рептилии 169, 172
ресурсы кормовые 270, 272, 274
— пищевые 288
Рио-Пара р. 313
ритм биологический миграционный 232
риф Австралийский барьерный 192
— Б. Барьерный 215, 216
рифообразователи 400
— коралловые 195, 394, 395
рифы коралловые 70, 146, 192, 247, 277,
278, 394, 395, 400
Ровиньо 288
родофиции — см. водоросли красные
Розас 250
Росков 194, 403
«Россинантэ» э. с. 19
Россия 193
рост животных 48, 313, 314, 324
— китов 324
рыбка-песчанка 328
рыборазведение 14
рыбы 14,40,69,145,147, 149. 162, 165, 168,
169, 180, 187—189, 200, 209, 220, 229—
231, 240, 251—253, 256, 258, 269—271,
275, 288, 295, 298, 307, 311, 313, 315,
320 и сл., 342, 353, 357, 371, 375, 378,
381, 384, 385, 391, 393, 396, 397, 401—
403, 409—416, 437
— бентосоядные 272, 334, 335
— глубоководные 71, 295—297, 401
— донные 190, 286, 295, 413
4«0
рыбы коралловые 395
— лазающие 395
— летучие 213, 401
— методы изучения питания 320—322
— массовая гибель 161, 125
— морские (происхождение) 168
— пелагические 295, 308, 336, 413, 414
— питание 320 и сл.
— планктоноядные 231, 235, 239
— пресноводные 168, 411, 439
— промысловые 48, 123, 246, 271, 410—
416
— проходные 411, 439
— рост 324
— светящиеся 295, 297
— семейство бычковых 315
-----камбаловых 410, 416
— — карповых 410
-----корюшковых 315
-----лососевых 315, 410, 414, 439
----- окуневых 411
-----осетровых 315, 411
-----сельдевых 315, 410, 414, 416
-----сиговых 315
-----скумбриевых 411, 416
-----тресковых 375, 410, 414, 416
----- тунцовых 411
-----угревых 411, 414
— сублиторальные 163
— теплолюбивые 125
— узко-пелагические 327—331
— хищные 425
— холоднолюбивые 125
— широко-пелагические 331—334
ряды пищевые 41
С
сагитта 209
сазан 419
сайка 413
Салернский залив 284
сальпы 189, 205, 329, 345, 369
Самоа острова 430
Сан-Диэго залив 226, 228, 303
Сан-Хуан-архипелаг 293
----- острова 195
----- пролив 291
Саргассово море — см. море
саргассы 257, 401
сардина 125, 162, 288, 385, 414
— питание 330
— тихоокеанская (иваси) 415, 416
Сарматский бассейн — см. бассейн
Сарматское море 176
Сахалин 297, 415, 432
сверхрассеяние 182
свет 61, 98 пел., 117, 157, 158, 334
— влияние на миграции планктона 231,
232
— лупный 194, 233
— небесного свода 98
— солнечный 93, 98
свечение моря 14, 208,434
свинец 170
«свинцовый метод» определения возраста
земли 170
связи биоценотические 34, 48
— пищевые 48, 271, 328, 331
— трофические — см. связи пищевые
связывание азота микроорганизмами 105
Севастополь 423, 434
Севастопольская бухта 32
Севан оз. 412
Северная Земля 75, 141
Северное море — см. море
Северный Ледовитый океан — см. океан
Северный полюс 209
«Северный полюс» дрейфующая станция
152
северобореальная область 404
севрюга 322
седиментаторы 320
седиментация 152, 294
сезонная смена планктона 200, 233, 404,
405
сезонные явления 233 и сл.
сезонный показатель биомассы 235, 236
сезоны биологические 233 исл.,237—240
— гидрологические 233
— климатические 233
сейвал 417
сейши 122
сельдевые 315, 410, 414, 416
сельдь 18, 40, 116, 125, 162, 232, 239,
240, 322, 331, 332, 342, 374, 413, 414
— атлантическая 327, 328
— каспийская 162
— питание 327—330, 333
семейства рыб — см. рыбы
Сенегал 390
Сен-Жуан залив 114
Сен-Мало 190
сера 304
сердцевидка 245, 270, 418, 419
серобактерии 96
сероводород 17, 61, 95—97, 105, 116,
117, 120, 149, 297, 405
сестон 189
сестоноеды 320
сеть планктонная 32, 200, 201, 340, 343,
344
Сиваш 64, 71, 432
сиги 439
сиговые 315
Сидней 195
сизигий 257, 260
сила Кориолиса 119
силикаты 108
силикофлагелляты 145
силур 168, 173, 174
силы космические 124, 125
сильт 144
симбиоз 181, 395
синовакула 418
синэкология 34, 35, 177, 339
сипункулиды 260, 334, 389
— питание 320
Сиракузы 103
Сирия 71
сифонофоры 161, 189, 209, 213, 220, 224,
251, 354, 372, 373
Сицилианский мост 405
Сицилия 85, 277
Скагерак 78, 263—265, 276
«Скагерак» э. с. 152
скалы 147
Скандинавия 141, 360, 363, 379
Скандинавская область 340
Скандинавские страны 193
скаты 190, 411
скелеты организмов ИЗ, 152, 242, 354
— агглютинирующие 71
— арагонитовые 70
— белково-апатитовые 177
— белковые 176, 177
— из BaSO4 71
— известковые 70, 71, 147, 176
— кремневые 70
— минеральные 176
— органические 71, 176
— планктонных организмов 70, 146
— хитиновые 70, 76
склон континентальный 195, 294
— материковый 54—56, 58, 89, 145
скопления водорослей 165, 251, 260
— макрофитов 165, 246
— птиц 307
— рыб промысловых 18
— филлофоры 260
Скоресби-Саунд фиорд 265, 266
скорость движения водных животных 188
---при миграциях 188, 228
— переваривания пищи рыбами 323
— передачи жизни 156
— передвижения водных масс 125, 126
— размножения растений 157
«Скриппс» э. с. 20
скумбриевые 411, 416
скумбрия 162, 288, 334
«слой компенсации» 204
— придонный 97
— «промежуточный» 130
— скачка 122, 125, 126, 134, 135, 218
— фотосинтеза 107
слон индийский 156
смена фауны 87, 363, 366, 403
сменность вод океанов и морей 119, 120
Совет Постоянный Международный по
изучению моря 22
«сокращение» вод 125
солевой состав крови 168
соленость 64, 71 и сл., 85, 89, 105, 117,
129, 130, 135, 136, 141, 143, 309—313,
316—318, 401
— влияние на рост 314
— мирового океана-происхождение 65—
67
— морей СССР 309
— распределение в мировом океане 71
соли минеральные 203
— питательные 38, 39, 110, ИЗ, 114, 117,
213, 235, 236, 401
солнечные лучи 302
— пятна 124
солнечный спектр 99, 101
солнце 155
Соловецкие острова 191
солоноватоводность 309
сообщество 179, 182, 264
состав биоценоза 181
— морских организмов химический 67,
69, 70, 176
— морской воды солевой 14, 62—65
• элементарный 62, 69
— органических веществ химический 68
— полостной жидкости химический 68
— речной .воды солевой 62, 63, 65
31 Зенкевич, т. I
481
состав скелетов химический 68
спектр солнечный 99, 101
спикулы губок 147
Сплит 114, 214
споры 155
среда абиотическая 34
— бескислородная 150
— воздушная 168, 169, 172
— пресноводная 168
Средиземное море — см. море
Средиземноморская область 340, 393
Средиземноморский бассейн — см. бас-
сейн
Средиземноморско-атлантическая область
402, 405
СССР 275, 309, 410—413, 419, 421—423,
427—432, 439
ставрида 288
стагнация 149
Станция биологическая в Аркашоне 22
----Вудсхоллская 23
- — Датская 28, 32, 37, 262, 271, 350
- — Ковденская 24
— в Мильпорте 28
----Мурманская 23, 24, 26
-- — Неаполитанская 23. 32
----в Одессе 24
----Плимутская 23, 28
----в Пюджет-Саунд 195, 291 , 293
----в Роскове 23
— — Севастопольская 23, 32
----Соловецкая 23
— зоологическая в Виллафранке 24, 28
— морская Бакинская 23
----рыбохозяйственная в Астрахани 23
— рыбохозяйственная Бергенская 28
ста филин иды 256
стации 178
стационарное число однородного живого
вещества 156
стенотермия 299, 368
степень дисперсности 182, 186
сток 41
— материковый 65
— речной 53
стратификация 117, 132
— газовая 97
— питательных солей ИЗ
— солевая 74
— температурная 132
стратосфера 135, 137
ступени гидробиологии 34, 35
ступень континентальная 294
Субантарктика 391
субантарктическая область 132, 362
субарктическая область — см. область
субдоминанты 183
субипфлюэнты 183
сублитораль 145, 191, 192, 195—197,199,
247, 249, 251, 252, 262, 283, 293, 294,
296, 300, 358, 360, 361, 363, 394, 400
судак 419
сукцессии 267
сульфаты 95, 304
сульфиды 304
супралитораль 191, 192, 194. 196, 247—
250, 256, 258—260, 290
суточный ритм питания 336
Суэцкий канал — см. капал
482
схема зоогеографического районирования
Ортманна 359
— — — Экмана 363
— фитогеографического районирования
Щаповой 363
сходство фаун 375
сцепление частиц грунта 256
США 195, 275, 349,'411, 4 12, 415, 419—
421, 423, 425, 426, 429,430,433,439
сырьевые запасы рыб 414
Съезд 2-й Всесоюзный гидрологический
196, 197
Т
талитриды 250
такое 418
тараканы морские 380
Тасмания 439
Татарский пролив 434
Тахо р. 284, 285
таяние льдов 71, 72
темп размножения 324, 325, 327
«темп растекания» живого вещества 156
темп роста сардины 330
— эволюции 172
температура воды 73—75, 80 и сл., 117,
129, 130, 133—136, 141, 143, 187, 188,
368, 379, 394, 401, 404
---глубинная 83—87
--- поверхностная 80—83, 88, 89
— воздуха 73, 74
— замерзания воды 316
— наибольшей плотности воды 316
Тенсберг-фиорд 247
«теория катастроф» 174
— Сейвелла 125
термическая неоднородность фауны 403,
404
термический режим грунта ваттенов 250
251
течение Английского канала 140
— Антильское 119, 140, 400
— Атлантическое 124, 139—141, 369, 400
— Босфорское 120
— Бразильское 369
— Восточно-Гренландское 72, 73, 139—
141, 369
— Восточно-Исландское 140
— Гренландское 369
— Гумбольдта 306, 307
— Западно-Грепландское 140, 141
— Канарское 400
— Куро-Сиво 87, 141. 369
— Лабродорское 87. 89, 139, 140, 366,
369
— Н о рвеж с кое 13 9— 1 \ 1
— Нордкапское 141
— Ока-Сиво 87
— Полярное 140
— южно-экваториалыюе 213
течения 73, 77, 117 и сл., 129, 133, 135,
138—143, 162, 368, 369
— аптициклонические 119. 121, 141, 400
— бароградиентпые 119
— вертикальные 117, 119
— в проливах 80
— глубинные 77, 79, 80, 119, 369
— горизонтальные 117, 119, 121
течения дрейфовые 119, 132, 369
— кольцевые 121, 132, 139—141, 400
— компенсационные 119
— конвекционные 117, 119
— морские (типы их) 117, 119
— низовые 74, 130
— океанические 118, 119, 121, 124
— пассатные экваториальные 121
— периодические 119
— поверхностные 77, 79, 94, 113, 119,
138, 143
— постоянные 119
— прибрежные 119
— приливно-отливные 119
— прямолинейные 119
— системы Гольфстрима 119
— скорость 118, 119, 141
— случайные 119
— смешанные 117
— солепые 79
— теплые 73, 80, 83, 87, 88, 119
— тропические 369
— Тэтиса 369
— холодные 73, 83, 87,119, 139, 141, 369
— циклопические 119, 121
— экваториальные 121, 135, 136
Тимаво 404
типы планктона 375
— формы тела и скелета 278
Тирренское море — см. море
Тирренский район 85
Титр 42, 43
Тихий океан — см. океан
ткани опорные белковые 176
-----хитиновые 177
токи конвекционные 120
токсины 302
торий 170
Тортугас остров 278
точка компенсации 103, 104, 204, 207,
208
— оптимального роста 207
трава морская 431, 432
трал 243, 244
траловый лов 295, 416
трансгрессии океанические 124, 125
трахимедузы 210
трепанг 395, 430
треска 18, 116, 125, 162, 268, 269, 272—
274,336, 342,345, 374, 414, 416, 419,
420, 436
—питание 331, 332, 335, 336
тресковые 375, 410, 414, 416
третичный период — см. период
триас 173
тридакна 354, 395
Триест 404
трилобиты 171
триптон 189
Тристан-да-Кунья 385
Тромсе 278, 279
Тротгеймфпорд 141
тропик Рака 409
тропосфера 127, 135—138
трофология 48
трубач 418
трубка для взятия грунта 152
тунец 125, 288,401
Тунис 430
тунцовые 411
турбеллярип 169, 170, 253, 283, ЗГ»
— питание 320
турбо 418, 421
турбулентное перемешивание 117, 141
тэтис — см. море
тюлени 48, 375, 385
тяготение земное 117
У
углеводороды нефти 304
углеводы 68, 116
углекислота 61, 95, 97, 176, 230, 314
углекислый кальций — см. кальций
углерод 111, 115, 116, 206, 207, 304
— органический 115,116
угорь 256, 399, 401
угревые 411, 414
удобрение морских водоемов 14
улптка наземная 185
улов 186, 187
— камбалы 274
— рыб 411 и сл.
— трески 274
ульва 257
умбеллула 354
Управление гидрографическое 145
уран 170
усвояемость пищи 324
Уссурийский залив 290
устрица «португальская» 439
— черноморская 423
— «японская» 439
устрицы 70, 102, 190, 270,320, 395,418—
420, 423—425, 437, 439
— условия нереста 439
устричные банки 423
— парки 102, 439
устье Ла-Платы 312
— Нила 284, 286
— Эльбы 245
устья рек 151, 169, 250, 284, 286, 313,
315
Утлюкский лиман 432
учет количественный 34, 35, 165, 185,
200, 242, 244, 245, 254, 262, 267, 275,
279, 281, 284, 288, 348, 350—352
— — содержимого кишечников 272
учреждения морские научные 21, 22,
25—28
Ф
фаза ксеротермическая 173
— ледниковая 173
— умеренная 173
фазы биологического продуцирования f»7
— развития акклиматизированных форм
438
фактор исторический 62
факторы внешней среды 61, 62
— воздействующие на соленость 71, 72
— обусловливающие развитие жизни в
море 61, 62
— определяющие распространение орга-
низмов 367—369
— роста 105
Фарерские острова — см. острова
483
Фарерский капал 221
фауна (определение понятия) 357
— абиссали 198, 296—300, 360
— австралийская 396
— амфиатлантическая 354, 382, 437
— амфибиальная 395
— амфибореальная 382, 437
— амфипацифическая 354, 382, 437
— антарктическая 354, 355, 363, 436
— антибореально-южноамериканская 363
— арктическая 160, 281, 295, 297, 354—
356, 360, 363, 368, 371,376, 436
— архибентальная 138
— атлантическая 363, 375, 376, 394, 400,
405, 436, 437
- батиали 294, 295
- батипелагическая 198
— Белого моря 369
— биполярная 390, 391
— бореальная 252, 281, 354, 360, 363,
371, 376, 384, 402
— вест-индская 401
— воздушная 355
- - воздушных почв 170
— восточно-американская 363
восточно-тихоокеанская 436
- - высокоарктическая 376, 381
гавайская 396
— глубоководная 219, 220, 269, 291,295—
300, 363, 368, 390, 401, 405
— — происхождение 297
- грунтовая 170, 252, 253
- дочерняя 436
западно-американская 363
- - западно-африканская 363
— западно-тихоокеанская 396, 436
- - индийская 402
— индо-вестпацифики 355, 363, 393, 395—
397, 400
— индопацифическая 363
— ископаемая четвертичная 390
— кергуэленская 363
— Красного моря 402
— литоральная 169, 170,183, 198, 248 и
сл., 257—259, 355, 360, 369, 395
— лузитанско-средиземноморская 403
— малайская 396
— мелководная 296, 376, 436
— наддонная 220, 225
— наземная 169, 361
— неритическая 160
— новозеландская 363
— потальная 354
— пелагическая 198
- перуанско-северочилийская 363
- - пещерная 239, 355, 368
— поверхностная 221
— подземных вод 299, 368
— пресноводная 66, 169, 250, 309, 310,
312, 315, 318, 355
— прибойной полосы 257
— прибрежная 296, 393, 394
— приполярных областей 390
— псевдоабиссали 29'«
— реликтовая 315, 318
— сарматская 363
— северо-тихоокеанская 363, 396, 437
— солоповатоводная 169, 309 и сл., 314,
315, 318, 319, 355
фауна Средиземного моря 401—405
— средиземноморско-атлантическая 363
— среднеглубинная 221
— стенобатная 223
— сублиторальная 295, 358 и сл.
— тепловодпая 354, 355, 368
— теплолюбивая 236, 379, 392
— тихоокеанская 257, 258, 371, 375, 376
394, 396, 436
— третичных морей 400
— тропическая 160, 354, 355, 363, 376,
392 и сл., 402, 436
— Тэтиса 297, 368, 393, 402, 403, 405,
436
— умеренных широт 160, 355, 363, 392
— холодноводная 354, 363, 368, 392
— холоднолюбивая 224
— эврибатная 223
— эпипелагиали 198, 363
— эритрейская 402
— юго-западноафриканская 363
— южно-австралийская 363
— южно-африканская 363
— южно-тихоокеанская 396
— южно-японская 396
фаунистика 33
фаунула 177, 178
фации 178, 193, 194, 258, 259
фаэтон 307
фекалии 36, 144, 205
фенология океанологическая 240
Феодосия 104
феоцистис 329
Филиппинские острова 75
филлофора 248, 260, 432
филлофорное поле 193
фильтраторы 14, 116, 320
фильтрационный аппарат 36
фильтрация воды животными 35
финвал 417
Финляндия 275, 349, 439
Финмаркен 366
Финский залив — см. залив
фиорд Гюльмарфиорд 102, 186, 197, 199,
243, 245, 266, 267, 275—278, 293
— Дибсофиорд 266, 267
— Лимфнорд 266, 267, 270, 271, 273—275,
322
— Осло-фиорд 207
— Рамфиорд 278, 279
— Ранде рс-фиорд 312
— Скоресби-Саунд 265, 266
— Тенсберг 247
— Тронтгеймфиорд 141
фиорды 94, 96, 123, 198, 199, 263
— Гренландии 275, 294
— Норвегии 61, 97, 123, 149
фитобентос 48, 246—248, 326
фитопланктон 47—49, 61, 94, 103, 105,
108, 112—114, 116, 120, 121, 123, 136,
157, 188, 200 и сл., 215, 218, 219, 230,
232, 235—237, 302, 304, 305, 320, 325,
326, 333, 334, 344, 345, 347, 401
флора донная 103, 391
— земноводная 174
— наземная 174
— неритическая 160
— псилофитовая 174
Флорида 87, 195
484
Флоридский пролив 119, 140
фораминиферы 70, 71, 145, 147, 152, 171,
190, 195, 208, 314, 354, 395
форель 439
«Форингеп» э. с. 17
формирование «центральных» водных масс
127
формы амфиамериканские 397
— амфиатлантические 377, 378
— амфибореальные 439
— антарктические 210, 387
— антарктическо-антибореальные 374
— арктические 210, 279, 295, 350, 366,
367, 369, 379, 381, 382, 384, 387
— арктическо-бореальные 210, 350, 362,
366, 374, 377, 384
— атлантические 369, 381, 404, 437
— батипланктонные 221
— биполярные 210, 385—391, 436, 439
— бореальные 210, 279, 350, 362, 378,
382, 384, 385, 387
— бореоарктические 362, 378
— «весенние» 234
— викариирующие 259, 284, 373, 397,
435, 436
— второстепенные 183—187
— высокоарктические 224, 362, 363, 378,
381, 385
— глубоководные 71, 210, 295, 381, 382,
404
— господствующие 178
— гости 183
— доминирующие 183, 185, 367
— индикаторы 14
— индо-вестпацифические 396
— индо-пацифические 402
— константы 185
— кормовые для рыб 272
— космополитные 373
— «летние» 234
— массовые 186, 187, 178, 244, 245,262,
269, 281, 367
— нижнеарктические 362, 363, 381
— нота л ыю-бореальные 402
— нотальные 210, 385, 387
— отступающие 183
— папарктические 362, 363, 374
— пелагические 369
— подчиненные 183
— посетители 183
— преобладающие 183
— пресноводные 319
— роющиеся в грунте 254
— руководящие 165, 181, 183—187, 194,
276
— свободно подвижные 187
— северные 363, 389, 390, 404
— северо-тихоокеанские 381
— сидячие 187
— случайные 183, 185
— с ограниченным ареалом 373
— сорняковые 274
— стенобитные 221
— стеногалинные 319
— средиземноморско-бореальные 384
— средиземпоморско-лузитанско-южно-
бореалыше 384
— субантарктические 210, 385, 387
— субарктические 138, 385, 387
формы субарктическо-бореальные 384
— субтропические 378
— тепловодные 87, 210, 382, 402
— тропические 378, 387, 404
-----материнские 389
— тропическо-эритрейские 402
— умеренно-северо-бореальные 366
— характерные 165, 184, 185, 187
— холодноводные 87, 369
— циркумарктические 363
— циркумполярные 363
— циркумтропические 393, 397, 402
— широкобореальные 366
— эврибатные 221, 297, 298
— эвригалинные 309, 310, 313, 314, 319,
437
— эвриоксибионтные 150
— эндемичные 373, 374, 387, 398, 400,
402—404, 406
— южно-бореальпые 366
— южного генезиса 391
— южные 363, 369, 389
фосфаты 47, 61, 107—ИЗ, 121, 135, 151 #
203, 204, 208, 215, 304
фосфор 15, 47, 105, 109—112, 114, 151,
169, 205, 215, 304
— концентрация в морских организмах
70
фосфоресценция 220
фотосинтез 40, 47, 98, 103, 204, 206
фототропизм 231, 233, 299
Фрайди бухта 114
«Фрам» э. с. 17
Франца Иосифа Земля — см. Земля
Франция 190, 191, 193, 194, 249, 360, 403,
412, 419, 420, 422, 426, 430, 439
фрегаты 307
фронт полярный 41, 118, 123, 138, 352
фукусовые 173, 246
фукусы 190, 191, 247, 248, 252, 253, 257,
260, 283, 284, 403
Фунафути атолл 195
Фунди залив 249.
X
халистазы 400
хамагури 418, 421
хамса 116, 162, 239, 288, 334, 403
химический состав организмов 67, 69, 70,
176
хитин 304
— переработка в шелк 425
хищники 48, 267, 320, 328, 425
хлорофил 157
хордовые 157, 168, 171, 172
хорология 34
хребет Уайвилл я Томсона 89, 90
хронология земли 172
ц
цвет воды 99, 100, 329
цветение диатомовых ИЗ
— моря 14, 40, 100, 235, 434
— фитопланктона 40, 94, 12f0, 157, 182
цветковые растения 246, 247
Цедар-Бог оз. 48, 49
цен 177
485
центры формирования фаун 348, 368
цепи пищевые 14, 49, 269^ 271, 325—327,
332—334
пестоды 327
Цехштейновое палеозойское море 65
«Цигольф» э. с. 17
циклические процессы в водоемах 39—41
циклы геологические 173
— развития планктона 346, 347
— физико-химические 121
ципк-накопление в теле кишечнополост-
ных 69
циркон -171
циркуляция вод Атлантического океана
132 и сл.
---вертикальная 110, 112, 123, 124,
129, 135, 136, 213, 236, 316
— воды в природе 393
циркумэкваториальность 393
цистозира 191, 248
цисты 155
цицинделлиды 250
Ч
чайка глупыш 307
частота встречаемости 183, 185, 245
---пищевых компонентов 321—323
Чезапик-бай залив 120, 421
«Челленджер» э. с. 16, 17, 32
человек доисторический 420
— палеолитический 420
черни 149, 168, 190, 220, 235, 242, 252—
254, 290, 292, 395, 417, 429, 430
— дождевые 255
— кольчатые 171
— корабельные 434
— малощетинковые 48, 169
— низшие 327
— пелагические хищные 328
— планктонные 209
черепаха каретта 393
черепахи морские 116, 354, 401
Черное море — см. море
четвертичный период 62
Чечень остров 432
Чили 431, 439
Чилька оз. 315
число видов макрофитов 165
---морской фауны 161—163
членистоногие 157, 158, 168, 177
Чукотская провинция 370
Чукотское море — см. море
Ш
Шаптарские острова — см. острова
Шавтарское море 194
Швеция 197, 275, 276, 349, 419, 422, 426
шельф 58, 151
Шотландия 123, 141
Шотландские острова 363
Шпицберген 19, 141, 308, 348, 350, 361,
369, 379, 434
шпрот 288
штормы 103
Щ
щебень 144
щелочной резерв 95
486
щетипкочелюстные 189, 209, 320
щетки мидий 293
— устриц 395
щупальщики 320
Э
эволюция абиссальной фауны 298
— животных 158, 171, 172
— растений 157, 158
эвритопность населения литорали 250
эвтрофия 41
эвфаузииды — см. ракообразные эвфау-
зиевые
Эгейское море — см. море
экология 33, 35, 48, 177, 339
экосистема 177
экспедиция Агассица 18
— адмирала Макарова 17
— на «Альбатросе» 17
— английская на Б. Барьерный риф
215
— Андрусова и Лебединцева 17
— на «Армауер Ганзен» 20
— на «Атлантис» 20
— Беринга 15
— Беллинсгаузена и Лазарева 15
— Билингса и Сарычева 15
— на «Блеке» 17
— Бэра 15
— Бэра и Данилевского 16
— на «Вальдивии» 18
— на «Дана» 19, 86, 136
— на «Дана II» 20
— Датская 1908—1910 гг. ИЗ, 214, 27 5,
284
— па «Дисковери» 109
— па «Дисковери II» 20
— «для научно-промысловых исследова-
ний у берегов Мурмана» 18
— Дригальского 18
— на «Ингольфе» 17
— Ионга 20
— на «Карнеги» 20, 135, 136
— Кука 15
— Литке 15
— на «Метеоре» 20, 111, 116, 345
— па «Михаэле Сарсе» 19, 31
— Нансена на «Веслемое» 139
— на «Национале» 17, 343. 344
— Норденшельда антарктическая 18
---на «Веге» 17, 192
— па «Персее» 20
— планктонная 1889 г. 213
— на «Поркьюпайн» 16
— принца Монакского 17
— Росса 16
— на «Снеллиусе» 20
— Тол л я на «Заре» 18
— на «Форингене» 17
— на «Челленджере» 17, 32, 217, 385
— Корейско-Сахалинская П. Шмидта 18
— на «Holsatia» 213
— па «Scripps» 20
элементы биогенные 47, 105 и сл., ИЗ, 120
— тяжелые 69
элитораль 191, 192, 196, 197, 199-
Эллиса острова 195
Эльба р. 245, 313
эндемизм 384, 400, 436
эндемизм фауны Средиземного моря 403,
405
эндемики — см. формы эндемичные
— амфиамернканские 397
— атлантические 376
— Белого моря 374
— Индо-ьестнацифики 396
— тихоокеанские 376
эндобиоз 177
энергетический баланс карпа 324
энергия 48
— - биохимическая 301
— геохимическая организмов 156
— космическая 155
— световая 101, 155
— солнечная 156
— тепловая 155, 156
— химическая 155, 156
энзимы 114
эоцен 376, 391
эпибиоз 177, 250, 276—278
эпипланктон 190, 217
эпифауна 148, 149, 190, 264, 276, 281, 287.
293
эпоха ледниковая 152, 282, 368, 376, 378,
390, 436
эпоха межледниковая 152
эпохи геологические 368, 391
Эри оз. 412
эстуарии р. Тахо 285
этапы изучения морской фауны 31—33
эхолот 56, 152
Ю
юра 173
Ютландия западная 253, 255
Ютландский полуостров 195, 249
Я
Ява 431
яйца 219. 224, 328, 330
— донных животных 208, 235
— рыб 208
Якшина губа 288
Ян-Майен 350
Япония 130, 163, 165, 395, 411, 412, 41
416, 419—421, 425, 426, 430, 431
Японские острова 297
Японское море — см. море
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИИ
А
Abatus (Ecninodermata Echinoidea) 406
Abra (Mollusca Lamellibranchiata) 265
— alba 261, 263, 266, 267
— longicallis 270
— nitida 263
— prismatica 270
Acanthephyridae (Crustacea Decapoda) 220
Acanthine (Mollusca Gastropoda) 260
Acanthodoris (Mollusca Gastropoda) 253
Acanthogorgia (Coelenterata Anthozoa) 295
Acartia (Crustacea Copepoda) 226
— clausi 221, 404
Acmea (Mollusca Gastropoda) 257, 258
— cassis 257, 292
— digitalis 257
— digitata 292
— testudinalis 253
Acrania 166
Acronida branchiata (E chi noderma ta
Ophiuroidea) 276, 277
Actiniae (Coelenterata Anthozoa) 258
Actinia equina (Coelenterata Anthozoa)
253, 403
Actinocyclus ehrenbergi (Algae Diatomeae)
175, 176
Aeolis (Mollusca Gastropoda) 253
Aglantha digitalis (Coelenterata Hydro-
zoa) 213, 220
Ahnpheltia (Algae Rhodophyceae) 247,431,
432
Alaria (Algae Phaeophyceae) 248
Alcyonaria (Coelenterata Anthozoa) 395,
396
Alcyonidium (Bryozoa) 253
— digitatum 242, 270
Alexia myosothis (Mollusca Gastropoda)
257
Allorchestes (Crustacea Amphipoda) 260
Alopias vulpes (Pisces) 387
Amage anops (Annelida Polycbaeta) 231
Ammodytes (Pisces) 328
Ampelisca (Crustacea Amphipoda) 292
— macrocephala 290
Amphibia 166
Amphilepis (Echinodermata Ophiuroidea)
261, 266
— norvegica 264
Amphineura (Mollusca) 164, 166
Amphiodia craterodmeta (Echinodermala
Ophiuroidea) 290
488
Amphioxus (Vertebrata Acrania) 288, 289
Amphiphelia (Coelenterata Anthozoa) 295
Amphipoda (Crustacea) 164, 165, 170, 209,
243, 249, 258, 313, 315, 320, 328, 330,
371
Amphiporus (Nemertini) 254
Amphissa (Mollusca Gastropoda) 258
Amphiura (Echinodermata Ophiuroidea)
261, 282
— chiajei 263, 266, 270, 276, 277, 286, 288
— filiformis 263, 276, 277, 281, 282, 286
Ancistrosyllis robusta (Annelida Poly-
chaeta) 290
Anguilla (Pisces) 399
— australis 332
Anomalocera (Crustacea Copepoda) 223
Anomalophora magna (Crustacea Copepoda)
220
Anomura (Crustacea Decapoda) 30, 296,
400
Antedon (Echinodermata Crinoidea) 284
Antedonidae (Echinodermata Crinoidea)
397
Anthoptillum (Coelenterata Anthozoa) 295
Anthozoa (Coelenterata) 163, 164, 166, 167,
315, 395
Anti patharia (Coelenterata Anthozoa) 296
Aphelasterias (Echinodermata Asteroidea)
258
Aphrodite aculeata (Annelida Polychaeta)
281, 295
Apogonops anomalus (Pisces) 332
Appendicularia (Tunicata) 166, 209, 210,
312
Arachnoidea 166
Area (Mollusca Lamellibranchiata) 285
— glacialis 283
— inflata 421
Archidoris britannica (Mollusca Gastro-
poda) 69
Arenicola (Annelida Polychaeta) 253, 255,
256
— claparedii 258, 292
— cristata 258
— grubei 403
— marina 254, 255, 259, 262, 263, 270, 403,
430
Argobuccinum oregonensis (Mollusca Gast-
ropoda) 292
Artediellus (Pisces) 381
— pacificus 381
— scaber 381
Artediellus uncinnatus 381
Artemisia (Crustacea Decapoda) 424
Arthropoda 69, 176
Asabellides sibirica (Annelida, Polychae-
ta) 290
Ascidia (Tunicata) 166, 285, 293
— obliqua 278
Ascophyllum (Algae Phaeophyceae) 246, 293
— nodosum 191, 253, 278
Astacura (Crustacea Decapoda) 296
Astacus leptodactylus (Crustacea Decapoda)
429
— pachypus 429
Astarte banksi (Mollusca Laniellibran-
chiata) 263, 283
— borealis 234, 261, 263, 269, 270, 276,
277, 283, 285
— crenata 282, 283, 285
— elliptica 234, 263, 283
— montagui 234, 276, 277
— sulcata 281
Asteras (Echi nodermata Asteroidea) 430
— rubens 253, 267
Asteroidea (Echinodermata) 165, 166
Asterionella (Algae Diatomeae) 375
Astrogonium pareli (Echinodermata Astero-
idea) 295
Astropecten (Echi node rmata Asteroidea)
396
— jonstoni 286
Asychis punctata (/\nnelida Poychaeta) 290
Atlanta (Mollusca Gastropoda) 147
Augaptilus (Crustacea Copepoda) 220
Aulographis pandora (Protozoa Radiola-
ria) 220
Aulospathis (Protozoa Radiolaria) 220
Aves 166
Avicennia (Gynoeciatae Angiospermae) 395
Axinopsis orbiculata (Mollusca Lamelli-
branchiata) 290
Axinus flexuosus (Mollusca Lamellibran-
chiata) 263, 264, 276, 277, 281, 283, 294
Azotobacter (Bacteria) 106
В
Balaenoptera borealis (Mammalia Cetacea)
334, 417
— musculus 417
— physalus 417
Balanidae (Crustace<i Cirripedia) 265
Balanus (Crustacea Cirripedia) 69, 249,
257—259, 265, 278
- balanoides 182, 253, 257 260, 293
— balanus 278
— cariosus 257, 291
- - crenatus 253
— glandula 257, 258, 291
— nobilis 258
— perforatus 403
— porcatus 389
— tintinnabulum californicum 258
Bangia (Algae Rhodophyceae) 257
Bankia setacea (Mollusca Lamellibran-
chiata) 434
— sibirica 434
Bathybiaster (Echinodermata Asteroidea)
295
— robustus 296
— vaxillifer 296
Bathycrinus (Echinoderma ta Crinoidea) 296
Bentneuphausia amblyops (Crustacea,
Euphausiacea) 222
Beroe cucumis (Coelenterata Ctenophora)
372
Bifurcaria tuberculata (Algae Phaeophy-
ceae) 191
Birgus latro (Crustacea Decapoda) 396
Bitnynia (Mollusca Gastropoda) 314
Bledius (Insecta) 256
Blepharipoda occidentalis (Crustacea Deca-
poda) 259
Bodotriidae (Crustacea Cumacea) 387, 388
Boleophthalmus (Pisces) 395
Boreomysis nobilis (Crustacea Mysidacea)
295
— scyphops 295
Bowerbankia (Bryozoa) 253
Brachiopoda 69, 71, 163, 166, 176
Brachiura (Crustecea Decapoda) 400
Brachynotus (Crustacea Decapoda) 258
Branchiomma vesiculosum (Annelida Po-
lychaeta) 288
Brissopsis (Echinodermata Echinoidea)
261, 265
— lyrifera 263, 266, 269, 270, 276, 277,
403
Brizaster fragilis (Echinodermata Echino-
idea) 295
Bryopsis plumosa (Algae Chlorophyceae)
366, 367
Bryozoa 163—167, 176
Buccinum (Mollusca, Gastropoda) 71, 267,
270
— groenlandicum 284
— hydrophanum 284
— undatum 403, 422
Byblis gaimardi (Crustacea Amphipoda)
290
Bythocaris leucopis (Crustacea Decapoda)
295
— payeri 295
C
Calanidae (Crustacea Copepoda) 209, 211,
212
Calanoida (Crustacea Copepoda) 211
Calanus (Crustacea Copepoda) 223, 226,
231, 328
— acutus 373
— finmarchicus 44, 92, 165, 188, 209, 220,
221, 224—233, 238, 239, 333, 334, 369,
372, 373, 401
— hyperboreus 219, 220, 224, 373
Calicophorae (Coelenterata Siphonophora)
210
Callinectes (Crustacea Decapoda) 424, 427
— sapidus 426
Calliossoma costatum (Mollusca Gastro-
poda) 292
Calocaris marandreae (Crustacea Decapoda)
270
Calyptosphaera oblonga (Protozoa Cocco-
litnopnorida) 165
Cambarus (Crustacea Decapoda) 424
Cancer (Crustacea Decapoda) 424, 426, 427
— amphioetus 377
Carcinoscorpius (Palaeostraca Xiphosura)
396
489
Careinus maenas (Crustacea Decapoda) 253.
288, 429
Cardiosoma (Crustacea Decapoda) 395
Cardium (Mollusca Lamellibranchiata) 253.
422
— californiense 423
— cilia turn 281, 283, 293
- - corbis 258, 292, 421
— edule 44, 182, 199, 253, 254, 259, 261,
263, 267, 270, 282, 294, 313, 314, 379,
421, 422
— groenlandicum 187
Caretta (Reptilia) 393
Carinaria (Mollusca Gastropoda) 147
Caryophyllia clavus (Coelenterata Antho-
zoa) 291
Caulerpa (Algae Chlorophyceae) 246—248
Cavolinia (Coelenterata llydrozoa) 147
Centoscus scolopax (Pisces) 332
Cenlropages (Crustacea Copepoda) 329
— hamatus 220
— typicus 213, 220, 404
Centropagidae (Crustacea Copepoda) 211,
212
Cephalopoda (Mollusca) 164, 166, 167, 209
Ceramium (Algae, Rhodophyceae) 246, 249,
257
— rubrum 366
Cerebratulus (Nemertini) 254
Cerithium (Mollusca Gastropoda) 395
Chaetoceros (Algae, Diatomeae) 176, 205,
235
Chaetognatha 163, 166, 209, 210, 212, 226,
373
Chaetopterus (Annelida Polychaeta) 278,
293
Challengeria (Protozoa, Radiolaria) 220
Charybdis (Crustacea Decapoda) 424, 427
— japonica 428
Cheiragonus (Crustacea Decapoda) 424, 427
Chelonia (Reptilia) 393
Chelura terebrans (Crustacea Amphipoda)
434
Chielostomata (Bryozoa) 315
Chione gallina (Mollusca Lamellibran-
chiata) 289
— verrucosa 288
Chionocoetes (Crustacea Decapoda) 382,
424, 427
— angulatus 382
— bairdi 382
— elongatus.bathyalis 291
— opilio 38'1 , 382, 428
— tahneri 382
Chiridius (Crustacea Copepoda) 220
— armatus 220
— obtusifrons 220
Chiridota discolor (Echinodermata Holo-
thurioidea) 258, 288
Chlamis sulcata (Mollusca) 286
Chlorophyceae (Algae) 201, 202, 246, 248
Chondrus (Algae Rhodophyceae) 246, 247
Chone cincta (Annelida Polychaeta) 290
Chorda filum (Algae Phaeophyceae) 191,
253, 278
Chordacea (Algae Phaeophyceae) 173
Chordaria flagelliformis (Algae Phaeophy-
ceae) 386
— magellanica 386
CMhamalus (Crustacea Cirripedia) 2'i9, 257
— challenger! 258
— dalli 257, 292
— stellatus 250, 257, 403
Cidaroidea (Echinodermata Echinoidea)
396
Ciliata (Protozoa Infusoria) 163, 166, 167
Cirolana (Crustacea Isopoda) 260
— harfordi 258
— linguiformis 259
Cirripedia (Crustacea) 163, 164, 209, 253,
257 329
Cladocera (Crustacea) 189, 209, 210, 226
Qadophora (Algae Chlorophyceae) 246—
248
— fracta 253
— gracilis 253
Clava (Coelenterata Hydrozoa) 253
Clio (Mollusca Gastropoda) 174, 375
Clione limacina (Mollusca Gastropoda)
401
Clostridium (Bacteria) 106
Clupea harengus (Pisces) 327
-----harengus 414
-----pallasi 414
Clymene gracilis (Annelida Polychaeta)
281, 282
Coccolithinae (Protozoa Flagellata) 201,
202
Coccolithophora fragilis (Protozoa Flagel-
lata) 165, 213
Coccolithophoridae (Protozoa Flagellata)
147
Codium (Algae Chlorophyceae) 246, 248
Coelenterata 69, 176, 212, 327
Coelopa (Insecta) 258
Coenobita (Crustacea Decapoda) 395
Collidae (Protozoa Radiolaria) 220
Collossendeis (Arthropoda Pantopoda) 295
Comatulida (Echinodermata Crinoidea) 396
Conchopis (Protozoa Radiolaria) 220
Copepoda (Crustacea) 69, 70, 163—165, 189,
205, 209—213, 219, 226, 235—239, 312,
313, 327—332, 342
Copilia (Crustacea Copepoda) 220
Corallium rubrum (Coelenterata Anthozoa)
430
Corallina (Algae Rhodophyceae) 146, 246,
247, 249, 257, 258
Corbula gibba (Mollusca Lamellibran-
chiata) 286, 288
Corbulomya maeotica (Mollusca Lamelli-
branchiata) 313
Cornacuspongida (Porifera) 396
Corophium (Crustacea Amphipoda) 253, 257
— volutator 253
Coscinosira polychorda (Algae Diatomeae)
207
Cosmetia (Coelenterata Scyphozoa) 223
Cosmetira pilosella (Coelenterata Scypho-
zoa) 225
Cottidae (Pisces) 296, 332
Crangon (Crustacea Decapoda) 424, 426
— crangon 256, 426
Crangonidae (Crustacea Decapoda) 223
Cribrina xanthogrammica (Coelenterata
Anthozoa) 257
- Crinoidea (Echinodermata) 1669 400
Crustacea 166, 167, 212, 336
490
Ctenodiscus crispatus (Echinodermata
Asteroide.a) 281, 282, 291
Ctenophora (Coelenterata) 163, 166
Cucumaria (Echinodermata Holothurio-
idea) 233
— elongata 288
— frondosa 430
Cultellus adriaticus (Mollusca Lamelli-
branchiata) 288
— pellucidus 286
Cumacea (Crustacea) 164, 165, 315, 331,
387, 388
Cyanophyceae (Algae) 201—203
Cyclonassa heritaea (Mollusca Gastropoda)
*286, 288
Cyclostomata 166
Cydaroidea (Echinodermata Echinoidea)
163
Cyprina islandica (Mollusca
branchi ata) 263, 270, 379, 403
Cystoseira (Algae Phaeophyceae)
Lamelli-
246, 247,
407
ericoides 191
— fibrosa 191
.— granulata 191
D
Dasycladus (Algae Chlorophyceae) 246, 248
Decapoda (Crustacea) 164, 209, 220, 320,
329, 331, 400
— Natantia 384
— Reptantia 384
Deima (Echinodermata Holothurioidea)
236
Delesseria (Algae Rhodophyceae) 246, 247
— middendorffii 258
Dendraster excentricus (Echinodermata
Echinoidea) 259
Dendronotus (Mollusca Gactropoda) 253
Dentalium (Mollusca Scaphopoda) 291
Deschampsietum (Gynoeciatae Angiosper-
mae) 182, 183
Desmarestia (Algae Phaeophyceae) 284—
286
Desmarestiacea (Algae Phaeophyceae) 173
Diastylidae (Crustacea Cumacea) 387
Diatomeae (Algae) 201, 202
Dictyosiphon (Algae Phaeophyceae) 253
Dicyemida (Mesozoa) 166
Dimophyes arctica (Coelenterata Siphono-
phora) 372
Diplosoma (Tunicata Ascidia) 401
Dipsacus lacinatus (Gynoeciatae Angio-
spermae) 184
Diptera (Insecta) 250
Disyringa (Spongia Tetraxonida) 296
Doliolidae (Tunicata Salpae) 210
Donax (Mollusca Lamellibranchiata) 281
— variabilis 422
Doris (Mollusca Gastropoda) 253
Dudresnaia (Algae Rhodophyceae) 248
Dynamena (Coelenterata Ilydrozoa) 253
E
Echinocardium (Echinodermata Echino-
idea) 261, 265, 279, 281, 292
— cordatum 263, 266, 270, 276, 277,
286
Echinocardium flavescens 270, 281, 282
— mediterraneum 288
Echinodermata 164, 167, 176, 314, 336, 400
Echinoidea (Echinodermata) 30, 166
Echinorachnius parma (Echinodermata
Echinoidea) 292
Echinothuriidae (Echinodermata Echino-
idea) 295, 296
Echinus esculentus (Echinodermata Echi-
noidea) 430
Echiurida (Annelida) 162, 163, 166, 315
Ecklogia (Algae Phaeophyceae) 246, 248
Eisenia (Algae Phaeophyceae) 384
— arborea 384
— bicyclus 384
— cookeri 384
— desmarestioides 384
— masonii 384
Elachista ficiola (Algae Phaeophyceae) 366
Elasipoda (Echinodermata Holothurio-
idea) 296
Elpidia glacialis (Echinodermata IIolo-
thurioidea) 235
Embiotocidae (Pisces) 377
Emerita (Crustacea Decapoda) 259
Empetrum nigrum (Archegoniatae Bryo-
phyta) 183
Engraulis (Pisces) 307
— encrasicholus 403
Ensis ensis (Mollusca Lamellibranchiata)
288
— silique 288
Enteromorpha (Algae Chlorophyceae) 246,
249, 253, 257
— compressa 366
Enteropneusta (Chordata Hemichordata)
166
Entoprocta 166
Epigeichthys atro-purpureus (Pisces) 258
Erimacrus (Crustacea Decapoda) 427
— isenbecki 428
Eriocheir (Crustacea Decapoda) 427, 438
— japonicus 428
— sinensis 429, 437
Eriphia spinifrons (Crustacea Decapoda)
429
Eucalanus attenuatus (Crustacea Copepoda)
221
Euchaeta glacialis (Crustacea Copepoda)
220
— norvegica 220, 221
Euchone papillosa (Annelida Polychaeta)
276
— rubrocincta 281
Eukrohnia hamata (Chaetognatha) 223, 373
Eumenia crassa (Annelida Polychaeta) 270,
276, 277
Eunephthya rubiformis (Coelenterata
Anthozoa) 379, 381
Eunice viridis (Annelida Polychaeta) 430
Euphausia hemigibba (Crustacea Euphau-
siacea) 222
— krohnii 222
— pseudogibba 222
— tenera 222
Euphausiacea (Crustacea) 209, 210, 220,
222, 226, 308, 330, 333
Euphausiidae (Crustacea Euphausiacea)
231
491
Euplectella (Porifera Triaxonida) 296
— speciosa 68
Eupyrgus (Echinodermata Holothurioidea)
292
— pacificus 290
Euryale (Echinodermata Ophiuroidea) 400
Isopoda)
Eurycope spinifrons (Crustacea
291, 297
Eurytemora (Crustacea Copepoda) 329
Eurytemora grimmi (Crustacea Copepoda)
228—230
— hirundoides 334
Euspongia officinalis (Porifera Cornacu-
spongida) 430
Evadne nordmanni (Crustacea Cladocera)
220, 389
F
Fabricia sabella (Annelida Polychaeta) 254,
259, 389
Filipenduletum (Gynoeciatae Angiosper-
mae) 185
Flabellum (Coelenterata Anthozoa) 295
Flagellata (Protozoa) 166, 167, 201, 202
Flustrella (Bryozoa) 253
Foraminifera (Protozoa Rhizopoda) 68, 71,
163—165, 283, 314, 327, 395
Forficulidae (Insecta) 258
Freyella (Echinodermata Asteroidea) 296
Fritillaria (Tunicata Appendiculariae) 220
Fucaceae (Algae Phaeopnyceae) 173, 259,
387
Fucus (Algae Phaeophyceae) 246, 285, 286,
293
— evanescens 258
— inflatus 247, 253
— platycarpus 191
— serratus 191, 247, 253, 259, 278
— spiralis 253
— vesiculosus 191, 253, 278
Fungia (Coelenterata Anthozoa) 396
Fungidae (Coelenterata Anthozoa) 195, 396
Furcellaria (Algae Rhodophyceae) 246, 247
Fusus (Mollusca Gastropoda) 267
G
Gadidae (Pisces) 391
Gadus morhua macrocephalus (Pisces) 414
-----morhua 414
— poutassou 336
— virens 402
Gaetanus (Crustacea Copepoda) 220
Gaidius (Crustacea Copepoda) 220
Galathea (Crustacea Decapoda) 223
Galaxiidae (Pisces) 391
Gammaridae (Crustacea Amphipoda) 210,
331, 332
Gammarus locusta (Crustacea Amphipoda)
253, 281, 403
— marinus 253
Gastropoda (Mollusca) 164—167, 253
Gastrotricha (Vermes) 166, 190
Gebia (Crustacea Decapoda) 223
Gelidiopsis (Algae Rhodophyceae) 246, 248
Gelidium (Algae Rhodophyceae) 247
Geortia (Pisces) 391
Gephyrea (Annelida) 164
Globigerina (Protozoa Foraminifera) 147,
208
— bulloides 165
— inflata 165
Gloiopeltis (Algae) 258
Glycera (Annelida Polychaeta) 430
Gnathia stygia (Crustacea Isopoda) 295
Gonothyrea (Coelenterata Hydrozoa) 253
Gordiacea (Nemathelminthes) 166
Gorgonaria (Coelenterata Anthozoa) 296,
400
Gorgonocephalus lincki (Echinodermata
Ophiuroidea) 295
Gracillaria (Algae Rhodophyceae) 246, 248
Grapsus grapsus (Crustacea Decapoda) 393
Graptolithoidea 176
• Gymnosomata (Bryozoa) 210
H
Halcompoides (Coelenterata Anthozoa) 283
Halichondria (Porifera Cornacuspongida)
258
Halicryptus spinulosus (Annelida Priapulo-
idea) 199, 254, 281
Ilalidrys siliquosa (Algae) 191, 278^
Halimeda (Algae Chlorophyceae) 195, 247,
395
Haliotis (Mollusca Gastropoda) 270, 422,
423
— giganteus 421
Halosydna insiguis 258
Haplochitonidae (Pisces) 391
Haploops (Crustacea Amphipoda) 261, 265
— tubicola 264, 266, 290
Haplophragmoides canariensis (Protozoa
Rhyzopoda) 291
Harmothoe derjugini (Annelida Polychaeta)
291, 292, 297
— imbricata 283
— impar 291, 297
Harpacticoida (Crustacea Copepoda) 190,
220
Heliometra (Echinodermata Crinoidea) 292
— glacialis 291
— maxima 291
Ileliopora coerulea (Coelenterata Antho-
zoa) 195
Ilelioporidae (Coelenterata Anthozoa) 396
Hemigrapsus nudus (Crustacea Decapoda)
258
Heterocontae (Algae) 201, 202
Heteromastus filiformis (Annelida Poly-
chaeta) 254
Heteropoda (Mollusca Gastropoda) 147,
209, 210
Heterorhabdus (Crustacea Copepoda) 220
Hexactinellidae (Porifera Triaxonida) 296
Hexapodinae (Crustacea Decapoda) 396
Himanthalia lorea (Algae Phaeophyceae)
191
Hippoglossus hippoglossus (Pisces) 376
------stenolepis 376
Hippopodius serratia (Coelenterata Sipho-
nophora) 373
Hippospongia canaliculata (Porifera .Cor-
nacuspongida) 430
— equina 430
Hirudinea (Annelida) 166
492
Holothuria atra (Echinodermata Holo-
thurioidea) 393
Ilolothurioidea (Echinodermata) 165, 166,
400
Homarus (Crustacea Decapoda) 68, 426
— americanus 425
— vulgaris 425, 426
Hormosira (Algae Phaeophyceae) 246, 248
Hvalinaecia fauveli (Annelida Polychaeta)
288
Hyalonema (Porifera Triaxonida) 296
Hyas araneus (Crustacea Decapoda) 253,
380—382
Hydrallmania (Coelenterata Hydrozoa) 430
Hydrobi a (Mollusca Gastropoda) 253
— ulvae 253, 254, 267
Hydrobiidae (Mollusca Gastropoda) 170
Hydrozoa (Coelenterata) 163—167, 395
Hyperammina fragilis (Protozoa Forami-
nifera) 291
Ilyperia galba (Crustacea Amphipoda) 373
Hyperiidae (Crustacea Amphipoda) 210,
328
Hypnea (Algae Rhodophyceae) 246, 248
laera marina (Crustacea Isopoda) 253
Idothea (Crustacea Isopoda) 265
— granulosa 253
Ilyocrinus carpenteri (Echinodermata Cri-
noidea) 295
Insecta 166, 169
Iridaea cordata (Algae Rhodophyceae) 390,
391
Isopoda (Crustacea) 164, 165, 170, 209, 315,
320, 371, 434
Isotomurus palustris (Insecta) 254
J
Jasmineira (Annelida Polychaeta) 291
— pacifica 291
К
Kinorhyncha (Vermes) 166
Krohnia hamata (Chaetognatha) 220
L
Labirinthula (Myxomycetes) 246
Laetmonice filicornis (Annelida Poly-
chaeta) 295
Lafoeina maxima (Coelenterata Hydrozoa)
291
Lamellibranchiata (Mollusca) 164—167,
242, 279
Lamellidoris (Mollusca Gastropoda) 253
Laminaria (Algae Phaeophyceae) 246, 286,
293, 431, 432
— bulbosa 191
— bullata 290
— clonstoni 248
— digitata 191, 248, 278, 432
— iberica 386
— japonica 290
— pallida 386
— saccharina 45, 191, 248, 290, 432
Laminariacea (Algae Phaeophyceae) 173,
387, 388
Laminariales (Algae Phaeophyceae) 375
Lampropidae (Crustacea Cumacea) 387
Laomedea (Coelenterata Hydrozoa) 253
Laurensia (Algae Rhodophyceae) 246—248,
' 258
Leander (Crustacea Decapoda) 424,426, 438
— adspersus 428
— squilla 428
Leda pernula (Mollusca Lamellibranchiata)
263, 277, 283
Lepas (Crustacea Cirripedia) 401
Lepeta coeca (Mollusca Gastropoda) 283
Leptasterias (Echinodermata Asteroidea)
260
Leptosynapta inhaerus (Echinodermata
Holothurioidea) 258, 292
Lessonia (Algae Phaeophyceae) 246—248
— laminariacoides 386, 391
— ovata 386
Leuconidae (Crustacea Cumacea) 387, 388
Leucosiinae (Crustacea Decapoda) 396
Ligia (Crustacea Isopoda) 249
— brandtii 257
— cinerescens 258
— italica 250
• — occidentalis 260
Lima (Mollusca Lamellibranchiata) 270
— fragilis 289
— inf lata 288
Limacina (Mollusca Gastropoda) 147
— helicina 389
Limapontia (Mollusca Gastropoda) 253
Limnoria iignorum (Crustacea Isopoda)
434
Limulus (Palaeostraca Xiphosura) 427
— molljicanus 395
Lineus (Nemertini) 254
Liparidae (Pisces) 296
Lithodes (Crustacea Decapoda) 427
Lithodidae (Crustacea Decapoda) 296
Lithoicea (Lichenes) 249
Lithophyllum antillarium (Algae Rhodo-
phyceae) 68
Lithothamnion (Algae Rhodophyceae) 146,
195, 246, 247, 395
— foecundum 366
Litiola (Mollusca Gastropoda) 401
Littorina (Mollusca Gastropoda) 249, 257,
265, 395, 422
— littorea 191, 253, 254, 282, 379, 422
— neritoides 191, 250, 257, 403
— obtusata 253
— palliata 253
— planaxis 260
— rudis 191, 253, 267
— scutulata 257, 292
— sitchana 257, 258, 292
— squalida 258
Loligo (Mollusca Cephalopoda) 422
— media 421
— opalescens 421
— pealei 421
— vulgaris 421
Lophophelia (Coelenterata Anthozoa) 270,
394
Lottia (Mollusca Gastropoda) 260
Loxechinus albus (Echinodermata Echino-
idea) 430
Lucernosa sant-hilairi (Coelenterata Scy-
phozoa) 374
493
Luidiaster (Echinodermata Asteroidea) 292
— tuberculatus 291
Lumbriconereis fragilis (Annelida Poly-
chaeta) 276, 281, 291, 295
Lycastis (Annelida Polychaeta) 258
Lycodes frigidus (Pisces) 295
Lygia italica (Crustacea Isopoda) 403
M
Macoma (Mollusca Lamellibranchiata) 253,
265, 282
— baltica 199, 253, 254, 258—263, 266,
269, 270, 282, 293, 313, 403
— calcarea 186, 187, 254, 261, 263, 270,
281—283, 290, 294
— inquianta 258, 292
— nasuta 258, 292
— secta 258, 292
Macrocallista chishimana (Mollusca Lamel-
libranchiata) 423
Macrocystis (Algae Phaeophyceae) 246—
248
— pyrifera 391
Macruridae (Pisces) 295, 296
Mactra (Mollusca Lamellibranchiata) 282,
422
— elliptica 270, 281
— stultorum 27 9—281
— subtruncata 270
Madrepora (Coelenterata Anthozoa) 195,
400
Madreporaria (Coelenterata Anthozoa) 195,
296
Mage lone longicornis (Annelida Polychae-
ta) 290
Maldane (Annelida Polychaeta) 282, 292
— globifex 288
— sarsi 276, 277, 282, 290, 389
Mallotus villosus (Pisces) 403
Mammalia 166, 167
Manayunkia (Annelida Polychaeta) 254
Margarita groenlandica (Mollusca Gastro-
poda) 284
— helicina 284
Marphysa bellii (Annelida Polychaeta) 288
Meganyctiphanes norvcgica (Crustacea Eu-
phausiacea) 402
Megaptera nodosa (Mammalia Cetacea) 417
Melinna cristata (Annelida Polychaeta)
276, 277, 281
Melitodidae (Coelenterata Anthozoa) 396
Melobesia (Algae Rhodophyceae) 246, 248
Membranipora (Bryozoa) 253, 401
Menippe (Crustacea Decapoda) 424, 426,
427
Mercenaria stimpsoni (Mollusca Lamelli-
branchiata) 423
Merethrix merethrix (Mollusca Lamellib-
ranchiata) 421
XIerluccius vulgaris (Pisces) 336
Merostomata (Arthropoda Chelicerata) 176
Mertensia ovum (Mollusca Lamellibranchi-
ata) 369
Mesidothea (Crustacea Isopoda) 380, 381
— entomon 380, 381, 429
— megalura 295, 380, 381
— sabini 380, 381
— sibirica 380
Metridia (CrustaceaCopepoda) 219, 220, 226
— longa 220, 221, 224, 227, 231, 389
— lucens 220
Metridium dianthus (Coelenterata Antho-
zoa) 253
Microcalanus pusillus (Crustacea Copepoda)
220
— pygmaeus 389
Microcosmus (Tunicata Ascidiae) 288, 430
— claudicans 373, 37 4
— exasperatus 373, 374
Microsetella atlantica (Crustacea Copepoda)
225
Microspira (Bacteria) 95
Milleporidae (Coelenterata Hydrozoa) 395
Mitella polymerus (Crustacea Cirripedia)
257, 260, 292
Modiola (Mollusca Lamellibranchiata) 282,
293
— barbata 286
— modiolus 261, 264, 266, 270, 281, 379
Modiolaria laevigata (Mollusca Lamelli-
branchiata) 284
Molgula (Tunicata Ascidiae) 258
Mollusca 69, 71, 212, 314, 336
Molva molva (Pisces) 402
Monorhaphis (Porifera Triaxonida) .296
Monostroma fuscum (Algae Chlorophyceae)
253
Mordacea (Pisces) 391
Mugil cephalus (Pisces) 393
Munida tenuimana (Crustacea Decapoda)
295
Muraenolepidae (Pisces) 391
Murex trunculus (Mollusca Lamellibran-
chiata) 286
Muricea (Coelenterata Anthozoa) 397
Mya (Mollusca Lamellibranchiata) 253
— arenaria 253, 254, 258, 262, 263, 270,
282, 313, 403, 422, 437
— truncata 281, 283, 284, 293
Myriapoda 166
Myriochele heeri (Annelida Polychaeta) 263,
281
Myriotrochus (Echinodermata Holothurio-
idea) 291
— mitsukui 290
Mvsidacea (Crustacea) 209, 226, 231, 315,
330—332
Mysis oculata relicta (Crustacea Mysidacea)
404
— relicta 404
Mytilaster (Mollusca Lamellibranchiata)
438
Mytilus (Mollusca Lamellibranchiata) 253,
*257, 278, 282, 293, 422
— californianus 257, 258, 260, 292, 421
— canaliculatus 421
— dunckeri 421—423
— edulis 182, 191, 199, 253, 254, 257—
259, 266, 267, 278, 281, 292, 313, 379,
389, 403, 421, 422
— galloprovincialis 257
— magellanicus 4 21
— undulatus 421
N
Nainereis (Annelida Polycnaeta) 254
Nannastacidae (Crustacea Cumacea) 387
494
Nassa (Mollusca Gastropoda) 267
— mammilla ta 286, 288
Natica alderi (Mollusca Gastropoda) 27 9,
281
Nautilus (Mollusca Cephalopoda) 396
Nobalia (Crustacea Leptostraca) 389
Nectocrangon dentata (Crustacea Decapoda)
291
Nemalion (Algae Rhodophyceae) 246
Nematoda (Nemathelminthes) 163—167
Nematoscelis megalops (Crustacea Euphau-
siacea) 222
— microps 222
— tenella 222
Nemertini 163—167
Neoplatycephalus macrodon (Pisces) 332,
333
Nephrops norvegicus (Crustacea Decapoda)
402, 426
Nephthys ciliata (Annelida Polychaeta)
281, 283
— coeca 279
— hombergi 259, 288
— incisa 276, 277
— longisetosa 281, 291
Nereis (Annelida Polychaeta) 258, 430
— diversicolor 254, 255, 267, 281
— pelagica 259
— virens 258, 292
— zonata 283
Nereocystis (Algae Phaeophyceae) 246—
248
Nitzschia closterium (Algae Diatomeae) 204,
334
Noctiluca miliaris (Protozoa Flagellata)
208
Notomastus latericeus (Annelida Polychae-
ta) 288
Nucula (Mollusca Lamellibranchiata) 265
— nitida 276, 277
— nucleus 276, 277, 286, 288
— sulcata 270
— tenuis 263, 276, 277, 283, 290
Nullipores (Algae Rhodophyceae) 395
О
Obelia (Coelenterata Hydrozoa) 292
Octopus (Mollusca Cephalopoda) 422
— hongkongensis 421
— punctatus 421
— tuberculatus 421
— vulgaris 421
Oculina (Coelenterata Anthozoa) 295
— diffusa 68
Odonthalia (Algae Rhodophyceae) 247
Oikopleura (Tunicata Appendiculariae)
220
Oithona atlantica (Crustacea Copepoda) 213
— plumifera 220
— similis 220, 225, 372
Oligochaeta (Annelida) 158, 166, 258, 259
Ommatostrephes (Mollusca Cephalopoda)
422
— sloanei pacificus 421
Oncaea conifera (Crustacea Copepoda) 220
— media 404
Oncorhynchus (Pisces) 438, 439
Oneirophanta (Echinodermata Ilolothuri
oidea) 296
Ophelia (Annelida Polychaeta) 254
Ophiactis abyssicola (Echinodermata Ophi-
uroidea) 295
Ophiocantha bidentata (Echinodermata
Ophiuroidea) 283, 381
Ophiocten (Echinodermata Ophiuroidea)
285
— sericeum 283, 294—296
Ophioglypha (Echinodermata Ophiuroidea)
261
— robusta 264
— sarsi 263
Ophiopholis aculeata (Echinodermata Ophi-
uroidea) 264, 270
Ophiopleura borealis (Echinodermata
Ophiuroidea) 283, 294
Ophiotrix fragilis (Echinodermata Ophi-
uroidea) 288
Ophipsila aranea (Echinodermata Ophiu-
roidea) 288
Ophiura albida (Echinodermata Ophiu-
roidea) 288
— leptoctenia 291
— robusta 284
— sarsi 291, 292
-----vadicola 290
Ophiuroidea (Echinodermata) 165, 166,
400
Orbitolites marginalis (Protozoa Foramini-
fera) 68
Orchestia (Crustacea Amphipoda) 258
— littorea 253
— mediterranea 403
Oregonia gracilis (Crustacea Decapoda) 382.
383
Oridia (Annelida Polychaeta) 258
Orthonectida (Mesozoa) 166
Osmeridae (Pisces) 391
Ostracoda (Crustacea) 163, 164, 190, 209.
220, 313, 330
Ostracodermi (Pisces) 176
Ostrea (Mollusca Lamellibranchiata) 422
— angulata 420, 439
— chiloensis 420
— cristata 420
— edulis 420, 439
— gigas 420, 439
— laperousi 420
— lurida 420
— plicata 420
— taurica 420
— virginica 420, 439
Oweniafusiforimis (Annelida Polvchaeta)
288
Oxyptila trux (Arachnoidea) 182
P
Pachvgrapsus marmoratus (Crustacea Deca-
poda) 250, 257, 403
Paguridae (Crustacea Decapoda) 223, 288
Pagurus bernhardus (Crustacea Decapoda)
253
— pubescens 253, 295
Palaemon (Crustacea Decapoda) 424, 426
Palaeostraca 166
Palinura (Crustacea Decapoda) 296
Palinurus (Crustacea Decapoda) 426
— argus 426
— interruptus 426
495"
Palinurus vulgaris 425
Panda lopsis (Crustacea Decapoda) 426
Pandalus (Crustacea Decapoda) 424, 426
— borealis 429
— hypsinotus 428
— kessleri 428
Panopea norvegica (Mollusca gastropoda)
403
Pan topoda 30, 164, 167
Paphia philippinarum (Mollusca Lamelli-
branchiata) 423
— staminea 258, 292
Paracalanus parvus (Crustacea Copepoda)
220, 225, 404
Paraeuchaeta glacialis (Crustacea Copepoda)
369
Paraliparis bathybii (Pisces) 295
Paralithodes camtschatica (Crustacea Deca-
poda) 419, 426
— platypus 426
Parapenaeus (Crustacea Decapoda) 426
Parasabella maculata (Annelida Polvchae-
ta) 258, 288, 293
Parengyroides (Tunicata Ascidiae) 292
— japonica 290
Patella (Mollusca Gastropoda) 249, 258
— lusitanica 250, 403
— pontica 257
— vulgata 253
Patiria (Porifera) 258
Pecten (Mollusca Lamellibranchiata) 261,
266, 420, 422, 423
— aequisulcatus 421
— frigidus 269, 270, 295, 296
— groenlandicus 283
— irradianus 421
— islandicus 284, 403
— jessoensis 421—423
— magellanicus 421
— maximus 421, 422
— randolfi 291, 292, 297
— septemradiatus 264
— undatus (fragilis) 296
— vitreus 264, 270
Pectinaria (Annelida Polychaeta) 285
— auricoma 276
— belgica 389
— granulata 283
— koreni 186, 276, 281
Pplagonemertes (Nemertini) 209
Pelonaja (Tunicata Ascidiae) 292
— corrugata 187, 290
Pelvetia canaliculata (Algae Phaeophyceae)
191, 253, 384, 385
— frisigiata 384
— galapagensis 384
— wrigntii 384
Penaeus (Crustacea Decapoda) 424, 426
Peniagone (Echinodermata Holothurio-
idea) 296
Pennatulacea (Coelenterata Anthozoa) 295
Pennatularia (Coelenterata Anthozoa) 190,
296
Pentacrinus (Echinodermata Crinoidea) 296
Peridineae (Protozoa Flagellata) 201, 202
Periophthalmus (Pisces) 395
Petrolisthes cinctipes (Coelenterata Antho-
zoa) 258
Petro my zonidae (Pisces) 391
Peysonella (Algae Rhodophyceae) 246—
248
Phacocystis (Protozoa Flagellata) 40, 322,
329, 330
— pouchetii 208, 329
Phaeophyceae (Algae) 246
Pharyngella (Protozoa Radiolaria) 220
Phascolosoma margaritaceum (Annelida
Sipunculida) 389
Pheronema carpenteri (Porifera Triaxonida)
295
Philine (Mollusca Gastropoda) 292
— aperta 242
Phocidae (Mammalia Pinnipedia) 375
Pholis gunellus (Pisces) 253
Phoronida 162, 163, 166
Phy Hi tis fascia (Algae Rhodophyceae) 366
Phyllophora (Algae Rhodophyceae) 246,
431
— nervosa 165
Phyllospadix (Gynoeciatae Angiospermae)
246, 431, 432
— scouleri 431
Physcosoma agassizi (Annelida Sipunculida)
258
Phvsophorae (Coelenterata Siphonophora)
210
Physeter macrocephalus (Mammalia Ceta-
cea) 417
Pisaster (Echinodermata Asteroidea) 260
— ochraceus 258
Pisces 164, 166, 167, 314
Plagusia depressa (Crustacea Decapoda) 393
Planes minutus (Crustacea Decapoda) 393,
401
Platycephalidae (Pisces) 332
Pleuromamma abdominalis (Crustacea Co-
pepoda) 221
Pleuronectes plalessa (Pisces) 402
Pododesmus macroschisma (Mollusca Gas-
tropoda) 292
Podon leukarti (Crustacea Cladocera) 220,
389
Pogonophora 163, 166
Pollicipes cornucopia (Crustacea Cirri-
pedia) 424
Polychaeta (Annelida) 163—167, 210, 212,
243, 331, 336
Polycirrus eous (Annelida Polychaeta) 291
Polyclada (Plathelminthes, Turbellaria)
315
Polydectinae (Crustacea Decapoda) 396
Polydora (Annelida Polychaeta) 258
— coeca 290
— quadrilobata 254
Polysiphonia (Algae Rhodophyceae)
246,
366
— violacea 367
Pontaster (Echinodermata Asteroidea) 295
— tenuispinus 296
Pontella atlantica (Crustacea Copepoda)
369
Pontellidae (Crustacea Copepoda) 212
. Pontophilus norwegicus (Crustacea Deca-
poda) 295
Pontoporeia (Crustacea Amphipoda). 282
— femorata 281, 282
Pontosphaera huxleyi (Protozoa Flagella-
ta) 165, 343, 344
496
Porcullanhlae (Crustacea Decapoda) 400
Porcupinia (Protozoa Radiolaria) 220
Porifera 69, 163—167, 176, 327
Pori les (Coelenterata Anthozoa) 195
Porospathis (Protozoa Radiolaria) 220
Porpliyra (Algae Rhodophyceae) 246, 247,
431, 432
Portlandia arctica (Mollusca Lamelli-
branchiata) 269,270, 283, 299, 368,378,
379
— frigida 283
— intermedia 283
— lenticula 187, 283
Portunus (Crustacea Decapoda) 401
— pelagic us 396
Posidonia (Algae Phaeophyceae) 403
Potamides (Mollusca Gastropoda) 395
Potamobius (Crustacea Decapoda) 424
Pourtalesia jeffreysi (Echinodermata Echi-
noidea) 295
Priapulida (Annelida) 162, 163, 166
Priapulus (Annelida Priapulida) 255, 387
— caudatus 199, 254, 283, 387, 389
— tuberculato-spi nosus 387
Primnoa (Coelenterata Anthozoa) 292
— resedaeformis pacifica 291
Prionospio malmgreni (Annelida Polychae-
ta) 281, 282
Protocystis harstoni (Protozoa Radiola-
ria) 389
— sloggetti 389
Protophyta 218
Prototroctes maraena (Pisces) 332
Protozoa 69, 164
Pro trachea ta 166
Psammobia faeroensis (Mollusca Lamelli-
branchiata) 270
— tellinella 281
Psammobiidae (Mollusca Lamellibranchi-
ata) 395
Pseudocalanus (Crustacea Copepoda) 226
— elongatus 220, 329
Pseudocucumis (Echinodermata Ilolo-
thurioidea) 399
Psilaster andromeda (Echinodermata Aste-
roidea) 295
Psolus chitionoides (Echinodermata llolo-
thurioidea) 292
Pte raster multipes (Echinodermata Aste-
roidea) 295
Pterobranchia 166
Pteroides griseus (Mollusca) 286
Pteropoda (Mollusca Gastropoda) 147, 209,
330, 333
Ptilota (Algae Rhodophyceae) 246, 247
Pulmonata (Mollusca Gastropoda) 169
Purpura (Mollusca Gastropoda) 257, 258
— lapillus 191, 253, 403
Pygospio (Annelida Polychaeta) 253, 257
— elegans 199, 254, 259
Pyrosomidae (Tunicata Ascidiae) 210
R
Radiolaria (Protozoa) 71, 163, 327
Ralfsia (Algae Rhodophyceae) 257
Reptilia 166
Retro pi nuidae (Pisces) 391
Rhabdammina (Protozoa Fora mi nif era) 291
Rhinocalanus (Crustacea Copepoda) 373
— cornutus L atlantica 373
— gigas 373
— nasutus 373
Rhizoclonium (Algae Chlorophyceae) 258
Rhizocrinus (Echinodermata Cri noidea)
296
— lofotensis 295
Rhizophora (Coelenterata Siphonophora)
395
Rhizopoda (Protozoa) 166, 167
Rhi zosolenia (Algae Diatomeae) 438
— calcar-avis 40, 176
— styliformis 329
Rhodichthys regina (Pisces) 295
Rhodine loveni (Annelida Polychaeta) 276,
277
Rhodophyceae (Algae) 246, 247, 260
Rhodimenia (Algae Rhodophyceae) 247
Rhopilema esculenta (Coelenterata Scy-
phozoa) 430
Rissoa (Mollusca Gastropoda) 191, 265
— aculeus 253
Rivularia (Algap Cyanophyceae) 258
Rossia macrosoma (Mollusca Cephalopoda)
421
Rotatoria 166
Ruppia (Gynoeciatae, Angiospermae) 246
S
Sagitta elegans (Chaetognatha) 220, 231
Salenia hastigera (Echinodermata Echino-
idea) 296
Salmonoidei (Pisces) 391
Salpa (Tunicata Salpae) 166
Salpae (Tunicata) 209
Salpidae (Tunicata Salpae) 210
Sardina (Pisces) 386
— pilchardus 386
Sardinops (Pisces) 386
— sagax 386, 414
-----melanostica 414
-----sagax 414
Sargassum (Alcrae Phaeophyceae) 246—
248, 376, 387, 401
Saxicava (Mollusca Lamellibranchiata) 293
— arctica 258, 283, 284, 293, 402
Scalibregma inf la turn (Annelida Poly-
chaeta) 186, 291
Scalpellum (Crustacea Cirripedia) 71
Scaphopoda (Mollusca) 166
Schizaster (Echinodermata Echinoidea)
289
— canaiiferus 288
— fragilis 187
Schizophyceae (Algae Cyanophyceae) 341
Schizopoda (Crustacea) 164, 307, 329
Scinaia furcelata (Algae Rhodophyceae)
366, 367
Sclerocrangon salebrosa (Crustacea Deca-
poda) 428
Scolecithrix (Crustacea Copepoda) 220
Scolecolepis (Annelida Polychaeta) 257
Scoloplos (Annelida Polychaeta) 257, 292
— armiger 186, 254, 259, 276, 277, 290
Scopimerinae (Crustacea Decapoda) 396
Scrobicularia (Mollusca Lamellibranchia-
ta) 253, 257
— plana 254
it
32 Зенкевич, т. 1
497
Scyphozoa (Coelenterata) 163, 164, 1GG
167, 313
Scytosiphon (Algae Phaeophyceae) 24 9,
257
Sepia officinalis (Mollusca Cephalopoda)
421
Sepiella japonjca (Mollusca Cephalopoda)
421
Sepiola rondeleti (Mollusca Cephalopoda)
421
Septibranchiata (Mollusca La me Hi branchi-
ata) 320
Sertularia (Coelenterata Ihdrozoa) 253,
430
Sesarma (Crustacea Decapoda) 395
Silicoflagellata (Protozoa Flagellata) 147,
201, 202
Siliqua pa tula (Mollusca La nielli branchi-
ate) 259, 422
Sipunculida (Annelida) 163, 166
Sinovacula constricta (Mollusca Lammeli-
branchiata) 421
Skenea planorbis (Mollusca Gastropoda) 253
Smittina foliacea (Mollusca Lamellibran-
chiata) 285
Solariella (Mollusca Gastropoda) 292
— varicosa 290
Solaster papposus (Echinodermata Astero-
idea) 284
Solen (Mollusca Lamellibranchiata) 37,
292
— pellucidus 267, 274
Solenidae (Mollusca Lamellibranchiata) 395
Solenogastres (Mollusca) 166
Sosane gracilis (Annelida Polychaeta) 299
Spatangus purpureus (Echinodermata
E chi noidea) 270, 276, 277
— rasehi 295
Sphagnum fuscum (Arch^goniata Brvophv-
ta) 183
Spiochaetopterus (Annelida Polychaeta) 2'i2
— typicus 187
Spirontocaris linguis (Crustacea Decapo-
da) 291
Spirorbis (Annelida Polychaeta) 242, 253,
401
Spisula elliptica (Mollusca Lamellibran-
chiata) 28 2
— grajana 4 23
— sachaJinensis 422, 4 23
— subtruncata 279- 281
Spongia (Porifera Cornacuspongida) 16G
— graminea 430
Squamaria (Algae Rhodophyceae) 246, 248
Squilla manthis (Crustacea Stomatopoda)
426
Staphylinidae (Insecta) 250, 258
Stegophiura (Echinoidea Ophiuroidea) 292
— brachiactis 290
— nodosa 290
Stephanasterias albula (Echinodermata As-
teroidea) 284
Stephanotrochus (Coelenterata Anthozoa)
295
Sternaspis scutata (Annelida Polychaeta)
288
Stevelina (Protozoa Rhyzopoda) 291
Stichopus (Echinodermata Holothurioidea)
395
Stichopus californiciN 292
— japonicus 430
Stoma to pod a (Crustacea) 320. 426
Strombus (Mollusca Gastropoda) 4 22
St rongylocentrotus droebachiensis (Echi-
nodermata Echinoidea) 283, 284, 292
— - luridus 4 30
— franciscarnis 292
Strophogorgia (Coelenterata Anthozoa) 295
Styela atlantica (Tunicata Ascidiae) 377
Stylocherion longicorne (Crustacea Euphau-
siacea) 222
— maximus 222
Sue to ria (Protozoa Infusoria) 166
Svndesmya (Mollusca Lamellibranchiata)
* 150
— alba 288
- fragilis 288
nitida 276, 286
- ovata 44, 294, 313
— prismatica 276, 277
T
Tachyplens (Palaeostraca Xiphosura) 396
Tali tri dae (Crustacea Amphipoda) 249,
' 250, 258
Talitrus locusta (Crustacea Amphipoda) 403
Tanaidacea (Crustacea) 297
Tapes auratus (Mollusca Lamellibranchiata)
286, 288
— decussa tus 421
— philippinarum 421, 4 25
Tardigrada 166
Teleostei (Pisces) 176
Те Ilina distorta (Mollusca Lamellibran-
chiata) 288, 289
— tabula 263. 270
Temora (Crustacea Copepoda) 328, 329
— longicornis 329
— sty I if era 404
Tere be Hides stroemi (Annelida Polvchae-
ta) 276, 277, 389
Terebra tulidae (Brachiopoda) 270
'I’erebratula coreanica (Brachiopoda) 291
Teredinidae (Mollusca Lamellibranchiata)
434
Teredo megotara (Mollusca Lamellibran-
chiata) 434
— navalis 434
— norvegica 434
— sinensis 434
- — utriculus 434
Thais (Mollusca Gastropoda) 258
— emarginata 258
Thalassema (Annelida Echiuroidea) 315
Thalassina (Crustacea Decapoda) 395
Thallophyta 157
.Thecosomata (Bryozoa) 210
Thelaceros rhizophorae (Coelenterata An-
thozoa) 395
Thelmessus cheiragonus (Crustacea Deca-
poda) 428
Themisto abyssorum (Crustacea Amphi-
poda) 382, 383
— compressa 382, 383
— libellula 382
Theodoxus (Mollusca Gastropoda) 314
Thoumatometra (Echinodermata Cri noidea)
292
498
Thoumatometra tenuis 29
Thyasira sarsi (Mollusca Lamellibranchiata)
186, 281
Thyone briarens (Echinodermata Holo-
thurioidea) 430
Thysanoessa gregaria (Crustacea Euphau-
siacea) 222, 389
— inermis 308
— longicaudata 222
— parva 222
Thysanopoda aequalis (Crustacea Euphau-
siacea) 222
— orientalis 222
Tintinnoidea (Protozoa Infusoria) 209
Tivela (Mollusca Lamellibrahciata) 422
— stultorium 259
Tmetonvx cicada (Crustacea Amphipoda)
291, 297
Todarodes sagittatus (Mollusca Cepha-
lopoda) 421
— verangi 421
Tomopteridae (Annelida Polychaeta) 210,
212
Tomopterisseptontrionales (Annelida Poly-
chaeta) 389
Travisia (Annelida Polychaeta) 254
— forbesi 290, 291
Triaxonida (Porifera) 71
Trichodesmium (Algae Cyanophyceae) 341
Trichotropis cancellata (Mollusca Gastro-
poda) 292
Triglidae (Pisces) 30
Trilobita 176
Trophonia plumosa (Annelida Polychaeta)
265
Tubifex (Annelida Oligochaeta) 48
Tubiporidae (Coelenterata Anthozoa) 395,
396
Tunicata 164, 167, 212
Turbo Ila ria (Plathelminthes) 160, 164,190
Turbinaria (Algae Phaeophyceae) 246—248
Turbo cornutus (Mollusca Gastropoda) 4 21
— sangarensis 258
Turitella (Mollusca Gastropoda) 265, 266,
289, 292
— communis 276—278
— fortilirata 290
— terebra 263, 286
Turris (Mollusca Gastropoda) 223
Tuscarora (Protozoa Radiolaria) 220
Tylaster (Echinodermata Asteroidea) 295
Tvphlotanais (Crustecea Isopoda) 291,297
U
Uca (Crustacea Decapoda) 395
Udotea (Algae Rhodophyceae) 246, 277
Ulva (Algae Chlorophyceae) 246, 248,
431
— lactuca 366, 432
Umbellula (Coelenterata Anthozoa) 295
— encrinus 295, 296
— lindahli 296
Undinella оblonga (Crustacea Copepoda)
220, 369
Upogebia littoralis (Crustacea Decapoda)
288
Urticina crassicornis (Coelenterata Antho-
zoa) 253
Vallonia pulchella (Mollusca Gastropoda)
185
Velella spirans (Coelenterata Siphonophora)
224
Venus (Mollusca Lamellibranchiata) 258,
261, 265, 281, 292, 422
— fluctuosa 283, 288, 290, 293
— gallina 263, 264, 266, 269, 270, 282,
286, 403
Vermes 69, 176
Verruca ria (Lichenes) 249
Vertebra ta 177
Verticardia nadina (Mollusca Lamelli-
branchiata) 291
Virgularia (Coelenterata Anthozoa) 315
W
Waldheimia septata (Brachiopoda) 295
X
Xenophyophora (Protozoa Rhizopoda) 71
Xiphister mucosus (Pisces) 258
Xiphozura (Palaeostraca) 162
Yoldia (Mollusca Lamellibranchiata) 282
— hyperborea 281, 282
— johanni 290
Yoldiella (Mollusca Lamellibranchiata) 292
— derjugini 290
Z
Zoarces viviparus (Pisces) 253, 272
Zoarcidae (Pisces) 296
Zembos arc tic us (Crustacea Amphipoda)
290
Zoroaster fulgens (Echinodermata Aster-
oidea) 295
Zostera (Gynoecia tae Angiospermae) 150,
246, 247, 292, 431
— marina 191, 246, 431
— nana 246, 731
32*
УКАЗАТЕЛЬ АВТОРОВ
Русски й
Агассиц 17, 18, 342
Агеноров 130
Аллен 208, 334
Альм 37
Андрияшев 317, 361, 370, 377, 378
Андрусов 17
Аппелеф 192, 195, 196, 295, 362, 363
Арнольди 249
Аткинс 47, 206
Аттербек 99
Аугенер 362
Ауривиллиус 192
Бальсе 402, 403
Баскина 182, 183, 185
Беклемишев 178, 181—183, 186
Беллинсгаузен 15
Бен-Дауэс 324
Берг 168, 355, 376, 378, 379, 385, 389—
391
Березкин 118
Беринг 15
Бертель 302
Биб 99, 277, 401
Бидже л оу 31
Билингс 15
Бирштейн 4, 65—67, 275
Блевад 242, 245, 270—275, 321—323
Блинов 65
Богоров4,42, 43, 200, 201, 224, 227—238,
240, 242, 275, 321, 323, 346, 347, 434
Бойсен-Иенсен 37, 38, 42, 43, 150, 241,
245, 246, 267, 271, 322
Бок 342
Бокова 324
Болдовский 240
Болтовской — см. Морду хай-Болтов-
ской
Борупкий 38, 43, 45
Брандт 37, 105—107, 200, 305, 318, 323
Бродский 38, 219
Брокман-Ерош 185
Брох 186, 187, 279, 379, 383
Броцкая 4, 34, 38, 39, 43, 44, 183, 184,
187, 243, 244, 323, 332, 335, 351,353, 357
Бруевич 47, 48, 152, 206
Брэди 259
Брюс 252
Буткевич 106, 305
Бьюкенен-Уолластон 27 9
Бэр 15, 16, 18
Вагнер 23
Ваксман 206, 301, 302
Вальтер 31, 32, 178, 321, 342
Варминг 195
Ватова 201, 284—288, 367
Ватсон 168
Баттенберг 93, 96, 106, 107,110,111,119,
125, 143
Ваттерман 226
Вебер 18
Вегенер 376
Ведрипский 431
Везенберг-Лунд 195, 254
Великангас 313, 318
Вержбинская 47, 206
Верилль 192
Вернадский 37, 62, 65, 67, 155—158, 171
Вильямс 37, 105
Винберг 38, 47, 206
Виноградов 62, 64, 67—71, 361, 428
Воган 31, 195
Водяпицкая-Морозова — см. Морозова-
Во дяницка я
Вознесенский 16
Воленберг 195, 254—256
Воробьев 42, 43, 45, 180, 184, 185, 314,
322, 348, 357, 423
Воронков 234
Воскресенский 36
Вэйан 191
Вюст 53, 71, 73, 76, 84, 130, 131
Гавеши 170
Гагмейер 177, 275
Гаевская 48
Гайл 234, 432
Гальцов 439
Гамулин 214, 215, 404
Гарвей 31, 105, 107, 112, 114, 204—207,
304, 334
Гарден 207
Гардер 102
Гарди 205, 230, 232, 240, 322, 327—329
Гардинер 114, 192, 228, 229
Гармс 260
Гарстанг 233
Гартмейер 196
500
Га узе 181
Гейльприп 191
Гек 177
Геккель 33, 34, 339
Гексли 28
Геллянд-Ганзен 124
Гемлей 191
Темп 431
Гензен 17, 32, 200, 213, 241, 245, 343, 345
Гентшель 111, 116, 143, 160, 203, 218,
345, 346, 348, 350, 363, 401
Гердман 245, 322
Герт 205
Герненштейн 191, 196, 197
Гессе 31, 340
Гессле 275
Гесснер 318
Гетчинсон 48, 49, 439
Гефлер 183
Гиклинг 322, 330, 336
1 ирмер 173
Гмслен 42, 43, 103, 177, 186, 197, 198,
244, 275, 277, 278
Горбунов 355
Гордеев 241, 243
Гордеева 357
Горшкова 152
Гофстен 19Г, 362, 370, 380, 381
Гран 47, 206, 207, 234,348, 375
Гримм 16
Гров 168, 245
Гукер 16, 385
Гунтер 205, 230, 240, 342
Гурвич 243
Гурьянова 30, 194, 295, 355, 356, 361,
370, 371, 380
Гуус 374, 377
Гюнтер 342, 375
Давидсон 439
Давиташвили 173, 174
Даль 217, 245, 340, 342
Дама 345
Дамас 219, 221, 34 5
Дана 192, 340, 341, 375, 385, 389
Данилевский 16
Данневич 116
Дарвин 15, 192, 255, 341, 375, 390, 436
Дебошам 194
Девенпор 192
Демоль 37, 38, 42—44, 47
Державин 322
Дерюгин 19, 28, 32, 79, 96, 123, 178, 179,
193—199, 257, 290—294, 297,362,385,
388—390
Дефапт 77, 124, 125, 127, 134, 135, 137,
139
Дехтерева 335
Джесперсен 214, 287
Джонсон 31, 188, 198 , 199
Дя$онстон 194, 195, 234, 245
Дикон 125, 133, 134
Докучаев 37
Долло. 389
Донати 32, 340
Дорн 23
Дригальский 18
Дрью 302
Дэвис 279—281
Дэкип 230, 232, 332
Дэли 36, 56
Дюри 183
Дьяконов 30, 355, 381
Ерцегович 114, 201, 214, 215
Жадин 38, 41, 42
Жеффрейз 16
Жуков 107
Закс 194, 250, 257, 288, 290, 293, 4 27
Зацепин 322
Зенге р 191
Зенкевич 34, 38, 39, 43, 44, 96, 149, 160,
161, 172, 183, 184, 187, 236, 243, 244 ,
265, 275, 288, 310, 322—326, 351—353,
367
Зернов 28, 32, 34, 35, 64, 179, 189, 191,
193, 196, 197, 217, 233, 250, 318
Знаменский 427
Зубов 64, 120—122, 131, 132, 236
Зюсс 155
Иванов, II. 243
Иванов, Л. 103, 104
Ивлев 48, 324
Идельсон 238
Йенсен 245, 295
Иогансен 245, 310, 312
Ионг 20, 320, 346
Порт 31, 89, 90, 333
Исаченко 305
Иселин 120
Иссель 214
Казаков 151
Калитин 104
Карзинкин 38, 39, 43, 44, 324
Карпевич 324
Карпентер 16
Карпов 423
Кашкаров 34, 178
Кетчум 204
Кимбел 99
Кинг 194
Кинкейд 431
Киреева 45
Кирсоп 243, 292
Кларк 89—92, 99—103, 226, 230, 231, 400
Клеве 348, 374, 375
Клементс 69, 183, 345
Кленова 145, 150, 151
Книпович 18, 191, 316—318, 429
Кнюдсен 62, 241—243
Кобякова 291
Ковальский 22, 23
Кожевников 191
Кольбе 313
Коротнев 28
Коссина 54, 55
Кост 22
Костычев 37
Коцебу 15
Крамп 278
Крашенинников 15
Крепе 47, 206
Кречман 234
Крог 114—116
Кросслепд 369
501
Крузенштерн 15
Ку 369
Куделин 30
Кук 15
Куличеико 357
Купер 111, 206
Курсанов 157
Кусморская 238
Куэнен 36, 56
Кэри 277
Кэфэд 219, 221
Лазарев 15
Лаказ-Дютье 23
Лами 194
Ларсен 245
Лаури 281
Леб 231
Лебединцев 17
Лебур 230, 327
Ле-Дануа 124—126
Леннберг 192
Либих 105
Ливит 219, 222
Лип деман 48, 4 9
Линдрот 243, 2'1 \
Линке 177, 183, 195. 250- 256
Линко 30
Лисянский 15
Литке 15
Личков 65, 173, 174
Ловен 17, 190, 382,. 384
Локар 296
Ломан 200, 208, 213, 218, 233, 234, 343,
344
Лоренц 32, 178, 191, 404
Лотка 53
Лунд 245
Лысенко 37, 233
Люкш 87
Л юн д бе к 37
Мадсен 281, 283
Майор 278
Макаров 17, 30, 120, 291
Малятский 188
Мантейфель 234, 239. 240
Марион 22, 191, 321
Марсильи 32, 340
Мартенс 296
Маршалл 206, 207, 216, 230, 232
Мебиус 31, 32, 178
Мейер 138, 139, 323, 432
Мейнардус 140
Мерк 15
Меррей 28, 31, 89,90, 145, 146,195,298,
389
Месяцев 19, 20, 243, 244
Миддендорф 16
Микимото 425
Миклуха-Маклай 23
Миланкович 124
Милашевич 30
Мильн-Эдвардс 17, 32, 190, 277, 340
Митчерлих 105
Михаельсен 296, 316
Мичурин 437
Мозер 372
Моландер 199, 275—277
Мордухай-Болтовской 184, 185, 243
Морозова-Водяницкая 45, 185
Мортенсен 195, 245, 254
Мосевич 378
Муравейский 34, 35, 38—41
Мэр 303
Мэрфи 306, 307
Мюллер 32, 340
Мюнстер-Стрём 37—40, 43, 44
Нансен 122, 124, 137, 139
Науман 37, 41, 42
Пид гем 169
Никитин 224, 233, 293, 423, 428, 433
Пикольс 227
Никольский 322, 357, 358
Нинбург 366, 367
Норденшельд 17, 18, 192
Ньюкомб 195
Овидий 433
Однер 195
Оду эн 17, 32, 190, 340
Окснер 391, 404
Окул 42
Оливер 195
Опарин 171
Остроумов 191
Орр 206, 207
Ортман 31, 296, 342, 347, 353, 359, 360—
362, 370, 390
Панникар 315
Паннинг 393
Паркер 188
Натон 232
Паульсен 245
Пеггот 152
Перселл 150
Персидский 175, 176
Песта 214
Петерсен 28, 32, 33, 37, 177, 241—243,
246, 261—272, 275, 322, 348—350,
353, 357
Петтерссон 102, 124, 152, 277
Петр Великий 15
Пирс 188
Попов 427
Поульсеп 332
Пратье 147
Пробатов 317
Прюво 32, 191, 193, 250, 403
Пу рта ле 17
Пфеффер 183, 342, 389
Пюттер 47, 116
Разин 422, 423
Расс 353, 409—411, 413, 415—418, 424
Редеке 318
Редикорцев 30
Редфильд 206
Ремане 310, 312, 314, 318, 350
Ренар 145
Рессел 194, 208, 210, 215, 221, 223, 225,
231—233, 346, 416
Робертсон 68
Роджерс 68 *
Ромер 168
Росс 15, 16, 385, 390
Россолимо 38
Рууд 333, 334, 346
502
Рэй-Ланкестер 28
Рэлей 206, 207
Рэтбеп 398, 400
Савилов 278
Самойлов 97, 150
Саразинов 316
Саре, Г. 17, 342, 402
Саре, М. 17, 32, 144, 190, 340
Сарычев 15
Саутерн 162, 194
Сведберг 182
Свердруп 3, 31, 56, 58, 64, 68, 77, 79,
83, 86, 101, 108, 114, 126—136, 139,
148, 188, 199, 204
Светловидов 30
Световидов 386
Сегерштрэле 275
Сейвелл 120, 125—127, 206
Семпер 342
Сернапдер 249
Скотсберг 391
Совинский 28
Сомова 290, 292
Соот-Риен 278, 279
Синее 308
Спэрк 102, 183, 185, 245, 275, 281—288,
349, 350, 353, 367
Стеггерда 188
Стеллер 15
Стефен 281
Стиаснп 389
Стукеборг 192
Сэведж 240, 322, 329
Съюэл 368, 369
Тамдруп 195, 245, 252, 254, 256
Таранец 419, 423, 428, 431, 432
Тарасов 433
Таулер 195
Тиле 296
Тиль 245, 389, 400
Тиман 312, 313
Тинеман 37, 38, 41
Тодд 334—336
Толль 18
Томпсон 114
Томсон 3, 16, 112, 296, 385
Торсон 245, 253, 265, 266, 275, 281,
283—288
Траск 150, 151
Тресслер 31
Трофимов 102
Туле 152
Тулин 275
Уда 218
Уимпенни 392
Ульянин 191
Уолластон 279
Уолтон 194
Усачев 43, 201, 202
Ушаков 194, 361, 370
Уэткин 281
Фельтем 304
Ферран 194
Ферсман 170
Фидлер 419
Филатова 185, 350, 353
Финк 195
Фифер 114
Флаттели 194
Флеминг 3,31, 111,
Ф01Л 190, 284
Фольц 86, 404
Форбс 17, 32, 178, 190, 320, 321, 340,
385, 403, 404
Форд 279
Фоулер 217, 221
Францев 150
Фрейман 375
Фридерикс 177
Фридман 182, 183, 185
Фрич 390
Хаджи 392
Хворостянский 191
Хедлей 195
Хун 18
Хьювотт 195
Циммер 387, 388
Цо-Белл 31, 301—305
Чельмап 191, 196, 197
Чугунов 313, 322
Чэпмен 248
Шелфорд 177, 183, 195, 291—293
Шепард 56, 151
Шименц 321
Шимкевич 30
Ширшов 47, 234, 240, 434
Шлыков 436
Шмидт 18, 173, 348, 355, 361, 376, 399
Шокальский 57, 73, 74, 81, 82
Шорыгин 42, 43, 320—323, 351
Шотт 72, 87, 88, 140, 151, 152, 400
Шпарлинский 423
Шренк 16
Штамме р 404
Штейер 31, 200, 214, 233, 347, 348, 353,
362, 369
Штейниц 402
Штот 308
Штудер 192
Шухерт 171, 394
Щапова 45, 163, 246, 361, 3G3, 371, 376,
384—388, 391
Экман 3, 31, 37, 73, 197, 241, 296—298,
340, 343, 353, 355, 363, 370, 372, 373,
376—378, 380, 382—385, 389, 391—394,
396—399, 402, 403, 405, 406
Элмхирст 194
Элтон 49, 177
Эмери 56, 151
Энгель 399
Энгельгардт 170
Эренберг 192
Эстерли 231—233
Эттингаузен 389
Яшнов 42, 43, 201, 219, 234, 238
503
Иностранный
Agassiz 17, 18, 342
Allen 208, 334
Alm 37
Alsterberg 255, 256
Appelloff 192, 195, 196, 295, 362, 363
Apstein 234
Atkins 47, 206
Audouin 17, 32, 190, 340
Aurivillius 192
Bacesco 315
Balss 402, 403
Beebe 99, 277, 401
Ben Dawes 324
Bertel 302
Bigelow 31
Blegvad 242, 270—275, 321—323
Boeck 342
Boysen-Jensen 37, 38, 42, 43, 150, 241,
245, 246, 267, 271, 322
Braarud 234
Brady 259
Brandt 37, 105—107, 200, 305, 318, 323
Broch 186, 187, 279, 379, 383
Brook 322
Buchanan — Wollaston 279
Caldervood 322
Carpenter 16
Cary 277
Chadwick 234
Chun 18
Clarke 89—92, 99—103, 226, 230, 231,400
Clements 69, 183, 345
Coe 369
Cooper 111, 206
Coste 22
Crossland 369
Dahl 217, 245, 340, 342
Dakin 230, 232, 332
Daly 36, 56
Damas 219, 221, 345
Dana 192, 340, 341, 375, 385, 389
Davenport 192
Davidson 439
Davis 69, 279—281
De-Beauchamp 134
Deichmann 400
Delff 323
Demoll 37, 38, 42—44, 47
Dietz 151
Doderlein 400
Dollfus 194
Dollo 389
Donati 32, 340
Dorn 23
Drew 302
Dury 183
Ehrenberg 192
Ekman 3, 31, 37, 73, 197, 241, 296—298,
340, 343, 353, 355, 363, 370, 372, 373,
376—378, 380, 382—385, 389, 391, 394,
396—399, 402, 403, 405, 406
Elmhirst 194
Elton 49, 177
Emery 56, 151
504
Ercegovic 114, 201, 214, 215
Esterly 231—233
Ettinhausen 389
Farran 194
Fiedler 419
Finck 195
Flattely 194
Fleming 3, 31, 111
Fogt 190, 284
Forbes 17, 32, 178, 190, 320, 321, 340,
385, 403, 404
Ford 279
Fowler 217, 221
Franz 231
Friederichs 177
Gaarder 102
Galtsoff 439
Gamulin 214, 215, 404
Gardiner 114, 192, 228, 229
Garstang 233
Geng 323
Gislen 42, 43, 103, 177, 186, 197, 198,
244, 275, 277, 278,
Gran 375
Grove 168, 245
Gunther 205, 230, 240, 342, 375
Hadzi 392
Haeckel 33, 34, 339
Haecker 220
Hagmeier 177, 275
Hanley 191
Hardy 205, 230, 232, 240, 322, 327—329
Harms 260
Hart 205
Harvey 31, 105, 107, 112, 114, 204—207,
304, 334
Heck 177
Hedley 195
Heilprin 191
Hensen 17, 32, 200, 213, 241, 245, 343,
345
Hentschel 111, 116, 143, 160, 203, 218,
345, 346, 348, 350, 363, 401
Hesse 31, 340
Hessle 275
Hewatt 195, 230
Hickling 322, 330, 336
Hjort 31, 195, 333
Hofler 183
llofsten 195, 362, 370, 380, 381
Hooker 16, 385
Hutchinson 48, 69, 439
Huus 374, 377
Huxley 28
Issel 214
Jeffreys 16
Jenkin 208
Jensen 24 5, 295
Jespersen 214, 287
Johansen 245, 310, 312
Johnson 31, 188, 198, 199
Johnstone 194, 195, 234, 245
Jowett 69
Ketchum 20
Kimball 99
King 194
Kirsop 2*3, 292
К jellman 191
Knauthe 323, 324
Knudsen 62, 241—243
Koefoed 219, 221
Kolbe 313
Kossina 54, 55
Kraeft 234
Kramp 278
Krogh 114—116
Kroyer 321
Kuenen 36, 56
Kiikenthal 192
Labbe 194
Lacaze-Duthier 23
Lami 194
Lankester 28
Lawrie 281
Lebour 230, 327
Lindeman 48, 49
Lindroth 243, 244
Linke 177, 183, 195, 250—256
Locard 296
Loeb 231
Lohmann 200, 208, 213, 218, 233, 234,
343, 344
Lonnberg 192
Lorenz 32, 178, 191, 404
Loven 17, 190, 382, 384
Lu nd bee к 37
Mac Grinitie 258
Madsen 281, 283
Mann 323
Mare 303
Marion 22, 191, 321
Marshall 206, 207, 216, 230, 232
Marsigli 32, 340
Martens 296
Masters 69
Mayor 278
McCance 69
Meyer 138, 139, 323, 432
Michaelsen 296, 316
Milne-Edwards 17, 32, 190, 277, 340
Moebius 31, 32, 178
Molander 199, 275—277
Mortensen 195, 245, 254
Muller 32, 340
Miinster-Srtom 37—40, 43, 44
Murray 28, 31, 195, 298, 389
Naumann 37, 41, 42
Nelson 268
Newcombe 195
Nicholls 227
Nidham 169
Nielsen 205
Odhner 195
Oliver 195
Orr 206, 207
Ortmann 31, 296, 342, 347, 353, 359,
360—362, 370, 390
Oxner 391, 404
Panikkar 315
Parker 188
Paton 232
Peggott 152
Pesta 214
Petersen 28, 32, 33, 37, 177, 241—243,
246, 261—272, 275, 322, 348—350, 353,
357
Pettersson 102, 124, 152, 277
Pfeffer 183, 342, 389
Pierse 188
Poulsen 332
Pourtales 17
Pruvot 32, 191, 193, 250, 403
Purcell 150
Putter 47, 116
Raben 323
Rathbun 398, 400
Remane 310, 312, 314, 318, 350
Riddel 322
Riley 206, 207
Roch 433
Rose 231
Route 71
Russel 194, 208, 210, 215, 221, 223, 225,
231—233, 346, 416,
Ruud 333, 334, 346
Sars, G. O. 17, 342, 402
Sars, M. 17, 32, 144, 190, 340
Sauvageau 194
Savage 240, 322, 329
Schiemenz 321
Schmarda 340, 404
Schott 72, 87, 88, 140, 151, 152, 400
Scott 234, 321, 322
Scottsberg 391
Segerstrale 275
Sei well 120, 125—127, 206
Semper 342
Sernander 249
Sewel 368, 369
Shelford 177, 183, 195, 291—293
Shepard 56, 151
Soot-Ryen 278, 279
Southern 162, 194
Sparck 102, 183, 185, 245, 275, 281—288,
349, 350, 353, 367
Spiess 308
Stammer 404
Steggerd 188
Steinitz 402
Stephen 281
Steuer 31, 200, 214, 233, 347, 348, 362,
369
Stiasny 389
Stott 308
Studer 192
Stuxberg 192
Svedberg 182
Sverdrup 3, 31, 56, 58, 64, 68, 77, 79,
83, 86, 101, 108, 114, 126—136, 139,
148, 188, 199, 204
Tester 323
Thamdrup 195, 245, 252, 254, 256
Theel 245, 389, 400
Thiele 296
505
Thiemann 312, 313 Thienemann 37, 38, 41 Thomson 3,. 16, 112, 296, 385 Thorson 245, 253, 265, 266, 275, 283—288 Thoulet 152 Thulin 275 Todd 334—336 Towle г 195 Tressler 31 Uda 218 Utterback 99 Vail lan t 191 Valikangas 313, 318 Vatova 201, 284—288, 367 Vaughan 31, 195 Verille 192 Vogt 190, 284 Volz 404 Waksman 206, 301. 302 Walther 31, 32, 178, 321, 342 Walton 194 281, Warming 195 Watkin 281 Wattenberg 93, 96, 106, 107, 110, 111, 119, 125, 143 Wesenberg-Lund 195, 254 Wimpenny 240, 392 Wohlenberg 195, 254—256 Wohlgemut 324 Wollaston 279 Wust 53, 71, 73, 76, 84, 130, 131 Yong 20, 320, 346 Zimmer 387, 388 Zo-Bell 31, 301—305 Zuntz 324
ОПЕЧАТКИ
Стр. Строка Напечатано Следует читать
31 13 сверху Derptus Depths
93 подпись под рис. 41 мг!л МЛ 1л
129 таблица (в мг на литр) мл/л
131 14 сверху и 9 млн. в 9 млн.
135 подпись под рис. 75,В О2 л3/л О2 мл/л
135 подпись под рис. 75,Г р атом!л атом р('л
253 подпись под рис. 129 (по Торсону) (по Тамдрупу)
256 подпись под рис. 132 поселения Volutator поселения с. volutator
341 3 сверху > 1,7° ! <1,7°
ТЛ-2. Зак. № 972
Редакторы: Я. Л. Бирштейп и В. А. Б p-о ц к а Я.
Подписано к печати 17/XI 1950 г. Тираж 1000 экз. Объем ЗГХ п. л., 48 уч.-изд. л.
Т-08788 Формат 70 X 108716. Заказ № 1253. Цена 35 руб.
Отпечатано с матриц типографии № 2 «Печатный двор» им. А. М.« Горького
в типографии № 2 Управления издательств и полиграфии Исполкома Ленгорсовета.
Ленинград, Социалистическая, 14. Заказ № 972.