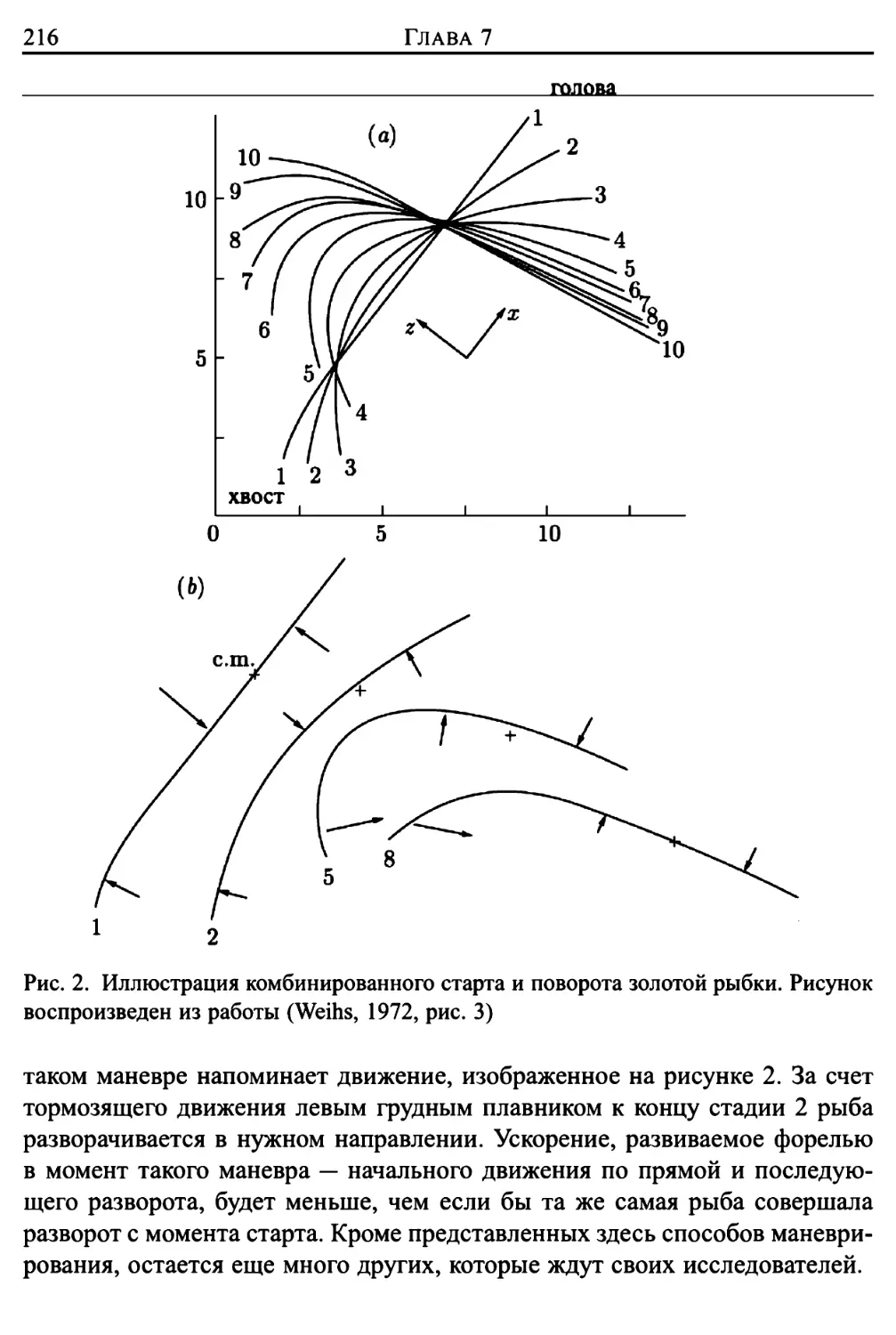
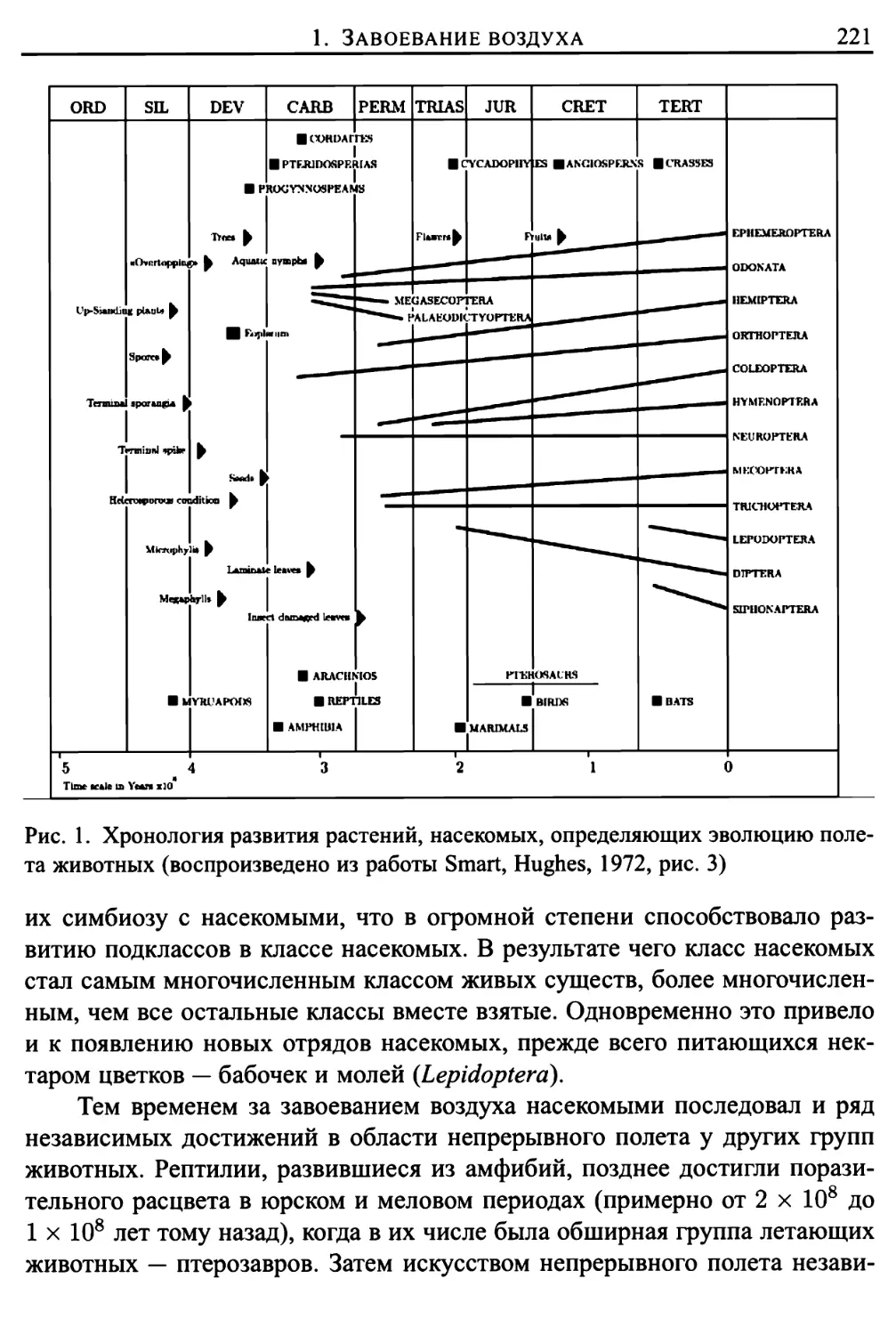
/

































































































































































































































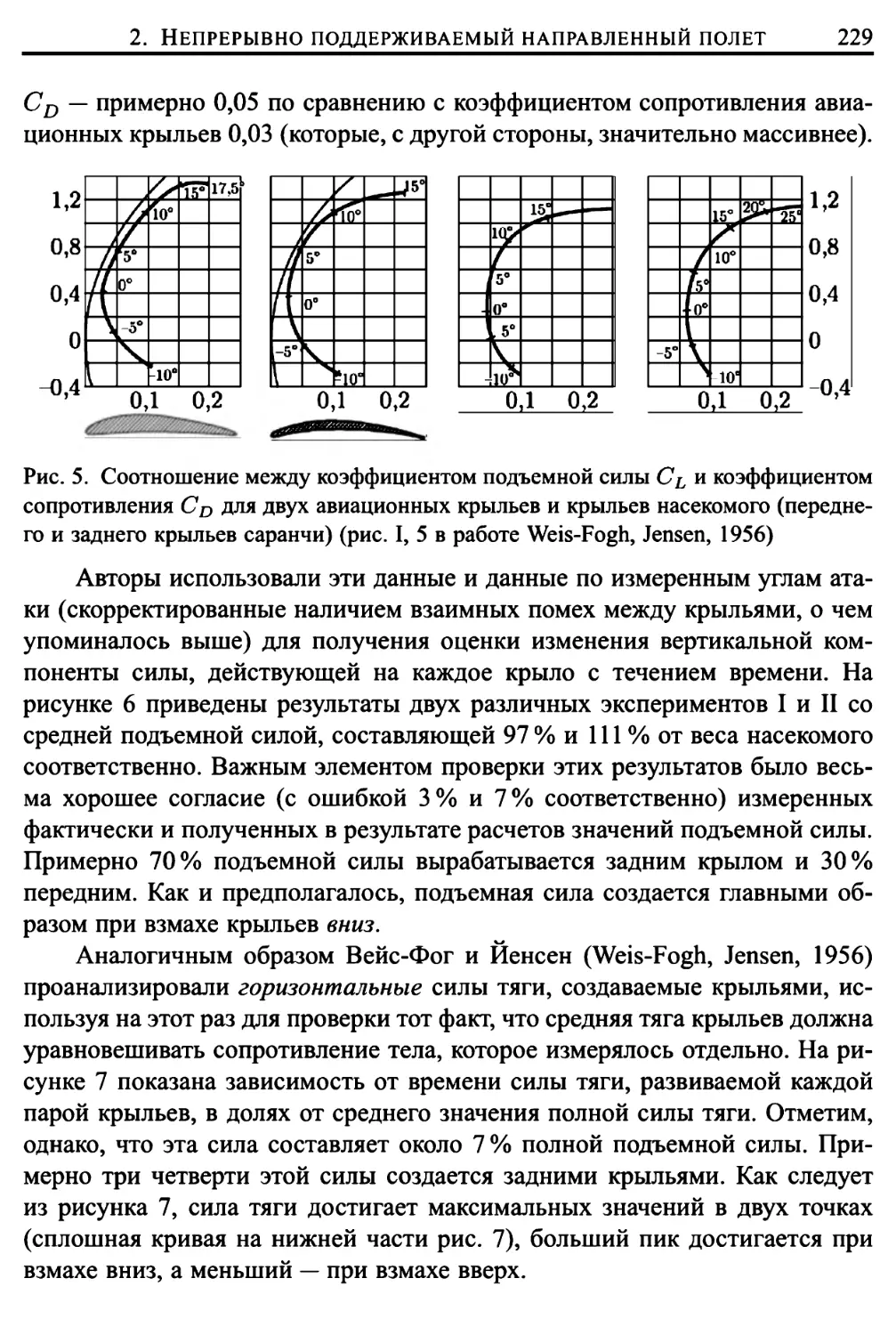

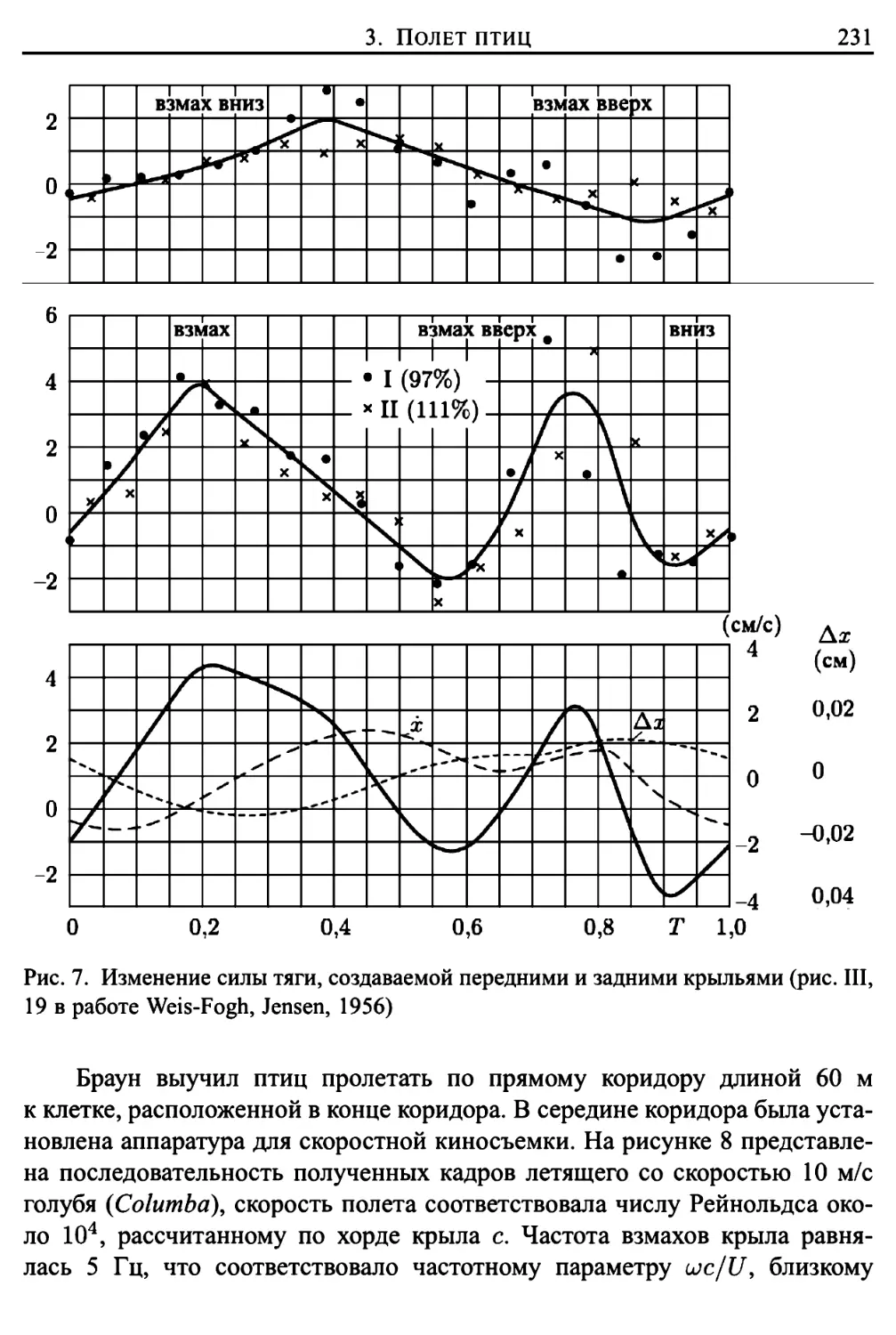
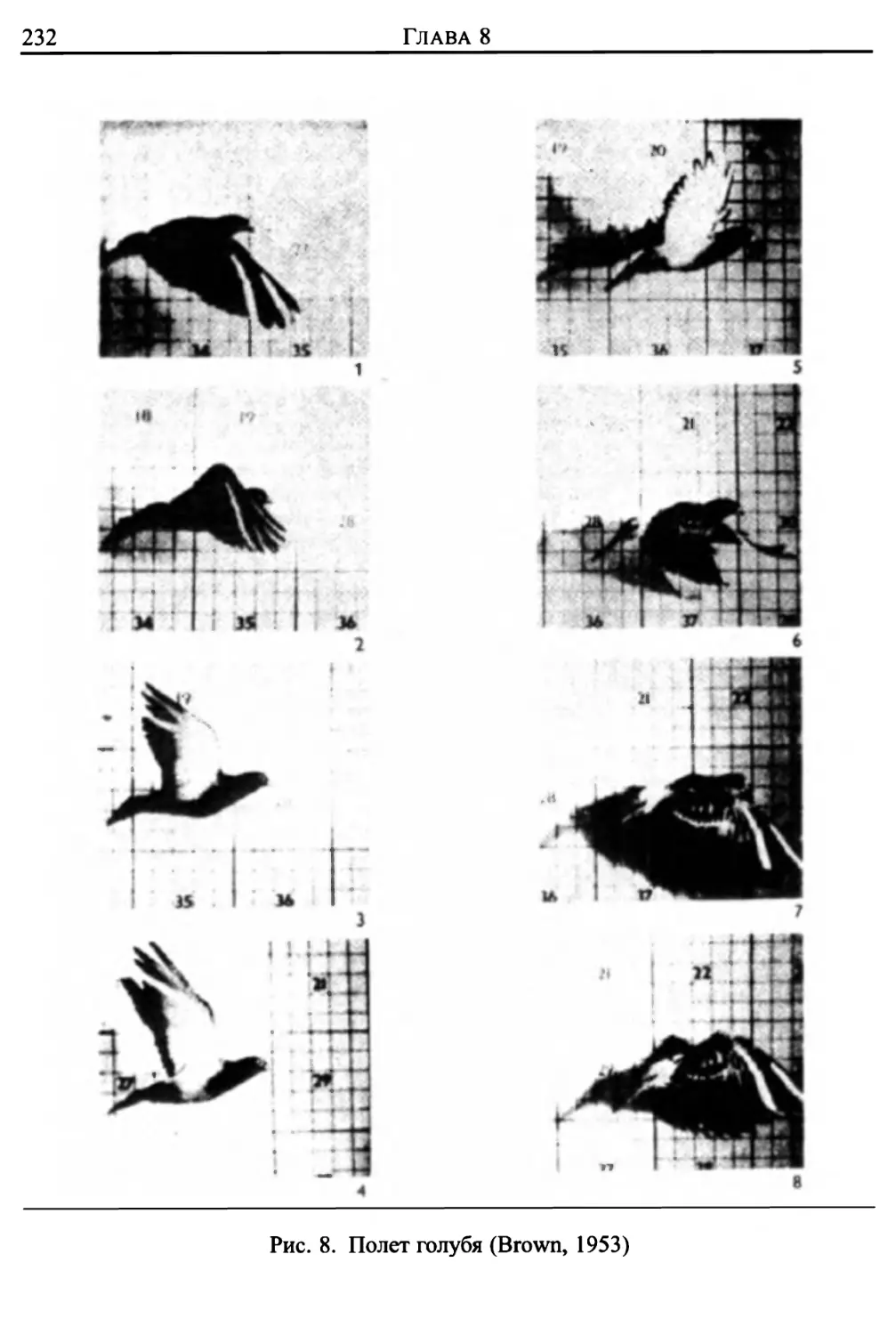




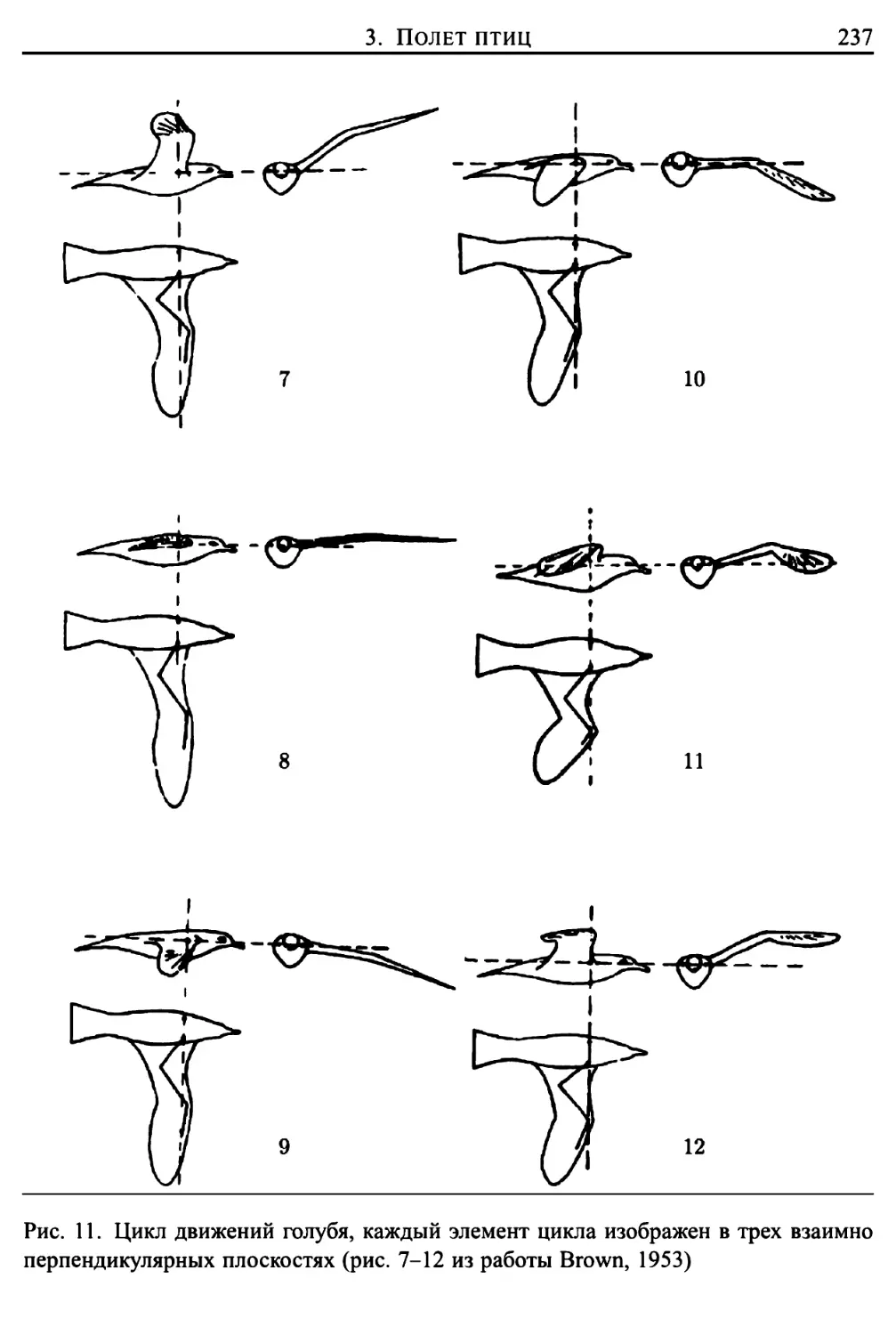











































































































































Автор: Лайтхилл Дж.
Теги: движение жидкостей гидродинамика механика медицина математика биология биомеханика математическая биогидродинамика
ISBN: 978-5-4344-0161-6
Год: 2014




Текст
т лл
|_
• •■"-tf .Ч/У- '.
Лаборатория нелинейного анализа
и конструирования новых средств передвижения
Лаборатория нелинейного анализа и конструирования новых средств
передвижения (http://lab.ics.org.ru) создана на базе Удмуртского
государственного университета в 2010 году благодаря гранту Правительства Российской
Федерации для государственной поддержки научных исследований,
проводимых под руководством ведущих ученых в российских образовательных
учреждениях высшего профессионального образования.
Важнейшей целью деятельности лаборатории являются поиск и реализация
новых способов передвижения на суше и в жидкости. С одной стороны,
актуальность таких исследований обусловлена растущей необходимостью
в увеличении надежности и маневренности существующих транспортных
средств. С другой стороны, не менее важной задачей является разработка
принципиально новых транспортных устройств для применения в
специфических условиях (например, в космосе, на дне океана). Совершенно
очевидно, что самые перспективные, действительно прорывные разработки
связаны с созданием мобильных роботов, использующих способы перемещения,
заимствованные из живой природы (механизмы полета птиц и насекомых,
плавания рыб и микроорганизмов, перемещения змей).
Эффективное решение поставленных задач возможно только при условии
глубокого понимания физических явлений, а также изучения и обобщения
передового мирового опыта в такой наукоемкой области, как
робототехника. Именно поэтому неотъемлемой частью работы лаборатории является
издание трудов по теоретической и прикладной механике, теории
управления, теории вычислительных систем, биомеханике и биомиметике. Большая
часть публикаций является переводами наиболее ценных зарубежных
материалов и фактически представляет собой прекрасную базу для
отечественных научно-исследовательских работ и постановки экспериментов.
Sir James Lighthill
University of Cambridge
Cambridge, England
Mathematical
Biofluiddynamics
A treatment in book form of the material in the lecture course delivered to the
"Mathematical Biofluiddynamics" Research Conference of the National Science Foundation
held from July 16-20,1973 at Rensselaer Polytechnic Institute, Troy, New York.
aiariL.
SOCIETY FOR INDUSTRIAL AND APPLIED MATHEMATICS
PHILADELPHIA
Джеймс Лайтхилл
МАТЕМАТИЧЕСКАЯ
БИОГИДРОДИНАМИКА
Перевод с английского
Т. В. Рамодановой
Под редакцией
К. В. Кошеля
1
7 ?
Москва ♦ Ижевск
2014
УДК 532.59
ББК 22.253.3
Л 185
Лайтхилл Дж.
Математическая биогидродинамика. — М.-Ижевск: Институт
компьютерных исследований, 2014. — 408 с.
В основу книги положены материалы лекций по математической
биогидродинамике, прочитанных автором на конференции Национального научного фонда (16-
29 июля 1973 года) в Политехническом институте Ренсселлера (Трои, Нью-Йорк).
Значительная часть материала была опубликована в таких ведущих журналах, как
Annual review of Fluid Mechanics и Journal of Fluid Mechanics.
В книге представлены методы и стиль исследований автора, который внес
значительный вклад в развитие этого направления в динамике в XX веке:
проанализированы различные механизмы достижения высоких скоростей и маневренности
(использование энергии волн, снижение сопротивления жидкости и оптимальные
режимы движения), а также вопросы внутренней биогидродинамики, связанные
с распространением волновых возмущений, ролью вихревых процессов и
эластичности стенок сосудов.
Книга будет полезна специалистам в области гидродинамики и биомеханики,
студентам соответствующих специальностей, а также всем, кто интересуется
механикой природных явлений. Особо следует отметить, что, по мнению автора, данная
книга чрезвычайно полезна для биологов и медиков.
ISBN 978-5-4344-0161-6 ББК 22.253.3
Mathematical Biofluiddynamics
© Society for Industrial and Applied Mathematics, 1975
Published by Izhevsk Institute of Computer Science with permission
© Ижевский институт компьютерных исследований,
перевод на русский язык, 2014
http://shop.rcd.ru
http://ics.org.ru
Оглавление
От редактора ix
Выражение признательности xi
Глава 1. Введение в биогидродинамику 1
1. Общие положения 1
2. Внешняя биогидродинамика 5
3. Внутренняя биогидродинамика 7
Часть I. Внешняя биогидродинамика 13
Глава 2. Гидромеханика самодвижения водных животных. Обзор 15
1. Область исследования 15
2. Самодвижение беспозвоночных в воде 16
3. Введение в кинематику плавания рыб 21
4. Плавание угреобразных 26
5. Плавание скомброидных 31
6. Лабораторные исследования скомброидного типа плавания . . 34
7. Скомброидный тип движения рыб с хвостом в виде полумесяца 37
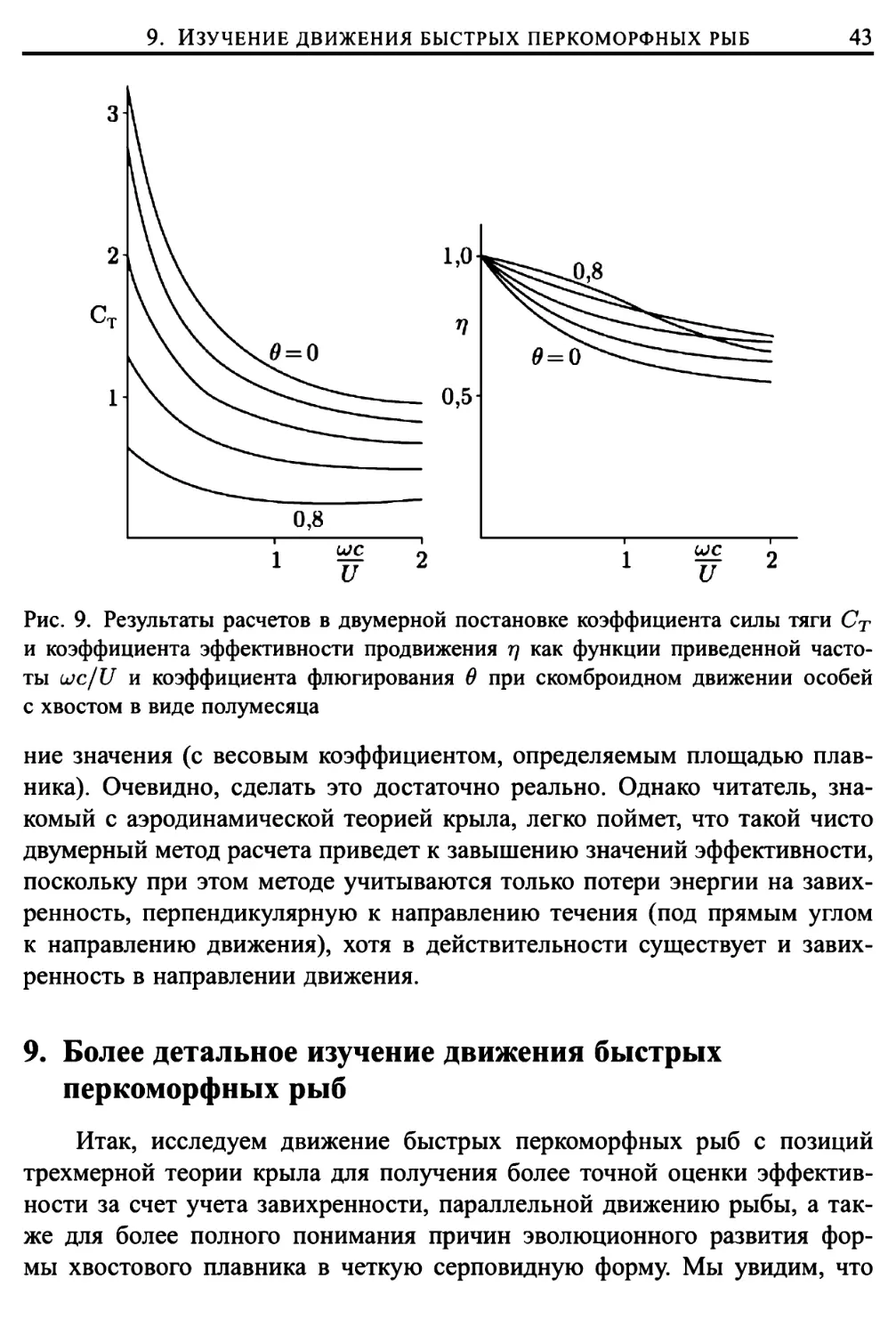
8. Двумерный анализ динамики серповидного хвоста 40
9. Более детальное изучение движения быстрых перкоморфных
рыб 43
10. Плавучесть рыб 46
11. Передвижение «панцирных» рыб 51
12. Использование конечностей для плавания: земноводные и
членистоногие 52
13. Перемещение в воде пресмыкающихся 54
14. Передвижение в воде млекопитающих 58
15. Заключение 60
Глава 3. Математические аспекты передвижения водных
животных при низких числах Рейнольдса 63
1. Введение 63
vi
Оглавление
2. Уравнения движения жидкости и их фундаментальные
сингулярные решения 64
3. Сингулярные решения на отрезке 69
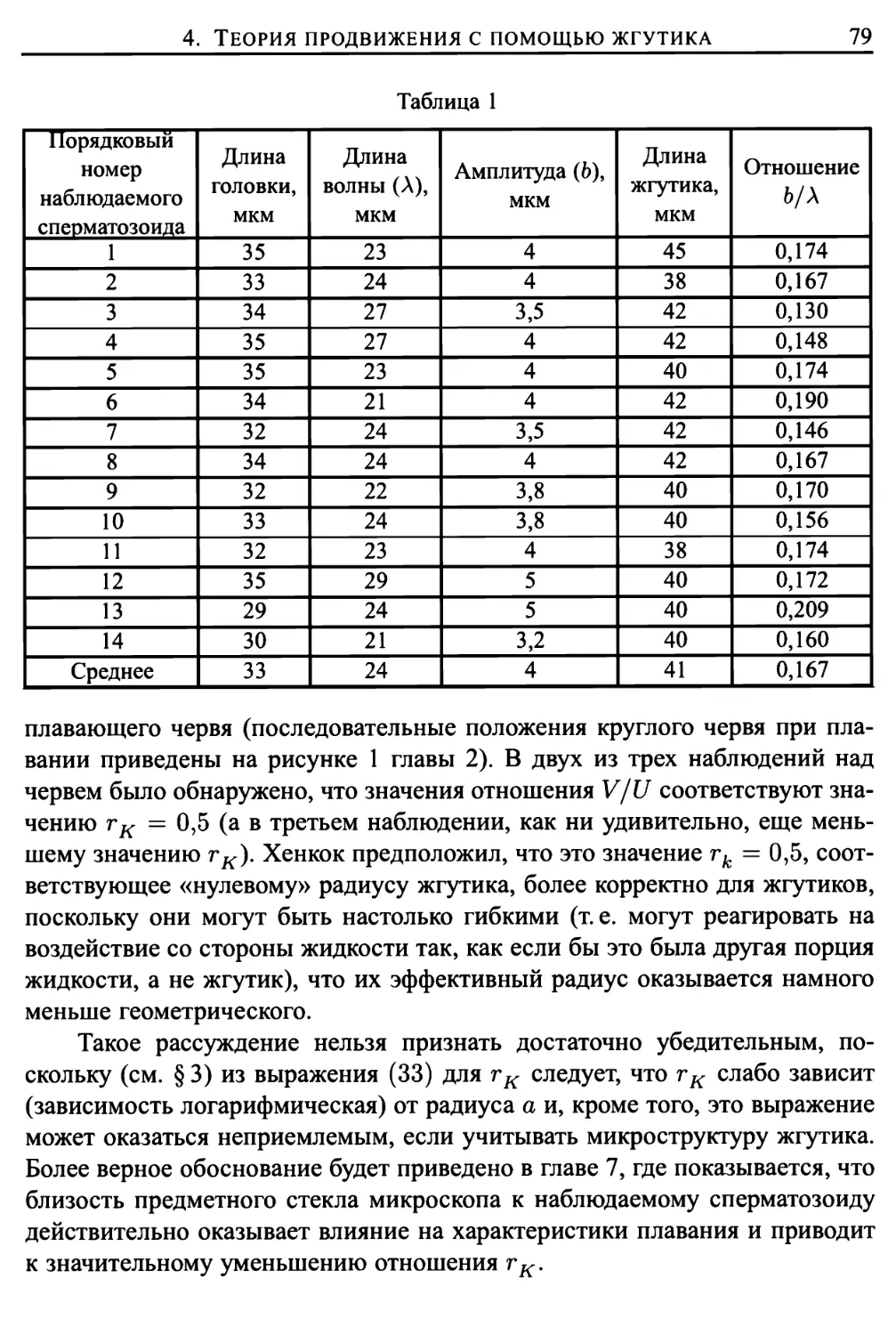
4. Теория продвижения с помощью жгутика, основанная на
силе сопротивления 74
5. Плавание с минимальной потерей энергии 82
6. Теория тонкого тела 86
Глава 4. Высокоэффективные с гидродинамической точки зрения
механизмы плавания животных 91
1. Введение 94
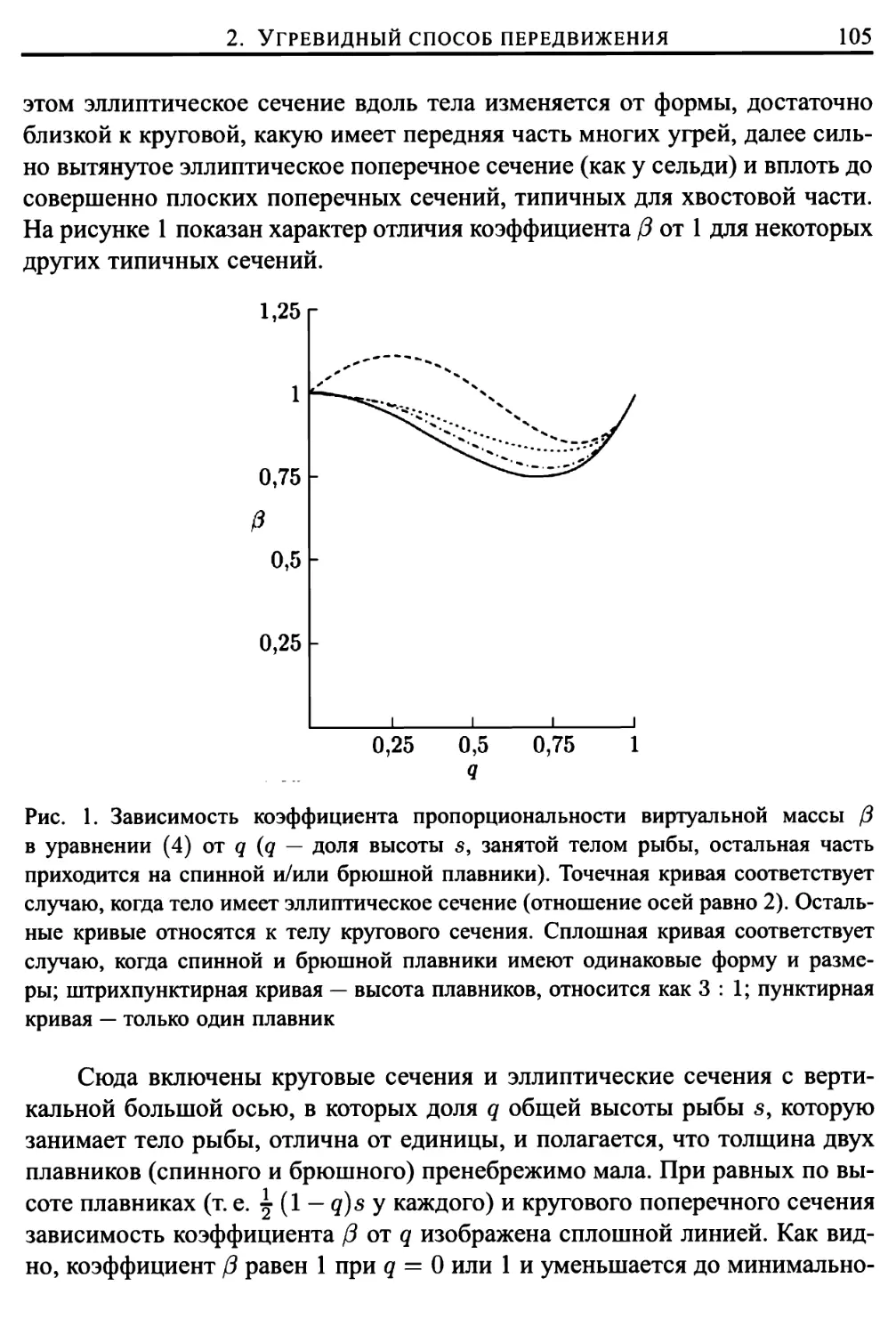
2. Угревидный способ передвижения 102
3. Вихревые пелены за плавниками 113
4. Механика скомброидного типа движения 122
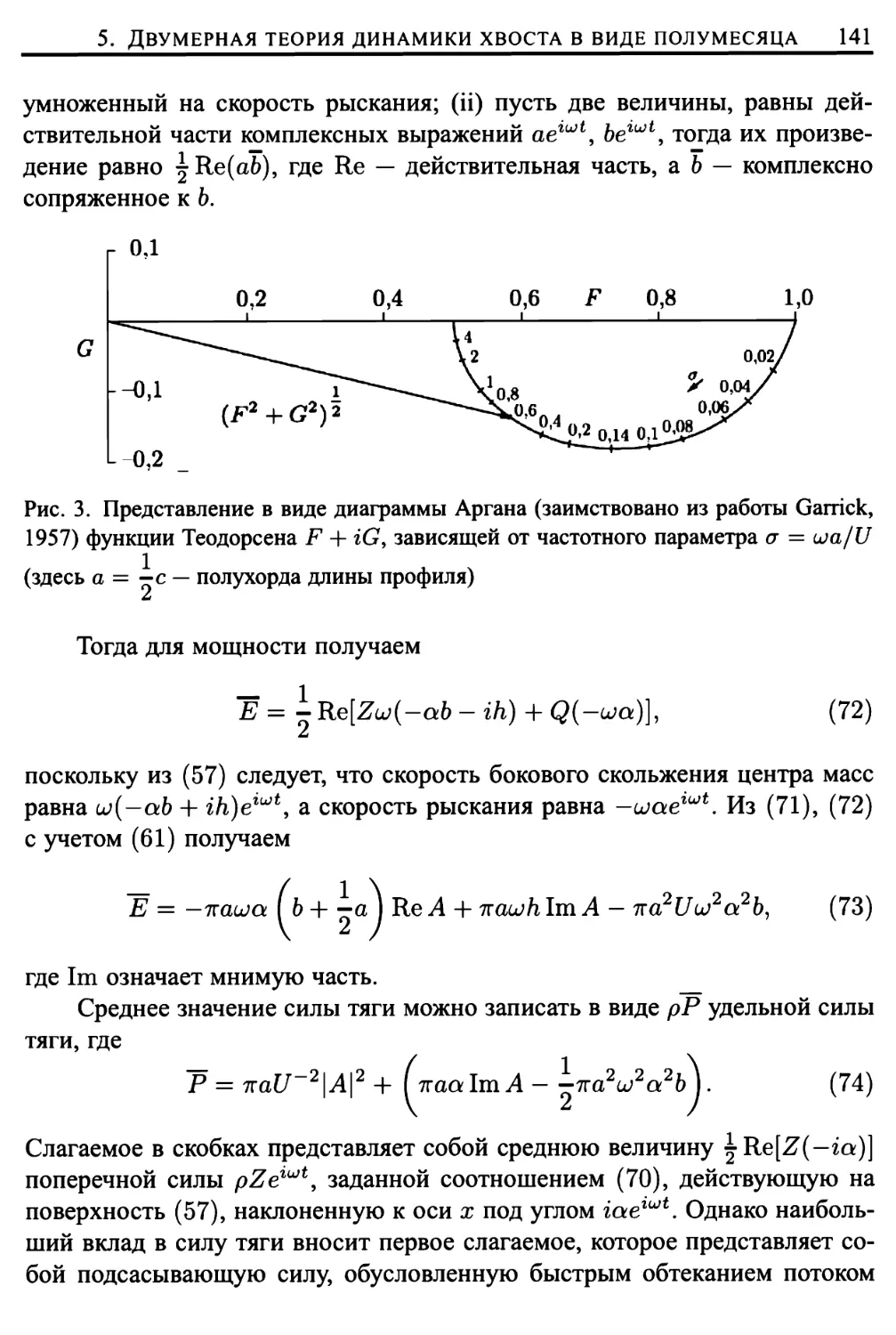
5. Двумерная теория динамики хвоста в виде полумесяца .... 134
Глава 5. Приложение теории тонкого тела к продвижению рыб
при волнообразном поперечном движении тела с большой
амплитудой 149
1. Введение 149
2. Расчет реактивной силы 154
3. Обсуждение баланса силы тяги и силы сопротивления на
материале наблюдений 161
Глава 6. Передвижение водных животных. Обзор новейших
теоретических исследований 171
1. Введение 171
2. Разделы гидродинамики, применяемые для изучения
передвижения животных в воде 174
3. Гидродинамика ресничных организмов 185
4. Теория вытянутого тела при широкоамплитудных
волнообразных поперечных движениях 192
5. Вихревые следы 200
Глава 7. Некоторые современные исследования перемещения
водных животных 205
1. Движение при малых числах Рейнольдса 205
2. Движение при больших числах Рейнольдса 210
Оглавление
vn
Глава 8. Аэродинамика полета животных 219
1. Завоевание воздуха 219
2. Непрерывно поддерживаемый направленный полет
насекомых 225
3. Полет птиц 230
4. Трепещущий полет 243
Глава 9. Механизм Вейс-Фога образования подъемной силы . . 255
1. Введение 256
2. Двумерная постановка задачи о движении крыльев
насекомого в невязком потоке 259
3. Двумерная постановка задачи о движении крыльев
насекомого с учетом вязкости 265
4. Заключение 272
Часть II. Внутренняя биогидродинамика 279
Глава 10. Физиологическая гидродинамика: общий обзор . . . 281
1. Введение 281
2. Установившиеся вторичные потоки 285
3. Входные зоны 286
4. Атероматозное поражение артерий 287
5. Распределение деформации сдвига в разветвленной
сосудистой системе 288
6. Сопротивление в ветвящихся системах 290
7. Бронхиальное сопротивление 291
8. Распределение скорости в пульсирующем потоке 292
9. Распространение пульсаций 293
10. Турбулентность в системе кровообращения 294
11. Мочевыводящие пути 295
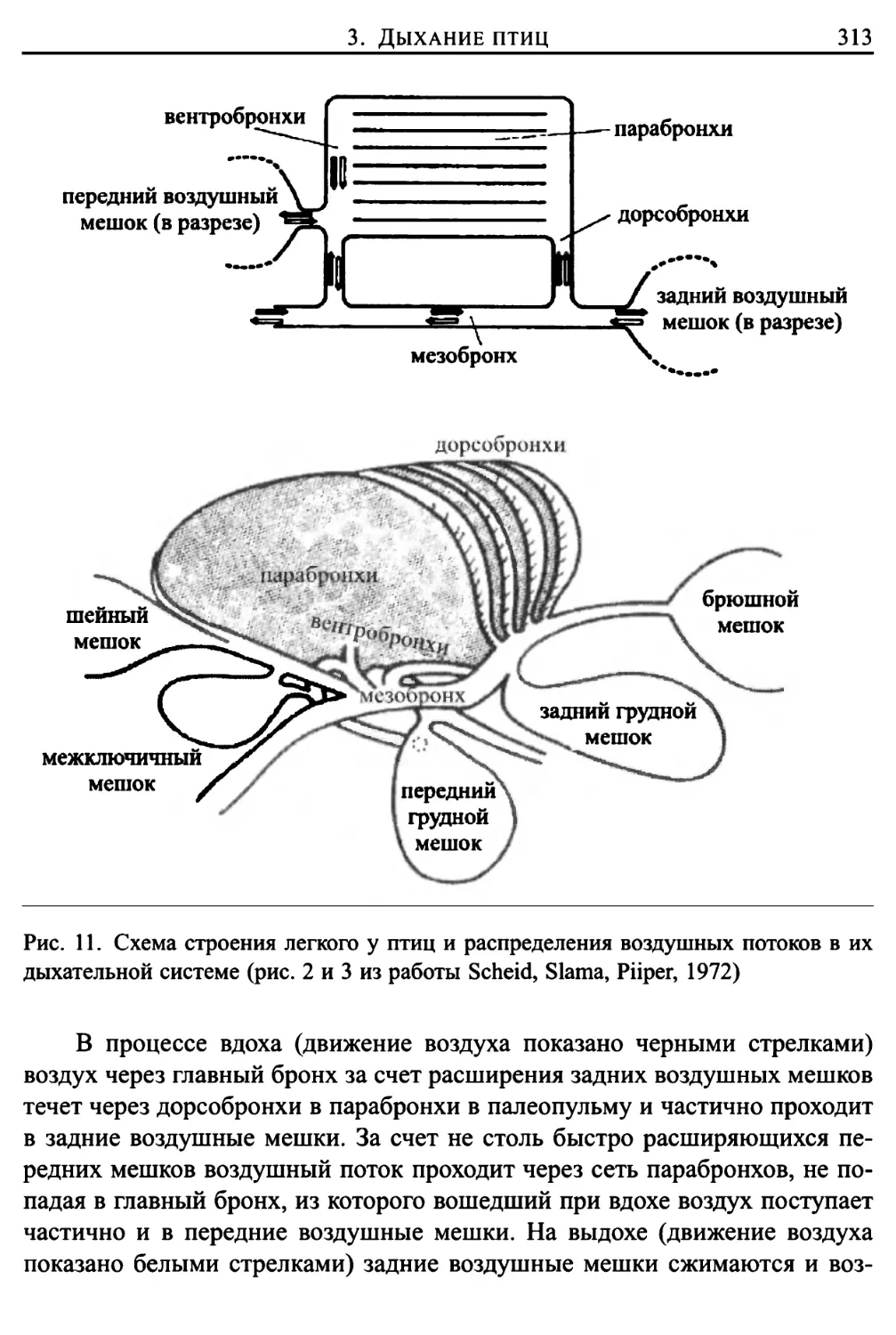
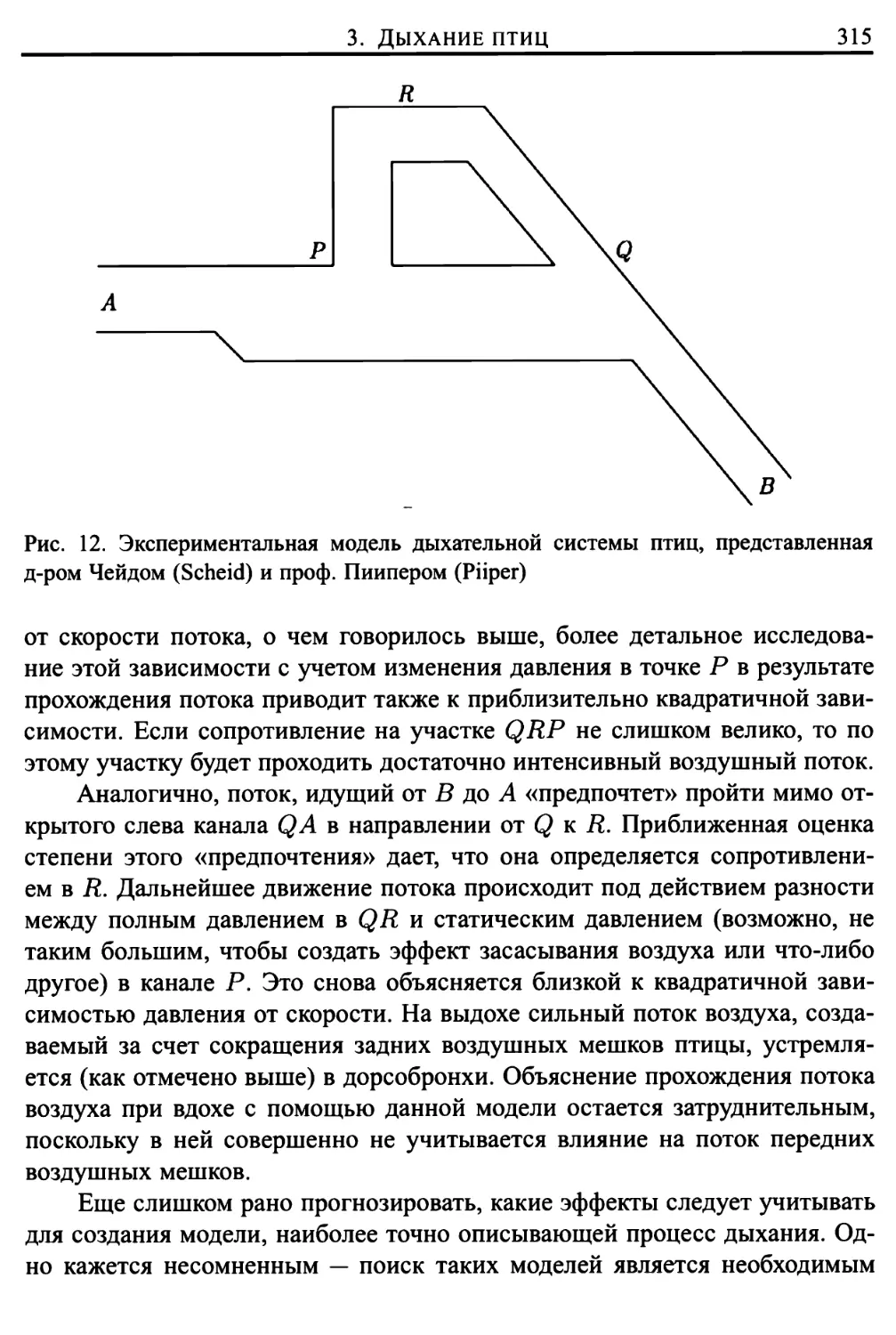
Глава 11. Воздушные потоки в дыхательной системе 297
1. Введение 297
2. Движение воздушных потоков в бронхах человека 302
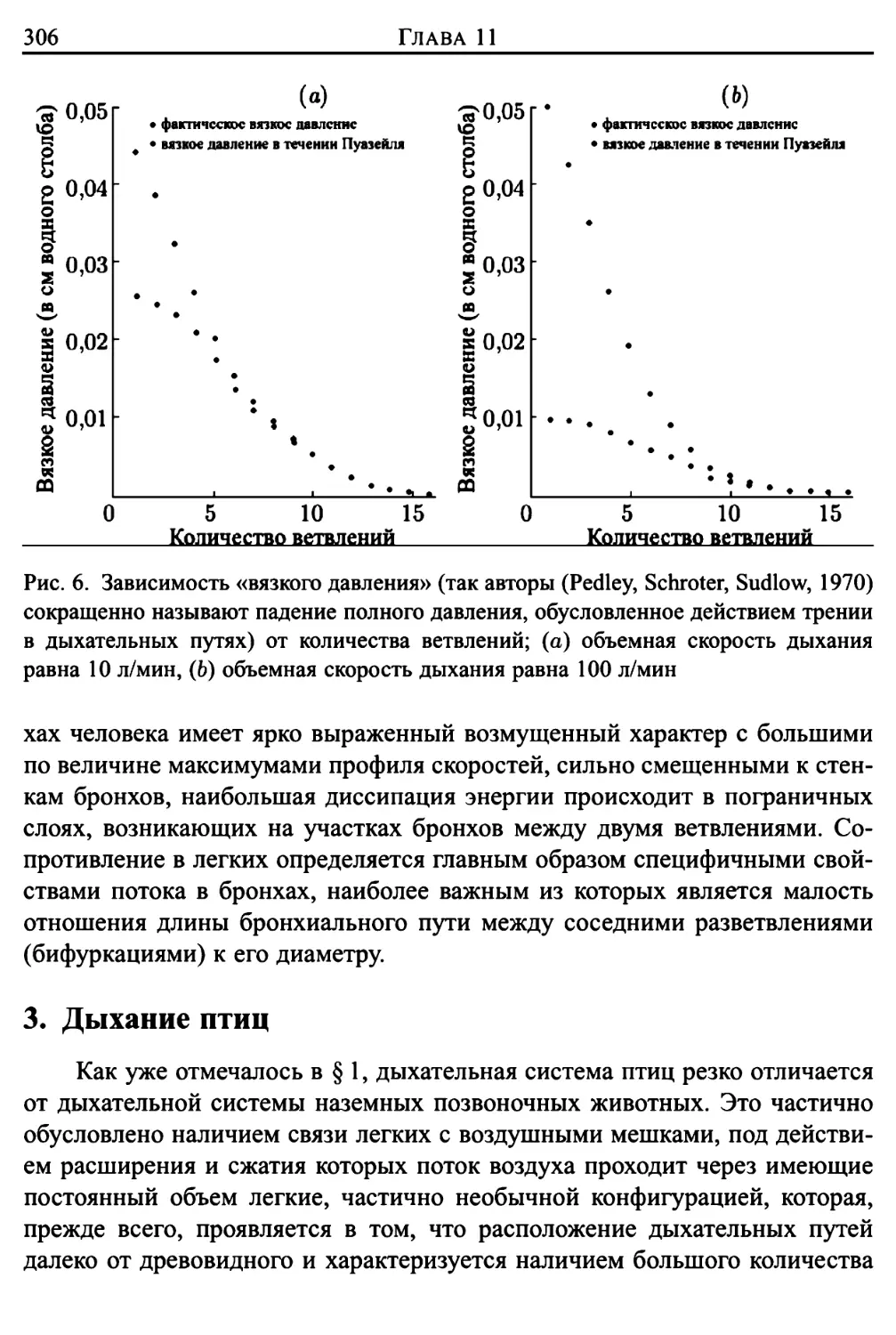
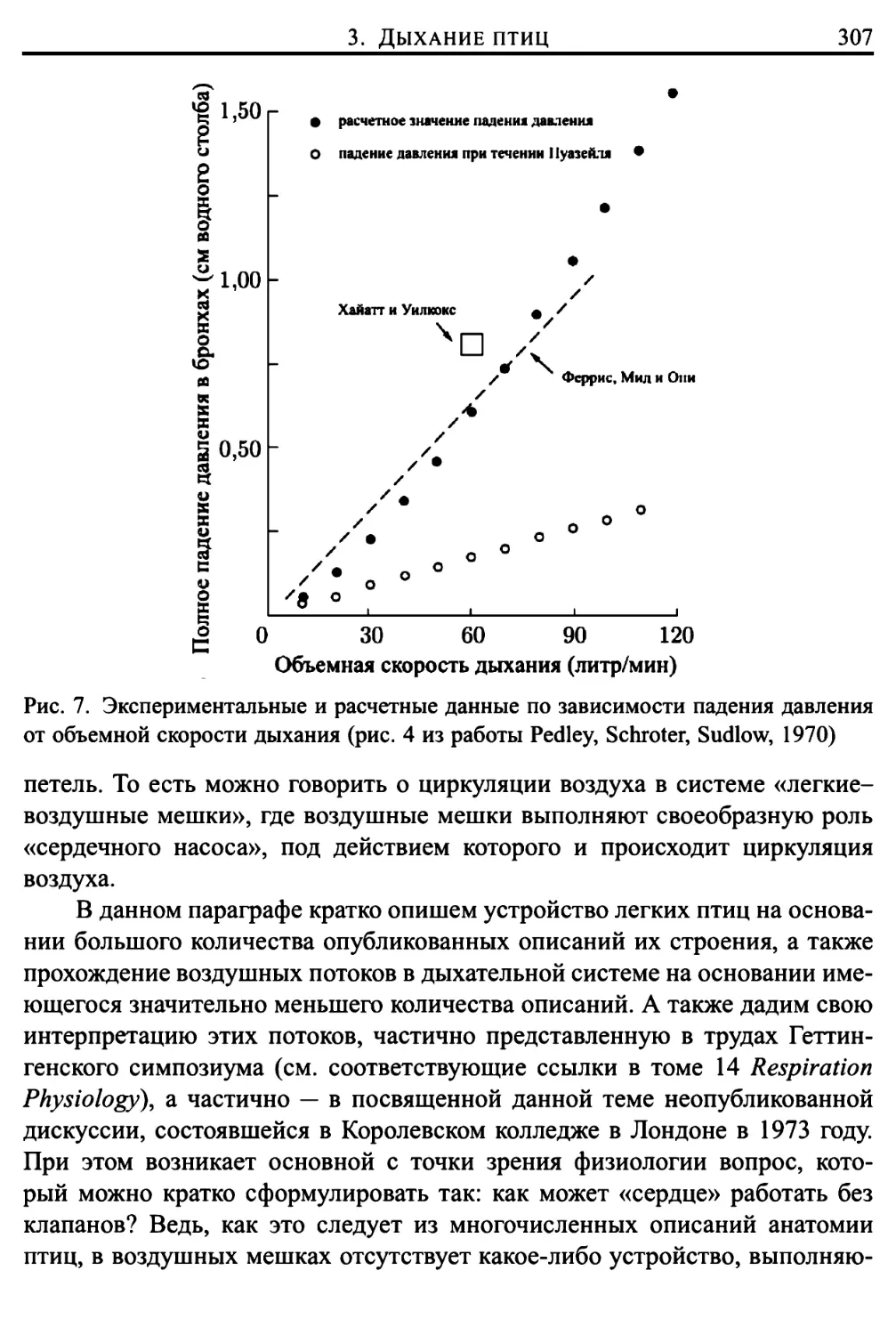
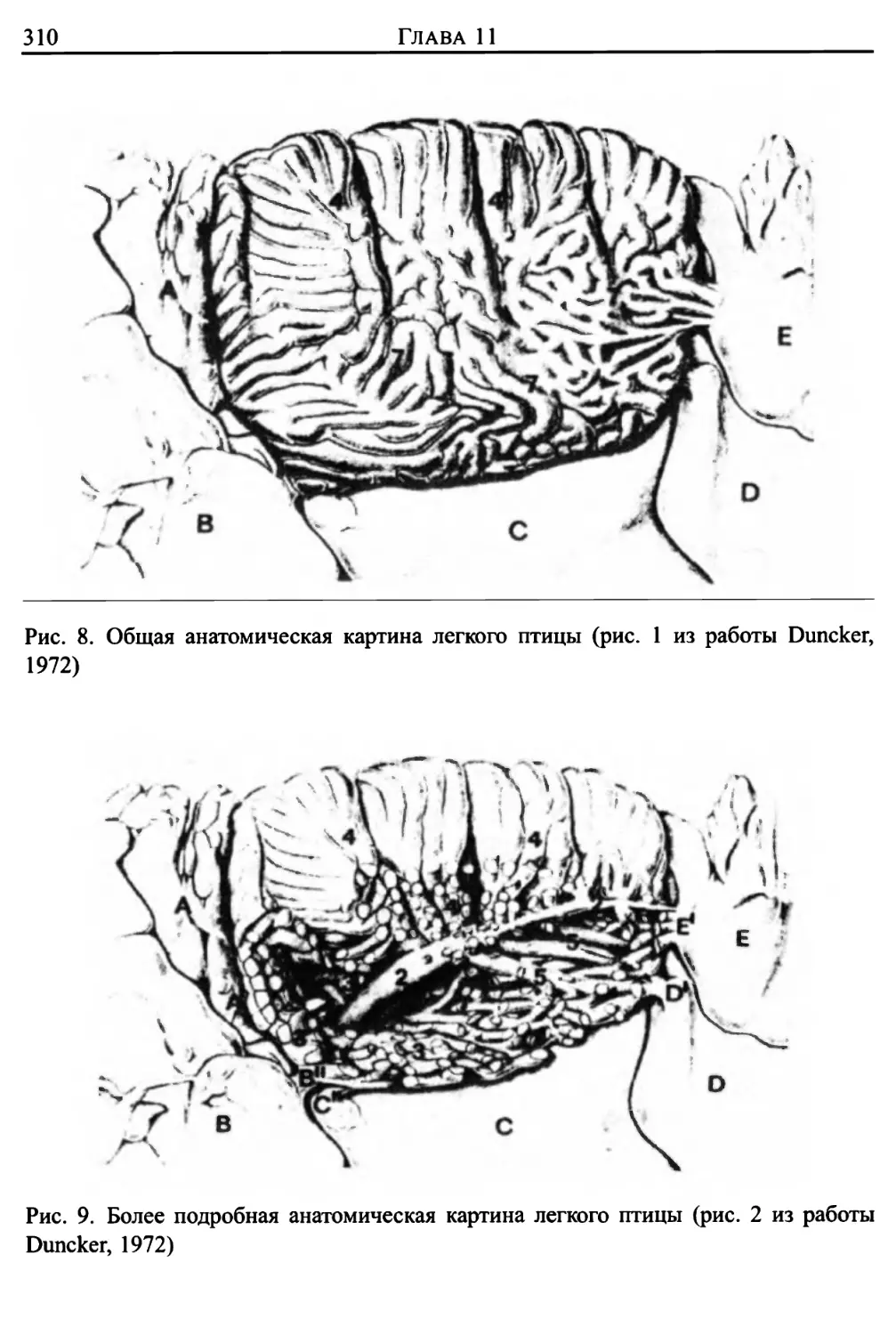
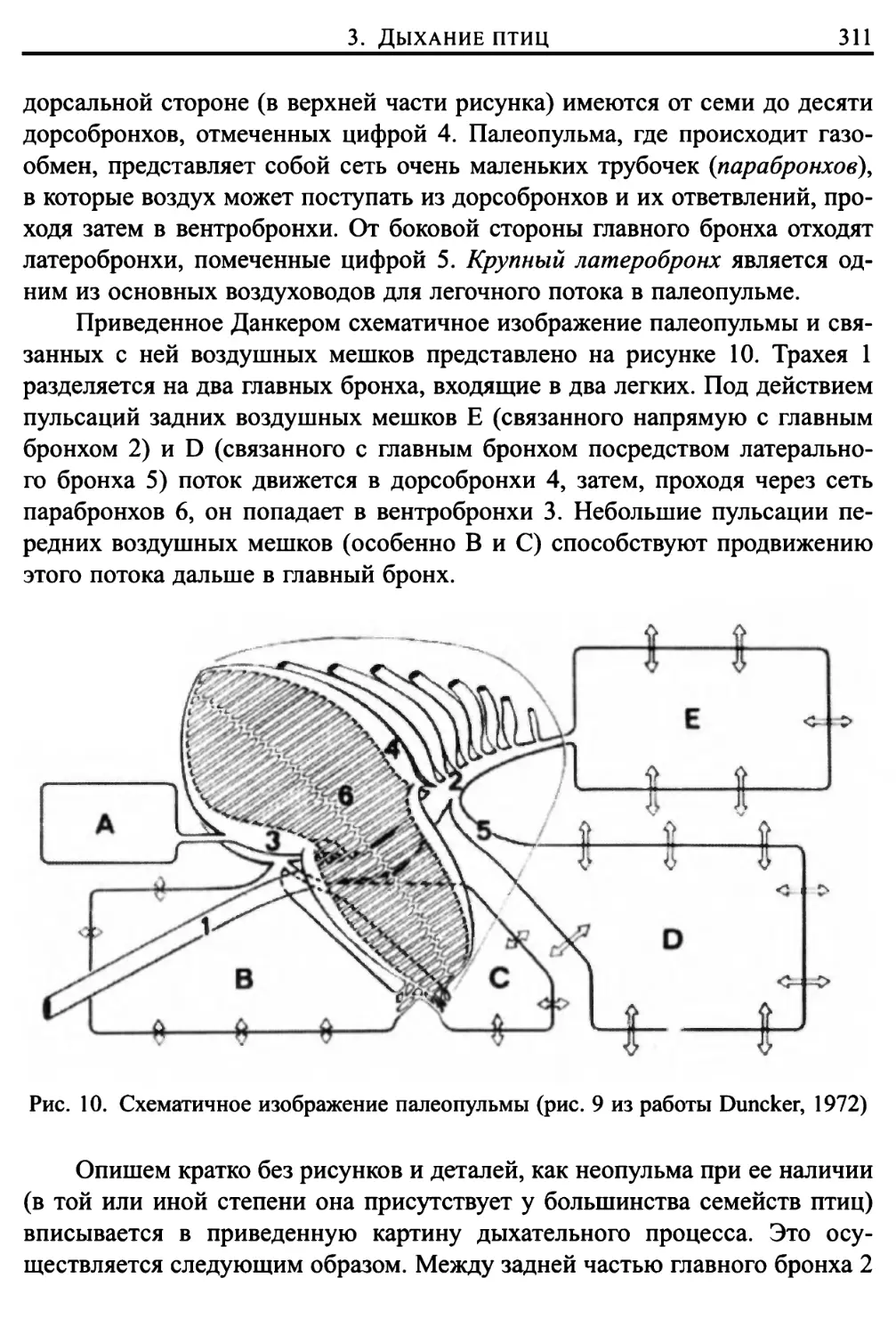
3. Дыхание птиц 306
Глава 12. Распространение пульсаций в кровеносной системе . 317
1. Введение 317
2. Распределение напряжений в стенках сосуда 320
viii Оглавление
3. Прохождение волн в местах ветвления артерий 325
4. Взаимодействие разветвлений в сосудистой системе 332
5. Изменение амплитуды волны в сосуде 337
6. Сравнение с экспериментальными данными 344
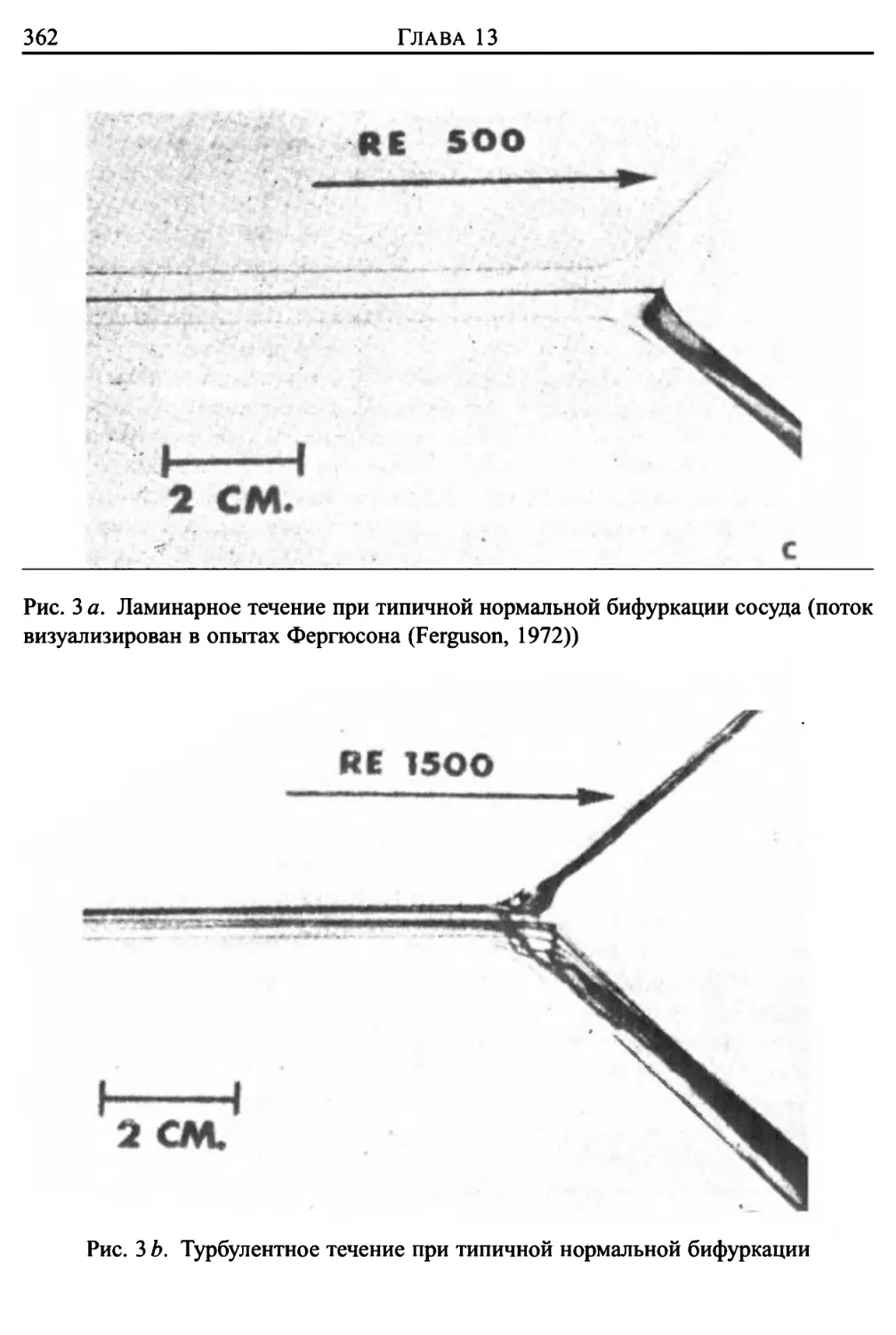
Глава 13. Кровоток и заболевание артерий 355
1. Турбулентность в потоке крови 355
2. Течение в области постстенотического расширения сосуда . . 358
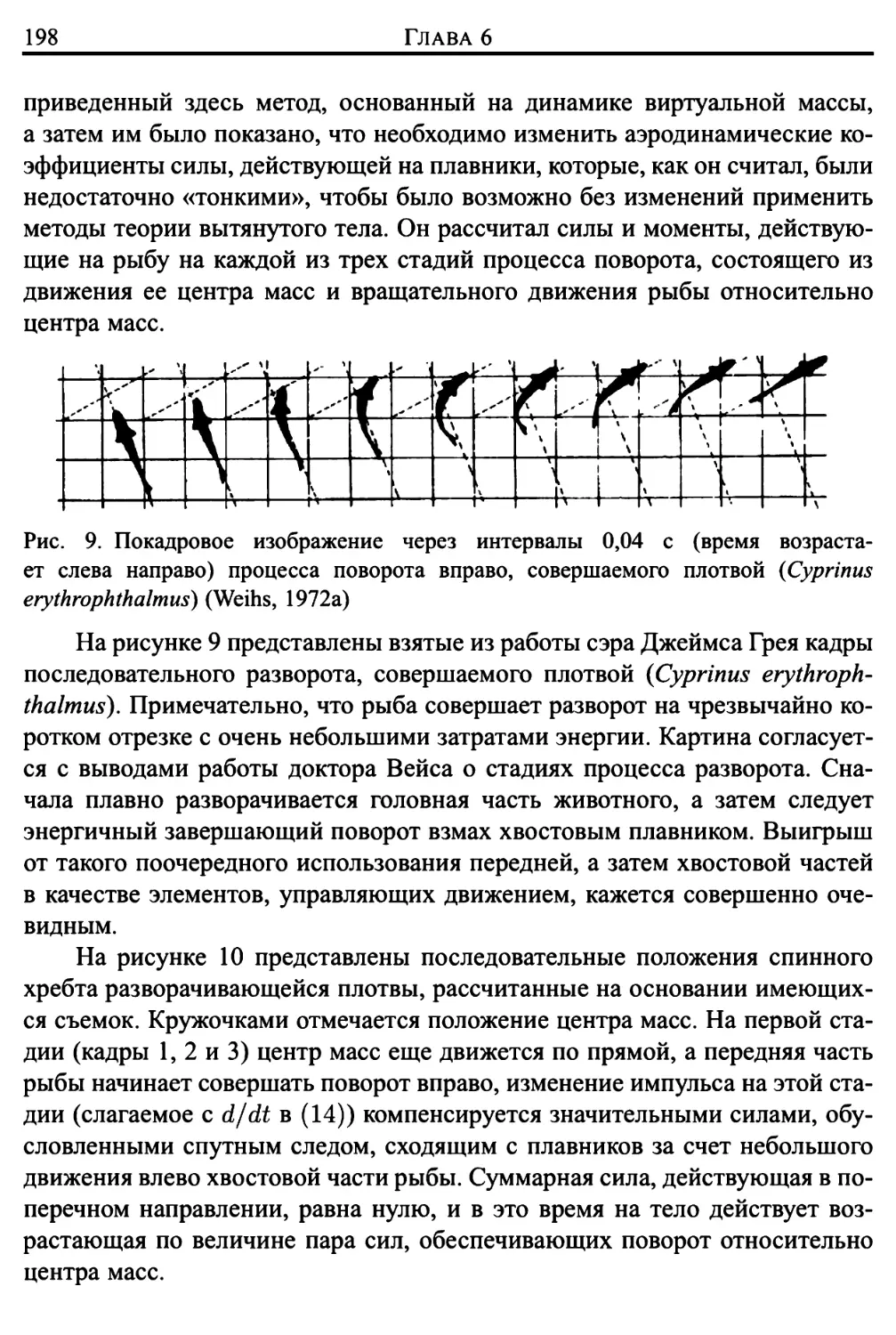
3. Аневризмы сосудов головного мозга 360
4. Развитие атеросклероза, обусловленное межклеточным
отложением жиров 364
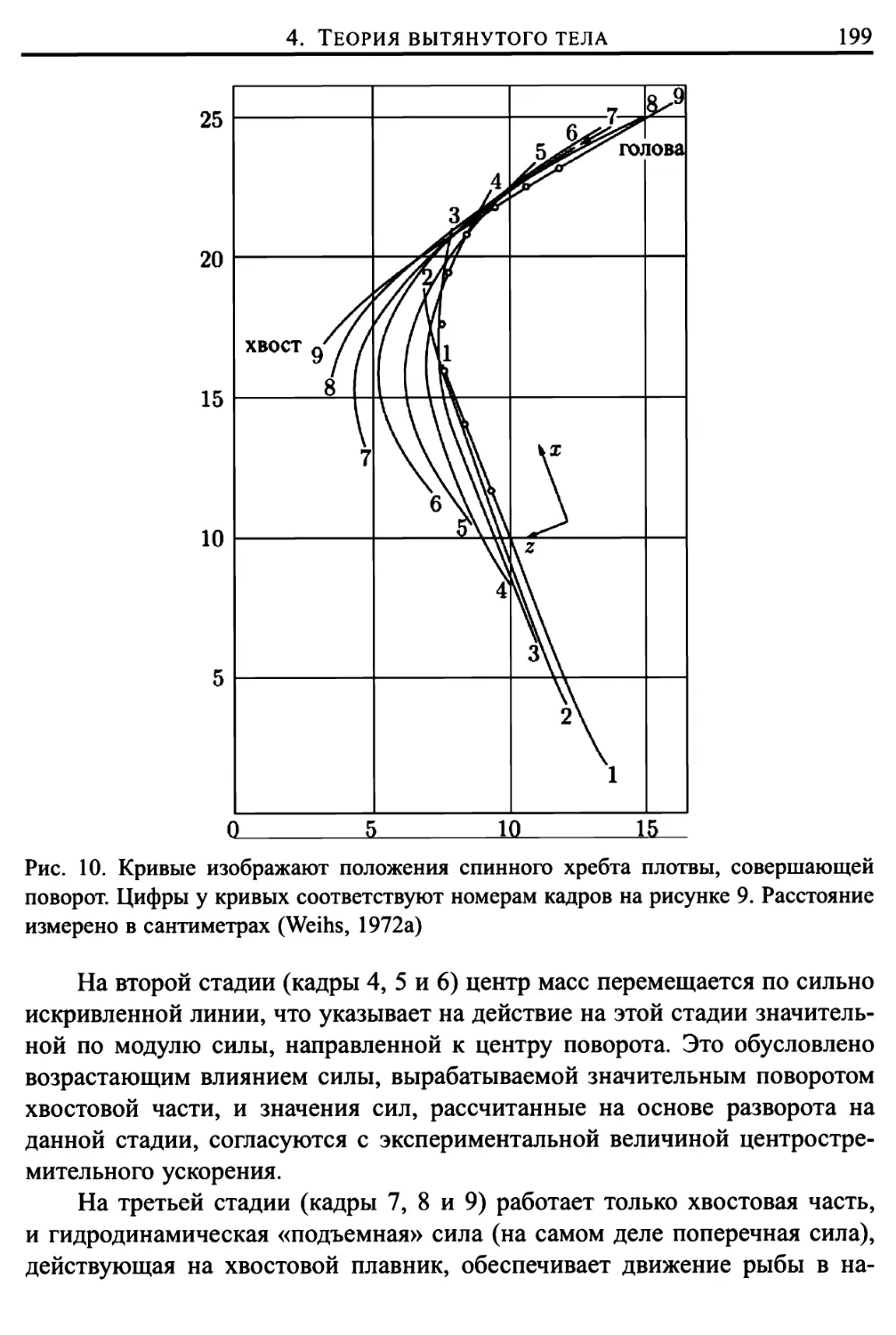
5. Внутриклеточное накопление липидов 370
Глава 14. Микроциркуляция 377
1. Введение 377
2. Вазомоторный контроль периферической перфузии 379
3. Легочная перфузия и вентиляция 381
4. Концентрация эритроцитов вдоль оси капилляров 384
5. Слой смазки в очень узких капиллярах 388
От редактора
Биогидромеханика, как и гидродинамика в целом, бурно развивалась
за прошедшие годы: достаточно просмотреть оглавление Annual Review
Fluid Mechanics, и мы обнаружим как минимум 1-2 обзора из этой области
в каждом выпуске. Как правило, это достаточно узкоспециализированные
обзоры, и «Математическая биогидродинамика» Дж. Лайтхилла, по сути,
представляет собой один из таких обзоров, подводя итог заметному
прорыву, достигнутому ко времени публикации оригинала книги (1975 год).
Прорыв этот сам по себе очень интересен и стал возможен во многом
благодаря взаимодействию биологов и специалистов по гидродинамике,
проводивших эксперименты, которые позволили понять некоторые физические
механизмы и заложить основы их математического описания. Вся эта
эпопея изложена в серии обзоров Дж. Лайтхилла, объединенных в одной
книге. Бесспорно, достойна восхищения и личность самого автора, внесшего
существенный вклад в описываемые достижения: Лайтхилл обладал
изумительной интуицией и способностью генерировать идеи, облекая их в
достаточно строгую математическую форму.
Биогидродинамика в общем сложнее, чем классическая
гидродинамика, а специальной литературы на русском языке не так уж много: например,
поиск в интернете на 90% выдает сборник под редакцией В.М. Ентова
(«Биогидродинамика плавания и полета», Москва: Мир, 1980). Указанный
сборник посвящен только внешней биогидродинамике и содержит всего
одну статью Дж. Лайтхилла, так что предлагаемая книга будет очень хорошим
дополнением ко все еще популярному сборнику. В части внутренней
биогидродинамики подобных изданий я не знаю. Конечно, работ очень много,
но видится перекос в сторону очень рафинированных упрощенных
моделей или мощного численного счета. Думается, что подход, представленный
в обзорах Дж. Лайтхилла, не потерял своей актуальности и по-прежнему
заслуживает внимания.
«Побольше смотрите на движения жидкости и изображения
движений жидкости» — известный совет начинающим исследователям. Чтение
этой книги, несомненно, позволит, в определенном смысле, «посмотреть»
X
ОТ РЕДАКТОРА
на многие типы движения так, как их видел выдающийся исследователь
Дж. Лайтхилл.
Все вышесказанное и служит основанием для публикации русского
перевода относительно старой книги, вполне доступной на языке оригинала.
Помимо прочего, заметим, что книги и статьи Дж. Лайтхилла очень
глубоки идейно, но не очень просты для восприятия. Поэтому будем надеяться,
что русский перевод облегчит чтение и понимание идей Дж. Лайтхилла,
многие из которых еще требуют осмысления и дальнейшего развития.
Выражение признательности
Хотелось бы поблагодарить многих моих коллег, в частности Ричарда
Бейнбриджа, Джона Блейка, Квентина Боуна, Колина Каро, Джима
Фитцджеральда, Джеймса Грея, Кена Макина, Нормана Маршалла, Тима Педли,
Марго Роуч, Боба Шрётера, Тони Сида, Даниеля Вейса и Торкела Вейс-
Фога, за их неоценимую помощь и плодотворное сотрудничество при
подготовке и подборке материала для данной работы, проводившихся в течение
пяти лет. Я также признателен Дику ДиПрима и Ле Рубенфельду за помощь
в компоновке материала книги «Математическая биогидродинамика».
Выражаю свою признательность Политехническому институту Ренсселлера,
а также Национальному научному фонду за поддержку и финансирование
проведенных исследований.
Джеймс Лайтхилл
Глава 1
Введение в биогидродинамику
1. Общие положения
Движущей силой большинства исследований в области гидродинамики
являются две наиболее важные практические задачи:
(i) усовершенствование способов передвижения в жидкой среде;
(ii) более глубокое осмысление процесса движения жидкости в
циркуляционных системах.
Научные исследования, проводимые в течение многих лет в соответствии
с этими целями, привели (i) к созданию эффективных транспортных
воздушных и водных средств перемещения и (ii) углубленному изучению
процесса передачи тепла или различных субстанций в виде растворов или
суспензий в больших циркуляционных системах (таких как атмосфера,
круговорот воды в природе, системы с циркулирующей жидкой средой на
промышленных предприятиях).
Подобным образом и научные исследования в биогидродинамике
стимулируются двумя различными практическими целями, что отражено в
схеме изложения материала в данной работе. Биогидродинамика плавания
и полета («внешняя» биогидродинамика) исследует перемещение живых
существ в жидкой среде (в воде или воздухе), а биогидродинамика жидких
сред организмов («внутренняя» биогидродинамика) исследует процессы
переноса веществ или тепла в сосудистой системе. Эти две ветви
биогидродинамики имеют одну общую черту, которая выделяет их из гидродинамики:
в обоих случаях движение жидкости может возникать за счет перемещений
поверхностей (внутренних или внешних) живого организма, некоторые
части которых чрезвычайно эластичны. Поскольку мы будем рассматривать
движение представителей только животного мира (так что, возможно,
более подходящим названием данной отрасли гидродинамики было бы «зо-
огидродинамика»), с характерной для них способностью к произвольному
2
Глава 1
движению (способностью совершать управляемое ими активное движение
в ответ на раздражающее воздействие), то в этом случае и возникает эта
характерная особенность, объединяющая обе указанные выше части
биогидродинамики.
В данной работе проблема в основном будет рассматриваться с
позиций биологической науки, включая фундаментальные и прикладные
аспекты. С позиций фундаментальной биологии задача состоит в изучении
не только функционирования животных, но и эволюции различных систем
организма, обеспечивающих самодвижение животных в окружающей их
жидкой среде, а также внутренний массо- и теплоперенос за счет движения
жидкости внутри организма животного. Свойства таких систем организма,
которые приводят к некоторому увеличению шанса животного на
выживание и воспроизведение, могут немного усилиться в следующем
поколении и возрасти еще больше в последующих, а в долгосрочной
перспективе могут ассимилироваться в видовые характеристики. Процесс развития,
в результате которого организм либо адаптируется к условиям окружающей
среды, либо находит экологическую нишу, может быть изучен с
привлечением сведений из общей биологии и палеонтологии, а также на основании
исследований биогидродинамики.
И хотя подобного рода исследования представляют интерес прежде
всего для биологов, тем не менее они будут интересны и для инженеров,
поскольку такое приспособление организма к окружающей внешней среде
можно интерпретировать как решение им биогридродинамических задач,
поставленных перед ним окружающей средой или экологией. Анализ
методов решения живыми организмами подобных задач часто приводит к
результатам, которые открывают перед инженерной наукой новые
возможности. Исследователи, занимающиеся прикладными биологическими
науками, также могут почерпнуть из данной работы полезные сведения. Так,
медицина нуждается в информации о внутренней биогидродинамике
человека (и о тех млекопитающих, которые используются в медицинских
исследованиях в качестве подопытных животных). А при решении проблем
рационального использования естественных биоресурсов необходимы
сведения из различных разделов биогидродинамики для оптимального ведения
рыбного промысла, борьбы с вредителями, а также для использования в
ветеринарии.
Решение сложных биогидродинамических задач возможно лишь при
тесном сотрудничестве специалистов из различных областей научного
знания — биологии, математики, инженерной науки. Сложные движения жид-
1. Общие положения
3
кости, обусловленные наличием подвижных поверхностей организма,
некоторые из которых эластичны, требуют для анализа профессионального
опыта в решении гидродинамических задач, основанного на методах
математики и инженерии. Однако результаты такого анализа должны быть
тщательно и детально интерпретированы в биологическом контексте. Для того
чтобы исследование проблемы методами биогидродинамики было
результативным, необходимы совместные усилия математиков и инженеров с одной
стороны и физиологов и зоологов с другой. Описание таких исследований
неизбежно является результатом тесного сотрудничества указанных
участников исследования. В настоящей работе с математической точки зрения
описываются проблемы, в решении которых автор, как математик,
принимал либо непосредственное участие, либо на уровне консультаций.
Важность любой биогидродинамической задачи, будь то задача о
самодвижении организма в окружающей жидкости или о движении
жидкости в его сосудистой системе для осознания и осмысления биологических
функций различных групп животных, едва ли может быть переоценена.
Такой систематический подход позволяет в полной степени ответить на
фундаментальные вопросы зоологии о том, как действуют законы
биогидродинамики в организме. Кроме того, этот подход является важным источником
для понимания первоочередности тех или иных практических задач
медицины, ветеринарии, охраны окружающей среды. Однако при описании
методов математической биогидродинамики следует соблюдать баланс между
зоологическим аспектом проблемы и аналитическим изучением движения
жидкостей.
В настоящей книге автор постарался соблюсти этот баланс, предваряя
описание первой из четырех основных рассмотренных тем, всесторонним
сравнительным обзором (глава 2). Систематический обзор проблемы
движения животного в водной среде (главы 3 и 4) проводится с целью
осветить ту обширную область, которая будет исследована с позиций
биогидродинамики. Этот обзор можно рассматривать как математическое
приложение, в котором изложены теоретические основы (для движений при малых
и больших числах Рейнольдса соответственно) тех прикладных проблем,
о которых говорилось в сравнительном обзоре. В главах 5, 6 и 7 описаны
последние достижения в области анализа передвижения организма в
водной среде.
Что касается последних тем, то для них столь подробный полный
обзор не проводится. Надеемся, что материал главы 2 достаточен для того,
чтобы читатель смог получить общее представление о том, какие биогид-
4
Глава 1
родинамические методы здесь следует привлечь. Обзор последних тем
проведен достаточно кратко, с тем чтобы сразу перейти к их анализу.
Поэтому в главе 8 представлен краткий обзор способов полета представителей
животного мира, преимущественно насекомых и птиц, и предшествующий
ему анализ биогидродинамических аспектов такого движения. В большей
части задач, рассмотренных в данной теме, для исследования
эффективности движения возможно применение методов классической аэродинамики.
Однако в некоторых случаях для решения поставленной задачи требуется
развитие совершенно новых методов анализа, пример такой задачи
подробно рассмотрен в главе 9.
Во второй части книги речь идет о внутренней биогидродинамике
дыхательной и сосудистой систем. Причем внимание уделяется в основном
только позвоночным животным. Предварительный обзор (глава 10) в
большей своей части посвящен сравнению потоков жидкости в различных
частях дыхательной и сердечно-сосудистой систем человека, а также их
сравнению с потоками жидкости в трубах в различных технических системах.
А в главе 11 проводится сравнение циркуляции жидкой среды в сосудистых
системах различных классов позвоночных, в качестве примера
рассматриваются дыхательные системы птиц и млекопитающих и показывается, что
для их исследования требуется совершенно различный теоретический
аппарат.
И наконец, в главах 12, 13 и 14 изучаются исключительно
сердечно-сосудистые системы млекопитающих. Здесь особое внимание уделяется
прикладному аспекту проблемы. В частности, в главе 13 рассматривается
влияние сосудистых заболеваний на кровоток и обратное влияние. И в связи
с этим постановка задач биогидродинамики дается в медицинском и
биохимическом контексте. В главах 12 и 14 изучаются потоки жидкости в
сосудистой системе в случае нормальной физиологии в крупных и малых
кровеносных сосудах соответственно.
Подготовка данной книги заняла пять лет. Некоторые части книги
(а именно, главы 2, 4, 5, 10 и 14) были ранее опубликованы в виде
отдельных статей задолго до того, как это было запланировано настоящей
научной конференцией. Материал в этих статьях был изложен так, чтобы его
можно было использовать в дальнейшем без дополнительной переработки
в качестве составной части более общего обзора. Автор выражает особую
благодарность журналу Annual Reviews Inc. за разрешение перепечатки
главы 2, а также Международному центру механических наук в Удине, Италия
(International Centre for Mechanical Sciences at Udine), за разрешение пе-
2. Внешняя биогидродинамика
5
репечатки глав 10 и 14, которые в исходном виде были представлены как
лекции для Международного центра.
При написании еще двух глав преследовались двойные цели. Одной из
целей было изложение на конференции и включение в книгу по мотивам
выступлений на конференции, также были другие цели. Так, глава 6
представляет собой материал обзорной лекции на XIII Международном
конгрессе теоретической и прикладной механики, а глава 9 является статьей,
опубликованной в Journal of Fluid Mechanics. Остальные семь глав (1,3,
7, 8, 11, 12 и 13) были написаны специально для Научной конференции,
чтобы данная монография в наиболее полном виде освещала современное
состояние математической биогидродинамики, как его представляет себе
автор.
На заключительном этапе работы над книгой особо рассматривалась
возможность внесения изменений в те главы, которые ранее были
опубликованы в виде отдельных работ. И было принято окончательное решение
о том, что это нежелательно и излишне. Нежелательно потому, что читателя
мог бы смутить тот факт, что одна и та же работа существует в двух пусть
немного, но различающихся между собой формах, а излишне потому, что
все, что хотелось бы еще внести в качестве дополнения, можно добавить
в новые главы.
2. Внешняя биогидродинамика
Необходимый математический аппарат, который используется при
решении задач внешней биогидродинамики, изложен в двух дополняющих
друг друга книгах под редакцией Твайтса (Thwaites, 1960) и Розенхеда
(Rosenhead, 1963) по основам движения несжимаемой жидкости.
Основные сведения по биологическим аспектам передвижения животных в воде
приведены в работе Грея (Gray, 1968).
Проблема исследования передвижения животных в воде включает в
себя вопросы о самодвижении и управлении движением. Общий обзор
проблемы самодвижения и связанных с этим задач гидродинамики изложен
в главе 2. Этот обзор был составлен автором в сотрудничестве с
учеными зоологического факультета Кэмбриджского университета,
Британского музея естествознания (Лондон) и Морской биологической ассоциации
(Плимут).
Теоретические методы исследования поставленных задач
биогидродинамики основаны на теории движения тонкого тела в жидкости и газе,
6
Глава 1
причем основным предположением является то, что взаимодействие
каждого поперечного сечения тонкого тела с окружающей средой
определяется его скоростью относительно невозмущенной среды. В случае движения
при низких числах Рейнольдса (главы 3 и 7) сила взаимодействия между
поперечным сечением и жидкой средой, а при больших числах
Рейнольдса (главы 4 и 5) момент количества движения, связанный с поперечным
сечением тела, зависят от относительной скорости, что приводит нас
соответственно к теориям «резистивного» и «реактивного» типов.
Другие сведения из гидродинамики движения тел при больших числах
Рейнольдса привлекаются при изучении продвижения тел с серповидным
хвостом, часто называемым хвостом в виде полумесяца, которым
обладают большинство наиболее быстрых морских животных (Marshall, 1965
и глава 2). Для описания самодвижения животных, обладающих крыльями
с относительно большим удлинением, используются методы теории крыла
(глава 4). Кроме того, различные методы аэродинамики весьма эффективны
при решении задач о движении морских животных тяжелее воды (глава 2).
К задачам о движении тела при низких числах Рейнольдса
приводит изучение продвижения реснитчатых организмов, которое
обеспечивается совместным согласованным колебательным движением большого числа
микроскопических вытянутых элементов, называемых ресничками,
расположенных на поверхности организма. Исследования кембриджского
математика Блейка (Blake, 1971, 1972), побудительной причиной которых
явились экспериментальные работы протозоологов Лондонского и
Бристольского университетов, привели к статистически верному описанию
движения жидкости вблизи поверхности организмов. Это были одни из
первых работ, которые можно считать исходными для исследований в области
внутренней биогидродинамики за счет возможности изучения на их основе
движения жидкостей в трубках, поверхность которых покрыта ресничками
(глава 7).
При изучении маневренного движения рыб большой интерес
представляют не только задачи самодвижения, но также и проблемы управления
движением при больших числах Рейнольдса (Marshall, 1965, Weighs, 1972,
а также главы 6 и 7). Однако об управлении движением одноклеточных
организмов известно весьма немного и было бы желательным в будущем
привлечь научный интерес протозоологов к изучению всех таких фактов,
которые помогли бы пролить свет на механизмы управления движением.
Поступательный полет насекомых (в силу непроницаемости их
крыльев к изучению их полета можно привлекать методы аэродинамики) был
3. Внутренняя биогидродинамика
7
подробно исследован в совместной работе зоолога и аэродинамика (Weis-
Fogh, Jensen, 1956). Ряд других интересных проблем возникают при
исследовании полета птиц с учетом оперенности крыльев, а также при
исследовании парящего полета за счет использования воздушных потоков
(см. Brown, 1948, 1953, Gray, 1968). В главе 8 описаны ставшие уже
классическими исследования, а также результаты некоторых новых работ,
которые выходят за пределы изучения исключительно полетов птиц и в
которых поставлены довольно интересные задачи, относящиеся к
гидродинамике.
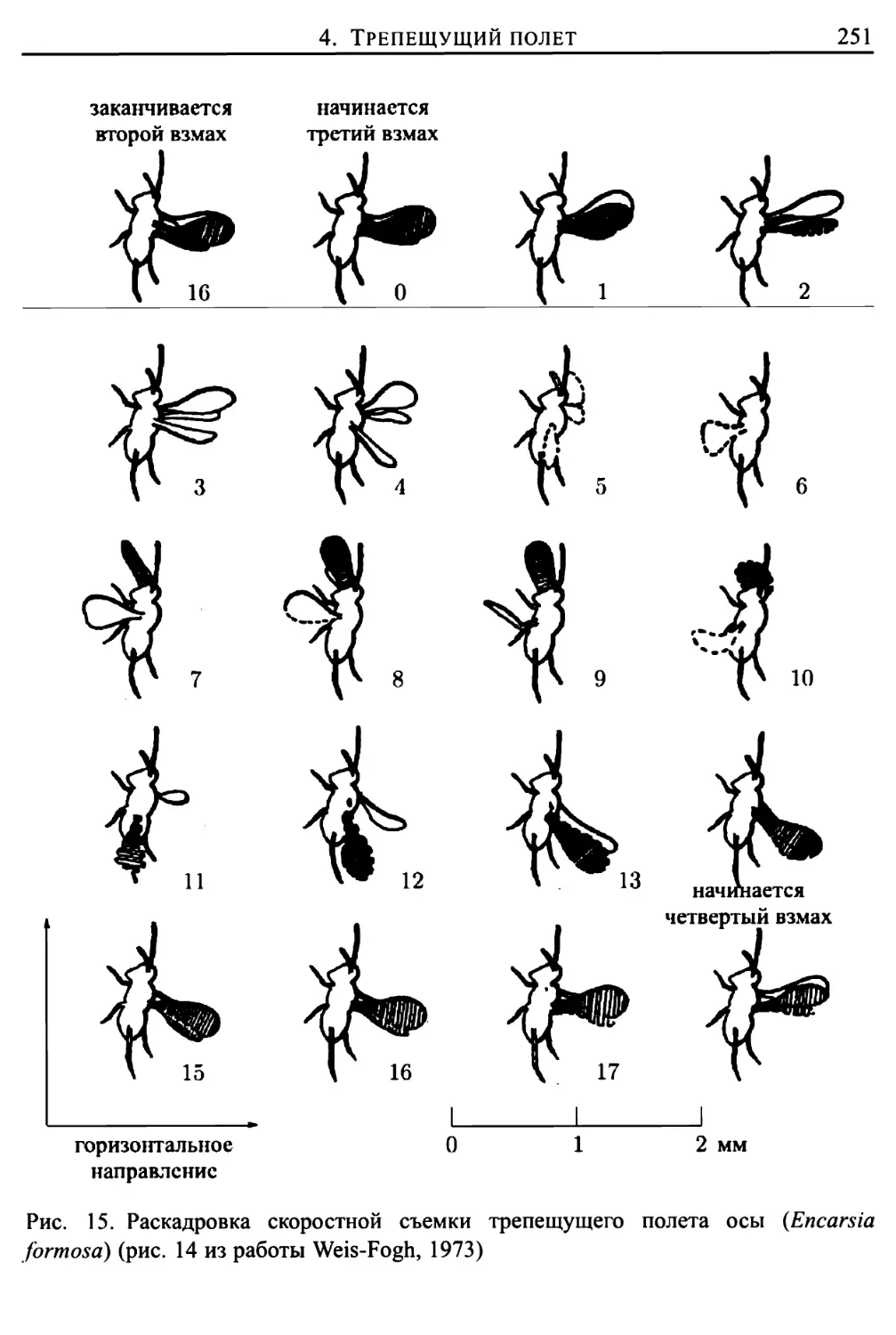
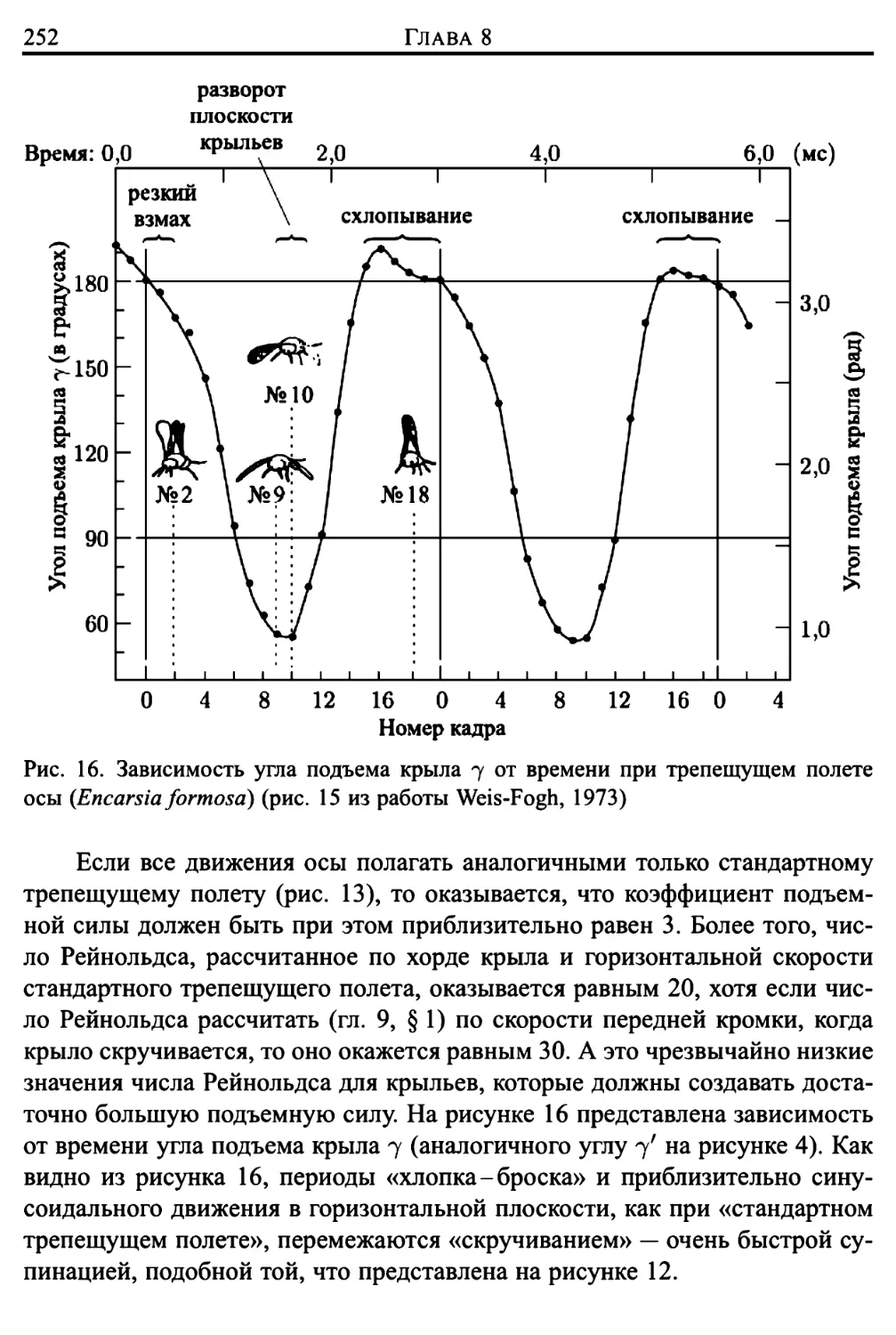
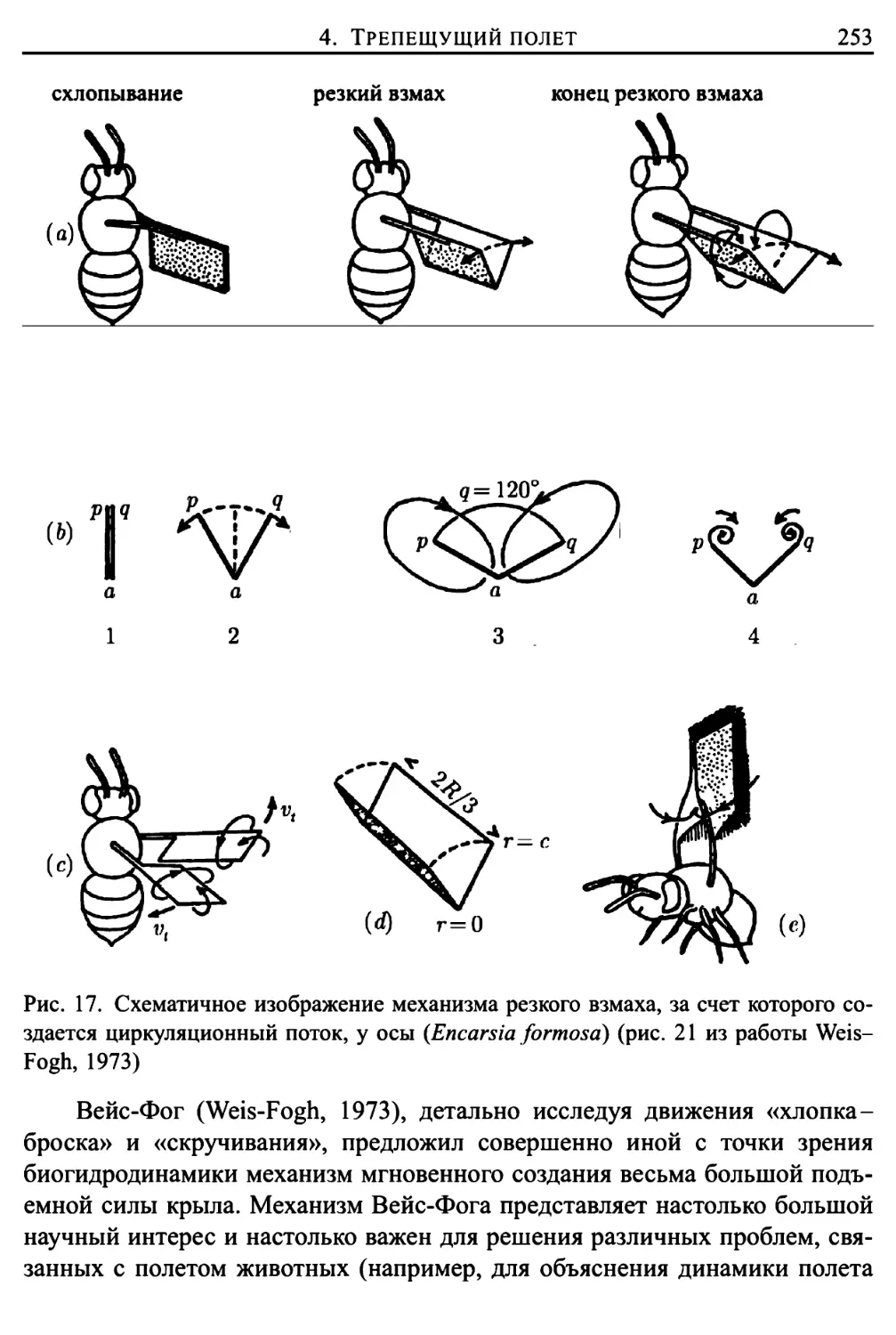
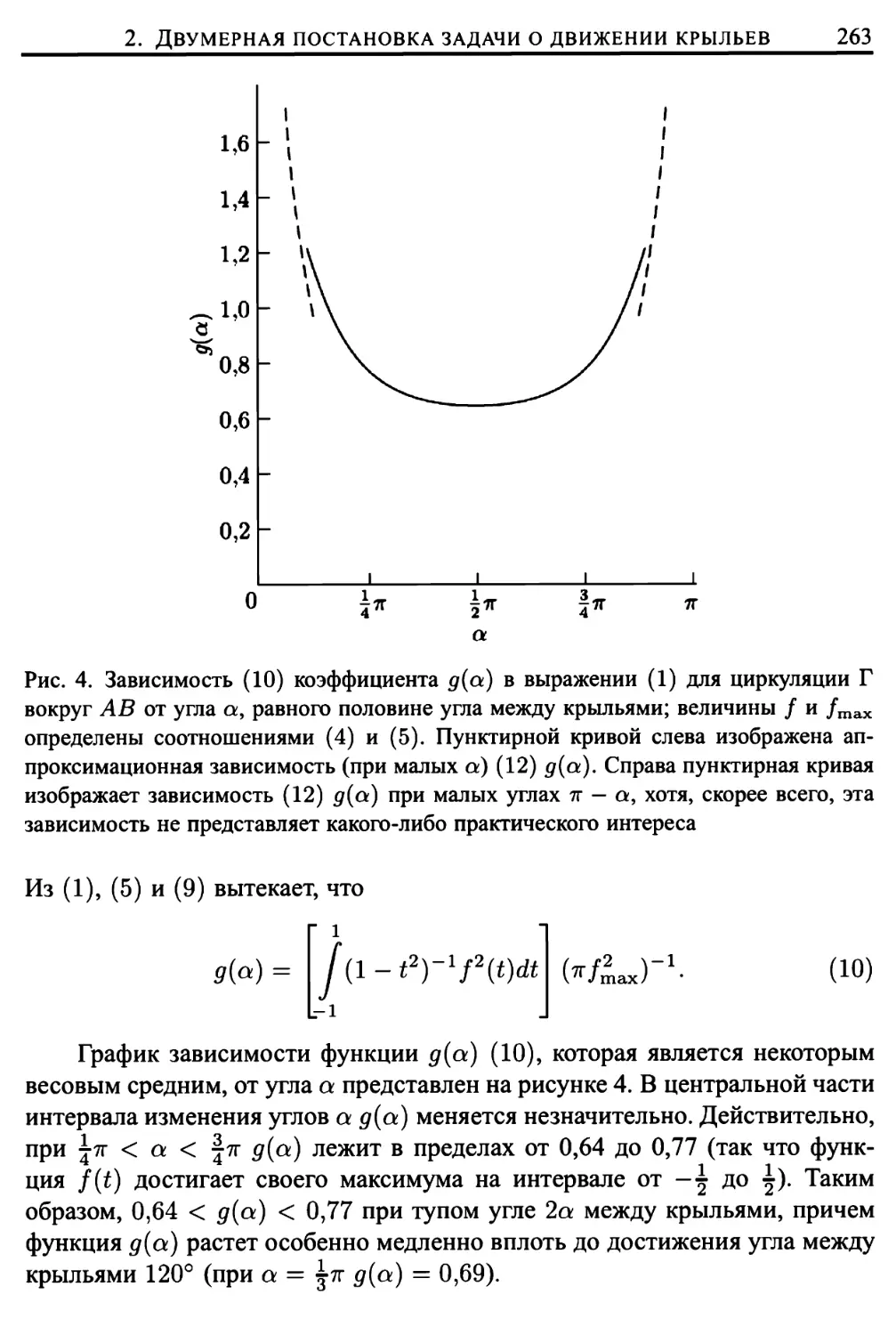
Здесь же представлено подробное исследование трепещущего полета
представителей животного мира (птиц, летучих мышей и многих отрядов
насекомых), проведенное Вайс-Фогом (Weis-Fogh, 1972, 1973) в
сотрудничестве с Лайтхиллом (математический аспект работы). Подавляющее
большинство животных в трепещущем полете поддерживают себя за счет
различных движений, «решая» при этом достаточно сложные задачи
классической аэродинамики. Однако имеются и весьма важные исключения, когда
полет более эффективно осуществляется за счет другого механизма
создания подъемной силы, техническая сторона которого еще не изучена
инженерами в должной степени (такой механизм использует, например,
крошечная оса Encarsia formosa). Подробное математическое описание подобного
механизма создания подъемной силы приведено в главе 9.
3. Внутренняя биогидродинамика
Внутренняя биогидродинамика человеческого организма подробно
описана в книге под редакцией Вольстенгольма и Найта (Wolstenholme,
Knight, 1969), а также ей частично посвящена и книга под редакцией Фанга,
Перрона и Анликера (Fung, Perrone, Anliker, 1972), в которой в основном
изложена «внутренняя биомеханика твердых биологических сред». Обе книги
представляют собой собрание докладов на междисциплинарных
симпозиумах с участием медиков, физиологов, инженеров и математиков. Вторая
часть этой книги описывает те области, которые автор разработал при
участии в работе отдела по исследованию физиологических потоков
Лондонского университета, возглавляемого доктором Каро.
Каро (Саго, 1966) подготовил к печати книгу, содержащую много
интересных работ, посвященных проблемам физиологии и механики дыхания
млекопитающих. Последние достижения в этой области ученых отдела по
исследованию физиологических потоков Лондонского университета описа-
8
Глава 1
ны в работах Педли и др. (Pedley et al., 1970, 1972). Главы 11 и 14
посвящены некоторым проблемам кровотока и газообмена в легких, которые по
возможности проанализированы с математической точки зрения.
Много интересных биогидродинамических проблем, связанных с
дыханием позвоночных, освещены в обзоре Пиипера (Piiper, 1972). В главе 11
обсуждается механизм жаберного дыхания рыб, исследуются воздушные
потоки в жабрах. Этот материал также можно рассматривать как
переходный к внешней гидродинамике движения рыб, поскольку воздушные
потоки в жабрах приводят к образованию пограничного слоя на поверхности
тела рыбы (глава 2). Разделы обзора Пиипера, посвященные дыхательной
системе птиц, особенно интересны с точки зрения биогидродинамики, а
именно процесс газообмена, обеспеченный однонаправленным потоком воздуха
в большей части легкого птицы. Это явление подробно обсуждается с
использованием материалов междисциплинарного симпозиума в 1973 году,
посвященных этой теме.
Наиболее значительные достижения в анализе сложного движения
в сердечно-сосудистой системе млекопитающих были описаны в
совместной работе Уомерсли (Womersley, 1955) и МакДональда (McDonald, 1960),
математика и физиолога, которые в тесном сотрудничестве работали над
этой проблемой. При анализе того, каким образом такая сложная по своему
составу жидкость, как кровь, движется под действием сердечных
сокращений по разветвленной системе эластичных трубок, встречается немало
трудностей. Уомерсли и МакДональд занимались в основном локальной
биогидродинамикой — движением в одном или двух сосудах и их
ответвлениях, в то время как их коллега Тейлор (Taylor, 1966) исследовал
кровеносную систему в целом (см. главу 12).
Во второй части книги приведены некоторые попытки продвинуться
далее в этих, ставших теперь классическими, исследованиях. Так, в
главе 10 в данной проблеме учитываются геометрические аспекты, что ранее
не делалось; например, можно учесть влияние кривизны (например, у
дуги аорты), используя теорию вторичного потока (McConalogue, 1970) или
различные эффекты, связанные с учетом длины аорты.
В главе 13 обсуждаются некоторые взаимосвязи между указанными
задачами биогидродинамики и проблемами возникновения
сердечно-сосудистых заболеваний. Так, атеросклероз тесно связан с перераспределением
потока крови. В работе Каро и др. (Саго et al., 1971) приводятся
некоторые идеи относительно таких взаимосвязей. Довольно подробно эти связи
освещены в главе 13 на основе материалов недавнего междисциплинарного
симпозиума под председательством автора.
3. Внутренняя биогидродинамика
9
И наконец, вновь возвращаемся (глава 14) к движению при низких
числах Рейнольдса в задачах, связанных как с микроциркуляцией
жидкости в системе кровообращения в легких (функционирование которой
связано с легочным дыханием), так и с системной микроциркуляцией. Такое
движение имеет место в тех местах, где становится значимым состав
крови, в этом случае кровь рассматривается как суспензия красных кровяных
телец и других составляющих. В сборниках статей под редакцией Коп-
ли (Copley, 1968) и Вольстенхольма и Найта (Wolstenholme, Knight, 1969)
имеется большое количество работ, в которых исследуется процесс
перераспределения частиц в потоке крови при ее движении по мелким
артериям (артериолам) и капиллярам. Кроме того, в этих работах
рассматриваются газообмен и обмен веществ в периферической системе
кровообращения, а также газообмен в легких. При движении крови в капиллярах,
сосудах весьма узких, красные кровяные тельца проходят через
капилляры одно за одним, часто при этом значительно деформируясь. Такой тип
движения при низких числах Рейнольдса рассматривался во многих
математических исследованиях (Fitz-Gerald, 1969, Aroesty, Gross, 1970). В
заключительной части работы приведен обзор последних достижений в этой
области.
Автор, посвятивший большую часть своей жизни гидродинамике,
попытался в данной книге показать, что в течение последних двух десятков
лет возник новый раздел гидродинамики, а именно биогидродинамика,
которая встала в один ряд с геофизической гидродинамикой, гидравликой и др.
В силу ограниченного объема данной книги автор смог только очертить
границы этой весьма важной новой области научного знания. Однако он
надеется, что ему все же удалось показать, сколь много нового и
захватывающе интересного откроется тому, кто готов углубиться в эту область.
Литература
[1] Aroesty, J. & Gross, J. 1970. Convention and Diffusion in the
Microcirculation, Rand Corp. Rep. RM-6214-NIH.
[2] Blake, J. R. 1971. A note on the image system for a Stokeslet in a no-slip
boundary, Proc. Camb. Phil. Soc. 70, 303-310.
[3] Blake, J. R. 1972. A model for the micro-structure in ciliated organisms,
J. Fluid Mech. 55, 1-23.
[4] Brown, R. H. J. 1948 & 1953. The flight of birds, J. Exptl. Biol. 25, 322-333
& 30, 90-103.
Ш Глава 1
Саго, C.G. (ed.) 1966. Advanced in Respiratory Physiology, Edward
Arnold, London.
Caro, C.G., Fitz-Gerald, J.M. & Schroter, R.C. 1971. Atheroma and
arterial wall shear, Proc. Roy. Soc. B, 177, 109-159.
Copley, A. L. (ed.) 1968. Hemorheology, Pergamon, London.
Fitz-Gerald, J. M. 1969. Mechanics of real-cell motion through very narrow
capillaries, Proc. Roy. Soc. B, 174, 193-227.
Fung, Y. C, Perrone, N. & Anliker, M. (ed.) 1972. Biomechanics: its
Foundations and Objectives, Prentice-Hall, Englewood Cliffs, N. J.
Gray, J. 1968. Animal Locomotion, Weidenfeld & Nicolson, London.
Marshall, N. B. 1965. The Life of Fishes, Weidenfeld & Nicolson, London.
McConalogue, D.J. 1970. The effects of secondary flow on the laminar
dispersion of an injected substance in a curved tube, Proc. Roy. Soc. A,
315,99-113.
McDonald, D. A. 1960. Blood Flow in Arteries, Edward Arnold, London.
Pedley, T.J., Schroter, R.C. & Sudlow, M.F. 1970. Energy losses and
pressure drop in models of human airways, Respiration Physiol. 9,
371-386.
Pedley, T. J., Sudlow, M.F. & Milic-Emili, J. 1972. A non-linear theory of
the distribution of pulmonary ventilation, Ibid. 15, 1-39.
Piiper, J. (ed.) 1972. Proceedings of the Symposium «Comparative
Physiology of Respiration in Vertebrates», Ibid. 14, 1-236.
Rosenhead, L. (ed.) 1963. Laminar Boundary Layers, Oxford University
Press.
Taylor, M. G. 1966. Wave transmission though an assembly of randomly
branching elastic tubes, Biophys. J. 6, 697-716.
Thwaites, B. (ed.) 1960. Incompressible Aerodynamics, Oxford University
Press.
Weighs, D. 1972. A hydrodynamical analysis offish turning manoeuvres,
Proc. Roy. Soc. B, 182, 59-72.
Weis-Fogh, T. & Jensen, M. 1956. Biology and physics of locust flight,
Phil. Trans. Roy. Soc. B, 239, 415-584.
Weis-Fogh, T. 1972. Energetics of hovering flight in hummingbirds and
drosophila, J. Exptl. Biol. 56, 79-104.
Weis-Fogh, T. 1973. Quick estimates of flight fitness in hovering animals,
including novel mechanisms for lift production, Ibid. 59, 169-230.
3. Внутренняя биогидродинамика
11
[24] Wolstenholme, G. E. & Knight, J. (ed.) 1969. Circulatory and Respiratory
Mass Transport, J. & A. Churchill, London.
[25] Womersley, J. R. 1955. Method for the calculation of velocity, rate of
flow and viscous drag in arteries when the pressure gradient is known.
J. Physiol. 127, 553-563.
Замечание, добавленное при чтении корректуры. Эдвард Арнольд
(Edward Arnold) опубликовал второе переработанное издание работы
D. A. McDonald, Blood Flow in Arteries в 1974 году, в это же время
была запланирована публикация в десятом юбилейном номере Bulletin
of the Institute of Mathematics and its Applications работы М. Дж. Лайт-
хилла Aerodynamic aspects of animal flight, где приводится значительно
более подробный сравнительный тематический обзор.
Часть I
Внешняя биогидродинамика
Глава 2
Гидромеханика самодвижения водных
животных. Обзор1
1. Область исследования
Основной целью данной работы является исследование самодвижения
водных животных с точки зрения гидромеханики. Здесь не будет
рассматриваться движение в воде представителей животного мира, которые не
живут в ней постоянно (т. е., к примеру, достаточно неумелое плавание Homo
sapiens). Кроме того, не будет изучаться движение водных животных в
среде, которая не является водной полностью или хотя бы частично, например
ползание по дну или зарывание в нору донных животных, или
передвижение по некоторой поверхности, или какая-либо другая ситуация, в которой
водное животное на время оставляет водную среду для перемещения по
суше, для совершения прыжка, скольжения по поверхности воды или для
полета.
Исследование передвижения с позиций биомеханики концентрируется
на водной среде, окружающей животное, и возмущениях, которые в ней
происходят при перемещении в ней животных (а где необходимо,
рассматривается движение водной среды и внутри животного). И в то же время
сравнительно мало сказано как о внутренней биомеханике животного, так
и об органах координации и побуждения к движениям, которые включают
органы чувств, нервную систему, а также мускулы и другие органы,
обеспечивающие движение животного. Более того, в области гидромеханики
мы будем рассматривать в большей степени проблемы самодвижения, а не
проблемы устойчивости и управления.
Даже при таких ограничениях область наших исследований будет
достаточно интересной и актуальной, поскольку в результате
эволюционного развития животных в водной среде в течение 109 лет закреплялись те
1 Перепечатывается с разрешения правообладателя из Annual Review of Fluid Mechanics.
Volume 1, pages 413-446. © 1969 by Annual Reviews Inc. Все права защищены.
16
Глава 2
специфические изменения в организме, которые повышали выживаемость
вида и его способность в воспроизводству. В результате чего с
неизбежностью возникали приспособления для осуществления быстрого движения
с наименьшей затратой энергии, которые заслуживают тщательного
изучения. И все это происходило потому, что в пищевой цепи или во
взаимоотношениях в связке «жертва-хищник» способность к быстрым перемещениям
жертвы или хищника или способность к быстрому преодолению
расстояния между жертвой и хищником является основным фактором,
обеспечивающим выживание вида.
Несомненно, что и другие факторы при некоторых воздействиях со
стороны окружающей среды оказываются более предпочтительными, и
эволюция может пойти в направлении уменьшения мобильности вида
(например, за счет образования брони у животного), приводящем к повышению
выживаемости. В данной работе рассмотрены отдельные подобные случаи
(см. раздел 11), но в основном мы сконцентрируемся на достаточно
большой группе представителей животного царства, которые способны
перемещаться в воде со скоростью, сравнимой с той, которую наземные
животные такой же массы могут развить и сохранить в течение некоторого
времени.
Вероятнее всего, большинство читателей обладают специальными
знаниями только в одной из областей — либо в области зоологии, либо в
области механики жидкости, а не одновременно в этих двух областях. Автор
постарался в меру своих сил подать материал так, чтобы он был доступен
и тем и другим.
2. Самодвижение беспозвоночных в воде
Из всего многообразия типов или главных подразделов животного
царства (большинство современных классификаций выделяют порядка 20
типов) мы рассмотрим самодвижение характерных представителей из 10
типов (см. таблицу 1), и в первую очередь представителей простейших. Из
этих одноклеточных организмов особое внимание будет уделено классу
жгутиковых, которые продвигаются за счет волнообразного движения
плетевидных жгутиков, строение которых подобно строению сперматозоидов
высших животных. Основным вопросом, который обсуждается в данном
исследовании, является эффективность волнообразного способа
продвижения и его различных модификаций, присущего практически всем наиболее
быстрым водным животным.
2. Самодвижение беспозвоночных в воде
17
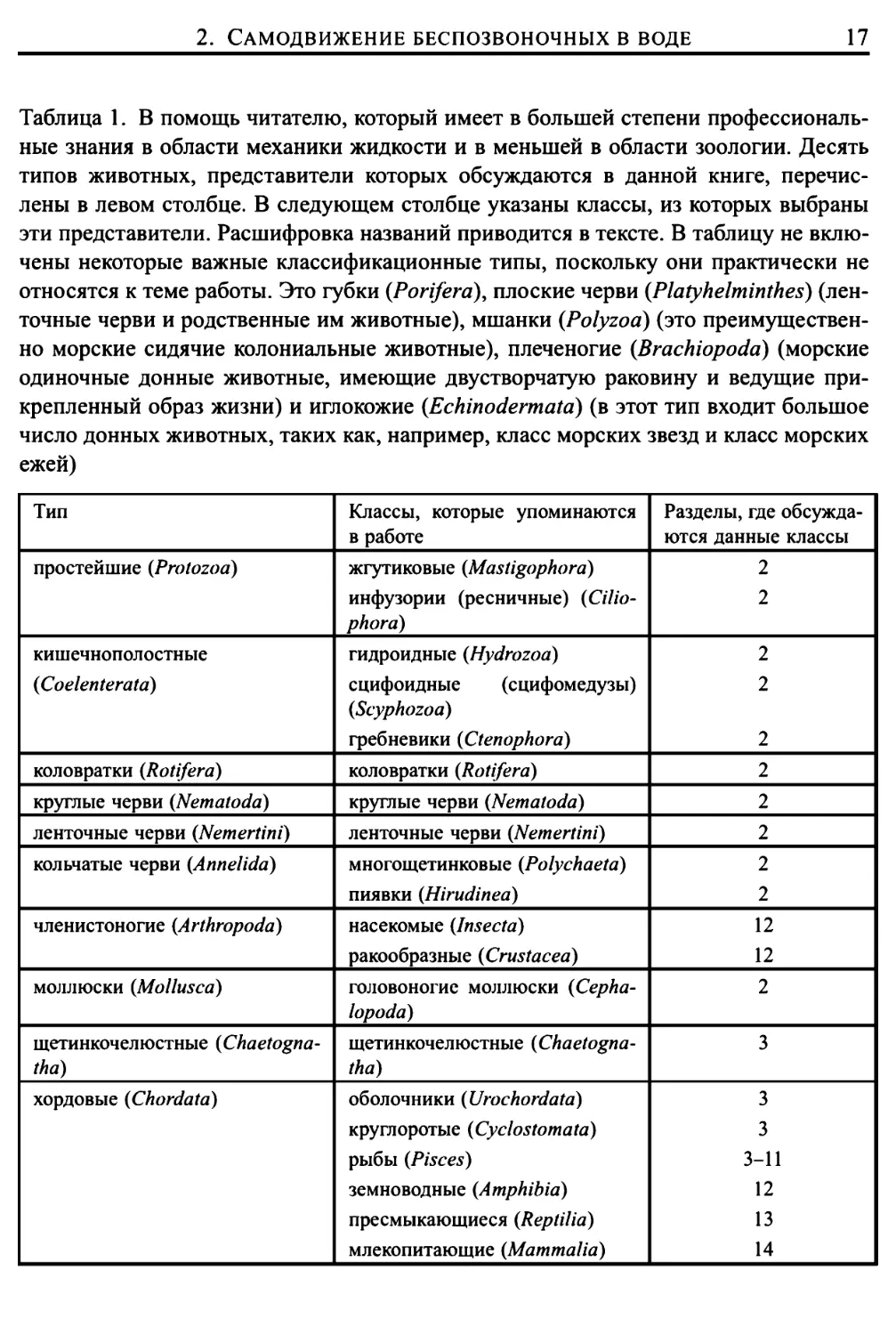
Таблица 1. В помощь читателю, который имеет в большей степени
профессиональные знания в области механики жидкости и в меньшей в области зоологии. Десять
типов животных, представители которых обсуждаются в данной книге,
перечислены в левом столбце. В следующем столбце указаны классы, из которых выбраны
эти представители. Расшифровка названий приводится в тексте. В таблицу не
включены некоторые важные классификационные типы, поскольку они практически не
относятся к теме работы. Это губки (Porifera), плоские черви (Platyhelminthes)
(ленточные черви и родственные им животные), мшанки (Polyzoa) (это
преимущественно морские сидячие колониальные животные), плеченогие (Brachiopoda) (морские
одиночные донные животные, имеющие двустворчатую раковину и ведущие
прикрепленный образ жизни) и иглокожие {Echinodermatd) (в этот тип входит большое
число донных животных, таких как, например, класс морских звезд и класс морских
ежей)
Тип
простейшие (Protozoa)
кишечнополостные
(Coelenterata)
\ коловратки (Rotifera)
\ круглые черви (Nematoda)
\ ленточные черви (Nemertini)
кольчатые черви (Annelida)
членистоногие (Arthropoda)
моллюски (Mollusca)
щетинкочелюстные (Chaetogna-
tha)
хордовые (Chordata)
Классы, которые упоминаются
в работе
жгутиковые (Mastigophora)
инфузории (ресничные) (ОПо-
phora)
гидроидные (Hydrozoa)
сцифоидные (сцифомедузы)
(Scyphozoa)
гребневики (Ctenophora)
коловратки (Rotifera)
круглые черви (Nematoda)
ленточные черви (Nemertini)
многощетинковые (Polychaeta)
пиявки (Hirudinea)
насекомые (Insecta)
ракообразные (Crustacea)
головоногие моллюски
(Cephalopoda)
щетинкочелюстные (Chaetogna-
tha)
оболочники (Urochordata)
круглоротые (Cyclostomata)
рыбы (Pisces)
земноводные (Amphibia)
пресмыкающиеся (Reptilia)
млекопитающие (Mammalia)
Разделы, где
обсуждаются данные классы |
2
2
2
2
2
2
2
2
2
2
12
12
2
3
3
3
3-11
12
13
14
18
Глава 2
Относительно менее эффективными в эволюционном смысле
(гидромеханические причины этого будут рассмотрены ниже) являются два
других основных способа перемещения водных животных, которые мы
рассмотрим. Один из них (этот способ в дальнейшем будет изучен с позиций
гидромеханики) представляет собой перемещение организма за счет
движения большого количества ресничек (см. главным образом работу (Gray,
1928)), как это происходит у представителей типа простейших из класса
инфузорий, или ресничных. Отметим, что в типе кишечнополостных
(который включает медуз и актиний) представители класса гребневиков
обладают хорошо развитыми восемью образованиями, которые получили
название гребных пластинок и которые обеспечивают самодвижение. Каждая
гребная пластинка состоит из сросшихся колеблющихся ресничек. А вот
большинство представителей типа коловраток продвигаются за счет двух
венчиков ресничек вокруг ротовой полости.
Другим альтернативным способом волнообразного перемещения в
воде, кроме упомянутого перемещения за счет согласованного колебания
ресничек, является реактивное движение. Медузоидные формы двух классов
кишечнополостных — гидроидные и сцифоидные (сцифомедузы) — имеют
характерный (как у медуз) зонтик, и при сокращении мышц нижней
полости зонтика из нее выбрасывается струя воды, за счет чего животное
медленно продвигается в противоположном направлении. Аналогичное
реактивное движение встречается и у представителей моллюсков — животных
довольно высокого уровня организации. У кальмаров (класс головоногих)
вода выталкивается через суженную воронку, в результате чего хорошо
обтекаемое тело кальмара может двигаться с достаточно большой скоростью
(от 2 до 4 м/с в зависимости от размера).
Большинство головоногих моллюсков (в частности, десятиногие
кальмары) имеют плавники, которые обеспечивают волнообразный тип
движения для маневрирования на относительно небольших скоростях, а
внезапное изменение направления движения обеспечивается за счет
выбрасывания струи воды из воронки в передней части тела кальмара. У некоторых
кальмаров обычным способом передвижения является периодическое
быстрое выталкивание через воронку воды, которая предварительно медленно
вбирается в мантийную полость. Сикменн (Siekmann, 1963) исследовал эту
форму движения с позиций гидромеханики. Ранее проведенные
исследования в рамках классической гидромеханики самодвижения показали, что
довольно малая часть затрачиваемой на создание струи энергии идет
непосредственно на совершение полезной работы по продвижению животного.
2. Самодвижение беспозвоночных в воде
19
Дополнительно отметим, что тип хордовые (Chordata), в который
входят все позвоночные и другие животные, имеющие внутренний скелетный
хрящ, представители класса оболочников (Urochordata) (асцидии и саль-
пы, безголовые, а также соответственно беспозвоночные и, конечно же, те
животные, у которых спинная струна (хорда) совершенно исчезает при
превращении их во взрослую особь), втягивают воду сквозь жаберные
отверстия и выталкивают ее из полости в области сердца. За счет этого
свободно плавающие животные из отряда пелагических оболочников (Thaliacea)
могут перемещаться в воде достаточно быстро. Среди позвоночных
реактивное движение встречается достаточно редко. Так, движение рыб в воде,
как это отмечается в разделе 9, не является реактивным движением, а
выталкивание рыбой воды через жаберные щели приводит в основном лишь
к уменьшению сопротивления окружающей жидкости за счет образования
пограничного слоя.
Итак, как уже отмечалось, волнообразное перемещение наряду с его
модификациями следует рассматривать как основной способ передвижения
водных животных. Одной из причин этого является то, что такой способ
слабо чувствителен к масштабам (размерам животного). На языке
механики жидкости это означает, что подобное движение может происходить при
самых различных числах Рейнольдса
R = Ul/v, (1)
где U и I — скорость (мм/с) и длина (мм) животного соответственно, a v —
кинематическая вязкость (вязкость, деленная на плотность), которая для
большинства встречающихся в природе водных сред лежит в пределах
от 1 до 2 мм2/с.
Таким образом, как правило, реактивное движение происходит при
достаточно больших числах Рейнольдса (в этом случае силы инерции
преобладают над силами вязкости), а передвижение в воде за счет согласованного
колебания ресничек может происходить только при довольно малых
числах Рейнольдса, в общем случае волнообразное движение животных в воде
осуществляется в достаточно широком диапазоне чисел Рейнольдса.
Поэтому волнообразное движение может осуществляться животными самых
разных размеров. Например, для жгутикового одноклеточного организма
число Рейнольдса имеет порядок 10~3. За счет поперечных колебаний
жгутика, распространяющихся от его основания со скоростью V и с
возрастающей амплитудой, организм движется вперед со скоростью О,2V (Gray,
Hancock, 1955).
20
Глава 2
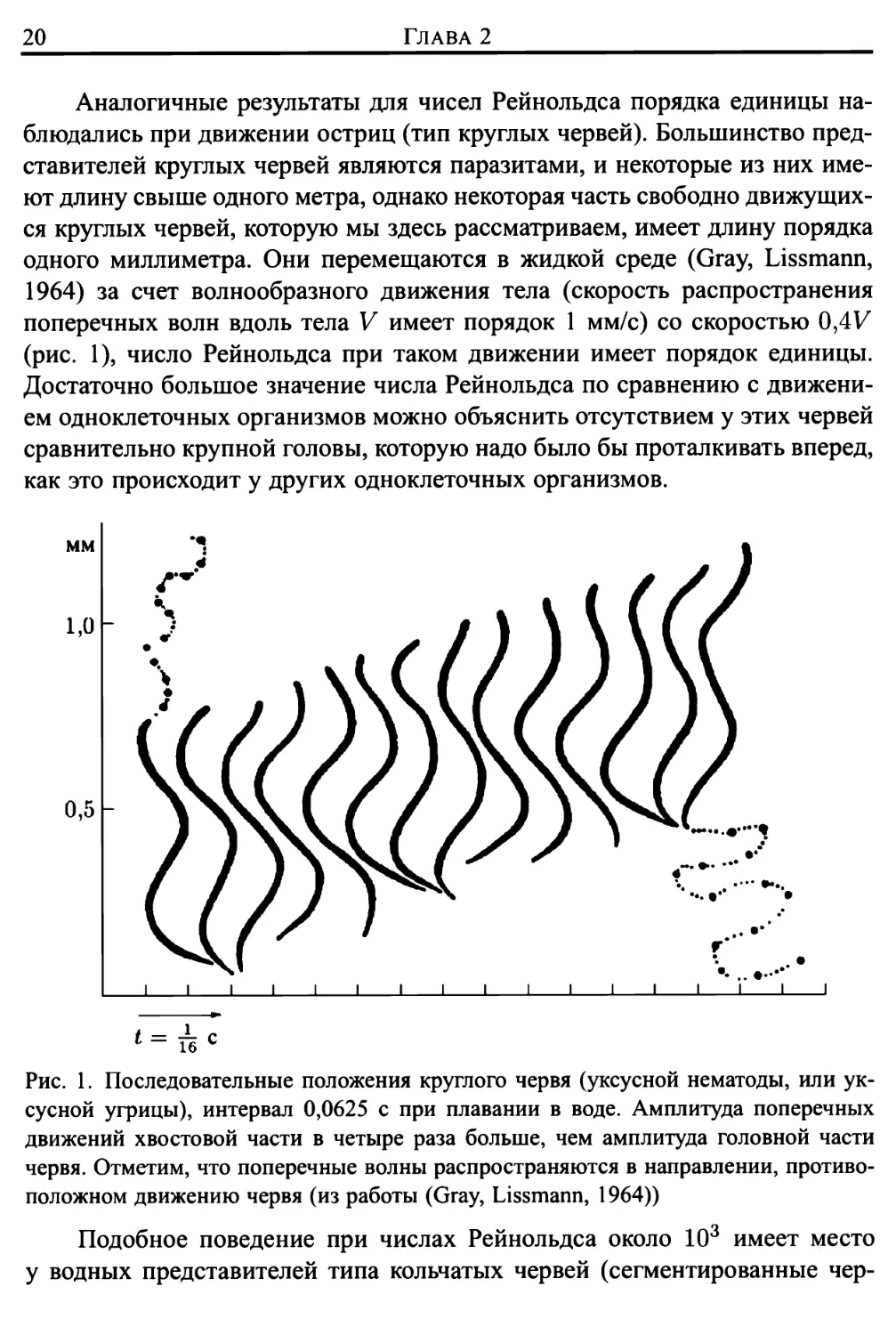
Аналогичные результаты для чисел Рейнольдса порядка единицы
наблюдались при движении остриц (тип круглых червей). Большинство
представителей круглых червей являются паразитами, и некоторые из них
имеют длину свыше одного метра, однако некоторая часть свободно
движущихся круглых червей, которую мы здесь рассматриваем, имеет длину порядка
одного миллиметра. Они перемещаются в жидкой среде (Gray, Lissmann,
1964) за счет волнообразного движения тела (скорость распространения
поперечных волн вдоль тела V имеет порядок 1 мм/с) со скоростью 0,4К
(рис. 1), число Рейнольдса при таком движении имеет порядок единицы.
Достаточно большое значение числа Рейнольдса по сравнению с
движением одноклеточных организмов можно объяснить отсутствием у этих червей
сравнительно крупной головы, которую надо было бы проталкивать вперед,
как это происходит у других одноклеточных организмов.
мм
1,0 h
0,5 Ь
1=Гь*
Рис. 1. Последовательные положения круглого червя (уксусной нематоды, или
уксусной угрицы), интервал 0,0625 с при плавании в воде. Амплитуда поперечных
движений хвостовой части в четыре раза больше, чем амплитуда головной части
червя. Отметим, что поперечные волны распространяются в направлении,
противоположном движению червя (из работы (Gray, Lissmann, 1964))
Подобное поведение при числах Рейнольдса около 103 имеет место
у водных представителей типа кольчатых червей (сегментированные чер-
3. Введение в кинематику плавания рыб
21
ви), имеющих длину порядка 100 мм и плавающих со скоростью
приблизительно 10 мм/с. Например, представители класса пиявок с гладкой
поверхностью тела при плавании волнообразно изгибают свое тело, и
амплитуда поперечных колебаний тела возрастает от головы к задней части
(Gray, 1939a), скорость продвижения пиявок в воде составляет 0,3V. Грей
отмечает такое же поведение у свободноплавающих представителей типа
ленточных червей (несегментированные черви с удлиненной головной
частью (хоботом), которая может втягиваться внутрь тела). Одна из наиболее
важных модификаций волнообразного способа продвижения была
обнаружена при исследовании водных представителей другого класса кольчатых
червей — класса многощетинковых.
Такие многощетинковые черви совершают реверсивные колебательные
движения (Gray, 1939b) — поперечные колебания распространяются вдоль
тела от задней части животного к его головной части с возрастающей
амплитудой. Животное движется в направлении распространения поперечных
волн. Отметим, что скорость перемещения животного в воде U составляет
около —0,2\Л Такое же реверсивное распространение поперечных
колебаний наблюдается у представителей рода многощетинковых червей
семейства нереид (Nereis) (рис. 2), у которых параподий (мускулистые выросты
на теле многощетинковых червей, расположенные попарно на каждом
сегменте туловища) необходимы главным образом в качестве органов
движения, причем, обычно они работают как «вёсла». Продвижение
осуществляется за счет волнообразного движения параподий с каждой стороны тела
червя, причем параподий удлиняются на пиках волн и укорачиваются во
впадинах. Реверсивное движение (от задней части к головной) также
наблюдается и у представителей рода нефтиды, у которых параподий
остаются пассивными. Тейлор (Taylor, 1952) первым исследовал механический
аспект реверсивных волн (см. раздел 13), образуемых движением параподий.
3. Введение в кинематику плавания рыб
Гидродинамический анализ волнообразного способа перемещения
в воде довольно гладких и имеющих вытянутую форму животных при
достаточно больших числах Рейнольдса (с использованием теории движения
тонких тел, см. раздел 4) показывает существенное преимущество
поперечного сжатия (уплощения) задней части тела животного, так что конец тела
приобретает форму широкого лезвия, расположенного под прямым углом
как к продольной оси животного, так и к направлению поперечных коле-
22
Глава 2
Рис. 2. Последовательные положения (интервал 0,05 с) многощетинкового червя
семейства нереид при движении влево. Числа над движущимися гребнями
нанесены для демонстрации того, что поперечные волны тела червя движутся в том же
направлении, что и сам червь, хотя и значительно быстрее (Gray, 1939b)
баний, обеспечивающих продвижение животного. Здесь говорится, скорее,
о «поперечном» уплощении, чем о «сжатии с боков» (которая встречается
у рыб, земноводных, премыкающихся), для того чтобы подчеркнуть, что
уплощенная часть расположена так, чтобы ее возвратно-поступательное
движение обеспечивало продвижение животного. Когда поперечное
движение происходит в дорсовентральном направлении (от спины к животу), т. е.
3. Введение в кинематику плавания рыб
23
в вертикальной плоскости (как это имеет место у китообразных
млекопитающих и некоторых ракообразных), то и уплощение задней части тела будет
также дорсовентральным (т.е. в направлении вертикали она будет более
плоской).
Уже у ранних форм позвоночных имеется поперечное уплощение
задней части тела, дающее им преимущество при передвижении по
сравнению с беспозвоночными (уплощение задней части тела достаточно
редко встречается среди беспозвоночных, хотя у небольшой изолированной
группы щетинкочелюстных (тип морских хищных беспозвоночных) —
морских стреловидных червей — такое уплощение имеет место). В частности,
единственным современным позвоночным классом бесчелюстных
являются круглоротые (Cyclostomatd) (миноги и миксины). Они обладают
вытянутыми вдоль всего тела спинным и грудным плавниками, площадь
поперечного сечения тела уменьшается к хвостовой части и становится
практически равной нулю, а совокупная высота тела и плавника остается постоянной
(как у морского угря, см. рис. 4). Преимущество такого строения тела
становится очевидным с точки зрения динамики движения в воде вытянутого
тела. Представители высшего класса позвоночных — класса рыб, как
правило с более сложным расположением плавников, используют
усовершенствованный способ плавания за счет усложненного строения хвостового
плавника.
Ранее способы плавания рыб кратко описывались в связи с
упоминанием других классов водных животных. Дадим теперь более подробное
описание наиболее характерных способов плавания представителей
некоторых отрядов класса рыб. Класс рыб насчитывает приблизительно 30
отрядов (Marshall, 1965). В данной работе ограничимся семнадцатью
отрядами (таблица 2). Сначала обсудим те отряды, у которых способы
плавания несильно отличаются от плавания жгутиковых, круглых червей, пиявок,
миног, а затем опишем модификации этих способов за счет: (а)
возрастания интенсивности движения хвостовой части, (Ь) изменения
геометрических размеров и пропорций хвостовой части, (с) специфических изменений
строения рыб, которые тяжелее воды, а также рыб, покрытых панцирем,
вследствие чего утративших присущую другим видам гибкость.
Остановимся сначала на типичном расположении плавников у
представителей класса рыб и ознакомимся с терминологией, используемой для
описания плавников (рис. 3). К плавникам, расположенным в вертикальной
плоскости симметрии, относятся: хвостовой плавник, который играет
наиболее важную роль в обеспечении продвижения рыбы; спинной плавник,
24
Глава 2
4» * • • •
ч
Sf
Рис. 3. Представитель отряда лососевых Salmo salar (солнечный лосось).
Схематичное изображения внешнего вида рыбы (Marshall, 1965)
за счет которого эффективно увеличивается размер рыбы в наивысшей
точке ее тела; и расположенные симметрично ему на нижней части тела один
или более плавников, называемых анальными, если они находятся близко
к хвостовому плавнику рыбы, или брюшными (вентральными), если они
находятся достаточно далеко от хвостового плавника. Согласованное
движение спинного и брюшных плавников с хвостовым обеспечивают
стабилизацию тела рыбы при резком изменении направления движения. Кроме
того, как это будет показано в разделе 5, за счет движения этих плавников
существенно повышается эффективность движения рыб в воде.
Вне вертикальной плоскости симметрии в передней части
расположены парные грудные плавники, играющие весьма важную роль в движении
рыбы, а ближе к хвостовой части могут быть (хотя и не всегда)
расположены играющие уже не столь значительную роль парные анальные
плавники. Возможно, наиболее простой функцией из всех, выполняемых парными
плавниками (Harris, 1938), является функция стабилизации тела при крене
и продольных наклонах. В дальнейшем будет дополнительно описано
множество других функций грудных плавников у различных групп рыб (и это
множество функций грудных плавников могло бы быть значительно более
обширным, если бы в нашу задачу входило исследование движения
животных не только в водной среде).
В качестве иллюстрации к сказанному отметим такие специфические
проблемы при движении, как резкая остановка и резкий старт (хотя в
дальнейшем будет рассматриваться только движение рыб с постоянной
скоростью). При низких числах Рейнольдса (менее единицы), когда преобладают
силы вязкости, эти проблемы не возникают: животное останавливается
сразу же, как только оно прекращает проталкивать себя, и при начале движения
3. Введение в кинематику плавания рыб
25
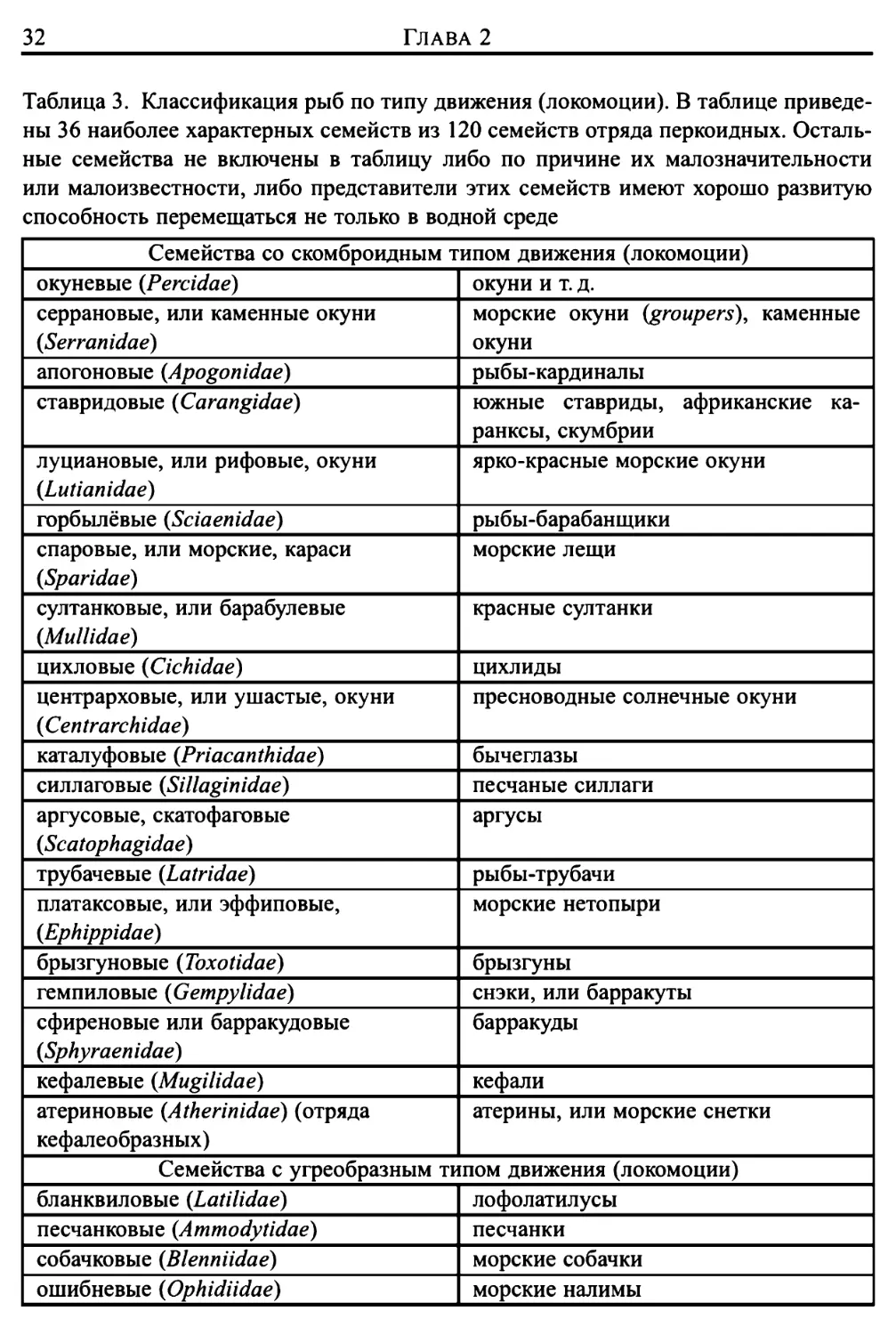
Таблица 2.
В таблице представлены отряды класса рыб, которые обсуждаются в тексте. Первые два
отряда принадлежат к древней группе — подклассу хрящевых рыб (Chondrichthyes) (скелет
состоит из хрящей). В подклассе костных рыб (Osteichthyes) (в скелете костных рыб
имеется костная ткань) отряд двоякодышащих {Dipnoi) относится к надотряду кистеперых рыб
(Crossopterygii), который дал начало классу земноводных (Amphibia), и отряд палеонискои-
дов (Palaeoniscoidea), которые представляют другую архаичную группу. Оставшиеся отряды
принадлежат к наиболее развитой группе рыб — надотряду костистые (Teleostei), характерной
чертой которых является наличие плавательного пузыря, выполняющего гидростатическую
функцию (за исключением тех рыб, у которых пузырь превратился в рудиментарный орган;
к таким рыбам относятся некоторые виды донных или быстро перемещающихся рыб).
Некоторые отряды класса рыб в данной работе не рассматриваются в силу того, что образ
жизни представителей этих отрядов выходит за пределы наших исследований. К ним относятся
донные отряда цельноголовых (Holocephali) (химеры и т. д.), представители отряда присоско-
образных (Xenopterygii) (другое его название Gobiesociformes) (семейство присосковых, или
морских уточек), представители отряда панцирнощеких (Scleroparei) (морские петухи, липа-
рисы и т. д.) и наконец, весьма специфичные отряды прилипалообразных (Discocephali)
(акулья ремора (акулья прилипала)) и рукоперых (Pediculati) (рыбы-удильщики и т.д.). Другие
отряды исключены как малоизвестные или не представляющие интереса с точки зрения их
способов передвижения в воде. Это отряд костных ганоидов (Holostei) (еще одна архаичная
группа), отряд сарганообразных (Synentognathi, или, что то же самое, Beloniformes) (среди
них наиболее интересными представителями являются летающие рыбы), отряд колюшкооб-
разных (Thoracostei) (среди них весьма интересна колюшка со свойствами, которые, однако,
не связаны со способом перемещения в воде), отряд бериксоидных (Berycomorphi) (для нас
интереса не представляет, поскольку представители этого отряда незначительно отличаются
от представителей многочисленного отряда перкоидных (Percomorphi)), и наконец, нами не
включены в обзор отряды карпозубообразных (Microcyprini), гигантохвостых (Giganturoidea),
отряды китовидкообразных (Cetunculi) и Miripinnati
1 Отряд
| акулы (Selachii)
| скаты (Batoidei)
| двоякодышащие (Dipnoi)
| палеонискоиды (Palaeoniscoidea)
| равнопозвонковые (Isospondyli)
| гаплоидные (Haplomi)
1 костнопузырные (Ostariophysi)
| безногие (Apodes)
1 многошиповые (лучеперые)
| (Heteromi)
| мешкоротообразные (Lyomeri)
1 опахообразные (Allotriognathi)
| мягкоперые (Anacanthini)
| пер ко иди ые (Percomorphi)
| солнечникообразные (Zeomorphi)
| сростночелюстные (Plectognathi)
| иглообразные (Solenichthyes)
\ камбалообразные (Heterosomata)
Представители
акула, морская собака (колючая акула)
ромбовый скат, скат, рыба-пила
двоякодышащие
осетр и др.
сельдь, сардина, семга, форель и др.
щука и т. д.
карп, зубастый карп, плотва, лещ, линь
и т. д.
угорь
колючий угорь
мешкоротый угорь
ремень-рыба (сельдяной король),
нарвал и т. д.
треска, хек, нитеперый налим и т. д.
(см. таблицу 3)
солнечник, рыба-кабан и т. д.
спинорог, фугу, рыба-шар и т. д.
морской конек
камбала
Разделы, в которых опи- 1
сывается данный отряд
10
10
4
10
6
9
6
4
4
4
4
4
7-9
11
11
" 1
10 |
26
Глава 2
достигает максимальной скорости за пренебрежимо малый отрезок
времени. При больших же числах Рейнольдса преобладают силы инерции, и для
того, чтобы рыба разогналась до максимальной скорости из состояния
покоя, требуется дополнительная энергия. А уже движущаяся рыба, если она
имеет обтекаемую форму, может проплыть достаточно большое расстояние
по инерции.
При разгоне из состояния покоя до максимальной скорости (Hertel,
1966, с. 160), а также при резком изменении направления движения (Gray,
1968, с. 76) за счет работы мышц рыба может на короткий промежуток
времени (несколько секунд) развить мощность, которая в несколько раз
(порядка 4) превышает мощность, развиваемую все остальное время
(вырабатываемая мощность определяется процессом снабжения мышц кислородом).
Эти маневры выполняются с помощью проталкивающих или рыскающих
движений со значительно увеличенной (в течение короткого промежутка
времени) силой. С резкой остановкой все не так просто, для остановки
рыба использует довольно гибкие грудные плавники, подобно тому как летчик
использует аэродинамический тормоз. Кроме того, некоторые рыбы,
сталкиваясь с необходимостью зависнуть в «нужном месте» при медленном
и часто неравномерно ускоренном течении воды и наличии неподвижных
препятствий разнообразной формы, с успехом используют грудные и
другие плавники для непрерывной корректировки своего движения вблизи
препятствий.
4. Плавание угреобразных
Начиная с (Breder, 1926), ихтиологи описывают как «угреподобный»
(по представителю рода речных угрей) волнообразный тип передвижения,
в котором участвует все тело (хотя амплитуда колебаний тела
увеличивается к хвосту). У типичного представителя отряда безногих (Apodes) —
угря — спинной и анальный плавники очень длинны и вместе с хвостовым
сливаются в один неразрывный плавник, окаймляющий всю заднюю часть
тела (так же как и у представителей уже упоминавшегося класса круглоро-
тых (Cyclostomata)). Тело угря сильно сужается к хвостовой части, однако
суммарный размер в вертикальном направлении задней части тела и
плавниковой ленты остается практически постоянным (рис. 4). Подобное
строение тела имеют и представители отряда двоякодышащих (Dipnoi); попутно
отметим, что двоякодыщащие рыбы имеют особенно интересное
эволюционное развитие.
4. Плавание угреобразных
27
Проведем расчет эффективности продвижения вперед за счет
волнового бокового смещения, распространяющегося со скоростью V вдоль тела,
(Lighthill, 1960). Если скорость хвостовой части в поперечном направлении
равна W, то вода вблизи тела будет двигаться со скоростью w, меньшей,
чем W:
w = W(V - U)/V < W, (2)
где U — скорость продвижения рыбы в воде (которая, как правило,
составляет 0,6V при угреподобном способе передвижения). Это
соотношение между скоростями иллюстрируется на рисунке 5, и можно заметить,
что скорость w становится все меньше при приближении скорости U к V,
в предельном случае при U = V рыба будет скользить в воде без
какого-либо бокового смещения.
ч
1
/|
•" %л ^.тщ
| 1 1 '
^ \ J 1 У
I 1 1 |_ ,
II
1 J i
1 к к L
> 1
> 1 1
1
1 1 1
8
10 11
12
Рис. 4. Вверху изображен речной угорь (Anguilla vulgaris). Внизу представлены
последовательные положения с интервалом 0,09 с тела молодой особи, плывущей в
воде (Gray, 1968). Сторона квадрата равна 25,4 мм. Положения максимального изгиба
тела отмечены точками и крестиками, это дает возможность увидеть, что
распространение поперечных волн происходит в направлении от головы к хвосту
При волнообразном поперечном движении рыба при продвижении
в воде вовлекает в поперечное движение прилегающий к телу
вертикальный тонкий слой воды (перпендикулярный направлению перемещения
рыбы). Скорость поперечного перемещения этого слоя воды у хвостовой части
28
Глава 2
рыбы равна w. Если размах хвостовой части тела рыбы достаточно велик,
то количество бокового движения (пропорциональное скорости w) также
будет достаточно большим по величине. Когда вертикальный слой воды
срывается с хвостовой части рыбы (срыв слоя происходит за счет
движения рыбы со скоростью U) в виде вихревой пелены, то сила реакции,
действующая на рыбу, равна скорости изменения количества движения воды
и пропорциональна Uw.
Сила, действующая со стороны хвоста рыбы, движущегося со
скоростью W, на воду, совершает работу с мощностью, пропорциональной
произведению UwW. Однако часть этой работы расходуется на «бесполезную»
энергию вихревой пелены, которая представляет собой энергию движения
со скоростью w, генерируемую на скорости продольного перемещения
рыбы [/, и, следовательно, пропорциональной2 величине U ( \w2).
Оставшаяся часть энергии рыбы (разность между указанными двумя величинами)
расходуется на продвижение, т. е. на создание движения в воде со
скоростью U с преодолением вязкого сопротивления, препятствующего этому
продвижению.
Для того чтобы произвести достаточно эффективную тягу, скорость
движения U при волнообразном способе продвижения должна составлять
значительную часть от V, поскольку в этом случае, как следует из
уравнения (2), скорость w будет значительно меньше W и, следовательно, потери
энергии на образование вихревой пелены, пропорциональные U(\w2 J,
будут малы по сравнению с общей затраченной энергией, пропорциональной
UwW. С другой стороны, величина скорости w не должна быть слишком
малой (т. е. скорость U не должна быть слишком близкой по величине к ско-
2 Слово «пропорционально» уже использовалось четыре раза, и читатель вправе задать
вопрос: чему же равен этот коэффициент пропорциональности. В каждом случае, если
использовать гидродинамическую терминологию, этот коэффициент равен «присоединенной массе
на единицу длины», которая с достаточной степенью точности может быть вычислена как
произведение плотности воды на площадь круга диаметром, равным вертикальному
размеру хвостовой части рыбы (Lighthill, 1960). Приблизительно, не вдаваясь в подробности, это
можно пояснить так: если тело, будучи в воде, внезапно придет в состояние движения, то
вода вблизи тела также мгновенно придет в движение (строго говоря, возмущения от тела
распространяются с конечной скоростью, равной скорости звука, но такая скорость в данной
постановке настолько велика, что распространение возмущений можно считать происходящим
с бесконечной скоростью) и, следовательно, количество движения тела, равное произведению
его скорости на массу тела, увеличится за счет «некоторой дополнительной массы» воды.
Важно отметить, что соответствующее количество движения изменяется при осциллирующем
движении иначе, чем при гидродинамических сопротивлениях другой природы, в частности,
происходит при фазе, меньшей 90°.
4. Плавание угреобразных
29
рости V), иначе поперечного размера хвостовой части рыбы (или
развиваемой рыбой энергии) будет недостаточно для преодоления вязкого
сопротивления. Очевидно, что это довольно приближенные рассуждения
(необходимо учитывать степень вытянутости тела рыбы, при которой
движение выделенных вертикальных слоев воды вдоль всего тела можно считать
независимым, поэтому для точного описания движения рыбы необходимо
привлечь методы теории обтекания вытянутых тел), однако уже эти
рассуждения дают возможность приблизиться к пониманию гидромеханики
движения угреобразных.
Угревидные рыбы отряда лучеперых, наряду с ремень-рыбой
(сельдяной король) и нарвалом отряда опахообразных, подобны
представителям отряда безногих, за исключением того, что вертикальный размер рыбы
(суммарная величина хвостовой части рыбы и ленты плавника) не
является постоянным вплоть до хвостовой части, а резко сужается к хвостовому
плавнику. Результатом такого строения является некоторая потеря
эффективности движения (сужение тела приводит в большей степени к
увеличению сопротивления, чем к увеличению развиваемой тяги). У некоторых
мешкоротых угрей отряда мешкоротообразных и двух семейств перкоид-
ных рыб (таблица 3) эта диспропорция между увеличением сопротивления
и увеличением силы тяги еще больше и, как следствие, еще больше потеря
эффективности движения.
На самом деле угри и подобные им рыбы используют угреподобный
метод продвижения только при больших скоростях движения, при более
медленном движении рыбы перемещаются за счет колебательных
движений спинных и анальных плавников. Медленное движение задним ходом
обеспечивается колебанием плавников в противоположном направлении.
С другой стороны, рыбы, у которых спинной и анальный плавники не
расположены вдоль тела в виде непрерывной ленты, также пользуются
способом продвижения, весьма напоминающим угреподобное движение. В
частности, представители отряда мягкоперых, такие как треска, хек, нитепе-
рый налим с несколькими отдельными спинными плавниками и большими
по величине анальным и хвостовым плавником, используют иногда способ
движения, похожий на угреподобный, практически с такой же
результативностью, несмотря на наличие промежутков между плавниками. С точки
зрения теории это объясняется тем, что вихревая поверхность,
образующаяся между плавниками, ведет себя как твердая поверхность, в
результате чего с некоторой степенью приближения при быстром движении рыбы
с несколькими отдельными плавниками можно считать, что строение ее
тела близко к строению угреподобных.
30
Глава 2
направление
движения рыбы,
Vr
01514 13
10 98
слои воды
задняя
кромка
. хвостового
D \ I. плавника
6 6789
плавник движется
вправо
10 15 14 13 12 11109
плавник движется
влево
(V Ц)т
Ut
Рис. 5. Движение хвостового плавника вправо (левая часть диаграммы) и затем
влево (правая часть диаграммы). Поперечная волна распространяется со скоростью V,
в то время как рыба продвигается со скоростью U. Период колебательного
движения хвоста (время, через которое хвост занимает исходное положение) равен 16т,
числа 1, 2, 3, ... на диаграмме соответствуют положениям задней кромки и
некоторой, находящейся перед ней, части хвостового плавника через моменты времени
т, 2т, Зт, ..., отсчитываемые от момента времени, который принимаем за нулевой.
За время т (одной шестнадцатой части периода) поперечная волна перемещается от
указанной предшествующей части хвостового плавника до его задней кромки,
поскольку расстояние между ними принимается равным Vr, следовательно,
количество позиций, занимаемых задней кромкой хвостового плавника, на единицу
меньше количества позиций предшествующей его части. Рассмотрим слой воды в
момент времени, скажем, 5т, он расположен относительно задней кромки хвостового
плавника рыбы на расстоянии [/т, этот слой в момент времени 6т будет совпадать
с задней кромкой (т. к. рыба продвигается вперед со скоростью £/). Следовательно,
расстояние, на которое переместится тело рыбы в поперечном направлении в слое
воды за время т, равно АВ. Из подобия треугольников следует, что AB/CD =
= PB/PD = (V — U)/V. Кроме того, CD = Wt9 где W — скорость бокового
перемещения задней кромки хвостового плавника. Следовательно, скорость w, с
которой рыба толкает слой воды, равна АВ/т = (CD/r)(V — U)/V = W(V — U)/V
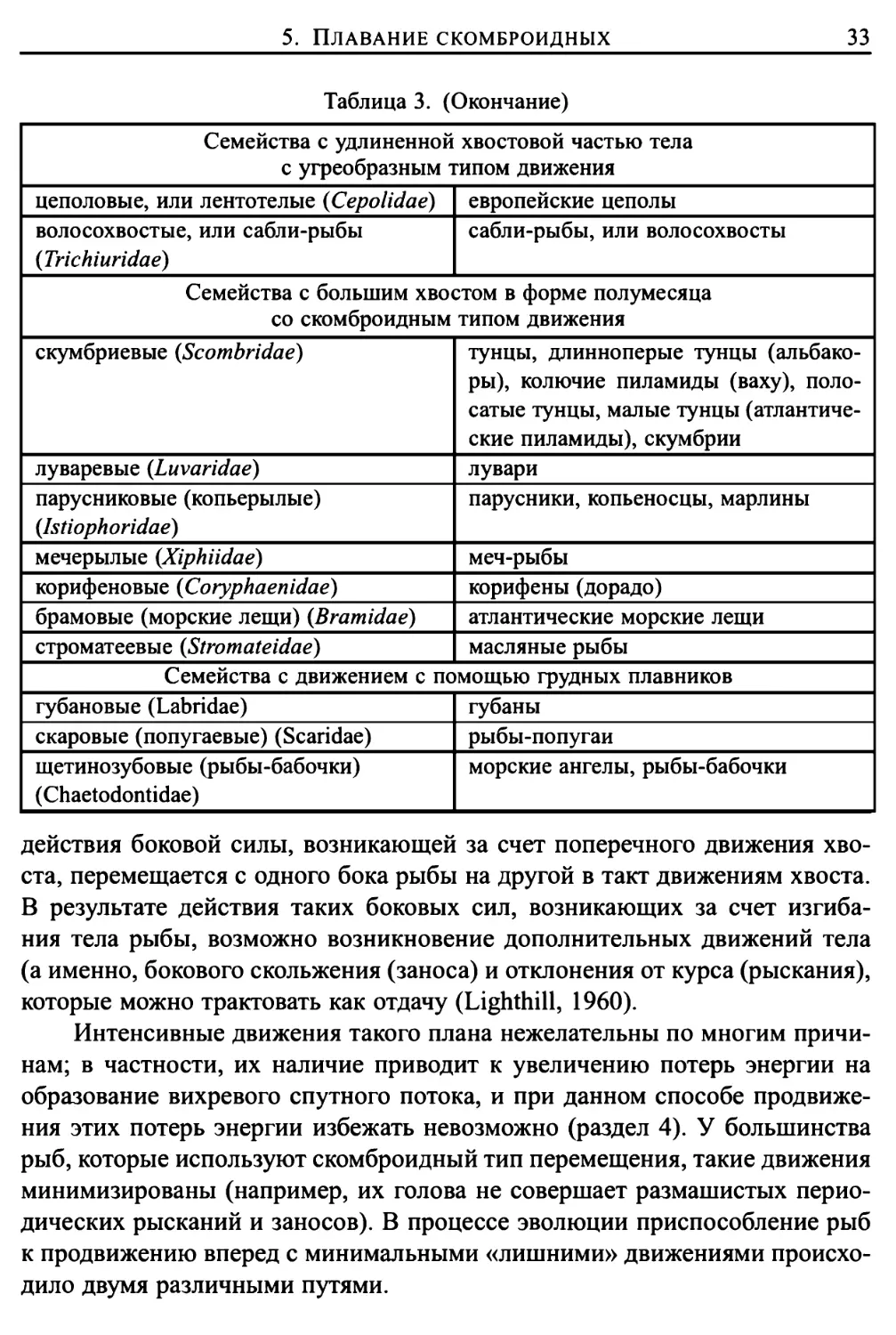
5. Плавание скомброидных
31
5. Плавание скомброидных
Рассмотрим теперь наиболее важную модификацию волнообразного
или угреподобного способа перемещения. А именно модификацию,
названную скомброидной (Breder, 1926) (по названию семейства скомброидных,
см. таблицу 3). Эволюцию скомброидного способа передвижения в воде из
угреподобного можно объяснить с точки зрения гидромеханики. При
больших числах Рейнольдса волнообразное движение всего тела практически не
происходит. Амплитуда поперечных колебаний передней части тела очень
мала или даже равна нулю и возрастает до сравнительно больших значений
по мере приближения к хвостовой части.
Если в волнообразном движении заметно участвует лишь задняя
половина тела или даже только треть, то в случае, когда амплитуда достигает
своего наибольшего значения вблизи задней кромки хвостового плавника
(см. рис. 5), количество движения и энергия, затраченная на образование
спутного потока, будут иметь такие же значения, как и при угреподобном
способе движения, а развиваемая сила тяги и эффективность движения,
скорее всего, не изменятся. Более того, эффективность может даже и возрасти,
поскольку слой воды в поперечном направлении движется в течение
значительно меньшего времени и, следовательно, потери энергии на образование
вихрей на кромках будут меньше.
При скомброидном способе рыбы плывут вперед за счет интенсивного
изгибания хвостового отдела тела. Волнообразные движения
производятся только этим отделом, и длина волны такого колебательного движения
превышает длину хвостовой части тела. Особенно важным является тот
факт, что эта часть тела рыбы постоянно совершает колебательные
движения в поперечном направлении. А это означает, что (см. рис. 5) хвостовой
плавник двигается из стороны в сторону таким образом, что его движение
всегда имеет возвратную составляющую (можно было бы предположить,
что сопротивление этому возвратному движению и обеспечивает тягу,
однако при таком подходе практически невозможно провести количественные
оценки или определить, какая часть вырабатываемой энергии теряется за
счет образования вихревого следа).
У рыб, приспособленных к скомброидному типу движения,
выработалась определенная геометрическая форма тела. В силу того, что длина
поперечной волны больше длины хвостовой части, боковые силы,
возникающие при поперечном колебании, не уравновешиваются, как это
имеет место при угревидном способе продвижения. В данном случае линия
32
Глава 2
Таблица 3. Классификация рыб по типу движения (локомоции). В таблице
приведены 36 наиболее характерных семейств из 120 семейств отряда перкоидных.
Остальные семейства не включены в таблицу либо по причине их малозначительности
или малоизвестности, либо представители этих семейств имеют хорошо развитую
способность перемещаться не только в водной среде
Семейства со скомброидным типом движения (локомоции)
окуневые (Percidae)
серрановые, или каменные окуни
{Serranidae)
апогоновые (Apogonidae)
ставридовые (Carangidae)
луциановые, или рифовые, окуни
(Lutianidae)
горбылёвые (Sciaenidae)
спаровые, или морские, караси
(Sparidae)
султанковые, или барабулевые
(Mullidae)
цихловые (Cichidae)
центрарховые, или ушастые, окуни
| {Centrarchidae)
| каталуфовые (Priacanthidae)
| силлаговые (Sillaginidae)
1 аргусовые, скатофаговые
| (Scatophagidae)
| трубачевые (Latridae)
платаксовые, или эффиповые,
| (Ephippidae)
| брызгуновые (Toxotidae)
| гемпиловые (Gempylidae)
сфиреновые или барракудовые
| (Sphyraenidae)
| кефалевые (Mugilidae)
1 атериновые (Atherinidae) (отряда
кефалеобразных)
окуни и т. д.
морские окуни (groupers), каменные
окуни
рыбы-кардиналы
южные ставриды, африканские ка-
ранксы, скумбрии
ярко-красные морские окуни
рыбы-барабанщики
морские лещи
красные султанки
цихлиды
пресноводные солнечные окуни
бычеглазы
песчаные силлаги
аргусы
рыбы-трубачи
морские нетопыри
брызгуны
снэки, или барракуты
барракуды
кефали
атерины, или морские снетки
| Семейства с угреобразным типом движения (локомоции)
| бланквиловые (Latilidae)
| песчанковые (Ammodytidae)
| собачковые (Blenniidae)
| ошибневые (Ophidiidae)
лофолатилусы
песчанки
морские собачки
морские налимы
5. Плавание скомброидных
33
Таблица 3. (Окончание)
Семейства с удлиненной хвостовой частью тела
с угреобразным типом движения
| цеполовые, или лентотелые (Cepolidae)
1 волосохвостые, или сабли-рыбы
| {Trichiuridae)
европейские цеполы
сабли-рыбы, или волосохвосты
Семейства с большим хвостом в форме полумесяца
со скомброидным типом движения
1 скумбриевые (Scombridae)
| луваревые (Luvaridae)
парусниковые (копьерылые)
| (Istiophoridae)
мечерылые (Xiphiidae)
корифеновые (Coryphaenidae)
брамовые (морские лещи) (Bramidae)
строматеевые (Stromateidae)
тунцы, длинноперые тунцы (альбако-
ры), колючие пиламиды (ваху),
полосатые тунцы, малые тунцы
(атлантические пиламиды), скумбрии
лувари
парусники, копьеносцы, марлины
меч-рыбы
корифены (дорадо)
атлантические морские лещи
масляные рыбы
| Семейства с движением с помощью грудных плавников
губановые (Labridae)
скаровые (попугаевые) (Scaridae)
1 щетинозубовые (рыбы-бабочки)
(Chaetodontidae)
губаны
рыбы-попугаи
морские ангелы, рыбы-бабочки
действия боковой силы, возникающей за счет поперечного движения
хвоста, перемещается с одного бока рыбы на другой в такт движениям хвоста.
В результате действия таких боковых сил, возникающих за счет
изгибания тела рыбы, возможно возникновение дополнительных движений тела
(а именно, бокового скольжения (заноса) и отклонения от курса (рыскания),
которые можно трактовать как отдачу (Lighthill, 1960).
Интенсивные движения такого плана нежелательны по многим
причинам; в частности, их наличие приводит к увеличению потерь энергии на
образование вихревого спутного потока, и при данном способе
продвижения этих потерь энергии избежать невозможно (раздел 4). У большинства
рыб, которые используют скомброидный тип перемещения, такие движения
минимизированы (например, их голова не совершает размашистых
периодических рысканий и заносов). В процессе эволюции приспособление рыб
к продвижению вперед с минимальными «лишними» движениями
происходило двумя различными путями.
34
Глава 2
Наиболее очевидный способ — это уменьшить величину самой
боковой силы. При скомброидном типе движения амплитуда колебаний вдоль
тела увеличивается от практически нулевого значения в передней части
до довольно больших значений в небольшой хвостовой части тела. В
области среднего участка тела рыбы, где происходит изменение амплитуды,
возникают резкие, большие по величине, изменения количества движения
прилегающего к нему слоя воды и, следовательно, возникают достаточно
большие боковые силы, за исключением того случая, когда этот участок
тела рыбы имеет достаточно малый размер и, значит, сможет приводить
в движение небольшое количество воды. Это явилось основанием для
сужения тела (локального уменьшения высоты тела рыбы) от среднего участка
вплоть до хвостового плавника. В этом случае рост амплитуды
поперечных колебаний происходит на суженном участке тела рыбы, а в области
хвостового плавника амплитуда бегущей волны изменяться уже не будет.
Конечно, боковая сила возникает за счет поперечных колебаний
и в этом случае, однако ее величина определяется скоростью изменения
количества движения спутного следа (эта скорость, как было указано ранее,
пропорциональна произведению Uw). Боковой занос и рыскание,
обусловленные этой силой, изменяют как W, так и w. Эти изменения определяются
лишь частью скорости w и не приводят к сколько-нибудь значительному
уменьшению эффективности при условии, что отношение w/W
остается достаточно малым. В небольшой степени отношение можно изменить
за счет изменения некоторых геометрических характеристик рыбы. Если
суммарная высота тела рыбы и плавников достаточно велика, то боковое
скольжение и рыскание вызывают ускорение значительных масс воды (см.
предыдущую сноску), в этом случае боковая сила дает лишь небольшой
вклад в занос и рыскание.
6. Лабораторные исследования скомброидного типа
плавания
Когда представители отряда равнопозвонковых (Isospondyli) (во
многих отношениях он является наиболее примитивным отрядом), приобрели
в процессе эволюции свою современную форму, то они смогли
использовать скомброидный тип плавания. Многие представители этого отряда
хорошо известны как физически сильные и активные пловцы.
Сужение тела вблизи хвостового плавника совместно с увеличением
высоты других отделов за счет спинных и анальных плавников (см. рис. 3)
6. Лабораторные исследования скомброидного типа плавания 35
привели к уменьшению эффектов бокового заноса и рыскания, а
гидродинамическое сопротивление, которое уравновешивается силой тяги,
уменьшилось за счет обтекаемой формы.
Большая часть представителей обширного отряда карпообразных
(Pstariophysi) (отряд, состоящий из более специализированных
пресноводных рыб, имеющий такие преимущества перед равнопозвонковыми, как
специализированные орган для восприятия акустических сигналов и более
гибкие грудные плавники) также освоила скомброидный тип плавания.
Исключением являются представители подотряда сомообразных (Siluroidea),
которые освоили практически угревидный способ перемещения в воде,
а также не похожий на других рыб электрический угорь {Gymnotus),
который обычно использует угревидный способ перемещения, поперечные
колебания распространяются вдоль его огромного анального плавника,
и (в отличие от безногих) его движение никак не скоординировано с
движениями тела.
Бейнбридж (Bainbridge, 1958; 1960; 1963) экспериментально
исследовал скомброидный тип движения у представителей отряда карпообразных
и равнопозвонковых на так называемом «рыбьем колесе». При
применении этой экспериментальной установки стало возможным провести более
точные измерения скорости, чем при наблюдении за рыбами в естественной
среде (где, например, достаточно часто неизвестна скорость течения), а
также стала возможна покадровая киносъемка волнообразных движений тела
рыбы. При наблюдении за несколькими видами рыб различных форм и
размеров (золотой рыбкой {Carassius auratus), плотвой {Leuciscus leuciscus),
форелью (Salmo irideus) и лещем (Abramis brama), которые имеют
значительно более тонкое тело, чем остальные) Бейнбридж установил, что длина
рыбы / является основным кинематическим параметром скомброидного
типа движения.
Бейнбриджем было также найдено, что максимальная амплитуда
движений хвостового плавника (т.е. максимальное отклонение хвостового
плавника от нейтрального положения в поперечном направлении)
составляет около 0,2/ и практически не зависит от каких-либо других параметров.
Значения важного гидромеханического параметра ul/U (угловая частота и9
умноженная на 27Г, измеряется в герцах) колеблются в окрестности десяти.
Имеющиеся экспериментальные данные из других источников
подтверждают полученные Бейнбриджем характерные значения параметров
скомброидного типа движения.
С увеличением размера особи максимальная частота колебаний ее
хвоста постепенно уменьшается, так что ее предельная крейсерская скорость,
36
Глава 2
приходящаяся на единицу длины тела, U/1 (ее типичные значения лежат
в пределах от 3 до 6 с-1) уменьшается, хотя сама скорость U
увеличивается. У самых маленьких особей в общем случае частота колебаний хвоста
наибольшая, и чрезвычайно забавно наблюдать за скомброидным
плаванием мелких представителей отряда карпообразных семейства харацино-
вых {Characidae), таких как тетры с их высокочастотными колебаниями
хвостов. Визуально практически невозможно увидеть распространение
поперечных колебаний вдоль хвостовой части таких особей, и только
киносъемка позволяет увидеть и изучить в подробностях плавание этих особей.
Именно таким способом Бейнбридж установил, что для золотых рыбок
отношение U/V приблизительно равно 0,7.
Большинство карпообразных обладают чрезвычайно гибким
хвостовым плавником, вследствие чего под действием боковой силы,
противодействующей поперечным перемещениям, хвостовой плавник при
движении принимает вогнутую форму. Такое явление приводит к нежелательным
гидромеханическим эффектам (например, к изменению количества
движения воды, обусловленного движением плавника). С другой стороны,
Бейнбридж (Bainbridge, 1963) показал, что изменения площади поверхности
хвостового плавника, происходящие при его растяжении и сжатии при
поперечном колебательном движении, дают рыбе дополнительные возможности
в управлении своим движением. Более того, с помощью вертикальных
колебаний хвостового плавника рыба может медленно смещаться в
вертикальной плоскости.
На основании проведенных измерений гидродинамического
сопротивления рыб, использующих скомброидный тип движения (сопротивление
определялось различными способами, например по ускорению торможения
при скольжении, по установившейся скорости движения застывшего тела
мертвой рыбы и т.д.), Грей (Gray, 1968, с. 52) получил, что
сопротивление тела рыбы близко к сопротивлению гладкого тела такой же, как у
рыбы, формы. Среднее значение коэффициента лобового сопротивления CD
на единицу площади максимального поперечного сечения, приблизительно
равное 0,25, немного уменьшается с ростом числа Рейнольдса. Грей
отмечает, что с увеличением размеров особи мощность, развиваемая ее мускулами,
пропорциональна объему, т.е. /3, а мощность, необходимая для
преодоления трения, равна ~pU3CDS9 где S (некоторая репрезентативная площадь
поперечного сечения) пропорциональна /2, тогда скорость движения U
равна (l/CD)1^. To есть зависимость скорости движения рыбы от ее длины /
имеет степенной характер, величина показателя степени определяется за-
7. СКОМБРОИДНЫЙ ТИП ДВИЖЕНИЯ РЫБ С ХВОСТОМ В ВИДЕ ПОЛУМЕСЯЦА 37
висимостью коэффициента CD от числа Рейнольдса R = Ul/v и лежит
в пределах от 0,33 до 0,60. Полученные таким образом зависимости очень
хорошо согласуются с экспериментальными данными.
7. Скомброидный тип движения рыб с хвостом в виде
полумесяца
Колючеперые представители большого отряда перкоморф (Percomorphi)
(см. таблицу 3), в целом приспособились к скомброидному типу
движения. Однако следует отметить три основных момента. Во-первых (не очень
существенно и об этом в дальнейшем упоминаться не будет), некоторые
семейства (см. таблицу 3) сохранили угревидный способ движения. А вот
второй момент является очень важным для нас, поскольку некоторые
семейства, модифицировав скомброидный тип движения, могут развивать
наибольшую наблюдаемую у рыб скорость. Так, Ферштайн и Уолтере (Fierstine,
Walters, 1968) провели тщательные измерения скорости рыб и получили,
что представители этих семейств могут развивать скорость от 10 до 20 м/с.
Здесь будут рассмотрены представители наиболее значимых семи семейств
данного отряда, включая, прежде всего, скумбриевых (Scombridae) (свое
название семейство получило по входящей в него скумбрии атлантической
(Scomber scombrus), однако с точки зрения быстроходности наиболее
яркими представителями этого семейства являются скорее тунцы, длинноперые
тунцы, колючие пиламиды, полосатые тунцы и малые тунцы).
Действительно, представители семейства скумбриевых имеют
большие размеры, чем представители других семейств, перечисленные в левом
столбце таблицы 3, и, подобно сельдям и другим представителям
подотряда сельдевых (Clupeoidea) отряда равнопозвонковых (Isospondyli)
имеют хвостовой плавник в форме латинской буквы V, или, говоря языком
аэродинамики, «стреловидное крыло». Угол между элементами V
составляет от 50° до 60°. Исследование эффективности продвижения методами
теории обтекания тонкого тела (а применение данной теории правомерно
при такой степени стреловидности) показало, что при движении
хвостовой V-образный плавник работает так же эффективно, как если бы выемка
отсутствовала (в силу свойств вихревой плены, заполняющей выемку).
Очевидно, что для увеличения силы тяги, с целью увеличения
развиваемой скорости, необходимо удлинение хвостового плавника. Однако в силу
морфологических особенностей рыб такое удлинение хвостового плавника
при сохранении данной степени стреловидности невозможно.
38
Глава 2
У одной группы рыб эволюционное развитие в борьбе за выживание
в верхних слоях океана, где преимущество получали более быстрые особи,
пошло по пути увеличения «относительного удлинения» хвостового
плавника (еще один аэродинамический термин, означающий отношение
квадрата размаха хвостового плавника к его площади). Изменения коснулись
прежде всего формы, и таким образом сформировался серповидный
хвостовой плавник (рис. 6), часто называемый хвостовым плавником в виде
полумесяца (Marshall, 1965, с. 10), хотя, как свидетельствуют фотосъемки
движения, проведенные Ферштайном и Уолтерсом (Fierstine, Walters, 1968),
эта группа рыб сохранила скомброидный тип движения.
Типичные представители семи семейств (возглавляемых семейством
скумбриевых (Scombridae)), приведенные в таблице 3, приобрели в
процессе эволюции серповидный хвост и мощную мускулатуру, способную
приводить его в движение. У них практически разделились структуры для
создания рывка при продвижении и для создания силы тяги. И такое разделение
было достигнуто, прежде всего, за счет изменения формы тела — сужения
хвостовой части к хвостовому плавнику. То есть произошло радикальное
уменьшение размаха хвостового плавника с одновременным увеличением
амплитуды поперечных колебаний хвоста (рис. 6). Таким образом,
развиваемая сила тяги осталась практически такой же, как у рыб с хвостовым
плавником с «большим относительным удлинением крыла», совершающим
колебательные движения, при движении в невозмущенной воде.
Поскольку здесь было использовано слово «крыло», то сразу возникает аналогия
с методом выработки силы тяги при наиболее простом способе птичьего
полета. И эта аналогия станет еще более ясной, если рисунке 5 повернуть
на 90°, он займет положение рисунка 7, иллюстрирующего полет птиц.
Надо сказать, что птице необходимо крыло с большим относительным
удлинением в каждом случае, когда требуется эффективная поддержка ее
тела, которую возможно обеспечить только при большом отношении
подъемной силы к силе лобового сопротивления (а следовательно, при малом
угле скольжения). Однако для того, чтобы при скомброидных
колебаниях движение было эквивалентным полету птицы, необходимо наложить на
колебания движение с постоянным по величине углом атаки, создающее
подъемную силу, необходимую для поддержания тела птицы.
Результат представляет собой комбинацию (Gray, 1968, с. 222) взмаха
по направлению вниз, где крыло создает силу, значительно большую, чем
вес птицы, и взмаха оперенного крыла вверх, при этом создается
значительно меньшая сила. Поэтому у птицы значительно более развиты мускулы,
7. СКОМБРОИДНЫЙ ТИП ДВИЖЕНИЯ РЫБ С ХВОСТОМ В ВИДЕ ПОЛУМЕСЯЦА 39
Рис. 6. Варианты серповидного хвоста у шести перкоидных рыб, двух акул, двух
китообразных млекопитающих и одной вымершей рептилии. Слева направо и
сверху вниз представлены: вид сбоку колючей пиламиды, тунца, полосатого марлина,
луваря, парусника и меч-рыбы (род ваху Acanthocybium, тунцов Thunnus, род
копьеносцев Teirapturus, род луварей Luvarus, род парусников Istiophorus и род
меч-рыбы Xiphius) из надотряда перкоидных (Percomorphi). Китовая акула и обыкновенная
сельдевая акула (Rhineodon и Lamnd) из отряда акул (Selachii). Вид снизу ивасевого
кита Boloenoptera и дельфина из рода клювоголовых дельфинов Cephalorhynchus
(сведения взяты из работы (Norman, Fraser, 1937)). Последнее необычное
изображение относится к ихтиозавру Ichthyosaurus (найден в Лейасе в Вюртемберге
(Woodward, 1910)), на рисунке ясно видна структура плавников
40
Глава 2
с помощью которых совершается взмах вниз, чем мускулы,
осуществляющие взмах вверх.
направление движения птиц ^ч.
Рис. 7. Диаграмма слева представляет вертикальные скомброидные колебания,
полученные поворотом диаграммы скомброидных колебаний на рисунке 5 на 90°.
Средняя часть диаграммы представляет движение с постоянным углом атаки,
создающее подъемную силу. Результат наложения двух типов движения показан на
диаграмме справа и представляет собой изображение типичного полета птиц
8. Двумерный анализ динамики серповидного хвоста
Из проведенной аналогии между плаванием рыб и полетом птиц
следует, что движение хвостового плавника с большим относительным
удлинением может быть изучено теми же методами, какими исследуется полет птиц
или насекомых или биение и закручивание крыла летательного аппарата,
при которых может развиться (что нежелательно) вибрация. Среди таких
аэродинамических методов исследования (Garrick, 1957) наиболее развита
так называемая двумерная теория осцилляции аэродинамических
поверхностей. В работе By (Wu, 1961) приведен подробный и в высшей степени
интересный обзор методов данной теории и особое внимание уделено
способам оценки эффективности создания силы тяги. Следует отметить, что
методами двумерной теории изучаются гидродинамические свойства
поверхностей с большим относительным удлинением (т. е. главным образом
8. Двумерный анализ динамики серповидного хвоста 41
серповидных хвостовых плавников и в некоторых случаях, как будет
видно позднее, грудных плавников). И неправомерно было бы применять ее
к исследованию собственно движения рыбы как целого объекта. Так,
рассчитанная на основании этой теории, приведенная частота крыла ujc/U (с —
хорда крыла) оказалась равной 10 (как ul/U; см. выше), а не меньшей или
равной единице.
При двумерном анализе (вертикальная составляющая не учитывается,
рассматриваются горизонтальные слои воды, которые приводятся в
движение за счет поперечных колебаний хвостового плавника) прежде всего
подробно исследуется баланс между (а) необходимостью избежать слишком
больших потерь энергии на вихревой след и (Ь) необходимостью затратить
энергию на создание вихревого следа, за счет которого и
вырабатывается сила тяги. Пояснения к затратам энергии (6), данные в работе Кармана
и Бюргерса (Karman, Burgers, 1934), приведены здесь в несколько
измененной форме на рисунке 8 (в их работе см. рис. 105).
Рис. 8. Изображены вихри, сходящие с задней кромки хвостового плавника
(рыба движется влево), а также линии тока (подобные линиям тока при реактивном
движении) течения воды, обусловленного этими вихрями. Это схематичное
изображение отражает наблюдавшуюся экспериментально структуру потока с данной
конфигурацией вихрей (Goldstein, 1938, вклейка 1)
На рисунке 8 изображен горизонтальный слой воды, который
приводится в движение хвостовым плавником, за счет чего рыба движется влево.
Совместное действие вихрей, вращающихся по часовой стрелке, которые
срываются с задней кромки хвостового плавника, когда он находится в
своем крайнем нижнем (на рисунке) положении, с вихрями, вращающимися
против часовой стрелки, которые срываются с задней кромки хвостового
плавника, когда он находится в своем крайнем верхнем (на рисунке)
положении, приводит к созданию течения, подобного реактивной струе. На
основании проведенных достаточно упрощенных рассуждений можно
сказать, что хвостовой плавник работает по реактивному принципу с хорошей
эффективностью за счет достаточно большого поперечного сечения
реактивной струи. (Отметим, что «дорожка Кармана», связанная с
торможением воздушного потока при обтекании тонкого цилиндра (Goldstein, 1938),
42
Глава 2
отличается от представленной на рисунке 8, связанной с созданием тяги.
С другой стороны, сопротивление тела рыбы определяется более сложным,
существенно трехмерным следом, в котором вода закручивается и движется
в противоположном направлении. Действие этого следа и вихревого следа,
создаваемого плавником, в общем случае не уравновешивается, за
исключением случая, когда их результирующая сила будет горизонтальной.)
Из теории обтекания тонкого тела следует, что предсказанные
потери энергии в вихревой пелене могут быть частично минимизированы за
счет флюгирования плавника во время движения при скомброидном типе
продвижения (рис. 5), поскольку сила тяги пропорциональна первой
степени интенсивности вихря (рис. 8), а потери энергии пропорциональны ее
квадрату. При расчете этих величин необходимо принимать во внимание
изменение эффективного угла атаки рыбы под действием движения воды
в вихревом следе. Подробности здесь рассматриваться не будут, лишь
только отметим, что математические методы теории обтекания тонких тел
существенно упрощают расчеты параметров течения (Wu, 1961). Расчет
распределения давления основан на том факте, что градиент давления в
направлении, перпендикулярном плавнику, равен скорости, с которой в этом
направлении изменяется количество движения воды.
На рисунке 9 приведены результаты некоторых расчетов.
Предполагается, что хвостовая часть совершает поперечные колебания с максимальной
скоростью W и максимальным углом атаки (достигающимся в той же
фазе, что и максимальная скорость) 0W/U. Таким образом, в не превышает
единицы и его можно рассматривать как коэффициент флюгирования, т. к.
в = 1 соответствует полному геометрическому флюгированию плавника.
Коэффициент силы тяги Ст равен силе тяги, приходящейся на единицу
площади плавника, деленной на величину ~pW2, а эффективность г\
равна доле мощности, расходуемой на создание силы тяги. Из приведенных
графиков ясно видно, что с ростом коэффициента флюгирования
эффективность растет, а коэффициент силы тяги убывает. Поэтому при плавании
рыба должна как-то выбирать оптимальное значение коэффициента
флюгирования.
Для того чтобы применить результаты, представленные на рисунке 9,
к оценке эффективности и силы тяги в случае хвостового плавника в виде
полумесяца с хордой, равной с (размер хвостового плавника в направлении
движения рыбы), уменьшающейся к кончикам плавника, необходимо было
бы использовать зависимость Ст и ту от ljc/U, соответствующую
истинному значению коэффициента флюгирования 0, для того чтобы оценить сред-
9. Изучение движения быстрых перкоморфных рыб 43
Рис. 9. Результаты расчетов в двумерной постановке коэффициента силы тяги Ст
и коэффициента эффективности продвижения rj как функции приведенной
частоты шс/и и коэффициента флюгирования в при скомброидном движении особей
с хвостом в виде полумесяца
ние значения (с весовым коэффициентом, определяемым площадью
плавника). Очевидно, сделать это достаточно реально. Однако читатель,
знакомый с аэродинамической теорией крыла, легко поймет, что такой чисто
двумерный метод расчета приведет к завышению значений эффективности,
поскольку при этом методе учитываются только потери энергии на
завихренность, перпендикулярную к направлению течения (под прямым углом
к направлению движения), хотя в действительности существует и
завихренность в направлении движения.
9. Более детальное изучение движения быстрых
перкоморфных рыб
Итак, исследуем движение быстрых перкоморфных рыб с позиций
трехмерной теории крыла для получения более точной оценки
эффективности за счет учета завихренности, параллельной движению рыбы, а
также для более полного понимания причин эволюционного развития
формы хвостового плавника в четкую серповидную форму. Мы увидим, что
44
Глава 2
в процессе эволюции животные совершенно различных семейств
приобрели серповидную форму хвостового плавника как средство, обеспечивающее
наиболее быстрое перемещение в воде. Это относится не только к тем семи
семействам, указанным в таблице 3, но также и к большой группе акул из
совершенно другого подкласса класса рыб (Pisces) и к (из совершенно
иного класса) большинству млекопитающих семейства китообразных (рис. 6).
К тому же хвосты всех указанных животных в поперечном сечении
приобрели профиль крыла; их плавная закругленная передняя кромка весьма
важна, поскольку как в двумерной теории крыла, так и в трехмерной
устанавливается, что подъемная сила, полученная на основании этих теорий,
образуется за счет подсасывания воды, движущейся вблизи такой передней
кромки (а из теории обтекания тонкого тела следует, что если хвостовой
плавник имеет резко выраженную стреловидную форму, то за счет наличия
закругленного переднего края плавника животное никакого преимущества
в скорости перемещения не получит).
Конечно, можно было бы предположить, что преимущества
серповидной формы определяются некоторой внутренней механикой, но
различная структура хвоста у различных быстро движущихся водных животных
(с костным скелетом у перкоморфных, с хрящевым скелетом у акул и с
полным отсутствием скелета у китообразных (Cetacea)) подтверждает тот факт,
что преимущества серповидной формы хвоста можно четко обосновать
с позиций гидромеханики. Возможная причина такого
гидромеханического преимущества может быть найдена (учитывая теорему (Goldstein, 1938)
о том, что вихревые линии замкнуты и не могут заканчиваться в жидкости),
при трехмерном анализе вихревой пелены или двумерном анализе слоя
воды, представленного на рисунке 8.
В силу того, что завихренность (перпендикулярная направлению
движения) пелены, сходящей со средней части хвостового плавника,
значительно больше завихренности пелены, сходящей с его заостренных концов,
вихревые линии пелены за центральной частью хвостового плавника
должны изгибаться вниз по потоку. Затем они должны заворачиваться внутрь и,
наконец, замыкаться, когда они встречаются с вихревыми линиями пелены
с противоположным направлением завихренности, сошедшей с хвоста на
половину периода раньше. Таким образом, можно представить, что спут-
ный след состоит из серии вертикальных вихревых колец, закрученных
поочередно вправо и влево (на рис. 8 представлено горизонтальное сечение
таких серий колец). Эти кольца были бы практически круговыми, если бы
приведенная частота ujI/U была бы приблизительно равна величине ivl/s
9. Изучение движения быстрых перкоморфных рыб 45
(где s — размах хвостового плавника), типичное значение которой равно 10,
как следует из данных измерений, представленных в небольшом числе
источников (см. главным образом у Ферштайна и Уолтерса (Fierstine, Walters,
1968), хотя, конечно, крайне необходимы в большом количестве надежные
измерения частоты у представителей всех семейств с серповидным
хвостом).
Возможно, что серповидная форма хвоста является наиболее
благоприятной, поскольку при частотах колебаний, соответствующих быстрому
движению, именно с хвостового плавника серповидной формы чаще всего
сходят вихревые кольца практически круговой формы. А для них
характерен перенос большего количества движения относительно их энергии, и
таким образом могла бы быть минимизирована доля скорости сброса энергии
со следом относительно эффективной мощности, равной W, умноженной
на максимальную скорость схождения количества движения в пелене.
Количественное обоснование этого утверждения, или, иначе, этой гипотезы,
с позиции гидромеханики будет проведено автором в рамках теории крыла,
разработанной для крыльев с искривленной центральной линией.
Быстрые перкоморфные рыбы семейства скумбриевых (Scombridae)
и других родственных им семейств развивают большие скорости не только
за счет большей силы тяги, но также за счет уменьшения трения.
Во-первых, суммарная высота спинного плавника, тела рыбы, и брюшного
плавника у всех этих рыб достаточно велика, так что боковая отдача тела рыбы
в ответ на флуктуации боковой силы, действующей на хвостовой плавник,
будет наименьшей за счет большой «присоединенной массы» воды,
которая также должна быть приведена в движение (раздел 4). Большой
суммарной высотой особенно отличаются парусники семейства парусниковых
(Istiophoridae). Во-вторых, тело быстрых рыб имеет исключительно
гладкую поверхность и хорошо обтекаемую форму, и, кроме того, у многих
видов для уменьшения гидродинамического сопротивления грудной и
анальный плавники при быстром плавании убираются в специальные щели.
На сегодняшний день трудно ответить положительно на вопрос, можно
ли уменьшить силу сопротивления за счет управления пограничным слоем
у тела рыбы. Однако кажется вполне вероятным, что во многих случаях
большое количество воды, которое выталкивает рыба через жаберные
щели, обладает достаточно большой кинетической энергией, чтобы избежать
отрыва пограничного слоя. В объемном труде Бредера (Breder, 1926),
посвященном исследованию плавания рыб (на которое, он считает, в
значительной степени влияет поток воды, прокачиваемой через жаберные щели,
46
Глава 2
хотя и не приводит обоснования этого утверждения с позиций
гидромеханики), приводится весьма интересный пример такого влияния. Совершая
поворот, рыба закрывает жаберные щели с внутренней стороны поворота,
и весь объем воды проталкивается через жаберные щели с другой стороны,
где возможность отрыва пограничного слоя наибольшая.
Ихтиологи считают, что рыба уменьшает гидродинамическое
сопротивление при движении за счет смазки тела, вырабатываемой ею
слизью. Действительно, слизь вырабатывается в большом количестве на коже
у большинства быстрых рыб. Надо сказать, что слизь выполняет и другие
функции, включая защиту от проникновения через кожу болезнетворных
бактерий, а вот ее гидродинамическая значимость кажется достаточно
сомнительной. Силу трения, обусловленную турбулентным пограничным
слоем, возможно уменьшить за счет введения в граничащий с телом слой воды
небольших количеств высокомолекулярных фракций или за счет введения
в воду вблизи тела рыбы вязкой жидкости, которая может двигаться в
полостях эластичной поверхности тела рыбы Эти процессы исключить в общем
случае нельзя. Однако такие эффекты наблюдались только в специально
организованных лабораторных условиях (Lumley, 1967; Kramer, 1960).
Невозможно закончить разговор об отряде перкоморфных (Percomorphi),
не упомянув кратко третью модификацию базового волнообразного способа
передвижения в воде, к которой пришли некоторые семейства этого отряда
(см. таблицу 3). Эта довольно радикальная модификация представляет
собой такой способ плавания, когда волнообразное движение осуществляется
только двумя грудными плавниками, их волнообразное движение в
вертикальной плоскости подобно горизонтальному волнообразному движению
хвоста при скомброидном типе перемещения (рис. 5). Само тело рыбы при
этом сравнительно негибкое. Семейства, у которых плавание происходит
за счет движения грудных плавников (см. таблицу 3), имеют другие общие
черты (Marshall, 1965, с. 210), а именно, им присущ осторожный
маневренный тип плавания. Такие пловцы являются постоянными обитателями
коралловых рифов, кроме того, это часто «чистильщики» других рыб.
Более того, некоторыми семействами, включая представителей отряда
гаплоидных (Haplomi\ таких как щука, плавание с помощью грудных плавников
используется в комбинации со скомброидным типом движения.
10. Плавучесть рыб
В данном обзоре остановимся в основном на представителях надотря-
да костистых рыб (Teleostei) (см. подпись к таблице 2), — костных рыбах,
10. Плавучесть рыб
47
которые в своем большинстве имеют гидростатический орган
(плавательный пузырь), с помощью которого они регулируют свою плавучесть. Этот
орган находится в рудиментарном состоянии или вовсе отсутствует
только у (а) некоторых донных костистых рыб, которые в своем
эволюционном развитии приобрели выгодную для них манеру поведения — в момент
неактивности опускаться на дно (где они часто защищаются от нападения,
приобретая защитную окраску), и у (Ь) некоторых семейств быстрых рыб,
таких как скумбриевые (Scornbridae), которые при наличии плавательного
пузыря не смогли бы достаточно быстро перемещаться в толще воды в
вертикальном направлении. А так они легко могут подняться в вышележащие
слои воды за счет силы тяги и за счет того, что они находятся в постоянном
движении (обеспечивая при этом возможность дыхания в силу
специфического устройства дыхательного механизма).
В случае когда рыбы не ведут придонный образ жизни или не столь
активны, как, например, скумбриевые, отсутствие плавательного пузыря
налагает дополнительные ограничения на стиль поведения рыбы. Например,
представители подкласса хрящевых рыб (Chondrichthyes) (см. таблицу 2),
у которых скелет состоит из хрящей, а плавательный пузырь отсутствует,
вынуждены поддерживать свою плавучесть на уровне 5% (Marshall, 1965,
с. 69), в некоторых случаях плавучесть повышается за счет запасов жира
в печени и в мышечной ткани. Подобная плавучесть наблюдается и у
семейства осетровых (Acipenseridae) (у осетров гидростатическая функция
плавательного пузыря недостаточна), принадлежащего древнейшему
отряду костных рыб — палеонискоидов (Palaeoniscoidea)
Как осетровые, так и большинство представителей отряда акул (Sela-
chii) (акулы и черные собачьи акулы), имеют гетероцеркальный хвостовой
плавник (плавник с увеличенной верхней лопастью, в которую
продолжается хвостовой отдел позвоночника), такой плавник не симметричен
относительно горизонтальной плоскости (см. рис. 10). Верхняя лопасть сильно
вытянута, имеет ярко выраженную форму стреловидного крыла,
отходящего вверх и назад. Нижняя лопасть имеет значительно менее выраженную
крыловидную форму, размах которой составляет от 0,1 до 0,5 размаха
верхней лопасти. Для рыб с гетероцеркальными хвостовыми плавниками
характерен угревидный тип движения, хотя при распространении поперечных
колебаний по телу у них наблюдается существенное увеличение
амплитуды колебаний хвостовой части тела. Движение верхней лопасти хвостового
плавника приводит к возникновению дополнительной силы тяги. Величину
этой силы можно найти на основании двумерной теории крыла (см.
раздел 8).
48
Глава 2
Очевидно, что при таком анализе необходимо рассматривать
движение стреловидной верхней лопасти плавника в воде как совокупность
движений, создающих тягу в направлении, перпендикулярном хорде лопасти,
(в этом направлении и создается наибольшая по величине составляющая
силы тяги), и в направлении, параллельном хорде (составляющая силы
тяги в этом направлении пренебрежимо мала). Вертикальная компонента
составляющей силы тяги, перпендикулярной верхней половине плавника,
представляет собой «подъемную силу». Эта сила совместно с
гидромеханической подъемной силой действует на грудные плавники, которые у этих
животных легко могут удерживаться в позиции положительного угла атаки
относительно продольной оси тела, в результате чего компенсируется
превышение веса тела над выталкивающей силой. Известно, что при
расположении грудных плавников немного впереди центра масс такая компенсация
будет наибольшей, однако без поддержки хвостового плавника движение
животного не будет устойчивым. Два источника возникновения подъемной
силы придают рыбе свойства, подобные свойствам аэроплана, она может
даже совершать петли в вертикальной плоскости (что достаточно часто
наблюдается, например, у представителей рода осетров Acipenser).
Представители многих семейств отряда акул (Selachii) имеют ярко
выраженный гетероцеркальный хвостовой плавник. Например, у семейств
песчаных (Odontaspidae), серых (Carcharinidae), колючих (Squalidae) и
рогатых (Heterodontidae) акул. У других же семейств хвостовой плавник вовсе
не заметен, например у таких, как ведущие донный образ жизни кошачьи
акулы семейства кошачьих акул {Scyliorhinidae). И наиболее интересным
исключением (хвостовой плавник не имеет ярко выраженной гетероцер-
кальной формы) являются быстрые акулы, включая таких представителей
семейства сельдевых акул (Lamnidae), как сельдевая акула, макрелевая
акула и акула-мако, или серо-голубые акулы, родов сельдевые акулы (Lamnd),
акулы-мако (hums) и большая белая акула (Carcharodon) (но не лисья
акула Alopias vulpes), а также представители семейства ковровых или усатых
акул (Orectolobidae), такие как китовая акула Rhineodon (но не
длиннохвостая зебровая акула Stegostomd).
Именно эти быстрые акулы, представленные выше на рисунке 6,
имеют хорошо развитый хвостовой плавник в форме полумесяца, практически
такой же формы, как и у наиболее быстрых костистых рыб. (По существу
же, такая форма плавника обусловлена тем, что нижняя беспозвоночная
лопасть приобрела похожую на верхнюю лопасть форму, и ее размах
составляет по меньшей мере 0,75 размаха верхней.) Более того, движения хвоста
10. Плавучесть рыб
49
i
, *.'
* ■>£
J
Рис. 10. Слева: голубая акула Prionace glauca семейства серых акул (Carcharinidae)
имеет ярко выраженный гетероцеркальный хвостовой плавник. Справа: спинорог
Xanthichthys, скалозуб или фугу Sphaeroides, рыба-луна Mola и морской конек
Hippocampus. Черным цветом отмечены плавники, обеспечивающие продвижение
рыбы, т.е. создающие тягу (движители) (Marshall, 1965)
у них практически такие же, как и при скомброидном типе движения. По
той же причине, что и скумбриевые, они не нуждаются в специальном
органе, изменяющем их плавучесть. То есть потому, что они являются такими
же быстрыми и ведущими активный образ жизни пловцами.
У многочисленного отряда скатов (Batoidei) класса хрящевых рыб
представители семейства пилорылых скатов (Pristidae) по внешней
форме, способу передвижения и обеспечению плавучести довольно похожи
на быстрых акул. Другие семейства этого отряда в результате длительного
процесса адаптации приспособили свою форму к придонному образу
жизни. Уже у семейства гитарных (Rhinobatidae) (рыбы-гитары) и гнюсовых
(Torpedinidae) (электрические скаты) имеются большие грудные плавники,
которые способны обеспечить плавучесть рыбы, хотя хвостовой плавник
еще выполняет функции движителя и совместно с колебательными
движениями грудных плавников обеспечивает продвижение рыбы в воде.
У других четырех семейств хвостовой плавник пренебрежимо мал или
вовсе отсутствует. Семейство скатовых (Raiidae) приспособилось к чрезвы-
50
Глава 2
чайно элегантному стилю плавания. Их огромные грудные плавники
сливаются с телом и вместе с ним составляют практически квадрат, при этом
тело ската располагается вдоль диагонали, и только тонкий хвост
выходит вдоль диагонали за границы квадрата. Перемещение ската происходит
за счет волнообразных движений, распространяющихся по грудным
плавникам в направлении противоположном движению, причем, причем длина
волны значительно меньше длины грудных плавников. Такой способ
плавания очень похож на тот, с описания которого начинался данный обзор
(только колебания плавников скатов происходят в вертикальной плоскости, в то
время как при угревидном способе плавания колебания распространяются
по телу в горизонтальной плоскости). Подобный способ плавания
используют и представители семейства хвостоколовых (Dasybatidae) (скаты-хво-
стоколы).
Движение скатов достаточно сложно проанализировать с точки зрения
гидромеханики, поскольку их тело имеет относительное удлинение,
равное 2. А для такого значения относительного удлинения не могут быть
применены ни теория обтекания тонкого тела, ни двумерная теория
крыла. Однако если скомбинировать результаты анализа движения с помощью
обоих подходов, то можно понять, что наибольшая эффективность
движения может быть достигнута в том случае, когда скорость распространения
волны лишь немного превышает скорость продвижения. Крылоподобная
форма тела животных с отрицательной плавучестью в состоянии покоя (за
исключением некоторых глубоководных форм) дает возможность
перемещаться с небольшим углом скольжения, когда они не находятся в состоянии
активного плавания.
У двух семейств: орляковых (Myliobatidae) и мантовых (Mobulidae), за
счет более поздней адаптации к свободному плаванию движения их
плавников еще более напоминают движение крыльев птиц, что даже отражено
в их названии — орляки. Действительно, их грудные плавники совместно
с телом ската образуют «ромб» с относительным удлинением, равным 4
(тело располагается вдоль меньшей диагонали ромба). Длина волны,
распространяющаяся по плавникам, превышает длину плавника, так что способ
движения этих скатов не отличается от способа, приведенного на рисунке 7
(хотя при этом добавляется постоянный угол атаки, величина этого угла
небольшая, поскольку подъемная сила должна уравновесить лишь
превышение веса рыбы над весом вытесненной воды, и, кроме того, кривизна
волнообразных движений у них несколько большая).
Интересным примером конвергентной эволюции является один отряд
костистых рыб, а именно отряд камбалообразных {Heterosomatd) (см. таб-
11. Передвижение «панцирных» рыб
51
лицу 2; знакомые представители этого отряда: камбала, морской язык,
палтус, калкан). В результате эволюции представители этого отряда приобрели
форму и выработали тип движения, похожие на их соседей, также
ведущих донный образ жизни, — ромбовых скатов (Raiidae). Так, камбала не
имеет плавательного пузыря, и на определенной стадии своей жизни она
поворачивается на бок, а глаз с нижней стороны постепенно перемещается
на верхнюю. Камбала перемещается, используя угревидный тип движения
(при активном плавании подобно угреобразным (Apodes) колебания
распространяются по всему телу рыбы, а при медленном плавании используются
только спинной и анальный плавники), конечно, после того, как она
переворачивается на один бок, движения ее тела происходят в вертикальной
плоскости. Крыловидное тело рыбы имеет форму ромба (вид сверху) с
относительным удлинением, близким к 1 (голова располагается по большей
диагонали).
11. Передвижение «панцирных» рыб
Представители оставшихся трех отрядов, которые предполагалось
обсудить в данной работе (см. таблицу 2), приобрели в процессе эволюции
защитную броню, что привело к уменьшению скорости их передвижения.
Не только их тело, но у некоторых семейств и хвост потеряли свою
гибкость до такой степени, что движение обеспечивается только за счет
плавников. Рыбы отряда солнечникообразных (Zeomorphi) имеют сильно сжатое
с боков высокое тело и продвигаются в воде за счет колебаний очень
гибких заднего спинного и анального плавников, практически таким же
способом плавают рыбы семейств единороговых (Monacanthidae) и спинорого-
вых (Balistidae) (например рыба-луна, см. рис. 10) отряда сростночелюст-
ных (Plectognathi)). Аналогичным способом плавают и имеющие вытянутое
сжатое с боков тело представители семейства кривохвостых (Centriscidae)
отряда иглообразных {Solenichthyes). В этом последнем отряде
представители семейства трубкоротых {Fistularidae) (рыбы-свистульки) и семейства
флейторылых (Aulostomatidae) в критических ситуациях для перемещения
могут также использовать и хвостовой плавник.
У семейства игловых (Syngnathidae) (морские коньки, морские иглы)
из всех иглообразных (Solenichthyes) панцирь наиболее жесткий, и эти
обитатели морских джунглей приспособились к весьма точному
маневрированию, значительно для них более важному, чем большая скорость
перемещения. Плавание происходит за счет колебаний, весьма небольших (для этой
52
Глава 2
цели), спинного и грудного плавников (см. рис. 10), для обеспечения
различных скоростей перемещения частота колебаний варьируется в весьма
широких пределах (зафиксирована частота колебаний в 70 герц). Гибкие
хвосты морских коньков, плавающих в вертикальном положении,
приспособлены в основном для прикрепления к стеблям подводных растений, но
иногда конек совершает колебательные движения хвостом для
перемещения в воде в вертикальном направлении. А перемещения в горизонтальном
направлении производятся за счет вибраций плавника.
Среди представителей отряда сростночелюстных (Plectognathi)
имеются обитатели коралловых рифов, одетые в исключительно мощную
броню, к ним относятся представители семейства ежи-рыбы (Diodontidae)
(рыбы-дикообразы), семейства иглобрюхих, или четырехзубых (Tetraodon-
tidae) (рыба-фугу, см. рис. 10), и семейства кузовковых (Ostraciontidae)
(кузовки). Эти рыбы перемещаются за счет колебательных движений
спинного и анального плавников, практически комбинируя способы плавания,
характерные для отряда солнечникообразных (Zeomorphi) и семейства иг-
ловых (Syngnathidae). С другой стороны, представители семейства рыб-лун
(Molidae) (обыкновенная рыба-луна) в процессе реадаптации приобрели
способность к пелагическому образу жизни. Из-за почти полного
отсутствия хвостового плавника им пришлость приобрести возможность
перемещения за счет колебательных движения спинного и грудного плавников
(рис. 10), относительное удлинение которых значительно возросло в
процессе приспособления к плаванию. Плавники у рыбы-луны работают
наподобие плавников семейства орляковых (Myliobatidae), но в отличие от орля-
ковых колебания плавников рыбы-луны происходят в вертикальной
плоскости. А тот факт, что пара разъединенных крыльев с высоким значением
относительного удлинения развернута в вертикальной плоскости, является
уникальным в животном царстве.
12. Использование конечностей для плавания:
земноводные и членистоногие
Проведенный довольно подробный обзор класса рыб дает возможность
лишь кратко осветить механизмы перемещения в воде представителей тех
классов позвоночных, которые имеют четыре конечности и приспособились
наряду с жизнью в воде и к жизни на суше. Большинство земноводных на
личиночной стадии развития унаследовало от своих предков-рыб сжатый
12. Использование конечностей для плавания 53
с боков хвост и угревидный способ перемещения в воде. Взрослых
особей, даже если у них, например как у представителей подкласса хвостатых
(Urodele), и сохранился хвост, можно лишь с натяжкой считать водными
животными. К таким земноводным относятся представители рода
саламандр {Salamandra), чей хвост стал непригоден для осуществления
перемещения в воде. Однако у большинства хвостатых земноводных (Urodeles),
от тритонов до исполинской саламандры (Megalobatrackus), сжатый с боков
хвост в значительной степени приспособлен для перемещения в воде.
Что касается бесхвостых земноводных (лягушки и жабы подкласса
бесхвостых (Апига)\ то во взрослом состоянии большинство из них
практически не ведут водный образ жизни. Нами будет рассмотрена с точки
зрения приспособления бесхвостых четырехлапых земноводных к
перемещению в воде та небольшая, но важная для нас часть бесхвостых
земноводных, вторично вернувшихся к водному образу жизни. Способ
перемещения в воде, который используют эти животные, не похож ни на один из
рассмотренных ранее способов плавания рыб (симметричное
использование плавников встречается только у тех рыб, которые перемещаются с
помощью грудных плавников). Эти бесхвостые земноводные имеют гибкие
задние конечности с перепончатыми стопами, при плавании их движения
напоминают движения человека, плывущего брассом, но значительно более
эффективны, поскольку конечности у них имеют более обтекаемую форму.
Вернемся ненадолго к тем беспозвоночным, чьи способы перемещения
в воде ранее не рассматривались, а именно к представителям большого типа
членистоногих {Arthropoda) (см. таблицу 1). Большинство членистоногих,
ведущих как водный, так и наземный образ жизни, прекрасно
приспособлены к плаванию. У представителей класса насекомых (Insecta)
приспособление к передвижению в воде произошло подобно земноводным за счет
изменения конечностей. Так, у многих семейств отряда жесткокрылых, или
жуков {Coleoptera\ изменилась форма задних и средних лапок, а у
плавунцов (Dytiscus) на этих лапках дополнительно появились плавательные
волоски. И в каждом случае лапки приобрели хорошо обтекаемую форму,
так чтобы развивать движущую силу с минимальными потерями энергии.
Однако их плавательные движения не симметричны, поскольку лапки на
каждой стороне тела жука гребут поочередно.
Из класса ракообразных (Crustacea) рассмотрим три отряда:
ротоногие ракообразные (Stomatopoda), эуфаузиевые (высшие ракообразные)
(Euphausiacea) и десятиногие ракообразные (Decapoda), в которые входят
все крупные животные этого класса (превосходный обзор класса ракообраз-
54
Глава 2
ных можно найти в работе Локхеда (Lochhead, 1961)). Тело этих животных
состоит из 19 сегментов (из 13 состоит головной и грудной отделы и из 6 —
брюшко), каждый сегмент снабжен парой конечностей, каждая из которых
может сильно отличаться от других или их рост может быть приостановлен,
за счет чего конечности животного могут принимать форму, немного
напоминающую плавники рыб. Из таких конечностей отметим, прежде всего,
тельсон, или эластичную хвостовую лопасть, сформированную из
отростков последнего брюшного сегмента. В процессе движения животное
совершает колебательные движения брюшком, эффективность которых
определяется главным образом значительным горизонтальным размахом тельсона.
Те представители рассматриваемых отрядов, которые сохранили другие
десять конечностей брюшка (или плеоподы), получили дополнительную
возможность плавать за счет их волнообразных движений. Эти
волнообразные движения у креветок и подобных им животных напоминают плавание
рыб с помощью грудных плавников. При медленном плавании десятиногие
ракообразные используют только эти десять конечностей, а для быстрого
плавания дополнительно подключается и движение тельсона.
А вот у крабов (подотряды крабоидов (ненастоящих крабов) (Апотига)
и настоящих крабов (короткохвостых раков) (Brachyura)) брюшные ножки
редуцированы. Эти крабы в основном ползают по дну и ведут роющий
образ жизни. Представители же отряда крабов-плавунцов (Portunidae)
являются отличными пловцами. Они плавают, отталкиваясь грудными
конечностями (особенно последней парой), имеющими крыловидное поперечное
сечение. Толчки, подобные взмахам крыльями, создают тягу, достаточную
для перемещения со скоростью до 1 м/с. После этого краткого обзора
способов перемещения некоторых членистоногих (Arthropoda) вернемся к
позвоночным.
13. Перемещение в воде пресмыкающихся
Если рассматривать животных с точки зрения степени
приспособленности к перемещению в воде, то происхождение современных
членов класса пресмыкающихся (Reptilia) от архаичных хвостатых
земноводных (Amphibia) особенно очевидно на примере отряда крокодилов
(Crocodilia) (крокодилы и аллигаторы), которые используют тип
движения, чрезвычайно сходный со скомброидным, совершая колебательные
движения своим высоким сжатым с боков хвостом. Отметим и другие, на
сегодняшний день самые многочисленные по количеству видов, отряды
13. Перемещение в воде пресмыкающихся
55
приспособленных к перемещению в воде пресмыкающихся — это отряд
чешуйчатых (Squamatd) (змеи, ящерицы и хамелеоны) и отряд черепах
(Chelonia) (пресноводные черепахи и сухопутные черепахи). Прежде чем
заняться их исследованием, вероятно, необходимо отметить тот
удивительный факт, что вымершая морская рептилия ихтиозавр (Ichthyosaurus) хотя
и произошла от сухопутных предков, но имела хвост в виде полумесяца,
как и у представителей отряда скумбриевых (Scombridae) (рис. 6).
«Змеевидный» способ (Gray, 1946) перемещения большинства змей по
земле (подотряд змеи (Ophidia)) напоминает угревидный способ
перемещения рыб, с той лишь разницей, что при движении змей выполняется
равенство U = V, т. е. при перемещении животного по земле по его телу в
направлении от головы к хвосту проходит поперечная волна с той же самой
скоростью, с какой животное в результате этого волнообразного движения
перемещается вперед. Подобное передвижение с U = V возможно
только при наличии неровностей, и в действительности за счет наличия этих
неровностей возникает «нормальная» сила (т. е. сила, действующая под
углом в 90° к оси тела) даже и тогда, когда нормальная компонента скорости
равна нулю, в то же время тангенциальная сила не зависит подобным
образом от тангенциальной скорости. Другими словами, в отсутствие движения
нормальное и тангенциальное сопротивление не являются
пропорциональными величинами.
Змеи, ведущие наземный образ жизни, несмотря на их «плохую» с
точки зрения гидромеханики форму — поперечное сечение близко к кругу, тело
к концу постепенно сужается, — в случае необходимости довольно быстро
плавают. Правда, при волнообразных движениях тела в воде, подобных тем,
которые они совершают на земле при ползании, скорость продвижения U
меньше, чем скорость распространения волны по телу V (так, у ужа
обыкновенного (Tropidonotus natrix), по наблюдениям Грея (Gray, 1939a), эти
скорости связаны соотношением U = 0,7V). Построим некоторую
упрощенную теорию продвижения за счет волнообразного движения тела,
имеющего «плохую» форму с точки зрения гидромеханики на основании работ
Тейлора (Taylor, 1952) и Грея и Хенкока (Gray, Hancock, 1955).
Предположим, что на рисунке 5 описывается движение некоторого короткого
сегмента тела животного, и примем, что для нормальной и тангенциальной
скорости выполняется линейный закон для силы сопротивления.
На рисунке 5 показан наклон сегмента (т. е. тангенс угла, который он
составляет с направлением движения), равный CD/PD = W/V в
произвольный момент времени. Считаем этот угол достаточно малым, чтобы его
56
Глава 2
косинус можно было заменить единицей, а нормальная скорость движения
сегмента относительно жидкости была бы равной w. Нормальное
сопротивление движению сегмента будет равно произведению этой скорости на
коэффициент KN. Произведение скорости W на тангенс угла наклона
сегмента W2 /V равно тангенциальной скорости. Тангенциальное
сопротивление движению сегмента равно произведению тангенциальной скорости на
коэффициент Кт. Здесь KN и Кт — коэффициенты пропорциональности
в выражении для нормального и тангенциального сопротивления движению
соответственно.
Теперь можно записать величину осевого усилия Р, действующего на
сегмент как разность нормального сопротивления, умноженного на тангенс
угла с направлением движения, и тангенциального сопротивления, т. е.
Р = (KNw)(W/V) - KT{W2/V). (3)
В то же время животное расходует энергию со скоростью Е = (KNw)W
при движении со скоростью W, преодолевая сопротивление KNw.
Совершенная полезная работа, или коэффициент полезного действия ту, равна
произведению UP, разделенному на Е, т. е.
П = UP/E = (U/V)[l - (KT/KN)(W/w)). (4)
Из уравнения (3) следует, что осевое усилие будет положительным, только
если отношение w/W лежит в пределах от г до единицы, где за г
обозначено отношение коэффициентов KT/KN. Из уравнений (2) и (4) следует,
что отношение скоростей и максимальный КПД равны
w/W = г1'2, U/V = l- г1'2, г) = (1 - г1/2)2. (5)
При движении змеи по земле отношение коэффициентов г будет равно
нулю, поскольку коэффициент пропорциональности для нормальной
компоненты силы сопротивления будет бесконечен, поскольку нормальная
скорость равна нулю. Тогда из уравнения (5) следует, что выполняется
равенство U = V, а механический КПД равен единице. С другой стороны,
при плавании ужа обыкновенного (Tropidonotus natrix) при числах Рей-
нольдса, рассчитанных по длине, имеющих порядок 106, значение г можно
приближенно принять равным 0,1, что дает отношение скоростей, равное
U/V = 0,7 с точностью до одной значащей цифры, и это совпадает с
экспериментальными данными, а для КПД ту получаем значение, немного
меньшее 0,5. С уменьшением числа Рейнольдса отношение г будет медленно
13. Перемещение в воде пресмыкающихся
57
возрастать, что приведет к уменьшению отношения скоростей U/V (до тех
пор, пока число Рейнольдса не станет слишком малым). Как следует из
расчетов, проведенных Хенкоком (Hancock, 1953), при г = 0,5 отношение
скоростей равно U/V = 0,3, а КПД равен 7? = 0,1.
Полученные результаты хорошо согласуются с многочисленными
наблюдениями над беспозвоночными, описанными ранее (раздел 2), и, кроме
того, эти результаты позволяют объяснить аномальное поведение много-
щетинковых кольчатых червей. Их длинные параподии при движении
испытывают сопротивление только в тангенциальном направлении, поэтому
величина г может превышать единицу, так что отношение U/V
становится отрицательным, что и наблюдается в природе. Греем и Хенкоком (Gray,
Hancock, 1955) (для низких чисел Рейнольдса) и Тейлором (Taylor, 1952)
(для больших чисел Рейнольдса) рассмотренная приближенная теория
была уточнена, что позволило применить ее для многих других типов
продвижения за счет волнообразного движения. Однако для «хороших» с точки
зрения гидродинамики форм тела, подобных формам тела рыбы, эта
теория дает совершенно неудовлетворительные результаты, поскольку в ней
не учитываются эффект наличия «присоединенной массы» (раздел 4) и
наличие дополнительной большой по величине силы тяги, возникающей за
счет движения высокого сжатого в вертикальной плоскости хвоста. Для
форм промежуточного вида было бы желательно, насколько это возможно,
объединить указанные два подхода.
Представители одного подсемейства отряда змей (Ophidia\ а
именно подсемейства ластохвостых морских змей (Hydrophinae), ведут
исключительно морской образ жизни, несмотря на наземных предков. Эти змеи
передвигаются в воде за счет волнообразных движений сплющенной с
боков задней части, заканчивающейся широким и плоским в вертикальной
плоскости ластообразным хвостом. Такая форма тела дает, по-видимому,
большой выигрыш в скорости. Прежде чем завершить обсуждение
класса пресмыкающихся (Reptilia) необходимо отметить, что в отряде черепахи
(Chelonia) наиболее сложный способ перемещения с точки зрения
гидромеханики используют представители семейства морских черепах (Chelonidae).
Они прекрасно решили задачу перемещения животных, имеющих четыре
конечности. В процессе адаптации к водному образу жизни их передние
конечности приняли форму ласт с большим относительным удлинением
и имеющих в поперечном сечении крыловидную форму. Движение ласт
морских черепах напоминает взмахи крыльев птиц (рис. 7) даже в большей
степени, чем движение плавников у скатов орляков.
58
Глава 2
14. Передвижение в воде млекопитающих
Среди млекопитающих (последний класс в таблице 1)
преимущественно водный образ жизни ведут представители только трех следующих
отрядов: хищные (Carnivora), китообразные (Cetacea), сирены (Sirenia),
причем в первом из указанных отрядов преимущественно водный образ
жизни ведут только тюлени (семейство тюленей настоящих (Phocidae),
семейство ушастых тюленей (Otariidae)) и моржи (семейство моржовых
(Odobenidae)). Эти четырехногие в процессе эволюции приспособили свои
конечности к возможности передвигаться в воде, и в то же время у них не
исчезла полностью возможность передвигаться на суше. Так, например,
передние конечности тюленей семейства настоящих тюленей (Phocidae)
(наружные ушные раковины у них отсутствуют, сюда входят все европейские
тюлени) служат опорой при передвижении по суше, а при плавании они
плотно прижимаются к телу, как бы встраиваясь в подмышечные
углубления (передние конечности в воде используются только при
маневрировании), при этом задние ласты совершают поочередные колебательные
движения вправо-влево, одновременно изгибаясь сверху вниз. Пятипалыми
задними конечностями тюлень сначала отталкивается назад, пальцы при
этом вытягиваются, затем он пальцы втягивает внутрь, далее его колени
сгибаются и пальцы ног сжимаются, ноги подтягиваются вперед, затем
снова следует сильный толчок назад (Backhouse, 1961). Способ плавания
тюленей напоминает в некоторой степени скомброидный тип движения.
Представители отряда китообразных (Cetacea) (киты и дельфины),
в отличие от тюленей и моржей, ведут исключительно водный образ
жизни, и их задние конечности полностью редуцировались. Вместо этого у них
на сужающейся задней части позвоночного столба образовался новый
орган в виде горизонтального хвоста в форме полумесяца, состоящий из двух
«камбалоподобных» лопастей (направленные в разные стороны
увеличенные кожные складки, изнутри поддерживаемые опорным каркасом из
хрящевой ткани). Хвост при движении совершает волнообразные движения
в вертикальной плоскости, создавая при этом необходимую для
поступательного движения в воде силу тяги. Плавники, развившиеся из передних
конечностей, используются животным для стабилизации тела в воде и
осуществления маневров. На целесообразность таких эволюционных
изменений конечностей указывает, в частности, тот факт, что животные
совершенно другого вида, а именно отряда сирен (Sirenia), состоящего на
сегодняшний день из двух родов — дюгоней (Halicore) и ламантинов (морских
14. Передвижение в воде млекопитающих
59
коров) {Trichechus), в процессе эволюции пришли практически к такому же
способу передвижения и к такому же преобразованию конечностей, вплоть
до потери задних конечностей и приобретения большого распластанного
в стороны хвоста, хотя только у китов хвостовой плавник приобрел ярко
выраженную форму полумесяца.
Из превосходных киносъемок, проведенных Парри (Рапу, 1949) и Лан-
гом (Lang, 1966), видно, что волнообразные движения в вертикальной
плоскости полулунного хвоста кита совершенно аналогичны волнообразным
движениям хвостового плавника у рыб в горизонтальной плоскости
(изображены на рис. 5). Отдача при таком строении задней части тела кита
(отдача действует в вертикальной плоскости и возникает в результате колебаний
величины выталкивающей силы, действующей на хвост) также
минимальна, главным образом за счет того, что в передней части хвостового
плавника тело имеет очень небольшой размер в горизонтальном направлении,
а скорость возрастания амплитуды колебаний хвоста здесь наибольшая.
Если кит плавает в условиях, когда выталкивающая сила превышает
его вес, то его плавники и хвост способны двигаться так, чтобы возникла
отрицательная подъемная сила. Лопасти хвоста, будучи расположенными
далеко от центра масс, вносят лишь небольшой вклад в создание подъемной
силы, которая, однако, для кита может оказаться достаточной в силу
большой площади хвоста. В этом случае (при положительной плавучести) ском-
броидный тип движения кита должен быть дополнен отрицательным углом
атаки, противоположным углу атаки, изображенному в средней части
диаграммы (рис. 7) схематичного представления типичного полета птиц.
Тогда наибольшее усилие необходимо приложить при движении хвоста вверх,
и интересно отметить, что у морской свиньи (Phocaena) (Gray, 1968, с. 289)
и у кита-полосатика (Bolaenoptera) (Kermack, 1948) мышцы,
обеспечивающие движение хвостового плавника вверх, значительно более развиты, чем
мышцы, обеспечивающие движение хвостового плавника вниз. И еще раз
отметим (как и для углов атаки) инверсный характер развития мышц
хвостового плавника у китовых и крыльев у птиц (раздел 7).
Поскольку большинство китообразных имеет гладкое хорошо
обтекаемое тело, а силу тяги вырабатывает только хвостовой плавник за счет
колебательных движений в вертикальной плоскости, то гидродинамическое
сопротивление, испытываемое животным, относительно невелико. Наличие
мощной хвостовой мускулатуры, создающей большую по величине силу
тяги, и относительно небольшое гидродинамическое сопротивление
приводят к тому, что животное может развивать достаточно большую скорость
60
Глава 2
(например, скорость U равна 8 м/с при длине животного / в 2 м). Однако,
так же как и в случае быстрых рыб, отсутствуют надежные доказательства
(см. (Bainbridge, 1961; Slijper, 1961; Lang, 1966)) того, что трение в
пограничном слое у тела животного может быть им сделано ниже того, которое
ему присуще при данной гладкости поверхности и при фиксированном
числе Рейнольдса (порядка 107 или больше). Заметим, что к многочисленным
сообщениям об исключительно высоких скоростях, развиваемых
дельфинами, следует относиться с осторожностью, поскольку возможно, что
животные двигались в струе морского течения, в спутном потоке или вместе
с волнами.
15. Заключение
Все наиболее быстрые морские животные, такие как млекопитающие,
хрящевые рыбы, костные рыбы, используют скомброидный тип плавания
с помощью большого полулунного хвоста. Крайне желательно в
дальнейшем было бы накопить большой экспериментальный материал по
передвижению этих животных в естественных условиях и на его основе провести
теоретическое изучение механизма их плавания.
Далее, следовало бы развить более совершенную теорию
передвижения удлиненного тела, в которой были бы учтены как эффект
присоединенной массы, так и наличие вязкого сопротивления при движении тела в
потоке. Такая теория была бы значительно более полезна при решении
различных прикладных задач. Для того чтобы в теории возможно было учесть
эти два эффекта, с нашей точки зрения, необходимо предварительно
изучить (а) колебательные движения одного плавника; (Ь) механизм создания
движущей силы ромбовых скатов, других скатов и рыб плоской формы; (с)
механизмы начала движения и маневров рыб; (d) механизмы передвижения
десятиногих ракообразных, подобных креветкам, роль плавательных ножек
в создании движущей силы (е) возможность применения методов
исследования Грея и Хенкока (Gray, Hancock, 1955) к изучению передвижения рес-
ничковых организмов. Несомненно, что вдумчивый читатель сможет
указать и множество других направлений исследований, связанных с темой
данного обзора.
Выражение признательности
С большим удовольствием выражаю свою благодарность доктору
Квентину Боуну (Quentin Bone), доктору Виктору Ховарту (Victor Howarth)
15. Заключение
61
и доктору Ричарду Бейнбриджу (Richard Bainbridge) за весьма ценные
дискуссии, а также Зоологическому обществу Лондона и Британскому музею
(Музею естественной истории) за те результаты исследований, которые
были получены при изучении их знаменитых коллекций. Моя глубокая
признательность сотруднику музея, доктору Н. Б. Маршаллу, совершенно ясно
следует из самого текста книги. И наконец, с самым теплым чувствам
признательности хотелось бы от своего имени и от имени своих
коллег-специалистов в области биогидродинамики поблагодарить сэра Джеймса Грея за
его работы, которые в течение последних пятидесяти лет являются мощным
стимулом развития этой области научного знания.
[
[19;
[20
Литература
Gray, J. 1928. Ciliary Movement. Cambridge Univ. Press, 162 p.
Siekmann, J. 1963. J. Fluid Mech., 15, 399^18.
Gray, J., Hancock, G.J. 1955. J. Exptl. Biol., 32, 802-14.
Gray, J., Lissmann, H. W. 1964. J. Exptl. Biol., 41, 135-54.
Gray, J. 1939a. Proc. Roy. Soc. (London) Sen B, 128, 28-62.
Gray, J. 1939b. J. Exptl. Biol., 16, 9-17.
Taylor, G. I. 1952. Proc. Roy. Soc. London, Ser. A, 214, 158-83.
Marshall, N. B. 1965. The Life of Fishes. Weidenfeld & Nicolson, London,
402 p.
Harris, J. E. 1938. J. Exptl. Biol, 15, 32-47.
Hertel, H. 1966. Structure, Form and Movement. Reinhold, New York,
251 p.
Gray, J. 1968. Animal Locomotion. Weidenfeld & Nicolson, London, 479 p.
Breder, CM. 1926. Zoologica, 4, 159-297.
Lighthill, M. J. 1960. J. Fluid Mech., 9, 305-17.
Bainbridge, R. 1958. J. Exptl. Biol., 35, 109-33.
Bainbridge, R. 1960. J. Exptl. Biol., 37, 129-53.
Bainbridge, R. 1963. J. Exptl. Biol., 40, 23-56.
Fierstine, H.L., Walters, V. 1968. Mem. S. Calif Acad. Sci., 6, 1-31.
Norman, J. R., Fraser, F. С 1937. Giant Fishes, Whales and Dolphins.
Putnam, London, 375 p.
Woodward, A. S. 1910. Encycl. Britann. (11th Ed.), 23, 141^17.
Garrick, I.E. 1957. Nonsteady Wing Characteristics. In Aerodynamic
Components of Aircraft at High Speeds, 658-793. (Donovan, A. F.,
Lawrence, H. R., eds.) Princeton Univ. Press, 845 p.
62
Глава 2
[21] Wu, Т. Y. 1961.J. Fluid Meek, 10, 321-44.
[22] von Karman, Т., Burgers, J. M. 1934. General Aerodynamic Theory:
Perfect Fluids. Vol. II (367 p.) of Aerodynamic Theory. (Durand, W. R,
ed.) Springer Verlag, Leipzig, vol. 6.
[23] Goldstein, S., ed. 1938. Modern Developments in Fluid Dynamics. Oxford
Univ. Press, 702 p.
[24] Lumley, J.L. 1967. Appl Meek Rev, 20, 1139-49.
[25] Kramer, M. O. 1960. J. Am. Soc. Naval Engn, 72, 25-34.
[26] Lochhead, J. H. 1961. Locomotion. In Physiology of the Crustacea, vol. 2,
313-64 (Waterman, Т. Н., ed.) Academic Press, New York.
[27] Gray, J. 1946. J. Exptl. Biol, 23, 101-20.
[28] Hancock, G.J. 1953. Proc. Roy. Soc. London. Ser. A, 111, 96-121.
[29] Backhouse, K.M. 1961. Symp. Zool. Soc. London, 5, 59-75.
[30] Parry, D. A. 1949. J. Exptl. Biol., 26, 24-34.
[31] Lang, T. G. 1966. Hydrodynamic analysis of cetacean performance. In
Whales, Dolphins and Porpoises: Proceedings of the First International
Symposium on Cetacean Research, 410-32 (Norris, K. S., ed.) Univ. of
California Press, 789 p.
[32] Kermack, K. A. 1948. J. Exptl. Biol., 25, 237^0.
[33] Bainbridge, R. 1961. Symp. Zool. Soc. London, 5, 13-32.
[34] Slijper, E.J. 1961. Symp. Zool. Soc. London, 5, 77-94.
Глава 3
Математические аспекты передвижения
водных животных при низких числах
Рейнольдса
1. Введение
Как отмечалось в главе 2, при размерах животного, не превышающих
1 мм, и при числе Рейнольдса меньше единицы силами инерции можно
пренебречь. В этой главе рассматриваются некоторые математические аспекты
движения при низких числах Рейнольдса.
При таком движении силы, действующие на каждый элемент
жидкости, находятся в равновесии, точно так же как в статике, поскольку
инерционные силы (произведение массы на ускорение), которые могли бы
нарушить это равновесие, пренебрежимо малы. Подобным образом и со
стороны жидкости на плывущее животное действует система сил, которая также
постоянно должна быть в состоянии статического равновесия, поскольку
скорость изменения количества движения животного или его момента
количества движения пренебрежимо мала по сравнению с величиной вязких
сил.
Эти статические соотношения дополним термодинамическими и
кинематическими соотношениями: скорость жидкости должна удовлетворять
уравнению неразрывности в предположении, что плотность жидкости есть
величина постоянная, для жидкости выполняется закон вязкости Ньютона.
Из кинематики следует, что движение тела животного является суммой
(i) движения относительно центра массы, связанного с изменением
формы тела и другими деформациями тела (включая растяжение или
расслабление), возникающими за счет подвижности (лабильности)
животного;
64
Глава 3
(ii) твердотельного движения, в общем случае представляющего собой
поступательное движение центра масс животного со скоростью U и
вращение с угловой скоростью ft относительно центра масс.
Пусть составляющая (i), определяемая подвижностью животного,
известна. В этом случае движение будет определено, если найдены
неизвестные векторы U и ft. Очевидно, что условие равенства нулю суммы всех
сил и моментов, действующих между жидкостью и животным, дает
нужное число уравнений (два векторных уравнения) для определения U и Q.
Отметим, что их мгновенные значения определяются только мгновенными
значениями параметров, характеризующих подвижность (лабильность)
животного, поскольку инерционные эффекты пренебрежимо малы. Уравнения
динамики в нашем случае сводятся к кинематическим и статическим
уравнениям, что в действительности является достаточно часто применяемым
упрощением.
2. Уравнения движения жидкости и их фундаментальные
сингулярные решения
Объемными силами, действующими на элемент ньютоновской
жидкости, являются градиенты давления —Vp и силы вязкости ^V2u, где р —
давление, р, — вязкость, и — вектор скорости жидкости. Условие
равновесия этих сил
-Vp + /j,V2u = 0 (1)
с уравнением неразрывности
V • и = 0 (2)
приводит к уравнению Лапласа для давления
V2p = 0. (3)
То есть давление является гармонической функцией в безынерционном
потоке. Из уравнений (1) и (3) следует, что вектор скорости и является би-
гармонической функцией (решением уравнения V4u = 0; на самом деле
векторным решением, для которого уравнение (2) дает дополнительную
информацию).
Для того чтобы представить движение жидкости, обусловленное
колебанием жгутика, различные участки которого действуют с различными
2. Уравнения движения жидкости
65
силами на жидкость, можно использовать уравнения движения
безынерционной жидкости под действием некоторой силы. Такую объемную силу
запишем в общем в виде
Щг). (4)
Здесь 6 — дельта-функция, г — вектор перемещения с началом в точке
приложения силы F. Условие равновесия (1) элементарного объема жидкости
при действии внешней силы (4) принимает вид
-Vp + AiV2u + F5(r) = 0. (5)
С учетом (2) получаем
V2p = V-[F*(r)]. (6)
Решением уравнения (6) в случае неограниченной жидкости является
классическое поле диполя
p=-V-[F/47rr] (7)
с интенсивностью, равной внешней силе F, приложенной к жидкости в
точке г = 0. Здесь интересно провести параллель с теорией звука, где
соответствующее поле скоростей, совершенно отличное от рассматриваемого
случая, находится приравниванием произведения —р du/dt (не к силе
вязкости ^V2u) к величине Vp.
Найдем поле скоростей в случае, когда сила
F = (F, 0, 0) (8)
направлена вдоль оси х. Тогда уравнение (7) принимает вид
р = Fx/Airr3. (9)
Из (5) вытекает, что в любой точке, за исключением г = 0, выполняется
„2 1 F ( 1 Зх2 Зху 3xz\
V2u = - Vp = — -j - -g-, f, r . 10
Воспользуемся известным способом нахождения бигармонической
функции, лапласиан которой равен заданной гармонической однородной
функции степени п. Для этого рассмотрим тождество
V2(r20) = r2V2<f> + 0V2(r2) + 2V0 • V(r2) =
= г2Ч2ф + 6ф + 4г- V<£.
66
Глава 3
Если V20 = 0, где ф — однородная функция n-го порядка, то правая часть
этого тождества принимает вид (6 + 4п)ф. В этом случае лапласиан от
функции (6 + 4п)~1г20 равен ф.
Правая часть уравнения (10) является однородной функцией
порядка -3, следовательно, частное решение уравнения (10) имеет вид
_ _1 2 ( F „ дЛ _ F (х2 -r2/3 xy_ xz\
6 \47ГД Г3/ 87ГД \ Г3 ' г3 ' г3 у
Эта функция не удовлетворяет уравнению неразрывности (2).
Действительно,
V • [r2V(x/r3)] = r2V2(x/r3) + V(r2) • V(x/r3) =
= 0 + 2r.V(x/r3). (12)
Поскольку функция x/r3 является однородной функцией порядка —2, то
-Ах/г3 = V-(4/r, 0, 0). (13)
Если добавить вектор (F/6717/r, 0, 0) в правую часть выражения (11), то
получившаяся функция будет удовлетворять уравнению неразрывности (2),
а ее лапласиан V2u не будет отличаться от (10), поскольку 1/г является
гармонической функцией.
Скорость и давление, обусловленные точечной силой (8), имеют вид
_ F (х2 + r2 xy xz\ _ Fx
U" 8^1 V г3 ' 73"' ~)' Р" 4^з' ( }
Радиальная компонента скорости равна
иг = (г • u)/r = (F/87r/i)(2cos^/r), (15)
где 9 — полярный угол и для него выполняется cos 9 = х/г. Тангенциальная
компонента скорости ив равна
ив = (—у2 — z2\ xy, xz) • u/r2sin0 = (F/8тг/л)(— sin9/r). (16)
На рисунке 1 приведены два характерных решения этих уравнений.
Если бы множитель 2 в (15) отсутствовал, то получилось бы поле
скоростей, сохраняющее постоянную величину на сфере радиусом г и
имеющее постоянное (вдоль оси х) направление. Отметим также подобие полю
скоростей V0, создаваемому диполем, с потенциалом скорости, равным
ф = Gx/Airr3. (17)
2. Уравнения движения жидкости
67
2cos0 F
г Ъттц
cosfl F
г 8тгц
2sin0 F
Snfi
(о) Stokeslet field
(b) Dipole field
Рис. 1. Изображены компоненты скорости в сферической системе координат: (а)
поле скоростей создано стокслетом интенсивности F; (Ь) поле скоростей создано
диполем интенсивности G
Такой диполь с моментом (G, О, 0) создает поле скоростей и давления
G (—-
U =
47Г
(18)
т. к. u = V0 удовлетворяет условию V2u = 0. Радиальная и трансверсаль-
ная компоненты этого поля скоростей имеют вид
ur = (-G/47r)(2cos0/r3), щ = (-G/47r)(sin0/r3). (19)
Это поле отличается от поля скоростей (15) и (16), обусловленного
действием точечной силы множителем 1/г3 вместо 1/г и знаком одной из
компонент.
68
Глава 3
Наиболее эффективным методом нахождения решения линейного
уравнения в частных производных, удовлетворяющего заданным
граничным условиям, является представление такого решения в виде комбинации
фундаментальных сингулярных решений. С этой точки зрения особенно
важными для уравнений движения Стокса (1) и (2) безынерционной
жидкости являются два сингулярных решения (14) и (18). В решении (14)
отражено влияние точечной силы (F, 0, 0), приложенной в начале координат,
точку приложения силы можно назвать стокслетом (stokeslet)
интенсивности (F, 0, 0). Поле скоростей (18) создается диполем с интенсивностью
(G, 0, 0) при отсутствии внешних сил, этот диполь иногда называют ди-
польным источником.
Простейшая иллюстрация этого метода состоит в конструировании
поля с постоянной по величине и направлению скоростью на границе сферы
радиуса г = а. Если мы возьмем линейную комбинацию поля стокселета
(рис. la) и поля точечного диполя (рис. 1 Ь), то мы можем добиться
требуемого результата за счет выбора момента диполя или амплитуды точечной
силы. Из рисунка 1 следует, что если момент диполя взять равным
G = Fa2 /6/х, (20)
то на сфере радиуса г — а скорость, обусловленная наложением диполя
и стокслета, будет иметь постоянную величину F/Qir/ia и иметь
направление, параллельное приложенной точечной силе, поскольку при этом
радиальные компоненты, содержащие множитель cos(0), сократятся, а транс-
версальные, содержащие множитель — sin(0), увеличатся аддитивно. Этот
же результат может быть получен аналитическим путем из равенств (14)
и (18).
Таким образом, мы пришли к формуле Стокса для силы сопротивления
действующей на движущуюся со скоростью U сферу, со стороны вязкой
жидкости
F = бтг/лаи. (21)
Мы можем в точности удовлетворить граничным условиям: на
поверхности тела скорость жидкости равна скорости тела, т.е. (С/, 0, 0) при г = а,
за счет того, что на поле скоростей, созданное стокслетом интенсивности
(F, 0, 0) при г = 0, который представляет собой внешнюю силу (F, 0, 0),
действующую на жидкость, накладывается поле скоростей, созданное
диполем интенсивности (20) при отсутствии внешней силы.
3. Сингулярные решения на отрезке
69
3. Сингулярные решения на отрезке
Рассмотрим теперь методы использования сингулярных решений (14)
и (18) для изучения движения микроскопических организмов,
перемещающихся за счет колебательных движений жгутиков (см. главу 2). Закон Сток-
са (21) возможно применять к таким организмам, у которых сила
сопротивления, действующая на микроорганизм, с формой, близкой к сферической,
уравновешивается силой тяги, создаваемой колебательными движениями
жгутика. Однако следует иметь дополнительную информацию
относительно силы, действующей между жидкостью и органеллой длинного
небольшого в поперечном сечении жгутика (т.е. здесь мы уже встречаемся не
с чем-то напоминающим по форме сферу, а скорее, с некоторым подобием
длинного цилиндра).
В этом случае следует помнить, что для бесконечно длинного
тонкого цилиндра не существует закона, подобного закону Стокса (21).
Известный парадокс Стокса утверждает, что уравнения движения и
неразрывности (1) и (2) безынерционной жидкости не имеют решения,
удовлетворяющего условиям прилипания на поверхности цилиндра. Это следует иметь
в виду при изучении движения жгутика, когда мы исследуем поле
скоростей жидкости, обсловленное стокслетами, распределенными на конечном
линейном промежутке.
Исследуем суммарный эффект от распределенных на отрезке стоксле-
тов (14), скажем, от Z = —Ь до Z = с с общей интенсивностью F на
интервале (Z, Z + dZ), равной (/dZ, 0, 0). То есть интенсивность стокслета
в направлении оси х имеет постоянную величину /, отнесенную к единице
длины. Поле скоростей найдем интегрированием (14) от Z = — Ь до Z = с,
заменив переменную z на z — Z, а величину F на fdZ.
Рассмотрим поле скоростей вокруг поперечного сечения z = 0
кругового цилиндра
х2 + у2 = а2. (22)
На оси цилиндра распределены стокслеты, радиус цилиндра а считаем
малым по сравнению с b и с. (Полагаем z = 0 в том поперечном сечении, для
которого определяется поле скоростей, и через Ь и с обозначим
расстояния от нулевого сечения до двух концов отрезка, на котором распределены
стокслеты.) В (14) при условии (22) и замены z на z — Z величина г2 при
z = 0 равна Z2 + а2. Тогда
fdZ 2 fzdZ л
-ъ -ъ
70
Глава 3
u = ^(^ + log^,^,0 . (25)
поскольку Ьис велики по сравнению с а, значения этих интегралов близки
к значениям несобственных сходящихся интегралов с бесконечными
пределами —оо и оо. Несобственный интеграл с бесконечными пределами от 1/г
расходится. Поэтому найдем точные значения определенных интегралов,
а затем запишем их аппроксимацию:
-ъ
Здесь log(2x) аппроксимирует обратный гиперболический синус sh_1 х
с погрешностью порядка х~2 при х —► оо. Используя приведенные в (14)
результаты, получим приближенное выражение для поля скоростей на кру-
ге х2 + у2 = a2, z = О в виде
/ (2х2 4cb 2ху
Полученное поле скоростей, обусловленное стокслетами,
распределенными на отрезке, непостоянно вдоль окружности х2 + у2 = a2, z = 0.
Так же как и в случае движения сферы, для компенсации этой
зависимости наложим на него поле, созданное расположенными на отрезке от —Ь
до с диполями с моментом (gdZ, О, 0) на каждом элементарном отрезке
(Z, Z + dZ). Проводя рассуждения, аналогичные случаю стокселетов, для
выражения (18) и используя дополнительно оценки интегралов
fdZ 4 [ZdZ л
/--з^' J—ф0' (26)
-ъ -ъ
с той же точностью получаем не зависящее от Ь и с поле скоростей,
индуцированное таким распределением диполей:
д ( 2 4х2 4ху
Выбрав
9 = fa2/Ар (28)
(отметим, что это соотношение отличается от соотношения (20), которое
возникло в задаче со сферой), получим, сложив (25) и (27),
результирующее поле:
87ф \ а?
« = £(^-^.-^.0. (27)
и = ^ (1 +log-J-, 0, 0 . (29)
3. Сингулярные решения на отрезке
71
Теперь скорость постоянна по величине вдоль окружности и во всех точках
окружности направлена вдоль оси цилиндра.
Здесь стокслеты интенсивности (/, 0, 0), рассчитанной на единицу
длины, соответствуют силам (/, 0, 0), приложенным к жидкости. Таким
образом, круговой цилиндр радиуса а и длины b + с » а действует с силой
(/, 0, 0), приходящейся на единицу длины цилиндра, на жидкость, когда
его фиксированное поперечное сечение (в данном случае сечение z = 0)
движется со скоростью (29), где b и с — расстояния от этого сечения до
концов цилиндра. Несомненно, что парадокс Стокса связан с
расходимостью интеграла (24) и со стремлением (29) к бесконечности, когда b или с
стремится к бесконечности.
Действительно, имеется слабая (логарифмическая) зависимость
линейной силы /, необходимой для приведения в движение цилиндра, от
отношения длины цилиндра к его радиусу. Более того, при постоянной
линейной силе / скорость (29) имеет слабую (логарифмическую) зависимость
от расстояний 6, с от рассматриваемого сечения цилиндра до его концов.
Скорость максимальна в середине цилиндра, где be/а2 принимает
наибольшее значение, что можно объяснить действием распределенных на обоих
концах цилиндра сил. Вблизи концов цилиндра скорость (29) довольно
мала по сравнению с ее значением в середине цилиндра, изменения скорости
вдоль длины цилиндра составляют чуть более 80% (расчеты для случая,
когда Ъ и с сравнимы с радиусом цилиндра а, не проводились).
Интересно сравнить полученные результаты для цилиндров,
движущихся в направлении, перпендикулярном оси, и вдоль оси (в
тангенциальном направлении). Для того чтобы рассчитать поле скоростей (14) в том
случае, когда вектор интенсивности стокслета направлен вдоль оси х,
примем, что элементарная интенсивность равна (fdX, 0, 0) на каждом
элементарном отрезке промежутка — 6 < X < с оси X. Заменим в (14) х на
х — X и проинтегрируем на круге y2 + z2 — а2, х = 0, используя (23) и (24),
заменив в них z на я, и в результате получим
f ( 4cb \
U=8^(-2 + 21Og^0'°> (30)
Множитель 2 в (30) появился за счет того, что в (14) числитель (х—Х)2+г2
при у2 + z2 = а2 равен — а2 + 2г2.
При распределении тангенциальных стокслетов полученная скорость
уже направлена вдоль оси х и имеет постоянную величину вдоль
окружности у2 + z2 = а2, и в отличие от рассмотренного выше случая дополни-
72
Глава 3
тельно вводить распределенные диполи не требуется. Отметим другие
качественные характеристики тангенциального движения цилиндра: созданная
распределенными стокслетами скорость (т. е. скорость, возникшая за счет
приложенной силы) имеет тангенциальное направление, а по величине
имеет слабую логарифмическую зависимость от отношения длины цилиндра
Ь + с к его радиусу а, кроме того, она имеет слабую логарифмическую
зависимость от расстояний b и с от рассматриваемого сечения до концов
цилиндра и достигает максимального значения, как и в ранее
рассмотренном случае в среднем сечении цилиндра.
Однако по величине скорости (29) и (30), созданные силами /,
приходящимися на единицу длины цилиндра, действующими в нормальном
и тангенциальном направлениях соответственно, значительно различаются.
При достаточном большом значении log(4c6/a2) их отношение становится
близким к 2, т. е. при действии тангенциальных сил, приводящих цилиндр
в движение, цилиндр движется почти в два раза быстрее, чем под
действием сил, направленных перпендикулярно к оси цилиндра.
Этот результат можно выразить в терминах сопротивления движению.
При движении цилиндра по нормали к своей оси отношение приложенной
силы /, приходящейся на единицу длины цилиндра, к скорости и
цилиндра называется коэффициентом сопротивления KN нормальному
движению, который равен
KN = Snfi^l + \og^y (31)
Это достаточно приближенная теория, ее приближенный характер
проявляется хотя бы уже в том, что величина скорости, обусловленной
действием постоянных распределенных сил, изменяется вдоль оси цилиндра (хотя
и не сильно). Тем не менее она может быть использована для
исследования движений микроорганизмов, скорости которых могут быть измерены
с небольшой степенью точности.
На основании равенства (30) введем соответствующий
тангенциальный коэффициент сопротивления (как отношение тангенциальной силы /,
приложенной к единице длины цилиндра, к локальной тангенциальной
скорости и движения поверхности цилиндра):
KT = 47Tfi Д-1 + log^V (32)
3. Сингулярные решения на отрезке
73
Значительное превышение сопротивления нормальному движению
цилиндра по сравнению с сопротивлением тангенциальному движению,
вытекающее из (31) и (32), помогает нам лучше понять проталкивающий эффект,
обусловленный волнообразным движением жгутика. При волнообразных
движениях жгутика скорость его элемента можно представить как
геометрическую сумму нормальной и тангенциальной скоростей, имеющих
отличные от нуля и противоположные по знаку проекции на направление
распространения волн. Сила, действующая со стороны жидкости на
элемент жгутика, в нормальном направлении превышает силу, действующую
со стороны жидкости на элемент жгутика в касательном направлении (это
следует и из равенств (31), (32)), и эти силы имеют противоположные по
знаку проекции на направление распространения волнообразных
движений. Результатом действия этих сил являются преодоление организмом
сопротивления и продвижение его в жидкости.
Поэтому важной характеристикой для рассматриваемого движения
является отношение коэффициента тангенциального сопротивления к
коэффициенту нормального сопротивления (о нем уже упоминалось в главе 2,
§ 13), которое определяет скорость плавания, обусловленного
волнообразным движением жгутика. Поэтому становится очевидным важность
отношения коэффициентов сопротивления для рассматриваемого движения:
г к = KT/KN = I (i + fog i£*) Д-1 + log ^). (33)
Отметим, что это отношение в небольшой степени зависит от отношений
Ъ/а и с/а. Так, его значение изменяется в пределах от 0,6 до 0,7 при
соответствующем изменении (сЬ)1/2/а от 120 до 10.
И все же желательно определить, какие значения (cfe)1/2 (а значит,
и какие значения гк) подходят для описания волнообразного движения
с длиной волны, равной Л. Поскольку линейная сила далека от
постоянной и ее изменение происходит с периодом Л, то значение (сб)1/2 для
фиксированного поперечного сечения жгутика, при котором ее можно считать
практически постоянной, скорее всего, составляет лишь малую часть от Л.
Действительно, из вычислений следует, что эта доля составляет 0,09Л для
колебаний с малой амплитудой. При проведении вычислений сила / в этом
сечении представляется в виде felkZ, где к = 27г/А иА>а. При этом
множитель elkZ не изменяет значения интегралов (23) и (26), а интеграл (24)
становится сходящимся, даже при интегрировании на (—ос, оо), и его зна-
74
Глава 3
чение равно
2К0{ка) = 2[log(2/fca) - 7], (34)
где К0 — модифицированная функция Бесселя, а 7 = 0,577 — эйлерова
константа. Это согласуется с (24) при
(сЬ)1/2 = е-^/k = А/(2тге7) = 0,09А. (35)
Проведенные расчеты дали достаточно малое значение (35), меньшее,
чем принято в большинстве случаев. Вычисления проводились в том
предположении, что при установившемся плавании при малой амплитуде
волнообразных движений интенсивность стокслета имеет очень малую по
величине компоненту в направлении перемещения организма и достаточно
большую по величине компоненту в перпендикулярном. Следовательно,
локальное действие компоненты стокслета в локальном нормальном (или
локальном тангенциальном) направлении можно описывать с достаточной
степенью достоверности приближенной формулой (31) (или (32)
соответственно), когда (сб)1/2 определена из (35). Даже в случае волнообразных
движений с большой амплитудой применение этих приближений
приемлемо для случая движения прямого цилиндра в жидкости, действующего на
нее с синусоидально изменяющейся силой, поскольку на расстоянии,
задаваемом величиной 0,09А от данного фиксированного сечения жгутика,
отклонение от прямолинейной формы еще невелико.
4. Теория продвижения с помощью жгутика,
основанная на силе сопротивления
Результаты § 3 могут быть применены к изучению продвижения
микроорганизма с помощью жгутика. При этом возможны два различных метода
исследования.
(i) Указанная аппроксимация для коэффициентов сопротивления (31)
и (32) применяется к каждому элементу жгутика для нормального
и тангенциального движения выделенного элемента при подходящем
подборе значения (cfe)1/2, таком, например, как (35). Такой метод
исследования был применен Греем и Хенкоком (Gray, Hancock, 1955),
который приводится в данном параграфе в дополненном виде.
(ii) Более точный, но значительно более сложный метод, разработанный
Хенкоком (Hancock, 1953), который в общих чертах будет описан в § 6.
4. Теория продвижения с помощью жгутика
75
Рассмотрим жгутик, длина которого в вытянутом виде равна L и
который может вращать, а может и не вращать само тело микроорганизма.
Пусть форма волны, проходящей по жгутику от начала к концу,
описывается уравнением
(х, у, z) = (X(s), Y(s), Z(s)), (36)
где s — параметр, равный расстоянию от начала жгутика до текущей точки,
так что выполняется
X'2{s) + Y'2{s) + Z'2{s) = l. (37)
Пусть функции в (36) имеют период Л, так что выполняются
Y{s + A) = Y{s), Z{s + A) = Z(s), но Х{з + Л) = Х{з) + А. (38)
То есть Л — длина волны, измеряемая вдоль искривленного жгутика, а Л =
= аЛ — длина волны, измеренная в направлении продвижения организма.
Здесь множитель а по величине меньше единицы и характеризует степень
кажущегося сжатия жгутика из-за принимаемой им в процессе движения
волнообразной формы.
В системе координат, связанной с волной, нерастяжимый жгутик
движется по касательной к кривой, описывающей форму волны (36), с
постоянной скоростью с. Так что в произвольный момент времени t выполняется
(ж, у, z) = {X{s - ct), Y{s - ct), Z{s - ct)). (39)
Отметим, что с — скорость, с которой волны распространяются вдоль
искривленного жгутика. Она отличается от V, определенной как скорость,
наблюдаемая в системе координат, где скорость продвижения организма равна
нулю. Действительно,
V = ас, (40)
поскольку период волнообразного движения может быть записан как Л/с
или X/V = aA/V.
Если скорость перемещения организма равна U (ее направление
противоположно направлению скорости распространения волны (40)), тогда
волна движется в противоположном направлении со скоростью V — U
относительно жидкости. Скорость относительно жидкости произвольного
элемента жгутика равна векторной сумме скорости с, направленной вдоль
касательной по ходу движения организма (как уже отмечалось, это скорость
76
Глава 3
в системе координат, связанной с волной), и скорости (V — С/, 0, 0). Ее
компонента вдоль касательной, наклоненной против хода движения
организма, равна
{V - U)X'{s - ct) - с. (41)
Компонента скорости в проекции на нормаль к касательной, наклоненной
против хода движения организма, равна
{V-U)[l-X'2{s-ct))1'2, (42)
где X' и (1 — X'2)1/2 — направляющие косинусы касательной и нормали
соответственно.
Общее осевое усилие Р, развиваемое жгутиком, может быть записано
как проекция на ось х суммы касательной силы сопротивления, равной
скорости (41), умноженной на коэффициент сопротивления КТ, и нормальной
силы сопротивления, равной скорости (42), умноженной на коэффициент
сопротивления KN. Умножая тангенциальную силу на X', а нормальную
силу сопротивления на (1 — X72)1/2, для осевого усилия получаем
L
Р = f{KT[(V - U)X'{s - ct) - c]X'(s - ct)+
0
+ KN(V - U)[l - X'2(s - ct)]}ds. (43)
Воспользуемся тем фактом, что
L
/ X'(s - ct)ds = l = aL = VL/c, (44)
о
где / — длина организма, совершающего волнообразное движение, так что
выполняется 1/L — а. Также мы определим
f X,2{s-ct)ds = PL, (45)
о
где /3 — средний квадрат направляющего косинуса касательной. С
использованием равенств (44) и (45) для силы (43) получаем
Р = KTL[(V - U)0 -V] + KNL{V - U){1 - /?). (46)
4. Теория продвижения с помощью жгутика
77
Если жгутик не вращает тело организма, то осевое усилие должно
уравновешивать силу сопротивления, действующую на голову
микроорганизма, плывущего со скоростью U. Запишем эту силу сопротивления как
KNLU69 где коэффициент 6 определяется как отношение
модуль силы сопротивления, действующей на голов-
г. _ ку микроорганизма при его передвижении вперед ,.-.
~~ модуль силы сопротивления, действующей на жгутик ' ^ '
при его движении в нормальном направлении
Для типичной длины жгутика коэффициент S довольно мал, как это
следует из сравнения силы сопротивления (21) при движении сферы радиуса а
и соотношения (31) для коэффициента KN.
Приравнивая (46) к силе сопротивления KNLU5, получаем для
отношения скорости перемещения организма к скорости распространения
волны
U = (1-(3)(1-гк)
V l-p + rKp + 5' K ;
Поскольку (3 (средний квадрат направляющего косинуса касательной)
уменьшается от 1 (организм не перемещается в жидкости) до нуля, то
отношение U/V возрастает от нуля до максимального значения
\ / max '
при J3 —> 0.
Эти результаты для случая гк = 0,5 (предельного значения гк для
бесконечно тонкого жгутика, см. (33)) были получены Греем и Хенкоком (Gray,
Hancock, 1955). В данной работе результат записан в общем виде для
произвольного значения тк главным образом потому, что даже при (cb)1^2/а =
= 120 рассчитанное значение коэффициента гк превышает 0,5,
предсказанное ими на 20%, кроме того, возможно оценить зависимость полученных
результатов от этого параметра. Грей и Хенкок (Gray, Hancock, 1955) в
своей работе привели данные (рис. 2), из которых следует, что у
сперматозоидов морского ежа {Psammechinus miliaris) волны, распространяющиеся
по жгутику, имеют синусоидальную форму с длиной волны Л = 24 мкм
и амплитудой b = 4 мкм, а радиус жгутика приблизительно равен 0,2 мкм.
Подставляя эти числовые данные в (31) и (35), получим для KN
г 67Г//Л
О =
х+ 21 (2) (0,09) (24 мкм)
87г/лЬ [ (0,2 мкм)
т- <50)
78
Глава 3
где А — радиус (или эффективный радиус) головки микроорганизма. Как
следует из данных, приведенных на рисунке 2, А = 1 мкм, L = 45 мкм,
что дает значение 5 = 0,11. Для коэффициента /?, рассчитанного Греем
и Хенкоком (Gray, Hancock, 1955) по приближенной формуле (3 = (1 +
+ 27г2Ь2/Л2)"1, было получено значение, близкое к 0,65.
А вот использованное ими значение гк = 0,5 сильно отличается от
гк = 0,7, полученного на основании равенств (33) и (35). Величина
отношения (48) существенно зависит от значения коэффициента гк. Так, для
гк = 0,5 это отношение равно 0,23 и лежит в середине множества
отношений скорости перемещения к скорости распространения волны, которые
наблюдались для этих сперматозоидов, тогда как отношение (48),
вычисленное при тк = 0,7, значительно более обоснованное с теоретической
точки зрения, в два раза меньше, чем при гк = 0,5, что далеко от значений
отношения, наблюдаемых в экспериментах. Грей и Хенкок знали об этом
противоречии, хотя они и использовали другой подход (о нем будет идти
речь ниже, в § 6) для того, чтобы провести более точные расчеты, а не
просто изменить значение тк. Отметим, что на наблюдения могло как-то
повлиять то, что исследуемые сперматозоиды перемещались слишком близко
к предметному стеклу микроскопа, поэтому Грей и Хенкок пришли к
заключению, что в силу баланса всех действующих факторов, скорее всего,
исследования с принятым значением тк = 0,5 наиболее корректно
описывают плавание сперматозоидов.
I А = 24 мкм Г
0 10 мкм
i i I I i i
Рис. 2. Таблица 1 и рисунок 4 из работы Грея и Хенкока (Gray, Hancock, 1955)
иллюстрируют методику расчета авторами основных характеристик синусоидальной
волны, распространяющейся вдоль жгутика
Хенкок (Hancock, 1953, с. 114) пришел к тому же парадоксальному
заключению при изучении плавания нематод (круглых червей) (Turbatrix
aceti). На рисунке 3 показан визуализированный поток жидкости вокруг
4. Теория продвижения с помощью жгутика 79
Таблица 1
1 Порядковый
номер
наблюдаемого
сперматозоида
1
2
3
4
5
6
7
8
9
10
11
12
13
14
| Среднее
Длина
головки,
мкм
35
33
34
35
35
34
32
34
32
33
32
35
29
30
33
Длина
волны (Л),
мкм
23
24
27
27
23
21
24
24
22
24
23
29
24
21
24
Амплитуда (6),
мкм
4
4
3,5
4
4
4
3,5
4
3,8
3,8
4
5
5
3,2
4
Длина
жгутика,
мкм
45
38
42
42
40
42
42
42
40
40
38
40
40
40
41
Отношение
Ь/Х
0,174
0,167
0,130
0,148
0,174
0,190
0,146
0,167
0,170
0,156
0,174
0,172
0,209
0,160
0,167 |
плавающего червя (последовательные положения круглого червя при
плавании приведены на рисунке 1 главы 2). В двух из трех наблюдений над
червем было обнаружено, что значения отношения V/U соответствуют
значению гк — 0,5 (а в третьем наблюдении, как ни удивительно, еще
меньшему значению гк). Хенкок предположил, что это значение rk = 0,5,
соответствующее «нулевому» радиусу жгутика, более корректно для жгутиков,
поскольку они могут быть настолько гибкими (т. е. могут реагировать на
воздействие со стороны жидкости так, как если бы это была другая порция
жидкости, а не жгутик), что их эффективный радиус оказывается намного
меньше геометрического.
Такое рассуждение нельзя признать достаточно убедительным,
поскольку (см. § 3) из выражения (33) для тк следует, что гк слабо зависит
(зависимость логарифмическая) от радиуса а и, кроме того, это выражение
может оказаться неприемлемым, если учитывать микроструктуру жгутика.
Более верное обоснование будет приведено в главе 7, где показывается, что
близость предметного стекла микроскопа к наблюдаемому сперматозоиду
действительно оказывает влияние на характеристики плавания и приводит
к значительному уменьшению отношения гк.
80
Глава 3
Рис. 3. Линии тока жидкости около плывущей нематоды (рис. 16.12 из работы Грея
(Gray, 1968))
Тейлором были проведены наблюдения над плаванием
самодвижущегося механического приборчика (достаточно большого размера, что дало
возможность получить точные экспериментальные данные) в большом
объеме жидкости, вязкость которой обеспечила очень низкие значения
числа Рейнольдса. В результате наблюдений было получено, что отношение
U/V существенно ниже тех значений, которые получались бы при тк =
= 0,5. Действительно (см. рис. 4), при А/а = 100 экспериментальные
значения отношения скоростей согласуются со значениями,
соответствующими гк = 0,7, как следует из (33) и (35). В этих экспериментах «головка
микроорганизма» содержит моторчик, который вращает с постоянной
скоростью проволочку-«хвост», имеющую спиральную форму. Такая спираль
составляет постоянный угол ф с направлением движения, так что направ-
4. Теория продвижения с помощью жгутика
81
ляющий косинус касательной X1 имеет постоянное значение cos^. В этом
случае (44) и (45) принимают вид
а = cos^, (3 = cos2 ф. (51)
При таком движении система сил, действующих на спираль (Кт,
умноженный на (41), плюс KN, умноженный на (42)), приводится не только
к равнодействующей силе (силе тяги), но также и к паре сил, момент
которой направлен вдоль оси спирали. Действие этой пары было
нейтрализовано Тейлором за счет перераспределения массы в «головке» приборчика.
Проведенные наблюдения в значительной степени прояснили
механизм движения микроорганизмов. Стало понятно, что продвижение
многих микроорганизмов происходит за счет спирального движения жгутика.
Однако вряд ли возможно значительно уменьшить величину момента
пары, возникающей при движении жгутика, за счет специальной
структуры клетки тела таких организмов. Ведь некоторые организмы (подобные
спирохетам) вообще не имеют тела как такового, а представляют собой
некоторое штопорообразное образование. В своей недавней работе Чанг
и By (Chwang, Wu, 1971) отметили, что во всех этих случаях
компенсирующая пара сил может возникнуть только за счет того, что они
назвали «вращением тела»: за счет вращения всего жгутика как твердого тела
(или за счет вращения головки, если таковая имеется) относительно
своей искривленной осевой линии. Скорость вращения должна быть такой,
чтобы возникающий момент скомпенсировал действие момента,
возникающего при волнообразном движении жгутика. И видимо, за счет
микроструктуры жгутика, который состоит из двух центральных волоконец,
обвитых девятью тонкими эластичными, способными растягиваться и
сжиматься волоконцами, возможно создание такого компенсирующего «вращения
тела».
При вращении кругового цилиндра радиуса а с угловой скоростью и
в вязкой жидкости поле скоростей будет таким, какое создает
распределенный вдоль оси цилиндра линейный вихрь интенсивностью 2тга2ш. Сила
вязкости, действующая на элемент жидкости, и градиент давления равны
нулю, а на поверхности цилиндра действует напряжение 2до;, приводящее
к возникновению пары сил 47га2/хи; на единицу длины.
Аналогично при спиральном движении жгутика можно предположить,
что любое самовращение не изменяет силы, действующие между жгутиком
и жидкостью, а вращению противодействует пара сил с моментом 47га2да;,
приходящимся на единицу длины. А это в результате приводит к паре сил
82
Глава 3
0,4
0,2
0_ 2 4
Рис. 4. Экспериментальная зависимость отношения U/V от 2тгЬ/\ (Ь —
амплитуда спирального движения) при движении спиральной проволочки,
исследованная Тейлором, сравнивается с теоретической зависимостью, полученной Хенкоком
(Hancock, 1953). Верхняя, средняя и нижняя кривые относятся к теоретическим
зависимостям, найденным для значений а/Х = 0, 0,01 и 0,02 соответственно.
Точками указаны экспериментальные значения U/V, цифры указывают соответствующие
величины а/Х
с моментом, действующим вдоль центральной линии жгутика, равным
47ra2LiJ,u)cosip. (52)
Значение и определяется так, что величина (52) совместно с парой сил,
препятствующих вращению головки (если головка у организма имеется1),
компенсирует момент касательной и нормальной сил, создаваемых
жгутиком. Однако вращение тела не влияет на сами эти силы, определяющие
осевое усилие, и поэтому они могут быть рассчитаны методами,
описанными в данном параграфе.
5. Плавание с минимальной потерей энергии
После того как в предыдущих параграфах с общих позиций была
описана теория плавания микроорганизмов за счет волнообразных движений
1 Здесь следует отметить, что вращению сферы радиуса А препятствует пара сил, момент
которой 87гЛ3да; значительно больше момента (52). И следовательно, при перемещении
микроорганизмов с большой головкой величина и должна оцениваться по моменту именно этой
пары.
5. Плавание с минимальной потерей энергии 83
жгутика при низких числах Рейнольдса, рассмотрим теперь некоторые
отдельные вопросы. В частности, рассмотрим наиболее эффективные
способы плавания. Об этом уже упоминалось в главе 2 при обсуждении плавания
на основании гораздо более грубого приближения (см. особенно
соотношения (5)). Найдем такие способы плавания микроорганизмов, при которых
для заданной скорости перемещения U потери энергии минимальны.
Для движения, описанного в §4, мощность, вырабатываемая
животным для преодоления различных сил сопротивления (при тангенциальном
движении — это сила (41), при нормальном движении это сила (42), а также
сопротивление, испытываемое головкой микроорганизма), равна
L
Е = f{KT[{V - U)X'(s - ct) - с]2+
о
+ KN(V - U)2[\ - Xf2{s - ct)]}ds + KNLU26. (53)
Для того чтобы получить, например первое слагаемое в суммарной
энергии, надо тангенциальную силу, приходящуюся на единицу длины (она
равна Кт, умноженному на (41)), умножить на тангенциальную скорость.
Аналогичным образом и в других слагаемых получаются квадраты
скоростей. Мощность (53) может быть вычислена в системе координат,
связанной с жидкостью, но поскольку результирующая сила, действующая на
животное, равна нулю, то значение мощности Е должно быть одним и тем
же во всех системах координат.
Так как в выражение для затрачиваемой в единицу времени энергии Е
входит квадрат скорости, то задачу об определении минимальной энергии
сведем к эквивалентной при заданной скорости U задаче определения
минимума отношения E/U2. Будем рассматривать безразмерную комбинацию
г)-1 = Е/{КТ + KNS)LU2, (54)
а именно отношение мощности, вырабатываемой животным при плавании
со скоростью U, к работе внешней силы, под действием которой животное,
преодолевая сопротивление, движется в воде со скоростью U с вытянутым
хвостом. То есть отношение г) сходно с числом Фруда.
С использованием равенств (44) и (45) интеграл (53) может быть
представлен в виде
Е = KTL[{V - U)2(3 - 2{V - U)ca + c2} +
+ KNL(V - U)2{\ - /?) + KNLU26. (55)
84
Глава 3
Отсюда, выражая с через V с помощью (40), а V через U с помощью (48),
получаем
Т1 = (1 - гк)"2(1 - РГ2[(гк + 6)(1 - /3 4- /?г*) -
- 2гк(1 - /3 + гк/3 + 5)] 4- 6(гк + б)-1 +
+ а~2гк(1 - тк)-2{\ - /3)2(гк + 6)-l(l -p + rKP + S)2. (56)
Здесь появились два параметра формы волны, а именно а и (3 —
средние значения направляющего косинуса касательной и его квадрата
соответственно. Найдем минимум (56) при заданном параметре (3.
Используем тот факт, что среднее значение квадрата произвольной
функции не меньше, чем квадрат ее среднего значения, причем равенство
достигается, только когда функция является константой (более строго, для
функций, которые отличны от константы на множестве меры нуль).
Применяя это «неравенство Коши-Буняковского-Шварца» к направляющему
косинусу касательной, получаем
а2 ^ /?, (57)
равенство выполняется только при постоянном направляющем косинусе.
Таким образом, при заданном (3 минимальное значение величины а~2
равно (3~1. А значит, минимальное значение (56) будет найдено, если в него
подставить (3~1 вместо а~2. Минимум отношения (56) для случая
постоянного направляющего косинуса касательной (за исключением множества
меры нуль) при заданном /? равен
mm(r) г) =
(1 - гк)2
гк(1 + 6)2 +гк+6
гк+д
(гк + 8)(3 \-(3
Минимального значения правая часть равенства (58) достигает при
_0_ _ rj/2(l-H)
Х-0- rK+S ■ (59)
Находя отсюда (3 и подставляя его в (58), получаем, что максимальная
эффективность г)т плавания микроорганизма равна
Vm = (l-r¥)2/(l + 6), (60)
5. Плавание с минимальной потерей энергии 85
а отношение скоростей соответственно равно
U/V = (l-r]{2)/(l + S). (61)
Интересно отметить, что максимальная эффективность (60) и
связанное с ней отношение скоростей (61) в случае 6 = 0, что соответствует
микроорганизму, не имеющему головки, в точности совпадает с полученными
в главе 2 результатами (см. соотношения (5)) при более приближенных
рассуждениях. Вероятно, это имеет место в силу принятого
предположения о том, что волнообразное движение жгутика является исключительно
трансверсальным. И это предположение корректно, если плавание
организма осуществляется при постоянном направляющем косинусе касательной.
Эти выводы подтверждают все замечания, сделанные в главе 2
относительно существенной зависимости г) и U/V от отношения коэффициентов
сопротивления гк (в главе 2 оно обозначено через г). Для жгутика,
продвигающего микроорганизм с небольшим значением параметра 6, как гу, так
uU/V уменьшаются в 1 + 6 раз.
Поскольку наибольшая эффективность достигается только при тех
способах плавания, когда направляющий косинус касательной сохраняет
постоянное значение везде (за исключением множества меры нуль), то угол ф
между касательной и направлением перемещения организма также
постоянен, причем выполняется а = cost/; и /? = cos2 ф (см. (51)). Из (59)
получаем, что оптимальное значение угла ф равно
ф0 = arctg[rj/4(l + бгк1)1'2^ + *Г1/2Ь (62)
Значение оптимального угла ф0 практически нечувствительно к изменению
как гк, так и 6. Действительно, расчеты показывают, что при
возрастании гк от 0,5 до 0,7 угол ф0 возрастает (1) от 40° до 42° при 6 = 0, (2) от
41° до 43° при 6 = ОД и (3) от 42° до 43° при 6 = 0,2. На основании чего
получаем, что в качестве среднего значения угла ф0 можно взять ф0 = 42°.
Волнообразные движения жгутика в радиальной плоскости (осевой
плоскости), при которых выполняется ф = ф0, имеют «пилообразный»
профиль. То есть оптимальные волнообразные колебания,
распространяющиеся в направлении от головной к хвостовой части организма, представляют
собой последовательность диагонально расположенных отрезков,
составляющих с направлением движения угол в 42°. Конечно, у реальных жгутиков
существует некоторый минимальный радиус кривизны при изгибе, из-за
86
Глава 3
чего жгутик не сможет принять строго пилообразную форму.
Пилообразная форма с закругленными вершинами пиков наблюдалась у некоторых
сперматозоидов (Brokaw, 1965) и, кроме того, нематоды в процессе
движения (глава 2, рис. 1) также принимают довольно близкую к пилообразной
форму.
С другой стороны, уже упоминавшееся трехмерное волнообразное
движение, при котором угол ф сохраняет постоянное значение и не
возникает требования малости радиуса кривизны, представляет собой
спиралевидное движение. Интересно отметить, что спиралевидная волна, движущаяся
в направлении, противоположном движению организма, у которой угол
наклона касательной составляет с направлением движения 42°, удовлетворяет
условиям оптимальности движения. И это было принято главным образом
со времени открытия Чангом и By (Chwang, Wu, 1971) того факта, что,
как уже отмечалось в конце § 4, вращающая пара сил, прежде считавшаяся
препятствием для оптимальности движения такого типа, может нейтрали-
зовываться организмом за счет вращения тела.
Примером спиралевидного движения жгутика при плавании
организма, является плавание эвглены зеленой (Euglena viridis); длина ее жгутика
составляет L = 100 мкм, длина волны равна Л = 35 мкм, амплитуда
волны — Ь = 6 мкм, частота колебаний — 12 Гц (Holwill, 1966). При таких
параметрах угол наклона касательной равен ф = 47°, однако амплитуда Ъ
определена с точностью только до одной значащей цифры, поэтому
отличие от теоретического оптимального значения ф0 = 42° можно отнести за
счет погрешности измерения. Это животное имеет достаточно большое
тело — около 60 мкм длины и 10 мкм в диаметре, и пара сил, препятствующая
вращению тела в силу вязкости жидкости, достаточно велика, в силу чего
это вращение происходит с небольшой скоростью — тело вращается с
частотой около 1 Гц. Холвилл (Holwill, 1966) проанализировал возможность
создания проталкивающего эффекта за счет наблюдаемых вращений такого
большого тела животного, однако пришел к заключению, что
проталкивающий эффект за счет колебаний жгутика все же на порядок больше.
6. Теория тонкого тела
Для биологических исследований приближенный теоретический
подход, развитый в §4 и §5, дает адекватные результаты в связи с тем, что
точность наблюдений недостаточно высока, а изменчивость
биологического материала весьма велика. Однако было бы интересно, по крайней ме-
6. Теория тонкого тела
87
ре, с математической точки зрения, развить теорию, в которой отсутствует
произвол в выборе таких параметров, как (сЪ)1/2/а и, как следствие,
отношения сил сопротивления гк. Начало подобной теории было положено
работой Хенкока (Hancock, 1953), хотя завершенная теория на сегодняшний
день все еще отсутствует.
Метод Хенкока основан на ряде идей, известных как «теория тонкого
тела» (см. также Batchelor, 1970, Сох, 1970). Рассмотрим тело (такое как
жгутик), радиус которого мал по сравнению с геометрическими размерами,
включая радиус кривизны при изгибе тела, течение жидкости вокруг
такого тела будем изучать в двумерной постановке — в плоскости поперечного
сечения тела. Причем при рассмотрении течения в плоскости данного
поперечного сечения будем учитывать, что движение тела происходит уже в
возмущенной жидкости, возмущение которой обусловлено действием других
поперечных сечений тела. В этом случае нельзя пренебречь действием
удаленных от данного поперечного сечения стокслетов, поскольку, как
следует из (14), скорость убывает пропорционально г-1 при г —► оо и такой
характер убывания приводит к расходимости интеграла (24). А влиянием
диполей возможно пренебречь, поскольку скорость течения, созданного
диполем, убывает пропорционально г~3.
Как было описано в § 1, движение жгутика можно представить в
виде суммы двух движений, а именно собственного движения жгутика
относительно центра масс, связанного с изменением формы тела и другими
деформациями тела (включая растяжение или релаксацию),
возникающими за счет его подвижности, обозначенной через (i), и его твердотельного
движения, обозначенного через (ii). Твердотельное движение, представляет
собой поступательное движение центра масс животного со скоростью U
и вращение с угловой скоростью ft относительно центра масс. Скорости U
и ft находятся из условия равновесия сил и моментов, действующих между
телом организма и жидкостью.
Для решения этой задачи Хенкок ввел некоторую длину 7, которая
достаточно велика по сравнению с радиусом жгутика а и достаточно мала по
сравнению с радиусом кривизны при изгибе. Хенкок представил поле
скоростей движений (i) и (ii) как обусловленное действием фундаментальных
сингулярностей, распределенных вдоль искривленной осевой линии
жгутика, а именно тангенциальных стокслетов, интенсивность которых является
неизвестной функцией от расстояния s, измеряемого вдоль оси жгутика,
нормальных стокслетов с отличной от тангенциальных стокслетов
интенсивностью, а также нормальных диполей с интенсивностью, определяемой
88
Глава 3
соотношением (28) (за счет такого выбора интенсивности диполя на
поверхности цилиндра выполняются граничные условия).
Поток вокруг фиксированного поперечного сечения в этом случае
может быть представлен как сумма:
(a) потока, созданного стокслетами и диполями, расположенными на
расстоянии не более чем 7 от данного поперечного сечения; поскольку 7
мало по сравнению с радиусом кривизны при изгибе осевой линии жгутика,
то можно считать, что стокслеты и диполи распределены на прямолинейном
отрезке и создают скорости на границе поперечного сечения, определяемые
выражениями (29) и (30), в которых следует положить Ь — с — 7,
-Lfl + logl^ или J-f-1 + loglV (63)
здесь f — интенсивность нормального или тангенциального (к осевой линии
жгутика) стокслетов;
(b) потока, созданного стокслетами, расположенными от данного
поперечного сечения на расстоянии, большем 7. Поскольку 7 3> «, то мы можем
считать, что скорость определяется стокслетами, расположенными в центре
поперечных сечений. По этой же причине (7 > а) влиянием диполей,
удаленных на расстояние, большее 7> можно пренебречь. Скорость потока (Ь)
может быть записана через определенный интеграл с пределами от 0 до
s — 7 и от s + 7 до L от полей, созданных стокслетами, распределенными
на указанных интервалах и расположенных вдоль осевой линии жгутика.
Отметим, что сумма потоков (а) и (Ь) не зависит от конкретного
значения расстояния 7- Например, если 7 уменьшить, то уменьшатся значения
логарифмов в (63), но при этом произойдут равные по величине и
противоположные по знаку изменения скоростей в потоке (Ь), поскольку при
уменьшении 7 в поток (Ь) добавятся скорости, созданные теми новыми
стокслетами, которые распределены на двух дополнительных (за счет уменьшения 7)
прямолинейных отрезках. Таким образом, с математической точки зрения
задача об определении нормальной и тангенциальной компонент
интенсивности f является корректной. Действительно, приравняв сумму скоростей
(а) и (Ь) к скорости, которая представляет собой сумму скоростей
движений (i) и (ii), получим интегральное уравнение Фредгольма второго рода
относительно неизвестной f. По найденной интенсивности можно далее
определить силы, приходящиеся на единицу длины, действующие между
жгутиком и жидкостью, а затем из условия равновесия найти скорость
перемещения организма U и угловую скорость ft.
6. Теория тонкого тела
89
Конечно, возникают трудности при нахождении аналитического
решения интегрального уравнения, полученного Хенкоком. Однако при
современном развитии вычислительной техники, возможно решить задачу
численно и найти все интересующие исследователя формы, которые может
принимать жгутик в процессе совершения движений, создающих силу
тяги, например подобные пилообразной форме, о которой говорилось в § 5,
а также сравнить результаты для различных значений угла ф с
результатами, которые дает теория продвижения с помощью жгутика, основанная на
силе сопротивления
Литература
[1] Batchelor, G.K. 1970. J. Fluid Mech. 44, 419-440.
[2] Brokaw, C.J. 1965. J. Exptl. Biol. 43, 155-169.
[3] Chwang, A.T. & Wu, T. Y. 1971. Proc. Roy. Soc. B, 178, 327-346.
[4] Cox, R.G. 1970. J. Fluid Mech. 44, 791-810.
[5] Gray, J. 1968. Animal Locomotion, Weidenfeld & Nicolson, London.
[6] Gray, J. & Hancock, G.J. 1955. J. Exptl. Biol. 32, 802-814.
[7] Hancock, G.J. 1953. Proc. Roy. Soc. A, 217, 96-121.
[8] Holwill, M.E.J. 1966. J. Exptl. Biol. 44, 579-588.
Глава 4
Высокоэффективные
с гидродинамической точки зрения
механизмы плавания животных1
В данной работе делается попытка (за образец взято серьезное
исследование Гольдштейна (Goldstein, 1929) «О вихревом следе за гребным
винтом» (On the vortex wake of a screw propeller)), разработать
динамическую теорию других более эффективных моделей движительных установок.
Способ перемещения животных воде за счет волнообразных движений
довольно распространен среди беспозвоночных, а нами будет рассмотрено,
как некоторые модификации подобного способа перемещений у
позвоночных (особенно у рыб) привели к увеличению скорости перемещения в воде
и повышению его гидромеханической эффективности.
Одной из наиболее существенных модификаций является
приобретенное рыбами в процессе эволюции сжатое с боков тело. Обратимся к
исследованию угревидного типа движения рыб, задняя часть тела которых сжата
с боков, а высота сужающегося к хвосту тела остается постоянной за счет
распределенных вдоль тела вплоть до вертикально расположенного хвоста
спинных и грудных плавников. Боковое движение каждого элемента тела
рыбы приводит к мгновенному обмену количеством движения с «присо-
единненной массой» воды, расположенной вблизи.
В § 2 приводятся подробное изложение и обоснование «теории тонкого
тела» (обобщение теории тонкого тела в невязкой жидкости) в приложении
к чисто угревидному типу движения. В предположении о том, что свойства
рыбы в поперечном сечении слабо изменяются вдоль тела, если за масштаб
взять высоту тела рыбы s (т. е., например, длины волн основных гармоник
превышают 5s), возможно, как и прежде, считать, что количество движения
1 Перепечатано из журнала J. Fluid Mech., 44 (1970), pp. 265-301.
92
Глава 4
воды в боковом направлении пропорционально квадрату высоты тела рыбы
в данном поперечном сечении.
При движении рыбы в результате волнообразных движений ее
хвоста возникает поперечная сила, действующая на жидкость, расположенную
вблизи тела, которая изменяет количество движения воды в поперечном
направлении. Средняя мощность, развиваемая рыбой, равная усредненному
произведению боковой силы на величину боковой скорости хвоста,
возрастает за счет эффекта «присоединенной массы», что обусловливает
тесную связь между боковыми движениями и величиной количества движения
жидкости. Частично эта мощность затрачивается на компенсацию потерь
энергии для создания поперечного импульса в вихревом следе. По
мощности, развиваемой рыбой, и потерям на вихреообразование можно
рассчитать силу тяги и эффективность самодвижения.
В § 3 изучается (все еще с позиции теории невязкой жидкости) влияние
наличия не одного непрерывного, а нескольких отделенных друг от друга
грудных и спинных плавников. Казалось бы, наличие нескольких спинных
(или грудных) плавников может сделать неверным предположение о
непрерывной и слабой зависимости свойств рыбы от расстояния, измеряемого
в продольном направлении, на чем основана теория тонкого тела. Однако
промежутки между плавниками заполняются вихревыми пеленами таким
образом, что при этом сила тяги практически не изменяется, а сила
сопротивления слегка уменьшается, так что с достаточной степенью точности
совокупность плавников и промежутков между ними, заполненных
вихрями, можно принять за один непрерывный плавник.
Еще одно преимущество некоторые рыбы приобрели (при сохранении
в целом угревидного типа движения) за счет того, что их передние
спинные и брюшные плавники вытянуты почти вдоль всего тела. Вихревые
пелены в промежутке между сильно вытянутыми плавниками и хвостовым
плавником поглощаются пограничными слоем около хвостового плавника,
и поэтому сколько-нибудь заметного роста потерь энергии не происходит.
Средняя мощность, развиваемая рыбой, увеличивается за счет работы
спинных и брюшных плавников, сдвиг фаз колебаний плавников и хвоста
препятствует корреляции бокового движения с импульсом вихревой пелены,
которая поглощается в пограничном слое.
В § 4 учитываются силы вязкости, действующие в поперечном
направлении. В этом случае увеличиваются количество движения и энергия
жидкости при ее движении в поперечном направлении, но в то же время
эффективность угревидного движения может уменьшиться, т. к. дополнитель-
Высокоэффективные механизмы плавания животных 93
ный импульс жидкости у хвостовой части, обусловленный действием сил,
развиваемых передней частью, приведет к затруднению боковых движений
хвоста в силу их несогласованности с возникновением дополнительного
импульса. Усвоение некоторыми видами рыб движения, подобного ском-
броидному, при котором амплитуда колебаний тела возрастает постепенно
от практически нулевого значения первой половины или даже двух
третей длины тела до весьма больших значений у хвостовой части, позволяет
избежать указанной трудности.
Любое движение, совершаемое рыбой, вследствие законов сохранения
количества движения и момента количества движения вызывает «отдачу»,
т. е. возникает дополнительное движение в виде смещения или некоторого
вращения. Подобные движения, приводящие к серьезным потерям в силе
тяги и эффективности движения при скомброидном типе движения,
рассмотрены в § 4. Там же показывается, как можно уменьшить подобные
потери за счет перераспределения совокупной инерционной массы (суммы
массы рыбы и виртуальной массы воды). По-видимому, неслучайно, что
скомброидным типом движения пользуются рыбы, у которых высота
передней части велика (имеет большой момент инерции), а хвостовая часть
сильно уменьшается по высоте к хвостовому плавнику.
В § 5 проведено теоретическое обоснование того, что уменьшение
площади хвостового плавника при той же высоте в результате преобразования
его к виду (напоминающему хвост селедки) «пары стреловидных крыльев»,
может уменьшить силу сопротивления без существенной потери силы тяги.
Подобный эффект можно ожидать (хотя в этом случае теория тонкого тела
не применима) и при увеличении ширины этой «пары крыльев» (при
уменьшении прямой стреловидности). Такое направление развития скомброид-
ного типа плавания у многих перкоидных (Percomorphi) привело к
развитию хвостового плавника в виде полумесяца, что дало наибольший прирост
в скорости и эффективности движения. Заметим, что такой же эффект
наблюдался у животных, которые шли совершенно другим эволюционным
путем.
Исследование преимуществ перечисленных модификаций проводится
в данной главе на основе двумерной линейной теории. Для различных
положений оси рыскания исследуются движения произвольного
горизонтального сечения хвостового плавника при изменениях угла рыскания,
совпадающих по фазе с боковой скоростью. Потери энергии в спутном потоке
будут наименьшими, если ось рыскания расположена на расстоянии три
четверти длины хвоста, а мощность возрастает, когда эта ось расположена
94
Глава 4
дальше чем на три четверти. Что-то похожее на оптимум эффективности,
силы тяги и доли силы тяги при подсасывании в передней части хвостового
плавника найдено при расположении оси рыскания на задней кромке.
На основе настоящего сверхупрощенного подхода мы приходим к
утверждению, что с точки зрения гидромеханики наилучшей конфигурацией тела
будет следующая: контур головной части изогнут, а хвостовая кромка
прямая. В конце главы приведено краткое обсуждение различных направлений
развития идей данной работы, например решение задачи в трехмерной
постановке, учет нелинейных эффектов, которые могли бы пояснить тот факт,
что у наиболее быстрых морских животных хвостовая кромка слегка
вогнута в направлении движения.
1. Введение
При составлении общего обзора по гидромеханике движения водных
животных Лайтхилл (Lighthill, 1969) воспользовался помощью своих
коллег-зоологов, для того чтобы, насколько это возможно, в равной степени
отразить для читателя зоологические и гидромеханические аспекты
проблемы. Данный обзор главным образом посвящен эффективности
самодвижения г) при движении водных животных за счет волнообразных
изгибающихся движений. Эффективность определяется подобно эффективности
Фруда для винта и равна
г) = UP/E, (1)
где U — скорость перемещения; Р — средняя сила тяги, необходимая для
преодоления вязкого сопротивления и поддержания скорости перемещения,
равной U, при твердотельном движении животного; Е — средняя мощность
изгибных движений, расходуемая на преодоление сопротивления
окружающей жидкости.
В обзоре проводится обсуждение способов перемещения в воде
представителей 12 классов животного мира, наибольшее внимание при этом
уделено классу рыб. Наиболее распространенным способом передвижения
является перемещение за счет волнообразных движений, при которых
поперечная волна проходит по телу в направлении от головы к хвосту.
Такой пропульсивный метод и его различные модификации с успехом
используются животными при движении как при высоких, так и при
низких числах Рейнольдса Я, рассчитанных по длине животного и скорости
перемещения. Альтернативные методы, такие как продвижение при низких
1. Введение
95
числах Рейнольдса за счет колебания ресничек или реактивное движение
при высоких числах Рейнольдса, встречаются в животном мире
значительно реже.
По эффективности способы перемещения можно разделить на две
категории: перемещения с высокой эффективностью и перемещения с
низкой эффективностью, границей раздела является значение
эффективности г) = 0,5. Фактически это деление относится только к перемещениям за
счет волнообразных движений тела (и их модификациям), поскольку
способы перемещения, встречающиеся только при низких числах
Рейнольдса, подобные перемещениям за счет колебательного движения ресничек,
с необходимостью происходят при низких значениях г) по указанному
критерию, в то время как реактивное движение при скорости струи Uj имеет
низкую эффективность Фруда, если Uj существенно больше С/, как это
обычно имеет место у водных животных.
В работе также исследуются различия для пропульсивного метода
перемещения при относительно высоких числах Рейнольдса между
«хорошими гидродинамическими формами» с высокой эффективностью rj и
«плохими» с низкой эффективностью. К типичным «плохим гидродинамическим
формам» при высоких числах Рейнольдса относятся формы тела
животного с почти круглым поперечным сечением (как у большинства наземных
змей), при умеренных числах Рейнольдса (не больше чем 103) все формы
тела животных являются «плохими» с точки зрения гидродинамики.
Среди серьезных научных исследований передвижения водных
животных за счет волнообразных движений тела, имеющего «плохую» с точки
зрения гидродинамики форму, следует отметить работы Тейлора (Taylor,
1952) и Грея и Хенкока (Gray, Hancock, 1955). В своей работе автор
данного обзора (Lighthill, 1969, § 13) резюмирует их результаты в краткой
упрощенной форме. Основное предположение, которое эти авторы используют,
состоит в том, что мгновенная сила, действующая между водой и
элементом тела животного, будет такой же, как если бы этот элемент равномерно
двигался в воде с таким же вектором относительной скорости. Это
предположение применимо для движений при низких числах Рейнольдса, когда
силы вязкости значительно больше сил инерции.
Очень часто принимается, что такая «квазистатическая» теория
волнообразного перемещения подходит и для больших чисел Рейнольдса. Однако
в общем случае это неверно, поскольку в ней не учитываются
дополнительные силы, которые при нестационарном движении ускоряют массу
воды, находящуюся вблизи тела (ее обычно называют «виртуальной массой).
96
Глава 4
Скорее всего, именно при «хороших гидродинамических формах»
возможно значительное увеличение эффективности продвижения за счет
использования эффекта виртуальной массы.
Для «тонких» животных (длина таких животных значительно
превышает остальные геометрические размеры) Лайтхилл (Lighthill, 1969),
используя теоретический метод, развитый в его работе (Lighthill, 1960),
показал, что именно те животные (практически все они позвоночные),
у которых хвостовая часть имеет поперечное сжатие и которые
используют эффект виртуальной массы воды, имеют высокую эффективность
самодвижения. «Сжатие» здесь означает уплощенность поперечного
сечения тела хвостового отдела или «задней части», а «поперечное
сжатие» означает, что сжатие происходит в поперечном направлении, в
котором происходит волнообразное изгибание тела, распространяющееся в
направлении от головы к хвосту. Именно поперечные (из стороны в
сторону) волнообразные движения типичны для водных позвоночных, включая
рыб, земноводных и пресмыкающихся, а характерное уплощение задней
части тела у большинства из них приводит к повышению
эффективности самодвижения. Однако когда волнообразные движения производятся
в спинно-брюшном (т. е. в вертикальном) направлении, как это имеет место
у китообразных млекопитающих, то тело уплощено в вертикальном
направлении.
Поперечное сжатие хвостовой части тела важно по той причине, что
такое строение позволяет хвостовой части приводить в движение большую
виртуальную массу воды без увеличения гидромеханического
сопротивления, которое имело бы место при отсутствии сжатия. Выигрыш от
увеличения величины виртуальной массы у хвостовой части тела объясняется
с механической точки зрения в §4 работы Лайтхилла (Lighthill, 1969),
однако в силу важности этого момента в данной работе его обоснование
приводится ниже. Отметим, что другие характерные черты различных типов
животных, кроме поперечного сжатия тела, выработанные в процессе
приспособления к перемещению в воде за счет волнообразных движений тела,
требуют дальнейшего изучения.
Характерные способы плавания большинства рыб (некоторые
наиболее важные исключения рассмотрены в работе Лайтхилла (Lighthill, 1969,
§§ 10 и 11) можно разделить на два класса: угревидный и скомброидный.
У рыб, использующих угревидный способ плавания, все тело гибкое, и
поперечные волны распространяются от головы до хвоста с довольно
большой амплитудой, немного возрастающей к хвостовой части. При скомбро-
1. Введение
97
идном типе плавания, напротив, амплитуда поперечной волны становится
заметной только во второй половине или даже в последней трети тела рыб,
другая часть тела относительно жесткая.
Чисто угревидное движение (волнообразное движение всего тела)
наблюдается у тех угрей (включая представителей рода речных угрей
(Anguilla), которые и дали название всему роду) и других рыб, у
которых спинные и брюшные плавники вытянуты вдоль всего тела и
высота тела (суммарный размера самого тела и обоих плавников в
вертикальном направлении) или увеличивается, или остается практически
постоянной (в вертикальном направлении) от головы до хвоста. С этой группы
животных начинается ниже изучение угревидного способа движения (§2).
Как и везде в данной статье, описание ограничивается теми случаями,
когда колебательные движения таковы, что возможно применить
упрощающее предположение Лайтхилла (Lighthill, 1960, 1969), при котором
движения тела рыбы считаются происходящими только в боковых направлениях.
Оцениваются не только средняя величина сил, возникающих при таких
колебательных движениях, но также и их флуктуации, обусловленные
поперечными движениями объемов воды, расположенных в непосредственной
близости от тела, и вихревым следом.
Угревидный способ движения наблюдается также и у некоторых
других групп рыб, таких как зубатки (см. в работе Лайтхилла (Lighthill, 1969)
полный список таких рыб), с обособленными спинными и брюшными
плавниками, за которыми общая поперечная высота значительно уменьшается
к хвосту, а затем снова возрастает за счет хвостового плавника практически
до прежней величины. Наличие вихревого спутного потока, который
развивается за спинным и грудным плавниками, приводит к изменению сил при
плавании таких рыб (это явление исследовано в § 3). Далее показано, что
при этом средняя сила тяги и эффективность самодвижения возрастают,
даже если вихри в виде отдельных образований движутся к хвостовому
плавнику и только за ним поглощаются вихревой пеленой.
Та же теория тонкого тела, методами которой изучается угревидное
движение, может быть использована при изучении той основной
модификации этого движения (когда поперечные колебания совершаются в
основном хвостовой частью), которую называют скомброидным движением. Этот
способ передвижения исследуется в основном в § 4, где будет показано, что
такой способ более предпочтителен с точки зрения эффективности
самодвижения.2
2 Частный случай скомброидного движения, к которому неприменима теория тонкого тела,
исследован в § 5.
98
Глава 4
Однако такое преимущество будет реализовано только в случае весьма
сильного уменьшения высоты (т. е. должно присутствовать «сужение»,
если смотреть на рыбу сбоку) непосредственно перед хвостовым плавником,
где амплитуда волнообразных движений сильно возрастает. А суммарная
высота спинного плавника, тела рыбы и брюшного плавника может и
действительно будет равна высоте хвостового плавника и остается
примерно постоянной практически на всей длине рыбы. Возникновение вихревой
пелены за спинным и брюшным плавниками не оказывает
существенного влияния на эффективность самодвижения, поскольку при скомброидном
способе передвижения роль плавников незначительна.
В §§2-4 проведены расчеты в предположении, что рыба движется по
прямой, а углы отклонения от прямого курса считаются достаточно
малыми (скажем, не превышающими 30°), так что с достаточной степенью
точности можно считать, что перемещение рыбы происходит за счет
колебательных движений тела в боковых направлениях. Одним из преимуществ
проведения расчетов, основанных на этом предположении, является то, что
их можно проводить или на основании точного, но достаточно сложного
разложения возмущения, в котором силы, действующие между водой и
телом рыбы, получены интегрированием по поверхности возмущений
давления, так как это изложено в приложении в работе Лайтхилла (Lighthill,
1960), или как сделано в данной работе — на основании закона
изменения количества движений и энергии в единице объема. Эти два подхода
дают идентичные результаты, что подтверждает правомерность обоих
подходов.
Желательно было бы снять некоторые ограничения, поскольку,
например, амплитуды колебаний тела рыбы при больших скоростях в
действительности достигают весьма большой величины, а хвостовой плавник
отклоняется от направления движения на угол порядка 60°. Более того,
хотелось бы, чтобы развитыми при этом теоретическими методами можно
было провести исследование маневренного движения. В связи с этим в
настоящее время автором разрабатывается новая теория тонкого тела,
которую предполагается в дальнейшем опубликовать. Эта теория основывается
на приведенном ниже методе исследования (надежность и достоверность
которого обоснована в данной работе), использующем законы сохранения
объемного количества движения и энергии, но при менее строгих
ограничениях на геометрическую форму тела рыбы.
С нашей точки зрения, слишком непродуманно и категорично
утверждение о том, что для «плохих гидродинамических форм» следует учи-
1. Введение
99
тывать только квазистатическую силу сопротивления движению элемента
тела (например у Тейлора (Taylor, 1952) и Грея и Хенкока (Gray, Hancock,
1955)), а для «хороших гидродинамических форм» при применении теории
тонкого тела влияние силы сопротивления считать пренебрежимо малым
по сравнению с влиянием виртуальной массы воды. Желательно
разработать теорию, в которой учитывались бы в равной степени оба эффекта.
В данной работе положено начало такой теории, в связи с этим особенно
следует уделить внимание § 4, где получила развитие идея автора (Lighthill,
1960) о совместном учете этих эффектов.
Последний раздел данного обзора посвящен исследованиям, в
основу которых легло заключение Лайтхилла (Lighthill, 1969) (обратите особое
внимание на рис. 6) о том, что все наиболее быстрые водные животные,
в частности некоторые представители класса костных рыб, куда входят
тунцы, некоторые самые быстрые акулы, а также почти все водные
млекопитающие, усвоили скомброидный способ плавания, а их хвостовые плавники
в процессе эволюции приобрели практически одинаковые формы. Эти
хвосты в форме полумесяца имеют большое относительное удлинение. Такие
хвостовые плавники часто называют серповидными независимо от того,
расположен ли он в вертикальной плоскости, как у рыб, или в
горизонтальной плоскости, как у китообразных. Утверждается, что эти быстрые
животные, принадлежащие к различным классам животного царства, в процессе
эволюции приобрели хвостовой плавник в форме полумесяца (см. также
Kramer, 1960, рис. 18), прежде всего, потому, что такой хвост обладает
преимуществом с точки зрения гидромеханики. В своей работе автор также
намечает пути исследования работы хвостового плавника с точки зрения
гидромеханики.
При изучении гидромеханики серповидного хвоста теория тонкого
тела неприменима, поскольку в теории тонкого тела предполагается, что
окружающая рыбу вода приводится в движение за счет воздействий со
стороны тела, распределенных вдоль направления движения, так что каждый
вертикальный слой воды, перпендикулярный этому направлению,
находится под воздействием главным образом со стороны элемента тела, близкого
к выделенному слою. А серповидный хвост у рыб при волнообразных
движениях отклоняется от направления движения в вертикальной плоскости
вплоть до угла в 90°. При таком большом отклонении хвоста от
вертикальной плоскости предположения теории тонкого тела несостоятельны,
поскольку весьма резкие воздействия со стороны хвоста на близлежащие
вертикальные слои уже не будут независимыми.
100
Глава 4
С другой стороны, действие серповидного хвоста при его боковом
движении на различные горизонтальные слои воды значительно меньше
зависит от расположения слоя, и, возможно, более приемлемым было бы
рассматривать эти воздействия в первом приближении как независимые. Это
говорит в пользу применения двумерной теории осциллирующего крыла
для изучения движения серповидного хвоста. В своих работах Лайтхилл
(Lighthill, 1969, §§8 и 9) предлагает в качестве первого приближения
подобный подход, учитывая поперечную компоненту вектора вихря в спутном
потоке (так, в случае плавания рыб учитывается вертикальная компонента),
хотя далее им предложен и более точный метод (но и более трудоемкий),
основанный на использовании подъемной силы, в котором учитывается
также и продольная компонента вектора вихря. Последняя более точная теория
дает более низкие значения эффективности за счет учета потерь энергии,
связанных с продольной составляющей вектора вихря.
Скорее всего, если двумерная теория осциллирующего крыла и
находит свое применение в качестве первого приближения к исследованию
гидродинамики рыб, то именно при исследовании колебательных
движений серповидного хвостового плавника. Напротив, любая попытка
изучения продвижения рыб с более округлым телом в двумерной постановке не
может дать хотя бы даже приближенно достоверные результаты, поскольку
основное предположение двумерной теории, состоящее в том, что тело
влияет на прилежащую жидкость в перпендикулярном направлении к
движению, для рыб с такой формой выполняться не будет. Следовательно,
аэродинамический частотный параметр ujc/U (рассчитанный по угловой
частоте и; и скорости продвижения С/), характеризующий передвижение рыбы,
должен включать в себя длину с, равную не длине тела рыбы /, а размеру
серповидного хвоста в направлении движения. Такие значения
параметра ujc/U, как правило, меньше единицы, хотя значения ujI/U колеблются
около 10.
Основной характеристикой скомброидного типа плавания (Lighthill,
1969, § 5) является совпадение фаз колебаний угла атаки хвостового
плавника и его боковой скорости. Лайтхилл провел расчеты основных
гидродинамических характеристик серповидного хвоста, результаты которых
представлены на рисунке 9 его работы. При расчетах предполагалось, что у
рассматриваемого профиля крыла осцилляция бокового скольжения
происходит со скоростью W cosuot, рыскание с углом атаки (0W/U) cos ut, и
фазы осцилляции рыскания и угла атаки совпадают. Параметр 9 называется
«коэффициентом флюгирования». Также было принято упрощающее пред-
1. Введение
101
положение о том, что ось осцилляции рыскания совпадает с центральной
осью (или полухордой) крыла. При в = 1 сила тяги отсутствует, и, хотя
сила тяги возрастает с уменьшением 0, наибольшая эффективность
создания силы тяги будет достигаться при значениях 9 не сильно отличающихся
от 1. При стремлении 0 к нулю эффективность уменьшается (эти результаты
дает даже двумерная теория, в которой пренебрегается продольной
компонентой вектора вихря, эффективность снижается до 0,65 при uc/U = 1),
а величина силы тяги при этом достигает максимума.
Случай в = 0 соответствует чистому боковому скольжению без
рыскания, когда сила тяги не может быть найдена как разность между
давлением с двух сторон плавника. В этом случае (сила тяги наибольшая при
небольшой эффективности) сила тяги возникает за счет подсасывания на
передней кромке, т. е. за счет уменьшения давления на передней
закругленной кромке при движении крыла в воде. При отличном от нуля в не вся, но
значительная часть силы тяги образуется за счет подсасывания. Эта сила
возникает при обтекании переднего края плавника, а именно такое
строение и имеют все водные животные, обладающие серповидным хвостовым
плавником.
В § 5 повторены вышеописанные расчеты, причем относительно оси
рыскания сделаны более общие предположения. Зависимость
эффективности от положения оси рыскания оказалась неожиданно значительной.
При заданных амплитудах потери энергии минимальны, когда ось
рыскания расположена на расстоянии в три четверти хорды (т. е. расстояние до
оси от передней кромки в три раза превышает расстояние до задней
кромки). Мощность возрастает при приближении оси рыскания к этой точке,
а оптимальное значение силы тяги и эффективности достигается, когда ось
расположена достаточно близко к задней кромке.
Изложенные выше рассуждения дают ответ на вопрос, почему же
серповидный хвост обладает наибольшей эффективностью. Если бы хвостовой
плавник рыскал как единое целое относительно единственной оси и если
бы возможно было применить двумерную теорию к каждому сечению
плавника, то мы смогли бы сделать вывод, что наибольшая сила тяги и
наибольшая эффективность имеют место в случае, когда задняя кромка совпадает
с осью рыскания. Для клиновидного плавника это означало бы, что задняя
кромка должна быть прямой, а передняя часть сильно изогнута в
направлении движения. Такое отклонение от «прямого крыла» (когда передняя часть
выгнута вперед, а задняя часть таким же образом вогнута) немного
улучшает гидродинамические характеристики. Возможно, более общий подход на
102
Глава 4
основании трехмерной теории позволил бы объяснить тот факт, что задняя
кромка даже слегка выгнута вперед.
Скорее всего, из всех подходов на основе трехмерных теорий более
всего подходит что-то наподобие теории подъемной силы, в которой
отдельно рассматривается двумерный поток относительно каждого
поперечного сечения и учитывается влияние угла атаки на всю структуру вихревого
следа. В § 6 обоснован выбор такой двумерной модели, чтобы результаты,
полученные на основании ее применения, могли быть использованы в
трехмерном случае.
Между тем общие выводы, сделанные относительно хвостовой части,
могут оказаться полезными и при рассмотрении других аспектов
продвижения животных. Например, при полете птиц машущие движения крыльев
можно исследовать с точки зрения двумерной теории как совокупность
колебательных ритмичных движений около поперечной оси (наложенных, см.
рис. 7, в работе (Lighthill, 1969) на движение с постоянным углом атаки,
необходимого для поддержания птицы в воздухе) и движения со
скоростью, колебания которой по фазе совпадают с колебаниями угла тангажа,
в этом случае эффективность полета будет наибольшей, при совпадении
оси тангажа с задней кромкой.
Параграфы 2-5 данной работы в какой-то степени представляют собой
математическое обоснование в целом нематематического введения и
обсуждения работы Лайтхилла (Lighthill, 1969).
2. Угревидный способ передвижения
Можно сказать, что рыбы приспособились к «чисто» угревидному
способу передвижения, когда подобно угрю обыкновенному {Anguilla vulgaris)
приобрели спинные и брюшные плавники, вытянутые вдоль всего тела.
Причем в поперечном направлении эти плавники таковы, что суммарный
размер по высоте плавников и тела рыбы вдоль тела рыбы остается
постоянным или даже немного увеличивается к хвостовой части. У
хвостовой части возможно настолько резкое сужение тела в боковом плане, что
можно считать, что тело рыбы имеет вертикальную заднюю кромку.
Такая рыба при небольшой скорости плавания часто перемещается за счет
колебательных движений плавников, а при высоких скоростях использует
способ перемещения, достаточно близкий к угревидному, т.е. совершает
волнообразные поперечные движения всем телом, включая плавники,
распространяющиеся вдоль тела в направлении от головы к хвостовой части.
2. Угревидный способ передвижения
103
Займемся анализом такого способа перемещения с использованием теории
тонкого тела (в авиации она называется теорией тонкого крыла).
Прежде всего, следуя Лайтхиллу (Lighthill, 1960), в качестве системы
координат выберем систему, движущуюся со скоростью рыбы [/, так что
в этой системе координат рыба совершает волнообразные движения для
того, чтобы остаться неподвижной в потоке воды, скорость которого
равна U и направлена вдоль оси х. Опишем волнообразные движения рыбы
относительно исходного нейтрального положения, когда тело совершенно
симметрично относительно вертикальной плоскости z = 0. Для того чтобы
удержать рыбу в потоке воды в нейтральном положении, потребуется сила,
равная силе вязкого сопротивления D (величину этой силы можно
определить на основании экспериментальных значений ускорения рыбы
известной массы, когда она скользит в воде как твердое тело). Исследуем, как без
приложения какой-либо внешней силы рыба может за счет волнообразных
поперечных колебаний тела оставаться неподвижной в потоке воды, т.е.
другими словами, как она создает силу тяги Р, равную по величине силе
сопротивления D.
В нейтральном положении поперечное сечение рыбы на расстоянии х
от головы обозначено в работе Лайтхилла (Lighthill, 1960) через 5Х, там
же в дальнейшем предполагается, что в процессе волнообразных движений
поперечное сечение смещается на величину h(x,i) в направлении оси z
(т.е. в боковом направлении). В работе Лайтхилла (Lightill, 1969) через W
обозначена боковая скорость dh/dt фиксированного поперечного сечения.
Здесь также будет использовано это обозначение. Кроме того, будем
использовать обозначение, принятое в работе Лайтхилла (Lighthill, 1969),
dh TT dh
w = Ti+u^ (2)
вместо скорости V из работы Лайтхилла (Lighthill, 1960). Здесь через V
обозначим скорость бокового движения вертикального слоя воды
(перпендикулярного оси х), обусловленную действием поперечных сечений, мимо
которых этот слой перемещается со скоростью U.
В работе Лайтхилла (Lighthill, 1969, §4) достаточно наглядно
объяснено, почему w значительно меньше, чем скорость W у хвостовой части,
если в ее окрестности волнообразное движение представляет собой бегущую
волну с постоянной амплитудой, движущуюся со скоростью V, ненамного
превышающей скорость U. Тогда выражение dh/dt+ V dh/dx равно нулю,
104
Глава 4
так что равенство (2) принимает вид
w = W{V - U)/V. (3)
Как было отмечено ранее, для того чтобы эффективность самодвижения
была достаточно высокой, отношение w/W должно быть относительно
небольшим, но все же таким, чтобы развиваемая при этом сила тяги смогла
уравновесить силу вязкого сопротивления D.
Рассмотрим количество движения в поперечном направлении, которое
приобретают вертикальные слои воды, под действием поперечных
колебаний тела. Когда начинается движение тела рыбы в боковом направлении, то
количество движения прилегающей к телу рыбы воды возрастает сразу же
на величину mw, приходящуюся на единицу длины (более строго,
количество движения сообщается воде за счет возмущения, распространяющегося
во скоростью звука, но в данной постановке задачи скорость
распространения возмущений бесконечна). Здесь т — виртуальная масса на единицу
длины рыбы. При последующем движении тела рыбы в боковом
направлении количество движения несколько возрастает за счет действия сил
вязкости, особенно за счет образования спутного вихревого потока в поперечном
направлении. Это явление исследовано в § 4, а в настоящий момент
предполагается, что подобный прирост количества движения равен нулю.
В теории тонкого тела, развитой в работе Лайтхилла (Lighthill, 1960),
предполагается, что изменения размеров поперечного сечения вдоль тела
рыбы малы и что волнообразные движения несильно изменяют
распределение виртуальной массы, так что изменение массы происходит
достаточно плавно, поэтому для каждого поперечного сечения Sx массу на единицу
длины (здесь она записывается как рА) можно рассчитать как массу
твердотельного цилиндра Сх с тем же поперечным сечением. Массу легко можно
рассчитать методами теории функций комплексного переменного для
двумерного безвихревого потока для различных форм поперечного сечения,
соответствующих реально существующим рыбам.
Из расчетов следует, что в первом приближении масса т зависит
только от высоты поперечного сечения s. Действительно, запишем
т= j7rs2pP, (4)
где р — плотность воды, а переменный безразмерный параметр (3 имеет
значения, близкие к 1. В случае эллиптического поперечного сечения
параметр (3 в точности равен единице для произвольного эксцентриситета. При
2. Угревидный способ передвижения
105
этом эллиптическое сечение вдоль тела изменяется от формы, достаточно
близкой к круговой, какую имеет передняя часть многих угрей, далее
сильно вытянутое эллиптическое поперечное сечение (как у сельди) и вплоть до
совершенно плоских поперечных сечений, типичных для хвостовой части.
На рисунке 1 показан характер отличия коэффициента (3 от 1 для некоторых
других типичных сечений.
1,25
1
0,75
0,5
0,25
0,25 0,5 0,75 1
Я
Рис. 1. Зависимость коэффициента пропорциональности виртуальной массы (3
в уравнении (4) от q (q — доля высоты s, занятой телом рыбы, остальная часть
приходится на спинной и/или брюшной плавники). Точечная кривая соответствует
случаю, когда тело имеет эллиптическое сечение (отношение осей равно 2).
Остальные кривые относятся к телу кругового сечения. Сплошная кривая соответствует
случаю, когда спинной и брюшной плавники имеют одинаковые форму и
размеры; штрихпунктирная кривая — высота плавников, относится как 3:1; пунктирная
кривая — только один плавник
Сюда включены круговые сечения и эллиптические сечения с
вертикальной большой осью, в которых доля q общей высоты рыбы s, которую
занимает тело рыбы, отлична от единицы, и полагается, что толщина двух
плавников (спинного и брюшного) пренебрежимо мала. При равных по
высоте плавниках (т. е. ^ (1 — q)s у каждого) и кругового поперечного сечения
зависимость коэффициента /? от q изображена сплошной линией. Как
видно, коэффициент /3 равен 1 при q = 0 или 1 и уменьшается до минимально-
106
Глава 4
го значения 0,75 при q = 0,7. В случае эллиптического поперечного сечения
и равных по высоте плавников отклонение (3 от единицы значительно
меньше, эта зависимость изображена точечной линией. Зависимость /? от q при
неравных по высоте плавниках ~(1 — q)s и ~(1 — q)s и при единственном
плавнике высоты (1—q)s изображается штрихпунктирной и пунктирной
линиями соответственно; как видно, даже при круговом поперечном сечении
отклонение коэффициента (3 от единицы меньше, чем для случая равных по
высоте плавников, в то время как для эллиптических поперечных сечений
(на рис. 1 не представлены) отклонение от единицы коэффициента /?, как
и следовало ожидать, еще меньше.
Поскольку достоверное знание величины виртуальной массы га имеет
большое значение, то необходимо провести дополнительное обоснование
заключения, сделанного на основании теории тонкого тела о том, что в
сечении Sx виртуальная масса такая же, как и для твердого цилиндра Сх
с тем же сечением. Для того чтобы понять, как велика должна быть
коррекция полученной ранее виртуальной массы, приходящейся на единицу
длины, необходимо дать ответ на следующие два вопроса. Каково влияние
(i) того, что тело рыбы эластично, а значит, различные поперечные
сечения приводят в движение воду с различными скоростями, и (ii) того, что
поперечные сечения не имеют постоянную форму? Довольно просто
ответы на поставленные вопросы могут быть получены для частного случая
кругового поперечного сечения тела рыбы. Полученные результаты
свидетельствуют о том, что коррекция не будет слишком велика при условии,
что длина волны Л главной гармоники поперечных колебаний тела должна
быть не меньше чем 5s.
Этот вывод сделан на основании исследования тех решений
записанного в цилиндрических координатах (я, г, в) уравнения Лапласа, которые
пропорциональны Kl(kr) cos в cos кх9 где к = 2п/\. Так, если, например,
круговой цилиндр толкает воду в направлении оси в = 0 (что соответствует
оси z декартовой системы координат) с изменяющейся как coskx
скоростью, то с использованием решений уравнения Лапласа можно получить,
что коэффициент (3 равен
0 = К1{*8/\)/[-(*8/\)К[(*8/\)] (5)
и по нему рассчитать виртуальную массу (4). Как видно из рисунка 2,
коэффициент (3 отклоняется от единицы не более чем на 25% при Л > 5s.
Аналогично проведенные расчеты для случая, когда высота поперечного
сечения изменяется по гармоническому закону, а каждое сечение толкает
2. Угревидный способ передвижения
107
0,5 \-
5
A/s
10
15
Рис. 2. Зависимость коэффициента виртуальной массы /3 от отношения Л/5 длины
волны к высоте рыбы в данном поперечном сечении (для кругового поперечного
сечения)
воду с одинаковой скоростью, показали, что наибольшее отклонение
коэффициента (3 от единицы будет таким же, какое дало бы уравнение (5),
а затем умноженным на отношение амплитуды к величине ^ s (это
отношение, очевидно, меньше 1).
Так как значения коэффициента /? не сильно отличаются от 1 для
кругового цилиндра Sx, то можно надеяться, что виртуальная масса,
рассчитанная на основании довольно грубой аппроксимации, основанной на том,
что масса будет такой же, как и при наличии твердого цилиндра Сх с таким
же поперечным сечением, в случае когда длина волны главной гармоники,
распространяющейся по течению, превышает величину 5s, вероятно, будет
такой же и для различных более уплощенных (т. е. сжатых с боков)
поперечных сечений. В этом случае прилегающая масса воды просто повторяет
боковое движение поперечного сечения. Следовательно, поскольку при
чисто угревидном перемещении характерная длина волнообразного движения
тела (см., например, рис. 4 в работе Lighthill, 1969) значительно
превышает 5s, то возможно использование теории тонкого тела, в которой принято,
что виртуальная масса рассчитывается на основании указанной выше
аппроксимации.
Компонента силы, направленная вдоль оси г, с которой рыба действует
на прилегающий к ней вертикальный слой воды единичной длины в направ-
108
Глава 4
лении оси х, должна быть равна скорости изменения z-компоненты
количества движения слоя воды, движущегося вдоль рыбы со скоростью U. То
есть
Z=(J-t+U-^j[m(x)w(x,t)l (6)
где га(х) — масса, рассчитанная по (4) и зависящая только от высоты s(x)
поперечного сечения Sx (поскольку J3 близок к единице); w(x, t)
определяется соотношением (2). В равенстве (6) отклонением х-компоненты
скорости движения вертикального слоя воды вдоль тела рыбы от
величины U можно пренебречь, поскольку оно имеет второй порядок малости при
волнообразном движении тела (порядок отклонения равен произведению
трансверсальнои скорости на локальные углы поворота тела относительно
оси х, направленной вдоль исходного положения тела рыбы).
Мощность Е9 которую развивает рыба при совершении трансверсаль-
ных движений со скоростью dh/dt (скорость dh/dt обеспечивается
действием силы Z, приходящейся на единицу длины), равна
J at
dx. (7)
Используя равенство (6) для силы Z и равенство (2) для w, для
мощности Е получаем
о о
д [ ( dh 1 2\ J Г дЪЛ
= тг / mw— -mw ) ах + и \ mw -т- . (8)
dtj \ at 2 ; [ at\x=l }
Внеинтегральный член в правой части равенства (8) (последний член в
правой части) содержит вклад в мощность только хвостовой части рыбы х = /,
поскольку при х = I масса т отлична от нуля за счет бокового сжатия,
а у передней части рыбы при х = 0 масса т обращается в нуль.
Последний член в (8) особенно важен, поскольку он равен средней
мощности. Отметим, что среднее по времени от первого слагаемого,
равного частной производной от изменяющейся периодически по времени, равно
нулю.
2. Угревидный способ передвижения
109
В своей работе Лайтхилл (Lighthill, 1969) приводит простую
механическую интерпретацию наиболее важного для нашей задачи последнего
слагаемого в (8). В использующихся здесь обозначениях оно равно UmwW.
Здесь mw — количество движения в боковом направлении, приходящееся
на единицу длины слоя воды, расположенного вблизи хвостовой части, и,
таким образом, Umw представляет собой скорость сброса этого
количества движения в вихревой след за единицу времени, которая фактически
равна силе, с которой хвостовая часть действует на след, совершая
работу в единицу времени, равную UmwW, т.к. боковая скорость хвостовой
части равна W. Как следует из (3), мощность положительна и ее среднее
значение пропорционально среднему значению квадрата боковой скорости
хвоста W.
Однако мы должны задаться вопросом о том, почему это простое
рассуждение приводит к корректному результату только в отношении средней
мощности, а не в отношении мгновенного значения, определяемого в
равенстве (8) первым членом в правой части. Причина состоит в том, что при
волнообразном движении рыбы мощность, развиваемая рыбой, расходуется
не только на создание спутного следа. Частично мощность идет и на обмен
энергией с той частью воды, которая еще не образовала спутный след.
Величина энергии такого объема воды перед задним фронтом колеблется, то
увеличиваясь, то уменьшаясь, так что мгновенная скорость подобного
изменения отлична от нуля, хотя средняя скорость за длительное время равна
нулю (за время, превышающее период осцилляции).
Такая интерпретация равенства (8) может быть подтверждена, если его
преобразовать, используя (2), к виду
Е - U[mwW]x=l = tj- f (\mw<1 - Umw |^) dx- (9)
о
Здесь слева записана мощность, развиваемая рыбой, минус мощность,
теряемая в спутном следе, а справа — скорость изменения энергии,
представленной в интегральной форме. Под знаком интеграла первое слагаемое
является кинетической энергией воды, движущейся в направлении осей у и z.
Кроме того, в используемой системе координат присутствуют флуктуации
энергии воды, движущейся в направлении оси х.
Более точно, когда скорость единицы массы воды меняется от U до
U + и, то в первом приближении энергия меняется на величину Uu.
Следовательно, когда большой объем воды, движущейся со скоростью [/, испы-
по
Глава 4
тывает некоторое непостоянное, небольшое изменение в скорости, то его
энергия изменяется на С/, умноженную на изменение величины количества
движения в направлении оси х. Количество движения воды, обусловленное
трансверсальным движением рыбы, равно mw, но это изменение
происходит не точно в направлении оси z9 поскольку тело рыбы наклонено под
углом dh/dx к оси х, тогда в первом приближении появляется х-компо-
нента количества движения, которая равна —mw dh/dx (по-прежнему на
единицу длины тела рыбы), а соответствующее изменение энергии равно
—Umw dh/dx, которое и равно второму члену в подынтегральном
выражении (9).
Здесь использовано важное обобщение основного утверждения о том,
что импульс воды, обусловленный движением поперечного сечения
рыбы Sx в направлении оси z со скоростью го, будет таким же, как если
бы этот импульс был обусловлен движением цилиндра Сх с таким же
поперечным сечением. Обобщенное утверждение состоит в том, что если
поперечное сечение Sx имеет отличный от нуля наклон к оси х, то
соответствующий цилиндр Сх должен быть также наклонен таким образом, что
связанный с ним импульс, перпендикулярный образующим цилиндра,
имеет отрицательную проекцию на ось z. Это утверждение полностью
согласуется с аналогичным утверждением, относящимся к виртуальной массе,
которое было приведено выше.
Мощность Е, выраженная через силу тяги Р (силу тяги мы и найдем,
сравнив два выражения для мощности Е), в системе координат, в которой
жидкость на бесконечности покоится, имеет вид
E = UP + U
1 2
—mw
2
я/й*
+ — / (-тю )dx- (10)
х—1
Здесь первое слагаемое справа представляет собой мощность рыбы,
когда она движется со скоростью U в направлении действия силы тяги Р.
Второе слагаемое представляет собой скорость поступления кинетической
энергии воды, а именно ^mw29 приходящейся на единицу длины рыбы,
в спутный поток у задней кромки тела рыбы. Этой «потерянной» энергии
Лайтхилл (Lighthill, 1969, §4) в своей работе придавал особое значение.
Третье слагаемое в (10) представляет собой скорость изменения
осциллирующей составляющей, а именно общей кинетической энергии (в новой
системе координат, где жидкость покоится на бесконечности) перед задней
кромкой.
2. Угревидный способ передвижения
111
Вычитая из (9) выражение (10), получаем для силы тяги
=l-^l{mw^)dx- (11)
р =
Соотношения (9) и (10) могут быть использованы для получения
гидромеханической эффективности самодвижения rj, введенной в равенстве (1) как
UP/E. Используя тот факт, что среднее значение производной по времени
от каждого интеграла равно нулю, получаем
v = 1 _ (Е- UT)/E= 1 - \[й2}х=1/\Ш}х=1. (12)
Используя соотношение (3) между скоростями w и W при х = /,
получаем, что 77 = 1 — \{V — U)/V. Это подтверждает аргументы, приведенные
в работе Лайтхилла (Lighthill, 1969), что эффективность будет высокой,
когда w значительно меньше, чем W (и следовательно, V не слишком велика
по сравнению с С/), хотя для значимой величины силы тяги, как следует
из (11), скорость w, конечно, не должна быть слишком маленькой.
В равенство (11) входит не только средняя, но и мгновенная
составляющая силы тяги Р. Эту составляющую легче выявить при вычитании
равенств (9) и (10), чем получить ее значение на основании физических
рассуждений. Последнее, конечно, также возможно. Для этого
преобразуем (11) к виду
Р +
dh 1 2
и raw — —mw
дх 2
J X — l
hl(-mwi£)dx- <13)
Как можно заметить, такой вид согласуется с выведенным ранее
равенством абсолютно независимым способом (за счет интегрирования
давления, распределенного на поверхности, обусловленного распространением
возмущений) в приложении (см. уравнения (А21), (А26) и (А27)) в работе
Лайтхилла (Lighthill, 1960).
Для того чтобы дать физическую интерпретацию равенству (13),
отметим, что правая часть представляет собой скорость изменения х-компо-
ненты количества движения воды перед плоскостью П, проходящей через
заднюю кромку тела рыбы перпендикулярно оси х. Эта компонента
количества движения изменяется частично за счет силы Р, действующей между
112
Глава 4
телом рыбы и окружающей его водой, а также частично за счет
переноса количества движения через плоскость П, это изменение представлено
слагаемым в квадратных скобках. В слагаемом в квадратных скобках
впервые появляются потери, обусловленные адвекцией со скоростью U
количества движения —mwdh/dx, приходящегося на единицу длины. Второе
слагаемое представляет собой потери, обусловленные результирующей
силой давления, действующей на плоскости П.
Непосредственный вывод величины силы Р на основании физических
рассуждений значительно более сложен за счет того, что перенос
количества движения происходит дополнительно за счет действия этой отличной
от нуля силы давления. Ее величина равна ~mw2 и выводится довольно
легко из нестационарной формы уравнения Бернулли, записанного в
системе координат, в которой жидкость на большом расстоянии от тела рыбы
имеет скорость, равную —w в направлении оси z. В этой системе
координат движение рыбы относительно воды у задней кромки равно нулю. Тогда
квадратичные члены в уравнении Бернулли дают давление
1
2Р
w2-
|'-(*)1
(9
=
-+w)
i
"2"
:»♦(*♦•:
(14)
отсюда результирующее давление можно выразить через полный
импульс mw и кинетическую энергию ~mw2 единицы длины для поля
скоростей (дф/ду, дф/dz + w) в виде
w(mw) — —mw2 = —mw2. (15)
(С другой стороны, вклад слагаемого —рдф/dt в результирующее давление
равен нулю, поскольку для симметричного относительно плоскости z — О
сечения потенциал ф является нечетной функцией относительно z.)
Присутствие такой результирующей силы давления, как ^mw2, на
одной стороне плоскости П снова приводит к необходимости рассмотрения
вихрей (см. § 3), включая различие в изменениях х-компоненты количества
движения в попутном потоке и при схождении потока с задней кромки.
В двух последующих параграфах проведена дальнейшая разработка
изложенной выше теории с целью исследования модификаций чисто
угревидного способа перемещения, возникших в процессе эволюции.
3. Вихревые пелены за плавниками
113
3. Вихревые пелены за плавниками
В данном параграфе, прежде чем переходить к исследованию вихре-
образования в окрестности плавников, обсудим форму потока воды
непосредственно за задней кромкой. Именно с этим потоком переносится то
количество движения и кинетической энергии, о котором говорилось в § 2
как о потерях в спутном потоке. Для некоторых читателей, особенно для
тех, кто знаком с аэродинамикой полета, практически очевидно, что такой
поток принимает форму вихревой пелены, а более точно (поскольку
рассматривается крыло небольшого удлинения) вихревой пелены с таким
распределением завихренности, которая соответствует «эллиптическому
распределению» потенциала скорости (минимизирующим на единицу длины
спутного следа, кинетическую энергию при заданном боковом импульсе
и размахе).
Для того чтобы проследить, как это вытекает из теории § 2, отметим,
что когда поперечное сечение Sx располагается в вертикальной задней
кромке соответствующий цилиндр Сх будет бесконечно длинным,
бесконечно близким по форме к тонкой пластинке шириной s. Потенциал
скорости ф при движении такой пластинки со скоростью w в направлении оси z
принимает значение
1/2
(16)
ф = ^fw
js2 -{у- t/o)2
на сторонах пластинки \у — у0| ^С ^s, z ^ 0 соответственно. Уже из
равенства (16) следует эллиптичность распределения потенциала скорости
в окрестности задней кромки. Это означает, что изменение вертикальной
компоненты скорости дф/ду равно
-1/2
А(дф/ду) = 2w(y - у0)
\s2 -(У- УоГ
(17)
между z < 0 и z > 0 сторонами хвостовой части.
Такой разрывный характер скорости указывает на наличие вихревой
пелены — тонкого слоя, в котором модуль х-компоненты завихренности
£ принимает большие значения, а интеграл от нее в направлении,
перпендикулярном плоскости пластинки от одной ее стороны до другой,
равен Е. Или поскольку в действительности на обеих сторонах пластинки
образуются пограничные слои, то интеграл от £ необходимо брать вдоль
пути, включающего пограничные слои. Отсюда вытекает, что может быть
выстроено совершенно непротиворечивое описание потока, для которого
114
Глава 4
равенство (16) определяет поле скоростей поперечного потока в
окрестности хвостовой части тела рыбы (более точно, в окрестности области,
занятой пограничными слоями, образовавшимися с обеих сторон хвостовой
части), в то время как за задней кромкой тела рыбы образуется свободная
вихревая пелена с той же завихренностью S, с тем же значением
потенциала (16) по обеим сторонам этой пелены и тем же самым наведенным
поперечным потоком в ее окрестности.
Эта модель непротиворечива, поскольку (i) такой непрерывный
переход на задней кромке от поперечного потока перед ней к поперечному
потоку за ней означает, что поперечный поток развивается постепенно и
непрерывно, так же как и в окрестности большей части тела рыбы, что
объясняет (см. § 2) возможность исследования поперечного потока в двумерной
постановке (как вокруг цилиндра Сх)\ (ii) непрерывность интегрируемой
х-компоненты завихренности Е на задней кромке согласуется с соленои-
дальным характером вихревого поля и с вмороженностью вихревых линий
в жидкость.
В системе координат, в которой невозмущенная жидкость покоится,
сошедшая в спутный поток система вихрей сохраняет постоянную
завихренность и продолжает двигаться со скоростью w в направлении оси z9
которая определяется значениями потенциала (16) на поверхности. В
каждой точке спутного следа локальное значение скорости w (которая связана
с локальным значением интенсивности вихревой пелены Е (17)) равно
тому значению поперечной скорости, которое она имела на задней кромке,
когда кромка проходила через рассматриваемую точку. Отсюда следует, что
величина w не будет постоянной вдоль спутного следа, что обусловлено
изменением по времени скорости w на задней кромке.
Вектор ротора в вихревой пелене направлен вдоль оси у в силу
изменения скорости w с изменением х, угловая скорость вращения, касательная
к вихревым линиям, равна —dw/dx. Вектор связанного с ними количества
движения mw на единицу длины также вращается. Вектор этого количества
движения достаточно близок по направлению к оси z9 но вращение
приводит к тому, что дополнительно появляется х-составляющая количества
движения со скоростью —mw dw/dx. В этом случае суммарная скорость
изменения количества движения в спутном потоке количества движения,
меняющаяся со скоростью, равна
ос
(18)
/
—mw——
дх )
1 dx =
(
1 2
—mw \
2
3. Вихревые пелены за плавниками
115
(жидкость на бесконечности покоится). Полученное в (18) значение
скорости изменения совпадает с полученным ранее в § 2 (см. (15)) на основании
совершенно других рассуждений, связанных с распределением давления).
На этом можно было бы закончить обсуждение структуры спутно-
го потока в случае чисто угревидного движения животных со спинными
и брюшными плавниками, непрерывно расположенными вдоль тела.
Однако следует привести еще некоторые соображения, аналогичные
указанным выше, для понимания спутного потока, обусловленного наличием
дискретно расположенных спинных и брюшных плавников, с задней кромкой,
не имеющей ярко выраженную стреловидность, при угревидном способе
перемещения. В окрестности поперечного сечения, где расположен такой
плавник, потенциал скорости принимает различные значения по обеим
сторонам плавника, как и в (16), что соответствует наличию вихревой пелены
интенсивности 5 = —А(дф/ду)9 как и в (17). Равенства (16) и (17) дают
значения величин, стоящих в левой части, с достаточно хорошим
приближением, только если толщина тела рыбы мала по сравнению с его высотой.
В случае округлых тел для получения подобных результатов следует
применять теорию функций комплексного переменного.
Сразу же за задней кромкой плавника должна образовываться вихревая
пелена с той же интенсивностью Е на основании совершенно аналогичных
приведенным выше рассуждений. В системе координат, где невозмущенная
вода покоится, эта интенсивность остается постоянной, и в любой точке
ее значение пропорционально той величине скорости w на задней
кромке плавника, когда кромка проходит через эту точку. В системе координат,
введенной в § 2, когда невозмущенная вода движется со скоростью [/,
интенсивность вихревой пелены соответственно является функцией от
переменных t — (x/U) и у.
Применим проведенные теоретические рассуждения к таким рыбам,
как треска, и к другим представителям семейства тресковых (Gadidae)
отряда мягкоперых костистых рыб (Anacanthini), которые имеют по несколько
спинных и брюшных плавников, расположенных достаточно близко друг
к другу, и плавают, используя угревидный способ перемещения. За
передним спинным плавником образуется вихревая пелена, заполняющая
пространство между ним и последующим плавником, в результате чего
обеспечивается непрерывность поперечного потока, значительно большая, чем
это можно было бы ожидать исходя из геометрических соображений.
У второго спинного плавника эта вихревая пелена смешивается с
пограничным слоем. Такая распространяющаяся вниз по потоку завихрен-
116
Глава 4
ность не окажет значительного влияния на поперечный поток или на общую
завихренность за этим плавником, поскольку указанные завихренности
однозначно определяются локальным значением скорости w. Таким образом,
присутствие отдельной обособленной вихревой пелены между двумя
плавниками можно рассматривать как исключительно локальное изменение,
которое окажет малое влияние на движущуюся вниз по потоку жидкость.
Подобным образом и другие промежутки между плавниками заполняются
вихрями.
Когда, как это имеет место у многих тресковых, эти промежутки
малы (случай больших промежутков между соседними спинными плавниками
будет рассмотрен ниже), то гидродинамический эффект от них практически
такой же, как если бы спинной плавник был непрерывным. Действительно,
некоторая разница в поперечном потоке вокруг сечения Sx, с учетом
поперечного размера вихревой пелены и вокруг того сечения, когда эта вихревая
пелена была бы заменена жестким плавником, может иметь место только
за счет (i) разницы между формой поперечного сечения тела, где имеется
вихревая пелена за плавником и формой поперечного сечения Sx9 или (ii)
разницы между величинами скорости w на этих участках. Если оба
различия отсутствуют, тогда интенсивность Н вихревой пелены будет такой
же, как и в случае отсутствия промежутка между плавниками, и на
основании теоремы единственности гидродинамики поперечный поток будет
таким же, как и при отсутствии промежутка между плавниками.
Для небольших промежутков между плавниками различия (i) и (ii)
незначительны. Заметим, что скорость распространения волны вдоль
тела V несильно отличается от скорости [/, скорость w сечения Sx только
немного опережает по фазе скорость у тех сечений, где имеется между
плавниками вихревая пелена, а ее амплитуда может немного возрастать
в хвостовой части. Как установлено в работе Лайтхилла (Lighthill, 1969,
§4) все рассуждения, проведенные для дискретных спинных плавников,
справедливы и для брюшных плавников, и в обоих случаях при
угревидном способе передвижения предполагается, что последовательность близко
расположенных плавников с динамической точки зрения эквивалентна
одному длинному непрерывному плавнику.
У некоторых же рыб, включая большинство сомов подотряда сомовых
(Siluroidea) (отряд костнопузырных (Ostariophysi)), плавающих угревидным
способом, расстояние между основным спинным и хвостовым
плавниками достаточно велико (второй спинной плавник сравнительно мал). При
анализе движения подобных рыб следует принимать во внимание нали-
3. Вихревые пелены за плавниками
117
чие большой вихревой области между спинным и хвостовым плавниками,
где перечисленные выше эффекты (i) и (и) могут играть достаточно
важную роль, и тогда поведение области между плавниками, занятой вихревой
пеленой, будет существенно отличаться от того, как если бы эта область
представляла бы собой продолжение спинного плавника.
Такие различия приводят к необходимости определить для каждого
поперечного сечения тела Sx виртуальную массу т(х) на единицу длины
рыбы при наличии вихревой пелены около тела. Для того чтобы это
сделать, не будем забывать, что значение потенциала скорости с обеих сторон
вихревой пелены пропорционально скорости
wF = w(xF, t - (х - xF)/U). (19)
Эта скорость равна скорости w конца плавника х = xF в тот момент
времени, когда вертикальный слой воды, находящийся в настоящий момент
времени у сечения Sx, находился у конца плавника. Коэффициент
пропорциональности зависит от формы поперечного сечения при х = xF и
задается равенством (16) с s = s(xF) для рыб, толщина которых значительно
меньше высоты.
Под массой т(х) по-прежнему будем понимать виртуальную массу на
единицу длины, обусловленную движением бесконечного твердого
цилиндра Сх с поперечным сечением Sx в направлении оси z при отсутствии
какой бы то ни было вихревой пелены. При движении цилиндра Сх со
скоростью w относительно воды потенциал поперечного потока ф
удовлетворяет условиям
— = wnz на Сж, ф = 0 на вихревой поверхности, (20)
где nz — z-компонента единичного вектора нормали к Sz. Количество
движения поперечного потока равно m(x)w.
Далее определим массу т(х) как виртуальную массу, которую
приводит в движение вихревая пелена, когда цилиндр Сх совершенно
неподвижен. Потенциал этого движения ф удовлетворяет условиям
— = 0 на Сж, ф = заданной величине на вихревой пелене, (21)
заданное значение пропорционально скорости wF (см. (19)). Количество
движения соответствующего потока равно m(x)wF на единицу длины.
118
Глава 4
Суммарный потенциал поперечного потока, возникшего при движении Sx
при наличии вихревой пелены, очевидно, равен сумме ф + ф, и,
следовательно, количество движения равно
m(x)w + fh(x)wF. (22)
Кинетическая энергия суммарного потока также имеет простую форму,
поскольку из теоремы Грина следует, что потенциалы фиф двух
поперечных потоков ортогональны в пространстве, где кинетическая энергия не
имеет особенностей. (Для этого интеграл J 0(<90/<9n)ds, взятый по
контуру, состоящему из Сх и двух сторон вихревой пелены, должен быть равен
нулю. Равенство нулю этого интеграла с очевидностью следует из равенств
(20) и (21).) Следовательно, кинетическая энергия равна
-m(x)w2 + —m(x)w2F. (23)
Zt Zt
Справедливость формул (22) и (23) легко можно подтвердить,
рассмотрев конкретные поперечные сечения. Так, для тонкого поперечного
сечения Sx получаем эти формулы непосредственным подсчетом для масс
га = —^(s2 — s2)p, га = -irs2p, (24)
где ? и s — локальные высоты поперечного сечения при наличии и
отсутствии вихревой пелены соответственно. Из (24) становится ясным, что
в точке х = xF (от этой точки начинается вихревая пелена) между т(х)
при х < xF и т(х) + т(х) при х > xF разрыва нет. Действительно, это
выполняется для всех форм поперечного сечения тела, поскольку при х =
= xF выполняется w = wF (см. (19)). Следовательно, из (22) вытекает, что
количество движения (которое, как говорилось выше, должно быть
непрерывным) равно [т(х) + m(x)]w.
Теперь можно применить полученные выше результаты к определению
сил, действующих между телом и окружающей его водой. Вместо
равенства (6) для силы Z, используя (19) и (20), запишем
V dt дхА
m(x)w(x, t) + m(x)w ( xF, t
X X F
и
§i + Ulb) H*M^ *)] + Um'(x)w (xF, t X JFj, (25)
3. Вихревые пелены за плавниками
119
причем, как мы помним, масса га в точке х = xF имеет скачок от 0 до
rh(xF), так что в га'(х) входит сомножитель, содержащий дельта-функцию
fh(xF)S(x — xF), а из предыдущего параграфа следует, что га'(х) содержит
равный по величине и противоположный по знаку член с дельта-функцией.
Равенство (7) для мощности Е, которую развивает рыба, совершая
трансверсальные движения со скоростью dh/dt и создавая силу Z на
единицу длины, может быть теперь преобразовано к виду, подобному (8)
с некоторыми дополнительными членами. Первое слагаемое в правой
части (25) преобразуется к такому же, как первое слагаемое в правой части
(8), хотя т(х) имеет разрыв при х = xF. Слагаемое, содержащее
множитель га'(х), дает два слагаемых в (7), первое за счет дельта-функции
и второе за счет остальной части га'(#), так что для мощности Е имеем
z. д Г ( dh 1 2\j тт
Е = — / mw— —mw ax + и
dt J V dt 2 J
dh
mw-rr-
dt
+
x—l
+ Ufh(xF)w{xF, t)—h(xF, t) +
i
+ J Um'{x)w (xF, t - X~rjXF) -q1 ^x' ^dx- (26)
X p
При записи мощности Е в другом виде (см. (10)), из которой
впоследствии можно найти силу тяги Р, находим скорость изменения энергии
в системе координат, в которой невозмущенная вода покоится. Используя
запись кинетической энергии в виде (32), получаем для Е:
E = UP + U
i
1 2 1~ 2
-mw +-mwF
I/&
+ ftJ [^mw2 + ^ihwl) dx. (27)
x~ о
У рыб, которые рассматриваются в данном параграфе, высота
хвостового плавника практически равна (или больше) высоте тела рыбы с
учетом спинного плавника. У таких рыб вихревая пелена, развивающаяся за
спинным плавником вплоть до хвостового, поглощается пограничным
слоем у хвостового плавника, так что в результате вихревая пелена сходит
с хвостового плавника. Она возникает за счет движения хвостового
плавника и практически не зависит от пелены, возникшей за спинным
плавником. На заднем фронте х = I не существует отдельной вихревой пелены
120
Глава 4
от спинного плавника, поэтому га = 0, и второй член в правой части (27)
имеет такую же форму, как и в (10). Другими словами, на величину потерь
энергии в спутном потоке наличие пелены между спинным и хвостовым
плавниками влияния не оказывает.
Надо сказать, что последнее слагаемое в правой части (27)
изменяется за счет этой вихревой пелены, но поскольку оно представляет собой
скорость изменения пульсирующей величины, то ее среднее значение
равно нулю. Следовательно, согласно принятой аппроксимации, наличие
вихревой пелены в промежутке между спинным и хвостовым плавником не
влияет на среднюю величину потерь Е — UP. В связи с этим было бы
интересно задаться вопросом о том, может ли возрасти общая средняя
мощность Е (26) за счет новых слагаемых, обусловленных наличием вихревой
пелены, поскольку в таком случае вся эта дополнительная мощность могла
бы эффективно расходоваться на продвижение и при этом произошло бы
увеличение как силы тяги, так и эффективности.
Такое кажется возможным, дадим этому физическую интерпретацию.
Среднее значение первого слагаемого в равенстве (26) равно нулю, а второе
слагаемое, равное UmwW (физический смысл которого был выяснен
выше), представляет собой скорость схода с хвостового плавника поперечной
компоненты количества движения, умноженной на поперечную скорость
задней кромки хвоста. Среднее значение этого слагаемого, как мы уже
упоминали, положительно. Подобный физический смысл имеет и третье
слагаемое, обусловленное потоком за спинным плавником. И следовательно, на
том же основании его среднее значение также будет (по существу, за счет
выполнения неравенства V > U) положительно.
Посмотрим, сократится ли последнее слагаемое в (26), в котором
множитель т'(х) будет в среднем отрицательным, поскольку га уменьшается
от положительного значения rh(xF) до нулевого га(/), с третьим
слагаемым, среднее значение которого положительно. Последнее в (26)
слагаемое можно рассматривать как отрицательную мощность, равную взятому со
знаком минус интегралу по передней кромке хвостового плавника от
произведения скорости поступления поперечной компоненты количества
движения из вихревой пелены U(—dm)wF на поперечную скорость передней
части хвостового плавника dh/dt. Последнее слагаемое будет
отрицательным, только если среднее значение множителя wF dh/dt положительно.
А это так и будет при V > С/, и, видимо, рассматриваемое среднее
значение wF dh/dt будет даже превышать среднее значение этой величины
на задней кромке спинного плавника, т. к. амплитуда поперечных колеба-
3. Вихревые пелены за плавниками
121
ний в направлении к хвосту возрастает. Однако если между wF и
скоростью dh/dt передней части хвостового плавника имеется существенный
сдвиг фаз, то это условие выполняться не будет. При сдвиге фаз,
превышающем 7г/2, среднее значение произведения этих скоростей может быть
отрицательным.
Разность фаз имеет среднее значение, равное
2"(xVXf)(f-')' (28)
где х — хс — среднее значения координаты передней части хвостового
плавника, Л — длина волны при волнообразном движении тела. Величина
27г(хс — xF)/\ представляет собой разность фаз скорости dh/dt в точках
х = xF и хс, в то время как разность фаз скорости wF, как следует из (19),
больше этой величины в V/U раз из-за того, что скорость адвекции
вихрей U меньше скорости V распространения поперечной волны вдоль тела.
В общем случае при длине промежутка (хс — xF)9 не превышающем
половины длины волны, и при значениях отношения скоростей U/V около 2/3
разность фаз (28) может принимать значения, равные или больше чем 7г/2.
Приведенные рассуждения показывают, что наличие достаточно
большого расстояния между спинным и хвостовым плавниками может
улучшить характеристики угревидного способа движения за счет того, что
суммарная выходная мощность возрастает без какого-либо увеличения потерь
мощности из-за возникновения вихревой пелены. Это объясняется
следующим. Движение спинного плавника обусловливает наличие
дополнительной мощности. Фаза изменения импульса сходящей со спинного плавника
воды совпадает с фазой его поперечного движения. У хвостового плавника
фаза импульса сошедшей со спинного плавника воды, движущейся со
скоростью U относительно рыбы, не будет совпадать с фазой поперечного
движения хвостового плавника (и может даже быть в противофазе), поскольку
скорость U меньше скорости распространения поперечной волны вдоль
тела V, в результате чего скорость поперечного движения воды, сошедшей
со спинного плавника, погашается и уменьшения выходной мощности не
происходит.
Возможно, приведенные в данном параграфе рассуждения в какой-то
степени могут объяснить появление в процессе эволюции дискретных
спинных плавников. Примитивные бесчелюстные позвоночные класса
круглоротых (Cyclostomata), такие как миноги и миксины, имеют
непрерывные спинные плавники. Думается, что их замена на некоторой стадии
122
Глава 4
эволюционного процесса развития рыб на ряд близко расположенных
отдельных плавников была благоприятной в отношении силы тяги, в то же
время такая замена могла привести к уменьшению силы сопротивления.
Упрочение в дальнейшем первого спинного плавника и уменьшение
остальных привели, прежде всего, к дополнительному (за счет фазового сдвига)
увеличению силы тяги, хотя преимущества такого расположения плавников
в отношении повышения устойчивости и возможности управления
рысканием проявились позднее.
4. Механика скомброидного типа движения
Дальнейшее увеличение силы тяги и повышение эффективности
движения произошли за счет постепенного перехода к скомброидному типу
плавания. При таком типе плавания со спинного уже не очень
длинного плавника сходит вихревая пелена, которая играет здесь другую, хотя
по-прежнему существенную роль. Рассмотрим с точки зрения механики
преимущества перехода к другому типу плавания. Как будет видно ниже,
при скомброидном способе перемещения волнообразные движения
осуществляются в основном хвостовой частью тела рыбы, в результате чего
увеличивается эффективность движения. Однако это увеличение
эффективности может быть сведено на нет за счет эффектов «отдачи», если переход
к скомброидному типу плавания не сопровождается специфическими
морфологическими изменениями, уменьшающими эту отдачу.
Для того чтобы понять преимущество перехода к скомброидному
способу плавания, нам необходимо в дальнейшем усовершенствовать
теоретические методы, изложенные в § 2, принимая во внимание тот факт
(отмеченный уже в тексте этого параграфа после равенства (3)), что при
проталкивании телом слоя воды виртуальной массы т этот слой воды не только
приобретает импульс mw, но этот импульс постепенно увеличивается по
мере продвижения слоя вдоль тела. Это связано с силой, действующей на
тело, возникновение которой является непосредственным результатом
образования вихрей около тела рыбы при проталкивании ею слоев воды. При
этом кинетическая энергия воды превышает величину кинетической
энергии при безвихревом обтекании ~mw2 (на величину, равную, как следует
из теоремы Кельвина о минимуме энергии, той энергии, которой обладала
бы система вихрей при неподвижном теле).
В работе Лайтхилла (Lighthill, 1960) было высказано предположение
о том, что теоретический подход, изложенный в §2, необходимо расши-
4. Механика скомброидного типа движения
123
рить, приняв во внимание отличия (если их возможно оценить
количественно) количества движения воды М и ее кинетической энергии Т на единицу
длины от величин mw и ^mw2. Отметим, что при обсуждении равенств (9)
и (10) в § 2 было высказано предположение о том, что те физические
положения, на основе которых были получены данные равенства, справедливы
и в более общем случае. И тогда для средних значений Р и Е получаем
E=U[MW]x=l, (29)
ТГ-u7=Ufflx=l. (зо)
Отличие в записи в равенствах определяется тем, что среднее значение
скорости изменения кинетической энергии движущейся вблизи тела воды
вычисляется в системе координат, в которой тело или вода соответственно
находятся в покое. Величина кинетической энергии воды, непосредственно
прилегающей к телу рыбы, изменяется по периодическому закону, так что
ее средняя скорость изменения по времени равна нулю.
Для среднего значения силы тяги и эффективности из
соотношений (29) и (30) получаем
Т = [MW - T\x=l = [{v/(v - и)}Ш; - т]х=ь (31)
r) = l-{{V- и)/У}Щх=1/[1Щх=1Г (32)
здесь скорости W и w связаны соотношением (3). Отсюда следует, что та
часть количества движения М на задней кромке х = /, которая слабо
связана с локальной скоростью w, практически не оказывает влияния на
величину силы тяги, в то время как соответствующая ей кинетическая энергия Т
может дать существенную прибавку к потерям энергии, обусловленным
наличием вихревого следа.
Количество движения М слоя воды, перпендикулярного телу,
изменяется при достижении слоем воды задней кромки хвостового плавника от
значения mw на величину, равную вихревой силе, действующей со
стороны тела рыбы по мере продвижения слоя воды вдоль тела,
проинтегрированной по времени. К сожалению, найти точное значение этой силы
невозможно, поскольку мы имеем дело с чрезвычайно неустойчивым процессом.
Слой воды последовательно проталкивается вдоль тела поперечными
сечениями переменной формы. Причем скорость прохождения слоя воды
изменяется при переходе от сечения к сечению. И еще тяжелее определить
приращение кинетической энергии ^mw2 к моменту прохождения слоем
124
Глава 4
воды задней кромки тела рыбы, т. к. трудно вычислить локальное значение
энергии системы вихрей. Однако даже на основании очень приближенных
расчетов можно показать, что за счет указанных эффектов происходит
существенное уменьшение эффективности плавания при угревидном способе
движения, и подобного уменьшения эффективности можно избежать при
переходе к скомброидному типу перемещения.
В данной работе проводится довольно приблизительная оценка
величины вихревой силы на единицу длины, действующей со стороны сечения
тела, движущегося в поперечном направлении с локальной скоростью w,
на прилежащий слой воды. При этом используются два упрощающих
предположения. Первое: сила сопротивления зависит только от скорости w9
высоты поперечного сечения s и от параметра стационарного движения
тонкой длинной полоски в направлении, перпендикулярном ее плоскости
(Flachsbart, 1935); т.е. равна psw2 sgnw, где sgnw (+1, если w > О, и -1,
если w < 0) отражает тот факт, что сила сопротивления совпадает по
направлению со скоростью w. Второе: используется линейное приближение
величины этой силы. Если скорость w изменяется между — w0 и +w0, то
абсолютная средняя квадратичная ошибка при замене величины w2 sgn w на
jw0w составит 0,1 Iwq. Подобная линеаризация приемлема в нашей
постановке, когда силы сопротивления при различных скоростях вносят вклад,
изменяющийся по линейному закону, в суммарное количество движения М
(так что учитываться будут только абсолютные ошибки, а не
относительные, которые становятся слишком большими при малых скоростях w).
В результате наших рассуждений получаем, что для вихревой силы
можно принять выражение jpsw0w. Вклад этой силы в суммарное
количество движения рассматривается наряду со вкладом от виртуальной
массы и отражает реакцию воды на движение поперечного сечения тела
рыбы, которая представляется в виде комбинации линейного сопротивления
и инерции. Избыток количества движения на задней кромке х = I в
момент времени tx можно теперь представить в виде интеграла по времени от
вихревой силы, пропорциональной произведению sw при х = I — U(t — £х):
t
[M]x=l = [mw]x=l + J jpw0[sw(l-U{t-t1),t1)]dt =
t-i/u
i
= [mw]x=l + / -pw0[sw(x, t-(l- x)U~l)]dx/U. (33)
о
4. Механика скомброидного типа движения
125
Можно сравнить соотношение фаз между М и w при х = I для
различных скоростей роста амплитуды волны в зависимости от переменной х.
Пусть
sw{x, t) = s0w0eK{x~l) cos[u;{t + {I - x)V~1)}, (34)
где V — скорость распространения волны относительно тела, к мало при
угревидном способе плавания и велико при скомброидном. Если массу т
записать в виде приближенного выражения ~ps2 (см. § 2), то с учетом (34)
выражение (33) принимает вид
3 2 х . IpsQwllKCosut + ujU-1 -V-^smut]
[M]x=i = -^pso^ocosujt + Ultf+LjtiU-i-V-1)2] ' ( }
здесь значением первообразной на нижнем пределе пренебрегается.
Следовательно, далее можем получить
И„, = f p« + cv + fe-'-K-MT <36'
Полученные результаты показывают, что для малых к отставание по
фазе по отношению к [w]x=l прироста количества движения [М]ж=/,
обусловленного перемещением воды относительно тела рыбы, совершающей
волнообразные движения, приводит к значительному уменьшению их
среднего произведения (36). Величина [Т?]ж=/ не столь чувствительна к
величине сдвига фаз. Часть, обусловленная действием вихревой силы,
может быть представлена в виде интеграла от скорости w, умноженной на
вихревую силу, относительно момента времени tv И кинетическую
энергию [Т]х=1 можно записать как
[Т)х=1 =
При
i
1
+
J X = l
—mw2
2
( jpw0[sw2{x, t-{l- x)U-l)]dx/U. (37)
о
sw2{x, t) = s0wleT{x~l) cos2[u{t + {I - x)V'% (38)
где s принимает свое наибольшее значение s0 на задней кромке
(подразумевается, что к ^ г ^ 2ас), равенство (37) принимает вид
ЕПх=1 = JHP*o*>l + IpsowlU-'T-1. (39)
126
Глава 4
Из (39) следует, что поток энергии, уходящий с вихревой пеленой,
возрастает за счет влияния вихревой силы на поперечные сечения тела
рыбы в 1 + 2w0/Uts0 раз. Здесь множитель 2w0/U имеет значения, близкие
к единице для большинства интересующих нас движений рыб, так что
возрастание потерь энергии будет значимым, если г не превышает s^1, что
имеет место при угревидном движении. Такое увеличение потерь энергии
не компенсируется соответствующим увеличением силы тяги, поскольку
для таких значений г (и соответствующих значений к менее половины),
как следует из (36), произведение [Mw]x=l незначительно возрастает3
потому, что второе слагаемое в выражении для [M]x=i слабо коррелирует
с [гу]ж_^ когда, как это и имеет место, величина uj(U~1 — V~l) в (36) имеет
порядок Sq1.
С другой стороны, при скомброидном движении амплитуда
поперечного движения тела нарастает у хвостовой части и г, относительная скорость
роста sw2, значительно превышает s^"1, в результате чего рост потерь
энергии будет значительно меньшим. Таким образом, переход к скомброидному
типу движения представляет собой логическое следствие процесса
бокового уплощения хвостовой части. Использование эффекта виртуальной массы
(«инерционности» воды, а не ее «сопротивляемости»), обусловленной
боковым уплощением, становится при скомброидном типе движения
практически единственным механизмом возникновения силы тяги.
Однако с переходом к скомброидному типу плавания возникает
возможность уменьшения силы тяги за счет недостаточного согласования меж-
ДУ [^1ж=1 и [W]x=j> a именно эффект «отдачи» (Lighthill, 1960). Это может
привести к возникновению дополнительной вихревой силы, а также к
нарушению соотношения (3), в котором предполагается, что фазы скоростей w
и W совпадают. Возникновение сдвига фаз между этими скоростями может
привести к уменьшению мощности, а уменьшение отношения их амплитуд
может к тому же привести и к возрастанию потерь энергии. При выводе
этого уравнения и других предполагается, что рыба может за счет мускульного
усилия совершать волновые движения в поперечном направлении,
распространяющиеся вдоль тела, которые можно описать функцией h(x, t). При
3Оценка (36) для E(V — U)/VU основана на принятом соотношении (29), которое
может дать завышенное значение степени влияния движений хвостовой части на вихревую силу
в спутном следе. Однако противоположное допущение о том, что возрастание Е9
обусловленное вихревыми силами, в точности равно дополнительной мощности, за счет толкания со
скоростью W9 привело бы к замене второго слагаемого в (36), равного TZ(V — U)/VU9
выражением ^ps0WqU~1t~1 и соответствующее увеличение мощности в этом случае составило
бы только половину от увеличения потерь энергии.
4. Механика скомброидного типа движения
127
этом не учитывается тот факт, что поперечные движения рыбы должны
подчиняться основным законам движения: (i) скорость изменения
количества движения в поперечном направлении должна быть равна суммарной
силе, действующей в поперечном направлении на тело рыбы со стороны
воды; (ii) скорость изменения момента количества движения относительно
фиксированной оси (в качестве которой нами принята ось х = /, z = 0)
должна быть равной моменту суммарной боковой силы, действующей на
тело рыбы со стороны воды.
Действительно, за счет мышечного усилия рыба может изменять свою
форму относительно центра тяжести, а перемещения и вращения тела,
сопровождающие эти изменения, подчиняются указанным выше законам (i)
и (ii). Например, попытка изменить свою форму по закону
h = H{x) cos[uj{t + {l- x)V~1)} (40)
на практике приводит к следующей зависимости формы тела от
переменных t и х:
h = Н(х) cos[u>(t+(l—x)V~1)] + [hl+h3(l—x)] cos cut+[h2+h4(l—x)] sinu;£,
(41)
где константы hl9 h2, h3 и /i4 определяются из условий удовлетворения
законам (i) и (ii) с последующим приравниванием коэффициентов при cos cut
и sin ut с обеих сторон.
Проведем эти выкладки, пренебрегая величиной вихревой силы.
Прежде всего получим «чистую» результирующую боковую силу (под «чистой»
силой понимается сила, когда дополнительно учитывается изменение
собственного количества движения тела рыбы), обеспечивающую изменение
формы тела рыбы по закону (40), т. е.
i
lZ + rrij -^-y \dx = Zx coscji + Z2 sinu;£, (42)
о
где mAx) — масса рыбы, приходящаяся на единицу длины тела.
Соответствующий «чистый» момент боковых сил относительно оси х = /, z = 0
(которая совпадает с положением задней кромки тела рыбы в
невозмущенном положении) равен
i
(I — х) lZ + rrif -jr-£ J dx = Z3 cos ut + Z4 sinu;£.
(43)
128
Глава 4
Далее определим, какова должна быть форма рыбы (41), чтобы
выполнялись законы движения (i) и (ii), а дополнительные сила и момент имели
вид (42) и (43). Линейные члены в (41) при этом должны быть такими,
чтобы интегралы (42) и (43), в которые подставлены только эти линейные
члены, имели равные и противоположные по знаку значения.
Расчеты проведены последовательно в два этапа: сначала
подставляется форма тела рыбы в виде (40), на втором этапе добавляются линейные
слагаемые, включенные в (41), поскольку в (42) и (43) одни слагаемые
существенно зависят от синусоидальных колебаний, а другие — от линейных
членов, входящих в (41). Например, синусоидальный член при скомброид-
ном типе плавания (т. е. при Н(х), имеющей достаточно большое значение
только вблизи х = I) соответствует пренебрежимо малому элементу массы
тела рыбы в (42) и (43), поскольку за счет имеющегося бокового сжатия
тела рыбы масса этого элемента тела рыбы пренебрежимо мала по
сравнению с виртуальной массой воды вблизи х = I. В то время как
соответствующие виртуальные массы воды определяются тем, что скорость w, связанная
с этим членом, значительно меньше скорости W (см. равенство (3)).
А линейные члены в (41) обусловливают элементы массы рыбы в (42)
и (43), в которых в полной мере учитывается инерция рыбы, равно как
и статический момент и момент инерции относительно оси х = I. Кроме
того, для этих линейных членов адвективные производные U д/дх в (2) и (6)
не столь важны, как локальные d/dt9 в силу использующихся значений
частот и (отношение ul/U приблизительно равно 10). В первом
приближении, пренебрегая производными U д/дх, получаем Z = m(x)d2h/dt2,
так что интегралы в равенствах (42) и (43) могут быть рассчитаны как для
твердого тела, на которое не действуют внешние силы и масса которого
равна т(х) + тЛх).
Это первое приближение, при котором реакция со стороны рыбы на
заданные поперечные силы принимает форму инерционного отклика, на
основе комбинации инерционности рыбы и воды, будет весьма полезна для
нас. Если ввести обозначения
/ / i
М = (т+т^)(1ху ML = / (/—x)(m+m^)dx,
0 0 0
(44)
где L — расстояние от центра масс до задней кромки, I — ML2 — момент
инерции относительно центра масс, то для определения [/i1? /i2, ^35 ^4] по"
4. Механика скомброидного типа движения
129
лучаем уравнение
( Ми? MLuj2\ (hx + ih2\ _ (Zx + iZ2\ ( .
\MLoj2 Iu2 J [h3 + ihj ~ \Z3 + iZ4) ' [ '
Решение уравнения (45) для [h1, h2, h3, h4] имеет вид
[(I/M)Z1 - LZ3, (I/M)Z2 - LZ^ Z3 - LZly Z4 - LZ2]
u2(I-ML2) ' [ }
Отметим, что можно записать матрицу в левой части (45) в точном виде
/ Ми2 + m(l)Uiu> MLu2 - MwUiu> + m(l)U2\ ( ,
\MLuj2 + MwUiuj IJ1 + MWU2 J ' [ }
i
где Mw = Jmdx — суммарная виртуальная масса (масса воды, которая
о
приводится в движение за счет поперечных движений рыбы как твердого
тела). Решение (46) приближенного уравнения (45) несильно отличается от
точного уравнения с матрицей (47).
Проведенные исследования силы реакции рыбы весьма важны для
скомброидного типа движения (когда амплитуда поперечных движений Н(х)
достигает больших значений только в хвостовой части тела рыбы),
поскольку поперечная сила (42) может быть достаточно большой. При /i,
определяемой равенством (40), скорость w, задаваемая уравнением (2), принимает
вид
w = UH\x)cos[u(t+{l-x)V-1)]-(V-U)V-1H{x)usm[u{t^
(48)
Из (48) видно, что небольшие значения w по сравнению с W (они связаны
соотношением (3)) сосредоточены на малом участке тела вблизи хвостовой
кромки, где амплитуда Н(х) волнообразного движения имеет практически
постоянное значение с пренебрежимо малой производной #'(х).
Достаточно большой величины скорость w достигает в предшествующей части тела
рыбы, где амплитуда Н(х) быстро нарастает.
Если виртуальная масса т(х) принимает в этой области большие
значения (как было показано в § 2, такие значения достигаются у хвостовой
части), то количество движения жидкости m(x)w(xJ t) имеет большие
амплитуды и, следовательно, соответствующая боковая сила (6) становится
130
Глава 4
очень большой. Действительно, ее суммарное значение
Z dx = — / mwdx + [Umw]x=l (49)
о о
состоит из двух слагаемых, одно представляет собой флуктуации
поперечного суммарного количества движения воды от передней части до хвоста,
а другое — количество движения, которое уходит в спутный след.
Последнее слагаемое играет большую роль при определении мощности (§ 2), а
первое слагаемое, если амплитуда колебаний mw велика, определяет сильно
пульсирующую поперечную силу.
Предположим, что рост Н(х) от нуля до его значения на хвостовой
кромке Н(1) сконцентрирован вблизи точки х = хг, тогда, заменяя в
подынтегральном выражении равенства (49) cos[uj(t + (I — x)V~x)] на cos[uj(t +
+ (I - xr)V~1)] и т(х) на m(xr), получим
f Zdx = -Um(xr)H(l)ujsm[u(t + (I - xT)V-1)]-
o
- Um(l)H(l)(V - и)У-гш8ти>г. (50)
Различие в фазах у этих двух слагаемых незначительно, первое
слагаемое вносит более существенный вклад в суммарный результат, поскольку
у него нет множителя (V — U)V~l. В случае угревидного движения
подобное увеличение суммарной поперечной силы отсутствует, т. к. производной
по переменной х уже пренебречь нельзя и наличие содержащего косинус
слагаемого в (48) приведет к значительному уменьшению флуктуационной
составляющей в интеграле (49).
Из проведенных рассуждений становится ясно, что только в том
случае, когда приспособление к скомброидному типу движения
сопровождается значительным уменьшением высоты (и, как следует из (4), с
соответствующим значительным уменьшением т(х)) в окрестности х = хТ,
где происходит быстрый рост амплитуды поперечных колебаний,
возможно значительное увеличение поперечной силы. Соответственно, как
следует из (46), амплитуды составляющих колебательных движений тела будут
также увеличиваться. Действительно, составляющие силы Zx и Z2 играют
более существенную роль в этом случае по сравнению с моментом
поперечной силы относительно прямой х = I (т. к. боковая сила имеет наибольшее
4. Механика скомброидного типа движения
131
значение вблизи х = I). Приблизительное значение поперечной отдачи
может быть получено умножением выражения (50) на величину
I/[M(I-ML2)u2]. (51)
Здесь I - ML2 — суммарный момент инерции рыбы и слоя воды
около нее относительно их центра масс, его можно записать в виде Mfc2,
где к — радиус инерции, величина которого, скорее всего, значительно
меньше, чем L. Амплитуда силы отдачи, соответствующая первому
слагаемому в выражении для суммарной поперечной силы (50), может быть
записана в виде
[(L2 + k2)/k2][m(xr)l/M](U/ul)H(l) (52)
и, как видно, является заметной (не пренебрежимой) функцией от Н(1).
Так как множитель U/ul имеет порядок ДО"1, то множитель (L2 + к2)/к2
во многих случаях принимает значения порядка 4 или 5, а множитель
т(хг)1/М приблизительно равен 1, если только величина т(хг)
каким-либо способом искусственно не уменьшена.
При движении возникает существенная по величине и угловая отдача
(определяется величинами h3 и /i4), в результате чего возникают
значительные движения в поперечном направлении вдоль всего тела рыбы. Это
сводит на нет все преимущество скомброидного типа движения, при
котором боковые движения тела совершаются хвостовой частью тела. Объем
воды, продвигаясь к хвостовой кромке, испытывает сопротивление,
обусловленное вихревым движением, не только в течение того промежутка
времени, когда вода движется вдоль достаточно короткой хвостовой части,
но и в течение значительно более длинного промежутка времени, в
результате чего жидкость испытывает дополнительные вихревые движения
со значительной энергией, импульс которых практически не коррелирован
со скоростью хвоста (как следует из (29) и (30), оба эффекта являются
нежелательными).
Поэтому неудивительно, что приспособление к скомброидному типу
движения происходит совместно (Lighthill, 1969) со значительным
уменьшением высоты тела и хвостового плавника в области резкого
возрастания амплитуды. Тогда вблизи х = хТ виртуальная масса т(х) будет
настолько малой, что поперечная составляющая количества движения будет
небольшой, а следовательно, небольшой будет и соответствующая
поперечная сила.
132
Глава 4
И все же некоторая боковая сила останется. Правда, уменьшенная за
счет множителя (V — U)V~l во втором слагаемом (50), но все еще
достаточная для создания наблюдаемой отдачи, поскольку величина т(1),
определяющая силу тяги, не должна быть слишком маленькой. Таким образом,
получаем, что величина множителя (51) должна быть маленькой, чтобы
оставшаяся боковая сила не привела к слишком большой отдаче. А это
означает, что передняя часть тела рыбы должна иметь достаточно большую
высоту.
Поясним сказанное. Предположим, что форма хвостового плавника
задана (главным образом форма хвостового плавника определяется
необходимой силой тяги) и его масса равна Мс. Рассмотрим различные
возможные формы передней части рыбы (пусть масса передней части равна Мл)9
центр масс лежит в х = I — 1Л, момент инерции относительно центра масс
равен МАк\. Тогда М = Мл + Мс и приближенно ML = МЛ1Л, I =
= МА{1\ + к2А). В этом случае множитель (51) можно записать в виде
ш-2[Мс + МАк2А/(12А + к2А)}-\ (53)
Для того чтобы этот множитель был небольшим по величине (поскольку к\
с необходимостью значительно меньше 12А), необходимо, чтобы часть тела
рыбы перед хвостовым плавником имела достаточно большой момент
инерции Мдк\ относительно центра масс (здесь учитывается как масса рыбы,
так и масса воды).
С точки зрения морфологии это означает следующее. Для того чтобы
переход к скомброидному типу передвижения был целесообразен,
необходимо, чтобы тело рыбы имело вытянутую переднюю часть, масса которой
распределена таким образом, чтобы масса воды, вовлеченной в поперечное
движение, была большой. В этом случае тело рыбы сжато с боков
преимущественно в его передней части. Кроме того, наиболее эффективной
с точки зрения вовлечения большой массы воды т(х) в боковое движение
является наибольшая суммарная высота s тела в области, занимаемой
спинным и брюшным плавниками (как следует из (4), эта масса
пропорциональна s2). Чем большая часть тела рыбы будет иметь указанную наибольшую
высоту, тем с большей эффективностью будет подавляться отдача.
Среди многих отрицательных эффектов, связанных с отдачей, особое
место занимает эффект, связанный с потерями, обусловленными действием
вихревой силы. Однако и любые другие эффекты могут уменьшить
эффективность передвижения животного в воде. При х = I из равенства (41)
4. Механика скомброидного типа движения
133
следует
W = — = — uj[H(1) + кг] smut + uh2 cosut. (54)
С/L
Из уравнений для коэффициентов отдачи следует, что коэффициент hx
отрицателен. Отрицательный коэффициент hx приводит к уменьшению
амплитуды скорости W хвостовой части рыбы (при заданном законе
изменения формы тела рыбы), в результате чего уменьшается и средняя сила тяги:
Р =
(55)
х=1
(Более точно, из (50) или любого другого приближения равенства (49)
следует, что не только коэффициент Z2 отрицателен и велик по модулю, но
также и коэффициент Zx при cosu;i отрицателен; моменты (43) меньше по
величине, а коэффициент Z3 оказывается положительным, вследствие чего
из (46) получаем, что hx (равно как и h2) меньше нуля. Если для
нахождения коэффициентов hl9 h2, h3, h4 использовать более точную матрицу (47),
то легко получить, что фаза /ix + ih2 немного уменьшится, так что
коэффициент hx также будет отрицательным.)
Далее, совпадение фаз скоростей w и W, которое отражено в
равенстве (3), в общем случае нарушается на задней кромке под влиянием
отдачи, что приводит к уменьшению средней силы тяги (55) и
эффективности (12). При х = / из (2) и (41) получаем
w = -[u{V-U)V-1H{l)+uh1 + Uh^]sinut+[UH\l)+uh2
(56)
При отсутствии отдачи величина #'(/) в (56) пренебрежимо мала (т.е. как
раз перед задней кромкой амплитуда колебаний прекращает расти), так что
разность фаз скоростей w и W будет невелика. Однако если величиной
#'(/) пренебречь при наличии отдачи (из-за отдачи как ujh2, так и — Uh3
отрицательны), то коэффициент при cos cut в выражении для скорости w
будет больше, чем для скорости W, в результате чего эффективность (12)
уменьшится. Если принять, что производная Н\1) отлична от нуля и
положительна и отдача имеет место, то коэффициент при cosu;i будет
отрицательным и небольшим по модулю (что приведет к уменьшению разности
фаз между w и W). Такое предположение могло бы привести к слишком
точному согласованию фаз, что делает это предположение практически
бесполезным при решении задачи о повышении эффективности движения.
134
Глава 4
Итак, подводя итоги, можно сказать, что при переходе к
скомброидному типу плавания рыбы приобретали морфологические признаки,
способствовавшие уменьшению эффектов отдачи, в частности, произошло
значительное уменьшение высоты тела вблизи хвостового плавника, где
амплитуда волнообразного движения быстро нарастает, с последующим
увеличением высоты хвостового плавника.
5. Двумерная теория динамики хвоста в виде полумесяца
Рассмотрим с позиции гидромеханики три различные стадии
перехода к скомброидному типу плавания у костных рыб, в результате которых
произошло повышение скорости и эффективности плавания. Во-первых,
«деградация» средней части хвостового плавника (как у сельди (Оиреа)),
в результате чего хвостовой плавник становится похожим на пару крыльев
очень большой стреловидности. Как было показано в § 3, такое
преобразование хвоста (при его движении как единого целого) не приводит к потере
в силе тяги, поскольку интенсивность вихревой пелены в стреловидной
выемке хвоста будет равна интенсивности у твердых частей хвостового
плавника. С другой стороны, уменьшение площади плавника может привести
к небольшому уменьшению сопротивления воды и соответственно к
выигрышу в скорости.
Во-вторых, уменьшение прямой стреловидности крыловидных частей
хвостового плавника (как у ставрид (Сагапх)). Это приводит к увеличению
размаха хвостового плавника s (и, как следует из (4) и (55), к
увеличению силы тяги) без увеличения площади его поверхности и, как результат,
к еще большему увеличению скорости. Если к тому же рыба может (как
это имеет место у ставрид) за счет мускульного усилия изменять степень
стреловидности хвостового плавника, то ее маневренность повышается.
При уменьшении стреловидности, начиная с некоторого ее значения,
для исследования динамики движения рыб использование методов теории
тонкого тела §§2-4 становится неправомерным (как это показано в § 1).
Действительно на третьей стадии, когда удлинение s2/5, где S — площадь
поверхности, достигает больших значений (что имеет место для хвоста
в виде полумесяца), речь о применении теории тонкого тела не идет.
Возможно, что для костных рыб достижение такого значения удлинения
является кульминационным моментом в процессе повышения скорости и
эффективности плавания. Высказанное предположение подтверждается и тем
5. Двумерная теория динамики хвоста в виде полумесяца 135
фактом, что подобные хвосты в виде полумесяца приобрели и самые
быстрые акулы, и морские млекопитающие (равно как и ихтиозавры) в процессе
совершенно различного эволюционного развития.
Не останавливаясь более подробно на обсуждении конвергентной
эволюции, в оставшейся части параграфа рассмотрим в первом приближении
скомброидный тип плавания животных, имеющих хвост в форме
полумесяца. Некоторые основания подобного приближенного подхода были
приведены в § 1. Будем рассматривать задачу в двумерной постановке. То есть
полагаем, что под действием хвостового плавника (здесь рассмотрим
только плавание рыб) горизонтальные слои воды движутся независимо один от
другого. Конечно, при таком подходе можно получить завышенное
значение эффективности, поскольку учитывается только поперечная компонента
завихренности. Здесь выкладки будут записаны в такой форме, которая
может быть использована и при дальнейшем уточнении постановки задачи —
при переходе к использованию методов теории, где учитывается подъемная
сила и, как следствие, учитывается компонента завихренности,
направленная вдоль потока жидкости.
В двумерной постановке полагаем, что сечение хвостового плавника
горизонтальной плоскостью у = const занимает интервал от х = —а до
х = +а (так что точке х = 0 соответствует середина хорды профиля),
а скорость невозмущенного потока жидкости равна U. Тогда зависимость
бокового смещения сечения от переменных х и t имеет вид
z = [h-ia(x-b)]eioj\ (57)
где действительные h и а — амплитуды бокового скольжения и рыскания
соответственно, г. х = b, z = 0 — положение оси рыскания. Разность фаз
между боковым скольжением и рысканием полагается равной 90°. Однако
если в действительности разность фаз будет отличной от 90°, то для
учета этого факта амплитуду h в (57) представим в виде комплексного числа,
что соответствует простому изменению положения оси рыскания
(изменению 6).
Граничные условия для потенциала скорости фегш1 возмущенного
потока жидкости в силу линейной постановки задачи ставятся на плоскости
2 = 0и имеют вид при —а<х<а
(Ё£\ = i(uh - Uot) + uja{x - 6), (58)
136
Глава 4
т. е. поперечная скорость жидкости равна скорости dz/dt+U dz/dx
бокового смещения (57) сечения относительно соответствующего
горизонтального слоя воды. Из соображений симметрии следует, что потенциал ф должен
быть нечетной функцией, и тогда на плоскости z = 0 потенциал ф
обращается в нуль при х < — а (вверх по течению от передней кромки). Однако
при х > а, т. е. вниз по течению от задней кромки сечения, это рассуждение
о равенстве нулю потенциала на плоскости z = 0 не проходит, поскольку
там, как это было показано в § 3 (но только при двумерной постановке,
когда учитывается лишь поперечная компонента завихренности), образуется
вихревая пелена. Напряженность вихревой пелены постоянна относительно
жидкости, так что на плоскости z = 0 при х > а величина
Ф = шф + и^- (59)
ох
непрерывна как нечетная функция относительно z, а следовательно,
обращается в нуль в этой области при z = 0.
Следуя Поссио (Possio, 1938) и By (Wu, 1961), в дальнейшем в качестве
новой искомой функции будем использовать Ф, поскольку она непрерывна
и обращается в нуль на плоскости z = 0 как при х < —а, так и при х > а.
Кроме того, она имеет и физическую интерпретацию, поскольку в линейной
постановке через нее можно выразить разность между текущим
давлением р и гидростатическим давлением р0 в виде
р-Ро = ~Р*еш. (60)
А это означает, что и давление является непрерывной функцией в
направлении, перпендикулярном вихревой пелене, что в дальнейшем и будет
использовано. Из (58) и (59) следует, что при —а<х<а имеем
( —) =В + Сх, где В = 2Uu>a-u2(h + iab), С = ш2а,(61)
уравнение, связывающее поперечный градиент давления с поперечным
ускорением элемента жидкости, обусловленным боковым смещением (57).
Решение уравнения Лапласа У2Ф = 0, имеющее вид (61), легко может
быть получено, однако необходимо рассмотреть и сингулярные решения,
удовлетворяющие поставленным граничным условиям. Как и во всех
задачах, связанных с установившимися колебательными процессами, удобно
представить частоту колебаний и в виде комплексного числа с небольшой
5. Двумерная теория динамики хвоста в виде полумесяца 137
по модулю мнимой частью (которую в конечном итоге возможно устремить
к нулю), что отражает медленное нарастание колебаний до конечного
установившегося значения. Тогда решение уравнения (59) относительно
потенциала ф имеет вид
— ос
здесь ф обращается в нуль на плоскости z = О при х < —а, если обращается
в нуль Ф.
Хорошо известны сингулярные решения для ф9 которые описывают
поток вблизи передней и задней кромок сечения. На на заостренном заднем
крае сечения х = а, где выполняются условия Кутта-Жуковского об
отсутствии потока вокруг кромки, потенциал ф имеет особенность степени 3/2
(т.е. (ф)г=0 имеет порядок (а — х)3/2). Из этого следует, что Ф и
давление (60) имеют особенность степени 1/2. Поток на передней кромке х =
= — а имеет хорошие гидродинамические характеристики, только если эта
кромка закруглена, такому обтеканию соответствуют особенность порядка
1/2 для ф (так что (ф)г=0 имеет порядок (х + а)1//2) и особенность порядка
обратного корня, квадратного для Ф. (Более точный анализ показывает, что
особенность располагается на небольшом расстоянии за передней кромкой,
в точке, равной половине радиуса кривизны сечения (Lighthill, 1951).
Однако, как следует из расчетов, если особенность расположить в х = —а, то
при этом получается достаточно хорошее приближение для расчета полной
силы.)
Из приведенных соображений следует, что решение уравнения У2Ф = 0,
удовлетворяющее условиям (61), следует искать в виде
Ф = АФ1 + ВФ2 + СФ3, (63)
где функции Фх, Ф2 и Ф3 определены в таблице 1. Здесь коэффициент А
произволен, слагаемое, содержащее Фх, включено для того, чтобы в
решении присутствовали все особенности, о которых говорилось выше.
Для определения А необходимо потребовать, чтобы величина ф,
определенная равенствами (62) и (63), удовлетворяла условиям (58),
выполнение которых, как было отмечено раньше, обеспечивает выполнение других
условий, накладываемых на ф. На основании данных таблицы 1 при |ж| < а
[Таблица 1. Приведены свойства трех решений ФьФ2иФ3 уравнения У2Ф = 0. Каждое из этих решений стремится
к нулю при х2 + z2 —► ос, и каждое из них представлено как комплекснозначная функция Q + гФ от переменной
г = х + iz9 регулярная вне разреза от С = —о, до С = о,. Здесь под корнем квадратным из (£ - а)/(С + а) и С2 ~
к- а2 понимаются те ветви, которые ведут себя подобно 1 и С соответственно при больших значениях |£|. Под корнем
радратным из действительного переменного х понимается арифметический корень. При переходе через разрез z = 0,
х\ < a Q непрерывна, а Ф терпит разрыв. Во всех точках выполняется
ЗФ _ сЮ
dz дх'
ф
ft + гФ
(п)„±0 {]*' iа-
\\х\ > а
( \х\ < а,
^ \х\ > а
Z = 2 /(Ф)г=_0<Ь
— а
Q = 2 /(-гг)(Ф)2=_о^х
— а
*i
1-[«-а)/(С + а)]1/2
1
\-[(х-а)/{х + а)}1'2
Т[(а-х)/(х + а)}1/2
0
27га
7га2
Ф2
С "(С2-а2)1/2
/ 2 2\1/2
ж — {х — а ) ' sgnx
=F(a2 - х2)1/2
0
7га2
0
Фз
|С2 - \«? - «2)1/2
1 2
1х* - 1\х\(х> - а*У/2
^(а2-*2)1/2
0
0
1 4
— —тга
8
5. Двумерная теория динамики хвоста в виде полумесяца 139
получаем
— а
jj-1 / €ш(х1-х)/и х
— ос
х < Аа(х1 +а)-1 + Вхх + С [х\ - -а2) \ (х\ - а2)"1/2 (1хг +
X
+ U~l I eM*i-*)/^(£ + Cx1)dx1 = i(uh - Ua) + иа(х - b). (64)
Здесь использовано равенство производных дФ/dz и дП/дх.
Несобственные интегралы рассматриваются как обобщенные функции (Lighthill, 1958)
или иначе сходящиеся в смысле Абеля, поскольку хх —> — оо (это
эквивалентно тому, что и должна иметь отрицательную мнимую часть, что
обеспечивает стремление подынтегрального выражения к нулю), и в
смысле Адамара, поскольку хг —► —а. Это обусловлено тем, что производная
дФг/дг равна производной от ft по переменной х, а в предел этой
производной при г —► 0 входит член — [(х — а)/(х + а)]1//2, имеющий особенности
при х < —а, но не имеющий особенности при — а < х < а. И, таким
образом, соответствующее интегральное слагаемое в (64) может быть корректно
вычислено по частям, т. к. проинтегрированное слагаемое при подстановке
пределов обращается в нуль.
Легко убедиться в том, что в (64) вторая строка, как и можно
было ожидать, представляет собой тождество, поскольку левая часть равна
{дф/дг)г={), что вытекает из (61), если выражение для (дФ/дг)2,=0
выполняется для всех х. Тогда выражение в первой строчке (64) обращается
в нуль. Разделив его на множитель и~1е~гшх^и, получим уравнение для
определения коэффициента А.
Если интегралы выразить через бесселевы функции от переменной
г = гау где а = — (65)
(предполагается, что переменная г имеет небольшую по величине
положительную действительную часть), то уравнение для А принимает вид
Аг[К0{г) + Кг{г)] - ВаКг(г) + ^Са2[К0{г) + 2Г-1К1{г)] = 0. (66)
140
Глава 4
Проведем вычисление интеграла-множителя при А (несобственный
интеграл с двойной особенностью). При замене хх = — achu этот интеграл
принимает вид
ос
1 -100
/.—(сЬ«-1Г'*.=
о
e-rchucth-u +
2 Jo
оо
+ r f e-rchu(l + chu)du. (67)
о
Проинтегрированное слагаемое в правой части (67), как уже отмечалось
выше, обращается в нуль, а интегральное слагаемое равно г[Я0(г)+.К'1(г)].
С использованием функции Теодорсена (Garrick, 1957)
F(a) + iG{a) = „ *l(><2 /• ч. (68)
v ' ; K0(^) + #iM v '
значения которой представлены на рисунке 3, из уравнения (66) с учетом
(62) и (65) получаем
A=-U
llujalb- -a) + i(Ua - ujK) (F + iG) + -шаа \
(69)
Запишем поперечную силу (в направлении оси z) на единицу размаха
хвостового плавника в виде pZe%UJt, момент рыскания на единицу
размаха относительно оси, проходящей через положение полухорды х = z = 0
в положительном направлении (т. е. этот момент стремится развернуть
переднюю кромку в направлении оси г), запишем в виде QeluJi. Как следует
из (60), (63) и таблицы 1, поперечная сила и момент рыскания равны
соответственно
а
Z = 2/(*)„_0(fa = 2^WB, (70)
—а
а
Q = 2 [(-х)(Ф)2=_0(1х = жа2А - ^жа4С. (71)
— а
Отсюда можно найти мощность рЕ, приходящуюся на единицу длины
размаха. Для этого будем опираться на два факта: (i) мощность с учетом
скольжения и рыскания равна поперечной силе, умноженной на скорость
скольжения центра масс, плюс момент рыскания относительно оси центра масс,
5. Двумерная теория динамики хвоста в виде полумесяца 141
умноженный на скорость рыскания; (ii) пусть две величины, равны
действительной части комплексных выражений аеги;19 ЬешЬ9 тогда их
произведение равно ^Re(ab), где Re — действительная часть, а Ь — комплексно
сопряженное к Ь.
г ОЛ
0,2
0,4
0,6
0,8
1,0
0,02>
L 0,2
.0,8
0,4
У 0,04,
0,06>
2'2 0Д4 0Д^2^
Рис. 3. Представление в виде диаграммы Аргана (заимствовано из работы Garrick,
1957) функции Теодорсена F + iG, зависящей от частотного параметра а — uja/U
(здесь а = —с — полухорда длины профиля)
Тогда для мощности получаем
Е = - Re[Zu;(-ab - ih) + Q(-ua)},
(72)
поскольку из (57) следует, что скорость бокового скольжения центра масс
равна uj(-ab + ih)elujt, а скорость рыскания равна -иаегшЬ. Из (71), (72)
с учетом (61) получаем
7? = -ттаиа I b + -а ) Re A + nauh Im A - тга2ии2а2Ь, (73)
где Im означает мнимую часть.
Среднее значение силы тяги можно записать в виде рР удельной силы
тяги, где
(•
Р = TraU~2\A\2 + ( TraalmA - -тга2ш2а2Ь
(74)
Слагаемое в скобках представляет собой среднюю величину ^Re[Z(—га)]
поперечной силы pZetuJt, заданной соотношением (70), действующую на
поверхность (57), наклоненную к оси х под углом iaeluJt. Однако
наибольший вклад в силу тяги вносит первое слагаемое, которое представляет
собой подсасывающую силу, обусловленную быстрым обтеканием потоком
142
Глава 4
передней кромки профиля. Подсасывающая сила в фиксированный момент
времени принимает значение, равное значению силы при стационарном
обтекании, полученному на основании теоремы Блазиуса (см., например,
работу Robinson, Laurmann, 1956, с. 126). Это связано только с квадратичными
членами —~р[(дф/дх)2 + (дф/dz)2], входящими в выражение для давления
(потому что линейный член (60), будучи нечетной функцией z9 имеет
нулевую результирующую). И среднее значение подсасывающей силы равно
^7г|/Г|2, где К(х + а)-1/2 представляет собой приближенное выражение,
равное значению производной (дф/дх)г=+0 при х [ —а, которая в
соответствии с (59) равна [/_1Ф (поскольку ф обращается в нуль при х = —а,
z = 0). В соответствии с (63) и таблицей 1 К равно -U~1A(2a)1/2,
подставляя это значение К в выражение для среднего значения
подсасывающей силы, получаем первое слагаемое в уравнении (74).
В результате алгебраических преобразований из (73), (74) с
учетом (69) получаем, что мощность потерь р(Е — UP) равна
~E-UT={U7ra)
Здесь каждое слагаемое положительно, средний сомножитель, как функция
от Ь, принимает наименьшее значение, когда ось рыскания х = Ь
расположена в точке Ь = \а, т. е. на расстоянии три четверти хорды от передней
кромки.
Формулу (75) для потерь можно было бы вывести и другим, более
трудоемким способом. Для этого необходимо было показать, что при х > а
(в области вихревой пелены) из равенств (62) и (63) при использовании
тождеств для функций Бесселя следует, что (0)2=±о — ТФт
w-iux/U^ где
= тгУ [uh -Ua + iua (b - \a)}
9w u[K0(ia) + K^ia)} ' [ }
Средняя энергия, приходящаяся на единицу длины, в вихревой пелене на
достаточно большом расстоянии от тела |х| < а равна \p{u/U)^w\2, а
скорость роста этой энергии равна \ри\ф^2. С использованием тождества
\K0(ia) + Ki(ia)|-2 = (2u>a/icU)(F - F2 - G2) (77)
мощность потерь \ри\Фы\2 можно записать в виде (75). Данный более
сложный способ вывода формулы для потерь энергии приведен здесь
с единственной целью — для контроля полученного ранее соотношения.
, 2 2
\U) OL
ь-\а.
+ (uh - Ua):
{F-F2-G2). (75)
5. Двумерная теория динамики хвоста в виде полумесяца 143
Получим выражение для эффективности 77, заданной
соотношением (1). В данном параграфе входящие в выражение для эффективности
величины Е и Р отнесены к единице размаха и разделены на плотность.
Из (69), (73), (75) получаем
1-4 = (Е- TfP)fE =
\u2a2{b-\a2f + (uh-Ua)2}(F-F2-G2)
~ {ша (Ь —\а) [а (Ъ +±а) F—hG- ±аа] + (u>h - Ua) [hF + a (b+ Ja) G]} ш'
(78)
Числитель в (78) пропорционален мощности потерь и как функция от
переменной Ъ имеет строгий минимум при Ъ = ^а. Причем чем больше
частотный параметр а = urn/U и параметр флюгерирования Ua/uh, тем
меньше по величине минимум (параметр Ua/uh в работе (Lighthill, 1969)
обозначен через 0, его значение при положительной силе тяги не превышает
единицу).
Наибольшее значение эффективности г/, как следует из знаменателя,
достигается при 6, немного превосходящем ^а. В окрестности Ь = ^а
стоящие в квадратных скобках множители при выражениях ujol {Ь — ^а)-и (uh —
— Uа) будут положительными, в силу чего суммарная мощность растет
с ростом разности (р — ~а). Следует отметить, что в большинстве случаев,
представляющих для нас интерес, первый множитель существенно меньше
второго, и значит, когда числитель достигает строгого минимума,
наибольшее значение эффективности не может достигаться в точках, слишком
далеко отстоящих от Ь = ~а. С другой стороны, из этого следует, что сила
тяги немного возрастает при значениях Ь, больших чем \а. И это
необходимо принимать во внимание при исследовании зависимости эффективности
от переменной Ь.
Указанные замечания поясняются рисунком 4. На нем приведены
рассчитанные в рамках двумерной теории зависимости силы тяги и
эффективности от переменных в и а для четырех положений оси рыскания (на
расстоянии полухорды, трех четвертей хорды, хорды (ось рыскания совпадает
с задней кромкой) и пяти четвертей хорды от передней кромки).
Наибольшее значение интенсивности достигается при втором и третьем
положениях, для большого значения частотного параметра uc/U = 2a,
рассчитанного по хорде с = 2а.
Безразмерный коэффициент силы тяги Ст равен силе тяги,
приходящейся на единицу площади плавника, разделенной на ^p(uh)29 т. е.
CT=T/uj2h2a. (79)
144
Глава 4
Рис. 4. Зависимость коэффициента силы тяги Ст и эффективности г) (в рамках
двумерной теории крыла) от переменной wc/U (равной 2а) для различных значений
параметра флюгерирования в = Ua/ush: 0, 0,2, 0,4, 0,6, 0,8 и различных положений
оси рыскания х — Ь (а именно, для полухорды 6 = 0, три четверти хорды Ь = ^а,
для хорды b = а и пяти четвертей хорды b = |а). Пунктирной линией изображена
зависимость доли коэффициента силы тяги, обусловленной подсасывающей силой,
при параметре в = 0,6, а штрихпунктирной линией — при параметре в = 0,8
Это соотношение связывает силу тяги с амплитудой uh бокового
движения хвостового плавника. Как следует из расчетов, коэффициент силы тяги
уменьшается с ростом частотного параметра uC/U, а это означает, что при
заданной частоте колебаний хвостового плавника сила тяги имеет меньшие
значения при меньших скоростях перемещения U животного.
Для больших значений параметра в (в нашем случае это 0,6, 0,8), при
которых эффективность принимает наибольшие значения, сила тяги тем
больше, чем дальше вниз по течению расположена ось рыскания, и
предпочтительными будут положения оси рыскания, расположенные дальше от
передней кромки (так, положение оси рыскания b = а предпочтительнее,
чем b = \a). В этом случае сила тяги будет иметь достаточно большую
величину и при небольших скоростях перемещения животного. Однако к
этому утверждению следует относиться осторожно. На рисунке 4 пунктирной
линией изображена зависимость части силы тяги, обусловленной эффек-
5. Двумерная теория динамики хвоста в виде полумесяца 145
том подсасывания у передней кромки, от переменной ujc/U при 9 = 0,6,
остальная часть силы тяги обусловлена действием направленной назад
компонентой поперечной силы, а штрихпунктирной линией изображена та же
зависимость при в = 0,8.4 (Отметим, что при в = 0 в отличие от других
значений в хвостовой плавник не разворачивается относительно оси х =
= Ь (т. к. а = 0) и вся сила тяги определяется эффектом подсасывания на
передней кромке.)
Очевидно, что рост силы тяги с ростом b при больших значениях
переменных в и ujc/U полностью определяется резким увеличением
подсасывающей силы на передней кромке, а влияние направленной назад компоненты
поперечной силы практически исчезает и даже может стать отрицательным.
Возможность отрицательного влияния поперечной силы на величину силы
тяги можно заметить из ее представления и в виде (74). Второе слагаемое
в скобках в (74), входящее в выражение для поперечной силы (70), (61),
растет с ростом Ь за счет наличия мнимой части выражения В, равной
—iuj2ab, что может привести к отрицательному значению силы тяги. Это
можно объяснить влиянием виртуальной массы на величину поперечной
силы. Поперечные колебательные движения хвостового плавника из
стороны в сторону приводят к изменению количества движения виртуальной
массы, так что инерционность жидкости с позиций теории тонкого тела
здесь играет важную роль. Чем дальше расположена ось рыскания за
центром масс профиля, тем менее вклад этого эффекта в боковую силу связан
со скошенностью плавника.
Тенденция уменьшения силы тяги за счет направленной назад
компоненты боковой силы при возрастании Ь не скомпенсируется ее увеличением
за счет роста подсасывающей силы, вклад которой в силу тяги определяется
величиной \А\2. Этот рост действительной части коэффициента А (см. (69))
с ростом 6, связанный с колебаниями боковой силы (70), не совпадает по
фазе с колебаниями угла атаки, обеспечивающего увеличение
подсасывающей силы.
Из проведенных рассуждений следует, что в рамках двумерной теории
крыла решение задачи об увеличении коэффициента силы тяги Ст за счет
расположения оси рыскания за задней кромкой профиля (например, в точке
Ь = |а на рис. 4), скорее всего, не должно проводиться без
одновременного решения задачи об увеличении эффективности. Без их одновременного
рассмотрения невозможно обойтись, когда решается проблема реализации
4Рисунок 4 заимствован из работы (Lighthill, 1969) (рис. 9), единственным отличием от
рисунка 9 являются результаты расчетов при 6 = 0.
146
Глава 4
большой подсасывающей силы на закругленном переднем крае профиля.
Если считать, что только эта сила обусловливает силу тяги, то в результате
значение тяги будет заниженным. Возможно, для получения наибольшего
значения силы тяги, а также и эффективности наиболее оптимальным
будет расположение оси рыскания где-то между точками Ь = \а и Ь = а, т. е.
практически весьма близко к задней кромке профиля.
Возможно, именно этим объясняется тот факт, что хвостовой плавник
в форме полумесяца наиболее эффективен с точки зрения гидродинамики.
Если бы хвостовой плавник имел как целое единственную ось рыскания,
причем колебания угла рыскания совпадали бы по фазе с поперечными
колебаниями скорости хвостового плавника, то в рамках двумерной теории,
примененной к каждому сечению, получили бы, что наилучшая
эффективность и наибольшая сила тяги могут быть достигнуты, когда ось
рыскания расположена очень близко к задней кромке каждого сечения. А отсюда
следует, что задняя кромка всего плавника должна практически совпадать
с осью рыскания.
Начиная от выгнутого вперед переднего края, плавник должен иметь
клиновидную форму. Степень выгнутости задней кромки должна быть мала
по сравнению с изогнутостью переднего края плавника. В процессе
рассуждений было установлено, что задняя кромка плавника должна быть
практически вертикальной, а колебания угла рыскания и боковой скорости
должны совпадать по фазе, что, несомненно, является идеализацией, о чем уже
упоминалось в § 2.
Такая степень отклонения от «прямого крыла» является действительно
оптимальной в рассматриваемой двумерной постановке (см. рис. 6 в
работе Lighthill, 1969), хотя, конечно, ее нельзя рассматривать как
окончательную. Возможно, в более полной трехмерной постановке можно было
бы объяснить тот факт, что в большинстве случаев серповидные хвосты
также имеют вогнутую вперед заднюю кромку, хотя и не в столь же
сильной степени, как передняя. В любом случае результаты решения задачи
об эффективности хвостового плавника в форме полумесяца в двумерной
постановке можно рассматривать в качестве исходных для уточнения их
при трехмерном подходе, когда возможно использовать понятие подъемной
силы.
Не будем проводить здесь дальнейшее обсуждение трехмерной
постановки. Завершим главу несколькими замечаниями касательно
правомерности линеаризации граничных условий в рамках двумерной модели,
которая была проведена в данном параграфе, а также и некоторых предыду-
5. Двумерная теория динамики хвоста в виде полумесяца 147
щих при использовании методов теории вытянутого тела. Как упоминалось
в § 1, применение методов теории вытянутого тела к движениям с большой
амплитудой кажется вполне возможным, однако даже в двумерном случае
полное использование методов этой теории еще далеко от завершения.
Нельзя сказать, что линейная теория бесполезна для описания
широкоамплитудных движений серповидного хвоста (Fierstine, Walters, 1968).
Достаточно часто аэродинамические характеристики потока при достаточно
больших амплитудах (но не таких, при которых возможно опрокидывание,
и которое, конечно, рыбы стараются предотвратить) можно получить
экстраполяцией характеристик при малых амплитудах. Тем не менее хотелось
бы понять тенденцию и определить степень отклонения истинных силы
тяги и эффективности движения от предсказанных линейной теорией.
Можно было бы попытаться исследовать поток в случае двумерного
крыла, которое легко может разворачиваться по течению (хорошо флюге-
рированное крыло), при его больших смещениях в комбинации с такими
изменениями угла атаки, что изменения скорости потока можно считать
достаточно малыми. В этом случае еще можно считать, что давление
'-«■-'(S^S) (80>
линейно зависит от потенциала скорости (как в (60) и (59), но теперь
потенциал ф не будет представлен в виде фегш1\ р удовлетворяет уравнению
Лапласа и непрерывно при переходе через вихревую пелену. Производная
от давления в направлении нормали во всех точках крыла в каждый
момент времени определяется заданной на границе нормальной скоростью.
Для плоского крыла давление р затем может быть получено, если его
представить в виде, подобном (63) и включающем произвольную константу А,
которая в данном случае является произвольной функцией от времени. Эта
функция может быть определена исходя из того, что решение ф
уравнения (80), полученное при интегрировании вдоль линий, где у, z и х — Ut
сохраняют постоянное значение, удовлетворяет граничным условиям.
Этот последний шаг решения, который приведет к соотношению,
являющемуся обобщением (66), весьма трудоемок вследствие крайне
громоздкого процесса интегрирования. В целом данный метод все же проще по
сравнению с процессом нахождения поля скоростей, обусловленного
синусоидальной вихревой пеленой с амплитудой, близкой к половине длины
волны (Fierstine, Walters, 1968). Однако последний метод все же
отбрасывать нельзя, поскольку в будущем он может быть применен к решению
148
Глава 4
нелинейной задачи в трехмерной постановке при использовании
положения, высказанного Лайтхиллом (LighthiU, 1969), о том, что серповидный
хвост может создавать вихревую пелену, состоящую из вихревых колец.
Но для решения такой задачи требуется гений, подобный Гольдштейну
(Goldstein, 1929), который исследовал развитие спиралевидной вихревой
пелены, сходящей с лопастей воздушного винта.
Автор благодарен семилетнему плодотворному периоду своей научной
деятельности, посвященному разработке методов, в основе которых лежат
идеи гениального Сидни Гольдштейна.
[1
[2
[3
[4
[5
[6
[7
[8
[9
[10
[Н
[12
[13
[14
Литература
Fierstine, H. L. & Walters, V. 1968. Mem. S. Calif. Acad. Sci. 6, 1.
Flachsbart, О. 1935. Z. angew. Math. Mech. 15, 32.
Garrick, I. E. 1957. In Aerodynamic Components of Aircraft at High Speeds
(A. F. Donovan & H. R. Lawrence eds.), pp. 658-793. Princeton University
Press.
Goldstein, S. 1929. Proc. Roy. Soc. A, 123, 440.
Gray, J. & Hancock, G.J. 1955. J. Exp. Biol. 41, 135.
Kramer, E. 1960. Z Wiss. Zool. 163, 1.
LighthiU, M.J. 1951. Aeron. Quart. 3, 193.
LighthiU, M.J. 1958. Fourier Analysis and Generalized Functions.
Cambridge University Press.
LighthiU, M.J. 1960. J. Fluid Mech. 9, 305.
LighthiU, M.J. 1969. Ann. Rev. Fluid Mech. 1, 413.
Possio, С 1938. Aerotecnica, 18, 441.
Robinson, A. & Laurmann, J. A. 1956. Wing Theory. Cambridge University
Press.
Taylor, G. I. 1952. Proc. Roy. Soc. A, 1U, 158.
Wu, T. Y. 1961. J. Fluid Mech. 10, 321.
Глава 5
Приложение теории тонкого тела
к продвижению рыб при волнообразном
поперечном движении тела с большой
амплитудой1
1. Введение
Большинство животных, обитающих в водной среде и имеющих
вытянутое тело, усвоили способ перемещения за счет волнообразных движений.
В данной главе задача о перемещении таких водных животных исследуется
методами двух наиболее часто применяемых теорий, определяется
величина потерь энергии, а также находятся силы, действующие между телом
животного и водной средой. Более ранняя теория — теория резистивности,
в которой сила взаимодействия между сечением тела животного и водой
рассматривается как сила сопротивления, зависящая исключительно, хотя
и не обязательно линейно, от мгновенной скорости перемещения этого
элемента тела относительно воды. Главное достоинство работ, в которых
авторы в своих исследованиях опирались на резистивную теорию (Taylor, 1952,
Hancock, 1953, Gray, Hancock, 1955), состоит в том, что методы этой теории
были разработаны для решения задач о волнообразном способе
перемещения с произвольно большими амплитудами.
Более позднюю теорию (Lighthill, 1960) можно назвать реактивной
теорией, поскольку в качестве основных сил взаимодействия между телом
рыбы и окружающей ее средой при движении рассматриваются реактивные
силы. Эти силы, обусловленные инертностью воды и пропорциональные
производной по времени от относительной скорости животного, в рези-
стивной теории не учитываются. Они могут играть достаточно важную
]В оригинале статья перепечатана из Proc. Roy. Soc. В., 179 (1971), pp. 125-138.
150
Глава 5
роль при таком строении тела животного, когда его поперечное сечение
имеет значительно меньший размер в направлении поперечных смещений,
осуществляемых животным для продвижения в водной среде, чем в
вертикальном направлении. И в этом случае «виртуальная масса» воды, которая
приобретает импульс за счет таких смещений, намного превосходит массу
тела животного.
В обзоре, посвященном механизму движения водных животных (Light-
hill, 1969), отмечалось, что механизм движения беспозвоночных с помощью
волнообразных движений тела хорошо изучен методами резистивной
теории. Это относится к большинству случаев движения при низких числах
Рейнольдса (инерционность водной массы не оказывает влияния на
движение), однако, вероятно, такой подход можно применить и при движении
при числах Рейнольдса порядка 103 тех червеобразных, форма поперечного
сечения тела которых такова, что эффектами наличия виртуальной массы
можно пренебречь.
В процессе эволюции некоторые позвоночные водные животные
приобретают боковое сжатие задней части тела. Преимущество такого
строения заключается в том, что при волнообразном извивающемся движении
возникают значительные реактивные силы, которые и составляют
существенную часть силы тяги. При этом величина потерь энергии на создание
вихревой пелены значительно меньше по сравнению с тем случаем, когда
основной движущей силой являются силы сопротивления. Способ
движения рыб за счет волнообразных движений всего тела назван угревидным.
Именно таким способом перемещаются угри Anguilla. При их способе
движения за счет волнообразных извивающихся движений всего тела потери
энергии наиболее низки, что для них существенно, поскольку в силу своего
образа жизни им приходится преодолевать огромные расстояния.
При угревидном движении рыб, имеющих вытянутое тело, сила тяги
определяется комбинацией сил сопротивления и реактивных сил (Lighthill,
1970). При другом наиболее распространенном способе продвижения
водных животных, также обеспечивающем достаточно большие скорости,
видимо, более важную роль играют реактивные силы. При переходе к
такому способу продвижения, названному скомброидным, передняя часть тела
рыбы потеряла свою гибкость, а извивающиеся движения стали
осуществляться задней половиной, или даже третьей частью, тела.
Характерная особенность угревидного передвижения, состоящая в том,
что фаза колебаний поперечного сечения в боковом направлении
запаздывает по отношению к фазе колебаний любого находящегося перед ним се-
1. Введение
151
чения, сохраняется и при скомброидном передвижении, которое является
результатом развития угревидного способа движения. С другой стороны,
скомброидный способ существенно отличается от угревидного: амплитуда
колебаний (при угревидном способе она постепенно нарастает от головы
к хвосту) резко возрастает в направлении к хвостовой части где-то в
последней трети тела рыбы, так что наблюдатель в фиксированный момент
времени не сможет увидеть что-нибудь подобное полному колебанию всего
тела. Он просто видит боковые колебательные движения хвостовой части,
так что колебание последующего сечения отстает по фазе от колебательных
движений предыдущих сечений.
При скомброидном способе перемещения преобладают реактивные
силы, поскольку ускорение водных объемов, продвигающихся относительно
тела рыбы, нарастает очень быстро — в течение отрезка времени,
необходимого для прохождения вдоль тела и сбрасывания объема воды,
расположенного у хвостовой части. То есть силы определяются мгновенным
действием виртуальной массы. И за этот небольшой отрезок времени в
результате схождения вихрей в поперечном потоке возникают силы сопротивления
(как правило, прилегающие к телу рыбы объемы воды приводятся в
движение только частью боковой поверхности сечения).
В работе Лайтхилла (Lighthill, 1970), которая, по существу, является
математическим дополнением к его обзору, изданному в 1969 году,
показано, что указанные особенности обеспечивают преимущество скомброидно-
го способа продвижения по сравнению с угревидным. И это преимущество
состоит в большем значении эффективности при условии, что скомброид-
ное движение сопровождается некоторыми морфологическими
изменениями (что постоянно встречается на практике), прежде всего уже
упоминавшимся сужением тела по высоте перед хвостовым плавником. Подобное
сужение минимизирует эффект возникающих движений за счет «отдачи»,
таких как рыскание и скольжение, которые обусловлены
несбалансированностью колебаний поперечной силы. Хотя в работе Лайтхилла (Lighthill,
1960) и предполагалось, что наличие такой отдачи приведет к слишком
большой потере эффективности, за исключением того случая, когда
длина колеблющейся части рыбы практически равна длине волны поперечных
колебаний. Автором (Lighthill, 1970) было показано, что локальное
уменьшение высоты тела приводит к значительному уменьшению колебаний
величины поперечной силы. В то же время если высота тела в центральной
части тела будет достаточно большой, то эффекты, обусловленные
рысканием и боковым скольжением, уменьшатся за счет вовлечения в боковое
движение большого объема водной среды.
152
Глава 5
Еще в работе Лайтхилла (Lighthill, 1960) были использованы методы
теории вытянутого тела (специалисты в области аэродинамики называют ее
теорией тонкого тела), которая, по существу, является реактивной теорией.
Методы этой теории могут быть применены при исследовании скомброид-
ного способа движения таких рыб, которые обладают «узким» хвостовым
плавником, в том смысле, что каждый луч плавника составляет
небольшой угол (не более 30°) с позвоночником. Несколько иные методы,
основанные на реактивной теории, следует использовать при исследовании
плавания тех немногочисленных представителей рыбного царства, которые
в процессе эволюции приобрели способность к более быстрому
передвижению за счет трансформации «узкого» хвостового плавника в плавник
большого удлинения в виде полумесяца, или серповидный плавник. Такие
методы должны основываться на теории крыла. Они были разработаны,
например, в работе By (Wu, 1961) и др., а затем применены (Lighthill, 1969,
1970) к исследованию движения рыб и водных млекопитающих с
хвостовым плавником в форме полумесяца. В данной работе основное внимание
будет уделено дальнейшей разработке теории тонкого тела применительно
к гидродинамике движения водных животных.
Основной целью данной работы является развитие тех методов
реактивной теории, применение которых дало бы возможность объяснить
результаты экспериментальных наблюдений за движением водных животных
с большой амплитудой волнообразных поперечных движений (как выше
было отмечено, подобная задача ранее была решена с использованием ре-
зистивной теории). Существующий в настоящее время подход основан на
разложении колебательного движения по степеням амплитудного
параметра б (Lighthill, 1960, Appendix), когда сила тяги и расход энергии
пропорциональны б2 (степенями порядка б4 пренебрегается). Однако такое
разложение не всегда правомерно, хотя бы, например, в силу того, что при
скомброидном способе перемещения скорость хвостового плавника в
поперечном направлении может почти в два раза превышать значение скорости
продвижения животного. Поэтому возникает необходимость разработки
такого подхода, при котором были бы учтены и значительные изгибы тела,
и большие значения скорости в поперечном направлении при
перемещении животного. Подобный подход оказался бы полезен также при изучении
механизмов маневрирования водных животных (хотя в данной работе эти
задачи не освещаются).
Одним из путей создания теории, учитывающей широкоамплитудные
движения в поперечном направлении, может быть использование довольно
1. Введение
153
сложной теории возмущений и трактовки ее положений и методов
применительно к рассматриваемым прикладным задачам. Такой подход был
осуществлен в работе Лайтхилла (Lighthill, 1960, Section 2), где
методами теории возмущений исследовались суммарная мощность и мощность
потерь энергии. Свое дальнейшее развитие этот подход получил в
работах Лайтхилла (Lighthill, 1969, 1970), однако применение его к изучению
продвижения животных при широкоамплитудных движениях в поперечном
направлении оказалось довольно затруднительным.
В конечном итоге Лайтхилл (Lighthill, 1970) пришел к
альтернативному, совершенно иному способу применения теории возмущений к решению
подобных задач, сформулировав ее положения применительно к силе тяги
(см. уравнения (13)-(15) цитируемой работы). В его работе полагалось, что
вода на достаточно большом расстоянии от рыбы находится в состоянии
покоя, и были сделаны следующие основные предположения.
(i) Количество движения воды вблизи поперечного сечения рыбы
перпендикулярно к спинному хребту и равно произведению виртуальной
массы т, приходящейся на единицу длины, на поперечную компоненту w
скорости движения рыбы.
(ii) Сила тяги определяется скоростью изменения количества движения
водного объема, находящегося в непосредственной близости от тела рыбы,
ограниченного в каждый момент времени плоскостью П,
перпендикулярной хвостовому плавнику и проходящей через его заднюю кромку.
(iii) При составлении уравнения баланса количества движения
необходимо учитывать перенос количества движения через плоскость П не только
за счет адвекции, но также за счет действия ^mw2 суммарного давления,
обусловленного движением в плоскости П.
Положения, сформулированные в пунктах (i)-(iii), можно сразу же
применить без дополнительных уточнений в реактивной теории для
широкоамплитудных движений. Это утверждение подкрепляется подробными
выкладками, проведенными в §2, но один из наиболее важных аспектов
следует привести здесь: даже если индивидуальные сечения тела рыбы
имеют большую по величине боковую скорость, пусть и превосходящую
скорость продвижения животного, поперечная компонента w
(перпендикулярная к направлению, задаваемому спинным хребтом рыбы), как
правило, намного меньше (что и имеет место при движениях с большой
эффективностью). Более того, следует отметить, что оценка отдачи, играющей
существенную роль при скомброидном продвижении, облегчается при
выбранном новом теоретическом подходе, который в конечном итоге можно
154
Глава 5
использовать также и для изучения механизма влияния всех видов
движений, совершаемых рыбой при движении вперед, на уменьшение разброса
значений силы тяги около ее среднего значения.
Разумеется, было бы желательно в конечном итоге разработать такую
комбинированную резистивно-реактивную теорию гидродинамики водных
животных для широкоамплитудных боковых движений, которую можно
было бы применить и к общему случаю движения водных животных,
имеющих удлиненное тело. Безусловно, такая задача весьма сложна, и в качестве
первого подхода к ее решению естественно исследовать движение рыб,
обладающих узким хвостовым плавником, с позиций только реактивной
теории, как это и сделано в данной работе.
2. Расчет реактивной силы
Как уже отмечалось выше, будем считать, что водная среда на
достаточно большом расстоянии от животного находится в состоянии покоя.
Предполагаем также, что движения рыбы, приводящие к возникновению
силы тяги, происходят в горизонтальной плоскости. Пусть ось у
направлена по вертикали, а взаимно перпендикулярные оси х и z — по горизонтали.
Направления этих осей можно выбрать произвольно, хотя при плавании
рыбы со средней постоянной скоростью удобнее в качестве положительного
направления оси х выбрать направление плавания рыбы.
Для описания широкоамплитудных движений позвоночного столба
животного в горизонтальной плоскости у = О выберем лагранжеву
координату а, которая принимает значения от нуля до / (длина недеформированного
тела рыбы) и равна расстоянию от произвольной точки позвоночника рыбы
до задней кромки ее хвостового плавника, измеренная вдоль позвоночника,
растяжимостью которого в силу ее малости пренебрегается. Эйлеровы
координаты этой точки (х, z) в момент времени t в горизонтальной плоскости
у = О являются функциями переменных а и £, т. е. х = х(а, t) и z = z(a^ t).
Произвольная фиксированная точка позвоночника перемещается (i) за счет
плавательных волнообразных движений, (ii) за счет результирующего
продвижения вперед самой рыбы, (iii) за счет любого движения в боковом
направлении, обусловленного явлением отдачи, которая носит случайный
характер (рис. 1).
Из предположения о нерастяжимости спинного хребта следует
(£)"♦ (©"-■
2. Расчет реактивной силы
155
В горизонтальной плоскости вектор скорости (дх/dt, dz/dt) имеет
компоненту
_ дх дх dz dz
u = ~dt~d^ + ~di^ (2)
направленную по касательной к позвоночному столбу в направлении
движения рыбы, и компоненту
dz дх дх dz
w =
dt да dt да'
(3)
перпендикулярную позвоночному столбу и направленную так, что (и, w)
составляют левую систему, если оси координат (х, z) направлены так, как
это указано на рисунке 2.
голова рыбы
7
хвост рыбы
*2
О
Рис. 1. Система координат, выбранная для описания конфигурации позвоночного
столба рыбы в плоскости у = О в момент времени t
О
касательная
Рис. 2. Конфигурация позвоночного столба рыбы при двух последовательных
моментах времени tx ut2. Стрелка, указывающая направление смещения
фиксированной произвольной точки позвоночника с момента времени t\ к моменту времени t2y
представляет собой вектор скорости точки, умноженный на длину отрезка
времени £2 — *i- На нижнем рисунке вектор скорости представлен в виде суммы двух
компонент — касательной и и перпендикулярной w к оси позвоночника
Основным характерным свойством удлиненного тела является то, что
виртуальная масса т, приходящаяся на единицу длины, образующаяся за
счет движений с поперечной скоростью w, достаточно велика, а при
движениях с продольной скоростью и (движение происходит вдоль касательной
156
Глава 5
>К
kz
х* "0
Рис. 3. Движение оси хвостового плавника. На верхнем рисунке показано
положение в два последовательных момента времени, и введены углы в и а. На нижнем
рисунке вектор скорости V конечной точки оси представлен как сумма двух
компонент — касательной и и перпендикулярной w к оси (вместо компоненты w можно
выбрать компоненту W, перпендикулярную направлению продвижения рыбы, см.
ниже уравнение (10))
к позвоночному столбу) соответствующей вовлеченной в тангенциальное
движение виртуальной массой можно пренебречь. Такие движения в
направлении касательной к позвоночному столбу вносят существенный вклад
в величину вязкого сопротивления, которое, как будет видно ниже,
уравновешивается силой тяги, возникшей в результате реактивного движения.
Будем полагать, что тангенциальные движения приводят к появлению сил
вязкого сопротивления, а движения в направлении нормали — к
возникновению реактивных сил. Перейдем к определению силы тяги и силы,
действующей в поперечном направлении. Для упрощения будем искать реактивную
силу, обусловленную импульсом движений со скоростью w, в
предположении отсутствия вязкости.
Импульс единицы длины тела рыбы представим в виде вектора
me (-£•£)• <4)
где множитель в скобках представляет собой единичный вектор в
направлении скорости w. В работе Лайтхилла (Lighthill, 1970) (изложена
выше в гл. 4) проводится подробное обсуждение2 правомерности
рассмотрения массы т для данного поперечного сечения как постоянной величины
2 Отметим, что в работе обсуждается также и влияние переменного размера поперечного
сечения. Если изменение размера поперечного сечения представить в виде синусоидальной
зависимости, то погрешность не превышает 25% для всех видов волнообразных движений
2. Расчет реактивной силы
157
близкой к jTrps2, где р—плотность воды, a s—высота поперечного сече
ния, т. е. возможности полагать, что т является известной функцией т(а)
от расстоянии а от хвоста рыбы. Однако в случаях, подобных описанным
Бейнбриджем (Bainbridge, 1963), когда рыбы, обладающие достаточно
гибким хвостовым плавником, способны менять высоту тела за один
плавательный цикл, необходимо рассматривать величину массы т как функцию
двух переменных га (a, t). Для общности в дальнейшем будем
предполагать, что масса зависит от а и времени.
Казалось бы, что для определения реактивных сил, действующих
между рыбой и прилегающей к ней водой, можно непосредственно
использовать импульс (4). Однако Лайтхилл в своей работе (Lighthill, 1970) показал,
что метод, основанный на определении скорости изменения импульса всей
массы воды вблизи рыбы, приводит к серьезным техническим трудностям,
и для решения поставленной задачи достаточно следить лишь за импульсом
в спутном следе (который представляет собой вихревую пелену, сходящую
с хвостового плавника рыбы). Но к рассматриваемым в данной главе
широкоамплитудным движениям приведенное им в своей работе приближенное
выражение (18) для импульса в спутном следе неприменимо.
Более приемлемый подход к изучению движения за счет
широкоамплитудных волнообразных движений тела основан на исследовании
скорости изменения импульса в некотором объеме V водной среды вне спутного
следа. Для определения объема V рассмотрим вертикальную плоскость П,
проходящую через концевую часть хвостового плавника. Эта плоскость
перпендикулярна конечной части позвоночного столба и при плавательных
движениях плавника поворачивается вместе с плавником. За искомый
объем V принимаем ту часть водной среды, которая в каждый момент времени
находится с той же стороны от плоскости П, что и рыба.
Импульс3 объема V представим в виде интеграла от (4) в пределах от 0
до / по переменной а. Изменение импульса по времени можно представить
в виде трех слагаемых: (i) изменение импульса за счет уноса импульса
из объема V через плоскость П; (и) изменение импульса, обусловленное
с длиной волны не менее 55. Следует также заметить, что из теории крыла вытекает, что
суммарная сила при малых углах атаки дельтавидного крыла (крыло имеет форму
равнобедренного треугольника, вершина которого обращена в сторону передней части тела рыбы, хвостовой
плавник исследуемых в работе рыб можно представить в виде дельтавидного крыла) может
быть выражена через такое приближенное значение га, если угол при вершине
равнобедренного треугольника не больше 60° и погрешность не превысит 25 %.
3Импульс рассчитывается на основании реактивной теории, т.е. в пренебрежении
какого-либо влияния на него пограничного слоя на поверхности тела рыбы, возникающего за
счет вязкости среды.
158
Глава 5
действием давления на плоскости П; (iii) действие реактивной силы (Р, Q)
на тело со стороны жидкости.
Для оценки слагаемого (i) заметим, что адвективный перенос
импульса через плоскость П незначителен и происходит исключительно за счет
движения жидкости со скоростью и перпендикулярно плоскости П, а
значимое изменение импульса жидкости происходит за счет движений хвоста
со скоростью w. Перенос через плоскость является результатом движений
самой плоскости, которые состоят из поступательного перемещения со
скоростью и (2) и вращения. Однако поскольку тело рыбы обладает
симметрией относительно вертикальной плоскости, проходящей через позвоночный
столб, то и импульс воды, приведенной в движение телом рыбы, обладает
такой же симметрией, а следовательно, вращение плоскости П не оказывает
влияния на величину импульса объема V.
Лайтхилл в (Lighthill, 1970, уравнения (14), (15)) привел простой
аргумент, непосредственно приложимый и к широкоамплитудным движениям,
доказывая, что суммарная сила давления в (ii), обусловленная движением
со скоростью w симметрично расположенных виртуальных масс ш,
приходящихся на единицу длины, равна ~mw2. Эта сила действует в
тангенциальном направлении, задаваемом единичным вектором (дх/да, dz/da). На
основании проведенных здесь рассуждений и с использованием результатов
двух предыдущих параграфов можем записать
d f ( dz дх\ _
dt J \ да' да J
о
/ dz дх\ 1 о f 9х dz\\ ,_ _ч . ч
~UmW Ы' Та) + 2mW [Га' -а)\а,о ~ (* Q)' (Ч
где слагаемые в правой части представляют собой указанные выше
изменения импульса (i), (ii), (iii) соответственно.
Выражение в квадратных скобках равенства (5) можно упростить.
Если бы множитель ^ отсутствовал, то, как следует из (1)-(3), выражение
в квадратных скобках было бы равно mw(dz/dt, —dx/dt). С учетом этого
получаем
<* Q) = |т„ | - ~ ) - ^ (|, %
d [ ( dz
а=0
(6)
2. Расчет реактивной силы
159
Уравнение (5) и его более простая форма записи (6) представляют собой
обобщение уравнений (13) и (И) из (Lighthill, 1970) и могут быть
использованы для исследования продвижения рыб за счет широкоамплитудных
поперечных волнообразных движений.
Заметим, что для периодически повторяющихся плавательных
движений, в результате которых происходит продвижение животного, скажем,
в положительном направлении оси х, производная по времени от интеграла
в (6) не оказывает влияние на среднее значение силы тяги Р. Для среднего
значения силы тяги на основании (6) можно записать
Р =
(dz 1 дх
raw ——w-
dt 2 да
(7)
а=0
Таким образом, как и в случае поперечных колебаний с малой амплитудой,
можно сделать вывод, что большая по величине сила тяги может
возникать, только если между величинами w и dz/dt существует тесная
положительная корреляционная связь, хотя последняя и имеет значительно
большую амплитуду осцилляции. Накладываемое на скорость w требование об
относительно небольшой амплитуде может быть ослаблено, если
флуктуации w2 будут отрицательно коррелированы с флуктуациями дх/да при
а = 0, что, как правило, имеет место при скомброидном типе движения.
Поскольку дх/да равен косинусу угла между хвостовым плавником и
направлением поступательного перемещения, этот угол имеет наибольшую
величину при больших значениях скорости w.
Следует отметить, что из векторного равенства (6) можно получить не
только среднюю силу тяги Р, но также и ее мгновенное значение Р, записав
его проекцию на ось х. Хотя выражение в квадратных скобках может
принимать большое положительное значение, однако его величина обращается
в нуль дважды за один цикл плавательного движения (когда скорость w
проходит через нуль), так что изменение силы тяги при этом носит ярко
выраженный скачкообразный характер. К счастью, при скомброидном типе
движения флуктуации второго слагаемого
i
d [ dz . /оЧ
— / mw-z-da (8)
dt J да v '
о
в выражении для силы тяги Р часто гасят друг друга, что приводит в
результате к значительному сглаживанию флуктуации силы тяги.
160
Глава 5
Для того чтобы это увидеть, прежде всего отметим, что при скомбро-
идном типе плавания амплитуда осцилляции z(a, £), равно как и
произведения mw, быстро убывает с увеличением параметра а от его наименьшего
нулевого значения. Интеграл (8) может быть приближенно представлен как
произведение величины [—z]a=0, на которую изменяется z между а = 0
и передней областью тела рыбы, и [^™^]а=0> которая представляет собой
довольно грубую оценку средней величины произведения mw в этой
области. Такое, правда, довольно грубое приближение амплитуды и
недооценка фазы усредненного произведения mw приводят к полному погашению
флуктуации силы тяги при условии, что z и mw при а = 0 изменяются по
синусоидальному закону:
z = z0 sin ut, mw = m0w0 cos cut. (9)
Тогда из (6) получаем
2 d f 1 . \ 1
P = m0w0z0uj cos ujt+— I — — m0w0z0 sm ut cos u;t I = — m0w0z0uj. (10)
at \ 2 у 2
Вероятно, подобное сглаживание флуктуации, хотя и не в столь полной
степени, может встретиться и на практике.
Кроме того, из равенства (6) можно получить и выражение для флукту-
ационных изменений боковой силы (—Q), действующей на рыбу, что
позволит нам оценить величину отдачи. На самом деле отличия,
касающиеся боковой силы, при широкоамплитудных движениях от случая движений
с малой амплитудой пока еще невелики. Из (6) следует
-Q =
дх 1 о dz
mw_+.mw _
+
а=0
-г / mw—- da. (11)
dtj да к J
Это выражение для боковой силы отличается от полученного ранее
выражения (49) в (Lighthill, 1970) для случая волнообразных поперечных
движений с малой амплитудой тем, что (i) средняя скорость U заменена на
переменную скорость dx/dt хвостового плавника, (и) включено слагаемое
-raw2 dz/da третьего порядка малости, (iii) в подынтегральное выражение
входит множитель дх/да.
Эти отличия невелики, и поэтому в данном случае также справедливо
заключение (Lighthill, 1970) о том, что достаточно большая высота
передней части тела рыбы с последующим сужением его к хвостовому плавнику
3. Обсуждение баланса силы тяги и силы сопротивления 161
минимизирует проявление отдачи. Для пояснения необходимости сужения
тела к хвостовому плавнику, обеспечивающего минимизацию величины
боковой силы отметим что если амплитуда осцилляции функции z(a, t)
быстро изменяется с изменением а, то второе слагаемое в (3) должно иметь
большое значение, которое не будет компенсироваться первым слагаемым.
Кроме того, величина флуктуации интеграла в (11) была бы достаточно
большой, если бы величина виртуальной массы вблизи хвостовой части не
уменьшилась бы.
И наконец, следует отметить, что приведенный метод расчета
реактивной силы может быть применен с небольшими изменениями к расчету
скорости, с которой происходит потеря энергии в спутном следе,
обусловленная этой силой. Без учета вязкости кинетическая энергия,
определяемая скоростью w, равна \mw2. Объем V теряет эту энергию за счет ее
адвекции через плоскость П, обусловленной поступательным движением
плоскости со скоростью и (2). (За счет вращения плоскости П потерь
энергии не происходит в силу симметричности распределения в направлении
оси z — в поперечном направлении. Потери энергии, обусловленные
действием давления на плоскости П, также невелики в силу пренебрежимой
малости нормальной к плоскости П компоненты скорости.) Следовательно,
средняя скорость потери энергии равна
2
|—mw2u\ . (12)
а=0
По сравнению со скоростью потерь энергии в случае волнообразных
поперечных движений с малой амплитудой скорость потерь энергии
увеличилась. Действительно, в выражении для потерь при малых амплитудах
поперечных движений вместо скорости и стоит средняя скорость
продвижения животного в воде:
L да1а=0
величина которой меньше и, поскольку дх/да < 1.
3. Обсуждение баланса силы тяги и силы сопротивления
на материале наблюдений
В данном параграфе попытаемся использовать методы теории § 2 для
анализа экспериментальных данных с целью определения средней силы тя-
162
Глава 5
ги и ее баланса с силой сопротивления среды при движениях животного
с постоянной скоростью. Будем исследовать реальный скомброидный тип
плавания рыб с узким хвостовым плавником и в рамках ограничений,
накладываемых на механизм возникновения силы тяги положениями
реактивной теории, рассмотрим баланс средней силы тяги, возникающей в
силу поперечных движений (перпендикулярно позвоночному столбу) со
скоростью w9 и средней силы сопротивления, обусловленной движениями со
скоростью и (в направлении касательной к позвоночному столбу).
Далее будет показано, что имеющихся экспериментальных данных
совершенно недостаточно для того, чтобы на их основании сделать
обоснованные заключения касательно баланса указанных выше сил. Однако
даже такое приближенное исследование позволит сделать выводы,
представляющие интерес с практической точки зрения и, несомненно, подчеркнет
необходимость проведения дальнейших экспериментальных исследований
с целью получения более полных данных. В идеале желательно провести
съемку процесса плавания в горизонтальной плоскости в невозмущенной
воде с помощью вертикальной кинокамеры, получая не меньше чем 16
кадров за один плавательный цикл. Причем наблюдения должны вестись над
одним и тем же экземпляром при различных скоростях плавания. Для
сравнения теоретической силы сопротивления, рассчитанной по движениям со
скоростью и, с ее экспериментальным значением было бы желательно
найти силу сопротивления на основании съемок в тех же условиях простого
скольжения той же самой рыбы без совершения ею волнообразных
поперечных движений.
Имеющиеся данные для одного экземпляра рыбы далеко не полны.
Бейнбридж (Bainbridge, 1958, 1960, 1963) сумел создать контролируемые
условия водной среды, создав так называемый «рыбный бассейн»
(резервуар кольцеобразной формы для рыб, снабженный механизмом для
предотвращения завихрения водного потока). Ниже будет проведен анализ
полученных Бейнбриджем с помощью киносъемки характеристик плавания
ельца (Leuciscus) длиной 0,3 м со скоростью 0,48 м/с. К сожалению,
такая скорость в несколько раз ниже максимальной скорости, развиваемой
водными животными, а данные наблюдений для других более высоких
скоростей плавания на настоящее время отсутствуют. Кроме того, на сегодня
отсутствуют данные и по измерению силы сопротивления. То же относится
и к полученным много раньше Греем (Gray, 1933) съемкам процесса
плавания различных особей. А некоторые экспериментаторы определяли силу
сопротивления в случае движения рыб как жесткого неупругого симметрич-
3. Обсуждение баланса силы тяги и силы сопротивления 163
ного тела. Например, Санднес (Sundnes, 1963) нашел величину силы
сопротивления при буксировке в водном потоке особей лососей и сельдей.
Полученный экспериментально коэффициент сопротивления4 CD равен
приблизительно 0,01 при числе Рейнольдса 106. Величина силы сопротивления
немного, но не чрезмерно, больше, чем при обтекании гладкого тела (см.
ниже, а также Hoerner, 1965, chapter 6). Возможно, что это превышение
обусловлено наличием буксировочной веревки. Однако, кроме буксировки
в воде, видеозаписи плавательных движений той же рыбы отсутствуют.
Лишь на небольшой части существующих записей плавательных
движений представлен скомброидный тип движения рыб с узким хвостовым
плавником. И эти наиболее качественные записи — записи плавания ельцов
сделаны Бейнбриджем (Bainbridge, 1963). На рисунке 1 в его работе
изображены положение и форма хвостового плавника в движении (вид сверху)
через каждые 0,02 с при средней скорости 0,48 м/с, которая достигается при
частоте колебаний хвостового плавника 2,5 Гц. На основании этих данных
и данных об изменении поперечного размера плавника, представленных на
рисунке 3 указанной работы, по формуле (7) возможно рассчитать среднюю
силу тяги Р.
В таблице 1 представлены результаты измерений некоторых
параметров, относящихся к движению хвостового плавника, в различные моменты
времени t плавательного цикла. Здесь V означает абсолютную величину
скорости рыбы относительно воды, угол 9 — угол наклона хвостового
плавника к направлению среднего движения, а угол а + в — угол наклона
траектории хвостового плавника к этому направлению. Указанные углы
положительны, если они отсчитываются вправо от направления движения рыбы.
Таким образом, угол а — угол атаки — будет положительным, если
боковая сила, действующая на плавник, направлена влево. Тогда справедливы
следующие соотношения:
Oz дх
w = Vsma, W = — = Vsm(a + 0), — =cos0. (14)
at да
Значения параметров, входящих в соотношения (14), приведены в
таблице 1. Там же приведены значения т = \-Kps2, полученные Бейнбриджем
(s — поперечный размер хвостового плавника).
На основании этих данных рассчитаны произведения mwW и ^raw2f^
и их разность. Значения этих произведений приведены в последних трех
43десь и далее коэффициент CD равен силе сопротивления, деленной на ^ри2^и на
площадь поверхности рыбы, соприкасающейся с водой.
Таблица 1. Значения параметров плавания ельца, полученные Бейнбриджем при съемке (данные заимствованы из
Bainbridge, 1963, рис. 1 и рис. 3)
(Примечание: 1 Н = 1 кг-м-с~2 = 105 дин)
Параметр
Единицы
измерения
t
с
0,04
0,08
0,12
0,16
0,20
0,24
0,28
0,32
0,36
0,40
V
м-с-1
0,50
0,60
0,74
0,68
0,52
0,62
0,87
0,67
0,52
0,48
а + в
градусы
17
45
60
48
13
-36
-60
-46
-25
-3
а
градусы
3
3
5
16
10
-8
-16
-10
-6
0
W
м-с-1
0,15
0,42
0,64
0,50
0,12
-0,36
-0,75
-0,48
-0,22
-0,02
w
м-с-1
0,03
0,03
0,06
0,19
0,09
-0,09
-0,24
-0,12
-0,05
0,00
дх
да
1
0,97
0,74
0,57
0,85
1,00
0,88
0,72
0,81
0,95
1,00
s
м
0,094
0,087
0,079
0,083
0,087
0,088
0,083
0,084
0,092
0,093
то
кг-м-1
6,9
5,9
4,9
5,4
5,9
6Д
5,4
5,5
6,6
6,8
mwW
Н
0,03
0,07
0,19
0,51
0,06
0,20
0,97
0,32
0,07
0,00
\mw2y.
v дх
_ да
н
0,00
0,00
0,01
0,08
0,02
0,02
0,11
0,03
0,01
0,00
IILLU VV —
- \vfiw1 х
xdZ
н
0,03
0,07
0,18
0,43
0,04
0,18
0,86
0,29
0,06
0,00
3. Обсуждение баланса силы тяги и силы сопротивления 165
столбцах таблицы 1. Второе произведение (определяемое действием силы
давления на плоскости П), как видно, составляет лишь небольшую часть
от первого и вносит малый вклад в величину их разности. Среднее
значение этой разности за один плавательный цикл равно средней силе тяги. Как
следует из таблицы, это среднее значение равно 0,2 Н. Точность
приведенных экспериментальных данных не позволяет получить значение силы тяги
с точностью более одной значащей цифры.
Подобное заключение представляет интерес с той точки зрения, что
дает возможность получить значения средней силы сопротивления,
связанной с движением тела в тангенциальном направлении, и эти значения
в несколько раз больше, чем сила сопротивления среды при скольжении
вытянутого тела, подобного ельцу, длиной 0,3 м со скоростью 0,48 м/с.
Площадь поверхности тела рыбы 5, соприкасающейся с водой, найденная по
фотографиям Бейнбриджа, равна приблизительно 0,04 м2, так что значение
произведения ^pU2S равно приблизительно 5 Н. При силе сопротивления
0,2 Н коэффициент сопротивления равен CD = 0,04, а это в четыре
раза больше, чем коэффициент силы сопротивления при числах Рейнольдса
R = ю5, отнесенных к длине.
Такое расхождение довольно велико и, возможно, объясняется
неточностью расчетов силы тяги, равно как и ненадежностью полученного
значения CD при скольжении. В силу сказанного оценим коэффициент CD на
основании достаточно хорошо известных соотношений для движения
вытянутых тел вращения. Максимальное поперечное сечение рыбы (см.
таблицу I у Бейнбриджа) представляет собой овал с большой и малой
осями, равными соответственно 0,07 м и 0,04 м, сечение при расчете силы
сопротивления можно заменить кругом диаметром приблизительно 0,06 м
(периметр поперечного сечения приблизительно равен длине тела рыбы).
Отношение длины эквивалентного тела вращения к его максимальной
толщине равно 5, если в качестве длины выбрать полную длину тела рыбы,
или 4, если за длину принять расстояние от головы до хвостового стебля
рыбы (где площадь поперечного сечения мала). На основании
проведенных расчетов Хёрнер (Hoerner, 1965, гл. 6, рис. 22) получил, что
значение коэффициента силы сопротивления CD приблизительно равно 0,01 для
отношения длины тела вращения к его максимальной толщине, лежащего
в пределах от 4 до 5 при числах Рейнольдса 105. (Можно было бы
предположить, что увеличение коэффициента CD обусловлено турбулентностью,
которая, скорее всего, присутствует в «рыбном бассейне» Бейнбриджа,
однако приведенные в работе Хёрнера данные (рис. 23, гл. 6) указывают на
166
Глава 5
то, что сила сопротивления возрастает на пренебрежимо малую величину
за счет турбулизации потока, возникающей при числах Рейнольдса 105.)
Очевидно, что необходимо исследовать возникшее различие в
значениях силы тяги и силы сопротивления (более чем в 4 раза), хотя смысл этого
расхождения не прояснится, пока подобные расчеты не будут проведены
на основании наблюдений над многими другими рыбами. А между тем
какие-то комментарии, касающиеся факторов, которые могли бы оказать или
не оказать влияние на такое расхождение, совершенно необходимы.
В качестве источников возможных ошибок в оценке силы тяги
наиболее очевидным представляется пренебрежение силами вязкости, но, скорее
всего, этот фактор следует исключить из рассмотрения как причину
возникновения такого расхождения, поскольку его учет приведет скорее к росту,
чем к уменьшению расхождения (Lighthill, 1970). В то же время ошибка
в оценке реактивной силы тяги сама по себе могла быть обусловлена
частично тем, что при расчете силы тяги предполагается наличие некоторого
небольшого вклада со стороны подсасывающей силы на передней кромке
хвостового плавника. Такой вклад в целом невелик (имеет порядок w2 по
сравнению с wW)9 а его учет является довольно сложной задачей из-за
разделения потока воды, обусловленного наличием относительно острой
передней кромки хвостового плавника. С другой стороны, возможно, с учетом
довольно заметной гибкости хвостового плавника, когда передняя кромка
достаточно сильно изгибается, разделение потоков происходит значительно
позже. Наблюдаемое Бейнбриджем (Bainbridge, 1963) изгибание хвостового
плавника подтверждает это предположение, но даже если разделение
потоков имеет место, то возникшая в силу этого ошибка слишком мала, чтобы
быть единственной причиной полученного расхождения.
Заметим, что в экспериментах Бейнбриджа исследуемая особь плавала
слишком близко ко дну бассейна. Конечно, это нельзя считать причиной
полученного расхождения между силой тяги и силой сопротивления, но,
скорее всего, за счет этого возможно увеличение силы тяги. Можно
провести оценку влияния близости дна при плавании рыбы, рассматривая дно
как плоскость симметрии, при этом эффективная высота хвоста рыбы и его
симметричного отражения относительно плоскости удваивается, что
приведет к учетверению виртуальной массы воды. Часть количества движения,
которая соответствует виртуальной массе, находящейся над плоскостью,
составляет половину от того учетверенного количества движения,
обусловленного действием сдвоенного плавника. Следовательно, могут иметь
место удвоение поперечного количества движения, обусловленного движени-
3. Обсуждение баланса силы тяги и силы сопротивления 167
ем хвостового плавника, когда рыба находится достаточно близко от дна,
и сопутствующее этому удвоение силы тяги.
Все подобные исследования возможных уточнений при расчете силы
тяги приводят к возрастающему неприятию мнения, высказанного автору
д-ром Боуном (Bone), о том, что сила вязкого сопротивления, действующая
на рыбу при ее движении в воде (т. е. та сила сопротивления, которая
необходима для уравновешивания реактивной силы), должна быть по
некоторым основаниям во много раз больше, чем та, которая определяется только
скольжением рыбы в воде. Этот вывод противоречит знаменитому
парадоксу Грея, однако необходимо подчеркнуть, что такое заключение сделано на
основании изучения движения совсем другой группы животных.
При формулировке этого вывода термин «сила вязкого сопротивления»
был использован с целью подчеркнуть то, что учитывается сопротивление
воды, связанное с движениями рыбы со скоростью и. При волнообразных
движениях направление вихревой силы, обусловленной поперечными
движениями (w-движения), должно совпадать с направлением силы тяги.
Поэтому, как уже упоминалось и ранее, действие любой подобной силы
приводит к увеличению силы тяги.
В некоторых других исследованиях показано, как при пассивном
движении рыбы в воде (при этом допустимы слабо выраженные
волнообразные движения тела и плавников) сопротивление превышает (в несколько
раз) сопротивление при скольжении рыбы как твердого тела. Этот эффект
был обнаружен Веббом (Webb, 1970, с. 98) в экспериментах по скольжению
парной мертвой форели в туннеле. Им было получено, что сопротивление
возрастает при возрастании степени допустимости волнообразных
движений тела мертвой форели. В случае практически полной невозможности
волнообразных движений было получено, что коэффициент
сопротивления CD приблизительно равен 0,015, что хорошо согласуется со значениями
коэффициента сопротивления при скольжении живой форели,
полученными Греем (Gray, 1957). В тех экспериментах, когда хвостовая часть мертвой
форели могла колебаться со значительной амплитудой, хотя ей и была
придана некоторая жесткость с помощью проволоки, Вебб получил значения
коэффициента сопротивления, лежащие в окрестности 0,03. Правда, точно
неизвестно, является ли это главным образом приростом вязкого
сопротивления, такого, как было постулировано в случае сопротивления,
обусловленного плавательными движениями, но из обсуждений условий
экспериментов и полученных результатов вытекает, что такое возможно.
Независимо от Вебба Смит (Smit, 1965) пришел к таким же
выводам о росте вязкого сопротивления при наличии плавательных движений.
168
Глава 5
Он оценивал мощность, развиваемую золотой рыбкой (Carassius auratus)
в аквариуме за счет мускульного усилия на основании расхода
кислорода, и получил (в предположении 80 % эффективности при движении), что
мощность превышает в 3,6 раза мощность, необходимую для преодоления
вязкого сопротивления при скольжении рыбки как твердого тела (оценка
проводилась указанным выше способом). Значение множителя 3,6,
полученное при проведении опытов в кольцеообразном аквариуме, где
имелась возможность достаточно точного измерения скорости рыбки,
вызывает большее доверие, чем полученное Смитом при проведении измерений
скорости рыбки в гидроканале, где за счет возможного движения в более
медленном потоке в пограничном слое вырабатываемая мощность могла
уменьшиться.
Доктор Боун предложил автору следующее объяснение прироста
вязкой силы сопротивления при возможности совершения животным
плавательных движений, которое на сегодняшний день видится наиболее
обоснованным. Его рассуждения основаны на том положении, что трение на
поверхности тела рыбы на большей части ее поверхности, возможно,
значительно больше, чем можно было бы ожидать для тела данной длины для
чисел Рейнольдса порядка 105, т.к. поперечные волнообразные движения
препятствуют росту толщины пограничного слоя и он не достигает той
толщины, которую имел бы на теле данной длины, если бы волнообразных
движений не было.
Возможность такого большого прироста силы сопротивления им было
подтверждено расчетом толщины пограничного слоя при сильных
упрощающих предположениях. Например, пограничный слой Блазиуса (при
постоянной скорости потока U вне пограничного слоя) имеет толщину слоя 6
(толщина определена так, что напряжение трения равно /if//5), равную
ZyJvxjU на расстоянии х от головы. Мы можем сравнить с этим значением
толщину пограничного слоя S для пластины толщиной s, которая движется
в направлении нормали к своей поверхности со скоростью w. Эта толщина
пограничного слоя равна 0,6yjvsjw^ s отсчитывается от края, обращенного
в сторону движения.
Очевидно, что указанные толщины различаются почти в 5 раз,
поскольку множители — квадратные корни при типичных параметрах
плавания рыбы — имеют достаточно близкие значения. На практике явление
отдачи приводит к значительным величинам скорости w вдоль всего тела
плывущей рыбы, а следовательно, толщина пограничного слоя на ее
поверхности может существенно уменьшиться по сравнению с толщиной по-
3. Обсуждение баланса силы тяги и силы сопротивления 169
граничного слоя, развивающегося на поверхности рыбы при ее скольжении
как твердого тела.
Согласно приведенным рассуждениям, именно баланс, существующий
между вязкой диффузией вихрей, сходящих с поверхности рыбы, и
адвекцией завихренности к поверхности за счет бокового движения тела рыбы,
обусловливает толщину пограничного слоя, а также и силу трения,
возникающую при тангенциальном движении со скоростью и. При продвижении
рыбы в воде на тех участках ее поверхности, которые движутся в
направлении скорости [/, возникающая сила трения может возрасти почти в 5 раз по
сравнению с силой трения, действующей при скольжении рыбы как
твердого тела. Подобное изменение может быть одним из объяснений различия
между значениями силы тяги и силы сопротивления.
Таким образом, присущее большинству рыб колебательное движение
хвостовой части с относительно большими амплитудами вызвано
необходимостью выработки достаточно большой силы тяги для компенсации
значительной силы вязкого сопротивления, которая возникает при движении
рыбы. По сравнению с плаванием рыб с постоянной скоростью за счет
небольших по амплитуде поперечных колебаний более эффективным
способом плавания с точки зрения выработки силы тяги является тот, который
обычно и можно увидеть при наблюдении за плаванием рыб в аквариумах,
когда интенсивные волнообразные движения хвостовой частью с большой
амплитудой, приводящие к движению с большой скоростью, сменяются
периодами пассивного скольжения.
И в заключение хотелось бы еще раз подчеркнуть гипотетический
характер выдвинутых предположений о возможных причинах расхождения
между величиной силы тяги и силы сопротивления и их обоснований. Для
более обоснованных заключений совершенно необходимы дополнительные
экспериментальные исследования.
С большим удовольствием выражаю признательность доктору
Квентину Боуну за плодотворные всесторонние обсуждения материала данной
главы, и особенно материала § 3.
Литература
[1] Bainbridge, R. 1958. J. Exp. Biol. 35, 109.
[2] Bainbridge, R. 1960. J. Exp. Biol. 37, 129.
[3] Bainbridge, R. 1963. J. Exp. Biol. 40, 23.
[4] Gray, J. 1933. J. Exp. Biol. 10, 88.
170
Глава 5
Gray, J. 1957. Scient. Am. 197 (Aug.), 48.
Gray, J. & Hancock, G.J. 1955. J. Exp. Biol. 32, 802.
Hancock, G.J. 1953. Proc. R. Soc. Lond. A, 217, 96.
Hoerner, S. F. 1965. Fluid-dynamic drag. Midland Park, N. J.: S. F. Hoerner.
Lighthill, M.J. 1960. J. Fluid Mech. 9, 305.
Lighthill, M.J. 1969. Ann. Rev. Fluid Mech. 1, 413.
Lighthill, M.J. 1970. J. Fluid Mech. 44, 265.
Smit, H. 1965. Can. J. Zool. 43, 632.
Sundnes, G. 1963. FiskDir. Skr. {Bergen), 13, 126.
Taylor, G.I. 1952. Proc. R. Soc. Lond. A, 214, 158.
Webb, P. 1970. Ph.D. thesis, University of Bristol.
Wu, T. Y. 1961. J. Fluid Mech., 10, 321.
Глава 6
Передвижение водных животных.
Обзор новейших теоретических
исследований1
1. Введение
В своем докладе конгрессу по теоретической и прикладной механике
хотелось бы сообщить об одном направлении прикладной зоомеханики —
теоретических аспектах механики взаимодействий между водной средой
и плывущим в ней животным.
Вообще говоря, прогресс в прикладной зоомеханике возможен
только при тесном сотрудничестве между зоологами и механиками. Животное
царство безгранично, и задача распознавания и описания типов движения
отдельных групп животных, прежде всего, с позиций эволюционного
развития является одной из наиболее важных задач, стоящих перед зоологами.
В то же время сложные взаимосвязи между такими движениями и
окружающей водной средой являются предметом изучения гидромеханики. Таким
образом, когда зоологи и гидромеханики хорошо знают друг друга и
готовы сотрудничать, то постепенно они способны настолько овладеть
профессиональным языком своих коллег, что становится возможным совместное
продвижение в направлении решения стоящих перед ними общих задач,
включая задачи исследования наиболее значимых, с точки зрения зоологов,
типов движения методами гидродинамики.
Особенно значимым для исследования передвижения животных было
сотрудничество, начавшееся в 50-х годах, сэра Джефри Тейлора (Geoffrey
Taylor), гидродинамика с огромным опытом, и сэра Джеймса Грея (James
Gray), зоологическая школа в Кембридже которого на протяжении более
20 лет являлась важнейшим источником сведений и исследований почти
1 Обзорная лекция на Международном конгрессе по теоретической и прикладной механике.
Москва, 1972.
172
Глава 6
всех аспектов проблемы перемещения животных (Gray, 1968). Совместное
обсуждение проблем перемещения водных животных привело к появлению
значимых работ Тейлора (Taylor, 1951, 1952а, 1952b) по перемещению как
при низких, так и при высоких числах Рейнольдса.
Все опубликованные работы Джефри Тейлора по гидродинамике
настолько интересны, что многие молодые ученые, ознакомившись с ними,
с энтузиазмом брались за дальнейшую разработку содержавшихся в них
идей. Одним из них был и автор доклада, который опубликовал работы
в 1952 году (Lighthill, 1952) и в 1953 году в соавторстве со своим учеником
Хенкоком (Hancock, 1953). Однако изложенные в них результаты нашей
работы в Манчестере по исследованию движений при очень низких
числах Рейнольдса остались незамеченными, поскольку в то время мы еще
не имели навыков общения с зоологами — нами не были поняты наиболее
значимые для зоологов проблемы, равно как и им были непонятны выводы,
сделанные исключительно на языке гидродинамики. К счастью, Д. Ж. Хен-
кок через некоторое время начал сотрудничать с сэром Джеймсом Греем,
и ими были опубликованы совместные работы (Gray, Hancock, 1955),
содержащие весьма значительные результаты.
Вплоть до 1960 года автор не был непосредственно знаком с
Джеймсом Греем, а в 1960 году ему посчастливилось провести незабываемый день
в обсуждении с ним проблемы передвижения водных животных при
высоких числах Рейнольдса. В результате проведенного обсуждения у меня
созрела мысль, что можно было бы применить методы гидродинамической
теории вытянутого тела к изучению механизма плавания рыб. Это было
сделано довольно поспешно и результаты были опубликованы в 1960 г.
(Lighthill, 1960). Однако в процессе работы не уделялось достаточного
внимания сотрудничеству с зоологами из Кембриджа, а это было совершенно
необходимо для понимания ими значимости работы. Отчасти такое
непонимание произошло из-за того, что руководитель Королевского авиационного
центра не ознакомился лично с работой, отчасти из-за того, что автор пока
не оценил всей важности изучения литературы по зоологии и овладения ее
терминологией для понимания существа проблем зоологии и для
возможности донести до зоологов смысл проводимых гидродинамических
исследований. Аналогичные гидродинамические задачи были блестяще решены
доктором By (Wu, 1961) на основе двумерной теории крыла, однако и в его
работах не были четко обозначены прикладной характер полученных
результатов и возможность их применения для решения зоологических проблем.
Много позже, в 1968 году, автор доклада вернулся к этой проблеме
в связи с предложением редакции Annual Reviews of Fluid Mechanics на-
1. Введение
173
писать обзорную статью по этому вопросу для первого тома журнала. При
работе над ней автором наконец было понято, насколько тесно должны
взаимодействовать биологи и гидродинамики при проведении совместных
исследований, например, в области динамики циркуляции физиологических
жидкостей и воздуха в организме. В кратком виде эти исследования
изложены в недавней работе автора (Lighthill, 1972). Приобретенный в процессе
совместной работы опыт, а также изучение литературы по движению
водных животных обнажили всю существующую на то время
несостоятельность сотрудничества между представителями двух областей знаний,
которая проявлялась в большинстве случаев при исследовании одних и тех же
задач. Все это утвердило автора в том, что для того, чтобы сделать
доступными для всех заинтересованных в этих проблемах специалистов в области
зоологии гидродинамические аспекты движения водных животных, следует
обсудить их с зоологами и постоянно обсуждать их с ними, ознакомиться
с их работами в этой области и постоянно читать их труды и следить за их
достижениями, изучить их коллекции (в музеях и аквариумах) и постоянно
продолжать изучать их все время!
В конечном счете при активном содействии сэра Джеймса Грея и его
коллеги доктора Бейнбриджа (R. Bainbridge), а также доктора Маршалла
(N.B.Marshall), сотрудника Британского музея естествознания, и
доктора Квентина Боуна (Quentin Bone) из Морской биологической ассоциации
в Плимуте, автором была написана (см. Lighthill, 1969 и главу 2 настоящей
книги) обзорная работа, в которой, в частности, имелись следующие слова:
«Скорее всего, читатели обладают специальными знаниями в области
зоологии или механики жидкости, но вряд ли они одинаково хорошо
ориентируются в обеих областях. Автор постарался изложить материал так, чтобы
он был доступен и тем и другим». Фактически, автор попытался, опираясь
на содержание бесед с зоологами, на материалы, почерпнутые из
литературы, на знания, полученные при изучении коллекций, провести
экскурсию в мир животных и осветить способы перемещения водных животных.
В обзоре приведены многочисленные данные зоологов, интерпретация этих
данных с точки зрения эволюционного развития и указаны работы
гидродинамиков, в которых соответствующие проблемы, представляющие интерес
для специалистов в области зоологии, решались с позиций гидродинамики.
А также была сделана попытка очертить области приложения имеющихся
на сегодняшний день теоретических работ гидродинамиков к различным
разделам биологии. Кроме того, в этом обзоре автор попытался также
выделить те области биологии, где применение методов гидродинамики было
бы наиболее полезным и эффективным.
174
Глава 6
Некоторые исследования, о которых шла речь в том обзоре, с тех пор
были завершены, и их результаты изложены в новых статьях (гл. 4 и 5
настоящей работы), в новой работе доктора By (Wu, 1971a, 1971b), а также
в работе моих новых коллег доктора Блейка (J.R.Blake) и доктора Вей-
са (D. Weihs). И именно эта работа составляет основной предмет данной
лекции. Было необыкновенно интересно наблюдать, насколько быстро мои
коллеги доктор Блейк и доктор Вейс, сумели освоить научный язык
зоологов и применять его в общении с ними, это моим коллегам удалось сделать
значительно быстрее, чем мне! В лекции приводятся выдержки из их
работы, а также с любезного разрешения авторов их еще не опубликованные
результаты.
В данной работе впервые с точки зрения гидродинамики
анализируются многочисленные данные наблюдений, собранные сэром Джеймсом
Греем и другими зоологами, за последние 40 лет. Вместе с тем и сами
зоологи в настоящее время начали публиковать новые материалы
наблюдений и самостоятельно анализировать их, используя аппарат гидродинамики
(см., например, Webb, 1971).
В качестве отправной точки, как уже упоминалось, в сегодняшнем
докладе используется материал обзорной статьи для журнала Annual Reviews
of Fluid Mechanics (глава 2 настоящей работы). Те, кому не довелось
прочитать эту статью, но хотел бы понимать рассуждения о приложениях
гидродинамики к задачам в области зоологии, могут найти полезную для себя
информацию по этому вопросу в доступных на сегодняшний день обзорных
статьях. Более того, автор даже настоятельно советует сделать это,
поскольку излагаемый здесь материал наиболее значим и важен именно в
зоологическом контексте. Такая рекомендация объясняется тем, что в сегодняшнем
докладе на 13-м Конгрессе по теоретической и прикладной механике
зоологические аспекты будут оставлены в стороне, иначе не останется времени
на изложение гидродинамических проблем, а также в силу того, что
основной целью доклада является обзор результатов исследований, проведенных
уже после 12-го конгресса. Другими словами, доклад посвящен тем
работам, которые были проведены после написания обзорной статьи.
2. Разделы гидродинамики, применяемые для изучения
передвижения животных в воде
Прежде чем переходить к подробному детализированному изложению,
хотелось бы остановиться на основных типах подразделений гидродина-
2. Разделы гидродинамики
175
мики в приложении к проблеме передвижения водных животных (рис. 1).
Наиболее очевидное и важное разграничение обусловлено величиной
числа Рейнольдса. Перемещения водных животных размером не более 1 мм
характерны тем, что влияние сил инерции на скорость перемещения
пренебрежимо мало, поскольку число Рейнольдса при таких перемещениях мало
(не превышает единицы). В этом случае давление и сила трения
удовлетворяют уравнениям равновесия и, как следствие, скорость является решением
бигармонического уравнения. Задача определения скорости перемещения
животных большего размера значительно более сложна, поскольку
существенную роль играют и силы инерции, и силы трения, а уравнения
движения становятся нелинейными.
При больших числах Рейнольдса возникает необходимость
дальнейшего подразделения гидродинамики. Выделяются разделы гидродинамики
применительно к способам движения водных животных (им
предполагается уделить более всего внимания в данном докладе), которые можно
рассматривать как различные модификации способов перемещения в воде при
низких числах Рейнольдса, а также к совершенно другим типам
перемещения — реактивному движению. Следует отметить немногочисленность
появившихся исследований механизмов реактивного движения с момента
выхода обзорной работы в 1969 году. Такой механизм движения
используют многие беспозвоночные при появлении опасности — за счет резкого
мышечного сокращения с большой скоростью выбрасывается струя воды,
и животное быстро перемещается в противоположную сторону.
Значительно меньшее число животных использует реактивное движение просто как
способ перемещения, и отметим, что к ним относятся не только
кальмары и животные, упомянутые в работе Лайтхилла (Lighthill, 1969), но
также и некоторые двустворчатые моллюски, которые демонстрируют
удивительный способ передвижения за счет повторяющихся резких схлопываний
створок раковины.
В рамках обсуждения способов перемещения и их последующих
модификаций в результате эволюционного развития заметим, что, прежде всего,
эти способы можно разделить на две основные группы — способы
перемещения при очень низких числах Рейнольдса и при высоких числах
Рейнольдса. Это разделение уже проводилось в работах, вышедших в начале
пятидесятых годов. Для исследования обоих типов перемещения
используются методы комплексной геометрии в четырехмерном пространстве — три
пространственные координаты и одна временная.
Так, Тейлор (Taylor, 1952a) при исследовании механизмов
передвижения микроскопических одноклеточных организмов за счет волнообразных
176
Глава 6
колебаний отростком или жгутиком использовал модель движения тела
вращения, усовершенствовав более раннюю (Taylor, 1951) довольно грубую
двумерную математическую модель, в которой направление
результирующего движения животного в качестве пространственной координаты не
рассматривалось. Последние теории либо основаны на двумерном анализе,
либо используют методы теории вытянутого тела. Методы теории вытянутого
тела, безусловно, предпочтительнее в том случае, когда исследуемый
организм имеет продолговатое тело и его движения носят пространственный
характер. Однако некоторые водные животные обладают (как будет
показано позже) поверхностями с весьма большим удлинением (подобно хвосту
марлина, см. рис. 1), и в этом случае возможно использование и двумерного
анализа.
Обе указанные выше ранние работы имели одну общую характерную
черту: сложные плавательные движения рассматривались как небольшие
возмущения стационарного движения вытянутого в струну тела. Позже
теория малых возмущений была применена и к движениям с большим числом
Рейнольдса Лайтхиллом (Lighthill, 1960) и By (Wu, 1961) при исследовании
движений с позиций теории вытянутого тела и методами двумерного
анализа соответственно. Однако очевидно, что такой подход неприемлем в тех
случаях, когда возмущения достаточно велики, что имеет место для многих
практически интересных плавательных движений.
Например, Тейлор получил, что скорость плавания сперматозоида
возрастает пропорционально квадрату амплитуды плавательных
движений (и следовательно, может возрастать неограниченно). Хенкок (Hancock,
1953) под научным руководством автора доклада использовал совершенно
отличный от метода малых возмущений подход. Поле скоростей в его
работе моделируется распределением особенностей, которые расположены на
искривленной осевой линии организма (линия проходит через центры
поперечных сечений тела). Эти наиболее важные сингулярности, которые
являются решениями уравнений Стокса при низких числах Рейнольдса
(впервые эти сингулярности были введены Стоксом в 1851 году), нами были
названы стокслетами.
Решения Хенкока включают стокслеты и диполи, расположенные на
мгновенной осевой центральной линии организма. Для определения их
интенсивности им было выведено интегральное уравнение, которое можно
было решить численным методом, хотя нахождение решения было очень
непростым (с доступными на то время вычислительными устройствами).
Им было получено, что скорость сперматозоида с ростом амплитуды пла-
2. Разделы гидродинамики
177
Д»1
(влияние сил инерции значительно)
Д<1
(влияние сил инерции пренебрежимо мало)
перемещение за счет волнообразного
поперечного движения и его модификации
сферическая геометрия / ДВУМСрНЫе Теории
извивающиеся сферические / ^^^"^ ||||| двумерные рыбы
ш __ „„ и
МаЛЫе (Блейк. 1971а) / (Блейк, 1971b)
возмущения
стационарного
состояния
теория вытянутого тела
теория малых
колебания
вытянутою тела
(Лайтхилл. 1970)
чГ^ш) —•—»—i
(By. 1961)
мосты ■ »цшс
атхуывема»
(ЛаЯпми. 1УШ>
вихревой след
(Вейс. 1972с)
/перемещение за счет
/выбрасывания
/ реактивной струи
<Е
малые
v возмущения
Х^Сихмсин. 1963) стационарного
состояния
возмущения,
распространяющиеся вдоль
осевой линии
организма
теория вытянутого
тела для широхо-
амллитудных движений
| (Лайтхилл, 1971)
маневрирование рыб (Вейс, 1972а)
старт рыб (Вейс, 1972b)
теория, учитывающая наличие сил сопротивления
\
сперматозоиды (Грен к Хснгох, 1955)
возмущения,
распространяющиеся вдоль
осевой линии
организма
(Тейлор. 1952b)
движение, обусловленное согласованными колебательными движениями частей организма
антнпдеатнтовос движение ресничек (Ьлснк, 1971а)
в> -£
образование косяков у рыб
(Вейс, неопубликованная
работа)
Рис. 1. Иллюстрация к подразделению гидродинамики применительно к
перемещению водных животных
вательных движений стремится к некоторой постоянной величине. Этим
же методом была решена задача о скорости движения круглых червей, два
представителя которых изображены на рисунке 1. Однако полный
математический анализ поставленной задачи был настолько сложным, что не было
никаких шансов привлечь к данному методу решения внимание зоологов.
Чрезмерная же громоздкость проведенных вычислений может быть
объяснена их слишком высокой точностью, которая значительно превышала
точность имеющихся экспериментальных данных.
И только при сотрудничестве доктора Хенкока с сэром Джеймсом
Греем в конце концов был успешно разработан метод, который давал количе-
178
Глава 6
ственные результаты с достаточной степенью точности и был понятен всем
заинтересованным в этой задаче специалистам (Gray, Hancock, 1955).
Принятые ими довольно грубые приближения при выводе интегрального
уравнения привели к тому, что интеграл из уравнения исчез, однако полученные
ими результаты достаточно хорошо согласовывались с имеющимися
экспериментальными данными наблюдений за передвижениями сперматозоидов.
Фактически эта аппроксимация означала, что рассматривалось локальное
взаимодействие между малым по длине сегментом вытянутого тела и
находящейся в непосредственной близости к нему жидкой среды.
Взаимодействие определяется только локальной силой сопротивления, зависящей
лишь от относительной скорости этих выделенных объемов и действующей
в направлении по нормали к осевой линии организма.
Ранее в подобной постановке Тейлором (Taylor, 1952b) была решена
задача о перемещении при высоких числах Реинольдса животных (таких как
черви и змеи), не очень хорошо приспособленных к плаванию в воде. Им
также было предложено использовать силы сопротивления с нормальной
или тангенциальной составляющей соответственно при нормальной или
тангенциальной относительной скорости. Единственное отличие состояло
в том, что зависимость между силой и скоростью полагалась линейной при
очень низких числах Реинольдса и квадратичной при высоких.
Возможно, что при средних числах Реинольдса следовало бы выбрать некоторую
промежуточную форму зависимости. Такие теории, где мгновенная сила
взаимодействия, приходящаяся на единицу длины тела животного, зависит
линейно или нелинейно от мгновенной скорости животного относительно
окружающей его водной среды, будем называть резистивными теориями.
Позднее Сэр Джеймс Грей показал в своей монографии
«Передвижение животных» (Gray, 1968), насколько широкий круг проблем может быть
решен методами резистивной теории. Сейчас хотелось бы уделить немного
внимания изложению этой теории для простого случая, когда сила
сопротивления (для любых чисел Реинольдса) линейно зависит от скорости и
косинус угла между касательной к осевой линии животного и направлением
перемещения животного равен единице.
Выделим небольшой участок ВЕС осевой линии животного,
имеющего вытянутую форму тела (рис. 2). Волнообразные движения из стороны
в сторону препятствуют перемещению тела назад при обтекании тела
потоком жидкости со скоростью U. (То есть U — средняя скорость перемещения
животного относительно потока.) Колебания распространяются вдоль тела
в направлении к хвостовой части со скоростью V. Поперечная скорость
2. Разделы гидродинамики
179
осевая линия
направление ^Tj
скорости
потока U
Рис. 2. Связь между поперечными скоростями W относительно животного и w
относительно воды при волнообразном движении
равна W', так что за время т точка осевой линии перемещается в
поперечном направлении на расстояние Wr из точки А в точку В. Волна при
этом пройдет вдоль тела на расстоянии W, переместившись из точки А
в точку С. То есть в точке С находилась точка А за время т до настоящего
момента. За временной интервал т вода переместится на расстояние Ur9
и относительно воды осевая линия переместится в поперечном
направлении на расстояние DE = wr, где w — нормальная скорость относительно
воды. Тогда
(см. уравнение (2) и рис. 5 в работе Lighthill, 1969). Это же уравнение
можно получить аналитическим путем, используя обозначение /i(x, i) для
описания бокового смещения осевой линии и соотношений (Lighthill, 1960)
dh dh TTdh л dh Trdh
w = Tv W=m+UT*< 0=m+vT* (2)
Силу сопротивления, приходящуюся на единицу длины, при
движении в поперечном направлении со скоростью w, обозначим через KNw.
Здесь KN — коэффициент сопротивления (для нормальной компоненты
силы сопротивления). Тогда сила тяги, действующая в продольном
направлении, равна произведению модуля силы сопротивления на тангенс угла
наклона АСВ W/V (см. рис. 2): KNw(W/V). За счет наличия наклона,
характеризующегося отношением W/V, появляется тангенциальная
компонента скорости W(W/V)9 при умножении которой на коэффициент Кт
получаем тангенциальную компоненту силы сопротивления KTW(W/V),
которая уменьшает силу тяги. При этом сила тяги имеет вид
Р = {KNw - KTW)(W/V). (3)
Сила тяги (3), осредненная вдоль осевой линии, уравновешивает
тормозящую силу KTU.
180
Глава 6
Используя равенство (3), можно получить два полезных соотношения.
Полезную мощность UP, определяемую силой тяги, сравним со скоростью
W(KNw) расхода механической энергии при поперечном движении,
когда преодолевается сопротивление среды, определяемое нормальной силой
сопротивления KNw. Отношение этих мощностей называется
эффективностью и равно
" = й^) = (1-^)(1-г^)' (4)
где
т=¥ (5)
— отношение тангенциального к нормальному коэффициенту
сопротивления. Максимальное значение эффективности достигается при
Z = r1/2, тогда £ = 1-^ и г? =(1-г1/2)2. (6)
W V
Эти формулы дают результаты, согласующиеся с имеющимися
наблюдениями — при числах Рейнольдса, не превышающих 1, и при значениях г,
близких к 0,5 (Gray, Hancock, 1955) отношение скорости плавания
животного к скорости распространения волны вдоль его тела U/V равно 0,3.
Отметим, что при таких числах Рейнольдса эффективность достаточно
мала (приблизительно равна 0,1). В случае движения ужа обыкновенного
{Tropidonotus natrix), которое характеризуется числами Рейнольдса
порядка 106, отношение г имеет порядок 0,1. Тогда из (6) следует, что отношение
скоростей U/V равно приблизительно 0,7, что также согласуется с
экспериментальными данными (Gray, 1939). А эффективность движения ужа
имеет большую величину по сравнению с движениями при низких числах
Рейнольдса и, как это следует из (6), равна приблизительно 0,7. Как
отмечал Тейлор (Taylor, 1952b) у многощетинковых червей с многочисленными
выростами-параподиями (рис. 3) тангенциальный коэффициент силы
сопротивления превышает нормальный коэффициент и их отношение может
быть равно г = 1,5. Тогда соотношение скоростей U/V принимает
значение —0,2, что также совпадает с экспериментальными данными. У
многощетинковых червей движение происходит за счет энергичных
волнообразных движений тела, хотя эффективность их движения не превышает 0,05.
Во всех этих случаях результирующую скорость плавания U можно
вывести из условия равновесия между силой тяги Р и силой лобового со-
2. Разделы гидродинамики
181
Рис. 3. Последовательные положения многощетинкового червя {Nereis diversicolor)
при его движении справа налево. Направление распространения поперечной волны
совпадает с направлением движения. На это указывает смещение влево
пронумерованных максимумов. Скорость распространения поперечной волны значительно
больше скорости результирующего движения червя (Gray, 1939)
противления KTU. Из условия равновесия получаем
V WJ\W J r
Максимальное значение скорости продвижения достигается при
1_г2
W 1 /л \
F=2(1 + r)'
и2
что дает —т
W
4г
(7)
(8)
Левая часть (8) изменяется от 2 при г = ОД (для ужа), затем 0,12 при г =
= 0,5 (для круглого червя) и до 0,04 при г = 1,5 (для многощетинкового
червя) и в целом согласуется с имеющимися данными отношения
квадрата скорости перемещения в воде к квадрату средней поперечной скорости.
Действительно, в условиях максимальной эффективности (6) скорость
плавания равна
U2 (1-г1/2)2
Л/2
(9)
182
Глава 6
Значение отношения квадратов скоростей немногим меньше, чем
полученное при расчете по формуле (8) даже при г = ОД, что, возможно, объясняет
тот факт, почему в подобном случае не наблюдаются более высокие
значения отношения w/W.
Эти довольно простые выводы, сделанные на основе резистивной
теории при принятых предположениях о небольшом отклонении формы
осевой линии от прямой (т. е. о равенстве 1 косинуса угла между касательной
к осевой линии и направлением движения животного), можно уточнить,
сняв предположение о равенстве косинуса единице либо о линейной
зависимости силы сопротивления от скорости либо сняв оба эти
предположения, а также проведя более подробные вычисления, которые при данной
постановке остаются все еще не слишком громоздкими. Именно этот
метод можно использовать при исследовании движения организмов, которые
двигаются в среде, затрачивая достаточно много энергии, т. е. с точки
зрения гидродинамики — для движений с достаточно низким числом Фруда,
о котором здесь уже упоминалось.
При появлении рыб и других позвоночных, имеющих широкую
уплощенную хвостовую часть, эффективность движения заметно повысилась.
Такие изменения накапливались отчасти за счет приспособления формы
тела к наилучшему взаимодействию с окружающей водной средой, и
подобное приспособление обусловлено главным образом не наличием силы
сопротивления, величина которой определяется скоростью относительно
воды, а скорее наличием реактивной силы, зависящей от изменения этой
относительной скорости. При высоких числах Рейнольдса сила реакции
возрастает при изменении импульса воды, обусловленного изменением
либо скорости, либо виртуальной массы воды, находящейся в
непосредственной близости от поверхности тела животного.
Сначала был достаточно распространен угревидный способ
передвижения, при котором животное плывет подобно угрю, сильно извиваясь всем
телом, причем расстояние от головы до хвоста значительно превышает
длину волны, распространяющейся вдоль тела (сила тяги при таком способе
в наибольшей степени обусловлена силами сопротивления). Если же
увеличение силы тяги происходит за счет сброса импульса в вихревую пелену,
что обусловлено движением сжатой с боков хвостовой частью, то такое
изменение силы тяги можно рассматривать как результат действия и
силы реакции, и силы сопротивления. Появление совершенно другого
способа плавания, скомброидного, которым пользуются многочисленные группы
рыб и других позвоночных животных, можно объяснить тем, что продвиже-
2. Разделы гидродинамики
183
ние в воде при таком способе плавания обусловлено исключительно силой
реакции, действие которой приводит, по существу, к повышению
быстроходности, характеризующейся большим числом Фруда.
При скомброидном способе плавания по телу животного проходят
поперечные колебания в направлении к хвостовой части, причем фаза
колебаний в поперечном направлении последующих сечений тела отстает от
фазы колебаний предыдущих. Амплитуда колебаний значительно
изменяется; так, на передней половине или даже двух третях длины тела
амплитуда очень мала и затем быстро возрастает по направлению к хвостовой
части. Использующие такой способ плавания рыбы способны внезапно
развить довольно большую скорость и совершать резкие изменения движения.
Результирующая сила тяги в основном обусловлена скоростью изменения
импульса.
Лайтхиллом (Lighthill, 1960, 1969, 1970) впервые был применен
подход к исследованию плавания животных, имеющих вытянутое тело, с
позиций реактивной теории (теории, в которой предполагается, что сила тяги
образуется только за счет действия сил реакции). Им была опубликована
серия работ, где движение животного представлялось в виде малых
возмущений стационарного состояния. Эта теория представляет собой развитие
аэродинамической теории тонкого крыла, однако формулировки и выводы
на основании теории вытянутого тела представляются более доступными
для понимания специалистами в области зоологии. С точки зрения
зоологии при таком типе движения возникает существенная черта, состоящая
в том, что любые некомпенсированные боковые силы, приводят к
потерям, связанным с так называемыми движениями отдачи: скольжения центра
масс рыбы и рыскания относительно центра масс. Лайтхиллом (Lighthill,
1969, 1970) было показано, что скомброидному типу движения
практически всегда сопутствуют некоторые морфологические особенности строения
тела животного, которые минимизируют нежелательные явления «отдачи».
Во-первых, высота рыбы увеличивается в достаточно большой
окрестности центра масс, так что при волнообразном движении тела в
поперечное движение вовлекается большая виртуальная масса воды, что приводит
к уменьшению амплитуды побочных движений, а во-вторых, за счет
того, что непосредственно перед хвостовым плавником, где поперечные силы
особенно велики за счет колебаний хвоста с большой амплитудой,
имеет место существенное сужение тела рыбы и, как следствие, уменьшается
и объем виртуальной массы.
Дальнейшее увеличение скорости движения и повышение
эффективности произошли при скомброидном типе плавания за счет того, что некото-
184
Глава 6
рые животные приобрели хвостовой плавник большого удлинения, в
частности, это были некоторые виды акул и китов. Серповидный хвостовой
плавник с его хорошо закругленным передним краем имеет много
замечательных качеств, присущих аэродинамическому крылу (у представителей
семейства китообразных — китов и дельфинов — хвост располагается в
горизонтальной плоскости и движется сверху вниз, а у большинства акул
и скомброидных рыб хвостовой плавник расположен в вертикальной
плоскости и совершает колебательные движения из стороны в сторону). Сила
тяги и эффективность движения таких животных может быть определена
методами двумерной теории (см. главу 2, § 8 настоящей работы).
В последних разделах своего доклада хотелось бы сделать
сообщение о некоторых моментах, связанных с дальнейшей разработкой теории
скомброидного движения. Во-первых, развит вариант реактивной теории
(глава 5), не использующий теорию возмущений, а по аналогии с ранее
развитой резистивной теорией использующий метод сингулярностей,
расположенных на осевой линии плывущего животного. Автором была разработана
теория движения вытянутого тела, совершающего широкоамплитудные
поперечные колебания, что дало возможность повысить точность получаемых
значений силы тяги при скомброидном способе плавания. Доктору Вейсу
(Weihs, 1972a, 1972b) в результате некоторых усовершенствований методов
этой теории удалось применить ее к решению задач, когда возможность
применения метода малых возмущений исключена, — задач
маневрирования и резкого старта. (На рис. 1 представлены 10 последовательных
положений стартующей и маневрирующей золотой рыбки {Cyprinus auratus).)
Действительно, при движении эти рыбы, практически не затрачивая
энергии, способны самым изящным образом развернуться, скажем, на 90° на
расстоянии менее половины длины своего тела. Было бы весьма
интересно определить, какие же движения они совершают и почему эти движения
столь эффективны с точки зрения затрат энергии.
И наконец, следует осветить некоторые последние результаты
приложения двумерной теории к исследованию движений серповидных
хвостовых плавников и вихревой пелены, образующейся за ними. К ним относятся
и разработки автора в этой области (глава 4), посвященные исследованию
вопроса, почему же такие разные водные животные, как киты, акулы и
тунцы, «пришли» в процессе поиска способов достижения более высоких
скоростей и большей эффективности движения именно к серповидной форме
хвостового плавника. В работах доктора Вейса (Weihs, 1972c)
исследуются вихри, образующиеся в процессе плавания отдельных рыб с серповид-
3. Гидродинамика ресничных организмов
185
ным хвостом, а также проблемы образования и перемещения косяков рыб
с тонким удлиненным телом. В подобных задачах методы теории движения
животных в воде при больших числах Рейнольдса используются для
изучения плавания большого числа тел, совершающих волнообразные движения
и взаимодействующих между собой.
Последняя проблема перекликается с тем материалом настоящей
лекции, где автором описаны наиболее важные недавние разработки доктора
Блейка, касающиеся методов теории движения отдельных жгутиков при
очень низких числах Рейнольдса, для применения их к решению задачи
о перемещении организма за счет согласованных колебательных движений
ресничек. Практически синхронные движения большого числа ресничек
приводят к перемещению организма в жидкой среде. Для изучения
подобных движений были разработаны два существенно различающихся между
собой подхода. Один из них основан на теории малых возмущений,
примененной к симлектическому движению ресничек у одноклеточных
организмов, подобных опалине (Opalina), а другой — на резистивной теории
широкоамплитудных волнообразных поперечных движений, примененной
к антиплектическому движению ресничек у организмов, подобных
туфелькам (Paramecium).
3. Гидродинамика ресничных организмов
Ресничные (Ciliophora) — это класс одноклеточных организмов,
которые перемещаются в окружающей водной среде за счет согласованного
биения большого числа волосковидных отростков на поверхности клетки. Эти
отростки, реснички, совершают не полностью синхронные колебательные
движения. Реснички движутся так, что фаза движения каждого слоя
ресничек запаздывает относительно фазы движения предыдущего слоя (так
называемые метахронные волны, метахронизм). Создается впечатление, что
по поверхности ресничек проходят волны. Движение отдельной реснички
представляет собой сначала быстрый резкий жесткий гребок в
направлении, противоположном направлению движения организма, при этом
ресничка практически не изгибается, а затем медленное плавное возвратное
движение уже относительно расслабленной реснички. Метахронизм
называется симплектическим, когда метахронная волна и гребной удар
одинаково направлены. Это напоминает механизм продвижения водных животных
за счет волнообразных движений тела, когда поперечная волна
распространяется вдоль тела в направлении, противоположном движению животного.
186
Глава 6
Типичным представителем простейших с симплектическим метахро-
низмом является опалина (Opalina) — простейшее дискообразной формы,
размером около 200 мкм ширины и 20 мкм толщины, в клетке которого
находится несколько ядер. Отдельная ресничка имеет длину порядка 10 мкм,
реснички расположены в одном ряду на расстоянии 0,3 мкм друг от друга,
ряды ресничек располагаются на расстоянии 3 мкм друг от друга. Кончики
ресничек расположены близко один от другого. Это объясняется тем, что
при быстром, резком гребке, в результате совершения которого ресничка
могла бы отделиться от соседних ресничек, она движется в направлении
к той соседней, которая несколько ранее по времени совершает это
движение, и от той соседней, циклическое движение которой запаздывает. В
результате их согласованного движения по кончикам ресничек, образующим
поверхность, проходит волна подобно волнам, пробегающим по полю
злаковых. Это похоже на то, как если бы дискообразное тело с очень гибкой
поверхностью совершало бы волнообразные движения, так что волны на
поверхности распространялись бы в направлении, обратном направлению
движения, в результате чего вырабатывалась сила тяги, необходимая для
перемещения организма в жидкости.
Интересно, что это была именно та гидродинамическая задача,
которую автор пытался решить 20 лет назад (Lighthill, 1952) методами
сферической геометрии. В то время автор в начале своей научной деятельности
рассуждал так: «Тейлор пытается найти ответ на вопрос, как клетки могут
продвигаться в окружающей среде за счет движения хлыста или жгутика,
а я займусь исследованием продвижения клеток без жгутика, которое
обусловлено какими-нибудь извивающимися движениями...», хотя, конечно,
следовало бы прежде всего обратиться к зоологам для выяснения
механизма продвижения простейших, не имеющих жгутиков. В процессе
написания автором обзора 1969 года стало очевидно, что этот метод анализа
применим именно к симплектическому движению ресничных (Ciliophora). И,
как следствие, первая часть работы, которую Блейк выполнил после того,
как автор посоветовал ему выбрать в качестве научной темы диссертации
на звание доктора философии гидродинамику продвижения ресничковых
организмов, была посвящена разработке и усовершенствованию методов
той самой сферической геометрии применительно к продвижению
опалины (Blake, 1971а).
Он достаточно быстро добился прогресса в этом направлении, его
теоретически полученные выводы о том, что скорость продвижения опалины
может достигать скорости около 100 мкм/с, подтверждались имеющимися
3. Гидродинамика ресничных организмов
187
экспериментальными данными. Интересно, оказалось, что найденное
значение потребной мощности, выраженной через эффективность, не
превышало 0,03, потребная мощность определялась при расчете движения
организма с такой же скоростью под действием постоянной внешней силы.
(Это соответствует значению 0,1 при низком числе Рейнольдса,
полученному в § 2 при перемещении вытянутого тела, обусловленном
волнообразными поперечными движениями.) Хотя были проведены расчеты далеко
не для всех форм волн, обеспечивающих большие скорости или даже
просто положительные, но совпадение результатов проведенных расчетов
формы волны, совпадение предсказанных теоретически расположений пиков
и ровных участков с имеющимися экспериментальными данными дает
возможность надеяться, что разработанный подход достаточно эффективен.
Другой подход, разработанный Блейком (Blake, 1971b) при
исследовании перемещения опалин состоял в том, что оболочка, образованная
кончиками ресничек, рассматривалась как плоская неограниченная поверхность.
Такой выбор модели он обосновывал тем, что дискообразное тело
занимает промежуточное положение между сферой и плоскостью. Исследуя
поведение простейших в свете выбора модели движения, Блейк пришел
к выводу, что «плоскостность оболочки значительно более важна, чем ее
конечность», показав, что колебательное движение ресничек,
прикрепленных к плоскости, параллельной направлению перемещения организма,
более точно моделирует истинное перемещение, чем движение ресничек,
прикрепленных по всей сферической поверхности. При наблюдении за
животными создается впечатление, что в природе у большинства организмов,
перемещающихся за счет движения ресничек, реснички расположены на
поверхностях, параллельных направлению движения. Отметим, что
предсказанная теорией скорость плавания не зависит от площади диска,
поскольку сила тяги и сила лобового сопротивления в равной степени возрастают
пропорционально росту площади поверхности диска, так что плоскостная
модель поверхности организма имеет право на существование и может дать
с приемлемой степенью точности верные результаты.
Плоскостная модель имеет и другие преимущества. Так, ее применение
дает возможность провести сравнение эффективности различных по форме
волнообразных движений, и на основании этой модели из теоретических
исследований вытекает, что с наименьшими энергетическими затратами
заданная скорость перемещения достигается при симплектическом движении
в том случае, когда кончики ресничек описывают траектории, подобные
траекториям частиц на поверхности волны на воде. И как раз у опалины
188
Глава 6
движение кончиков ресничек очень напоминает движение волн на воде.
Другим достоинством плоскостной модели является то, что ее
применение дает возможность в методе малых возмущений использовать
разложение решения по малому параметру до степеней высокого порядка. Блейк
(Blake, 1971b, 1972b) смог показать, используя метод малых возмущений,
что даже при волнах максимально возможной амплитуды с заостренными
пиками отношение скорости плавания к скорости распространения волны
вдоль организма (то, что в настоящей работе, в § 2, обозначено через U/V)
не может превышать 0,5. А это согласуется с большим числом наблюдений
за симплектическим движением.
И теперь остановимся кратко на третьем подходе, который
использовал Блейк (Blake, 1971b), моделируя перемещение организма с
антиплектическим движением тела вращения. Этот подход совершенно отличен от
других им разработанных и основан на положении, что скорость плавания
тесно связана со скоростью распространения волны вдоль организма, либо
с мощностью (более точно, с корнем квадратным из мощности) либо с
обеими вместе. Прежде всего, это происходит потому, что отдельная ресничка
имеет больше свободы движения за время совершения резкого гребка при
антиплектическом движении по сравнению с симплектическим.
Действительно, во время резкого гребка ресничка движется в сторону соседней
реснички с опережающей фазой совершения гребка, которая уже
отклонилась от нее, а другая соседняя ресничка с запаздывающей фазой
цикла плавательного движения еще отклонена от совершающей резкий гребок
реснички в противоположном направлении. Таким образом, самый важный
такт движения каждой реснички, за счет которого и происходит
продвижение организма, выполняется ею без влияния со стороны соседних
ресничек. И следовательно, при антиплектическом движении моделирование
геометрических мест кончиков ресничек непрерывной гибкой
поверхностью невозможно. Одним из представителей простейших с
антиплектическим метахронизмом является инфузория туфелька (Paramecium). Это
многоядерное одноклеточное простейшее имеет форму сплюснутого
вытянутого сфероида длиной 250 мкм, длина ресничек составляет 10 мкм, а длина
метахрональной волны равна 20 мкм. Инфузория туфелька способна
передвигаться со скоростью до 1000 мкм/с, что на порядок превышает скорость
опалины (Opalina) (а отношение U/V даже превышает единицу).
Блейк (Blake, 1972a) разработал подход к решению задачи о
движении простейших с антиплектическим движением ресничек, основанный на
фундаменте, заложенном Греем и Хенкоком (Gray, Hancock, 1955). Он рас-
3. Гидродинамика ресничных организмов
189
сматривал каждую ресничку как некоторое распределение стокслетов на
ее осевой линии, обусловленных силами взаимодействия между ресничкой
и окружающей ее жидкостью. В его модели имелись два существенных
отличия от модели Грея и Хенкока.
Во-первых, Блейк учел наличие поверхности, к которой прикреплена
ресничка. Он рассматривал стокслет вблизи бесконечной плоской стенки
и показал, что (Blake, 1971c) решение уравнений Стокса в этом случае
может быть выражено через систему изображений. Каждая система
изображений определяется компонентой силы — перпендикулярной или
параллельной границе (см. рис. 4 и 5). В первом случае система состоит из равных по
величине и противоположных по направлению стокслетов, плюс стокс-ди-
поль (состоит из двух равных по величине и противоположно
ориентированных стокслетов), плюс источник-диполь. Поле на достаточном удалении
от стенки представляется как комбинация стоке-диполей со скоростями,
убывающими как г~2. Во втором случае стокс-диполь уже другой и
уравновешивает эквивалентный стокс-диполь, обусловленный исходным стокс-
летом и его противоположно направленным изображением. В этом случае
поле на достаточном удалении от стенки уже значительно слабее и убывает
со скоростью г~3. Для продвижения реснитчатого организма это означает,
что наибольшее влияние на относительное движение воды и стенки
оказывают силы, параллельные границе, а не перпендикулярные ей. Позднее
Блейк обнаружил подобную систему изображений у выдающегося ученого
прошлого столетия Лоренца (1896 г.), однако до Блейка столь эффективно
эта система в прикладных задачах не использовалась.
Во-вторых, он учел тот факт, что ресничка совершает гребки в среде,
возмущенной плавательными движениями других ресничек. Именно этот
учет влияния большого числа ресничек и представляет наибольшую
сложность.
Выбранный метод решения аналогичен тому, который используется
в ядерной физике при изучении структуры атомов с большим атомным
числом, т. е. содержащих большое число электронов. Используется так
называемая модель самосогласованного поля, когда взаимодействие между
электронами определяется усредненным радиальным электростатическим
полем. Одночастичные волновые функции каждой частицы удовлетворяют
уравнению Шрёдингера и должны быть согласованными со средним
потенциалом введенного электростатического поля, в котором движутся эти
электроны. Для решения задачи, поставленной Блейком, подобный метод
подходил как нельзя лучше.
190
Глава 6
Ч
поле на достаточном ш
удалении от стенки: I • |
стокс-диполь
стокслет (F)
У,-'''
V- +J.f + . .
система стокслет стоке- источник-
изображений: ( F) диполь диполь
(2hF) (-2hF)
Рис. 4. Система изображений для стокслета с интенсивностью F, направленной
параллельно плоской твердой стенке х3 = 0. Стокслет находится на расстоянии h
от стенки (Blake, 1971c)
*з
стокслет (F)
У У
поле на достаточном
удалении от стенки:
стокс-диполь,
источник-диполь
У \
V
+
система стокслет стоке- источник-
изображений: (-F) диполь диполь
(2hF) (-2h2F)
Рис. 5. Система изображений для стокслета с интенсивностью F, направленной
перпендикулярно твердой стенке х3 = 0. Стокслет находится на расстоянии h от
стенки (Blake, 1971c)
Компонента скорости, параллельная стенке, зависит от расстояния от
стенки и обозначается через U(x3). При х3 —> оо эта скорость равна
скорости перемещения организма, при стремлении х3 к нулю скорость умень-
3. Гидродинамика ресничных организмов
191
шается до нуля, что соответствует тормозящему свойству подслоя,
образованного ресничками. Функция U(x3) определяется из условий
самосогласованности следующим образом. Грей и Хенкок (Gray, Hancock, 1955)
определяют силу, с которой каждая ресничка действует на жидкость, через
скорость реснички относительно жидкости (U(x3), О, 0). Такая сила
представляется стокслетом и его системой изображений, о которой говорилось
выше. Создаваемое каждым элементом системы поле скоростей
записывается и затем суммируется. Полученная сумма упрощается с использованием
формулы Пуассона. Из условий самосогласования следует, что эта сумма
есть искомая скорость. Выведенное интегральное уравнение относительно
скорости U(x3) решается численным методом.
На рисунке 6 представлены результаты расчетов скоростей для трех
животных: инфузории туфельки, опалины и гребневика плевробрахии. По
оси х откладывается безразмерное расстояние до стенки (х3 делится на
длину реснички L), а по оси у — безразмерная скорость (скорость U
делится на произведение длины L и частоты <т, измеряемой в рад/с). Расчеты
были проведены для различных значений частотного параметра к9 который
равен отношению <тЬ к скорости волны с, и различных значений 7,
равного отношению коэффициента нормальной составляющей силы к
коэффициенту тангенциальной составляющей силы, который обратно
пропорционален величине г (см. § 2). Отметим, что скорость перемещения
инфузории туфельки (Paramecium) (антиплектическая метахрония) значительно
выше, чем у опалины (Opalina) с ее симлектической метахронией. Третье
животное гребневик плевробрахия (Pleurobrachia) — организм,ё подобный
медузам, имеет студенистое тело, перемещается при помощи специальных
гребных пластинок — склеенных между собой пучков ресничек с антиплек-
тическими метахрональными волнами.
В своих расчетах Блейк (Blake, 1972a) использовал еще один
параметр т, пропорциональный плотности реснички на единицу площади.
Результаты, приведенные на рисунке 6, получены при т = 150. На рисунке 7
представлена зависимость скорости U°° (скорости жидкости относительно
организма вне слоя ресничек) от параметра т. Ограниченное время доклада
не позволяет подробно остановиться на многих других полученных
результатах, которые представляют несомненный интерес для зоологов: а именно,
как силы и изгибающие моменты, прикладываемые к ресничкам со
стороны животного, изменяются в течение одного плавательного цикла, а
также как изменяется величина развиваемой организмом средней мощности.
И, несмотря на недостаток времени, все же отметим, что Блейком (Blake,
192
Глава 6
1972b) было получено хорошее согласие (только для симлектического ме-
тахронизма) между оболочковой моделью и приведенным здесь
значительно более полным анализом.
О 0,5 1,0 1,5 0 0,5 1,0 0 0,2 0,4
U/al U/al U/al
Рис. 6. Распределение скорости U(x3) в подслое ресничек. L — длина реснички,
а — частота в радианах, г = 150 (Blake, 1972a)
4. Теория вытянутого тела при широкоамплитудных
волнообразных поперечных движениях
А сейчас следует вернуться к движениям при больших числах Рей-
нольдса и методам, основанным на учете силы реакции (методы резистив-
ных теорий), исследования которых приведены в последней работе автора
(Lighthill, 1971). Резистивная теория доработана таким образом, чтобы ее
методы можно было применять для исследования движений организмов,
имеющих вытянутое тело и совершающих поперечные волнообразные
движения с произвольной амплитудой.
Так же как и для движений при низких числах Рейнольдса, на осевой
линии животного распределяются определенные сингулярности, в данном
случае это диполи, обусловленные переносом импульса (источник-дублет).
Поскольку в дальнейшем речь пойдет только о рыбах, то центральную ось
4. Теория вытянутого тела
193
1,6
300
I - для гребневика плевробрахии
II - для инфузории-туфельки
III - для опалины
Рис. 7. Зависимость скорости передвижения организма относительно жидкости на
достаточно большом расстоянии от поверхности тела организма, т. е. lim U(x3) =
х3—оо
= U°°9 в зависимости от параметра т (Blake, 1972a)
симметрии животного будем называть позвоночным столбом, или
позвоночником.
В результате долгих поисков правильного подхода к проблеме автор
в конце концов пришел к заключению, что теоретический подход должен
основываться на следующих трех положениях.
(i) Импульс воды вблизи тела рыбы направлен перпендикулярно
позвоночнику, и величина импульса равна произведению виртуальной
массы га, приходящейся на единицу длины, на компоненту w скорости
рыбы в этом направлении,
(ii) Сила тяги определяется скоростью изменения импульса в объеме V,
охватывающем рыбу и с одной стороны ограниченном плоскостью П,
которая проходит через заднюю кромку хвостового плавника и
перпендикулярна ему.
194
Глава 6
(iii) В уравнении для количества движения учитывается перенос
импульса через плоскость П не только за счет адвекции, но и за счет
действия суммарного давления ~mw2, обусловленного движением в
плоскости П.
Так как в выбранный объем V вихревая пелена за хвостовым
плавником не включается, то при составлении уравнений движения особенности
потока в спутном следе рассматриваться не будут. И поэтому
результирующее давление на плоскости П, обусловленное движением на плоскости,
равно ~mw2.
Для описания движения выберем систему координат следующим
образом (см. рис. 8). Лагранжева координата а произвольной точки
позвоночника равна расстоянию от хвоста рыбы до этой точки, измеренному вдоль
позвоночника. Ось у направлена вертикально вверх, а взаимно
перпендикулярные оси х и z расположены в горизонтальной плоскости и в общем
случае развернуты произвольно. При стационарном движении рыбы
направление оси х совпадает с направлением скорости перемещения и.
Компонента скорости, перпендикулярная к позвоночнику равна w, а тангенциальная
компонента скорости равна и.
голова рыбы
((*(М),*(М)))
хвост рыбы
((x(a,t2),z(a,t2)))
((x(Mi),z(a,ti)))
х
касательная
Рис. 8. Обозначения, принятые в работе Лайтхилла (Lighthill, 1971) для
описания широкоамплитудных поперечных движений плывущих рыб. Система координат
(х, у, z) неподвижна относительно жидкости на бесконечности
4. Теория вытянутого тела
195
Условие нерастяжимости позвоночника запишем в виде
(10)
Единичные векторы в направлении нормали и касательной к позвоночнику
в горизонтальной плоскости у = 0 имеют соответственно координаты
(§•§) и (-£•©• (п)
Откуда получаем
_ дх дх dz dz _ dz дх дх dz .
dt да dt да' dt да dt да'
Теперь запишем уравнение количества движения для определения
силы реакции (Р, Q), возникающей при поперечном движении тела рыбы.
Эта сила реакции (Р, Q) приводит к возникновению силы тяги,
необходимой для преодоления действия вязкой силы сопротивления при движении
рыбы в тангенциальном направлении. Другими словами, основное
положение данного метода состоит в том, что поперечному движению
препятствует сила реакции, а тангенциальному — сила сопротивления.
Сила реакции (Р, Q), представляющая собой часть общей суммарной
силы, удовлетворяет уравнению
d Г ( dz дх\ _
dt J \ да' да J
Г / dz дх\ 1 2 (дх дг^
V да" да) 2 V да' да
-(P,Q). (13)
J а=0
Слева стоит изменение импульса, приходящегося на единицу длины, в
объеме V, охватывающем рыбу без вихревой пелены, которая развивается за
ней в процессе плавания. Величина импульса равна проекции mw на
единичный вектор, перпендикулярный к позвоночнику (см. (11)). В правой
части (13) три слагаемых соответственно означают (i) потери импульса за
счет адвекции со скоростью и через плоскость П; (ii) прирост за счет
действия силы давления ^rnw2, действующей в направлении нормали к
плоскости П; (Hi) потери импульса за счет воздействия на рыбу со стороны
жидкости с силой (Р, Q).
196
Глава 6
С использованием (12) равенство (13) принимает вид
(Р, Q) =
dz дх
1
mw
Tt^Tt~2mW
(дх_ dz\\
Уда' да)\а=0
i
-11mw (-£• ё)da- (14)
С учетом того, что среднее значение производной по времени равно нулю,
для средней силы тяги Р получаем
Р-=
mw
dt
— —mw'
2
>дх
да
(15)
Здесь черточка сверху означает осреднение. Производная по времени в (14)
для силы тяги играет существенную роль, поскольку иначе колебание в
значении силы тяги было бы очень велико, обращаясь в нуль всякий раз, когда
w = 0 при а = 0. Лайтхилл (Lighthill, 1971) показал, что слагаемое,
содержащее производную по времени, сглаживает флуктуации силы тяги. Вторая
компонента Q силы реакции играет существенную роль в процессах,
которые названы отдачей (§ 2).
Проводя аналогичные рассуждения, получим среднюю скорость
потерь энергии:
Т =
-mw2u
(16)
которые обусловлены сходом со скоростью и через плоскость П
кинетической энергии ^mw2, приходящейся на единицу длины. Тогда
эффективность по Фруду следующая:
т? =
UP
UP + T'
(17)
в которую входят полученные средняя сила тяги, средняя скорость потерь
энергии и средняя скорость плавания
U =
дх
ъ
да
(18)
а=0
которая может быть выражена через величины на задней кромке
хвостового плавника. Это позволит понять, почему эффективность по Фруду равна
4. Теория вытянутого тела
197
приблизительно 80%, что сильно отличается от значений эффективности
в 50 % (§2) при плавании при больших числах Рейнольдса, когда в
качестве сил, препятствующих поперечному волнообразному движению тела,
рассматриваются силы сопротивления (подход с позиций резистивной
теории).
При анализе возможности за счет силы тяги, обусловленной силами
реакции при поперечном волнообразном движении, преодолеть
сопротивление движению в тангенциальном направлении (такой анализ был
проведен Лайтхиллом (Lighthill, 1971) при исследовании плавания плотвы
(Leuciscus)) следует учитывать, что толщина пограничного слоя,
развивающегося на теле плывущей рыбы, мала. При боковых движениях тело рыбы
вступает в контакт каждый раз со свежей порцией воды, следовательно,
сила сопротивления имеет большую величину при плавании рыбы, чем при ее
скольжении. Относительно большие амплитуды волнообразного движения
хвостового плавника наблюдаются у большинства плывущих рыб, и такой
способ плавания можно объяснить в терминах производства силы тяги. При
больших амплитудах движения хвостовой части тела за счет сил реакции со
стороны жидкости возникает сила тяги, достаточная для преодоления
животным значительной по величине силы вязкости, действие которой
проявляется с момента начала волнообразного движения. Равномерное движение
с малыми по амплитуде поперечными колебаниями хвостового плавника
может быть довольно энергозатратным и значительно менее эффективным
по сравнению с неравномерным движением, когда широкоамплитудные
резкие движения хвостового плавника, за счет которого достигается довольно
большая скорость, перемежаются периодами пассивного скольжения. Такое
неравномерное движение довольно часто можно увидеть при наблюдении
за плаванием рыб в аквариумах.
Рассмотренный здесь теоретический подход был использован Вейсом
(Weihs, 1972a, 1972b) при исследовании маневрирования и начальной фазы
движения (старта) рыб. Опишем здесь его исследование процесса
поворота — изменения направления движения. В 1933 году сэром Джеймсом
Греем были получены прекрасные кадры, где был запечатлен процесс
поворота рыбы при изменении курса. До работ Вейса анализ маневрирования
рыб проводился исключительно с позиций гидродинамики. Начиная с
момента сотрудничества в Кембридже с автором данного доклада (с 1971 г.),
Вейс внес дополнительные усовершенствования в представленный выше
метод, связанные с детальным рассмотрением движения плавников при
маневрировании рыбы. Для сил, действующих на тело рыбы, он использовал
198
Глава 6
приведенный здесь метод, основанный на динамике виртуальной массы,
а затем им было показано, что необходимо изменить аэродинамические
коэффициенты силы, действующей на плавники, которые, как он считал, были
недостаточно «тонкими», чтобы было возможно без изменений применить
методы теории вытянутого тела. Он рассчитал силы и моменты,
действующие на рыбу на каждой из трех стадий процесса поворота, состоящего из
движения ее центра масс и вращательного движения рыбы относительно
центра масс.
Рис. 9. Покадровое изображение через интервалы 0,04 с (время
возрастает слева направо) процесса поворота вправо, совершаемого плотвой (Cyprinus
erythrophthalmus) (Weihs, 1972a)
На рисунке 9 представлены взятые из работы сэра Джеймса Грея кадры
последовательного разворота, совершаемого плотвой {Cyprinus
erythrophthalmus). Примечательно, что рыба совершает разворот на чрезвычайно
коротком отрезке с очень небольшими затратами энергии. Картина
согласуется с выводами работы доктора Вейса о стадиях процесса разворота.
Сначала плавно разворачивается головная часть животного, а затем следует
энергичный завершающий поворот взмах хвостовым плавником. Выигрыш
от такого поочередного использования передней, а затем хвостовой частей
в качестве элементов, управляющих движением, кажется совершенно
очевидным.
На рисунке 10 представлены последовательные положения спинного
хребта разворачивающейся плотвы, рассчитанные на основании
имеющихся съемок. Кружочками отмечается положение центра масс. На первой
стадии (кадры 1, 2 и 3) центр масс еще движется по прямой, а передняя часть
рыбы начинает совершать поворот вправо, изменение импульса на этой
стадии (слагаемое с d/dt в (14)) компенсируется значительными силами,
обусловленными спутным следом, сходящим с плавников за счет небольшого
движения влево хвостовой части рыбы. Суммарная сила, действующая в
поперечном направлении, равна нулю, и в это время на тело действует
возрастающая по величине пара сил, обеспечивающих поворот относительно
центра масс.
4. Теория вытянутого тела 199
25
20
15
10
5
0 5 Ш 15_
Рис. 10. Кривые изображают положения спинного хребта плотвы, совершающей
поворот. Цифры у кривых соответствуют номерам кадров на рисунке 9. Расстояние
измерено в сантиметрах (Weihs, 1972a)
На второй стадии (кадры 4, 5 и 6) центр масс перемещается по сильно
искривленной линии, что указывает на действие на этой стадии
значительной по модулю силы, направленной к центру поворота. Это обусловлено
возрастающим влиянием силы, вырабатываемой значительным поворотом
хвостовой части, и значения сил, рассчитанные на основе разворота на
данной стадии, согласуются с экспериментальной величиной
центростремительного ускорения.
На третьей стадии (кадры 7, 8 и 9) работает только хвостовая часть,
и гидродинамическая «подъемная» сила (на самом деле поперечная сила),
действующая на хвостовой плавник, обеспечивает движение рыбы в на-
200
Глава 6
правлении уже нового курса движения с ускорением, наличие которого
сбалансирует все потери импульса за счет трения в процессе разворота.
Момент этой силы относительно центра масс таков, что предотвращается
дальнейший разворот рыбы на третьей стадии, и при завершении поворота
происходит движение только по прямой.
Изложенный анализ подтверждается расчетами сил и моментов,
проведенными доктором Вейсом, за исключением того, что на второй и третьей
стадии под действием рассчитанных моментов поворот должен был
происходить со значительно меньшей скоростью, чем следовало из наблюдений.
Затем Вейс обратил внимание на одну важную деталь, ранее не
замеченную им на кинокадрах. А именно, непосредственно перед началом второй
стадии рыба расправляет и затем поворачивает правый грудной плавник,
за счет чего скорость прямолинейного движения уменьшается и
увеличивается скорость поворота. Этим объясняются довольно большая скорость
поворота в начале второй стадии и тот факт, что вращающие моменты
имеют большие значения, чем при отсутствии дополнительного движения
правого грудного плавника. Незначительное уменьшение скорости движения
вдоль кривой уменьшает радиус поворота, происходящего под действием
заданной боковой силы, пропорционально корню квадратному из
величины снижения скорости. А затем в результате завершающего резкого взмаха
хвостового плавника скорость движения довольно легко увеличивается до
прежнего значения! Действие грудного плавника подобно
аэродинамическому тормозу, который расположен с той стороны, в которую рыба
поворачивает, и проявляется в резком гребке в момент между стадией 1 (когда
головная часть только начинает разворот) и второй стадией, когда хвостовая
часть вступает в движение, сильно изгибаясь, а головная часть уже
движется по прямой. И наконец, третья стадия — резкое выпрямление хвостовой
части завершает — процесс поворота. Вот интересно, сможет ли
когда-нибудь так же эффективно развернуться корабль?
5. Вихревые следы
Время позволяет лишь кратко коснуться последней темы — вихревых
следов, — которая является последней потому, что именно их оставляет
плывущая рыба за собой. Прежде всего, с огромным удовольствием
хотелось бы начать с вклада Кармана и Бюргерса (Karman, Burgers, 1934).
Рисунок 11 заимствован из их знаменитой совместной работы в серии
«Аэродинамика».
5. Вихревые следы
201
Рис. 11. Иллюстрация к процессу сбрасывания вихрей с хвостового плавника
при движении рыбы влево и струйного течения, индуцированного этими вихрями
(Lighthill, 1969 из работы Karman, Burgers, 1934)
В своей работе Карман и Бюргере подчеркивают, что знаменитая
дорожка Кармана может давать и обратный эффект, а именно если
направление всех вихрей изменится, то дорожка Кармана будет способствовать
возникновению силы тяги, а не приводить к увеличению сопротивления
движению. При изменении направления вращения вихрей созданное ими
струйное течение переносит импульс, который обусловливает силу тяги
рыбы. И возникает значительно более тесная взаимосвязь, чем это может
показаться на первый взгляд, между волнообразным способом перемещения
и реактивным движением.
Следует упомянуть о нескольких способах усовершенствования
этого упрощенного представления о течении за телом и о двух приложениях,
интересных для зоологов. Картину вихревого течения за телом рыбы,
представленную на рисунке 11, можно рассматривать как иллюстрацию чисто
двумерного течения в спутном следе, скорее всего, за рыбами,
обладающими серповидным хвостовым плавником большого удлинения (см. §2).
Доктор Вейс предположил, что такая двумерная картина реализуется
реально при коллективном движении рыб, расположение которых одна над
другой позволяет достичь высокого отношения вертикального масштаба к
горизонтальному.
Кроме того, доктор Вейс (Weihs, 1971c) изучил в двумерной
постановке поведение этих ранее не рассматривавшихся вихревых дорожек, которые
порождают силу тяги, и обнаружил интересное поведение вихрей с
удалением от животного. Оказалось, что вниз по течению дорожка становится
шире где-то в 1,5 раза и расстояние между вихрями изменяется.
Конечно, где-нибудь далеко от рыбы такое распределение вихрей
в спутном следе будет видоизменяться в результате тормозящего действия
окружающей среды. Отметим, что для рыб с серповидным хвостом вихри
будут более сконцетрированы и иначе расположены (в окрестности
центра масс плавника). На некотором расстоянии вниз по течению изменения
в распределении завихренности должны привести к уменьшению до нуля
202
Глава 6
импульса в следе, поскольку средние значения силы тяги и сопротивления
там обращаются в нуль.
И теперь хотелось бы остановиться на трехмерных эффектах и
возможном объяснении на основании этих эффектов того факта, почему наиболее
быстрые морские животные совершенно неродственных групп в процессе
эволюции приобрели серповидный хвостовой плавник. Детальный
математический анализ в двумерной постановке положительных качеств
хвостового плавника, проведенный автором (глава 4), показал, что оптимальным
будет плавник с прямой задней кромкой, более точно — передняя кромка
выпукла вперед, а задняя кромка прямая. А из этого следует, что любое
преимущество, которое дает искривление задней кромки в сторону движения
животного, необходимо обосновывать исходя из трехмерной постановки.
В трехмерной постановке при рассмотрении вихрей, сходящих с
серповидного плавника конечного размаха, следует также учитывать
завихренность и в поперечном к потоку направлении (показано на рис. 11).
Схематичная картина распределения вихрей за рыбой, изображенная на
рисунке 11, при трехмерном подходе представляет собой горизонтальное сечение
серии вихревых колец, которые образуются поочередно то с левой, то с
правой стороны животного за счет биения хвостовым плавником. При наиболее
типичной частоте колебательных движений плавника срывающиеся вихри
имеют форму практически круговых колец, и, очевидно, именно
искривленная задняя кромка плавника способствует образованию таких круговых
колец, которые в общем случае являются лучшим механизмом переноса
наибольшего по величине импульса при заданной энергии.
Сначала вихревые кольца движутся назад в диагональном
направлении относительно направления движения животного, а затем в результате
тормозящего действия силы сопротивления движение их будет происходить
только в поперечном направлении. Доктор Маккатчен (McCutcheon) в
частной беседе с автором упомянул, что он совсем недавно наблюдал подобное
распространение вихревых колец в поперечном направлении в вихревом
следе за рыбой, использующей скомброидный способ плавания. С нашей
точки зрения кажется наиболее убедительным то, что именно серповидным
хвостом определяется образование круговых вихрей в следе за животным,
и в своих дальнейших исследованиях автор продолжит попытки более
удовлетворительно обосновать это утверждение.
В заключение перейдем к процессу образования косяков из рыб. Как
уже упоминалось, схематичное изображение спутного следа за плывущей
рыбой на рисунке 11 подобно двумерному изображению движения несколь-
5. Вихревые следы
203
ких особей, плывущих одна над другой. Такой способ передвижения
группы рыб дает им выигрыш в мощности при обеспечении движения с
заданной скоростью. Понятно также, что плывущей рыбе помогают продвигаться
вперед вихри в спутном следе за плывущей впереди нее по диагонали
другой рыбой. Именно этим можно объяснить ромбовидное расположение рыб
(рис. 12), при котором на движение с заданной скоростью отдельная особь
затрачивает меньше энергии. Доктор Вейс в неопубликованной работе
рассчитал реальный выигрыш в мощности при ромбовидном размещении.
Рис. 12. Ромбовидное расположение рыб в косяке
Имеется достаточно большое число наблюдений, указывающих на то,
что подобное ромбовидное решетчатое распределение рыб действительно
имеет место в случае перемещения косяка с большой скоростью. И
возникает вопрос: за счет ли реакций связи (пассивных сил) поддерживается
такое размещение или существуют некоторые другие сложные
управляющие процессы, действующие в косяке для поддержания подобного
размещения. Доктору Вейсу удалось выяснить некоторые механизмы достаточно
эффективного воздействия пассивных сил. В шутку автор предположил,
что в целом данный феномен подобен тому процессу, когда с
понижением температуры молекулы жидкости формируют решетчатую
кристаллическую структуру. Другими словами, при росте скорости косяка выигрыш
в энергии при перестраивании рыб в решетчатую ромбовидную структуру
предотвращает потери энтропии.
204 Глава 6
Литература
Blake, J.R. 1971a. J. Fluid Mech. 46, 199-208.
Blake, J.R. 1971b. Ibid. 49, 209-22.
Blake, J.R. 1971c. Proc. Camb. Phil. Soc. 70, 303-310.
Blake, J.R. 1972a. J. Fluid Mech. 55, 1-23.
Blake, J.R. 1972b. Ph.D. thesis, University of Cambridge.
Gray, J. 1939. J. Exptl. Biol. 16, 9-17.
Gray, J. 1968. Animal Locomotion, Weidenfeld & Nicolson, London.
Gray, J. & Hancock, G.J. 1955. J. Exptl. Biol. 32, 802-814.
Hancock, G.J. 1953. Proc. Roy. Soc. A, 217, 96-121.
Karman T. von & Burgers, J. M. 1934. General Aerodynamic Theory:
Perfect Fluids, vol. 2 of Aerodynamic Theory (ed. Durand, W. F.), Springer,
Leipzig.
Lighthill, M.J. 1952. Comm. Pure Appl. Math. 5, 109-118.
Lighthill, M.J. 1960. J. Fluid Mech., 9, 305-317.
Lighthill, M.J. 1969. Ann. Rev. Fluid Mech., 1, 413-446. (Глава 2.)
Lighthill, M.J. 1970. J. Fluid Mech. 44, 265-301. (Глава 4.)
Lighthill, M.J. 1971. Proc. Roy. Soc. B, 179, 125-138. (Глава 5.)
Lighthill, M.J. 1972. J. Fluid Mech. 52, 475^97. (Главы 10 и 14.)
Lorentz, H.A. 1896. Zittingsverlag Akad. v. Wet. 5, 168-187.
Siekmann, J. 1963. J. Fluid Mech. 15, 399-418.
Stokes, G.G. 1851. Trans. Camb. Phil. Soc. 9, pt. II, 8-106.
Taylor, G. I. 1951. Proc. Roy. Soc. A, 209, 447^61.
Taylor, G.I. 1952a. Ibid. 211, 225-239.
Taylor, G.I. 1952b. Ibid. 214, 158-183.
Webb, P. W. 1971. J. Exptl. Biol. 55, 489-540.
Weihs, D. 1972a. Proc. Roy. Soc. B, 182, 59-72.
Weihs, D. 1972b. Submitted to 1st Internat. Congr. Biorheology. (В итоге
опубликовано как последняя ссылка в библиографии главы 7.)
Weihs, D. 1972c. J. Fluid Mech. 54, 679-691.
Wu, T. Y. 1961. Ibid. 10, 321-344.
Wu, Т. Y. 1971a. Ibid. 46, 337-355.
Wu, T. Y 1971b. Ibid. 46, 521-568.
Глава 7
Некоторые современные исследования
перемещения водных животных
1. Движение при малых числах Рейнольдса
В завершение описания различных способов продвижения водных
животных изложим кратко некоторые современные исследования, которые
являются естественным продолжением работ, освещенных в обзоре главы 6.
И начнем с исследований продвижения организмов при низких числах
Рейнольдса, основанных на рассматриваемой ниже работе доктора Блейка.
Прежде всего, отметим, что некоторые последние работы доктора
Блейка (Blake, 1973a, Blake, Sleigh, 1974) являются связующим звеном
между внешней и внутренней биогидродинамикой. В своих
исследованиях Блейк использовал модель ресничного подслоя для анализа движений
жидкости по каналам в организме, происходящих под действием колебаний
ресничек, которыми выстлана внутренняя поверхность канала. Движение
жидкости в каналах, поддерживаемое ресничками, широко распространено
в животном царстве. В данной главе рассмотрим только два вида, играющих
наиболее важную роль в физиологии человека: движение слизи в
воздушных путях легких и движение спермы в проводящих канальцах мужской
половой системы.
Поскольку метахрональная координация в обоих случаях, думается,
является антиплектической, то единственной подходящей
гидродинамической моделью на сегодняшний день для исследования указанных процессов
является модель ресничного подслоя, разработанная Блейком. На
рисунке 1 представлены результаты применения этой модели при расчете
скорости спермы в протоках при j = lfi (соответствует значению 2/3 для
отношения коэффициента тангенциального сопротивления к коэффициенту
нормального сопротивления тк в главе 3) и при различных значениях к =
= aL/c (где а — частота в радианах, L — длина реснички, а с — скорость
метахрональной волны).
206
Глава 7
ось трубы
Рис. 1. Расчеты Блейка скорости спермы Us в семенных протоках
Задача о движении слизи в воздушных путях легких совершенно
другая, поскольку слизь полностью канал не заполняет и, кроме того, слизь
является тиксотропной жидкостью. Другими словами, ее длинноцепочечные
мукополисахаридные молекулы в состоянии покоя образуют такую
структуру, при которой вязкость жидкости становится очень большой, а в случае
больших скоростей сдвига вязкость существенно уменьшается за счет
разрушения образовавшейся в состоянии покоя крупномасштабной структуры.
Проведенные рассуждения указывают на особое значение
распределения скорости, представленного на рисунке 1 (или рис. 6 в главе 6) с его
областью большой сдвиговой скорости в наружной части подслоя
ресничек. Расчеты Блейка для антиплектического метахронизма показали, что
максимальное значение скорости в этой области имеет порядок 2а (где а —
частота колебаний реснички, выраженная в радианах). Это соответствует
(Blake, 1973b) максимальному сдвигу 200 с-1 для слизи в воздушных путях
легких, величина которого достаточна для того, чтобы слизь в наружной
части подслоя ресничек приобрела «водную» консистенцию (со значительно
меньшей вязкостью), позволяя «густой» (с очень большой вязкостью)
слизи, находящейся вне этого маловязкого слоя, относительно легко скользить
по нему. Многочисленные экспериментальные наблюдения подтверждают
такой механизм движения слизи.
Возвращаясь к внешней биогидродинамике, рассмотрим еще одно
применение системы изображений, построенной Блейком, соответствующей
стокслету на плоской твердой границе. Покажем в общих чертах, что эта
1. Движение при малых числах Рейнольдса
207
система может быть применена для описания движения жгутика в
присутствии твердой плоской границы, такой, например, как предметное стекло
микроскопа.
Рассмотрим плывущий организм, будем считать, что его центральная
линия (продольная ось) остается в плоскости х3 = h, параллельной твердой
плоской стенке х3 = 0. Поток вокруг организма (как и в главе 3) обусловлен
нормальными и тангенциальными стокслетами, распределенными вдоль его
осевой линии. Граничные условия будут удовлетворены при выполнении
следующих положений:
(i) нормальным стокслетам сопутствуют диполи, удовлетворяющие
уравнению (28) главы 3;
(ii) для нормальных и тангенциальных стокслетов, расположенных
параллельно стенке, система изображений будет такой же, как и на рисунке 4
главы 6 так, чтобы граничные условия на стенке были удовлетворены;
(iii) с другой стороны, эти граничные условия имеют пренебрежимо
малые возмущения со стороны диполей (i), влияние которых на стенке
незначительно при условии, что отношение h/a достаточно велико;
(iv) при этом же условии характеристики поля системы изображений (ii)
в каждом поперечном сечении изменяются незначительно и
приблизительно равны своим значениям на осевой центральной линии.
Вычисления, проведенные в главе 3, дают поле скоростей (29) для
нормальных стокслетов и сопутствующих им диполей и поле скоростей (30)
для тангенциальных стокслетов. Полученные результаты необходимо
немного изменить за счет учета эффекта наличия системы изображений, который
рассчитывается в соответствии с (iv). Этот эффект проявляется в том, что
уменьшается скорость обоих полей, поскольку основной элемент в
системе изображений, построенной для стокслета (это единственный элемент,
в котором поле на достаточном удалении убывает со скоростью 1/г, после
его интегрирования (24) главы 3 получаем логарифмическую зависимость),
является равным по величине и противоположно направленным стоксле-
том. Далее, меньшая скорость (29) уменьшается сильнее, чем большая
скорость (30). Это означает, что отношение коэффициентов сил
сопротивления гк уменьшается при движениях жгутика, происходящих в плоскостях,
параллельных стенке. И, как следует из рассуждений, проведенных в
главах 2 и 3, отношение скоростей U/V при этом должно возрасти.
Проведя приближенные расчеты, покажем, что указанный эффект
достаточно существен. Полагаем, что жгутик можно заменить прямым цилин-
208
Глава 7
дром длиной Ъ + с, причем выполняется неравенство Ъ + с > 2h9 где 2h —
расстояние до его системы изображений.
Приближенное значение интеграла, взятого от этой системы
изображений по переменной 1/г до произвольной точки на жгутике, может быть
найдено аналогично (24) главы 3. При этом в соотношении (24) величину а
надо заменить на 2h (расстояние по нормали к стенке до системы
изображений), и логарифм log(4c6/a2) заменится на \og(cb/h2). В главе 3 для
стокслетов напряженности f на единицу длины были рассчитаны скорости
для нормального и тангенциального стокслетов соответственно
— (l + log^-j или _^2 + 21og^-j. (1)
Проведя указанные выше замены, получаем
JL(1 + Iogg) или JL(-2 + 2]og£). (2)
Здесь логарифмическое слагаемое в первом случае и все второе
соотношение обусловлены наличием изображения стокслета. В случае нормального
стокслета изображение-стокслет-дублет приводит к появлению этого
дополнительного логарифмического слагаемого, умноженного на 2. Однако
этот множитель становится равным 1 в силу взаимодействия с
источником-дублетом, или диполем, интенсивности 2/i2f j\x (который, конечно, дает
значительно большую скорость на достаточно большом удалении порядка h
или 2h9 чем скорость, обусловленную диполем интенсивности fa2/4^x,
сопутствующим каждому нормальному стокслету). Заметим, что скоростью,
обусловленной диполем, мы пренебрегаем.
Отношение коэффициентов сил гк равно отношению первой
скорости в (1) ко второй в (1) (т.е. отношению нормальной скорости,
обусловленной действием заданной нормальной силы, к тангенциальной скорости,
обусловленной эквивалентной тангенциальной силой). Как следует из
проводимого анализа (рассматриваются скорости (2), полученные
преобразованием (1)), отношение коэффициентов сил уменьшается от 0,7, о чем
говорилось в главе 3, для случая движения жгутика в неограниченной
жидкости до значения 0,5, которое и использовалось в классической работе Грея
и Хенкока (Gray, Hancock, 1955). Очевидно, что это лишь грубая оценка,
которая тем не менее все же указывает на то, что значение тк для
жгутиков может значительно уменьшаться при наличии близко расположенной
стенки.
1. Движение при малых числах Рейнольдса
209
Вышеприведенные выводы сделаны при условии, что движение
жгутика происходит в плоскости, параллельной твердой стенке, т. е. выполняются
два основных предположения: а) все точки центральной осевой линии
находятся на одном и том же расстоянии h от стенки; Ь) используется система
изображений, представленная на рисунке 4 главы 6. Анализ более общих
движений будет значительно более сложным за счет необходимости
введения переменной величины h9 изменяющейся вдоль оси жгутика, а также за
счет необходимости введения стокслетов, перпендикулярных твердой
стенке, и соответствующей им системы изображений (рис. 5 главы 6).
Отметим здесь только одну характерную особенность применения
более общего вида движения. Как вытекает из исследования системы
изображений для перпендикулярного к стенке стокслета, скорость жгутика (1)
уменьшается в результате действия системы образов и становится равной
(что превышает значение скорости (2)). Надо сказать, что это довольно
грубая аппроксимация, как и ранее основанная на изображениях,
распределенных вдоль прямого цилиндра длиной с + 6, параллельного стенке,
не подходит для случая, когда движение жгутика происходит в плоскости,
наклонной к стенке. И это наводит на мысль, что необходимо учитывать
следующий факт. При некоторых способах перемещения жгутика вдоль
плоской твердой стенки в результате движений в направлении нормали
к центральной осевой линии возникают нормальные к стенке
компоненты, значительно большие по величине, чем в результате тангенциального
движения. И в этом случае существенное уменьшение скорости жгутика
под действием нормальной компоненты силы обусловлено в большей
степени нормальными, чем тангенциальными, движениями и приводит к еще
более значительному уменьшению отношения гк.
Отмеченной особенностью можно объяснить, почему в выражении
в скобках (3) присутствует дополнительное слагаемое, равное 2, по
сравнению с первым выражением в скобках (2). Дополнительное слагаемое
полностью обусловлено более сильным влиянием изображения стокслета. Как
показано в главе 3, § 1, стокслет создает на заданном расстоянии г
большую скорость в направлении от стокслета, чем в перпендикулярном
направлении. Именно этим эффектом объясняется превышение скорости (3)
над скоростью (2), а влияние стокса-дублета и источника-дублета остается
прежним. Для произвольного стокслета, как параллельного, так и перпен-
210
Глава 7
дикулярного к стенке, скорость жгутика уменьшается главным образом за
счет наличия равного по величине и противоположно направленного стокс-
лета. Преобладание влияния именно этого стокслета может быть выявлено
в дальнейших исследованиях движения жгутика при более подробном
рассмотрении геометрических аспектов движения.
2. Движение при больших числах Рейнольдса
Обращение к задачам движения водных животных при больших
числах Рейнольдса стимулирует новые разработки в области математической
биогидродинамики. В данном параграфе отметим некоторые перспективы
развития биогидродинамики в связи с новыми задачами, а затем
расскажем о возможных направлениях исследований, вытекающих из материала,
изложенного в главе 6.
Одна особенность движения водных животных при высоких числах
Рейнольдса исследована значительно меньше, чем в случае низких чисел
Рейнольдса. Об этом уже упоминалось в главе 3, § 1, а именно, о том, что
передвижение животного определялось движением тела относительно его
центра масс, в то время как общее движение представляет собой
комбинацию перемещения центра масс и вращения относительно центра масс.
При высоких числах Рейнольдса инерционные силы играют существенную
роль, и при определении скорости U центра масс и угловой скорости
вращения ft относительно центра масс инерционные силы уже не
приравниваются к нулю. В этом случае результирующая сила равна скорости
изменения импульса, а результирующая пара — скорости изменения момента
импульса.
Эти принципы были использованы в нескольких работах,
посвященных плаванию животных с учетом осциллирующих компонент
результирующей силы и пары. Одно из таких исследований для скомброидного
движения с точки зрения «реактивной» теории (учитывающей силы реакции)
в случае поперечных волнообразных движений с малой амплитудой было
проведено в главе 4, § 4. Это довольно исчерпывающее исследование,
основанное на представлении движения в поперечном направлении в виде (40),
являет собой пример методов оценки влияния различных морфологических
особенностей строения удлиненного тела рыбы на характеристики
скольжения и рыскания при скомброидном способе плавания, оба движения
изучаются для случая малых амплитуд поперечных движений и затем результаты
обобщаются на случай широкоамплитудных движений.
2. Движение при больших числах Рейнольдса 211
В начале §4 главы 4 представлен метод исследования, требующий
дальнейшей основательной доработки: учет сил реакции и сил
сопротивления в комбинированной теории, методами которой можно было бы
описать плавательные движения при угревидном способе плавания. В работе
рассматриваются две предельных теории, использующие линейный подход
даже для широкоамплитудных движений. Одна теория учитывает только
силы сопротивления при движении при низких числах Рейнольдса и
приводит к бигармоническому уравнению, полученному на основании линейной
комбинации полей, обусловленных стокслетами, распределенными вдоль
осевой центральной линии (произвольной формы) животного. Другая
учитывает только силы реакции при движении при высоких числах
Рейнольдса. Применение этой теории приводит к уравнению Лапласа, полученному
на основании аналогичного распределения диполей. Между этими
значительно различающимися теориями лежит некая промежуточная
нелинейная теория, использование которой необходимо, поскольку исследуемое
явление существенно нелинейно. В §4 главы 4 эта существенная
нелинейность была обойдена за счет линеаризации выражения для силы
сопротивления (выражение psw2sgnw было заменено на ^psw0w на интервале
-w0 < w < w0).
Другие сложности, упомянутые в главе 4 в сноске, возникают в
результате неясности, как на вихревую силу в спутном потоке, возникшем
за передней частью тела, влияет последующий спутный поток.
Желательно разработать теории, в которых можно было бы обойти те модификации,
которые указаны в той сноске. Здесь следовало бы учесть действие
локальной силы сопротивления psw2 sgniu, приходящейся на единицу длины, при
нормальном движении со скоростью w, а для тангенциального движения
учесть силы реакции, методы расчета которых изложены в главе 4 или для
широкоамплитудных волнообразных движений, в главе 5.
Аналогичные неясности возникают и при рассмотрении другого типа
вихрей, распространяющихся вдоль направления плавания, а именно
вихревой пелены, сбегающей с передних плавников. В главе 4 часть § 3,
начиная с (19), посвящена анализу этих вихрей методом малых возмущений
в предположении, что вихревая пелена сходит с плавников
перпендикулярно позвоночнику, и после схода ее характеристики не изменяются. Из
расчетов следовало, что передний плавник участвует в образовании силы
тяги, и при выполнении определенных условий для фаз дополнительных
потерь энергии не происходит. То есть изменение импульса (или
импульса «бокового потока») вихревой пелены, сбегающей с переднего плавника
212
Глава 7
в поперечном направлении, по крайней мере на 90° расходится по фазе
с движением хвостового плавника, где происходит ее поглощение. Так как
волна вдоль тела распространяется со скоростью V, а вихри перемещаются
относительно рыбы со скоростью плавания U = | V, то разность фаз между
ними может достигать 90°, если расстояние между плавниками превышает
половину длины волны (глава 4, соотношение (28)).
Ньюмен и By (Newman, Wu, 1972, Newman, 1973) далее развили эту
теорию. Они учли изменение вихревой пелены, сходящей с передних
плавников, за счет адвекции в направлении течения в потоке вдоль вытянутого
тела рыбы. Такой подход важен в случае перемещения животных, по форме
похожих на субмарину. Такие объекты имеют тело вращения, жесткие
плавники, а размах хвостового плавника значительно меньше, чем у передних,
и движутся подобные объекты с постоянным углом атаки. А вот тело рыбы
с его сильным боковым сжатием не может внести столь существенные
изменения в движение вихрей, сошедших с грудного и спинного плавников.
Более того, взаимодействие вихрей с потоком, созданным хвостовым
плавником, приводит к их значительному ослаблению не за счет постоянного
угла атаки, а за счет того, что волнообразное движение тела создает
вышеупомянутую разность фаз. Ньюмен и By указывают и на другое возможное
обобщение — учет скошенности задней кромки плавников.
В § 5 главы 4 динамика серповидного хвостового плавника
исследована методами двумерной теории малых возмущений. Доктор М. Дж. Чоп-
ра (Chopra, 1974) исследовал эту задачу в трехмерной постановке, однако
остался в рамках теории малых возмущений. Конечно, было бы очень
интересно для исследования динамики хвостового плавника разработать
наиболее общую теорию широкоамплитудного движения, подобно подходу к
решению данной задачи, осуществленному в главе 5 на основании теории
вытянутого тела.
Значительный пробел в сегодняшнем понимании движения вытянутого
тела отмечается в главе 5, § 3: изучение плавания рыб носит односторонний
характер, поскольку большее внимание уделено процессу выработки силы
тяги по сравнению с процессом образования силы сопротивления. В
главе 3 отмечалось различие теоретических подходов для движений при
низких и высоких числах Рейнольдса. В этой главе для движений при низких
числах Рейнольдса были определены как сила сопротивления,
обусловленная тангенциальным движением жгутика, так и сила тяги, обусловленная
нормальным движением жгутика, и условия равновесия этих сил. Однако
при больших числах Рейнольдса в силу существенной нелинейности урав-
2. Движение при больших числах Рейнольдса 213
нений Навье-Стокса подобное решение задачи — единый подход к
определению и силы тяги, и силы сопротивления затруднено. Наоборот, считается,
что сила сопротивления является результатом действия вязкости,
тормозящей продвижение рыбы в воде в тангенциальном направлении, когда на
поверхности тела развивается тонкий ламинарный или турбулентный
пограничный слой, а сила тяги вырабатывается за счет движений тела рыбы
в нормальном направлении и определяется импульсом объема воды,
расположенного главным образом вне пограничного слоя.
При скольжении рыбы по прямой на нее действует только сила
сопротивления, определяемая условием стационарного скольжения, и величина
этой силы равна скорости потери импульса животным. Очевидно, что для
того, чтобы поддерживать постоянную скорость передвижения, рыба
должна совершать плавательные волнообразные движения, приводящие к
возникновению силы тяги за счет реакции. Интенсивность этих движений
должна быть такой, чтобы по меньшей мере компенсировать силу
сопротивления при стационарном движении. Однако в главе 5 высказывалась мысль
о том, что на самом деле требуемая для продвижения сила тяги должна
быть значительно больше, поскольку необходимо преодолевать и силу
сопротивления окружающей воды при поперечном движении тела, которая
для рыб, имеющих длинное вытянутое тело, может значительно превышать
силу сопротивления при равномерном скольжении. (Естественно, что эти
рассуждения не относятся к животным с серповидным хвостом, включая
китообразных, относительно которых Грей пришел к совершенно другим
выводам в своей знаменитой работе.)
Описанный в главе 5, § 3, эффект нашел подтверждение в работах Веб-
ба (Webb, 1971), особенно в той части его исследований, где проводилась
оценка вырабатываемой форелью мощности на основе определения
потребленного ею кислорода. Как следует из его работы, сила сопротивления,
обусловленная поперечными движениями тела рыбы, приблизительно в 3 раза
превышает силу сопротивления при стационарном скольжении. Это в
целом согласуется с полученным в главе 5 результатом на основании изучения
плавания форели о том, что эти силы сопротивления различаются в 4 раза.
Такое расхождение заставляет более внимательно подойти к оценке силы
тяги, обусловленной силами реакции, на основании теории вытянутого
тела. Возможно, следует учесть увеличение впятеро силы трения с той
стороны тела рыбы, к которой направлена поперечная скорость w, что
объясняется уменьшением в 5 раз толщины пограничного слоя с этой стороны
рыбы за счет поступления к телу при движении все новых порций воды.
214
Глава 7
Увеличение приблизительно в три раза отношения сил сопротивления
доктор Вейс попытался обосновать с точки зрения биологии.
Характерной особенностью рыб, плотность которых равна плотности воды, является
то, что когда они проплывают фиксированное расстояние, чередуя период
скольжения как твердого тела с периодом энергичного продвижения за счет
волнообразных движений тела, то затраты энергии в два раза меньше, чем
при преодолении этого же расстояния с постоянной скоростью. Подобный
обнадеживающий подход и сравнение полученных с его помощью
результатов с данными наблюдений находятся в стадии разработки. Этим
исследованиям предшествовала работа Вейса (Weihs, 1973a), посвященная
плаванию рыб, плотность которых выше плотности воды. В ней было отмечено,
что такие рыбы затрачивают в два раза меньше энергии, если их движение
представляет собой чередование подъема вверх и скольжения вниз.
Кроме того, разработанный им подход доктор Вейс применил к
изучению разворота рыб (Weihs, 1972). Эти исследования, а именно результаты
по старту и одновременному развороту золотой рыбки (Carassius auratus),
в главе 6 подробно не освещались. Между результатами этой работы и
описанным ранее в главе 6 процессом поворота уже плывущей красноперой
плотвы просматривается интересная связь: в обоих случаях поворот
осуществляется на 90° и сам процесс можно разделить на три стадии.
На рисунке 2, воспроизведенном из работы (Weihs, 1972),
представлена разработанная им схема комбинированного процесса старта и разворота.
Так же как и в случае движения красноперки, на первой стадии процесса
(кадры 1, 2 и 3) никакие силы на рыбу не действуют, центр масс рыбы
остается на месте (в отличие от плывущей красноперки, центр масс
которой на этой стадии движется по прямой). За счет мускульного усилия
рыба изгибает тело, действующие силы равны изменению импульса
жидкости в направлении нормали к телу. Эти силы указаны на кривых 1 и 2
в нижней части рисунка 2. На данном этапе силы находятся в статическом
равновесии и вращения рыбы относительно центра масс не происходит. На
этапе, изображенном кривой 3, начинают действовать силы, равные
изменению импульса жидкости, сходящей с хвостового плавника, в то время
как силы, действующие на этапах 1 и 2, ослабевают и начинается поворот
рыбы по часовой стрелке. Вращающая пара действует на рыбу за счет того,
что хвостовой плавник перемещается из позиции 3 в позицию 4. Действие
вращающей пары, обусловленной перемещением хвостового плавника из
положения 3 в положение 4, аналогично приводящему к развороту (и
одновременному необходимому торможению) действию грудного плавника
плывущей красноперки.
2. Движение при больших числах Рейнольдса 215
На второй стадии (так же как и у красноперки) поворот продолжается
и начинает действовать большая по величине сила, под действием
которой происходит необходимое изменение импульса. В позиции 4 хвостовой
плавник меняет знак своего угла атаки и под действием сил,
обусловленных изменением импульса срывающейся с плавника жидкости, центр масс
начинает двигаться со значительной скоростью в направлении своего
результирующего движения. В это время еще не возникает сколько-нибудь
значительного момента, тормозящего вращение относительно центра масс,
и еще действуют значительные по величине силы на переднюю часть тела
(см. кривую 5 на нижней части рис. 2). На третьей стадии движения, так же
как и у красноперки, преобладающее влияние имеют силы, обусловленные
перемещением хвостового плавника. В результате действия этих сил
завершается ускорение центра масс и предотвращается вращение тела рыбы
относительно него.
Проводимые в настоящее время исследования доктора Вейса
свидетельствуют о том, что многие рыбы при быстром повороте (как из
положения покоя, так и из состояния движения) совершают поворот именно
на 90°. Например, съемки поворота на 180° показали, что он
совершается в два этапа — поворота на 90°, а между этими этапами рыба движется
по прямой. Недавняя работа доктора Вейса (Weihs, 1973b) посвящена
исследованию быстрого старта таких рыб, как щука (Esox lucius), которые,
как правило, во время охоты сначала затаиваются и выслеживают жертву,
а затем внезапно бросаются на добычу, развивая ускорение до 50 м/с2.
Интересно, что резкий бросок таких рыб часто сопровождается разворотом
почти на 90°, а динамика движения при этом подобна представленной на
рисунке 2. Доктор Вейс высказал предположение, что единственной силой,
способной обеспечить резкий рывок, является сила, обусловленная
движением хвостового плавника. Сначала рыба изгибается так, как схематически
изображено на рисунке 2, и только после этого хвостовой плавник
совершает резкие движения в направлении, перпендикулярном движению рыбы при
умеренных углах атаки. Бросок с разворотом происходит в целом по схеме
рисунка 2. Способность щуки к особенно быстрым броскам обусловлена
строением ее тела. Ее спинной и брюшной плавники расположены очень
близко к хвостовому, и при рывке их движения придают дополнительное
ускорение хищнице.
А вот форель при нападении на добычу стартует сначала в том
направлении, в каком было расположено ее тела до броска, и лишь затем,
совершая движения грудными плавниками, совершает разворот. Ее движение при
216
Глава 7
голова
Рис. 2. Иллюстрация комбинированного старта и поворота золотой рыбки. Рисунок
воспроизведен из работы (Weihs, 1972, рис. 3)
таком маневре напоминает движение, изображенное на рисунке 2. За счет
тормозящего движения левым грудным плавником к концу стадии 2 рыба
разворачивается в нужном направлении. Ускорение, развиваемое форелью
в момент такого маневра — начального движения по прямой и
последующего разворота, будет меньше, чем если бы та же самая рыба совершала
разворот с момента старта. Кроме представленных здесь способов
маневрирования, остается еще много других, которые ждут своих исследователей.
2. Движение при больших числах Рейнольдса 217
Литература
1] Blake, J. R. 1973a. Flow in tubules due to ciliary activity, Bull. Math. Biol.,
35,513-523.
[2] Blake, J. R. 1973b. A note on mucus shear rates, Resp. Physiol., 17,
394-399.
[3] Blake, J. R. & Sleigh, M. A. 1974. Mechanics of ciliary locomotion, Biol.
Reviews, 49, 85-125.
[4] Chopra, M. G. 1974. Hydromechanics of lunate-tail swimming propulsion,
J. Fluid Mech., 64, 375-391.
[5] Gray, J. & Hancock, G. J. 1955. The propulsion of sea-urchin spermatozoa,
J. Exptl. Biol., 32, 802-814.
[6] Newman, J.N. 1973. The force on a slender fish-like body, J. Fluid Mech.,
58, 689-702.
[7] Newman, J.N. & Wu, T. Y. 1972. A generalized slender-body theory for
fish-like forms, Ibid., 57, 673-693.
[8] Weihs, D. 1972. A hydrodynamical analysis offish turning manoeuvres,
Proc. Roy. Soc. B, 182, 59-72.
[9] Weihs, D. 1973a. Mechanically efficient swimming techniques for fish with
negative buoyancy, J. Marine Res., 31, 194-209.
10] Weihs, D. 1973b. The mechanics of rapid starting of slender fish,
Biorheology, 10, 343-350.
Глава 8
Аэродинамика полета животных
1. Завоевание воздуха
В области внешней биогидродинамики перемещение животных в воде,
чему посвящены главы с 1 по 7, представляет собой первые достижения
на пути эволюции передвижения животных. Не менее интересны и более
поздние способы перемещения: по многочисленным окаменел остям можно
проследить эволюцию морских животных на протяжении по крайней
мере 6 х 108 лет с начала кембрийского периода и до настоящего времени.
В противоположность этому перемещение животных в воздухе,
составляющее аэродинамический раздел внешней биогидродинамики (ему будут
посвящены главы 8 и 9), которое можно было бы назвать завоеванием воздуха,
представляет собой событие, значительно более недавнее, нежели
плавание.
Тем не менее история полета животных более давняя, и эпоху
завоевания воздуха приходится поместить раньше, чем многие могли бы
вообразить. Уже 3 х 108 лет назад в середине каменноугольного периода
проблемы длительного активного полета были решены насекомыми. Прекрасное
описание некоторых процессов эволюции полета дано Смартом и Хьюзом
(Smart, Hughes, 1972), схематичное изображение процесса эволюции
воспроизведено здесь на рисунке 1.
Согласно их описанию, завоевание суши растениями и животными,
в том числе примитивными членистоногими {Arthropodd), такими как
тысяченожки, произошло в силурийском периоде, немногим более 4 х 108 лет
назад. После этого в девонском периоде эволюция членистоногих на суше
привела к появлению шестиногих форм, характерных для насекомых, до
какого-либо развития крыльев. Однако уже к середине каменноугольного
периода появляются многочисленные крылатые насекомые (о чем
свидетельствуют многочисленные ископаемые останки этих насекомых) с
замечательным разнообразием их форм: например, уже произошло отделение
220
Глава 8
древнекрылых, таких как стрекозы (Odonata), которые не могут, отдыхая,
складывать крылья над брюшком, и новокрылых, включавших тогда
кузнечиков (Orthoptera), которые уже были способны это сделать. Имеются
достоверные доказательства, что тогда уже существовали насекомые с
полным превращением (в стадиях личинки и имаго (взрослой особи),
разделявшихся стадией куколки, насекомые сильно различались по внешнему
виду).
Смарт и Хьюз (Smart, Hughes, 1972) связывают явление развития
полета насекомых в весьма разнообразных формах в течение первой половины
каменноугольного периода с появлением в то время в изобилии высоких
наземных растений, образовавших густые «леса», остатки которых,
спрессовавшись, превратились в пласты угля. Формы растительной жизни в
каменноугольном периоде сильно отличались от типичной для третичной эпохи
покрытосемянной (цветковой) растительности. Однако тогда существовали
«деревья» высотой до 30 м: плауны, каламиты и древние голосемянные,
в том числе и деревья с развитыми листовыми пластинками, и многие
другие источники пищи для ползающих насекомых.
Таковы условия, которые могли благоприятствовать развитию
аэродинамических поверхностей, первоначально для того, чтобы
способствовать перемещению в горизонтальных плоскостях за счет планирования
с высоты. В дальнейшем преимущества летающих животных в
расселении и отыскании пищи могли нарастать благодаря постепенному
развитию суставов на аэродинамических поверхностях, проходящему ряд стадий
и в конечном итоге приведшему к осуществлению непрерывного крылатого
полета.
Дальнейшая эволюция могла пойти по пути выделения, например,
хищных форм, таких как стрекозы (Odonata), а затем последующее
развитие полета в различных группах насекомых могло определяться не только
преимуществами в расселении и способах отыскания пищи, но и влиянием
со стороны хищников-насекомых, а также хищников-ненасекомых,
например амфибий (которые в этот период стали первыми наземными
позвоночными) и появившихся к тому времени представителей класса
членистоногих — паукообразных (Arachnida).
После каменноугольного периода разнообразие классов насекомых
продолжало нарастать, так что многочисленные разновидности, типичные
для большинства ныне существующих отрядов, присутствовали уже в
триасовом периоде, задолго до появления цветковых растений. Много позже
развитие покрытосеменных растений привело к чрезвычайно успешному
1. Завоевание воздуха
221
CARB
PERM
■ (ЛЖРАГГКЧ
I
| PTFJUDOSPERIAS
I PROGYNNUSPEAMS
■СЬпПоррИцг» Ь Aquatic пушрЫ ^
Up-S»iMiJUK pt&ub* Ш
I Sparc* ^
| ftiplwiim
SmkIi ^
ЕМспмрогаж caaditioa b
Рис. 1. Хронология развития растений, насекомых, определяющих эволюцию
полета животных (воспроизведено из работы Smart, Hughes, 1972, рис. 3)
их симбиозу с насекомыми, что в огромной степени способствовало
развитию подклассов в классе насекомых. В результате чего класс насекомых
стал самым многочисленным классом живых существ, более
многочисленным, чем все остальные классы вместе взятые. Одновременно это привело
и к появлению новых отрядов насекомых, прежде всего питающихся
нектаром цветков — бабочек и молей (Lepidoptera).
Тем временем за завоеванием воздуха насекомыми последовал и ряд
независимых достижений в области непрерывного полета у других групп
животных. Рептилии, развившиеся из амфибий, позднее достигли
поразительного расцвета в юрском и меловом периодах (примерно от 2 х 108 до
1 х 108 лет тому назад), когда в их числе была обширная группа летающих
животных — птерозавров. Затем искусством непрерывного полета незави-
222
Глава 8
симо от них овладели птицы (развившиеся из бескрылых форм). В другом
классе позвоночных — млекопитающих — еще позднее, примерно 0,5 х 108
лет тому назад, появились летучие мыши.
Как говорилось в главе 1, в задачи данной книги не входит полный
сравнительный обзор наиболее важных областей научных изысканий, как
это было сделано в главе 2, относительно перемещения животных в
воде. Поэтому главы 8 и 9 посвящены почти полностью аэродинамическому
исследованию движений, обеспечивающих полет, у отдельных
представителей животного мира. В оставшейся части первого параграфа приведем
еще несколько комментариев относительно эволюционного развития
механизмов полета.
Способные к непрерывному полету животные должны приобрести
разнообразные способности. Так, для поддержания состояния непрерывного
полета требуется, чтобы движение крыльев приводило к созданию как
силы тяги, так и подъемной силы. В главе 2 приведено схематичное
изображение движения (рис. 7) морских животных тяжелее воды с конечностями,
подобными крыльям, таких как черепахи (§ 13). В § 2 данной главы
описывается соответствующее движение типичных представителей класса
насекомых с четырьмя крыльями и способных к полету с достаточно большой
скоростью (пустынной саранчи (Schistocerca gregarid) отряда прямокрылых
(Orthoptera)), а в § 3 проведены аналогичные исследования для
представителей двух видов птиц (чайки и голубя). Несмотря на очевидные различия,
характер движения крыла птицы имеет много общего с описанным
схематично в главе 2 (рис. 7) скомброидным движением водных животных.
Машущее движение крыла можно рассматривать как суперпозицию ском-
броидного движения, приводящего к созданию силы тяги, и движения с
постоянным положительным углом атаки.
Большинство летающих животных способны к парящему полету либо
в течение длительных периодов, либо по крайней мере в моменты взлета
и посадки.1 Очевидно, что проблемы поддержания тяжелого тела
требуют совершенно другого подхода при парящем полете — в отсутствие
какого-либо относительного движения между воздухом и поверхностью
животного.
Серьезное сравнительное исследование парящего полета насекомых,
птиц и летучих мышей провел недавно Вейс-Фог (Weis-Fogh, 1973). Он
установил, что широко распространен полет, названный им «нормальным
1 Если использовать термины авиации, то можно сказать, что летающие животные подобны
самолетам с вертикальным взлетом и посадкой (СВВП), за исключением некоторых
водоплавающих птиц, которые используют водную поверхность как взлетно-посадочную полосу.
1. Завоевание воздуха
223
парящим полетом». При «нормальном парящем полете» тело и голова
животного находятся практически на одной прямой (т. е. голова не
наклонена). Для создания необходимой подъемной силы крылья совершают взмахи
назад и вперед в горизонтальной плоскости, причем каждый взмах крыла
приводит к образованию воздушного потока, за счет чего птица летит под
постоянным углом атаки. В §4 приводятся основные выводы, к которым
пришел Вейс-Фог при изучении парящего полета. Кроме того,
описывается его важное открытие в области биогидродинамики, к которому он
пришел, исследуя виды летающих животных с необычным парящим полетом,
а именно к механизму создания подъемной силы, который будет
проанализирован в главе 9.
Между двумя крайне различающимися способностями — осуществлять
непрерывный полет с постоянной скоростью и парением в спокойном
воздухе — существуют и другие типы полетов, о которых будет кратко
упомянуто, и обсуждены преимущества, которые получают животные, пользуясь
таким способом. Так, один из способов, при котором используются
восходящие потоки воздуха, описан в § 3 на примере полета птиц. Кроме того,
у многих летающих животных замечательно развиты также способности
к маневрированию и управлению полетом.
Интересно отметить, что непрерывный полет двукрылых насекомых,
птиц и летучих мышей осуществляется без какого-либо дополнительного
стабилизирующего действия задних аэродинамических поверхностей.
Например, хвостовые перья у птиц при быстром полете находятся, как
правило, в нерасправленном состоянии и составляют конусовидную
конфигурацию, не способствующую возникновению подъемной силы. Животные,
крылья которых способны совершать сложные трехмерные движения (со
сложной угловой ориентацией), могут активно управлять точкой
приложения подъемной силы, что помогает им избегать появления нежелательных
продольных моментов. При других видах полета, включая резкие маневры,
парение и скольжение, птицы расправляют хвостовое оперение.
При описании этих вопросов особо обратим внимание на тот факт,
что при систематизации полетов по маневренности, будь то животное или
некий созданный человеком механизм, вопросы устойчивости не
рассматриваются. Отметим также, что был бы очень интересен общий
сравнительный обзор с точки зрения биогидродинамики способов управления
полетом.
А теперь зададимся вопросом: какова же характерная особенность
полета птиц и насекомых, присущая именно этим двум классам, освоившим
224
Глава 8
наиболее эффективные способы полета? Вероятно, лучше всего для
ответа обратиться к структурным аспектам, поскольку подход с позиции
биогидродинамики приведет к характерным особенностям, присущим многим
классам летающих животных.
Представители класса птиц обладают многими специфическими
анатомическими чертами, весьма полезными для осуществления полета. Так,
например, соответствующее распределение воздушных мешков, кроме
того, что делает дыхательную систему птиц весьма эффективной (глава 11),
помогает также сочетать легкий вес с достаточно большой жесткостью
корпуса. Ну и самое главное, наличие перьев приводит к уменьшению нагрузки
на костную систему крыла.
Кроме того, наличие перьев улучшает некоторые гидродинамические
характеристики полета. В отличие от плотных крыльев насекомых,
птерозавров и летучих мышей, крыло птицы с его проницаемостью,
обусловленной перьями, позволяет совершать полет с лучшими аэродинамическими
характеристиками при высоких числах Рейнольдса (§ 3) за счет управления
толщиной пограничного слоя подобно тому, как это делается в авиации при
выдуве через систему щелей тонкой струи воздуха.
Рассматривая особенности строения насекомых, обеспечивающие
возможность полета, коснемся и их способности совершать крыльями
высокочастотные колебания. При таких колебаниях нет необходимости
производить отдельные мускульные усилия при каждом взмахе крыла для
преодоления инерции. У насекомых инерция крыла преодолевается за счет сил
упругости, а мускульное усилие требуется им для аэродинамического
сопротивления.
Это сделалось возможным, когда в процессе эволюции насекомые
приобрели внешний скелет, состоящий из прочной эластичной кутикулы, а
также из соединительной ткани, в которую входит обладающий упругими
свойствами белок резилин (Weis-Fogh, 1960). Свойства такой
соединительной ткани напоминают свойства вулканизированной резины. Резилин
обладает очень высокой упругостью и мгновенно реагирует с очень низкой
потерей энергии на широкоамплитудные колебания с частотой от 20 до
500 Гц, характерной для колебательных движений крыльев насекомых.
Позвоночные, при всех их многочисленных других преимуществах,
не могли использовать выскочастотные колебательные движения в
качестве механизма передвижения, поскольку их скелет недостаточно
эластичен, и любой эластомер в их тканях даже отдаленно не напоминает
резилин. Основным эластомером, присутствующим в эластичных волокнах
2. Непрерывно поддерживаемый направленный полет 225
соединительной ткани позвоночных, является эластин, наличие которого
придает упругость артериям млекопитающих. Как мы увидим в главах 12
и 13, его эластичные свойства достаточны для колебаний с частотой
сердечного пульса, однако при частотах около 100 Гц этот белок не успевает
их проявлять.
Часть энергии мышечных усилий позвоночные вынуждены
расходовать на преодоление инерции конечностей при движении, что также
препятствует развитию способов передвижения с использованием
высокочастотных колебательных движений. Хотя некоторые отдельные виды
позвоночных, таких как колибри (§ 4) или морские коньки (глава 2, § 11), успешно
преодолели эти трудности и приобрели способность совершать крыльями
или плавниками колебательные движения с частотой порядка 50 Гц.
Возможность совершать высокочастотные колебательные движения за счет
эластичных систем организма, чьи свойства определяются свойствами
кутикулы или резилина, способствовала развитию замечательных техник полета
и скольжения у насекомых.
2. Непрерывно поддерживаемый направленный полет
насекомых
Приведем здесь результаты наблюдений над движением крыла
пустынной саранчи (Schistocerca gregoria) в полете и подробный анализ
экспериментальных данных из работы Вейса-Фога и Йенсена (зоолога и
аэродинамика, их совместная работа была чрезвычайно плодотворной) (Weis-Fogh,
Jensen, 1956). Аэродинамическая труба, в которой они проводили
исследования, оказалась таким же бесценным оборудованием в этой области, как
и в области аэродинамики.
Исследователи подвешивали исследуемое насекомое в
аэродинамической трубе на весах таким образом, чтобы исключить помехи в движении
грудного отдела (поскольку именно там располагались четыре крыла).
Система регулирования с обратной связью, управляющая скоростью вращения
вентилятора аэродинамической трубы, позволяла воспроизводить условия
свободного равномерного полета: скорость воздушного потока, при
условии, что насекомое машет регулярно, автоматически подстраивалась таким
образом, чтобы полная горизонтальная сила, измеряемая с помощью
весов, обращалась в нуль. При этом гидродинамическое сопротивление и тяга
уравновешивали друг друга как в свободном полете.
В каждом проводимом эксперименте по показаниям весов
отслеживалось среднее значение вертикальной силы. В тех случаях, когда она была
226
Глава 8
равна нулю, реализовывались условия стационарного горизонтального
полета, когда подъемная сила, создаваемая крыльями, уравновешивала вес
насекомого. Другие интересные исследования проводились в условиях,
когда средняя сила, направленная по вертикали вниз или вверх, составляла
приблизительно 70% от веса насекомого, что соответствовало движению
с вертикальным ускорением, достигающим OJg. Кроме указанных, можно
было создать и другие условия полета насекомого (например, таких, как
нагруженный полет).
Типичное исследуемое насекомое имело массу 2 г и крылья длиной 4
см. Типичная скорость полета U равнялась 4 м/с, что соответствовало
числу Рейнольдса Uc/u, рассчитанному по хорде с переднего крыла, близкому
к 2000. Типичная частота взмахов крыла была равна 20 Гц, так что
частотный параметр uc/U (см. главу 4) принимал очень небольшое значение,
близкое к 1/4.
Вообще, для данных, имеющихся по непрерывному направленному
полету как насекомых, так и птиц, частотный параметр, рассчитанный по
угловой скорости и9 хорде крыла с и скорости полета /7, принимает значения,
значительно меньшие 1 — типичного частотного параметра при движении
морских животных с серповидным хвостовым плавником (главы 2 и 4).
Как видно из рисунка 2, на котором представлено движение в воздухе
сечения переднего крыла (срединного сечения), частотный параметр принимает
небольшие значения. При таких значениях частотного параметра
аэродинамические силы можно рассматривать как квазистационарные.
Рис. 2. Движение относительно воздуха среднего сечения переднего крыла
пустынной саранчи (Schistocerca gregoria) (рис. Ill, 8 в работе Weis-Fogh, Jensen, 1956)
Отметим, в частности, долгий взмах вниз, при котором хорда крыла
почти параллельна направлению движения (что очень похоже на схему рис. 7
главы 2). На протяжении этого взмаха имеется достаточно времени для
2. Непрерывно поддерживаемый направленный полет 227
того, чтобы на крыле установилась стационарная аэродинамическая сила,
направленная под прямым углом к направлению движения крыла в воздухе
и поэтому имеющая компоненты как силы тяги, так и подъемной силы.
После быстрого разворота крыла, называемого пронацией (в авиационном
деле ей соответствует термин «пикирование»), непосредственно перед
взмахом крыла вниз квазистационарные значения составляющих указанных сил
достигаются уже на расстоянии нескольких длин хорды.
Быстрая супинация (соответствует авиационному термину
«кабрирование») указывает на переход к взмаху вверх с большим положительным
углом тангажа относительно направления среднего движения (вновь как на
рис. 7 главы 2). Отметим, что в это же время угол атаки относительно
направления мгновенного движения имеет отрицательное значение, причем
степень отклонения от условий квазистационарности на этой фазе
движения увеличивается и действующая сила увеличивается за счет возрастания
силы тяги.
На рисунке 3 представлено закручивание крыла при пронации и
супинации. Здесь дано также использованное Вейсом-Фогом и Иенсеном (Weis-
Fogh, Jensen, 1956) определение угла атаки как угла между направлением
крыла на задней кромке и направлением движения воздуха относительно
крыла.
направление полета
^основание крыла крылец
а
конец крыла ' направление
пронация нескрученное супинация движения воздуха
крыло относительно крыла
Рис. 3. Закручивание крыла при пронации и супинации. Определение угла атаки а
(рис. I, 2 и I, 3 из работы Weis-Fogh, Jensen, 1956)
На рисунке 4 представлена зависимость угла подъема крыла У
передних и задних крыльев при стандартном взмахе крыльев пустынной саранчи
(Schistocerca gregorid). Здесь 7' — угол между (i) линией, направленной
вдоль крыла, и (ii) плоскостью симметрии насекомого, тупой или острый
в зависимости от расположения крыла относительно своего среднего
положения выше или ниже соответственно.
228
Глава 8
i ■ 1 ■ 1 ■ 1 ■ 1 ■ г
Г ■ i ■ i . i . i . i .11 . I
0 20 40 60
Время, отсчитываемое от того момента,
когда задние крылья выполнили половину взмаха вверх (мс)
Рис. 4. Взмахи заднего и переднего крылев при полете пустынной саранчи
(Schistocerca gregoria) (рис. II, 20 в работе Weis-Fogh, Jensen, 1956)
Отметим, что отставание по фазе движения передних крыльев
относительно задних характерно для четырехкрылых насекомых. Вейс-Фог
и Йенсен (Weis-Fogh, Jensen, 1956) считали, что между двумя
крыльями возникает достаточно тесное аэродинамическое взаимодействие, однако
оно не играет главной роли в процессе полета. Эффективный угол атаки
заднего крыла уменьшается примерно на 2° благодаря отклонению вниз
потока за передним крылом, а эффективный угол атаки переднего крыла
увеличивается на ту же величину за счет направленного вверх потока
перед задним крылом.
Вейс-Фог и Йенсен (Weis-Fogh, Jensen, 1956) провели отдельные
измерения коэффициентов подъемной силы и силы сопротивления при
различных углах атаки в условиях стационарного относительного движения
воздуха. Результаты этих измерений представлены на рисунке 5. Здесь же для
сравнения приведены соответствующие кривые для двух стандартных
профилей крыла, используемых в авиации. Главный недостаток крыла
насекомого — более высокое минимальное значение коэффициента сопротивления
2. Непрерывно поддерживаемый направленный полет 229
CD — примерно 0,05 по сравнению с коэффициентом сопротивления
авиационных крыльев 0,03 (которые, с другой стороны, значительно массивнее).
1,2
0,8
0,4
0
-0,4
/
1
1
[
L
А
Л
г/
ж
f5°
0°
-5°
^
1о°
-10й
IF
17,5h
0,1 0,2
-5е
L
//
7
5°
0°
/
Ж
*ж
Jb°\
0,1 0,2
1
_
10р
г
0е
5°
к
ho*
15J
ОД 0,2
А1 0,2
Рис. 5. Соотношение между коэффициентом подъемной силы CL и коэффициентом
сопротивления CD для двух авиационных крыльев и крыльев насекомого
(переднего и заднего крыльев саранчи) (рис. I, 5 в работе Weis-Fogh, Jensen, 1956)
Авторы использовали эти данные и данные по измеренным углам
атаки (скорректированные наличием взаимных помех между крыльями, о чем
упоминалось выше) для получения оценки изменения вертикальной
компоненты силы, действующей на каждое крыло с течением времени. На
рисунке 6 приведены результаты двух различных экспериментов I и II со
средней подъемной силой, составляющей 97 % и 111 % от веса насекомого
соответственно. Важным элементом проверки этих результатов было
весьма хорошее согласие (с ошибкой 3% и 7% соответственно) измеренных
фактически и полученных в результате расчетов значений подъемной силы.
Примерно 70% подъемной силы вырабатывается задним крылом и 30%
передним. Как и предполагалось, подъемная сила создается главными
образом при взмахе крыльев вниз.
Аналогичным образом Вейс-Фог и Йенсен (Weis-Fogh, Jensen, 1956)
проанализировали горизонтальные силы тяги, создаваемые крыльями,
используя на этот раз для проверки тот факт, что средняя тяга крыльев должна
уравновешивать сопротивление тела, которое измерялось отдельно. На
рисунке 7 показана зависимость от времени силы тяги, развиваемой каждой
парой крыльев, в долях от среднего значения полной силы тяги. Отметим,
однако, что эта сила составляет около 7 % полной подъемной силы.
Примерно три четверти этой силы создается задними крыльями. Как следует
из рисунка 7, сила тяги достигает максимальных значений в двух точках
(сплошная кривая на нижней части рис. 7), больший пик достигается при
взмахе вниз, а меньший — при взмахе вверх.
230
Глава 8
0,8
0,4
(
0
■^h-
\jum
взмах вниз
к
i
>
-^
.^—
л
взмах вверх
•^J!
•
щ
fci
^
z,u
1,6
1,2
0,8
0,4
0
(
\
)
X
f
>
•
ВЗ
>
t
0
мах
•
К^
,2
•
X
*
0,
4
—^.
4^J
^—
1 —I Г 1
взмах вверх
>
о,
1 1 1 1
• I (97%) -
>гч
6
к
X
t
I
X
•
о,
-to»
ft
8
— i i
ВНИЗ
^ 1 1 1
•
х
Т
"йт
1,0
Рис. 6. Изменение со временем подъемной силы, создаваемой передними и задними
крыльями (рис. III, 16 в работе Weis-Fogh, Jensen, 1956)
В данном параграфе были приведены в кратком виде наиболее важные
с аэродинамической точки зрения результаты работы Вейс-Фога и
Иенсена (Weis-Fogh, Jensen, 1956), их работу можно рассматривать как пример
наиболее полного, методического и детального исследования проблем пе-
редвижения животных. Основной вывод Вейс-Фога и Иенсена заключался
в том, что полет насекомых происходит при значениях частотного
параметра, слишком малых для того, чтобы возможно было применить
квазистационарный подход, и этот вывод относительно насекомых в целом был
подтвержден другими исследованиями.
3. Полет птиц
В данном параграфе остановимся, прежде всего, на классических
наблюдениях за полетом чайки и голубя (Brown, 1953) и покажем, что в
биогидродинамике механизма непрерывного полета птиц и насекомых
гораздо больше сходства, чем различия. Затем рассмотрим проблемы расхода
энергии крупными птицами и связанный с этим вопрос об освоенных ими
интересных способах экономии энергии.
3. Полет птиц
231
?,
о,
2
*^
взмах вниз
V
х
•
X
•^J-
*
9
1 1 1 1
взмах вверх
•
•
^х
•
f
•
X
ш
"к
-2
♦
» J
т X
ц/>
вза
•
iax
* •
*
X
•
X
\
1 Т
взмах Bi
1
л у%/ i /v/
х ТТ М11<ЙЛ
N
W )
<
:
> >,
X
;epj
• /
X
с.
* X
л
*
*
к
ВНИЗ
JC^
x/i
4
2
0
2
У-
^^«»
—
*^.
.
^ ■
>""
**
у
,
,-'•
- ^.
Jp
^.
-s.
VI
^
Г-
л
"-~-
V4
Да
.
\
V
"S ^
V
"*-J
'^J
(см/с)
4
2
0
-2
-4
Да:
(см)
0,02
О
-0,02
0,04
0,2
0,4
0,6
0,8 Т 1,0
Рис. 7. Изменение силы тяги, создаваемой передними и задними крыльями (рис. III,
19 в работе Weis-Fogh, Jensen, 1956)
Браун выучил птиц пролетать по прямому коридору длиной 60 м
к клетке, расположенной в конце коридора. В середине коридора была
установлена аппаратура для скоростной киносъемки. На рисунке 8
представлена последовательность полученных кадров летящего со скоростью 10 м/с
голубя (Columba), скорость полета соответствовала числу Рейнольдса
около 104, рассчитанному по хорде крыла с. Частота взмахов крыла
равнялась 5 Гц, что соответствовало частотному параметру ujc/U, близкому
232
Глава 8
Рис. 8. Полет голубя (Brown, 1953)
3. Полет птиц
233
к 1/2. Подобные значения наблюдались также и при экспериментальных
исследованиях Пенникуика (Pennycuick, 1968) полета голубя в
аэродинамической трубе (методика подобного исследования впервые была разработана
Такером (Tucker, 1968)). При таком полете аэродинамические силы
отклоняются от квазистационарных значений в большей степени, чем при полете
саранчи, хотя отклонение все еще остается небольшим.
На рисунке 9 приведено схематичное изображение голубя в трех
взаимно перпендикулярных плоскостях для шести последовательных фаз цикла
взмаха крыла, начиная с наивысшего положения крыла. На этом
рисунке ясно просматривается сходство между взмахом вниз, когда хорда крыла
остается практически горизонтальной, с картиной взмаха крыла саранчи
(рис. 2) или со схемой машущего «полета», представленной на рисунке 7
главы 2. Быстрая супинация в конце взмаха вниз для обеспечения хорошо
обтекаемого взмаха вверх (с большим положительным углом наклона к
направлению среднего движения) происходит в точности так же, как показано
на рисунке 2.
С другой стороны, новым обстоятельством является изгиб лучезапяст-
ного сустава, сопровождающий движение крыла голубя вверх. Вероятно,
это способствует поддержанию значительного тягового усилия на крыле
в середине взмаха вверх, когда маховые перья (расположенные вблизи
конца крыла) сильно отгибаются назад.
С другой стороны, подъемная сила обеспечивается, вероятно, главным
образом при взмахе вниз. Браун установил, что расхождение маховых
перьев при взмахе вниз (что видно на кадре 5 рис. 8) обеспечивает увеличение
подъемной силы. Он провел в аэродинамической трубе опыты с различным
расположением перьев в жесткой держалке и показал, что такое разделение
маховых перьев (что напоминает эффект увеличения подъемной силы от
наличия щелей в передней части авиационного разрезного крыла) позволяет
получить увеличение максимальной подъемной силы на 60 % по сравнению
с «жестким» фиксированным расположением перьев.
Хотя именно голуби обычно применяют методы увеличения тягового
усилия и подъемной силы, о которых было упомянуто в последних двух
параграфах, следует отметить, что похожие методы используются и
другими птицами. Так, Браун (Brown, 1953) в своей работе приводит
рисунки последовательного взмаха крыльев у чаек (Larus). На рисунке 10
можно видеть значительно большее расхождение маховых крыльев при взмахе
вниз (кадры с 1 по 5), чем при взмахе вверх (кадры с 6 по 10). На
рисунке 11 представлено расположение при полете крыльев в трех взаимно
234
Глава 8
^С &\
КЗ
f? ^
J$? ff\
^^ J^ ft-"4
Рис. 9. Схематичное изображение голубя в трех взаимно перпендикулярных
плоскостях для шести последовательных фаз цикла взмаха крыла (рис. 1-6 из работы
Brown, 1953)
3. Полет птиц
235
Рис. 10. Полет чайки (Brown, 1953)
236
Глава 8
перпендикулярных плоскостях (сравните с рис. 9). Снова можно увидеть,
как и на рисунке 9 (хотя и не столь отчетливый), изгиб лучезапястного
сустава при взмахе вверх, в результате которого происходит направленное
назад движение маховых перьев.
Однако более важным, чем небольшое различие в
биогидродинамических параметрах между непрерывным полетом птиц и насекомых, является
довольно существенное различие в развиваемой мощности,
приходящейся на единицу массы тела. Пенникуик (Pennycuick, 1968) провел оценку
мощности, развиваемой голубем при непрерывном полете, и получил
приближенное значение 20 Вт/кг, составляющее 96 % от энергии,
вырабатываемой в процессе метаболизма. Если рассматривать затраты энергии с
точки зрения потребления кислорода, то для обеспечения непрерывного
полета потребление кислорода составляет до 130 мл/мин, а при состоянии
покоя — только 5 мл/мин. С другой стороны, при изучении непрерывного
полета чайки Вейс-Фог и Йенсен (Weis-Fogh, Jensen, 1956) получили, что
для обеспечения непрерывного полета чайке требуется только около трети
мощности, вырабатываемой мышцами птицы на единицу массы.
Отложим до главы 11 обсуждение внутренней биогидродинамики
специальных приспособлений в дыхательной системе, которые значительно
усиливают вентилирование легких. Что касается ранее упоминавшегося
различия птиц и насекомых, то в § 1 все уже практически было сказано
о роли в экономии мощности кутикулы и резилина, каучукоподобного
белка, за счет которых обеспечиваются высокочастотные колебания крыльев
насекомых. Рассмотрим с позиций биогидродинамики влияние размера
животного на потребление мощности при полете.
Пенникуик нашел (Pennycuick, 1968), что для птиц минимальная сила
тяги, необходимая для осуществления полета животного массой га, равна
весу тд, деленному на максимальное отношение коэффициентов
подъемной силы и силы сопротивления, которое может быть достигнуто при
заданной форме тела и его аэродинамических характеристиках (это также
знакомо и авиационным инженерам). Минимальная мощность, необходимая
для выработки силы тяги, равна силе тяги, умноженной на скорость,
которая растет пропорционально массе тела га в степени более высокой, чем
сила тяги. Это происходит потому, что скорость /7, при которой
выполняется условие максимума отношения подъемной силы к силе сопротивления
и через которую определяется коэффициент подъемной силы
тд
°ь " \pU*S* U)
3. Полет птиц
237
(j 7 (/ 10
т. т
Рис. 11. Цикл движений голубя, каждый элемент цикла изображен в трех взаимно
перпендикулярных плоскостях (рис. 7-12 из работы Brown, 1953)
238
Глава 8
изменяется пропорционально нагрузке крыла mg/S (здесь S — площадь
крыла). Для животных, имеющих подобные размеры, при изменении
длины / нагрузка крыла меняется пропорционально первой степени /,
поскольку нагрузка крыла меняется пропорционально /3, а площадь крыла S —
пропорционально I2. Следовательно, скорость U меняется
пропорционально /1//2, и такую же зависимость от длины имеет и минимальная мощность
на единицу массы тела птицы, требуемая для осуществления полета.
Приближенные вычисления (если не принимать во внимание
возможную экономию расхода мощности за счет эластичной системы у насекомых
или хороших аэродинамических характеристик крыла птиц) показывают,
что у голубя и саранчи, отношение полуразмахов крыльев которых равно
приблизительно 6 (25 см у голубя и 4 см у саранчи), а отношение масс
равно приблизительно б3 (450 г у голубя и 2 г у саранчи), отношение
характерных скоростей равно б1/2 (скажем, 10 м/с у голубя и 4 м/с у саранчи),
и отношение расхода мощности в целом имеет такое же значение. Однако
такие расчеты представляют еще больший интерес для случая полета
больших птиц.
Действительно, максимальная мощность на единицу массы
животного определяется фундаментальными свойствами мышечной системы. Так
что если минимальное значение мощности, требуемой для осуществления
полета, возрастает пропорционально /1//2 (или, что то же самое,
пропорционально га1/6), то поддержание тела в воздухе при непрерывном полете с
какого-то момента становится невозможным при росте линейного размера /
(или массы животного). Начиная с некоторого размера, задача поддержания
тела в полете станет невыполнимой.
Из проведенных наблюдений следует, что максимальная масса
животного, при которой возможно вести такой образ жизни, когда птица
временами совершает непрерывный полет, равна приблизительно 12 кг.
Такой массой обладает большинство представителей четырех отрядов (со-
колообразные, или дневные хищные птицы (Falconiformes), гусеобразные,
или пластинчатоклювые (Anseriforrnes), пеликанообразные, или веслоногие
птицы (Pelecaniformes), журавлеобразные {Gruiformes)). С другой стороны,
эта масса, значительно превышает максимальную массу, при которой
возможен продолжительный парящий полет, равную 20 г (см. § 4).
Возрастание при длительном полете затрат на поддержку тела в
воздухе с ростом размеров птицы объясняет тот факт, что большие птицы
приобрели замечательную способность экономить расход энергии за счет
использования естественных восходящих воздушных потоков или за счет
3. Полет птиц
239
использования разности скорости ветра на разных высотах (см. Gray, 1968
и ссылки в этой работе). Заканчивая этот раздел, остановимся кратко на
биогидродинамике планирования и полета ввысь.
Пусть планер массы га, животное ли это или аппарат, созданный
человеком, планирует, спукаясь в спокойном воздухе под углом в к горизонту,
сила сопротивления D направлена против движения, а подъемная сила L
перпендикулярна направлению движения, тогда
L = m<7cos0, D = mgsinO, в = arctg(L)/L). (2)
Угол в принимает наименьшее значение при тех же условиях, когда
отношение подъемной силы к силе сопротивления принимает наибольшее
значение, а именно при минимальной силе тяги в горизонтальном полете.
Далее будет изложена аэродинамика парящего полета, значительно более
простая, чем аэродинамика машущего полета. И результаты проведенного
исследования можно будет использовать не только для описания парящего
движения в спокойном воздухе, но также и в случае, когда скорость
парения представляет собой векторную сумму скорости движения вниз под
углом в к горизонту и скорости ветра.
Осмысление механизма планирования приводит к необходимости
учета двух составляющих силы сопротивления D, когда на тело действует
подъемная сила L: часть силы лобового сопротивления, обтекаемого
потоком тела, которая возникает вследствие трения воздуха о поверхность
тела Dj и приводит к возникновению спутной струи за телом,
обусловленной трением, и индуктивного сопротивления Di9 генерирующего вихревой
жгут, в котором воздух движется перпендикулярно направлению движения
тела, а скорость изменения количества движения равна перпендикулярной
силе L. Можно ожидать, что для больших птиц при числах Рейнольдса,
превышающих 104, составляющая силы лобового сопротивления,
обусловленная трением, пропорциональна скорости в степени, немного меньшей 2,
и, следовательно, выражение ^pU2SCDj9 где CDj — коэффициент
составляющей суммарной силы лобового сопротивления, обусловленной трением,
изменяется с ростом скорости очень медленно. Импульс присоединенного
вихревого жгута в расчете на единицу длины равен L/C/, а масса
присоединенного вихревого жгута на единицу длины пропорциональна pb29 где Ъ —
полуразмах крыльев, который приблизительно равен радиусу жгута.
Кинетическая энергия на единицу длины следа D{ изменяется пропорционально
квадрату импульса (L/C/)2, деленному на массу, так что Di можно записать
240
Глава 8
B^&Hfffi^fCL2 / ( ^рЦ^Ь^ч^-rpfi-K пра^^чееюмоет^нная
величина—Важно отметить обратную зависимость не только от скорости t/, но также и от
полуразмаха Ь. В процессе планирования птицы могут менять полуразмах
крыла. Как следует из проведенных измерений для различных профилей
крыла значение константы К колеблется около 0,10, что несколько больше
теоретического значения, равного (1/47г).
Для анализа планирования при постоянном угле в подъемную силу L
можно считать постоянной, поскольку она уравновешивает компоненту
силы тяжести тд cos 9. Тогда при заданном размахе b суммарная сила
лобового сопротивления
D = \PU*SCDf + KL>/(±pUW) (3)
зависит от скорости U. Причем сила сопротивления велика при
достаточно больших или достаточно малых значениях U и принимает минимальное
значение при некоторой промежуточной скорости Umd. Так, если
пренебречь изменением величин CDf и К, то Umd равна
и ( 4К L2\l/4 и\
\P2CDfSb2) • [)
При этом значении скорости индуктивное сопротивление равно
сопротивлению трения, т. е. в (3) оба слагаемых принимают одинаковые значения.
При U > Umd сила лобового сопротивления растет с ростом U (в этом
случае сопротивление трения превосходит индуктивное сопротивление).
Устойчивое планирование в этом случае возможно, поскольку любое
возрастание скорости приведет к немедленному увеличению силы
сопротивления. А при U < Umd возрастание скорости приведет к уменьшению силы
сопротивления (в этом случае индуктивное сопротивление превосходит
сопротивление трения), так что устойчивое планирование при этом условии
невозможно.
Значит, следует ожидать, что устойчивое планирование у птиц
осуществляется при скоростях полета, удовлетворяющих условию
U > Umd. (5)
При приближенном значении скорости Umd (4) (вычисленном в
предположении о постоянстве CDj и К) условие устойчивости (5) эквивалентно
3. Полет птиц
241
тому, что индуктивное сопротивление не должно превышать половину сум-
марной силы лобового сопротивления. То есть полуразмах крыла Ъ должен
быть достаточно большим, а именно, удовлетворять неравенству
При уменьшении скорости планирования U (скорость U должна, конечно,
превышать скорость сваливания, при которой коэффициент подъемной
силы L I l~pU2S) достигает своего максимально возможного значения)
птица расправляет крылья, так чтобы условие (6) продолжало выполняться.
При планировании птицы массой т под углом 9 под действием
силы сопротивления mg sin 9 происходит подрегулировка величины
полуразмаха крыльев Ь так, чтобы условие (6) не было нарушено. При
устойчивом планировании отношение полной силы сопротивления к силе трения
\pU2SCDf лежит в пределах от 1 до 2, а следовательно, скорость /7, при
которой возможно устойчивое планирование, должна лежать в пределах
/ \ 1/2 / \ 1/2
/ та sin 9 \ I 2mg sin 9 \ . ч
г также превышать скорость сваливания. Очевидно, что пределы
изменения скорости U расширяются с ростом угла 9, кроме того, эти пределы
птица может расширить, увеличивая коэффициент CDj (подобно тому как
это делает пилот, используя механизмы самолета для уменьшения скорости
самолета при пикировании).
Многим большим птицами необходимо достаточно долго пребывать на
довольно большой высоте над землей для того, чтобы высмотреть добычу,
затрачивая при этом как можно меньше энергии. Планирование в
наибольшей степени соответствует этой цели. Например, с наветренной стороны
скал или холмов птицы могут найти воздушные потоки, восходящие вверх
под углом 9 к горизонту, и, входя в этот поток со скоростью /7, птица
может планировать, спускаясь вниз под тем же углом 9 и со скоростью U
относительно воздуха, таким образом, оставаясь на постоянной высоте
относительно земли.
В жарких странах не только пилоты, но и крупные птицы пользуются
«термиками» — областями восходящих конвективных потоков. Например,
242
Глава 8
грифы (Aegypiinae) способны совершать планирование, удерживаясь в
центральной части термика за счёт кружения на месте. Если их скорость и угол
спуска, соответствующий скорости восходящего потока v, рассматривать
как скорость U и угол 0, то sin в = v/U и условие (7) будет удовлетворено,
если скорость U лежит в пределах
от -е^Г- ДО -W7T- • (8)
При возрастании скорости v кружение на месте становится возможным, как
только скорости в (8) превысят скорость сваливания.
Наконец, проанализируем знаменитое парящее движение альбатроса
(Diomedea). Альбатросы активно используют довольно заметную разность
скорости ветра на разных высотах над океаном. В главных чертах анализ
полета альбатроса был проведен Релеем (Rayleigh, 1883), однако здесь
будет приведено его изложение в современных терминах, предложенное
профессором С. Коррсином (S. Corrsin).
Пусть средняя скорость ветра направлена вдоль оси х, равная TT(z),
изменяется от высоты над океаном z, и пусть мгновенная скорость альбатроса
равна
fu{z) + u\v\w'). (9)
Для того чтобы избежать сваливания, кинетическая энергия
lm(u'2 + v'2 + w'2) (10)
движения альбатроса относительно воздушного потока должна
постоянно превышать диссипацию энергии за счет аэродинамического трения. Как
следует из теории турбулентности, при произвольном возмущенном
движении со скоростью (9) (вихрей или альбатроса) средняя скорость изменения
полной энергии (кинетическая и потенциальная) равна
du
—mvJw1—-, (11)
dz
где черточка над скоростями означает осреднение.2 Если множитель ufwf
отрицательный и большой по модулю, тогда (11) будет положительной
2По существу, выражение (11) представляет собой мощность инерционной силы, равной
(—ш), умноженной на ускорение (wf du/dz, 0, 0), в системе координат, где скорость
альбатроса равна (г*7, vf, w').
4. Трепещущий полет
243
и большой по величине, обеспечивая, таким образом, необходимое
среднее кинетической энергии (10). Отрицательность множителя
обеспечивается движением альбатроса, когда он, двигаясь против ветра, набирает высоту
и опускается, двигаясь по ветру Если проследить за движением
альбатроса, то можно увидеть, что его траектория движения похожа на спираль, ось
которой совпадает с направлением движения ветра и витки которой сильно
отклонены назад, так что движения, связанные с набором высоты,
происходят против ветра.
4. Трепещущий полет
Ранее было отмечено, что при полете животных со скоростями
(относительно воздуха), превышающими скорость сваливания, они вполне
справляются с задачей удержания себя в воздухе, хотя крупные
животные тратят достаточно много энергии на единицу массы для полета,
когда они не используют дополнительно энергию восходящих воздушных
потоков или потоков, образующихся из-за разности скорости ветра. А для
осуществления трепещущего полета, т. е. для такого движения, когда
животное удерживает себя в воздухе, не совершая сколько-нибудь значимых
движений в горизонтальной плоскости (относительно воздуха), требуется
очень большая энергия. Действительно, довольно длительный трепещущий
полет, о котором речь пойдет в этом параграфе, может быть осуществлен
только небольшими животными, большее из которых (например, такое, как
исполинский колибри (Patagona gigas)) имеет массу 20 г. Исследование
трепещущего полета преследует значительно более общие цели, чем просто
изучение движения мелких животных, — многие крупные животные могут
анаэробным способом выработать энергию, достаточную для
осуществления трепещущего полета в течение короткого времени движения, например
при посадке или взлете.
В своем общем обзоре, посвященном длительному трепещущему
полету, Вейс-Фог (Weis-Fogh, 1973), выделил общую схему полета, которую он
назвал «стандартным зависанием», характерную для колибри (Trochilidae)
и насекомых — различных представителей одиннадцати отрядов. Ниже
речь пойдет, о представителях четырех отрядов (жуки, или жесткокрылые
(Coleoptera), бабочки, мотыльки, моли, или чешуекрылые (Lepidoptera),
перепончатокрылые насекомые (Hymenoptera) и двукрылые (Diptera)). С
точки зрения биогидродинамики движения стандартного зависания
представляют собой приспособление тех машуших движений, которые используют-
244
Глава 8
ся при прямолинейном полете. Для создания подъемной силы,
удерживающей тело в воздухе, необходимы движения крыльев относительно воздуха
в горизонтальной плоскости следовательно, при отсутствии перемещения
животного в горизонтальной плоскости, крылья должны совершать
горизонтальные биения назад и вперед. Очевидно, что при прямолинейном
полете крылья совершают махи в плоскости, перпендикулярной
горизонтальной оси тела, если же ось тела сделать вертикальной, машущий полет
можно легко перевести в трепещущий полет.
На рисунке 12 изображен стандартный трепещущий полет бражника
табачного {Manduca sexta) (вид сверху). Как только что было сказано,
положение оси тела близко к вертикальному, вследствие чего махи крыльев
происходят практически в горизонтальной плоскости. Для того чтобы
обеспечить движение передней кромки крыла вперед при угле атаки,
обеспечивающем благоприятное соотношение между подъемной силой и
сопротивлением, поворот крыла при машущем полете, представленный на рисунке 3,
должен быть достаточно большим. Пронация крыла в начале взмаха вниз
значительно превышает обычную пронацию при машущем полете
(кадры (а) и (6)), в результате чего крылья движутся в горизонтальной
плоскости (кадры с (с) до (g)\ а плоскость крыльев развернута под углом атаки
менее 30°. (Фактический угол атаки еще меньше, поскольку крылья
движутся в общем нисходящем потоке, обусловленном трепещущими движениями
животного.) Затем супинация в начале взмаха вверх значительно (кадры (Л),
(/)) усиливается, так что крыло движется по тому же горизонтальному пути
в обратном направлении приблизительно с тем же углом атаки.
Этот широко распространенный «стандартный трепещущий полет»
является логическим завершением процесса адаптации движения крыльев
при машущем полете к движениям при трепещущем полете. Переход от
одной формы полета к другой можно наблюдать у крупных птиц при
приближении к земле. Тело птицы занимает почти вертикальное положение,
она расправляет хвостовые перья для торможения, разворот крыла
увеличивается, так что маховые движения происходят уже не в вертикальной,
а в горизонтальной плоскости, и при посадке вес птицы
уравновешивается подъемной силой, создаваемой машущими в горизонтальной
плоскости крыльями. При взлете, когда требуется существенная сила тяги для
создания ускорения в горизонтальном направлении, стандартный
трепещущий полет почти сразу сменяется некоторой переходной формой
машущего полета: плоскость махов крыльев расположена наклонно, так что даже
при небольшой скорости продвижения вперед крыло может создавать тя-
4. Трепещущий полет
245
К, У. S? V.
(a) -V^ (Ь) -^ (С) -^ (rf)
"^ ^^ ^Ач
(е) (У) ^Ы *S ^(fc)
А А А ^^
(m) (n) (о) (р)
I .... I
Q Zlqm
Рис. 12. Стандартный трепещущий полет бражника табачного (Manduca sexto)
(рис. 8 из работы Weis-Fogh, 1973)
гу и подъемную силу, достаточные для взлета. Особенно четкую картину
медленного полета вперед (промежуточную фазу машущего полета) можно
наблюдать при полете голубя (описанном Брауном (Brown, 1948)).
В оставшейся части главы хотелось бы подробнее остановиться на
способах полета, представляющих собой промежуточные фазы между
машущим полетом при движении вперед и трепещущим полетом. Не будем
останавливаться на искусной компоновке костно-мышечной системы,
позволяющей осуществлять сложные движения при трепещущем полете, как
те, что изображенные на рисунке 12, когда крылья совершают колебания
с частотой 30 Гц, например у мотыльков или колибри, или с частотой на
порядок больше у небольших перепончатокрылых (Hymenoptera) и
двукрылых (Diptera). Сконцентрируем внимание только на биогидродинамических
проблемах — механизме поддержания тела животного в воздухе при
стандартном трепещущем полете или его модификациях.
В 1973 году Вейс-Фог (Weis-Fogh, 1973) опубликовал обширный
обзор имеющихся на то время наблюдений за животными, способными
осуществлять трепещущий полет в течение продолжительного времени. И он
246
Глава 8
пришел к заключению, что колибри и большинство насекомых совершают
движения, обеспечивающие трепещущий полет, как те, что изображены на
рисунке 12. Однако существуют и весьма важные следующие исключения:
(i) некоторые очень маленькие насекомые (наибольшее число
задокументированных наблюдений касается осы {Encarsia formosa)) в процессе
полета, представляющего некоторую модификацию «стандартного
трепещущего полета», складывают крылья за спиной в течение одного
махового цикла; этот механизм движения более подробно будет описан
в главе 9;
(ii) представители семейства дневных бабочек (Papilionoidea) из отряда
бабочек (Lepidoptera) совершают в полете движения, включая
складывание крыльев (как за спиной, так и перед грудью), которые
представляются еще более искусными и сложными;
(iii) стрекозы (Odonata) в трепещущем полете (продольная ось тела при
этом практически горизонтальна) совершают сложные хорошо
скоординированные движения своими четырьмя крыльями;
(iv) представители подсемейства журчалок (Syrphinae) отряда двукрылых
(Diptera) или «цветочниц» обитают там же, где и стрекозы, и обладают
похожей способностью к трепещущему полету с почти
горизонтальным расположением тела, но с совершенно другими движениями их
единственной «стреловидной» пары крыльев.
Среди большого количества данных наблюдений,
проанализированных и обобщенных Вейс-Фогом (Weis-Fogh, 1973), наличие
кинодокументов по движениям при трепещущем полете предоставило дополнительные
возможности и для теоретического исследования полета. Были
рассчитаны значения среднего коэффициента подъемной силы CL, достаточной для
поддержания веса животного при трепещущем полете, обусловленной
взмахами крыльев с частотой, измеренной акустическими методами и заметной
для невооруженного глаза амплитудой. На рисунке 13 представлены
результаты расчетов коэффициента подъемной силы. Звездочки означают
насекомых, относящихся к указанным ранее группам исключений (i), (ii) (летучие
мыши в данном параграфе не рассматриваются), для которых к расчетам,
проведенным в предположении «стандартного трепещущего полета»,
следует относиться с осторожностью. Расчетные данные для этих групп
насекомых приведены в скобках.
Данные рисунке 13 (за исключением групп, отмеченных звездочкой)
подтверждают тот взгляд на движения при «стандартном трепещущем по-
4. Трепещущий полет
247
Летучие мыши: * Plecotus auritus
Птицы: hummingbirds, Trochili
Archilochus colubris
Amazilia Jimbriata fluviatilis
Patagona gigas
Coleoptera: жуки
Melolontha vulgaris
Amphimallon solstitialis
Heliocopris sp.
H. sp
Cetonia aurata
Cerambycid species
Lepidoptera: бабочки, мотыльки, моли
Pieris napi * *
P. brassicae * *
Sphinx ligustri
Manduca sexta
A/, sexta
Macroglossum stellatarum
Amathes bicolorago
Hymenoptera: осы, пчелы
Vespa crabro
V. vulgaris
Bombus terrestris
B. lapidarius
Apis mellifica
Encarsia formosa (Chalcid wasp) * * *
E. formosa (Chalcid wasp) * * *
Diptera: комары-долгоножки, комары и мухи
Jipula sp.
Theobaldia annulata
Aedes aegypti
Eristalis tenax
Calliphora erythrocephala
Drosophila virilis * * *
Средний
коэффициент
подъемной
силы С L
(1,3)
1,9
2,0
1,8
0,6
0,7
0,6
0,5
0,5
1,1
(2,2)
(0,9)
1,2
1,2
0,9
1,1
1,6
0,8
0,8
1,2
0,9
0,8
(2,4)
(3,2)
0,8
0,7
0,6
0,9
1,3
1,0
Число
Рейнольдса
(Re)
14000
6400
7500
15 000
4700
3000
17000
23 000
4300
1600
1400
4000
6300
6700
6100
2800
1600
4200
1600
4500
3700
1900
18
15
770
480
170
2000
1000
210
Рис. 13. Значения среднего коэффициента СL подъемной силы для различных
представителей мира летающих животных, необходимой для поддержания тела
животного при стандартном трепещущем полете, и чисел Рейнольдса, рассчитанных по
хорде крыла, для соответствующих горизонтальных движений крыла (Weis-Fogh,
1973)
248
Глава 8
лете», что в этом случае подъемная сила, создаваемая при движении
крыльев, способна поддерживать тело животного, а значения коэффициента
подъемной силы не являются неправдоподобно большими. Для насекомых,
даже при их плотных крыльях, значения среднего коэффициента подъемной
силы вполне приемлемы. Так, например, при полете табачного бражника
(Manduca sextet) (схематически его полет изображен на рис. 12) средний
коэффициент подъемной силы принимает значение 1,2. Такое же значение
имеет коэффициент подъемной силы при стандартном трепещущем
полете большого земляного шмеля (Bombus terrestris) (масса особей этого вида
составляет в среднем 0,9 г). Следует отметить, что некоторые авторы
сомневались, что трепещущий полет можно описать на основании
аэродинамических принципов. Довольно маленькие значения коэффициента у
представителей отряда жесткокрылых, жуков (Coleoptera) (всего 0,5 для
самого большого навозного жука из рода пластинчатоусых жуков (Heliocopris)
весом 13 г), а также для представителей отряда двукрылых (Diptera)
(находящихся на другом конце шкалы по массе тела, коэффициент С L равен
только 0,6 при полете желтолихорадочного комара (Aedes aegypti), вес
которого составляет 1 мг). Для осуществления трепещущего полета колибри
требуется значительно большая подъемная сила — среднее значение
коэффициента для этих птиц равно 1,9. Заметим, однако, что устройство их
первостепенных маховых перьев исключительно подходит для достижения
того увеличения максимального значения подъемной силы, о котором шла
речь в § 3. При просмотре в замедленном режиме высокоскоростной
съемки полета было обнаружено, что колибри расправляет первостепенные
маховые перья, в результате чего происходит увеличение коэффициента CL
на 60%-с 1,2 до 1,9. _
При расчете значений коэффициента CL, приведенных на рисунке 13,
для каждой птицы потребовалось определить величину, которая здесь
обозначена через 52, равную второму моменту поверхности крыла
относительно оси шарнира. Действительно, при взмахе крыла и заданной угловой
скорости П создаваемую на единицу площади подъемную силу можно
записать в виде ^pU2CL, где U представляет собой скорость крыла в воздухе,
равную произведению Q на расстояние до оси шарнира. Тогда подъемная
сила, действующая на все крыло, может быть записана в виде ^pfi252CL,
здесь С L — взвешенное среднее коэффициента CL подъемной силы,
действующей в поперечном сечении крыла. При заданной частоте и угловой
амплитуде взмахов крыла птицы может быть легко вычислено осреднен-
ное по времени значение Л в предположении, что крыло описывает си-
4. Трепещущий полет
249
нусоидальную траекторию с течением времени. Тогда после осреднения по
времени угловой скорости (I коэффициент ~СL может быть найден из урав-
нения баланса сил
\p&2S2CL = mg. (12)
Оставим в стороне другие аспекты «стандартного трепещущего
полета», подробно изученные Вейс-Фогом (Weis-Fogh, 1973), включая
особенности строения крыла, обусловленные действием изгибающих моментов
у основания, и подробное сравнительное изучение расхода мощности при
полете. Завершая эту главу, опишем некоторые модификации стандартного
трепещущего полета, о которых говорилось выше в пунктах (i)-(iv). Вейс-
Фогу удалось прояснить роль «хлопка-броска» (т. е. быстрого складывания
насекомым крыльев за спиной с последующим резким их раскрытием) на
примере полета осы (Encarsia formosa). И это открыло путь к исследованию
биогидродинамики бабочек, поскольку бабочки во время отдыха
складывают крылья вместе за спиной, а при полете совершают крыльями движения
типа «хлопок-бросок», за счет чего возникает существенная по величине
подъемная сила. Так, согласно данным рисунка 13, белянка брюквенная
(Pieris napi) может создавать подъемную силу, коэффициент С L которой
достигает значения 2,2.
Приблизительная оценка коэффициента СL для сильно
различающихся трепещущих полетов журчалок (Syrphinae) и стрекоз (Odonata)
показала, что эти коэффициенты достаточно велики. К очень интересным
выводам пришел Вейс-Фог (Weis-Fogh, 1973), исследуя специальный механизм
увеличения подъемной силы у журчалок, включая постулированный
механизм «разворота плоскости крыла», напоминающий механизм резких схло-
пывающих движений осы (Encarsia formosa). Дальнейшее изучение
полета журчалок представляет несомненный интерес. Отдельно следует сказать
и о многообещающей области исследования полета стрекоз с их
доведенным до совершенства взаимодействием четырех крыльев, что, вероятно,
потребует более глубокого и серьезного подхода к этой задаче с позиций
биогидромеханики.
На рисунке 14 представлено общее строение маленькой
паразитической осы (Encarsia formosa), использующейся как средство биологического
контроля над размножением растительной тли. Размах крыльев осы очень
небольшой и составляет всего 0,7 мм. «Крючочки» обеспечивают
подвижную связь между передним и задним крыльями, за счет чего оба крыла
движутся как единое целое. Щетинки на крыльях, возможно, предотвраща-
250
Глава 8
Рис. 14. Строение осы (Encarsia formosd) (рис. 13(A) из работы Weis-Fogh, 1973)
ют слипание крыльев при их соединении за спиной, возможно, они также
предотвращают отрыв пограничного слоя. О значимости этого явления еще
будет упомянуто в главе 9, § 3.
Выполненная профессором Вейс-Фогом высокотехничная скоростная
съемка трепещушего полета осы {Encarsia formosa) (со скоростью 7000
кадров в секунду) показала, что цикл движений ее крыльев значительно
более сложный, нежели представленный на рисунке 12, — ей удается
совершать до 400 взмахов в секунду На рисунке 15 приведено покадровое
представление полета этого насекомого. Отметим еще раз почти
вертикальное расположение тела насекомого при трепещущем полете. Обратим также
внимание на положение ножек осы. Такое положение указывает на то, что
на этих кадрах спинка осы (за спинкой схлопываются крылья) все время
повернута вправо.
Цикл движений начинается (кадры 16 и 0) с момента, когда крылья
сложены вместе и неподвижны. Затем происходит резкое движение крыльев
(кадры 1 и 2): быстрый разворот каждой пары как одного целого (см. выше)
относительно их общей оси вращения, расположенной вдоль задней
кромки крыла. На остальных кадрах (кадры с 3 по 13) происходит движение
крыльев такое же, как и при стандартном трепещущем полете (см. рис. 12)
до следующего схлопывания крыльев за спиной (кадры с 14 по 16).
4. Трепещущий полет
251
заканчивается начинается
второй взмах третий взмах
Q
6
10
11
12
13
15
горизонтальное
направление
начинается
четвертый взмах
2 мм
Рис. 15. Раскадровка скоростной съемки трепещущего полета осы (Encarsia
formosd) (рис. 14 из работы Weis-Fogh, 1973)
252
Глава 8
Время: 0,0
разворот
плоскости
крыльев
6,0 (мс)
12 16 0 4
Номер кадра
12 16 0
Рис. 16. Зависимость угла подъема крыла 7 от времени при трепещущем полете
осы {Encarsia formosa) (рис. 15 из работы Weis-Fogh, 1973)
Если все движения осы полагать аналогичными только стандартному
трепещущему полету (рис. 13), то оказывается, что коэффициент
подъемной силы должен быть при этом приблизительно равен 3. Более того,
число Рейнольдса, рассчитанное по хорде крыла и горизонтальной скорости
стандартного трепещущего полета, оказывается равным 20, хотя если
число Рейнольдса рассчитать (гл. 9, § 1) по скорости передней кромки, когда
крыло скручивается, то оно окажется равным 30. А это чрезвычайно низкие
значения числа Рейнольдса для крыльев, которые должны создавать
достаточно большую подъемную силу. На рисунке 16 представлена зависимость
от времени угла подъема крыла 7 (аналогичного углу 7' на рисунке 4). Как
видно из рисунка 16, периоды «хлопка-броска» и приблизительно
синусоидального движения в горизонтальной плоскости, как при «стандартном
трепещущем полете», перемежаются «скручиванием» — очень быстрой
супинацией, подобной той, что представлена на рисунке 12.
4. Трепещущий полет
253
схлопывание резкий взмах конец резкого взмаха
Рис. 17. Схематичное изображение механизма резкого взмаха, за счет которого
создается циркуляционный поток, у осы (Encarsia formosa) (рис. 21 из работы Weis-
Fogh, 1973)
Вейс-Фог (Weis-Fogh, 1973), детально исследуя движения «хлопка-
броска» и «скручивания», предложил совершенно иной с точки зрения
биогидродинамики механизм мгновенного создания весьма большой
подъемной силы крыла. Механизм Вейс-Фога представляет настолько большой
научный интерес и настолько важен для решения различных проблем,
связанных с полетом животных (например, для объяснения динамики полета
254
Глава 8
бабочек), что следующая глава 9 полностью посвящена анализу
предложенного им механизма с точки зрения аэродинамики. И в завершение главы 8
приведем схему, составленную Вейс-Фогом (рис. 17), на которой
проиллюстрировано возникновение большой по величине циркуляции, за счет чего
и появляется большая подъемная сила — по существу, за счет резкого
ускоренного движения воздуха в раскрывающиеся крылья.
Литература
[1] Brown, R.H.J. 1948. J. Exptl. Biol. 25, 322-333.
[2] Brown, R.H.J. 1953. Ibid. 30, 90-103.
[3] Gray, J. 1968. Animal Locomotion, Weidenfeld & Nicolson, London.
[4] Pennycuick, C.J. 1968. J. Exptl. Biol. 49, 527-555.
[5] Rayleigh, Lord. 1883. Nature, 27, 534-535.
[6] Smart, J. & Hughes, N.F. 1972. In Insect/Plant Relationships: Sympos. R.
Entomol. Soc. Lond. no. 6, 143-155.
[7] Tucker, V. A. 1958. J. Explt. Biol. 48, 67-87.
[8] Weis-Fogh, T. 1960. Ibid. 37, 889-907.
[9] Weis-Fogh, T. 1973. Ibid. 59, 169-230.
[10] Weis-Fogh, T. & Jensen, M. 1956. Proc. Roy. Soc. B, 239, 415-584.
Глава 9
Механизм Вейс-Фога образования
подъемной силы1
Вейс-Фог (Weis-Fogh, 1973) предложил новый механизм образования
подъемной силы, представляющий большой научный интерес.
Поразительно, что этим механизмом можно объяснить возникновение подъемной силы
даже при двумерном описании движения, начавшегося из состояния покоя.
Из теоремы Кельвина следует, что для таких движений циркуляция по
контуру, охватывающему тело, равна нулю. Однако если тело состоит из двух
не связанных между собой областей, то циркуляции по контурам,
охватывающим каждую область, могут быть отличны от нуля, равны по величине
и противоположны по знаку. Каждая область может создавать достаточную
для ее поддержания подъемную силу В полете при резком разведении
насекомым двух крыльев с хордой с (рис. 1) с угловой скоростью ft между
раскрывающимися лопастями крыльев в результате засасывания воздуха
генерируется безвихревое движение, причем, как показано в § 2, циркуляции
по контурам, охватывающим каждое крыло, равны —0,690с2 и +0,69Пс2.
В момент, когда задние кромки крыльев разъединяются, в безвихревом
движении возникают точки застоя (позиция (/)). Под действием сходящих
вихрей в безвихревом потоке возникают вязкие силы, которые
генерируют (§3) сход с передней кромки замкнутого объема воздуха, что
приводит к некоторому увеличению суммарной завихренности. В § 4 освещается
роль вихрей в действии механизма Вейс-Фога генерирования подъемной
силы в трехмерной постановке. Кроме того, в этом параграфе
рассматривается образование сходящих с концов крыльев вихрей, в частности, и для
случая трепещущего полета подобных осе (Encarsia formosa) насекомых,
крылья которых при трепещущем полете описывают круговые траектории.
В конце главы отмечается, что поле скоростей воздуха на достаточно
большом расстоянии от таких очень маленьких летящих насекомых является
Перепечатано из журанала J. Fluid Mech., 60 (1973), pp. 1-17.
256
Глава 9
точным решением уравнений Навье-Стокса (Landau, 1944; Squire, 1951).
При этом считается, что точечная сила (равная весу тд насекомого)
действует на жидкость плотностью рис кинематической вязкостью v и
отношение тд/pv2 достаточно мало, так что возникшее струйное течение будет
устойчивым.
1. Введение
Вейс-Фог (Weis-Fogh, 1973) при анализе трепещущего полета халь-
циды-осы (Encarsia formosa) (эта оса используется для биологического
контроля над распространением тли и является важным с точки зрения
экономики паразитирующим насекомым) пришел к выводу, что по сравнению
с большинством других насекомых она создает намного большую
подъемную силу. Это происходит в силу использования ею особого способа
движения крыльями (особого механизма создания циркуляционных потоков),
который до сих пор не был знаком специалистам в области аэродинамики,
так что он представляет значительный научный интерес. При обычном
трепещущем полете крылья животного быстро движутся назад и вперед в
горизонтальной плоскости, создавая при каждом движении подъемную силу
за счет разворота крыльев, вследствие которого одна и та же кромка крыла
обращена в сторону его движения под углом атаки, обеспечивающим
наибольшую подъемную силу. Однако увеличение коэффициента подъемной
силы замедляется в силу эффекта Вагнера: для схождения вихрей с задней
кромки крыла, обусловливающих возникновение необходимых
циркуляционных потоков вокруг крыла, требуется определенное время (Wagner, 1925).
Оса же {Encarsia formosa) каждое движение крыла в горизонтальном
направлении предваряет специальным движением («хлопок-бросок»),
которое, как считает Вейс-Фог, приводит к практически немедленному
возникновению циркуляционных потоков, за счет чего без какой-либо временной
задержки создается максимальная подъемная сила. Для выявления техники
полета этого насекомого потребовалась высокоскоростная съемка,
поскольку частота биений ее крылышек составляет 400 Гц.
Механизм Вейс-Фога подобно эффекту Вагнера может быть описан
с позиций двумерной теории. Приводимое ниже описание корректно для
тех животных, у которых удлинение крыла приблизительно равно 5. В
данной главе приводится количественный анализ механизма Вейс-Фога в
двумерной постановке. Некоторые комментарии относительно трехмерной
постановки изложены в заключительном § 4.
1. Введение
257
Особенно примечательно, что, основываясь на предложенном им
механизме, Вейс-Фог (Weis-Fogh, 1973) смог описать процесс образования
подъемной силы, используя теорию течения невязкой жидкости. Анализ
невязкого двумерного потока приведен в §2, а в §3 при учете вязкости
потока внесены некоторые корректирующие поправки в результаты § 2.
Отметим, что механизм образования подъемной силы в двумерном невязком
потоке жидкости был открыт в последующие шесть десятилетий после
работ Прандтля, Жуковского, Кутта и Ланчестера (Lanchester).
Кроме того, механизм Вейс-Фога срабатывает и в жидкости с нулевой
вязкостью, а не просто в предельном случае исчезающе малой вязкости,
когда тонкие пограничные слои Прандтля у острых задних кромок, где
выполняются условия Кутта-Жуковского, переходят в вихревой след — вихри
Ланчестера. При невязком двумерном потоке из теорем Гельмгольца, Сток-
са, Кельвина следует, что если тело начинает движение в жидкости из
состояния покоя, то циркуляция потока относительно тела равна нулю во все
время движения, в силу чего образование подъемной силы (или
какой-либо другой силы, за исключением сил, обусловленных наличием
виртуальных масс) невозможно. Этим теоремам совершенно не противоречит
возможность появления циркуляции, когда тело распадается на две несвязные
части, вокруг каждой части равных по величине и противоположных по
знаку. Каждый возникший циркуляционный поток воздуха приводит к
появлению подъемной силы, действующей на каждую часть, которая и
обеспечивает движение частей.
На рисунке 1 схематично изображены последовательные движения
крыльев, включающие те движения (механизм Вейс-Фога), которые
приводят к образованию подъемной силы. Тело животного, совершающего
трепещущий полет, расположено вертикально головой вверх. Вертикальная
плоскость проходит со стороны спинки животного, посередине между телом
животного и концами его крыльев. На рисунке показаны последовательные
положения сечений крыльев АВ и CD вертикальной плоскостью,
проходящей через середину крыльев.
Последние три позиции (/), (д) и (h) на рисунке 1 представляют
обычный трепещущий полет, когда оба крыла движутся в горизонтальной
плоскости с положительными углами атаки. Позднее будет пояснено, почему
в процессе этого движения с предшествующим «хлопком-броском», с
самого начала образуется максимально возможная подъемная сила. Прежде
чем это описывать, отметим, что трепещущие в горизонтальной плоскости
крылья при обычном трепещущем полете изображены в виде прямолиней-
258
Глава 9
B„D B„D В, ,D Вч ,D
)L V
AC AC AC AC
(a) (b) (c) (d)
Вк /D В D В D В D
AC AC AC AC
(e) {f) _ (9) (h)
Рис. 1. Последовательность движений сечений крыльев осы (Encarsiaformosa).
Сечения лежат в вертикальной плоскости, проведенной со стороны спинки
вертикально расположенного насекомого, посередине между телом насекомого и концами его
крыльев. Позиции (а) и (6) — крылья находятся в состоянии мгновенного отдыха
после «схлопывания»; позиции (&)-(/) — крылья резко раздвигаются; в позиции (/)
крылья отъединяются друг от друга; позиции (/), (д) и (h) — крылья совершают
обычные движения трепещущего полета
ных движений в позициях (/), (д) и (/i), хотя в действительности крылья
описывают круги в горизонтальной плоскости вокруг вертикально
расположенного тела. Такое движение можно было бы изобразить более точно,
если бы движение крыльев представлялось бы не в сечении их вертикальной
плоскостью, а при сечении цилиндрической поверхностью, ось симметрии
которой совпадает с осью вертикально расположенного насекомого.
В определенный момент цикла машущего движения насекомое
совершает «хлопок» крыльями. Его крылья схлопываются за спиной, соединяясь
так, как показано на первых двух позициях рисунка 1, передние кромки
крыльев занимают вертикальное положение и располагаются выше задних
кромок. В позиции (6) окружающая среда уже находится в невозмущенном
состоянии. Этот момент можно рассматривать как начало действия
механизма Вейс-Фога. Поскольку к этому времени оставшиеся вихри, которые
образовались в процессе схлопывания крыльев, унесены с потоком воздуха
достаточно далеко от крыльев, и их влияние на дальнейший процесс можно
не учитывать.
В позициях (6) -(g) на рисунке 1 показаны положения крыльев АВ
и CD, когда они еще, как и в четырех первых позициях, касаются друг
друга (точки А и С совпадают) и составляют единое целое. В момент (/)
крылья начинают разъединяться и полностью разделяются, занимая
положение (д). Механизм Вейс-Фога образования подъемной силы состоит
в следующем. Вращательные движения хорды АВ относительно точки А
2. Двумерная постановка задачи о движении крыльев 259
и хорды CD относительно точки С (т.е. «бросок» крыльев) (позиции (&)-
(/)) порождают в момент (/), когда крылья начинают разъединяться,
циркуляционные потоки воздуха вокруг них с равными по величине и
противоположно направленными циркуляциями — отрицательными вокруг
хорды АВ и положительными вокруг CD. Причем величина циркуляции
достаточна для образования максимально возможной подъемной силы сразу
же, как только крылья начинают двигаться в горизонтальной плоскости.
В § 2 приводится математическая двумерная модель механизма Вейс-Фога
образования подъемной силы в случае невязкой жидкости. А в § 3 вносятся
поправки, связанные с учетом вязкости окружающей среды. Необходимость
учета вязкости становится понятной, если учесть, что речь идет о движении
при низких числах Рейнольдса — порядка 30. Число Рейнольдса
рассчитано по хорде крыла, равной 0,22 мм, и скорости движения передней кромки
2,2 м/с. И в конце главы в § 4 приводятся некоторые соображения
относительно трехмерной постановки задачи о движении крыльев насекомого.
2. Двумерная постановка задачи о движении крыльев
насекомого в невязком потоке
На рисунке 2 представлена двумерная модель процесса «броска»
крыльев. Пунктирная линия EF — ось симметрии, отрезки АВ и CD
длиной с — поперечные хорды крыла. Каждая хорда составляет с прямой EF
угол а. В процессе «броска» угол а возрастает от нуля до некоторого
определенного угла а0 (порядка 7г/3), начиная с которого совпадавшие до этого
точки А и С начинают расходиться.
Вследствие отсутствия вязкости возникающий поток воздуха при
движении крыльев с угловой скоростью fi = da/dt является безвихревым.
В данном параграфе будут рассчитаны параметры этого потока. Отметим
для начала некоторые его характерные черты. В потоке имеются на обеих
сторонах угла точки, где скорость потока равна нулю (точки торможения),
а циркуляция потока вокруг каждого крыла равна
Г - Пс2д(а) (1)
(отрицательная вокруг крыла АВ и положительная вокруг CD). Здесь
функция д(а) подлежит определению.
С достаточной степенью точности можно считать, что в тот момент,
когда крылья развернулись относительно прямой EF на угол а = а0, угловая
260
Глава 9
F
сЧ | Ус
А]С
L
J?
Рис. 2. Двумерная модель процесса резкого разведения крыльев насекомого.
Крылья АВ и CD поворачиваются относительно совпадающих точек А и С с угловой
скоростью f2 = da/dt. Движение воздуха симметрично относительно прямой EF
скорость крыльев достигла значения £] = О0, и начинается их раздвиже-
ние (т. е. точки А и С начинают расходиться), поток воздуха не меняется.
В этом случае циркуляция потока воздуха относительно каждого крыла
равна Q0c2g(a0) и вследствие предположения о невязкости потока будет
оставаться постоянной при последующем движении. Тогда, согласно теории
неустановившегося обтекания крыла, возникает подъемная сила,
действующая на каждое крыло, и при отсутствии сходящих вихрей. Отложим до § 3
исследование возможной коррекции описания процесса «броска» крыльев
на основе предположения о двумерности и невязкости потока за счет учета
сходящей как с передней кромки, так и с задней кромки после раздвижения
крыльев вихревой пелены. И перейдем к определению функции д(а).
В потоке, обусловленном движением крыльев, изображенным на
рисунке 2, линия симметрии EF является линией тока. В силу этого будем
рассматривать движение воздуха вокруг крыла АВ с левой стороны от EF
и рассчитаем результирующую циркуляцию Г. Введем комплексные пере-
2. Двумерная постановка задачи о движении крыльев 261
z плоскость
Е
(«)
Zплоскость
Ах В А2
Е г •—г Е
„-1 (») '
Рис. 3. Комплексные плоскости, которые используются для описания потока
воздуха вокруг крыла АВ: (а) на плоскости z9 представлена левая область потока (рис. 2),
развернутая на 90°; (6) на верхнюю полуплоскость Z конформно отображена
плоскость z
менные. Исследование будет проводиться (рис. 3) на верхней
полуплоскости комплексной плоскости z. Пусть действительная ось совпадает с
прямой EF. На плоскости имеется разрез от начальной точки А до точки В,
где z = сега. Обозначим через Ах и А2 вершину угла с разных сторон от
разреза.
Отобразим верхнюю полуплоскость с разрезом плоскости z на
верхнюю полуплоскость плоскости Z с разрезом на действительной оси,
используя конформное отображение Шварца-Кристофеля
а/7Г Z - а
Z + 1
(2)
Аргумент комплексного числа в скобках равен нулю при Z = сю.
Бесконечно удаленная точка отображается в бесконечно удаленную, точки Аг и А2
отображаются в точки Z = — 1 и +1 соответственно. Так как на плоскости z
точки Ах и А2 совпадают, то следовательно, интеграл (2) с пределами
интегрирования от Z = —1 до Z = +1 обращается в нуль, что дает значение
а = 1-(2а/тг), (3)
которое определяет координату точки В на плоскости Z. Переменная ze~~za
на крыле АВ определяет расстояние от точки А до произвольной точки
262
Глава 9
крыла, может быть записана в виде
а/тг
ze-ia = Kf(Z), где f(Z) = j (l-J^*'" ^dZ. (4)
-1
Переменная ze~ia принимает значение с в точке В (где Z = а). Тогда К,
определяемая как
K = cfmL, W /max = /(<*)> (5)
равна максимальному значению /(Z) на интервале — 1 < Z < 1.
Функция тока ^ (мнимая часть комплексного потенциала w) равна
нулю на линиях тока ЕАХ и A2F и отлична от нуля на крыле АВ9 каждая
точка которого имеет линейную нормальную скорость, равную расстоянию
ze~lCL от точки А, умноженному на угловую скорость Q. Зависимость
функции тока ф от расстояния на крыле А В может быть записана в виде
ЫМ = ф = -1п(*е"'в)2, (6)
что является граничным условием для функции тока как на АХВ, так
и на А2В.
Соответствующее граничное условие на плоскости Z для
действительных Z имеет вид
ым=< - ?«*№ (-1 < * < I», (7)
О (иначе).
Комплексный потенциал w(Z), удовлетворяющий этим граничным
условиям, имеет вид
ПК2 } f2{t)dt
W =
2тг
-1
т-
Циркуляция Г вокруг крыла АВ в отрицательном направлении равна
изменению потенциала скорости ф при обходе вокруг крыла от точки Ах к
точке Л2, что соответствует движению на комплексной плоскости Z от — 1
до +1. Тогда
1
'2
r = ^-J(l-t2)-'f(t)dt. (9)
-1
2. Двумерная постановка задачи о движении крыльев 263
1,6
1,4
1,2
^1,0
0,8
0,6
0,4 |-
0,2
0
J*
о;
!«
7Г
Рис. 4. Зависимость (10) коэффициента д(а) в выражении (1) для циркуляции Г
вокруг АВ от угла а, равного половине угла между крыльями; величины / и /тах
определены соотношениями (4) и (5). Пунктирной кривой слева изображена ап-
проксимационная зависимость (при малых а) (12) д(а). Справа пунктирная кривая
изображает зависимость (12) д(а) при малых углах 7г — а, хотя, скорее всего, эта
зависимость не представляет какого-либо практического интереса
Из (1), (5) и (9) вытекает, что
9(<*) =
1
j{\-t2)-'f{t)dt
L-l
2 \-1
(""/max)
(10)
График зависимости функции д(а) (10), которая является некоторым
весовым средним, от угла а представлен на рисунке 4. В центральной части
интервала изменения углов а д(а) меняется незначительно. Действительно,
при j7r < а < f 7г д(а) лежит в пределах от 0,64 до 0,77 (так что
функция /(£) достигает своего максимума на интервале от — ^ до ^). Таким
образом, 0,64 < д(а) < 0,77 при тупом угле 2а между крыльями, причем
функция д(а) растет особенно медленно вплоть до достижения угла между
крыльями 120° (при а = ^тг д(а) = 0,69).
264
Глава 9
Этот факт представляется достаточно важным не только потому, что
при нахождении циркуляции g(a0)Q0c2 оказывается не обязательно знать
точное значение угла а0, при котором крылья начинают разъединяться, но
также и потому, что перепадом давлений в образующемся между крыльями
промежутке можно пренебречь. Перепад давлений между точками
торможения А1 и А2 равен производной —рдф/dt, величина которой равна
т. к. Г равна разности значения потенциала скорости в точках Ах и А2.
Поскольку в средней части изменения угла а функция д(а) меняется
незначительно, то величина (11) должна быть небольшой.
Отметим, что для малых значений угла а функцию д(а) можно
представить приближенно в виде
9{a)~h+hx<- (12)
На рисунке 4 эта зависимость изображена пунктирной линией. Такое ап-
проксимационное поведение функции д(а) можно получить аналитически
из (10). Дадим зависимости (12) физическую интерпретацию.
В начальной стадии процесса «броска» крыльев поток окружающего
воздуха определяется практически полностью тем, что воздух
устремляется в образующийся промежуток между крыльями. Объем воздуха,
приходящийся на единицу длины поперечной хорды крыла, на расстоянии от
точки Л, меньшем, чем г, равен от2. Скорость его роста
Пг2 (13)
определяется (рис. 5) втеканием потока через дугу длиной 2аг со
средней скоростью Qr/2a. Интеграл от этой скорости Qr/2a по переменной г
в пределах от 0 до с равен Пс2/4а, что и объясняет порядок главного члена
разложения (12). Главный член разложения (12) вносит наибольший вклад
в циркуляцию и определяется интегралом от скорости в области между
крыльями, где входящий поток (13) проходит через небольшую по длине
дугу2. Вне стекающего между крыльями потока воздух движется со всех
сторон к дуге BD, скорость изменения объема воздуха, приходящегося на
2Исследование процесса «броска» крыльев с точки зрения энергетики животного можно
найти в работе Вейс-Фога (Weis-Fogh, 1973). В общих чертах это значит, что мышечная си-
3. Двумерная постановка задачи о движении крыльев 265
единицу длины поперечной хорды крыла, равна Qc2. На расстоянии с от
точки А в направлении к В отнесенная к единице длины поперечной
хорды площадь, занимаемая движущимся воздухом, изменяется с 2тгс до 2ас
(рис. 5), и соответствующее изменение потенциала скорости равно
ПС2 л 7Г
это выражение соответствует второму слагаемому в (12).
Строго говоря, вопросы о том, какой поток возникает и какова при
этом циркуляция при малых углах а, не столь важны в невязком течении.
Поскольку при таком движении циркуляция в момент времени, когда угол а
достигает значения а — а0, определяется только движением границ и
состоянием потока в этот момент времени. Однако здесь следует об этом
упомянуть, т. к. влияние вязкости на поток, исследованное в § 3, подобно
«памяти» потока. Действительно, на него влияют конвекция и диффузия
вихрей с момента их отрыва от границы со скоростью, зависящей от того,
как граница движется в данный момент времени.
3* Двумерная постановка задачи о движении крыльев
насекомого с учетом вязкости
Из исследований § 2, проведенных на основании теории невязкого
потока, вытекает, что после того, как крылья разъединились, поток воздуха
остается безвихревым (т. е. с нулевой суммарной завихренностью) с
циркуляцией П0с2д(а0) вокруг АВ и CD в отрицательном и положительном
направлениях соответственно. В данном параграфе введем некоторые
уточнения этого результата, оставаясь в рамках двумерной модели и учитывая
влияние на поток сходящих с крыльев вихрей до и после разъединения
крыльев.
Если вихри сходят с границы, подобной изображенной на рисунке 2,
то поток воздуха можно представить состоящим из (i) безвихревого потока
(рассчитанного в § 2), когда на границе движение происходит в направлении
нормали, и (ii) потока, обусловленного вихрями, в предположении, что
граница остается неподвижной. Исследование потока (ii) можно проводить
методом изображений. Действительно, когда система вихрей отображается на
стема в совокупности с эластичными элементами насекомого действует таким образом, что
«броску» предшествует большое потребление энергии, что обеспечивает возможность
создания скорости Qr/2a при малых углах а (который за счет наличия волосков не может стать
в точности равным нулю).
266
Глава 9
движение,
обусловленное
стоком
Пс/2тгс
окружность
27ГС
А С
движение,
обусловленное
стоком
Пс/2тгс
Рис. 5. Начальная стадия резкого раздвижения крыльев. Площадь от2 сектора
радиусом г и с углом раствора а возрастает со скоростью fir2, за счет чего поток
втекает через дугу длиной 2аг со средней скоростью Г£г/2а. Внешнее (вне
сектора) движение представляет собой течение в окрестности стока мощностью £2с2,
обусловливающее поток жидкости через дугу BD
плоскость Z (см. рис. 3) с бесконечной прямолинейной границей,
соответствующий поток представляет собой систему вихрей плюс
зеркально-симметричная система вихрей с такой же по величине интенсивностью и
противоположной по знаку.
Движение системы вихрей в двумерном потоке определяется на
основании двух положений: (i) имеет место конвекция вихрей за счет движе-
3. Двумерная постановка задачи о движении крыльев 267
ния жидкости и диффузия с коэффициентом диффузии v (кинематической
вязкостью); (ii) границы потока являются источником образования вихрей
(Lighthill, 1963), скорость движения которых определяется условием
отсутствия проскальзывания на границе. Например, для движения,
представленного на рисунке 2, изменение потока происходят за короткое время, когда
условие равенства нормальной составляющей скорости потока и
нормальной скорости стенки не нарушается. При новом значении угла а и угловой
скорости П крыла безвихревая компонента потока изменяется тотчас же,
а перераспределение системы вихрей и их изображений и образование
новой их конфигурации происходит вследствие адвекции и диффузии в
течение достаточно длительного времени, так что условие равенства
нормальных составляющих скорости на границе не нарушается. А тангенциальная
составляющая скорости на границе изменяется, так что условие отсутствия
проскальзывания может выполняться только в том случае, если новые
вихри возникают вблизи границы в виде вихревой пелены, интенсивность
которой равна возмущенной тангенциальной скорости — вихревая пелена
возникает, а затем происходят ее конвективное движение и диффузия.
При определенных условиях конвекция может происходить
исключительно в тангенциальном направлении, так что возникшие вихри остаются
вблизи границы, образуя через время t пограничный слой, толщина
которого пропорциональна диффузионной длине (vt)1/2. В этом случае
безвихревая составляющая потока движется ускоренно вблизи границы, так что
завихренность имеет один и тот же знак, в результате чего происходят
движение вихрей поперек пограничного слоя и их увеличение. Например,
такое ускоренное движение было обнаружено на нижних поверхностях
крыльев (рис. 2). В начальный момент времени, когда угол а мал (см. рис. 5),
ускоряющийся поток устремляется в раскрывающееся пространство через
дугу BD. Однако для всех а скорость, рассчитанная на основании
предположения о невязком потоке (§ 2), возрастает у нижней поверхности каждого
крыла даже при постоянной угловой скорости £}, хотя ускорение воздуха
значительно более заметно, когда угловая скорость растет с течением
времени.
Это является достаточно важным фактом, т.к., когда а
увеличивается до значения 7г/3, крылья начинают разъединяться и около половины от
суммарной циркуляции Пс2д(а), найденной в §2 (изменение функции д(а)
описывается почти горизонтальным участком на рис. 4), для каждого крыла
обеспечивается за счет циркуляционного потока, возникшего у нижней
части крыльев. Учет вязкости не приведет к значительному изменению вклада
268
Глава 9
этого течения в циркуляцию, создаваемую потоком вокруг крыла, и
возникающего при этом пограничного слоя.
А невязкая компонента потока будет замедляться вблизи
пограничного слоя, величина завихренности, за счет которой возможно перемещение
вихрей поперек пограничного слоя, уменьшается, и в результате на
границе возникает система вихрей с противоположной по знаку завихренностью.
Когда в результате образования новой системы вихрей диффузия
первоначально существующих вихрей к стенке будет уменьшаться, в области
ускоренного движения возникает обратный поток и воздух начинает двигаться
от поверхности. Такой отходящий от стенки поток может переносить вихри
за фиксированное время t на значительно большее расстояние от нее, чем
при чисто диффузионном процессе.
Безвихревой поток, составляющий значительную часть потока вокруг
передней острой или имеющей маленький радиус кривизны кромки крыла,
является классическим примером того случая, когда тангенциальная
скорость потока увеличивается до достижения им кромки, а затем
уменьшается. Это приводит к возникновению возвратного потока на некотором
расстоянии от кромки. В этом случае может произойти срыв потока с кромки.
Возникающая конвекция вихрей называется схождением вихревой пелены
с передней кромки. Когда такая система вихрей продвигается на достаточно
большое расстояние от поверхности, то происходит полный отрыв потока
от передней кромки (как и при обтекании авиационного крыла). В
других случаях поток может снова прижаться к верхней поверхности, образуя
«пузырь у передней кромки», представляющий собой небольшую область,
занятую сошедшим с передней кромки потоком. Такое может произойти
в стационарном потоке, или в потоке, возникающем при начале движения
крыла, на расстояниях, меньших его хорды.
Пузырь у передней кромки, скорее всего, появится и в потоке,
образующемся при разведении крыльев (рис. 2). На передней кромке каждого
крыла тангенциальная скорость возрастает до своего максимального
значения, а затем резко уменьшается, в результате чего отрыв потока
становится практически неизбежным. В начальной стадии процесса разъединения
крыльев (рис. 5) в силу уравнения неразрывности поток воздуха
втягивается в образующийся промежуток между крыльями, перенося с собой
вихри. Подобный перенос вихрей существенно ограничивает возможность для
вихревой пелены отойти на большое расстояние от границы. В то же самое
время диффузия в этой раскрывающейся области может привести к
взаимному погашению завихренностей противоположных знаков справа и слева
от прямой симметрии за счет диффузионного потока через нее.
3. Двумерная постановка задачи о движении крыльев 269
При разъединении крыльев передние кромки каждого крыла
перемещаются в воздухе на расстояние одной хорды крыла, и вихри,
образовавшиеся на начальной стадии, втягиваются в образующееся между крыльями
пространство и распадаются в результате диффузии. Возможно, поэтому
большинство оставшихся вихрей сосредотачиваются в отрывном пузыре
у передней кромки, представляющем собой небольшую область с
возвратным потоком. При полете осы (Encarsia formosd) низкие числа Рейнольдса
(порядка 30) способствуют тому, что пузырь прижимается к крылу за счет
возрастающего влияния диффузионных явлений. Известно, что при
больших числах Рейнольдса отошедшие от передних кромок крыльев области
ламинарных течений могут снова прижаться к стенке при переходе к
турбулентному течению в отделившемся пограничном слое, главным образом
потому, что такой переход усиливает диффузионные процессы. Аналогичное
усиление влияния диффузионных процессов при небольших числах
Рейнольдса (порядка 30), возможно, приведет к подобному эффекту.
Замкнутая область возвратного движения, образовавшаяся у передней
кромки, не является помехой для создания подъемной силы за счет
механизма Вейс-Фога. Когда крылья разъединяются (рис. 1, кадры (/), (д) и (/i)),
то образованию большой подъемной силы дополнительно способствует
закругление передней кромки крыла (как у сечения авиационного крыла),
обусловленное наличием отрывной области (отрывного пузыря).
Более того, циркуляция вокруг такого профиля, состоящего из крыла
и области возвратного движения, будет более интенсивной по сравнению
с тем случаем, когда такая область отсутствует. На рисунке 6
схематически изображена возможная форма (неважно, это абсолютно точное
соответствие действительной форме или нет) вихревой пелены вокруг отрывной
зоны в комплексных плоскостях z и Z (см. рис. 3). На плоскости Z, кроме
того, показаны изображения вихрей относительно прямолинейной границы,
имеющие равные по величине и противоположные по знаку завихренности.
Комплексный потенциал w в этой плоскости равен сумме комплексного
потенциала безвихревого потока (8) и потенциала, обусловленного вихрями
и их изображениями. Циркуляция вокруг крыла и отрывной зоны в
направлении от точки А г к точке А2 равна изменению потенциала скорости ф
при обходе по контуру, охватывающему все вихри от точки Z = —1 до
точки Z = +1. Величина циркуляции превышает значение для
безвихревого потока (1) за счет немного большего вклада вихрей с отрицательной
завихренностью в верхней половине комплексной плоскости по сравнению
с вкладом их изображений, имеющих положительную завихренность и
расположенных в нижней половине комплексной плоскости.
270
Глава 9
Z ПЛОСКОСТЬ
Е-
(«О
Z плоскость
4* *&4<
-\ ■ >. i
1 +К*) +1
1*1
Рис. 6. Возможная форма вихревой пелены, огибающей отрывную зону у передней
кромки, (а) на плоскости z и (Ь) на плоскости Z (см. рис. 3). В плоскости Z указаны
положения осей вихря и его изображения, а также направления их вращений, что
дает возможность оценить влияние вихря на циркуляцию Г
Для пояснения этого довольно слабого эффекта (если он имеет место)
рассмотрим точечный вихрь интенсивности —К, расположенный в точке
Z = I + im (см. рис. 6). Циркуляция скорости безвихревого потока при
— 1 < Z < 1 равна Г/2, поскольку Г равна интегралу от скорости от — 1
до 1. Так что точечный вихрь, расположенный в точке Z = I + гт, и его
изображение, расположенное в точке Z = I — im, имеют завихренности,
равные завихренности возвратного потока, если К/тгт = Г/2.
Дополнительная циркуляция при обходе от —1 до 1 по контуру, охватывающему
вихрь, равна
к
7Г
arctg |
( т \
Kl-l)
Тт2
+ arctg
(ш)
(14)
и имеет порядок
! _ /2 « Г- (15)
К аналогичному результату можно прийти, рассматривая завихренность
вихревой пелены, охватывающей отрывную зону. Здесь просматривается
аналогия с классическим прогнозированием некоторого увеличения
циркуляции вокруг двумерного крыла в стационарном потоке при заданном угле
атаки за счет возрастания толщины крыла. На практике такое увеличение
циркуляции часто уравновешивается другими эффектами.
3. Двумерная постановка задачи о движении крыльев 271
Рассмотрим вихревую пелену, образующуюся после разъединения
крыльев. Прежде всего, отметим, что схождение с задней кромки крыла
вихревой пелены в теории крыла объясняется разностью давлений на
задней кромке. Сначала при разведении крыльев (рис. 1, (/)) давление с обеих
сторон крыльев при их начинающемся разъединении приблизительно
одинаково, и, значит, на этом этапе движения крыльев вихревой пелены не
образуется.
Следовательно, в начальный момент поток на верхней стороне
крыла АВ движется вдоль линий, изображенных на рисунке 6 а, т. е.
происходит обтекание крыла при наличии на передней кромке отрывного пузыря.
Последующее присоединение потока, движущегося вдоль верхней части
крыла, происходит в том числе и за счет возникновения ускорения
поступательного движения крыла. Примечательно, что прекращение вращения
крыла и переход к чисто поступательному движению приводят к тому, что
тангенциальное движение у нижней части происходит в направлении,
противоположном внешнему потоку. При этом у нижней части образуется
новый пограничный слой. Его положительная завихренность должна
превышать уже существующую отрицательную завихренность в ранее
образовавшемся пограничном слое за счет обтекания крыла внешним потоком (§ 2)
настолько, чтобы суммарная завихренность стала положительной, в
результате чего и возникнет необходимое проскальзывание между нижней частью
крыла и внешним потоком. На самом деле пограничные слои образуются
на стенках и в том случае, когда стенка движется относительно жидкости
и может изменять направление тангенциального движения (для сравнения
можно вспомнить хорошо известные пограничные слои Стокса на
движущейся стенке, периодически изменяющей направление движения). За счет
диффузии происходит постепенное уменьшение по величине как
положительной, так и отрицательной завихренности, но в каждый момент времени
в любом случае суммарная завихренность существенно влияет на внешний
поток.
Пусть скорость поступательного движения равна U. Циркуляция Г
может изменяться вследствие схождения вихревой пелены с задней кромки,
но она останется постоянной, если выполняется равенство Г = ~UcCL,
где CL — коэффициент подъемной силы, возникающей при движении
крыла в окружающей среде под углом атаки3, при котором суммарное вихре-
3 Эффективный угол атаки в действительности будет меньше, чем геометрический угол
(который составляет каждое крыло с горизонтальным направлением), поскольку (§ 4) каждое
крыло движется в поле скоса потока другого крыла.
272
Глава 9
образование равно нулю (обобщение условий Жуковского-Кутта на случай
вязкого потока). Отсюда следует ограничение на скорость U. Она должна
быть такой, чтобы суммарная подъемная сила, приходящаяся на единицу
размаха
pUT = \PU2cCL, (16)
могла образоваться с начала движения. Далее, в § 4, будет исследованы
полученные условия применительно к вопросу о возможности полного
отсутствия эффекта Вагнера.
4. Заключение
Вейс-Фог (Weis-Fogh, 1973) описал действие открытого им
механизма образования подъемной силы в животном мире. В данной работе были
рассмотрены аэродинамические аспекты этого механизма. И на основании
проведенного исследования можно сделать вывод, что предложенный им
механизм весьма перспективен с точки зрения аэродинамики.
При двумерной постановке задачи (рис. 1) «бросок» крыльев после их
резкого разведения, когда точки А и С еще совпадают, приводит к
возникновению довольно близкой по величине к максимальному значению
подъемной силы потому, что на языке теории Прандтля-Вагнера каждое крыло
действует на другое как «начальный вихрь»: интенсивность каждого вихря,
скорее, больше половины интенсивности классического начального вихря
Вагнера (Wagner, 1925). Группировка вихрей в виде двух систем
присоединенных вихрей (это вихри на крыльях, зона возвратного движения у
передней кромки крыла и т.д., все они вносят вклад в увеличение циркуляции
вокруг крыла) способствует тому, что импульс жидкости направлен вниз
(он рассчитывается через интеграл с подынтегральной функцией,
подобной моменту от горизонтально направленной завихренности) и движение
происходит с постоянно возрастающей скоростью, что приводит к
возникновению постоянной подъемной силы.
Задача перехода от двумерной модели, исследованной в §§ 2 и 3, к
полноценной трехмерной модели потока вокруг реального крыла конечного
размаха может быть решена с использованием теории крыла конечного
размаха, представленной в завершенном виде Прандтлем в 1918 году.
Рассмотрим, прежде всего, задачу в постановке Прандтля, о прямолинейном
движении в противоположных направлениях двух крыльев конечного размаха,
продольное сечение которых представлено на рисунке 1.
4. Заключение
273
(а)
Рис. 7. Механизм Вейс-Фога образования подъемной силы для крыльев конечного
размаха при прямолинейном движении. В момент «броска» мгновенно возникает
циркуляция Г вокруг каждого крыла, что сопровождается (а) возникновением
вихрей у концов крыльев с той же самой циркуляцией Г. При дальнейшем движении
крыльев концевые вихри образуют вихревую пелену (Ь)
Циркуляция вокруг продольного сечения каждого крыла (механизм
возникновения циркуляции изучен в §§2 и 3) приводит к появлению
систем присоединенных вихрей с завихренностью противоположных знаков
на двух крыльях (рис. 1): на левом крыле вращение потока происходит
в направлении в плоскость чертежа, а на правом — из плоскости чертежа.
В трехмерном пространстве это вихревое поле является соленоидальным,
а следовательно, вихревые линии должны быть замкнутыми. Тогда,
согласно теории Прандтля, должны возникать вихри на концах крыльев (рис. 7).
И их циркуляция должна быть достаточной для того, чтобы произошел
разворот завихренности на 180° при переходе от левого крыла к правому.
Направление циркуляции сформировавшегося потока вблизи концов
крыльев (векторов завихренности) представлено на рисунке 7 а. Поток воз-
274
Глава 9
духа, находящегося между крыльями, направлен от концов крыльев в
область, расположенную над крыльями, и поток вне крыльев направляется
в область, расположенную под ними. Поток внутри может рассматриваться
на ранних стадиях «броска» как поток, устремляющийся в открывающуюся
щель. Быстрее всего воздух втягивается между срединными поперечными
сечениями (при расположении крыльев, представленном на рис. 5). Вблизи
концов крыльев всасывание воздуха может привести к образованию
концевых вихрей.
На рисунке 7 Ъ показано, как на крыльях, двигающихся в направлении
друг от друга, присоединенные вихри на каждом их них соединяются за
счет наличия концевых вихрей в такую вихревую систему, что
результирующее вихревое поле является соленоидальным. Они удлиняются в результате
действия обычного механизма, который заставляет поднимающиеся крылья
сбрасывать концевые вихри: превышение давления с нижней стороны
выталкивает воздух вовне вблизи концов, в то время как меньшее давление
с верхней стороны втягивает воздух внутрь, происходит завихрение потока
вблизи концов крыла в направлении движения крыла. И направленный вниз
импульс жидкости (он рассчитывается через интеграл с подынтегральной
функцией, подобной моменту от горизонтально направленной
завихренности; для уединенного замкнутого вихря завихренность равна произведению
циркуляции на площадь) увеличивается с постоянно возрастающей
скоростью, что приводит к возникновению постоянной подъемной силы.
Трехмерная картина течения, изображенная на рисунке 7, может быть
описана двумерным течением вокруг срединного сечения крыла с учетом
тех же положений, что и в теории Прандтля. А именно, что
результирующее движение каждого сечения осуществляется не в невозмущенной среде,
а в среде, на движение которой влияет движение нисходящего потока или
скос потока, определяемого трехмерной вихревой структурой. При учете
этих факторов эффективный угол атаки каждого сечения будет меньше
геометрического на величину отношения скоса потока к скорости крыла.
Подобная поправка на скос потока в нестационарных
аэродинамических задачах (включая и рассматриваемую в данной работе) необходима
даже и при двумерной постановке: например, эффект Вагнера можно
представить себе происходящим за счет уменьшения эффективного угла атаки
вследствие скоса потока, обусловленного стартовым вихрем. В механизме
Вейс-Фога присутствует такое уменьшение, как отмечено в сноске в
конце § 3, потому что каждое крыло движется в поле скоростей, обусловленном
скосом потока, возникшим за счет присоединенного вихря на другом крыле.
4. Заключение
275
Однако результирующая подъемная сила в последнем случае не
изменяется, поскольку циркуляция вокруг сечения крыла не зависит от «броска».
Уменьшение означает только, что относительно большие геометрические
углы атаки (например, 30° согласно рис. 1) достигаются лишь в
начальной стадии движения крыльев в направлении друг от друга (причем срыва
потока при этих значениях угла не происходит). Отметим далее, что
трехмерная вихревая структура на рисунке 7, действующая, возможно,
совместно с некоторой системой вихрей, возникшей при предшествующих взмахах
крыла, немного усиливает тот наведенный в данном сечении нисходящий
поток, обусловленный действием сечений, расположенных выше, по
сравнению с его значением в двумерной модели.
Крылья осы (Encarsia formosa) не могут совершать такое
прямолинейное движение, которое изображено на рисунке 7: концы ее крыльев
описывают, как это пояснено в § 1, дуги окружности вокруг вертикально
расположенного тела. В результате чего на концах крыльев возникают концевые
вихри с вихревыми линиями в виде дуги окружности. На рисунке 8
показан рост площади, охватываемой концевыми вихрями и, как следствие,
рост направленного вниз импульса вихревой системы, соответствующего,
как и ранее, постоянной подъемной силе. Величина подъемной силы не
зависит от индуцированных скосов потока, которые влияют только на
величину эффективных локальных углов атаки.
Конечно, требует доказательства такое утверждение, что наиболее
полное использование эффекта от механизма Вейс-Фога можно осуществить
в том случае, когда животное могло бы продолжать полет при движении
обоих крыльев по окружности, как это показано на рисунке 8 так,
чтобы крылья описывали угол в 180°, совершая «хлопок-бросок» дважды за
один цикл махового движения крыльев, вместо одного «хлопка-броска»,
как это имеет место у осы {Encarsia formosa). Когда круговой концевой
вихрь (рис. 8) увеличивается и перерастает в нисходящее вниз вихревое
кольцо, то «хлопок» завершается, сброшенные концы крыльев
соединяются ниже тела4 и образуется замкнутое вихревое кольцо (чья
диаметрально противоположная сторона начинает опускаться). Затем случается другой
«бросок» — резкое разъединение крыльев, генерируя новую с противопо-
43аметим, что движение воздуха в процессе «хлопка» не является реверсивным
относительно движения воздуха при «броске» — резком разъединении крыльев, внешний поток от
закрывающейся щели не будет представлять из себя поток как в окрестности источника
(течение, обратное по направлению, по сравнению с течением в окрестности стока — рис. 5).
При полном развороте крыльев возникает течение, подобное истечению из отверстия в виде
сужающейся устремленной вниз струи.
276
Глава 9
Рис. 8. Схематичное изображение наружного и внутреннего концевых вихрей,
возникающих при трепещущем полете насекомого. Движение крыльев происходит по
окружности в горизонтальной плоскости вокруг вертикально расположенного тела
ложным знаком циркуляцию вокруг крыльев (поскольку крылья
движутся в противоположном направлении), и в дальнейшем создается вихревое
кольцо с той же самой циркуляцией, как и при первом «хлопке-броске».
Картина «идеального» использования механизма Вейс-Фога,
приводящего к поддержке животного в воздухе за счет генерирования импульса
за один мах крыла в виде двух последовательностей нисходящих
круговых вихревых колец, представляет теоретический интерес, поскольку за
счет круговой формы вихревых линий возникает наибольший по величине
импульс (пропорциональный охватываемой вихрями площади) на
единицу кинетической энергии. Попутно можно отметить, что на достаточно
большом расстоянии от животного диссипация энергии за счет вязкости
без изменения количества движения потока приведет к постепенной
перестройке потока в классическую «ламинарную круглую струю», которая
является решением уравнений движения (Landau, 1944; Squire, 1951) при
действии точечного источника в вязкой жидкости. Несомненно, течение на
достаточно большом расстоянии вниз от совершающего трепещущий полет
небольшого насекомого массой т должно принимать форму
автомодельного решения. Это автомодельное решение зависит только от одного
параметра — отношения между силой тяжести тд, с которой насекомое действует
на окружающую его среду, и имеющей ту же размерность величиной pi/2,
4. Заключение
277
Рис. 9. Линии тока на достаточно большом удалении от насекомого. Течение
описывается уравнением количества движения воздуха плотности р и кинематической
вязкости v при действии силы тяжести тд на единицу времени, приложенной в
отмеченной точке (в точке помещено насекомое). Отношение тд/ри2 равно 900.
Линии тока изображены для равноотстоящих значений функции тока Стокса
где р — плотность окружающей среды, v — кинематическая вязкость
среды. Для осы {Encarsia formosd) (массой 0,025 мг) это отношение (подобное
квадрату числа Рейнольдса) имеет порядок 900 — слишком мало для того,
чтобы уходящая вниз струя воздуха на большом расстоянии от насекомого
сохранила устойчивость.
Известно, что оса {Encarsia formosd) не может развести крылья так,
чтобы они составляли развернутый угол, что необходимо для совершения
двух циклов «хлопок-бросок» за одно маховое движение крыльев.
Крылья осы в процессе «хлопка» расходятся не более чем на 130°. При таком
278
Глава 9
ограничении требуется дополнительное обоснование того, что описанное
выше движение весьма близко к имеющему место в природе движению
крыльев осы. Крылья этого насекомого совершают полноценный «хлопок-
бросок» во время каждого махового движения, а затем нечто подобное
хлесткому движению без последующего сведения крыльев вместе (при той
же величине угловой скорости крыльев), это движение трудно поддается
наблюдению. Комбинация таких приемов может дать движение, близкое
к идеальному.
Литература
[1] Landau, L.D. 1944. Dokl. Akad. Nauk SSSR, 43, 286.
[2] Lighthill, M.J. 1963. In Laminar Boundary Layers (ed. L.Rosenhead),
chap. 2, § 1.7. Oxford University Press.
[3] Prandtl, L. 1918. Tragflugeltheorie. Nachr. Ges. Wiss. Gottengen, pp. 107,
451.
[4] Squire, H.B. 1951. Quart J. Mech. Appl. Math. 4, 321.
[5] Wagner, H. 1925. Z. angew. Math. Meek 5, 17.
[6] Weis-Fogh, T. 1973. J. Exp. Biol, (to appear).
Часть II
Внутренняя биогидродинамика
Глава 10
Физиологическая гидродинамика:
общий обзор1
1. Введение
Для успешной работы в области физиологической гидродинамики
необходимо тесное сотрудничество между специалистами в области
физиологии и гидродинамики. При решении смежных прикладных задач
сотрудничеству между специалистами в разных областях должен предшествовать
достаточно длительный процесс взаимного ознакомления с терминологией
каждой области научного знания для адекватного понимания научного
языка, описания фактов и утверждений, используемых сотрудничающей
стороной. Понимание этих фактов дает возможность смежной дисциплине
развивать весьма обширные и сложные области своего научного знания, и когда
возникает необходимость, использовать разработанные в данной области
методы для решения прикладных задач. И после такого предварительного
знакомства со смежной дисциплиной возможно действенное, плодотворное
сотрудничество между представителями различных областей научных
знаний, которое может привести к существенному продвижению в решении
поставленных задач.
В настоящей лекции автор хотел бы остановиться на некоторых
деталях совместного исследования, с которыми пришлось столкнуться автору
как специалисту в области гидродинамики, а именно в течение 6 лет, когда
приглашенный к сотрудничеству автор работал с коллективом,
возглавляемым со стороны физиологов доктором Колином Каро (Colin Саго).
Первое, что хотелось бы подчеркнуть, это наличие исключительно обширной
области физиологии, где оказалось возможным применить методы
гидромеханики. Думается, что это не удивит любого человека, который хотя бы
Секция (1971) в Международном центре механических наук в Удине (Италия)
(International Centre for Mechanical Sciences at Udine).
282
Глава 10
элементарно знаком со сложным устройством человеческого тела, и в
особенности с дыхательной системой и системой кровообращения, не говоря
уже о лимфатической или мочевыделительной системах. Несомненно, что
профессиональное знание обширных областей гидродинамики и богатый
опыт работы в этой области помогут внести существенный вклад в решение
физиологических проблем, для чего конечно овладеть основами анатомии
и физиологии, что бы понимать коллег, которые формулируют
соответствующие (физиологические) задачи.
Второй характерной чертой совместной работы специалистов в
разных научных областях, с точки зрения автора, является особенность, прямо
противоположная первой. Большинство наиболее значимых проблем в
научной области, если взять отдельно, например, гидродинамику, встречается
в тех случаях (как вытекает из всей предыдущей истории развития
гидродинамики), когда поднимается вопрос, который до сих пор перед наукой не
ставился. Просто отметим, что часто проблемы гидродинамики, встающие
перед ней при исследовании задач в области физиологии, оказывались
проблемами, которые никто не пытался исследовать в течение всего времени
развития гидромеханики, поскольку ее методы в основном использовались
в прикладных технических задачах. С другой стороны, такие
возникающие на стыке разных наук проблемы обогащает гидродинамику, и это
ясно любому специалисту в области динамики жидкости. Очевидно, что ему
следует постоянно проводить новые исследования, касающиеся основных
проблем течения жидкости, и многие из них так же интересны, как и
большинство не менее интересных исследований, подсказанных технической
мыслью.
Здесь может возникнуть вопрос: почему же задачи, связанные с
потоками жидкости в человеческом организме, так уж сильно отличаются от
того огромного числа задач, связанных с потоками жидкости в
инженерных системах? Попытаемся ответить на него, перечислив основные пункты
отличия.
(i) Непривычный диапазон чисел Рейнольдса. Как в крупных
дыхательных путях, так и в крупных кровеносных сосудах число Рейнольдса
принимает значения от нескольких сотен до нескольких тысяч, и для такого
диапазона изучение внутренних потоков практически полностью
игнорировалось, в частности, при высоких числах Рейнольдса не рассматривался
переход ламинарного течения в турбулентное. Инженеры в основном
решали задачи, связанные с полностью сформировавшимися турбулентными
потоками (за исключением рассмотрения чрезвычайно вязких жидкостей,
1. Введение
283
когда числа Рейнольдса были очень небольшими). А в человеческом теле,
напротив, только в отдельных случаях или в очень ограниченных областях
возможно возникновение турбулентности (§ 10). С другой стороны,
ламинарный внутренний поток имеет очень сложную структуру при данных
числах Рейнольдса, потому что огромная по протяженности так называемая
«входная зона» требуется для того, чтобы установилось некоторое
достаточно простое течение, подобное течению Пуазейля в прямой трубе (§ 3).
А также потому, что центростремительные силы, обусловленные
кривизной сосудов, приводят к появлению достаточно интенсивных вторичных
потоков (см. § 2).
(п) Необычайнаяразветвленность системы сосудов. Как дыхательная,
так и кровеносная система представляют собой крайне сложную и
запутанную разветвленную сеть сосудов. Поток воздуха, входящий через
трахею (дыхательное горло), или поток крови, вытекающий из сердца в аорту,
пройдя от 20 до 30 точек ветвления, разделяются на огромное число (сотни
миллионов) небольших отдельных потоков — потоки воздуха в альвеолах,
диаметр которых составляет несколько сотен микрон, или потоки крови
в капиллярах, диаметр которых не превышает 10 микрон. На ранних
стадиях процесса разветвления поток испытывает серьезное изменение при
каждом разделении на два потока, от которого по причине, указанной в
пункте (i), он не успевает восстановиться, прежде чем произойдет следующее
разделение (§ 7). На поздних стадиях, когда число Рейнольдса становится
слишком малым, для того чтобы преодолеть возникающие сложности при
очередном делении потока, характер потока становится сложным, и анализ
такого течения затруднен из-за необозримого числа сосудов, по которым
проходят потоки, и нехватки информации о длинах и диаметрах этих
сосудов.
(iii) Необычная растяжимость сосудов. Сети сосудов, в которых
текут воздушные потоки и поток крови, сложным образом
взаимодействуют между собой за счет растяжимости сосудов. Например, при
мускульном усилии, приводящем к расширению грудной клетки, объем различных
участков легких изменяется по-разному в зависимости от того, как каждый
элемент этой чрезвычайно гибкой структуры деформируется под
действием силы тяжести. А гибкость артерий играет существенную роль в
согласованной работе сердца и непрерывного протекания крови в периферических
капиллярах. К тому же это усложняется реакцией стенок сосудов,
обладающих вязкоупругими свойствами, играющими значительную роль в
сглаживании пульсирующего характера потока, который он приобретает при
284
Глава 10
поступлении из сердца в аорту. Существенное влияние на поток оказывают
также различные нелинейные эффекты, обусловленные наличием в стенках
сосудов волокон из двух совершенно различных по своим свойствам
материалов (эластин и коллаген), оказывающих сопротивление растяжению
артерий, не говоря уже о влиянии гладкой мускулатуры стенок сосудов,
осуществляющей различные управляющие функции, в частности
(относительно артериол см. главу 14) управление скоростью перфузии через стенки
периферических сосудов.
(iv) Необычные свойства жидких сред организма. Это особенно
касается крови, которая представляет собой суспензию. От 40 % до 50 % ее
объема занимают маленькие деформируемые клетки, главным образом красные
кровяные тельца, которые представляют собой очень гибкие дискообразные
образования диаметром около 8 микрон. Клетки крови находятся во
взвешенном состоянии в бесцветной жидкости — плазме крови. Хотя плазма
сама по себе имеет свойства, близкие к свойствам ньютоновской
жидкости, измерение вязкости всей крови показывает, что значения так
называемой эффективной вязкости существенно возрастают с уменьшением
скорости деформации. Это происходит за счет образования эритроцитами при
небольших скоростях деформации различных пространственных структур.
Как следует из экспериментальных данных, скорости деформации в
артериях достаточно велики, поэтому предположение о том, что кровь является
ньютоновской жидкостью, не приведет к серьезным ошибкам. В настоящее
время проводятся серьезные исследования свойств течения суспензии, хотя
это и не убеждает нас в необходимости рассматривать кровь как жидкость
с необычной реологией, но заставляет обратить внимание на тот факт, что
в малых сосудах с диаметром в несколько диаметров красных кровяных
телец концентрация эритроцитов в поперечном сечении сосуда может быть
очень неравномерной, что влияет на сопротивление току крови (см.
главу 14). Кстати и воздух, которым мы дышим, также, к нашему
сожалению, представляет собой суспензию с мелкими частичками пыли. Одной
из ключевых проблем в области биогидродинамики, связанной с
движением воздушных потоков в дыхательной системе, является определение доли
твердых частиц различных размеров в воздушном потоке, осаждающихся
в различных отделах бронхиальной системы.
(v) Наличие необычной пулъсационной составляющей. Регулярное
изменение направления потоков в легких при вдохе и выдохе требует
тщательного изучения, а именно, следует определить те отклонения от строго
реверсивного движения в различных отделах, за счет которых и возможно
2. Установившиеся вторичные потоки
285
отложение твердых частиц в дыхательной системе. В кровеносной
системе клапаны предотвращают движение крови в обратном направлении, но
у стенок артерий возможно существование обратных токов. Более того,
изменение общего тока с течением времени в восходящей части аорты, куда
кровь поступает из левого желудочка сердца, принимает форму большой
мощной волны за время, меньшее, чем половина цикла, после чего в
оставшееся время интенсивность потока уменьшается. Если это импульсное
движение крови представить в виде ряда Фурье, то окажется, что амплитуда
нескольких первых гармоник сравнима с величиной установившейся
компоненты. Было бы интересно отдельно изучить распространение
различных гармоник и, возможно, убедиться в том, что их можно представить
в виде линейной суперпозиции, однако на практике наблюдаются
существенно нелинейные взаимодействия этих гармонических составляющих.
Нелинейные взаимодействия могут возникнуть за счет феноменов
«входной зоны» и «нелинейной упругости», упоминавшихся в пунктах (i) и (ш)
соответственно. С другой стороны, ослабление пульсирующей волны
делает наличие пульсирующей составляющей в потоке, значительно менее
существенном при движении крови по капиллярам и венам.
Ниже, в главе 10, будут обозначены некоторые направления
исследований потоков сред в человеческом организме с учетом указанных выше
свойств этих сред, привлекшие научный интерес докладчика.
2. Установившиеся вторичные потоки
Центробежные силы, появление которых вызвано кривизной линий
тока «первичного потока», приводят к появлению «вторичного потока»,
линии тока которого перпендикулярны исходному При этом прямой поток
оттесняется к наружной стенке изгиба, а у внутренней стенки изгиба
возникает обратный поток. Влияние искривления и разветвления сосудов при
больших числах Рейнольдса на ламинарное течение очень мало изучено.
В настоящее время известно (Ваша, 1963; McConalogue, Srivastava, 1968),
что при течении в искривленной трубке в таком режиме под действием
градиента давления присутствует практически равномерное движение в
направлении наружной стенки изгиба, тогда как обратный поток тормозится
под действием вязких сил и не выходит за пределы тонкого пограничного
слоя. Это, возможно, объясняет (Lighthill, 1970, с. 114), почему
критическое число Рейнольдса перехода к турбулентному режиму течения
возрастает по сравнению со средним значением 2000 в случае прямых трубок до
286
Глава 10
среднего значения в 6000 для искривленных трубок. Эти числа Рейнольд-
са рассчитаны по диаметру трубки, а числа Рейнольдса, рассчитанные по
толщине пограничного слоя, различаются значительно меньше. Как следует
из экспериментальных исследований Каро (Саго, 1966), вторичные потоки
действуют таким образом, что способствуют перемешиванию субстанций
в поперечном направлении. Причем перемешивание потока при низком
коэффициенте диффузии в отсутствие турбулентности было бы значительно
менее интенсивным.
3* Входные зоны
Как хорошо известно, при установившемся течении потока в прямом
канале при больших числах Рейнольдса (см. прекрасный недавний обзор
в работе Fargie, Martin, 1971) длина входной зоны, необходимой для того,
чтобы приблизительно постоянная начальная скорость получила
распределение по сечению, отличающееся от распределения скорости при течении
Пуазейля не более чем на 5 %, равна (0,03R)d. Здесь R — число
Рейнольдса, рассчитанное по диаметру d трубы, и U0 — начальная скорость потока.
Отсюда следует, что в некоторых крупных артериях входные зоны могут
занимать по длине весь сосуд, и при исследовании течения в них
использование закона Пуазейля (хотя он и был физиологом!) уже будет
некорректным. Заметим, что при стационарном течении перепад давлений во
входной зоне превышает это значение в течении Пуазейля на величину, которую
можно назвать поправочной кинетической энергией. При вхождении потока
в сосуд требуется ускорить основной поток так, чтобы профиль скоростей
стал более заостренным, как при течении Пуазейля, со средней
кинетической энергией, превышающей значение \рЩ. Общая поправка к
кинетической энергии, учитывающая дополнительное сопротивление в тонком
вблизи входа пограничном слое, имеет порядок 0,67рС/о. При исследовании
результирующего падения давления во всей системе с ее
многочисленными разветвлениями (подобной кровеносной системе) оказывается, что даже
если поправка в одном сосуде и мала по сравнению с указанной общей
поправкой, то в сумме по всем сосудам они могут дать значение, которым
пренебречь нельзя.
Работа по совместному учету последних двух указанных эффектов
в настоящее время ведется главным образом потому, что самая первая
входная зона в большом круге кровообращения — восходящая часть аорты —
является сама по себе сильно искривленным каналом. Не следует ожидать,
4. АТЕРОМАТОЗНОЕ ПОРАЖЕНИЕ АРТЕРИЙ
287
что в этой входной зоне полностью разовьется вторичный поток. Если
аортальный клапан открывается должным образом, то в этой зоне кровь имеет
небольшую завихренность, и завихренность основного и вторичного
потоков может быть ограничена только пограничным слоем. Центральный
поток будет безвихревым на внутренней стенке изгиба (поскольку вторичный
поток еще не полностью развит). Вторичные течения, зарождающиеся в
пограничном слое, развившись, могли бы привести к тому, что наибольшая
скорость сдвига была бы ближе к наружной стенке изгиба. В результате
совместного обсуждения докладчиком и доктором Каро (и особенно
доктором Скартоном (Scarton) с позиций теоретического подхода и доктором
Сидом (Seed) с позиций экспериментальной науки) это довольно простое
описание было признано вполне приемлемым.
4. Атероматозное поражение артерий
Можно задаться вопросом о том, почему так много внимания уделяется
точному определению скорости течения или скорости сдвига и массопере-
носу. На это имеется несколько оснований. Но наиболее важным является
то, что связано с атеромой — дегенеративным изменением стенок артерий.
Действительно, напряжение трения и массоперенос на стенке имеют в
общем похожие распределения в системах, где происходит течение
жидкости. Поэтому можно выделить несколько интересных направлений
исследования связи этих величин с возникновением атеромы. К сожалению, два
направления исследования, описанные Фраем (Fry, 1968) и Каро,
Фитцджеральдом и Шрётером (Саго, Fitz-Gerald, Schroter, 1971), прямо
противоречат друг другу. На симпозиуме, организованном Ciba Foundation в 1972
году, где автор был председателем оргкомитета, была предпринята попытка
прийти к единой оценке значимости этих двух представленных работ, не
вынося заранее суждение о них до подробного рассмотрения каждой
работы. С одной стороны, в работе Фрая (Fry, 1968) показано, что необычно
большие скорости сдвига способны привести к механическому
повреждению эндотелиального пласта на стенках артерии и что это может
инициировать процесс образования атеромы. С другой стороны Каро, Фитцджеральд
и Шрётер (Саго, Fitz-Gerald, Schroter, 1971) утверждают, что такие
большие скорости сдвига, которые могут привести к механическим
повреждениям стенок сосудов, в реальных системах кровообращения не
наблюдаются. Они изучали распределение локализации атеромы на начальной стадии
болезни у большого числа людей и представителей семейства псовых и вы-
288
Глава 10
явили тесную связь между этими локализациями и областями, где имеют
место небольшое значение скорости сдвига и массоперенос, т.е. перенос
некоторой субстанции в направлении от стенок артерии за счет
захватывания субстанции протекающим потоком жидкости. Они представили
довольно сложное объяснение возникновения атеромы в такой «стоячей воде»
и появления областей с низкими значениями скорости сдвига,
невозможностью кровотоку достаточно быстро унести от стенок артерии имеющийся
в крови холестерин. Не ссылаясь на чье-либо мнение, нам хотелось бы
заметить, что поскольку атерома сосудов является одной из самых
распространенных смертельных болезней современного человеческого общества,
то обе работы весьма важны, несмотря на их различные пути исследования,
и они показывают нам, насколько необходимы достоверные знания о
процессах массопереноса и скоростях сдвига в сосудистой системе.
5. Распределение деформации сдвига в разветвленной
сосудистой системе
Еще немного внимания уделим гидродинамике установившихся
потоков в разветвленной сосудистой системе. На основании таких исследований
можно указать типичные распределения скоростей в реальной
сердечно-сосудистой и дыхательной системах, когда скорость потока максимальна,
а влияние скорости изменения потока наименьшее. Кроме того, для потока
крови можно получить распределение установившейся компоненты
скорости (только для тех случаев, когда скорость можно представить в виде ряда
Фурье).
В соответствии с работой Каро, Фитцджеральда и Шрётера (Саго, Fitz-
Gerald, Schroter, 1971) скорость сдвига в отдельной точке отдельного сосуда
разветвленной системы при стационарном потоке зависит от двух
факторов: (i) фактор, определяемый параметрами потока, а именно расходом Q,
в этой отдельной трубе; этот фактор равен 32Q/7rd3 и характеризует сдвиг
на стенке для течения Пуазейля при расходе Q в круглой трубе
диаметром d\ (ii) геометрический фактор, который характеризует локальное
отклонение скорости сдвига от скорости сдвига при течении Пуазейля и зависит
от расположения сечения в трубе. Причины и величины этих отклонений
уже обсуждались. Так, в любой входной зоне (§ 3) геометрический фактор
значительно больше единицы вверх по течению и уменьшается вниз по
течению. В искривленной трубе (§ 2) он имеет большую величину на
наружной стенке изгиба сосуда, чем на внутренней. При ветвлении сосудов гео-
5. Распределение деформации сдвига
289
метрический множитель достаточно велик в потоке у делителя потока, где
на стенках дочерних сосудов начинает образовываться новый пограничный
слой, и достаточно мал на противоположных стенках трубы (внутренние
стенки изгиба дочерних сосудов) для потока, устремляющегося в дочерние
сосуды.
К сказанному добавим несколько кратких замечаний касательно того,
как геометрический фактор изменяется в ветвящейся системе. Для этого
введем коэффициент (3 — отношение площадей при разветвлении. Здесь (3
равен отношению суммы площадей в дочерних сосудах к площади
«материнского» сосуда. Эта характеристика достаточно важна как для
сердечно-сосудистой, так и для дыхательной системы, поскольку суммарная
площадь поперечных сечений сосудов должна быть как можно большей.
Этот параметр увеличивается на два порядка при прохождении по системе
от аорты до капилляров и еще больше при прохождении по дыхательной
системе от трахеи до альвеол.
Специалист в области гидродинамики определенно сказал бы, что
параметр (3 должен превышать единицу при каждом ветвлении, однако он
поостерегся бы придать этому параметру значение, существенно
превышающее единицу при произвольном количестве произошедших перед этим
ветвлений. И все это потому, что скорость потока при каждом ветвлении
должна делиться на (39 а такое замедление потока может привести к его
срыву при значениях (3 больших, чем 1,2. Явление срыва потока
нежелательно по двум причинам: (i) это приводит к возрастанию сопротивления,
(ii) образование застойной зоны на стенках сосуда может привести к
пагубным последствиям.
Однако на ранних стадиях ветвления параметр (3 может быть, как
показывают измерения, и небольшим: меньше единицы при первом ветвлении
аорты (Саго, Fitz-Gerald, Schroter, 1971), но не более 1,2 при последующих
ветвлениях. Размеры параметра (3 приведены здесь для случая самого
простого дихотомического ветвления, когда в каждый дочерний сосуд проходит
ровно половина основного подходящего к точке ветвления потока.
В этом случае при каждом ветвлении скорость основного потока умно-
-1 (л \1/2
жается на (3 , диаметр сосуда — на ( ^/Я , а число Рейнольдса — на
(2/5)_1/2. Факт уменьшения числа Рейнольдса значим, поскольку при
достаточно большом числе ветвлений оно может уменьшиться менее чем
до 10 и срыв потока не сможет произойти даже при больших (3. А
следовательно, такие значения могут иметь место. И наконец, фактор, опре-
290
Глава 10
деляемый параметрами потока, а именно величина 32ф/яч23, умножается
на (2//33)1/2. Поскольку при начальных ветвлениях значения параметра /3
превышают единицу, то фактор, определяемый параметрами потока,
уменьшается вниз по течению. В общем случае скорость сдвига имеет
наименьшее значение в проксимальных (ближайших к сердцу) артериях, т. е. в тех
артериях, где геометрический фактор наименьший.
6. Сопротивление в ветвящихся системах
Рассмотрим, как влияет падение давления в различных участках
ветвящейся системы на полное падение давления во всей системе. Для того,
чтобы провести такой анализ, сделаем достаточно сильное упрощающее
предположение. Будем считать, что сосуды с меньшим диаметром имеют
и пропорционально меньшую длину. Такое утверждение в общих чертах
является правильным.
Падение давления при течении Пуазейля имеет довольно простую
зависимость: для того чтобы получить падение давления в дочерних
сосудах, необходимо умножить падение давления в исходном сосуде, ветвление
которого мы рассматриваем, на величину (2//33)1/2, такую же, как и для
сдвига. Необходимо помнить, что при высоких числах Рейнольдса
истинное падение давления превышает падение давления при течении Пуазейля
во входных зонах и по другим причинам (см. ниже § 7, а также
вышеназванную причину под пунктом (i)). Однако можно видеть, что в то время как
скорость сдвига увеличивается в направлении от центра (т. е. к периферии,
и, следовательно, значение параметра (3 не должно значительно превышать
единицу), степень приближения падения давления к падению при течении
Пуазейля также будет увеличиваться.
Сердечно-сосудистая и дыхательная системы в этом отношении
различаются. Оказывается, что в первой системе увеличение значений /3
замедляется с ростом числа ветвлений, особенно когда диаметр сосудов становится
меньше 0,1 мм. Это согласуется с законами гидродинамики — числа
Рейнольдса должны сначала уменьшиться настолько, чтобы при дальнейшем
росте J3 не произошло срыва потока. В дыхательной системе отношение
(2//?3)1/2 принимает значения, меньшие единицы, уже после первых пяти
ветвлений, когда число Рейнольдса еще имеет порядок нескольких сотен.
Отрыв потока при следующих ветвлениях становится весьма вероятным,
но в силу различных причин отрыв потока не будет иметь столь серьезные
последствия, как в кровеносной системе.
7. Бронхиальное сопротивление
291
В кровеносной системе наибольшее падение давления происходит
в небольших сосудах диаметром менее 0,1 мм (некоторые свойства этих
сосудов будут рассмотрены в главе 14), а в дыхательной системе
значительное падение давления происходит уже после десяти ветвлений в
сосудах, диаметр которых превышает 1 мм (Macklem, Mead, 1966). Или, говоря
по-другому, задача нахождения сопротивления в артериях связана с
движением при низких числах Рейнольдса, а нахождение сопротивления в
дыхательной системе связано с движением при высоких числах Рейнольдса.
7. Бронхиальное сопротивление
В соответствии со сказанным выше в главе 10 при определении
бронхиального сопротивления и оценки превышения падения давления в
бронхах над падением давления в случае течения Пуазейля необходимо
рассматривать течение при высоких числах Рейнольдса. Руководствуясь этим
положением, Педли, Шрётер и Садлоу (Pedley, Schroter, Sudlow, 1970)
исследовали бронхиальное сопротивление в системе после десяти ветвлений
дыхательных путей и пришли к некоторым весьма интересным выводам.
Ими было показано, что вторичные потоки, образующиеся при
дихотомическом ветвлении, настолько мощны, что исходное распределение
скоростей в разветвленных сосудах сильно искажается. Распределение
скорости по сечению имеет вид островершинной кривой, максимум которой
расположен вблизи внутренней стенки изгиба, причем искажение, внесенное
вторичными потоками, слегка размывается вниз по течению. И, как
следствие, отношение вязкой диссипации в потоке к вязкой диссипации в
течении Пуазейля значительно больше по сравнению с подобным отношением
во входной зоне, в которую входит поток с равномерным распределением
скорости по сечению. Это отношение равно 0,33(i?d//)1/2, где R — число
Рейнольдса, рассчитанное по диаметру d и средней скорости, а / — длина
сосуда.
Это означает, что вклад в падение давления нескольких первых
ветвлений, где число Рейнольдса R наибольшее, достаточно велик. Более того,
полное падение давления при таких больших числах Рейнольдса в
ламинарном потоке пропорционально скорости потока в степени 3/2.
Действительно, поток воздуха в трахее является турбулентным, а это имеет место при
числах Рейнольдса порядка 10000, где закон турбулентного сопротивления
близок к степенному с показателем степени 3/2. Педли, Шрётер и
Садлоу (Pedley, Schroter, Sudlow, 1970) приходят к заключению, что и падение
292
Глава 10
давления при прохождении воздуха через всю дыхательную систему имеет
такую же степенную зависимость, что подтверждается приводимыми ими
экспериментальными данными.
8- Распределение скорости в пульсирующем потоке
Вернемся к рассмотрению влияния пульсаций на кровоток в
артериях. Сначала проведем исследование без учета эластичности стенок
артерий. Возможно, это и не так уж плохо в качестве первого приближения,
если нас интересует только связь между распределением пульсирующей
скорости по сечению сосуда и пульсационным изменением градиента
давления. (Заметим, что характерные изменения диаметра артерии при
пульсациях составляют ±2%. Отметим также, что скорость распространения
пульсационнои волны по крайней мере в 5 раз больше максимальной
скорости течения крови, так что если эластичностью сосуда пренебречь, т. е.
считать, что изменения давления в данном сечении под действием
пульсаций происходят бесконечно быстро, то это не будет сильным
преувеличением!)
Распределение скорости, определяемой главной компонентой в
разложении пульсационнои волны в ряд Фурье с угловой частотой а;, зависит от
величины, которую Уомерсли (Womersley, 1955) обозначил через а.
Параметр а равен отношению радиуса сосуда а = ^d к осциллирующей
толщине пограничного слоя (iz/o;)1/2, где v — кинематическая вязкость (для
кровотока в больших артериях она равна 4 мм2/с). При ш = 8 с-1 (для
человеческого организма это типичное значение угловой частоты первой
компоненты ряда Фурье) толщина пограничного слоя равна 0,7 мм, а для п-й
компоненты ряда Фурье эту величину необходимо умножить на п-1/2.
В больших артериях а равна 10, и при синусоидальном градиенте
давления скорость по поперечному сечению сохраняет постоянное значение,
за исключением пограничного слоя толщиной, составляющей 10% радиуса
сосуда. Почти весь перепад давления расходуется на преодоление инерции
жидкости, а не на преодоление вязкого сопротивления, так что поток
запаздывает по фазе почти на 90° по отношению к фазе градиента давления2,
а по отношению к фазе скорости сдвига на стенке — на 45°.
2 Это выполняется даже в том случае, когда сила трения превышает трение в течении Пу-
азейля в два раза (за счет тонкого пограничного слоя) для а = 10. Только для а < 3 силы
инерции становятся менее значимыми по сравнению с силами вязкости. Напряжение трения
для таких сосудов (диаметр артерий не превышает 4 мм) определяется законом Пуазейля.
9. Распространение пульсаций
293
9. Распространение пульсаций
В классическом случае скорость распространении волны в эластичном
сосуде с тонкими стенками равна (Eh/2pa)1/2, где Е — модуль упругости
Юнга, h — толщина стенки. В большинстве работ оценивается отклонение
от этого значения за счет влияния вязкости или за счет учета свойств
стенок (толщина стенок, сжимаемость, вязкоупругие свойства, наличие
ограничений перемещений стенок, нелинейность), при этом получается, что для
реальных артерий это отклонение невелико. Скорее всего, учет только
вязкости жидкости даст заниженное значение степени ослабления волны.
Существенную роль в этом процессе играют, конечно, и вязкоупругие
свойства стенок сосудов. Скорость волны, наверное, уменьшится, если, кроме
главной компоненты пульсационной составляющей (коэффициент,
зависящий от инерции, при главной гармонике тем больше, чем больше толщина
пограничного слоя), дополнительно учесть гармоники высших порядков.
Как следует из проведенных измерений, скорость волны возрастает
от значения в аорте 5 м/с до значений 10 м/с в периферической
сосудистой системе в силу значительно меньшего модуля упругости стенок аорты.
Измерения проводились при наблюдении за распространением с течением
времени искусственно созданного возмущения давления.
В кровеносной системе человека пульсации, распространяющиеся
в направлении от сердца, нельзя представить только в виде бегущей
волны. В формировании этой волны участвуют и многочисленные отраженные
волны, возникающие при каждом ветвлении сосудов. Как следует из теории
отражения волны, при ветвлении скорость волны зависит от полной
проводимости (адмитанса) Y — величины, обратной импедансу (полному
сопротивлению), и представляет собой отношение амплитуды объемных
флуктуации потока к амплитуде флуктуации давления. Для бегущей волны в трубе
с площадью поперечного сечения А величина Y равна Y = А/рс. Волна
сжатия приводит к возникновению отраженных волн сжатия (с меньшей
амплитудой) при каждом ветвлении, где сумма полных проводимостей
дочерних сосудов меньше проводимости разветвляющегося сосуда. Это имеет
место и при первом ветвлении аорты (суммарная площадь после ветвления
уменьшается, а скорость волны с увеличивается), и при некоторых первых
ветвлениях артериального дерева.
Вот почему волны в аорте и некоторых других крупных артериях
включают в себя отраженные волны сжатия и представляют собой комбинацию
бегущей и стоячей волн. Из приведенных рассуждений следует, что круп-
294
Глава 10
ные артерии, в частности аорта, действуют подобно так называемому ре-
сиверу, где скапливаются порции крови, поступающие в них в
результате сердечного сокращения, для последующего продвижения с практически
постоянной скоростью в периферической системе кровообращения (см.,
например, работу Taylor, 1966).
Кроме того, возникновением отраженных волн при каждом ветвлении
объясняется тот факт, что амплитуда давления в одной из артерий нижней
конечности может превышать амплитуду давления в аорте за счет
уменьшения проводимости. Сглаженную форму кривой зависимости давления от
времени в таком месте можно объяснить влиянием гармоник более
высокого порядка. Другая причина, приводящая к подобной форме
(опережение фазы второй гармоники, приводящее к более заметному пику давления
при диастоле), в настоящее время обсуждается. Отметим, что к
указанному выше эффекту могут приводить либо нелинейные эффекты, либо малая
скорость первой гармоники ряда Фурье.
10. Турбулентность в системе кровообращения
Неясность в вопросе о существовании в аорте турбулентности в
настоящее время разрешена в результате экспериментальных исследований
скорости с помощью ее измерения высокочувствительными зондами (работа
доктора Линя (Ling) в Вашингтоне, доктора Шульца (Schultz) в Оксфорде,
доктора Сида (Seed) в Лондоне совместно с коллегами). Наибольшее
значение числа Рейнольдса таково, что при установившемся режиме вторичный
(ретроградный) поток в дуге аорты отодвигает начало возниковения
турбулентности (см. §2). Фактически при каждом сердечном цикле возникают
малые области турбулентности в момент уменьшения скорости (см.,
например, Seed, Wood, 1971).
Автор склоняется к мысли, что турбулентность обусловливается
изменением профиля скоростей в результате реакции на запаздывающий
градиент давления. Изменение профиля скоростей вблизи стенки опережает по
фазе течение в центре сосуда на 45° (см. § 8). Профиль скоростей
вблизи стенки имеет точку перегиба, наличие которой, как хорошо известно,
является наиболее существенной причиной неустойчивости. Зоны
турбулентности возникают сразу же, как только такая точка перегиба возникает.
Однако при изменении условий эти области турбулентности могут
довольно быстро исчезнуть (см., например, Lighthill, 1970).
Турбулентность в системе кровообращения, как правило, не
возникает, за исключением тех случаев, когда поток приобретает форму струи.
11. МОЧЕВЫВОДЯЩИЕ ПУТИ
295
Например, это может происходить при стенозе аортального клапана
аорты (полного открытия клапана не происходит), что представляет серьезную
медицинскую проблему. При обычном способе измерения давления с
использованием нарукавной манжеты, в которую накачивается воздух, при
сдувании манжеты медик слушает хорошо известные тоны Короткова —
в момент появления первого тона определяется систолическое давление,
а в момент перехода громких тонов в тихие — диастолическое. Однако эти
звуки более звонкие, чем звуки, обусловленные наличием турбулентности.
Недавно причина появления тонов Короткова была довольно ясно
описана в прекрасной работе, сочетающей теоретические и
экспериментальные методы исследования (Conrad, 1969; Katz et al., 1969). Давление
можно определить по довольно устойчивым нелинейным колебаниям, которые
возникают в сосуде, когда он достаточно сильно сжимается. Упрощенно
можно представить, что механизм действия сжимающего давления состоит
в следующем. Степень сжатия сосуда увеличивают до момента, когда
поток временно останавливается или даже имеет обратную скорость, затем
сжатие ослабляется и скорость движения крови начинает снова возрастать,
поскольку внешнее давление еще довольно значительно.
11. Мочевыводящие пути
Кроме движения крови в кровеносной системе и воздуха в
дыхательной, в человеческом организме существуют и другие сосудистые системы,
по которым движется жидкая среда. К ним относятся, например,
система мочевыводящих путей. В почке, удивительном механизме, включающем
органы, обеспечивающие обратное всасывание, формируется моча из
фракций плазмы крови. Моча продвигается в мочевой пузырь по мочеточнику
(каждая почка имеет один мочеточник) за счет перистальтического
сокращения его гладких мышц. Перистальтический механизм мочевыводящих
путей еще требует детального изучения по многим основаниям.
Превосходный обзор по проблемам перистальтики имеется в недавно вышедшей
работе Джаффрина и Шапиро (Jaffrin, Shapiro, 1971).
Литература
[1] Ваша, S.N. 1963. Quart. J. Mech. Appl. Math. 16, 61.
[2] Саго, C.G. 1966. J. Physiol. 185, 501.
[3] Caro, C.G., Fitz-Gerald, J.M. & Schroter, R.C. 1971. Proc. Roy. Soc. B,
177, 109.
296
Глава 10
[4] Conrad, W. A. 1969. IEEE Trans. Bio-Med. Eng. BME-16, 284.
[5] Fargie, D. & Martin, B. W. 1971. Proc. Roy. Soc. A, 321, 461.
[6] Fry, D.L. 1968. Circulation Res. 22, 165.
[7] Jaffrin, M. Y. & Shapiro, A. H. 1971. Ann. Rev. Fluid Mech. 3, 13.
[8] Katz, A. I., Chen, Y. & Moreno, A.H. 1969. Biophys. J. 9, 1261.
[9] Lighthill, M.J. 1970. Turbulence. In Osborne Reynolds and Engineering
Science Today (ed. McDowell, D. M. & Jackson, J. D.), pp. 83-146,
Manchester University Press.
[10] Macklem, P.T. & Mead, J. 1966. J. Appl. Physiol. 22, 395.
[11] McConalogue, D.J. & Srivastava, R.S. 1968. Proc. Roy. Soc. A, 307, 37.
[12] Pedley, T. J., Schroter, R.C. & Sudlow, M.F. 1970. Pespiration Physiol. 9,
371.
[13] Seed, W. A. & Wood, N. B. 1971. Cardiovascular Res. 5, 319.
[14] Taylor, M.G. 1966. Biophys. J. 6, 697.
[15] Womersley, J.R. 1955. J. Physiol. 127, 553.
Глава 11
Воздушные потоки
в дыхательной системе
1. Введение
В данной главе продолжим (см. § 2) начатое ранее, в главе 10, § 7,
описание движения воздуха в крупных дыхательных путях, которые
обеспечивают связь окружающей среды с главными органами дыхательной системы
человека — легкими. В процессе движения воздуха в дыхательных путях,
происходящего при высоких числах Рейнольдса, перенос кислорода и
углекислого газа осуществляется за счет адвекции. Исследование движения
жидкой среды в альвеолах и легочных капиллярах при низких числах
Рейнольдса и главным образом процесса газообмена между атмосферным
воздухом, достигшим легочных альвеол, и кровью, протекающей по легочным
капиллярам, проводится в главе 14.
Кроме того, представляет интерес сравнительный анализ дыхательной
системы млекопитающих с дыхательной системой других классов
животных. В данной работе для сравнения выбраны два класса, о которых шла
речь в первой части книги, а именно птицы и рыбы. Дыхательная система
насекомых в дальнейшем упоминаться не будет главным образом в силу
того, что поступление кислорода в дыхательную систему и удаление из нее
углекислого газа происходят, по существу, только путем диффузии через
постоянно открытые дыхальца.
В трудах Геттингенского симпозиума по сравнительной физиологии
дыхательной системы позвоночных, опубликованных в томе 14 Respiration
Physilogy, подробно изложены различные типы потоков жидких сред,
связанных с дыхательным процессом. Характер биогидродинамических
процессов сильно варьируется от класса к классу (и в меньшей степени внутри
самого класса). И их основное назначение — приведение в
непосредственное соприкосновение красных кровяных телец с кислородом или водой —
298
Глава 11
реализуется в большинстве случаев за счет сходного пространственного
расположения элементов дыхательной и кровеносной систем. Так,
например, (Weibel, 1972; Hughes, 1972) на рисунке 1 показано расположение
эритроцита в межальвеолярной перегородке легкого человека, и подобная же
конфигурация имеет место в жабрах рыбы (рис. 2, электронная
микрофотография вторичной ламеллы линя (Tinea tinea)). А вот толщина барьера
между клетками крови и молекулами воздуха у млекопитающих очень невелика
(от 0,3 до 0,5 мкм) по сравнению с таким же барьером у рыб (от 2 до 3 мкм
у линя, до 10 мкм у морской собаки, а у рыб, ведущих очень активный
образ жизни, таких как представители семейства скумбриевых, толщина
барьера снижается до 0,6 мкм).
В своем большинстве рыбы для вентиляции жабер используют
прокачивание в два этапа. Они набирают воду в ротовую полость, после чего
ротовая полость закрывается и за счет мышечного сокращения вода
выталкивается через жаберные щели. Но они частично сглаживают прерывистое
действие «ротового» насоса последующим несинфазным всасыванием воды
за счет расширения полостей в жаберных крышках. Однако имеется и
довольно много исключений (см. довольно обширный обзор в работе Marshall,
1965, глава 6). Так, скумбриевые рот не закрывают, и вода проталкивается
сквозь жабры при их быстром поступательном движении в соответствии
с законом Бернулли; у угрей отсутствует активное проталкивающее воду
через жаберные щели движение и т. д. В отличие от легких у
млекопитающих жабры у рыб в основном непрерывно омываются проточной водой.
Потоки в жабрах обладают еще одной характерной чертой, дающей
дополнительное преимущество при газообмене. Кровь во вторичных ламел-
лах течет в направлении, противоположном направлению движения воды,
проходящей по поверхностям ламелл. Такая созданная природой система
«противотока» чрезвычайно увеличивает эффективность газообмена и
взята на вооружение инженерами-химиками. По существу, вся поверхность,
через которую проходят потоки, участвует в газообмене. Действительно,
между первой порцией воды с большим содержанием кислорода и кровью
в ламеллах происходит газообмен, и кровь частично обогащается
кислородом, затем последующая порция воды, потерявшая часть кислорода,
обменивается с кровью, уже получившей некоторое количество кислорода,
и обогащенный кислородом поток крови втекает в капиллярную сеть.
Большое количество семейств рыб различных отрядов приобрели
дополнительную способность дышать атмосферным воздухом. Подобной
способностью обладают рыбы, живущие в стоячих тропических водоемах, ко-
1. Введение
299
IN
v
\
0,35pm
»
■i:
м
• J
1
ЕР
ВМ
EN
л .
\
. -N
V % •
Рис. 1. Расположение эритроцита в межальвеолярной перегородке легкого птицы
(рис. 3 из работы Weibel, 1972)
300
Глава 11
Ч
Рис. 2. Расположение красных кровяных телец во вторичной ламелле жабр рыбы
(рис. 12 из работы Hughes, 1972)
торые могут время от времени высыхать или в них может уменьшаться
количество кислорода в силу тех или иных причин. И рыбы
приспособились к таким условиям за счет изменения жабер или плавательного пузыря
(или даже кишечного тракта).
У большого числа двоякодышащих рыб с плавательным пузырем,
приобретшим ячеистое строение или преобразовавшимся в своеобразные
легкие, существовал такой тип дыхательной системы, из которой в
каменноугольном периоде развилась дыхательная система земноводных (из группы
вымерших к настоящему времени кистеперых рыб). В течение всего
развития легочной системы земноводных, рептилий и млекопитающих
газообмен остается локализованным на поверхности концевых воздуховодных
канальцев, наименьшими из которых являются альвеолы. У всех
четвероногих позвоночных газообмен в легких (в противоположность газообмену
через кожу животного) носит ярко выраженный ритмический характер.
1. Введение
301
А у птиц в процессе эволюции образовались дополнительные
насосные блоки, называемые воздушными мешками, наличие которых
позволило легким развиться в органы, представляющие собой небольшие плотные
губчатые тела постоянного объема. Воздушные мешки, сжимаясь и
расширяясь, прогоняют воздух в область, где происходит газообмен,
представляющую собой тонкие жесткие воздуховоды диаметром от 3 до 10 мкм
(Duncker, 1972), расположенные в непосредственной близости от
капилляров кровеносной системы: воздуховоды и капилляры расположены в
поперечном направлении к их границе раздела (рис. 3). То, что легкие
имеют постоянные объем, является существенным преимуществом. Например,
площадь поверхности, через которую совершается газообмен, составляет
250 мм-1 на единицу объема, что на порядок больше, чем у легких
млекопитающих (расширенные альвеолы имеют диаметры на порядок больше,
чем у млекопитающих). Воздушные мешки играют важную роль в
улучшении характеристик полета за счет повышения качества дыхательного акта
и за счет уменьшения удельного веса птицы.
легкие
жабры рыбы легкие птицы млекопитающего
встречное движение движение в поперечном вспомогательная
направлении емкость
Рис. 3. Схема газообмена у рыб, птиц и млекопитающих (рис. 1 в работе Piiper,
Scheid, 1972)
Более подробное описание дыхательной системы птиц приведено в § 3.
Предварительно обрисуем только интересующую нас проблему, а именно
как накачивание и выталкивание воздуха в передних и задних мешках
могут обеспечивать прохождение воздуха по всей дыхательной системе при
отсутствии каких-либо клапанов. Жаберные крышки, выполняющие роль
насосов, имеют клапан и участвуют в работе ротового насоса, где рот
играет роль клапана. У птиц происходящее всегда в одном направлении
движение воздуха через легкие обусловлено расширением и сокращением перед-
302
Глава 11
них и задних воздушных мешков, которые не имеют приспособлений для
перекрытия воздушного потока при его прохождении либо через выходной
канал при расширении, либо через входной канал при сжатии. Глава
завершается обсуждением особенностей пространственной структуры легких
и воздушных мешков, наличие которых позволяет осуществлять подобное
движение.
2. Движение воздушных потоков в бронхах человека
Шрётер и Садлоу (Schroter, Sudlow, 1969) в силу серьезных
затруднений, связанных с измерением поля скоростей в легочной системе в
процессе дыхания в естественных условиях, создали из плексигласа модель
ветвящегося воздухопровода с первыми десятью точками ветвления,
подобную сети сосудов в человеческих легких. На рисунке 4 представлено
последовательное разделение сосуда на две части, расходящиеся под углом
70° с отношением диаметров, равным 0,78 (соответствующее отношению
площадей /3 = 1,22, определенное в главе 10). Отметим важную деталь:
типичное для бронхов отношение длины трубки к ее диаметру составляет
3,5, так что скорость потока, возмущенного в момент разделения сосуда,
устанавливается фактически на небольшом расстоянии от места ветвления
перед тем, как поток снова будет возмущен очередным ветвлением.
Действительно, при числах Рейнольдса, характерных для этих
воздушных путей, длина «входной зоны», на которой (глава 10, § 3) входное
течение становится достаточно близким к течению Пуазейля с тем же расходом,
на порядок больше, чем длина участка провода до следующего ветвления.
И длина зоны при входном потоке, близком к пуазейлевскому, больше, чем
длина входной зоны, при равномерном по всему сечению входном потоке.
Это частично объясняется тем, что при разделении потока образуется
новый пограничный слой, где скорость вне пограничного слоя близка к
максимальному значению скорости в исходном (материнском) трубопроводе,
а частично тем, что образуются вторичные потоки, обусловленные
искривлением движения потока, после его вхождения в разветвившиеся дочерние
участки. Максимум скорости сдвигается к внешней стенке трубопровода,
недалеко от точки разделения потока.
На рисунке 5 представлены профили скоростей в дочерних
трубопроводах на расстоянии одного, двух и трех диаметров от точки ветвления при
числе Рейнольдса, равном 700, и при условии, что в материнском
трубопроводе имеет место течение Пуазейля. Отметим сильное отклонение профиля
2. Движение воздушных потоков в бронхах человека 303
третий
участок
первый
участок
второй
участок
Рис. 4. Схема воздухопровода из плексигласа, который использовался в
экспериментальных исследованиях (рис. 2 из работы Schroter, Sudlow, 1970)
скорости от профиля при течении Пуазейля, такой резкий пик в профиле
скоростей возникает недалеко от точки ветвления и сохраняется на
протяжении всего движения по этой трубе.
На основании полученных профилей скоростей (рис. 5) Педли, Шрётер
и Садлоу (Pedley, Schroter, Sudlow, 1970) провели расчеты диссипации
энергии, обусловленной наличием вязкости в трубе. Расчеты проводились для
широкого диапазона чисел Рейнольдса для потоков после первого и после
второго ветвления. Было получено, что отношение скорости диссипации
энергии в реальном течении к скорости диссипации энергии в течении
Пуазейля зависит главным образом от числа Рейнольдса R, рассчитанного по
диаметру трубы d и средней скорости U.
Зависимость от числа Рейнольдса можно понимать следующим
образом. Диссипация энергии в потоке при наличии максимума профиля
скорости, расположенного вблизи стенки, происходит главным образом в тонком
пограничном слое (толщиной S) в области между стенкой трубы и пиком
профиля скорости. Градиент скорости в этой области приблизительно
равен (С//5), тогда скорость диссипации энергии на единицу объема в
пограничном слое с площадью поперечного сечения d8 имеет порядок fi(U/8)2.
Скорость диссипации энергии на единицу длины трубы равна fiU2(d/d)
и превышает скорость диссипации энергии в течении Пуазейля в (d/S) раз.
304
Глава 11
1 DIA на расстоянии одного диаметра
2 DIA - на расстоянии двух диаметров
3 DIА - на расстоянии трех диаметров
1,00,75 0,50,25 0 0,250,5 0,751,0
Безразмерное расстояние
в поперечном направлении
в долях от радиуса
1,00,75 0,50,25 0 0,250,5 0,751,0
Безразмерное расстояние
в поперечном направлении
в долях от радиуса
Рис. 5. Профили скоростей, полученные Педли, Шрётером и Садлоу (рис. 1 в работе
Pedley Schroter, Sudlow, 1970)
Отношение (d/S) изменяется пропорционально R1/2. Если длина трубы /
кратна d9 то d/S изменяется пропорционально (Rd/l)1/2.
Как следует из расчетов проведенных при шестидесяти различных
наборах параметров (Pedley, Schroter, Sudlow, 1970), отношение скорости
диссипации энергии рассматриваемого течения к скорости диссипации энергии
при течении Пуазейля довольно близко к величине
0,33(it!d/Z)1/2,
(1)
что подтверждает проведенные выше рассуждения. Смысл подобной
зависимости от числа Рейнольдса может быть понят исходя из того факта, что
число Рейнольдса в состоянии покоя (объемная скорость дыхания 10 л/мин)
изменяется от 800 в трахее до 100 в дыхательных путях на пятом ветвлении
и до 10 в дыхательных путях на десятом ветвлении. При интенсивном
дыхании (объемная скорость достигает 100 л/мин) эти значения увеличиваются
в 10 раз.
Проведенные рассуждения подчеркивают значимость первых
ветвлений дыхательных путей как основного фактора, приводящего к падению
давления в дыхательной системе. По существу, получаем, что сопротив-
2. Движение воздушных потоков в бронхах человека 305
ление пропорционально степени 1/2 скорости потока и падение полного
давления (равное сопротивлению, умноженному на скорость)
пропорционально степени 3/2 скорости. (Здесь под полным давлением понимается
сумма кинетической и потенциальной энергии жидкости на единицу
объема и «статического» давления.)
При интенсивном дыхании числа Рейнольдса превышают критическое
значение (2000) еще на первых трех или четырех ветвлениях дыхательных
путей, и возникает вопрос о том, как следует преобразовать полученные
выше зависимости при меньших значениях чисел Рейнольдса, если возможен
переход к турбулентному режиму движения воздуха. Педли, Шрётер и Сад-
лоу (Pedley, Schroter, Sudlow, 1970) утверждают, что на образование у
разделителя потока нового тонкого вязкого пограничного слоя возникающая
при данных числах Рейнольдса крупномасштабная турбулентность не будет
оказывать существенного влияния. В силу чего диссипация энергии в
основном будет происходить в пограничном слое, так что зависимость
полного давления от скорости будет все еще степенной с показателем степени
3/2. В любом случае в этом интервале чисел Рейнольдса имеет место
степенная зависимость падения давления, обусловленного турбулентностью.
На рисунке 6 представлены зависимости полного давления в
предположении, что его отношение к полному давлению при течении Пуазейля
принимается равным выражению (1), где это отношение превышает
единицу, и в других случаях (т. е. при числах Рейнольдса не больше 40) полагаем
его равным 1. Отметим, насколько значительным является прогнозируемый
прирост полного давления по сравнению с его значением при течении
Пуазейля даже в случае дыхания в состоянии покоя (при объемной скорости
дыхания 10 л/мин), а уж в случае интенсивного дыхания (рис. 6 6), при
объемной скорости дыхания 100 л/мин, превышение становится чрезвычайно
большим. Это еще раз подчеркивает, что полное давление существенным
образом меняется при первых десяти ветвлениях системы.
На рисунке 7 представлена зависимость падения полного давления во
всей дыхательной системе от объемной скорости дыхания. Сплошными
кружочками отмечено падение давления, полученное на основании
экспериментальных данных; незакрашенными кружочками — при течении
Пуазейля. Отметим, что экспериментальные данные, полученные в работах
Хайатта и Уилкокса (Hyatt, Wilcox, 1963) и Ферриса, Мида и Опи (Ferris,
Mead, Opie, 1964), хорошо ложатся на прогнозируемую линейную прямую.
Как вытекает из исследований, проведенных Педли, Шрётером и Сад-
лоу (Pedley, Schroter, Sudlow, 1970), движение воздушного потока в брон-
306
Глава 11
Я 0,05г
ю
5
g 0,04
о
X
э
2 0,03 h
о
4>
0,02 [
0,01 h
(«О
■ фактическое вязкое давление
> вязкое давление в течении Пуазейля
• 1 ш
0 5 10 15
Количество ветвлений
^О.Об
§
§
g0,04
о
s
5
"0,03
о
ffi
J0.02
?0,01
Вязко
Р ф V /
• фактическое вязкое давление
• вязкое давление в течении Пуазейля
ф
ф
ф
ф
ф
" • • • ф
ф
ф ф
ф
• ф
0 5 10 15
Количество ветвлений
Рис. 6. Зависимость «вязкого давления» (так авторы (Pedley, Schroter, Sudlow, 1970)
сокращенно называют падение полного давления, обусловленное действием трении
в дыхательных путях) от количества ветвлений; (а) объемная скорость дыхания
равна 10 л/мин, (Ь) объемная скорость дыхания равна 100 л/мин
хах человека имеет ярко выраженный возмущенный характер с большими
по величине максимумами профиля скоростей, сильно смещенными к
стенкам бронхов, наибольшая диссипация энергии происходит в пограничных
слоях, возникающих на участках бронхов между двумя ветвлениями.
Сопротивление в легких определяется главным образом специфичными
свойствами потока в бронхах, наиболее важным из которых является малость
отношения длины бронхиального пути между соседними разветвлениями
(бифуркациями) к его диаметру.
Зф Дыхание птиц
Как уже отмечалось в § 1, дыхательная система птиц резко отличается
от дыхательной системы наземных позвоночных животных. Это частично
обусловлено наличием связи легких с воздушными мешками, под
действием расширения и сжатия которых поток воздуха проходит через имеющие
постоянный объем легкие, частично необычной конфигурацией, которая,
прежде всего, проявляется в том, что расположение дыхательных путей
далеко от древовидного и характеризуется наличием большого количества
3. Дыхание птиц
307
4 1,50 г
2
о
о
со
S
S
1,00 h
о
о,
ю
со
ОС
§0,50
§
S
с
и
о
о
расчетное значение падения давления
падение давления при течении Иуазейля
Хайатт и Уилкокс
\
п
,'\
Фсррис, Мил и Они
/ •
'8
о
С
0 30 60 90 120
Объемная скорость дыхания (литр/мин)
Рис. 7. Экспериментальные и расчетные данные по зависимости падения давления
от объемной скорости дыхания (рис. 4 из работы Pedley, Schroter, Sudlow, 1970)
петель. То есть можно говорить о циркуляции воздуха в системе «легкие-
воздушные мешки», где воздушные мешки выполняют своеобразную роль
«сердечного насоса», под действием которого и происходит циркуляция
воздуха.
В данном параграфе кратко опишем устройство легких птиц на
основании большого количества опубликованных описаний их строения, а также
прохождение воздушных потоков в дыхательной системе на основании
имеющегося значительно меньшего количества описаний. А также дадим свою
интерпретацию этих потоков, частично представленную в трудах Геттин-
генского симпозиума (см. соответствующие ссылки в томе 14 Respiration
Physiology), а частично — в посвященной данной теме неопубликованной
дискуссии, состоявшейся в Королевском колледже в Лондоне в 1973 году.
При этом возникает основной с точки зрения физиологии вопрос,
который можно кратко сформулировать так: как может «сердце» работать без
клапанов? Ведь, как это следует из многочисленных описаний анатомии
птиц, в воздушных мешках отсутствует какое-либо устройство, выполняю-
308
Глава 11
щее функции клапана, а за счет последовательных их сжатий и расширений
поток воздуха в легких проходит только в одном направлении. Специалист
в области биогидродинамики (Hazelhoff, 1943) задается вопросом: что же
выполняет роль аэродинамического клапана! И приступает к исследованию
этого вопроса, занимается толкованием имеющихся экспериментальных
наблюдений, которые связаны с выдвигаемой им гипотезой о существовании
аэродинамического клапана.
Данкер (Duncker, 1972), изготавливая слепки дыхательной системы,
изучил и описал анатомическое строение дыхательной системы 155 особей
47 различных семейств, по одному из 15 семейств и более чем по одному из
остальных. В его работе область, где происходит газообмен, разделяется на
две части. Одна из них, которую он называет палеопульмой,
присутствует и имеет удивительно схожее строение у представителей всех
исследованных им птиц, а другая область, названная им неопульмой, отсутствует
у большинства примитивных семейств, таких как пингвины (Spheniscidae),
семейство эму (Dromiceiidae). У таких семейств, как аисты (Ciconiidae),
голубиные (Columbidae), утиные (Anatidae) и чайковые (Laridae), неопуль-
ма появляется в некоторой переходной форме, а у семейств фазановые
(Phasianidae), дроздовые (Turdidae) неопульма принимает уже развитую
современную форму.
Палеопульма, которая изначально присутствовала в дыхательной
системе птиц, представляет собой некоторую структуру, обладающую
замечательным свойством, а именно расширением и сжатием воздушных мешков,
не имеющих клапанов, приводит к созданию однонаправленного потока
через ее газообменные конструкции — парабронхи — тонкие трубочки
диаметром около 1 мм, — «воздуховодные капилляры», о которых говорилось в § 1.
В парабронхах неопульмы, у тех видов, где она имеется, воздух движется
в прямом, а затем и в обратном направлениях.
Перед описанием анатомии хотелось бы провести некоторые
рассуждения математического характера. Реакция потока в неопульме на
пульсации воздуха в воздушных мешках носит линейный характер, тогда как
в палеопульме такая реакция существенно нелинейная, что объясняется
необходимостью сгенерировать процесс прохождения потока через легкие
только в одном направлении. К примеру, наличие квадратичной
нелинейности было бы достаточным для этой цели. Ниже будут описаны
анатомические структуры, обеспечивающие нелинейную (квадратичную) реакцию
потока на пульсации воздуха. Квадратичная зависимость вентиляции
воздуха в воздушных капиллярах от амплитуды дыхательного движения подошла
3. Дыхание птиц
309
бы тем птицам, у которых в силу образа их жизни в определенные моменты
резко возрастает потребность в кислороде. Однако это приводит и к
некоторым сложностям, поскольку и в условиях покоя, когда амплитуда
дыхательных движений невелика, действие такого механизма приводит к
ненужному с энергетической точки зрения усиленному потреблению кислорода.
Данкер считает, что неопульма, отвечающая за линейную реакцию на
амплитуду пульсаций воздушных мешков, обеспечивает потребность птицы
в кислороде, когда она находится в состоянии покоя и амплитуда
пульсаций минимальна, а за счет движения воздуха в палеопульме формируется
быстрый отклик на сильное возрастание амплитуды пульсаций, что
обеспечивает полноценное дыхание (удовлетворяется резко возросшая
потребность в кислороде) птицы при полете.
Опишем кратко изложенную в работе Данкера анатомию дыхательной
системы птицы, сосредоточив основное внимание на палеопульме в свете
решения задачи о механизме газообмена в этой области. На рисунке 8
изображен гипсовый слепок левого легкого и связанных с ним пяти воздушных
мешков петуха (Gallus domesticus). Задние мешки (задний грудной
воздушный мешок D и брюшной мешок Е) — в них поступает вдыхаемый воздух
при их расширении и сжатии за счет скоординированных мускульных
усилий мышц грудины и задних ребер. Передние мешки (шейный воздушный
мешок А, межключичный воздушный мешок В и передний грудной
воздушный мешок С) — расширение и сжатие этих мешков происходит в
значительно меньшей степени при тех же мускульных усилиях, а движение
некоторой другой группы мышц происходит в противофазе к сокращениям
вышеупомянутых групп мышц. Мускульные усилия в противофазе
поддерживают неизменным объем легких (см. § 1).
Для лучшего понимания внутренней структуры легкого, и особенно
палеопульмы, наряду с общим схематичным изображением легкого на
рисунке 8, на другом рисунке, рисунке 9 показано то же легкое с удаленной
неопульмой. Воздухопровод 2 представляет собой главный бронх, через
который воздух попадает в легкое. В передней части (слева на рисунке) от
него расходятся четыре вентробронха, отмеченные цифрой 4, через вен-
тробронхи собирается воздушный поток, прошедший через палеопульму,
и возвращается в главный бронх. Каждый из передних мешков А, В и С
связан с одним из вентробронхов. Расширение и сжатие передних мешков
обеспечивает возвращение воздушного потока в главный бронх.
От другого конца главного бронха (справа на рисунке) до его входа
в брюшной мешок Е отходит большое число воздуховодных трубок. На
310
Глава 11
>
V)
) '
V
Рис. 8. Общая анатомическая картина легкого птицы (рис. 1 из работы Duncker,
1972)
ч.
в
ч
Рис. 9. Более подробная анатомическая картина легкого птицы (рис. 2 из работы
Duncker, 1972)
3. Дыхание птиц
311
дорсальной стороне (в верхней части рисунка) имеются от семи до десяти
дорсобронхов, отмеченных цифрой 4. Палеопульма, где происходит
газообмен, представляет собой сеть очень маленьких трубочек (парабронхов),
в которые воздух может поступать из дорсобронхов и их ответвлений,
проходя затем в вентробронхи. От боковой стороны главного бронха отходят
латеробронхи, помеченные цифрой 5. Крупный латеробронх является
одним из основных воздуховодов для легочного потока в палеопульме.
Приведенное Данкером схематичное изображение палеопульмы и
связанных с ней воздушных мешков представлено на рисунке 10. Трахея 1
разделяется на два главных бронха, входящие в два легких. Под действием
пульсаций задних воздушных мешков Е (связанного напрямую с главным
бронхом 2) и D (связанного с главным бронхом посредством
латерального бронха 5) поток движется в дорсобронхи 4, затем, проходя через сеть
парабронхов 6, он попадает в вентробронхи 3. Небольшие пульсации
передних воздушных мешков (особенно В и С) способствуют продвижению
этого потока дальше в главный бронх.
с4с>
Рис. 10. Схематичное изображение палеопульмы (рис. 9 из работы Duncker, 1972)
Опишем кратко без рисунков и деталей, как неопульма при ее наличии
(в той или иной степени она присутствует у большинства семейств птиц)
вписывается в приведенную картину дыхательного процесса. Это
осуществляется следующим образом. Между задней частью главного бронха 2
312
Глава 11
и задними воздушными мешками D и Е располагается дополнительная па-
рабронхиальная ткань. Через эту область в прямом и обратном
направлениях проходят воздушные потоки, обусловленные пульсацией задних
воздушных мешков. (Здесь не рассматривается картина прохождения воздушных
потоков в легких представителей тех семейств, у которых в результате
эволюционного развития возникла связь неопульмы и с передними мешками.)
В работе Данкера подчеркнута одна геометрическая особенность
устройства легких большинства птиц, которая не отражена в блок-схеме
рисунка 10, но может быть довольно значимой для обеспечения
одностороннего движения воздушного потока через дорсобронхи (а следовательно,
и через сеть парабронхов палеопульмы). Входные участки задних дорсо-
бронхов обращены в сторону брюшного воздушного мешка Е, а входные
участки передних дорсобронход, расположенных напротив латеробронха 5,
обращены в его сторону. Возможно, эти две особенности способствуют
тому, что в большей части дорсобронхов прохождение потока воздуха
происходит при сжатии задних воздушных мешков D и Е.
Отметим, что в отличие от подробнейших анатомических описаний
легких птиц в работе Данкера (Duncker, 1972), описание прохождения
воздушных потоков в дыхательной системе птиц весьма ограничено. Так,
только для одной особи (утка-кряква (Anas platyrhynchos)) была
проведена экспериментальная демонстрация одностороннего потока в дорсоброн-
хах. В независимо проведенных опытах на легких утки Чейдом и Пиипе-
ром (Scheid, Piiper, 1971) и Бретцем и Шмидтом-Нельсеном (Bretz, Schmidt-
Nielsen, 1971), а также Чейдом, Сламой и Пиипером (Scheid, Slama, Piiper,
1972) было получено значительное количество экспериментальных данных,
а в последней работе не только описаны результаты экспериментов, но
и приведено их подробное обсуждение.
В верхней части рисунка 11 (рисунок взят из работы Scheid, Slama,
Piiper, 1972) приводится схематичное изображение палеопульмы, а в
нижней части палеопульма изображается с той же степенью подробности, как
и на рисунке 10 (только главный бронх здесь назван мезобронхом). Верхняя
часть представляет собой блок-схему дыхательного процесса, многие
несущественные для общего описания процесса детали опущены. Так, два
задних воздушных мешка (D и Е на рис. 10) объединены в один «хвостовой»
мешок, а вместо трех передних воздушных мешков изображен один
«головной» воздушный мешок. Кроме того, дорсобронхи и вентробронхи
изображены схематично в одном блоке. Экспериментально определенное
направление всех воздушных потоков указано на диаграмме вверху стрелочками.
3. Дыхание птиц
313
вентробронхи
передний воздушный
мешок (в разрезе)
парабронхи
дорсобронхи
задний воздушный
мешок (в разрезе)
мезобронх
дорсобронхи
шеиньга
мешок
-J---T-/ *-.- .„- :--.•»- ^\ -;г , t I
t' ?.* -;•** параб " ахи ... * '-*.
\
межключичныи
мешок
1СЗОО ОНХ
передний
грудной
мешок
брюшной
мешок
Рис. 11. Схема строения легкого у птиц и распределения воздушных потоков в их
дыхательной системе (рис. 2 и 3 из работы Scheid, Slama, Piiper, 1972)
В процессе вдоха (движение воздуха показано черными стрелками)
воздух через главный бронх за счет расширения задних воздушных мешков
течет через дорсобронхи в парабронхи в палеопульму и частично проходит
в задние воздушные мешки. За счет не столь быстро расширяющихся
передних мешков воздушный поток проходит через сеть парабронхов, не
попадая в главный бронх, из которого вошедший при вдохе воздух поступает
частично и в передние воздушные мешки. На выдохе (движение воздуха
показано белыми стрелками) задние воздушные мешки сжимаются и воз-
314
Глава 11
душный поток частично проходит через дорсобронхи, а частично
непосредственно через главный бронх. Воздух, прошедший через парабронхи, идет
в вентробронхи и выводится вместе с потоком, поступающим из передних
мешков в главный бронх.
Хотя в работе и приведено большое количество экспериментальных
данных, касающихся направлений воздушных потоков в дыхательной
системе, однако количественные данные относительно объемов проходящих
потоков и их зависимости от амплитуды пульсации воздушных мешков
в работе представлены в недостаточном количестве. Конечно, получение
таких данных экспериментальным путем представляет значительную
сложность. Желательно создать гидродинамическую модель на основании этих
ограниченных данных, с помощью которой можно было бы описывать
процессы в дыхательной системе птиц без привлечения дополнительных
экспериментальных сведений.
Кратко остановимся на той теории, которая должна объяснить факты,
о которых говорилось в данном параграфе. Конечно, она должна
отличаться от общепринятых теорий, связанных с внутренней биогидродинамикой
в силу отсутствия древовидного строения бронхов. С другой стороны, она
должна включать в себя идеи теории струйного течения, где развит подход
к решению задач о распространении потоков, имеющих тенденцию к
образованию струй при истечении из щели, и механизме подсасывания потока
для изменения его направления.
Простая экспериментальная модель, учитывающая наличие
струйного течения, в которой поток через легкие проходит всегда только в одном
направлении, была представлена профессором Пиипером на симпозиуме
в Имперском колледже науки, техники и медицины. На рисунке 12
схематически изображены воздушные пути дыхательной системы. Поток воздуха
проходит от точки Q к точке Я, а затем к точке Р всегда в одном
направлении под действием осциллирующего с нулевым средним значением
перепада давления между А и В. (Здесь В представляет собой задний воздушный
мешок, а схематическое изображение переднего воздушного мешка на
рисунке и в модели отсутствует.)
В первом приближении однонаправленность потока в легких можно
пояснить так. Поток, двигаясь от А к В, проходит мимо Р и, замедляясь,
движется до точки Q, останавливаясь в ней, в результате чего
устанавливается такая разность между полным давлением в точке Q и статическим
давлением в точке Р, что становится возможным движение потока от Q
к R и затем к Р. Эта разность давлений имеет квадратичную зависимость
3. Дыхание птиц
315
Я
Р_
А
\
Рис. 12. Экспериментальная модель дыхательной системы птиц, представленная
д-ром Чейдом (Scheid) и проф. Пиипером (Piiper)
от скорости потока, о чем говорилось выше, более детальное
исследование этой зависимости с учетом изменения давления в точке Р в результате
прохождения потока приводит также к приблизительно квадратичной
зависимости. Если сопротивление на участке QRP не слишком велико, то по
этому участку будет проходить достаточно интенсивный воздушный поток.
Аналогично, поток, идущий от В до А «предпочтет» пройти мимо
открытого слева канала QA в направлении от Q к R. Приближенная оценка
степени этого «предпочтения» дает, что она определяется
сопротивлением в R. Дальнейшее движение потока происходит под действием разности
между полным давлением в QR и статическим давлением (возможно, не
таким большим, чтобы создать эффект засасывания воздуха или что-либо
другое) в канале Р. Это снова объясняется близкой к квадратичной
зависимостью давления от скорости. На выдохе сильный поток воздуха,
создаваемый за счет сокращения задних воздушных мешков птицы,
устремляется (как отмечено выше) в дорсобронхи. Объяснение прохождения потока
воздуха при вдохе с помощью данной модели остается затруднительным,
поскольку в ней совершенно не учитывается влияние на поток передних
воздушных мешков.
Еще слишком рано прогнозировать, какие эффекты следует учитывать
для создания модели, наиболее точно описывающей процесс дыхания.
Одно кажется несомненным — поиск таких моделей является необходимым
316
Глава 11
и полезным для решения многих задач в новых областях
биогидродинамики, и, несомненно, их следует разрабатывать при сотрудничестве зоологов
и специалистов в области ветеринарии с одной стороны и
экспериментаторов и ученых-теоретиков в области гидродинамики с другой стороны.
Литература
Bretz, W.L. & Schmidt-Nielsen, К. 1971. J. Exptl. Biol. 54, 103-118.
Duncker, H.R. 1972. Respiration Physiol. 14, 44-63.
Ferris, B.G., Mead, J. & Opie, L.H. 1964. J. Appl. Physiol. 19, 611-622.
Hazelhoff, E. H. 1943. Versl. gewone Vergad. Afd. Natuurk. Kon. Ned.
Akad. Wet. 52, 391^00. English translation (1951): Poultry Sci. 30, 3-10.
Hughes, G.M. 1972. Respiration Physiol. 14, 1-25.
Hyatt, R.E. & Wilcox, R.E. 1963. J. Clin. Invest. 42, 29-39.
Marshall, N.B. 1965. The Life of Fishes, Weidenfeld & Nicolson, London.
Pedley, T. J., Schroter, R.C. & Sudlow, M.F. 1970. Respiration Physiol. 9,
371^105.
Piiper, J. & Scheid, P. 1972. Ibid. 14, 115-124.
Scheid, P. & Piiper, J. 1971. Ibid. 11, 308-314.
Schied, P., Slama, H. & Piiper, J. 1972. Ibid. 14, 83-95.
Schroter, R.C. & Sudlow, M.F. 1969. Ibid. 7, 341-355.
Weibel, E.R. 1972. Ibid. 14, 26^13.
Глава 12
Распространение пульсаций
в кровеносной системе
1. Введение
Теория распространения пульсаций в артериях является частным
случаем более общей теории распространения одномерных волн в жидкости.
В ней локальному скоплению жидкости препятствуют либо эластичные
стенки сосуда в случае пульсовых волн, либо сила тяжести в случае
длинноволновых колебаний жидкости в открытых каналах, либо сжимаемость
среды в случае звуковых волн. В любом случае характер
распространения волны определяется балансом сил сопротивления и инерции жидкости.
В данной главе теория будет применена к интересному с точки зрения
физиологии случаю распространения пульсаций при движении несжимаемой
жидкости (крови) в эластичных сосудах. В конце главы кратко будет
отмечено, как, внеся небольшие изменения, можно использовать рассмотренные
методы для изучения других сосудистых систем.
Будет рассмотрено движение крови постоянной плотности р в
кровеносной системе животного, которое находится в состоянии покоя.
Обозначим гидростатическое давление крови через р0. Это давление
устанавливается в отсутствие сердечного импульса и равно атмосферному давлению
на уровне аортального клапана, гидростатическое давление растет со
скоростью рд на единицу длины при превышении этого уровня. Разность
давлений
Ре=Р~Ро (1)
в каждой точке системы обусловливается работой сердца.
Порядок превышения давления в данной точке над гидростатическим
в артериях млекопитающих составляет 104 Н/м (или 10"1 от стандартного
атмосферного давления). Рост давления при нормальных физиологических
318
Глава 12
условиях приводит к изменению просвета сосуда (внутренней площади
поперечного сечения) А приблизительно на 10%. Растяжимость D сосуда,
определенная как скорость изменения относительной площади
поперечного сечения при изменении давления, вычисляется по формуле
и имеет порядок 10~5 м2/Н. Хотя ее действительная величина различна
для различных сосудов. В аорте она принимает наибольшее значение, а
затем в других сосудах уменьшается — чем больше удаленность сосуда от
сердца, тем меньше ее значение. Более того, в силу сложной композитной
структуры стенок артерии, состоящих из волокон эластомера, эластина,
достижение растяжимостью среднего значения возможно уже при достаточно
умеренных значениях избыточного давления. При избыточном давлении,
близком к максимальному, жесткие коллагеновые и гладкомышечные
волокна существенно ограничивают прирост площади поперечного сечения
сосуда. Характер распространения пульсовой волны в отдельном сосуде
зависит от наличия ветвления, закона зависимости растяжимости D от
давления ре как для растяжимых сосудов, так и для сосудов с постоянной
площадью поперечного сечения.
В данном параграфе рассмотрим задачу распространения пульсаций
в линейной постановке без учета вязкости в одной артерии с площадью
поперечного сечения в невозмущенном состоянии, равной постоянной
величине А. Полагаем, что пульсации достаточно слабые и растяжимость сосуда
можно считать постоянной величиной, пренебрежем адвективными
членами и • Vu в выражении для ускорения жидкости. Случай ветвления сосуда
рассмотрен в § 3 и § 4, зависимость напряжения в стенках от
растяжимости сосудов исследована в § 2. Влияние изменения параметров сосуда вдоль
его продольной оси, а также влияние вязкости исследуются в § 5, а эффекту
нелинейности посвящен § 6.
Пусть х — продольная координата (вдоль осевой линии артерии), и —
продольная скорость. Более точно скорость и можно определить в
каждый момент времени как осредненное по поперечному сечению значение
продольной скорости, хотя заметим, что при наличии вязкости скорость
существенным образом изменяется в плоскости поперечного сечения. Этот
эффект будет учтен только в § 5.
Из второго закона Ньютона следует, что продольная сила равна
произведению массы на продольное ускорение. Это произведение при уело-
1. Введение
319
вии, что адвективными членами ускорения можно пренебречь, принимает
вид pdu/dt. Поскольку вязкими эффектами в первом приближении пре-
небрегается, то продольная сила, приходящаяся на единицу объема, равна
—дре/дх. Здесь в выражение для силы входит только избыточное
давление (1), поскольку градиент гидростатического давления уравновешивается
силой тяжести, действующей на жидкость. Таким образом, линейное
уравнение количества движения имеет вид
рж = ~' (3>
к нему следует добавить уравнение неразрывности
Здесь с левой стороны стоит относительная скорость изменения
площади поперечного сечения A~l dA/dt (см. соотношение (2) для
растяжимости D), которая для несжимаемой жидкости равна взятой с
противоположным знаком средней относительной скорости расширения ди/дх в
направлении, перпендикулярном поперечному сечению.
Исключая скорость и из уравнений (3) и (4), получаем волновое
уравнение
аЧ=о2|&, (5)
где скорость с равна
dt2 дх2 '
c=(pD)~1/2. (6)
Скорость с при р = 103 кг/м3 и D = 10~5 м2/Н равна 10 м/с, что
согласуется со многими экспериментальными данными. С другой
стороны, в здоровой грудной аорте растяжимость может достигать значений
4 • 10~5 м2/Н (при искусственно созданном импульсе давления), а
измеренная при этом скорость распространения волны уменьшается до значения
5 м/с (McDonald, 1960).
Заметим, что сформулированная выше постановка может быть без
изменений применена к задаче распространения длинных гравитационных
волн в открытых каналах с постоянной площадью поперечного сечения А
и длиной свободной поверхности в этом сечении, равной В. Избыточное
давление ре приводит к повышению уровня воды на величину ре/рд, а
площадь поперечного сечения увеличивается на Вре/рд. Тогда в соответствии
320
Глава 12
с (2) и (6) можно получить
D = ^A И C=(9A/B)V\ (7)
известную скорость распространения длинных тяжелых волн. Отметим
также, что если принять во внимание сжимаемость жидкости К, то
растяжимость D в левой части (4) увеличится на эту величину, и для скорости
с из (6) получим с = [p(D + K)]~1/2. Конечно, сжимаемость жидкостей,
включая кровь, не превышает 10~9 м2/Н, так что поправка, внесенная
учетом сжимаемости, в сердечно-сосудистой системе пренебрежимо мала. Для
жидкостей, текущих в металлических трубках, величина D значительно
меньше сжимаемости К, и в таком случае скорость распространения
возмущений определяется, как и в теории звука, величиной с = (рК)~1/2.
Во всех случаях решение уравнения (5) относительно давления ре
представляет собой распространяющуюся волну давления в направлении
возрастания х. Это решение, а также связанное с ним решение
уравнения (3) имеют вид
Pe = f(t-c-lx), u = (pc)-lf(t-c-lx). (8)
Соответственно распространение волны давления в направлении убывания
переменной х описывается соотношениями
Ре = 9(* + с^х), и = -{pc)-xg{t + c-lx). (9)
Разные знаки у решения для скорости и в равенствах (8) и (9) означает,
что волны распространяются в обоих направлениях, и в каждой точке
скорость и равна избыточному давлению, умноженному на величину (рс)~1.
Общее решение уравнений (4) и (3) представляет собой сумму
решений (8), (9). И в этом случае соотношение между скоростью и избыточным
давлением будет иметь более сложный характер. Вывод этих соотношений
будет приведен в § 3.
1. Распределение напряжений в стенках сосуда
Обсудим кратко силы, действующие со стороны сосуда на поток, при
распространении в нем пульсовой волны, т. е. силы, обусловленные
растяжимостью сосуда. Это обращение к биомеханике твердого тела не вызвано
2. Распределение напряжений в стенках сосуда 321
необходимостью определить величину растяжимости стенок (она
достаточно легко измеряется) на основании других констант,
характеризующих свойства материала стенок (величины, которые возможно значительно
сложнее измерить). Главной целью этого обращения к механике твердого
тела является выявление факторов, которые могут привести к
повреждению сосуда или, напротив, помогут сохранить его здоровым при
протекании крови. Об этих факторах уже упоминалось в главе 10 и более
подробно будет обсуждаться в главе 13. Обсуждение реакции стенок сосуда на
распространение в нем пульсовой волны совершенно необходимо для
более полного понимания процесса ее распространения. В силу чего сначала
и займемся исследованием реакции стенок.
Отношение h/a0 толщины стенки к радиусу а0 сосуда, находящегося
в невозмущенном состоянии, колеблется в пределах от 0,1 до 0,2, что дает
возможность воспользоваться методами теории течения жидкости в
трубах с тонкими стенками, причем мы не будем делать неподходящего для
данной задачи предположения об изотропной упругости материала
стенки. Что в конечном итоге нам потребуется, так это утверждение,
известное физиологам как закон Лапласа — термин, который специалисты
используют в теоретической механике при исследовании совершенно других
фундаментальных явлений, описываемых в трудах Лапласа. Специалисты
в области теоретической механики могут удивиться такому
использованию термина, но, возможно, им следовало бы повнимательней прочесть
его труды.
Закон Лапласа утверждает, что давление ре внутри трубы с
внутренним радиусом а0 приводит к возникновению1 окружного (направленного
по касательной к окружности) напряжения а0ре на единице длины сосуда.
Конечно, это не может вызвать удивления, однако для того, чтобы лучше
понять механизм возникновения окружного напряжения, представим, что
сосуд единичной длины разделен на две равные части с сечениями в виде
половины круга. Пусть на обе части действует растягивающее внутреннее
давление ре на площади 2а0, т.е. результирующая сила равна 2а0ре,
которая уравновешивается окружным напряжением, действующим в плоскости
соединения этих половинок.
Это окружное напряжение а0ре действует на площади разреза
половинок, равной толщине стенок h. Тогда осредненное по толщине стенок
окружное напряжение растяжения гх (действует на плоскости разделения
Напряжения, вызванные действием давления ре, равны напряжениям, которые возникают
при действии давления ре минус напряжения при его нулевом значении.
322
Глава 12
половинок перпендикулярно окружной силе) равно
т, = 2йЬ. (Ю)
В артериях, как и в других тонкостенных трубах, отношение h/a0 мало.
Среднее окружное напряжение растяжения (10) значительно превышает
величину радиального напряжения (которое изменяется от значения ре на
внутренней стенке до 0 на внешней стенке).
Из проведенных рассуждений следует, что в окружном напряжении,
обусловленном внутренним давлением, превалирующее значение имеет
окружное напряжение растяжения (иногда называемое кольцевым
напряжением). К этому предположению мы вернемся немного позже. Сейчас кратко
рассмотрим то утверждение, что напряжение в стенке артерии, как и при
простом испытании на разрыв, направлено по окружности. Если
внутренний радиус изменяется от а0 до а и изменения толщины стенки
пренебрежимо малы (см. ниже), то соответствующее удлинение вдоль окружности
имеет вид
?—^. (П)
Следовательно, если модуль Юнга при испытании на разрыв,
пересчитанный для деформации вдоль окружности, равен Е, то ^ = Е(а — а0)/а0
представляет собой напряжение растяжения в каждой точке, а Тг = Е(а —
- а0)/а0, где Е — средний по толщине модуль Юнга £*, равный среднему
напряжению растяжения (10).
Тогда с учетом (10) получаем
а0 Eh' K )
Поскольку относительное изменение площади внутренней поверхности
сосуда в два раза больше величины (а — а0)/а0, то на основании (2)
растяжимость сосуда принимает вид
D = ж- <13>
а скорость распространения пульсовой волны (6) соответственно равна
с = (Eh/2pa0)1/2. (14)
2. Распределение напряжений в стенках сосуда 323
Это согласуется с классической формулой Моенса -Кортевега для скорости
распространения пульсовой волны, если в ней модуль упругости Е для
однородного и изотропного материала стенок заменить на осредненный по
толщине стенки сосуда модуль упругости.
Рассмотренное выше приближение для напряжения в стенках сосуда
и вытекающие из него следствия правомерны для многих задач. Читатель,
знакомый с теорией упругости, может заметить, что до сих пор ни разу не
упоминался один хорошо известный факт, а именно: растяжение в
направлении действия силы должно сопровождаться сжатием в
перпендикулярном направлении. Коэффициент пропорциональности между поперечной
и продольной деформациями в случае материала с изотропными упругими
свойствами называется коэффициентом Пуассона, который обозначается
через а. Рассмотрим такое сжатие, не предполагая изотропности упругих
свойств материала стенки.
Суммарное сжатие в радиальном направлении, в которое входит
меньшее по величине, чем окружное растяжение (11), отрицательное
радиальное растяжение, играет в рассматриваемом случае весьма существенную
роль. За счет него происходит изменение толщины стенок, которое по
величине меньше произведения h[(a — а0)/а0] двух малых величин. Это
подтверждает сделанное ранее предположение о том, что окружное
растяжение (11) есть величина приблизительно постоянная, поскольку оно
означает, например, что относительные изменения внутреннего и внешнего
радиусов трубы отличаются только на множитель 1 + O(h/a0).
Более интересные проблемы возникают при продольном сжатии
стенок артерии, которое сопровождает возникновение окружного напряжения.
Например, действительно ли это является единственной реакцией на
изменение внутреннего давления. Пусть индекс 1 относится к окружному
напряжению и удлинению, а индекс 2 — для продольного напряжения и
деформации. Далее используем обозначения Лява (Love, 1927, гл. 3).
Рассмотрим матрицу элементов Cij9 где г-тые компоненты деформации
обусловлены бесконечно малым приращением j-той компоненты напряжения. Так,
Си представляет собой окружную деформацию, приходящуюся на
единицу окружного напряжения, и в наших ранее использованных
обозначениях она равна Е~1. С другой стороны, элементы С12 и С21, равенство
которых обусловлено законом сохранения энергии, представляют окружную
деформацию, приходящуюся на единицу продольного напряжения и
продольную деформацию, приходящуюся на единицу окружного напряжения
соответственно. Для изотропных материалов эти элементы равны —аЕ~1.
324
Глава 12
Элемент С22 представляет собой продольную деформацию, приходящуюся
на единицу продольного напряжения, и в общем случае необязательно этот
элемент равен Сп = Е~1, хотя для жесткого материала должно
выполняться неравенство
СцС22 > ^12- (15)
При простом окружном растяжении под действием окружного
напряжения тг продольная деформация в стенке артерии удовлетворяет
соотношению
Сит, = СпЕ ^ = \СиЕ ±^Ь.. (16)
По сравнению с деформацией в жидкости —(А — А0)/А0 деформация
в стенке артерии уменьшается в (—^C12Ej раз. Множитель (— ^C12Ej
принимает значение 0,2 для типичных для несжимаемых биологических
материалов значений отношения С12/Си. С полученным результатом,
утверждающим, что продольные деформации в стенке сосуда составляют
0,2 от продольных деформаций в жидкости, очень трудно согласиться.
В действительности волокна в артериях хотя и достаточно эластичны
и их реакция на пульсационное изменение давления внутри сосуда слаба
(как предполагалось при выводе закона Лапласа), но, вероятно, все же
достаточна для предотвращения продольных смещений стенки сосуда. А
если это так, то ограничивающее действие упругих волокон стенок артерии
должно привести к возникновению деформации —С12т19 равной по
величине и противоположной по знаку деформации (16). Это происходит за счет
возникновения напряжений, противодействующих продольному
напряжению, величина которых равна
^22
(17)
поскольку С22 представляет собой продольную деформацию,
приходящуюся на единицу продольного напряжения. В силу действия напряжения (17)
возникает дополнительная окружная деформация равная С12, умноженному
на (17), которую следует добавить к окружной деформации Cnrl9
обусловленной действием напряжения тг. Результирующая окружная деформация
принимает вид
^ = С..(1-с^-)г,. (18)
а0 \ °11°22/
Таким образом, возникающая как реакция на изменение давления в
потоке продольная деформация стенки приводит к возрастанию окружного
3. Прохождение волн в местах ветвления артерий 325
напряжения тх от значения Е(а — а0)/а0 (где Е = Сп ) до значения Е'(а —
- а0)/ао> где
1 ^19
^11^22
— эффективный модуль упругости Юнга. Он отличается от модуля
Юнга Е положительным (в силу неравенства (15)) множителем и превышает
его приблизительно на 20 %. И значит, средний модуль упругости Е в
равенствах (12), (13), (14) должен быть заменен на Е . В результате такой
реакции стенок артерий скорость потока хотя и увеличивается при
прохождении пульсовой волны, но увеличение не превышает 10%.
Расчетные данные, полученные на основании (13) и (14), согласуются
с имеющимися экспериментальными данными (McDonald, 1960). Однако
это согласование нельзя признать удовлетворительным, т. к. с той же
степенью точности согласуются с этими экспериментальными данными и
расчетные данные (13) и (14), если в них подставить Е вместо Е.
Вышеприведенная дискуссия проведена не с целью ввести небольшие изменения
в формулах, а хотелось бы указать на все возникающие напряжения в
стенках артерий, связанные с прохождением пульсовой волны при наличии
избыточного давления ре. Это не только среднее окружное напряжение (10),
но также и пропорциональное увеличение продольного напряжения (17),
обусловленное реакцией стенок сосудов, препятствующей возникновению
существенных продольных деформаций.
3. Прохождение волн в местах ветвления артерий
Выше, в § 1 и § 2, были рассмотрены вопросы, связанные с
распространением пульсовой волны в одной артерии. А сейчас исследуем
прохождение волны в составных сосудах. Затем результаты проведенного здесь
исследования будут использованы при изучении прохождения волн через
ветвящуюся артериальную систему.
Как отмечал М. Дж. Тейлор (Taylor, 1965, 1966), методы, которые здесь
могут быть применены, очень схожи с методами расчета тока в
электрической цепи. Прохождение тока подчиняется законам Кирхгофа: в любом узле
сумма токов, направленных к узлу, равна сумме токов, направленных от
узла, и алгебраическая сумма падений напряжения вдоль любого замкнутого
контура электрической цепи равна нулю. Соответственно для
сердечно-сосудистой системы примем правила прохождения потока крови при наличии
ветвления: избыток давления не испытывает скачка при прохождении точки
326
Глава 12
сочленения, а суммарный расход крови, входящей в сосуд, равен
суммарному расходу крови в сосудах после ветвления.
Оценим относительную ошибку, которая определяется этими
предположениями, выразив ее через длину / — расстояние от входа в сосуд до
его первого ветвления и радианную частоту и; сердечного пульса. Градиент
избыточного давления, определяемого уравнением (3), приравняем к
величине рии, где скорость жидкости и, как следует из (8), равна ре/рс.
Изменение избыточного давления ре равно произведению / на градиент давления
или (ul/c), умноженному на ре. Аналогично, разность между входящим
в сосуд и выходящим за точкой ветвления потоком жидкости равна скорости
изменения расхода жидкости в области ветвления и может быть рассчитана
как произведение / на скорость изменения площади поперечного сечения
ADdpe/dt (см. определение (2) растяжимости D). Приравнивая скорость
dpe/dt произведению и;реи и используя равенство (6), заменяем
растяжимость D на р~1с~2 и запишем разность между втекающим и вытекающим
объемом жидкости как произведение (ljI/c) на расход жидкости Аи.
Относительная ошибка в обоих случаях равна ul/c и является малой
для ветвлений, размер которых удовлетворяет следующему условию:
/ мало по сравнению с с /и. (20)
При скорости с = 10 м/с (§ 1) и частоте и = 10 с-1 (частота в герцах,
умноженная на 27г) длина с/и равна 1 м, а типичное значение / имеет порядок
10~2 м, следовательно, условие (20) выполняется. Значит, с достаточной
степенью точности можно принять правила прохождения крови по
ветвящейся системе сосудов, аналогичные законам Кирхгофа для электрических
цепей.
Прежде чем исследовать реальную ветвящуюся систему,
предварительно ответим на следующий вопрос. Что произойдет с пульсовой волной,
распространяющейся вдоль одной трубы, при ее переходе в трубу с другим
поперечным сечением и другой растяжимостью? Пусть ось координат х
выбрана так, что соединение этих труб происходит при х = 0. Исследуем
возможность возникновения отраженных волн, которые пойдут в
противоположном направлении, при прохождении пульсовой волны через точку
х = 0 в другой сосуд, в котором за счет другой растяжимости скорость
проходящей волны изменится.
Изменение избыточного давления ре и расхода жидкости J = Аи
описывается волнами, бегущими вперед, (8), при х = 0 эти величины равны
Ре = /(*), J = 17(0,
(21)
3. Прохождение волн в местах ветвления артерий 327
где величину
Y = — (22)
рс
определим как адмитанс (проводимость) сосуда, т.е. отношение расхода
текущей жидкости к избыточному давлению для волны, бегущей в том
направлении, в котором распространяется волна изменения скорости.
Подразумевается, что равенства (21) справедливы при условии, что
распространяющиеся волны в двух трубках удовлетворяют (i) условию непрерывности
избыточного давления в сечении х = О, которое выполняется, если и только
если форма волн f(t) одинакова, и (и) условию баланса количества крови,
втекающего в сечение, где соединяются два сосуда, и вытекающего из него,
которое выполняется, если и только если трубки имеют один и тот же
адмитанс Y. Такое выравнивание адмитансов (при увеличении скорости
волны площадь поперечного сечения возрастает и одновременно согласованно
с этим уменьшается растяжимость) является необходимым и достаточным
условием отсутствия отраженных волн при прохождении пульсовой волной
области соединения сосудов.
В общем случае, когда адмитансы не согласованы, будем записывать
характеристики волны при х < О с нижним индексом 1 (поперечное
сечение Al9 скорость распространения волны сх и адмитанс А1/рс1 = Yx) и
соответственно характеристики волны при х > 0 — с нижним индексом 2.
Общее решение уравнения (5), допускающее существование отраженных
волн, является суммой волн (8) и (9), при х < О имеет вид
Ре = /(* " С11х) + 9(* + С11х)> ( ,
J = Y1[f(t-c^1x)-g(t + c-1x)}.
Жидкость в трубке х > 0 находится в покое до тех пор, пока пришедшая
пульсовая волна не достигнет точки соединения сосудов, волна давления
может проходить по покоящейся жидкости только в направлении
возрастания х (отсчитывается от соединения) и удовлетворяет соотношениям
ре = h(t - с^х), J = Y2h(t - c^x). (24)
Из условия непрерывности давления и баланса расхода жидкости в области
ветвления следует
f(t) + g(t) = h(t), Y1[f(t)-g(t)] = Y2h(t). (25)
328
Глава 12
Эти соотношения могут быть использованы в течение всего времени, пока
волна (24) не достигнет дальнего конца трубки при х > О и возникшая
у дальнего конца трубки отраженная волна не вернется к месту
соединения, что приведет к изменению правой части уравнений. К исследованию
множественных отражений вернемся позднее.
Разделив первое уравнение в (25) на второе, получим
у Я*) ~ 9(t) _ у т)
Yif(t) + 9(t)-Y>- (26)
Отсюда для отношения g/f получаем
9(t) Y1 - Y2
f(t) Yx + Y2
Из (27) и второго уравнения (25) следует
h(t) 2YX
f(t)-Y1+Y2
(27)
(28)
Левая часть (26) представляет собой эффективный адмитанс при х = О для
трубки х < О, т. е. отношение расхода жидкости к избыточному давлению.
Его следует принимать в расчет при использовании общего решения (23),
в котором присутствует отраженная волна. Соотношение (26) представляет
собой условие согласования эффективного адмитанса при х < О с адмитан-
сом У2 трубки х > О в отсутствие какой-либо отраженной волны.
При Y2 < Yx из равенства (27) следует, что при наличии
согласованных адмитансов имеет место положительное отражение (т.е. функция д
имеет такой же знак, как и функция /). В предельном случае, когда
трубка х > 0 имеет закрытый конец (У2 = 0)> выполняется равенство д = /,
т. е. имеет место полное положительное отражение. С другой стороны, при
Y2 > Yx (может быть за счет большего поперечного сечения трубки х > О
или большей растяжимости) из (26) вытекает, что возникает
отрицательное отражение, т. е. выполняется g/f<0.
В случае любой распространяющейся волны поток энергии
достаточно просто связан с адмитансом. Скорость переноса энергии в направлении
распространения волны равна скорости жидкости, умноженной на силу,
которая равна произведению избыточного давления на площадь поперечного
сечения. То есть равна объемной скорости расхода жидкости, умноженной
3. Прохождение волн в местах ветвления артерий 329
на избыточное давление, которое, в свою очередь, равно адмитансу,
умноженному на квадрат избыточного давления. Например, в подходящей к
месту соединения волне скорость переноса энергии равна У1/2(^-с]~1ж),
аналогично для отраженной волны имеем Yi#2(£ + с^1^), а для проходящей —
Y2h2(t-c2lx).
Теперь можно записать на основании (27) и (28) условие сохранения
энергии при х = 0. Доля энергии, переносимая отраженной волной, от
энергии падающей волны равна
92(t) = (Yi ~ Y2)2
Pit) (П+П)2' К }
а доля энергии, переносимой прошедшей волной, равна
Y2h2(t) _ 4YlY2
YjHt)- (Y1 + Y2r {Щ
Эти доли энергии больше или равны нулю и в сумме составляют единицу,
т. е. вся энергия, которая не ушла с отраженной волной, уносится
прошедшей волной.
Заметим, что если условие согласования адмитансов выполняется не
точно, а только приблизительно (т.е. отношение Y2/Yl близко к единице,
но не равно ей), тогда при прохождении волны через сечение, где
сосуды соединяются, можно считать, что энергия передается дальше почти без
потерь. Доля энергии, ушедшей с отраженной волной (29), в этом случае
равна \(У2/Уг — I)2. Так, при рассогласовании адмитансов в 5% эта доля
принимает значение 0,0006. При последовательном соединении нескольких
сосудов с относительно небольшим рассогласованием адмитансов при
каждом соединении с отраженной волной уходит незначительная доля энергии
пульсовой волны (пропорциональная квадрату величины рассогласования),
так что можно приближенно считать, что при прохождении соединений
потерь энергии не происходит. Это предположение будет использовано в § 5
при исследовании случая постепенно и непрерывно меняющегося адмитан-
са трубы.
В другом достаточно интересном случае, когда трубка х > 0 хотя и не
закрыта на другом конце, однако ее адмитанс У2 очень мал по сравнению
с Уг, из (27) следует, что отношение g/f близко к единице (к значению
этого отношения при наличии закрытого конца у трубки х > 0, ведь в
таком случае выполняется условие Y2 = 0)- А из (30) вытекает, что
отношение h/f близко к 2, что довольно удивительно. Надо сказать, что в силу
330
Глава 12
малости Y2 поток энергии Y2h2 при х > 0 невелик. Действительно,
отношение (30) проходящего потока энергии к потоку энергии, поступающему
с падающей волной, приблизительно равно A(Y2/Yl). Тем не менее
амплитуда давления в проходящей волне в два раза больше, чем в падающей,
главным образом потому, что при наличии почти полностью закрытого
конца у трубки х < 0 (т. е. соединение эквивалентно почти полностью
закрытому концу) происходят практически полное положительное отражение и,
как следствие, удвоение амплитуды давления. Удвоение амплитуды
колебаний давления в трубке х > 0 является следствием удвоения амплитуды
давления вблизи почти полностью закрытого конца трубки х > 0.
Немного видоизменив проведенный выше анализ, рассмотрим теперь
случай прохождения пульсовой волны области ветвления, когда
проксимальная артерия (расположенная ближе к сердцу) разветвляется на две или
три дистальные артерии (расположены дальше от сердца). Пусть ось х
направлена в сторону удаления от сердца. Абсцисса сечения, где происходит
разветвление, равна нулю. Индекс 1, как и раньше, относится к
характеристикам неразветвленной проксимальной трубки х < 0, а индексы от 2 до N
относятся к характеристикам трубок х > 0, образовавшихся при
ветвлении (см. рис. 1, где изображены типичные случая ветвления одного сосуда
при N, равном 3 или 4). Равенства (23) справедливы относительно
движения в трубке х < 0 (включая как падающую так и отраженную волну), но
равенства (24) для проходящей волны следует заменить несколькими:
Ре = h(t - c~lx), J = Yrh(t - c~lx), x > 0, (31)
для каждой дистальной трубки г ^ 2. Из непрерывности давления в
сечении ветвления х = 0 вытекает сохранение формы h(t) в каждой трубке,
и выполняется, как и в (25), условие
f(t)+g(t) = h(t). (32)
Приравнивая расход жидкости, входящей в сечение х = 0, равный
(23), если в нем подставить х = 0, сумме расходов жидкости в каждой
дистальной трубке, выходящей из сечения х = 0 (в соотношении (31) следует
подставить х = 0), получаем
YAf(t)-9(t)]=nTYr)h(t). (33)
3. Прохождение волн в местах ветвления артерий 331
Следует отметить, что равенства (32) и (33) совпадают по форме с (25),
если в нем адмитанс Y2 трубки х > О заменить на сумму адмитансов всех
N
дистальных трубок х > О, равную ^ Уг- ^ак же как и в электрических це-
г=2
пях адмитансы параллельно соединенных трубок складываются, и область,
где сосуд разветвляется на несколько, ведет себя так, как будто в этом
сечении произошло соединение проксимальной трубки с одной дистальной
N
х > О с адмитансом, равным ^ Уг-
г=2
В этом случае соотношения (27) и (28) для проходящей и отраженной
волн принимают вид
N
д(Я Yl~^2Yr ОД = 2Y,
f(t) А ' f(t) А ' [
г=2 г=2
Можно также показать, что энергия, уносимая от х = 0 всеми проходящи-
ми волнами (31), равна сумме ( ^ Yr)h2(t). Для расчета в соответствии
с законом сохранения энергии доли энергии, переносимой прошедшими
в трубки волнами, и доли энергии, переносимой отраженной волной, от
энергии падающей волны необходимо в (29) и (30) заменить адмитанс У2
N
на £ Yr.
г=2
Ветвлением с наибольшим углом расхождения сосудов в
сердечно-сосудистой системе является подвздошная бифуркация, когда аорта, пройдя
в брюшную полость, разделяется на две подвздошные артерии.
Рассмотрим кратко, какое влияние на пульсовую волну, идущую от сердца,
оказывает одна эта бифуркация. Конечно, при подобном рассмотрении не
учитывается вся сложность структуры сердечно-сосудистой системы (включая
большое число взаимодействующих между собой разветвлений и
изменение свойств стенок артерий между этими ветвлениями). Однако
результаты такого исследования в целом адекватно отражают поведение пульсовой
волны в системе, поскольку рассматриваемая бифуркация является
наиболее существенной.
В этом случае N = 3 и сумма адмитансов Y2 и Y3 подвздошных
артерий оказывается меньше, чем адмитанс Ух аорты, частично за счет того, что
суммарная площадь их поперечных сечений меньше площади поперечного
332
Глава 12
сечения аорты (приблизительно на 20 %, хотя для каждого конкретного
индивидуума это отношение может сильно отклоняться от среднего значения),
и частично за счет большей растяжимости стенок аорты, а следовательно
и меньшей скорости пульсовой волны с, по сравнению с подвздошными
и другими периферическими артериями. Тогда из уравнения (34) следует,
что отражение будет положительным, т. е. g/f > 0.
Экспериментальные данные относительно флуктуации давления в
аорте подтверждают наличие положительной отраженной волны. С точки
зрения физиологии увеличение давления в относительно растяжимой аорте
при поступлении в нее крови за счет сокращения сердечной мышцы
необходимо для обеспечения кровотока с относительно постоянной скоростью
в периферической системе кровообращения.
Более того, в подвздошной аорте имеется большое число различных
видов ветвлений сосудов, среди которых можно указать ответвление двух
узких межреберных артерий, отходящих от аорты, через них идет
небольшой кровоток, снабжающий кислородом межреберные мышцы. При таком
ветвлении (рис. 1 при N — 4) адмитансы Yx и Y2 проксимального и ди-
стального относительно данного ветвления отделов аорты практически
равны и велики по сравнению с адмитансами У3 и У4 межреберных артерий.
N
Тогда величина суммы ^ Уг близка к Yx, а значит, происходит практически
г=2
полное без отражений прохождение пульсовой волны. Кроме отсутствия
отраженных волн, что неудивительно, следует отметить, что в межреберных
артериях пульсация давления имеет такой же характер, как и в самой аорте.
4. Взаимодействие разветвлений в сосудистой системе
Кажется совершенно невозможным проанализировать
распространение пульсовой волны в чрезвычайно сложной ветвящейся системе
артерий человека, поскольку имеющиеся в огромном количестве ответвления
оказывают влияние на весь кровоток. Это явление в § 3 совершенно не
учитывалось. И все же в данном параграфе попытаемся рассмотреть влияние
ветвлений на примере взаимного воздействия двух ветвлений в системе.
Как будет видно, даже наличие только двух ветвлений значительно
усложняет анализ кровотока, в частности хотя бы в силу сильной зависимости
процесса от частоты колебаний. К счастью оказывается, что методы
исследования взаимного воздействия двух ветвлений могут быть
непосредственно применены к сложной ветвящейся системе (см. § 6).
4. Взаимодействие разветвлений в сосудистой системе 333
х=
2Г<0
х>0
х=0
х<0
х>0
х>0
Рис. 1. Схематичное изображение различных видов ветвления в
сердечно-сосудистой системе
Для изучения взаимодействия двух ветвлений рассмотрим (рис. 2)
ветвление в сечении В, в точности такое же, как изображено на
рисунке 1, только трубка х < О длиной / сама является одним из разветвившихся
в сечении А сосудов, и в ней распространяется волна, прошедшая
проксимальное разветвление в сечении А, я-координата которого равна х = — I.
Полагаем, что отношение g{t)/f(t) отраженной и падающей волны в
сечении В (х = 0) удовлетворяет соотношению (34).
Рассчитаем эффективный адмитанс Y^ трубки х < 0. Индекс 1
относится к характеристикам на участке АВ. Для адмитанса, равного отно-
334
Глава 12
х=0
х<0
Рис. 2. Взаимное влияние двух разветвлений сосудов в сечениях А и В
шению расхода J к избыточному давлению ре, с учетом прохождения волн
в обоих направлениях (23) для трубки х < О имеем
-1;
Yf* = Yx
f(t + c;ll)-g(t-c;4)
-1;
/(г + с^О + ^г-с]-1/)
(35)
Если один раз рассчитать отношение Y*ff объемного расхода к
избыточному давлению для ветвления в сечении А для трубки АВ с учетом наличия
ветвления в сечении В, то анализ прохождения волны в сечении А
может быть проведен в точном соответствии с исследованиями прохождения
волны в месте ветвления, когда это ветвление единственно (как это было
рассмотрено в § 3), с заменой адмитанса Yx на эффективный адмитанс Y^
в сумме адмитансов всех трубок, через которые проходит пульсовая волна,
прошедшая ветвление А. Затем этот же алгоритм может быть
последовательно применен ко всей ветвящейся системе.
Качественно новым здесь является появление временного
интервала с^11 между волнами в сечении А и В, т.е. между волнами / (см. (35)),
распространяющимися от А к В и волнами д, распространяющимися от В
к А. Этот момент достаточно важен, поскольку при его наличии
невозможно непосредственное применение соотношения (34) для g{t)/f(t) для
расчета (35).
Представление волны в комплексной форме, содержащей
множитель elujt, и применение анализа Фурье позволят избежать возникшей
трудности. В линейной теории любое внешнее воздействие, влияющее на рас-
4. Взаимодействие разветвлений в сосудистой системе 335
пространение волн, может быть представлено в виде линейной
комбинации величин егш1 для различных частот и9 и если реакция системы на это
воздействие может быть рассчитана отдельно для каждой гармоники, то
суммарная реакция системы на все внешнее воздействие равно той же
линейной комбинации реакций на каждую гармонику. Так, например,
периодическое, но не синусоидальное внешнее воздействие, такое как изменение
давления в аортальном клапане, может быть представлено в виде ряда
Фурье по егш19 где и принимает целые значения, умноженные на 27г/Т, где Т —
период.
Тот факт, что в линейной теории внешнее воздействие,
пропорциональное eluJt9 вызывает реакцию тоже пропорциональную eluJt во всей системе,
означает, что соответствующие значения f(t) и g(t) в соотношениях (34)
и (35) также пропорциональны величине егшЬ. Отсюда следует, что
(y1 + Е Yr\ е^Г1' - (Vi - Е Yr) еГ^1{
veff _ v Л г=2 ' Л г=2 ' (ia\
1 *? * \ ; 1 * \ г* ( j
( У\ + Е Yr\e™i 1 + i Yx - Е Yr ) e-*"ci«
Отражение и прохождение в сечении А волн, пропорциональных егшЬ9
должны рассчитываться с учетом наличия ветвления в сечении Б, и ад-
митанс трубки АВ представляет собой комплексное число (36), зависящее
от и. Таким образом, использование комплексных адмитансов, зависящих
от частоты, позволяет применять методы, аналогичные методам расчета
электрических цепей.
Равенство (36) для комплексного эффективного адмитанса трубки АВ
в сечении А при наличии ветвления в сечении В может быть представлено
в более удобной для исследования форме:
En + ^itgCwcj-1/)
yiCff = П Г=2 /дг ч • (37)
n + '^EnJtg^cr1*)
Хотя в § 5 в равенства (36) и (37) будут внесены поправки, учитывающие
уменьшение давления за счет вязкости (главным образом путем
добавления мнимой части к величине с]-1), полученное в настоящий момент более
простое выражение (37) для эффективного адмитанса дает возможность
336
Глава 12
выделить многие характерные моменты взаимного влияния ветвлений в
сечениях А и В при не очень большой длине / трубки АВ.
Если длина / мала по сравнению с сх/и, то равенство (37) может быть
записано в виде
N
yf = T,yr- (38)
Это в точности совпадает с тем случаем, как если бы трубка АВ
совершенно отсутствовала бы или, более строго, как если бы она была бы одним из
сосудов, разветвившихся в В. Это согласуется со сделанным ранее выводом
о том, что давление при прохождении волны через ветвление изменяется
непрерывно при условии выполнения (20).
Отметим, что та же «прозрачность» трубки АВ наблюдается при
прохождении волны, частота и которой такова, что выражение шс±11 близко
к 7г, т. е. трубка АВ имеет длину, равную половине длины волны, в этом
случае волна, прошедшая по трубке АВ к сечению В не претерпевает
никаких изменений, т. к. ее фаза изменяется на 7г. Если длина трубки I равна
целому числу полуволн, то также сохранится свойство «прозрачности»,
хотя из-за большого пройденного волной расстояния в силу тех или иных
причин будет происходить ее затухание.
В промежуточном случае, когда значения tjcj"1/, лежащие между
крайними значениями, обеспечивающими «прозрачность», т. е. близкими либо
к 0, либо к 7г, равны ^7г, длина трубки АВ приблизительно равна четверти
длины волны, и ее адмитанс примет значение
Yf = Y?/(fy\ (39)
TV
Таким образом, если Yx больше суммы ^ Уг, тогда Y*^ все еще будет
г=2
N
превышать значение Ух; если же Yx меньше суммы ^ Уг, тогда Y^ будет
г=2
меньше, чем Y1.
Для основной частоты сокращения сердечной мышцы длина аорты
у человека близка к четверти длины волны. При такой частоте эффективный
адмитанс аорты в районе аортального клапана имеет наибольшую
величину. С точки зрения физиологии этот момент достаточно важен, поскольку
в этом случае главная составляющая колебания объемного расхода при про-
5. Изменение амплитуды волны в сосуде
337
хождении через клапан определяется только умеренными по величине
колебаниями избыточного давления, обусловленными сердечными
сокращениями. Отметим также, что длины подвздошных артерий приблизительно
равны четверти длины высокочастотной составляющей пульсовой волны,
что приводит к уменьшению адмитанса и к возникновению
положительного отражения в месте ветвления подвздошных артерий. Более подробно
этот вопрос будет освещен в § 6.
5. Изменение амплитуды волны в сосуде
При изучении сосудистой системы в § 4 предполагалось, что площадь
поперечного сечения каждого отдельного сосуда постоянна и что
амплитуда волны, распространяющейся в нем в направлении возрастания х или
в противоположном направлении, сохраняет постоянное значение. В
данном параграфе, оставаясь в рамках линейной постановки, исследуем
изменение амплитуды при распространении волны в сосуде, которое может
быть обусловлено изменением площади поперечного сечения сосуда либо
диссипацией энергии из-за наличия вязкости.
Постепенное изменение растяжимости стенок сосуда и площади
поперечного сечения при отсутствии диссипации энергии можно исследовать
методами, изложенными в §3, а изменение амплитуды будет
описываться соотношениями, подобными (29). Трубку с меняющимися непрерывно
свойствами представим в виде последовательно соединенных трубок,
свойства которых постоянны и несильно отличаются от соседних. Бегущую
волну в сосуде с переменным поперечным сечением и изменяющейся
растяжимостью будем рассматривать как волну, распространяющуюся со
скоростью с, которая меняется при переходе из одного сосуда с постоянными
свойствами в другой в соответствии с равенством (6). При этом амплитуда
волны изменяется таким образом, что поток энергии Ypl (где адмитанс Y
также меняется от одного сосуда к другому) остается постоянным при
распространении волны в исходном сосуде. Отсутствие диссипации энергии
можно обосновать следующим образом. У каждого из п сосудов адмитанс
отличается от соседних на малую величину, имеющую порядок п"1, тогда
энергия отраженной в каждом соединении волны пропорциональна п-2.
Поскольку соединений п, то суммарная энергия отраженной волны
пропорциональна п(п~2) = п"1. Отсюда видно, что суммарная энергия
отраженной волны стремится к нулю при п —> оо.
Таким образом, в пределе избыточное давление ре и расход J
для бегущей в направлении увеличения х волны могут быть записаны
338
Глава 12
(согласно Taylor, 1965) в виде
Ре =
Y(x)
Y(0)\
1-1/2
/
X
-/■
dx
J = Y(x)Pe,
(40)
где f(t) — форма волны при х = 0, а /с-1 dx — время, необходимое для
о
прохождения волной расстояния от сечения х = 0 до сечения х.
Соответственно для бегущей в сторону убывания х волны имеем
Ре =
Y(x)
Г(0)
1-1/2
9\t +
X
I
-1
dx
J = -Y(x)Pe,
(41)
где g(t) — форма волны при х = 0. В каждом случае получаем, что если
волна распространяется, например, по сосуду с убывающим адмитансом,
то давление возрастает, хотя соответствующий расход будет убывать, а их
произведение остается постоянным.
Естественно задаться вопросом о том, каким образом должны
изменяться скорость с и адмитанс Y с изменением х, чтобы соотношения (40)
и (41) наилучшим образом описывали решения линейных уравнений
движения. Ответим на этот вопрос очень кратко. Линеаризованные уравнения
движения (обобщение уравнений (3) и (4)) могут быть записаны в виде
dt dx ' dt
е = -cY-1 —.
дх
(42)
Оценим точность, например, представлений решений в виде (40) для ре.
Подставим соотношение (40) для избыточного давления в (42), исключив
предварительно расход J. В результате находим, что с достаточной
степенью точности для с и У, а также dY/dx изменения должны быть малы
на расстоянии с/и (это же расстояние уже возникало в исследованиях при
совсем других условиях (20)). Квадрат малого изменения является мерой
расхождения между двумя слагаемыми, составляющими избыточное
давление^.
Для сердечно-сосудистой системы это означает, что между двумя
соседними соединениями сосудов имеет место только небольшое, очень
постепенное изменение свойств (относительное изменение, приходящееся на
5. Изменение амплитуды волны в сосуде
339
единицу длины сосуд, мало), которое может быть изучено на основании
равенств (40) и (41), когда отраженной волной можно пренебречь, а
давление ре и расход J изменяются пропорционально У-1/2 и У1/2
соответственно. Правда, обоснование постепенности изменения следует провести
дополнительно.
Заметим, при другом крайнем случае, который соответствует настолько
малой частоте, что отношение с/и значительно превышает длины сосудов,
правая часть (42) может быть приравнена к нулю. При таких малых
частотах снова приходим к общему положению относительно длины сосуда
и отношения с/и в виде соотношения (20), когда отражение может иметь
место, при ре и J остающихся постоянными. Конечно, между указанными
крайними случаями будут и промежуточные. Например, в аорте происходит
постепенное уменьшение адмитанса вдоль ее довольно значительной
длины. И все же, думается, можно считать, что здесь возможно применение
приближения, принятого в первом крайнем случае.
В любой ветвящейся системе равенство (37) для эффективного
адмитанса в сечении А (где х = — I) трубки АВ, когда учитываются отражения
от места ветвления в сечении В (где х = 0), может быть легко обобщено
на случай постепенного изменения свойств трубки АВ. Это может быть
проведено по той причине, что отношение [Y(x)/Y(0)]~1^2 в волне,
бегущей вперед (40), и в отраженной волне (41) сокращается в выражении (35).
Действительно, из равенства Y^ отношению расхода J в сечении А к
избыточному давлению для пульсовой волны, представленной в виде суммы (40)
или (41), получаем
\^eff _ \sA /(* + сАв0 ~ 9(t ~ сАв0 (Лп\
1 ~~ ! 777"!—-1 ;\ ,—и ^ГТТ' \*6'
f{t + cAlBl)+g{t-cAlBl)
где У^4 — адмитанс Y(—l) трубки АВ в сечении А и с~АВ — величина,
обратная средней скорости распространения волны в АВ, определяемая
соотношением
о
сав1= /c_1dx. (44)
-/
С другой стороны, в первом соотношении в (34), определяющем
отношение отраженной волны g(t) к падающей f(t) при х = 0 (соединение
в сечении Б), адмитанс Yx принимается равным адмитансу Yf* = Y(0)
340
Глава 12
трубки АВ в сечении В. Тогда получаем соотношение, подобное (37),
£ Yr + iY* tg{uc?Bl)
Yf = Yf Г=2 ,N v • (45)
Это соотношение можно использовать для расчета свойств ветвящейся
системы, состоящей из трубок, вдоль которых их свойства меняются
постепенно, аналогично тому, как используется равенство (37) для трубок с
постоянными свойствами. Например, в случае резонанса, когда длина
сосуда равна четверти длины волны (условия этого резонанса можно записать
в виде шсд1^ = ^7г), резонансное значение эффективного адмитанса
(подобно (39)) равно YAYB/(Y: Yr).
4=2 '
Другими факторами ослабления амплитуды пульсовой волны
являются вязкость крови и вязкоэластичные свойства стенок сосуда. Рассмотрим
влияние вязкости крови и кратко опишем способ учета вязкоэластичных
свойств стенки трубы.
Скорость деформации сдвига крови в артериях при нормальных
физиологических условиях не ниже 102 с-1. Хотя эксперименты по
определению вязкости крови указывают на совершенно аномальные свойства
крови при значительно меньших скоростях деформации сдвига (при которых
наблюдается агрегация (склеивание) клеток крови), из экспериментов,
проводимых при скоростях деформации сдвига порядка 102 с-1, следует, что
с достаточной степенью точности кровь может рассматриваться как
ньютоновская жидкость с вязкостью /i, приблизительно равной 4 • 10~3 Нс/м2
(что в два с половиной раза превышает вязкость плазмы и в четыре раза
вязкость воды). Соответствующая кинематическая вязкость или
коэффициент диффузии импульса равен
^ = ^ = 4.1СГ6м2/с. (46)
Р
При пульсации крови в артерии вязкость приводит к такому профилю
скорости в сечении трубы, когда отсутствует проскальзывание жидкости
на стенке и распределение скорости по сечению становится
неравномерным. Перенос импульса с коэффициентом диффузии v приводит к обмену
количества движения между кровью, находящейся в состоянии покоя при
контакте со стенками сосуда и движущейся с постоянной скоростью вдали
5. Изменение амплитуды волны в сосуде
341
от них. Область, где жидкость движется с промежуточными скоростями,
называется пограничным слоем.
Это является еще одной причиной, по которой распространение
пульсовой волны в рамках линейной теории лучше всего исследовать в
зависимости от каждой гармоники волны, пропорциональной eluJt. Пограничный
слой на стенке, определяемый одной гармоникой, имеет толщину,
пропорциональную (v/u)1/2, которая определяется как расстояние на которое про-
диффундирует импульс при коэффициенте диффузии v на половину
периода 7г/ал Распределение скорости в пограничном слое, найденное Стоксом
(см. Rosenhead, 1964, глава 7), определяется при условии, что радиус
трубки а (для некруговых сечений радиус кривизны стенок) велик по сравнению
с толщиной пограничного слоя, т. е. при условии, что параметр Уомерсли
а = а{и/и)1/2 (47)
достаточно велик (на практике достаточно, чтобы он был больше 4).
Типичное значение параметра Уомерсли для большинства
млекопитающих удовлетворяет этому условию. Так, при частоте сердечных
сокращений, типичной для каждого вида, параметр Уомерсли а равен 20 для волов,
15 для человека, 10 для собак, 4 для кроликов (McDonald, 1960, гл. 5). Как
следует из (47), с ростом частоты параметр а возрастает. Однако в
небольших артериях параметр Уомерсли а принимает небольшие значения (так,
этот параметр равен 3 для бедренной артерии человека) там, где вязкость
крови играет существенную роль, при этом пограничный слой занимает
практически весь просвет сосуда.
Рассмотрим случай больших значений параметра Уомерсли а (а
затем укажем, что следует изменить в анализе, чтобы исследовать процесс
при малых значения параметра Уомерсли а). Скорость в пограничном
слое Стокса, возникшем при прохождении пульсовой волны с
осциллирующим градиентом дре/дх избыточного давления, пропорционального
величине eluJt, равна
и = (piuj)-l(-dpjdx){\ - exp[-z(zu;/^)1/2]}, (48)
где z — расстояние от стенки. На больших расстояниях от стенки z
скорость и стремится к значению
u = {piu)-l{-dpjdx), (49)
которое можно рассматривать как чисто реактивный ответ (т. е.
величина скорости обусловливается только инерцией и запаздывание по фазе
равно 90°) на действующую силу (—дре/дх), приходящуюся на единицу
342
Глава 12
объема. При малых z (в области вязкого пограничного слоя) в
выражение для скорости (48) входит не только чисто мнимый множитель при
(—дре/дх)9 но также и реальный положительный множитель, который
можно рассматривать как резистивный ответ, обусловленный силами
сопротивления (запаздывание по фазе отсутствует), на действующую силу,
приходящуюся на единицу объема.
Расход J, связанный с равномерным по сечению площадью А
распределением скорости (49), удовлетворяет уравнению
| = Л^ = -Л,-'^. (50)
Такое уравнение уже было ранее записано (см. первое уравнение в (42)).
Эффекты вязкости при больших значениях параметра а проявляются в
довольно тонком пограничном слое Стокса, тогда с учетом (48)
уравнение (50) принимает вид
где s — периметр поперечного сечения. Здесь второе слагаемое в фигурных
скобках представляет собой «дефект» расхода в пограничном слое,
обусловленный как ростом влияния инерции, так и эффектом наличия сил
сопротивления. Для кругового поперечного сечения радиусом а отношение A/s
равно ~а9 так что множитель в правой части (51) больше множителя в (50)
{1-2Г1/2а~1} (52)
раз.
Хотя наличие пограничного слоя приводит к изменению множителя
в правой части первого уравнения в (42), вид второго уравнения не
изменяется. Оно просто определяет скорость изменения объема жидкости,
приходящегося на единицу длины, а именно величины dA/dt, которую можно
записать как AD dpe/dt, равную градиенту расхода, взятому с
противоположным знаком. Отметим, что из (6) и (22) следует, что произведение AD
равно c~lY.
Ранее уже было показано, что если скорость с и адмитанс Y
изменяются достаточно постепенно, частные решения уравнений (42) можно
записать в виде (40) и (41) и, сложив их, получить общее решение. В
рассматриваемом случае в первом уравнении (42) правая часть имеет множитель (52),
а второе уравнение (42) не изменилось. Тогда можно представить, что
уравнения остались по форме те же, а скорость с и адмитанс Y умножены на
5. Изменение амплитуды волны в сосуде
343
корень квадратный из выражения (52). И решения уравнений в случае учета
вязкости будут иметь вид (40) и (41), только в них как скорость с, так и ад-
митанс Y необходимо умножить на корень квадратный из выражения (52).
Действительно, малые изменения адмитанса Y приводят к малым
изменениям самих решений, а вот малые изменения величины с"1, которая
входит в аргумент функций f(t) или g(t) (здесь имеющих множитель e2u;t),
могут привести к значительным изменениям. Воспользовавшись
приближенным представлением выражения, обратного квадратному корню из (52),
для больших значений а можем в решениях (40) и (41) величину с"1
заменить на
С-1(1 + Г1/2а-1) = с-1
1 +
W2,
(53)
И тогда, например, множитель
ехр
ги
-1
dx
(54)
в волне (40), распространяющейся в направлении возрастания х, после
такой замены принимает вид
ехр
iujt — iu
(55)
В (55) второе слагаемое определяет уменьшение фазы скорости,
обусловленное возрастанием влияния инерции, а третье слагаемое определяет
уменьшение амплитуды за счет влияния сил сопротивления. Относительно
небольшое уменьшение фазы будет наибольшим для основной гармоники
и меньшим для гармоник более высокого порядка при больших
значениях а. С другой стороны, скорость uj/cay/2 экспоненциального уменьшения
амплитуды с ростом расстояния будет наименьшей для основной
гармоники (в силу равенства (47)) и будет возрастать как о;1/2 для более высоких
гармоник пульсации крови.
Все результаты, полученные ранее в этом параграфе при отсутствии
вязкости, могут быть уточнены в связи с учетом наличия тонкого
пограничного слоя, если величину с-1 (см. уравнение (44)) заменить на (53).
Тот факт, что с~^в представляет собой теперь комплексное число,
приводит, конечно, к более заостренному пику при резонансе, когда длина сосуда
кратна четверти длины волны.
344
Глава 12
Подобные результаты были получены Уомерсли (Womersley, 1957) при
произвольном значении его параметра а на основе анализа, начинающегося
с того, что скорость (48) заменяется выражением, содержащим бесселевы
функции. В пределе при малых а распределение скорости по сечению
становится параболическим (течение Пуазейля) и (51) заменяется равенством
4dJ 8v T . 1 дре . „ч
ш+^^-Ар •£• (56)
где 4/3 является инерционным множителем, учитывающим превышение
кинетической энергии при течении Пуазейля по сравнению с течением,
когда скорость распределена равномерно по сечению при одном и том же
расходе J. Величина (8и/а2) представляет собой обычное гидродинамическое
сопротивление Пуазейля. При использовании равенства (56) величину с"1
следует заменить на
(57)
и У"1 следует умножить на такое же выражение. Фактически
использование выражения (53) при а > 4 и выражения (57) при а < 4 дает решения,
хорошо согласующиеся с общим решением Уомерсли.
Завершим этот параграф кратким обсуждением вязкоупругих свойств
стенок артерий. Не будем углубляться в проблемы биомеханики твердого
тела более, чем это было сделано в §2, заметим только, что учет вязко-
упругих свойств приводит к представлению растяжимости D в виде
комплексного числа, зависящего от частоты. Измерения Бергеля (Bergel, 1961)
в артериях показали, что аргумент растяжимости равен от 5° до 10°. Тогда
из равенства (6) следует, что с-1 необходимо умножить на новый
множитель — с отрицательной мнимой частью, как в (53), что приводит к
затуханию волны, подобному (55). А при учете и вязкости крови новое
выражение, которым заменяется с-1, следует умножить еще и на множитель
в выражении (53).
6. Сравнение с экспериментальными данными
В данном разделе объясним некоторые экспериментальные
результаты с точки зрения теоретических исследований распространения пульсовой
волны с учетом вязкости крови и вязкоупругих свойств стенок сосуда и без
их учета.
6. Сравнение с экспериментальными данными 345
Профессор М. Дж. Тейлор проводил теоретические и
экспериментальные исследования, касающиеся входного адмитанса аорты
непосредственно у аортального клапана. Он указал (Taylor, 1966) на возможность расчета
входного адмитанса для сложной ветвящейся системы, какой является
артериальное дерево, путем последовательного применения алгоритма расчета
адмитанса по формулам (37) или (45), когда в качестве исходного
эффективного адмитанса выбирается дистальный. При постепенном
продвижении к началу сосудистой системы, выполняя на каждом этапе расчетов
эффективного адмитанса в конечном итоге им был найден входной адмитанс.
Характер зависимости входного адмитанса от частоты и представляет
для исследователей несомненный интерес. Это частично объясняется тем,
что основная частота биений сердца не является постоянной величиной,
а особенно тем, что амплитуды гармоник более высокого порядка могут
достигать довольно больших значений. В конце § 4 было отмечено, что длина
аорты приблизительно в четыре раза превосходит длину волны основной
гармоники, что приводит к резонансу, а следовательно, и к возрастанию
входного адмитанса. Что касается гармоник более высокого порядка, то
возникновение связанных с ними резонансов приводит к уменьшению
эффективных адмитансов, к примеру, в подвздошных артериях и может
приводить к увеличению эффективного адмитанса аорты.
В работе Тейлора отмечалось, что большое разнообразие длин труб
между двумя последовательными ветвлениями в артериальном дереве
должно было бы сгладить все резонансные пики, не относящиеся к
основной частоте, и допустить согласованный с этим высокий уровень входного
адмитанса, чтобы он сохранился и при больших частотах. Тейлор
продемонстрировал это (Taylor, 1966) на многочисленных моделях ветвящейся
системы, в частности такой, которая представлена на рисунке 3,
включающей семь ветвлений, когда суммарная площадь поперечных сечений
отходящих сосудов меньше площади поперечного сечения материнского
сосуда. При этом скорость распространения волны увеличивалась. Сосудистая
система заканчивалась сетью артериол с их общим заданным
периферическим сопротивлением.
На рисунке 4 представлена зависимость входного импеданса этой
системы (величины, обратной адмитансу Z = У"1) от частоты. Понятие
импеданса является важной физиологической характеристикой,
определяющей требуемое избыточное давление, которое необходимо создать
сердечной мышце для обеспечения заданного расхода крови при
определенных частотах. Отметим, что очень большой импеданс, соответствующий
346
Глава 12
случаю и = О, представляет собой чистое сопротивление системы
проходящему по ней осредненному потоку. Такое большое значение импеданса при
и — О определяется тем, что при типичном значении основной частоты
сердечных сокращений имеет место очень небольшой минимум импеданса
(резонансный пик адмитанса). Отметим также, что при всех частотах, больших
или достаточно близких к ней (все частоты сердечных сокращений),
импеданс остается меньшим по сравнению с его значением при нулевой частоте.
-, 1
II \ , 1 1
Hri—utf
i "-+
hV
g
—i—i—i
Г 1
i * '
—п
1 ~l—
ilH 1 I
и \
1 1 *
1 T&
II '1
■ t
1И l
J_
1
— I —
Рис. З. Схематичное представление рассматриваемой в данном параграфе
ветвящейся системы (рис. 1 из работы Taylor, 1966)
Такой низкий уровень значений импеданса для всех частот, за
исключением самых низких, является характерной чертой данной модели.
Тейлор (Taylor, 1964, 1965) обратил внимание на тот факт, что для реальных
артериальных систем подобное явление имеет место. Им было найдено,
что работа, совершаемая сердечной мышцей для создания пульсирующего
6. Сравнение с экспериментальными данными 347
6 L
о
2
4
3
Л 2
5
а 1
0)
О
7Г
2тг
Зтг
47Г
Рис. 4. Зависимость модуля и фазы входного импеданса модели артериального
дерева от частоты (рис. 9 из работы Taylor, 1966)
348
Глава 12
кровотока, на 5 %-15 % превышает работу, которую ей пришлось бы
совершить для создания среднего объемного потока крови, движущегося с
постоянной скоростью. С другой стороны, при исследовании упрощенных
моделей, в которых, естественно, не учтено все многообразие длин сосудов
(рис. 3), получены значительно большие по величине резонансные пики
импеданса при частотах, близких к основной частоте сердечных сокращений.
На рисунке 4 представлена зависимость модуля и фазового угла
импеданса, равного фазовому углу адмитанса У, взятому с противоположным
знаком, от частоты. Положительные величины фазового угла адмитанса
возрастают с ростом частоты и (точно так же, как и емкость С в
выражении для адмитанса Y = Ciu) при низких частотах. Это отражает
такое свойство аорты, как ее емкость — накопительное свойство растяжимой
эластичной камеры (Windkessel) (или гофрированной оболочки). Для
вычисления и дальнейшего исследования эффективного входного адмитанса
в выражение (37) подставим в качестве Yx аортальный адмитанс. Величи-
N
на его значительно больше, чем комбинация дистальных адмитансов ^ Уг,
поэтому при низких частотах и положительная мнимая часть эффективного
адмитанса Y*ff будет пропорциональна частоте и9 а при резонансной
четвертой частоте мнимая часть приближается к нулю. Однако, как видно из
рисунка 4, при больших частотах размах значений фазового угла входного
импеданса значительно менее выражен.
Сплошной линией на рисунке 4 изображены расчетные зависимости
модуля и фазы импеданса при течении вязкой крови в жесткой трубке (т. е.
не учитываются вязкоупругие свойства стенок артерий). Прерывистыми
линиями изображены расчетные кривые для входного импеданса с учетом
свойств материала стенок. Учет свойств материала стенок сосуда
проводился введением 10° запаздывания по фазе растяжимости стенок. Как вытекает
из § 5, такой учет приводит к небольшим изменениям при низких частотах
и к существенным — при больших, что сглаживает колебания импеданса
после достижения им первого ярко выраженного минимума.
Интересно рассмотреть форму пульсовой волны, зарегистрированной
одновременно в различных участках артериального дерева. На рисунке 5
представлены кривые давления в различных участках аорты собаки (рис.
заимствован из работы McDonald, 1960). Заметим, что Ремингтон и Вуд
(Remington, Wood, 1956) получили аналогичные результаты для артерий
человека. Наиболее удивительным является факт возрастания амплитуды
давления по мере удаления от сердца.
6. Сравнение с экспериментальными данными 349
Так, среднее значение квадратного корня из вариации давления
увеличивается на 50 % при распространении волны от проксимальной
восходящей аорты (кривая 1) до дистальной брюшной аорты (кривая 4), и
затем происходит дальнейший рост в дочерних сосудах, таких как большая
подкожная артерия. Такое поведение амплитуды объясняется (Taylor, 1965)
главным образом за счет уменьшения периферийного адмитанса.
Постепенное уменьшение адмитанса (связанное с постепенным уменьшением как
растяжимости сосуда, так и площади его поперечного сечения при
удалении от сердца), как было показано в § 5, приводит к постепенному
увеличению амплитуды давления и соответствующему уменьшению амплитуды
скорости потока крови. На рисунке 6 (рисунок также заимствован из работы
McDonald, 1960) показаны кривые скорости потока, измеренные в
различных точках кровеносной системы, амплитуды которых, как и
предсказывали теоретические исследования, уменьшаются. Скачкообразное изменение
адмитанса при переходе потока через место соединения сосудов приводит
к увеличению амплитуды давления в дочерних сосудах, что объясняется
с точки зрения теории на основании второго уравнения (34).
Еще одной интересной особенностью формы волны, представленной
на рисунке 5, является небольшая по величине иррегулярность
проксимального профиля, которая довольно быстро сглаживается при перемещении
волны дальше от сердца. Согласно теоретическим исследованиям,
проведенным в §5, такой иррегулярный «зубец» может появиться на профиле
волны за счет влияния вязкости крови, возможно совместного с влиянием
вязкоупругости стенок сосуда.
Кроме того, следует отметить появление на периферии диастоличе-
ского пика давления, т.е. небольшого по величине второстепенного
максимума через половину периода после основного (систолического) пика.
При исследовании процесса распространения пульсовой волны с
использованием анализа Фурье необходимо определить сдвиг фаз второстепенного
максимума относительно основного. С точки зрения линейной теории
возникновение подобного запаздывания можно объяснить несколькими
причинами. Например, параметр Уомерсли а для основной частоты меньше,
чем у других гармоник волны, тогда при учете вязкости фазовая скорость
(определяемая соотношением (55)) основной гармоники будет меньше
других фазовых скоростей. Кроме того, форма проходящей волны
определяется эффективными адмитансами (вычисляемыми по формулам (37)),
которые и сами по себе являются комплексными числами, зависящими от
частоты.
350
Глава 12
Рис. 5. Форма пульсовой волны давления в разных участках артериального русла
(рис. 12.4 из работы McDonald, 1960)
Казалось бы, что уже можно и не искать объяснение изменения
формы волны за счет нелинейных эффектов. Тем не менее и нелинейные
эффекты влияют на нее аналогичным образом. В рамках нелинейной теории
участки волны, соответствующие нарастанию давления, распространяются
с несколько большей скоростью (главным образом за счет того, что при
нарастании давления скорость крови увеличивается, что приводит к
увеличению скорости распространения этой части волны), чем участки,
соответствующие падению давления (в это время скорость крови уменьшается,
а следовательно, уменьшается и скорость распространения этой части
волны). В результате энергия переносится не с основной гармоникой волны,
а с ее первой гармоникой с запаздыванием по фазе на 90°, что и объясняет
вышеуказанный эффект.
В работе Линга и Атабека (Ling, Atabek, 1972) проведен подробный
обзор нелинейных теорий распространения пульсовых волн, в которых учте-
6. Сравнение с экспериментальными данными 351
g 100
i 80
60 h
140 h
о 100
^ 60
20
-20 h
аорта
восходящая брюшная
аорта грудная а°рта бедренная
аорта артерия
большая
подкожная
артерия
Рис. 6. Сопоставление кривых давления и скорости крови в разных участках
артериального русла. Амплитуда давления уменьшается, а амплитуда скорости
возрастает при распространении волн в направлении к периферии (рис. 12.6 из работы
McDonald, 1960)
но распределение скорости по поперечному сечению, обусловленное
вязкостью крови. Авторы сравнили результаты теоретических исследований
с профилями скоростей, измеренными в модели аорты собаки. И это
довольно хорошее совпадение экспериментальных и теоретических данных
представлено на рисунке 7.
При обсуждении столь хорошего совпадения экспериментальных и
теоретических данных авторы, возможно, переоценили нелинейный аспект
вычислений. Они сравнили данные наблюдений с теоретическими расчетами
только для частного случая линейной постановки, что действительно дало
довольно плохое совпадение результатов, полученных на основании
линейной теории, с экспериментами. И не учли тот факт, что достаточно было
слегка изменить теоретическую модель течения, оставаясь в рамках
линейной теории, чтобы согласие с экспериментальными данными значительно
улучшилось бы.
352
Глава 12
1,0
zLO
Рис. 7. Экспериментальные и теоретические профили продольной скорости w в
модели аорты собаки в различные моменты времени одного цикла пульсовой волны.
Измерения проведены Лингом и Атабеком (Ling, Atabek, 1972). Период пульсовой
волны равен 0,8 с. Максимальное значение скорости wm = 0,77 м/с. Теоретические
кривые изображены сплошными линиями, экспериментальные данные — точками
Для пояснения отметим, что учет вязкости крови в § 5 применялся
только для случая распространения синусоидальных компонент пульсовой
волны. Такой подход корректен для самого волнового процесса и
составляющих его гармоник, однако применение подобного подхода для
осредненного потока требует дополнительных обоснований. Довольно
неоднозначный подход к изучению осредненного потока применялся в большинстве
ранних теорий распространения пульсовых волн. Действительно, в этих
теориях допускалась серьезная неточность, отмеченная в главе 10, §3,
которая возникает, если не учитывать входной эффект для осредненного
потока и если поток в трубки считать пуазейлевским. И именно такая
теоретическая модель была принята Атабеком и Лингом для сравнения
экспериментальных данных и теоретических результатов. Однако если учесть,
что длина аорты равна длине входного участка для осредненного потока
6. Сравнение с экспериментальными данными 353
и что толщина пограничного слоя в нем незначительно превышает
толщину пограничного слоя, определяемого компонентами ряда Фурье волнового
профиля, то, оставаясь в рамках линейной теории, можно получить
практически столь же хорошее совпадение теоретических расчетов с
экспериментальными данными, представленными на рисунке 7, какое получили
Линг и Атабек, используя вышеуказанную нелинейную постановку
задачи. С другой стороны, Анликером (Anliker, 1972) были приведены
интересные данные, касающиеся нелинейных эффектов. В нормальных
физиологических условиях их влияние практически незаметно. Однако влияние
таких эффектов становится весьма существенным при некоторых патало-
гиях, таких как недостаточность аортального клапана, которые могут
привести к гипертрофии сердца и, как следствие, к увеличению объема крови,
выброшенной в кровяное русло за одно сердечное сокращение.
Литература
[1] Anliker, M. 1972. In Biomechanics: its Foundations and Objectives (ed.
Fung, Y. C, Perrone, N. & Anliker, M.), Prentice-Hall, Englewood Cliffs,
N.J.
[2] Bergel, D.H. 1961. J. Physiol. 156, 458-469.
[3] Ling, S.C. & Atabek, H.B. 1972. J. Fluid Mech. 55, 493-511.
[4] Love, A. E. H. 1927. A Treatise on the Mathematical Theory of Elasticity,
4th Edition, Cambridge University Press.
[5] McDonald, D. A. 1960. Blood Flow in Arteries, Edward Arnold, London.
[6] Remington, J. W. & Wood, E.H. 1956. J. Appl. Physiol. 9, 433-442.
[7] Rosenhead, L. (ed.) 1964. Laminar Boundary Layers, Oxford University
Press.
[8] Taylor, M. G. 1964. In Pulsatile Blood Flow (ed. Attinger, E. O.), McGraw-
Hill, New York.
[9] Taylor, M.G. 1965. Phys. Med. Biol. 10, 539-550.
[10] Taylor, M.G. 1966. Biophys. J. 6, 29-51.
[11] Womersley, J.R. 1957. Phys. Med. Biol. 2, 178-187.
Глава 13
Кровоток и заболевание артерий
1. Турбулентность в потоке крови
В этой главе проиллюстрируем применение методов внутренней
биогидродинамики к исследованию связи некоторых особенностей потока
крови с начальной стадией заболевания артерий. В двух последних параграфах
будет более подробно освещена проблема начинающейся атеросклеротиче-
ской болезни, о которой уже говорилось в общих чертах в §4 главы 10.
Вопросы, которые поднимаются в § 8 и § 9, предварительно обсуждались
в главе 12. Начнем с проблем, обсуждавшихся в главе 10, § 10,
касающихся турбулентности в кровотоке. В последних параграфах данной главы
более подробно обсудим диагностику паталогических процессов, о начале
которых свидетельствует наличие турбулентности в кровотоке (например,
процессов, связанных с церебральной геморрагией).
В работе Сида и Вуда (Seed, Wood, 1971) отмечается наличие
коротких всплесков (см. рис. 1, кровоток в аорте собаки) турбулентных
флуктуации скорости на заднем фронте пульсовой волны кровотока, когда скорость
крови уменьшается. Их исследования затем были продолжены и
расширены в работах Нерема и Сида (Nerem, Seed, 1972), а также Паркера и
Сида (Parker, Seed, 1973). Ими были развиты технические методы получения
с использованием датчика пленочного анемометра «среднего по ансамблю»
значения скорости на основании около 500 пульсирующих профилей
скорости в зависимости от времени. За начальный момент принимался момент
достижения возрастающей с течением времени скорости некоторого
положительного значения. Турбулентные компоненты скорости при каждой
пульсации определялись по отличию скорости от ее среднего по ансамблю
значения.
В аорте млекопитающих различают три режима течения крови:
(i) ламинарный — линии тока одинаковы при всех пульсациях, и,
следовательно, в этом случае «турбулентные» составляющие скорости
пренебрежимо малы;
356
Глава 13
ECG
\
I 1 1 1
0 0,5 1,0 1,5
Время (с)
Рис. 1. Волна скорости в аорте собаки, полученная на основании ЭКГ. В начальной
фазе сокращения сердечной мышцы возникает турбулентность (Seed, Wood, 1971)
(ii) переходный — имеют место существующие в течение короткого
времени значительные по размаху флуктуации, которые практически сразу
сменяются достаточно длительными по времени пульсовыми
колебаниями средней скорости;
(iii) турбулентный — в потоке возникают длительные по времени и
достаточно большие по размаху флуктуации скорости со
среднеквадратичным значением, составляющим 10% от максимальной скорости,
которые распространяются вниз по потоку на диастоле.
Эксперименты, проведенные при различных условиях на добровольцах и на
животных, показали, что о характере течения можно судить по числу
Рейнольдса Д, рассчитанному по максимальному значению скорости и
диаметру сосуда. Ламинарный режим течения имеет место, когда число
Рейнольдса не превышает 4000; турбулентное течение возникает при числах
Рейнольдса, превышающих 8000; а промежуточный режим — при числах
Рейнольдса порядка 6000. Числа Рейнольдса в аорте человека изменяются
во всем указанном диапазоне, и, значит, в аорте имеют место все три
режима течения. У собак числа Рейнольдса меньше по величине, кровоток,
как правило, ламинарный, однако возникают и переходные течения (рис. 1).
Числа Рейнольдса у лошади более высокие, чем у человека, и поток в аорте,
как правило, турбулентный.
1
I
и
J.UUH
50 J
°1
1. Турбулентность в потоке крови
357
На режим течения оказывает влияние также и параметр Уомерсли а =
= aiyjjv)1!2 (равенство (47), глава 12). С ростом а (например, когда
возрастает частота сердечного пульса ш) критическое число Рейнольдса Rc
для переходного режима немного возрастает. Это связано с тем, что, когда
значения Rc при любом течении в сосуде, отличном от течения Пуазей-
ля, значительно превышает 2000 (критическое значение числа Рейнольдса
для течения Пуазейля), завихренность развивается в тонком пограничном
слое, и именно число Рейнольдса, рассчитанное по толщине пограничного
слоя, определяет переходный режим. Эта точка зрения уже высказывалась
в § 2 главы 10 для стационарных вторичных потоков. Аналогичные
рассуждения можно применить к пограничному слою Стокса, толщина которого
пропорциональна величине (^/и;)1/2, или к тонкому пограничному слою на
входном участке. Нерем и Сид (Nerem, Seed, 1972), а также Паркер и Сид
(Parker, Seed, 1973) получили, что значения числа Рейнольдса,
рассчитанного по толщине пограничного слоя (v/u)1/2, для переходного режима имеют
порядок 100. Кроме того, Коллинз (Collins, 1963), проводя наблюдения над
визуализированным потоком в пограничном слое Стокса на дне водного
русла при течении с небольшими поверхностными волнами, получил такое
же значение переходного числа Рейнольдса.
Механизм возмущения ламинарного потока состоит в быстром росте
небольших возмущений, которые всегда в силу тех или иных причин
присутствуют в потоке крови. Возмущения нарастают по величине, как только
поток начинает замедляться и на профиле скоростей появляется точка
перегиба. В 1970 г. Лайтхилл (Lighthill, 1970) подвел итог проводившимся
исследованиям и обосновал эффективность указанного механизма
возникновения турбулентности при не очень низких числах Рейнольдса и
механизма генерирования крупномасштабной турбулентности далеко от области
возникновения перегиба в профиле скоростей.
Описанная выше турбулентность, которая возникает в аорте
человека в период уменьшения скорости (на диастоле) перед моментом закрытия
аортального клапана, как правило, плохо прослушивается с помощью
стетоскопа. Хотелось бы знать, почему это происходит, ведь турбулентность,
которая возникает за счет образования струйного течения при стенозе
аортального клапана (клапан полностью не раскрывается), прослушивается
совершенно отчетливо. Паркер и Сид (Parker, Seed, 1973) при измерении
скорости течения крови с помощью катетера для определения
турбулентности за суженным аортальным клапаном получили, что сренеквадратичная
скорость в этой области в два раза больше, чем скорость турбулентного те-
358
Глава 13
чения при здоровом клапане. Соответствующие амплитуды давления могут
в четыре раза превышать обычные значения, в результате создается шум, на
12 дБ превышающий шум при обычном течении, при условии, что в обоих
случаях действуют одинаковые механизмы, ослабляющие звук при его
прохождении через стенки артерии и через мягкие ткани, окружающие сосуд.
Наличие клапана нарушает непрерывность характеристик материала стенок
с разных сторон от него, что, возможно, способствует прохождению
флуктуации давления, а колебание струи, обусловленное периодическим
функционированием стенозированного клапана, приводит к усилению шума до
20 дБ, что и объясняет различие между ясно слышимым турбулентным
течением и не различимым на слух.
Ранее измерение турбулентности в кровотоке было довольно
затруднительным и требовало большого мастерства. В настоящее время находят
широкое применение пленочные анемометры. Методы измерения
турбулентности с помощью пленочного анемометра в живом организме (in vivo) дадут
возможность получить обширный материал о распределении турбулентных
областей в здоровых и пораженных сосудах.
2. Течение в области постстенотического расширения
сосуда
Неблагоприятное влияние турбулентности на течение в области,
расположенной за сужением сосуда, впервые было продемонстрировано Роуч
(Roach, 1963a, 1963b) при изучении избыточного расширения артерии,
наблюдающегося сразу за стенозированным аортальным клапаном, или
клапаном легочной артерии, или за другими суженными участками артерий
(также называемыми стенозированными, даже если это сужение не связано со
стенозом клапана). Такое постстенотическое расширение рассматривалось
как «гемодинамический парадокс». Обширные экспериментальные
исследования стационарных и пульсирующих потоков в жестких и эластичных
сосудах, в изолированных аортах и аортах в живом организме, проведенные
Де Фризом и ван дер Бергом (de Vries, van den Berg, 1958), подтвердили тот
факт, что давление в области за суженным сосудом в норме меньше, чем
проксимальное давление, и никогда его не превышает. Следовательно,
расширение сосуда за областью сужения должно обусловливаться локальным
увеличением растяжимости стенок сосуда.
Роуч (Roach, 1963a) провела более 50 экспериментов. Измерения
проводились на бедренной и /или сонной артериях в живом организме, сосуды
2. Течение в области постстенотического расширения сосуда 359
пережимались наложением нейлонных бандажей. Как оказалось,
постстенозное расширение образовывалось в том и только в том случае, когда за
сужением турбулентное течение становилось слышимым, что
обнаруживалось с помощью микрофона, приложенного к сосуду. Турбулентное
течение прослушивалось в 39 случаях пережатия сосудов, когда просвет сосуда
уменьшался от 23 % до 76 %. За сужением диаметр сосуда в среднем
увеличивался на 20 %. Частота звуковых сигналов, поступавших с микрофона,
лежала в пределах от 50 до 500 Гц, при этом соответствующие сигналы
воспринимались как «вибрация» или «гул».
А в 7 артериях постстенозного расширения обнаружено не было.
Степень стеноза этих артерий была таковой, что за сужением не
обнаруживались признаки наличия турбулентности. Кроме того, в 4 из них при
сужении просвета более чем на 85 % и при последующем образовании медленно
текущей тонкой струи вновь не было обнаружено с помощью микрофона
каких бы то ни было признаков возникновения турбулентности.
Роуч (Roach, 1963b) экспериментально изучала причины
возникновения турбулентности и ее влияние на кровоток. Автором было обнаружено,
что значительные изменения эластичных свойств сегмента артерии
происходят тогда и только тогда, когда в сосуде или возникает турбулентное
течение, или же в нем создаются с помощью наложенного бандажа
вибрации с теми же частотами, что и турбулентное течение. Природа изменений
была такой, что, вероятнее всего, именно эластиновые (в отличие от
коллагена) волокна изменяли свои упругие свойства. То есть возникновение
высокочастотных флуктуации давления и, как следствие, высокочастотных
напряжений на стенке приводило к явлению, напоминающему в общих
чертах усталость материала. Кроме того, флуктуации могут ослаблять связи
в матриксе, состоящем из коллагеновых и эластиновых волокон. В более
поздней работе Роуч (Roach, 1970) было показано, что при нормальных
условиях восстановительные процессы в артериях собаки приводят к
восстановлению диаметра и упругих свойств расширенной артерии сразу же
после того, как сужение сосуда устранено.
Ответ на вопрос о том, может ли низкий уровень турбулентности,
описанный в § 1, связанный не с образованием струи за сужением артерии,
а с замедлением потока при высоких числах Рейнольдса в грудной
аорте, оказать влияние на упругие свойства стенки аорты, в настоящее время
еще окончательно не получен. Пока еще неизвестно, какое влияние
оказывают возникающие флуктуации давления (о которых говорилось в § 1), на
долю которых приходится до четверти всех возмущений, воздействующих
360
Глава 13
на течение за сужением, достаточны ли они для оказания хотя бы слабого
влияния на поток. Но следует иметь в виду, что такое влияние возможно.
3. Аневризмы сосудов головного мозга
В данном параграфе опишем результаты исследований Фергюсона
(Ferguson, 1972), занимавшегося изучением ослабления стенок артерий под
воздействием флуктуации давления, обусловленных турбулентностью
потока (см. § 2). Такое воздействие становится патологическим фактором,
приводящим к серьезным последствиям. В частности, к возникновению
мешковидной аневризмы, которая, как правило, образуется (рис. 2) в месте
бифуркации артерий, чаще всего, артерий головного мозга.
угол бифуркации
разветвившийся
сосуд
вершина
бифуркации
угол отхождения
сосуда
основной
сосудистый ствол
дно (купол)
полость
мешок (тело)
шейка аневризмы
Рис. 2. Схематичное изображение типичной мешковидной аневризмы (рис. 1 в
работе Ferguson, 1972)
Исследования шумов в артериях с помощью микрофона были
предприняты Фергюсоном (Ferguson, 1972) для установления связи
турбулентности с мешковидными аневризмами сосудов головного мозга. У некоторых
пациентов при трепанации черепа удалось записать звуковые сигналы,
исходящие от стенок артерий и аневризм. У четырех пациентов (аневризмы
отсутствовали) какие-либо звуковые сигналы при прохождении потока че-
3. Аневризмы сосудов головного мозга
361
рез бифуркации или другие участки артерий записать не удалось. А у 12 из
19 больных с аневризмами сосудов оказалось возможным записать шумы
с частотой от 200 до 700 Гц, причем распределение амплитуды звуковых
сигналов ясно указывало на то, что источник шума находился в самой
аневризме. У 19 пациентов шум отсутствовал в тех случаях, когда среднее
артериальное давление под действием лекарственных препаратов
уменьшалось до очень низких значений (около 60 мб или ниже), в случае же более
близкого к норме низкого давления — в пределах 100 мб — шум
прослушивался достаточно отчетливо.
Фергюсон (Ferguson, 1972) провел серию модельных экспериментов
с целью объяснения полученных им ранее результатов и выяснения роли
турбулентности в разрыве аневризмы. Тщательные наблюдения дали
возможность установить характер потока при прохождении бифуркации, когда
дистальные сосуды расходились под углом 90°. Вихревые линии,
параллельные плоскости симметрии, при прохождении бифуркации разделялись,
что приводило к возмущениям потока, обусловленным локальным
увеличением завихренности в области разделения потока. На рисунке 3 а приведено
движение подцвеченной жидкости под влиянием возникающей
завихренности в стационарном потоке при числе Рейнольдса 500, а на рисунке 3 Ъ
представлен сложный характер линий тока под действием турбулентности,
возникающей после прохождения точки бифуркации при числах
Рейнольдса порядка 1500. Переходное число Рейнольдса оказалось равным 1300 для
стационарного потока и 900 для пульсирующего.
Другие модельные эксперименты, включая модели с мешковидными
аневризмами в месте разветвления сосудов, показали, что имеет место
взаимосвязь между интенсивной завихренностью в области бифуркации и
полостью, обусловливающей возникновение сильной турбулентности в
аневризме даже при числах Рейнольдса порядка 500 (рис. 3 с). Среднее
переходное число Рейнольдса равнялось 500 для стационарных потоков и 400
для пульсирующих.
Фергюсон (Ferguson, 1972) привел доводы в пользу того, что влияние
флуктуации давления в месте соединения сосудов — в точке бифуркации —
будет значительно большим в сосудах головного мозга, чем где-либо еще,
по трем причинам. У артерий головного мозга отношение толщины стенки
сосуда к его диаметру меньше, чем у других артерий. Кроме того, стенки
этих артерий содержат значительно меньше эластина и не столь надежно
поддерживаются окружающей сосуд тканью. Принимая все это во
внимание, Фергюсон представлял себе, что инициация и рост внутричерепных
362
Глава 13
Рис. 3 а. Ламинарное течение при типичной нормальной бифуркации сосуда (поток
визуализирован в опытах Ферпосона (Ferguson, 1972))
Рис. 3 Ъ. Турбулентное течение при типичной нормальной бифуркации
3. Аневризмы сосудов головного мозга
363
*
N
Рис. 3 с. Мешковидная аневризма, турбулентность возникает уже при числах Рей-
нольдса, равных 500
мешковидных аневризм происходит под действием патологического
влияния турбулентности на эластиновые волокна и на целостность матрикса
волокон в стенке артерии.
Локализованное нарушение внутренней эластичной мембраны в
точках бифуркаций большинства артерий головного мозга может являться
следствием гидродинамических напряжений различного вида, включая
пики систолического давления, а также напряжения сдвига, связанного с
интенсификацией завихренности совместно с действием случайно
возникающих областей турбулентности при переходном режиме течения при
сердечном выбросе крови, когда число Рейнольдса течения может
достигать 1000. Воздействие пульсового гидростатического давления при систоле
на непрочные стенки в области бифуркации может привести к
выпячиванию стенки в месте разделения сосудов, что является начальной стадией
развития аневризмы.
Нарастание и взаимное усиление действия отрицательных факторов
являются результатом последовательно усиливающейся турбулизации
потока за бифуркацией. Возникновение турбулентности обусловлено
понижением критического числа Рейнольдса с 900 до 400 из-за наличия аневриз-
364
Глава 13
мы. Проведенное Фергюсоном тестирование на упругость ткани аневризмы
показало, что под действием турбулентности эластин теряет свои упругие
свойства. Кроме того, могут быть ослаблены связи коллагеновых волокон
в матриксе стенки сосуда, что также способствует прогрессивному росту
аневризмы. Возможность разрыва аневризмы возрастает по мере ее роста,
т.к. максимальное напряжение в стенках аневризмы равно ~ape/h, где а —
радиус, ре — избыточное давлениие на систоле, h — минимальная толщина
стенки. Когда радиус а становится достаточно большим (при этом
соответственно уменьшается толщина стенки /i), то при разрыве, приводящем
к мозговому кровоизлиянию, в любом случае возникает необычно большое
системное артериальное давление.
Приведенное здесь краткое описание развития аневризмы является
ключом к проблемам диагностики и оценке необходимости хирургического
вмешательства — трепанации черепа для вскрытия и клипирования
аневризмы.
4. Развитие атеросклероза, обусловленное межклеточным
отложением жиров
Среди всех заболеваний артерий, проявляющихся в потере
эластичности или склерозировании стенок, одно заболевание является наиболее
распространенным — это атеросклероз. Название болезни происходит от
греческого слова авероя — «кашица». При этой болезни происходит
разрушение стенок артерии, когда в интиме (ближайший к просвету сосуда
слой материала стенки) накапливаются липиды. Липиды — это
обобщающее название всех жирных кислот, стеринов и сложных эфиров. На
начальной стадии заболевания на стенках сосудов возникают кашеобразные
или желатиноподобные образования, что и определило название болезни.
На более поздней стадии происходит разрастание волокнистой ткани,
содержащей стерины, эта часть стенки отверждается в результате локального
нарушения процесса растворения фибринового сгустка, что может
привести к росту прилипания тромбоцитов, образованию фибриновой бляшки,
кальцинированию и образованию тромбов.
В данной работе не будем рассматривать серьезные последствия
поздних стадий атеросклероза: не только образования тромбов, но и
образования атеросклеротических бляшек, приводящих к сужению просвета сосуда
и уменьшению растяжимости стенок. Отметим лишь, что развитие этих
процессов приводит к чрезмерной работе сердечной мышцы. В настоящей
4. Развитие атеросклероза
365
работе исследуем развитие атеросклероза с точки зрения
биогидродинамики. Рассмотрим начальную стадию этого заболевания, атеросклеротическое
повреждение и его расположение на стенках, обусловленное протеканием
крови в сосудах.
На симпозиуме, организованном фондом Ciba Foundation,
посвященном болезни века — атеросклерозу, Лайтхилл (Lighthill, 1973)
председательствовал на двух секциях, где обсуждались проблемы старта заболевания,
вызванного межклеточным отложением липидов (липиды откладываются
в межклеточном пространстве), и механизм возникновения заболевания,
в котором основную роль играли процессы внутриклеточного накопления
липидов (избыточное поступление липидов в клетки). Эти два механизма
возникновения болезни изложены соответственно в § 4 и § 5, однако не
стоит их очень жестко разграничивать. Ниже будет отмечено, что связь между
этими двумя процессами отложения липидов окончательно не установлена.
Одним из наиболее существенных отличий этих процессов является
меньшая степень обратимости процесса межклеточного накоплении липидов по
сравнению с внутриклеточным. Однако их взаимное влияние и
приоритетность в инициировании заболевания до сих пор являются предметом
обсуждения, и на этот счет единое мнение отсутствует.
Подобного рода неопределенность становится понятной, поскольку
исчерпывающий анализ артерий пациента возможен лишь после его смерти.
Когда атеросклероз приводит к смерти больного, то болезнь обычно заходит
уже слишком далеко, так что посмертное исследование не слишком
проясняет механизм развития атеросклероза. Ранние стадии отложения липидов
могут быть изучены только в том случае, когда смерть пациента наступила
в силу каких-то других причин, однако в этом случае, как правило,
отсутствует информация о прижизненном процессе повреждения сосудов.
В обоих процессах накопления липидов из всех групп липидов, таких
как свободный холестерин, триглицериды или фосфолипиды, в значительно
большей степени участвуют эфиры холестерина. Среди них следует
особенно выделить эфир линолевой кислоты, который образуется именно на
ранних стадиях развития фиброзных бляшек. Так, Смит и Слейтер (Smith,
Slater, 1973) считают этот факт основополагающим в развитии
атеросклероза, вызванного межклеточным отложением липидов, поскольку из
наблюдений следует, что отложение липидов происходит вдоль волокон матрикса
артериального коллагена и эластина. Как следует из проведенных авторами
наблюдений, среди липидов, отложившихся в фиброзной ткани,
преобладают эфиры линолевой кислоты. А вот эфиры олеатовой кислоты преоб-
366
Глава 13
ладают в процессе внутриклеточного отложения липидов, что, возможно,
является результатом этерификации внутри клетки.
Подобно большинству липидов холестерин и его эфиры
нерастворимы, и они не могут перемещаться в организме в виде растворов. Однако
они легко образуют соединение с белками — липопротеины, которые
обладают достаточно хорошей растворимостью и присутствуют в виде
раствора в плазме крови. Накопление липидов в стенке артерии происходит за
счет проникновения липопротеинов в интиму и последующей диссоциации
на отдельные компоненты (диссоциировавшийся белковый компонент
называется апопротеином). На скорость диссоциации липопротеинов влияют
следующие факторы:
(i) средняя концентрация липопротеинов в плазме крови;
(ii) факторы, которые определяют величину равновесных концентраций
липопротеина в интиме и в плазме крови;
(iii) проницаемость эндотелиального слоя стенки артерии, что определяет
скорость диффузии липопротеина из плазмы, когда концентрация в
интиме меньше ее равновесного значения (или диффузию липопротеинов
в плазму, если концентрация липопротеина в интиме превышает
равновесное значение);
(iv) скорость диссоциации при проникновении липидов через клеточную
мембрану (процесс, при котором апопротеин может оставаться вне
клетки, а липидный компонент липопротеина соединяется с другим
апопротеином внутри клетки);
(v) скорость диссоциации при осаждении липидного компонента вдоль
фиброзного волокна.
Последний тип диссоциации, в результате которой происходит
внеклеточное отложение липидов, рассматривается как потенциально
саморазгоняющийся процесс в течение достаточно длительного периода времени
(Smith, Slater, 1973). Накопление липидов в виде фибриновых отложений
через определенный период времени приводит к образованию рыхлых,
загустевших, довольно прозрачных желатиноподобных структур в интиме, за
счет чего равновесная концентрация липопротеинов в интиме в два и более
раз превышает равновесную концентрацию в плазме.
Очевидно, что эти желатинообразные структуры являются
прототипами атеросклеротических бляшек. Развитие атеросклеротических бляшек
часто происходит в результате формирования фиброзной ткани в участках
отложения липидов, т. е. желатинообразная структура является как бы цен-
4. Развитие атеросклероза
367
тром роста бляшки. Концентрация липопротеина в желатинообразной
области увеличивается более чем в 4 раза по сравнению с его концентрацией
в нормальной, неповрежденной интиме, что, вероятно, ускоряет
дальнейшее накопление липидов. Развившуюся бляшку можно достаточно легко
обнаружить при пальпации.
Изучение физико-химических процессов (Iverius, 1973) показало, что
наиболее значимым в начальной стадии развития атеросклероза является
процесс накопления мукополисахаридов, называемых также гликозамино-
гликанами, в стенках артерии. Эти линейные полимеры вытесняют другие
длинноцепочечные молекулы, включая липопротеины, в результате чего их
излишек оседает на стенках сосудов. Рыхлые структуры липопротеинов (их
молекулы имеют размер порядка 0,02 мкм и называются многими
биохимиками ЛНП, или липопротеинами низкой плотности, и именно ЛНП
в наибольшей степени ответственны за перенос эфира холестерина, т. е.
являются особо атеромагенными), как показано Ивериусом, довольно легко
соединяются с гликозаминогликанами, такими как дерматансульфат с
сульфатными и карбоксильными группами, имеющими отрицательный заряд.
А именно, гликозаминогликаны участвуют в формировании волокнистых
структур соединительной коллагеновой ткани.
Биогидродинамические факторы, влияющие на развитие
атеросклероза, изучались Фраем (Fry, 1973). Им было отмечено, что эти факторы могут
вызывать изменение структуры стенок артерий таким образом, что
значительно облегчается процесс проникновения липопротеина в стенки и
усиливается его межклеточное отложение. Некоторые из этих факторов уже
знакомы читателю из главы 12 и некоторых предыдущих параграфов
настоящей главы.
Проведение дискуссии на эту тему не имеет смысла до уточнения
терминологии, поскольку некоторые термины допускают двоякое толкование.
Так, например, термин «стресс» используется в совершенно разных
значениях. В общей биологии (включая медицину) он означает реакцию
организма на предъявление к нему со стороны окружающей среды аномальных
требований, а в механике твердого тела стресс — напряжение, означает
силу, приходящуюся на единицу площади. В медицинском значении стресс
может привести к тому, что стенки артерий становятся более
подверженными отложению на них липидов, и, как следствие, стресс может
явиться причиной развития атеросклероза. Подобное же влияние стресса может
определяться, как можно будет видеть ниже, и биогидродинамическими
факторами. Однако большое число причин возникновения атеросклероза,
368
Глава 13
таких как, например, биохимические процессы в организме, останется вне
сферы нашего внимания.
С этого момента будем понимать стресс как в механике твердого тела —
напряжение, действующее на стенках артерий (как в §2 главы 12). Фрай
экспериментально изучил реакции стенок артерий на различные виды
механических напряжений, действие некоторых из них приводит к изменению
проницаемости стенок, об этом факторе говорилось выше, в пункте (Ш).
Это представляет особый интерес в свете возникновения
турбулентности, описанной в § 2 и § 3, поскольку при наблюдении над живым
организмом Фраем была обнаружена хорошая согласованность между
локализацией интенсивного потока протеина через эндотелий и локализацией выскоча-
стотных турбулентных напряжений. Биогидродинамик мог бы склониться
к постулированию связи между интенсивным потоком и уменьшением
сопротивления диффузии поперек пограничного слоя, что, возможно, имеет
место при наличии турбулентности. Однако Фрай показал, что
сопротивление диффузионным процессам, обусловленное развитием пограничного
слоя, даже в случае ламинарного пограничного слоя пренебрежимо мало
по сравнению с сопротивлением процессу проникновения протеинов
через эндотелиальный слой стенок артерии. При этом он имел в виду, что
изменение в стенках сосудов, обусловленное турбулентностью, в общем
виде описанное в наиболее экстремальных ситуациях в § 2 и § 3, оказывает
наибольшее влияние на проникновение протеинов через эндотелиальный
слой.
Фрай показал, что среди различных видов распределений напряжения
только те напряжения, которые вызывают удлинение вырезанного
участка артерии, приводят к увеличению проницаемости стенки для протеинов.
Как показано в главе 12, § 2, расширение сосуда в радиальном направлении
обусловлено повышением давления, а следовательно, интенсивность
проникновения протеинов в стенки может значительно повышаться всякий раз,
когда давление достигает своего наибольшего значения, особенно у
пациентов с повышенным давлением (и значит, такой пациент предрасположен
к атеросклерозу). Из других экспериментов in vitro (проведенных над
выделенными из организма сосудами), следовало, что в отсутствие расширения
повышение трансмурального давления оказывает на проницаемость стенок
пренебрежимо малое влияние. Эти наблюдения сводят на нет концепцию
о повышении проницаемости за счет повышения давления и подчеркивают
диффузионную природу этого феномена (проникновения липидов в стенки
сосудов).
4. Развитие атеросклероза
369
Керью было показано (Carew, 1971) в экспериментах in vitro, что
проницаемость протеинов возрастает пропорционально квадрату напряжения
сдвига. Такая практически функциональная зависимость от квадрата
векторной величины напряжения сдвига предполагает независимость от его
направленности. При очень большом напряжении сдвига, порядка 40 Н/м2,
значительно большем, чем физиологические значения напряжения сдвига,
проявляется свойство текучести эндотелия в техническом значении. Это то,
что доктор Фрай называл напряжением текучести, при котором происходит
серьезное повреждение стенки. Это наводит на размышление, что
исключительно большие напряжения сдвига в пределах, допустимых в
человеческом организме, могут в конечном счете привести к изменениям в
эндотелии, вызывающим атеросклероз. Такие изменения возникали в
экспериментах, проводимых Каро и Неремом (Саго, Nerem, 1973), и описаны
в § 5. Уместно рассмотреть период времени, в течение которого приложено
напряжение, или некоторый цикл напряжений, поскольку в экспериментах
доктора Фрая по определению предела текучести напряжения
прикладывались в течение совершенно короткого промежутка времени, а также
поскольку приложение высокочастотных напряжений сдвига за малое время,
как это показано в § 2 и § 3, способно вызвать значительные повреждения.
В некоторых экспериментах над собаками Фрай с коллегами создавали
шунт между артерией и веной в одной или нескольких подвздошных
артериях и получили, что число Рейнольдса увеличивается с 70 в контрольной
до 800 в шунтированной артериях. В шунтированной артерии происходила
значительная эрозия эндотелия. На основании наблюдаемой скорости в
такой артерии в предположении о течении Пуазейля было найдено, что
напряжение сдвига имеет порядок 3 Н/м2. С другой стороны, бифуркация сосуда
может привести к увеличению напряжения сдвига, описанному в главе 11,
§ 2, или даже к турбулизации потока (см. § 3), что может иметь
разрушительные последствия.
Конечно, в стенках сосудов постоянно идет компенсационный
процесс, препятствующий действию напряжения сдвига, износу и
возможному разрыву. Фраем была экспериментально продемонстрирована
восстановительная реакция со стороны стенок сосуда при направленном действии
напряжения сдвига, которое прилагалось к сосуду в течение нескольких
недель. Реакция сосуда на приложенное напряжение проявлялась, в
частности, в переориентации ядер клеток в направлении действия напряжения,
что, возможно, усиливало сопротивление эндотелия действующему
напряжению сдвига. Кроме того, существенное изменение скорости кровотока
370
Глава 13
в периферийных областях может привести к изменению долей потока
крови, расходящихся в дочерние сосуды после разветвления, что может
вызвать новые сдвиговые напряжения, достаточно большие для ослабленных
участков эндотелия.
Реакция сосудов на большое по величине механическое напряжение
может также проявляться в виде утолщения интимы вследствие
избыточного роста фиброзно-мышечной ткани. Такая реакция наблюдается
достаточно часто и провоцирует отложение липидов. Для получения реакции
такого типа в ответ на возрастающие напряжения сдвига Фраем был
проведен еще один эксперимент с шунтом между артерией и веной — на этот
раз была зашунтирована сонная артерия собаки.
Здесь уже было уделено много внимания вопросу о том, какая
структура потока может спровоцировать возникновение атеросклероза. Однако
окончательное строгое соответствие до сих пор не установлено. Частично
потому, что экспериментальные данные о структуре потока, а также о
механизме отложения липидов недостаточны, поэтому требуется более
детальное изучение обеих указанных причин атеросклероза как по отдельности,
так и в их взаимосвязи.
5. Внутриклеточное накопление липидов
Заканчивая эту главу, остановимся на некоторых факторах, которые,
как было показано, влияют на накопление липидов внутри клеток вообще
и в клетках интимы в частности при нормальном и патологическом
протекании данного процесса. Затем рассмотрим природу связей между подобным
накоплением липидов и начальной стадией атеросклероза.
Клетки испытывают потребность в липидах, в частности, они входят
в состав митохондрий и клеточных мембран. Бейли (Bailey, 1973) описал
в общих чертах процесс регулирования количества липидов в клетках.
Используя радиоактивный метод, он показал, что если клетка находится в
состоянии равновесия с сывороткой крови, где содержатся протеины
сыворотки и липопротеины, то происходит постоянное выделение липидов,
которые, связываясь с протеинами сыворотки, такими как альфа-глобулины,
образуют липопротеины, и одновременно происходит процесс поглощения
липидов, при этом за пределами клетки остаются альфа-глобулины как
диссоциировавшие апопротеины. Он указал на высокую степень обратимости
механизма (в деталях этот механизм в работе не описан) переноса липидов
через мембрану.
5. Внутриклеточное накопление липидов
371
Если липидов в окружающей среде нет, то они синтезируются внутри
клетки (Bailey, 1973) из глюкозы и других простых
молекул-предшественников. При избыточном количестве липидов в окружающей среде синтез
липидов в клетке практически полностью прекращается после того, как из
окружающей среды в клетку поступит необходимое количество липидов.
Таким образом, имеется по крайней мере два механизма управления
содержанием липидов в клетке. Биосинтез липидов в клетке прекращается по
достижении некоторого порогового уровня, а при его превышении вступает
в действие механизм выделения излишнего количества липидов.
Некоторые типы клеток, включая эндотелиальные клетки в стенках
артерий, имеют везикулы, которые, как было доказано, способны участвовать
в процессе «пиноцитоза» для захватывания даже таких больших молекул,
как липопротеины, и транспортировки их внутрь клетки. Штейн и Штейн
(Stein, Stein, 1973) рассматривали это как дополнительный механизм,
возможно наиболее значимый, аккумуляции липидов клетками интимы, хотя
Бейли (Bailey, 1973) высказывал противоположные взгляды. Он считал, что
такие клетки накапливают липиды только из липопротеинов, проникающих
во внутриклеточное пространство за счет такой транспортировки, при
которой они освобождаются от апопротеинов.
Какой бы ни был механизм синтеза при поглощении или выделении
липидов клетками интимы, одно совершенно очевидно: при слишком
высокой концентрации липидов в окружающей среде согласованное течение
процессов управления содержанием липидов в этих клетках может быть
нарушено. В этом случае липиды аккумулируются в других местах клетки
(Peters, Takano, Duve, 1973), в результате возникают клетки, описанные как
жировые клетки (содержат аномальное количество липидов).
В артериях молодых совершеннолетних людей, погибших в
результате несчастного случая, обычно находят значительное количество жировых
полосок — областей на стенках артерий, где содержится большое число
жировых клеток.
С одной стороны, Митчелл и Шварц (Mitchell, Schwartz, 1965) изучали
расположение жировых полосок и атеросклеротических бляшек в аортах
600 человек, умерших вследствие каких-либо других причин. У молодых
людей наблюдалось относительно небольшое количество жировых полосок
в брюшной аорте, значительно большее — в грудной, на боковых и передних
стенках их практически не было, и относительно чистыми были устья
межреберных артерий. У людей преклонного возраста со сформировавшимися
атеросклеросклеротическими бляшками в брюшной аорте их значительно
372
Глава 13
больше чем в грудной, в которой боковые и передние стенки, а также устья
межреберных артерий поражены этими образованиями. Общая площадь
поверхности, занятой жировыми полосками, не коррелирует с возрастом или
полом, а общая площадь бляшек увеличивается с возрастом, кроме того,
у мужчин она значительно больше, чем у женщин (и, кстати, коррелирует
с высоким кровяным давлением).
Митчелл и Шварц (Mitchell, Schwartz, 1965) рассматривали эти данные
по локации как опровержение утверждения о том, что жировые полоски
перерождаются в фиброзные бляшки. О других свидетельствах, приведенных
Смитом и Слейтером (Smith, Slater, 1973), речь шла в §4. Там отмечалось,
что эфир линолевой кислоты преобладает в фиброзных бляшках и в их «же-
латинообразных» предшественниках, в то время как эфир олеатовой
кислоты преобладает в жировых клетках.
С другой стороны, Флаерти, Ферранс, Пирс, Керью и Фрай (Flaherty,
Ferrans, Pierce, Carew, Fry, 1972) придавали особое значение взаимосвязям
между жировыми полосками и фиброзными бляшками, хотя и не
отрицали возможность перерождения одних в другие под воздействием
определенных сильных факторов, таких как генетические, влияние
окружающей среды или структурные или биогидродинамические параметры
различных участков стенок артерий. Уолквист, Дей и Тьюм (Wahlqvist, Day,
Tume, 1969) изучали пенистые клетки (жировые клетки с большим
содержанием жировых капель) в пораженных артериях человека и
показали, что изолированные пенистые клетки поглощают олеиновую кислоту из
окружающей их среды таким образом, что при этом в них накапливается
в большей степени олеат холестерина. Пенистые клетки ответственны за
процесс перехода липидов, накопленных внутри клетки, в межклеточное
пространство и обратно. Переход липидов из клетки в межклеточное
пространство может происходить за счет разрушения пенистых клеток
вследствие их крайней хрупкости. При этом выделяющиеся липиды вызывают
различные болезненные явления в стенках артерии (включая
воспалительные реакции). Выделение липидов из пенистых клеток и вызванные ими
болезненные явления способствуют развитию атеросклеротической
бляшки. И обратно, липиды из межклеточного пространства могут переходить
в клетку. Это происходит следующим образом. Окисление находящихся
на стенке артерии липидов представляет угрозу для организма.
Иммунная система активизируется, в опасную зону поступают клетки,
заглатывающие липиды, на данном этапе эти макрофаги называются пенистыми
клетками.
5. Внутриклеточное накопление липидов
373
Каро, Фицжеральд и Шрётер (Саго, Fitz-Gerald, Schroter, 1971)
подчеркивали, что различные биогидродинамические факторы могут
оказывать разнонаправленные воздействия на расположение мест повреждения
стенки. Например, как уже говорилось, процесс внутриклеточного
отложения липидов на ранних стадиях развития заполненной жиром клетки может
быть обратимым и жировые полоски могут полностью исчезать при
диете с пониженным содержанием жиров, что подтверждалось исследованием
сосудов у жертв концентрационных лагерей. Каро, Фицжеральд и Шрётер
показали, что любым реверсивным типом разрушения стенки можно
управлять в том смысле, что факторы, приводящие к росту проницаемости
протеинового окружения липидов, о чем шла речь в § 4, могут также и уменьшать
ее. Это возможно, если сопротивление выходу липидов, отделившихся от
связанных с ними носителей протеинов, больше, чем сопротивление
соединению липидов с различными видами носителей протеинов (или,
возможно, даже более простых молекул, из которых клетки синтезируют лепиды),
или если пиноцитоз является основным механизмом, поддерживающим
поступление липидов, за счет их экскреции.
Среди факторов, определяющих просачиваемость протеинов (§ 4),
Каро, Фицжеральд и Шрётер (Саго, Fitz-Gerald, Schroter, 1971) особо
отмечают роль напряжения сдвига на стенке. В своей работе Каро (Саго, 1973)
проводя исследования с использованием меченых липопротеинов в своей
лаборатории и подводя итоги исследований Каро и Нерема (Саго, Nerem,
1973), отметил, что все однозначно указывает на возрастание
проницаемости с ростом напряжения сдвига, об этом же свидетельствуют
независимые эксперименты под руководством доктора Фрая (§ 4). Кроме того, обе
группы ученых пришли к выводу, что наблюдаемая зависимость
проницаемости от напряжения сдвига не может быть объяснена только изменением
скорости диффузии протеинов поперек пограничного слоя. Скорее всего,
непосредственное влияние сдвигового напряжения на стенку артерии или,
возможно, на структуру адсорбционного слоя (Miller, Graet, Frei, 1973)
оказывает значительно большее влияние на увеличение скорости
транспортировки липидов.
Один из выводов, к которым пришли исследователи, состоит в том, что
эпидемиологические данные свидетельствуют о связи низкого уровня
физической активности с атеросклерозом, что можно объяснить небольшими
сдвиговыми напряжениям, которые недостаточны для вымывания липидов
на ранней стадии, когда еще накопление липидов носит обратимый
характер. Возможно также, что обратимые повреждения, как правило имеющие
374
Глава 13
место в юном возрасте, могут образовываться в областях, где сдвиговые
напряжения малы, а другие биогидродинамические факторы, которые могли
бы ускорить процесс вымывания липидов, отсутствуют. Это не исключает
утверждения о том, что дальнейшее развитие уже сформировавшихся ате-
росклеротических бляшек является в значительно меньшей степени
обратимым процессом, который может ускоряться за счет абсорбции липопротеи-
нов (§ 4) и может развиваться в основном в тех местах стенки сосуда, где
биогидродинамические сдвиги приводят к повышению проницаемости, как
и при больших сдвиговых напряжениях. Этим можно обосновать и данные,
представленные в работе Митчелла и Шварца (Mitchell, Schwartz, 1965),
о различных локализациях различных видов повреждения сосудов, что,
однако, не исключает возможность перерождения одного вида повреждения
в другой.
Полученные после смерти пациентов дополнительные
экспериментальные данные о расположении повреждений в аортах у подростков,
представленные в работе Каро, Фицжеральда и Шрётера (Саго, Fitz-Gerald,
Schroter, 1971), подтверждают и уточняют выводы Митчелла и Шварца
(Mitchell, Schwartz, 1965). Экспериментальные исследования подтвердили
предположение о том, что образование жировых полосок происходит в
основном в зонах низких сдвиговых напряжений. К таким зонам
относится, в частности, и область грудной аорты, где в силу образования
вторичных потоков возникает достаточно большой по толщине пограничный слой,
а также в большинстве периферических артерий. При этом области
высоких сдвиговых напряжений оказываются небольшими по размеру, включая
область разделения потока в результате бифуркации. В этой связи Фрай
(Fry, 1973) отмечает тот факт, что площадь стенок артерии с
наибольшим сдвиговым напряжением в местах разделения потока имеет
тенденцию к уменьшению, хотя он связывает это уменьшение с усилением
защитной реакции (включая разрастание соединительной ткани) на столь высокие
сдвиговые напряжения.
Каро, Фицжеральд и Шрётер (Саго, Fitz-Gerald, Schroter, 1971)
высказали предположение, что в плазме крови должна быть крайне высокая
концентрация липопротеинов, при которой процесс повреждения стенки
сосуда, сопровождающийся выделением липидов, становится процессом,
сопровождающимся поглощением липидов. Они утверждают, что
искусственно вызванная начальная стадия атеросклероза (атерогенез) у
подопытных животных за счет диеты с повышенным содержанием жиров
определяется процессом именно такого типа. Жировые полоски в этом случае
5. Внутриклеточное накопление липидов 375
возникали в совершенно других местах, чем у молодых людей,
соблюдающих обычную диету. Липиды довольно быстро накапливались главным
образом в областях высокого сдвигового напряжения, в частности у
разделителей потока. Такая интерпретация возникновения жировых полосок
у животных как процесса, связанного с поглощением липидов,
противоположна интерпретации процесса образования жировых полосок у молодых
людей. Вследствие чего изучение атерогенеза у людей на основании
данных, полученных при исследовании сосудов животных, следует проводить
крайне осторожно. Можно только отметить, что, видимо, на эти
экспериментальные данные можно опираться только для выяснения
справедливости утверждения о преобразовании жировых полосок в атеросклеротиче-
ские бляшки.
Главной целью этой главы было представить читателю все
разнообразие (скорее всего, далеко не полное) факторов, не связанных с
биогидродинамикой, которые значительно усложняют взаимосвязь между
протеканием крови в сосудах и их болезнями. Очевидно, что многочисленные
практические исследования биологической стороны этой взаимосвязи,
проводившиеся отдельно медиками, физиологами и биохимиками, должны быть
освещены с единой точки зрения. Для этого следует создать одну
группу, которая сосредоточит совместные усилия на решении подобных
проблем. Другая сложность данной проблемы проистекает из необходимости
владеть теоретическими и практическими знаниями в областях
биогидродинамики, связанных с рассматриваемыми задачами. Поэтому в созданной
команде ученых обязательно должен быть и биогидродинамик!
Возможно, представителям различных областей знаний весьма сложно объединить
усилия, чтобы решать задачи на стыке нескольких наук, в частности,
досконально исследовать болезни артерий, обусловленных гемодинамикой.
Однако решение такой задачи чрезвычайно важно для человечества и
попытки ее решить должны быть предприняты. Об этом следует всегда помнить
и прежде всего следует заниматься тем, что в наибольшей степени важно
для человечества.
Литература
(Замечание. В приводимом ниже списке литературы использовано
общепринятое сокращение Ath. для Atherogenesis.)
[1] Bailey, J.M. 1973. Ath., 63-88.
376
Глава 13
[2] Carew, Т.Е. 1971. Mechano-chemical response of canine aortic
endothelium to elevated shear stress in vitro, PhD thesis, The Catholic
University of America, Washington, DC.
[3] Caro, C.G. 1973. Ath., 127-149.
[4] Caro, C.G., Fitz-Gerald, J.M. & Schroter, R.C. 1971. Proc. Roy. Soc. B,
177, 109-159.
[5] Caro, C.G. & Nerem, R.M. 1973. Circulation Res. 32, 187-205.
[6] Collins, J.I. 1963. J. Geophys. Res. 68, 6007-6014.
[7] Ferguson, G.G. 1972. J. Neurosurgery, 37, 666-677.
[8] Flaherty, J.Т., Ferrans, V.J., Pierce, J.E., Carew, Т.Е. & Fry, D.L. 1972.
Localizing factors in experimental atherosclerosis. In Atherosclerosis and
Coronary Heart Disease (ed. Likoff, W. et al.) Grune & Stratton, New
York.
[9] Fry, D.L. 1973.93-120.
[10] Iverius, P.H. 1973. Ath., 185-193.
[11] Lighthill, M.J. 1970. Turbulence. In Osborne Reynolds and Engineering
Science Today (ed. McDowell, D.M. & Jackson, J.D.) Manchester
University Press.
[12] Lighthill, M.J. 1973. Ath., 277-282.
[13] Miller, I.R., Graet, H. & Frei, Y.F. 1973. Ath., 251-262.
[14] Mitchell, J.R. & Schwartz, C.J. 1965. Arterial Disease, Blackwell
Scientific Publication, Oxford.
[15] Nerem, R.M. & Seed, W.A. 1972. Cardiovascular Res. 6, 1-14.
[16] Parker, К.Н. & Seed, W.A. 1973. Turbulence in the Aorta, unpublished
lecture at Euromech Colloquium 32, Imperial College, London.
[17] Peters, T. J., Takano, T. & de Duve, С 1973. Ath., 197-214.
[18] Roach, M.R. 1963a. Circulation Res. 13, 537-551.
[19] Roach, M.R. 1963b. Amer. J. Cardiol. 12, 802-815.
[20] Roach, M.R. 1970. Circulation Res. 27, 985-993.
[21] Seed, W.A. & Wood, N.B. 1971. Cardiovascular Res. 5, 319-330.
[22] Smith, E. B. & Slater, R. S. 1973. Ath., 39-52.
[23] Stein, Y. & Stein, O. 1973. Ath., 165-179.
[24] de Vries, H. & van den Berg, J. 1958. Cardiologia, 33, 195-211.
[25] Wahlqvist, M.L., Day, A.J. & Tume, R.K. 1969. Circulation Res. 24,
123-130.
Глава 14
Микроциркуляция'
1. Введение
В заключительной главе рассматривается кровоток в сосудах, диаметр
которых не достигает 0,1 мм (т.е. 100 мкм), в частности в артериолах,
капиллярах и венулах диаметром 5-50 мкм. В этой периферической части
циркуляции, называемой микроциркуляцией, которая уже рассматривалась
в главе 10 (§ 6), возникает максимальный перепад давления между
артериями и венами, у здоровых людей достигающий 100-200 мбар.
В главе 14 рассматриваются количественные аспекты
периферического сопротивления, в частности зависимость перепада давления от скорости
потока, которая становится нелинейной. Периферическое сопротивление
имеет большое значение, поскольку оно определяет необходимую
интенсивность работы сердца и поскольку необходимое для тела большое
разнообразие скоростей перфузии его различных периферических частей (где
скорости потока уменьшаются на порядок) обусловлено силой гладкомы-
шечного слоя артериол, обеспечивающего расширение и сжатие артериол,
что вызывает значительные изменения периферического сопротивления.
Второй важнейшей характеристикой микроциркуляции является
функция капилляров (сосудов диаметром менее 10 мкм, не имеющих гладко-
мышечного слоя) при различных обменных процессах. Капилляры,
находящиеся в легких, отделены от альвеол чрезвычайно тонкими мембранами,
допускающими обмен 02 и С02 между воздухом, находящимся в
раздутых альвеолах, и кровью в перфузируемых капиллярах. В самой крови
подобные обменные процессы происходят между плазмой и гемоглобином,
находящимся, конечно, внутри эритроцитов, окруженных тончайшей
клеточной мембраной. В капиллярах протекают такие же обменные процессы
(только противоположно направленные), дополняемые обменом различных
Лекция в Международном центре механики в Удине, Италия, 1971.
378
Глава 14
питательных веществ между плазмой и окружающими тканями. Одни из
этих процессов обусловлены диффузией, другие — конвекцией мелких
порций плазмы (не достигающих 1 % общего потока), проникающих через
щели (шириной менее 0,1 мкм) в эндотелиальной стенке капилляра (Guyton,
1966). Многие из этих фракций плазмы в конечном счете всасываются
обратно за счет низкого давления, создающегося в конце того же капилляра,
но остальные отводятся по лимфатической системе и попадают в русло
одной из крупных вен.
Течение жидкости в микроциркуляторном русле заметно отличается от
течения в более крупных кровеносных сосудах (см. главы 10 и 12), а
также от большинства других привычных потоков, поскольку инерция потока
пренебрежимо мала. Здесь будут рассматриваться потоки, чьи параметры
определяются исключительно уравновешивающими друг друга градиентом
давления и силой вязкости, поскольку инерционные эффекты (ускорение,
умноженное на плотность) являются столь малыми, что совсем не
нарушают это равновесие. В частности, это объясняется тем, что число Рейнольдса
в сосудах диаметром менее 0,1 мм всегда значительно меньше 1 и
снижается до 0,001 в самых узких капиллярах, а параметр Уомерсли а (гл. 10,
§ 8), характеризующий соотношение инерционных эффектов,
обусловленных пульсацией, с вязкостными эффектами, значительно меньше 1. В мире
микроциркуляции, т. е. в мире сверхнизких чисел Рейнольдса, где инерция
пренебрежимо мала, следует забыть большую часть того, что было
найдено при изучении крупных сосудов, и сконцентрироваться на совершенно
других положениях.
Перечислим пункты, о которых надо постараться забыть в условиях,
когда градиенты давления полностью уравновешены силами вязкости.
Надо забыть об уравнении Бернулли, поскольку нет заметной разницы между
статическим и динамическим давлениями. Следует забыть о центробежных
силах: жидкость может преодолевать крутые повороты без малейших
трудностей и без формирования каких бы то ни было вторичных потоков. Иначе
говоря, движение жидкости фактически обладает меньшей зависимостью
от геометрии сосуда и практически не проявляет тенденции к разделению
потоков. Действительно, в некоторых случаях (выполняется закон вязкости
Ньютона, жидкость течет в сосуде с твердыми стенками) поток жидкости
является полностью «обратимым».
Нечувствительность к геометрическим деталям в дальнейшем
проявляется в виртуальном отсутствии «входной зоны». Например, если
жидкость, удовлетворяющая закону вязкости Ньютона, входит в трубу под боль-
2. Вазомоторный контроль периферической перфузии 379
шим углом к оси, то в одном и том же потоке разница между реальным
течением и течением Пуазейля практически исчезает на расстоянии,
равном половине радиуса (Fitz-Gerald, 1971): исключительно быстрый
переход к течению Пуазейля. Благодаря таким упрощениям и возможности
использования идей, касающихся динамики жидкости с низким числом Рей-
нольдса, разработанных в прикладных науках, в частности в теории
гидродинамики смазочных материалов, изучение микроциркуляции значительно
упрощается.
Однако, несмотря на эти положительные моменты, существуют и
достаточно большие трудности, упомянутые во введении в главе 10, в
подпункте (iv), — необходимо принимать во внимание взаимодействие между
эритроцитами и стенками каждого из таких капилляров, диаметр которых
равен одному или двум диаметрам эритроцитов. Соответственно, большая
часть этой главы (начиная с § 4) посвящена особенностям
микроциркуляции, обусловленным тем, что кровь представляет собой
высококонцентрированную суспензию эритроцитов (здесь не учитываются другие
компоненты, которые значительно меньше влияют на гемодинамику) в плазме.
2. Вазомоторный контроль периферической перфузии
Характерной чертой системы кровообращения, прокомментированной
в главе 10, является наличие вазомоторного контроля периферической
перфузии, заключающегося в том, что гладкая мускулатура меняет диаметр
мелких сосудов, изменяя на порядок скорость потока крови в отдельном
участке микроциркуляторного русла. Хотя мышечный слой отсутствует
в стенках самих капилляров, существует масса доказательств того, что
вазомоторный контроль осуществляется в мельчайших прекапиллярах, диаметр
которых не превышает 0,1 мм.
Было бы естественно предположить, что в сложной разветвленной
системе контроль локального сопротивления должен осуществляться в той
группе сосудов, которые вносят максимальный вклад в общее
сопротивление, и имеется серьезное подтверждение этого предположения. В главе 10,
в подпункте (ii), было отмечено, что набор длин и диаметров сосудов
ограничен, а в §6 указан способ оценки распределения перепадов давления
в разветвленной системе в предположении выполнения закона Пуазейля.
Расчеты, проведенные Кремером (Kraemer, 1967) показали, что при вазоди-
латации около половины общего перепада давления в циркуляторном русле
возникает в прекапиллярах диаметром менее 50 мкм (которые по его клас-
380
Глава 14
сификации относятся к группе артериол и мельчайших артерий), четверть —
в более крупных сосудах и около четверти — в капиллярах. (Заметим, что
при таком разделении под термином «давление» подразумевается давление,
откорректированное только для гидростатического компонента таким
образом, чтобы оно стало эквивалентным «давлению на высоте сердца».)
Из этого можно сделать неожиданный вывод, что даже до вазокон-
стрикции около половины общего сопротивления сердечно-сосудистой
системы обеспечивается группой сосудов малого размера (15-50 мкм), тех
самых сосудов, с помощью которых осуществляется вазомоторный контроль
периферической перфузии. Хотя Кремер должен был принять, что
вазомоторное сопротивление этих сосудов очень велико, для того чтобы объяснить
наблюдаемые большие изменения скорости перфузии, но все же половина
невозмущенного общего сопротивления осталась необъясненной.
Естественно попытаться устранить возникшее затруднение путем
учета отклонения течения от пуазейлевого. В главе 10 было исследовано
отклонение течения от течения Пуазейля при высоких числах Рейнольдса.
Отклонение давления от случая течения Пуазейля было невелико, поскольку
отклонение от течения Пуазейля существенно только в достаточно широких
трубках, где перепад давления мал. В микрососудах отклонение давления
от пуазейлевого объясняется эффектом Фареуса-Линдквиста (Fahraeus-
Lindquist) (см. ниже §4), который, согласно расчетам Кремера,
обусловливает 40 %-ное падение давления ниже давления Пуазейля в трубках
диаметром менее 25 мкм. В последнее время было выдвинуто предположение
Лайтхилла (Lighthill, 1968, с. 116) о том, что нелинейность сопротивления
микроциркуляторного русла, обусловленная особенностями кровотока в
самых узких капиллярах, может увеличивать эффективность вазомоторного
контроля; подробно этот эффект описан в § 5.
Не будем обсуждать здесь различные типы раздражителей,
вызывающих вазомоторную реакцию; большинство из них (например, температура,
вызывающая вазодилатацию, или широкий спектр химических стимулов)
мало связаны с гидродинамикой. Отметим только, что Каро, Фоли и Садлоу
(Саго, Foley, Sudlow, 1970) было доказано наличие вазомоторных реакций
на быстрое изменение внутреннего давления жидкостей. Их исследование
позволяет предположить, что быстрое снижение давления, обусловленное
спадением манжеты, повышающей кровенаполнение вен предплечья,
приводящее к повышению венозного давления до 40 мб, может спустя 2-3
секунды вызвать внезапное, едва ли не трехкратное вазомоторное повышение
кровотока в предплечье по сравнению с состоянием покоя; хотя во время
3. Легочная перфузия и вентиляция
381
короткой задержки перед возникновением вазомоторной реакции скорость
кровотока превышает величину, характерную для состояния покоя, только
на 25 %. Этот пример активной реакции на изменения давления
подчеркивает необходимость для специалиста по гидромеханике и течению
жидкости в пассивно деформирующихся трубках тщательно исследовать и
учитывать наличие физиологических феноменов при решении задач в области
биогидродинамики.
3. Легочная перфузия и вентиляция
Микроциркуляция в легких имеет несколько особенностей, которые
сильно отличаются от особенностей микроциркуляции, перечисленных
в §2. В частности, ни в легких, ни в прекапиллярах, ни где-либо еще
не наблюдается вазомоторной активности и большая часть сопротивления
обеспечивается самими капиллярами (Fowler, West, Pain, 1966). Кроме того,
в разных частях легкого перфузия капилляров может быть крайне
неравномерной, хотя в дальнейшем будет видно, что эта неравномерность является
главным образом пассивной и ведущую роль в ней играет сила тяжести.
Эффективность легочной газообменной системы зависит от взаимного
соответствия распределения перфузии и вентиляции в различных частях
легкого (Farhi, 1966), и из наблюдений следует, что сила тяжести также
вызывает неравномерность вентиляции некоторых (хотя и не всех) альвеол,
совпадающую с неравномерностью перфузии (West, 1966).
Типичные капилляры в легких проходят через тонкую перегородку,
разделяющую две альвеолы, и отделяются от альвеолярного газа очень
тонкой легочной мембраной. Поэтому они очень легко деформируются, когда
альвеолярное давление рА превышает давление крови в соседнем
капилляре. Эта деформация, обнаруженная Уэстом, Глейзером, Хьюгом и Мелони
(West, Glazier, Hughes, Maloney, 1969), значительно уменьшает среднюю
ширину сосуда, с 6,5 до 2,3 мкм, и даже для ньютоновской жидкости
изменение ширины капилляра вызывает аномальное повышение сопротивления
и, как следствие, снижение кровотока. Для реальной крови в силу
достаточно больших размеров отдельных эритроцитов (см. § 5) повышение
сопротивления приводит к полной остановке кровотока.
Поясним значение двух сравниваемых здесь давлений, начиная с
альвеолярного давления рА. При раздувании легких происходит расширение
сотен миллионов альвеол до диаметра, достигающего нескольких сотен
микрон, при этом значительно увеличивается площадь влажной поверхно-
382
Глава 14
сти. Этот процесс подобен вспениванию! Поэтому для раздувания альвеол
требуется очень большое усилие, поскольку жидкость, увлажняющая
поверхность, представляет собой воду, обладающую большим поверхностным
натяжением. К счастью, в этой жидкости присутствуют некоторые белки,
играющие роль детергентов и облегчающие образование пены; они
значительно снижают поверхностное натяжение (Pattle, 1966). Это также
означает, что давление рА газа в альвеолах приводит к закрытию соседних
капилляров, а поправка, вносимая силой поверхностного натяжения, большого
значения не имеет.
Что же касается давления крови в соседних капиллярах, то его
величина имеет промежуточное значение между величинами ра и ру давлений
на артериальной и венозной сторонах легочной циркуляции; перепад
давления между ними уравновешивается сопротивлением капилляров.
Сердечный насос создает избыточное давление в легочных артериях, не
достигающее величины давления в аорте и в покое приблизительно равное 12 мбар,
но при физической нагрузке его величина легко удваивается. Такая картина
сравнима с разницей гидростатического давления между верхушкой и
нижней частью легкого, и здесь нельзя просто игнорировать гидростатический
компонент, как это имело место в § 2, поскольку величины давления в цир-
куляторном русле сравнимы не только друг с другом, но и с давлением рА
газа в альвеолах.
Давления ра и ру отражают падение гидростатического давления с
высотой на 1 мбар на см. Фактически альвеолярное давление рА тоже падает
с высотой приблизительно на 0,3 мбар на см. В целом связь этих данных
с механикой легких, подвешенных под воздействием силы тяжести и
удерживаемых только тонким слоем плевральной жидкости, требует
длительного обсуждения, не являющегося предметом обсуждения в гидродинамике,
но все же можно сделать вывод, что легко деформирующаяся структура
легких обладает некоторыми общими свойствами с жидкостью, в их число
входит, в частности, градиент внутреннего давления, который при величине
удельного веса легочной ткани, равной 0,3, близок к гидростатической
величине. Уэст (West, 1966) показал, как градиент влияет на вентиляцию,
которая изменяется, с высотой так же, как и большая часть обсуждаемых
изменений перфузии.
Мы видели, что по сравнению с рА давления ра и ру наиболее
значительно изменяются по вертикали. В зависимости от соотношения
величин рА, ра, ру можно выделить три режима (или три зоны) кровотока,
определяемые Уэстом (West, 1966) следующим образом.
3. Легочная перфузия и вентиляция
383
Зона 1 — зона самого высокого давления в альвеолах, где ра < рА.
Зона 2 — зона давления в альвеолах принимает промежуточное
значение: ру < рА < ра.
Зона 3 — зона самого низкого альвеолярного давления, где рА < pv.
В зоне 1, как экспериментально доказали Уэст, Глейзер, Хьюг и Мело-
ни (West, Glazier, Hughes, Maloney, 1969), любой отрезок капилляра
(давление в котором имеет промежуточное значение между величинами ра и ру)
сжимается при альвеолярном давлении рА, и кровоток в нем прекращается.
Как было обнаружено, в зоне 3 полностью открытые трубки слегка
раздуваются; механизм этого явления, обсуждающийся в § 5, связан с чрезвычайно
низким падением давления ра — ру.
Возможно, достаточно было бы сказать, что зона 2 представляет
собой переходную зону, но ее поведение вызывает определенный
теоретический интерес и поэтому привлекает внимание. У тонких сжимающихся
трубок, по которым гомогенная жидкость перемещается между резервуарами
под давлением ра и pv, промежуточное давление рА, действующее на
трубки снаружи, вызывает частичное сужение дистального сосудистого русла,
результатом которого является высокое местное сопротивление,
уравновешивающее внезапное падение давления в жидкости от рА до pv (Permutt,
Bromberger-Barnea, Bane, 1962). Co временем нормальное сопротивление
главной трубки становится равным падению давления ра — рА вплоть до ди-
стальной констрикции. Следствием этого является свойство потока, часто
описываемое как резистор Старлинга (более правильное обозначение, чем
данное вышеупомянутыми авторами название «водопад», подчеркивающее
наличие инерционной составляющей): скорость течения пропорциональна
разности ра — pv, пока pv остается ниже рА,ив дальнейшем становится
пропорциональной ра — рА. Для гомогенной жидкости частичный дистальный
коллапс неизбежен, поскольку поток в сосуде, заполняющий все его
сечение, может привести к схлопыванию сосуда, когда давление падает ниже
рА, но при полном схлопывании проксимальное артериальное давление рй
может распространиться по всему сосуду и привести к его открытию.
Ситуация еще сильнее осложняется в случае цельной крови,
поскольку кровоток может прекращаться в не полностью закрытых каналах и
поддерживать градиент давления за счет сухого трения эритроцитов о
стенки (см. §5). Наблюдения показывают, что взаимосвязь между давлением
и потоком обусловливается действием механизма резистора Старлинга, но
в несколько измененном виде (West, Glazier, Hughes, Maloney, 1969). Если
говорить коротко, то этот механизм действует следующим образом. Если
384
Глава 14
давление в венах ру падает ниже альвеолярного, то число схлопнувшихся
протоков увеличивается, а при вновь возрастающем давлении в венах pv
число открытых протоков нарастает.
Отметим, что при физической нагрузке ра возрастает, так что зона 1
(где нет кровотока) может полностью отсутствовать, поскольку при
возросшем артериальном давлении ра границы всех зон смещаются вверх.
Заметим также, что в приведенном кратком изложении принципов легочной
циркуляции не подразумевалось, что легочные капиллярные каналы имеют
такую же геометрию, как и капилляры большого круга кровообращения.
Недавно появилась тенденция (см., например, Fung, Sobin, 1968)
представлять себе распределение капиллярных каналов в перегородках подобным
распределению путей для прохода через зал, тесно заставленный
толстыми колоннами! В силу такого распределения большая часть легочных
капилляров представляет собой каналы, поперечное сечение которых меньше
диаметра эритроцита в недеформированном состоянии, равного 8 мкм.
Чтобы пройти через узкие капилляры, эритроциты должны деформироваться.
Такое «протискивание» эритроцитов через легочные капилляры облегчает
перенос газов между легочной мембраной и гемоглобином, однако здесь
возникают проблемы в продвижении эритроцитов, обусловленные
созданием смазочного слоя и наличием сопротивления. Движение в очень узких
капиллярах можно проанализировать методами теории смазки, о чем
более подробно говорится в § 5, а предварительно в § 4 исследуется течение
в более крупных сосудах.
4. Концентрация эритроцитов вдоль оси капилляров
Для завершения обсуждения отклонений от закона Пуазейля в крупных
сосудах, рассмотренных в предыдущих главах, оставшаяся часть главы 14
будет посвящена отклонениям в крайне мелких сосудах и их роли в
микроциркуляции. Такие отклонения наблюдаются в каналах, диаметр
которых сравним с диаметром недеформированного эритроцита, равным 8 мкм,
и принимают достаточно неожиданные формы; в частности, когда диаметр
трубки падает ниже 30 мкм, возникающий в нем перепад давления
значительно меньше, чем перепад давления при течении Пуазейля, а вязкость
такая же, как и в больших трубках. Этот феномен, обнаруженный Фаре-
усом и Линдквистом (Fahraeus, Lindquist, 1931), обсуждается в §4. Однако
в самых узких капиллярах шириной менее 6 мкм обнаруживается обратная
тенденция (рассматривается ниже, в § 5). Таким образом, «кажущаяся вяз-
4. Концентрация эритроцитов вдоль оси капилляров 385
кость», определяемая законом Пуазейля, при уменьшении диаметра
трубки сначала падает (примерно на 50 %), достигает минимума при диаметре
трубки, приблизительно равном 12 мкм, а при дальнейшем уменьшении
диаметра повышается, превышая величину кажущейся вязкости в трубках
большого диаметра.
Эти явления практически не зависят от любых изменений вязкости
цельной крови и являются функцией скорости сдвига, как это упоминалось
в пункте (iv) главы 10. Действительно, в микроциркуляторном русле, как
и в крупных сосудах, скорости сдвига слишком велики для таких
изменений, чтобы быть единственной причиной изменения кажущейся вязкости.
В §4 изучается взаимодействие суспензированных эритроцитов со
стенками сосудов, вызывающее эффект Фареуса-Линдквиста. А рассмотрение
соображений, касающихся эффекта возвратного движения, при движении
в настолько узких каналах, что эритроциты подвергаются значительной
деформации даже для того, чтобы проникнуть в них, отложено до § 5.
Фареус и Линдквист (Fahraeus, Lindquist, 1931) выявили
статистическую закономерность в поведении эритроцитов — они отходят от стенок
и в большей степени концентрируются вблизи оси сосуда. Чуть позже
обсудим этот наблюдаемый феномен «аксиальной концентрации» с позиции
механики текучих сред, а также покажем, как этот феномен приводит к
снижению кажущейся вязкости.
Кинематическая вязкость плазмы крови при нормальной температуре
тела постоянна и приблизительно равна 1,5 мм2/с, а вязкость цельной
крови возрастает при увеличении гематокрита (объемной концентрации
эритроцитов) сначала медленно, а затем скорость роста увеличивается. То есть
кривая зависимости кинематической вязкости от гематокрита вогнута. При
нормальном гематокрите, равном 0,4-0,5, вязкость равна 4, а затем
возрастает, достигая больших значений. При аксиальной концентрации
эритроцитов вязкость значительно изменяется в радиальном направлении, начиная
от 1,5 в свободной от клеток области, в нескольких микронах от стенки, до
весьма больших величин в окрестности оси.
С другой стороны, из теории осесимметричного ламинарного потока
следует, что если вязкость /л зависит от расстояния от оси г, то при
заданном градиенте давления при стационарном движении в круглой трубе
скорость сдвига в сечении должна изменяться как /i"1/-. При кровотоке, когда
эритроциты концентрируются вблизи оси, совместное действие обоих
факторов — уменьшение вязкости с ростом радиуса и пропорциональность
скорости сдвига величине ^~гг — приводит к тому, что в области стенок сдвиг
386
Глава 14
увеличивается сильнее, чем в каком-либо другом месте. Расход жидкости
через поперечное сечение равен интегралу по г от произведения тгг2 на
удвоенную скорость сдвига, которая пропорциональна /х-1 г3. Около
стенки (при максимальном г) множитель г3 значительно больше величины /х-1.
Таким образом величина, обратная кажущейся вязкости ц^1
(пропорциональная скорости сдвига, разделенной на градиент давления), представляет
собой среднее взвешенное значение /х-1 по г, где весом является г3.
Наличие такого большого весового коэффициента при больших
значениях /i"1 в тонком пристеночном слое, где отсутствуют эритроциты,
объясняет тот факт, что кажущаяся низкая вязкость, равная 2-2,5 мм2/с, обычно
наблюдается в трубках диаметром 10-30 мкм. Уитмор (Whitmore, 1968)
отмечает, что даже в предположении о бесконечно большой вязкости при г,
превышающих 80 % от радиуса трубки, вязкость в этой части сечения и
постоянная конечная вязкость в остальной части поперечного сечения дают
в совокупности кажущуюся вязкость, только в 1,7 раза превышающую
постоянную величину. Согласно такой модели, кровь с высоким гематокритом
заполняет трубку диаметром 30 мкм, за исключением бесклеточного слоя
толщиной в 3 мкм вблизи стенки. В другом случае, при диаметре трубки
в 10 мкм, эритроциты при прохождении через трубку, отклоняясь от
стенок и смещаясь к оси, создают так называемую осевую цепочку диаметром
8 мкм, препятствующую возникновению сдвига слоев плазмы.
Откладывая до § 5 вопрос о том, что происходит при меньших
диаметрах трубок, в которых толщина плазменного слоя еще меньше,
упомянем, помимо эффекта Фареуса-Линдквиста, другие последствия
аксиальной концентрации. Во-первых, имеется в виду, что в сосудах диаметром
10-30 мкм «средняя скорость эритроцитов» превышает «среднюю скорость
плазмы». Следовательно, в среднем эритроцит затрачивает меньше времени
на прохождение этих сосудов, чем средняя частица плазмы! Из этого
следует (Whitmore, 1968), что гематокрит внутри таких сосудов значительно
ниже, чем в теле в целом. Такое снижение гематокрита может иметь
важное значение в органах, подобных почкам, которые перерабатывают только
плазму (гл. 10, § 11). Действительно, в почечном циркуляторном русле
происходит дальнейшее снижение гематокрита (Pappenheimer, Kinter, 1956) за
счет оттеснения плазмы из области стенок артериол и попадания ее в сеть
мелких капилляров, по которой потом скользят имеющиеся в этой сети
эритроциты.
В оставшейся части данного параграфа попытаемся ответить на
непростой вопрос, почему «эритроциты избегают стенок», т. е. рассмотреть ме-
4. Концентрация эритроцитов вдоль оси капилляров 387
ханизм аксиальной концентрации эритроцитов с позиций гидродинамики.
Некоторые неверные объяснения следует сразу исключить: например,
такие толкования, которые основываются на инерциальных эффектах
(включающих в себя «эффект Магнуса»), они в действительности совершенно
незначительны, как упоминалось в § 1; пояснения, правомерные только для
разбавленных растворов, и пояснения, игнорирующие форму эритроцитов
и их способность к деформации.
Мейсон и Голдсмит (Mason, Goldsmith, 1969) проводят обзор гипотез
механизма аксиальной концентрации эритроцитов и указывают на их
несостоятельность, хотя и отмечают, что до сегодняшнего дня не предложено
удовлетворительного объяснения данного явления. В разбавленных
суспензиях при небольших числах Рейнольдса подобные эритроцитам дисковид-
ные тела большую часть своего времени перемещаются в сдвиговом потоке
таким образом, что практически все время продольная ось диска почти
совпадает с направлением потока (т. е. угол атаки мал), но через определенные
интервалы времени, приблизительно равные обратной величине скорости
сдвига, они «переворачиваются» (так что в направлении движения потока
обращен уже противоположный край диска). При переворачивании
эритроцитов вблизи стенки столкновения их со стенкой не происходит, поскольку
за счет высокого давления в тонком пристеночном слое плазмы,
выполняющем роль смазки при движении эритроцитов в капиллярах (см. ниже, § 5),
клетки от стенки оттесняются.
В случае движения недеформируемых дисковидных клеток аксиальная
концентрация отсутствует и течение суспензии происходит таким образом,
что столкновения дисков со стенкой не происходит. Но если частицы
способны к деформации, то в разбавленных суспензиях имеет место
дополнительный механизм их аксиальной концентрации. В процессе
«переворачивания» деформируемый диск пересекает слои жидкости с разной скоростью
сдвига, там, где скорость велика, жидкость воздействует на диск,
вытягивая его в направлении дистального края, а в тех слоях, где скорость мала,
жидкость сжимает диск. Возникающее при этом радиальное смещение
поверхности диска, направленное внутрь, превышает смещение его наружу,
и происходит миграция диска в направлении оси.
Однако совершенно неясно, как реализуется такой механизм при
нормальном гематокрите, поскольку в данном случае каждая клетка все
время сталкивается с соседними. Большинство экспериментаторов наблюдали
некоторую степень аксиальной концентрации клеток в сочетании с
наличием относительно свободного от клеток слоя толщиной в несколько мик-
388
Глава 14
ронов, существование которого обусловлено оттеснением клеток от стенок,
но причина такого течения суспензий оставалась неизвестной. Возможно,
что статистически среднее поведение красных клеток под действием их
хаотических столкновений с соседними можно лучше понять, если для начала
рассмотреть равновесную ситуацию (в статистическом смысле). Действие
соседних клеток с той стороны, где сдвиговые скорости больше (где, скорее
всего, в среднем ротация клеток происходит быстрее либо «перевороты»
возникают более часто), уравновешивается большей плотностью соседних
клеток, находящихся с той стороны, где сдвиговые скорости ниже. Этим
гипотетическим рассуждением мы и закончим обсуждение вопроса о
механизме аксиальной концентрации.
5. Слой смазки в очень узких капиллярах
Завершим главу 14 обсуждением прохождения крови через столь узкие
капилляры, что форма эритроцитов должна значительно измениться,
чтобы клетки смогли пройти через них. Предварительно рассмотрим немного
подробнее форму недеформированного эритроцита и его способность к
деформации.
Эритроцит представляет собой необычно простую клетку. Эта
клетка не имеет ядра и содержит жидковатую массу гемоглобина, окруженную
тонкой гибкой клеточной мембраной, пассивно реагирующей на
механические воздействия (в отличие от фагоцитов, больших белых кровяных
клеток, имеющих ядро и активно реагирующих на воздействия, в частности, за
счет чрезвычайно сильного изменения формы). Недеформированные
эритроциты, описанные ранее как диски диаметром около 8 мкм, на самом
деле имеют еще с обеих сторон в центре диска углубления, т.е.
эритроциты вогнуты с обеих сторон диска. Здесь мы не будем подробно обсуждать
выпукло-вогнутую форму недеформированного эритроцита, лишь отметим,
что клетки, имеющие такую форму, значительно легче деформируются без
изменения объема и площади поверхности, чем клетки просто выпуклой
формы, что облегчает вытягивание и деформирование клетки для
прохождения сквозь крайне узкие капилляры.
И все-таки для деформирования исходно недеформированной клетки
требуется некоторое внешнее напряжение. В соответствии с данными,
полученными Рендом и Бартоном (Rand, Burton, 1964) и Фангом (Fung, 1966),
величина нагрузки, вызывающей достаточно сильную деформацию,
сравнимую с размерами клетки, составляет 0,5 мбар (нагрузка может представ-
5. Слой смазки в очень узких капиллярах 389
лять собой, например, разницу величин внешнего давления, приложенных
к разным частями поверхности клетки). Отметим, что эта нагрузка
значительно меньше, чем избыточное давление, необходимое для растяжения
капилляра. В легких, согласно исследованиям Уэста, Глейзера, Хьюга и Мело-
ни (West, Glazier, Hughes, Maloney, 1969), расхождение в соответствующих
давлениях на два порядка выше, а в капиллярах эта разница еще
значительнее (Fung, 1966).
При исследовании под микроскопом потока крови в очень узком
капилляре можно увидеть то, что и ожидается. Эритроциты проходят по
сосуду по одному, и прохождение клеток происходит не за счет растягивания
стенок сосуда, а за счет деформации самих эритроцитов. В
экспериментальных исследованиях наблюдалась значительная деформация
эритроцитов. Причем в не слишком узких капиллярах эритроциты принимали
форму «парашютов» (представляющих собой асимметричную конфигурацию,
выпуклую в направлении движения), а в очень узких капиллярах
эритроциты изгибались и даже складывались по диаметру как тонкие блинчики.
Аналогичные результаты наблюдались при протекании крови в
стеклянных трубках такого же диаметра (Hochmuth, Marple, Sutera, 1970).
Необыкновенное разнообразие форм деформированных эритроцитов наблюдается
в потоке крови, протекающем через капиллярную сеть легких, описанную
в конце §3. Во всех этих случаях нагрузка порядка 0,5 мбар вызывает,
как правило, достаточно сильную деформацию (и в более редких случаях
небольшую деформацию).
В своих работах Лайтхилл (Lighthill, 1968, 1969) отмечал связь течения
кровотока в узких капиллярах с гидродинамикой движения смазки,
разработанной для задач, поставленных промышленностью. В ней
рассматривается скольжение одной поверхности по другой, когда между ними имеется
очень тонкая масляная пленка, за счет наличия которой от одной
поверхности к другой передается значительная сила. В прикладных инженерных
задачах теории смазки высокие скорости развиваются в условиях высокой
вязкости масляной пленки, так что имеет место движение с низким числом
Рейнольдса (как и в микроциркуляторном русле). Действительно, именно за
счет распределения вязкого напряжения в пленке передается действие
одной поверхности на другую и толщина масляной пленки пассивно
подстраивается под характер передаваемого действия. В тех случаях, когда такая
«требуемая толщина» в некотором смысле слишком мала (так что может
возникнуть сцепление неровностей поверхностей или могут нарушиться
какие-либо гидродинамические свойства масла), то в отдельных случаях
390
Глава 14
возникает «заедание» — прерывистое скольжение, когда
«гидродинамическая смазка» заменяется «сухим трением».
Лайтхилл привел обоснование того, что воздействие со стороны стенки
очень узкого капилляра на проходящий сквозь капилляр эритроцит
приводит к деформации клетки, достаточной для ее скольжения в капилляре.
Воздействие передается посредством тонкого смазочный слоя плазмы, толщина
которого обычно пассивно регулируется в соответствии с величиной
воздействия. В том случае, если необходимая толщина этого слоя в некотором
отношении «слишком мала», то может произойти прекращение скольжения.
Исследования, проведенные Лайтхиллом, получили дальнейшее развитие
и были существенно дополнены в работах Фитц-Джеральда (Fitz-Gerald,
1969а, 1969b).
Необходимую для прохождения через очень узкий капилляр
деформацию клетки в основном вызывает радиальная компонента напряжения,
которая может передаваться через смазочный слой за счет действия в нем
избыточного давления (давление связано с толщиной смазочного слоя).
Хотя в случае движения крови в не столь узких капиллярах клетки немного
деформируются за счет продольной компоненты вязкого напряжения и
принимают форму парашюта (Fitz-Gerald, 1969a). При наличии трения
давление изменяется вдоль смазочного слоя. Градиент давления направлен вдоль
стенки и из соображений размерности (это согласуется с теорией смазки)
и пропорционален /ipU/h2, т.е. прямо пропорционален вязкости /хр плазмы
и скорости скольжения U и обратно пропорционален квадрату толщины h
пленки плазмы. Давление в пленке толщиной в несколько десятков микрон
может обеспечить необходимую деформацию эритроцитов при типичной
скорости потока в капилляре 0,5 мм/с. Наиболее существенным выводом,
вытекающим из теоретических рассуждений, является то, что толщина h
пленки должна быть пропорциональной U1/2 (так как величину fxpU/h2
можно считать в первом приближении постоянной).
Действительно, при малой толщине h пленки сила вязкости,
препятствующая движению эритроцитов в соответствии с законом вязкости
Ньютона, пропорциональна fipU/h. Отсюда следует, что если h
пропорциональна то падение избыточного давления в капилляре также
пропорционально U1/2. Из нелинейной зависимости толщины пленки и падения
избыточного давления от скорости U (они пропорциональны С/1/2) следует
достаточно интересный вывод. Например, падение давления в нелинейном
случае при достаточно малых скоростях U превышает падение давления
в линейном случае (пропорциональное U). При разработке математических
5. СЛОЙ СМАЗКИ В ОЧЕНЬ УЗКИХ КАПИЛЛЯРАХ 391
моделей протекания крови в капиллярах Лайтхтилл и Фитц-Джеральд
показали, что эти модели могут быть построены на основании теории
гидродинамической смазки при ряде дополнительных предположений (например,
принятие в расчет отклонения от осесимметричности движения и потери
жидкости через щели в капиллярной стенке). Хотя на настоящий момент
нет ни одной модели этого процесса, столь же подробно и адекватно его
описывающей, как, например, разработанная детальная механика
деформации эритроцитов.
Поскольку обычные микроскопические исследования движения
эритроцитов в капиллярах организма не позволяют определять толщину
смазочной пленки плазмы, Хочмат, Марпл и Сатера (Hochmuth, Marple, Sutera,
1970) провели тщательное измерение параметров кровотока в стеклянных
микрокапиллярах. Полученная ими в экспериментальных исследованиях
толщина пленки плазмы подтвердила правомерность применения теории
гидродинамической смазки к прохождению эритроцитов в очень узких
капиллярах. Так, например, на рисунке 5 в их работе представлена
зависимость толщины пленки плазмы от скорости U. Откуда следует, что в
трубках диаметром (а) 4,5, (Ь) 6,7 и (с) 8 мкм зависимость толщины пленки при
низких скоростях U хорошо описывается степенной зависимостью от U1/2
(если U измеряется в мм/с, то полученные зависимости необходимо
умножить на (а) 0,6, (Ь), 1,0 (с) 1,3 мкм соответственно). При превышении
скорости значения U = 1 мм/с кривая становится более пологой, т. е. толщина
пленки остается практически постоянной (возможно, что дальнейшему
нарастанию толщины пленки препятствует значительное увеличение
сопротивления клеток сильным деформациям).
Эти результаты свидетельствуют о необходимости дальнейших
исследований периферического сопротивления in vivo и нахождения некоторой
составляющей общего падения давления, пропорциональной квадратному
корню из скорости. Наличие такой составляющей давления объяснит
прохождение крови через наиболее узкие капилляры с очень небольшой
скоростью. Эта составляющая дополнит главную составляющую падения
давления, прямо пропорциональную скорости потока (дополнит, возможно,
наряду с другой составляющей, пропорциональной квадрату потока и
связанной с эффектами входной зоны, роль которой становится значительной при
больших скоростях). В настоящее время имеются многочисленные
опубликованные экспериментальные данные, связанные с периферическим
сопротивлением, в которых кривая «давление-скорость» потока вблизи
начала координат имеет параболическую зависимость и имеет при больших U
392
Глава 14
вертикальную касательную (Green, Rapela, Conrad, 1963). В прошлом эти
данные интерпретировались по-разному (например, такая зависимость
объяснялась расширением микроциркуляторного русла при повышении перфу-
зионного давления). Однако утверждение о том, что значительное падение
давления при малой величине потока обусловлено уменьшением толщины
смазочной плазменной пленки в самых узких капиллярах, кажется
довольно сомнительным.
Поэтому можно предположить, что при снижении диаметра трубки
эффект Фареуса-Линдквиста будет проявляться только в виде местного
снижения кажущейся вязкости (см. § 4), за которым следует повышение вплоть
до превышения нормальной вязкости цельной крови, характерной для
трубок малого диаметра. Такие же результаты были получены Динтенфассом
(Dintenfass, 1968) при исследовании протекания крови с гематокритом,
равным 0,49, через крайне узкую щель между параллельными пластинами
(см. рис. 7 в его работе).
Кровоток в легочных капиллярах (см. конец §3), возможно,
является промежуточным между током крови в трубках и между параллельными
пластинами, и несомненно, что многие его параметры могут быть
интерпретированы в соответствии с теорией гидродинамической смазки, в том
числе и прекращение тока в отдельных каналах при разрушении
гидродинамической смазки. Недавно доктор Дж. Б. Уэст (J. B. West) разработал
компьютерную модель капиллярной сети (не опубликовано). И им был сделан
вывод о том, что неравномерное заполнение различных каналов легочного
циркуляторного русла, наблюдаемое в экспериментах им и его коллегами
(см. § 3), предположительно играет очень заметную роль при нелинейном
законе сопротивления.
В неопубликованных экспериментах доктора Реджинальда Грина (Dr.
Reginald Greene) получено некоторое подтверждение того, что взаимосвязь
давления и потока в легких непосредственно зависит от способности
эритроцитов к деформации. Проводя эксперименты с эритроцитами,
способность которых к деформации была значительно снижена путем тепловой
обработки, он показал, что составляющая общего падения давления,
пропорциональная квадратному корню из скорости потока, присутствует при
движении обычных клеток и имеет значительно большую величину при
движении клеток, подвергнутых обработке. Эти результаты согласуются
и с теоретическими исследованиями течения методами теории
гидродинамической смазки, в соответствии с которыми было получено, что для
обеспечения продвижения клеток, менее способных к деформации, требуется
значительно больший градиент давления fipU/h2.
5. Слой смазки в очень узких капиллярах 393
В системе кровообращения наличие нелинейного сопротивления
в очень узких капиллярах может привести к повышению эффективности
вазомоторного контроля периферической перфузии (см. §2). Так как
повышение сопротивления артериол снижает скорость потока, что
автоматически увеличивает капиллярный компонент сопротивления. Отметим, что
при этом в капиллярах наблюдается некоторое «слипание» эритроцитов.
Это может быть объяснено наличием небольших колебаний размеров
эритроцитов или степени их сжимаемости (см. Whitmore, 1968), что приводит
к незначительным колебаниям толщины плазменной пленки и, как
следствие, к возникновению обратного движения плазмы в пленке. В этих
случаях плазма может накапливаться позади любой необычно маленькой или
легко сжимаемой клетки.
Ароести и Гросс (Aroesty, Gross, 1970) и Фитц-Джеральд (Fitz-Gerald,
1971) смоделировали линии тока плазмы между соседними
эритроцитами в очень узких капиллярах. И полученные результаты использовали для
оценки совместного влияния адвекции и диффузии на состояние
отдельных растворов (см. § 1). Авторы пришли к заключению о том, что
диффузия преобладает в случае растворенных газов вроде 02 и С02, а
конвекция, обусловленная тороидальной циркуляцией в «болюсе» (как его назвали
Протеро и Бартон (Prothero, Burton, 1961)) вязкой жидкости между двумя
эритроцитами и обратным движением плазмы в смазочной пленке, может
играть достаточно заметную роль в переносе слабо диффундирующих
макромолекул в системе кровообращения.
Все исследования, касающиеся проблем образования и воздействия на
кровоток смазочной пленки плазмы в микроциркуляторном русле,
находятся еще на очень ранней стадии развития. Однако несомненно, что через
некоторое время на их основе будут решены многие важные прикладные
физиологические проблемы.
Литература
[1] Aroesty, J. & Gross, J. 1970. Convection and Diffusion in the
Microcirculation, Rep. RM-6214-NIH, The Rand Corporation, Santa
Monica, California.
[2] Caro, C.G., Foley, Т.Н. & Sudlow, M.F. 1970. J. Physiol. 207, 257.
[3] Dintenfass, L. 1968. The viscosity of blood ... In Hemorheology
(ed. Copley, A. L.), pp. 197-210, Pergamon, London.
[4] Fahraeus, R. & Lindquist, T. 1931. Amer. J. Physiol. 96, 562.
394
Глава 14
[5] Farhi, L. E. 1966. Ventilation-perfusion relationship ant its role in alveolar
gas exchange. In Advances in Respiratory Physiology (ed. Саго, С. G.),
pp. 148-197, Edward Arnold, London.
[6] Fitz-Gerald, J.M. 1969a. Proc. Roy. Soc. B, 147, 193.
[7] Fitz-Gerald, J.M. 1969b. J. Appl. Physiol. 27, 921.
[8] Fitz-Gerald, J.M. 1971. J. Fluid Mech. 51, 463.
[9] Fowler, K.T., West, J.B. & Pain, M.C.F. 1966. Resp. Physiol. 1, 88.
[10] Fung, Y.C. 1966. Federation Proc. 25, 1761.
[11] Fung, Y.C. & Sobin, S.S. 1968. Ibid. 27, 578.
[12] Green, H.D., Rapela, C.E. & Conrad, M.C. 1963. Resistance and
capacitance phenomena in terminal vascular beds. In Handbook of
Physiology, Section 2: Circulation (ed. Hamilton, W. F. & Dow, P.),
pp. 935-960, American Physiological Society, Washington.
[13] Guyton, A.C. 1966. Textbook of Medical Physiology, W.D.Saunders,
London.
[14] Hochmuth, R. M., Marple, R. N. & Sutera, S. P. 1970. Microvascular Res. 2,
409.
[15] Kraemer, K. 1967. Arch. f. Kreislaufforschung, 52, 79.
[16] LighthiU, M.J. 1968. J. Fluid Mech. 34, 113.
[17] LighthiU, M.J. 1969. Motion in narrow capillaries from the standpoint
of lubrication theory. In Circulatory and Respiratory Mass Transport
(ed. Wolstenholme, G.E. W. & Knight, J.), pp. 85-104, J. & A. Churchill,
London.
[18] Mason, S.G. & Goldsmith, H.L. 1969. The flow behaviour of
particulate suspensions. In Circulatory and Respiratory Mass Transport
(ed. Wolstenholme, G.E. & Knight, J.), pp. 105-129, J. & A. Churchill,
London.
[19] Pappenheimer, J.R. & Kinter, W.B. 1956. Amer. J. Physiol. 185, 377.
[20] Pattle, R. E. 1966. Surface tension and the lining of the lung alveoli. In
Advances in Respiratory Physiology (ed. Саго, С G.), pp. 83-105, Edward
Arnold, London.
[21] Prothero, J. & Burton, A. C. 1961. Biophys. J. 1, 565.
[22] Rand, R.P. & Burton, A. C. 1964. Ibid. 4, 115.
[23] West, J.B. 1966. Regional differences in blood flow and ventilation in the
lung. In Advances in Respiratory Physiology (ed. Саго, С G.), pp. 198-254,
Edward Arnold, London.
5. Слой смазки в очень узких капиллярах 395
[24] West, J.В., Glazier, J.В., Hughes, J.M.B. & Maloney, J.E. 1969.
Pulmonary capillary flow, diffusion, ventilation and gas exchange. In
Circulatory and Respiratory Mass Transport (ed. Wolstenholme, G. E. &
Knight, J.), pp. 256-276, J. & A. Churchill, London.
[25] Whitmore, R. L. 1968. The dynamics of blood flow in capillaries. In
Hemorheology (ed. Copley, A. L.), pp. 77-87, Pergamon, London.
Полный ассортимент литературы издательств
«Институт компьютерных селе ований>
и «Регулярная и хаотическая динамика»
по самьм доступны * ценам преде авлен
в отделах прямы продаж:
/Й.
Рос , io
Институт ашиноведени им. А. А. Бла онравова РА
ул. Бс рдина . 4, корп. 3, к. 415 (м. Ленинский пр-т)
тел.: +7 (925) 280-78-9 , +7 (499) 135-54-37
e-mail: rhd m@mail.ru
Росс , е с
У муртский государст енныи университет
ул. Университетская, д. 1 корп. 4, оф. 201 а 207
телефакс: +7 (3412) 50-02-95
е mail: subscribe@rcd.ru
нт рн т- г зи
Ш55
http: /shop.rcd.
■ Отпрао а заказоа осуществляется
ПОЧТОЙ РФ И Г. ИЖ "ИСК
• Цены на cai> re уклзамы
без чегастоиуюст < с иики
Кни
о о приобрест так. е:
Московский дом книги
1осква, ул. Новый Арбат, д. 8 (м. .Арбатская»)
Тел.:+ (495) 89-35-91
Дом технической книги
М к а, Ленин к просп чет, д. 40
(м. le инский Проспект»)
Тел.: + ( 9)137 0-19
ь*ые оск ООО «Ар нт»
Москва, Ленинский проспе. , д. 65
(м. !енински~1 Проспект») Главное здание
РГУ нефти и газа им. И. М. Губкина
ООО «Санкт-Петербургская книготорговая
о п н
Санкт-Петербург, ул. капитана Воронина, д. 8
Тел.: 7(812)295-0 -57
ООО «К >
Самара, ул. Ново Садовая, д. 381, й эта —
«ТЦн Н о-С оной
Толь1Тти,у .Дзержин кого, 21, нус 1- т ,
секция 803а — «ТЦ Капитал
000«П рмкн . а»
Перь ь, > 1. Лодыгина, д.
Тел.: +7 (342) 42- 90 242- 2-
ОООиИзд те ьст о« нфр -И н »
Вологда, ул. М аши ноет рои тел ьнан д. 19, о . 238
Тел.: 7(911)512 8-48
ОООкВЕЛЕС
Омск
тел/ 7(381 )4 -31-12,46-31 1
Джеймс Лайтхилл
Математическая биогидродинамика
Дизайнер А. А. Гурьянова
Технический редактор А. В. Бакиев
Компьютерный набор и верстка А. В. Моторин
Корректор Е. В. Огородникова
Подписано в печать 31.01.2014. Формат 60 х 84 У16.
Печать цифровая. Усл. печ.л. 23,72. Уч. изд. л. 25,41.
Гарнитура Тайме. Бумага Xerox. Заказ № 14-07.
АНО «Ижевский институт компьютерных исследований»
426034, г. Ижевск, ул. Кооперативная, 5.
http://shop.rcd.ru E-mail: mail@rcd.ru Тел./факс: +7 (3412) 50-02-95
т лл
|_
• •■"-tf .Ч/У- '.