/
























































































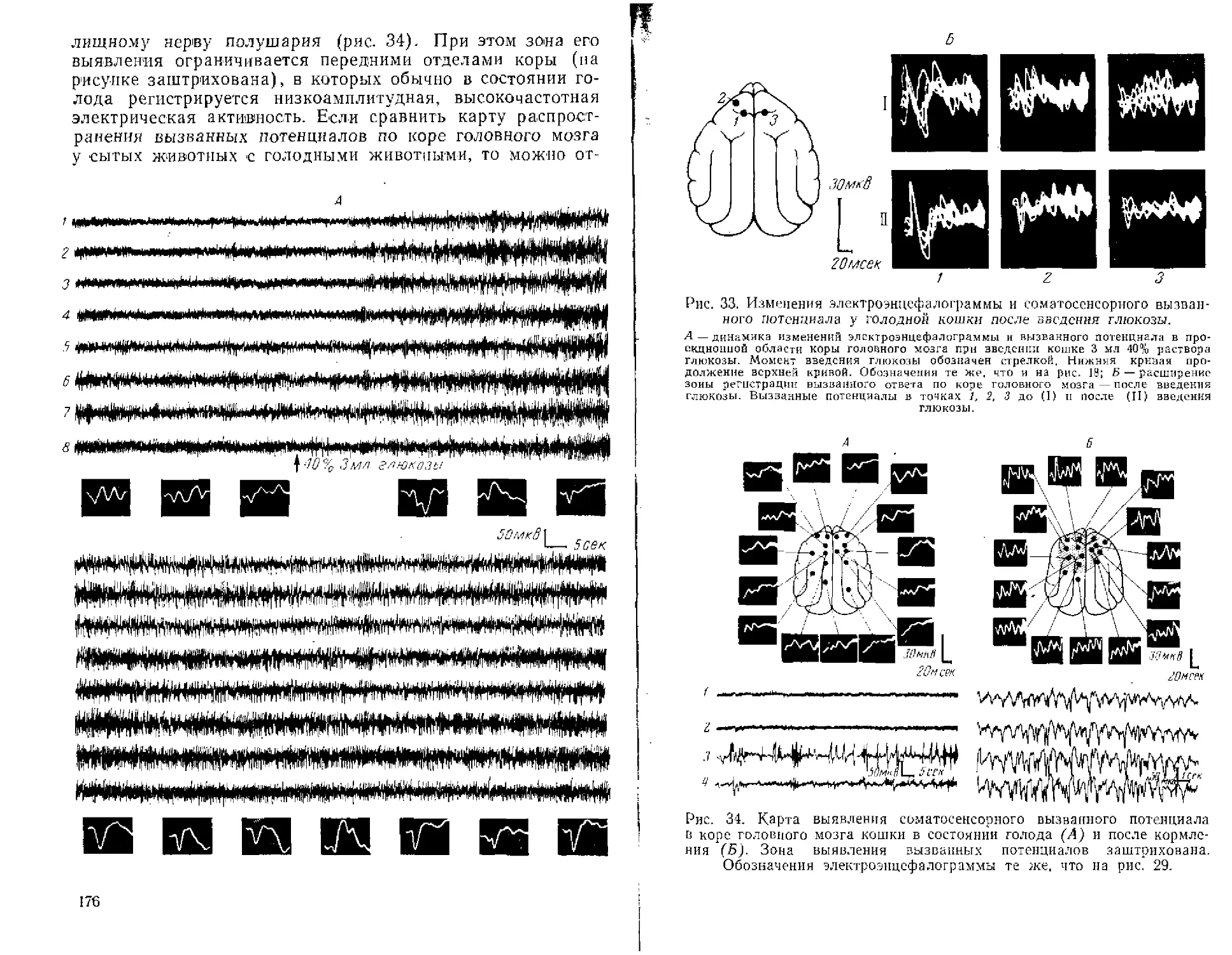

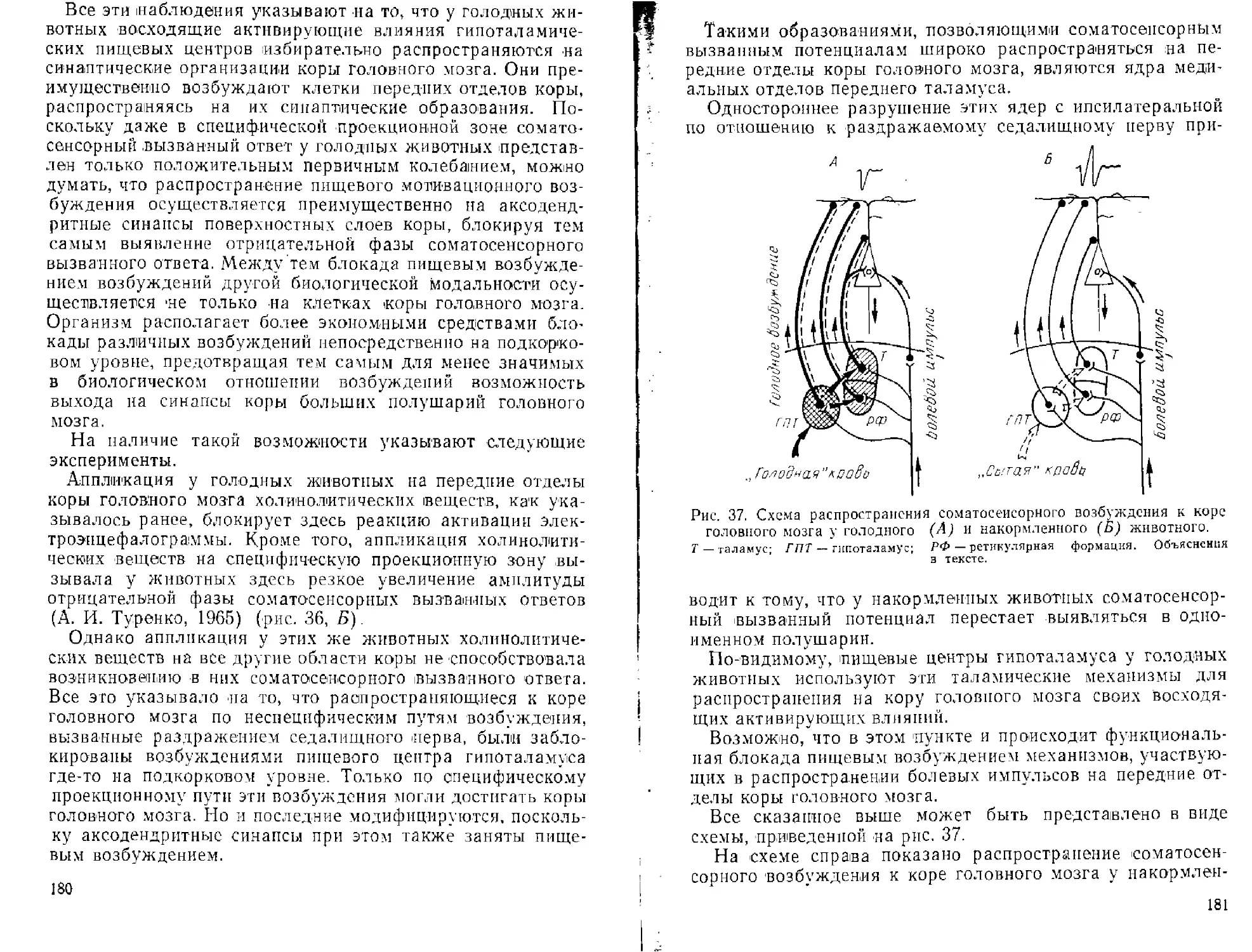


































































Текст
К. В. СУДАКОВ
БИОЛОГИЧЕСКИЕ
МОТИВАЦИИ
Издательство «Медицина»
Мо сква—1971
УДК 612.825.1
АННОТАЦИЯ
Книга «Биологические мотивации» посвящена вопросу
о природе таких эмоционально окрашенных состояний ор-
ганизма, как голод, жажда, страх, половое возбуждение,
определяющих целенаправленную деятельность животных
и человека. В работе показана роль мотивации в целост-
ных приспособительных реакциях организма, представлены
современные теории и нейрофизиологические механизмы мо-
тиваций и их подкрепления, роль эмоций в мотивационном
возбуждении, патология и психофармакология мотиваций.
Настоящая монография является первой попыткой в Со-
ветском Союзе дать систематическое изложение указанных
выше вопросов. Книга представит интерес не только для
физиологов, психологов и медиков, но и для инженеров,
занимающихся проблемами бионики.
Судаков Константин Викторович
биологические мотивации
Редактор Е. А, Колпакова
Техн, редактор Н. И. Людковская Корректор Т. П. Осокина
Художественный редактор Ф. К. Мороз
Переплет художника В. Г. Германа
Сдано в набор 17/IV 1971 г. Подписано к печати 25/VIII 1971 г.
Формат бумаги 84XI08‘/л 9,50 печ. л. (условных 15,96 л.) 17,66
уч.-изд. л. Бум. тип. № 2. Тираж 3000 экз. Т-13282. МН-71.
Издательство «Медицина», Москва, Петроверигский пер., 6/8.
Тип. изд-ва «Горьковская правда», г. Горький, ул. Фнгиер, 32.
Заказ 5074. Цена 1 р. 88 к.
5-3-1
71—56
ПРЕДИСЛОВИЕ
Развитие нейрофизиологии в последние годы характери-
зуется настойчивой тенденцией к дискретизации сложных
физиологических деятельностей, обеспечивающих гармони-
ческое приспособительное поведение животных и человека.
Считается, что выделение какого-то качественно очерченно-
го свойства, механизма или комплекса механизмов будет
способствовать более успешному пониманию целостной дея-
тельности организмов.
На основе этой тенденции в виде самостоятельных функ-
ций были выделены память, мотивации, эмоции и т. д. Они
стали объектом изолированных исследований, а часто да-
же и номенклатурной принадлежности ученого. Мы очень
часто говорим: «Он специалист по памяти»... Или: «Он фи-
зиолог эмоций»...
В результате такого дробления и вырывания отдельных
компонентов из системы и сам исследовательский процесс
по изучению этих объектов часто теряет в своем познава-
тельном значении, а иногда принимает и уродливые формы.
Примером может служить хотя бы изучение памяти.
Хорошо известно, что подавляющее большинство исследо-
ваний в этой области направлено в сторону механизмов
фиксации пережитого организмом опыта. Наоборот, почти
не пользуется вниманием другая не менее важная сторона
памяти — извлечение пережитого опыта из памяти в момен-
ты необходимости.
Как известно, наше поведение развивается в форме кон-
тинуума отдельных действий и отдельных результатов, пе-
реходящих одно в другое. А это значит, что все мы живем,
действуем, говорим и читаем, практически непрерывно (!!),
3
извлекая из памяти сегодня то, что было в ней зафиксиро-
вано вчера и на протяжении всей жизни. Поражает и явля-
ется крайне парадоксальной та легкость, с которой происхо-
дит это моментальное извлечение, если сопоставить его
с экспериментами, в которых ни охлаждение, ни кардио-
зольный шок не могли разрушить фиксацию в памяти не-
давних событий.
Извлечение из прошлого опыта идет в таком строгом по-
веденческом континууме, что хочется сказать: «да уж и есть
ли на самом деле „память” как нечто самостоятельное?»
Не является ли она органической составной частью какого-
то целого комплекса и потому, естественно, не может быть
понята без этого комплекса. Действительно, все попытки
понять «фиксацию» в памяти без одновременного исследо-
вания «извлечения» из памяти являются малоутешитель-
ными. Очевидно, оба процесса должны быть изучаемы
вместе, поскольку извлечение из памяти идет по путям, ко-
торые в прошлом послужили фиксации в ней непрерывного
континуума внешних событий.
Становится все более и более очевидным, что наступило
время, когда должен быть начат обратный процесс — про-
цесс возвращения изолированных и специфически очерчен-
ных функций в лоно целостной деятельности, т. е. в то син-
тетическое целое, которому они служат и в пределах кото-
рого они только и могут осуществить свои функциональные
особенности.
Развившийся в последние годы системный подход, так же
как «общая теория систем», не дал каких-либо конкретных
сведений о природе системы и о ее функциональной архи-
тектуре. Следовательно, исследователь опять-таки не может
определить те узловые 'Механизмы, в которых память, эмо-
ция и мотивация работают как неотделимая часть целого.
Совершенно очевидно, что память и мотивация как фи-
зиологические явления могут быть только тогда продуктив-
но исследованы, когда будут вскрыты их динамические
взаимодействия и органическая связь с другими деятельно-
стями организма (афферентация, подкрепление и др.).
4
Значительное облегчение для включения памяти и мо-
тивации в систему, я бы сказал для «натурализации» приве-
денных выше деятельностей, дала нам концепция о функ-
циональной системе (см. П. К. Анохин «Успехи физиологи-
ческих наук», 1970, № 1).
Именно благодаря тому, что функциональная система
имеет четко очерченную физиологическую архитектуру и фи-
зиологически идентифицированные конкретные механизмы,
она позволяет наметить точное место для каждой из упомя^
нутых выше деятельностей и показать их органическую
связь друг с другом в пределах функциональной системы
как интегративной единицы. .
В свете всего вышесказанного монография профессора
К. В. Судакова представляет собой несомненно оригиналь-
ное явление в физиологической литературе. Автор не толь-
ко свел воедино весь имеющийся материал по мотивациям
и эмоциям, но и сделал существенный шаг для включения
этих явлений именно в те узловые механизмы системы, ко-
торые в совокупности поддерживают системную организа-
цию до получения качественно очерченных результатов.
Такой подход к проблеме мотиваций вообще и к биоло-
гическим мотивациям в частности, несомненно, более про-
грессивен, чем рассмотрение мотиваций в изолированном
виде, в отрыве от органического единства функциональной
системы. Достаточно указать, что только при таком подхо-
де стала ясной универсальная роль конвергенции различ-
ных возбуждений на одном и том же нейроне.
Успех такого подхода в монографии К. В. Судакова, не-
сомненно, облегчился еще и тем, что сам автор в течение
ряда лет, анализируя на тонком нейрональном уровне
мотивационное поведение, не отрывал его от большой функ-
циональной системы. Это обстоятельство вносит несомнен-
ный вклад в современную нейрофизиологическую литера-
туру и будет способствовать развитию более широкого под-
хода к проблеме мотиваций. Особенно важно указать, что
«подкрепление» условного рефлекса и принцип доминанты
5
А. А, Ухтомского приведены в историческую связь с пробле-
мой мотивации.
Кроме того, К- В. Судаков ib течение длительного времени
работал в лаборатории одного из выдающихся исследова-
телей мотивационного поведения и физиологии лимбиче-
ской системы, именно в лаборатории МакЛейна (США, Бе-
тезда), что позволило ему сопоставить достоинства систем-
ного анализа мотиваций с традиционным физиологическим
подходом к этой проблеме.
Можно быть уверенным, что книга К- В. Судакова послу-
жит отправным пунктом для практического применения
теории функциональной системы и к другим конкретным
проблемам синтетической нейрофизиологии, развивавшимся
до сих пор в традициях классической нейрофизиологии.
Академик АН и АМН СССР
П. К. АНОХИН
ВВЕДЕНИЕ
«...Вся наша жизнь состоит из непрерывной
цепи формирующихся целей и их достижений.
Такие цели могут быть большими, малыми,
а иногда и просто пустяковыми, однако только
поставив перед собой такую цель, человек мо-
жет в дальнейшем формировать гармониче-
ское, а не хаотическое поведение. Сомнитель-
но, чтобы вообще можно было представить
себе какой-либо поведенческий акт человека
или даже высших животных, который мог бы
реализоваться без предварительного формиро-
вания самой цели или намерения к соверше-
нию данного действия».
(77. К- Анохин, 19626, стр, 7).
Проблема формирования целенаправленного поведения
занимает одно из важнейших мест среди многочисленных
проблем современной физиологии и биологии.
Интерес к этой проблеме не случаен.
Она является одной из самых старых и тем не менее до
настоящего времени окончательно не решенной проблемой
физиологии и медицины. В течение ряда столетий
проблема целенаправленного поведения была окутана тума-
ном мистицизма и агностицизма, чему в значительной сте-
пени способствовали безраздельно господствовавшие в эпо-
ху Средневековья религиозные теологические представле-
ния. Их влияние еще долго и в последующие этапы истории
общества и науки накладывало свой отпечаток на мировоз-
зрение многих даже очень авторитетных естествоиспытате-
лей.
Именно признанием сверхъестественных божественных
сил объясняли все акты животного и человеческого орга-
низма, имеющие хотя сколько-нибудь выраженный целесо-
образный характер. Однако пытливый ум человека, преодо-
левая пути религиозного миропонимания, настойчиво искал
материальные причины этих реакций.
Историческое развитие физиологии сложилось таким об-
разом, что первоначально наибольшее развитие получило
:7
изучение более простых реакций, в отношении которых бы-
ло установлено, что они развиваются по принципу рефлек-
са, открытому Р. Декартом. Сложные же формы целена-
правленного поведения в силу господства идеалистических
представлений вплоть до недавного времени оставались
почти совершенно не изученными.
Здесь прочно господствовали различные идеалистиче-
ские концепции, содержание которых сводилось к одному:
все формы целенаправленной деятельности определяются
и направляются действием различных божественных
сил и поэтому недоступны познанию человеческим ра-
зумом.
Такое положение объективно было обусловлено целым
рядом причин. Главная из них заключалась в том, что от-
крытый Декартом принцип рефлекса ходом исторического
развития аналитической физиологии был связан прежде
всего с различными формами спинальной деятельности. Для
западной физиологии вплоть до последнего времени голов-
ной мозг оставался сферой действия духовных сил. Это на-
шло выражение в работах даже таких крупных нейрофи-
зиологов, как Sherrington и Eccles,
Только И. М. Сеченов и особенно И. П. Павло;в рефлек-
торный принцип прочно распространили на деятельность
головного мозга и поведение.
Между тем рефлекторный принцип, как правило, не мог
объяснить многих форм' целенаправленной деятельности
живых существ.
В самом деле, нередко животные без видимой внешней
причины начинают совершать активную деятельность во
внешней среде. Они активно исследуют различные объекты
окружающего их внешнего мира и, только обнаружив оп-
ределенные из них, вступают с ними в различные взаимодей-
ствия. Целенаправленное поведение таким образом часто
возникает без видимого внешнего стимула, который, соглас-
но классической рефлекторной теории, является главным
условием для формирования ответной реакции организма.
Как же увязать такую форму поведения с классической схе-
мой рефлекса «стимул — реакция?»
Отсутствие конкретных данных о физиологических меха-
низмах этих явлений снова давало основание для различ-
ных идеалистических допущений.
Это особенно отчетливо проявилось у ряда таких крупных
Исследователей нового научного направления — этологии,
как Lorenz, Tinbergen и др. Авторы, будучи крупными нату-
8
радистами, собрали огромной важности ценнейший науч-
ный материал о поведении живых существ, об их взаимоот-
ношениях во внешней среде. Однако в объяснении внутрен-
них механизмов целенаправленной деятельности животных
авторы, как правило, апеллируют к таким неопределенным
понятиям, как «спонтанные реакции», «вакуум реакции»
И пр.
Второй причиной задержки исследований целенаправ-
ленной деятельности явилось следующее обстоятельство.
Личный опыт человеческих ощущений подсказывал, что лю-
бая целенаправленная деятельность связана с определен-
ным качеством эмоционального переживания. Однако
объективные методы выявления эмоциональных пережива-
ний у животных длительное время отсутствовали. Это яви-
лось причиной развития двух крайних дискредитировавших
себя направлений. Одно из них — антропоморфизм, т. е.
стремление исследователей наделить животных субъектив-
ными переживаниями, сходными с эмоциональными пере-
живаниями человека.
Другое направление вообще полностью отрицало нали-
чие субъективного мира у животных. Авторы этого направ-
ления настаивали на изучении только физиологических ме-
ханизмов поведения.
Известно отношение к этому вопросу И. П. Павлова.
На одной из «сред» в 1934 г. он говорил: «Глупо было бы
отрицать субъективный мир. Само собой разумеется, он, ко-
нечно, есть. Психология как формулировка явлений нашего
субъективного мира совершенно законная вещь. И нелепо
было бы с этим спорить. На этой основе мы действуем, на
этом складывается вся социальная и личная жизнь... Речь
заключается в анализе этого субъективного мира» (Пав-
ловские среды, 1949, т. 2, стр. 41'5).
К сожалению, эти замечательные мысли И. П. Павлова
не были подвергнуты глубокому экспериментальному ана-
лизу.
Таким образом, проблема целенаправленного поведения
во всех случаях упиралась в неразрешенные механизмы
внутреннего состояния животных.
Каковы же те внутренние причины, приводящие живот-
ных и человека к целенаправленному поведению? Что же
это за состояние, вызывающее целенаправленное 'поведение
животного? Каковы нейрофизиологические механизмы этих
состояний?
На все эти вопросы мы и постараемся ответить ниже.
9
ИСТОРИЯ ВОПРОСА
Рассмотрим прежде всего, как складывались научные
представления о внутренних состояниях, вызывающих у жи-
вотных целенаправленную деятельность.
Еще в конце прошлого столетия американский исследо-
ватель Sully (1884) впервые заявил, что «желания, которые
предшествуют действию и определяют его, могут быть на-
званы как движущие силы или мотив». «Избранное же-
лание становится мотивом»,— писал Dewey в 1886 г.
Так возникло представление о мотивациях как об особых
состояниях организма, которые заставляют его действовать
в определенном плане.
Термин «мотивация», как указывает Lorenz (1950), про-
исходит от латинского movere — двигаться, что в широком
смысле слова означает движение.
McDougall (1908) предположил, что первичным мотивом
всей активности человека являются инстинкты. Однако он
распространил понятие мотивации и на произвольное пове-
дение.
Отсутствие конкретных физиологических данных явилось,
по-видимому, причиной того, что проблема мотиваций ста-
ла развиваться по особому руслу. Ее развитие сначала по-
шло в основном в плане общих рассуждений, выяснения ро-
ли и места мотивации в целенаправленном поведении. По
существу только с 30-х годов нашего столетия проблема мо-
тиваций встала на научную основу и прочно заняла свое
место среди основных проблем экспериментальной психо-
логии.
В разработке этой проблемы приняли участие многие из-
вестные американские психологи (Troland, 1928; Warden,
'1931; Tolman, 1932; Joung, 1936, и др.). И только в послед-
ние годы появились экспериментальные работы, посвящен-
ные анализу нейрофизиологических механизмов мотиваций.
Эта проблема в России развивалась самобытным путем.
В отечественной литературе мотивационные состояния ор-
ганизма больше известны как влечения или побуждения.
Расстройства в сфере влечения .составляли предмет спе-
циального изучения знаменитого русского психиатра
С. С. Корсакова (1916).
Проблеме влечений посвящены специальные работы
И. М. Сеченова и И. П.'Павлова.
Рассмотрим прежде всего отношение И. М. Сеченова
к этой проблеме. В «Рефлексах головного мозга» И. М. Се-
10
ченов подробно разбирает особый класс психических явле-
ний, в которых, как он пишет, аппаратом, усиливающим
рефлексы, является «чувственное наслаждение в обширном
смысле слова».
И. М. Сеченов считает, что невольные движения, вытека-
ющие из чувственного наслаждения, зависят от физиологи-
ческого состояния нервного центра.
Согласно И. М. Сеченову, страх или отвращение, с одной
стороны, и наслаждение — с другой, есть причина целена-
правленной деятельности. Животное стремится найти
и удержать чувственное возбуждение, если оно приятно,
и, напротив, удалиться и избавиться от действия раздражи-
теля, вызывающего неприятные ощущения.
Все эти рассуждения приводят И. М. Сеченова к призна-
нию особой формы целенаправленной деятельности «психи-
ческих рефлексов с усиленным концом» или «страстных
рефлексов».
«Страстные психические рефлексы», согласно представ-
лениям И. М. Сеченова, основываются на таких эмоцио-
нальных возбуждениях, как голод, жажда, страх, половое
чувство и т. п.
И. М. Сеченов вводит категорию «желаний», которые от-
личаются от действительных чувственных наслаждений.
К таким желаниям он относит голод, жажду. Он объединяет
их термином «тоскливые ощущения», под которыми по су-
ществу понимает инстинктивные влечения. Различные же-
лания, пишет И. М. Сеченов, будучи столько же томитель-
ными, как голод и жажда, должны вызывать при долгом
неудовлетворении ту же реакцию, как и последние.
Желание (тоскливое ощущение) составляет, по И. М. Се-
ченову, основу страстного психического акта, его первые
две трети. В томительной стороне желания он усматривает
источник страсти. По его мнению, страстные психические
рефлексы лежат в основе нашего страстного поклонения
Добродетелям и отвращении от порока... Все это в наибо-
лее яркой форме излагается И. М. Сеченовым на примере
формирования любви к женщине.
«В любви к женщине,— пишет И. М. Сеченов,— есть
инстинктивная сторона'—половое стремление. Это ее нача-
ло, потому что любовь начинается, как известно, в мальчике
лишь во время созревания половых органов. Вопрос, ассо-
циирует ли мальчик уже первые половые ощущения с обра-
зом женщины невольно или эта ассоциация подготовлена
знанием наперед, решить я не берусь. Известно только, что
при нашем воспитании детей последнее случается наверно
у девяти десятых всех мальчиков...
Когда же, наконец, идеал более или менее определился,
и мальчику случалось встретить женщину, похожую по его
мысли на этот идеал, то он, как говорится, переносит свою
мечту на эту женщину и начинает ее любить в ней. По-на-
шему, он ассоциировал свой страстный идеал с реальным
образом. Это и есть так называемая платоническая любовь.
В ней половой характер чрезвычайно бледен на том основа-
нии, что рядом с яркими, следовательно, страстными, зри-
тельными и слуховыми ощущениями лежат неопределив-
шиеся, еще темные половые желания.
Но вот мужчина начинает обладать своим идеалом.
Страсть его вспыхивает еще живее, ярче, потому что место
темных, неопределенных половых стремлений заступают
теперь яркие, трепетные ощущения любви, да и сама жен-
щина является,-в небывалом дотоле блеске. Проходят меся-
цы, год, много два, и обыкновенно страсть уже потухла да-
же в тех счастливых случаях, когда с обеих сторон действи-
тельность соответствовала идеалам. Отчего это? Да на
основании закона, по которому яркость страсти поддержи-
вается лишь изменчивостью страстного образа. В год, в два,
при жизни, очень близкой друг к другу, сумма возможных
перемен и с той и с другой стороны дивным давно исчерпа-
лась, и яркость страсти исчезла. Любовь, однако, не уничто-
жилась: от частого повторения рефлекса, в котором психи-
ческим содержанием является представление любовницы
с теми или другими, или со всеми ее свойствами, образ ее
сочетается, так сказать, со всеми движениями души любов-
ника, и она стала действительно половиной его самого. Это
любовь по привычке — дружба.
Человек, раз переживший все эти натуральные фазы
полной любви, едва ли может любить страстно во второй
раз. Повторные страсти признак неудовлетворенности
предшествовавшими» (И. М. Сеченов. Избранные произве-
дения, 1953, стр. 107).
Этот пример является классическим для характеристики
формирования полового влечения и его удовлетворения,
и, как мы увидим ниже, полностью отражает многие совре-
менные взгляды на природу мотивационного возбуждения.
Особенно ценным у И. М. Сеченова является то, что он
не только обратил внимание на важность «страстных реф-
лексов», но и определил их роль в поведении. Желание как
ощущение,. пишет И. М. Сеченов, имеет всегда более или ме-
12
нее томительный, отрицательный характер; напротив, ощу-
щения, сопровождающие поступок, т. е. удовлетворение
страстного желания, имеют всегда яркий положительный
характер. 'I
И. М. Сеченов разбирает, почему желания могут прояв-
ляться или же быть подавлены. Он по существу первый об-
ратил внимание на взаимодействие различных влечений
и указал на чисто материальные процессы, лежащие в их
основе. «Бесстрастное хотение, каким бы независимым от
внешних явлений оно ни казалось, в сущности столько же
зависит от них, как любое ощущение».
Всеми своими блестящими примерами И. М. Сеченов до-
казывает, что «причина всякого человеческого действия ле-
жит вне его». Однако в данной работе И. М. Сеченов, как
он сам это пишет, разобрал только внешнюю сторону пси-
хических рефлексов, «о сущности самого процесса нет и по-
мина».
Тем не менее это был безусловно важный этап развития
нейрофизиологии. И. М. Сеченов окончательно нанес удар
по различным идеалистическим представлениям, пытавшим-
ся целенаправленную деятельность животных и человека
свести только к действию сил «божественного разума».
Большое значение исследованию «основных влечений»
организма придавал И. П. Павлов.
На III съезде по экспериментальной педагогике в Петро-
граде в 1916 г. в докладе на тему «Рефлекс цели» он указы-
вал на огромное значение «основных влечений» в генезе
«рефлекса цели». «Рефлекс цели,— говорил И. П. Павлов,—
имеет огромное жизненное значение, он есть основная фор-
ма жизненной энергии каждого из нас... Вся жизнь, все ее
улучшения, вся ее культура делается рефлексом цели, де-
лается только людьми, стремящимися к той или другой по-
ставленной ими себе в жизни цели... Наоборот, жизнь
перестает привязывать к себе, как только исчезает цель...
До какой степени у нас отсутствуют практические сведения
относительно такого важного фактора жизни, как рефлекс
цели. А эти сведения нужны во всех областях жизни, начи-
ная с капитальнейшей области — воспитания» (И. П. Пав-
лов, 1951, стр. 199).
«Рефлекс цели» И. П. Павлов понимал таким образом,
как стремление к действующему на организм объекту,
стремление к обладанию этим раздражающим объектом.
Он подчеркивал, что «рефлекс цели» формируется на базе
«основных влечений» организма, таких, как голод, страх, по-
13
ловое возбуждение и др. И. П. Павлов считал, что именно
эти влечения являются (первым толчком для целенаправ-
ленной деятельности.
В качестве примера И. П. Павлов разбирает пищевое
влечение. Он пишет: «каждый день мы стремимся к извест-
ному веществу, .необходимому нам как материал для совер-
шения нашего жизненного химического процесса, вводим
его в себя, временно успокаиваемся, останавливаемся, что-
бы через несколько часов или завтра снова стремиться за-
хватить новую порцию этого материала — пищи. Вместе
с тем ежеминутно 'всякий новый раздражитель, падающий
на нас, вызывает соответствующее движение с нашей сторо-
ны, чтобы лучше, полнее осведомиться относительно этого
раздражителя» (И. П. Павлов, 1951, стр. 198).
И. П. Павлов показывает, что на основе влечения разви-
вается мощная исследовательская деятельность («хвата-
тельный рефлекс»). С захватыванием .необходимого объек-
та развивается успокоение и равнодушие. Он указывает ина
другую важную сторону рефлекса цели —его периодич-
ность.
Говоря о том, что рефлекс цели есть универсальное яв-
ление природы, И. П. Павлов показывает пути, ,по которым
должно строиться его воспитание.
Замечательны в этом отношении следующие слова И. П.
Павлова: «Правильным пищевым режимом — соответству-
ющей массой еды и правильной периодичностью в приеме
пищи — обеспечивается всегда здоровый сильный аппетит,
нормальной пищевой рефлекс, а за ним и нормальное пи-
тание.
...Для полного правильного, плодотворного проявления
рефлекса цели требуется известное его напряжение. Англо-
сакс, высшее воплощение этого рефлекса, хорошо знает это,
и вот почему на вопрос: какое главное условие достиже-
ния цели?— он отвечает неожиданным, невероятным для
русского глаза и уха образом: существование препятст-
вий...
При продолжительном ограничении в удовлетворении ос-
новных влечений, при постоянном сокращении работы ос-
новных рефлексов падает даже инстинкт жизни, привязан-
ность к жизни...
Если каждый из нас будет лелеять этот рефлекс в себе
как драгоценнейшую часть своего существа, если родители
и все учительство всех рангов сделает своей главной зада-
чей укрепление и развитие этого рефлекса в опекаемой мас-
14
се, если наши общественность и государственность откроют
широкие возможности для 'практики этого рефлекса, то мы
сделаемся тем, что мы должны и можем быть, судя по мно-
гим эпизодам нашей исторической жизни и по некоторым
взмахам нашей творческой силы» (И. П. Павлов, 1951,
стр. 200).
К сожалению, эти замечательные мысли И. П. Павлова
не нашли широкого развития в трудах его учеников и по-
следователей.
Формулируя понятие о «рефлексе цели», И. П. Павлов ис-
пытывал, ino-видимому, определенные трудности. Действи-
тельно, все теоретические концепции И. П. Павлова строи-
лись на традиционной схеме рефлекторной деятельности,
в которой, как указывалось ранее, ведущая роль принадле-
жала внешним стимулам.
«Рефлекс цели» как стремление к объекту потребовал су-
щественного изменения этой схемы. Необходимо было при-
нять новую трактовку приспособительной деятельности,
в которой цель опережала бы действие. Это требовало изме-
нения традиционных представлений о рефлекторной дуге.
Однако, сформулировав понятие о «рефлексе цели», И. П.
Павлов уже тем самым поставил изучение проблемы цели
на строго материальную основу. А это в свою очередь по-
служило мощным толчком к дальнейшим научным откры-
тиям.
Проблема целенаправленного поведения получила новую
интерпретацию в связи с открытием динамических органи-
заций— функциональных систем организма (П. К- Анохин,
1935, 1949).
Только с позиций представлений о функциональных си-
стемах организма оказалось возможным с материалисти-
ческих позиций исследовать такое веками освещенное тео-
логами понятие, каким является «цель» .к действию.
Как же интерпретирует «цель к действию» теория функ-
циональных систем организма?
Рассмотрим, что такое функциональные системы орга-
низма.
Согласно П. К- Анохину, функциональные системы пред-
ставляют собой динамические организации, деятельность
которых направлена на обеспечение тех или иных полезных
для организма приспособительных результатов.
Такими полезными приспособительными результатами
могут быть определенные константы внутренней среды, та-
кие, например, как уровень питательных веществ, кисло-
15
рода, углекислого газа, реакции и осмотического давления
крови и др. В то же время в качестве полезных приспосо-
бительных результатов выступают все те результаты пове-
денческой деятельности организма, направленные на удов-
летворение его внутренних потребностей, т. е. на обеспече-
ние в конечном счете выживания данной особи или
продление ее рода.
Рис. 1. Схема саморегуляторного механизма функциональной системы
(по II. К. Анохину, 1968).
Так, например, вся деятельность функциональной системы
питания направлена на поддержание постоянного уровня
питательных веществ в организме, обеспечивающего его
нормальное существование. Точно так же функциональная
система дыхания поддерживает постоянный жизненно важ-
ный уровень содержания кислорода и углекислого газа
в крови. Аналогичным образом любой сложный поведенче-
ский акт, направленный на удовлетворение той или иной
потребности организма, использует множество крайне ди-
намичных функциональных систем, которые складываются
для обеспечения только какого-либо определенного приспо-
собительного этапа целостной поведенческой реакции и не-
медленно распадаются при успешном его совершении.
Таким образом, полезный приспособительный эффект вы-
ступает как центральное звено в динамической организа-
ции любой функциональной системы организма.
16
Каждая такая функциональная система работает по
принципу саморегуляции. Это означает, что любое отклоне-
ние полезного для организма приспособительного результа-
та от уровня, обеспечивающего его нормальную жизнедея-
тельность, является стимулом для включения в действие
всех аппаратов соответствующей функциональной системы,
следствием чего является возвращение данного приспособи-
тельного эффекта к исходному уровню (рис. 1). Все функ-
циональные системы имеют принципиально общие узловые
пункты. Это — конечный приспособительный эффект, спе-
цифический рецептор, воспринимающий этот эффект, об-
ратная афферентация, поступающая в центр о приспособи-
тельном эффекте, центральные воспринимающие и испол-
нительные аппараты. Последние вместе с соответствующими
исполнительными приборами (эффекторами) обеспечивают
регуляцию приспособительного эффекта, т. е. удержание его
в измененном состоянии определенное время, необходимое
для выполнения тех или иных приспособительных функций
организма в особых условиях.
Деятельность каждой такой функциональной системы,
обеспечивающей свой частный приспособительный резуль-
тат, в большинстве случаев начинается с изменения соот-
ветствующей константы крови. Эти изменения немедленно
воспринимаются соответствующими рецепторами, метабо-
лизм которых избирательно чувствителен к изменению со-
держания только данного вещества. Затем посредством
нервных и гуморальных сигнализаций возбуждаются спе-
циальные нервные центры. Возбуждения последних дина-
мически распространяются на соответствующие эффекторы,
которых может быть несколько и степень участия каждого
из них может изменяться по мере необходимости. Послед-
ние в конце концов восстанавливают исходную константу
крови. При этом уровень ее восстановления всегда контроли-
руется рецепторами, посылающими обратную афферента-
цию в центр о степени достаточности и полноценности восста-
новления данного полезного приспособительного результата.
Циклическая саморегулирующая организация каждой
функциональной системы всегда строится не на анатомиче-
ском принципе, а на основе того, чтобы осуществить наилуч-
шее, самое быстрое, наиболее экономное обеспечение той
или иной приспособительной функции организма. Вовлече-
ние различных исполнительных компонентов в деятельность
функциональной системы не является жестоко детерминиро-
ванным. В каждом случае это могут быть различные ком-
17
бинации органов и центрально-периферических приборов
нервной системы, но, как правило, все эти компоненты объе-
динены избирательной взаимозависимостью на основе их
избирательного тонического напряжения и пластично под-
держивают друг друга на путях получения конечного при-
способительного результата. Нередко различные функцио-
нальные системы могут использовать одни и те же компо-
ненты для достижения различных приспособительных
результатов.
Так, работа сердца может быть использована как для
поддержания постоянного уровня артериального давления,
так и для обеспечения газообмена и т. д.
Такая пластичность каждой функциональной системы оп-
ределяется гибкими функциональными перестройками их
центральных аппаратов.
Внутри каждой функциональной системы имеется возмож-
ность чрезвычайной взаимозаменяемости, взаимо,компенса-
ции их эффекторных механизмов. Вот почему при выходе
из строя одного или нескольких исполнительных компонен-
тов такой функциональной системы обеспечение ее конечно-
го приспособительного эффекта может осуществляться дру-
гими входящими в нее компонентами.
Так, для выправления экстренного изменения концентра-
ции СО2—О2 в крови имеются десятки различных механиз-
мов в обширной функциональной системе дыхания.
Этот эффект может быть достигнут различными путями —
углублением каждого вдоха, изменением частоты дыхания,
увеличением ударного объема сердца, учащением сердечных
сокращений, перестройкой буферных свойств крови, увели-
чением кислородной емкости крови, количества гемоглоби-
на, ускорением эритропоэза, увеличением легочной венти-
ляции и т. д. При этом, несмотря на перестройку различных
компонентов функциональной системы дыхания, конечный
приспособительный результат в виде необходимого жизнен-
но важного уровня концентрации кислорода и углекислоты
в крови остается постоянным.
Представления о функциональных системах организма
потребовали заменить старые представления о линейном
характере распространения возбуждения по центральной
нервной системе представлениями о циклических формах
взаимосвязи различных уровней центральной нервной си-
стемы.
Следовательно, динамическая организация функциональ-
ной системы приобретает уже не линейный и поступатель-
18
ный характер движения возбуждения по дуге, а вертикаль-
ную, крайне пластичную организацию, в которой наблюда-
ется неравномерное возбуждение различных входящих
в нее структур.
Факт вертикальной организации любой функциональной
системы заставляет рассматривать функциональную орга-
низацию головного мозга как совокупность .множества
пронизывающих друг друга вертикально организованных
центральных аппаратов различных функциональных си-
стем. При этом механизмы саморегуляции различных функ-
циональных систем могут замыкаться на различных уров-
нях.
Все это определяет чрезвычайную сложность динамичес-
кой организации головного мозга.
Особенно сложную организацию функциональные систе-
мы приобретают в том случае, когда в них для обеспечения
конечного жизненно важного результата включаются пове-
денческие акты.
Целостный поведенческий акт таким образом может вы-
ступать как компонент той или иной функциональной систе-
мы, поддерживающей определенную внутреннюю констан-
ту. В то же время он может формировать самостоятельные
динамические функциональные системы, обеспечивающие
выполнение определенных этапов на пути удовлетворения
основной исходной потребности организма.
Функциональные системы, обеспечивающие тот или иной
приспособительный акт организма, складываются в каждом
случае в зависимости от внешней ситуации, исходного со-
стояния организма и предшествующего опыта данного жи-
вотного. Для того чтобы понять, каким же образом в цент-
ральной нервной системе формируется состояние, опреде-
ляющее цель к действию, необходимо рассмотреть механизм
центральной архитектуры функциональных систем орга-
низма.
Центральные аппараты функциональных систем, по П. К.
Анохину, также имеют принципиально общую для каждой
из них архитектуру (рис. 2).
Первой и наиболее ответственной стадией центральной
архитектуры функциональной системы любой сложности
является стадия афферентного синтеза. Всякое отклонение
той или иной внутренней константы организма от уровня,
обеспечивающего нормальную жизнедеятельность, пред-
ставляющее собой его внутреннюю потребность, немедлен-
но через возбуждение специальных рецепторов направляет
19
в центральную нервную -систему мощные потоки афферент-
ных импульсов. Эти импульсы нередко вместе с прямым
гуморальным действием возбуждают -специальные цент-
ральные чувствительные аппараты соответствующей функ-
циональной системы. На этой основе, как мы увидим ниже,
и формируется мотивационное возбуждение.
Однако каждый организм живет во внешней среде. По-
этому наряду с воздействиями со стороны внутренней среды
О Atptp
ОЛ
ПА
ОА
Рис. 2. Принципиальная схема центральной архитектуры функциональ-
ной системы (по П. К. Анохину, 1968).
Л! — доминирующая мотивация; Z7 —память; ОД — обстановочная аффер&нтация;
ПА — пусковая афферентацяя; ПР — принятие решения; ПД — программа дейст-
вия; АРД —акцептор результатов действия; ЭВ — эфферентные возбуждения;
Д —действие; Рез.—результат; Пар. Рез.— параметры результата; ОАфф — обрат-
ная афферентация.
он подвергается постоянным внешним раздражениям. По-
этому возникшее под влиянием той или иной внутренней
потребности возбуждение на клетках центральной нерв-
ной системы (особенно коры головного мозга) па стадии
афферентного синтеза взаимодействует с возбужде-
ниями, обусловленными действием на организм специаль-
ных обстановочных и пусковых раздражителей внешней
среды.
В свою очередь возбуждения, обусловленные действием
на организм внешних и внутренних раздражителей, взаимо-
действуют, кроме того, с аппаратами памяти, т. е. прошлым
опытом данного индивидуума по удовлетворению соответ-
ствующей потребности.
20
Эту функцию взаимодействия возбуждений, вызванных
действием иа организм внешних и внутренних раздражите-
лей в каждой функциональной системе, в значительной сте-
пени берут на себя высшие отделы центральной нервной
системы— головной мозг и особенно его кора. Для построе-
ния приспособительном деятельности клетки коры головно-
го мозга должны, таким образом, «выбрать» из массы воз-
буждений, обусловленных действием на организм внешних
и внутренних раздражителей, главное и наиболее важное
для удовлетворения соответствующей его потребности
в каждый данный момент времени.
На стадии афферентного синтеза таким образом решает-
ся несколько вопросов: «что делать» (на основе сопоставле-
ния внешних и внутренних раздражителей), «как делать»
(на основе памяти) и «когда делать» (на основе действия
специальных пусковых раздражителей).
Стадия афферентного синтеза заканчивается важным мо-
ментом— принятием решения к действию. Принципиальное
значение этой стадии чрезвычайно важно, так как она огра-
ничивает организм от многочисленных «степеней свободы»
и заставляет выполнять только одну конкретную форму по-
ведения, соответствующую его внутренней потребности
и окружающей обстановке.
Именно на этой стадии сложного приспособительного
акта и формируется цель к действию. Вот здесь-то мы
и встречаемся с важным моментом работы функциональных
систем--формированием цели к действию. Это связано
с деятельностью специального аппарата, который в лабо-
ратории П. К- Анохина был назван сначала «акцептором
действия», а затем «акцептором результатов действия»
(П. К. Анохин, 1945, 1949, 1968).
Акцептор результатов действия возникает всякий раз,
как только принимается соответствующее решение к дейст-
вию и формируется исполнительное действие. По своей глу-
бокой сути акцептор результатов действия является аппара-
том, программирующим результаты будущих событий. На
основе врожденного и индивидуального опыта данного жи-
вотного акцептор результатов действия заключает в себе
свойства внешних раздражителей, необходимых для удов-
летворения соответствующих внутренних потребностей ор-
ганизма. Благодаря этому организм имеет возможность
оценивать поступающую в центральную нервную систему
животного афферентацию о результатах и параметрах со-
вершенного действия, сравнивать ее со свойствами удовлет-
21
воряющего данную потребность раздражителя. Таким обра-
зом, этот аппарат в подлинном смысле слова является
аппаратом предвидения и оценки результатов деятель-
ности.
Такое предвидение результатов деятельности и особен-
но свойств специальных раздражителей, удовлетворяющих
ту или иную внутреннюю потребность организма, и являет-
ся по существу материальным процессом постановки опре-
деленной цели к действию.
Следовательно, «цель» включается в деятельность любой
функциональной системы, в которой для удовлетворения
той или иной внутренней потребности необходим поиск спе-
циального раздражителя во внешней среде.
Принятие таких представлений о цели к действию по су-
ществу меняет традиционное мышление с позиций рефлек-
са. Согласно классическим представлениям, весь процесс
в рефлексе развертывается линейно и идет последователь-
но от начала до конца, от стимула к действию. Понятие це-
ли, естественно, требует признания такой формы реагирова-
ния, когда сама «цель» опережает реальные события.
Таким образом, «цель» по самой своей глубокой сути
есть опережение реальных событий действительности.
Каждая цель реализуется в действие. При этом реализа-
ция цели в действие также представляет собой сложный
интегративный процесс. В каждом случае он избирательно
объединяет специальный комплекс исполнительных аппара-
тов, соматические и вегетативные компоненты («эфферент-
ный интеграл» по П. К. Анохину).
В результате всех этих 'Процессов формируется соответ-
ствующее целенаправленное поведение, которое сопровож-
дается специальными вегетативными реакциями.
Это целенаправленное поведение строится на основе по-
стоянной оценки организмом результатов, совершенных во
внешней среде действий, сравнения их со свойствами акцеп-
тора результатов действия.
Результат любой деятельности организма прежде всего
оценивается с той точки зрения, в какой степени он удов-
етворяет исходную потребность организма.
Итак, постановка цели к действию вытекает из самой
сущности динамической саморегулирующейся организации
функциональных систем организма. Какое же место зани-
мают мотивации в формировании целенаправленной дея-
тельности?
I. ПРОБЛЕМА МОТИВАЦИИ
КЛАССИФИКАЦИЯ мотивации
В целенаправленной деятельности животных и человека
чрезвычайно важную роль играют такие эмоционально
окрашенные внутренние состояния, как голод, жажда,
страх, агрессия, половое возбуждение и пр. Именно они й
составляют наследственно обусловленный фонд основных
биологических мотиваций.
Действительно, все животные рождаются с примитивными
биологическими влечениями голода, страха, жажды. В даль-
нейшем к ним присоединяются также возникающие на осно-
ве наследственных механизмов половые влечения.
Именно эти биологические мотивации прежде всего и
определяют целенаправленную деятельность животных и че-
ловека, поскольку они первостепенно важны для сохранения
жизни индивидуума и продолжения его рода. В самом деле,
голодное животное упорно и настойчиво ищет пищу, живот-
ное в состоянии полового возбуждения — особь противопо-
ложного пола, а страх гонит животное на большие расстояния
И все это во имя сохранения жизни животного и продле-
ния его рода. Эти мотивации обладают высокой энергети-
ческой силой, которая заставляет животных активно дейст-
вовать во внешней среде.
Между тем побудительные мотивы значительно усложня-
ются в процессе индивидуальной жизни животного, в про-
цессе его обучения.
Woodworth (1918) первый выдвинул представление о том,
что привычки могут стать побуждениями, и тем самым вы-
ступил против теории McDongal (1908), который связывал
мотивации только с инстинктами.
В дальнейшем многие исследователи подтвердили эти
представления. Было установлено, что в процессе обучения
влечения могут возникать и под влиянием специальных фак-
торов внешней среды.
Между тем совершенно ясно, что такие мотивации также
формируются на основе врожденных «основных влечений».
23
Однако, каковы взаимоотношения между ними, каким обра-
зом определенные внешние раздражители приобретают са-
мостоятельное побудительное значение, остается почти не
выясненным.
В конечном счете вся наша жизнь состоит из непрерывной
цепи формирующихся целей и их достижений: ищет ли
голодный пищу, а трус бежит, гонимый страхом, эгоист стре-
мится к достижению личного благополучия, а человек с вы-
сокими моральными идеалами — к работе на благо общест-
ва, ученый — к познанию новых тайн природы, поэт — к пи-
санию стихов и т. д. Все это различные примеры мотиваций.
Даже этот незначительный перечень примеров мотиваций
указывает на многообразие их форм.
В свое время известный русский психиатр С. С. Корса-
ков (1916) пытался классифицировать различные мотива-
ции. Однако эта классификация, несмотря на ее глубокий
смысл, не является четкой и достаточно удобной, поскольку
в ней смешаны психологические и физиологические понятия.
Согласно С. С. Корсакову, «первую, наиболее низшую
группу мотивов, составляют побуждения, вытекающие из не-
посредственного стремления к достижению того, что прият-
но самому себе и что удовлетворяет животным инстинктам
человека. Вторую — составляют побуждения, хотя тоже вы-
текающие из непосредственного стремления к тому, что при-
ятно самому себе, но вытекающие из примитивных интеллек-
туальных стремлений—пытливости, любознательности,
развлечения (игры, занятия и пр.), подражательности,
стремления к красивому. Третью группу составляют по-
буждения, вытекающие из непосредственного стремления
доставить приятное другим или удалить от них неприятное
(сочувствие, любовь).
Четвертую группу составляет более сложный ряд моти-
вов— мотивов пользы и вреда (себе или другим людям)
от данного поступка. Пятая группа — мотивы обязанности,
долга.
Наконец, высшую группу мотивов составляют те, которые
обусловливаются побуждениями не только непосредственно-
го удовлетворения душевных интересов (притом не для себя"
лично, но и для других), не только правильным рассужде-
нием о пользе и вреде поступков и не только непосредст-
венным чувством долга и обязанности, но и разумом и со-
знанием теснейшей связи всех явлений и своего личного я
со всем строем жизни, стремлением быть в полной гармо-
нии с высшими идеалами разума, любви, труда, самопо-
24
жертвования и мудрого подчинения всех своих побуждений,
даже и таких, которые кажутся происходящими из беско-
рыстных источников,— высшему идеалу соединенного доб-
ра, разума, любви и энергии (С. С. Корсаков, 1954, стр.
108—109).
Более простая классификация мотиваций предложена
Н. Миллером (1960). Он подразделяет мотивации на низ-
шие, или первичные, и высшие, или вторичные. К низшим он
относит все формы врожденных биологических влечений.
Иначе он их называет инстинктивными, или висцеральными.
Основное биологическое значение первичных мотиваций со-
стоит в обеспечении нормальных условий для обмена ве-
ществ в организме. Они же являются исходными реакция-
ми организма, приводящими к продлению его рода.
Именно эти состояния организма И. П. Павлов (1916)
рассматривал как «основные влечения» и относил их к раз-
ряду сложнейших безусловных рефлексов.
Характеризуя сложные безусловные рефлексы как фор-
му приспособления организма, И. П. Павлов в статье
«Условный рефлекс» писал: «Первое обеспечение уравнове-
шенности, а следовательно, и целостности отдельного орга-
низма, как и его вида, составляют безусловные рефлексы
как самые простые (например, кашель, при попадании по-
сторонних тел в дыхательное горло), так и сложнейшие,
обыкновенно называемые инстинктами — пищевой, оборони-
тельный, половой и др. Эти рефлексы возбуждаются как
внутренними агентами, возникающими в самом организме,
так и внешними, что и обусловливает совершенство уравно-
вешенности» (И. П. Павлов, 1951, стр. 450).
К «основным влечениям» организма можно отнести сле-
дующие: влечение к пище (голод), к воде (жажда), к воз-
духу, к избеганию повреждения (страх), к поддержанию
оптимальной температуры, к отдыху (после продолжитель-
ной деятельности), ко сну (после длительного бодрствова-
ния) , к активности (после бездействия).
Все они формируются по единому трафарету на основе
врожденных наследственных механизмов.
С известной долей осторожности в эту же группу могут
быть отнесены и наиболее примитивные мотивации — позы-
вы, которые предшествуют таким актам организма, как мо-
чеиспускание, дефекация, кашель и чиханье.
Нетрудно заметить, что все первичные влечения связаны
с поддержанием постоянства внутренней среды (или гомео-
стаза по Кеннону).
25
Они характеризуются рядом особенностей. Любое такое
влечение формируется под влиянием весьма определенных
раздражителей внутренней среды. Так, пищевое влечение
возникает при истощении питательных запасов организма,
половое — в связи с накоплением половых гормонов и т. п.
Такие влечения присущи как человеку, так и всем формам
живых существ.
При всех первичных мотивациях исходные изменения во
внутренней среде организма вызывают целую цепь взаимо-
обусловленных процессов, которые можно подразделить на
внутренние и внешние. Внутренние процессы основываются
целиком на вегетативных механизмах. Внешние механиз-
мы мотивации включают соответствующее поведение, на-
правленное на добычу необходимых веществ при освобож-
дении организма от продуктов жизнедеятельности, или из-
бавление его от неблагоприятных условий жизни.
К наиболее сложным врожденным формам первичных мо-
тиваций относятся исследовательские и половые влечения.
Вторую группу мотиваций составляют высшие мотивации.
Они приобретаются в течение индивидуальной жизни и стро-
ятся на основе первичных мотиваций и накопленного опыта.
У животных к группе высших мотиваций относятся раз-
нообразные целенаправленные сложные реакции, связанные
с деятельностью во внешней среде, основу которой состав-
ляет условнорефлекторная деятельность.
В процессе обучения животных и человека сигналы, ра-
нее индифферентные, после неоднократного сочетания с тем
или иным подкреплением (например, пища, вода) приобре-
тают способность вызывать соответствующую мотивацию.
Как возникают такие реакции, отчетливо показывает сле-
дующий эксперимент (Miller, 1960).
Крыс помещали в специальную камеру, состоящую из
двух отделений. В одном из отделений, окрашенном в белый
цвет, через решетку пола животные получали раздражение
электрическим током. Однако они могли избежать этого
раздражения, если перебегали во второе отделение, окра-
шенное в черный цвет, В последнее вела специальная дверь,
открыть которую крыса могла, только повернув специаль-
ное колесо или нажав на специальный засов.
Опыт проводили таким образом, что сначала при полу-
чении тока в белом отделении крысам позволяли беспрепят-
ственно перебегать в черное отделение через открытую
дверь. Затем условия опыта были изменены. Крыс помеща-
ли в белое отделение, но дверь, ведущую из него во второе
26
отделение, закрывали. Оказывалось, что даже в случае, ес-
ли животные не получали электрического раздражения в бе-
лом отделении клетки, они проявляли подчеркнутые при-
знаки страха, выражающиеся в беспокойстве, мочеиспуска-
нии, испражнении, попытках притаиться. Характерно, что
многие животные настойчиво стремились проникнуть через
дверь в черное отделение и после ряда проб обучались от-
крывать дверь.
Таким образом, прежде нейтральные раздражители бе-
лого отделения (общий вид, запах и пр.) приобрели значе-
ние мощных стимулов, формирующих мотивацию страха.
Перебегание из белого в черное отделение приобретало
свойства подкрепления.
Нетрудно заметить, что главное отличие мотивации, воз-
никающей в данном случае, заключается в том, что она сти-
мулируется определенными внешними, ранее индифферент-
ными раздражителями. Эти формы реакции строятся по ти-
пу классического условного рефлекса И. П. Павлова.
Высшие мотивации человека крайне разнообразны и ка-
чественно отличаются от таковых у животных. Они опреде-
ляются прежде всего наличием у человека второй сигналь-
ной системы и абстрактного мышления. Социальная среда
формирует у человека такие мотивации, как долг перед Ро-
диной, политические убеждения, нравственные и эстетиче-
ские идеалы, творческие устремления, спортивные увлече-
ния, страсть к коллекционированию и т. д., превращаю-
щиеся в форму постоянного побуждения часто целой
жизни.
Несмотря на то что в основе высших мотиваций, как пра-
вило, лежат механизмы «основных влечений», высшие моти-
вации могут значительно изменять последние. Достаточно
сопоставить самопожертвование патриота со страхом смер-
ти, любовь — с половым инстинктом, вкус к определенной
пище, воспитанный национальными обычаями и личными
привычками,— с голодом.
По-видимому, первичные мотивации в их «чистом» виде
в реальной действительности могут наблюдаться только в
первые часы или дни после рождения животного. В процес-
се его индивидуальной жизни е каждым днем механизмы
мотиваций все в большей степени усложняются и приобре-
тают новые многогранные качества.
В современной психологии проблема мотиваций имеет,
как указывают Bindra и Steward (1966), три главных ас-
пекта.
27
1. Механизмы влечения (drive). Этот аспект
проблемы связан с необходимостью раскрытия природы ос- [
новных влечений организма, тех внутренних механизмов, I
которые заставляют животных и человека проявлять настой-
чивое целенаправленное поведение.
2. Целенаправленное поведение. Здесь пер- ,
воочередным является необходимость ответа на вопросы:
что заставляет животных достигать определенных целей
и какие механизмы определяют целенаправленность пове-
дения.
3. Природа п одкрепления (удовлетворение вле- t
чения). Здесь возникают такие вопросы, как: почему живот- '
ные неоднократно повторяют одни формы деятельности и
стремятся избегать другие и какова природа положительных
и отрицательных эмоций в этом механизме.
ОСОБЕННОСТИ МОТИВАЦИОННОГО ВОЗБУЖДЕНИЯ
Отличительной особенностью мотивационных реакций ,
является то, что они возникают без видимого внешнего сти-
мула и, наоборот, определяют активное воздействие живот-
ного на окружающую среду (П. К. Анохин, 19626).
Они являются теми исходными состояниями организма,
которые заставляют животных и человека активно искать 1
или избегать определенных раздражителей внешнего мира.
Такая особенность мотиваций коренным образом отлича-
ется от реакций организма, совершаемых непосредственно
под влиянием раздражителей внешней среды и развиваю-
щихся по типу «стимул — реакция».
В физиологии уже давно накапливались факты, показы-
вающие, что обычная схема «стимул — реакция» не всегда
может быть применена к объяснению различных поведенче-
ских реакций целого организма. По существу учение о мо-
тивации, включающей врожденные и приобретенные меха-
низмы поведения, возникло как бы в противоположность
бихевиористскому направлению, которое рассматривало по-
поведение только согласно схеме «стимул — реакция».
В зависимости от исходной мотивации может значительно
изменяться отношение животных к одним и тем же раздра-
жителям внешней среды.
Общеизвестен факт, неоднократно наблюдавшийся в пав-
ловской лаборатории, что условные пищевые рефлексы в
ответ на предъявление условных сигналов хорошо проявля-
ются у голодных и совершенно исчезают у накормленных
28
животных (С. Б. Хазен, 1908; П. Н. Никифоровский, 1910;
И. П. Павлов, 1933, и др.).
Активная роль мотиваций отчетливо выражается и в об-
щеповеденческих реакциях животных. Первым признаком
развивающегося мотивационного возбуждения всегда явля-
ется подчеркнутая ориентировочно-исследовательская реак-
ция (животные настораживаются, прислушиваются, приню-
хиваются, присматриваются к окружающим их предметам).
Вслед за этим начинается активный поиск определенных ве-
ществ во внешней среде. Все это указывает на то, что внеш-
ние раздражители не всегда определяют характер и особен-
ности поведения. Сплошь и рядом мы видим примеры того,
что поведение обусловливается внутренними факторами и
развивается по типу «организм — реакция, направленная
на внешнюю среду и активный поиск специальных раздра-
жителей внешней среды». Именно такая принципиальная
схема поведения наблюдается при мотивациях.
Однако принятие такого принципа организации поведе-
ния требует определенной осторожности. В самом деле, ес-
ли признается, что внутренние факторы поведения являют-
ся в организме индетерминированными, или «спонтанными»,
то такая позиция является бесспорно идеалистической.
Именно таких воззрений, как уже указывалось ранее, при-
держиваются некоторые представители этологического на-
правления (Lorenz, Tinbergen и др.). Напротив, с точки
зрения материалистической методологии, внутренние фак-
торы, организующие целенаправленное поведение, являются
материальными факторами, связанными прежде всего с из-
менениями во внутренней среде организма. Именно эти
материальные факторы запускают специальные механизмы
мотиваций внутри организма, которые и формируют целе-
направленные акты без непосредственного действия на
организм объекта поиска. И если первая из упомянутых
выше точек зрения разоружает исследователя, заранее
предопределяя его беспомощность, то вторая точка зрения
побуждает исследователя настойчиво искать особенности
внутренних механизмов, определяющих целенаправленные
реакции живых существ, тех механизмов, которые и состав-
ляют физиологическую сущность мотивации.
Естественно возникает вопрос: какие же факторы опре-
деляют столь активный характер поведения, обусловленного
исходной мотивацией, и каковы нейрофизиологические ме-
ханизмы мотивационного возбуждения?
29
Но прежде чем перейти к этим вопросам, рассмотрим, ка-
кими методами современная психология и нейрофизиология
изучают различные стороны мотивационного возбуждения.
ОБЪЕКТИВНЫЕ МЕТОДЫ ИЗУЧЕНИЯ МОТИВАЦИЙ
Мотивации являются определенным внутренним состояни-
ем организма. Будучи полностью сформированными, они
приводят к целенаправленным поведенческим реакциям. По-
этому вполне естественно, что целенаправленные поведенче-
ские реакции являются одним из основных внешних прояв-
лений мотиваций. Таким образом, поведение животного мо-
жет характеризовать наличие у него определенной мотива-
ции. В самом деле, когда мы видим, что спокойно лежавшее
животное вдруг встает и начинает направленно совершать
поиск чего-либо, то совершенно определенно мы имеем ос-
нование полагать, что у такого животного возникла какая-то
мотивация. И если это поведение животного направлено на
поиски пищи, то можно говорить о пищевой мотивации; если
животное ищет воду,—о питьевой мотивации, и т. п. При этом
такое целенаправленное поведение у любого животного вы-
ражается в весьма специфической форме, в результате чего
правильно охарактеризовать его не представляет никакого
труда даже ребенку.
Обширный экспериментальный материал, посвященный
изучению механизмов свободного целенаправленного пове-
дения животных, представлен работами отечественных
физиологов (И. С. Беритов, П. С. Купалов, П. К. Анохин
и др.).
П. К. Анохин (1932) предложил оригинальную методику
свободного выбора. Она состоит в том, что животному на че-
тырехстороннем станке предлагается на выбор несколько
сортов пищи (например, мясо, сухари, молоко, солевой рас-
твор и т, д.), находящейся на разных сторонах станка. Ес-
тественно, что животное в этих условиях проявляет направ-
ленный интерес только по отношению к тому веществу, ко-
торое соответствует его исходной мотивации.
Многочисленные опыты показали, что мотивации обла-
дают высокой энергетической силой. Животное, реализуя
исходную мотивацию, двигаясь к потребному ему веществу,
способно преодолевать сложные препятствия (барьеры, за-
городки и даже площадки, через которые пропускается
электрический ток). Варьируя подобные препятствия, мож-
30
но определить и измерить силу исходного побуждения жи-
вотного к тому или иному веществу.
Изучение мотивации производится также путем наблюде-
ния за целенаправленным поведением животного при спе-
циальных воздействиях на интерорецепторы его внутренних
органов. В качестве примера могут служить эксперименты
А. М. Уголева и В. Н. Черниговского (1959). Авторы показа-
ли, что можно искусственно получить целенаправленные ре-
акции животного к определенному сорту пищи при раздра-
жении слизистой оболочки желудка различными пищевыми
и химическими веществами.
Так, при введении в желудок раствора хлористого натрия
собака предпочитает пить воду и избегает концентрирован-
ных растворов хлористого натрия.
Другим примером изучения мотиваций являются исследо-
вания поведения животных при удалении половых желез
(М. К. Петрова, 1936). Клиническая практика знает много
случаев избирательных влечений, которые появляются при
нарушениях гормональных функций.
Каким же образом поведение может характеризовать ис-
ходное мотивационное возбуждение?
Показателем мотивационного возбуждения может слу-
жить измерение общей двигательной активности животного
в специальных камерах. Wang (1923) показал, например,
что у самок крыс двигательная активность возрастает в со-
ответствии с нарастанием уровня эстрогенов в крови и по-
вышением в связи с этим половой возбудимости. При сниже-
нии уровня эстрогенов двигательная активность падает.
Двигательная активность животных возрастает также при
лишении их пищи, воды, витаминов, изменении окружаю-
щей температуры и т. д.
Кроме того, степень мотивационного возбуждения может
определяться объемом потребляемого фактора. Так, на-
пример, чем более голодно животное, тем большее количе-
чество пищи оно способно принять за одно потребление.
Крысы, например, при лишении их воды вплоть до 8-го
дня пьют все возрастающее количество воды (Stellar, Hill,
1952). В случае полового возбуждения это может быть бы-
строта возникновения эякуляции и т. д.
Применение стереотаксической техники исследования и
усовершенствование методов раздражения головного мозга
позволили активно вмешиваться в работу различных обра-
зований центральной нервной системы и следить за изме-
нением соответствующего поведения животных.
31
Так, например, было показано, что раздражение электри-
ческим током области латерального гипоталамуса приводит
к возникновению целенаправленного пищевого поведения
и гиперфагии. Разрушение этих отделов головного мозга,
наоборот, приводит к устранению пищевых побуждений.
Голодные животные с такой операцией не едят даже при-
влекательную для них пищу и погибают от истощения
(Anand, Brobeck, 1951; Teitelbaum, Stellar, 1954; Anand,
Dua, 1955, и др.).
Противоположный эффект, т. е. отказ животных от пред-
лагаемой пищи или значительное уменьшение потребления
пищи, наблюдался при раздражении медиальной гипотала-
мической области (Anand, Dua, 1955).
Разрушение медиальных отделов гипоталамуса приводи-
ло к увеличению приема пищи и ожирению подопытных жи-
вотных (Hetherington, 1943; Brobeck, 1943; Anand, Brobeck,
1951; Teitelbaum, Campbell, 1958).
Изменение поведения при искусственном раздражении
определенных подкорковых структур наблюдал А. Б. Коган
(1949).
Между тем, как показывает опыт, целенаправленное по-
ведение животных не всегда может в достаточной степени
верно указывать на наличие той или иной мотивации. В этих
случаях всегда имеется опасность антропоморфической ин-
терпретации поведения, т. е. приписывание животному эмо-
циональных переживаний, свойственных только человеку.
Точно так же отказ животного от совершения той или иной
поведенческой реакции (например, еды) еще не является
показателем отсутствия мотивации.
Поэтому для объективного изучения поведенческих про-
явлений мотивации применяются специальные методики.
Самыми распространенными являются специальные инстру-
ментальные методики (Skinner, 1938; Miller, 1960). Они по-
зволяют судить о наличии у животного той или иной моти-
вации по специальной реакции (нажатие на рычаг, повора-
чивание специальной вертушки, дергание за кольцо и т. п.),
вызывающей соответствующее подкрепление. Так, если жи-
вотное успешно нажимает на рычаг, вызывающий появле-
ние пищи, можно говорить о наличии у него голодной мо-
тивации. Аналогично о мотивации жажды можно говорить
в том случае, когда животное нажимает на рычаг, чтобы
получить воду, и т. д.
Такие же эксперименты (многочисленные) были проведе-
ны Miller (1960, 1961). Он отметил, что у животных, обучен-
32
ных нажимать для получения порции пищи на специальный
рычаг, по мере голодания наблюдалось возрастающее уве-
личение скорости и частоты нажатия на этот рычаг. В ана-
логичном эксперименте состояние жажды также сопро-
вождалось увеличением скорости и частоты нажатия на
рычаг.
В специальных экспериментах было показано, что пище-
вые побуждения (скорость нажатия на рычаг) снижаются
как при введении пищи в ротовую полость, так и в желудок.
При этом введение пищи в ротовую полость вызывает более
сильное ослабление побуждений (Miller, Kessen, 1952). Все
это указывает на то, что интенсивность проявления у живот-
ного усвоенной им в результате обучения инструментальной
реакции может характеризовать интенсивность исходного
влечения.
Особенно успешно изучение мотиваций в условиях инстру-
ментальных методик развернулось, когда экспериментаторы
стали исследовать инструментальные реакции одновременно
с активным воздействием (удаление, раздражение) на раз-
личные структуры головного мозга.
В результате этого с начала 1950 г. в изучении проблемы
мотиваций начался новый этап. Накопились новые экспери-
ментальные факты, и проблема мотивации прочно заняла
свое место среди основных проблем нейрофизиологии.
Многочисленные эксперименты показали, что раздраже-
нием определенных нервных центров можно вызвать не
только первичные мотивации, на что указывалось выше, но
и определенные навыки, ведущие к удовлетворению данной
мотивации.
Так, например, если у крыс был выработан определенный
навык в виде нажатия на рычаг, за которым следовало пи-
щевое подкрепление, то при раздражении латеральной гипо-
таламической области даже после кормления такие крысы
снова начинали нажимать на рычаг (Miller, I960). Такие
же реакции нажатия на рычаг для получения воды наблю-
дались у кошек при инъекции в область гипоталамуса ги-
пертонического раствора хлористого натрия.
Появление ранее заученной реакции нажатия на рычаг
наблюдалось при раздражении латерального гипоталамуса
адренергическими и холинергическими веществами (Gros-
sman, 1960, 1966).
Специальными экспериментами Miller (1960) показал так-
же, что при искусственном раздражении головного мозга,
вызывающем реакцию страха, животные быстро и успешно
2 Заказ Б074
33
образуют соответствующие навыки. Кошки, которые ранее
были обучены вращать специальное колесо, чтобы выклю-
чать болевое раздражение, в ответ на раздражение этих
структур головного мозга немедленно вращали это колесо.
Аналогичные наблюдения были сделаны Delgado (1959)
и Lilli (1960). На основе электрического раздражения опре-
деленных структур промежуточного мозга у кошек и обезьян
авторам удалось образовать двигательный оборонитель-
ный навык в виде вращения колеса, выключавшего раздра-
жение. В дальнейших экспериментах удалось вызвать реак-
цию вращения колеса на предварительно включавшийся
звук.
На основании этих данных можно думать, что при искус-
ственном воздействии на определенные структуры головно-
го мозга удается вызвать поведенческую реакцию, которая
внешне соответствует различным внутренним побуждени-
ям. Однако естественно поставить вопрос, является ли
состояние животного, которое наблюдается при искусствен-
ном раздражении его мозговых структур, именно тем состоя-
нием, которое возникает при натуральном побуждении.
Эксперименты Miller показывают, что для исследования
физиологических механизмов различных мотиваций изуче-
ние только поведения экспериментального животного не яв-
ляется достаточным.
Скорость нажатия на рычаг не всегда является
мерой исходной мотивации. Если у крыс осуществля-
лось двустороннее разрушение медиальных отделов гипота-
ламуса, то такие животные съедали пищи гораздо больше
контрольных, однако скорость нажатия на рычаг была у них
значительно меньше, чем у контрольных. Резкое возраста-
ние скорости нажатия на рычаг также отмечалось при дли-
тельном голодании, однако количество принятой пищи не
превышало того, которое крыса потребляла после 24-часо-
вого голодания.*
Это обстоятельство заставило предположить, что слож-
ные навыки, образованные на основе различных первичных
мотиваций, могут определяться другими структурами го-
ловного мозга. Такого же мнения придерживается Morgane
(1961, 1966), который считает, что латеральный гипоталами-
ческий отдел пищевого центра состоит из двух функцио-
нально различных частей. Одна из них (передне-боковая
часть) определяет пищевые поведенческие реакции, а вторая
(средне-боковая) — пищевое мотивационное возбуждение.
Все это показывает, что поведение не всегда может дать
34
правильное представление о возникновении пли исчезнове-
нии биологической мотивации.
Например, если животное отказывается от еды, это еще
не критерий его «сытого» состояния. Отказ от еды в дан-
ном случае может быть вызван самыми разнообразными
причинами (страх и другие биологические состояния).
В связи с этим следует высказать критические замечания по
следующему вопросу.
Некоторые авторы (А. И. Баранов, 1951; Paintai, 1954,
и др.) используют механическое раздражение желудка для
изучения механизмов насыщения. Это делается на том осно-
вании, что раздувание резинового баллона в желудке часто
приводит к отказу животного от еды.
Специальные опыты показали, однако, что раздувание
баллона в желудке вызывает в центральной нервной систе-
ме изменения, противоположные тем, которые возникают
при натуральном пищевом раздражении (Н. В. Асмаян,
К. В. Судаков, 1964; К. В. Судаков, Л. С. Мнасин, 1965).
Если воздействие пищи па рецепторы желудка приводит к
снижению активности подкорковых и корковых отделов пи-
щевого центра, то механическое раздражение желудка, на-
оборот, вызывает значительное повышение активности тех
же структур.
Таким образом, несмотря на то что при натуральном на-
сыщении и при воздействии на желудок резинового баллона
наблюдается с внешней стороны один и тот же эффект—от-
каз животного от еды, однако причины, его вызывающие,
различны, различно и функциональное состояние пищевого
центра. При механическом раздражении желудка, как по-
казывают наши наблюдения, отказ животного от еды вызы-
вает в центральной нервной системе неприятное состояние,
близкое к тошнотному.
К такому же заключению приходит Miller (1957). В его
опытах, проведенных на крысах в Т-образном лабиринте,
животные избегали той стороны, где им производилось ме-
ханическое раздражение желудка. Тем не менее скорость
ранее выработанной реакции нажатия на рычаг, за которым
следовало пищевое подкрепление у этих животных, при раз-
дувании баллона в желудке понижалась примерно так же,
как у контрольных крыс, которым в желудок вводили мо-
локо.
Все это заставляет очень осторожно относиться к оценке
поведенческого показателя в изучении механизма мотива-
ции.
2*
35
Особый интерес для изучения мотиваций представляют
эксперименты (Olds, Milner, 1954; Olds, 1956) по изучению
феномена «самораздражения» у крыс. Оказалось, что раз-
дражение определенных структур головного мозга (гипота-
ламус, перегородка и др.) через вживленные электроды при-
водит к тому, что животное впоследствии начинает само
включать ток, чтобы наносить себе раздражение. Таким об-
разом, в этих экспериментах стимулом к соответствующему
поведению становится искусственное раздражение мозговых
образований. И эти мотивации, как показали авторы, так
же как и естественные влечения, обладают значительной
энергетической силой, поскольку для того, чтобы получить
подкрепление в виде электрического раздражения, крысы
могут осуществлять весьма сложные поведенческие акты.
Экспериментам по самораздражению мы уделим специаль-
ное внимание ниже.
В последнее время одно из основных мест среди других
методов изучения мотиваций заняли электрофизиологиче-
ские методы исследования. С помощью электрофизиологиче-
ских исследований удается обнаружить конкретные нервные
структуры и механизмы, лежащие в основе мотивационных
реакций. Особенно плодотворной оказалась в этом отноше-
нии методика телеметрии (Delgado, 1960, 1965).
В хроническом опыте электроэнцефалографические изме-
нения в коре и различных подкорковых структурах сопо-
ставляются, коррелируются с поведением, и с определен-
ным состоянием животного (голод, жажда, страх) и с удов-
летворением его потребностей.
Надо заметить, что и данные электроэнцефалограммы
также не всегда являются специфическим показателем той
или иной мотивации, так как одни и те же электрические
феномены, например реакция десинхронизации, проявляют-
ся при различных эмоциональных состояниях (П. К. Ано-
хин, 1964а). Критически оценивая каждый из методов объ-
ективного изучения мотиваций, нельзя не прийти к выводу,
что успешное изучение их физиологических механизмов
возможно только при комплексном изучении поведенческих
реакций, вегетативных проявлений этих реакций, биоэлект-
рической активности различных структур головного мозга
в сочетании с направленным воздействием на специальные
мотивационные аппараты головного мозга.
В связи с тем что мотивации заняли почетное место среди
основных проблем нейрофизиологии, интерес к этой пробле-
ме значительно усилился.
36
Однако для того, чтобы понять природу мотиваций, не-
обходимо совершенно точно определить их место в целена-
правленных приспособительных реакциях организма.
Ниже будут изложены основные теории мотиваций,
ТЕОРИИ МОТИВАЦИЙ
Как указывалось ранее, проблема мотиваций историче-
ски возникла в психологических лабораториях.
Это определило и то обстоятельство, что большинство
теорий мотиваций носит чисто описательный характер и по
существу в большинстве своем не идет дальше общих рас-
суждений, не имеющих в основе конкретных физиологиче-
ских механизмов. Эти теории пытаются объяснить мотива-
ции как общебиологическое явление, показать их место в це-
ленаправленной деятельности животных и человека. Ниже
будут рассмотрены только некоторые наиболее существен-
ные, на наш взгляд, общепсихологические теории мотива-
ций. Не претендуя на всестороннее и глубокое освещение
этого вопроса, мы рекомендуем интересующимся читателям
познакомиться с подробными монографиями следующих ав-
торов: Bindra (1959), Brown (1961), Hall (1961), Joung
(1961), Cofer и Appley (1964), Atkinson (1964), Bindra, Ste-
ward (1966), Bartoshuk (1966) и др.
Основная цель настоящей работы заключается в том,
чтобы показать нейрофизиологические механизмы биологи-
ческих мотиваций. Поэтому нейрофизиологическим теориям
мотиваций, естественно, будет уделено большее внимание.
Общие теории мотиваций
Многие психологические теории мотиваций возникли как
своего рода протест против традиционной манеры мышле-
ния американских бихевиористов, рассматривающих все
формы поведения с позиции трафаретной схемы «стимул —
реакция».
Важные критические замечания в адрес бихевиористов
сделал Craig (1918). Он одним из первых указал на то, что
имеется множество примеров поведения животных при от-
сутствии непосредственно объекта, на который в конечном
счете направлено воздействие данного животного и которые
не укладываются в обычную схему «стимул — реакция».
Такие формы поведения были названы Craig «аппетентны-
ми». К ним Craig относил поиск пищи, особи противополож-
37
ного пола, партнеров по стаду и стае, материала для по-
стройки гнезда и т. д.
Именно этот центральный момент — отсутствие явного
внешнего стимула в ряде целенаправленных реакций, явил-
ся решающим в построении теорий мотиваций. Условно су-
ществующие общие теории мотиваций можно подразделить
на две группы.
Одна группа теорий мотивации посвящена энергетической
стороне мотивации, ее внутренним факторам, тому, что у
психологов получило название собственно влечения (drive).
Согласно этой точке зрения, влечение является своеобраз-
ной «внутренней готовностью, определяющей целенаправ-
ленные реакции животных».
Другая группа теорий в происхождении мотиваций значи-
тельную роль уделяет специальным направляющим факто-
рам внешней среды.
Рассмотрим обе эти точки зрения.
Концепция об энергетической роли влечений берет свое
начало от McDougall (1908), который считал, что источни-
ком энергии (первичными двигателями) всех разнообраз-
ных форм произвольной деятельности являются врожденные
механизмы инстинкта. Инстинкты, по его мнению, определя-
ют характер восприятия и реагирования животных на мно-
гочисленные окружающие его раздражители определенным
способом.
Такой же по существу точки зрения придерживался Fre-
ud (1925), а также в своих ранних работах по этологии
Lorenz (1950) и Tinbergen (1951).
Согласно Freud, высшие психические функции определя-
ют две группы первичных инстинктов: инстинкты самосохра-
нения (ego-instincts) и инстинкты сохранения рода (sexu-
al- instincts). Выдвигая эти две формы инстинктивной дея-
тельности как источник психической деятельности, Freud
в то же время совершенно ошибочно, па наш взгляд, пола-
гал, что энергетическая сила этой деятельности определяет-
ся своеобразным конфликтом между этими двумя формами
инстинктов.
По Lorenz (1953), целенаправленное поведение зависит от
непрерывного накопления энергии в нервных центрах. Эта
энергия освобождается при совершении животным опреде-
ленных действий при наличии специальных раздражителей
внешней среды. Однако опа может выявляться и при их
отсутствии. Часто животные в неволе, лишенные всех
естественных соответствующих раздражителей (releasing
38
situation), продолжают совершать врожденные реакции со-
вершенно так же, как они совершали бы их на свободе, в ес-
тественных условиях жизни. Так, голодный скворец, по опи-
санию Lorenz, без всякого раздражения из внешней среды
часто фиксирует глазами невидимую добычу и хватает ее.
Такие реакции Lorenz назвал реакциями в пустоте (vacuum
reaction). Эти реакции, по мнению Lorenz, определяются са-
мопроизвольным освобождением энергии, накопленной в
нервных центрах.
Все указанные выше авторы исходят из одного общего по-
ложения: энергия поведения освобождается и убывает по
мере его выполнения. Такую точку зрения характерно выра-
зил Mac Dougall, когда он писал об «открывании инстинк-
том двери для получения специфических раздражителей».
Представление об энергетическом значении влечений в
возникновении целенаправленных действий и их обеспечении
было развито Woodworth (1918). Однако в отличие от ука-
занных выше авторов Woodworth считал, что энергия влече-
ния, возникая в определенных нервных центрах, затем под-
держивается энергией нервов и мышц, участвующих в ре-
акциях.
Woodworth, а вслед за ним Lashley (1938) и Hebb (1955)
выдвинули представление о том, что энергетическую основу
мотивации составляет неспецифическое генерализованное
состояние возбуждения (своего рода внимание). Это, как
будет видно ниже, нашло подтверждение в работах некото-
рых нейрофизиологов (Fisher, 1956; Morgan, 1957). Однако
эта теория не нашла достаточного распространения. По-ви-
димому, вследствие того что ее авторы утверждали наличие
неспецифического механизма мотиваций. Ведь для каждого
исследователя было очевидно, что мотивация всегда на-
правлена на определенные специфические раздражители
внешней среды.
Концепция об исключительно внутренних энергетических
факторах мотивации была подвергнута справедливой крити-
ке еще и в связи с тем, что естествоиспытатели накаплива-
ли все большее число факторов, показывающих, что в мо-
тивациях примают существенное участие определенные фак-
торы внешней среды. Так, например, материнские влечения
у крысы, выражающиеся в уходе за потомством, можно под-
держивать вместо обычных 15—20 дней вплоть до одного
года. Для этого необходимо только подкладывать все новых
новорожденных животных. Этот пример показывает, что
мотивации определяются не только аккумулированной энер-
39
гией, а определенными специальными направляющими фак-
торами внешней среды.
Точка зрения об освобождении инстинктивной энергии
влечения была также подвергнута критике Nissen (1953)
и Hinde (1960). Авторы выдвинули представление о том,
что не только освобождающаяся энергия врожденных ин-
стинктов является причиной целенаправленной деятельности
животных. Влечение определяется, по их мнению, опреде-
ленным действием. Это могут быть приближение или удале-
ние от необходимого объекта, определенная поза, исследо-
вательская деятельность и т. д. На основании этого Nissen
формулирует, теорию множественности мотиваций. Он
утверждает, что каждое незначительное движение имеет
свою собственную мотивацию.
Nissen (1951) считает, что в основе целенаправленной
деятельности лежит некий «сенсибилизирующий компо-
нент». Эта сенсибилизация является избирательной и опре-
деляется внутренними потребностями организма. Наряду
с этим фактором, по Nissen, существуют «направляющие
факторы» внешней среды, определяющие поведение живот-
ного даже в случае отсутствия явных внутренних потребнос-
тей. Как подтверждение своей точки зрения автор приводит
следующий пример. Взрослый самец шимпанзе может спо-
койно отдыхать и даже спагь в тот момент, как в клетку
вносят еду или впускают охочую самку. Незамедлительно
вслед за этим проявляются явные признаки пищевого или
полового возбуждения и, если позволяет обстановка, живот-
ные едят или спариваются.
Beach (1947) подтвердил эту точку зрения. Он показал,
что восприимчивость к половому возбуждению у низших жи-
вотных сильно зависит от половых гормонов. Однако у бо-
лее высокоорганизованных видов, например человекообраз-
ных обезьян и человека, половое влечение может возник-
нуть без внутренних гормональных факторов, под влиянием
только внешних раздражителей.
Все это явилось причиной того, что наряду с указанной
выше возникла и другая точка зрения, согласно которой
влечения стали рассматривать как результат влияния опре-
деленных направляющих факторов внешней среды.
Эта точка зрения исходит из того положения, что мотива-
ции определяются специальными внешними факторами
(«разрешающей ситуацией»), к которым животное стремит-
ся в процессе своей целенаправленной деятельности и при
наличии которых они могут совершать определенные физио-
40
логические реакции. Эти реакции получили специальное
название «завершающих» (consummatory responses).
Так, если голодное животное стремится к пище, то его
стремление к ней, согласно данной точке зрения, определя-
ется реакцией на пищу как на «завершающий фактор».
Аналогично «завершающие факторы» направляют живот-
ных к поиску воды, особи противоположного пола и т. д.
Такая точка зрения была обоснована Craig (1918), который
полагал, что все доведение животных складывается из ап-
ветентного поведения и завершающего акта.
Значение «завершающих реакций» отмечено уже в рабо-
тах Lorenz и Tinbergen.
Так, Lorenz пишет о «врожденном высвобождающем ме-
ханизме». Последний представляет собой определенную про-
грамму, которая развертывается только при наличии соот-
ветствующих факторов внешней и внутренней среды; по-
следние являются своеобразными «ключевыми» раздражи-
телями.
Tinbergen (1951) развивает эту точку зрения в своеобраз-
ную концепцию об «иерархии поведения». Наиболее образно
описание такой иерархии дается им в отношении репродук-
тивного поведения самца колюшки. Созревание половых
гормонов, а также определенная длительность дня застав-
ляют самца колюшки в определенный период жизни мигри-
ровать в мелкую воду. Так у него формируется общий мотив
(инстинкт воспроизведения по Тинбергену). Однако этот
мотив реализуется через ряд промежуточных. Сначала са-
мец выбирает собственную территорию и яростно защищает
ее от притязаний других самцов, затем строит гнездо (ко-
пает, ищет материалы, склеивает их и т. д.). Только после
этого самец допускает самку к гнезду и спаривается с ней,
после чего вся цепь поведенческих реакций завершается
своеобразной заботой о потомстве. Таким образом, Tinber-
gen расчленяет цикл половой деятельности на более мелкие,
дробные элементы, которые и составляют своеобразную ие-
рархию инстинкта.
Все эти элементы, по мнению Tinbergen, представляют
собой своеобразные «завершающие акты». Последние ха-
рактеризуются строго стереотипным двигательным выраже-
нием. Так, «завершающие акты» в нападении у самца ко-
люшки заключаются в преследовании, кусании, угрозе
и т. д. «Завершающие акты» при спаривании заключаются
в зигзагообразном танце, направлении самки к гнезду, по-
казывании входа, дрожаний, оплодотворении икры и т. д.
41
Как указывает Tinbergen, «завершающие акты» стано- I
вятся менее подвижными по мере приближения к концу со- |
ответствующей иерархии инстинкта. !
Характерно, что для выполнения любого такого «завер- |
шающего акта» необходимо наличие своеобразных раздра- |
жителей внешней среды. Так, например, «завершающие ак- |
ты» при спаривании возникают только при наличии самки;
нападение проявляется только при наличии рыбы соответ- г-
ствующей окраски — другого самца и т. д. <
Сам Tnibergen считает, что выявление каждой последую- (
щей ступени иерархии связано со снятием определенных |
тормозящих влияний. (-
Однако нередко мотивационные реакции происходят и в 5
отсутствие соответствующих внешних раздражителей, как бы
«в запас». При этом в своей деятельности животное руко- *
водствуется только определенными «моделями» будущей
«завершающей реакции».
Bailliey и Morrison (1963) показали, что при разрушении -i
латеральной гипоталамической области, несмотря на то,
что животные отказываются самостоятельно есть, они про-
должают успешно нажимать на рычаг в случае, если вслед
за этой процедурой пища инъецируется им через фистулу
в желудок, Следовательно, как полагают авторы, и в этом
случае при невозможности самостоятельного потребления
пищи у животных формируется определенная пищевая «мо-
дель».
Удаление половых желез у высших млекопитающих и че-
ловека также не расстраивает их половые влечения и соот- -
ветствующее поведение во внешней среде.
Young (1961) считает, что половые гормоны только акти-
визируют определенную врожденную модель поведения, ко-
торая может усложняться индивидуальным опытом.
Все эти примеры указывают на то, что направляющими
факторами мотивации могут быть определенные внутренние
врожденные и приобретенные в индивидуальной жизни мо-
дели поведения.
В формировании таких внутренних моделей поведения
у человека значительная роль принадлежит прежде всего
социальным факторам. Это особенно отчетливо заметно на
примере формирования у человека полового влечения.
По сути дела «образ» полового партнера формируется
у ребенка задолго до полового созревания. Этому способст-
вуют личные наблюдения за окружающими, разговоры'
взрослых, книги и т. д. Половое созревание только активи-
зирует эти представления. Еще более характерный пример
зависимости половых влечений от социальных факторов при-
водят Kinsey и др. (1953).
На основании статистических исследований американские
авторы указывают на то, что женщины проявляют повы-
шенную сексуальность преимущественно в пре- и постмен-
струальный период. В то же время в середине менструаль-
ного цикла, т. е. в момент овуляции и повышенного содер-
жания в организме половых гормонов, их половое влечение
минимально.
С точки зрения биологии человека такое явление пара-
доксально. Казалось бы, что половое влечение должно быть
максимальным в период, когда условия для продления рода
наиболее благоприятные. Вместе с тем, как полагают авто-
ры, снижение полового влечения в этот период определяет-
ся прежде всего таким социальным фактором, как'боязнью
беременности.
Имеется множество примеров несоответствия гормональ-
ного состояния человека его половой активности. Так, жен-
щины в период менопаузы, как правило, сохраняют обычное
для них половое влечение.
Young (1961) описывает случай, когда гермофродит муж-
ского пола при медицинском обследовании ошибочно был
принят за женщину. Этого оказалось достаточно для того,
чтобы во всей своей жизни (в одежде, привычках и т. д.)
он в дальнейшем проявлял поведение женщины.
К этой же группе теорий мотивации тесно примыкают тео-
рии, постулирующие, что влечение определяется положи-
тельными эмоциями подкрепления.
Так, Harlow (1959), изучая поведение новорожденных
обезьян, наблюдал у них поразительное влечение к опреде-
ленной ткани. Такие животные предпочитали макет «мате-
ри» из мягкой одежды проволочному, несмотря на то, что
они, как правило, кормились на последнем.
Harlow и др. (1950) считают также, что влечения к ма-
нипулированию или любопытство, которое проявляется у
обезьян в стремлении исследовать окружающее без специ-
ального вознаграждения, также разыгрывается на основе
положительных эмоциональных ощущений.
Все эти влечения рассматриваются автором как стремле-
ние к получению определенного гедонического аспекта за-
вершающего ответа.
Наибольшее распространение получила теория, согласно
которой мотивации обусловлены стремлением животных к
;42
43
подкреплению, поскольку соответствующее подкрепление
ослабляет силу исходного влечения (drive reduction) (Hull,
1943; Miller, Dollard, 1941).
Согласно этой теории, голодное животное стремится к пи-,
ще, поскольку пища снижает тоническую силу исходного
влечения голода. Так же животное, испытывающее жажду,
стремится к воде, поскольку последняя снижает неприятные
эмоциональные ощущения жажды, и т. д. Miller считает, что
всякое снижение исходной мотивации всегда является дос-
таточным условием для подкрепления, на основе которого
и происходит целенаправленное поведение и обучение.
Skinner (1953), Brown (1953), Spence (1956) и Estes
(1958) вообще полагают, что все мотивации направляются
только определенными внутренними и внешними (ключевы-
ми— key) стимулами, и выступили вообще против общей
теории генерализованного влечения.
Однако в последнее время большинство исследователей
стали склоняться к компромиссной точке зрения (Hull, 1943;
Smith, 1966; Karli, 1968; Morgan, 1969, и др.), согласно ко-
торой любая мотивациия имеет два аспекта. Один из них
чисто энергетический, побуждающий фактор, а другой —
фактор, направляющий животное к цели.
Эти два разных аспекта определили по существу и два ас-
пекта физиологических теорий мотивации. Между тем, если
в отношении энергетического аспекта мотивации имеются
определенные нейрофизиологические подтверждения, второй
аспект по-прежнему выглядит довольно тенденциозным.
Это со своей стороны снова оставляет окрытой дверь раз-
личным теологического смысла токованиям.
Физиологические теории мотиваций
Изучение механизмов мотивации с физиологических пози-
ций значительно задержалось. Это объясняется тем, что
для их исследования потребовалась разработка специаль-
ных объективных методов.
Вместе с тем подходу с физиологических позиций к про-
блеме мотиваций, так же как и к проблеме эмоций, препят-
ствовала предубежденность многих, даже очень крупных
исследователей, которые в последние годы пытались вообще
отрицать наличие у животных внутренних субъективных
состояний.
Естественный ход развития науки все более определенно
указывает на то , что основу любого субъективного состоя-
44
ния организма составляют физиологические механизмы. По-
этому задача современных исследований состоит не в отри-
цании субъективного мира, а прежде всего в том, чтобы по-
казать, как и когда на основе физиологических механизмов
формируются субъективные эмоциональные состояния
и какую роль они играют в приспособительных поведенче-
ских реакциях. Таким образом, проблема мотивации явля-
ется одновременно психологической и физиологической
проблемой. И. М. Сеченов очень точно определил эти состо-
яния организма как «психические рефлексы с усиленным
концом», или «страстные психические рефлексы»;
Рассмотрим теперь различные представления о физио-
логических механизмах мотиваций.
Периферические теории
Одной- из первых теорий, пытающихся объяснить физио-
логические механизмы влечений, явилась теория Кэннона
(1932, 1934). Автор полагал, что различные влечения воз-
никают в результате специфической деятельности опреде-
ленных периферических органов, что определило название
«периферическая теория» мотиваций. Так, возникновение
жажды Cannon рассматривал как результат сухости в ро-
товой полости за счет снижения секреции слюны; возникно-
вение голода — как результат влияний из пустого желудка
и т. п.
По существу Cannon сформулировал унитарную теорию
мотиваций. Возникновение мотивации, так же как и ее
прекращение, согласно этой теории, связано с действием на
одни и те же периферические органы. Так, жажда возникает
при сухости ротовой полости и исчезает при смачивании тех
же рецепторов водой. При этом подкрепление выступает в
своеобразной роли «отмерителя» силы исходного влечения.
Заслуга Cannon состоит в том, что ои первый перешел от
изучения чисто поведенческой стороны мотиваций к изуче-
нию их внутренних механизмов. Его теория вскрывает один
из возможных компонентов мотивационного возбуждения—
импульсацию от периферических органов.
В начале XX века «периферическая теория», казалось бы,
получила определенное экспериментальное подтверждение.
Так, после открытия моторной периодической деятельности
желудка в состоянии голода (В. Н. Болдырев, 1904) ряд ав-
торов, и прежде всего Cannon и Washborn (1912), Carlson
(1919), Cannon (1934) и др., пытались связать ощущения
45
голода с этими периодическими сокращениями пустого же-
лудка. На этом основании Carlson создал специальную тео-
рию регуляции аппетита. Он-полагал, что весь комплекс ощу-
щений голода определяется наличием или отсутствием ак-
тивности мускулатуры желудка.
Эти представления были поддержаны рядом отечествен-
ных исследований.
Так, Д. Ф. Филимонов и Б. X. Чирейкина (1929) рассмат-
ривали «голодную» периодическую деятельность желудка
как важный стимул, заставляющий животных устремляться
на поиски пищи.
Однако уже наблюдения С. В. Аничкова (1924) заставили
усомниться в правильности этих представлений.
С. В. Аничков в экспериментах, проведенных на самом се-
бе, не смог обнаружить зависимости между приступами же-
лудочных сокращений и ощущениями голода. Он отметил,
что чувство голода нередко возникало и в период полного
покоя желудка. В то же время желудочные сокращения не-
редко вызывали такое состояние, которое можно было ско-
рее квалифицировать как «тошнотное».
К аналогичным выводам пришел и В. И. Иванов (1926),
обследовавший 24 здоровых человека и обнаруживший, что
периодические сокращения желудка часто не сопровожда-
лись у них субъективными ощущениями голода.
Р. И. Гаврилов и А. А. Значкова (1933), изучая длитель-
ное голодание у собак, также не смогли установить прямой
зависимости между реакцией на пищу и характером «голод-
ных» желудочных сокращений. На 3—4-й день голодания
пищевая реакция животных была значительно выражена,
а моторика желудка угнетена и, наоборот, в более поздние
сроки голодания пищевая реакция отсутствовала, несмотря
на усиленную моторную деятельность желудка.
В 1900 г. Sherrington обратил внимание на то, что живот-
ные даже после полного удаления желудка в состоянии го-
лода проявляют отчетливое стремление к пище. Этот факт
никак не мог быть объяснен с точки зрения теории Кэннона
и Карлсона. В дальнейшем целой серией убедительных экс-
периментальных доказательств было показано, что как при
полной ваготомии, так и при спланхникотомии, при которых
полностью устранялись «голодные» сокращения желудка, ,
реакции животных к пище сохранялись полностью (Bash,
1939; Grossman a. oth., 1947; Harris a. oth., 1947, и др.).
Аналогичные наблюдения были сделаны и при операци-
ях на людях. При этом также было выяснено, что после
46
полного удаления желудка или его денервации голодные
ощущения продолжали проявляться в совершенно отчетли-
вой форме.
Сохранение стремления к пище у голодных собак с пол-
ной перерезкой спинного мозга на грудном уровне и обоих
блуждающих нервов наблюдал также Н. Ф. Попов (1950).
Все эти опыты отчетливо указывали на то, что централь-
ный нервный пункт регуляции пищевого поведения в со-
стоянии голода находится вне пищеварительного тракта
и что «голодная» периодическая деятельность желудка не
является единственным источником голодных побуждений.
Близка к этой другая теория мотивации, выдвинутая
Hebb (1949, 1953). Эта теория непосредственно вытекает из
концепции «drive reduction». Hebb в отличие от Cannon
в центре внимания ставит эмоциональный компонент. Hebb
считает, что все мотивации определяются стремлением ор-
ганизма избежать неприятных, нередко болевого характера
эмоциональных ощущений («страданий»), которые могут
возникать при раздражении различных органов. Согласно
этой теории, голодное влечение рассматривается как стрем-
ление избавиться от «голодных болей», возникающих в эпи-
гастральной области при сокращениях пустого желудка,
жажда — как стремление избавиться от неприятных ощу-
щений сухости в полости рта и глотке. Таким же образом
даже материнские влечения связываются со стремлением
избавиться от неприятных, ощущений, связанных с растя-
жением молочной железы накапливающимся в ней мо-
локом.
Нетрудно заметить, что хотя в концепции Hebb также
отводится важная роль импульсациям, идущим от перифе-
рических органов, она принципиально отличается от приве-
денной выше концепции Cannon, поскольку в ней мотива-
ции определяются не прямыми влияниями от перифериче-
ских органов, а, наоборот, стремлением животных избежать
этих раздражений.
Теория Hebb, так же как и теория Cannon, справедливо
указывает на значение в формировании мотиваций состоя-
ния различных периферических органов как возможных
провоцирующих факторов. Однако сохранение влечения
к пище после удаления всего желудка и материнских влече-
ний после удаления грудных желез делает теорию Hebb
также не совсем убедительной.
Все это, естественно, направило .исследовательскую
мысль в сторону более настойчивого выяснения значения
47
роли внутренней среды организма и тех гуморальных ве-
ществ крови, которые могли бы определять мотивационное
возбуждение.
Так, для объяснения голодных мотиваций возникла тео-
рия «голодной» крови. Согласно этой теории, причиной воз-
никновения голодных мотиваций считали раздражающее
действ.ие крови с измененным содержанием питательных ве-
ществ на определенные нервные центры (И. П. Павлов,
1911).
Эта теория заставила Bulatao и Carlson (1924) пересмот-
реть раннюю теорию голода, постулированную Cannon.
Вместо этого они выдвинули представление о том, что со-
стояние голода определяется гипогликемическими условия-
ми. Основаниями для этого послужили следующие наблю-
дения. Авторы заметили, что «голодные» сокращения
желудка так же, как и ощущения голода, тормозились по-
сле внутривенного введения глюкозы, в го время как введе-
ние инсулина, снижая уровень глюкозы в крови, наоборот,
усиливало «голодные» сокращения желудка и ощущения
голода. Исходя из этого, они предположили, что возникно-
вение периодических «голодных» сокращений желудка,
определяющих чувство голода, обусловливается первичным
снижением уровня глюкозы в крови.
Однако и эти представления не нашли в дальнейшем
экспериментального подтверждения.
Было установлено, что ощущения голода не совпадают
с самым низким уровнем сахара в крови и проявляются да-
же в том случае, когда уровень сахара в крови возрастает
(Scott a. oth., 1938; Grossman, Stain, 1948; Janowitz, Ivy,
1949).
Тем не менее в последние годы Mayer и др. (1951), Van
Itallie и др. (1953), Stunkard и др. (1958) и другие авторы
убедительно показали, что в возникновении голодных моти-
ваций первоочередное значение имеет не абсолютный уро-
вень глюкозы в крови, а артерио-венозная разница ее содер-
жания— так называемая дельта-глюкоза.
Установлено, что голодные мотивации возникают только
тогда, когда артерио-венозная разница глюкозы стремится
к нулю. Именно это состояние обычно совпадает с периодом
«голодных» сокращений желудка.
Исходя из концепции К. Бернара о постоянстве внутрен- .
ней среды и Cannon о гомеостазе, Richter (1942, 1947)
сформулировал представления, согласно которым мотива-
ционное возбуждение является следствием определенных
48
физиологических потребностей. Он указывал >на то, что-воз-
никающее под влиянием мотивации поведение направлено
на восстановление постоянства внутренней среды. Своими
экспериментами он отчетливо показал, что крысы обычно
всегда предпочитают тот сорт пищи, компоненты которой
отсутствуют в питании. Так, если в ежедневном рационе жи-
вотных отсутствует соль, они предпочитают употреблять со-
леную пищу. При отсутствии в питании определенного ви-
тамина они закономерно избирают только ту пищу, в кото-
рой содержится этот витамин. Особое значение
в формировании мотиваций Richter придавал изменению
деятельности желез внутренней секреции. Он показал, что
удаление надпочечников у крыс вызывает возникновение
подчеркнутого влечения к употреблению поваренной соли.
Такие животные поддерживают постоянный уровень нат-
рия в организме путем употребления больших количеств
растворов хлористого натрия. Однако, будучи убежденным
бихевиористом, Richter объяснял мотивации, возникающие
при определенных изменениях внутренней среды организма,
как следствие возрастания чувствительности соответствую-
щих периферических рецепторных аппаратов к внешним
раздражителям. Так, он полагал, что удаление надпочечни-
ков повышает чувствительность вкусовых рецепторов к со-
ли. В подтверждение такой точки зрения Richter произвел
специальный эксперимент, показавший, что перерезка вку-
совых чувствительных нервов у крыс с удаленными надпо-
чечниками расстраивала у них избирательное отношение
к солевым растворам.
Исследования ряда авторов показывают, что гумораль-
ные факторы участвуют в формировании и других моти-
ваций.
Так, Falk.считает, что исходным стимулом в формирова-
нии питьевой мотивации является изменение осмотического
давления плазмы или снижения внеклеточной воды
в тканях.
Beach (1948) рассматривает половое влечение как ре-
зультат нарастания в крови уровня половых гормонов.
У низших животных удаление половых желез немедленно
разрушает половое поведение. Любопытны в этом отноше-
нии эксперименты Green, Clemente и de Groot (1957).
Авторы показали, что введение неполовозрелым котам те-
стостерона вызывает появление в их поведении отчетливой
сексуальной активности. То же наблюдается при введении
неполовозрелым кошкам эстрогенов.
49
Другие наблюдения показывают, что кастрированные сам-
цы крыс прекращают реагировать на охочую самку в со-
стоянии течки. Однако их половая активность немедленно
восстанавливается после введения половых гормонов.
Тем не менее, несмотря на убедительные данные, показы-
вающие значение гуморальных изменений в генезе мотива-
ционного возбуждения, ряд фактов не укладывается в эти
представления. Мы уже указывали ранее на то, что у выс-
ших обезьян и у человека половое влечение может возни-
кать и при отсутствии гормональных влияний.
Наиболее убедительные в этом отношении данные были
получены Т. Т. Алексеевой (1958) в результате ее система-
тических наблюдений, проведенных па неразделившихся
близнецах. Было исследовано две пары таких сросшихся
близнецов, сакропагов—Ира—Галя и Маша—Даша.
Особенность строения и физиологии этих близнецов со-
стояла в том, что у них было общее кровообращение и со-
вершенно раздельные головной мозг, спинной мозг и иннер-
вация периферических органов.
Глюкоза, введенная в вену руки одной из девочек, уже
через 2 минуты переходила в кровь другой, а через несколь-
ко минут концентрация сахара в крови у обеих девочек
почти уравновешивалась. Однако при раздражении легким
уколом кожи на 1 см вправо от средней линии (условной
линии разделения их тел) вызывало реакцию только соот-
ветствующей головки. Другая девочка оставалась к этому
раздражению безучастной. Аналогичный результат полу-
чался и при обратных 'соотношениях.
Обе пары близнецов имели раздельную верхнюю часть
пищеварительного тракта. Оказалось, что кормление одной
из девочек и, следовательно, поступление питательных ве-
ществ в кровь другой никогда не приводило к развитию на-
сыщения у последней. У другой девочки длительное время
обнаруживалась потребность в пище, несмотря на то что ее
кровь и ткани получили достаточное количество питатель-
ных веществ.
Все это указывало на то, что гуморальные факторы не
являются ведущими в механизме голода и насыщения, что
состояние пищеварительного тракта имеет определенное
влияние на эти процессы. Подтверждением этого являются
и многочисленные клинические наблюдения, которые пока-
зывают, что полноценное насыщение никогда не развивается
после внутривенного введения голодным больным раствора
глюкозы, при наличии у них пустого желудка.
50
Janowitz и Grossman (1949) установили, что введение
в кровь животных глюкозы или гидролизата белка в коли-
честве, даже значительно превышающем калорийную зна-
чимость обычно потребляемой пищи, не вызывает тормо-
жения пищевой реакции к последующим кормлениям.
Все эти факты совершенно определенно указывают на то,
что гуморальные факторы сами по себе не играют решаю-
щей роли в механизме возникновения мотиваций.
Попытки все более углубленного анализа мотивационных
возбуждений привели к .необходимости каким-то образом
объединить нервные и гуморальные механизмы, участвую-
щие в их возникновении, в единый нейро-гуморальный
механизм.
Вот почему многие авторы в настоящее время стали при-
держиваться той точки зрения, что мотивации и их удовлет-
ворение определяются множественными факторами, одни
из которых могут быть ведущими, другие заменять ведущие
в случае их повреждения. Так, возникли концепции мно-
жественного генеза мотиваций. Такая точка зрения была
подробно развита в исследованиях Beach (1948), Stellar
(1954), Morgan (1959).
Beach, например, считает, что для мотивационного поло-
вого возбуждения необходим определенный минимум сен-
сорной или гуморальной информации, а также предшест-
вующее обучение.
Множественные факторы определяют и удовлетворение
мотиваций. Это отчетливо можно видеть на примере пище-
вого насыщения.
Наиболее существенным фактором насыщения является
сигнализация от рецепторов, ротовой полости (Berkun
a. oth., 1952; Miller a. oth.; 19’52).
Наряду с этим, согласно Janowitz и Grossman (1949),
в приеме пищи существенная роль принадлежит фактору
растяжения желудка. По данным этих авторов, животные
при мнимом кормлении съедают значительно больше пищи,
чем нормальные животные.
Показано, что прием пищи сопровождается мощной сим-
патической реакцией, в результате которой происходит не-
медленный выброс глюкозы из печени (К- М. Быков, 1947).
Значение глюкозы как фактора насыщения постулировано
в работе ряда авторов (Van Italie a. oth., 1953; Mayer,
1955),
Brobeck (1943), Strominger (1953) и Brobeck (1953), кро-
ме того, показали значение температурного фактора в прие-
51
ме пищи. Насыщение, по мнению этих авторов, связано
с повышением температуры тела и крови.
Kennedy (1953) полагает, что насыщение определяется
метаболическими процессами, связанными с изменениями
обмена жира. Эта концепция нашла подтверждение в на-
блюдениях над парабиотическими крысами (Hervey, 1959).
Разрушение медиального гипоталамуса у одной из таких
крыс вызывает у нее гиперфапию и ожирение на фоне резко-
го исхудания другой крысы.
Все данные постулируют, таким образом, множествен-
ность пусковых факторов, участвующих 'в формировании
мотивационного возбуждения и в его удовлетворении. Если
теперь дать общую оценку всех рассмотренных выше тео-
рий мотиваций, то можно отметить следующее.
По существу все эти теории посвящены только вопросу
о пусковых факторах мотиваций. Одни авторы (Cannon,
Carlson, Hebb) полагают, что мотивации определяются со-
стоянием определенных периферических органов, другие
(И. П. Павлов, Richter, Mayer и др.) усматривают причину
мотивационного возбуждения в определенных изменениях
внутренней среды организма и, в частности, крови. И, на-
конец, существует теория множественного генеза мотива-
ций (Beach, Stellar, Richter). Безусловно, все эти факторы
участвуют в формировании мотивационного возбуждения
и каждый из упомянутых выше авторов прав по-своему,.
Однако вопрос заключается в следующем. Если мотива-
ции определяются множественными нервными и гумораль-
ными факторами, то где же происходит интеграция этих
влияний, порождающая мотивационное возбуждение?
Современная нейрофизиология представляет все большее
количество экспериментальных доказательств того, что мо-
тивационное возбуждение складывается в центральной
нервной системе.
Центральные теории мотиваций
Lashley (1938) в противоположность Cannon и Richter
предположил, что мотивация возникает как результат воз-
буждения сенсорными и гуморальными факторами опреде-
ленных нервных центров. Хотя Lashley и не определил ло-
кализацию этих центров, его общие представления об
участии центральной нервной системы в формировании мо-
тиваций дали мощный толчок развитию центральных теорий
мотиваций. Эти представления в дальнейшем были развиты
Morgan (1959).
52
Morgan по аналогии с «центральным возбудительным со-
стоянием» Шеррингтона сформулировал понятие о цент-
ральном мотивационном состоянии. Такое понятие было
выдвинуто автором для того, чтобы подчеркнуть локализа-
цию мотивационного возбуждения в центральной нервной
оистеме и тем самым несколько приуменьшить значение
периферических теорий мотиваций. Morgan считает, что все
специфические влечения могут возникнуть только при опре-
деленном уровне центрального мотивационного состояния.
Развивая эту точку зрения, Beach (1958) пришел к выво-
ду, что такое центральное возбуждение в случае половых
мотиваций определяется двумя факторами. С одной сторо-
ны, это половые гормоны, которые действуют на подкорко-
вые центры, с другой — он предполагает существование
коркового механизма активации, который может обуслов-
ливать половое поведение и в отсутствие половых гор-
монов.
В соответствии с этими представлениями Dell (1958) вы-
двинул теорию, согласно которой в любом инстинктивном
поведении имеется неспецифичесний компонент — повышен-
ный уровень сенсорного и моторного возбуждения, который
и определяет мотивацию. Этот неспецифический компонент
мотивационных реакций связывался Dell с неспецифичёски-
ми восходящими активирующими влияниями ретикулярной
формации.
Dell представил обзор основных механизмов, посредством
которых органические потребности организма трансформи-
руются в акты поведения.
Так, в случае кислородного голодания низкое парциаль-
ное давление кислорода и высокое парциальное давление
углекислоты являются сильными раздражителями ретику-
лярной формации головного мозга. На этой основе строится
соответствующее приспособительное поведение, например
избегание душного помещения.
Пищедобывательное поведение, согласно Dell, также
определяется восходящими активирующими влияниями ре-
тикулярной формации. Возбуждение последней в данном
случае определяется адреналином, выделяющимся у голод-
ных животных под влиянием гипогликемии.
Dell считает, что все формы инстинктивного поведения
начинаются с обменных сдвигов, которые все более откло-
няют состав внутренней среды от нормы и это отклонение
воздействует на активность ретикулярной формации пря-
мым или опосредствованным путем.
53
Ретикулярные влияния, как он полагает, создают один из
главных механизмов трансформации потребностей организ-
ма в поведение. Независимо от природы потребности все
влечения, по его мнению, имеют общий неспецкфичесюий
компонент—изменение активности или степени «бдитель-
ности».
Эта точка зрения по существу является развитием пред-
ставлений Lindsley (1951), который предложил известную
«теорию активации эмоций». Согласно этой теории, веду-
щая роль в эмоциональных состояниях также приписыва-
лась ретикулярной формации.
Stellar в 1954 г. выдвинул гипоталамическую теорию, ко-
торая связывает формирование мотиваций с деятельностью
гипоталамических центров головного мозга. Согласно этой
теории, «центральное мотивационное состояние» поддержи-
вается возбуждением специальных гипоталамических цент-
ров, на которые оказывают влияние кортикальные импуль-
сы, тормозные гипоталамические центры, чувствительные
стимулы, гуморальные факторы и др. (рис. 3).
Таким образом, эта теория утверждает, что гипоталамус
является местонахождением «центрального мотивационного
состояния», о котором писал Morgan.
Stellar считает, что побуждение основывается на актив-
ности возбуждающегося гипоталамического механизма,
а насыщение, или удовлетворение зависит от гипоталамиче-
ского тормозного механизма. При этом ои полагает, что
действие обоих этих механизмов контролируется соответ-
ствующими сенсорными раздражителями, изменением внут-
ренних состояний и влиянием коры головного мозга.
В отличие от этого К. Лишак и Э. Эндрёци (1965) пола-
гают, что мотивационное возбуждение определяется мезо-
диэнцефалическим кругом. Основу этого круга составляют
мезенцефаличеокая ретикулярная формация, ядра средней
линии таламуса, стриопаллидарные связи через нижние
ножки таламуса, базальное поле перегородки, включая ди-
агональный пучок Брока, медиальный пучок переднего
мозга и, наконец, как отправная, так и конечная точки акти-
вирующего круга — снова ретикулярная формация ствола.
Разрушение этого круга в любой точке вызывает, по мне-
нию этих авторов, нарушение влечения и способности к обу-
чению.
Представление о неспецифических механизмах мотивации
было в значительной степени поколеблено опытами Olds и
Milner (1954), Olds (1960) с самор аздражением у животных.
54
Olds наблюдал, что раздражение определенных подкор-
ковых областей имитирует в одних случаях пищевое под-
крепление (частота самораздражения увеличивается у го-
лодных животных). В других случаях скорость реакции
самораздражения зависит от уровня половых гормонов.
Она как бы имитирует половое подкрепление. На основа-
нии этих экспериментов он выдвинул предположение о на-
личии в головном мозге специфических мотивационных
механизмов.
Рис. 3. Схема взаимодействия нервных и гуморальных механизмов в ре-
гуляции мотивационного поведения. Нейтральный пункт мотивации —
гипоталамическая область, возбуждающий и тормозящий отделы кото-
рой находятся под влиянием периферических и центральных нервных
импульсов, а также различных химических агентов (по Stellar, 1954).
/ — внутренние химические и физические факторы (гормоны, температура крови,
осмотическое давление, лекарственные вещества); 2 — регуляция постоянства внут-
ренней среды; 3 — обратная информация об успешности поведения; 4 — сенсорные
раздражители (условные и безусловные); 5 — пути формирования соответствую-
щего поведения.
55
Все это позволило Gellhorn и Loofbourrow (1963) гово-
рить об общем и специфическом влечении. Под общим вле-
чением они понимают определенное внутреннее состояние
животного, его готовность к действию. Специфические вле-'
чения изменяют акты поведения в определенном направле-
нии, модулируя своеобразным путем общее влечение.
Авторы считают, что специфические 'влечения возникают
при действии нервных и гуморальных факторов на механиз-
мы «поощрения» и «наказания».
Близка к этим представлениям точка зрения Smith
(1966). Автор считает, что мотивационное возбуждение
обеспечивается двумя системами, определяющими ее два
разные аспекта: интенсивность и направленность. Первый
аспект мотивационного возбуждения определяется восходя-
щей ретикулярной активирующей системой. Второй аспект
определяется экстраталамической системой, осуществляю-
щей активацию коры через базальный диэнцефалон.
Вслед за Olds, который экспериментально показал нали-
чие в головном мозге специальных центров подкрепле-
ния— «вознаграждения» и «наказания», в нейрофизиологии
по существу сложилось две точки зрения.
Сторонники одной из них считают, что мотивация и ее
прекращение связаны с деятельностью одних и тех же
нервных центров. Считается, что .подкрепление в этом, 'слу-
чае вызыв-ает только снижение исходного влечения (drive
reduction). Согласно второй точке зрения, в головном мозге
имеются две реципрокные системы: наказания и поощрения,
и мотивация определяется их взаимодействием. Субстра-
том поощряющего механизма сторонники этой точки зрения
считают медиальный пучок переднего мозга, а субстратом
«наказания» — перивентрикулярные структуры (рис. 4).
Lilly (1960) выдвигает -весьма дискуссионное утвержде-
ние о том, что любая двигательная активность сама по себе
может вызывать либо внутреннее поощрение («старт-эф-
фект»), либо внутреннее-наказание («стоп-эффект»), кото-
рые формируются в виде определенной активности головно-
го мозга.
Последняя точка зрения оспаривается Mac Lean (1955),
который утверждает, что все приспособительные реакции
следует рассматривать с позиции сохранения особи или ви-
да. Их отрицательный и положительный характер зависит
от того, несут ли они угрозу существованию или устранение
этой угрозы. Все приведенные выше теории показывают,
что природа мотивационного возбуждения все еще остается
56
недостаточно ясной. Нейрофизиологические механизмы мо-
тиваций экспериментально разработаны неудовлетворитель-
но. Это находит свое выражение и в отсутствии четких
определений понятия мотивации. Различные авторы дают
различные определе-
ния мотиваций. Од-
ни авторы понимают
под мотивацией им-
пульс к действию,
другие — само целе-
направленное пове-
дение. Сплошь да
рядом можно встре-
тить смешение поня-
тий «инстинкт», «вле-
чение», «мотивация»,
«эмоциональное по-
ведение», «потреб-
ность», «инстинк-
тивное поведение»
и т. д.
Так, Gellhorn и
Loofbourrow (1966)
пишут: «В самом ши-
роком смысле моти-
вация включает все
факторы, которые
вызывают и направ-
ляют акты поведе-
ния». Авторы пола-
гают, что «основные
влечения» (голод,
жажда, половое вле-
чение) составляют
только основу моти-
вации .в широком
Рис. 4. Локализация механизмов «награды»
(Л) и «наказания» (Б) в головном мозге
млекопитающих (по Le Gross Clark, 1938),
Механизмы «награды» связаны с медиаль-
ным пучком переднего мозга, механизмы
«наказания» — с перивентрикулярными об-
разованиями.
/“Передняя комиссура; 2“ ядро диагонального
пучка; 3 — мамяллярное тело; ^—перегородка;
5 — передний гипоталамус; 6 — таламус; 7 — зад-
ний гипоталамус; 8 — покрышка среднего мозга.
смысле слова
Bindra (1959) пишет о «мотивационной активности», та-
кой, как поиск пищи, воды, половые реакции и т. д.
Сюда же некоторые авторы (Bartoshuk, 1966) включают
все разновидности инструментальной активности живот-
ных, а также деятельность человека, направленную на до-
стижение специальных социальных раздражителей, таких,
как деньги, положение в обществе и т. п. Таким образом,
57
мотивация включается этими авторами в само целенаправ-
ленное поведение, обусловливая его направленность.
В отличие от этих авторов Smith (1966) считает, что мо-
тивация имеет два аспекта. Один связан с энергетической
силой поведения. Это — влечение (drive). Другой аспект ка-
сается направленности поведения.
Ruch (1960) отождествляет мотивацию и влечение
и определяет их как нервные процессы, толкающие орга-
низм к определенным действиям или цели, достижение ко-
торой снижает силу исходного влечения.
В. Детьер и Э. Стеллар (1967) пишут: «Побуждение или
влечение является стремлением к определенной цели... Оно
выражается в повышенной активности, желании действо-
вать для преодоления препятствий к достижению цели
и в обучении новым навыкам... Мотивация, таким образом,
является побуждением или влечением, которое ведет к це-
ленаправленному поведению и насыщению».
П. В. Симонов (1966) определяет влечение как целена-
правленную потребность, обращенную к объектам внеш-
ней среды, которые могут обеспечить се удовлетворение.
Дж. Миллер, Е. Галантер и К. Прибрам (1965) пишут
о программирующем свойстве мотиваций, о создании ими
определенных врожденных программ поведения. Эта точка
зрения весьма интересна. Однако авторы оставляют в сто-
роне вопрос о природе мотивационного возбуждения.
Karli (1968) считает, что в любой мотивации необходимо
различать две составляющие: энергетическое влияние (ди-
намическая составляющая) и направляющее влияние, ко-
торые отражают семантическое содержание мотивации.
Если сопоставить все эти высказывания, можно совер-
шенно определенно отметить, что удовлетворительного
определения мотивации не существует.
Все указанные.выше теории трактуют проблему мотива-
ций слишком обще и не учитывают то важное обстоятель-
ство, что каждая мотивация имеет специальные внутренние
раздражители и подчеркнутое биологическое значение.
Именно этот критерий является наиболее важным в меха-
низме мотиваций. Исходя из этого критерия, можно видеть,
что каждая мотивация представляет собой основу для спе-
цифической функциональной системы организма. Эти сис-
темы формируются из нервных элементов различных уров-
ней, по всегда находятся в энергетической зависимости от
исходного пункта — рецептора гуморальных изменений
(П. К. Анохин, 1962).
58
Теория функциональных систем совершенно четко увязы-
вает периферические и центральные теории мотиваций. Кро-
ме того, она показывает место мотивации в целостных при-
способительных актах, а также с материалистических пози-
ций объясняет энергетическую и направляющую роль моти-
ваций, о которой столь неопределенно пишут психологи.
Каково же место мотивации в функциональных системах
организма?
МОТИВАЦИИ И ФУНКЦИОНАЛЬНЫЕ СИСТЕМЫ
Как указывалось выше, в каждой функциональной систе-
ме организма центральным звеном является тот или иной
полезный приспособительный результат. Последний удер-
живается па уровне, обеспечивающим нормальную жизне-
деятельность на основе принципа саморегуляции, т. е. само
отклонение от этого приспособительного результата явля-
ется стимулом к его восстановлению. Для поддержания
полезных приспособительных эффектов организма все фун-
кциональные системы избирательно объединяют централь-
ные и периферические аппараты.
Мотивационное возбуждение возникает только в тех
функциональных системах, в которых полезный приспосо-
бительный результат не может быть обеспечен только внут-
ренними механизмами и в которых для поддержания по-
лезного приспособительного результата включается внеш-
нее звено — соответствующее поведение во внешней среде
(рис. 5).
Рис. 5. Место мотивационного возбуждения в функциональной системе.
X— константа крови; Л'(, 2G — отклонение константы от жизненно важного уровня
через возбуждение специальных рецепторов или гуморальным путем вызывает в
Центральной нервной системе мотивационное возбуждение. Последнее приводит
к целенаправленному поведению и удовлетворению исходной потребности организма.
59
Таким образом, мотивация возникает всякий раз, как
тот или иной полезный приспособительный результат функ-
циональной системы изменяется и не может быть восстанов-
лен только внутренними резервами организма. Только на-
личие мотивационного возбуждения в функциональных
системах приводит организм к строго специфическому це-
ленаправленному поведению, которое определяется физио-
логическими особенностями исходной потребности орга-
низма.
Из этого следует, что мотивация исчезает только в том
случае, когда организм достигает во внешней среде успеш-
ного результата, удовлетворяющего его исходную потреб-
ность.
Поскольку мотивации являются составной частью многих
функциональных систем, имеющих принципиально одина-
ковую организацию, принципиальная архитектура различ-
ных мотиваций также является аналогичной.
Если теперь снова вернуться к центральной архитектуре
функциональной системы, о которой мы писали ранее, то
можно заметить, что мотивационное возбуждение, обуслов-
ленное изменениями во внутренней среде организма, явля-
ется существенным компонентом стадии афферентного син-
теза.
Именно мотивационное возбуждение на стадии аффе-
рентного синтеза определяет, каким образом животное
будет реагировать на окружающие его раздражители.
Только при наличии мотивации организм, испытывающий
определенную потребность, будет настойчиво совершать
деятельность, направленную на удовлетворение этой по-
требности.
Мотивационное возбуждение крайне важно и в таком
важном компоненте афферентного синтеза, как механизмы
памяти. В настоящее время можно с полным основанием
утверждать, что только на основе мотивационного возбуж-
дения происходит извлечение следов памяти для построе-
ния целенаправленного поведения.
Стадия афферентного синтеза позволяет также понять,
почему в определенной (для человека прежде всего в соци-
альной) обстановке тс или иные биологические мотивации
могут совсем не проявиться. В этом случае обстановочные
(социальные) раздражители на основе механизма памяти
формируют адекватную себе высшую мотивацию, которая
определяется опытом и воспитанием данного индивидуума
и которая вытормаживает биологическую мотивацию.
60
Включаясь в деятельность различных функциональных
систем, каждая из которых поддерживает на постоянном
уровне только определенный жизненно важный эффект ор-
ганизма, любое мотивационное возбуждение является под-
черкнуто специфичным.
Возникает естественный вопрос, каким образом в цен-
тральной нервной системе при наличии нескольких потреб-
ностей одновременно реализуется несколько мотиваций?
Формирование мотивационного возбуждения в централь-
ной нервной системе подчиняется принципу доминанты,
описанному А. А. Ухтомским (1923, 1927, 1928). Это зна-
чит, что в каждом случае поведение определяется домини-
рующей мотивацией, которая завладевает всей деятель-
ностью организма. И только после удовлетворения этой
доминирующей мотивации может доминировать следую-
щая по биологической зависимости мотивация и т. д.
Рассмотрим несколько подробнее представления
А. А. Ухтомского о доминанте. В статье «Инстинкт и доми-
нанта» (1923) А. А. Ухтомский определяет принцип доми-
нанты следующим образом: «Доминанта как общий рабочий
принцип центров заключается в том, что частью под влия-
нием внутренних гормонов организма, частью под рефлек-
торным влиянием совне в нервной системе образуются оча-
ги повышенной возбудимости, которые именно в силу своей
повышенной возбудимости будут облегченным образом
реагировать на самые разнообразные и отдаленные раздра-
жения, прилагающиеся к организму. Сама по себе доми-
нанта представляется как временно переживаемая, доста-
точно инертная констелляция центров с повышенной воз-
будимостью в разнообразных этажах головного и спинного
мозга, а также в автономной системе. Когда доминанта за-
ключается в ускоренном акте глотания воды, которая вли-
вается в рот, отдаленные раздражения, не имеющие прямо-
го отношения к акту глотания, стимулируют глотание, не
производя реакций на своих ближайших путях. Когда до-
минанта заключается на весенней лягушке в рефлексе об-
нимания, отдаленные болевые раздражения, например,
в задних конечностях, дают явные усиления обнимания при
затруднении других ближайших рефлексов. Когда доми-
нанта заключается в повышенной возбудимости чувствую-
щих центров в поясничном отделе спинного мозга лягушки,
то самые отдаленные раздражения кожи животного вызы-
вают в задней лапке рефлекс потирания, направленный так,
как будто раздражитель действовал на кожу поясничных
61
дерматомеров» (А. А. Ухтомский. Собрание сочинений, 1950, j
стр. 186). По А. А. Ухтомскому, представление о том, что
в основе инстинктов лежит анатомическое возбуждение
определенных центров внутренними химическими (гормо-
нальными) влияниями, на которое уже вторично накладыва-
ются рефлекторные влияния, близко к понятию доминанты. /
В высшей психической жизни инертность господствующе-
го возбуждения, т. е. доминанты переживаемого момента,
как полагает А. А. Ухтомский, может служить источником
«предубеждения», «навязчивых образов», «галлюцинаций»,.
по она же дает ученому то маховое колесо, «руководящую j
идею», «основную гипотезу», которые избавляют мысль от
толчков и пестроты и содействуют сцеплению фактов в еди- ,
ный опыт.
В статье «Доминанта как фактор поведения» (1927) i
А. А. Ухтомский развивает эти мысли следующим образом:
«В конце концов не прирожденное наследие рефлексов
и инстинктов, по борьба текущих конкретных доминант
с унаследованным и привычным поведением приводит J
к оплодотворению всей работы, к поднятию достижений
высшей центральной нервной системы. Симптомокомплекс
доминанты заключается в том, что определенная централь-
ная группа, в данный момент особенно впечатлительная и ,
возбудимая, в первую голову принимает на себя текущие
импульсы, но это связано с торможением в других централь-
ных областях, т. е. с угнетением специфических рефлексов
на адекватные раздражители в других центральных обла- >
стях, и тогда множество данных из среды, которые должны
были бы вызвать соответствующие рефлексы, если бы при-
шли к нам в другое время, остаются теперь без прежнего
эффекта, а лишь усиливают текущую доминанту (действуют !
на руку текущего поведения)» (А. А. Ухтомский. Собрание '
сочинений,1950, стр. 309).
Наконец, в статье «О доминанте» (1928) он высказал
очень важную идею.
«В понятии доминанты скрывается та мысль, что орга- ’
низм человека представляет из себя более или менее опре- ’
деленный энергетический фонд, который расходуется ;
в каждое мгновение преимущественно по определенному •
вектору и тем самым снимаются с очереди другие возмож- J
ные работы» (А. А. Ухтомский. Собрание сочинений, 1950,
стр. 317).
Все изложенное выше определенным образом указывает
на чрезвычайную близость понятий «доминанта» и «моти-
62
вация». Однако А. А. Ухтомский, указав на чрезвычайно
важное значение принципа доминанты для целенаправлен-
ной деятельности, не вскрыл природу того, каким образом
доминирующее возбуждение складывается в центральной
нервной системе и каковы его конкретные механизмы.
Он писал: «Доминанта как общая формула еще ничего
не обещает. Нужно знать ее содержание и конкретные
условия возникновения...»'
Ответ на этот вопрос в совершенно отчетливой форме
дает функциональная система П. К. Анохина. Благодаря
доминирующей мотивации в каждый данный момент вре-
мени в центральной нервной системе может доминировать
только одна функциональная система.
В чем же заключается энергетическая сила доминирую-
щей мотивации? Безусловно, что она связана с деятельно-
стью определенных нервных элементов, поскольку мотива-
ционное возбуждение складывается в центральной! нервной
системе.
Каков же нервный субстрат мотиваций?
П. НЕРВНЫЙ СУБСТРАТ БИОЛОГИЧЕСКИХ
МОТИВАЦИИ
Современная нейрофизиология убедительно показывает,
что биологические мотивации животных и человека опреде-
ляются практически всеми структурами головного мозга,
начиная с его низших, стволовых отделов и кончая корой
больших полушарий. Вместе с тем теперь уже совершенно
определенно можно утверждать, что каждая структура го-
ловного мозга вносит нечто свое, специфическое в мотива-
ционное возбуждение.
Многочисленные примеры участия различных структур
головного мозга в формировании мотиваций представила
клиническая практика, убедительно показавшая, что раз-
личные формы нарушения биологических влечений наблю-
даются при поражениях как подкорковых так и некоторых
образований коры головного мозга. Особенно ярко выра-
жены эти расстройства при поражениях гипоталамической
области, при которых наблюдался комплекс различных па-
тологических влечений (гиперфагия, гиперсексуализм, ги-
пердипсия и т. п.) (А. М. Гринштейн, 1925; Е, А. Попов,
1927; Н. Н. Бурденко, Б. Н. Могильницкий— Burdenko, Мо-
gilnitzkij, 1926, и др.). Аналогичные неврологические рас-
стройства наряду с изменением в сфере высших мотиваций
наблюдались при удалении лобных отделов коры головного
мозга (Fulton 1949; А. Р. Лурия, 1963, и др.). Penfild и Jas-
per (1954) провели многочисленные клинические наблюде-
ния над расстройствами мотиваций при повреждениях ви-
сочной доли коры головного мозга, амигдалярной области
и гиппокампа.
Вслед за клиническими наблюдениями начались интен-
сивные экспериментальные исследования роли различных
структур головного мозга в мотивационных реакциях жи-
вотных.
В 1937 г. Keuver и Вису описали характерные изменения
в поведении обезьян после удаления височных долей и при-
лежащих областей палеокортекса, включая амигдалу и
энторинальную кору и вентральную часть гиппокампа.
64
Эти изменения проявляются ё виде характерного синдро-
ма. Животные становятся ручными и теряют страх, Они
проявляют отчетливую тенденцию повторно обследовать
предметы визуально, тактильно и особенно орально («ги-
перметаморфозис»). Кроме того, они начинают без разбо-
ра захватывать в рот не только пищу, но и различные не-
съедобные предметы («зрительная агнозия»). Животные
становятся гиперсексуальными и проявляют тенденцию к
спариванию с особями даже одного пола и даже с живот-
ными другого вида.
Таким образом, обезьяны после подобных поражений
проявляют расстройства почти всех биологических мотива-
ций и теряют способность адекватно оценивать свое со-
стояние соответственно окружающим их предметам.
. В последнее десятилетие началось интенсивное исследо-
вание нервного субстрата различных мотиваций с исполь-
зованием техники прямого вмешательства в различные под-
корковые образования. Экспериментальные исследования
подтвердили многие клинические наблюдения и наряду с
этим значительно расширили представления о локализации
в головном мозге нервных образований, принимающих уча-
стие в формировании биологических мотиваций. Оказа-
лось, что нервные структуры, раздражение которых вызы-
вает целенаправленные реакции, широко представлены под-
корковыми образованиями.
В течение двух последних десятилетий многочисленные
эксперименты показали, что ограниченные двусторонние
повреждения гипоталамуса могут оказывать сильное влия-
ние на такие основные биологические мотивации, как го-
лод, жажда, сон, страх, агрессия и половое влечение. Среди
прочих мозговых структур наиболее тесное отношение к мо-
тивационным реакциям имеют нервные образования, со-
ставляющие так называемую лимбическую систему голов-
ного мозга (Mac Lean, 1954; Brady, 1958, и др.). Последняя
представляет взаимосвязанную группу структур • голов-
ного мозга, локализованную преимущественно в области
риненцефалона. Сюда входят лобно-теменная кора, поясная
извилина, область перегородки, хвостатое ядро, амигдала,
гиппокамп, энторинальная и инсулярная кора.
К лимбическим структурам относят ограниченные области
переднего таламуса и гипоталамуса, находящиеся в тесных
функциональных связях с риненцефалоном. Nauta (1960)
относит к лимбической системе и некоторые образования
ретикулярной формации среднего мозга.,
3 Заказ 5074
65
Все упомянутые выше структуры находятся между собой
в тесных морфологических и функциональных взаимоотно-
шениях.
Настоящая работа не претендует на всестороннее изло-
жение вопроса о морфологических и функциональных вза-
имоотношениях внутри лимбической системы. По этим во-
просам имеется обширная литература (Э. Ш. Айрапетянц,
Т. С. Сотниченко, 1967; Т. А. Леонтович, 1968; К- В. Суда-
Рис. 6. Функциональные
взаимосвязи лимбических
образований головного
мозга.
a — круг Пейпса, схема стро-
ения (заштрихованы лимби-
ческие образования): 1 — по-
ясная извилина; 2 — обоня-
тельная область; 3 — гиппо-
кампальная извилина; 4 —
мамиллярное тело; 5— перед-
ние ядра таламуса (по Гель-
горну н Луфборроу, 1966);
б — схема взаимодействия
структур лимбической систе-
мы на примере круга Пейп-
са (обозначено черными
стрелками, белыми стрелка-
ми обозначены связи, не вхо-
дящие в круг Пейпса): 7 —
миндалина; 2— обонятельная
система; 3 — перегородка;
4— свод.; J —поясная изви-
лина; 6— гиппокамп; 7 — пе-
реднее ядро таламуса; S —
гипоталамус; 9 — энториналь-
няя кора (по Cleary и Moore, 1965); в — круговые взаимосвязи внутри лимбиче-
ских образований: /-—миндалина; 2 — латеральная обонятельная полоска; 3 —
•диагональный пучок (Брока); 4 — обонятельный бугорок; 5 — обонятельная лу-
ковица; 6 — медиальная обонятельная полоска; 7 — перегородка; 8— лобная до-
ля; 9 — мозолистое тело; 10— надмозолистые полоски; 11 ~ свод; 12 — мозговая
полоска; /3 —передние ядра таламуса; 14 — уздечка; 15 —* ствол головного мозга;
16 — конечная полоска; 17 — медиальный пучок переднего мозга; 18 — интерпедун-
кулярное ядро; 19— зубчатая извилина; 20 — гиппокамп; 21 — мамиллярное тело.
66
ков, Е. М. Богомолова, 1969; Brady, 1958; Nauta, 1960,
и др.).
Здесь нам хотелось бы отметить только некоторые осо-
бенности функциональной организации лимбических обра-
зований, которые важны для понимания природы биологи-
ческих мотиваций.
Впервые Papez (1937) обратил внимание на то, что лим-
бические структуры находятся в циклических взаимоотно-
шениях. Он показал, что нервные импульсы благодаря мор-
фофункциональным взаимодействиям различных структур
этой системы могут циркулировать по кругу, состоящему
из гиппокампа, свода, мамиллярных тел гипоталамуса, пе-
редних ядер таламуса, поясной извилины и снова гиппо-
кампа (рис. 6 а, б). Papez предложил рассматривать эти
структуры как нервный субстрат эмоций.
Такие же циклические взаимоотношения существуют меж-
ду другими лимбическими образованиями (рис. 6, в), на-
пример, между амигдалой, концевой полоской, перегород-
кой, диагональным пучком и амигдалой, затем между пе-
регородкой, мозговой полоской, уздечкой, интерпедункуляр-
ным ядром, медиальным пучком переднего мозга, перего-
родкой. Имеется и другой «септальный круг» (так называе-
мый круг Наута) —перегородка, супракаллозальные по-
лоски, гиппокамп, форникс, перегородка.
Если, кроме того, учесть, что амигдала, септум и гиппо-
камп непосредственно связаны и между собой, можно ви-
деть, какие широкие возможности циклических взаимодей-
ствий представляют эти структуры.
Для того чтобы проследить, как различные отделы лим-
бической системы участвуют в мотивационном возбужде-
нии, рассмотрим их значение по отношению к отдельным
биологическим мотивациям,
ПИЩЕВЫЕ МОТИВАЦИИ
Мы рассмотрим влияние разрушения и раздражения раз-
личных структур головного мозга на такие аспекты пище-
вого поведения, как поиск и поглощение пищи, которые мо-
гут в какой-то степени характеризовать пищевые мотива-
ции.
Многочисленные эксперименты последнего времени убе-
дительно показали, что основная роль в формировании пи-
щевых мотиваций принадлежит гипоталамической области.
Увеличение потребления пищи при повреждениях гипота-
67
ламуса было описано во многих работах (Keller, Noble,
1935; Keller, Witt, Batsel, 1959, и др.).
В то же время сообщалось, что у некоторых животных
* при повреждении гипоталамуса, наоборот, уменьшался или
полностью прекращался прием пищи (Ingram, Barris, Ran-
son, 1936; Ranson, 1939; Hetherington a. Ranson, 1940 и др.).
Серия последующих экспериментальных исследований
была посвящена детальному выяснению участия гипота-
ламических образований в регуляции приема пищи.
" Briigger (1943) установил, что раздражение окрестностей
мамиллоталамического тракта у кошек вызывает булимию.
При искусственном раздражении этой области животные
ели не только пищу, но и несъедобные вещества.
Значение вентро-медиальных ядер гипоталамуса в раз-
витии гиперфагии и ожирения было показано в работе May-
er и Barrnett (1955). Авторы отметили, что у некоторых
крыс гиперфагия развивалась даже после односторонних
повреждений вентро-медиальных ядер гипоталамуса, хотя
при этом ожирение было приблизительно в 2 раза меньше,
чем при двусторонних повреждениях.
Ruch, Maire и Patton (1956) обнаружили, что прием пи-
щи изменяется при раздражении премамиллярной области
и передних ядер таламуса.
Morrisson и Мауег (1957) наблюдали возникновение ожи-
рения и афагии у крыс также и после разрушения латераль-
ных отделов субталамуса.
В других работах было описано появление ожирения
у собак при повреждении заднего гипоталамуса и областей,
расположенных каудальнее паравентрикулярных ядер.
У обезьян ожирение развивалось при нарушении более кау-
дальных областей в вентральной части таламуса и рост-
ральной части покрышки среднего мозга.
Все эти наблюдения дали основание Brobeck (1946) вы-
сказать предположение о том, что аксоны клеток вентро-ме-
диальных ядер распространяются к области, лежащей вы-
ше и латеральнее сосцевидных тел, и оттуда — внутрь по-
крышки среднего мозга. Вот почему поражение этих воло-
кон на любом уровне может вызвать гиперфагию и ожире-
ние. Вместе с тем Anand и Brobeck (1951) наблюдали, что
гиперфагию и ожирение вызывает также двустороннее
поражение области, расположенной латеральнее вентро-ме-
диальных ядер. Это дало им основание считать, что аксоны
клеток вентро-медиальных ядер проецируются также и в
латеральную гипоталамическую область.
68
Наоборот, двустороннее повреждение самых боковых ча-
стей латерального гипоталамуса в том же рострокаудаль-
ном плане, в котором расположены вентро-медиальные яд-
ра, вызывало полную афагию и гибель животного от исто-
щения даже в случае наличия около него избыточного ко-
личества пищи.
Этот факт латеральных гипоталамических повреждений
отчетливо проявлялся у животных не только при интактной,
но и при поврежденной вентро-медиальной области. Жи-
вотные, у которых в результате повреждения медиальных
отделов гипоталамуса наблюдалась гиперфагия, после по-
вреждений латерального гипоталамуса немедленно пол-
ностью прекращали всякие пищевые реакции. Все это ука-
зывает на то, что в цищевых реакциях латеральная гипо-
таламическая область функционально преобладает над
медиальными его отделами. В противоположность медиаль-
ным поражениям односторонние повреждения латерально-
го гипоталамуса не вызывают гипофагии.
Все это позволило Anand и Brobeck (1951) сформулиро-
вать представления о наличии в области латерального ги-
поталамуса, раздражение которого вызывает увеличение
приема пищи, а разрушение — отказ от еды, специального
«центра питания» («feeding center»). В противоположность
этому медиальные отделы гипоталамуса, ограничивающие
прием пищи и способствующие отложению питательных ве-
ществ в организме, получили название «центра насыщения»
(«satiety center»).
Согласно Anand и Brobeck, передней границей «центра
питания» латерального гипоталамуса являются паравентри-
кулярные и супраоптические ядра. Задняя граница опре-
деляется премамиллярными ядрами и дугообразным
ядром.
Наряду с этими наблюдениями Reynolds (1963), исполь-
зуя методику разрушения подкорковых структур радиоча-.
стотным облучением, не обнаружил у животных возникно-
вения гиперфагии после разрушения медиальных отделов
гипоталамуса. Он считает, что гиперфагия, наблюдаемая
в опытах других авторов, при электролитическом повреж-
дении медиальных отделов гипоталамуса могла быть след-
ствием раздражения, вызванного кровоизлияниями и отло-
жением тяжелых металлов.
Эти и другие эксперименты позволили Reynolds выска-
зать сомнение в отношении существования центра насыще-
ния в медиальной гипоталамической области.
69
Однако в последнее время Hoebel (1965) не смог подтвер-
дить данные Reynolds.
Подтверждение наличия в гипоталамусе центров «пита-
ния» и «насыщения» было получено в опытах, в которых
у неанестезированных животных производилось искусствен-
ное раздражение этих отделов гипоталамуса через по-
гружные вживленные электроды.
Так, Delgado и Anand (1953), Anand и Dua (1955) заме-
тили, что электрические раздражения импульсным током
в течение часа латеральной гипоталамической области у ко-
шек, хотя и не вызывали пищевой реакции в момент раздра-
жения, тем не менее значительно увеличивали потребление
пищи в экспериментальные и последующие дни. Кроме того,'
это раздражение вызывало у животных увеличение содер-
жания сахара в крови.
При раздражении медиальных отделов гипоталамуса
у некоторых животных, наоборот, отмечалось угнетение пи-
щевой реакции в дни раздражения и падение содержания
сахара в крови. Аналогичные данные приводит Smith
(1961).
Larsson (1954) в опытах, проведенных на овцах и козах,
получил гиперфагию при электрическом раздражении ла-
терального гипоталамуса, а также при инъекциях в эту об-
ласть гипертонического раствора хлористого натрия. Разд-
ражение. медиальных отделов гипоталамуса в его опытах
вызывало у животных только облизывание, жевание и не-
которое уменьшение ежедневного приема пищи.
Все эти опыты указывают на то, что латеральные и ме-
диальные отделы гипоталамуса имеют прямое отношение
к механизму приема пищи.
Наиболее отчетливые изменения пищевого поведения
удалось наблюдать при применении методов микроинъек-
ции разных химических веществ непосредственно в струк-
туры гипоталамической области.
При использовании этого метода введение химических
и фармакологических веществ в типоталамическую область
осуществляется через специальные тонкие канюли — иглы,
которые вводят в головной мозг при помощи стереотакси-
ческой техники. Впервые такая методика микроинъекций
в гипоталамическую область растворов хлористого натрия
была осуществлена Andersson (1953) на овцах. Вслед за
ним Larsson (1954), инъецируя в область между сводом
и мамиллоталамическим трактом 0,012 мл 3—5% раство-
ра хлористого натрия, наблюдал у овец в течение 40 минут-
70
резко выраженную гиперфагию. Такая процедура приводи-
ла у животных также к падению уровня сахара в крови.
Вместе с тем гипертонические растворы других веществ
в соответствующей концентрации (глюкоза, сахароза, ин-
сулин) не вызывали специфической пищевой реакции.
Позднее Grossman (1960, 1964, 1969) изучал изменения
пищевого поведения у крыс путем введения им в гипотала-
мус микрокристаллов холинергетических и адренергических
веществ. Введение в его опытах накормленным крысам
в область латерального гипоталамуса адренергических ве-
ществ (эпинефрина, допамина и норэпинефрина) вызывало
у них снова в течение 20—40 минут реакцию потребления
пищи. Однако холинергические вещества (ацетилхолин, ди-
метиламиноэтанол, карбохол) не вызывали пищевой реак-
ции. Вслед за их введением в область латерального гипо-
таламуса, как правило, возникала питьевая реакция.
Раздражение адренергическими и холинергическими ве-
ществами латеральной гипоталамической . области наряду
с возникновением пищевых и питьевых реакций нередко
приводило к появлению ранее заученных инструменталь-
ных реакций (например, реакции нажатия на рычаг, кото-
рая обычно приводила к получению пищи или воды). При
этом введение в латеральные отделы гипоталамуса адре-
нергических веществ вызывало у животных нажатие на
рычаг, который обеспечивал подачу пищи. Наоборот, вве-
дение в область латерального гипоталамуса холинергиче-
ческих веществ вызывало нажатие на тот рычаг, который
обеспечивал подачу воды. . .
У голодных животных введение в латеральный гипота-
ламус холинергических веществ тормозило пищевые реак-
ции. Точно так же тормозились питьевые реакции у живот-
ных, содержавшихся длительное время без воды при вве-
дении им в эту область адренергических веществ. Вместе,
с тем введение в область латерального гипоталамуса хло-
ристого натрия, хлористого бария и экстракта задней доли
гипофиза в опытах Grossman не вызвало изменений в. пи-
щевом и питьевом поведении.
Введение в область латерального гипоталамуса атропи-.
на не подавляло пищевой, но значительно снижало питье-
вую реакцию.
Аналогичные реакции при введении карбохола и.норад-
реналина в область переднего гипоталамуса, особенно в об-
ласть вентро-медиальных ядер, наблюдали Miller, Gottes-
man, Emery (1964). Fisher (1964) также наблюдал резкое
TH
увеличение приема воды при введении в область переднего
гипоталамуса ацетилхолина и мускарина.
Используя метод микроионофореза таких веществ, как
ацетилхолин, норадреналин и серотонин, и изучая характер
ответной реакции нейронов гипоталамуса при помощи ми-
кроэлектродов, Bloom, Oliver, Salmoirachi (1963) пришли
к выводу, что клетки, чувствительные к этим веществам,
диффузно представлены по всему гипоталамусу. Вместе
с тем наиболее чувствительными к ним являлись зоны па-
равентрикулярных и вентро-медиальных ядер.
Epstein (1960) отметил отчетливую реципрокность пище-
вых реакций, возникающих при введении одних и тех же
химических веществ в медиальные и латеральные отделы
гипоталамуса. Так, в его опытах введение крысам через
вживленные канюли в медиальный гипоталамус хлористо-
водородного прокаина вызывало значительное увеличение
приема пищи даже у накормленных животных. Хлористый
натрий, введенный в эту же область гипоталамуса, наобо-
рот, подавлял пищевые реакции, что было особенно замет-
но у голодных животных. В то же время введение хлори-
стоводородного прокаина в область латерального гипота-
ламуса тормозило пищевые реакции, а хлористый натрий
их стимулировал. Delgado (1965) наблюдал изменение пи-
щевых реакций животных при введении различных химиче-
ских веществ в головной мозг благодаря применению мето-
да телемикроинъекций. При этом введение в головной мозг
различных веществ осуществлялось посредством специаль-
ного «хемотрода» — электрода, позволяющего вводить в го-
ловной мозг микродозы химических веществ при управле-
нии на расстоянии.
В последние годы для изучения центральных механизмов
голода и насыщения все в большей степени привлекаются
электрофизиологические методы.
Так, Anand, Dua, Singh (1961), Anand, China, Singh
(1962) у бодрствующих кошек й обезьян зарегистрировали
электрическую активность латеральных («центр питания»)
и медиальных («центр насыщения») отделов гипоталамуса.
После внутривенного введения этим животным раствора
глюкозы авторы наблюдали увеличение частоты электри-
ческой активности в медиальных отделах гипоталамуса.
В то же время в латеральных отделах гипоталамуса отме-
чалось уменьшение амплитуды электрических колебаний.
Аналогичные изменения электрической картины центров
гипоталамуса наблюдали у голодных животных после корм-
72
дения (Anand, 1962). В противоположность этому гипогли-
кемия, которая достигалась введением инсулина, наоборот,
урежала частоту и увеличивала амплитуду биопотенциа-
лов в медиальных отделах гипоталамуса, в то время как
в латеральных его отделах частота электрических колеба-
ний увеличивалась.
Н. Н. Любимов (1958) и Л. Г. Трофимов (I960), регист-
рируя одновременно с поверхностной электроэнцефало-
граммой электрическую активность латеральных и меди-
альных отделов гипоталамуса цри выработке пищевого
условного рефлекса, отметили, что вид пищи так же, как
и условные пищевые раздражители, вызывает в этих струк-
турах возрастание частоты электрических колебаний. При
еде в этих структурах появляются медленные колебания.
Появление медленных высокоамплитудных колебаний в
пищевых центрах гипоталамуса во время кормления и как
реакцию на условные пищевые раздражители отмечают
и другие авторы (Clemente a. oth., 1964; Wyrwicka, 1964,
и др.).
В последнее время появился ряд работ, авторы которых
изучали электрическую активность отдельных нейронов пи-
щевого центра гипоталамуса с помощью микроэлектродно-
го метода. Так, В. А. Заболотных (1963) на кошках под
хлоралозным наркозом обнаружил в гипоталамусе раз-
личные нейроны. Одни из них реагировали преимуществен-
но на механическое растяжение желудка, другие — на вве-
дение инсулина или глюкозы. На основании своих опытов
автор высказывает предположение о сложной «мозаичной»
картине «центра питания» и «насыщения» в гипоталамусе.
Он также обнаружил, что у голодных животных количество
активных нейронов в гипоталамусе значительно выше, чем
у животных накормленных. В работе японских авторов
(Oomura, Kimura, Oojama, Масло, Iki, Kuniyoshi, 1964; Ma-
fasaburo, 1964) у наркотизированных кошек регистрирова-
лась активность нейронов латерального и вентро-медиаль-
ного гипоталамуса. Им удалось отметить строго рецип-
рокные соотношения между этими отделами гипоталамуса.
Так, раздражение латерального гипоталамуса в их опы-
тах, увеличивая активность расположенных здесь нейро-
нов, в то же время тормозило разряды в клетках вентро-
медиальных ядер. Наоборот, раздражение вентро-медиаль-
ных ядер гипоталамуса, увеличивая активность нейронов
этой области, в то же время прекращало активность ней-
ронов латерального гипоталамуса. Внутривенное введение
73
глюкозы уменьшало частоту разрядов в клетках латераль-
ного гипоталамуса и резко увеличивало активность клеток:
вентро-медиальных гипоталамических ядер. Такая подчерк-
нутая реакция вентро-медиальных ядер гипоталамуса на1,
введение глюкозы является, по мнению авторов, доказа-
тельством их высокой избирательной к ней чувствительно-
сти. Аналогичные данные были получены Anand и др..
(1964), а также Ю. Л. Пинесом (1965).
Все приведенные выше эксперименты в целом подтверж-
дают наличие в гипоталамусе центров «питания» и «насы-
щения». Вместе с тем в настоящее время возникла тенден-
ция более тонко дифференцировать пищевые центры гипо-
таламуса— делить его на специальные подотделы.
Так, Morgane (1961а, б) на основании собственных эк-
спериментов с применением инструментальных методов
приходит к заключению, что в латеральном гипоталамусе
следует выделить две части. Средняя его часть определяет
исходные побуждения к пище. Более латеральные его от-
делы обеспечивают регуляцию приема пищи.
Такое предположение Morgane доказывает следующими
экспериментами. Он проводил раздражение различных от-
делов латерального гипоталамуса у крыс, обученных для-
получения пищевого подкрепления преодолевать специаль-
ный электрический барьер и после этого нажимать на ры-
чаг. При этом преодоление электрического барьера харак-
теризовало исходную пищевую мотивацию животных.
Его опыты показали, что раздражение более медиальной
части латерального гипоталамуса вызывало преодоление
животным электрического барьера, подбегание к рычагу
и нажатие на него. Раздражение же латеральной части ги-
поталамуса никогда не приводило к подбеганию и нажатию
на рычаг, но вызывало непосредственный акт еды.
Как уже указывалось ранее, Miller, Bailey, Stevenson
(1950), а также Anliker, Mayer (1957) показали, что гипер-
фагия, возникающая после разрушения у животных вентро-
медиальных ядер гипоталамуса, не сопровождается возра-
станием голодных побуждений. Оказалось, что такие крысы
меньше нажимают на рычаг, чтобы получить пищевое под-
крепление, чем контрольные. Это же наблюдал в своих
экспериментах Teitelbaum (1962).
Весьма интересные факты приводят Baillie и Morrisson
(1963). В своих опытах они использовали инструменталь-
ную методику нажатия на рычаг, вслед за чем обычно жи-
вотное получало пищу. Затем они производили у животных
74
двустороннее разрушение латеральных отделов гипотала-
муса. Авторы установили, что в первые 1—2 дня, когда
у животных наблюдалась полная афагия, они также не про-
являли реакцию нажатия на рычаг. Затем наблюдалось
восстановление спонтанной реакции нажатия на рычаг,
однако животные по-прежнему не принимали пищу. Реак-
ция нажатия на рычаг не исчезала даже в том случае, ког-
да животным через фистулу в желудок вкладывали пищу.
На основании этих экспериментов авторы приходят к вы-
воду, что разрушение латеральных отделов гипоталамуса
вызывает только расстройство двигательных механизмов
пищевого акта, в то время как пищевая мотивационная ре-
акция при этом мало изменяется.
Все эти данные свидетельствуют о том, насколько слож-
на дифференцировка в гипоталамических отделах пищево-
го центра. Вместе с тем этот вопрос, безусловно, требует
дальнейшего, более конкретного экспериментального обос-
нования.
Все приведенные выше экспериментальные данные со-
вершенно отчетливо указывают на центральную роль гипо-
таламических образований в формировании пищевых реак-
ций. Тем не менее возникает совершенно закономерный
вопрос, в каких взаимоотношениях находятся между собой
латеральные и медиальные отделы гипоталамуса? Каковы
их взаимоотношения с другими отделами головного мозга?
Вполне правомерным является и вопрос о том, в какой
степени другие структуры головного мозга участвуют в фор-
мировании пищевых реакций и, в частности, пищевого по-
ведения?
Anand и Brobeck первые обратили внимание на следую-
щий факт. Животные проявляли повышенную «прожорли-
вость» как в случае повреждения медиальных отделов ги-
поталамуса, так и при раздражении латеральных его от-
делов. Это позволило авторам высказать предположение,
которое затем они сформулировали как постулат о том,
что «центр питания» латерального гипоталамуса и «центр
насыщения» медиального гипоталамуса по аналогии с цент-
ром вдоха и выдоха находятся в реципрокных соотноше-
ниях.
Реакция потребления пищи, по их мнению, определяется
тормозными влияниями латеральной области на медиаль-
ную. И, наоборот, явления гиперфагии при разрушении ме-
диальных отделов гипоталамуса они рассматривают как
результат снижения тормозящих влияний медиальной об-
75
ласти на латеральную. Среди этих двух центров «центр пй-
тания», по мнению Anand и Brobeck, является доминирую-
щим.
Исходя из этих представлений, Anand и Brobeck считают,
что латеральные гипоталамические «центры питания» сле-
дует рассматривать как облегчающие, а медиальные отде-
лы — как тормозящие стволовые пищевые рефлексы.
К последним относятся такие формы пищевой деятельно-
сти, как жевание, глотание, лизание, рвота и т. д., которые
нередко называют «пищевыми автоматизмами». Извест-
но, что «пищевые автоматизмы» отчетливо выступают у де-
церебрированных животных, что является полным доказа-
тельством их формирования на уровне ствола головного
мозга. Оказалось, что такие пищевые автоматизмы в форме
облизывания, жевания, глотания и т. и. часто наблюдаются
в дополнение к гиперфагии при раздражении области ла-
терального гипоталамуса (Delgado, Anand, 1953; Larsson,
1954).
Раздражение обонятельного бугорка и амигдалы (Мас
Lean, Delgado, 1953) также вызывает у животного жева-
тельные и глотательные движения. Уже эти данные ука-
зывают на то, что гипоталамические структуры не являются
единственной основой пищевого поведения.
Ствол головного мозга, так же как и спинной мозг, со-
держит специальные проводящие пути, необходимые для
пищевой деятельности, раздражение которых способно вы-
зывать пищевые реакции.
Подтверждением этого являются наблюдения Miller и
Sherrington (1916), Baset и Penfild (1922), которые на де-
церебрированных кошках показали возможность пищевых
ответов и рефлексов отвергания определенных веществ.
Так, у накормленных децеребрированных кошек было наме-
чено появление мурлыканья, свидетельствующее о состоя-
нии насыщения. Кроме того, Larsson (1954) получил по-
лифагию у коз при электрическом раздражении ствола
головного мозга в области дорсального моторного ядра
вагуса. Andersson, Kitchell и Person (1958) при раздражении
близлежащих областей также наблюдали появление у коз
реакции жевания. В дополнение к этому авторы установи-
ли, что гиперфагия у коз развивается и При раздражении
областей, расположенных латеральнее и сзади от сосцевид-
ных тел.
В опытах Miles и Thomas (1962) гиперфагия наблюда-
лась у кошек после электролитических повреждений пери-
76
акведуктуального серого вещества на уровне верхних бу-
горков четверохолмия. Maire и Patton (1954) нашли, что
прием пищи увеличивался у крыс при раздражении меди-
альных мамиллярных тел, а также переднего таламуса.
После открытия восходящих активирующих влияний ре-
тикулярной формации на кору головного мозга Dell (1958)
первый высказал предположение об участии ретикулярной
формации в исследовательском пищедобывательном по-
ведении.
Вместе с тем оказалось возможным вызвать пищевое
поведение у накормленной кошки, раздражая область вент-
ральной покрышки Тцая или вентро-латеральную границу
центрального серого вещества (Wyrwicka, Doty, 1966). -
Kesner, Fiedler, Thomas (1967), Parker, Feldman (1967)
показали, что повреждение мезэнцефалической ретикуляр-
ной формации у крыс приводит к длительной афагии и адип-
сии.
Участие структур ретикулярной формации в пищевом по-
ведении кролика было показано прямыми экспериментами
(В. Гавличек, 1958; Ю. А. Макаров, 1960; А. И. Шумили-
на, 1961, и др.).
В настоящее время установлено, что ретикулярная фор-
мация среднего и продолговатого мозга имеет богатые мор-
фологические связи с гипоталамусом (Magoun, Ranson,
Hetherington, 1938; Wang, Ranson, 1939; Wang, Borisson,
1950; - Thompson, Bach, 1950). Показано, что нисходящие
связи гипоталамуса широко, диффузно представлены в раз-
ных отделах продолговатого и среднего мозга. Вместе
с тем, согласно данным Magoun, Ranson, Hetherington,
основные проекции из гипоталамуса распространяются пре-
имущественно в латеральную часть ствола головного мозга.
Проекции от вентро-медиальных ядер гипоталамуса в ре-
тикулярную формацию среднего мозга описывает также
Feldman (1963). Задний гипоталамус, по его данным, свя-
зан преимущественно с медиальной петлей.
Функциональные влияния мезэнцефалической ретику-
лярной формации на вентро-медиальные ядра гипоталамуса
обнаружены Tsubokawa и Sutin (1963).
Все эти и другие факты позволяют предполагать, что
влияния пищевых центров гипоталамуса могут распростра-
няться и на ретикулярную формацию ствола головного
мозга. Однако прямых доказательств этого не имеется.
Говоря о связях гипоталамуса, следует указать на связь
медиальных его отделов с передней долей гипофиза. Вот
77
почему при изменении функции гипофиза немедленно нару-
шаются взаимоурегулированные отношения между медиаль-
ными и латеральными отделами гипоталамуса и может раз-
виваться ожирение (синдром Фрейлиха) или кахексия
(синдром Симондса).
Гипоталамус, согласно Бробеку, действует наподобие
«тормоза и акселератора автомобиля», облегчая или подав-
ляя рефлекторную деятельность более низших уровней
центральной нервной системы.
Brobeck (1957) подчеркивает большое сходство между
регуляцией приема пищи и регуляцией дыхания, И потреб-
ление пищи, и дыхание являются периодическими и ритми-
ческими явлениями, подчиненными произвольному контро-
лю. Количественные регуляции — минутный объем и по-
требление пищи соответственно осуществляются в каждом
случае изменением частоты. Дыхательный минутный объ-
ем зависит от частоты дыхания и количества вдыхаемого
воздуха, тогда как количество потребляемой пищи опреде-
ляется частотой и объемом приемов пищи. Можно устано-
вить определенную аналогию между центрами вдоха и ап-
петита и центрами выдоха и насыщения. Обе эти функции
связаны соответственно с болезненными ощущениями го-
лода и воздушной недостаточности и т. д.
Наряду с этим регуляция приема пищи имеет много об-
щего с нервной регуляцией большинства других внутренних
и автономных деятельностей, поддерживающих постоянство
внутренней среды организма.
Anand (1961, 1963) выдвинул представление о том, что
следующим за гипоталамусом уровнем регуляции приема
пищи является лимбическая система головного мозга. Со-
гласно этому предположению, лимбические структуры по-
добно ассоциативным областям коры больших полушарий
производят интеграцию и переработку всех афферентных
импульсов, идущих из внутренней среды организма. Толь-
ко после этого лимбическая система осуществляет свои ре-
гуляторные влияния.
Эти регуляторные влияния осуществляются преимущест-
венно через гипоталамус, который, по мнению Anand, по-
добен моторным областям головного мозга. Он осуществ-
ляет облегчающий и тормозящий контроль над автономны-
ми процессами стволового и спинального уровня, а также
регулирует секреторную деятельность гипофиза.
Основным коллектором, связывающим лимбическую си-
стему с пищевыми центрами гипоталамуса, является так
78
называемый медиальный пучок переднего мозга (Morgane,
1961, 1962).
Имеется множество научных фактов, подтверждающих
участие лимбической системы, особенно миндалевидного
комплекса, в регуляции приема пищи. Так, показано (Fon-
berg, Delgado, 1961; Morgane, 1962; Э. Фонберг, 1965;
S. Grossman, L. Grossman, 1963, и др.), что раздражение
латеральных базальных и вентро-медиальных ядер амигда-
лы тормозит естественные пищевые побуждения и инстру-
ментальные пищевые реакции животных, а их разрушение
ведет к развитию гиперфагии и ожирения. При этом Fon-
berg считает, что торможение пищевых реакций в данном
случае происходит в результате возникновения какого-то
другого биологического качества положительной эмоции.
Различными исследованиями показано, что результатом
разрушения миндалевидного комплекса, особенно его ба-
золатеральной части, является гиперфагия (Green, Clemen-
te, de Groot, 1957; Morgane, Kosman, 1959).
Brutkowski, Fonberg, Mempel (1960) показали, что уда-
ление миндалевидного комплекса, особенно его базолате-
ральной части, не нарушало проявления пищевых инстру-
ментальных реакций. В то же время у животных резко
расстраивались реакции на пищевые дифференцировочные
сигналы.
Раздражение базолатеральной части миндалин вызыва-
ет у голодных животных прекращение приема пищи и вос-
произведение предварительно выработанных пищевых ин-
струментальных реакций (Fonberg, Delgado, 1961; Э. Фон-
берг, 1965).
По аналогии с этим Schwartzbaum (1961) считает, что
гиперфагия, возникающая при разрушении амигдалы, свя-
зана не с усилением голодных ощущений, а с уменьшением
чувства насыщения.
Такое предположение может быть вполне обоснованным,
поскольку Niemer, Goodfellow, Speaker (1963) показали,
что именно базальная часть миндалевидного комплекса
наиболее тесно связана с вентральным гипоталамусом.
Ряд авторов отмечают возникновение гиперфагии при
прямом раздражении миндалевидного комплекса (Kaada,
1951; Fuller, Green a. oth., 1957; Wood, 1958; Allikmets, Dit-
rich, 1965). В противоположность этому Anand (1961),
Brutkowski и др. (1962), Fonberg (1966), В. А. Черкес
(1967) после полного удаления миндалевидного комплекса
наблюдали возникновение афагии.
79
Химическое раздражение амигдалЫ позволяет выявить
в ней две различные области. Одна из них холинергическая,
другая—-адренергическая. Холинергическое раздражение
вентральной амигдалы вызывает уменьшение приема пищи,
а адренергическое раздражение той же области, наобо-
рот,— повышение потребления пищи (Grossman, 1964).
Ряд исследователей показали функциональные взаимо-
связи миндалевидного комплекса с медиальными отделами
гипоталамуса.
Так, японские авторы (Masaichi, Naoshige, Taigi, Shizuo,
1959) описывают влияния амигдалярных ядер на клеточную
активность вентро-медиальных ядер гипоталамуса.
Or den и Sutin (1963) наблюдали увеличение амплитуды
вызванных потенциалов в перегородке и уменьшение их
амплитуды в миндалевидном ядре в ответ на раздражение
вентро-медиальных ядер гипоталамуса вслед за апплика-
цией в эту область или введением в кровь норадреналина.
Ryoichi и др. (1959) также отметили, что раздражение
медиальной части базальных ядер миндалины вызывало
изменения электрической активности только в вентро-ме-
диальных ядрах гипоталамуса. Авторы наблюдали, что
раздражение латеральной части базального ядра миндали-
ны вызывало преимущественные изменения в латеральном
ядре гипоталамуса.
Nauta (1960) сообщает о прямых проекционных связях
миндалевидного комплекса с перегородкой и гипоталаму-
сом. Он указывает, что, кроме волокон stria terminalis,
оканчивающихся в медиальных группах клеток преоптиче-
ской и гипоталамической области, от миндалевидного ядра
отходит толстый пучок проекционных волокон, который
идет медиально и рострально и оканчивается в латераль-
ной части преоптической и гипоталамической области и
в перегородке.
Кроме миндалевидного комплекса, другие отделы лимби-
ческой системы также участвуют в регуляции пищевого по-
ведения.
В последнее время все большее количество авторов ука-
зывают на участие гиппокампа в формировании пищевых
условных рефлексов (А. И. Шумилина, 1965; Grastiyan а.
oth., 1959).
Эксперименты Лищщак и Эндреци (1967) убедительно
показывают, что гиппокамп имеет преимущественное отно-
шение к формированию некоторых механизмов памяти, свя-
занных с пищевым подкреплением
80
Что касается повреждений перегородки, то эффекты этих
операций на пищевое поведение противоречивы.
Raphelson и др. (1961) и Wolfe (1967) указывают, что
крысы с повреждением перегородки предпочитают в лаби-
ринте сторону пищевого подкрепления подкреплению водой
и бегут в этом отсеке значительно быстрее.
Delgado (1960) указывает, что у голодных обезьян при
раздражении области перегородки значительно утрачива-
ется интерес к пище.
Наличие богатых связей внутри лимбической системы
позволило Morgane (1962) рассматривать подкорковые
структуры, включающие большую часть среднего гипота-
ламуса и распространяющиеся по краю третьего желудочка
латерально к оптическому тракту и внутренней капсуле
посредством медиального пучка переднего мозга, как свое-
образный «аппестат» — аппарат, регулирующий постоян-
ство питательных веществ в организме. По его мнению, эта
система включает реципрокные «старт» и «стоп» механиз-
мы. Она -испытывает влияния со стороны активирующих
аппаратов ретикулярной формации, а также риненцефалона
и коры головного мозга.
Имеются все основания считать, что именно в лимбиче-
ской системе головного мозга, включающих пищевые цент-
ры гипоталамуса, происходит точное сопоставление уровня
обмена питательных веществ в организме с соответствую-
щей потребностью в них.
Вместе с тем «лимбический» уровень регуляции вместе
с гипоталамическими центрами принимает также участие
и в формировании соответствующего пищевого поведения.
Уже давно было замечено, что голодное животное прояв-
ляет значительное беспокойство, двигательную активность,
выражающуюся в активном поиске пищевых веществ. На-
оборот, накормленные животные обычно мало подвижны.
Hetherington (1943) наблюдал, что двигательная актив-
ность голодных крыс резко уменьшалась при повреждении
медиальных отделов гипоталамуса. Аналогичные данные
приводят Miller, Bailey, Stevenson (1950). Они показали,
что животные после поражений медиальных отделов гипо-
таламуса часто становятся вялыми и не склонны к инстру-
ментальной деятельности по добыванию пищи.
Вместе с тем Mendelson и Chorover (1965) показали, что
раздражение латеральных отделов гипоталамуса стимули-
рует пищедобывательное поведение даже у накормленных
животных и способствует их лучшему обучению;
81
Наряду с этим показано, что после удаления определен'
ных частей лобных долей коры и базальных ганглиев (Rich-
ter, Hawkes, 1939; Ruch, Shenkin, 1943), так же как и после
повреждений в переднем гипоталамусе (Maire Patton,
1954), наблюдается увеличение двигательной активности
животных. На основании этого Brobeck (1955) высказал
предположение, что гипоталамические механизмы, опреде-
ляющие пищевые движения животного, значительно сдер-
живаются влияниями, идущими со стороны передних отде-
лов коры головного мозга и хвостатого ядра.
В настоящее время имеется множество доказательств то-
го, что регуляция приема пищи и пищевого поведения в
значительной степени осуществляется высшими отделами
головного мозга—деятельностью коры и ближайших под-
корковых образований. Так, Morgane (1961) наблюдал, что
у крыс после разрушения вентральной части бледного шара
развиваются афагия и адипсия.
Rubinstein и Delgado (1962) отмечают, что раздражение
у обезьян передней части головки хвостатого ядра тормо-
зит пищевые мотивации.
Перерезка паллидофугальных волокон, направляющихся
к латеральному гипоталамусу и среднему мозгу, вызывает
афагию (Morgane, 1961; Gold, 1967).
Раздражение областей паллидума, связанных с покрыш-
кой среднего мозга, наоборот, «запускает» пищевое пове-
дение (Wyrwiska, Doty, 1966).
Еще давно было замечено (Goltz, 1892), что животные,
лишенные полушарий головного мозга, не проявляют сколь-
ко-нибудь выраженных целенаправленных действий. Извест-
но, что «бесполушарное» животное, будучи голодным, тем
не менее не отправляется на поиск пищи и потребляет ее
только в том случае, когда она вкладывается ему непо-
средственно в ротовую полость.
Как показали опыты Ц. А. Орджоникидзе и М. А. Нуцу-
бидзе (1961), кошки после удаления у них коры больших
полушарий отчетливо проявляют чувство голода и насыще-
ния. В голодном состоянии при отсутствии пищедобыва-
тельной деятельности такие животные становятся беспо-
койными, мяукают и даже неистово кричат. При насыще-
нии они успокаиваются и перестают кричать.
Все это заставляет думать, что гипоталамический и ре-
тикулярный уровни лимбической системы обеспечивают
только примитивно организованные, относительно индиффе-
рентные, т. е. лишенные выраженной целенаправленности,
82
типы пищевых мотиваций. Эти мотивации не могут идти ни
в какое сравнение с мотивациями нормальных животных.
Н. А. Рожанский (1957) у декортицированных собак
также не наблюдал целенаправленного поиска пищи. Он
полагает, что пищенаправленные реакции связаны щ дея-
тельностью полосатого тела. Такие реакции были получены
им при раздражении ряда подкорковых образований (хво-
статое ядро, передние ядра таламуса, центральное ядро
таламуса).
Методом экстирпации различных отделов коры больших
полушарий головного мозга установлено, что регуляция
пищевых мотиваций, а также пищевых условных рефлексов
осуществляется преимущественно фронтальными и височ-
ными отделами, которые относятся к лимбической системе
(А. И. Шумилина, 1949; А. Н. Советов и В. Н. Чернийов-
ский, I960; Ю. М. Конорский, 1962; М. А. Нуцубидзе, 1962;
В. Н. Коваленко, 1962, 1963; Pribram, Bagshaw, 1953;
Andersson, Larsson, 1956; Soulairac, 1958, 1962; Anand, Dua,
Chine, 1958; Robinson, Mishkin, 1962; Szwejkowska, Kreiner,
Sychova, 1963; Brutkowski, Dabrowska, 1963; Brutkowski,
Mishkin, Rosvold, 1963, и др.).
Brown и Schafer еще в 1888 г. сообщили о гиперфагии
у двух обезьян, которая развилась у них после височной
лоботомии. То же самое обнаружили несколько лет спустя
Pribram и Bagshow (1953), Fulier, Rosvold, Pribram (1957)
после хирургических повреждений, включающих миндале-
видную и гиппокампову области. Green, Clemente, de Lroot
(1957), Wood (1958), Morgane и Kosman (1958) также на-
блюдали увеличение потребления пищи у кошек после по-
вреждений миндалевидного комплекса, особенно если эти
разрушения производились в базальном и латеральном мин-
далевидных ядрах, центральном и медиальном ядрах или
по всей грушевидной доле.
Височная лоботомия у человека также приводит к неко-
торой гиперфагии (Sawa a. oth., 1954; Tersian a. Ore, 1955,
и др.).
При поражениях структур во фронтальных долях коры
головного мозга также наблюдаются изменения в потребле-
нии пищи. СооЬ (1944) показал, что разрушение связей
между передними долями коры головного мозга и таламу-
сом нередка приводит к увеличению аппетита у человека.
Andersson и Larsson (1956) наблюдали уменьшение по-
требления пищи у собак после полного отсечения передне-
го мозга, включающего и перерезку обонятельных трактов.
83
Наиболее систематичные эксперименты по изучению ро-
ли лимбической-коры были проведены Anand, Dua и China
(1958). Авторы производили билатеральные повреждения
различных частей фронтальной доли коры и лимбической
системы у обезьян и кошек и изучали соответствующие из-
менения в потреблений пищи. Как они сообщают, повреж-
дения передней доли коры головного мозга, включающие
или ограниченные задней орбитальной корой, приводят к
уменьшению потребления пищи. Вместе с тем повреждения
только фронтальных кончиков и прилегающего к ним не-
большого участка задней орбитальной коры ведет к увели-
чению потребления пищи. Повреждения только передней
поясной извилины в их опытах не вызывали изменений по-
требления пищи. Повреждения, ограниченные миндалевид-
ным ядром и околоминдалевидной областью, вызывали вре-
менную афагию или гиперфагию, после чего обычно потреб-
ление пищи у животных возвращалось к норме. Обширные
повреждения височной доли коры, которые не затрагивали
миндалевидного комплекса, сопровождались, как правило,
гипофагией. Когда же эти повреждения включали также
и миндалевидный комплекс, заметная исходная гипофагия
сменялась в конечном счете гиперфагией. На основании
этих экспериментов авторы считают, что влияние передних
отделов коры на пищевое поведение не является однотип-
ным. Орбитальная поверхность фронтальной коры и минда-
левидного комплекса облегчает пищевые реакции, в то вре-
мя как дорзальные, фронтальные и височные области ока-
зывают на них тормозные влияния.
Одновременно с этим авторы обнаружили, что изменения
в потреблении пищи после повреждения лимбической об-
ласти были более заметны у обезьян, чем у кошек, и со-
вершенно не обнаруживались у крыс. Эти наблюдения го-
ворят о процессе «энцефализации» у высших животных да-
же на лимбическом уровне.
Изменения в потреблении пищи после повреждений лим-
бической области оказались менее выраженными, когда
они сочетались с повреждениями гипоталамуса.
После повреждений лимбической коры у кошек наруша-
лись «спонтанные» ритмы потребления пищи, которые на-
блюдались до операции. Обезьяны после повреждений ви-
сочной доли не различали съедобные и несъедобные веще-
ства и проявляли синдром «психической слепоты» Клювер
Бьюси. Интересно отметить, что такие повреждения у обезь-
ян вызывали преимущественное потребление овощной пищи
84
fid сравнению с крупой и орехами, содержащими большое
количество калорий. На основании этих наблюдений Anand
и др. заключили, что, вероятно, лимбические структуры
передней и височной областей коры головного мозга изме-
няют потребление пищи благодаря «дискриминационному
механизму», осуществляющему оценку вкуса. Эти механиз-
мы они назвали «аппетитом», в то время как такие прими-
тивные побуждения, как «голод» и «насыщение», они от-
несли к гипоталамическому уровню.
При раздражении различных лимбических структур из-
менений в потреблении пищи не наблюдается (Anand, Dua,
1956). При этом часто возникает жевание, глотание, при-
нюхивание, открывание и закрывание рта с высовыванием
языка и даже слюноотделение, т. е. все те реакции, которые
известны как «пищевые автоматизмы». Однако при этом ни-
когда не возникает целенаправленного пищевого поведения.
Все эти эксперименты позволяют высказать предположе-
ние, что лимбические образования высшего уровня, бес-
спорно, участвуют в механизме приема пищи. Вместе с тем
их роль, по-видимому, больше сводится к оценке количест-
ва и качества принимаемой пищи. Формирование же пище-
вых побуждений происходит на более низком гипоталами-
ческом уровне. Вопрос же о том, как формируется пищедо-
бывательное поведение, остается неясным. Ясно, что одного
лимбического уровня головного мозга для формирования
пищедобывательного поведения недостаточно.
Все высшие формы пищевого поведения — привычки, на-
циональные обычаи, предубеждения, выбор или предпочте-
ние одной пищи другой и т. д. — составляют неотъемлемые
атрибуты человеческого общества. Некоторые из этих ре-
акций проявляются и у высших животных. Каждый из нас
сознает, что на наше пищевое поведение сильно влияют со-
знательные, волевые действия. Бесспорно, что вся эта дея-
тельность определяется корой больших полушарий его.
Значительный материал в этом отношении представлен
опытами, в которых производилось изучение влияний по-
вреждений различных участков коры головного мозга на
потребление пищи.
Так, Anand, Dua и China (1961) исследовали влияние дву-
сторонних повреждений некоторых областей коры больших
полушарий головного мозга на потребление пищи у кошек
и обезьян. Они наблюдали заметное уменьшение потребле-
ния пищи после повреждений передне-латеральных отделов
коры головного мозга, включающих также хвостатое ядро.
85
В то же время они отметйлй, что повреждения в области
латеральных теменных, латеральных затылочных и задне-
латеральных височных отделов коры больших полушарий
не приводили к каким-либо изменениям в потреблении пи-
щи. Повреждения передней височной коры, как уже указы-
валось выше, увеличивали потребление пищи.
Meyer (1962) установил, что экстирпация передних отде-
лов коры головного мозга у крыс вызывает нарушение при-
ема пищи. Однако равные по объему разрушения в заты-
лочных отделах коры головного мозга не вызывают подоб-
ного эффекта.
Применение электроэнцефалографических методов позво-
лило исследовать участие коры головного мозга в пищевых
реакциях по изменению ее электрической активности. Впер-
вые Hess и др. (1953) отметили появление у кошек в коре
головного мозга при питье молока высоковольтных коле-
баний (5—8 кол/сек), которые были более отчетливо выра-
жены в париентальной и затылочных областях.
А. И. Шумилина (1958, 1961) наблюдала у кролика при
приеме пищи появление в электроэнцефалограмме высоко-
амплитудных колебаний. Позднее такие же изменения
в электрической активности коры головного мозга во время
кормления и после насыщения были отмечены рядом авто-
ров (Sadowski, Longo, 1962; К. В. Шулейкина, 1963, 1964;
Clemente, Sterman, Wyrwicka, 1964; Buchwald a. oth., 1964;
Roth, Sterman, Clemente, 1967; Marczynski a. oth., 1968).
Появление медленных, высокоамплитудных волн на элект-
роэнцефалограмме после введения в желудок собаки рас-
твора глюкозы наблюдали в своих опытах И. А. Булыгин
и Ю. Н. Дорожкин (1964). Такую же реакцию в коре го-
ловного мозга наблюдали Е. Вассар, И. Лайднаи П. И. Кин-
гисепп (1961) после введения глюкозы.
В состоянии голода у животных и человека ряд авторов,
наоборот, зарегистрировали высокочастотную, низкоампли-
тудную активность электроэнцефалограммы (А. М. Мариц,
1962; Morgane, 1961; И. И. Гохблит, 1961; Sadowski, Longo,
1962; Hockman, 1964).
Однако специального анализа нейрофизиологических ме-
ханизмов различной электроэнцефалографической карти-
ны голодных и накормленных животных ни один из упомя-
нутых авторов не производил.
Специальное исследование активности отдельных нейро-
нов передних отделов коры головного мозга при голоде и
после приема пищи было проведено Ю. А. Фадеевым (1963,
86
1964). Он обнаружил, что количество активных нейронов
в сенсомоторных отделах коры головного мозга у голодных
животных значительно выше, чем у накормленных живот-
ных.
Аналогичный эффект снижения активности корковых ней-
ронов у голодных животных в его опытах наблюдался и по-
сле внутривенного введения глюкозы.
Наиболее значительные и неопровержимые доказательст-
ва участия коры больших полушарий головного мозга в ре-
гуляции пищевого поведения представлены многочислен-
ными экспериментами с условными пищевыми рефлексами,
проведенными в школе И. П. Павлова, а также его учени-
ками и последователями.
Все полученные в этом отношении экспериментальные
данные указывают на то, что роль коры головного мозга
особенно отчетливо проявляется в таких реакциях, как на-
хождение пищи, ее оценка и выбор, а также в формирова-
нии аппетита.
Вместе с тем вопрос о том, каким образом происходит
вовлечение корковых элементов в деятельность гипотала-
мических отделов пищевого центра, остается соверешенно
не изученным.
Впервые И. А. Черешнев (1950, 1960) применил методику
локального раздражения и разрушения гипоталамических
образований под контролем рентгеновых лучей для изуче-
ния их роли в высшей нервной деятельности. Ему удалось
показать, что раздражение и разрушение структур заднего
гипоталамуса вызывали усиление условных оборонитель-
ных реакций и одновременное торможение условных секре-
торно-двигательных пищевых реакций. При этом наблюда-
лась отчетливая диссоциация двигательных и секреторных
компонентов условных реакций.
Изменения условнорефлекторной деятельности при вве-
дении различных химических веществ в область заднего
гипоталамуса отметил Л. В. Калюжный (1962, 1963, 1964).
Введение адренергических веществ в его опытах усиливало
условные оборонительные реакции и тормозило пищевые.
Инъекции холинергических веществ вызывали противопо-
ложный эффект.
Многочисленные эксперименты по изучению роли пище-
вых центров гипоталамуса в формировании условных и ин-
струментальных пищевых реакций были проведены рядом
авторов (Wyrwicka, Dobrzecka, Tarnecki, 1953, 1960; Wyr-
wicka, Dobr?ecka, 1960; Balinska, Lewipska, Romanjuk, Wyr-
87
wicka, 1961; Matsumoto a. oth., 1963; Balinska, 1963, 1966;
Baiinska, Brutkowski, 1964, и др.).
Было показано, что раздражение «центра питания» лате-
рального гипоталамуса, а также разрушение медиального
«центра насыщения» вызывали усиление пищевых услов-
ных и инструментальных реакций.
Разрушение латеральной гипоталамической области, на-
оборот, вызывало исчезновение ранее выработанной инстру-
ментальной пищевой реакции (Balinska, 1963; Mishkin,
1954).
Mishkin и Pribram (1954) установили, что удаление вент-
рального отдела височной области коры головного мозга
ухудшает зрительные инструментальные дифференцировки,
основанные на пищевом подкреплении.
С. В. Брутковский (1965) показал, что условнорефлек-
торные пищевые дифференцировки у собак расстраиваются
при повреждениях медиальной области коры лобных долей,
а также «area gennuaiis» передней части поясной извилины
и базолатеральных областей миндалевидного ядра. Он
считает, что именно эти отделы лимбической системы участ-
вуют в процессах торможения пищевой деятельности.
Оценивая в целом весь приведенный выше эксперимен-
тальный материал, можно видеть, как обширно представле-
ны образования пищевого центра в головном мозге.
При этом более высокоорганизованные отделы головного
мозга участвуют в осуществлении все более сложных пище-
вых реакций.
Если на уровне продолговатого и спинного мозга осуще-
ствляются только такие простые врожденные рефлексы, как
глотание, жевание, рвота, то уже следующий, более высо-
кий уровень центральной нервной системы—гипоталамус—
приобретает свойства интеграции соматических и вегета-
тивных функций в регуляции приема пищи. Однако за счет
только гипоталамических структур целостный пищедобыва-
тельный акт не может быть сформирован. Для его отчетли-
вого проявления необходимы другие, более высокие орга-
низации — лимбические образования и кора головного моз-
га. Только объединенное участие гипоталамуса, лимбиче-
ских образований и коры головного мозга формирует це-
лостные пищевые реакции животных.
Все это подтверждает представления И. П. Павлова о
сложной динамической организации пищевого центра («дис-
танция огромного размера»), находящегося на различных
уровнях центральной нервной системы.
88
Насколько широки функциональные взаимосвязи струк-
тур, определяющих пищевую деятельность организма, по-
казывают эксперименты Robinson и Mishkin (1962). Авторы
установили, что прием пищи у накормленных животных
может быть вызван раздражением 139 пунктов гипотала-
муса и 99 пунктов, расположенных вне его. Пищевое пове-
дение в их опытах вызывалось раздражением латерально-
го гипоталамуса, задней орбитофронтальной коры, обоня-
тельного бугорка, subst. innominata, диагонального пучка
Брока, амигдалы, перегородки, преоптического поля и пе-
редней поясной извилины.
Morgane (1969) выдвинул представления, согласно кото-
рым регуляция приема пищи осуществляется комплексом
функционального объединения структур, расположенных
в лимбической области, гипоталамусе и среднем мозге.
В связи с этим, естественно, возникает вопрос: каким же
образом происходит функциональное объединение различ-
ных отделов пищевого центра в единую функциональную
систему? Какие конкретные нейрофизиологические меха-
низмы производят это объединение? В результате чего
возникает целенаправленная пищедобывательная деятель-
ность?. Но ответа на эти вопросы литературные источники не
дают. Аналогичная функциональная организация определяет
контроль, поиск и поглощение воды. В области переднего
латерального гипоталамуса расположен центр жажды.
В вентро-медиальной области гипоталамуса расположен
центр питьевого насыщения (Smith, McCann, 1962; Steven-
son, 1964; Fitzsimoz, 1966; Mogenson, Stevenson, 1967, и др.).
Впервые Andersson (1952, 1953) при электрическом и хи-
мическом раздражении гипоталамуса получил у коз уси-
ленные питьевые реакции. Зона раздражения охватывала
область воронки гипоталамуса между сводом и мамиллота-
ламическим трактом, расположенную латерально к пери-
вентрикулярному ядру.
Подобные наблюдения были проведены и на крысах (Gre-
er, 1955; Montemurro a. Stevenson, 1957). Montemurro и
Stevenson (1957) установили, что у крыс питьевые реакции
могут возникать при раздражении области латерального
гипоталамуса. Двустороннее разрушение этой области ве-
дет к комбинированной афагии и адипсии (Tinbergen, 1951).
Удаление латеральной гипоталамической области у собак
также вызывало адипсию (Andersson, McCann, 1956).
Питьевые реакции у коз возникают при электрическом
или химическом раздражении области между форниксом
89
и мамиллоталамическим трактом (Andersson, McCann,
1955).
Fisher и Coury (1964) произвели детальное исследование
влияний холинергического раздражения различных пунктов
головного мозга крысы на питьевое поведение. Они показа-
ли, что увеличение потребления воды возникает при разд-
ражении различных структур круга Пейпса, перегородки,
диагонального пучка переднего мозга. Из структур гиппо-
кампа только его дорсо-медиальная область при ее хими-
ческом раздражении вызывает у животных питьевые реак-
ции. Таким образом, питьевые мотивации также определя-
ются целым комплексом функционально объединенных
образований головного мозга.
МОТИВАЦИИ СТРАХА И АГРЕССИИ
Изучение центральных механизмов защитных мотива-
ций по существу началось с классических экспериментов
Bard (1928, 1934). Он впервые показал, что удаление коры
головного мозга у кошек и собак приводит к развитию ре-
акции «ложной ярости». В ответ на раздражение перифе-
рических рецепторов у таких животных легко возникает
приступ ярости. У таких бесполушарных кошек обнаружи-
ваются реакции страха и в той ситуации, где они предвари-
тельно подвергались болевому раздражению (И. С. Бери-
ташвили, 1961).
Bard полагал, что такая повышенная возбудимость яв-
ляется следствием высвобождения функций гипоталамуса
из-под контроля коры и первый высказал предположение
о.тормозной роли коры головного мозга в развитии защит-
ных эмоциональных реакций.
Однако впоследствии Bard и Mountcastle (1947) устано-
вили, что такая же реакция у кошек наблюдается и после
удаления части риненцефалона, особенно амигдалы и ме-
диальной коры. Вместе с тем было обнаружено, что удале-
ние только коры больших полушарий, приводит, наоборот,
к развитию у животных покорности.
В других исследованиях на обезьянах и кошках (Kliiver,
Вису, 1939; Schreiner a. Kling, 1953) было показано, что
удаление верхушки височной доли и гиппокампа также ча-
ще приводит к развитию покорности, чем ярости и агрес-
сии.
Spiegel, Miller, Oppenheimer (1940) в опытах на кошках
заметили, что разрушение обонятельных луковиц, миндале-
90
видного ядра или гиппокампосводовых трактов такЖё мен
жет вызвать реакцию «ложной ярости». Между тем этого
эффекта не наблюдалось при разрушении фронтальных или
других отделов коры больших полушарий или грушевидной
области обонятельного мозга.
Дополнительно к этому Kiiiver и Вису (1938, 1939) от-
метили, что двустороннее разрушение височной области
у обезьян делает их необычайно спокойными. Животные ут-
рачивают страх. Они смело и неоднократно схватывают
зажженную спичку. Одна из таких обезьян даже смело ли-
зала ядовитые зубы змеи. Эта реакция является крайне по-
разительной, поскольку обычно такие обезьяны испытыва-
ют панический ужас при одном только виде змеи.
В более поздних экспериментах Bard и Mauntcastle
(1948) удаляли у кошек большие полушария, оставляя при
этом интактными обонятельный отдел и поясную извилину.
Удаление всего неокортекса при сохранении лимбических
структур делает животное явно апатичным и эмоционально
ареактивным. Однако самым удивительным является тот
факт, что в результате такой операции у животных в ответ
на внешние раздражители не проявлялась реакция «лож-
ной ярости». Только после того, как у этих животных были
дополнительно удалены миндалины, грушевидная область
и гиппокамп, у них отчетливо проявилась реакция «ложной
ярости». Все это позволило Bard и Mauntcastle высказать
предположение о том, что сдерживающее влияние на низ-
шие центры ствола, определяющие агрессивные мотивации,
оказывает не только кора больших полушарий, но и древ-
няя лимбическая кора.
Авторы предполагают, что все тормозные влияния коры
больших полушарий на функции гипоталамуса осуществля-
ются через миндалевидное ядро. Это, как видно, противо-
речит данным Kluver и-Вису, так как в их опытах разруше-
ние миндалевидных ядер у обезьян не вызывало реакции
«ложной ярости».
Fulton и Ingram1 (1929) описали подобные реакции гне-
ва у кошек после двусторонних разрушений прехиазмаль-
ной области в основании головного мозга. Эти данные были
позднее подтверждены Bradey и Nauta (1953, 1955), Har-
rison-и Lyon (1957) и King (1958).
Однако Heath (1954) при разрушении у кошек области
перегородки, хвостатого ядра и обонятельных бугорков, на-
оборот, наблюдал снижение эмоциональной реактивности
и апатию.
91
Smith (1950) показал, что когда экспериментальные по-
вреждения у обезьян были ограничены лишь грушевидной
долей, миндалевидным комплексом и гиппокампом при
сохранении большей части коры больших полушарий, жи-
вотные становились покорными и ручными.
Thompson и Walker (1951) в опытах на обезьянах с дву-
сторонним повреждением медиальной поверхности височ-
ной доли также получили эффекты «укрощения» при раз-
рушениях только миндалевидного комплекса и гиппокампа.
Как видно, данные о роли различных лимбических обра-
зований в проявлении мотивации страха и агрессии весьма
противоречивы.
Центральная роль в формировании оборонительных
и защитных реакций принадлежит гипоталамусу.
По существу Hess (1930) первый продемонстрировал,
что агрессивные и защитные реакции у кошек возникают
при раздражении окрестностей гипоталамической области.
Hess и Briigger (1961) путем раздражения различных
точек гипоталамуса составили карту, показывающую, что
зона, раздражение которой наиболее отчетливо вызывает
защитные реакции у кошек, расположена вокруг свода в об-
ласти заднего гипоталамуса.
Возникновение реакции ярости и атаки при раздраже-
нии задней гипоталамической области наблюдали также
Ranson и др. (1935), Stachnic и др. (1966).
Ranson (1936) отмечал разнообразные проявления яро-
сти при раздражении медиального пучка переднего мозга
в латеральной части гипоталамуса. Аналогичный эффект
наблюдался при раздражении свода в том месте, где он
входит в латеральную область гипоталамуса, а также зо-
ны, расположенной между латеральным гипоталамусом
и центральным серым веществом.
Оборонительные реакции удается вызвать при раздра-
жении участков гипоталамуса, расположенных вблизи сво-
да в преоптической области, в вентральной части перего-
родки и в центральном сером веществе (Hess, 1949; Hess а.
Akert, 1955).Повреждение заднего гипоталамуса снижает
агрессивность и приводит к развитию у животных явлений
покорности (Ranson, 1939).
Наоборот, после разрушения вентро-медиальных ядер ги-
поталамуса дружелюбное животное становилось исключи-
тельно злобным (Wheatley, 1944; Ingram, 1962).
Если теперь сопоставить данные приведенных выше авто-
ров, можно легко заметить, что зона, вызывающая защит-
92
ные реакции животных, представляет собой узкую полоску
не более 1—1,5.мм длины, распространяющуюся от перед-
него мозга через вентральный диэнцефалон к среднему
мозгу. Между тем перифорникальная область заднего ги-
поталамуса является своего рода центром этой зоны. За-
щитные реакции возникают из этой области при наимень-
шей силе электрического раздражения.
De Molina и Hunsperger (1959) опубликовали специаль-
ный.обзор, в котором они приводят карту расположения в го-
ловном мозге зон, раздражение которых вызывает различ-
ные проявления эффективных реакций (рис. 7).
Рис. 7. Схематическое изображение областей переднего и среднего моз-
га кошки, при раздражении которых возникают аффективные реакции
(по Hunsperger, 1956).
На рисунке нанесены изображения сагиттальных срезов на двух уров-
нях. Активная область гипоталамуса и среднего мозга состоит из «внут-
ренней зоны» (черное), раздражением которой можно вызвать реакцию
шипения, и «внешней зоны» (штриховка в клеточку), при раздражении
которой возникает реакция бегства. Активная область миндалевидного
ядра, Str. terminalis и ложе Str. terminalis (поперечная штриховка)
Вызывает при раздражении реакцию рычания.
/—миндалевидное ядро; 2—передняя комиссура; 5—ложе Str. terminalis; 4—нис-
ходящее колено свода; 5 — Str. terminalis; 5 — fibria; 7 —гиппокамп; 8 централь-
ное серое вещество среднего мозга; 9 — гипоталамус.
93
Рис. 7 показывает, что реакция ворчания возникает при
раздражении миндалевидного ядра, конечных полосоки их
ложа.
Этой реакции обычно предшествует поворот головы
кошки в сторону наблюдателя, расширение глазной щели
и зрачков и усиление дыхания. За первым тихим ворчанием
следовали более громкие звуки; при этом у Животного были
расширены зрачки, прижаты уши, опущена голова,
а шерсть на спине и хвосте стояла дыбом.
Таким образом, налицо были все признаки оборонитель-
ного состояния.
Авторы полагают, что в гипоталамусе и центральном се-
ром веществе среднего мозга находится «внутренняя зона»,
раздражением которой можно вызвать «реакцию шипения».
Вокруг нее располагается «внешняя зона». При ее раздра-
жении животное начинает искать пути к спасению и нахо-
дит его в бегстве.
Если теперь обратиться к морфологическим связям ги-
поталамуса, то нетрудно заметить, что все упомянутые на
рис. 7 области, имеющие отношение к формированию за-
щитных реакций, тесно связаны друг с другом.
Именно наличие таких тесных функциональных взаимо-
отношений в передних отделах головного мозга определяет
то обстоятельство, что оборонительные реакции определя-
ются многими структурами головного мозга.
При повреждении ряда мозговых структур нарушается
своеобразный баланс спокойствия и раздражительности,
адекватного реагирования на внешние раздражители.
Так, после удаления коры больших полушарий или разру-
шения области перегородки или вентро-медиальных ядер
гипоталамуса животные на слабые раздражители начина-
ют проявлять бурную реакцию ярости.
В формировании мотиваций страха и агрессии значитель- .
ная роль принадлежит амигдалярной области. Однако эф-
фекты влияния удаления миндалины на проявление оборо-
нительных реакций противоречивы.
Bard и Mountcastle (1948), Bunnel (1966), Wood (1958),
Spiegel n др. (1940), В. А. Черкес (1967) после таких опе-
раций наблюдали у животных нарастание страха и агрес-
сии. Другие авторы (Schreiner, Kling, 1953; Fuller a. oth.,
1957) отмечали прямо противоположные эффекты. .
Karli (1956) и Woods (1956) сообщают, что двусторон-
нее разрушение амигдальных ядер делает животных уди-
вительно спокойными.
94
Обезьяны, обычно панически боящиеся змей, После дву-
сторонней амигдалотомии спокойно берут их в руки и даже
едят.
' Удаление базолатеральных отделов миндалины наруша-
ет проявление классических условных оборонительных ре-
акций, не оказывая в то же время заметного влияния на
проявление ранее выработанных инструментальных реак-
ций избегания (Fonberg, Delgado, 1961; Э. Фонберг,
1965).
При раздражении медиальных отделов амигдалы наблю-
даются такие реакции, как шипение и рычание. Delgado
(1959) при раздражении миндалевидного ядра у кошек вы-
зывал отчетливые реакции ярости.
Электрическое раздражение дорсо-медиальной области
миндалины вызывает у собак возникновение страха (убе-
гания), попытки вырваться из экспериментальной обста-
новки, скуление, лай. Эти реакции очень напоминают реак-
ции, возникающие при раздражении задних отделов гипо-
таламуса.
Однако эффекты раздражения миндалины имеют
больший латентный период и большую продолжительность.
На основе раздражения этих структур собаки легко выра-
батывают инструментальные оборонительные реакции на
условные раздражители.
Эффект стимуляции базолатеральной части противополо-
жен эффекту раздражения дорсо-медиальной части
(Э' Фонберг, 1965). Раздражение этой области угнетает со-
стояние страха, вызванное как ноцицептивными раздраже-
ниями, так и прямым раздражением гипоталамуса. Однако
оно не тормозит реакцию избегания, развивающуюся в от-
вет на действие условных раздражителей (Fonberg, Delga-
do, 1961).
Egger и Flynn (1963, 1967) убедительными опытами пока-
зывают, что раздражение амигдалы у кошек может как уси-
лить, так и ослабить агрессивную реакцию атаки,' вызван-
ную раздражением задней гипоталамической области. Эф-
фект торможения вызывался из крупноклеточной части ба-
зальных миндалевидных ядер и из передней и медиальной
частей латерального миндалевидного ядра. Облегчение вы-
зывалось при раздражении дорсо-латеральной области зад-
ней части латерального ядра.
De Molina и Hunsperger (1959) показали, что только раз-
рушение оборонительных центров гипоталамуса устраняет
оборонительные реакции, возникающие при раздражении
95
ймигдал. Наоборот, разрушение аМигДалярной области не
устраняет оборонительные реакции, вызванные раздраже-
нием гипоталамических центров.
Аналогичным образом спокойствие, вызванное амигдал-
эктомией, снова сменяется повышенной яростью после до-
полнительного повреждения вентро-медиальных ядер гипо-
таламуса (Schreiner a, Kling, 1953).
Разрушения латерального гипоталамуса понижают вы-
раженность реакции ярости при раздражении амигдалы.
Все это указывает на то, что в формировании оборони-
тельных реакций ведущая роль все же принадлежит гипота-
ламическим центрам.
На участке других отделов лимбической системы в фор-
мировании мотиваций страха и агрессии указывают следую-
щие наблюдения. Повреждения области перегородки вызы-
вают у крыс появление ярко выраженных неадекватных
эмоциональных реакций на тактильные раздражения (Bra-
dy, 1955).
Heath и Mickle (1960) описывают, что у больных раздра-
жение области перегородки значительно снижает болевые
ощущения.
Л. Алликметс и И. Лапин (1965) и Bunnel (1966) указы-
вают на то, что разрушение перегородки повышает агрес-
сивные реакции у крыс.
Повышение агрессивности наблюдается при разруше-
нии срединных и передних ядер таламуса (Lindsley a. oth.,
1949; Jasper, 1954; Schreiner a. oth., 1953; Magoun 1954).
Раздражение передней части поясной извилины вызыва-
ет разнообразные проявления ярости (Anand a. Dua, 1955).
Наоборот, удаление поясной извилины и фронтальной коры
снижает агрессивность животных и превращает диких
животных в ручных (Pribram1 a. oth., 1954, 1957).
Adey (1962) у австралийского фалангера наблюдал уст-
ранение агрессивности после удаления энторинальной коры.
Животные после такой операции охотно шли на руки, по-
зволяли ласкать себя, а также проявляли большое любо-
пытство к другим живым существам, обнюхивали их и т. д.
Такие же реакции защиты и нападения наблюдали
у обезьян при раздражении гиппокампа (Mac Lean, Delgado,
1953; Brady, Hunt, 1955).
Разрушением гиппокампа, как указывают Green и др.
(1957), у животных можно вызвать резкое усиление страха.
Оперированные таким образом коты забиваются в угол при
появлении других животных или экспериментатора. При
96
попытке дотронуться до них они шипят и проявляют агрес-
сивность.
Вместе с тем другие авторы указывают на то, что агрес-
сивные реакции у обезьян отчетливо снижаются при дву-
стороннем повреждении гиппокампа (Shapito, 1965), а так-
же после инъекции новокаина в эту область (Delgado,
Kitahata, 1967).
Реакция страха и ярости наблюдается и при раздражении
поясной извилины (М. А. Нуцубидзе, 1963).
Целая серия опытов (Baird a. oth., 1951; Schreiner a. oth.,
1953) показывает, что разрушение интраламинарных ядер
таламуса приводит к сонливости и понижению эмоциональ-
ной реактивности.
Противоречивые данные получены при удалении орби-
тальной области. Одни исследователи (Kennard, 1945) при
этом наблюдали усиление реакции ярости, другие, наобо-
рот, отмечали, что животные становятся ручными (Living-
ston a. oth., 1948). Последующие исследования показали,
что эффекты зависели от того, в какой степени при этой
операции были повреждены такие лимбические структуры,
как миндалевидное ядро, гиппокамп и пириформная кора.
В том случае, когда эти структуры оставались интактны-
ми, животные проявляли спокойствие.
Stellar (1954) высказывает предположение, что в избега-
нии боли решающую роль играет лобная кора.
Двусторонние разрушения у кошек базальной части пере-
городки или передне-латерального гипоталамуса с повреж-
дением преоптического поля и медиального пучка передне-
го мозга приводят к появлению на электроэнцефалограм-
ме постоянной медленной высокоамплитудной активности,
развитию состояния гиперсомнии и отсутствию реакций на
внешние стимулы, особенно на сигналы опасности (К. Лиш-
шак и Э. Эндреци, 1965). Подобные картины наблюдаются
в тех случаях, когда разрушения затрагивают вентро-меди-
альную часть бледного шара, включая нижние ножки тала-
муса.
Стимуляция ретикулярной формации у таких кошек не
вызывает пи электроэнцефалографической, ни поведенче-
ской реакции пробуждения. У таких животных утрачивается
способность к обучению и исчезает ранее выработанные ус-
ловные рефлексы. У крыс с двусторонними повреждениями
базальной перегородки или медиального пучка среднего
мозга, а также и в стриопаллидарной области полностью
исчезают ранее выработанные условные рефлексы избега-
4 Заказ 5074
97
ния; животные совершенно перестают реагировать нй ус-
ловный сигнал; на болевое электрическое раздражение от-
вечают нецеленаправленной реакцией.
Brady и др. (1954) указывают на то, что после двусторон-
него удаления миндалевидного комплекса у кошек значи-
тельно медленнее вырабатываются условнорефлекторные
реакции избегания. Между тем эта операция практически
не оказывает влияния на уже выработанные реакции избе-
гания.
Weiskrantz и Pribram (1956) наблюдали на приматах
значительное уменьшение условной реакции избегания
после двустороннего повреждения как миндалевидного
комплекса, так и лобно-височной коры и поясной изви-
лины.
Brady и Nauta (1953) обнаружили, что условнорефлек-
торная реакция беспокойства при последующем болевом
подкреплении значительно снижалась после перерезки
свода и повреждения перегородки. Такую же реакцию вы-
зывает повреждение и раздражение гиппокампа (Koranyi,
Endroczi, 1965).
Итак, в формировании оборонительных и агрессивных
мотиваций также участвуют обширные, функционально объ-
единенные участки головного мозга. Эти мотивации объеди-
няют многие лимбические структуры, включая и кору боль-
ших полушарий.
Разные отделы лимбической системы имеют отношение
к разным сторонам проявления оборонительных и агрессив-
ных мотиваций.
ПОЛОВЫЕ МОТИВАЦИИ
Практически почти все уровни центральной нервной си-
стемы также участвуют в интеграции полового поведения.
Так, за счет спинальных механизмов у самцов обеспечива-
ется эрекция полового члена и эякуляция. Самки с сохран-
ным средним мозгом могут принимать соответствующие по-
зы при раздражении влагалища.
Но не только низшие центры нервной системы обусловли-
вают половое поведение животных. Известно, что оператив-
ное удаление матки и влагалища или их деафферентация
не препятствует копулятивной деятельности самок крыс и
кроликов (Bali, 1934; Brooks, 1937). После разрушения кре-
стцового отдела спинного мозга коты проявляют агрессив-
ное половое поведение, несмотря на то что область таза,
98
в том числе половые органы, у них были полностью пара-
лизованы (Ruch, Bard, 1947), Половая деятельность таких
животных отчетливо проявляется, несмотря даже на невоз-
можную эрекцию полового члена. Безусловно, что афферен-
тация, идущая от половых органов, весьма существенна
в построении полового возбуждения и особенно полового
акта. Вместе с тем половое влечение и соответствующее
поведение формируются другими механизмами. Наиболее
тесное отношение к формированию половых влечений при-
надлежит гипоталамической области (Sowyer, Robinson,
1956).
Разрушение переднего и заднего гипоталамуса приводит
к устранению полового поведения и изменениям половых же-
лез и полового аппарата.
Fisher, Magoun, Ranson (1938) первыми выявили, что пос-
ле повреждения передней части гипоталамуса у кошки по-
ловое поведение прекращается. То же самое наблюдали
в своих экспериментах Dempsey и Rioch (1939) и Bard
(1940).
Введение таким животным женских половых гормонов не
восстанавливает половое поведение. Это, следовательно,
указывает на то, что половое поведение возникает как след-
ствие действия половых гормонов на передние отделы гипо-
таламуса.
Повреждения, расположенные между вентро-медиальны-
ми ядрами и мамиллярными телами гипоталамуса, вызыва-
ют атрофию яичников и также нарушают половое поведе-
ние животных. Однако половое поведение при этих пора-
жениях головного мозга восстанавливается при введении
животным эстрогенных гормонов. Все это дает основание
думать, что в данном случае поражается в основном гона-
дотропная функция гипофиза.
Другие авторы (Harris, 1948; Markee a. oth., 1946) пока-
зали, что раздражение вентро-медиальных ядер гипотала-
муса вызывает овуляцию.
У кошек в отсутствие эструса раздражение дорсо-меди-
ального ядра гипоталамуса стимулирует секрецию эстро-
генов.
Раздражение гипоталамической области также влияет на
половое поведение самцов. Так, раздражение туберальной
области гипоталамуса у самцов крыс вызывает эякуляцию.
Раздражение медиального пучка переднего мозга в области
латерального гипоталамуса у самцов вызывает эрекцию по-
лового члена и соответствующие эмоциональные проявле-
4:
99
ния. Раздражение у крыс переднего гипоталамуса понижа-
ет половые влечения (Mac Lean a. oth., 1960).
О чувствительности гипоталамуса к эстрогенам свидетель-
ствует ряд опытов, в ходе которых в различные участки ги-
поталамуса подсаживались кристаллы стероидов. Lisk
(1960) у крыс под влиянием эстрадиола, подсаженно-
го в вещество дугообразного ядра, наблюдал прекращение
полового цикла и появление синдрома «кастрационной
матки».
Davidson и Sawyer (1961) под влиянием эстрадиола, под-
саженного в область заднего серого бугра и срединного
возвышения, наблюдали у кроликов торможение ову-
ляции, вызванной копуляцией, и атрофию матки. Har-
ris, Michael, Scott (1958) под влиянием стильбэстрола, под-
саженного в область бугрово-соскового поля, отмечали
у кастрированных кошек резко выраженную реакцию поло-
вого влечения.
Palva и Sawyer (1966) также вызывали резкое возраста-
ние половой активности у самок овариэктомированных кро-
ликов после имплантации им в премамиллярную вентро-ме-
диальную область гипоталамуса эстрадиола.
Предположение, согласно которому область переднего ги-
поталамуса содержит чувствительные к эстрогенам струк-
туры, было подтверждено опытами Donovan и Vander Werff
Ten Bosch (1956), которые путем разрушения у крыс перед-
него гипоталамуса вызывали наступление преждевременно-
го полового созревания.
Разрушение области, расположенной около мамиллярно-
го тела, приводит у самцов крыс к повышению половой ак-
тивности. При этом наблюдается раннее возникновение
эякуляции, укорочение фазы очередной эрекции, отсутствие
полового насыщения (Lisk, 1966).
Инъекции стильбэстрола или эстрадиола животным до
наступления половой зрелости способны вызвать поведение,
характерное для взрослой самки. Инъекции тестостерона
неполовозрелым животным вызывают возникновение у них
агрессивного поведения, драку с другими животными и т. п.
Путем чередования инъекций неполовозрелым животным
стильбэстрола и тестостерона можно наблюдать у них смену
двчх типов поведения.
Д. У. Гарис (1962) показал, что имплантация стильбэст-
рола в область мамиллярных тел гипоталамуса приводит
у овариэктомированной кошки к возникновению полной кар-
тины полового поведения,
100 .
Ч. X. Сойер (1962) наблюдал у крольчих овуляцию при
электрическом раздражении базальной области заднего ги-
поталамуса, расположенной между серым бугром и мамил-
лярными телами.
Fisher (1961) приводит данные о том, что инъекция поло-
вых гормонов в латеральную преоптическую область у крыс
обоего пола вызывает отчетливо выраженное половое по-
ведение.
При введении половых гормонов самцам и самкам в зад-
ний край перегородки или медиальную преоптическую об-
ласть животные проявляют выраженные материнские
инстинкты в поведении. Разрушение этой области, наобо-
рот, приводит к исчезновению всех инстинктов материнства
у самок крыс по отношению к детенышам.
Двустороннее повреждение вентральной части переднего
гипоталамуса между зрительным перекрестком и ножкой
гипофиза подавляло половое поведение у самки морской
свинки. В этих случаях даже инъекция половых гормонов
не могла вызвать полового поведения Brookhart a. oth., 1940;
Day a, oth., 1940).
Подобные данные были получены на самцах морских сви-
нок (Brookhart, 1941) и на крысах обоего пола Clark,
1942).
Раздражением латерального преоптического поля у крыс
обоего пола путем инъекций незначительных количеств со-
ответствующих половых гормонов можно вызвать продол-
жительное половое возбуждение (Fisher, 1956).
Все эти эксперименты свидетельствуют о наличии в гипо-
таламической области специальных хеморецепторов, изби-
рательно чувствительных к уровню циркулирующих поло-
вых гормонов и участвующих в формировании полового
влечения животных.
Деятельность гипоталамических центров, кроме того,
в значительной степени определяет функцию тесно связан-
ного с ним гипофиза.
Точная локализация в гипоталамусе центров, контроли-
рующих выделение адренокортикотропного (АКТГ), тирео-
тропного (ТТГ) и гонадотропного (ГТГ) гормонов гипофи-
за, представлена Д. М. Хьюмом (1962). Он установил, что
клеточные тела, регулирующие выделения АКТГ и ТТГ,
у собаки расположены в заднем отделе надоптического яд-
ра задней оптической области и в переднем отделе средин-
ного возвышения. Клетки, осуществляющие контроль над
выделением ГТГ, локализуются в заднебугровой области.
101
Вопрос об участии высщих отделов головного мозга в по-
ловых мотивациях оказался более сложным. Установлено,
что степень их участия в этих реакциях зависит от ряда
фактов: пола животных и уровня их эволюционного разви-
тия.
Удаление коры больших полушарий, особенно ее фрон-
тальных отделов, тормозит половое поведение у самцов
(Beach, 1940). При этом, как указывает Beach, степень вы-
раженности данных реакций зависит от общей массы пора-
жения коры головного мозга. Двустороннее повреждение
коры головного мозга у самцов крыс, превышающее 60%
общей массы коркового вещества, вызывает у животных
полную импотенцию.
Любопытно, что эти же животные могут спариваться пос-
ле инъекции большого количества мужского полового гор-
мона. На основании этих опытов Beach (1950) считает, что
кора головного мозга непосредственно не участвует в ор-
ганизации половых реакций самцов крыс, а лишь поддер-
живает возбудимость низших центров половой мотивации.
Для самок крыс, по мнению этого автора, нисходящие кор-
ковые влияния на половые подкорковые центры менее су-
щественны.
Особенно важными отделами для формирования поло-
вых реакций у самцов кошек Bard считает передние отделы
коры головного мозга.
Полное удаление коры головного мозга, гиппокамповой
области и полосатого тела не уничтожает половой активно-
сти у самок морских свинок, кроликов, крыс и кошек (Bard,
1936; Brooks, 1937; Davis, 1939; Dempsey a. Rioch, 1933).
Как указывает Beach (1944), после удаления коры боль-
ших полушарий у самок крыс часто обнаруживается даже
повышенная половая возбудимость. Вместе с тем у них
Наблюдаются некоторые недостатки в поведении. Так, на-
пример, такие животные перестают обращать свои призыв-
ные сигналы к самцу.
В противоположность эффектам удаления коры больших
полушарий удаление лимбических структур, таких, как пи-
риформная кора и амигдал, вызывают отчетливые изменения
Половых влечений. Наиболее характерно, что при таких по-
вреждениях животные утрачивают способность различать
и оценивать половой объект и проявляют попытки спари-
ваться с любым движущимся предметом.
Значение архикортекса и архистриатума (миндалевид-
ного ядра) в организации полового поведения было пока-
102
зано Kliiver и Вису (1939) впервые при синдроме пораже-
ния височной доли. В их опытах на обезьянах, как указыва-
лось выше, после удаления височной доли наблюдалась ги-
персексуальность. Аналогично выраженную гиперсексуаль-
ность после удаления миндалевидной ядерной группы у сам-
цов. кошек и обезьян наблюдали Schreiner и Kling (1953).
Вместе с тем Green и др. (1959), подробно исследуя участие
миндалевидного тела в половых реакциях у кошек после
амигдалэктомии, не отмечают развития гиперсексуальнос-
ти. Такая реакция наблюдалась только у котов. Поврежде-
ния амигдалярной и околоамигдалярной области у самок
в отличие от самцов не вызывает у них гиперсексуальности.
Еще более выраженные гиперсексуальные реакции, по дан-
ным этих авторов, наблюдаются при изолированном разру-
шении грушевидной коры.
Гиперсексуальные реакции у этих животных значитель-
но подавлялись кастрацией.
Разрушение амигдалы приводит к тому, что у самца сни-
жается активность по преодолению электрического раздра-
жения, для удовлетворения полового влечения (Rasmus-
sen a. oth., 1960).
Такие же гиперсексуальные реакции наблюдаются при
удалении энторинальной коры. Раздражение этой области
у самок вызывает овуляцию, движения матки и реакцию
полового поведения (Koikegami, Yamada a. Usui, 1950;
Chritchlow, 1957; Endroczi a. Lissak, 1961). У кошек и обезь-
ян при двустороннем повреждении пириформной коры так-
же наблюдается значительное повышение половой активно-
сти. Такие животные проявляют попытки копулировать да-
же с особями одного пола или с неодушевленными предме-
тами. '
У неполовозрелых животных кортико-медиальная часть
миндалины оказывает тормозящее влияние на половое со-
зревание. Двустороннее разрушение этой области приводит
к раннему половому созреванию (Elwers a. Critchlow,
1960). Такой же эффект вызывает двусторонняя перерезка
концевых полосок. Наоборот, ритмические повторные элек-
трические раздражения этой области вызывают замедление
полового созревания (Bar-Sela a. Critchlow, 1966). Такое
же замедление полового созревания наблюдается при пере-
резке обонятельных трактов у кроликов (Franck, 1965).
Имплантация в область латерального гипоталамуса тесте-
рона приводит к развитию у самок поведения, характерного
для самцов.
103
Электрическое и химическое (путем инъекций карбахола)
раздражение гиппокампа, поясной извилины и перегород-
ки вызывает различные проявления половых реакций—уха-
живание, лизание гениталий, эрекцию полового члена и др.
У котов с разрушенным гиппокампом также обнаружи-
ваются гиперсексуальные реакции, и они пытаются произво-
дить половой акт не только с кошками, но и с целым рядом
различных предметов (Mac Lean, 1954, 1955, 1958),
Эрекция полового члена вызывается раздражением септо-
преоптической медиальной области и медиальной части дор-
со-медиального ядра таламуса (Mac Lean, 1966).
Изучение электрической активности головного мозга во
время копуляции и в период после нее показало, что во вре-
мя оргазма в области переднемозгового пучка и прехиаз-
мальной области возникает активность 8—14 имп/сек
с большой амплитудой (Porter, Cavanaugh, Critchlow а.
Sawyer, 1957). Подобные изменения электрической актив-
ности в этих структурах головного мозга Lissak и Endroczi
(1962) наблюдали у кошек в период полового возбуждения.
У кролика в период после копуляции в коре головного моз-
га преобладает медленная электрическая активность, про-
являющаяся в образовании веретен 8—12 в секунду. Затем
животное впадает как бы в коматозное, сноподобное состоя-
ние. В это время появляется внезапно наступающая элект-
роэнцефалографическая реакция пробуждения, хотя пове-
дение животного скорее напоминает более глубокий сон,
чем в период медленной синхронной электрической актив-
ности. Sawyer рассмативает это состояние как «парадок-
сальный сон».
По наблюдениям Kawakami и Sawyer, такая же элект-
роэнцефалографическая активность может быть вызвана
как раздражением влагалища, так и электрической стиму-
ляцией вентро-медиального гипоталамуса.
Тщательное изучение половых реакций у древесных ля-
гушек (Aronson a. Noble 1945) показывает, что различные
отделы головного мозга участвуют в формировании целост-
ного полового акта.
Разрушение спинного мозга устраняет только реакцию
схватывания самки, однако другие компоненты полового
возбуждения при этом сохраняются в отчетливой форме.
Призывные реакции квакания самцов связаны со сред-
ним мозгом и с внутренними бугорками четверохолмия. На
движения самца, которые он производит всякий раз, когда
самка выделяет порцию яиц, не влияет удаление переднего
104
мозга. Они сохраняются при наличии неповрежденного
среднего и заднего мозга. Лягушки с повреждением зри-
тельного бугра не отпускают самку после оплодотворения.
Целенаправленные реакции приближения партнеров друг
к другу определяются передним мозгом.
Все эти наблюдения показывают, что половые мотивации
и их удовлетворение представляют собой сложные интегра-
тивные акты с участием различных отделов нервной систе-
мы, расположенных на ее различных уровнях.
ИСКУССТВЕННЫЕ МОТИВАЦИИ. САМОРАЗДРАЖБНИЕ
В 1953 г. Olds в лаборатории Milner обнаружил, что кры-
са, раздражаемая электрическим током через электроды,
копчики которых находились в области передней комиссу-
ры, стремится к повторному раздражению. Это проявлялось
в том, что животное предпочитало то место, где ему произ-
водилось электрическое раздражение головного мозга.
В дальнейшем техника эксперимента была усовершенство-
вана. Животное помещали в камеру Скиннера, в которой
ему предоставляли возможность наносить самораздраже-
ние. Принципиально это достигалось следующим образом.
Нажимая на рычаг, имеющийся в камере, животное тем са-
мым замыкало цепь электрического тока, идущего от сти-
мулятора к электродам, вживленным в его головной мозг.
Раздражение прекращалось через полсекунды после вклю-
чения тока даже в том случае, если крыса продолжала на-
жимать на педаль. Поэтому для повторного раздражения
крыса должна была снова нажать на педаль.
Оказалось, что в таких условиях животные самостоятель-
но производили раздражение собственного мозга, нажимая
на рычаг и тем самым замыкая цепь электрического тока,
раздражающего мозг.
С помощью этой методики Olds удалось исследовать ре-
акцию животного на раздражение многих пунктов мозга
(Olds, Milner, 1954). В результате было установлено, что
раздражение ряда пунктов вызывало самостоятельную по-
вторную реакцию нажатия крысой на рычаг. При этом час-
тота повторных раздражений варьировала в разных точках,
Достигая в некоторых из них (при локализации, например,
кончика электрода в дорсальном гипоталамусе) 5000—6000
раздражений в час. Эти точки были названы Olds положи-
тельными зонами самораздражения. В отличие от них раз-
дражение других пунктов головного мозга вызывало резко
10S
выраженные реакции отрицательного эмоционального ха-
рактера. Получив однажды раздражение подобного пунк-
та, животные никогда больше не осуществляли повторное
раздражение этого пункта. Мало того, как показали экспе-
рименты Lilli (1960), животные производили сложные опе-
рации для того, чтобы покинуть помещение, где осуществ-
лялось это раздражение. Эти области получили название
отрицательных зон самораздражения. Наряду с этим были
найдены пункты мозга, раздражение которых не вызыва-
ло заметных реакций животного. Они получили название
нейтральных зон. Olds составил соответствующие карты
локализации положительных, отрицательных и нейтраль-
ных зон самораздражения в головном мозге крыс (Olds,
1956, 1957,1960).
Положительные зоны самораздражения у крысы состав-
ляют около 35%, отрицательные — около 5% и нейтраль-
ные— около 60% всей массы головного мозга крысы.
Характерно, что положительный эффект самораздражения
может быть вызван практически из любой точки лимбиче-
ской системы. Между тем внутри этой системы имеется оп-
ределенный градиент самораздражения. Эффект самораз-
дражения, измеряемый частотой реакции, уменьшается по
мере продвижения раздражающих электродов к коре. При
раздражении в области интерпедункулярного ядра покрыш-
ки частота самораздражения достигает 7000 в час, при
раздражении задней области гипоталамуса — около 5000
в час, при раздражении базальных ядер — 2000—3000
в час и при раздражении коры обонятельного мозга — око-
ло 200 в час.
В целом можно сказать, что точки, вызывающие само-
раздражение, расположены в медиальном пучке мозговой
ткани, простирающемся от миндалины через гипоталамус
к покрышке среднего мозга. Наибольшая скорость само-
раздражения наблюдается при раздражении перегородки,
заднего гипоталамуса и покрышки.
Brady (1958) подтвердил эти наблюдения Olds и также
показал, что животные многих видов охотно наносят само-
раздражение через электроды, вживленные в различные
участки лимбической системы и связанных с нею структур.
По данным Lilli (1960), положительные реакции самораз-
дражения у обезьян отчетливо наблюдаются при нахождении
кончика электрода в вентро-медиальном ядре гипоталаму-
са. Более слабые реакции (в порядке уменьшения эффек-
тивности) получаются при раздражении интраламинарных
106
ядер таламуса, перегородки, верхней части свода, головкй
хвостатого ядра и скорлупы,
Lilli у обезьян обнаружил следующую иерархию (по сте-
пени уменьшения эффекта) зон самораздражения: базаль-
ная покрышка среднего мозга, вентро-медиальное ядро ги-
поталамуса, интраламинарные ядра таламуса, перегородка,
верхний отдел свода, головка хвостатого ядра и скорлупа.
Точки, раздражение которых вызывают реакцию избегания,
расположены в латеральной части заднего гипоталамуса,
дорсальной части среднего мозга и в эиторинальной коре.
Delgado (I960) показал, что раздражение некоторых то-
чек гиппокампа, задне-вентрального ядра таламуса, по-
крышки и центрального серого вещества у обезьян также
вызывает отрицательные эмоциональные реакции, которые
могут быть рассматриваемы как «наказание». После несколь-
ких опытов с раздражением этих областей у животных об-
наруживаются признаки беспокойства, когда они попадают
в помещение, где ставился опыт.
Как показал Lilli, животные при этом быстро производи-
ли обученные инструментальные реакции, позволяющие им
покинуть «неприятную» обстановку опыта.
Прямое раздражение этих областей головного мозга не-
редко вызывает аффективные реакции, такие, как агрессия,
ярость, ужас, страх, нападение или защита и т. п., внешне
напоминающие соответствующие естественные отрицатель-
ные мотивации.
Если теперь сравнить общий объем мозговых структур,
раздражение которых вызывает положительную реакцию
самораздражения, с объемом структур, дающих отрицатель-
ную реакцию, можно совершенно определенно сказать, что
положительные эмоциональные центры представлены зна-
чительно шире. Такое более обширное представительство
положительных эмоциональных центров имеет, по-видимо-
му, важное биологическое значение—положительная эмо-
ция подкрепления должна обладать значительной силой,
чтобы подавить отрицательные эмоции любой мотивации.
Реакция самораздражения и естественные
влечения животных
Возникает вопрос, какова природа реакции самораздра-
жения. Связано ли электрическое раздражение определен-
ных пунктов головного мозга с механизмами естественных
влечений, а если связано, то каким образом.
107
Уже в экспериментах Olds было обращено внимание на
то, что стремление крысы к рычагу, с помощью которого
осуществляется самораздражение, обладает большой при-
тягательной силой. Чтобы получить такое электрическое
«вознаграждение», крысы могут решать сложные задачи
в лабиринте, а также преодолевать значительные препятст-
вия в виде сетки, на 'Которую подается сильный электриче-
ский ток.
Все это указывает на то, что возникающая у животных
в условиях самораздражения «тяга» к рычагу обладает
такой ясе высокой энергетической силой, как и естествен-
ные мотивации.
Позволительно спросить, что же является более сильным
по своей побудительной силе—естественные влечения, обус-
ловленные внутренними потребностями организма, или же
искусственное влечение, вызванное электрическим раздра-
жением головного мозга.
Оказалось, что «тяга» животных к самораздражению зна-
чительно сильнее, чем, например, стремление голодного
животного к пище.
Опыты Olds (1960) показали, что обучение ясивотных,
подкрепляемых пищей или электрическим раздражением
положительных зон головного мозга, в лабиринте происхо-
дит примерно с одинаковой скоростью, и даже быстрее
в случае электрического подкрепления. Мало того, для по-
лучения электрического раздражения головного мозга кры-
сы преодолевали более сильное болевое раздражение, по-
дававшееся как препятствие на пути, чем голодавшие в те-
чение суток животные. Голодные крысы для того, чтобы
получить пищевое вознаграждение, пересекали решетку, на
которую подавался ток не выше 70 мка. В то же время
для того, чтобы получить раздражение положительной зо-
ны головного мозга, те же животные преодолевали решет-
ку с током 300 мка и более.
Наиболее демонстративны опыты Spies (1965). В этих
опытах крысам предоставлялся выбор двух рычагов. Один
из них обеспечивал подачу пищи, другой — раздражение го-
ловного. мозга.
Опыты показали, что крысы, у которых электрический ток
производил раздражение области латерального гипотала-
муса, медиального пучка переднего мозга, преоптической
области, покрышки среднего мозга, ядер уздечки, красных
ядер и др., значительно предпочитали самораздражение пи-
щевому подкреплению.
108
Такие же реакции наблюдали Endrozi и Lissak (1966)
при раздражении у кошек медиального пучка переднего
мозга.
Все это указывает на то, что искусственное влечение при
самораздражении внешне очень напоминает естественные
влечения.
В подтверждение этого эксперименты Ю. А. Макаренко
(1964) показали, что при реакции самораздражения наблю-
дается такая же генерализованная активация электроэнце-
фалограммы, как и при естественных мотивационных реак-
циях. После самораздражения на электроэнцефалограмме
появляются медленные высокоамплитудные волны.
Эти наблюдения в дальнейшем были подтверждены New-
man (1967) и Н. Г. Михайловой (1968).
Рассмотрим теперь вопрос о связи искусственных влече-
ний в условиях самораздражения с естественными влече-
ниями.
Показано, что при голодании животных частота самораз-
дражения увеличивается (Brady, 1955). Учащение реакции
самораздражения при голодании животных особенно отчет-
ливо наблюдается при раздражении заднего отдела гипота-
ламуса. При большей силе раздражающего тока эту реак-
цию можно наблюдать и при раздражении вентро-медиаль-
ной области гипоталамуса, а также перегородки (Olds,
1956, 1957). При локализации электродов в переднем гипо-
таламусе подобный эффект не обнаруживается.
Olds показал также, что при нахождении кончика элект-
рода в области латерального гипоталамуса, раздражение
которой у накормленных животных вызывает дополнитель-
ный прием пищи, у животных обнаруживается высокая ча-
стота самораздражения. Wilkinson и Peele (1962), Margu-
les и Olds (1962), Coons и Cruce (1967) наблюдали, что
у голодных крыс значительно увеличивается самораздра-
жение из зоны «центра питания» в латеральном гипотала-
мусе.
Аналогично этому Hoebel и Teutelbaum (1962) и Morgane
(1964) нашли, что раздражение вентро-медиального ядра
у крыс, так же как и введение пищи в желудок, прекраща-
ло прием пищи и угнетало реакцию самораздражения. На-
оборот, раздражение латерального гипоталамуса усилива-
ло прием пищи и реакцию самораздражения.
Разрушение вентро-медиальных ядер гипоталамуса уси-
ливало как реакцию самораздражения, так и прием пищи
(Hoebel, 1965).
109
В. И. Вадиков (1968) наблюдал у нескольких кроликов
с электродами, вживленными в область латерального гипо-
таламуса, следующую реакцию. Предварительно накорм-
ленный перед опытом кролик с помощью специального кон-
тактного устройства осуществлял реакцию самораздраже-
ния. Однако после 3—4 «сеансов» электрического саморазд-
ражения области латерального гипоталамуса кролик с
жадностью набрасывался на лежащую перед ним капусту
и начинал ее усиленно поглощать. После приема определен-
ного количества пищи животное снова производило само-
раздражение, потом снова принимало пищу и т. д.
Все эти наблюдения показывают, что реакция самораз-
дражения из указанных выше областей гипоталамуса нахо-
дится в тесной функциональной зависимости от состояния
пищевого возбуждения животного. Вместе с тем самор аз-
дражение этих центров оказывает значительное влияние на
пищевые мотивации.
В других экспериментах Olds (1960) показал связь реак-
ции самораздражения с половыми влечениями животных.
При локализации кончиков электродов в дорсо-медиаль-
ной части хвостатого ядра кастрация животных резко сни-
жала у них частоту самораздражения. В то же время вве-
дение кастрированным животным тестостерона снова зна-
чительно повышало у них частоту реакции самораздраже-
ния из этих пунктов головного мозга.
Такие же эффекты наблюдались при раздражении неко-
торых точек латерального гипоталамуса и перегородки.
Характерно, что эти зоны, реакция самораздражения из
которых значительно зависела от содержания в организме
андрогенов, имеют тесное отношение к половым реакциям.
Как уже указывалось выше, Porter и др. (1937) обнару-
жили изменения электрической активности в передне-ла-
теральном гипоталамусе при раздражении влагалища у кош-
ки в период течки. Sawyer и Kawakami (1959) зарегистри-
ровали у самки (кролика изменения электрической актив-
ности в латеральном гипоталамусе и перегородке при
искусственном раздражении влагалища, а также во время
и после покрытия.
Следует отметить, что частота реакции самораздражения
из указанных выше точек изменялась только при измене-
нии содержания андрогенов в крови. В то же время на нее
не оказывала влияния степень голодания животного. Все
это позволило Olds говорить о дифференцировании зон под-
крепления, об их подчеркнутой специфичности.
ПО
На основании всёх свойх экспериментов Olds сформули-
ровал представления, что реакция самораздражения обла-
дает выраженными свойствами биологического подкрепле-
ния. Зоны, вызывающие у животных самораздражение, рас-
сматривались им как положительное, а зоны избегания как
отрицательное подкрепление.
Развивая эти представления, Bovard (1961) выдвинул ги-
потезу о наличии в головном мозге животных и человека
двух систем, связанных с отрицательными и положительны-
ми подкреплениями. Первая представлена главным образом
в задних и медиальных отделах гипоталамуса, вторая —
в передних и латеральных его отделах. Обе эти системы
охватывают, как полагает Bovard, лобную, височную и те-
менную области коры больших полушарий, а также гип-
покамп и амигдалу. Однако в этих структурах трудно раз-
граничить локализацию этих систем; нередко рядом распо-
ложенные участки дают как однонаправленные, так и
разнонаправленные по эмоциональному знаку реакции само-
раздраження. Автор связывает первую систему с симпати-
ческими, а вторую—с парасимпатическими образованиями
(Bovard, 1962).
И все же, несмотря на наличие тесной функциональной
зависимости реакции самораздражения от естественных мо-
тиваций, до настоящего времени трудно утверждать, что
стремление животного к раздражению собственного голов-
ного мозга представляет собой мотивацию, разыгрываю-
щуюся по механизму естественных биологических влечений.
В отличие от Olds, который считает, что самораздраже-
ние наблюдается в результате получения животным опреде-
ленного «эмоционального» удовольствия, в последние годы
появились работы, убедительно опровергающие эту точку
зрения.
Авторы этих работ настойчиво утверждают, что стрем-
ление животного к раздражению определенных структур
головного мозга связано скорее не с положительным, а с
неприятным эмоциональным состоянием, возникающим
у них после раздражения. Таким образом, самораздраже-
ние, согласно этой точке зрения, есть избавление от непри-
ятного последействия раздражения мозговых структур. Эта
точка зрения основана на ряде убедительных фактов.
Так, Ball и Adams (1965) в опытах, проведенных на кры-
сах, показали, что реакция самораздражения зависит от
длительности интервала между повторными раздражения-
ми. При малых интервалах самораздражения из медиально-
1Н
го пучка переднего мозга частота самораздражения возра-
стает, при длительных интервалах — падает.
Кроме того, было обнаружено, что в специальном скин-
неровском ящике животные предпочитают находиться на
той половине, где последующие раздражения головного
мозга подаются в более частом ритме.
На основании этих наблюдений авторы полагают, что
каждое раздражение головного мозга оставляет после се-
бя отрицательное эмоциональное последействие и повторным
раздражением животные как бы нивелируют это последей-
ствие. При длительных интервалах самораздражения это по-
следействие успевает завершиться, в результате чего, как
считают авторы, и падает скорость самораздражения.
В более поздних работах Olds (1962), вслед за ним
Deutsch и Howarth (1963) выдвинули представление о том,
что при самораздражении одновременно возбуждаются мо-
тивационные и подкрепляющие системы. Самораздражение
как бы снижает мотивацию, вызванную предыдущим раз-
дражением, оказывая тем самым подкрепляющее действие.
Близка к этим представлениям точка зрения Grastian
и др. (1966). Авторы считают, что слабое раздражение ла-
теральных отделов гипоталамуса на фоне тета-ритма вы-
зывает положительные реакции приближения. Сильное
раздражение этих центров, приводя к десинхронизации
электрической активности гиппокампа, вызывает реакции
выключения раздражения (избегание).
Авторы обнаружили, что реакция самораздражения пред-
ставляет собой последовательную смену тета-ритма в гип-
покампе реакций десинхронизации. Первое состояние вы-
зывает самораздражение, второе — прекращает его.
Все эти наблюдения показывают, что в своей глубокой
сущности природа искусственных мотиваций все еще оста-
ется не изученной.
НЕКОТОРЫЕ ЗАМЕЧАНИЯ
Мы привели лишь относительно небольшое число приме-
ров об участии различных отделов центральной нервной си-
стемы в формировании мотивационного возбуждения.
Они отчетливо показывают, что мотивации преимущест-
венно связаны с деятельностью лимбико-ретикулярного ком-
плекса. Однако образования и различные уровни старой
и новой коры играют различную роль в формировании мо-
тивационного возбуждения.
112
На основании экспериментов с раздражением и Экстир-
пацией таких подкорковых образований, как гипоталамус,
таламус, средний мозг и т. д., можно определенно сказать,
что эти образования имеют наиболее тесное отношение
к возникновению мотивационного возбуждения любого био-
логического качества.
В отличие от этого образования древней, старой и новой
коры производят преимущественно оценку мотивационного
состояния и в соответствии с действием на животное внеш-
них раздражителей формируют на этом основании целена-
правленное поведение во внешней среде. Такая точка зре-
ния находит подтверждение в тех многочисленных экспе-
риментах, которые показывают, что как удаление, так
и раздражение этих образований, как правило, не уничто-
жает исходного влечения, а только значительно изменяет
его в сторону ослабления или, наоборот, усиления. Наи-
более отчетливо это наблюдается при синдроме Клювера —
Бюси.
Среди подкорковых образований центральная роль в фор-
мировании мотиваций принадлежит, по-видимому, гипота-
ламическим центрам. Различные другие лимбические обра-
зования оказывают на них модулирующее влияние, усили-
вая или ослабляя эффекты их возбуждения.
Однако на основании литературных данных мы все еще
не можем определенно сказать, какие отделы головного
мозга несут воспринимающую, а какие исполнительную
функцию в формировании мотиваций. Некоторые исследо-
ватели, например Anand (1963), рассматривают лимбиче-
скую систему как чисто афферентный интегративный аппа-
рат мотивации. Эффекторные проявления мотиваций, по их
мнению, складываются на гипоталамическом уровне. Мож-
но, однако, предположить, что и в гипоталамусе должны су-
ществовать чувствительные и моторные элементы мотива-
ционного возбуждения.
Как было показано выше, в формировании мотиваций
различного биологического качества участвуют практиче-
ски одни и те же лимбические структуры. Спрашивается,
каким же образом каждое мотивационное возбуждение
охватывает эти структуры для формирования специального
целенаправленного поведения? Rosvold, Mirsky, Pribram
(1954) считают, что эффекты удаления лимбических струк-
тур во многом зависят от обстановки, в которую попадает
животное после операции. Они провели наблюдения за по-
ведением восьми самцов макак резус, помещенных в отдель-
на
Hbte и групповые клетки до и после удаления миндале-
видного комплекса. Оказалось, что в отдельных клетках
обезьяны после операции проявляли повышенную агрессив-
ность. Вместе с тем в групповой клетке они проявляли под-
черкнутую трусливость, и обезьяны, ранее занимавшие гос-
подствующее положение, теперь попадали в подчинение.
Kling и Lewartz (1961), Kling (1962) считают, что эф-
фекты удаления лимбических структур определяются так-
же возрастом и индивидуальным развитием животного.
Так, у животных в раннем возрасте удаление амигдалярной
области обычно не вызывает таких изменений в пищевом
поведении, как у взрослого животного. Таким образом,
можно думать, что амигдалярная область включается в пи-
щедобывательную деятельность только в процессе индиви-
дуального обучения особи.
В отличие от этих точек зрения Mac Lean (1959) полага-
ет, что внутри лимбической системы имеется отчетливая
функциональная дифференциация. Одни ее структуры обес-
печивают реакции самосохранения данной особи, такие, на-
пример, как прием и поиск пищи и различные оборонитель-
ные и защитные реакции. Эти структуры, по мнению Мас
Lean, охватывают «лимбическую среднемозговую область
Наута», куда входят парамедиальная ретикулярная форма-
ция и центральное серое вещество вместе с ядром Гуддена.
Отсюда имеются восходящие волокна к различным частям
гипоталамуса и от него через медиальный пучок среднего
мозга латерально к амигдалярной области. К ним же от-
носятся волокна латерального обонятельного тракта, под-
ходящие к амигдале. Из амигдалы волокна распространя-
ются к лимбической коре лобно-височной области. Таким
образом, все эти структуры представляют функционально
объединенный комплекс, обеспечивающий различные сто-
роны пищевых и оборонительных реакций, включая слож-
ные формы целенаправленного поведения.
Другой функциональный комплекс лимбической системы,
по мнению Mac Lean, обеспечивает животным реакции, на-
правленные на сохранение рода и, в частности, половые
реакции. Он объединяет гипоталамус, медиальный пучок
переднего мозга, перегородку, куда также конвергируют
волокна медиального обонятельного тракта. Из перегородки
волокна распространяются через цингулум и свод на пояс-
ную извилину и гиппокамп.
Таким образом, согласно Mac Lean, каждая мотивация
должна строиться на своей специфической структурной ос-
114
пове. Тогда спрашивается, почему же часто повреждения
одних и тех же лимбических образований (например, ви-
сочной области) вызывают одновременно расстройство не-
скольких мотиваций? Ответа на этот вопрос литературные
данные не дают. По-видимому, чем более высоко развито
животное и чем в большей степени оно обучено, тем в боль-
шей мере мотивационное возбуждение охватывает высшие
отделы головного мозга, включая кору больших полушарий,
которая может в равной степени участвовать в различных
мотивациях.
Учитывая, что наиболее значительные нарушения поло-
вого, пищевого и оборонительного мотивационного поведе-
ния и особенно целенаправленных реакций наблюдаются
при поврежденных лобных отделов коры головного мозга
(Ю, М. Коморский, 1962; Л. С. Гамбарян, 1962; Anand,
1961; Pribram, Bagshaw, 1953), можно думать, что именно
лобные отделы коры являются высшим интегративным от-
делом мотиваций.
Все указанные выше экспериментальные данные не ос-
тавляют сомнения в том, что мотивации возникают на
структурной основе лимбических образований головного
мозга. Однако все же остается открытым вопрос о том, как
осуществляется интеграция различных структур головного
мозга в формировании тех или иных мотиваций, а также,
что определяет вовлечение различных структур головного
мозга в каждое мотивационное возбуждение? Все это обус-
ловлено тем, что имеющиеся литературные данные о меха-
низмах мотиваций являются крайне разрозненными. Только
с позиции теории функциональных систем организма этот
вопрос приобретает определенную ясность. Учитывая это,
все последующее изложение будет посвящено вопросу
о том, как складывается мотивационное возбуждение
в функциональных системах организма.
III. ПРИРОДА МОТИВАЦИОННОГО ВОЗБУЖДЕНИЯ
Главной особенностью каждой мотивации, как следует из
всего изложенного выше, является ее подчеркнутая специ-
фичность. Эта специфичность наиболее отчетливо проявля-
ется в поведении, которое возникает на ее основе. Это пове-
дение всегда избирательно направлено только на специаль-
ный раздражитель внешней среды. Так, животное в состоя-
нии питьевого возбуждения ищет только воду, половые
мотивации направляют животных только на поиск особи
противоположного пола и т. д.
Естественно поставить вопрос: что же определяет такую
подчеркнутую специфику каждой мотивации? Почему при
каждом мотивационном возбуждении животное настойчи-
во ищет только специальные раздражители в окружающей
его среде? Чтобы ответить на эти вопросы, мы должны те-
перь рассмотреть те внутренние механизмы, которые приво-
дят к возникновению мотивационного возбуждения в цент-
ральной нервной системе, а также особенности нейрофизио-
логических механизмов мотиваций.
ОСОБЕННОСТИ ФОРМИРОВАНИЯ МОТИВАЦИОННОГО
ВОЗБУЖДЕНИЯ
В ФУНКЦИОНАЛЬНЫХ СИСТЕМАХ ОРГАНИЗМА
Специфика мотивационного возбуждения вытекает преж-
де всего из природы различных функциональных систем,
в которые оно, как уже указывалось ранее, входит как со-
ставной компонент. Последние же, обеспечивая каждая тот
или иной полезный приспособительный эффект организма,
по своей природе являются подчеркнуто специфичными.
Мотивационное возбуждение, как указывалось выше, воз-
никает не во всех функциональных системах. Оно склады-
вается лишь в тех случаях, когда та или иная функциональ-
ная система организма для обеспечения своего конечного
приспособительного результата не ограничивается процес-
сами, происходящими внутри организма, а обязательно
включает внешний компонент — специальное пеленаправ-
116
ленное поведение. При этом мотивационное возбуждение
возникает в центральном звене саморегуляторных меха-
низмов данной функциональной системы, т. е. в головном
мозге. В то же время каждое мотивационное возбуждение
отражает в себе все свойства конечного приспособительного
результата — деятельности данной функциональной систе-
мы, поскольку оно возникает только в случае отклонения
этого результата от уровня, обеспечивающего нормальную
жизнедеятельность организма.
Каким же образом различные отклонения конечных при-
способительных результатов деятельности разных функцио-
нальных систем формируют мотивационное возбуждение?
ПОСТОЯНСТВО И ИЗМЕНЧИВОСТЬ ВНУТРЕННЕЙ СРЕДЫ.
ВНУТРЕННИЕ «ПОТРЕБНОСТИ» ОРГАНИЗМА
Жизнь характеризуется непрерывно идущим в организме
обменом веществ, постоянным потреблением, накоплением,
и разрушением определенных продуктов жизнедеятельности.
Понятно, что нормальный (оптимальный) уровень обмена
веществ в организме может быть достигнут только при оп-
ределенном содержании специальных веществ внутри орга-
низма.
К. Бернар по этому поводу писал; «... постоянство внут-
ренней среды организма есть условие его свободной жиз-
ни»1. Вслед за ним Cannon провозгласил понятие о гомео-
стазисе как поддержании устойчивости внутренней среды.
В действительности Же внутренняя среда организма все
время изменяется и ни секунду не может быть постоянной.
Ее постоянство значило бы смерть для организма, прекра-
щение его обмена веществ.
Итак, с одной стороны, постоянная изменчивость внутрен-
ней среды, а с другой — жизненная необходимость ее по-
стоянства.
Что же в таком случае определяет стабильность внутрен-
ней среды?
Вот здесь-то на помощь организму и приходят аппараты
саморегуляции. Любое отклонение того или иного показа-
теля внутренней среды от уровня, обеспечивающего нор-
мальную жизнедеятельность организма, немедленно вызы-
вает цепь процессов, направленных на восстановление ее
относительного постоянства.
1 С. Bernard. Lecons sur les propietes physiologiques et les alte-
rations pathologtques des llquldes de I’organtsme, 1859, 1.
117
Таким образом, наиболее Ёажйые приспособительное
эффекты организма в первую очередь связаны с поддержа-
нием определенных параметров внутренней среды на относи-
тельно постоянном уровне, что создает благоприятные усло-
вия для обменных процессов.
Понятно, что эти приспособительные эффекты организма
не являются строго фиксированными. Они имеют опреде-
ленную степень «разброса», вследствие постоянно изменяю-
щихся условий внешней и внутренней среды организма.
Однако эта флюктуация всегда происходит около опреде-
ленного уровня.
Именно эту роль поддержания содержания определенных
веществ внутри организма на жизненно важном уровне бе-
рут на себя различные функциональные системы организма.
Так, например, функциональная система дыхания поддер-
живает оптимальный уровень СОг—О2 в организме,
функциональная система питания — уровень питательных
веществ и т. д.
Принцип саморегуляции является центральным в под-
держании постоянного уровня содержания веществ внутри
организма, и любое отклонение содержания, того или иного
жизненно важного вещества от уровня, обеспечивающего
нормальную жизнедеятельность организма, приводит в дей-
ствие все аппараты обеспечивающей его функциональной
системы, в результате чего происходит возвращение его
содержания к исходному уровню.
Это, по мнению П. К- Анохина, является «золотым прави-
лом» саморегуляции. Отклонение от результата само
является стимулом к возвращению разультата.
Итак, мы подошли к важному пункту изложения. Несмот-
ря на постоянно идущий обмен веществ в организме, не-
смотря на постоянное колебание в содержании тех или иных
жизненно важных веществ внутри организма, деятельность
функциональных систем поддерживает их содержа-
ние на постоянном уровне.
При этом важно отметить следующее. Поддержание по-
стоянного уровня тех или иных жизненно важных веществ,
необходимых для нормального течения обменных процессов
в тканях, обеспечивается уже в кровяном русле. Вот почему
мы можем говорить об определенных ко нстантах крови.
К таким константам крови относятся: постоянство осмо-
тического давления, реакция (ph) крови, ее температура,
содержание в крови определенных количеств воды, пита-
тельных веществ, солей, витаминов, газов и др.
118
к
s.
Поддержание состояния определенных жизненно важных
веществ на константном уровне уже в кровеносном русле
является, по-видимому, большим достижением эволюцион-
ного развития. Это серьезно предохраняет тканевый обмен
веществ от всех возможных неполадок—«ударов судьбы»,
связанных с перебоями в снабжении или, наоборот, удале-
нии определенных продуктов.
В этом факте поддержания постоянства определенных
констант крови мы имеем поразительный пример защитных
предупредительных реакций организма, поскольку вся регу-
ляция внутренних констант осуществляется еще задолго до
реальных изменений в обмене веществ в организме. В ре-
зультате этого дело обстоит таким образом, что каждому
конечному приспособительному эффекту может предшество-
вать несколько стадий так называемого предконечного
эффекта, которые, поддерживаясь на постоянном уровне,
уже сами по себе могут определять все условия конечного
эффекта.
Создав в эволюции механизмы поддержания уже «пред-
эффектов» на константном уровне, организм тем самым
обеспечил себе строгую гарантию постоянства обменных
процессов.
Отсюда понятно, почему, например, в организме имеются
механизмы саморегуляции постоянства артериального дав-
ления. Само артериальное давление не является, конечно,
фактором, участвующим непосредственно в обмене веществ,
но оно косвенным образом определяет нормальные условия
для его обеспечения.
Точно таю же поддерживается постоянство питательных
веществ в крови, парциальное напряжение кислорода и угле-
кислого газа и др. Поддержание постоянного уровня опре-
деленных констант крови есть условие для нормального
течения обменных процессов в тканях организма.
Итак, различные функциональные системы своей деятель-
ностью в условиях непрерывно изменяющейся внутренней
среды организма поддерживают жизненно важные констан-
ты крови на относительно постоянном уровне. Именно эти
механизмы и обеспечивают то «постоянство внутренней сре-
ды», о котором писал К. Бернар, и «гомеостазис» У. Кен-
нона.
Таким образом, функциональные системы являются
аппаратами саморегуляции и гомеостазиса.
Следует отметить, что ритмы работы разных функцио-
нальным систем раздицны. Это определяет их важное
119
свойство: как быстро будет восстановлена та или иная
константа крови в случае ее изменения. В свою очередь
ритм работы каждой функциональной системы определяет-
ся тем, насколько большие и опасные для жизни изменения
вызывают отклонения той константы, которую она поддер-
живает на постоянном уровне.
В организме имеются такие константы, отклонение
которых на непродолжительное время вызывает резчайшие
нарушения обмена веществ и критическое состояние орга-
низма. Примером служит уровень сахара в крови, осмоти-
ческое давление и др. Это так называемые жесткие кон-
станты организма (П. К. Анохин, 1962).
Наряду с этим есть такие константы крови, значительное
отклонение которых в ту или иную сторону не приводит
к существенному нарушению жизнедеятельности организма.
Примером служит снижение уровня питательных веществ
в крови, которое определенное время может поддерживаться
за счет депонирующих механизмов или перераспределения
питательных веществ внутри организма.
Примером другой смещаемой в довольно широких пре-
делах константы является артериальное давление. Оно
может значительно повышаться и длительно удерживаться
на высоком уровне при физических или эмоциональных
напряжениях организма, обеспечивая тем самым лучшее
кровоснабжение работающих с большим напряжением
органов. Примерно таким же свойством обладают и другие
константы крови, такие, как содержание воды, температура
и т. д.
В отличие от жестких последние названы в нашей лабо-
ратории пластичными константами.
Понятно, что в результате непрерывно идущего обмена
веществ практически каждая константа организма непре-
рывно смещается. Быстрота же поддержания ее на постоян-
ном уровне таким образом определяется скоростью процес-
сов саморегуляции.
Малейшее изменение жестких констант организма приво-
дит в действие аппараты саморегуляции, которые должны
обеспечить возвращение их к исходному уровню почти
немедленно. В случае пластичных констант аппараты само-
регуляции могут довольно длительное время не обеспечи-
вать возвращение их к норме и жизнедеятельность организ-
ма при этом не будет значительно нарушена. Более того,
столь длительное смещение некоторых внутренних констант
нередко позволяет организму совершать напряженную
120 :
деятельность, направленную на внешнюю среду и связанную
с сохранением его вида или рода.
Это особенно отчетливо наблюдается в случае израсходо-
вания в организме питательных веществ или накопления
половых гормонов.
Однако, несмотря на то что отклонение таких констант
может быть компенсировано внутренними возможностями,
эти возможности, естественно, не безграничны. Вот почему
в организме немедленно начинают формироваться процессы,
направленные на потребление данного вещества из внеш-
ней среды, т. е. соответствующая мотивация. Именно этот
момент, характеризующийся изменением той или иной жиз-
ненно важной константы в крови, может быть расценен как
возникновение определенной потребности организма.
Таким образом, обособившись в процессе эволюции от
неживой природы, живые организмы оказались неспособны-
ми существовать только за счет своих внутренних ресурсов.
Они вынуждены все время апеллировать к окружающей их
среде как в смысле потребления из нее определенных про-
дуктов, так и выделения в нее продуктов своей жизне-
деятельности. В этом заключается теснейшая взаимосвязь
живых организмов и окружающей их природы.
Сколько же потребностей у организма? По-видимому, их
бесчисленное множество. Любое отклонение содержания
того или иного вещества в организме от константного уровня
может быть рассматриваемо как потребность. Однако не
все потребности сразу реализуются в соответствующие мо-
тивации. Но об этом ниже.
Итак, мотивация возникает на основе отклонения той
или иной жизненно важной константы организма от уровня,
обеспечивающего его нормальную жизнедеятельность, т. е.
на основе определенной потребности. Именно потребность
является первым толчком в цепи процессов саморегуляции,
направленных на поддержание той или иной константы
внутренней среды на постоянном уровне.
Возникает следующий вопрос: каким образом организм
воспринимает каждую потребность?
МЕХАНИЗМЫ ВОСПРИЯТИЯ ПОТРЕБНОСТИ
Любое незначительное отклонение жизненно важной,
константы от уровня, обеспечивающего нормальную жизне-
деятельность, еще даже не опасное для метаболизма тканей,'
немедленно воспринимается рецепторными аппаратами.
121
Такие рецепторы в изобилии представлены на внутренней
поверхности кровеносного русла. Их особенно много
в специальных чувствительных зонах сосудов — аортальной
и синокаротидной. Известно, что именно в этих областях
в изобилии находятся многочисленные хеморецепторы,
воспринимающие все параметры химических изменений
крови, а также механо- и барорецепторы.
В последние годы аналогичные рецепторы описаны вну-
три самой центральной нервной системы. Особенно богаты
ими некоторые области ретикулярной формации ствола го-
ловного мозга и гипоталамуса. Эти зоны центральной нерв-
ной системы, обладающие рецепторными свойствами, полу-
чили специальное название — «хеморецепторные триггер-
ные зоны». Ниже мы покажем, почему в их название
включено техническое понятие «триггер».
Рассмотрим прежде всего локализацию центральных хе-
морецепторов. В области ретикулярной формации продолго-
ватого мозга описаны рецепторы, воспринимающие измене-
ние уровня О2—СОг в организме (Dell, Bonvallet, 1954),
а также рецепторы, воспринимающие химические вещества
и токсины, вызывающие рвоту (Wanf, Borisson, 1952),
В передних отделах гипоталамической области в преде-
лах супраоптического ядра Javell (1953) и Verney (1953)
обнаружили особые нервные клетки, способные восприни-
мать изменения осмотического давления крови. Любопытно,
что конструкция этих клеток способствует восприятию ос-
мотического давления. Каждая такая клетка имеет специ-
альную вакуоль. Эта вакуоль сморщивается при увеличении
осмотического давления крови за счет отдачи воды в окру-
жающую среду и набухает при его снижении за счет вхож-
дения воды в клетку. Все эти процессы являются специ-
альными раздражителями данных клеток.
В вентро-медиальных ядрах гипоталамуса Мауег и др.
(1951, 1952) предполагают наличие рецепторов, чувствитель-
ных к уровню глюкозы в крови. Эти рецепторы были назва-
ны «глюкорецепторами». Существование таких рецепторов
доказывается на основании экспериментов с разрушением
данной области, после чего животные становятся гиперфа-
гичными.
Показано, что нейроны вентро-медиального и латераль-
ного поля гипоталамуса избирательно чувствительны к из-
менению концентрации глюкозы в крови (Б. И. Котляр,
Т. М. Ярошенко, 1969; Б. В. Журавлев, 1969; Anand a. oth.,
1964; Oomura a. oth., 1964),
122
При введении крысам соединения глюкозы с серой (аурб-
тиоглюкозы) Marshall и др. (1954, 1955), Мауег и Marshall
(1956) и Liebelt и др. (1961) обнаружили, что у крыс разви-
ваются явления чрезвычайной прожорливости и ожирения.
При гистологических исследованиях у таких животных
в вентро-медиальной области гипоталамуса были обнаруже-
ны отеки, пикнозы и дегенерация нервных клеток. Наблю-
даемое избирательное поражение именно клеток вентро-
медиальных отделов гипоталамуса связано с тем, что эти
клетки, избирательно чувствительные к глюкозе, концентри-
руют в себе ауротиоглюкозу, которая и вызывает их деге-
нерацию.
Наиболее убедительные данные о существовании глюко-
зочувствительных нейронов в гипоталамусе представлены
Оошига, Опо, Ооуаша и Wayner (1969). Используя методи-
ку микроинофореза, авторы обнаружили, что глюкозочув-
ствительные нейроны располагаются как в вентро-медиаль-
ном ядре, так и в латеральной гипоталамической области.
При этом большинство нейронов вентро-медиального гипо-
таламуса отвечало на инъекцию глюкозы учащением спон-
танной активности, в то время как нейроны латерального
гипоталамуса проявляли как возбуждающие, так и тормоз-
ные реакции.
В передней гипоталамической области (супраоптическая
область) описаны рецепторы, воспринимающие повышение
температуры крови, а в области заднего гипоталамуса
рецепторы, возбуждающиеся при снижении температуры
крови (Andersson a. Larsson, 1961).
Michael (1962) указывает на наличие в гипоталамусе
клеток, избирательно чувствительных к половым гормонам.
Dell и Bonvallet (1954) в ростральном отделе ретикуляр-
ной формации обнаружили клетки, чувствительные к гормо-
ну — адреналину.
Таким образом, хеморецепторные зоны широко представ-
лены в центральной нервной системе. Следует, однако, об-
ратить внимание на одно важное обстоятельство. Все цент-
ральные хеморецепторные зоны расположены в области
богатой васкуляризации. Такая их локализация вполне по-
нятна, если вспомнить их основное назначение — восприни-
мать изменения содержания тех или иных констант крови.
Итак, потребности организма прежде всего воспринима-
ются хеморецепторами кровеносного русла и специальными
центральными хеморецепторами. Следует подчеркнуть, что
при этом воспринимается не истинная потребность тканей,
123
а только изменения предконечного приспособительного
результата. Однако это надежно гарантирует организм
от возможных резких колебаний конечного полезного при-
способительного эффекта.
Наряду с периферическими и центральными хеморецеп-
торами у высших животных и человека имеются и другие
I
Рис. 8. Афферентная импульсация в желудочной веточке блуждающего
нерва у кошки под уретановым наркозом. У накормленного животного
(А) и после суточного голодания (Б). В — веточки блуждающего нер-
ва на желудке, от которых отводили электрическую активность (пока-
заны стрелкой).
124
предупредительные механизмы восприятия потребности.
Это — рецепторы, расположенные в определенных внутрен-
них органах. Сигнализация от этих рецепторов начинается
еще задолго до изменения основных жизненно важных
констант крови и на этой основе формирует соответствую-
щую мотивацию.
ice*
Г |)4*МММ1|ММЙ1
Рис. 9. Афферентная активность большого чревного нерва собаки в со-
стоянии голода и после приема пищи (по Л. И, Стадниченко, L969).
Рассмотрим несколько примеров. По мере эвакуации
питательных веществ из желудка в кишечник и в кровь
желудок постепенно запустевает. Тонус его мускалатуры
увеличивается. При этом, как показали наши наблюдения
( Н. В. Асмаян и К. В. Судаков, 1961; К. В. Судаков и С, К.
Рогачева, 1962), нарастает уровень афферентной импульса-
ции, идущей от желудка по волокнам блуждающего нерва
(рис. 8). Это в свою очередь приводит к возрастанию тонуса
ядра блуждающего нерва и усилению его восходящих влия-
ний на другие образования центральной нервной системы.
Аналогичные изменения электрической активности в волок-
нах большого чревного нерва у бодрствующих животных
наблюдала Л. И. Стадниченко (рис. 9). Все это чисто
нервным путем формирует мотивационное возбужде-
125
ние и на его основе — поиск пищи. Заметим, что в крови при
этом еще нет значительных изменений уровня питательных
веществ. Здесь мы снова встречаемся с предупредительным
механизмом.
Точно так же при наполнении мочевого пузыря постепен-
но нарастает уровень афферентной сигнализации в тазовом
нерве (Iggo, 1955). Возникающая на этой основе мотива-
ция также формируется только нервными механизмами.
Потребность может восприниматься и непосредственно
тканями. Так, Russek и др. (Rogriguez-Zendejas, Vega, Soto-
Mora, Russek, 1968; Russek, Rodriguez-Zendejas a. Pina,
1968) показали наличие специальных глюкорецепторов
в печени.
Все изложенное выше показывает, что восприятие потреб-
ности организма и формирование на ее основе мотивацион-
ного возбуждения осуществляются многими рецепторами
в несколько этапов. Эти рецепторы расположены как в спе-
циальных внутренних органах, так внутри сосудистой стенки
и, наконец, непосредственно в центральной нервной системе.
При этом каждый предшествующий этап формирования
потребности выступает в предупредительной роли по отно-
шению к последующему.
Рассмотрим теперь основные физиологические свойства
рецепторов, воспринимающих различные потребности орга-
низма и участвующих в последующем в формировании
мотивационного возбуждения.
Главным свойством этих рецепторных образований явля-
ется их подчеркнутая специфичность. Эта специ-
фика рецепторов непосредственно вытекает из специфичес-
ких свойств каждой потребности, которую они воспринимают.
Так, изменения артериального давления воспринимают
специальные барорецепторы, расположенные по всему кро-
веносному руслу и особенно в аортальной и синокаротидной
области. Изменения напряжения Ог—СО2 в крови воспри-
нимаются хеморецепторами кровяного русла и т. д. Благо-
даря такой подчеркнутой специфике рецепторов обеспе-
чивается строгое «наблюдение» за каждой жизненно важной
константой организма и всеми ее параметрами. Рецептор
приобретает таким образом способность воспринимать ма-
лейшие ее изменения. При этом включение рецептора в вос-
приятие каждой потребности происходит постепенно. Это
значит, что возбуждение рецептора прямо пропорционально
степени отклонения соответствующей константы от уровня,
обеспечивающего нормальную жизнедеятельность.
126
Таким образом, Соотношения рецёптора и полезного
приспособительнного эффекта в каждой функциональной
системе организма являются очень тесными. Они строятся
по принципу комплементных взаимосвязей. А это значит,
что любое отклонение того или иного полезного приспособи-
тельного эффекта в организме от уровня, обеспечивающего
нормальную жизнедеятельность, немедленно воспринимает-
ся соответствующим рецептором. Следовательно, соотноше-
ние «полезный приспособительный эффект — рецептор» яв-
ляется центральным звеном в каждой саморегулирующейся
функциональной системе организма.
Какие же механизмы определяют столь интимные взаимо-
отношения рецептора с конечным приспособительным эф-
фектом? Почему каждый рецептор «следит» за определен-
ной константной крови и усиливает свою деятельность во
всех случаях ее изменения?
Такая тонкая специфика рецепторов внутренней среды
организма, воспринимающих только специальные изменения
ее внутренних констант, основана на их подчеркнутой изби-
рательной химической чувствительности к данной константе.
Для каждого из этих рецепторов константное содержание
того или иного вещества внутри организма определяет
нормальное течение его метаболизма, т. е. это вещество
входит в интимные биохимические процессы его жизнедея-
тельности. Отсюда понятно, что любое изменение содержа-
ния того или иного вещества внутри организма немедленно
возбуждает именно те рецепторные окончания, которые ис-
пользуют это вещество в своем метаболизме.
Включение определенных веществ в метаболизм рецеп-
торов делает их крайне консервативными и заставляет
«служить» только одной потребности.
Работа периферического рецептора в каждой функцио-
нальной системе подчинена следующим закономерностям.
Как уже указывалось, меньшее отклонение функции от
константного уровня вызывает меньшее возбуждение рецеп-
торов, а это в свою очередь порождает соответственно
меньшую афферентацию, поступающую к центральным
аппаратам данной функциональной системы. Наоборот,
большее отклонение константы от жизненно важного уровня
вызывает наиболее сильное возбуждение этих рецепторов
н интенсивную нервную сигнализацию в центральную нер-
вную систему. Для того чтобы понять эти закономерности,
рассмотрим поведение барорецептора при изменениях
уровня артериального давления.
Опыты П. к. Анохина и А. И. Шумилиной (Anochin,
Schumilina, 1962) показали, что количество и продолжитель-
ность импульсаций при каждом сердцебиении, регистрируе-
мых в одиночном нервном волокне депрессорного нерва,
идущего от аортальной области, прямо пропорциональны
величине артериального давления (рис. 10). При артериаль-
ном давлении 110 мм рт. ст. в каждом залпе регистрируется
3 импульса, а при увеличении давления до 180 мм рт. ст.
А
рост импульсаций резко возрастает. Эта же
ке (Б) в сравнении /г1 и h2. На осн ординат
цисс — вртернальное давление в миллиметрах
хину и А. И. Шумилиной, 1962),
Рис. 10. Изменения импуль-
сации одиночного барорецеп-
тора дуги аорты кролика
при повышении артериаль-
ного давления,
Д — импульсация барорецептора
при увеличении артериального
давления от ПО до 180 мм рт.
ст. (/); электрокардиограмма,
отметка времени 0,02 секунды
(2). Прирост импульсаций на
каждые 10 мм увеличения дав-
ления (обозначено стрелками)
непропорционален. При боль-
ших величинах давления при-
зависнмость приведена на графи-
— число импульсаций, на оси абс-
ртутиого столба (по П. К- Ано-
каждый залп импульсов состоит уже из 65—70 импульсов.
Это связано с тем, что каждое рецепторное образование
аортальной области отвечает только на определенные изме-
нения артериального давления. Барорецептор строго следит
за каждым изменением артериального давления. Точно так
же работают и другие рецепторы.
Таким образом, в условиях постоянно изменяющейся внут-
ренней среды организма консерватизм рецептора за счет
различной интенсивности возбуждения механизмов саморе-
гуляции обеспечивает важное свойство—поддержание той
или иной жизненно важной функции на константном уровне.
Соотношение «полезный приспособительный результат —
рецептор» — это основной «узел» саморегуляции, Без этого
128
«узла» нельзя представить саморегуляцию. Рецептор в своей
эволюции, в своем метаболизме, твердо законсервировал
свойства конечного приспособительного эффекта, на стра-
же которого он находится.
Если вернуться к знаменитому выражению К- Бернара
о том, что «постоянство внутренней среды есть условие сво-
бодной жизни организма», то можно заметить в нем некото-
рые противоречия. Условием свободной жизни организма
является по сути дела не постоянство внутренней среды,
которая постоянно изменяется, а своеобразный консерва-
тизм рецепторных образований, возбуждение которых через
саморегуляцию приводит к стабилизации внутренней среды.
Именно консерватизм рецепторов является ограничителем
пластических изменений внутренних констант организма.
Консервативный рецептор во всех случаях устанавливает
нужный уровень приспособительного эффекта. В случае
же длительного отклонения последнего от консервативного
уровня рецептор начинает непрерывно сигнализировать
в центральную нервную систему, создавая тем самым усло-
вия для возникновения мотивации.
Включение только определенных веществ в метаболизм
каждого рецептора определяет и другое их важное свойст-
во — избирательную чувствительность толь-
ко к определенной потребности.
Такое избирательное отношение рецепторов только к оп-
ределенной потребности определяет в дальнейшем важное
свойство мотивационного возбуждения — его подчеркнутую
нейрофизиологическую специфику. Каким образом прояв-
ляется эта подчеркнутая избирательная чувствительность
рецепторных образований в формировании мотивации, мы
рассмотрим в последующем изложении при разборе свойств
гипотамических мотивационных центров.
Сейчас перейдем к ответу на вопрос, каким образом воз-
бужденный соответствующей потребностью рецептор приво-
дит к формированию мотивационного возбуждения в цен-
тральной нервной системе.
НЕРВНЫЙ И ГУМОРАЛЬНЫЙ МЕХАНИЗМ ФОРМИРОВАНИЯ
МОТИВАЦИОННОГО ВОЗБУЖДЕНИЯ
Истинная потребность организма всегда возникает в его
тканях. Однако сигнализация о потребности организма
в том 'Пли ином веществе начинается задолго до существен-
ных изменений в тканях.
5 Заказ 5074
129
Как указывалось выше, мотивационное возбуждение на-
чинает формироваться прежде всего за счет возбуждения
специальных рецепторов, находящихся в определенных
внутренних органах (желудок, печень, мочевой пузырь, по-
ловые органы и т, д.), т. е, на основе чисто нер-
вных механизмов. Рассмотрим теперь этот механизм
более подробно на примере формирования мотивации го-
лода.
По мере эвакуации пищи из желудка последний все в боль-
шей степени запустевает. В свою очередь отсутствие пищи
А Б
Рис. 11. Схема операции
анастомозов блуждающего
нерва у собаки
А — анастомоз блуждающего
нерва со смешанным нервом яе-
родней конечности и Б — блуж-
дающего нерва с барабанной
струной: I — g. nodosum; 2—ваго-
снмпатический ствол; 3— ана-
стомоз; 4 — слюнная железа;
5 — барабанная струна; 6 — сме-
шанный нерв передней конечно-
сти; 7 — верхний шейный симпа-
тический узел.
в желудке является мощным раздражителем рецепторов его
слизистой и мышечной оболочек, что, как уже указывалось
ранее, приводит к значительному возрастанию афферент-
ной сигнализации, идущей в центральную нервную систему
по волокнам блуждающего нерва и симпатических нервов.
Эта импульсация, поступая в центр блуждающего нерва,
расположенный на уровне продолговатого мозга, начинает
вызывать в нем определенные изменения. Эти изменения
хорошо могут быть прослежены в случае создания операции
искусственного анастомоза центрального конца одного из
стволов блуждающего нерва с периферическим концом
перерезанного нерва передней конечности или с нервом
барабанной струны (П. К. Анохин, 1935; А. П. Анохина, 1946;
К. В. Судаков, 1960; Н. В. Асмаян, 1961) (рис. 11).
В результате такой операции эфферентные волокна блу-
ждающего нерва прорастают и устанавливают функцио-
130
нальный контакт с поперечнополосатыми мышцами перед-
ней конечности (в случае анастомоза Vagus-Radialis) или
с подчелюстной слюнной железой (в случае анастомоза
vagus-chorda tympani). Создаются условия, позволяющие
по изменениям в деятельности этих органов, получивших
искусственную иннервацию от блуждающего нерва, следить
за состоянием его центра.
Я
Рис, 12. Сопряженные периодические изменения деятельности органов
собаки, естественно и искусственно связанных с ядром блуждающего
нерва.
— после 20-часозого голодания и Б— после кормления (момент кормления обо-
значен стрелкой): а — электрическая активность поперечнополосатой мышцы, ин-
нервируемой блуждающим нервом; б —моторика желудка; & — количество слюны,
выделяемой подчелюстной железой, связанной с блуждающим нервом в условных
баллах (0, 2, d, 6); г — отметка времени 15 минут.
Проведенные в этом направлении опыты показали прежде
всего, что уровень функциональной активности ядра блу-
ждающего нерва у накормленных животных очень низок.
Активность его возрастает по мере голодания и эвакуации
пищи из желудка. Эту закономерность можно проследить
как в изменении электрической активности мышц перед-
ней конечности, иннервированных блуждающим нервом,
так и секреторной деятельности слюнной подчелюстной
железы в случае анастомоза vagus-chorda tympani (рис. 12),
Все это указывает па то, что нарастающая по мере голо-
дания животного от желудка импульсация приводит к воз-
5*
131
растанию функциональной активности центров блуждаю-
щего нерва. Что это действительно так, подтверждают
специальные эксперименты (К. В. Судаков, 1959), в которых
при наличии анастомоза одного из блуждающих нервов осу-
ществлялась механическая или химическая блокада прове-
дения возбуждения по афферентным волокнам другого ин-
тактного выведенного в кожную манжету блуждающего
нерва. Во всех этих случаях возрастания активности ядра
блуждающего нерва не наблюдалось.
Наблюдения за деятельностью периферических органов
в условиях анастомоза блуждающего нерва показали также,
что при голодании в ядре блуждающего нерва, кроме обще-
го возрастания активности, периодически наблюдается еще
большее ее возрастание. Эти периодические изменения ак-
тивности ядра блуждающего нерва у голодных животных
точно соответствуют ритмике «голодной» моторной перио-
дической деятельности желудка.
Максимальное нарастание активности ядра блуждающего
нерва наблюдается в период моторики желудка и особенно
к его концу. После завершения «периода работы» эта
активность падает до минимального уровня и постепенно
нарастает к следующему «периоду работы».
Какие же механизмы обусловливают периодические
«взрывы» возбуждения в ядре блуждающих нервов у голод-
ных животных?
Специальные опыты (К. В, Судаков, 1960) показали, что
взрывы возбуждения в ядре блуждающего нерва у голод-
ных животных происходят па основе периодического изме-
нения возбудимости составляющих их нервных элементов
в так называемый период покоя за счет афферентной им-
пульсации, поступающей от желудка.
Это доказывается следующим экспериментом.
В процессе регенерации ствола блуждающего нерва
в ствол сшитого с ним нерва передней конечности, например
лучевого нерва, регенерируют не только эфферентные, как
указывалось выше, но и афферентные волокна. Благодаря
этому они устанавливают функциональная контакт с рецеп-
торами кожи. Последнее обстоятельство позволяет в экспе-
рименте искусственно раздражать эти искусственные рецеп-
торные образования и по изменениям, например, моторной
деятельности пустого желудка следить за возбудимостью
ядра блуждающего нерва.
Опыты показали, что при приложении к соответствующе-
му участку кожи, предварительно десквамированному под-
132
бриванием, раствора хлористого натрия определенной кон-
центрации удается полностью воспроизвести внеочередной
период сокращений желудка.
При использовании пороговых раздражений рецепторов
блуждающего нерва в коже удалось показать, что возбу-
димость ядра блуждающего нерва в течение «периода по-
Рис, 13. Моторная реакция желудка у голодной собаки в ответ на
раздражение хлористым натрием рецепторов кожи, искусственно ин-
нервированных блуждающим нервом.
.4 — в начале периода покоя; Б —в конце периода покоя; В — последовательное
раздражение в течение одного периода покоя; Г — подпороговое раздражение на
фоне введения в желудок 5% раствора хлористого натрия. Отметка времени
5 мгшут.
коя» периодически нарастает. В начале «периода по-
коя» пороговое раздражение 5% хлористым натрием совсем
не вызывает реакции. В середине «периода покоя» в ответ
на такое же раздражение возникает несколько единичных
желудочных сокращений, В конце «периода покоя» то же
раздражение приводит к возникновению полноценного пе-
риода желудочных сокращений (рис. 13).
Наблюдаемые изменения возбудимости в ядре блуждаю-
щего нерва в «период покоя» моторной деятельности желуд-
ка в свою очередь зависят от афферентных импульсаций,
133
непрерывно поступающих в центр по блуждающему нерву.
Блокада интактного блуждающего нерва в условиях ана-
стомоза второго нерва, вызванная механическим или хими-
ческим путем, приводит к тому, что в период блокады не
наблюдается возрастания возбудимости ядра блуждающего
нерва и очередной «период работы» отставляется точно на
интервал времени, равный времени блокады.
Аналогичным образом во всех случаях, когда удается
искусственно за счет раздражения рецепторов блуждающего
нерва в коже вызвать внеочередной полноценный приступ
моторной деятельности желудка, последующий приступ
развивается только через обычный интервал «периода по-
коя» после искусственно вызванного приступа.
Все это показывает, что у голодных животных за счет
постоянной сигнализации, идущей от пустого желудка, про-
исходит периодическое изменение возбудимости центров
блуждающего нерва. При достижении возбудимости крити-
ческого уровня в ядре под влиянием той же афферентной
импульсации возникает взрывной процесс возбуждения, ко-
торый по соответствующим эффекторным каналам распро-
страняется на периферию и вызывает приступ так называе-
мой голодной периодической деятельности.
Одновременно с распространением в нисходящем направ-
лении к пищеварительным органам ритмически возникаю-
щие в ядре блуждающего нерва возбуждения распростра-
няются, по-видимому, и в восходящем направлении
К расположенным выше отделам пищевого центра и, в част-
ности, в гипоталамус, принимая участие в формировании
мотивационного возбуждения. Такое предположение весьма
согласуется с данными Bronk, Pitts и Larrable (1940), пока-
завшими, что вагусные импульсы достигают гипоталамуса.
Естественно поставить вопрос: какой биологический смысл
такой ритмической деятельности ядра блуждающего нерва
у голодных животных?
Если проанализировать работу других функциональных
систем организма, то мы легко обнаружим у них наличие
подобной ритмической деятельности.
Так, в функциональной системе дыхания — это ритмиче-
ская смена вдоха выдоха; в половой функциональной систе-
ме— циклические процессы, связанные с созреванием яйце-
вой клетки и'т. д.
По-видимому, основной биологический смысл таких рит-
мических процессов в каждой функциональной системе
заключается в периодическом стимулировании мотивацион-
134
них центров во избежание их адаптации. Эти ритмические
процессы важны и в другом отношении. Они определяют
периодическое распределение соответствующих веществ
в организме, благодаря чему соответствующие жизненно
важные константы внутренней среды поддерживаются
некоторое время на относительно постоянном уровне за счет
внутренних резервов организма.
Известно, что в период голодной периодической деятель-
ности наблюдаются не только сокращения желудка и верх-
ней части кишечника. Одновременно с этим происходят
изменения дыхания, состава крови, а также выброс желчи
и панкреатического сока в просвет двенадцатиперстной
кишки. Этот сок богат белковыми и ферментными про-
дуктами и, как считает В. Н. Болдырев (1914), является
источником питательных веществ для организма при голо-
дании. На этом основании В. Н. Болдырев полагает, что
голодная периодическая деятельность обеспечивает пере-
распределение питательных веществ внутри организма у го-
лодных животных. Возможно, что именно посредством этих
периодических процессов происходит и перераспределение
содержания питательных веществ в различных органах при
голодании, открытое И. П. Разенковым (1949), при котором
белок из органов, менее значимых для сохранения жизни,
поступает к таким жизненно важным органам, как нервная
система, сердце и железы внутренней секреции.
Итак, ритмические изменения в определенных нервных
центрах, возникающие под влиянием сигнализации, идущей
от рецепторов соответствующих периферических органов,
являются первым этапом формирования мотивационного
возбуждения.
Иногда уже этих чисто нервных процессов бывает доста-
точно для того, чтобы возникло возбуждение соответствую-
щих отделов центральной нервной системы и возникли
процессы, направленные на перераспределение определен-
ных веществ в организме и поиск соответствующего веще-
ства во внешней среде.
Однако, как правило, этим дело не ограничивается.
Под влиянием нервной сигнализации, идущей от рецепто-
ров соответствующих периферических органов в центральную
нервную систему, формируются соответствующие изменения
в крови. В одних случаях (например, при голодании) орга-
низм начинает «прятать» необходимые для его метаболизма
вещества в депо в качестве резервов. В других случаях
(например, при половом возбуждении), наоборот, выбрасы-
135
вает определенные вещества в кровь. Таким образом, всту-
пают в действие раздражающие факторы самой крови. Они
в свою очередь приводят в возбуждение специальные рецеп-
торы, которые обильно представлены в стенке сосудистого
русла (В. Ф. Маркелова и А. М. Уголев, 1960), а также не-
посредственно в центральной нервной системе.
Следует отметить, что изменения в крови снова выступают
в предупредительной роли, поскольку уровень тех или иных
веществ в организме, его депо, может еще вполне обеспечи-
вать недостаток содержания этих веществ в крови.
Возникает вопрос, каким же образом на гуморальной ос-
нове происходит раздражение рецепторов сосудистого русла
и специальных центральных рецепторов. Здесь возмож-
ны принципиально два механизма: либо эти рецепторы
возбуждаются периодически, т. е. при изменении той или
иной константы крови, и прекращают возбуждаться по
мере того, как эта константа снова временно восстанавли-
вается за счет работы депонирующих механизмов. Есть все
основания думать, что периодический выброс питательных
веществ из депо происходит в конце каждого «периода
работы». На это указывает резкое снижение тонуса блу-
ждающего нерва в данный период, совершенно аналогичное
тому, которое наблюдается непосредственно после кормле-
ния животного.
Второе предположение, которое нам кажется наиболее
вероятным, заключается в том, что по мере увеличения сиг-
нализации из внутренних органов соответствующие констан-
ты крови (за счет депонирования или, наоборот, выброса
определенных веществ в кровь) приходят в состояние дли-
тельного отклонения, на фоне которого периодические выб-
росы веществ из депо производят только незначительную
временную нормализацию той или иной физиологической
функции. Именно такое устойчивое отклонение константы
от уровня, обеспечивающего нормальную жизнедеятельность,
является мощным раздражителем мотивационных центров.
Оба эти предположения, безусловно, нуждаются в даль-
нейшей экспериментальной проверке.
Следует отметить, что возбуждения сосудистых рецепто-
ров также распространяются в центральные нервные обра-
зования нервным путем.
Таким образом, мотивационное возбуждение формирует-
ся на основе внутренней потребности тремя принципиально
различными нейро-гуморальными механизмами: 1) нервной
сигнализацией, идущей от соответствующих внутренних ор-
136
ганов, обеспечивающих тесную связь организма с внешней
средой его существования; 2) нервной сигнализацией от
рецепторов сосудов; 3) непосредственным действием гумо-
ральных факторов на центральные хеморецепторы.
Значение всех этих факторов в формировании мотиваци-
онного возбуждения велико. Весь процесс формирования
мотивации развертывается во времени с определенным
опережением одними событиями других. Нервный механизм
сигнализации от рецепторов внутренних органов определяет
формирование мотивационного возбуждения задолго до
того, как содержание определенных веществ существенно
изменится не только в тканях организма, но и в кровеносном
pi еле. Однако это воздействие не стабильно, оно периодиче-
ское и, формируясь на более низших уровнях центральной
нервной системы, может быть подавлено другими, более
сильными центральными возбуждениями. Последующее
включение в формирование данного мотивационного воз-
буждения гуморальных факторов обеспечивает устойчивое
и длительное раздражение, которое, как мы увидим ниже,
приобретает свойства доминирующего для данного живот-
ного возбуждения.
Все изложенное выше показывает, что нервные и гумо-
ральные механизмы в формировании мотивационного воз-
буждения тесно взаимосвязаны.
Если вспомнить наблюдения Sherrington и Н. Ф. Попова
о том, что люди с удаленным желудком продолжают по-
прежнему испытывать голодное возбуждение, то становится
совершенно очевидной роль гуморальных факторов в этом
процессе. Если же вспомнить наблюдения Т, Т. Алексеевой
над сросшимися близнецами, становится ясным и тот факт,
что, несмотря на общее кровообращение, обе девочки по-
разному испытывали чувство голода и насыщения, незави-
симо друг от друга. В этом случае на первое место высту-
пает значение нервного фактора мотивационного возбуж-
дения.
По-видимому, для естественного формирования мотива-
ционного возбуждения необходимо, чтобы соответствующие
центры получили раздражение сначала нервным, а затем
гуморальным путем. Любое изменение естественной после-
довательности раздражения нарушает нормальный меха-
низм возбуждения этих центров.
Наконец, следует отметить, что в формировании различ-
ных мотивационных возбуждений значение нервных и гумо-
ральных факторов и их взаимозаменяемость различны. Так,
137
в формировании половых мотиваций большее значение име-
ет гуморальный фактор, накопление в крови половых гормо-
нов, а в мотивации, связанной с опорожнением мочевого
пузыря, преобладает чисто нервный механизм.
Однако, какой бы из перечисленных выше механизмов ни
преобладал в формировании любого мотивационного воз-
буждения, в нем при ближайшем исследовании всегда
можно обнаружить взаимодействие нервных и гуморальных
факторов.
Возникает вопрос, к каким отделам центральной нервной
системы адресуются нервные и гуморальные влияния, вы-
званные раздражением рецепторов, воспринимающих ту
или иную внутреннюю потребность организма?
Исследования последних лет показывают, что такой обла-
стью является гипоталамус. Гипоталамические отделы,
к которым адресуются возбуждения, вызванные отклонения-
ми тех или иных констант крови от уровня, обеспечивающего
нормальную жизнедеятельность организма, получили назва-
ние гипоталамических мотивационных центров.
К рассмотрению физиологических свойств этих центров
и вопроса о том, как эти центры порождают мотивационное
возбуждение, мы и переходим в последующем изложении.
ГИПОТАЛАМУС КАК ЦЕНТРАЛЬНОЕ ЗВЕНО
ВОСПРИЯТИЯ ВНУТРЕННИХ ПОТРЕБНОСТЕЙ
ОРГАНИЗМА
Трудно говорить об изолированных свойствах гипотала-
мических центров. В настоящее время показано, что гипо-
таламус имеет тесные функциональные связи с ретикуляр-
ной формацией ствола головного мозга, с лимбическими
образованиями, а также с корой головного мозга. Между
тем участие гипоталамических центров в мотивационном
возбуждении не подлежит сомнению. Можно указать на
некоторые особенности функционирования мотивационных
центров гипоталамуса.
Прежде всего следует отметить, что именно гипоталами-
ческие образования обладают свойством воспринимать из-
менения внутренней среды, а также смещения констант
крови, т. е. они обладают центральной рецепторной функци-
ей. Главная же особенность гипоталамических центров
состоит в том, что они не только воспринимают эти измене-
ния, но трансформируют их в специальный нервный про-
цесс, завершающийся формированием поведения во внешней
138
Среде, направленного на удовлетворение внутренних потреб-
ностей. Именно поэтому гипоталамус является централь-
ным звеном в формировании мотивационного возбуждения.
Рассмотрим теперь вопрос о том, как же внутренние
потребности организма оказывают свое действие на гипо-
таламические центры. Возбуждение этих центров, как уже
указывалось выше, осуществляется двояким — нервным
и гуморальным путем.
Как же эти влияния сказываются на деятельности гипо-
таламических центров?
Рис, 14. Схема избирательной химической чувствительности рецептор-
ных клеток гипоталамической области,
Каждая из клеток А Б, 5, Г реагирует только на изменение специфического
фактора крови. На схеме показано, что при изменении в крови фактора а воз'
буждэется только клетка Л. При изменении фактора е? — только клетка 5.
Как мы уже говорили выше, именно в гипоталамической
области имеется ряд зон, метаболизм клеток которых изби-
рательно чувствителен к изменениям той или иной констан-
ты крови. Из этого следует, что каждая такая клетка начи-
нает возбуждаться только при изменении содержания
определенного вещества крови. Так, при изменении осмоти-
ческого давления крови возбуждаются только клетки, мета-
болизм которых высокочувствителен к изменению ионного
или водного состава плазмы крови. Однако при этом не
возбуждаются клетки, метаболизм которых чувствителен,
например, к определенному содержанию глюкозы в крови.
Последние возбуждаются только при изменении в крови
содержания глюкозы (рис. 14).
Все это определяет резко выраженную избирательность
возбуждения гипоталамических центров при различных
изменениях состава крови.
139
Другой особенностью гипоталамических центров является
то, что их клетки, избирательно чувствительные к изменени-
ям определенной константы крови, в то же время избира-
тельно возбуждаются нервными импульсациями, идущими
500мкв
1 сек
Рис. 15. Реакция одной и
той же клетки латераль-
ного гипоталамуса кро-
лика на пищевые раздра-
жения (по Б. В. Журав-
леву, 1969).
/—фон; 2 —орошение ротовой
полости (первая пунктирная
линия) и введение морковно-
го сока в желудок (вторая
пунктирная линия); 3 — 4 —
введение в сонную артерию
5 мл 5%' раствора глюкозы
(момент введения обозначен
пунктирной линией); 5— кон-
троль: введение физиологи-
ческого раствора в сонную
артерию (момент введения
обозначен пунктирной ли-
нией); б — локализация кон-
чика микроэлектрода {пока-
зано стрелкой).
140
из соответствующих, связанных с данной потребностью
органов.
Так, экспериментами нашего сотрудника Б. В. Журавлева
(1969) с помощью микроэлектродных исследований показа-
но, что одна и та же клетка латеральной области гипотала-
муса у кролика реагирует как на введение глюкозы в кровь,
так и на введение питательных веществ непосредственно
в желудок (рис. 15).
Рис. 16. Принципиальная схема триггерного механизма работы клеток
гипоталамических мотивационных центров.
а — изменения соответствующей константы крови; б — изменения возбудимости
клеток гипоталамуса; / — возникновение потребности; fl — удовлетворение потреб
пости. При изменении константы крови от К до 0 изменяется возбудимость клеток
гипоталамуса. Когда нх возбудимость достигает критического уровня 4 (точка /),
они приходят в состояние возбуждения и порождают соответствующую импульса-
цию. При удовлетворении потребности н восстановлении константы крови (точки
2, 3, 4) уровень возбудимости этих клеток снижается и их импульсация прекра-
щается. Активность этих клеток восстанавливается только в том случае, если под
влиянием изменения константы крови или других факторов уровень их возбудимо-
сти снова достигает критического порога (точка 5).
Интересен механизм возбуждения этих центров гипота-
ламуса. Все его клетки, обладающие избирателыгой рецеп-
торной функцией, работают по триггерному типу. Это зна-
чит, что возбуждение в этих клетках возникает не сразу
при изменении соответствующей константы крови или при
поступлении к ней нервных импульсаций, а периодически,
только тогда, когда изменения их возбудимости достигают
определенного критического уровня (рис. 16).
При слабых степенях раздражения происходит лишь из-
менение возбудимости этих центров без взрывных деполя-
ризационных процессов. Взрывной процесс возбуждения
возникает в клетках мотивационных центров гипоталамуса
только тогда, когда изменения их возбудимости под влияни-
141
ем раздражающего фактора достигнут определенного крити-
ческого уровня.
Только при достижении порога этого критического уровня
клетки мотивационных центров гипоталамуса начинают
генерировать разряды нервных импульсов. Отсюда стано-
j вится ясным, почему в формировании мотивационного воз-
Г буждения участвуют нервные и гуморальные механизмы.
В том случае, когда чувствительные клетки гипоталами-
ческих центров подвергаются действию только нервных
импульсаций со стороны периодически возбуждаемых аф-
ферентными импульсациями, идущими от периферических
органов нижележащих центров, возбуждение в них возни-
кает также периодически. Каждый взрыв возбуждения
в этих центрах сменяется последующим падением возбуди-
мости до исходного низкого уровня, и для того, чтобы воз-
никло следующее возбуждение, нервные импульсации дол-
жны снова поднять возбудимость этих’ клеток до критиче-
ского порогового уровня.
Другое явление наблюдается в том случае, когда в про-
цесс включаются гуморальные факторы. В этом случае
длительное и устойчивое изменение содержания того или
иного фактора в крови немедленно поднимает возбудимость
воспринимающих этот фактор клеток гипоталамуса до по-
рогового уровня и поддерживает ее на этом высоком уровне
все время, пока существует соответствующая потребность.
Благодаря этому клетки начинают генерировать непрерыв-
ную импульсацию. И эта импульсация продолжается до тех
пор пока не будет устранено порождающее ее раздражение,
т. е. не будет восстановлено нормальное содержание в крови
определенного вещества, являющегося для нее специфиче-
ским раздражителем. Подтверждение этой мысли дают не-
которые общебиологические наблюдения.
У крысы и человека семенная железа, например, функцио-
нирует постоянно. Это определяет у них постоянность поло-
вого влечения. У некоторых животных в деятельности этой
железы наблюдается сезонная цикличность. Так, у хорька,
бурундука и оленя активный сперматогенез и отчетливое
половое влечение наблюдаются лишь во время сезонного
брачного периода. В стимуляции этого состояния у этих
животных большую роль играют некоторые факторы внеш-
ней среды — яркий свет, тепло и т. д.
Подчеркнутая цикличность половой активности, связан-
ная с периодическим усилением деятельности половых же-
лез, наблюдается у самок многих млекопитающих.
142
Триггерные механизмы— определяют важное свойство
гипоталамических мотивационных центров — периодичность
их работы.
Характерной особенностью возуждеиия мотивационных
центров гипоталамуса является то, что их триггерный ме-
ханизм значительно растянут во времени^ Это определяет
довольна/ замедленную ритмику мотивационных возбужде-
ний. Некоторые из них возникают через интервалы в не-
сколько часов (например, голодное возбуждение), другие—
через несколько суток и даже месяцев (половое возбуж-
дение).
Особенностью мотивационных центров гипоталамуса яв-
ляется и то, что изменения возбудимости до порогового
уровня, порождающего развитие взрывного процесса воз-
буждения, могут происходить в них и без участия нервных
и гуморальных факторов, т. е. автоматически, за счет внут-
ренних метаболических процессов:—Вот почему иногда
мотивационное вбзЗуХдепие может рождаться в самом
нервном центре.
Эта способность некоторых клеток центральной нервной
системы периодически «самовозбуждаться» под влиянием
метаболических процессов, нервных и гуморальных фак-
торов в последнее время приобрела специальное название;
«биологические ритмы», или «биологические часы» (Harker,
1964).
По-видимому, именно эти клеточные процессы, объеди-
няясь далее в ансамбли синхронно работающих клеток,
и составляют ту наследственную основу мотивационного
возбуждения, которая у взрослых животных значительно
модулируется нервными и гуморальными влияниями.
По триггерному типу формируются и искусственные мо-
тивации у животных, вызванные электрическим или хими-
ческим раздражением этих центров. Рассмотрим случай,
когда предварительно накормленное животное при раздра-
жении латеральной гипоталамической области снова начи-
нает есть. В желудке и крови этого животного имеется
достаточное количество питательных веществ. Следователь-
но, исходный уровень возбудимости клеток латеральной гипо-
таламической области, воспринимающих изменения содер-
жания питательных веществ в крови, низок. Однако
электрическое раздражение этих клеток поднимает их
возбудимость допорогового уровня. Вследствие этого клетки
приходят в состояние возбуждения и возникает пищевая
мотивация.
143
Рис, 17. Схема генерализованного
распространения, возникшего на гу-
моральной основе возбуждения из
гипоталамических центров на другие
отделы головного мозга (по П. К.
Анохину, 1962). Схематически пока-
зано формирование одного из силь-
нейших влечений человеческого орга-
низма — жажды.
Гипертоническая кровь через капиллярную
стенку Л раздражает осморецептор ОР,
находящийся в гипоталамусе Г; /СР —кора;
ПК — подкорка; РФ — ретикулярная форма-
ция.
Наконец, отметим пос-
леднюю особенность моти-
вационных центров гипо-
таламуса. Чувствительные
клетки гипоталамуса име-
ют обширные связи с дру-
гими клетками головного
мозга, в частности с ре-
тикулярной формацией
среднего мозга и лимби-
ческими образованиями
переднего мозга. Эти свя-
зи, как указывалось вы-
ше, осуществляются пре-
имущественно через jn.e-
диальный„.тгу1цж передне-
го мозга и другие обра-
зования. Благодаря таким
обширным связям клеток
гипоталамуса с другими
отделами головного моз-
га, возбуждение, возник-
шее в них, способно ши-
роко распространяться и
возбуждать целый комп-
лекс корково-подкорковых
аппаратов (рис. 17). Это
в свою очередь определяет наиболее важное свойство мо-
тивационных центров гипоталамуса, их восходящие акти-
вирующие влияния на другие отделы головного мозга,
ВОСХОДЯЩИЕ АКТИВИРУЮЩИЕ ВЛИЯНИЯ
ГИПОТАЛАМИЧЕСКИХ ЦЕНТРОВ КАК ОСНОВА
МОТИВАЦИОННОГО ВОЗБУЖДЕНИЯ
Как известно, впервые Magoun и Moruzzi в 1949 г. уста-
новили, что ретикулярная формация ствола головного мозга
обладает свойством оказывать на кору головного мозга
восходящие активирующие влияния. Только при наличии
этих влияний клетки коры головного мозга оказываются
способными устанавливать активные связи с внешним ми-
ром и осуществлять свою аналитико-синтетическую дея-
тельность. Разрушение ретикулярной формации, так же как
и подавление ее функции различными фармакологическими
144
и особенно наркотическими средствами, приводит к потере
сознания и состоянию беспробудного сна.
Восходящие активирующие влияния ретикулярной фор-
мации на кору головного мозга имеют характерное выра-
жение в электроэнцефалограмме. Обычно они проявляются
в форме низкоамплитудных, высокочастотных, электриче-
ских колебаний. Если такая активность возникает на фоне
медленных высокоамплитудных колебаний, она носит на-
звание реакции «пробуждения», «активации», «десинхрони-
зации» и т. д. Характерно, что почти во всех случаях восхо-
дящих влияний ретикулярной формации на кору головного
мозга наблюдается однотипная электроэнцефалографиче-
ская картина. Это дало основание ряду исследователей
(Magoun, Moruzzi a. oth.) говорить о неспецифических влия-
ниях ретикулярной формации на кору головного мозга.
Между тем в лаборатории, руководимой П. К. Анохиным
(1952 в), была выдвинута другая точка зрения. Было пока-
зано, что, несмотря на однотипность электроэнцефалогра-
фических изменений, восходящие активирующие влияния
ретикулярной формации на кору головного мозга неодно-
родны. Они являются специфичными при реакциях различ-
ного биологического качества. Неоднородность восхо-
дящих активирующих влияний ретикулярной формации
на кору головного мозга отчетливо проявляется по ее отно-
шению к различным наркотическим веществам. Так, напри-
мер, оказалось, что под уретановым наркозом, несмотря на
сон животного, блокаду его активных связей с внешним ми-
ром и восходящих влияний ретикулярной формации на кору,
наблюдаемых в бодрствующем состоянии, болевое раздра-
жение седалищного нерва вызывает отчетливую реакцию
десинхронизации электроэнцефалограммы (В. Г, Агафонов,
1956).
Другим примером диссоциации восходящих ретикулярных
влияний являются опыты с применением аминазина. В экс-
периментах А. И. Шумилиной (1956), В. А. Гавличека
(1958), Ю. А. Макарова (1960) было показано, что амина-
зин избирательно устраняет активацию коры головного моз-
га, вызванную оборонительным напряжением животного,
но не снимает восходящие активирующие влияния, обуслов-
ленные пищевым возбуждением животного.
Итак, восходящие активирующие влияния ретикулярной
формации па кору головного мозга_ неоднородны и _специ-~
фичны при реакциях различного биологического качества,
145
Какую долю в эти влияния вносят мотивационные центры
гипоталамуса, возбуждение которых специальными раздра-
жителями внутренней среды формирует биологическую
окраску поведенческих реакций?
Эксперименты, проведенные в нашей лаборатории (К. В.
Судаков, 1962, 1963; В. И. Вадиков и К- В. Судаков, 1968),
убедительно показывают, что любая мотивация строится на
основе восходящих активирующих влияний мотивационных
центров гипоталамуса на кору головного мозга.лЭти влия-
ния имеют характерное электроэнцефалографическое выра-
жение. ’ --------------------
А 5
2
4
6
I 50мк8
8
,г.рк I 50 mkS
Рис. 18. Избирательная активация элект-
роэнцефалограммы передних отделов ко-
ры головного мозга после суточного го-
лодания (Д) и у накормленной кошки
(Б). Уретановый наркоз.
/ — лобная правая; 2 — лобная левая; 3— сен-
сомоторная правая; 4 — сенсомоторная левая;
5 -^-теменная правая; 6 — теменная левая;
7—затылочная правая; 8 — затылочная левая область. На схеме (В) точками
обозначена зона регистрации высокочастотной низкоамплитудной электрической
активности в коре головного мозга у кошки после суточного голодания.
{46
Так, установлено, что у кошек после 1—2-суточного голо-
дания в передних отделах коры головного мозга даже под
уретановым наркозом наблюдается регионарная реакция ак-
тивации электроэнцефалограммы (рис. 18). Эта реакция
проявляется на фоне медленных высокоамплитудных коле-
баний в затылочной области, Принадлежность этой реакции
к пищевому возбуждению доказывают эксперименты с ис-
кусственным Кормлением таких животных путем введения
в ротовую полость и желудок жидких пищевых веществ или
раствора глюкозы непосредственно в кровь. Во всех этих
*
z
>8
f | ’ IWwf
^iV.wA^JVt1
/{Л
„—............
Рис. 19. Изменения электроэнцефалограммы голодной кошки после
внутриартериального введения глюкозы,
Введение 1 мл 40% раствора глюкозы обозначено стрелками, /—3 — то же, что на
рис, 18; 9 — правый медиальный гипоталамус,- /г? — левый медиальный глпотала*
Мус. Нижняя кривая — продолжение верхней.
147
случаях исходная активация коры головного мозга исчезает
и заменяется медленной высокоамплитудной активностью
(рис. 19).
Характерно, что у накормленных животных во всех отде-
лах коры головного мозга наблюдается медленная высоко-
амплитудная активность (см. рис. 17, Б). Аналогичные на-
блюдения были сделаны впоследствии и другими авторами
(А. Н. Бакурадзе, 1963; К. В. Шулейкина, 1967; В. А. Забо-
лотных, 1964; С. А. Чхенкели, 1963, 1968; Hockman, 1964;
Clemente a. oth., 1964).
Аналогичную активацию лобных отделов коры головного
мозга наблюдал А. И. Лакомкин (1966) и при питьевом воз-
буждении животных.
При более длительном голодании животного (например,
у кошек при 4-суточном голодании) наблюдается генерали-
зованная активация электроэнцефалограммы (А. А. Панфи-
лов и Т. Н. Лосева, 1966) (рис. 20, а).
Эти влияния избирательно блокируются введением ами-
назина, который, как указывает ряд авторов (Rothballer,
1956; В. Г. Агафонов, 1956; П. К. Анохин, 1962), преимуще-
ственно действует на активирующие аппараты ретикулярном
формации ствола головного мозга. Характерно, что регио-
нарные активирующие влияния на передние отделы коры
головного мозга после введения аминазина проявляются в
достаточно отчетливом степени (рис. 20, б).
Все это указывает на то, что генерализованные влияния
на кору головного мозга у животных при длительном голо-
дании определяются восходящими активирующими влияни-
ями ретикулярной формации ствола головного мозга.
При кратковременном голодании (сутки и менее) актива-
ция передних отделов коры головного мозга у кошек неред-
ко не выявляется. Однако она проявляется в своеобразной
форме в активности лимбических образований (амигдала,
гиппокамп). В амигдалярных образованиях при этом на-
блюдаются высокочастотные веретена, а в гиппокампе —
медленный упорядоченный ритм 4—6 колебаний в секунду.
Все эти особенности распространения пищевого возбуж-
дения на кору головного мозга отчетливо выявляются в
эксперименте с искусственным раздражением пищевого мо-
тивационного центра латерального гипоталамуса у кролика
через вживленные электроды.
Опыт состоял в том, что у накормленного кролика предва-
рительным раздражением латерального гипоталамуса полу-
чалась направленная пищевая реакция. После этого голову
148
кролика укрепляли в стереотаксическом аппарате и по ме-
ре постепенного увеличения силы раздражения латерально-
го гипоталамуса исследовали изменения электроэнцефало-
граммы лимбических образований, ретикулярной формаций
и коры головного мозга.
Рис. 20. Электроэнцефалограмма кошки после 4-суточного голодания
(а) и ее изменения после введения аминазина (б) (по А, А. Панфило-
ву и Т. Н. Лосевой, 1966).
1—8 то же, что на рис. 18; 9 — схема- в — восходящие активирующие влияния ги-
поталамических отделов пищевого центра на передние отделы коры головного
мозга и генерализованные влияния ретикулярной формации на кору головного
мозга у длительно голодавших животных; -з — устранение генерализованных восхо-
дящих влияний ретикулярной формации после введения аминазина.
На рис. 21 представлена динамика изменений электроэн-
цефалограммы у кролика в процессе увеличения силы раз-
дражения «центра голода» латерального гипоталамуса. Как
149
можно видеть, на кривой при раздражении латерального ги-
поталамуса при силе раздражающего тока 0,5 мка не отме-
чается изменений электрической активности в подкорковых
структурах и в коре головного мозга. При увеличении силы
раздражающего тока до 1 мка наблюдаются изменения
электрической активности типа десинхронизации в медиаль-
гб ГО.Змкд.! -22б(1^к.а)
A iV Ч Wa А V Л-V f/ jJ
7
^WjW.w*«*wwti^4
Tf iW^WW^Ww ^MWvMSK'^v4"***-
i ,(W»!WW®W:» ww^»V~AL^Wwim» ^ijVViANWWWyiW»'
Рис. 21, Динамика изменений электроэнцефалограммы у кролика при
увеличении силы раздражения пищевого центра латерального гипота-
ламуса.
/ — отметка времени — 1 секунда; 2 — величина раздражения (мка); 3 — сенсомо-
торная правая; 4 — сенсомоторная левая; 5 — затылочная правая; б — затылочная
левая; 7 — перегородка левая; 8 — амигдала левая; Р — гиппокамп левый; 10 — ре-
тикулярная формация среднего мозга левая. Раздражение гипоталамуса показано
толстой линией во II отведении; II — дыхание; 12 — реконструкция локализации
кончиков подкорковых электродов в данном опыте.
ной области перегородки и в амигдалярной области, упоря-
доченный ритм в гиппокампе и ретикулярной формации
среднего мозга, незначительные изменения электроэнцефа-
лографической активности в левой и правой сенсомоторной
области. Изменяется дыхание. При увеличении силы раздра-
жающего тока до 6 мка наблюдаются генерализованные из-
менения электроэнцефалограммы во всех представленных
структурах.
Таким образом, эти эксперименты указывают на то, что
при низких порогах раздражения латерального гипоталаму-
са изменения электрической активности прежде всего на-
блюдаются в подкорковых образованиях и особенно в лим-
бических структурах. Только при дальнейшем увеличении
150
силы раздражения центров латерального гипоталамуса из-
меняется электрическая активность и коры головного моз-
га. Эти изменения могут носить локальный, наблюдаясь
преимущественно в передних отделах коры головного мозга,
или генерализованный характер в зависимости от силы раз-
дражения пищевого центра гипоталамуса,
Наиболее интересным оказался тот факт, что именно та
сила электрического тока, которая при предварительном
тестировании вызывала у кролика целенаправленную пи-
щевую реакцию, вызывает изменения электрической актив-
ности ретикулярной формации среднего мозга и коры боль-
ших полушарий. Эта величина раздражающего тока уди-
вительно точно совпадала у каждого кролика, Этот факт
указывает, по-видимому, на то, что для возникновения на-
правленной пищевой деятельности необходима вполне опре-
деленная интенсивность восходящих активирующих влияний
пищевого центра гипоталамуса, при которой они распростра-
няются на кору головного мозга кролика.
Все эти наблюдения показывают, что мотивационное воз-
буждение строится на основе восходящих активирующих
влияний гипоталамических центров на лимбическую и но-
вую кору головного мозга. При этом зона распространения
этих вдаяний-^авдодт от силы исходного, даздражения-ЕИШъ
таламических центров внутренней.потребнортью организма.
Наблюдается, таким образом, градуальное включение все
I большего количества лимбических и корковых элементов
> в пищевую мотивацию в зависимости от степени исходного
голодания животного.
Первой инстанцией, куда адресуется возбуждение любо-
го мотивационного центра гипоталамуса, являются лимби-
ческие образования головного мозга. Последние объединя-
ют такие морфологические образования, как перегородку,
миндалину, гиппокамп, поясную извилину, свод головного
мозга и мамиллярные тела. Выход гипоталамических воз-
буждений на эти структуры головного мозга осуществляется
преимущественно через медиальный пучок переднего мозга,
t/ При усилении гипоталамического возбуждения последнее
через передние ядра таламуса начинает распространяться
и на передние отделы коры больших полушарий. Это
проявляется в виде десинхронизации спонтанной электро-
энцефалограммы в этих отделах коры головного мозга.
Такая активация коры головного мозга, вызванная голод-
ным возбуждением животного, выступает в наиболее отчет-
ливой форме у животных, находящихся под уретановым
151
наркозом, т. е, при блокаде активирующих влияний на кору
головного мозга, связанных с состоянием бодрствования.
При еще большем увеличении силы возбуждения гипота-
ламических центров (например, при длительном голодании)
возбуждение последних начинает вовлекать в свою дея-
тельность (по-видимому) через гипоталамо-тегмептальные
связи, активирующие аппараты ретикулярной формации
среднего мозга (рис. 22).
Рис. 22. Последовательные стадии вовлечения различных подкорковых
образований и коры головного мозга в восходящие влияния пищевого
центра латерального гипоталамуса при постепенном нарастании его
возбуждения.
I — при слабых степенях возбуждения латерального гипоталамуса его влияния
распространяются только на область перегородки; II—при увеличении силы
возбуждения латерального гипоталамуса его влияния распространяются на гиппо-
кампальную область; Ш — возбуждение охватывает все лимбические образования
и формирует ориентировочно-исследовательскую (О/5) реакцию животного;
IV — возбуждение выходит на кору больших полушарий, формирует пищевую мо-
тивацию и пищедобывательиую реакцию (ПР) животного, / — латеральный гипо-
таламус; 2 — перегородка; 3 — гиппокамп; 4 — миндалина; 5 — ретикулярная фор-
мация среднего мозга.
^Последняя оказывает на кору больших полушарий вос-
ходящие генерализованные активирующие влияния, форми-
руя тем самым состояние эмоционального напряжения.
Итак, мотивационное возбуждение характеризуется нали-
чием восходящих активирующих влияний на кору головного
мозгд^ Именно эти влияния и являются энергетической ос-
152
новой формирования целенаправленного поведения для
удовлетворения исходных потребностей организма.
Какие подкорковые образования участвуют в осущест-
влении этих влияний? Осуществляются ли они непосредст-
венно самими мотивационными центрами гипоталамуса или
опосредованно, через активирующие влияния ретикулярной
формации ствола головного мозга и таламуса. Наконец,
где ведущий пункт этих влияний?
ПЕЙЦМЕКЕРНАЯ ТЕОРИЯ МОТИВАЦИОННОГО
ВОЗБУЖДЕНИЯ
Исследования, проведенные на голодных животных, по-
казывают, что у таких животных наряду с активацией коры
головного мозга наблюдается аналогичное возбуждение в
лимбических структурах, в латеральных и медиальных отде-
лах гипоталамуса, медиальных ядрах таламуса, а также
в структурах ретикулярной формации ствола головного моз-
га (рис. 23, А),
Электрическая активность этих подкорковых структур
почти точно соответствует активности передних отделов ко-
ры головного мозга. Все это указывает на то, что голодное
мотивационное возбуждение охватывает большой комплекс
корково-подкорковых образований. После кормления эта
активация исчезает на всех уровнях (рис. 23. Б).
Для того чтобы выяснить, какие подкорковые образова-
ния являются ведущими в механизме восходящих активиру-
ющих влияний на кору головного мозга у голодных живот-
ных, в специальной серии экспериментов производилась
функциональная блокада различных подкорковых образова-
ний с помощью анода постоянного тока. Известно, что мест-
ное приложение анода к нервной ткани вызывает снижение
ее возбудимости и проводимости,— явление, описанное под
названием анэлектротона.
Естественно, что прежде всего было испробовано действие
анода постоянного тока на пищевые мотивационные цент-
ры гипоталамуса, т. е. на «центр голода» в латеральной ги-
поталамической области и «центр насыщения» в области
вентро-медиальных ядер гипоталамуса. Выбор этих образо-
ваний был не случайным, поскольку, как уже указывалось
выше, именно в этих отделах гипоталамуса расположены
клетки, чувствительные к изменению содержания глюкозы
в крови, и именно эти центры при их электрическом раздра-
жении у кроликов вызывали активацию лимбических обра-
зований и коры головного мозга.
153
154
5
ляркая формация; // — правый
Рис, 23, Электрическая, активность
коры головного мозга, гипотала-
муса, таламуса и ретикулярной
формации среднего мозга
А — у голодной и Б — у накормленной
кошки под уретановым наркозом; / —
лобная правая; 2 — лобная леная; 3 —
сенсомоторная правая; 4 — сенсомотор-
ная левая; 5 —затылочная правая; 6 —
затылочная левая область; 7 —правый
медиальный таламус; 3 — левый лате-
ральный гипоталамус; 9 — правая рети-
кулярная формация; 10—левая ретнку-
гипоталамус. На препаратах стрел-
медиальный
ками показана локализация кончиков электродов.
155
Опыты показали, что такое'действие анодом постоянного
тока у голодных животных на латеральный гипоталамус или
же особенно на область вентро-медиальных ядер устраняло
активацию в подкорковых образованиях (передние ядра та-
ламуса, ретикулярная формация ствола головного мозга),
а также в коре головного мозга ипсилатерального по от-
ношению к пункту аподизации полушария. При двусторон-
ней аподизации одноименных отделов пищевых центров ги-
Д Б
Рис. 24. Изменения
электрической актив-
ности коры головного
мозга голодной кош-
ки после поляризации
медиальных отделов
гипоталамуса анодом
постоянного тока.
/— лобная правая; 2 —
лобная левая; 3 — сенсо-
моторная правая; 4 •—
сенсомоторная левая; 5 —
теменная прааая; б—Те*
менная левая; 7 — За-
тылочная правая; & —
затылочная. левая; 9 —
правый медиальный ги-
поталамус; 10 — левый
медиальный гипотала-
мус; Д — фон: Б — пос-
ле поляризации обоих медиальных гипоталамусов анодом постоянного тока
(0,5 ма—30 сек,); В — локализация кончиков электродов,
156
поталамуса эффект проявляется в обоих полушариях. Во
всех этих отделах вслед за анодной поляризацией гипотала-
муса возникала медленная высокоамплитудная электриче-
ская активность (рис. 24). Таким образом, локальным дей-
ствием анода постоянного тока на пищевые центры гипота-
ламуса удавалось устранить пищевую активацию у голод-
ных животных на всех уровнях головного мозга.
То, что такое воздействие было локальным, подтверждают
контрольные эксперименты, в которых аналогичное действие
анода постоянного тока той же силы на соседние области
гипоталамуса не приводило к устранению «голодной» акти-
вации коры головного мозга.
Поляризация медиальных отделов переднего таламуса
у голодных животных устраняла только активацию перед-
них отделов коры ипсилатерального полушария головного
мозга. В гипоталамических центрах и в ретикулярной фор-
мации при этом сохранялась реакция активации электро-
энцефалограммы.
Последующее действие анода постоянного тока на меди-
альные отделы гипоталамуса немедленно устраняло актива-
цию во всех отделах головного мозга (рис. 25).
Аноднческая поляризация ретикулярной формации не
сказывалась на электрической активности гипоталамуса, та-
ламуса и передних отделов коры головного мозга у живот-
ных после 1—2-суточного голодания. У длительно голодав-
ших животных такое воздействие вызывало только устране-
ние генерализованной активации коры в теменно-затылоч-
ных ее отделах.
Все эти опыты указывают на ведущую роль гипоталами-
ческих пищевых центров в механизме формирования восхо-
дящих активирующих влияний на кору головного мозга у го-
лодных животных.
Наряду с восходящими активирующими влияниями на ко-
ру головного мозга эти же центры оказывают активирующее
влияние на лимбические структуры и ретикулярную фор-
мацию ствола головного мозга. Степень этих влияний раз-
лична у животных с разной степенью выраженности пище-
вой мотивации. При кратковременном голодании возбужде-
ние гипоталамических пищевых центров распространяется
на лимбическую структуру, а при длительном голодании —
и на ретикулярную формацию ствола головного мозга.
В зависимости от силы этих влияний последние структуры
проявляют свои специфические восходящие активирующие
влияния на кору головного мозга.
157
/ ........ ..................................I
7 ,w~—.........................»,ч.>>1м|>|1>.и1.—i,.,w^wl .,i4l,|^y.
3 ..-.^V>miii«ii«MMi4iii hi,I ^^>ii»y»iitRw>iw»t*>wiW*'«|i<i
4 Ml. «^^МЙММММ***Д| II ,,НМ| I
7 *Л**Л6*
ч АЛллУ1^к(^(пМ1|Л0'#*»«|**ЧМ«^|*«‘М**МЛ*Ч1^
| Ann:) hn А ил Г MH |
, r— |f;iYjy:nra^pWBeBl.dLiiiii^^^^^^^^
a *----^д|11И111НЦаММ11М11М1М||1Ш1М*11««
|^1чм4»>|****11^1' *'i '** >i*Mi*i"
*11 I < Pl *** l«* *|*,„||, ч* 1*4*1 —
'чННН“4НН^***‘
| Анод //nA на Г МЛ
|j'4***
рм^4г*4^1ПГ4л)^м4<1йн44'4**^^
..............................4rrt 'W*H ^,^!>
\50mhS
Hy 1 '\ Т" lri,’'i'1 "Hilf v -
>win«»!>!>,,l>^r.>4.».im*n,M|W*W‘l УНЕЙР1*
Sl'Ia
'<• л iiwaiwi 4r**i n»' i' 4w i"4lfp'‘Ччч
Рис. 25. Динамика изменений электрической активности коры головного
мозга и различных подкорковых структур кошки после коагуляции ядер
правого медиального таламуса и левого медиального гипоталамуса (мо-
мент коагуляции показан стрелками).
f, 2, 3, 4, 7, 8—то же, что на рис.ь 24; 5 —правый медиальный таламус;
6 — левый медиальный гипоталамус; 9 — правая ретикулярная формация; // — ле-
вая ретикулярная формация; 12 — локализация копчиков электродов (показана
стрелками). Средняя и нижняя кривые — продолжение верхней кривой.
Есть все основания полагать (Kowamura a. oth., 1961;
Kawamura, Oshima, 1962), что активация передних отделов
коры головного мозга у голодных животных осуществляется
преимущественно через лимбические образования переднего
мозга. Вот почему эти влияния не снимаются после выклю-
чения аминазином адренергического субстрата ретикуляр-
ной формации ствола головного мозга и устранения ее гене-
рализованных восходящих активирующих влияний на кору-
головного мозга.
По-видимому, аналогичны механизмы формирования и
других биологических мотиваций.
Все изложенное выше заставляет думать о том, что моти-
вационным центрам гипоталамуса принадлежит особая, ве-
дущая роль в формировании корково-подкорковых комплек-
сов мотивационных возбуждений (рис. 26).
В общем виде механизм формирования мотивации очень
напоминает возникновение возбуждения в сердечной мыш-
це, где в синусовом узле имеется специальный «задаватель
ритма» сердечных сокращений — «pacemaker».
159
Аналогичным образом центры гипоталамуса являются
своеобразными «задавателями ритма» («пейцмекерами»)
каждого мотивационного возбуждения, возникающего на ос-
нове специфической .внутренней потребности организма.
Разрушение или функциональное выключение этих центров
ликвидирует соответствующее мотивационное возбуждение
на любом уровне головного мозга и тем самым прекращает
целенаправленное поведение. И это действительно наблю-
дается, если вспомнить классические эксперименты Brobeck
Рис. 26. Схема восходящих активирующих влияний гипоталамических
отделов пищевого центра на кору головного мозга у голодных живот-
ных (а) и их устранение после кормления (б).
/ — действие «голодной» крови; 2 — действие афферентных импульсаций; 3 —ла-
теральный гипоталамус; 4 — таламус; 5 — ретикулярная формация среднего мозга.
и Anand с двусторонним разрушением у животных лате-
ральных областей гипоталамуса. Такие животные при об-
щем внешнем благополучии даже после длительного голо-
дания совершенно утрачивали всякий интерес к пище.
Такой «пейцмекер» каждой мотивации в высшей степени
чувствителен к только определенной форме нервного и гу-
морального раздражения и возбуждается при изменении
уровня в крови только того химического вещества, которое
оказывает специфическое влияние на его метаболизм. По
мере того, как содержание этого химического вещества
в крови снова восстанавливается в пределах физиологиче-
ской нормы, возбуждение гипоталамического «пейцмекера»
мотивации прекращается.
Все это позволило П. К- Анохину (1968) сформулировать
«пейцмекерную» теорию мотивации.
Согласно этой теории, именно гипоталамические отделы
головного мозга приобретают значение «пейцмекеров» мо-
160
тивациоиных возбуждений, поскольку к ним адресуется
нервная и гуморальная сигнализация о различных внутрен-
них потребностях организма.
Все это определяет важнейшие свойства этих центров,
их чрезвычайную специфичность и высо-
кий уровень метаболизма по сравнению с други-
ми компонентами центрального мотивационного возбуж-
дения.
Эта специфика определяется уже тем, что каждая моти-
вация определяется специфическими изменениями в содер-
жании того или иного жизненно важного вещества в орга-
низме. Кроме того, как указывалось выше, клетки каждого
мотивационного центра гипоталамуса используют в своем
метаболизме только специальные вещества. Вот почему
это возбуждение не блокируется наркотиком уретаном и спе-
цифическим адренолитическим веществом — аминазином.
Отсюда становится понятным, что каждое мотивационное
возбуждение, несмотря на принципиальную общность их
нейрофизиологических механизмов, строится на основе
специфических восходящих активирующих
влияний гипоталамических центров на дру-
гие подкорковые образования и на кору го-
ловного мозга.
В этом мы имеем еще один пример, подтверждающий
концепцию П. К. Анохина о множественном и специфиче-
ском характере восходящих активирующих влияний подкор-
ковых образований па кору головного мозга.
Именно эти восходящие активирующие влияния возбуж-
денных определенной внутренней потребностью организма
гипоталамических центров на старую и новую кору голов-
ного мозга и определяют по существу возникновение состо-
яния центрального мотивационного возбуждения.
Таким образом, мотивации представляют собой такие со-
стояния организма, при которых за счет восходящих акти-
вирующих влияний избирательно чувствительных к той или
иной потребности гипоталамических центров организуются
системные объединения корково-подкорковых аппаратов,
направляющие животное на удовлетворение этой потребно-
сти.
Из этого следует, что главным свойством мотивационного
возбуждения является его высокая побудительная сила. По
сути дела любая мотивация определенным образом «заря-
жает» организм, направляя его поведение на удовлетворе-
ние возникшей потребности (рис. 27).
6 Заказ 5074
161
Каждое мотивационное возбуждение любого биологиче-
ского качества характеризуется такой же системой избира-
тельно объединенных корково-подкорковых элементов.
Однако в каждом мотивационном возбуждении ведущая
роль принадлежит своим специфическим центрам гипотала-
мической области. Периодически возбуждаясь под влиянием
специфических нервных и гуморальных раздражителей, эти
центры через свои обширные морфологические связи пропз-
Рис. 27. Формирующиеся на основе гуморальных (1) и нервных (2) фак-
торов восходящие активирующие влияния мотивационных гипоталами-
ческих центров на ретикулярную формацию и кору головного мозга
создают основу целенаправленного поведения, приводящего к удовлетво-
рению исходной потребности.
водят в каждом случае избирательное вовлечение подкорко-
вых и корковых образований для построения специальных
форм поведения.
Естественно спросить, как же многочисленные специфиче-
ские влияния мотивационных центров гипоталамуса распре-
деляются на клетки коры головного мозга? Иными слова-
ми, каким образом корковые клетки перерабатывают каж-
дое приходящее к ним из подкорковой области мотива-
ционное возбуждение и в этом потоке множественных при-
ходящих к ним возбуждений строят соответствующей
направленности поведение?
Однако прежде чем ответить на эти вопросы, разберем,
в чем же проявляется специфичность восходящих активиру-
ющих влияний, лежащих в основе любого мотивационного
возбуждения, и чем она определяется.
162
ХИМИЧЕСКАЯ СПЕЦИФИКА МОТИВАЦИОННОГО
ВОЗБУЖДЕНИЯ.
ИЗБИРАТЕЛЬНОЕ ОБЪЕДИНЕНИЕ МОТИВАЦИЕЙ
КОРКОВО-ПОДКОРКОВЫХ АППАРАТОВ
В последние годы все больше экспериментов указывает
на то, что различные мотивации животных избирательно по-
ражаются при введении им раличных фармакологических
веществ, особенно веществ с так называемым психотроп-
ным действием. Было обнаружено, что некоторые наркоти-
ческие вещества искусственно устраняли различные моти-
вации, приводя примерно к тому же эффекту, как и нату-
ральное подкрепление. Congen (1949) обнаружил снижение
голода и страха у крыс под действием алкоголя.
Подобный эффект наблюдали Masserman и Jum (1946),
которые выявили, что в оборонительной ситуации кошки
предпочитают молоко с алкоголем обычному молоку. Gantt
(1940) наблюдал, что алкоголь снижал у собак половые мо-
тивации. Примерно такое же действие оказывает морфин.
Seevers (1936), Tatum и др. (1920), И. В. Стрельчук (1949)
обнаружили тормозящее действие морфина на половые вле-
чения, голод и жажду. Такая избирательная чувствитель-
ность различных мотиваций к определенным фармакологи-
ческим веществам открывает простор для исследований в об-
ласти экспериментальной психофармакологии различных
мотиваций. Уже сейчас мы можем наблюдать случаи изби-
рательной блокады оборонительных мотиваций у животных
трансквилизаторами типа аминазина (В. Гавличек, А. И.
Шумилина, Ю. А. Макаров). О том же свидетельствует и
практика применения нейролептических веществ в психиа-
трической клинике. Все более интенсивно разрабатываются
препараты, избирательно устраняющие мотивации голода.
Все эти наблюдения послужили для нас исходной предпо-
сылкой для исследования химических механизмов восходя-
щих активирующих влияний подкорковых мотивационных
центров на кору головного мозга, лежащих в основе различ-
ных мотиваций.
Опыты проводились следующим образом.
У голодных кошек под уретановым наркозом на фоне ре-
гистрации высокочастотной низкоамплитудной электри-
ческой активности в передних отделах коры головного моз-
га наносилось ритмическое раздражение электрическим то-
ком седалищного нерва (К. В. Судаков и Ю. А. Фадеев,
1963). Это вызывало генерализованную активацию всей ко-
6*
163
ры головного мозга, включая и передние ее отделы, уже акти-
вированные голодным состоянием. Затем животным инъеци-
ровали аминазин в дозе не выше 5 мг/кг. Через некоторое
время снова осуществляли раздражение седалищного нер-
ва. Однако получить генерализованную активацию коры го-
ловного мозга в этих условиях при той же силе раздраже-
ния уже оказалось невозможным. Вместе с тем «голодная»
активация передних отделов коры головного мозга у этих
животных сохранялась в прежнем виде (рис. 28). Все это
*''*** .........................
>*>«мм*ммим*ммр>
7 ................ -..............
..1 i.....................................।......।...............
J в 50 гц.......................3650 гц
Рис. 28. Изменения электроэнцефалограммы голодной кошки под
уретановым наркозом при раздражении седалищного нерва.
Л — фон; Б — после введения аминазина (5 мг/кг). Электрическое раздражение
седалищного нерва показано стрелками; /—8 — то же, что и на рис. 18; 9 — отмет-
ка времени 1 секунда; а — два восходящих активирующих влияния на кору го-
ловного мозга. Толстые стрелки — восходящие активирующие влияния пищевых
центров гипоталамуса на передние отделы головного мозга, обусловливающие «го-
лодную» активацию. Тонкие стрелки — генерализованные восходящие активирую-
щие влияния ретикулярной формации, обусловливающие «болевую» активацию;
б — избирательное устранение «болевой» активации аминазином; ГК — «голодная»
кровь; БВ — болевое возбуждение.
указывало на то, что химические механизмы восходящих
активирующих влияний в случае этих двух разных по био-
логическому качеству состояний различны. Аминазин как
адренолитическое вещество избирательно блокировал меха-
низмы «болевой» активации и практически не оказал дей-
164
ствие на активацию передних отделов коры, обусловленную
состоянием голода.
На основании этих экспериментов возник естественный
вопрос: каковы химические механизмы голодного мотива-
ционного возбуждения?
Специальные эксперименты (К. В. Судаков, 1966) показа-
ли, что активация передних отделов коры головного мозга
2""
б
| 2мл 0,1% раствор атропина.
3
4
I Зв 80гц 0,1 мсек 1 ООмн.Ь
5 'vmnrmvwtf¥infuWinnr^^ д,л^д'пгт"зтлтлгпгп"л!"Г1Р.'ги "i« гглг."^."лттт’Ж"^гтпя^
Рис. 29. Изменения электроэнцефалограммы голодной кошки после
введения атропина.
Л “ исходный фон; Б — после подкожного введения 2 мл 0,1% раствора атропина.
В правой части рисунка показан эффект раздражения седалищного нерва на фоне
действия атропина. Момент раздражевия— прекращение отметки времени, / — сен-
сомоторная правая область; 2 — сенсомоторная левая область; 3 — затылочная
правая область; 4 — затылочная левая область; 5 — отметка времени г секунда.
На схемах: восходящие активирующие влияния на кору головного мозга, а — «го-
лодная» активация; б — устранение «голодной» активации атропином и сохранение
«болевой» активации.
у голодных животных устраняется после введения холино-
литических веществ, таких, как атропин и амизил (рис. 29).
После внутримышечного введения этих веществ в электро-
энцефалограмме голодных животных появляются медлен-
ные высокоамплитудные колебания. Однако раздражение
седалищного нерва на этом фоне по-прежнему вызывает ге-
нерализованную активацию коры головного мозга.
165
Все это указывало на холинергическую природу восхо-
дящих активирующих влияний на кору головного мозга у
голодных животных.
Однако на основании этих опытов еще нельзя было ска-
зать, на какие звенья корково-подкоркового комплекса го-
лодного мотивационного возбуждения оказывали действие
холинолитические вещества. Для выяснения этого вопроса
Рис. 30. Устранение активации коры головного мозга у голодной кошки
после локальной аппликации 0,5 % раствора амизила на левую сенсомо-
торную область (по А. И. Туре.чхо, 1965).
/1 — фоновая активность: Б — возрастание амплитуды в левой сенсомоторной об-
ласти через 18 минут после аппликации амизнла. Раздражение седалищного
нерва на этом фоне вызывает генерализованную активацию всей коры головного
мозга; В — электроэнцефалограмма через 23 минуты после аппликации; отведения:
J —севсомоторная левая область; 2 — сенсомоторвая правая область; 3~ темен-
ная левая область; 4 — теменная правая область; 5 —затылочная левая область;
6 — затылочная правая область. На схеме ввизу черные точки — распространение
«голодной» активации по коре (а). Она отсутствует в пувкте аппликации ами-
зила (б). Крестики — «болевая* активация коры головного мозга. Она проявляет-
ся в пункте аппликации амизила (в).
в экспериментах А. И, Туренко (1964) осуществлялась ап-
пликация холинолитических веществ непосредственно на пе-
редние отделы коры головного мозга, активированные го-
лодным состоянием.
Опыты показали, что аппликация на передние отделы ко-
ры одного из полушарий, активированного состоянием го-
166
лода, фильтровальной бумажки размером 2X2 мм, смочен-
ной 0,5% раствором амизила или 0,1% раствором атропина,
вызывала в пункте аппликации устранение этой активации.
В этом пункте коры появлялась медленная высокоампли-
тудная активность (рис. 30).
Характерно, что в ответ на раздражение у этого же жи-
вотного седалищного нерва во всей коре головного мозга,
включая пункт аппликации холинолитических веществ, на-
блюдалась генерализованная активация электроэнцефало-
граммы. Все это указывало на то, что холинолитические ве-
щества избирательно блокировали корковые механизмы «го-
лодной» активации н не оказали заметного влияния на ме-
ханизмы болевой активации коры головного мозга.
Таким образом, было установлено, что голодное мотива-
ционное возбуждение при средних сроках голодания, при
которых наблюдается активация только передних отделов
коры головного мозга, избирательно строится на основе хо-
линергических химических механизмов. Это же отмечает
в своих опытах Marczyncki (1969).
Следующие эксперименты (А. А. Панфилов , 1966) пока-
зали, что и гипоталамические пищевые центры в построении
своих восходящих активирующих влияний на кору головно-
го мозга у голодных животных используют также холинер-
гические механизмы.
Наиболее показателен в этом отношении следующий
эксперимент. Кошке в область латерального гипоталамуса
вживлялась специальная канюля, через которую экспери-
ментатор в любой момент имел возможность с помощью спе-
циального устройства ИЕЕъецировать малое количество хими-
ческого вещества. Конструкция канюли позволяла избежать
механического эффекта инъекции, так как канюля состояла
из двух вставленных одна в другую полых игл. Инъекция
производилась через внутреннюю иглу, копчик которой рас-
полагался на 0,1 мм выше кончика наружной иглы. Таким
образом, инъецируемое вещество как бы апплицировалось
к тем отделам головного мозга, в которых располагался
кончик наружной иглы. Инъекция через такую кянюлео
голодным кошкам, находящимся под уретановым наркозом,
в область литерального гипоталамуса 0,00025% раствора
амизила или атропина устраняла в ипсилатеральном полу-
шарии передних отделов коры ранее наблюдавшуюся там
реакцию активации электроэнцефалограммы (рис. 31).
Характерно, что и в этом случае действие холинолити-
ческих веществ непосредственно на гипоталамус не препят-
167
ствует реакции десинхронизации, возникающей у тех же жи-
вотных в ответ на ноцицептивное раздражение. Крайне важ-
но отметить, что эффект устранения «голодной» активации
передних отделов коры головного мозга наблюдался только
А 6 В
/ — 60гц,
Рис. 31. Устранение восходящих активирующих влияний на кору го-
ловного мозга у голодной кошки после локальной инъекции 0,03 мл
0,00025% раствора амизила в область латерального гипоталамуса (по
А. А. Панфилову, 1966).
Д — исходная электроэнцефалограмма кошки после суточного голодания; Б из-
менение электроэнцефалограммы преимущественно в ипсилатеральном полуша-
рии после инъекции амизила в левый латеральный гипоталамус. На этом фоне
раздражение седалищного нерва вызывает обычную генерализованную активацию
коры головного мозга; В — электроэнцефалограмма через 10 минут после инъек-
ции амизила в гипоталамус, / — отметка раздражения; 2 — Сенсомоторная левая
область; 3 — сенсомоторная правая область; 4 — теменная левая область; 5 — те-
менная правая область: в — затылочная левая область; 7 — затылочная правая
область; 8 — латеральный гипоталамус; 9 — отметка времени I секунда; Г — лока-
лизация кончика канюли в области левого латерального гипоталамуса.
168
при инъекции холинолитических веществ в пищевые центры
латерального или вентро-медиального отделов гипоталаму-
са. Инъекция этих же веществ в другие отделы гипотала-
мической области не вызывала подобного эффекта.
Все это указывало на то, что только пищевое мотивацион-
ное возбуждение избирательно строится преимущественно
на холинергических механизмах.
Дело, однако, существенно изменяется при увеличении
сроков голодания животного. У длительно голодавших жи-
вотных активация коры головного мозга уже не устраняет-
ся аппликацией на кору холинолитических веществ. В этом
случае вовлечение корковых нейронов в мотивационное воз-
буждение происходит уже не только на основе холинергиче-
ских механизмов, но к этому все в большей степени присо-
единяются другие (по-видимому, адренергические) не-
блокируемые холинолитическими веществами механизмы.
Последние, по-видимому, определяются восходящими гене-
рализованными активирующими влияниями стволовой ре-
тикулярной формации на кору головного мозга.
Такую же особенность перемены химического знака кор-
ково-подкорковых взаимоотношений при длительном голо-
дании у голубей на преимущественно адренергический отме-
тили в своих исследованиях Г. Г. Гасанов и В В. Рубцова
(1969).
Следует обратить внимание еще па одно обстоятельство.
Химическая чувствительность в различных частях корково-
подкоркового комплекса мотивационного пищевого возбуж-
дения различна. Наибольшей чувствительностью к соответ-
ствующему раздражителю обладают центры гипоталамуса.
Для того чтобы устранить восходящие активирующие влия-
ния гипоталамических центров на кору головного мозга
у голодных животных, необходимо инъецировать в эту об-
ласть раствор амизила в концентрации в 10 000 раз ниже,
чем при прямой аппликации этого вещества непосредственно
на кору головного мозга.
Все это, по-видимому, связано с тем, что интенсивность
метаболических процессов в данных центрах наиболее вы-
сокая.
Итак, пищевое мотивационное возбуждение голодных жи-
вотных, выражающееся в регионарной активации передних
отделов коры головного мозга, на всех уровнях центральной
нервной системы избирательно строится на основе специфи-
ческих холинергических механизмов. Такое избирательное
химическое объединение корково-подкорковых комплексов
169
степени специфическим химизмом пищевых центров гипота-
ламуса. Именно особенность метаболизма этих центров оп-
ределяет качество химических процессов, используемых
всеми другими отделами центральной нервной системы, объ-
единенными в мотивационное возбуждение данной биологи-
ческой модальности. Разрушение или химическая блокада
этих центров гипоталамуса является причиной прекращения
соответствующих химических реакций всей системы моти-
вационного возбуждения данной биологической модаль-
ности.
Каждый мотивационный центр гипоталамуса имеет свою
подчеркнутую химическую специфику, которая определяет-
ся прежде всего его избирательным использованием в сво-
ем метаболизме специальных химических веществ. И эта
подчеркнутая химическая специфика мотивационных цент-
ров гипоталамуса сохраняется на всех уровнях, куда адре-
суются его восходящие активирующие влияния.
Поскольку химизм каждого мотивационного центра гипо-
таламуса чрезвычайно специфичен, из этого следует, что
каждое мотивационное возбуждение избирательно строится
на основе специфических химических механизмов. Это же
определяет и специфичность восходящих активирующих
влияний подкорковых мотивационных центров на кору го-
ловного мозга при мотивациях различного биологического
качества.
Это положение является чрезвычайно важным. Оно позво-
ляет понять механизм участия клеток коры головного мозга
в различных мотивационных возбуждениях. В самом деле,
даже одна и та же нервная клетка коры головного мозга
способна участвовать в различных реакциях за счет различ-
ных химических механизмов. И не только участвовать, но и
формировать соответствующую форму поведения животного.
ОРГАНИЗАЦИЯ ВОСХОДЯЩЕГО МОТИВАЦИОННОГО
АКТИВИРУЮЩЕГО ВЛИЯНИЯ НА КОРУ
ГОЛОВНОГО МОЗГА
На какой же основе происходит вовлечение корковых
клеток в мотивационное возбуждение?
Такое функциональное объединение происходит строго
избирательно на основе специфических механизмов
восходящих активирующих влияний мотивационных цент-
ров гипоталамической области,
(70
Это было показано следующими экспериментами (К- В.
Судаков, 1966; К. В. Судаков и Э. М. Набиль, 1966). Для
того чтобы исследовать, как и к каким синаптическим обра-
зованиям коры головного мозга адресуются восходящие по-
токи возбуждений гипоталамических пищевых центров у го-
лодных животных, в качестве показателя был использован
соматосенсорный вызванный потенциал. Этот вызванный
потенциал регистрировали в коре головного мозга в ответ на
одиночное раздражение седалищного нерва электрическим
током. Фокус максимальной активности вызванного потенци-
ала находился в области переднего края задней крестовид-
ной извилины. Опыты проводили на кошках, находящихся
под уретановым наркозом.
В настоящее время большинство исследователей считают,
что положительная фаза первичного ответа вызванного по-
тенциала отражает возбуждение аксосоматических синап-
сов III—IV слоя коры. Считается, что мозговая ткань пред-
ставляет собой объемный проводник второго рода, причем
верхние слои коры рассматриваются как правильно ориен-
тированные дипольные организации. Вот почему возбужде-
ние, всякий раз возникающее у одного конца диполей в III—
IV слое коры, создавая здесь относительную электроотрица-
тельность, регистрируется у другого их конца на поверхно-
сти коры как противоположное по знаку положительное ко-
лебание потенциала (Eccles, 1951).
Однако по поводу механизма происхождения отрицатель-
ной фазы первичного ответа вызванного поверхностного по-
тенциала коры до настоящего времени не существует единого
мнения. Некоторые исследователи (Bischop a. Clare, 1953;
Li a. oth., 1956; Chang, 1959) рассматривают отрицательную
фазу первичного ответа, регистрируемую с поверхности ко-
ры, как результат электротонического распространения воз-
буждений, первично возникших в клетках III—IV слоя коры,
по их дендритам до поверхностного плексиморфного слоя.
Согласно этой точке зрения, отрицательная фаза первично-
го ответа генетически связана с положительной фазой.
Другие авторы считают, что поверхностный отрицательный
потенциал образуется благодаря приходу к поверхностным
слоям коры возбуждений по коллатералям аксонов уже воз-
бужденных клеток III—IV слоев коры через цепь вставоч-
ных нейронов (Marshall a. oth., 1941; Eccles, 1951; А. И.
Ройтбак, 1955, и др.).
Это общепринятое представление о природе первичного
ответа вызванных потенциалов в коре головного мозга со-
171
держит в себе два исходных принципиально важных поло-
жения; 1) афферентное возбуждение приходит в кору го-
ловного мозга по одному и тому же структурному пути и
является качественно гомогенным; 2) отрицательная фаза
вызванного ответа является электрическим следствием то-
го же процесса постсинаптического-возбуждения в клетках
III—IV слоя, который еще раньше был причиной возникно-
вения положительной фазы вызванного потенциала.
Наряду с этим в последнее время появилось принципи-
ально новое представление о происхождении в коре голов-
ного мозга вызванного потенциала. Это представление было
выдвинуто и экспериментально обосновано в лаборатории
П. К. Анохина.
Исследования этой лаборатории показали, что вызванный
потенциал, регистрируемый с поверхности коры головного
мозга, представляет собой результат множественных несин-
хронных восходящих влияний, поступающих к синапсам
корковых клеток по различным каналам от различных под-
корковых образований.
Представление П. К- Анохина о множественном характе-
ре восходящих активирующих влияний подкорковых обра-
зований на кору головного мозга при реакции различного
биологического качества позволяет с принципиально новых
позиций рассматривать происхождение различных фаз кор-
ковых вызванных потенциалов. Опыты Ф. Ата-Мурадовой
(1960, 1963), Лю Джуан-Гуй (1960), В. А. Полянцева и
М. В. Сербиненко (1962), Сунь-Вень (1964) и др. показали,
что различные фазы поверхностного вызванного потенциа-
ла определяются возбуждением различных корковых и под-
корковых образований. Таким образом, корковый вызван-
ный потенциал всегда является результатом взаимодействия
разнородных по качеству н происхождению восходящих
влияний на кору головного мозга.
Опыты Ф. Ата-Мурадовой убедительно показали, что от-
рицательная фаза первичного ответа в своем происхождении
не зависит от возбуждений, формирующих положительную
фазу. Так, у новорожденных кроликов до 6—8-го дня онто-
генетического развития в коре головного мозга в ответ на
раздражение седалищного нерва вообще не удается зареги-
стрировать первичноположительную фазу вызванного отве-
та, в то время как первичноотрицательная фаза выражена
отчетливо.
Морфологические наблюдения (Ф. Ата-Мурадова и И. А.
Чернышевская, 1961, 1966) показывают, что в это время
172
в коре созрели только аксодендритные синапсы плексиморф-
ного слоя.
Только с б—8-го дня онтогенетического развития в ответ
на раздражение седалищного нерва в коре появляется
и первичноположительная фаза вызванного ответа. Морфо-
логически при этом отмечается созревание аксосоматиче-
ских синапсов в IV слое коры.
Наряду с этим было обнаружено, что даже положитель-
ная и отрицательная фазы первичного ответа скрывают за
собой еще несколько различных возбуждений, которые мо-
гут быть выявлены при специальных химических воздейст-
виях.
Так, в опытах Ф. Ата-Мурадовой (1963) было показано,
что при аппликации на кору специальных веществ (ГАМК
и др.) удается расщепить обычно регистрируемую первую
отрицательную фазу и выявить одновременно с ней проте-
кающую вторую отрицательную фазу.
На основании этого было выдвинуто представление, со-
гласно которому отрицательная фаза и вторичные ответы
вызванного потенциала обусловливаются в коре множест-
венными восходящими влияниями подкорковых образова-
ний, которые адресуются к аксодендритическим синапсам
поверхностного плексиморфного слоя. В то же время по-
ложительная фаза первичного ответа определяется возбуж-
дениями аксосоматических синапсов на нейронах IV слоя
коры вследствие прихода возбуждений по классическим
лемнисковым путям (IL K- Анохин, 19626; Ф. Ата-Мурадо-
ва, 1960).
Убедительным доказательством этого положения явились
еще одни эксперименты Ф. Ата-Мурадовой (1966). Исполь-
зуя метод локального послойного обогревания коры голов-
ного мозга, ей удалось обнаружить следующие интересные
факты.
По мере того как под нембуталовым наркозом прогре-
вание распространялось от поверхностных в более глубо-
кие слои коры, наблюдалась характерная динамика изме-
нения различных фаз вызванного потенциала, возникающе-
го в ответ на раздражение седалищного нерва. Прежде все-
го исчезала отрицательная фаза первичного ответа, затем
деформировался и исчезал вторичный положительный ответ
Форбса и, наконец, позднее всего исчезал первичный поло-
жительный потенциал. После прекращения обогревания ко-
ры восстановление фаз вызванного потенциала шло в об-
ратном порядке.
173
Все эти эксперименты в совершенно отчетливой форме по-
казывают, что формирование различных фаз коркового
вызванного потенциала определяется возбуждением различ-
ных специфических синаптических организаций различных
слоев коры головного мозга. По мере того как прогревание
распространяет свое действие от поверхностных к более глу-
боким слоям коры головного мозга, происходит функцио-
нальная блокада различных синаптических организаций.
Естественно, что прежде всего блокируются аксодендритиче-
ские синапсы поверхностного плексиморфного слоя коры и
их блокада и определяет исчезновение первичной отрица-
тельной фазы вызванных потенциалов. Исчезновение вто-
ричного положительного разряда Форбса, по-видимому, свя-
зано с блокадой еще ниже расположенных (возможно, на
середине дендритов клеток III—IV слоя) синапсов. Нако-
нец, при распространении тепловой блокады на III—IV
слой коры и блокаде аксосоматических синапсов этих слоев
исчезает первичноположительный потенциал.
В связи со всеми изложенными выше исследованиями ста-
новится ясным, что в настоящее время открывается широ-
кая перспектива изучения синаптических организаций коры
головного мозга по изменению конфигурации корковых вы-
званных потенциалов.
Замысел эиопериментов, о которых говорилось выше, со-
стоял в следующем.
Состояние голода на основе восходящих активирующих
влияний подкорковых образований мобилизует определенные
корковые синаптические организации. В этом состоянии лю-
бое возбуждение биологической модальности, поступающее
в данный момент к коре головного мозга, должно пройти
через «фильтр» системы пищевого возбуждения и, естест-
венно, претерпеть определенные изменения. Особенно отчет-
ливо эти изменения должны быть выражены у соматосен-
сорного вызванного ответа, поскольку формирующее его
возбуждение адресуется к тем же передним отделам коры,
куда поступает возбуждение пищевых мотивационных цент-
ров гипоталамуса. В то же время после приема пищи, ког-
да устраняются восходящие активирующие влияния под-
корковых отделов пищевого центра на кору, соматосенсор-
ный вызванный потенциал должен проявиться в своей
классической форме. Опыты подтвердили такое предполо-
жение. У голодных животных на фоне активации электро-
энцефалограммы в передних отделах соматосенсорный вы-
званный потенциал плохо выявляется даже в специфической
174
проекционной зоне коры. Как правило, у него наблюдалась
только положительная фаза первичного ответа. Иногда же
вызванный потенциал совсем нельзя было зарегистриро-
вать (рис. 32, А).
Иная картина наблюдалась у животных, предварительно
накормленных до эксперимента. У таких животных на фоне
медленной вы со ко амплитудной электрической активности
коры головного мозга в ответ на одиночное раздражение
30w.fi
А
ЗОмсек
ЗОмкв
Б
20мсек
Рис. 32. Соматосенсорный вызванный потенциал в проекционной обла-
сти коры у голодной (Л) и накормленной (5) кошки.
Точкой на поверхности головного мозга обозначен пункт отведения
вызванных потенциалов.
седалищного нерва в сенсомоторной области всегда реги-
стрировался вызванный потенциал, который имел хорошо
выраженное первичноположительное колебание, большую
первичную отрицательную фазу и выраженный вторичный
положительный потенциал (рис. 32, 5).
Аналогичные изменения наблюдаются и после введения
в кровь голодным животным раствора глюкозы (рис. 33).
В этом случае изменения соматосенсорного вызванного по-
тенциала в коре, увеличение его различных фаз начинали
выявляться по мере возрастания амплитуды электрических
колебаний электроэнцефалограммы в передних отделах ко-
ры головного мозга.
Обращает на себя внимание и тот факт, что у накормлен-
ных животных соматосенсорный вызванный потенциал реги-
стрируется широко по поверхности не только контра-, но
и ипсилатерального по отношению к раздражаемому седа-
175
лищному нер'ву полушария (рис. 34). При этом зона его
выявления ограничивается передними отделами коры (на
рисунке заштрихована), в которых обычно в состоянии го-
лода регистрируется низкоамплитудная, высокочастотная
электрическая активность. Если сравнить карту распрост-
ранения вызванных потенциалов по коре головного мозга
у сытых животных с голодными животными, то можно от-
.*....
1^0%, Змл глюкозы
176
Рис. 33. Изменения электроэнцефалограммы и соматосенсорного вызван-
ного потенциала у голодной кошки после введения глюкозы.
>4 — динамика изменений электроэнцефалограммы и вызванного потенциала в про-
екционной области коры головного мозга при введении кошке 3 мл 40% раствора
глюкозы. Момент введения глюкозы обозначен стрелкой. Нижняя кривая про-
должение верхней кривой. Обозначения те же. что и на рис. 13; Б — расширение
зоны регистрации вызванного ответа по коре головного мозга—после введения
глюкозы. Вызванные потенциалы в точках 1, 2, 3 до (I) и после (II) введения
глюкозы.
Рис. 34. Карта выявления соматосенсорного вызванного потенциала
в коре головного мозга кошки в состоянии голода (А) и после кормле-
ния (Б). Зона выявления вызванных потенциалов заштрихована.
Обозначения электроэнцефалограммы те же. что на рис. 29.
метить, что у последних они выявляются только в фокусе
максимальной активности.
Все это также указывает на то, что трудность выявления
соматосенсорных вызванных потенциалов в коре головного
мозга у голодных животных связана с распространением на
передние отделы коры восходящих активирующих влиянии
пищевых центров гипоталамуса.
Рис. 35. Изменение характера и распространения вызванного ответа
по коре головного мозга у голодной кошки после коагуляции вентро-
медиальных отделов гипоталамуса и субталамуса.
I — исходный соматосенсорный вызванный потенциал в точках /, 2, 3; II —потен-
циал в тех же точках после коагуляции медиальных отделов гипоталамуса и суб-
таламуса; III —зона коагуляция.
[ESSES
7 Z J
Прямым подтверждением этого положения явились сле-
дующие эксперименты.
У голодных кошек, у которых соматосенсорный вызван-
ный ответ выявлялся только в специфической проекционной
зоне и притом в форме только первичноположительного
колебания, производилась коагуляция вентромедиальных
отделов гипоталамуса и таламуса с целью устранения вос-
ходящих активирующих влияний гипоталамических цент-
ров на кору головного мозга.
Вслед за такой операцией вызванный соматосенсорный
ответ претерпевал в коре головного мозга характерные из-
менения.
В фокусе его максимальной активности начинала отчет-
ливо регистрироваться отрицательная фаза. При этом зна-
чительно расширялась зона регистрации вызванного ответа.
Теперь его можно было зарегистрировать не только
в контра-, но испилатеральном по отношению к раздра-
жаемому седалищному нерву полушарий (рис. 35).
178
Аналогичные изменения соматосенсорного вызванного по-
тенциала у голодных животных наблюдаются и после внут-
римышечного введения или аппликации непосредственно на
проекционную зону коры холинолитических веществ (атро-
пин, амизил), которые, как указывалось ранее, избиратель-
но блокируют корково-подкорковые механизмы пищевого
возбуждения (рис. 36, Л).
После введения холинолитических веществ наблюдается
резкое возрастание первичноотрицательной фазы вызван-
ного ответа и появляются вторичные потенциалы.
Рис. 36, Изменения соматосенсорно вызванного потенциала в проек-
ционной зоне коры головного мозга.
А — после внутримышечного введения атропина; Б — после аппликации атропина
на кору. Слева каждого опыта представлены фоновые вызванные потенциалы;
В — схематическое обозначение коркового пункта отведения вызванных потенциа-
лов; Г — схема распространения восходящего пищевого и болевого возбуждений
на корковый нейрон (а). Атропин избирательно устраняет только пищевое воз-
буждение (б). ПВ — пищевое возбуждение; БВ — болевое возбуждение.
179
Все эти наблюдения указывают на то, что у голодных жи-
вотных восходящие активирующие влияния гипоталамиче-
ских пищевых центров избирательно распространяются на
синаптические организации коры головного мозга. Они пре-
имущественно возбуждают клетки передних отделов коры,
распространяясь на их синаптические образования. По-
скольку даже в специфической проекционной зоне сомато-
сенсорный .вызванный ответ у голодных животных представ-
лен только положительным первичным колебанием, можно
думать, что распространение пищевого мотивационного воз-
буждения осуществляется преимущественно на аксоденд-
ритные синапсы поверхностных слоев коры, блокируя тем
самым выявление отрицательной фазы соматосенсорного
вызванного ответа. Между тем блокада пищевым возбужде-
нием возбуждений другой биологической модальности осу-
ществляется не только на клетках коры головного мозга.
Организм располагает более экономными средствами бло-
кады различных возбуждений непосредственно на подкорко-
вом уровне, предотвращая тем самым для менее значимых
в биологическом отношении возбуждений возможность
выхода на синапсы коры больших полушарий головного
мозга.
На наличие такой возможности указывают следующие
эксперименты.
Аппликация у голодных животных на передние отделы
коры головного мозга холинолитических веществ, как ука-
зывалось ранее, блокирует здесь реакцию активации элек-
троэнцефалограммы. Кроме того, аппликация холинолити-
чесюих веществ на специфическую проекционную зону вы-
зывала у животных здесь резкое увеличение амплитуды
отрицательной фазы соматосенсорных вызванных ответов
(А. И. Туренко, 1965) (рис. 36, 5).
Однако аппликация у этих же животных холинолитиче-
ских веществ на все другие области коры не способствовала
возникновению в них соматосенсорного вызванного ответа.
Все это укалывало па то, что распространяющиеся к коре
головного мозга по неспецифическим путям возбуждения,
вызванные раздражением седалищного нерва, были забло-
кированы возбуждениями пищевого центра гипоталамуса
где-то на подкорковом уровне. Только по специфическому
проекционному пути эти возбуждения могли достигать коры
головного мозга. Но и последние модифицируются, посколь-
ку аксодендритныс синапсы при этом также заняты пище-
вым возбуждением.
180
Такими образованиями, позволяющими соматосенсорным
вызванным потенциалам широко распространяться на пе-
редние отделы коры головного мозга, являются ядра меди-
альных отделов переднего таламуса.
Одностороннее разрушение этих ядер с ипсилатеральной
по отношению к раздражаемому седалищному нерву при-
Рис. 37. Схема распространения соматосенсорного возбуждения к коре
головного мозга у голодного (А) и накормленного (Б) животного.
Г —таламус; ГПТ — гипоталамус; РФ — ретикулярная формация. Объяснения
в тексте.
водит к тому, что у накормленных животных соматосенсор-
ный вызванный потенциал перестает выявляться в одно-
именном полушарии.
По-видимому, пищевые центры гипоталамуса у голодных
животных используют эти таламические механизмы для
распространения на кору головного мозга своих восходя-
щих активирующих влияний.
Возможно, что в этом пункте и происходит функциональ-
ная блокада пищевым возбуждением механизмов, участвую-
щих в распространении болевых импульсов на передние от-
делы коры головного мозга.
Все сказанное выше может быть представлено в виде
схемы, приведенной на рис. 37.
На схеме справа показано распространение соматосен-
сорного возбуждения к коре головного мозга у накормлеи-
181
ного Животного, т. е. при отсутствии пищевой мотивации.
При этом болевой импульс свободно распространяется на
корковую клетку двумя путями: 1) по лемнисковому пути,
достигая специфической проекционной зоны коры; 2) по кол-
латералям от лемнискового пути через структуры ретику-
лярной формации, гипоталамуса и таламуса, широко гене-
рализуясь по передним отделам коры головного мозга.
При этом в фокусе максимальной активности регистрирует-
ся .соматосенсорный вызванный потенциал с характерными
фазами первичного и вторичного ответов.
На схеме слева показаны функциональные взаимоотноше-
ния у голодного животного. У голодного животного под
влиянием пищевой потребности приходят в возбуждение
соответствующие центры гипоталамуса. Последние оказы-
вают восходящее активирующее влияние на передние отде-
лы таламуса, а через них генерализованно на передние от-
делы коры головного мозга. Одновременно с этим возбужде-
ние распространяется и на ретикулярную формацию
среднего мозга (показано штриховкой).
Вследствие этого пищевое возбуждение распространяется
непосредственно на аксодендрптные синапсы коры головно-
го мозга и на указанные выше подкорковые образования.
В результате этого происходят блокада генерализованного
распространения соматосенсорных потенциалов к передним
отделам коры и изменение их формы в специфической про-
екционной зоне, т. е. блокада здесь отрицательной фазы
первичного ответа. При усилении пищевой мотивации, на-
пример при длительном голодании, восходящие активирую-
щие (влияния гипоталамических центров начинают распрост-
раняться и на аксосоматические синапсы III—IV слоя, бло-
кируя тем самым полностью выявление соматосенсорного
вызванного потенциала в коре головного мозга.
Таким образом, на основании сказанного выше можно ви-
деть, что мотивационное возбуждение избирательно
распространяется на клетки коры головного
мозга. Тем самым производится как бы своеобразная
«мобилизация» определенных корковых синапсов из общей
их массы для построения той или иной формы целенаправ-
ленного поведения.
Благодаря этому любое мотивационное возбуждение вы-
ступает в роли своеобразного активного «фильтра» на пути
распространения возбуждений различной сенсорной и био-
логической модальности от подкорковых образований к ко-
ре головного мозга. При наличии мотивации эти возбужде-
182
ния могут быть избирательно заблокированы или, 'наоборот,
облегчены. А это все определяет то важное свойство моти-
вации, на которое мы уже указывали ранее,— активное ис-
следование и подбор специальных раздражителей внешнего
мира.
Эти же динамические процессы лежат в основе чрезвы-
чайной изменчивости условных рефлексов в зависимости от
исходных состояний животного. В самом деле, в каждом
случае возбуждения, обусловленные действием на организм
условных раздражителей, должны «просочиться» до клеток
коры головного мозга через сложившиеся мотивационные
системы возбуждений, обусловленные исходным состоянием
животного. В конечном счете результат действия условных
раздражителей на организм всегда определяется тем, в ка-
кой .степени корковые синапсы свободны и не заблокирова-
ны 'возбуждениями другой биологической модальности.
Вместе с тем действие условного сигнала резко облегчается
при наличии соответствующей ему мотивации. Так,
А. И. Шумилина (1966) показала, что если вспышка света
неоднократно подкрепляется кормлением, то в голодном со-
стоянии величина вызванного потенциала на свет резко
возрастает.
Многочисленные наблюдения показывают изменчивость
характера вызванных ответов в зависимости от исходного
состояния подопытных животных. Впервые Bremer (1962),
наблюдая блокаду вызванных потенциалов в коре при нали-
чии восходящих активирующих влияний ретикулярной фор-
мации на кору головного мозга, назвал это явление феноме-
ном «маскировки» вызванного ответа. А. И. Шумилина по-
казала, что обстановочная оборонительная доминанта резко
усиливает величину вторичных колебаний вызванного отве-
та у кролика па световой условный раздражитель, ранее
подкрепляющийся болью. Аналогично этому Galambos и
Sheatz (1962) обнаружили увеличение амплитуды вызван-
ных ответов при ориентировочно-исследовательской и обо-
ронительной реакции подопытных животных. Freeman (1962)
в хронических опытах на кошках по мере их голодания
наблюдал в грушевидной извилине коры в ответ на аффе-
рентные раздражения увеличение амплитуды вызванных
ответов. Наоборот, насыщение и условные пищевые раз-
дражители, по его данным, снижают амплитуду вызванных
ответов.
Аналогично нашим данным Kitai и др. (1965) наблюдали
блокаду слухового вызванного ответа в слуховой коре у го-
183
лодных животных. Точно так же снижает амплитуду слухо-
вого вызванного потенциала световой условный раздражи-
тель.
Marczynski и др. (1968, 1969) наблюдали у голодных ко-
шек отсутствие слуховых вызванных потенциалов и их рез-
кое возрастание по амплитуде во время приема пищи.
Все эти наблюдения позволяют говорить не просто
о блокаде вызванных ответов, а об активной фильтрации
сенсорных возбуждений исходными мотивационными воз-
буждениями.
Итак, мотивационное возбуждение, избирательно объеди-
няя различные подкорковые образования, избирательно рас-
пространяется на кору головного мозга. Именно такие спе-
цифические корково-подкоркозые комплексы и характеризу-
ют центральное мотивационное состояние, описанное
Morgan.
Возникает естественный вопрос, каким образом клетки
коры головного мозга воспринимают различные мотиваци-
онные восходящие возбуждения и па этой основе формиру-
ют поисковую активность животного?
ОБРАБОТКА КОРОЙ ГОЛОВНОГО МОЗГА
ВОСХОДЯЩИХ МОТИВАЦИОННЫХ ВОЗБУЖДЕНИЙ
Концепция П. К- Анохина о множественных восходящих
активирующих влияниях подкорковых центров на кору го-
ловного мозга, о которой мы писали выше, позволяет с но-
вых позиций представить вопрос о функционировании от-
дельной нервной клетки коры головного мозга в интегра-
тивной деятельности целого организма.
В самом деле она заставляет принять, что на одну и ту
же клетку'коры головного мозга могут поступать возбужде-
ния различной сенсорной и биологической модальности.
Конвергенция на одну и ту же клетку возбуждений, вызван-
ных действием на организм раздражителей различной сен-
сорной модальности (например; звуковых, световых, так-
тильных и т. п.), была обнаружена Fessard (1958) и Jung
(1961) и названа соответственно «мультисенсорной» кон-
вергенцией.
Ю. А. Фадеев (1964) в лаборатории, руководимой
П. К. Анохиным, обнаружил, что к одной и той же клетке
сенсомоторной области кошки адресуются возбуждения раз-
личной биологической модальности. Опыты, обнаружившие
эту закономерность, состояли в следующем, У голодной
184
кошки под уретановым наркозом в сенсомоторной области
регистрировали реакции отдельных иейронов на раздраже-
ние седалищного «ерва и введение глюкозы в кровь. Опыты
показали, что из 108 обследованных нейронов коры 40 отве-
чали только на раздражение седалищного нерва, 15 — толь-
ко на введение глюкозы. Наряду с этим было зарегистриро-
5
t Зв
!lll!lll!lll|l!lll IIIIIIIIIIIIIIIII
£
f ЗВ
Рис, 38. Конвергенция возбуждений двух биологических модальностей
на одном и том же корковом нейроне (по Ю. А. Фадееву, 1964).
4 — разражение седалищного нерва током напряжением 3 в на фоне регулярных
разрядов нейрона у голодной кошки; Б — фоновая активность того же нейрона
до введения глюкозы; В, Г, Д — последовательные стадии устранения разрядов
В периоды 15—23—28 секунд после введения глюкозы; Е—реакция «замолчавшего»
пищевого нейрона на раздражение током седалищного нерва напряжением 3 в.
Видна отчетливая реакция на болевое раздражение, как и в случае Л, хотя пи-
щевая возбудимость была устранена путем инъекции глюкозы.
вано 45 нейронов, реагирующих как на раздражение седа-
лищного 1нерва, так и на введение глюкозы (рис. 38). Как
правило, ®а введение глюкозы реакция большинства нейро-
нов в передних отделах коры выражалась в урежении или
подавлении его исходной активности, хотя наряду с этим
было зарегистрировано некоторое количество нейронов, ко-
торые после введения глюкозы, наоборот, увеличивали час-
тоту разрядов
185
Все эти опыты показывают, что наряду со специализиро-
ванными нейронами в передних отделах коры головного
мозга имеются нейроны, на которые адресуются возбужде-
ния различной биологической модальности. На этом осно-
вании п. к. Анохин сформулировал представление о «муль-
тибиологнческой конвергенции» возбуждений на нейронах
коры головного мозга.
Дальнейшие опыты Ю. А. Фадеева показали, что в перед-
них отделах коры головного мозга имеются нейроны, на
которые поступают возбуждения сенсорной (например, 'Све-
товые или звуковые) и биологической модальности (напри-
мер, болевое возбуждение). Все это дало основание гово-
рить о «сенсорно-биологической конвергенции» возбуждений
на нейронах коры головного мозга.
В последние годы представления о конвергенции возбуж-
дений на нейронах коры головного мозга значительно рас-
ширились. В. А. Правдивцев (1967) и В. В. Синичкин (1967)
в лаборатории, руководимой П. К. Анохиным, обнаружили,
что при антидромном раздражении пирамидного тракта
у кошек удается зарегистрировать реакцию вставочных ней-
ронов не только в сенсомоторной, но даже в зрительной об-
ласти коры. Оказалось, что эти же нейроны могут отвечать
не только на антидромное раздражение пирамидного трак-
та, но и .на различные сенсорные раздражения. Было сдела-
но предположение, что реакция этих нейронов при анти-
дромном раздражении определялась распространением
к ним возбуждений по коллатералям аксонов пирамидного
тракта (П. К- Анохин, 1968). В связи с этим такая форма
конвергенции возбуждений на нейронах коры головного
мозга была названа «аксонально-сенсорной конвергенцией».
На рис. 39 схематически представлены различные формы
конвергенции возбуждений на нейронах коры головного
мозга.
Все это показывает, что один и тот же корковый нейрон
принципиально может получать на себя множественную ин-
формацию. Она определяется восходящими влияниями мо-
тивационных центров гипоталамуса, действием различных
раздражителей внешней среды на разные органы чувств
и, наконец, возбуждениями, ответвляющимися от пирамид-
ных трактов при распространении то ним потоков эффе-
рентных возбуждений. Все эти процессы и составляют ту
стадию целостного поведенческого акта, которая получила
название в лаборатории П. К. Анохина «стадии афферент-
ного синтеза».
186
Спрашивается, как же клетка перерабатывает такую мно-
жественную поступающую к ней информацию и «строит»
адекватную исходному состоянию животного и действию
внешних раздражителей ответную реакцию?
Многочисленные эксперименты показывают, что возбуж-
дения различной биологической и сенсорной модальности на
одной и той же корковой клетке определяются различными
Рис. 39. Различные виды конвергенции восходящих возбуждений на
нейронах коры головного мозга.
А — мультисенсорная конвергенция; Б — мультибиодогическая конвергенция;
В — сенсорно-биологическая конвергенция: Г — аксонально-сенсорная конверген-
ция: / — свет; 2 —звук; 3— боль; 4 — голод; 5 — ориентировочная реакция,
специфичными химическими механизмами (Ф. А. Ата-Му-
радова, 1967, 1968; Сунь-Вень, 1964; К- М. Катраманов,
1956; Бадам-Ханд, 1965; Л. М. Чулпина, 1967).
Наибольший интерес в этом отношении представляют эк-
сперименты Л. М. Чуппиной (1967). Они показывают, что
различные фазы вызванных потенциалов, регистрируемых
в одном и том же пункте коры головного мозга в ответ на
раздражение контралатерального седалищного нерва
и транскаллозально на раздражение симметричного
коркового пункта притивоположного полушария, имеют раз-
ное отношение к аппликации различных химических ве-
ществ.
Так, наложение на пункт регистрации вызванных потен-
циалов ГАМК почти сразу же блокирует отрицательный
компонент потенциала, вызванного раздражением седалищ-
ного нерва, и почти не изменяет такого же отрицательного
компонента транскаллозально вызванного ответа. Наобо-
рот, новокаин преимущественно блокирует отрицательный
187
компонент транскаллозально вызванного потенциала.
Стрихнин усиливает отрицательную фазу обоих вызванных
ответов.
Если учесть, что в этом эксперименте разнородные вы-
званные потенциалы регистрировались с одного 'и того же
пункта .коры головного мозга, то различное их отношение
к разным химическим веществам может быть объяснено
только различием химизма, синаптических организаций, фор-
мирующих оба эти процесса возбуждения. Наиболее пока-
зательно это химическое многообразие корковых синапсов
было выявлено в опытах А. И. Туренко (1965).
Как указывалось ранее в опытах, аппликация на сенсомо-
торную кору у голодных животных холинолитических ве-
ществ способствовала выявлению в пункте аппликации от-
рицательной фазы соматосенсорного вызванного ответа.
Все эти факты совершенно понятны, если исходить из кон-
цепции о множественном характере распространения вос-
ходящих возбуждений на нейроны коры головного мозга.
Так, в опытах А. И. Туренко холинолитические вещества
блокируют распространение пищевого .возбуждения из гипо-
таламических центров на нейроны коры и тем самым предо-
ставляют возможность соматосенсорным имлульсациям
формировать с помощью этих же клеток характерный для
них вызванный потенциал.
Точно так же н в экспериментах Л- М. Чуппиной разные
химические вещества в различной степени блокируют в од-
ном случае проведение возбуждения к клеткам через синап-
сы, участвующие в соматосенсорном, а в другом случае —
в транскалло'зальном возбуждении.
Возникает вопрос, с чем же связана химическая специфи-
ка различных восходящих возбуждений, адресующихся
к одному и тому же нейрону коры головного мозга?
В объяснении этих данных имеются две возможности:
1) для каждого возбуждения специфической сенсорной или
биологической модальности имеются специфические нейро-
ны коры головного мозга различной химической чувстви-
тельности. При этом в таком случае определенной модаль-
ности возбуждения из подкорки распространяются на свои
специфические нейроны; 2) многочисленные подкорковые
возбуждения разной модальности распространяются на
один и тот же нейрон коры головного мозга. При этом хи-
мическая специфика подкорковых возбуждений может быть
связана со спецификой химического передатчика в синапсах
или специфическими процессами, которые развертываются
188
в протоплазме клетки после распространения возбуждения
на постсинаптическую мембрану (П. К. Анохин, 1968).
Наши данные позволяют думать, что корковые клетки ис-
пользуют все указанные выше механизмы. Подтверждением
этого являются 'следующие наблюдения.
—IWIHHI1IIIII-
2в h# 66
4В
66
Рис. 40. Мультихимичсская чувствительность нейрона сенсомоторной
коры головного мозга (по А. В. Котову, 1967).
А — реакция нейрона на раздражение седалищного нерва электрическим током;
Б — отсутствие реакции поел? аппликации амизила; S — восстановление реакции
на раздражение седалищного нерва После отмывания амизила', Г — отсутствие
реакции после аппликации аминазина; Д — восстановление реакции после отмы-
вания аминазина,
В опытах А. В. Котова (1967) было показано, что реакции
нейронов сенсомоторной области коры головного мозга на
- раздражение седалищного черва имеют разное отношение
к аппликации на кору холино- и адренолитических веществ.
Одни нейроны прекращают реагировать .на ноцицептивное
раздражение при аппликации амизила, другие — только при
аппликации аминазина. Вместе с тем обнаружена специаль-
у пая группа «мультихимических» нейронов, прекращавших
реакцию как при аппликации холино-, так и адренолитиче-
у ских веществ (рис. 40).
Г 189
й‘
4
Эти опыты показывают, что реакция определенного био-
логического качества осуществляется как специализирован-
ными нейронами, имеющими подчеркнутую химическую спе-
цифичность, так и нейронами с по.тихимическими свойст-
вами.
Точно так же могут быть объяснены и опыты А. И. Ту-
ренко, которые показывают, что при блокаде аппликацией
холинолитических веществ в коре пищевого возбуждения
болевое -возбуждение по-прежнему вызывает характерную
для него реакцию генерализованной активации электроэн-
цефалограммы, которая развивается ив пункте аппликации.
Возможно, что в этом случае болевое возбуждение исполь-
зует иной химической чувствительности корковые клетки, не
заблокированные холинолитическим веществом, или же
в условиях блокады холинолитическим веществом на кор-
ковой клетке одних синаптических (пищевых) образований
другие синаптические образования могут проводить боле-
вое возбуждение. Не исключено и другое объяснение, что
холинолитическое вещество блокирует определенные хими-
ческие реакции в постсинаптической мембране, которые
обычно используются пищевым возбуждением. Ответ на все
эти вопросы, безусловно, может быть получен только в том
случае, когда будет поставлен специальный эксперимент
с регистрацией реакции микроэлектродным методом одного
и того же коркового нейрона на «голодное» и «болевое»
возбуждение на фоне аппликации холинолитического ве-
щества непосредственно на кору головного мозга.
В настоящее время можно определенно сказать только
следующее. Корковая клетка обрабатывает поступающую
к .ней по различным каналам многочисленную информацию
на основе подчеркнутой химической специфики различных
подкорковых возбуждений.
Что же создает клетке особую «химическую» настроен-
ность, на основе которой она подбирает или отвергает
поступающую к ней информацию? Опыты показывают, что
эту исходную настроенность клетки и создают приходящие
к ней из -подкорки мотивационные возбуждения. Именно ис-
ходная мотивация создает тот фон, на котором к клеткам
коры головного мозга поступают возбуждения различноной
модальности. Она определяет избирательное отношение
этих клеток к действию раздражителей различной сенсорной
и биологической модальности, способствуя или, наоборот,
препятствуя конвергенции на них определенных возбужде-
ний.
190
Подтверждением этого являются эксперименты, проведен-
ные С. Н. Хаклиным (1969) в лаборатории, руководимой
П. К. Анохиным. Опыты проводились следующим образом.
У кролика определялась реакция нейрона зрительной коры
на вспышку света определенной частоты. После этого раз-
дражали область латерального гипоталамуса, вызывавшую
обычно у данного кролика при предварительном тестирова-
нии пищевую реакцию, и регистр провал и реакцию указан-
ного выше нейрона на это раздражение. Такое раздраже-
ние, как правило, вызывало несколько отличную от светово-
I
j~4--II Ц i| 1 Д !---1---г P I 1 l-l i;> | ’I |t а ц i| । i..u. ip । ii
1 сек
_ -H-f-t-t IP-н P I I! 1 I I II I I1 I1 P I 1 III mu II H I ! ' Н 4 >
Рис. 41. Изменение реакции нейрона зрительной области коры головного
мозга кролика на ритмическое световое раздражение до и после
раздражения центра пищевой мотивации в латеральном гипоталамусе
(по С. Н. Хаютину, 1969).
Момент раздражения гипоталамуса обозначен стрелкой, / — электроэнцефалограм-
ма; 2 — активность нейрона; 3 — отметка вспышек света.
го раздражения реакцию нейрона. После раздражения
гипоталамуса снова исследовали реакцию данного нейрона
на ритмическое действие света. Оказалось, что после моти-
вационного раздражения нейрон приобретал свойство реа-
гировать на более высокую частоту световых вспышек
(рис. 41). Повышалась дискриминационная способность
нейрона.
Все это указывает на то, что мотивационное возбуждение
определенным образом изменяет реакцию нейрона на сен-
сорный раздражитель.
Близки к этому и эксперименты нашего сотрудника
А. Сосновского (1969). Он показал, что слабое раздражение
мотивационного пищевого центра у кролика, вызывающее
только ориентировочно-исследовательскую реакцию кроли-
ка, значительно снижает порог восприятия клеткой светово-
го раздражения.
Особая форма мотивационных влияний на корковый ней-
рон показана В. А. Макаровым (1969). Он обнаружил, что
раздражение мотивационных центров амигдалярной облас-
ти значительно повышает способность корковых нейронов
191
к конвергенции возбуждений различной биологической мо-
дальности.
Все эти примеры показывают, что мотивационное возбуж-
дение значительно изменяет реакции корковых клеток на
раздражители внешнего мира, а также определяет конвер-
генцию на них возбуждений различной биолопической и сен-
сорной модальности. Исходная мотивация, таким образом,
специфически завладевает корковыми нейронами, направляя
их деятельность на удовлетворение исходной потребности
организма.
НИСХОДЯЩИЕ ВЛИЯНИЯ КОРЫ ГОЛОВНОГО МОЗГА
НА МОТИВАЦИОННЫЕ ЦЕНТРЫ ГИПОТАЛАМУСА
Наше -изложение роли корковых нейронов в мотивацион-
ном -возбуждении было бы неполным, если бы мы не косну-
лись одного важного вопроса. Корковые нейроны наряду
с получением и обработкой -информации об исходном состоя-
нии организма и окружающей организм внешней среды са-
ми оказывают нисходящие влияния на инициативные моти-
вационные центры гипоталамуса, регулируя тем самым силу
тонического напряжения последних. Подтверждением этого
являются эксперименты, проведенные в нашей лаборатории
А. И. Туренко (1965).
Опыты проводились на голодных кошках, находившихся
под уретановым -наркозом, У животных регистрировалась
электрическая активность коры головного мозга и с по-
мощью двух погружных электродов — электрическая актив-
ность правого и левого пищевых центров латерального ги-
поталамуса.
Ка-к уже указывалось ранее, аппликация на передние от-
делы коры одного из полушарий головного мозга фильтро-
вальной бумажки, смоченной раствором х-ол-иполитического
вещества (амизил), через некоторое время устраняет ис-
ходную «голодную» активацию в пункте аппликации. Здесь
появляются медленные вы-сокоамплитудные волны. Одно-
временно с этим, а в ряде опытов с некоторым запозданием
такая же активность возникает в латеральном гипоталамусе
одноименной стороны. Спустя некоторое время такая же вы-
сокоампл-итудная активность появляется -в латеральном ги-
поталамусе -контралатеральной по отношению к пункту
аппликации холинолитического вещества -стороны и затем—
в передних отделах коры головного мозга этой стороны
(рис. 42).
192
Ёсе это указывает на то, что при аппликации холинолиФй-
ческого вещества на передние отделы коры головного мозга
одного из полушарий животного, устранение голодного мо-
тивационного возбуждения происходит в определенной по-
следовательности. Сначала устраняется возбуждение в пунк-
те аппликации. Это в свою очередь подавляет возбуждение
в латеральном гипоталамусе одноименной стороны. Послед-
ний оказывает однотипное влияние на латеральный гипота-
5 Г
1
. !о/о Qbu:„l.‘l т
з
7 ... - . . -1 - VsAAV V-sV.1,*- V--—'
lc£n IJ.U1 /J, a v
Рис. 42. Динамика изменений электрической активности коры головного
мозга и латерального гипоталамуса после аппликации на правую сенсо-
моторную область 1% раствора амизнла.
А — фон; Б — через 17 минут; В — через 52 минуты; Г — через 74 минуты после
аппликации; / — сенсомоторная левая область; 2 — сенсомоторная правая область;
3—теменная левая область; 4 — теменная правая область; 5 — затылочная левая
область; (? — затылочная правая область; 7 — правый лагеральяый гипоталамус.
ламус противоположной стороны, ,в результате чего послед-
ний прекращает свои восходящие активирующие влияния
на соответствующее полушарие.
Таким образом, мотивационное возбуждение характери-
зуется постоянными циклическими реверберациями возбуж-
дений между инициативными центрами гипоталамуса и ко-
рой головного мозга.
Мысль о реверберационных циклах возбуждения между
корой и подкорковыми образованиями была сформулирована
Dusser de Barrenne, Mac Culloch (1938). По их мнению,
каждое поле коры может функционировать лишь в том слу-
чае, если связано с соответствующим ядром таламуса или
метаталамуса.
Chang (19'50) удалось показать, что посредством кор-
тико-таламического отраженного механизма одиночный
залп афферентного импульса может производить изменения
7 Заказ 5071
193
в головном мозге, продолжающиеся гораздо дольше, чем про-
должительность самого первичного стимула. Представления
о кортико-таламическом реверберационном цикле теперь
значительно расширились. Существование таких цикличе-
ских взаимоотношений находит подтверждение в двусторон-
нем характере анатомических связей как «специфических»,
так и «неспецифических» ядер таламуса с корой головного
мозга (В. М. Бехтерев, 1896; Ramon у Cajal, 1902; Stratford,
1951; Jasper, Ajmone-Marson, Stoll, 4952; Nauta a. oth.,
1954, и др.).
Весьма возможно, что реализация циклического движения
возбуждения, столь важного для корково-подкорковой интег-
рации, может обеспечиваться различными, тесно связанными
друг с другом корково-подкорковыми системами возбужде-
ний: |рети;куло^кор’викальнЫ1М1и (Bonvailet, Dell, Hiebel,
1954), гипоталамо-кортикальпыми (Papez, 1937; Mac Lean,
1955; Gellhorn a. Loofbourow, 1963), таламокортикальными
(Chang, 1950, 1959).
Это находит подтверждение при изучении корково-под-
корковой реверберации возбуждений в условнорефлектор-
ной деятельности. Так, А. И. Шумилина (1966) установила,
что после безусловного болевого подкрепления происходит
удвоение или утроение медленного колебания вызванного
потенциала в ответ на вспышку света, применяемую в ка-
честве условного раздражителя. Такая реакция постепенно
распространяется .из коры головного мозга ;в подкорковые
образования п снова ограничивается корой.
По данным Bishop и O’Leary (1950), удвоение медленно-
го колебания вызванного потенциала служит проявлением
таламо-кортикальной реверберации возбуждении, которая
возникает благодаря повторному возбуждению клеток лате-
рального колончатого тела под влиянием импульсов, рас-
пространяющихся по нисходящим аксонам таламо-корти-
кальных нейронов. Однако, как полагают авторы, в меха-
низм возникающей реакции реверберации возбуждений при
этом включаются также ретикулярная формация и гипота-
ламус.
Таким образом, механизмы корково-подкорковой ревер-
берации возбуждений определяются не только таламо-кор-
тикальными связями, но и более обширной системой под-
корковых образований, подобной, например, гиппокампову
кругу (Papez, 1937).
Существуют и такие фюрмы корково-подкорковой ревер-
берации возбуждения, когда обратные,влияния подкорковых
194
образований на кору носят генерализованный характер.
Эта форма реверберирующих влияний была установлена
В. Н. Шелиховым (1960) в лаборатории, руководимой
П. К. Анохиным, и названа возратной генерализацией воз-
буждений. Такая реакция отчетливо возникает, если на
определенный пункт .коры наложить бумажку, смоченную
стрихнином. Через несколько минут будут заметны парок-
сизмальные и эпилептиформные разряды, возникающие
с одним и тем же латентным 'периодом по всей коре голов-
ного мозга, независимо от расстояния до первичного пункта
наложения стрихнина. Перерезка мозолистого тела не ис-
ключает этих генерализованных (влияний. Они исчезают
только при разрушении неспец,ифичеокого комплекса ядер
таламуса и субталамуса.
Все это указывает на то, что возбуждение, локализован-
ное вначале в коре головного мозга, 'распространяясь в по-
следующем в кортико-фугальном направлении, создает
мощное возбуждение в подкорковых аппаратах. В свою оче-
редь субкортикальное возбуждение оказывает возвратное
генерализованное влияние на кору больших полушарий. Об
этом свидетельствует такой факт: если охладить первичный
стрихнинный пункт и подавить в нем всякую электрическую
активность, то 'пароксизмальные разряды ® других частях
головного мозга все-таки сохраняются. Такая же реакция
наблюдается и при локальной механической травме коры
головного мозга (С. К- Рогачева, I960).
Физиологическое значение корково-подкорковых меха-
низмов реверберации заключается в следующем.
1. Корково-подкорковая реверберация за счет (включения
в реакцию активирующих влияний подкорковых образова-
ний способствует длительному тоническому напряжению
корковых элементов. Это особенно важно при таких биоло-
гических состояниях, как страх, голод, ориентировочно-ис-
следовательская реакция. Можно предполагать, что между
подкорковыми образованиями и корой головного мозга при
реакциях определенного биологического качества всегда
устанавливаются специфические циклические взаимодей-
ствия.
Важно подчеркнуть, что инициативный источник этих воз-
буждений всегда лежит в подкорковых образованиях, обла-
дающих более высокой чувствительностью к действию спе-
циальных гуморальных и нервных раздражений.
2. В последнее время реверберирующим взаимодействиям
в коре и между корой и подкорковыми образованиями при-
7*
195
дается все большее значение в механизме формирования
акцептора результатов действия (П. К- Анохин, 1968). Со-
гласно этим представлениям, в момент формирования
любого исполнительного акта и выхода эффекторных воз-
буждений по аксонам корковых пирамидных клеток импуль-
сы от последних (определенная «команда») направляются
не только к исполнительным органам; по коллатералям
аксона копии этих команд одновременно «раздаются» спе-
циальным группам чувствительных клеток, которые могут
располагаться как в коре, гак и в подкорковых образова-
ниях.
В момент совершения действия и достижения определен-
ного результата эти клетки производят сравнение информа-
ций о достигнутом приспособительном эффекте с ранее по-
данной командой. Впоследствии они мобилизуются всякий
раз, когда организм принимает решение к совершению дан-
ного действия.
При неоднократном совершении определенных действий
в системе этих клеток устанавливаются реверберирующие
взаимоотношения, составляющие основу механизма памяти
и способствующие быстрому и надежному достижению по-
лезного приспособительного результата.
Это положение, как мы увидим ниже, очень важно для
характеристики мотивационного возбуждения.
МОТИВАЦИИ И АФФЕРЕНТНЫЙ СИНТЕЗ.
ПРОБЛЕМА ДОМИНИРОВАНИЯ МОТИВАЦИЙ
Как указывалось ранее, согласно представлениям
П. К. Анохина, отправным пунктом поведенческого акта лю-
бой степени сложности является стадия афферентного син-
теза.
Принципиальное значение этой стадии заключается в том,
что во время ее формирования на нейронах центральной
нервной системы и особенно коры больших полушарий на
основе избирательных восходящих подкорковых активирую-
щих влияний происходит взаимодействие возбуждений, обу-
словленных внутренним состоянием организма и воздействи-
ем на него различных факторов окружающей среды. При
этом также происходит сопоставление этих -возбуждений
с аппаратами памяти животного. Именно на этой стадии фор-
мирования целостного поведенческого акта мотивационное
возбуждение, обусловленное той или иной внутренней по-
требностью организма, выступает как главный фактор, объ-
единяющий корково-подкорковые аппараты в единую систр-
196
Му для активного исследования внешних раздражителей
и подбора специальных из них, удовлетворяющих исходную
потребность организма.
Из этого следует, что мотивационное возбуждение
является наиболее важным и решающим компонентом
афферентного синтеза. Его значение состоит в сле-
дующем.
Во-первых, каждое мотивационное возбуждение отражает
собой жизненно важную потребность организма. Во-вторых,
формируясь на основе внутренних механизмов, мотивацион-
ное возбуждение определяет активное отношение животно-
го к раздражителям внешнего мира. В-третьих, оно приво-
дит в действие прежний опыт животного по удовлетворению
соответствующей потребности, как бы оживляет соответ-
ствующие аппараты памяти и тем самым способствует наи-
более оптимальному удовлетворению лежащей в его основе
жизненно важной потребности.
Благодаря всему этому .каждая функциональная .система
обладает по существу всеми необходимыми аппаратами для
достижения своего конечного приспособительного резуль-
тата.
Так, аппарат афферентного синтеза на основе исходного
мотивационного возбуждения и действия обстановочных
раздражителей определяет, какой результат должен
быть получен. Специальные пусковые (условные) раздра-
жители обусловливают время, когда этот результат дол-
жен быть получен. Аппараты памяти и акцептора резуль-
тата действия определяют, каким образом он должен
быть получен. Имея свойства конечного результата в аппа-
рате акцептора результата действия, организм всегда, та-
ким образом, имеет возможность активно сравнивать с ним
результаты своей деятельности во внешней среде.
На основании всех этих процессов, которые, как указыва-
лось выше, в большей степени свойственны клеткам коры
головного мозга, происходит оценка внутреннего состояния
животного. Кроме того, последнее сопоставляется с внеш-
ними условиями, в которых данное животное в данный мо-
мент находится, т. е. внешний мир также оценивается через
призму внутреннего состояния. Отсюда становятся понят-
ными эксперименты Goltz и Bard с удалением коры, когда
животное приобретает неадеквантную форму реагирования
иа внешние раздражители. То же самое наблюдается® опы-
тах Kluver и Вису с удалением височной коры и амигдаляр-
ной области.
197
Во всех этих случаях страдает аппарат оценки исходного
состояния животного и результатов его деятельности во
внешней среде.
Однако не только корковые элементы обладают этой
функцией оценки внутреннего состояния и результатов дея-
тельности организма во внешней среде. Она присуща и не-
которым лимбическим образованиям (старая 'И древняя
кора). Вот почему раздражение или разрушение различных
лимбических образований приводит к появлению неадекват-
ных исходной потребности организма мотиваций, которые
могут быть как резко подавленными, так и, наоборот, резко
усиленными.
На основе процессов афферентного синтеза организм при-
нимает решение к построению определенного действия или
поведенческого акта (т. е. где, когда и как действовать).
Одним словом, ставит перед собой определенную цель
Нужно отметить, что значение обстановочных раздражи-
телей в построении цели к действию все в большей степени
возрастает у высших животных и особенно у человека. Про-
стейшие животные строят свою целенаправленную дея-
тельность преимущественно на основе своих биологических
мотивационных возбуждений, обусловленных их внутренни-
ми потребностями.
У высших животных и человека на мотивационные воз-
буждения, обусловленные внутренними потребностями,
оказывает значительное влияние среда существования,
а у человека —социальная среда. Поэтому цель к действию
строится у них не только биологическими мотивациями.
Возникающие на основе биологических мотиваций и обста-
новочных раздражителей возбуждения и являются по су-
ществу теми высшими формами мотиваций, о которых
сообщалось ранее.
Рассмотрим теперь следующий вопрос.
Животное может иметь одновременно несколько потреб-
ностей, каждая из которых, возбуждая определенные цент-
ры гипоталамуса, способна привести в действие соответ-
ствующую мотивацию. Как же строится поведение в этом
случае?
Опыт показывает, что в условиях наличия в организме
нескольких потребностей, мотивационное возбуждение фор-
мируется всегда по доминирующему типу, т. е. сначала за-
владевает всей деятельностью головного мозга, и удовлетво-
ряется первостепенная по своей биологической важности
мотивация. Только после удовлетворения ведущей моти-
198
вации деятельностью организма может завладеть следую-
щее по своей биологической значимости мотивационное
возбуждение и т. д.
Что же определяет доминирование той или иной мотива-
ции? Оно вытекает из того обстоятельства, насколько та
или иная потребность первостепенно жизненно важна для
организма ,в данный момент времени, насколько отклонение
той или иной жизненно важной константы крови опасно в
данный момент времени для жизнедеятельности организма.
На этой основе доминирующая мотивация практически
завладевает всей деятельностью организма на данный мо-
мент. В результате этого практически любая внешняя ин-
формация, попадающая в центральную нервную систему
животного, неизбежно сопоставляется н оценивается на ве-
сах этой доминирующей в данный момент мотивации. Таким
образом, становится понятным то фильтрирующее свойство
мотивационного возбуждения, о котором мы писали ранее.
Как полагал А. А. Ухтомский, доминирующее возбуж-
дение отбирает необходимое и отбрасывает ненужное в дан-
ный момент для организма.
На основании всего сказанного выше становится понят-
ным, как на основе доминирующей мотивации происходит
этот отбор. Восходящие активирующие влияния мотиваци-
онных центров гипоталамуса, избирательно завладевая
клетками коры головного мозга, значительно меняют харак-
тер их реакций на раздражения различной сенсорной мо-
дальности. Если вспомнить эксперименты С. Н. Хаютина,
А. С. Сосновского и В. А. Макарова, то становится ясно,
что при наличии мотивационного возбуждения клетки коры
приобретают качественно иную форму реагирования на сен-
сорные раздражители. Именно на этой основе организм,
испытывающий ту или иную жизненно важную потребность,
смотрит на внешний мир и оценивает его раздражители
только через призму этой доминирующей мотивации.
Доминанта, как пишет П. К. Анохин (1958), является об-
щим принципом функционирования. Если распространить
понятие доминанты к мотивационному возбуждению, то мы
должны совершенно определенно заявить, что каждая
мот и в ац настроится по принципу домин ан ТЬЕ.
Как же складываются >в Центральной нервной системе
процессы, обусловливающие доминирование той или другой
мотивации?
Такое доминирование складывается, по-видимому, двумя
путями: сильное мотивационное возбуждение на основе сво-
199
их специфических химических избирательностей оккупирует
почти все синаптические образования коры головного мозга.
Кроме того, а это, по-видимому, является наиболее важным
Рис. 43. Схематическое изображе-
ние механизма завладевания до-
минирующей мотивацией корковы-
ми синаптическими образованиями.
Схема показывает, что под влия-
нием биологически более важной
потребности (Л) происходит силь-
ное возбуждение определенных
гипоталамических центров. По-
следние тормозят возбуждение
других центров, обусловленное
менее важной потребностью (Б),
и распространяют свои активирую-
щие влияния на клетки коры го-
ловного мозга.
и экономичным меха-
низмом, сильное моти-
вационное возбуждение
уже на уровне пейцме-
керпых механизмов ги-
поталамуса может тор-
мозить другие возбуж-
дения, обусловленные
другой внутренней по-
требностью, подавляя
тем самым их восходя-
щие активирующие
влияния на кору голов-
ного мозга в самом кор-
не (рис. 43).
В результате всех
этих процессов домини-
рующее мотивационное
возбуждение избира-
тельно завладевает си-
наптическими образо-
ваниями коры головно-
го мозга, активирует их
и создает тем самым
условия для построения
адекватной, направлен-
ной на удовлетворение
только данной потреб-
ности поведенческой
реакции.
Именно эти процес-
сы лежат в основе под-
бора текущей информа-
ции в интересах доми-
нирующей мотивации. Вместе с тем это определяет и другое
важное свойство доминирующей мотивации.
Доминирующая мотивация на основе своих специфиче-
ских нейрофизиологических механизмов осуществляет
активный подбор внешней информации. Отражая потреб-
ность организма на данный момент времени, она производит
своеобразный «перебор информации» и выделяет полезный
200
сигнал из шума. Такой «перебор информации» идет в на-
правлении выбора наиболее пригодной информации для
удовлетворения лежащей в основе данной мотивации по-
требности. Одновременно с этим происходит отметание всей
информации, мешающей удовлетворению этой потребности.
Даже активный подбор внешней информации с помощью
ориентировочно-исследовательской реакции происходит
только на основе доминирующего мотивационного возбуж-
дения.
Все эти процессы ограничивают многочисленные избыточ-
ные степени свободы деятельности организма. Они обеспе-
чивают наиболее экономную форму жизнедеятельности в из-
меняющихся условиях существования.
Доминирующая мотивация, занимая главенствующие
позиции на корковых клетках, определяет таким образом
для животного выбор в соответствующих условиях только
основной линии поведения, создавая тем самым начальные
условия к целенаправленному поиску соответствующих ис-
ходной потребности раздражителей во внешней среде.
Наиболее характерным примером в данном смысле может
служить явление, описанное И. А. Аршавским (1956), как
«доминанта беременности». По существу состояние беремен-
ности на основе нейро-гуморальных сдвигов в организме со-
здает постоянную специфическую мотивацию. Биологиче-
ский смысл ее совершенно понятен. Она направлена на со-
хранение жизни будущего потомка. Эта мотивация меняет
все отношение беременной женской особи к раздражителям
внешней среды. Здесь проявляется и раздражительность,
и избирательное отношение к некоторым пищевым ве-
ществам и т. д. -
С точки зрения доминирования- мотиваций может быть
с новых позиций рассмотрен вопрос о влиянии коры на
подкорковую деятельность в целенаправленной деятель-
ности.
Иногда в научной литературе высказывается мнение
о том, что корковые процессы в построении целенаправлен-
ных реакций могут подавлять подкорковую деятельность.
Особенно распространено это мнение по отношению к ин-
стинктивной деятельности. Такая концепция, например, от-
четливо выступает у Барда.
На основании всего сказанного выше мы можем утверж-
дать, что любое вытормаживаиие одной целенаправленной
деятельностью другой может происходить только путем
создания другой доминирующей мотивации. Последняя
,20.1
и вытормаживает предшествующую деятельность. Это вы-
тормаживание, безусловно, происходит, как указывалось
выше, прежде всего на подкорковом уровне. Тормозятся
прежде всего те пейцмекерные центры, которые определяют
всю корково-подкорковую интеграцию предыдущей мотива-
ции. Тем самым разрушаются ее восходящие активирующие
влияния на кору головного мозга и создаются все условия,
позволяющие новому мотивационному возбуждению завла-
деть корковой деятельностью. Трудно представить себе этот
механизм иначе. В самом деле, как было бы энергетически
неэкономно, если бы вторая мотивация тормозила первую
непосредственно на корковом уровне при наличии активного
подкоркового пейцмекера!
До сих пор мы рассматривали энергетическую сторону
мотивационного возбуждения, т. е. все те нейрофизиологи-
ческие процессы, которые лежат в основе построения орга-
низмом целенаправленной деятельности, связанной с удов-
летворением исходной потребности. Мы показали, что
энергетическую силу мотиваций определяет избирательная
активация со стороны подкорковых образований на кору
головного мозга. Именно эти механизмы обусловливают ту
силу, которая заставляет животных, испытывающих потреб-
ность, активно преодолевать препятствия.
Наряду с этим мотивации характеризуются
своими архитектурными особ е н но с тя ми, т. е.
содержанием. Каждое мотивационное возбуждение за-
ключает в себе определенную программу действия по удов-
летворению соответствующей потребности. Мотивации
представляют таким образом запрограмми-
рованное возбуждение. Это свойство мотиваций
определяет многие особенности поведения животных во
внешней среде, направленного на удовлетворение их исход-
ных потребностей.
Рассмотрим, как же складывается программа мотиваци-
онного возбуждения и какие факторы ее определяют?
МОТИВАЦИИ И ПРОГРАММИРОВАНИЕ ПОВЕДЕНИЯ
Для естествоиспытателей давно являлось загадкой, поче-
му животные осуществляют столь направленные действия
во внешней среде. Почему, например, голодное животное
ищет только пищу, а испытывающее жажду животное
ищет только воду, животное в состоянии полового возбуж-
дения =только особь противоположного пола? Даже ново-
202
рождённое млекопитающее животное, будучи голодный,
неуверенно переползая, в конце концов находит сосок ма-
тери?
Объяснение этому становится понятным из представле-
ний, которые сложились в лаборатории, руководимой
Эфферентные Возбуждения
Обратная афферентация
Эфферентные Возбуждения
Эфферентные Возбуждения
Рис. 44. Схема построения целенаправленного поведенческого акта
(по П. К. Анохину, 1968).
ОА — обстановочная афферентация; ПА — пусковая афферентация; М — мотива-
ция; П — память, ОР — ориентировочная реакция. Объяснения в тексте.
203
П. К. Анохиным, о наличии в каждой функциональной си-
стеме аппарата — акцептора результатов действия. Этот ап-
парат формируется всякий раз, когда на основе афферент-
ного синтеза в центральной нервной системе развертывают-
ся процессы принятия решения к действию (рис. 44, А).
В этом аппарате заключен как врожденный, так и приобре-
тенный опыт, а самое главное— параметры удовлетворяю-
щих исходную доминирующую мотивацию раздражителей.
Этот аппарат формируется всякий раз, когда возникает та
или иная внутренняя потребность, и, заключая в себе все
свойства удовлетворяющего данную потребность раздражи-
теля, способствует тем самым оценке результатов будущего
действия по удовлетворению исходной потребности. Только
при (Наличии такого аппарата оргамизм обладает способ-
ностью оценить действие на него раздражителей внешнего
мира. Поскольку внешние раздражители действуют только
на рецепторы животного, оценка результатов действия идет
за счет поступления в центральную нервную систему аффе-
рентной нервной ммпульсации. Эта афферентация о резуль-
тате действия получила в лаборатории П. К. Анохина назва-
ние обратной афферентации.
Однако аппарат акцептора результатов действия не толь-
ко оценивает результат совершенного действия, но оценива-
ет его активно, своеобразно «одобряя» или «не одобряя».
В случае, если сигнализация о параметрах совершенного
результата действия полностью соответствует заготовлен-
ным свойствам акцептора результатов действия, то поиско-
вая реакция заканчивается. Соответствующая потребность
удовлетворяется. Мотивация исчезает и животное при от-
сутствии других мотиваций успокаивается (рис. 44, Б).
В том же случае, когда результат совершенного действия
и сигнализация о его 'параметрах не соответствуют свой-
ствам акцептора результатов действия, последний немедлен-
но включает ряд аппаратов (ориентировочно-исследователь-
скую деятельность, отрицательную эмоцию), которые ведут
к соответствующей «перестройке» афферентного синтеза
(рис. 44, В). На основе этого принимается новое решение
к действию, строится новая программа действия и действие
таким образом будет продолжаться до тех пор, пока его ре-
зультаты не будут соответствовать свойствам акцептора
результатов действия.
Возникающие на основе соответствующей внутренней по-
требности и действия определенных раздражителей внешней
среды мотивационное возбуждение в своей архитектуре уже
204
заключает свойства тех раздражителей, которые приводят
к удовлетворению данной потребности (рис. 45). Действи-
тельно, в каждом мотивационном возбуждении уже заклю-
чены определенные параметры будущего подкрепления, с ко-
торыми в процессе поиска все время осуществляется сравне-
ние свойств внешних раздражителей. Если же к тому же
Рис. 45. Схема демонстрирует наличие акцептора результатов действия
в механизме биологических мотиваций.
Возникая на гуморальной основе, акцептор результатов действия (А РД)
производит постоянное сравнение параметров результатов (Рр Р-А
поведенческой деятельности (ПД) с исходной потребностью и тем са-
мым направляет животное на поиск удовлетворяющего потребность
подкрепления. Целенаправленный поиск прекращается только в том
случае, когда параметры подкрепления (П) будут полностью соответ-
ствовать параметрам акцептора результатов действия.
учесть, что большинство мотиваций формируется на основе
врожденных механизмов, то нетрудно заметить, что свой-
ства подкрепляющего мотивацию агента запрограммирова-
ны генетически в возбуждении специальных нервных аппа-
ратов. Действительно, даже .новорожденное животное, ис-
пытывая голодное возбуждение первый раз в жизни, уже
направленно ищет только пищевые и немедленно отвергает
все непищевые вещества. На основе мотивации создается,
таким образом, такое возбуждение центральной нервной си-
стемы, .в котором параметры будущего результата, удовлет-
воряющего данную потребность, уже готовы и организм
ищет только адекватный раздражитель во внешней среде.
305
Итак, программирование мотивационного .возбуждения
заключается в создании определенного аппарата, отражаю-
щего все параметры внешнего раздражителя, удовлетворя-
ющего лежащую в ее основе потребность— аппарата акцеп-
тора результатов действия.
Такая же архитектура наблюдается в любых формах мо-
тивационного возбуждения, возникает ли мотивация только
под влиянием раздражителей .внутренней среды или в ее
формировании участвуют внешние факторы. Только благо-
даря акцептору результатов действия мотивационное воз-
буждение направляет поведение животного к специальным
внешним раздражителям. Наличие в любом мотивационном
возбуждении аппарата — акцептора результата действия
определяет программирование целенаправленного поведе-
ния на удовлетворение соответствующей потребности.
Такое программирование наблюдается даже у новорож-
денных животных. Оно происходит на основе врожденных
связей.
В том 'Случае, когда животное неоднократно удовлетворя-
ет свои потребности, соответствующие аппараты акцепто-
ра результатов действия мотивации все более обогащаются,
становятся более разветвленными, обеспечивая тем самым
наиболее успешное и экономное целенаправленное поведе-
ние.
Если сравнить поведение взрослого и новорожденного
животного по удовлетворению одной и той же потребности,
можно отметить их заметное различие. Соответствующий
поиск новорожденного животного неуверен. В то же время
взрослое животное немедленно бежит к тому месту, где оно
ранее неоднократно удовлетворяло данную потребность.
Что же заставляет животных, кроме исходного 'мотиваци-
онного возбуждения, устремляться к цели? Какие факторы
внешней среды способствуют этому?
IV. МОТИВАЦИИ И ЦЕЛЕНАПРАВЛЕННОЕ
ПОВЕДЕНИЕ
Само по себе мотивационное возбуждение не способно
привести животное к соответствующей цели. Это только
внутренняя неопределенная, как писал И. П. Павлов, «тем-
ная» сила. Для ее реализации необходимы специальные
внешние раздражители,
«Голод,— как пишет И. М. Сеченов,—способен поднять
животное на ноги, способен придать поискам более или ме-
нее страстный характер, но в нем нет никаких элементов,
чтобы направить движение в ту пли другую сторону и видо-
изменять его сообразно требованиям местности и случайных
встреч1.
Эту мысль И. М. Сеченова подтверждают многие психо-
логические наблюдения. В качестве примера, показывающе-
го значение внешних факторов для целенаправленной дея-
тельности человека, рассмотрим наблюдения, приводимые
Baxton, Neron и Scott (1954).
Авторы описывают следующий эксперимент, проведенный
на людях.
Обследуемые за щедрое денежное вознаграждение долж-
ны были находиться в специальных условиях, в которых они
не должны были ничего делать, видеть, слушать и не иметь
даже достаточного объема тактильных раздражений. При
этом обследуемых кормили по требованию и не подвергали
никаким неприятным раздражениям. Однако, несмотря на
наличие сильной мотивации (денежное вознаграждение и
научный интерес к эксперименту), обследуемые не смогли
находиться в указанных выше условиях более 8 часов.
Еще более показателен случай, описанный Nissen (19'53).
Автор наблюдал поведение пяти молодых пар самцов и са-
мок шимпанзе, воспитанных в изоляции. В период полового
созревания животные были разделены и снова допущены
друг к другу после начала менструаций у самок, т. е. в пе-
1 И. М. Сеченов. Первая лекция в Московском университете. Избран-
ные произведения. М., 1952, стр. 581.
207
риод их половой готовности. Однако автору в бтих условиях
не удалось наблюдать ни одного случая полового акта. Во
всех случаях поведение животных близко напоминало сек-
суальное поведение и выражалось только во взаимном уха-
живании, но не завершалось половым актом.
Близки к этому наблюдения И. С. Цитовича (1911) в ла-
боратории И. П. Павлова. Он показал, что голодные щенки,
никогда в своей жизни не питавшиеся мясом, не проявляют
никакой реакции па его запах до тех пор, пока они хотя бы
раз его ни попробуют.
Все это показывает, что, для того чтобы созревшее моти-
вационное возбуждение завершилось конечным приспособи-
тельным результатом, необходимо наличие специальных
раздражителей внешней среды, направляющих животных
к подкреплению.
Что же это за раздражители? Как организм различает
нх? Здесь мы должны более подробно остановиться на сле-
дующем вопросе,
НЕКОТОРЫЕ ОСОБЕННОСТИ ЭВОЛЮЦИОННОГО
РАЗВИТИЯ
Возраст земной коры определяется приблизительно
в 4,5 млрд. лет. Жизнь в ее примитивных формах возникла
на 0,5—1 млрд, лет позже. Все это обусловило то обстоя-
тельство, что живые существа, как указывает П. К- Анохин,
возникнув как более позднее явление эволюции, неизбежно
«вписались» в рамки существующего неорганического мира.
Явления и предметы внешнего мира 'Служили ареной, на ко-
торой развертывалась эволюция органического мира. Жи-
вые существа встретили жесткие границы, имевшие опреде-
ленную пространственно-временную конфигурацию, строгие
температурные, барометрические, гидрологические и другие
параметры. Одни группы явлений окружающего мира чере-
довались (закономерно и периодически повторялись), дру-
гие— характеризовались стабильностью, т. е. держались
неизмененными на протяжении целых геологических эпох,
третьи — раз'возникнув, никогда больше не повторялись,
и т. д. Естественно, что биологическая эволюция вошла
в соприкосновение как со стабильными (или четко периоди-
ческими) явлениями, так и с неожиданными и апериодичны-
ми событиями внешнего мира.
Последующее усложнение во взаимоотношениях организ-
мов с внешней средой было внесено эволюцией жизни. Жи-
208
йЫе существа (индивидуумы) вынуждены вступать в кон-
такты между собой (внутривидовые и межвидовые взаимо-
отношения), что особенно расширило круг неожиданных
и апериодичных событий. Тем не менее такое разнообразие
внешних по отношению к индивидууму событий не приоста-
новило поступательного хода эволюции, но потребовало
развития определенных приспособительных реакций.
Все многообразие форм приспособительных реакций всех
без исключения живых существ, когда-либо населявших
землю или населяющих ее сейчас, можно разделить на две
основные группы. К первой группе относятся инстинк-
тивные формы поведения, т. е. реакции, осуществляе-
мые по жесткой, наследственно запрограммированной схе-
ме— как форма приспособления к стабиль-
ным и четко периодичным явлениям
окружающего мира. Ко второй группе адаптивных
реакций относятся формы поведения, приобретенные в про-
цессе индивидуальной жизни; поведение, которое
развивается и совершенствуется на протя-
жении жизни одного поколения. Эта группа
поведенческих реакций обеспечивает при-
способление организма (в конечном счете выжива-
ние индивидуума и воспроизведение потомства) к неожи-
данным (экстренно изменяющимся) и апериодичным
событиям окружающего мира. Именно эта форма
приспособительного поведения была названа И. П. Павло-
вым условным рефлексом.
Следует отметить, что обе формы приспособительной дея-
тельности имеют ряд общих принципиально важных черт.
S Инициативным моментом к целенаправленной деятель-
ности во всех случаях является формирование соответствую-
щей внутренней потребности и на ее основе—мотивации.
Благодаря мотивации организм все время исследует окру-
жающие его предметы .и постоянно получает обратную ин-
формацию об их параметрах. Параметры раздражения
окружающими объектами органов чувств животного посто-
янно сравниваются с аппаратом акцептора результатов
действия, который, как мы указывали ранее, формируется
при возникновении всякой мотивации.
Таким образом, при наличии соответствующей мотивации
центральная нервная система все время программирует ко-
нечный результат, т. е. подкрепление данной мотивации со
всеми его параметрами, которые определяются как врож-
денным, так и приобретенным опытом данного животного.
209
На основе всех этих механизмов возникает специальной
поведение, которое приводит к удовлетворению соответст-
вующей потребности, лежащей в основе данной мотивации.
Итак, каждая приспособительная реакция животного может
быть представлена в 'виде следующей схемы. Начало реак-
ции определяется соответствующей внутренней потреб-
ностью организма, его мотивацией с определенной програм-
мой поведения (акцептором результатов действия). Ее за-
вершение связано с удовлетворением этой потребности,
т, е. получением полезного приспособительного результата,
удовлетворяющего данную потребность. Между этими край-
ними состояниями включено поведение и его промежуточ-
ные результаты, направленные па получение конечного при-
способительного результата. Таким образом, вся деятель-
ность организма строится на основе непрерывно идущего
сравнения свойств внешних раздражителей со свойствами
запрограммированного конечного приспособительного ре-
зультата, имеющегося в аппарате акцептора результатов
действия. Однако реализация программы поведения у раз-
ных живых существ осуществляется по-разному.
ЖЕСТКОЕ ПРОГРАММИРОВАНИЕ ПОВЕДЕНИЯ
Все формы приспособительного поведения, обеспечиваю-
щие выживание индивидуума в относительно неизмененных
условиях существования и направленного на удовлетворе-
ние его внутренних потребностей, можно с достаточной сте-
пенью условности подразделить на ряд этапов, каждый из
которых заключает определенный биологический смысл —
удовлетворение какой-либо из жизненно важных потребно-
стей, обеспечивающих сохранение индивидуума или вида.
В основе каждого такого отрезка деятельности лежит соот-
ветствующая потребность.
За примером обратимся к трудам выдающегося натура-
листа-энтомолога Ж- Фабра. Рассмотрим ряд этапов гене-
ративного поведения желтокрылого сфекса. Этот этап его
поведения может 'служить типичным примером любой фор-
мы инстинктивной деятельности.
На определенном этапе развития у осы сфекс под влия-
нием внутренних гормональных изменений и факторов
-внешней среды (температура, влажность, продолжитель-
ность дня и т, д.) возникает половая деятельность. Через
некоторое время она завершается созреванием яиц и воз-
никновением потребности в их откладывании.
210
Рассмотрим, какие процессы вызывает эта потребность.
Данный фрагмент деятельности оса начинает с того, что
в определенном месте роет всегда определенной формы нор-
ку. Затем она улетает на охоту, находит и парализует поле-
вого сверчка, нанося ему три жалящих удара в его главные
нервные узлы. Подтащив сверчка к норке, оса оставляет его
около норки на поверхности, а сама, возможно, с целью
проверки залезает в норку. После этого оса втаскивает в
норку сверчка и откладывает на его грудку яйцо. Таким же
образом она втаскивает в норку еще одного или двух сверч-
ков. Затем замуровывает вход в норку и улетает с тем, что-
бы больше сюда никогда не вернуться.
Если внимательно рассмотреть все этапы поведения осы,
нетрудно заметить, что эта деятельность развертывается
по определенной программе (рис. 46, Л). При этом вся прог-
рамма действия сфекса во внешней среде определяется ко-
нечным приспособительным жизненно важным результатом
(в данном случае кладка яйца).
По отношению к этому конечному результату деятель-
ности все предшествующие формы поведения выстраивают-
ся в определенный ряд. При этом каждый предшествующий
этап деятельности является своеобразным сигналом для
развертывания последующего.
Если воспрепятствовать совершению осой того или иного
этапа деятельности, ведущего к откладыванию яйца, она не
может перейти к дальнейшим этапам -и повторяет неудав-
шийся акт множество раз. Фабр описывает случай, когда он
неоднократно отодвигал сверчка, оставленного сфекоом
у входа во время «проверки» порки. В этом случае, выбрав-
шись из норки и обнаружив добычу слишком далеко, оса
хватает ее, подтаскивает к входу, а затем спускается в нор-
ку, но снова одна. Исследователь 40 раз отодвигал добычу
и каждый раз оса подтаскивала добычу к входу, но обяза-
тельно, оставив сверчка у входа, лезла «проверять» норку
и лишь затем возвращалась за сверчком.
Точно так же, если препятствовать выполнению любого
другого промежуточного этапа деятельности осы, например
строительству норки, охоте за сверчком и т. д., оса не при-
ступит к следующему этапу своей деятельности, пока не
совершит предыдущей и не получит полноценной информа-
ции о результате совершенной деятельности.
Таким образом, очевидно, что у осы имеется определен-
ная жестко фиксированная программа поведения, направ-
ленная на достижения конечного жизненно важного резуль-
211
Рис. 46. Разные формы
программирования пове-
дения.
Л — поведение по врожден*
ной жесткой программе.
Очередной этап деятельно-
сти развивается только пос-
ле совершения предыдущего
и получения информации Об
успехе его совершения; / —
оса роет норку, 2 — летит
на охоту, 3—парализует
сверчка, 4 — тащит его К
норке, 5 — проверяет норку,
6— откладывает яйца 7—за-
муровывает норку; .Б —прин-
ципиальная схема формиро-
вания акцептора результатов
действия и поведения при
жестком программировании.
Pi. Р2. Pi — этапные резуль-
таты деятельности; П — под-
крепление; В — поведение по
приобретенной динамической
программе. Обученное пове-
дение осуществляется с про-
пуском промежуточных ре-
зультатов действия. Объяс-
нения в тексте;
Г — принципиальная схема
формирования акцептора ре-
зультатов действия н поведе-
ния при динамическом про-
граммировании. Наиболее су*
щественный результат пове-
дения (Pi) немедленно вы-
зывает действие, направлен-
ное иа конечное подкрепле-
ние (77).
212
тэта, Эта программа,
возникая на основе
внутренней потребнос-
ти, определяется конеч-
ным приспособитель-
ным эффектом. Что это
так, указывает следую-
щий факт. После того
как оса отложила по-
следнее яйцо и замуро-
вала вход в гнездо,
можно спокойно разру-
шить всю норку па гла-
Рис. 46. (продолжение).
зах у осы вместе с от-
ложенными яйцами, и оса не проявит больше никакой реак-
ции. Она свою программу полностью выполнила. Анало-
гично реализуются и другие программы инстинктивной
деятельности, возникающие на основе других пищевых, по-
ловых и защитных потребностей.
Подобные программы в случае инстинктивной деятель-
ности определяются наследственными механизмами. Они
проявляются у таких животных, у которых потомки никогда
не встречаются с родителями. Однако реализация этих на-
следственных механизмов в действие происходит только при
наличии определенных для каждого последующего поколе-
ния факторов внешней среды, имеющих, таким образом, для
них сигнальное значение. Именно эти факторы, получившие
в зарубежной научной литературе название ключевых, и на-
правляют животное к конечному приспособительному ре-
зультату, вызывающему удовлетворение его исходных по-
требностей. При их отсутствии (например, в указанном
выше случае отсутствие в данной местности соответствую-
щей температуры и мягкой почвы), даже при наличии соот-
ветствующей мотивации, выполнение всей цепи действий,
направленных на достижение конечного результата, окажет-
ся невозможным и вид в данной местности обречен на вы-
мирание.
Имеется только очень небольшой процент сфексов, кото-
рые, как указывает Фабр, способны при неоднократном уда-
лении сверчка прекратить деятельность в этом месте и уле-
теть, чтобы снова начать ее в другом месте. Большинство
же ‘Их осуществляет многочисленные повторные и безуспеш-
ные действия. Таковы особенности программирования и со-
вершения инстинктивной деятельности.
213
Рассмотрим теперь, как осуществляется программирова-
ние и достижение конечного приспособительного эффекта
у животных, приспособившихся к жизни в изменяющихся
условиях внешней среды.
ДИНАМИЧЕСКОЕ ПРОГРАММИРОВАНИЕ ПОВЕДЕНИЯ
В изменяющихся условиях внешней среды целенаправлен-
ное поведение также периодически формируется на основе
внутренних потребностей, возникающих при изменениях во
внутренней среде организма. Эти потребности также форми-
руют системные комплексы мотивационных возбуждений,
программирующие конечный, удовлетворяющий данную мо-
тивацию результат.
Однако отличительной особенностью программ, возни-
кающих ,на основе внутренних потребностей у данных жи-
вотных, является то, что только ничтожная их часть, своего
рода «скелет», формируется наследственными механизмами.
Такие довольно примитивные, основанные на врожденных
механизмах программы по удовлетворению исходных по-
требностей наблюдаются у таких животных только сразу
после рождения и очень ненадежно сами по себе обеспечи-
вают их приспособление к окружающей среде.
Главное отличие целенаправленного поведения у живот-
ных, живущих в изменяющихся условиях среды, заключает-
ся в том, что в построении программ поведения, направлен-
ных на удовлетворение их жизненно важных потребностей,
у таких животных значительная роль принадлежит индиви-
дуальному обучению. Обучение таких животных, как прави-
ло, сначала осуществляется родителями и только затем
происходит самостоятельно. Без помощи родителей такие
животные обречены на верную гибель. Характерно, что чем
более высокоорганизовано существо (например, обезьяна
и человек), тем беспомощнее при рождении его потомок и
тем длиннее период обучения.
В результате индивидуального обучения животные начи-
нают выделять из огромного разнообразия изменяющихся
раздражителей специальные ключевые или условные раз-
дражители, надежно сигнализирующие о возможности удов-
летворения их потребностей. Параметры этих раздражите-
лей на основе механизмов памяти включаются во все более
усложняющиеся мотивационные программы поведения и на-
правляют животных на удовлетворение их потребностей.
С помощью этих направляющих сигнальных раздражителей
214
такие животные в каждом последующем случае удовлетво-
рения той или иной внутренней потребности не совершают
все те этапы, которые приводили к ее удовлетворению
в прошлом.
Следовательно, реализация программы по удовлетворе-
нию потребности идет в этом случае не по жесткому типу,
когда каждый предшествующий этап строго обязателен для
последующей деятельности, а более успешно, с устранением
промежуточных несущественных этапов. Отличительной
особенностью динамических программ является их изменя-
емость, совершенствование и экстренная перестройка в со-
ответствии с требованиями окружающей среды.
В тех случаях, когда специальные раздражители по ка-
ким-либо причинам утрачивают свое сигнальное по отноше-
нию к удовлетворению потребности значение, они устраня-
ются из программ, и их место занимают другие, более на-
дежные раздражители.
Этот .процесс выделения во внешней среде только специ-
альных раздражителей, по отношению ж которым только
и развертывается целенаправленная деятельность, происхо-
дит на всех этапах индивидуальной жизни таких животных.
Нетрудно заметить, что это выделение животными специ-
альных сигналов из массы изменчивых раздражителей
внешнего мира и построение на их основе поведения для
удовлетворения различных исходных потребностей, являет-
ся той формой поведения, которая была названа И. П. Пав-
ловым условным рефлексом.
Как строится подобный процесс, можно посмотреть на
примере поведения новорожденного голодного щенка. Вы-
пущенный в незнакомую комнату, щенок активно исследует
все окружающие его предметы. Его поведение неорганизо-
ванно. Затем в определенном месте он обнаруживает вкус-
ную для него пищу и с жадностью ее съедает. Теперь при
повторном посещении данной комнаты поведение щенка ра-
дикально изменяется. Попав в то же помещение, он сразу
устремляется к месту, где в прошлый раз обнаружил пищу.
В отличие от сфекса он не воспроизводит все элементы пре-
дыдущего поискового поведения. Он немедленно реагирует
только на те раздражители, которые непосредственно свя-
заны с пищей (в данном случае на комнату и положение пи-
щи в ней). Его поведение можно изложить в виде следую-
щих схем (см. рис. 46, В).
Следовательно, для приспособительной деятельности
щенка, направленной на удовлетворение пищевой потреб-
215
ности, из всего комплекса внешних раздражителей приобре-
тают особое значение только определенные раздражители,
которые наиболее надежно связаны с пищевым подкрепле-
нием и которые немедленно направляют его к этому под-
креплению. Если теперь мы приведем щенка в другую не-
знакомую для него комнату, то весь процесс обучения по-
вторится сначала. Однако если часто изменять комнаты,
в одних из которых животное получает пищу, а в других —
нет, то комната как сигнальный раздражитель утрачивает
свое значение. Животное в построении своего пищедобыва-
тельного поведения будет ориентироваться на другие, более
надежные раздражители.
Таким образом, поведение, организованное по динамиче-
ской программе, включает два обязательных момента.
а) Выделение из множества раздражителей внешнего ми-
ра тех, которые связываются с последующей жизненно важ-
ной деятельностью, т, е. образование условных рефлексов.
б) В случае, если эти раздражители не обеспечивают
удовлетворения потребностей животного, они теряют свое
сигнальное значение.
Все эти программы строятся на связях, как на это ука-
зывал И. П. Павлов, которые в подлинном смысле этого
слова имеют характер «временных».
Итак, одни программы по удовлетворению внутренних
потребностей организма строятся по жесткому типу на ос-
нове врожденных механизмов. В этом случае для того, что-
бы удовлетворить исходную потребность, животное обяза-
тельно должно произвести определенную серию этапных
действий и получить информацию об успешном выполнении
каждого этапа предыдущей деятельности. Другие програм-
мы динамичные, складываются в процессе индивидуального
обучения. Они не требуют для достижения конечного при-
способительного эффекта осуществления всех предшеству-
ющих этапов деятельности, а развертываются немедленно
при наличии определенных сигнальных раздражителей
внешнего мира со значительным опережением реальных
событий.
Следует отметить, что все эти формы программирования
имеют ряд общих моментов. Во всех случаях поведение
строится на основе предварительного программирования
свойств конечного удовлетворяющего данную потребность
полезного приспособительного результата, а в процессе
деятельности — на получении животным постоянной инфор-
мации об этапных результатах совершенного действия,
210
Только в случае динамического программирования дости-
жение конечного результата всегда происходит значительно
быстрее за счет пропускания ряда несущественных проме-
жуточных этапов поведения.
В каждой такой цепочке приспособительной деятельности,
направленной на удовлетворение той или иной внутренней
потребности организма, можно выделить конечный по отно-
шению к удовлетворению этой потребности полезный при-
способительный эффект и этапные, ведущие к нему резуль-
таты деятельности.
Наличие этапных результатов в случае динамического
программирования мотивационного возбуждения, естествен-
но, зависит от обученности животного. Чем более обучено
животное, тем меньше промежуточных этапных действий
оно совершает на пути удовлетворения своих потребностей.
Таким образом, в любом мотивационном возбуждении
запрограммирован окончательный результат — параметры
удовлетворяющего исходную потребность раздражителей,
т. е. подкрепления. Кроме того, запрограммированы менее
главные цели — параметры тех сигнальных раздражителей,
наличие которых уже тесно связано с последующим под-
креплением. Все это, естественно, находит свое отражение
в составе аппарата акцептора результатов действия, про-
граммирующего такое поведение.
В каждом акцепторе результатов действия, возникающем
на основе той или иной мотивации, в специальных нервных
процессах «закодированы» все ’параметры удовлетворяюще-
го исходную потребность подкрепления. В этом же аппара-
те зафиксирован опыт данного животного по удовлетворе-
нию этой потребности в прошлом.
В случае, если этот опыт фиксирован жесткими наследст-
венно предопределенными связями, то возникающее на
основе соответствующей мотивации поведение при наличии
определенных внешних раздражителей осуществляется ав-
томатически. Достижение конечного приспособительного
результата (удовлетворение данной потребности) возможно
в этом случае только по мере получения всего комплекса
афферентных возбуждений от всех этапных результатов
действия (см. рис. 46, Б).
При отсутствии же соответствующих внешних раздражи-
телей достижение конечного результата в данном случае
совсем невозможно.
В случае динамического программирования индивидуаль-
ный опыт животного фиксируется временными связями.
217
Удовлетворение основных биологических потребностей про-
исходит да этой основе значительно быстрее, так как живот-
ное в каждом последующем случае не совершает всех про-
межуточных этапов поведения, которые оно совершало
ранее для удовлетворения данной потребности. При наличии
специальных раздражителей внешней среды в данном слу-
чае животное немедленно устремляется к подкрепляющему
агенту. При этом достижение конечного результата дейст-
вия, удовлетворяющего ту или иную потребность, возможно
даже в том случае, когда отсутствуют .некоторые этапные
направляющие раздражители внешней среды (см.
рис. 46, Г).
При помощи аппарата акцептора результатов действия
каждое животное способно в деятельности своего мозга как
бы опережать реальные события действительности.
Возникает вопрос, откуда же взялась эта способность
нервной системы животных к программированию последую-
щих результатов деятельности.
Такое опережение живыми организмами явлений окру-
жающей их действительности, как указывает П. К- Анохин
(1962а), является следствием определенной направленности
эволюционного развития. Вписавшись в определенную по-
следовательность ритмических событий внешнего мира, жи-
вые клетки благодаря развитию у них 'Специальных фермен-
тативных реакций приобрели способность в ответ на дейст-
вие раздражителей, занимающих в последующей цепи
воздействия на данный организм первое место, быстро от-
вечать реакцией, характерной для действия последующих
раздражителей.
Это свойство опережения реальных событий в деятельно-
сти живой клетки явилось универсальным и получило наи-
более отчетливое выражение у клеток центральной нервной
системы, особенно в аппарате акцептора результатов дейст-
вия.
«Аппарат акцептора действия, опережающий своим по-
явлением осуществление самого действия и его конечных
результатов, и"есть то последнее звено, которое когда-то
закономерно создавалось последовательно
развивающимися событиями внешней дей-
ствительности и фиксацией в нервной системе аффе-
рентных возбуждений от конечного результата
этого ряда действий — результата, адекватно по-
ставленной для этого цели к действию» (П. К- Анохин,
1962а).
218
В функциональных системах это свойство 'наиболее отчет-
ливо выражено при формировании мотивационного возбуж-
дения. Это и понятно, поскольку из поколения в поколение
каждая мотивация имеет однотипное подкрепление. Наряду
с этим у высших животных ,и человека опережающее воз-
буждение формируется в процессе индивидуальной жизни
по отношению к действию специальных сигнальных раздра-
жителей (по типу условного рефлекса).
Значение акцептора результатов действия в целенаправ-
ленном поведении чрезвычайно велико.
Именно этот аппарат наряду с инициативной, тонизирую-
щей силой восходящих активирующих влияний на кору го-
ловного мозга подкорковых мотивационных центров направ-
ляет животное на активный поиск потребного вещества. Его
роль в этом смысле заключается прежде псего в том, что он
определяет целенаправленное поведение животного по свое-
образной «модели», постоянно осуществляя сравнение
с ней параметров воздействия внешних факторов. Близкую
точку зрения отстаивает И. С. Бериташвили (1961, 1966).
Согласно И. С. Бериташвили, целенаправленная деятель-
ность животных и человека направляется образами опреде-
ленных раздражителей внешнего мира. Формирование та-
ких образов зависит от обучения и памяти животных.
Понятно, что чем более обучен организм, тем богаче его
аппарат акцептора результатов действия, тем успешнее
происходит удовлетворение возникшей потребности. Необу-
ченный организм, .как правило,'направляется к цели только
несовершенным аппаратом акцептора результатов действия,
заложенным генотипически .в соответствующем мотивацион-
ном возбуждении.
Каким же образом организм оценивает результат совер-
шенного действия? В этом смысле значительная роль при-
надлежит аппарату эмоций.
МОТИВАЦИИ И ЭМОЦИИ
Согласно биологической теории эмоций П. К- Анохина
(1949, 19646), последние возникли в эволюции как средство
для обеспечения наиболее успешного и быстрого удовлетво-
рения потребностей организма. Основное биологическое
значение эмоционального переживания заключается в том,
что по существу только оно позволяет животному и челове-
ку быстро оценить свое внутреннее состояние, свою возник-
шую потребность и быстро построить адекватную форму
219
реагирования: будь то примитивное влечение или сознатель-
ная социальная деятельность. Наряду с этим эмоции явля-
ются и главным средством оценки удовлетворения потреб-
ности.
В самом деле, как животные, так и человек, например,
никогда де оценивают истинную пищевую 'потребность в ко-
личестве белков, жиров, углеводов, витаминов и т. п. Одно
только возникновение эмоции голода достаточно для оцен-
ки возникшей потребности в пище. Аналогичным образом
потребление пищи также оценивается только эмоцией насы-
щения, а не тонким учетом того количества различных пи-
тательных веществ, которое поступает с пищей. При отсут-
ствии эмоции голода животные, естественно, могли бы по-
гибнуть в окружении питательных веществ. Не менее пе-
чальный результат мог бы быть и при отсутствии эмоции
насыщения.
Таким образом, в процессе эволюционного развития эмо-
ции заняли крайние позиции в ряду потребностей и их удов-
летворения.
Как правило, эмоции, сопровождающие любое мотива-
ционное возбуждение, относят к эмоциям отрицательного
характера. Они субъективно неприятны. Эти сопровождаю-
щие мотивации, такие отрицательные эмоциональные пере-
живания, как чувство голода, жажды, страха и т. п., опре-
деляются прежде всего «рассогласованием» во внутренней
среде, объективным выражением чего является (возникнове-
ние лежащей в основе мотивации потребности. Отрицатель-
ная эмоция, сопровождающая мотивацию, имеет важное
биологическое значение. Она мобилизует усилия животного
на удовлетворение возникшей потребности.
Эти неприятные эмоциональные переживания усиливают-
ся во всех тех случаях,когда поведение животного во внеш-
ней среде не ведет к удовлетворению возникшей потребно-
сти. т. е. к нахождению соответствующего подкрепления.
Из этого следует, что сопровождающие мотивационное
возбуждение отрицательные тягостные эмоциональные ощу-
щения способствуют тому, чтобы подкрепление было осу-
ществлено более оптимальным и ускоренным образом.
Вместе с тем удовлетворение потребности (действие под-
крепляющего раздражителя на организм), наоборот, (всегда
связано с положительными эмоциональными пережива-
ниями.
Все это показывает, что положительная эмоция возника-
ет во всех случаях, когда результаты совершенного дейсл-
220
вия соответствуют свойствам акцептора результатов дейст-
вия данной мотивации. Во всех случаях несоответствия
формируется отрицательная эмоция.
Биологическое значение положительной эмоции при удов-
летворении потребностей понятно, поскольку ома как бы
санкционирует успех поиска. Однако только этим ее значе-
ние не ограничивается. Она фиксируется в памяти и впо-
следствии как своеобразное «представление» о будущем воз-
никает всякий раз при возникновении соответствующей
мотивации. Таким образом, положительные эмоции включа-
ются в аппарат акцептора результатов действия соответст-
вующей мотивации, а это в свою очередь определяет важное
свойство мотивационного возбуждения. Обученный неодно-
кратным удовлетворением своих потребностей организм
впоследствии стимулируется к целенаправленной деятель-
ности не только отрицательной эмоцией мотивационного
состояния, но и представлением о той положительной эмо-
ции, которая связана с будущим подкреплением. Именно
на этой основе происходит формирование того состояния,
которое iB обыденной жизни носит название аппетита. Та-
ким образом, аппетит есть не что иное, как представление
о положительной эмоции, связанной с удовлетворением
данной потребности. Механизм аппетита является вторым
стимулирующим моментом целенаправленного мотивацион-
ного поведения. Он в большей степени выражен у обучен-
ных животных и может отсутствовать у новорожденных.
Итак, эмоции не только заняли в эволюции важные клю-
чевые позиции между потребностью и ее удовлетворением,
но непосредственно включились в аппарат акцептора ре-
зультатов действия соответствующей мотивации.
Во всех случаях возникновения потребности как отраже-
ния определенных изменений во внутренней среде организ-
ма появляется отрицательная эмоция неприятного характе-
ра. Эта эмоция способствует активной целенаправленной
деятельности животного при .наличии у него внутренней по-
требности. Она усиливается всякий раз, когда результаты
деятельности организма во внешней среде не удовлетворя-
ют данную исходную потребность. Наоборот, .во всех случа-
ях, когда происходит удовлетворение потребности, т. е.
организм получает адекватное исходной потребности под-
крепление, возникает положительная эмоция. При этом чем
более обучен организм, чем большее число раз в прошлом
он соответствующим образом удовлетворял данную потреб-
ность, тем в большей степени положительный эмоциональ-
221
ный компонент подкрепления представлен в акцепторе ре-
зультатов действия, формирующемся при данной мотивации.
Как видно, эта точка зрения несколько отличается от
представлений П. В. Симонова (1966), который считает, что
отрицательные эмоции возникают в результате недостатка
информации в случае невозможности удовлетворения по-
требностей, а положительные эмоции только в результате
избыточной информации. Трудно согласиться с П. В. Симо-
новым, который утверждает, что чем более обучен организм,
тем менее участвуют в его деятельности эмоциональные
ощущения. Наши наблюдения, наоборот, показывают, что
чем более обучен организм, тем в большей степени при воз-
никновении у пего соответствующей мотивации наряду с от-
рицательной представлена положительная эмоция будуще-
го подкрепления.
Поскольку, как было показано выше, эмоциональные ап-
параты тесно связаны с деятельностью лимбических образо-
ваний головного мозга, можно думать, что эмоциональный
компонент мотивационного возбуждения, особенно связан-
ный с оценкой внутренних потребностей и их удовлетворе-
ния, также локализуется в этих структурах.
Вместе с тем есть, по-видимому, все основания считать,
что эмоциональную окраску возникающей на основе моти-
вации поведенческой деятельности определяют и элементы
коры больших полушарий.
Удаление коры больших полушарий, как это наблюдалось
в .случае Goltz и Bard, вызывает у животных потерю спо-
собности адекватно оценивать внешнюю обстановку и соот-
ветственным образом реагировать на внешние стимулы,
вследствие чего и возникает реакция «ложной ярости».
Аналогично этому синдром Клювера Бюси, наблюдаю-
щийся при удалении височных и амигдальных областей ко-
ры, есть, по-видимому, не что иное, как потеря животным
способности адекватно оценивать результат совершенного
действия и внутреннего эмоционального переживания.
В процессе целенаправленного поведения немаловажную
роль играет еще один механизм.
ОРИЕНТИРОВОЧНО-ИССЛЕДОВАТЕЛЬСКАЯ ДЕЯТЕЛЬНОСТЬ
Мы уже отмечали ранее, что любая мотивация, лежащая
в основе афферентного синтеза, приводит к активному от-
ношению животных и человека к раздражителям окружаю-
щей среды.
222
Это активное отношение к внешней среде строится с nd-
мощью подчеркнутой ориентировочно-исследовательской
реакции. Последняя подобно отрицательной эмоции возни-
кает как следствие «рассогласования» во внутренней среде
организма и возникновения соответствующей потребности.
Это рассогласование обусловлено изменением тех или иных
внутренних констант организма, отклонением их от уровня,
обеспечивающего нормальную жизнедеятельность. В ре-
зультате именно эти процессы и приводят к тому, что жи-
вотные и человек начинают активно исследовать все окру-
жающие их раздражители внешней среды и выбирать толь-
ко те из них, которые удовлетворяют их исходную потреб-
ность.
Когда мы голодны, мы ищем только пищу, когда испыты-
ваем жажду,— воду и т. д. Таким образом, целенаправ-
ленный поиск всегда происходит в интересах доминирую-
щей мотивации. Вместе с тем поиск соответствующего ве-
щества во внешней среде происходит с активным исследо-
ванием всего окружающего. Организм ведет такое исследо-
вание всеми своими средствами: обнюхиванием, при-
сматриванием, прислушиванием и даже опробованием
«на зуб».
Исследования последних лет показывают, что каждое
мотивационное возбуждение не только распространяется
на определенные моторные элементы, но и приводит к по-
вышению чувствительности некоторых, в каждом случае
специальных периферических рецепторов. Так, у голодного
животного и человека резко повышается чувствительность
обонятельных и вкусовых рецепторов. В то же время она
значительно снижается после приема пищи (Н. С. Зайко,
1956; А. И. Есаков, 1961). По-видимому, именно этот меха-
низм позволяет голодному более направленно и «обострен-
но» искать пищу.
Подобные наблюдения были сделаны Richter (1947) и
Young (1941). Авторы искусственно повышали у крыс по-
требность к определенным веществам. Это достигалось
путем лишения животных определенного вида пищи, мине-
ральных веществ или витаминов, а также экстирпацией
некоторых эндокринных желез. Такие процедуры резко
повышали предпочтение животных только к тем веществам,
которых вследствие экспериментальных приемов не хвата-
ло в организме. После перерезки вкусовых нервных волокон
эти животные утрачивали избирательное отношение к этим
веществам. Здесь снова отчетливо выступает биологическое
223
Значение повышения чувствительности пищевых рецепторов
для активного поиска пищи во внешней среде.
Однотипные изменения чувствительности происходят в
рецепторах глотки и в случае мотивации жажды. Это на-
ходит свое выраженное изменение в субъективном ощуще-
нии «сухости в горле». Аналогично повышается чувстви-
тельность рецепторов половых органов при половом воз-
буждении.
Указанные выше наблюдения находят подтверждение в эк-
спериментах с искусственным вызыванием мотивации при
раздражении определенных центров гипоталамуса. Мас
Donne! и Flynn (1966), раздражая у кошек через вживлен-
ные электроды область заднего гипоталамуса, получали
реакцию атаки на крысу. При этом резко возрастала чув-
ствительность рецепторов кожи, расположенных около ро-
товой полости.
Н. А. Рожанский (1957) указывает на то, что проявление
пиломоторной реакции на спине при вспышке ярости созда-
ет особую повышенную чувствительность волосковых ре-
цепторов. У таких животных легкое прикосновение даже
к отдельному волоску вызывает вспышку реакции ярости.
По-видимому, такая реакция является биологически оправ-
данной, она повышает возможность животного на выжива-
ние, создавая ему наиболее выгодные условия для мгновен-
ного реагирования на опасность.
Таким образом, мотивационное возбуждение в своей глу-
бокой природе за счет распространения на определенные
периферические рецепторы создает условия наиболее на-
правленного поиска потребного вещества.
Следует подчеркнуть, что ориентировочно-исследователь-
ская реакция является постоянным атрибутом любой моти-
вации. Она тесно связана с ней даже своими нейрофизиоло-
гическими механизмами.
Эксперименты показывают, что при слабом раздражении
мотивационных центров возникает ориентировочно-иссле-
довательская реакция соответствующей биологической на-
правленности. Показателен в этом отношении эксперимент,
проведенный в нашей лаборатории В. И. Вадиковым (В. И.
Вадиков, К. В. Судаков, 1968). Кролику в область лате-
рального гипоталамуса вживлялся биполярный электрод.
После операции раздражение этой области электрическим
током вызывало у накормленного животного дополнитель-
ный прием пищи. Во время раздражения кролик набрасы-
вался на капусту и поедал ее с большой жадностью. Одна-
224
ко, если исследователь уменьшал силу раздражения, кро-
лик проявлял только следующую реакцию. Он обнюхивал
окружающие предметы, поднимал уши, прислушивался,
т. е. проявлял отчетливое беспокойство и поиск.
Как же объяснить приведенный выше эксперимент?
По-видимому, раздражая пищевой центр гипоталамуса
и повышая тем самым его возбудимость, мы, естественно,
вызываем своего рода внутреннее «рассогласование» в ор-
ганизме кролика. Животное накормлено, его кровь богата
питательными веществами, и обычно это снижает актив-
ность пищевого центра до минимального уровня. И вдруг
на этом фоне активность этого центра резко возрастает.
Именно это «внутреннее рассогласование» и вызывает ори-
ентировочно-исследовательскую, а затем и поисковую ре-
акцию животного. Сначала поисковая реакция животного
неопределенна, но при усилении раздражения, при дости-
жении определенного порога, когда возбуждения пищевого
центра начинают распространяться на другие отделы голов-
ного мозга и, в частности, на его кору, возникает направ-
ленная пищевая реакция.
Итак, ориентировочно-исследовательская реакция всегда
сопровождает мотивационное возбуждение.
Именно с помощью ориентировочно-исследовательской
реакции животное при наличии соответствующей мотива-
ции целенаправленно осуществляет поиск раздражителя,
удовлетворяющего его исходную потребность. Мотивации,
таким образом, представляют собой только основу, с по-
мощью которой разыгрываются активные исследователь-
ские реакции животного во внешней среде. Но мотивации
при этом играют ведущую роль. Достаточно вспомнить на-
блюдения Beach, показывающие, что самки обезьян начи-
нают избирательно принимать самцов только на высоте
астрального цикла.
В осуществлении мотивационной ориентировочно-иссле-
довательской деятельности важная роль принадлежит так-
же аппарату акцептора результатов действия. Именно с
этим аппаратом происходит постоянное сравнение резуль-
татов совершенного действия.
Ориентировочно-исследовательская реакция возникает и
усиливается во всех случаях, когда результат совершенного
действия не соответствует свойствам сформированного на
основе афферентного синтеза акцептора результатов дей-
ствия, т. е. во всех случаях, когда возникает «рассогласо-
вание» в поведенческой деятельности.
8 Заказ 5074
225
В результате включения ориентировочно-исследователь-
ской реакции немедленно перестраивается афферентный
синтез, принимается новое решение, строится новая про-
грамма действия и поиск продолжается в новом направле-
нии.
Таким образом, ориентировочно-исследовательские реак-
ции активно включаются не только в процесс формирова-
ния мотивации, но и в другое звено центральной архитек-
туры функциональных систем, а именно в тот пункт пове-
денческого акта, где включена эмоция и происходит оцен-
ка результатов совершенных действий.
Столь важная интегрирующая роль ориентировочно-ис-
следовательской реакции в приспособительных реакциях
имеет под собой определенный нейрофизиологический ме-
ханизм.
Известно, что любая ориентировочно-исследовательская
деятельность строится на основе в высшей степени энерге-
тически богатых генерализованных восходящих активирую-
щих влияний ретикулярной формации на кору головного
мозга (Е. Н. Соколов, 1959; О. С. Виноградова, 1961,
и др.).
Некоторые авторы (Mac Dougall, 1923) пишут о специ-
альном исследовательском влечении или инстинкте любо-
пытства.
У животных, стоящих ближе всего к человеку, напри-
мер у обезьян, любопытство выражено, как известно, очень
сильно. Оно проявляется в стремлении не только прибли-
зиться к определенному объекту и сосредоточить на нем
деятельность всех своих органов чувств, но и активно ощу-
пать его руками.
Tolman (1932) считает, что любопытство так же, как
и стадность, является вторичной мотивацией, формирую-
щейся на основе обучения. Вместе с тем эту точку зрения
не разделяют другие авторы, которые считают, что иссле-
довательские влечения являются врожденными первичны-
ми мотивациями и относят их к разряду «основных влече-
ний» организма.
В то же время признание врожденности исследователь-
ского влечения ни в коей степени не должно означать от-
рицания влияния факторов внешней среды на его форми-
рование. Постоянное восприятие внешней среды и выбор из
нее специальных раздражителей, удовлетворяющих внут-
ренние потребности организма, значительно усложняет
функциональную организацию этого влечения.
226
Будучи на первых порах жизни животного тесно связан-
ной с другими основными влечениями, ориентировочно-ис-
следовательская реакция в дальнейшем может приобрести
и самостоятельное биологическое значение.
И еще одно замечание. Мотивационное возбуждение,
формируясь в определенных нервных центрах, только пос-
ле этого выходит на соответствующие периферические ре-
цепторы. Повышение чувствительности определенных ре-
цепторов является, таким образом, только вспомогатель-
ным механизмом в целенаправленной деятельности по
удовлетворению той или иной внутренней потребности.
А это в свою очередь значит, что соответствующее поведе-
ние будет проявляться и после повреждения этих рецеп-
торов, Beach (1945) и Reynnard (1938) подтвердили это
специальными опытами. Они удаляли у крыс и мышей сра-
зу же после рождения матку и влагалище, и тем не менее
у таких животных все же обнаруживалось отчетливое по-
ловое поведение. Существует, по-видимому, несколько ка-
налов распространения мотивационного возбуждения,
сформированного в нервных центрах, сразу на несколько
различных рецепторных образований, из которых одни яв-
ляются ведущими, а другие второстепенными в восприятии
результатов поведения, направленного на удовлетворение
данной внутренней потребности. Удаление ведущих рецеп-
торов приводит к компенсаторной перестройке дополнитель-
ных рецепторов, на которые и начинает усиленно выходить
данное мотивационное возбуждение и которые берут на
себя функцию оценки результатов соответствующей дея-
тельности.
Итак, все указанные выше механизмы (акцептор резуль-
татов действия, эмоции, ориентировочно-исследовательская
деятельность) направляют животное на поиск специально-
го раздражителя, удовлетворяющего исходную потреб-
ность, лежащую в основе данной мотивации. Эти раздра-
жители получили название специальных подкрепляющих
раздражителей, поскольку при их действии возникают осо-
бые процессы в организме, получившие название «под-
крепление». Естественно спросить, что же приносит под-
крепление в деятельность возбужденных мотиваций нерв-
ных центров? Каковы механизмы устранения мотивации
при подкреплении?
V. МЕХАНИЗМЫ ПОДКРЕПЛЕНИЯ
По существу подкрепление представляет собой финаль-
ную стадию всех процессов, которые приводятся в действие
каждой мотивацией.
Прежде чем перейти к изложению механизма подкреп-
ления, разберем, что же представляет собой «подкрепляю-
щий агент».
Как уже неоднократно указывалось выше, каждая моти-
вация направляет животное на ряд действий, ведущих к по-
иску специальных потребных организму веществ или, на-
оборот, удалению некоторых избыточно накапливающихся
в нем продуктов, т. е. к полезному приспособительному ре-
зультату.
Таким образом, среди многочисленных раздражителей
внешней среды животное, испытывающее ту или иную по-
требность, целенаправленно ищет только специальные. Осо-
бенность этих раздражителей состоит в том, что они по
своим физико-химическим параметрам полностью удовлет-
воряют всем «требованиям» сформированного па основе
данной потребности аппарата акцептора результатов дей-
ствия. Именно эти соответствующие свойствам сформиро-
ванного на основе мотивации акцептора результатов дей-
ствия полезные приспособительные результаты действия и
являются «подкреплением». В соответствии подкрепления
параметрам акцептора результатов действия данной ис-
ходной мотивации и заключаются подкрепляющие свойст-
ва определенных внешних раздражителей.
Таким образом, аппарат акцептора результатов действия
в мотивационном возбуждении и возбуждение, обусловлен-
ное действием на организм подкрепления (всех его пара-
метров), взаимодействует на нейронах центральной нерв-
ной системы наподобие комплементных отношений по типу
«ключ—замок». Каждая мотивационная система с акцепто-
ром результатов действия в ее составе представляет собой
специфический рецептор многопараметренных результатов
поведения животного во внешней среде (рис. 47).
228
Между мотивационным возбуждением и соответствую-
щим подкреплением всегда включено ориентировочно-ис-
следовательское целенаправленное поведение животного.
Естественно, что в случае адекватного подкрепления, ког-
да его параметры полностью совпадут с параметрами воз-
никшей на основе мотивационного возбуждения модели
акцептора результатов действия, поиск заканчивается.
В противном случае поиск продолжается.
Рис. 47. Схема взаимодействия мотивационного и подкрепляющего дан-
ную мотивацию возбуждений по комплементному типу.
М — мотивация; П — подкрепление; / — гуморальные раздражители; 2 — нервные
раздражители.
Если учесть, что и рецепторы внутренней среды, находя-
щиеся внутри кровеносных сосудов, взаимодействуют
с соответствующими константами крови по такому же прин-
ципу жестких комплементных взаимоотношений, то аппа-
рат акцептора результатов действия может быть рассмат-
риваем как многопараметренный рецептор мотивации, на-
правленный на внешнюю среду. Отличительной особен-
ностью этого рецептора является лишь то, что его свойства
определяются не только генетически, но и обучением жи-
вотного.
Важной особенностью действия на организм подкрепле-
ния в любой его форме (прием пищи, воды, избегание опас-
229
ности, половой акт и пр.) является снижение активности
всей системы инициативного мотивационного возбуждения.
Это находит свое выражение и в характере электрической
активности головного мозга.
Так, у кошки, погруженной в уретановый наркоз, как
указывалось выше, после ее кормления во всех отделах
коры головного мозга и в подкорковых пищевых центрах
регистрируется медленная высокоамплитудная активность,
характерная для состояния покоя и сна.
Такая же активность наблюдается после приема воды,
полового акта и т. д.
Как правило, действие подкрепляющего раздражителя
на организм, испытывающий мотивационное возбуждение,
сопровождается эмоцией положительного акта. Это эмо-
циональное возбуждение определяет то состояние, которое
в широком смысле слова носит название насыщения. Такое
эмоциональное насыщение наблюдается при всех формах
подкрепления: при приеме пищи, при совершении полового
акта, утолении жажды, избегании опасности и т. п.
Насыщением в широком смысле слова можно назвать
даже и такой удовлетворяющий исходную потребность
организма акт, как набирание определенного количества
воздуха в легкие и т. п.
Между тем, как правило, мы наблюдаем, что при одном
и том же мотивационном возбуждении даже одного биоло-
гического качества, скажем при голоде, разные животные,
а иногда одно и то же животное в разные периоды своей
жизни, ищут различные пищевые вещества. Из этого следу-
ет, что свойства подкрепляющего акта даже при мотиваци-
онном возбуждении одного биологического качества всегда
избирательно различны.
Что же определяет такую избирательность подкрепле-
ния? Здесь мы встречаемся с тем состоянием, которое в обы-
денной жизни, а также и в научной литературе получило
название аппетита.
ПРОБЛЕМА АППЕТИТА
Прежде всего рассмотрим вопрос о происхождении ап-
петита.
Из всего сказанного выше ясно, что аппетит связан преж-
де всего со свойствами аппарата акцептора результатов
действия, возникающего в каждом мотивационном воз-
буждении.
230
Поскольку каждая мотивация возникает на основе внут-
ренней потребности, этот аппарат прежде всего отражает
в себе свойства именно этой потребности. В формировании
сложных биологически окрашенных мотиваций (таких, на-
пример, как мотивация жажды, голода и половая) внут-
ренние потребности организма многопараметренны.
Так, у голодного животного наблюдаются изменения ком-
плекса питательных веществ внутри организма. Это — бел-
ки, жиры, углеводы, соли, витамины и т. п. Понятно, что
при возникновении пищевой мотивации пропорции в по-
требности в тех или иных питательных веществ у разных
животных, так же как и у одного и того же животного,
в разных случаях будут различны.
Все это, безусловно, находит свое отражение в свойствах
возникающего на основе данной потребности акцептора
результатов действия. А это в свою очередь и определяет
характер и направленность возникающего на основе дан-
ной мотивации поведения. Отражая специфику потребно-
сти, такое поведение направлено поэтому всегда на спе-
циальные раздражители. Характерен в этом отношении при-
мер, который приводят Wilkins и Richter (1940). Авторы
наблюдали мальчика с врожденной опухолью надпочечни-
ков, организм которого вследствие Этого недуга не был
способен удерживать соли натрия, С самого рождения этот
ребенок проявлял постоянное беспокойство и не успокаи-
вался после приема обычной пищи. В дальнейшем он начал
отдавать предпочтение соленой пище и быстро научился
просить соль жестами. Характерно, что первым произне-
сенным словом было слово «соль». Соленая пища и все,
связанное с солью, стали для этого ребенка главной целью
всей его короткой жизни. Это и понятно, поскольку возни-
кающий на основе его постоянной солевой недостаточности
акцептор результатов действия отражал в себе все пара-
метры солевого подкрепления.
Показателен в этом отношении и другой эксперимент,
разработанный в лаборатории П. К- Анохина. Опыт состо-
ит в том, что одна группа крыс лишается на 1-—2 суток
пищи, другая группа (на этот же срок)—воды, третья
группа кормится соленой пищей и, наконец, у животных
четвертой группы удаляются надпочечники. После такой
предварительной подготовки животных пускают в клетку,
где им предоставляется выбор пищи, воды и соли. Через
некоторое время все животные занимают место соответст-
венно своей доминирующей мотивации, т. е. там, где они
231
имеют возможность удовлетворить свою исходную по-
требность. Крысы первой группы сосредоточиваются у кор-
мушки с едой, второй группы — у воды, третьей группы —-
только у пресной воды и четвертой группы — у соли.
Таким образом, исходная потребность совершенно точно
определяет поведение каждого животного во внешней
среде.
Однако не только внутренние потребности определяют
избирательную направленность возникающего на основе
мотивации поведения. Этот механизм, как правило, наблю-
дается только у новорожденных высших животных, а так-
же у животных, поведение которых строится по жесткой
(инстинктивной) программе. У большинства высших жи-
вотных аппетит и его избирательность в значительной сте-
пени определяются условиями жизни индивидуума, его
приобретенными навыками. У этих животных соответству-
ющий «аппетит» может возникнуть и без исходной потреб-
ности, при действии только определенных обстановочных
раздражителей. Это особенно характерно для человека.
«Люди,— пишет Н. Миллер (1960),— не рождаются
с жаждой наживы, со стремлением открывать научные исти-
ны или занимать высокое общественное положение. Подоб-
ные побудительные мотивы усваиваются в процессе жизни
в обществе. Многие из них в значительной степени различ-
ны в различных обществах... Даже первичные побуждения
сами по себе могут видоизменяться посредством научения;
так, например, голод может стать потребностью в опреде-
ленном виде пищи, аппетитно приготовленной. Известно,
что многие французы питают отвращение к сладкой куку-
рузе и любят улиток».
Однако характерно, что такое избирательное отношение
животного к определенным факторам внешней среды в боль-
шинстве случаев строится только на основе эмоциональных
ощущений, без сознательной оценки истинной потребности.
Аппетит, возникающий только на основе внутренней по-
требности, как правило, представлен эмоциональными пере-
живаниями неприятного характера. Между тем аппетит обу-
ченного животного и человека характеризуется эмоциями
приятного характера, представлением о тех положительных
эмоциях, которые обычно сопровождает тот или иной
процесс насыщения. При одной и той же внутренней
потребности произошла своеобразная перемена знака
эмоции!
В чем же здесь дело?
232
Все становится ясным, если мы снова обратимся к мате-
риалу, изложенному в главе IV.
По мере обучения животного, неоднократно удовлетво-
рения соответствующей его внутренней потребности свойст-
ва будущего подкрепления так же, как и сопровождающая
их положительная эмоция, входят обязательными компонен-
тами в состав акцептора результатов действия данной мо-
тивации. Вследствие этого при возникновении соответствую-
щей потребности организм начинает уже испытывать пред-
ставление о той будущей положительной эмоции, которая
возникает у него при потреблении соответствующего веще-
ства, удовлетворяющего по своим параметрам исходную
потребность.
Аналогично формируется и реализуется мотивация любо-
го другого биологического качества.
Таким образом, избирательность поведения, возникающе-
го на основе каждой мотивации, определяется, с одной сто-
роны, свойствами внутренней потребности, а с другой—
свойствами, возникающими на основе данной потребности
акцептора результатов действия, в котором отража-
ется не только генетический, но и индивидуальный
опыт данного животного, а также положительные эмо-
циональные переживания, связанные с будущим под-
креплением.
Итак, исходная мотивация и формирующийся на ее осно-
ве акцептор результатов действия представляют собой еди-
ный комплекс, определяющий характер избирательного по-
ведения животного во внешней среде. В случае, если раздра-
житель внешней среды соответствует свойствам акцептора
результатов действия данной мотивации, возникает положи-
тельная эмоция и мотивация прекращается. В противном
случае за счет ориентировочно-исследовательской реакции
она еще более усиливается и заставляет животное более
активно действовать для того, чтобы удовлетворить свои
внутренние потребности.
Следует отметить, что в том случае, когда параметры
подкрепляющего раздражителя по своему жизненно важно-
му значению превышают ожидаемые на основе аппарата
акцептора результатов действия, положительная эмоция
также может усиливаться.
Какие же процессы разыгрываются в организме при дей-
ствии на него подкрепляющих раздражителей? В чем состо-
ит «подкрепляющий» смысл удовлетворяющих исходную
потребность организма раздражителей?
233
НЕЙРО-ГУМОРАЛЬНЫЕ МЕХАНИЗМЫ ПОДКРЕПЛЕНИЯ
Непосредственное взаимодействие организма с окружа-
ющим его миром осуществляется через органы чувств. Сле-
довательно, сличение свойств имеющегося в каждом моти-
вационном возбуждении акцептора результатов действия
и раздражителей внешней среды происходит прежде всего
чисто нервным путем.
Любой, в том числе и подкрепляющий, агент при дейст-
вии на организм оказывает на его рецепторы комплексное
воздействие. Это воздействие осуществляется как через ди-
стантные (глаза, уши, нос), так и контактные (кожа, слизи-
стые оболочки) рецепторы.
Таким образом, действие внешних раздражителей вызы-
вает всегда многопараметренное комплексное возбуждение,
поступающее в центральную нервную систему, где оно и со-
поставляется с акцептором результатов действия исходного
мотивационного возбуждения.
Если параметры возбуждения, обусловленного действием
внешних раздражителей, не соответствуют параметрам име-
ющегося в мотивационном возбуждении акцептора резуль-
татов действия, возникает своеобразное «рассогласование».
В результате этого усиливается ориентировочно-исследова-
тельская реакция и поиск продолжается.
Совсем другие процессы разыгрпваются при действии на
организм раздражителя, свойства которого удовлетворяют
исходную доминирующую потребность организма.
Рассмотрим все эти процессы на примере действия пищи
на организм голодного животного.
СЕНСОРНОЕ НАСЫЩЕНИЕ
Как уже указывалось выше, состояние голода у кошек
после 1—2-суточного голодания даже под уретановым нарко-
зом характеризуется наличием восходящих активирующих
влияний гипоталамических центров на кору головного моз-
га. Эти влияния проявляются в форме регионарной актива-
ции передних ее отделов.
Специальные опыты показали, что под уретановым нар-
козом животные сохраняют способность при орошении мо-
локом ротовой полости, особенно корня языка, заглатывать
молоко в желудок. Таким образом, под уретановым нарко-
зом сохранялись незаблокированными не только централь-
ные механизмы «голодного» возбуждения, но и исполнитель-
234
ные аппараты акта глотания. Этот пример является типич-
ным для характеристики избирательного действия наркоти-
ческих веществ на центральную нервную систему. Харак-
терно, что и другие наркотики с резко выраженным блоки-
рующим действием на механизмы корково-подкорковых
взаимоотношений, например нембутал, оставляют неблоки-
рованными такие стволовые реакции, как например, ды-
хание.
Факт наличия акта глотания у животных, находящихся
под уретановым наркозом, позволил нам поставить ряд спе-
циальных опытов по изучению центральных механизмов
действия на голодное животное подкрепляющих пищевых
факторов.
Опыты показали, что введение таким животным теплого
молока в ротовую полость, т. е. только орошение молоком
рецепторов языка и ротовой полости, без заглатывания мо-
лока в желудок нередко уже само по себе вызывало отчет-
ливые изменения на электроэнцефалограмме голодных жи-
вотных. Эти изменения проявлялись в двух формах. В одних
опытах вслед за орошением ротовой полости молоком на-
блюдалась генерализованная активация электроэнцефало-
грамм во всех отделах коры головного мозга. Эта актива-
ция впоследствии сменялась высокоамплитудной медленной
активностью. При этом высокоамплитудные волны нередко
группировались в «пачки» или «веретена».
В других опытах такая высокоамплитудная активность
возникала в ответ на пищевое раздражение сразу без фазы ге-
нерализованной активации. Однако в обоих случаях фаза
медленной высокоамплитудной активности была довольно
кратковременной, не продолжаясь обычно более 2—3 ми-
нут при орошении ротовой полости 20 мл молока. После
этого в передних отделах коры снова появлялась низкоам-
плитудная, высокочастотная активность.
Все эти опыты указывали на то, что устранение актива-
ции коры головного мозга, связанной с голодным состояни-
ем животного, уже происходит при действии подкрепляю-
щего пищевого раздражителя на рецепторы ротовой поло-
сти. Однако эффект такого раздражения не является дли-
тельным.
Последующие наблюдения показали, что если молоко
вслед за действием на рецепторы ротовой полости загла-
тывалось затем животным в желудок, эффект устранения
«голодной активации» в коре головного мозга был выражен
более отчетливо. После заглатывания животным в желудок
235
60—80 мл молока в ряде опытов можно было наблюдать
проявление медленной высокоамплитудной активности во
всех отделах коры головного мозга в течение длительного
времени (рис. 48J.
’*<'IA|r У-Л vA^AV'-v^T/^|М
ww L 1СГН
Рис. 48. Изменения электроэнцефалограммы голодной кошки под
уретановым наркозом после раздражения пищей рецепторов ротовой
полости и заглатывания 20 мл молока в желудок (сенсорное насыще-
ние). Обозначения электроэнцефалограммы те же, что на рис. 29.
Момент раздражения обозначен линией. Средняя и нижняя кривые —
продолжение верхней кривой.
Как же трактовать полученные данные?
Пища в приведенных выше экспериментах оказывала раз-
дражающее действие только на соответствующие рецепторы
пищеварительного тракта. При этом возникал только чисто
нервный процесс возбуждения, распространяющийся в
центральную нервную систему. Однако этого нервного
возбуждения оказывалось достаточно для того, чтобы затор-
236
мозить возбужденный «голодной» кровью пищевой центр
и подавить восходящие активирующие влияния пищевых
центров гипоталамуса на кору головного мозга.
Значит, подкрепляющее действие пищи на организм го-
лодного животного связано прежде всего с нервными
процессами. Вот почему эта фаза насыщения была названа
в нашей лаборатории фазой «сенсорного насыщения»
(П. К- Анохин, 19626). Аналогично Pfaffman (1960) называ-
ет эти процессы сенсорным подкреплением.
Значение этой фазы в биологическом смысле очень велико.
Ведь каждый из нас действительно насыщается уже за обе-
денным столом, когда пища поступает еще только в же-
лудок.
В механизме первичного насыщения заключается глубо-
кий смысл в высшей степени совершенного приспособления
животных к внешнему миру. Благодаря этому механизму
значительно сокращается тот длительный интервал времени,
который необходим для химического преобразования добы-
тых питательных веществ и поступления их в кровь. Он
позволяет закончить акт насыщения в короткий интервал
времени, несмотря на то что питательный материал нахо-
дится только в желудке и еще, естественно, не поступил в
кровь.
Как же можно представить себе механизм первичного
насыщения?
Как уже указывалось выше, пищевое возбуждение у го-
лодных животных всегда представляет собой комплекс функ-
циональных объединенных корково-подкорковых механиз-
мов. Это возбуждение, будучи основой пищевой мотивации,
вызывающей целенаправленное пищедобывательное пове-
дение, всегда имеет в себе параметры будущего необходи-
мого для организма пищевого подкрепления. При попада-
нии соответствующей пищи в ротовую полость и затем в же-
лудок происходит специфическое и избирательное возбуж-
дение тех многочисленных нервных рецепторов, которые
в обильном количестве находятся в верхней части пищевари-
тельного тракта и чувствительность которых к соответству-
ющим раздражителям, как указывалось выше, значительно
повышена.
Следствием этого является формирование специального
комплекса афферентных возбуждений, поступающих в пи-
щевой центр.
Если этот комплекс афферентных возбуждений соответ-
ствует ранее заготовленным параметрам пищевого подкреп-
237
ления в акцепторе результатов действия, имеющемся у
голодного животного, то состояние насыщения может возник-
нуть немедленно на основе чисто нервных механизмов. Про-
исходит это вследствие того, что данные афферентные нерв-
ные импульсации, поступающие от рецепторов ротовой по-
лости и желудка, могут оказаться достаточно сильными,
чтобы затормозить возбужденные «голодной» кровью «цент-
ры голода» латерального гипоталамуса.
Следствием этого является то, что система «голодного»
возбуждения в центральной нервной системе распадается,
снижаются восходящие активирующие влияния гипоталами-
ческих центров на кору головного мозга и прекращается
прием пищи.
Кроме того, в механизм подавления возбужденного пище-
вого центра импульсациями, идущими от рецепторов рото-
вой полости и желудка, может быть включен еще один до-
полнительный механизм. Он заключается в рефлекторном
выбрасывании в кровь тех питательных веществ, например
глюкозы, которые находились до этого в депонированном
состоянии.
В физиологическом смысле мы должны допустить, что по-
токи афферентных возбуждений, возникающих в рецепторах
пищеварительного тракта, должны быть всегда достаточны
по количеству, чтобы затормозить возникшее на гумораль-
ной основе «голодное» возбуждение пищевых центров гипо-
таламуса.
Естественно, что полное удовлетворение и распад системы
пищевого возбуждения, сформированного голодным состоя-
нием, происходит только в случае полного совпадения всего
комплекса афферентных нервных возбуждений, поступаю-
щих от пищеварительного канала в пищевой центр, с ранее
заготовленными свойствами акцептора результатов дейст-
вия. В случае, если интегративный комплекс возбуждения
при приеме пищи в какой-то степени не соответствует заго-
товленному голодным состоянием комплексу возбуждения,
поиск и прием пищи будут продолжаться до тех пор, пока
не будет употреблено нужное для организма количество пи-
тательных веществ.
К каким же отделам пищевого центра адресуется тот ком-
плекс афферентных нервных возбуждений, который возни-
кает в связи с приемом пищи?
Естественно предположить, что это возбуждение прежде
всего адресуется именно к тем образованиям медиальной
гипоталамической области, которые согласно литературным
238
данным в реципрокных взаимоотношениях с латеральными
отделами гипоталамуса.
В состоянии голода афферентные импультации от пище-
варительных органов вместе с действием «голодной» крови
возбуждают латеральный гипоталамус и формируют восхо-
дящие активирующие влияния на кору головного мозга. При
Рис. 49. Схематические механизмы сен-
сорного насыщения (по П. К. Анохину,
1962).
А—'раздражение пищей рецепторов ротовой
полости и желудка приводит к торможению
активированного «голодной» кровью пищевого
центра латеральной гипоталамической области.
Эти процессы в дальнейшем «подкрепляются»
поступлением питательных веществ в кровь:
ГХ — «голодная» кровь; ПА — пищевая аффе-
рентация; ПЦ — пищедобывательное поведе-
ние; Г—гипоталамус' РФ — ретикулярная фор-
мация; Б — функциональные взаимоотношения
медиального (2) и латерального (/) отделов
гипоталамуса в состоянии голода (/) и после
приема пищи (//); 3 — афферентные импуль-
сации от языка; 4 — афферентные импульса-
цни от желудка; 5 — гуморальные влияния
через гипоталамический капилляр. Объясне-
ния в тексте.
239
приеме пищи афферентные импульсации от пищеваритель-
ного тракта, возбуждая вентро-медиальное ядро гипотала-
муса, тормозят тем самым латеральный центр голода и
прекращают прием пищи.
IlfflIIHi'WHIHI MlllllllWIIIh
/------------------------
| 2--------
Рие. 50. Реакции нейронов гипоталамической области кролика на
раздражение пищей рецепторов ротовой полости и желудка (но
Б. В. Журавлеву, 1969).
/ — ответы двух нейронов (а и б) вентро-медиальной области на введение мор-
ковного сока в ротовую полость и желудок. Момент введения пищи обозначен ли-
ниямн. 1 — раздражение пищей рецепторов ротовой полости; 2 — раздражение же-
лудка. //—соотношение реакций нейронов латеральной (4) И медиальной (5)
области гипоталамуса на раздражение пищей рецепторов ротовой полости и же-
лудка; /—торможение; 2—учащение исходной активности; 3—арейктивные нейроны.
Возбуждение клеток медиального гипоталамуса должно
быть, таким образом, достаточно сильным, чтобы в свою
очередь полностью затормозить возбуждение латеральных
его отделов, определяющих всю систему центрального воз-
буждения у голодных животных (рис. 49). В этом процессе
вспомогательную роль играют активирующие механизмы той
положительной эмоции, которая сопровождает прием пищи.
240
Специальные опыты, проведенные в нашей лаборатории
Б. В. Журавлевым, подтвердили такое предположение.
С помощью микроэлектродов автор регистрировал актив-
ность различных нейронов латерального и медиального от-
делов гипоталамуса у кроликов. Он обнаружил, что в ответ
на раздражение пищей рецепторов ротовой полости или не-
посредственно желудка (последнее достигалось введением
морковного сока в желудок через зонд) преимущественно
реагировали нейроны медиальной гипоталамической обла-
сти. Характерно, что эти же нейроны отвечали и на введение
глюкозы в кровь (рис. 50).
При использовании двух микроэлектродов можно было
зарегистрировать реципрокные реакции нейронов латераль-
ного и медиального гипоталамуса. Аналогичные данные
приводят Oomura и др. (1964) и Anand и др. (1964).
На основании этих опытов можно было бы думать о прос-
тых реципрокных отношениях между медиальным «цент-
ром насыщения» и латеральным «центром голода». Такой
точки зрения, как известно, придерживаются большинство
авторов (Brobeck, 1957; Anand, 1961; Oomura a. oth., 1964,
и др.). Согласно этой точке зрения, в состоянии голода воз-
буждаются «центры голода» латеральной гипоталамиче-
ской области и последние в свою очередь оказывают тор-
мозное действие на «центры насыщения», расположенные
в области медиального гипоталамуса. Наоборот, при приеме
пищи возбуждаются «центры насыщения» и тормозятся
«центры голода».
Опыты, однако, показывают, что функциональные взаимо-
отношения этих центров гипоталамуса более сложные.
Уже тот факт, что у голодных животных, а также у жи-
вотных после приема пищи под уретановым наркозом в ла-
теральных и медиальных отделах гипоталамуса регистриру-
ется однотипная электрическая активность (К. В- Судаков,
1963; К. В. Шулейкина, 1967; С. А. Чхенкели, 1968, и др.),
заставляет выступить против признания существования
между этими центрами только реципрокных взаимоотноше-
ний.
Представления о наличии аппарата акцептора результа-
тов действия в любом мотивационном возбуждении позво-
ляет представить механизм насыщения с совершенно иной
точки зрения.
В самом деле, если в мотивационном возбуждении имеет-
ся аппарат, ожидающий результат подкрепления, следова-
тельно, в центральной нервной системе должны быть клетки,
10 Заказ 5071
241
которые возбуждаются голодным состоянием и опреде^
ленным образом «ожидают» прихода возбуждения, вызван-
ного действием пищи на рецепторы пищеварительного
тракта. Только на этой основе происходит оценка подкреп-
ления. Где же такие клетки расположены?
Согласно экспериментам Б. В. Журавлева, которые были
приведены выше, такими и являются клетки медиальной ги-
поталамической области. Благодаря конвергенции на них
всех афферентных возбуждений, идущих от пищеваритель-
ного тракта, эти клетки и производят «оценку» параметров
подкрепляющих раздражений. Наряду с этим возбуждение
клеток вентро-медиальных отделов гипоталамуса определя-
ется активностью нейронов латеральной гипоталамической
области. Снижение активности нейронов латерального гипо-
таламуса с помощью слабой (25—30 мка) анодной поляри-
зации приводит к тому, что нейроны медиальной гипотала-
мической области изменяют или полностью прекращают
свои реакции на раздражение пищей рецепторов пищевари-
тельного тракта. Следовательно, у голодных животных воз-
бужденные клетки латеральной гипоталамической области
возможно через коллатерали своих аксонов соответствен-
ным образом «настраивают» клетки вентро-медиального от-
дела на подкрепляющий раздражитель. В результате этого
такие клетки приобретают способность оценивать качество
подкрепляющего раздражения.
Специальные эксперименты, проведенные нашим сотруд-
ником А. В. Котовым (К. В; Судаков, А. В. Котов, 1969),
показали, кроме того, что такие же клетки располагаются
и в коре головного мозга, особенно в ее лобных и орбиталь-
ных отделах.
Эксперимент, подтверждающий это положение, состоял
в следующем. Кролику вживляли биполярный нихромовый
изолированный, за исключением кончика, электрод в об-
ласть «центра голода» латерального гипоталамуса. Локали-
зация кончика этого электрода именно в центре «голода»
подтверждалась тем, что электрическое раздражение, по-
даваемое на этот электрод, вызывало у предварительно на-
кормленного животного выраженную пищевую реакцию.
После тестирования локализации кончика электрода в пи-
щевом центре гипоталамуса кролика фиксировали в стерео-
таксическом аппарате (без наркоза). Под новокаиновой
блокадой у него производилась трепанация черепа и в сенсо-
моторную или в орбитальную область коры опускался мик-
роэлектрод. Замысел экспериментов состоял в следующем.
242
Если мотивационное возбуждение действительно формиру-
ет аппарат акцептора результата действия, клетки которого
«ожидают» подкрепления, значит на одних и тех же клетках
коры должна наблюдаться конвергенция мотивационных
и подкрепляющих возбуждений. Было интересно выяснить,
имеются ли такие клетки в сенсомоторной и орбитальной
области коры. Для этой.цели исследовалась реакция нейро-
А
имни
н
Рис. 51. Реакция одного и того же
нейрона сенсомоторной области коры
головного мозга на мотивационное
(А) раздражение центра «голода»
в латеральном гипоталамусе и под-
крепляющее (Б) (введение пищи в
ротовую полость) раздражение; В —
схема функциональных взаимоотно-
шений мотивационного (1) и под-
крепляющего (2) возбуждений на
одном нейроне коры головного мозга.
(По А. В. Котову, 1969).
нов этих областей код»ы на искусственное раздражение
«центров голода» гипоталамуса (т. е. при создании пище-
вой мотивации) и па раздражение рецепторов пищевари-
тельного тракта.
Опыты показали, что в сенсомоторной и орбитальной коре
кролика можно было зарегистрировать нейроны, реагирую-
щие только па раздражение пищевого центра латерального
гипоталамуса или на раздражение рецепторов ротовой по-
лости или желудка пищей. Эти реакции были самыми разно-
образными. В одном случае наблюдалось торможение спон-
танной активности, в других — ее учащение. Вместе с тем
в указанных выше областях коры были зарегистрированы
нейроны, реагирующие как на раздражение центра голода
1Q*
243
гипоталамуса, так и на раздражение рецепторов ротовой по-
лости или желудка пищей (рис. 51).
Характерно, что реакции на мотивационное и подкреп-
ляющее раздражения по направленности изменений фоно-
вой активности нейрона, как правило, были противопо-
ложны.
Если на раздражение гипоталамуса наблюдалось учаще-
ние нейронной активности, пищевое раздражение рецепто-
ров пищеварительного тракта тормозило ее. Наоборот если
раздражение гипоталамуса тормозило спонтанную актив-
ность нейрона, пищевое раздражение рецепторов пи-
щеварительного тракта активировало ее.
Характерно, что после поляризации анодом постоянного
тока области латерального гипоталамуса, т. е. разрушения
тем самым, как указывалось ранее, всего комплекса «голод-
ного» возбуждения, те же клетки коры головного мозга, ко-
торые ранее отчетливо реагировали на действие пищи, пе-
реставали на нее реагировать. Все это указывало на то, что
способность этих клеток реагировать на подкрепляющее
раздражение определялась восходящими активирующими
влияниями мотивационных центров гипоталамической об-
ласти. Последние таким образом своеобразно заряжают
клетки коры, создавая тем самым для них возможность
«ожидания» будущего подкрепления и его оценки. Благода-
ря этим влияниям создаются комплексы возбуждения мо-
торных клеток. Последние наряду с распространением сво-
их эффекторных импульсаций по пирамидному тракту к со-
ответствующему двигательному аппарату по коллатералям,
отходящим от пирамидного тракта, и возбуждают, как по-
лагает П. К. Анохин, клетки, составляющие аппарат акцеп-
тора результатов действия.
Последующие эксперименты (В. В. Синичкин, 1967) под-
твердили правомерность такого предположения. Оказалось,
что некоторые нейроны в сенсомоторной и особенно в орби-
тальной коре головного мозга у кошек и кроликов отчетливо
реагировали как на прямое раздражение «центра голода»
латерального гипоталамуса, так и на антидромное раздра-
жение пирамидного тракта (рис. 52).
Естественно, что при таком антидромном раздражении
возбуждение могло распространяться на кору головного
мозга только через коллатерали, отходящие от пирамидного
тракта.
Таким образом, антидромное раздражение пирамидного
тракта выявляет через его коллатерали все те вставочные
244
нейроны, которые имеют непосредственное отношение к ап-
парату акцептора результатов действия, созданному исход-
ной мотивацией.
Следовательно, аппарат акцептора результатов действия
при пищевой мотивации широко представлен в коре голов-
ного мозга. Есть все основания думать, что он не менее ши-
роко представлен во многих лимбических структурах, а так-
же, как указывалось выше, непосредственно на уровне гипо-
таламических пищевых центров.
Рис. 52. Реакция одного н того же нейрона орбитальной коры на
раздражение латеральной гипоталамической области и антидромное
раздражение пирамидного тракта (А) (по В. В. Синичкину, 1969).
1 реакция на антидромное раздражение пирамидного тракта; 2 —через 2 мину-
ты после раздражения; 3 — реакция нейрона на раздражение латерального гипо-
таламуса; 4— повторное раздражение латерального гипоталамуса; Б — локали-
зация кончика микроэлектрода в орбитальной коре (обозначена точкой); В — схема
конвергенции антидромного и гипоталамического возбуждения на нейроне орби-
тальной коры: ( — микроэлектрод; 2 — пункт раздражения латерального гипотала-
муса; 3 — пункт антидромного раздражения пирамидного тракта въчме места ста
перерезки.
245
Можно представить механизм формирования голодной
мотивации и насыщения следующим образом. Под влияни-
ем пищевой потребности (действия соответствующей и.м-
пульсацни от пищеварительного тракта и «голодной крови»)
возбуждаются соответствующие центры латерального гипо-
таламуса. Благодаря своим восходящим активирующим вли-
яниям на кору и подкорковые образования они наряду с воз-
буждением моторных элементов формируют «ожидающие»
соответствующего подкрепления комплексы возбуждения,
широко разветвленные по коре и подкорковым образованиям,
включая и клетки вентро-медиального отдела гипоталамуса.
Характерно, что разрушение всех этих отделов аппарата ак-
цептора результатов действия вызывает нарушение только
одной функции — способности животных адекватно оцени-
вать свое внутреннее состояние (соответствующую потреб-
ность) и количество принятой пищи. Вот почему после по-
вреждения этих отделов наблюдаются различные формы
гиперфагии или гипофагии. Аналогично гиперфагия наблю-
дается при разрушении вентро-медиальных отделов гипота-
ламуса.
Только при разрушении пейцмекера пищевой мотива-
ции— латеральной гипоталамической области — наблюда-
ется полная афагия, в основе которой лежит полное разру-
шение всей архитектуры пищевого мотивационного возбуж-
дения.
Итак, в отличие от Ананда и других авторов мы склонны
рассматривать область вентро-медиального гипоталамуса
не как центр насыщения в прямом смысле слова, а как
фрагмент акцептора результатов действия, широко пред-
ставленного при пищевой мотивации в структурах централь-
ной нервной системы.
Подтверждением такой точки зрения является и то, что
ни один автор не наблюдал, чтобы разражением вентро-ме-
диального гипоталамуса можно было вызвать натуральное
пищевое насыщение и появление на электроэнцефалограм-
ме медленной высокоамплитудной активности, характерной
для сытого состояния.
Как правило, раздражение этих отделов гипотала-
муса вызывает возникновение ориентировочной исследо-
вательской реакции или даже реакции ярости. С нашей
точки зрения, объяснение этих реакций достаточно
просто.
Раздражение вентро-медиального гипоталамуса вызывает
неадекватное подкрепление, вследствие чего возникает внут-
246
реннее рассогласование, которое при его усилении, естест-
венно, завершается реакцией ярости.
Таким образом, пищевой центр гипоталамуса устроен до-
вольно сложно. Вместе с тем в нем, безусловно, могут отме-
И!!! 1111)1 lliill^ir^l’llllthd
rill 1‘ !* 3 4 * 6 *Г|ННГН| ! II!'q!i• НИ II Н811И<Г:)Г!'ЗД
f мл
। н I и iiii^iwirrs
< Zv-'l
4 Mil
'I!' 'IBH H"111 > ! I- Ml
j 6 MB
Рис. 53. Градуальный ответ
нейрона вентро-медиальной
области гипоталамуса на
все возрастающее раздра-
жение рецепторов ротовой
полости морковным соком у
голодного кролика (по
Б. В. Журавлеву, 1970).
/ — фон; 2 — раздражение / мл;
3 — раздражение 2 мл; 4 —
раздражение 4 мл; 5 — раздра*
женне 6 мл морковного сока.
Момент орошения ротовой по-
лости обозначен утолщенной
линией внизу каждой кривой;
6 — локализация кончика мик-
роэлектрода.
247.
чаться и реакции реципрокного типа, которые наблюдались
и в экспериментах Б. В, Журавлева.
Следует подчеркнуть еще одну особенность процесса на-
сыщения.
Насыщение всегда происходит постепенно. При этом гра-
дуально нарастает возбудимость «центров насыщения» ме-
диального гипоталамуса и градуально тормозится деятель-
ность «центра питания» латеральных его отделов. Только
в том случае, когда возбуждение «центров насыщения» пол-
ностью превысит активность центров питания, происходит
окончание акта еды.
Наличие такого градуального механизма у нейронов ме-
диальной гипоталамической области показано Б. В. Журав-
левым. На рис. 53 показана реакция одного из нейронов
этой области на все увеличивающееся по продолжительно-
сти раздражение рецепторов ротовой полости пищей. За-
метно, что на каждое удлинение раздражения нейрон так-
же увеличивает продолжительность своей ответной реакции.
Именно эти механизмы определяют, почему при разной
степени мотивационного возбуждения (например, при раз-
личной длительности голодания) насыщение определяется
разным количеством принятой пищи. В каждом случае про-
цессы насыщения должны подавить различной степени
возбужденный пищевой центр латерального гипоталамуса.
Все эти закономерности отчетливо выявляются и при ис-
следовании электрической активности коры головного мозга
у голодных животных в процессе их насыщения.
Как уже указывалось выше, орошение молоком ротовой
полости у голодных животных вызывает только кратко-
временное устранение электроэнцефалографической десин-
хронизации в передних отделах коры головного мозга. Од-
нако продолжительность и величина этой реакции увеличи-
ваются при введении пищи в желудок.
Таким образом, можно совершенно произвольно удлинять
или укорачивать электроэнцефалографический эффект на-
сыщения, производя соответственно более продолжительное
или укороченное раздражение рецепторов ротовой полости
пищей. Аналогичные эффекты наблюдаются и при введении
разных количеств пищи в желудок.
Следует отметить качественное отличие механизма под-
крепления от механизма формирования мотивации. Как ука-
зывалось ранее, мотивационное возбуждение формируется
на основе триггерных механизмов. Насыщение же разверты-
вается градуально. Такая градуальность определяет важное
248
свойство процесса насыщения — точное отмеривание количе-
ства пищи, необходимого для удовлетворения исходной го-
лодной мотивации.
Иначе говоря, эфферентные возбуждения, сопровождаю-
щие акт приема пищи, тончайшим образом дозируются по
отношению к исходной возбудимости пищевых центров ги-
поталамуса, но непременно могут подавить их возбуждение,
каким бы сильным оно ни было.
На основании приведенных выше экспериментов и рас-
суждений можно заключить, что сенсорное насыщение на-
ступает только в том случае, когда общая сумма афферент-
ных нервных импульсаций, идущих от возбужденных пищей
рецепторов пищеварительного тракта, является достаточ-
ной, чтобы через возбуждения клеток вентро-медиальных
ядер затормозить возбужденные «голодной» кровью и соот-
ветствующими импульсациями от пищеварительного тракта
клетки латеральной гипоталамической области. Вот почему
небольшое количество пищи не снимает полностью чувство
голода. Оно снова нарастает через некоторое время, по-
скольку кровь при этом в значительной степени сохраняет
свойства «голодной».
Надо иметь в виду, что такой нервный механизм на-
сыщения становится более устойчивым в случае поступле-
ния пищи в желудок, чем при раздражении рецепторов
только ротовой полости. Все это еще раз указывает на то,
что в механизме торможения мотивационного центра голода
латерального гипоталамуса важное значение принадлежит
общей сумме импульсаций от рецепторов всего пищевари-
тельного тракта.
Конечно, наиболее устойчивое насыщение развивается
только тогда, когда питательные вещества из пищевари-
тельного тракта поступают в кровь. При этом развивается
истинное насыщение тканей. На смену сенсорному насыще-
нию приходит обменное, метаболическое насыщение.
Сенсорное насыщение широко представлено в различных
функциональных системах и является общебиологическим
явлением. Оно наиболее отчетливо проявляется в случае
утоления жажды. Общеизвестно, что при приеме воды жаж-
да исчезает почти мгновенно, когда вода еще только посту-
пает в желудок. Используя этот механизм, некоторые люди
нередко подавляют неприятные ощущения жажды просто
путем полоскания водой ротовой полости и глотки.
Половой акт также представляет собой типичный пример
сенсорного насыщения. Он разыгрывается полностью на ос-
249
нове чисто нервных механизмов и только после завершения
полового акта в случае оплодотворения вступают в дейст-
вие метаболические процессы.
Даже в дыхательной функциональной системе можно на-
блюдать фазу «сенсорного насыщения». Ведь вдох прекра-
щается чисто нервным путем за счет импульсаций от меха-
норецепторов альвеол еще задолго до того, как кислород по-
ступит в кровяное русло.
По существу, даже речевая и мыслительная деятельность
человека часто строится по типу сенсорного насыщения.
В различных функциональных системах интервал между
сенсорным и метаболическим насыщением может быть очень
различным. Если в дыхательной функциональной системе
он составляет секунды, то в функциональной системе пита-
ния уже десятки минут (в пределах часа). В половой функ-
циональной системе от полового акта, разыгрывающегося
на чисто нервной основе, до момента оплодотворения и рож-
дения потомства проходит несколько часов и месяцев.
Каково же биологическое значение сенсорного насыще-
ния? Рассмотрим этот вопрос на примере пищевого под-
крепления. Афферентные возбуждения от акта еды высту-
пают, как можно видеть из предыдущего изложения, в ка-
честве сигналов истинного обменного насыщения. Они ока-
зываются способными затормозить возникшую на гумораль-
ной основе пищевую мотивацию.
Каков же смысл этих процессов? Он заключается в сле-
дующем. Действие пищи на рецепторы ротовой полости и
желудка является уже в достаточной степени надежным мо-
ментом для того, чтобы организм начал расходовать имею-
щиеся у него еще запасы питательных веществ. При дейст-
вии пищи на рецепторы пищеварительного тракта по суще-
ству в организме начинают разыгрываться все те процессы,
которые характеризуют истинно метаболические процессы
насыщения. Как показывают исследования К. М. Быкова
(1947), при действии пищи на рецепторы ротовой полости
происходит увеличение содержания глюкозы в крови. Глю-
коза, как думает автор, выбрасывается из депонированного
состояния, хотя поступления питательных веществ в кровь
еще не наблюдается. Это предположение было подтвержде-
но Steffens (1969), а также С. М. Будылиной и Л. Д. Рез-
цовой (1969). Таким образом, действие пищи на рецепторы
пищеварительного тракта является для организма уже на-
дежной гарантией того, что питательные вещества в буду-
щем поступят в кровь. Такая гарантия отработана всем хо-
250
ДоМ эволюционного развития. Ведь в природе никогда не
наблюдалось случая, чтобы съеденная пища была извлечена
из желудка животного и впоследствии не поступила бы в
кровь. Только наличие из поколения в поколение такой
жесткой последовательности физиологических процессов
насыщения позволило организму использовать каждый
предшествующий этап этого сложного процесса в ка-
честве надежного сигнала для последующей деятель-
ности.
Вот почему афферентная сигнализация, идущая от ре-
цепторов пищеварительного тракта при действии на них
пищи, тормозя пищевую мотивацию, уже сама по себе при-
водит к метаболическим изменениям в организме.
Предшествуя у многих миллионов поколений живых су-
ществ истинному удовлетворению потребности, фаза сенсор-
ного насыщения приобрела по отношению к ней сигнальное
значение и закрепилась эволюцией в соответствующих мор-
фофизиологических врожденных связях.
В результате этого каждый предыдущий этап прохожде-
ния пищи по пищеварительному каналу вызывает соответ-
ствующие изменения в пищевом центре, которые как бы про-
граммируют последующие процессы. Здесь мы снова встре-
чаемся с тем типом жесткого программирования, который
наблюдается в любой инстинктивной деятельности: после-
дующий этап деятельности развертывается сразу же после
того, как центром получена соответствующая информация
о результате совершения предыдущего этапа. При этом, чем
ближе соответствующий процесс к конечному приспособи-
тельному эффекту, тем более отчетливое влияние он оказы-
вает на пищевой центр. Каждый последующий этап насыще-
ния в свою очередь усиливает и удлиняет эффект предыду-
щего этапа. Так, влияние пищи из желудка на пищевой
центр значительно сильнее, чем изолированное раздражение
пищей рецепторов ротовой полости. Введение питательных
веществ в кровь значительно усиливает эффект сенсорного
насыщения.
Здесь снова проявляется общебиологическая закономер-
ность, описанная в лаборатории, руководимой П. К. Анохи-
ным. Наибольшая сила «замыкания» в цепи потребности и
ее удовлетворения имеется между теми раздражителями,
которые наиболее близко расположены к реальному под-
креплению.
Однако и предыдущий этап оказывает заметное влияние
на последующий. Так, без предварительного раздражения
251
пищей рецепторов ротовой полости влияние на пищевой
центр от рецепторов желудка резко понижается.
Таким образом, подкрепление представляет собой после-
довательно развертывающуюся цепь событий. При этом
каждое предшествующее событие имеет сигнальное значе-
ние по отношению к последующему. Каждое же последую-
щее событие обладает по отношению к предшествующему
подкрепляющим свойством. Это подкрепляющее значение
все более нарастает по мере приближения к исходному ко-
нечному приспособительному эффекту данной функциональ-
ной системы.
Только непосредственное влияние на этот конечный при-
способительный эффект по существу может быть рассмат-
риваемо как истинное или метаболическое насыщение.
Рассмотрим теперь особенности метаболического насы-
щения.
МЕТАБОЛИЧЕСКОЕ НАСЫЩЕНИЕ
Как уже указывалось выше, раздражение пищей рецепто-
ров пищеварительного тракта тормозит активность гипота-
ламических центров пищевой мотивации. Но при отсутствии
последующего поступления питательных веществ в кровь
это торможение недлительное. Обычно через 15—20 минут
после этого снова наблюдается возрастание активности пи-
щевых центров гипоталамуса и формируются их восходя-
щие активирующие влияния на кору головного мозга. Это
наблюдается, например, в том случае, когда пища из желуд-
ка не поступает в кишечник, а выпадает через фистульное
отверстие наружу. Однако устойчивое и длительное устране-
ние «голодной» активации коры головного мозга наступает
только после введения этим животным глюкозы в кровь.
Таким образом, устойчивое насыщение определяется толь-
ко поступлением питательных веществ в кровь. Это находит
свое Подтверждение и в специальных экспериментах. Так,
устойчивое устранение исходной электроэнцефалографиче-
ской активации передних отделов коры головного мозга у
голодных животных наблюдается только через 40—60 ми-
йут после поступления пищи в желудок. После этого мед-
ленная высокоамплитудная активность сохраняется во всех
отделах коры головного мозга животного в течение несколь-
ких часов.
При анализе механизма метаболического насыщения надо
иметь в виду следующее.
252
Устойчивое насыщение (по различным показателям) раз-
вивается только в том случае, если питательные вещества
перед поступлением в кровь оказали свое действие на ре-
цепторы пищеварительного тракта. Иными словами, обмен-
ному насыщению обязательно должна предшествовать фаза
сенсорного насыщения. На это указывают следующие опыты.
Введение 2—3 мл 20% раствора глюкозы в кровь голод-
ным животным при наличии у них пустого желудка приво-
дит к устранению электроэнцефалографической активации
только кратковременно, не более чем на 15 минут. По-види-
мому, такое кратковременное действие глюкозы связано
с ее быстрой утилизацией голодным организмом при нали-
чии постоянно идущей сигнализации от пустого желудка.
Этот факт объясняет отсутствие чувства сытости у больных
при парентеральном введении питательных веществ при пус-
том желудке. По той же причине не происходит насыщения
у одного из сросшихся близнецов, с общим кровообраще-
нием. По-видимому, во всех этих случаях стимулирующее
влияние на пищевой центр оказывает афферентная сигна-
лизация от пустого желудка, которая, несмотря на времен-
ное «насыщение» крови, снова приводит животное в состоя-
ние голода.
Таким образом, на клетках гипоталамуса все время про-
исходит интеграция нервных и гуморальных факторов насы-
щения. Устойчивое насыщение развивается только в том
случае, если нервные и гуморальные факторы насыщения
действуют вместе. При этом они должны действовать в опре-
деленной последовательности: питательные вещества, посту-
пившие в кровь, должны действовать па пищевой центр толь-
ко после действия пищи на рецепторы пищеварительного
тракта.
При этом так же, как и при формировании мотивацион-
ного возбуждения, мы снова встречаемся с определенной
последовательностью событий. Сначала действуют нервные
факторы, затем их действие подкрепляется гуморальными
факторами. Любое нарушение естественной последователь-
ности поступления питательных веществ в организм приво-
дит у голодных животных только к кратковременному сни-
жению функциональной активности пищевого центра. Все
это указывает на то, что у животных в процессе эволюции
сложились тесные пространственно-временные взаимоотно-
шения между различными этапами поступления питатель-
ных веществ в организм и соответствующими изменениями
в пищевом центре.
253
Окончательное насыщение возникает только в том случае,
если пищевой центр получает весь комплекс возбуждений
как от пищеварительного тракта, так и со стороны крови, по-
полненной питательными веществами, т. е. процесс насыще-
ния идет в обычном порядке: рот — пищевод — желудок —
кишечник — кровь.
Таким образом, развертывание процесса насыщения про-
исходит по определенному закону. Каждый предшествую-
щий этап насыщения по сигнальному типу вызывает сле-
дующий.
ЭМОЦИОНАЛЬНЫЕ МЕХАНИЗМЫ ПОДКРЕПЛЕНИЯ-
ЗНАЧЕНИЕ ВОЗБУЖДЕНИЯ
ПОДКРЕПЛЯЮЩИХ ПОДКОРКОВЫХ ЦЕНТРОВ
ДЛЯ ДЕЯТЕЛЬНОСТИ КОРЫ БОЛЬШИХ ПОЛУШАРИЙ
В механизме подкрепления существенная роль принадле-
жит эмоциональному возбуждению. Как правило, любое
подкрепление сопровождается эмоцией приятного характе-
ра. Биологический смысл этой эмоции состоит в том, что
она как бы санкционирует успех поиска и позволяет оце-
нить степень насыщения, т. е. момент удовлетворения исход-
ной потребности организма.
Кроме того, как указывалось ранее, биологическое значе-
ние положительной эмоции, входящей в аппарат акцептора
результатов действия каждой мотивации, заключается в
обеспечении наиболее оптимальных путем удовлетворения
внутренних потребностей организма. В связи с этим особой
важности вопрос заключается в том, каким образом поло-
жительное эмоциональное возбуждение формируется в цент-
ральной нервной системе и как оно влияет на аппарат моти-
вационного возбуждения. У нас имеются все основания пола-
гать, что положительное эмоциональное ощущение возни-
кает при снижении функциональной активности центров,
определяющих исходное мотивационное возбуждение. Это
подтверждает ряд факторов.
Процесс потребления пищи голодным животным связан
с появлением в коре головного мозга медленных высокоам-
плитудных электрических колебаний (А. И. Шумилина,
1961; К- В. Судаков, 1962; А. Н. Бакурадзе, 1963; В. А. За-
болотных, 1964; К. В. Шулейкина, 1967; Hockman, 1964;
Clemente a. oth., 1964; Morgane, 1966). Такая же электри-
ческая активность появляется в коре и при половом акте
(И. В. Орлов, 1961; К. Лишшак и Э. Эндреци, 1967;
Kawakami a. Sayer, 1959). То же самое наблюдается у жи-
254
вотных после реакции самораздражения (Ю. А. Макарен-
ко, 1964, 1966; Н. Г. Михайлова, 1968; Newman, 1967).
Между тем не исключено, что положительные эмоции на-
сыщения связаны с возбуждением и специального аппарата,
отличающегося от аппарата исходной мотивации.
На справедливость подобного предположения указывают
наблюдения Ю. А. Фадеева и А. В. Котова, которые обнару-
жили при действии пищи на рецепторы пищеварительного
тракта, а также при введении глюкозы голодным животным
в сенсомоторной коре наряду с нейронами, снижающими
свою исходную функциональную активность, нейроны, отве-
чающие усилением своей активности.
За такое предположение высказывается и Stein (1964),
который считает, что в центральной нервной системе имеют-
ся две различные системы. Одна обеспечивает положитель-
ные, другая — отрицательные эмоции.
В лаборатории, руководимой П. К. Анохиным, В. И. Ва-
диков (1969) произвел специальные эксперименты с целью
определить, как скажется в деятельности различных нейро-
нов коры головного мозга у кролика раздражение центров
гипоталамуса, дающих при их предварительном тестирова-
нии реакцию самораздражения или избегания. Опыты по-
казали, что как зоны самораздражения, так и зоны избега-
ния при их раздражении оказывают отчетливое влияние на
деятельность отдельных нейронов коры больших полуша-
рий (рис. 54, А). Такие влияния проявлялись как в учаще-
нии, так и в урежении спонтанной активности этих нейро-
нов. Больше всего на данные раздражения реагировали
нейроны сенсомоторной области, затем орбитальной, височ-
ной и слабее всего реакция проявлялась у нейронов заты-
лочной области. Все это еще раз подтверждает ранее вы-
сказанное нами положение о преимущественном распрост-
ранении мотивационного возбуждения па нейроны передних
отделов коры головного мозга.
Характерно, что процент нейронов коры, реагирующих на
раздражение зон избегания, был значительно выше.
Раздражение указанных выше зон гипоталамуса оказы-
вало, кроме того, заметное влияние и на реакцию корковых
нейронов на специфические сенсорные раздражители. На
рис. 54, Б показано изменение реакции нейрона затылочной
области на свет при одновременном раздражении «поло-
жительной зоны» гипоталамуса.
Интересным оказался тот факт, что в передних отделах
коры головного мозга были обнаружены нейроны, которые
255
отвечали как на раздражение положительной, так и отрица-
тельной эмоциональной зоны гипоталамуса. Это еще более
подтверждает точку зрения о том, что каждое различное по
биологическому знаку (положительное и отрицательное)
эмоциональное состояние может определяться разными си-
латерального гипоталамуса
переднего гипоталамуса
Рис. 54. Реакции корковых нейронов на
раздражение эмоциональных центров гипо-
таламуса (по В. И. Вадикову, 1969).
Л—реакция одного и того же нейрона сенсомо-
торной области коры на раздражение положитель-
ной и отрицательной зоны самораздражения в ги-
поталамусе; / — раздражение положительной зоны
(3 в, 48 гц); 2 — раздражение отрицательной зоны
(2,5 в, 48 гц); Б — реакция одного и того же нейрона затылочной области коры
на световое раздражение (1), на раздражение положительной зоны самораздра-
жения латерального гипоталамуса (2) н на совместное разраженнс (3). Световое
раздражение обозначено точками. Раздражение гипоталамуса — вертикальными
линиями и артефактом раздражения; В — схема конвергенции на одном и том
же нейроне коры сенсорного (3), отрицательного (2) и положительного (1) эмо-
циональных возбуждений.
стемами корково-подкорковых элементов, проявляясь раз-
личным образом в деятельности даже одних и тех же нейро-
нов коры.
Таким образом, возбуждение эмоциональных центров ги-
поталамуса оказывает заметное влияние на деятельность
клеток коры головного мозга. Именно эти процессы акти-
вирующих влияний подкорковых эмоциональных центров
на кору головного мозга, по-видимому, обусловливают эмо-
циональный компонент мотиваций и их удовлетворения.
Как это происходит? Можно думать, что сначала клетки
коры головного мозга получают сигнализацию от возбуж-
денных соответствующей потребностью мотивационных
центров гипоталамуса. После этого вторичным путем они
получают сигнализацию от соответствующих подкорковых
эмоциональных центров, порождающих отрицательную эмо-
256
цию. Точно так же и при подкреплении эти клетки сначала
получают афферентацию о действии соответствующего
подкрепляющего раздражителя на организм и после этого
санкционирующую афферентацию от эмоциональных цент-
ров, порождающих эмоцию положительного знака.
Важно подчеркнуть, что к одной и той же клетке коры
головного мозга принципиально могут поступать различные
потоки восходящих возбуждений. С одной стороны, это —
восходящие возбуждения мотивационных гипоталамических
центров и центров соответствующей отрицательной эмоции,
с другой — подкрепляющие возбуждения. Таким образом,
определенные клетки коры как бы находятся в тисках мо-
тивационного и подкрепляющего возбуждения.
Если учесть, что эти же клетки могут вовлекаться в воз-
буждения различной сенсорной модальности, например при
действии на организм света или звука, становится ясным,
что здесь происходит взаимодействие мотивационных, сен-
сорных и подкрепляющих возбуждений в построении пове-
денческих актов. Эти же процессы в коре головного мозга
определяют и механизмы образования условного рефлекса.
Согласно представлениям П. К. Анохина (1968), услов-
ный рефлекс образуется в результате конвергенции возбуж-
дений различной сенсорной и биологической модальности
на одни и те же нейроны коры головного мозга. Именно
это и послужило исходной предпосылкой для постулирова-
ния «конгенвертной теории» замыкания условного рефлек-
са (П. К. Анохин, 1968).
Вопрос состоит в том, какие процессы разыгрываются
в самой нервной клетке коры головного мозга при поступле-
нии к ней мотивационных, сенсорных и подкрепляющих
возбуждений? В чем истинный смысл подкрепления? Поче-
му при неоднократном удовлетворении потребности эти
клетки программируют всю цепь событий, ведущих к под-
креплению? Для ответа на эти вопросы необходимы даль-
нейшие эксперименты .
ОБЩИЕ ЗАМЕЧАНИЯ
Итак, мы рассмотрели вопрос, какое место занимают мо-
тивации в деятельности функциональных систем организма,
а также каким образом строится в этих системах мотива-
ционное возбуждение. Подведем некоторые итоги.
ЭНЕРГЕТИЧЕСКАЯ ОСНОВА МОТИВАЦИЙ
Если рассматривать мотивации с физиологической точки
зрения, то мотивационное возбуждение — это прежде всего
организованное на уровне подкорковых образований и коры
возбуждение. Это возбуждение строится на основе внутрен-
них потребностей и избирательно охватывает элементы
различного уровня центральной нервной системы. При этом
центральным пунктом любого мотивационного возбуждения
является гипоталамическая область. Здесь сосредоточены
клетки, метаболизм которых избирательно чувствителен
К той или иной внутренней потребности. Под влиянием этой
потребности гипоталамические центры строят специфичес-
кие для каждой мотивации восходящие активирующие
влияния на другие отделы головного мозга (лимбические
образования, ретикулярная формация среднего мозга
и, наконец, кора больших полушарий).
Эти специфические восходящие влияния при разных моти-
вациях объединяют корковые элементы всегда избиратель-
но на основе их специфической химической чувствительно-
сти. Именно на основе такого избирательного объединения
корково-подкорковых элементов при каждой мотивации
возникает целенаправленное поведение, ведущее к удовлет-
ворению исходной потребности организма. Наряду с восхо-
дящими активирующими влияниями подкорковых мотива-
ционных центров на кору головного мозга клетки послед-
ней в свою очередь оказывают нисходящее влияние на эти
центры. Таким образом между подкорковыми мотивацион-
ными центрами и корой головного мозга устанавливаются
циклические, реверберирующие взаимоотношения. Эти про-
цессы, взаимоусиливая друг друга, и составляют энергети-
258
ческий субстрат мотиваций, активно направляющий живот-
ных к удовлетворению внутренних потребностей. Таким об-
разом, та «слепая сила» подкорки, о которой писал
И. П. Павлов, приобретает конкретное нейрофизиологи-
ческое выражение.
Мотивации формируются именно на основе таких восхо-
дящих активирующих влияний подкорковых образований
на кору головного мозга. Благодаря этим механизмам весь-
ма малое количество начальной энергии, которое образует-
ся при возбуждении центральных рецепторов, может через
мобилизацию корковых элементов привести последователь-
но к появлению огромной энергии. Упомянутые механизмы
обеспечивают протекание этих реакций по типу доминант-
ных реакций А. А. Ухтомского.
Следует подчеркнуть, что формирующиеся из нервных
элементов различных уровней центральной нервной систе-
мы мотивационные возбуждения находятся всегда в зави-
симости от исходного пункта — рецептора гуморальных
изменений. Этот пункт является своеобразным пейцмеке-
ром мотивационного возбуждения.
Мотивационное возбуждение, таким образом, является
активным компонентом различных функциональных систем
организма, деятельность которых направлена на удовлет-
ворение внутренних потребностей. Каждая потребность ор-
ганизма формирует специфическую мотивацию, которая че-
рез соответствующее целенаправленное поведение и приво-
дит организм к удовлетворению соответствующей потреб-
ности. Из этого явствует, что значение мотивационного воз-
буждения в деятельности различных функциональных сис-
тем заключается прежде всего в мобилизации организма
на активное поведение во внешней среде.
Наряду с этим мотивационное возбуждение «оживляет»
и мобилизует на удовлетворение соответствующей потреб-
ности как врожденный, так и приобретенный опыт дан-
ного индивидуума. Но не только «оживляет». На основе
мотивации в свою очередь происходит активное обучение,
животные и человек строят свой индивидуальный опыт и в
конце концов ' свою личность. Вспомним пример Richter
с мальчиком, у которого было врожденное поражение над-
почечников. Ведь первым его словом было «соль»!
По-впдимому, в зависимости от доминирования той или
иной врожденной мотивации происходит формирование
различных типов психической деятельности животных
и человека. Конечно, при этом, безусловно, огромное влия-
259
ние оказывает внешняя, а у человека социальная среда
и соответствующее воспитание. Но тем не менее, учитывая,
что именно мотивации определяют активное отношение
животных и человека к внешним раздражителям, можно
думать, что исходная доминирующая биологическая моти-
вация, безусловно, должна сказываться на формировании
личности.
МОТИВАЦИИ и эмоции
В формировании целенаправленного поведения, вызванно-
го исходной мотивацией, как указывалось выше, важная
роль принадлежит эмоциональному компоненту.
В процессе эволюционного развития эмоции заняли клю-
чевые 'Позиции в целенаправленном поведении между
потребностью и ее удовлетворением. Потребность сопровож-
дается эмоциями неприятного качества, 'в то время как ее
удовлетворение сопровождается приятными эмоциональны-
ми переживаниями.
Эмоции, сопутствующие мотивационным реакциям,
возникли в эволюции как одно из средств наиболее быстрого
и надежного обеспечения приспособительных функций.
И. П. Павлов по этому поводу писал: «Кто отделил бы
в безусловных сложнейших рефлексах (инстинктах) физио-
логическое соматическое от психического, т. е. от пережива-
ний, могучих эмоций голода, полового влечения, гнева и т. д.?
Наши чувства приятного, неприятного, легкости, трудности,
радости, мучения, торжества, отчаяния и т. д. связаны то
с переходом сильнейших инстинктов и их раздражителей
в соответствующие эффекторные акты, то с их задержани-
ем...» (Условный рефлекс. Большая медицинская энцикло-
педия.)
Эмоции голода, жажды и какие-либо иные неприятные тя-
гостные эмоции стимулируют возникновение соответствую-
щих мотиваций, т. е. способствуют оценке организмом по-
требности и более успешной мобилизации организма на
поиск необходимого раздражителя. Наоборот, любое под-
крепление (удовлетворение) основных влечений всегда
связано с положительными эмоциями, которые определяют
удовлетворение потребности, и, как правило, завершается
общим успокоением и даже сном.
Эмоции, сопровождающие различные мотивации у ряда
животных, имеют внешнее проявление в виде специальных
эффекторных реакций, а также в форме разнообразных
260
Звуковых сигналов. Каждому хорошо известны звуковые ре-
акции различных животных, сопровождающие состояние
голода, половое возбуждение и т. д. Точно также и положи-
тельные эмоции, сопровождающие удовлетворение мотива-
ции, сопровождаются у некоторых животных специальными
звуками. Это «звуковое оформление» мотивации обладает
такой выразительностью и специфичностью, что вместе с це-
ленаправленным поведением представляет собой весьма
отчетливую форму передачи информации. Участие эмоцио-
нальных компонентов определяет и другую сторону моти-
вационного возбуждения. Наряду с формированием целе-
направленного поведения любая мотивация всегда специ-
фически сказывается на работе внутренних органов.
Вегетативное обеспечение каждой мотивации является край-
не специфичным и весьма тонко отражает сопровождающее
ее эмоциональное состояние.
мотивации и Акцептор результатов действия
Наследственные механизмы, а также индивидуальный
опыт животного формируют в каждом мотивационном
возбуждении аппарат, ожидающий результаты будущего
подкрепления (акцептор результатов действия). Во всех
случаях целенаправленного поведения, обусловленного ис-
ходной мотивацией, результаты поведения всегда контроли-
руются и сравниваются со свойствами этого аппарата,
который, по-видимому, и представляет ту констелляцию
возбужденных центров, о которой писал А. А. Ухтомский-
Акцептор результатов действия в каждом мотивационном
возбуждении в своей сути всегда заключает все необходимые
свойства потребного вещества. Таким образом, этот аппарат
как бы программирует все свойства адекватного данной
мотивации подкрепления и, представляя своего рода «рецеп-
тор», следящий за поведением животного во внешней среде,
активно оценивает результаты поведенческой деятельности.
По мере неоднократного подкрепления каждой мотивации
механизмы акцептора результатов Действия отшлифовы-
ваются, автоматизируются и начинают проявлять свое по-
буждающее действие всякий раз в связи с возникновением
той или иной мотивации.
Следует отметить, что На основе обучения аппарат акцеп-
тора результатов действия может возникнуть и действовать
И без наличия непосредственно первичной мотивации, а
только под влиянием внешних, а у человека словесных
261
раздражителей, которые могут быть связаны с «основными
влечениями» только самым отдаленным образом.
Аппарат акцептора результатов действия оценивает как
исходную мотивацию, так и удовлетворяющие ее факторы и
широко представлен в центральной нервной системе в коре
и в подкорковых аппаратах. Эта оценка, как правило, преж-
де всего осуществляется с помощью эмоциональных меха-
низмов. Именно эмоции у большинства животных являются
единственным аппаратом оценки мотивационного возбуж-
дения и его удовлетворения. На основе литературных дан-
ных (Mac Lean, 1958, И. С. Бериташвили, 1961) и наших
собственных исследований есть все основания предполагать,
что субъективная оценка мотивационного возбуждения и не-
которых основных параметров будущего подкрепления свя-
зана прежде всего с деятельностью лимбических образова-
ний головного мозга. В тс же время аппарат акцептора ре-
зультатов действия в каждом мотивационном возбуждении
(т. е. ожидание определенных свойств будущего подкрепле-
ния), связанный с приобретенным опытом данного индиви-
дуума, безусловно, наряду с лимбическими образованиями
определяется деятельностью и коры больших полушарий.
Однако все эти аппараты функционируют только при на-
личии исходного энергетического компонента мотивацион-
ного возбуждения — восходящих активирующих влияний
гипоталамуса и ретикулярной формации на кору головного
мозга.
ОПРЕДЕЛЕНИЕ МОТИВАЦИИ
Как же на основании всего сказанного выше дать опре-
деление понятия «биологические мотивации»?
Как мы могли видеть из всего приведенного материала,
«биологические мотивации» (основные влечения организ-
ма) возникают на основе определенных, в каждом случае
специальных внутренних потребностей организма. Эти по-
требности приводят в возбуждение специальные подкорко-
вые центры, особенно гипоталамической области, которые
за счет тонических восходящих активирующих влияний па
кору головного мозга избирательно охватывают корково-
подкорковые аппараты и заставляют организм активно вы-
бирать из внешней среды раздражители, удовлетворяющие
его исходную потребность.
Таким образом, мотивации представляют собой возни-
кающее на основе внутренней потребности специально ор-
ганизованное возбуждение корково-подкорковых аппара-
262
тов, вызывающее целенаправленную деятельность живых
существ.
Любая мотивация высокоактивна. Такое «заряженное»
мотивационное состояние организма определяется избира-
тельной активацией со стороны подкорковых образований
па кору головного мозга.
Наряду с энергетической стороной мотивация характе-
ризуется содержанием, определенной программой, направ-
ляющей поведение животного только на определенные
внешние раздражители. Программа мотивационного воз-
буждения определяется как врожденными механизмами,
так и приобретенным опытом (памятью) по удовлетворе-
нию соответствующей потребности.
Направленность мотивации определяется наличием в
структуре мотивационного возбуждения аппарата, заклю-
чающего в себе свойства подкрепления (акцептор резуль-
татов действия). ?1менно этот аппарат и определяет на-
правленность мотивации, поскольку в своей поисковой
деятельности животные постоянно производят сравнение
параметров внешних раздражителей с исходными свой-
ствами этого аппарата.
Кроме того, каждое мотивационное возбуждение так же,
как и его удовлетворение, характеризуется определенными
эмоциональными ощущениями.
Таким образом, мотивации представляют со-
бой возникающие под влиянием первичных
изменений во внутренней среде эмоцио-
нально окрашенные состояния организма,
характеризующиеся избирательными акти-
вирующими влияниями специальных под-
корковых аппаратов на кору головного
мозга и направляющие животное на удов-
летворение исходной потребности.
Как правило, все мотивации формируются по доми-
нантному принципу, т. е. сначала формируется и удовлет-
воряется та мотивация, в основе которой лежит наиболее
существенная для жизни организма потребность.
В отличие от реакций, развивающихся по принципу
«стимул — реакция», в случае мотивационного возбужде-
ния происходит активный подбор раздражителей внешней
среды. Это происходит потому, что в любом мотивационном
возбуждении есть свойства потребного вещества. ВнеЩние
раздражители, таким образом, могут только вскрыть ис-
ходное мотивационное возбуждение. Такой активный под-
263
бор специальных раздражителей внешней среды происхо-
дит с помощью постоянно сопутствующей мотивационному
возбуждению ориентировочно-исследовательской реакции.
Как было показано, при мотивационном возбуждении про-
исходит избирательное повышение чувствительности опре-
деленных специальных рецепторов. Поиск заканчивается
только в том случае, когда параметры внешнего раздражи-
теля будут полностью соответствовать исходным програм-
мирующим свойствам мотивационного возбуждения.
Изучение мотиваций заставляет с иных позиций рассмат-
ривать такие, казалось бы, уже сложившиеся понятия,
как безусловный и условный рефлекс.
МОТИВАЦИИ И ИНСТИНКТ
Понимаемая в обычном смысле слова безусловнорефлек-
торная реакция обозначает чаще всего эффекторный ответ
организма на действие того или иного внешнего раздражи-
теля.
Однако при исследовании целостной приспособительной
деятельности животных дело значительно усложняется.
Внешние раздражители всегда действуют на организм,
находящийся в различных исходных состояниях в след-
ствие процессов, происходящих внутри него. Таким образом,
Bice сложное поведение невозможно сводить к действию
только внешних раздражителей. Любая безусловнорефлек-
торная поведенческая реакция целого организма обязатель-
но включает в себя определенную мотивацию, обусловлен-
ную первичными изменениями во внутренней среде. И эта
мотивация, выступая в роли своеобразного «фильтра»,
в значительной степени определяет характер реагирования
организма на внешние раздражители.
Точно так же строятся сложные формы безусловнореф-
лекторной деятельности — инстинкты. В этом случае
на основе своих внутренних потребностей животные также
активно относятся к раздражителям внешней среды. Однако
их жизнедеятельность жестко детерминирована наличием
в окружающей среде определенных раздражителей, только
при наличии которых они оказываются способными удовлет-
ворить свои потребности. При отсутствии таких раздражи-
телей приспособительная деятельность этих животных
становится невозможной.
Так, новорожденный котенок при возникновении у него го-
лодного возбуждения успешно найдет сосок матери только
264
в том случае, если он находится в непосредственной бли-
зости к ней и имеет возможность ориентироваться по харак-
теру ее шерстного покрова. Будучи, однако, отнесен на
значительное расстояние, он может погибнуть, если его
крик не будет услышан матерью.
Такое направленное отношение животных к определенным
раздражителям окружающей среды в случае инстинктив-
ной деятельности определяют жесткие программы поведе-
Рис. 55. Схема циклической саморегуляции инстинктив-
ной деятельности. Объяснения в тексте.
ния, которые являются наследственно предопределенными и
формируются в каждом случае возникновения соответству-
ющего мотивационного возбуждения.
Успешная реализация этих программ возможна у таких
животных только при наличии определенных раздражите-
лей внешней среды.
Таким образом, мотивации являются важным звеном
врожденной инстинктивной деятельности. По существу вся
инстинктивная деятельность совершенно определенно рас-
кладывается на отдельные этапы возникновения соответ-
ствующих потребностей, мотивации и их удовлетворения.
Отличительной особенностью этой формы приспособитель-
ной деятельности является наличие наследственно предошре-
деленнных программ поведения, направленных на удовле-
творение соответствующей потребности организма. Любая
сложная инстинктивная деятельность в конечном счете
265
развертывается на основе двух механизмов: механизмов
влечения и подкрепления, между которыми включается
(именуемое иногда собственно инстинктом) поведение.
Любую инстинктивную деятельность, следовательно, можно
рассматривать как циклическую, саморегулирующуюся
организацию, которая принципиально состоит из механиз-
мов «основных влечений», целенаправленного поведения
и, наконец, механизмов подкрепления (рис. 55).
Именно биологические мотивации направляют в данном
случае животных к совершению полезной для них приспосо-
бительной деятельности. Удовлетворение же лежащих в
основе этих мотиваций потребностей происходит через по-
стоянное сравнение и оценку результатов совершенного
действия с запрограммированными в каждой мотивации
свойствами соответствующего подкрепления.
Врожденные механизмы мотивации могут обеспечить це-
ленаправленную деятельность, по-видимому, только при на-
личии во внешней среде специальных раздражителей. У выс-
щих млекопитающих животных и человека врожденных
механизмов мотивации недостаточно для совершения при-
способительной деятельности. Здесь все в большей степени
включается фактор обучения.
МОТИВАЦИИ И УСЛОВНЫЙ РЕФЛЕКС
Работы школы И. П. Павлова убедительно показали,
что условные рефлексы образуются только на базе слож-
ных безусловных рефлексов, которые возникают при под-
креплении. Считается, например, что условный пищевой
рефлекс возникает на основе подкрепления пищей, оборо-
нительный — при болевом подкреплении и т. п. Однако,
как уже указывалось выше, у сытых животных очень
трудно образовать пищевой условный рефлекс. Исследова-
ния М. К. Петровой показали, что кастрация также значи-
тельно понижает способность к образованию половых и
других условных рефлексов. Все эти и многие другие экспе-
риментальные данные позволяют думать, что основой
успешного образования любого условного рефлекса явля-
ется, кроме безусловного подкрепления, то первичное вле-
чение, которое определяет исходное состояние центральной
нервной системы.
Значение мотивации для образования различных навы-
ков подробно исследовано Miller (1957, 1960). Используя
болевое раздражение в качестве подкрепления, Miller вы-
266
рабатываЛ у крыс в определенной обстановке возникнове-
ние состояния «страха». И па этой основе ему удалось
образовать сложные поведенческие навыки.
Другим примером, показывающим, как сложные навыки
формируются под влиянием оборонительной доминанты
и влечения, обусловленного страхом, являются экспери-
менты Д. С. Фурсикова (1921). На основе подкрепления
электрическим током ему удалось образовать у собаки ус-
ловный рефлекс даже третьего порядка. Таким образом,
оборонительное состояние обеспечивает наиболее быстрое
образование условных рефлексов. Не меньшее значение в
механизме образования различных навыков имеют голод,
жажда и другие мотивации. Как было показано выше, ис-
следованиями Lilli (1960) установлено, что на основе вле-
чений голода можно обучать обезьян совершать сложные
операции: включать и выключать различные рычаги, аппа-
ратуру и т.п. Miller (1963) показал, что у животных с силь-
ной жаждой (при лишении воды за 22 часа до эксперимен-
та) условная реакция па свет с подкреплением водой
вырабатывается чрезвычайно быстро. Однако если в желу-
док таких животных через фистульное отверстие предвари-
тельно вводили большое количество воды, условного
рефлекса у них не образовывалось.
Многочисленными экспериментами было показано, что
раздражение различных структур гипоталамической об-
ласти электрическим током или химическими веществами,
которые обычно вызывают определенные мотивации, спо-
собствует образованию и лучшему выявлению различных
условных рефлексов (Н. А. Костепецкая, 1963; Г. П. Уда-
лова, 1968; Л. В. Калюжный, 1962; Delgado, 1965; Wyr-
wicka a. oth., 1959; Nakao, 1958). В опытах Л. В. Калюж-
ного, например, инъекция адренергетическнх веществ в
область заднего гипоталамуса улучшала проявление
оборонительного условного рефлекса. Инъекция в эту об-
ласть холинергических веществ усиливала пищевые услов-
ные рефлексы. К- Лишшак и Э. Эндреци (1965) считают,
что облегчение условнорефлекторных реакций при электри-
ческом раздражении подкорковых структур является
результатом искусственного усиления влечения. Тем не
менее Masserman (1943) полагал, что, хотя раздражение
определенных отделов гипоталамуса вызывало реакцию
ярости у кошек, на основе этого раздражения нельзя вы-
работать условного рефлекса. Иного мнения придержи-
вается Nakao (1958). Ему удалось выработать условные
бборонительныё реакции у кошек на раздражение перед-
него и среднего отделов гипоталамуса.
Мотивации, таким образом, являются мощной энергети-
ческой основой, совершенно необходимой для образования
условных рефлексов и сложных навыков. Как уже указы-
валось, основу первичных мотиваций составляют механиз-
мы активирующих влияний со стороны подкорковых обра-
зований на кору головного мозга. Они же определяют ту
доминантную деятельность, благодаря которой организм
активно воздействует на раздражители внешнего мира.
Поэтому все условные раздражители прежде всего связы-
ваются с механизмами «основных влечений» организма.
Таким образом, механизмы влечения лежат в основе обра-
зования условных рефлексов.
Однако условные раздражители сами могут стимулиро-
вать различные мотивации, вскрывать ее скрытые меха-
низмы. Голодное животное может спать или спокойно
сидеть в тот момент, когда к нему подносят пищу. Однако
запах пищи сразу же пробуждает животное, запах и вид
пищи оказывает на него выраженное активирующее влия-
ние и животное с жадностью набрасывается на еду.
Условные обстановочные раздражители в данном случае
приобретают пусковое значение в выявлении скрытого мо-
тивационного возбуждения и возникновении целенаправ-
ленного поведения.
Все специальные пусковые раздражители вызывают це-
ленаправленную реакцию только за счет активирования ап-
паратов мотивации. Другой пример этого представлен в экс-
периментах нашего сотрудника А. С. Сосновского. У кроли-
ка вырабатывается условная пищевая реакция. В качестве
условного раздражителя используется свет, который под-
крепляется пищей. После выработки и упрочения условной
реакции кролику вживляется электрод в область центра
голода латерального гипоталамуса. Теперь в ответ на разд-
ражение этого центра, как указано выше, у накормленного
кролика возникала пищевая реакция. Если же этот центр
раздражаем током подпороговой силы, то у животного на-
блюдается только ориентировочно-исследовательская реак-
ция. Эксперимент проводился следующим образом. На-
кормленного кролика помещали в обычную эксперимен-
тальную обстановку. В ответ на вспышку света он не про-
являл никакой пищевой реакции. Затем ему одновремен-
но с вспышками света наносилось слабое подпороговое раз-
дражение центра голода латерального гипоталамуса, кото-
268
рое ранее вызывало только ориентировочно-исследователь-
скую реакцию животного. Животное немедленно проявляло
пищевую реакцию. Раздражение, вызванное светом, в дан-
ном опыте усилило возбуждение центра голода латерально-
го гипоталамуса и сформировало у данного животного пол-
ноценную пищевую мотивацию.
В опытах И. С. Бериташвили (1966) наблюдался сле-
дующий факт. Если накормленной собаке показать пищу,
то при отсутствии соответствующей мотивации живот-
ное не устремляется к ней. Пищу на глазах животного пря-
чут в определенное место. Теперь, будучи приведенным
в это же помещение голодным, животное немедленно уст-
ремляется к тому месту, где ранее была спрятана пища.
По-видимому, даже при отсутствии соответствующей моти-
вации в ответ на действие пищевого раздражения и всей
массы сигналов, связанных с ним, последние фиксируются
аппаратом афферентного синтеза и включаются в аппарат
акцептора результатов действия. Как только усиливаются
мотивационные влияния, процессы памяти немедленно
оживляются и животное устремляется к тому месту, где
спрятана пища. Интересны в этом отношении эксперименты
А. В. Котова, выполненные в хронических опытах на кро-
ликах. У кроликов вырабатывался инструментальный услов-
ный рефлекс. На вспышку света кролик обучался нажимать
на площадку. При нажатии на эту площадку открывалась
кормушка с пищей. После анадической поляризации цент-
ра голода латерального гипоталамуса голодный кролик при
зажигании лампочки по-прежнему нажимал на площадку.
Однако он отказывался брать пищу из кормушки даже
в том случае, если подходил к ней.
Эти эксперименты указывают на то, что комплекс кор-
ковых элементов, вовлекаемых в ту или иную мотивацию,
иногда может возбуждаться внешними стимулами и без
наличия соответствующей потребности.
Близко в этим наблюдениям примыкают опыты Н. В.
Асмаяна, проведенные в лаборатории П. К. Анохина. Он
вырабатывал у собаки инструментальную реакцию нажа-
тия на рычаг, которая приводила к подаче пищи. Экспери-
мент обычно проводился таким образом, что сначала вклю-
чался условный сигнал (например, свет), затем животное
должно было нажать на рычаг и получало вслед за этим
пищу. После определенного срока тренировки эта реакция
была успешно выработана и отчетливо проявлялась у го-
лодного животного. Однако затем оказалось, что и сытно
269
накормленное животное при действии условного раздражи-
теля в данной обстановке нажимает на рычаг, хотя при этом
не потребляет подающуюся пищу. Значит, условный сигнал
активирует только этапную часть всего комплекса пищевого
возбуждения, связанную с инструментальной реакцией.
Попять этот факт можно, по-видимому, следующим об-
разом.
Если тот или иной раздражитель внешнего мира длитель-
ное время или на протяжении даже всей жизни данного ин-
дивидуума успешно сигнализирует животному о последую-
щем подкреплении, то в этом случае происходит процесс
автоматизации этой деятельности. Соответствующий внеш-
ний раздражитель заставляет животное принудительно со-
вершать определенную деятельность даже при отсутствии
соответствующей потребности.
Однако подчеркнем, что и в этих случаях, если соответст-
вующий раздражитель внешнего мира перестает по каким-
либо причинам сигнализировать о последующей жизненно
важной деятельности, он утрачивает свое сигнальное зна-
чение, т. е. теряет свои связи с подкреплением.
Таким образом, мотивационное возбуждение является
обязательным компонентом классического условного реф-
лекса. В отечественной и иностранной научной литературе
до последнего времени усиленно дискутируется вопрос иден-
тичности и различия павловского условного рефлекса и ин-
струментальных реакций, широко применяемых американ-
скими нейропсихологами.
Если подойти к этому вопросу с позиций мотивационного
возбуждения, то можно отметить следующее.
Как условный рефлекс, так и инструментальные реакции
отчетливо проявляются только при наличии соответствую-
щей мотивации. И в том и в другом случае имеется под-
крепление, удовлетворяющее данную мотивацию. Все раз-
личие заключается в том, каким образом осуществляется
в каждой из этих реакций подкрепление. В случае условно-
го рефлекса оно пассивно предоставляется животному экс-
периментатором вслед за условным сигналом. В инструмен-
тальной реакции животное для того, чтобы получить под-
крепление, должно активно совершить ряд предварительных
действий (нажать на рычаг, толкнуть дверцу и т. д.), т. е.
получить предварительный этапный результат и активно
оценить его.
В выработанном условном рефлексе тот или иной сигнал
уже заключает в себе надежную гарантию будущего под-
270
крепления, которое осуществляет экспериментатор. Для то-
го чтобы получить его, животное не должно осуществлять
никакой деятельности. Таким образом, акцептор результа-
тов действия в случае условного рефлекса заключает в себе
только параметры следующего за сигналом подкрепления.
Совершенно другая картина наблюдается в случае инстру-
ментальной реакции. Здесь для того, чтобы получить соот-
Рис. 56, Схема соотношения мотивационных и условнорефлекторных
механизмов (по П. К. Анохину. 1968). Объяснения в тексте.
ЛД — акцептор результатов действия.
ветствующее подкрепление, животное должно еще путем
проб и ошибок совершить этапные действия, оценить их
и получить адекватное подкрепление. Следовательно, аппа-
рат акцептора результатов действия в этом случае более
сложен. Он включает в себе не только свойства будущего
подкрепления, но и параметры результатов тех этапных дей-
ствий, которые ведут к этому подкреплению (рис. 56).
Итак, различие павловского условного рефлекса и инст-
рументальных реакций состоит в различной сложности фор-
мирующихся при наличии одинаковой мотивации аппара-
тов акцептора результатов действия, направляющего жи-
вотных на подкрепление соответствующей мотивации. В свя-
зи с этим нет никаких оснований для противопоставления
271
‘Гих реакций. Обе эти формы деятельности являются сиг-
'альными реакциями, возникающими на основе определен-
ых доминирующих мотиваций. В то же время инструмен-
тальные реакции представляют пример более сложной дея-
ельности, которую некоторые авторы (Миллер, 1966) рас-
Матривают как высшие мотивации.
Если рассматривать классический условный рефлекс
* инструментальные реакции с точки зрения центральной
фхитектуры поведенческого акта по П. К- Анохину (см.
kic. 2), то можно отчетливо заметить, что условный стимул
1:ключается в аппарат афферентного синтеза, в то время
’’ак при инструментальных реакциях активная деятельность
фоисходит за счет постоянной оценки параметров внешних
Раздражителей, которые включаются в другой узловой ме-
}анизм этой архитектуры в пункт, где происходит сравнение
*араметров полученного результата со свойствами акцеп-
тора результатов действия.
ВЫСШИЕ МОТИВАЦИИ
Как правило, высшие мотивации возникают на основе ме-
ханизмов низших биологических мотиваций. В процессе ин-
дивидуальной жизни для удовлетворения «основных вле-
чений» организм все в большей мере «знакомится» с внеш-
ним миром. На этой основе в центральной нервной системе
'Ьрмируется специальный аппарат афферентного синтеза.
Н процессе обучения в результате установления связей
Нежду механизмами «основных влечений» и внешними воз-
действиями роль мотивационного стимула может перейти
1же к любому внешнему раздражителю (обстановочному
Или специальному). Таким образом, в формировании выс-
Ьих мотиваций прежде всего участвуют условнорефлектор-
Ные механизмы.
Интересно, что в формировании высших мотиваций роль
Подкрепляющего фактора может перейти от реально удов-
летворяющего данную потребность раздражителя к разд-
ражителю, который сам по себе не удовлетворяет ее, но
Находится в таких тесных взаимоотношениях с реальным
Подкреплением, что надежно гарантирует последующее под-
крепление. Наблюдается своего рода «абстрагирование»
Подкрепления. Животные начинают совершать целенаправ-
ленную деятельность ради определенных раздражителей,
которые сами еще являются только сигналами будущего
Подкрепления. Формируются специальные комплексы моти-
ва
НйЦиойных возбуждений, которые лишь отдаленным спосо-
бом связаны с биологическими потребностями.
Такая форма деятельности впервые наблюдалась в лабо-
ратории И. П. Павлова при выработке условных рефлек-
сов второго и третьего порядка. Однако наиболее характер-
но она проявляется в свободном поведении животных.
Интересен в этом отношении эксперимент Cowless (1937).
Автор тренировал голодных шимпанзе вставлять жетоны
в особый аппарат, который автоматически за каждый жетон
выдавал определенную порцию изюма. Но для того чтобы
получить жетон, обезьяны должны были совершить опреде-
ленную работу, например повернуть определенную рукоят-
ку против ее тяжести.
Оказалось, что животные легко обучались проводить эту
инструментальную реакцию для того, чтобы получить же-
тоны. Таким образом, жетон превратился у этих животных
в определенное подкрепление. Обращает на себя внимание
тот факт, что значение жетона у обезьян значительно осла-
бевает, если подкрепление в опытах отсрочено. Здесь,
по-видимому, включаются механизмы угасательного тормо-
жения, открытого И. П. Павловым и его школой,
У шимпанзе обнаруживается способность различать раз-
нообразные виды жетонов в зависимости от их значения для
удовлетворения исходных мотиваций.
Такие реакции наблюдали Wolfe (1936) и А. И. Счастный
(1963, 1968). В опытах А. И. Снастного шимпанзе совер-
шенно отчетливо выбирали ту или иную форму жетона для
удовлетворения своей доминирующей мотивации. Так, в го-
лодном состоянии животные выбирали из серин самых раз-
нообразных жетонов только те, на предъявление которых
экспериментатору они получали пищу. Только удовлетво-
рив пищевую мотивацию, животные начинали выбирать
другие жетоны, предъявление которых обеспечивало полу-
чение воды, игрушки.
Эти эксперименты отчетливо показывают значение доми-
нирующей мотивации для реакции животного на внешние
раздражители. Они указывают на то, что сигналы внешнего
мира получают свое подкрепляющее значение только при
наличии соответствующей биологической мотивации.
Отсюда становится совершенно очевидной важная роль
биологических мотиваций для всех форм сложной поведен-
ческой деятельности животных и человека.
Проблема формирования сложных мотиваций может быть
решена только при подходе к пей, во-первых, с эволюцион-
11 Заказ 5074
273
ных позиций (филогенез) и, во-вторых, в аспекте индиви-
дуального развития данной особи (данной личности, если
речь идет о человеке). Вопрос о том, как формируются
высшие мотивации у человека, в настоящее время еще мало
изучен.
По существу вся сложная психическая деятельность че-
ловека связана в конечном счете со стремлением наилуч-
шим и более успешным способом удовлетворять различ-
ные его потребности. Создание машин, заводов, т. е. весь
прогресс промышленного производства, направлено на то,
чтобы лучше обеспечить человека питанием, одеждой, жиль-
ем и т. д. Здесь уже проблема мотиваций начинает взаимо-
действовать с социальными и классовыми вопросами.
Проявление мотиваций у человека имеет качественные
особенности. У человека мы постоянно наблюдаем не толь-
ко эмоциональное, но и словесное выражение различных
мотиваций, которые выступают в форме самых разнообраз-
ных желаний и намерений, отражающих внутреннее состоя-
ние организма. Таков результат абстрактно-логического
мышления человека. И эта способность человека на основе
второй сигнальной системы оценивать свое собственное со-
стояние является важным пунктом всей целенаправленной
человеческой деятельности. Осознав свои потребности, че-
ловек не только приспосабливает их к окружающей среде,
но и активно изменяет внешнюю среду сообразно своим
потребностям. Качественное своеобразие мотиваций челове-
ка и особенности их выявления являются предметом пси-
хологии.
Здесь мы остановимся только на следующем вопросе. Ес-
ли биологические мотивации имеют столь важное значение
для образования различных навыков, каково их значение
для восприятия вообще?
МОТИВАЦИИ И ВОСПИТАНИЕ
Как уже указывалось ранее, животные сначала с по-
мощью родителей, а затем самостоятельно начинают выде-
лять в окружающем их мире специальные сигналы, которые
способствуют удовлетворению их основных биологических
потребностей. В этом умении выделить из массы окружаю-
щих раздражителей специальные, способствующие или, на-
оборот, препятствующие удовлетворению биологических по-
требностей раздражители собственно и заключается глав-
ный смысл обучения.
274
В случае, если потомство не встречается с родителями
(как это наблюдается у большинства насекомых), этот про-
цесс выделения определенных направляющих факторов
внешней среды происходит благодаря наследственно пред-
определенным жестким программам поведения. Большинст-
во высших животных приобретает эту способность в инди-
видуальной жизни. Однако везде успешному достижению
полезного результата, удовлетворению потребности способ-
ствуют определенного качества эмоциональные пережива-
ния. При возникновении мотивации это, как правило, ощу-
щения неприятного характера. При совпадении результата
действия с ожидаемым, запрограммированным в соответст-
вующей мотивации, возникает эмоция положительного
знака.
Этот механизм является настолько распространенным,
что мы вряд ли можем отрицать его наличие и у разных
насекомых и животных, хотя, безусловно, у разных живот-
ных он имеет различные качественные особенности.
Особую важность в этом смысле приобретают положи-
тельные эмоции, которые, возникая при удовлетворении той
или иной потребности, тем самым как бы санкционируют ус-
пех поиска. Наоборот, если результат поведения не при-
водит к удовлетворению исходной потребности организма,
усиливается противоположного знака тенденция — усили-
вается отрицательная эмоция. Из этого становится ясным,
что воспитание всегда должно строиться по этой отработан-
ной эволюцией схеме: потребность — целенаправленное по-
ведение — удовлетворение потребности.
Если мы проследим любую деятельность человека, на-
правленную на достижение какой-либо цели, то отчетливо
увидим, что этот процесс всегда происходит через ряд по-
следовательных смен состояний эмоционального неудовлет-
ворения положительными эмоциями радости и удовольст-
вия. И самое глубокое удовольствие человек испытывает
в том случае, когда достигнута главная цель, решена основ-
ная поставленная перед ним задача.
Этот процесс смены эмоционального состояния в процес-
се целенаправленной деятельности более отчетливо наблю-
дается у ребенка. Даже новорожденный ребенок выражает
свои голодные побуждения криком и плачем, а насыще-
ние— блаженной улыбкой.
То же самое наблюдается, если ребенок поставил перед
собой какую-нибудь задачу, например построить пирамиду
из кубиков. Какое неудовольствие, выражающееся иногда
||*
§75
и в форме плача, он проявляет, если ему это не удается по
каким-либо причинам сделать. И наоборот, улыбка и все его
поведение выражают большую радость, когда поставлен-
ная цель достигнута.
Аналогичная смена эмоциональных состояний в процессе
целенаправленной деятельности наблюдается и у животных.
Так, служебная собака, если не может перепрыгнуть барь-
ер, всем своим видом выражает неудовольствие, вину. А вот
барьер взят и животное оживленно виляет хвостом, броса-
ется к хозяину. Одним словом, оно своим поведением так-
же выражает удовлетворение.
И точно так, пишет ли писатель увлекательный роман,
пытается ли ученый проникнуть в тайны своей науки, взби-
рается ли альпинист на вершину Памира — всюду достиже-
ние цели — это всегда эмоция радости и удовольствия!
Совершенно иная картина наблюдается, если воспитание
происходит вопреки этой отработанной эволюцией схеме.
Это чаще всего наблюдается в человеческом обществе, под
влиянием социальных факторов.
Приведем пример. Кормление голодного ребенка вызыва-
ет у него чувство удовлетворения, положительные эмоции,
так как пища по всем своим параметрам удовлетворяет
требованиям голодного мотивационного возбуждения.
Теперь представьте такой случай, что ребенка, не испы-
тывающего голода, при отсутствии потребности в питатель-
ных веществах пытаются кормить насильно. Что же про-
изойдет в этом случае? Насильственное введение питатель-
ных веществ в организм при отсутствии пищевой потреб-
ности вызывает в нем вместо положительной эмоции насы-
щения отрицательную эмоцию рассогласования. Таким об-
разом, грубо нарушена естественная последовательность
событий — от отрицательной эмоции мотивации к положи-
тельной эмоции подкрепления! При постоянном насильном
кормлении может произойти большая опасность. Неприят-
ная отрицательная эмоция, каждый раз возникающая при
насильственном кормлении, может включиться в аппарат
акцептора результатов действия соответствующей голодной
мотивации и тогда человек на всю жизнь утрачивает инте-
рес к пище.
Это опасно еще и потому, что именно на основе пищевой
потребности и ее удовлетворения у человека по существу
строятся все формы целенаправленной деятельности. Ведь
пищевая мотивация созревает у новорожденного одной из
первых.
276
Этот пример показывает, что все воспитание ребенка
должно проводиться в направлении естественного удовлет-
ворения сначала биологических, а затем и социальных
потребностей. Отсюда ясно, что если ребенок поставил
перед собой даже пустяковую задачу, важно, чтобы дело
было доведено до определенного положительного резуль-
тата. Это вызывает у него положительную эмоцию радости.
Только такой путь создаст в дальнейшем стремление к тру-
ду, искусству и науке!
Это и понятно, поскольку положительные эмоции входят
как обязательный компонент в аппарат акцептора резуль-
татов действия и по мере обучения возникают затем всякий
раз при формировании той или иной потребности. Стрем-
ление к положительным эмоциям создает в жизни челове-
ка преобладание оптимистического, радостного начала. На
этой основе человек станет стремиться к высоким идеалам,
к тому, чтобы приносить как можно больше пользы людям.
И наоборот. Если у людей отсутствует стремление к ра-
дости жизни, творчеству и борьбе, их постоянно разъедает
яд неудовлетворенности. Они становятся скептиками и пес-
симистами.
Особое значение в процессе воспитания имеет игра.
Главное же биологическое значение игры заключа-
ется в тренировке естественных механизмов целена-
правленной деятельности, т. е. умения при наличии той или
иной мотивации достичь соответствующего подкрепления.
МОТИВАЦИИ И ВОСПРИЯТИЕ
Другая проблема, которая непосредственно вытекает из
проблемы мотиваций,— это восприятие.
Как мы указывали ранее, доминирующая мотивация рез-
ко изменяет отношение субъекта к раздражителям внешне-
го мира. С помощью ориентировочно-исследовательской ре-
акции организм активно подбирает только те раздражители,
которые ликвидируют лежащую в основе данной мотиваций
потребность.
Если стоять на позиции классической Схемы «стимул —
реакция», то может оказаться неясным, почему при дейст-
вии некоторых внешних факторов на организм иногда име-
ется, а в других случаях отсутствует его реакция на эти
раздражители. Именно доминирующая мотивация выступа-
ет в роли «активного фильтра», который определяет отно-
шение животного к окружающим его раздражителям. В свя-
277
зй с этим наибольший интерес представляет тот факт, что
каждая мотивация избирательно повышает чувствитель-
ность именно тех рецепторов, которые принимают участие
в ее удовлетворении, тем самым как бы еще более «обост-
ряя» отношение животного - к подкрепляющим факторам
внешней среды. При устранении мотивации чувствитель-
ность рецепторов к соответствующим раздражителям па-
дает. Наряду с этим в механизме восприятия существенная
роль принадлежит аппарату памяти.
МОТИВАЦИИ И ПАМЯТЬ
Мы уже указывали на то, что в афферентном синтезе на-
ряду с мотивационным возбуждением принимает участие
механизм памяти. Память, как указывает П. К- Анохин
(1968), есть свойство только организованных функциональ-
ных систем. В связи с этим все попытки найти субстрат па-
мяти в изолированных нервных элементах хотя и представ-
ляют интерес для объяснения поведенческой деятельности,
по-видимому, не будут иметь большого значения. Естест-
венно поставить вопрос; каковы взаимоотношения между
мотивацией и памятью?
Мы должны совершенно определенно подчеркнуть, что
именно доминирующая мотивация является тем механиз-
мом, который позволяет организму извлекать из памяти на-
копленный (генетический и индивидуальный) опыт для
построения целенаправленных реакций. Вместе с тем все
успешные акты и их результаты, направленные на удовлет-
ворение каждой потребности, немедленно фиксируются в па-
мяти и «оживляются» всякий раз при возникновении соот-
ветствующей мотивации.
Итак, проблема мотивации заставляет с принципиально
новых позиций рассматривать многие актуальные вопросы
современной нейрофизиологии (инстинкт, условный реф-
лекс, восприятие, память).
ПАТОЛОГИЧЕСКИЕ МОТИВАЦИИ
Наряду с естественными нормальными мотивациями кли-
ническая практика имеет множество случаев их патологи-
ческих расстройств. Большинство патологических расст-
ройств мотиваций связано с нарушением внутренней среды
организма и особенной деятельности желез внутренней сек-
реции. Это и понятно, поскольку железы внутренней секре-
278
ции выделяют свои гормоны непосредственно в кровь. Пос-
ледняя же оказывает непосредственное действие на моти-
вационные центры гипоталамической области.
Расстройства мотиваций наблюдаются при многих фор-
мах патологии желез внутренней секреции. Мы уже упомина-
ли случай, описанный Wilkins и Richter, когда у мальчика
с опухолью надпочечников было патологическое влечение
к соли. Это связывалось с постоянной потерей соли через
почки в связи с отсутствием гормона надпочечника—аль-
достерона.
Повреждение надпочечников также иногда приводит
у мужчин к развитию импотенции. При опухолях надпочеч-
ников у мужчин известны случаи феминизации. Патологи-
ческие мотивации наблюдаются при измененной функции
щитовидной железы. Нередко при гипертиреодозе наблю-
дается булимия. Гиперфункция щитовидной железы увели-
чивает также прием воды (Н. А. Рожанский, 1957).
Дисфункция половых желез определяет различные расст-
ройства половых влечений от импотенции до гиперсексуаль-
ности. Наиболее богатая картина расстройства влечений
наблюдается при поражении гипофиза. При этом могут
наблюдаться различные нарушения пищевых, питьевых и
половых влечений, поскольку деятельность гипофиза тесно
связана с работой других желез внутренней секреции, име-
ющих прямое отношение к этим функциям.
Такое непосредственное участие гормонов желез внутрен-
ней секреции в формировании основных биологических вле-
чений показывает, насколько важны пропорциональные со-
отношения этих гормонов во внутренней среде организма для
формирования нормальной целенаправленной деятельности.
Значительная группа патологии мотиваций наблюдается
при поражении гипоталамуса, его центров, которые непо-
средственно определяют мотивационное возбуждение, а так-
же лимбических образований на различных уровнях. При-
меры этого были описаны нами ранее. Важно подчеркнуть
одно обстоятельство. Все мотивации формируются по пейц-
мекерному типу с последующим распространением моти-
вационного возбуждения вплоть до коры головного мозга.
Из этого совершенно очевидно, что любое изменение актив-
ности данного «пейцмекера» (при действии факторов крови,
структурные изменения и т, д.) вызывает значительное из-
менение деятельности всей системы такой модальности.
А это в свою очередь может явиться основой формирова-
ния патологической личности.
279
НАРКОМАНИИ
Наряду с указанными выше формами патологии мотива-
ций особую группу составляют искусственные мотивации,
которые формируются путем введения в организм опреде-
ленных химических веществ (никотина, морфина, алкоголя
и др.) и проявляются в том, что после неоднократного при-
менения этих веществ у некоторых людей развивается не-
преодолимая тяга к их употреблению (наркомании).
На основании приведенного выше материала о механиз-
ме мотиваций мы можем выдвинуть следующую гипотезу
формирования наркоманий.
По-видимому, упомянутые выше химические вещества
обладают резко выраженным свойством включаться в мета-
болизм мотивационных центров гипоталамуса. Как это про-
исходит в деталях, мы, к сожалению, еще не знаем. Воз-
можно, здесь играют важную роль врожденные особенности
организации этих центров, поскольку наркомании наблю-
даются не у всех лиц.
Неоднократное применение подобных веществ приводит
к тому, что их отсутствие резко расстраивает метаболизм
соответствующих центров гипоталамуса. А это в свою оче-
редь формирует мотивационное возбуждение, направленное
на поиск соответствующего вещества во внешней среде.
Формирование подобной мотивации подчинено всем законо-
мерностям, присущим любому мотивационному возбужде-
нию, а указанные выше химические вещества приобретают
свойства подкрепления.
Так формируются влечения к курению, алкоголю, нарко-
тикам. Прием каждого из этих веществ устраняет исходную
мотивацию. Кроме того, он сопровождается положительны-
ми эмоциональными переживаниями, которые впоследствии
включаются в аппарат акцептора результатов действия. По-
следний фактор значительно усиливает тягу к этим вещест-
вам.
Поскольку эти вещества включаются в метаболизм кле-
ток центральной нервной системы, все попытки лечения этих
влечений различными средствами (гипнотерапия, голода-
ние и пр.) являются безуспешными. Лечение будет успеш-
ным только в том случае, если удастся «выбить» это веще-
ство из порочного круга метаболических процессов.
Другая опасность подобных патологических мотиваций
заключается еще и в том, что, становясь при отсутствии со-
ответствующего подкрепления доминирующими, они завла-
280
девают всем сознанием больного. Вот почему, образно го-
воря, алкоголик смотрит на жизнь через призму водочной
бутылки, а наркоман оценивает все окружающие его явле-
ния с точки зрения возможности или невозможности доста-
вания соответствующего наркотика.
ФАРМАКОЛОГИЯ МОТИВАЦИИ
Избирательная чувствительность различных мотиваций
к химическим веществам послужила основой для разработ-
ки специальных фармакологических веществ, избирательно
действующих только на определенную мотивацию. Посколь-
ку, устраняя мотивацию, эти вещества действуют опреде-
ленным образом и на психику, они получили название
психофармакологических веществ. Возникло новое направ-
ление— психофармакология мотиваций.
Еще в 1943 г. Masserman обнаружил, что алкоголь и мор-
фин понижают страх животного в обстановке, в которой ему
предварительно наносилось болевое раздражение.
Congen (1949) подтвердил эти наблюдения и показал,
что после инъекции алкоголя крысы утрачивали страх и бе-
жали к пище, расположенной у того конца прохода, на пути
к которому обычно подавался электрический ток.
Н. Миллер (1966) установил, что страх у животных ус-
пешно может быть подавлен барбитуратами (например,
амобарбиталом). Это вещество оказалось очень эффектив-
ным для снятия страха в следующем эксперименте.
Голодные крысы обучались нажатию на рычаг для полу-
чения пищи. После закрепления навыка нажатие на рычаг
однажды было подкреплено сильным ударом электрическо-
го тока. Этого оказалось достаточным, чтобы крысы, даже
будучи сильно голодными, перестали нажимать на рычаг.
Однако после инъекции амобарбитала животные совершен-
но спокойно нажимали на рычаг и получали вслед за этим
пищу.
Аналогичного типа наблюдения были сделаны В. А. Гав-
личеком (1958) и Ю. А. Макаровым (1960), которые показа-
ли, что таким же свойством устранения болевой доминан-
ты обладает аминазин. После введения аминазина живот-
ное, которое ранее в обстановке, в которой ему наносилось
болевое раздражение, отказывалось от еды, теперь охотно
ее поедало.
В других опытах Davis и Miller (1963) показали, что
в безысходной обстановке болевого подкрепления крысы
успешно обучались инъецировать себе через специальный
281
катетер, введенный в яремную вену, раствор амобарбитала
или гексобарбитала.
Olds (I960) показал, что введение крысам хлорпромази-
на (аминазина) в дозе 2 мг/кг полностью блокирует эффект
самораздражения при локализации электродов в вентраль-
ной области заднего гипоталамуса, В то же время хлор-
промазин оказывает меньшее влияние на реакцию самораз-
дражения, возникающую при раздражении средних отделов
гипоталамуса. А при локализации электродов в переднем
отделе гипоталамуса влияние хлорпромазина на реакцию
самораздражения вообще не проявляется. Это влияние так-
же мало выражено при локализации электродов в передних
участках преоптической области и снова резко усиливается
при локализации электродов в области перегородки.
Точно так же резерпин угнетает резекцию самораздраже-
ния при нахождении электродов в заднем отделе гипотала-
муса и не оказывает заметного влияния на эту реакцию при
локализации электродов в переднем мозге (Olds, Killam,
Bach-y-Rita, 1956).
Все эти эксперименты показывают, что в пределах раз-
личных частей, подкрепляющих мотивации систем, имеется
отчетливая химическая специфика.
Приведенные выше наши экспериментальные данные по-
казывают, что каждое мотивационное возбуждение строит-
ся на основе специфических химических механизмов акти-
вирующих влияний подкорковых мотивационных центров
на кору головного мозга. Так, введением адренолитических
веществ (аминазина) удается избирательно заблокировать
механизмы активации коры головного мозга при ноцицеп-
тивном раздражении. Активация коры головного мозга при
пищевом возбуждении голодных животных избирательно
блокируется холинолитическими веществами.
Однако во всех случаях применения психофармакологи-
ческих веществ надо иметь в виду следующее. Все эти ве-
щества преимущественно влияют на пейцмекерные центры
каждой мотивации. Это происходит, по-видимому, благода-
ря наиболее интенсивному обмену веществ в этой области
и повышению ее химической чувствительности.
Однако уже преимущественное действие химических ве-
ществ на эти отделы приводит к подавлению всей системы
мотивационного возбуждения данной модальности на лю-
бом уровне и соответствующему нарушению психической
деятельности.
-В вопросе широкого использования психофармакологи-
282
веских средств для избирательного действия на мотиваци-
онные возбуждения всегда надо иметь в виду и еще одно
обстоятельство.
Как мы указывали выше, биологическое значение моти-
ваций и сопутствующих им отрицательных эмоций заклю-
чается в обеспечивании наиболее быстрого удовлетворения
потребности. Поэтому всякое снижение мотивации расстраи-
вает эту функцию, в результате чего удовлетворение со-
ответствующей потребности становится невозможным. Осо-
бенно это следует учитывать при химических воздействиях
на мотивацию страха. Подавление мотивации страха, на-
пример, у спортсменов, может привести к нежелательному
эффекту — снижению спортивных результатов.
МОТИВАЦИИ И МОДЕЛИРОВАНИЕ ПОВЕДЕНИЯ
Исключительно четкая организация различных мотива-
ций и их место в саморегулирующихся аппаратах различ-
ных функциональных системах, принципиальная схема ко-
торых обрисована выше, позволили в настоящее время при-
ступить к искусственному их моделированию. Этот вопрос
имеет особо важное значение для создания систем, способ-
ных совершать целенаправленные «запрограммированные»
реакции.
К сожалению, многие имеющиеся в настоящее время ис-
кусственные модели не имеют в своей принципиальной схе-
ме аналога мотивационного возбуждения.
ЗАКЛЮЧЕНИЕ
Внелогические мотивации, как это следует из всего ска-
занного выше, играют, таким образом, весьма существен-
ную роль в целенаправленной деятельности животных и че-
ловека.
Поистине, как писал И. П. Павлов, это «основная форма
жизненной энергии каждого из нас».
Не будь мотивационное возбуждение столь сильно по
своей побуждающей силе, животные и человек не смогли
бы преодолеть ни одного, даже малейшего препятствия на
пути к удовлетворению своих внутренних потребностей.
Эта энергетическая сила мотивации возникает благодаря
восходящим активирующим влияниям специальных подкор-
ковых центров на кору головного мозга, в результате чего
создается своеобразная внутренняя «заряженность» живот-
ного, испытывающего ту или иную потребность. Вместе
с тем наличие в каждом мотивационном возбуждении ап-
парата акцептора результатов действия со всеми парамет-
рами будущего подкрепления активно направляет живот-
ное на поиск соответствующих раздражений внешней среды.
Благодаря этому создается направленная «заряженность»
животного. Такая двойственная природа мотивационного
возбуждения вполне примиряет крайние точки зрения отно-
сительно энергетических и направляющих факторов моти-
вации.
Проблема мотивации значительно меняет многие наши
привычные представления. Она заставляет прежде всего
по-новому рассматривать значение эмоций в приспособи-
тельной деятельности животных.
Такие практически важные проблемы, как инстинкт, обу-
чение, восприятие, воспитание, условный рефлекс и память,
Также не могут изучаться без глубокого применения теории
мотивационного возбуждения.
В свою очередь проблема мотиваций и их место в целост-
ном приспособительном поведении может быть понята
только в свете общей теории П. К- Анохина о функциональ-
ных системах организма.
284
Как указывалось ранее, мотивационное возбуждение яв-
ляется одним из компонентов стадии афферентного синте-
за различных функциональных систем, которые для обеспе-
чения своих полезных для организма приспособительных
результатов включают поведенческую деятельность во
внешней среде.
Только рассматривая мотивационное возбуждение с по-
зиции теории функциональных систем организма, можно
изучать его целиком с материалистической точки зрения.
Все иные попытки изучения проблемы мотиваций всегда
связаны с опасностью идеалистического толкования.
Проблема мотивации заставляет по-новому осмыслить
сложные формы приспособления живых существ к окру-
жающей их среде. Взамен принципа «стимул — реакция»
она провозглашает принцип активного подбора внешних
раздражителей на основе постоянной оценки результатов
совершенной деятельности и сравнения их с запрограмми-
рованными в каждой мотивации свойствами аппарата'ак-
цептора результатов действия.
Мы рассмотрели в основном нейрофизиологические ме-
ханизмы только биологических мотиваций («основных
влечений»). Бесспорно, что в психической деятельности че-
ловека все большую роль играют приобретенные и социаль-
ные мотивы. Изучение этих высших мотиваций одна из важ-
нейших задач психологии. Важно только подчеркнуть, что,
по-видимому, и высшие мотивации человека строятся по об-
щим законам, отработанным эволюцией по отношению
к биологическим мотивациям, с использованием тех же уз-
ловых механизмов, которые имеет каждая биологическая
функциональная система,
Предлагаемая читателю книга является первой попыткой
в Советском Союзе представить нейрофизиологические ме-
ханизмы основных биологических мотиваций организма и
понять их место и значение для целостной приспособитель-
ной деятельности животных и человека.
Мы пытались сделать некоторые обобщения и выдви-
нуть ряд новых гипотез. Возможно, что некоторые стороны
проблемы остались изложенными недостаточно глубоко.
Конечно, в этой области имеется еще много неясных во-
просов. Тем не менее уже сама возможность объективного
исследования конкретных физиологических механизмов мо-
тиваций делает эти исследования весьма перспективными.
Мы с большим вниманием готовы изучить все критиче-
ские замечания.
ЛИТЕРАТУРА
Агафонов В. Г. Журн. певропатол. и психиатр., 1956, 56, 94.
Айрапетяну Э. Ш., Сотниченко Т. С. Лимбика. Л., 1967.
Алексеева Т. Т. Физиол. журн. СССР, 1958, 44, 4, 295.
Аничков С. В. Р,усск. физиол. журн., 1924, 6, 4, 84.
Анохан П. Ж Нижегородский детрн. клинич. и эксперим. мед.. 1932, 7—8,
42—52.
Анохин П. К. Проблема центра и периферии в современной физиологии
нервной деятельности. В сб.: Проблема центра и периферии физио-
логии нервной деятельности. Горький, 1935, с, 9—71.
Анохин И. К. Ученые записки МГУ, 1945, 2, 3, 40.
Анохин П, К, Физиол. журн. СССР, 1949, 35, 5, 491.
Анохин П. К. Проблемы высшей нервной деятельности. М., 1949.
Анохин П. К. Внутреннее торможение как проблема физиологии. М.,
1958.
Анохин П. К. Вопросы философии, 1962а, 7, 97.
Анохин П. К. Журн. высш. нерв, деят., 19626, 12, 1, 7.
Анохин П. К. Электроэнцефалографические исследования высшей нерв-
ной деятельности. М., 1962, с, 241,
Анохин П. К. В сб.: Современные проблемы электрофизиологических ис-
следований нервной системы. М., 1964а, с. 132.
Анохин П. К. Эмоции. БМЭ. Изд. 2-е, 19646, 36, с. 339.
Анохин П. К. Биология и нейрофизиология условного рефлекса. М.,
1968.
Анохина А. П. Физиол. журн. СССР, 1946, 32, 1, 120.
Аршавский И. А- Труды Научно-исследовательского института биологии
и биол. ф-та Харьковского государственного ун-та. Харьков, 1956,
24, с. 161.
Аслйяя И. В. В сб.: Проблемы общей нейрофизиологии и высшей нерв-
ной деятельности. ЛА, 1961, с. 53.
Асмаян Н. В., Судаков К. В. физиол. журн. СССР, 1961, 47, 5, 605.
Асмаян Н. В., Судаков К. В. Физиол. журн. СССР, 1964, 50, 7, 861.
Ата-Мурадова Ф, Материалы первой научной конференции, посвящен-
ной проблемам физиологии, морфологии, фармакологии и клиники
ретикулярной формация головного мозга. М-, 1960, с. 14.
Ата-Мурадоеа Ф. Физиол. журн СССР, 1963, 49, 7, 781.
Ата-Мурадова Ф. Физиол. жури. СССР, 1966, 6, 696.
Ата-Мурадова Ф. Принципы онтогенетического развития восходящих
систем коры мозга. Докт. дисс. М., 1968.
Ата-Мурадова Ф., Чернышевская И. А. III научное совещание по эво-
люционной физиологии, посвященной памяти акад. Л. А. Орбели. Л..
1961, с. 13.
Ата-Мурадова Ф., Чернышевская И. А. Физиол. журн. СССР, 1986, 12,
1410.
Бадам-Ханд Л. С. Электрофизиологический анализ гипоталамо-кортн-
кальных восходящих влиянии. Дисс. канд. М., 1965.
286
Бадикдв В. И. Тезисы и рефераты докладов XXII совещания по пробле-
мам высшей нервной деятельности. Рязань, 1969, с. 18.
Бодиков В. И., Судаков К. В. Общая нейрофизиология и эксперимен-
тальная патология нервной системы. Труды Института нормальной
и патологической физиологии АМН СССР. М., 1968, 11, с. 9.
Бакурадзе А. Н. XX совещание по проблемам высшей нервной деятель-
ности. Л., 1963, с. 18.
Баранов А. И. Бюлл. экспер. биол. и мед., 1951, 33, 12, 423.
Бериташвили И. С. Труды Института физиологии АН Груз. ССР, 1961,
12, с. 51.
Бериташвили И. С. Об образной психонервной деятельности животных.
М„ 1966.
Беритов И. С. Нервные механизмы поведения высших позвоночных жи-
вотных. М., 1961.
Бехтерев В. М. Проводящие пути спинного и головного мозга. СПб,,
1896—1898, т. 1 и 2.
Болдырев В. Н. Периодическая работа пищеварительного аппарата при
пустом желудке. Дисс. СПб., 1904.
Болдырев В. Н. Русек, врач., 1914, 45—49.
Бремер Ф. (Bremer Г.). В кн.; Электроэнцефалографическое исследо-
вание высшей нервной деятельности. М., 1962, 5.
Брутковский С. В кн.: Рефлексы головного мозга. М., 1965, с. 326.
Будылина С. М., Резцова Л. Д. Билл, экспер. биол. и мед., 1969,
7, 11.
Булыгин И. А., Дорожкин К). И. Докл. АН Белорусск. ССР, 1964, 8,
1, 63.
Быков К. М. Кора п внутренние органы. М.—Л., 1947.
Вассар Е., Лайдна И,, Кингисепп П. Н. Ученые записки Тартуского ун-
та, 1961, 112, 160.
Виноградова О. С. Ориентировочный рефлекс и его нейрофизиологиче-
ские механизмы. М., 196Т.
Гавличек В. Физиол. жури. СССР, 1958, 44, 4, 305.
Гамбарян Л. С. Вопросы физиологии двигательного анализатора. М„
1962.
Гарис Дж. У. В кн.: Ретикулярная формация мозга. М., 1962, 19].
Гасанов Г. Г., Рубцова В. В. Изв. АН Азербайджана:. ССР. Серия биол.,
1969, 2, 96.
Гохблит И. И. Бюлл, экспер. биол. и мед., 1961, 52, 8, 12.
Гринлитейн А. М. Врач, дело, 1925, 24—26.
Грин Дж. В сб.: Ретикулярная формация мозга. М,, 1962, с. 533.
Детьер В., Стеллар Э. Поведение животных. Л., 1967.
Есаков А. И. Бюлл. экспер. биол. и мед., 1961, 3, 3.
Журавлев Б. В. Труды Института нормальной и патологической Физио-
логии АМН СССР. М., 1969, 12, 194,
Заболотных В, А. Докл. АН СССР, 196'3, 153, 6, 145$
Заболотных В. А. Тезисы X Всесоюзного съезда физиологов. Ереван
1964, с. 300.
Зайко Н. С. Бюлл. эксп. биол. и мед,, 1956, 1, 19.
Иванов В. Н. Врач, дело, 1926, 20, 1593.
Инграм У. (Ingram К'. R.). В кн.: Ретикулярная формация мозга. М.,
1962, с. 471.
Каграманов К. М. О холинергических и адренергических механизмах
деятельности головного мозга. Дисс. докт. М., 1965.
Калюжный Л. В. /Кури. высш. вера, деят., 1962, 12, 2, 318.
Калюжный Л. В, Успехи совр. био.т., 1964, 57, 2, 232.
287
Кассиль В. Г., Уголев А. М., Черниговский В. Н. Докл. АН СССР, 1959,
126, 3, 692.
Коваленко В. Н. Журн. высш. нерв, деят., 1962, 12, 3, 503.
Коваленко В. Н. В сб.: 5-я научная конференция Кемеровского мед. ин-
та. Кемерово, 1963, с. 76.
Коган А. Б. Электрофизиологические исследования центральных меха-
низмов некоторых сложных рефлексов. М., 1949,
Конорский Ю, М. В сб.: Электроэнцефалографическое исследование выс-
шей нервной деятельности, М., 196'2, с. 79.
Корсаков С. С. (1916). Избранные произведения. М„ 1954, с. 108.
Костенецкая Н. А. Ежегодн. ИЭМ АМН СССР за 1961—1962 гг. Л.,
1963, 42,
Красусский В. К. Труды физиологической лаборатории. М.—Л., 1949,
16, 145.
Котляр Б. И., Ерошенко Т. .41. Жури. высш. нерв, деят., 1969, 19, 5, 882.
Лакомкин А. И. В сб.: Электрофизиология нервной системы. Ростов-на-
Дону, 1963, с. 218.
Леоитович Т. А. Успехи совр. биол., 1968, 65, 34.
Линдслей Д. Б. В сб.: Экспериментальная психология. М., 1960, с. 629.
Лишак К , Эндреци Э. В кн.: Рефлексы головного мозга. М., 1965, с. 352.
Лишак К., Эндреци Э. Нейро-эндокринная регуляция адаптационной
деятельности. Будапешт, 1967.
Лурия А. Р. Мозг человека и психические процессы. М., 1963.
Любимов Н. Н. Журн. высш. нерв, деят., 1958, 8, 4, 560.
Лю Джуан-Гуй. Электрофизиологический анализ механизмов генерали-
зации возбуждений в коре больших полушарий головного мозга.
Дисс. канд. М., 1960.
Макаров В, А. Тезисы ХХШ совещания по проблеме высшей нервной
деятельности. Рязань, 1969, с. 157.
Макаров Ю. А. Физиологические механизмы взаимодействия условных
реакций разного биологического качества. Дисс. канд. М., I960.
Макаренко Ю. А. Анализ центральных механизмов реакции саморездра-
жения. Дисс. канд, М„ 1964.
Макаренко Ю. А. В сб.: Физиология и патология гипоталамуса. М„ 1966,
с. 100.
Мариц А. М. Физиол. журн. СССР, 1962, 48, 8, 889.
Маркелова В. Ф., Уголев А. М, Бюлл. экспер. биол. и мед., 1960, 50, 9,24.
Миллер Н. Экспериментальная психология. Под ред. Стивенса. М.,
1960.
Миллер Н. В сб.: Высшая нервная деятельность. М., 1963, с. 23.
Миллер Н. Экспериментальные исследования по теории обучения и пси-
хопатологии. Лекция на 18-м Международном психологическом кон-
грессе. М., 1966.
Миллер Дж., Галантер Е. и Прибрам К- Планы н структура поведения.
М„ 1965.
Михайлова И, Г, В сб.: Физиология и патология лимбико-ретикулярно-
го комплекса. М., 1968, с. 58.
Никифоровский П, Н, Труды Общества русских врачей в СПб., 1910, Т. 77.
Нуцубидзе М. А. Сообщение АН Груз. ССР, 1962, 28, 6, 717.
Нуцубидзе М. А. Труды Института физиологии АН Груз. ССР, 1963,
3, 103.
Орджоникидзе Ц. А. и Нуцубидзе М. А. Труды Института физиологии
АН Груз. ССР, 1961, 12, 85.
Орлов И. В. Сборник научных трудов Института акушерства и гинеко-
логии. М„ 1961, с. 56.
288
Павлов И. П. Труды Общества русских врачей, 1911, т. 78, с. 31.
Павлов И. П. (1916). Рефлекс цели. Поли. собр. сочим., 1949, г. 3, с. 243.
Павлов И. П. (1933). О повышении пищевой возбудимости В кн.; Пав-
ловские среды М.—Л., 1949, т. 1, с. 306.
Павлов И. П. Двадцатилетний опыт объективного изучения высшей нерв-
ной деятельности (поведения) животных. М„ 1951.
Панфилов А. А., Лосева Т. И. Физиол. журн. СССР, 1966, 52, 5, 447.
Пенфилд У., Джаспер Г. Эпилепсия и функциональная анатомия голов-
ного мозга человека. М., 1958.
Петрова М. Д. Труды физиологических лабораторий акад, И. П. Пав-
лова. Л„ 1936, 6, 1, 5.
Пинес Ю. Л. Докл. АН СССР, 1965, 160, 2, 482.
Полянцев В. А., Сербиненко М. В. В сб.: Вопросы физиологии и патоло-
гии нервной системы, 1962, 6, с. 8.
Попов £. А. Современная психоневрология, 1927, 5, 6—8, 76.
Попов П. Ф. Изв. АН СССР, 1950, 6, 124.
Правдивцев В. А. В сб.: Интегративная деятельность мозга. М„ 1967,
с. 67.
Разенков И. П. В кн,: Проблемы современной физиологии, биохимии и
фармакологии. Под ред. Л. А. Орбели. М., 1949, кн. 1, 13.
Рогачева С, Д. Первая научная конференция, посвященная проблемам
физиологии, морфологии, фармакологии и клиники ретикулярной фор-
мации головного мозга. М., 1960, с. 95.
Ройтбак А. И. Биоэлектрические явления в коре больших полушарий.
Изд-во АН Груз. ССР. Тбилиси, 1955.
Рожанский И. А. Очерки по физиологии нервной деятельности. Медгиз.
Л., 1957.
Сеченов И. М. Рефлексы головного мозга (186'6). Избранные произведе-
ния. М„ 1953.
Симонов Н. В. Что такое эмоция? М., 1966.
Синичкин В. В. В сб.: Интегративная деятельность мозга. М-, 1967,
с. 72—74.
Советов А. Н., Черниговский В. Н. Бюлл. экспер. биол. и мед., 1960, 50,
9, 16.
Сойер Ч. X. В кн.: Ретикулярная формация мозга. М., 1962, с. 204-
Соколов Е. Н. В кн.: Ориентировочный рефлекс и вопросы высшей нерв-
ной деятельности. М., 1959, с. 5.
Стадниченко Л. И. Физиол. журн. СССР, 1969, 55, 4, 461.
Стрельчук И. В. Клиника и лечение наркоманий. М., 1949.
Судаков К. В. О рефлекторном возбуждении ядра блуждающего нерва
в условиях относительного голода. Дисс. канд. М., 1959.
Судаков Д. В. Бюлл. экспер. биол. и мед,, I960, 50, 11, 23.
Судаков Д. В. В сб.: Вопросы физиологии и патологии органов пищева-
рения. Труды, 1 Моск. мед. ин-та им. И. М. Сеченова, 1960, 8, 91.
Судаков Д. В. Физиол. журн. СССР, 1962, 48, 2, 150,
Судаков Д. В. Физиол. журн. СССР, 1963, 49, 8, 901,
Судаков К. В. Мотивации. БМЭ. Изд. 2-е, 1964, т. 36, с. 750.
Судаков Д. В. В сб.: Функциональная нейрохимия. Баку, 1966, с. 163.
Судаков Д. В. В сб.: Физиология и патология гипоталамуса. М., 1966,
с. 114.
Судаков Д. В., Богомолова Е. А1. Лимбическая система (Обзор). БМЭ.
Ежегодник, 1969, № 2, с. 506.
Судаков Д. В., Мнасин Л. С. Бюлл. экспер. биол. и мед., 1965, 4, 8,
Судаков Д. В., Набиль Э. М. Физиол. журн. СССР, 1966, 52, 7, 785.
Судаков Д. В., Рогачева С, Д. Физиол, жури СССР, 1962, 48, 6, 728.
289
Судаков К- В., Фадеев Ю. А. Физиол. жури. СССР, 1963, 49, 11, 1310.
Судаков К. В., Котов А. В. Материалы симпозиума XXII совещания по
проблемам высшей нервной деятельности. Л., 1909, с. 20.
Сунь-Вень. Цит. по статье II. К. Анохина. Физиол. журн. СССР, 1964,
50, 6, 773.
Снастный А. И. Докл, АН СССР, 1963, 149, 4, 995.
Снастный А. И. Физиологическое изучение сложных форм поведения че-
ловекообразных обезьян (шимпанзе). Докт. дисс. Л., 1968.
Трофимов Л. Г. В кн.: Вопросы электрофизиологии и электроэнцефало-
графии. М.—Л., Изд. АН СССР, I960, с. 344.
Туренко А. И. Труды Института физиологии АМН СССР, 1964, т. 7, с. 100.
Туренко А. И. Нейрофизиологический анализ восходящих активаций раз-
личного биологического качества. Дисс. канд. М., 1965,
Уголев А. М., Черниговский В. Н. Докл. АН СССР, 1959, 126, 2, 450.
Уголев А. М., Кассиль В. Г. Успехи совр. биол., 1961, 51, 3, 352.
Удалова Г. П. Журн. высш. нерв, деят., 1968, 58, 6, 1066'.
Ухтомский А. А. (1923). Инстинкт и доминанта. Собр, соч. Л., 1950, т. 1,
с. 186,
Ухтомский А. А. (1927). Доминанта как фактор поведения. Собр. соч.,
1950, 1, с. 293.
Ухтомский А. А. О доминанте (1928). Собр. соч. Л., 1950, т. 1, с, 316.
Фадеев Ю. А. Рефераты работ VIII конференции молодых ученых Ин-
ститута нормальной и патологической физиологии. М., 1963, с. 107.
Фадеев Ю, А, Анализ активности корковых нейронов в условиях восхо-
дящих влияний различного биологического качества. Дисс. каид.
М„ 1964.
Филимонов Д. В., Чирейкина Б. X. Врач, дело, 1929, 2, 107.
Фонберг Е. В. В ки,: Рефлексы головного мозга. М., 1965, с. 382.
Фурсиков Д. С. Русск. физиол. журн,, 1921, 4, 257.
Хазен С. В. О соотношении размеров безусловного и условного слюно-
отделительных рефлексов. Дисс. СПб., 1908.
Хаютин С. В. Материалы XXII совещания по проблемам высшей нервной
деятельности, Рязань, 1969, 23.
Хьюм Д. М. В кн.: Ретикулярная формация мозге. М„ 1962, с. 211.
Цитовия И. С. Происхождение и образование натуральных условных реф-
лексов. Дисс. СПб., 1911.
Черешнев И. А. Бюлл. экспер. биол. и мед., 1950, 29, 6, 429,
Черешнев И, А. Журн. высш. нерв, деят., 1960, 10, 6, 896.
Черкес В. А. Журн. высш. нерв, деят., 1967, 17, 70.
Черниговский В. Н. Жури. высш. нерв, деят., 1960, 10, 3, 313.
Черниговский В. Н. Значение интероцептивной сигнализации в пищевом
поведении животных. М.—Л., 1962.
Чуппина Л. М. Физиол. журн. СССР, 1967, 53, 4, 385.
Чхенкели С. А. Сообщение АН Груз. ССР, 1963, 3, 699.
Чхенкели С. А. О физиологических механизмах голода и насыщения.
Дисс. канд. Тбилиси, 1968,
Шелихов В. Н. Жури, невропатол. и психиатр,, 1960, 60, 145,
Шулейкина К- В. В сб.: Материалы VI научной конференции по вопро-
сам возрастной морфологии, физиологии и биохимии. М„ 1963, с. 506.
Шулейкина К- В. Физиол. журн. СССР, 1964, 50, 8, 1025.
Шулейкина К. В. Функциональная организация пищевого поведения в
раннем онтогенезе. Дисс. докт. М., 1967.
Шумилина А. И. В сб.: Проблемы высшей нервной деятельности. М.,
1949, с. 561.
Шумилина А. И, Журн. невропатол. и психиатр., 1956, 56, 116.
290
Шумилина А. И. Конференция по электрофизиологии центральной нерв-
ной системы. М., 1958, с. 4.
Шумилина А. И. Физиол. журн. СССР, 1961, 47, 1, 3.
Шумилина А. И. В сб/. Современные проблемы физиологии и патологии
нервной снтемы. М., 1965, с. 240.
Эйди У. (Adey W7. R.). В ки.: Ретикулярная формация мозга. М., 1962,
с. 544.
Allikments L. И., Dltrich .И. Е. Zh. vysh. пег/. Deyat. Pavlova, 1965
15, 86.
Allikmets L. H.a. lapin J. (Алликметс Л.и Лапин И.) Actlv. nervosa
sup. (Praha), 1966 , 8, 2. 129.
Akeri K. In; Electrical Stimulation of the Brain, 1961, 288.
Anabd О. К. Physiol. Rev., 1961, 41, 4, 677.
Anand В. X. 22 Intern. Congress of Physiol. Sci. Amsterdam, 1962, 1,
p, 2, 680.
Anand В. K. Ind. J. Med. Res., 1963, 51, 175.
Anand В, К., Brobeck J. R. Yale J. Biol. Med., 1951, 24, 123.
Anand В, K., Chhina (i. S.. Singh B. 22 Int. Congr. of Physiol. Sci,
Leiden, 1962, v. 1, p. 682.
Anand B. X., Chhina (i. S’., Sharma K. N., Dua S., Singh B. Am.
J. Physiol., 1964, 207, 5, 1146.
Anand В. K., Rua S. Ind. J. Med. Research, 1955, 43, 1, 113.
Anand В. X., Rua S. Science, 1955, 122, 1139.
Anand В. X., Dua S. Ind. J. Med. Res., 1956, 44, 107.
Anand В. X., Dua S., Chhina G. S. Ind. J. Med. Res., 1958, 46, 2,
277.
Anand B. A’., Dua S., Chhina G. S. Ind. J. Med. Res., 1961, 49 , 4,
491.
Anand В. X., Dua S., Singh B. Electroenceph. Clin. Neurophysiol.,
1961b, 13, 54.
Andersson В Experientia, 1952, 8, 157.
Andersson B. Acta physiol. Scand., 1953, 28, 188.
Andersson B., Me Cann S. M. Acta Physiol. Scand., 1955 , 33 , 333.
Andersson B., Me Cann S. M. Acta Physiol. Scand., 1956, 35, 312.
Andersson B., Kitchell R. L. a. Person N. Acta Physiol. Scand., 1958,
45, 319.
Andersson B., Larsson B. Acta Physiol. Scand., 1961, 52, 75.
Anliker J., Mayer J. Am. J. clin. Nutrition, 1957, 5, 148.
Anochin P. K., Schumilina A. I. (Анохин П. К., Шумилина А. И.).
Arch. Int. be Pharmacodyn. et de Therapie, 1962, 160, fasc, I—11, 1.
Anochin P. X., Sudakov i\. J. Proceedings of 7th Int. Congress of
Nutrition, 111. Hamburg, 1966.
Aronson L. R., Noble G. X. Bull. Am. Mus. Nat. Hist., 1945 , 86 , 87.
Atkinson J. IF. An Introduction to motivation. Princeton D. Von Nost-
randeo Inc., 1964.
Bailley P., Morrison S. D. j Physiol. (LonJ.), 1963, 165, 2, 227.
Baird H., Guidetti B., Reyes V , IFyc/s H. T., Spiegel £. A. Fed.
Proc., 1951, 10, 8.
Balinska H. Acta biol. Exp. (Warsaw) 1963 23 , 2, 115.
Balinska H. Acta biol. Exp. (Aarsaiv), 1963, 23, 1, 33,
Balinska H. Rev. roumaine blol. Ser. Zool., 1966, 11, 4, 243.
Balinska H., Brufkrwski S, Acta blol, Exp., 1964, 24, 4, 213.
Balinska. H., Lewinska K., Romaniuk A., Wyrwicka IF. Acta blol.
Exp., 1961, 21, 189,
291
Ball J. J. Comp. Psychol., 1934, 18, 419.
Ball G. G., Adams D. IV. Psychon. Scl., 1965 , 3 , 39.
Bar-Sela Л1. E., Critchlow 7, Am. J. Physiol., 1966, 211, 1103.
Bard P. Am. J. Physiol., 1928 , 84, 490.
Bard P. Psychol. Rev., 1934 , 41, 309 , 429.
Bard P. Am. J. Physiol., 1936, 116, 4.
Bard P. The hypothalamus and central levels of Autonomic function.
Baltimore, Williams and Wilkins, 1940.
Bard P., Mountcastle V. B. A. Res. Nerv. and Ment. Dis. Proc., 1947,
27, 362.
Bard P., Mountcastle V. B. Res. Publ. Nerv. Ment. Dis., 1948 , 27,
362.
Bartoshuk A. K., Motivation IT. C. Brown Company publ. Iowa, 1966.
Bask K. W. J. Comp., Psychol., 1939. 28, 109.
Baxton W. H., Heron 17., Scott T. H. Canad. J, Physiol., 1954, 8,
70.
Bazett H. C., Penfield W. G. Brain, 1922, 45, 185.
Beach F. A. J. Comp. Psychol., 1940 , 29, 193.
Beach F. A. Psychosom. Med., 1944, 6, 40.
Beach F. A. Anat. Rec., 1945 , 92, 289.
Beach F. A. Psychol. Rev., 1947, 54, 297.
Beach F. A. Hormones and Behavior. New York, Paul B. Hoeber Inc,
1948.
Beach F. A. The Harvey Lectures 1947—1948. Springfield 111, Thomas,
Ed. 1950 254.
Beach F. A. J. Comp, physiol. Psychol., 1958, 51, 37.
Beach F. A. In: Biological and Biochemical Bases of Behavior Harlow.
H. F. a. Woolsey C. N. Madison Uni. Wise. Press, 1958.
Berkun M. M., Hessen M. L., Miller N. E. J. Comp. Physiol. Psy-
chol., 1952, 45, 550.
Bernard C. Lecons sur les proprietes physiologlques et les alterations
pathologlques des llquldes de Ibrgauisme, Paris Balliere, 1859,
1, 2.
Blndra D. Motivation: A Systematic reinterpretation. New York, Ronald
Press, 1959.
Blndra D., Stewart J. .Motivation (Selected Readings) Penguin modern
Psychol., 1966.
Bischop G. H., Clare M. H. J. Neurophysiol., 1953, 16, 1.
Bishop G. H. OLeary Electroenceph. clin. Neurophysiol., 1950, 2, 401.
Bloom F. E., Oliver A. P., Salmolrachi G. C. Int. J. Neuropharma-
coi. 1963, 2, 3—4, 181.
Bonvallet M., Dell P., Hiebel G., Electroenceph, clin. Neurophysil,
1954, 6, 119.
Bovard E. TP. Perspectives in Biology and Medicine, 1961. 5, 1.
Bovard E. 17. Perspectives in Biology and Medicine, 1962 , 6, 1.
Brady J, V. Am. Psychologist, 1955, 10 , 369 (abstr).
Brady J. 7. In: Biol, and Biochem. Bases of Behavior. (Ed. H. F. Har-
low and C. N. Woolsey), 1958, 193.
Brady J. 7., Hunt F. J. Psychol., 1955, 40, 313.
Brady J. 7., Nauta 17. J. H. J. Comp. Physiol. Psychol., 1953 , 46,
339
Brady J. 7., Naufa IF. J. H. J. Comp. Physiol. Psychol., 1955 , 48,
412.
292
4 Brady J. V., Shreiiier L., Geller I., Kling A. J. Comp. Physiol. Psy-
: chol., 1954, 47, 179.
r Brobeck J. R. Yale J. Biol. Med., 1943, 20, 545.
: Brobeckfj.“R. Physiol. Rev., 1946 , 26, 541.
; Brobeck J.R. Am. New York Acad. Scl., 1955 , 63, 44.
Brobeck J. R. Gastroenterology, 1957, 32, 2, 169.
>' Bronk D. W., Pitts R.F., Larrabee M.G, Res. Publ, Ass. nerv. ment.
i Dis., 1940, 20, 323.
[ Brookhart J. Л1., Dey F. L. Am. J. Physiol., 1941, 133, 551.
; Brooks C. Me. C, Am. J. Physiol., 1937, 120, 544.
: Browman L. G. J. Comp. Psychol., 1943, 36, 33.
; Brown J. S. Nebraska Symp. on Motivation. Univ, of Nebraska Pres.
i Lincoln, 1953, 1.
‘ Brown J. S. The motivation of behablor, New York, McGraw hill.,
' 1961.
Brown G, IP., Cohen B. D. Am. J. Physiol., 1959, 197, 153.
f Brutkowski S., Dabrowska J. Science, 1963, 139 , 3554, 505.
,s Brutkowski S., Fonberg E., Mempel E. Acta blol. Exp. (Warsaw),
i 1960, 20, 263.
Brutkowski S., Fohberg E., KreinerJ., Mempel E., Sychowa B. Acta
blol. Exp. (Warsaw), 1962, 221, 143.
i Brutkowski S., Mishkin M., Rosvold H. E. In: Central and Peripher.
Meehan, of motor functions. Prague, 1963, 133.
BrUgger M. Helv. physiol, pharmacol. Acta, 1943, 1, 183.
Buchwald N. A.. Horlath F. S., Wyers E. S., Wakefield C. Nature
(Engl.), 1964, 201, 4921, 830.
Bulatao E., Carlson A. Am. J. Physiol., 1924, 59, 107.
Bunnell B. N. Abctr. XXIII Int. Congr. Psychol. Moscow, 1966, 1, 314.
Burdenko N. N., Mogilnitzklj B. N. Z. Neurol. Psychol., 1926, 2,
1-2, 42.
: Cannon W. B. Bodily Changes in Pain, Hunger, Fear and Rage 2nd.
ed. New York, Appleton, 1929.
Cannon IP. S. The Wisdom of the body. New York, Norton, 1932.
1 Cannon IP. B. Hunger and thrlst. A Handbook of gen. exp. Physiology,
ed. by C.Murchison, 1934, 247, Worcester, Mass.—London.
Cannon VP. B., Washdorn A. L. Am. J. Physiol., 1912, 295, 441.
Carlson A. J, The control of hunger in health and disease. Chicago,
1919, 176.
I Chang H. T. J. Neurophysiol., 1950, 13, 235.
Chang H. G. Handbook pf Physiolog, 1959, Sect. 1, 1, Neurophys.,
299, Washington.
Clark G. Amer. J. Physiol., 1942, 137, 746.
Clemente C. D., Sterman M. B,t Wyrwicka IP. Electroenceph. Clin.
Neurophysiol., 1964, 16, 4, 355.
Cobb S. Med. Clin. N. Amer., 1944, 28, 1210.
Cofer C. N., Appleu M. H. Motivation: theory and research. New Ork,
Wiley, 1964.
Congen J. J. An analysis of the effect of alcohol on conflict behavior
in the albino rat. Ph. D. Dissertation, Yale Univ., 1949.
Coons E. E., Cruce J. A. F. Science, 1967, 159, 1117.
Cowless I. T. Comp. Psychol. Monogr., 1937, 14, 5.
Craig TP. Biol. Bull., 1918, 34, 91.
Criiclow V. B. Anat. Rec., 1957, 127, 283.
Critchlow E. Am. J. Physiol., 1960, 198 , 381.
293
Davidson j. AL, Sawyer С. H. Acta endocr. (Kbh.), 1961, 37, 185.
Davis C. D. Am. J. Physiol., 1939, 127, 374.
Davis J. D., Miller Л. E. Science, 1963. 141, 1286.
Delgado J. M. R. Trans. New York Acad. Sci, 1959, 21, 689.
Delgado J. M. R. Psychiat, Res. Reports. Am. Psychiat. Ass., I960,
12, 259.
Delgado J. M. R. Evolution of Physical control of the brain. The amer.
Museum of Nature history. New York, 1965.
Delgado J. M. R., Anand В. R. Am, J. Physiol., 1953, 172, 162.
Delgado J. M. B., Ritahata L. M. Electroenceph. Ciln. Neurophysiol.,
1967, 22, 453.
Dell P. C. Ciba. Found. Symp. .Neurological Basis of Behavior", Bos-
ton, L. Brown, 1958, 187.
Dell P., Bonvallet M. C. R. Soc. Biol. Paris, 1954, 148, 855.
De Molina F. A., Hunsperger R. IF. J. Physiol., 1959, 145, 251.
Dempsey E.IF., Rioch D. Me. R. J. Neurophysiol., 1933, 11, 9.
Deutsch J. A. J. Theoret. Biol., 1963, 4, 2, 193.
Deutsch J. A., Howarth С. I., Ball G. C., Deutsch D. J. Nature,
1962, 196, N 4855, 699-700.
Deutsch J. A.. Howarth С. I. Psychol. Rev., 1963 , 70, 444.
Dewey J. Psychology. Harper. New York, 1886.
Dey F, L., Fisher C., Berry С. M., Ranson. Amer. J. Physiol., 1940,
129 , 39.
Donovan В. T., Van der Werff Ten Bosch J. Physiol., 1956, 132 , 57.
Dusser de Barenne J. G,,Mc Culloch IF.S.J. Neurophysiol.,1938,1,176.
Eccles J. C. EEG a.clin. Neurophys., 1951, 3, 449.
Egger M. D., Flynn J. P. J. Neurophysiol., 1963 , 26 , 705.
Egger M. D., Flunn J. P. progress in Brain Research. Structure and
Function of the Limbic System, 1967, 27, 165.
Ebdroczl E., Llssak K. Acta physiol. Acad. Sci. Hung., 1961, 18, 301.
Endroczi E., Llssak R. Phys, and behavior, 1966, 1, 3, 233.
Epstein A. N. Am. J. Physiol., 1960, 199, 6 , 969,
Epstein A, N., Teitelbaum P. J. Comp. Physiol. Psychol., 1962 , 55,753.
Estes TF. R. In: Nebraska Sumposium on Motivation. Univ, of Nebraska
Press. Lincoln, 1958, 35.
Falk J. R. In: Nebraska Symposium on motivation. Л1. R. Jones ed.
Lincoln, Neb, Univ of Nebraska Press, 1961.
Feldman S. EEQ a. clln. Neurophys., 1963, 15, 4, 672.
Fessard A. The Moscow Colloquium, on EEC of higher nervous acti-
vity. Eds. H. Jasper a. Q. Smirnow. Moscow, 1958.
Fisher A. E. Science, 1956, 124, 228.
Fisher A. E. Jn: Current trends in psychological theory. Pittsburgh,
Univ, of Pittsburgh Press, 1961, 70.
Fisher A. E. Scienl, Am., 1964, 210, 6, 60.
Fisher A. E., Coury J. N. In M. J. Wayner, Thirst, 1st Symp. on
thirst in the regulation of body water. Perganion Press, 1964.
Fisher C., Magoun H. IF., Ranson S. IP. Am. J. Obstet. Gynec.,
1938, 36, 1.
Fitzsimonz J, T. In: Recent Studies on the Hypothalamus. Brit. med.
Bull., 1966, 22, 232.
Fonberg E. Bull. Acad. Polon Sci., 1966, 14 , 719.
Fonberg E., Delgado J. M. R. J. Neurophysiol., 1951, 24, 651.
Franck H. Soc. Biol. Paris, 1965, 160, 389.
Freeman IF. J. Exp. Neurol., 1962, 6, 1, 12.
294
Freud S. Instincts and their vicissitudes. Collected Papers. Hogarta.
Г onion, 1925.
Fuller J. L., Rosvold H. E., Pribram К. H. J. Comp. Physiol. Psy-
chol.. 1957, 50, 89.
Fulton J. F. Functional localization in the frontal lobes and cerebel-
lum. Oxford, Clarenion Press, 1949.
Fulton J. F., Ingram F. F>. J. Physiol,, 1929 , 67, 67.
Galamhos P. Federal. Proc., 1961, 20, 2, 1, 603.
Galamhos P., Sheatz G. C. Am. J. Physiol., 1962, 203, 1, 173.
Gantt IF. H. Am. J. Physiol., 1940, 129 , 360.
Gavrilov R. J., Znachkova А. А. (Гаврилов P. И.. Значкова A. A.).
Zeitschs. f. d. ges. exp. Med. Bd, 1933, 89, 183.
Gellhorn E., Eoofhourrow G. N. N. Y. Ehanston, L., 1963.
Gold P. <W, Physiol. Behaviour, 1967, 2, 211.
Goltz F. Pfliig. Arch. ges. Physiol., 1892, 51, 570.
Grastyan F., llssak., Madarasz I., Donhoffer H. Electroenceph. clin.
Neurophysiol., 1959, 11,4.
Grastyan F., Earmos G., Vereczkey L, J. Cropt. Acad, of Sci. Hava-
na, Cuba, 1966, 155.
Grastyan F., Karmos G., Vereczkey I., Kellenji L. EEG and Clin.
Neurophys., 1966 , 21, 34.
Green J. Г)., Clemente C. D., De Groot J. J. Comp. Neurol., 1957,
108, 505.
Greer M. A. Proc. Soc. exp. Biol (N. Y.), 1955 , 89, 59.
Gross K. The play of animals. New York, Appleton-Century Crofts,
Grossman W. I,, Cummins G. M,, Ivy A. C. Am. J. Physiol., 1947,
149, 100.
Grossman M. I., Stein I. F. J, Appl. Physiol., 1948, 1, 263,
Grossman S. P. Science, 1960, 132 , 3422 , 301.
Grossman S. P. Int. J. Neuropharmacol., 1964 , 3 , 45.
Grossman S. Discovery, 1966 , 27, 5, 19.
Grossman S. P. Ann. New York Acad. Sci, 1969, 157, 902.
Grossman S. P., Grossman I. Am. J. Physiol., 1963 , 205 , 4 , 761.
Hatl J. F. Psychology of Motivation. Lippincott. New York, 1961.
Harker J. E. The physiology of diurnal rhythms. Cambridge, Univ.
Press, 1964.
Harlow H. F. Sci, Am., 1959, 200, 68.
Harlow 4. F., Harlow M. K., Mayer D. L. J. exp. Physiol., 1950,
40, 228.
Harris G. IF. J. Physiol., 1948, 107, 418.
Harris G, IF., Levine S. J. Physiol, (bond.), 1962, 163 , 42P.
Harris G. IF., Michael L. P., Scott P. P. Clba Found. Coll, on Endo-
crinol., 1958, 236.
Harris S. C., Ivy A. C., Searle L. M. J.A.M.A,. 1947, 134, 1468.
Harrison J.. M, Lyon M. J. Comp. Neurol., 1957, 108, 1, 121.
Heath R. J In: R.G. Heath et at. Studies in Schizophrenia. Cambridge
Mass. Harvard Univ. Press, 1954, 83.
Heath R. G,, Mickle IF. A. In: Electrical Studies on the Unanestheti-
zed Brain. New Yorkt, Hoeber, 1960, 214,
Hebb D, O. Ordanizatlon of behavior. New York, Wiley, 1949.
Hebb D, O. Canad. J. Psychol., 1953, 7, 99.
Hebb D. 0. Psychol, Rev., 1955 , 62 , 243.
295
Helnbecker P.t White H. L. a. Rolf D. Am, J. Physiol,, 1944, 141,
54g______5g5
Herberg L. J. Drive and the brain. Discovery, 1965, 26, 2, 8.
Hervey G. R. J. Physiol., 1959, 145 , 2, 336.
Hess W. R. Die ReguHerung des Blutkrelslaufes. Leipzig, 1930.
Hess W. R. Das Zwischenhlrn, Basel, Schwabe, 1949,
Hess VP. R. Diencephalon: Autonomic and Extrapyramidal Functions,
New York, 1954.
Hess IP. R., Akeri K. Arch. Neurol. Psychiat. (Chlcado), 1955 , 73,
127—129.
Hess W. R., Brugger At., Akert K. In: Electrical Stimulation of the
Brain, 1961, 288.
Hess R. W., Koella IP. P., Akert K. Electroenceph. clin. Neurophysl-
ol., 1953 , 5 , 75.
Hetherington A. W. Am. J. Physiol., 1943, 140 , 89.
Hetherington A. W., Ranson S. W. Anat. Rec., 1940, 78, 149.
Helnd R. A. Symp. of the Society for Experimental Biology. Cambridge
Univ. Press. Cambridge, 1960, 14, 199.
Hockman С. H. Electroenceph, Clin. Neurophysiol., 1964, 17 , 420.
Hoebel B. G., Teitelbaum P. H. Science, 1962, 135, 3501, 375.
Hoebel B. G. Science, 1965, 149, 3682 , 452.
Hull C. L. Principles of Behavior. Appl-Century-Crofts. New York.
1943.
Iggo A. J. Physiol., 1955, 128, 593.
Ingram W. R., Barris K. W,, Ranson S. IP. J.M.A. Arch. Neurol.
Psychiat. (Chicago), 1936 , 35, 1175.
Janowitz H. D., Grossman M. I. Am. J. Physiol., 1949, 159, 143.
Janowitz H. D., Grossman M. I. Am, J. Physiol., 1951, 161, 182.
Janowitz H. D., Ivy A. C. J. Aopl. Physiol., 1949, 1, 643.
Jasper H. H. In: Brain Mechanisms and Consciousness. Ed. by J. F.
Delafresnaye, Springfield. Thomas, 1954 , 374,
Jasper H, H., AJmone-Marsan C., Stoll J. Arch. Neurol. Psychiat.,
1952, 67, 155.
Javell В. P. J. Physiol., 1953, 121, 1, 167.
Jung R. in: Sensory communication. Ed. Rosenbtith, New. York—Lon-
don, 1961, 627.
Kaada B. R, Acta Physiol. Scand., 1951, 24, 285, Suppl. 83.
Karli P. Behavior, 1956, 10, 81.
Karli P. J. Physiol. (Paris), 1968, 60, 3, Suppl. 1.
Kawakami M., Sawyer С. H. Endocrinology, 1959, 65, 652.
Kawamura H., Nakamura Y., Toklzane T. The Japan Physiol. (Lond.),
1961 П 5 564.
Kawamura H., Oshima K. The Japan. J. Physiol. (Lond.), 1962, 12 , 3,
Keller A. D., Noble TV. Am J. Physiol., 1935, 113, 79.
Keller A. D., Witt D. M., Batsel H. L. Fed. Proc., 1959, 18 , 80.
Kennard M. A. J. Neuropath, exp. Neurol., 1945 , 4, 295.
Kennedy G. C. Proc. roy. Soc. Biol., 1953, 140, 578.
Keener R. P., Fiedler P., Thomas G. S. J. Comp. Physiol. Psychol.,
1967 , 63 , 452.
King F. A. A. nerv. ment. Dis., 1958, 126, 1, 57,
Kinsey A. C., Pomerov TV. B., Matrin C. F., Gebhard P. H. Sexual
behavior In the human female. Philadelfia W. B. Saunders. Co.,
1953.
К It al S. T., Morin F, Nature (Engl.), 1965, 206, 4991, 1375.
296
Kltai S. T., Cohen B., Morin F. Electroenceph. clln. Neurophyslol.,
10RK 1Я 4 444
Kling A. Science, 1962, 137 , 429.
Kling A., Schwartz N. B. Fed. Proc., 1961, 2Q, 335.
KHiver H., Вису P. C. Am. J. physiol,, 1937, 119, 352.
Kliiver H., Вису P. C. J. Psychol., 1938, 5, 33.
KHiver H., Васу P. С. A.M.A. Arch. Neurol. Psychiat. (Chicago),
1939 , 42, 979.
Koikegaml H., Yamada T. Med. J. Osaka Univ., 1950 , 2, 1.
Koranyt L., Endroczi E. Acta physiol. Acad, Sci. hung,, 1965, 28,
339.
Kotov A. V. (Котов A. B.). Wlssenschafl. Z. De K. Marx Univ. Leip-
zig 1967, 441.
Kubie L. S. J. Psychosom., 1948, 10, 15.
Larsson S., Forssberg A. Acta physiol, scand., 1954, 32, 1—2, Suppl.
115.
Lashley K. S. Psychol. Rev., 1938 , 45 , 445.
Li C. L., Cullen C., Jasper H. H, J. Neurophyslol., 1956, 19, 131.
Liebelt R. A., Dear IV., Guillemin ,R. Proc. Soc. exp. Blol. (N. Y.),
1961, 108, 2, 377.
Lilli J. C. In: Electrical Studies of the unanesthetized brain', 1960, 78.
Lindsley D, B. In: Handbook of experimental Psychology Ed. S. S. Ste-
vens. New York, 1951, 473.
Lindsley D. B., Bowden J. IV., Magoun H. IV. Etectroenceph. clln.
Neurophyslol., 1949, 1, 475.
Lisk R. D. J. exp. Zool., i960, 145, 197.
Lisk R. D. Science, 1966, 152, 669.
Lissak K-, Endrdczi E. In: Physiol, de IHippocampe. CNRC Symp.
Montpellier, CNRC. Parts, 1962.
Livingston R. B., Fulton J.F., Delgado J.M. R., Sachs E, jr., Brend-
ler S. J., Davis G. D. Res. Publ. Nerv. Ment. Dis., 1948, 27, 5,
405.
Lorenz K- L, Symp. Soc. exp. Blol., 1950, 4, 221.
Lorenz K. L. Zoolog. Anzelg. Suppl., 1953, 17, 36.
Mac Donnel M. F,, Fiynn J, P, Science, 1966, 152 , 3727, 1406.
Mac Dougall И7. An Introduction to Social Psychology. Methnen, Lon-
don, 1908.
Mac Dougall W. An Introduction to Social Psychology. Boston Luce,
1923.
Mac Lean P. D, J. Neurosurg., 1954, 11, 29.
Mac. Lean P. D. Arch. Neurol. Psychiat., 1955, 73, 130.
Mac Lean P. D. J. nerw. ment. Dis., 1958, 127.
Mac Lean P. D. In; The Central Nervous System and behavior. Ed. M.
Brazier, 1959, 31.
Mac Lean P. D. Brain and behavior. In; Brain and Gonadal function,
1966.
Mac Lean P. D., Delgado J. M. R. Electroenceph. clln. Neurophyslol.,
1953 5 91
Mac Lean P. D., Ploog P. IV., Robinson B. W. Physiol. Rev., 1960,
40 (Suppl. 4), 105.
Magoun H. TV. In: Brain Mechanisms and Consciousness. Ed. by J. F.
Delafresnaye. Springfield: Thomas. 1954, 1.
Magoun H. W., Ranson S. W., Hetherington A. Arch. Neurol. Psy-
chiat. (Chicago), 1938 , 39, 1127.
297
Malre F. W., Patton H. D. Am. J. Physiol., 1954, 178, 315.
Marczynskl T. J., Rosen A. J., Hackett J. T. Electroenceph. clin.
Neurophysiol., 1968, 24, 227.
Marczynskl T. J., Hackett J. T, Electroenceph. clin. Neurophysiol.,
1969, 26, 41.
Marczynskl T. J. Fed. Proc., 1969, 28, 1, 132,
Margules D. L., Olds J. Science, 1962, 135, 3501, 374.
Markee J. E., Sayer С. H., Hollinshead 11'. H. Endocrinology, 1946,
38 345.
Marshall N. B., Mayer J, Am. J. Physiol., 1954, 178, 271.
Marshall N.B., Barrnett R.J., Mayer J. Proc. Soc. exp. Blol. (N.Y.),
1955 90 240.
Marshall В7. H., WoolseyC.N., Bard Ph.]. Neurophysiol., 1941, 4, 1.
Masalchi S., Naoshlge M., Talgl H., Shlzuo K. Folia psychlatr. et
neurol. Japan, 1959, 13, 3, 253.
Massertnan J. H. Behavior and neurosis. Chicago, Univ, of Chicago,
Press, 1943.
Masserman J. H., Jam K. S. Psychosom.med., 1946 , 8 , 36.
Mafasaburo J. Acta med. Univ. Kagoshimaensls, 1964 , 6 , 2, 155.
Matsumota E, Kijono S., Ide K. Med. journ. Osaka University, 1963,
13 325
Mayer J. Bull. New. Engl. med. Cent., 1952, 14, 43.
Mayer J. Ann. New York Acad. Sc., 1955, 63, 15.
Mayer J., Barrnett R. J. Science, 1955, 121, 599-
Mayer J., Marshall N. B. Nature, 1956, 178, 1399-
Mayer J. J., Vitale J., Bates M. 11 . Nature (London), 1951, 167, 562-
Mendelson J., Chorover S. L. Science, 1965, 149, 3683, 559.
Michael R. P. Science, 1962, 136, 322.
Miles R. C. Comp, and Physiol. Psychol,, 1962, 55, 3, 358.
Miles S. F., Thomas G. M. Neurology, 1962, 12, 6 , 394.
Miller F. R., SherrlngtonC. S. Quart. J. exp. Physiol., 1916,9,2, 147
Miller N. E. Science, 1957, 126, 1271.
Miller N. E. Fed. Proc,, i960, 19, 846.
Miller N. E. Ann. New York Acad. Sci., 1961, 92, 3, 830.
Miller N. In: Electrical stimulation of the brain, 1961, 387.
Miller N. E. Ann. New York Acad . Sci., 1961, 92, 3, 830. Discuss.
890-897.
Miller N. E. Electroenceph. clin. Neurophysiol., 1963, Suppl. 24.
Miller N, E., Bailey C. J., Stevenson J.A.F. Science, 1950, 112 , 256.
Miller N. E., Dollard J. Social learning and imitation New Haven. Yale
Univ. Press, 1941.
Miller N. E., Gottesman K. S., Emery N. Am J. Physiol., 1964 , 206,
6, 1384.
Miller N. E., Hessen M. L, J. Comp. Physiol., Psychol., 1952, 45, 555.
Mishkin M. J. Comp. Physiol. Psychol., 1954, 47, 187.
Mishkin M., Pribram R. H. J. Comp. Physiol. Psychol., 1954, 47, 14.
Mogenson C. J., Stevenson J.A.F. Exp. Neurol., 1967, 17, 119.
Monfemurro D. G., Stevenson J. A. F. Canad. J. Blochem, Physiol.,
1957, 35, 31.
Monfemurro D. G., Stevenson J. A. F. Canad. J. Biochem., 1957, 191,
248.
Morgan С. T. Physiological psychology. New York, Me. Qraw HUI,
1943.
Morgan С. T. In: Nebraska Syntpos. on Motivation.Univ, of Nebraska
Press. Lincoln, 1957, 1.
298
Morgan С. T. Physiological theory of drive In Psychology: A <)tudv
of a Science (Sensory, perceptual and physiological formulations?
Koch S. ed. 1, New fork, McGraw, 1959, 64-5. ’
Morgane P. J. Science, 1961a, 133, 3456, 887.
Morgane P. J. J. Comp. Neurol,, 1961b, 117, 1, 1.
Morgane P. Intern. Congress of Physiol. Sci. Amsterdam, 1962,
Morgane P. J. Ann. New York. Acad. Sci., 1969, 157, 80S.
Morgan M. Motivation „Cambr. Res"., 1969, 5, 3, 11.
Morgane P. J. In: M. J. Wagner, „Thrist", 1 st Int. Symp, on thirst
in the regulation of body water, 1964.
Morgane P. J. In: Proceedings of the 7th Int. Congress of Nutrition
Hamburg, 1966, 1—5, 1. ’
Morgane P. J., Kosman A. J. Am. J. Physiol,, 1959, 197, 158.
Morrison S. D., Mayer J. Am. J. Physiol., 1957, 191, 248.
Moruzzi Magoun H. IF. Electroenceph. clin. Neurophysiol., 194g
1, 455. ’
Nakao 1!. Am. J. Physiol., 1958, 194, 411.
Nauta Th. J. H. In: Electrical Studies on the unanesthetized Brain
P. B. Hoeber Inc., i960, 1,
Nauta IF. J. H., Whitlock. D, G. Jn: Brain Mechanisms and Conscious-
ness. Oxford, 1954 , 81.
Newman B. L. J. Comp. Physiol. Psychol., 1967, 63, 3, 516.
Nlemer IF. T., Goodfellow E. F., Speaker G. Electroenceph. clin. Neu-
rophysiol., 1963, 15, 5, 827.
Nissen H. In: Handbook of experimental psychology. E’d by Stivens,
1951, 458.
Nissen H. W. In: IF. C. Allee, H. IF. Nissen, M. F. Nimkoff. A re-
examination of the concept Of instinct, Psychological Rewlew, 1953,
60, 287,
Olds J. J, Comp, Physiol, Psychol., 1956, 49, 281.
Olds J, In: Electrical Stimulation of the Brain. Ed. D. E. Sheer.
Houston, Univ, Houston and the Hogg Foundation Symposium on
Brain Stimulation, 1957.
Olds J. In: Electrical Studies on the Unanesthetized Brain. E. R. Ro-
mey, D. C. O'Ooherty Eds. New York Hoeber, 1960, 17.
Olds J. Physiol. Rev., 1962. 42, 554.
Olds J., Killam K. F., Bach-Y. Rita P. Science, 1956, 124, 265.
Olds J., Milner P. J. Comp. Physiol. Psychol., 1954 , 47, 419.
Olds M. E., Olds J. J. Comp. Neurol., 1963, 12, 2, 259.
Oomura Y., Kimura K. Science, 1964, 143, 3505, 484.
Oomura Y., Ono T.,. Ooyama H,, Wagner M. J. Nature, 1969, 222,
5190, 282.
Orden L. S., Satin J. Electroenceph. clin. Neurophysiol., 1963, 15, 5,
796.
Paintai A. S. J. Physiol., 1954, 126, 255.
Palka V. S., Sawyer С. H. J. Physiol., 1986, 185, 251.
Papes J. M. Arch. Neurol. Psychiat. (Chicago), 1937, 38, 725.
Parker S. IF., Feldman S. W. Exp. Neurol., 1957, 17, 313.
Penfield IF., Jasper H. Epilepsy and the functional anatomy of the hu-
man brain. Boston, 1954.
Pfafman C. Psychol. Rev., I960 , 67 , 253.
Porter R. IF-, Cavanaugh E. B., Critchlow В. V., Sawyer С. H- Am.
J. Phoysil., 1957, 189, 145.
299
Pribram К, H., Bagshaw М. J. Comp. Neurol., 1953, 99, 347.
Pribram R. H„ Fulton J. F. Brain, 1954 , 77 , 34.
Pribram R. H., Welskrantz L. J. Comp. Physiol. Psychol., 1957, 50,
Ramon у Cajal (1902). Studies of the cerebral cortex (limbic Stmctu-
res). London, 1955.
Ranson S. IV. In: Harvey Lectures. Baltimore Williams and Wilkins,
1936, 92.
Ranson S. W. A. M.A. Arch. Neurol. Psychat (Chicago), 1939 , 41, 1.
Ranson S. W., Rabat H., Magoun H. W. A. M. A. Arch. Ne urol. Psy-
chiat (Chicago) 1935, 33,. 467,
Raphelson A. C., Isaacson R. L., Douglas R. J. Neuropsychologla,
1966, 4, 253.
Rasmussen E. W., Raada B. R., Bruland H. Acta physiol, scand.,
1960, 50, suppl. 175, 126.
Reynolds R. J. Comp. Physiol. Psychol., 1963, 56, 6 , 965.
Reynnard A. C. R. Soc. Biol. Paris, 1938, 127, 903.
Richter С. P. Harvey Lectures, 1943 - 43, 38, 63.
Richter С. P, J. Comp. Physiol. Psychol., 1947, 40, 129.
Robinson В. H7., Mishkin M. Science, 1962, 136, 260.
Rodrlguez-Zendejas A. M., Yega G., Soto-Mora L. M., Russek R. M.
Physiol, and Behavior, 1968 , 3 , 259.
Rosvold H. E., Mirsky A. F., Pribram R. H. J. Comp. Physiol. Psy-
chol., 1954, 47, 173.
Roth S. R., Sterman M. B., Clemente C. D. Electroenceph. clin. Neu-
rophysiol., 1967 , 23 , 509.
Rothballer A. B. Electroenceph. clin. Neurophysiol., 1956, 8, 603.
Rubinstein H., Delgado J.M.R.22 Int. Congress of Physiol. Sci., Free,
Comm., 1962 2, 113, 366.
Ruch С. M., Bard P. Res. Nerv. Ment. Dis., 1947, 27, 420.
Ruch T. C. In: Medical physiology and biophysics. Saunders Company,
1960, p. 483.
Ruch T. C., Maire F. W., Pation H. D. Abst, Comm. Congr. Int. Phy-
siol., 1956, 20, 788.
Ruch T. C., Shenkin H. A. J. Neurophysiol., 1943, 6, 349.
Russek M. Nature (EnglA 1963, 197, 4862, 73.
Russek M., Rodrlguez-Zendejas A. M., Pina S. Physiol. Behavior,
1968, 3, 249.
Ryorchl Y., Ryotchlro I., Shlgetosfii O., Minoru M. J. Physiol. Soc.
Japan, 1959, 21, 5, 556.
Sadowski B., Longo V7. C. Electroenceph. clin. Neurophys io I., 1962,
14, 465.
Sawa M., Heki Y., Arita M., HaradaT. Folia psychiat. neurol. Jap.,
1954, 7, 309.
Sawyer С. H., Rawakami M. Endocrinology, 1959, 65, 622.
Sawyer C. H., Robinson B. J. clin. Endocr., 1956, 16, 601.
Schreiner L., Rling A, J. Neurophysiol., 1953, 16, 643.
Schreiner L. D., Rlochy Me. R., Pechtel C., Masserman J. H. J.
Neurophyslol,, 1953, 16, 234.
Schwartzbaum J. S. Am. J. Psychol., 1961, 74, 252.
Scott W. W., Scott С. C., Luckhabdt A. B. Am. J. Physiol., 1938,
123, 243.
Seevers M. H. J. Pharmacol., 1936 , 56, 157.
300
Shapiro M. M. Jel A., Kel/away Р. Exp. Neurol., 1965, 13, 128.
Sherrington C. S. Proc. roy. Soc. B., 1900, 66, 390.
Skinner B. F, The behavior of organisms. New York Appl. Centurico.
Inc., 1938.
Skinner B. F. Science and Human Behavior. New York, 1953.
Smith 0. A. In: Electrical stimulation of the brain, 1961, 367.
Smith O. A. Physiologic Basic of Motivation. Neurophysiology, 1966,
494. Sanders К. B. Co. Philadelphia—London.
Smith IF. K. Fed. Proc., 1950, 9, 118.
Smith R. W., McCann S. M. Am. J. Physiol., 1962, 203 , 2 , 366.
Soulalrac A. J. Physiol. Paris. 1958, 50, 663.
Soulairac A. (Discussion). 22 Int. Congress Physiol., 1962, 6, 678.
Spence R. M. Behavior Theory and Conditioning. Yale Univ. Press, 1956.
Spiegel E. A., Miller H. R., Oppenheimer. M. J. J. Neurophysiol.,
1940, 3.
Spies G. J. Comp. Physiol. Psychol., 1965, 60, 2, 153.
Stachnik T. I., Ulrich R., Mabry I. H. Am. Zoologost, 1966, 6, 663.
Steffens A. B. Physiol. Behav., 1969, 4, 215.
Stein L. In: The role of pleasure in Behavior, 1964, 113.
Stellar E. Psychoi. Rev., 1954, 61, 5.
Stellar E., Hill J. H. J. Comp. Physiol. Psychol., 1952, 45, 96.
Stevenson J. A. F. In: M. J. Wayner. .Thrlst", 1st Int, Symp. on
thirst in the regulation of body water, 1964.
Stratford J. G. A thesis presented to the Faculty of the Graduate
Sch. of Art. and Sci of McGill Univ., 1951, 63.
Strominger J. L., Brobeck J. Yale J. Biol. Med., 1953, 25, 383.
Stunkard A. I., Wolff H. G. Psychosom Med., 1958, 20, 17.
Sully J. Outlines of Psychology. D. Appleton and Co. New York, 1884.
Szwejkowska G., Kreiner J., Sychowa B. Acta Biol. exp.. 1963 , 23,
3, 81.
Tatum A. L., Seevers M. H., Collins К. H. J. Pharmacol, exp. Ther.,
1929, 36, 447.
Teitelbaum P. 22 Int. Congress of Physiol. Sci., Amsterdam, 1962, 1,
2, 697.
Teitelbaum P., Campbell B. J. Comp. Physiol. Psychol., 1958, 51, 2,
135.
Teitelbaum P., Stellar E. Science, 1954, 120, 894.
Persian H., Ore D. Neurol., 1955, 5, 373.
Thompson A. F.t Walken A. E. Arch. Neurol. Psychiat (Chicago),
1951, 65, 251.
Thompson И'. C., Bach. L. M. N. J. Neurophysiol., 1950, 13, 455,
Thorpe If'. H. Learning and Instinct in Animals. Cambridge, Harv.
Univ. Press, 1956.
Tinbergen N. The Study of Instict, Oxford, Oxford. Univ Press, 1951.
Tolman E. C. Purposive Behavior in Animals and Men. Century, New
York., 1932.
Troland L. T. The Fundamentals of Human Motivation, Van Nostrand.
Princeton, 1928.
Tsubokawa T., Sutin J. Electroenceph. clin. Neurophysiol., 1963, 15,
5, 804.
Van Itallie T. B., Beaudoin R., Mayer J. J. clin. Nutrition, 1953, 1,
208.
Verney E. B. Proc. Roy. Soc. (Lond.), B., 1947, 135, 25.
Wang С. H. Comp. Psychol. Monogr., 1923 , 2, N 6.
301
JVfZflg' S. C,, Borison H. L. Arch. Neurol. Psychiat. (Chicago), 1950,
63, 928.
Wang S. C., Borisson H. L. Gastroenterol., 1952, 22, 1.
Wang S. C. Ranson S. IF. J. Comp. Neurol., 1939, 71, 457.
Warden C. J. Animal Motivation: Experimental Studies on the Albino
Rat. Golumb. Univ. Press. New York, 1931.
Weiskrantz L., Pribram К. H. J. Comp. Physiol. Psychol., 1956 , 49,
4, 381.
JFAeafley Л7. D. A.M.A. Arch. Neurol, and Psychiat., 1944, 52, 296.
Wilkins L., Richter С. P. J.A.M.A., 1940, 114, 866.
Wilkinson H. A., Peelle T. L. Am. J. Physiol., 1962, 203, 3, 537.
Wolfe J. B. Comp. Psychol. Monogr., 1936, 12, 60.
Wolfe J. IF., Lubar J. F., Ison J. R. Physiology and Beha tor,
1967, 2, 239.
IT’ooR C. D. Neurology, 1958, 8, 215.
Woorfs J. IF. Nature (Lond.), 1956, 178, 869.
Woodworth R. S. Dynamic Psychology. Columbia Univ, Press. New
York, 1918.
Wyrwicka IF. Electroenceph. din. Neurophysiol., 1964, 17, 164.
Wyrwicka IF., Dobrzecka C., Tarnecki R. Science, 1959, 130, 3371,
336.
Wyrwicka IF., Dobrzecka С., Tarneckl R. Acta biol. exp,, i960, 20,
121.
Wyrwicka IF., Dobrzecka C. Science, 1960, 132, 3430, 805.
Wlrwycka IF., Doty R. IF. Exp. Brain Res., 1966, 1, 2, 152.
Young P. T. Motivation of Behavior. The Fundamental Determinants of
human and animal activity. Wiley, New York, 1936.
Young P. T. The experimental analysis of appetite. Psychol. Bull.,
1941, 38, 123.
Young P. T. Motivation and Emotion, J. Wiley, New York, 1961.
Young IF. C. Chap. 19. In: Sex and internal secretion, 1961, 2, Balti-
more, Williams and Wilkins Co.
СОДЕРЖАНИЕ
ВВЕДЕНИЕ..................................................... 7
История вопроса...............................................Ю
1. Проблема мотиваций 23
Классификация мотиваций ...................................23
Особенности мотивационного возбуждения.....................28
Объективные методы изучения мотиваций .................... 30
Теории мотиваций...........................................37
Общие теории мотиваций .................................37
Физиологические теории мотиваций........................44
Мотивации и функциональные системы.........................59
11. Нервный субстрат биологических мотиваций.................64
Пищевые мотивации .... ................67
Мотивации страха и агрессии................................91
Половые мотивации..........................................98
Искусственные мотивации. Самораздражение...............105
Реакция самораздражения и естественные влечения живот-
ных .............................................107
Некоторые замечания.....................................112
III. Природа мотивационного возбуждения...................116й
Особенности формирования мотивационного возбуждения в
функциональных системах организма..........................116 -
Постоянство и изменчивость внутренней среды. Внутренние
«потребности» организма.............................117
Механизмы восприятия потребности......................... 121
Нервный и гуморальный механизм формирования мотивацион-
ного возбуждения...................................Д29.
Гипоталамус как центральное звено восприятия внутренних *
«потребности» организма ............................... 138
Восходящие активирующие влияния гипоталамических цен-
тров как основа мотивационного возбуждения . 144
Пейцмекерная теория мотивационного возбуждения . . 153
Химическая специфика мотивационного возбуждения. Избира-
тельное объединение мотивацией корково-подкорковых ап-
паратов ...............................................163
Организация восходящего мотивационного активирующего
влияния на кору головного мозга ...................... 170
Обработка корой головного мозга восходящих мотивационных
возбуждений.........................................184
Нисходящие влияния коры головного мозга на мотивационные
центры гипоталамуса ............................... 192
303
Мотивации и афферентный синтез. Проблема доминирования
мотиваций...........................................196
Мотивации и программирование поведения....................203
IV. Мотивации и целенаправленное поведение ................ 207
Некоторые особенности эволюционного развития .... 208
Жесткое программирование поведения........................210
Динамическое программирование поведения...................214
Мотивации и эмоции........................................219
Ориентировочно-исследовательская деятельность .... 222
V. Механизмы подкрепления...................................228
Проблема аппетита.........................................230
Нейро-гуморальные механизмы подкрепления..................234
Сенсорное насыщение.......................................234
Метаболическое насыщение..................................252
Эмоциональные механизмы подкрепления. Значение возбужде-
ния подкрепляющих подкорковых центров для деятельности
коры больших полушарий....................................254
Общие замечания 258
Энергетическая основа мотиваций...........................258
Мотивации и эмоции..........................260
Мотивации и акцептор результатов действия..............261
Определение и мотивации ..................................262
Мотивации и инстинкт.........................264
Мотивации и условный рефлекс.....................266
Высшие мотивации.........................................2Z2
Мотивации и воспитание........................274
Мотивации и восприятие........................277
Мотивации и память.......................................278
Патологические мотивации............................... 278
Наркомании ..............................................280
Фармакология мотивации...................................281
Мотивации и моделирование поведения......................283
Заключение..................................................284
к. в. судаков
биологические
мотивации