/










































































Текст
Д. М. ЗАЛЕССКИМ
(ijuioi//
ЛЕНИНГРАДСКИЙ ОРДЕНА ЛЕНИНА
И ОРДЕНА ТРУДОВОГО КРАСНОГО ЗНАМЕНИ
ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ имени А. А. ЖДАНОВА
Д. М. ЗАЛЕССКИЙ
СЕНПОЛИИ
ИХ ДИКОРАСТУЩИЕ
ВИДЫ
И ВОПРОСЫ
культуры
ЛЕНИНГРАД
ИЗДАТЕЛЬСТВО ЛЕНИНГРАДСКОГО УНИВЕРСИТЕТА
1983
Печатается по постановлению
Редакционно-издательского совета
Ленинградского университета
УДК 582.952 : 001.4+633/635(213)
Залесский Д. М. Сенполии: Их дикорастущие виды
и вопросы культуры. — Л.: Изд-во Ленингр. ун-та, 1983.—
144 с.
В книге подробно характеризуются условия обитания и
морфологические особенности дикорастущих видов рода Сен-
полия, введенных в культуру и пользующихся популярностью
в промышленном и комнатном садоводстве в качестве деко-
ративных растений. Дается их систематическое описание, ха-
рактеризуются биологические особенности, географическое
распространение. Даются советы по выращиванию сенполий
в комнатных условиях. Научные данные изложены в доступ-
ной форме.
Предназначена для работников ботанических садов и лю-
бителей комнатного садоводства. Библиогр. 51 назв. Ил. 60.
Рецензенты: д-р биол.'наук Н. А. Миняев (Ленингр.
ун-т), д-р биол. наук Г. И. Родионенко (БИН АН СССР).
ИБ № 1768
Дмитрий Михайлович Залесский
СЕНПОЛИИ
Их дикорастущие виды и вопросы культуры
Редактор И. П. Дубровская
Художественный редактор А. Г. Голубев
Технический редактор Г. М. Матвеева
Корректоры С. К- Школьникова, В. Л. Латыгина
Сдано в набор 24.06.83. Подписано в печать 04.11.83. М-19306. Формат бум. 60Х90'Л«.
Бумага тип. № 2. Гарнитура литературная. Печать высокая. Усл. печ. л. 9,0.
Усл. кр.-отт. 9,19. Уч.-изд. л. 9,02. Тираж 11626 экз. Заказ № 265. Цена 55 к.
Издательство ЛГУ имени А. А. Жданова. 199164, Ленинград, Уннверснтетская наб., 7/9.
Типография Изд-ва ЛГУ нм. А. А. Жданова. 199164. Ленинград, Университетская наб., 7/9.
2004000000—181
076(021-83
109—83
Издательство
Ленинградского
университета, 1983 1.
Светлой памяти родителей
Александры Романовны Залесской
и Михаила Дмитриевича Залесского
посвящает автор
ПРЕДИСЛОВИЕ
В ходе проверки определений оранжерейных растений кол-
лекции Ботанического сада Ленинградского университета мною
обрабатывались различные группы растений и в их числе род
Saintpaulia A. Wendl. из семейства Gesneriaceae (Геснериевых),
Это привело к установлению того факта, что в наших коллек-
циях представлены растения только одного вида Saintpaulia
ionantha Н. Wendl. или его производные, в то время как в ли-
тературе было известно упоминание о других видах, которых
в СССР не оказалось.
Установление личных связей с отдельными зарубежными
коллегами дало возможность постепенно привлечь для изуче-
ния не только известные из литературы и описанные до 1945 г.
Ниды, но и ряд других, открытых и описанных в последующие
годы. Так, английским исследователем Б. Л. Бэрттом, работаю-
щим над изучением Геснериевых Старого Света в Эдинбурге,
описан ряд новых видов, большая часть которых была введена
в культуру. Материал этих видов (в виде листовых черенков)
был получен автором в.. 1959 г. через хранителя Эдинбургского
ботанического сада X. Р. Флетчера (Н. R. Fletcher), а вскоре
нвтор описаний Б. Л. Бэртт (В. L. Burtt) прислал мне оттиск
своей работы с ботаническими описаниями всех этих видов.
Описания еще двух видов, сделанные американским бота-
ником И. Робертсом (Ev. Roberts) в 1950 г. и опубликованные
В журнале «African Violet Magazine» в США, были получены
от Альмы Райт (Alma Wriht).
Позже, в 1960 и 1962 гг., были получены из США от Альмы
Райт и из Эдинбурга от Б. Л. Бэртта черенки еще трех видов—
Одного, описанного Энглером еще в 1901 г., в виде стеблевых
черенков (Saintpaulia goetzeana A. Engl.), отсутствующего в
Культуре в Англии и Европе, и в виде листовых черенков двух,
Описанных Бэрттом в 1961 г. (S. rupicola и S. brevipilosa
3
В. L. Burtt) и введенных в культуру. В результате в нашем
распоряжении оказались все виды этого рода, которые были
введены в культуру к настоящему времени.
Это позволило сделать ряд наблюдений и зарисовок по
мере развития растений из черенков и подробно изучить осо-
бенности их морфологии, в том числе морфологии их плодов;
они были изучены, описаны и изображены. Наряду с этим были
высеяны семена различных культиваров этого растения, полу-
ченных в США путем свободного опыления. Выращенные ра-
стения послужили материалом для широкого изучения вопросов
культуры и отбора среди сеянцев различных новых вариаций.
Иллюстрирующие текст рисунки сделаны мной с натуры.
В заключение считаю своим долгом выразить X. Р. Флет-
черу, Б. Л. Бэртту и Альме Райт искреннюю благодарность за
их любезность и неоценимую помощь.
Глава 1
ОБЩИЕ СВЕДЕНИЯ
КРАТКИЙ ОЧЕРК ОТКРЫТИЯ И ОПИСАНИЯ ВИДОВ
РОДА SAINTPAULIA Н. WENDL.
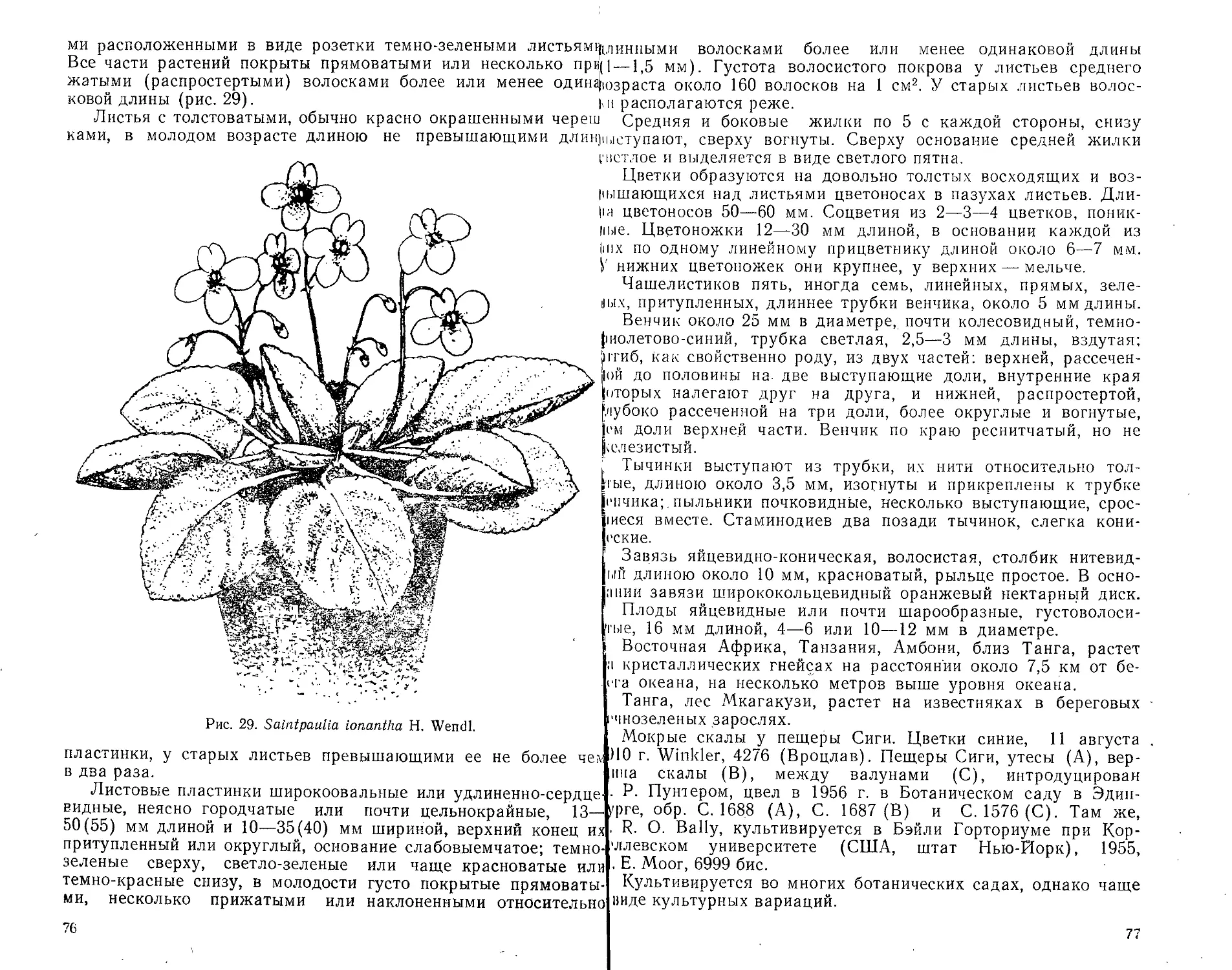
В настоящее время насчитываются двадцать один вид и две
разновидности рода Сенполий. Из них вид Saintpaulia ionantha
Н. Wendl. (сенполия фиалкоцветковая), описанный в 1893 г.,
широко известен и распространен в культуре в виде различных
садовых вариаций, часто неудачно называемых африканской
или усамбарской фиалкой, хотя с настоящими фиалками они
ничего общего не имеют. Упомянутый первый представитель
сенполий был открыт в 1892 г. Вальтером Сен-Полем (Albert
Emile Walter Radcliff le Таппёе" von Saint Paul Hillaire).
В. Сен-Поль переслал собранные им в Восточной Африке семе-
на своему отцу — президенту Германского дендрологического
общества Ульриху фон Сен-Полю в Фйшбах в Силезии, кото-
рый в свою очередь передал их Германну Вендланду, руково-
дителю Ботанического сада в Геренгаузене близ Ганновера.
Вендланд высеял полученные семена, и в 1893 г. растения
зацвели. Изучая эти растения, Вендланд пришел к заключению,
что они не могут быть отнесены ни к одному из известных в то
время представителей семейства геснериевых (Gesneriaceae),
и описал их как новый род, назвав его по имени сына и отца
Сен-Полей Saintpaulia (Сенполйя). Видовое название ionantha
(фиалкоцветковая) было дано ввиду некоторого (в общих чер-
тах) сходства цветков сенполий с цветками фиалок (Viola).
В том же году растения этого вида экспонировались на Меж-
дународной выставке садоводства в Генте (Бельгия), а бота-
ническое описание и изображение опубликовано в журнале
«Гартенфлора» (Gartenflora, 1893, Вd 42, S. 321—324, t. 1391,
abb. 66).
После выставки право промышленного разведения сенполий
было приобретено Эрфуртской садоводческой фирмой Эрнста
Бенари. Вскоре описания этого растения были повторены в Ита-
лии, Бельгии и Англии [Grilli 1894; Neubert Gart Mag, 1894;
Pynaert, 1894; Hooker, 1895]. Через некоторое время сенполий
распространились во многие ботанические сады Европы и Анг-
лии и другие собрания. В связи с введением в культуру сенпо-
лий Ульрих Сен-Поль писал в Англию Джозефу Гуккеру: «Сен-
полия открыта моим сыном, живущим в Восточной Африке.
Растения были найдены в двух пунктах. Один близ меридиана
У Танги в лесистом месте, в трещинах известковых скал, в ко-
торых в изобилии накопляется растительный перегной, не бо-
лее чем на 50—150 футов выше уровня моря. Второй — девст-
венные леса Усамбары, также в тенистом месте, но на гранит-
ных скалах, на высоте 2500 футов над уровнем моря. Она более
обильна в первом месте». Эрфуртский садовод Э. Бенари в
письме в Англию в Кью д-ру Максвеллу отмечал: «Кажется,
у этих растений имеется особенность образовывать семенные
коробочки двух различных форм: удлиненной и округлой».
Эти сообщения явились основанием для предположения о
наличии двух типов сенполий, отличающихся по форме плодов.
В конце концов выяснилось, что в культуру одновременно были
введены два различных вида: один — из прибрежных районов
Восточной тропической Африки, другой — с гор Усамбара и
Что описание Г. Вендланда относится лишь к одному из них.
Спустя 8 лет, в 1901 г. А. Энглер описал еще два вида сен-
полий, происходивших с гор Улугуру. Один из них представлял
собой растение того же типа, что и известная уже сснполия
фиалкоцветковая с розеточным расположением листьев на ко-
ротком стебле. Он был назван за свою миниатюрность Saint-
paulia pusilia A. Engl, (сенполия крошечная). Второй вид Отли-
чался удлиненными ползучими стеблями и супротивными пар-
ными листьями и был назван по имени открывшего его Гётце
(Gotze)'—Saintpaulia goetzeana A. Engl, (сенполия Гётца).
В 1906 г. Кларке сделал, как он предполагал, описание вто-
рого вида из первоначально введенных в культуру видов сенпо-
лий, происходившего из береговых районов Восточной тропиче-
ской Африки, считая, что описание Вендланда относится к об-
разцам, собранным в Усамбарских горах. Он назвал вид Saint-
paulia kewensis С. В. Clarke (сенполия кьюская)—по назва-
нию местечка Кью, в котором находится Лондонский ботаниче-
ский сад. Однако, как выяснилось позже, Кларке ошибочно
описал то же растение, которое было описано Г. Вендландом,
а второе растение — с Усамбарских гор — так и осталось не-
описанным.
Выяснению ошибки Кларке способствовали большие сборы
живого материала, произведенные супругами Моро. Это дало
возможность в 1947 г. Б. Л. Бэртту описать и назвать упомя-
нутый выше вид. Он был назван по характерному двойствен-
ному покрову листьев из очень коротких и длинных волосков—
Saintpaulia diplotricha В. L. Burtt (сенполия двояковолоско-
вая). Этот вид был основан на гербарном образце и отождест-
влен с живыми растениями, распространенными в культуре на-
ряду с сенполией фиалкоцветковой (S. ionantha). Как выясни-
лось только в 1956 г., после повторной интродукции растения,
соответствовавшего гербарному образцу, послужившему типом
для описания названного выше растения, этот вид оказался не
тождественным культивируемым растениям с Усамбарских гор,
введенным в культуру в 1893 г. В результате это последнее
растение получило название Saintpaulia confusa В. L. Burtt
(сенполия спутанная).
Благодаря селекционной работе кроме двух введенных в
культуру в 1893 г. видов сенполий, описанных как один вид,
уже в 1910 г. были известны три садовые вариации, отличав-
шиеся от типичных образцов размерами и окраской цветков:
var. purpureas (с очень темными фиолетовыми цветками), var.
grandiflora violacea (с темноокрашенными крупными цветка-
ми) и var. albescens (с белыми цветками, которые иногда имели
либо синеватый, либо розоватый оттенок). Кроме того, еще в
1898 г. фирмой )5енари была выведена разновидность rubra
(с красными цветками).
В 1922 г. А. Энглер описал еще один вид сенполий с Усам-
барских гор, по облику сходный с сенполией Гётца, т. е. имею-
щий удлиненный ползучий стебель. Этот вид был назван по
имени собравшего его в природе Гроте (Grote)—Saintpaulia
grotei Pc. Engl, (сенполия Гроте).
В 1947 г. одновременно с описанием упомянутого выше вида
S. diplotricha Б. Л. Бэртт дал краткие диагнозы еще двух ви-
дов— одного, интродуцированного в 1938 г. Р. Е. Моро и про-
исходящего из Западных Усамбарских гор — Saintpaulia orbi-
cularis В. L. Burtt (сенполия округлая), и второго — с г. Тонгве
близ Пангани, названного Saintpaulia tongwenis В. L. Burtt
(сенполия тонгвейская).
В 1950 г. в США И. Робертс [Roberts, 1950а, 19506] описал
еще два близких друг к другу вида из Восточных Усамбарских
гор, отличающихся несколько удлиненными лежачими стебля-
ми, наподобие стеблей у сенполий Гётца и Гроте. Эти виды по-
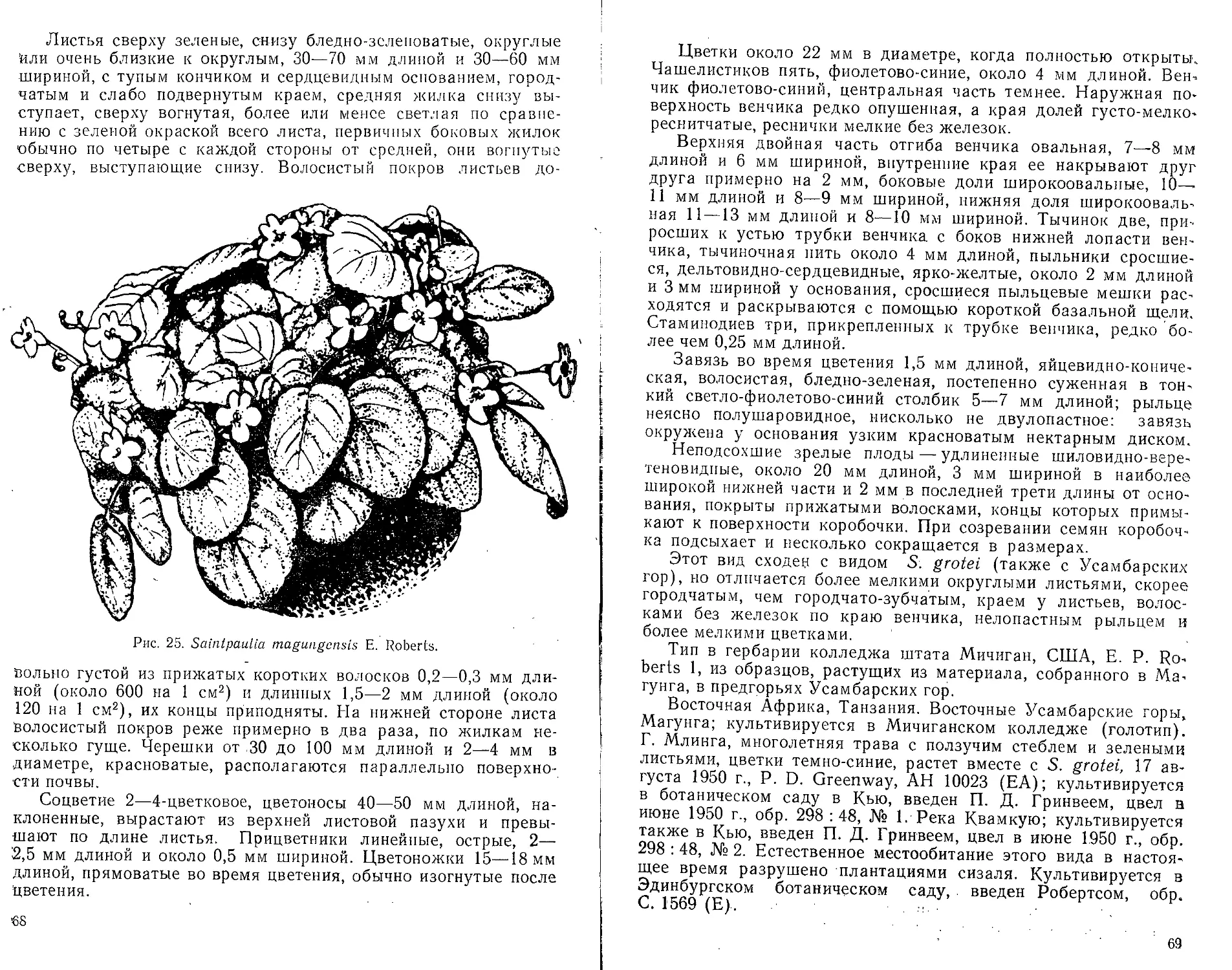
лучили свои названия по населенным, пунктам Амани и Магун-
ге, вблизи которых они были найдены, Saintpaulia amaniensis
Roberts (сенполия аманийская) и S. magungensis Roberts (сен-
полия магунгейская). В 1955 г. Б. Л. Бэртт описал вид из За-
падных Усамбарских гор, отличающийся изящным обликом и
относительно малыми размерами. Этот вид получил название
по населенному пункту Шуме, вблизи которого он был найден,
Saintpaulia shumensis В. L. Burtt (сенполия шумейская).
В 1958 г. Бэрттом были описаны еще восемь видов: один
с холмов Тейте в Кении — Saintpaulia teitensis В. L. Burtt (сен-
полия теитейская), один с гор Нгуру (Унгуу) — Saintpaulia
nitida В. L. Burtt (сенполия блестящая), один с гор Улугуру—
Saintpaulia inconspicua В. L. Burtt (сенполия малозаметная)
6
7
и пять из Усамбарских гор, из них два — Saintpaulia grandi-
folla В. L. Burtt (сенполия крупнолистная) и Saintpaulia velu-
tina В. L. Burtt (сенполия бархатная) — из западной части
этих гор, все с укороченными стеблями и розеточным располо-
жением листьев, и три — из восточной: Saintpaulia difficilis
В. L. Burtt (сенполия трудная) с укороченным стеблем, Saini-
paulia pendula В. L. Burtt (сенполия повислая) с удлиненным
стелющимся стеблем и Saintpaulia intermedia В. L. Burtt (сен-
полия промежуточная) тоже с удлиненным (но в значительно
меньшей степени, чем это свойственно всем остальным извест-
ным видам) стеблем. Этот вид представляет собой переходный
тип между типичными стелющимися стеблевыми и так назы-
ваемыми розеточными, короткостебельчатыми видами сенполий.
Одновременно с описаниями указанных видов Бэрттом был
упомянут гербарный образец, собранный еще в 1887 г. Тейло-
ром с холмов Тсимба (Шимба) и Гирияма в Кении (между
Момбасой и границей Танзании), но из-за плохой сохранности
не описанный.
Наконец, в 1964 г. тем же Бэрттом были опубликованы опи-
сания еще двух видов — Saintpaulia brevipilosa В. L. Burtt
(сенполия коротковолосковая) с гор Нгуру и Saintpaulia ги-
picola В. L. Burtt (сенполия скальная) с холмов Тсимба и Мбо-
лоло из Кении. Последний из них, по-видимому, соответствует
упомянутому выше образцу, собранному Тейлором. Одновремен-
но Бэртт привел описания двух выделенных им разновидностей
уже известных видов: S. orbicularis var. purpurea В. L. Burtt
(сенполия округлая, разновидн. пурпурная), отличающейся
главным образом окраской цветков (сплошь фиолетово-синяя)
и S. pendula var. kizarae В. L. Burtt (сенполия повислая, раз-
новидн. кизарская), отличающейся несколькими признаками.
Он высказал мнение, что описанные И. Робертсом сенполии
аманийская и магунгейская являются разновидностями одного
вида,* и предложил называть их соответственно Saintpaulia.
magungensis var. minima В. L. Burtt (сенполия магунгейская,
разновидн. наименьшая) и S. magungensis var. occidentalis
В. L. Burtt (сенполия магунгейская, разновидн. западная),
а также сделал некоторые дополнения к ранее описанным ви-
дам 5. teitensis и S. velutina.
Таким образом, в настоящее время описан 21 вид, из кото-
рых 19 введены в культуру. Отсутствуют пока в культуре два
вида и очень редок один. Это соответственно Saintpaulia pusila,
S. inconspicua и S. goetzeana.
В нашем распоряжении в настоящее время имеются 15 ви-
* С мнением Бэртта нельзя согласиться, так как между этими видами
много существенных различий, что дает право оассматривать их как отдель-
ные виды, хотя и близкие между собой. Поэтому мы приводим их описания
раздельно, обращая внимание на их различия.
8
дов и две разновидности, хотя в течение нескольких лет у нас
были еще три вида (ныне утраченные) и мы имели возмож-
ность их изучить.
С момента введения в культуру первых сенполий в Европе-
и Англии было получено значительное количество садовых ва-
риаций. В 1927 г. сенполии из Германии были завезены в США,
где за короткий срок приобрели большую популярность как
комнатные растения. В США в 1936 г. было 10 садовых вариа-
ций сенполий, в 1949 г.—уже более 100, а в настоящее время
число их превысило 1,3 тыс.
ФИЗИКО-ГЕОГРАФИЧЕСКАЯ ХАРАКТЕРИСТИКА
ОБЛАСТИ РАСПРОСТРАНЕНИЯ РОДА
SAINTPAULIA Н. WENDL. В ВОСТОЧНОЙ АФРИКЕ
Область Восточной Африки, откуда происходят известные
виды сенполий, включает в себя часть территории Объединен-
ной Республики Танзания (собственно Танганьика) и неболь-
шую часть примыкающей к ней с северо-востока территории
Республики Кения. На рис. 1 схематически изображена инте-
ресующая нас часть Восточной тропической Африки, включаю-
щая весь ареал рода Saintpaulia Н. WendL
Восточная Африка, в том числе упомянутые части респуб-
лик Танзания и Кения, характеризуется следующими чертами.
В целом — это возвышенная страна типа плоскогорья, от внут-
ренних районов постепенно спускающаяся к Индийскому океа-
ну. Непосредственно к нему она выходит по большей части в-
виде крутых берегов высотой от 10 до 40 м. Местами этот
уступ, сложенный из коралловых известняков третичного воз-
раста и песчаников, не доходит до океана, и тогда берега
отлоги.
Среди довольно обширной ровной страны местами возвыша-
ются горные группы сбросового происхождения, отличающиеся
от остальной территории лучшим увлажнением. В направлении
с востока на запад в глубь материка, примерно от районов
прибрежных населенных пунктов Танга и Пангани, возвыша-
ются группы гор, тянущиеся с некоторыми перерывами почти
до г. Килиманджаро, отделенной от них низменностью (рис. 1).
Между реками Умба на северо-востоке и Пангани на юго-
западе лежат горы Усамбара, занимающие площадь около
10 тыс. кв. км, с высотами, достигающими более 2 тыс. м над
уровнем моря. Они довольно резко отграничены со всех сторон
и распадаются на несколько групп гор.
Усамбара, как и многие другие Горные области Восточной
Африки, представляет собою сбросовый выступ, круто спускаю-
щийся к линиям трещин почти во все стороны, но особенно
круто (несколькими уступами) к долине р. Пангани. Во многих
местах благодаря незначительному наклону пластов Усамбара
9
носит характер плато, лишь местами поднимаясь в виде мощ-
ных скалистых вершин, округленных вымыванием и выветрива-
нием.
Рис. 1. Ареал рода сенполии (схема).
J — S. ionantha; 2 — S. teltensls; 3 — S. rupicola; 4 - S. shumensis; 5 — S. confusa:
°S.pendula; 7 — S. amanlensis; 3 — S. grandlfolia; 9 — S. brevlptlosa; 10— S. nltida-
11 s- goetzeana; 13 — S. tongwensls 14 - S. diplot rlctia; 15— S. velutl-
na; 16 — S. dtfjtails', ,17 - S. orbicularis; IS — S. magungensis', 19 — S.grotel: 20 —S.
inconspicua; 21 — S. intermedia.
10
Вдоль р. Пангани, по ее левому берегу, почти до Килиманд-
жаро тянется система возвышенностей такого же характера,
как и Усамбара. Это — горы Паре, состоящие из нескольких
обособленных более мелких групп гор.
Южнее р. Пангани горные острова, выступающие среди рав-
нины, не сходны с горами Усамбара и Паре. Горы местностей
Нгуру, Улугуру (Уками) и Узегуга, хотя также сложены глав-
ным образом из кристаллических сланцев, отличаются от
гор Усамбары более сложным строением. Горы Нгуру с высо-
тами, достигающими 1,5 тыс. м, находятся почти на одной дол-
готе с Усамбарскими, примерно на половине расстояния от них
до гор Улугуру. Горы Улугуру представляют собой гнейсовый
массив, почти непосредственно выступающий из равнины и до-
стигающий высот 2,4—2,8 тыс. м. Они обильно увлажнены и
питают водой несколько небольших рек. Значительное степное
пространство отделяет их от гор Усагара, с возвышенностями,
достигающими 1,5—2,1 тыс. м.
Возраст горных пород, слагающих перечисленные возвышен-
ности, докембрийский, в отдельных точках древнепалеозойский.
С севера, за долиной р. Умбы к системе Усамбарских гор,
примыкают холмы Тейте, Тсимба и Гирияма, находящиеся уже
в Кении и представляющие собою возвышенности, не превы-
шающие нескольких сот метров.
Севернее и северо-западнее перечисленных горных областей
расположены во многом (по высотам, климату) имеющие об-
щие черты с ними горы Меру и Килиманджаро, но это горы
вулканического происхождения, значительно более молодого
третичного геологического возраста.
С гор к Индийскому океану течет несколько рек, большая
часть из них порожистые: это — небольшие реки Умба и Зиги,
крупная р. Пангани с притоками, берущая свое начало в горах
Килиманджаро, меньшая по величине р. Вами с главными при-
токами, впадающими в нее с пространств, лежащих между
горами Улугуру, Нгуру и Усагара, р. Руе (Руву) с притоками,
берущая свое начало в горах Улугуру и в своем нижнем тече-
нии называемая Кингани, реки Руфиджи, Руага и другие.
Климат интересующей нас страны на большей площади (как
и в значительной части восточно-африканского побережья) но-
сит ярко выраженный муссонный характер. С декабря господ-
ствует северо-восточный муссон, а юго-западный начинается в
конце февраля — начале марта и ослабевает в июле или в ав-
густе, затем снова усиливается до конца сентября. С октября
и в особенности в ноябре бывают затишья, после которых опять
дует северо-восточный муссон. Периоды дождей повторяются
здесь два раза в год: приблизительно во время равноденст-
вия — в апреле или -мае и в ноябре, но они сравнительно
коротки.
11
В прибрежной полосе годовое количество осадков превы-
шает 1000 мм; так, например, в районе Танга норма годовых
осадков равна 1586 мм, для Дар-эс-Салама, находящегося юж-
нее по побережью, она равна 1145 мм.
В глубь страны количество осадков заметно снижается, при-
мерно до 900 мм. У подножья г. Килиманджаро с середины
марта до середины апреля почти ежедневно идут дожди. Ино-
гда это короткие ливни, а иногда обложные дожди, продолжаю-
щиеся 20 и более часов подряд. С июня до октября господ-
ствует сухая жаркая погода. В сухой период на равнинах вре-
менами выпадают дожди. Особо важное значение для снабже-
ния растений влагой в сухой сезон имеют ночные росы, быстро
испаряющиеся с наступлением утра. В этот же период на рав-
нинах резкие различия ночных и дневных температур в период
с мая до августа иногда по утрам вызывают образование инея.
Горы Усамбара расположены гораздо ближе к океану, по-
этому здесь образуется резко выраженный климатический
базис с пассатными дождями и отсутствием полностью бездо-
ждных периодов. Годовой уровень осадков здесь, однако, срав-
нительно мало отличается от такового на побережье и бывает
близок к 1500 мм.
Восточная тропическая Африка в общем теплее Западной
на той же широте, но для внутренней возвышенной части тро-
пической Африки существует много исключений из этого пра-
вила и даже прибрежные районы не следует считать особо жар-
кими. В силу этого в Восточной Африке, по существу, очень
немного мест, в которых всегда тепло. Так, например, даже в
прибрежной Танге по утрам бывает сравнительно прохладно,
пока поднявшееся солнце не согреет воздух.
Среднегодовая температура береговой зоны близка к +26 °C
с амплитудой колебаний от -j-18 до +33°C.
В глубине страны, за береговой зоной, в местах более сухих
и возвышенных над уровнем океана, суточные температуры от-
личаются большими колебаниями. Так, например, на высоте
1120 м амплитуда температур в дождливый период приближа-
ется к 11°, а в сухой — к 20°. На отдельных пространствах тем-
пература в сухое время может снижаться до +4 °C. В горных
районах с более влажным климатом на той же высоте столь
резкие колебания температуры не наблюдаются. Для этих рай-
онов абсолютный минимум близок к +13 °C, а максимум редко
превышает +25 °C.
КРАТКАЯ ХАРАКТЕРИСТИКА
РАСТИТЕЛЬНЫХ ФОРМАЦИЙ ВОСТОЧНОЙ
ТРОПИЧЕСКОЙ АФРИКИ
В общих чертах растительные формации, свойственные этой
части Африки, таковы: по берегам Индийского океана, в устьях
12
рек развиты однообразные темно-зеленые мангровые заросли,
сходные по характеру с таковыми в тропической Азии, но более
бедные по видовому составу. Там, где этих зарослей пет, берег
голый, плоский.
В глубь страны идет пояс так называемых криков, состав-
ленный частично из травянистых растений, над которыми воз-
вышаются небольшие кустарники или одинокие баобабы (Adan-
sonia digitata L.), а кое-где пальмы дум (Hyphaene thebaica
Mart.), частично из вечнозеленых кустарников, переплетенных
лианами, с преобладанием представителей семейства бобовых,
акокантеры (Acokanthera abyssinica К. Schum) и др. и частич-
но из колючих листопадных кустарников на твердой красной
почве. По берегам крупных рек к этим формациям присоединя-
ются аллювиальные леса, характерным представителем кото-
рых является распространенное в Австралии миртоподобное
дерево с кожистыми листьями и длинными свешивающимися
кистями цветков — баррингтония (Barringtonia racemosa Roxb.,
Lecydiaccae).
До высоты 125 м над уровнем океана развита флора ниж-
ней кустарниковой области, для которой характерны акации и
небольшие полусухие кустарники.
Вдоль рек тянутся галерейные леса с зарослями африкан-
ских бамбуков (Oxythenanthera) и с крупными кустарниками,
чередующиеся с однообразными участками древесно-кустарни-
ковой степи с одинокими пальмами борассус (Borassus flabelli-
Ior mis Murr.).
Далее, в глубь материка простираются восточно-африкан-
ские степи. Степь местами поднимается до значительной высо-
ты, проникая по сухим склонам в горные области, и на различ-
ных участках имеет различный характер. Либо она сочетается
с чащами степных листопадных кустарников, либо образует
так называемую степь марио с кустарниками акокантеры, по-
дымающуюся до высоты 1,9 тыс. м. На твердых латеритовых
почвах развиты молочайные степи с кактусоподобными кустар-
никовидными или крупными (достигающими 20 м высоты) дре-
вовидными представителями молочаев (Euphorbia), переме-
жающихся с колючими кустарниками, либо по латериту раз-
бросаны отдельные деревья и кустарники с множеством расте-
ний двух видов сансевьерий (Sansevieria) и других суккулен-
тов. Наконец, встречаются травяные степи в различных вариан-
тах с преобладанием злака сетария (Setaria aurea Hochst. ex
А. Вг.), то чисто травяные, то с одиночными деревьями и кус-
тарниками, среди которых преобладают баобабы и акации.
Эти степные формации подходят к склонам горных местно-
стей и при слиянии с горными формациями образуют смешан-
ные группировки, переходящие в настоящий горный тропиче-
ский лес, поднимающийся до высоты 1,5 тыс. м, а местами и
выше. В теплых сырых ущельях, где в изобилии накапливается
13
растительный перегной, развивается роскошная тропическая
растительность с мощными деревьями, обильным подлеском и
множеством эпифитов. Такие леса лучше всего развиты в
обильных водой долинах и ущельях Усамбары, но встречаются
и в других горных странах.
В горах выше тропического лесного пояса, на высотах до
1,7 тыс. м, лес приобретает иной характер; это уже густые ку-
старниковые массивы с отдельными группами деревьев, преры-
ваемые вторичными зарослями папоротника-орляка и сухими
или сырыми луговинами. Начиная с высоты 1,7 тыс. м прости-
раются леса высоких гор, состоящие из невысоких толстых, до-
вольно редко расположенных деревьев с редким подлеском,
с лианами по северным склонам, папоротниками и другими
травянистыми растениями по ручьям.
Выше 2,5 тыс. м появляются горные луга, преобладающие
с высотой и переходящие далее в пояс подальпийских кустар-
ников, а на г. Килиманджаро, начиная с высоты 4 тыс. м, —
в небогаты?! по своему составу альпийский пояс, выше сменяю-
щийся лишайниковым.
В предгорьях, на холмах и в долинах рек значительные про-
странства заняты окультуренными участками, используемыми
под различные плантации и поля.
ХАРАКТЕРИСТИКА МЕСТ ОБИТАНИЯ СЕНПОЛИЙ
Сенполии встречаются в названных выше горных областях
на высотах от 0,6 до 2 тыс. м за исключением единственного
вида Saintpaulia ionantha, растущего в прибрежных районах
в зоне вечнозеленых кустарниковых зарослей до высоты 125 м
над уровнем моря. Их распространение связано с зонами тро-
пических лесов, кустарниковых зарослей с одиночными деревья-
ми и высокогорных лесов. Более высокогорные виды сенполий
живут в условиях более сухих и прохладных дождевых лесов,
состоящих из различных представителей вечнозеленых и листо-
падных деревьев и кустарников, часто эндемичных: хвойных,
маслинных, мирсиновых, флякуртиевых, розоцветных (./uni-
perus, Podocarpus, Olea, Myrsine, Cassearia, Cossipurea и др.).
В таких лесах обитают Saintpaulia shumensis, S. orbicularis,
S. velutina, S. pusilia, S. goetzeana, S. brevipilosa, S. nitida,
S. inconspicua.
Виды сенполий, растущие на меньших высотах и составляю-
щие большинство, являются обитателями более влажных лесов,
основную массу которых составляют вечнозеленые растения.
Эти леса образованы главным образом из деревьев, относя-
щихся к семействам бобовых (Isoberlinia, Piptadenia, Cinomet-
га), сапотовых (Allophyllus, Chrisophyllum), розоцветных (Pa~
rinari), флякуртиевых (Flacurtia, Zahna), бигнониевых (круп-
14
ный кустарник Dolichondrone zanzibarica К. Schum.), марено-
вых (Tricalysia), липовых (Mystroxylon, Elaeodendron), черно-
устовых (Memecylon), анноновых (например, Polyalthia olivieri
Engl., по облику напоминающая некоторые виды фикусов), лав-
ровых (Ocotea), зверобойных • (Allanblackia), миристиковых
(Cephalosphaera usumbariensis) и т. д. Им сопутствуют многие
травянистые растения, эпифитные и наземные, из которых мо-
гут быть упомянуты различные, в том числе древовидные, па-
поротники, дикие бананы, хлорофитумы и мн. др.
Наряду с перечисленными деревьями и кустарниками, расту-
щими на более влажных местах, в непосредственном соседстве
с ними на более сухих склонах и скалах можно встретить ти-
пичные суккуленты, такие, как гигантские кактусоподобные
молочаи (Euphorbia ingens Е. Меу), достигающие 20 м высоты
и кустообразные молочаи (Euphorbia wekefieldii Engl.), виды
алоэ (Aloe secundiflora Engl., A. lateritia Engl.), виды сансевье-
рнй (Sansevieria) и перемежающиеся с ними цикадовики
(Encephalartos) и драцены (Dracaena').
Низинные леса, произрастающие в высотных пределах до
800 м, по поясной схеме, предложенной Р. Е. Моро, по мнению
П. У. Ричардса [1961], основанному на изучении их видового
состава, должны быть отнесены к лесам вторичного происхож-
дения как в горах Усамбара, так и в других горах Восточной
тропической Африки.
В древостое таких лесов преобладают Antiaris usambariensis
Engl., Trema guinensis Priemer (Urticaceae), Chlorophora excel-
sa Aubl., различные виды Ficus (Moraceae), Sterculia appendl-
culata K- Schum. (Sterculiaceae), различные виды Albizzia (Le-
guminosae). В подлеске большое число колючих кустарников,
главным образом Acacia pennata Willd. (Leguminosae), Harri-
sonia abyssinica Oliver (Simarubaceae). Злак Olyra latifolia L.
образует густые трудно проходимые заросли высотою до 2,5 м.
Других травянистых растений мало. Лианы многочисленны,;
эпифиты редки, в их числе встречается папоротник Platycerium.
angolense Welw.
Изолированные участки леса на высотах от 800 до 1400 м,
сохранившиеся в настоящее время главным образом на восточ-
ных склонах Усамбары, в горах Нгуру и Улугуру и в общей
сложности занимающие не более 1000 кв. км, похожи по,
своему характеру на тропический дождевой лес. Такой лес,
растущий в горах Усамбара, по мнению его исследователя
Р. Е. Моро, является самым пышным среди лесов Восточной
Африки и в предложенной им схеме поясов отнесен к проме-
жуточному (среднему) поясу горных лесов этой страны. Число
видов деревьев здесь очень велико, многие из них являются
единственными представителями родов и эндемичны. Среди до-
минирующих могут быть отмечены Piptadenia buchanonit
15
Backer, Isoberlinia scheffleri (Harms) Greenway (Leguminosae),
виды Parinari AubL (Rosaceae), Macarango usambarica Pax et
R. Hoffm. (Euphorbiaceae), Allanblackia stuhlmanii Engl.,
A. ulugurensis Engl. (Hyperricaceae), большое число древовид-
ных папоротников, лиан, фикусов-душителей, видов и индиви-
дуумов различных эпифитов, на лучше освещенных местах
очень разнообразный и богатый травяной покров.
Флора и фауна этих лесов, хоть и состоит из эндемичных
видов, обнаруживает, по свидетельству П. У. Ричардса [1961J,
автора известной книги «Тропический дождевой лес», порази-
тельное сходство с флорой и фауной тропического дождевого
леса в Западной и Центральной Африке. Исследовавший эти
леса Р. Е. Моро (R. Е. Moreau) на основании изучения глав-
ным образом птиц делает интересный вывод, что эти восточно-
африканские леса представляют собой реликты обширного лес-
ного массива, который когда-то составлял единое целое с леса-
ми Центральной Африки, а другой исследователь, Д. П. Чепин
(J. Р. Chapin) также на основании изучения птиц высказывает
предположение, что помимо Африки леса такого типа во время
влажного периода в плейстоцене занимали значительно боль-
шую территорию и простирались через Аравию до влажных
тропических районов Азии.
Третий тип леса, на высотах свыше 1400 м в горных странах
Восточной Африки, в горах Усамбара достигает полного раз-
вития только в западной их части. Этот тип назван по схеме
поясов Р. Е. Моро нагорным и по облику несколько напоми-
нает леса промежуточного типа, расположенные на меньших
высотах, хотя и отличается от них по своему флористическому
составу. Среди лесообразующих деревьев преобладают виды
Podocarpus (Podocarpaceae), Ocotea usumbariensis Engl. (Lau-
raceae). Здесь же встречаются заросли можжевельника (Juni-
perus procera Hochst.). По мнению P. E. Моро, эти леса могут
быть отнесены к горным дождевым лесам.
Каждому из перечисленных типов леса присущи свои виды
сенполий, со свойственными им морфологическими особенностя-
ми, обусловленными степенью и периодичностью увлажнения.
В большинстве случаев сенполии растут во влажных в той
или иной степени лесах на скалистых поверхностях отдельных
утесов, выступах скал, скалистых террасах, голых и обомше-
лых, известковых или гнейсовых, между валунами, в трещинах,
иногда на глине, обычно в тени, которая может быть названа
густой, среди мхов и папоротников, иногда эпифитно на де-
ревьях. Отдельные виды встречаются в менее густой тени, часто
вблизи водопадов, где воздух насыщен брызгами падающей
воды и потому очень влажный, или на выступах скал, увлаж-
няемых просачивающейся сквозь них водой. Температура ко-
леблется здесь от +13 до +22—25 °C. Их корневая система
развивается либо в трещинах скал, где в изобилии накопляется
16
перегной, либо на обомшелых поверхностях камней, либо в ка-
менистой почве или глине.
Если корни так или иначе получают воду, растения благо-
денствуют, однако в дневные часы, в период наиболее интен-
сивного солнечного освещения и нагрева они могут несколько
подвядать под влиянием превосходящего испарения надземны-
ми частями, оправляясь лишь к вечеру и в ночные часы.
Интенсивный солнечный свет в течение сравнительно корот-
кого сухого сезона и связанная с этим сезоном несколько раз-
реженная тень за счет листопадных деревьев и кустарников,
приводят также к сильному подвяданию растений (что особен-
но заметно на высотах ниже 1,2 тыс. м), способствуя одновре-
менно их интенсивному цветению. В такой обстановке обычно
небольшие и раскидистые растения (особенно видов, обладаю-
щих удлиненными стеблями) проявляют тенденцию стелиться
и укореняются на различных участках своих стеблей; отдель-
ные обламывающиеся листья или участки стеблей во время
дождя смываются водой по склонам и задерживаются в какой-
либо трещине, и в месте, где всегда имеется в изобилии вода,
укореняются и воспроизводят новые растения.
То, что некоторые сенполии встречаются растущими на де-
ревьях, как настоящие эпифиты, и все они обитают на склонах
или обомшелых камнях с малым количеством почвенного суб-
страта, позволяет всех представителей этого рода считать
растениями полуэпифитного (протоэпифиты *) образа жизни.
С этой точки зрения становятся понятными особенности облика
сенполий, которые им присущи, — сочность и мясистость стеб-
лей и листьев, волосистый покров всех частей и способность
из частей тела воспроизводить целые растения, т. е. черты, ха-
рактерные для некоторых эпифитных и подавляющего боль-
шинства суккулентных растений.
ОСОБЕННОСТИ ГЕОГРАФИИ
ПРЕДСТАВИТЕЛЕЙ РОДА SAINTPAULIA Н. WENDL.,
АРЕАЛ РОДА И ОТДЕЛЬНЫХ ВИДОВ
Одной из особенностей географии рода сенполий является
ограниченность распространения сравнительно небольшим рай-
оном Восточной Африки, за пределами которого они нигде на
земном шаре не обнаружены. Самым южным районом нахож-
дения сенполий являются горы Улугуру, расположенные, при-
* Протоэпифиты — растения, не имеющие специальных приспособлений
для накопления воды или почвы и не спускающие в почву воздушных кор-
ней. Оии представляют собой наименее специализированную группу и в
очень малой степени защищены от влияния периодических засух и недостат-
ка почвы. Например: виды пеперомий, многие папоротники, растения с водо-
удерживающими органами различного типа (суккулентные листья, псевдо-
бульбы орхидей и т. п.). Многие из них отличаются ксероморфной струк-
турой.
2 Д. М. ЗАЛЕССКИЙ
17
близительно, на 7° ю. ш. и 37° в. д.. Отсюда известны три свое-
образных вида: S. pusilia, S, goetzeana и 3. inconspicua. Горь:
Улугуру отделены от других горных областей, где найдень:
сенполии, значительными степными пространствами.
Наибольшее количество видов известно в различных частям
гор Усамбара, расположенных примерно на 5° ю. ш. и 38° в. д
В западной части этой области, где высоты превышают 2 тыс. м
обитают 4 вида: S. grandifolia, S. orbicularis, S. shumensis v
S. velutina, в восточной части, с высотами, не достигающими
2 тыс. м, известно 9 видов: S. grotei, S, magungensis, S. ата
niensis, S. intermedia, S. pendula, S. difficilis, S. diplotricha,
3. confusa и 3. tongwensis. Сюда же примыкает один вид из
прибрежного района, выходящего к Индийскому океану на вы-
соте, не превышающей 100—150 м, — 3. ionantha Н. WendL.
описанный первым в конце прошлого столетия.
В горах Нгуру, отделенных от Усамбарских гор долиной
Пангани и степным пространством, известны два вида —
3. nitida и 3. brevipilosa.
Самым северным местонахождением сенполий являются
холмы Тейте, Тсимба и Гирияма. Здесь найдены два вида:
3. teitensis и 3. rupicola.
Таким образом, сенполии образуют разорванный ареал, со-
стоящий из четырех обособленных субареалов, внутри которых
различаются локальные ареалы каждого из известных видов.
Исключение составляют только два вида — S. grotei и 3. ma-
gungensis, ареалы которых соприкасаются. Судя по наиболь-
шему (14) числу видов общего ареала рода, его центр — горы
Усамбара, а остальные части общего ареала являются перифе-
рическими.
Все виды центральной части ареала, как выше упомянуто,
образуют две группы видов — восточную, с девятью горными
видами и одним прибрежным, и западную, из четырех видов.
Все четыре вида западной группы, или, иначе, западно-усам-
барского субареала, хорошо различимы между собою по мор-
фологическим признакам. Морфологические различия десяти ви-
дов восточной группы, или восточно-усамбарского субареала,
менее явственны, что привело автора их описаний Б. Л. Бэртта
к поискам новых, более мелких признаков их различия; таким
признаком он счел волосистый покров листовых пластинок,
различный для каждого из видов. Однако, как нам кажется,
этот признак может быть использован только в дополнение ко
всему комплексу отличающих признаков. Так же как и виды
западной группы, каждый из видов восточной группы образует
свой обособленный локальный ареал с одним уже упомянутым
исключением для двух видов, ареалы которых соприкасаются.
Однако это соприкосновение не приводит к образованию про-
межуточных форм, и каждый из видов сохраняет свою само-
стоятельность. В более южных нгурском (с двумя видами) и
18
улугурском (с тремя видами) субареалах, в первом из которых
виды встречаются раздельно, а во втором совместно, и в север-
ном кенийском субареале (с двумя видами) каждый из видов
также образует свой обособленный ареал.
Второй существенной особенностью географии сенполий яв-
ляется факт сосредоточенности относительно большого количе-
ства видов на сравнительно малой площади.
Нельзя не обратить внимание и на третью особенность гео-
графии этих растений, а именно на их отсутствие в ряде гор-
ных районов, прилегающих к тем горным районам, где они
обитают, и обладающих теми же климатическими чертами.
Сенполии отсутствуют в горах Паре, находящихся на расстоя-
нии 22 км степного пространства от Усамбарских гор и 37 км
от ближайшего пункта нахождения сенполий (S. shumensis).
Горы Меру и Килиманджаро, расположенные несколько запад-
нее Усамбарских гор и отделенные от них расстояниями, не
большими, чем расстояния между горными странами, где оби-
тают сенполии, и сходные по климатическим характеристикам.,
также лишены этих растений.
Единственной особенностью, которая нам представляется
заслуживающей внимания (помимо особенностей рельефа и ха-
рактера растительности, играющих определенную роль в рассе-
лении растений), является геологический возраст всех упомя-
нутых горных стран. Все близлежащие горные" страны, где не
найдены сенполии, начинают свою историю с третичного вре-
мени, тогда как страны, в которых они обитают, — с докемб-
рийского. В горах древнего происхождения сохранились релик-
товые островки, где уцелёли эти растения и откуда они из-за
особенности рельефа и других естественных преград не смогли
распространиться за пределы своих местообитаний.
Особенность распространения видов сенполий, ограничен-
ность его незначительной областью в целом и характер их
современного распространения позволяют считать этот род эн-
демичным для Восточной тропической Африки, а отдельные
виды — эндемичными..для занятого ими ареала наряду с други-
ми эндемичными растениями этого района. Приуроченность
распространения сенполий к горным странам древнего геологи-
ческого возраста позволяет, по нашему мнению, расценивать
рассматриваемый род, как реликтовый.
Что касается вопроса локализации ареалов отдельных ви-
дов, свойственной подавляющему большинству сенполий, то
она может быть легко объяснена особенностями строения релье-
фа и характером распределения растений, создающими в сово-
купности естественные преграды, исключающие возможность
миграции видов за пределы их ограниченных ареалов. Кроме
того, по-видимому, этому способствуют некоторые морфолого-
биологические черты сенполий, на которых мы остановимся
ниже.
2*
19
Глава 2
СИСТЕМАТИКА И МОРФОЛОГИЯ
СИСТЕМАТИЧЕСКОЕ ПОЛОЖЕНИЕ
РОДА SAINTPAULIA Н.WENDL.
Род Saintpaulia относится к семейству Геснериевых (Gesne-
п’асеае), представленных в подавляющем большинстве травя-
нистыми многолетними растениями, распространенными пре-
имущественно в тропических районах Азии, Африки и Америки,
® также небольшим числом представителей в субтропических
дайонах этих материков и в Южной Европе. Это — многолет-
ние, редко однолетние травы, иногда с клубневидно утолщен-
ными или чешуйчатыми корневищами; полукустарники, кустар-
ники, лианы, редко небольшие деревца (чаще наземные, но не-
редко и эпифитные) часто с сочными стеблями и листьями.
Семейство Геснериевых помещается в порядок Scrophula-
riales (в понимании А. Л. Тахтаджяна). Оно включает около
120 родов с 1800 видами, распространенными в тропиках Цент-
ральной и Южной Америки, Восточной и Юго-Восточной Азии,
в Новой Гвинее, в Австралии, Экваториальной и Южной Аф-
рике, на Мадагаскаре и в Южной Европе. В Америке Гесне-
риевые распространены в тропической зоне, заходят местами в
южную часть умеренной зоны, в Азии они распространены от
Северо-Восточного Китая по всему юго-востоку материка.
Семейство разделяется на три подсемейства: Cyrtandroideae
е 9 трибами, Episcioideae с 2 трибами и Gesnerioideae с 6 три-
бами.
Род Saintpaulia включается в одну из девяти триб подсе-
мейства Cyrtandroideae, выделенную на основе карпологиче-
ских данных Л. И. Иваниной в 1965 г. и названную ею Saint-
paulieae, с четырьмя родами: Saintpaulia, Caralofritshia, Асап-
thonema и Linnaeopsis.
Представители подсемейства Cyrtandroideae распространены
в основном в тропической и субтропической зонах Старого
Света. Среди них триба Saintpaulieae является наиболее древ-
ней и обладает наряду с трибой Ramondeae наиболее прими-
тивными признаками, такими как пятичленный отгиб венчика,
зигоморфия и т. д.
20
ОБЩАЯ ХАРАКТЕРИСТИКА
РОДА SAINTPAULIA Н. WENDL.
Как уже указывалось, представители рода сенполий — это
многолетние травы. Стебли их либо довольно короткие и тол-
стые с густой кроной листьев на концах, либо лежачие, укоре-
няющиеся, с удлиненными междоузлиями, на узлах которых
расположены попарно листья.
Листья чередующиеся, черешчатые, довольно мясистые, оде-
тые бесцветными волосками, покрывающими и остальные части
растений. Листовые пластинки от почти округлых до эллипти-
ческих. Цветоносы пазушные, одно-, многоцветковые; цветки
вырастают цимозно, распускаются сериально на каждой дихо-
томии соцветия. Часто один из цветков дихотомии не разви-
вается, в этом случае в его пазухе возникает продолжение оси
цветоноса, несущее следующую дихотомическую пару цветков
и т. д. При таком развитии образуется нечетное число цветков.
В случае нечетного числа нормально развивается только по-
следняя дихотомическая пара цветков.
Прицветники маленькие, обычно линейные или узколанцет-
ные, смещенные под углом относительно осей цветоножек.
Чашечка разделена почти до основания на пять ланцетных
сегментов, они чередуются с долями венчика, и обычно два
верхних сегмента чуть длиннее остальных трех. Венчик с ко-
роткой бочонковидной трубкой и характерным пятилопастным
отгибом, отходящим от трубки под тупым углом, реже под пря-
мым. Отгиб состоит из двух частей: верхней — двулопастной
(верхние лопасти или доли чаще всего короче остальных трех)
и нижней, из трех долей — двух боковых и одной средней.
Доли венчика гладкие изнутри, волосистые снаружи и реснит-
чатые или железисто-реснитчатые по краю. Плодущих тычинок
две, нити их располагаются против синусов, образуемых боко-
выми и средней долями нижней части отгиба венчика, прира-
стая уплощенными основаниями к внутренней стороне трубки
венчика. Стаминодиев два-три, расположены они соответствен-
но синусам, образуемым долями венчика верхней части отгиба.
В случае двух стаминодиев третий, всегда средний, —• редуци-
рованный. Пыльники (два соответственно двум тычинкам)
крупные, двухгнездные, ярко-желтые, подпирающие устье труб-
ки венчика. На каждом пыльнике на стороне, обращенной к
соседнему пыльнику, намечаются две щели, образующие в со-
вокупности дугообразную линию. С той же стороны, где распо4
лагаются щели, пыльники срастаются между собой, препятст-
вуя этим вскрытию щелей и высвобождению пыльцы при её
созревании. Завязь верхняя короткоконическая, волосистая,
постепенно утончающаяся и переходящая в гладкий (за исклю-
чением волосистого его основания) столбик, превосходящий ее
по длине примерно в три раза. Плоды нераскрывающиеся
28
коробочки от почти округлой, яйцевидной, до шиловидной фор-
мы, характерные для каждого вида. Семена мелкие.
Корневая система функционирует кратковременно и затем
заменяется новой, развивающейся за счет придаточных корней,
образующихся в любой точке поверхности стеблей и черешков
листьев.
Стебель. По облику растений обычно
(иногда противопоставляя их др
стеблевые и розеточные. Однако
различают два их типа
их друг другу), обозначаемые как
.... _д’.шкэ при сравнительном изучении
всех известных видов намечаются и переходные типы, связы-
вающие их в единый ряд.
У так называемого стеблевого типа листья отделены друг
©т друга хорошо заметными междоузлиями, более длинными
у оснований стебля и более короткими к их вершине. Длина
стеблей у различных видов достигает 80—300 мм, иногда и бо-
22
лее. Благодаря удлиненности, относительно небольшой толщи-
не и мягкости стебли по мере роста растений и увеличения
числа междоузлий становятся распростертыми и ползучими.
Вопреки мнению некоторых авторов, в том числе автора
большинства видов Б. Л. Бэртта, ошибочно указывающих на
то, что придаточные корни образуются в узлах, на самом деле
они образуются в любой точке поверхности стебля. С отмира-
нием главного стебля при его старении, обычно начинающимся
от основания или от его участков в основании боковых ответ-
влений, образующихся в пазухах листьев, растение распадается
на несколько отдельных особей, и их корневая система разви-
вается за счет ранее возникших придаточных корней.
У другой части видов, относимых к розеточному типу, стеб-
ли сильно укорочены и утолщены, расстояния между листьями
Рис. 3. Схема вегетативного размножения путем деления стебля у
растений с укороченным стеблем.
. почти отсутствуют. В силу этого стебли растений данного типа
; прямостоячие и лишь с возрастом несколько удлиняющиеся и
полегающие, причем к этому времени их основания оголяются,
! так как листья нижних ярусов отмирают, и поверхность стебля
становится бугристой от листовых рубцов. По мере старения и
полегания у всех сенполий основания стеблей вместе с базаль-
, ными корнями отмирают, а взамен отмерших корней начинают
функционировать корни, образующиеся как придаточные на
; местах соприкосновения полегшего стебля с субстратом. Эти
придаточные корни, зачатки которых в виде небольших бугор-
ков заметны на поверхности стебля между листовыми рубцами,
так же как и у стеблевых видов в виде небольших бугорков,
возникают в любом месте стебля.
У видов розеточного типа стебель сначала не разветвлен,
по рано или поздно в пазухах нижних листьев или рубцов от-
павших нижних листьев образуются боковые укороченные вет-
ви, воспринимаемые обычно как отпрыски. Они сосредоточены
больше близ основания старого стебля и со временем форми-
руют за счет придаточных корней самостоятельную корневую
i систему и обособляются от материнского растения так же, как
! и ветви растений стеблевого типа (рис. 2, 3 и 4). Эту способ-
ность растений сенполий расчленяться на отдельные участки
стеблей или ветви-отпрыски с образованием самостоятельной
корневой системы следует, по-видимому, рассматривать как
23
неясно,
нижней
бурова-
—ные на поверхности окрашенного стебля. Их значение
Часто у стареющих растений поверхность стеблей в
кор- части покрывается пробковым слоем, придающим им
густым тый оттенок.
Как уже было упомянуто, среди видов сенполий мы
наблю-
один из способов вегетативного размножения и заселения близ-.
лежащего пространства.
Помимо упомянутых бугорков (зачатков придаточных
ней) поверхность стеблей сенполий одета относительно гуимя
ВОЛОСИСТЫМ покровом (в ОСНОВНЫХ чертах соответствующим ха- х\ак уже ОЫЛО упомянуто, среди ВИДОВ сспиилии мш navvnv-
рактеру волосистого покрова, присущего тому или иному виду), даем, с одной стороны, растения-с ярко выраженными стеоляш
(то более удлиненными, то более короткими), с другой сторо-
соп-
ны, такие, у которых стебель удлиняется только в зрелом
расте (S. intermedia и в известной степени S. shumensis, S.
Рис. 5. Попарное слегка смещающееся расположение листьев у сен-
полий с укороченным стеблем.
Рис. 4. Корневищеподобный стебель S. rupicola и его
ветвление (схема).
Его образуют торчащие, разной длины или более или менее
одинаковые, прозрачные или в редких случаях буроватые мно-
гоклеточные волоски, иногда в той или иной степени прижатые
или распростертые, длинные или короткие.
Стебли либо зеленоватые, либо, чаще, окрашенные в крас-
новатый или темно-красно-бурачный цвет, соответствуют по
окраске черешкам листьев и в известной степени нижней сто-
роне листовых пластинок. У отдельных стеблевых видов (S. gro-
tei, S, magungensis и S. pendula) наблюдаются своеобразные
продольные светлые черточки, сравнительно редко расположен- ।
fusa) либо, наоборот, только в юном возрасте (S. nitida, S. or-
bicularis, S. teitensis, в известной мере S. grandifolia). Впослед-
ствии стебель становится незаметным, и растения приобретают
облик розеточных, правда с менее утолщенными стеблями по,
сравнению с типичными розеточными видами.
В результате по характеру стеблей все известные виды сен-
полий могут быть расположены в последовательный ряд от
типично стеблевых до типично розеточных, где переходные
виды связывают эти два противоположных типа. Этот ряд,
в который виды, известные нам только по описаниям и изобра-
жениям, включены лишь ориентировочно, выглядит так:
S. grotei, S. pendula, S. goetzeana, S. magungensis, S. amanien-
sis, S. intermedia, S. rupicola, S. inconspicua, S. nitida, S. orbi-
cularis, S. grandifolia, S. teitensis, S. confusa, S. shumensis,
24
25
При развитии молодых растений из семян появляющиеся
через 10—14 дней всходы имеют две одинаковые семядоли,
одна из которых достигает 3 мм, а вторая вырастает до 14 мм
(черта, характерная для всех Геснериевых). На кончиках сред-
них жилок семядолей имеются по одному, иногда по два апи-
кальных устьица. На нижней стороне семядолей волоски слабо-
головчатые, по краям палицеобразные, обычные волоски, за-
остренные на концах, появляются несколько позже. После
окончательного сформирования семядолей начинают последо-
вательно появляться настоящие листья: первый лист, вскоре
супротивно второй лист, через некоторый промежуток, после
незначительного вытягивания стебля, перекрестно первой паре
появляется вторая пара листьев и т. д. При развитии растений
стеблевого облика междоузлия удлиненные. У растений с уко-
роченным стеблем при нормальных условиях освещения после
образования первой пары листьев следующая за нею пара в
силу укорочения стебля несколько смещается, не достигая пря-
мого угла, относительно первой пары листьев и т. д. Это в
дальнейшем создает впечатление смещенного по кругу листо-
расположения, образующего в совокупности розетку.
В условиях недостаточного освещения, когда в силу этиоля-
ции междоузлия удлиняются, это впечатление исчезает, и листо-
расположение оказывается в общих чертах сходным для всех
сенполий, а именно супротивно-парным. Все прочие упомяну-
'тые типы листорасположения представляют собой результат
приспособительных модификаций исходного перекрестно-пар-
[пого листорасположения, которое среди представителей рода
Рис. 6. Супротивное расположение
листьев и удлиненные междоузлия у ли-
стовых черенков при недостаточном осве-
щении.
секции
Saint-
сделал
1893-
время,
были
как это
[Fritsch,
еще в то
известны
2 вида сенполий,
1958].
Листорасположение
и лист. Листья на стеб-
лях у части сенполий рас-
полагаются перекрестно-
парно, у других — пооче-
редно по одному листу
на узле (второй лист не
развивается), у третьих
они смещены и распола-
гаются на верхнем участ-
ке укороченного стебля
в виде розетки (рис. 5, 6). У первой группы междоузлия в
основании стеблей более длинные, к их концам укороченные;
у второй группы нижние листья отмирают относительно рано,
-а функционирующие листья сосредоточены на концах стеблей,
где междоузлия особенно короткие, и их облиственные вершины
выглядят как розетки; у третьей группы междоузлия очень ко-
ротки, и листья смещаются, образуя розетку на укороченном
стебле.
26
<S. brevipilosa, S. pusilia, S. difficilis, S. velutina, S. diplotricha,
S. ionantha, S. tongwensis.
Учитывая то, что укорочение стеблей вместе с изменением
характера листорасположения (см. ниже) является результатом
определенной специали-
зации в связи со специфи-
кой условий окружаю-
щей среды, следует счи-
тать растения, имеющие
развитые в длину стебли,
более древним типом, а
розеточные — более моло-
дым и специализирован-
ным. Однако разбивать
эти типы на противостоя-
щие группы или
Archisaintpaulia и
paulia,
Фрич
1894]
когда
только
вряд ли целесообразно,
хотя бы из-за наличия
упомянутых растений пе-
реходного типа. Эта же
точка зрения высказана
Б. Л. Бэрттом [Burtt, является и более древним.
Характер строения листьев сенполий в общих чертах более
или менее однотипен (исключение составляют многие культи-
вары). Черешки по длине превышают (в развитом состоянии)
листовую пластинку. Край листа городчатый или зубчатый в
(разной степени у разных видов или, в отдельных случаях, почти
(цельный, или цельный. Однако если мы обратимся к деталям,
-то неизбежно находим немало существенных отличий, харак-
теризующих тот или иной вид.
! По очертанию различаются листовые пластинки овальные,
широкоовальные и почти округлые. Верхушки их заострены,
широко заострены или тупые, почти округлые. Основания серд-
I цевидные, сердцевидно-лопастные с налегающими друг на дру-
га лопастями либо не налегающими или почти щитковидные
(во всяком случае на определенной стадии развития листа).
Края приподнятые, слегка подогнутые вниз или почти плоские.
Очертание листьев различных видов сенполий и характер их
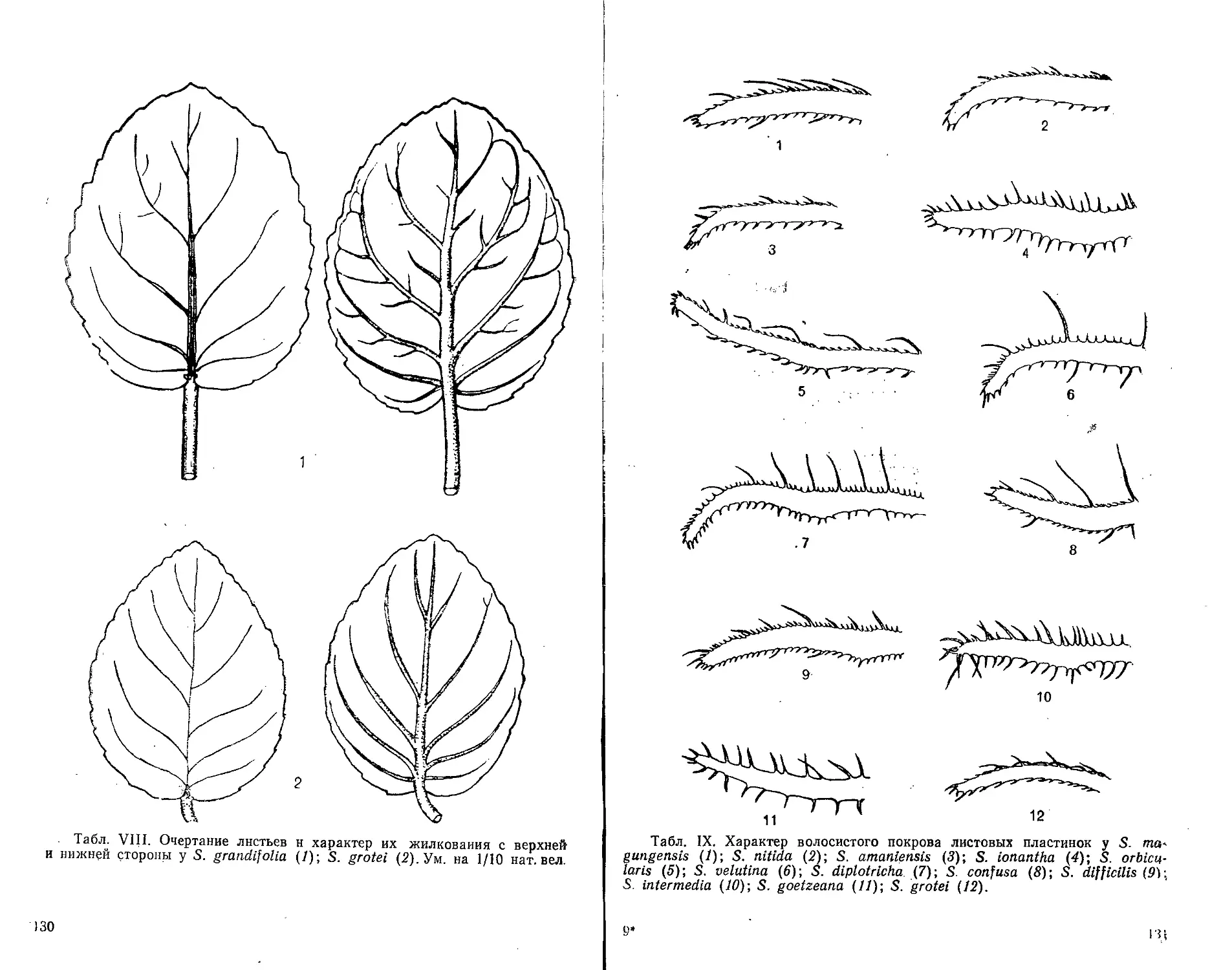
жилкования с верхней и нижней стороны см. табл. I—VIII.
Средняя и боковые жилки листа хорошо заметны сверху.
Большинство жилок мало заметно. Во многих случаях жилки
сверху светлее всей листовой пластинки либо они одной с нею
। 27
Рис. 7. Различные по длине и наличию на концах же-
лезок типы волосков на листе S. ionantha.
окраски. Сама окраска листовой пластинки различна у разны^3?31^16? волосистого покрова и окраска с нижней стороны
видов. То она темная, то светлая, правда, интенсивность окрасг'1ИС13, „ .
ки в известной степени зависит от возраста листьев, условий Волосистый покров способствует у нь р >
освещения и питательности субстрата, на котором развиваютс?ащищает поверхность листьев от „ отрицательного в здс
растения. ультрафиолетовых лучей, с другой стороны, он предохраняет
С нижней стороны листовой пластинки сеть жилок, как прасе от чрезмерного смачивания при попадании воды, что при
вило, выступает, но в одних случаях очень сильно, в други)<'войственной этим листьям мясистости может угрожать загни-
слабее. Жилки или светлые или розово- и красноокрашенные|1анием’ Красноватая или
Нижняя сторона листовой пластинки у одних видов беловато
зеленая, у других светло- или темно-красноокрашенная с жил
ками, не отличающимися по окраске или более светлыми.
Листовая пластинка у одной группы видов тонковатая
у другой утолщенная. Толщина пластинки обычно меньше )
стеблевых и высокогорных видов, у розеточных видов пластин
ка всегда более мясистая и хрупкая.
Существенные различия наблюдаются в характере волосис-
того покрова. Общей чертой почти всех видов является .нали-
чие, помимо хорошо заметного волосистого покрова, густоп
покрова из короткостебельчатых или сидячих железок, разли-
чимых только при рассматривании в сильную лупу и при опре
деленном освещении. Густота расположения этих железок раз-
лична. Волосистый покров, оказывается, может служить важ-
ным признаком (наряду с другими) для характеристики мно-
гих видов. Он состоит, как правило, из многоклеточных волос;
ков, прямостоячих или распростертых, прижатых, приподни-
мающихся на концах или концами направленных к поверхност!
пластинки, более или менее однородных по длине или переме
шанных длинных и коротких или очень коротких, прямых ил;
прижатых (см. табл. IX и рис. 7). Степень густоты покров;
различна у разных видов и зависит от возраста отдельны)
листьев. Молодые листья всегда имеют более густой волосистьй
покров.
Размеры листовых пластинок (если они вполне развить?
у разных видов значительно отличаются. На размеры листье;
оказывают известное влияние условия роста растений и их воз
раст.
Вышеперечисленные признаки строения листьев сенполий
по-видимому, являются отражением определенного комплекс;
условий окружающей среды, в которых формировались те ил;
иные виды.
Степень толщины листовой пластинки и ее сочность (как
впрочем, и остальных частей растений) определенно связанг
с приспособленностью отдельных видов к существованию в ус
ловиях различной степени и периодичности увлажнения. Боле
толстые и мясистые листья свойственны видам, обитающим j
условиях неравномерного увлажнения и обычно встречаю
щимся на скалистом субстрате, способном в известные периодь
перегреваться. С последним обстоятельством тесно связаны ;
красная окраска нижней стороны
противодействует перегреву листьев от нагревшегося камени-
стого субстрата.
Более тонкие и соответственно менее сочные листья свойст-
венны видам, обитающим в условиях более равномерного
увлажнения и поселяющимся на менее нагреваемом субстрате,
например одетом моховым покровом. Для видов, имеющих тон-
кие листья, обычно характерна меньшая густота волосистого
покрова, его прижатость к поверхности листьев, а также слабо
выраженная или вовсе отсутствующая красноватая окраска
нижней стороны листьев.
Интенсивность зеленой окраски верхней стороны листьев,
по-видимому, связана со степенью затененности местообитаний
отдельных видов. Темноокрашенные листья имеют обитатели
смешанных вечнозеленых тропических лесов, тогда как светлая
29
окраска свойственна более высокогорным видам, обитающим в
смешанных вечнозеленых и листопадных лесах, листопадные
элементы которых (в известные периоды сбрасывающие листья)
разрежают тень.
Для распознавания отдельных видов сенполий в живом со-
стоянии в большинстве случаев оказывается достаточным руко-
водствоваться общим обликом растения, очертанием листовой!
пластинки и особенностью ее края, а также характером воло-
систого покрова.
Цветоносы и цветки. Цветоносные побеги у сенполий обра-
зуются, как правило, в пазухах листьев, чаще всего до образо-
вания боковых стеблевых побегов на главном стебле. У одних
видов они в той или иной степени восходящие, у других — от-
клоненные и никогда не восходящие, почти касательные к
листьям, разные по длине: то не поднимающиеся выше листьев,
то более или менее равные им по длине, то возвышающиеся
над ними.
Цветоносы образуют цимозные соцветия из попарно после-
довательно ответвляющихся цветоножек с цветками; число по-
следних кратно двум, нечетное число—результат недоразви-
тия одного из цветков пары. Изображения цветоносов боль-
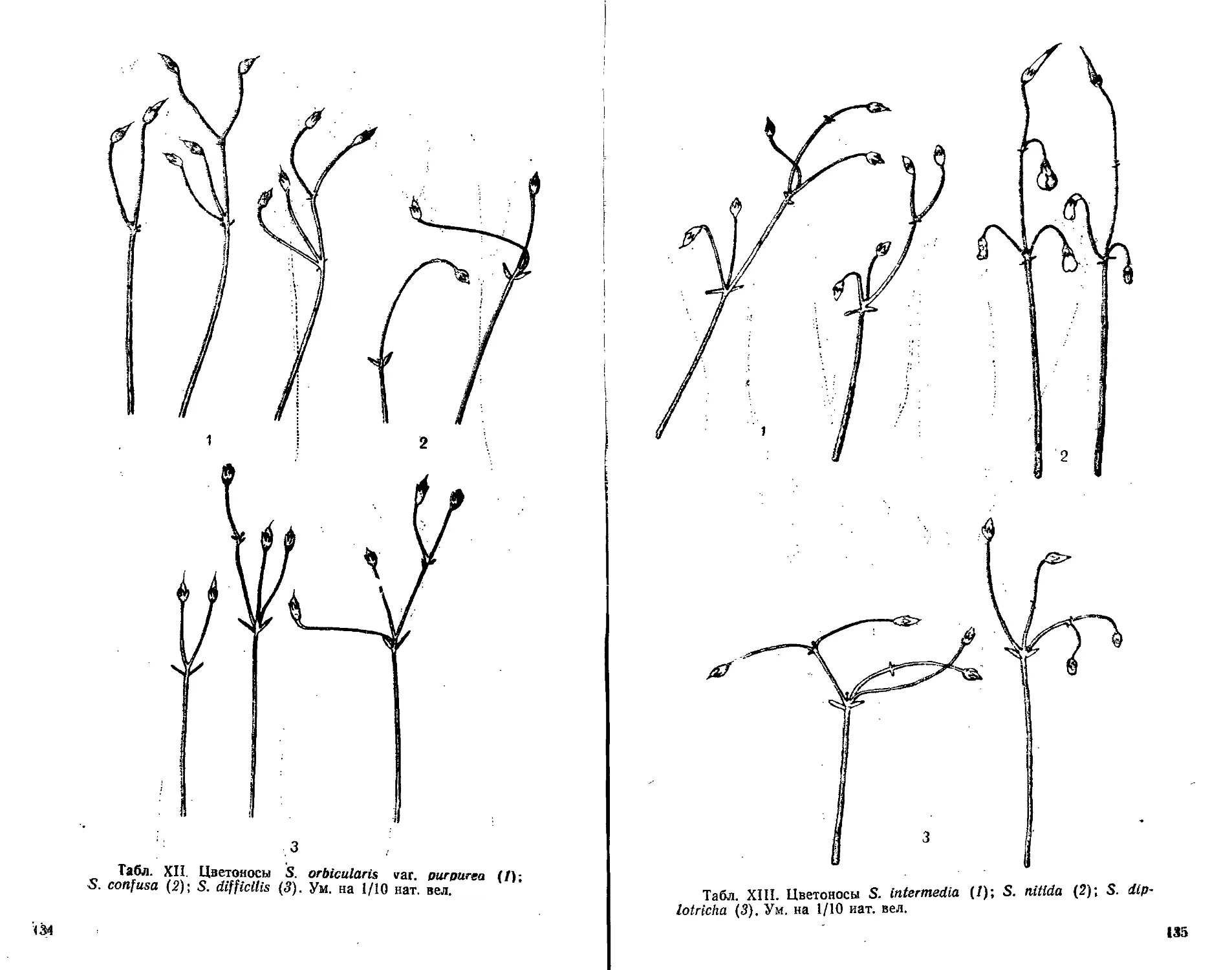
шинства видов сенполий на табл. XI—XVI дают ясное пред-
ставление о различиях, свойственных каждому из видов.
Длина цветоножек в зависимости от вида равна или немного
превышает диаметр венчика. До цветения цветоножки крючко-
образно или спирально изогнуты. Перед распусканием цветков
они несколько распрямляются до слабой дуги и после цвете-
ния, при развитии семенных коробочек, вновь принимают пер-
воначальное положение.
В основании каждой цветоножки располагается по одному,
обычно смещенному почти под прямым углом или расположен-
ному с одной стороны цветоносов (табл. XVI, 1, 2) прицветнику,
чаще всего узколанцетной формы, больших размеров в нижней
части соцветия и меньших — к вершине (табл. XI—XVI).
В случае недоразвития одного из цветков пары в основании
цветоножки образуются два прицветника, противостоящие друг
другу. У культиваров прйцветцики часто крупнее и имеют ли-
стообразную форму.
Главная ось соцветия после образования первой дихотоми-
ческой пары цветоножек продолжается из пазухи одного из
прицветников; то же повторяется и в последующих парах цве-
тоножек.
Чашечка несколько асимметричная из пяти сросшихся в осно-
вании узколанцетных долей, из которых две верхние, противо-
стоящие долям верхней части отгиба венчика, несколько длин-
нее остальных трех. Доли чашечки плотно прилегают к трубке
венчика, резко изгибаясь у его отгиба, и несколько оттопыри-
30
ваются кончиками. После опадения венчика доли чашечки плот-
но прилегают к завязи или оттопыриваются.
Нормальные цветки сенполий имеют неправильный пяти-
членный отгиб венчика, соединенный с помощью короткой бо-
чонкообразной трубки и состоящий из двух частей, неточно
называемых многими авторами губами (labium): верхней (pars
superior), составленной из двух наполовину от основания срос-
шихся долей, и нижней (pars inferior), состоящей из трех глу-
боко рассеченных долей. Эти рассечения образуют синусы,
у одних видов открытые, у других в той или иной мере прикры-
тые боковыми лопастями, образующимися в основании средней
доли, нижней доли или одной из долей верхней части отгиба
венчика.
Верхняя часть отгиба венчика противостоит средней доле
нижней части отгиба, а две боковые доли нижней части отги-
ба — друг другу, причем у одних видов их оси образуют пря-
мую линию, у других же — изломанную в результате некото-
рого опускания этих долей, тогда как оси верхней части отгиба
и средней доли нижней части всегда образуют прямую линию.
Многие авторы называют отгиб венчика сенполий колесовид-
ным или почти колесовидным, т. е. подразумевают положение
долей венчика относительно трубки равным или близким к
прямому углу. На деле же плоскости верхней части отгиба, так
же как и нижней, всегда образуют с трубкой угол, превышаю-
щий прямой у одних видов на большую величину, у других —
на меньшую. Обе части отгиба, верхняя и нижняя, между собою
также образуют угол, величина которого меняется в зависимо-
сти от видов и от возраста отдельного цветка, но он всегда
является тупым (см. сравнительное изображение разных видов
сенполий на табл. XVII—XIX, где можно заметить характерные
для каждого вида очертания венчика и отношения верхних и
нижних его долей).
В андроцее имеются две плодущие тычинки и два или три,
в зависимости от вида, стаминодия. Иногда наблюдаются цвет-
ки с 3—4—5 плодущими тычинками (атавизм).
Уплощенные тычиночные нити прирастают изнутри к трубке
венчика в местах, противостоящих синусам, образуемым сред-
ней долей нижней части отгиба и ее боковыми долями. Выходя
вначале параллельно стенкам трубки, тычиночные нити обра-
зуют (в направлении одна к другой) изгиб и заканчиваются
двугнездными пыльниками. Форма пыльников, при взгляде на
них сбоку, может быть названа сердцевидной, состоящей из
двух сложенных вместе почковидных частей, соответствующих
двум гнездам пыльника. В основании каждой половины пыль-
ника снизу наблюдаются мешковидные выступы, представляю-
щие собою камеры для накопления зрелой пыльцы, поступаю-
щей из пыльцевых мешков, с которыми эти камеры сообщают-
ся. Поэтому при рассматривании сросшихся пыльников двух
31
'тычинок снизу видны четыре основания этих камер и вокруг
них четыре основания пыльцевых мешков. Сверху же видны
только вершины четырех пыльцевых мешков. Срастание дву-
Рис. 8. Сросшиеся пыльники у сенполий (Х80).
1 — вид от основания (5. Ttipicola); 2 — искусственно отделенный пыльник (S.dlf-
ficllis}; 3— вид сросшихся пыльников сверху; 4— искусственно разделенные пыльники
Л. difftclis’, 5 — поперечный разрез пыльника S. rupicola с гнездами и камерами, где
накапливается зрелая пыльца; 6 — схема этого разреза.
гнездных пыльников двух тычинок происходит в верхней трети
их высоты сторонами, обращенными друг к другу (рис. 8). Это
срастание в большинстве случаев оказывается настолько проч-
ным, что препятствует вскрытию пыльников при созревании
пыльцы и ее высвобождению.
32
У культивируемых растений вскрытие щелей происходит
поздно, когда венчики с приросшими к их трубкам тычинками
отпадают и пыльца бесполезно высыпается. Как происходит
вскрытие щели в природе, неизвестно.
Стаминодии расположены, как и основания тычиночных
нитей, внутри трубки, два из них противостоят выемкам (си-
нусам) между боковыми долями нижней части отгиба и дву-
лопастной верхней части отгиба, а при наличии третьего стами-
нодия он расположен между двумя первыми и противостоит
рассечению долей верхней части отгиба. Во всех случаях ста-
минодии представляют собою различно развитые сосочковид-
ные образования, причем средний, если он есть, всегда развит
слабее остальных двух, принимая часто вид небольшого бугор-
ка (рис. 9).
Как уже сказано, двугнездные пыльники каждой тычинки
срастаются между собой теми же сторонами, с которых про-
исходит их вскрытие при созревании пыльцы. При вскрытии
Рис. 9. Два типа стаминодиев в цветках сенпо-
лий из двух и трех выростов.
образуются щели в виде неправильной линии (соответственно
каждому их гнезду), сливающиеся в одну, приобретающую
форму Дуги или линии, сходной с перевернутыми латинскими
буквами V или U (рис. 8).
Гинецей состоит, из верхней довольно однотипной, по форме
конической, неполностью двугнездной завязи (рис. 10), перехо-
дящей в длинный ..столбик, заканчивающийся различными по
форме у разных видов'рыльцами (рис. 11): либо булавовидным
округловатым, либо коническим уплощенным или относительно
слаборасширенным, дву- или трехлопастным, причем в случае
двулопастности одна из лопастей всегда меньшего размера.
Положение столбика относительно оси завязи у подавляю-
щего большинства видов искривленное то вправо, то влево.
Лишь у S. velutina, как мы наблюдали неоднократно, положе-
ние столбика часто соответствует продолжению оси цветка.
Пыльники у этого вида часто не срастаются, и столбик поэтому
прямой. Это, по-видимому, дает возможность поставить искрив-
ление столбика в зависимость от срастания пыльников, проис-
ходящего задолго до развития столбиков.
Иногда у некоторых культивируемых растений разных видов
сенполий можно наблюдать ненормально правильные цветки,
имеющие по пять лопастей отгиба с пятью плодущими, тычин-
Л. М. ЗАЛЕССКИЙ
3.3-
ками, либо цветки с шестью долями, тремя или четырьмя пло-
дущими тычинками, с завязью или без завязи.
Размеры цветков различных видов сенполий колеблются в,
пределах от 8 до 30—35 мм в диаметре. У некоторых культи-
варов диаметр цветков достигает 50—7'5 мм. Следует, однако,
Рис. 10. Продольный (а) и поперечный (б) разрезы завязи
сенполий.
1 — у вершины; 2 — в средней части; 3 — у основания.
отметить, что диаметр цветков может несколько меняться в
пределах одного и того же вида в зависимости от условий,
в которых развиваются растения, а также — у культивируемых
растений — при поражении болезнями и вредителями. Указы-
ваемые размеры соответствуют оптимальным условиям разви-
тия. Измерения диаметра цветков следует производить у впол-
34
не сформировавшихся растений и не ранее чем на третий день
после начала распускания цветков, так как до этого они про-
должают расти.
У здоровых, растений отдельные цветки сохраняются в тече-
ние 6—7—14 дней, после чего отпадают. Опадение цветков
ранее этого срока свидетельствует о каких-то болезненных
явлениях.
Окраска цветков сенполий различная.у разных видов: or
голубовато- или розовато-белой с более темным пятном в обла-
сти расположения пыльников, особенно в верхней части отгиба,
до разных оттенков сине-фиолетовой и пурпурной (красно-фио-
летовой).. В пределах одного и того же вида в некоторых слу-
1 2 3 4 5 6 7
Рис. 11. Различные типы рылец у сенполий:
J “ 5. grandifolia; 2 — S. difficilis; .3 — 5. shtimensis, .S'. atnanten.sis; 4 — S. bred-
pilosa; .5 — S. ionantha, S. confusa, S. intermedia, S. magungensis, S. rupicola, S. orbicu-
laris', 6 — 5. nitida; 7 — S. velutina.
чаях наблюдаются клопы с различными по оттенку цветками,
ио каждый вид обладает достаточно характерной окраской
цветков, которая, если и повторяется у некоторых видов, то
сочетается с теми или иными отличительными особенностями
по другим признакам.
У культиваров окраска цветков значительно разнообразнее:
от чисто-белой, желтоватой, бледно-розовой, розовой разных
оттенков и интенсивности, пурпурной и почти красной до голу-
бой, синей и темно-синей различных оттенков. Кроме того, из-
вестны культивары с различными (фестончатыми, волнистыми
и т. д.) краями долей венчика, а также с махровыми цветками..
Цветоносы, цветоножки, прицветники, доли чашечки и за-
вязь покрыты, так же как листья и стебли, многоклеточными
различной длины волосками, а также рассеянными между ними
шарообразными железками на коротких стебельках. Волоси-
стый покров наблюдается у. основания; тычиночных нитей вну?
три трубки венчика, в месте их прикрепления. Волоски одина-
ковой длины имеются на наружной стороне лопастей отгиба
венчика, а также по их краям; у некоторых видов добавляются
такой же длины железистые волоски, имеющие на концах бу-
лавовидные утолщения и образующие так - называемый ресниц
у»
35
чатый'край (рис. 12). На внутренней стороне отгиба венчика
волосистый покров отсутствует, наблюдаются лишь разбросан-
ные железки на стебельках (такие же, как между волосками,
покрывающими листья). На внутренней стороне долей чашечки,
у основания столбика, в том месте, где кончается волосистость
завязи/. И; между покрывающими ее волосками имеются такие
же короткостебельчатые железки. В основании завязи, в месте
прикрепления чашечки, у всех видов наблюдается четко выра-
женный, обычно оранжевый или желтовато-коричневый коль-
цевидный нектарник.
Корневая система. При описании видов сенполий их корне-
вой системе обычно не уделяется внимания. По-видимому, это
' ?ис. 12. Различные, типы волосков по краю венчика.
'. / — реснитчатые; 2 — железистые.
ввязано- с- относительным ее однообразием. Однако при более
внимательном рассмотрении оказывается; что корневая система
сенполий’ обладает некоторыми особенностями, на которые
нельзя не обратить внимания. .
В общих чертах корневая система сенполий, формирующая-
ся при развитии растений из семян, представляет собою базаль
•ную мочку относительно слабо разветвленных тонких п неж-
ных корней. По мере роста растений, удлинения стеблей и ста-
рения наблюдается постепенное отмирание этих корней и об-
разование им на смену придаточных корней на стеблях, кото-
рые к этому времени оказываются лежащими на земле. Одно-
временно происходит постепенное отмирание базальной части
ё^ебля.
Таким образом, корневая система, образующаяся первона-
чально в базальной части растения, функционирует относитель-
но, короткое время, затем отмирает и заменяется придаточны- .
й-и- корням®; возникающими на стеблях между листовыми руб-
3&. ' I
i вами, остающимися на месте отмирающих листьев. Эти прида-
точные корни после выполнения своих функций вновь заменя-
ются придаточными корнями, вырастающими на более молодых
частях стеблей, и т. д. Иначе говоря, через определенный про-
межуток времени происходит полное обновление корневой си-
стемы, способствующее развитию всех наземных частей расте-
ния. Благодаря этой особенности в результате формирования
боковых ответвлений стебля и образования на них придаточ-
ных корней происходит вегетативное размножение растений,
приводящее, при благоприятных почвенных условиях, к возник-
новению колонии.
У некоторых видов стебли первоначально возникших розе-
точных растений с базальной мочкой корней ведут себя, как
корневище (S. rupicola). Они, не принимая вертикального на-
ложения, сразу же, по мере удлинения, образуют придаточные
корни, и развивающаяся разветвленная особь представляет со-
бою нерасчлененную на части колонию или дерновину (см,
рис. 4).
У некоторых видов корневая система сначала развита отно-
сительно слабее, чем свойственно большинству. Это виды, ко-
торые ведут себя в природе, как эпифиты. К числу их относит-
ся S. goetzeana (см. рис. 2, 3, 4).
Плоды сенполий — почти округлая, яйцевидная или удлй-
L ценная коробочка, образующаяся из . синкарпного гинецея из
двух плодолистиков. В основании плода и в верхней части об-
ё разуются два гнезда, сообщающиеся в центральной части, об-
разуя общую полость. В сечении плоды слегка четырехгранны
или почти округлы, сужены на базальном конце довольно
резко и округло, на апикальном — в.верхней трети длины всего
плода — постепенно сужаются и заостряются, заканчиваясь
обычно долго сохраняющимся подсохшим столбиком пестика
(рис. 13).
. На так называемой брюшной и спинной сторонах плода име-
ются (не у всех видов) четко заметные продольные светлые,
слегка выпуклые линии — «швы», представляющие собою сред-
ние жилки двух плодолистиков, в завязях образующих гинецей,
в состоянии плодов — их наружные оболочки, а внутри плода-—
плаценту, на которой располагаются семена. С боков плода ино-
гда заметны две (по одной с каждой стороны) беловатые ли-
нии, .соответствующие, по-видимому, местам срастания двух
плодолистиков, уходящих краями внутрь плода, В поперечном
разрезе неподсохшей коробочки внутри стенки плода, в местах,
соответствующих средним жилкам плодолистиков, наблюдаются
V-образные выемки (рис. 10,6).
При характеристике формы плодов некоторые коллекторы,
собиравшие растения в природе, а также автор описаний боль-
шинства видов Б. Л. Бэртт,' называют плоды цилиндрическими.
Мы не можем согласиться с тем, что плоды-коробочкй какого-
3?
Рис. 13. Типы плодов культурных форм S. ionantha (cv).
либо из известных видов сенполий можно назвать цилиндриче-
скими. Та форма плодов, которая свойственна части видов сен-
полий и обычно называется цилиндрической, по нашему мне-
нию, должна характеризоваться как веретеновидная или, в бт-
33
Рис. 13. (продолжение).
дельных случаях, шиловидная, у другой части видов форма пло-
дов яйцевидная или удлиненно-яйцевидная. .
Вообще же форма коробочек варьирует в зависимости от
принадлежности к тому или иному виду от широкояйцевидной,
почти шарообразной, яйцевидной (S. ionantha, S. iong-wensis),
удлиненно-яйцевидной (S. pendula, S. velutina, S. intermedia)
до веретеновидной и шиловидной (S. leitensis и все остальные
виды). Коробочки то более или менее симметричны во всех
плоскостях, то относительно утолщены с брюшной стороны,
прямые или в той или иной степени крючкообразно изогнутые.
Они покрыты бесцветными прямыми, более или менее торча-
щими или прижатыми, иногда волнистыми многоклеточными
волосками различной длины и разной степени густоты обычно
такого же типа, как и волоски листьев и прочих частей ра-
стений.
В неподсохшем состоянии поверхность коробочек зеленая,
или красновато окрашенная сплошь, или по зеленому фону идут
короткие продольные красноватые черточки. После подсыхания
плоды коричневато-серые, либо красновато-сетчатые, либо
серые.
Ниже мы приводим общую характеристику плодов различ-
ных видов, которые при искусственном опылении давали в на-
39
ших условиях плоды. На табл. XX показаны очертания пло-
дов в двух плоскостях: с так называемой спинки (сверху) и
сбоку, что дает возможность их сравнения.* В эту же таблицу
мы включили очертания плодов, воспроизведенные на основа-
нии кратких литературных сведений, поэтому они могут ока-
заться недостаточно точными (S. pusilia, S. goetzeana, S. incon-
spicua, S. teitensis).
В связи с тем что в форме коробочек существует много пе-
реходов, мы при определении их формы используем отношение
их длины (от основания до верхнего конца) к их наибольшему
диаметру в нижней трети длины. При таком подходе к опреде-
лению формы коробочек можно выделить три группы плодов:
яйцевидные с отношением длины к диаметру от 2 : 1 дэ
3:1, веретеновидные с отношением от 4 : 1 до 5 : 1 и
шиловидные с отношением от 6:1 до 9:1 (плоды тех
видов, форма коробочек которых воспроизводится на основа-
нии литературных сообщений, мы пока не включили ни в одну
из перечисленных трех групп).
В первую группу, с яйцевидными плодами, входят пять ви-
дов: S. ionantha, S. pendula, S. velutia, S. intermedia, S. tong-
wensis; во вторую, с веретеновидными плодами, — 8 видов:
X. brevipilosa, S. shumensis, S. confusa, S. difficilis, S. rupicola,
S. grandifolia, S. amaniensis, S. magungensis (последние два
вида Б. Л. Бэртт рассматривает сейчас как разновидности
одного, что, на наш взгляд, недостаточно оправдано см. с. S
и примечание); в третью, с шиловидными плодами, — 4 вида:
X. orbicularis, X. nitida, X. diplotricha, X. teitensis. Вне этих
групп остались плоды четырех видов, известных нам только по
литературе: 1) X. inconspicua с плодами (незрелыми) около
10 мм длиной, часто изогнутыми; 2) X. goetzeana с плодами'
м.енее 10 мм длиной; 3) X. pusilia с удлиненными плодами око-
ло 10 мм длиной и около 2 мм шириной (изображение А. Энг-
лера), на конце крючковидно оттянутыми; 4) X. teitensis
с удлиненными плодами (по нашей номенклатуре — шиловид-
ные) .
О вскрытии коробочек и освобождении семян в природе в
литературе никаких указаний не имеется. Известно лишь пред-
положение, высказанное Л. И. Иваниной в ее двух работах:
«Вскрывание большей частью позднее, вероятно, дорсальное,
створки не закручиваются» (1965, с. 31), коробочка «вскрываю-
щаяся поздно, дорсально (возможно, не вскрывающаяся,
* Эта таблица, составленная нами в ходе работы с сенполиями, была
опубликована Л. И. Иваниной. К сожалению, при перерисовке с нашего
оригинала отдельных рисунков и их перераспределении были допущены, до-
садные ошибки. В результате подписи к рисункам оказались перепутанными
и появились подписи, которых не было в оригинале.
и сгнивающая в почве)» (1967, с. 26),* «в трибе Saintpaulieae
имеют место базикарпия и геокарпия» и далее «цветоножки
иидов Saintpaulia в культуре по мере созревания плодов изги-
баются й опускаются вниз, так что зрелые коробочки оказы-
ваются под листьями розетки; по литературным данным, плоды
Saintpaulia зарываются в землю (геокарпия)» (1967, с. 113).**
По нашим наблюдениям на культивируемых растениях раз-
личных видов, после отцветания и образования плодов цвето-
ножка и позже цветонос вместе с развивающимися плодами,
иначе говоря, все соцветие, подгибаются вниз и в сторону, в на-
правлении стебля. При этом цветоножки, становящиеся к этому
времени плодоножками (они обычно и до цветения часто крюч-
кообразно или даже кольцеобразно изогнуты, во время цвете-
ния несколько выпрямляются), изгибаются таким образом, что
коробочки располагаются параллельно или почти параллельно
поверхности нижнего яруса листьев. Созревание плодов продол-
жается долго.*** На первых порах они растут довольно мед-
ленно. После того как коробочки перестают увеличиваться в
размере, цветонос начинает постепенно подсыхать, а затем уже
подсыхают и коробочки. К этому времени растущее растение
развивает несколько ярусов нижних листьев,' так что созревшая
коробочка бывает основательно прикрыта ими, оказываясь в
условиях полного затенения, повышенной влажности и незна-
: чптельной циркуляции воздуха; в это время и происходит ее
вскрытие и освобождение семян.
1 В действительности вскрытие происходит не так, как можно
: было бы предполагать, а путем постепенного разрушения обо-
лочек плода, созревающего как в условиях повышенной влаж-
ности, так и в условиях относительной сухости. Створки же
остаются сросшимися. Последовательность и характер разруше-
ния оболочек коробочки можно видеть на рис. 14. О геокарпии
же применительно к плодам сенполий вряд ли можно говорить,
>отя бы потому, что семена у них мелкие и в тех условиях,
и которых обитают растения в природе, не требуют для своего
прорастания попадания под землю.
Семена сенполий очень мелкие: от 0,4—0,7 мм длиной и
0,15—0,30 мм шириной, яйцевидные или удлиненно-яйцевидные
1с шестью продольными валиками-ребрами, в сечении почти
। округлые или слегка сплющенные, беспорядочно почти сплошь
мелкобугорчатые,**** рыжеватые, темно-коричневые или почти
* Данное предположение, правда, без уточнения о сгнивании в почве
было высказано автором этих строк в личной беседе с Л. И. Иваниной
и то время, когда мы производили эксперименты, отрицавшие возможность
вскрывания створками.
** Без ссылки на литературные источники.
*** При опылении весной — около 6, при опылении зймой — до 9 ме-
сяцев.
**** Бугорки, вероятно, служат для прикрепления рассеивающихся се-
мян к тем или иным предметам.
40
।
41
Рис. 14. Стадии
разрушения коробочки
у S. shumensis.
I и 2 — Х2; 3 и 4 — в нату
ральную величину; 5 — ни
чальная стадия; 6 — коней
ная стадия (увеличено).
черные. Во впадинах между округлыми ребрами иногда наблю-
дается поперечная полосатость из расположенных в ряд воз-
вышающихся бугорков, ничем не отличающихся от остальных
бугорков, покрывающих всю поверхность семени. Участки, об-
разованные продольными валиками-ребрами и рядами возвы-
шенных бугорков, выглядят вроде «ячеек». Мы наблюдали по-
добные «ячейки» чаще Всего в одной из шести впадин между
2
Рис. 15. Сравнительное изображение'семян сенполий.
1 — S. shumensis, вид сбоку и со стороны основания; 2— S. difficilis, вид
сбоку и со стороны основания; 3 — S. tongwensis, вид сбоку; 4 — 6. ionantha, вид сбоку^
валиками-ребрами, обращенной в сторону прикрепления семян
к плаценте.
При сравнении семян различных видов заметных особенно-
стей мы не обнаружили (рис. 15).
Семена различных видов отличаются по длине и диаметру
и, может быть, по интенсивности коричневой окраски их бугор-
чатой поверхности, то более светлой, то темной.
НЕКОТОРЫЕ БИОЛОГИЧЕСКИЕ ОСОБЕННОСТИ
СЕНПОЛИЙ И ПОПЫТКА ИХ ОБЪЯСНЕНИЯ
Как уже было сказано, растения сенполий в природе растут
в расселинах скал на- каменистом субстрате и на обомшелых
43
камнях в качестве полуэпифитов или, иногда, на деревьях как;
эпифиты во влажной атмосфере горных районов Восточной
Африки. Эти условия,, по-видимому, служат причиной появле-
ния своеобразных черт строения, что выражается прежде всего»
в мясистости стеблей и листьев, покрытых прозрачными много-
клеточными волосками. Таким же покровом одеты обычно и
остальные части растений, за исключением внутренних сторон
долей венчика, на наружной стороне и по'краям которых на-
блюдаются простые или железистые волоски.
Мясистость надземных органов у различных видов сенполий
выражена неодинаково и коррелирует с разной степенью уко-
рочевности стеблей. То же можно сказать о густоте и харак-
тере их волосистого покрова. Это приводит к формированию-
двух типов растений, связанных между собой многими перехо-
дами, характеризующими те или иные виды. Один тип — с силь-
но укороченными и соответственно утолщенными стеблями,,
с листьями, располагающимися (в результате сокращения дли-
ны междоузлий) в виде розетки на укороченных узлах. Дру-
гой тип — с удлиненными стеблями, менее утолщенными, с не-
сколько смещенным, почти супротивно-парным расположением
листьев. Обычно в прямой связи со степенью утолщенное™
стеблей находится и утолщенность или мясистость листовых:
пластинок, и густота и характер волосистого покрова. У расте-
ний с укороченными стеблями листья наиболее мясисты,, хруп-
ки, волосисты'.
Черешки листьев всех без исключения видов сенполий об-
ладают способностью образовывать (в соответствующих усло-
виях) придаточные корни и у подавляющего числа видов —
адвентивные ростовые почки, способные воспроизвести целые
растения. Способность эта проявляется не только при черенко-
вании листьями, но и независимо от него — на черешках и в
основании пластинки листа на средней жилке, как показано на>
рис. 16, 17. При образовании адвентивных почек в основании
черешка листа молодые корешки формируются не из каллуса,,
а у основания развивающихся почек. Верхняя почка побега,,
находящаяся в менее благоприятных условиях увлажнения, от-
стает в развитии и корешков не образует.
Таким образом, сенполии обладают той же способностью к
вегетативному размножению, которая характерна для многих
суккулентных и некоторых эпифитных растений. Эта способ-
ность у различных видов проявляется различно. Так, например,
на листьях улугурского вида S. goetzeana, обладающего удли-
ненным и сравнительно мало утолщенным стеблем и живущего
часто эпифитно на скалах и деревьях, адвентивные почки ни-
когда не образуются, хотя придаточные корни формируются^
Иначе говоря, растения этого вида к вегетативному размноже-
нию листьями, свойственному всем остальным видам, не спо-
собны. Все же остальные виды обладают этой способностью,
44
юднако, по нашим наблюдениям, и среди них имеются виды
(S. tongwensis, S. pendula из Восточной Усамбары), у которых
эта способность проявляется в меньшей степени и выражается
в значительно меньшем количестве образующихся адвентивных
почек на черешках листьев при их черенковании. Первый вид
•относится к розеточному типу, второй имеет удлиненные, но
утолщенные лежачие и укореняющиеся стебли.
Следует отметить также часто наблюдающуюся при черен
ковании листьями зависимость образования адвентивных почек
от того или иного участка черешка. В основании черешка их
•образуется больше, чем на конце, ближе к пластинке листа.
Рис.' 16. Образование адвентивных почек на листьях и черешке S. niti-
da (а) и на средней жилке<S. orbicularis (б).
В процессе развития растений в пазухах опавших листьев,
а у S. pendula и S. goetzeana в пазухах живых листьев обра-
зуются боковые ответвления, которые после образования на
них придаточных корней обособляются от материнского расте-
ния. Такое обособление дочерних растений наблюдается у ра-
стений как с удлиненным, так и с укороченным стеблем, только
у последних это часто внешне выглядит, как кущение.
Исходя из сказанного мы можем констатировать, что у ви-
дов сенполий, за исключением S. goetzeana, отчетливо выра-
жена способность к вегетативному размножению, проявляю-
щаяся в двух формах: с помощью обламывающихся листьев
и в виде обособления боковых ответвлений.
Как уже упоминалось, при характеристике цветков сенполий
обращает на себя внимание искривленное положение столбика
пестика относительно оси цветка (на одних цветках он искрив-
лен вправо, на других — влево), что послужило поводом для
неоправдавшегося предположения Джона Вилсона о связи это»
особенности с перекрестным опылением [Wilson, 1898].
В строении цветков сенполий обнаруживается еще одна ха-
рактерная особенность. Это — срастание пыльников двух пло-
дущих тычинок теми сторонами, где расположены щели, пост
Рис. 17. Образование адвентивных почек в основании черешка
листа при черенковании.
вскрытия которых происходит высвобождение зрелой пыльцы|
из пыльцевых камер. При наблюдавшемся нами у S. velutinat
частом отсутствии указанного срастания пыльников столбик'
пестика всегда занимает вертикальное положение, что позво-
ляет понять причину его искривления у других видов. При раз-
витии отдельных частей цветка столбик искривляется в ту или
другую сторону в результате срастания пыльников, происходя-
щего уже на ранних стадиях развития цветка. Сросшиеся пыль-
ники мешают развивающемуся столбику занять положение, со-
ответствующее оси цветка.
46
У двух видов' сенполий, а именно у S, orbicularis и S. shu-
mensis, наблюдается образование большого количества плодов.
У этих видов срастание пыльников столь, незначительно, что
не мешает вскрытию щелей й высвобождению пыльцы до опа-
дения венчика. Опыление здесь происходит в момент опадения
венчика с приросшими к его трубке тычинками. Опавший вен-
чик постепенно сползает и неизбежно касается пыльниками
рыльца. При этом при малейшем сотрясении можно наблюдать
облачка пыльцы, часть которой попадает на рыльце. Таким об-
разом, у этих видов имеет место самоопыление.
Известно, что первый из введенных в культуру видов —-
S. ionantha — в результате применения искусственного пере-
крестного опыления дал большое число различных вариаций*
морфологически существенно отклоняющихся от исходного
типа. Это, по-видимому, говорит о накоплении внутри большин-
ства видов сенполий и в первую очередь у S. ionantha больших
потенциальных генетических возможностей, которые проявля-
ются только при перекрестном опылении. Однако перекрестно-
му опылению препятствует указанное выше срастание пыльни-
ков, исключающее возможность высвобождения из пыльцевых
камер зрелых пыльцевых зерен. Через некоторый промежуток
времени после отцветания венчик вместе с приросшими к его-
трубке тычинками опадает, и весь запас пыльцы погибает.
Наряду с примитивными чертами, характерными для пред-
; ггавителей р. Saintpaulia, такими, как архаичные морфологиче-
L ские признаки (пятичленная чашечка, почти колесовидный от-
1 гиб венчика, не вполне редуцированный андроцей), они обла-
f дают признаками, свидетельствующими об известной специали-
зации, что говорит об известной эволюционной продвинутое™
этих растений. Это такие признаки, как укороченные и утол-
щенные стебли, способность образовывать адвентивные почки*
воспроизводящие целые растения, срастание пыльников, ограни-
чивающие пли почти исключающие семенное размножение.
Сочетание архаичных признаков с явной специализацией
черт растений с учетом особенностей окружающей растительно-
сти и рельефа местности позволяет: 1) понять особенности гео-
графического распространения представителей р. Saintpaulia,.
ограниченного небольшим районом, и приуроченность их обита-
ний к древним геологическим образованиям; 2) считать, что-
p. Saintpaulia обладает огромными нереализованными потен-
циальными генетическими возможностями, что подтверждается
изобилием форм, получаемых при искусственном опылении;
3) заключить, что особенности, присущие р. Saintpaulia, позво-
ляют отнести его представителей к числу растений, находящих-
ся на грани вымирания, а также, что этот род является пред-
ставителем древней флоры, сохранившимся в Восточной Аф-
рике со времени не позднее плейстоцена, наиболее благоприят-
ного Климатически для формирования тропической флоры.
47
Кариологическйй анализ многих видов сенполий показал,
что исследованные виды генетически однотипны — диплоидное
число их хромосом равно 30, поэтому тот факт, что в резуль-
тате межвидовых скрещиваний образуются плоды с жизнеспо-
собными семенами, вполне понятен и объясним (рис. 18).
Межвидовые скрещивания сенполий, произведенные Д. Клей-
бергом [Clayberg, 1961], С. Ридом, а также автором этих строг
для пяти видов, которые не были использованы упомянутыми
ным ареал, он был представлен большим числом видов, харак-
теризующихся дихогамией. В силу изменившихся условий, вы-
звавших гибель части видов и изоляцию друг от друга уцелев-
ших, приспособившихся к новым условиям, произошли какие-то
изменения, приведшие к срастанию пыльников, исключившему
перекрестное опыление. У одних видов это привело к появле-
нию облигатной дихогамии, у других — к самоопылению, пол-
ностью или частично заменившему нормальное перекрестное
авторами, почти во всех случаях дали жизнеспособные гибри-
ды. Те исключения, которые не дали гибридов, по-видимому,
следует считать случайными, так как в некоторых из повторен-
опыление.
Наличие суккулентности привело к распространению как
главного вегетативного способа размножения, в то время как
срастание пыльников, обусловившее очень позднее вскрытие их,
почти исключало возможность опыления, причем большая часть
пыльцы погибала вместе с опадающим венчиком и опыление
сделалось явлением чрезвычайно редким. Таким образом, на-
капливаемые растениями генетические возможности оказыва-
лись нереализуемыми.
НЕКОТОРЫЕ СООБРАЖЕНИЯ
О ВЗАИМООТНОШЕНИЯХ ВНУТРИ РОДА
SAINTPAULIA Н. WENDL.
Очевидно, что группу всех сенполий, обладающих наиболь-
?шим количеством примитивных признаков строения (стебель
(с удлиненными междоузлиями, хорошо выраженное супротив-
' но-парное листорасположение, а также минимально редуциро-
ванный андроцей, т. е. .две плодущие тычинки и три стамино-
дия), следует считать наиболее древней и исходной.
Им противостоят виды, обладающие признаками, отражаю-
f щими их приспособительную эволюцию, такими, как укорочен-
ii пый и утолщенный стебель с розеточным листорасположением,
। густой волосистый покров, различная толщина листовых пла-
М’тинок и различная степень редукции элементов андроцея,
j Характер листорасположения, который мы наблюдаем у
тыне живущих видов и по которому они разделяются на две
(Связанные между собою переходами группы, на деле оказы-
вается однотипным, так как при развитии молодых растений
из семян в условиях различного по интенсивности освещения
у сеянцев розеточной группы проявляются при ослабленном
освещении признаки атавистического характера, незаметные
у взрослых растений, — удлинение стебля и супротивно-парное
листорасположение. Это, по нашему мнению, подтверждает тот
факт, что признаки, наблюдаемые у так называемого розеточ-
ного типа, появились позже в результате воздействия условий
окружающей среды.
Рис. 18. Кариотипы некоторых видов сенполий.
1 — S. amaniensis (2п); 2 — S. confusa (п); 3 - S, magun-
gensis (2п)\ 4 — S. orbicularis (л); 5 — S. shumensis (п).
ных позже тех же комбинаций скрещиваний гибриды были по-
лучены [Reed, 1954, 1961, 1962].
Особенностью вегетативных частей сенполий является и раз-
лично выраженная или отсутствующая антоциановая окрашен-1
кость стеблей, цветоносов, черешков, а главное, нижней сторо-1
ны листовых пластинок. Эту особенность, по-видимому, следует
связывать с характером местообитаний отдельных видов — на;
каменистом нагреваемом солнцем субстрате, на обомшелых
камнях, либо в качестве эпифитов на деревьях. Те из них, ко-,
торые приспособлены к жизни на нагреваемых солнцем поверх-
ностях, имеют красноватую окраску, защищающую их от отри-
цательного влияния отраженных от субстрата тепловых лучей.
В ту пору, когда род сенполий был молод (в плейстоцене
или ранее) и занимал значительный по сравнению с современ-
48
Д. М. ЗАЛЕССКИЙ
49
Однако названные нами примитивные признаки друг с дру-
гом не коррелируют. В результате обнаруживаются виды, у ко-
торых сохраняется только какой-либо один из упомянутых при-
митивных признаков, а остальные оказываются исчезнувшими
или явно прогрессивными. Так, например, среди трех видов улу-
гурского субареала р. Saintpaulia два вида обладают удлинен-
ными стеблями и лишь один — укороченным, с соответствую-
щим листорасположением. Среди 14 видов усамбарского суб-
ареала различаются 6 с удлиненными стеблями и 8 с укорочен-
ными, тогда как среди этих же видов обладают слабо редуци-
Улугуру Нгуру Зап. Усамбара Вост. Усамбара’ Теите.Тсимба
О Б Ш, И Е ПРЕДКИ
S.goetzeana
S.incon-
spicua. (?)
S.pusilla.
(?)
S. brevipt-
losa. (3)
Рис. 19.
сенполий.
S.grandifolia,
S.nitida
(2)
S.diplotri-I
cha (3) /
S.velutina
(3)
S. grot ее
S.shumen- /
sis (3) /
S.intermedia
(3)
S.dtfflciUs
(3)
S.orbiculans
(2)
S.pendula.
v.KizaraeiZ}
S. pendula
(3)
S.magun-
gensis
S. ton-
gwensis
S.amanien:
sis(2)
S. confute (i)
S.ionantha A
(2) /\
/ S.ruptcola
S.teitensis (2)
V
Схема предполагаемых родственных отношений внутри рода
Цифры в скобках означают количество стаминодиев.
рованным андроцеем (две тычинки и три стаминодия), соот-
ветственно 3 вида и 5.
Нгурский субареал, представленный двумя видами с укоро-
ченным стеблем, содержит один вид с тремя стаминодиями.
Северный субареал включает два вида с укороченными стебля-
ми, один из которых с двумя стаминодиями.
Руководствуясь упомянутыми примитивными признаками,
мы сделали попытку составить схему родственных взаимоотно-
шений среди известных в наше время видов (рис. 19).
50
Мы исходим из того, что каждая из географически разоб-
щенных групп видов, образующих упомянутые субареалы и
связанных в прошлом какими-то общими предками, представ-
лена лишь небольшой частью видов, приспособившихся к изме-
нившимся условиям и сохранившихся до нашего времени. Меж-
ду собой они непосредственно не связаны.
Таким образом, схема включает связанные в прошлом об-
щими предками линии развития, наиболее примитивными пред-
ставителями которых оказываются два вида: S. goetzeana ив
улугурского субареала и S. grotei из усамбарского. В отдель-
ных субареалах, по-видимому, исходные типы не .сохранились.
Глава 3
ОПИСАНИЕ ОТДЕЛЬНЫХ ВИДОВ
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ
РОЛА SAINTPAULIA Н. WENDL.
(для культивируемых растений *)
1. Стебель развивается только ниже собранного на его верх-
нем конце пучка листьев в виде розетки, обычно очень уко-
рочен и утолщен, лишь у старых растений он несколько удли-;
нен и тогда полегает. !
А. Волосистый покров верхней стороны листовых пласти-j
нок состоит из однотипных относительно ДЛИННЫХ ВО-г
лосков разной густоты (в зависимости от вида), не-
значительно различающихся по длине, более или менее
прямых или согнутых по всей длине (не на концах) !
и прижатых к поверхности пластинки. В промежутках
между волосками неравномерно распределенные по по-
верхности очень короткостебельчатые железки, свой-
ственные всем известным видам.
1. Цветки почти белые с фиолетово-синим пятном в
центре, не более 20 мм в диаметре.
а. Листья эллиптические, почти цельнокрайные;
изящное маленькое растение с тонким стеб-
лем. Плоды около 10 мм длиной. В культуре
не известно .... 8. S. pusilia A. Engl,
аа. Листья округловатые, городчатые по краю,
редко-длинноволосистые (не считая упомяну-
тых стебельчатых железок). Миниатюрное
растение, в природе часто образующее круп-
ные дернины. Стебли около 5 мм в диаметре.
Плоды веретеновидные до 20 мм длиной.
.....................9. S. shumensis В. L. Burtt
ааа. Листовые пластинки почти щитковидные и
почти цельнокрайные, соцветие короче, чем
листья. Волосистый покров листьев составлен
* Сюда включены и виды, отсутствующие в культуре.
S2
из прижатых изогнутых волосков. Плоды ши-
ловидные, до 25 мм длиной ....................
. . . ' . . . . 10. S. teitensis В. L. Burtt
2. Цветки однородной окраски, нормально не менее
25 мм в диаметре. Листовые пластинки у основания
сердцевидно вырезанные, городчатые по краю. Плоды
не более 20 мм длиной.
-J-. Листья темно-зеленые, снизу багряные, плотные,
толстоватые, до 80 мм длиной и 60 мм шириной,
б. Листья округловатые или широкоовальные,
темные, со светлым основанием средней жилки.
Цветки темно-фиолетово-синие или бледно-
красновато-фиолетовые (пурпурные), в этом
случае более или менее прозрачные. Плоды
яйцевидные, около 15 мм длиной . . . . ,
.........................11. S. ionantha Н. Wendl.
бб. Листья эллиптические, более светлой окраски,
с заметно выделяющимися светлыми жилками.
Цветки бледно-лиловато-голубые, но не про-
зрачные. Плоды до 14 мм длиной . . . . ,
12. S. tongwensis В. L. Burtt
++. Листья светло-зеленые, более тонкие, до 100 мм
длиной и 80 мм шириной. Цветки фиолетово-
синие. Плоды веретеновидпые, до 16 мм длиной
...........................13. S. grandifolia В. L. Burtt
Б. Волосистый покров верхней стороны листовых пласти-
нок составлен из относительно длинных прямых или
наклоненных в одном направлении (распростертых)
волосков или только из коротких (за исключением са-
мого края листовой пластинки) ...........................
3. Листья покрыты длинными волосками, в промежут-
ках между ними очень короткие волоски длиной не
более 1/10 длинных. Края листьев часто приподняты.
в. Цветки мелкие, около 20 мм в диаметре, фио-
летово-синие, у отдельных клонов часто со
светлыми краями. Листья тонковатые, изум-
рудно-зеленые, бархатистые, по жилкам более
светлые, снизу красноватые. Жилки слабо вы-
деляются сверху, снизу выступают. Плоды яй-
цевидные, 11—12 мм длиной........................
......................14. S. velutina В. L. Burtt
вв. Цветки крупнее, около 25 мм в диаметре, от-
носительно темно-фиолетово-синие. Листья
сверху темно-зеленые, бархатистые, с ясно вы-
раженной сверху разветвленной сетью жилок,
снизу красноватые. Плоды шиловидные, до
17—20 мм длиной...................................
.................15. S. diplotricha В. L. Burtt
53
4. Листья сверху покрыты либо перемешанными отно-
сительно короткими и более длинными волосками,
среди которых выделяются еще более длинные, снаб-
женные на концах головчатыми железками, либо
только короткими волосками (за исключением края
пластинки).
г. Листья яйцевидные, постепенно заостренные к
концу. Цветки фиолетово-синие. Листовые пла-
стинки покрыты перемешанными волосками
разной длины. Стебли у старых растений ле-
жачие, разветвленные, несущие несколько вер-
шин. Плоды веретеновидные, до 20 мм длиной
........................16. S. rupicola В. L. Burtt
гг. Листья почти округлые, сверху покрыты очень
короткими волосками и только по самому краю
пластинки имеются более длинные волоски
........................17. S. brevipilosa В. L. Burtt
В. Покров из волосков на верхней стороне листовых пла-
стинок всегда составлен из многих коротких, иногда
очень коротких или более длинных прижатых волосков;
длинные волоски (если имеются) в отдельных случаях
распростертые.
5. Волоски на верхней стороне листьев короткие, отно-
сительно редко расположенные, малозаметные, поэто-
му листья выглядят неволосистыми и блестящими.
Листья темно-зеленые сверху, снизу светлые, главные
жилки снизу красновато окрашенные. Цветки темно-
фиолетово-синие (самые темные среди других видов).
Плоды тонкие шиловидные, около 13—15 мм длиной
и 1 —1,5 мм в диаметре . . 18. S. nitida В. L. Burtt
5. Волоски на верхней стороне листовых пластинок раз-
ной длины, прижатые или распростертые. Листья
явно волосистые, относительно светло-зеленые с явно
заметной сверху сетью жилок.
д. Цветки почти белые с фиолетово-синим пят-
ном в основании долей венчика, особенно в
основании двух верхних; иногда цветки фио-
летово-синие (var. purpurea В. L. Burtt).
Листья более или менее округлые, относитель-
но тонкие, с заметной сверху сетью жилок,
светло-зеленые, снизу светлые, с красноваты-
ми жилками, прижатоволосистые. Плоды об-
разуются в большом количестве, тонкие шило-
видные, около 18 мм длиной, часто изогнутые
. . . . 19. S. orbicularis В. L. Burtt, typica
дд. Цветки фиолетово-синие. Листья овальные,
длина их заметно превышает ширину, с ясно
заметной сверху сетью жилок, никогда не
54!
окрашены снизу, сверху сравнительно светло-
зеленые; волоски более или менее прямые или
распростертые. Плоды веретеновидные, около
14 мм длиной, прижатоволосистые . . . .
................................20. S. difficilis В. L. Burtt
ддд. Цветки относительно темно-фиолетово-синие
или пурпурные (красновато-фиолетовые).
Длинные волоски на верхней стороне листо-
вой пластинки прижатые, достигают 3 мм
длины. Листья овальные или более или менее
округлые, с постепенно заостренным верхним
концом, сверху светло-зеленые с очень слабо
заметной сетью жилок, снизу обычно светлые.
Плоды веретеновидные, часто изогнутые, до
18 мм длиной . . 21. S. confusa В. L. Burtt
И. Растения с явно выраженным удлиненным (в той или иной
степени) стеблем, слабо утолщенным, с удлиненными меж-
доузлиями. Листья расположены на стеблях либо супротив-
но-парно, либо спирально (в этом случае растения в моло-
дом возрасте производят впечатление розеточных), или же
листья расположены на концах удлиненных стеблей в виде
концентрического пучка.
Па. Стебли удлиненные только при полном развитии расте-
ний, в молодом возрасте они незаметны или заметны
слабо.
Г. Листья волосистые только по краям пластинки и на
жилках, снизу и в остальных частях без волосков.
Цветки мелкие, около 12,5 мм в диаметре, светлые,
с темным пятном вокруг тычинок. Плоды около 10 мм
длиной. В культуре неизвестно ........................
..........................1. S. inconspicua В. L. Burtt
Д. Покров на верхней стороне листовых пластинок состав-
лен из более или менее однородных по длине волосков,
почти прямых или полуприжатых, расположенных гус-
то или сравнительно редко.
7. Листья по краю в зрелом состоянии городчатые, в мо-
лодом возрасте не всегда явственно; листовые пла-
стинки не менее 40—50 мм длиной. Цветки фиолето-
во-синие.
е. Листья светло-зеленые (несколько более тем-
ные со светлыми жилками у var. kizarae
В. L. Burtt), черешки подкрашены буровато-
красным. Цветоносы дву-, многоцветковые,
сегменты чашечки более узколанцетные. Пло-
ды более тонкие (var. kizarae). (Листья у типа
не окрашены.) Цветоносы двухцветковые.
Плоды яйцевидные, 15 мм длиной и 6—7 мм
в диаметре . . . 2. S. pendula В. L. Burtt
ее. Листья те.мно-серовато-зеленые (сероватый от-
тенок придают густо покрывающие поверх-
ность листа волоски), снизу багряные или
красноватые. Листорасположение не супро-
тивное. Цветоносы многоцветковые. Плоды
удлиненно-яйцевидные, 12 мм длиной и 4 мм
J в диаметре . . . 3. S. intermedia В. A. Engl.
8. Листовая пластинка от 20 до 30 мм длиной с почти
цельным краем; покров верхней стороны листьев из
довольно длинных, более или менее одинаковых по
длине, почти прямых волосков. Цветки белые с лило-
вато-розовым оттенком на двух верхних долях отгиба
венчика. Плоды менее 10 мм длиной, толстоватые.
В культуре встречается очень редко ...................
.........................4. S. goetzeana A. Engl.
116. Стебли явно удлиненные, их удлиненность заметна и у
относительно молодых растений.
Е. Покров на верхней стороне листьев из перемешанных
более длинных и немного более коротких прижатых
или распростертых волосков, в промежутках между
ними имеются значительно более короткие прижатые
волоски.
9. Черешки листьев 70—100 мм длиной, в культуре
иногда длиннее. Листья тонковатые, отчетливо го-
родчатые, цравнительно крупные, зеленые сверху,
бледные снизу. Цветки фиолетово-синие. Плоды ве-
ретеновидные 16—17 мм длиной и 4 мм в диаметре
...............................5. S. grotei A. Engl.
10. Черешки листьев около 50 мм длиной. Листья не-
крупные, городчатые по краю. Цветки фиолетово-
синие (пурпурные).
ж. Листья эллиптические, постепенно заострен-
ные на верхнем конце. Цветки фиолетово-си-
ние или пурпурные (non vidil), доли венчика
по краю железисто-реснитчатые, рыльце сла-
бодвулопастное, стаминодиев 2. Плоды шило-
видные, 12—13 мм длиной и 3 мм в диаметре,
покрывающие их волоски концами торчат
кверху .... 6. S. aminiensis Е. Roberts
жж. Листья округлые с подогнутыми вниз краями,
на концах притупленные. Цветки только фио-
летово-синие, доли венчика по краю не желе-
зисто-реснитчатые, волоски простые, рыльце
расширенное, не двулопастное, стаминодиев 3.
Плоды шиловидные, 18—20 мм длиной и 3 мм
в диаметре; волоски, их покрывающие, кон-
цами достигают поверхности плодов . . .
...................7. S. magungensis Е. Roberts
56
1. SAINTPAULIA 1NC0NSPICUA В. L. Burtt
Notes Roy. Bot. Gard. Edinb., 1958, XXII, 6: 557.
Вид, близкий к Saintpaulia pusilia A. Engler, с удлиненными
стеблями, отличается отсутствием * волосистого покрова па
верхней стороне (исключая край) листовой пластинки, воло-
систостью на нижней стороне (только по жилкам) и мелкими
цветками.
Слабое стебельчатое травянистое растение с прижатоволо-
систым стеблем до 150 мм длиной. Листья с волосистыми че-
решками, варьирующими по длине до 20—35 мм; листовая пла-
стинка более или менее эллиптическая длиной около 40—50 мм
и шириной 20—30 мм, закругленной вершиной или притуплен-
ная, у основания более или менее округленная (и часто нерав-
нобокая). Пластинка сверху гладкая, за исключением редко-
волосистых краев, снизу редковолосисто опушенная (по жил-
кам), бледная, с пятью выступающими снизу боковыми жилка-
ми с каждой стороны.
Соцветие пазушное, 1—6-цветковое, цветоножки короче
листьев или чуть длиннее, редковолос'истые. Прицветники ли-
нейные, волосистые, до 3 мм длиной.
Цветоножки 5—7 мм (у плода 10 мм) длиной; цветки мел-
кие, 12,5 мм в диаметре, белые с голубым в центре. Чашечка
из пяти линейных сегментов длиной 3 мм, с немного отогнутыми
кончиками, полуприжато опушенная. Трубка венчика 2 мм дли-
ной; отгиб из двух частей: верхней, длиной 5,5 мм, тупой и дву-
лопастной, и нижней, длиной 6 мм, притупленной и трехлопаст-
ной. Тычинки 2,5 мм длиной; пыльники 1 мм длиной и шири-
ной. Завязь 2,5 мм длиной, волосистая, со столбиком тоже
2,5 мм длиной, у основания волосистым, кверху гладким. Плоды
(незрелые) около 10 мм длиной, часто слегка изогнутые.
Восточная Африка, Танзания, Горы Улугуру, Морогоро, Ки-
заки, 1370 м, трава 15 см, белые цветки с синим пятном, изредка
на влажной почве в лесу, 18 декабря 1934, Е. М. Bruce 328 (го-
лотип К); северо-западная сторона, 1400 м, девственный лес,
разреженная заросль, цветки белые, голубоватые, 10 декабря
1932, Schlieben 3068/В. В культуре неизвестна.
2. SAINTPAULIA PENDULA В. L. Burtt
Notes Roy. Bot. Gard. Edinb., 1958, XXII, 6: 561.
По мнению Б. Л. Бэртта, растения несколько сходны с ра-
стениями Saintpaulia intermedia, от которых отличаются более
удлиненными лежачими стеблями, менее густо облиственными,
* Исключая разбросанные короткостебельчатые железки, имеющиеся у
большинства видов.
57
светло-зеленой окраской листьев, покрытых сверху почти пря-
мыми волосками, и обычно одноцветковыми цветоносами с бо-
лее крупными плодами (рис. 20). Нельзя не согласиться с мне-
нием о наличии у этих видов некоторых общих черт в строении
их отдельных частей, однако отличий оказывается гораздо
больше, так что черты сходства теряют свое значение.
Распростертое ползучее растение с удлиненным, явно воло-
систым разветвляющимся стеблем длиною около 120—150 мм,
укореняющимся при соприкосновении с землей и испещренным
Рис. 20. Saintpaulia pendula В. L. Burtt.
продольными светлыми черточками (ср. с S. grotei). Междо-
узлия около 30 мм, листорасположение супротивно-парное, че-
редующееся.
Листья широкоовальные или почти округлые, по краю город-
чатые, 33—46 мм длиной и 30—45 мм шириной, на вершине
притупленные, у основания почти сердцевидные. Черешки
листьев около 20—50 мм длиной. Верхняя сторона листовой
пластинки светло-зеленая, довольно густо * (500 на 1 см2) по-
крыта почти прямыми, длинными (1—2 мм) волосками, ниж-
няя сторона беловато-зеленая, покрыта волосками более ко-
роткими, до 1 мм длины.
* Во всех случаях подсчет количества волосков производился визуально
на вырезанном квадратном участке листа через бинокулярную лупу.
58
Цветоносы около 5 см длиной, волосистые, большей частью
одноцветковые, иногда двухцветковые, наклоненные. Цветонож-
ки около 2 см длиной, при бутонах и плодах часто спирально
изогнутые. У основания цветоножки расположено по два узко-
ланцетных прицветника до 3 мм длиной.
Чашечка от основания рассеченная, из пяти ланцетных
сегментов 3 мм длиной, коротковолосистая. Трубка венчика
2 мм длиной, отгиб 28—30 мм в диаметре, верхняя его часть
до половины рассеченная, 7—10 мм длиной и шириной, боковые
лопасти нижней части 8—12 мм длиной и 9—13 мм шириной,
средняя 8—10 мм длиной и 10—12 мм шириной, железисто-рес-
нитчатые по краю. Окраска венчика относительно темно-фио-
летово-синяя, как у S. confusa. Тычиночные нити 4 мм длиной,
гладкие, пыльники 2,5 мм длиной и 3,5 мм шириной. Стамино-
диев два. Цветоносы, прицветники, черешки листьев и чаше-
листики в той или иной степени красновато окрашенные.
Завязь коническая, длиной 2 мм, густо-длинноволосистая,
столбик 7 мм длиной, гладкий, пурпурный. Плоды яйцевидные,
густо-длинноволосистые, длиной 16 мм и шириной 6,5 мм в
основании, до 2 мм на его верхнем конце.
Восточная Африка, Танзания, Восточные Усамбарские горы,
гора Мтаи, на скале, сборы В. Р. Пунтера, образец U, культи-
вируется в Эдинбургском ботаническом саду, С. 1686 (голо-
тип Е).
Культивируется в Ботаническом саду Ленинградского уни-
верситета с 1959 г., куда поступил в виде листовых черенков
из Эдинбургского ботанического сада с индексом «Punter U».
2а. SAINTPAULIA PENDULA var. KIZARAE В. L. Burtt
Notes Roy. Bot. Gard. Edinb., 1964, XXV, 3; repr. Gesn.
Sain. News, 1964, Nov. — Dec.: 46—47.
В 1964 г. Б. Л. Бэртт опубликовал краткое описание выде-
ленной им разновидности,* названной по месту ее обитания
S. р. var. kizarae. Она отличается от типа количеством (от двух
и более) цветков, образующихся на цветоносах, а также более
узкими сегментами чашечки, более тонкими и темными листья-
ми с более светлыми жилками и более узкими плодами.
Восточная Африка, Танзания, северо-восточная Усамбара,
Кизара, сборы Пунтера, 1958, № 59/3589, культивируется в
Эдинбургском ботаническом саду, С. 3066 (голотип Е); там же,
* Эту разновидность, по нашему мнению, по совокупности признаков и
общему облику растения вполне возможно рассматривать как особый вид,
который мог бы быть в этом случае назван Saintpaulia kizarae (В. L. Burtt)
D. Zalessky.
59
сборы Пунтера, 1959, «Kizara II», № 59/4354, культивируется
в Эдинбургском ботаническом саду, С. 3725.
Культивируется в Ботаническом саду Ленинградского уни-
верситета, куда получена в виде листовых черенков в 1966 г.
из Эдинбургского ботанического сада.
3. SAINTPAULIA INTERMEDIA В. L. Burtt
Notes Roy. Bot. Gard. Edinb., 195,8, XXII, 6: 562.
Растения с удлиненными, ползучими, укореняющимися стеб-
лями со сближенными листьями и небольшими, редко дости-
гающими 20 мм длины междоузлиями (рис. 21). Листья темно-
Рис. 21. Saintpaulia intermedia В. L. Burtt.
зеленые, из-за характера волосистого покрова выглядят серо-
ватыми, снизу красновато-пурпурные. Черешки листьев 35—
50 мм и до 80—90 мм длиной, явноволосистые, окрашенные.
Листорасположение перекрестно-парное. Листовые пластинки
обратно округло-овальные, овальные или овально-округлые,
35—55 мм длиной и 32—47 мм шириной, на вершинах округ-1
ленные, у оснований округлые или слегка сердцевидные с го-
родчато-зубчатыми краями, сверху покрытые густоватыми по-
луприжатыми (около 150 на 1 см2) волосками 1—2 мм длиной,
направленными от середины к краям пластинок; снизу воло-
еистый покров состоит из более коротких волосков, не превы-
шающих в длину 1 мм.
Цветоносы многоцветковые, большей частью по 4 цветка,
волосистые, от 40 до 60 мм длиной, наклоненные. Прицветники
туполанцетные, от 3 до 5 мм длиной и 1 мм шириной. Цвето-
ножки от 20 до 30 мм длиной, как и прицветники, волосистые.
Чашечки, от основания разделенные на ланцетные сегменты,
4 мм длиной и до 1,5 мм шириной, волосистые. Трубки венчи-
ков до 2,5 мм длиной, отгиб около 25 мм в диаметре; лопасти
верхней части 6 мм длиной и 5 мм шириной, боковые лопасти
нижней части 7 мм длиной и 9 мм шириной и средняя 9 мм
длиной и 10 мм шириной, по краям железисто-реснитчатые,
светло-фиолетово-синие. Тычиночные нити 4 мм длиной, глад-
кие; пыльники 2,5 мм длиной и 3,5 мм шириной. Стаминодиев
два.
Завязь коническая, до 4 мм длиной и 2 мм в диаметре, во-
лосистая, столбик 7 мм длиной, гладкий, пурпурный, рыльце
слегка расширенное (рис. 12).
Плоды несколько удлиненно-яйцевидные, асимметричные,
12 мм длиной и 4 мм в диаметре.
Восточная Африка, Танзания, Восточные Усамбарские горы,
Кигонгои, на скалах, сбор В. Р. Пунтера, образец V, культиви-
руется в Эдинбургском ботаническом саду, С. 2007 (голотип Е);
зам же, образец W, С. 1580 (голотип Е).
Культивируется в Ботаническом саду Ленинградского уни-
верситета, куда получен в 1959 г. из Эдинбурга в виде листо-
вых черенков с индексом «Punter V».
4. SAINTPAULIA GOETZEANA A. Engler
Engl. Jahrb., 1901, XXVIII: 481, t VI.
Травянистое многолетнее, довольно густо-беловолосистое
растение, разветвленное от основания, с ползучим, удлиненным
до 120—150 мм стеблем, прирастающим к субстрату и обра-
зующим восходящие побеги, несущие несколько пар почти су-
противных листьев. Листья расположены на узлах, междоузлия
в нижней и средней частях побегов равны 15—20 мм, в верхней
части короче (рис. 22).
Листовые пластинки мясистые, почти округлые или коротко-
овальные, с обоих концов притупленные, цельнокрайные, сверху
темно-зеленые, снизу пурпурные, 20—30 мм длины и почти та-
кой же ширины, беловолосистые. Листорасположение перекре-
стно-парное.
Волосистый покров из более или менее однородных волосков
1,5—1,7 мм длиной, около 500 на 1 см2 с верхней стороны,
с нижней волоски короче и реже. Черешки менее волосистые,
равны пластинке или в 1,5—2 раза длиннее ее. Жилки снизу
' 61
60
слабо выступают, сверху не выступают и почти незаметны.*
Цветоносы тонкие, длиной 30—35 мм, большей частью двуцвет-
ковые. Прицветники острые ланцетные, по одному у основании
каждой цветоножки; цветоножки 6—8 мм длины.
Доли пятилопастной чашечки — 3,2 мм. Венчик из двух со-
единенных трубкой частей: верхней лиловатой, овальной, глубо-
кодвулопастной, в полтора раза короче (до 0,67 мм и длиннее)
и уже, чем три белые доли нижней части (non vidi!). Доли
Рис. 22. Saintpaulia goetzeana A. Engier.
нижней части неодинаковые: боковые 7—8 мм длиной и 7 мм
шириной и средняя 9—10 мм длиной и 7—8 мм шириной. Тычи-
нок две длиною 3,2 мм, с широкими сердцевидными выступаю-
щими пыльниками шириною 2 мм. Стаминодиев три, маленькие.
Завязь короткоконусовидная 1,2 мм длиной, с кольцевидным
нектарным диском у основания, оттянута в косой столбик дли
ной 2—2,5 мм. Рыльце очень с^абодвулопастное. Плоды яйце-
видные (non vidi!), менее 10 мм длиной. В культуре очень ред-
кий вид.
* В пазухах живых листьев образуются боковые вегетативные побеги.
Влага осаждается капельками на волосках листьев и других частей растения.
62
Восточная Африка, Танзания. Горы Улугуру, в соседстве
с S. pusilla, центральная часть Улугурских гор, в горном лесу
южнее плато Луквангуле на высоте 1300 и 2000 м, на скалах,
покрытых мхом, 1898, Goetze, 245 (В).
Лупанга, 1800—1950 м, ползучая трава на поверхности
обомшелых скал во влажном лесу, цветки голубые с белым,
выше отсутствуют, 23 декабря 1933 г., Michelmore, 861 (К).
Лупанга, 1950 м, скалы в лесу, мясистые декоративные
листья, красные с изнанки, изредка, 7 апреля 1935 г., Е. М. Bru-
ce (К).
Край плато Луквангуле, 2000 м, на скалах — в густой тени
в лесу, на склоне холма — во влажном нагорном лесу, ползу-
чее и укореняющееся в узлах растение, венчик белый с незна-
чительным розовато-лиловым оттенком на двух верхних долях,
17 марта 1953 г., Drumond et Hemsley, 1656 (К, Е).
Северо-западный склон Паратапасс, 1910 м, трава на дере-
ве, одиночно, цветки белые с лилово-голубым, 18 октября
1932 г., Schlieben, 2829 (BR).
5. SAINTPAULIA GROTEI A. Engler
Bot. Jahrb., 1921, Ivii: 202—203; Cox H., Roberts E.
Afr. Viol. Mag., 1950, iii, 4: 5; Moore H. E., Afr. Viol,
etc., 1957: 151.
Травянистое многолетнее растение с относительно толстым
и мясистым стеблем до 50 мм толщиной и до 100—150 мм дли-
ной, с междоузлиями 10—15 мм длиной на концах стеблей и до
30—40 мм у их основания (рис. 23). Стебель ползучий, укоре-
няющийся в местах соприкосновения его с субстратом, покры-
тый, как и все вегетативные части растения, сильно прижатыми
волосками длиной до 1,5 мм, все волоски многоклеточные, бе-
ловатые или на черешках, а также снизу на жилках листьев
светло-буроватые. Стебель зеленый или красноватый, на нем
редкие желтоватые или желтовато-зеленые продольные черточ-
ки длиной от 1 до 1,5 мм и бугорки — зачатки придаточных
корней. Листорасположение перекрестно-парное. Листовая пла-
стинка слегка толстоватая, 0,6—1 мм толщиной, овальная,
у основания слабосердцевидная, 40—60 мм длиной и 30—50 мм
шириной, светло-зеленая, иногда пурпурная (non vidi!), с ниж-
ней стороны светлее, короткозубчатая по краю. Черешки 40—
75 мм длиной, в отдельных случаях более длинные. Густота
покрывающих листья длинных прижатых волосков около 250
па 1 см2, длина около 1,5 мм, густота коротких волосков около
500 на 1 см2, длина 0,3—0,4 мм.
Цветоносы тонкие, 1—2-цветковые (или более), длиной 50—
60 мм (иногда более), возникают в пазухах верхних листьев.
Цветоножки короткие, от 5 до 10 мм длиной, очень коротко-
63
волосистые. Прицветники линейные, 2—3 мм длиной, до 1 мм
шириной.
Ланцетные доли чашечки рассечены почти до основания.
Трубка венчика около 3 мм длиной. Отгиб венчика из двух ча-
стей: верхней двулопастной, 15 мм длиной и 6—7 мм шириной
и нижней, рассеченной на три доли овальной формы с тупова-
тыми концами: средняя 5—6 мм, боковые — 6—7 мм шириной
и около 20 мм длиной. Венчик по краю не железисто-реснитча-
тый, реснички простые, однако автор вида Энглер в своем ори-
гинальном описании говорит, что края долей венчика слегка
железисто-волосистые. Венчик имеет светловатую фиолетово-
синюю окраску, такую же, как у S. grandifolia, или близкую
к ней.
Рис. 23. Saintpaulia grotei A. Engler.
Тычиночные нити 3—3,5 мм длиной, изогнутые, пыльники
желтые, 2—2,5 мм длиной и 2,5—3 мм шириной. Стаминодиев
три. Завязь удлиненная коническая, 6 мм длиной, густоволо-
систая, столбик 19 мм длиной, розоватый, рыльце двулопастное
или неясно трехлопастное.
Плоды веретеновидные, слабоасимметричные, при рассмот-
рении сбоку с явно заметными светловатыми «швами» створок.
Они покрыты относительно длинными прижатыми волосками,
более короткими, чем волоски на цветоносах и чашечках.
Сквозь поверхность неподсохших коробочек просвечивают се-
мена. Длина плодов 16—17 мм, диаметр в наиболее широкой
нижней трети 4 мм и на верхнем конце 2 мм.
По зубчатым овальным листьям на длинных черешках и от-
носительно длинным стеблям со сравнительно длинными междо-
64
алиями этот вид выделяется среди других видов, имеющих
развитые стебли. Тип хранится в гербарии в Кью.
Восточная Африка, Танзания. Восточная часть Усамбарских
юр близ Амани, Grote, 3708. В плохой сохранности. Амани, на
скалах близ водопада и культурно, 2 февраля 1939 г. Moreau,
1>(К). Г. Млинга, 1080 м, многолетняя ползучая трава со сплошь
пурпурными листьями и бледно-лиловыми цветками, август
1950 г., Р. D. Greenway, АН 10022. Естественное местообитание
уничтожено плантациями сизаля. Культивируется в Эдинбург-
।ком ботаническом саду.
Культивируется в Ботаническом саду Ленинградского уни-
нгрситета, куда был получен в 1959 г. из Эдинбурга в виде ли-
। |овых черенков с индексом «Barker Е».
6. SAINTPAULIA AMANIENSIS Е. Roberts
Afr. Viol. Mag., 1950, 4, 2: 7.
Стебель лежачий, ветвящийся, укореняющийся в местах со-
прикосновения с субстратом (в культуре часто ниспадающий),
•Медно-зеленый в более старых частях, красноватый в более
молодых, до 100 мм длиной, 4—5 мм в диаметре, с междоузлия-
ми 2—15 мм длиной (рис. 24).
Вегетативные части растения покрыты шелковистым опуше-
нием из длинных, и коротких, перемешанных между собою во-
нп'ков; короткие волоски прижатые, длинные распростертые
(кроме листьев) и в 3—6 раз длиннее коротких.
Листья зеленые сверху, бледно-зеленоватые снизу, овальные
к» овально-эллиптических, 25—55 мм длиной и 20—35 мм ши-
риной, с заостренным или почти заостренным кончиком, не-
>колько сердцевидным основанием и городчато-зубчатым ' или
>убчатым, заметно подвернутым книзу краем. Средняя и боко-
нЫе жилки снизу выступают, сверху вдавленные, несколько бо-
НЧ' бледные, чем зеленая поверхность листа. Первичные боко-
вые жилки обычно по 4, иногда 3 с каждой стороны от средней.
Волосистый покров верхней стороны листьев составлен из при-
битых волосков, более длинных, 1—1,5 мм длиной (около 120
ни I см2) и коротких — вполовину (или менее) короче длинных
(около 600 на 1 см2). С нижней стороны волосистый покров
примерно в два раза реже, по жилкам несколько гуще. Череш-
• II 40—90 мм длиной, 20—30 мм в диаметре, почти цилиндри-
ческие, суживающиеся кверху, красновато окрашенные, при-
поднимающиеся.
Соцветие двухцветковое, иногда одноцветковое, цветоносы
•II 40 мм длиной, поднимающиеся от верхней листовой пазухи
•• протягивающиеся почти касательно листьям, прицветники
•Инейные, острые, 1—2 мм длиной, около 0,3 мм шириной, цве-
Ч Л, М. ЗАЛЕССКИЙ Q5
тоножки 1,5—2 мм длиной, прямые во время цветения и обычнс
изогнутые после него.
Цветки около 23 мм в диаметре (когда совершенно откры
тые), чашелистиков 5, темновато-пурпурных, ланцетных, 2,5—
3 мм длиной. ~
часть темнее, с
долей, густо
Отгиб венчика фиолетово-синий, центральна;
редко опушенной верхней поверхностью и краям!
покрытыми мелкими железистыми волосками
Рис. 24. Saintpaulia amaniensis Е. Roberts.
Верхняя двулопастная часть отгиба венчика овальная, 7—10 mi
длиной и 4—5 мм шириной, внутренние края ее долей налегаю'
друг на друга примерно на 4 мм в длину, боковые доли оваль
ные, 10—12 мм длиной и 7—10 мм шириной, нижняя широко
овальная, 12—15 мм длиной и 8—10 мм шириной. Тычинок две
в устье трубки венчика с боков нижней его части. Тычиночньк
нити 2,5—3 мм длиной, пыльники сросшиеся, дельтовидно-серд
невидные, ярко-желтые, около 1,5—2 мм длиной и 2 мм шири’
ной у основания, сросшиеся пыльцевые мешки расходятся I1
вскрываются в виде широкой базальной щели. Стаминодие!
два, сосочковидных, расположенных на трубке венчика, редко
более 0,1 мм длиной.
66
Завязь во время цветения 15 мм длиной, яйцевидно-кониче-
ская, прижатоволосистая, бледно-зеленая, постепенно заострен-
ная в тонкий бледно-фиолетово-синий столбик длиной 5—7 мм,
в нижней трети коротко опушенный; рыльце слабодвулопаст-
ное; завязь у основания окружена узким красноватым нектар-
ным диском.
Плоды шиловидно-веретеновидные 12—13 мм длиной и 3 мм
диаметром в наиболее широкой нижней части и 2,5 мм в верх-
ней трети, покрыты прижатыми волосками, не касающимися
поверхности коробочки.
Вид очень похожий на S. magun.gen.sis, также из Усамбар-
ских гор, от которого отличается очертанием листьев и воло-
систым покровом, двумя стаминодиями (вместо трех) и желе-
зистыми волосками по краю венчика, а также Соцветием, имею-
щим меньшее число цветков (1—2). Многие общие черты меж-
ду S. magungensis и S. amaniensis послужили основанием для
Б. Л. Бэртта рассматривать эти виды как разновидности одного
[Burtt, 1964]. Так как наряду со сходством существуют значи-
тельные различия, мы предпочитаем рассматривать эти виды
как близкие, но различные.
Тип в гербарии колледжа штата Мичиган, США, Е. Р. Ro-
berts 2, из растений, культивируемых из материала, собранного
в предгорьях Усамбарских гор, близ Амани, Танзания, Восточ-
ная Африка.
Восточная Африка, Танзания. Восточные Усамбарские горы,
близ Амани. Культивируется в Эдинбургском ботаническом
саду, исходный образец Е. Р. Roberts, С. 1568 (Е).
Культивируется в Ботаническом саду Ленинградского уни-
верситета, куда был получен в виде листовых черенков из Эдин-
бурга с индексом «Michigan».
7. SAINTPAULIA MAGUNGENSIS Е. Roberts
Afr. Viol. Mag., 1950, 3, 1: 6—7.
Стебель ветвящийся, ползучий, укореняющийся при сопри-
косновении с субстратом (в культуре часто ниспадающий),
бледно-зеленоватый, в более молодом возрасте на концах побе-
гов красноватый, с развитой темноватой пробкой наружного
слоя в старости, 60—140 мм длиной, 30—70 мм в диаметре,
с междоузлиями 3—30 мм длиной (рис. 25).
Вегетативные части растения покрыты шелковистыми длин-
ными и короткими, перемешанными между собою волосками;
короткие волоски прижатые, длинные распростертые везде, кро-
ме листьев. Длинные волоски в 5—8 раз превышают по длит?
короткие. Листья располагаются попарно, но в последующих
парах смещаются.
5’ (•?
Листья сверху зеленые, снизу бледно-зеленоватые, округлые
’или очень близкие к округлым, 30—70 мм длиной и 30—60 мм
шириной, с тупым кончиком и сердцевидным основанием, город-
чатым и слабо подвернутым краем, средняя жилка снизу вы-
ступает, сверху вогнутая, более или менее светлая по сравне-
нию с зеленой окраской всего листа, первичных боковых жилок
обычно по четыре с каждой стороны от средней, они вогнутые
сверху, выступающие снизу. Волосистый покров листьев до-
Рис. 25. Saintpaulia magungensis Е. Roberts.
Вольно густой из прижатых коротких волосков 0,2—0,3 мм дли-
ной (около 600 на 1 см2) и длинных 1,5—2 мм длиной (около
120 на 1 см2), их концы приподняты. На нижней стороне листа
Волосистый покров реже примерно в два раза, по жилкам не-
сколько гуще. Черешки от .30 до 100 мм длиной и 2—4 мм в
диаметре, красноватые, располагаются параллельно поверхно-
сти почвы.
Соцветие 2—4-цветковое, цветоносы 40—50 мм длиной, на-
клоненные, вырастают из верхней листовой пазухи и превы-
шают по длине листья. Прицветники линейные, острые, 2—
2,5 мм длиной и около 0,5 мм шириной. Цветоножки 15—18 мм
длиной, прямоватые во время цветения, обычно изогнутые после
Цветения.
€8
Цветки около 22 мм в диаметре, когда полностью открыты.
Чашелистиков пять, фиолетово-синие, около 4 мм длиной. Вен-
чик фиолетово-синий, центральная часть темнее. Наружная по-
верхность венчика редко опушенная, а края долей густо-мелко-
реснитчатые, реснички мелкие без железок.
Верхняя двойная часть отгиба венчика овальная, 7—8 мм
длиной и 6 мм шириной, внутренние края ее накрывают друг
друга примерно на 2 мм, боковые доли широкоовальные, 10—
11 мм длиной и 8—9 мм шириной, нижняя доля широкооваль-
ная 11 —13 мм длиной и 8—10 мм шириной. Тычинок две, при-
росших к устью трубки венчика с боков нижней лопасти вен-
чика, тычиночная нить около 4 мм длиной, пыльники сросшие-
ся, дельтовидно-сердцевидные, ярко-желтые, около 2 мм длиной
и 3 мм шириной у основания, сросшиеся пыльцевые мешки рас-
ходятся и раскрываются с помощью короткой базальной щели.
Стаминодиев три, прикрепленных к трубке венчика, редко бо-
лее чем 0,25 мм длиной.
Завязь во время цветения 1,5 мм длиной, яйцевидно-кониче-
ская, волосистая, бледно-зеленая, постепенно суженная в тон-
кий светло-фиолетово-синий столбик 5—7 мм длиной; рыльце
неясно полушаровидное, нисколько не двулопастное: завязь
окружена у основания узким красноватым нектарным диском.
Неподсохшие зрелые плоды — удлиненные шиловидно-вере-
теновидпые, около 20 мм длиной, 3 мм шириной в наиболее
широкой нижней части и 2 мм в последней трети длины от осно-
вания, покрыты прижатыми волосками, концы которых примы-
кают к поверхности коробочки. При созревании семян коробоч-
ка подсыхает и несколько сокращается в размерах.
Этот вид сходен с видом S. grotei (также с Усамбарских
гор), но отличается более мелкими округлыми листьями, скорее
городчатым, чем городчато-зубчатым, краем у листьев, волос-
ками без железок по краю венчика, нелопастным рыльцем и
более мелкими цветками.
Тип в гербарии колледжа штата Мичиган, США, Е. Р. Ro-
berts 1, из образцов, растущих из материала, собранного в Ма-
гунга, в предгорьях Усамбарских гор.
Восточная Африка, Танзания. Восточные Усамбарские горы,
Магунга; культивируется в Мичиганском колледже (голотип).
Г. Млинга, многолетняя трава с ползучим стеблем и зелеными
листьями, цветки темно-синие, растет вместе с S. grotei, 17 ав-
густа 1950 г., Р. D. Greenway, АН 10023 (ЕА); культивируется
в ботаническом саду в Кью, введен П. Д. Тринвеем, цвел в
июне 1950 г., обр. 298 : 48, № 1. Река Квамкую; культивируется
также в Кью, введен П. Д. Тринвеем, цвел в июне 1950 г., обр.
298: 48, № 2. Естественное местообитание этого вида в настоя-
щее время разрушено плантациями сизаля. Культивируется в
Эдинбургском ботаническом саду, введен Робертсом, обр.
С. 1569 (Е) .
69
Культивируется в Ботаническом саду Ленинградского уни-
верситета, куда получен в виде листовых черенков из Эдин-
бурга с индексом «Michigan».
8. SAINTPAULIA PUSILLA A. Engler
Engl. Bot. Jahrb., 1901, XXVIII: 481—482; Clarke.
Oliver, Fl. Trop. Afr„ 1906, IV: 501.
Небольшое розеточное травянистое растение, по облику
сходное с 3. ionantha, но отличающееся от него меньшими раз-
мерами всех частей и многочисленными цветками, верхняя
часть венчика которых окрашена в голубоватый цвет, а ниж-
няя— белая (рис. 26). Растение толстоватое, все части редко-
волосистые.
Рис. 26. Saintpaulia pusilia A. Engler.
Листовая пластинка короткоовальная, с обоих концов при*
тупленная, цельнокрайная, 25—35 мм длиной и 17—25 мм ши-
риной, пурпурная снизу. Черешки равны по длине пластинке,
несколько короче или иногда длиннее, 15—35 мм длиной.
Цветоносы тонкие восходящие, одинаковой длины с череш-
ками или несколько длиннее (но не длиннее 30 мм). Соцветие
немногоцветковое, от 1 до 3 цветков. Соцветий обычно много.
Цветоножки тонкие, почти равные по длине цветкам, 5—7 мм
длиной, прицветники маленькие ланцетные.
70
Чашелистики удлиненно-треугольные, у основания срос-
шиеся, 2,5—3 мм длиной. Венчик редко-тонковолосистый, около
8,5 мм диаметром; трубка венчика короткая, до 3 мм. Две доли
верхней части отгиба голубые, более овальные, чем доли ниж-
ней, и в полтора раза короче их, около 3 мм длиной, три доли
нижней части венчика белые, около 4 мм длиной.
Тычинок две по бокам завязи, прикрепленных к основанию
двух боковых долей нижней части отгиба; тычиночные нити,
почти равные трубке венчика, изогнутые; пыльники яйцевидные
с коротко растрескивающимися швами с внутренней стороны;
стаминодии крупные. Автор описания Энглер не называет их
числа, а па его рисунках, иллюстрирующих текст, они не изо-
бражены.
Завязь около 1 мм длиной, овальная, у основания окружен-
ная кольцеобразным нектарным диском и переходящая в стол-
бик почти вдвое большей длины, заканчивающийся обратно-
яйцевидным рыльцем, достигающим 2 мм ширины.
Плоды удлиненные, около 10 мм длиной и 2 мм в диаметре,
па конце крючковидно оттянутые.
. В культуру вид пока не введен.
Восточная Африка, Танзания, центральная часть гор Улу-
гуру. На юго-западном склоне плато Луквангуле, на высоте
1200—1800 м, в девственном лесу, па скалах между мхом,
ноябрь 1898 г. 205 (голотип).
Северо-западный склон пика Бондуа, туманный лес, 1650 м,
одиночная трава па мокрых скалах между мхом и папоротни-
ками, цветки белые с лиловым пятном, 24 ноября 1933 г., Schi-
lieben, ЗОЮ (ВМ, BP, М). Пик Бондуа, близ восточного скло-
на, 1680 м, на скалистой поверхности между нежными папорот-
никами под нависающими склонами, в месте, не доступном пря-
мому дождю, цветки белые, исключая две маленькие верхние
доли, которые лиловаты. Семенная коробочка удлиненная,
листья багряные снизу, январь 1953 г., W. J. Eggeling 6443 (К).
Там же, ноябрь 1953 г., W. J. Eggeling, 6712(К). Там же, лес,
1680 м, 25 ноября 1932, G. В. Wallace, 492В (голотип К сме-
шан с S. goetzeana).
9. SAINTPAULIA SHUMENSIS В. L. Burtt
Notes Roy. Bot. Gard. Edinb., 1955, XXI, 5: 238;
Moore II. E„ Afr. Viol, etc., 1957: 153.
Многолетнее миниатюрное растение с небольшими округло-
ватыми листьями, расположенными розеткой, около 12 см диа-
метром, па укороченном стебле. Листья сверху покрытьгпрямо-
.ватыми редкими, довольно длинными волосками, между кото-
рыми встречаются мельчайшие густые трудноразличимые во-
71
доски. Цветки беловатые, многочисленные, пыльники часто сво
бедные, столбик светлый, рыльце темноватое (рис. 27).
Компактное растение, стебель укороченный, около 1—2 сг.
длиной, незначительно утолщенный.
Листовые пластинки тонковатые, широкоовальные или почт,
округлые, 25—35 мм длиной и 29—30 мм шириной, у основанш
почти ушасто-сердцевидные, по краям городчато-зубчатые, вер
хушечный зубчик всегда подвернут вниз (у молодых листьер
Рис. 27. Saintpaulia shumensis В. L. Burtt.
края пильчато-зубчатые, у зрелых — городчатые и подвернуты
вниз). Верхняя сторона светлая, от желто-зеленого до относи-
тельно темного оливково-зеленого цвета, покрыта редкими (око-
ло 25 на 1 см2) длинными (до 3 мм) волосками, прямыми или
от основания наклонными по направлению от середины к краям.
Промежутки между длинными редкими волосками густо покры-
ты мельчайшими волосками-железками, свойственными другим
видам, длина которых менее 1/10 длины длинных волосков.
Нижняя сторона пластинок бледная, иногда чуть розоватая
вдоль розоватых слабо выступающих средней и боковых жилок,
покрыта по жилкам довольно длинными прижатыми волосками,
между жилок — очень короткими наклонными волосками около
0,3 мм длиной.
Черешки красноватые, редковолосистые, по длине равные
пластинкам либо несколько длиннее или короче их (20—40 мм),
толщиной около 1,5—2 мм.
Цветоносы восходящие, 1—2-цветковые, иногда 5-цветковые,
около 50—60 мм длиной, как и черешки, красноватые и покры-
72
тые волосками разной длины. Прицветники линейные зеленые,
1,5—5 мм длиной, около 0,3—0,5 мм шириной, тупые, волосис-
тые. Цветоножки 15—20 мм длиной, волосистые, при плодах
крючкообразно изогнутые.
Чашечка глубоко рассеченная, из пяти долей, чашелистики
5 мм длиной, у основания 1 мм шириной, верхушки их притуп-
ленные, 0,5 мм шириной, волосистые. Трубка венчика 3 мм дли-
ной; отгиб почти под прямым углом к трубке, очень бледный,
от середины к периферии голубовато-белый. Верхняя часть от-
гиба несколько глубже половины длины, двулопастная, от се-
редины к основанию сине-фиолетовая, 9—10 мм длиной и 14—
15 мм шириной. Нижняя часть глубокотрехлопастная, в общем
очертании почти округлая, 15 мм длиной и 27 мм шириной.
Лопасти нижней части удлиненно-округленные, средняя 9 мм
длиной и 7 мм шириной, боковые 10 мм длиной и 8 мм шири-
ной, в основании белые, слегка подкрашенные сине-фиолето-
вым.
Тычиночные нити 3 мм длиной, пыльники почти округлые,
1 мм длиной, прикрепленные к нитям спинками, не сросшиеся
или едва сросшиеся. Стаминодиев три, в виде одинаковых не-
больших сосочков.
Завязь коническая с волнистым оранжевым' нектарным дис-
ком 3 мм длиной, прижато-волосистая (волоски по направле-
нию к верхушке короче нижних), зеленая, переходит в равный
по длине гладкий столбик, очень бледно-лиловоокрашенный,
рыльце явно двулопастное, интенсивно фиолетовое.
Плоды асимметричные, в той или иной степени крючковидно
изогнутые, шиловидные, веретеновидные или веретеновидно-
шиловидные, 15 мм длиной и 3 мм шириной, красноватые ог
красноватых небольших черточек вдоль плодов, коротковоло-
систые. Б. Л. Бэртт приводит сообщение В. М. Моро, что расте-
ние обтает в более прохладных лесах, чем большинство других
видов, и обладает чувствительными плодами, не сообщая, как
чувствительность проявляется.
Восточная Африка, Танзания, Западные Усамбарские горы.
Обитает в более прохладных лесах, чем большинство других
видов, W. М. Moreau, Шуме, Ворлдс Вью, 1900—1950 м, 8 мар-
ча 1947 г., Greenway, 7934 (ЕА, К), культивируется в Ботани-
ческом саду в Эдинбурге от этого номера (голотип К, Е).
W. М. Moreau, Аманский гербарий, № 9840 (ЕА). 4 июня
1953 г., Drummond Hemsley, 2852А (К, Е). Маноло до Магам-
ба, 1750 м, 31 мая 1914 г., Peter 0 I 126 (В). Младо, вечнозеле-
ный дождевой лес, 1400—1600 м, 7 октября 1902 г., Engler,
1397 (В).
По сообщению Б. Л. Бэртта, в горах Нгуру в 1933 г. Шли-
беном был собран образец, близкий к этому виду (вероятно,
S. brevipilosa — предположение автора). Культивируется в
Эдинбургском ботаническом саду.
73
Культивировался в Ботаническом саду Ленинградского уни-
верситета, куда был получен из Эдинбургского ботанического
сада в 1959 г. в виде листовых черенков с индексом «Barker N».
10. SAINTPAULIA TEITENSIS В. L Burtt
Notes Roy. Bot. Gard. Edinb., 1958, XXII, 6: 559.
Растения с укороченным стеблем, длинночерешчатыми ли-
стьями и почти щитковидной листовой пластинкой, с более ко-
роткими, чем листья, соцветиями и удлиненными плодами (упо-
мянутая щитковидность оснований листовых пластинок отно-
сится только к старым листьям, у молодых они сердцевидные)
(рис. 28).
Рис. 28. Saintpaulia teitensis В. L. Burtt.
Стебель неразветвленный, до 100 мм в длину и до 10 мм
в толщину, на вершине в виде плотной розетки расположены
листья. Все вегетативные части растений густо покрыты отно-
сительно длинными прижатыми волосками.
Листовые пластинки широкоэллиптические, широкоовальные
или почти округлые, часто ложкообразные вогнутые, сверху
изумрудно-зеленые, 55—75 мм длиной и 45—55 мм шириной,
на вершине широко заостренные или, реже, туповатые, у осно-
вания— от сердцевидных до почти щитковидных, выемчатых
с неясно городчатым краем, снизу красновато-пурпурные. Сред-
няя и боковые (по 4—5 с каждой стороны) жилки снизу (лишь
в основании) выступающие, сверху слабо вогнутые, светлые.
74
Листья сверху густовато прижато-волосистые (около 150 волос-
ков на 1 см2), снизу умеренно волосистые. Сверху волоски до
1,5—2 мм длиной. Черешки до 150 мм длиной, красноватые.
Соцветия короче листьев, густоволосистые; цветоносы до
50 мм длиной, прицветники 3—8 мм длиной, линейные; цвето-
ножки около 15 мм длиной. Чашечка от основания разделена
на 5 линейных долей длиной 7 мм.
Трубка венчика 2 мм длиной; верхняя часть отгиба венчика
почти до основания рассечена на две доли, ее длина — 13 мм,
нижняя часть-—15 мм длиной и 20 мм шириной, образует три
доли. По сообщению Лайтбури [Lightbourn, 1964], цветки сред-
не сине-фиолетовые, с темным глазком в основании долей око-
лоцветника (наше изображение взято с фотографии из этой
статьи). Тычинки 4 мм длиной, пыльники 2,5 мм длиной и
3,5 мм шириной. Завязь 3 мм длиной, густоволосистая, сверху
гладкая, утоньчающаяся. Плоды шиловидные, на конце сужен-
ные, 25 мм длиной и 3 мм в диаметре, умеренно волосистые.
Характеристика генеративных частей приведена нами по дан-
ным оригинального описания, когда в распоряжении автора
вида имелся только гербарный материал. В настоящее время
этот вид интродуцирован, но описание генеративных частей по
живым образцам отсутствует. Наши экземпляры не цвели.
Автор вида Б. Л. Бэртт считает этот вид более близким к
береговым видам сенполий из Танги по сравнению с видами из
западной Усамбары.
Восточная Африка, Кения, холмы Тейте, холм Мбололо,
в лесу близ воды, сентябрь-октябрь 1938 г., Boy Joanna (Со-
ryndon Mus. No. 8982 — ЕА, К).
Растение культивируется в Эдинбургском ботаническом саду,
по-видимому, с 1960 или 1961 г. Тип хранится в гербарии
в Кью.
Культивировался в Ботаническом саду Ленинградского уни-
верситета с 1962 по 1965 г., куда был получен в виде листовых
черенков из Эдинбурга, в настоящее время отсутствует.
11. SAINTPAULIA IONANTHA Н. Wendl.
Gartenflora, 1893, XLIII: 321, tab. 1391 et 323, fig. 66;
I- r i t s c h. Engler u. Prantl. Pflanzenfam, 1893—94,
IV, 3. B. 185; Bull. Soc. Tosc. ort., 1894, XIX; 13, fig. 1;
Rev. Hort. Beige, 1894, XX: 109, cum ic.; Neubert
Gart. Mag., 1894: 362, fig. 97; Hooker Bot. Mag.,
1895, 51, t. 7408; Engler Pflanzen. Ost. afr.: 362.
Syn: Petrocosmea ionantha (H. Wendl.) Rodi-
g a s. Ill. Hortic., 1895, xlii: 108.—Saint-
paulia kewensis С. B. Clarke. Oliver, Fl.
Trop. Afr., 1906, iv (2): 501.
Растения с укороченным стеблем в молодости до 10 мм и в
старости до 150 мм длиной и 12—15 мм толщиной, с мясисты-
75
ми расположенными в виде розетки темно-зелеными листьям^,линными волосками более или менее одинаковой длины
Все части растений покрыты прямоватыми или несколько при(1 —1,5 мм). Густота волосистого покрова у листьев среднего
жатыми (распростертыми) волосками более или менее одина^озраста около 160 волосков на 1 см2. У старых листьев волос-
ковой длины (рис. 29). ),ц располагаются реже.
Листья с толстоватыми, обычно красно окрашенными череш Средняя и боковые жилки по 5 с каждой стороны, снизу
ками, в молодом возрасте длиною не превышающими длин)1ыступают, сверху вогнуты. Сверху основание средней жилки
Рис. 29. Saintpaulia ionantha Н. Wendl.
пластинки, у старых листьев превышающими ее не более чем
в два раза.
Листовые пластинки широкоовальные или удлиненно-сердце-
видные, неясно городчатые или почти цельнокрайные, 13—
50(55) мм длиной и 10—35(40) мм шириной, верхний конец их
притупленный или округлый, основание слабовыемчатое; темно-
зеленые сверху, светло-зеленые или чаще красноватые или
темно-красные снизу, в молодости густо покрытые прямоваты-
ми, несколько прижатыми или наклоненными относительно
зстлое и выделяется в виде светлого пятна.
Цветки образуются на довольно толстых восходящих и воз-
ышающихся над листьями цветоносах в пазухах листьев. Дли-
а цветоносов 50—60 мм. Соцветия из 2—3—4 цветков, поник-
ые. Цветоножки 12—30 мм длиной, в основании каждой из
их по одному линейному прицветнику длиной около 6—7 мм.
' нижних цветоножек они крупнее, у верхних — мельче.
Чашелистиков пять, иногда семь, линейных, прямых, зеле-
ы.х, притупленных, длиннее трубки венчика, около 5 мм длины.
Венчик около 25 мм в диаметре, почти колесовидный, темно-
шолетово-синий, трубка светлая, 2,5—3 мм длины, вздутая;
1тиб, как свойственно роду, из двух частей: верхней, рассечен-
ой до половины на. две выступающие доли, внутренние края
нторых налегают друг на друга, и нижней, распростертой,
лубоко рассеченной на три доли, более округлые и вогнутые,
|см доли верхней части. Венчик по краю реснитчатый, но не
Железистый.
Тычинки выступают из трубки, их нити относительно тол-
тые, длиною около 3,5 мм, изогнуты и прикреплены к трубке
и'пчика;. пыльники почковидные, несколько выступающие, срос-
шеся вместе. Стаминодиев два позади тычинок, слегка кони-
еские.
Завязь яйцевидно-коническая, волосистая, столбик нитевид-
ен длиною около 10 мм, красноватый, рыльце простое. В осно-
ании завязи ширококольцевидный оранжевый нектарный диск.
Плоды яйцевидные или почти шарообразные, густоволоси-
тые, 16 мм длиной, 4—6 или 10—12 мм в диаметре.
Восточная Африка, Танзания, Амбони, близ Танга, растет
а кристаллических гнейсах на расстоянии около 7,5 км от бе-
та океана, на несколько метров выше уровня океана.
Танга, лес Мкагакузи, растет на известняках в береговых -
чнозеленых зарослях.
Мокрые скалы у пещеры Сиги. Цветки синие, 11 августа .
)10 г. Winkler, 4276 (Вроцлав). Пещеры Сиги, утесы (А), вер-
ина скалы (В), между валунами (С), интродуцирован
. Р. Пунтером, цвел в 1956 г. в Ботаническом саду в Эдип-
/рге, обр. С. 1688 (А), С. 1687 (B) и С. 1576 (C). Там же,
. R. О. Bally, культивируется в Бэйли Горториуме при Кор-
/ллевском университете (США, штат Нью-Йорк), 1955,
, Е. Moor, 6999 бис.
Культивируется во многих ботанических садах, однако чаще
виде культурных вариаций.
76
77
12. SAINTPAULIA TONGWENSIS В. L. Burtt
Gard. Chron., 1947, cxxii: 23; Bot. Mag., 1948, nov.
ser., clxv, pt. I, tab. 11; Cox et Roberts. Afr.
Viol. Mag., 1950, 3, 4: 5; Moore H. E., Afr. Viol, etc.,
1957: 153; Burtt B. L. Notes Roy. Bot. Gard. Edinb.,
1958, XXII, 6: 556.
Растения с укороченными стеблями, до 30 мм длиной и обыч
но 10—i2 мм в диаметре, в старости диаметр достигает 20 mi
относительно крупные, с длинночерешковыми листьями, расш
ложенными в виде розетки. Все вегетативные части доводы;
густо покрыты наклоненными волосками, различной длин:
(рис. 30).
Листовые пластинки эллиптические, овально-эллиптически-
или яйцевидные, тупо заостренные на верхушках и в завпсимл
сти от возраста неясно сердцевидные, сердцевидные или глубе
ковыемчатые, слегка неравнобокие у основания, 45—78 мм д.и)
ной и 30—50 мм шириной, городчатые и в молодости круши;
волнистые, слегка подогнутые по краю, относительно мясист^
С верхней стороны ярко-зеленые с более светлыми полосам!
по жилкам, снизу светло-зеленые или красновато окрашенны!
в юности густоволосистые (около 400 волосков на 1 сан), бел®
шелковистые от мало различающихся по длине наклоненны|
длинноватых (от 1 до 1,5 мм длиной) волосков. Снизу волос;
более короткие. Средняя и боковые первичные жилки, по 5
каждой стороны, снизу выступающие, сверху расположен;;;
почти в одной плоскости с поверхностью листа, светлые. Чере;
ки до 150 мм длиной.
Цветоносы восходящие, около 70 мм длиной, 4—5-цветк
вые; цветоножки 15—25 мм длиной, парные; прицветники ла
цетные или широколанцетные, к основанию суженные напод
бие черешка и согнутые книзу, нижние почти 12 мм длипо
верхние короче, как и цветоносы, красновато окрашенные.
Чашелистики линейные, около 6 мм длиной, две верхние к
роче трех нижних, красновато окрашенные, волосистые. Тру
ка венчика 3 мм длиной, отгиб (около 30 мм в диаметре) поч'
под прямым углом к трубке, светло-фиолетово-голубой, бл,
трубки более темный, доли по краю железисто-волосистые.
Две верхние доли отгиба венчика до половины сросшиес
12—15 мм длиной, 9—12 мм шириной, боковые доли 12—17 а
длиной и 11—15 мм шириной, средняя 12—15 мм ширине
с угловатыми лопастями у основания, налегающая на боков;
доли. Тычиночные нити 2,5 мм длиной, изогнутые; пыльни
почковидные, сросшиеся, около 2 мм в диаметре, стаминоди
два.
Завязь конусовидная, очень густоволосистая, 2 мм длине
резко суженная в длинный, гладкий бледно-лиловый столб
до 7 мм длиной. Рыльце двулопастное.
78
Плоды яйцевидные, слегка асимметричные, в неподсохшем
состоянии 12—14 мм длиной и 3—4 мм в диаметре, густо покры-
тые длинными волнистыми волосками.
Этот вид, помимо морфологических отличий, характеризует-
ся также особенностью очень редко образовывать боковые от-
ветвления, расцениваемые обычно как отпрыски, и слабо выра-
женной способностью к размножению листьями, в случае уко-
. Рис. 30. Saintpaulia tongwensis В. L. Burtt,
ренения которых образуется не более одной-двух ростовых-
почек.
Для полноты характеристики этого вида приведем описание-
условий обитания в природе, помещенное Херви Коксом и Ро-
бертсом в их статье: «Для того чтобы достигнуть горы Тонгве,
необходимо преодолеть значительное пространство, покрытое
высокой травой, достигающей 1,5 м и более в вышину. Этот
пояс, покрытый травой, эффективно препятствует проникнове^
нию туда других видов извне. В дополнение к этому травяному
поясу местообитание описываемого вида на вершине горы огра-
ничено пластами прочной скалы, достигающей 45 м длины»
(Сох, Roberts, 1950, р. 5). В настоящее время местообитание
79
вида в природе разрушено в связи с проводившимися археоло-
гическими раскопками.
Восточная Африка, Танзания. Восточные Усамбарские горьи
Известен в единственном местообитании. Г. Тонгве близ Пан-
гани, местообитание ограничено выходами гнейса в 45 м от
вершины на высоте 690—720 м над уровнем моря (Gerring).
Там же, сбор R. Е. Moreau, культивируется в Ботаническом
саду в Кью (голотип К). Там же, 600 м, многолетняя трава,
иногда с плотным полуодревесневшим стеблем и листьями в ро-
зетках; листья темные и светло-зеленые сверху, часто малино-
вые снизу, цветков помногу, в цимозных соцветиях, лавандово
окрашенных; очень редко растет на гнейсовых скалах вместе
с Encephalarios в лоскутном лесу из Isoberlinia, Mystroxylon.
Flacurtia, Chrysophyllum, Allophyllus, Tricalysia, Memecylon на
скалистом горном пике, 22 сентября 1940 г. Тринвей (Green-
way), 6024 (ЕА). Культивируется в Эдинбургском ботаническом
саду, стебель с возрастом достигает толщины большого пальца,
июнь 1954 г., образец С. 1559 (Е); сбор (W. R. Punter, образец
С. 1574 (Е). Тип хранится в Кыо.
Культивируется в Ботаническом саду Ленинградского уни-
верситета с 1962 г., куда был получен в виде листовых черен-
ков из Эдинбургского ботанического сада с индексом «from
Greenway», позже погибших. Современные растения воспроиз-
ведены из семян, собранных- с погибших растений.
13. SAINTPAULIA GRANDIFOLIA В. L. Burtt
Notes Roy. Bot. Gard. Edinb., 1958, XXII, 6: 560.
От остальных видов отличается крупными светло-зелеными
листьями широко овально-эллиптической формы с округленной
вершиной и сердцевидным ушастым основанием. Волоски на
верхней стороне листовой пластинки почти одинаковой длины
и относительно прямые; по этому признаку описываемый вид
может быть сравним с 3. ionantha и, в некоторой степени, с
3. grotei, от которых он отличается более тонкими, мягкими и
крупными листьями и их светлой окраской (рис. 31).
Стебли укороченные, листья расположены в виде розетки.
Листовая пластинка широко овально-эллиптическая, 65—
100 мм длиной и 55—80 мм шириной, на вершине округленная,
у основания сердцевидная с приподнятыми или выгнутыми
кверху лопастями, по краю явно городчато-зубчатая. Средняя
и боковые первичные жилки на верхней стороне не выступают
и почти незаметны, снизу выпуклые. Верхняя сторона пластинки
относительно густо покрыта (около 500 на 1 см2) прямоватыми
волосками от 1 до 2 мм длиной, светло-зеленая, нижняя — бе-
ловато-зеленая с более короткими волосками (0,7—1,5 мм дли-
ной).
80
Черешки 60—100 мм длиной и 3—5 мм толщиной, покрыты
многочисленными длинными прямыми волосками, перемешан-
ными с менее многочисленными более короткими.
Цветоносы около 70 мм длиной, большею частью трехцвет-
ковые, волосистые, восходящие, прицветники линейные 6 мм
длиной, также волосистые; цветоножки около 15 мм длиной.
Чашечка разделена на 5 линейных сегментов, длинноволо-
систая.
Трубка венчика 2,5 мм длиной, отгиб почти под прямым
углом около 25 мм в диаметре, светло-фиолетово-синий, верх-
ние доли 9 мм длиной и 6 мм шириной, боковые 10 мм длиной
Рис. 31. Saintpaulia granclifolia В. L. Burtt.
и 9 мм шириной, средняя доля 12 мм длиной и 9 мм шириной,
все по краю железисто-реснитчатые. Тычиночные нити 4 мм
длиной, пыльники 2,5 мм длиной и 3 мм шириной. Стаминодиев
три, из них средний едва заметен.
Завязь коническая, 3 мм длиной, густоволосистая с оттяну-
тым голым столбиком, длиной 5 мм. Рыльце дву- или неясно
трехлопастное. Плоды веретеновидные, асимметричные, около
13—14 мм длиной и 2—4 мм шириной, волосистые.
Восточная Африка, Танзания, Западные Усамбарские горы,
Лутинди, в крутой долине, на глине, сбор W. R. Punter, обра-
зец S. Культивируется в Эдинбургском ботаническом саду.
С. 1575 (голотип Е).
Культивируется в Ботаническом саду Ленинградского уни-
верситета с 1959 г., куда был получен в виде листовых черен-
ков из Эдинбургского ботанического сада с индексом «Punter S».
(j д. М. ЗАЛЕССКИЙ
81
14. SAINTPAULIA VELUT1NA В L. Burtt
Notes Roy. Bot. Gard. Edinb., 1958, XXII, 6: 563.
Этот вид отличается красивыми темно-зелеными бархатис-
тыми листьями с более светлыми жилками с верхней стороны
и светлыми розоватыми жилками снизу, а также сравнительно
мелкими фиолетово-синими, часто по краям долей более свет-
лыми цветками. Внешне растения этого вида слегка напоми-
нают растение S. diplotricha, отличаясь от них окраской цвет-
ков, более темными листьями с менее выраженной и мелкой
пильчатостыо по краям, а также более миниатюрными разме-
рами растений. Все вегетативные части белово.тосистые
(рис. 32).
Стебель укороченный, толстоватый, до 15 мм длиной и / мм
в диаметре, с густой розеткой листьев на вершине. Черешки
листьев 40—90 мм длиной. Листовые пластинки почти округ-
лые, 40—50 мм длиной и шириной, на вершине почти округлые-
или тупо заостренные, всегда приподнятые у сердцевидного
основания. По краю молодые листья явно зазубренно-пильча-
тые, более старые неясно пильчато-городчатые, края пластинки
часто приподнятые или, в старости, подогнутые. Листья покры-
ты относительно редко (около 25 на 1 см2) длинными, от 1,5
до 4 мм длиной, наклоненными волосками; между ними густо
(около 800 на 1 см2) располагаются очень короткие прямые
волоски, около 0,2 мм длиной, как у S. diplotricha.
Длинные волоски покрывают пространство между развет-
влениями жилок и наклоненные от центра пластинки к ее пери-
ферии, на самих же жилках они почти отсутствуют пли рас-
положены очень редко. Это выделяет светлые жилки на темном
бархатистом фоне всего листа. Боковых жилок по пять с каж-
дой стороны от средней, снизу они светлые, выступающие,,
сверху слабовогнутые.
Цветоносы 50—70 мм длиной, многоцветковые, от 2 до 5
цветков, наклоненные. Прицветники узколинейные до 2 мм дли-
ной, цветоножки 15—20 мм длиной. Чашечка от основания раз-
делена на ланцетные сегменты около 3 мм длиной. Трубка вен-
чика 2 мм длиной. Верхняя двулопастная часть отгиба венчика
4 мм длиной, 5 мм шириной, боковые и средняя доли нижней
части отгиба 8 мм длиной и 9 мм шириной. Отгиб 20—25 мм
диаметром, фиолетово-синий, верхняя часть его темнее, доли
одноцветные или по краям беловатые, железисто-реснитчатые.
Тычиночные нити 3 мм длиной, пыльники часто несросшиеся„
2,5 мм длиной, 3 мм шириной. Стаминодиев три, два боковых
в виде сосочков и средний в виде бугорка.
Завязь 2,5—3 мм длиной, волосистая, столбик бледно-розо-
вый или беловатый, почти до половины опушенный прямыми
мелкими волосками, сверху гладкий, часто прямой (когда пыль-
82
ники несросшиеся), расположенный между пыльниками; рыль-
це крупное двулопастное, белое или розовое.
Плоды яйцевидной формы, красноватые, на кончике белова-
тые, покрыты густыми короткими, почти прямыми волосками.
Длина коробочек в неподсохшсм состоянии 11 —12 мм, диаметр
4 мм в наиболее широкой нижней трети их длины и 3,5 мм
в верхней более узкой части. Плодоножки часто спирально или
дугообразно изогнуты.
Как отмечает Б. Л. Бэртт, растение в сухом виде легко мо-
жет быть спутано с 3. shumensis. Их различает лишь характер
Рис. 32. Saintpaulia velutina В. L. Burtt.
волосистого покрова листьев. Действительно, между этими ви-
дами много общих черт, в том числе очень сходны очертания
и размеры их венчиков, в известной мере темная у обоих верх-
няя часть отгиба венчика, относительно небольшие размеры
растений, но их легко различить по характеру и форме семей-
ных коробочек, если обратить внимание на их детали.
У этого вида на наших живых образцах наблюдались откло-
нения от нормального строения цветка: цветки с четырьмя раз-
витыми тычинками и четырьмя долями венчика и без столбика;
без тычинок с шестью долями венчика — тремя в верхней части
и тремя в нижней; с тремя тычинками и с шестью долями вен-
чика— двумя сверху и четырьмя снизу; с нормально развитыми
тычинками и пестиком, но с семью долями венчика — двумя
сверху и пятью снизу. На аналогичные отклонения в условиях
культуры указывает и Б. Л. Бэртт.
6* 83
Растение было открыто и собрано еще в 1916 г., но описано
не было до 1958 г.
Восточная Африка, Танзания, Западные Усамбарские горы.
Девственный лес близ Балангаи, 1280 м, бледно-лилово-синяя,
10 марта 1916 г., Peter 0 IV 29 (В). Балангаи, в 5 милях от
Саккаре, W. R. Punter, образец D культивируется в Ботаниче-
ском саду в Эдинбурге, С. 1579 (голотип Е). Балангаи, 900м,
на увлажняемых скалистых, террасах в тенистом лесу, малень*
кое розеточное растение с бледно-лиловыми цветками, с листья-
ми с обратной стороны от телесно-розового до светло-кашта-
нового цвета, 23 января 1953 г. Faulkner, 1126 (К).
Культивируется в Ботаническом саду Ленинградского уни-
верситета с 1959 г., куда получен в виде листовых черенков из
Эдинбургского ботанического сада с индексом «Barker».
15. SAINTPAULIA DIPLOTRICHA В. L. Burtt
Gard. Chron. 1947, 3 ser., cxxii: p. 23 — excl. pl. culta;
Baileya, 1956, 4: 164; Moore H. E., Afr. Viol, etc.,
1957: 150.
Растения с укороченным стеблем длиной около 20—30 мм
И диаметром около 6 мм, красноокрашенным, с пучком распо-
ложенных розеткой темных листьев (рис. 33).
Листья с длинными (до 100 мм) красноокрашениыми че-
решками. Листовая пластинка около 0,5 мм толщиной, широко-
овальная, овальная с почти округлой или слегка заостренной
Вершиной, темно-зеленая сверху, красно-пурпурная снизу, края
слегка приподнятые, пильчато-зубчатые. Основание листовой
пластинки почти сердцевидное, с налегающими друг на друга
лопастями, всегда приподнятое. Листья покрыты длинными рас-
простертыми волосками, между которыми имеется густой по-
кров из едва заметных очень коротких прямых волосков. При
рассматривании' сверху этот покров увидеть трудно, так как
Волоски сильно просвечивают; их лучше видно, если разрезать
лист И смотреть в лупу со стороны разреза. Длинные волоски
имеют размеры до 5 мм в длину, их около 50 на 1 см2. Корот-
кие не более 0,4 мм в длину, их около 2000 на 1 см2. Сеть жи-
лок листа сильно выступает снизу, а сверху сильно вогнута,
поэтому промежутки между жилками сильно возвышаются.
Такой характер жилкования (в отличие от большинства ви-
дов) имеется еще только у двух видов — S. difficilis и S. orbi-
cularis.
Цветоносы не длиннее листьев, обычно четырехцветковые,
восходящие, красно окрашенные; прицветники линейные, около
2 мм длиной и 0,5 мм шириной у основания каждой цветонож-
ки; цветоножки около 25 мм длиной, изогнутые.
84
Чашечка рассечена до основания на пять красновато окра*
шенных долей.
Отгиб венчика около 25 мм в диаметре, темновато-фиолето-
во-синий, по краю железисто-реснитчатый. Тычинки 4 мм дли-
ной, изогнутые, с почковидными сросшимися или несросшимися
желтыми пыльниками не более 2 мм шириной.
Стаминодиев три в виде небольших светлых сосочков, при-
крепленных к трубке венчика и противостоящих верхней части
отгиба.
Рис. 33. Saintpaulia diplotricha В. L. Burtt.
Завязь коническая, густо прижато-волосистая, переходящая
в фиолетовый, в нижней части коротко опушенный столбик, за-
канчивающийся едва заметно двулопастным рыльцем. Нектар-
ный диск у основания завязи желтый.
Плоды до 17—20 мм длиной, шиловидные, . крючковидно
изогнутые, около 2,5 мм в диаметре в наиболее широкой части.
Автор вида Бэртт указывает на то, что нижняя часть отгиба
венчика образует угол около 135° с прямостоячей верхней
частью, и считает это примечательной особенностью описывае-
мого вида в отличие от других видов, у которых, как он счи-
тает, отгиб венчика плоский. Однако это мнение Бэртта после
открытия многих новых видов оказалось не соответствующим
действительности, тем более что практически ни один ..из опи-
санных видов не имеет плоского (под прямым углом), ртгиба
85
и у всех видов между верхней и нижней частями отгиба обра-
зуется тупой угол, но различной величины, меняющийся в зави-
симости от возраста цветка.
Тип хранится в гербарии в Кью, голо- и изотип — там же.
Образец, являющийся типом, Кларк (Clarke) как раз и по-
считал настоящей S. ionantha, что привело его к ошибочному
описанию истинной 5. ionantha под другим, теперь уже синони-
мичным названием 5. kewensis.
Восточная Африка, Танзания, Восточные Усамбарские горы,
на высоте 1000 м над уровнем моря, 26 ноября 1895 г., Buch-
wald, 149 (голотип К, изотип ВМ, BR).. Мавени, Танга, на ска-
лах, W. R. Punter, образец Т, культивируется в Ботаническом
саду Эдинбурга, С. 1571 (Е). Долина Мкулумуци, W. R. Punter,
образец О, культивируется в Ботаническом саду Эдинбурга,
С. 1573 (Е). Там же, образец Р, С. 1572 (Е). Канге, W. R. Pun-
ter, культивируется в Ботаническом саду Эдинбурга, С.1577 (Е).
Канге, узкий известковый проход на пластах скалы в темном,
мрачном, сухом вечнозеленом лесу, листья темно-зеленые свер-
ху, бледнее снизу, цветки очень бледно-лиловые, 13 ноября
1956 г. Milne Redhead et Taylor, образец 7287 (К). Культиви-
руется в Эдинбургском ботаническом саду образец из Мавени,
введенный Пунтером.
Культивировался в Ботаническом саду Ленинградского уни-
верситета с 1962 г., куда был получен из Эдинбурга в виде ли-
стовых черенков с индексом «from W. R. Punter». В настоящее
время отсутствует.
16 SAINTPAULIA RUP1COLA В. L. Burtt
Notes Roy. Bot. Gard. Edinb., 1964, XXX, 3: 46.
По мнению Б. Л. Бэртта, данный вид близок к S. diplotricha
и S. ionantha, от которых отличается волосистым покровом
верхней стороны ярко-зеленых листовых пластинок, состоящим
из очень коротких волосков, смешанных с неодинаковыми по
длине прямыми волосками, и определенно удлиненными листья-
ми (рис. 34). Этот вид также сходен с 5. brevipilosa — видом
с листовыми пластинками, покрытыми сверху одинаковой дли-
ны короткими волосками. В юности розеточное травянистое
растение; стебли рано полегают и, разветвляясь, образуют по-
добие корневища. Черешки до 90 мм длиной, коротко- и откло-
ненно-волосистые, с немногочисленными более длинными волос-
ками. Пластинка яйцевидная, около 65 мм длиной и 45 мм ши-
риной, у основания слегка открыто-сердцевидная, на конце при-
тупленно заостренная, по краю вначале заметно, затем неясно
городчато-зазубренная, сверху ярко-зеленая, снизу красновато
окрашенная, негусто покрыта сверху короткими прямыми во-
лосками, железисто-головчатыми и более длинными, снизу ко-
ротко отклоненью опушенная; густота волосистого покрова
1600 волосков на 1 см2, из них 460 длинных, коротких — 1140.
Боковых жилок с каждой стороны около 5 (3 суббазальные),
восходящих, сверху незаметных, снизу выступающих. Цветоно-
сы 3—6-цвстковые, равные по длине листовым черешкам и бо-
лее коротко отклоненпо опушенные, с немногочисленными длин-
ными волосками. Прицветники ланцетные, 1—2 мм длиной, все
почти одного размера. Чашечка отклоненно-волосистая, почти
до основания рассеченная на ланцетные узкие сегменты длиной
3 мм. Трубка венчика 3 мм длиной, фиолетово-синяя, отгиб
Рис. 34. Saintpaulia rupicola В. L. Burtt.
около 20—25 мм в диаметре, его верхняя часть, как и у всех
видов рода, двулопастная, нижняя трехлопастная. Тычиночные
нити 4 мм длиной, сверху желтые, пыльники интенсивно жел-
тые, 1,5 на 2 мм. Стаминодиев два. Завязь коническая, 2,5 мм
длиной, мелко опушенная, на столбике гладкая, слегка пурпур-
ная, резко суженная. Нектарный диск бледно-желто-зеленый.
Рыльце слегка расширенное и слегка коническое. Плоды вере-
теновидные, 15—18 мм длиной, опушенные, швы створок мало-
заметны, при созревании не раскрываются.
Юго-восточная Кения, Калолени, между Мариакари и Ка-
лифи, около 25 миль северо-западнее Момбасы, кол. R. Bayliss,
передан W. R. Punter за № 8, от 30 октября 1958 г., интрод.
№ 58/3591. Культивируется в Ботаническом саду в Эдинбурге,
С. 3065 (голотип Е).
87
Культивируется в Ботаническом саду Ленинградского уни-j
верситета с 1966 г., куда был получен из Эдинбурга в. виде ли г
стовых черенков. |
Примечание автора вида Б. Л. Бэртта при его описании: |
«Скрытая общность характера между этим видом и S. brevi-.
pilosa с гор Нгуру (также и других крайних в ареале видов
рода) весьма примечательна. Этот вид не кажется очень близко I
родственным другому виду из Кении — S. tietensis. Они оби- •
тают в расселинах на пластах скал и кажутся более приуро-
ченными к скальным местообитаниям по сравнению с другими.
видами, об обитании которых мне сообщили» (Burtt, 1964).
17. SAINTPAULIA BREVIPILOSA В. L. Burtt
Notes Roy. Bot. Gard. Edinb., 1964, XXV, 3: 47.
По мнению Б. Л. Бэртта, вид близок к 5. nitida и S. rupi-
cola; отличается от них почти округлыми, сверху ярко-зелены-
ми листьями, покрытыми очень короткими прямыми волосками ,
(рис. 35).
Розеточное травянистое растение. Черешки до 4 см длиной, г
покрыты явственными густо расположенными волосками. Пла- I
стинка почти округлая, около 40 мм длиной и шириной, на [
вершине округлая, у основания сердцевидная, с городчатым 1
краем, сверху ярко-зеленая, густо покрытая очень короткими,
трудно различимыми прямыми волосками. Густота волосков
около 8000 на 1 см2; снизу также явно коротковолосистая; жил-
ки снизу немного выступающие, сверху слегка вогнутые, боко-
вых жилок по 5—6 с каждой стороны. Соцветия пазушные,
1—2 (6)-цветковые, цветоносы около 6 см длиной, как и цвето-
ножки, покрыты более длинными и более короткими волосками.
Прицветники 2—3 мм длиной, ланцетные. Чашечка явно воло-
систая, рассеченная почти до основания на 5 ланцетных чаше-
листиков 2,5 мм длиной. Венчик светловато-фиолетово-синий,
с трубкой, у основания белой, 2 мм длиной. Доли венчика по '
краю железисто-волосистые, реснитчатые. Верхняя часть вен-
чика, 10 на 13 мм, более чем до середины двулопастная; ниж-
няя— 15 на 25 мм, более чем до середины трехлопастная. Ты-
чинки расположены в устье трубки венчика, тычиночные нити
4 мм длиной, белые или слегка пурпурноватые, бородавчато-
сосочковые; стаминодиев три, средний в виде небольшого со-
сочка, остальные два на небольших стебельках; пыльники жел-
тые, 2 мм длиной и шириной, сросшиеся между собою; завязь
коническая, около 2,5 мм длиной, густо покрыта жесткими
острыми волосками; нектарный диск кольцевидный, оранже-
вый; столбик у основания волосистый, выше гладкий, 6,5 мм
длиной, резко суженный, рыльце бледно-фиолетовое, слабодву-
лопастное.
в
Плоды удлиненно-веретеновидные, 16 мм длиной и 2—3 мм
диаметре.
Восточная Африка, Танзания, Горы Нгуру, Лулага; передан
автору вида В. Р. Пунтером в сентябре 1959 г. По мнению
Бэртта, в культуре является слаборастущим видом с не очень
отчетливыми чертами строения.
Рис. 35. Saintpaulia brevipilosa В. L. Burtt
Культивируется в
Культивируется в
верситета с 1966 г.,
листовых черепков.
Эдинбургском ботаническом саду.
Ботаническом саду Ленинградского уни-
куда был получен из Эдинбурга в виде
18. SAINTPAULIA NITIDA В. L. Burtt
Notes Roy. Bot. Gard. Edinb., 1958, XXII, 6: 564.
Вид отличается почти округлыми блестящими темно-зелены-
ми листьями, покрытыми короткими, довольно густо располо-
женными прижатыми волосками, с почти цельным или неясно-
городчатым краем и тонкими, довольно длинными плодами
I (рис. 36).
i Травянистое растение с укороченным (до 20 мм длиной)
стеблем, простым или снизу компактно разветвленным. Листья
собраны в розетку.
Листовая пластинка широкоовальная или почти округлая,
45—55 мм длиной и 40—50 мм шириной, с округлой вершиной
и слабо сердцевидным основанием, по краю сглаженно-город-
88
чатая, сверху темно-зеленая (у молодых листьев ярко-зеленая)^ нии BeDXHeg части отгиба, позади тычинок. Завязь кониче-
™ИЛУ П0КРыта, короткими, сильно прижа-f 2 Рм длиной> ко.ротКоволосистая, столбик 5 мм длиной,
XhoZcOtCoK7okoS ?200Мня расположе™ЫМЙ СР*В- т выступающего основания гладкий. Рыльце булавовидно утол-
тельно густо (около 1200 на 1 см2). Волосистый покров ниж-Г,рннп/неяРН0 ЛВулопастное.
ней стороны состоит из более коротких (около 0,2 мм длиной),; Плоды шиловидные 13—14 мм длиной в наиболее широкой
hLTr бопТп™Х волосков’ Реже расположенных и приурочен- * 2 диаметром, красновато окрашены и довольно
ных в большей мере к жилкам. Черешки от 50 до 90 мм дли-^™ помыты прижатыми короткими волосками.
нои, коротко прижато-волосистые. Цветоносы незначительно1^ у ЭТОГР ввда Рна наших образцах наблюдалось образование
1в отдельных случаях адвентивных ростовых почек и побегов
на черешках листьев (см. рис. 16, а).
Восточная Африка, Танзания. Горы Нгуру, Мкоове, северо-
восточная сторона близ Туриани, около 1000 м, на скалах по
берегу лесного ручья, 29 марта 1953 г., Drummond et Hensley,
Рис. 36. Saintpaulia nitida В. L. Burtt.
берегу лесного ручья, 29 марта 1953 г.,
1862 (К, E).
Колугамба, Туриани, ноябрь 1953 г. S. R. Semsei, 1463
(ЕА, К).
Там же, декабрь 1953 г., S. R. Semsei for Eggeling
t (E. 6776—К). Культивируется в Ботаническом саду в Эдинбур-
Iге растение, полученное через Harvey Сох, образец А (С. 1557—
голотип Е).
I Культивируется в Ботаническом саду Ленинградского уни-
верситета с 1959 г.; исходный образец получен из Эдинбург-
ского ботанического сада в виде листовых черенков с индексом
«Harvey Сох А1».
19. SAINTPAULIA ORBICULARIS В. L. Burtt
Gard. Chron., 3 ser., cxxii, 1947: 22—23, in adnot;
Moore H. E. Afr. Viol, etc., 1957: 153; Notes Roy.
Bot. Gard. Edinb., 1958, XXII, 6: 564.
Очень краткое описание, сделанное Б. Л. Бэрттом [Burtt,
1947], и еще более краткое, сделанное Г. Е. Муром [Moore,
1957], являются единственными описаниями, в которых сообща-
отличаются по длине или равны черешкам, 4—5-цветковые; по-
крыты, так же, как и черешки, короткими прижатыми волоска-
ми, полувосходящие; прицветники линейные, 4—5 мм длиной,
цветоножки около 25 мм длиной, чашечка из пяти рассечен-
ных до основания сегментов 6 мм длиной, сегменты при созре-
вании плодов прижаты к ним; венчик интенсивно-фиолетово-
синий, по краю без железистых волосков, его трубка 2 мм дли-
ной, верхние, сросшиеся до половины длины доли отгиба 7 мм
длиной и 8 мм шириной, боковые доли 8 мм длиной и 9 мм ши-
риной, средняя нижняя доля 10 мм длиной и шириной.
Тычиночные нити 4 мм длиной, пыльники 1,5 мм длиной и
2 мм шириной.
Стаминодиев два, удлиненно-сосочковидной формы в осно-
90
ются самые общие сведения для характеристики этого вида
(рис. 37). Более подробное описание сделано нами по живым
растениям, культивируемым в Ботаническом саду Ленинград-
ского университета. Они были получены в виде листовых черен-
ков из Эдинбургского ботанического сада.
Прежде чем привести наше описание, приведем перевод ав-
торского диагноза: «Растение по облику сходно с S. ionantha.
Отличается округлыми листьями с сердцевидным основанием.
Соцветие многоцветковое, пониклое, цветки мельче, чем у 5.
ionantha, бледные; внутренняя сторона венчика почти чисто-бе-
лая, но с темно-лиловым ободком вокруг тычиночных камер,
наружная поверхность долей венчика белая, со светло-малино-
вой средней жилкой, края долей обмыты светло-малиновым.
Розеточное растение с широкоовальными или почти округлыми
91
светло-зелеными листьями, с многочисленными голубовато-бе-
лыми цветками, верхняя часть отгиба венчика которых в осно-(
вании своем окрашена в фиолетово-синий цвет, а нижняя —'
беловатая». Г
Стебель укороченный, около 50 мм длиной и до 5 мм в дна-!1
метре, с междоузлиями, достигающими 8 мм в основании стеб-!,
ля и кверху уменьшающимися. В отличие от других розеточных!1
видов листья располагаются от оснований до вершины укоро-?
ченного стебля и не сосредоточены на его верхушечной части.
юнцами. Листья, особенно молодые, из-за прижатости волос^
<ов кажутся почти гладкими и глянцевитыми. С нижней сторо-
ны более длинные волоски расположены только по жилкам,
Промежутки между ними покрыты только короткими волосками.
J Цветоносы тонкие, длиной около 100 мм, многоцветковые
,1(5—7), восходящие и возвышающиеся над листьями. Цвето-
ножки тонкие, около 25 мм длиной, красновато окрашенные;
прицветники ланцетные.
.Чашелистики, сросшиеся у основания чашечки, удлиненно
Грехугольно-ланцетные, около 3 мм длиной. Трубка венчика
Ч мм длиной, отгиб около 25 мм в диаметре, редковолосистый,
верхняя его часть до половины сросшаяся, синевато-белая,
основания фиолетово-синяя, 12—13 мм длиной и шириной,
гри доли нижней части отгиба 8—10 мм длиной и шириной
каждая. Края лопастей не железисто-волосистые. Тычинок две,
iix пыльники, как и у большинства видов, сросшиеся. Стамино-
диев два, в виде небольших бугорков на границах верхней и
|шжней частей отгиба венчика. Завязь удлиненно-коническая,
[юлосистая, до 3 мм длиной, в ее основании коричневатый сла-
ю развитый нектарный диск. Столбик длинный (до 5 мм), за-
учивается слегка расширенным и на копчике конически за-
остренным рыльцем.
’ Плоды шиловидные, притупленные у основания, 12—18 мм
шлиной и около 2 мм шириной в наиболее широкой нижней
|юловине.
На отдельных наших экземплярах этого вида наблюдалось
>бразование адвентивных ростовых почек у основания листовой
1ластинки, как это наблюдается у бегоний (см. рис. 16, б).
Вид открыт в 1916 г. Петером, интродуцирован в культуру
। 1938 г. Моро.
Восточная Африка, Танзания. Западные Усамбарские горы,
>унгу до Амбангулу, 1060 м, 29 февраля 1916 г., A. Peter,
I IV 16 (В). Сакарре, Амбангулу, 1200 м, 4 августа 1938 г.,
(. Е. Moreau (голотип К); культивируется в Ботаническом саду
Кью под этим номером (К). Культивируется в Эдинбургском
ютаническом саду от W. R. Punter (С. 1689—Е) и Harvey Сох
С. 1561—Е). Тип хранится в Кью.
Культивируется в Ботаническом саду Ленинградского уни-
ерситета, куда был получен в виде листовых черенков изЭдип-
ургского
ег».
Рис. 37. Saintpaulia orbicularis В. L. Burtt.
Листовая пластинка широкоовальная или почти округлая,
с тупо заостренной вершиной и ушастым или сердцевидным
неравнобоким основанием, лопасти ушастого основания часто
налегают одна на другую, по краю пластинка неясно городча-
тая, края ее, особенно в основании, приподняты. Размеры пла-
стинки 40—60 мм длиной и 38—50(55) мм шириной, черешки
красновато окрашенные, по длине равны либо незначительно
длиннее или короче пластинок; красновато окрашены также
средняя и боковые жилки с нижней стороны листовой пластин-
ки. Жилки снизу выступают, сверху заметно вдавлены. Верхняя
сторона пластинки относительно густо (около 400 на 1 см2) по-
крыта прижатыми к поверхности волосками длиною около 1 мм
и. равномерно расположенными между ними более короткими
.волосками длиною не более 0,3 мм с изогнутыми к поверхности
.92
ботанического сада в 1959 г. с индексом «from Pun-
19 а. SAINTPAULIA ORBICULARIS,
var. PURPUREA В. L. Burtt
Notes Roy. Bot. Gard. Edinb., 1964, XXV, 3.
Л. рБэртт-у отличается главным образом сплошь ин-
” отличительные
По Б.
енсивно фирлеторотсиними цветками. Другие
93
черты следующие: волоски черешков листьев и цветоносов от-
клоненные, листовые пластинки более или менее овальные,,
у основания открыто-сердцевидные, доли чашечки удлиненные
(слегка удлиненно-обратноовальные).
По нашему мнению, общий облик растений вариации очень
близок или почти не отличим от типичной S. orbicularis. Те же
красноватоскрашенные черешки листьев и цветоносы, однако
при более детальном рассмотрении растения вариации отлича-
ются, помимо окраски цветков, более овальными листовыми
пластинками с сердцевидным основанием (не асимметрично ло-
пастным, как у типа) и очень четко выраженной вогнутостью
жилок на верхней стороне листовых пластинок.
Восточная Африка, Танзания. Западные Усамбарские горы,
Амбангулу, сбор W. R. Punter, № 3, октябрь 1958 г. Поступле-
ние № 58/3586. Культивируется в Эдинбургском ботаническом
саду, С.2959 (голотип Е).
Культивируется в Ботаническом саду Ленинградского уни-
верситета, куда было получено в виде живого растения из Эдии-|
бургского ботанического сада.
20. SAINTPAULIA DIFFICILIS В. L. Burtt
Notes Roy. Bot. Gard. Edinb., 1958, XXII, 6: 565.
Розеточные растения относительно мощного облика, с эл-
липтическими городчато-зубчатыми листовыми пластинками на
длинных черешках, с резко вогнутым рисунком жилок. На верх-
ней стороне листовые пластинки сравнительно редко покрыты
длинными и густо малозаметными очень короткими прижатыми
волосками (рис. 38). Цветки фиолетово-синие, относительно
крупные, сходные с цветками S. confusa. v
Травянистое растение, с укороченным толстоватым стеблем
длиной 20 мм у молодых растений и до 50 мм — у более ста-
рых, с розеткой листьев на вершине. Черевйки листьев 50—
120 мм длиной, листовая пластинка светло-зеленая, яйцевид-
ная, крупно городчато-зубчатая по краю, 50—75 мм длиной и
40—55 мм шириной, с сердцевидным основанием. Вершина ту-
повато заостренная. Верхняя сторона пластинки покрыта длин-
ными, сравнительно редко расположенными (около 150 на
1 см2) почти прямыми или полураспростертыми волосками дли-
ной до 2—7 мм, перемешанными с значительно более коротки-
ми (в 3—5 раз короче) прижатыми волосками, расположеиньь
ми более густо (3500 на 1 см2). Наклонены волоски в направ-
лении от средней жилки к периферии пластинки параллельно
боковым жилкам. Нижняя более светло-зеленая сторона листо-
вой пластинки покрыта примерно однородными короткими
(0,5 мм длиной) волосками, расположенными довольно густо
(около 2500 на 1 см2), несколько гуще по жилкам. Все жилки
сильно вдавлены с верхней стороны и выпуклы с нижнеи, что
отличает листья этого вида от большинства других видов; про-
странства между жилками благодаря этому выпуклы, что осо-
бенно подчеркивает всю сеть жилок.
Цветоносы многоцветковые (от 2—4 и более), полувосходя-
щие, 50—70 мм длиной, также покрыты длинными и короткими,
полураспростертыми волосками; прицветники линейные, до 8 мм,
длиной и около 0,5 мм шириной. Цветоножки 20—30 мм дли-
Рпс. 38. Saintpaulia clifficilis В. L. Burtt.
ной. Чашечка разделена на узкие ланцетные прижато-волоси-
стые сегменты 3 мм длиной.
Венчик фиолетово-синий, с простыми волосками по краю,,
по своему характеру очень сходен с венчиком S. confusa, труб-
ка 2 мм длиной, отгиб около 25—30 мм в диаметре; верхняя
лопасть отгиба 8—10 мм длиной и 5—6 мм шириной рассечена
до половины длины, боковые и средняя — около 10—12 мм дли-
ной и 8 мм шириной. Тычиночные нити 4 мм длиной, пыльники
1,75 мм длиной и 2,5 мм шириной. Стаминодиев три, средний
из них развит слабее.
Завязь коническая, 2 мм длиной, прижато-волосистая; стол-
94
95-
рик § Jim длиной, у основания волосистый, далее гладкий, свет-
ло-ф'ио^товый. Рыльце слегка двулопастное.
Плоды веретеновидные, до 14 мм длиной и 3,5 мм в диа-
.метре, асимметричные, густо прижато-волосистые.
Восточная Африка, Танзания. Восточные Усамбарские горы,
.в .истоке р. Сиги выше Монга, на скалах в дождевом лесу, сбор
•W. R. Punter. Образец культивируется в Эдинбургском ботани-
ческом саду, С. 1578 (голотип Е). Монга, на скалах в дожде-
ром лесу, 900 м, колл. Р. J. Greenway, 1939, сбор R. Е. Moreau,
7(К). Верховья р. Сиги, Убири до Мгамбо, 1050 м, очень ло-
кально, преобладает на скалах берегов ручья, в сухом долип-
,но,м печнозеленом лесу из Parinari, Piptadenia, Cynometra,
<22 цюдя 1940 г., сбор Р. J. Greenway, 5974 (ЕА, К). Истоки
р~ Сиги, 1050 м, очень локально, преобладает на мокрых скалах
!р pvc.ie ручья в вечнозеленом дождевом лесу из Piptadenia,
'Parinari, Allanblackia, на крутом склоне, 24 октября 1945 г.,
-Р. J. Greenway, 7551 (ЕА, К). Истоки р. Сиги, Убири — Монга,
'очень локально, преобладает на поверхности гнейсовых скал и
,в листовом перегное на поверхности скал в дождевом лесу из
Parinari, Piptadenia, Zahna, Isoberinia, 24 мая 1950 г., Green-
way et (Verdcourt, 8454 (ЕА, К)- Сбор W. R. Punter, образец L
.культивируется в Эдинбургском ботаническом саду (С.1945—Е).
'Культивируется в Эдинбургском ботаническом саду, сбор Наг-
Аёу Сох, 1954 г., образец С (С. 1570—Е).
В Ботаническом саду Ленинградского университета культи-
вируется образец, полученный из Эдинбургского ботанического
'^ада в 1959 г. с индексом «Punter Q».
,21. SAINTPAULIA CONFUSA В. L. Burtt
Baileya, 1947, 4: 163—164.
,Растения с укороченным полулежачим стеблем и листьями,
.расположенными розеткой * (рис. 39).
' Стебель не более 20—30 мм длиной и 10 мм толщиной.
Листья расположены попарно (заметно на верхушках побе-
гов), затем смещаются. Листовая пластинка эллиптическая до
щирокоовальной,’ закругленная на вершине, сердцевидно-выем-
чатая у основания, по’краю явно городчато- или пильчато-зуб-
чатая, тонковатая (менее 1 мм толщиной), но прочная, 37—
50 мм длиной и 32—37—45 мм шириной. Края пластинки у мо-
лодых листьев приподнятые, у старых листьев узко подогнутые.
Нерешки длиннее пластинок (у зрелых листьев), до 75 мм
длиной.
* Растения этого вида в молодом возрасте образуют правильную ро-
зетку, которая с возрастом .становится как бы растрепанной 'и однобокой
(из-за раннего полегания‘стебля).
₽6
Волосистый покров листьев состоит из прижатых волосков
двух типов: длинных — до 3—4 мм (около 50 волосков на Гем2)
и коротких — до 0,2—0,3 мм (около 900 волосков на 1 см2),
причем короткие прижаты так, что их кончики касаются или
почти касаются поверхности листа.
Цветоносы полувосходящие, слегка красноватые или зеле-
ные, около 50 мм длиной, не длиннее чем листья, 2—4-цветко-
вые. Цветоножки 25—30 мм длиной, слабоизогнутые.
Прицветники у основания каждой цветоножки линейные,
около 7 мм длиной и 1 мм шириной, иногда меньше.
Рис. 39. Saintpaulia confusa В. L. Burtt.
Цветки 25—32 мм в диаметре, у одних клонов фиолетово-
синие, у других фиолетовые (non vidi!), в центре несколько бо-
лее темные, по краю с волосками без железок. Отгиб плоскова-
тый, трубка менее 3 мм. Стаминодиев два.
Завязь округловато-коническая около 2,5 мм длиной, пере-
ходящая в окрашенный столбик около 5 мм длиной. Завязь и
столбик до половины его длины волосистые; волоски прямые
длинные, густые на завязи и короткие и редкие на столбике.
Рыльце слабодвулопастное.
Неподсохшие зрелые плоды бледно-зеленые, довольно густо
и длинно прижато-волосистые, веретеновидные, в той или иной
мере изогнутые, длиной 13—18 мм и диаметром (в наиболее
широкой нижней трети) 2,35—2,5 мм. Подсохшие плоды корич-
невато-серые, несколько меньших размеров.
Тип культивируется в Ботаническом саду в Кью с 1947 г.,
7 Д. М. ЗАЛЕССКИЙ
97
голотип — в Ботаническом саду в Эдинбурге, изотип — в Бэйли
Горториуме при Корнеллевском университете (США, штат
Нью-Йорк).
Восточная Африка, Танзания. Восточная часть Усамбарских
гор, Нгуа, дорога Потве, близ Амани, 900 м над уровнем моря,
1938, сборы R. Е. Moreau, № 1 (К). Культивируется в Ботани-
ческом саду Кью, 1947 (голотип К, Е). Квамкоро-Потве, 1050 м
над уровнем моря, очень локально, обычно на увлажненных
гнейсовых скалах в руслах ручьев, в вечнозеленых дождевых)
лесах, составленных из Cephalosphaera, Piptadenia, Ocotecp
Polyalihia oliveri, 3 ноября 1940 г., Greenway, 6050 (EA, K).|
Культивируется в Кыо, в Эдинбурге, в Итаке. I
Культивируется в Ботаническом саду Ленинградского уни-i
верситета, куда получен в виде листовых черенков из Эдин-)
бурга с индексом «Michigan».
Глава 4
ВОПРОСЫ КУЛЬТУРЫ
Мы посвятили все предшествующее изложение исключитель-
но дикорастущим видам сенполий и дали возможно более пол-
ное их описание совершенно сознательно. По нашему глубоко-
му убеждению только ясное представление о диких видах, осо-
бенностях их строения, связанных с условиями существования
в природе, и ряде других особенностей, их характеризующих,
дает возможность правильно, осмысленно подойти к вопросам
их культуры. Это мы сделали и потому, что до сих пор боль-
шинство авторов, говоря о культуре тех или иных растений,
предлагают читателям готовые рецепты, не объясняя, на чем
основаны их рекомендации. Предлагаемые рецепты чаще всего
созданы в результате длительных практических исканий от-
дельных культиваторов, авторы рецептов в той или иной степени
учитывают требования растений и поэтому их рекомендации
правильны, однако их изобилие, а подчас и противоречивость,
нередко ставят в тупик начинающего культиватора.
Сенполии как декоративные и продолжительно цветущие
растения выгодно отличаются от многих других культивируе-
мых растений прежде всего тем, что могут цвести в любое вре-
мя года и этим восполнять тот пробел, который ощущается зи-
мой в удовлетворении спроса на цветущие растения. Кроме
того, большое количество культурных вариаций, которые спо-
собны давать сенполии, может удовлетворить самые разнооб-
разные вкусы. Строение и окраска их цветков с множеством
оттенков — от белого до розового и почти красного и от фиоле-
тового, голубого до бархатисто-синего, махровость и бахром-
чатость цветков, расположение соцветий, различная форма ли-
стьев, форма розеток и т. п. — все это позволяет видеть в сен-
полиях одно из весьма интересных растений для использования
во внутрикомнатном озеленении. Поэтому эти растения, несом-
ненно, заслуживают широкого внедрения в промышленное садо-
водство. Для любителей выращивать растения в комнатных
7* 99
условиях сенполии представляют также самый разнообразный
интерес. Они не занимают много места, отличаются изящест-
вом и миниатюрностью, способностью хорошо адаптироваться
к свойственной жилым помещениям обстановке.
Помимо тех качеств, которые перечислены выше, эти расте-
ния обладают сильно выраженной способностью образовывать
при семенном размножении самые неожиданные отклонения от
исходного типа, позволяющие создавать разнообразнейшие и
все новые формы растений. В США за относительно короткий
отрезок времени (около 50 лет с момента завоза растений в эту
страну из Европы) было выведено свыше 1300 сортов сенполий.
И это не предел. Могут быть получены все новые и новые ва-
риации и сорта, особенно если к работе по их созданию будут
привлечены ставшие известными к настоящему времени дикие
виды.
Несмотря на то, что многие культурные растения в ходе
культивирования значительно уклонились по внешним призна-
кам от диких исходных форм (это имеет место и у сенполий),
все же они в большинстве случаев сохраняют основные черты
своих диких родичей, в особенности те, которые сформирова-
лись в ходе эволюции под воздействием условий окружающей
среды. Именно поэтому знание условий существования расте-
ний в природе позволяет выбирать такие приемы культуры, ко-
торые наилучшим образом помогают решить поставленную за-
дачу. Излагаемые здесь рекомендации по культуре сенполий
каждый раз будут основываться на приведенных выше сведе-
ниях об особенностях их жизни в природе и на нашем опыте
работы с сенполиями.
Как уже было сказано, все виды сенполий происходят из
Восточной Африки и их распространение связано с определен-
ными условиями, среди которых важнейшими являются особен-
ности освещения, температурный режим, водоснабжение и ха-
рактер субстрата, в котором и на котором развивается корне-
вая система растений.
УСЛОВИЯ ОСВЕЩЕНИЯ
Как известно, в субэкваториальном поясе продолжитель-
ность освещения в течение суток составляет примерно 12 ч.
Интенсивность падающего на поверхность Земли света в целом
высокая и меняется в зависимости от высоты, наличия или от-
сутствия и степени облачности, от особенностей мест произра-
стания растений на открытых местах или под пологом других
растений и т. п.
На тех высотах, на которых обитает большинство видов сен-
нолий, интенсивность света высокая, однако присущая горным
и прибрежным районам восточно-африканского плоскогорья
значительная степень облачности в дневные часы ослабляет ее.
100
Этому же способствует тень, падающая от деревьев и кустарни-
ков, под пологом которых обитают сенполии. Таким образом,
при общей высокой интенсивности падающего света растения
сенполий получают его в четыре раза меньше по сравнению
с растениями открытых мест. На основании этого мы можем
рассматривать сенполии как теневые растения, для которых
необходимо продолжительное, но не очень интенсивное и не
прямое солнечное освещение. Чрезмерное освещение действует
на сенполии отрицательно, что сказывается в заметном сокра-
щении энергии роста, образовании соцветий и в частичном раз-
рушении зеленого аппарата листьев: листья бледнеют, при дли-
тельном освещении даже желтеют, в конечном итоге растения
перестают цвести.
Особенности солнечного освещения на той или иной широте
в разное время года требуют для сенполий различных условий
при их культуре как в оранжереях, так и в жилых помещениях.
В южных районах нашей страны в осенне-зимний период коли-
чество света (продолжительность и его интенсивность) оказы-
вается достаточным для нормального развития растений.
В центральных районах зимой продолжительность освещения
I даже при достаточной интенсивности оказывается недостаточ-
ной. Хотя рост и цветение сенполий продолжается, однако эти
процессы в значительной мере замедлены. В северных районах
нашей страны зимой растения сенполий обычно прекращают
рост и цветение, возобновляя их лишь с улучшением освещен-
ности ближе к весне.
Применение искусственных источников света позволяет вы-
ращивать сенполии круглый год. В качестве искусственных ис-
точников света можно использовать обычные осветительные
лампы накаливания, или дающие лучшие результаты люминес-
центные лампы, или же (самое лучшее) — комбинацию тех и
других.
В наших опытах по применению искусственного освещения
растений в течение круглого года мы применяли 12-часовое
освещение люминесцентными лампами типа ЛДЦ-30 на рас-
стоянии около 40 см от растений при температуре около 25—
26 °C и относительной влажности воздуха 15—50%. Итоги этих
опытов можно суммировать следующим образом: минимальная
освещенность, при которой идет развитие вегетативных частей
растений без резко заметных изменений в их морфологическом
строении, равна 800 лк. Однако при таком освещении цветение
долго не наступает. Оптимальная освещенность, при которой
наблюдается хорошее развитие вегетативных частей, сопровож-
дающееся рано наступающим и интенсивным цветением, оказа-
лась равной 2500 лк. При максимальном освещении в 5000 лк
наблюдались различные отрицательные явления, приводящие
к сокращению, а в отдельных случаях к прекращению роста и,
соответственно, цветения.
юа
Оптимальную освещенность давали 5 ламп мощностью в
30 Вт, подвешенные параллельно на расстоянии от 5 до 10 см
одна от другой на высоте 40—50 см над растениями. Иначе
говоря, это количество ламп освещало участок примерно 0,6 м2,
на котором помещается 15 нормально развитых растений с диа-
метром розетки листьев около 20—25 см.
При выращивании растений в условиях освещения, свойст-
венного Ленинграду, при температуре 18—20°C в качестве ис-
точника дополнительного освещения использовались обычные
лампы накаливания. При освещении такой же площади лампой
мощностью 200 Вт на расстоянии 70 см от растения в течение
5—6 ч наблюдается рост растений и их цветение, но интенсив-
ность того и другого оказывается сравнительно слабой.
Приведенные выше значения освещенности соответствуют,
по-видимому, потребностям растений сенполий в свете, которые
сложились у них в естественной обстановке и которые отража-
ют их приспособленность к жизни в условиях затенения де-
ревьями и кустарниками. Однако диапазон освещенности, при
которой растения сенполий могут существовать без угрозы для
жизни, на основании этих же данных следует считать довольно
значительным.
В весенне-летний период применение искусственных источ-
ников света как в центральных, так и в северных районах ис-
ключается, если только растения не содержатся в темном по-
мещении. Однако в этот период приходится применять различ-
ные способы сокращения интенсивности освещения путем при-
тенения растений, что не требует таких затрат, как искусствен-
ное подсвечивание в осенне-зимний период.
ТЕМПЕРАТУРНЫЙ РЕЖИМ
Температурные условия мест обитания сенполий в природе
обусловлены прежде всего тропическим поясом и в известной
мере высотой над уровнем океана. Среднегодовая температура
близка к 26°C, однако разница между максимальной и мини-
мальной температурами в известные периоды может достигать
значительной величины. Минимальная температура мест обита-
ния сенполий близка к 13°С, при этом растения не испытывают
каких-либо отрицательных воздействий, максимальная же тем-
пература достигает 30 °C.
Исходя из этих данных, а также из наших наблюдений за
состоянием растений при культивировании их в различных
температурных условиях мы можем сказать, что температур-
ный минимум, который сенполии способны выдержать непро-
должительное время без вреда, даже ниже минимума, свойст-
венного их родине, он достигает 8—10 °C.
В Англии (51—56° с. ш.) садоводы обычно круглый год со-
держат сенполии в теплых орхидных оранжереях с температу-
102
рой 26—30°C. По их наблюдениям, в таких условиях растения
цветут обильнее, чем при более низких температурах. Немец-
кие садоводы (48—54° с. ш.) рекомендуют для зимнего содер-
жания сенполий температуру от 16 до 20 °C, а летом — более
высокую. Американские садоводы, которые располагают наи-
более обширным опытом культуры сенполий, рекомендуют для
содержания растений температуру 22—23°C.
По нашим наблюдениям, рост растений идет наиболее ин-
тенсивно при температуре, близкой к 25°С днем и 20° ночью.
Однако надо учитывать характер освещения в наших широтах
(Ленинград) в различное время года (если исключить искус-
ственное дополнительное освещение в осенне-зимний период).
Мы можем рекомендовать содержать растения зимой при тем-
пературе 18—20 °C. В этих условиях наблюдается слабый рост
и цветение отдельных экземпляров, усиливающиеся с улучше-
нием освещения к весне.
ЗЕМЛЯ
В природной обстановке сенполии развивают свою корневую
систему в очень незначительном рыхлом объеме субстрата, яв-
ляющегося продуктом разложения растительных и частично
животных остатков, накапливающихся в трещинах известковых
и гранитных скал. В отдельных случаях сенполии поселяются
на поверхностях обомшелых камней вместе с папоротниками.
Подавляющее большинство, если не все виды, сенполий пред-
ставляют собою полуэпифитные растения, развивающиеся в та-
ких условиях, когда их корневая система размещается в рых-
лом, влажном и хорошо проветриваемом субстрате. Этим по-
следним обстоятельством, по-видимому, и обусловливается
характерная особенность сенполий — мясистость стеблей и
листьев в сочетании с волосистым покровом, защищающим их
поверхность от чрезмерного испарения, особенность, позволяю-
щая им в случае подсыхания субстрата некоторое время суще-
ствовать за счет накопленной в теле влаги.
Поэтому земля, приготовляемая для сенполий, должна от-
вечать следующим условиям: быть питательной (т. е. содержать
достаточное количество органических и минеральных частиц),
в известной мере рыхлой, хорошо проветриваемой и одновре-
менно влагоемкой. Состав земли, применяемой для посадки
сенполий, может быть самым разнообразным.*
Предлагаются различные составы смесей. Приведем неко^
торые из них:
1. Волокнистая глинистая дерновая земля — 2 части, грубый
торф — 1 часть, крупнозернистый песок с добавлением толче-
ного на кусочки древесного угля — 1 часть.
* О стерилизации земли см. с. 116—117.
103
2. Листовой перегной — 4 части, резаный сфагновый мох —
1 часть, крупнотолченый уголь — 1 часть, песок — 1 часть, тол-
ченый доломит (известняк) — 1 часть. 1
3. Листовой перегной — 2 части, навозный перегной — i
1 часть, песок — 1 часть. . I
4. Дерновая земля — 1 часть, подстилочный торф — 2 части,
листовая земля — 1 часть, перегной — 1 часть (или: перегной— )
2 части, листовая земля 3 части), песок— 1 часть. J
5. Хорошая черная земля — 3 части, песок — 2 части, торф— 1
1 часть, компост (ок. 50% удобрений) — ! часть, перегной— I
1 часть. I
На каждые 2 ведра любой из этих смесей следует добавить •
объем горшка 15 см диаметром суперфосфата и 4 л древесной !
золы с кусочками угля. Из перечисленных смесей мы отдаем I
предпочтение четвертой, составляемой нами. Пятую смесь ре-
комендует автор популярной в США книги, посвященной куль- i
туре сенполий, Ван-Пельт Вилсон [Wilson Н. v. Pelt, 1953J. !
Однако мы ее не применяли. '
ОТНОШЕНИЕ К ВЛАГЕ
(поливка)
Как говорилось выше, в природе сенполии селятся на каме- {
нистых склонах вблизи воды или в местах, где постоянно
сквозь трещины сочится вода, или же на поверхности камней,
покрытых хорошо задерживающим воду мхом. Окружающий
их воздух в постоянно увлажняемых местах всегда насыщен
влагой. Иначе говоря, корневая система сенполий требует ре-
гулярного снабжения водой.
Эти сведения помогают правильно решить вопрос о поливке
сенполий при содержании их в культуре. Поливать сенполии
необходимо регулярно, не допуская пересыхания земляного
кома. Частота поливки зависит от температурных условий,
в которых содержатся растения, от интенсивности освещения,
степени влажности воздуха и, естественно, от характера земель-
ной смеси и размеров посуды. При более низкой температуре
и слабом освещении поливать приходится реже, чем при более
высоких температуре и интенсивности освещения. Если земель-
ная смесь содержит достаточное количество элементов, облада-
ющих выраженной влагоудерживающей способностью, и расте-
ния выращиваются в крупных горшках, нужна более редкая
поливка, тогда как при использовании рыхлой песчанистой или
торфяной земли и при малых размерах посуды она должна
быть чаще.
Общей рекомендации относительно поливки дать невозмож-
но, и каждый культиватор исходя из особенностей содержания
растений в данных конкретных условиях должен сам устано-
вить для себя необходимые сроки поливки.
104
Следует отметить, что существенное значение имеет темпе-
ратура воды, применяемой для поливки. При использовании
слишком холодной или слишком теплой воды в случае ее попа-
дания на листья поверхность их покрывается рыжевато-желты-
ми пятнами неправильной формы, которые не исчезают. Тем-
пература поливочной воды должна быть всегда на 2—3° (и не
более чем на 5°) выше температуры воздуха в помещении,
в котором содержатся растения. Всякий застой воды при полив-
ке представляет опасность для растений, поэтому дренаж в
горшках должен быть хорошим (из черепков и из крупнозер-
нистого песка). Поливать можно разными способами: либо с
поддонов, либо с поверхности земли.
В заключение следует сказать несколько слов о других усло-
виях содержания сенполий (помимо освещения, температуры и
поливки). Растения должны располагаться в хорошо вентили-
руемых помещениях, причем не следует допускать непосредст-
венного тока воздуха на растения, сквозняков и резких коле-
баний температуры при вентиляции. Размещать их следует та-
ким образом, чтобы они не соприкасались листьями. Растения
должны содержаться в постоянной чистоте. Отмершие листья,
опавшие цветки и процветшие соцветия нужно регулярно уда-
лять; накапливающуюся на листьях пыль время от времени счи-
щать с помощью мягкой кисточки, а все заболевшие или подо-
зрительные растения изымать и изолировать или уничтожать.
РАЗВИТИЕ КОРНЕВОЙ СИСТЕМЫ
И РАЗМЕРЫ ПОСУДЫ
Как уже было сказано, корневая система сенполий в длину
развита сравнительно слабо и функционирует относительно не-
долго, заменяясь по мере роста растения новой, образующейся
на стеблях после отмирания нижних ярусов листьев. Поэтому
применение для посадки сенполий горшков большого размера
исключается.
В зависимости от возраста растений и их размеров исполь-
зуются горшки диаметром от 5—7 до 12—13 см (в очень ред-
ких случаях для особенно мощно развитых растений — до 15 см).
Развитые цветущие растения лучше всего сажать в горшки диа-
метром 10—11 см.
Кратковременное существование корневой системы застав-
ляет постоянно следить за тем, чтобы длина оголенной части
стеблей не превышала 5—6 см. При большем оголении стебля
необходимо растения пересаживать или переваливать, углуб-
ляя при этом оголенную часть стебля под землю (при пересад-
ке — с освобождением верхнего слоя земли, при перевалке —
105
срезая ножом слой нижней части земляного кома на величину
предполагаемого углубления стебля).
При регулярной пересадке с углублением растения форми-
руют правильно развитые симметричные розетки; в ряде слу-
чаев на стеблях после пересадки образуются боковые ответвле-
ния— дочерние розетки, сосуществующие с материнским ра-
стением. Их можно оставлять, не отсаживая в отдельные горш-
ки. Правда, при этом симметричность растений нарушается,
и в целом они приобретают неряшливый вид. Для сохранения
розеточного облика растений и придания им аккуратного вида
следует боковые ответвления отделять от материнского расте-
ния и сажать отдельно. Это вызывает, естественно, более ча-
стую пересадку растений, что сказывается на них положитель-
но. Можно пользоваться и третьим путем — вырывать или вы-
резать боковые розетки, сажать их в песок для окоренения и
только после этого высаживать в отдельные горшки в земель-
ную смесь.
В условиях нашего Ботанического сада мы практикуем пе-
ресадку не реже двух раз в год, а отдельные экземпляры пере-
саживаем до четырех, а иногда и до шести раз в год; это осо-
бенно относится к молодым и быстрорастущим экземплярам.
Между этими пересадками в случае необходимости применя-
ется перевалка (увеличение объема посуды с добавлением зем-
ли без разрушения старого кома).
После всякой пересадки, с которой связано неизбежное
частичное повреждение корневой системы, проходит некоторый
промежуток времени, необходимый для ее восстановления, —
обычно не более 10—15 дней. По истечении этого срока наблю-
дается интенсивный рост листьев, а за ним — образование цве-
тоносов.
РАЗМНОЖЕНИЕ
Вегетативное, размножение сенполий в условиях
культуры является главным способом их размножения. Оно
происходит путем отделения боковых ветвей, образующих свои
корни, иначе говоря, путем деления, или путем черенкования,
т. е. отделением тех же боковых ответвлений, но до образова-
ния ими своих корней, или же, при необходимости получения
большого числа новых растений, путем черенкования листьями.
- Отделение боковых ответвлений производится во время пе-
ресадок, причем отделению подлежат абсолютно все ответвле-
ния независимо от их размеров и развития самостоятельной
корневой системы. Отдельные ветви, которые имеют достаточно
развитую корневую систему, сажаются в горшки, размеры ко-
торых должны соответствовать степени развития корней. Ответ-
вления, не имеющие корней, используются как черенки и раз-
106
мещаются для укоренения в парник (на укоренение уходит
обычно около 25 дней).
Следует отметить, что сохранение признаков, характеризую-
щих ту или иную культурную форму, возможно только при ве-
гетативном размножении. Поэтому черенкование растений ли-
стьями оказывается единственным путем, с помощью которого
можно получить одновременно большое количество молодых
растений одного варианта. С момента черенкования листьями
до начала цветения обычно проходит срок от 6 до 9 месяцев.
Для черенкования листьями следует брать крупные и хоро-
шо развитые листья — наиболее старые в розетке, т. е. находя-
щиеся в нижних ее ярусах. Листья, имеющие желтоватую
окраску или признаки подсыхания края, использовать не сле-
дует, они редко успевают образовать корни и отмирают. Уста-
новлена следующая зависимость: чем длиннее отрезан черешок,
тем большее количество почек и соответственно молодых расте-
ний формируется на нем (иногда 5—7). По мере срезания че-
решка ближе к пластинке число почек сокращается. В произ-
водственных условиях черенкование лучше всего осуществлять
в разводочном парнике, располагая черенки на расстоянии
около 2—3 см друг от друга в слегка наклонном положении.
Окоренившиеся листовые черенки можно считать готовыми .
к высадке в горшки после того, как листовые пластинки моло-
дых растений, возникших у основания черешка, достигнут не
менее одной трети длины пластинки маточного листа. Окоре-
ненный листовой черенок следует сажать вместе с образовав-
шимися на нем молодыми растеньицами. Маточные листья мож-
но срезать с возможно более длинным черешком и использо-
вать для повторного черенкования.
Если черенкование произведено в весенние месяцы, то выса-
женные группой молодые растения через 3—4 месяца следует
рассадить по одному. Операцию разделения необходимо произ-
водить по возможности осторожно, чтобы повреждение корней
рассаживаемых растений было минимальным. Растения следует
сажать в горшки диаметром 5 см, в рыхлую земельную смесь,
слегка обжимая ее по краю горшка. Рассаженные таким обра-
зом молодые растения обычно через 2—3 месяца начинают
зацветать. В случае же более позднего (летнего и, тем более,
осеннего) черенкования разделение молодых растений можно
осуществлять только весной или летом следующего года. Зимой
при содержании растений в условиях дополнительного освеще-
ния или полностью в условиях искусственного света и ровной
высокой температуры, а также на юге рассаживание растений
можно производить через 2—3 месяца независимо от времени
черенкования. Рассаживание молодых растений при их разделе-
нии рационально (во избежание лишней перевалки) произво-
дить в горшки довольна большого диаметра (около 10 см),
обжимая землю только по краю горшка.
107
Неоднократное черенкование одного листа, взятого с длин-
ным черешком, позволяет получить до 25 молодых растений,
причем каждая из пересаженных групп растений отличается по
возрасту примерно на 1 месяц и достигает цветения почти
одновременно или вскоре одна вслед за другой.
При черенковании небольших количеств листовых черенков
вместо разводочного парника можно воспользоваться обыкно-
венным горшком или иным сосудом, заполненным на 2/3 пес-
ком, который потом накрывается стеклом. Лучше всего брать
два горшка, один большего диаметра, а второй — меньшего.
Большой нужно заполнить промытым песком, а малый углубить
в песок в центре большого. Черенки следует разместить в про-
странстве между краями двух горшков. Увлажнение посажен-
ных в песок черенков производится через малый горшок — пу-
тем вливания в него воды. Такой способ увлажнения песка
исключает смачивание поверхности листовых черенков, что при
густой посадке и содержании под стеклом может привести к
их подгниванию.
Можно производить укоренение листовых черенков и в воде.
В этом случае поверхность сосуда, заполненного водой, затя-
гивается вощеной бумагой или полиэтиленом и обвязывается
ниткой или тонкой резинкой, в бумаге (полиэтилене) делаются
отверстия, соответствующие диаметру черешков черенкуемых
листьев. Через эти отверстия черешки опускают на 1 см в воду.
Обычно воды в сосуде оказывается достаточно до момента пе-
ресадки окорененных черенков в горшки. В случае необходи-
мости добавить воду, бумагу или полиэтилен слегка припод-
нимают вместе с укрепленными в отверстиях черенками.
При таком способе черенкования окорененные черенки могут
считаться готовыми для посадки в горшки только тогда, когда
помимо хорошо сформировавшихся корней разовьются молодые
растения с двумя-тремя парами листьев. Вполне окоренившие-
ся черенки с образовавшимися молодыми растениями перед
посадкой в землю следует аккуратно вынуть из воды вместе
с бумагой или полиэтиленом, разрезать бумагу и извлечь из
нее черенки или извлекать их по одному и сажать в горшки.
Посадку необходимо производить в рыхлую землю с обжима-
нием ее только по краю горшка и затем обильно поливать че-
рез мелкое ситечко тепловатой водой. В дальнейшем поливку
производить, как описано выше, с поддона или сверху, но не
смачивая листьев. Размещать посаженные растения необходимо
на светлом, но защищенном от прямого солнца месте. В неко-
торых случаях, особенно на первых порах, растения приходит-
ся притенять. Цветение растений при таком способе черенкова-
ния наступает обычно спустя 8—9 месяцев, иногда раньше (но
не ранее 6 месяцев).
Семенное размножение — это более кропотливый
по сравнению с вегетативным, требующий большего времени,
108
но и более интересный способ размножения. Он (прежде всего
при работе с культурными формами) дает возможность полу-
чать путем отбора новые формы, отличающиеся от родитель-
ских, и выделять среди них кандидатов в новые сорта.
В плодах, завязывающих семена большей частью только
после искусственного опыления, образуется большое количество
семян. Они очень мелкие, и поэтому требуется соблюдение со-
ответствующих условий для успешного получения всходов.
Посев семян сенполий производится на подготовленную вы-
ровненную и несколько уплотненную, политую поверхность про-
сеянной через мелкое сито земли. После завершения посева
сосуд снабжается этикеткой. Она должна быть таких размеров,
чтобы не мешала покрыть стеклом сосуд с посевом. В этикетке
указываются название растения или его номер и дата посева.
Одновременно аналогичная (но более подробная) запись де-
лается в специальном журнале посевов. Затем посев закрыва-
ется стеклом и ставится в теплое, светлое место (не на солнце!).
Поверхность земли с посевом должна быть всегда влажной.
Увлажнение осуществляется с помощью пульверизатора мелко
распыленной водой (но не струйками). При употреблении для
этого лейки, даже с очень мелким ситом, всегда имеется опас-
ность смыть семена.
Через 10—14 дней, а иногда (при содержании посева в не-
благоприятных температурных условиях) и позже начинают
появляться всходы. Обычно появление всходов растягивается
на довольно продолжительный срок и завершается более чем
через месяц со дня посева.
Всходы очень мелкие, состоят из двух одинаковой величины
семядолей диаметром менее 1 мм. Вскоре один из семядольных
листиков начинает заметно увеличиваться в размерах, тогда
как второй остается неизменным (особенность, присущая всем
растениям семейства Геснериевых). Через некоторое время по-
являются первые два настоящих листа размером около 2—3 мм.
За ними следует вторая пара, располагающаяся перпендику-
лярно первой, и т. д. Поливку всходов следует производить
теплой водой сначала пульверизатором, когда же они образуют
не менее двух пар настоящих листьев — через ситку лейки.
Пикировку всходов можно производить только тогда, когда
появятся не менее 3—4 пар настоящих листьев. Первая пики-
ровка обычно преследует цель разредить всходы, поэтому на-
добность в ней возникает только в том случае, когда при посе-
ве семена были рассыпаны неравномерно. При равномерном
расположении всходов рационально дождаться смыкания их
друг с другом и только тогда рассадить на расстояние около
2 см. Если же надобность в пикировке обнаруживается рань-
ше, то рассаживание надо производить дважды: сначала на
2 см друг от друга, а потом (через 1,5—2 месяца) на 5 см.
В этом возрасте молодые растения можно сажать и в неболь-
109
шие горшки, однако они занимают больше места и в них труд-
нее поддерживать равномерную влажность, поэтому следует
отдать предпочтение плоским ящикам или глиняным плошкам.
По истечении 7—8 недель рассаженные в ящиках растения
разрастаются настолько, что вновь смыкаются своими листья-
ми. В это время растения нужно рассадить в мелкие горшки
диаметром около 7 см.
Через 24—25 недель после посева первые растения начина-
ют зацветать. Первое цветение обычно непродолжительно,
и спустя 3—4 недели процветшие растения надо перевалить в
горшки диаметром 10 см, в которых они остаются до пересадки
(примерно через год).
Во все время развития сеянцев следует их регулярно поли-
вать, удалять отмирающие нижние листья, время от времени
опрыскивать мелко распыленной теплой водой, одновременно
следя за проветриванием помещения, где производится выра-
щивание растений.
БОЛЕЗНИ И ВРЕДИТЕЛИ.
МЕРЫ БОРЬБЫ С НИМИ
Сенполии, как и другие растения, подвержены поражению
различными болезнями и вредителями. Поэтому мы кратко*
остановимся на наиболее часто встречающихся болезнях и вре-
дителях этих растений и основных мерах борьбы с ними.
Из заболеваний или поражений, не связанных с нападением
тех или иных вредителей, мы в первую очередь укажем на
гниль и желто-коричневую пятнистость листьев. Эти заболева-
ния наиболее распространены у неопытных культиваторов.
Гниль. Загнивание стеблей может возникнуть либо после
разделения растений (операции, требующей большой осторож-
ности и чистоты), либо вследствие заболевания корней, пора-
женных вредителями, либо в результате неравномерной полив-
ки (чрезмерная подсушка с последующей обильной поливкой,
отчего отмирает часть жизнедеятельных корней и на их месте
появляются ранки, через которые проникают гнилостные микро-
организмы) .
На первых стадиях загнивание проявляется в потере листья-
ми тургора, они становятся мягкими и слабыми, теряют нор-
мальную окраску и растение подвядает. Такое состояние расте-
ния чаще всего вызывается проникновением в ткани гнилост-
ных микроорганизмов и требует оперативного вмешательства,
т. е. срезания нижней части растения с корнем и помещения
вершины в парник для укоренения.
Для предотвращения загнивания при делении растений их
необходимо слегка подсушить, чтобы пораненные поверхности
обветрились и обсохли (на 1—2 ч), затем посадить их в землю,
ПО
если при делении сохранилось много корней, или, если корней
мало, — в песок в парник. Еще лучше (кроме того) разделен-
ные растения в местах поранений обсыпать либо порошком,
составленным из равных частей соли муравьиной кислоты и
серы, либо толченым древесным углем. Уже пораженные
гнилью места при обнаружении следует срезать острым ножом
и места срезов посыпать одним из вышеназванных порошков.
При загнивании стеблей в результате избыточной поливки
нарушается нормальное снабжение земли воздухом, а с ним и
дыхание корней. Корни частично отмирают, и гнилостные ми-
кроорганизмы, всегда имеющиеся в плодородной земле, легко
проникают в ткани корней и далее, поэтому всякое нездоровое
состояние растений может закончиться подгниванием.
Иногда черешки старых нижних листьев становятся студне-
образными, касаются краев горшка и в этих местах поражают-
ся гнилью. Во избежание этого под черешки следует подстав-
лять подпорки или периодически удалять стареющие листья.
При сильно увлажненном воздухе, особенно в парниках для
черенкования, при касании черенков, при тесном размещении
для окоренения развитых растений, при обильном опрыскива-
нии их, а также когда отпадающие отцветшие цветки своевре-
менно не удаляются, могут подгнивать отдельные листья и,
если их не удалить, гниль может охватить все растение.
Вообще же правильная культура, равномерная поливка,
опрыскивание, поддержание чистоты и соблюдение мер пред-
осторожности, о которых сказано выше, позволят полностью
избежать появления этого неприятного заболевания. Следует
также избегать применения слишком жирных земельных
смесей.
Пятнистость листьев характеризуется желтыми или
коричневатыми, чаще неправильной формы округловатыми пят-
нами на верхней стороне темно-зеленых листьев. Они появля-
ются в результате слишком сильного прямого солнечного осве-
щения или забрызгивания листьев водой переменной темпера-
туры. Пятнистости легко избежать, поливая и опрыскивая ра-
стения водой, температура которой равна температуре воздуха
помещения или не более чем на 3—5° превышает ее, и не под-
вергая их освещению прямым солнечным светом.
Задержка роста — это заболевание сенполий, природа
которого неизвестна. Внешне оно сходно с тем, что мы наблю-
даем при заражении цикламенным. клещом. Листья становятся
короче и шире, чем обычно. Они выглядят более толстыми, как
бы вспухшими и хрупкими. Краевая городчатость листьев сгла-
живается, края подымаются кверху. Эти изменения особенно
заметны на молодых листьях, они делаются блестящими из-за
уменьшения в 2—3 раза длины волосков, черешки укорачива-
ются. При размножении листьями, взятыми с больного расте-
ния, эти особенности больного растения сохраняются. Обработ-
111
ка какими-либо контактными ядами или горячей водой резуль-
татов не дает. Высказываются предположения о вирусном про-
исхождении этого заболевания. При обнаружении этого забо-
левания пораженные растения следует сжигать.
Из поражающих сенполии вредителей наиболее распростра-
ненными являются тли, трипсы, клещи, червецы, поселяющиеся
на листьях и стеблях, и нематоды, поражающие ткани корней,
реже стеблей и листьев.*
Тли обычно группируются на молодых растущих частях
растений. Для своевременного обнаружения тлей и других со-
сущих вредителей необходимо время от времени производить
осмотр и мытье листьев с помощью мягкой кисточки. В каче-
стве хорошей профилактической меры следует практиковать
регулярные опрыскивания тепловатой водой.
Растения, пораженные тлями, нужно изолировать от осталь-
ных растений и подвергнуть опылению водой с боков и снизу,
поворачивая их постепенно вокруг своей оси так, чтобы струйки
воды омывали все пораженные места. В случае же сильного
заражения растений тлями следует опрыскивать их одним из
доступных препаратов для борьбы с сосущими насекомыми
(никотин-сульфат, анабазин-сульфат, тиофос, цветофос, карбо-
фос и т. п.). Этими же препаратами можно пользоваться и для
борьбы с клещиками и отчасти с червецами. При применении
препарата в течение 10 дней можно полностью избавиться от
тлей.
Трипсы. На сенполиях присутствие трипса можно обна-
ружить по белым жилкам на долях околоцветника и в виде
белых пятен или неправильных колец на листьях. Благодаря
применению в последние годы высокоэффективных фосфорорга-
нических препаратов (тиофос, карбофос и рагор) поражение
трипсом в наших условиях стало редким явлением.
При обнаружении трипсов следует применить контактную
обработку растений путем опрыскивания эмульсией карбофоса
(0,1—0,2% по тех. препарату) или тиофоса (0,05—0,1% потех,
препарату).
Трипсов иногда можно обнаружить в личиночной стадии на
цветках сенполий, особенно на пыльниках и внутри них, среди
пыльцы. При этом наблюдается преждевременное опадение
цветков.
Клещи. Землянично-цикламенного клеща скорее можно
обнаружить по его деятельности, нежели заметить при обыч-
ном осмотре растений, так как видеть его можно только с по-
* Кроме перечисленных вредителей иногда у сенполий наблюдается за-
болевание мучнистой росой. Оно проявляется в появлении на поверхности
листьев серых пятен, состоящих из пылящих нитей. Такие растения обычно
оказываются обреченными и их следует сжигать.
112
мощью сильной лупы. Поражает клещик обычно верхнюю сто-
! роиу листьев и, кроме того, конечные почки.
Растения, пораженные цикламенным клещиком, проявляют
признаки заболевания уже на ранних стадиях. Рост таких ра-
стений — карликовый, молодые растущие части (листья) обыч-
но деформируются и уменьшаются в размерах. Волосистость
у больных растений более явственная, чем у здоровых, листья
приобретают чашеобразную форму и становятся очень хрупки-
ми. Fla пораженных растениях могут образовываться только
уродливые цветки или серии бутонов, которые начинают рас-
пускаться и преждевременно опадают.
При подозрении на пораженность клещом следует прежде
всего изолировать заболевшие растения, поливку их произво-
дить с поддона и перед работой со здоровыми растениями обя-
зательно тщательно мыть руки, так как клещи распространя-
ются путем контакта.'
Простейшим способом уничтожения клещей является фуми-
гация, повторяемая каждые 10—12 дней (промежутки между
кладками яиц), при этом уничтожаются появляющиеся молодые
особи. Повторность обработки необходима для полного уничто-
жения всех клещей. Подвергающиеся фумигации растения по-
мещаются в герметическую камеру или герметически закрываю-
щийся ящик, куда кладется порция нафталина (лучше в виде
шариков). В результате нагревания ящика нафталин образует
пары, убивающие клещей.
Цикламенный, или земляничный, клещ, обычно активный в
холодную погоду, нападает на растения зимой или ранней вес-
ной, поэтому наиболее надежными оказываются предупреди-
тельные меры.
Широкий клещ. Если цикламенный, или земляничный,
клещ в большинстве случаев поражает поверхность листьев,
то широкий клещ поселяется на их нижней стороне. Его, так
же как й цикламенного клещика, удается обнаружить только
с помощью сильной лупы, причем приходится раздвигать острым
предметом волоски, покрывающие листья. Пораженные широ-
ким клещом растения образуют деформированные листья с бо-
лее редкой, чем обычно, волосистостью, края которых подгиба-
ются вниз. Особенно много вреда широкий клещ причиняет в
жаркую погоду, продуцируя новые поколения через каждые
4—5 дней. Однако уничтожать его значительно легче, нежели
цикламенного, так как он откладывает свои яйца на поверхно-
сти нижней стороны листа, а не в разных углублениях и щелях,
как это делает цикламенный клещ.
При установлении факта поражения растений широким кле-
щом первой простейшей мерой является опыление нижней сто-
роны листьев мелкой серной пудрой (серным цветом), повто-
ряемое через 4—5 дней до полного уничтожения клеща.
Очень хорошие результаты дает обработка пораженных ра-
8 д. М. ЗАЛЕССКИЙ 113
стений тиофосом, повторяемая через 7—10 дней, или, еще луч-
ше, одно-, двукратное опрыскивание в течение месяца кельта-
ном (4—5 капель на 1 л воды).
Ниже мы приводим один из способов борьбы с клещами,
рекомендуемый многими американскими культиваторами сен-
полий. Его мы проверяли на практике и в общем оцениваем
положительно. Это — тепловой способ. Он заключается в сле-
дующем. Пораженные растения полностью погружают на
15 мин в воду с температурой 44,5 °C. Для этого воду, нагретую
до температуры 45°, наливают в деревянный или металлический
сосуд. Кроме того, нужно приготовить некоторый запас воды,
нагретой до 50 °C. Растение, подлежащее обработке, предвари-
тельно подготавливают, расправив его центральные листья, и
по возможности освобождают поверхность земли для обеспече-
ния глубокого проникновения тепла. Затем, проверив с по-
мощью водяного термометра температуру приготовленной воды
и убедившись, что она равна 44,5°, полностью погрузить горшок
с растением в сосуд и по мере остывания воды добавлять в со-
суд более горячую воду, помешивая деревянной мешалкой и
поддерживая температуру на уровне 44,5°C. При добавлении
воды во избежание ожогов и механических повреждений ли-
стьев вливать ее так, чтобы она не попадала на листья, и разме-
шивать ее, не задевая листьев. Поэтому при выборе посуды для
тепловой обработки необходимо иметь в виду, что ее диаметр
должен превышать диаметр розетки растения не менее, чем в
полтора раза. После 15-минутной обработки растение следует
обмыть, опрыскивая его тепловатой водой и одновременно очи-
щая листья от комочков земли. Затем в течение нескольких
дней растение нужно защищать от света и только после этого
его можно поместить на освещенное место.
По поводу приведенного теплового способа борьбы с клеща-
ми существуют разноречивые мнения. Одни считают, что этот
метод вполне удовлетворителен, другие — что одновременно с
уничтожением клещей он вредно сказывается на состоянии ра-
стений, ослабляет их. Успех обработки растений этим спосо-
бом, конечно, зависит от состояния обрабатываемого растения.
Сильно пораженные клещом растения, ослабленные в резуль-
тате поражения, обрабатывать этим способом несколько риско-
ванно, но это отнюдь не значит, что этот способ следует ис-
ключить. При достаточно внимательном отношении к проведе-
нию обработки, точном соблюдении температуры и выборе не
слишком ослабленных растений он вполне себя оправдывает.
Червецы группируются в розетках растений и на нижней
поверхности листьев. Они разрушают ткани листа, высасывая
клеточный сок.
Простейшим способом уничтожения червецов, конечно, при
сравнительно небольшом количестве пораженных листьев, яв-
ляется поштучное их собирание путем накалывания на иглу,
114
тонко заостренную палочку и т. п. Этим путем в течение не-,
скольких недель можно полностью очистить растения от вре-
дителя.
Можно также применять смачивание отдельных особей чер-
веца спиртом, используя для этого небольшую кисточку. Одна-
ко при этом можно повредить стебли и листья. После обработ-
ки растения необходимо обмыть теплой водой с помощью мелко
распыляющего опрыскивателя, положив их для этого на бок.
Очень сильно пораженные червецом растения целесообраз-
нее всего уничтожать, так как усилия, необходимые для окон-
чательного истребления этих вредителей, оказываются неоправ-
данными.
Практика обработки растений уже упомянутыми контактны-
ми ядами дает положительный результат только в том случае,
если применяется в начальных стадиях поражения. Имеются
сообщения о хороших результатах обработки зараженных ра-
стений раствором рагора, однако этой обработки мы не приме-
няли. Появление на растениях червецов лучше всего предупре-
ждать путем их регулярного опрыскивания тепловатой водой.
Нематоды представляют собой одного из самых опас-
ных паразитов сенполий, так же, впрочем, как и всех геснери-
евых и ряда других растений. Они внедряются в живую ткань
растений и высасывают содержимое клеток, выделяя при этом
ядовитые для тканей растений вещества. В зависимости от ви-
дов нематод это внедрение может быть частичным (эктонемато-
ды) или полным (эндонематоды). Жизнедеятельность нематод
внутри ткани растения вызывает ненормальное деление клеток,
приводящее к нарушению функций пораженных органов.
Наиболее распространенными на сенполиях нематодами яв-
ляются так называемые галловые (Meloidogyne), значительно
реже встречаются листовые нематоды.
Галловые нематоды вызывают большие повреждения корней
и оснований стеблей, иногда даже черешков листьев, образуя
на них вздутия. Эти нематоды обнаруживаются по узелкам
(галлам) на корнях, разрастаниям и ранам у основания стеб-
лей. На ранних стадиях поражения растения выглядят нор-
мальными, позже появляются признаки угнетения: подвядание,
усиленное образование у основания стеблей молодых боковых
побегов, ненормальное утолщение основания стебля и возникно-
вение на нем трещин и ран, способствующих проникновению
внутрь растения гнилостных микроорганизмов. Пораженное не-
матодами растение, осложненное гнилью, через некоторое вре-
мя погибает.
Листовые нематоды проникают внутрь ткани листа, где ак-
тивно передвигаются в межклеточных пространствах. На
листьях образуются темные пятна, увеличивающиеся в разме-
рах, в результате чего пораженный лист постепенно отмирает,
Как и во всех случаях паразитарных заболеваний растений.
8* 115
основное внимание в борьбе с нематодами на сенполиях сле-
дует уделить профилактическим мероприятиям. Источником
заражения нематодами является земля, так как многие виды
нематод паразитируют на растениях открытого грунта. Поэто-
му первой профилактической мерой является стерилизация зем-
ли и посуды, используемых для посадки растений.
При обнаружении (или даже при подозрении) заболевания
растение необходимо изолировать от здоровых. Кроме того, при
работе с растениями необходимо соблюдать максимальную чи-
стоту, постоянно мыть с мылом руки после соприкосновения
с землей и пораженными (или подозрительными) растениями.
Следует иметь в виду, что многие нематоды и их личинки
Рис. 40. Схема простейшего
устройства для пропаривания
способны передвигаться по сла-
бо увлажненным поверхностям,
по песку, гравию, по рассыпан-
ной из горшка или вымытой'при
поливке земле. Если поражен-
ные растения не представляют
собой большой ценности, реко-
мендуется уничтожать их путем
сжигания; в случае же ценности
того или иного образца могут
быть использованы для черенко-
вания только более молодые
листья, находящиеся в верхних
частях стебля.
Наконец, все поступающие
извне растения до размещения
их рядом с остальными необхо-
димо в течение 6—8 недель со-
держать в изолированном месте,
земли. лучше всего в специальном изо-
ляторе. Стерилизация земли и
Посуды может производиться путем пропаривания при темпе-
ратуре выше 100°C в течение 15—20 мин. При. температуре
Нара от 80 до 100° требуется более продолжительное время
Пропаривания — от 1 до 2 часов. Следует иметь в виду, что
равномерность пропаривания зависит от толщины стерилизуе-
мого слоя земли. Она не должна превышать 25—30 см. При
более мощном слое земли требуется больше времени на пропа-
ривание.
В производственных условиях стерилизации земли лучше
Нсего пользоваться специальными почвопропаривающими уста-
новками, состоящими из парогенератора, нагрев которого про-
изводится топливом или электричеством, и бака-пароприемни-
ка, куда загружается земля. Для пропаривания 1 м3 земли
Требуется около 100 кг пара при давлении 0,3 атм, на что рас-
ходуется 15—20 кг твердого топлива или 30—60 кВт/ч электро-
не
энергии. Небольшой слой (около 4 см) земли можно стерили-
зовать, проливая его кипятком. Однако при этом не всегда
имеется гарантия, что вся земля подверглась горячей обра-
ботке.
В качестве доступного всем способа пропаривания земли мы
рекомендуем применяемый нами. Для этого используется ци-
линдрический металлический бак с крышкой (см. рис. 40).
В бак на высоте около 12 см над дном вставляется на круго-
вой опоре второе дно, имеющее по всей поверхности отверстия
диаметром около 3—4 см и обтянутое мелкой (с ячейками до
5 мм) металлической сеткой. В пространство между первым и
вторым дном вливается горячая вода, затем поверх сетки засы-
пается земля, бак закрывается' крышкой и ставится па огонь.
Кипящая на дне бачка вода образует пар, который проникает
сквозь слой земли и стерилизует ее. При этом земля не должна
быть слишком сухой или мокрой. С момента начала кипения
воды в бачке до окончания обработки земли проходит 2—3 ч.
Объем земли, обрабатываемый за один прием, равен объему
земли двух пикировочных ящиков (60X40X10).
Посуда, употребляемая для посадки растений, стерилизуется
путем пропаривания или кипячения в воде в течение 10—15 мин.
Одним из термических способов стерилизации земли явля-
ется прокаливание ее на неглубоких противнях в течение 30—
40 мин при помешивании на огне при температуре около 90°C,
После остывания земли ее следует проветрить, несколько раз
перемешав. Спустя три дня землю можно использовать для по-
садки. Опасение, что при такой обработке будет уничтожена
вся микрофлора, имеющая значение для жизни растений, обыч-
но не оправдывается, так как вскоре после остывания микро-
флора в земле восстанавливается.
Что касается химических методов борьбы с нематодами, то
применительно к горшечным растениям они недостаточно раз-
работаны. Имеются указания о благоприятных результатах об-
работки земли фосфорорганическими препаратами типа тиофо-
са, рагора и др. (фосфаты легко поглощаются растениями).
ПРИЛОЖЕНИЯ
I. ВНЕШНИЕ ПРИЗНАКИ ЗАБОЛЕВАНИЙ СЕНПОЛИЙ
И ВОЗМОЖНЫЕ ИХ ПРИЧИНЫ
Признаки
Растем и я:
подвядают
Неплотно держатся в горшках
Подвядают, не реагируют на
поливку при здоровых корнях
появление большого числа
Молодых боковых ответвле-
ний у основания стеблей, ча-
сто ненормально утолщенных
Листья:
искривленные
пятнистые от округлых жел-
товатых или коричневатых
разводов и пятен
темно-коричневые пятна на
нижней стороне листьев, уве-
личивающиеся и проявляю-
щиеся на их верхней стороне
бледные зеленые с подверну-
тым вниз краем, часто бле-
стящие
бледно-зеленые, мелкие и ис-
кривленные, с краями, подня-
тыми вверх
скопление белого пуха, а так-
же часто подвижных, напо-
минающих мокриц, белова-
тых насекомых, при раздав-
ливании красных, на нижней
(18
Возможная причина
Подгнивание корней, стеблей
как следствие избыточной по-
ливки или обильной поливки
после пересушки, заражение
нематодами
Нематоды или гниль корней
как следствие поражения и
нарушения режима поливки
Стеблевая гниль
Ранние стадии заражения не-
матодами, рано или поздно
завершающегося гибелью ра-
стения
Тли
Повреждение водой низкой
или высокой температуры или
содержание на сильно осве-
щенных солнцем местах
Листовая нематода
Широкий клещ
Цикламенный клещ
Червец
стороне и у оснований ли-
стьев
утолщенные, блестящие, мел-
кие, хрупкие, края подняты
кверху
Цветки и бутоны:
искривленные, мелкие, блед-
ные, распускаются и сразу
опадают
бледные, коричневатые или
почти сухие, не успевающие
распускаться и опадающие
серая плесень на бутонах
Остановка роста
(возможно, вирус)
Ц и к л а м е 11 н ы й клещ
Трипс, клещи
Корни отмирают
Бутонная гниль как резуль-
тат плохой вентиляции при
высокой влажности воздуха
Корневая гниль как резуль-
тат избыточной поливки или
поражения нематодами, или
плотной плохо проветривае-
мой земельной смеси, обиль-
ной поливки после пересушки,
особенно при использовании
сильноторфянистой земельной
смеси, которая после пересы-
хания плохо смачивается
2. НЕКОТОРЫЕ ОБЩИЕ РЕКОМЕНДАЦИИ
ДЛЯ УСПЕШНОЙ КУЛЬТУРЫ СЕНПОЛИЙ
1. Размещать растения в местах, не доступных прямому сол-
нечному освещению,, в особенности полуденному. На солнце на
листьях появляются непроходящие желтоватые пятна, портя-
щие вид растений, или они желтеют целиком.
2. Летом размещать растения на не освещаемых солнцем
местах в хорошо проветриваемом, но не прохладном помеще-
нии, на юге после постепенного приучения — на подоконниках
открытых окон.
3. Содержать растения при температуре от +22 до +25 °C
с понижением ночью не более чем на 5°.
4. Оберегать растения, от резких охлаждений, в том числе
при проветривании помещения.
5. Поливать растения водой, температура'которой равна или
не более чем на 5° выше или ниже температуры воздуха в по-
мещении, в котором они содержатся.
119
6. Увлажнять помещения, где содержатся растения, если
окружающий воздух заметно сух или если температура воздуха
превышает +25°C. Увлажнение осуществлять с помощью пуль-
веризатора или иного мелко распыляющего опрыскивателя
теплой водой ( + 27—30°С в зависимости от степени повыше-
ния температуры воздуха). Еженедельное опрыскивание расте-
ний мелко распыленной теплой водой способствует росту ли-
стьев. Употребление холодной или слишком горячей воды при-
водит к появлению на листьях портящих вид желтых или- ко-
ричневых пятен, которые не удаляются.
' 7. Удалять опавшие цветки, процветшие цветоносы, подсы-
хающие с краев и желтеющие листья нижних ярусов.
8. Стерилизовать землю, употребляемую для посадки расте-
ний, а также посуду, в которую их сажают. Это обезопасит
растения от заражения нематодами. Стерилизовать следует
также полки и стеллажи, на которых размещаются растения,
ошпаривая их два-три раза кипятком.
9. Изолировать все больные, подозреваемые и поступающие
извне растения на срок 2 месяца, в течение которого проверить
их на зараженность и в случае ее обнаружения принять необ-
ходимые меры. При обнаружении сильного заражения расте-
ния сжечь.
10. Для уничтожения клещей, трипсов, тлей, червецов при-
менять опрыскивание (опыление) теплым раствором тиофоса,
карбофоса, кельтана (от клещей), препаратом Цветофос или
делать горячие ванны (против клещей). При этом строго со-
блюдать пропорции и правила пользования препаратами, пом-
ня, что все они ядовиты для человека и животных.
11. Во избежание заражения и его распространения строго
соблюдать чистоту, при соприкосновении с непроверенными,
подозреваемыми или больными растениями тщательно мыть
руки с мылом.
12. Не применять крупной посуды. Молодые растения са-
жать в сосуды диаметром 7—10 см, старые — в сосуды диамет-
ром 12—13 см.
13. Поливать растения равномерно, не пересушивать и не
переливать, соблюдать первое правило поливки — поливать
так, чтобы вода смочила весь земляной ком и прошла насквозь.
При поливке к каждому растению подходить индивидуально,
учитывая его состояние, размеры посуды, характер земельной
смеси, примененной для посадки, и т. п.
14. К работе с растениями подходить внимательно и вдум-
чиво, какая бы черновая она ни была. Работая, наблюдать. Не-
внимательный подход приводит к отрицательным результатам.
•120
3. к ВОПРОСУ О КЛАССИФИКАЦИИ
КУЛЬТУРНЫХ ФОРМ СЕНПОЛИЙ
Многие любители культурных форм сенполий в СССР пы-
таются отыскать и установить те основные признаки, ио кото-
рым можно было бы различать и классифицировать эти расте-
ния. Необходимо сразу же предупредить, что задача эта крайне
трудная, так как исходные растения попали к нам в страну в
большинстве своем с утраченными названиями сортов и полу-
чили новые имена, данные им их обладателями по своему вкусу
и разумению.
Эти исходные формы послужили материалом для получения
при выращивании из семян многих отклоняющихся от материн-
ских растений вариаций, которые распространяются также под
произвольно данными им названиями.
Многие из таких вариаций представляют собою повторение
уже известных и зарегистрированных сортов и поэтому давае-
мые им произвольные названия являются ненужными синони-
мами. Чтобы выяснить, можно ли их считать сортами, их нужно
подвергнуть проверке и сортоиспытанию, которое у нас для
сенполий не организовано. Распространяемые же под видом
сортов вариации могут до этого называться только сеянцами,
полученными тем или иным автором, и числиться под тем или
иным номером, с указанием родительских растений или того,
что они получены в результате свободного опыления.
Для облегчения задачи классификации сеянцев, выращивае-
мых любителями в СССР, позволю себе кратко ознакомить чи-
тателей с принципами, которые положены в основу классифи-
кации сеянцев и новых сортов в США, где в конце 40-х годов
было организовано специальное Общество Африканских фиа-
лок Америки (African Violet Society of America, Inc.).
Тогда же несколько первоначально выведенных сортов были
зарегистрированы специальным классификационным комитетом,
позже получившим наименование Номенклатурного комитета
(Nominating Committee) в составе упомянутого Общества.
Комитет различает растения сенполий по четырем основным
признакам:
1) окраске цветков, 2) типу листьев, 3) типу цветков,
4) размерам растений.
В первой группе различают девять окрасок цветков, обозна-
чая каждую из них той или иной буквой: В — голубая, синяя
(Blue), С-—разноцветная (Multicolor), Р — ярко-розовая или
темно-розовая (Pink or Rose), О — орхидейная, розово-лило-
вая, лавандовая, т. е. бледно-лиловая (Orchid, Mauve, Leven-
der), P — красная, бурачно-каштановая, сливовая или цвета
бургундского вина, винная (Red, Maraon, Plum, Burgundy),
V — фиолетовая или пурпурная (Violet or Purple), W — белая,
кремовая, или подрумяненная, т. е. слегка розовая (White,
121
Creamy or Blush), X — двуцветная (Bicolor) и Y — желтая, no
существу белая с желтоватым оттенком (Yellow).
Во второй группе признаков различают также девять типов
листьев, давая каждому типу свой номер: 1—длиннолистные
или паукообразные (Longifolia or Spider), 2 — обычные, пло-
ские простые (Plain), 3 — стеганные (Quilted), жилки этого
типа листьев настолько углублены, что получается впечатление
простеганной поверхности, 4 — типа Girl, Semi-Girl (форма
листьев одного из первых сортов с этим именем)—обратно-
овальные, несколько сложенные по средней жилке, в виде со-
вочка, 5 — гофрированные, бахромчатые, волнистые, выемча-
тые или гребенчатые (Ruffled, Fringed, Wavy, Fluted or Scalo-
ped), 6 — типа Supreme Amazon or Du Pont — крупные, как
у сортов с этими названиями (группу Дю-Понт представляют
собою полиплоидные растения, названные по фамилии получив-
шего их автора. Они характеризуются крупными мясистыми
листьями светло-зеленой окраски, густо покрыты волосками и
имеют синие, лиловато-розовые или лиловые цветки), 7 — раз-
ноцветные, пестрые (Variegated), 8 — ложковидные, овальные,
чашеобразные (Spooned, Ovate, Cupped up), 9 — заостренные,
остроконечные (Pointed).
В третьей группе признаков различают цветки: s — простые
(одинакие) (Single), d — махровые и полумахровые (Double
or Semidouble), с — звездообразные (Star shaped), f — гофри-
рованные или бахромчатые (Fringed or Ruffled).
Четвертую группу признаков составляют растения, разли-
чаемые по размерам: М — миниатюрные или почти миниатюр-
ные (Miniature or Semiminiature), S —средние, стандартного
размера (Standard size), L — очень крупные (Very large).
Каждый из сортов после проверки получает регистрацион-
ный номер с датой регистрации и код, составленный из тех букв
и цифр, которые характеризуют его черты.
Так, например, V. 6.3.S.L — фиолетовая, Амазонская, сте-
ганная, цветки одинакие, очень крупная — соответствует сорту
«Асе of Cedar Rapids», №834—1956, S. A. Nemec (оригинатор).
P.6.S.L — розовая, Амазонская, цветки одинакие, очень
крупная — сорт «Amason Pink Delight», № 10—1953, автор Mrs.
George Vallina (оригинатор).
Т АБЛИЦЫ
Табл. I. Очертание листьев и характер их жилкования с верх-
ней и нижней стороны у S. orbicularis (1), S. orbicularis, var. pur-
purea (2). Ум. иа 1/10 иат. вел.
123
Табл. II. Очертание листьев и характер их жил-
кования с верхней и нижней стороны у S. niti-
da (1)', S. ionantha (2); S. tongwensis (3). Ум. на
1/10 нат. вел.
Табл. Ш. Очертание листьев и характер их
жилкования с верхней и нижней стороны у S. tei-
tensis (/); S confusa (2); S. pendula (3). Ум. на
1/10 нат. вел.
125
Табл. IV. Очертание листьев и характер их жилкования-
с верхней и нижней стороны у S. pendula, var kizarae (/);
S. magungensis (2); S. amaniensis (3). Ум. на 1/10 нат. вел.
126
Табл. V. Очертание листьев и характер их жилкования
с верхней и ннжней стороны у S. brevipilosa (1); S. veluti-
па (2); S. diplotricha (3). Ум. на 1/10 нат. вел.
127
Табл. VI. Очертание листьев и характер их
жилкования с верхней и нижней стороны у S. in-
termedia (/); S. goetzeana (2); S. shumensis (3).
Ум. на 1/10 нат. вел.
128
Табл VII. Очертание листьев и характер их жилкования с верхней
и нижней стороны у S. difficilis (/); S. rupicola (2). Ум. иа 1/10 нат.вел.
9 Д. М. ЗАЛЕССКИЙ
139
Табл. VIII. Очертание листьев н характер их жилкования с верхней
и нижней стороны у S. grandifolia (/); S. grotei (2). Ум. на 1/10 нат. вел.
Табл. IX. Характер волосистого покрова листовых пластинок у S. та
gungensis (/); S. nitida (2); S. amaniensis (3); S. ionantha (4); S. orbicu
laris (5); S. velutina (6); S. diplotricha (7); S. confusa (3); S. difficilis (9}
S. intermedia (10); S. goetzeana (11); S. grotei (12).
)30
ГЦ
7
Табл. X. Характер волосистого покрова листовых пластинок у S. rupl-
cola (/); S. brevipilosa (2); S. tongwensis (3); S. shumensis (4); S. grand/.-
folia (5); S. teitensis (ff); S. pendula (7).
Табл. XI. Цветоносы S. grandifolia (/); S. orbicularis (2);
S. shumensis (3). Ум. иа 1/10 нат. вел.
!32
133
3
Табл. XII. Цветоносы 5. orbicularis var. purpurea in-
S. confusa (2); S. difficllis (3). Ум. на 1/10 нат. вел.
Табл. ХШ. Цветоносы S. Intermedia (/); S. nitlda (2); S. dtp-
lotricha (3). Ум. на 1/10 иат. вел.
135
136
137
В“етоносы S- ruPicola (') = 5. velutina (2); S. ionantha
'.LVJ W. Ум. на 1/10 нат. вел.
138
Табл. XVII. Очертание цветков и вид сбоку S. tongwensis (/);
5 pendula (2); S. Ionantha (3); S. magungensis (4)| S. veliM-
na (5); S. amaniensis (3). Ум. на 1/10 нат. вел.
139
G
Табл. XVIII. Очертание цветков и вид сбоку S. grotei (1);
Л. teitensis (2); S. diplotricha (3); S. shumensis (4); S. difficilis
(5); S. intermedia (6). Ум. иа 1/10 нат. вел.
140
Табл. XIX. Очертание цветков и вид сбоку S. confusa (/);
S. rupicola (2); S. nitida (5); S. brevipilosa (#); S. grandifolia (5);
S. orbicularis (6); S. goetzeana (7). Ум. на 1/10 наг. вел.
141
Таблица XX. Плоды (с двух сторон) S. ionantha (/); S. pendula (2);
S. velutina (3); S. intermedia (4); S. tongwensis (5); S. brevipilosa (6);
S. shumensis (7); S. confusa (8); S. difficilis (9); S. rupicola (10); S. gran-
difolia (11); S. amaniensis (12); S. magungensis (13); S. grotei (14);
S. pusilia (15); S. diplotricha (16); S. orbicularis (17); S. nitida (18); S.
teitensis (19); S. goetzeana (20); S. inconspicua (21). (puc. 15, 20 и 21
воспроизведены по описаниям и неточным изображениям). Ум. на 1/10 нат. вел.
142
ЛИТЕРАТУРА
Берг Л. С. Основы климатологии. Л, 1938. 456 с.
Воейков А. И. Климаты земного шара. — В кн.: Воейков А. И. Из-
бранные сочинения. М.; Л.: Изд-во АН СССР, 1948, т. I, с. 163—677.
Ган Ф. Африка. СПб., Просвещение, 1903. 600 с.
Залесский Д. М. Сенполии. — Цветоводство, 1965, № 10, с. 18—20.
Иванина А. И. Применение карпологического метода в систематике
сем. Gesneriaceae Dum. — Бот. журн., 1965, т. 50, № 1, с. 29—43.
Иваиина Л. И. Семейство геснериевых: Карпологпческий обзор. Л.:
Наука, 1967, с. 3—115.
Ричардс П. У. Тропический дождевой лес / Нер. с англ. М., 196Е
448 с.
Тахтаджян А. Л. Морфологическая эволюция покрытосеменных.
М., 1948. 301 с.
Тахтаджян А. Л. Основы эволюционной морфологии покрытосемен-
ных. М.; Л., 1964. 236 с.
Толмачев А. И. Основы учения об ареале. Л., 1962. 100 с.
Bald J. С., Chandler Ph. A., Bowmann Kenneth. Stunt disease
of African violets. — Gesn. Saintp. News, Jan. — Febr., 1964, p. 45—47.
Beddome G. H. Gesneriaceae: with annotated list of the genera and
species which have been introduced to cultivation.— J. Roy. Hort. Soc., 1908,
vol. XXXIII, p. 74—100.
Burtt B. L. Plants new noteworthy species of Saintpaulia. — Gard.
Chron., 3 ser., 1947, vol. СХХП, p. 22—23.
Burtt B. L. Plants new noteworthy species of Saintpaulia. — Curtis Bot..
Mag., 1948, nov. ser., vol. CLXV, pt. I, tab. 11.
В u r 11 B. L. A confusion in African violets. — Baileya, 1956, vol. 4,
p. 163—164. -
Burtt B. L. Studies in Gesneriaceae of the old world: the genus Saint-
paulia.— Notes Roy. Bot. Gard. Edinburgh, 1958, vol. XXII, N 6, p. 547—568.
Burtt B. L. Studies in Gesneriaceae of the old world: Additional notes
on Saintpaulia. — Notes Roy. Bot. Gard. Edinburgh, 1964, vol. XXV, N 3,
p. 46—47.
Burtt B. L. Studies in Gesneriaceae of the old world: Additional notes
on Saintpaulia. — Gesn. Saintp., News, Nov. — Dec., 1964, p. 46—47.
Clayberg D. Hybridizing with African violet species. — Afr. VioL
Mag., 1961, vol. 15, N 1, p. 105—107.
Clayberg D. Hybridizing result with Petrocosmea. — Afr. Viol. Mag.,
1962, vol. 16, N 1, p. 107.
143
Clarcke С. В. — In: Oliver, Fl. trop. Afr., 1906, IV (2), p. 501.
Cox Harvey, Roberts E. Saintpaulia tongwensis. — Afr. Viol. Mag.,
1950, vol. 3, N 4, p. 5.
Engler A. Die von W. Goetze u. Dr. Stukmann in Uluguru gebierge...
des Pflanzen. — Bot. Jahrbiicher, 1901, N 28, S. 481—482.
Engler A. Gesneriaceae africanae. — Bot. Jahrbiicher, 1921, N 57.
Fritsch K. Gesneriaceae. — In: Engler A. u. Prantl K. Die natiirliche
Pflanzenfamilien, 1893—1894, Bd IV, Abt. 36.
F u s s e 1 С. P. Chromosome number in the Gesneriaceae. — Baileya, 1958,
vol. 6, N 2, p. 117.
F u s s e 1 С. P. Chromosome number in the Gesneriaceae: Saintpaulia. —
Afr. Viol. Mag., 1961, vol. 14, N 3, p. 107.
G r i 11 i M. Nuove specie et varieta di plante da fioritura. — Bull. Soc.
Toscano Ortic., 1894, Anno XIX, A 13, fig. 1.
Ha ar er A. E. Saintpaulias in the wild. — Garden Chron., 1955,
vol. 138, p. 172.
Hooker J. D. Saintpaulia ionantha H. Wendl. — Curtis Bot. Mag., 1895,
vol. 51, t. 7408.
Howard E. Cytological studies in Saintpaulia Wendl.— Amer. J. Bot.,
1958, vol. 45, N 8.
Huber E. Beitrag zur anatomischen Untersuchung der antheren von
Saintpaulia. — Sitzungsber. Osterr. Akad. Wiss., Math. Naturw. К.1., 1953,
Abt. i, p. 162.
Lawrence G. H. M. Discussion on botanical names of cultural plant.—
In Gents Herb., 1949, vol. VIII, fasc. 1, p. 68—69.
Lightbourn R. E. The Saintpaulia species. — Gesn. Saintp. News,
1964, Nov. — Dec., p. 28—31.
Moore H. African violets, Gloxinias and their relatives. New York, 1957.
Naylor E. E., Johnson B. A histological study of vegetative repro-
duction in Saintpaulia.—Amer. J. Bot., 1937, vol. XXIV, p. 673.
Naylor E. E. African violets for the window garden. A note on their
culture and propagation. — J. New York Bot. Gard., 1940, vol. 41, N 487,
p. 166—168.
Neubert Gart. Mag. Saintpaulia ionantha H. Wendl., 1894, p. 362,
fig- 97.
Neubert Gart. Mag. Saintpaulia ionantha H. Wendl., 1898, vol. LI,
p. 193.
Pynaert E. Saintpaulia ionantha H. Wendl. — Rev. Hort. Beige, 1894,
t. XX, p. 109—110, cum ic.
Rector С. K. Some notes on Saintpaulia species. — Afr. Viol. Mag.,
1961, vol. 14, N 3, p. 87.
Reed S. C. Genetic sheck-list.—Afr. Viol. Mag., 1961, vol. 14, N 3,
p. 104—106.
Reed S. C. Research report.— Afr. Viol. Mag., 1962, vol. 16, N 1,
p. 105—106.
Reed S. C. African violet genetics.—J. Hered., 1954, vol. XLII, p. 225—
230.
Roberts E. A new species of Saintpaulia. — Afr. Viol. Mag., 1950a,
vol. 3, N 4, p. 6—7.
Roberts E. Saintpaulia amaniensis a new species of Saintpaulia.—
Afr. Viol. Mag., 1950b, vol. 4, N 2, p. 6—7.
Rogers О. M. Some chromosome counts in the Gesneriaceae. — Baileya,
1954, vol. 2, N 1, p. 14—18.
Saunders E. R. Floral anatomy and its morphological interpretation.—
New Phytologist, 1934, vol. XXXIII, p. 127—170. (Saintpaulia — p. 153).
Wendl and H. Saintpaulia ionantha. — Gartenflora, 1893, vol. XLII,
p. 321—324, tab. 1391, fig. 66.
Wilson J. H. Observations on the flowers, fruit and seedlings of Saint-
paulia ionantha H. Wendl. — Bot. Jaarboek, 1898, p. 86.
Wilson H. v. Pelt. The complete book of African violets. New York,
1953.
144
Оглавление
Предисловие....................................................... 3
Глава 1. Общие сведения.............................................5
Краткий очерк открытия и описания видов рода Saintpaulia
Н. Wendl. (5). Физико-географическая характеристика области
распространения рода Saintpaulia Н. Wendl. в Восточной Афри-
ке (9). Краткая характеристика растительных формаций Восточ-
ной тропической 'Африки (12). Характеристика мест обитания
сенполий (14). Особенности географии представителей рода
Saintpaulia Н. Wendl., ареал рода и отдельных видов (17)
Глава 2. Систематика и морфология..................................20
Систематическое положение рода Saintpaulia Н. Wendl. (20). Об-
щая характеристика рода Saintpaulia Н, Wendl. (21). Некоторые
биологические особенности сенполий и попытка их объясне-
ния (43). Некоторые соображения о взаимоотношениях внутри
рода Saintpaulia Н. Wendl. (49)...........................
Глава 3. Описание отдельных видов..................................52
Таблица для определения видов рода Saintpaulia Н. Wendl. (для
культивируемых растений) (52). 1. Saintpaulia inconspicua
В. L. Burtt (57). 2. Saintpaulia pendula В. L. Burtt (57). 2a. Sa-
intpaulia pendula var. kizarae B. L. Burtt (59). 3. Saintpaulia
intermedia B. L. Burtt (60). 4. Saintpaulia goetzeana A. Engler
(61). 5. Saintpaulia grotei A. Engler (63). 6. Saintpaulia ama
niensis E. Roberts (65). 7. Saintpaulia magungensis E, Roberts
(67). 8. Saintpaulia pusilia A. Engler (70). 9. Saintpaulia sliu-
mensis B. L. Burtt (71). 10. Saintpaulia teitensis B. L. Burtt (74).
11. Saintpaulia ionantha H. Wendl. (75). 12. Saintpaulia tongwen-
sis B. L. Burtt (78). 13. Saintpaulia grandifolia B. L. Burtt (80).
14, Saintpaulia velutina B. L. Burtt (82). 15. Saintpaulia diplo-
tricha B. L. Burtt (84). 16. Saintpaulia rupicola B. L. Burtt (86).
17. Saintpaulia brevipilosa B. L. Burtt (88). 18. Saintpaulia niti-
da B. L. Burtt (89). 19. Saintpaulia orbicularis B. L. Burtt (91).
19a. Saintpaulia orbicularis, var. purpurea B. L. Burtt (93).
20. Saintpaulia difficilis B. L. Burtt (94). 21. Saintpaulia confusa
B. L. Burtt (96). i t
Глава 4. Вопросы культуры..........................................99
Условия освещения (100). Температурный режим (102). Зем-
ля (103). Отношение к влаге (поливка) (104). Развитие корне-
вой системы и размеры посуды (105). Размножение (106). Бо-
лезни и вредители. Меры борьбы с ними (ИО)................
Приложения........................................., , ,1:18
1. Внешние признаки заболеваний сенполий и возможные их
причины (118). 2. Некоторые общие рекомендации для успешной
культуры сенполий (119). 3. К вопросу о классификации куль-
турных форм сенполий (121)................................
Таблицы......................................................... 123
Литература . , ............................... 143