/






















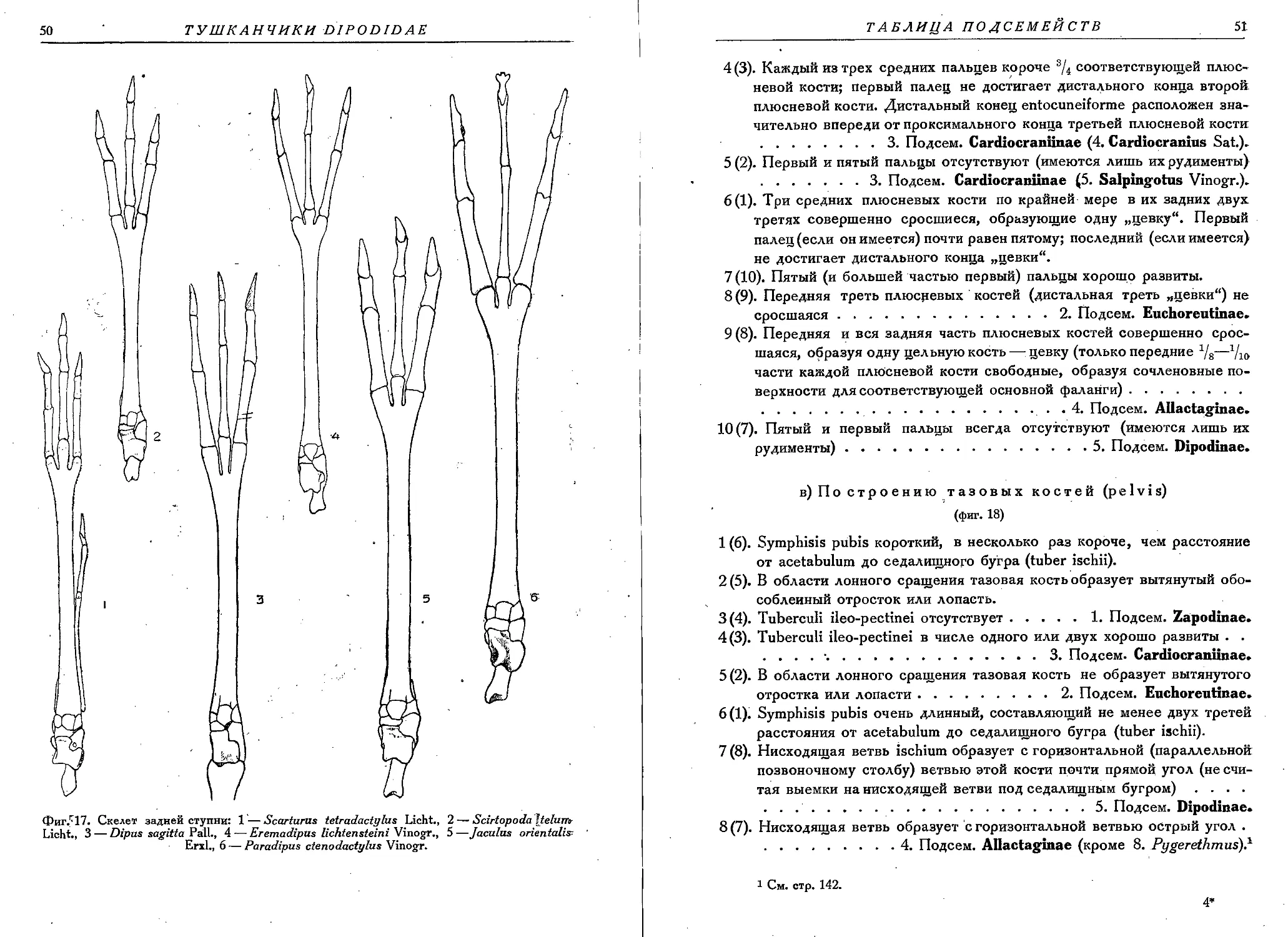






































































Текст
JNSTITUT ZOOLOGIQUE DE L’ACADEMIE DES SCIENCES DE L’URSS
NOUVELLE SERIE n° 13
FAUNE DE L’URSS
Sous la direction de S. A. SERNOV, de I*Academic
Redige par A. A. STACKELBERG
M|AMMIFfiRES
Vol. Ill, n° 4
Fam. DIPODIDAE
par B. S. VINOGRADOV
EDITION DE I’ACADEMIE DES SCIENCES DE L’URSS
MOSCOU • 1937 • LENINGRAD
ЗООЛОГИЧЕСКИЙ ИНСТИТУТ АКАДЕМИИ НАУК СССР
НОВАЯ СЕРИЯ №13
ФАУНА СССР
Главный редактор акад. С. А. ЗЕРНОВ
Редактор А. А. ШТАКЕЛЬБЕРГ
МЛЕКОПИТАЮЩИЕ
Том Ш, вып, 4
Б. С. ВИНОГРАДОВ
ТУШКАНЧИКИ
ИЗДАТЕЛЬСТВО АКАДЕМИИ НАУК СССР
МОСКВА . 1937 • ЛЕНИНГРАД
Напечатано по распоряжению Академия Наук СССР
Июль 1937 г.
Директор Зоологического Института акад. С. А. Зерноа
Редактор издания А. А. Штакельберг
Технический редактор н ученый корректор Н. Г. Редько.
Сдано в набор 25 декабря 1936 г. — Подписано к печати 14 июля 1937 г.
VIII -г- 198 стр.
Формат бум. 72 X 1Ю см- — 13 печ. л. — 15.62 уч.-авт. л. — 48055 тип. зн. — Тираж 1500
Ленгорлит № 3321. — РИСО № 264. — АНИ № 1485. — Заказ № 15
Типография Академии Наук СССР. Ленинград, В. О., 9 линия, 12
ПРЕДИСЛОВИЕ
В течение более десятка лет, на ряду с другими группами мелких мле-
копитающих, мною было уделено значительное внимание изучению одного,
из наиболее своеобразных семейств грызунов — тушканчиков (Dipodidae)\
эта группа с давнего времени привлекала внимание многих исследовате-
лей своими своеобразными чертами организации, резко выраженными
приспособлениями к обитанию в пустынях и чрезвычайно интересными,
биологическими особенностями. Эта группа оказалась также весьма инте-
ресной в систематическом отношении; несмотря на чрезвычайно резко
выраженные признаки отличия между отдельными видами, родами и под-
семействами, общая классификация современных представителей этой
группы оставалась в весьма неудовлетворительном состоянии; еще более
неясным оставался вопрос о взаимоотношениях между известными
в настоящее время ископаемыми представителями этой группы. Следует
также отметить, что эта группа в систематическом отношении была столь
слабо изучена, что до недавнего времени оставался неизвестным не
только ряд резко обособленных видов, но даже родов. Лично мне за
последнее десятилетие удалось установить, кроме нескольких хороша
обособленных новых видов, три своеобразных новых рода и одно под-
семейство; из них два рода и три вида принадлежат к фауне нашей
страны.
Я далек от мысли считать свои исследования по систематике, мор-
фологии и отчасти экологии этой группы законченными, но все же надеюсь,
что некоторые из результатов этой работы могут способствовать даль-
нейшему углубленному изучению этой группы, в чем я предполагаю при-
нять также посильное участие.
В течение моих работ над Dipodidae имелся ряд благоприятных
условий, которыми я старался воспользоваться с возможной полнотой.
Среди этих условий особенно следует отметить наличие в Зоологическом
Институте Академии Наук СССР богатейшей коллекции по рассматри-
ваемой группе, особенно обильных спиртовых материалов, позволивших
выйти за пределы обычно применяемых систематиками стандартных
признаков, наблюдаемых на шкурках и черепах, и использовать ряд при-
знаков внутренней организации этих животных. Исключительно благо-
приятным для этой работы условием была также возможность личного
VI
ТУШКАНЧИКИ DIPODIDAE
просмотра всех материалов по Dipodidae не только в важнейших совет-
ских музеях, но также в крупнейших музеях Западной Европы — Есте-
ственно-Историческом Музее в Берлине и Британском в Лондоне; осо-
бенно ценным было ознакомление в этих музеях с многочисленными
оригинальными экземплярами, по которым были установлены различными
авторами многие новые роды и виды, а также с экземплярами из тех
•стран, откуда в советских музеях материалы были слишком скудны или
отсутствовали вовсе, как, например, Малая Азия, Аравия, Месопотамия,
Иран, Афганистан, Белуджистан, Сахара и пр.
В своих исследованиях по рассматриваемой группе я обращал наи-
большее внимание на изучение взаимоотношений между более крупными
систематическими подразделениями—подсемействами, родами и видами,
полагая, что установление низших таксономических групп в рассматривае-
мом семействе еще не может быть осуществлено в настоящее время
с достаточной точностью в виду недостатка для этой цели коллекционного
материала в количественном отношении и в отношении совершенства его
сохранности, а также в связи с отсутствием общепринятого взгляда
по вопросу о низших таксономических единицах; однако, где возможно,
-я делал попытки дать некоторое освещение общим закономерностям гео-
графической внутривидовой изменчивости в пределах этой группы, считая,
что подобные попытки могут в дальнейшем способствовать более деталь-
ному изучению низших таксономических единиц.
В настоящем обзоре, как и в предыдущих своих работах, я недо-
статочно строго следовал правилам номенклатуры, приоритета и пр., не
считая во многих случаях эти вопросы существенными; однако, в отдель-
ных случаях, где это вызывалось необходимостью, мне пришлось сделать
-некоторые изменения в общепринятой номенклатуре во избежание могу-
щей возникнуть в дальнейшем путаницы (Sicista subtilis вместо S. nord-
manni и Sicista betulina вместо S. montana).
Помещенные в работе карты представляют ареалы распростра-
нения отдельных видов и групп тушканчиков в весьма схематическом
виде, вследствие недостаточности имеющихся фактических данных, т. е.
точно проверенных нахождений отдельных видов (особенно вне пределов
СССР); по указанной причине эти карты являются лишь ориентировочными
>и предварительными.
Большинство иллюстраций, помещенных в настоящей работе, .выпол-
-иено автором, за исключением нескольких рисунков живых зверьков
А. Н. Формозова, а также нескольких фотографий А. И. Аргиропуло и
М. К. Серебренникова. За предоставление указанных иллюстраций автор
Яриносит названным лицам сердечную благодарность.
Б. Виноградов
Зоологический Институт
Академии Наук СССР
Май 1936 г.
СИСТЕМАТИЧЕСКИМ УКАЗАТЕЛЬ ВИДОВ
Стр»
Сем. Dipodidae
1. Подсем. Zapodinae
Род 1. Sicisia Gray
1. S. subtilis Pallas................................................... 67
2. S. betulina Pallas................................................... 72,
3. S. napaea Hollister.................................................... 75
4. S. tianschanica Salensky................................................76
5. S. caucasica Vinogradov ........................ ..................... 77
6. S. caudata Thomas..................................................... 78
Род 2. Zapus Coues
1. Zapus (Eozapus) setchuanus Pousargues.................................. 82
2. Подсем. Euchoreutinae
Род 3. Euchoreutes W. L. Sdater
1. E. naso W. L. Sclater.................................................. 86
3. Подсем. Cardiocraniinae
Род 4. Cardiocranius Satunin
1. C. paradoxus Satunin................................................. 93
Род 5. Salpingotus Vinogradov
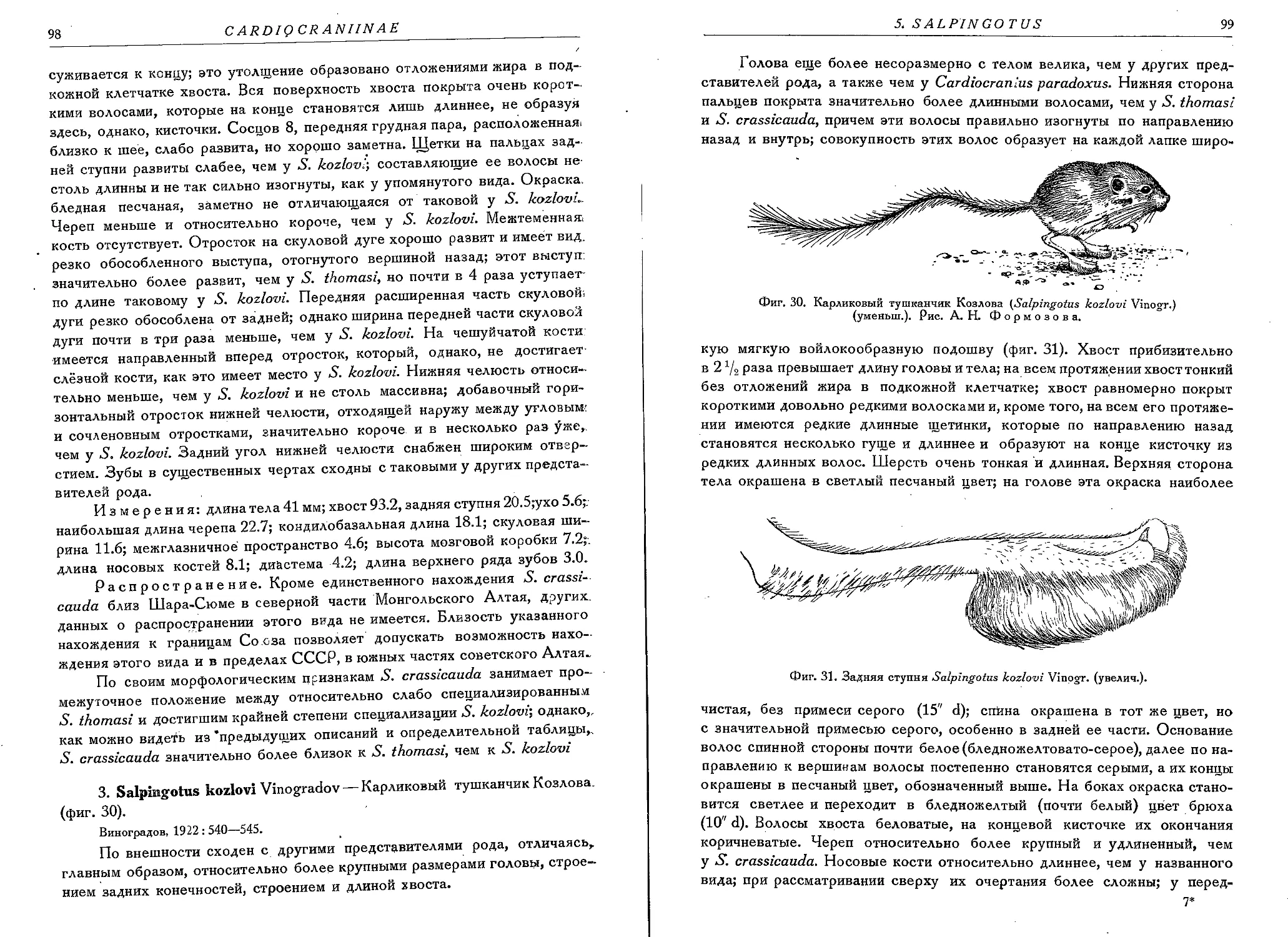
1. S. thomasi Vinogradov.................................................. 96
2. S. crassicauda Vinogradov.............................................. 97
3. S. kozlovi Vinogradov................................................. 98.
4. Подсем. Allactaginae
Род 6. Allactaga F. Cuv.
1. A. jaculus Pallas..................................................... Ill
2. A. severtzovi Vinogradov.............................................. 117
3. A. elater Lichtenstein............................................... 119
4. A. williamsi Thomas................................................... 125
5. A. euphratica Thomas................................................. 127
6. A. hotsoni Thomas ................................................... 128.
7. A. bullata Glover Allen............................................... 128
8. A. saltator Eversmann................................................ 129
VIII
ТУШКАНЧИКИ DIPODIDAE
Стр.
Род 7. Alactagulus Nehring
1. A. acontion Pallas ............................................................ 134
8. Род Pygsrethmus Gloger
1. P. platyurus Lichtenstein ... ............... ............................. 143
2. P. zhitkovi Kuznetzov.......................................................... 144
Род 9. Scarturus Gloger
1. S. tetradactylus Lichtenstein................................................ 149
5. Подсем. Dipodinae
Род 10. Dipus Gmelin
1. D. sagitta Pallas.............................................................. 155
Род 11. Scirtopoda Brandt
1. S. telum Lichtenstein..................................................... . 168
2. S. andrewsi Glover Allen..................................................... 172
Род 12. Paradipus Vinogradov
1. P. ctenodactylus Vinogradov.................................................... 174
Род 13. Eremodipus Vinogradov
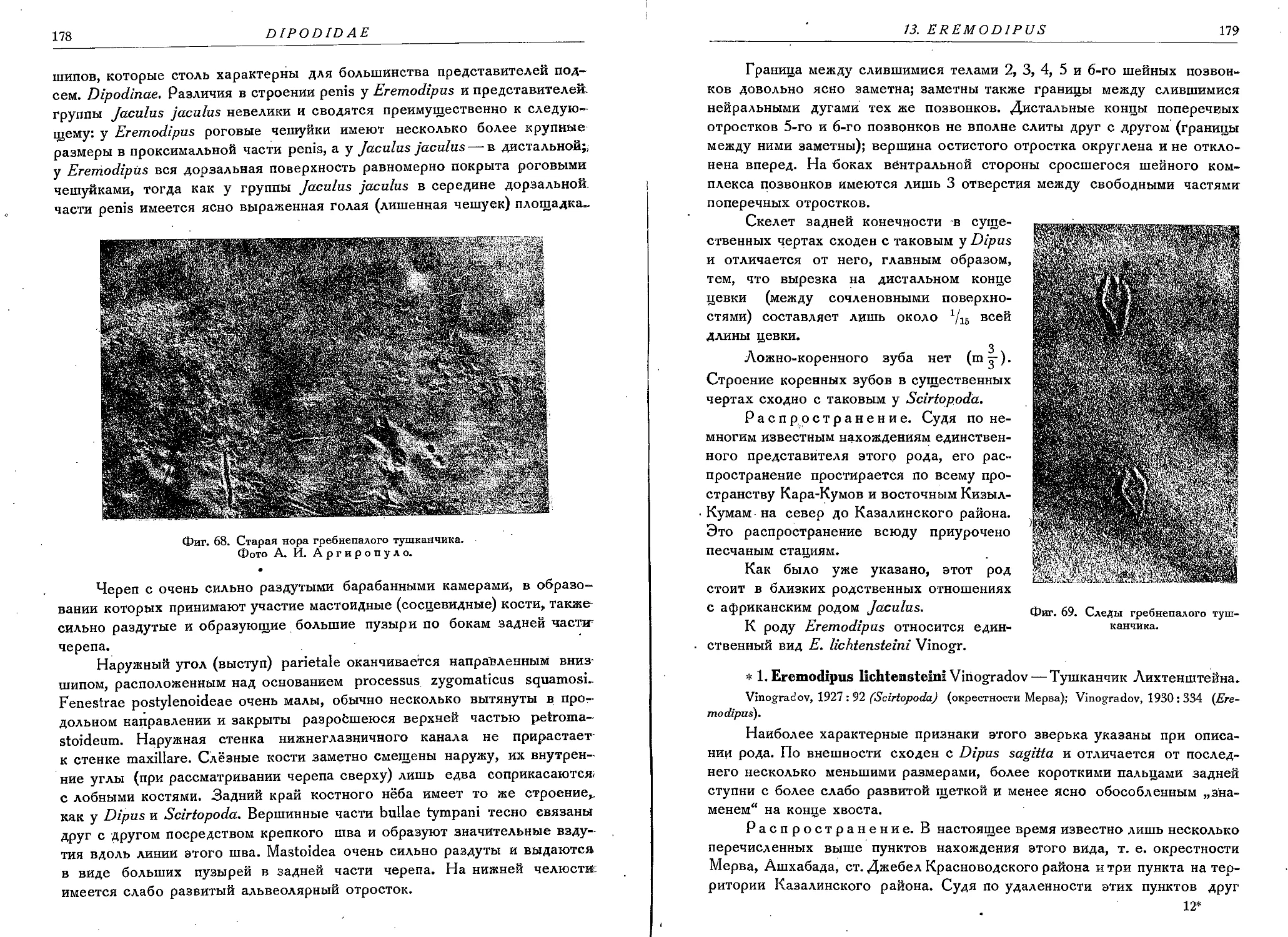
3. E. lichtensteini Vinogradov............................................... . 179
Род 14. Jaculus Erxleben
1. J. jaculus Linne............................................................. 181
ВВЕДЕНИЕ
КРАТКИЙ ОЧЕРК ИСТОРИИ ИЗУЧЕНИЯ СИСТЕМАТИКИ ГРУППЫ
Из числа наиболее ранних капитальных исследований по тушканчикам
га мышевкам следует отметить работы Палласа, особенно его известную
.монографию „Novae species quadrupedum е glirium ordinae" (1778), содер-
жащую описание ряда видов этой группы, данные по их анатомии, рас-
пространению и биологии. Паллас не делал, однако, попыток дать клас-
сификацию всей группы и причислил тушканчиков вместе с большинством
известных ему грызунов к роду Mus, причем отнес тушканчиков и мыше-
вок вместе с сонями, песчанками и кафрским долгоногом к группе „mures
iethargici".
Более полную классификацию тушканчиков дал в 1825 году Lichten-
stein, обработавший сборы Эверсмана из Казахстана и другие имевшиеся
у него материалы и установивший ряд неизвестных до того времени форм.
Общая система классификации тушканчиков и взаимоотношения их родов
более полно и точно была разработана Ф. Брандтом (1843), включившим
в семейство ,,Dipodau также роды Zapus и Pedetes. Брандтом была дана
следующая классификация рассматриваемой группы:
Fam. Dipoda s. Macropoda Wagn.
subf. I. Dipodina s. Thylarodactyli.
sect. A. Dipodes genuini.
1 gen. Dipus Fr. Cuv. (1836), Jarocki (1821)
subg. 1. Scirtopoda Br.
sect. 1. Halticus Br.
sect. 2. Haltomys Br.
subg. 2. Dipus Br.
sect. B. Scirtetides.
2 gen. Scirtetes Wagn. s. Allactaga Fr. Cuv.
subg. 1. Scirtomys Br.
subg. 2. Scirteta Br.
3 gen. Platycercomys Br.
Фауна СССР, Виноградов
1
2
ТУШКАНЧИКИ DIPODIDAE
subf. II. Merionina
1 gen. Meriones Fr. Cuv.
subf. III. Pedetina
1 gen. Pedetes Illig.
В группу Haltomys включены африканские трехпалые тушканчики,
к группе Halticus причислен азиатский Н. telum Licht. В подрод Scirtomys
выделен четырехпалый тушканчик S. tetradactylus Licht. Подрод Scirteta
содержит виды, причисляемые ныне к родам Allactaga и Alactagalus. Под-
семейство Merionina с'родом Meriones заключает виды относящиеся совре-
менными систематиками к роду Zapus (автор полагает, что по правилам
приоритета, название рода Meriones не может применяться в системе песча-
нок, а должен быть оставлен для групп африканских мышевок).
В 1876 году Alston отнес тушканчиков к sectio Myomorpha и под-
разделил семейство Dipodidae на три следующих подсемейства:
1) Jaculinae (с родом Jaculus, соответствующим Zapus по современ-
ной номенклатуре);
2) Dipodinae (роды Dipus, Allactaga, Plaiycercomys)}
3) Pedetinae (род Pedetes).
За период времени с 1878 г. до начала девятисотых годов A. Mering
опубликовал ряд работ по систематике и остеологии тушканчиков, при-
чем этим автором уделено также значительное внимание изучению остат-
ков четвертичных тушканчиков.
В 1882 г. Dobson, на основании изучения мускулатуры задней
конечности тушканчиков, пришел к заключению, что это семейство имеет
сходство с группой Hystricomorpha, с которым его сближает также устрой-
ство нижне-глазничного отверстия, скуловой дуги, отсутствие бугорков
на зубах, наличие роговых чешуй и игол на glans penis и т. д.
В 1887 г. Winge дал следующую схему классификации рассматри-
ваемой группы:
Dipodidae
Eomyini: Eomys
Dipodini: Sminthus, Jaculus, Scirtetes, Dipus
Spalacini: Spalax.
Из приведенной схемы можно видеть, что Winge причислил к семей-
ству Dipodidae олигоценового .’грызуна Eomys, а также слепышей (род
Spalax), относимых в настоящее время к семейству Spalacidae.
В 1890 г. W. Sclater’oM был описан своеобразный тушканчик
Euchoreutes naso из Китайского Туркестана. В 1896 г. Pousargues впервые
установил нахождение одного из представителей Zapus на материке-Азии,
описав Eozapus setchuanus из Сычуани.
ВВЕДЕНИЕ
3
В 1896 г. О. Thomas причислил тушканчиков к группе Myomorpha
и разделил их на следующие подсемейства.
. A. Sminthinae: Sminthus Keys & Blas.
В. Zapodinae: Zapus Coues.
C. Dipodinae: Dipus Gmel., Allactaga F. Cuv.,
Platycercomys Br., Euchoreutes W. Scl.
В 1899 г. была опубликована обширная и чрезвычайно детальная
монография грызунов Т. Tullberg’a, содержащая ценные данные по анатомии
и систематике различных представителей этого отряда, в частности рас-
сматриваемого семейства. Семейство Dipodidae причислено Tullberg’oM
к subsectio Dipodiformes и sectio Myoidei с родами Sminthus, Zapus,
Dipus, Alactaga, Euchoreutes, Platycercomys и Eomys (последний род
причислен к этому семейству предположительно).
В 1899 г. Preble опубликовал сводку по систематике рода Zapus.
В 1901 г. М. Lyon опубликовал ценную работу по остеологии тушкан-
чиков и их классификации. В 1914 г. появилась детально разработанная
монография европейских мышевок (Sicistinae) L. Mehely; следует отметить,,
что при разработке систематики этой группы Mehely удачно использовал
особенности строения наружных genitalia.
В 1918 г. Miller и Gidley причислили тушканчиков и мышевок к над-
семейству (superfamily) Dipodoidae и разделили его следующим образом:
Zapodidae.
Theridomyinae: Sciuroides, Theriodomys, Issiodoromys etc.
Sicistinae: Sicista ?Eomys.
Zapodinae: Eozapus, Zapus, Napaeozapus.
Dipodidae.
Protoptychinae: Prdloptychus.
Dipodinae.
Кроме семейства Zapodidae и Dipodidae, авторы причисляют также
к надсемейству Dipodoidae следующие семейства: Paramyidae, Graphiuri~
dae, Allomyidae, Aplodontiidae, Cylindrodontidae („position doubtful")^
Pseudosciuridae, Mylagaulidae, Anomaluridae, Idiuridae, Sciuravidae, Cteno-
dactylidae и Pedetidae.
В 1922 г. было опубликовано ценное исследование по систематике
тушканчиков, преимущественно на' основании изучения их genitalia
R. Pocock’a.
В работе Н. Winge, опубликованной в 1924 г., автор более детально
разработал данную им прежде классификацию тушканчиков и признал
в качестве родоначальной для них группы, семейство Anomaluridae.
Из исследований последнего периода следует указать также значи-
тельное количество статей О. Thomas’a, преимущественно с описаниями
1*
4
ТУШКАНЧИКИ DIPODIDAE
новых видов и подвидов азиатских и африканских тушканчиков и ряд
работ К. А. Сатунина, особенно его посмертную монографию 1920 г.
„Грызуны Кавказа", т. И, содержащую ряд данных по тушканчикам СССР.
С 1922 по 1933 г. ряд работ по систематике, морфологии и биологии
тушканчиков был опубликован Б. С. Виноградовым, которым в частности
было установлено несколько новых родов рассматриваемой группы, как
Salpingotus (с тремя видами), Paradipus и Eremodipus. В работе 1925 г.
Б. С. Виноградов дал общую классификацию Семейства на основании
строения genitalia, а в работе 1933 г. — на основании строения черепа,
коренных зубов и слуховых косточек; эта классификация с небольшими
изменениями положена в основу настоящей монографии.
В последние годы (1932) два американских исследователя — R. Hatt
и В. Howell опубликовали две чрезвычайно ценные работы по анатомии
Dipodidae и других прыгающих грызунов и выяснению приспособлений
в их организации в связи со способом передвижения этих зверьков
прыжками.
Перечисленные выше исследования посвящены главным образом
выяснению вопросов систематики рассматриваемого семейства. Свое-
образные экологические особенности этой группы изучены очень слабо
до настоящего времени, хотя в последний период наблюдается значитель-
ный сдвиг в этом отношении.
Наблюдения над образом жизни этих животных имеются в работах
Сатунина (1920 и др.), Огнева (1924), Фенюка (1928,1929), Беме и Крассов-
ского (1931), Колесникова (1934), Виноградова и Аргиропуло (1931)
и других авторов.
ОБЩАЯ МОРФОЛОГИЧЕСКАЯ ХАРАКТЕРИСТИКА СЕМЕЙСТВА
, Форма тела у наиболее примитивных представителей мышеобразная,
характеризующаяся относительно острой мордой, средней величины ушами
и глазами, относительно длинными задними конечностями, значительно
превосходящими по длине передние, с относительно длинной задней ступ-
ней и длинным хвостом, превышающим длину тела; тело покрыто большей
частью относительно грубой шерстью. У более специализированных форм
на ряде переходных стадий наблюдается приобретение специфического
„тушканчикообразного" облика, характеризующегося следующими внеш-
ними признаками: морда в большей или меньшей степени укорочена и на
ее конце обычно образуется „пятачок", т. е. притупленный и расширенный
rhinarium; голова в ее задней части сильно расширена, глаза значительно
увеличены в размерах, причем разрез век имеет значительный наклон
вперед по отношению к продольной оси головы; края ушной раковины
обыкновенно срастаются своими основными частями и все наружное ухо
принимает в связи с этим трубчатую форму; иногда ушная раковина
достигает значительной длины (фиг. 14_6); задние конечности сильно типе-
ВВЕДЕНИЕ
5
трофированы при одновременной значительной редукции передних; один
или оба боковых пальца (1-й и 5-й) более или менее редуцированы, нередко
до полного исчезновения видимых снаружи частей, но при сохранении
остатков их костных элементов; на концах пальцев образуются дольчатые
эластические подушечки, на нижней поверхности пальцев иногда имеется
щетка из упругих волос; хвост обычно превышает длину тела и часто
снабжен на конце расположенной горизонтально, плоской, ланцетообраз-
ной кисточкой („знамя"); лишь у немногих представителей хвост равен
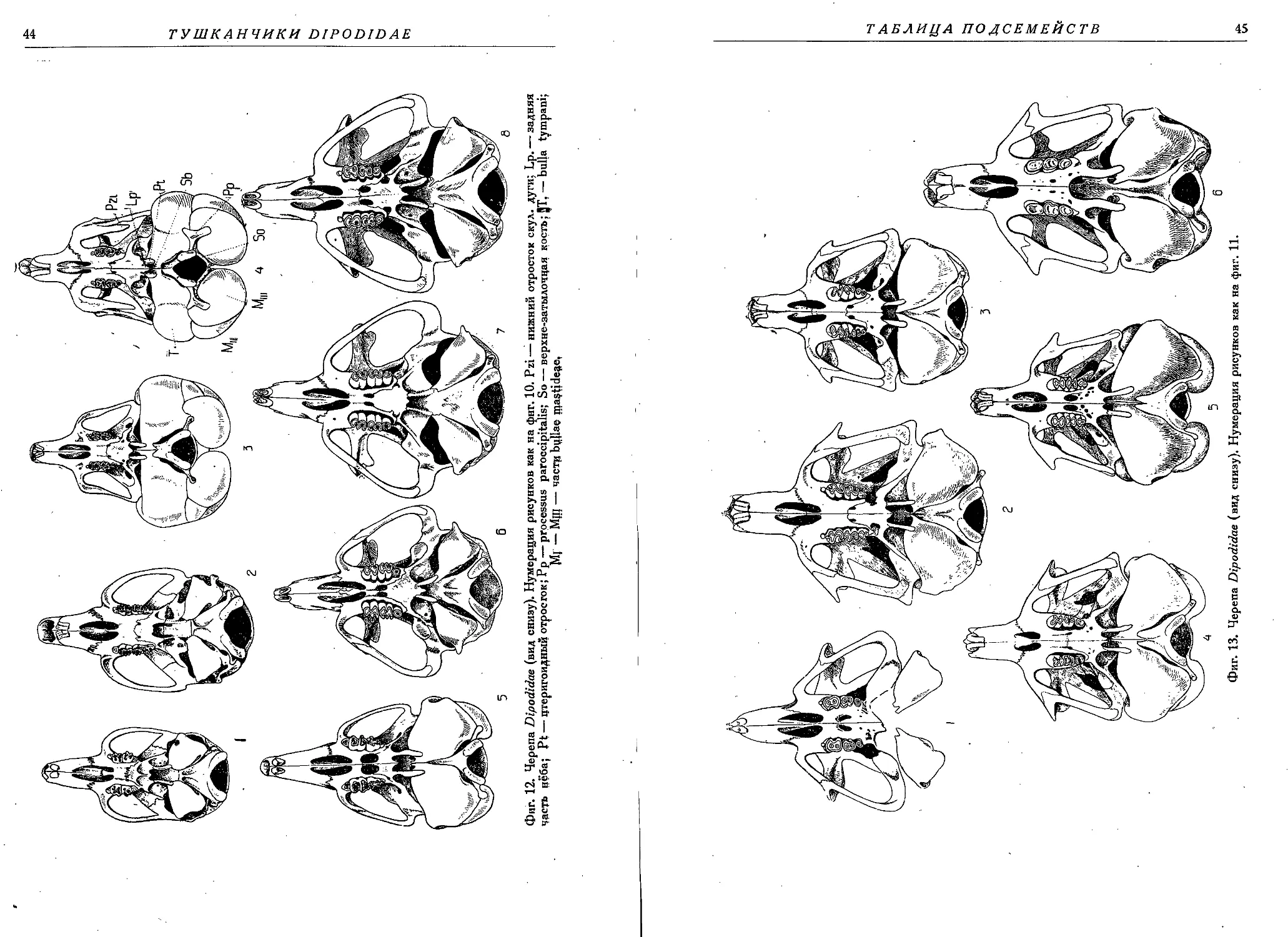
Фиг. 1. Головы тушканчиков: 1 —Sicista betulina Pall., 2— Cardiocranius paradoxus Satun.,
3 — Salpingotus kozlovi Vinorg., 4 — Euchoreutes naso Sclat., 5 — Allactaga elater Licht.,
6 — Alactagulus acontion Pall.
или короче длины тела и в таких случаях он обычно утолщен в связи
с образованием значительного жирового слоя в подкожной клетчатке;
шерсть обычно длинная и тонкая. Окраска шерсти у наиболее примитив-
ных форм одноцветная или с образованием на поверхности спины темного
рисунка, состоящего из одной или трех темных продольных полос или
широкой буроватой мантии; у более специализированных форм окраска
верха довольно однообразная, типа „пустынной" окраски, т. е. светлая
с преобладанием песчаных тонов; нижняя поверхность тела чисто белая;
в задней части тела на бедрах обычно имеется с каждой стороны резко
очерченная, широкая, поперечная белая полоса, идущая к основанию
хвоста. „Знамя" в его основной части обычно окрашено в черный цвет
и в конечной — в белый.
6
ТУШКАНЧИКИ DIPODIDAE
Клитор расположен внутри переднего края vulva и urethra, открывается
в vagina. Glans penis покрыт роговыми шипиками или бляшками, причем
нередко отдельные шипики достигают значительной длины и имеют вид
острых конусов или стилетов. Форма glans penis, а также форма и распо-
ложение его роговых образований дают ряд резко выраженных и посто-
янных признаков для характеристики подсемейств, родов и видов рассма-
триваемой группы.
Общая конфигурация черепа у наиболее примитивных представителей
мышеобразная, т. е. с округлой мозговой коробкой, почти овальной при
рассматривании сверху и дугообразными скуловыми дугами; область
чешуйчатой кости гладкая, без выростов или гребней; межглазничное
сужение приблизительно совпадает с серединой длины лобных костей
и расположено значительно сзади от слёзных костей; у более специализи-
рованных форм мозговая коробка приобретает более или менее трапецио-
дальную форму И сильно расширена в затылочной области с более или
менее прямолинейными скуловыми дугами, сильно разведенными сзади
и резко сходящимися по направлению вперед; на поверхности чешуйчатой
кости образуется округлая выпуклость, или резко обозначенный гребень
или шип. Наиболее узкая часть лобных костей (межглазничное сужение)
расположено непосредственнЬ за слёзными костями.
Слёзные кости у наиболее примитивных форм маленькие и округлые;
у более специализированных наблюдается тенденция к значительному
увеличению их размеров, особенно в поперечном направлении.
Для всех представителей семейства характерно Наличие широкого
нижнеглазничного отверстия с отдельным каналом на его внутренней
стенке для прохождения ветви тройничного нерва; стенки этого канала
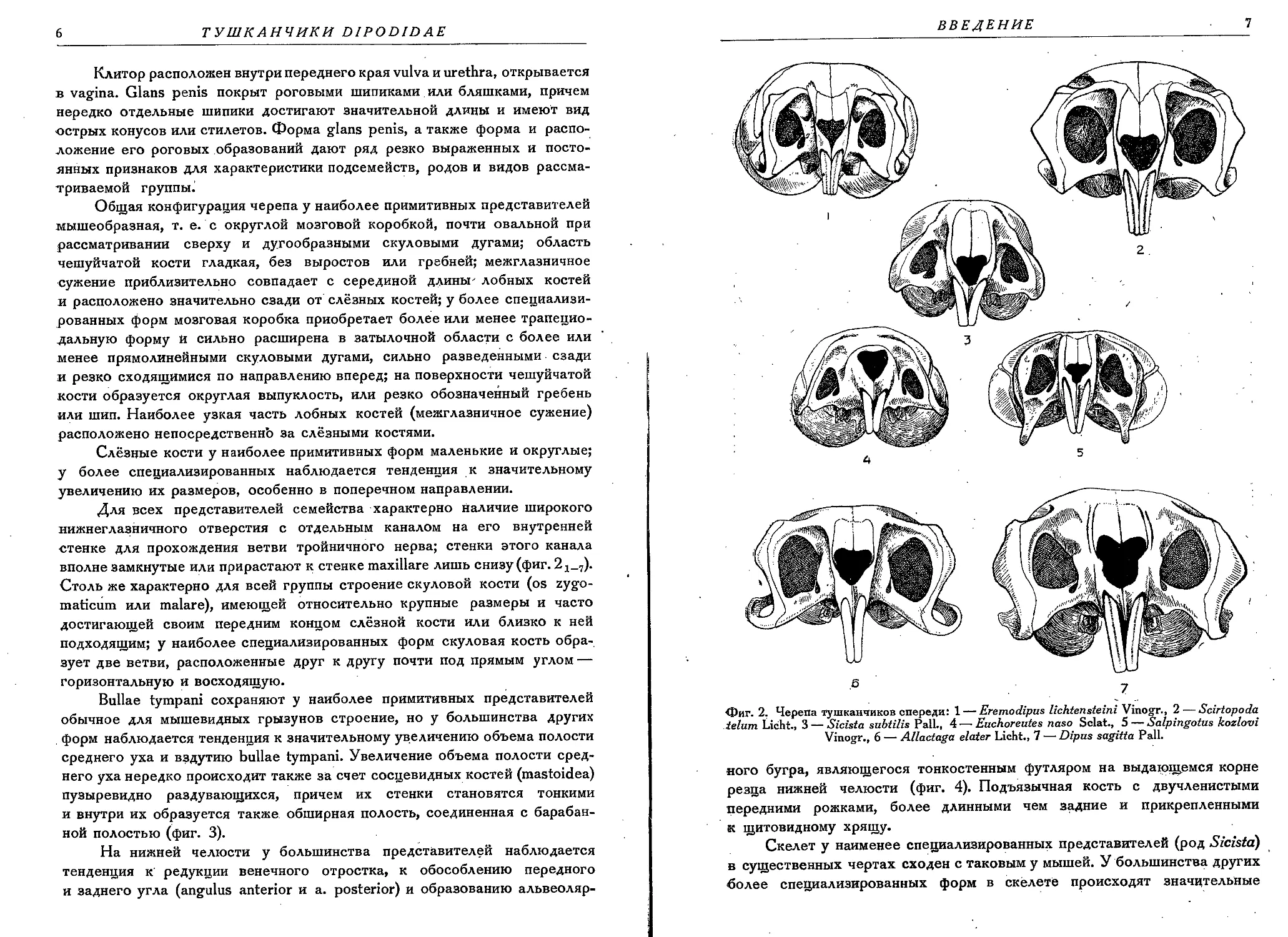
вполне замкнутые или прирастают к стенке maxillare лишь снизу (фиг. 2
Столь же характерно для всей группы строение скуловой кости (os zygo-
maticum или malare), имеющей относительно крупные размеры и часто
достигающей своим передним концом слёзной кости или близко к ней
подходящим; у наиболее специализированных форм скуловая кость обра-
зует две ветви, расположенные друг к другу почти под прямым углом —
горизонтальную и восходящую.
Bullae tympani сохраняют у наиболее примитивных представителей
обычное для мышевидных грызунов строение, но у большинства других
форм наблюдается тенденция к значительному увеличению объема полости
среднего уха и вздутию bullae tympani. Увеличение объема полости сред-
него уха нередко происходит также за счет сосцевидных костей (mastoidea)
пузыревидно раздувающихся, причем их стенки становятся тонкими
и внутри их образуется также обширная полость, соединенная с барабан-
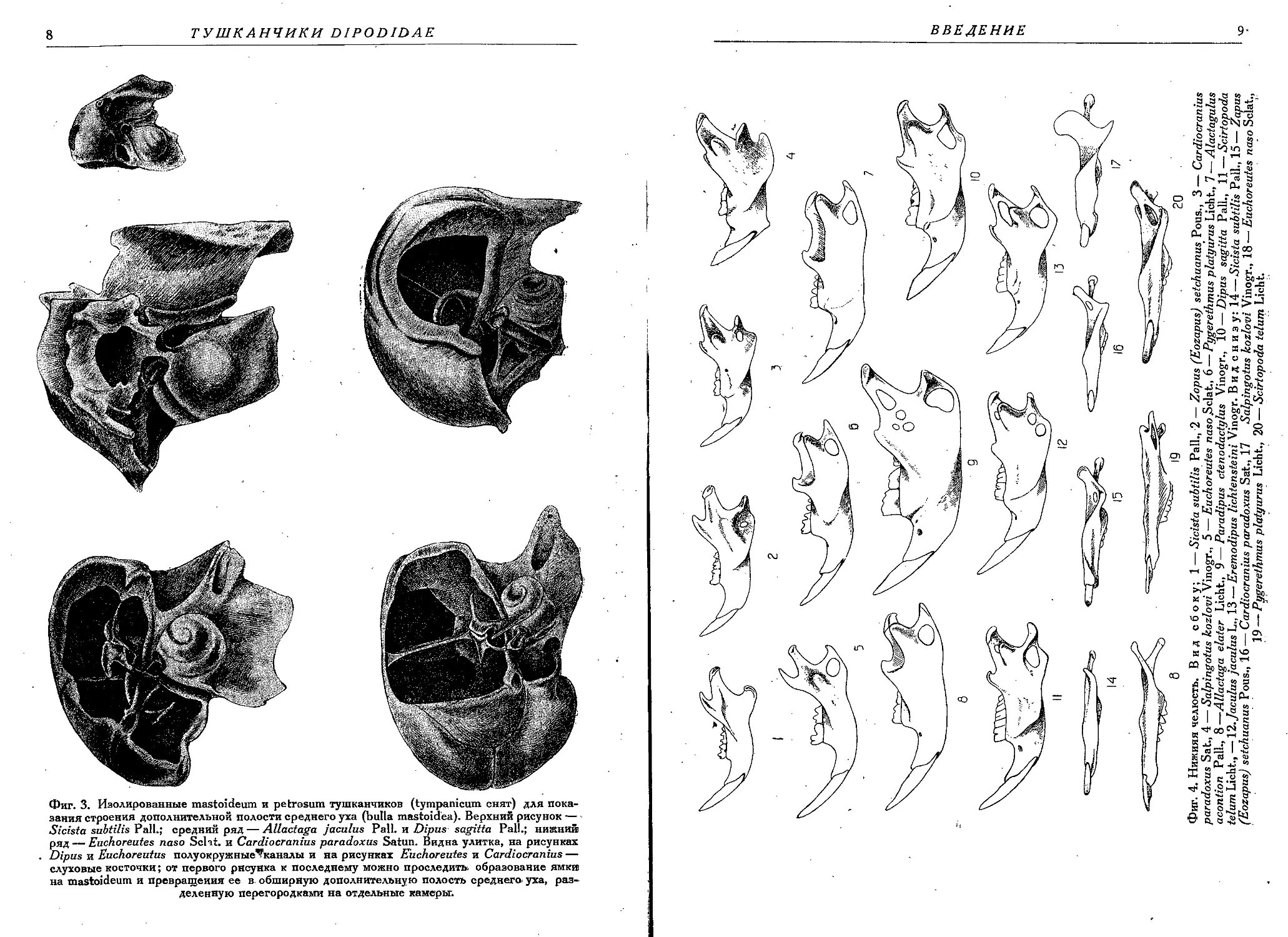
ной полостью (фиг. 3).
На нижней челюсти у большинства представителей наблюдается
тенденция к редукции венечного отростка, к обособлению переднего
и заднего угла (angulus anterior и a. posterior) и образованию альвеоляр-
ВВЕДЕНИЕ
7
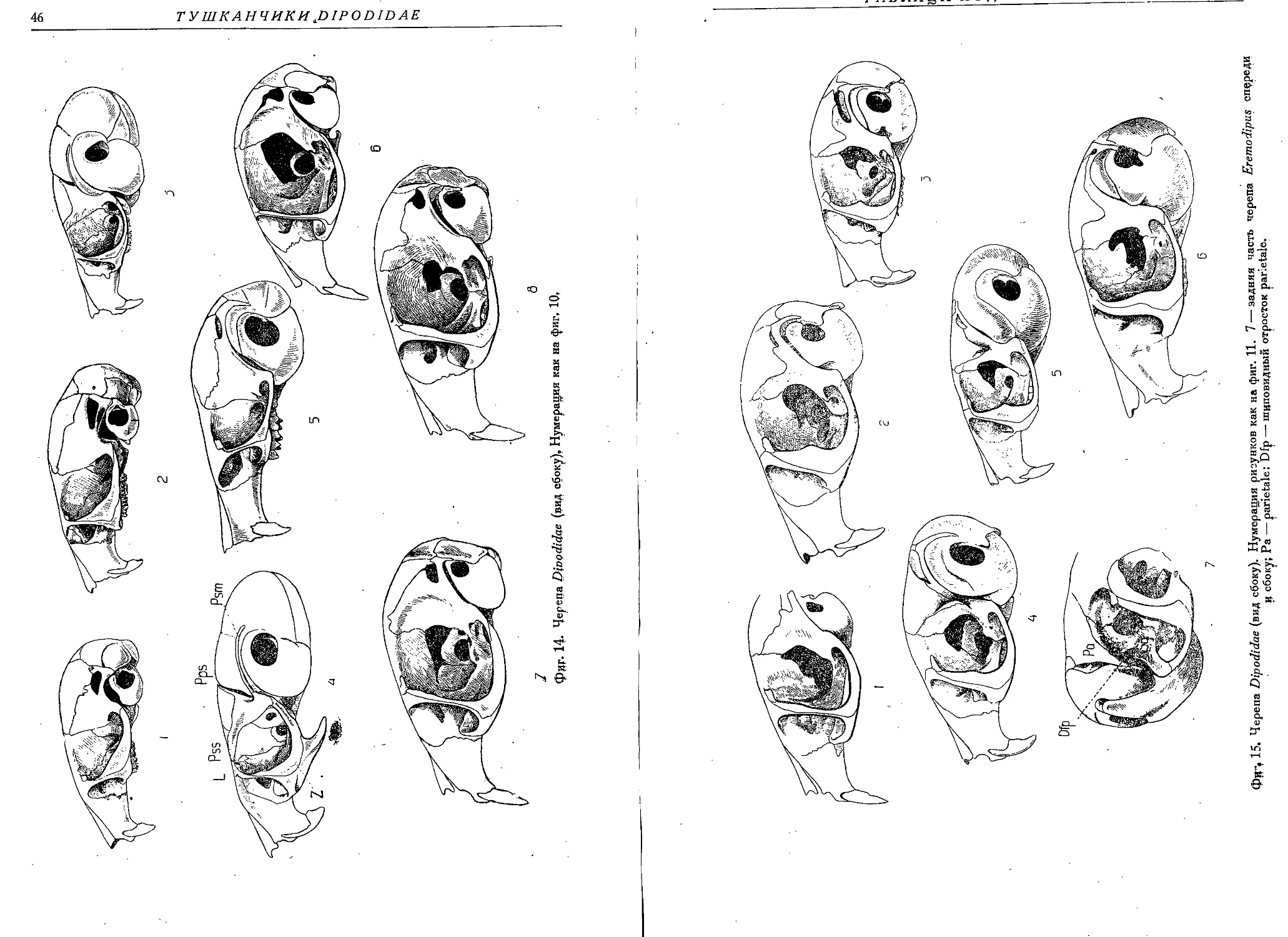
Фиг. 2, Черепа тушканчиков спереди: 1 — Eremodipus lichtensteini Vinogr., 2 — Scirtopoda
ielum Licht., 3 — Sicista subtilis Pall., 4 — Euchoreutes naso Sclat., 5 — Salpingotus kozlovi
Vinogr., 6 — Allactaga elater Licht., 7 — Dipus sagitta Pall.
ного бугра, являющегося тонкостенным футляром на выдающемся корне
резца нижней челюсти (фиг. 4). Подъязычная кость с двучленистыми
передними рожками, более длинными чем задние и прикрепленными
к щитовидному хрящу.
Скелет у наименее специализированных представителей (род Sicista)
в существенных чертах сходен с таковым у мышей. У большинства других
более специализированных форм в скелете происходят значительные
8
ТУШКАНЧИКИ DIPODIDAE
Фиг. 3. Изолированные mastoideum и petrosum тушканчиков (tympanicum снят) для пока-
зания строения дополнительной полости среднего уха (bulla mastoidea). Верхний рисунок —
Sicista subtilis Pall.; средний ряд — Allactaga jaculus Pall, и Dipus sagitta Pall.; нижний
ряд — Euchoreutes naso Sclat. и Cardiocranius paradoxus Satun. Видна улитка, на рисунках
Dipus и Euchoreutus полуокружные^каналы и на рисунках Euchoreutes и Cardiocranius —
слуховые косточки; от первого рисунка к последнему можно проследит!» образование ямки
на mastoideum и превращения ее в обширную дополнительную полость среднего, уха, раз-
деленную перегородками на отдельные камеры.
ВВЕДЕНИЕ
9
“10
ТУШКАНЧИКИ DIPODIDAE
изменения, в частности следующие: кости задних конечностей чрезвы-
чайно удлинены, особенно в дистальном отделе (кости голени и ступни),
при одновременной редукции передних конечностей.
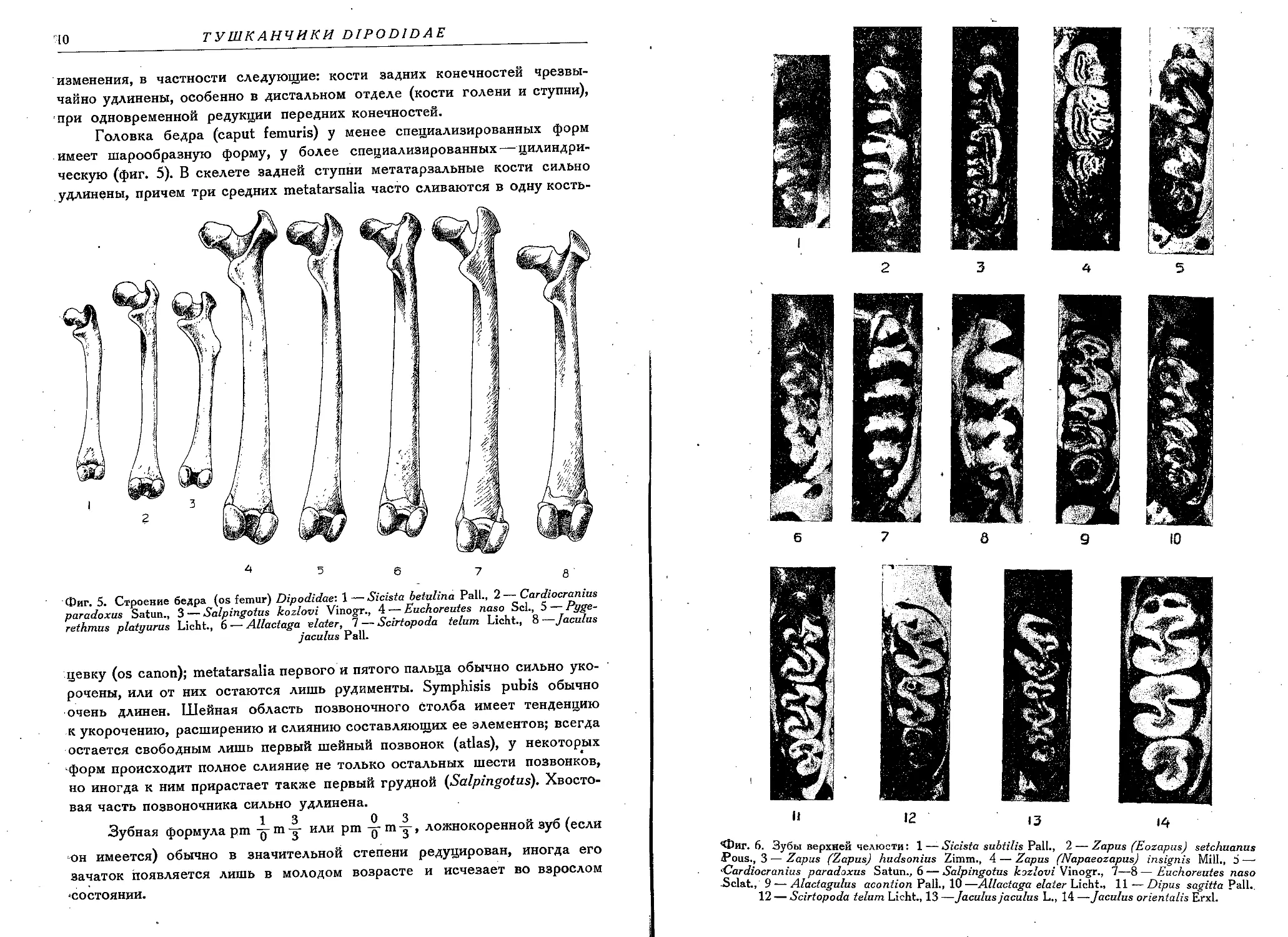
Головка бедра (caput femuris) у менее специализированных форм
имеет шарообразную форму, у более специализированных — цилиндри-
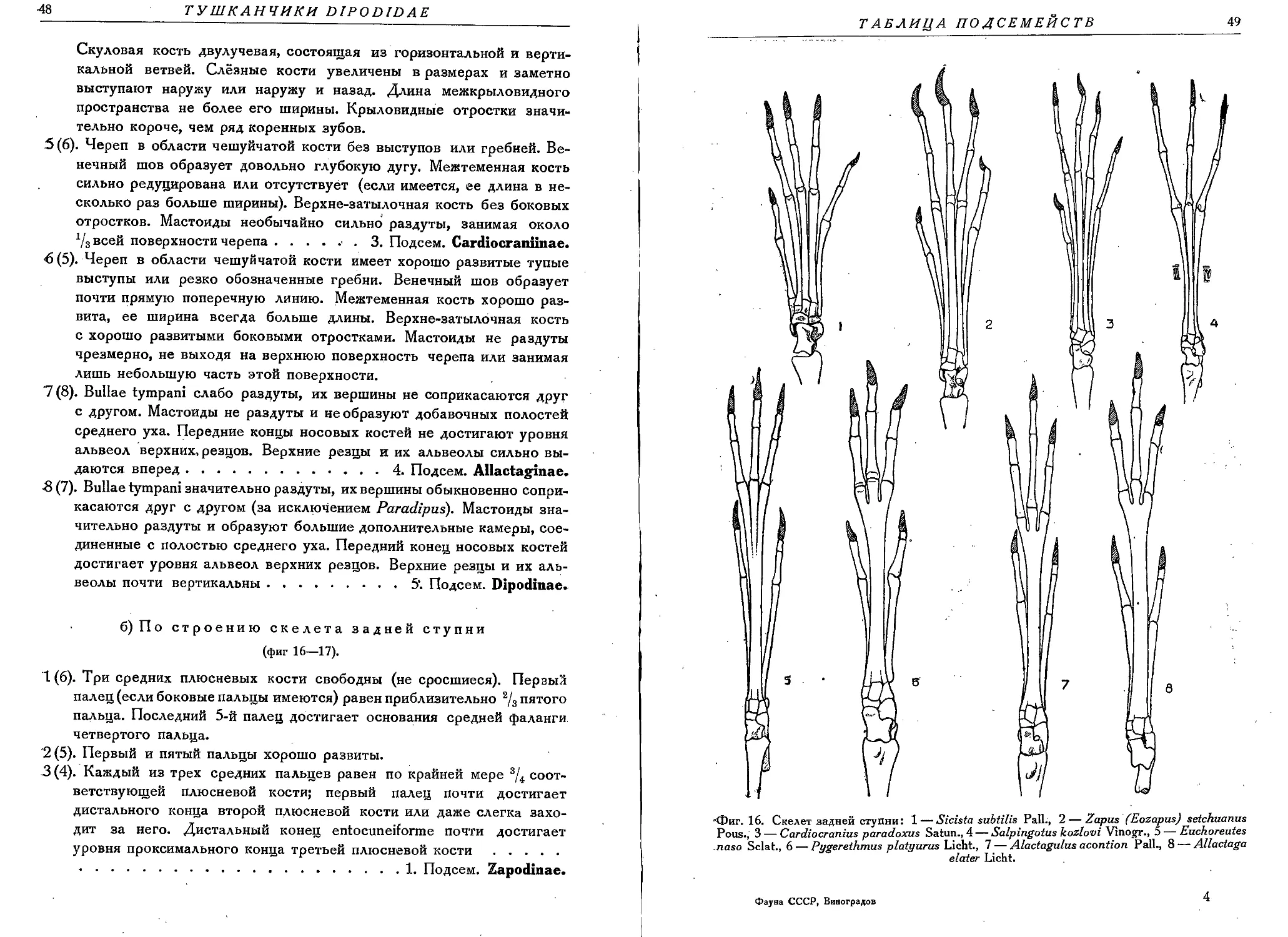
ческую (фиг. 5). В скелете задней ступни метатарзальные кости сильно
удлинены, причем три средних metatarsalia часто сливаются в одну кость-
Фиг. 5. Строение бедра (os femur) Dipodidae-. 1 — Sicista betulina Pall., 2 — Cardiocranius
paradoxus Satun., 3 — Salpingotus kozlovi Vinogr., 4 — Euchoreutes naso Scl., 5 — Pygc-
rethmus platyurus Licht., 6 — Allactaga elater, 7 — Scirtopoda telum Licht., 8 —Jaculus
jaculus Pall.
цевку (os canon); metatarsalia первого и пятого пальца обычно сильно уко-
рочены, или от них остаются лишь рудименты. Symphisis pubis обычно
очень длинен. Шейная область позвоночного Столба имеет тенденцию
к укорочению, расширению и слиянию составляющих ее элементов; всегда
остается свободным лишь первый шейный позвонок (atlas), у некоторых
форм происходит полное слияние не только остальных шести позвонков,
но иногда к ним прирастает также первый грудной (Salpingotus). Хвосто-
вая часть позвоночника сильно удлинена.
о , 13 03 „ е ,
Зубная формула pm у m у или pm у m у, ложнокоренной зуб (если
он имеется) обычно в значительной степени редуцирован, иногда его
зачаток появляется лишь в молодом возрасте и исчезает во взрослом
состоянии.
2 3 4 5
6 7 a g 10
'Фиг. 6. Зубы верхней челюсти: 1—-Sicista subtilis Pall., 2 — Zapus (Eozapus) setckuanus
Pous., 3 — Zapus (Zapus) hudsonius Zimm., 4 — Zapus (Napaeozapus) insignis Mill., 5 —
‘Cardiocranius paradoxus Satun., 6 — Salpingotus kozlovi Vinogr., 7—8 — Euchoreutes naso
-Sclat., 9 — Alactagulus acontion Pall., 10—Allactaga elater Licht., 11—Dipus sagitta Pall.,
12 — Scirtopoda telum Licht., 13 —Jaculus jaculus L., 14 —Jaculus orientalis Erxl.
12
ТУШКАНЧИКИ DIPODIDAE
Коренные зубы Dipodidae (фиг. 6) значительно различаются по строе-
нию и сложности в пределах различных подсемейств и родов, но вполне
очевидно, что они развились из четырехбугорчатой структуры; эта структура
обычно замаскирована уплощением бугорков и развитием складок в раннем
возрасте. Четыре главных бугорка замаскированы вторичными или доба-
вочными бугорками или складками, которые занимают определенное поло-
жение на коронке: на верхних коренных зубах первый и второй добавочный
бугорки (или складки) расположены сзади от первого и второго главных
бугорков (или складок) последовательно; третий добавочный бугорок
расположен на переднем краю коронки в средней части (как например
у Sicista) или сдвинут наружу (как у многих других представителей
семейства). Подобное же строение наблюдается на нижних коренных
зубах, но с обратным расположением элементов, т. е. два добавочных
бугорка расположены здесь на внутренней стороне коронки, а передний
бугорок (или складка) имеет тенденцию к смещению на внутреннюю
сторону.
Язык с тремя papillae circumvallatae. Желудок без внутреннего рого-
вого слоя. Слепая кишка имеется. Дистальная часть толстой кишки
образует непосредственно около слепой кишки более или менее изогнутую
парацекальную извилину.
Слизистая оболочка проксимальной части толстой кишки с косыми
сходящимися вдоль шовной линии складками.
ОСНОВНЫЕ НАПРАВЛЕНИЯ СПЕЦИАЛИЗАЦИИ СЕМ. DIPODIDAE
Наиболее обобщенный мышеобразный тип представляет организация
наиболее примитивных представителей семейства — Sicista и Zapus, раз-
личные виды которых приспособлены к разнообразным комплексам жиз-
ненных условий, встречающимся в различных широтах от северной границы
леса до зоны полупустыни включительно и от высокогорных лугов до
равнины; однако уже у названных представителей намечаются некото-
рые основные пути, по которым направлена адаптация у большинства
других более специализированных представителей семейства, а именно,
удлинение задних конечностей и удлинение хвоста, что связано с выра-
боткой у этих животных способности к передвижению на поверхности
прыжками при помощи задних конечностей. Вместе с тем у некоторых:
представителей рассматриваемой группы наблюдаются также слабо выра-
женные приспособления к лазанью.
Более резко выражены признаки специализации у большинства других
представителей семейства, связанные с приспособлением этих животных
к обитанию в условиях открытого и большей частью равнинного ланд-
шафта степей и пустынь с разреженным растительным покровом, где для
существования мелких животных, не обладающих активными средствами
сопротивления врагам, необходимы весьма совершенные средства пассивной
ВВЕДЕНИЕ
13
защиты. Эволюция тушканчиков в указанном направлении шла по одному
из немногих возможных в данном случае путей, а именно по пути выра-
ботки способности стремительного бега на задних конечностях, а также
способности рыть особого устройства норы с замаскированными запас-
ными выходами, позволяющими животному неожиданно и часто, незаметно
для хищника, раскапывающего нору выскакивать из норы и быстро скры-
ваться из его поля зрения. Несмотря на кажущуюся несовместимость
в одном и том же организме приспособлений к прыганию и в то же время
к рытью, у тушканчиков эти два направления специализации находятся
в гармоничном сочетании, причем доминирующее значение во всем ком-
плексе морфологических признаков имеют изменения, связанные с выра-
боткой способности к прыганью („рикошетированию") на задних конеч-
ностях, что определяет существование следующих наиболее характерных
черт рассматриваемого семейства.
Значительное удлинение задних конечностей, особенно в их дисталь-
ных частях, причем у некоторых представителей длина ступни может
достигать 50% длины тела (Salpingotus)-, удлинение задних конечностей
увеличивает размах „пружины" при прыжках и ослабляет толчки при сопри-
косновении с землей. В связи с указанной функцией достигают также
значительного развития мышцы задних конечностей и таза.
Тенденция к слиянию трех средних плюсневых костей в одну кость —
„цевку" при редукции боковых metatarsalia, тенденция к одновременной
редукции боковых пальцев и перенесению опоры тела при прыжках на
дистальные концы трех средних пальцев, исе эти изменения в значитель-
ной степени напоминают аналогичные изменения в конечностях копытных
и биологическая целесообразность этих приспособлений заключается,
как у упомянутой группы, в создании необходимой компактности и проч-
ности дистального отдела конечностей и выработке Способности наиболее
совершенного механизма бега „на цыпочках" (при использовании при
беге пятки и всей ступни бег не может быть эластичным, т. е. в этом
случае он неизбежно был бы сопряжен со значительными толчками).
С указанной функцией задних конечностей связано также наличие на
нижней Поверхности пальцев (особенно на их дистальных концах, на
которые опирается тело при прыжках), эластических кожистых подушечек
{при передвижении на твердом субстрате) или твердых торчащих щетин
(при передвижении на песке), еще более способствующих смягчению
толчков при прыжках. При однообразии движений задних конечностей
в одной плоскости у наиболее специализированных форм вырабатывается
более или менее цилиндрическая форма сочленовной головки бедра (Dipo-
dinae, Allactaginae).
Необходимое при прыжках на задних конечностях балансирование
тела достигается при помощи сильно удлиненного хвоста, превышаю-
щего длину тела и обычно снабженного на конце ланцетообразной, распо-
ложенной в горизонтальной плоскости кисточкой из волос; у некоторых
14
ТУШКАНЧИКИ DIPODIDAE
представителей имеющих более короткий хвост, значение этого органа,,
как баланса, компенсируется его более значительной толщиной, зависящей
от отложения в его подкожной клетчатке толстого жирового слоя (Pyge-
rethmus, Cardiocranius, Salpingotus crassicauda).
При прыжках хвост имеет также значение руля, помогающего живот-
ному делать резкие крутые повороты и двигаться зигзагами, что делает
зверька во время его бега почти неуловимым; в этом отношении имеет
также большое значение упомянутая плоская кисточка из волос на конце:
хвоста, играющая роль лопасти этого воздушного руля. Отчасти хвост
помогает также животному при первом прыжке, давая опору зверьку
и действуя в этот момент подобно пружине (при дальнейших прыжках,
хвост не касается земли, что подтверждается непосредственными наблю-
дениями в природе). Описанный выше способ передвижения тушканчиков
прыжками на задних конечностях делает бесполезными в качестве органов
движения передние конечности, подвергающиеся в связи с этим значи-
тельной редукции. Основной функцией передних конечностей при этих
условиях является рытье и поддерживание пищи во время еды; при сла-
бости передних конечностей их значение, как органов рытья, усиливается,
описанными ниже приспособлениями в строении нижних резцов, челюстного
аппарата и мягких частей рта, в связи с тем, что основная работа при
процессе рытья у этих зверьков падает на резцы (см. ниже).
Наблюдающаяся у многих представителей семейства тенденция,
к укорочению шейной части позвоночника, а также частичное или полное
слияние шейных позвонков, расположенных сзади от атланта, вероятно
следует также поставить в связь с передвижением тушканчиков прыжками,,
так как это способствует большей компактности переднего отдела тела,
и предохраняет голову от резких колебаний при прыжках (Hatt, 1932).
Характерная для тушканчиков форма черепа, сильно расширенного
сзади, связана, главным образом, со значительным или иногда чрезмерным
развитием bullae tympani и mastoidei, что вызывает также ряд глубоких
изменений в костях задней части мозговой коробки, в частности в тенден-
ции к редукции interparietale и supraoccipitale. В свою очередь сильное
развитие bullae вероятно связано какими-то косвенными и неизученными
в настоящее время путями с передвижением при помощи прыжков на зад-
них конечностях и с обитанием в пустынях (если оба названных фактора
сочетаются вместе), как об этом можно судить по аналогии с многими
другими пустынными прыгающими или вообще быстро передвигающимися,
млекопитающими (Heteromyidae, а также некоторые Gerbillinae, Leporidae,
Felidae и даже Macroscelididae). Обычно у тушканчиков чрезмерное раз-
витие bullae tympani коррелятивно связано с редукцией наружного уха
и наоборот слабое развитие bullae часто связано с значительным увели-
чением размеров наружного уха (примером первого случая могут служить
Salpingotus, Cardiocranius, Eremodipus и другие и для второго — Allac-
tagd). Следует, однако, отметить, что подобная закономерность не является:
ВВЕДЕНИЕ
15.
безусловной и имеет некоторые исключения (длинное ухо у Euchoreutes
при сильно раздутых bullae, длинное ухо у Allactaga hotsoni при относи-
тельно более крупных bullae tympani, чем у других представителей назван- •
ного рода). Биологическая целесообразность указанных приспособлений
очевидно связана с значительным увеличением остроты слуха (Howell,.
1932), но пока остается совершенно неясной связь этих приспособлений
с быстротой передвижения на поверхности, а подобная связь, повидимому,
существует, что подтверждается также и на обратных примерах, т. е. на
наличии в самых суровых пустынях целого ряда млекопитающих с слабо
развитыми bullae tympani и ведущих мало подвижной образ жизни (хомячки,
слепушенка, некоторые полевки и др.).
Другое направление специализации в пределах семейства, т. е. при-
способления к рытью, в значительной степени подавлено и замаскировано
приспособлениями к прыганью. При наличии слабых передних конечностей,.
редуцированных вместе с гипертрофией задних, их значение, как роющего
органа, усиливается отчасти развитием острых серпообразных когтей
{Salpingotus, Dipus и др.), но основной роющий механизм переносится,
повидимому, на челюстной аппарат, во всяком случае, иначе нельзя объяс-
нить чрезмерное удлинение нижних резцов, выступающих своими зад-
ними концами у многих представителей на боковой поверхности нижней
челюсти в виде альвеолярных бугров, срастания в области нёба верхне-
губных лопастей и пр. В указанных отношениях тушканчики обнаруживают
определенное сходство с такими высоко специализированными роющими
грызунами с слабыми передними конечностями как Ellobius, Spalax, Geo-
rhychus, Heterocephalus и др. (Виноградов, 1926). Непосредственные наблю-
дения над процессом рытья у тушканчиков также указывают на то, что
главным орудием разрыхления почвы у этих зверьков являются резцы,,
а передние и задние конечности служат главным образом для отбрасы-
вания уже разрыхленной земли, в чем помогают также движения головы
и груди, иногда животное поворачивается в норе и выталкивает скопляю-
щуюся землю наружу. При всей кажущейся слабости резцов и челюстного
аппарата тушканчиков, они нередко роют норы в чрезвычайно твердом
глинистом грунте „такыров", при раскопке которого человек должен при-
менять лом или топор (Alactagulus acontion Pall.).
Из других признаков специализации тушканчиков обращает внимание
у большинства видов значительное увеличение в размерах глазного яблока,,
что является характерным признаком для многих ночных животных;
совершенно неясной, однако, является целесообразность характерного
для тушканчиков косого разреза век, резко наклоненного по отношению
к продольной оси головы.
' Чрезвычайно характерны для всех видов тушканчиков весьма совер-
шенная „пустынная" окраска. Я не имею здесь возможности останавли-
ваться на выяснении сложного и спорного вопроса о биологической
целесообразности этого типа окраски, столь распространенного у живот-
16
ТУШКАНЧИКИ DIPODIDAE
*ных пустынь; по этому вопросу существуют различные взгляды, нашедшие
отражение в литературе (Benson, Формозов, Бобринский и др.), отмечу
однако, что мне пришлось убедиться на личном опыте, что положенная
на песок набитая шкурка тушканчика в сумерки становится для челове-
ческого глаза невидимой уже на расстоянии 5—7 шагов; в песках Средней
-Азии живые тушканчики на свободе абсолютно невидимы даже в светлые
лунные ночи при наблюдениях на близких расстояниях, как это показали
наблюдения сделанные мною и А. И. Аргиропуло (см. описание образа
жизни Dipus sagitta).
Как можно видеть из предыдущего краткого очерка и более деталь-
ных описаний морфологических особенностей отдельных групп и видов
"тушканчиков в специальной части этой работы, в пределах этого семейства
явления морфологической адаптации достигают чрезвычайно сильного
i развития; среди других групп наших грызунов эти явления в не менее
, резкой форме выражены лишь у некоторых роющих форм, но имеют здесь
‘-совершенно иной характер.
КРАТКАЯ ЭКОЛОГИЧЕСКАЯ ХАРАКТЕРИСТИКА СЕМЕЙСТВА DIPODIDAE
По условиям обитания, как было уже упомянуто выше, наибольшее
разнообразие представляет группа Zapodinae, характеризующаяся в то же
время отсутствием ясно выраженных признаков приспособления к какому
-либо определенному комплексу жизненных условий. Различные виды этого
подсемейства обитают в лесах различного типа, на лугах, в различных
зонах гор до альпийских лугов включительно, в степи и полупустыне.
Высоко специализированные формы, к которым принадлежит наиболее
-значительная часть семейства, являются, за очень немногими исключе-
ниями, характерными обитателями зоны пустынь и полупустынь, причем
для глинистых пустынь являются особенно характерными представители
пятипалых тушканчиков (подсемейство Allactaginae), а для песчаных —
трехпалые тушканчики (подсемейство Dipodinae)-, один из представителей
первой группы {Allactaga jaculus) на севере ареала своего обитания встре-
чается также в песках и проникает далеко на север и запад в степную
зону, достигая даже лесостепи. Единственный представитель второй
группы — Scirtopoda telum, в противоположность своим родичам, является
обитателем глинистых пустынь и полупустынь, отчасти проникая также
в степную зону, где он нередко селится также в песках. Преимущественно
-обитателями песков являются вероятно также представители подсемейства
Euchoreutinae и Cardiocraniinae (из последних особенно характерным для
барханных песков несомненно является Salpingotus kozlovi). Некоторые
виды встречаются также в щебнистых пустынях {Allactaga elater, Scirtopoda
telum), хотя более характерны они для различного типа глинистых пустынь.
Большинство видов тушканчиков являются обитателями равнин и
лишь в виде исключения некоторые виды проникают в горы, как, напри-
ВВЕДЕНИЕ
17
•мер, Allactaga williamsi, встречающийся на Арарате на высоте 2500 метров,
или A. saltator, найденный в Тянь-шане (долина р. Аксая) на высоте
около 3000 метров.
Все виды тушканчиков принадлежат к числу характерных ночных
животных, выходящих на поверхность с наступлением темноты и уходящих
в норы до рассвета. Общими чертами строения нор тушканчиков является
то, что зверьки обычно забивают изнутри отверстие главного входа,
•изнутри же вырывают вверх один или несколько ходов, доходящих почти
до поверхности или открывающихся наружу отверстием закупориваемым
на день пробочкой из земли (это образование обычно носит название
„копеечки" или „пупочка"); в тех случаях, когда неожиданная опасность
-застигает зверька в норе, он выскакивает наружу через один из упомя-
нутых запасных ходов, разрушая земляную пробочку или тонкий слой
-земли над слепым окончанием запасного хода. Кроме различного типа
.жилых нор, тушканчики обычно роют значительное количество временных
нор с открытыми отверстиями; эти норы каждую ночь посещаются зверь-
ками, как об этом можно следить по свежим следам у этих нор, а также
путем расстановки у нх отверстий ловушек.
Для всех представителей семейства характерна продолжительная и
• глубокая зимняя спячка; о том, что эта спячка не прерывается до весны,
можно судить по отсутствию в норах этих зверьков зимних запасов пищи.
Спячка у некоторых видов начинается лишь поздней осенью с наступлением
-заморозков (Фенюк 1928,1929); иногда даже наблюдали следы тушканчиков
на снегу (Формозов, 1929). Приведенные примеры свидетельствуют о том,
что не все представители семейства столь чувствительны к понижению
температуры, как это вообще характерно для большинства видов этой
группы. Следует, однако, отметить, что тушканчики не выносят и высокой
температуры, в чем можно убедиться на основании самых элементарных
опытов (например, подвергая клетку со зверьком действию солнечных лучей
в жаркую погоду). Таким образом подобно многим другим обитателям жар-
ких пустынь, тушканчики способны жить здесь лишь избегая влияния высо-
кой температуры, чему способствует их ночной образ жизни н наличие
:нор, в которых зверьки скрываются днем.
Для большинства представителей семейства характерно наличие растя-
нутого периода размножения, т. е. рождение детенышей у различных
особен обычно происходит в различные сроки, начиная с весны и до
позднего лета. Количество детенышей в помете обычно колеблется
•от 2 до 5.
Основной пищей тушканчиков служат различные растительные
вещества и, кроме того, некоторые виды отчасти-питаются также насеко-
мыми; можно предполагать, что насекомые имеют относительно большее
значение в пищевом режиме тех видов, у которых в большей степени
•сохраняется бугорчатая структура коренных зубов, как например у Sicista
или Euchoreutes', относительно некоторых представителей первого рода это
Фауна СССР, Виноградов 2
18
ТУШКАНЧИКИ DIPODIDAE
доказано и непосредственными наблюдениями. Из растительных веществ,
тушканчики наиболее охотно поедают семена различных растений, а также
луковицы некоторых лилейных. При поедании даже мелких семян тушкан-
чики очищают их от оболочки. При поедании луковиц тушканчики выры-
вают характерные ямкн, так называемые „копанки", которые встречаются,
в большом числе в местах их обитаний и в которых можно найти
шелуху от съеденных зверьками луковиц.
На большинство тушканчиков сильно действует искусственный свет,,
причем попадая в полосу сильного света (например фонарей автомобиля),
зверьки теряют ориентировку и становятся беспомощными.
В пределах зоны пустынь н полупустынь тушканчики являются
существенными членами соответствующих биоценозов, нередко преобладая:
по численности над другими представителями класса млекопитающих,,
а иногда являясь почти единственными представителями отряда грызунов-
в некоторых местностях. Этн животные в соответствующих стациях оказы-
вают значительное влияние на растительный покров н почвы и во многих,
случаях являются основными ресурсами питания для некоторых хищных,
птиц.
КРАТКИЙ ОЧЕРК ГЕОЛОГИЧЕСКОЙ ИСТОРИИ СЕМЕЙСТВА DIPODIDAE И.
НЕКОТОРЫЕ ДАННЫЕ О ЕГО ЭВОЛЮЦИИ
Происхождение семейства Dipodidae и его родственные связи с дру-
гими группами грызунов остаются до настоящего времени неясными. Ряд
авторов до недавнего времени считал за исходную форму для всего
семейства род Eomys, описанный Schlosser’oM (1885) из олигоценовых.
фосфоритов Франции {Eomys zitteli Schloss.); однако сам названный
автор отнес эту форму к Myomorpha и поставил ее в системе рядом,
с родом Cricetodon. Eomys zitteli известен по одному фрагменту нижней
челюсти с 4 нижними коренными зубами (1 ложнокоренной и 3 настоя-
щих коренных), имеющими четырехбугорчатое строение. Оба наружных,
бугорка соединены угловато изогнутым гребнем, отсылающим от каждого-
угла отросток к середине зуба, от каждого наружного бугорка отходит
также по одному гребню к соответствующим внутренним бугоркам.
Впоследствии Н. Winge (1887) признал Eomys за. общую родоначаль-
ную форму для всего семейства Dipodidae, что было принято также рядом
других авторов. Следует, однако, отметить, что такой выдающийся иссле-
дователь как Т. Tullberg (1899) высказал некоторое сомнение относительно
родства Eomys и Dipodidae, указав на необходимость дальнейшего
изучения этого вопроса. Подобные же сомнения были высказаны также
самим Schlosser’oM в его более поздней работе (1924), который, однако,
считал, что род Eomys может пролить некоторый свет на происхождение
Sicista и Sminthoides. Можно считать не лишенным вероятия, что-
строение зубов Eomys в основных чертах сходно с исходным примитив-
ВВЕДЕНИЕ
19
ным для Dipodidae строением; однако, отсутствие данных о других
признаках Eomys не позволяет пока считать достоверным наличие род-
ственных отношений между этим родом и семейством Dipodidae.
К семейству Eomyidae были предположительно отнесены Matthew и
Granger’oM (1923) описанные этими авторами из олигоцена Монголии
(Hsanda Gol, Loh) роды Tataromys и Karakoromys. Это предположение
было поддержано также Young’oM (1927). Следует, однако, отметить, что
описания этих родов у названных авторов столь кратки, а известные остатки
этих животных столь скудны, что в настоящее время не может быть уверен-
ности в принадлежности названных родов к семейству Eomyidae, хотя
возможность этого и не исключена, поскольку общий план строения зубов
этих животных напоминает таковой у Eomys.
Так как история рассматриваемого семейства в Европе и Азии
значительно разнится между собой, здесь излагаются сначала краткие
данные по истории европейских представителей семейства и затем данные:
по истории азиатских форм по материалам из Китая и Монголии.
Наиболее древние достоверные остатки Dipodidae принадлежат
олигоцену Европы и относятся к представителям подсемейства Zapodinae
(— Sicistinae auct.). Чрезвычайный интерес представляет описанный Viret
(1926) род Plesiosminthus (фиг. 73_5), представители которого известны
из плиоцена Западной Европы от верхней границы стампийского (Stam-
pien) до верхне-аквитанского яруса. В настоящее время известны 3 сле-
дующих вида этого рода:
1) Р. promyarion Schaub из верхне-стампийского яруса Puy de
Montdoury и Rickenbach; 2) Р. schaubi Viret из того же яруса, найденный
в Braussat; 3) Р. myarion Schaub, из аквитанского яруса, найденный
в Paulhiac, Tommerdingen и Chavroches. S. Schaub (1930), посвятивший
специальную работу описанию остатков ископаемых Zapodinae (=Sicisti-
п'ае) Европы сравнивает этот род с представителями рода Sicista, упоми-
ная лишь, что по наличию бороздок на передней поверхности коренных
зубов этот род сходен с представителями Zapus. Однако, если сравнивать
общую структуру зубов Plesiosminthus (фиг. 7 3_5) с современными пред-
ставителями подсемейства, то обращает на себя внимание сходство не
только в строении резцов, но и в расположении бугорков и гребней на
коренных зубах Plesiosminthus в гораздо большей степени с Zapus, чем
с Sicista', особенно велико сходство в строении коренных зубов Plesiosmin-
thus и современного сычуанского прыгунчика (Eozapus setchuanus), как
это можно видеть на прилагаемых схематических рисунках (фиг. 72).
Указанное обстоятельство позволяет предполагать, что Plesiosminthus
является родственным современному Eozapus и что этот последний
является реликтом близким к представителям древней плиоценовой фауны
Евразии. Наряду с этим обращает на себя внимание тот факт, что род
Sicista, являющийся по своим морфологическим признакам более прими-
тивным, чем род Zapus, однако, появляется значительно позже, чем
2*
20
ТУШКАНЧИКИ DIPODIDAE
представители рода Plesiosminthus (первые представители рода Sicista
известны лишь с начала четвертичного времени, как зто будет ука-
зано ниже).
В древнейшем квартере Западной Европы начинают встречаться
остатки Sicista. В частности Th. Kormos (1903) описал мышевку из долед-
никовой фауны Somlyoberg близ Piispokfiirdo в Венгрии под названием
Sicista praeoloriger. S. Schaub’oM были найдены остатки мышевки, отнесен-
ные им к тому же виду в Nagyharsanyberg близ Villany. Относящиеся
ж более поздним эпохам квартера остатки Sicista были найдены в лёссе
Nussdorf близ Вены (Nehring 1879), в пещере Dobsina (Nehring 1890),
в мадлене Puskaporos в Венгрии (Kormos, 1911), в мадлене Thierstein и
Etinger в Швейцарии (Stehlin, 1921) и в мадлене Petersfeld (Е. Mandach,
1930). Эти остатки обычно относились к .S', montana или S. loriger, но как
показывает изучение современного материала, точно различать виды
мышевок по их черепам и тем более по их фрагментам не представляется
возможным, вследствие чего я не останавливаюсь здесь на видовой
принадлежности четвертичных мышевок. Интересным является тот факт,
что в квартере мышевки были распространены в Европе значительно
далее на запад, чем в настоящее время.
В квартере в Западную Европу проникает вместе с другими степными
животными и единственный представитель настоящих тушканчиков —
Allactaga jaculus-, в послеледниковую эпоху распространение этого вида
на запад достигло Брауншвейга (Thiede и др.), Тюрингского леса (Saalfeld),
северной Баварии (Wurzburg) и южной Богемии (Zuzlawitz) (A. Nehring,
1890). В современную эпоху этот вид распространен к западу лишь до
Днепра (в немногих пунктах он встречается на правом берегу названной
реки). Вероятно в ту же эпоху проникли в Крым Allactaga elater и Scirto oda
telum-, последний вид сохранился в виде реликта в материковой части
Крыма (Алешкинские пески) и в настоящее время.
Из кратко изложенных здесь данных по истории европейских Dipo-
didae можно видеть, что здесь первые достоверные остатки представите-
лей этой группы найдены в олигоцене (верхне-'стампийский ярус), причем,
как было уже отмечено выше, древнейший европейский род Plesiosminthus
является, повидимому, ближайшим родичем современного рода Zapus и
в частности его подрода Eozapus—рецентного представителя фауны Сы-
чуани и Гансу; начиная с раннего квартера появляются остатки предста-
вителей современного рода Sicista; таким образом в течение третичного
времени в Европе известен лишь один род, относящийся к подсемейству
Zapodinae; в квартере он сменяется другим родом того же подсемейства
Sicista. Кроме того, в квартере в Западную Европу проникает с востока
единственный представитель настоящих тушканчиков — Allactaga jaculus,
а в восточную Европу (Крым), кроме того, еще Scirtopoda telum и Allac-
taga elater.
Иначе протекает история этой группы в Китае и Монголии.
ВВЕДЕНИЕ
21
Фиг. 7. Строение зубов у некоторых современных и ископаемых Dipodidae'. 1 — Sicista
subfilis Pall., 2 — Zapus (Eozapus) setchuanus Pous., 3 — Plesiosminthus schaubi Viret.
(no Schaub), 4—5 — Plesiosminthus myarion Schaub (no Schaub), 6 — Paralactaga andersoni
Young (no Ch. Ch. Young), 7 — Protalactaga grabaui Young (no Ch. Ch. Young); 8 — Plesiodipus
ZeeZz Young (no Ch. Ch. Young), 9. Allactaga jaculus Рай., 10 — Alactagulus acontion Pall.
22
ТУШКАНЧИКИ DIPODIDAE
Как было уже упомянуто, некоторые авторы предположительно
относят описанных из олигоцена Монголии Karakoromys и Tataromys
к семейству Eomyidae, которое столь же предположительно считается
родоначальным для Dipodidae. В виду спорности этого вопроса здесь изла-
гаются лишь данные относительно остатков достоверных представителей
семейства Dipodidae.
Наиболее древние остатки тушканчиков известны в Китае и Монголии
лишь с нижнего плиоцена (понтийский ярус). М. Schlosser’oM (1924) описаны
из Ertemte и Olan Chorea в Монголии по нескольким остаткам из понтий-
ского яруса нижнего плиоцена два интересных представителя семейства
Dipodidae — Sminthoides fraudator и Allactaga wimani. S. Schaub (1930), ис-
следовавший оригинальные экземпляры Schlosser’a, пришел к заключению,
что типом S. fraudator следует считать фрагмент верхней челюсти и
отдельные верхние коренные зубы, изображенные Schlo&ser’oM на таблице
III фиг. 2 и’З и что изображенные тем же автором зубы нижней челюсти
принадлежат другому животному, которого Schaub относит к хомякам н
выделяет в новый род и вид — Sinocricetus zdanskii. Schlosser сближает
описанного им грызуна с родом Sicista, но указывает, что описанная им
форма все же значительно разнится от мышевок по расстановке бугров
на жевательной поверхности коренных зубов и по размерам. S. Schaub
(1930) пришел к выводу, что Sminthoides не принадлежит к подсемейству
Sicistinaevi что он имеет некоторые признаки более сходные с представите-
лями настоящих тушканчиков. Насколько можно судить по рисункам Schlos-
ser’a и его описанию, я склонен предполагать, что из современных форм
Sminthoides стоит по устройству коренных зубов ближе всего к Cardiocra-
nius и вероятно должен быть отнесен к подсемейству Cardiocraniinae, что
может быть точно установлено лишь после того, как будут найдены более
полные остатки этого животного; в частности сходство между названными
родами выражается в общей конфигурации жевательной поверхности и
в расположении бугорков; следует отметить, что у Sminthoides, как и у
Cardiocranius бугорки правой и левой стороны чередуются друг с другом,
тогда как у Sicista они расположены друг против друга. Необходимо
также указать, что род Sminthoides обнаруживает также определенное
сходство с описанным Young-’ом ископаемым родом Paralactaga, как это
будет изложено ниже.
Из того же понтийского яруса и из той же местности М. Schlosser’oM
(1924) описан второй интересный представитель семейства, причисленный
автором к роду Allactaga и названный им A. wimani. Schlosser причислил
это животное к названному роду на основании сходного строения корен-
ных зубов, головки бедра и некоторых других признаков; насколько
можно судить по описанию автора и тщательно выполненным рисункам,
по упомянутым признакам этот тушканчик действительно имеет значитель-
ное сходство с родом Allactaga', однако, по некоторым другим признакам
это животное резко отличается от представителей рода Allactaga, в част-
ВВЕДЕНИЕ
23
:ности по следующим: верхние резцы у A. wimani имеют значительно
большую кривизну, чем это имеет место у Allactaga', metatarsus у A. wi-
mani обнаруживает иное строение, чем у представителей названного рода,
а именно у A. wimani при наличии цевки сохраняются явные следы
-срастания трех средних metatarsalia в виде резко обозначенных борозд
на значительном пространстве дистального конца цевки, особенно на ее
.дорзальной поверхности; следует также отметить, что судя по рисунку,
петли на нижних коренных зубах у A. wimani имеют, повидимому, несколько
•более упрощенное строение, чем у настоящих Allactaga. Даже в том случае,
если Schlosser ошибочно причислил к описанному животному верхние резцы
от другого грызуна, все же остается несколько признаков, несвойственных
•современным представителям подсемейства Allactaginae. Указанные обстоя-
тельства не дают возможности в настоящее время выяснить точное систе-
матическое положение A. wimani, но все же следует считать вполне
.достоверным тот факт, что A. wimani должен быть отнесен к подсемейству
Allactaginae, и что таким образом один из представителей этой высоко
специализированной группы существовал уже в нижнем плиоцене Монголии.
Несколько своеобразных третичных тушканчиков были описаны
также Young’oM (1927) из Гансу, найденных в нескольких пунктах близ
•города Lanchow; эти находки автор относит к плиоцену или верхнему
миоцену. В виду скудости найденных остатков точное систематическое
положение этих форм, как и предыдущих, в настоящее время не вполне
:ясно, вследствие чего высказываемые ниже предположения носят лишь
весьма предварительный и ориентировочный характер. Упомянутый автор
•описал из Hsien-Shui-Ho в Ping-Fang-Hsien близ Lanchow Protalactaga
grabaui (фиг. 7 7); наличие лишь скудных остатков этого животного, краткое
их описание с иллюстрациями позволяют пока сделать лишь следующие
заключения: по строению бугорчатых зубов в нижней челюсти Protalactaga
имеет сходство с представителями Zapodinae или Cardiocraniinae, однако
по наличию трех сросшихся средних metatarsalia этот род имеет черты
более высокой специализации, чем перечисленные роды. Впрочем следует
•отметить, что цевка Protalactaga вряд ли была сходной с таковой у Eucho-
.reutes и Allactaga, так как боковые пальцы (I и У) не были так редуцированы
и так обособлены, как у названных родов, да и слияние трех средних meta-
tarsalia вероятно не было столь полным, как у названных родов. Комбина-
ция признаков примитивного строения зубной системы и наличие трех слив-
шихся metatarsalia или тенденции к их слиянию позволяет искать для
рода Protalactaga промежуточное место между Cardiocraniinae и Dipodi-
nae. Так как у упомянутого, выше Sminthoides неизвестно строение задних
конечностей, в настоящее время не представляется возможным судить об
отношении Protalactaga к этому роду, хотя близость обоих родов между
собой кажется вполне возможной.
Ch. Ch. Joung’oM (1927) в той же работе был описан род Plesiodipus
t(P. leeli) (фиг. 7 8) из того же района и горизонта, как и предыдущий род.
24
ТУШКАНЧИКИ DIPODIDAE
Чтобы судить о положении рода Plesiodipus в системе Dipodidae, необхо-
димо отметить следующие его признаки: 1) зубы верхней челюсти, как ука-
зывает автор, до некоторой степени действительно напоминают таковые:
у Dipus, но не менее сходны они с таковыми у Scirtopoda, а быть может
в еще большей степени—у Cardiocranius-, 2) зубы нижней челюсти
имеют некоторые черты сходства даже с Eozapus, особенно по малой
степени редукции М3 и по наличию маленьких дополнительных петель на
внутренней стороне; 3) головка бедра круглая, как у всех Zapodinae и
Cardiocraniinae, что хорошо видно на рисунке в цитированной работе;;
4) чрезвычайно характерно строение metatarsus, причем сам автор указы-
вает, что три средних metatarsalia слабо слиты; на тщательно выполнен-
ном рисунке автора ясно видно, что это слияние даже в проксимальной
части цевки было очень слабым или даже отсутствовало вовсе (в подоб-
ных случаях это не легко решить даже на рецентном материале). Сово-
купность указанных признаков позволяет предполагать, вопреки мнению
Young’a, что Plesiodipus должен быть поставлен в системе ближе к под-
семейству Cardiocraniinae или отчасти даже к Zapodinae. Мнение назван-
ного автора, что Plesiodipus связывает роды Allactaga и Dipus, не может
быть поддерживаемо. Ch. Ch. Young'(1927) описал кроме того из Гансу
еще один род — Paralactaga с двумя видами — Р. major и Р. anderssoni
(фиг. 7 е); позже Teilhard de Chardin и Ch. Ch. Young описали еще один;
вид, относящийся к тому же роду, Р. suni. Остатки первых двух упо-
мянутых видов найдены в Ching-Chuan-Hsien в плиоцене или верхнем
миоцене, остатки последнего в красной понтийской глине Shemnu в Шенси:
(плиоцен).
Род Paralactaga является несомненно представителем подсемейства.
Allactaginae и близок по своим признакам к роду Allactaga, в частности;
по устройству коренных зубов, степени редукции М3 и пр. Расположение
эмалевых петель сходно с таковым у Allactaga^ быть может одним из,
наиболее существенных признаков различия является слабое развитие
первой наружной добавочной петли у Paralactaga-, зубы нижней челюсти
также сходны, но М3 у Paralactaga менее редуцирован, чем у Allactaga
(этот зуб у Paralactaga почти равен по длине коронки предыдущему М2).
Цевка характерного для Allactaga строения. Головка бедра имеет харак-
терную для Allactaga цилиндрическую форму. Таким образом, указанна»
находка, наряду с упомянутой выше находкой Allactaga wimani, подтвер-
ждает существование подсемейства Allactaginae в нижнем плиоцене Мон-
голии и Китая.
Имеющиеся в настоящее время скудные и отрывочные данные
о геологической истории рассматриваемого семейства и сравнительное
изучение морфологических признаков современных его представителей,,
дают возможность составить лишь в чрезвычайно общем виде возможную,
схему эволюции этой группы; подобная попытка в графическом виде
сделана на прилагаемой таблице (фиг. 8).
Фиг. 8. Схема взаимоотношений родов Dipodidae и возможные пути их эволюций.
onwanadgoj
HdWMOWJlJ
нанолиц
nation^
26
ТУШКАНЧИКИ DIPODIDAE
Как было указано выше, уже в плиоцене имеется ряд резко диффе-
ренцированных форм тушканчиков, вероятно укладывающихся в рамки
большинства современных подсемейств, следует поэтому думать, что
расщепление основного родоначального ствола этой группы произошло
по крайней мере в миоцене или даже ранее..
Выше было уже указано, что Н. Winge (1887) и за ним некоторые
другие авторы считали одной из родоначальных для семейства тушканчи-
ков форм олигоценовый род Eomys, что, однако, в настоящее время
нельзя считать доказанным, особенно имея в виду, что род Eomys изве-
стен лишь по строению коренных зубов и другие его морфологические
признаки остались пока совершенно неизученными; однако следует отме-
тить, что в строении и расположении бугорков на коренных зубах имеется
значительное сходство также между представителями Dipodidae и пред-
ставителями вымершего семейства Cricetodontidae‘, указанное сходство
послужило даже причиной существенных разногласий в определении
некоторых третичных грызунов Китая крупными специалистами-палеонто-
логами, как это было указано выше (Sminthoides fraudator Schlosser
и Sinocricetus zdanskii Schaub.). Впрочем сам Schlosser, описавший Eomys,
вероятно был более прав, чем Winge и другие авторы, так как он относил
установленный им род Eomys к Myomorpha и сближал его с родом Cri-
cetodon. Таким образом, не исключена возможность, что семейство Dipo-
didae, имеющее несомненную близость к надсемейству Myomorpha, связано
с последней группой именно через Cricetodontidae. Повидимому, более
отдаленной является связь родоначальных для Dipodidae форм с семей-
ством Muscardinidae, которая признается некоторыми исследователями
(Tullberg, 1891).
В настоящее время не имеется палеонтологических данных для дока-
зательства связи сем. Dipodidae также с сем. Spalacidae-, на родственные
отношения между названными семействами было впервые обращено вни-
мание таким выдающимся исследователем как Н. Winge; вероятность
подобного предположения согласуется с некоторыми фактами.
Наиболее рано отошедшей от мышеобразных предков рассматривае-
мого семейства ветвью следует считать подсемейство Zapodinae, сохра-
нившее по сравнению с другими тушканчиками наибольшее количество
примитивных мышеобразных черт организации, как это указывается
в соответствующих главах (внешний облик, строение конечностей, очер-
тания черепа, форма слуховых косточек, строение коренных зубов и пр.).
К рассматриваемой ветви следует также причислить плиоценовый евро-
пейский род Plesiosminthus, имевший, повидимому, значительное сходство
с современным Eozapus. Остается, однако, неясной эволюция рода
Sicista — наиболее примитивного в морфологическом отношении рода
рассматриваемого подсемейства, так как остатки Sicista известны лишь
с начала четвертичного периода, тогда как более специализированный
род Plesiosminthus известен уже в плиоцене.
ВВЕДЕНИЕ
27
Древние корни происхождения имеет вероятно также современное
подсемейство Euchoreutinae, сохранившее ряд примитивных признаков
при общей высокой специализации свего организма (см. описание назван-
ного подсемейства). Наиболее вероятно считать эту группу рано ответ-
вившейся от родоначального ствола Dipodidae и развивавшейся до изве-
стной степени параллельно с подсемейством Allactaginae, не имея, однако,
с ним прямых родственных связей.
Одной из наиболее древних ветвей родоначального ствола Dipodidae
является вероятно также своеобразное подсемейство Cardiocraniinae,
сохранившее при наличии значительных изменений во всем организме
ряд признаков примитивной организации. Вполне возможно, что к этой же
ветви должны быть причислены найденные в плиоцене Монголии и Гансу
роды Sminthoides и Plesiodipus. Более спорным является происхождение
рода Protalactaga, быть может относящегося к той же ветви или же свя-
зывающего эту ветвь с родоначальной ветвью подсемейства Dipodinae.
Интересно отметить, что подобно тому, как Euchoreutinae развивались
до известной степени параллельно Allactaginae, так и рассматриваемая
ветвь Cardiocraniinae развивалась до некоторой степени параллельно
с Dipodinae (трехпалая задняя конечность у Salpingotus, наличие щетки
на пальцах, строение верхних резцов, строение наружного уха и пр.).
Из современных родов этого подсемейства относительно более
примитивные черты организации сохранили представители рода Cardiocra-
nius, а наиболее измененными морфологическими особенностями характе-
ризуется род Salpingotus (особенно S. kozlovi Vinogr.).
Вероятно позже других ветвей отходят от основного ствола две
последних ветви семейства, соответствующие подсемействам Allactaginae
и Dipodinae.
В пределах подсемейства Allactaginae пока достоверно известен
лишь один вымерший плиоценовый род—Paralactaga. Древние корни
имеет также современный род Allactaga, один из представителей кото-
рого (A. wimani) известен нз понтийского яруса (нижний плиоцен) Монго-
лии. Дальнейшие стадии эволюции наблюдаются у современных родов
Alactagulus и Pygerethmus (в частности по утрате бугорчатого строения
коренных зубов и редукции pm); оба названных рода близко родственны
между собой и их обособление не вполне закончилось еще и в настоя-
щее время (наличие формы промежуточного характера в виде Pygerethmus
zhitkovi). Наиболее уклоняющийся от других современных представите-
лей рассматриваемого подсемейства род Scarturus (редукция одного
пальца задней конечности) вероятно раньше других названных родов
обособился от основной ветви подсемейства Allactaginae.
Среди представителей подсемейства Dipodinae не известно вымер-
ших родов (я склонен придавать роду Plesiodipus иное систематическое
значение, чем это делал Young, как это было уже указано выше). Род
Dipus известен с плейстоцена, а другие роды — лишь в современном
28
ТУШКАНЧИКИ DIPODIDAE
состоянии. На основании изучения современного материала можно считать,,
что представители рода Dipus имеют по сравнению с другими родами
этой группы более примитивные черты организации (наличие pm, бугор-
чатое строение зубов, слабо раздутые mastoidea и пр.). Остальные роды
рассматриваемого подсемейства, несмотря на значительные различия
между ними, имеют более близкое, сходство между собой, чем с родом
Dipus. Из родов относящихся к этой группе (Scirtopoda, Paradipus,.
Eremodipus и Jaculus) более низкую эволюционную ступень занимает род
Scirtopoda (отсутствие „знамени" на хвосте, наличие в молодом или взро-
слом состоянии рудимента pm, относительно слабо раздутые mastoidea
и пр.); более измененной является организация Paradipus и в еще боль-
шей степени у Eremodipus и Jaculus', последние два рода близки друг
к другу и вероятно обособились относительно недавно; из них несколько
более примитивные особенности сохранил Eremodipus (отсутствие гребня
на squamosum, слабо развитое знамя и пр.). Указанные отношения между
двумя последними родами свидетельствуют о том, что родину для афри-
канских Jaculus вероятно следует искать в пустынях Средней Азии
и, быть может, Ирана, где сохранился наиболее примитивный представи-
тель этой группы. Более подробные данные о возможных очагах, в кото-
рых развивались отдельные группы тушканчиков приводятся в главе
о географическом распространении этого семейства.
ОБЩИЙ ОЧЕРК ГЕОГРАФИЧЕСКОГО РАСПРОСТРАНЕНИЯ
Все семейство в целом свойственно исключительно Голарктической
области, причем в Неарктике распространены представители лишь одного,
подсемейства Zapodinae, относящиеся к одному роду Zapus. Значитель-
ное большинство палеарктических представителей семейства связано
в своем распространении с пустынно-степной зоной, за исключением рода
Sicista, охватывающего своим распространением и значительную часть,
лесной зоны (одним из представителей этого рода — «S', montana местами
достигает к северу даже северной границы лесной зоны) (фиг. 9).
В Палеарктике граница распространения семейства простирается
к западу до Венгрии и Дании; северная граница достигает Норвегии,
северного Урала, Томска, побережья Байкала и Сахалина (до указанных
пределов достигает распространение лишь представителей рода Sicista)..
К югу представители семейства распространены до Румынии, побе-
режья Черного моря, Малой Азии, Северной Аравии, северной Африки,
Ирана, Белуджистана, Афганистана, Кашмира, северной окраины Тибет-
ского нагорья, Сычуани, Гансу, Шенси и Чили.
Если выделить из рассматриваемого семейства примитивный и резко
отличный в экологическом отношении род Sicista и рассматривать рас-
пространение остальных групп семейства, т. е. собственно тушканчиков,
то очерченная выше северная граница распространения семейства должна
ВВЕДЕНИЕ
29
Фиг. 9. Карта распространения сем. Dipodidae в Палеарктике: 1. Распространение рода Sicista, 2. Распространение других родов.
От Гринича
30
ТУШКАНЧИКИ DIPODIDAE
быть значительно отодвинута к югу, а именно до следующих пределов^
р. Ока, южные районы Горьковского края, южный Урал, Омск, Барнаул,
Танну-Тувинская республика, южное Забайкалье, Манчжурия. Следует,
однако, отметить, что и до этой границы доходит распространение лишь
двух наиболее северных видов одного рода Allactaga (Л. jaculus и A. sal-
tator), границы же распространения большинства других видов лежат еще
южнее, как это будет указано ниже, охватывая преимущественно зону
полупустыни и пустыни.
Рассматривая состав фауны семейства Dipodidae в различных частях
Палеарктики, здесь можно наметить три основных территории, имеющие
свои специфические особенности в рассматриваемом отношении.
Первой из них следует признать территорию средне-азиатской части
СССР (Туркестана) и южного Казахстана, где наблюдается наибольшее
богатство форм рассматриваемой группы представленных в числе по
крайней мере 11 видов, относящихся к 7 родам, как это можно видеть из
следующего списка.
Allactaga jaculus
„ severtzovi
„ saltator
„ elater
Alactagulus acontion
Pygerethmus platyurus
Pygerethmus zhitkovi
Dipus sagitta
Scirtopoda telum
Eremodipus lichtensteini
Paradipus ctenodactylus
Из приведенного списка являются эндемичными для рассматриваемой
территории следующие: Allactaga severtzovi, Pygerethmus platyurus,
P. zhitkovi, Eremodipus lichtensteini, Paradipus ctenodactylus, что соста-
вляет 3 эндемичных рода и 5 видов. Общими с описываемой ниже
монгольской фауной тушканчиков являются лишь Allactaga saltator,.
Alactagulus acontion и Dipus sagitta, а с иранской фауной — лишь Allac-
taga elater и Dipus sagitta.
Средне-азиатская фауна тушканчиков проникает довольно далеко на
запад в пределы европейской части Союза, обнаруживая постепенное
обеднение видового состава: до побережья Каспийского моря распростра-
нены Allactaga severtzovi, Paradipus ctenodactylus и Eremodipus lichten-
steini, до низовий p. Урала — Allactaga saltator и Pygerethmus platyurus, до
Калмыцкой области и северо-восточного Предкавказья — Allactaga elater,
Alactagulus acontion и Dipus sagitta и наконец на Украину проникают
лишь два вида — Allactaga jaculus и Scirtopoda telum, причем первый из
них доходит на запад до Днепра, а второй имеет островное реликтовое
распространение в песках нижнего течения Днепра (Алешкинские пески).
Некоторые из свойственных Средней Азии видов тушканчиков про-
никают также в пустыни Ирана и Афганистана, как A. elater- однако
в Иранские пустыни проникают также некоторые африканские или близ-
кие к ним виды тушканчиков (род Jaculus), как это будет указано ниже.
ВВЕДЕНИЕ
зт.
Euchoreutes naso
Cardiocranius paradoxus
Salpingotus kozlovi
„ crassicauda
Вторым очагом распространения тушканчиков следует признать тер-
риторию Монголии. Здесь известно нахождение 9-ти следующих видов,
относящихся к 7-ми родам:
Allactaga saltator
„ bullata
Alactagulus acontion
Dipus sagitta
Scirtopoda andrewsi
Процент эндемичных видов здесь также очень высок, считая эндемич-
ными для этой территории следующие виды: Allactaga bullata, Scirtopoda
andrewsi, Cardiocranius paradoxus, Salpingotus kozlovi, S. crassicauda
и в значительной степени Euchoreutes naso, основной ареал которого
лежит в пределах Монголии, но захватывет также часть территории Синц-
зяна (Китайского Туркестана); таким образом из 9-ти монгольских видов 5
или 6 являются эндемичными. Особую своеобразность монгольской фауны
тушканчиков придает наличие здесь таких замечательных родов, как Car-
diocranius, Salpingotus и Euchoreutes. Как было уже указано, лишь 3 сле-
дующих вида являются общими для Монголии и Советской Средней Азии:
Allactaga saltator, Alactagulus acontion и Dipus sagitta.
Третьей, характерной по фауне тушканчиков, территорией является
территория Ирана, Аравни и Северо-африканских пустынь. Для этой
территории характерно присутствие представителей рода Jaculus (Jaculus
loftusi, J. blanfordi в Иране, J. orientalis и J. jaculus с ближайшими фор-
мами— в Африке); кроме того, здесь представлен род Scarturus с одним
видом — 6”. tetradactylus, найденным в Ливийской пустыне и Египте, A. hot-
soni (Белуджистан), A. euphratica (Месопотамия) и под сомнением
Salpingotus thomasi, точное нахождение которого неизвестно, но предпо-
ложительно относится к южному Афганистану, что, однако, внушает
сомнения (см. ниже). Таким образом для Иранско-африканских пустынь
характерно присутствие 4 родов с 8 или более видами, которые являются
эндемичными для рассматриваемой территории. Единственным отголоском
рода Jaculus в более северных пустынях является родственный род Eremo-
dipus, распространенный в пустынях Средней Азии. Близкий к A. euphra-
tica вид — A. williamsi распространен в Малой Азии и Закавказья.
Если сравнивать фауну тушканчиков азиатских и африканских пу-
стынь, то обращает на себя внимание значительно большее богатство и
разнообразие в этом отношении азиатских пустынь, что позволяет счи-
тать основной родиной рассматриваемой группы Азию, откуда проникли
в Африку лишь немногие представители этой группы, заняв здесь лишь
самую северную часть материка от Египта до Марокко. В этом отноше-
нии представляет полную противоположность другая характерная группа
пустынных грызунов — песчанки (Gerbillinae), представленная на всем про-
тяжении африканского материка большим количеством родов и видов
и проникшая в азиатские пустыни в виде лишь 2—4 родов, представлен-
32
ТУШКАНЧИКИ DIPODIDAE
ных лишь небольшим количеством видов. Таким образом можно предполо-
гать, что основной путь эволюции тушканчиков связан с азиатскими
пустынями, а песчанок — с африканскими. Первое предположение подтвер-
ждается и палеонтологическими находками, как это указывается в главе
по истории фауны тушканчиков.
КЛАССИФИКАЦИЯ
В настоящем обзоре для классификации семейства Dipodidae приме-
нена с немногими дополнениями та же система, которую я применял
в предыдущих своих работах (особенно в работе 1925 года); эта система
является коренной переработкой системы R. Рососк’а (1922) и других
исследователей; основанием для этой переработки послужило углубленно
сравнительное изучение морфологических признаков современных туш-
канчиков и открытие нескольких своеобразных новых родов этого
семейства (особенно рода Salpingotus с тремя видами).
Принятое до последнего времени большинством исследователей
разделение тушканчиков на два самостоятельных семейства Zapodidae
(мышевки и прыгунчики) и Dipodidae (настоящие тушканчики) в настоящее
время не может быть сохранено, так как открытие рода Salpingotus и более
детальное исследование морфологических признаков Cardiocranius и Eucho-
reutes, обладающих рядом признаков промежуточного характера между
названными группами, дало возможность заполнить пропасть между мышев-
ками и тушканчиками и сделало необходимым объединить всех известных
мышевок, прыгунчиков и настоящих тушканчиков в одно семейство —
Dipodidae с пятью подсемействами — Zapodinae (мышевки и прыгунчики),
Euchoreutinae (единственный род и вид — Euchoreutes naso), Cardiocraniinae
(Cardiocranius и Salpingotus), Allactaginae (пятипалые настоящие тушкан-
чики) и Dipodidae (трехпалые настоящие тушканчики). Взаимоотношения
этих групп и их возможные родственные связи с ископаемыми формами
кратко изложены выше в обзоре геологической истории и вероятного
. хода эволюции всего семейства; детальные диагнозы названных под-
семейств приводятся ниже в соответствующих местах систематической
части и в определительных таблицах. В тех же частях обзора даны
диагнозы родов, составляющих названные подсемейства и освещены
взаимоотношения между отдельными родами.
Принимаемые здесь подсемейства и роды располагаются в следую-
щую систему:
Подсемейство 1. Zapodinae
Род Sicista Gray.
„ Zapus Cones.
Подсемейство 2. Euchoreutinae
Род Euchoreutes Sclat.
Мышевки и прыгунчики.
, Мышевки.
Прыгунчики.
Длинноухие тушканчики.
ВВЕДЕНИЕ
33
Подсемейство 3. Cardiocraniinae
Род Cardiocranius Satun.
„ Salpingotus Vinogr.
Подсемейство 4. Allactaginae
Род Allactaga F. Cuv.
„ Alactagulus Nehr.
„ Pygerethmus Gloger.
„ Scarturus Gloger.
Подсемейство 5. Dipodinae
Род Dipus Gmel.
„ Scirtopoda Br.
„ Paradipus Vinogr.
„ Eremodipus Vinogr.
„ Jaculus Erxleb.
Карликовые тушканчики.
Пятипалые карликовые
тушканчики.
Трубкоухие или трехпалые
карликовые тушканчики.
Пятипалые и четырехпалые
настоящие тушканчики.
Земляные зайцы.
Земляные зайчики
или тарбаганчики.
Толстохвостые тушканчики.
Четырехпалые тушканчики.
Трехпалые настоящие тушкан-
чики.
Мохноногие тушканчики.
Емураичики.
Гребнепалые тушканчики.
Пустынные тушканчики.
Африканские трехпалые
тушканчики.
В настоящем обзоре я не всегда строго следовал формальным прави-
лам номенклатуры и в частности, так называемому, „правилу приоритета",
ме считая целесообразным загромождать настоящую работу второстепен-
ными или излишними данными по синонимике и пр.; однако в наиболее
необходимых случаях, где это требовала ясность изложения, сделаны
«соответствующие изменения номенклатуры с краткими пояснениями.
Во всех других случаях я твердо стоял на этой позиции, что для зоолога
более целесообразно заниматься непосредственным изучением самих
животных, чем данных им в различное время различными авторами
названий.
«СТАТИСТИЧЕСКИЕ ДАННЫЕ О СИСТЕМАТИЧЕСКОМ СОСТАВЕ СЕМЕЙСТВА
На основании современных данных можно считать, что семейство
Dipodidae в современном его объеме содержит 4 подсемейства с 14 родами
и 50 видами. Подсемейства можно считать, повидимому, более или менее
установленными; почти то же относится и к родам, за исключением
.вероятно родов Alactagulus и Pygerethmus, которые, быть может, после
«более детального исследования будут объединены вместе; однако не
исключена еще возможность установления новых родов, как об этом
Фауна СССР, Виноградов 3
34
ТУШКАНЧИКИ DIPODIDAE
можно судить по имевшим место еще недавно фактам описания трех
резко дифференцированных родов (Salpingotus, Paradipus и Eremodipus^
за период времени с 1923 по 1930 г.
Указанное количество видов является приближенным, так как пока
не установлено еще точное систематическое положение некоторых видов-
мышевок (род Sicista), относящихся к группе одноцветных мышевок
(5- Jlava, S. leathemi и др.), равным образом, недостаточно точно выяснены
еще видовые взаимоотношения между некоторыми американскими предста-
вителями рода Zapus и африканско-аравийскими представителями рода.
Jaculus. Во всех указанных случаях при дальнейших исследованиях
возможно ожидать некоторого сокращения количества описанных до
настоящего времени форм в качестве самостоятельных видов; однако
возможно в дальнейшем и установление новых хорошо дифференцирован-
ных видов (за последнее десятилетие было установлено 10 несомненно
новых видов и из них 5—с территории Союза).
Подсемейство Zapodinae содержит 2 рода — Sicista и Zapus с 22 ви-
дами, из которых к первому роду относятся 9 видов и к второму—13.
Подсемейство Euchoreutinae содержит один род и вид (Euchoreu-
tes naso).
Подсемейство Cardiocraniinae составлено 2 родами: Cardiocranius
и Salpingotus, причем первый род имеет лишь один вид, а второй —
3 вида.
Подсемейство Allactaginae содержит 4 рода: Allactaga с 8 видами,.
Alactagulus с 1 видом, Pygerethmus с двумя и Scarturus — с одним.
В подсемействе Dipodinae в настоящее время известно 5 родов::
JJipus, Paradipus, Eremodipus, Scirtopoda и Jaculus-, первые 3 рода имеют
по одному виду, род Scirtopoda — 2 вида и род Jaculus предположительно
4 вида, так как большое количество форм, описанных из северной
Африки и Аравии столь сходны во всех существенных признаках, что
вероятно в дальнейшем будут сведены к указанному количеству видов.
(в настоящем обзоре количество этих видов предположительно прини-
мается за 4).
Из пределов Палеарктики в настоящее время известно 38 видов,,
относящихся к 14 родам (с указанными выше оговорками), в том числе из
пределов СССР 18 видов, составляющих 8 родов.
По количеству видов сем. Dipodidae занимает в фауне грызунов
Союза 3-е место (на первом месте Muridae с 73 видами, на втором
Sciuridae с 22 видами).
Численность особей отдельных видов в некоторых пустынных частях
Союза чрезвычайно велика, составляя местами преобладающую в коли-
чественном отношении часть всего животного населечния, относящуюся
к классу млекопитающих. Уже по указанной причине тушканчики играют
важную роль в биоценозах пустыни, как это описывается в соответ-
ствующей главе.
ВВЕДЕНИЕ
35
ЗНАЧЕНИЕ ПРЕДСТАВИТЕЛЕЙ СЕМЕЙСТВА DIPODIDAE ДЛЯ ЧЕЛОВЕКА
В настоящее время при скудности имеющихся данных по биологии
тушканчиков, не представляется возможным дать в достаточной мере
полную оценку экономического значения этой группы в целом или даже
отдельных ее представителей. Однако при значительной численности
этих животных или даже преобладании их среди других видов млекопи-
тающих в некоторых районах Союза, тушканчики являются здесь важным
компонентом пустынных биоценозов и уже по этой причине они прямым
или косвенным образом входят в сферу хозяйственных интересов человека.
Положительная хозяйственная роль тушканчиков заключается прежде
всего в том, что некоторые виды этого семейства в настоящее время
являются объектами промыслового использования. В этом отношении
наибольшее значение имеет большой тушканчик {Allactaga jaculus),
шкурки которого на ряду с другими второстепенными объектами пушнины
стали заготовляться с 1925 г. и с тех пор их заготовка начала значи-
тельно расти, достигнув в 1929/30 г. количества около 32000 штук. Про-
должающийся дальнейший рост заготовок еще далеко не использовал
всех имеющихся возможностей, т. к. некоторые виды, мало уступающие
по величине большому тушканчику {A. severtzovi, A. saltator и др.), до
настоящего времени почти еще не заготовлялись.
Другая сторона положительного хозяйственного значения тушкан-
чиков заключается в том, что эти животные в некоторых местах состав-
ляют существенную часть кормовой базы для некоторых ценных промыс-
ловых видов, как лисица, корсак, степной хорек, что также следует
учитывать при промысловом использовании территории пустынь.
Отрицательная хозяйственная роль представителей рассматриваемой
группы выражается в наибольшей степени в области сельского хозяйства.
Особенно это резко сказывается в период освоения отдельных частей
территории пустынь или полупустынь в целях их сельско-хозяйственного
использования. Вредоносная деятельность тушканчиков отмечена на бах-
чевых, зерновых и технических культурах, где выражается более всего
в поедании посеянных в землю семян культурных растений.
Вред тушканчиков на бахчевых и зерновых культурах наиболее
изучен в Сталинградском крае (Фенюк, 1928,1929), на технических куль-
турах—в пределах Аулие-атинского и Туркестанского районов Казах-
стана (Колесников, 1934); в частности последним из упомянутых авторов
установлено, что на плантациях ценного каучуконоса тау-сагыза в окре-
стностях кишлака им. Атабаева в предгорьях Кара-тау процент повре-
ждения площади питомников тау-сагыза тушканчиками выражался в цифре
около 32%. Впрочем следует отметить, что общий удельный вес тушкан-
чиков среди других вредных в сельско-хозяйственном отношении грызу-
нов относительно невелик и причиняемые им повреждения являются
3*
36
ТУШКАНЧИКИ DIPODIDAE
скорее исключением, чем правилом, что, однако, не умаляет наличия их
значительной вредоносной деятельности для отдельных районов и отдель-
ных культур. Некоторый вред тушканчики причиняют также растительности
насаждаемой на песках с целью их закрепления, что особенно относится
к кустарникам — саксаулу (Arthrophyton haloxylon), черкезу (Salsola
Richteri) и кандыму (Calligonum caput-medusae и др.); в этом случае вред
выражается также преимущественно в поедании семян после их посева.
Пока совершенно не выяснена роль тушканчиков на пастбищах, хотя
не исключено их отрицательное влияние на растительность этих угодий,
особенно принимая во внимание пристрастие тушканчиков к луковичным
растениям, среди которых имеются ценные в кормовом отношении виды.
Слабо изучено также значение тушканчиков, как переносчиков пара-
зитов и болезней человека и домашних животных; однако считается, что
эти животные наряду с другими грызунами, могут являться резервуарами
вируса, например, чумы, как это установлено для некоторых районов
южного Заволжья.
СПИСОК ЦИТИРОВАННОЙ ЛИТЕРАТУРЫ
Allen, Glover, М. Gerboas from Mongolia. Amer. Mus. Novitates, n° 161. 1925.
Alston, E. K. On the classification of the order Glires. Proc. Zool. Soc. bond. 1876. 61.
Anderson, John and Winton, W. E. Zoology of Egypt, Mammalia, bond. 1902.
Barre t-Hamilton. Proc. Zool. Soc. bond. (Dipus deasyi.)
[В e 1 j a e v A. M.] Беляев, A. M. Список грызунов Казакстана. Tp. Зоол. Инет. Акад.
Наук, 1933, т. 2 : 27.
(В i 1 j s k i j] Б i л ь с к и й. Поширення великого земляного зайца (Alactaga jaculus Pall.)
на Правобережной Украине. Тр. физ. мат. отд. Укр. Акад. Наук. XIII, I, 1929: 145.
Blanford, W. Т. Eastern Persia, bondon, 1876: 75.
Brandt, F. Remarques sur la classification des Gerboises etc. Bull. Cl. Phys.-Math. Acad.
Sciences St. Petersb. II, 1844 : 209.
Cha w о r t h-M usters. A new Species of the genus Sicista. Ann. Mag. Nat. Hist, XIX,
1927: 542 (Sicista).
Cuvier, F. Memoires sur les Gerboises et les Gerbilles. Transact. Zool. Soc. bondon, II,
1841: 131.
Dobs о n, G. E. On the natural position of the family Dipodidae. Proc. Zool. Soc. bondon,
1882: 640.
Eversmann, E. Einige Beitrage zur Mammalogie und Ornithologie des Russischen Reiches.
Bull. Soc. Nat. Moscou, 1848, I: 188. (Allactaga saltator).
'[F a u s s e k, V. А.] Ф а у с с e к, В. А. Биологические исследования в Закаспийской обла-
сти. Зап. Русск. Геогр. общ., 1906: 1.
[F е n j и к, В. К.] Ф е н ю к, Б. К. К биологии тушканчиков, Матер, к позн. фауны Ниж-
него Поволжья, III, изд. Научно-Иссл. лаб. Отр. Вещ. Саратов, 1928.
[F е n j u к, В. К.] Фенюк, Б. А. Еще о биологии тушканчиков и о мерах борьбы с ними.
Матер, к позн. фауны Нижнего Поволжья. III, Саратов, 1929.
F or mo so v, A. N. Ueber die Saugetiere der Nordlichen Mongolei. „Folia Zool. et Hybro-
biol.“ Riga III, n° I, 1930: 79 (Sicista montana strandi).
[F orm os о v, A. N.] Ф о p м о з о в, A. H. Млекопитающие Северной Монголии. Пред-
варительный отчет о Зоолог, экспедиции в Сев. Монголию за 1926 г. изд. Акад.
Наук, 1929.
ВВЕДЕНИЕ
37
Granger, W. and Matthew, W. Nine new Rodents from the Oligocene of Mongolia.
Amer. Mus. Novitates, 1923, n° 102 :5 ( Tataromys, Karakoromys).
Hatt, R. G. The vertebral columns of ricochetal Rodents. Bull. Amer. Mus. Nat. Hist.,.
LXIII, 1932.
H e p t n e r, W. G. Systematische und tiergeographische Notizen liber einige russische Sauger
„Folia Zool. et Hydrobiol. “ Riga, VI, n° 1934 :17 (Allactaga elater и Scirtopoda telum).
Hollister, N. New Mammals from the Highlands of Siberia. Smithsonian Mascell. Collec-
tions, 1912, n° 60, n° 14. (Allactaga grisescens, Sicista napaea).
Howell, A. B. New Chinese Mammals. Proc. Biol. Soc. Washington, 1928, 41: 41 (Eucho-
reutes naso alashanicus).
Howell, A. B. The saltatorial rodent Dipodomys-, the functional and comparative anatomy
of its muscular and osseous systems. Proc. Amer. Acad. Arts and Sci., 1932, 67,
n° 10.
Young, Chun g-C h i e n. Fossile Nagetiere aus Nord-China. Palaeontologia Sinica. 1927.
Ser. C.
[Кар lan о v, A. u Rajevsky, V.] Капланов, А. и Раевский, В. Материалы
по фауне млекопитающих центрально-промышленной области. Тр. Гос. Музея Центр,
промышленной области. Москва, 1928, стр. 17 (наблюдения над Sicista montana).
[Kolesnikov, I. I.] Колесников, И. И. Вредные грызуны каучоконоса тау-сагыза
САОГИЗ, Москва—Ташкент, 1934.
Kormos, Th. Die pleistocane Saugetierfauna der Felsnische Purkaporos bei Hamor. Mittel.
aus d. Jahrb. d. konigl. Ungar, geol. Reichsanst. XIX : 191 (Sicista).
Kormos, Th. Beitrage zur Praglazialfauna des Somblyoberges bei Piisporkiido. Allatani
Kozlemenyek, XXVII, 1930 (Zapodinae).
[Kuznetzov, В. А.] Кузнецов, Б. А. Доклады Акад. Наук СССР, 1933 (Pygerethmus
schitkovi).
Lie h t e n s t e i n, M. H. K. Ueber die Springmause, oder die Arten der Gattung Dipus.
Berl. 1825 (1828).
Lyon, Marcus. A comparison of the osteology of the Jerboas and Jumping-mice. Proc.
Unit States Nat. Museum., 1901, XXIII: 659.
M a n d a c h, К. V. Die kleinere Wirbeltiere der prahistorischen Stationen Bilstockfels und
Peterfels, in Peters E. Die alsteinzeitliche kulturstatte Peterfels, Augsburg, 1930
(Zapodinae).
M e h e 1 y, L. Streifenmause (Sicistinae) Europas. Ann. Mus. Nat. Hungarici, XI, 1913 : 220.
Miller, G. S. Four new Chinese Mammals. Proc. Biol. Soc. Washington, 1911 :53 (Alla-
ctaga mongolica longior).
Miller, G. S. List of American recent mammals. Unit. Stat. Nat. Museum. Bull. 128,
1924,: 432 (список Zapus).
Miller, G. S. and G i d 1 e у, I. W. Synopsis of the supergeneric groups of Rodents.
Journ. Washingt. Acad. Sci., VIII, n° 13 : 431.
N e h r i n g, A. Fossilreste kleiner Saugetiere aus dem Dilluvium von Nussdorf bei Wien.
Jahrb. k. geol. Reichsanstalt, 29, 1879 (Sicista).
N e h r i n g, A. Ueber Tundren und Steppen der Jetzt - und . Vorzeit, 1890 (Dipodidae).
N e h r i n g, A. Ueber Gebiss-und Schadelunterscheide von Alactaga elater Licht, und
A. acontion Pall. Sitzt.-Ber. Gesellsch. Naturforsch. Freunde Berlin, 1897, n°
9:151.
N e h r i n g, A. Ueber Alactaga saliens fossilis Nehr. Neues Jahrb. fiir Mineralogie etc. 1898,
Bd. II: 1.
N e h r i n g, A. Ueber Schadel, Gebiss- und Schwanzbildung bei Platycercomys platyurus Lich-
tenstein. Zool. Anzeiger, XXIII, n° 619, 1900 : 361.
Milne-Edwards, A. Recherches pour servir a 1’histoire natur. des Mammiferes, 1868—
74:146.
Nordmann, A. Voyage de Demidoff d. 1. Russie meridionale, III, 1840.
38
ТУШКАНЧИКИ DIPODIDAE
[Ognev, S. I.] Огнев, С. И. Млекопитающие Таврической губ. I, 1916:49.
[О g n e v, S. I.] Огнев, С. И. Грызуны Северного Кавказа, Ростов-Дон, 1924.
[О g n е v, S. I.] Огнев, С. И. Uebersicht der in der russischen Fauna einheimischen Spring-
mause der Gattung Dipodipus. Zool. Anzeiger, I, Heft 5/8, 1933 : 207 (Dipus sagitta Pall.).
Pallas, P. S. Novae species Quadrupedum Glirium Ordinae, 1778.
Pallas, P. S. Zoographia Rosso-Asiatica, 1781.
P e t e n у i. Termeszetrajzi Fuzetek, 1832, V : 18 (биологические наблюдения над Sicista).
P о с о c k, R. The External Characters of Scarturas and other Jehrboas, compared with those
of Zapus and Pedetes. Proc. Zool. Soc. of London 1922, part. Ill: 659 (наружные при-
знаки Dipodidae и строение их genitalia)..'
Р о m е 1, A. Catalogue methodique et descriptif des vertebres fpssiles decouverts dans le
bassin hydrographic superieur de la Loir etc., Paris 1853 (Zapodinae).
Pousargues, M. E. Sur la faune mammalogique du Setchuan et sur uns espece asiatique
du genere Zapus. Bull. Mus. d’Hist. Nat., Paris, t. 2, 1896: II (Zapus seichuanus).
Preble, E. Revision of the Jumping Mice of the genus Zapus. „North-Amer. Fauna",
n° 15, 1899.
Rad de, G. Neue Sagethier-Arten aus Ost-Sibirien, Mel. Biol.FAcad. Petersb., Ill, 1861 : 680
(Allactaga mongolica).
[Rajevskij] Раевский „Московский Краевед" n° 4 (12). Москва 1929:33 (Северная
граница распространения Allactaga jaculus в Московской области).
S а 1 е n s к у, W. Ueber eine neue Sminthus-art aus dem Tian-schan. VIII, 1903, p. 17 (Sicista
tianschanica).
(S a t u n i n, К. А.] С а т у н н н, К. А. Млекопитающие Кавказа, т. II, Тифлис, 1920.
[S a t u n i п, К. А.] С а т у н и н, К. А. Обзор исследования млекопитающих Кавказского
края. Записки кавказского отдела русского геогр. общ. 1903, XXIV: 60.
S a t u n i п, К. Eine neue Springmauu aus der Kirgisen-Steppe (Allactaga suschkini). Zool.
Anzeiger XXIII, 1900, n° 610:137.
Satunin, K. Neue Nagetiere aus Centralasien. Ежегод. Зоол. Музея Акад. Наук, VII,
1902 (1903) : 36 (Cardiocranius paradoxus).
Schapiro, В. Das Verhaltnis der Gattung Dipus zu den Myomorphen. Morphol. Jahrb.
Leipzig, 1913: 209 (строение мускулатуры задней конечности у Myomorpha).
Schaub, S. Fossile Sicistinae Europas. Eclogae geolog. Helvetiae, 23 (2), 1930.
Schaub, S. Die hamsterartigen Nagetiere des Tertiars und ihre lebende Verwandten.
Abhandl. Schweiz, palaeont. Gesellschaft, XLV, 1925.
Schlosser, M. Tertiary vertebrates from Mongolia „Palaeont. Sinica", ser. с. I, 1929.
Schlosser, M. Palaeontographica, Bd. 31, 1835 :102 (Eomys zitteli.)
Schumann, A. Das Skelett der Hinterertremitat von Dipus aegyptius. Morphol. Jahrb.
Leipzig, 1904, XXXII: 233 (остеология задних конечностей).
S elate r, W. One new Genus and Species of Rodents of the Family Dipodidae from
Central Asia. Proc. Zool. Soc. Lodon, 1890:610.
[S h e p e, А. К.] Шепе, А. К. Збирник праць Зоол. Музею (Укр. Акад. Наук) п° 13,
1934, стр. 159 (вред Allactaga jaculus на плантациях тау-сагыза).
{Shnitnikov, V. N.] Шнитников, В. Н. Фауна мелких млекопитающих дол. р.
Алматннки в Семиречьи „Защит, раст. от вредит.", 1931.
S о w е г b у, А. С. A new Three-toed Jahrboa from China. Ann. Mag. Nat. Hist. Ser. 9, vol.
V, 1920 : 219 (Dipus halli).
S t e h 1 i n, H. G. Sicista spec, im schweizerlichen Pleistocan. Eclogae geol. Helveciae, XVI, 1922.
Thomas, O. On Mammals from Central Asia, collected by Mr Douglas Carruthers. Ann.
Mag. Nat. Hist., ser. 8, 1912, v. IX: 406 (Allactaga elater dzungariae).
Thomas, O. List of small Mammals from the Islands of Saghalien and Hokkaido. Proc.
Zool. Loud., 1907: 413 (Sicista caudata).
Thomas, O. On two new Rodents from Van, Kurdistan. Ann. Mag. Nat. Hist., ser. 6,
IX, 1897:309.
ВВЕДЕНИЕ
39
Thomas, О. Ann. Mag. Nat. Hist., v. VII, 1831 :15 {Allactaga euphratica).
Thomas, O. On small Mammals from Djarkent, Central Asia. 1914, v. 8, XIII: 571
(Allactaga riickbeili).
Thomas, O. On the genera of Rodents. Proc. Zool. Soc. London, 1896:1012.
T u 11 b e r g, T. Ueber das System der Nagetiere. Uppsala, 1899.
JT u г о v, S. S.] Туров, С. С. Опыт систематического обзора млекопитающих Осетии.
Учен. Зап. Сев.-Кавказ. Общ. Краевед.: 1,1926 : 321 (Sicista caucasica).
Vinogradov, В. S. On the classification of Dipodidae (Rodentia) I. Cranial and dental
characters. Изд. Акад. Наук СССР. 1930 :331—350, 453—466.
Vinogradov, В. S. On the mechanism of gnawing and mastication in some fossorial
Rodents. Ежегодн. Зоол. Муз. Акад. Наук, 1926: 275.
Vinogradov, В. S. Notes on some Gerboas from Mongolia. Докл. Акад. Наук СССР,
1926 : 232.
Vinogradov, В. S. On the structure of external genitalia in Dipodidae and Zapodidae
as a classificatory character. Proc. Zool. Soc. Lond., 1925 : 577.
Vinogradov, B. S. A Second interesting Species of Mongolian Gerboa of the Genus
Salpingotus Vinogr. Zool. Anzeiger, LXI, 1924 (Salpingotus crassicauda).
Vinogradov, B. S. A Third Species of Dwarf Jahrboa. Ann. Mag. Nat. Hist. ser. 10,
v. 1:372 (Salpingotus ihomasi).
Vinogradov, B. S. und Argyropulo, A. J. Zur Biologie der turkestanischen Spring-
mause. Zeitschr. fur Saugetierkunde, 6, Heft 4, 1931 :164.
Виноградов, Б. С. Млекопитающие СССР. Грызуны (определитель). Изд. Акад. Наук
СССР, 1933.
Виноградов, Б. С. О новом виде тушканчика. Доклады Акад. Наук СССР, 1929 :248
(Paradipus ctenodactglus).
Виноградов, Б. С. Заметки о своеобразном новом роде и виде тушканчика из Хара-
хото (Монголия); в книге II. К. Козлова — „Монголия и Амдо“, Петерб. 1922,
стр. 540 (описание рода Salpingotus и вида У. kozlovi).
Виноградов, Б. С. и Аргиропуло, А. И. Биологические наблюдения над туш-
канчиками. Ежегодн. Зоологии. Музея Акад. Наук, т. XXXII, 1931 :135.
Vinogradov, В. S. Ueber eine neue Springmaus aus der Karakum Wiiste, Russisch-
Turkestan. Zeitschr. fur Saugetierkunde, В. II Heft I, 1927 :92.
Winge, H. Pattedyr-SIaegter, II Rodentia, Kjobenhavn, 1924.
Winje, H. Jordfundene og nulevende Gnavere, 1887.
Z d a n s k у, O. Die alttertiaren Saugetiere Chinas nebst stratigraphischen Bemerkungen
„Palaeontologa Sinica", ser. С, VI, 1930.
СПЕЦИАЛЬНАЯ ЧАСТЬ
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ПОДСЕМЕЙСТВ
а) По краниологическим признакам
(фиг. 10—15)
1 (4). Наиболее узкая часть лобных костей совпадает с их серединой на
значительном расстоянии сзади от слёзных костей. Оба корня скуло-
вых отростков верхней челюсти отходят один над другим (на одной
вертикальной линии). Скуловая кость (jugale) постепенно опускается
сверху (от lacrymale) вниз и назад, не образуя резко обособленных
горизонтальной и вертикальной ветвей. Слёзные кости очень малень-
кие, не выдающиеся заметно наружу. Межкрыловидное пространство
приблизительно в 2 раза больше в длину, чем в ширину. Крыло-
видные отростки почти равны по длине ряду коренных зубов
(без РМ).
2(3). Мозговая коробка при рассматривании сверху почти овальной формы.
Скуловые дуги дугообразны, не сходящиеся или едва сходящиеся
по направлению вперед. Птеригоидные ямки мелкие, не расширен-
ные заметно сзади. Барабанные камеры слабо раздуты, их вершины
не соприкасаются друг с другом по средней линии. Сосцевидные
кости (mastoidea) не раздуты и не образуют мастоидных пузырей
(bullae mastoideae). Задние нёбные отверстия малой величины . . .
......................................1. Под сем. Zapodinae.
3(2). Мозговая коробка почти трапециоидальная. Скуловые дуги почти
прямолинейные при рассматривании сверху, значительно сходящиеся
по направлению вперед. Птеригоидные ямки глубокие и сильно рас-
ширяющиеся кзади. Bullae tympani значительно раздуты, их вершины
соприкасаются друг с другом по средней линии. Мастоиды сильно
раздуты, образуя большие камеры, соединенные с полостью bullae
tympani. Задние нёбные отверстия очень большие..............
...................................2. Поде ем. Euchoreutinae.
4(1). Наиболее узкая часть лобных костей расположена непосредственно
сзади от слёзных костей. Верхние корни скуловых отростков мак-
силлы отходят на значительном расстоянии сзади от нижних корней.
Фиг. 10. Черепа Dipodidae (сверху): 1 — Sicista subiilis Pall., 2 —Zapus (Eozapus) setchuanus Pous., 3 — Cardriocranius
paradoxus Satun., 4 — Salpingotus kozloviVinogr., 5 — Euchoreutes naso Sclat., 6 — Pygerethmus platyurus Licht., 7 — Alacta-
gulus acontion Pall., 8 — Allactaga elater Licht.
ТАБЛИЦА ПОДСЕМЕЙСТВ
Фиг. И. Черепа Dipodidae (сверху): 1 — Scarturus tetradactylus Licht., 2 - Dipus sagitta Pall., 3 — Scirtopoda telum
Licht., 4—Jaculus jaculus L., 5 — Eremodipus lichtensteini Vinogr., 6 — Paradipus ctenodactylus Vinogr.
CO
Фиг. 12. Черепа Dipodidae (вид снизу). Нумерация рисунков как на фнг. 10. Pzi — нижний отросток скул, дуги; Lp. — задняя
часть нёба; Pt — птеригоидный отросток; Рр — processus paroccipitalis; So — верхне-затылочцая кость;JT,— bulla tympani;
Ml — Мщ — частд Ьц|1ае nia$tideae,
Фиг. 13. Черепа Dipodidae (вид снизу). Нумерация рисунков как на фиг. 11.
ТУШКАНЧИКИ DIPODIDAE _______________ТАБЛИЦА ПОДСЕМЕИС ТВ
Фиг. 14. Черепа Dinodidae (вид сбоку). Нумерация как на фиг. 10,
ТУШКАНЧИКИ ^DIPODIDAE
фи-, 15. Черепа Dipodidae (вид сбоку). Нумерация рисунков как на фиг. 11. 7 — задняя часть черепа Eremodipus спереди
jj сбоку; Ра — parietale; Dfp — шиповидный отросток panetale.
48
ТУШКАНЧИКИ DIPODIDAE
Скуловая кость двулучевая, состоящая из горизонтальной и верти-
кальной ветвей. Слёзные кости увеличены в размерах и заметно
выступают наружу или наружу и назад. Длина межкрыловидного
пространства не более его ширины. Крыловидные отростки значи-
тельно короче, чем ряд коренных зубов.
5(6). Череп в области чешуйчатой кости без выступов или гребней. Ве-
нечный шов образует довольно глубокую дугу. Межтеменная кость
сильно редуцирована или отсутствует (если имеется, ее длина в не-
сколько раз больше ширины). Верхне-затылочная кость без боковых
отростков. Мастоиды необычайно сильно раздуты, занимая около
% всей поверхности черепа............ . 3. Подсем. Cardiocraniinae.
*6 (5). Череп в области чешуйчатой кости имеет хорошо развитые тупые
выступы или резко обозначенные гребни. Венечный шов образует
почти прямую поперечную линию. Межтеменная кость хорошо раз-
вита, ее ширина всегда больше длины. Верхне-затылочная кость
с хорошо развитыми боковыми отростками. Мастоиды не раздуты
чрезмерно, не выходя на верхнюю поверхность черепа или занимая
лишь небольшую часть этой поверхности.
7 (8). Bullae tympani слабо раздуты, их вершины не соприкасаются друг
с другом. Мастоиды не раздуты и не образуют добавочных полостей
среднего уха. Передние концы носовых костей не достигают уровня
альвеол верхних, резцов. Верхние резцы и их альвеолы сильно вы-
даются вперед.............................4. Подсем. Allactaginae.
3 (7). Bullae tympani значительно раздуты, их вершины обыкновенно сопри-
касаются друг с другом (за исключением Paradipus). Мастоиды зна-
чительно раздуты и образуют большие дополнительные камеры, сое-
диненные с полостью среднего уха. Передний конец носовых костей
достигает уровня альвеол верхних резцов. Верхние резцы и их аль-
веолы почти вертикальны.....................5*. Подсем. Dipodinae.
б) По строению скелета задней ступни
(фиг 16—17).
1 (6). Три средних плюсневых кости свободны (не сросшиеся). Первый
палец (если боковые пальцы имеются) равен приблизительно 2/з пятого
пальца. Последний 5-й палец достигает основания средней фаланги
четвертого пальца.
2(5). Первый и пятый пальцы хорошо развиты.
3 (4). Каждый из трех средних пальцев равен по крайней мере 3/4 соот-
ветствующей плюсневой кости; первый палец почти достигает
дистального конца второй плюсневой кости или даже слегка захо-
дит за него. Дистальный конец entocuneiforme почти достигает
уровня проксимального конца третьей плюсневой кости..............
........................................1. Подсем. Zapodinae.
ТАБЛИЦА ПОДСЕМЕЙСТВ
49
пФиг. 16. Скелет задней ступни: 1 — Sicista subtilis Pall., 2-—Zapus (Eozapus) setchuanus
Pous., 3 — Cardiocranius paradoxus Satun., 4 — Salpingotus kozlovi Vinogr., 5 — Euchoreutes
..naso Sclat., 6 — Pygerethmus platyurus Licht., 7 — Alactagulus acontion Pall., 8 Allactaga
elater Licht.
Фауна СССР, Виноградов
4
50
ТУШКАНЧИКИ DIPODIDAE
Фиг517. Скелет задней ступни: 1 — Scarturus tetradactylus Licht., 2 — Scirtopoda [telum
Licht., 3 — Dipus sagitta Pall., 4 — Eremadipus lichtensteini Vinogr., 5 —Jaculus orientalis;
Erxl., 6 — Paradipus ctenodactylus Vinog-r.
ТАБЛИЦА ПОДСЕМЕЙСТВ 51
4 (3). Каждый из трех средних пальцев короче 3/4 соответствующей плюс-
невой кости; первый палец не достигает дистального конца второй
плюсневой кости. Дистальный конец entocuneiforme расположен зна-
чительно впереди от проксимального конца третьей плюсневой кости
....................3. Подсем. Cardiocraniinae (4. Cardiocranius Sat.),
5 (2). Первый и пятый пальцы отсутствуют (имеются лишь их рудименты)
........................3. Подсем. Cardiocraniinae (5. Salpingotus Vinogr.),
6(1). Три средних плюсневых кости по крайней мере в их задних двух
третях совершенно сросшиеся, образующие одну „цевку". Первый
палец (если он имеется) почти равен пятому; последний (если имеется)
не достигает дистального конца „цевки".
7 (10). Пятый (и большей частью первый) пальцы хорошо развиты.
8 (9). Передняя треть плюсревых костей (дистальная треть „цевки") не
сросшаяся.................................2. Подсем. Euchoreutinae.
9 (8). Передняя и вся задняя часть плюсневых костей совершенно срос-
шаяся, образуя одну цельную кость — цевку (только передние х/8—^io
части каждой плюсневой кости свободные, образуя сочленовные по-
верхности для соответствующей основной фаланги)...................
................................... . 4. Подсем. Allactaginae.
10(7). Пятый и первый пальцы всегда отсутствуют (имеются лишь их
рудименты).....................................5. Подсем. Dipodinae.
в)Построению тазовых костей (pelvis)
(фиг. 18)
1 (6). Symphisis pubis короткий, в несколько раз короче, чем расстояние
от acetabulum до седалищного бугра (tuber ischii).
2 (5). В области лонного сращения тазовая кость образует вытянутый обо-
собленный отросток или лопасть.
3(4). Tuberculi ileo-pectinei отсутствует.....1. Подсем. Zapodinae.
4(3). Tuberculi ileo-pectinei в числе одного или двух хорошо развиты . .
.....................................3. Подсем. Cardiocraniinae.
5(2). В области лонного сращения тазовая кость не образует вытянутого
отростка или лопасти......................2. Подсем. Euchoreutinae.
6(1). Symphisis pubis очень длинный, составляющий не менее двух третей
расстояния от acetabulum до седалищного бугра (tuber ischii).
7 (8). Нисходящая ветвь ischium образует с горизонтальной (параллельной
позвоночному столбу) ветвью этой кости почти прямой угол (не счи-
тая выемки на нисходящей ветви под седалищным бугром) ....
.........................................5. Подсем. Dipodinae.
8 (7). Нисходящая ветвь образует с горизонтальной ветвью острый угол .
......................4. Подсем. Allactaginae (кроме 8. Pggerethmus)?
1 См. стр. 142.
4’
52
ТУШКАНЧИКИ DIPODIDAE
Фиг. 18. Таз (pelvis) тушканчиков: 1 — Sicista betulina Pall., 2 — Euchoreutes naso Sclat.,
3 — Cardiocranius paradoxus Satun., 4 — Salpingotus kozlovi Vinogr., 5 — Salpingotus crassi-
cauda Vinogr., 6 — Pygerethmus platyurus Licht., 7 — Allactaga saltator Evers., 8 — Allac-
taga elater Licht., 9 — Alactagulus acontion Pall., 10 — Dipus sagitta Pall., 11—Jaculus
orientalis Erxl.
ТАБЛИЦА ПОДСЕМЕЙСТВ 53
г) По строению шейных позвонков
(фиг. 19)
1 (6). Позвонки не сросшиеся.
2(5). Поперечные отростки короткие, непревышающие подлине половину
ширины тела соответствующих позвонков.
3(4). Вершины поперечных отростков 5-го, 6-го и 7-го шейных позвон-
ков не сближены, поперечные отростки почти параллельны друг
другу..........................................1. Подсем. Zapodinae.
4 (3). Вершины поперечных отростков 5-го, 6-го и 7-го шейных позвонков •
соприкасаются друг с другом...............2. Подсем. Euchoreutinae.
5 (2). Поперечные отростки относительно длинные, превышающие по длине
половину ширины тела соответствующих позвонков.....................
....................................4. Подсем. Allactaginae.
6 (1). Позвонки сросшиеся (лишь 7-й позвонок часто остается свободным).
7 (8). Седьмой позвонок свободен. На вентральной стороне сросшегося
комплекса шейных позвонков имеется 3 или 4 отверстия...............
......................Г ....... . 5. Подсем. Dipodinae.
8(7). Седьмой позвонок прирастает к 6-му. На боках шейного комплекса
позвонков имеется лишь 1 отверстие, в котором можно видеть тон-
чайшие костные нити — остатки редуцированных поперечных отрост-
ков ......................................3. Подсем. Cardiocraniinae.
д) По строению наружных genitalia самцов
(фиг. 20—22)
1 (2). Os penis всегда имеется; его основная часть без срединного выдаю-
щегося гребня..................................1. Подсем. Zapodinae.
2(1). Os penis имеется или отсутствует; если имеется, его основная часть
всегда снабжена срединным выдающимся гребнем.
3 (6). Penis тонкий, Постепенно суживающийся к вершине или уплощенный
с боков; на его поверхности нет ясйо выраженных роговых шипиков
или чешуек (могут иметься лишь очень мелкие роговые зернышки).
4 (5). Os penis имеется; его вершина покрытая эпителием резко загнута
кверху. Penis цилиндрический или уплощенный с боков................
....................................2. Подсем. Euchorentinae.
5 (4). Os penis отсутствует; penis тонкий, постепенно суживающийся к вер-
шине и лишь слегка уплощенный с боков вблизи вершины ....
..................................3. Подсем. Cardiocraniinae.
6(3). Penis толстый с хорошо развитыми роговыми шипиками или чешуй-
ками, большей частью направленными назад.
7 (8). Os penis отсутствует. Вершина penis простая, без боковых складок
(карманов); на нижней поверхности penis нет глубоких продольных
борозд. Форма penis (при рассматривании сверху) овальная,
54
ТУШКАНЧИКИ DIPODIDAE
2
Фиг. 19. Шейные позвонки тушканчиков на фиг. 1, 8, 9 и 10 первый шейный позвонок
(atlas) удален. 1 — Sicista betulina Pall., 2 — Allactaga saltator Eversm., 3 — Alactagulus
acontion Pall., 4 — Euchoreutes naso Sclat., 5 — Dipus sagitta Pall., 6 — Eremodipus lichten-
steini Vinogr., 7 — Scirtopoda telum Licht., 8—Jaculus orientalis Erxl., 9 — Salpingotus
crassicauda Vinogr., 10 — Cardiocranius paradoxus Satun., 11 — Paradipus ctenodactylus
Vinogr.
ТАБЛИЦА ПОДСЕМЕЙСТВ
55
•Фиг. 20. Наружное строение penis: 1 — Sicista subtilis Pall., 2 — Sicista betulina Pall., 3—
-4 — Sicista subtilis Pall, (вскрытая часть penis над семенной щелью для показания строе-
ния шипа и расположенной над ним бляшки), 5—6 — Sicista параеа Hallist (спереди
-.и сбоку), 7 — Sicista caucasica Vinogr., 8 — Sicista caudata Thos., 9 — Zapus (Eozapus)
.setchuanus Pous., 10—11 — Euchoreutes naso Sclat. (спереди и сбоку), 12—13 — Salpingo-
tus kozlovi Vinogr. (спереди и сбоку), 14 — Scarturus tetradactylus Licht, (no R. Pocock).
56
ТУШКАНЧИКИ DIPODIDAE
Фиг. 21. Наружное строение penis: 1 — Allactaga jaculus Pall., 2 — A. severtzovi Vinogr.,.
3 — A. saltator Eversm., 4 — A. elater Licht., 5 — Д. williamsi Thos., 6 — A. euphratica
Thos., 7 — Alactagulus acontion Pall., 8 — Pygerethmus platyurus Licht., 9 — Paradipus:
ctenodactylus Vinogr., 10—Jaculus orientalis Erxl., 11 — Dipus sagitta Pall., 12 — Scirtopodaz
telum Licht., 13—Jaculus jaculus L., 14 — Eremodipus lichtensteini Vinogr.
ТАБЛИЦА ПОДСЕМЕЙСТВ
Фиг. 22. Os penis мышевок и тушканчиков: 1—2 — Sicista subtilis Pall., 3—4 — S. betu-
lina Pall., 5—6 — napaea Hollist., 7—8 — S. caucasica Vinogr., 9—10 — S. caudata Thos.;
11—12 — Zapus (Eozapus) setchuanus Pous., 13—14— Euchoreutes naso Sclat., 15—16 —
Scirtopoda telum Licht., 17—18 — Dipus sagitta Pall., 19—20—Jaculus jaculus Pall., 21—
22 — Jaculus orientalis Duv.
58
ТУШКАНЧИКИ DIPODIDAE
сердцевидная или ланцетовидная; дорзальная поверхность покрыта
мелкими шипами (длинные шипы всегда отсутствуют)..........
. .. . ..............................4. Подсем. Allactaginae.
8(7). Os penis всегда имеется. Вершина penis большей частью снабжена
глубокими складками (карманами) с каждой стороны наружного отвер-
стия urethra; на нижней стороне penis всегда имеется глубокая про-
дольная щель; с каждой стороны от нее может быть еще по одной
более короткой щели. Penis овальный или с параллельными краями.
Кроме мелких шипов большей частью имеются две длинных роговых
иглы........................................ 5. Подсем. Dipodinae.
1. Подсем. ZAPODINAE — МЫШЕВКИ И ПРЫГУНЧИКИ
Внешность мышеобразная. Голова характерной для мышей формы,
-с короткими ушами, причем каждая ушная раковина имеет свободные
жрая, не сросшиеся при основании, в противоположность настоящим
тушканчикам. Глаза относительно маленькие, разрез век расположен
параллельно продольной оси головы. Верхняя губа не имеет продольного
разреза; rhinarium не расширен и окружен лишь короткими прилегающими
и относительно редкими волосами. Нижняя губа не окружает оснований
нижних резцов. Вибриссы относительно короткие, лишь едва превышаю-
щие длину головы. На первом (внутреннем) пальце передней конечности
имеется короткий ноготь с округленным свободным краем. Задняя ступня
относительно длинная, имеющая характерное для мышеобразных грызунов
строение (Sicista) или значительно более удлиненная (Zapus). Хвост длин-
ный с едва удлиненными на конце волосами, не образующими обособлен-
ной кисточки „знамени".—-Строение penis имеет свои характерные
особенности в пределах обоих родов подсемейства и у каждого вида.
Общими для всего подсемейства признаками является постоянное присут-
ствие os penis, которая не имеет выдающегося гребня в ее основной
части. Поверхн'ость penis покрыта мелкими шипиками или бляшками, среди
которых у некоторых форм обособляются отдельные крупные конические
или стилевидные шипы. Череп мышеобразного типа, т. е. с округленной
мозговой коробкой, имеющей при рассматривании сверху почти овальную
форму; бока мозговой коробки округлы, без образования резко обозначен-
ных выпуклостей; по направлению вперед боковые края мозговой коробки
постепенно сближаются в области межглазничного пространства и опять
несколько расходятся при приближении к слёзным костям; таким образом
межглазничное сужение (довольно слабо выраженное) находится значи-
тельно кзади от линии проведенной через задние края слёзных костей.
Верхние корни скуловых отростков максиллы лишь немного отклонены
назад от перпендикулярной линии, проведенной к сагитальной линии
черепа. Zygomata дугообразны, они наиболее широко расставлены в их
«средней или задней части; в последнем случае .они лишь немного сходятся
ZAPODINAE
59
кпереди. Скуловой отросток чешуйчатой кости резко отогнут вниз; при
рассматривании сверху он имеет косое направление, отчасти совпадающее
с общим направлением дуги. Нижний корень скулового отростка максиллы
лежит над верхним и поэтому при рассматривании черепа сверху совер-
шенно не выступает вперед из под верхнего корня. Скуловая кость не резко
разделена на горизонтальную и восходящую ветвь и постепенно изгибается
по направлению вперед и вверх, где она оканчивается острым концом вблизи
от слёзной кости или соприкасается с последней. Слёзные кости малы
и не вытянуты в поперечном направлении. Rostrum относительно длинный,
превосходящий по длине расстояние от его основания до заднего края нёба
(считая от переднего края основанйя наружных ветвей скуловых отростков
maxillare до gnathion). Венечный шов образует довольно глубокий обра-
щенный назад угол или глубокую дугу. Межтеменная кость хорошо
развитая, постепенно суживающаяся к внешним краям и заканчивающаяся
по бокам заостренными углами. Внутренняя стенка глазницы вполне
окостеневшая, без образования затянутого перепонкой отверстия или
редко лишь с слабыми следами последнего. Межкрыловидное простран-
ство наполовину больше в длину, чем в ширину, с почти параллельными
краями. Крыловидные отростки относительно длинны, равные или почти
равные по длине зубному ряду (без pm); hamuli касаются передне-внутрен-
них углов bullae tympani. Крыловидные ямки узкие, длинные и почти плоские.
Bullae tympani малы и мало раздуты (их наибольший поперечный диаметр
лишь немного более длины зубного ряда); их передне-внутренние углы
далеко отстоят друг от друга и почти не налегают на basis phenoideum.
Mastoidea не раздуты и не принимают участия в образовании bullae;
внутренняя их часть почти целиком занята полостью fossa floccularis
мозжечка. Нижняя челюсть в задней части высокая — в 2 или почти
в 2 раза выше, чем на уровне Мх. Венечный отросток хорошо развит,
будучи лишь немного короче,- чем сочленовный отросток; вершина венеч-
ного отростка почти достигает уровня сочленовной головки. Выступ
на боковой поверхности челюсти (альвеолярный бугор), соответствующий
корню нижнего резца, едва намечен. Задний угол нижней челюсти отвер-
нут наружу. Строение слуховых косточек резко отличается от такового
у всех других подсемейств Dipodidae (фиг. 23 14—15). Incus очень малень-
кий, в 3 или 4 раза меньше, чем corpus mallei. Corpus mallei уплощен и
образует очень тонкую пластинку; manubrium mallei резко повернут вперед
и идет почти параллельно длинной оси corpus mallei и corpus incudis.Описан-
ные отношения не наблюдаются более ни в одной группе Dipodidae, но
являются очень характерными для представителей семейства Muridae. Все
семь шейных позвонков свободны, шея относительно длинная, поперечные
отростки короткие, не превышающие по длине половину ширины тела
соответствующих позвонков. Скелет задней ступни имеет нормальное
„мышеобразное" строение, т. е. сохраняются все 5 пальцев при наличии
5 свободных (несросшихся) плюсневых костей, причем entocuneiforme
60
ТУШКАНЧИКИ DIPODIDAE
Фиг. 23. Слуховые косточки Dipodidae — malleus и uncus: 1 — Allactaga jaculus Pall., 2 —
A. bullata Gl. Allen., 3— A. saltator Eversm., 4— A. elater Licht., 5 — Alactagulus acontion
Pall., 6 — Pygerethus platyurus Licht., 7 — Dipus sagitta Pall., 8 — Scirtopoda telum Licht.,
9—Jaculus orientalis Erxl., 10—J. jaculus L., 11 — Euchoreutes naso Sclat., 12—Cardio-
cranius paradoxus Sat., 13 — Zapus hudsonius Zimm., 15 — Sicista subtilis Pall., 16 — Rattus
norvegicus Erxl.
Z APODIN АЕ
61
‘(т. e. первая tarsale) не удлинена, как у настоящих тушканчиков, и мало
разнится от этой косточки у Muridae. Плюсневые кости относительно
более коротки, чем у Cardiocranius, в связи с чем каждый из трех сред-
них пальцев равен по крайней мере 3/4 длины соответствующей плюсневой
кости. Пятый палец относительно очень сильно развит, достигая своим
концом до основания средней фаланги 4-го пальца. Строение таза харак-
теризуется следующими признаками: crista iliaca отсутствует или развита
лишь близ переднего конца ileum; tuber ischii слабо обособлен или почти
отсутствует вовсе; symphisis pubis очень короток, он образован обосо-
бленными треугольными отростками ischio-pubis. Tuberculi-ileo-pectinei —
отсутствует. Головка бедра шарообразная, без седловины на верхней
поверхности в противоположность настоящим тушканчикам. Ложно-корен-
ные зубы имеются почти у всех представителей подсемейства (за исклю-
чением подрода Napeozapus). У представителей Zapodinae коронка корен-
ных зубов имеет довольно хорошо развитое бугорчатое строение, когда
зубы еще не сношены (кроме представителей подрода Napaeozapus)
•с 4 главными и 3 добавочными бугорками.
Представители Zapodinae сохраняют наиболее примитивные для
Dipodidae признаки, унаследованные от мышеобразных предков рассма-
триваемого семейства. Наличие у мышевок и прыгунчиков признаков
характерных для Dipodidae, как строение коренных зубов, скуловой дуги,
нижнеглазничного отверстия, penis и пр. делает несомненной принадлеж-
ность этой группы к сем. Dipodidae. Вместе с тем признаки различия
между мышевками и прыгунчиками с одной стороны и настоящими туш-
канчиками с другой столь велики, что еще недавно эти группы рассмат-
ривались как отдельные семейства; однако, в настоящее время, после
нахождения связующих звеньев между обеими этими группами в виде
Euchoreutes, Cardiocranius, Salpingotus и ряда ископаемых форм, наиболее
правильно рассматривать мышевок и прыгунчиков, как особое подсемей-
ство {Zapodinae) семейства Dipodidae. Современные представители рас-
сматриваемого подсемейства имеют близкое отношение к ряду ископаемых
форм, как это было подробно изложено в главе о геологической истории
«семейства. #
Распространение подсемейства Zapodinae охватывает значительное
пространство Европейско-азитского материка и Северной Америки.
В Европе и Азии представители этой группы встречаются на запад в Вен-
грии и Дании и на восток до Сахалина и Сычуани; к северу представители
рассматриваемого подсемейства встречаются до Норвегии, северного
Урала, окрестностей Томска, Прибайкалья и Сахалина, а к югу до Закав-
казья, Тянь-Шаня, Западных Гималаев, Алтая, Саян, Гансу и Сычуани.
.Для Европы и Азии характерен род Sicista и лишь в пределах Сычуани
и Гансу встречается представитель рода Zapus {Eozapus) setchuanus
Pousargues, вероятно являющийся здесь реликтом плиоценовой фауны
(см. главу о геологической истории Dipodidae)’, в Северной Америке широко
62
ZAPODINAE
распространены исключительно представители рода Zapus (подроды Zapus
s. str. и Napaeozapus).
В экологическом отношении представители подсемейства Zapodinae
характеризуются широкой приспособляемостью к разнообразным жизнен-
ным условиям, встречающимся от субарктической зоны до полупустыни
включительно и от равнины до альпийской зоны, что связано с относи-
тельно слабо выраженной специализацией организма рассматриваемых
животных по отношению к какому-либо определенному комплексу жизнен-
ных условий. Однако при всей широкой амплитуде приспособляемости этих
животных они не смогли приспособиться к условиям пустыни в противо-
положность всем другим группам семейства.
В экономическом отношении представители рассматриваемой группы
не имеют сколько нйбудь серьезного значения.
Подсемейство Zapodinae составлено двумя родами — Sicista и Zapus;
к этому же подсемейству вероятно должны быть отнесены несколько
ископаемых родов (во всяком случае род Plesiosminthus, из плиоцена
Западной Европы).
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ И ПОДРОДОВ
1 (2). Передний конец скуловой кости не достигает слёзной. Нижне-глаз-
ничный канал неполный, его наружная стенка не касается своим
краем максиллярной кости. Межтеменная кость в поперечном напра-
влении приблизительно в два раза больше, чем в продольном. Задний
край костного нёба значительно выдается назад от уровня заднего-
края М3, нёбо оканчивается хорошо развитым, заостренным выступом.
В задней части нёба имеются два неглубоких, резко ограниченных
спереди вдавления. Передний угол нижней челюсти (angulus anterior)
не ясно отделен от заднего угла и не резко повернут внутрь . . .
.............................................1. Sicista Gray.
2 (1). Передний конец скуловой кости достигает слёзной. Нижне-глазнич-
ный канал полный, его наружная пластинка у взрослых экземпляров
большей частью сливается своим верхний краем с maxillare. Меж-
теменная кость в поперечном направлении почти в три или четыре
раза больше, чем в продольном. Задний край костного нёба выдается
назад не дальше уровня заднего края М3; задний край нёба без
заостренного выступа, большей частью даже вогнутый; в задней
части нёба вдавленных площадок нет. Передний угол (angulus ante-
rior) нижней челюсти ясно отделен от заднего (angulus posterior)
и резко отогнут внутрь ............. 2. Zapus Coues.
3 (4). Ложно-коренной зуб (Р4) верхней челюсти отсутствует. Все бугорки
на жевательной поверхности коренных зубов исчезают в раннем
возрасте и поверхность зубов становится плоской..................
.....................Подрод Napaeozapus Preble (Сев. Америка).
1. SICISTA
63-
4 (3). Ложно-коренной (P4) верхней челюсти имеется. Бугорчатая структура
жевательной поверхности коренных зубов сохраняется во взрослом:
состоянии.
5 (6). Бугорки на коренных зубах сильно выдающиеся и разделенные глу-
бокими впадинами. Наружные добавочные бугорки на зубах верхней
челюсти соединены поперечными гребнями с внутренними главными
бугорками...........................Подрод Eozapus Preble (Вост. Китай).
6 (5). Бугорки на коренных зубах менее выдающиеся и разделенные менее
глубокими впадинами. Наружные добавочные бугорки изолированы^
т. е. не соединены поперечными гребнями с внутренними....................
........................ . Подрод Zapus Cones (Сев. Америка)..
1. Род SICISTA GRAY (1827) — МЫШЕВКИ
Gray, in Griffith’s Cuvier Anim. Kingd., V, 1827: 227.
Из внешних признаков, по сравнению с Zapus, для представителей
рассматриваемого рода характерны относительно более короткие задние
конечности и особенно задняя ступня, более острая морда, более короткие
вибрисы, а также отсутствие темной резко ограниченной мантии на спине
(темный рисунок на спине отсутствует или представлен в виде одной или
трех продольных полос).
Дистальная часть penis на верхней поверхности снабжена продоль-
ной, большой частью очень глубокой щелью („Samenfalte" Mehely); в конце
этой щели заметен выдающийся бугорок, образованный вершиной os penis.
Вся поверхность penis покрыта мелкими роговыми шипами и чешуйками,,
среди которых у некоторых форм имеются более крупные шипы. Os penis
более или менее дугообразная. Скуловые дуги наиболее широко расста-
влены в их средней (или иногда даже передней) части. Скуловая кость
(zygomaticum) оканчивается острым концом вблизи от lacrymale и лишь-
в очень редких случаях соприкасается с последней. Canalis infraorbitalis.
образует лишь открытый сверху желобок, так как ограничивающая его
снаружи костная пластинка не касается стенки maxillare. Nasalia своими
передними концами доходят лишь до вертикальной линии, проведенной
через gnathion. Поперечный диаметр interparietale приблизительно'
в 1 у/2—2V2 Раза больше продольного. Теменные кости касаются своими
задними наружными углами fenestra postglenoidea; fenestra postglenoidea
не разделены продольным костным мостиком на два отверстия. Задний
край нёба расположен далеко позади линии, соединяющей задние корен-
ные зубы, нёбо заканчивается ясно выраженным шипом. Задняя половина,
или треть нёбных костей образует два. широких плоских вдавления, раз-
деленных низким продольным гребнем. Нижняя челюсть относительно'
более тонкая, чему у Zapus, ее передний угол (angulus anterior) ясно не
отделен от заднего угла и не отогнут резко внутрь. Строение слуховых
косточек сходно с таковыми у Zapus, но у Sicista processus brevis incudis.
64
Z APO DIN АЕ
короче, чем processus long-us (у Zapus оба отростка почти равны); incus отно-
сительно меньше, чем у Zapus (фиг. 14). Остистый отросток второго шейного
позвонка слабо развит. Относительная длина ступни меньше, чем у Zapus.
Первый палец заходит своим концом за дистальный конец 2-й плюсневой
кости; пятый палец заходит своим концом за дистальный конец основной
фаланги 4-го пальца. Когтевые фаланги почти прямые. Коренные зубы
имеют бугорчатое строение, но у Sicista бугорки низкие и складки эмали
выражены не столь резко, как у Eozapus. Третий (передний) бугорок (или
складка) довольно короткий и расположен на сагиттальной линии
(у других Zapodinae он более удлинен и смещен в наружном направлении).
Как можно видеть из предыдущего описания, род Sicista является
наиболее примитивной группой среди рассматриваемого семейства, сохра-
няя ряд мышеобразных признаков, как общее строение и пропорции тела
и’ скелета, общая форма черепа, строение слуховых косточек и пр. Как
будет указано ниже, наиболее близкий к Sicista род Zapus характеризуется
рядом признаков более высокой специализации в направлении настоящих
тушканчиков. Из всех известных видов Sicista наиболее примитивный,
т. е. наиболее удаленный от настоящих тушканчиков формой, является
. 5. nordmanni, обладающая наиболее коротким хвостом и задней ступней;
дальнейшую стадию специализации в этом направлении представляет
S. montana и особенно группа одноцветных мышевок, приближающихся
по длине ступни и хвоста до некоторой степени к роду Zapus\ следует
также отметить, что строение penis самцов у некоторых представителей
последней группы (напр. S. caucasica, S. caudata) также несколько при-
ближается в сторону тех признаков, которые имеют место у Zapus.
Виды рода Sicista представляют подобно мышам средний „генерали-
зованный" тип специализации; с этим находится в соответствии
обитание этих зверьков в весьма разнообразной жизненной обстановке:
на окраинах тундры нижней Печоры и Северного Урала, в хвойных лесах
европейской части Союза и Сибири, в лиственных лесах Европы, лесо-
степи, степи, полупустыне, в горах Кавказа, Тянь-Шаня, западных Гима-
лаев, Алтая, Сихотэ-Алина, Сахалина и пр. В противоположность своим
узко специализированным родичам — настоящим тушканчикам, мышевки
не приспособились к условиям жизни в пустыне и в своем распростране-
нии к югу достигли лишь северных пределов этой зоны, местами про-
никая, однако, по горным хребтам с более обильной влажностью и более
богатой растительностью дальше к югу (Тянь-Шань, Зап. Гималаи),
подобно целому ряду других более влаголюбивых грызунов (лесные мыши,
большинство полевок и пр.).
Различные виды мышевок характерны для различных мест обитания,
в частности S. betulina характерна для различного типа лесов, subtilis —
для степи и полупустыни; остальные виды — преимущественно для раз-
-личного типа горных ландшафтов иногда с выходом из горных местностей
:в равнину (S. параеа), как это будет указано при описании последнего вида.
I
/. SICISTA
Фиг. 24. Распространение видов рода Sicista. 1. 5. ketulina Pall., 3. Изолированные нахождения вне очерченного ареала.
2. 5. subtilis Pall., 4. S. caucasica Vinogr., 5. S. tianschanica Salensky, 6. 5. flava True (leathemi Thos.), 7. 5. napaea Hollist,
8. Изолированные нахождения вне очерченного ареала, 9. S. concolor Buchn, 10. S. caudata Thos.
Фауна СССР, Виноградов
5
66
ZAPODINAE
Биология мышевок известна весьма слабо и те немногие отрывочные
данные, которые имеются относительно некоторых видов, излагаются
в сжатом виде в соответствующих местах.- Своеобразной особенностью
5. montana, а может быть и некоторых других видов, является наличие
цепкого хвоста, что указывает на способность этих зверьков к лазанию.
Род Sicista в противоположность Zapus, свойственен исключительно
Палеарктике. Общее распространение этого рода охватывает обширную
территорию от Венгрии, Дании и Норвегии на восток до Сахалина и
Восточного Китая включительно. К северу представители рода распро-
странены до Норвегии, Северного Урала, бассейна р. Таза, Томска,.
Байкала, северной части хребта Сихотэ-Алин в ДВК и острова Сахалина;,
к югу до главного Кавказского хребта и Малого Кавказа, низовий Эмбы,.
Семиречья, Кашмира, Алтая, Саян и Китая (Гансу) (фиг. 24).
Всего известно 9 видов этого рода, из которых 6 видов обитают
в пределах Союза. Всюду, где встречаются эти зверьки, их относитель-
ная численность не велика или даже ничтожна; по указанной причине эти
зверьки не играют заметной роли в соответствующих биоценозах и не
имеют сколько-нибудь существенного экономического значения, даже
в тех случаях, когда они встречаются среди культурных растений
(см. S. subtilis). Следует вместе с тем отметить, что эта группа чрезвы-
чайно интересна в теоретическом отношении, давая ключ к пониманию
возможных путей эволюции всего семейства Dipodidae и направления
специализации последних.
Соотношения между видами рассматриваемого рода не являются еще
вполне ясными, что особенно относится к группе, так называемых одно-
цветных мышевок (без полоски на спине), в частности к -S', tianschanica,.
кашмирским S. flava и S. leathemi и восточно-китайскому 5. concolor;.
у указанных видов не известно строение penis, которое дает в пределах
рассматриваемого рода наиболее существенные и постоянные видовые
признаки. Я считал также преждевременным разделение отдельных видов
мышевок на подвиды, даже 5. subtilis, у которой наблюдаются наиболее
ясные изменения в окраске экземпляров из различных частей ареала;
поэтому я ограничился лишь изучением общей географической изменчи-
вости отдельных видов, насколько это позволил наличный материал.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДО'В^
1 (4). На спине имеется черная продольная полоска, проходящая по хребту.
2 (3). Хвост приблизительно на одну треть длиннее тела (длина хвоста
не свыше 90 мм). Задняя ступня относительно короткая, не более
1 Вследствие малой изученности, встречающихся вне пределов Союза, видов (<£. con-
color, S. flava, S. leathemi), последние не включены в таблицу. Из видов распространенных
в Союзе не включена в таблицу S. tianschanica, т. к. до сего времени не известно строение
penis у представителей этого вида, что имеет решающее значение при определении группы,
одноцветных мышевок.
1. SICISTA 67
15.7 мм. Цвет спины серый с охристо-желтым налетом. По обеим
сторонам от темной продольной полоски на спине имеются узкие
продольные светложелтоватые участки и кнаружи от них обычно
расположены темные или почти черные продольные участки, особенно
в задней части тела. • На верхней поверхности penis имеется лишь
один длинный шип (кроме мелких шипиков); перед его основанием
имеется вогнутая роговая чешуйка различной формы. От основания
срединной щели отходят две боковых складки, расходящихся в виде
фигуры V и не переходящих на боковую поверхность. Penis не раз-
делен на две лопасти.........................* 1. S. subtilis Pall.
3 (2). Хвост по крайней мере наполовину превышает длину тела (длина
хвоста обыкновенно от 90 до 102 мм); задняя ступня длинная:
от 15.8 до 17.7 мм. Окраска верха равномерная, желтовато-коричне-
вая, без образования светлых и темных полос по бокам от средин-
ной полосы. На верхней поверхности penis имеется два длинных
шипа; перед их основанием имеются лишь мелкие многочисленные
роговые чешуйки. Две боковых щели, отходящих от основания
срединной щели, расходятся W-образно, переходя на боковую
поверхность penis и разделяя penis на две лопасти (верхнюю и
нижнюю)...........................................* 2. S. betulina Pall.
4 (1). На спине черной продольной полоски нет.
5 (6). Penis кроме мелких шипиков имеет на боковой поверхности два
больших толстых шипа до 2-х мм длины; нижняя поверхность воору-
жена меньшего размера шипами, из которых 6 наиболее наружных
расположены в два продольных ряда.................* 3. S. napaea Holl.
6 (5). Penis покрыт мелкими шипиками, без отдельных крупных шипов.
7 (8). Penis длинный и тонкий, слегка булавообразной формы, покрытый
мелкими шипиками. Семенная щель занимает около трети длины
penis. Основная пластинка os penis очень узкая и заметно выпуклая,
постепенно переходящая в стержень . . . * 5. S. caucasica Vinogr.
8 (7). Penis толстый и относительно короткий. Семенная щель занимает
около половины всей длины penis. Основание os penis несколько
более расширенное, заметно вогнутое и более резко обособленное
от стержня....................................* 6. S. caudata Thomas.
* 1. Sicista subtilis Pallas — Степная мышевка (фиг. 25 — 26J.
Pallas, 1778: 327 (Mus); Mehely, 1913 : 237; Огнев, 1916 : 49. — vagus Pallas, 1781:169
(Mus).—nordmanni Keyserling- und Blasius, 1840, X 38: (окрестности Одессы) [.SmznfAus]. —
loriger Nordmann, 1840 : 49 (Sminthus).
Длина хвоста не свыше 90 мм (обыкновенно менее); хвост приблизи-
тельно на Уз превышает длину тела. Задняя ступня не более 15.7 мм.
Задний внутренний бугорок на подошве задней ступни овальный, мало
вытянутый в продольном направлении. Характерным типом окраски при
наибольшем развитии темных пигментов можно .считать следующую:
5*
68
ZAPODINAE
основания волос темно аспидно-серые (blackish slate), предконцевая зона
бледно охристо-желтая, концы волос черновато-бурые. В результате
смешения этих оттенков, общий тон спины кажется серым с охристо-
желтым налетом. Окраска спины более или менее резко ограничена от
цвета боков. Начиная от темени или от конца носа и до основания хвоста
вдоль спины проходит черная полоса, более резко выраженная в задней
части спины; по бокам к ней примыкают желтоватые продольные полосы,
более светлые, чем остальная поверхность спины; кнаружи от них обычно
имеются темные широкие участки (особенно в задней части спины). Уши
темнобурые или почти черные с светлой желтоватой каймой. Брюшная
Фиг. 25. Степная мышевка Sicista subtilis Pall. Фото М. К. Серебренникова.
поверхность тела светлосерая с слабым желтоватым налетом.1 От опи-
санного типа окр’аски встречается ряд уклонений в сторону общего
посветления, а также в сторону развития большей или меньшей примеси
охристых тонов, как это описывается ниже в обзоре географической
изменчивости вида. Penis с почти параллельными или немного расходя-
щимися назад краями, с закругленной или заостренной вершиной, вся его
поверхность усажена мелкими роговыми шипиками. В задней трети penis
имеется ограниченная глубокой полулунной складкой площадка, пересе-
ченная в медиальном направлении семенною щелью; эта щель ограничена
по бокам довольно слабо выпуклыми губкообразными складками. Из
переднего конца семенной щели выдается заостренный, слабо изогнутый
шип, Достигающий у взрослых экземпляров 2 мм длины, большей частью
видимый снаружи без препаровки; впереди от основания этого шипа
расположена роговая чешуя изменчивой формы, имеющая вид или оваль-
ной слабо вогнутой пластинки, полукруга или полулуния. На конце семен-
ной щели выдается бугорок, образованный верхушкой os penis, одетой
1 Описание окраски сделано по экземпляру № 13232 из окр. г. Змиева на Донце.
1. SICISTA
69
вполне надежных краниологических
Фиг. 26. Задняя ступня мышевок. 1 — Si-
cista subtilis Pall., 2 — S. caucasica Vinogr.
слизистой оболочкой. Os penis заметно изогнута; базальная пластинка
почти треугольной формы; стержень относительно длинный, окаймленный
тонкими пластинчатыми краями; на конце os penis желобчатого расшире-
ния нет. По G. Millery и L. Mehely (I. с.) наименьшая высота rostrum
позади резцов у S. subtilis почти равна ширине rostrum в той же области,
в отличие от «S', betulina, имеющей более высокий rostrum; уже последний
из упомянутых авторов указывает на непостоянство этого признака, в чем
я мог также достаточно убедиться при просмотре серий черепов обоих
видов. Таким образом мы не имеем
отличий между обоими видами, также
как и отличий в строении коренных
зубов; впрочем один из признаков,
кажущихся наименее существенным,
а именно — отсутствие у -S', subtilis
скоплений темного пигмента в глу-
бине ямок между бугорками коренных
зубов—является, повидимому, вполне
реальным, тем более что у S. betulina
эти скопления наблюдаются с доста-
точным постоянством.
Как было уже упомянуто выше,
S. subtilis по своим морфологическим
признакам является среди прочих
видов рода наименее специализиро-
ванной формой, по крайней мере
по короткой задней ступне и хвосту.
5. subtilis, как и другие представи-
тели рода, является хорошо диффе-
ренцированным видом, что особенно резко сказывается на строении наруж-
ных genitalia и своеобразной окраске. В общем ряду видов рода «S’, sub-
tilis более близка к 5. betulina по пропорциям частей тела (ступня, хвост),
по окраске (наличие полоски на спине).
Распространение. В своем распространении 5. subtilis связана
с зоной степи и отчасти полупустыни. На западе распространение этого
вида начинается в степях Венгрии и Румынии и простирается по степям
и- полупустыням европейской части Союза, Казахстана и южной Сибири
на восток до юго-западного побережья Байкала; в Восточной Сибири
распространение этого вида является островным, соответственно наличию
здесь островных степей, вкрапленных в зону тайги. Южная граница
распространения этого вида достигает Добруджи, побережья Черного
и Азовского морей, северных предгорий Кавказского хребта, северного
побережья Каспийского моря, озера Кургальджин в Казахстане, равнинной
части Семиречья, оз. Зайсана, Алтая и Саян. Северная граница распро-
странения вида точно не известна, но имеющиеся данные позволяют
70
ZAPODIN AE
утверждать, что она нигде не заходит далее северных пределов лесостеп-
ной зоны европейской части Союза, Казахстана и Сибири.
Географическая изменчивость. Географическая изменчи-
вость рассматриваемого вида выражена ясно и значительно в большей
степени, чем у других мышевок; однако, на основании имеющихся мате-
риалов можно подметить определенную изменчивость лишь в окраске
шерсти при относительном постоянстве других морфологических призна-
ков. Изменения окраски, как и в большинстве, других случаев, имеют
определенное направление и выражаются в более значительном развитии
темных пигментов у особей в более северных и более влажных частях
ареала вида и в постепенном посветлении окраски по направлению на юг и
на юго-восток в сторону более сухих и пустынных частей ареала. Таким
образом, наиболее темная окраска наблюдается у особей из лесостепной
зоны, средней интенсивности окраска — из южных частей зоны степей
и наиболее бледная — из зоны полупустыни. Указанная закономерность
подтверждается приводимыми ниже примерами. 1) Наиболее темная
окраска с различными вариациями оттенков наблюдается (по имею-
щейся серии) на экземплярах из следующих местностей: северная Украина
(окр. г. Змиева) и из западной части Харьковской области, Среднее Поволжье
(б. Бузулукский округ), Приалтайская степь (б. Рубцовский округ), б. Мину-
синский округ (с. Белоярское). Большинство указанных местонахождений
относятся к зоне лесостепи, не исключая Минусинского округа, где экзем-
пляры мышевок добыты в Тубинской лесостепи. Окраска серий из указан-
ных местностей несколько отличается между собой по оттенку, в частности
Бузулукские экземпляры несколько отличаются от Змиевских большей
примесью на спине охристых тонов, особенно в области светлого продоль-
ного поля кнаружи от черной продольной полоски. С последней серией
в существенных чертах сходны экземпляры из Рубцовского округа (с. Но-
венькое); серия из Минусинского округа несколько темнее, чем Рубцовские
и Бузулукские экземпляры и характеризуется меньшим развитием охристых
тонов, наиболее приближаясь по окраске к упомянутым выше змиевским
экземплярам. 2) Окраска промежуточного типа, т. е. более
светлая, чем описанная выше, но более темная, чем у следующей группы,
наблюдается у экземпляров из более сухих, преимущественно более южных
районов степной зоны. В частности экземпляры из б. Херсонского округа
и с Северного Кавказа значительно светлее, чем Змиевские и Полтавские
экземпляры, из Сарепты и Актюбинска — светлее Бузулукских, экземпляры
из Павлодара — светлее приалтийских; к этому же типу окраски можно
отнести экземпляры из Аларской степи б. Иркутского округа. 3) Окраска
третьего типа характеризуется крайней степенью
побледнения меха; она свойственна экземплярам из наиболее южных
частей ареала, лежащих в пределах полупустыни и северной окраины
пустынной зоны. Из имеющейся серии особенно характерна окраска экзем-
пляров с низовий р. Эмбы, у которых общая бледная окраска меха зави-
1. SICISTA
71
сит от значительного побледнения предконцевых зон отделочных волос,
укорочения темной концевой зоны до полной утраты темных окончаний
у многих волос. Редукция темного пигмента наблюдается и на других
частях волосяного покрова и выражается в побледнении брюшной стороны,
замене черного пигмента на ушах и спинной полоске коричневым, посвет-
лении хвоста и т. д. Редукция пигмента на спинной полоске выражается
также у некоторых экземпляров в полной утрате этой полоски в передней
половине спины. Кроме экземпляров с Эмбы, подобный же тип окраски
имеют экземпляры из северо-восточного Прибалхашья (Лепса, Лягуз), из
окрестностей Семипалатинска и побережья оз. Зайсана. Между описан-
ными тремя типами окраски имеется постепенный ряд переходов, вслед-
ствие чего при настоящей степени изученности географической изменчи-
вости этого вида не является целесообразным разделять его на подвиды
или иные мелкие систематические группы.1
Попытки установления подвидов .S’, subtilis на основании иных мор-
фологических признаков до сего времени также не увенчались успехом;
в частности восстановленная L. Mehely (1. с.) форма S. trizona Petenyi,
характеризующаяся, по мнению упомянутого автора, некоторыми особен-
ностями строения penis, вряд-ли существует в природе, так как даже на
•серии экземпляров из одной местностй можно наблюдать признаки, кото-
рые Mehely считал характерными с одной стороны для S', subtilis и с дру-
гой — S. trizona (параллельные или расходящиеся края penis, остроугольное
или тупоугольное его окончание, короткий или длинный при основании
семенной щели и пр.).
Биология. Биология степной мышевки очень слабо изучена. Как
было уже указано, характерным местообитанием этого вида является
степь, лесостепь и отчасти полупустыня.
Несколько интересных наблюдений над этими зверьками были сде-
ланы еще Палласом, однако, этот автор, повидимому, не всегда строго
разграничивал оба описанных им вида мышевок („Mus subtilis" или
„vagus" и „М. betulinus") и поэтому очень трудно выбрать из этих наблю-
дий те данные, которые относятся именно к рассматриваемому виду. По
наблюдениям Petenyi (1882) (цитируется по Mehely 1913) в Венгрии, степ-
ная мышевка (S', loriger trizona у названного автора) относится к числу
наиболее слабых и нежных мышевидных грызунов; уже ничтожное давле-
ние, холод, даже сырость и загрязнение действуют на зверька роковым
образом. В неволе мышевка не проявляет по отношению к человеку боязни
и является доверчивым зверьком, беря пищу из рук ухаживающего за ней
человека, не кусаясь и т. д. В неволе мышевка поедала зерна проса,
конопли, пшеницы, ячменя, овса и семена тыквы; мышевка охотно поедала
1 После сдачи настоящей работы в печать С. И. Огневым (Бюлл. Н.-исслед. Инет.
Зоологии МГУ, № 2, 1935) был опубликован систематический обзор Sicista СССР, где
автор высказывает иную точку зрения на рассматриваемый вопрос и устанавливает
.5 подвидов X subtilis.
12
ZAPODINAE
также сырое мясо. Mehely (I. с.) предполагает, что на воле мышевка
питается преимущественно животной пищей, так как автор находил во
вскрытых им желудках мышевок хитиновые части насекомых и остатки:
гусениц.
Некоторые данные об экономическом значении степной
мышевки на Украине были опубликованы А. А. Танджием (1932), который;
нашел несколько мышевок в защищенных канавках около свекловичных
плантаций. Наблюдения над этим зверьком в неволе показали, что
мышевка поедала в больших количествах различных насекомых, в част-
ности свекловичных долгоносиков (Bothynoderes punctiventris Germ.), хлеб-
ных жуков (Anisoplia), жужелиц и др., причем за 26 дней мышевка съела
2154 вредных насекомых. Зверек в условиях садка совершенно не трогал
предлагавшиеся ему зерна пшеницы, овса и гречихи. Упомянутый автор
считает степную мышевку полезным для полевых культур зверьком,,
истребляющим значительное количество вредных насекомых. Однако,
принимая во внимание малую численность или даже редкость этого зверька,,
следует думать что практическое значение этого зверька в указанном,
отношении вряд ли существенно.
* 2. Sicista betulina Pallas — Лесная мышевка (фиг. 27).
Pallas, 1775 : 332; (Mus); 1811 :169 (Mus). — montana Mehely, 1913 : 246. — norvegicar
Chaworth-Musters, 1927 : 542. — montana strandi Formosov, 1930 : 79.
S. betulina хорошо отличается от S. subtilis большой длиной хвоста
и ступни, окраской и характерным устройством penis.
Длина хвоста обыкновенно от 90 до 102 мм (редко немного менее),,
хвост по крайней мере наполовину превышает длину тела. Задний вну-
тренний бугорок задней ступни заметно более вытянут в продольном,
направлении, чем у S. subtilis. Мех несколько более грубый чём
у S. subtilis.
Общий тон верха желтовато-коричневый (между 15/z i и 17'" i Ридж-
вея) с примесью черных волос. Основания волос аспидно-серые, немногие
остевые волосы с черной вершрной, большая часть других — с оранжево-
желтым концевым поясом, некоторые волосы с узким желтоватым пред-
концевым пояском и черной верхушкой. По средине спины проходит
черная полоса, начинающаяся большей частью на голове. Окраска спины
постепенно переходит в желтовато-серый цвет брюха (21"" d). Уши темно-
коричневого цвета с слабо выраженной желтоватой каймой. Хвост дву-
цветный, верхняя и нижняя его поверхности окрашены сходно со спинной
и брюшной сторонами тела. В строении penis характерны следующие,
признаки: в глубине продольной щели penis имеются два длинных рого-
вых шипа; перед их основанием, а также по всей остальной поверхности
penis расположены мелкие роговые чешуйки. От вершины срединной щели
отходят косо Две боковые щели, образующие W-образную фигуру, пере-
ходящие на боковую поверхность и разделяющие penis на две лопасти.
1. SICISTA
73-
(дорзальную и вентральную). Базальная пластинка os penis ромбоидальной
формы и значительных размеров. Стержень относительно короток без
пластинчатой каймы. На вершине os penis имеется желобообразное расши-
рение.
Выше уже было указано, что .S', betulina по более удлиненному хвосту
и задней ступне является -несколько более специализированной в напра-
влении тушканчикообразного типа формой, чем S. subtilis. Как и последний;
вид, S. betulina является хорошо дифференцированным видом, особенно-
резко отличающимся от других представителей рода строением penis
и расцветкой меха.
Распространение. В своем распространении S. betulina связана
с лесной зоной, от хвойных лесов на севере ареала до широколиствен-
ных— на юге. На западе распространение этого вида начинается в север-
ной Венгрии,. Польше (Силезия), Дании (Ютландия), Восточной Пруссии
(Кенигсберг) и Норвегии и простирается на восток до Байкала, причем
этот вид отчасти проникает даже в затадные районы Забайкалья (окр..
Верхнеудинска, Като-Кель). Наиболе северными нахождениями этого
вида являются Дания, Вост. Пруссия (Кенигсберг), Центральная Норвегия,,
Финляндия, окр. Ленинграда, низовья реки Печоры, б. Архангельский у.,
74
ZAPODINAE
Северный Урал, верховья р. Таза (устье р. Мотльки), откуда граница
опускается к юго-востоку, охватывая Алтай, Саяны и южное Прибайкалье.
Южная граница слабо известна: кроме нескольких пунктов Московской
области, этот вид найден в лесистых балках близ Сарепты и даже в лесах
Карачая на северном склоне Главного Кавказского хребта. На востоке
ареала южную границу составляют леса Башкирии, Алтая, Саян и южного
Прибайкалья.
Географическая изменчивость у рассматриваемого вида
слабо выражена. Экземпляры из различных районов европейской части
Союза и Сибири, поскольку позволяют судить имеющиеся скудные кол-
лекционные материалы, не имеют ясно выраженных отличий. За послед-
нее время была сделана одна попытка выделить особую расу (подвид)
лесной мышевки А. Н. Формозовым (1. с.), который установил подвид
S. Ъ. strandi из Карачая (Сев. Кавказ) на основании некоторых отличий
в оттенке меха.
S. betulina селится в довольно разнообразной обстановке: среди
хвойного или лиственного леса, в кустарниках, на полянах, лугах, в гор-
ных местностях, местами до альпийской зоны включительно.
Образ жизни этого зверька известен лишь по немногим отры-
вочным наблюдениям некоторых авторов, в частности по наблюдениям
Капланова и Раевского (1928) в Московской и Калининской областях.
Лесная мышевка, пр наблюдениям названных авторов, селится в разно-
образных стациях. Этот вид был находим в молодом березовом лесу
с примесью дуба, осины и липы, кроме того этот зверек был добыт
в сосновом лесу с примесью ели в густых зарослях черники; авторы наблю-
дали также эту мышевку на поляне среди соснового леса и в дубовом
лесу. Об образе жизни этого животного авторы сообщают следующее:
„все три случая встречи с Sicista на свободе происходили днем. Однако,
живя в садке, мышевки возились и ели всегда только ночью и только
в исключительных случаях проявляли деятельность днем, под утро, как
указывалось оанее другими авторами для близкого вида; лесная мышевка
засыпает крепким сном в характерной позе спящего грызуна, производя
впечатление совсем умирающего зверька, но после первых же попыток
•согреть его в руках, мышевка открывает рот, как бы продолжительно
зевая и издает тихий, мелодичный звук, похожий на свист. В неволе
любимой пищей мышевки являются фрукты и ягоды, в особенности зем-
ляника и черника. Двигаясь по клетке, или посаженная на руку, мышевка
все время старается обхватить своим длинным хвостом какой-нибудь
предмет, причем если хвост обвивается вокруг пальца, на последнем
ощущается заметное давление. Виснуть на хвосте мышевка не может.
С конца сентября, несмотря на постоянную комнатную температуру, оба
зверька стали впадать в спячку, не проявляя признаков жизни в течение
пяти-шести дней. Прожив более трех месяцев в неволе зверьки погибли
ло недосмотру от перегрева террариума, поставленного на горячий кало-
1. SICISTA
75
рифер“ (стр. 17). По наблюдениям А. Косуап (1887) (цитировано по Mehely
1913) S. betulina селится в Венгрии на южных склонах гор и встречается
преимущественно на овсяных полях, заросших кустарниками пашнях
и около засохших пней. На поверхности земли этот зверек очень прово-
рен, но будучи испуган он никогда не бросается в свою нору, но быстро
вбегает на камни или пни и старается спрятаться под корой. Зверек ведет
ночной образ жизни и зимой впадает в продолжительную спячку. Содер-
жащиеся в неволе мышевки много спали даже летом, когда температура
опускалась до 10° С. Некоторые зверьки были очень дики, другие напро-
тив позволяли брать себя в руки и бегали вечером вокруг открытой клетки,
не убегая прочь. Если приближали к зверьку палец или стержень птичьего
пера, он поднимал свой длинный, тонкий хвост и обвивал его вокруг,
но не схватывал этот предмет. К этим наблюдениям Mehely добавляет,
что зверек должен считаться всеядным, так как автор находил во вскры-
тых им желудках хитиновые части насекомых, непереваренные остатки
гусениц и отчасти размельченные семена растений. Паллас (1778) указы-
вает, что эта мышевка {„Mus betulinus“) хорошо лазит, пользуясь при
этом своим хвостом, обхватывающим ветви и травы.
* 3. Sicista napaea Hollister — Алтайская мышевка
Hollister, 1912: 2 (с. Топучая, Семинский хребет, Алтай); Vinogradov, 1925: 581.
Довольно своеобразный вид, по окраске меха сходный с представи-
телями группы одноцветных мышевок {„concolor“ s. 1.), а по длине хвоста
и задней ступни занимающий промежуточное положение между -S', subtilis
и S. betulina и по устройству penis стоящий совершенно особняком среди
всех других видов рода.
Длина хвоста не более 98 мм (78—98); задняя ступня не более 17 мм
(14.6—17). Спинная сторона тела окрашена в светлокоричневый цвет,
сходно с S. betulina, но с. большей примесью оливкового тона (между
Saccardo’s umber a. tawny olive) и без черной продольной полосы. Уши
коричневые с слабой желтоватой каймой по краю. Окраска брюшной
стороны желтовато-серая. Хвост двуцветный, коричневый сверху и светло-
серый снизу. Наиболее характерно строение penis. Наружные края penis
почти параллельны, вершина остооугольная. Вентральная поверхность
покрыта довольно равномерно мелкими шипиками. От верхнего угла
половой щели, как у S. betulina, расходятся в стороны, борозды, которые,
однако, здесь очень неглубоки и проходят лишь на короткое расстояние,
быстро сглаживаясь и не разделяя penis на две лопасти, как у S. betulina.
Шип при основании половой щели отсутствует. Особенно своеобразно
устройство боковой и дорзальной поверхности penis: на боковой поверх-
ности, в нижней ее трети, сидят почти на одной продольной линии два
толстых роговых конических шипа длиною около двух миллиметров,
направленных вершинами назад; немного кверху от них и ближе к концу
penis сидит небольшой шип сходного устройства, выделяющийся среди
76
Z APODINАЕ
окружающих его мелких шипиков. Вся дорзальная поверхность усеяна,
крупными толстыми шипами, из которых особенно выделяются шесть-
шипов, расположенных в два продольных ряда, все они обращены верши-
нами назад. Os penis слабо изогнута. Базальная пластинка хорошо развита.
Распространение. Известные нахождения этого вида ограни-
чиваются несколькими пунктами Алтая и прилегающих степных районов.
В частности эта мышевка найдена в окрестностях сел. Топучая (Семин-
ский хребет), сел. Онгудай, Усть-Кан (верховья р. Чарыша), Чинеты.
на р. Ине (приток р. Чарыша), Колывань (Рубцовского окр.), Фунтиков-
ское (Барнаульского окр.); особенно интересными являются нахождения
этого вида в Кипринском районе Каменского окр., далеко к северу
от предгорий Алтая. Остается невыясненным вопрос о нахождении этого
вида на южном Алтае, в частности имевшиеся у меня одноцветные
мышевки из долины Бухтармы (Катон-Карагай) являлись полувзрослыми
и имели недостаточно удовлетворительную сохранность, что не дало воз-
можности их точного определения. Добытая Б. А. Кузнецовым серия
мышевок из Риддерского окр. (с. Пихтовка) отнесена этим автором
к виду — У. tianschanica Salensky (см. ниже).
Биологических данных об алтайской мышевке не имеется. Мною был
добыт один экземпляр этого вида среди кустарника в долине горной
речки близ с. Усть-Кан на Алтае.
* 4. Sicista tianschanica Salensky — Тян-шанская мышевка
Salensky, 1903:17 (р. Чапчагай и р. Цанма на южных склонах Тан-шаня).
Наиболее крупная из всех мышевок: длина хвоста до 114 мм,
задняя ступня до 19.3 мм, кондилобазальная длина черепа до 20 мм.
Задний внутренний бугорок задней ступни вытянут в продольном напра-
влении, но несколько более короток, чём у У. concolor и S. caucasica
и не искривлен, как у упомянутых видов.
Окраска верха одноцветная, серовато-коричневая (близкая к 17’i),
затемненная многочисленными отдельными черными волосами. На боках
цвет спины переходит в сероватую с бледножелтым налетом окраску брюш-
ной стороны. Строение penis неизвестно. Вероятно строение penis у этого
вида в существенных чертах сходно с таковым у S. caucasica и У. caudata,
как об этом можно судить по единственному бывшему у меня экзем-
пляру с Зайсана (р. Базариха), предположительно относимому к S. tianscha-
nica. У этого экземпляра penis сходен с таковым S. caucasica; он имеет
булавовидную форму, но несколько более укорочен, чем у -S', caucasica,
семенная щель не достигает половины длины penis, в дистальной части
верхняя поверхность penis покрыта относительно крупными шипиками,
направленными назад и несколько наружу; в проксимальной части имеются
лишь очень мелкие, направленные остриями назад шипики.
Отношение S. tianschanica к другим видам рода не может быть
выяснено в настоящее время с достаточной точностью вследствие скудности
1. SICISTA
77
коллекционного материала по рассматриваемому виду и особенно вслед-
ствие того, что до сего времени точно неизвестен признак, являющийся
(Наиболее характерным для одноцветных мышевок, т. е. строение penis.
Судя по значительной длине хвоста и задней ступни, S. tianschanica
наиболее близка к S. caucasica, S. caudata, S. concolor и S.flava (= leathemi),
т. e. к группе настоящих одноцветных мышевок, но не является родствен-
ной алтайской мышевке (.S', параеа). Косвенным подтверждением выска-
занного предположения служит нахождение на Зайсане (р. Базариха)
одноцветной мышевки, сходной по наружным признакам с S. tianschanica
и с характерным для группы настоящих одноцветных мышевок строением
penis, как это было указано выше. Вполне вероятно, что S. tianschanica
наиболее близка к кашмирской S. flava (— leathemi) или быть может обе
эти формы принадлежат даже к одному и тому же виду.
Распространение. Данные о географическом распространении
тянь-шанской мышевки скудны. Это? вид найден в следующих пунктах
Тянь-Шаня: р. Цанма, р. Капсилан, Кунгес, Заилийский Алатау (близ
г. Алма-Ата), р. Куелю (б. Пржевальский у.); можно предполагать, что
S. tianschanica распространена на восток до Зайсана (см. изложенные
выше данные о экземплярах с р. Базарихи) или даже до южного Алтая,
если правильно определение Б. А. Кузнецова (1931), указывающего этот
вид для Риддерского округа (с. Пихтовка); предположение упомянутого
автора подтверждается относительно крупными измерениями добытых
им зверьков.
Данные по биологии. Е. М. Вакуленко (in litt.), наблюдавшая
тяньшанскую мышевку в Заилийском Алатау, считает этого зверька харак-
терным обитателем зоны лиственного леса; один экземпляр этого зверька
дсбыт автором в осиннике на высоте 1500 метров. (Мною здесь же был
добыт этот зверек у нижней границы елового леса в кустарнике). В. Н. Шнит-
ников (1931) добывал этих зверьков в Заилийском Алатау в зоне яблони
на границе с елью, среди густой травянистой растительности, под кустами
шиповника, на открытых, лишенных древесной растительности, местах;
под снопами пшеницы, в посевах подсолнечника, в высокой травянистой
растительности и т. д. На южном Алтае Б. А. Кузнецов (1931) находил
тянь-шанскую мышевку чаще всего под снопами на сжатых полосах хлеба,
расположенных среди леса. Значительно реже эти зверьки встречались
в зарослях кустов по долинам и логам. Желудки добытых на полях экзем-
пляров были набиты мелко пережеванными зернами пшеницы; изредка
в них попадались остатки насекомых.
* 5. Sicista caucasica Vinogradov — Кавказская мышевка
Vinogradov, 1925: 584 (Майкопский окр.); Туров, 1926: 321; Туров, 1928 :25; Сатунин,
1903: 60 (Sminthus) concolor; Огнев, 1916; 52.
По внешнему облику и размерам сходна с другими представите-
лями группы concolor, характеризуясь, главным образом, особенностями
78
ZAPODINAE
строения penis. Длина хвоста 103—115 мм; длина ступни 16.3—18 мм.
Хвост, в отличие от S. concolor и S. caudata, двуцветный. Окраскам
спины несколько краснее и желтее, чем у других одноцветных мышевок
(17'i — 17'k Риджвея). Нижняя поверхность тела светлоохристожелтая
(15'd). У основания уха имеется рыжеватое пятно. На верхней поверх-
ности хвоста узкая полоска из черных волос. Череп со значительно
более коротким и низким rostrum, чем у S. concolor, nasalia длиннее, чем
у последнего. Penis тонкий и длинный, слегка булавовидной формы, покры-
тый тонкими острыми шипиками, направленными вершинами назад. Продоль-
ная щель занимает около трети длины, penis. Основная пластинка os penis
очень узкая и заметно выпуклая, постепенно переходящая в стержень.
Распространение. S. caucasica распространена преимуще-
ственно в западной половине Главного Кавказского хребта от верховий.
Терека до территории Кавказского заповедника включительно; вероятно'
к этому же виду относится недавнее нахождение одноцветной мышевки
в Закавказьи близ Караклиса в Армении (сообщение Аргиропуло) и еще
непроверенное указание о нахождении одноцветной мышевки в горном
Дагестане. На Главном хребте нахождение этого вида известно для сле-
дующих пунктов: аул Тепи в истоках Терека (С. С. Туров, 1926), Кавказ-
ский заповедник (Лаганаки близ г. Фишт) (С. С. Туров, 1928), б. Майкоп-
ский окр. (без точного местонахождения), р. Доута (приток Кубани}
(С. И. Огнев, 1916), г. Шугус (западная часть Главного хребта) (С. И.
Огнев, 1916), окр. С. Псху, Сухумского окр. (экз. Зоологического Инет.),.
Сванетия (К. А. Сатунин).
* 6. Sicista caudata Thomas — Хвостовая мышевка
О. Thomas, 1907: 413 (описана по экземпляру из окрестностей Корсаковского поста
на Сахалине); Vinogradov, 1925 : 581.
По длине хвоста и ступни в существенных чертах сходна с другими
представителями группы concolor (хвост 110—115 мм, ступня 16.8—18 мм)..
Задний внутренний бугорок задней ступни вытянут в длину, как у S. топ-
tana, но слабее, чем у 5. concolor и S. caucasica. Окраска верха сходна
с S. tianschanica, но с несколько более рыжеватым оттенком (близкая к clay
color Риджвея); бока тела и низ окрашены несколько светлее, чем
у S. tianschanica, хвост одноцветный, темный сверху и снизу, как у S. con-
color Biichn. Penis узкий и длинный (длина 3 мм), от основания слегка
расширяющийся к концу и покрытый на всем протяжении мелкими рого-
выми шипиками. Семенная щель узкая и длинная, доходящая до половины
длины penis; на конце семенная щель расходится и из нее выдается конец
os penis, одетый слизистой оболочкой. Os penis с довольно узкой основ-
ной пластинкой, но не в такой степени редуцированной, как у S. caucasica;
поверхность пластинки заметно вогнута.
Из всех одноцветных мышевок S. caudata наиболее близка к S. con-
color Biichn., описанной из Гансу; не лишено вероятности даже, что оба
1. SICISTA
791
эти вида являются тождественными, что удастся установить лишь после
того, когда будет известно строение penis у concolor (единственный
спиртовой экземпляр этого вида хранящийся в коллекциях Зоологического
Института является самкой).
Распространение. Сахалин, хребет Сихотэ-Алин, Уссурий-
ская область. Известные нахождения: пост Корсаковский на Сахалине,,
р. Ботча в северной части Сихотэ-Алина, долина реки Одарка (бассейн
оз. Ханка), долина р. Сучана близ Владивостока, долина реки Колумбе.
Заметки о видах Sicista, не встречающихся в преде-
лах СССР.
Известно лишь 3 следующих азиатских вида Sicista, не встречаю-
щихся в пределах СССР: S. flava True — Центральный Кашмир; S. leat-
hemi Thomas — Krishnye valley, Wardwan, Кашмир, найденная также
в горах Kagnag в Кашмире; S. concolor— Гансу близ г. Синина (Гуй-дуй-оха).
Последний вид найден также в Манчжурии, на севере Гиринской провин-
ции (Sowerby, 1923) и в юго-западной части названной провинции. Систе-
матическое положение этих форм в настоящее время не может быть
выяснено с достаточной точностью, особенно вследствие того, что у этих
видов неизвестно строение penis. Просмотренные мною два экземпляра
S. leathemi в Британском музее кажутся в существенных чертах сходными
с S. tianschanica. Вполне возможно также, что обе кашмирских формы
(«S', flava и S. leathemi) в видовом отношении являются тождественными.
Третий вид, т., е. S. concolor, как было уже указано, кажется близким
к S. caudata, что, однако, остается под вопросом до тех пор, пока не
будет известно строение penis у S. concolor.
2. Род ZAPUS COUES (1875) —ПРЫГУНЧИКИ
Coues, Bull. U. S. Geol. Survey Terr, 5, 1875:255.
По внешности представители этого рода несколько уклоняются
от примитивного мышеобразного внешнего облика характерного для
Sicista в сторону настоящих тушканчиков, особенно по сильно удлинен-
ной задней ступне (фиг. 28) и длинному хвосту.
Для всех представителей рода характерна своеобразная трехцветная
окраска: вся средняя часть спины, включая темя и лоб, окрашена в виде
резко ограниченной мантии в буровато-серый цвет (близкий к Saccarod’s
umber); бока имеют светлую желтовато-оранжевую окраску (между zinc
orange и ochraceous buff); вся брюшная сторона чисто белого цвета; окраска
низа резко ограничена от окраски боков. Для Eozapus характерно при-
сутствие на брюшке узкой продольной срединной полоски бурого цвета.
У многих представителей рода конец хвоста белый. Penis длинный цилин-
дрический. На верхней поверхности его дистальной части не имеется про-
дольной щели. Выдающийся бугорок, образованный вершиной os penis,
80
ZAPODINAE
Фиг. 28. Задняя
ступня сычуанского
прыгунчика (Eoza-
pus setchuanus
Pous.).
выходит из колокольчатого дистального края penis. Поверхность penis,
за исключением его основной части, покрыта овальными бляшками. Os
penis слабо изогнута, причем ее изгиб направлен в противоположную
сторону по сравнению с родом Sicista', только вершина os penis загнута
кверху, как у Sicista. Череп во многих существенных признаках сходен
с черепом Sicista. Главнейшие отличия выражаются в следующем: скуловые
дуги обычно расставлены в задней части немного шире,
чем в средней, поэтому наибольшая скуловая ширина
обыкновенно падает на заднюю часть скуловых дуг;
впрочем и здесь этот признак не выражен столь резко,
как у настоящих тушканчиков. Скуловая кость почти
всегда достигает передним концом lacrymale и тесно
соприкасается с последней. Canalis infraorbitalis вполне
замкнут, т. к. край его наружной стенки достигает,
maxilare, и у взрослых экземпляров срастается с по-
следней. Nasalia заходят далеко от вертикальной линии,
проведенной через gnathion. Поперечный диаметр inter-
parietale приблизительно в 3—4т/2 раза больше его про-
дольного диаметра. Задний, наружный угол parietale
не доходит до fenestra postglenoidea, которое целиком
окружено чешуйчатой костью; fenestra postglenoidea
разделены узким продольным костным мостиком, отхо-
дящим от основания скулового отростка, на два отвер-
стия. Задний край нёба доходит назад не далее линии,
соединяющей задние края коренных зубов или чаще
даже не достигает этой линии; нёбо оканчивается
назади выемкой или редко очень слабо обособленным
выступом; задняя часть нёба плоская, без вдавлен-
ных площадок. Отросток на передне-внутреннем углу
bullae tympani относительно шире и короче, чем
у Sicista. Нижняя челюсть более массивна, чём у Si-
cista, ее передний угол (angulus anterior) ясно обо-
соблен от заднего (angulus posterior) и резко повернут
внутрь. Слуховые косточки сходны с таковыми у Sicista
и вместе с тем более сходны по строению с слухо-
выми косточками Muridae, чем Dipodidae. Processus
brevis incudis почти равен по длине processus longus (в отличие от
. Sicista); incus относительно крупнее, чем у Sicista. Описанный отросток
на нейральной дуге второго шейного позвонка развит сильнее, чем
у Sicista. Ступня значительно длиннее, чем у Sicista. Первый палец
задней конечности не заходит за дистальный конец второй плюсневой
кости. Когтевые фаланги трех средних пальцев заметно изогнуты.
Пятый палец не выдается за дистальный конец основной фаланги четвер-
того пальца. Ложно-коренной зуб имеется, за исключением подрода
2. ZAPUS
81
.Napaeozapus. Коренные зубы имеют бугорчатую жевательную поверхность,
за исключением подрода Napaeozapus, у которого жевательная поверх-
ность плоская. Третий (передний) бугорок или складка удлинены и сме-
щены наружу. У Eozapus все бугорки сильно выдающиеся и отделены
.друг от друга глубокими впадинами; у подрода Zapus (с не сношенными
коронками) наружные добавочные бугорки изолированы, т. е. не соеди-
нены с главными бугорками поперечными складками; у Napaeozapus все
бугорки исчезают в раннем возрасте и жевательная поверхность стано-
вится плоской, но очертания эмалевых складок позволяют найти здесь
то же самое расположение составных элементов, как и у других предста-
вителей рода Zapus.
Распространение. Представители рода Zapus распространены
в Северной Америке, за исключением одного азиатского вида {Eozapus
:setchuanus Pous.), распространенного в Китае (Сычуань, Гансу). В Соеди-
ненных Штатах и Канаде представители этого рода (подроды Zapus
и Napaeozapus) распространены к северу до Лабрадора, Гудзонова залива,
Большого Невольничьего озера и реки Юкона на Аляске, к югу — до
Северной Каролины, Новой Мексики и Калифорнии. Распространение
сычуанского прыгунчика (подрод Eozapus) слабо выяснено и до сего
времени он найден лишь в некоторых пунктах Сычуани и Гансу.
Б'и'ологические данные. Данные об образе жизни имеются
.лишь относительно американских видов. В Северной Америке (Preble,
1899) прыгунчики селятся на лугах, заросших кустарниками полях, на
опушках лесов, на берегах лесных ручьев и пр. и предпочитают более
влажные места обитания. Будучи потревожены, прыгунчики передвигаются
•большими прыжками до 6—8 футов; при прыжках зверьки пользуются
хвостом как балансом. Рождение детенышей в числе 5—6 происходит
обыкновенно в подземных гнездах, чаще всего в мае или июне, но иногда
наблюдались пометы даже в сентябре, что объясняется наличием второго
помета или растягиванием сезона размножения на все лето. В конце
лета, по окончании сезона размножения, прыгунчики делают в густой
траве или мелком кустарнике шарообразные гнезда из травы с малень-
•кими отверстиями сбоку; обыкновенно это гнездо занимается двумя
зверьками.
Прыгунчики всех видов впадают в зимнюю спячку, хотя у некоторых
эта спячка не полная и зверьки выходят зимой в теплую погоду на поверх-
ность. Хотя зверьки часто делают летом запасы пищи в гнездах или норах,
неизвестно употребляют ли они эту пищу зимой. Спящие зверьки нахо-
дятся по одному или парами в гнездах, в норах. Иногда зверьки залегают
® спячку в гнездах на поверхности земли. Образ жизни сычуанского
тушканчика не изучен.
Видовой состав. В настоящее время американские зоологи
(G. Miller, 1824) насчитывают 13 видов этого рода, из которых 11 принадлежат
•подроду Zapus Coues (1875), 1 вид — подроду Napaeozapus Preble (1899),
Фауна СССР, Виноградов 6
82
EUCHOREUTINAE
и 1 вид — подроду Eozapus Preble (1899). Ниже излагаются краткие сведения
о единственном палеарктическом представителе этого рода — Zapus
(Eozapus) setchuanus Pousargues.
1. Подрод Eozapus Preble (1899).
Preble, N.-Amer. Fauna, 15, 1899 : 37.
Характеризуется, по сравнению с другими подродами Zapus, сильно'
выдающимися бугорками на жевательной поверхности коренных зубов,
причем эти бугорки отделены друг от друга глубокими впадинами. Наруж-
ные добавочные бугорки на зубах верхней челюсти соединены попереч-
ными гребнями с внутренними главными бугорками. Ложно-коренной зуб'
имеется. На брюшной стороне тела имеется ясно обозначенная бурая
полоска. Хвост с белым кончиком. Окраска верха как у других предста-
вителей рода. ’
Из современных представителей рода Eozapus по устройству корен-
ных зубов стоит ближе к подроду Zapus чем к Napaeozapus, сохраняя,,
однако, более примитивное строение коронки, чем у первого из названных,
родов. Еще более близок Eozapus к плиоценовому европейскому Plesio-
sminthus, как это было уже указано в главе о геологической истории:
тушканчиков. Таким образом Eozapus может считаться одним из релик-
тов плиоценовой европейско-азиатской фауны. К рассматриваемому под-
роду относится единственный указываемый ниже вид, который в то же
время является единственным современным представителем рода Zapus на.
материке Азии.
1. Zapus (Eozapus) setchuanus Pousargues.
Pousargues, 1896 : 220 (Zapus) [описан из Та-цен-лоу (Ta-tsien-lou) в Сычуани].
Основные признаки этого вида были указаны при характеристике
подрода. Кроме указанного выше нахождения, этот вид найден в настоя-
щее время в немногих других пунктах Сычуани, а также в Гансу.
2. Подсем. EUCHOREUTINAE —ДЛИННОУХИЕ ТУШКАНЧИКИ
Голова необычайной для тушканчиков удлиненной конической формы
с сильно выдающимся вперед вытянутым и притупленным на конце свинооб-
разным рылом (фиг. 1у). Относительные размеры глаз значительно меньше,,
чем у Allactaginae и Dipodinae", ось разреза век заметно наклонена по
отношению к продольной оси головы, но значительно в меньшей степени,
чем у упомянутых выше групп. Ушные раковины громадного размера,,
достигающие своими вершинами крестцовой области. Вибриссы длинные,
почти достигающие своими концами основания хвоста. Внутренний палец
передней конечности относительно хорошо развит и снабжен длинным
изогнутым когтем с острой вершиной. Задние конечности в существенных
ОПИСАНИЕ ПОДСЕМЕЙСТВА
83
чертах сходятся с таковыми у Allactaginae, т. е. с тремя длинными средними
пальцами и двумя значительно более короткими боковыми, но средний
палец лишь едва выдается за концы соседних пальцев (2-го и 4-го); поду-
шечки на концах пальцев сильно развиты и разделены на дольки, как у
Allactaginae. При основании боковых пальцев (1-го и 5-го) на нижней
поверхности имеются маленькие бугорки („мозоли")- Кисточка на конце
хвоста при основании окрашена в черный цвет и в дистальной части
в белый, но она не уплощена, как у Allactaginae. Penis удлиненной формы,
почти цилиндрический. На его конце из вертикальйого широкого, щеле-
видного отверстия далеко выдается крючкообразно изогнутый кверху
конец os penis, одетый слизистой оболочкой. Поверхность penis гладкая
без шипиков или чешуек. Os penis длинная и тонкая, почти на всем про-
тяжении прямая, кроме крючкообразно изогнутого кверху дистального ее
конца. Базальная пластинка маленькая, треугольной формы; гребень на
базальной пластинке сильно развит. Мозговая коробка имеет наибольшую
ширину в задней своей части, начиная отсюда она довольно постепенно
суживается по направлению вперед; в области межглазничного простран-
ства образуется перетяжка, гораздо более ясно выраженная, чем у Sicista
и Zapus', начиная от этой перетяжки и до слёзных костей боковые края
мозговой коробки опять расходятся: таким образом межглазничное суже-
ние лежит далеко позади линии преходящей через задние края слёзных
костей (в противоположность другим тушканчикам и сходно с Zapodinae).
Скуловые дуги нииболее широко расставлены в их задней части и сильно
сходятся кпереди, имея почти прямолинейное направление. Скуловой
отросток чешуйчатой кости сильно отогнут вниз; при рассматривании
черепа сверху он имеет почти перпендикулярное направление к продоль-
ной оси черепа и образует с скуловой дугой резкий угол. Нижний корень
скулового отростка максиллы лежит под верхним корнем, так что при
рассматривани черепа сверху корень совершенно закрыт верхним и не
выступает вперед от последнего, как это имеет место у других тушкан-
чиков. Верхний корень скулового отростка максиллы образует с сагит-
тальной осью черепа необычайно острый угол (тогда как у настоящих
тушканчиков, даже отчасти у Zapus и Sicista, этот угол приближается
к прямому). Передняя часть скуловой дуги, образующая наружную стенку
foramen infraorbitale, идет (при рассматривании черепа сбоку) косо сверху
вниз и назад в противоположность всем остальным тушканчикам, даже
Zapus, у которых эта часть или вертикальна или же идет сверху вниз
и вперед (лишь у Sicista имеется состояние сходное с Euchoreutes, но-
у первого отклонение назад более слабо выражено). Canalis infraorbitalij
.хорошо развит, причем его внешняя пластинка доходит или почти дохо-
дит до стенки maxillare, но не срастается с последней. Os zygomaticum
передним концом обыкновенно сочленяется с lacrymale. Nasalia своими
передними концами заходят вперед за вертикальную линию, проведенную
через gnathion. Lacrymalia очень маленькие, не вытянутые в поперечном
6*
®4
EUCHOREUTINAE
направлении и более сходны по форме с Zapus и Sicista, чем с настоящими
тушканчиками. Rostrum очень длинен, почти равен по длине расстоянию
ют его основания до задних концов processus pterygoidei; при рассматри-
вании сбоку rostrum имеет более или менее горизонтальное направление,
лишь едва отклоняющееся от продольной оси черепа. Венечный шов
образует довольно глубокую дугу, круто опускающуюся по бокам и вы-
прямленную в средней части. Межтеменная кость крупная, обыкновенно
с почти параллельными передними и задними краями и неправильно за-
кругленная на наружных краях. Fenestra postglenoidea маленькие, за-
крытые снизу частью bullae tympani. Внутренняя стенка глазницы с боль-
шим не окостеневшим отверстием затянутым перепонкой. Задний край
foramina incisiva заходит назад за линию проведенную через передний
край настоящих коренных зубов. Нёбные отверстия очень крупны, их
.длина приблизительно равна длине коронки 1-го коренного зуба; оба
отверстия отделены друг от друга очень узким, костным мостиком. Задний
край нёба расположен довольно далеко позади линии, соединяющей зад-
ние края последних коренных зубов; нёбо заканчивается довольно длин-
ным направленным назад шипом. Мезоптеригоидное пространство при-
близительно наполовину больше в длину, чем в ширину, с параллельными
краями. Птеригоидные отростки длинные, далеко заходящие на bullae
tympani; птеригоидные ямки широкие и глубокие. Bullae tympani очень
сильно раздуты и соприкосаются друг с другом своими передне-внутрен-
ними углами, не вступая, однако, в столь тесную связь друг с другом, как
это имеет место у Jaculus, Cardiocranius и Salpingotus’, bullae mastoideae
очень сильно развиты (приблизительно в такой же степени, как у Jaculus,
но значительно слабее, чем у Cardiocranius и Salpingotus)', внутренняя
полость каждой из bulla mastoidea разделена перегородками на три камеры,
соединенные с полостью bulla tympani. Нижняя челюсть в задней части
высокая, приблизительно в 2 раза выше, чем на уровне Mv Венечный
отросток необыкновенно длинный, не короче чем сочленовный отросток;
его вершина выдается выше уровня сочленовной головки. На боковой
поверхности челюсти имеется лишь слабая выпуклость соответствующая
корню нижнего резца. Задний угол (angulus posterior) челюсти отогнут на-
ружу; передний угол (angulus anterior) не ясно отделен от заднего угла
и не отогнут значительно внутрь. Incus лишь немного меньше, чем
corpus mallei. Manubrium mallei не отогнут вперед; близ основания manu-
brium выступа нет. Обе слуховые косточки относительно велики (при
сравнении с общими размерами тела животного) и по абсолютным разме-
рам крупнее, чем даже у Allactaga jaculus. Corpus incudis и mallei значи-
тельно удлинены вдоль линии соединяющей proc, brevis incudis и proc,
anterior mallei. Первый из названных отростков почти такой же длины,
как processus longus. Шейные позвонки свободные, их поперечные отростки
короткие, как у Zapodinae, но в отличие от этой группы у Euchoreutinae,
вершины поперечных отростков 5-го, 6-го и 7-го позвонков соприкасаются
ОПИСАНИЕ ПОДСЕМЕЙСТВА 85
друг с другом (как у Allactaginae). Скелет задней ступни сходен с тако-
вым у представителей подсемейства Allactaginae, т. е. имеет три срос-
шихся средних плюсневых кости, образующих „цевку", сильно укорочен-
ные боковые пальцы, почти равные по длине, не достигающие своими
концами до дистального конца „цевки". Однако, в отличие от Allactaginae-
цевка у Euchoreutinae неполная и в ее дистальной трети плюсневые
кости остаются не слившимися. Строение тазовых костей характеризуется
следующими признаками: crista iliaca хорошо развита; tuber ischii имеется,,
но довольно слабо обособлен; symphisis pubis (сравнительно с Allactaginae
и Dipodinae) короток, но значительно длинней, чем у Zapodinae и Cardio-
craniinae’, tuberculi ileo-pectinei — отсутствуют. Головка бедра цилин-
дрическая, с ясно выраженной седловиной на верхней поверхности (как.
у Allactaginae и Dipodinae)’, trochanter major сильно отклонен от оси
бедра наружу, расстояние от его дистального конца до основания головки
больше длины головки. Ложно-коренной зуб относительно более крупен,,
чем у других Dipodidae’, бугорки коренных зубов значительно более:
высокие и острые, чем у других представителей семейства. Бугорки
наружного и внутреннего ряда расположены друг против друга (не чере-
дуются, как у некоторых других представителей семейства); передняя
и задняя пары бугорков каждого зуба разделены друг от друга глубокими
впадинами; на коронке можно видеть маленькую, слабо развитую добавоч-
ную складку, которая обычно несколько смещена наружу; на несношен-
ных зубах можно также видеть две слабо развитые наружные добавочные
складки.
Единственный представитель рассматриваемой группы, Euchoreutes
naso, обладает столь своеобразными особенностями организации, что
заслуживает выделения не только в отдельный род, но и в отдельное
подсемейство, как это указывалось мною еще ранее (Виноградов, 1925).
Из описанных выше морфологических признаков рассматриваемой группы:
особенно обращает на себя внимание следующее: при общем сходстве
внешних признаков Euchoreutes с таковыми у представителей Allactaginae,
для рассматриваемой группы характерно наличие необычайно вытянутой
морды, очень крупных ушей и описанного выше строения penis. Как было
уже упомянуто, череп имеет очень своеобразное строение, характеризуясь
сильно вытянутым rostrum, сильно раздутыми bullae, особенностями строе-
ния скуловой дуги, слёзных костей твердого нёба, чрезвычайно прими-
тивным для тушканчикоЬ устройством нижней челюсти, резко выраженной
бугорчатой структурой коренных зубов и пр. В устройстве скелета обра-
щает внимание слияние трех средних плюсневых костей (обычно не столь
полное как у Allactaginae), строение таза, цилиндрическая форма сочленов-
ной головки бедра и пр.
Таким образом, подсемейство Euchoreutinae, резко отличаясь по
совокупности своих признаков от Zapodinae и представляя высокую
степень специализации, приближается в некоторых отношениях к под-
86
EUCHOREUTINAE
семейству Allactaginae', однако при всем этом, как можно видеть из преды-
дущего описания подсемейства, Euchoreutincue сохраняет удивительным
образом ряд примитивных признаков характерных для подсемейства
Zapodinae, как строение нижней челюсти, слёзных костей, скуловой дуги,
зубной системы и пр. По указанной причине, несмотря на резкие различия
между подсем. Zapodinae и Euchoreutinae, эти подсемейства помещены
здесь рядом, так как ни одно из других подсемейств тушканчиков не сох-
ранило столь значительного количества признаков примитивного строения
характерного для Zapodinae, как подсемейство Euchoreutinae.
Распространение. До недавнего времени единственный пред-
ставитель рассматриваемого подсемейства—Euchoreutes naso Scl. считался
характерным эндемиком пустынь Синцзяня (Китайского Туркестана);
однако сравнительно недавно Euchoreutes был найден также в Алашани
(Виноградов, 1926, Howell, 1928), что позволяет предполагать, что этот
зверек кроме Синцзяня широко распространен также в пустынях южной
Монголии, представляя в этом отношении аналогию с некоторыми другими
видами, как например с песчанкой Пржевальского ( Meriones przewalskii
Biichn.), найденной в настоящее время кроме Синцзяня, также в песках
Центральной Гоби (Эцзин-гол).
3. Род EUCHOREUTES SCLATER
W. L. Sclater, Proc. Zool. Soc. London, 1890: 610.
К подсемейству Euchoreutinae относится лишь единственный род
Euchoreutes Sclater (1890) с единственным видом Е. naso Sclater; характе-
ристика названного рода дана выше в диагнозе подсемейства; ниже сооб-
щаются лишь некоторые дополнительные данные о названном виде.
1. Euchoreutes naso W. Sclater — Длинноухий тушканчик
Sclater W., 1890 : 610 (без точного местонахождения, предположительно окрестности
Яркенда); Vinogradov, 1926; 1931. — naso alashanicus Howell, 1928 (Алашань 100 миль к СВ
от Ningsia в Гансу).
Описание наиболее существенных признаков этого вида было при-
ведено выше при характеристике подсемейства, вследствие чего здесь
указываются лишь некоторые дополнительные признаки.
Размеры средние; длина тела 90—80 мм; хвост 157—162; ступня
44—46 мм; ухо относительно более крупное, чем у всех других предста-
вителей семейства (37—43 мм). Кондилобазальная длина черепа 26—28;
скуловая ширина 16—16.5; межглазничное пространство 7—7.2; максил-
лярный ряд зубов 5—5.2. Окраска верха серая с примесью рыжеватого,
особенно близ основания хвоста; бока и низ белые. Кисточка из длин-
ных волос на конце хвоста хорошо развита, но длинные волосы
растут кругом стержня хвоста не образуя плоского „знамени"; кисточка
при основании белая, в средней части черная и ее конец белый.
3. EUCHOREUTES
87
Распространение этого вида было указано при характеристике
подсемейства; здесь приводятся следущие известные нахождения рассма-
триваемого вида: окрестности Яркенда, дол. р. Тарима, Ак-Лянгар (дорога
Яркенд-Хотан), Chipon Jagh в 30 милях от Яркенда (экземпляры Британ-
ского Музея), Хами, Люкчун, Ончи-Гургуль на сев. границе Алашани
(экземпляры Зоологического Инет. Академии Наук), Алашань в 100 милях
к ССВ от Ningsia в Гансу (Howell, 1928).
В 1928 г. Howell описал особый подвид длинноухого тушканчика —
Е. п. alashanicus из Алашани (см. выше); по мнению названного автора,
эта форма отличается от типичной более охристой и бледной окраской
и более крупными bullae. Мною (Виноградов, 1926) было также обращено
внимание на некоторые отличия алашанского длинноухого тушканчика от
синцзянского, в частности следующие: мозговая коробка у алашанских
.экземпляров сзади шире чем у типичных и bullae tympani несколько
крупнее; волосы на нижней поверхности пальцев несколько длиннее; однако
я не считаю рациональным выделить эту форму, так как имеющийся
.в музеях материал по „типичной" форме слишком недостаточен для точного
ее сравнения с алашанскими экземплярами.
Образ жизни этого замечательного зверька почти неизвестен.
Е. naso является вероятно обитателем песчаных пустынь, как об
этом свидетельствуют наблюдения Козакевича (Виноградов, 1926) и ука-
зания Sclater’a (1890), хотя по внешности этого зверька и особенно по
устройству его задней ступни можно было бы принять его за обитателя
глинистых или щебнистых пустынь. По наблюдениям Козакевича в Мон-
голии (Виноградов, 1926), этот тушканчик селится в песчаных долинах,
заросших низкими кустами саксаула. Названный исследователь наблюдал
их у костра на расстоянии ближе 4-х метров и находил их в юртах кочев-
ников. Один экземпляр пойманный в юрте, прожил в неволе 4 дня и обна-
руживал сильное стремление кусаться.
3. Подсем. CARDIOCRANIINAE —КАРЛИКОВЫЕ ТУШКАНЧИКИ
Мелкие тушканчики, не превышающие по длине тела домашнюю
мышь. Голова по отношению к размерам туловища очень крупная, значи-
тельно крупнее, чем у,всех других тушканчиков. Уши маленькие, трубчатые,
т. е. внутренние края ушной раковины при ее основании сросшиеся.
Глаза относительно крупнее, чем у Zapodinae, но значительно меньше,
•чем у Allactaginae и Dipodinae, с ясно выраженным наклоном разреза век
по отношению к оси головы. Задняя конечность с пятью или тремя паль-
цами с развитой в большей или меньшей степени щеткой из волос на
подошве. Хвост длинный и тонкий или относительно короткий и толстый
•с отложениями жира в подкожной клетчатке; кисточка на конце хвоста
отсутствует или образована редкими длинными волосками, не образующими
..„знамени".
88
CARD IOCR ANIINAE
Строение genitalia известно лишь у Salpinogotus kozlovi Vinogr.,,
у которого penis резко отличается от такового у представителей всех дру-
гих подсемейств, характеризуясь тонкой вытянутой и слегка суживающейся
к концу формой и отсутствием os penis; кроме того на его поверхности'
не имеется роговых шипиков или чешуй, которые заменены чрезвычайно
мелкими слабо заметными зернышками в дистальной части penis. Череп,
тушканчикообразного типа, т. е. с сильно расширенной сзади мозговой
коробкой, имеющей сверху трапециодальную форму; резко обозначенных
выпуклостей на боках мозговой коробки нет; по направлению вперед
боковые края мозговой коробки быстро сближаются, образуя почти пря-
мые линии и достигая наибольшего сближения непосредственно позади от’
слёзных костей, таким образом межглазничное сужение резко выражено,
так как непосредственно впереди от этого сужения слёзные кости сильно
выдаются в стороны. Верхние корни скуловых отростков верхне-челюст-
ной кости немного отклонены вперед от перпендикулярной линии прове-
денной к сагиттальной оси черепа. Скуловые дуги почти прямолинейны,
наиболее сильно расставлены в задней их части, откуда они идут почти
прямолинейно вперед, быстро сближаясь друг с другом до соединения
с верхними и нижними скуловыми отростками максиллы. Скуловой отросток
чешуйчатой кости сильно отклонен вниз; при рассматривании сверху он
отходит почти перпендикулярно к сагиттальной оси черепа или даж&
несколько отклонен назад и образует с общим направлением скуловой дуги:
резко обозначенный прямой или острый угол. Нижний корень скулового'
отростка максиллы лежит значительно впереди от верхнего и при рас-
сматривании черепа сверху далеко выступает вперед от последнего. Ску-
ловая кость ясно разделена на горизонтальную и восходящую ветвь,
но обе эти ветви с постепенным закруглением переходят друг в друга,
не образуя резкого угла как у настоящих тушканчиков; своим передним1
концом скуловая кость не достигает lacrymale. У представителей рода.
Salpingotus (см. ниже) скуловая дуга имеет резко выраженный отросток,
отходящий от ее средины косо назад и вниз. Слёзные кости несколько
увеличены в размерах и вытянуты, но не в поперечном направлении, как
у настоящих тушканчиков, а в косом. Rostrum относительно короткий,
приблизительно равный по длине линии проведенной от его основания до-
заднего края зубного ряда (длина rostrum условно считается от переднего
края основания нижних ветвей скуловых отростков maxillare до gnathion).
Венечный шов образует довольно глубокую, обращенную выпуклостью,
назад дугу. Межтеменная кость очень сильно редуцирована, она представ-
лена лишь очень узкой маленькой косточкой, вытянутой в продольном
направлении, или она отсутствует вовсе. Supraoccipitale в связи с чрез-
мерно увеличенными bullae mastoidea представлена лишь узкой полоской
зажатой между bullae. Внутренняя стенка глазницы вполне окостеневшая,
без образования затянутых перепонкой отверстий. Межкрыловидное про-
странство очень короткое, его длина не более его ширины'(обыкновенно<
ОПИСАНИЕ ПОДСЕМЕЙСТВА
89’
значительно короче). Птеригоидные отростки относительно короткие,,
заметно короче длины зубного ряда (без pm’) или даже короче половины
его длины; hamuli заходит далеко назад за переднюю границу bullae.
Птеригоидные ямки очень сильно расширены по направлению назад и глу-
бокие; их дно продырявлено довольно крупными отверстиями. Bullae
tympani очень крупны и сильно раздуты; их передне-внутренние углы
тесно соприкасаются друг с другом, образуя в месте соприкосновения
дискообразное расширение и закрывая собой всю заднюю часть basisphe-
noideum. Mastoidea необычайно раздуты, образуя огромные пузыри, полость-
которых соединена с полостью bullae tympani; bullae mastoidea превосхо-
дит по объему несколько раз bullae tympani; они выдаются в виде огром-
ных пузырей далеко назад от задних краев condyli occipitales и выходят-
также на верхнюю поверхность черепа, составляя около половины всей
этой поверхности и почти соприкасаясь своими внутренними краями.
Нижняя челюсть в задней части высока, как у Zapodinae и Euchoreutinae..
Венечный отросток едва развит, его вершина не достигает уровня сочле-
новной головки челюсти. На боковой поверхности челюсти имеется
выпуклость, образованная корнем нижнего резца, но эта выпуклость,
развита значительно слабее чем Allactaginae и Dipodinae. Задний угол,
(angulus posterior) нижней челюсти резко повернут наружу, образуя
хорошо обособленную горизонтальную пластинку. Передний угол резко-
выражен и значительно повернут внутрь. Строение слуховых косточек
сходно с таковым у Euchoreutinae, но у Cardiocraniinae corpus incudis-
и особенно corpus mallei более узкие и более удлиненные. По строению<
шейных позвонков эта группа обнаруживает наивысшую степень специа-
лизации по сравнению со всеми другими представителями семейства.
Шейные позвонки у Cardiocraniinae чрезвычайно укороченные и сливаются,
друг с другом, причем срастаются не только 2, 3, 4, 5 и-6-й позвонки, но '
к ним вполне прирастают также и 7-й -позвонок, чего не наблюдается
у других групп семейства (более того, у представителей рода Salpingotus
к шейному комплексу целиком прирастает также 1-й грудной позвонок)..
Кроме того, у представителей подсемейства Cardiocraniinae проис-
ходит более полная редукция свободных частей поперечных отростков,
чем у Dipodinae (см. ниже), в связи с чем на боках сросшегося комплекса,
шейных позвонков имеется лишь по одному большому отверстию, в кото-
ром можно видеть тончайшие костные нити — остатки средних частей
редуцированных поперечных отростков. По строению скелета задней сту-
пни роды Cardiocranius и Salpingotus столь резко разнятся между собой,,
что не представляется возможным дать детальный диагноз всего подсе-
мейства на основании этого признака. Общим признаком для обоих родов
является полное отсутствие срастания плюсневых костей, что сближает по
этому признаку эту группу с подсемейством Zapodinae.
Строение тазовых костей характеризуется сочетанием следующих
признаков: crista iliaca хорошо развита; tuber ischii хорошо обособлен-
90
CARDIOCRANIINAE
и заметно загнут в дорзальном направлении. Symphisis pubis очень
короток; он образован лопастевидными отростками ischio-pubis (Cardiocra-
nius) или чрезвычайно тонкими и длинными отростками (Salpingotus)',
Tuberculi ileo-pectinei хорошо выражены. Головка бедра шарообразная,
без седловины на верхней поверхности. Ложно-коренной зуб имеется.
Строение коронки первого верхнего коренного зуба, по сравнению с Zapo-
dinae и Euchoreutinae, значительно упрощено; передняя добавочная
складка имеется; две наружные добавочные складки отсутствуют.
Распространение рассматриваемой группы слабо изучено,
вследствие редкости нахождения составляющих ее видов; насколько можно
•судить по немногим известным нахождениям представителей этой группы,
распространение подсемейства Cardiocraniinae охватывает Монголию от
северных районов Монгольского Алтая на севере до Нань-Шаня на юге.
Остается под сомнением нахождение одного из представителей рассма-
триваемого подсемейства (Salpingotus thomasi Vinogr.) в южном Афгани-
-стане, как это более подробно излагается в соответствующем месте.
Подсемейство Cardiocraniinae является вероятно древней группой,
рано ответвившейся от основного ствола семейства Dipodinae', во всяком
случае есть основания предполагать, что некоторые ископаемые тушкан-
чики из миоцена и плиоцена Китая и Монголии принадлежат к этому
подсемейству или родственным ему группам. Выше в главе о геологической
истории семейства подобное предположение было высказано относительно
Sminthoides fraudator Schlosser (1924) из нижнего плиоцена (понтийского
яруса) из Ertemte и Olan Chorea в Монголии, а также относительно Plesiodi-
pus leeli Young (1927) из плиоцена или верхнего миоцена Гансу (близ г. Lan-
chow), менее ясным является систематическое положение Protalactaga
grabaui Young (1927) из той же местности и того же горизонта; по изве-
стным немногим остаткам этого зверька можно предполагать, что ои
имеет черты сходства с Cardiocraniinae и в то же время с Dipodinae или
быть может занимает до некоторой степени промежуточное положение
между этими группами.
Подсемейство Cardiocraniinae является чрезвычайно своеобразной
труппой соединяющей в себе признаки высокой специализации с призна-
ками примитивности унаследованными от мышеобразных предков семей-
ства и в наибольшей степени сохранившимися у представителей подсемей-
ства Zapodinae. Из признаков примитивной для рассматриваемого семейства
организации сохранившихся у Cardiocraniiae следует отметить наличие
несросшихся metatarsalia, круглой головки бедра, отсутствие выпуклостей
или гребней на краях parietale, дугообразную форму венечного шва, вполне
окостеневшую стенку глазницы, относительно высокую нижнюю челюсть
в задней ее части и пр. На ряду с указанными признаками рассматривае-
мое подсемейство характеризуется некоторыми особенностями высокой
специализации, не повторяющимися в других подсемействах Dipodidae, как
.например, описанное выше своеобразное строение шейных позвонков,
4. CARDIOCRANIUS
91
таза, penis, редукция или полное отсутствие межтеменной кости, отсут-
ствие processus lateralis на supraoccipitalae, гигантская величина bullae
и особенно их части образованной раздутой сосцевидной костью и пр.
Третья группа признаков, характерных для Cardiocraniinae, сближает это
подсемейство с Allactaginae и в еще большей степени с Dipodidae, как
трапециодальная форма мозговой коробки, очертания скуловой дуги,
:ясное разделение zygomaticum на горизонтальную и восходящую ветвь,
короткий rostrum, короткие крыловидные отростки, глубокие и широкие
крыловидные ямки, редукция венечного отростка нижней челюсти и пр.
Как было указано уже выше, установление подсемейства Cardiocra-
niinae и составляющих его родов стирает предполагавшуюся ранее рез-
кую грань между Zapodinae с одной стороны и настоящими тушканчиками
с другой.
В настоящее время известны два рода, составляющие подсемейство
Cardiocraniinae с четырьмя видами.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ
1 (2). Задняя конечность с 5-ю пальцами. Скуловая дуга без
отростка отходящего от ее середины вниз. Первый грудной позвонок
не прирастает к последнему шейному. Лонное сращение образовано
•относительно короткими и широкими лопастевидными отростками ischio-
pubis. Бугорчатая структура коренных зубов сохраняется более или
менее ясно у взрослых особей................... . 4. Cardiocranius Satun.
2 (1). Задняя конечность с 3-мя пальцами. Скуловая дуга с хорошо
)развитым угловатым выступом или саблевидным отростком, отходящим
от ее середины вниз. Первый грудной позвонок прирастает к последнему
шейному. Лонное сращение образовано узким и чрезвычайно вытянутым
отростком ischio-pubis. Бугорчатая структура коренных зубов сохра-
няется у молодых и полувзрослых особей...........5. Salpingotus Vinogr.
4. Род Cardiocranius Satunin (1902) — Карликовые пятипалые тушканчики
. Satunin, Ann. Mus. Zool. Acad. Sci. Petersb., VII, 1902 : 582.
Голова по отношению к размерам тела значительно крупнее, чем
у Zapodinae. Глаза меньше, чем у Salpingotus', ось разреза век имеет ясный
наклон. Вибрисы вокруг rhinarium значительно более длинные и более
густо расположенные, чем у Zapodinae, но уступают в обоих отношениях
таковым у Salpingotus. Наиболее длинные вибрисы достигают половины
.длины туловища. Уши трубкообразные, маленькие; их размеры приблизи-
тельно в 2 раза меньше, чем у Salpingotus. Коготь на первом (внутрен-
нем) пальце передних конечностей имеет вид длинного изогнутого ногтя
с закругленным свободным краем. Задняя конечность с 5 пальцами;
на концах пальцев имеются разделенные на дольки эластические поду-
92
CARDIOCRANIINAE
шечки, сходные с Allactaginae. Щетка на нижней стороне пальцев задней
конечности развита значительно слабее, чем у Salpingotus. Canalis,
infraorbitalis не вполне замкнут, так как верхний край его наружной стенки
немного не доходит по стенки maxillarae. Расширенная передняя половина
скуловой дуги постепенно переходит в узкую заднюю часть; никаких,
следов отростка, отходящего от середины скуловой дуги вниз не имеется..
Nasalia доходят своими передними концами лишь до вертикальной линии,,
проведенной через gnathion. Профиль расширеннной части черепа почти
прямой (т. е. мало изогнутый вниз). Задний край неба простирается назад
за линию, соединяющую задние края molares, на расстоянии приблизи-
тельно в 2 раза меньше чем длина зубного ряда (без pm'). Птеригоидные
отростки заметно короче длины зубного ряда, но превышают половину
его длины. Нижняя челюсть более тонкая (менее массивная), чем.
у Salpingotus; ее диастемная часть более удлинена и без угловатого-
отростка, отходящего вниз и назад от основания нижних резцов. Первый
грудной позвонок не прирастает к последнему шейному; следы срастания
2—7 шейных позвонков слабо заметны. Крючкообразные отростки на
боках вентральной стороны б-го позвонка хорошо развиты. Задняя ступня
с хорошо раззитыми пятью пальцами и пятью несросшимися плюсневыми
костями. В отличие от пятипалых Zapodinae с несросшимися плюсневыми
костями, у Cardiocranius каждый из трех средних пальцев короче трех
четвертей соответствующих плюсневых костей; первый палец не дости-
гает дистального конца второй плюсневой кости. Дистальный конец
entocuneiforme расположен значительно впереди от проксимального конца
третьей плюсневой кости. В строении таза характерны следующие приз-
наки; symphisis pubis образован относительно короткими и широкими
лопастевидными отростками ischio-pubis. Tuberculi ileo-pectinei двойные..
На бедренной кости trochanter tertius отсутствует. Бедро относительно
длинное. На коренных зубах бугорчатая стуктура коронки сохраняется,
более или менее ясно у взрослых особей. Передняя добавочная складка,
значительно смещена наружу, образуя добавочный (третий) выдающийся
угол на наружной стороне коронки.СкладКи эмали образуют замкнутые
чередующиеся петли.
Род Cardiocranius, обладающий рядом признаков сходных с Salpin-
gotus, в то же время характеризуется весьма существенными и своеобраз-
ными особенностями организации, что придает ему значение резко очер-
ченной систематической группы. При общей высокой специализации орга-
низма Cardiocranius все же значительно уступает в этом отношении роду
Salpingotus (строение задних конечностей, таза, шейных позвонков, скуло-
вой дуги, жевательной поверхности коренных зубов и пр.). Среди современ-
ных родов Dipodidae. Cardiocranius, как и Salpingotus, занимают изолиро-
ванное положение. Как было уже указано в главе о геологической истории
тушканчиков, оба названных рода, повидимому, имеют наиболее близкие
родственные отношения к плиоценовым родам Plesiodipus и Sminthoides*
4. CARDIOCRANIUS
93
'что окажется возможным установить точно лишь по?ле нахождения более
полных остатков упомянутых вымерших родов.
Отмеченная выше для подсем. Cardiocraniinae тенденция к образова-
нию групп до известной степени аналогичных (параллельных) пятипалым
и трехпалым тушканчикам {Allactaginae и Dipodinae) выражается у рода
Cardiocranius в приобретении [внешнего облика до некоторой степени
соответствующего таковому у рода Pygerethmus (короткий жирный
хвост, пятипалая ко-
нечность).
Наличие у Car-
diocranius пятипалых
задних конечностей
с относительно слабо
-развитой щеткой из
волос, по аналогии
•с другими тушканчи-
ками, позволяет счи-
тать этого зверька
преимущественно
обитателем твердых
-субстратов глинистых
или щебнистых пу-
стынь; этому предпо-
ложению быть может
не противоречит на- . .
ЦЛиг. 2У. 1 олова Cardiocranius paradoxus oatun. (увелич.).
хождение Козакеви-
чем (Виноградов, 1926) двух экземпляров Cardiocranius среди песчаных
барханов в северной Гоби, так как многие обитатели глинистых пустынь
в северных частях ареала распространения становятся обитателями песча-
ных субстратов {Scirtopoda telum, Citellus fulvus и др.).
Распространение рода в настоящее время является слабо выя-
сненным в связи с редкостью нахождения этого зверька. В настоящее время
известны лишь два далеко отстоящих друг от друга нахождения этого
зверька — Нань-Шань (место описания первого экземпляра) и северная
Гоби (Алгай-Хайрхан, около 500 км к юго-западу от Улан-Батора), что
указывает на широкое распространение этого зверька в пустянях Монголии.
В настоящее время известен лишь один вид.
1. Cardiocranius paradoxus Satunin — Карликовый пятипалый туш-
канчик (фиг. 29).
Satunin, 1903 : 36 (Шарогол-джин в Нань-шане); Виноградов, 1929 : 223; Vinogradov,
1930: 331, 453 и др.
Наиболее характерные признаки этого вида указаны в диагнозе
рода. В дополнение к приведенному описанию здесь приводятся лишь
94
C ARDIOCRANIIN АЕ
следующие признаки. Размеры мелкие: длина тела 50—54 мм, хвост 74—78,.
ступня 25—26; ухо 4, 5—6.0. Череп относительно крупный: наибольшая
длина черепа — 23.0—24.0, кондилобазальная длина черепа 17—18.2,.
скуловая ширина 11.4—12.0, межглазничное пространство 4.2—4.3. Окраска
верха серовато-коричневая; вся нижняя сторона чисто белая. Наружная
сторона конечностей и узкая пограничная зона между окраской верха
и брюшной стороны светлоржаво-желтая. Сосков — 8.
Распространение. Кроме Нань-шаня (Шарагол-джин) этот вил
найден также в северной Гоби (Алгай-хайран, около 500 км к юго-западу
от г. Улан-батора).
5. Род SALPINGOTUS VINOGR.— ТРУБКОУХИЕ ИЛИ КАРЛИКОВЫЕ
ТРЕХПАЛЫЕ ТУШКАНЧИКИ
Vinogradov, 1922: 540.
Голова относительно крупная, как у Cardiocranius или крупнее?
глаза более крупные. Уши такого же строения, как у Cardiocranius, но
значительно больших размеров. Rhinarium более уплощен, чем у назван-
ного рода; окружность rhinarium и конец морды густо покрыты твердыми
длинными волосами; наиболее длинные выбрисы почти достигают своими
концами основания хвоста. Когти на передних конечностях еще более
длинные, чем у Cardiocranius и еще более изогнуты. Коготь первого
внутреннего пальца с острой вершиной. Задняя конечность с тремя
пальцами. Подушечки на конце пальцев слабо развиты, маленькие и не
разделенные на дольки. Щетка на задней ступне необычайно сильно,
развита и образована длинными прямыми или правильно изогнутыми
волосами. Строение penis известно лишь у 6". kozlovi и было описано
выше при характеристике подсемейства. Нижнеглазничный канал полный;,
верхний край его наружной пластинки соприкасается с шахШаге, ио не
срастается с ней. Передняя часть скуловой дуги образует расширенную
пластинку, резко отделенную от ее тонкой задней части; от середины
каждой скуловой дуги отходит отросток, направленный косо вниз и назад;
этот отросток образован выростом нижнего корня скулового отростка
верхне-челюстной кости. Передняя часть носовых костей выдается вперед,
значительно далее альвеолярных концов межчелюстных костей. Rostrum,
отогнут вниз. Нёбные кости значительно длиннее, чем верхний зубной
ряд и необычайно далеко выдаются назад. Крыловидные отростки очень
коротки, короче половины длины верхнего зубного ряда. Нижняя челюсть
более масивная, чем у Cardiocranius, ее диастомная часть более укорочена,
чем у представителей предыдущего рода и снабжена угловатым выступом,
отходящим вниз и назад от основания альвеол нижних резцов. Первый
грудной позвонок совершенно слит с последним шейным (об этом можно-
судить по местам прикрепления 1-й и 2-й пары ребер). Границы между
слившимися шейными позвонками, а также между 7-м шейным позвонком
5. SALPINGOTUS
95-'
и 1-м грудным совершенно не заметны. Крючкообразные отростки на 6-м
позвонке совершенно редуцированы. Задняя ступня имеет лишь 3 пальца;,
от двух боковых пальцев (1-го и 5-го) сохраняются лишь рудименты, как
у представителей подсем. Dipodinae. Три средних плюсневых кости
свободны, т. е. не сросшиеся и не образующие цевки. В отличие от всех
Dipodinae у Salpingotus фаланги имеют большую относительную длину,,
а плюсневые кости меньшую, в связи с чем длина каждой фаланги
у Salpingotus почти равна длине соответствующей metatarsale, тогда как.
у всех Dipodinae фаланги составляют менее 2/8 длины цевки. В строении
таза характерно то, что symphisis pubis образован узким и чрезвычайно-
вытянутым отростком ischiopubis, а также тем, что имеется лишь один
хорошо развитый tuberculins ileo-pectineus. На бедренной кости trochanter
tertius хорошо развит; бедренная кость относительно более короткая, чем
у Cardiocranius. Бугорчатая структура на жевательной поверхности корен-
ных зубов сохраняется лишь у молодых и полувзрослых особей. Перед-
няя добавочная складка расположена на срединной линии зуба (не смещена
наружу). Все эмалевые петли на жевательной поверхности не замкнуты
и их наружные и внутренние углы расположены друг против друга,
(не чередующиеся).
Как это было указано в главе о Cardiocranius, оба рода слагающих,
подсем. Cardiocraniinae являются высоко специализированными, но в этом
отношении Salpingotus значительно превосходит род Cardiocranius, обладая
трехпалой задней конечностью и сильно развитой щеткой из волос, свое-
образным строением скуловой дуги, шейных позвонков, таза, утратой
бугорчатого строения коренных зубов и пр. Род Salpingotus, как и Cardio-
cranius занимают изолированное положение среди современных предста-
вителей семейства. Как уже было указано выше, наиболее близкими к ним.
группами вероятно являются плиоценовые роды Plesiodipus и Sminthoides.
По своим морфологическим признакам представители рода Salpin-
gotus представляют крайнюю степень приспособления к обитанию в пусты-
нях, в частности в песчаных пустынях; по устройству задней конечности
(трехпалость, сильно развитая щетка) представители рода Salpingotus
обнаруживают хорошо выраженный параллелизм с другими обитателями
песков (роды Dipus, Eremodipus, Jaculus, Paradipus).
Биологические данные о представителях рода Salpingotus отсут-
ствуют.
О географическом распространении рода Salpingotus можно судить
лишь по нахождениям четырех известных экземпляров Salpingotus, отно-
сящихся к трем видам: 2 известных экземпляра 6". kozlovi добыты в песках
окрестностей Хоро-Хото в Центральной Гоби, 1 экземпляр S. crassicauda —
в северной части Монгольского Алтая, близ Шара-Сюме; точное нахож-
дение третьего вида — S. thomasi неизвестно; судя по маршрутам добыв-
шего его путешественника Griffith, этот экземпляр добыт в южном Афга-
нистане или южном Тибете (см. ниже). Таким образом, можно предположить,.
'96
CARDIO C RAN I IN AE
что представители рода Salpingotus широко распространены в Центральной
„Азии от северной части Монгольского Алтая через пустыню Гоби на юг
может быть до южного Тибета (нахождение в Афганистане менее ве-
роятно).
Как было уже указано, в настоящее время известны 3 вида рода
Salpingotus'. S. kozlovi Vinogr., -S. crassicauda Vinogr. и «S', thomasi Vinogr.
Интересно отметить, что эти удивительные животные стали известны лишь
сравнительно недавно: род Salpingotus с единственным тогда видом
,S. kozlovi был описан Виноградовым в 1922 г., тот же автор описал
в 1924 г. второй резко дифференцированный вид — 6". crassicauda и им же
был описан в 1928 г. по экземпляру Британского музея третий вид —
S. thomasi.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
?1 (4). Хвост в передней половине или передней трети утолщенный с отло-
жениями жира в подкожном слое. На всем протяжении хвост покрыт
короткими волосами, которые становятся лишь едва длиннее на конце
хвоста, не образуя, однако, заметной кисточки. Отросток на скуло-
вой дуге короткий, значительно короче половины задней части
скуловой дуги.................................................. 2.
2 (3). Длина хвоста более 100 мм. Передняя расширенная часть скуловой
дуги не резко отделена от задней тонкой. На нижнем краю скуловой
дуги в ее середине имеется лишь слабо выраженный угловатый
отросток, не отогнутый назад...................1. S. thomasi Vinogr.
-3 (2). Длина хвоста менее 100 мм. Передняя расширенная часть скуловой
дуги резко отделена от задней тонкой. На нижнем краю скуловой
дуги в середине имеется хорошо обособленный (хотя и короткий)
отросток ясно отогнутый назад...............2. S. crassicauda Vinogr.
4 (1). Хвост тонкий без отложений жира в подкожном слое. На всем про-
тяжении, кроме коротких прилегающих волос, хвост покрыт редкими
длинными щетинками, которые на конце становятся заметно длиннее,
образуя редкую кисточку на конце. Отросток на скуловой дуге
длинный, саблевидный, заметно превышающий половину длины задней
части скуловой дуги............................3. S. kozlovi Vinogr.
1. Salpingotus thomasi Vinogradov — Карликовый тушканчик
Томаса.
Vinogradov, 1928: 372 (описан по экземпляру Британского музея без точного место-
нахождения, добытому W. Griffith в южном Тибете или южном Афганистане).
Хвост около 105 мм, значительно более длинный, чем у .S', crassicauda,
но более короткий чем у S. kozlovi. В передней половине хвост несколько
утолщен с отложениями жира в подкожной клетчатке. На всем протяжении
хвост покрыт очень короткими волосками, которые лишь едва длиннее на
5. SALPINGO TUS
97
конце. Задняя ступня несколько длиннее, чем у »S. crassicauda. Отросток
отходящий от середины скуловой дуги слабо развит и образует только
маленький угловатый выступ, не обособленный от скуловой дуги и не
отогнутый назад как у .S’, crassicauda. Расширенная передняя половина
скуловой дуги не резко отделена от задней тонкой ее части: начиная от
вершины упомянутого отростка скуловая дуга постепенно становится
более тонкой. Окраска S. thomasi не может быть точно описана,
так как единственный известный экземпляр этого вида сохранялся в кол-
лекции до его описания около 90 лет и в значительной степени утратил
свою естественную окраску.
Измерения: длина тела 57 мм (приблизительно); хвост 105 мм;
задняя ступня 23.3 мм; ухо — 9 мм (приблизительно); межглазничное про-
странство 4.5; носовые кости 8.0; максиллярный ряд зубов 3.0 мм.
Недостаточная удовлетворительная сохранность экземпляра, послу-
жившего для описания S. thomasi, не дает возможности точно выяснить
отношения этого вида к другим представителям рода. Однако, уже
на основании имеющихся данных можно утверждать, что S. thomasi
.является среди других представителей рода наименее специализированной
формой, что особенно ясно выражается в описанном выше строении ску-
ловой дуги. По своим признакам S. thomasi стоит значительно ближе
к S. crassicauda, чем к »S. kozlovi, на что указывает описанное строение
скуловой дуги, наличие жирного хвоста, покрытого короткими волосками,
относительно слабо развитая щетка на задней ступне и пр.
Распространение S. thomasi, как было уже отмечено, неиз-
вестно. Экземпляр этого зверька первоначально сохранялся в музее Ост-
Индской компании с этикеткой: „Афганистан, W. Griffith" и впоследствии
был передан в Британский музей. Насколько можно судить по опублико-
ванному W. Griffith „Journal of Travels", vol. i-ii, Calcutta, 1847, он путе-
шествовал и коллекционировал также в некоторых пограничных с Тибетом
частях Гималаев (верхний Ассам и Бутан) и на территории Индостанского
полуострова. Судя по общему распространению рода Salpingotus трудно
.думать, что это животное действительно добыто на территории Афгани-
стана, характеризующегося совершенно иными элементами фауны. Скорее
□можно предполагать, что этот зверек добыт где-нибудь в пограничных
с Индией частях южного Тибета.1
2. Salpingotus crassicauda Vinogradov — Жирнохвостый карликовый
тушканчик
Vinogradov, 1924 :150 (описан по экземпляру из Шара-Сюме в Гобийском Алтае,
около 160 км к югу от границы Монголии и СССР).
По внешности сходен с другими представителями рода, но хвост
.значительно короче, чем у S. thomasi и 5. kozlovi (93.3 мм); в передней
^четверти хвоста имеется значительное утолщение, которое постепенно
1 Оригинальной этикетки коллектора при этом экземпляре не было.
Фауна СССР, Виноградов 7
98
CARDIO CRANIINAE
суживается к концу; это утолщение образовано отложениями жира в под-
кожной клетчатке хвоста. Вся поверхность хвоста покрыта очень корот-
кими волосами, которые на конце становятся лишь длиннее, не образуя
здесь, однако, кисточки. Сосцов 8, передняя грудная пара, расположенная,
близко к шее, слабо развита, но хорошо заметна. Щетки на пальцах зад-
ней ступни развиты слабее, чем у -S', kozlovi', составляющие ее волосы не-
столь длинны и не так сильно изогнуты, как у упомянутого вида. Окраска,
бледная песчаная, заметно не отличающаяся от таковой у S. kozlovi..
Череп меньше и относительно короче, чем у 5. kozlovi. Межтеменная,
кость отсутствует. Отросток на скуловой дуге хорошо развит и имеет вид.
резко обособленного выступа, отогнутого вершиной назад; этот выступ:
значительно более развит, чем у S. thomasi, но почти в 4 раза уступает-
по длине таковому у S. kozlovi. Передняя расширенная часть скуловой;
дуги резко обособлена от задней; однако ширина передней части скуловой
дуги почти в три раза меньше, чем у «S’, kozlovi. На чешуйчатой кости
имеется направленный вперед отросток, который, однако, не достигает
слёзной кости, как это имеет место у S. kozlovi. Нижняя челюсть относи-
тельно меньше, чем у S. kozlovi и не столь массивна; добавочный гори-
зонтальный отросток нижней челюсти, отходящей наружу между угловым:
и сочленовным отростками, значительно короче и в несколько раз уже,,
чем у S. kozlovi. Задний угол нижней челюсти снабжен широким отвер-
стием. Зубы в существенных чертах сходны с таковыми у других предста-
вителей рода.
Измерения: длина тела 41 мм; хвост 93.2, задняя ступня 20.5;ухо 5.6;.
наибольшая длина черепа 22.7; кондилобазальная длина 18.1; скуловая ши-
рина 11.6; межглазничноё пространство 4.6; высота мозговой коробки 7.2;.
длина носовых костей 8.1; диастема 4.2; длина верхнего ряда зубов 3.0.
Распространение. Кроме единственного нахождения S. crassi-
cauda близ Шара-Сюме в северной части Монгольского Алтая, других,
данных о распространении этого вида не имеется. Близость указанного
нахождения к границам Со.-оза позволяет допускать возможность нахо-
ждения этого вида и в пределах СССР, в южных частях советского Алтая.,
По своим морфологическим признакам S. crassicauda занимает про-
межуточное положение между относительно слабо специализированным
S. thomasi и достигшим крайней степени специализации S. kozlovi', однако,,
как можно видеть из ’предыдущих описаний и определительной таблицы,.
S. crassicauda значительно более близок к S. thomasi, чем к •S', kozlovi
3. Salpingotus kozlovi Vinogradov — Карликовый тушканчик Козлова,
(фиг. 30).
Виноградов, 1922:540—545.
По внешности сходен с другими представителями рода, отличаясь,,
главным образом, относительно более крупными размерами головы, строе-
нием задних конечностей, строением и длиной хвоста.
5. SALPINGOTUS
99
Голова еще более несоразмерно с телом велика, чем у других пред-
ставителей рода, а также чем у Cardiocranius paradoxus. Нижняя сторона
пальцев покрыта значительно более длинными волосами, чем у S. thomasi
и aS. crassicauda, причем эти волосы правильно изогнуты по направлению
назад и внутрь; совокупность этих волос образует на каждой лапке широ-
Фиг. 30. Карликовый тушканчик Козлова (Salpingotus kozlovi Vinogr.)
(уменьш.). Рис. А. Н. Формозова.
кую мягкую войлокообразную подошву (фиг. 31). Хвост прибизительно
в 21/2 раза превышает длину головы и тела; на всем протяжении хвост тонкий
без отложений жира в подкожной клетчатке; хвост равномерно покрыт
короткими довольно редкими волосками и, кроме того, на всем его протяже-
нии имеются редкие длинные щетинки, которые по направлению назад
становятся несколько гуще и длиннее и образуют на конце кисточку из
редких длинных волос. Шерсть очень тонкая и длинная. Верхняя сторона
тела окрашена в светлый песчаный цвет; на голове эта окраска наиболее
Фиг. 31. Задняя ступня Salpingotus kozlovi Vinogr. (увелич.).
чистая, без примеси серого (15" d); спйна окрашена в тот же цвет, но
с значительной примесью серого, особенно в задней ее части. Основание
волос спинной стороны почти белое (бледножелтовато-серое), далее по на-
правлению к вершинам волосы постепенно становятся серыми, а их концы
окрашены в песчаный цвет, обозначенный выше. На боках окраска стано-
вится светлее и переходит в бледножелтый (почти белый) цвет брюха
(10" d). Волосы хвоста беловатые, на концевой кисточке их окончания
коричневатые. Череп относительно более крупный и удлиненный, чем
у «S', crassicauda. Носовые кости относительно длиннее, чем у названного
вида; при рассматривании сверху их очертания более сложны; у перед-
7*
100
CARDIOCRANIINAE
него конца они сильно стянуты, в средней их части имеется вторая не-
сколько более слабая перетяжка; на заднем конце носовых костей имеется
значительно более глубокая вырезка, чем у S. crassicauda. Лобные
кости вполне сросшиеся, следы сагиттального шва остаются лишь на
очень небольшом протяжении близ переднего их края; в передней их
части имеется глубокое продольное вдавление, постепенно переходящее
вперед в желобообразную заднюю часть носовых костей. Задний край
лобных костей образует обращенную выпуклостью назад дугу, на вершине
которой имеется вырезка; вдоль верхнего края орбит лобные кости
покрыты сверху длинными направленными вперед отростками чешуйчатой
кости, достигающими своими передними концами слёзной кости. Меж-
теменная кость имеется, хотя она сильно редуцирована и сохраняется
лишь в виде узкой продольной пластинки зажатой между близко сходящи-
мися мастоидальными камерами. Строение скуловой дуги еще более свое-
образно, чем у 5. thomasi и S. crassicauda-. ее передняя половина необы-
чайно широкая, пластинчатая, вся нижняя часть ее в этой половине
образована чрезвычайно сильно увеличенным продолжением processus
zygomatici ossis maxillaris, который продолжаясь далее образует длинный
саблевидный отросток, отходящий от скуловой дуги по направлению назад
и вниз; начиная от места отхождения этого отростка скуловая дуга резко
суживается, будучи образована здесь одной скуловой костью (jugale).
Чешуйчатая кость еще более изменена, чем у 6”. crassicauda, от нее отхо-
дит по направлению вперед длинный отросток с почти параллельными
краями, идущий по верхнему краю глазницы и достигающий слёзной
кости; этот отросток отчасти покрывает наружный край теменной кости
и налегает на лобную кость разделяя ее две кажущихся обособленными
части — подглазничную и глазничную; кроме того, чешуйчатая кость дает
тонкий отросток, налегающий на задний край теменной кости и доходящий до
половины ее ширины (фиг. 10 Pps); processus posttympanicus (Psm) развит
в виде необычайно тонкой и длинной слегка изогнутой костной палочки,
идущей над верхним краем tympanicum и доходящей немного далее заднего
края meatus auditorius; наконец, squamosum дает еще один отросток, напра-
вленный вниз и налегающий на задний край alisphenoideum (Pas). Нижняя
челюсть по отношению к размерам черепа маленькая, но весьма массив-
ная. Венечный отросток (Рс) выражен лишь в виде небольшого зубца;
сочленовный отросток (Par) широкий и слегка дугообразно изогнуты!;
угловой отросток несколько загнут внутрь. Между сочленовным и угло-
вым отростками имеется большой добавочный отросток (Ptr) в виде гори-
зонтальной пластинки, имеющей значительно большие размеры, чем
у kS. crassicauda.
По строению penis «S', kozlovi резко отличается от всех других изве-
стных представителей сем. Dipodidae.1 Penis длинный и относительно
1 Строение penis у других видов Salpingotus, а также у Card'.ocranius paradoxus
остается пока неизвестным.
5. SALPINGOTUS
101
тонкий, постепенно суживающийся к концу, на котором находится отвер-
стие семенной щели, окруженное складками слизистой оболочки. На вен-
тральной стороне penis в передней его половине проходит неглубокая
продольная бороздка. Вся поверхность penis гладкая, лишь в дистальной
его трети едва заметны очень мелкие роговидные зернышки.
Измерения: голова и тело—51; хвост—126; ступня — 25.2; ухо (от
основания) —11.8; наружное отверстие слуховой раковины — 7.3; кисточка
хвоста—18. Наибольшая длина черепа — 27.4; кондилобазальная длина —
22.0; скуловая ширина (сзади)—13.5. Межглазничное пространство — 7.9;
nasalia 10.8; диастема—4.8; нижняя челюсть —12.1; максилярный ряд
зубов — 3.8; мандибулярный ряд зубов — 3.2.
Распространение. Оба известных экземпляра .этого редкого
вида добыты экспедициями П. К. Козлова близ развалин города Хара-
хото в Центральной Гоби. Других данных о распространении S. kozlovi
не имеется. Упомянутые экземпляры были добыты в песчаной пустыне.
О стационарной принадлежности этого вида пескам свидетельствует
такое устройство задней ступни этого зверька, снабженной мощно разви-
той щеткой на пальцах задней конечности.
Как можно видеть из предыдущего описания, а также описания рода,
S. kozlovi является одним из наиболее высоко специализированных туш-
канчиков, что особенно резко выражается в удивительных особенностях
строения его черепа и скелета. В то же время У. kozlovi, как и другие
представители этого замечательного рода, сохраняют ряд примитивных
черт организации, напоминающих таковые у представителей подсем. Zapo-
dinae, как это уже отмечалось выше.
4. Подсем. ALLACTAGINAE — ЗЕМЛЯНЫЕ ЗАЙЦЫ
Голова широкая и толстая, резко суживающаяся вперед и заканчиваю-
щаяся коротким притупленным спереди свинообразным рылом. Вибрисы
длинные. Глаза очень крупные с косым разрезом век (передняя часть про-
дольной оси разреза сильно поднята вверх, а задняя опущена). Уши длинные,
будучи отогнуты вперед у немногих форм не достающие носа, у большин-
ства же форм заходящие за него. Передние конечности с рудиментарным
первым пальцем, снабженным ногтем с округлым свободным краем. Задние
конечности обычно с 5-ю пальцами и только у одного представителя (Scartu-
rus) имеется лишь 4 пальца, вследствие редукции первого (внутреннего);
боковые (1-й и 5-й) пальцы короткие и не достигают своими концами
оснований трех средних пальцев (фиг. 32 А). На конце каждого из трех
средних пальцев имеется сильно выдающаяся, сжатая с боков, эластическая
подушечка, разделенная глубокими бороздками на вееробразно располо-
женные дольки; подушечки на боковых пальцах имеют более простое
устройство. Вся подошва и нижняя поверхность пальцев голая. У основа-
ния трех средних пальцев имеется сильно выдающийся конусовидный
102
ALLACTAGINAE
бугорок. Когти на всех пальцах короткие и тупые. Хвост разнообразного
строения: он длинный и тонкий или заметно укороченный и утолщенный
с отложениями жира в подкожной клетчатке, принимая иногда сильно раз-
дутую колбасообразную форму {Pygerethmus)’, „знамя" может совершенно
отсутствовать {Pygerethmus platyurus) или оно едва намечено {Pygereth-
mus zhitkovi), или развито достаточно ясно {Alactagulus), или, наконец,
оно может иметь вид резко обособленной плоской ланцетовидной кисточки
из длинных, растущих на боках концевой части хвоста волос {Allactaga).
В строении penis для рассматриваемого подсемейства характерны следую-
щие признаки: os penis совершенно отсутствует. Вершина penis простая,
Фиг. 32. Задняя ступня пятипалых и трехпалых тушканчиков: А — Allactaga
elate' Licht., Б — Scirtopoda telum Licht., В — Dipus sagitta Pall.
без образования боковых складок („карманов"): на нижней поверхности
penis не имеется глубокой продольной бороздки. При рассматривании
сверху, penis имеет овальную, сердцевидную или ланцетообразную форму;
его поверхность покрыта мелкими шипиками (длинные отдельные шипы
всегда отсутствуют). Череп резко выраженного тушканчикообразного
типа, т. е. укороченный и с сильно расширенной сзади мозговой коробкой,
имеющей трапециодальную форму; на боках задней части мозговой коробки
имеются резко выраженные выпуклости. По направлению вперед боковые
края мозговой коробки быстро сближаются, образуя почти прямые линии
и достигая наибольшего сближения непосредственно позади от слёзных
костей; таКим образом, межглазничное сужение очень резко выражено,
так как непосредственно впереди от этого сужения слёзные кости сильно
выдаются в стороны. Верхние корни скуловых отростков максиллы почти
совпадают с перпендикулярной линией, проведенной к сагиттальной линии
черепа. Скуловые дуги почти прямолинейны; они наиболее широко рас-
ставлены в их средней части, откуда они идут почти прямолинейно вперед,
ОПИСАНИЕ ПОДСЕМЕЙСТВА
103
быстро сближаются друг с другом до соединения с верхними и нижними
•скуловыми отростками максиллы. Скуловой отросток чешуйчатой кости
имеет почти горизонтальное направление или лишь едва наклонен вниз;
при рассматривании сверху, он направлен почти перпендикулярно к сагит-
тальной оси черепа; он образует с общим направлением дуги резко обозна-
ченный, почти прямой угол. Нижний корень скулового отростка максиллы
лежит далеко впереди от верхнего и поэтому, при рассматривании черепа
сверху, далеко выступает вперед от последнего. Передняя часть скуловой
дуги (мостик между foramen infraorbitale и глазницей) имеет вертикальное
^направление. Скуловая кость резко разделена на горизонтальную и восхо-
дящую ветвь, образующие друг с другом почти прямой угол; передний
конец jugale тесно соприкасается с lacrymale. Слёзные кости очень круп-
ные, сильно вытянутые в поперечном направлении, их поперечный диа-
метр лишь в 2—2.5 раза меньше межглазничного сужения. Rostrum корот-
кий, далеко не достигающий по длине расстояния от его основания до
заднего края нёба (не далее заднего края М2). Передние концы nasalia
.далеко не достигают перпендикулярной линии, проведенной через gnathion.
Венечный шов образует почти прямую поперечную линию. Межтеменная
кость довольно крупная, постепенно суживающаяся к бокам, заостренная
или закругленная на своих наружных концах. Внутренняя стенка глаз-
ницы с большими неокостевшими отверстиями, затянутыми перепонкой.
Мезотеригоидное пространство приблизительно одинаково в длину и ши-
рину, трапецеидальной формы или с почти параллельными боковыми краями.
Птеригоидные отростки короткие, заметно короче длины зубного ряда;
bamuli оканчиваются против передних внутренних углов bullae, птеригоид-
яые ямки глубокие, сильно расширенные кзади, отверстия в их дне нет.
Bullae tympani обычно слабо раздутые, при рассматривании снизу почти
треугольные; их передние внутренние углы не сближены друг с другом
и почти не налегают на basis phenoideum (за исключением немногих форм).
Mastoidea не раздутые и не образуют bullae mastoideae; в верхней их
части имеется отверстие, затянутое перепонкой. Ossa mastoidea не высту-
пают назад за задний край condyli и не выходят на верхнюю поверхность
черепа. Нижняя челюсть в задней части довольно низка, приблизительно
.лишь в 1.5 раза выше, чем на уровне М1. Венечный отросток значительно
редуцирован и его вершина достигает лишь уровня сочленовной головки
челюсти. Корень нижнего резца образует на наружной стороне челюсти
•сильно выдающийся альвеолярный отросток. Передний и задний углы
челюсти (angulus anterior и a. posterior) не обособлены друг от друга и не
•отвернуты наружу или внутрь, в связи с чем нижний край челюсти (при
рассматривании снизу) является почти прямолинейным. Слуховые косточки
характеризуются относительно малым incus (хотя не столь малым как
у Zapodinae) и укороченным processus brevis mallei, который значительно
короче, чем processus longus. Corpus mallei и incudis не столь удлинены
как у других Dipodidae (за исключением Zapodinae) и шире и толще, чем
104
ALLA СТА GINAE
у других групп. Все семь шейных позвонков свободны, но в отличие от
Zapodinae и Euchoreutinae, поперечные отростки значительно более-
длинны и превышают по длине половину ширины тела соответствующих,
позвонков; вершины поперечных отростков 5-го, б-го и 7-го позвонков
еще более сближены, чем у Euchoreutinae', остистый отросток на втором:
шейном позвонке (epistropheus) значительно более развит, чем у Zapodi-
nae и Echoreutinae. Задняя ступня с 5-ю пальцами (кроме Scarturus)’
и с вполне развитой цевкой, образованной полным срастанием трех сред-
них metatarsalia. Боковые пальцы (1-й и 5-й) укорочены; они имеют почти:
одинаковую длину и далеко не достают своими концами дистального-
конца цевки. (У Scarturus 1-й, т. е. внутренний палец редуцирован и от
него остается entocuneiforme, с сидящей на его конце очень маленькой,
косточкой, являющейся рудиментом metatarsale 1-го пальца). В строении,
таза для представителей Allactaginae характерно сочетание следующих,
признаков: crista iliaca хорошо развита. Tuber ischii хорошо развит"
и вместе с прилежащей частью седалищной кости вытянут в направлен-
ный назад клювовидный отросток. Symphisis pubis очень длинен: его длина,
составляет не менее двух третей расстояния от acetabulum до седалищ-
ного бугра. Вентральный край ischio-pubicum, образующий сращение,,
переходит в задний свободный край этой кости более или менее посте-
пенно, не образуя резко выраженного закругления или излома. Tubercu-
lum ileo-pectineum присутствует и не раздвоен, как у Dipodinae. Нисходя-
щая ветвь ischium образует с ее горизонтальной (параллельной позвоноч-
нику) ветвью острый угол. От описанного типа строения таза предста-
вляют значительные уклонения представители рода Pygerethmus, как это
будет указано ниже. Головка бедра цилиндрическая с ясно выраженной,
седловиной на верхней поверхности. Trochanter major слабо отклонен от-
оси бедра наружу; расстояние от его верхнего (дистального) конца до
основания головки меньше длины головки. Наружный угол дистального*
конца trochanter major округлый или слабо угловатый. Ложно-коренной,
зуб имеется или редуцирован. Коронка коренных зубов плоская или почти:
плоская с тремя выдающимися углами на наружной стороне (некоторые
из этих углов могут быть двулопастными, как это будет указано при опи-
сании родов). Можно предположить, что первый из этих углов соответ-
ствует передней добавочной петле, которая сильно увеличена в размерах
и значительно смещена наружу, что подтверждается при сравнительном
изучении этой петли у Sicista, Zapus {Eozapus) и Cardiocranius. Если
это предположение правильно, то можно считать, что второй и тре-
тий выдающиеся углы соответствуют первому и второму главным,
бугоркам (внутренние главные бугорки образуют два внутренних выдаю-
щихся, угла).
Географическое распространение подсем. Allactaginae чрезвычайно
обширно: оно охватывает большую часть степной и пустынной зоны Европы
и Азии от бассейна Днепра на западе до Манчжурии на востоке. К северу
ОПИСАНИЕ ПОДСЕМЕЙСТВА 105
представители этого подсемейства распространяются до Десны, Оки, верх-
ней Волги в пределах Татарской АССР, Камы, Белой, северного Казах-
стана, Барабинской степи, Танну-Тувинской республики, южного Забай-
калья и северной Манчжурии. К югу представители этой группы распро-
страняются до побережья Черного и Азовского морей, Закавказья, Малой
Азии, Сирии, Палестины, Северной Аравии, Северо-западной Африки
(Ливийская пустыня), Южного Ирана, Белуджистана, Южного Афгани-
стана, Северного Таджикистана, Синцзяня, южной Гоби и провинции Чили
в Китае.
Представители рассматриваемого подсемейства известны с плиоцена
(роды Paralactaga и Allactaga). Подсем. Allactaginae представляет наиболее
цельную и однородную среди всех других Dipodidae группу и содержит
представителей с весьма сходной во всех существенных чертах организа-
цией. Подсем. Allactaginae и еще в большей степени Dipodinae содержат
специализированных представителей и наиболее далеко уклонившихся
по своей организации от первоначального плана строения сем. Dipodidae,
в наибольшей степени сохранившегося у мышевок и прыгунчиков
(подсем. Zapodinae), как это было уже указано. Следует, однако, отме-
тить, что в пределах подсемейства Allactaginae все же сохранилось
несколько больше примитивных черт строения, чем у Dipodinae (на-
личие 1-го и 5-го пальцев задней конечности и соответствующих
плюсневых костей, отсутствие срастания элементов в шейной части по-
звоночника, большая степень сохранности бугорчатого строения коренных
зубов и пр.).
Из характерных для подсем. Allactaginae биологических особенностей
прежде всего следует отметить, что его представители являются по преи-
муществу обитателями твердых субстратов (степи, глинистые и щебнистые
пустыни), в противоположность Dipodinae, обитающим по преимуществу на
песчаных почвах. Для Allactaginae характерна также относительно широ-
кая климатическая приспособленность: представители этой группы обитают
от условий климата лесостепи (Allactaga jaculus) до климата жарких пустынь
(A. elater и др.); некоторые виды приспособились даже к обитанию на зна-
чительных высотах (до 2000 м), как A. williamsi и A. saltator; некоторые
из представителей этой группы переносят значительные понижения темпе-
ратуры (Формозов, 1929, наблюдал в Монголии следы A. saltator даже на
снегу).
Нередко представители этой группы селятся на совершенно бес-
плодных участках, как например на голых лишенных растительности
участках — „такырах". Норы у большинства представителей устроены
по одному плану, характеризуясь наличием одного или нескольких запасных
выходов, не доведенных до поверхности земли или закрываемых земляной
пробочкой („копеечка").
Рассматриваемая группа представлена 4-мя довольно близкими
между собой родами, содержащими не менее 12 видов.
Г,б
ALL ACT AGIN АЕ
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ
1 (6). Задняя конечность с 5-ю пальцами.
2 (3). Ложно-коренной зуб в верхней челюсти имеется. На наружной
стороне жевательной поверхности первого коренного зуба (на несно-
шенных зубах) 4 или 5 выдающихся эмалевых петель. На краю
чешуйчатой кости, граничащем с теменной костью, имеется большая
вырезка соответствующая выступу теменной кости. Шипики на верх-
ней поверхности penis направлены назад (не отогнуты внутрь) . .
............................................6. Allactaga Cuv.
3 (2). Ложно-коренной зуб в верхней челюсти у взрослых особей отсут-
ствует. На наружной стороне жевательной поверхности первого
коренного зуба имеется лишь 3 выдающихся эмалевых петли (угла).
На краю чешуйчатой кости, граничащем с теменной костью, вырезки
нет. Шипики на верхней поверхности penis близ его основания
отогнуты внутрь.
4 (5). Хвост тонкий и длинный, без заметных отложений жира в подкожной
клетчатке; на конце хвоста имеется плоская кисточка из расчесанных
на две стороны волос („знамя'*)- Лонное сращение длинное (как
у Allactaga). В середине верхней поверхности penis имеется два
слабых вдавления, не ограниченных от остальной поверхности.
Шипики близ-основания penis слабо отогнуты внутрь.................
..........................................7. Alactagulus Nehr.
5 (4). Хвост обычно сильно утолщенный и относительно короткий, обычно
со значительными отложениями жира в подкожной клетчатке.
На конце хвоста вместо „знамени** могут быть лишь удлиненные
концевые волосы, не образующие плоской расчесанной на две стороны
кисточки. В середине верхней поверхности penis имеется резко
ограниченное, узкое, глубокое продольное вдавление. Шипики близ
основания penis сильно отогнуты внутрь . . 6. Pygerethmus Glog.
6 (1). Задняя конечность с 4-мя пальцами (первый палец отсутствует) —
..........................................9. Scarturus Glog.
Род ALLACTAGA F. CUVIER (1836)— ЗЕМЛЯНЫЕ ЗАЙЦЫ
F. Cuvier, Proc. Zool. Soc. bond., 1836 :141.
Уши длинные, будучи отогнуты вперед, обычно достигают конца
носа или заходят за него. Хвост длинный и тонкий, значительно превосхо-
дящий длину тела, без отложений жира в подкожной клетчатке: на конце
хвоста имеется резко обособленное „знамя** из растущих на боках хвосто-
вого стержня длинных волос. Penis сердцевидной, овальной или яйцевид-
ной формы. Верхняя поверхность penis равномерно покрыта мелкими
шипиками, которые могут отсутствовать лишь у основания и вершины
penis (голой площадки в средней части penis не имеется), все шипики
6. ALLACTAGA
107
•обращены остриями назад (не загнуты внутрь, как у Allactagulusw. Pyge-
.rethmus). Межглазничное пространство черепа приблизительно в 3 раза
меньше наибольшей длины черепа. Верхняя поверхность мозговой коробки
более или менее округло переходит в внутреннюю стенку глазницы,
т. е. лобная и глазничные пластинки frontale не образуют резкого пере-
лома при переходе одна в другую; боковые линии мозговой коробки
от межглазничного сужения до середины наружного края parietale (при
рассматривании черепа сверху) заметно дугообразные (не прямолинейны).
Костный мостик над foramen infraorbitale (вертикальная передняя часть
•скуловой дуги) имеет почти ту же ширину, как горизонтальная часть
•скуловой дуги. Париетальный край squamosum образует глубокую выемку,
в которую входит соответствующий лопастевидный выступ parietale.
Foramina incisiva доходят назад не далее линии, проведенной через перед-
ние края 1-го коренного зуба. Первый палец на задней конечности хорошо
развит и он немного превышает по длине пятый палец; основная фаланга
второго пальца приблизительно в три раза длиннее средней фаланги
того же пальца. Ложнокоренной зуб имеется (сохраняется во взрослом
состоянии); по сравнению со всеми другими представителями подсемейства
Allactaginae коренные зубы Allactaga удерживают наиболее примитивные
признаки, имея почти все компоненты, встречающиеся у менее специализи-
рованных групп; таким образом, у 'Allactaga имеется сзади от второго
наружного выдающегося угла (соответствующего первому наружному глав-
ному бугорку)маленький выступ, который можно считать за рудимент первой
наружной добавочной петли и сзади от третьего угла иногда имеется другой
маленький выступ, соответствующий второй добавочной наружной петле.
Представители рода Allactaga сохраняют некоторые признаки более
примитивной организации, чем это имеет место у Alactagulus и Pygerethmus,
особенно по наличию ложнокоренного зуба и большей степени сохранности
бугорчатой структуры коренных зубов. Впрочем, по некоторым наружным
признакам род Allactaga является более специализованным, чем названные
другие роды, что особенно относится к резкому обособлению на конце
хвоста „знамени" и удлинению ушей.
Ископаемые остатки представителей рода Allactaga известны с плио-
цена. Современное распространение рода охватывает обширный ареал,
совпадающий с таковыми для всего подсемейства (кроме Египта и Ливии),
как это было указано выше (фиг. 33, 34).
На протяжении этого ареала, представители рассматриваемого рода
обитают в довольно разнообразной обстановке от лесостепи на севере
до глинистых и щебнистых пустынь на юге; кроме того, некоторые виды
этого рода имеют значительное вертикальное распространение, проникая
в горные местности до высоты свыше 2000 метров (Л. williamsi и
A. saltator).
В четвертичное время, в течение ксеротермического периода, один
из представителей рода (A. jaculus) был распространен далеко на запад
фиг, 33. Распространение видов рода Allactaga-. 1, A. jaculus Pall., 2. Д. saltator Eversm., 3. A. severtzovi Vinogr,
Фиг. 34. Распространение видов рода Allactaga: 1. A. williamsi Thos., 2. A. euphratica Thos., ч. A. elater Licht., 4. A. bullata Gl. A len.
Ю8 ALLACTAGINAE 6. ALLACTAGA
110
ALLACTAGINAE
в пределы Германии и Чехии; границы распространения некоторых других:
видов (например: A. elater) были также значительно сдвинуты к западу.
Род Allactaga содержит 8 видов, из которых 5 представлены в фауне:
Союза.
Виды рода Allactaga можно разделить на следующие 3 группы, беря
в основу этого разделения особенности строения их genitalia: 1) группа
jaculus-elater с расходящимися бороздками на верхней поверхности
penis; 2) группа saltator— без средней бороздки на верхней поверхности
penis; 3) группа williamsi-euphratica с более или менее ясной средней
бороздкой, проходящей по всей длине дорзальной поверхности penis;
к последней группе по строению penis очень близко стоит также Scartarus
tetradactylus.
Подобное разделение в значительной степени согласуется также
с географическим распространением указанных групп: 1) jaculus, severtzovi,.
elater — Туранская низменность, Казахстан, юго-восток и юг европейской
части СССР; 2) saltator—-Монголия,северная полоса Туранской низменно-
сти; 3) williamsi, euphratica—Закавказье, Малая Азия, Месопотамия, Сирия.
Составляя во многих степных и пустынных районах Союза суще-
ственную часть соответствующих биоценозов, представители этого рода,
оказывают значительное влияние на растительность и почвы и являются
в некоторых местах заметными вредителями сельско-хозяйственных
культур (зерновые культуры, огороды, бахчи, технические культуры);,
вместе с тем некоторые крупные виды (A. jaculus и др.) в последнее
время имеют значение как объекты пушных заготовок.
Следует также учитывать, что при значительной численности некото-
рых видов Allactaga местами они составляют существенную часть кормовой
базы некоторых хищных млекопитающих и птиц.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (14). Ложнокоренной зуб в 2—3 раза меньше, чем задний коренной..
Передние части скуловых дуг (при рассматривании черепа сверху)-
отходят почти перпендикулярно к продольной оси черепа. Penis
с продольной бороздкой на дорзальной поверхности.
2 (11). Bullae tympani слабо раздуты и не сближены своими передними
концами.
3 (6). Длина задней ступни взрослых экземпляров не менее 70 мм. Конди-
лобазальная длина черепа взрослых экземпляров не менее 35 мм..
Срединная бороздка на дорзальной поверхности penis доходит при-
близительно лишь до половины длины penis и здесь разделяется на
две расходящихся под косым углом бороздки.
4 (5). Черная часть знамени на нижней поверхности обычно не прервана
в середине (вдоль стержня хвоста) белой полосой. На дорзальной
поверхности penis имеется около 60 шипиков . * 1. A. jaculus Pall.
6. ALLACTAGA
111
5 (4). Черная часть знамени на нижней поверхности прервана светлой
полосой вдоль стержня хвоста (часто на нижней поверхности светлого'
стержня хвоста по срединной линии в свою очередь проходит узкая
полоска из темных волос). На дорзальной поверхности penis имеется
около 30 шипиков.......................... * 2. A. severtzovi Vinogr..
6 (3). Длина задней ступни взрослых экземпляров менее 70 мм, кондилаба-
зальная длина черепа взрослых экземпляров менее 33 мм. Срединная
бороздка на дорзальной поверхности penis доходит по крайней мере
до дистальной четверти penis или достигает его вершины.
7 (8). Размеры мелкие: кондилобазальная длина черепа взрослых экзем-
пляров не свыше 27.5 мм, диастема не более 8.7 мм; максиллярный
ряд зубов не более 5.5 мм. Срединная бороздка на дорзальной,
поверхности penis немного не доходит до его вершины и разделяется
на две коротких расходящихся бороздки . . . . * 3. A. elater Licht..
8 (7). Размеры средние: кондилобазальная длина черепа свыше 27.5 мм;,
диастема обыкновенно более 8.7 мм; максиллярный ряд зубов больше
5.5 мм. Срединная бороздка на дорзальной поверхности penis дости-
гает его вершины и не разделяется на две бороздки.
9 (10). Задняя ступня не менее 62 мм (62—68); кондилобазальная длина,
не менее 30 мм (30—32).........................* 4. A. wllliamsi Thos.
10 (9). Задняя ступня менее 62 мм (52—58); кондилобазальная длина
не более 30 мм (28—30) . ...................... 5. A. euphratica Thos-
11 (2). Bullae tympani сильно раздуты и сближены своими передними
концами почти до взаимного соприкосновения.
12 (13). Длина уха не свыше 36 мм (обыкновенно короче). Межглазничное
пространство не менее 10 мм. Верхний ряд зубов свыше 5 мм (5.8—
6.7). Foramina incisiva относительно узкие . . 6. A. bullata Gl. Allen-
13 (12). Длина уха более 36 мм (у единственного известного экземпляра
41 мм). Межглазничное пространство около 9 мм; верхний ряд зубов,
около 5 мм. Foramina incisiva сильно расширенные ..................
........................................... 7. A. hotsoni Thos..
14 (1). Ложнокоренной зуб почти нравен заднему коренному или лишь
немного более последнего. Передние части скуловых дуг (при рас-
сматривании черепа сверху) отходят косо к продольной оси черепа.
Penis без продольной бороздки на дорзальной поверхности ....
........................................ * 8. A. saltator Eversm.,
* 1. Allactaga jaculus Pall.—Большой тушканчик (фиг. 35, 36).
Pallas, 1778: 275 (Mus); 1781 : 181 (Dipus); Сатунин, 1920:172; Огнев, 1924; Фенюк,.
1928, 1929 (биология).
Наиболее крупный представитель подсемейства земляных зайцев
(Allactaginae); длина тела 190—250 мм, длина задней ступни 85—93 мм;
кондилобазальная длина черепа 40—47 мм. Голова более короткая и широ-
кая, че.м у большинства других видов рода. Впереди от черной частж
112
ALLA СТА GINAE
знамени белого кольца нет; черная часть знамени на нижней поверхности
не прервана вдоль стержйя хвоста белой полосой. Передний (ложнокорен-
ной) зуб в 2—3 раза меньше, чем последний коренной. Передние части
скуловых дуг (при рассматривании черепа сверху) отходят почти перпен-
дикулярно к продольной оси черепа. Продольная бороздка на верхней
поверхности penis достигает лишь его середины и разветвляется на две
расходящихся под углом бороздки; на верхней поверхности penis обыкно-
венно около 60 шипов. Общий тон окраски спины колеблется у различных
географических форм от буровато-серой до бледной песчано-серой; брюш-
ная сторона и внутренняя сторона ног белые; в задней части тела на бедрах
Фиг. 35. Большой тушканчик (Allactaga jaculus Pall.). Фото М. К. Серебренникова.
имеется с нижней стороны резко очерченная широкая белая полоса, идущая
к основанию хвоста. Бедра с наружной стороны ржаво-желтые. Края
подошвы задней ступни покрыты черно-бурыми волосами. Хвост до основ-
ной части знамени светлый ржаво-бурый; основная часть знамени черная,
концевая белая.
Распространение. Лесостепи, степи и полупустыни южной
и средней полосы Европейской части СССР, Западной Сибири и Казах-
стана. В европейской части СССР распространен к западу до Криворож-
ского района включительно, к северу до реки Десны, низовья Сейма
до реки Оки, частично заходя местами к северу по этой реке в пределах
Московской области (Раевский, 1929), реки Камы и Белой; далее на восток
северная граница выяснена слабо: в частности этот вид найден у Зла-
тоуста, в южной части Шадринского округа, близ г. Ишима, в Барабин-
ской степи и в Ординском районе Ново-Сибирского округа; к югу распро-
странение этого вида простирается до нижнего Днепра, степей Крыма,
побережья Азовского моря, предгорий Главного Кавказского хребта,
северного берега Каспийского моря, Северного Усть-урта (включительно),
6. ALLACTAGA
113
^Семиречья, юзера Зайсана и Приалтайских степей. На восток этот вид
«распространен не далее долины Оби в пределах Новосибирского и Бар-
наульского округов.
Географическая изменчивость: хотя изменчивость в пре-
делах рассматриваемого вида чрезвычайно невелика, ряд прежних и сов-
ременных авторов признавали и признают существование нескольких
«подвидов A. jaculus в пределах занятого этим видом ареала; в частности
'Фиг. 36. Большой тушканчик (Allactaga jaculus Pall.). Фото М. К. Серебренникова.
С. И. Огнев (1924), изучавший этот вопрос, признает существование
следующих подвидов:
1) A. j. jaculus Pallas (1778:285); описан из Крымских степей;
по мнению С. И. Огнева, характеризуется относительно тусклой серо-
желтоватой окраской с серо-желтовато-пепельным тоном верхней части
головы и некоторыми особенностями строения черепа (относительно
широкая расстановка скуловых дуг, длинное и широкое предглазничное
кольцо, более широкое .межглазничное пространство, чем у предкавказ-
ского подвида), а также относительно крупными резцами. По мнению
названного автора эта форма распространена в Крыму, на Украине,
Воронежской области, на Дону, в северной части .Куйбышевской обла-
сти и пр.
2) A. j. fuscus Ognev (1924:8); описан из окрестностей^ Тушиловки
Кизлярского округа Северо-Кавказскбго края. По мнению описавшего
.эту форму автора, легко отличается от типичных A. jaculus (из Крымских
Фауна СССР, Виноградов 8
114
ALLACTAGINAE
степей) более насыщенной красновато-рыжеватой окраской спины и верхом
головы с ясным ржаво-серо-желтоватым оттенком. Череп этого подвида
имеет относительно не очень широкую расстановку скул,, более узкое
в поперечнике и менее длинное в отвесном направлении переднеглазнич-
ное кольцо, несколько более суженный глазничный промежуток. Резцы
меньше и слабее, чем у типичной формы.
3) A. j. decumanus Lichtenstein (1825:154); описан из окрестностей
Златоуста. Этот подвид был восстановлен В. Э. Мартино (1912:107), кото-
Фиг. 37. Головы тушканчиков. 1 — Allactaga jaculus Pall., 2 — A. saltator Eversm., 3 —
Dipus sagitta Pall.
рый указывает, что для него характерна серая окраска Лба и обилие
черных волос на спине, которые, группируясь, образуют у некоторых
экземпляров неясные черные полоски. С. И. Огнев предполагает, что
эта форма обитает в степных райнах Башкирии и в южной части
б. Пермской губернии.
4) A. j. vexillarius Eversmann (1840:42); описан с Усть-Урта. По мне-
нию С. И. Огнева, эта форма характеризуется бледной песчано-желтова-
той окраской и светлым пепельно-желтоватым верхом головы и лбом..
Названный автор причисляет к этой форме также экземпляр с, реки
Урала.
5) A. j. spiculum Lichtenstein (1825:143); описан из Западной Сибири.
Относительно тусклая и серая окраска без песчаных и желтоватых оттен-
6. ALLACTAGA
115
ков; лоб и верх головы не разнятся заметно от окраски спины. Уши отно-
сительно более короткие, чем у других форм. Кроме перечисленных форм,
указываемых С. И. Огневым, позже была описана В. Э. Мартино еще
одна следующая форма:
б) A. j. chachlavi Martino (1924) (описан с оз. Зайсана). Мартино
считает, что зайсанские тушканчики отличаются от тушканчиков Западного
Казахстана более длинными ушами, и более серым цветом спины.
Проверка приведенных выше признаков на основании просмотра
значительной серии шкурок большого тушканчика в коллекциях Зоологи-
ческого Института, заставляет сомневаться в реальности существования
приведенной выше схемы. Амплитуда цветовой изменчивости у этого вида
не велика; окраска верха колеблется у различных особей от буровато-
серой до более бледной песчано-серой, причем эти колебания наблюдаются
даже у экземпляров из одной и той же местности. Однако, как и у боль-
шинства других млекопитающих, экземпляры из более южных и более
пустынных районов в общем характеризуются несколько более бледной
окраской, чем из северных частей ареала; так экземпляры из Северного-
Казахстана имеют несколько более темную окраску, чем экземпляры
с Усть-урта; сходное соотношение существует между экземплярами
из Западной Сибири и с Зайсана и т. д. Другие указываемые авторами
признаки, как окраска лба, также значительно колеблются у особей
из одной и той же местности, но здесь не наблюдается определенной
направленности изменчивости этого признака. Можно также сомневаться
в правильности указываемых различными авторами краниологических
отличий предполагаемых рас большого тушканчика; во всяком случае
я на основании просмотренного материала не мог найти подтверждения
реальности этих признаков. Нельзя отрицать, что при улучшении методики
исследования и накоплении более обильного и более совершенно сохра-
ненного материала, удастся расчленить рассматриваемый вид на ряд хорошо
характеризующихся рас, но в настоящее время вряд-ли целесообразно
принимать существование перечисленных выше форм.
Данные по биологии. Большой тушканчик в пределах своего-
распространения, встречается в разнообразных стациях от луговой степи
на севере до окраинных частей глинистой пустыни на юге; в более север-
ных частях своего ареала, большой тушканчик иногда встречается также
в песках. Кроме того, этот вид встречается на полях, бахчах и других
возделанных местах.
Наиболее детальными исследованиями по биологии тушканчика,,
являются работы Фенюка (1928,1929), которые составляют главную основу
кратко излагаемых ниже данных. Норы земляного зайца имеют тот же тип
строения, который характеризует всю группу. Запасных выходов может
быть одцн или два, причем один из них часто не доводится на 2—5 см
до поверхности почвы и над его концом остается таким образом тонкий
слой земли, который легко проламывается тушканчиком и зверек выска-
8*
116
ALL А СТА GINAE
кивает через образовавшееся отверстие в случае опасности. Норы туш-
канчиков упомянутый автор делит на следующие категории: 1) постоянные
норы, характеризующиеся наличием камеры, присутствием забитого
землею хода и одного или двух дополнительных ходов, закрытых земляной
пробочкой (или один из этих ходов может быть не доведен до поверх-
ности земли, как это было описано выше); кроме главных камер, могут
быть несколько расширений на месте отхождения отдельных колен или
отнорков. 2) Временные дневные норы характеризуются более простым
устройством и меньшей глубиной, не превышающей 20—35 мм, отсутствием
камеры и забитой землей начальной частью хода. 3) Временные ночные
норы имеют еще^более простое устройство и обычно представляют про-
стой короткий прямой канал, отверстие которого остается открытым.
4) Зимние норы отличаются от других типов главным образом более
значительной глубиной (до двух метров и более). Выход большого
тушканчика на поверхность наблюдается относительно рано, через 30—
40 минут после захода солнца. Вышедшие на поверхность зверьки начи-
нают прежде всего утолять голод и после насыщения начинают бегать
и играть. Тушканчики уходят в норы, в зависимости от времени года,
за 20 минут — 1 ч. 30 мин. до восхода солнца. В Нижнем Поволжье
залегание большого тушканчика в спячку наблюдается при наступлении
первых ночных заморозков, пробуждение от спячки — во второй половине
марта или в начале апреля. Детеныши — в количестве 1—4 в каждом
помете появляются на свет в конце апреля и переходят к самостоятель-
ному образу жизни около конца июня; у отдельных особей нередко
наблюдается второй помет в конце июня или в июле. Большой туш-
канчик питается степными и некоторыми культурными растениями,
особенно их семенами, корнями, луковицами и клубнями; при поедании
семян он очищает их от оболочки, даже тогда, когда семена очень мелкие,
как, например, просяные. При выкапывании луковиц и других подземных
частей растений, тушканчики оставляют в этих местах характерные ямки
(„копанцы"), в которых, или около которых можно найти шелуху от луко-
виц и другие остатки съеденной пищи. Из культурных растений, тушкан-
чики поедают семена тыквы, арбуза, дыни, семена хлебных злаков (просо,
пшеница, рожь, ячмень, овес, кукуруза). Иногда эти зверьки частично
питаются также насекомыми.
В некоторых районах большой тушканчик должен считаться одним
из вредителей сельского хозяйства, особенно в Нижнем Поволжье
и в земледельческих районах Казахстана. Весной тушканчик вредит
на бахчах поеданием посеянных семян арбузов, дынь и тыкв; после созре-
вания хлебов он приносит вред поеданием зерен перечисленных выше
хлебных злаков, а также вредит отчасти подсолнуху, гороху и чечевице.
В последнее время наблюдениями И. И. Колесникова (1934) в Казах-
стане установлено, что большой тушканчик наряду с малым (A. elater)
является таг же существенным вредителем ценного каучуконосного расте-
6. ALLACTAGA
117
ния тау-сагыза (Scorzonera tau-saghyz). Большой тушканчик наносит куль-
турам тау-сагыза такие же повреждения, как и малый, т. е. выедает
посеянные семена; как только семена начинают прорастать, угроза повре-
ждений отпадает. На полях тау-сагыза в окрестностях кишлака имени
Атабаева Туркестанского района большим и малым тушканчиками из 11 га
рядового посева полностью было уничтожено 1.59 га, а из 10 га гнездового
— 5 га. При поздних осенних посевах тушканчики, повидимому, не трогают
семян тау-сагыза. При культивировании тау-сагыза на Украине, наблю-
дались также случаи повреждения этого растения тушканчиком (большим),
как об этом сообщает А. К. Шепе (1934). Здесь повреждения носили
иной характер и заключались в объедании зеленых частей растений
у корневой шейки.
За последнее время (с 1925 г.) большой тушканчик приобрел зна-
чение промыслового вида. Запасы этого тушканчика во многих районах
достаточно велики и имеются основания ожидать дальнейшего расшире-
ния этого промысла. Наибольшего развития этот промысел достиг
в Казахстане.
* 2. Allactaga severtzovi Vinogradov — Тушканчик Северцова
Vinogradov (1925 : 583) (Тамар-уткуль, б. Копальского у., Семиречье).
По строению тела и черепа этот вид сходен с A. jaculus, но отли-
чается от последнего заметно меньшей величиной, распределением белого
и черного цвета на нижней поверхности хвостового знамени и строением
мужского наружного полового органа. В частности одним из наиболее
наглядных наружных признаков отличия A. severtzovi от A. jaculus является
то, что черная часть „знамени" на нижней поверхности прервана светлой
полосой вдоль стержневой части хвоста, причем часто на этом светлом
стержне проходит в свою очередь узкая темная полоска. Окраска верхней
поверхности тела бледнее и серее, чем у A. jaculus, сходная с A. saltator
riickbeili Thos. Задняя ступня обыкновенно 70—80 мм (редко больше),,
кондилобазальная длина черепа обычно не более 41 мм. Верхняя поверх-
ность penis имеет около 30 шипов (у A. jaculus — около 60); основная
раздутая часть penis относительно меньше, чем у A. jaculus; длина penis,
около 6.3 мм.
Измерения: длина тела 140—170; хвост 212—227; задняя ступня
71—81; ухо 45—60; кондилобазальная длина черепа 37.3—40.7; скуловая
ширина 27—29; межглазничное пространство —11.0—11.5.
A. severtzovi имеет ряд сходных признаков с A. jaculus вследствие-
чего лишь при более внимательном исследовании признаков того или
другого вида удалось обосновать видовую самостоятельность тушканчика
Северцова. Интересно отметить, что необходимость выделения тушкан-
чика Северцова в особый вид возникла впервые при сравнительном изу-
чении строения половых органов у представителей семейства тушкан-
чиков, причем были найдены вполне определенные отличия в строении
118
ALL ACT A GINAE
указанных органов у обоих видов; эти отличия были подтверждены также
другими морфологическими признаками, указанными выше (Vinogradov,
1925).
Распространение. Распространение этого вида охватывает
пустыни Средней Азии и Казахстана, кроме песчаных пустынь, от южного
Усть-Урта, Приаральских пустынь, нижнего течения Сыр-Дарьи и побе-
режья Балхаша на юг до Копет-Дага,1 долины верхнего течения Аму-
Дарьи (по крайней мере до Термеза), предгорий Гиссарского хребта,
хребтов Ферганы и хребтов системы Тянь-Шаня в пределах юго-восточного
Казахстана. Таким образом, этот вид заменяет в Средней Азии большого
тушканчика (Л. jaculus), который там не встречается; ареалы обоих
названных видов, повидимому, частично заходят друг за друга лишь
на Усть-урте, в приаральских пустынях, в долине Сыр-Дарьи и Прибал-
хашья. Так как в настоящее время известно лишь небольшое число точно
проверенных пунктов нахождения этого вида, здесь приводятся все эти
местонахождения: Моюн-кумы (уроч. Базоба в дол. р. Чу), долина Сыр-
Дарьи (ст. Берказан близ г. Кзыл-орда), Фергана (Кара-тау, Мусульм-
куль) б. Кашка-дарьинский окр. (г. Бек-буди), б. Сурхана-дарьинский окр.
(окрестности Термеза, Джар-кургана и ст. Болдыр), Западная Туркмения
(Туаркырский район, южный Усть-урт), центр. Кзыл-кумы(ур. Тамды).
Данные по биологии. Тушканчик Северцова является обита-
телем различного типа глинистых пустынь. Об образе жизни этого зверька
имеется лишь несколько отрывочных наблюдений, в частности В. С. Вино-
градовым и А. И. Аргиропуло (1931) исследовано несколько нор этого
тушканчика в окрестностях Термеза, а также близ сТ. Болдыр (в 63 км
к западу от этого города); в последнем пункте норы этого тушканчика
встречаются на открытых голых солонцеватых площадках вместе с норами
Alactagulus, а также среди зарослей солянок (Salsola), верблюжьей колючки
(Alhagi) и других травянистых растений. Норы этого тушканчика хорошо
•отличаются от нор Alactagulus и Allactaga elater уже по внешнему виду:
кучка у норы A. severtzovi более - крупна и более вытянута в длину, чем
у других упомянутых видов и поперечное сечение норы имеет форму
•более вытянутого овала. Раскопанная названными авторами нора тушкан-
чика имела следующее устройство: начальная часть хода была забита
землей на расстоянии метра; далее нора на протяжении одного метра
круто углублялась и переходила в небольшую камеру, за которой
ход углублялся опять на протяжении метра и вновь образовывал камеру,
за которой следовал опять ход длиной около г/г метра, заканчивающийся
большой камерой, в которой находился зверек. Другая раскопанная нора
была построена по тому же плану, т. е. представляла собою ход, с чере-
дующимися друг за другом камерами, но в этом случае конечная часть
1 В Туркмении, кроме южного Усть-урта и Прикарабугазья, известно лишь одно
нахождение этого вида в окрестностях ст. Джебел; по этой причине распространение этого
вида до предгорий Копет-дага указывается здесь предположительно.
6. ALLACTAGA
119
хода поднималась постепенно вверх ^заканчивалась в 2—3-х сантиметрах
от поверхности, образуя запасный выход, незаметный снаружи и сходный
с этим образованием в норе Allactaga elater как это было описано выше.
В настоящее время не имеется данных об экономическом значении
тушканчика Северцова; мне удалось лишь получить, путем распросов,
•сведения о повреждениях этого тушканчика, наблюдавшихся на вновь
освоенных землях в совхозах Джизакского и других районов Узбекистана.
•*’ 3. Allactaga elater Lichtenstein — Малый тушканчик (фиг. 38).
'Lichtenstein 1825: 23, (Dipus) (описан по экземплярам из Западного Казахстана);
dray, 1842:262; Сатунин, 1920:185; Heptner, 1934:17 (подвиды); Vinogradov u. Argyropulo
1931:173 (биол.), Виноградов и Аргиропуло 1931:153 (биол.); Колесников 1934:15 (биол.,
вред, меры борьбы).
По наружным признакам в общих чертах сходен с A. jaculus, отличаясь
несколько более узкой и вытянутой головой и значительно меньшей вели-
чиной; по размерам приблизительно в 2 раза меньше, чем A. jaculus',
дл.' тела 95—115 мм, длина хвоста 150—180, дл. ступни 48—56, высота
уха 34—42, кондилобазальная дл. черепа 25—27, скуловая шир. 19.2—
21.7, максиллярный ряд зубов 4.8—5.5.1
По строению penis этот вид характеризуется тем, что продольная
бороздка на его дорзальной поверхности почти доходит до вершины
и здесь лишь едва разветвляется на две коротких боковых бороздки;
основная треть или четверть penis голая (без шипиков), вершина penis
заостренная.
Окраска у экземпляров из различных местностей колеблется от отно-
сительно темной серовато-ржаво-бурой до более светлой песчаного тона
с особенно значительным побледнением на боках.
Как уже было указано при характеристике подсемейства, A. elater
по своей организации стоит ближе к A. jaculus и A. severtzovi, чем к дру-
гим представителям подсемейства. Кроме сходства наружных признаков
и устройства черепа, названные три вида обнаруживают сходное в основ-
ных чертах устройство penis, хорошо отличаясь по этому признаку от
других видов подсемейства Allactaginae.
Распространение. A. elater является одним из наиболее харак-
терных представителей фауны пустынь (кроме песчаных) Средней и Запад-
ной Азии, проникая к северу в зону полупустыни. На северо-западе ареал
распространения A. elater начинается в полупустынях северо-восточного
Предкавказья в пределах б. Кизлярского округа и охватывает восточную
(приморскую) часть Калмыцкой АССР на север до Бешкуля и Калмыц-
кого Базара (к с.-з. от Астрахани), далее в пределах Волжско-Уральских
1 В приводимые здесь пределы колебаний наружных измерений черепа не включены
соответствующие величины описываемой ниже формы из Аулие-атинского района, так как
•систематическое положение этой формы еще недостаточно выяснено и остается еще недо-
казанной даже ее принадлежность к рассматриваемому виду A. elater (см. ниже).
120
ALLACTAGINAE
полупустынь северная граница этого вида, за отсутствием коллекционных
материалов, не выяснена, но известно, что в долине р.. Урала этот вид.
достигает с. Калмыкова, далее северная граница распространения этого-
вида идет через г. Иргиз и далее на восток до Зайсанской котловины не-
заходя на север выше 48—49° с. ш. (Беляев, 1933), от Зайсана иАлакуля
малый тушканчик распространен еще далее на восток в пределы Джун-
гарии, где он найден восточнее Гучена.
Южная граница известна менее определенно. В пределах Закавказья,,
малый тушканчик распространен в Куро-Аракской низменности, достигая
по Куре с. Варвара Ганжинского округа и по Араксу — поста Шах-Чинар-
(Сатунин 1920); кроме того имеется отдельное нахождение этого вида,
в Аралыхской равнине у подножья Арарата; далее на юг этот вид прони-
кает в Иран, где он найден у Тегерана, в округе Kain в пределах Хора-
сана; самыми крайними юго-восточными известными пунктами нахождения-
малого тушканчика является Кандагар в Афганистане и Сураб в Белуд-
жистане; отсюда граница идет к соверо-востоку, достигая верхнего тече-
ния Аму-Дарьи (Термез) и далее по равнинам вдоль предгорий хребтов.
Узбекистана и Казахстана (в пределах Семиречья), достигая алакульской
и зайсанской равнины и выклиниваясь в Джунгарии, как это. было ука-
зано выше.
Географическая изменчивость. До настоящего времени,
было описано 7 географических форм A. elater, из которых б считались-
описавшими их авторами за подвиды и 1 за самостоятельный вид (A. indica:.
Gray).
Здесь приводится краткий обзор этих форм с критическим пересмо-
тром их признаков, где это возможно.
1) А. е. kizljaricus Satunin 1907 (Кизлярский окр.) характеризуется
относительно темной окраской, значительно более темной, чем у закав-
казских форм; белого кольца перед черным цветом знамени нет, как
и у других кавказских форм. Известно лишь 2 экземпляра из Кизлярского-
округа (непосредственно исследовать эту форму мне не удалось);
2) Л. е. caucasicus Nehring 1900 (Куро-Аракская низменность восточ-
ного Закавказья). По мнению Неринга эта форма отличается от А. е. ela-
ter, описанной из Казахстана, тем, что у первой череп несколько ниже
и костный мостик над foramen infraorbitale шире; эти признаки, однако*
не подтверждаются на просмотренном мною материале и вероятно- един-
ственным реальным отличием А. е. caucasicus от северо-кавказской формы,
является несколько более светлая окраска. Эта окраска заметно не отли-
чается от таковой у типичной формы из Казахстана, как на это- указывает-
и Сатунин (1903).
3) А. е. aralychensis Satunin, 1901 (Аралых у подножья Арарата
и другие районы Армении). По мнению Сатунина, главным отличием этой
формы является „чрезвычайная яркость" окраски летней шерсти;, другими
отличительными признаки этой формы, по мнению названного автора*
6. ALL ACT A GA
121
является большая ширина черепа, чем у А. е. caucasicus (особенно меж-
глазничная ширина), более длинные уши и более короткая ступня. При
просмотре двух имеющихся в Зоологическом Институте экземпляров
Сатунина из Аралыха и серии экземпляров из других местностей Армении
перечисленные признаки не подтверждаются, в частности окраска этих
экземпляров в существенных чертах сходна с таковой у А. е. elater
нисколько не выделяется своей яркостью.
4) А. е. elater Lichtenstein, 1825 (Западный Казахстан) характери-
зуется относительно ярким охристо-желтым тоном основной окраски:
и относительно слабым развитием темного оттенка на спине, зависящего
Фиг. 38. Малый тушканчик (Allactaga elater Licht).
от степени развития остевых волос с темными окончаниями. Экземпляры,
имеющие подобный тип окраски, распространены от низовий р. Урала
до побережья Аральского моря или еще далее на восток.
5) А. е. subsp.? Гептнер (1934) предположительно выделяе'т экзем-
пляры из восточного Семиречья (Алакульская равнина) в особый подвид,,
характеризующийся более значительным развитием темного оттенка на
спине, чем у предыдущей формы. Это подтверждается также и на про-
смотренных мною экземплярах с Алакуля, из Прибалхашья, из Аулие-
атинского округа.
Следует отметить, однако, что экземпляры из Аулие-атинского
округа, отличаются кроме того, исключительно крупными для этого вида
размерами, в частности у промеренных мною экземпляров из окрестностей
с. Ровное и Бурное были получены следующие размеры черепа: кондило-
базальная длина черепа до 29.7 мм (у других форм до 27.5 мм); скуловая
ширина до 22.5 (у др. форм до 21.8), диастема до 9.6 (у др. форм до 9.1),
максиллярный ряд зубов до 6.4 (у др. форм до 5.8). Отсутствие доста-
точного количества материала из упомянутой местности и особенно.'
122
ALL А СТА GINAE
отсутствие данных о строении penis не дает возможности выяснить точное
систематическое положение этого тушканчика в настоящее время, несмотря
на то, что указанные признаки являются весьма значительными. Череп
аулие-атинской формы весьма сильно отличается по измерениям от всех
известных форм A. elater и приближается по своим размерам к черепу
A. euphratica Thos.
6) А. е. strandi Hepther 1934 (окрестности Мерва). Эта форма отли-
чается от более северной А. е. elater несколькоболее бледным песчаным
тоном основной окраски верха, светлая окраска особенно заметна на
боках тела, голове и бедрах. Подобного типа окраска характерна для
экземпляров из южной Туркмении и южного Узбекистана.
7) А. е. dzungariae Thomas (Джунгария, „100 миль от Гучена").
Относительно темная форма, сходная с семиреченской или несколько
темнее.
8) А. е. indica Gray 1842 (Афганистан, окрестности Канда-гара). Эта
форма многократно упоминалась в литературе в качестве самостоятельного
вида. Мною был исследован в Британском музее один из оригинальных
экземпляров Грея (лектотип) и ряд других экземпляров из Афганистана,
Белуджистана и Ирана, причем я не нашел достаточно существенных отли-
чий между A. indica и A. elater и поэтому считаю их принадлежащими
к одному виду. А. е. indica должен считаться одним из наиболее ясно
дифференцированных подвидов рассматриваемой группы. При окраске
шерсти, сходной с таковой у среднеазиатского А. е. strandi Hept, харак-
теризуется А. е. indica несколько более крупными размерами черепа, чем
упомянутая форма (кондилобазальная длина у А. е. indica до 27.5 мм)
и особенно более значительным межглазничным пространством, дости-
гающим 9.8 мм (8.3—9.8), причем у большинства экземпляров, эта вели-
чина составляет не менее 9 мм. Из всех известных форм A. elater, лишь
описанная выше форма из Аулие-атинского округа превышает по разме-
рам А. е. indica, кроме межглазничной ширины. Кроме Кандагара, экзем-
пляры А. е. indica в Британском музее имеются из других местностей
Афганистана, из Белуджистана (Surab) и из Ирана (округ Kain в Хорас-
сане и окрестности Тегерана).
Из перечисленных форм наиболее ясно дифференцированными
являются три: А. е. indica Gray, описанная выше крупная форма из Аулие-
атинского округа и совокупность перечисленных выше форм, которые
примыкают к А. е. elater и вероятно могут быть объединены с ней под
одним подвидовым названием (А. е. kizljaricus, А. е. caucasicus, А. е. araly-
chensis, А. е. strandi, А. е. dzungariae и А. е. elater). Последняя группа форм
разнится друг от друга в сущности лишь некоторыми особенностями
окраски, причем изменение этой окраски подчиняется обычной зональной
закономерности, т. е. в более северных или на предгорных частях ареала
окраска более темная и постепенно светлеет по направлению к южным
-или пустынным частям ареала; в частности северо-кавказская форма
6. ALLACTAGA
123
(Л. e. kizljaricus) темнее чем обе закавказских (А. в. caucasicus и А. е. ага-
lychensi), которые возможно являются идентичными; казахстанская форма
(Л. е. elater Licht.) несколько темнее, чем более южная средне-азиатская
(Л. е. strandi Hept.) и чем афгано-иранская форма (Л. е. indica), темная
форма из Аулие-атинского округа связана в своем распространении
с предгорными районами хребта Кара-тау и пр.
Из всех перечисленных форм наиболее сильно уклоняющиеся от обыч-
ного для Л. elater стандарта, является форма, распространенная в Аулие-
атинском округе; недостаточное количество материала и отсутствие данных
о строении genitalia пока не позволяют дать точный диагноз этой формы
и даже удостовериться в ее принадлежности к рассматриваемому виду.
Данные по биологии. Биологические наблюдения над малым
тушканчиком довольно отрывочны и немногочисленны. В частности Вино-
градов и Аргиропуло (1931) наблюдали этого тушканчика в ближайших
окрестностях г. Термеза, где он обитает на самых безотрадных участках
глинистой степи, покрытых скудной растительностью; местами этот зверек
селится даже в черте города на пустырях и некоторых площадях, заро-
сших верблюжьей колючкой (Alhagi camelorum); по наблюдениям названных
авторов, строение нор у A. elater заметно отличается от такового
I у A. acontion; особенно это заметно во внешнем устройстве. Начальная
часть хода в жилой норе A. elater всегда забита землей, но эта пробка
никогда не достигает значительной длины и обычно ход открывается уже
после 2—3 ударов лопатой; нужно отметить в связи с этим, что около
Термеза этот тушканчик устраивает свои норы в очень твердом грунте.
Кучка нарытой земли у норки бывает обычно очень мала и слабо заметна
на окружающем фоне, нередко она не превышает размеров кучки у норки
копра (Scarabaeus sacer L.). Форма нарытой кучки разнообразна, но чаще
неправильной округлой формы. A. elater часто вырывает свои норки
в небольших углублениях почвы, в колеях дорог и т. д.; в этих случаях
нарытая земля заполняет эти углубления и тогда нахождение норки ста-
новится еще более затруднительным; трудность нахождения норки Л. elater
увеличивается еще тем, что нередко эти норки располагаются под кусти-
ками растений или среди них. Другой отличительной особенностью норки
Л. elater от Alactagulus является отсутствие заметного снаружи окошечка.
Нам пришлось разрыть несколько десятков нор Л. elater и убедиться
в том, что боковой ход, ведущий к поверхности, имеется всегда, но он не
доводится до самой поверхности всего лишь на 1—2 см. При раскопке
норы, если заранее не были приняты соответствующие меры, тушканчик
-проламывал тонкую корку над концом бокового хода и убегал через
открывшееся отверстие. Глинистая почва у Термеза, в которой мы нахо-
дили норы Л. elater, оченб плотна и с трудом поддается раскопке, в связи
с чем все найденные нами норы были коротки и имели малую глубину;
общее протяжение главного хода от входного отверстия до конца камеры
не превышало одного метра, чаще же не достигало и этой длины.
124
ALL ACTA GINAE
Выпугнутый из норы раскопкой A. elater стремительно убегает на.
большое расстояние и затем пытается укрыться в первую попавшуюся
открытую норку, а также в норку песчанки или другого зверька. Как и дру-
гие пятипалые тушканчики A. elater имеет обыкновение бегать и играть,
по дорогам, и утром в дорожной пыли всегда можно найти в большом коли-
честве его следы, а также отпечатки тела и хвоста. Мы неоднократно слы-
шали рассказы о том, как при свете автомобильных фонарей удается
наблюдать на близком расстоянии скачки тушканчиков по дорогам. Нам
также удалось однажды видеть эту картину, при быстром ночном переезде
на автомобиле из Термеза в Ширабад; вскоре после наступления темноты,
когда были зажжены фонари, на дороге стали мелькать фигуры тушкан-
чиков, причем кроме A. elater здесь встречались также экземпляры
A. severtzovi, часто в одном месте попадалось сразу 2—3 экземпляра;
некоторые тушканчики быстро скакали, большинство же встречавшихся
зверьков, попадая в полосу света, останавливались на месте и не убегали
с дороги даже тогда, когда колеса автомобиля проезжали на расстоянии
не более метра от них; по словам шоффера, ему нередко приходилось
давить колесами этих зверьков во время ночных поездок. Получалось
впечатление, что в этом случае тушканчики не прибегали на свет, а будучи
неожиданно застигнуты на дороге снопом ослепительного света, были им
ошеломлены и находились в полной растерянности. Как нам сообщали,,
в этих случаях не составляет никакого труда поймать сачком даже такого
быстрого тушканчика, как A. severtzovi.
В последнее время более детальные исследования по биологии ма-
лого тушканчика, были произведены Колесниковым (1934) в связи с вредо-
носной деятельностью этого зверька на плантациях тау-сагыза (Scorzo-
nera tau-saghyz) в Туркестанском районе южного Казахстана. Названный
автор различает три типа строения нор этого зверька: к первому типу он
относит норы, имеющие снаружи одну или ‘ две пробочки (чаще одну),
один или три идущих к поверхности и заканчивающихся слепо (не дохо-
дящих на один—два сантиметра до поверхности) тупика, одну галлерею,
ведущую от пробочки вглубь земли к гнездовой шарообразной камере.
Второй тип норы имеет снаружи одну пробочку, от которой идет один
ход вглубь, заканчивающийся гнездовой камерой также шарообразного
вида. Третий тип напоминает в общих чертах второй, но отличается от
последнего присутствием особых расширений, расположенных по ходу
галлереи; количество этих расширений, на'зываемых автором дополнитель-
ными камерами, колеблется от 1 до 3 штук в норе; эти камеры ие имеют
гнезд и лежат на различной глубине от поверхности земли. Последнего-
типа норы служат, по наблюдениям автора, зимними норами, в которых
зверьки проводят спячку.
Автор таким образом описывает способ рытья норы тушканчиком:
„при копании он применяет в дело передние лапки и зубы. По мере нара-
стания кучки нагребаемой земли, тушканчик отгребал ее или задними
6. ALLACTAGA
125
лапками, или же прижимаясь туловищем к разрыхленной кучке земли,
движением вперед отгребал ее дальше. Работая таким образом и повторяя
описанные движения несколько раз, он постепенно углублял норку. При-
мерно через 30 минут после начала работы, мы увидели, что тушканчик
начинает забивать вход. Проделывал это он оригинальным способом:
подводя рыхлую и небольшую кучку земли к выходу, частым движением
головы, равномерно распределяя частицы у самого края входа, грызун
постепенно образовывал пробочку. На всю эту работу, как было отмечено,
ушло 40 минут".
Автор предполагает, что у малого тушканчика имеются три помета:
„дружный" весенний и осенний и слабо выраженный летний. Повидимому
более вероятным было бы предположить, что у этого вида, как и боль-
шого тушканчика, имеется растянутый период размножения, т. е. что
у отдельных особей спаривание и беременность наступают в различное
время, начиная с весны и до осени. Количество детенышей в помете
колеблется от 2 до б (в среднем 5); период спаривания носит затяжной
характер, он, видимо, наступает вскоре после пробуждения. Автор пред-
полагает, что появление молодняка первого помета падает на конец апреля
и начало мая. По предположению автора, период активной жизнедеятель-
ности этих грызунов продолжается приблизительно 8 месяцев — с половины
марта до середины ноября.
Автором сделан ряд наблюдений в естественных условиях и в неволе
над влиянием низких температур на жизнедеятельность тушканчиков, при-
чем в частности было выяснено, что в садке уже при температуре ниже
-+-10° С тушканчики впадают в оцепенение.
Врагами малого тушканчика являются: пустынный сыч, болотная
сова, степной хорек, однако эти животные в обследованном автором районе
немногочисленны; главными врагами' малого тушканчика в этом районе
автор считает степного удава и полоза Ревергиера.
Экономическое значение. В большинстве случаев малый
тушканчик живет на непригодных для земледелия участках и не является
вредителем сельского хозяйства. Однако в некоторых случаях, при освое-
нии отдельных частей пустыни, этот зверек может стать существенным
вредителем, как это имело например место при культивировании тау-сагыза.
Сведения о повреждениях тау-сагыза малым тушканчиком были приведены
в главе о большом тушканчике (наблюдения Колесникова).
*4. Allactaga williamsi Thomas— Малоазийский тушканчик.
Thomas, 1897 : 309. — williamsi latieps Nehring, 1903 : 357. — w. schmidti Сатунин,
1920:180. Вид описан по экземпляру из Вана в с-в. Турции; A. w. laticeps Nehring —
из Koktschi-Kissik, с-з. часть Малой Азии; A. w. schmidti Satun. — из восточного Закав-
казья (пустынные степи южных склонов восточной оконечности Главного Кавказского
хребта).
Ухо относительно короче, чем у других представителей рода (будучи
отогнуто вперед ухо не достигает конца морды или лишь едва достигает
126
ALLAСТА GINAE
его). Другие внешние признаки сходны с таковыми у других „широко-
головых" представителей рода, но голова* не столь широка и коротка, как
у A. jaculus. Размеры средние между A. jaculus и A. elater, задняя ступня
62—68 мм; кондилобазальная длина черепа 30.2—31.7; диастема—9.3—
10.5; максиллярный ряд зубов — 6.3—6.9. Характерными особенностями
строения penis, в отличие от ранее рассмотренных видов, является нали-
чие на дорзальной его поверхности сплошной продольной бороздки, про-
ходящей от его основания и доходящей до вершины; вершина penis обычно
притуплена.
Малоазийский тушканчик вместе с близкими к нему евфратским
(A. euphratica) составляют довольно хорошо обособленную группу среди
других представителей рода, особенно по описанному выше строению
genitalia. Интересно отметить, что по тем же признакам с этой же группой
имеет значительное сходство своеобразный представитель подсемейства
Allactaginae — четырехпалый тушканчик (Scarturus tetradactylus}. Морфо-
логическое сходство между названными формами согласуется также с их.
географическим распространением, как это будет указано ниже. Отноше-
ния A. williamsi к A. euphratica рассматриваются ниже, при описании,
последнего вида.
Распространение. A. williamsi ограничено Малой Азией на вос-
ток до оз. Ван и на север до южных районов Закавказья включительно.
На территории Закавказья этот вид распространен на Апшеронском полу-
острове и прилегающих с запада низменностях б. Гекчайского, Шемахин-
ского и Кубинского уездов, далее на запад он найден на Арарате, в Сардар-
Абадской степи и на Карском плоскогорье. Из пределов Малой Азии, на
основании просмотренных мною в Берлинском и Лондонском музеях
экземплярах, известны следующие немногочисленные нахождения: 1) Ван?.
2) Eregli (к востоку от Босфора); 3) Кападокия (Utsch-Kopu-Dagh между Nigde
и Berekezli Maaden); 4) Kokcoschi Kissik (с.-з. часть Малой Азии — (оригин.
экз. A. w. laticeps Неринга). Что касается указываемого К. А. Сатуниным
(1920) экземпляра Бланфорда из Буширд на Персидском заливе, то по
местонахождению здесь скорее можно было бы ожидать A. euphratica
(см. ниже), совпадение измерений задней ступни с таковыми у A. wil-
liamsi быть может объясняется неточностью измерений у названного автора
(у A. euphratica ступня короче, чем у рассматриваемого вида).
Географическая изменчивость: имеющиеся в настоящее
время коллекционные материалы по малоазийскому тушканчику нельзя
признать достаточными для изучения его географическей изменчивости
и особенно для установления подвидов, как это было сделано Нерингом
(1903) и Сатуниным (1920), тем более, что оба названных автора основы-
вались лишь на разнице в скуловой ширине по сравнению с впервые опи-
санной формой, не зная точные пределы вариации этого признака у этой
формы. Нерингом был описан подвид •—A. w. laticeps из северо-запад-
ной части Малой Азии (Koktschi— Kissik), отличающейся, по мнению
6. ALLACTAGA
127
автора, большей скуловой шириной (24.5), чем у описанной Томасом формы;;
Сатуниным был установлен еще подвид — A. w. schmidti из восточного
Закавказья (б. Кубинский, Шемахинский и Гекчайский уезды), характе- -
ризующийся еще большей скуловой шириной (25—26 мм).
Данные по биологии. Малоазийский тушканчик, в противопо-
ложность большинству видов этого рода, имеет значительное вертикаль-
ное распространение, встречаясь почти от уровня моря в восточном За-
кавказья до значительных высот в горных местностях; в частности на.
Арарате он найден Сатуниным на высоте около 2500 метров. В этом отно-
шении единственным известным аналогом этого вида является A. saltator
(см. ниже). Характерным местообитанием этого зверька Сатунин считает
предгорья и горы с ксерофильной растительностью. Образ жизни этого
животного известен очень мало; по наблюдениям К. А. Сатунина, в неволе -
этот тушканчик охотнее, чем другие виды, ест сухие семена. Летом люби-
мую пищу этого зверька составляли зеленые колосья диких и культурных
злаков; он охотно ел также морковь и разные семена, особенно подсол-
нуха. Экземпляр этого зверька прожил у К. А. Сатунина целую зиму,
не обнаруживая склонности к спячке; днем он был неподвижен, а вечером
становился резв и деятелен.
Экономическое значение этого вида не выяснено, хотя возможно,
что местами горный тушканчик приносит некоторый вред посевам пше-
ницы, о чем свидетельствуют наблюдавшиеся К. А. Сатуниным, перета-
скивания колосьев пшеницы этим зверьком в норки.
5. Allactaga euphratica Thomas. — Евфратский тушканчик
О. Thomas, 1881:15 (описан из Месопотамии без точного указания местности).
Сходен с A. williamsi, отличаясь от него несколько меньшей вели-
чиной, особенно более короткой ступней и меньшей длиной черепа: длина
задней ступни 52—58 мм, кондилобазальная длина черепа 28.3—30; ску-
ловая ширина — 21.7—23 мм, диастема 8.6—9.4; максиллярный ряд зубов
6.0—6.6.
Penis сходен по строению с таковым у A. williamsi, но его вершина
более заостренная и при его основании имеются несколько более крупных
шипов, чем на остальной части дорзальной поверхности (обычно особенно
выделяются по величине 2 шипа).
Евфратский тушканчик по своим признакам близок к малоазийскому
A. williamsi. Возможно, что при накоплении более обильного коллекцион-
ного материала по обеим формам и при более детальном их исследова-
нии, они будут соединены в один вид и A. williamsi будет рассматри-
ваться в этом случае как более крупная северная форма A. euphratica.1
Распространение. В настоящее время на основании просмо-
тренных мною экземпляров этого вида в музеях Берлина и Лондона, можно
1 В качестве видового названия в гаком случае должно быть сохранено название.
A. euphratica, как данное рацее, чем A. williamsi.
128
ALLACTAGINAE
'считать, что этот тушканчик распространен, кроме Месопотамии, также
в Сирии. В частности в Месопотамии он найден в Chabur и других пунктах
' (без точных указаний в этикетках); из пределов Сирии он известен из
Алеппо, из Caryatein и других местностей (без точных указаний на эти-
. кетках).
6. Allactaga hotsoni Thomas — Тушканчик Хотсона
Thomas, 1920:936 (Белуджистан, Кант в 20 милях к юго-западу от г. Сибб).
Размеры средние (крупнее, чем A. elater, но несколько меньше, чем
- A. williamsi). Ухо относительно длиннее, достигая у единственного изве-
стного экземпляра 41 мм. Общая окраска верха тусклая, серовато-желтая,
« приблизительно, как у A. elater. Стержень хвоста тускло-буроватого
цвета. Череп крупнее, чем у A. elater, но меньше чем у A. williamsi.
Мозговая коробка относительно крупная и округлая; ее наиболее широкая
часть расположена дальше вперед, чем у других видов (у A. hotsoni наи-
более широкая часть мозговой коробки расположена почти на уровне зад-
“ них корней скуловых дуг, тогда как у A. williamsi и A. elater эта часть
лежит несколько назад от указанного уровня). Foramina incisiva относи-
тельно шире, чем у A. elater', наибольшая ширина этих отверстий падает
. на их заднюю часть. Bullae tympani по крайней мере в 2 раза крупнее, чем
у A. elater и по крайней мере в I1/2 раза крупнее, чем у имеющего более
значительные размеры тела A. williamsi', передние заостренные концы
обеих bullae почти соприкасаются друг с другом в области передней части
основной затылочной кости. Foramina postglenoidea узкие и вытянутые
в продольном направлении.
Измерения (единственного известного экземпляра): длина тела 121 мм;
длина хвоста 215; длина ступни 57.2; длина уха — 41; кондилобазальная
длина черепа 29.0; скуловая ширина 21.1; затылочная ширина 14.6; заты-
лочная высота 10.6; межглазничное пространство 9.2; максиллярный ряд
зубов 5.3; носовые кости 11.5; диастема 9.5.
Распространение. A.. hostoni известен лишь по одному экзем-
пляру, добытому в Белуджистане близ Кант, в 20 милях к юго-западу
от Сибб, и хранящемуся в Британском музее.
A. hotsoni по своим морфологическим признакам хорошо отличается
от рассмотренных выше видов и приближается лишь к A. bullata GI.
- Allen; при описании последнего вида, указываются признаки его различия
• от A. hotsoni.
7. Allactaga bullata Glover Allen — Гобийский тушканчик
Glover Allen, 1925 : 2 (Цаган-нор в Монголии); Vinogradov, 1926 : 233.
По внешности сходен с обитающим в тех же районах A. saltator, но
несколько меньшего размера и с значительно более крупными bullae tym-
pani, которые почти соприкасаются своими передними концами. Окраска
весьма сходна с таковой у A. saltator. Основной тон окраски верхней
6. ALLACTAGA
129
•стороны тела между avellaneous и vinaceous fawn (Ridgeway, X. L.). Стер-
жень хвоста имеет несколько более яркую окраску, чем у A. saltator,
с большей примесью рыжевато-желтых тонов. Череп имеет значительное
сходство с таковым у A. hotsoni, характеризуясь относительно крупной
мозговой коробкой с наиболее широкой частью расположенной несколько
далее вперед, чем у других видов, а также крупными.bullae tympani, почти
соприкасающимися своими передними концами в области передней части
основной затылочной кости. Foramina postglenoidea сходны по очертаниям
•и размерам с таковыми у A. hotsoni.
Измерения: длина тела 110—132; ступня 57—64; ухо 27—36;
кондилобазальная длина черепа 30—32.5; скуловая ширина 22.5—?23.5; меж-
глазничное пространство —9.9—10.5; верхний ряд зубов — 5.8—6.7.
По краниологическим признакам, как было уже указано, A. bullata
стоит близко к A. hotsoni. Точного взаимоотношения между названными
видами в настоящее время не может быть выяснено в связи с тем, что
•строение genitalia у обоих видов пока не известно. Из наиболее ясных
•отличий между этими видами нужно указать следующие: ухо у A. bullata
значительно короче, чем у A. hotsoni, межглазничное пространство шире,
•верхний ряд зубов длиннее, как это можно видеть при сравнении приве-
денных для обоих видов измерений; кроме того, у A. bullata foramina
incisiva заметно уже, чем у A. hotsoni.
Распространение. Большинство известных нахождений A. bul-
lata приурочено к сравнительно узкой полосе Гоби от р. Туин-гол и озера
Цаган-нор на юго-восток вдоль окраинных хребтов Монгольского Алтая
до северной Алашани и центральной Гоби (Саир-усу, Ончи-гургуль,
Холоин-гашун); кроме того имеется также нахождение этого вида в вос-
точной Гоби близ Ирен-дабасу (Эрлиен) (Gl. Allen, 1925). Таким образом,
можно предполагать, что этот вид широко распространен в подходящих
•стациях на всем пространстве Гоби. Об образе жизни этого зверька дан-
ных не имеется.
*8. Allactaga saltator Eversmann — Тушканчик-прыгун или монголь-
ский тушканчик.
Eversmann, 1848:188 (Dipus) [описан из Чуйской степи Алтай]; Виноградов, 1933.—
mongolica Radde, 1861 : 680. — annulate Milne-Edwards, 1867 : 376. — suschkini Satunin,
1900:137. — riickbeili Thomas, 1914: 571. — mongolica longior Miller, 1911: 53. — Формозов,
1929: 7778 (A mongolica и saltator).
Размеры значительно меньше, чем у A. jaculus и A. severtzovi. Длина
тела 130—150 мм; хвост 190—220 мм; задняя ступня 68.75 мм; ухо 43—
52 мм; кондилобазальная длина черепа 35—38 мм; впереди от черной части
знамени имеется белое кольцо (стержень хвоста на протяжении 30 мм или
более покрыт светлыми волосами). Голова значительно более узкая и
.вытянутая, чем у предыдущих форм, с более узким и вытянутым рылом.
Передний (ложнокоренной) зуб почти равен по размерам заднему корен-
Фауна СССР, Виноградов 9
130
ALLA СТА GINAE
Ному или немного превышает его. Передние части скуловых дуг (при
рассматривании черепа сверху) отходят косо к продольной оси черепа.
Окраска у различных географических форм вариирует от относительно
темной и насыщенной (Л. s. mongolica) до светлой серовато-желтоватой
(A. s. saltator и A. s. riickbeili).
Penis имеет ббченкообразную форму; средня» продольная бороздка,
на его верхней поверхности отсутствует.
Под видовым названием — A. saltator здесь соединено несколько*
форм, считавшихся в недавнее время за самостоятельные виды, а именно*
следующие: A. saltator Eversmann, A. mongolica Radde, A. annulafa Milne-
Edwards, A. suschkini Satunin и A. riickbeili Thomas. После накопления
достаточного количества коллекционного материала но названным формам
и детального его исследования стало совершенно очевидно, что все наз-
ванные формы должны быть объединены под общим видовым названием
A. saltator, так как это название было дано Эверсманном р^нее других
авторов.
A. saltator занимает между другими представителями рода довольно
изолированное положение, хорошо отличаясь от других видов по
описанным выше особенностям строения черепа, коренных зубов,
и penis.
Распространение. Казахстан от северо-восточного побережья.
Каспийского моря на восток до Зайсана, Чуйской степи (на Алтае) и
Семиречья включительно, вся Монголия, южное Забайкалье (Кяхтинский
и Борзинский окр.), Манчжурия до юго-заподного Хейлунгкианга и вероятно
северо-западного Гирина и северный Китай в пределах провинций Чили„
Шанси и Гансу. В пределах СССР рассматриваемый вид распространен
к западу до низовий р. Урала, северо-восточного побережья Аральского
моря и полуострова Мангышлака. Северная граница распростронения
этого вида идет от низовий р. Урала, приблизительно через Актюбинск,,
Тургай, Каркаралинск и Семипалатинск; далее на восток в пределах СССР
имеется изолированное нахождение этого вида в юго-восточном Алтае
(Чуйская степь) и еще далее к востоку ареал распространения A. saltator
близко подходит к границам Союза (не переходя их) в пределах Танну-
Тувинской республики; далее на восток на территории Союза этот вид,
появляется снова вместе с рядом других элементов монгольской фауны
в пределах южного Забайкалья (Кяхтинский и Борзинский окр.). Южная
граница распространения тушканчика-прыгуна в пределах СССР изучена
более слабо: от южного чинка Усть-урта она простирается к северному
и северо-восточному побережью Аральского моря; далее на восток она
идет от низовий р. Сыр-дарья приблизительно по направлению р. Чу до
южной окраины Бетпак-дала и Чу-Илийских гор до хребтов системы Тянь-
шана в юго-восточном Семиречье, местами проникая здесь в горы, где
наиболее южным пунктом нахождения этого вида является Аксайское
плато в Киргизии.
6. ALLACTAGA
131
Географическая изменчивость. A. saltator на протяжении
обширного ареала своего распространения образует несколько хорошо
дифференцированных форм, считавшихся до последеего время самостоя-
тельными видами: этими формами являются следующие.
1. A. s. suschkini Satunin, 1900 (пустыня Сара-копа к югу от г. Ир-
гиза б. Тургайск. обл.). Окраска верха охристо-буроватая с сероватым
оттенком. Бока серовато-белые с охристым оттенком. Голова почти серая,
окрашенная сходно с передней частью спины, но серее. Задняя часть
бедер сероватая или охристо-сероватая. Верхняя поверхность хвоста
охристо-серая. Размеры сходны с таковыми у других форм, за исключе-.
нием более крупного mongolica. К этой же форме, повидимому, следует
причислить riickbeili Thomas (1914) из горных районов Семиречья; по край-
ней мере, имевшиеся у меня несколько экземпляров этой формы (в том
числе топотип из окрестностей Джаркента) не отличимы от A. s. su-
schkini по окраске и другим признакам: кондилобазальная длина черепа
34.5—37 мм; скуловая ширина 23.5—25.5 мм; максиллярный ряд зубов
6.7—7.5 мм.
Распространение A. suschkini охватывает значительную часть Казах-
стана от низовий р. Урала и северо-восточного побережья Каспийского
моря до Семиречья включительно, где местами этот тушканчик встречается
и в горных местностях. Северная и южная границы распространения этой
формы были указаны при характеристике вида.
2. A. s. saltator Eversmann (1848, Чуйская степь на Алтае). Наиболее
светло окрашенная форма. Общий тон верха желтовато-серый с легким
буроватым оттенком. Бока серовато-белые, почти без примеси охристых
тонов. Голова серая. Задняя часть бедер серая (от светлосерого до темно-
серого цвета). Хвост сверху светлый, охристо-сероватый. Размеры как
у A. s. suschkini hjm несколько меньше.
Кроме Чуйской степи, рассматриваемая форма широко распростра-
нена в Монголии, за исключением ее северо-восточных частей (от Улан-
Батора и долины р. Керулен на север до границ СССР, где распро-
странена другая форма — A. s. mongolica Radde).
3. A. s. mongolica Radde, 1861 (с. Кулуссутай на оз. Tарей-нор, в ю.-в.
Забайкалья). Наиболее темно окрашенная и наиболее крупная форма.
Окраска верха охристо-бурая с темнооливковым оттенком. На боках
головы, позади глаз, имеется рыжеватое пятно. Бока охристо-буроватые,
с ясно заметной охристой каймой, вдоль границы с светлой окраской
нижней стороны тела. Лоб окрашен сходно с передней частью спины,
но рыжее. Задняя сторона бедер бурая или черно-бурая. Хвост сверху
охристо-буроватый. Размеры более крупные, чем у всех других форм:
кондилобазальная длина черепа 36.0—39 мм; скуловая ширина 25.5—27.8;
максиллярный ряд зубов 7.0—8.8.
Распространение этой формы охватывает южное Забайкалье
(Кяхтинский и Борзинский окр.) и северо-восточную Монголию на юг
9*
132
ALLACTAGINAE
приблизительно до Улан-Батора и р. Керулена, как об этом можно судить
по просмотренным мною нескольким экземплярам из этих местностей.
4. A. s. annulata Milne-Edwards, 1867 (Северный Китай, вероятно
к западу от Бейпина). Окраска во всех существенных признаках сходна
с таковой у A. s. mongolica, но несколько более тусклая. Размеры заметно
меньше, чем у предудущей формы.
A. s. annulata распространен в северном Китае, пункты нахождения
этой формы указаны выше в очерке распространения вида.
Данные по биологии. По наблюдениям Формозова (1929)
в Монголии, этот вид встречается в довольно разнообразных стациях:
наиболее многочисленным этот зверек в глинистой и щебнистой пустыне
с редкой растительностью с преобладанием лука(А1шш polyrhizum и др.).
Довольно многочисленен этот зверек также на участках полупустынного
характера и несколько реже в высоких степях с преобладанием тырсы
(Stipa capillata); местами вместе со степью он поднимается очень высоко,
в частности его присутствие было обнаружено в горах Чаган-арзирга на
высоте до 2000 м. A, saltator довольно постоянно встречается также
в котловинах озер у солончаков и может считаться характерным для
речных долин, где он селится в ближайшей к воде полосе и где лишенные
растительности глинистые или галечные пространства чередуются с лужай-
ками или гривами заросшими караганой и другими кустарниками; по реч-
ным долинам A. saltator заходит вглубь гористых местностей. По наблюде-
ниям того же автора этот тушканчик имеет по нескольку нор, из которых
одна является жилой, а остальные временными. Раскопанные автором
норы достигали 5 метров длины и имели до трех отнорков; обычно
у места расхождения отнорков находилась гнездовая камера, один из ходов
круто опускался вниз, где иногда имелось второе гнездо, другой развет-
вляясь шел до самой поверхности почвы, давал расширения и оканчивался
незаметной снаружи отдушиной, через которую выскакивал потревожен-
ный тушканчик. Автор наблюдал выход наружу тушканчиков и добывал
их ловушками, еще 7—8 сентяря, когда по утрам были заморозки и выпа-
дал иней. Тот же автор наблюдал в конце сентября следы тушканчика
на довольно глубоком снегу при морозе около —17° С; автор предпола-
гает, что эти следы принадлежали A. s., mongolica. В Казахстане A. s. susch-
kini является преимуществённо обитателем глинистой и щебнистой
пустыни, хотя он встречается и в других стациях, в частности П. П.
Сушкин добыл экземпляр, послуживший К. А. Сатунину оригиналом для
описания названного подвида, в приаральских песках; мною на побережьи
Балахаша был добыт экземпляр этого тушканчика на пухлом солончаке на
месте прежнего залива Балахаша. В. Н. Шнитниковым получен экземпляр
этого тушканчика с Аксайского плато в Киргизии, где он был добыт на
высоте около 2000 м.
Экономическое значение A. saltator не выяснено. По наблюдениям
Е, П. Лупповой (личное сообщение) в песках Барсуки этот тушканчик вре-
7. AL ACT A GUL US
133
дит посадкам каучуконоса хондриллы. Следует предполагать, что этот вид
может служить объектом промысла, так как его шкурка имеет достаточно
крупные размеры, а его мех заметно не отличается по качеству от заго-
товляемого большого тушканчика (A. jaculus).
7. ALACTAGULUS NEHRING (1897) - ЗЕМЛЯНЫЕ ЗАЙЧИКИ
Nehring, Sitz.-Berichte Naturf. Freunde Berl., 1879 : 151.
По внешним признакам сходны с представителями рода Allactaga,
уши более короткие, отогнутые вперед, значительно не достигающие носа;
голова очень широкая и короткая; в теменной области она значительно
шире, чем у сходного по размерам Allactaga elater. „Знамя" на конце
хвоста более слабо развито, чем у представителей рода Allactaga. Penis
ланцетовидной формы; шипики покрывают его верхнюю поверхность
лишь на внешних краях и близ вершины, оставляя в средней и основной
части голую площадку; в области этой площадки имеется два слабых
продольных вдавления. Шипики, расположенные близ боковых частей
основания penis, слегка отогнуты по направлению внутрь (в сторону средин-
ной линии). Строение черепа в основных чертах сходно с таковыми у Alla-
ctaga, но имеет следующие характерные особенности. Верхняя поверхность
мозговой коробки более или менее резко переходит в внутреннюю стенку
глазницы, т. е. лобная и глазничная пластинка frontale образует ясный
излом или грань при переходе одна в другую; боковые линии мозговой
коробки от межглазничного сужения до середины наружного края parietale
(при рассматривании черепа сверху) почти прямолинейные. Костный
мостик над foramen infraorbitale (вертикальная часть скуловой дуги) очень
узок: его наименьшая ширина приблизительно вдвое или почти вдвое уже
горизонтальной части скуловой дуги. Париетальный край squamosum не
образует выемки,, поэтому сквамозо-париетальный шов почти прямой.
Foramina incisiva доходит назад не далее линии, проведенной через перед-
ние края 1-го коренного зуба. Первый палец на задней конечности
хорошо развит и он немного превышает по длине пятый палец; основная
фаланга второго пальца приблизительно вдвое длиннее средней фаланги
того же пальца. Ложно-коренной зуб в верхней челюсти у взрослых
особей отсутствует, но иногда в виде рудимента он сохраняется некоторое
время у молодых экземпляров. Жевательная поверхность коренных зубов
более упрощена, чем у Allactaga, и обе добавочные наружные петли совер-
шенно атрофированы, т. е. с наружной стороны на жевательной поверхности
имеется лишь 3 выдающихся нераздвоенных угла; выдающиеся углы более
заострены, чем у Allactaga.
Некоторые характерные особенности рода Alactagulus указывают
на то, что этот род более значительно уклонился от основного плана
строения более примитивных представителей подсемейства, чем род Alla-
ctaga, в частности по редукции РМ, по утере следов бугорчатого строения
134
ALLACTAGINAE
жевательной поверхности коренных зубов и упрощению их петель,
а также по строению penis. По совокупности признаков рассматриваемый
род стоит значительно ближе к роду Pygerethmus, чем к Allactaga, как
будет указано ниже при описании первого из названных родов.
Род Alactagulus заключает лишь один вид A. acontion, при описа-
нии которого сообщаются также данные о его географическом распро-
странении, образе жизни и пр.
Фиг. 39. Голова тарбаганчика (Alactagulus acontion Pall.) (увелич.).
*1. Alactagulus acontion Pallas — Земляной зайчик или тарбаганчик
(фиг. 39—42).
Pallas, 1878: 284, 295 (описан из западного Казахстана); Nehring, 1900: 63; Сатунин,
1920:196.
Основные признаки указаны в характеристике рода. Окраска верха
у различных географических форм колеблется от относительно темной
оливково-буроватой на северо-западе ареала (Предкавказье) до чрезвы-
чайно бледной песчано-серой на юге ареала (южная Туркмения и южный
Узбекистан). Размеры сходны с таковыми у Allactaga elater.
Распространение (фиг. 43). Зона полынковых полупустынь
и полынковосолянковых пустынь; от восточного Предкавказья на восток
до Алакуля и Кульджи; как указывается ниже, отдельные нахождения
имеются и значительно далее на восток в пределах внутренней Монголии.
К северу приблизительно до 50° с. ш., к югу до южного Усть-урта, низо-
7. ALACTAGULUS
135
®ий Аму-дарьи, побережья Аральского моря и вероятно долины Сыр-дарьи;
отдельными островками распространен далеко на юг по пятнам солянковых
пустынь (такыров) до южной Туркмении (найден близ ст. Ахча-куйма
к западу от Кзыл-Арвата,
близ ст. Артык к востоку
от Ашхабада и ст. Болдыр
к западу от Термеза).
;В Предкавказьи на запад
до Ставропольской возвы-
шенности, далее на север
встречается в Калмыцкой
АССР и Сталинградском
округе (ст. Иловлинская на
Дону); приблизительно от
‘Сталинграда граница идет
по левому берегу Волги до
.широты Вольска; следую-
Фиг. 40. Голова тарбаганчика (Alactagulus acontion
Pall.).
щими наиболее северными
известными нахождениями
являются поселок Ново-
российский Актюбинского окр., окрестности Тургая и долина реки Токрау
в северном Прибалхашье. Отдельные оторванные нахождения известны
далеко на востоке в пределах Алашаня и Ордоса (внутренняя Монголия).
Г еографиче-
Фиг. 41. Тарбаганчик (Alactagulus acontion Pall.).
ская изменчи-
вость в пределах
рассматриваемого ви-
да выражена до вольно
ясно, хотя она отно-
сится преимуществен-
но лишь к измене-
ниям окраски у осо-
бей из различных
частей ареала, причем
эти изменения подчи-
няются общей законо-
мерности, отмеченной
многократно выше и
заключающейся в по-
степенном побледне-
нии окраски от северо-западных и северных частей ареала по направле-
нию на юг и юго-восток. Наиболее темно окрашенной формой является
A. a. dinniki из восточного Предкавказья и наиболее светлой — форма
из юго-восточного Узбекистана (окрестности Термеза); разница между
136
ALLA СТА GINAE
этими крайними формами весьма значительна, но между ними имеется
ряд постепенных переходов, как это указывается ниже.
1. Экземпляры из восточного Предкавказья (A. a. din-
niki Satunin, 1920). Общий тон окраски верхней поверхности тела отно-
сительно темный, оливково-буроватый. Окраска головы довольно темная,
но несколько светлее спины. На боках тела и на брюхе относительно
сильно развит рыжеватый налет. На бедрах имеются темные (почти черные
пятна). Беловатый цвет на верхней стороне хвоста впереди от черной части,
„знамени" отсутствует или лишь едва намечен.
Фиг. 42. Мертвый тарбаганчик на такыре.
2. Экземпляры из западного Казахстана (Актюбинск,.
Эмба, низовья реки Урала). Общий тон окраски верха охристо-сероватый,,
сходный с A. a. dinniki, но бледнее. Темные окончания волос на голове
менее развиты.
3. Юго-западное Прибалхашье (серия из сел. Буру-Байтал)..
Общий тон окраски верха сходен с таковым у экземпляров из западного
Казахстана, но значительно серее и с более ясно выраженными мелкими
темными пестринками. Голова светлее, чем у предыдущей серии. Ржавые-
тона светлее. Брюшная сторона чисто белая. Белая окраска на хвосте перед
черной частью „знамени" развита сильнее.
4. Северное побережье Аральского моря. В основных
чертах по окраске сходны с балахашскими экземплярами, но охристые
тона слабее, переходящие в песчаные. Серый оттенок,более ясно выражен..
Бока тела сероватые. Черный цвет на „знамени" занимает меньшую
поверхность, чем у предыдущих.
5. Западная Туркмения (Южный Усть-урт, Прикарабугазье*.
ст. Ахча-куйма). Сходны с аральскими, но значительно бледнее и серее.
Черный цвет окончаний волос заменен черно-бурым. Охристые тона, почти.
7. ALACTAGULUS
137
Фиг. 43. Распространение Alactagulus и Pygerethmus: 1. A. acontion Pall., 2. P. platyurus
Licht, 3. P. zhitkovi Kuzn.
138
ALLA СТА GINAE
Фиг. 44. Тарбаганчик. Рис. А. Н. Формозова.
7. ALACTAGULUS
139
отсутствуют. Белая окраска на хвосте перед черной частью „знамени"
развита еще сильнее. На бедрах сильно развиты серые пятна. Черная
часть „знамени" значительно сокращена в размерах.
6. Юго-восточный Узбекистан (ст. Болдыр близ Термеза),
Окраска верха чрезвычайно бледная, бледнее чем у всех других форм.
Общий тон верха песчано-серый, голова почти белая с слабым песчаным
налетом. Пятна на бедрах светлосерые или почти отсутствуют. На хвосте
впереди от черной^части „знамени" сильно развитое белое кольцо. Черная
Фиг. 45. Внешний вид норы тарбаганчика.
часть „знамени" сильно сокращена в размерах. Внутренняя поверхность
ушей голая, не покрытая волосками; небольшая голая площадка имеется,
кроме того, на наружной поверхности уха близ его края.
7. Ордос и Алашань (A. a. potanini Vinogradov, 1926). Уста-
новленное мною нахождение A. acontion в Ордосе и Алашани расширяет
очень далеко на юго-восток известные границы ареала этого вида; до сего
времени на громадном пространстве между Семиречьем и Алашанью
не установлено ни одного нахождения этого вида, и поэтому до настоящего
времени не известно, является ли A. a. potanini в современную эпоху
совершенно изолированной от основного ареала вида формой или же
представители этого вида встречаются также на указанном выше обшир-
ном промежуточном пространстве. Рассматриваемая форма отличается
от указанных выше более значительными размерами тела и несколько
более раздутыми и округленными bullae tympani. У единственного, вполне
взрослого известного экземпляра A. a. potanini наиболее характерны
140
ALLACTAGINAE
следующие измерения; задняя ступня — 52 мм; кондилобазальная длина!
черепа 26.5 мм; затылочная ширина 18.0; длина нижней челюсти 17.6;
носовые кости— 10.6; диастема 9.3. Точная окраска не известна, так как.
эта форма была описана по экземпляру изменившему от продолжительного
хранения в спирту естественную окраску.
Данные по биологии. Мы наблюдали земляного зайчика,
в окрестностях ст. Болдыр (56 км к западу от Термеза), где он селится
преимущественно на совершенно плоских солонцеватых площадках, почти
лишенных растительности. Норки этих зверьков, как у большинства!
других тушканчиков, имеют закрытое днем входное отверстие, которое
можно найти по плоской насыпи из выкопанной земли, закрывающей это
отверстие (фиг. 45); засыпанный вход в нору обычно находится где-нибудь
на краю кучки, где, при некотором навыке, можно заметить несколько отли-
чающихся от остальной масс насыпи зернистую пробочку из комочков земли,
впрочем нередко эта пробочка совершенно сливается с общим фоном
насыпи. Часто на краях кучки имеются 2—3 неглубоких ямки („копанцы"),
производящих впечатление начальной части хода. Начальная часть хода,
забитая землей, идет на некотором протяжении почти горизонтально в рых-
лом поверхностном слое солончака непосредственно под наружной коркой
и заполняющая ход масса земли почти не отличается по окраске и струк-
туре от окружающего грунта, вследствие чего ход легко теряется при рас-
копке. Заполненная землей часть норы иногда имеет очень небольшое
протяжение (5—10), но чаще она тянется на гораздо большее расстояние,
иногда достигая 2—4 метров. Свободная от заполняющей земли часть '
норы обычно вскоре делится на два выхода: одна начинает быстро углу-
бляться от места отхождения второго хода и оканчивается расширенной
яйцевидной камерой; вторая ветвь отходит в сторону от главного хода
и поднимается вверх до самой или почти до самой поверхности земли;,
в первом случае на поверхности земли образуется маленькое отверстие
аккуратно закупоренное в жилых норах в дневное время пробочкой из.
небольшого количества маленьких комочков земли, хорошо заметное
снаружи („окошечко" или „копеечка") (фиг. 46); если боковой ход не
достигает поверхности земли, над его слепым концом, остается тонкий
свод в 1—2 см толщины и в таком случае „окошечко" незаметно. Раскопку
норы удобнее начинать с этого „окошечка" (если оно заметно снаружи),
т. к. здесь ход норы открывается сейчас же после удаления пробочки,
закупоривающей это отверстие. Если раскопку вести со стороны главного
хода и заранее не заложить „окошечко" каким-нибудь предметом, тушканчик,
стремительно выбегает из этого отверстия и быстро скрывается с глаз.
Мы изгоняли земляных зайцев из нор при помощи небольшой дозы хлор-
пикрина, закладываемого со стороны вскрытого „окошечка" или главного'
входа, этот газ действует на тушканчика необычайно сильно и часто уже
через несколько секунд после закладки хлорпикрина в нору,тушканчик
выбегает наружу. В окрестностях ст. Болдар земляной зайчик преобладает
7. ALA СТА GULUS
141
по численности перед другими видами обитающих здесь тушканчиков,
в частности над Allactaga elater, который здесь довольно редок, тогда
как в окрестностях Термеза последний вид встречается в изобилии, а
A. acontion здесь не был найден нами вовсе. Относительно размножения
A. acontion мы располагаем следующими данными: 16—18 июня мы нахо-
дили в окрестностях Болдыр беременных самок с маленькими еще не
вполне развитыми эмбрионами (в двух вскрытых самках было до 3 эмбриона);
в то же время в одной из нор была самка вместе с 5-ю детенышами, дости-
гшими уже приблизительно половины роста взрослой особи. Приведенные
Фиг. 46. „Копеечка" тарбаганчика (запасный выход, закрытый изнутри
пробкой из комочков земли).
данные позволяют предполагать, что здесь имеют место два помета — один
весенний и один летний. Аналогичные наблюдения были опубликованы
Б. К. Фенюком1 относительно Allactaga jaculus и Scirtopoda telum, причем
автор пришел к выводу, что вторичное метание детенышей не носит такого
массового характера, как весеннее.
Экономическое значение. По наблюдениям Б. К. Фенюка
в Нижнем Поволжьи, земляной зайчик вместе с другими обитающими
здесь видами считается вредителем сельского хозяйства, особенно весной
на бахчах, где тушканчики выкапывают из земли и поедают посеянные
семена арбузов, дынь и тыкв; тушканчики. вредят здесь также пшенице
и другим зерновым хлебам поеданием зелени, особенно после выхода
пшеницы в трубку и еще более они вредят выеданием зерен из колосьев
1 Б. К. Ф енюк. „К биологии тушканчиков". Матер, к позн. фауны Нижн. Поволжья,
Саратов, 1928.
142
ALLACT A GINAE
после их созревания (Фенюк, 1928:33—35; 1929:36—28). За отсутствием:
соответствующих наблюдений экономическое значение этого вида в Сред-
ней Азии пока остается неизвестным.
8. Род PYGERETHMUS GLOGER (1841) — ТОЛСТОХВОСТЫЕ ТУШКАНЧИКИ1
Gloger, Hand- und Hilfsbuch Naturg., 1, 1841:106.
По наружным признакам сходны с Allactaga и Alactagulus, но хвост
имеет иное строение; на его конце имеются лишь короткие волосы,,
не образующие совершенно „знамени" (Р. platyurus) или же длинные
волосы, не образующие, однако, плоской кисточки, как это имеет место-
у названных родов (Р. zhitkovl)', конец хвоста темный или с примесью
лишь небольшого количества белых волос. Обычно в подкожной клетчатке
хвоста имеются значительные отложения жира, в связи с чем нередко-
хвост сильно раздут и имеет вальковатую форму, достигая иногда в попе-
речнике 12 см (уР. zhitkovl). Задняя ступня относительно более короткая^,
чем у Allactaga и Alactagulus. Первый палец на задней конечности немного
короче, чем пятый; оба боковых пальца несколько более редуцированы,,
чем у Allactaga и Alactagulus. Penis сходен по своему строению с таковым,
у Alactagulus, но шипики на его дорзальной поверхности еще более резко
отогнуты внутрь и вдоль его срединной линии имеется резко ограниченная
узкая вдавленная площадка. Межглазничное пространство почти в 4 раза,
меньше наибольшей длины черепа. Наружные края мозговой коробки еще
более резко обособлены и более прямолинейны, чем у Alactagulus. Костный,
мостик над foramen infraorbitale столь же узок, как у Alactagulus. Парие-
тальный край squamosum имеет те же особенности как у Alactagulus..
По совокупности признаков череп Pygerethmus имеет значительное
сходство с Alactagulus. По строению тазовых костей представители,
этого рода резко отличаются от Allactaga, Alactagulus и Scarturus чрезвы-
чайно коротким symphisis pubis, напоминая в этом отношении представи-
телей подсемейства Zapodinae и Cardiocraniinae', это сходство усиливается
тем, что в области сращения на ischio-pubicum образуются довольно ясно
обособленные отростки. Однако, в отличие от Zapodinae и Cardiocranii-
nae, нисходящая ветвь ischium образует с горизонтальной (параллельной:
позвоночному столбу) частью этой кости острый угол, сохраняя в этом,
отношении те признаки, которые характерны для представителей подсе-
мейства Allactaginae. Ложно-коренной зуб в верхней челюсти у взрослых
особей отсутствует. Строение коренных зубов совершенно сходно с тако-
выми у Alactagulus (см. выше).
1 Название „плоскохвостый1' тушканчик, данное Лихтенштейном описанному им:
представителю этой группы (Р. platyurus), основано на недоразумении, так как названный
автор описал строение хвоста этого животного по уродливо приготовленному чучелу
хранящемуся в Берлинском Зоологическом музее; у этого экземпляра хвост был сделан,
препаратором плоским и ланцетовидным.
8. PYGERETHMUS
143..
По своим признакам род Pygerethmus стоит очень близко к роду
Alactagulus, особенно по отсутствию ложно-коренных зубов, строению
эмалевых петель коренных зубов и строению черепа. Сходство указанных
признаков столь велико, что руководствуясь только ими, следовало бы
объединить обе эти группы в один род; однако наличие некоторых,
других признаков, особенно своеобразное строение тазовых костей
У Pygerethmus делает необходимым сохранение родовой самостоятель-
ности толстохвостых тушканчиков. Как будет указано ниже, особенно
приближается по своим признакам к Alactagulus недавно описанный
Р. zhitkovi.
Географическое распространение рассматриваемого рода выяснено.
недостаточно. На основании немногих известных нахождений представи-
телей Pygerethmus ареал этого рода представляется разорванным: первая.,
часть этого ареала охватывает пространство от низовий р. Урала до северо-
восточного побережья Аральского моря (Куван-дарья) и к югу до южного
Усть-урта (Р. platyurus}’, вторая часть ареала охватывает Аулие-атинский
округ, Прибалхашье и Алакульскую равнину (Р. zhitkovi).
В настоящее время известны два хорошо дифференцированных,
вида этого рода.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ
1 (2). Концевые волосы хвоста белые; длина концевых волос, приблизи-
тельно 13—15 см. Нижняя поверхность хвоста в предконцевой части
(как и на всем протяжении), светлая. Длина хвоста 95—150 см.
Задняя ступня 40—45 мм........................* 2. Р. zhitkovi Kuzn.
2 (1). Концевые волосы хвоста темные (в более редких случаях среди
них могут присутствовать до десятка белых волос); длина концевых
волос хвоста не более 6—8 мм; нижняя поверхность хвоста в пред-
концевой части темная. Длина хвоста 80—90 мм; задняя ступня
-32—35 мм................................ * 1. Р. platyurus Licht.,
* 1. Pygerethmus platyurus Licht.—Толстохвостый тушканчик.
Lichtenstein, 1821 :23 (описан по экземпляру с р. Куван-дарья близ побережья >
Аральского моря); Nehring, 1900 : 361.
Маленький тушканчик, имеющий заметно меньшие размеры, чем
Pygerethmus zhitkovi (длина тела 80—90 мм). Длина задней ступни 32—
35 мм. Хвост приблизительно равен длине тела и сильно утолщен,,
с отложениями жира на позвонках, „знамя" на конце хвоста отсутствует
вовсе; концевые волосы хвоста лишь едва удлинены, достигая в длину
6—8 мм, они окрашены в темный цвет или в более редких случаях среди
них могут присутствовать лишь единичные белые волосы. Задняя ступня.,
коротка, достигая в длину 32—35 мм. Ухо короткое 17—20 мм. Череп
с относительно узким межглазничным пространством (более узким чем..
144
ALLACTAGINAE
у Alactagulus acontion). Наружные края мозговой коробки (при рассматри-
вании черепа сверху) более угловатые и более резко обозначенные чем
у Alactagulus. Вертикальная передняя часть скуловой дуги узкая, как
у Alactagulus или еще уже. По размерам череп значительно меньше, чем
у Р. zhitkovi и Alactagulus. Корённых зубов у; строение их вполне сходно
с Р. zhitkovi и A. acontion. Penis на верхней поверхности близ его
основания с загнутыми внутрь шипами; в середине верхней поверх-
ности имеется резко очерченное узкое, голое, лишенное шипов, про-
странство.
Измерения. Длина тела 80—90 мм; хвост 80—90 мм; задняя
ступня* 32—35 мм; ухо 17—20 мм.
Толстохвостый тушканчик по строению черепа, зубов и penis имеет
значительное сходство с тушканчиком Житкова, от которого он отличается
более короткими ушами, отсутствием на хвосте „знамени" и значительно
меньшими размерами.
Распространение. Этот редкий в коллекциях зверек имеет, неви-
димому, ограниченное распространение — долина р. Урала и Эмбы и приле-
жащие районы (Индерск, Чингильды б. Гурьевского у., Манайли, Сор-куль
б. Лбищенского у.), а также в б. Казалинском у. (Куван-дарья близ
впадения в Аральское море). Недавно один экземпляр этого вида найден
также на южном Усть-урте. Таким образом, на основании имеющихся
немногих данных можно считать, что ареал распространения этого вида
охватывает полупустыни и пустыни от нижнего течения р. Урала на восток
до северо-восточного побережья Аральского моря и на юг до южного
Усть-урта.
* 2. Pygerethmus zhitkovi Kuznetzov — Тушканчик Житкова
(фиг. 47)
schitlcovi (Alactagulus) Кузнецов, 1930:623 (описан по экземплярам с оз. Ала-куль
в восточном Семиречьи); Виноградов, 1930 : 23.
Хвост относительно короткий, обычно не превышающий длину тела
или лишь немного превосходящий последнюю; обыкновенно хвост заметно
утолщен вследствие развития жирового слоя на хвостовых позвонках,
причем у некоторых экземпляров этот жировой слой достигает значитель-
ной толщины и хвост становится в этих случаях очень сильно раздутым,
достигая иногда 12 мм в поперечнике. Концевые волосы хвоста не обра-
зуют настоящего „знамени", но они заметно удлинены, причем почти
вся кисточка окрашена в черный или темный цвет и лишь очень маленький
участок на ее конце может быть белым, вместо которого нередко имеются
лишь одиночные белые волосы среди черных на конце кисточки. Зад-
няя ступня, по сравнению с Alactagulus acontion относительно короткая,
достигая обычно 40—43 мм в длину. Уши длиннее, чем у A. acontion
(30—40 мм). Окраска верха желтовато-серая, с темными остевыми воло-
сами. Брюшная сторона желтоватого цвета; хвост сверху желтовато-
8. PYGERETHMUS
145
серый с рассеянными отдельными черными волосами; кисточка на конце
хвоста черная с небольшой примесью белых волос на заднем ее конце.
Череп сходен по своему строению с таковым Alactagulus acontion, но
несколько крупнее и с более ясно обозначенными надглазничными греб-
нями. Зубная формула у; коренные зубы имеют такое же строение, как
у Alactagulus acontion и Pygerethmus platyurus. Строение penis в основных
чертах сходно с. Pygerethmus platyurus, особенно по сильно отогнутым
внутрь роговым шипам близ его основания и по наличию на дорзальной
поверхности резко очерченной узкой продольной углубленной площадки.
Фиг. 47. Голова тушканчика Житкова (Pygerethmus zhitkovi Kuzn.).
Из особенностей строения скелета для Pygerethmus zhitkoviособенно
характерно строение таза, в значительной степени сходное с таковым
У Pygerethmus platyurus, особенно по очень короткому лонному сращению
и по тонким ветвям лобковой кости, сходящимися в области сращения
под острым углом.
Измерения: длина тела 97—122; хвост 101—128; ступня 38—43;
ухо 30—34; кондилобазальная длина черепа 27—28 мм; скуловая ширина
20—23; межглазничное пространство 7—7.3.
По своим систематическим признакам тушканчик Житкова занимает
до некоторой степени промежуточное положение между Pygerethmus
platyurus и Alactagulus acontion в частности по длине хвоста, развитию
его концевой кисточки, длине ступни и т. п. То же самое следует сказать
относительно строения тазовых костей, которое является одним из наиболее
крупных признаков различия между родами Alactagulus и Pygerethmus.
После того, как стал известен тушканчик Житкова, возникли вполне
определенные сомнения о целесообразности сохранения отдельных родов
Фауна СССР, Виноградов 10
146
ALLACTAGINAE
Alactagulus и Pygerethmus. До окончательного выяснения этого вопроса
мы причисляем здесь тушканчика Житкова к роду Pygerethmus на том
основании, что более внимательное изучение его признаков после получе-
ния нового и более совершенного сохраняемого материала, чем тот,,
который имелся в сборах Б. А. Кузнецова, дало возможность установить
большую близость этой формы к роду Pygerethmus, чем к Alactagulus*
в противоположность поддержанному мной первоначальному мнению
Б. А. Кузнецова, основанному лишь на исследовании 3-х шкурок с черепами..
Распространение. В настоящее время Р. zhitkovi известен
лишь из немногих районов, в частности следующих: Алакульская равнина,,
р. Каратал, юго-западное побережье Балхаша (Буру-байтал), хребет
Кан-тау (Чу-Илийские горы), Аулие-атинский округ (хребет Джамбул,,
долина Кара-джар). Таким образом, по имеющимся в настоящее время,
немногим данным можно считать, что тушканчик Житкова распространен
от озера Ала-куль и равнин Прибалахашья на юго-восток до Аулие-
атинского округа; можно предполагать также, что этот вид распространен,
и в пустыне Бедпак-дала, по крайней мере в ее восточной части.
Данные по биологии. По наблюдениям Б. А. Кузнецова,,
на побережья Ала-куля этот вид обитает в глинистой полупустыне, где-
он встречается гораздо реже других тушканчиков, и что этот зверек
роет неглубокие норы, которые были найдены по краям дороги близ-
ст. Рыбальное. На этикетках П. Окунева, добывшего этого тушканчика
в Аулие-атинском районе, указывается, что этот вид добыт в полынной
степи. В 1934 г. мне удалось собрать значительную серию тушканчика
Житкова на юго-западном побережья Балахаша близ поселка Буру-Байтал.
Здесь эти зверьки встречались исключительно на обширных низинах,
образовавшихся на месте прежних заливов Балахаша, • с мягкой почвой,
типа пухлых солончаков и довольно обильным и высоким растительным!
покровом, преимущественно из солянок и отчасти из мелких кустов тама-
риска и небольших зарослей тростника. Основной фон растительности
здесь составлен следующими видами: Salsola brachiata Pall., S. crassa MB,.
S. kali L., Sueda setigera ;M. Tand., S. dendroides L., Petrosimonia bra-
chiata Bg., Kalidium foliatum M. Tand., Artemisia terrae-albae H. Krasch.,
Atriplex tatarica L., Polygonum argyroleucum Send, и др.
Из всех мелких зверьков тушканчик Житкова был здесь наиболее
многочисленным. Здесь всюду имелись в большом количестве норы
Р. zhitkovi и других видов тушканчиков, их копанки, „купальные ямки"'
(„купанье" в пыли), многочисленные следы на наиболее рыхлых частях,
солончака; во многих местах имелись длинные протоптанные тушканчиками
Тропинки, часто усеянные отгрызанными веточками солянок, которые
вероятно являются здесь основной пищей тушканчиков. Следует отметить,
что при наличии чрезвычайно рыхлого с поверхности грунта здесь очень
трудно было находить закопавшихся в норках зверьков, так как заполнен-
ная землею часть норы, не отличающаяся по окраске и по консистенции
9. SC ART UR US
147
от окружающего грунта, быстро терялась при раскопке. Большин-
ство добытых здесь экземпляров Р. zhitkovi было поймано в кап-
каны у открытых нежилых нор, которые обычно посещаются зверьками
по ночам.
9. Род SCARTURUS GLOGER — ЧЕТЫРЕХПАЛЫЕ ТУШКАНЧИКИ
Glgoer, Hand- und Hilfsb. Naturg. 1, 1841 : XXXI et 106.
По наружным признакам сходны с представителями рода Allactaga,
но на задней конечности имеются лишь четыре пальца (внутренний палец
отсутствует). Penis в существенных чертах сходен с таковым у Allactaga’,
срединная бороздка на его дорзальной поверхности доходит до вершины
penis, как у A. williamsi и A. elater.
Череп Scarturus очень сходен с таковым у представителей рода
Allactaga, особенно представителей группы A. euphratica; сходство может
быть обнаружено не только в общих очертаниях, но даже в размерах;
однако, межглазничное пространство у Scarturus значительно шире, чем
у A. euphratica’, обыкновенно у последнего вида межглазничное простран-
ство составляет не более 8.6 мм, а у Scarturus эта величина обычно
равна 10—И мм. Лобно-теменной шов чешуйчатой кости не имеет вырезки,
которая характерна для всех представителей рода Allactaga. Foramina
incisiva у Scarturus несколько более широки и более Открыты, чем
у Allactaga. Однако не представляется возможным найти другие более
важные краниологические различия между обоими родами. Задняя ступня
Scarturus представляет собой значительное уклонение от строения этой
части скелета у всех других Allactaginae, в связи с отсутствием первого
(внутреннего) пальца, от которого остается лишь рудимент, в виде удли-
ненного entocumiforme и сидящей на его дистальном конце очень маленькой
косточки, являющейся рудиментом плюсневой кости первого пальца,
(фиг. 48 Б). В остальных признаках ступня Scarturus сходна с другими
представителями подсемейства Allactaginae. Ложно-коренной зуб имеется
у взрослых особей, его размеры (поперечник) очень маленькие. Коренные
зубы по строению совершенно сходны с таковыми у Allactaga, т. е.
с раздвоенными выдающимися углами на наружной стороне.
По морфологическим признакам род Scarturus стоит очень близко
к роду Allactaga, отличаясь от последнего лишь редукцией внутреннего
пальца на задней конечности; если не принимать во внимание указанный
признак, то сходство между родами Scarturus и Allactaga является более
значительным, чем между Allactaga и Pygerethmus. Выше было указано,
что особенно значительным является сходство в наружных признаках,
строении черепа и в размерах между Scarturus и группой Allactaga
williamsi—euphratica.
Распространение. S. tetradactylus является редким объектом
в коллекциях и поэтому в настоящее время имеется лишь небольшое
10*
148
DIPODINAE
фиг. 48. Редукция боковых пальцев задней конечности у трехпалых тушканчиков (А. — Dipus
sagitta) и четырехпалого (Scarturus tetradactylus). 1 и 4 — внутренняя сторона; 2 и 5 перед-
няя, 3 и 6 — наружная. Astr. — astragalus. Calc. — calcaneus, Cbd. — cuboideum, Ectcn. —
ectocuneiforme, Mscn. — mesocuneiforme, navic — naviculare, Pis — pisiforme, Ses — sesamo-
ideum, X — рудименты 5-го пальца, II—IV metatarsalia (,,os canon”), V—5 — metatarsalc.
ОПИСАНИЕ ПОДСЕМЕЙСТВА
149’
количество пунктов нахождения этого зверька, находящихся на терри-
тории Египта и Ливии.
К рассмотренному роду относится лишь один вид, описанный по экзем-
пляру добытому в Ливийской пустыне.
1. Scarturus tetradactylus Licht.
Lichtenstein, 1825; Pocock, 1922.
Этот вид был описан по экземпляру из Ливийской пустыни. Наи-
более характерные признаки этого животного описаны при характери-
стике рода.
Подсем. DIPODINAE —ТРЕХПАЛЫЕ ТУШКАНЧИКИ
Форма головы в существенных чертах сходна с таковой у Allacta-
ginae. Уши короткие, при основании трубчатые, будучи отогнуты вперед
обычно достигают лишь глаза. Когти на передних конечностях более
длинные и более изогнутые, чем у Allactaginae. Задние конечности с тремя
пальцами; нижняя поверхность пальцев покрыта длинными щетинистыми
волосами, образующими щетку; на концах пальцев имеются маленькие,
сжатые с боков зазубренные подушечки, значительно уступающие по раз-
мерам таковым у Allactaginae. Os penis имеется. Вершина penis большей
частью снабжена глубокими складками („карманами") с каждой стороны
наружного отверстия уретры; на нижней поверхности penis всегда имеется
глубокая продольная щель; с каждой стороны от этой щели может быть,
еще одна более короткая щель. Penis овальный или с почти парал-
лельными краями. Кроме мелких шипиков на поверхности penis, часто
имеется пара длинных стилевидных шипов. В средней части верхней
поверхности penis большей частью имеется голая, хорошо обособленная,
площадка (эта площадка расположена над срединным гребнем os penis).
Череп по основному плану строения сходен с таковым у Allactaginae.
Главнейшими признаками отличия являются следующие. Межчелюстная
кость не вытянута так сильно вперед, как у Allactaginae, в связи с чем
ее передний край лежит на одной вертикальной линии с передним краем
носовых костей. Передняя восходящая часть скуловой дуги относительно
шире, в связи с чем foramen infraorbitale относительно уже, чем у Allacta-
ginae. Lacrymalia крупные, но относительно меньшей величины, чем
у Allactaginae, их наибольший диаметр в 2% или почти в 3 раза меньше
межглазничного сужения. Hamuli птеригоидных отростков в большей или
меньшей степени заходят за передние края bullae. Bullae tympani сильно
раздуты, своими передне-внутренними углами они налегают на basis phe-
noideum й соприкасаются друг с другом, иногда приходя в тесную взаим-
ную связь с образованием диска в месте соприкосновения. Mastoidea
большей частью очень сильно раздуты, как ни у одного из представи-
телей Allactaginae', даже при относительно слабом их развитии, как у рода
150
DIPODINAE
Dipus-, их внутреннее строение сходно с таковым у Cardiocraniinae и Eucho-
reutinae mastoidea частично выходят на верхнюю поверхность черепа
(преимущественно через расширенные fenestra postglenoida), Нижняя
челюсть довольно низкая, как у Allactaginae, но задний угол (angulus
posterior) резко повернут наружу и хорошо развитый передний угол
(angulus anterior) заметно повернут внутрь. Венечный отросток сильно
редуцирован, как у Allactaginae или у Cardiocraniinae. Некоторые при-
знаки строения слуховых' косточек у представителей подсемейства
Dipodinae, как напр. относительно крупный incus и довольно длин-
ный processus brevis, сходны с таковыми у Cardiocraniinae-, однако у Dipo-
dinae corpus mallei и incudis не столь узкие и не столь вытянутые,
как у Euchoreutinae и Cardiocraniinae. Шейные позвонки у всех
представителей Dipodinae сросшиеся, за исключением первого (атланта)
и седьмого; срастаются не только тела позвонков, но также нейраль-
ные дуги и дистальные части поперечных отростков. Между свободными
проксимальными частями поперечных отростков имеются 4 или редко 3
отверстия. Весь комплекс шейных позвонков укорочен и расширен;
максимальное укорочение шеи имеет место у представителей рода Jaculus.
В строении задних конечностей характерно полное слияние трех средних
плюсневых костей, образующих „цевку“, а также редукция первого и чет-
вертого пальцев, от которых остаются лишь ничтожные рудименты; инте-
ресно отметить, что характерная для всех настоящих тушканчиков сильно
удлиненная entocuneiforme, несмотря на редукцию первого пальца,
остается хорошо развитой (рис. 48), от первого пальца сохраняется лишь
ничтожный остаток в виде маленькой косточки, сидящей на дистальном
конце entocuneiforme. Рудименты 5-го пальца обычно сохраняются в виде
трех маленьких косточек (рис. 45), гомологию которых не представляется
возможным выяснить точно без специального исследования этих признаков
на постэмбриональных стадиях. В строении тазовых костей у Dipodinae
имеют сходство с Allactaginae, особенно по значительной длине лонного
сращения; главнейшими отличительными признаками между названными
подсемействами является указываемая ниже , постоянная разница в строе-
нии задней части таза и наличие Dipodinae раздвоенного tuberculum
ileo-pectineum. Таким образом строение таза у Dipodinae характеризуется
сочетанием следующих признаков: crista iliaca хорошо развита. Tuber
ischii сильно развит и вместе с прилежащей частью ischium заметно
отогнут в дорзальном направлении. Symphisis pubis очень длинен (как
у Allactaginae)-, его длина составляет не менее двух третей расстояния
от acetabulum до седалищного бугра. Вентральный край ischio-pubicum,
образующий сращение, переходит в задний свободный край этой кости
более или менее резким закруглением .или изломом. Tuberculum ileo-
pectineum присутствует и ясно разделен на два отдельных бугра. Нисходя-
щая ветвь ischium образует с ее горизонтальной (параллельной позвоноч-
нику) ветвью почти прямой угол. Ложно-коренной зуб имеется во взрослом
ОПИСАНИЕ ПОДСЕМЕЙСТВА
151
•состоянии или сохраняется лишь у молодых особей, или отсутствует
вовсе. Жевательная поверхность первого коренного зуба, плоская или
почти плоская и строение эмалевых складок упрощено; сохраняется лишь
по два выдающихся угла на наружной и внутренней стороне: передняя
добавочная складка недоразвита или отсутствует вовсе; если она имеется,
•она не смещена на наружную сторону.
Распространение рассматриваемой группы охватывает пре-
имущественно песчаные пустыни юго-восточной Европы, Азии и Север-
ной Африки (лишь представители рода Scirtopoda встречаются в степях,
полупустынях и глинистых пустынях). В Европе распространение этой
.группы достигает к западу до нижнего течения Днепра (изолированное
реликтовое нахождение); далее представители этой группы распространены
ют северо-восточного Предкавказья на север до широты Камышина;
к востоку от Волги северная граница рассматриваемого подсемейства
в пределах Казахстана и Западной Сибири достигает приблизительно 50°
и в немногих местах даже 52° с. ш.; далее на восток северная граница
распространения подсемейства лежит в пределах Северной Монголии
и Манчжурии. На юг представители рассматриваемой группы распростра-
нены в Иране, Месопотамии, Сирии, Северной Аравии и Северной Африке
(Сахаре), на запад до Алжира и Туниса; далее на восток южная граница
лежит в пределах южного Ирана и вероятно Афганистана, южного Синц-
зяня (Китайского Туркестана), Цайдама, Нань-шаня, Южного Алашаня
и Северного Китая (Шенси и Чили).
Достоверные ископаемые остатки рассматриваемого подсемейства
известны лишь с плейстоцена (род Dipus). Как уже было указано в главе
о геологической истории семейства, описанный Ch. Young’oM (1927) из плио-
цена или верхнего миоцена род Plesiodipus, занимающий, по мнению наз-
ванного автора, промежуточное положение между Allactaga и Dipus,
в действительности вероятно является представителем подсемейства Car-
diocraniinae.
Подсемейство Dipodinae является довольно резко дифференцирован-
ной группой. Эта группа имеет небольшее число сходных признаков с под-
семейством Allactaginae', Dipodinae имеют также некоторые черты сход-
ства с Cardiocraniinae (форма и размеры уха, щетка на задней ступне,
сильно раздутые mastoidea, бороздки на резцах и пр.); возможность
последней связи до некоторой степени подтверждается, повидимому,
и на палеонтологическом материале; во всяком случае, возможно, что
описанный Young’oM (1927) из плиоцена или верхнего миоцена Ганьсу
Protalactaga grabaui, имеющий примитивное строение зубов сходное
с Cardiocranius и сросшиеся 3 средних плюсневых кости, занимает проме-
жуточное положение между Cardiocraniinae и Dipodinae.
В экологическом отношении рассматриваемая группа характеризуется
тем, что почти все ее представители обитают в песчаных пустынях;
•единственным исключением в этом отношении является вероятно лишь
152
DIPODINAE
Scirtopodaielum, приуроченный к твердым субстратам, но иногда встречаю-
щийся также и в песках. Наиболее очевидным приспособлением к рых-
лому песчаному субстрату у Dipodinae является наличие у них на задней
ступне щетки из твердых прямых волос.
Экономическое значение рассматриваемой группы невелико и выра-
жается в наблюдающихся иногда повреждениях посадок, служащих для
закрепления песков; положительная экономическая роль этих грызунов,
заключается в том, что местами они являются вместе с некоторыми
другими грызунами основной кормовой базой для промысловых хищных
млекопитающих песчаных пустынь (пустынные кошки, лисицы, перевязки
и др.).
Подсемейство Dipodinae состоит из 5 хорошо дифференцирован-
ных родов, большинство которых является монотипическими, т. е. содер-
жат по одному виду. Из этих родов относительно более примитивное
положение 'занимает Dipus (наличие РМ, бугорчатое строение коренных
зубов, относительно слабо раздутые mastoidea и пр.); все другие роды
более близки друг к другу, чем к Dipus. Из этих родов относительно
более низкую эволюционную ступень занимает Scirtopoda (отсутствие
дифференцированного „знамени", наличие в молодом или взрослом состоя-
нии рудимента РМ, относительно более слабо раздутые mastoidea, чем
у упомянутых выше родов); на следующем месте должен быть поставлен
Paradipus, отличающийся от Scirtopoda своеобразным строением щетки
на задней ступне, полным исчезнованием РМ, снабженной двумя лопастями:,
когтевой фалангой среднего пальца задней конечности; наиболее изме-
ненными следует считать близкие роды Eremodipus и Jaculus, имеющие
своеобразное строение черепа с чрезвычайно сильно раздутыми mastoi-
dea, с сильно развитым гребнем или шипом на чешуйчатой кости и т. д..
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ
1 (2). Сосцевидные кости не вздуты и не выдаются при рассматривании
черепа сверху в виде округлых выступов по бокам затылка. Постгле-
новидные отверстия открываются внутрь мозговой полости. Ложно-
коренной зуб относительно хорошо развит. Верхние резцы желтые.
.............................................10. Dipus Gmel.
2 (1). Сосцевидные кости значительно вздуты и выдаются при рассматри-
вании черепа сверху в виде округлых выпуклостей по бокам затылка..
Постгленоидные отверстия частично или целиком закрыты и внутри
частью petromastoideum. Ложно-коренной зуб во взрослом состоя-
нии обычно отсутствует и в виде слабого рудимента сохраняется
лишь у одного вида (Scirtopoda andrewsi Н. АП.). Верхние резцы
белые.
3 (6). Височная область черепа без гребня или зубца, образованных
выступом теменной кости, входящим в вырезку чешуйчатой. Вершины
ОПИСАНИЕ ПОДСЕМЕЙСТВ А
155
bullae tympani не соединены друг с другом швом или не сопри-
касаются вовсе. Сосцевидные кости значительно раздуты, но не
образуют больших пузырей.
4 (5). На хвосте нет резко обособленного „знамени" (длинные волосы
концевой части хвоста, постепенно укорачиваясь достигают почти
середины хвостового стержня. Щетка на подошве задней ступни
одноярусная, образованная темными изогнутыми волосами). Задний
край костного нёба оканчивается выступом значительно позади от
уровня заднего края m3. Fenestrae postglenoideae образуют довольно
широкую щель частично закрытую изнутри частью petromastoideum.
Слёзные кости не смещены значительно наружу, их внутренние
вершины соприкасаются с лобной костью. Нижняя челюсть с хо-
рошо развитым альвеолярным отростком...............................
.......... ............................11. Scirtopoda Brandt.
5 (4). На конце хвоста имеется хорошо обособленное „знамя". Щетка на
подошве задней ступни двухярусная, т. е. в ней, кроме тонких и длин-
ных волос, имеются более короткие и толстые щетинки. Задний край
костного нёба вогнутый и расположен на уровне середины m3. Fene-
strae postglenoideae очень маленькие и округлые, целиком закрытые
изнутри частью petromastoideum. Слёзные кости значительно сме-
щены наружу и причленяются к узкому поперечному отростку лобной
кости. Нижняя челюсть без альвеолярного отростка или бугра. . .
........................................ . 12. Paradipus Vinogr.
6 (3). Височная область черепа с резко обособленным гребнем или зубцом,,
образованным боковым отростком теменной кости, входящим в узкую
глубокую вырезку чешуйчатой кости. Вершины bullae tympani соеди-
нены друг с другом швом или слиты друг с другом (в обоих слу-
чаях в месте соединения обеих вершин образуется дискообразное
расширение).
7 (8). Боковые отростки каждой теменной кости снабжены резко обособлен-
ным зубцом или шипом. Постгленоидные отверстия очень маленькие,
обыкновенно несколько вытянутые. Верхний край наружной стенки,
нижне-глазничного канала не слит со стенкой максиллярной кости.
Корень нижнего резца образует слабо развитый альвеолярный
отросток....................................13. Eremodipus Vinogr..
8 (7). Боковые отростки каждой теменной кости снабжены резко развитым
гребнем, причем основание этого гребня образовано прилежащими
частями чешуйчатой кости. Постгленоидные отверстия очень широ-
кие и образуют почти равнобедренные треугольники. Верхний край
наружной стенки нижнеглазничного канала у взрослых экземпляров
совершенно прирастает к стенке верхнечелюстной кости. Корень
нижних резцов образует сильно выдающийся альвеолярный отросток
(бугор).............................................14. Jaculus ErxL
154
DIPODINAE
10. Род DIPUS GMELIN (1788) — МОХНОНОГИЕ ТУШКАНЧИКИ
Gmelin, Linne Syst. Nat., 1778.
Из наружных признаков для рода Dipus характерны короткие труб-
чатые уши, наличие хорошо развитой щетки из длинных прямых волос на
нижней поверхности пальцев задней ступни, довольно хорошо обособлен-
ное „знамя" на конце хвоста и желтая окраска передней поверхности
резцов (у всех других представителей подсемейства резцы белые). Сре-
динный гребень os penis расположен на ее стержне (не на базальной
пластинке) в его основной части. Передне-наружные углы базальной
пластинки не выражены и округлены. На нижней поверхности penis сре-
динная продольная щель начинается лишь от середины длины penis;
с каждой стороны от нее имеется еще по одной глубокой щели, начинаю-
щихся от основания и доходящих почти до его вершины. Два длинных
стилевидных шипа хорошо развиты. Выступ на наружной стороне parietale
(над основанием processus zygomaticus ossis squamosi) не образует острого
гребня или шипа, а лишь широкую округлую выпуклость. Fenestrae post-
g’lenoideae имеют вид довольно узкой щели, которая открывается
в внутреннюю полость мозговой коробки (т. е. не закрыта снизу разрос-
шеюся верхнею частью petromastoideum). Наружная стенка нйжне-глаз-
ничного канала не прирастает к стенке maxillare. Слёзные кости не сме-
щены наружу, их вершины лежат в переднем верхнем углу орбиты и
сочленяются с frontale. Задний край костного нёба образует направленный
назад угловатый выступ, оканчивающийся одной или двумя вершинами;
задняя часть нёба простирается значительно назад от уровня заднего
края т3. Вершины обоих bullae tympani соприкасаются друг с другом, не
образуя, однако, между собою тесного соединения. Mastoidea слабо раздуты,
при рассматривании черепа сверху они не образуют выступов по бокам
задней части черепа. Корень нижнего резца образует на поверхности
нижней челюсти хорошо развитый выступ (альвеолярный отросток), отсут-
ствующий лишь у Paradipus. В устройстве шейных позвонков характерны
следующие признаки: границы между слившимися телами 2—6-го шейных
позвонков довольно ясно заметны; равным образом заметны границы между
слившимися нейральными дугами тех же позвонков. Дистальные концы
поперечных отростков 5-го и 6-го позвонков совершенно слиты друг
с другом; вершина остистого отростка заострена и несколько отклонена
вперед. На боках вентральной стороны сросшегося шейного комплекса
позвонков имеются 4 отверстия между свободными частями поперечных
отростков. Цевка более чем вдвое длиннее среднего пальца. Основная
фаланга среднего пальца почти вдвое длиннее второй фаланги этого
пальца. Вырезки между сочленовными поверхностями на дистальном конце
цевки составляют около х/7 длины всей цевки. Зубовидный отросток на
нижней поверхности каждой когтевой фаланги слабо развит. Ложно-
10. DIPUS
155
коренной зуб хорошо развит и сохраняется во взрослом состоянии.
Передняя добавочная складка на первом коренном зубе, по сравнению
•с другими Dipodinae, обыкновенно более развита; иногда эта складка
заменена маленьким изолированным стилевидным бугорком.
Распространение. Распространение рассматриваемого рода,
представленного лишь одним видом (£>. sagitta) охватывает пески крайнего
юго-востока Европы, южного Казахстана, Средней Азии, вероятно, Север-
ного Ирана, Синцзяня, Монголии и Северного Китая. Подробности распро-
странения указываются при характеристике вида (D. sagitta). Распро-
странение мохноногих тушканчиков приурочено исключительно песчаным
массивам и поэтому общий ареал рода имеет прерывистый островной
характер. Представители этого рода обитают в разнообразного типа
песках от голых или.едва заросших барханов, до бугристых песков, покры-
тых кустарниками и даже иногда заросших сосновым бором.
К роду Dipus прежние исследователи относили несколько видов
(D. sagitta Pall., D. lagopus, D. nogai Satun., S. deasyi В.-Ham., D. sower-
byi Thos., D. halli Sowerby); в настоящее время при более тщательном
изучении морфологических признаков этих форм, является совершенно
очевидным, что этим формам нельзя придавать видового значения и что
они могут быть рассматриваемы лишь в качестве подвидов. Таким обра-
зом, род Dipus составлен единственным видом D. sagitta Pall.
*1, Dipus sagitta Pall. — Мохноногий тушканчик (фиг. 49—51).,
Pallas, 1773 (Mus). [пески по Иртышу близ станции Ямышевской]; Trouessart,
1910 : 207 (Dipodipus); Ognev, 1930: 207 (подвиды); Бёме и Крассовский, 1931 (биология);
-Виноградов и Аргиропуло, 1931 (биология). — lagopus Lichtenstein, 1828:121. — nogai
Satunin, 1907 :127; 1920: 203.
Наиболее существенные признаки рассматриваемого вида указаны
выше в диагнозе рода Dipus. Окраска у различных географических
•форм колеблется по относительно темной охристо-коричневатой или
буровато-серой до бледно-охристо-песчаной.
Измерения. Длина тела 105—130 мм; дл. хвоста 135—150 мм;
дл. ступни 60—65 мм; выс. уха 15—20 мм. Кондилобазальная длина
черепа 30.3—34.3 мм; скуловая ширина 21.0—24.5; межглазничная ширина
9.7—12.1; дл. носовых костей 12.2—15.5; диастема 8—9.5, максиллярный
ряд зубов 5.3—6.8
Распространение (фиг. 52). Общий ареал вида был кратко
охарактеризован при описании рода. Здесь приводятся следующие подроб-
ности распространения D. sagitta: на северо-западе распространение этого
вида достигает Прикумских песков в восточном Предкавказьи и Волжско-
Донских песков в пределах Сталинградского и Усть-Медведицкого
округов до устья р. Медведицы; в Волжско-Уральских песках этот вид
распространен до широты г. Урды и по р. Уралу до окрестностей
Индерска; далее на восток он встречен в песках Кок-джида б. Темирского у.
156
DIPODINAE
и на северном побережья Аральского моря, откуда он, вероятно, рас-
пространен несколько севернее на территории песков Большие Барсуки
и Приаральские Кара-кумы; далее на восток этот вид найден на терри-
тории Мугон-кумы; в пределах Семиречья этот вид найден в ряде
пунктов б. Лепсинского и Копальского уездов; по р. Иртышу D. sagitta
распространен до с. Семиярского (50°30" с. ш.) и далее на восток найден
в песках ленточных боров Приалтайской степи (с. Новенькое и Локоть-
Фиг. 49. Мохноногий тушканчик (Dipus sagitta Pall.), роющий нору.
Фото А. И. Аргиропуло.
Рубцовского окр.); далее северная граница распространения этого вида,
выходит за пределы Союза, следуя по отдельным песчаным массивам
пустыни Гоби. На восток распространен приблизительно до Манчжурской
границы (в Манчжурии не найден). К югу этот вид распространен вероятно-
в северном Иране и северном Афганистане, найден в нескольких пунктах
Синцзяня (Нура, Карашар, Хотан, Хами, Куфи), достигает северных
окраин Тибета (Цайдам), Нань-шаня, Южного Алашаня и Северного
Китая, где он найден в провинции Шенси (Yun-li, Yu- lin-fu) и в северной
части провинции Чили (Chile- feng). D. sagitta является одним из немно-
гих видов тушканчиков, образующих ясные географические формы, боль-
шинству которых можно придавать подвидовое значение; эти формы раз-
личаются между собой не только окраской меха, но и некоторыми
краниологическими признаками. До настоящего времени известны следую-
щие формы D. sagitta.
10. DI I'US
157
Фиг. 5C. Мохноногий тушканчик, съедающий ветку саксаула. Фото А. И. Аргиропуло.
Фиг. 51. Мохноногий тушканчик.
158
DIPODINAE
1. D. s. nogai Satunin, 1907; Ognev, 1930. Описан из Прикумских
песков Восточного Предкавказья. Отличается от других форм преобла-
данием серого цвета в окраске. Спинная сторона окрашена в тусклый
буровато-серый цвет с легкой примесью охристых или рыжеватых тонов.
D. s. nogai является одной из наиболее крупных форм рассматриваемого
вида: наибольшая длина черепа вполне взрослых экземпляров 35—
Зб.З мм; кондилобазальная длина 32.4—34.3; затылочная высота более
значительна, чем у всех других форм, колеблясь у вполне взрослых
экземпляров от 13.4 до 14.7 мм.
Распространение. Пески восточного Предкавказья (долина
р. Кумы и Кизлярский окр.).
2. D. s. innae Ognev (1930) (описан из Енотаевского окр.). Отличается
от других форм более ярким охристо-рыжеватым тоном окраски верхней
стороны тела. Измерения заметно не отличаются от таковых у описывае-
мого ниже D. s. lagopus Licht.
3. D. s. lagopus Lichtenstein (1823); Ognev (1930) (сев.-вост. побережье
Аральского моря в долине Сыр-Дарьи близ оз. Камышлы). Характери-
зуется бледной охристо-песчаной окраской верха. Верхняя поверхность
стержня хвоста несколько бледнее и серее, чем у D. s. innae Ogn. По
измерениям занимает среднее положение между рассматриваемыми фор-
мами: наибольшая дл. черепа 32—34.7 мм, кондилобазальная длина 30.2—
32.8; скуловая ширина 21.7—24.0; межглазничное пространство относи-
тельно широкое, достигая иногда 12.1 мм.
Распространение. Пески средней Азии и Южного Казахстана
К описываемой форме близко примыкает форма, обитающая в Муюн-
Кумах, отличающаяся, однако, от типичных D. s. lagopus несколько более
темной окраской и большей примесью бурых тонов.
4. D. s. deasyi Barret-Hamilton (1900) (Синьцзянь, Нура). Рассматри-
ваемая форма близка к D. s. lagopus, но некоторые экземпляры отли-
чаются еще более бледной окраской и имеют кремовый оттенок вместо
охристо-рыжеватого у предыдущих форм. По измерениям, повидимому,
является наиболее мелкой из всех известных форм рассматривае-
мого вида.1
Распространение. Пески Синцзяна (Китайского Туркестана).
5. D. s. sagitta Pallas (1773); Ognev (1930) (пески долины р. Иртыша
близ ст. Ямышевской). Характеризуется относительно темной окраской.
Окраска верха охристо-коричневатая с оливковым оттенком. Размеры
относительно крупные: наибольшая длина черепа до 36.2 мм, кондило-
базальная длина до 34 мм; скуловая ширина до 24.5 мм.
Распространение. Пески Западной Сибири и Восточного
Казахстана: долина р. Иртыша от Ямышевской до Семипалатинска;
ленточные боры Приалтайской степи (с. Локоть Рубцовского окр.).
1 Цифры не приводятся, в виду наличия измерений лишь немногих взрослых экзем-
пляров.
10. DIPUS
159
Фиг. 52. Распространение Dipas sagitta Pall. (1); основные массивы песчаных пустынь
в пределах СССР, где известны нахождения D. sagitta (2).
160
DIPODINAE
6..D. s. sowerbyi, Thomas (1908) (провинция Шенси, Youn-lin).
По окраске сходен с D. s. sagitta Pall., но с несколько более рыжим оттен-
ком. Размеры меньше, чем у предыдущей формы: наибольшая длина
черепа 32.2—34.2 мм: кондилобазальная длина 30.3—31.8; скуловая
ширина 21.1—22.8.
Кроме провинции Шенси, широко распространен в пустынях
Монголии.
7. D. s. halli Sowerby (1920) (сев.-вост. часть провинции Чили, Китай).
Окраска сходна с таковой у D. s. sowerbyi, но с меньшей примесью
желтоватых тонов и более темная. Размеры значительно крупнее, чем
у названной формы: наибольшая длина черепа до 36.6 мм; кондилобазаль-
ная длина до 33.9 мм.
Найден лишь в северо-восточной части провинции Чили в Китае,
близ границы Монголии.
Д а н н ы е п о биологии. Мохноногий тушканчик является харак-
терным обитателем барханных песков, но в отличие от гребнепалого
тушканчика он селится не только на голых барханах, но также и на
заросших кустарниками песках. Особой своеобразной стацией мохноногого
тушканчика являются пески, заросшие сосновым бором в окрестностях
Семипалатинска и с. Новенького, как это было упомянуто выше. Мохно-
ногий тушканчик закупоривает свою нору на день; в том месте, где
.закопался тушканчик, можно видеть небольшую истоптанную площадку,
имеющую большей частью вид треугольника или полуовала (фиг. 57);
пробка в начальной части норы бывает невелика, не превышая в толщину
нескольких сантиметров, нора углубляется под очень острым углом косо
и вниз и обычно имеет в наиболее глубокой части камеру, от которой ход
поднимается кверху, оканчиваясь под самой поверхностью песка („око-
шечко", или „копеечка"); часто это окошечко снаружи совершенно неза-
метно, иногда же здесь образуется небольшая рыхлая пробочка, заметная
снаружи (пока ветер не заметет все видимые следы ночной деятельности
зверьков на поверхности песка); общее протяжение обоих ходов в норах
самцов обычно не превышает 1—2 метров, а наибольшая глубина х/2 метра.
Норы самок, по нашим наблюдениям, имеют гораздо большее протяжение
и значительно более сложное строение, причем главный ход имеет
в длину до 5 метров и от него отходит несколько отнорков, достигающих
в длину до 1 метра и более. Когда раскопка норы тушканчика бывает
доведена до наиболее глубокой части и начинает раскапываться ход,
ведущий к „окошечку", неожиданно окошечко открывается и оттуда вместе
с кучей песка выскакивает тушканчик, который стремительно убегает
и скрывается из глаз; впрочем, в этом случае зверек обычно не убегает
далеко, а прячется где-нибудь под кустами. Следы мохноногого тушкан-
чика (фиг. 59) имеют характерный вид: длина каждого следа колеблется
в пределах 23—26 мм; форма следа, как и у других трехпалых тушкан-
чиков, более или менее треугольна, с закругленной передней частью;
10. DIPUS
161
-Фиг. 53.
Пески с зарослями черкеза (Salsola Richteri) близ города Каракуля
Характерные места обитания мохноногого тушканчика! ₽ У
Фиг. 54. Пески с зарослями селина (Aristida pennata) близ
Характерные места обитания мохноногого тушканчика.
Каракуль.
'Фауна СССР, Виноградов
11
162
dipodinae
Фиг 55. Бугристые пески близ ст. Хаджа-Давлет Каракульского района.
Характерные места обитания мохноногого тушканчика.
Фиг. 56. Бугристые пески близ ст. Джебел Красноводского района. Характерные
места обитания мохноногого тушканчика.
10. DI PUS
163
Фиг. 57. Закопанная на день нора мохноногого тушканчика.
Фиг. 58. Раскопанная нора мохноногого тушканчика.
11*
164
DIPODINAE
в отличие от гребнепалого тушканчика, следы D. sagitta имеют менее
вдавленные боковые края и очень слабо выраженную перетяжку в средней
их части. В песках Средней Азии мохноногий тушканчик ведет очень
Скрытный образ жизни. Мы неоднократно пытались сделать наблюдения
над тушканчиками в сумерки по выходе их из нор и для этой цели дежу-
рили на барханах целыми вечерами и несколько раз оставались там
до рассвета, и нам ни разу не
удалось видеть живого тушкан-
чика на воле, хотя во время
этих дежурств тушканчики были
рядом, на что указывали их
свежие следы на песке. Впро-
чем, однажды мне удалось ви-
деть занятого рытьем мохноно-
гого тушканчика на поверхности
песка при необычайных обстоя-
тельствах— поздно утром, при
ослепительном солнечном свете.
Относительно размножения D.
sagitta мы имеем лишь следую-
щие наблюдения: 15 мая нами
была вскрыта, добытая близ
ст. Ходжа-Давлет, самка с че-
тырьмя вполне развитыми дете-
нышами 20 мм длины (от темени
до крестцовой области); 25 июня
близ ст. Бургучи в одной норе
вместе с самкой были добыты
Фиг. 59. Следы мохноногого тушканчика.
особи 'и в другой норе три детеныша
полагаем точными данными о составе
два детеныша, достигших бо-
. лее половины роста взрослой
того же возраста. Мы не рас-
пищи мохноногого тушканчика
и можем лишь отметить, что в неволе тушканчик охотно поедал зеленые
веточки саксаула, кроме того, при раскопке норы мы находили внутри
ходов остатки поеденных тушканчиком плодов кандыма (Calligonum).
Опубликованные Л. Б. Бёме и Д. Б. Крассовским наблюдения (1931) над
ногайским тушканчиком (D. s. nogai Satun.) в песках северо-восточного
Предкавказья в некоторых отношениях расходятся с нашими наблюде-
ниями над среднеазиатским мохноногим тушканчиком (D. s. lagopus
Licht.), как это было уже подробно рассмотренно в статье Б. С. Виногра-
дова и А. И. Аргиропуло (1931) по биологии средне-азиатских тушкан-
чиков, что позволяет предполагать, что образ жизни различных подвидов
мохноногого тушканчика имеет свои характерные особенности: в част-
ности, по наблюдениям упомянуых авторов, норы ногайских тушканчиков
10. DIP US
165
хорошо заметны издали по присутствию характерного выброса песка,
тогда как норы среднеазиатского тушканчика в большинстве случаев
хорошо замаскированы. По наблюдениям тех же авторов, главный ход
ногайского тушканчика
в 40—60°, тогда как
у среднеазиатского туш-
канчика мы находили
почти исключительно
очень полого спускаю-
щийся вниз главный ход.
Бёме и Крассовский
неоднократно наблю-
дали ногайских тушкан-
чиков по вечерам на
воле и даже добывали
их стрельбой, наши же
неоднократные попытки
в этом этошении имели
лишь отрицательные ре-
зультаты. Упомянутые
авторы указывают, что
ногайские тушканчики
в дождливую и ненаст-
ную погоду зарываются
гораздо позднее, чем
в ясную, и поэтому во
время дождя их иногда
удается видеть даже
утром. Нам удалось на-
блюдать в песках Бу-
харского округа совер-
шенно противополож-
ную картину, когда
после дождя на уплот-
ненной поверхности смо-
ченного песка нельзя
было обнаружить ника-
идет в глубину почти отвесно, образуя угол
Фиг, 60. Копанка мохноногого тушканчика.
Фото А. И. Аргиропуло.
ких следов роющей деятельности тушканчиков и других ночных живот-
ных. По наблюдениям тех же авторов, ногайские тушканчики держатся
в норах всегда отдельно друг от друга, кроме кормящих самок с дете-
нышами; однако, в Средней Азии мы неоднократно находили в одной
норе самку с несколькими детенышами, достигшими уже более поло-
вины роста матери и способными самостоятельно питаться и быстро,
бегать по барханам. В дополнение к приведенным фактам, свидетель-
166
DIPODINAE
ствующим о наличии биологических отличий у различных рас D. sagitta,
следует указать, что западно-сибирская раса этого вида (D. s. sagitta)
имеет несомненно также свои биологические особенности; во всяком
случае этот тушканчик нередко встречается в совершенно необычай-
ной для этого вида обстановке — на песках, заросших сосновым бором
Экономическое значение. Мохноногий тушканчик обитает
только в песчаных пустынях и не встречается среди возделанных участков.
Однако нам пришлось слышать в некоторых пунктах жалобы на вред,
причиняемый этим зверьком молодым посадкам кустарников, насаждаемых
для закрепления барханных песков; особенно заметен этот вред после
посадки семян этих кустарников в лунки, откуда значительное их количе-
ство выкапывается и поедается не только тонкопалым сусликом и песчан-
ками, но также и тушканчиками.
11. Род SCIRTOPODA BRANDT (1844) — ЕМУРАНЧИКИ
Brandt, Bull. Class. Phys.-Math. Acad. Sc. St. Peterb., II, 1844: 212.
Из внешних признаков для представителей этого рода наиболее
. характерны следующие: хвост не имеет настоящего „знамени", т. е. удли-
ненные волосы покрывают почти всю заднюю половину хвоста, начинаясь
приблизительно от середины хвоста и постепенно удлиняясь по направле-
нию назад; концевые волосы хвоста окрашены в темный цвет и не имеют
белого кончика, характерного для большинства других туншканчиков.
Нижняя сторона пальцев задней ступни и большая часть подошвы покрыты
более короткими волосами, чем у других трехпалых тушканчиков, причем
эти волосы прилегают к подошве и не образуют прямо стоящей щетки как
у Dipus sagitta и других трехпалых тушканчиков; характерным признаком
представителей этого рода является также то, что волосы на его подошве
окрашены в темный цвет, а не в белый, как у других трехпалых тушкан-
чиков. Срединный гребень на os penis расположен на базальной пластинке;
передне-наружные углы базальной пластинки резко обозначенные, острые.
На нижней поверхности penis срединная продольная щель начинается от
основной части penis, с каждой стороны от этой щели могут быть лишь
неглубокие борозды. Стержень os penis не расширен и не имеет вида пла-
стинки и он не отделен перетяжкой от основной пластинки. Средний гре-
бень при основании os penis с остроугольным краем. На верхней поверх-
ности penis имеются два длинных шипа, которые, однако, не заходят за
вершину penis и не лежат в углублении на верхней поверхности penis,
как это имеет место у Jaculus. Наружный выступ parietale без острого
гребня или шипа. Fenestrae postglenoideae образуют более широкую щель,
чем у Dipus, причем эта щель почти наполовину закрыта частью petroma-
stoideum. Наружная стенка нижне-глазничного канала не прирастает
к шахШаге* Слёзные кости как у Dipus, или несколько смещены наружу
(в последнем случае их вершины касаются frontale). Задний край костного
77. SCIR TOPODA
167
иёба как у Dipus. Вершины bullae tympani сходны по строению с тако-
выми у Dipus. Mastoidea довольно сильно раздутые, при рассматривании
черепа сверху они образуют хорошо заметные округлые выступы на боках
задней части черепа. Нижняя челюсть с довольно хорошо развитыми pro-
cessus alveolares, но несколько более слабым, чем у Dipus. В устройстве
шейных позвонков характерны следующие признаки: границы между слив-
шимися телами 2—6-го шейных позвонков совершенно незаметны, или от
них остаются местами лишь неясные следы. На боках вентральной сто-
роны сросшегося комплекса шейных позвонков имеются 4 отверстия между
свободными частями поперечных отростков. (Указанные сочетания при-
знаков свойственны также Jaculus). В отличие от этого рода остистый
отросток на 2-м шейном позвонке развит слабее, чем у Jaculus и имеет
тупую вершину; на нейральных дугах имеются слабо выраженные гра-
ницы. срастания отдельных элементов. Скелет задней конечности отли-
чается от такового у других родов подсемейства тем, что средний палец
лишь немного тоньше каждого из боковых пальцев и когтевая фаланга
среднего пальца лишь немного меньше, чем когтевые фаланги боковых
пальцев. Ложно-коренной зуб сохраняется в виде рудимента лишь у моло-
дых особей (У. telum) или остается и во взрослом состоянии (S. andrewsi).
На первом коренном зубе верхней челюсти имеется с внешней и внутрен-
ней стороны лишь по два выдающихся угла.
Распространение этого рода начинается на западе изолиро-
ванным небольшим островком в низовьях Днепра (Алешкинские пески);
после значительного перерыва распространение рода Scirtopoda начи-
нается в северо-восточном Предкавказьи, в Калмыцкой АССР и Нижнем
Поволжья, достигая к северу до Камышина, далее на восток этот род
распространен в Казахстане до Зайсанской котловины включительно
местами достигая к северу 52° с. ш. (Кустанайский окр.) и к югу до Ман-
гышлака, северной окраины Кара-кумов и Семиречья;-вне пределов СССР
один представитель этого рода найден в нескольких пунктах Монголии
на восток до Калганского тракта.
Условия обитания. Представители этого рода; встречаются
в разнообразных стациях от ковыльных степей и песков на западе
и севере ареала до глинистых и щебнистых пустынь на юге и востоке.
Среди подсемейства Dipodinae этот род является единственным, не свя-
занным в своем распространении исключительно с песками.
Род Scirtopoda составлен двумя хорошо отличимыми видами У. telum
Licht, и У. andrewsi Gl. Allen.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (2). У взрослых экземпляров ложнокоренной зуб отсутствует. Длина
задней ступни 46—51 мм, кондилобазальная длина черепа 25—28 мм.
.........................................* 1. S. telum Licht.
168
DIPODINAE
2(1). У взрослых экземпляров ложнокоренной зуб имеется. Длина задней
ступни около 55 мм, кондилобазальная длина черепа около 30 мм
...................................... 2. S. andrewsi Gl. Allen.
*1. Scirtopoda telum Lichtenstein — Емуранчик.
Lichtenstein 1823:120 (Dipus) [Аральское море]; Сатунин, 1920:214; Виноградов,
1930:347, 454 и др.; Фенюк, 1928, 1929 (биология и сельско-хозяйственное значение). —
falz-feini Браунер, 1913 :1. — turovi Гентнер, 1934.
В отличие от У. andrewsi рассматриваемый вид характеризуется:
отсутствием во взрослом состоянии ложно-коренного зуба в верхней
челюсти (у молодых особей нередко сохраняется рудимент этого зуба
в виде тонкой палочки) и более мелкими размерами; дл. тела 90—125 мм, дл.
хвоста 133—-160; дл. ступни 46—51; вне. уха 14—18; кондилобазальная.
дл. черепа 25—28 мм.
Окраска у более северных и западных форм относительно темная,,
буровато-серая, у более южных более светлая, охристо-буроватая.
Распространение. Крайним западным пунктом, куда доходит'
распространение -S’, telum являются Алешкинские пески в низовьях:
р. Днепра, где распространение этого тушканчика занимает небольшую'
площадь, оторванную от основного ареала вида. Далее на восток этот
вид встречается в северо-восточном Предкавказьи, в Калмыцких степях,
доходя на север по степям между Волгой и Доном до широты
Камышина. Далее на восток этот вид широко распространен в Казах-
стане до Зайсанской котловины включительно; точные границы
распространения на этом пространстве неизвестны. Наиболее северными
пунктами нахождения здесь являются окрестности г. Темира и с. Семи-
озерного Кустанайского окр.; к югу распространение этого вида прости-
рается до Мангышлака, приаральских пустынь, северной окраины Кара-
Кумов и Семиречья. Таким образом, ареал распространения У. telum
представляется в виде относительно узкой, местами прерванной полосы
от низовий р. Днепра до оз. Зайсана.
Географическая изменчивость рассматриваемого вида
слабо изучена и таксономическое значение нескольких описанных до на-
стоящего времени географических рас S. telum, считающихся за подвиды,
остается еще неясным. На просмотренном мною сравнительно небольшом
материале можно подметить две основных градации в изменении окраски —
более темной у более северных и западных форм и несколько более
светлой — у южных. К более темной группе относятся следующие 3 формы
и к более светлой — четвертая.
1) Низовья Днепра (Алешкинские пески) (У. t. falz-feini Brauner,
1913). Экземпляры из указанной местности мною не были исследованы.
По описанию Браунера эта форма отличается от аральской („типичной")
более темной окраской. Общая окраска верха „палево-охристая или авел-
лановая, испещренная довольно густо темносерым".
11. SC1R TOPOD A
169
2) Донские степи (Федосеевка в 20 км к СВ от Торговой)
(.S’. t. turovi Heptner, 1934). Экземпляры мною не были исследованы. По
описанию Гептнера, эта форма отличается от живущих к востоку от
р. Урала более темным и тусклым тоном верхней стороны тела и относи-
тельно мелкими bullae tympani. Автор предполагает, что эта форма темнее,
чем У. t. falz-feini и имеет меньшие размеры.
Фиг. 61. Щебнистая солянковая пустыня на ю.-з. побережья Балхаша (одно
из типичных мест обитания Scirtopoda telum Licht.).
Распространение этой формы, по мнению автора, охватывает степи
между нижним течением Волги и Доном к северу до Камышина и степи
северо-восточного Предкавказья.
3) Северный Казахстан (экземпляры из окрестностей Темира,
Наурзумского заповедника Кустанайского окр., Семипалатинска, Зайсана
(<У. t. birulae Martino). Общий тон окраски относительно темный, буровато-
серый с оливковым оттенком. Окончания волос черные. Окраска головы
сходна с окраской спины. Внешняя сторона бедер темная. Семипалатин-
ские экземпляры в весеннем меху имеют более тусклую окраску.
4) Южный Казахстан (юг Темирского окр. низовья р. Урала,
Эмба, северные или приаральские Кара-Кумы) (У. t. telum Lichtenstein,
1823). Перечисленные серии относятся к более светлой группе. Общий
тон окраски охристо-буроватый, более светлый, чем у предыдущих форм.
Окончания волос бурые. Внешняя сторона бедер светлая.
Данные по биологи и. S. telum является, повидимому, единствен-
ным представителем трехпалых тушканчиков, селящимся в довольно раз-
170
DIPODINAE
нообразных стациях и не связанным в своем распространении исключи-
тельно с песками, как это имеет место у представителей рода Dipus,
Eremodipus, Paradipus. По наблюдениям А. А. Браунера в б. Днепровском
у. б. Таврической губ. S. telum falz-fein является „жителем ковыльной
степи, густо покрытой травой или песков, хорошо поросших раститель-
ностью" (1. с. стр. 10). В северо-восточном Предкавказьи (долина р. Кумы)
S. telum был найден в пескгх, а в Калмыцких степях — в степи с супес-
чаной почвой (К. А. Сатунин стр. 221). По наблюдениям Е. И. Орлова 1
и Б. К. Фенюка2 и на осно-
вании исследований, произве-
денных совместно обоими ука-
занными авторами3 можно счи-
и в Н. Поволжья (б. Сталин-
градская губ.) емуранчик встре-
чается в степях с суглинистой
почвой в бугристой степи,
в песках, на бахчах и пр.; по
сообщению М. К. Серебренни-
кова, в окрестностях г. Темира
(Казахстан) емуранчик селится
исключительно в глинистой
степи. На юго-западном побе-
режья Балхаша я наблюдал
емуранчика в значительном ко-
личестве в разнообразных ста-
циях, в частности на пухлых
солончаках, заросших солянками, в щебнистой пустыне (фиг. 61—62) и на
высоких холмах, сложенных из кварцита, где его приходилось добывать
даже в кучах камней (фиг. 63); на щебнистых склонах нередко этот зверек
рыл норы в чрезвычайно плотном щебне, который почти не поддавался
раскопке при помощи большого крепкого ножа.
В связи с тем, что S’. telum не придерживается исключительно
песчаных стаций, следует отметить, что у этого вида в строении задней
ступни имеются значительные отличия от обитающих исключительно
в песках других трехпалых тушканчиков, как Dipus sagitta, Eremodipus
Фиг. 62. Копанка емуранчика на щебне (побе-
режье Балхаша).
тать, что в Калмыцкой области
1 О р л о в, Е. И. Матер, к позн. фауны наземн. позв. Калм. обл. (Матер, к позн.
фауны Нижн. Поволжья, вып. III), Саратов, 1928.
2 Ф енюк, Б. К. К биоло.ии тушканчиков (Матер, к позн. фауны Нижн. Поволжья,
вып. II) Саратов, 1928.
Ф е н юк, Б. К Еще о биологии тушканчиков (Матер, к позн. фауныНижн. Поволжья,
вып. III) Саратов, 1929.
3 О р л о в, Е. И. и Ф е н ю к, Б. К. Матер, к позн. фауны назем, позв. Приморск,
полосы Калм. обл. (Матер, к позн. фауны Нижн. Поволжья, вып. I) Саратов, 1927.
11. SCIRTOPODA
171
lichtensteini и Paradipus ctenodactylus', в частности у емуранчика (как было
указано выше) на задней ступне имеется слабо развитая щетка из волос,
не столь совершенно приспособленная для передвижения животного по
рыхлому песку, как у перечисленных других трехпалых тушканчиков.
Сведения об образе жизни емуранчика очень скудны. За последнее
время они несколько пополнились наблюдениями Б. К. Фенюка, опубли-
кованными в цитированных выше его работах. По наблюдениям этого
автора, норы -S', telum имеют более сложное строение, чем у A. jaculus.
Фиг. 63. Нора емуранчика под кучей камней (отверстие норы
указано стрелкой).
Постоянные норы этого зверька характеризуются присутствием гнезда
и сильно ветвящимися ходами; количество выходных отверстий колеблется
от 1 до б; выход, которым пользуется тушканчик, закрывается на день
земляной пробочкой („копеечка"), остальные частично или полностью
забиваются землей. Временные норы емуранчика сходны с таковыми
у Alactaga jaculus и имеют вид прямого канала без камеры. Выход зверь-
ков из нор происходит через полчаса или более (до 1 часа) после захода
солнца. Находясь на поверхности земли емуранчик издает характерный
двукратный свист, по которому можно узнать зверька в темноте. Питаются
емуранчики различными степными растениями, в том числе луковичками
лилейных и полынью. Пробуждение от зимней спячки в Нижнем Поволжье
наблюдается в конце марта или начале апреля. Детеныши появляются
в период времени от апреля до конца июня; в каждом помете рождаются
3—4 детеныша. Залегание в спячку емуранчиков (как и земляных зайцев)
происходит вероятно поздней осенью, когда уже начинаются заморозки.
Осенью емуранчики сильно жиреют, причем у некоторых экземпляров
жир откладывается также в хвосте, отчего хвост сильно вздувается.
172
DIPODINAE
Экономическое значение. Емуранчик, подобно некоторым
другим тушканчикам, повидимому, повреждает бахчевые растения; однако,,
по наблюдениям Б. К. Фенюка в Н. Поволжья (1. с.), этот тушканчик,
приносит здесь значительно меньший вред, чем A. jaculus и A. acontion.
2. Scirtopoda andrewsi Glover M. Allen. — Монгольский емуранчик.
Allen, Glover 1925 : 4. (Stylodipus) [списан по экземпляру из Уссук, вероятно пра-
вильнее Ускук, к северу от оз. Цаган-нор].
По внешности сходен с «S’. telum', ложнокоренной зуб имеется у взрос-
лых экземпляров в виде тонкой палочки; размеры крупнее, чем у S. telumz.
задняя ступня около 55, кондилобазальная длина черепа около 30.5 мм.
Вся верхняя поверхность окрашена в светлый песчано-серый цвет,,
затемненный примесью волос с темными окончаниями. Над глазом имеется,
неясное беловатое пятно и несколько более бледное пространство ниже
глаза. Вся брюшная поверхность, передние конечности, внутренние сто-
роны задних конечностей белые. Подошвы задних конечностей покрыты,
темными волосами. Хвост окрашен сходно с спиной; череп по общим,
очертаниям сходен с таковым у «S'. telum\ rostrum более удлинен, носовые
кости относительно длиннее. Mastoidea земетно более раздуты, чем'
у «S', telum и более сильно выдаются по бокам затылка при рассматривании
черепа сверху; bullae tympani более тесно соприкасаются своими верши-
нами в области basisphenoideum. Fossae pterygoidei более вытянуты:
в продольном направлении.
. Этот вид был выделен описавшим его G1. АНеп’ом в особый род
Stylodipus. Детальное изучение признаков этого животного и непосред-
ственное его сравнение с S. telum 1 не оставляет сомнений в принадлеж-
ности обоих этих видов к одному и тому же роду, т. е. к ранее описан-
ному Брандтом роду Scirtopoda.
Распространение. Нахождение S. andrewsi пока известно лишь
для нескольких пунктов Монголии, где он был добыт экспедицией
Р. Эндрюса; пункты эти следующие: Уссук, Цаган-нор, Лох и Эрлиен
(Ирен-дабасу).
Условия обитания и образ жизни этого животного неизвестны.
12. Род PARADIPUS VINOGR. — ГРЕБНЕПАЛЫЕ ТУШКАНЧИКИ
Vinogradov, Изв. Акад. Наук, VII, 1930: 333.
Задняя ступня с тремя длинными пальцами почти одинаковой длины;
нижняя поверхность боковых (2-го и 4-го) пальцев покрыта с внутренней
стороны щеткой, состоящей из мягких длинных волос и с наружной сто-
роны снабженная гребнем из утолщенных щетинок, которые у старых экзем-
1 В коллекциях Зоологического Института Академии Наук СССР имеется один иа
оригинальных экземпляров S. andrewsi, полученный от G1. АПеп’а.
12. PARADIPUS
173.'
пляров почти в два раза короче, чем волосы на наружной стороне (фиг. 64).
Эти щетинки особенно хорошо заметны на концах пальцев, куда не дохо-
дят щетки из длинных волос. У более молодых экземпляров та и другая
категории волос не столь резко дифференцированы. На среднем пальце
твердые щетинки сидят в два продольных ряда по бокам нижней поверх-
ности пальца; между этими рядами располагаются тонкие длинные подош-
венные волосы; на конце пальца оба ряда Щетинок удаляются друг от
друга, расходясь веерообразно вперед и в стороны. Уши значительна
длиннее, чем у других представителей Dipodinae. Череп характеризуется
следующими особенно-
«стями. Строение на-
ружных выступов parie-
tale как у предыдущих
групп. Fenestrae post-
glenoideae очень малы
и округлы, целиком
закрытые разросши-
мися частями petro-
mastoideum. Наружная
стенка нижнеглазнич-
ного канала не при-
растает к тахШаге.
Слёзные кости значи-
тельно смещены наружу
И Прикреплены К уз фиг, 64, Щетка на пальцах ступни гребиепалого тушканчика,
кому отростку frontale,
идущему в поперечном направлении вдоль верхнего корня скуловой дуги.
Задний. край костного нёба вогнутый, с едва заметным шипиком посре-
дине; этот край расположен на уровне центра т3. Вершины bullae tym-
pani не соприкасаются друг с другом и не покрывают собою basisphenoi-
deum. Mastoidea относительно сильно раздуты и при рассматривании
черепа сверху образуют заметные выступы на боках задней части черепа.
Корень нижнего резца не образует на поверхности нижней челюсти
выступа (альвеолярного бугра).
Шейная часть позвоночника наиболее сильно изменена по сравнению
со всеми другими Dipodinae', кроме значительной укороченности шеи
и полного слияния 2, 3, 4, 5 и 6-го позвонков, боковые части позвонков
более изменены, чем у других представителей подсемейства, причем
в этой области у Paradipus остается лишь три неправильной формы
отверстия.
Скелет задней ступни отличается от других представителей под-
семейства тем, что у Paradipus пальцы относительно более длинны, чем
у других Dipodinae, и средний палец еще более тонок; чрезвычайно харак-
терно также для Paradipus наличие на когтевой фаланге среднего пальца
'174
DIPODINAE
двух боковых лопастей, служащих опорой для двух рядов крепких щетинок
на боковых частях концов пальцев.
Ложно-коренного зуба нет (ту). Строение коренных зубов в суще-
ственных чертах сходно с таковым у Scirtopoda.
Распространение. Распространение рода Paradipus, судя по
немногим известным нахождениям единственного его представителя,
охватывает пространство Кизыл-кумов и Кара-кумов.
Условия обитания и образ жизни. Род Paradipus харак-
терен для голых барханных песков, как это подробно указывается ниже
при характеристике вида. Это подтверждается также своеобразным строек
нием задних конечностей с длинными пальцами и двухярусной щеткой,,
что является весьма совершенным приспособлением для передвижения;
по особенно рыхлой поверхности песка. В отличие от других трехпалых
тушканчиков, норы Paradipus не всегда забиваются их обладателями на;
день песком и часто располагаются под подветренными склонами бар-
ханов, дающими им надежную естественную защиту.
К роду Paradipus принадлежит единственный вид Р. ctenodactylus-
Vinogr.
*1. Paradipus ctenodactylus Vinogr. — Гребнепалый ^тушканчик
(фиг. 59—69).
Виноградов, 1929:243 (Scirtopoda) [ст. Репетек Закасп. ж. д.]; Виноградов,
1930:333 (Paradipus).
В дополнение к признакам, указанным в характеристике рода, здесь
сообщаются следующие:
Верхние резцы окрашены в белый цвет, их передняя поверхность
3
гладкая, без продольной бороздки. Коренных зубов -у(ложнокоренной зуб
отсутствует).
Измерения оригинального экземпляра. Задняя ступня
78:6 мм, ухо 30 мм; наибольшая длина черепа 39.5; кондилобазальная
длина 35.0; скулов.ая ширина (сзади) 22.5 мм, межглазничное пространство
10.7 мм; длина носовых костей 15.3; диастема 9:5; максиллярный ряд.
зубов 6.4 мм.
Заметки по систематике. При первом описании гребнепалого
тушканчика Б. С. Виноградов предположительно отнес его к роду Scirto-
poda, рассматривая этот род в широком смысле (1. с.). После дальнейших,
более детальных исследований тот же автор выделил этот вид в особый
новый род Paradipus; основанием для этого послужило наличие у Р. cte-
nodactylus целого ряда своеобразных признаков, не встречающихся
у других известных представителей трехпалого тушканчика; наиболее
существенные из этих признаков были описаны выше.
Распространение. В настоящее время нахождение Р. ctenodacty-
lus точно известно для трех указанных выше пунктов, т. е. ст. Репетек.
12. PARADIPUS
175
и Ходжа-давлет Ср.-Аз. ж. д. и окрестности кишлака Богоабзаль близ,
ст. Шафрикан на северной окраине Бухарского оазиса; кроме того, мы
видели следы этого тушканчика в песках близ ст. Кара-куль (к востоку
от ст. Каган) и обнаружили остатки этого зворька в погадках птиц близ
ст. Джебел Красноводского района.
Данные по б и о ло г и и. Гребнепалый тушканчик является харак-
терным обитателем голых сыпучих барханов и селится в наиболее без-.
Фиг. 65. Гребнепалый тушканчик' (Paradipus ctenodactylus).
Фото М. К. Серебренникова.
отрадных районах песчаных пустынь. На приспособленность этого живот-
ного к передвижению на рыхлом песке указывают некоторые из описан-.
ных выше морфологических особенностей гребнепалого тушканчика,
в частности сильно удлиненные пальцы задней конечности, наличие длин-
ной щетки из прямо стоящих волос на нижней поверхности пальцев зад-
ней ступни, и присутствие дополнительной щетки из более коротких
и значительно более толстых щетинок.
Нора гребнепалого тушканчика в большинстве случаев располагается
на подветренном склоне бархана, причем часто она имеет откры-
тое выходное отверстие, или слегка засыпанное сползающим по склону
бархана песком. Раскопка норы гребнепалого тушканчика представляет
большие трудности, т. к. сползающие при раскопке по склону бархана
массы сухого песка устремляются в выкопанную яму и нередко уничто-
жают всю проделанную работу. Ход норы гребнепалого тушканчика имеет
176
DIPODINAE
до 10 сантиметров в диаметре; длина норы достигает двух метров и более,
причем нора обычно не образует ответвлений и не имеет расширений или
камер. Одним из существенных отличий нор гребнепалого тушканчика от
мохноногого является отсутствие в первых из них запасного выхода, окан-
чивающегося „окошечком" или „копеечкой"
Фиг. 66. Одно из характерных мест обитания Paradipus
ctenodactylus Vinogr. (окр. ст. Ходжа-давлет).
Фото А. И. Аргиропуло.
(см. описание норы мохно-
ногого тушканчика); таким
образом, нора гребнепа-
лого тушканчика имеет
единственное выходное от-
верстие, через которое
зверек прячется в нору и
выходит вечером на по-
верхность} как это нами
было установлено по сле-
дам. Обычно уже в самом
начале раскопки жилой
норы гребнепалого туш-
канчика можно слышать,
приложив ухо к песку,
характерное постукивание
в виде правильно чередую-
щихся двойных ударов,
которые производит зве-
рек, пытающийся уйти от
преследования вглубь бар-
хана.
Следы гребнепалого
тушканчика на песке
(фиг. 69) очень характерны
и хорошо отличаются от
следов мохноногого туш-
канчика; особенно харак-
терна значительно боль-
шая их величина, чем у по-
следнего вида, перетяжка
в средней их части и резко вдавленные боковые линии; кроме того,
в отличие от мохноногого тушканчика, следы Р. ctenodactylus располага-
ются не попарно, а большей частью наискось, причем отпечатки пальцев
правой и левой ноги отстоят друг от друга на значительное расстояние,
что указывает на наличие у этого зверька совершенно своеобразного
характера прыжков.
Гребнепалый тушканчик, как и мохноногий, часто „играет" на песке,
истаптывая здесь своими следами обширные площадки, причем тушканчики
валяются также на песке, оставляя здесь отпечатки своего тела и хвоста.
13. EREMODIPUS
177
Нам удавалось непосредственно наблюдать гребнепалого тушкан-
чика на воле, так как эти зверки нередко выходят из нор уже в су-
мерки.
Экономическое значение. 'Данных об экономическом значе-
нии этого вида в настоящее время не имеется; можно лишь предполагать,
что этот тушканчик в тех
случаях, когда он встре-
чается в районах песко-
укрепительных работ
(как это имеет место
во всех перечисленных
выше пунктах), отчасти
приносит вред вместе
•с мохноногим тущканчи-
.ком, песчанками и тон-
копалым сусликом по-
садкам саксаула и дру-
гих кустарников; во
.всяком случае, живший
у нас в неволе гребне-
палый тушканчик охотно
ел предлагавшиеся ему
молодые веточки сак-
саула.
13. Род EREMODIPUS VINO-
GRADOV (1930) — СРЕДНЕ-
АЗИАТСКИЕ ПУСТЫН-
НЫЕ ТУШКАНЧИКИ
Vinogradov, Изв. Акад.
Наук, VII, 1930:334.
По внешности СХОД- Фиг. 67. Вновь вырытая нора гребнепалого тушканчика,
ны с мохноногим туш- Фото А. И. А рги pony ло.
жанчиком, но меньше
последнего по размерам, с более короткими пальцами на задней ступне,
снабженными более редкой щеткой из белых волос; „знамя" на конце
хвоста более слабо развито и его белый кончик очень маленький или
отсутствует вовсе.
Penis Eremodipus по своим признакам наиболее сходен с таковым
у африканских тушканчиков, относящихся к роду Jaculus и особенно
к группе Jaculus jaculus. Penis Eremodipus имеет почти цилиндрическую
форму и покрыт на всем протяжении лишь мелкими роговыми чешуйками;
:на дорзальной поверхности penis не имеется никаких следов двух длинных
Фауна СССР, Виноградов 12
178
DIPODIDAE
шипов, которые столь характерны для большинства представителей под-
сем. Dipodinae. Различия в строении penis у Eremodipus и представителей,
группы Jaculus jaculus невелики и сводятся преимущественно к следую-
щему: у Eremodipus роговые чешуйки имеют несколько более крупные
размеры в проксимальной части penis, а у Jaculus jaculus — в дистальной;,
у Eremodipus вся дорзальная поверхность равномерно покрыта роговыми
чешуйками, тогда как у группы Jaculus jaculus в середине дорзальной,
части penis имеется ясно выраженная голая (лишенная чешуек) площадка..
Фиг. 68. Старая нора гребнепалого тушканчика.
Фото А. И. Аргиропуло.
Череп с очень сильно раздутыми барабанными камерами, в образо-
вании которых принимают участие мастоидные (сосцевидные) кости, также-
сильно раздутые и образующие большие пузыри по бокам задней частиг
черепа.
Наружный угол (выступ) parietale оканчивается направленным вниз-
шипом, расположенным над основанием processus zygomaticus squamosi-
Fenestrae postylenoideae очень малы, обычно несколько вытянуты в про-
дольном направлении и закрыты разробшеюся верхней частью petroma-
stoideum. Наружная стенка нижнеглазничного канала не прирастает
к стенке maxillare. Слёзные кости заметно смещены наружу, их внутрен-
ние углы (при рассматривании черепа сверху) лишь едва соприкасаются;
с лобными костями. Задний край костного нёба имеет то же строение,,
как у Dipus и Scirtopoda. Вершинные части bullae tympani тесно связаны
друг с другом посредством крепкого шва и образуют значительные взду-
тия вдоль линии этого шва. Mastoidea очень сильно раздуты и выдаются
в виде больших пузырей в задней части черепа. На нижней челюсти:
имеется слабо развитый альвеолярный отросток.
13. EREMODIPUS
179
Граница между слившимися телами 2, 3, 4, 5 и 6-го шейных позвон-
ков довольно ясно заметна; заметны также границы между слившимися
нейральными дугами тех же позвонков. Дистальные концы поперечных
отростков 5-го и 6-го позвонков не вполне слиты друг с другом (границы
между ними заметны); вершина остистого отростка округлена и не откло-
нена вперед. На боках вентральной стороны сросшегося шейного ком-
плекса позвонков имеются лишь 3 отверстия между свободными частями
поперечных отростков.
Скелет задней конечности в суще-
ственных чертах сходен с таковым у Dipus
и отличается от него, главным образом,
тем, что вырезка на дистальном конце
цевки (между сочленовными поверхно-
стями) составляет лишь около Vis всей
длины цевки.
3
Ложно-коренного зуба нет (ту).
Строение коренных зубов в существенных
чертах сходно с таковым у Scirtopoda.
Распространение. Судя по не-
многим известным нахождениям единствен-
ного представителя этого рода, его рас-
пространение простирается по всему про-
странству Кара-Кумов и восточным Кизыл-
Кумам на север до Казалинского района.
Это распространение всюду приурочено
песчаным стациям.
Как было уже указано, этот род
стоит в близких родственных отношениях
с африканским родом Jaculus.
К роду Eremodipus относится един-
ственный вид Е. lichtensteini Vinogr.
Фиг. 69. Следы гребнепалого туш-
канчика.
* 1. Eremodipus lichtensteini Vinogradov — Тушканчик Лихтенштейна.
Vinogradov, 1927 : 92 (Scirtopoda) (окрестности Мерва); Vinogradov, 1930; 334 (Ere-
modipus).
Наиболее характерные признаки этого зверька указаны при описа-
ния рода. По внешности сходен с Dipus sagitta и отличается от послед-
него несколько меньшими размерами, более короткими пальцами задней
ступни с более слабо развитой щеткой и менее ясно обособленным „зна-
менем" на конце хвоста.
Распространение. В настоящее время известно лишь несколько
перечисленных выше пунктов нахождения этого вида, т. е. окрестности
Мерва, Ашхабада, ст. Джебел Красноводского района и три пункта на тер-
ритории Казалинского района. Судя по удаленности этих пунктов друг
12*
180
DIPODINAE
от друга можно предполагать, что тушканчик Лихтенштейна распростра-
нен на всем обширном пространстве Кара-кумов от Мерва, Ашхабада
и Красноводска до северного побережья Аральского моря.
Образ жизни тушканчика Лихтенштейна неизвестен. На основании
некоторых морфологических особенностей этого вида (щетка на задней
ступне, сильно раздутые мастоидальные и барабанные камеры) по анало-
гии с другими видами, следует считать этого тушканчика за характерного
обитателя песчаных пустынь. Это предположение было подтверждено
нами и путем непосредственных исследований в песках Красноводского
района в окрестностях ст. Джебел.
14. Род JACULUS ERXLEBEN (1777) - АФРИКАНСКИЕ ТРЕХПАЛЫЕ ТУШКАНЧИКИ
Erxleben, Syst. Reg-n. Anim., 1, 1777 :404.
По наружным^признакам в существенных чертах сходны с Eremodi-
pus и Dipus. '
Genitalia самца в пределах рода Jaculus представляет два типа
строения. У представителей группы Jaculus orientalis penis снабжен двумя
длинными шипами, выдающимися за его конец; поверхность penis покрыта
хорошо развитыми короткими шипами, которые имеют наибольшую длину
близ основания двух упомянутых выше длинных шипов. В середине дор-
зальной поверхности имеется резко ограниченная голая площадка. Penis
широкий, несколько сплюснутый в дорзовентральном направлении, оваль-
ной формы (при рассматривании сверху). Срединный гребень на основной
пластинке os penis резко выдающийся и имеет широкий округленный
край (при рассматривании сбоку). Стержень os penis превращен в тонкую
широкую пластинку, отделенную от основной пластинки ясно выраженным
перехватом. Второй тип строения наблюдается у представителей Jaculus
jaculus, у которых два длинных шипа на penis совершенно отсутствуют;
поверхность penis покрыта очень мелкими чешуйками или шипиками,
несколько более крупными в дистальной части верхней поверхности penis.
Penis большей частью имеет цилиндрическую форму. Средний гребень
на основной пластинке os penis слабо развит. Последний тип строения
в значительной мере напоминает строение penis у Eremodipus; отличия
между обеими формами были указаны при описании penis у Eremodipus.
Череп по своему строению в значительной степени сходен с таковым
у Eremodipus, отличаясь, однако, от последнего некоторыми существен-
ными признаками. Наружный выступ parietale вместе с прилегающими
частями squamosum образует резко обособленный гребень. Fenestrae
postglenoideae очень велики, образуя почти равносторонние треугольники;
оба отверстия вполне закрыты разросшимися petromastoidea. Наружная
стенка нижнеглазничного канала у взрослых экземпляров прирастает
своим верхним краем к стенке maxillare. Слёзные кости и задний край
костного нёба имеют то же строение как у Dipus и Scirtopoda. Mastoidea
74. JACULUS
181
очень сильно раздуты, как у Eremodipus. Вершинные части bullae tympani
тесно связаны друг с другом посредством крепкого шва и образуют
значительное вздутие вдоль линии этого шва. Нижняя челюсть образует
хорошо развитой альвеолярный отросток.
Строение шейных позвонков сходно с таковым у Scirtopoda, но ости-
стый отросток на втором шейном позвонке хорошо развит и имеет за-
остренную вершину и границы между сросшимися нейральными дугами
2, 3, 4, 5 и 6-го позвонков, незаметны.
Скелет задней ступни сходен по строению с таковым у Dipus
и Eremodipus по следующим признакам: средний палец значительно
тоньше, чем каждый из боковых пальцев и его когтевая фаланга почти
в 2 раза меньше, чем на боковых пальцах. Отличиями в устройстве ске-
лета задней ступни от упомянутых родов у Jaculus являются следующие:
цевка не более чем в два раза длиннее, чем средний палец. Основная,
фаланга среднего пальца почти в три раза длиннее второй фаланги этого
пальца. , 1
Ложно-коренной зуб отсутствует (ш -|). Строение коренных зубов
в существенных чертах сходно с таковым у Scirtopoda, Paradipus и Ere-
modipus.
По совокупности признаков род Jaculus имеет наибольшее сходство
с родом Eremodipus', однако обе названных группы имеют ряд существен-
ных различий, свидетельствующих об их родовой самостоятельности;
в частности различия в строении mastoidea, fenestra postlamdoidea нижне-
глазничного отверстия и канала, слёзных костей, общей конфигурации
черепа, наличие гребня в височной области И пр.
Распространение. Представители рода Jaculus широко распро-
странены в северной Африке, Аравии, Сирии, Палестине и южном Иране.
В Африке их распространение не выходит за пределы Сахары, причем на
запад представители этого рода распространены здесь до Алжира и Туниса
и на восток до Египта и на юг до Аира (близ границы Судана) и верх-
него течения Нила (Кордофан); в Иране представители этого рода най-
дены на север до пунктов между Исфаганью и Тегераном и на восток
до Сеистана.
Краткие данные о видовом составе. До настоящего вре-
мени описано не менее двух десятков видов рода Jaculus', однако при
ближайшем рассмотрении этих форм они ясно группируются около двух
видов —J. jaculus и J. orientalis (—mauritanicus) и быть может являются
даже лишь географическими расами двух названных видов, или в некото-
рых случаях полными их синонимами. Первый из этих видов (или групп)
характеризуется относительно малыми размерами и устройством penis, не
имеющего на своей поверхности длинных стилевидных шипов; второй вид
(или группа) отличается более крупными размерами тела и наличием на
дорзальной поверхности penis двух длинных стилевидных шипов.
182
DIPODINAE
Распространенный в Иране J. loftusi Blanford заметно не отличается
от представителей первой группы (т. е. J. jaculits), a J. blanfordi вероятно
близок к второй группе (т. е. J. orientalis).1
Более детальное рассмотрение систематики видов рода Jaculus не
входит в задачу настоящего обзора.
1 Последний вопрос ие может быть разрешен точно, так как строение penis J. blan-
fordi неизвестно.
SUMMARY
Jumping’ mice and Gerboas belong to the most interesting and the most
peculiar groups of mammals of Palaearctic in morphological, systematical and
oecological senses. The author after detailed study of morphological characters
of all known representatives of the named groups comes to the conclusion
that Jumping mice (Sicista and Zapus}, which are considered by modern
authors as a separate family Zapodidae, must be united with true Gerboas
into single family Dipodidae', this opinion is confirmed by the presence of
some groups with intermediate or mixed characters, as Euchoreutes, Cardio-
cranius and Salpingotus.
After giving morphological characteristics of the family Dipodidae the
author explains essential adaptations of their organism in connection with
their environment, i. e. desert and steppe landscape; saltatorial adaptations
are prevailing and producing stricking modifications in many organs; fossorial
adaptations are feebly developed but rather evident. Saltatorial capacity may
Ъе considered as an important adaptation to avoid enemies in open land;
this adaptation is correlated with peculiar structure of burrows of these ani-
mals. The author describes further the most important oecological characters
.of this family.
Giving a short synopsis of palaeontological data on Dipodidae, the
author comes to the conclusion that Plesiosminthus from pliocen or Europe
may be considered as nearest relative of recent Zapus (especially Eozapus
from Szechwan and Kansu). The author considers provisionally fossil forms
from Mongolia and China — Sminthoides (lower pliocen) and Plesiodipus
(pliocen or upper miocen), as members of the subfamily Cardiocraniinae.
The position of Protalactaga (pliocen or upper miocen) is not clear, but it is
possible to suppose it has some mixed characters of Cardiocraniinae and
Dipodinae. The author gives further a hypothetical scheme of evolution of
«different genera and subfamilies of this group (fig. 8) and describes their
interrelationships.
In the chapter on the geographical distribution of Dipodidae the author
•considers this group as having Asiatic origin, quite differently with other
desert Rodents Gerhillinae, which may be considered as mainly African
group. The author describes also general outlines of the recent geographical
distribution of Dipodidae, especially on the Asiatic continent.
184
DIPODINAE
On the base of his recent and previous studies the author adopts in the-
present paper the following’ classification of subfamilies and genera:
Subf. 1. Zapodinae— genera: Sicista Gray, Zapus Coues.
„ 2. Euchoreutinae— „ • „ 3. Cardiocraniinae — „ „ 4. Allactaginae — „ Euchoreutes Sclat. Cardiocranius Satun. Salpingotus Vinogr. Allactaga F. Cuv. Alactagulus Nehr. Pygerethmus Glog.
„ 5. Dipodinae— „ Scarturus Glog. Dipus Gmel. Scirtopoda Br. Paradipus Vinogr. Eremodipus Vinogr. Jaculus Erxl.
All family contains 4 subfamilies, 14 genera and about 50 species; the-
exact number of species cannot be fixed now, as systematic position of some
Sicista {„concolor11 group), American Zapus and African Jaculus is not clear».
Some distinguishing characters of subfamilies, genera and species are
shown in the keys as follows.
Key to subfamilies
a) based on cranial characters
1 (4). Frontals with narrowest part in their middle at a considerable distance
posterior to lacrymals. Both roots of zygomatic process of maxilla arise
one directly above another. Malar (jugal) bone slopes from above down-
wards posteriorly, forming no sharply separated horizontal and vertical
branches. Lacrymals are very small, not expanded externally. Mesoptery-
goid space about twice as long as broad. Pterygoid processes about
as long as molar series (without PM).
2 (3). Braincase nearly oval, when viewed.from above. Zygomata bowshaped,,
not converging or slightly converging anteriorly. Pterygoid fossae
shallow, not conspicuosly broadened posteriorly. Tympanic bullae feebly
inflated, their apices are not in contact with each other. Mastoids are
not inflated, forming no mastoid bullae. Posterior palatine foramina,
small. ..........................................1. Subf. Zapodinae.
3 (2). Braincase nearly trapezoidal. Zygomata nearly rectilinear when viewed
from above, considerably converging anteriorly. Pterygoid fossae are
deep and considerably broadened posteriarly. Tympanic bullae much,
inflated, their apices being in contact with each other. Mastoids greatly
inflated, forming large chambers connected with the cavity of tympanic
bullae. Posterior palatine foramina very large. . 2. Subf. Euchoreutinae..
KEY TO SUBFAMILIES
185-
4 (1). Frontals with narrowest part approached close to lacrymals. Upper
roots of zygomatic processes come off at a considerable distance poste-
rior to lower roots. Malar bone biradiate, consisting of horizontal and
vertical branches. Lacrymals are enlarged and considerably project
outwards or in posteroexternal direction. Mesopterygoid space is not
longer than broad. Pterygoid processes considerably shorter than molar
series.
5 (6). Squamosal region without any protuberances or ridges. Coronoid
suture forms a rather deep arch. Interparietal much reduced or wanting;,
when present its longitudinal dimension in many times greater than its
transverse diameter. Supraoccipital without lateral processes. Mastoids-
enormously inflated, occupying about ond third of the whole surface ot
skull.................................... 3. Subf. Cardiocraniinae.
6 (5). Squamosal region with well developed rounded prominences or
sharply defined ridges. Coronoid suture forms nearly straight trans-
verse line. Interparietal is well developed, its transverse diameter
always greater than longitudinal dimension. Supraoccipital with well
developed lateral processes. Mastoids are not inflated enormously, not
reaching the upper surface of skull or occupying there only a small
part of it.
7 (8). Tympanic bullae feebly inflated, their apices are not in contact with
each other. Mastoids are not inflated and forming no additional cham-
bers of the middle ear. Anterior ends of nasals not reaching the level
of upper incisors. Upper incisors and their alveoli are much protrud ng
forwards...................................... 4.. Subf. Allactaginae.
8 (7). Tympanic bullae considerably inflated, their apices usually being in
contact with each other (with the exception of Paradipus). Mastoids
are much inflated and forming large additional chambers connected
with the cavity of middle ear. Anterior end of nasals reaching the level,
of alveoli of upper incisors. Upper incisors and their alveoli are nearly
vertical............................................5. Subf. Dipodinae.
Key to subfamilies
(based on the structure of theskeleton of hind foot).
1 (6). Three middle metatarsals are free (not ankylosed). The first toe (ifll
lateral toes are present) is equal about two thirds of fifth one. The
last toe is reaching the base of the middle phalanx of fourth toe.
2 (5). First and fifth toes are well developed.
3 (4). Each of three middle toes is equal at last three quarters of correspon-
ding metatarsal; first toe is projecting nearly to the distal end of the
second metatarsal or slightly surpassing it. The distal end of entocunei-
form is reaching about the same level as the proximal end of third,
metatarsal....................• •...................1. Subf. Zapodinae..
185
DIPODINAE
4 (3). Each of three middle toes is shorter than three quarters of correspon-
ding- metatarsal. The distal end of the entocuneiform is situated consi-
derably forwards of the proximal end of third metatarsal. ...........
•....................3. Subf. Cardiocraniinae (gen. Cardiocranius).
5 (2). First and fifth toes are wanting- (only their rudiments are present). . .
.....................3. Subf. Cardiocraniinae (g-en. Salpingotus).
6 (1). Three middle toes at last on their posterior two thirds are perfectly anky-
losed forming a „canon-bone“. The first toe (when present) is nearly
equal to the fifth toe; last toe (if present) is not reaching the distal end
of the ,,canon-bone“.
7 (10). Fifth (and mostly first) toe is well developed.
8 (9). The anterior third of metatarsals (distal third of canon-bone) is not
ankylosed.....................................2. Subf. Euchoreutinae.
9 (8). The anterior third of metatarsal as well as all posterior parts of it is
perfectly ankylosed forming a complete canon-bone (only the anterior
Vs—Vio th part of each metatarsal is free forming an articulating surface
for corresponding basal phalanx).................4. Subf. Allactaginae.
10 (7). Fifth and first toes are always absent (only their rudiments present) .
..............................................5. Subf. Dipodinae.
Key to subfamilies
(based on the structure of pelvis)
1 (6). Symphisis pubis short, sometimes shorter than distance from acetabulum
to ishiatic tuber.
2 (5). Pelvic bone forms near symphisis a long process or lobe.
3 (4). Ileo-pectineal tubercles are wanting . . •....!. Subf. Zapodinae.
4 (3). Ileo-pectineal tubercles (one or two) are well developed ......
.........................................2. Subf. Cardiocraniinae.
5 (2). Pelvic bone forms near symphisis no process or lobe..........
...........................................3. Subf. Euchoreutinae.
6 (1). Symphisis pubis very long, not shorter than two thirds of the distance
from acetabulum to ishiatic tuber.
7 (8). Ramus descendens of ishium forms with horizontal branch (parallel to
vertebral column) of this bone nearly right angle (without an incisure
below the ishiatic tuber)...........................5. Subf. Dipodinae.
8 (7). Ramus descendens of ishium forms with horizontal branch of this bone
an acute angle.......................................................
, 4. Subf. Aliactaginae (with the exception of the genus Pygerethmus).
Key to subfamilies
(based on the structure of cervical vertebrae)
1 (6). Vertebrae are free (not ankylosed).
2 (5). Transversal processes short, not longer than a half width of corpora
of corresponding vertebrae
KEY TO SUBFAMILIES
187
3
4
5
6
7
3
1
.2
3
4
.5
6
7
<8
(4). Apices of transversal processes of 5th, 6th and 7th vertebrae are
not in contact with each other; transversal processes are nearly parrallel
to each other.....................................1. Subf. Zapodinae.
(3). Apices of transversal processes of 5th, 6th and 7th vertebrae are in
contact with each other . . -................2. Subf. Euchoreutinae.
(2). Transversal processes relatively long, surpassing a half width of corre-
sponding vertebrae..............................4. Subf. Allactaginae.
(1). Vertebrae are ankylosed (only the 7th vertebra is sometimes
free).
(8). Seventh vertebra is free. There are on the ventral surface of ankylosed
complex of vertebrae 3 or 4 foramina..............5. Subf. Dipodinae.
(7). Seventh vertebra is ankylosed with 6th one. There is on the lateral
side of the ankylosed complex of vertebrae only one foramen wit-
some very thin bony tubercles corresponding to rudiments of reduce 1
transversal processes........................3. Subf. Cardiocraniinae.
Key to subfamilies
(based on the structure of genitalia)
(2). Os penis is always present; its basal part without median projecting
ridge.............................................1. Subf. Zapodinae.
(1). Os penis is present or wanting; if present its basal part is always fur-
nished with a median projecting ridge.
(6). Penis thin, gradually becomes thinner to the tip or flattened laterally;
there is on its surface no distinct horny spines or scales (on its surface
may be seen only very minute grains).
(5) . Os penis present; its tip, covered with epithelium, strongly curved
upwards. Penis cylindrical or somewhat flattened laterally............
..........................................2. Subf. Euchoreutinae.
(4) . Os penis absent; penis thin, gradually becomes thinner to its tip and
somewhat flattened laterally near the tip; the tip not curved upwards .
..........................................3. Subf. Cardiocraniinae.
(3). Penis stout with well developed horny spines or scales mostly dire-
cted backwards.
(8). Os penis absent. The tip of penis simple, without lateral folds
(„pouches"); on the lower surface of penis no deep longitudinal furrow.
Penis ovate cordate or lanceolate, covered with small spines (long spi-
kes always absent)........................... 4. Subf. Allactaginae.
(7). Os penis present. The tip of penis mostly provided with deep folds
(„poushes") on each side of external orifice of urethra; on the lower
surface of penis a deep longitudinal crease always present; on each side
of it may be another crease. Penis ovate or parallelsided. In addition
to the small spines a pair of long styliform spikes mostly present. . .
..............................................5. Subf. Dipodinae.
188 ZAPODINAE, EUCHOREUTINAE, CARDIOCRANIINAE
1. Subf. ZAPODINAE
1 (2). Anterior end of malar does not reaching’ the lacrymal. Interorbital
canal incomplete, its external wall not being in contact with maxilla..
Transverse dimension of interparietal about two times greater than its-
longitudinal diameter. Posterior edge of bony palate projects conside-
rably backwards, from the level of posterior margin of M3; palate ter-
minating with a well developed spinous process. Two shallow impres-
sions are present on the posterior part of the palate . . . .1. Sicista.
2 (1). Anterior end of malar is reaching lacrymal. Interorbital canal is com-
plete, its external plate being mostly fused to maxilla in adult specimens..
Transverse dimension of interparietal about three of four times greater
than its longitudinal diameter. Posterior edge of bony palate projects,
backwards not further than posterior margin of M3; posterior edge of
palate without a spinous process, usua ly being concave; no impressions
on the posterior part of the palate...............2. Zapus Coues..
3 (4). Premolar (P4) wanting. All tubercles on upper molars disappear in
early age and their crown surface becomes flat.................
. . . ................ subg. Napeozapus Preble
4 (3). Premolar (P4) present. Tubercular structure of crown surface of
upper molars remains in adult specimens.
5 (6) Tubercis of upper molars much projecting and separated by deep valleys.
External additional tubercles joined by transverse crests with interior
main tubercles...................•..........subg. Eozapus Preble
6 (5). Tubercles of upper molars not so projecting and separated by shallow
valleys. External additional tubercles are isolated, i. g. not connected!
by crests with interior tubercles.....• subg. Zapus Coues (s. str.).
1. Gen. SICISTA
1 (4). A well defined black stripe on the back is present.
2 (3). Tail is about one third longer than head and body (tail measurement is not
surpassing 90 mm). Hind foot relatively short, not more than 15.7 mm.
Colour of the dorsum grayish with ochraceous suffusion. Light narrow
bands present on both sides of dorsal stripes and externally dark longi-
tudinal spots or stripes are usually developed, especially on the poste-
rior parts of the dorsum. Upper surface of penis with only one long
spine (in addition to minute spines); before its base a separate concave
horny scale is situated of variable shape. Two lateral slits, rising
from median slit diverging V-shaped, not passing into lateral surface.
Penis is not divided in two lobes. — Steppe- and partly semidesert zone
from E. Ungaria to Baikal Sea. ................................
............1. S. subtilis Pall. (—S. nordmanni Keys, and Blas.).
3 (2). Tail at least at half longer than head and body (tail measurement
usually 90—102 mm); hind foot relatively long, usually more than.
/. SICISTA, 2. ZAPUS, 3. EUCHOREUTES
189
15.7 mm (15.8—17.7). Colour of the dorsum uniform, yellowish-brown,
without light and dark bands. Upper surface of penis furnished with
two long spines; before its base are situated only minute numerous
horny scales. Two lateral slits, rising from the median slit diverging
W-shaped, passing on the lateral surface and dividing penis in two
lobes — superior and inferior. — Forest zone from Norway, Danemark and
N. Ungaria to Baikal Sea . 2. S. betuiina Pall. (= S. montana Mehely).
4 (1). Black stripe on the dorsum is wanting.
5 (6). Penis in addition to minute spines is furnished with two stout spines
about 2 mm long; ventral surface of penis formed with less enlarged
spines; six of them, more prominent, are situated nearly in two longi-
tudinal rows. — Altai Mts.......................3. S. napaea Hollister.
6 (5). In addition to minute spines and scales there are on the penis no sepa-
rate prominent spines.
7 (8). Penis long and thin, somewhat clavate, covered with minute spines.
Longitudinal slit occupies about one third of total length of penis. The
base of os penis is feebly enlarged, rather convex ang gradually pas-
sing into the style. W part of Caucasus Range, mountains of Armenian
SSR........................................... 6. S. caucasica Vinogr.
8 (7). Penis rather and stout and short. Longitudinal slit occupies about a half
of total length of penis. The base of os penis somewhat more enlarged,
rather concave and more abruptly separated from the style. Saghalien,
Ussuri Province . ..............................4. S. caudata Thomas.
Notice: Other species of „concolor group" are not included into
the present key, as the characters of their genitalia are not known; other
characters as cranial, dental, external etc. are not sufficiently clear to use
them in the key. These species are as follows: 5. б', tianschanica Salensky,
7. S', concolor Biichn., 9. S. flava True and 8. S', leathemi Thos.
2. GEN. ZAPUS COUES.
Only one species — Z. {Eozapus) setchuanus Pous. — Szechwan,
C. Kansu.
2. Subf. EUCHOREUTINAE
Only one genus and species — Euchoreutes naso Sclat. — Sintsien,
Mongolia.
3. Subf. CARDIOCRANIINAE
1 (2). Hind foot with 5 toes. Zygoma without any process rising from its
middle. First thoracal vertebra is not ankylosed to posterior cervical
one. Symphisis pubis is formed by rather short and broad lobular
processes of ishio-pubis. Tubercular structure of molars is more or less
clearly visible in adult specimens..............4. Cardiocranius Satun.
190 CARDIOCRANIINAE, ALL ACT AGIN AE
2 (1). Hind foot with only 3 toes. Zygoma with well developed angular pro-
jection or sabre-shaped process rising from its middle and directed,
downwards and backwards. First thoracal vertebra is ankylosed to pos-
terior cervical one. Symphisis pubis is formed by narrow and very long
processes of ishio-pubis. Tubercular structure of molars is visible only
in young and subadult specimens................5. Salpingotus Vinogr..
4. Gen. CARDIOCRANIUS SATUN.
Only one species — C. paradoxus Satun. — Nan-shan, N. Gobi desert..
5. Gen. SALPINGOTUS VINOGR.
1 (4). Tail in its proximal half or third thick with fat covering on vertebrae..
All surface of tail covered with short hairs which are only a little
longer on the tip, not forming, however, a conspicuous pencil.
2 (3). Tail length more than 100 mm. Anterior broadened part of zygoma
not sharply separated from its posterior thin part. Lower margin of
zygoma in its middle is furnished only with feebly defined angular
process, not bent backwards. — Locality not exactly known (probably
Afghanistan near Kandagar)....................... 2. S. thomasi Vinogr.
3 (2). Tail length less than 100 mm. Anterior broadened part of zygoma
sharply separated from its posterior thin part. Lower margin of zygoma,
in its middle is furnished with well defined process (it is, however,,
short) clearly bent backwards. — N. part of Gobi-Altai (Shara-sume) .
........................................ 1. S. crassicauda Vinogr..
4 (1). Tail thin, without fat covering on vertebrae. All surface of tail is cove-
red not only with short hairs, but in addition to them there are long
projecting separate hairs which are on its tip still longer and forming
a long thin pencil. — Central Gobi (Khara-khoto)......................
.............................................3. S. kozlovi Vinogr.
4. Subf. ALLACTAGINAE
1 (6). Hind foot with 5 toes.
2 (3). Upper Pm is present. External surface of grinding surface of M1 with.
4 or 5 enamel angles. Margin of squamosal with large incisure corres-
ponding to a lobular process of parietal. Spines on dersal surface of
penis directed backwards (not bent inwards) .... 6. Allactaga Cuv.
3 (2). Upper Pm is wanting in adult specimens. External surface of M1 with
only 3 enamel angles. Margin of squamosal on squamoso-parietal suture
without any incisure. Spines on dorsal surface of penis are somewhat,
bent inwards.
4 (5). Tail thin and long, without conspicuous fat covering on vertebrae; tail,
tip is furnished with distichous hair pencil. Symphisis pubis long (as iit
Allactaga). Middle area of dorsal surface of penis with two feeble, not
4. CARDIOCRANIUS, 5. SALPINGOTUS, 6. ALLACTAGA 191.
sharply defined, depressions. Spines near the base of penis feebly bent
inwards.......................................7. Alactagulus Nehr.
5 (4). Taill usually much thickened and relatively short, usually with conside-
rable fat covering on vertebrae; tail tip without conspicuous distichous
flat pencil, sometimes only with rather long simple pencil, Symphisis
pubis short. Middle area of dorsal surface of penis with sharply
marked narrow deep impression. Spines near base of penis sharply
bent inwards...................................8. Pygerethmus Gloger.
6 (1). Hind foot with 4 toes (first toe wanting) .... 9. Scarturus Glov.
6. Gen. ALLACTAGA CUV.
1 (14). Pm twice or three times smaller than last molar. Anterior parts of zygc-
mata (as seeing from above) directed transversally. Penis furnished
with longitudinal furrow on its dorsal surface.
2 (11). Tympanic bullae feebly inflated and their apices not approached to
each other.
3 (6). Hind foot in adult specimens at last 70 mm (usually more). Condylo-
basal length in adult specimens at least 35 mm. Median furrov on the-
dorsal surface of penis reaches about centrum of its surface and then
diverging in two oblique furrows.
4 (5). Black proximal part of distichous tail pencil usually not interrupted
along vertebrae with white stripe. Dorsal surface of penis with about
60 spines. — Steppe- and semidesert zone of European part of USSR.
W. Siberia and Kazakhstan...........................1. A. jaculus Pallas.
5 (4). Black proximal part of distichous tail-pencil is interrupted along verte-
brae with white stripe (usually there is on this stripe a median line of black
hairs). Dorsal surface of penis usually with about 30 spines. Clay deserts
of Soviet Central Asia and Kazakhstan. . . . 2. A. severtzovi Vinogr..
6 (3). Hind foot in adult specimens shorter than 70 mm; condylobasal length,
in adult specimens less than 33 mm. Median furrow on the dorsal
surface of penis reaching at least to distal quarter of penis or going as
far as its. tip.
7 (8). Size small: condylobasal length in adult specimens not surpassing
27.5 mm, diastema not more than $.7 mm; maxillary tooth row not more
than 5.5 mm. Median furrow on the dorsal surface of penis not reaching ,
a little its tip and diverging in two short furrows.—-Semidesert and
desert zone from N. E. Ciscaucasia and E. Transcaucasia to Saissan
Lake, N. W. Dzungaria, Soviet Central Asia, Afghanistan, Baluchistan,
N. Iran...............................................3. A. elater Licht.
8 (7). Size medium: Condylobasal length more than 27.5 mm; diastema usually
more than 8.7 mm; maxillary tooth row more than 5.5 mm. Median furrow
on the dorsal surface of penis reaches its tip and not diverging.
9 (10). Hind foot at least 62 mm (62—68 mm); condylobasal length at least
30 mm (30—32). — Asia Minor, S. Transcaucasia . 4. A. williamsi Thos..
192
ALLACTAGINAE, DIPODINAE
10 (9). Hind foot less than 62 mm (52—58); condylobasal length usually less
than 30 mm (28—30). — Mesopotamia, Syria . 5. A. euphratica Thos.
11 (2). Tympanic bullae much inflated and their apices approached to each
other and sometimes nearly touching each other.
12 (13). Ear-conch not more than 36 mm (usually shorter). Interorbital constric-
tion not less than 10 mm. Upper tooth-row more than 5 mm (5.8—6.7).
Incisive foramina relatively narrow. — Gobi Desert......................
.......................................... 6. A. bullata Gl. Allen.
(1 3) 12. Ear-conch more than 36 mm (in only one known specimen it is 41 mm).
Interorbital constriction about 9 mm: upper tooth-row about 5 mm.
Incisive foramina considerably broadened. — Baluchistan.................
................................................7. A. hotsoni Thos.
14 (1). Premolar is nearly equal to posterior molar or somewhat surpassing it.
Anterior parts of zygomata (as seen fiom above) directed obliquely
(externally and backwards). Longitudinal furrow on the dorsal surface
of penis wanting. — Kazakhstan from E. shore of Caspian sea to S. Altai,
Mongolia, S. Transbaikalia, W. Manchuria, N. China (Cansu, Shansi,
Chili).............................................8. A. saltator Eversm.
7. Gen. ALACTAGULUS NEHR.
Only one species. Zone of semideserts and clay deserts; from E. Ciscau-
casia to Kuldja, southwards to Kopet-Dagh and Termez; found also in
Ordos and Alashan.................................A. acontian Pall.
8. Gen. PYGERETHMUS GLOG.
1 (2). Terminal hairs on the tail white; length of terminal hairs about 13—
15 mm. Under surface of tail in subterminal part dark. Tail length
95—150 mm. Hind foot 40—45 mm...................1, P. zhitkovi Kuznezov.
2 (1). Terminal hairs of the tail dark (in rare specimens among them few
separate white hairs can present); length of terminal hairs nor more
than 6—8 mm; under surface of tail in subterminal part and through
its whole length light. Tail length 80—90 mm; hind foot 32—35 mm.
Semideserts of lower course of Ural River to N. shore of Aral Sea and
to S. Ust-Urt plateau. . . . . t...................2. P. platyurus Licht.
9. Gen. SCARTURUS GLOVER
Only one species. S. tetradactylus Licht. — Lybian desert.
5. Subf. DIPODINAE
1(2). Mastoids as seen from above do not project on the lateral sides of the
posterior part of braincase. Postglenoid fenestrae open into internal
cavity of braincase. Pm well developed (but small). Upper incisors
yellow.................................................10. Dipus Gmel.
7. ALACTAGULUS, 8. PYGERETHMUS, 9. SCARTURUS 193
2 (1). Mastoids as seen from above form conspicuous rounded projections
on the lateral sides of the posterior part of braincase. Postglenoid fene-
strae are partly or entirely closed from inside by a portion of petro-
mastoideum. Pm in adult specimens usually wanting or retaining
as a minute rudiment only in one species {Scirtopoda andrewsi). Upper
incisors white.
3 (6). Temporal region of skull without a crest or prong formed by a reent-
rant lateral process of parietal. The apices of tympanic bullae are not
connected with each other by a suture. Mastoids are considerably
inflated but forming no large bladders.
-4 (5). Distichous tail pencil wanting (long hairs on the distal part of the tail
occupy nearly all distal part of the tail). Hair brush on hind foor simple
(without admixture of thick bristles), formed with curved hairs. Posterior
margin of bony palate projects backwards considerably further than
the level of the posterior side of Ms; this margin forms a salient angle
directed backwards. Postglenoid fenestrae form a rather broad slit
partly closed from inside by a portion of petromastoideum. Lacrymals
are not considerably displaced externally, their interior apices are in
contact with frontal bone. Mandible with well developed alveolar pro-
cess .................................................11. Scirtopoda Brandt.
5 (4). Distichous tail pencil well developed and occupies only terminal part
of vertebrae. Hair brush on hind foot consisting of two kinds of hairs:
in addition to long and rather thin hairs there are shorter and thicker
bristles. Posterior margin of bony palate situated on the level of the
centrum of M3; this margin is concave. Postglenoid fenestrae very small
and rounded, entirely closed from inside by a portion of petromastoi-
deum. Lacrymals considerably displaced externally and articulating with
a’narrow transversal process of frontal bone. Mandible without alveolar
process or tubercle................................. 12. Paradipus Vinogr.
6 (3). Temporal region with sharply ridge or prong formed by a reentrant
angle of lateral process of parietal. The apices of tympanic bullae are
connected with each other by a suture or ankylosed. Mastoids are greatly
inflated and forming large bladders.
7 (8). Lateral process of each parietal bone is furnished with a sharply deve-
loped conical prong directed externally and downwards. Postglenoid
fenestrae Very small, usually somewhat elongated. Superior margin
of external wall of infraorbital canal is not ankylosed to the wall of
maxilla. The root of the lower incisor forms a feebly developed alveolar
process..............................................13. Eremodipus Vinogr.
.8 (7). Lateral process of each parietal bone is furnished with a sharply deve-
loped crista, its base being formed by surrounding parts ot the squamo-
sal. Postglenoid fenestrae are greatly enlarged and form nearly equila-
teral triangles. The superior margin of the external wall of infraorbital
canal is completely fused to the wall of maxilla in adult specimens. The
Фауна СССР, Виноградов 13
194
DIPODINAE
root of the lower incisor forms a prominent alveolar process ....
.............................*............... . 14. Jaculus ErxL
10. Gen. DIPUS GMEL.
Only one species D. sagitta Pall, (including/), sowerbyi Thos., D. hall!
Sowerby, D. deasyi Barr.-Ham and D, lagopus Licht.; the named forms must
be considered as subspecies of D. sagitta, as it was shown above).—
Sable deserts from E. Ciscaucasia, Kazakhstan, Soviet Central Asia, Sintsien,,
to Mongolia and N. China.
11. Gen. SCIRTOPODA BRANDT
1 (2). Pm is wanting in adult specimens. Hind foot 46—51 mm, condylobasal
length 25-—28 mm. — Lower course of Dnieper, E. Ciscaucasia, lower
Volga, Kazakhstan Eastward to Saissan..................1. S. telum Licht.
2 (1). Pm is present in adult specimens. Hind foot about 55 mm, condylobasal
length about 30 mm. — Mongolia..................2. S. andrewsi Gl. Allen.
12. Gen. PARADIPUS VINOGR.
Only one species — Kara-Kum and Kizul-Kum Deserts.................
.......................................1. P. ctenodactylus Vinogr
13. Gen. EREMODIPUS VINOGR.
Only one species — Kara-Kum and Kizyl-Kum Deserts.................
..........................................1. E. lichtensteini Vinogrt
14. Gen. JACULUS ERXLEBEN
Systematic interrelationships^of the described forms are not clear at pre-
sent time. It is possible that all forms described from N. Africa and Arabia
may be united under two species —J. jaculus L. (small forms, penis without
large spines) and J. orientalis Duv. (large forms, penis with two long spines).
Iranian/, loftusi is similar to / jaculus and perhaps identical with it. Other
species from Iran —J. blanfordi differs from /. loftusi by larger dimensions;
its genitalia are unknown and therefore the true position of this species is not
clear.
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
acontion, Alactagulus — 134, 139, 141, 144,145,
192.
Alactagulus —102, 106, 107, 118, 123, 133,
134, 142, 143, 144, 145, 146, 184,191, 192.
alashanicus, Euchoreutes naso — 87.
Allactaga — 102, 105, 106, 107, 110, 13 \ 134,
142, 147,151, 184, 190.
Allactaginae — 48, 51, 53, 58, 82, 83, 85, 86,
87, 89, 91, 92, 93, 101, 104, 105, 107, 111,
119, 126, 142,147,149, 150, 151, 184, 186,
187.
annulata, Allactaga saltator — 129, 130, 132.
andrewsi, Scitropoda —152, 167, 168, 172, 193,
194.
aralychensis, Allactaga elater — 120, 122, 123.
betulina, Sicista — 64, 67,69,72, 73, 74, 75,189.
birulae, Scirtopoda telum — 169.
blanfordr, Jaculus — 182, 194.
bullata, Allactaga —11, 128, 129, 192.
Cardiocraniinae — 48, 51, 53, 85, 87, 88, 89,
90, 91, 93, 95, 142, 150, 151,183, 184,185.
186, 189.
Cardiocranius — 51, 61, 84, 89, 90, 91, 92, 93,
94, 95, 104, 183, 184, 186, 189,190.
chachlovi, Alactaga jaculus — 115.
caudata, Sicista — 64, 67, 76, 77, 78, 79, 189.
caucasicus, Allactaga elater —120, 121, 122,
123.
cancasica, Sicista — 64, 67, 76, 77, 78, 189.
concolor, Sicista — 66, 76, 77, 78, 79, 189.
crassicauda, Salpingotus — 95, 96, 97, 98, 99,
100, 190.
ctenodactylus, Paradipus — 171, 174,176,194.
decamanus, Allactaga jaculhs — 114.
deasyi, Dipus sagitta — 158.
dinniki, Alactagulns acontion 135, 136.
Dipodidae — 59, 61, 66, 80, 85, 90, 91, 92, 100,
103, 105, 183.
Dipodinae —48, 51, 53, 58, 82, 85, 87, 89, 90,
93, 95, 104, 105, 149, 150, 151, 152, 155,
173, 178,183,184, 185, 186,187, 192.
Dipus —95, 150, 151, 152, 154, 155, 166, 167,
170,178, 179, 180, 181, 184, 192, 194.
dzungariae, Allactaga elater— 122.
elater, Allactaga elater — 120, 121, 122, 123.
elater, Allactaga — 105, 110, 111, 116, 118,119,
120,122, 123, 124, 126, 127,133, 134, 141,
147, 191.
Eozapus —61, 63, 64, 79, 81, 82,104, 183, 188.
Eremodipus —95, 152, 153, 170, 177, 178, 179,
180,181, 184, 193, 194.
Euchoreutes —61, 83, 85, 86, 87, 183, 184,189.
Euchoreutinae — 42, 51, 53, 82, 83, 84, 85, 86,
89, 90, 104, 150, 184,186, 187, 189.
euphratica, Allactaga — 110, 111, 122, 126,127,
147, 192.
falz-feini, Scirtopoda telum — 168, 169, 170.
flava, Sicista — 66, 77, 79, 189.
fraudator, Sminthoides — 90.
fulvus, Citellus — 93.
fuscus, Allactaga jaculus — 113.
Gerbillinae —184.
grabaui, Protalactaga — 90, 151.
halli, Dipus sagitta—155, 160, 194.
hotsoni, Allactaga— 111, 128, 129, 192.
inna'e, Dipus sagitta — 158.
indica, Allactaga elater —• 120, 122, 123.
jaculus, Allactaga — 84, 95, 105, 107, 110, 111,
113, 117, 118, 119, 126, 129, 133, 150,152,
153,166, 167, 171, 172, 177, 179, 180, 181,
182, 184, 191, 194.
jaculus, Allactaga jaculus — 113, 177, 178, 180,
182, 194.
kizljaricus, Allactaga elater — 120, 122, 123.
kozlovi, Salpinogotus — 88, 94, 95, 96, 97, 98,
100,101, 190.
lagopus, Dipus sagitta — 155, 158, 164.
laticeps, Allactaga williamsi — 126.
leathemi, Sicista — 66, 79, 189.
leeli, Plestodipus — 90.
lichtensteini, Eremodipus—-171, 179, 194.
loftusi, Jaculus — 182, 194.
13*
196
УКАЗА ТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
loriger, Sicista — 67.
mongolica, Allactaga —129.
mongolica longior, Allactaga — 129.
mongolica, Allactaga saltator—130, 131, 132.
montana, Sicista — 64, 72, 78, 189.
montana strandi, Sicista — 72.
Muridae — 59, 61, 80.
napaea, Sicista — 64, 67, 75, 77, 189.
Napaeozapus — 61, 62, 81, 82, 188.
naso, Euchoreutes — 85, 86, 189.
nogai, Dipus sagitta — 158, 164.
nordmanni, Sicista — 64, 67, 188.
norvegica, Sicista — 72.
orientalis, Jaculus — 180, 181, 182, 194.
Paradipus — 95, 152, 153, 154, 170, 172, 173,
174, 181, 184, 193, 194.
Paralactaga —105.
paradoxus, Cardiocranius — 93, 99, 190.
platyurus, Pygerethmus —102, 142, 143, 145,
192.
Plesiodipus — 92, 95, 151, 183.
Plesiosminthus — 62, 82.
potanini, Alactagulus acontion — 139.
Protalactaga —183.
Pygerethmus — 51, 93, 102,104, 106, 107, 134,
142,143, 145, 146, 147,184, 186, 191,192.
riickbeili, Allactaga —129.
rilckbeili, Allactaga saltator — 130, 131.
sagitta, Dipus —155,156,164,166,170,179,194.
sagitta, Dipus sagita — 158, 160, 166.
Salpingotus — 51,61, 84, 88, 89, 90, 91, 92, 94,
95, 96, 97,183, 184, 186,190.
saltator, Allactaga —105, 107, 110, 111, 117,
127,128,129, 130,131, 132, 192.
saltator, Allactaga saltator — 130, 131.
Scarturus —101, 104, 106, 142, 147, 184, 191,
192.
Scirtopoda —151,152, 153, 166,167, 172,174,
178,179, 180, 181,184,193, 194.
severzovi, Allactaga —11, 117, 118, 119,124,
129.
setchuanus, Zapus — 82, 189.
schitkovi, Pygerethmus —144.
schmidti, Allactaga williamsi — 125, 127.
Sicista — 58, 61, 62, 63, 64, 66, 67, 68, 69, 70,.
71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 83, 84,.
104, 183,184,188.
Sminthoides — 92, 183.
sowerbyi, Dipus —155.
sowerbyi, Dipus, sagitta — 160.
spiculum Allactaga jaculus — 114.
strandi, Allactaga elater — 122.
Stylodipus —172.
subtilis, Sicista — 64, 66, 67, 69, 71, 72, 73, 75,.
188.
sushkini, Allactaga saltator—129, 130, 131,
132.
telum, Scirtopoda — 93, 141, 151, 167,168,169,.
170,171,172,194.
telum, Scirtopoda telum — 169.
thomasi, Salpingotus!—90, 95, 96, 97,98,99,.
100, 191.
tetradactylus, Scarturus — 110, 126, 147, 149,.
192.
tianschanica, Sicista — 66, 76, 77, 78, 79, 189.
trizona, Sicista — 71.
turovi, Scirtopoda telum — 168, 169.
vagus, Sicista — 67.
vexillarius, Allactaga jaculus — 114.
williamsi, Allactaga —105, 107, 110,111, 125,.
126, 127, 128,147,191.
Zapodinae —42, 48, 51, 53, 58, 61, 62, 64, 83,
84, 85, 86, 87, 89, 90, 91, 92, 101, 103, 104,.
105,142,183,184,185,186,187, 188.
Zapus —58, 61, 62, 63, 64,66,79,80,81,82,.
83, 84,104, 183,184,188,189.
zhitkovi, Pygerethmus — 102, 142, 143, 144,„
145, 146, 147, 192.
СОДЕРЖАНИЕ
Стр.
Предисловие................................................... V
Систематический указатель видов.............................. VII
Введение
Краткий очерк истории изучения систематики группы.............. 1
Общая морфологическая характеристика семейства................. 4
Основные направления специализации сем. Dipodidae ............ 12
Краткая экологическая характеристика сем.-Dipodidae.......... 16
Краткий очерк геологической истории сем. Dipodidae............ 18
Общий очерк географического распространения................. 28
Классификация ............................................ . . 32
Статистические данные о систематическом составе семейства..... 38
Значение представителей сем; Dipodidae для человека........... 35
Список цитированной литературы............................. 36
Специальная часть
Таблица для определения подсемейств........................... 41
1. Подсем. Zapodinae ...................... .58
2. Подсем. Euchoreutinae .......' .............. 82
3. Подсем. Cardiocraniinae ........................ ... 87
4. Подсем. Allactaginae ........ .... .............. 101
5. Подсем. Dipodinae ................... .... 149
Резюме....................................................... 183
Указатель латинских названий................................. 194
ФАУНА СССР
(новая серия)
Млекопитающие (Mammalia)
Том I. Вып. 1. Млекопитающие: введение; вып. 2. Олени и кабарги; вып. 3.
Полорогие (*быки, ^антилопы, козы, бараиы); вып. 4. Верблюды, свиньи,
лошади, ослы.
Том II. Вып. 1. Китообразные и сиреневые; вып. 2. Ластоногие; вып. 3.
Хищники; кошки; вып. 4. *Куньи; вып. 5. Гиены, собаки, медведи.
Том III. Грызуны. Вып. 1. Введение; вып. 2. *Беличьи; вып. 3. Бобры, сони,
дикобразы, слепыши; вып. 4. Тушканчики; вып. 5. Мыши; вып. 6.
Хомяки; вып. 7. ^Песчанки; вып. 8. Полевки; вып. 9. *Пищухи; вып. 10.
Зайцы.
Том IV. Вып. 1. ^Рукокрылые; вып. 2. ^Насекомоядные.
* Отмечены группы, находящиеся в работе, ---- законченные обработ-
кой, ===== •— напечатанные.
ФАУНА СССР
(новая серия)
Вышли из печати:
Млекопитающие т. III, вып.4—Тушканчики (Б.С.Виноградов), 1937, цена 10 руб.
Рыбы т. VI, вып. 1 — Triglidae (А. Н. Световидов), 1936, цена Гр. 25 к.
Насекомые двукрылые т. III, вып. 2 — Москиты (П. П. Перфильев), 1937,
цена 7 р.; т. VII, вып. 2 — Слепни (Н. Г. Олсуфьев), 1937, цена 17 р.; т. XIX,
вып. 1 —Сем. Sarcophagidae, ч. 1 (Б. Б. Родендорф), 1937, цена 20 р.
Насекомые перепончатокрылые т. II, вып. 1 — Пилильщики Tenthredinodea,
ч. 1 (В. В. Гуссаковский), 1935, цена 22 р. 75 к.; т. V, вып. 2 — сем.
Braconidae, ч. 1 (Н. А. Теленга), 1936, цена 20 р. 75 к.
Насекомые жесткокрылые т. XXI — Дровосеки, ч. 1 (Н. Н. Плавильщиков),
1936, цена 22 р.; т. XXVI, вып. 1 — Листоеды Galenzcinae (Д. А. Огло-
блин), 1936, цена 17 р.
Насекомые кожистокрылые (Г. Я. Бей-Биенко), 1936, цена 14 р.
Ракообразные т. VII, вып. 3—Равноногие Дальневосточных морей (Е. Ф. Гурья-
нова), 1936, цена 15 р. 50 к.
Губки т. II, вып. 2—Пресноводные губки (П. Д. Резвой), 1936, цена 6 р. 60 к.
Находятся в печати:
Птицы т. I, вып. 5 — Дневные хищники (Б. К. Штегман).
Насекомые двукрылые т. III, вып. 5 — Кровососущие комары Culicinae (А. А.
Штакельберг).
Насекомые чешуекрылые или бабочки т. XIII, вып. 3 — Совки Agrotinae
(И. В. Кожанчиков).
Подготовлены к печати:
Млекопитающие т. 1, вып. 2 — Олени (К. К. Флеров); т. III, вып. 1 — Мыши
и крысы (А. И. Аргиропуло).
Рыбы т. VII, вып. 4 — Трескообразные (А. Н. Световидов).
Насекомые перепончатокрылые т. II, вып. 2 — Пилильщики, ч. 2 (В. В. Гус- -
саковский); т. V, вып. 3 — Braconidae, ч. 2 (Н. А. Теленга); т. XIV,
вып. 1 — Муравьи, ч. 1 (К. В. Арнольди, В. А. Караваев).
Насекомые двукрылые т. VI, вып. 6 — Мошки Simuliidae (И. А. Рубцов);
т. IX, вып. 2 — Жужжала, ч. 1 (С. Я. Парамонов).
Насекомые жесткокрылые или жуки т. V, вып. 3 — Жуки-карапузики, ч. 1
(А. Н. Рейхардт); т. XIV, вып. 2 — Лжекороеды Bostrychidae (В. Н.
Старк); т. XXII — Дровосеки, ч. 2 (Н. Н. Плавильщиков).
Насекомые сосущие т. VI, вып. 3 — Тли. Триба Eriosomini (А. К. Мордвилко).
Паукообразные т. I, вып. 3 — Сольпуги (А. А. Бялыницкий-Бируля); т. V,
вып. 2 — Водные клещи Hydracarina (И. И. Соколов).
Ракообразные т. II, вып. 1 — Ракушковые Ostracoda континентальных водое-
мов (3. С. Бронштейн); т. VI, вып. 1 — Усоногие Cirrhipedia (Н. И.
Тарасов).
Моллюски т. III, вып. 1 — Unionidae (В. И. Жадин).