/






























































































































































































































































































































































Текст
c/l.^C.c/lun U H
ПРЕСНЫЕ ВОДЫ И ИХ жизнь
УЧ П ЕД ГИ 3 1 9 5 О
Нроф. А. И. липин
ПРЕСНЫЕ ВОДЫ и II X ЖИЗНЬ
Издание третье переработанное
Ч/Х/Ч ЧЧЧ VW4ZA444 KWV ЧЧЧ ЧАХЛ ХЧХЧЛЧЧ-» WV»V% VAX VA'VX'W "4ZV4.
ГОСУДАРСТВЕННОЕ УЧЕБНО-НЕДАГОГИЧЕСКОЕ ИЗДАТЕЛЬСТВО МИНИСТЕРСТВА ПРОСВЕЩЕНИЯ РСФСР МОСКВА * 1 V 6 О
В настоящем издании по сравнению с предыдущим книга проф. А. Н. Липина значительно переработана и дополнена.
В общей части дано подробное описание типов пресных водоёмов и их населения, растительного и животного. Подчёркнуто хозяйственное значение тех или иных групп водных обитателей и их роль в преобразовании природы нашей страны.
Во второй части даны таблицы для определения важнейших групп водных беспозвоночных животных и водных растений.
Книга может служить пособием для учителей средней школы и студентов педвузов.
ПРЕДИСЛОВИЕ
Послевоенные годы характеризуются чрезвычайно энергичным развитием всех областей нашего социалистического хозяйства. В связи с этим в сильнейшей степени возрастает значение активного воздействия человека на природу с целью получения от неё возможно большего количества материальных и энергетических ресурсов, необходимых для дальнейшею подъёма промышленности и сельского хозяйства нашего отечества.
Среди природных источников, из которых техника может черпать необходимые для неё сырьё и энергию, очень большое место занимает вода. Многообразие форм использования воды человеком обусловливает и большое различие типов водного хозяйства. Партия и правительство нашей страны придают огромное значение внутренним (пресноводным) водоёмам и в ряде постановлений довоенного и послевоенного периодов дали твёрдые указания относительно использования этих водоёмов для нужд народного хозяйства. Эти указания не ограничиваются только проблемой освоения существующих водоёмов, но и делают упор на необходимость устройства новых.
Постановление Совета Министров Союза ССР и ЦК ВКП(б) «О плане полезащитных лесонасаждений, внедрения травопольных севооборотов, строительства прудов и водоёмов для обеспечения высоких и устойчивых урожаев в степных и лесостепных районах Европейской части СССР» ясно подчёркивает теснейшую связь растения с водой. Планирование насаждений различных растительных культур (полевых, луговых, лесных и пр.) на огромных площадях в целях достаточного их орошения нераздельно связано с планированием строительства новых водоёмов.
Для вновь строящихся водоёмов, помимо основной их функции — накопления воды в количествах, достаточных для орошения проектируемых посевных площадей и лесных массивов, — планом предусматривается и другая форма их использования: разведение в них рыбы. Это сильно повышает хозяйственное значение этих водоёмов как источников дополнительного питания населения СССР.
Что же нужно, чтобы выполнить задания этого величественного плана в той его части, которая касается строительства и освоения искусственных водоёмов? Нужны организованные и объединённые усилия громадного числа работников физического и умственного труда. В разрешение вопросов, связанных с строительством новых водоёмов, включаются Академия наук СССР с её филиалами, республиканские академии наук, факультеты университетов, научно-исследовательские институты различных специальностей, большое количество работников министерств лесного хозяйства и земледелия и т. д. Очевидно, что по мере развёртывания работы в неё вольются школьные работники, краеведческие организации, кружки по изучению природы (школьные, студенческие), юннаты, наконец, просто отдельные лица, воодушевлённые желанием внести и свой труд в это великое, общенародное дело преобразования природы нашей родины.
Несомненно, значительная часть учителей-биологов будет участвовать в работах по строительству водоёмов и их освоению для рыборазведения. Понятно, что чем лучше учителя будут подготовлены к работе, тем сознательнее и успешнее они будут её выполнять. Частично эта подготовка заклю-
* 3
чается в приобретении знаний о внутренних водоёмах и о жизни их обитателей. Существует значительное количество книг, которые могут оказать помощь желающим ознакомиться с основами учения о пресных водах и жизни населяющих их существ. Автор надеется (и опыт двух первых изданий поддерживает в нём эту надежду), что и выпускаемая 3-м изданием его книга «Пресные воды и их жизнь» принесёт посильную пользу и оправдает своё появление в свет.
В настоящее (3-е) издание, по сравнению со 2-м, введены значительные дополнения. В обшей части книги некоторые главы («Пруд», «Болото») написаны заново; глава «Река» расширена введением более подробного описания генезиса и формирования реки (речных долип, русла, поймы), а также более детального описания речных биоценозов. Во 2-ю часть введены новые определительные таблицы — по гидре, коловраткам, олигохетам (сост. И. И. Малевич), мшанкам (сост. Г. Г. Абрикосов); более широко развёрнуты введением дополнительных объектов другие таблицы как по растениям, так и по животным (например, по листоногим и ветвистоусым рачкам, по некоторым отрядам насекомых и пр.).
Помимо указанных крупных добавлений к книге, была проведена общая переработка её в соответствии с появлением в литературе новых научных данных за период, протекший со времени выпуска 2-го издания.
Считаю своим долгом выразить искреннюю благодарность проф. С. Д. Муравейскому, проф. | Д. А. ЛасточкинуТ] от которых я получил целый ряд очень ценных указаний, а также И. .И. Малевичу и Г. Г. Абрикосову за их любезное согласие написать для моей книги определительные таблицы по олигохетам и мшанкам, и другу, доктору биологических наук Н. Н. Липиной.
Проф. А. Липин.
ВВЕДЕНИЕ
Вода па земном шаре занимает более*2/* его поверхности, и только около */3 приходится на долю суши. Почти вся эта огромная масса воды сосредоточена в океанах и морях и представляет негодную для питья солёную воду. Но, кроме этой морской воды, существует, как всем известно, вода, которую принято называть пресной. В то время как морская вода, за очень лишь немногими исключениями, омывает сушу снаружи, разделяя её на материки и острова, пресная вода, напротив, в громадном большинстве случаев сосредоточена внутри материков, причём общая площадь её относится к площади морской воды приблизительно как 1 : 20. Вот этими-то пресными водами нам и предстоит заняться в нашей книге.
Пресная вода находится н а поверхности земли или под ней в виде отдельных, в той или иной степени ограниченных сушей скоплений, обозначаемых общим названием водоёмов. Такое более или менее полное окружение пресных водоёмов сушей дало повод ещё к другому названию их: их нередко называют также внутренними водоёмами, противопоставляя внешним водоёмам, к которым относятся моря и океаны.
Из двух групп внутренних водоёмов — наземных и подземных (так называемых грунтовых) — нас здесь интересует лишь первая группа, и потому мы на ней и остановим своё внимание.
Из предыдущего ясно, что для возникновения на поверхности земли внутреннего водоёма необходима прежде всего наличность углубления в суше и заполняющей его воды. В результате вза имодействия воды и суши и образуется водоём. Это взаимодействие может быть очень разнообразным, отчего зависит разнообразие самих водоёмов. Попадая тем или иным путём на поверхность земли, вода в силу собственной тяжести стремится занять наиболее низкие места, в которые и стекает с более высоких уровней. Скорость течения зависит от величины и формы склона. Если бы сопоставить все внутренние водоёмы земной поверхности в нисходящий по скорости течения воды ряд, то мы получили бы постепенный переход от водопадов до водоёмов, совсем или почти совсем лишённых постоянного и определённо направленного стока воды. По этому признаку обычно и делят внутренние водоёмы на две группы: текучие — водопады, реки, ручьи и ключи (родники, источники) и стоячие — озёра, пруды, лужи и болота.
Не нужно, однако, думать, что во всех так называемых стоячих водоёмах всегда отсутствует течение воды; среди озёр и прудов немало существует проточных, в которых, с одной стороны, вода притекает через впадающие в них речки и ручьи, а с другой — вытекает таким же путём; этим вызывается ток воды, хотя слабый и медленный, но постоянный. Таким образом, можно различать три группы водоёмов: текучие, очень медленно текучие и стоячие. Современные учёные предпочитают первую из этих групп называть водоёмами быстрого стока, а вторую и третью группы объединять под названием водоёмов замедленного стока.
Давая термину «сток» широкое толкование, мы включаем в это понятие не только самую воду, но и всё, что стекающая вода несёт в себе: механически взвешенный материал как минерального, так и органического про-
5
исхождения (частицы размельчённого грунта и почвы, живые организмы, их трупы и остатки в различной степени разложения и т. п.) и растворённые химические вещества — газы и соли. Всё это вместе со скоростью течения воды представляет совокупность основных факторов стока. Различными комбинациями этих факторов, а также формой, величиной и геологическим строением русла стока и определяется всё разнообразие внутренних водоёмов.
Так как вода является одной из наиболее мощных производительных сил природы, то понятно, что внутренние водоёмы должны иметь и действительно имеют громадное хозяйственное значение. Формы и возможности их использования в промышленности и сельском хозяйстве зависят от типа, характера и размеров самих водоёмов, а также от их местоположения. Отсюда вытекает необходимость широкого и глубокого исследования внутренних водоёмов.
Все главнейшие составные части внутренних водоёмов — вода, многое из того, что в ней содержится (как живое, так и неживое) и, наконец, дно водоёма — могут быть использованы в народном хозяйстве. Прилагаемая схема представляет попытку сопоставить главные виды этого использования.
1. Использование воды водоёмов
1. Механическая энергия воды:
а) водные пути сообщения (судоходство, сплав); б) двигательные гидроустановки на основе использования движения воды; в) водоснабжение и канализация.
2. Тепловая энергия воды:
а) охладительные установки промышленных и сельскохозяйственных предприятий; б) согревательные установки — паровые, отопительные и т.п.
3. Химическая энергия воды:
а) орошение и удобрение; б) физиологические процессы в живых организмах; в) лечение, санитария и пр.
11. Использование содержащегося в воде водоёмов сырья
1. Минеральное сырьё (поваренная соль, сульфаты, железо и др.).
2. Органическое сырьё:
а) растительное — применение водной растительности в промышленности и сельском хозяйстве; б) животное — использование водных животных в пищевой промышленности (рыба, раки), в пуговичной (моллюски), в пушной (бобр, выдра, выхухоль, кутора, нутрия и др.). .
III. Использование донных отложений водоёмов
1. Сапропель.
2. Озёрная руда.
Эта схема ясно показывает, насколько широко и разносторонне могут быть использованы внутренние водоёмы в народном хозяйстве.
В дореволюционный период главнейшими формами хозяйственного использования внутренних водоёмов были судоходство и рыболовство. Второстепенную роль играло добывание, кроме рыбы, других видов животного сырья (раки, моллюски). В настоящее время у нас в СССР это использование ведётся не только по линии интенсивного развития рыбной промышленности и широкого развития судоходства, но и по линии всемерного использования энергетических ресурсов внутренних водоёмов. В этих целях на базе большинства крупных внутренних водоёмов СССР развернулось грандиозное строительство энергетических установок, мощных водных каналов, громадных водохранилищ, представляющих совершенно новые искусственные водоёмы озёрного типа, и пр. Весь этот гигантский размах хозяйственного использования внутренних водоёмов на пути социалистического строительства в нашем Союзе требует всестороннего изучения этих водоёмов, в результате которого наука могла бы играть роль мощного двигателя в техническом осуществлении этого строительства.
ГЛАВА ПЕРВАЯ
ОЗЕРО
1. ФИЗИКО-ГЕОГРАФИЧЕСКАЯ ХАРАКТЕРИСТИКА Озёрное ложе
ЧТО МЫ НАЗЫВАЕМ ОЗЕРОМ
Всякий, конечно, видал озёра, по далеко не каждый, если его спросить, сумеет точно определить это понятие. Научное определение озера одним учёным было сформулировано следующим образом: озеро представляет собой довольно значительную водную массу, постоянно покоящуюся в естественном замкнутом углублении на поверхности суши, или медленно текущую в нём и не имеющую близкого, непосредственного сообщения с морем.
Из этого определения видно, что озером мы можем называть далеко не всякое скопление воды, а только «довольно значительное», этим озеро отличается от более мелких водоёмов — прудов, болот, луж. Далее, водная масса озера всегда находится в замкнутом углублении, т. е. окружена со всех сторон сушей, в противоположность текучим водам, которые характеризуются, во-первых, тем, что на всём протяжении они ограничены сушей лишь с двух взаимно противоположных сторон, а во-вторых, более или менее значительной и вполне заметной скоростью течения. Наконец, если озеро находится в сообщении с морем посредством, например, реки, то это ещё не лишает его права называться озером; водоём же, находящийся в близком и непосредственном, т. е. открытом и широком соединении с морем, уже не может называться озером.
Всякий водоём, в частности и озеро, состоит из двух главнейших частей: углубления в суше (впадины, или котловины), называемого ложем, и заполняющей это углубление воды.
КЛАССИФИКАЦИЯ ОЗЁР ПО ПРОИСХОЖДЕНИЮ
Когда говорят о происхождении озера, то имеют в виду не столько способ заполнения его ложа водой, сколько историю образования самого ложа. А для этого необходимо знать строение земной коры в окружающей озеро местности. Если мы будем знать это строение, то увидим, какие геологические изменения земная кора претерпела в районе данного озера и, следовательно, как в результате этих изменений образовалась его впадина.
По способу происхождения ложа озёра могут быть разделены на несколько типов, из которых мы укажем лишь на главнейшие и наиболее обычные.
Запрудные, или плотинные, озёра образуются, как указывает самое название, путём запруды, т. е. полного или частичного преграждения течения реки. Такое преграждение может произойти от разных причин, например, от обвала или осыпи прилегающих гор, от наносов самой реки, от вмеша
тельства человека (искусственные озёра) и пр. Озёра с естественными запрудами наибольшее распространение имеют в горных местностях, например, в горах Средней Азии; запруды образуются здесь в результате горных обвалов, вследствие чего эти озёра называются ещё обвальными.
Запрудные озёра распространены в бассейнах крупных рек, например, на территории СССР — в долинах рек Волги, Оби, Амура и др. За последние годы этот тип озёр получил в нашем Союзе значительное распространение в связи с гигантским гидростроительством на наиболее мощных речных артериях: течение воды рек и каналов искусственно преграждается и задерживается мощными плотинами, перед которыми образуются благодаря этому водохранилища — громадные искусственные озёра: Московское море, ряд волжских водохранилищ (Рыбинское, Ярославское, Горьковское, Куйбышевское и др.), Днепропетровское, Магнитогорское (Урал), Мингечаурское (Кура) и многие другие.
П ровальные, или карстовые, озёра образуются в том случае, когда текущая подземная вода встречает 119 своём пути легко растворимые породы и, как говорят, выщелачивает их, т. е. растворяет и уносит с собой; естественно, в таких местах происходят пустоты — пещеры, которые с течением времени могут достичь такой величины, что прикрывающий их слой не выдерживает и проваливается; таким способом может образоваться достигающая иногда очень значительных размеров котловина, которая при заполнении её водой и образует озеро.
На территории СССР провальные озёра найдены в автономной Башкирской республике, в Горьковской области, на Урале и др.
Ледниковые, или моренные, озёра, как показывает самое название, происходят в результате деятельности ледников. Ледник при своём движении прокладывает себе более пли менее углублённое ложе, или русло; при этом он захватывает на своём пути камни, достигающие иногда значительной величины (валуны); перекатывая их по дну русла, он располагает их в виде широкой (в ширину самого ледника) гряды, носящей название донной морены. Кроме донной морены, существуют ещё так называемые боковые морены, представляющие гряды камней по боковым краям ледника; они образуются вследствие обвала камней с боковых стенок ледникового ложа, представляющих, так сказать, берега ледника. Наконец, известны и конечные морены; они происходят оттого, что ледник при своём движении толкает встречающиеся на пути камни; последние, накопляясь на его переднем конце, располагаются в виде поперечной гряды, которая и называется конечной мореной. Когда ледник начинает отступать, т. е. передний конец его начинает таять, то образующаяся при этом вода заполняет вместо ледника его ложе; склоны этого ложа образованы запрудой (конечной мореной) и окружающими горными породами; на дне ложа и отчасти по бокам его остаются камни донной и боковых морен, по присутствию и расположению которых и можно узнать о бывшем здесь леднике. Озёра этого типа имеют очень большое распространение. Их много и на территории РСФСР, например, в областях Ленинградской, Калининской, Ярославской и др.
Остаточные, или реликтовые, озёра представляют собой остатки моря, которые могут образоваться двояким способом: или простым отделением части моря, например, морского залива, посредством пересыпи (песчаного наноса) от самого моря, или путём сохранения остатков моря, отступившего из данной местности. При этом, если через такое озеро со временем устанавливается проток пресной воды, то оно постепенно превращается в пресноводное озеро; в противном случае вода его сохраняет свойства солёной воды, и тогда его называют солёным озером.
К озёрам морского происхождения относятся, например, Чудское, Каспийское, Аральское и др.
Относительно термина «реликтовые озёра» нужно, впрочем, заметить, что он применяется также и к предыдущей группе озёр, т. е. кледнико-s
вым озёрам. Одним из отличительных признаков реликтовых озёр является присутствие в них реликтовых организмов, представляющих остатки или от населения ледниковой эпохи, или от морской фауны и флоры; по присутствию этих организмов до известной степени можно судить и о происхождении реликтовых озёр — ледниковом или морском.
Упомяну также группу озёр с котловинами древнего тектонического происхождения. Эти котловины появились в процессе горообразования как результат имевших в то время место сдвигов, сбросов и складок земной коры. При заполнении таких котловин водой образовывались крупные горные озёра, примерами которых могут служить: Байкал в Сибири, Иссык-Куль в Тянь-Шане, Севан (Гокча) в Армении.
ВЕЛИЧИНА. ОЗЁР
Величина озёр складывается из различных элементов, главнейшими из которых являются длина, ширина, площадь и глубина озера, а также объём его водной массы.
Из всех названных понятий площадь и объём водной массы наиболее просты и не нуждаются в объяснениях; остальные же в применении к озеру оказываются более сложными, поэтому о них необходимо сказать несколько
слов.
Несмотря на кажущуюся простоту понятия длины озера, оно оказалось в действительности одним из наиболее трудных для определения вследствие большого разнообразия форм озёр. Из
многих определений этого понятия, данных разными авторами, мы приведём определение Г. Ю. Верещагина, наиболее, по нашему мнению, удовлетворительное : длиной озера является кратчайшее расстояние между двумя наиболее удалёнными друг от друга точками его береговой линии, считая по поверхности водоёма. Прямой линией длина озера будет лишь в случаях простой формы его, т. е. такой, при которой береговая линия не делает более или менее глубоких выступов внутрь водоёма. При сложной форме, когда берег образует сильные изгибы, далеко заходящие в озеро, длина последнего будет представлять изогнутую линию, состоящую из отрезков прямых, концы которых соединяются кривыми — касательными к главным выступам берега. Для наглядности приводим рис. 1,
взятый из книги Верещагина. Рис. 1. Проведение
Что касается ширины озера, то различают линии длины озера, наибольшую и среднюю ширину. Хотя данное Верещагиным определение максимальной ширины, по его собствен-
ному признанию, страдает в некоторой степени несовершенством и услов-
ностью, тем не менее, по сравнению с другими определениями, оно всё же является наиболее удачным: наибольшая ширина озера есть наибольшее расстояние между противоположными берегами озера в направлении, перпендикулярном к линии длины озера.
В определении понятия средней ширины нет разногласий между авторами — все они принимают за среднюю ширину озера отношение площади озера к его длине.
Величина площади различных озёр чрезвычайно разнообразна, начиная с небольших озёр, поверхность которых не достигает даже одного квадратного километра, и кончая озёрами с площадью в сотни тысяч квадратных километров. Если такие озёра обладают к тому же солёной водой, то
они обычно называются морями, как, например, Каспийское море и другие. Следует отметить, что значительная часть наибольших по площади озёр всего земного шара расположена на территории СССР; в качестве примеров можно указать самое большое из всех озёр — Каспийское море, площадь которого занимает 438 700 км2, озеро Байкал с поверхностью в 34 200 км2, Ладожское — наибольшее из европейских озёр, занимающее площадь в 18 000 км2, и др.
Что касается глубины, то она не стоит в каком-либо определённом соотношении с величиной поверхности озера: и крупные и малые по площади озёра могут быть как глубокими, так и мелкими. Глубина озера зависит скорей от рельефа окружающей местности: горные озёра большей частью бывают относительно (т. е. по отношению к площади озера) более глубокими, чем равнинные. Если же сравнивать по абсолютной глубине, то в общем можно сказать, что крупные озёра глубже маленьких. Глубочайшим озером на земле является Байкал, наибольшая глубина которого определена приблизительно в 1 745 м; Каспийское море имеет наибольшую глубину приблизительно в один километр.
Подобно ширине, и глубину различают также максимальную и среднюю. Первая получается в результате непосредственных промеров глубин, вторая определяется как отношение объёма водной массы озера к его площади.
О способах измерения и вычисления глубин, площади и объёма озера более подробно будет сказано дальше.
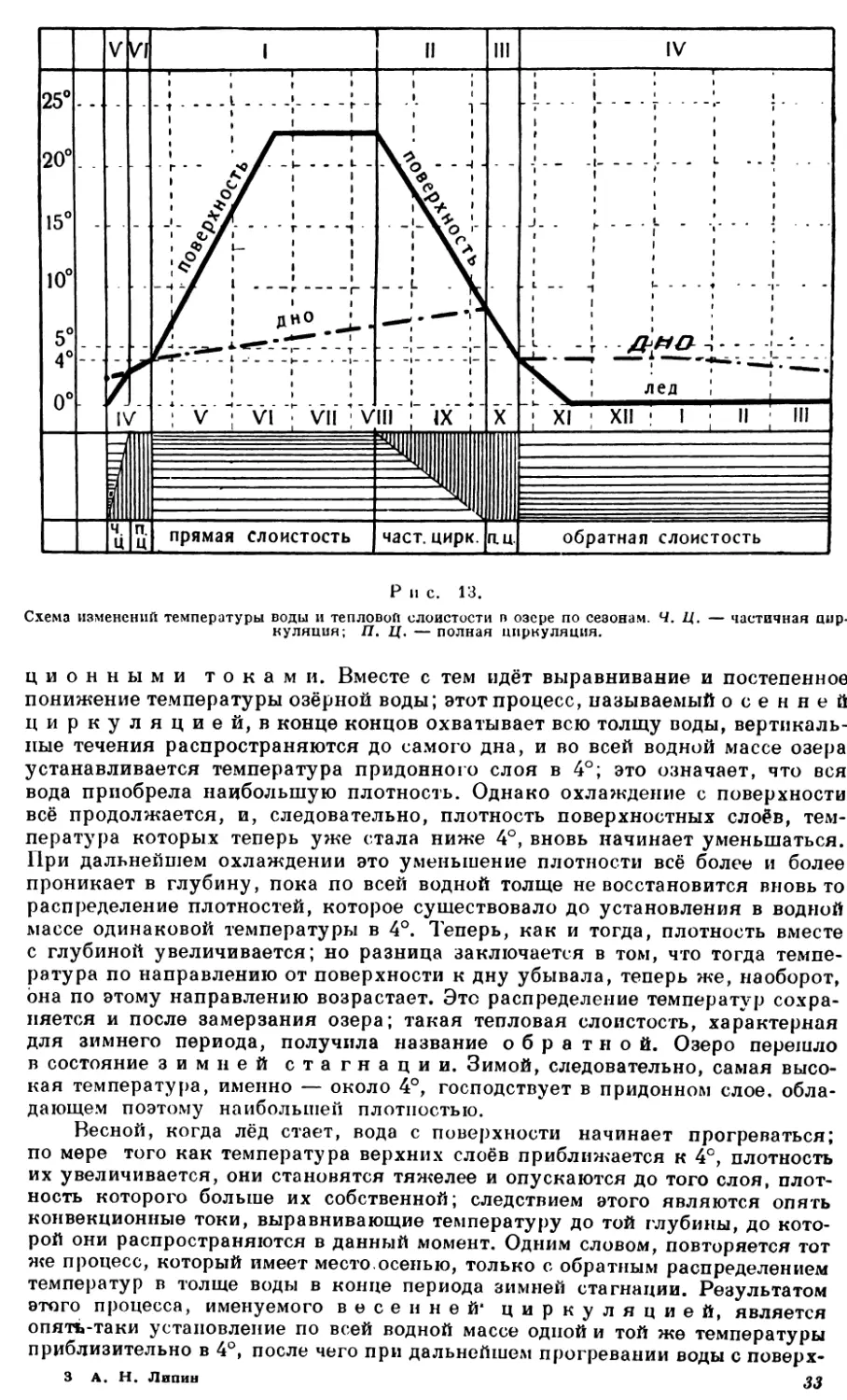
ЦИКЛ ЖИЗНИ И ВОЗРАСТНЫЕ ПЕРИОДЫ ОЗЕРА
Как только впадина суши заполнилась водой, так последняя уже начинает оказывать значительное воздействие на неё. С этого, так сказать, младенческого состояния начинается жизнь озера; под влиянием взаимодействия воды и суши водоём мало-помалу развивается в настоящее типичное озеро.
Пусть линия АБВГД (рис. 2) изображает первоначальный склон стенок впадины. Действие воды на эти стенки начинается с того, что возникающие под влиянием ветра прибойные волны, ударяясь в них и отрывая от них более или менее крупные частицы, постепенно разрушают и подмывают их.
уровень воды
3 -
Рис. 2. Схема вертикального разреза через прибрежную часть озера.
В результате этого у верхнего края впадины образуется выбоина, выемка АЕЖ, в которой более крутая часть ЛЕ называется береговым откосом, а остальная, более пологая часть ЕЖ — побережьем. Но куда же девается тот материал, который образовался от размыва верхнего края впадины? Он и дальше продолжает разрушаться от действия волн. Крупные обломки распадаются на более мелкие, которые постепенно обкатываются, закругляются, образуя разных размеров гальку и гравий. Отчасти от дальнейшего разрушения этого материала, отчасти от выветривания окружающих горных пород образуется п е с о к, а наряду с ним в качестве конечного продукта разрушения получаются мельчайшие частицы, совокупность которых даст уже более или менее тонкий и л. Весь этот материал 10
распределяется в воде сообразно своей величине и весу и в зависимости от двигательной силы действующих на него волн. Чем частицы мельче, тем они относительно легче и тем. следовательно, дольше могут держаться в воде и медленнее погружаться на дно. Отхлынувшая от берега волна захватывает с собой часть разрушенного материала, и вот тут-то и сказывается влияние величины и веса составляющих его частей: очевидно, что чем крупнее и, следовательно, тяжелее эти части, тем скорее они осядут на дно (более крупные даже совсем не поднимутся со дна и — самое большее — только перекатятся по нему на то или другое расстояние) и тем, значит, ближе к берегу они окажутся; наоборот, чем частицы мельче, тем дальше они будут унесены от берега. В результате всего этого процесса размыва и осаждения на дно разрушенного материала у берега под водой образуется нанос, имеющий форму уступа или обрыва (рис. 2 ВЗГ), крутая часть которого ЗГ носит название подводного откоса; часть ЖВЗ, расположенная между берегом и подводным откосом, называется береговой мелью; сумма же обеих этих частей, т. е. область ЖВЗГ, получила наименование п р и-брежной, или литоральной, области. Вся средняя центральная часть водоёма, начиная от нижней границы подводного откоса (на рис. 2 ГД и дальше), в отношении водной массы называется глубоководной, или пелагической, областью, пелагиалью, а в отношении дна — профундальной областью, профундалью. Таким образом, профундаль есть дно пелагиали. Из рис. 2 ясно, что прибрежная и глубоководная области резко отличаются друг от друга глубиной.
Итак, благодаря размывающему действию воды на стенки заполняемой ею впадины суши образуются три части водоёма, три непосредственно граничащие одна с другой области его: 1)береговая (от А до Ж), 2) п р и-брежная, или литораль (от Ж до Г) и 3) глубоководна я, или пелагиаль — профундаль (от Г в глубь и к центру)1.
Но на этом деятельность воды не останавливается. Наносная часть береговой мели образуется сначала из грубого, тяжёлого материала, к которому с течением времени прибавляются продукты его дальнейшего разрушения. В озёрах, имеющих один или несколько притоков в виде хотя бы небольших речек или ручьёв, к наносам, образуемым деятельностью самой озёрной воды, прибавляются ещё отложепия того материала, который приносится этими притоками. Эти отложения, имеющие обычно конусовидную форму, образуются у самого устья притока, причём в случае значительного накопления материала часть такого конуса может выступать над поверхностью воды. Что касается тонкого материала, состоящего из очень мелких частиц, которые вследствие своей лёгкости долго носятся в воде и оседают на дно медленно, то такой материал относится волнами далеко от берега. В конце концов и он осаждается, постепенно заполняя углубления дна и таким образом выравнивая его. И опять-таки в озёрах с притоками к этому илу, образовавшемуся в самом озере, присоединяется такой же ил, приносимый притоками; он, следовательно, вместе с озёрным илом участвует в выравнивании и заполнении дна, оседая преимущественно на самую глубокую центральную часть впадины. Наконец, следует упомянуть и о той пыли, которая при сильных ветрах несётся с окружающей озеро суши и, осаждаясь на воду, опускается затем на дно.
Но и этими источниками так называемого неорганического (так как в нём не участвуют организмы) происхождения озёрного ила не ограничивается его образование. Вскоре после возникновения озера в нём начинает развиваться жизнь. Конечно, жизнь при господствующих ныне на земле условиях не может зародиться из неживой материи — она приходит
1 Различают ещё одну область, найденную в очень многих озёрах и пограничную между литоральной и пелагической областями, так называемую сублиторальную. Подробная характеристика её будет дана ниже.
в воду откуда-то извне. Если наше озеро имеет сообщение с каким-нибудь другим водоёмом, например, если в него впадает река, то такое проникание жизни понятно само собой. Но и в случае полной обособленности водоёма от другой воды, уже ранее населённой, жизнь приносится в него тысячами путей. Очень многие водные организмы обладают способностью сохранять более или менее долго свою жизнь и вне воды; особенно развита эта способность у тех мельчайших, микроскопических растений и животных, которые в громадных количествах населяют водную массу озёр, прудов, рек и пр.
Способы для этого в природе есть разные: одни из этих микроскопических организмов целиком одеваются твёрдой, не допускающей испарения оболочкой, как говорят, «инцистируются», другие, не способные сами переносить высыхание, производят внутри своего тела яйца или споры, обладающие этой выносливостью. Под защитой оболочки такие «дремлющие» организмы или их зародыши лежат по берегам, заброшенные туда набежавшей волной, или остаются на надводных частях растений и при понижении уровня воды попадают то на перья плавающих птиц, то на шерсть пришедших пить животных. Порывом ветра они могут быть подняты с земли, с растений и переброшены во вновь народившееся озеро или могут попасть в него вместе с птицами и млекопитающими.
Но не для всех попавших в воду водных обитателей условия новой жизни окажутся приемлемыми; очень многие являются требовательными по отношению к физико-химическим условиям и к питательности воды: для них необходима вода, в которой ранее уже жили долгое время организмы и обогатили её питательными веществами. Вспомним, что из упавших на почву семян далеко не все могут произрасти, а лишь те, для которых эта почва будет подходящей; так и для водных организмов вода должна быть подходящей, т. е. опа должна содержать в растворе определённые вещества. Поэтому в молодом озере поселяются лишь определённые организмы, количество которых бывает сравнительно невелико.
Далее, в такую воду, уже «удобренную» развившимися в ней организмами, попадают япца более крупных животных, семена водных растений, а также споры и зародыши других, более прихотливых микроскопических животных и растений; из последних есть такие, которые способны в некоторые моменты жизни размножаться в громадном количестве, так что -даже вызывают окрашивание воды в тот или другой цвет (так называемое «цветение»). »
Семена крупных растений, упавшие на дно, развиваются лишь в том случае, если они попали на подходящий грунт и притом в мелкие места, куда проникают в достаточной мере солнечные лучи, так как без солнца жизнь растений невозможна. Мы уже знаем такие места: это береговая мель, образующая часть прибрежной области.
Водные растения, почти без исключения, обладают очень развитой способностью размножаться при помощи корневищ, стеблей, оторвавшихся частей и т. п. Попавшее даже в единичном числе семечко вскоре даёт целую заросль, быстро расходящуюся во все стороны, если не встретится ей на пути сильный конкурент в виде заросли другого растения. Часто два-три растения завладевают одним участком, перепутываясь своими подземными частями и образуя на поверхности воды и над водой причудливый рисунок из разнообразных листьев и стеблей.
Таким образом, вдоль берегов озера появляется пояс прибрежных растений.
Отмершие тела мельчайших организмов в сильнейшей степени обогащают воду мелкими взвешенными в ней частицами, совокупность которых называется детритом; эти частицы, падая на дно и примешиваясь к озёрному илу, сильно увеличивают его количество; трупы более крупных животных и остатки крупной водной растительности, падающие на дно, гниют там, разлагаются и распадаются на всё более и более мелкие части, внося этим свою лепту на пополнение массы покрывающих дно отложений. п
Но и это ещё не всё. Весь этот материал так называемого органического происхождения (потому что он происходит от организмов) образуется внутри самого озера. Между тем весьма даже немалое количество такого же материала попадает в озеро извне, отчасти теми же путями, как и неорганический материал, т. е. притоками и ветром (последний, например, приносит пыльпу наземных растений, листья и т. п.), отчасти — другими способами, например, падающие из воздуха трупы насекомых, сваливающиеся с берега в воду растения, опадающая листва, помёт приходящего на водопой скота, если поблизости есть селение, и т. д. Всё это, спустя то или иное время, претерпевает одну общую участь — оседает на дно озера, увеличивая слой уже находящихся там отложений.
Каким бы путём эти отложения ни образовались, все они с течением времени очень сильно изменяют дно и стенки первоначальной впадины, превращая её в озёрное ложе.
Легко понять, каков будет результат этого непрерывного, происходящего в течение долгих лет накопления на дне озера громадных масс осаждающегося материала: ясно, что дно озера должно постепенно повышаться, озеро должно год за годом мелеть. Так оно и происходит в действительности. Центральная яма котловины всё более заполняется, дно, повышаясь, становится всё более плоским, растения прибрежной полосы получают возможность распространяться всё дальше и дальше по направлению к центру водоёма, пока, наконец, обмеление озера не достигнет такой степени, при которой глубина его по всей площади дна становится равной глубине прибрежной области. Последняя как бы распространяется по всему озеру; поверхность его покрывается плавающими листьями водяной лилии (кувшинки) и других растений. Население самой водной толщи мало-помалу снова меняет свой состав. Этот момент необходимо отметить, так как он означает превращение озера в пруд. Следовательно, различие между озером и естественным прудом с физико-географической точки зрения сводится главным образом к разной глубине их.
Эта основная разница влечёт за собой, как следствие, различия физико-химического характера, например, различие в температуре (скорость прогревания воды), в силе освещения, в содержании кислорода и прочее; эти различия, в свою очередь, вызывают биологические изменения — в составе и распределении прежде всего водной флоры (растительности), а через неё и водной фауны (животных). Из всех этих изменений мы отметим лишь одно, как резко заметное и потому легко обнаруживаемое, — это распределение водной растительности. В типичном озере распространение надводной и подводной флоры ограничено исключительно лишь прибрежной областью, потому что здесь только она находит для себя подходящие условия жизни, главным образом освещение. Когда же в известной стадии обмеления озера глубина прибрежной области, а следовательно, и характерные для неё условия жизни распространяются по всей площади водоёма, то естественно, что и водная растительность распространяется тоже по всему водоёму, однако не вся, а только лишь подводная и с плавающими листьями; надводная же попрежнему остаётся ограниченной в своём местопребывании прибрежной полосой, так как для неё глубина средней части пруда всё ещё велика *.
Этот именно признак и является наиболее крупной, резко бросающейся в глаза биологической отличительной чертой между озером и прудом: в первом распространение растений, не возвышающихся над водой, ограничено только прибрежной областью, во втором же они могут расселяться по всему водоёму.
1 Подводными, или погружёнными, растениями в дальнейшем изложении мы будем называть такие, которые, за исключением иногда лишь нериода цветения, проводят всю свою жизнь под водой; растениями надводными будут обозначаться такие, которые выставляют над поверхностью воды свои листья или стебли. Термин с плавающими листьями понятен сам собой.
J3
Но кончается ли на этом обмеление озера? Нет, оно продолжается дальше. Когда растения завладели всей водной поверхностью, процесс заполнения котловины ускоряется благодаря обилию падающего на дно растительного материала. Количество животного населения также значительно увеличивается в зависимости от большего распространения высших растений, дающих прекрасные условия для жизни. Трупы их падают обильным дождём на дно пруда; последнее повышается всё быстрее, так что на нём могут уже укореняться надводные растения, и пруд превращается в болото. Следовательно, разница между прудом и болотом заключается в распределении надводной растительности: в первом она ограничена в своём распространении литоралью, во втором — расселяется по всей площади водоёма.
Но и это ещё не конец. При дальнейшем обмелении и разрастании болотной флоры, которая, становясь гуще и гуще, всё более вытесняет воду, последняя в конце концов совсем исчезает, и болото превращается в луг. Озеро умерло.
Швейцарский исследователь Форель различал в жизни озера 5 следующих «возрастных» периодов.
1. Юность. Наносы и отложения на дне водоёма ещё настолько незначительны, что не оказывают заметного влияния на конструкцию и форму дна и стенок первоначальной котловины.
2. Зрелость. Озёрные отложения образовали прибрежную область; приносимый притоками грубый материал отложился в их устьях в виде конусовидных наносов; несмотря на значительное уже количество осевшего речного ила, дно первоначального углубления всё же ещё не совсем выравнено им, не все его неровности скрыты, и в некоторых местах через тонкий покров ила выступает первоначальное строение дна впадины.
3. Старость. Дно и стенки первоначальной котловины на всём протяжении покрыты более или менее мощным слоем ила, совершенно скрывающим первоначальную форму дна и стенок; впадина вполне превратилась в озёрное ложе, представляющее собой большую ровную площадь, окружённую склонами подводных откосов литорали и речных наносных конусов.
4. Дряхлость. Плоская центральная часть ложа по своей глубине сравнялась с прибрежной областью; озеро превратилось в пруд.
5. Умирание. Дно поднялось ещё выше; толщина слоя воды настолько уменьшилась, что погружённая растительность в ней уже не может жить, и потому опа вытесняется надводной флорой; пруд превратился в болото.
Однако изображённая нами картина возникновения, развития и исчезновения озера есть лишь голая схема, которая приложима далеко не к каждому озеру. Нередки случаи, когда озеро, вступившее уже в период «дряхлости», получив значительный приток воды (например, в результате изменения сухого климата во влажный), становится глубже, теряет растительность в центральной своей части, оставляя её лишь у берегов, и вновь превращается, таким образом, из пруда в типичное озеро. Недавние исследования стратификации (слоистости) донных отложений некоторых уральских озёр показали, что в истории жизни последних были такие периоды, когда они были уже в стадии болот, так сказать, в предсмертной агонии, но по каким-то, пока ещё не выясненным причинам, переживали эту стадию и продолжали жить в состоянии нормальных озёр, давая наслоение донных отложений.
Есть и другой путь естественного умирания озера: зарастание его так называемой сплавиной с образованием торфяного болота, но об этом речь будет дальше.
ТЕМЫ ДЛЯ РАБОТ
Для того чтобы вполне ясно представить себе все особенности ложа взятого для изучения водоёма, необходимо произвести ряд работ.
Прежде всего нужно составить понятие о величине озера, о его очертаниях, о рельефе дна, о характере донных отложений. При исследовании последних интересно отметить границу распространения водной растительности, без чего не может быть решён вопрос 14
о том, к какой категории водоёмов — к прудам или озёрам — может быть отнесён интересующий нас водоём.
Затем следует попытаться разрешить вопрос о происхождении озера, что может быть сделано путём ориентировки в характере горных пород, окружающих озеро, и в установлении присутствия или отсутствия следов ледникового происхождения (стр. 8).
1. Физико-географическая характеристика водоёма
1. Установить контур (очертания) водоёма: съёмка плана водоёма.
2. Определить приблизительный характер окружающей водоём местности: съёмка плана этой местности (стр. 19).
3. Вычислить по плану площадь водной поверхности (стр. 21).
4. Вычислить длину и развитие береговой линии (стр. 21).
5. Произвести, поскольку есть возможность, ряд промеров глубин и, в завпсимостп от этого, вычертить профиль дна (стр. 19).
6. Определить характер донных отложений в разных пунктах водоёма.
II. Возраст озера (озеро или пруд?)
Об этом могут дать представление уже пункты 5 и 6 предыдущего отдела, так как по ним можно судить, есть ли или пет разделение па пелагическую и прибрежную области и насколько резко это разделение выражено. В дополнение к этому можно добавить ещё определение границы распространения погружённых растений.
III. Происхождение озёрного ложа
1. Установить характер горных пород в окружающей местности.
2. Обратить внимание, нет ли следов ледникового происхождения: присутствие валунов по берегам или на дне и характер их расположения.
МЕТОДИКА
Съёмка плана водоёма. Инструментарий.
1. Буссоль или компас для самодельной буссоли (при отсутствии того и другого — обыкновенная линейка; описание инструментов см. ниже).
2. Подставка для буссоли.
3. Рулетка или верёвка 20 м длины.
4. Несколько кольев.
5. Несколько шестов.
При всякой съёмке местности необходимо уметь определять две основные величины: 1) расстояние между двумя точками и 2) угол между двумя пересекающимися линиями. Первую величину получают прямым измерением посредством рулетки или, за неимением её,простой верёвкой,заранее вымеренной; для получения второй величины при--------------------------------------
точных съёмках пользуются особыми угло- /*"7\ м
мерными снарядами 1, но при менее точной съёмке такой снаряд может быть с успехом ХС-Х
заменён буссолью или даже простым __________________________________________
компасом. Посмотрим, как с помощью г .
лишь рулетки и компаса произвести съёмку Рис. 3. Самодельная буссоль (ориг.). водоёма.
Прежде всего из компаса нам необходимо устроить самодельную буссоль. Для этой пели (рис. 3) берём линейку и проводим на ней (хотя бы карандашом) среднюю продольную липию MHt в центре линейки прикрепляем к ней компас, причём располагаем последний так, чтобы его северо-южная линия NS по направлению вполне совпадала с линией МН\ теперь нам остаётся воткнуть в линейку в точках М и Н по булавке (которые должны стоять строго перпендикулярно к поверхности линейки), и паша буссоль готова. Воткнутые таким образом булавки называются диоптрами. Для того чтобы придать буссоли необходимую устойчивость, её во время работы следует класть на какую-нибудь подставку вроде, например, треножника от фотографического аппарата.
Приступаем к съёмке. Пусть перед нами какой-нибудь водоём (рис. 4) 2. Базой своей работы выбираем наиболее оголённый, т. е. лишённый растительности и притом, по возможности, ровный участок берега. Прежде всего мы должны отметить на этом берегу вер-
1 Точные съёмки производятся обычно при научных исследованиях (географических, геологических и пр.) или для технических целей (возведение построек, гидротехнических сооружений и т. п.). Для получения первого ориентировочного представления о плане местности или водоёма можно ограничиться менее точной съёмкой.
2 Само собой разумеется, что таким упрощенным методом можно делать съёмку лишь небольших сравнительно водоёмов; для съёмки же крупных водоёмов нужны более точные приёмы п усовершенствованные аппараты.
тикальными вехами-шестами две точки А и Б, расстояние между которыми должно быть таково, чтобы наблюдатель из точки А хорошо видел веху, поставленную в точке В, и наоборот. Длина линии АБ, называемой базисной линией, или просто базой, может быть непосредственно измерена рулеткой(или отмеренным куском верёвки). В этой работе должны участвовать по крайней мере два человека. Чтобы при измерении не отклоняться в стороны,аидти всё время по прямому направлению от А до В,поступают следующим образом. Первый экскурсант становится в одной из этих точек (отвернувшись от солнца), например, в точке А и устанавливает диоптры буссоли так, чтобы ближайшая к глазу булавка закрывала от него как другую булавку, так и шест Б. Второй экскурсант закрепляет на шесте Б конец лепты рулетки или 20-метровой верёвки 1 и затем, взяв в руки кол, с лентой (верёвкой) в руке идет прямо па первого экскурсанта. Пройдя 20 м (что отмечено будет рулеткой или концом верёвки), он останавливается и ставит кол вертикально на землю, ещё не втыкая его; первый экскурсант смотрит, правильно ли поставлен кол: при правильном положении ближняя к глазу булавка буссоли должна закрывать и вторую булавку и кол; когда последний будет правильно поставлен, второй экскурсант достаточно прочно втыкает кол в землю как раз на прямой АБ в 20 м от Б (при этом лента рулетки или измерительная верёвка должны быть хорошо натянуты и не провисать). После этого второй экскурсант с другим таким же колом продолжает свой путь к точке А и описанным способом ставит вторую веху, затем третью и т. д. Таким образом всю прямую линию АБ размечают целым рядом вех, стоящих в 20 м одна от другой. Сумма этих промежутков даст расстояние между точками А и Б.
Теперь нам нужно узнать направление линии АБ относительно стран света. Это мы можем сделать, определяя при помощи нашей буссоли угол между прямой АБ и линией, соединяющей север и юг, направление которой всегда указывается стрелкой компаса. Для этого устанавливают линию МН буссоли по направлению прямой АБ. Если только последняя не идёт прямо (^севера на юг, то очевидно, что при установке буссоли в положении АБ стрелка" компаса, всегда сохраняющая одно и то же положение (север — юг), теперь будет составлять некоторый угол с линией NS компаса, всегда совпадающей на нашей буссоли с линией МН, а в данном случае, следовательно, с линией АБ. Для опреде
ления величины этого угла нужно только посмотреть,на каком делении компаса (вся окружность последнего разделена на 360 градусов) стоит ближайший к 7V, т. е. к нулю, конец стрелки и в какую сторону от него он отклонён. Теперь остаётся лишь нанести линию АБ на бумагу. На экскурсии* в записной тетрадке делается лишь приблизительный чертёж, на котором отмечаются величины углов и расстояний, настоящий же план делается дома на основании этих данных. Прежде всего в экскурсионной тетрадке проводят прямую линию, один конец которой обозначают как северный 2V, другой — как южный S (рис. 4). Эта линия NS изображает направление компасной стрелки. Если буссоль показала нам, что линия АБ составляет со стрелкой, положим, угол в 32° и отклоняется от неё вправо, то на основании этих данных нам уже ничего не стоит при помощи транспортира нанести линию АБ на наш чертёж. Таким образом, полученные нами на экскурсии результаты измерений мы тут же заносим в записную тетрадь в виде двух пересекающихся прямых NS и АБ, обозначив на чертеже величину угла — в данном примере 32° — и расстояние между точками А и Б (рис. 4).
Теперь уже нетрудно посредством буссоли нанести на чертёж контур водоёма^ Отметим на противоположном берегу какую-нибудь точку В; за неимением естественной вехи, вроде дерева, небольшого кустарника, какой-нибудь постройки и т. и., можно поставить искусственную, например, воткнутый в землю шест с флагом хотя бы из бумаги и т. п., лишь бы она была хорошо видна из А и В. Направляя буссоль из точек А и Б в точку В, мы определяем описанным уже способом утлы, составляемые линиями АВ и БВ с линией NS\ отложив на чертеже углы от линии NS в точках Л и В, проводим прямые, пересечение которых и даёт нам точку В.
Таким образом, одна точка береговой линии уже найдена. Совершенно тем же способом мы находим и другие точки противоположного берега — Г, Д, Е и т. д. Соединяя эти точки между собой, мы получаем искомый контур. Чтобы получить линию ближнего берега, пользуемся методом расстояний.Берём какую-нибудь точку М и определяем угол линии AM с NS, а затем измеряем расстояние между точками А и М; этих данных вполне
1 Лучше, если в работе будет принимать участие ещё третий экскурсант, чтобы, стоя у шеста В, удерживать его от наклонения.
достаточно, чтобы нанести на чертёж точку Л/. Подобным же образом находим п другие точки — Н, О, П и пр. Соединением этих точек получаем линию ближнего берега.
Имея в экскурсионной тетради черновой чертёж с записями углов и расстояний, мы на основании их легко уже можем по возвращении с экскурсии начертить в определённом масштабе настоящий план водоёма, вооружившись для этого линейкой, циркулем л транспортиром (подробнее об этом — дальше).
За отсутствием компаса можно произвести съёмку ещё более простым способом. Но это можно сделать не на всех водоёмах, а лишь на тех. которые кругом открыты, т. е. берега которых доступны на всём своём протяжении. Для съёмки такого водоёма достаточно вооружиться рулеткой или, за её отсутствием, опять-таки размеренной верёвкой, затем прямоугольным угольником и, пожалуй, ещё кольями для расстановки вех. Угольник (рис. 5) легко сделать из двух половинок перереза иной поперёк линейки, которые своими концами скрепляются под прямым углом; проведя затем па линейках продольные средние линии МН и МО, втыкаем в линейки булавки в точках М, Н и О, и наш угольник готов; при этом необходимо озаботиться, чтобы угол между линиями МН и МО действительно и точно равнялся 90°. Для этого угольника так же, как и для буссоли, следует иметь подставку, па которой его можно было бы укрепить так, чтобы обе линейки
находились в горизонтальном положении.
При отсутствии компаса определить направление стран света можно только в солнечный день или с помощью вертикально стоящей палки, наиболее длинная тень от которой в 12 часов дня даёт нам северо-южное направление (линию NS), или по карманным
часам, причём в этом случае поступают следующим образом: ставят часы в такое положение, при котором часовая стрелка указывала бы на солпце,—тогда радиус цифер-
Р и с. 6. Схема съёмки плана озера с помощью угольника (ориг.).
Р и с. 5. Угольник для измерения прямых углов (ориг.).
блата, делящий пополам дугу между часовой стрелкой и 12 часами, будет представлять как раз линию NS.Сделав это, проводим на берегу прямую линию АВ, совпадающую по направлению с линией TVS’(рис. 6). Способ этого проведения изложен уже выше, причём диоптрами в данном случае будут служить булавки одной из линеек нашего угольника. В точках А и Б ставим вехи. Затем, поставив угольник в точке А так, чтобы одна из его линий, например МО, совпадала по направлению с АБ, проводим по другой линии МН прямую АГ, которая, очевидно, будет перпендикулярна к АБ. В точке Г ставим опять веху. Точно так же, поставив угольник в точках Б и Г, проводим прямую БВ, перпендикулярную к АБ, и прямую ГВ, перпендикулярную к АГ. Точку пересечения В прямых БВ и Г £ отмечаем тоже вехой. При этом положение точек А, Б, В и Г или, другими словами, длину линий АБ., БВ, ВГ и АГ нужно выбирать так, чтобы ни одна из этих линий не пересекала самый водоём, а все они прошли на берегу. Таким образом, мы вписали наш водоём в прямоугольник АБВГ. Теперь разделим каждую из 4 линий посредством втыкания кольев на несколько равных отрезков. Так, на линии АБ будут воткнуты колья, скажем, в точках а, 6, в, е, д, на линии Б В — в точках е, ж, <?, и, кит. д., причём все эти точки лучше расположить на одном и том же расстоянии друг от друга, например, 20 м. После этого ставим в точке а угольник в таком положении, чтобы одна из его сторон пришлась по линии АБ; тогда по другой стороне угольника проводим линию al до самой воды и здесь ставим веху, а затем измеряем расстояние между точками а и 1. Таким способом находим первую точку береговой линии. Переставляя угольник последовательно в точки 6, в, е, д, совершенно таким же образом определяем точки 2, 3, 4 и 5 береговой линии. То же самое проделываем и на остальных трёх сторонах прямоугольника БВ, ВГ и АГ. Соединив полученные точки между собой, получим более или менее точный план водоёма. Само собой разумеется, что чем большее количество точек мы возьмём на сторонах прямоугольника АБВГ (т, е. точек а, б, в, г,... и т. д.), тем меньшие расстояния получатся между точками 1,2,3, 4, 5 и т. д. и, следовательно, тем точнее будет произведена съёмка. При этом контур береговой линии между найденными точками её (1, 2, 3,...) зарисовывают гут же па глаз, стараясь, по возможности, точно воспроизвести, если не все, то, по крайней мере, главные излучины, изгибы и зигзаги береговой линии.
Итак, мы сняли контур водоёма. Все записи у пас в экскурсионной тетради. Возвратившись домой, мы должны сделать настоящий чертёж. Для этого* берём лист клетчатой или, по крайней мере, линованной бумаги и, принимая линии за направление меридиана (т. е. северо-южное —NS), выбираем для плана такой масштаб, чтобы водоём вполне поместился на листе. Отметив теперь в соответственном месте последнего на одной из линий NS точку Л, строим при ней посредством транспортира вправо от линии JVS угол в 32° (рис. 4); для этого накладываем транспортир на бумагу так, чтобы его центр совпал с точкой Л, а прямая JVS прошла бы через его н^ль;затем отмечаем на бумаге точку против 32-го деления транспортира и, убрав последний, проводим через эту точку и точку Л прямую линию. На этой новой прямой откладываем от точки А в принятом нами масштабе расстояние АБ. Таким путём находим на плане вторую точку базисной линии В. Если эта точка окажется не па меридиональной линии (2VW), необходимо провести через неё такую линию. Теперь при точках А и В мы должны построить целый ряд углов. Построив углы N АВ и IVБВ и продолжив прямые АВ в БВ до взаимного пересечения, находим как точку этого пересечения первый пункт береговой линии В. Затем построением углов NАГ и ЯБГ получаем вторую точку берега Г и таким же способом — остальные точки Д, Е, Ж... и т. д. Для получения контура ближнего берега опять-таки строим при А и В записанные в экскурсионной тетради углы и на соответственных линиях откладываем опять-таки в принятом масштабе расстояние ЛЛ/, АН, БО, БП... и т. д. Соединяя теперь
площади озера (ориг.).
в принятом масштабе расстояние AM, АН, БО, БП... и т. д. Соединяя теперь все полученные нами точки береговой линии — при этом необходимо всё время сверяться с зарисовкой контура водоёма, произведенной на экскурсии с натуры, — мы получаем план береговой линии нашего водоёма.
При втором способе съёмки (без буссоли) вычерчивание облегчается — нужно лишь иметь разграфлённую на квадратные клетки бумагу. Масштаб выорать лучше всего в зависимости от размера клеток, чтобы сторона клетки (квадратика) представляла целое число единиц размера, например, 1, 2, 3 м и т. п. Затем в принятом масштабе зачерчивают прямоугольник АБВГ (рис. 6), на сторонах которого отмечают по записям экскурсионной тетради точки а, б, в, а... и т. д. и, проведя линии al, 62, е<3..., находят точки /, 3, 4... кон-
тура водоёма.
Умея зарисовать на плане очертания водоёма, мы должны теперь научиться определять положение любого пункта на его площади. Это нужно для будущих экскурсий, когда придётся вести исследование с лодки на самой воде, чтобы знать и отметить на плане пункты этих исследований, или так называемые станции. Для этого выбирают на берегу три точки — А, 2>.2?(рис. 7) и отмечают их естественными или искусственными вехами 1. Допустим, что нам надо определить на водоёме положение пункта О, в котором находится в данный момент лодка. Сидя в последней, определяем буссолью углы, составляемые прямыми ОА, ОБ и О В с меридианом NS. Само собой понятно, что это возможно лишь при спокойном стоянии лодки, т. е. в тихую, беэветреную погоду; при сильном ветре вообще нельзя производить никаких исследований, а при слабом лодка должна быть так или иначе укреплена, например, посредством длинного шеста, воткнутого в дно, если последнее достаточно мягко, а водоём неглубок, или посредством двух якорей, которые, в случае их отсутствия, могут быть заменены крупными камнями. Затем наносим точки А, Б и В па план и, взяв кусок прозрачной бумаги достаточной величины, вычерчиваем на нём прямые ОА, ОБ и ОВ, углы между которыми нам уже известны. Водя прозрачной бумагой по плану, три точки которого А, Б и В через неё просвечивают, мы найдём, наконец, такое положение, при котором все три линии ОА, ОБ и ОВ прозрачной бумаги пройдут через соответствующие точки А, Б и В плана. Тогда протыкаем точку О булавкой, след которой на плане и будет искомым пунктом.
Съёмка окружающей водоём местности. Нанести на бумагу план водоёма ещё не значит сделать настоящий план его. Так как всякий водоём находится в тесной связи с окружающей его местностью, испытывая влияние с её стороны и, в свою очередь, влияя на неё сам, то естественно, что для более полного познания efro жизни необходимо изучить и эту местность, а следовательно, необходимо и её нанести на план. Это возможно лишь в том случае, если мы имеем хоть несколько опорных пунктов, по которым сможем любой пункт данной местности отмстить на плане или, наоборот, любую точку плана найти на местности. Лучше всего в качестве таких опорных пунктов иметь какие-нибудь естественные предметы, например, вершины холмов или бугров, выдающиеся чем-нибудь деревья и т. п. или же какие-либо постройки, и только за отсутствием подобных предметов прибегать к установке искусственных вех.
Как при помощи буссоли и рулетки отметить положение опорных пунктов, ясно из предыдущего изложения, и потому здесь мы останавливаться на этом не будем.
1 Для экономии работы желательно не ставить новых вех, а воспользоваться которыми-нибудь из старых, установленных уже при съёмке самого водоёма. 18
Однако, если мы хотим знать как следует окружающую водоём местность, то мы, конечно, не можем ограничиться нанесением па план одних лишь опорных пунктов, которые служат только для ориентировки. Мы должны произвести настоящую съёмку агой местности. Это значит, что на план должны быть нанесены, с соблюдением правильных отношений, различные по своему физико-географическому характеру участки этой местности. Так, например, необходимо отметить местоположение и площадь растительности (а также и её характер — лес, луг. поле, огород), обнажённой почвы с её морфологической характеристикой (глина, песок, чернозём и т. п.), положение близлежащих селений, направление рек и ручьёв, водопой скота, мойку белья и пр.Теперь нам уже известен метод съёмки контура водоёма. Этот метод может быть применён к съёмке контура, ограничивающего и любой участок суши, например, участок поля, луга и т. д., если только этот контур достаточно резко выражен и вехи вполне отчётливо видны. При этом если площадь участка невелика, он может быть нанесён на план целиком; если же он очень велик, то можно ограничиться той его частью, которая обращена к водоёму. Прп достаточном числе экскурсантов такую съёмку произвести нетрудно, тем более что здесь уже не требуется такой точности, как при съёмке самого водоёма: например, расстояние можно мерять не рулеткой, а просто ровными шагами, средняя величина которых должна быть извесша (проходится определённое расстояние, например, 40—50 м, п при этом сосчитывается число шагов, па которое затем и делится это расстояние).
Но если обстоятельства не позволяют произвести настоящую съёмку окружающей водоём местпости, то опа, по крайней мере приблизительно, на глаз, должна быть занесена на план, опять-таки, разумеется, с обозначением характера каждого участка. Однако если у водоёма имеются притоки и стоки, то необходимо но возможности точно заснять их положение и направление на некотором протяжении от водоёма (хотя бы на несколько десятков метров) и особенно места их соединения с водоёмом.
Глубина и рельеф дна. Инструментарий.
1. Л о т для измерения глубины: к концу верёвки длиной в несколько метров прикрепляется гирька или камень, верёвка размечается на метры и полуметры посредством нанесения на ней каких-нибудь значков масляной краской, завязывания цветных ленточек и т. и., причём метры отмечаются одним цветом, а полуметры другим. Такая размеченная верёвка называется ли нём; нижнюю поверхность гирьки или камня следует намазать салом, к которому должны пристать частицы грунта, когда лот упадёт па дно.
2. Б у с с о л ь; её устройство см. в главе о съёмке плана.
3. Кусок сала.
Летнее измерение глубин. Чтобы получить хотя какое-нибудь представление о рельефе дна, необходимо сделать несколько промеров глубины, которые производятся не где попало, а в каких-нибудь определённых пунктах водоёма, расположенных в известном порядке. Обычно для этой цели выбирают систему взаимно параллельных прямых линий, пересекающих водоём в поперечном направлении; каждую из этих лппий делят на несколько равных отрезков точками, которые и служат пунктами промеров. Вычерченный по таким промерам для каждой поперечной линии контур дна называется разрезом, или п рофи ле м, дна в данном месте. Совокупность всех профилей даёт представление об общем рельефе дна. Ясно, что чем больше будет промеров, тем более точную карту рельефа дна мы сможем составить.
Однако не на всякой экскурсии условия работы позволяют удовлетворить такие требования. Здесь волей-неволей приходится ограничиваться минимальным количеством промеров, и потому достаточно взять лишь одну поперечную линию, именно — среднюю. Таким образом, мы получим только один профиль.
Для того чтобы лодка шла прямо по промерной линии, не уклоняясь в стороны, поступаем так. На глаз намечаем направление профиля и у концов его на противоположных берегах ставим вехи, назовём их А и Б. Затем на одном из берегов, например, на том, где находится веха В, ставим третью веху В так, чтобы она стояла на одной прямой линии с А и Б 1. При таких условиях лодка должна двигаться от Л кБ, причём рулевой обязан непрерывно и строго следить за тем, чтобы веха Б всё время закрывала от него веху В. Только это даёт гарантию того, что лодка идёт правильно по линии разреза. Для большего удобства и ускорения работы желательно присутствие в лодке не менее 3 человек: гребца, рулевого и лотового, т. е. лица, производящего промеры.
Первое измерение глубины можно произвести на расстоянии 3—4 м от берега (это расстояние можно измерить непосредственно рулеткой). Следующий промер — через 10 гребков, причём грести нужно ровно, спокойно, слегка лишь задевая вёслами воду. Считая гребки вслух, гребец должен прекратить греблю на 9-м гребке, при 10-м же, так как лодка продолжает двигаться по инерции, нужно лишь опустить вёсла в воду и держать их неподвижно, чтобы остановить лодку, удерживая во всё время промера. Как только лодка остановилась, лотовой опускает лот с такой скоростью, чтобы успеть во время спуска отсчитать марки (значки) линя, наблюдая при этом, чтобы линь был в вертикальном положении, что и будет означать спокойное стояние лодки. Отметив глубину, лотовой произносит её вслух, и это является для гребца сигналом для возобновления гребли. Пока лодка идёт до следующего пункта промера, лотовой вытаскивает лот, отмечает по пристав-
1 Как это сделать, см. главу о съёмке водоёма. *
Рис. 8. Измерение глубин зимой (ориг.).
шим к сальной поверхности его частицам характер грунта (ил, песок, крупные растительные остатки и г. п.), смывает этот грунт с лота и записывает глубину. Таким же способом производятся и все следующие промеры, причем положение последнего промера—так же как и первого — определяется измерением его расстояния от берега. Ввиду того, что слой сала постепенно стирается с лота, последний время от времени приходится смазывать вновь.
Однако само’собой понятно, что определение положения лодки на водоёме посредством одного только счёта гребков есть способ довольно приблизительный и неточный, не гарантирующий от более или мепее грубых ошибок. Поэтому желательно время от времени прибегать к более точному способу с помощью буссоли. Этот способ описан нами в конце главы о съёмке (стр. 18), поэтому здесь останавливаться на нём нет надобпости.
Что касается числа и частоты этих определений, то в этом отношении руководствуются следующим правилом: пункты определений располагают так, чтобы между двумя соседними пунктами заключалось 5—10 промерных точек, найденных по числу гребков. При этом в записях, разумеется, особо отмечают те промеры, которые произведены в буссольных пунктах.
Теперь мы должны полученные глубины па нести на план. Прежде всего прочерчиваем на плане промерную линию, конечные точки которой А и Б мы можем найти на плане по опорным пунктам (см. главу о съёмке). Если у нас ни одним точки промера не определено по буссоли, а всё только по гребкам, то,.отметив па плане (по известному нам расстоянию от точек .4 и Б) положение первого и последнего промеров, делим промежуток между ними на общее число промеров без единицы (например, при общем количестве промеров 10 делим на 9); таким способом мы мо!кем получить на плане расположение промерных пунктов на линии разреза. Если же имеется несколько точек, определённых по буссоли, то, нанеся эти точки на план, делим расстояние между каждыми двумя соседними точками на число промеров, сделанных между ними по гребкам, плюс единица.
Остаётся лишь вычертить профиль дна водоёма. Переносим нашу промерную линию со всеми промерными пунктами на другой лист бумаги (лучше в том же масштабе). Затем в каждом промерном пункте проводим вниз перпендикуляр и откладываем на нём соответствующую данному пункту глубину. Полученные на перпендикулярах точки соединяем линией — и профиль готов. Выбор масштаба для нанесения глубины на план профиля дна зависит от размеров самого озера, именно — от отношения его ширины к наибольшей глубине. Понятно, что при очень большой величине этого отношения, например, при ширине озера в 2 км и максимальной глубине 4 м (в этом случае отношение, следовательно, равно 2 000 : 4 = 500), на небольшом, сравнительно, листе бумаги, на который наносится план профиля, даже максимальная глубина выразится, самое большее, линией в 1 мм. Разумеется, при таких условиях говорить о каких-либо деталях профиля не приходится. В таких случаях выбирают для
ширины и глубины разные масштабы, именно такие, при которых получилась бы вполне ясная картина профиля.
Если имеется возможность произвести измерения глубины по нескольким поперечным линиям, то, получив для каждой из них соответствующий профиль, мы можем по ряду профилей составить себе некоторое представление о рельефе дна.
Наконец, соединяя на плане пункты с одинаковой глубиной, получаем замкнутые кривые, так называемые изобаты.
Зимнее измерение глубин. Зимой промеры глубин можно сделать точнее, чем летом, так как благодаря большей сравнительно с лодкой устойчивости работы на льду уточняется как самое измерение глубины, так и расположение промерных пунктов.
Зимнее измерение лучше всего производить в начале зимы, когда, с одной стороны, лёд уже настолько крепок, что на нём можно свободно ходить и работать, а с другой стороны, он ещё не успел покрыться толстым слоем снега, сильно затрудняющим ходьбу по водоёму и нахождение границы между водой (вернее льдом) и берегом.
Прежде всего, конечно, нужно наметить промерные пункты. Для этого (рис. 8) проводим сначала среднюю линию АБ вдоль водоёма (мы теперь уже знаем, как это сделать) и делим её на несколько равных частей в зависимости от предполагаемого числа профилей или поперечных промерных линий (если, например, мы намерены сделать 3 профиля, то нужно делить на 4 части, если 5, то на 6 и т. д.); В найденных при таком делении точках Б, Г, Д восстанавливаем к продольной линии перпендикуляры, которые и будут » поперечными промерными линиями. По этим перпендикулярам, начиная от средней продольной линии, отмеряем по направлению к двум противоположным берегам последовательные вплоть до берега отрезки а, б, в, г... длиной в 10—20 м. Точки I, 2, 3, 4 и т. д., отделяющие эти отрезки друг от друга, и будут промерными пунктами. Отметив их вехами, приступаем затем к пробивке прорубей в этих пунктах и к измерению в них глубин 20
или посредством лота, или посредством размеченного па метры и полуметры шеста, если глубина водоёма вообще певелика. При этом начинать лучше со среднего профиля.
При большом числе экскурсантов и достаточной налаженности и соргапизованности работы это дело может пойти довольно быстро.
Ещё одно практическое замечание: зимние работы на водоёме следует производить отнюдь пе в валенках, а в непромокаемых сапогах, чтобы не промочить ноги выступающей из-подо льда или из-под снега водой.
Работа по плану. Вычисление длины и развития береговой линии. Чем более изрезан берег, тем извилистее будет контур водоёма, или, как говорят, тем более развита береговая л и п и я. Д л и н у последней можно измерить по плану циркулем, ножки которого раздвигаются на 1—2 мм (чем развитее береговая линия, тем меньший нужно брать раствор циркуля) :изполученной на плане длины получают затем по масштабу истинную длину береговой линии.
Чтобы определить развитие береговой линии, т. е. степень изрезанности берегов водоёма, нужно её длину разделить па окружность круга, площадь которого равна площади этого водоёма. Это отношение выражается формулой:
I
2|/ г.а
Где а — площадь водоёма, а I — длина береговой линии;
а = иг2, откуда г = ; окружность круга 2пг = 2- j/"= 2 j/"= 2 j/"~а.
Вычисление площади водоёма. Опишем три способа определения площади озера.
При первом способе пользуются особым инструментом — планиметром (продаётся в магазинах лабораторного оборудования).
В случае невозможности иметь планиметр — инструмент не дешёвый —для определения площади водоёма пользуются более доступным способом палетки, которую можно купить или изготовить самому. Палетка представляет собой прозрачный лист бумаги (или какого-либо другого материала—стекла, целлулоида и т. п.), расчерченный на квадраты. Её накладывают на план озера, контур которого через неё я<що виден, и подсчитывают число целых квадратов, охваченных контуром водоёма. Части квадрату пересечённых контуром, определяются или на глаз, или оолее точно, наложением мелкой палетки, на которой квадраты большой палетки разграфлены на мелкие квадратпкп (например, если сторона квадрата большой палетки равна 1 см, то сторона мелкого квадратика равна 1 мм)\ подсчитывая число мелких квадратиков, расположенных внутри контура озера, мы получаем площадь частей больших квадратов, прорезанных этим контуром. Теперь остаётся только суммировать площадь больших квадратов и их частей внутри контура, и мы получим всю плошадь озера на плане, а помножив её на масштаб плана, будем иметь реальную площадь водоёма, которую можно выразить в любых единицах квадратных мер — квадратных метрах, километрах или гектарах.
Третий способ — самый простой, лёгкий и скорый и, следовательно, самый удобный— применяется в тех случаях, когда не гонятся за особенно большой точностью. Он заключается в следующем. На листе хорошей бумаги, имеющей везде одинаковую толщину, очерчивают, хотя бы карандашом, контур водоёма в определённом масштабе. Определяют возможно точнее площадь этого листа и затем тоже возможно точнее взвешивают его. Потом вырезывают фигуру водоёма в точности по контуру и опять взвешивают её. Если взятый лист бумаги имеет действительно одинаковую по всей его площади толщину, то площадь и вес как всего листа, так и любой его части будут пропорциональны между собой. Поэтому, обозначая площадь вырезанной фигуры водоёма через х, будем иметь пропорцию:
х вес вырезанной фигуры
площадь всего листа вес всего листа ’
откуда определяется площадь х, из которой, путём умножения её па принятый масштаб, получают реальную плошадь водоёма.
Вычисление объёма видной массы. Объём водной массы озера вычисляется или способом конусов, или способом призм. Мы приведём здесь последний способ, как более простой. Он может быть применён в том случае, если имеется карта изобат. Йа плане изобаты представляют замкнутые кривые, проходящие через точки одинаковой глубины. Если мы мысленно проведём по контурам изобат вертикальные поверхности до пересечения с дном озера, то вся водная масса последнего разобьётся по вертикали па ряд горизонтальных слоёв, или зон, расположенных одна под другой.
Объём всей водной массы представляет сумму объёмов всехэтих зон. Так как с боков слои ограничены не вертикальными, а косыми поверхностями (склони дна водоёма), то объём каждого слоя будет равняться произведению его высоты на среднюю арифметическую из площадей, которыми ограничен этот слой сверху и снизу, т. е. па половину суммы этих площадей.
Площади, ограниченные изобатами, мы можем определить одним из способов» описанных выше для вычисления площади озера, а высота каждого слоя, или зоны,
есть разпость двух смежных глубпп, ограничивающих эту зону. Принимая контур самого озера за нулевую изобату (с глубиной в 0 м) и обозначая его площадь через а0> обозначим площади, ограниченные остальными изобатами, соответственно alf а2, а3,...ап (эти плотади по величине последовательно уменьшаются по направлению от берега к центру озера). Высоты слоёв, или зон, обозначим ссответственно через Ль Л2, Л3,...ЛЛ. Если зоны пронумеруем последовательно сверху вниз цифрами 7, /7, 777,...н, то получим следующую серию их объёмов:
Зоны их объёмы
Г («о + Д1) hl
2
ц _(^i 4“ Д2)
о
1П (я. + g3) h3
о
й (ал-1 ~Ь °л) .
2
если площадь срединной зоны ничтожна, так что может быть принята за нуль (ап = 0), то ооъем зоны будет---- ’
Суммируя объёмы всех зон, получим объём всей водной массы озера:
17 __ (gO 4~ 4“ (Д) 4" а1) ^2 4" (а2 4" аз) 4“ ал-1 hn
2
Если глубины, за исключением последней (Ал), равны между собой, т. е. = h2 = h9.. ... = то, обозначая эту одинаковую для всех слоёв глубину через h и вынося за скобки, получаем для объёма водной массы озера более простое выражение:
2 (ао 4- *Д) 4- 4" 2Дз 4- ••• 4- an-i) 4" у
Напоминаем, что здесь а0, аь а2....ап_} — площади, ограниченные последовательными изобатами, h — высота слоя, т. е. вертикальное расстояние между горизонтальными площадями, соответствующими двум соседним изобатам, a hn — высота последнего (самого нижнего) слоя.
Водная масса озера, главнейшие физико-химические свойства озера
До сих пор наше внимание было посвящено тем процессам, которые связаны с образованием озёрного ложа, или бассейна. Теперь мы обратимся к свойствам наполняющей ложе водной массы и к выяснению тех особенностей в жизни обитателей озера, которые связаны с этими свойствами.
Очевидно, что все наиболее характерные черты озера и его жизни, которые обусловливаются присутствием водной массы, должны зависеть от общих и основных физических и химических свойств воды. Поэтому они могут быть названы физико-химическими свойствами озера, важнейшие из которых мы и рассмотрим в связи с основными свойствами воды как физического тела.
СВОЙСТВА ВОДЫ КАК ЖИДКОГО ТЕЛА
Сила сцепления и плотность воды. Одним из основных физических свойств воды является её способность при тех колебаниях температуры, которые наблюдаются в нашем климате, принимать или все три состояния — твёрдое, жидкое и газообразное (зимой), или только два последних (летом). Важнейшим из этих состояний нужно считать, несомненно, жидкое, так как именно в этом состоянии вода представляет один из основных жизненных факторов и является тем веществом, без которого немыслима никакая жизнь на земле. Жидкая вода входит в состав всех живых организмов в таком количестве, которое в несколько раз превышает количество твёрдых веществ.
оо
Для интересующих нас организмов она составляет ту среду, в которой они проводят всю свою жизнь или, по крайней мере, часть её и которая постоянно окружает их со всех сторон. Что касаетс я озера, то мы уже видели, какую громадную работу проделывает там вода в жидком состоянии и какие изменения она производит в собственном ложе, на котором покоится. Всё это обязывает нас внимательнее присмотреться к свойствам жидкой воды, которые помогут нам объяснить многие явления в жизни водоёма.
Сущность жидкого состояния заключается в слабом сцеплении частиц жидкости между собой; из этого видно, чем отличается жидкое состояние от двух других: в газообразном состояв.ih частицы не только не сцеплены друг с другом, но даже, наоборот, взаимно отталкиваются и потому всегда находятся на некоторОхМ расстоянии одна от другой, между тем как в жидкости они соединены или сцеплены между собой; но это сцепление чрезвычайно слабое, настолько слабое, что достаточно применить самую ничтожную силу, чтобы разъединить частицы; этим жидкое тело отличается от твёрдого, в котором частицы, напротив, соединены плотно и прочно, так что для их разобщения (для раскалывания твёрдого тела) приходится употреблять иногда очень большую силу. Этим объясняется, что жидкость не имеет собственной формы, а принимает форму того сосуда, в который она налита; кроме того, если жидкость находится в покое и не подвергается действию никаких сил, кроме силы тяжести, то её свободная открытая поверхность всегда горизонтальна. Это, разумеется, относится ко всякому сосуду, в том числе и к тому, который мы называем озёрным ложем.
Влияние силы сцепления и вязкости воды на водные организмы. Если мы будем сравнивать с указанной точки зрения две основные среды, в которых проходит жизнь живых существ, — воду и воздух, то мы увидим здесь такое громадное различие, которое не может не отразиться на строении и жизненных проявлениях организмов. Но здесь необходимо вспомнить ещё одно обстоятельство: всякое погружённое в воду тело становится значительно легче, именно на столько, сколько весит равная ему по объёму часть воды, так что если взять предмет, вес которого равен весу такого же объёма воды, или, как говорят, удельный вес которого равняется 1, то тело будет держаться, не падая на дно и не всплывая на поверхность. Как уже указывалось, во всяком живом организме, а особенно^ растении, значительную часть составляет вода (в растении иногда 90%); ясно поэтому, что вес организмов большей частью довольно близок к весу воды; они, следовательно, становятся в воде почти невесомыми, и опускание их на дно должно происходить медленно. Но быстрота опускания зависит не только от веса тела, но и от его формы. При падении тело должно раздвигать встречающиеся ему на пути частицы. Даже в воздухе, частички которого расступаются легко, лист бумаги будет опускаться заметно на глаз медленнее, чем тот же лист, смятый в плотный комок. Частицы воды в силу их сцепления расступаются гораздо труднее; поэтому ясно, что здесь форма имеет ещё большее значение для замедления падения. Известно, что если тонкую металлическую пластинку плашмя положить на воду и отпустить, то она будет погружаться значительно медленнее, чем в случае падения ребром. Следовательно, для скорости падения большую роль играют величина и форма той поверхности тела, которая обращена вниз и должна раздвигать частицы воды: чем больше эта поверхность, тем медленнее будет погружаться тело. Это явление называется «сопротивлением формы» тела.
Наконец, для скорости погружения имеют значение не только свойства погружающегося тела, а также и некоторые свойства воды, а именно — так называемая вязкость её, под которой подразумевается внутреннее трение водных частиц друг о друга; чем больше последнее, тем медленнее погружается тело в воде. Эта вязкость стоит в обратной зависимости от температуры (в тёплой воде опа меньше, в холодной — больше) и д прямой — от количества растворённых в воде солей, а так как эти факторы в озёрной 23
воде оказываются изменчивыми, то и вязкость её в течение года подвержена значительным изменениям и потому отражается на жизни её обитателей.
Итак, скорость опускания тела в воде является прямо пропорциональной его удельному весу и обратно пропорциональной вязкости воды и сопротивлению формы тела. Поэтому одновременное влияние всех этих трёх факторов может быть выражено следующей формулой: скорость погружения равна: остаточный вес
сопротивление формы х вязкость воды ’
где под остаточным весом подразумевается вес тела в воде, т. е. разница между его весом в воздухе и весом вытесненной им воды.
Однако эта формула, выведенная для физических тел, приложима лишь для тех живых организмов, которые, подобно неживым телам, совершенно пассивны, т. е. не могут самостоятельно, активно передвигаться в воде, как, например, многочисленные взвешенные в воде мельчайшие растительные организмы. Но все животные, за немногими лишь исключениями, обладают способностью к активному движению, которое помогает животному преодолевать влекущую его вниз силу тяжести и держаться в воде. Ясно, что влияние такого важного фактора не может быть игнорировано, и он был введён в формулу, после чего она получила следующий вид: скорость, погружения равна:
остаточный вес х активное движение и руление вниз сопротивление трения X активное движение и руление вверх где сопротивление трения = сопротивление формы х вязкость воды, а под рулением подразумевается направление движения..
Другое изменение, введённое в первую формулу, состоит в том, что «сопротивление формы» заменено в ней «удельной поверхностью», т. е. отношением поверхности тела к его объёму.
Из всего этого ясно, в каких направлениях должно было идти приспособление к условиям существования тех организмов, для которых вода, постоянно или временно, оказывается единственной опорой: оно должно было, по возможности, уменьшать влияние тех факторов, которые стоят в числителе приведённой формулы, и увеличивать влияние факторов, стоящих в знаменателе. Той или иной комбинацией этих приспособлений и определяется степень способности всякого водного организма держаться в воде без помощи твёрдого субстрата. Рассмотрим несколько примеров.
Выше уже было указано, что у всех истинно водных организмов в тканях тела содержится громадный процент воды — иногда свыше 90, благодаря чему их удельный вес более или менее близок к единице. Но он все же не единица, и вот эта-то разность с последней, влекущая тело организма вниз, и является стимулом для развития таких приспособлений, которые компенсировали бы её влияние. Одним из таких приспособлений является дальнейшее уменьшение остаточного веса путём, например, выделения водными организмами в своих тканях веществ, обладающих удельным весом, значительно меньшим единицы, и потому в свободном состоянии легко всплывающих на поверхность воды, например, газы и жиры; однако эти вещества у производящих их живых организмов не выходят наружу, а остаются в теле и своим присутствием, естественно, облегчают его, уменьшают его остаточный вес. Всем известно, что у большинства рыб имеется наполненный газами плавательный пузырь; увеличение или уменьшение содержания газов в пузыре соответственно уменьшает или увеличивает остаточный вес тела рыбы и тем способствует его поднятию или погружению в воде. Действие этого аппарата настолько совершенно, что оно преодолевает даже отрицательное влияние другого фактора — сопротивление формы: у громадного большинства рыб наибольшая и наиболее плоская поверхность тела обращена 24
не вниз, а в боковые стороны, что связано уже с хорошо развитой у рыб способностью к активному движению. Подобными же плавательными пузырьками обладают личинки комаров Chaoborinae (рис. 434), у которых их две пары: одна — вблизи переднего конца, другая — вблизи заднего. В обоих этих случаях — у рыб и у личинок комаров — газовый аппарат настолько хорошо действует, что сводит остаточный вес животного к нулю и потому делает излишними все остальные приспособления.
11 среди водных растений известны примеры развития в их тканях воздухоносных полостей, но здесь к этому часто прибавляется ещё увеличение горизонтальных поверхностей (увеличение знаменателя формулы), как это мы видим на листьях кувшинки (рис. 98).
Уменьшение остаточного веса достигается выделением во внутренние полости тела не только газов, но и жировых веществ — масла. Это наблюдается, например, у некоторых микроскопических организмов —у диатомовых водорослей, у коловраток и рачков (особенно, веслоногих). Однако в последнем случае это приспособление оказывается не всегда достаточным для поддержания тела во взвешенном состоянии, и на помощь ему является активное движение животных: у коловраток — посредством коловратного аппарата (рис. 27, 28), у рачков — посредством плавательных усиков (рис. 241); например, дафнии из ветвистоусых рачков в спокойном состоянии медленно опускаются в воде и для избежания погружения на дно вынуждены время от времени ударять, как вёслами, второй парой усиков — плавательных. У многих крупных водных жуков тело сплющено в спинно-брюшном направлении, этим у них осуществляется «сопротивление формы», которое, однако, тоже недостаточно и дополняется способностью к активному перемещению (в неподвижном состоянии эти животные довольно быстро падают) в воде.
Увеличение поверхности как фактор, замедляющий или уничтожающий погружение тела в воде, имеет особенно большое значение для мелких организмов, свободно носящихся в открытой воде, т. е. для планктона1. Дело в том, что при этом играет роль не увеличение поверхности вообще, а увеличение её по отношению к объёму того же тела, т. е. увеличение так называемой удельной поверхности, равной отношению абсолютной поверхности к объёму тела.
Чем больше поверхность при одном и том же объёме или чем меньше объём при одной и той же поверхности, тем более будет замедление погружения тела. Вот почему увеличение поверхности играет у мелких организмов гораздо большую роль, чем у крупных. У многих микроскопических растений и животных развиваются отростки различной величины и формы. Так, среди планктических водорослей, коловраток и рачков очень распространены игловидные и щетинкообразные выросты тела, незначительно лишь изменяющие их объём, но сильно увеличивающие поверхность. Для примера можно указать на следующие формы: из коловраток — Кега-tella quadrata (Anuraea aculeate), Notholca longispina (рис. 27), из ракообразных — большинство ветвистоусых (Cladocera) и веслоногих (Copepoda) рачков, конечности которых — усики, ножки, хвостовая вилка — обычно густо усажены большим количеством более или менее длинных шипов и щетинок, причём многие из последних (перистые щетинки) несут на себе ещё вторичные ряды более мелких щетинок.
Итак, вода, благодаря силе сцепления частиц, прекрасно может поддерживать находящиеся в ней предметы, но зато эта же сила сцепления оказывает, по сравнению с воздухом, очень сильное препятствие движению. С о-противление движению, как и в случае погружения, зависит
1 Планктоном называют совокупность микроскопических растений и животных, которые находятся в виде во взвешенном состоянии (т. е. не связаны ни с каким твёрдым субстратом, как опорой для их тела) и которые не в состоянии противостоять даже слабым течениям благодаря или полному отсутствию способности к активному плаванию, или лишь слабому развитию этой способности.
в очень большой мере от фор-м ы предмета: чем больше раздвигающая воду поверхность, тем больше частиц должны будут перед ней расступиться и тем большее сопротивление сцепляющей их силы придётся ей преодолеть. Поэтому нежным, сильно раскидистым и лишённым внутренней опоры телом могут обладать лишь неподвижные организмы, как высшие растения и некоторые прикреплённые животные, например, гидра (.рис. 9). Те животные, которые в погоне за подвижной животной пищей совершают скачки в воде или быстро плавают, должны иметь тело, достаточно твёрдое для рассечения водной толщи и приспособленное к преодоле-
нию сцепления частиц: оно дол-Р и с. 9. Гидра. жн0 быть заострено с концов
и обладать или веретеновидной, или сплюснутой формой. Обоим требованиям прекрасно удовлетворяет строение тела, например, водных хищных жуков: они плоски и прикрыты очень твёрдым покровом. Подавляющее большинство рыб также имеют уплощённое тело.
Что касается перемещения тела в самой воде, то оно может совершаться очень разнообразными способами. Вертикальное перемещение в воде возможно и совсем без специальных органов. Вспомним, что вес животного в воде вообще невелик, а если тело станет ещё легче, то всплывание вверх должно произойти само собой. Для облегчения иногда достаточно небольшого скопления воздуха. У некоторых водных жуиов и клопов выпущенный из дыхалец отработанный, ненужный им воздух задерживается между телом и надкрыльями благодаря присутствию там несмачиваемых водой волосков и легко и быстро поднимает на поверхность часто очень крупное тело, например, плавунца (рис. 21); здесь насекомое выпускает этот воздух в атмосферу и после этого получает возможность опуститься снова вниз; если не дать ему выпустить воздух, то жук не может погрузиться в воду. Рыбы, регулируя количество газов в плавательном пузыре, могут то подниматься, то опускаться.
Произвольное перемещение тела в воде в различных направлениях тоже иногда совершается без специальных органов движения. Пиявки, поочерёдно сгибаясь и выпрямляясь, производят целый ряд отталкивательных движений, в результате которых происходит перемещение тела (рис. 10). Так же передвигаются в воде и некоторые другие черви, например, малощетинковые (рис. 128 и 129). Совсем своеобразно перемещается личинка коромысла: она забирает в свою очень объёмистую заднюю кишку большое количество воды, удерживая её плотно захлопывающимися клапанами; при нужде в перемещении она быстро выбрасывает эту воду и ею отталкивается от окружающей воды (так называемое реактивное движение).
В большинстве случаев водные животные перемещаются в воде при помощи особых, специально" для передвижения предназначенных органов. Некоторые микроскопические организмы, например, инфузории (рис. 11), коловратки (рис. 27, 28), движутся при помощи колеблющихся мельчайших ресничек; таким же способом перемещаются и ресничные плоские черви — 26
планарии (рис. 20); брюхоногие моллюски (например, прудовики, катушки) ползают ногой по подводным предметам или скользят по нижней стороне плёнки поверхностного натяжения воды (рис. 21); в последнем случае телу, свешивающемуся вниз и находящемуся в воде, помогает держаться у самой поверхности воды воздух, заключённый в дыхательной полости моллюска.
Р и с. 10. Европейские пиявки:
налево—плавает рйбьн — Piscicola; на камне — две малых ложноконских — Herpobdella; на дне заглатывает дождевого червя ложноконская— Haemopis; направо—плывёт ложноконская; на растениях и на дне— улитковая — Glussiphonia.
Рис. И. Туфелька — Paramaeci am.
Все хорошие пловцы, как жуки, клопы, личинки многочисленных насекомых, обладают ножками, состоящими из отдельных члеников; механизм их действия подобен механизму вёсел: сильными ударами широкой поверхности органа животное отталкивается от воды, а затем возвращает ножку в первоначальное положение. Возвращение лодочного весла происходит в воздухе и не требует от гребца почти никаких усилий; если бы он стал возвращать вёсла в воде, лодка его не ушла бы далеко. Нашим пловцам приходится возвращать свои конечности в воде, и для облегчения их движения в этом направлении есть целый ряд приспособлений: например, ножки ракообразных сгибаются в сочленениях только назад, поэтому при движении ножек вперёд они идут согнутыми и легко врезаются углом в воду, а при ударе
назад механически распрямляются водой; ножки клопа гладыша и жука плавунца снабжены длинными густыми щетинками, которые могут прилегать к ножке или распрямляться (рис. 12).
Необходимо, однако, добавить, что некоторые жуки движутся в воде обыкновенными ходильными ножками, которые остаются у них совершенно неизменёнными.
Все указанные активные способы передвижения осуществляются при помощи удара о воду и отталкивания от неё, они основаны на сопротивлении воды движущемуся телу и, следовательно, на силе сцепления её частиц.
Рис. 12. Ножка жука плавупца при движении его вперёд (правая) и назад.
Приход п расход воды в озере. Одним из следствий слабого сцепления частиц жидкой воды друг с другом является то, что при соприкосновении воды с атмосферой её частицы, расположенные на свободной водной поверхности, более или менее легко отрываются и улетают в воздух. Этот процесс, который мы называем испарением, играет в жизни озера огромную роль.
Испарение, как известно, зависит от температуры: с повышением её оно увеличивается, с понижением — уменьшается, но никогда не прекращается, так как даже в твёрдом состоянии (снег, лёд) вода всё-таки испаряется, хотя и очень медленно. Ввиду того, что испарение происходит только на открытой, соприкасающейся с воздухом поверхности воды, величина его зависит от размеров этой поверхности; поэтому из двух озёр, водная масса которых имеет один и тот же объём, но различные площади, испарение будет сильнее на том, площадь которого больше. Следовательно, для понимания этой стороны жизни озера необходимо учитывать обе указанные величины — объём и площадь водной массы.
Для более или менее продолжительного существования озера испаряющаяся вода должна как-нибудь пополняться, т. е. озеро должно, как говорят, питаться. Откуда же озеро берёт для этого воду? В конечном счёте, несомненно, из атмосферы — в виде атмосферных осадков (дождя и снега). Здесь мы встречаемся с новым свойством воды, прямо противоположным испарению, именно — со сгущением водяного пара в жидкую воду.
Убыль воды из озера в форме испарения и прибыль её в форме атмосферных осадков происходят во всех озёрах. Но во многих из них общая потеря воды, называемая её расходом, совершается ещё другим путём, кроме испарения, именно — посредством стоков; точно так же и общее пополнение озёрной воды, или так называемый приход её, может происходить, кроме атмосферных осадков, через притоки — реки, ручьи, ключи. Вот почему для учёта общего результата прихода и расхода воды в озере, пли так называемого водного баланса, нужно знать не только объём и площадь водной массы самого озера, но и количество притоков и стоков и их приблизительную мощность.
Итак, наряду с отдачей воды в атмосферу в форме испарения мы имеем возврат её из атмосферы в озеро в форме осадков; наряду с расходом воды в виде стоков имеется приход её в виде притоков. Ясно, что водный баланс озера определяется той или иной комбинацией всех этих четырёх процессов. Из этих процессов испарение и осадки стоят в прямой зависимости от климата данной местности, а притоки — в косвенной. Во влажном климате расход воды через испарение будет небольшой, а приход через притоки — значительный, и поэтому, если нет стока, уровень озера будет повышаться до тех пор, пока вода не найдёт себе какого-нибудь стока; в умеренном климате озеро в отношении водного баланса будет находиться в приблизительном равновесии; и, наконец, в жарких и сухих местностях во время бездождья вода будет всё время убывать и при очень продолжительной засухе может совсем испариться.
По отсутствию или присутствию стока озёра делятся на бессточные1 или конечные, и сточные1; если кроме стока имеется хотя бы один приток, озеро называется проточным. Бессточные озёра встречаются большей частью в сухих местностях, в пустынях и имеют в большинстве случаев солёную воду; к ним относятся, например, Каспийское и Аральское моря. Сточные озёра, наоборот, свойственны влажному климату; таковы озёра Ладожское, Онежское, Ильмень и многие другие.
Вести исследования по вопросу о балансе воды в озере непосильно для отдельных лиц или даже для группы лиц, не связанных с каким-либо спе-
1 Здесь слова «сток» и «сточный» применяются в более узком смысле,чем во введении (стр. 5), а именно — для обозначения в ы т е к а го ni и х из озера вод, в противоиолож-ность притокам, через которые вода втекает в озеро.
циальпо оборудованным учреждением, и потому здесь методика «того не даётся.
Движение воды. Течения. Течение воды, определяемое силой тяжести, имеет место только тогда, когда по крайней мере в двух пунктах уровень её будет различный; в таком случае вода будет течь по направлению от более высокого уровня к более низкому. Очевидно, что чем больше разность уровней, тем быстрее будет течение. В проточных озёрах разность уровней создаётся в местах притоков и стоков: у первых уровень воды несколько повышен (в форме очень плоского конуса), у вторых — несколько понижен (в форме очень плоской воронки). Однако течение воды, зависящее от этой разности уровней, ясно заметно лишь в устьях притоков и в начале стоков; в самом озере его можно обнаружить только в сравнительно небольших озёрах; но чем больше площадь озера, тем более слабым и менее заметным будет течение воды в нём. Кроме такого течения, на озере бывают и другие, вызываемые иными причинами. Для примера укажем на ветровые течения: ветер, дующий в течение продолжительного времени в одном направлении, вследствие трения движущегося воздуха о поверхность воды увлекает за собой последнюю и постепенно сгоняет её от одного берега к другому, противоположному; таким образом возникает течение, которое лишь с большим трудом и медленно проникает в глубину и потому остаётся более или менее поверхностным; благодаря этому течению уровень воды у первого берега понижается, у второго повышается; создаётся разность уровней, вызывающая течение от более высокого к более низкому, т. е. обратное по отношению к поверхностному. Так как на поверхности этому течению препятствует ветер, то оно опускается на глубину; происходит это так, что накопившаяся у одного берега вода погружается здесь на глубину, создавая нисходящий ток, у противоположного же берега, где уровень понижен, она, наоборот, образует восходящий ток. Таким образом в озере возникает движение, называемое циркуляцией воды: поверхностный слой последней гонится ветром к одному берегу, здесь опускается на глубину, затем течёт к противоположному берегу, где опять поднимается на поверхность, снова движется по ветру и т. д. Очевидно, что чем дольше дует ветер, тем большей и большей толщины слой воды будет захватываться циркуляцией; если озеро не слишком глубоко, то при достаточно!! продолжительности ветра циркуляция воды может углубиться до самого дна и захватить всю водную массу.
Кроме рассмотренных нами горизонтальных течений, в водной массе озера могут.возникать вертикальные перемещения, вызываемые уже не разностью уровней, а другими причинами, например, изменением плотности верхних и нижних слоёв воды; если плотность верхних слоёв стала вследствие каких-либо причин больше плотности нижних, то первые окажутся тяжелее нижних и будут опускаться, нижние же, наоборот, подниматься. Так образуются вертикальные, так называемые, конвекционные токи. Но о них мы будем говорить подробнее в другом месте (стр. 32), здесь же ограничиваемся лишь сказанным.
Волнения. Движение воды в озере не ограничивается одной только формой — течением, но принимает и другие формы, главнейшей из которых является волнообразное движение. Это движение вызывается разными причинами: движущимся судном, падающим на воду предметом, дождевыми каплями, ветром и т. п. Наиболее важной из них является, несомненно, последняя причина, и потому в дальнейшем мы опишем лишь ветровые волнения.
Падение на воду предмета вызывает, как известно, круговые волны, концентрически расходящиеся от места падения, как от центра. Ветер же обусловливает возникновение прямолинейных волн, перпендикулярных направлению ветра, но движущихся в ту же сторону. Непрерывно дующий потер всё время образует новые и новые волны, которые, накладываясь друг на друга, слагаются в более и более мощные волны, бегущие одна за другой в направлении ветра. Величина волн зависит, кроме силы и про-
должптельности ветра, также от площади и глубины водоёма, вследствие чего на больших и глубоких озёрах волнение бывает сильнее, чем на мелких.
Когда волны достигают суши, оказывающей непреодолимое препятствие их дальнейшему движению вперёд, то они откатываются назад. При встреч© их с волнами, бегущими на берег, вода рассыпается на мелкие капли (брызги), воздух, смешиваясь с водой, также разбивается ею на маленькие пузырьки, одетые тонкой водной оболочкой, и получается то, что мы называем пеной» Весь этот процесс набегания волн на берег называется прибоем. С разрушающим и размывающим действием прибойных волн на берег мы познакомились уже раньше — в главе о цикле жизни и возрастных периодах озера. Здесь лишь добавим следующее: та часть берега, которая своим склоном обращена навстречу господствующему в данной местности направлению ветров, подвергается, разумеется, в большей степени, чем другие части, их действию, больше размывается: вследствие этою береговой откос становится круче, и берег, называемый в этом случае прибойным приобретает характер обрывистого.
Несколько иной характер приобретает прибой в том случае, если береговая мель покрыта растительностью. Само собой разумеется, что последняя представляет для волн известный барьер, оказывающий им то или иное сопротивление. Волны уже здесь разбиваются, и если полоса зарослей достаточно широка и густа, то она в состоянии совершенно заглушить волны; тогда волнение прекращается, ещё не достигнув суши.
Влияние движения воды на водное население. Очевидно, что если прибрежная растительность оказывает определённое действие на волны, то, наоборот, и волны, конечно, так или иначе влияют на растения, о которые они ударяются. Это влиянйе отражается на организации растения.
Движение воды угрожает растениям в двух направлениях: с одной стороны, отрывом от грунта, в котором они укрепляются, с другой — разрывом тканей самого растения.
Опасности почти не существует для растений, не поднимающихся над водной поверхностью: ведь волнение не всегда доходит до придонных слоёв, а если и доходит, то уже в большой мере ослабленным. Растения с плавающими листьями обладают стеблями, способными к сильному вытягиванию, в очень хорошо противостоящими разрыву. Попробуйте разорвать стебель водяной лилии, и вы в этом сейчас же убедитесь. Растения с поднимающимися в воздух частями подвергаются риску быть вытянутыми с корнем из грунта или слом а н н ы м и напором волн, а также при порыве- ветра. Защитой против этой опасности служит прежде всего та же очень большая эластичность, способность сильно вытягиваться или сгибаться и снова приходить к первоначальной длине и форме по миновании действия ветра, а с другой стороны, некоторые особенности формы и строения. Прежде всего почпг все надводные растения снабжены узкими стеблями и листьями, оказывающими ветру малое сопротивление; затем у некоторых ра-’ стений (тростник) влагалища листьев могут поворачиваться вокруг стебля и этим направлять их пластинки в сторону от ветра. Если вы попадёте на озеро в ветреный день, то обратите внимание на то, как аккуратно расчёсаны в одну сторону листья тростника.
Сильной защитой для водных растений служит ещё и то, что они почтп всегда растут не в одиночку, а образуют более или менее густые заросли, где быстро глохнут и ветер и волнение.
Заросли высших растений, тсрмозя движение воды, тем самым сильно облегчают поселяющимся среди них животным борьбу с вечным колебанием окружающей водной массы; помимо того, они дают животным опору для постоянного или временного прикрепления. Постоянное прикрепление, т. е. сидячий образ жизни, встречается среди обитателей пресных вод не часто (губки, мшанки), но свободно передвигающиеся животные постоянно пользуются стебельками или листьями водных растений (для подстерегания добычи, для удержания у поверхности воды при воздушном дыхании и т. д.). 30
Помимо того, большое число свободно живущих животных прикрепляют к растениям свои яйца, то приклеивая их (моллюски, некоторые рыбы), то погружая в самую ткань растения (некоторые жуки и клопы).
В тех местах, где отсутствуют растения, условия жизни, благодаря сильному движению воды, так затруднительны, что лишь для немногих обитателей воды, обладающих особыми приспособлениями, возможна жизнь. В таких условиях могут существовать, например, пиявки, способные очень сильно держаться своими присосками за камни или другие погружённые предметы (рис. 10), некоторые личинки насекомых, живущие в твёрдых домиках, плотно приклеенных к чему-нибудь (Trichoptera, рис. 57—58, 59), улитки, нога которых может прочно присасываться к подводным предметам.
ТЕПЛОВЫЕ СВОЙСТВА
Приход и расход тепла в озере. Главнейшим и неисчерпаемым источником тепла для воды служит солнце, которое, как известно, не только светит, но и греет, т. е. посылает на землю, кроме световых лучей, также и тепловые. Вода, наряду со многими другими прозрачными предметами, до известной степени пропускает через себя световые лучи; однако она не только пропускает, но и отражает их, доказательством чего служит то, что на спокойной, гладкой поверхности озера или реки можно, как в зеркале, хорошо видеть окружающую природу: небо с облаками и светилами, горы, деревья и т. д.
Этими же свойствами — прозрачностью и отражательной способностью— вода обладает также и по отношению к тепловым солнечным лучам. Таким образом, последние, падая на поверхность озера, частью отражаются от неё, частью проходят в воду, проникая более или менее глубоко в зависимости от степени её теплопрозрачности, которая, подобно световой прозрачности, увеличивается вместе с чистотой воды и уменьшается с помутнением её. Этим свойством вода отличается от суши, у которой солнечными лучами нагревается лишь поверхностный слой. Вследствие этого поверхность суши и нагревается и охлаждается гораздо быстрее, чем поверхность воды. Но как у той, так и у другой наиболее нагретым будет самый поверхностный слей, с глубиной же температура убывает—у суши быстрее, у воды медленнее.
От теплопрозрачности нужно отличать теплопроводность воды. Благодаря первому свойству солнечные лучи проникают на ту или иную глубину и здесь, отдавая воде свою тепловую энергию, непосредственно нагревают её; благодаря второму свойству теплота, воспринятая верхними слоями воды, передаётся нижележащим слоям и нагревает их, хотя бы действие непосредственного источники тепла — солнца — в это время уже прекратилось. Теплопроводность воды чрезвычайно мала и в сравнении с теплопрозрачностью играет ничтожную роль в нагревании глубоких слоёв воды.
Наконец, следует отметить ещё одно свойство воды, которое имеет большое значение для её нагревания — это теплоёмкость, т. е. способность поглощать теплоту. Она измеряется особыми единицами, так называемыми калориями — большими или малыми, причём под большой калорией понимают количество тепла, которое необходимо затратить для нагревания одного килограмма (под малой калорией — одного грамма) чистой воды на один градус Цельсия. Измерения теплоёмкости воды показали, что она очень значительна и во всяком случае превосходит теплоёмкость суши.
Все указанные свойства воды — теплопрозрачность, теплопроводность и теплоёмкость — способствуют накоплению в ней теплоты, т. е. являются факторами прихода тепла в воде. Кроме непосредственного действия солнечных лучей, есть и другие источники тепла для озёрной воды, из которых на первом месте следует поставить передачу тепла воде от окружающей озеро суши и от озёрного дна, а также и от биохимических процессов, главным образом от процессов разложения и гниения органических веществ как в самой воде, так особенно и в донных отложениях.
ЗТ
Ясно, что вода не только получает, а также и отдаёт свою теплоту воздуху. Следовательно, на поверхности воды совершается, кроме прихода, также и расход тепла, главнейшей формой которого является лучеиспускание, или излучение тепла водой; этим свойством вода обладает в сильной степени. Таким образом, на поверхности озера происходит усиленный теплообмен между водой и воздухом, результатом которого является тепловой баланс озера.
Температурные колебания. Чередование нагревания и охлаждения верхних слоёв воды может иметь нерегулярный характер; но оно может быть вызвано и постоянными, регулярно действующими причинами, и в таком случае оно становится явлением правильным, повторяющимся через определённые промежутки времени. — оно делается, как говорят, периодическим. Такие периодические колебания температуры озёрной воды проявляются в двух формах: в виде суточныхив виде годовых колебаний. Суточные колебания обусловливаются сменой дня и ночи, причём наибольшая температура бывает ближе к вечеру, а наименьшая — ближе к утру. При годовых колебаниях, как известно, наибольшее нагревание имеет место летом, наибольшее охлаждение — зимой. Ясно, что годовые колебания, как гораздо более длительные, чем суточные, успевают распространиться на значительно большую глубину.
Тепловая слоистость. Посмотрим теперь, как проникшая в глубину теплота распространяется в толще водной массы озера и какие температурные отношения господствуют в различных её слоях. К решению этого вопроса мы должны привлечь ещё одно свойство воды — её плотность ив особенности отношение последней к температуре. Известно, что пресная вода обладает наибольшей плотностью при 4°, вверх же и вниз от этой температуры плотность убывает. Такое свойство воды очень сильно отражается на жизни озера, так как им определяется одно из чрезвычайно важных явлений, а именно: так называемая те п л о в а я слоистость водной массы. Сущность этого явления заключается в том, что различные слои озёрной воды, лежащие один над другим, всегда располагаются соответственно своей плотности и, следовательно, своей тяжести: само собой понятно, что более лёгкие слои всегда будут находиться над более тяжёлыми. Таким образом, на поверхности будет расположен слой наименьшей плотности, у дна, наоборот, — слой наиболее плотный. В местностях умеренного пояса при достаточной глубине озера придонный слой будет иметь плот-но<*гь, равную наибольшей плотности воды, а это значит, что его температура должна быть равна приблизительно 4°. Какова же будет температура вышележащих слоёв? Очевидно, что она может быть как выше, так и ниже 4°, или же равна 4°. В озёрах умеренных стран в течение года осуществляются все три случая: первый имеет место летом, второй — зимой, третий — весной и осенью. Происходит эта смена следующим образом (рис. 13).
Летом температура водной толщи по направлению от дна к поверхности возрастает, так как верхние слои прогреваются солнцем; такая тепловая слоистость, типичная для лета, называется прямо й; её отношение к плотности характеризуется тем, что с глубиной температура убывает, а плотность возрастает. Такое состояние озера носит название летней стагнации. Осенью тепловая энергия солнечных лучей уменьшается; вода ещё хранит в себе значительные запасы летней теплоты, но воздух охлаждается и начинает отнимать тепло у воды, которая, таким образом, тоже начинает с поверхности охлаждаться. Как только температура поверхностного слоя стала хотя бы немного ниже температуры непосредственно под ним лежащего слоя, первый, сделавшись плотнее и тяжелее, погружается и вытесняет последний на своё место, т. е. на поверхность, причём температура обоих слоёв сравнивается. При дальнейшем охлаждении повторяется то же самое, но теперь перемешивание слоёв распространяется уже на большую глубину; так возникают в водной массе озера вертикальные перемещения — холодных слоёв сверху вниз, а тёплых — снизу вверх; эти течения называются к он век-Я9
Схема изменений температуры воды и тепловой слоистости в озере по сезонам. Ч. Ц. — частичная циркуляция; П. Ц. — полная циркуляция.
ционными токами. Вместе с тем идёт выравнивание и постепенное понижение температуры озёрной воды; этот процесс, называемый осенней ц и р к у л я ц и е й, в конце концов охватывает всю толщу воды, вертикальные течения распространяются до самого дна, и во всей водной массе озера устанавливается температура придонного слоя в 4°; это означает, что вся вода приобрела наибольшую плотность. Однако охлаждение с поверхности всё продолжается, и, следовательно, плотность поверхностных слоёв, температура которых теперь уже стала ниже 4°, вновь начинает уменьшаться. При дальнейшем охлаждении это уменьшение плотности всё более и более проникает в глубину, пока по всей водной толще не восстановится вновь то распределение плотностей, которое существовало до установления в водной массе одинаковой температуры в 4°. Теперь, как и тогда, плотность вместе с глубиной увеличивается; но разница заключается в том, что тогда температура по направлению от поверхности к дну убывала, теперь же, наоборот, она по этому направлению возрастает. Это распределение температур сохраняется и после замерзания озера; такая тепловая слоистость, характерная для зимнего периода, получила название обратной. Озеро перешло в состояние зимней стагнации. Зимой, следовательно, самая высокая температура, именно — около 4°, господствует в придонном слое, обладающем поэтому наибольшей плотностью.
Весной, когда лёд стает, вода с поверхности начинает прогреваться; по мере того как температура верхних слоёв приближается к 4°, плотность их увеличивается, они становятся тяжелее и опускаются до того слоя, плотность которого больше их собственной; следствием этого являются опять конвекционные токи, выравнивающие температуру до той глубины, до которой они распространяются в данный момент. Одним словом, повторяется тот же процесс, который имеет место.осенью, только с обратным распределением температур в толще воды в конце периода зимней стагнации. Результатом этого процесса, именуемого весенней* циркуляцией, является опять-таки установление по всей водной массе одной и той же температуры приблизительно в 4°, после чего при дальнейшем прогревании воды с поверх-
3 А. Н. Липин
ности озеро переходит к летней, т. е. прямой тепловой слоистости, к летней стагнации.
При описании явления тепловой слоистости мы исходили из предположения, что лежащий на самом дне слой воды имеет температуру 4°. Однако прямые наблюдения далеко не всегда оправдывают это предположение: часто температура этого слоя уклоняется от Iе — летом в сторону повышения, а зимой в сторону понижения. Причиной этого является та циркуляция воды, которая возбуждается деятельностью ветра и о которой говорилось в главе о течениях. Мы уже знаем, что при достаточной силе и продолжительности ветра циркуляционные токи могут захватить всю водную массу; при этом, конечно, нагретые поверхностные слои, попадая на дно, нагревают нижние слои, температура последних становится несколько выше 4° и остаётся таковой и по прекращении вызванной ветром циркуляции; вследствие этого летом температура придонного слоя обычно выше 4е. Перед замерзанием озера та же самая причина — ветровая циркуляция воды, во время которой более холодные, в данном случае поверхностные слои, опускаясь, охлаждают нижние слои, — вызывают обратное явление, именно: падение температуры придонного слоя ниже 4°, остающееся на всю зиму.
Само собой разумеется, что если даже в озёрах значительной глубины могут иметь место описанные изменения температуры того слоя, который лежит у самого дна, то тем более они допустимы в прудах и мелких озёрах, где идущее от поверхности нагревание и охлаждение легко распространяется до самого дна и без всякой циркуляции может изменить температуру придонного слоя.
Температурный скачок. Но циркуляция воды может вызвать и ещё одно очень важное явление, с которым нам необходимо познакомиться, — это так называемый температурный скачок. Так как нагревание и охлаждение воды распространяются от поверхности в глубину медленно, то столь же медленно и постепенно происходит и выравнивание температуры конвекционными токами и проникновение последних в глубину. Ветровая циркуляция, перемешивая слои, несомненно, способствует выравниванию их температуры и ускоряет его. Вызванная ветром циркуляция воды захватывает сначала верхний слой сравнительно небольшой толщины; в этом слое температура выравнивается довольно быстро, между тем как нижележащие слои продолжают сохранять прежнюю температуру; тогда на нижней границе указанного верхнего слоя окажется более или менее резкий переход температуры сверху вниз; это явление и носит название температурного скачка. Таким образом, вводной толще озера появляется такой слой, в котором температура обнаруживает резкое изменение, в то время как выше и ниже этого слоя она изменяется очень постепенно. Слой воды, лежащий выше скачка, называется эпилимнионом, слой самого скачка — металимнионом, а слой, расположенный под скачком, — гиполимнионом. Раз появившись, — а это бывает весной по вскрытии льда, когда поверхность воды становится доступной как для нагревания солнцем, так и для деятельности ветра, — температурный скачок уже не исчезает, а остаётся на целое лето; в течение лета он постепенно спускается от поверхности ко дну, причём нередко ширина его слоя постепенно уменьшается, и, наконец, осенью он совершенно исчезает. Очевидно, что его существование тесно связано с тепловой слоистостью; поэтому он появляется и исчезает вместе с последней, т. е. имеет место летом и зимой и отсутствует весной и осенью.
Всё это можно проследить на рисунке 14, представляющем ход изменений температурного скачка на озере Глубоком в течение лета 1907 г. В мае скачок находится почти у самой поверхности (меньше 3 арш. глубины), в июле он несколько глубже (3—6 арш.), в августе — на глубине 7—8 арш., в сентябре спускается уже до 12—14 арш.: так же постепенно идёт на протяжении лета убывание ширины слоя скачка, пока он не исчезает окончательно (в октябре).
ял
Рис. 14. Температурный скачок в озере Глубоком в 1907 г. (горизонтальный ряд цифр обозначает глубины в аршинах, вертикальный — температуры в градусах Ц).
Замерзание озёр. Мы уже знаем, что перед замерзанием озера в нём устанавливается обратная тепловая слоистость, при которой самый тёплый слой воды находится на дне, а самый холодный на поверхности. При непрерывно продолжающемся охлаждении поверхностного слоя его температура спускается, наконец, до 0°; при этой температуре вода, как известно, превращается в лёд. Если озеро спокойно, то первая ледяная плёнка может покрыть всю его поверхность в течение одной ночи. В следующую ночь эта плёнка, если только не успела растаять за день, с нижней стороны увеличивается в толщину; при достаточно низкой температуре двух первых ночей обычно можно считать, что лёд стал уже окончательно. Дальнейший процесс замерзания заключается в постепенном утолщении ледяной коры, которая всё время нарастает с нижней поверхности; сначала это нарастание » идёт более или менее быстро, но затем мало-помалу замедляется и, наконец, совсем прекращается; это объясняется плохой теплопроводностью льда: хотя наружная поверхностьледяногопокрова непрерывно охлаждается, но чем толще он становится, тем медленнее проникает через него это охлаждение в глубину, и, наконец, наступает момент, когда оно уже* не может дойти до находящейся подо льдом воды, замерзание которой поэтому прекращается. Таким образом, водная масса озера не промерзает вся до дна, а остаётся в течение зимы в жидком виде и только сверху покрывается более или менее тол стымледянымпокровом; верхний, непосредственнопримыкающий снизу ко льду слой воды имеет температуру лишь немного выше 0°, но с увеличением глубины температура возрастает, пока, наконец, в придонном слое не достигнет 4° или немного ниже.
Всякий крупный водоём — озеро, пруд, река — замерзает только сверху. Когда весной начинается ледоход, то расколовшийся на отдельные глыбы лёд не тонет, а плавает по воде. Очевидно, он легче воды; другими словами, плотность его меньше, чем у воды. Чем это объясняется? Вода, как известно, при замерзании кристаллизуется, т. е. из неё образуются отдельные ледяные иголочки (ледяные кристаллы); эти иголочки смерзаются между собой в разных положениях, причём между ними остаются мельчайшие невидимые промежутки; таким образом, лёд пмеет не сплошное, а рыхлое строение, и если взять одинаковые по весу количества воды и льда, то последний займёт больший объём, чем первая; следовательно, вес одинаковых объёмов воды и льда будет тоже различный: лёд оказывается легче воды при той же температуре, т. е. при 0°.
Влияние тепловых свойств воды наорганизиы.Температураокружающей среды является одним из важнейших условий в жизни животных и растений. Каждый организм может жить лишь в пределах некоторых вполне опреде-
*
лённых температур минимальной и максимальной. Если температура перейдёт за эти границы, организм гибнет, если она подходит к ним близко — развивается плохо; в пределах этих границ есть для каждого организма такая температура, которая является для его процветания наиболее подходящей, — это так называемая оптимальная температура.
Пределы температур, при которых жизнь ещё возможна, различны для разных организмов; поэтому каждому климату свойственна своя характерная картина жизни. Как уже говорилось выше, температура воды значительно менее резко отвечает на колебания количества солнечного тепла, чем температура суши, — ясно, что и состав водных растений и животных не так зависит от климата, как состав наземных.
Для озёрных организмов нашего климата верхний предел возможной для жизни температуры1 не имеет особого значения, так как вода в природе никогда не прогревается выше этого
Рис. 15. Прорастание турионов:
А — телореза — Stratlotes aloldes; Б — урути — Myriophyllum; В — пузырчатки — Utrtcularta; Г — рдеста — Potamogeton.
стительной, а также и для животной — имеют низкие температуры. В этом отношении чрезвычайно важную и благотворную роль играет то обстоятельство, что озеро не промерзает до самого дна, а покрывается только сверху ледяной корой; вода подо льдом, сохраняющая благодаря его защите своё жидкое состояние, сохраняет вместе с тем для обитающих в ней живых существ и более или менее благоприятные условия жизни. Тем не менее, её охлаждение в нашем климате довольно часто достигает тех температур, которые для жизни некоторых водных организмов являются предельными, т. е. ниже которых они уже погибают. Долгие и холодные зимы ставят очень и очень многим из них предел для обычного хода их жизненных процессов.
Обратимся прежде к растениям. Надводные их части оказываются в таком же положении, как и сухопутные растения, т. е. в температуре гораздо более низкой, чем температура воды, и жизнедеятельность их должна прекращаться одновременно с гибелью травянистой наземной растительности. Погружённые растения — в случае, если они находятся на такой глубине, что им не грозит гибель от вмерзания в лед, — находятся в лучших температурных условиях, потому что, как уже указывалось, водная масса отстаёт от непосредственно над ней лежащего воздуха в быстроте охлаждения. Но тут жизни растения угрожает другая опасность. Снежный покров не даёт воз-
Известны водоросли, живущие в горячих источниках при температуре свыше
можности проникнуть в лежащую под ним водную массу солнечным лучам, и растения, за некоторыми исключениями, лишаются возможности питания, связанного для них с солнечным светом. Жизнь замирает; она хранится или в корнях и корневищах, или в опустившихся на дно частях стеблей, у различных растений различно устроенных, но большей частью состоящих из укороченного побега с плотно прилегающими листьями. Так, некоторые водные растения, как, например, рдесты, уруть, пузырчатка и другие, осенью образуют особые укороченные побеги с тесно сидящими листьями, так называемые турионы (рис. 15), — это зимующие почки, снабжённые большим количеством запасных питательных веществ. Перед наступлением зимы само растение погибает, а турионы падают на дно, перезимовывают и весной дают новые побеги.
Наряду с цельными растениями хранятся зимой под защитой плотных оболочек семена и споры. Вспомним, что размножение семенами у водных растений вообще отступает на второй план.
Из животных, связанных с водой, лишь очень немногие проводят зиму вне водоёма, например, зарывающиеся иногда на зимовку в мох или поры
тритоны или в сухие листья водомерки, а также комары, ищущие убежища от холода в людском жилье или в дуплах деревьев. Большинство из тех, которые вообще покидают в какой-либо момент своей жизни воду, делают это в тёплое время года и перед своей гибелью отдают вновь под защиту воды своё потомство или яйца; так делают стрекозы и вообще очень многие из насекомых. Животные, вся жизнь которых протекает в воде, в весьма различной степени выносливы к понижению температуры. Немногие гибнут, оставляя или яйца, или, подобно некото
Р и с. 16. Сёдлышко (ephippium) дафнии.
рым растениям, отделившиеся части своего
тела (зимующие почки), так или иначе приспособленные к перенесению холода (рис. 16). Большинство проводят зиму в подлёдных слоях воды или в илу; одни вполне деятельны, разве лишь немного менее подвижны, чем летом, как, например, ползающие по нижней поверхности льда улитки, роющиеся в илу олигохеты и личинки Tendipes—«мотыль», некоторые виды план-ктических животных (из коловраток и ракообразных), среди которых встречаются даже исключительно зимние виды, и пр.; другие уходят в ил, затихают и впадают в оцепенение (например, некоторые личинки насекомых, пиявки).
Само собой понятно, что активная жизнь под водой возможна лишь в тех местах водоёма, которые достаточно глубоки для того, чтобы промерзание не достигло дна. Но там, где остаётся небольшой слой подлёдной воды, её температура ниже, чем в придонных слоях более глубоких мест. Естественно, что с наступлением холода животные покидают прибрежную мелкую зону и стягиваются в более глубокие места, чтобы весной снова двинуться в обратный путь в поисках благоприятных условий, предоставляемых им летом растительной полосой.
Однако надо заметить, что даже многие вмёрзшие в лёд животные не погибают окончательно: у них прекращаются лишь активные проявления жизни, но в скрытом виде последняя сохраняется и при оттаивании вновь переходит в активную форму. Таких животных, перезимовывающих во льду, немало известно среди простейших, коловраток, червей, моллюсков, ракообразных и личинок насекомых.
В связи с сезонной сменой температуры стоит ещё одно интересное биологическое явление — это сезонные изменен ияв жизни планктона: от лета к зиме, и наоборот, меняется количество планктона и его видовой состав, а у круглогодичных видов — форма и строение тела. Подробнее об этом будет сказано дальше.
ТЕМЫ ДЛЯ РАБОТ
Исследователю, пришедшему на озеро с одним только термометром в руках, может найтись очень интересная и важная для понимания жизненных явлений в водоёме работа. В некоторых отношениях можно ограничиться однократным измерением температуры, но в большинстве случаев важно производить систематические измерения её на протяжении более или менее длительного периода времени.
При возможности лишь однократного исследования следует установить температурную слоистость. Как самостоятельное задание при этих же условиях может быть выполнено сравнительное исследование температур в различных участках озера.
При возможности работы на водоёме более долгое время или при условии возвращения на него вновь и вновь, очень интересно заняться изучением колебания температуры во времени. При ещё более длительных наблюдениях можно установить годовые колебания или хотя бы колебания на протяжении одного сезона — от момента вскрытия водоёма до момента замерзания.
1. Непериодические измерения температуры
1. Измерить температуру одновременно в пелагической и в прибрежной области на одной и той же глубине.
2. Измерить температуру по возможности одновременно на поверхности и у дна в нескольких пунктах прибрежной зоны — в зарослях разного типа.
3. Установить на самом глубоком месте или по крайней мере в центре водоёма наличность или отсутствие тепловой слоистости и температурного скачка (стр. 32, 34).
П. Периодические измерения температуры
1. Установить наличность суточных колебаний температуры в пелагической области: измерять температуру на поверхности и на какой-либо определённой глубине (но каждый раз на одной и той же) в 7 часов утра, в 1 час дня и в 7 часов вечера в течение нескольких дней, например, недели.
2. Сделать то же самое в различных пунктах прибрежной области в связи с присутствием и отсутствием растительности, с зарослями разного типа и т. п.
3. Установить ход сезонных изменении температуры: измерить температуру в одном или, если можно, в нескольких пунктах водоёма (в таком случае обязательно в пелагической области и в литорали) на поверхности и на какой-либо определённой глубине (одной и той же для всех пунктов) в конце февраля или в начале марта (перед вскрытием водоёма, в проруби), в мае, в августе и в ноябре. Пункты для измерения температуры зафиксировать па водоёме посредством воткнутых в дно кольев или шестов, или же буйков.
МЕТОДИКА
Инструментарий. Термометр. Если есть возможность, то лучше употреблять термометр в металлической оправе, на шкале которого нанесены не только целые градусы, но и их десятые доли или по крайней мере полуградусы; и только в случае полной невозможности иметь такой термометр можно ограничиться обыкновенным водным термометром в деревянной оправе, употребляемым для ванн. К нижнему концу термометра нужно привязать небольшую стеклянную или, в крайнем случае, жестяную банку (например, из-под консервов) так, чтобы оправа термометра упиралась в дно банки; снизу к этому дну (в случае деревянной оправы) прикрепляется груз, тяжесть которого должна погружать термометр в воду; чтобы оправа не стукалась о стенки баночки, между ними можно вставить в трёх местах кусочки пробки. Назначение этой баночки состоит в следующем: если опустить термометр, не снабжённый банкой, на какую-нибудь глубину, а затем через некоторое время его вытащить, то пока при вытаскивании он будет проходить через верхние, более тёплые слои воды и потом через воздух, он успеет хотя бы отчасти воспринять температуру этих слоёв, и показание его, следовательно, изменится; банка же предохраняет от такого изменения, потому что в ней будет находиться вода, взятая с глубины и приблизительно сохраняющая свою температуру, которую, конечно, сохранит и погружённый в эту воду термометр >. К верхнему концу термометра привязывают линь, т. е. верёвку, размеченную на метры и полуметры (подробнее об этом см главу об измерении глубины); размечать линь нужно начиная от ртутного шарика, т. е. принимать в расчёт длину самого термометра.
Измерение. Рекомендуется всегда делать два параллельных измерения температуры — сначала в воздухе, затем в воде — и производить их в затенённом месте, избегая действия на ртутпый шарик прямых солнечных лучей. В воде следует делать по крайней мере тоже два измерения: под самой поверхностью и около дна. При поверхностном измерении вводу погружается лишь нижний конец термометра (с ртутным шариком),
1 Конечно, такой способ измерения температуры груб и не точен и может дать результат с точностью лишь не более целого градуса. Более же точное измерение требует применения более совершенного и точного термометра. г 7
90
шкалу же держат над водной поверхностью, следя по пей до того момепта, когда температура перестанет изменяться и конец ртутного столбика остановится па одной точке, тогда записывают указываемую термометром температуру. Для измерения температуры у дна термометр опускают па нужную глубину и держат там минуты 3, чтобы банка с водой и находящийся в ней ртутный шарик успели воспринять господствующую на этой глубине температуру; затем быстро вытаскивают термометр и быстро же смотрят по шкале его показания, причём, если шкала с десятыми долями градуса, то смотрят сначала их, а потом уже целое число градусов.
Если исследование ведётся на озере, то хотя бы в одном пункте его, именно — в пункте наибольшей глубины, если она известна, или по крайпей мере в середине озера, желательно произвести более детальное измерение температуры по всей вертикали — от поверхности до дна, например, через каждые полметра: сначала измеряют под самой поверхностью, потом на глубине 1/2, затем 1, Р/г» 2 м и т. д. до конца; при этом необходимо соблюдать указанные выше два правила: выдерживание термометра на каждой глубине в течение достаточного времени и быстрое вытаскивание его и констатирование температуры. Такое детальное измерение нужно для того, чтобы узнать, есть ли температурная слоистость и температурный скачок.
ОПТИЧЕСКИЕ СВОЙСТВА ВОДЫ
Сюда относятся два явления, играющие большую роль в жизни озера, — это прозрачность воды и её цвет, окраска.
Прозрачность. Даже самая чистая, т. е. лишённая всяких примесей, вода не абсолютно прозрачна и в слое достаточно большой толщины уже не пропускает через себя свет, что зависит от способности воды поглощать световые лучи. Но в озёрах вода никогда не бывает совершенно чистой, она всегда содержит то или иное количество взвешенного материала — или неживого в виде детрита, или живого в виде планктона; этот материал, поглощая и отражая свет, сильно задерживает его, так что в более глубокие слои воды его проходит уже значительно меньше.
Из двух составных частей взвешенного материала — планктона и детрита — первого больше всего бывает в наших озёрах летом (с июня по август), второго — во время осеннего и в особенности весеннего паводков; наименьшее количество того и другого бывает зимой, подо льдом. Особенно большие скопления взвешенных частиц в большинстве наших озёр бывают летом в виде «цветения», вызываемого колоссальным развитием тех или иных представителей растительного планктона (фитопланктона). Эта сезонность в накоплении взвешенного материала в воде обусловливает и соответствующую сезонность в прозрачности, которая имеет минимальное значение обычно летом, а максимальное — зимой. Так, например, в одном из подмосковных косинских озёр — в озере Белом — наименьшая прозрачность 0,6 м констатирована в июле, а наибольшая 3,3 м — в декабре; в Переславском озере минимум прозрачности 2,(Ы наблюдался в августе,максимум 8,75л< — в марте.
Прозрачность воды в водоёмах измеряется погружением в неё белого диска; та глубина, на которой последний исчезает из вида, и считается мерилом прозрачности \
Наибольшее из полученных до сих пор таким способом значений для прозрачности пресной воды выражается числом 40 м (в оз. Байкал) — это довольно значительная цифра, приближающаяся к числам, характеризующим прозрачность воды некоторых океанов (Индийского 50м, Тихого 59 .и). В большинстве озёр прозрачность много меньше; в некоторых — там, где летом развивается обильное цветение, — она может доходить лишь до нескольких десятков сантиметров.
В связи с той громадной ролью, какую свет играет в жизни организмов, и прозрачность воды, конечно, приобретает столь же огромное значение для обитающих в ней растений и животных. Особенно это относится к растениям,
1 Подробнее об этом см. в «Методике». Этот способ — довольно грубый и приближённый. Гораздо более точен фотографический метод: погружают в воду кассету, которая посредством особых приспособлений может быть па любой глубине открыта и вновь закрыта; та глубина, на которой фотографическая пластинка перестаёт чернеть, и означает предел проникания света в воду.
которым свет необходим для питания. Ясно, что степень проникания света в воду должна в той или иной мере отражаться на распределении водной флоры. Это влияние прежде всего сказывается в том, что высшая растительность озера ограничена в своём распространении лишь прибрежнойобластью, в которой вследствие её небольшой глубины она находит для себя наиболее благоприятные условия освещения. Точно так же и низшая флора в лице водорослей и растительного планктона (фитопланктона) в своём вертикальном распределении в значительной степени зависит от прозрачности воды; так как последняя с глубиной быстро убывает, то и наибольшая масса растительных планктических организмов обычно сосредоточена в верхних слоях воды; это, однако, не мешает некоторым видам, довольствующимся минимальным количеством света, существовать на более или менее значительных глубинах, где они находят себе в других отношениях наилучшие условия жизни. Кроме того, нужно иметь ещё в виду, что вода не одинаково поглощает различные цвета спектра, вследствие чего лучи разной преломляемости проникают на разную глубину. Преломляемость возрастает по направлению от красной части спектра к фиолетовой, поэтому прежде всего погасают красные и жёлтые лучи, следовательно, самые яркие, наибольшей же глубины достигают лучи наименьшей яркости — голубые и синие. Это обстоятельство имеет значение для вертикального распространения отдельных видов, входящих в состав планктона.
Зимой, как мы уже сказали, в большинстве озёр прозрачность больше, чем летом, благодаря гораздо меньшему количеству планктона и детрита. Но зато роль последних, в смысле поглощения света, играет снежный покров озера. Вследствие этого подо льдом господствует тоже более или менее густой мрак, но не абсолютная темнота, не полное отсутствие света, которого остаётся ещё достаточно, чтобы обеспечить существование некоторых форм из фитопланктона.
Цвет воды в озере есть явление сложное и зависит от разнообразных причин. Цвет совершенно чистой воды, так же как и чистого воздуха, голубой. Но присутствие в озёрной воде растворённых органических веществ — главным образом гумусовых кислот—изменяет эту окраску, причём с возрастанием их количества цвет воды меняется в таком порядке: голубой, голубоватозелёный, жёлтозелёный, тёмножёлтый, коричневый; первая окраска свойственна некоторым альпийским озёрам, последняя — гумусовым озёрам, большинство же озёр по цвету своей воды располагаются в том или ином месте указанного ряда. Цвет воды влияет и на её прозрачность, которая уменьшается по направлению от голубого к коричневому цвету.
Для определения цветности озёрной воды существует прибор — так называемая шкала Ф oj) е л я-У л е. Эта шкала состоит из целого ряда (21) переходных между голубым и коричневым тонов, причём каждый тон отмечен своим номером. Сравнивая с ними пробу воды из данного озера, мы находим наиболее близко к ней подходящий тон и обозначаем её соответствующим номером.
На окраску воды оказывает влияние ещё присутствие в ней взвешенных частиц. Поэтому планктон при сильном его развитии способен в значительной степени затемнять основной собственный цвет воды, сообщая ей свою специфическую окраску. Особенно это проявляется во время массового размножения каких-нибудь видов растительного планктона, т. е. во время «цветения» воды; так, в тех озёрах, в которых весной и осенью диатомовые водоросли достигают своего максимума, вода принимает в эти периоды желтоватый оттенок, летом же цвет воды обусловливается максимальным развитием других представителей фитопланктона: например, Ceratium (рис. 32, Б) сообщает ей такую же желтоватую окраску, как и диатомовые, а цветение синезелёными водорослями окрашивает её в более или менее интенсивный зелёный цвет.
Таким образом, водные организмы в лице некоторых планктических водорослей оказывают сильное влияние на цвет воды. Но нельзя того же сказать об обратном влиянии. Сама по себе собственная окраска воды, обусловленная *40
физико-химическими факторами, невидимому, не играет роли в жизни обитателей озера, она имеет лишь косвенное значение, именно постольку, поскольку она связана, как уже было указано, с другими оптическими свойствами воды — с её прозрачностью, о влиянии которой говорилось уже выше.
ТЕМЫ ДЛЯ РАБОТ
По отношению к исследованию оптических свойств, или, точнее, прозрачности, так как лишь это исследование возможно без особо сложных приспособлений, можно повторить почти то, что было сказано по отношению к исследованию температуры; наибольший интерес представляют сравнительные исследования на протяжении оолее или менее длительного периода времени. Изменения, которые могут быть подмечены по сезонам, указывались выше.
При однократном экскурсионном обследовании необходимо произвести ряд сравнительных определений прозрачности в тех пунктах, где можно ждать резких отличий, в местах впадения притоков и вдали от пих, в различных участках берега и в открытой части озера, вблизи и вдали от мест загрязнения скотом и т. п.
*
I. Непериодические измерения прозрачности
1. Определить прозрачность в пелагической области и в нескольких пунктах прибрежной, сравнить полученные данные и постараться подметить причины различий, если таковые обнаружатся (планктон и детрит, механическое взмучивание, вызванное ветром на мелких местах, вхождением в воду скота на водопое и т. д.).
2. Сравнить прозрачность воды перед бурей и после неё.
II. Периодические определения прозрачности
Установить сезонные изменения прозрачности: измерить последнюю в феврале (в проруби), в апреле, в июле и в октябре.
МЕТОДИКА
Инструментарий. Диск Секки. Это цинковая круглая пластинка величиной с крупную тарелку, хорошо прокрашенная белилами в чистый белый цвет и подвешенная на лине. Её можно с успехом заменить белой эмалированной или лучше фаянсовой тарелкой, вделанной в проволочный каркас (проволока должна быть достаточно прочной); к этому каркасу в трёх точках его окружности прикрепляются короткие шнуры, пирамидой сходящиеся над центром тарелки, которые в точке их схождения связываются как между собой, так и с концом линя; размечать линь на метры и полуметры начинают от самой поверхности диска.
Производство работ. Определение прозрачности воды нужно производить при тихой погоде, так как волнение мешает видеть диск с достаточной ясностью. С затенённой стороны лодки опускаем диск в воду и погружаем его до тех пор, пока он не исчезнет из вида; замечаем глубину, соответствующую этому исчезновению. Затем, продержав диск 1—2 минуты ниже предела видимости (чтобы дать утомлённому видом диска глазу отдохнуть), начинаем его вновь медленно поднимать и замечаем глубину в тот момент, когда глаз опять увидит его. Из двух наблюдённых глубин берут арифметическую среднюю, которую и записывают как показатель прозрачности. Иногда прозрачность выражают числом, вдвое большим наблюдённой глубины: если, например, диск исчезает из вида на глубине Р/2 м. то показателем прозрачности считают 3 м. Делается это на том основании, что световой луч проходит расстояние, равное двойной глубине исчезновения диска из вида: от поверхности воды до диска и, отразившись от него, обратно—от диска до поверхности, после чего и попадает в наш глаз; это значит, что лучу приходится преодолеть, так сказать, световое сопротивление воды на двойном против наблюдённой глубины расстоянии. Если бы на пути луча не было диска, то он свой второй, т. е. обратный путь (от диска в глаз наблюдателя), прошёл бы в глубину и потух бы на глубине вдвое большей, чем наблюдённая нами. Если бы мы поместили свой глаз у самого диска в воде (или вместо диска взяли бы какой-либо яркий источник света, например, электрическую лампочку), то заметили бы его исчезновение именно па этой двойной глубине. Поэтому глубина исчезновения диска из вида может быть названа скорей границей видимости; пределом же собственно прозрачности воды является двойная глубина видимости.
ХИМИЧЕСКИЕ СВОЙСТВА ВОДЫ
Одним из самых важных свойств воды является её способность растворять в себе многие другие вещества. С явлением растворения в воде твёрдых веществ всякий знаком хотя бы на примерах соли и сахара. Но эта способность не ограничивается лишь твёрдыми телами, а распространяется также
на жидкие и газообразные. Если мы нальём в один сосуд две жидкости, которые, как говорят, смешиваются между собой, например, воду и спирт, то всякая граница между ними в конце концов исчезает, и в сосуде оказывается столь же однородная жидкость, как и каждая из двух смешиваемых в отдельности; с другой стороны, если мы нальём в сосуд две несмешиваю-щпеся жидкости, как вода и масло, то между ними всегда можно заметить более или менее резкую границу, причём жидкости располагаются в зависимости от своей плотности и удельного веса — более лёгкая (масло) наверху, более тяжёлая (вода) внизу. О первой паре жидкостей мы говорим, что они растворяют друг друга, причём каждая из них может быть как растворителем, так и растворимым веществом, смотря по количеству той и другой. Накопец, многие жидкости способны растворять в той или иной степени и некоторые газы. Само собой разумеется, что растворимость различных веществ в одной и той же жидкости при одинаковых условиях будет различна; она измеряется количеством объёмных единиц вещества, способных растворяться при данных условиях в 100 таких же единицах жидкости. Когда какое-нибудь вещество растворяется в жидкости, то составляющие его частицы, или молекулы, разъединяются и размещаются в промежутках между частицами растворителя, раздвигая их и тоже отделяя друг от друга. Мы уже знаем, что повышение температуры способствует взаимному разъединению молекул вещества и, преодолевая их сцепление, может в конце концов обратить со- < стояние этого вещества из твёрдого в жидкое, а из жидкого в газообразное— в пар; понижение температуры действует в противоположном направлении — оно содействует большему сцеплению частиц между собой, результатом чего является, наконец, превращение пара в жидкость, а последней — в твёрдое тело. Из этого ясно, как должна влиять температура на процесс растворения: так как при растворении какого-либо вещества в жидкости оно само переходит в жидкое состояние, то очевидно, что растворению твёрдого тела должно способствовать повышение температуры, а растворению газа — понижение её, и, наоборот, повышение температуры должно содействовать выделению из жидкости растворённого в ней газа, а понижение — выделению растворённого в ней твёрдого тела. Так это и бывает в действительности: всякому известно, что в горячей воде сахар растворяется значительно быстрее, чеАм в холодной, а с другой стороны, если принести холодную воду в тёплое помещение, то через некоторое время, когда она согреется, из неё начинают выделяться пузырьки воздуха — это выходит тот излишек воздуха, который в холодной воде был растворён, а в тёплой уже не может раствориться.
На процесс растворения оказывает влияние, кроме температуры, ещё другой фактор — давление. Физика учит нас, что газы, в отличие от несжимаемых твёрдых тел и почти несжимаемых жидких, в сильной степени поддаются действию внешнего давления, причём их сжатие, т. е. уменьшение объёма, при одной и той же температуре прямо пропорционально этому давлению. Поэтому — как на процесс превращения тела из твёрдого состояния в жидкое или обратно, так и на процесс растворения твёрдого тела в жидкости или, наоборот, выделения из последней уже растворённого тела — изменение давления не оказывает никакого влияния. Но совсем другое дело по отношению к газам: всякое повышение давления способствует, с одной стороны, сгущению пара или газа в жидкое состояние, с другой — растворимости газа в жидкости, а всякое понижение давления действует на оба эти процесса в обратном направлении. Если по обе стороны поверхности жидкости, т. е. внутри последней и вне её, давление газа будет неодинаково, то он направится в ту сторону, где давление меньше, пока оба давления не сравняются и не наступит состояние равновесия. Этим отчасти может быть объяснено то, что налитая из водопровода в стакан вода выделяет на его стенках пузырьки воздуха, который был растворён в ней при большом давлении, господствующем в водопроводе, а при переходе к обыкновенному атмосферному давлению выходит из воды; на этом же основано приготовление различных шипучих вин и вод, насыщаемых при большом давлении углекислым газом. 42
Вода всех открытых водоёмов на земной поверхности всегда находится в соприкосновении с атмосферой, а воздух, как известно, представляет собой смесь трёх главнейших газов — азота, кислорода и углекислого газа — в определённой объёмной пропорции с примесью того или иного количества водяного пара и ничтожного количества так называемых «благородных» газов (гелий, аргон, криптон и др.). Поэтому чрезвычайно важно и необходимо знать, каким правилам подчиняется растворение газовой смеси.
Если над поверхностью жидкости одновременно находятся два газа, то каждый из них производит на неё своё самостоятельное и независимое от другого газа давление; другими словами, каждый газ давит на жидкость так, как если бы он один только присутствовал над жидкостью, другого же газа совсем нет. Такое отдельное для каждого газа давление называется, как известно, парциальным давлением. Таким образом, общее давление газовой смеси будет равно сумме парциальных давлений всех составляющих её газов.
Вода, соприкасающаяся с атмосферным воздухом, естественно, будет содержать в растворе все входящие в состав его газы, причём относительное количество этих растворённых газов должно регулироваться их различной растворимостью. Воздух, как известно, содержит по объёму, не считая водяных паров, приблизительно 79% азота, около 21% кислорода и не более 0,05% углекислого газа. Но при поглощении воздуха водой это отношение благодаря различной растворимости газов нарушается; лучше всего растворяется в воде углекислый газ, за ним — кислород и хуже всего — азот. Поэтому в воде, сравнительно с воздухом, процентное содержание азота уменьшается, а кислорода и особенно углекислого газа увеличивается, так что состав растворённого в воде воздуха оказывается следующий: 62% азота, 34% кислорода и 4% углекислого газа.
Газовый режим озера. Рассмотрим теперь с точки зрения указанных выше основных законов, которым подчиняются растворы газов в жидкости, приход и расход их в массе озёрной воды, т. е. газовый баланс озера. Этот вопрос имеет огромную важность потому, что кислород и углекислый газ именно в растворённом состоянии принимают чрезвычайно деятельное участие в обмене веществ водных организмов; лишь немногие из водных растений и животных пользуются, подобно наземным организмам, для питания (растений) и дыхания атмосферным воздухом. В этом отношении особенно большое значение приобретает тот отмеченный уже выше факт, что растворённый в воде воздух отличается по своему составу от атмосферного: по сравнению с последним в нём возрастает процентное содержание как раз тех газов, которые играют громадную роль в жизни водных организмов (кислород — для дыхания растений и особенно животных, углекислый газ — для питания растений).
Ареной более или менее интенсивного газообмена между атмосферой и водной массой озера является поверхность последнего; здесь именно происходит поглощение водой атмосферного воздуха, причём количество растворённого воздуха в поверхностном слое воды зависит от комбинации в данный момент температуры и атмосферного давления: при понижении первой и повышении последнего верхний слой воды обогащается газами, в обратном случае, напротив, беднеет ими; когда же и температура и давление одновременно или повышаются, или понижаются, т. е. действуют на процесс растворения воздуха во взаимно противоположных направлениях, то температурные влияния берут верх над барометрическими. Поверхностный слой воды поглощает газы из атмосферы обыкновенно до полного насыщения, при котором их парциальное давление в атмосфере и в воде уравнивается. По отношению к кислороду в озёрах часто наблюдается даже временное состояние пересыщения, характеризующееся тем, что количество растворённого газа оказывается при этом большим, иногда значительно большим, чем то, которое соответствует температуре и давлению в данный момент; причиной этого явления служит усиленное выделение кислорода скопляющимися в верх-
них слоях воды зелёными водорослями, а также высшими водными растениями, как результат питания тех и других. Но этому состоянию пересыщения в значительной мере препятствуют, во-первых, дыхательные процессы животных и бактерий, во-вторых, процессы разложения органического материала (например, отмершего планктона), которые, утилизируя кислород, уменьшают его содержание в воде.
Если на поверхности вода оказывается насыщенной, а иногда и пересыщенной газами, то какое же количество их растворено в самой водной толще озера и как оно там распределено? Главнейшим источником снабжения воды газами является атмосфера. Следовательно, глубокие слои воды могут получить газы только от насыщенного поверхностного слоя, от которого они могут распространяться в глубь двумя способами; или — при спокойном состоянии воды — посредством диффузии, идущей по направлению от большего давления газа к меньшему, или же путём конвекционных токов — температурных или ветровых; ясно, что последний способ является гораздо более верным и быстрым, чем первый. Рассмотрим вертикальное распределение в озёрной воде кислорода, который в этом отношении изучен лучше, чем другие газы.
Полная циркуляция воды в озере весной и осенью переносит богатые кислородом верхние слои воды в глубину и этим содействует кислородному обогащению нижних слоёв. Изменения, которые испытывает количество кислорода на глубине, зависят от фотосинтеза глубоководных растений, от дыхания живых организмов, но больше всего от процессов разложения трупов водных организмов и детрита, а следовательно, и от количества того и другого. В этом отношении различают разные категории озёр. Одну категорию составляют такие озёра, в которых как летом, так и зимой в металим-нионе (в слое температурного скачка) не происходит более или менее резкого падения содержания кислорода, т. е., иными словами, температурному скачку здесь не соответствует никакого кислородного скачка (рис. 17). В таких озёрах та часть водной массы, которая находится под температурным скачком (гиполимнион), содержит ещё довольно большое количество кислорода, именно — не менее 70% насыщения, и это количество сохраняется также и подо льдом. Озёра этого типа называются субальпийскими; типичными их представителями являются горные озёра, по большей части довольно глубокие, отличающиеся значительным богатством кислорода.
Озёра другой категории, называемые балтийскими, обнаруживают в смысле содержания и распределения кислорода совершенно иные отношения. Сюда относятся равнинные озёра средней полосы РСФСР, т. е. большинство наших русских озёр. При сравнительно небольшой в общем глубине они характеризуются обилием летнего планктона, чем и определяется типичное для них
содержание кислорода
Рис. 47.
Вертикальное распределение температуры и кислорода в процентах от состояния насыщения в ов. Сенека субальпийского типа (Сев. Америка) летом 1010 г. На кривой температуры — ясный скачок, на кривой кислорода — почти никакого.
распределение кислорода. Верхние слои воды, расположенные в эпилимнионе, т. е. над слоем температурного скачка, могут быть, как мы уже указывали ранее, не только насыщены, но даже пересыщены кислородом благодаря изобилию в планктоне зелёны х водорослей; это изобилие обусловлено большим запасом в верхних слоях углекислого газа и достаточным количеством света, так как тот и другой необходимы для питания водорослей. Пышный расцвет последних, или так называемое «цветение», создаёт благоприятные условия для
содержание кислорода Рис. 18.
Вертикальное распределение температуры и кислорода в процентах от состояния насыщения в Плёнском озере (Германия) балтийского типа летом 1916 г. Обе кривые обнаруживают параллельный скачок.
усиленного развития зоопланктона. Отмирая,
планктические организмы падают и уже во время этого падения начинают разлагаться. Когда они доходят до слоя температурного скачка, то
скорость их погружения уменьшается, и потому в этом слое они задерживаются дольше, чем в верхних слоях; происходит это оттого, что при быстром понижении температуры в металимнионе (в слое скачка) значительно возрастает вязкость, или внутреннее трение воды, которое и замедляет опускание планктических трупов. На процессы разложения, так же как и на процессы дыхания, потребляется кислород. В насыщенных или пересыщенных этим газом верхних слоях воды его хватает и на живой, и на мёртвый планктон, и здесь он потребляется в столь большом количестве, что до слоя температурного скачка его доходит лишь незначительная часть; кроме того, находящийся в эпилимнионе густой слой планктона, сильно уменьшая прозрачность воды, пропускает мало света в металимнион, а это, в свою очередь, препятствует интенсивному развитию зелёных водорослей, которые процессом своего питания могли бы восполнить недостаток кислорода в этом слое; между тем остающиеся здесь трупы планктических организмов продолжают разлагаться и, следовательно, потреблять кислород. Всё это ведёт к тому, что в слое температурного скачка количество кислорода быстро и более или менее резко падает; другими словами, в этих озёрах температурному скачку соответствует параллельный ему кислородный скачок (рис. 18 и 19). Естественно, что в гиполимнионе, т. е. в глубоких частях водной толщи, находящихся под слоем скачка, содержание кислорода оказывается совсем ничтожным, или даже он совершенно исчезает. Таков кислородный режим балтийских озёр в период летней, т. е. прямой тепловой слоистости (летней стагнации).
Каков же он бывает в озёрах в зимний период? В этом отношении различают в свою очередь две группы озёр. К первой относятся преимущественно более глубокие озёра (субальпийского типа), в которых установившийся после полной осенней циркуляции кислородный баланс со значительным содержанием этого газа даже на больших глубинах остаётся на всю зиму. Ко второй группе принадлежат, напротив, мелкие плоские озёра балтийского типа, в которых в начале зимы имеется по всей водной* толще ещё значительный запас кислорода, распределённый более или менее равномерно осенней полной циркуляцией. Но в течение зимы водное население продолжает потреблять кислород, а процессы разложения мёртвых организмов и химические реакции, сопровождающиеся потреблением кислорода, идут своим чередом, пополне
ния этого газа нет ни из атмосферы благодаря присутствию ледяного покрова, ни от зелёных водорослей, которые не могут хорошо развиваться вследствие господствующего подо льдом мрака. В результате этого — в особенности если озеро зимой имеет слабую проточность или совершенно лишено её — вода обнаруживает в конце зимнего периода, перед вскрытием льда (а иногда и раньше), чрезвычайно низкое содержание кислорода или даже полное его отсутствие. Этим объясняется тот факт, что в таких озёрах нередко имеет место так называемый «замор», т. е. гибель в конце зимы рыбы и многих других обитателей озера.
Наконец, существует ещё третья категория озёр, в которых летом наблюдается тоже кислородная слоистость, но здесь она обусловлена не разложением отмершего планктона, а другими процессами. Дело в том, что такие озёра бедны планктоном, но зато богаты детритом, принесённым в озеро извне;
вот разложением этого-то детрита, заменяющего в этом отношении планктон, и вызывается кислородная слоистость в данном типе озёр. Вода таких озёр богата растворёнными в ней органическими веществами, так называемыми гуминовыми кислотами, которые сообщают ей более или менее густую коричневую окраску. Озёра этой категории называются гумусовыми.
Что касается другого газа, очень важного для организмов, в особенности
для растительных, именно углекислого, то он в своих превращениях и количественных изменениях тесно связан с кислородом, причём в озёрах балтийского типа оба газа находятся в отношении обратной зависимости друг от друга: при питании зелёных растений количество углекислого газа уменьшается, количество кислорода увеличивается, при дыхании и разложении организмов — наоборот. Поэтому все характерные черты распределения
кислорода в озере сохраняют свою силу и для углекислого газа, только в обратном отношении, так что кривые распределения обоих этих газов представляют приблизительно зеркальные изображения друг друга (рис. 19). Таким образом, в воде, как и в атмосфере, постоянно происходит взаимный кругооборот кислорода и углекислого газа, причём две группы живых существ — растения и животные — дополняют одна другую и взаимно поддер-
Рис. 19.
Вертикальное распределение температуры (Т), кислорода (О) и углекислоты — свободной (С) и связанной (Св) — в оз. Отмско (Сев. Америка) балтийского типа летом 1910 г. Кривые кислорода и углекислоты взаимно симметричны.
живают существование ДРУГ друга.
На азоте мы останавливаться не будем, так как, с одной стороны, его количественные колебания в озере ещё недостаточно хорошо изучены, а с другой — этот газ в свободном состоянии не играет, собственно, никакой роли в жизни водных организмов и является для них совершенно индиферентным.
Итак, основным источником обогащения водной массы озера газами является атмосфера, из которой вода получает главным образом кислород, углекислый газ (углекислоту) п азот. Но, кроме атмосферы, есть и другие внешние источники снабжения
озёрной воды этими газами, именно притоки: реки, ручьи, ключи. Впадающая в озёра вода притоков с тем или иным количественным и качественным газовым составом изменяет газовый состав озёрной воды в более или менее значительной степени, в зависимости от количества приносимой притоками воды.
Однако газы могут поступать в озёрную воду не только извне, но и из внутренних по отношению к озеру источников. Так, выше уже указывалось, что кислород поступает в воду в процессе ассимиляции (питания) водных растении, а углекислота, или углекислый газ, — в процессе дыхания растений и животных, а также в процессах брожения и гниения. Кроме того, при гниении в условиях незначительного доступа кислорода, или, как говорят, в анаэробных условиях, которые нередко осуществляются на дне наших озёр, образуются газы: сероводород при разложении белков и м е-т а н, или болотный газ, при разложении растительной клетчатки. Материал для образования этих газов может приноситься также притоками. Таким образом, притоки могут способствовать, как улучшению, так и ухудшению газового режима озера, в зависимости от того, какой материал они несут в него.
Солевой режим. Озёрная вода всегда содержит в себе то или иное количество растворённых в ней солей. Обычно это количество колеблется от 0,01 г до 0,2 г на 1 л воды (в противоположность морской воде, которая содержит в среднем 35 г на 1 л воды, а в некоторых морях даже до 47 г). Эти соли приносятся в озеро извне притоками, ключами, ручьями, дождевыми потоками и т. п., вода которых, протекая по различным горным породам, растворяет многие из них в большем или меньшем количестве. Попав в озеро, они подвергаются здесь разным химическим превращениям, характер которых зависит от того, с какими веществами они здесь встречаются. Таким образом, солевой состав озёрной воды зависит от геологического строения окружающей местности и должен быть различен в разных озёрах. Но даже в одном и том же озере он не остаётся постоянным, а изменяется в зависимости от климатических условий, влияющих на водный баланс озера. Мы уже знаем, что приход воды совершается через притоки и атмосферные осадки, расход же — через стоки и испарение. Главнейшим источником прихода солей является питающая озеро вода (отчасти — процессы минерализации органических веществ в самом озере), а расхода — стоки (и процесс питания водных растений). Очевидно, что той или другой комбинацией этих процессов и будет определяться солевой состав озёрной воды во всякий данный момент. Так, во время сильных и продолжительных дождей озеро должно опресняться, так как, с одной стороны, дождь приносит чистую воду х, а притоки благодаря тем же дождям — опреснённую, с другой же стороны, из двух факторов расхода — испарение, способствующее осолонению воды, отсутствует, а сток воды уносит вместе с водой и соли; напротив, в жаркую и сухую погоду раствор солей в озёрной воде, вследствие сильного испарения, сгущается.
Далее, состав растворённых в озёрной воде солей меняется в зависимости от изменений её температуры, повышение которой усиливает растворимость солей, а понижение, наоборот, ослабляет. Большую роль в процессе растворения некоторых углекислых соединений (например, кальция, магния и др.) играет количество растворённого в воде углекислого газа, который способствует растворению этих соединений (подробнее об этом будет сказано дальше).
В самом же озере постоянно происходят химические реакции, при которых сложные вещества распадаются на более простые, а эти, в свою очередь соединяясь, дают другие сложные вещества. Особенно значительные изменения химический состав озёрной воды испытывает со стороны населяющего
1 Строго говоря, даже дождевая вода не является совершенно чистой, т. е. абсолютно свободной от растворённых в ней веществ, так как, проходи через атмосферу, она, хотя и в ничтожном количестве, поглощает из воздуха некоторые газы.
её органического мира, так как жизнь организмов поддерживается постоянным химическим обменом веществ между ними и окружающей их водой. Но, влияя на солевой и газовый состав воды, организмы и сами испытывают с её стороны определённые химические воздействия.
Живые организмы в процессе своей жизнедеятельности, с одной стороны, поглощают различные растворённые в воде вещества, с другой — выделяют в воду отработанные вещества. Так, растения в процессе питания поглощают некоторые соли и углекислый газ и выделяют кислород; при дыхании как растения, так и животные, напротив, поглощают кислород и выделяют углекислоту; животные при питании поглощают различные органические вещества и выделяют другие органические вещества и т. п. После смерти растительные и животные организмы подвергаются процессам разложения, при которых сложные органические соединения, входящие в состав тела живого организма, постепенно распадаются на всё более простые, а эти в процессе дальнейшего распада минерализуются, т. е. превращаются уже в неорганические соединения — в минеральные соли. Таким путём водные организмы возвращают воде те минеральные вещества, которые они берут из неё для построения своего тела. Всякий, вероятно, по личному опыту знает, что такое «жёсткая» и «мягкая» вода. Химическое исследование показывает, что жёсткая вода содержит в растворённом состоянии значительно больше солей, чем мягкая; на первом месте из них по количеству следует поставить известковые соли. Это они именно отлагаются на волосах в виде лёгкого белого налёта после мытья их жёсткой водой и сообщают им, в свою очередь, жёсткость; это они образуют на стенках внутри самовара или металлического чайника слой так называемой накипи, становящийся со временем всё толще и толще. Известь, или окись кальция СаО, в свободном состоянии в природе не встречается, так как чрезвычайно жадно соединяется с различными кислотами, образуя с ними соли. Наиболее частыми и обыкновенными из них являются углекислая и сернокислая соли, из которых первая пгра°т в жизни озера очень важную роль. Она представляет собой соединение извести с углекислым газом (СаО ф СО2 = СаСО3) и встречается в природе в виде целых пород — известняков (грубый известняк, мел, мрамор и др.). Чистая вода, хотя и в чрезвычайно слабой степени, всё же растворяет в себе эту соль; но гораздо лучше углекислая известь растворяется в воде, насыщенной углекислым газом, потому что в этом случае она переходит в более растворимую, так называемую двууглекислую известь; так, в воде, лишённой углекислоты, углекислой извести растворяется 13 мг на 1 л воды, а в воде, содержащей углекислый газ,— 64,6 мг\ а так как этот газ содержится во всех естественных водах, то очевидно, что вода, приносимая притоками в озеро, протекая через известковые породы, растворяет их в той или иной степени (в зависимости от количества в ней углекислого газа) и доставляет в озеро углеизвестковые соли. Естественно, что в местностях, богатых известняками, и озёра будут содержать много извести, вода в них будет жёсткая, в местностях же, где известковых пород мало, озёра должны иметь мягкую воду.
Значительные изменения в солевом режиме озера, происходящие, например, вследствие усиления (при интенсивном испарении) или ослабления (при большом притоке дождевых вод) общей концентрации солевого раствора или вследствие изменения качественного состава солей (при усиленном притоке извне каких-либо определённых солей), ведут к более или менее значительному изменению состава населения озера: некоторые группы организмов при этом совсем выпадают или уменьшаются в своём развитии, другие, напротив, развиваются за этот счёт гораздо сильнее. Так происходит смена одних видов водных организмов другими х.
1 Одним из важнейших факторов такой смены являются ещё, как мы видели, колебания температуры, которые действуют в этом направлении отчасти непосредственно, отчасти косвенно — через вызываемые ими изменения концентрации солевого раствора. 48
С этой точки зрения большое значение имеют некоторые законы, которые первоначально были установлены в агрономии и затем перенесены в область гидробиологии. В половине XIX в. был установлен так называемый закон минимума, по которому урожай определяется тем фактором, который находится в наименьшем количестве и может быть увеличен только путём усиления этого фактора, причём величина остальных факторов, если ни один из них не находится в минимуме, не имеет никакого значения. Но в недавнее время этот закон был исправлен и дополнен в том смысле, что наряду со значением фактора, находящегося в минимуме, выдвигается значение и других факторов, вследствие чего этот закон получил название закона совокупного действия факторов. Согласно этому закону, каждый фактор вносит в обеспечение урожая свою долю влияния соответственно его относительному значению.
Этот закон даёт возможность человеку воздействовать в желательном для него направлении на те или иные факторы или их комплексы, чтобы этим путём гарантировать максимум урожая. Поэтому от правильного и целесообразного применения его к озёрам можно ожидать очень плодотворных результатов в смысле обслуживания некоторых видов озёрного хозяйства, в первую очередь — рыбного.
После этих общих замечаний перейдём к рассмотрению важнейших для водных организмов солей, растворённых в озёрной воде, и их биологического значения.
Следует заметить, что чистая вода растворяет лишь хлористые и сернокислые соединения и щелочные карбонаты, а вода с углекислотой растворяет все карбонаты и разлагает силикаты.
В большинстве наших озёр соли кальция — и особенно углекислая соль — по своему количеству стоят на первом месте и значительно превышают в этом отношении все другие соли. Этому соответствует и та большая биологическая роль, какую кальций играет в жизнедеятельности водных организмов. Яснее всего эта роль обнаруживается у тех организмов, в теле которых происходит отложение в том или ином виде углекислой извести, например, образование раковин некоторых простейших животных и моллюсков, обызвествление скелета у рыб и т. п. Вот почему все такие организмы находят для себя благоприятные условия в озёрах с жёсткой водой и очень немногочисленны или даже совсем отсутствуют в озёрах с мягкой водой; особенно наглядно это можно видеть на примере моллюсков, которые в озёрах с пониженным содержанием углекислой извести или совсем отсутствуют, или, если имеются, обладают слабо развитой тонкой раковиной.
Таким образом, некоторые растения и животные для своих потребностей извлекают углекислую известь из воды. Но она может быть выделена из раствора и другими способами, например, уменьшением количества растворённого в воде углекислого газа: нам уже известно, что чем меньше вода содержит этого газа, тем меньше она может растворить в себе углекислой извести, вследствие чего излишек последней выпадает в виде нерастворённого, опускающегося на дно озера осадка, если только понизится содержание вводе углекислого газа. Очевидно, что распределение извести, растворённой в озёрной воде, зависит от распределения в ней углекислого газа и изменяется в том же направлении, как и последнее. Этим определяются годовые колебания в количестве углекислой извести, растворённой в различных слоях водной массы озера. Весной и осенью, благодаря перемешивающему действию конвекционных токов, эта соль распределяется более или менее равномерно. Летом же присутствие температурного скачка нарушает эту равномерность, причём в верхних слоях воды, именно — в эпилимнионе, вследствие поглощения углекислого газа планктическими водорослями для целей питания, выделяется твёрдый осадок углекислого кальция (это доказано на одном из швейцарских озёр, именно — на Цюрихском); опускаясь на дно, он испытывает в гиполимнионе, под слоем температурного скачка, различную участь в зависимости опять-таки от количества растворённого в воде углекислого
4 А. Н. Липин 49
газа: при недостатке последнего он осаждается на дно озера, при избытке — снова растворяется. Подобно фитопланктону и в том же направлении действует на раствор кальция в воде, конечно, и прибрежная высшая растительность: извлекая из воды в процессе ассимиляции углекислый газ, она вызывает осаждение известковых солей из воды, которые или осаждаются на дно, или оседают на поверхности самих растений, образуя на них известковые корочки, известные под названием инкрустаций. Такие инкрустации нередко встречаются, например, на водяной чуме — Elodea, телорезе — Stra-tiotes. рдесте — Potamogeton, а больше всего на лучице — Chara.
Такое явление выпадения кальция из раствора, благодаря ассимилирующей деятельности зелёных растений, получило название биогенной декальцинации (обескальцивания).
Другой биологически важной составной частью растворённых в озёрной воде твёрдых веществ является кремнезём, который служит для постройки скелетных образований в теле некоторых водных организмов, как, например, па мцыри диатомовых водорослей или скелетные иглы губок. Несмотря на незначительное содержание и малую растворимость в озёрной воде кремневой кислоты, количество последней оказывается всё же достаточным для развития многочисленных и разнообразных видов диатомовых водорослей. Это возможно благодаря известного рода биологическому круговороту, в который вступает кремнезём; панцыри отмерших диатомовых, находящихся ещё долгое время во взвешенном состоянии, могут — особенно в воде, богатой известью, — постепенно вновь раствориться и превратиться таким путём в строительный материал для панцырей будущих поколений тех же водорослей.
Соединения железа, растворённые в озёрной воде, играют важнейшую роль в жизнедеятельности и распределении водорослей. Количество их в разных слоях воды во время летней стагнации далеко не одинаково: например, в Переславском озере в 1923 г. в области эпилимниона и металимниона оно было не более 1,2 мг/л, между тем как вблизи дна оно доходило до И мгл. Растворимыми в воде являются не всякие соединения железа; в озёрной воде обычно растворена двууглекислая закись железа — Fe(HCO3)2, которая приносится водой притоков. Но эта соль непрочна, так как легко отдаёт углекислоту, выделяющуюся в воздух или поглощаемую зелёными растениями, и, присоединяя кислород, переходит в форму окиси железа — Fe(OH); последняя, благодаря своей нерастворимости, выпадает из раствора и оседает па дно, на камни, на растения и т. п. в виде хлопьевидного бурожёлтого налёта.
Так как предельным для жизни многих водорослей раствором железа в воде является раствор в 5—6 мг/л, а содержание его в озёрной воде может доходить до десятков миллиграммлитров, то ясно, что во многих случаях этот элемент может лимитировать и регулировать развитие водорослей; поэтому среди остальных факторов он является в этом отношении первенствующим. Различные роды и виды пресноводных водорослей могут быть расположены в определённый ряд в зависимости от количества растворённого железа, при котором они могут жить.
Однако есть организмы, приспособленные к жизни в воде с довольно сильной концентрацией раствора железа, это так называемые железобактерии, например, Crenothrix, Gallionella и др. Если в воде водопроводных труб имеется растворённое железо, то это создаёт благоприятные условия, с одной стороны, для осаждения большого количества окиси железа, с другой — для массового развития железобактерий; всё это нередко приводит к закупориванию водопроводных труб.
Наличие в питьевой воде только 1,5 мг железа на 1 л воды уже сообщает ей характерный чернильный вкус.
Из других минеральных веществ, играющих для органического мира ту или иную роль, можно отметить, папример, соединения серы, имеющие значение для развития и жизни соответствующих бактерий, как 60
Reggiatoa, или серобактерии. Сернистые соединения могут быть или принесены притоками, особенно если в озеро впадают серные источники, или фабричные стоки, или же могут образоваться в самом озере; так, летом в глубоких озёрах при длительном застое придонных частей водной массы последние сильно беднеют кислородом, а это создаёт благоприятные условия для развития процессов гнилостного разложения, вызываемых жизнедеятельностью серных бактерий; при этом образуется сероводород, благодаря гибельному действию которого на подавляющее большинство живых организмов глубины таких озёр становятся почти необитаемыми.
Наконец, необходимо остановиться ещё па органических веществах.
Откуда берутся в воде органические соединения? Уже по самому названию их нетрудно догадаться, что они происходят от организмов, независимо от того, приносятся ли они извне притоками или образуются в самом озере.
Тело растений и животных состоит из многих, чрезвычайно разнообразных веществ как минеральных (вода, различные соли, газы), так и органических. Присутствие последних именно и отличает в отношении химического состава организмы от тел неживой природы. Это не значит, что между минеральными (неорганическими) и органическими веществами лежит непроходимая пропасть: в настоящее время многие органические вещества получаются в лабораториях искусственно — путём соединения неорганических веществ.
Органические вещества представляют гораздо более сложные соединения, чем минеральные соли, и делятся на три основные группы — белки, жиры и у г л е в о д ы. Из них самой необходимой, неотъемлемой составной частью, имеющейся в теле всех населяющих землю растений и животных, являются белки, между тем как присутствие жиров и углеводов необязательно. Белки же имеют и наиболее сложный химический состав: в то время как жиры и углеводы состоят в основном из трёх элементов — углерода, водорода и кислорода—в различных количественных сочетаниях, в состав белков, даже наиболеепростых, входит, кроме этих элементов, ещё азот, а более сложные белки характеризуются обязательным присутствием фосфора, серы и железа или порознь, или в разных комбинациях. Таким образом, белки резко отличаются по химическому составу от жиров и углеводов, и притом — не только количественно (числом атомов входящих в их состав элементов), но — что главное — качественно (присутствием в них таких элементов, которых нет в жирах и углеводах).
Так как белки, являющиеся главнейшей составной частью организмов, характеризуются присутствием азота и во многих случаях фосфора, то этим самым значение этих двух элементов для всех организмов — в том числе и водных — выдвигается на первый план. Поэтому, чтобы лучше понять это значение, следует, хотя бы вкратце, остановиться на круговороте азота в водоёме (этот элемент лучше всего изучен в отношении цикла превращений его соединений в озёрах).
Когда какой-либо водный организм, растительный или животный, умирает, его тело начинает разлагаться. Этот процесс происходит благодаря деятельности разнообразных бактерий. При этом белки последовательно распадаются па целый ряд всё более и более простых органических соединений 1, которые, в свою очередь, распадаются на неорганические, или минеральные соли, пока, наконец, процесс разложения не дойдёт до образования аммиачных солей (аммиак представляет соединение одного атома азота с тремя атомами водорода — NH3; это — газ с очень крепким, характерным запахом, знакомым каждому по нашатырному спирту, который является раствором аммиака в воде). Но аммиачные соли сами по себе мало пригодны для использования их растениями в целях питания. Азот усваивается водными расте
1 Органические соединении, в противоположность неорганическим, или минеральным, характеризуются одновременным присутствием в них двух элементов: углерода С и водорода Н. К ним могут присоединяться также и другие элементы, всего чаще — кислород Оа.
*
ниями лучше всего в форме азотнокислых солей, или так называемых н и-тр а то в. Поэтому для лучшего усвоения азота растениями аммиачные соли должны быть переведены в нитраты. Этот перевод осуществляется не сразу, а последовательно: сначала аммиачные соли окисляются до азотистокислых солей (соли азотистой кислоты), так называемых нитритов, а последние затем окисляются в азотнокислые соли (соли азотной кислоты) — нитраты. Эти два процесса выполняются двумя специфическими родами бактерий: перевод аммиачных солей в нитриты производится нитритными бактериями Nitrosomonas, перевод нитритов в нитраты — нитратными бактериями Nitrobacter. II те и другие бактерии вместе называются нитрифицирующими бактериями.
Таким образом, растения не способны усваивать непосредственно свободный (газообразный) азот, они могут получать его лишь путём всасывания раствора солей азотной кислоты. Принятый таким путём азот идёт на дальнейшие процессы построения тела. Животные же получают его в растительной и животной пище. Так совершается кругооборот азота в водоёме: азот, находящийся в воде или на дне озера в виде минеральных солей, поступает в процессе ассимиляции растений в тело последних, здесь идёт на образование белков, а белки в виде растительной пищи поступают в тело животных; растения и животные при жизни выделяют из тела отработанные продукты, а после смерти разлагаются, причём продуктами распада белков является целый ряд последовательно расщепляющихся соединений, сначала органических, но в конце концов доходящих до полной минерализации, чем и заканчивается кругооборот, а затем при помощи нитрифицирующих бактерий начинается новый круг.
Есть, однако, некоторые роды грибков и бактерий, которые способны усваивать и свободный газообразный азот. С другой стороны, есть бактерии, которые не окисляют, а, напротив, восстанавливают азотные соединения. Они, следовательно, действуют в противоположном сравнительно с нитрифицирующими бактериями направлении, и потому их называют денитрифицирующими бактериями.
Итак, в озёрной воде всегда находится в растворе то или иное количество азота в трёх состояниях: в молекулярном (растворённый газообразный азот), в виде минеральных солей (нитритов и нитратов, причём вторых всегда больше, чем первых) и в виде органических солей.
Среди органических соединений, лишённых азота, выделяются, по своему значению для жизни озера, так называемые гуминовые вещества. Некоторые из них имеют характер кислот, что и дало повод назвать их г у-мусовыми, или гуминовыми кислотами. Эти кислоты, если они присутствуют в большом количестве, придают водоёму совершенно особый характер: они сообщают воде своеобразную жёлтобурую окраску и делают её мягкой. Это явление накладывает особый отпечаток на весь водоём: бедность минеральными и богатство растворёнными органическими веществами препятствуют сильному развитию флоры, а это влечет за собой и скудость фауны. Поэтому в гумусовых озёрах обычно мало развита органическая жизнь. Только зоопланктон (животный планктон) может в них развиваться хорошо в том случае, если имеется в воде много детрита, приносимого извне притоками.
Присутствие в озере в умеренном количестве органических веществ, имеющих питательное значение, ведёт к пышному расцвету тех или иных групп водных обитателей: подобное явление часто можно наблюдать даже в отдельных, особо благоприятных в этом отношении участках водоёма, например, вблизи пастбища, в местах водопоя скота и т. п. Но сильное за грязнепие водоёма органическими веществами, напротив, вызывает гибель большинства живых организмов вследствие того, что происходящие при этом усиленные процессы гниения в большом количестве потребляют кислород, необходимый для дыхания. При полном гниении сложные органические соединения распадаются на всё более и более простые, пока, наконец, дело не дойдёт до 62
образования конечных продуктов распада — различных минеральных веществ, которые таким путём возвращаются в воду и могут быть снова использованы живыми существами.
Так совершается в озере, биологический круговорот веществ: простые минеральные соединения идут на питание растительных организмов, которыми, в свою очередь, питаются животные, а те и другие, разлагаясь после смерти, вновь превращаются в исходные минеральные вещества. Такой круговорот даёт право назвать озеро в условном смысле микрокосмосом — малым миром, который, благодаря значительной самостоятельности совершающихся в нём явлений, имеет более или менее резко выраженный индивидуальный характер.
Использование организмами растворённых в воде веществ. (Обмен веществ между организмами и средой.) Растворённым в воде кислородом большинство пресноводных организмов как растительных, так и животных пользуется для дыхания. В тех случаях, когда тело снабжено нежными покровами, оно большей частью не имеет специальных приспособлений для дыхания— последнее совершается всей поверхностью тела. Ясно, что дыхание будет происходить тем полнее, чем больше поверхность тела: всякие выступы, бахромки, рассечение на мелкие доли и т. д., увеличивая поверхность, тем самым способствуют наилучшему использованию кислорода. Вспомним, что расчленённость листьев и одновременно нежность их покровов, позволяющая газам проникать внутрь растения, возможны только в воде и что, кроме дыхания, это важно для растения и в смысле сохранения его целости. В этих случаях мы можем говорить о типично водном дыхании; но многие растения, спустившись с суши в воду, рядом с водным сохранили воздушный тип дыхания. Таковы растения, которые всегда или по временам поднимают некоторые свои части над поверхностью воды.
Известно, что у наземных растений воздух проникает внутрь тканей через устьица, расположенные преимущественно на нижней стороне листьев; у водных растений на поверхностях, погружённых в воду, устьица совершенно исчезают, но они остаются и действуют как дыхательные и питательные органы на поднимающихся над водой частях. Так, у плавающих на воде листьев, например, кувшинки они располагаются только на верхней, обращённой в воздух стороне.
И среди животных наблюдаются оба типа дыхания: типично водный и связанный с атмосферным воздухом. В первом случае кислород из воды поступает через тонкие стенки тела или дыхательных органов (жабры) в жидкость организма, и обратным путём из жидкости тела в воду поступает углекислый газ. Для успешности процесса необходимо, чтобы около тела происходило непрерывное обновление воды, чтобы вода, использованная для дыхания, возможно быстрее отбрасывалась от поверхности. Это достигается различного рода движениями, то связанными, то не связанными с перемещением тела.
Движения в целях дыхания иногда совершаются всем телом: так, например, мелкий червь трубочник (Tubifex, рис. 20 и 34) передним концом зарывается в ил, задний же конец выставляет вертикально в воду и почти непрерывно машет им из стороны в сторону. Личинки комаров тендипедид, живущие на дне в трубках, склеенных из ила, почти безостановочно колеблют своё тело внутри трубки, вызывая постоянное прохождение через трубку струи свежей воды. Иногда движется не всё тело целиком, а лишь какой-нибудь его орган.
Во многих случаях органы, приводящие в движение окружающую воду, как, например, реснички коловратки (рис. 27, 28) или ножки ракообразного, являются одновременно и органами дыхания, хотя бы они выполняли и другую работу.
Специальными приспособлепиями для обмена газов между водой и жидкостями тела являются жабры. Жабры представляют собой особые выпячивания тела, большей частью защищённые от повреждений, имеющие чрезвычайно нежные покровы и находящиеся на таком месте, где постоянно
Р II с. 20.
В лепим верхнем углу под| есилась задним концом к поверхностной плёнке воды личинка мухи львинки — Stratiomyia; направо от н< ё вверху и внизу клоп гладыш — Notonecta с длинными раскинутыми задними ножками; в середине рисунка плывущий водпной ослик — Asellus; вверху и направо от него ползущая по стеблю гребенчатая улитка — Valvala cristata; под ней на листе личинка стрекозы красотки — Agrion (б. Calopteryx), схватывающая червя; направо от улитки ползущая по стеблю личинка подёнки с боковыми трахейными жабрами; крайний направо на стебле клоп водяной скорпион— Nepa; у основания растений колония червей олигохет — Tubifcx; под ними на камне ресничный червь — Planaria; налево от него ползущая пи дну личинка тритона.
Р и с. 21.
В левом верхнем углу самка жука плавунца — byiiscus, набирающая воздух в трахеи; в середине внизу самец того же жука, схвативший личинку веснянки—Perla; между ними плывущий водяной паук — Argyroneta, несущий на брюшке слой воздуха в свой подводный воздушный колокол (в левом нижнем углу); направо от самки плавунца ползущая по нижней стироне поверхностной плёнки воды улитка прудовик — Limnaea; другой прудовик ползёт по стеблю, на котором ниже прудовика его икра в виде колбаски слизи с заключёнными в ней яйцами; в правом верхнем углу жук водолюб — Hydrophilus
происходит смена воды, например, у рака — на органах движения, у рыб — по бокам головы, стенки которой в этих местах пронизаны жаберными отверстиями, через которые проходит ток воды.
У многих насекомых, перешедших к жизни под водой от наземного образа жизни, сохранилось в органах дыхания иное приспособление, напоминающее об их наземном происхождении. Тело наземных насекомых и их личинок пронизано сетью трубочек — трахей, наполняющихся воздухом из атмосферы; между воздухом трахей и кровью и происходит обмен газов.
Этот принцип обмена газами с атмосферным воздухом сохранился у многих водных насекомых и их личинок. У некоторых животных обмен происходит непосредственно с атмосферой, и, следовательно, им приходится подниматься к поверхности воды. Выше (стр. 26) уже был описан способ поднятия жука плавунца (рис. 21) и то, как для этого поднятия за свежим воздухом используется воздух, уже отработанный; через дыхальца, на ходящиеся на конце тела, жук забирает свежий воздух в свои трахеи в таком количестве, какое позволяет ему погружаться на более или менее долгий срок под воду. По использовании всего кислорода из забранного в трахеи воздуха жук снова должен подняться кверху. Также и прудовик (рис. 21) периодически подвешивается к поверхности воды и открывает вход в большую дыхательную полость,
в которой он держит запас воздуха на время погружения в воду.
В некоторых случаях животные с воздушным типом дыхания используют пузырьки газообразного кислорода, выделяемого подводными частями
растений, или воздуха, выделенного в воде при изменении температуры, что нередко бывает зимой подо льдом; тогда мы можем сказать, что животное дышит кислородом в газообразном состоянии, хотя оно и не поднимается до атмосферы; пример подобного дыхания даёт нам зимой плавунец. Эти жуки при полном отсутствии подо льдом газообразного воздуха переходят к использованию воздуха, растворённого в воде: выделяемый из тела, лишённый кислорода и насыщенный углекислым газом воздух собирается на конце брюшка в виде пузырька, одетого тонкой плёнкой, через которую и совершается обмен газов с окружающей водой (рис. 22).
Но другие организмы, а именно личинки многих насекомых (подёнок, стрекоз и др.), сохранив орган воздушного дыхания — трахеи, совсем утратили связь с атмосферным воздухом: они остаются всё время под водой, и
воздух их трахей черпает нужный кислород непосредственно из воды и сюда же выделяет углекислый газ. При этом необходимо, конечно, чтобы в каком-
нибудь участке тела трахеи находились в возможно более тесном соприкосновении с водой, чтобы стенки тела в этих местах были тонкими й нежными и чтобы вода здесь возможно часто сменялась. Это достигается различными путями. В большинстве случаев образуются выпячивания тела, построенные по тому же принципу, как и настоящие жабры (трахейные жабры), и также находящиеся в движении ради облегчения смены воды. Прекрасный пример трахейных жабер дают личинки подёнок (рис. 64), где трахейные жабры в виде листочков помещаются по бокам тела и почти непрерывно колеблются, словно крошечные веера. У личинок коромысла (стрекоза) обновление газового состава крови совершается в задней кишке, пронизанной по стенкам густым сплетением трахей: личинка втягивает через заднепроходное отверстие в заднюю кишку свежую воду, которая, соприкасаясь с тонкими стен-
ками трахей, отдаёт в трахеи кислород и принимает из них углекислоту и затем выталки-
Р и с. 22. Дыхание плавунца зимой.
вается обратно наружу. Если выталкивание совершается с большой силой, то тело личинки скачкообразно перемещается вперёд (реактивное движение).
Во всех только что описанных случаях дыхание происходит исключительно за счёт растворённого в воде кислорода, т. е. так же, как и у животных с типично водным (жаберным) дыханием, о которых говорилось выше: рыбы, червь-трубочник, личинки тендипедид и прочие. Но разница между обеими описанными группами животных сводится к тому, что у животных с типично водным способом дыхания этот способ является основным, первичным, между тем как у личинок насекомых, снабжённых трахейными жабрами, первичным является воздушное дыхание, водное же носит уже характер вторичного приспособления.
Посмотрим теперь, как растворённые в воде вещества используются водными организмами для питания.
Свойство воды растворять углекислый газ играет громадную роль в жизни водных организмов, особенно растений. Очень многие растения совсем не поднимаются над уровнем воды, и, следовательно, весь нужный им для питания и построения тела углекислый газ черпается непосредственно из воды. Так же непосредственно из воды черпают они и минеральные вещества. Корни погружённых растений выполняют двоякую функцию: с одной стороны, они являются органами укрепления растений на дне, с другой стороны, они, как это доказано в последнее время, функционируют как органы питания, всасывая из грунта необходимые растворы солей. Растворённые в воде газы поступают в ткани растения непосредственно через стенки тела. У тех растений, где некоторые части выдвигаются над поверхностью, устьица их забирают углекислый газ, как и кислород для дыхания, из атмосферного воздуха.
Что касается питания минеральными солями у растений с плавающими листьями и у надводных, то у первых оно происходит в значительной мере, а у вторых, вероятно, исключительно путём всасывания из грунта корнями.
И среди животных возможно, невидимому, питание через стенки тела за счёт растворённых в воде органических соединений; конечно, это возможно лишь при очень большой нежности покровов, как, например, у простейших (одноклеточных) животных. Но подавляющее большинство животных питается, главным образом, твёрдой, так или иначе оформленной пищей, которая предварительно измельчается, обрабатывается пищеварительными соками и частично превращается в жидкость; только в жидком виде она всасывается специальными органам^, расположенными почти у всех многоклеточных животных внутри тела.
Взвешенное состояние мелких частичек (планктон или детрит) и перенос их движущейся водой определяют способы питания весьма многих организмов. Так, у некоторых животных органы принятия пищи построены по типу сита; для того чтобы отцедить достаточное количество пищевых частичек, приходится пропускать через это сито значительное количество воды. Здесь, как и в дыхании, мы сталкиваемся с необходимостью постоянного тока. Иногда ток воды достигается движением самого животного, как это имеет место у рыб. Рыба плывёт, то открывая, то закрывая рот, и при своём движении вбирает всё время новые порции воды в ротовую полость; отсюда вода устремляется в щели в глотке, ведущие прямо наружу, но на пути своём встречает фильтрующий аппарат (сито из мелких, плотно стоящих тычинок), не пропускающий содержащихся в воде частичек. Отфильтрованная вода омывает по дороге жабры и служит для дыхания; у планктоноядных рыб застрявшие на тычинках мелкие частички — в том числе планктон — поступают в кишечник. Таким образом, здесь аппарат питания тесно связан с аппаратом дыхания.
В других случаях животное остаётся само неподвижн ы.м, но особые приспособления — колеблющиеся реснички — производят вблизи ротового отверстия водоворот, которым находящиеся в воде частички загоняются в рот. Лакой водоворот имеется у большинства коловраток. 6t>
Рис. 23. Часть тёрки прудовика — Limnaea.
Из крупных водных животных типично растительноядных немного, да и то некоторые из них при случае могут переходить на мясной стол (как это делают прудовики). Интересно отметить, что растительноядные животные в природных условиях редко трогают для целей питания ткани высших растений, а большей частью соскабливают лишь с листьев и стебельков всегда очень обильный нарост водорослей; так, например, делают улитки (Limnaea и др.) при помощи своей «тёрки» (рис. 23). Но иногда животные въедаются внутрь растений, прогрызая себе в листьях или стеблях длинные коридоры (этих животных принято называть «минирующими»); такие ходы постоянно приходится наблюдать, например,, на листьях кувшинки, в стеблях или листьях камыша и (рис. 24).
Среди водных хищников, захватывающих более крупных животных, мы находим не только животных, но и растения. У распространённой в наших стоячих водах пу зырчатки (рис. 106) имеются на веточках мелкие пу
т. п.
зырьки, в которые попадают проплывающие мимо мелкие животные и перевариваются там, благодаря выделению внутри
Р п с. 24. Ходы личинок Gricotopus (из Tendipedidae) в листе телореза — Strati о tes.
В колах видны личинки, совершающие колебательные движения в це-члях дыхания, причём смена воды происходит черев отверстия на концах ходов.
Оказывается, что не
пузырька пищеварительного сока.
Способы завладения добычей у водных хищников довольно мало отличаются от таковых же у наземных. Здесь, как и на суше, пускаются в ход внезапные прыжки с довольно значительного расстояния (личинки коромысла, рис. 25), преследование добычи на ходу (жуки и клопы), подстерегание добычи в укромном уголке, где хищник незаметен (водяной скорпион), захватывание в специально построенные тенёта (водяной паук, рис. 21, личинки некоторых ручейников, рис. 26) и т. д. Пойманная добыча в большинстве случаев измельчается челюстями и отправляется в рот мелкими кусочками; но иногда она пронзается острым хоботком, через который всасываются её соки (клопы), или челюстями, но с каналом внутри, по которому поступает в цельное, неразмельчённое тело добычи сок, превращающий все мягкие ткани её в жидкость, которая затем и высасывается (личинки плавунца, рис. 373,Д).
Активная реакция среды. Мы должны отметить ещё один фактор, имеющий большое значение в жизни организмов, населяющих водоём. Этот фактор носит название активной реакции среды. Что под этим подразумевается, мы сейчас узнаем.
Известно, что химическая формула воды — Н2О; это означает, что молекула (частица) воды сострит из двух атомов водорода (Н) и одного атома кислорода (О). Так как атомный вес водорода равен 1, а кислорода 16, то молекулярный вес воды будет 2 -j- 16 = 18. все частицы воды находятся в состоянии полного соеди-
нения атомов водорода и кислорода, соответствующего формуле воды И2О. Некоторое число частиц распадается, но не на Н2 и О, а на Н и ОН. Такой распад молекул воды называется диссоциацией1, а ого
1 Явление диссоциации существует не только в чистой воде, но и в растворах различных солей, кислот и щелочей.
Рис. 25.
Налево две личинни коромысла—Aeschna, направо две личинки плавунца — Dy Use us, нижняя личинка коромысла (на растении) схватила своей маской пиявку, верхняя охотится эа водяным осликом — Asellus; выбрасывая в то же время иэ задней кишки струю воды; верхняя личинка плавунца — в состоянии дыхания, нижняя схватила челюстями головастика.
Рис. 26. Ловчие сети ручейников:
Вверху — Neureclipsls Ы-maculata, внизу — Poly-centropus tlavomacula-tus.
продукты — Ни ОН — носят название ионов. Ионы характеризуются тем, что они несут на себе электрические заряды, причём атом водорода Н заряжен положительно, что обозначается через Н’, а группа ОН, называемая гидрокислом, заряжена отрицательно и обозначается через ОН'. Таким образом, процесс диссоциации может быть символически представлен так: НиО = Н - Чем большее количество молекул воды диссоциировано (разложено) на ионы, тем большей проводимостью электрического тока обладает вода.
Кроме физического различия — противоположности электрических зарядов между ионами Н* и ОН’, существует также и химическая разница: водородные ионы придают воде кислотные свойства, а гидроксильные ионы — щелочные свойства. Когда те и другие ионы присутствуют в воде в оди-наковом числе, то вода имеет нейтральную реакцию; с увеличением количества ионов Н* и уменьшением числа ионов ОН' повышается кислотность и понижается щёлочность жидкости и наоборот. При этом во сколько раз увеличивается число водородных ионов, во столько же раз уменьшается число гидроксильных ионов; во сколько раз увеличивается число последних, во столько уменьшается число первых (произведение чисел тех и других ионов остаётся всегда постоянным).
В химически чистой нейтральной воде диссоциация её частиц чрезвычайно мала: из всего количества молекул воды на ионы распадается лишь 0,0000001 часть, т. е. на каждые 10 000 000, или на 107 частиц воды приходится только 1 диссоциированная частица, дающая при своём распадении один Н’-ион и один ОН -ион. Следовательно, сколько молекул воды с нейтральной реакцией подвергнется диссоциации, столько получится и водородных и гидроксильных ионов. Если мы к нейтральной воде прибавим некоторое количество кислоты, то число водородных ионов в определённое число раз увеличится, а число гидроксильных ионов во столько же раз уменьшится. Прибавление щёлочи вызовет обратное явление — уменьшение числа водородных ионов и увеличение числа гидроксильных.
Таким образом, по числу ионов можно судить не только о характере реакции жидкости — нейтральном, кислом или щелочном, но и о степени кислотности или щёлочности, т. е. числом ионов можно измерять последние. Для такого измерения служит число водородных ионов, которое и является выражением активной реакции жидкости (или среды). Если это число равно 1/107 всего количества молекул жидкости, заключающихся в данном её объёме, то это значит, что жидкость имеет нейтральную реакцию; если это число больше 1/107, например '/ю5, то реакция будет кислой, если меньше — допустим Vio9 — то щелочной. Следовательно, усиление активной реакции соответствует увеличению кислотности, а ослабление её — увеличению щёлочности. Для удобства активную реакцию принято выражать латинскими буквами pH, а число водородных ионов, вместо дроби, — только показателем степени при 10. Поэтому, выражение pH = 7 означает, что жидкость имеет нейтральную активную реакцию; чем кислее делается жидкость, тем меньше становится pH (в сравнении с 7), тем более усиливается её активная реакция; чем щелочнее делается жидкость, тем больше становится pH, тем более ослабляется активная реакция.
Из предыдущего ясно, что для определения активной реакции среды, т. е. для определения pH, необходимо уметь определять концентрацию водородных ионов в этой среде, т. е. число свободных ионов Н' в ней. Наиболее точно это число определяется электрометрическим способом — посредством особого сложного и дорогостоящего электрического прибора. Для измерения же активной реакции пресных вод в условиях полевых работ пользуются сравнительно дешёвым, простым и удобным колориметрическим методом, основанным на принципе сравнения окраски проб исследуемой воды (после прибавления к ним определённого реактива) с заранее заготовленными, различно окрашенными образцами жидкостей, в каждом из которых определённому тону окраски соответствует вполне определённое, заранее вычисленное значение pH. Такие образцы существуют в продаже (в магазинах лабораторного оборудования), причём к ним прилагается подробная инструкция, как нужно ими пользоваться для определения pH той воды, которую мы хотим исследовать на активную реакцию.
Понятно, что pH воды зависит от общего её химизма, т. е. от совокупности растворённых в ней веществ и от характера и количества каждого из этих веществ. Поэтому активная реакция должна изменяться вместе с изменениями общего химизма воды. И, действительно, она неодинакова как в разных водоёмах, так и в различные моменты времени в одном и том же водоёме.
В пресноводных водоёмах pH колеблется в пределах приблизительно от 3,5 до 9,5—10. Знакомые уже нам три типа озера в отношении pH могут быть охарактеризованы следующим образом: в субальпийских озёрах pH воды колеблется около 7 (немного меньше или немного больше), т. е. лежит в пределах нейтральной реакции, в балтийских — pH обычно выше 7 (во время цветения может доходить до 9,5—10), следовательно, обнаруживает щелочную реакцию воды, и, наконец, в гумусовых озёрах pH всегда ниже 7. значит их вода имеет кислую реакцию. В сфагновых болотах pH понижается иногда до 3,5.
Даже в одном и том же водоёме — в различных его участках и пунктах— pH может быть разным, например, в пелагической области озера и в литорали, в разных местах последней, в разных точках по вертикали (по глубине) и т. д. Значительные колебания этот фактор испытывает также во времени; эти колебания могут быть суточными, сезонными и годовыми.
Каковы же основные причины изменений pH в пресных водах? На первом месте среди этих причин нужно поставить жизнедеятельность водных организмов. Два главнейших фактора этой жизнедеятельности — кислород и углекислый газ — оказывают очень большое влияние на pH воды; всё, что способствует увеличению растворённой в воде углекислоты 59
и уменьшению растворённого кислорода (дыхание водных организмов, про-цессы гниения), содействует вместе с тем понижению pH, т. е. усилению кислой реакции; наоборот, всё, что вызывает накопление кислорода в воде и убыль углекислоты (питание зелёных растений), обусловливает повышение pH, т. е. подщелачивание воды. Отсюда понятны суточные и годовые изменения pH; последнее повышается при усилении солнечного освещения и понижается при ослаблении его, вследствие чего днём pH, выше, чем ночью, и летом выше, чем зимой. Далее, процессы обмена веществ водных организмов, выражающиеся в поглощении ими из воды одних веществ и в выделении в неё других, изменяя общий химизм воды, этим самым вызывают и колебания pH. Так, накопление органических продуктов выделения в условиях небольшого количества воды и растворённого в ней кислорода обусловливает понижение pH, так как разложение этих продуктов содействует уменьшению кислорода и увеличению углекислоты в воде.
Однако водные организмы не только воздействуют на pH, но и сами находятся под его влиянием. Для каждого организма существуют пределы изменений активной реакции, между которыми он вообще может жить. В этом отношении водные организмы могут быть разделены на две основные группы. Организмы одной из них характеризуются широкой приспособленностью к концентрации водородных ионов, т. е. могут существовать и в кислых, и в нейтральных, и в щелочных водах, вследствие чего они получили название евриионных; к другой группе принадлежат так называемые с т э нойон н ы е организмы (т. е. организмы с узкой приспособленностью к условиям активной реакции), существование которых возможно только в сравнительно узких пределах pH, именно: или в кислых водах (с pH, приблизительно, не выше 4,5), или в нейтрально-щелочных (pH = 7 и выше).
Наилучшей средой для развития и жизни громадного большинства водных организмов является всё же нейтрально-щелочная вода. Интересно отметить, что водная растительность, воздействуя на pH окружающей воды, сама себе подготовляет наилучшую среду: поглощая из воды для питания углекислоту, растения делают воду более щелочной, т. е. более подходящей для их существования. Однако известны случаи, когда вызванное растением перещелачиванне среды делает последнюю уже вредной для него; так, водоросль Pandorina начинает испытывать уже вредные влияния со стороны окружающей воды, когда её pH становится близким к 9.
Из всего сказанного об активной реакции среды ясно, что этот фактор играет очень большую роль в жизни водного населения.
Как и на суше, единственными производителями органических веществ в воде являются зелёные растения; всё же животное население питается или непосредственно ими (особенно, как уже указывалось, мелкими), или поедая растительноядных животных. С этой точки зрения водные организмы делятся на две крупные категории: продуценты (производители) и консументы (потребители).
Методику исследования химизма воды и темы для работы в этой области мы не считаем нужным приводить ввиду сложности этой методики и необходимости дорогостоящего оборудования.
Для лиц, желающих более подробно ознакомиться с методикой физико-географических и химических исследований озёр, рекомендуем следующие кпиги:
1. Программа предварительного исследования озера, составленная членами постоянной комиссии по изучению озёр России при Русском географическом обществе. Выл. I, Л. 1918; вып. II, Л. 1924.
2. Верещагин Г. 10., Методы морфометрической характеристики озёр. Труды Олонецкой научной экспедиции, ч. 2, География, вып. I, 1930, изд. Гос. гидрол. ин-та. Л.
3. Верещагин Г. Ю., Аничкова Н. И. и Форш Т. Б., Методы полевого гидрохимического анализа в их применении к гидрологической практике. Л.
4. Малинина В. С., Руководство для практических занятий по исследованию воды пресноводных водоёмов (физико-химическое исследование), М.—Л. 1933.
П. КАЧЕСТВЕННАЯ БИОЛОГИЧЕСКАЯ ХАРАКТЕРИСТИКА ОЗЕРА. БИОТОПЫ И БИОЦЕНОЗЫ
Разбирая в предыдущем изложении различные свойства воды, мы видели, что не во всём озере в одинаковой мере проявляются те или иные из этих свойств. Проникание света до дна в мелких и глубоких частях, движение воды у берегов, подверженных и не подверженных воздействию ветров, температурный режим и кислородный баланс у берегов и в глубоких частях и прочее и прочее — всё даёт нам несомненные доказательства того, что на различных участках озёрного ложа свойства воды будут проявляться не в равной мере. С другой стороны, и образование озёрного бассейна в некоторых местах подвергается особому воздействию воды, отличному от общего воздействия всей водной массы — хотя бы у мест впадения притоков или выхода стоков.
В результате этих сложных воздействий неживой природы создаются в озере области с весьма различными условиями для развития жизни и растительной и животной. Различные живые существа в свою очередь воздействуют и на озёрное ложе и на свойства непосредственно окружающей их воды различно, видоизменяя их в ту или иную сторону; этим они вносят ещё больше разнообразия в общую картину различных частей водоёма.
Таким образом в озере создаются области, носящие особые черты с физико-географической и с физико-химической точки зрения. Однородные условия существования в пределах каждой области влекут за собой определённую группировку организмов. Эти однородные области получили название «биотопов», а группировки организмов — «биоценозов». Следовательно, каждому биотопу соответствует свой биоценоз. Являясь частью одного целого — озера, каждый биотоп вместе со своим биоценозом живёт своеобразной жизнью, но оказывает прямое или косвенное влияние на другие биотопы, подвергаясь, со своей стороны, их постоянному воздействию. »
Попробуем теперь кратко обобщить особенности наиболее типичных биотопов и биоценозов озера. Не будем только забывать, что в природе ни те, ни другие не являются резко ограниченными и связаны постепенными переходами.
Прежде всего мы имеем в озере две принципиально различные по всему комплексу условий области — водную массу и дно. Но каждая из этих областей опять-таки приобретает очень резкие отличия в зависимости от своего положения в озёрном ложе и в первую очередь от глубины. Глубина влияет на прогреваемость, на освещение, на возможность заселения высшими растениями, а последнее обстоятельство привносит в свою очередь ряд новых черт в общую картину данного участка — и дна и воды, изменяя очень существенно все физико-химические и биологические условия. Так как именно заселение высшими растениями является штрихом, наиболее резко меняющим картину, то принято делить озеро, как это указано в начале книги, на крупные области: пелагиаль с про фу н д а л ь ю, где не могут произрастать высшие растения в силу неблагоприятных условий освещения у дна, и литораль, где освещение у дна делает возможным существование там высшей растительности. В большинстве случаев литораль является действительной областью прибрежных растений, но иногда другие условия — свойства грунта или прибой — лишают эту область растительности, и тогда граница может быть проведена лишь на основании донных отложений.
Таким образом, по горизонтали мы различаем два крупных биотопа: 1) пелагическую область и 2) литоральную область; в некоторых случаях различают ещё расположенную между ними узкую полосу — 3) сублитораль. Познакомимся сначала с пелагической областью, именно — с её водной массой (короче — пелагиалью) и её дном — профундалью.
Пелагическая область
ВОДНАЯ МАССА КАК СРЕДА
Водная масса пелагической области озера, в отличие от дна и прибрежной области с её зарослями, представляет комплекс специфических и совершенно своеобразных условий существования для обитающих в ней организмов. Эти условия и их влияние на строение и жизнь организмов были очерчены нами в общей форме уже ранее— в разделе о физико-химических свойствах озера. Поэтому здесь мы остановимся лишь на некоторых частных вопросах, дополняющих и освещающих картину жизни тех организмов, которые населяют самую воду озера.
Самой главной и существенной особенностью жизни в толще воды, имеющей для обитателей последней первостепенное значение, нужно считать отсутствие в ней всякого твёрдого субстрата, на котором могли бы укрепиться, постоянно или временно, организмы. Исключение составляют разве лишь некоторые сидячие формы из простейших животных да паразиты, укрепляющиеся на теле свободно живущих животных. Ясно, что у организмов, которые всё время проводят в толще водной массы л для которых она является единственной опорой, должны вырабатываться специальные приспособления к таким условиям жизни; главнейшие из них были нами рассмотрены уже выше. Присутствие или отсутствие твёрдой опоры имеет в этом случае настолько большое значение, что иногда представители одной и той же систематической группы животных, живущие в пелагической и в прибрежной области, имеют разную форму и разное строение. Это, конечно, не значит, что не существует форм, которые являются общими для обеих областей озера; но наряду с ними есть и такие, которые принадлежат исключительно какой-нибудь одной из них. Для примера достаточно указать на Gladocera, среди которых чисто пелагические формы обладают гораздо более тонкой и нежной раковиной (например, Leptodora), чем чисто прибрежные, снабжённые, напротив, большей частью довольно грубой и жёсткой раковиной, которая служит достаточно надёжной защитой для слабого и нежного тела.
Но отсутствие твёрдой опоры в водной массе пелагической области означает вместе с тем и отсутствие преград для передвижения, каковыми в литорали являются заросли. Таким образом, отсутствие в воде твёрдых субстратов играет двойную роль: вода как единственная опора для организмов должна была способствовать развитию у них путём естественного отбора таких приспособлений, которые помогали бы им использовать это свойство воды и в той или иной степени обеспечивали бы их телу положение равновесия, следовательно, состояние относительного покоя; вода как простор для движения должна, напротив, вырабатывать пловцов, т. е. вызывать развитие органов активного движения, единственной формой которого является в данном случае плавание. Чем большее сопротивление вода будет оказывать погружению тела (пассивная форма движения), тем больший толчок она будет давать развитию таких приспособлений, которые необходимы для преодоления этого сопротивления при активном плавании, тем более, что вода при этом играет опять-таки роль опоры для органов плавания. Следовательно, одно и то же свойство воды — вязкость —г- действует одновременно в двух разных направлениях, обусловливая две группы приспособлений. По существу обе эти группы однородны, так как у тех и других приспособлений в основу конструкции положен один и тот же принцип — увеличение рабочей поверхности; разница сводится лишь к тому, что к первой категории приспособлений относятся неподвижные органы или части тела, ко второй — подвижные. Иногда один и тот же орган может выполнять обе функции, смотря по тому, в каком состоянии он находится — покоя или деятельности (например, вторая пара усиков у Cladocera, рис.- 241). Первая группа приспособлений для поддержания тела в воде развита, хотя и не в одинаковой степени, но всё же довольно хорошо у всех населяющих воду планктических организмов; вторая группа — 62
приспособлений для активного плавания, напротив, развита у этих организмов чрезвычайно различно, вследствие чего и самые организмы образуют в этом отношении ряд градаций, начиная с совершенно пассивных (полное отсутствие приспособлений для активного движения) и кончая прекрасными пловцами. Так как с уменьшением размеров тела объём его убывает в третьей степени, а поверхность — только во второй, то чем мельче организм, тем относительно больше его удельная поверхность, и. следовательно, тем лучше он — уже по самым своим размерам — приспособлен для поддержания в воде во взвешенном состоянии; с другой стороны, у крупных организмов большие размеры их тела позволяют иметь и более сильные органы плавания, для чего необходимо наличие хорошо развитой системы некоторых частей и тканей тела, как, например, части скелета, мускулатуры и т. п. Таким образом, ясно обособляются две крайние группы организмов: мелкие (большей частью микроскопической величины) — совершенно пассивные или обладающие лишь незначительной способностью к самостоятельному перемещению в воде, и крупные — хорошо приспособленные к активному плаванию. Первые способны к самостоятельному перемещению лишь на небольшие расстояния, измеряемые самое большее метрами, и не могут противостоять даже небольшим течениям, которыми они пассивно переносятся из одного места в другое; вторые, напротив, обладают способностью активно перемещаться на десятки километров — даже против значительного по силе течения. Организмы первой группы объединяются под общим названием планктон, организмы второй — под названием нектон.
Как уже указано, в отношении способности к активному плаванию организмы образуют ряд переходов, и потому между планктоном и нектоном резкой грани провести нельзя. Бесспорными в этом смысле являются, как и всегда в подобных случаях, лишь крайние группы, а именно: в планктоне — громадное большинство растительных форм, или фитопланктона, в нектоне — рыбы; что же касается планктических животных, или зоопланктон а, то среди них встречаются уже такие формы, которые вызывают в этом отношении сомнение и лишь с большей или меньшей натяжкой могут быть отнесены к планктону, образуя скорей переход или к нектону (некоторые из более крупных ракообразных, мальки многих рыб), или к организмам прибрежной области (например, некоторые клещи, коловратки и др.), пли, наконец, к донной фауне (как некоторые из ракообразных).
Ввиду того, что единственной безусловно нектической группой организмов пелагической области являются рыбы, которых мы вообще оставляем в стороне, мы, в дальнейшем, совершенно не будем говорить о нектоне и ограничимся лишь рассмотрением планктона.
ПЛАНКТОН
Принесём домой в наполненной водой банке из-под варенья наловленный в озере или пруде планктон, пропустим всю эту воду через планктиче-скую сетку (см. дальше — «Методика») или через куоок материи, из которой она делается, и профильтрованный планктон смоем в стеклянную чашечку или в стакан с небольшим количеством воды: затем перенесём отсюда пипеткой несколько капель в часовое стёклышко, поместим его под микроскоп и будем наблюдать при слабом увеличении. Нашему взору представится довольно любопытная картина, в которой с первого взгляда разобраться нелегко: глаза разбегаются, не успевая следить за снующими туда и сюда организмами, взгляд не может долго останавливаться на одном организме и, привлекаемый каким-либо другим движущимся существом, перебегает на него, с'него перебрасывается на третий организм и т. д. И неопытный наблюдатель долго ещё не может сосредоточить своё внимание на каком-нибудь одном планкти-ческом растении или животном, чтобы лучше рассмотреть его и последить за его поведением. Но мало-помалу глаз привыкает и научается различать и разную форму организмов, и различные способы их движения.
подвижном состоянии, но затем —
Рис. 27. Коловратки — Rotatoria.
Слева направо: верхний ряд — Asplanchna priodonta, Filinia longiseta, Keratella quadrata, Keratella cochlea-rts; нижний ряд — Notbolca longispina, Trichocerca capucina, Brachionus urceus, Schizocerca diversicornis.
Сильными ударами своих передних нитевидных усиков производят прыжки веслоногие рачки (Copepoda): циклопы и диаптомусы (рис. 275); сделав такой прыжок, рачок находится некоторое время в не-удар усиками, новый скачок — и рачок опять неподвижен. Подобные же, но более слабые и притом подряд следующие один за другим прыжки производят своими разветвлёнными усиками ветвистоусые рачки (Cladocera): Daphnia (рис. 241), Bosmina (рис. 253), Chydorus (рис. 271), Geriodaphnia (рис. 247) и др.; особенно выделяется по своим размерам самый крупный не только из ветвистоусых, но и вообще из планктических пресноводных рачков — Lepto-dora, — который, однако, несмотря на большую величину (достигает иногда 1 см), принадлежит, особенно в неподвижном состоянии, к наиболее незаметным организмам; такой его делает чрезвычайно большая прозрачность, благодаря которой он становится видимым только во время движения. Совсем по-другому двигаются коловратки Rotatoria (рис. 27, 28), которые значительно мельче ракообразных: в то время как большинство последних имеют
размеры от 1 до нескольких миллиметров, величина первых измеряется долями миллиметра. Коловратки плавают не толчками, а плавно, в то же время вращаясь вокруг оси тела; такой способ передвижения обусловливается присутствием на переднем конце тела коловратного аппарата, состоящего из расположенных в круговой ряд ресничек, которые своими согласованными движениями и вызывают плавное перемещение животного в воде. В планктоне нам может встретиться, например, самая крупная из всех коловраток — мешковидная Asplanchna (рис. 27), интересная своей прозрачностью, затем — снабжённые игловидными щетинками Filinia (Triar-thra), Polyarthra (рис. 28); далее—одетые в панцырь коловратки: Trichocerca (Rattulus, рис. 27), цилиндрическое и немного изогнутое тело которой на конце снабжено очень маленькой, почти незаметной «ногой», несущей длинный игловидный «палец»; Brachionus, панцырь которого на переднем конце несёт несколько (2 или 4, или 6) шипов, на заднем же по большей части лишён их; Schizocerca (рис. 27)
у которой на переднем конце панциря 4 шипа (2 крайних длинных и 2 средних коротких), а на заднем 2 (или одинаковой, или различной длины); Keratella (Anuraea) с 6 передними шипами, из которых два средних обычно длиннее остальных, на заднем же конце у одного вида — Keratella quadrata (Anuraea aculeata) (рис. 27) — 2.шипа, а у другого — Keratella cochlearis
и c. 28. Коловратка Polyarthra trigla.
Рис. 29. Синезелёыые водоросли—Cyanophyceae:
А — Gloeocapsa (налево — колония клеток, одетых слизью); Б — часть разреза через колонию Nostoc; В — Anabaena (налево — колония, направо — часть нити, в середине — спора); Г — Aphanizomenon (налево — колония параллельно расположенных нитей, направо — отдельная нить); Д — Clathrocystla (налево вверху — отдельные клетки, остальное — разные формы слизистых колоний); Е — Oscllla-• toria; Ж — Gloeotrictiia.
(рис. 27) — 1 шип, а иногда и совсем не бывает; у одного из видов No-tholca панцырь снабжён игловидными отростками.
Все приведённые нами в качестве примеров планктические организмы, составляющие лишь очень малую часть из встречающихся вообще в планктоне, относятся к животному планктону, или к зоопланктону. Планктические животные обращают на себя внимание как более крупными размерами своего тела, так и своим движением. Но значительно труднее рассмотреть ф и т о п л а н к т о и, т. е. совокупность растительных организмов планктона, или планктические водоросли, вследствие их мелкости и неподвижности многих из них. Из подвижных планктических водорослей нам могут встретиться многие жгутиконосные водоросли (рис. 32), обладающие очень тонкими, у большинства живых организмов почти или совсем незаметными нитевидными органами плавания — жгутиками; к таким водорослям принадлежат, например, колония Volvox (рис. 30 Л), плавающая в виде зелёных, сравнительно крупных (1—1,5 мм) шариков; Dinobryon (рис. 32 Л) — тоже колония, но только очень изящной разветвлённой формы, причём каждая клетка заключена в тонкий прозрачный бокальчик; одиночная (не колониальная) мелкая водоросль Euglena, тело которой имеет веретеновидную форму; Ceratium (рис. 325), одетый в панцырь, несущий 3 или 4 рога, и др.
& А. Н. Липпа 65
Рис. 30. Зелёные водоросли—Chlorophyceae:
А__ Micrastertas; Б — Cosmart um; В — Xanthidtum; Г — Desmldium; Д — Staurastrum (вверху —
вид сверху внизу — вид сбоку); Е — Cladophora; Ж — Scenedesmus; 3 — Eudorina; И — Pandortna; К — Gunium Л — Volvox (налево — с дочерними колониями внутри, направо — с яйцами и спермато-воидами); М — Pcdlastrum.
Что касается малоподвижных или совсем неподвижных водорослей, то из них мы можем встретить, например, диатомовые (рис. 31), характеризующиеся присутствием кремнёвого панцыря, состоящего из двух створок, из которых одна закрывает другую, как крышка коробку; в наших озёрах часто встречаются следующие диатомовые водоросли: Melosira (рис. 31 А) — 66
Рис. 31. Диатомовые водоросли — Diatomeae:
А — Melosira: Б — Pinnularia (вверху — вид со стороны створки, внизу — вид со стороны пояска); В — Asterionella; Г — Surlrella; Д — Closterium; Е — Fragilaria; Ж — Gomphonema; 3 — Pleuro-sigma.
в виде нитей из расположенных в один ряд клеток; Fragilaria (рис. 31Е), узкие и удлинённые клетки которой соединены своими срединалми в колонии, имеющие вид длинных столбиков; Asterionella (рис. 31В), представляющая тонкую изящную 8-лучевую звёздочку, и др. Большинству береговых и донных диатомовых водорослей свойственна своеобразная форма передвижения в виде отдельных толчков, причиной которых является быстрое выбрасывание из клетки через находящуюся в её створке щель слизистого вещества, в результате чего клетка отталкивается в противоположную сторону (реактивное движение). Совершенно лишены способности к активному движению многочисленные представители синезелёных (рис. 29) и зелёных (рис. 30) водорослей, как, например, Anabaena (рис. 29В), которая представляет собой правильно или неправильно извитую нить, заключённую в слизь и состоящую из более или менее округлых клеток; Aphanizo-menon (рис. 29 Г) — в виде пучков небольших нитей, параллельно соединённых слизью и состоящих из удлинённых клеток; Pediastrum (рис. ЗОЛ/), имеющий форму округлённой пластинки, сложенной из плоских многогранных клеток, и многие другие (см. рис. 29—30).
Если тот планктон, который мы рассматриваем под микроскопом, наловлен лишь в одной единственной экскурсии в одном пункте водоёма, то он может дать нам лишь самое общее представление о родовом или видовом составе населяющих этот водоём планктических организмов в тот день, когда совершена экскурсия. Но он ничего не может сказать ни о том, претерпевает
ли планктон какие-либо изменения в течение, например, года и каковы эти изменения, ни о том, как распределён он в данном водоёме, т. е. одинаков он во всех пунктах последнего или неодинаков. Эти вопросы могут быть разрешены лишь специально поставленными исследованиями, при которых для решения первого вопроса планктон собирают через определённые промежутки времени (например, раз или два каждый месяц), для решения же второго вопроса сбор производят одновременно на разных пунктах или на различных глубинах водоёма. В сущности на оба эти вопроса нетрудно уже заранее предугадать ответ, если вспомнить, что планктон, как и всякий вообще биоценоз, зависит прежде всего от своего биотопа, т. е. от совокупности господствующих в воде физических и химических условий, в которых приходится жить планк-тическим организмам. А мы уже видели, что эти условия не остаются постоянными ни во времени, ни в пространстве. Ясно, следовательно, что вместе
с ними должен меняться и
Рис. 32. Жгутиковые водоросли — Flagella ta :
А — Dinobryon; Б — CeraUum.
изменений температуры в
планктон — как по количественному, так и по качественному своему составу. Результаты многочисленных исследований вполне подтверждают такое предположение.
Общий характер сезонных изменений всей совокупности физических условий жизни можно формулировать так: от зимы к лету эти условия для большинства организмов улучшаются, от лета к зиме ухудшаются. Соответственно этому и планктон наиболее богат во 2-й половине лета, наиболее беден зимой; и это касается как его общего количества, так и числа входящих в состав его видов; при этом необходимо отметить, что в озёрах умеренного пояса одни виды являются типично летними и на зиму выпадают из планктона, например, водоросль Aphanizomenon, другие, наоборот, встречаются только зимой, как один из видов циклопов —Cyclops strenuus, третьи, наконец, существуют в течение всего года, примером чего может служить коловратка Keratella (Anuraea) cochlearis.
Таким сезонным колебаниям подвержены многие планктические организмы. Так как при смене сезонов на первый план выступает теснейшая зависимость этой смены от определённых течение года, то и сезонные колебания
планктона можно поставить в связь (прямую и косвенную) с теми же
температурными изменениями, что и было действительно сделано некоторыми исследователями. Для многих видов планктических организмов существуют оптимальные температуры, способствующие наиболее пышному их развитию; чем больше температура удаляется в ту или другую сторону от этого оптимального значения, тем хуже развивается данный вид,
и при некоторых предельных температурах его представители уже не могут существовать и погибают. Таким образом, при сезонных сменах температур каждый такой вид при оптимальной для него температуре проходит через максимум своего развития, который характеризуется наибольшим его расцветом и преобладающим его значением в планктоне. Так, для большинства диатомовых водорослей оптимальная температура 15—16°, и потому их максимум падает то на весну, то на осень (впрочем, некоторые формы, как Fra-gilaria, имеют и тот и другой максимум); синезелёные водоросли, например, знакомые уже нам Anabaena и Aphanizomenon, имеют более
высокую оптимальную температуру, и потому период их максимального развития падает на разгар лета. Такие максимумы размножения той или иной водоросли часто вызывают так называемое цветение воды, причём вода ба
приобретает специфическую окраску в зависимости от той водоросли, которой она цветёт: диатомовые сообщают воде буроватый оттенок, жгутиковые — Ceralium и Dinobryon — зеленоватожёлтый, синезелёпые вызывают зелёную окраску с голубоватым оттенком и т. п.
Таким образом, в озере в течение года происходит смена одних планктических форм другими. Объяснить это исключительно лишь одними температурными влияниями едва ли возможно; здесь, несомненно, действует комбинация различных факторов, из которых одни находятся в более, другие в менее тесной связи с температурой. В отношении зоопланктона из таких факторов на первый план выдвигается питание: чем больше развивается в планктоне тех организмов, которые для представителей данного вида служат пищей, тем лучше расцветает и этот вид, если тому не препятствуют другие условия; наблюдения, произведённые в Любичанковском пруде Московской области, показали, что коловратка Asplanchna, питающаяся другими коловратками из рода Keratella, размножалась в большом количестве, пока было много Keratella; но в середине июля последние сильно уменьшились в числе, а за ними и Asplanchna; в августе количество Keratella вновь сильно возросло, и это повлекло за собой новую вспышку размножения Asplanchna.
Влияние сезонности, т. е. смены времён года, обнаруживается ешё в явлении так называемого цикломорфоза. Этим термином обозначают сезонную смену взрослых форм одного и того же рода и вида, которые в разные времена года имеют различный внешний облик и которые в этом случае называются сезонными вариациями. Такая смена известна у многих видов планктических организмов — как растительных, так и животных — и выражается в изменениях размеров или наружной формы тела, например, в появлении и исчезновении различных выростов, или же в одновременных изменениях и размеров и формы. Так, жгутиковая водоросль Ceratium hirundinella летом имеет 4 рога, а зимой превращается в 3-рогую форму. У некоторых дафний летом вырастает на голове более или мепее длинный шип — так называемый шлем, зимой же он исчезает. У Bosmina на нижней стороне головы имеется раздвоённый хоботок, который у летних форм значительно длиннее, чем у зимних. Таким образом, повышенная температура (летом) способствует развитию выступов и выростов, пониженная, наоборот, затрудняет это развитие.
Однако влияние температуры на цикломорфоз обнаруживается не только при таких резких сменах её, как от лета к зиме и наоборот, но и при более постепенных переходах. В зависимости от этого крайние (летние и зимние) сезонные вариации в некоторых случаях связываются целым рядом промежуточных переходных форм (например, у дафний и у Bosmina), которые создают большое многообразие форм. Благодаря этому раньше, когда цикломорфоз был ещё неизвестен, эти разнообразные формы описывались как самостоятельные виды. После изучения сезонных вариаций, в котором большую роль сыграли экспериментальные исследования, сотня прежних видов свелась к одному-двум видам.
Причиной цикломорфоза, кроме температуры, могут быть и другие факторы, среди которых на первом месте следует поставить питание. Это доказано, например, для коловратки Brachionus. Однако необходимо подчеркнуть, что цикломорфоз — явление очень сложное, обусловленное, вероятно, комбинированным действием нескольких факторов, детали которого далеко ещё не выяснены.
Что касается пространственного распределения планктона, то различают два его вида; вертикальное и горизонтальное распределение. Мы уже знаем, что в достаточно глубоких озёрах по вертикальному направлению господствуют далеко не однородные условия: достаточно вспомнить существование здесь температурного, а за ним и газового скачка и существование температурной слоистости вообще. Совокупность подобных различий всех условий по вертикали — тем-
пературных, химических, оптических, вязкости воды и т. д. — вызывает, конечно, соответствующие различия в количественном и качественном распре-делении планктона, как, например, сосредоточение фитопланктона в верхних, наиболее освещённых слоях воды, разница в содержании планктона над слоем скачка и под ним, богатство планктическими организмами более тёплых слоёв воды и бедность ими более холодных и т. п. Естественно, что для вертикального распределения растительных организмов определяющими причинами являются указанные физические факторы; но в распределении зоопланктона наряду с этими первичными факторами играют роль и вторичные, в виде зависимости животных организмов от растительных, хищников — от их добычи и т. д. Совокупное действие всех этих причин вызывает в озере ту сложность и то разнообразие в вертикальном распределении планктона, которые уже многократно были констатированы различными исследователями и с каждым разом получают всё новые и новые подтверждения.
Но вертикальное распределение физических условий не остаётся постоянным даже в течение суток: оно постепенно меняется от дня к ночи и от ночи к дню; другими словами, оно испытывает суточные колебания. И вот в зависимости от этих колебаний и распределение планктона претерпевает соответствующие изменения; многие организмы, днём обитающие на глубинах, ночью поднимаются в верхние слои воды, чтобы с рассветом вновь начать опускаться. Такие перемещения планктических организмов по вертикальному направлению в течение суток называются суточными вертикальными миграциями и обусловливаются совокупным действием нескольких факторов, главным образом, степенью освещения, температурой и вязкостью воды.
Обычно в летнее время днём планктон, особенно растительный, сосредоточивается главным образом в верхних слоях; зоопланктон распространяется иногда значительно дальше в глубину, чем фитопланктон, причём количество первого уменьшается медленнее, чем второго; в наиболее глубоких слоях, до которых простирается планктон, большей частью находим только животных, преимущественно веслоногих рачков (Copepoda). Ночью эти рачки поднимаются ближе к поверхности.
Что касается горизонтального распределения планктона, то в пелагической области больших и глубоких озёр оно, как показали исследования, носит равномерный характер, что и понятно, так как и физико-химические условия жизни здесь повсюду очень сходны между собой. Другое дело — мелкие озёра и пруды: здесь слой воды имеет сравнительно столь незначительную толщину, что равномерное распределение физико-химических и биологических факторов в большей или меньшей степени уже нарушается. Близость дна с его неровностями, различия в прогреваемости водного слоя, в прозрачности, в степени освещения, в распределении погружённой растительности и т. п. — всё это может вызвать и действительно вызывает неравномерное распределение планктона в горизонтальном направлении. Наблюдения показывают, например, что при таких условиях нередко образуются так называемые «рои» или «тучи» планктических организмов, состоящие иногда лишь из одного рода или даже вида (дафнии); иногда —из нескольких (коловратки).
ТЕМЫ ДЛЯ РАБОТ
Работы по планктону могут быть произведены, конечно, только при наличии микроскопа. Микроскопический мир живых существ настолько разнообразен и интересен сам по себе, что наблюдения над ним имеют значение даже и без отношения к какой-либо специальной теме. Мы не будем здесь касаться всех тех общих биологических вопросов, которые могут разрабатываться на основании изучения планктических форм, и лишь укажем на тетемы, которые стоят в связи с изучением водоёма как целого и взаимоотношений его частей. Проведённые по этим темам исследования могут дать также и полезные практические сведения для рыбного хозяйства, поскольку планктон является пищей для мальков почти всех пресноводных рыб, а у некоторых пород рыб — также и для взрослых особей.
7U
I. Непериодические наблюдения над планктоном
1. Первоначальное знакомство.
2. Сравнить планктон пелагической и прибрежной области.
3. Сравнить планктон в нескольких участках пелагической области.
4. Сравнить планктон в нескольких участках литорали в связи с различными типами зарослей.
5. Сравнить планктон у поверхности и, по возможности, вблизи дна.
II. Периодические наблюдения над планктоном
1. Установить суточные изменения планктона: сравнить поверхностный планктон среди дня, часа в 3, и ночью, часов в 12.
2. Установить сезонные изменения планктона: сравнить поверхностный планктон зимний (февраль — пропустить через планктическую сетку несколько вёдер воды из проруби), весенний (апрель, по вскрытии льда), летний (июль) и осенний (октябрь).
ME ТОДИКА
А
идет (за границей) под номером 25. А
50гл
б
50 см
8cw
2 см
Рис. 33. Выкройка для планктиче-ской сетки (ириг.).
к У-
Оборудование. 1. Качественная планктическая сетка делается из шёлка, употребляемого для изготовления мельничных сит, через которые просеивается мука, и потому называемого мельничным газом; для получения разных сортов муки служат сита различной частоты, т. е. с различными размерами ячей; все эти разнообразные сорта сит известны каждый под своим номером, причём с увеличением частоты мельппчпого газа, т. е. с уменьшением его ячей, возрастает и его номер, вследствие чего высшие сорта муки получаются соответственно из высших номеров газа.
В дореволюционной России высшие номера мельничных сит не выделывались, а выписывались из-за границы. Для планктических сеток употребляется самый густой газ, который раньше обозначался номером 20, а теперь идёт (за границей) под номером 25. За последние годы, однако, и у нас, в СССР, начали выделывать высшие номера мельничного газа, причём нумерация у нас другая — более дробная, чем за границей, и охватывающая свыше 70 номеров.
Мы опишем здесь так называемую к а-ч ветвенную планктическую сет-Она состоит из металлического кольца диаметром в 25 см, согнутого из толстой медной проволоки (толщиной в 3,5—4 мм), и из конуса, сшитого из мельничного газа. Для экономии шёлковой материи конус можно сшить из двух половинок, выкройки которых можно сделать по рисунку 33. Как видно из пего, на изготовление одной сетки нужно 1/4 м2 материи, поэтому достаточно купить последней полмотра и от этой полосы отрезать квадрат со стороной в 50 см. На этом квадрате проводим карандашом диагональ, из концов которой радиусом, равным стороне квадрата, проводим дуги АВ от двух других углов до пересечения с диагональю. Затем разрезаем квадрат по диагонали, а каждый из получившихся треугольников — по д^ге. Прежде чем сшивать полученные половинки копуса, необходимо по этой же выкройке вырезать из какой-либо другой материи (из бязи, холста ит. п.) дуговые полоски а и б и нашить их на половинки конуса из шёлка в том положении, как это указано па рисунке. Обрезав вершины обеих половинок конуса по самой короткой дуге, сшиваем эти половипки боковыми краями в целый конус, основанием которого (нашитой полоской из грубой материи) обшпваем медное кольцо. Вершина копуса прикрепляется к металлическому (лучше всего тоже медному, а если железному, то, по крайней мере, лужёному) стаканчику без дна длиной 6—7 см, диаметром 4 см Верхний конец стаканчика вставляется в узкое (вершинное) отверстие конической сетки, которое по размерам должно быт^ пригнано к стаканчику, и по кольцу из грубой материи его плотно завязывают прочной ниткой или шнурком. Для того чтобы стаканчик пе мог выскользнуть из сотки, лучше на его верхнем крае сделать кольцевое утолщение, под которым и завязывать нижний конец сетки.
Нижний конец стакапчика затягивается кусочком того же мельничного газа, из которого сделана сама сетка; при выкройке последней остаются обрезки, из которых вырезается кружок диаметром на 11/а—2 см больше, чем диаметр стаканчика, затем этот кружок накладывается на отверстие стаканчика, излишек его краёв загибается на боковую стенку стаканчика и завязывается так же, как сетка на верхнем его конце. И здесь очень целесообразно сделать утолщение по нижнему краю стаканчика, чтобы привязанное донышко из газа держалось крепче. Донышко должно быть натянуто ровно, без складок.
Для того чтобы удобно и быстро стаканчик снимать и снова надевать на сетку, что бывает необходимо при работе, его верхнее кольцо может быть соединено с нижним кольцом сетки двояким ооразом: или одно из колец навинчивается на другое, или вставляется одно в другое и закрепляется посредством вдвигания штифтика, укреплённого на внутреннем кольце, в прямоугольный вырез на краю наружного кольца. При этом, конечно, как оба кольца, так и штифтик с вырезом должны быть хорошо пригнаны друг к другу.
Как видно из описания, изготовление стаканчика далеко не всякому под силу — в большинстве случаев оно должно быть поручено опытному слесарю. Ио стаканчик можно заменить таким приспособлением, которое может сделать любой школьник. Для этого берут воронку (стеклянную или, лучше, металлическую, например, керосиновую), подгоняют диаметры верхнего (широкого) конца воронки и нижнего (узкого) конца сетки так, чтобы последний можно было надеть на первый, прочно завязывают, а на трубочку воронки надевают подходящий по диаметру кусочек каучуковой (резиновой) трубки длиной приблизительно в 7—8 см, которую тоже крепко привязывают к трубочке воронки, и зажимают каучуковую трубку зажимом Мора (можно купить в аптеках или магазинах лабораторного снабжения). Теперь остаётся только прикрепить линь, в качестве какового может послужить верёвочный шнур диаметром в 4—5 мм. Он прикрепляется посредством «уздечки», состоящей из 3 коротких шнурков, сходящихся над центром отверстия сетки в одной точке, в которой они скрепляются как между собой, так и с линем. Для привязывания этих шнурков к кольцу в материи сетки непосредственно под кольцом вырезаются на равном расстоянии друг от друга три отверстия такой величины, чтобы можно было продёрнуть через них шнуры, концы которых и завязываются вокруг кольца. Сетка готова.
2. Несколько широкогорлых склянок (приблизительно на 200 куб. см) с пробками.
IIроизводство работ. Работа с планктической сеткой основана на принципе фильтрации: при движении сетки в воде последняя проходит через ячеи сетки, а те планктические организмы, которые по своей величине не могут пройти через эти поры, задерживаются сеткой и таким образом улавливаются.
Так как главнейшая масса настоящего планктона обитает в пелагической области, то естественно, что для его лова необходимо иметь в своём распоряжении лодку. При сильном волнении собирать планктон не рекомендуется. Сбор производится следующим образом.
Планктонист помешается на корме лодки и во время остановки последней опускает сетку вод поверхность воды. Затем он отдаёт гребцу команду дать лодке ход и вместе с её движением и по мере его начинает травить линь сетки, который, однако, в то же время должен быть натянут настолько, чтобы не дать сетке погрузиться на дно; самое лучшее держать сетку в это время у самой поверхности, чтобы она, просвечивая через воду, была всё время видна. Вытравив метров 10 линя, планктонист задерживает его в руке и при продолжающемся движении лодки начинает тянуть сетку за собой. Движение лодки должно быть равномерным, а скорость его — средней. Только при таких условиях сетка будет двигаться тоже равномерно и в горизонтальном направлении; в противном случае — при слишком быстром ходе лодки — сетка будет вылезать на поверхность воды, а при слишком медленном, напротив, погружаться на дно, при соприкосновении с которым может зачерпнуть ил и этим испортить весь улов. Достаточно небольшого навыка, чтобы по степени натяжения линя в руках приспособиться к требуемой скорости движения, которую планктонист регулирует соответственной командой гребцу. Таким способом ловят в течение 3—5 минут. Затем лодку останавливают, и планктонист, ни на один момент не ослабляя натяжения линя и сохраняя этим прежнюю скорость движения сетки, постепенно выбирает линь; когда сетка достигнет лодки, планктонист придаёт ей вертикальное положение и вынимает из воды. Если материя на сетке не очень редкая, то по поднятии её на воздух в ней остаётся ещё более или менее порядочное количество воды, которая сначала стекает довольно быстро, но потом всё медленнее и медленнее, так как, чем плотнее материя сетки, тем хуже она пропускает через себя воду, особенно благодаря закупорке её отверстий планктическими организмами (иногда случается, что вода совершенно перестаёт вытекать из сетки}; в таком случае для ускорения фильтрации сетку нужно свободной рукой слегка потряхивать и поглаживать до тех пор, пока вся материя её не освободится от воды, которая вместе с пойманным планктоном останется лишь в привязанной к нижнему концу сетки баночке или воронке.
После этого отвинчивают нижнюю часть стаканчика и опрокидывают её пад широко-гордым сосудом, входное отверстие которого должно быть шире стаканчика, чтобы последний свободно входил в него. Чтобы смыть с сеточки, которой затянуто дно стаканчика, приставший к ней планктон, нужно поливать на неё сверху из баночки зачерпнутой в озере водой, поворачивая в то же время нижнюю часть стаканчика во все стороны так, чтобы стекающая через сетку внутрь его вода омывала бы вместе с тем и внутреннюю поверхность боковой стенки стаканчика. Удостовериться в том, достаточно ли промыта сетка я не осталось ли на ней организмов, можно просматриванием её на свет с помощью ручной лупы (при некотором опыте и навыке можно это делать и простым глазом). Если вместо стаканчика на нижнем конце планктической сетки прикреплена воронка, то стоит лишь открыть зажим трубки чтобы перелить воду с планктоном из воронки в склянку. Не следует ограничиваться только одним ловом. Так как на различной глубине состав планктона обычно бывает неодинаков, то для того, чтобы иметь более или менее полное представление о планктоне, нужно производить ловы в разных слоях воды, — во всяком 12
случае, не менее чем в двух: у самой поверхности и на какой-нибудь средней глубине, стараясь лишь, как указано выше, не допустить прикосновения сетки ко дну.
После каждого лова сетку необходимо споласкивать, что производится следующим образом. Сетка с открытым пижним концом осторожно опускается в воду до верхнего края — так, чтобы не зачерпнуть воды через этот край и чтобы вода профильтровывалась только через сетку снаружи внутрь. Затем сетку снова вынимают из воды. Проделав такие вертикальные движения сеткой поочерёдно вверх и вниз (не опуская верхний край её в воду) несколько раз, можно быть уверенным, что сетка достаточно промыта. Тогда закрывают нижний конец сетки, как полагается в зависимости от устройства (стаканчика или воронки), и она готова для следующего лова.
В зависимости от того, для какой цели собирают планктон, с уловами (пробами) последнего поступают различным образом. Если хотят ограничиться лишь просмотром под микроскопом живого планктона, то собранный планктической сеткой материал сливают в крупный по объёму сосуд (не менее литра, лучше — больше), чтобы привезти планктон, по возможности, в неповрежденном виде. По возвращении с экскурсии воду с планктоном вновь пропускают через сетку, чтобы сгустить всю пробу в небольшом количестве воды, берут отсюда пипеткои две-три капли на часовое или предметное стекло и помещают под микроскоп. Если же ставят себе целью определить хотя бы некоторые из плапктиче-ских организмов, то собранный плапктоп необходимо сохранить в неповреждённом состоянии в течение более или менее длительного периода времени, для чего его фиксируют 2— 3-процентным формалином. Для этого в баночку, в которой находится сконцентрированный в небольшом количестве воды планктон, прибавляют на глаз (Vu—1/ц этого количества) крепкий, 40-процеятный (такую именно крепость имеет продажный раствор) формалин.
В каждую склянку с материалом нужно положить этикетку с номером лова и в экскурсионной тетради против каждого номера записать следующие сведения:
1) Место лова.
2) Время лова (месяц, число и время дня).
3) Глубина водоёма в данном месте.
4) Глубина лова.
Условия жизни на дне
Дно (пелагиали) слагается из оседающего из воды материала. В более старых озёрах первоначально слагавшие его минеральные элементы, происшедшие от размывания его дна, лежат уже под слоями отложений, отчасти выпадающих из самой озёрной воды, отчасти приносимых с суши в виде минеральной или органической пыли, или остатков наземных растений.
После смерти планктических организмов их трупы начинают разрушаться и погружаться. Вместе с ними погружаются и остатки наземных организмов, принесённые в озеро с берега. Вся эта масса органических остатков падает на дно и присоединяется к разрушающимся трупам организмов, населяющим самое дно озера. В озёрах с малой производительностью живых организмов процесс накопления иловых отложений на дне идёт очень медленно не только вследствие малого количества организмов, но и вследствие того, что у подавляющего большинства их разложение трупов, благодаря значительному содержанию кислорода в окружающей воде, доходит до конца, т. е. до полной минерализации органических частиц; в результате этого органический материал вновь переходит в минеральные вещества, идущие на питание растений; круговорот начинается снова.
Но во многих озёрах дно впадины покрыто уже более или менее мощным слоем ила. Придонный слой воды в таких озёрах чрезвычайно беден кислородом, а нередко и совсем лишён его. Такой ил очень богат органическими веществами, которые накопляются в нём вследствие того, что падающие на дно трупы организмов или их остатки в условиях кислородного дефицита подвергаются гниению, причём они не разлагаются до конца. Такие иловые отложения, содержащие большое количество остатков планктона и бентоса, называются гиттией, или сап ропелем. Они представляют собой маслянистую, тонкозернистую массу. В той или иной мере к сапропелю (гиттии) примешиваются остатки растений литоральной зоны, особенно после отмирания летней растительности.
В озёрах, воды которых богаты гумусовыми кислотами, последние оседают на дно в виде студнеобразной бурой массы и, примешавшись к илу, придают ему коричневый оттенок. Такие илы носят название д ю, или д ь ю.
Необходимо вспомнить, что богатая гумусовыми кислотами вода имеет небогатое население и потому даёт мало сапропеля.
Таким образом, можно отличить три основных типа донных отложений, связанных, конечно, рядом переходов: 1) и лы, бедныеорганиче-скими веществами, оседающие в озёрах с чистой, богатой до дна кислородом и бедной планктоном водой; 2) илы, богатые сапропелем, оседающие в водоёмах, бедных у дна кислородом и богатых планктоном, и 3) илы, богатые дью, оседающие в водоёмах, вода которых содержит много гумусовых кислот и бедна кислородом и планктоном.
На дне глубоких озёр царит чрезвычайно большое однообразие в условиях жизни во многих отношениях в течение всех сезонов: однообразие температуры, постоянное отсутствие волнений, отсутствие растений во все сезоны. Это отражается и на населении дна: например, типичные донные обитатели не впадают в зимнюю спячку. Интересно, что на раковинах моллюсков со дна глубоких озёр нельзя видеть концентрических полос, которые вызываются у других моллюсков чередованием сезонов с богатым питанием и сезонов с приостановкой питания.
БЕНТОС(НАСЕЛЕНИЕ ДНА)
Фауна названных типов илов различна по составу своих представителей, по видам и иногда родам, но характерные для неё животные принадлежат к немногим группам. Фауна эта населяет верхние слои ила обычно приблизительно на 10—20 см глубины. Она состоит из организмов, приспособленных к своеобразным условиям жизни в илу на глубине. Прежде-всего, конечно, из состава населения исключаются все те формы, которые связаны с высшими растениями, а затем и те, которые требовательны по отношению к кислороду.
Нельзя ожидать встретить здесь, конечно, формы прикреплённые, так как нет твёрдых предметов, а также формы, дышащие атмосферным воздухом, так как расстояние до него слишком велико.
Рис. 34.
А — червь трубочник — Tublfex; Б—мотыль личинка Ten di pes plain оз us.
Пища обитателей дна состоит главным образом из разлагающихся органических остатков ила. Прошедший через кишечный канал ил выходит оттуда в виде шариков, комочков, содержащих уже немного органических веществ. Есть здесь и небольшое количество хищников: личинка двукрылых насекомых Chaoborus (Corethra), питающаяся ракообразными, и Tanypus, пишей которой служат олигохеты. Так как конкуренции здесь мало, то обитатели ила почти всегда развиваются массами.
Наиболее типичными группами бентоса надо считать личинок комаров-дергунов из родов Tendipes (Chironomus, рис 34 Z>), Tanypus и Tany-tarsus, личинку комара Chaoborus и червей из олигохет (малощетинковых), например, Tubifex (рис. 34 А). Наиболее распространёнными наиболее известным из всех этих организмов является личинка Tendipes plumosus, называемая в продаже мотылём; личинка эта содержит в крови гемоглобин, связывающий, подобно гемоглобину человеческой крови, ки-74
слород и отдающий его очень постепенно, почему опа может жить и при очень низком содержании кислорода. Личинка Chaoborus (Corethra)1 тоже способна к пребыванию в бескислородной среде, но у неё это обусловливается тем, что в ночное время личинка всплывает к поверхности и здесь забирает свежий воздух в пузырьки, расположенные вблизи переднего и заднего концов тела.
Типичными формами для сапропелевого ила являются: из насекомых — личинки Tendipes, из групп Plumosus и Bathophilus, и Chaoborus; изчервей — Ilyodrilus hammoniensis. Количество особей всегда очень большое.
Типичными формами для д ь ю являются тоже Tendipes (группы Plumosus) и Chaoborus, но к ним обычно присоединяется в большом количестве Culicoides. Количественно фауна всегда очень бедна.
Формы бедного органическими остатками ила иные: здесь господствуют личинки Tanytarsariae (главным образом Eutany-tarsus), которые более требовательны к кислороду, но зато менее требовательны к пище, чем личинки Tendipes из группы Plumosus и личинки Chaoborus (ни тех, ни других в типичных озёрах с бедным илом и большим содержанием кислорода мы не находим). Из олигохет здесь живёт Peloscolex ferox. Количество видов иных, менее распространённых организмов в этих озёрах много больше, чем в озёрах первых двух типов, так как здесь условия жизни менее отличаются от условий жизни в литорали, если не считать отсутствия высших растений, и потому некоторые литоральные формы могут опускаться в глубину, примешиваясь к истинно глубинному населению. Количественное богатство фауны различно — большей частью оно не велико; иногда, благодаря прозрачности воды и прониканию в глубину света, хотя и слабого, на дне развивается зелёный покров из некоторых водорослей, за счёт которых и существует богатая фауна.
Кроме организмов, доступных наблюдению простым глазом, дно даёт приют богатой и разнообразной микрофлоре в лице главным образохм бактерий и водорослей и микрофауне в лице простейших, коловраток и ракообразных. Но на них мы не будем останавливаться ввиду трудностей работ над ними.
ТЕМЫ ДЛЯ РАБОТ
Исследование профундали должно захватывать донные отложения (биотопы) и их обитателей (биоценозы).
Приборы, служащие для добывания образцов 1уунта, по своей сложности и дороговизне доступны лишь для специальных учреждении. Поэтому о характере грунта приходится судить на основании того материала, который захватывается драгой и скребком. Конечно, надо постараться выяснить состав оиоценозов различных донных отложений, а также различных глубин, так как иногда одинаковые отложения дают на разных глубинах различные пробы.
Затем очень интересно изучить население тех участков водоёма, куда достигает влияние человека, — загрязнение отбросами хозяйства, мытьём белья, пастьбой скота, фабричными отбросами и т. п. Также интересно узнать население тех мест, куда сносится притоками чуждая данному водоёму донная фауна, — выживают ли новые пришельцы и какие именно.
I. Непериодические наблюдения над бентосом профундали
1. Первоначальное знакомство с бентосом.
2. Взять драгой бентос с различных глубин и грунтов и сопоставить но глубинам (с однородным грунтом) и по грунтам (с одинаковых глубин).
3. Выяснить особенности грунта и бентоса у впадения притоков.
4. Выяснить особенности грунта и бентоса в местах загрязнения.
II. Периодические наблюдения
Наблюдения по сезонам могут дать интересные результаты в вопросе о развитии личинок двукрылых (моменты появления молоди, окукливания); такие наблюдения особенио
1 Нужно, впрочем, заметить, что личинка Chaoborus (Corethra) не может считаться, строго говоря, настоящим донным организмом, потому что живёт не в грунте и даже не на грунте, а в воде, плавая лишь над самым дном, вследствие чего она попадается как в донные орудия.так и в планктическую сетку; поэтому её следует скорее называть не донным, а придонным или наддонным организмом.
важны в практическом отношении, так как бентос является главной пищей большинства пресноводных взрослых рыб.
Пробы бентоса следует брать периодически — не реже двух раз в месяц (в течение летнего сезона); весной (в"течение первого месяца по вскрытии льда) и осенью (в августе в в течение последнего месяца перед первым появлением льда) желательно брать пробы чаще — не менее трёх раз в месяц.
ME ТО ДИКА
Оборудование. Драга. Это орудие лова употребляется для собирания донных животных в центральной глубоководной части водоёма и состоит из трёх- или четырёхугольной металлической рамы, к которой по одну сторону прикрепляется мешок из конгресс-канвы, а по другую — верёвка, так называемый трос, за который тянут драгу по дну. Драги бывают разной конструкции и величины. Мы опишем здесь наиболее простое устройство этого инструмента.
Железная полоса толщиной миллиметра в 2, шириной в 30—35 мм и длиной в 1 м пли немного более сгибается в виде равносторонней треугольной рамы (рис. 35 А, Б), свободные концы её скрепляются на одной из вершин треугольника. Как видно из рисунка, с одной стороны к этой раме прикрепляется верёвочный трос, а с другой — мешок, так называемая «мотня». Со стороны троса края рамы немного отгибаются наружу и заостряются, что ясно видно на рисунке35Б, это делается для того, чтобы драга при Движении по дну водоёма, врезаясь в грунт, лучше захватывала его. Вблизи противоположных краёв рамы на всех трёх сторонах её делается ряд отверстий миллиметра в 2 диаметром на рас-
стоянии приблизительно 15 мм друг от друга; эти отверстия служат для прикрепления к раме мотни посредством продеваемого в них шнура. Мотня представляет собой мешок, несколько суживающийся по направлению от рамы и слепо замкнутый на конце; он делается из конгресс-канвы, но при отсутствии последней она может быть заменена достаточно прочной, но редкой тканью, ячеи которой должны иметь диаметр не менее 1 мм (например, мешочный холст для овсяных мешков); длина мотни раза в полтора больше, чем длина каждой стороны рамы. Для прикрепления троса к раме в углах последней вделываются металлические кольца, которые могут быть сделаны из достаточно прочной проволоки, свитой в два-ijpH раза, или даже из толстой верёвки. К этим кольцам крепко привязываются небольшом длины верёвки, которые другими своими концами скрепляются над центром рамы как между собой, так и с концом троса; это скрепление можпо осуществить или тоже посредством кольца, или же, за неимением его, — непосредственным связыванием, причём проще всего это сделать следующим образом: вместо трёх отдельных коротких верёвок берём верёвку вдвое большей длины и концы её привязываем к двум кольцам рамы; к третьему же кольцу привязываем непосредственно конец троса, которым затем перевязываем середину первой верёвки так, чтобы все три верёвки, ооразуюгаие рёбра трёхгранпой пирамиды, основанием которой служит рама, были одинаковой длины. Верёвка для троса должна быть не тоньше, а лучше даже толще той, какая употребляется для вожжей; длина же троса берётся в зависимости от глубины водоёма — от нескольких метров до нескольких десятков метров.
Производетво работ. Для драгирования необходима лодка, так как работы производятся посредине водоёма. Перед началом работы лодку останавливают, и драгировщик, помещающийся на корме, спускает драгу. Тот момент, когда драга достигла дна, отмечается исчезновением ощущения тяжести в руках лрагировшнка и ослаблением троса. С этого момента лодке дают ход, а драгировщик начинает травить трос, свободно пропуская его через рукп. Это продолжается до тех пор, пока трос не примет такое направление, при котором он будет составлять с поверхностью воды угол меньше 45° (это легко определить на глаз). Тогда драгировщик перестаёт травить и, удерживая трос в руках, начинает тянуть драгу. При этом гребцу приходится, конечно,* несколько приналечь на вёсла, чтобы преодолеть сопротивление драги движению лодки. Драгировщик, руководствуясь натяжением троса (должно быть ощущение среднего натяжения — не слишком сильного и пе очень слабого), регулирует движение лодки, отдавая гребцу соответственную команду: «тише», «быстрее» и т. п.
Лов производится в течение трёх-пяти минут. После этого гребеп прекращает свою работу, а драгировщик начинает вытаскивать драгу. Когда последняя покажется из воды, 76
on берёт её эа раму и, держа всё время мотню в воде, начинает двигать ею по все стороны; это делается с целью промывки принесённого драгой материала, во время которой через ячеи мешка уходят из пего все мелкие частицы (ил), более же крупные, в том числе и многие из живущих па дне животных, задерживаются внутри драги и таким образом попадают в улов. По окончании промывки (этот момент обозначается прекращением выхождсния при промывании из драги в окружающую воду мути) драга вытаскивается на корму лодки, и содержимое её вываливается в таз с водой.
Здесь прежде всего необходимо записать грунт, причём нужно пометить, много ли при промывке ушло из драги ила. Дальнейшая процедура заключается в выборке животных из грунта, с которым они смешаны. Выбирать приходится, конечно, лишь тех животных, которые видпы простым, невооружённым глазом. Непосредственно в тазу это делать неудобно, так как животные зарываютсяв гущу грунта и стараются с поверхности скрыться в глубь его. Поэтому содержимое таза нужно просматривать отдельными небольшими порциями. Для этого обыкновенным чайным блюдцем или мелкой тарелкой зачерпываем небольшое количество грунта из таза и разбавляем его водой; тогда попавшие сюда животные становятся гораздо виднее, особенно благодаря своему движению; немногие лишь животные остаются неподвижными, как, например, моллюски, скрытые в своих раковинках, но они легко замечаются по этим раковинкам, и потому для собирания не представляют затруднений. Выбираем животных из блюдца пинцетом, стараясь захватывать их как можно осторожнее, чтобы не повредить, и складываем в склянку с 4-процентным формалином г. В каждую склянку необходимо бросить небольшой клочок бумаги с написанным на нём простым карандашом номером лова, а в записной тетради для каждого номера должны быть отмечены следующие сведения:
1) Место лова.
2) Время лова (год, месяц, число и час).
3) Глубина лова.
4) Грунт.
Зафиксированные животные определяются по приведённым во второй части книги определительным таблицам.
Литоральная область. Общая характеристика
Литоральная область, по сравнению с открытой частью озера, обладает многими чрезвычайно типичными особенностями.
Прежде всего она даёт гораздо более разнообразия в отдельных своих участках, нежели открытая часть; вызывается это разнообразие теми геологическими породами, которые подходят к берегу, той или иной крутизной склона, той или иной доступностью для ветров и, следовательно, волнений, большей или меньшей степенью инсоляции и пр., а в связи со всем этим — и тем или иным населением.
Но многие черты являются общими для всей прибрежной области, независимо от характера участка. Наиболее важной особенностью является её температурный режим. Небольшой слой воды днём легко прогревается солнцем — гораздо легче, чем мощный слой центральной части1 2, но зато за ночь мелкий слой воды успевает охладиться сильнее, чем толща центральной части. Таким образом, в литорали получаются более сильные, чем в открытой части, суточные колебания температуры. То же явление, но в более крупном масштабе, наблюдается и по отношению к сезонным колебаниям: зимой прибрежная область иногда совершенно промерзает, а если и не промерзает, то имеет в небольшом слое воды, лежащем непосредственно подо льдом, температуру, близкую к 0°, тогда как в более глубоких частях озера последняя держится выше — около 4°. Когда ранней весной начнёт пригревать солнце, свободная поверхность озера, покрытая ослепительно белой пеленой снега, отражает в большей мере солнечные лучи и прогревается значительно медленнее, чем литораль, где почти всюду есть тёмные группы прибрежных кустов, остатки надводных частей растений и т. п., поглощающие свет и тепло. Когда по берегам озера образуются закраины, вода в них может достигать высоких температур, тогда как вода подо льдом, конечно, хранит
1 Исключение составляют моллюски, раковины которых в формалине размягчаются, вследствие чего моллюсков после их выборки из грунта следует фиксировать разведённым (приблизительно 70-градусным) спиртом.
2 В одном озере констатирована разница в температуре воды в солнечный день между прибрежной и открытой частью в 15<
прежнюю низкую температуру. Всё лето дневная температура в прибрежной области довольно значительно выше, чем в открытой части озера. Такое непостоянство температуры позволяет жить в литорали только тем животным, которые способны переносить её резкие колебания.
Малой толщиной водного слоя обусловливается также значительно большее количество кислорода даже в тех частях прибрежной области, где это не связано с присутствием производящих кислород зелёных растений. Та же незначительная глубина вызывает проникание света до самого дна, что и влечёт за собой возможность поселения зелёных растений.
Но как уже указывалось, не во всех местах литоральной области присутствуют растения; свободные от растительности участки имеют настолько своеобразный характер, что более подробное описание их в дальнейшем изложении будет выделено в самостоятельный раздел.
УЧАСТКИ ЛИТОРАЛИ, ПОКРЫТЫЕ РАСТИТЕЛЬНОСТЬЮ
Растения прибрежной области можно разбить на биологические группы, в зависимости от их отношения, во-первых, ко дну и, во-вторых, к водной поверхности. Прежде всего громадное количество их укореняется в грунте и сравнительно незначительная часть живёт свободными странниками в водной массе. Эта вторая группа растений живёт лишь в таких местах, где вода относительно спокойна, так что перенос их на далёкие расстояния, за редкими исключениями, не имеет места. Напомним, что и многие из тех, которые связаны с грунтом, используют его, главным образом, лишь как опору. Поэтому отношение к водной поверхности сказывается на организации и биологии растений значительно резче, чем отношение к дну.
По отношению к водной поверхности растения могут быть разбиты на несколько групп. К первой группе относятся такие, вся жизнь которых проходит в толще водной массы, вплоть до размножения и появления нового поколения. Представители этой группы занимают сравнительно глубокие места прибрежной области, спускаются вниз вплоть до той границы, где глубина позволяет дойти до дна достаточному количеству солнечных лучей, необходимых для питания растений. Из представителей этой группы в наших водах чаще всего встречаются бесцветковые: лучица — Chara, Nitella (рис. 36), мхи — Calliergon (pHc.69),Fontinalis (рис. 37 А) и одно единственное цветковое растение роголистник — Ceratophyllum (рис. 110); возможность перекрёстного опыления у этого растения обусловлена тем, что пыльца, отрывающаяся от цветов, имеет удельный вес, равный единице, и потому свободно носится в водной толще, пока не прикоснётся к рыльцу.
Вторую группу водных растений составляют те, которые проводят под водой свою индивидуальную жизнь, но выдвигают на воздух цветы, чтобы предоставить заботу об их опылении ветру или перелетающему с цветка на цветок воздушному насекомому. Эта группа чрезвычайно обширна: она охватывает прежде всего (за единственным исключением) всю обширную семью рдестов — Potamogeton, имеющих очень сильное распространение в наших водах, и чуму наших вод — элодею — Elodea (рис. 38 А), иногда разрастающуюся на громадных пространствах; помимо того, сюда входят весьма многие, хотя и не образующие очень больших скоплений, но сильно распространённые растения: пузырчатка — Utricularia (рис. 106), лютик — Ranunculus (рис. 107, 108), уруть — Myriophyllum (рис. 111), турча— Hottonia (рис. 109) и др.
Так как растения первой и второй групп своими вегетативными частями всегда погружены в воду, то они и получили название погружённых, или подводных, растений.
Третья группа охватывает растения, поднимающие до границы воздуха свои листья, как всем известная кувшинка, земноводная гречиха и другие, или листовидно расширенные стебли, как тоже всем известная мелкая ряска, иногда затягивающая сплошным ковром небольшие 78
Рис. 37.
А — МОХ FOQtinalid; Б — расходник — leogtes.
тихие водоёмы. Цветы этих растений или также плавают по воде, или поднимают свои головки в воздух. Эти растения отчасти используют воздух и для целей питания, так как они имеют на верхней стороне листьев устьица. Представители этой группы носят название растений с плавающими листьями.
Наконец, четвёртая группа охватывает растения, уже на половину связанные с воздухом, выставляющие над по-
верхностью воды большую или меньшую часть своих зелёных стеблей и листьев, использующих газы воздуха через устьица уже в гораздо большем масштабе. Они как будто спустились с суши, погрузив в воду свои нижние части; они и держатся всегда ближе к берегам, чем все ранее рассмотренные растения. Сюда относятся камыши, тростники, хвощи и т. п. Живя по краям зеркала воды, эти полуназемные растения подвергаются действию более сильных, чем на суше, ветров, вследствие чего им должна быть свойственна очень большая устойчивость по -отношению к ветрам; чем это достигается, уже было указано ранее. Очень большую роль для устойчивости играет их способность к сильному вегетативному размножению, благодаря которой они образуют сплошные заросли очень плотно друг к другу стоящих стеблей. Растения четвёртой группы называются надводными.
Прибрежные заросли одевают широкой полосой берега наших озёр большей частью совсем без перерывов. Лишь очень открытые берега подветренной стороны бывают лишены крупной растительности. Обычно различные типы растений—поднимающиеся над водой, с плавающими листьями, погружённые, располагаются отдельными полосами, группируясь главным образом в зависимости от глубины. У самого берега прежде всего тянутся заросли тростников, камышей, хвощей и т. п., образуя над поверхностью воды густую щетину из узких, тесно стоящих высоких стеблей и линейных листьев. Далее к центру озера камыши уступают своё место целому ряду растений, не под-
мени, на протяжении ряда лет он оолее или
Рис. 38.
А — водяная чума — Elodea; Б — водяная сосенка — Hippurib vulgaris; В — лягушатник — Hydrocharis morsus ranae.
нпмающихся выше уровня воды, за исключением лишь периода цветения, но или распластывающих свои листья по воде (кувшинка), или поднимающих их почти до поверхности, так что они прекрасно видны через тонкий слой воды (рдесты). Далее уже идут те растения, которые близко жмутся ко дну и обнаружить которые, наклонившись над водой, трудно. Часто, однако, за-росли различных типов заходят одна в другую и дают смешанные растительные сообщества, а в связи с этим и смешанные биоценозы.
Кроме того, видовой состав зарослей не остаётся одним и тем же во вре-менее значительно меняется. Происходит это оттого, что, во-первых, растения истощают почву, высасывая из неё необходимые им соли, а во-вторых, растения некоторых видов выделяют в грунт (дно водоёма), в котором они укрепляются, вредные для себя вещества и этим сами прекращают своё дальнейшее развитие и погибают. На их месте развиваются другие виды, для которых эти вещества не являются вредными. Так происходит с м е-н а видового состава зарослей в водоёме вследствие тех изменений окружающей среды, которые вызываются жизнедеятельностью самих же растений (сукцессии).
В дальнейшем мы всё же охарактеризуем отдельно условия, господствующие в участках литорали, занятых определёнными типами растений, а теперь рассмотрим вкратце дно зоны зарослей.
При отмирании высших зелёных растений части их падают в воду; сначала, ввиду богатства растительных тканей воздухоносными путями, обрывки эти плавают на поверхности воды, а затем, по мере пропитывания водой,
начинают опускаться на дно. Какова же их судьба? Когда растительные трупы лежат на поверхности земли, судьба их зависит от того, попадут ли они в условия обильного доступа кислорода, или кислорода будет недостаточно. В первом случае они просто истлеют, оставив после себя незначительное количество неорганического вещества (золы), во втором они перегниют и дадут богатый углеродом перегной. Пока отмершие части водных растений плавают на поверхности, они находятся в условиях, вызывающих тление, но, очутившись ближе ко дну, они попадают или в среду, богатую кислородом, и тогда тление продолжается, или в среду, сравнительно бедную кислородом, если дело происходит в густых, мощных зарослях, не допускающих волнений в не пропускающих достаточного количества света и, следовательно, препятствующих развитию в воде хлорофилле
поносных растений и их частей; в последнем случае растительные остатки не могут уже разложиться до образования неорганических соединений и остаются в виде твёрдого, ещё богатого углеродом вещества, которое называет’ ся т о р ф о м. К торфу примешивается сапропель, и иловые отложения прибрежной области носят то более торфяной, то более сапропелевый характер, в зависимости от господства тех или иных отложений.
Фауна прибрежной полосы зарослей является теснейшим образом связанной с растительностью. Присутствие крупных растений задерживает колебание воды и создаёт вблизи берегов тихие, спокойные уголки, в которых указанные выше особенности температурного, кислородного и светового режима создают чрезвычайно благоприятные условия для существования животных в летнее время: высокая температура, отсутствие волнений (по крайней мере, сильных), богатство кислородом и растительный покров, оказывающий животным громадные услуги. Значение растений для животных в воде заключается, конечно, прежде всего в том, что растения в живом виде или после смерти дают животным пищу. Вспомним, что животные не могут быть производителями (продуцентами) органического вещества, они только — потребители (консументы) того запаса, который собирают растения. Водные животные редко питаются крупными растениями, но они соскабливают с пих налёты водорослей или роются на дне, поглощая разлагающиеся остатки. Количество разлагающегося органического вещества, поставляемое прибрежными растениями, громадно, и часть его, как мы уже видели, поступает и в пользование донных животных глубинной области.
Второе очень важное значение растений для животного мира заключается в обогащении воды кислородом.
Помимо того, растения самым своим присутствием создают благоприятные условия для животных: среди их стеблей и листьев животные могут прятаться от врага и, с другой стороны, могут использовать те же растения как прикрытие на охоте. На стебельках и листьях или внутри них всегда можно найти или самих животных, или их яйца, приклеенные или спрятанные в самую ткань; кроме того, стебли выступающих над водой растений используются как лестницы, по которым дышащие атмосферным воздухом животные добираются до поверхности воды.
По образу жизни представителей фауны зарослей можно разбить на несколько групп, в зависимости от места их обитания. Конечно, свободно живущие формы могут встречаться среди представителей то той, то другой группы, но мы будем рассматривать таких животных в той группе, для которой они наиболее типичны.
1. Среди надводных частей растений (животные, не водные в настоящем смысле этого слова, но так или иначе связанные с водой).
2. На поверхности воды.
3. На плавающих листьях или в их тканях.
4. Внутри растительных тканей, погружённых в воду.
5. Прикреплённые (постоянно или временно) на подводных частях растений.
6. В толще воды среди растений.
7. На дне.
Из животных, живущих среди надводных частей растений, мы упомянем лишь тех, которые имеют непосредственную связь с водой в некоторые периоды их жизни; к таковым относятся многочисленные стаи тех взрослых насекомых, личинки которых живут в воде. Более всего из них бросаются в глаза быстро снующие в погоне за более мелкими насекомыми хищницы - стрекозы (рис. 39, 541), лишь изредка присаживающиеся на стебельке, уцепившись своими нежными ножками, почти не приспособленными для ходьбы. Очень часто встречаются сидящие неподвижно невзрачные, большей частью тёмпосерые ручейники (рис. 536), сложив крышо-образно свои нежные, покрытые волосками крылышки. Сходны с ними по первому впечатлению вислокрылки (Sialis, рис. 40, 41). Так же не-
6 А. Н. Линии
подвижно, приподняв кверху совершенно прозрачные крылышки, сидят и изящные подёнки (рис. 42), прозванные так за свой недолгий век. Несколько реже можно встретить небольших, довольно невзрачных б а б о-
Рис. 39. Схема личин- Рис. 40. Личинка
ки стрекозы из Aesch- вислокрыл ни — Sia-
nidae. lis.
чек, гусеницы которых живут в воде. Большими роями толкутся в воздухе над прибрежными растениями, над водой или вблизи неё комары-дергуны — Tendipedidae (Chironomidae, рис. 532 Б), с довольно крупным телом и очень длинными ножками, сразу позволяющими отличить их от надоедливых кусающих комаров, распевающих тут же свои назойливые песенки. На стеблях иногда удаётся найти комки слизи, заключающие в себе яички некоторых ручейников; когда из яичек разо
вьются личинки, слизь эта разжижается и падает по капелькам в воду, увлекая за собой туда и личинок. Очень часто на листьях тростника и других растений попадаются плотные комочки паутины, в которых находятся яйца пауков, снующих тут же на листьях.
На поверхности воды, на самой границе воды и воздуха, быстро, как конькобежцы, скользят тонконогие длинные клопы водомерки (Hydro-
Р и с. 41. Вислокрылка — Sialis lutaria: взрослые насекомые и яйца (на растениях), личинка (в воде) и куколка (на берегу).
metridae); в солнечный день можно заметить, что отбрасываемая от каждой ножки тень утолщена на конце; это оттого, что тонкие лапки, не смачиваемые водой, вследствие покрывающего их слоя жира, вдавливают немного вниз поверхностную плёнку, не прорывая её, однако (рис. 43). В быстроте бега с ними конкурируют блестящие мелкие жуки вертячки — Gyrinus (рис. 367). Как водомерки, так и вертячки — опасные хищники, причём вертячки могут быстро нырять за своей добычей в глубь; они интересны, между прочим, устройством своих глаз (рис. 368): каждый из них разделён на две части, из которых одна смотрит вверх, а другая под воду. Водомерки вонзают в пойманную добычу свой хобот и высасывают из неё соки, оставляя иногда лишь пустую шкурку. Ловко прыгают тут же, отталкиваясь от поде
верхностной плёнки вилочкой заднего конца тела, лишённые крыльев мелкие подуры — Podura (рис. 281). Это всё постоянные обитатели поверхности, лишь изредка покидающие её.
Но здесь есть ещё и ряд таких организмов, которые поднимаются сюда
лишь по временам: это те насекомые и их личинки и те моллюски, которые, перейдя к водному образу жизни от наземного, сохранили дыхание атмосферным воздухом, за которым они и всплывают. Свесившись вниз раковиной,
скользят по плёнке поверхностного натяжения воды прудовики — Limnaea (рис. 21), время от времени открывая круглое дыхательное отверстие. На очень короткое время поднимаются многочисленные жуки, выставляя для дыхания то передний конец тела,как водолюб — Hyd-
Рис. 43. Клопы — Hemiptera (Rhynchota):
А — водомерка на поверхности воды; Б — личинка Gerris.
Р и с. 4?. Подёнки — Ephemeroptera:
А — на дне — личинка, налево - вылупление нимфы из личинки, направо — взрослые насекомые; Б — личинка Ba£tis; В—личинка Cloeon dipterum; Г—личинка Ephemcrella Ignita; Д — личинка Ordclla macrura; Е — личинка Paraleptophlebia submarginata.
rophilus (рис. 21), то задний, как плавунец —Dytiscus (рис. 21); различные личинки кусающихся комаров—Culex и Anopheles (рис. 434 Б,В) — подвешиваются на минутку у поверхности; личинки мухи ль винки — Stratiomyia — раскидывают здесь же звёздочку волосков, обнажая среди них дыхательное отверстие (рис. 20). Здесь же совершается превращение во взрослых насекомых плавающих в воде куколок тех же комаров, а также подёнок — Ephe-meridae (рис. 42); после вылупления их пустые шкурки ещё долго носятся по поверхности воды. Временами удаётся наблюдать плавающую куколку ручейника, освободившуюся уже из домика, поднявшуюся вверх и носящуюся по волнам, пока не прибьёт её к какому-нибудь растению, за-
*
83
целившись за которое, она сбросит свою шкурку и взлетит на воздух. Тут же плавают и яички комаров кусающихся и комаров-дергунов; яйца тендипедид опускаются через некоторое время, когда набухнет окружающая их слизь, на дно, а из яиц комаров кусающихся выскальзывают тут же в воду молодые личинки.
Часто на поверхности плавают зеленоватые или бурые лепёшки, то, что обычно называют тиной; это поднявшиеся со дна скопления колониальных нитчатых водорослей, всплывшие на поверхность благодаря пузырькам кислорода, которые накопляются среди запутанной массы нитей, составляющих колонию; пузырьки эти появляются в результате процесса питания этих водорослей.
Плавающие листья. Плавающие крупные зелёные островки листьев кувшинки — Nymphaea (рис. 98) и кубышки — Nuphar (рис. 99) и более мелкие — листьев л я гу ш а т н и к а — Hydrocharis morsus гапае (рис. 94), пластинки ряски — Lemna (рис. 96)и Spirodela (рис. 95)и прочих растений
Рис. 44. Мшанка — Plu-matella repens на листе кувшинки.
Рис. 45. Кладки яиц моллюсков:
А— прудовика — Limnaea; Б—катушки — Planorbis; В — битинии — Bithynia.
прежде всего используются некоторыми неводными животными: частенько можно видеть па них сидящих комаров, жуков и т. п.; часто встречаются здесь жуки Galerucella и Donacia, питающиеся тканью листьев. Водные животные не держатся, само собой понятно, на верхней поверхности, так как на ней нет воды, которая легко скатывается с этой поверхности, потому что последняя покрыта восковым налётом. Но зато нижняя поверхность плавающих пластинок — целый мир разнообразных существ. Некоторые из них используют листья как постоянную опору: охотно поселяются здесь мелкие, едва заметные простым глазом, полупрозрачные гидры — Hydra (рис. 9, 141,142), залавливающие своими щупальцами проплывающие мимо микроскопические организмы. Селятся на поверхности листьев и изящные веточки мшанки — Plumatelia repens (рис. 44). Очень многие организмы откладывают на нижнюю поверхность листьев свои яйца, то заключённые лишь в слизь (бабочка огнёвка-кувшин н и ц а — Nymphula nymphaeata, моллюски, рис. 45, жук-радужница—Donacia, ручейники, рис. 46), то прикрытые коконом (пиявка улитковая), то погружённые в самую ткань растения (стрекоза красотка — Agri on).
Медленно ползают по нижней поверхности листа более или менее часто встречающиеся здесь моллюски: прудовик — Limnaea (рис. 200), физа —Physa (рис. 197), чашечка —Ancylus (рис. 196) и др., соскабливая налёты водорослей. Плотным маленьким прозрачным комочком сидит иногда массами мелкий рачок сида — Sida crystallina (рис. 236), ползают мелкие ресничные черви — Turbellaria (рис. 20). Тут же вьются в прозрачных слизистых трубочках личинки мелких тендипедид из Orthocladiinae.
Рис. 46. Кладки яиц ручейников — Trichoptera; А — Limnophllus; Б— Phryganea; В — Triaenodca.
Рис. 47. Кладки яиц стрекоз — Odonata:
А — стрелки — Coenagrion pulchellum на нижней стороне листа; Б — лютни — Lestes на стебле частухн подорожника (налево вид снаружи, направо в разрезе); В — дедки — Gomphus; Г — стрекозы — Libellula; Д — бабки — Cordulia аепеа на лучице; Е— Sympetrum на водяном мху; Ж—Epitheca blmaculata на элодее.
Ткань листьев, особенно кувшинок, кубышек и рдеста, служит убежищем для некоторых форм: здесь прокладывают себе узкие и длинные коридоры целый ряд минирующих личинок Tendipedidao (рис. 24) и прогрызают ходы молодые гусенички — Nymphula nymphaeata; в более старшем возрасте эти гусеницы отгрызают целые куски листьев и приклеивают их к нижней стороне листа, устроив защитную крышу; ещё позднее гранича-
щпй с этой крышкой кусок листа выгрызается нацело, так что получается свободный домик, плавающий часто тут же между листьями (рис. 426), Подводные части растений. Внутри растительных
тканей—в черешках листьев кувшинковых и рдестов, в стеблях камыша и в других сочных подводных частях — почти всегда можно видеть просвечивающий сквозь тонкий слой кожицы коридор, внутри которого вьётся красноватая личинка из Tendipedidae; ра
Рис. 49. Кладка яиц жука плавунца — Dytiscus.
Р и с. 48. Кладки яиц клопов — Hemiptera:
А — гребняка — Corixa; Б — плавта — Naucoris; В — водомерки — Gerris.
найти и мелкую
у неё нет, как
недоступно, так как
яиц клещей — Hydracarina:
Рис. 50. Кладка
А — Plona; Б — Hydrophantes; В — Llmnochares; верхний ряд — кладки на растениях, нижний — отдельно (у велич.).
о о
□ о
о. о
зорвав такой стебелёк, часто в том же ходе можно олигохету, пользующуюся готовым помещением, сделать которое ей у личинки тендипедид, твёрдых челюстей. Внутри подводных растительных частей очень многие насекомые откладывают свои яйца: стрекозы — Erythrom-ma, Aeschna, Lestes (рис. 47) и др., клопы (рис. 48) Noto-necta, Naucoris, Nepa, водомерки семейства Gerridae, очень многие жуки (рис. 49). Ещё большее количество животных приклеивают свои яйца на поверхности подводных частей: таковы планарии, из стрекоз— Libellula (рис. 47 Р), из клопов —ко ри к сы (рис.48А), из моллюсков — прудовики, катушки, б и т и и и и (рис. 45), из пиявок —Nephelis, наконец, клещи (рис. 50).
Стебли крупных растений часто обрастают зеленоватым налётом нитей колониальных водорослей, образующих причудливые нежные кустики, просвечивающие через воду или склеенные студенистым вегце-
О
а Р б'о
ством в общую массу, в виде комочков шарообразной пли неправильной формы. Как кустики, так и комки водорослей являются излюбленной пищей для очень многих личинок насекомых; укажем для примера, что комки водорослей составляют преимущественную пищу личинок малярийного комара — Anopheles.
Часто подводные части растений служат опорой для прикреплённых сидячих коловраток, гидр (рис. 9), губок и мшанок (рис. 163 А). Обе последние формы образуют наросты различного вида, представляющие собой колонии мелких отдельных индивидуумов; питаются, как и гидра, микроскопическими организмами. Отличить их по форме, в особенности на воздухе (т. е. вынув из воды), для начинающих исследователей трудно, так как и те и другие могут образовать как сплошные наросты, так и свешивающиеся лопастные кустики; когда же они находятся в воде и притом в спокойном состоянии,они становятся несколько различимы по внешнему виду: губка остаётся неизменной, а мшанка покрывается белым пушистым колеблющимся налётом; это выставляются прятавшиеся на воздухе отдельные особи в виде звёздочек. Впрочем, есть и такие мшанки, которые могут быть узнаны сразу по форме своих тонкоразветвлённых колоний: это Plumatella repens (рис. 44). Колонии губок и мшанок дают приют другим животным: личинкам Tendipedidae, червям из Oligochaeta, а иногда и личинкам ручейников. Так как для указанных форм растения являются лишь опорой, то с таким же успехом все эти формы селятся и на мёртвых растениях, находящихся под водой, на сваях купален, мостков и т. п.
На нижних частях стеблей некоторых растений, выносящих свои верхние концы в воздух, висят куколки бабочки огнёвки Nymphula в своих листовых домиках и куколки жука радужницы Donacia в плотных коричневых коконах: обе куколки дышат атмосферным воздухом и пользуются воздухоносными тканями растения как вентиляционными воздушными трубами, соединив себя с ними посредством отверстий.
По стеблям ползают различные моллюски, личинки ручейников, гусеницы, личинки подёнок — в поисках пищи преимущественно из налётов водорослей, а иногда и из самой растительной ткани; цепляясь за растения, подстерегают добычу пиявки (рис. 10), личинки вислокрылки — Sialis (рис. 40, 41), водяной с к о р п и о н — Nepa (рис. 20), Ranalra и другие хищники, а среди подводных листьев прикрепляет колокол из выделений паутинных желез на конце брюшка единственный из истинно водных пауков наших мест серебрянка — Argyroneta (рис.21); построив колокол, паук наполняет его атмосферным воздухом и затем пользуется им как жилищем для себя и гнездом для молоди.
Кроме отдельных видов животных как одиночных, так и колониальных, временно или постоянно живущих на подводных частях растений, здесь встречаются ещё особые образования, так называемые на росты: это хлопьевидные или войлокообразиые покровы на подводных стеблях, занимающие более или менее значительную часть последних. Наросты состоят обычно из детрита и различных мелких организмов как растительного, так и животного происхождения, которые находят себе здесь пищу. Таким образом, наросты представляют собой своеобразные биоценозы, в состав которых в разных сочетаниях входят водоросли (в особенности — зелёные и диатомовые) и мелкие животные, например, ресничные и круглые черви, ракообразные, личинки тендипедиди некоторых других насекомых.
От наростов отличают обросты — подобные же биоценозы на неживых подводных предметах: камнях, сваях, корягах и т. п.
В толще воды между зарослями снуют, временно задерживаясь на растениях, разнообразные хищники; жуки и их личинки, к л о п ы и их личин-,ки, клещи, серебрян-ка, некоторые личинки подёнок, некоторые личинки стрекоз (с тонкими жаберными листочками на конце тела,
рис. 285). Тут же плавают и микроскопические животные планктона из коловраток (рис. 27, 28) и р а ч к о в.
На дне между растительными остатками население тоже достаточно богато. Двустворчатые моллюски: беззубки—Anodonta (рис. 212), шаровки — Sphaerium (рис. 215) сидят, приоткрыв свою раковину и пропуская через себя воду со взвешенными в ней частицами, служащими им пищей; бегает по дну водяной ослик — Asellus (рис. 221); там и сям ползают личинки вислокрылки и ручейников, роясь в растительных частичках; здесь же сидит и целый ряд хищников, замаскировавшись тёмной окраской своего тела среди грубых растительных остатков: это клопы плавты — Naucoris (рис. 349) и личинки стрекоз— Libellula (рис. 298), Cordulia, Epitheca (рис. 295) и др.
Рассмотрим теперь отдельные типы зарослей как биотопы с характерными для них биоценозами.
Область поднимающихся над водой растений. Заросли, образующие эту область, слагаются главным образом из камыша — Scirpus (рис. 70), тростника — Phragmites (рис. 75), трезубки — Scolochloa (рис. 77), рогоза — Typha (рис. 84, 85) и х в о щ а — Equisetum (рис. 71, 72).
Все эти растения обладают мощными многолетними корневищами; стебли и листья их отмирают осенью, но весной снова появляются чрезвычайно густой щёткой. Большая стойкость тростников и других живущих здесь растений против действия волн позволяет этому поясу занимать иногда и подветренные берега, где невозможно присутствие зарослей рдестов — растений, гораздо менее устойчивых против волн.
Большая густота в типично выраженных участках оказывает значительное влияние на физические и химические условия в воде, мешая, с одной стороны, прониканию в воду солнечного света и, следовательно, сильному развитию фитопланктона, с другой стороны, препятствуя освежению воздуха над поверхностью воды и, следовательно, освежению газового состава. Недостаток кислорода создаёт условия для торфообразования, а мощная растительность даёт для этого богатый материал.
Сами растения живут, в сущности, почти наземной жизнью, заимствуя газы для дыхания и питания и солнечный свет непосредственно из атмосферы.
Для животных в густых зарослях создаются неблагоприятные условия. Поэтому животные избегают таких зарослей: среди них живёт небольшое количество форм и в небольшом количестве особей; здесь селятся преимущественно моллюски, водяной ослик — Asellus и личинки ручейников— Trichoptera.
В менее густых зарослях, среди торчащих над водой стеблей, распластывают по воде своп листья кувшинки — Nymphaea, лягушатник — Hydrocharis и другие подобные им растения; в таких местах создаются тихие и хорошо освещённые и прогретые уголки, где находит приют разнообразная и богатая фауна.
Иногда описанные заросли образуют пояс на некотором расстоянии от берега, так называемый «барьер»; между ними и берегом образуются тихие, хорошо освещаемые и прогреваемые участки воды, представляющие очень благоприятные условия для развития фауны и для выкормки мальков рыб.
Заросли растений с плавающими по поверхности листьями. Растения с плавающими листьями часто занимают в озере очень большие пространства. Чаще всего они образуют бордюры по краям тростника и камыша, а иногда в тихих заливчиках — целые зелёные ковры. Распластанные по поверхности воды цельные, большей частью крупные листья умеряют движение воды, гасят возникающее волнение; в ветреный день ещё издали бросается в глаза занятая ими площадь, как тихий островок среди изборождённой волнами поверхности; только встают по всему островку зелёные гребешки приподнятых ветром краёв листьев кувшинки или кубышки. Редко заросли подобного 88
Рис. 51. Ходы личинки мухи Hydromyza livens в листьях кувшинковых.
типа бывают вполне однородными: большей частью тут господствуют Nym-phaea, Nuphar или Polygonum (рис. 100); к ним в большей или меньшей мере присоединяются: лягушатник — Hydrocharis morsus гапае (рис. 94), п а в у н — Limnanthemum, ряска — Lemna (рис. 96), рдест п л а-вающий — Potamogeton natans (рис. 101).
О фауне подобных зарослей уже говорилось па стр. 84. В дополнение к этому укажем здесь лишь на ходы в листьях кувшинки, прогрызенные личинкой мухи Hydromyza livens (рис. 51).
Заросли рдестов. Самым характерным для рдестов является весьма сильное отложение на их листьях извести, образующей часто целые корочки — так называемые инкрустации. Причина этого кроется в способности рдестов разлагать растворённый в воде двууглекислый кальций, отнимая от него углекислый газ; по отнятии газа остаётся одноуглекислый кальций, который растворяется в воде с большим трудом и потому тут же, на месте своего образования, т. е. на листьях, осаждается в виде мелких чешуек. Конечно, этого явления нельзя наблюдать в озёрах с малым содержанием извести в воде. Понятно, что и дно среди зарослей рдеста осенью покрывается известью от опавших листьев.
В озёрах, вода которых богата солями железа, на рдестах откладываются инкрустации этих Солей.
Рдесты подвергаются сильным колебаниям от волнений и потому представляют не очень благоприятные условия для обитания: они стоят на границе между биотопом прибойной зоны и биотопом тихих уголков среди поднимающихся над водой или стелющихся по её поверхности растений.
Часто встречается здесь ветвистоусый рачок — Sida (рис. 236) и очень мелкая личинка из Теп-
dipedidae — Corynoneura. Известковые инкрустации дают приют некоторым мелким личинкам тендипедид (особенно Cricotopus) и олигохетам — Nais (рис 165 б).
Заросли погружённых растений. Погружённые растения ниже всех прочих спускаются по дну озера; они являются последними членами растительной семьи, попадающимися на пути от берега к глубинам. Главными представителями здесь являются: у р у т ь — Myriophyllum (рис. 111), р о го-л и с т н и к —Ceratophyllum (рис. НО), э л о дея — Elodea (рис. 38 Л, ИЗ), мх и — Fontinalis (рис. 37 А), Calliergon (рис. 69) и другие и, наконец, глубже всех заходящие л у ч и ц ы — Chara, Nitella (рис. 36). Часто среди этих растений внедряются скопления водорослей: кустики Cladophora, зелёные комки Nostoc и некоторые другие. Погружённые растения нередко покрываются наростами микроскопических зелёных и диатомовых водорослей.
Население этой области очень богато и в количественном, и в качественном отношениях. Как характерную черту фауны можно отметить лишь отсутствие тех форм, которые нуждаются для дыхания в атмосферном воздухе.
Погружённые растения могут отчасти оставаться и зимой подо льдом зелёными и создавать около себя большое количество кислорода; вследствие этого такие участки служат местами скопления животных, прибывающих сюда из мелких участков литорали и спасающихся этим от замерзания.
В этой зоне было бы очень интересно поставить ряд наблюдений по сезонам, отмечая наплыв пришельцев из других областей к зиме и обратное передвижение их к берегам с наступлением тёплого времени года. На русских озёрах подобных наблюдений ешё не производилось.
Участки прибоя. В тех местах, где незащищённые берега подвергаются действию порывов ветра, постоянное движение воды не даёт возможности укорениться растениям. Если грунт такого места состоит из мелких отдельностей, последние, постоянно передвигаясь доходящим до дна волнением, создают такое неустойчивое дно, из которого корни растений будут постоянно вымываться. Более крупные камни, сдвинуть которые волнение не в состоянии, могут давать опору лишь для низших растений, использующих подвергшийся разрушению верхний слой камня.
Поселяющиеся здесь животные должны обладать таким строением тела и таким образом жизни, который позволял бы им бороться с волнами, которые в любой момент могут выбросить их на берег.
Условия существования здесь оказываются, таким образом, чрезвычайно сходными с теми; которые имеют место в ручьях или в прибрежье рек.
Первым средством борьбы с движением воды является, конечно, способность задерживаться на одном месте: 1) путём прикрепления к камням; 2) путём зарывания в песок или подрывания под камни; 3) путём отягощения тела.
Вторым средством является приобретение такой формы, которая предоставляла бы малую поверхность сопротивления движению воды (поверхность, перпендикулярную к их направлению); это достигается возможно большим уплощением тела.
Прекрасное освежение газового состава воды вплоть до дна, благодаря постоянному перемешиванию, и невозможность спокойного парения у поверхности обусловливают то, что фауна прибойной зоны слагается исключительно из животных, не нуждающихся в атмосферном воздухе.
Питание происходит здесь главным образом за счёт мелких растений или растительных остатков, часто принесённых в озеро извне и оседающих на камнях или песке тонким налётом.
Камни. Скудная растительность поверхности камней состоит чаще всего из мха Fontinalis и из налётов водорослей.
Животные сидят главным образом на камнях или под ними. Возможность удерживаться здесь и противостоять смывающему действию токов воды осуществляется различными приспособлениями: то плоской формой тела, то прикреплением его к камням посредством слизи (чашечка — Ancylus, рис. 196), крючков и зацепок (личинки подёнок), присосок (п и-явка— Herpobdella), то приклеиванием чехликов личинок к камням посредством нитей (личинки мошки Melusina (Simulium), рис. 437 и комаров-дергунов), то отягощением тела путём приклеивания к чехликам тяжёлых посторонних частиц, например, песчинок (личинки ручейников Goera, рис. 405 2? и Silo, рис. 57—58ZJ).
Защитой против напора волн, как уже указывалось, служит между прочим сильное уплощение тела или заключающего его домика, что характерно для всей фауны камней. Прекрасный пример подобного уплощения дают личинки подёнок — Ecdyonurus и Heptagenia (рис, 308).
Уплощение тела достигается не только сжатием ёго и ног в спинно-брюш-пом направлении, но и расположением ног в плоскости тела; иногда такое уплощение ещё усиливается разными боковыми выростами и каёмками из волосков. Нижняя поверхность тела у жителей камней обычно совершенно плоская, а выпуклость спины слаба и очень постепенно сходит на нет на краях.
Песок. Песчаная область прибоя возникает лишь на очень пологих участках берега. При волнении набегающая на берег вода трением о поверхностный слой песка перекатывает песчинки этого слоя на берег, а спадающая волна 90
обладает слишком малой скоростью, чтобы перекатить эти песчинки обратно в водоём. Таким путём толщина слоя песка на берегу постепенно нарастает. При малой глубине набегающей на берег волны температура воды в ней быстро выравнивается и повышается благодаря сильному прогреванию. Мелкие песчинки, постоянно перекатывающиеся с места на место, не дают возможности удержаться здесь даже низшим растениям. Отсутствие высших растений исключает возможность заселения подобных берегов растительноядными животными; лишь пожиратели разлагающихся частиц, оседающих на дно, находят здесь для себя пищу, но и этой пищи немного, так как источником её является лишь планктон. Конечно, могут жить здесь и питающиеся планктоном формы, а за ними следуют и хищники. Население этой области качественно всегда бедно ввиду того, что лишь немногие, весьма своеобразно организованные животные способны мириться с указанными условиями. В большинстве случаев обитатели песка прокладывают себе в грунте ходы; зарывшись в песок, они выставляют часть своего тела для захвата пищи и для дыхания.
Наиболее характерными обитателями являются: личинка стрекозы— Gomphus (рис. 289 Б), личинки ручейника — Molanna (рис. 403) и несколько личинок тендипедид; тут же можно встретить и рачка бокоплава — Gammarus, прозванного так за оригинальный способ плавания (на боку), к которому его тело хорошо приспособлено: с боков сплюснуто и, кроме того, в плоскости сжатия изогнуто дугой; встречаются здесь также двустворчатый моллюск горошинка — Pisidium, некоторые виды о л и г о х е т (из родов Limnodrilus, Chaetoga-ster, Nais, Paranais, Psammoryctes) и др.
Сублитораль
Сублитораль как область переходная между литоралью и глубинной областью обладает промежуточным режимом. Это — область начала падения кислорода, средних между прибрежной и глубинной температур и сред-него освещения. По кислороду она находится в лучших, нежели пелагическая область, условиях благодаря тому, что освежение газового состава происходит легче и ил на дне, перемешанный с раковинами, лежит более рыхло. В температурном отношении сублитораль находится большей частью в слое температурного скачка или даже несколько выше его.
Часто её фауну составляют выходцы из литорали, как, например, личинки вислокрылки — Sialis, моланны, подёнок; но здесь есть и свои специфические, только сублиторали свойственные организмы из тендипедид и олигохет. Из моллюсков здесь обитают горошинка — Pisidium (рис. 216), дрейссена — Dreis-sena (рис. 211), беззубка — Anodonta (рис. 212), затворка— Valvata, битиния — Bithynia (рис. 208).
Одной из характернейших и интересных черт сублиторали является массовое скопление пустых раковин моллюсков, носящее название ракушника. На первый взгляд это явление могло бы быть объяснено столь же массовым распространением в этой области соответственных видов живых моллюсков; однако это не так, что доказывается непосредственным наблюдением. Общепринятого объяснения этого факта пока ещё не существует; повидимому, снесение сюда масс пустых раковин обусловлено циркуляционными токами воды.
ТЕМЫ ДЛЯ РАБОТ
Литоральная область с её громадным разнообразием жизненных условий даёт неисчерпаемый материал для изучения, причём требует самого примитивного оборудования. В тёплое летнее время можно часто обойтись даже и без лодки, если не бояться войти поглубже в воду. Простой самодельный сачок из любой редкой материи, размеренная па дециметры палка (для измерения ширины зарослей и глубины), бумага (для сохранения
растений) и банка (для сбора животных) — вот и весь несложный инструментарий, при котором может быть произведён целый ряд ценных псследовапий и интереснейших наблюдении. Для более полного изучения литорали необходимо учесть и микроскопический мир, для чего необходимо иметь уже планктический сачок.
Прежде всего, конечно, все те разнообразные биотопы с их биоценозами, о которых говорилось выше, могут явиться темой для самостоятельного изучения. Листья одной только кувшинки или кубышки с их черешками дают приют четырём различным комбинациям сообществ животных: на верхней поверхности листа, на нижней, на черешке и в самой ткани; сопоставить эти сообщества животных друг с другом и провести различие между формами, общими всем им и специфическими лишь для некоторых из них, — задача нетрудная, но очень благодарная.
Очень мало затронуты изучением плавающие комки водорослей, а они имеют своеобразную фауну, равно как и почти неизученные корочки извести на погружённых растениях. Не менее интересны и сообщества животных, живущих в колониях мшанок, или те биоцепозы-обросты, которые приурочены к погружённым в воду деревянным предметам — мосткам, сваям купален и т. п.
Своеобразную группу образуют также скользящие по поверхности воды животные, отношение которых к прочим обитателям и связь с внешними условиями ещё далеко не выяснены; в теме, касающейся жизни поверхности воды, могут быть затронуты и те животные, которые не живут здесь постоянно, но обязательно должны появляться время от времени в целях обновления газов для дыхания.
Позволяем себе указать ещё на ряд вопросов, которые могли бы быть взяты для изучения: сравнение фауны пояса определённых зарослей у берегового конца и у пелагического; смена поясов растительности от берега к центру и ширина и последовательность этих поясов в различных типичных участках; минирующая фауна; кладка яиц различных животных и первые этапы их развития (наблюдения необходимо перенести в аквариум).
Большую ценность имеют наблюдения, проведённые на протяжении нескольких, хотя бы двух, сезонов. Здесь можно выяснить ещё почти не затронутый для наших озёр вопрос о переселении литоральной фауны перед наступлением холодов в более глубокие места и возвращение вновь к берегам весной, причём может быть учтён и момент появления той или иной животной формы, а также и момент начала прорастания водных растений.
Граница между растительностью и лишёнными последней более глубокими местами на наших озёрах мало изучена: есть или нет в изучаемом водоёме то, что мы назвали выше «сублитораль», и какова её физиономия — вот вопросы, на которые следует попытаться ответить.
Изучение литорали имеет также и большую хозяйственную ценность; так, надводная растительность озёр (тростник, камыш, рогоз) идёт — особенно в степных, безлесных районах — как строительный материал и как топливо; кроме того, она употребляется для выделки картона и бумаги. В рыбохозяйственном отношении водные заросли имеют значение как места нагула многих взрослых рыб и выкормки рыбьей молоди, а также как места нереста. Таково же значение их и для ракового промысла. Для охотничьего промысла литораль играет большую роль, поскольку она даёт приют водоплавающей дичи. Однако литораль может играть, кроме положительной, также и отрицательную роль. Так, с точки зрения народного здравоохранения прибрежная область озёр имеет значение как возможный очаг малярии, вследствие чего очень важным здесь является изучение фауны комаров. Сильное зарастание водоёмов (как естественных, так и искусственных — прудов, каналов, водохранилищ и т. д.) может явиться большим препятствием для рыболовства и судоходства.
L Непериодические наблюдения над прибрежной флорой и фауной
1. Первоначальное знакомство: сбор растений и животных и их определение.
2. Установить распространение прибрежной растительности вообще: определить ширину занятой ею полосы воды по всему берегу озера и нанести на план.
3. Установить типы зарослей с условным обозначением их на плане и с указанием: состава каждого типа, занятой им площади (приблизительно), грунта и глубины водоёма у ближней и дальней по отношению к берегам границы его распространения.
4. Установить состав фауны в различных типах зарослей, сравнить полученные данные и попытаться отметить животных, как общих для всех типов зарослей, так п живущих отдельно среди растений надводных, погружённых и с плавающими листьями.
5. Обследовать животное население однородной заросли какого-либо вида растений, например, тростника, плавающих листьев кувшинки, плавающего рдеста ит. п., включая минирующую фауну.
6* . Обследовать биоценозы различных подводных деревянных предметов: сваи, коряг, затонувших стволов деревьев и пр. (обростов).
7. Познакомиться с биоценозами участков прибрежной эоны, лишённых растительности: население камней, песка.
8. Установить состав планктона прибрежной области и сравнить его с планктоном пелагической области.
9. Сравнить бентос различных типов зарослей.
IL Периодические наблюдения над населением литоральной зоны
1, Установить хотя бы в одном-двух пунктах сезонные изменения прибрежной флоры и фауны по сборам в феврале (перед вскрытием льда — сбор сачком и скребком в проруби), мае, июле и октябре.
МЕТОДИКА
Флора (растительность). Для обследования высшей флоры водоёма очень желательно иметь в своём распоряжении лодку, чтобы объехать кругом озера вдоль прибрежной полосы растений 1 2.
Подъехав к какой-нибудь заросли, нужно взять несколько листьев с надводных и плавающих растений,составляющих данную заросль, и вытащцть чем-нибудь, например, граблями, погружённые в воду растения. По этим листьям тут же определяем растения, пользуясь прилагаемым определителем, который должен быть взят на экскурсию. Этот определитель представляет то удобство, что по нему можно определять растения в течение всего периода, пока на них имеются листья. Записав название растений, измеряем и отмечаем ширину заросли, т. е. расстояние начала и конца её от берега. Затем необходимо определить её д л и н у, т. е. начало и конец заросли вдоль берега, чтобы потом нанести площадь, занимаемую данной зарослью, на карту водоёма. Вместе с тем определяем и местонахождение заросли. Для большей точности определения следует производить его буссольным способом (см. конец главы о съёмке плана). Но ввиду кропотливости этого способа и значительной потери времени, которой он потребует в данном случае, можно лишь приблизительно, на глаз, отметить положение пограничных пунктов заросли по каким-нибудь береговым вехам и потом приблизительно же нанести её на план. Далее следует отметить сравнительную густоту зарастания каждого растения, т. е. записать, как оно растёт — в одиночку или образуя скопления той или иной густоты.
Наконец, отмечаем глубину и характер донных отложений у ближней и дальней по отношению к берегу границы заросли. Для каждой заросли должен быть заполнен отдельный листок со следующими сведениями:
1. Дата обследования (число, месяц и год).
2. Видовой состав заросли.
3. Приблизительная площадь заросли.
4. Местонахождение заросли.
5. Приблизительное соотношение различных видов растений в заросли по занимаемой ими плогаади и по густоте зарастания.
6. Глубина в начале и конце заросли по ширине её.
7. Грунт.
Если при определении по листьям встретится какое-нибудь затруднение или сомнение, то лучше прибегнуть к более точному способу определения, именно — по цветам: их можно собирать, конечно, лишь в известное время — в период цветения, который у большинства водных растений наступает в июле.
Книги для такого определения смотри в прилагаемом списке. Тем, кто пожелал бы запяться водной флорой более тщательно и подробно, советуем составить гербарий, для которого сбор растений производится более тщательно — так, чтобы были налицо все важные и необходимые части и органы растения: корень, стебель, лист и цветок или плод (для крестоцветных последний необходим) и притом в расправленном состоянии. Для этого необходимо иметь с собой па экскурсии походную папку с сушильной бумагой, в качестве каковой лучше всего употреблять белую фильтровальную бумагу; по при отсутствии последней её можно заменить другой, например, серой обёрточной, газетной и т. п., вообще — непроклеенпой бумагой. Самое собирание растенпй проф. Л. Иванов рекомендует производить следующим способом:
«Вып^чз из воды и обмыв корни, листья и стебли от приставшего к ним ила, слизняков и слизистых масс с их яичками, обтирают корпи досуха и помещают в обыкновенную бумагу для просушки растений. Нежные водяные растения трудно расправлять на бумаге, поэтому лист проклеенной (писчая, восковая) бумаги подводят под растение, лежащее в воде, и, расправив его, осторожно вынимают лист с растением, тогда последнее остаётся расправленным на листе. Дав сбежать воде, лист с лежащим на нём растением кладут в пропускную бумагу и сушат обыкновенным способом». Собрав таким образом в папку нужное количество растений, разделённых между собой пачками сушильной бумаги в несколько листов каждая, привозим их в таком виде с экскурсии и сменяем сушильную бумагу на свежую, стараясь как можно осторожнее обращаться с растениями 3; затем подкладываем под общую пачку с растениями продырявленную доску, положенную на два кирпича, кладём на пачку такую же доску с равномерно распределённым на ней грузом из камней или кирпичей и оставляем сохнуть при средней ровной температуре и при
1 За неимением лодки можно сделать обход озера по берегу, но при этом придётся, конечно, насколько позволит глубина, влезать в воду и ходить вброд.
2 Тот лист, в котором лежит растение, сменять не нужно до полного высыхания
последнего.
достаточной вентиляции Не позднее чем через полсуток необходимо посмотреть растения и, если они не высохли, ещё раз сменить бумагу. В случае надобности следует такую смену производить несколько раз через 8—12 часов, стараясь делать это каждый раз осторожно, чтобы не повредить растения. По окончании просушки каждое растение прикрепляется посредством бумажных полосок к свежему листу бумаги, и на последнем паклеивается этикетка, на которой должны быть помечены следующие касающиеся данного растения сведения:
1. Название растения.
2. Дата сбора.
3. Название водоёма.
4. Пункт сбора.
5. Глубпна.
6. Грунт.
7. Характер зарастания.
8. В сообществе с какими растениями произрастает.
Интересующихся специально вопросами гербаризации и методикой исследования водной растительности отсылаем за подробностями к специальным книгам, как, например:
С ю з е в, Гербарий. Руководство к собиранию и засушиванию растений для гербария и составлению флористических коллекций. С.-Петербург. 1912.
Рис. 53. Сачок: вверху — в цельном виде, внизу — кольцо (ориг.).
Л е п и л о в а Г. К.. Инструкция для полевого исследования высшей водной растительности. Инструкции по биологическим исследованиям вод, ч. II. Биология материковых вод. Разд. А. —Исследование бентоса, вып. 5, 1934, изд. ГГИ, Л.
Рычин Ю. В. Флора гигрофптов. Изд. «Сов. наука», М., 1948.
Фауна. Оборудование, 1. Скребок употребляется для сбора донных животных прибрежной области, где растительность препятствует пользованию драгой. Общий вид этого инструмента представлен на рисунке 52. Существенную часть его представляет изображённая на рисунке железная вилка, к концам которой прикреплена железная же пластинка толщиной в 1,5—2 мм. Как и у драги, один край этой пластинки заострён, а около другого — ряд отверстий для прикрепления мешка. На противоположном конце вилка расширяется в муфту с двумя отверстиями для надевания на деревянную палку. Мешок делается из той же материи, что и мотня драги, и прикрепляется к вилке путём обшивания её, а к пластинке — посредством продетой в отверстия крепкой питки или шнура. Палка должна быть прочная, диаметром сантиметра 3, а длиной метра 2. В муфте имеются 2 отверстия для прикрепления её гвоздями или винтами к концу палки.
2. Сачок употребляется для сбора всех плавающих среди прибрежной растительности животных. Он состоит из 3 частей (рис. 53): металлического круга, палки и
1 Хорошо также стянуть верёвками пачку между двумя продырявленными досками или, ещё лучше, между вделанными в рамку металлическими сетками и повесить в таком месте, где свободно циркулирует воздух.
мешка. Круг, диаметром приблизительно в 35 см,сгибается из круглого железпого стержня приблизительно в 5 мм шириной; концы этого круга, пе доходя друг до друга немного менее чем на ширину палки, отгибаются кнаружи и обрезаются на разной длине, причем самые кончики их заостряются и загибаются под прямым углом. Палка берётся такая же, как и для скребка; на одном конце её с противоположных сторон делается по желобку, в которые и вкладываются оттянутые концы круга, острия которых вколачиваются в палку; затем около самого круга конец палки плотно обхватывается железным кольцом или, проще, туго обматывается проволокой. Круг обшивается свободным краем мешка, длина которого равна приблизительно 40 см и который делается из такой же материи, как в драге и скребке; конец этого мешка лучше закруглить.
3. Планктическая сетка или планктический сачок. Если есть в распоряжении уже готовая планктическая сетка, то лучше воспользоваться ею; в противном случае её можно заменить планктическим сачком. Этот сачок представляет собой, собственно говоря, планктическую сетку, надетую на палку. Зная из предыдущего изложения, как сделать сетку и сачок, нетрудно уже скомбинировать эти два инструмента в планктический сачок.
4. Два тазика: побольше — для скребка и поменьше — для сачка.
5. Широкогорлые склянки, как для драги, — несколько штук.
6. Пинцеты.
7. Пипетки разных калибров.
8. Ведро.
Производство работ. — Работу скребком можно производить как с берега, так и с лодки. Держа скребок обеими руками возможно дальше от себя, погружаем противоположный конец до самого дна в таком положении, чтобы острый край пластинки был направлен вниз, а мешок — вверх. Затем, нажимая как можно сильнее на дно, отдельными короткими толчкообразными движениями подтягиваем скребок к себе и загребаем таким образом грунт в мешок. По положению собственного тела и палки работающий сумеет определить тот момент, когда ему нужно прекратить соскребание грунта со дна; тогда он, не прерывая работы, быстро повёртывает в руках палку такпм образом, чтобы самый скребок принял противоположное прежнему направление, т. е. мешком вниз, и в то же время вытаскивает его на поверхность воды. Здесь, так же как и при драгировке, производится энергичными движениями из стороны в сторону промывка материала до тех пор, пока не прекратится выступление мути из мешка в окружающую воду; при этом бывает полезно время от времени, взяв мешок в руку и не вынимая его из воды, осторожно производить им различные перебалтывающие движения, чтобы способствовать более сильному перемешиванию его содержимого. По окончании промывки это содержимое вываливается в таз с водой, и дальше с ним поступают так же, как с материалом, принесённым драгой.
Ловить сачком — дело настолько незамысловатое, что оно знакомо почти каждому. Тем не менее для большей успешности лова нужна некоторая сноровка, на которую мы и считаем необходимым указать. Чтобы наловить возможно большее количество животных, недостаточно один раз провести сачком в воде; отведя сачок как можно дальше в сторону от себя (допустим, вправо), опускаем его в воду ребром так, чтобы отверстие его было направлено влево, и проводим им равномерно, со средней скоростью, на такое же расстояние от себя влево; здесь быстро повёртываем сачок на 180°, чтобы отверстие eix) оказалось направленным вправо, проводим вновь до начального пункта, опять быстро поворачиваем на 180°, доводим снова до крайней точки противоположной стороны и т. д.; проведя так раз десять из стороны в сторону (5 раз в одну и 5 раз в другую), делаем последний поворот не на 180°, а только на 90° (разумеется, отверстием сачка вверх), и вынимаем сачок из воды. Ещё проще описывать сачком в воде лежащую восьмёрку оо, причём па концах восьмёрки сачок должен подниматься, а не опускаться. Дав воде стечь, выворачиваем мешок в тазик с водой, в которой его споласкиваем; по и после споласкивания на мешке остаются ещё некоторые животные, которых осторожно снимаем пинцетом. Затем из тазика пинцетом же вылавливаем всех животных и бросаем их в склянки с водой или с 4-процентным формалином, если их нужно зафиксировать (или с 70-градусным спиртом для моллюсков). Однако среди них встречаются такие, которых даже при большом навыке и искусстве не удаётся схватить пинцетом, например, мелкие жучки или водяные клещи, которые благодаря своей малой величине, шарообразной форме и довольно быстрому плаванию легко ускользают от пинцета; в таком случае лучше прибегнуть к пипетке, посредством которой их гораздо легче поймать. Попадутся и такие животные, которых вообще трудно схватить чем бы то ни было: оставаясь значительное время в совершенном покое, они вводят нас в заблуждение своей неподвижностью, и как только мы, хотя бы со всей осторожностью, приближаемся к ним со своим инструментом, они с молниеносной быстротой делают прыжок в сторону, и мы ловим пустое место; к таким принадлежат, например, личинки подёнок, некоторых стрекоз и др. Для их вылова можно рекомендовать следующий метод, который, впрочем, если уже вводить его в употребление, годится и для всех вообще плавающих животных. Маленькое кухонное ситечко погружаем в тазик с водой, где находятся пойманные нами животные, затем осторожно, чтобы не спугнуть их, подводим под них ситечко и так же осторожно вынимаем его из воды; как только верхние края ситечка достигли поверхности воды, попавшие в него животные уже не могут выбраться из него, и, когда из вынутого ситечка стечёт вода, они окажутся на дне в беспомощном состоянии, в котором их легко можно взять пинцетом.
Для сбора животных, сидящих па растениях, последних вынимают из воды, осматривают и снимают или пинцетом же, или даже просто рукой (например, более крупных животных, вроде прудовика). Впрочем, для некоторых животных, как гидра, даже пинцет оказывается слишком грубым инструментом; то же относится к икре некоторых животных, прикреплённой к растениям в виде разной величины и формы прозрачных комочков слизи, содержащих внутри более или менее многочисленные яйца (икра прудовиков в виде небольших студенистых колбасок, икра стрекоз, двукрылых, ручейников и пр.). Такие нежные объекты рискованно снимать пинцетом, которым легко может пх повредить; поэтому их совсем не отделяют от растения, а вырезают вокруг них кусочки последнего (листа, стебля), которые и бросают в склянку с водой. Рекомендуется также вынутые из воды стебли и листья растений ополаскивать в тазу с водой, из которой затем указанными выше способами вылавливать животных; для вылавливания можно применять также пипетку.
Лов планктических организмов в литорали можно производить разными способами, смотря по тому, какое орудие лова имеется в нашем распоряжении. Ловить планктон сеткой так, как мы это делаем в глубоководной области, здесь невозможно, так как мешают заросли. Поэтому сетка может получить здесь несколько иное применение. Если есть лодка, то в тех же участках прибрежной области, в которых производят сбор животных скребком и сачком, зачерпывают ведром воду и выливают её в сетку; пропустив через последнюю несколько вёдер, обычным способом сливают улов в склянку. При отсутствии лодки эта же операция производится с каких-нибудь мостков, подходящих свай, камней и т. д.; в этом случае приходится, разумеется, ограничиться ловом лишь в тех пунктах, где условия позволяют произвести таковой.
Лов планктическим сачком производится совершенно так же, как и обыкновенным, и поэтому в повторном описании не нуждается; разница, как это само собой понятно, сводится лишь к способу перевода собранного материала из сачка в склянку.
Бросив в каждую склянку номер лова, нужно для каждого из номеров записать в тетради следующие сведения:
1. Место лова.
2. Время лова (месяц, число и время дня).
8. Глубина вэдоёма в данном месте.
4. Расстояние от берега.
5. Способ лова.
6. Какая заросль.
7. Температура воды (желательно).
Сбор воздушных насекомых, цикл жизни которых имеет отношение к воде. Оборудование. 1. Воздушный сачок, который делается так же, как и водный, только легче; в связи с этим все твёрдые части сачка (круг, палка) делаются тоньше, а мешок, который должен быть длиннее диаметра круга по крайней мере в два раза, можно сшить из более тонкой и лёгкой материи — из кисеи или марли.
2. Ши^окогорлая склянка с обыкновенной пробкой, к нижней стороне которой прикрепляется булавкой клочок ваты, смоченный эфиром; дно склянки внутри покрывается фильтровальной бумагой, а на него насыпаются узкие полоски этой же бумаги.
3. Пузырёк с эфиром.
Производство работ. Часто насекомые, летая над водой, садятся на самих работающих, на лодку, вёсла и т. д.; тогда их можно ловить обычно прямо руками; так же можно собирать и насекомых, сидящих на надводных растениях; однако лучше ловить их, проводя воздушным сачком по этим растениям. Летающих над берегом насекомых, понятно, нужно ловить тоже сачком. Стрекоз рекомендуется ловить на заре, когда они ещё с "отягчёнными крыльями сидят на растениях, с которых их можпо обирать руками.
Пойманных так или иначе насекомых бросают тотчас же в склянку с памоченной эфиром ватой, стараясь как можно меньше держать склянку открытой. Дальнейшие операции относятся не только к воздушным, но и к постоянно живущим в воде насекомым, именно — к водяным жукам и водяным клопам. По возвращении с экскурсии нужно поскорей, пока ещё насекомые не высохли и сохраняют гибкость тела, наколоть их на булавки; булавка протыкается со спинной стороны в грудной отдел насекомого, которое располагается приблизительно посередине длины булавки; это необходимо для того, чтобы, с одной стороны, оно не касалось той поверхности, в которую воткнута булавка (торфяная пластинка, пробка, просто липовая или вообще из мягкого дерева доска), а, с другой стороны, над насекомым оставался бы ещё достаточно длинный кусок булавки, чтобы его можно было держать в руке, не задевая и не повреждая насекомого. Затем у насекомого (кроме жуков и клопов) нужно расправить брюшко,'усики, крылья и ножки и оставить на булавке, где они и засыхают в этом состоянии. У высохших стрекоз брюшко часто отпадает; для предупреждения этого в брюшко только что наколотой на булавку свежей стрекозы втыкается с заднего копца длинная энтомологическая булавка или тонкая деревянная палочка, несколько превосходящая длиной брюшко, чтобы её передний конец прошёл в грудной отдел; с таким приспособлением брюшко хорошо держится и после высыхания.
Тем, кто хочет запяться специально собиранием коллекций по водным и имеющим к воде отношение воздушным насекомым, рекомендуем следующие книги.
Кожевников Г. А., под ред., Руководство к зоологическим экскурсиям и собиранию зоологических коллекций, М. 1912.
Павловиче. А., Составление коллекций по естествознанию. Учпедгиз, 1938.
Программы и наставления для наблюдения и собирания коллекций. Изд. О-ва естествоиспытателей при Петербургском ун-те, 1913.
ПБ <1914° Р И К ° В А ’ Инструкция для коллеитЬвирования пресноводной фауны Обработка собрапного материала. Консервировка и определение. С точки зрения пашей непосредственной задачи — обследования водоёма как целого и единого физико-биологического комплекса — обитающие в нём животные важны и интересны для нас постольку, поскольку они служат для его биологической характеристики, поскольку они входят в число его составных частей и явлений, из которых в общем складывается жизнь водоёма. Очевидно, что для этой цели животные прежде всего должны быть зарегистрированы, т. е. должен быть определён их качественный состав, по крайней мере до родов включительно. Такое определение может быть произведено по прилагаемым таблицам, которые составлены с таким расчётом, чтобы каждое животное находить возможно кратчайшим и удобнейшим способом, пользуясь наиболее легко различимыми признаками.
Чтобы определить животных, необходимо их тщательно рассмотреть. Но быстрота движения многих животных часто не позволяет этого сделать. В таких случаях прибегают или к наркотизации животных посредством, например, кокаина, хлорал-гидрата и т. п., или, чаще всего, просто к умерщвлению их особыми жидкостями, которые не только убивают животных, но и сохраняют их от гниения и разложения в течение очень долгого времени (целые годы); такие жидкости, как говорят, фиксируют, или консервируют, животных. Наиболее обыкновенными фиксаторами, пли консервирующими жидкостями, являются 70—80-градусный спирт (можно денатурированный) и 3—4-процентный формалин. Ввиду малой доступности первого пользуются чаще всего вторым. Продажный формалин имеет обычно 40-процентпую крепость. Чтобы из такого формалина получить 4-процентный, пужно его разбавить в 10 раз, т. е. на 1 часть 40-процентного формалина надо брать 9 частей воды. Консервировка собранного на экскурсии материала производится обыкновенно тут же, на месте. При этом на экскурсию лучше брать не разведённый (4%) формалин, а крепкий (40%). Так как приходится фиксировать пробы уже содержащие значительное количество воды, то рекомендуется просто к пробе прибавлять одну десятую часть её (на глаз) 40-процентного формалина; тогда и получим приблизительно 4-процентный формалин.
Живой материал. Однако консервирование тоже имеет свою отрицательную сторону: животные при этом делаются менее прозрачными, некоторые из них, именно — нежные, незащищённые более или менее твёрдыми покровами, настолько сильно сжимаются, сокращаются, деформируются, что становятся подчас совершенно неузнаваемыми (например, гидра, некоторые из планктических животных и др.); наконец, даже среди более крупных животных есть такие, у которых в фиксированном состоянии отпадают некоторые нежные части и органы и этим затрудняют их определение (это относится, например, к личипкам некоторых насекомых, как подёнки, некоторые стрекозы п пр.). Поэтому не мешает определение, произведённое на консервированном материале, проверить на живых животных. Кроме того, раз в руках уже имеется живой материал, то вполне естественно заинтересоваться им как таковым и познакомиться с биологией мпогих водных животных. Для этого их нужно в живом виде доставить домой и устроить для них хотя бы самые простые аквариумы, например, в банках из-под варенья, в глубоких тарелках и т. н. При этом, начиная с момента самого лова и кончая водворением в аквариум, необходимо помнить следующее: ни в коем случае нельзя сажать вместе хищников (например, клопы, большинство жуков и др.) с теми животными, которые служат им пищей, иначе от последних ничего не останется. В аквариуме можно сделать много очень интересных наблюдений над жизпью и поведением водных животных; но в нашу задачу не входит излагать способы и правила устройства и содержания аквариума, и потому для интересующихся этим мы указываем книги, специально посвящённые этому предмету.
Емельяненко, Школьный аквариум, его устройство и уход за пим, СПБ 1912.
Золотницкий Н. Ф., Аквариум любителя, 1916.
Его ж е, В мире пресных вод. Наблюдения над жизнью животных и растений рек, прудов, озёр я луж в воде и в аквариумах (Библиотека И. Горбунова-Посадова для детей и юношества), Москва 1915.
Его же, Живая природа в школе, 1915.
Как устроить аквариум и террариум. Для начинающих любителей, СПБ 1914, пзд. 2-е, «Аквариум».
1 Пробой в данном случае мы называем совокупность наловленных в склянку организмов.
97
7 А. Н. Липин
Набатов А. Л., Устройство пресноводных аквариумов и уход за ними. изд. 2-е, Москва 1914.
Его же, Комнатный пресноводный аквариум и его население, СПБ 1914, изд. зоолог, маг. «Аквариум».
Школьный аквариум и террариум как пособие при изучении природоведения, Москва 1912.
Мы изложили так называемую качественную методику сбора и обработки озёрной флоры п фауньГ; целью этой методики является регистрация родов или видов живущих в озере растений и животных, т. е. определение, как говорят, ка мест не и по го состава их. Но паряду с качественной существует ещё к о л и ч е-ствеппая методика, преследующая количественный учёт водных организмов по их числу, весу или объёму. Для производства этого учёта организмы должны быть и собраны количественным способом, который осуществляется при помощи особо выработанного инструментария. Так, для количественного сбора донных животных глубоководной области употребляются так называемые дночерпатели, вырезаю-щпе и захватывающие групт вместе с живущими в нём животными с вполне определённой площади дна, причём сосчитывается число этих животных, отдельно для каждого рода и вида, и обычно перечисляется па 1 кв. м дна; количественный лов планктона производится тоже особыми количественными планктическими сетками, позволяющими облавливать определённый объём воды, в котором затем сосчитывают число пойманных планктических растений и животных и перечисляют на 1 куб. м воды или на столб воды с поперечным сечением в 1 кв. м, заключённый между поверхностью воды и дном водоёма.
Такие методы исследования нашли себе применение в первую очередь в открытых, т. е. но занятых растительностью местах водоёмов, именно в пелагической области и в профундали озёр, где ничто не мешает работе с количественным инструментарием. И здесь, действительно, количественные исследования ведутся уже не один десяток лет и дали много интересного материала и важных научных обобщений.
Совсем другое дело в литорали, занятой различными зарослями. Мы уже видели, что даже качественные исследования там вести труднее, чем в открытом плёсе озера. Применение же количественных орудий сбора фауны, особенно в густых зарослях, наталкивается на очень большие трудности, так как растительность мешает опуститься инструментам на должную глубину. Этим объясняется, что, несмотря на глубокий научный интерес и важное практическое значение (в особенности для рыбного хозяйства) количественного изучения зоны зарослей, такое изучение и до сих пор продвинулось вперёд ешё чрезвычайно слабо. А между тем здесь имеет громадное значение количественное исследование не только фауны, но и самой растительности.
В последние 20—25 лет были отдельные попытки как за границей, так и у нас ввести количественную методику в изучение литорали, однако до сих пор ни одна из этих попыток не получила всеобщего признания и универсального применения. В СССР проблема количественного исследования зоны зарослей с каждым годом всё настойчивее и настойчивее ставится на очередь требованиями со стороны различных хозяйственных организации и промышленных предприятий, главным образом рыбного хозяйства. Поэтому за последние годы у нас в этом направлении заметны некоторые сдвиги: некоторые исследователи пытаются разными путями подойти к разработке методики количественного учёта флоры и фауны литорали, стараясь обеспечить, по возможности, полный сбор с определённой площади воды и дна имеющихся там растительных и животных организмов. Но в общем этот вопрос ещё довольно далёк от окончательного решения, которое могло бы считаться удовлетворительным как с научной, так и с практической точки зрения.
Ввиду назначения данной книги оказать начинающим помощь в первоначальном ознакомлении с основными сведениями о пресных водах и их жизни, мы не можем вдаваться здесь в дальнейшие детали методики количественных исследований водного населения и давать описание количественного инструментария и способов работы с ними с той подробностью, с какой мы описывали качественную методику. Приобретение сложного и дорогостоящего оборудования для количественных исследований и большая затрата времени на производство последних делают их доступными лишь для специальных учреждений, организаций и научных работников, деятельность которых посвяшена исследованию водоёмов. Если же кто-нибудь заинтересуется этой областью работы, то может получить подробные сведения о ней в следующих книгах (в них же можно получить и более подробные указания по методике качественного биологического исследования пресных вод).
Инструкции по биологическим исследованиям вод, под ред. К. М. Дерюгина, ч. II, Биология материковых вод, разд. А.—Исследование бентоса, вып. 1—4, 1931, изд. ГГИ, Л. (статьи С. Г. Л е и н е в о й, И. И. Соколова, А. А. Чернов-с к о г о).
*Т о же, разд. Б. — Исследование планктона, вып. 1 (В. М. Р ы л о в).
Р ы л о в В. М., Краткое руководство к исследованию пресноводного планктона, Саратов 1926.
III. ЗНАЧЕНИЕ КОЛИЧЕСТВЕННОГО УЧЁТА НАСЕЛЕНИЯ ОЗЕРА.
Понятие о продуктивности. Классификация озёр.
Значение количественных исследований. На предыдущих страницах книги мы дали краткую качественную характеристику главнейших биоценозов озера и их взаимоотношений с биотопами. Качественное исследование кончается определением родов и видов водных растений и животных и, следовательно, установлением родового и видового состава водной флоры и фауны. Но достаточно ли этого для более глубокого и точного познания тех жизненных явлений, которые протекают в водоёме? Разумеется, недостаточно. Для пас важно и интересно зпать не только то, из каких форм складывается тот или другой биоценоз, но и количественные отношения между этими формами, а также и между самими биоценозами. Если мы не будем этого знать, то не сможем объяснить многих явлений в жизни озера, не сможем или не во всех случаях сможем ответить на самые существенные вопросы, как, например: остаются ли биоценозы количественно постоянными или изменяются во времени и пространстве, каковы характер и последовательность этих изменений, каково их направление, от чего они зависят и т. д. Разве можно достаточно глубоко и широко охватить изучением такое, например, частое в наших озёрах явление, как «цветение», если не ввести в это изучение количественных методов исследования? Можно ли без применения этих методов составить себе полное представление о жизнедеятельности растений и животных литорали и их взаимоотношениях или, например, о жизни бентоса, в которой такую громадную роль играют личинки Tendipedidae, в известный период исчезающие из водоёма вследствие окукливания и последующего вылета в воздух, чтобы потом, после кладки яиц, вновь появиться на дне озера? Таких вопросов можно было бы поставить много, но мы обратим внимание ещё только на один вопрос — вопрос основной, всеобъемлющий, имеющий важнейшее значение для всей жизни водоёма, — это вопрос о круговороте веществ в водоёме: едва ли есть надобность убеждать в том, что при решении этого вопроса шагу нельзя ступить без количественных исследований водного населения.
Но не в одном лишь чисто научном, теоретическом интересе заключается значение количественного изучения флоры и фауны водоёмов. Ещё большее значение имеет это для практических потребностей различных видов народного хозяйства, в первую очередь — для рыбного. Каждый, конечно, знает, что рыба питается теми организмами, которые она находит в водоёме. При этом для каждой породы рыб различают три рода пищи: главная пища — это та, которую данная порода рыб предпочитает всем остальным родам пищи, затем заменяющая, которую рыба поглощает лишь при недостатке или отсутствии главной, и, наконец, вынужденная, которой рыба питается лишь в крайнем случае, за неимением главной и заменяющей. Для разных пород рыб существуют и разные виды главной пищи: для хищников (щука, судак, окунь и др.) такой пищей являются другие рыбы, для большинства так называемых мирных пород (лещ, сазан, карась, карп и др.) — донные животные (зообентос), некоторые породы (например, плотва) питаются преимущественно растениями и только немногие породы (сиг, ряпушка) предпочитают плапктическую пищу. Этим разнообразием вкусов в значительной степени уменьшается конкуренция между различными породами рыб, но всё же она, разумеется, не устраняется нацело. В молодом возрасте между разными породами рыб господствует гораздо большее единообразие в отношении пищи, так как мальки почти всех пород в течение более или менее длительного периода жизни питаются планктоном.
Ввиду того что рыба вылавливается человеком для своего питания, ее называют конечной продукцией водоёма. Те животные, которыми питается рыба (зообентос, зоопланктон, прибрежная фауна), получили
ны е
рыба I конечная продукция
название промежуточной продукции. И, наконец, водная растительность, как высшая, так и низшая (водоросли, фитопланктон), питание которой происходит за счёт неорганических веществ (углекислота, минеральные соли), но которая сама производит органическое вещество, идущее на питание водной фауны (промежуточной продукции), называется первичной продукцией. Таким образом, всё водное население (флора и фауна, включая рыб) с точки зрения пищевых отношений разбивается на три основные группы:
водныерастения водные живот
(внешне и низшие) зообентос, зоопланктон,
| литоральная фауна
первичная продукция |
промежуточная продукция
Если мы каждую из этих трёх групп разобьём на более мелкие систематические группы или на отдельные роды и даже виды, то из полученных мелких подразделений можно составить так называемые пищевые цепи, причём эти цепи (в которых каждый предыдущий член является объектом питания следующего члена) могут состоять из разного числа членов. Пример пищевой цепи: водоросли — мелкие ракообразные — рыба — человек.
Из всего сказанного о питании рыб и о пищевых цепях совершенно ясно, что для того, чтобы сознательно, с открытыми глазами подойти к организации рыбного хозяйства, чтобы правильно и целесообразно вести его, мы должны подвергнуть учёту не только самую рыбу, но и её пищу; мы должны знать, сколько рыбы может прокормить тот или иной водоём, т. е. какими кормовыми запасами он обладает, как изменяются эти запасы во времени, должны научиться регулировать эти запасы, чтобы этим путём управлять самой рыбой в интересах хозяйства. Вполне понятно, что для этого необходимо владеть методами количественного учёта водной флоры и фауны. Что этот учёт имеет громадное хозяйственное значение, доказывает сама история его возникновения: первоначальным импульсом для введения количественного метода исследования планктона явилась потребность определить среднее число плавающих в море яиц камбалы и трески, приходящееся на каждый квадратный метр поверхности моря.
Акад. С. А. Зернов в своём классическом учебнике «Общая гидробиология» пишет:
«Все другие вопросы изучения и использования рыбных богатств — подсчёт пищевых запасов для рыбы, изменение рыбных запасов и рыбного населения в желательном направлении, акклиматизация той или иной породы, изучение естественно-исторических причин уменьшения рыболовства и пр.— всё это упирается в материалы, даваемые гидробиологией, которая может понять и подсчитать весь баланс жизни водоёма».
Итак, необходимость введения в гидробиологическую практику количественных методов исследования не подлежит сомнению. И эта необходимость особенно чувствуется в настоящее время у нас, в СССР, где рыбохозяйственная практика развивается такими невиданными в истории темпами, что гидробиологии необходимо двигаться вперёд ещё быстрее, чтобы не только не отставать, но, наоборот, встать во главе практики, играть для неё ведущую роль.
Понятие о продуктивности водоёмов. На основе, с одной стороны, требований рыбохозяйственной практики, с другой — теоретических обобщений того фактического материала, который получен в результате применения количественных методов к изучению кормовой базы рыб, в последние годы у гидробиологов возникла потребность выделить совокупность относящихся сюда многочисленных вопросов большой принципиальной и практической важности в особую проблему — так называемую проблему биологической продуктивности водоёмов. Попробуем вкратце познакомиться с этой проблемой.
Для того чтобы было вполне ясно, о чём идёт речь, мы должны начать с выяснения тех терминов и понятий, которые лежат в основе проблемы продуктивности. Термины продуктивность, продукт, продукция происходят от слова «продуцировать» (латинского происхождения), которое на русский язык точнее всего может быть переведено словом производить. Отсюда продуцирование означает производств о; продукт есть то, что продуцируется, производится, т. е. результат производства; продуктивность значит производительность. Поясним эту терминологию примерами применения её на практике. Продуктом производства на промышленных предприятиях разного типа (на различных заводах и фабриках) являются полуфабрикаты, фабрикаты, промышленные товары, как чугун, сталь, автомобили, электролампы, текстиль, консервы и т. д. Продуктом производства па сельскохозяйственных предприятиях являются семена и плоды разных зерновых культур, текстильное сырьё, молоко, шерсть и пр. Продуцирование есть самый процесс производства, т. е. процесс изготовления продукта. Продукцией называется количество продукта, произведённое предприятием за какой-либо (любой) промежуток времени. Другими словами, продукция есть прирост количества продукта за какой-либо период времени. В земледелии продукцию называют обычно урожаем. Продуктивностью мы называем продукцию (количество произведённого продукта) за условно выбранную единицу времени (час, день, месяц, год и т. п.). Такая продукция может служить для сравнения мощности однородных (по сорту выпускаемого продукта) предприятий между собой.
Обратимся теперь к главному вопросу: как применить все эти понятия и термины к водоёмам. Так как нашей задачей здесь является дать понятие о биологической продуктивности водоёма, то из всего круговорота веществ в нём нас в данном случае интересует та часть его, которая касается живых организмов. Они и являются тем продуктом, производителем которого является водоём. С точки зрения пищевых цепей, т. е. с точки зрения кормового использования одних организмов другими (в частности, рыбами), важно иметь в руках весовое выражение продукта, а живой вес той или иной группы организмов даёт представление об их массе. Поэтому вместо термина «продукт» среди гидробиологов вошёл в употребление другой термин — биомасса, что значит в переводе «живая масса», или, точнее, «масса (количество вещества) живых организмов». Продукция в данном случае есть биомасса, произведённая за определённый период времени, или, другими словами, прирост биомассы за этот период. Продуктивностью водоёма по какому-либо виду организмов мы называем прирост общей биомассы представителей этого вида за единицу времени.
Таким образом, учение о биологической продуктивности водоёмов, которое теоретически начало разрабатываться только в последние 10—15 лет, строится в настоящее время на трёх основных понятиях: биомасса, продукция и продуктивность водоёма. Эти три понятия, в свою очередь, могут быть в конечном счёте сведены к одному, именно — к первому (биомасса), как это будет видно из дальнейшего.
Биомасса, как выше указано, — это то количество вещества, которое находится в телах живых организмов. Она выражается обычно в единицах веса (живого) или — условного объёма. Мы можем учитывать биомассу организмов не только одного вида, но и какой-либо группы видов, биомассу определённого биоценоза, наконец, биомассу всех организмов водоёма. Биомассу водных организмов можно относить к единице поверхности воды или дна, например, к 1 кв. м или к 1 га и т. п., или же к единице объёма воды, например, к 1 куб. м \ но можно её вычислить и для всей площади озера или для объёма всей его водной массы. При каждом количественном исследовании любой группы живого населения водоёма, если только результаты этого исследования мы выражаем в весовых единицах, мы определяем
биомассу этой группы организмов. Таким образом, методами определения биомассы водных организмов гидробиологи владеют уже сейчас: это методы количественных исследований водного населения с весовым выражением их результатов. Если мы производим такое исследование только один раз, мы получаем разовую биомассу тех групп населения, которые охвачены этим исследованием. При многократных исследованиях в течение какого-либо промежутка времени мы можем констатировать колебания биомассы за этот промежуток. Если такие многократные исследования распространить на длительный период времени, например, на сезон или даже на целый год, то это позволит нам установить максимум, минимум и среднее значение биомассы за исследованный период (максимальная, минимальная и средняя сезонная или годовая биомасса). Ясно, что чем чаще будут производиться исследования, тем точнее будут выражены показатели биомассы.
Но сколько бы раз мы ни производили исследование биомассы, мы каждый раз будем получать лишь разовую биомассу, которую можно сравнить с моментальным фотографическим снимком. Такие «снимки» отдельных моментов, разумеется, говорят нам многое: при повторном их проведении они дают, некоторое представление о непрерывном процессе изменения биомассы, и чем чаще эти «снимки» производятся, тем больше их совокупность приближается к «кинематографической» картине, тем полнее, яснее и точнее становится наше представление о колебаниях биомассы во времени. И, несмотря на это, такая полная и точная картина всё же не может дать нам понятия о тех биологических процессах, результатом которых является биомасса того или иного момента. Она показывает нам лишь окончательный эффект действия этих процессов, но что это за процессы, каковы их качественные и количественные выражения, в каких взаимоотношениях они находятся между собой и пр. — всё это вопросы, ответы на которые не может дать одно лишь изучение биомассы. Так как эти биологические явления в своей взаимосвязи и комбинированном действии приводят к образованию той или иной биомассы, то совокупность этих явлений мы вправе назвать процессом производства или продуцирования биомассы. А это вплотную подводит нас к понятию продукции, как это легко видеть из приведённого выше краткого определения этого понятия.
П родукция есть результат накопления биомассы. Отсюда вытекают два следствия: во-первых, в основе понятия «продукция» лежит понятие «биомасса»; во-вторых, различие между этими двумя понятиями заключается в том, что первое из них есть понятие, как говорят, д и-намическое, а второе — статическое; это значит, что продукция связана с определённым процессом, именно процессом производства, увеличения, нарастания биомассы и, следовательно, с определённым периодом времени, в течение которого происходил этот процесс (всякий процесс протекает во времени), между тем как биомасса сама по себе ни с каким течением времени не связана — она представляет лишь то, что мы можем установить в водоёме в каждый отдельный момент.
Какие же биологические процессы определяют продукцию, накопление биомассы? Это два основных процесса: индивидуальный рост и размножение, или воспроизведение себе подобных, которое можно назвать ростом вида Как каждая из этих двух категорий роста (индивидуальный и видовой), так и их совокупность обусловливают рост, или увеличение биомассы, т. е. продукцию. Это, следовательно, процесс созидательны й, п о л ожительпы й.
По всякий хорошо знает, что наряду с этим явлением нарождения организмов и их роста существует другое явление, по своему характеру прямо ему противоположное, — это явление отхода, удаления организмов вследствие многих и разнообразных причин, как, например: естественная смерть, ги-
1 По выражению одного учёного (К. Бэр), размножение есть рост за пределы индивидуума.
бель от поедания другими организмами, от ухудшения условий существования, от вылова человеком и т. п. Это явление, обусловливающее уменьшение, убыль биомассы, мы можем назвать, следовательно, разрушительным, отрицательным процессом.
Взаимодействием этих двух основных взаимно противоположных процессов и определяется та или иная наличность биомассы в водоёме в любой момент времени. Смотря по тому, какой из этих двух процессов на данном отрезке времени берёт верх, в водоёме будет происходить или накопление биомассы (продукция), или временное состояние динамического (подвижного) равновесия её (постоянство биомассы), или же уменьшение, убыль биомассы. Таким путём продукция и биомасса количественно связываются между собой, и потому эту связь можно выразить математически. Одной из простейших форм этой связи будет следующая.
Предположим, что мы, произведя количественное исследование одного какого-либо вида организмов в водоёме, нашли в нём биомассу, которую обозначим через Второе исследование, произведённое спустя более или менее длительный промежуток времени, дало нам, допустим, биомассу В*. За этот промежуток времени одновременно протекали два указанных выше взаимно противоположных процесса — накопление биомассы путём роста и размножения организмов и убыль её путём их гибели. Обозначим результат первого процесса через Р (продукция), а результат второго — через N. Ясно, что в течение взятого нами периода времени произошло, с одной стороны, увеличение первоначальной биомассы на величину Р и уменьшение на величину 7V. В результате этих двух взаимно противоположных изменений получилась биомасса В^. Таким образом, все 4 величины связываются следующим уравнением :
B14-P-2V = 2?2.......................(1)
В этом уравнении величины Вг и В2 нам известны, так как мы получаем их непосредственно, как результат исследования. Другие же две величины Р и N неизвестны. Если бы мы получили каким-нибудь способом одну из этих величин, то другую могли бы определить из нашего уравнения, а именно:
Р = Вг - Вг 4- N.........................................(2)
или
7V = Вг + Р - 5,.........................................(3)
К сожалению, для громадного большинства видов организмов, населяющих наши водоёмы, мы ещё не обладаем достаточно разработанной методикой более или менее точного учёта нарастания биомассы в результате роста и размножения организмов и её убыли в результате гибели организмов. Но в некоторых случаях человек сам в хозяйственных целях, путём искусственных мероприятий, так организует режим водоёма, что гибель организмов, которые служат для него объектом хозяйства, или сводится к нулю, или может быть точно учтена. Примером такого хозяйства является рыбоводство, где человек может для любого момента определить величины отхода рыбы и её прироста, т. е. величины N и Р в приведённых выше уравнениях. Но в условиях искусственных (рыбоводных) прудов имеется возможность, хотя бы с грубым приближением, определить продукцию не только самой рыбы, но и той группы донных ркивотных, которая является главной и излюбленной пищей карпа, именно — личинок Tendipedidae («мотыль»). Существует дна главнейших фактора убыли этих личинок в пруде — это поедание их рыбой и превращение во взрослых насекомых (imago), вылетающих из водоёма. Первый фактор можно устранить, оставив пруд незарыбленным. Второй фактор начинает действовать только тогда, когда личинки вырастают настолько, что становятся готовыми к окукливанию. Таким образом, до периода созревания личинок последние почти не подвергаются гибели (если, конечно, в пруде юз
не произойдёт какое-либо резкое ухудшение условий их жизни) В таком случае величину N мы можем принять равной нулю, и продукция, в пределах периода существования личинок в пруде, легко определяется из уравнения (2).
Наибольшие трудности для определения продукции представляют мелкие организмы, например, планктические, у которых мы пока ещё не можем разделить процессы увеличения и уменьшения биомассы, протекающие здесь одновременно и параллельно. Только путём подробного и тщательного изучения биологии этих организмов — главным образом, явлений их роста и размножения — можно получить материал для вычисления величины Р, а затем по уравнению (3) определить и величину N.
Уравнению (1) можно придать ещё один вид, а именно:
P-N^B.-B,................................ (4)
Это уравнение особенно наглядно демонстрируёт связь между продукцией и биомассой.
Но математической связи этих двух величин можно дать и другие выражения. Так, один наш учёный (Л. А. Зенкевич) предложил пользоваться отношением годовой продукции к средней годовой биомассе — так называв-р
мым — коэфициентом. Этот коэфициент будет тем больше, чем больше Р и чем меньше В, и, наоборот, тем меньше, чем меньше Р и чем больше В. Первый случай осуществляется тогда, когда велико накопление биомассы, но зато велика и убыль её; во втором случае, напротив, то и другое выражаются малыми величинами. Эти отношения вытекают из нашего равенства (4), говорящего о том, что биомасса, которую мы констатируем в любой момент в водоёме, принципиально есть остаток от продукции в результате гибели части этой продукции.
В отношении хозяйственно ценных организмов водоёма задача человека заключается, с одной стороны, в содействии увеличению продукции (Р), с другой — в уменьшении убыли биомассы (N) от всяких естественных факторов и в хозяйственном использовании сохраняющейся таким путём биомассы (В).
Наконец, что касается термина продуктивность, то мы уже видели, что это понятие определяется через понятие продукции. Это значит, что первое понятие, подобно второму, имеет количественный характер, что играет большую практическую роль, так как даёт возможность сравнивать разные водоёмы по продуктивности. Для этого нужно иметь лишь меру сравнения. За такую меру можно принять продукцию, которую даёт водоём в течение такого периода времени, который мы могли бы принять за единицу времени, например, месяц, сезон, год и т. п. (в зависимости от цикла жизни данного вида организмов). Таким образом, продукция может быть отнесена к любому промежутку времени и измеряется приростом биомассы за этот промежуток, продуктивность же измеряется продукцией за период времени, принятый за е д и н и.ц у.
Из всего сказанного ясно, что продукция определяется через биомассу, а продуктивность — через продукцию, т. е. в конечном счёте тоже через биомассу.
Классификация озёр. Если мы расширим понятие продуктивности, включив в него также и неорганические вещества, имеющиеся в водоёме, то мы восстановим полный круговорот веществ, состоящий из двух основных комплексов — минерального и органического. Главнейшие этапы этого круговорота сводятся к следующему.
Из минерального комплекса зелёные водные растения — продуценты (производители), черпают необходимые для построения их тела вещества, газообразные и твёрдые (растворённые в воде минеральные соли). Растения
‘ Гибель их от поедания различными хищниками в хорошо содержащихся рыбоводных прудах мы считаем незначительной, так что этой величиной можно пренебречь. 104
являются, следовательно, лабораториями, где неорганические вещества перерабатываются в органические. Многие из растений в качестве пищи поглощаются животными; другие животные питаются не растительной, а животной же пищей. Животные, следовательно, принципиально отличаются от растений гем, что неспособны питаться непосредственно минеральными веществами, а только органическими, почему и называются консументами (потребителями). Как растения, так и животные в процессе обмена веществ выделяют в воду некоторые органические вещества. После отмирания тех и других тела их разлагаются, постепенно распадаясь тоже на всё более и более простые органические соединения. При дальнейшем процессе гниения органические вещества распадаются на минеральные соли, которые вновь вступают в круговорот как пища растений и т. д. Это, разумеется, самое общее и схематическое представление о круговороте веществ в водоеме. В природе этот процессуслож-няется многими частными деталями, среди которых одной из главнейших является то, что не все вещества и не всегда совершают полный круговорот: некоторые выходят из него й благодаря этому накопляются в водоёме. Так происходит, например, с многочисленными трупами многих планктических организмов, которые, не доходя в процессе гниения до полной минерализации, падают на дно и отлагаются здесь.
Мы видели, что основу учения о продуктивности составляют три основных вида биологических процессов: рост организмов, их размножение и их отмирание. Первый из этих процессов зиждется на процессе питания. Следовательно, процессы питания, или, как их называют, трофические процессы, образуют один из базисов продуктивности. С другой стороны, сама продуктивность лежит в основе ещё более широкого комплекса явлений в водоёме — в основе круговорота вещества, в который, кроме продуктивности, входит ещё сложная система химических превращений веществ. Таким образом, получается ряд: трофические процессы — продуктивность — круговорот веществ — с восходящей степенью сложности явлений, ряд, познание которого ведёт к высшему синтезу наших знаний о водоёме. Знание последнего члена этого ряда дало бы нам в руки исчерпывающий материал для полной, всесторонней и подробной классификации водоёмов.
Однако научный интерес исследователей и требования рыбохозяйственной практики не могут мириться с очень долгим ожиданием полного разрешения указанных выше проблем и с откладыванием в долгий ящик решения первоочередных и насущнейших вопросов. Поэтому учёные попытались уже теперь, на основании частичных, неполных знаний о трофических процессах, продуктивности и круговороте веществ в водоёме, создать хотя бы самую общую, схематическую классификацию водоёмов того типа, который лучше всего изучен, именно — озёр.
В основу современной классификации озёр положен не один какой-либо признак, а целый ряд признаков, или факторов, характеризующих озёра как с физико-химической, так и с биологической стороны. Основная связь между этими факторами идёт по линии трофичности и продуктивности озёр. Все озёра умеренного пояса по этой классификации делятся на 3 главнейших типа, с характеристикой которых по одному признаку — кислородной слоистости — мы познакомились уже выше, где эти типы назывались субальпийским, балтийским и гумусовым. В настоящее время озёра делятся на эти же три типа, но уже на основе целого комплекса факторов, причём этим типам присвоены другие названия, именно: вместо субальпийский — олиготрофный, т. е. малопитательный, вместо балтийский — эвтрофный, или хорошо питательный, вместо гумусовый — д и строф н ы й, или непитательный. Термин «питательный» относится здесь к количеству содержащихся в озере минеральных солей — так называемых биогенных веществ, которые идут на питание зелёных (автотрофных) растений и за счёт которых, следовательно, образуется первичная продукция озера.
Три основных типа озёр характеризуются следующими комплексами признаков.
I. Олиготрофный (субальпийский) тип. Относительная бедность воды растворёнными питательными для растений веществами; отсюда — бедность фитопланктоном и детритом внутреннего (т. е. в самом озере) происхождения; вследствие этого большая прозрачность воды и бесцветность её; как зимой, так и летом — отсутствие'резко выраженной кислородной слоистости; рН^7. 11л на дне беден органическими веществами, поэтому гниющего ила нет.
II. Эвтрофный (балтийский) тип. Богатство воды питательными для растений веществами, поэтому богатство фитопланктоном и детритом; отсюда малая прозрачность воды; последняя почти бесцветна, лишь с лёгким желтоватым оттенком ь, в озёрах со значительной глубиной летом ясно выраженная параллельная слоистость по температуре и кислороду; зимой в мелких озёрах исчезновение кислорода у дна. pH 7. Донные отложения богаты органическими остатками и представляют гниющий ил (гиттия, сапропель).
III. Дистрофный (гумусовый) тип. Бедность воды питательными для растений веществами и богатство её растворёнными гуминовыми веществами и принесённым в озеро извне детритом; цвет воды — от жёлтого до коричневого; малая прозрачность её; зимой и летом резко выраженная температурная и кислородная слоистость. pH 7. Донные отложения — дью или торф.
Само собой понятно, что в приведённых характеристиках трёх основных типов озёр свойственные каждому типу комплексы признаков взяты в их наиболее законченном и резком выражении. В природе встречается много переходных случаев, т. е. озёр с промежуточными степенями выражения признаков, характеризующих тот или иной тип, что, конечно, может затруднить отнесение озера к одному из них.
Считаем необходимым подчеркнуть, что приведённая классификация относится только к озёрам умеренного пояса, к озёрам же более высоких и низких широт она неприменима. Но и для озёр средних широт она даже в настоящее время становится недостаточной, так как встречаются озёра, которые очень трудно в неё втиснуть. Это ведёт к её постепенному расширению и углублению. В этом направлении многое делается нашими советскими гидробиологами. Правильная классификация водоёмов должна представлять собой прочную основу для их хозяйственного использования.
Кроме описанных трёх типов озёр, некоторые исследователи различают ещё 4 типа: алкалитрофные озёра с большим количеством растворённого кальция, ацидотрофные — с кислой реакцией (pH = 5,5 и ниже), сидеротрофные — с большим количеством железа, и а р-гиллотрофные — со значительным количеством глинистых частиц. В подробную характеристику этих четырёх типов мы входить не будем.
В приведённые выше краткие характеристики основных типов озёр входят факторы почти исключительно небиологического порядка. Из биологических факторов (живые организмы) там фигурирует только фитопланктон в качестве первичной продукции. Но совокупность всех этих факторов образует определённый биотоп или определённую комбинацию биотопов, которая в свою очередь определяет то или иное сочетание определённых биоценозов. Ясно, что эти биоценозы должны состоять не только из растительных, но и из животных комплексов. Поэтому и среди животных должны быть такие, которые, будучи приспособлены к определённым условиям жизни, могут быть характерными для того или иного типа озёр, так что по их присутствию или отсутствию или по их количеству можно судить о типе озера. И действительно, в более подробную и развёрнутую характеристику основных типов озёр входят животные, характерные для этих типов; это известные уже нам личинки комаров Tany tarsus, Tendipes и Chaoborus. Для олиготрофного типа озёр характерно присутствие Tanytarsus и отсутствиеChaoborus; для эвтрофного — присутствие Tendipes plumosus, Tendipes batophilus и Glyptotendi-pes polytomus, а также Chaoborus ( Гапу tarsus встречается только в прибреж-
1 Вода принимает более или менее густую окраску (зелёную, синезелёную) лишь при её «цветении» во время вспышек развития различных видов водорослей.
ной зоне, где он находит для себя достаточное количество кислорода); для дистрофного — присутствие Chaoborus, а из видов рода Tendipes — только plumosuB. Из рыб такой показательной породой служит глубоководный сиг, нуждающийся для дыхания в большом количестве кислорода: в олиготрофных озёрах он встречается часто, в эвтрофных — редко, а в дистрофных — никогда. Такие организмы, как только что перечисленные, называются п о-казателями, или индикаторами, так как они своим присутствием, отсутствием и степенью развития указывают на определённый комплекс условий существования, на то или иное состояние водоёма. Прилагаемая таблица (стр. 108) даёт представление о классификации озёр.
Особенно большую известность, как показатели состояния среды, приобрели те организмы, которые служат индикаторами на степень загрязнения водоёмов, или, как говорят, на степень их сапробност и. Это явление, как мы уже знаем, состоит в чрезмерном накоплении внутри водоёма органических веществ, поступающих в него (извне при водопое скота, с водой притоков, несупщх отбросы домашнего хозяйства или отходы промышленных и сельскохозяйственных предприятий, с канализационными стоками и т. п.) или происходящих в результате естественных процессов внутри самого водоёма, как, например, в рыбоводных прудах при нагуле карпа в условиях сильно уплотнённой его посадки (особенно при одновременном нагуле уток). Для разных степеней загрязнения характерны различные физико-химические свойства и комплексы ортанических веществ воды, создающие для организмов разные условия существования, следовательно, разные биотопы, которые, в свою очередь, определяют и различные биоценозы. Поэтому, если весь процесс возрастающего загрязнения — от чистой воды до самой загрязнённой — разделить на некоторое число степеней, то и все организмы можно соответственно разбить на такое же число групп, из которых каждая будет приурочена к определённой степени сапробности. Так именно и была создана для водных организмов система их сапробности, которая первоначально состояла из трёх групп: организмы, живущие при самой сильной степени загрязнения, или п о л и с а п р о б н ы е; организмы, живущие при среднем загрязнении, или мезосапробные; и организмы, живущие только в почти чистой воде, или олигосапробные;но затем средняя группа была разбита на две: а (альфа) — мезосапробвая, более близкая к по-лисапробной, и р (бета) — мезосапробная, более близкая к олигосапробной. Таким образом, различают четыре степени сапробности водных организмов, которые сокращённо обозначаются первыми буквами их латинских названий: р (полисапробные организмы, или полисапробы), ат (альфамезосапробы), (бетамезосапробы) и о (олигосапробы).
Со времени первого появления системы сапробности водных организмов прошло 40 лет, в течение которых эта система исправлялась и дополнялась. Значительная работа в этом направлении проделана советскими учёными *.
С возрастанием загрязнённости уменьшается число групп организмов, способных выносить её, причём выпадают более высоко организованные, а остаются всё менее и менее высоко организованные существа, так что высшую степень сапробности могут выносить только организмы, стоящие на самой низкой ступени организации. Усиление загрязнения вызывает интенсивное развитие и количественный рост бактерий, и это является чрезвычайно важным и ценным с точки зрения хозяйственных интересов моментом, потому что именно наличность бактерий обусловливает в явлении загрязнения воды поворотный пункт: они являются очистителями воды, так как, питаясь органическими веществами, бактерии разрушают и при благоприятных условиях (при достаточном притоке кислорода) переводят их в минеральные соли, уничтожая таким путём загрязнение воды. Особенно ясно это яв-
1 Синеок организмов и распределение их по степеням («зонам») сэнробногти см.: Долгов Г, И. и Никитинский Я. Я., Гидробиологические методы исследования. В книге «Стандартные методы исследования питьевых и сточных вод», изд. Постоянною Бюро всесоюзных водопроводных и санитарных технических съездов. Москва 1927.
Характеристика трёх основных типов озёр по различным признакам 1
Признаки Олиготрофный тип Эвтрофный тип Дистрофный тип
Морфометрия озёр Количество растворённого в воде Оа летом Причина кислородного дефицита Активная реакция воды Взвешенный материал Биогенные минеральные вещества (питательные для автотрофных растений соли) Органические вещества Донный ил Прозрачность воды Цветность воды Прибрежная водная растительность Планктон Личинки Tendipe-didae Личинка Chaoborus При зарастании превращаются Глубокие, с узкой прибрежной зоной; водная масса гиполимниона по отношению к таковой же эпилимниона велика Большое; в гиполимнионе до 60— 70% насыщения От кислой до щелочной (рН^7) Очень мало Немного Немного Мало Большая Переход от голубой до зелёной Мало Огносительно мало, но распределён по всей глубине Почти все Tanytar-sariae, часто Dia-mesini Отсутствует В эвтрофные водоёмы Не глубокие, с широкой прибрежной зоной; водная масса гиполимниона по отношению к таковой же эпилимниона мала Эпилимнион может быть богат О2(до состояния насыщения и даже пересыщения), гиполимнион беден (у дна до 0% насыщения) Летом разложение планктона и донпого ила, зимой только ила Нейтральная или щелочная (pH 2^7) Много, главным образом планктон в живом и отмершем состоянии Много Много Много; сильно преобладает автохтонный разлагающийся органический материал Малая вследствие большого скопления взвесей Переход от зелёной через жёлтую до бурой Много В эпилимнионе много, в гиполимнионе мало Tendipes plurnoeus, Т. batbophilus; Chaoborus, часто Glptotendipes poly tom us. Почти всегда есть В пруды и болота (верховые и визовые) Различной глубины, в болотных районах или в областях изверженных пород Как в эвтрофных озёрах (иногда как в олиготрофных) Разложение взвешенного и донного детрита Кислая (рН<7) в некоторых озёрах даже рН<5) Много, главным образом аллохтонного происхождения (приносимый в водоём извне) Мало Много (гумус) Много; преобладает аллохтонный гумусовый материал Малая вследствие присутствия аллохтонных взвесей в воде и тёмной окраски последней Переход от жёлтой через бурую до коричневой Мало, большейчастью болотного характера Фитопланктона мало, зоопланктона может быть много Как в эвтрофных озёрах В верховые (торфяные) болота
1 Таблица в сокращённом и несколько изменённом виде взята из «Общей гидробиологии» акад. С. А. Зернова.
лепие биологического самоочищения воды обнаруживается в реках, где оно находит для себя наиболее благоприятные условия: движение воды, перемешивание её слоев, большое количество кислорода и пр. «
Изучение явления сапробности водоёмов, несомненно, имеет не только большой научный интерес, но и громадное практическое хозяйственное значение (прежде всего для народного здравоохранения, поскольку грязная вода является поставщиком и разносчиком эпидемических болезней, а также для рыбного хозяйства). Это изучение ведёт к познанию тех организмов, которые служат индикаторами на степень загрязнения, и тех способов борьбы с ними, которые сама природа даёт в руки человека в виде способа биологической очистки воды.
ГЛАВА ВТОРАЯ
ПРУД
Название «пруд» прилагается к водоёмам различного рода. По своему этимологическому значению втот термин должен обозначать такой мелкий водоём, который образуется посредством запруды, т. е. устройства преграды, задерживающей течение воды, например, плотины в виде котя бы самой простой земляной насыпи. Такие пруды, как обязанные своим происхождением вмешательству человека, носят название искусственных. Но мы уже видели, что название «пруд» может быть отнесено и к таким водоёмам, которые образовались в природе сами по себе, без воздействия на них человека, например, как одна из стадий угасания озера. Такие пруды называются естественными. Они могут образоваться и вне всякой зависимости от озера — допустим, путём заполнения водой какого-нибудь небольшого углубления суши.
Познакомимся несколько ближе с обеими группами прудов, причём начнём с естественных.
ЕСТЕСТВЕННЫЕ ПРУДЫ
Среди естественных прудов можно также различать две группы: о с-военные человеком в целях их хозяйственного или промышленного использования ине освоенные, иначе говоря, «дики е». Существование второй группы прудов может быть обусловлено разными причинами: большой отдалённостью от человеческого жилья, недоступностью вследствие, например, нахождения прудов в неудобных для их использования местностях (лесистых или болотистых районах) и т. п.
Общей для всех прудов чертой является, как уже говорилось, их н е-знач и тельная глубина. Это основной фактор, обусловливающий появление и развитие в прудах таких характерных для них признаков и свойств, общая совокупность которых заставляет выделить пруды в особый тип водоёмов. Прежде всего следует отметить, что небольшая толща воды позволяет солнечным лучам проникать до самого дна, освещая и нагревая его, благодаря чему создаются условия для развития и распространения по всему пруду как растительного, так и животного населения. Гидрохимический режим прудов изменяется — также в зависимости от мелкости этих водоёмов — в сторону накопления в них биогенных элементов и, следовательно, создания больших запасов питательных веществ как для макро-, так и для микрофлоры. Таким путём создаётся широкая база для развития в прудах богатой гидрофауны. Всё это указывает на то. что общий комплекс факторов, господствующих в прудах, сильно интенсифицирует процессы обмена веществ в них и повышает трофику прудов, по сравнению с озёрами.
По общему режиму как жизненная среда для растительных и животных организмов пруды обнаруживают очень близкое сходство с прибрежной областью озёр. В сущности пруд — это лишённое пелагической области озеро, в котором литораль распространилась по всей его площади. Отсюда ясно, 110
что и животное население прудов в общем очень сходно с населением литоральной области озёр. Так, в связи с чрезвычайно разнообразными условиями среды в прудах пышно развивается фауна водных насекомых — одни лишь в стадии личинок (ручейники, стрекозы, подёнки, комары и пр.), другие — в стадии как личинок, так и имаго (жуки, клопы). Однако животное население прудов обнаруживает и некоторые особенности. Например, личинки иногда несколько отличаются по своему строению от тех же форм в озёрах (это указывается, например, для Chaoborus).
В планктоне прудов и прибрежной области озёр также встречается много общих форм, но вместе с тем имеются и различия, касающиеся присутствия или отсутствия некоторых видов организмов, а также количества представителей общих видов. Так, коловратки в прудах находят для себя лучшие условия жизни, чем в озёрах, и потому развиваются пышнее. Среди них имеются даже такие, которые можно назвать типично прудовыми формами, например, некоторые виды Erachionus и Schizocerca (рис. 27). Из ракообразных в прудах развивается в большем, чем в озёрах, количестве, ветвистоусый рачок Chydorus (рис. 271). С другой стороны, есть и такие рачки, которые, наоборот, живут лишь в озёрах, в типичных же прудах отсутствуют, например, Leptodora (тоже из Cladocera).
До сих пор шла речь о «диких», т. е. не освоенных человеком естественных прудах. Освоенные естественные пруды также могут быть разбиты па группы в зависимости от тех целей, которые ставит себе человек при их хозяйственном или промышленном использовании. Это использование очень часто заключается в обслуживании бытовых нужд домашнего хозяйства, как это имеет место в колхозных прудах: водопой скота, наличие домашней водной птицы, мытье белья, приём сточных вод со скотных дворов и т. п. Все эти процессы в той или иной мере содействуют накоплению органических веществ в пруде. Ускорение этого накопления и составляет наиболее существенное и характерное отличие этих прудов от «диких». Минеральными веществами такие пруды, наоборот, бедны, и в этом заключается причина отсутствия пли, по крайней мере, очень слабого развития в них крупной растительности. Но и микроскопическая флора оказывается здесь чрезвычайно скудной по числу видов, так как лишь немногие группы родов и видов водорослей — преимущественно синезелёпые — способны жить в таких условиях; зато они при отсутствии конкурентов размножаются в громадных количествах и заполняют водоём; примером может служить водоросль Aphanizonienon (рис. 29Г), часто вызывающая цветение в таких прудах. Само собой разумеется, что в связи с изменениямпхарактера среды и животный мпр колхозных прудов претерпевает определённые изменения: в нём получают преобладание формы, более приспособленные к существованию в условиях обильного содержания органических веществ в воде, как, например, некоторые виды рода Brachionus (В. urceus, В. angularis) из коловраток или наиболее крупные виды рода Daphnia (D. magna, D. pulex).
Скорость накопления органических веществ в колхозном пруде зависит, с одной стороны, от объёма водной массы пруда, с другой — от количества органических веществ, поступающих в пруд в единицу времени (другими словами, от количества людей и домашних животных в пользующемся этим прудом населённом пункте). При значительных размерах пруда, следовательно, при значительном количестве воды в нём и вместе с тем при малой плотности людского населения на берегу пруда масса растворённого в воде кислорода достаточна для разложения и минерализации поступающих в пруд сравнительно в небольшом количестве органических соединений; это способствует4 поддержанию его в хорошем, чистом состоянии. В этот период органические вещества играют положительную роль, так как служат удобрением пруда, повышающим его продуктивность. Но при увеличении поступления органической массы в этот пруд может наступить такой момент, когда кислорода в воде будет едва хватать на разложение этой массы. Этот момент является для пруда критическим, поворотным, так как после него, в процессе дальней-
шего нарастания поступления в пруд органических веществ, последние уже не успевают разложиться нацело; начинается их накопление в пруде, ведущее к его загрязнению. Загрязнение колхозных прудов наступает в тех случаях, когда они перегружены поступающими в них органическими веществами, количество которых несоразмерно велико по отношению к объёму водной массы пруда, и последний загнивает: вода его портится, как говорят, закисает вследствие накопления в ней различных кислот, содержание кислорода снижается до минимума, иногда до полного исчезновения, появляются дурно пахнущие вещества, как сероводород и др. В этих и подобных им явлениях и заключается сущность процесса загрязнения прудов.
Но как только в той или иной мере ослабляется или, тем более, совсем прекращается пользование прудом для нужд хозяйства, а следовательно, и искусственное загрязнение его, так, будучи предоставлен естественному ходу событий, он начинает постепенно очищаться; это проявляется в изменении химического состава воды, а в связи с этим и состава живого населения: формы, типичные для искусственно загрязнённого пруда, исчезают и заменяются организмами, свойственными нормальному пруду. Такое самоочищение воды является следствием деятельности различных гнилостных бактерий, которые при большом доступе кислорода, обусловленном перемешиванием воды в пруде от действия ветра, вызывают интенсивные процессы тления, т. е. распада органических веществ и их минерализации, чем и достигается очистка как воды, так и грунта.
В настоящее время — в период восстановления хозяйства и промышленности в нашем Союзе и реализации сталинского плана преобразования природы — выдвигается для колхозных прудов новая, очень почётная задача, которая должна поднять их значение в народном хозяйстве на ббльшую высоту: это участие их в процессе разведения и выращивания рыбы. Само собой разумеется, что эта задача наилучшим образом может быть выполнена лишь в тех случаях, когда в колхозных прудах не только не будет допускаться загрязнение, но, наоборот, будут поддерживаться чистота и порядок, так как в грязных и засорённых водоёмах рыба плохо себя чувствует, хуже питается и потому хуже растёт. Для опрятного содержания прудов их необходимо время от времени чистить.
Вторую группу освоенных человеком естественных прудов составляют фабрично-заводские пруды, выполняющие задачу, аналогичную задаче колхозных прудов: как эти последние принимают в себя отходы и отбросы домашнего хозяйства, так фабрично-заводские пруды принимают отработанные сточные воды от расположенных вблизи их фабрик и заводов. Ясно, что пруды этой группы могут загрязняться не только не в меньшей, но часто даже в большей степени, чем колхозные, причём загрязнение фабрично-заводских прудов может происходить от поступающих в них как минеральных, так и органическйх (в зависимости от характера производства) соединений — различных кислот, щелочей и солей, среди которых часто бывают вредные для организмов вещества, вызывающие гибель живого населения пруда.
Однако использование человеком естественных прудов не всегда сопровождается приведением их в негодное состояние. Так, в случаях использования механической энергии воды для приведения в действие каких-либо силовых установок пруду не наносится никакого вреда. Пример — мельнич н ы е пруды.
Из этой очень краткой характеристики естественных прудов видно, что в полной мере термин «естественные» приложим только к «д и к и м» прудам, которых, так сказать, не касалась человеческая рука. Уже в колхозных прудах самый факт их использования так или иначе изменяет их режим и этим вносит в них некоторый элемент искусственности. Ещё резче этот элемент выражен в фабрично-заводских прудах, где приходится устраивать кое-какие гидротехнические приспособления с целью усовершенствования процесса их использования. Но всё же у всех этих прудов имеется одно общее 112
свойство: это именно естественные, т. е. образованные самой природой, водоёмы, и влияние на них человека проявляется лишь в стремлении утилизировать их полезные для него природные свойства без особенной заботы об улучшении самих прудов. Вот почему я объединяю их в одну категорию естественных прудов. Теперь познакомимся с другой категорией — с искусственными прудами.
ИСКУССТВЕННЫЕ ПРУДЫ
Если не единственную, то во всяком случае обширнейшую группу искусственных прудов составляют рыбоводные пруды, играющие огромную роль в народном хозяйстве нашего отечества как поставщики одного из основных продуктов народного питания — рыбы. Искусственный характер этих прудов целиком определяется их назначением — разводить и выращивать культурные породы рыб. Всё устройство рыбоводных прудов — их грунт, водный режим, размеры (глубина, площадь), гидротехнические сооружения и пр. — должно быть приспособлено к тому, чтобы обеспечить выращиваемой породе рыб наиболее благоприятные условия её существования. Для этого необходимо хорошо знать биологию этих рыб, образ жизни, их разнообразные физиологические потребности, чтобы иметь возможность удовлетворять их путём регулирования режима прудов.
В СССР существуют два типа прудового рыбного хозяйства — холодно-водное и тепловодное. Первый тип распространён сравнительно мало, второй же — очень широко. В прудах первого типа выращиваются холодолюбивые породы рыб — лососевые (главным образом форель), в прудах второго типа — такие теплолюбивые породы, как карповые (почти исключительно карп). Форелевые пруды характеризуются большей глубиной, чистой, прозрачной водой, сравнительно низкой летней температурой её, высоким содержанием растворённого в воде кислорода, слабым развитием флоры и фауны; карповые пруды, наоборот, мелкие, хорошо прогреваемые водоёмы с малой прозрачностью воды, с пониженным количеством кислорода в ней, с более или менее развитой растительностью и богатым животным населением, следовательно, с более пли менее обеспеченной кормовой базой для рыб. В дальнейшем речь будет идти лишь о карповых прудах, как наиболее распространённых в наше.м Союзе.
При господствующей у нас системе карпового хозяйства с двухлетним оборотом товарная (столовая) рыба выпускается хозяйством в возрасте около полутора лет (так называемый двухлеток, так как он прожил в прудах два летних сезона). В течение этого времени карп проходит несколько возрастных периодов, которым рыбоводы присвоили специальные названия; для выращивания карпа в течение каждого из этих периодов устраиваются специальные пруды. Соответственно этому весь производственный процесс вырашиваипя товарного карпа в полном хозяйстве с двухлетним оборотом складывается из нескольких частей, а именно:
_ _ , Категории прудов, воярастпые периоды
Приблизительные сроки . карпа и рыбоводные операции.
1) Приблизительно середина мая (при Посадка производителей (самок и наступлении тёплой погоды). самцов карпа) в нерестовые пруды; процесс нереста (оплодотворение выметанной самками икры молоками самцов); развитие в икринках личинок карпа; выдерживание в нерестовых прудах вылупившихся из икры мальков в течение 10—15 дпей.
2) Вторая половина мая или начало Пересадка мальков из цересговых июня. в выростные пруды.
1 Для центральных районов СССР,
3) Май (пли июнь) — октябрь. Выращивание мальков в вырост -
1 ных прудах до стадии сеголетков;
пересадка сеголетков в зимовальные пруды.
4) Октябрь — апрель. Зимовка сеголетков.
5) Конец апреля. Пересадка годовиков (перезимо-
вавших сеголетков) из зимовальных в нагульные пруды.
6) Май — октябрь. Нагул годовиков в течение второго
лета их жизни.
7) Октябрь или ноябрь. Окончательный облов нагульных
прудов; полный вылов двухлетков — товарных (столовых) карпов.
Отсюда видно, что успех всего рыбоводного процесса в целом обеспечивается правильным проведением двух главнейших операций: 1) выращиванием карпа до стадии товарной рыбы и 2) полным выловом последней из прудов. С целью осуществления последней операции рыбоводные пруды делаются спускными: в них устраиваются особые гидротехнические сооружения — водоспуски (так называемые «монахи»), позволяющие спускать из прудов всю воду. Эти приспособления в наибольшей степени придают рыбоводным прудам искусственный характер и являются самой характерной чертой их отличия от естественных прудов.
Приведённая очень краткая характеристика искусственных рыбоводных прудов показывает, что успешные результаты ведения прудового рыбного хозяйства могут быть обеспечены лишь путём специального устройства прудов и поддержания в них определённого режима. Эти именно черты главным образом и придают рыбоводным прудам характер искусственных водоёмов, противопоставляя их прудам естественным, по отношению к которым, как сказано выше, человек не предпринимает каких-либо мер, направленных к их упорядочению и улучшению.
Само собой понятно, что такое энергичное вмешательство человека в жизнь прудов, какое имеет место в прудовом рыбном хозяйстве, должно в сильной степени изменить общий режим рыбоводных прудов. Ясно, что оно должно отразиться и на живом населении их, которое человек изменяет качественно и количественно в таком направлении, чтобы оно наилучшим образом отвечало его хозяйственным потребностям. Эта цель осуществляется двумя главными путями: 1)содействием размножению и развитию организмов, полезных для рыбного хозяйства; 2) борьбой с организмами, которые наносят хозяйству какой-либо вред.
К первой группе относятся в первую очередь организмы, идущие рыбе в пищу, главнейшими из которых являются в бентосе личинки разных насекомых, особенно личинки Tendipedidae (Chironomidae), а в планктоне — ракообразные. Затем в эту же группу входят те организмы, которые являются кормом для пищевых организмов рыбы, как, например, зелёные водоросли для ракообразных. Эти же зелёные водоросли и вообще зелёные растения полезны ещё тем, что в процессе фотосинтеза снабжают воду пруда кислородом.
Ко второй группе — врагов рыбного хозяйства — относятся организмы, наносящие вред как самой рыбе (хищные личинки некоторых насекомых — жуков плавунцов, стрекоз, клопов и др., нападающие главным образом на рыбью молодь, затем эктопаразиты, как Argulus foliaceus — рыбья вошь, наконец, различные болезнетворные микроорганизмы), так и организмам, составляющим кормовую базу рыбы.
Всё сказанное о прудах свидетельствует о том, что между естественными и искусственными прудами существует резкая разница, которую в общем можно охарактеризовать следующим образом: в естественных прудах явления протекают стихийно, а в искусственных — организованно, так как регулируются человеком в определённом направлении.
ТЕМЫ ДЛЯ РАБОТ
Ввиду того, что в отношении как тем, так и методики исследований почти всё сказанное для озера относится и к пруду, мы не будем повторяться и отсылаем к соответствующим страницам первой главы.
Заметим лишь, что с практической точки зрения важно исследовать колхозные п р у Д ы с целью разведения в них рыбы. Как использовать полученные в результате этих исследований сведения, а также как организовать и технически осуществить самое разведение рыбы, обо всём этом можно получить сведения из учебников и популярных книг по рыбоводству, например:
Арнольди Й. Н., Как использовать озёра и пруды в колхозах, Ленинград 1931,
Е л е о н с к и й А. Н., Рыбоводство, Москва 1933.
Елеонский А. Н., Прудовое рыбоводство, М. 1946.
Е л е о п с к и й А. Н. и Ч е р ф а с Б. И., Что надо знать колхознику о рыбе и рыбоводство, Москва 1933.
Протасов А. С., Прудовое рыбоводство в совхозах и колхозах, Москва, Снаб-техиздат, 1935.
Суховерхое Ф. М., Прудовое рыбоводство в совхозах, колхозах, при фабриках и заводах, Москва 1933.
Гримм О. А., Рыбоводство, под ред. И. В. Кучина, М.—Л. 1931.
Исаев А. И., Организация рыбного хозяйства па местных водоёмах, М. 1934.
Исаев А. И. и Дор ох о в С. М.. Рыбоводство в колхозах, М. 1946.
Мовчан В. А., Короп, вирощування в ставках, КиТв 1947.
Мартышев Ф. Г., Соснина Е. И., Каспии Б. А., Прудовое рыбоводство в колхозе, М. 1949.
ГЛАВА ТРЕТЬЯ
БОЛОТО
Известно несколько способов образования болот в природе, но все эти способы могут быть сведены к двум основным: 1) превращение какого-либо водоёма со стоячей или медленно текущей водой (озера, пруда, ручья и пр.) в болото путём постепенного разрастания водной флоры и вытеснения ею воды и 2) происхождение болота на сухом месте путём скопления на нём воды с одновременным развитием в ней характерных и типичных для болот растительных ассоциаций.
Первый из этих двух главнейших способов распадается в свою очередь на два: зарастание водоёма водной флорой и нарастание, или надвигание, последней на водоём. Процесс зарастания озера, обмелевшего вследствие непрерывного накопления в нём донных отложений, описан в начале этой книги. Мы видели, что этот процесс означает наступление заключительного периода жизни озера и представляет собой расширение зоны прибрежных зарослей путём распространения её от периферии (от берегов) к центру водоёма. Таким образом, сначала озеро, постепенно зарастая, превращается в пруд; затем при дальнейшем продолжении зарастания площадь водной поверхности всё более сокращается, и пруд переходит в болото; наконец, после полного высыхания болота на его месте образуется луг. Эта генетическая связь болота и луга дала повод назвать происходящие путём зарастания водоёмов болота луговыми или травяными болотами. У этих болот есть ещё третье название — низинные, обусловленное расположением водоёмов, давших начало этим болотам, в низко ле
жащих местностях.
Характерным признаком описываемой группы болот является то, что в процессе их образования водная растительность по мере повышения дна,
на котором она укореняется, тоже поднимается, постепенно вытесняя воду. Ежегодно отмирая, она покрывает дно своими остатками, совокупность ко-
торых образует торф, отчего и самый процесс возникновения болота назы-
квпавТростниковый и ИЯдИИ камышовый тирф
Рис. 54. Схема зарастания озера.
вается ещё заторфо-в ы в а и и е м водоёма (рис. 54).
Второй путь превращения озера в болото заключается в следующем. В природе нередки случаи, когда берег озера где-либо в одном месте на том или ином протяжении представляет собой сплош-
ной зелёный ковёр, который колеблется под
ногами, хотя хорошо вы-
держи вает тяжесть человеческого тела. Ясно, что этот ковёр лежит на какой-то зыбкой основе, — основой этой является вода. Если подъехать к подобному берегу на лодке, то можно легко убедиться, что подведенное под него весло без сопротивле
ния уходит Довольно Рис. 55. Схема нарастания сплавины на озеро, глубоко. Растительный
ковёр свободно лежит на воде в пловучем состоянии и только одним, обращённым к берегу краем прикреплён к нему (рис. 55). Начало образованию такого ковра, или так называемой сплавины, даёт растительность самого берега на топ стороне его, откуда дуют на водоём господствующие ветры и где, следовательно, вода, защищённая берегом от действия этих ветров, находится в наиболее спокойном состоянии \
Процесс образования сплавины начинается с того, что некоторые береговые растения, расположенные у самого уреза воды,—такими растениями обычно являются трифоль — Menyanthes, сабельник — Comarum, белокрыльник — Calla — разрастаются от берега в сторону открытой воды, причём их горизонтально растущие по направлению к середине озера стебли стелются по водной поверхности, а отходящие от их междоузлий корни свободно свешиваются вниз, в толщу воды. По мере своего роста эти стебли всё теснее переплетаются друг с другом, образуя таким путём основу, на которой может произрастать целое сообщество других растений, например, разные виды осоки — Carex, хвощ — Equisetum, стрелолист — Sagittaria, частуха — Alisma и др.
Корневища этих растений, сплетаясь, постепенно уплотняют сплавину, а отмершие части растений, оставаясь в сплавине, утолщают её. С течением времени её нижние слои под действием собственной тяжести отваливаются и, падая на дно, повышают его 2. Так происходит постепенное сближение сплавины с дном п вытеснение из промежутка между ними воды, пока, наконец, образовавшийся торф не заполнит всё пространство между дном и поверхностным слоем растительности. Тем временем окаймляющая свободный край сплавины полоса трифоли и сабельника продолжает надвигаться на открытую поверхность воды, всё более сокращая её площадь, а расположенная по направлению к берегу от неё растительная ассоциация постепенно погибает и сменяется повой, в которой господствуют или осоки, или мхи — Hypnum и Sphagnum (рис. 56а). В результате этих процессов на месте водоёма образуется болото или осоковое, или моховое. В последнем случае гипиовое или сфагновое: первое, когда вода содержит известь, второе — когда извести в ней нет. Характерным обитателем сфагновых болот является растение росянка — Drosera, обладающая способностью переваривать мелких насекомых, попадающих на её листочки (рис. 56в).
Бывают, однако, и такие случаи, когда сплавина обязана своим происхождением надвиганию на поверхность воды непосредственно мхов Hypnum или Sphagnum. Образуемый их сплетением ковёр вначале настолько нежен и слаб, что не в состоянии выдержать даже небольшого волнения. Но с течением времени прочность его возрастает, и тогда на нём поселяются другие растения, например, осоки, которые своими корневищами ещё более его
1 На противоположном берегу, на который дует ветер, условия для развития растительности неблагоприятны, так как вызываемые ветром заплески и волны не дают возможности укрепиться па берегу растениям.*
8 Такие оторвавшиеся от сплавины и лежащие па дно пласты торфа носят название «мутта».
укрепляют — настолько, что он может служить почвой для новых растительных ассоциаций. Нередки случаи, когда первыми надвигаются Hypnum, за ними идут осоки, а за осоками Sphagnum.
Таким образом, процесс заторфовывания водоёмов происходит двумя основными путями и приводит соответственно к возникновению болот двоякого рода: путём зарастания — травяных (луговых) и путём нарастания сплавин — моховых (преимущественно гипповых).
Переходим теперь к рассмотрению второй главной группы болот — к болотам, образующимся на сухом месте. Здесь также известно несколько различных типов происхождения болот, главнейшие из которых мы и рассмотрим. Общее всем им явление заключается в том, что образование болот в этих случаях сопровождается процессом, обратным тому, который описан выше для случаев превращения водоёмов в болота: в то время как там болото происходит путём постепенного усыхания водоёма, здесь оно происходит, напротив, путём увлажнения первоначально сухой почвы и постепенного накопления воды в её верхних слоях, другими словами, путём её заболачи-
вания.
Заболачиванию могут подвергаться разнообразные сухие местности, как покрытые
Р и с. 56.
а — мох Sphagnum; б — мох кукушкин леи — Polytrichum; в—росянка — Drosera.
растительностью (лес, луг), так и лишённые её (лесные вырубки и пожарища).
Естественно возникает вопрос: откуда в сухих местах берётся вода и при каких условиях она накопляется? Увлажнение почвы может быть вызвано различными факторами, но их можно свести к двум основным источникам: 1) грунтовые воды и 2) атмосферные осадки. Соответственно этому различают и болота двоякого рода: 1) болота грунтового и 2) болота атмосферного питания. Но появление влаги в почве само по себе ещё не служит признаком её заболачивания: как известно, дождь иногда обильно смачивает почву, которая тем не менее от этого не превращается в болото, так как после дождя часть воды испаряется, часть уходит в глубоко лежащие слои почвы. Для заболачивания последней необходимо, чтобы вода в ней застаивалась, а это может быть в тех случаях, когда почва лежит на водонепроницаемом грунте. Рассмотрим в качестве примеров образования болот на сухих местах два случая: 1) заболачивание леса и 2) заболачивание луга. Из разных типов леса наиболее благоприятные условия для заболачивания представляет хвойный лес. При густоте елового или соснового леса, не позволяющей проникать до поверхности земли ни солнечным лучам, ни ветру, испарение почвы сильно задерживается, воздух застаивается, что содействует накоплению влаги как в нижних слоях атмосферы, так и в верхних слоях почвы. В таких условиях большей частью начинает разрастаться Polytrichum — к укушкин лён (рис. 566), а потом на образовавшемся из него торфе поселяется Sphagnum (рис. 56а) — несомненный предвестник заболачивания леса.
Этот способ заболачивания наблюдается в тех случаях, когда почва, на которой оно начиналось, первоначально была более или менее богата минеральными веществами; на таких почвах Sphagnum не может расти, другие ?ке мхи могут. Развивающийся в данном случае кукушкин лён в процессе своего роста выкачивает из почвы минеральные соли и при своём отмирании образует слой торфа, в котором эти соли почти совершенно отсутствуют; на этом торфе и развивается сфагнум.
Но существует другой путь заболачивания леса, при котором почва с самого начала чрезвычайно бедна минеральными веществами, и тогда на ней начинает произрастать сразу сфагнум, минуя стадию кукушкина льна.
В обоих этих случаях необходимое для процесса заболачивания накопление влаги в почве происходит благодаря тому, что Polytrichum и особенно Sphagnum обладают большой влагоёмкостыо, сильно развитой способностью поглощать значительные количества воды, поступающей главным образом в виде дождя. При непрерывном нарастании мха вверх и одновременном отмирании его нижних частей получается всё утолщающаяся, выпуклая в середине подушка, не пропускающая в почву атмосферные осадки и хранящая в себе большие запасы воды. Так образуется при заболачивании хвойного деса моховое болото атмосферного питания, называемое благодаря его форме верховым или возвышенным.
При большом скоплении воды в верхних слоях почвы возвышенных болот глубоко лежащие слои почти совсем лишены её, что ведёт к постепенному отмиранию древесной растительности, начиная с пород, наиболее чувствительных к недостатку влаги.
В наших хвойных лесах первыми гибнут ели, а способные выносить засуху сосны некоторое время ещё сопротивляются; по затем и они в конце концов отмирают, а появившаяся уже на болоте молодая сосновая поросль принимает очень жалкий вид: корявые сосенки стоят поодиночке и не поднимаются высоко над землёй.
Следует отметить ещё один тип заболачивания леса — это заболачивание елового или смешанно-елового леса, на месте которого возникает болото не моховое, а травяное (луговое) с характерной для него растительностью: осоками, злаками, сабельником и др. Такое заболачивание может происходить лишь на почвах с обильным содержанием как воды, так и растворённых в ней минеральных солей. На таких болотах может дольше держаться ель, а также могут расти и некоторые представители лиственных пород, например, берёза, осина.
Перейдём теперь к вопросу о заболачивании сухих участков земли. Явление это изучено ещё очень слабо, и среди специалистов по этому вопросу существует немало разногласий. Основная причина заболачивания сухих мест (в том числе и лугов) заключается в поднятии грунтовых вод до уровня, близкого к поверхности почвы. Это явление может быть обусловлено разнообразными факторами. Так, если в данной местности климат становится более влажным, то интенсивность испарения понижается, влага в почве задерживается, грунтовые воды в верхних слоях земли накопляются, и таким путём создаются условия для заболачивания. Далее, болотам свойственна тенденция распространяться и наступать на граничащие с ними сухие участки земли; этим объясняется, что площадь поля или луга, по соседству с которой находится болото, постепенно сокращается под влиянием заболачивания, пока вся не превратится в болото. Сухопутная растительность — как древесная, так и луговая (травянистая) — является чрезвычайно сильным испарителем, не позволяющим накопляться в почве излишкам влаги. Но если покрытый лесом участок земли обнажается вследствие пожара или вырубки леса, то этим создаётся возможность при определённых условиях (высокий уровень грунтовых вод) превращения этого участка в болото.
Если основной фактор заболачивания — накопление грунтовых вод в поверхностных слоях земли — тем или иным путём осуществляется, то самый процесс заболачивания начинается с замены обычной травяной расти-
тельности влаголюбивой флорой — прежде всего мхами (кукушкин лён, гипнум и в особенности сфагнум), которые являются пионерами заболачивания. За мхами могут последовать и некоторые травянистые влаголюбивые растения, например, пушица (Eriophorum), осоки и др. При отмирании этой растительности происходит накопление её остатков в верхнем почвенном слое; так образуется постепенно нарастающий в толщину слой торфа, свидетельствующий о превращении сухого прежде участка земли в болото.
В нашем Союзе больше всего насыщена болотами его северная полоса, что вполне понятно, потому что эта полоса характеризуется большей влажностью климата и, следовательно, меньшей испаряемостью, чем в южных районах. Благодаря этому на севере сосредоточено наибольшее количество водоёмов различного типа, в том числе и болот.
По мере накопления знаний о болотах у исследователей явилась потребность систематизировать результаты их изучения с целью создать классификацию болот. Первая классификация делила болота на две главнейших группы — «подводные», т. е. лежащие ниже уровня стоячих и текучих вод (соответствуют плоским, или низинным по современной терминологии), и «надводные», лежащие выше указанного уровня (соответствуют выпуклым, или возвышенным, или верховым). Эта классификация, получившая общее признание, в дальнейшем дополнялась, детализировалась, но её основной принцип — деление болот на две большие группы — сохранялся. Однако продолжающиеся исследования показали недостаточность этого деления. Оказалось, что существуют болота, занимающие промежуточное положение. С этим фактом также согласны все исследователи, расхождение во мнениях существует лишь по вопросу о том, считать ли эту промежуточную группу совершенно самостоятельной, равноценной двум другим группам, или включить её в качестве подгруппы в первую группу. Среди наших отечественных болотоведов преобладает второе мнение. В виде окончательной классификации болот я привожу, по акад. Сукачеву, ту, которая предложена у нас так называемой Гумусовой комиссией.
1. Плоские болота. Воды их богаты питательными веществами. Располагаются они в понижениях. Поверхность их более или менее плоская или слабо вогнутая. Они подразделяются на:
1. Низинные болота. Сфагнум почти вовсе отсутствует. Воды значительно богаты питательными веществами. Сюда относятся тростниковые болота, болота с зарослями высших растений (злаки, осоки), отчасти лесные болота, ольшаники.
2. Переходные болота. Характерно участие сфагнума. Питательными веществами воды менее богаты. Лесные болота имеют характер берёзовых или смешанных лесов на болоте.
И. Возвышенные болота. Они развиваются при очень бедной питательными веществами воде, развиваются исключительно за счёт атмосферных осадков. Господствуют виды Sphagnum. Поверхность их выпукла. Древесная растительность представлена бывает лишь в виде корявой сосны, не образующей леса.
По типу питания болота разделяются следующим образом: низинные и переходные относятся к болотам грунтового питания, возвышенные — к болотам атмосферного питания.
Мне остаётся лишь сказать несколько слов о животном населении болот. Подходя к этому вопросу с точки зрения влияния на организмы окружающей их среды, мы прежде всего должны обратить внимание на характерные черты отличия болота как биотопа от других водных биотопов.
Из всех внешних факторов, действующих на водную фауну болот, наиболее сильное влияние оказывает, пожалуй, активная реакция воды. Именно этот фактор кладёт самую резкую грань, отделяющую болота от других типов пресных вод (озеро, пруд, река): в то время как в последних господствующей реакцией является нейтрально-щелочная (pH 7), в болотах она кислая 120
(pH 5) благодаря присутствию там гумусовых кислот. Ясно, что это обстоятельство должно наложить значительный отпечаток на животный мир болот. И действительно, есть такие виды животных, которые не выносят жизни в нейтрально-щелочных водах и приспособлены к существованию в воде с низким pH. К таким относятся, например, Hyalosphenia из простейших животных, Elosa worallii из коловраток и др. Они населяют центральные районы сфагновых болот, где pH доходит до 3,8.
Однако в болотах встречается целый ряд эвриионных организмов, живущих в водоёмах с широкими пределами колебаний pH. Так, из ракообразных сюда относятся веслоногий рачок — Cyclops languidus, ветвистоусый рачок — Chydorus ovalis.
Отметим некоторое своеобразие фауны в краевой зоне верховых болот. Здесь встречаются характерные виды: Cladocera — из родов Scapholeberis, Chydorus, Streblocerus; Copepoda — из родов Cyclops, Bryocamptus, Moraria и др.; Oligochaeta из родов Vejdovskyella, Pristina, Marionina; некоторые Tendipedidae; из моллюсков — Bithynia tentaculata (рис. 208 А) и др.
ТЕМЫ ДЛЯ РАБОТ
При исследовании луговых болот могут быть выдвинуты многие из тех тем, которые уже рассматривались при описании работ на озере.
Что касается торфяных болот, то здесь уже выступают на первый план специфические темы, связанные с возникновением сплавив и заболачиванием леса
При исследовании сплавины следует всегда отметить: 1) толщину сплавины; 2) глубину озера у конца сплавины; 3) растения авангарда; 4) растения основного ковра; 5) состояние древесной растительности.
Интересно отметить разницу в температуре поверхностного влажного слоя мха и более глубокого слоя.
Чтобы изучить микроскопическое население мха, необходимо выжать из него достаточное количество воды и пропустить её через планктическую сетку. Сгустив микроскопическое население мха в стаканчике сетки в небольшом количестве воды, выпустить последнюю в баночку или чашечку, из которой при помощи пипетки перенести каплю на предметное стекло, накрыть покровным и рассмотреть под микроскопом.Рассматривать можно или живые организмы, или предварительно зафиксированные небольшим количеством формалина.
ГЛАВА ЧЕТВЁРТАЯ
ВРЕМЕННЫЕ ВОДОЁМЫ
ХАРАКТЕРИСТИКА
Под временными водоёмами мы будем здесь понимать незначительные скопления воды, которые возникают периодически и сравнительно быстро исчезают. Весной во время таяния снега там и здесь в низинах блестят мелкие лужицы. Вода их холодна; однообразие температуры объясняется тем, что тающий снег всё время подбавляет новые порции холодной воды. В прозрачной воде весенней лужи мелькает богатое население. Как же попали сюда живые водные организмы? Несомненно, зачатки этой жизни могли или находиться тут же на месте, в том грунте, который пропитала весенняя вода, или могли быть занесены из воздуха. Окружающие водоёмы стоят скованные льдом, жизнь в них ещё скрыта от глаз. Когда окончится таяние снега, вода начнёт быстро прогреваться, испаряться и, наконец, исчезнет, чтобы появиться здесь снова, быть может, только через год, когда опять начнётся таяние снега. С исчезновением воды живые существа тоже должны замирать или покинуть водоём до следующей весны.
Коренное население подобных луж, не связанных с другими биотопами, должно состоять из таких организмов, которые могут переносить полное исчезновение влаги летом и промерзание зимой; одни организмы закрываются целиком твёрдыми и непроницаемыми оболочками — цистами, другие прекращают с высыханивхМ воды своё индивидуальное существование и оставляют яйца или зимующие почки. Все эти образования лежат в грунте и при пропитывании его весенней водой снова освобождаются из своих непроницаемых оболочек и вновь начинают жить и развиваться. Покоящиеся яйца не только прекрасно переносят засуху и промерзание, но в некоторых случаях даже требуют их как необходимые условия для своего развития.
К самым типичным формам весенних луж относятся несколько ракообразных: щптень — Lepidurus (рис. 231), жаброног — Branchipus (рис. 226), Pristicephalus (рис. 229), Lynceus. Кроме того, всегда встречаются некоторые впды циклопов и два вида Diaptomus, некоторые Ostracoda.
Вторую группу организмов, населяющих весенние лужи, составляют те насекомые, которые при высыхании луж перелетают во вновь образовавшиеся лужи (жуки, клопы). Наконец, к третьей группе относятся комары; зимующие самки откладывают в первые появившиеся лужицы свои яички, из которых вылупляются личинки, успевающие проделать все свои превращения вплоть до летающих взрослых кома ров за короткий срок существования лужи. Такова же причина появления здесь личинок к о м а р о в-де р г у-н о в и др. За кратковременное существование весенней лужи комары иногда успевают дать несколько поколений: быстрота развития является необходимым условием жизни в периодических водоёмах.
Помимо указанных групп животных, в весенние лужи попадают й в ременные обитатели, обладающие свободой передвижения по воздуху,— жуки и клопы.
Интересно отметить, что в л е с у, где испарение идёт много медленнее и где характер дна иной, нежели на открытом месте, и население луж тоже несколько иное, рядом с видами, характерными для луж открытых мест, здесь встречаются и другие виды тех же родов или даже другие роды. Наиболее частыми являются здесь Branchipus, Diaptomus, Cyclops (рис. 275) и Candona.
Есть ещё другой тип временных весенних водоёмов — это лужи, остающиеся после спада весенних высоких вод или рек. Их фауна заключает в себе, помимо указанных элементов, отчасти и животных, оставшихся от фауны основного водоёма.
Временные водоёмы могут возникать и от дождевых вод в самых разнообразных местах, лишь бы вода встретила здесь преграду для своего стока просачивания вглубь. Самые мелкие лужицы, вроде заполненной водой колеи колеса, банки от консервов, углубления в старом пне и т. п., оказываются заселёнными не только микроскопическим миром, но и личинками комаров Culex (рис. 434) и Tendipes (из группы Thummi). Даже лужицы на пнях имеют довольно богатое население из целого ряда личинок разнообразных комаров и крыски — личинки мухи Eri-stalis (рис. 444); известен и один жук — Prionocyphon; такая лужица по многу раз в лето высыхает и наполняется вновь, а в более глубоких частях пня истлевшая в труху древесина хранит остатки влаги.
ТЕМЫ ДЛЯ РАБОТ
Исследование весенней лужи, связанное с временем первого пробуждения жизни организмов от зимнего сна, может дать много интересного материала для наблюдений. Конечно, полное изучение этого чрезвычайно интересного биотопа возможно лишь при наличии микроскопа, но даже работа с лупой может дать достаточный материал для наблюдений.
1. Так как жизненные процессы развиваются здесь с большой быстротой, то желательно вести наблюдепия систематически — если возможно, ежедневно или через небольшие промежутки времени. Инструментарий должен состоять из планктического сачка и скребка, по возможности на очень длинных палках, чтобы можно было ими работать с берега, так как входить в ледяную воду опасно. Параллельно с исследованием н а-селения необходимо проследить и изменения температуры. Интересно подметить момент первого появления живых существ из грунта во вновь образовавшемся водоёме после таяния снега и льда; желательно отметить также появление обитателей, перелетевших из каких-нибудь соседних водоёмов (жуки, клопы).
2. Второй, не менее интересный, этап жизни весенней лужи — конец её существования; здесь придётся порыться в хранящем влагу грунте, куда уходят некоторые животные в попытках избежать засухи.
Интереспо учесть время развития тех насекомых, которые во взрослом состоянии покидают воду.
При исследовании так называемых полойных или п о й м е н н ы х, т. е. оставшихся после разлива рек луж, интересным вопросом для изучения является следующий: какая судьба постигает в оставшемся от текучих вод мелком стоячем водоёме те формы, которые остались здесь вместе с водой реки? Конечно, для этого нужно сравнительное изучение населения самой реки и населения отделившейся лужи в момент отделения и через некоторые промежутки времени; параллельно следует учитывать и ту разницу в температуре, которая выявляется для коренной реки и лужи после отделения последней.
ГЛАВА ПЯТАЯ
КЛЮЧ И РУЧЕЙ
ХАРАКТЕРИСТИКА КЛЮЧА
Ключами называются воды, пробивающиеся из-под земли. Иногда ключ выходит на поверхность земли под водой, впадая в озеро или реку, и тогда он, конечно, остаётся вне сферы непосредственного наблюдения.
Но часть ключа бьёт на поверхности земли, и тогда его воды дают интересный материал для наблюдения. По способу выхождения воды различают три типа ключей. Одни — так называемые реокрены — бьют на к р у т о м с к л о н е, и воды их непосредственно ниспадают вниз; быстро ниспадающие воды размывают встречающиеся на пути породы, и выход ключа обычно покрыт песком или камнями и мало заселён растениями. Другие — лимно-крены — выбиваются на дне небольшого бассейна, и вода, наполняя бассейн до краёв, выливается из него через край; дно такого бассейна большей частью илистое или песчаное и иногда густо заселено растениями. Третьи — гелокрены — просачиваются через более или менее толстый слой земли, превращая этот слой в род болота. Пробиваясь среди наземных растений, разливаясь кое-где лишь слабым слоем воды на растительном ковре, а кое-где образуя довольно большие скопления воды, ключ даёт место для обитания весьма разнообразному населению — от чисто наземного, случайно захваченного струёй, до чисто водного, включая в состав своих биоценозов формы, живущие в более или менее влажной среде.
Годовые и суточные колебания температуры очень малы. Годовые имеют размах всего 1—2°; это само собой понятно, так как вода ключа есть вода, только что покинувшая земные недра.
По химизму ключевые воды весьма различны. В зависимости от геологического строения местности, от горных пород, по которым протекал под землёй поток, содержание солей будет тем или иным. Содержание кислорода по выходе из земли низкое, но оно быстро возрастает за счёт кислорода воздуха, особенно если вода разливается сразу тонким слоем по сравнительно большой площади.
Ключи интересны тем, что на незначительном пространстве в них могут быть сосредоточены разнообразные условия существования, что вызывает возможность нахождения здесь весьма разнообразной флоры и фауны.
Растительное население почти отсутствует в ключах, где вода ниспадает сверху, но в ключах других типов оно может быть довольно богато. В связи с этим и пищевые запасы для животных в различных ключах будут неодинаковы; запасы то велики и состоят из типично водных свежих и перегнивающих растений, то очень незначительны — лишь в виде попавших случайно в воду частей наземных растений (главным образом листьев). Но большинство ключевых форм всё же является потребителем перегнивающего растительного вещества или живёт хищнически; очень немногие питаются живыми растениями.
Фауна ключей может быть разбита на ряд групп: прежде всего в месте выхода воды из-под земли можно ожидать появления на земную поверхность 124
тех форм, которые населяют подземные грунтовые воды. Все эти обитатели характеризуются отсутствием глаз и окраски, что стоит в связи с полной тьмой, господствующей в месте их коренного обитания (из ракообразных : слепой ослик — Asellus cavaticus — и колодезный рачок — Niphargus puteanus; из моллюсков: улитка — Lartetia и др.
В тихих местах, где вода почти стоит, всегда можно найти многих пред-о д, но, конечно, только из тех предста-
ставителей фауны стоячих
в
Рис. 57, 58.
Рис. 59. Чехлики ручейников:
А — Hydropsyche на камне; Б — Holoceatropus на листе; В — чехликп Hydroptila: налево — личинки, направо — куколки.
Чехлики личинок ручейников Trichoptera:
А — Rhyacophlla; Б — Silo; В — РШосо-lepus.
вителей, которые способны выносить низкие температуры: некоторые личинки мух и комаров, некоторые жуки, ручейники, веснянки, ракушковые раки, олигохеты и др.
По границе воды и суши скопляются обитатели влажной почвы: дождевой червь, личинки мух, пауки, мокрицы и пр.
В быстро низвергающихся ключах господствует своеобразная фауна потоков, для которой наиболее типичными явгляются некоторые личинки ручейников из родов Agapetus и др. (рис. 59), Plectrocnemia, Rhyacophila (рис. 57, 58), Apatania и бокоплав — Gammarus pulex (рис. 220).
Рис. 60. Куколки Blepharoceridae па камне.
Рис. 61. Личинка ди-ксы Dixa.
Перечисленные выше формы всё же не являются представителями типично ключевой фауны; к последним нужно отнести своеобразные формы, характерными для которых являются малые размеры и появление во многих случаях приспособлений, приближающих их к наземным формам; эти приспособления объясняются тем, что формы эти всегда рискуют попасть в условия мелководья. Так, у личинок ручейников редуцируются плавательные волоски на ногах, жабры, боковая линия.
Очень интересная и своеобразная фауна появляется на тех местах, где вода омывает очень тонким слоем камни (рис. 60) или листья, — здесь господствуют резкие колебания температуры и быстрые токи воды. В подобных условиях живёт, например, обычно встречающаяся в ключах личинка диксы — Dixa maculata (рис. 61); на нижней поверхности тела личинки имеются крючки, направленные в передней половине тела кзади, а в задней половине — кпереди. При движении личинка всегда изгибается U-образно — средней, перегнутой частью вперёд — и попеременно передвигает то одну, то другую половину своего тела; своеобразное расположение крючков понятно при таком способе передвижения. Чрезвычайно интересным приспособлением к жизни в ключе — то глубже в воде, то на мелком месте, где слой воды не толще тела самого животного, — обладает личинка мухи пер и ко мы — Pericoma (рис. 433 В, Г): попав на мелкое место, личинка дышит атмосферным воздухом, забирая его через стигмы на конце тела, а попав в более глубокое место, выпячивает из заднего прохода беловатые мешки — жабры, и производит обмен газов между заполняющей жабры жидкостью и водой. К типично ключевым формам, кроме только что описанных личинок и перикомы диксы, принадлежат ещё личинки ручейника — Crunoecia и личинки мух — Pedizia, Tipulidae (рис. 431), Orphnephila (рис. 439), Охусега (Hermione) и др.
ХАРАКТЕРИСТИКА РУЧЬЯ
Характер ручья в очень большой мере зависит от того, откуда берёт он своё начало и на каком расстоянии от истока мы его наблюдаем. Если ручей вытекает из озера, он обычно имеет сильное течение, всюду богат кислородом и имеет с самого начала много вынесенного из озера планктона. Если ручей является продолжением ключа, его воды будут у начала совсем бедны и кислородом и планктоном; лишь по мере удаления от истока будет происходить насыщение его атмосферным кислородом и обогащение планктоном, развивающимся в более тихих местах.
Часто на небольшом протяжении ручей много раз меняет свой характер, то разливаясь и замедляя своё течение, то суживаясь и быстро пробегая по узкому ложу. Если кислородный режим вследствие малой глубины и постоянного перемешивания остаётся постоянным, то температурный режим, наоборот, меняется резко. Смена температуры, скорость течения, глубина в различных местах, т. е. различие биотопов, вызывают и различие биоценозов. В ручьях можно различить 5 типичных биотопов: 1) камни на дне, 2) заросли мхов,3) тихие заводи, 4) свободная вода, 5) дно, лишённое камней.
Биоценоз камней слагается из форм, обладающих приспособлениями для удержания против течения. Приспособления эти уже были разобраны при описании биоценоза прибойной каменистой зоны; биоценозы камней в прибойной зоне и в ключах очень близки и имеют много сходных и даже тождественных форм. Наиболее типичными для ручьёв являются личинки ручейника Silo (рис. 57—58 В), некоторые виды Rhyacophila (рис. 57—58А), Hydropsyche (рис. 59А) и др., личинки и куколки двукрылых: Melusina (рис. 437), Blepharoceridae (рис. 60), Rheotanytarsus и др.
Биоценоз за рослей состоит не только из представителей самих мхов Fontinalis и др., но и из населяющих их животных, преимущественно из класса насекомых. Все животные обладают очень маленькими размерами, позволяющими им укрываться между мелкими листиками мхов, и снабжены крючками и зацепками, при помощи которых они противостоят течению воды. Наиболее частыми представителями фауны этого биоценоза являются некоторые жуки (Helmis, Hydraena), Pericoma, личинка ручейника Ptilocolepus (рис. 57—58В) и очень маленькая личинка Tendipedidae (Thienemaniella); очень часто попадается рачок Candona (из Oslracoda). 126
Заводи имеют обычную фауну прудов, и свободная вода в них заключает в себе значительное микроскопическое население. Как на характерную для ручья форму можно указать здесь на роющихся в опавших листьях бокоплавов — Gammarus (рис. 220).
Дно, не покрытое камнями, всегда бедно растениями, так как ил постоянно сносится, а песчинки подвергаются перекатыванию и не позволяют укрепиться растениям. Бедность пищей ведёт к бедности животного населения; только перловица — Unio (рис. 214) раскрывает здесь свои створки навстречу току мелких частиц ила, да прижимается ко дну сплющенный с боков бокоплав — Gammarus; здесь же можно часто найти чрезвычайно интересную личинку ручейника — Hydropsyche (рис. 59), сплетающую тонкую ловчую сеть, улавливающую, как в планктическую сетку, проплывающие мимо маленькие питательные частицы.
ТЕМЫ ДЛЯ РАБОТ
Исследование ручья всего целиком, включая сюда его исток (ключ, если ручей не озёрного происхождения), может дать богатый материал. При значительной длине ручья такое обследование, конечно, может вестись лишь экскурсионным путём, причём большим плюсом, в сравнении с более крупными водоёмами, является возможность производить работу без лодки и даже почти без инструментов: малая глубина позволяет просто войти в ручей вброд, измерить палкой глубину, верёвкой ширину и доставать руками со дна камни, пригоршни опавших гниюших листьев, дерновинки мха, затопленные коряги и пр., чтобы подвергнуть потом захваченные организмы разборке просто в каком-ниоудь тазике или тарелке; так же примитивно можно обставить и изучение растительности и отчасти её животного населения, подходя к растениям вплотную и вырывая с корнем руками. Если к этому прибавить ешё измерение температуры, то можно получить много ценного, так как в литературе данных по нашим ключам и ручьям мало. Очень желательно прибавить ещё некоторый инструментарий, например, компас, скребок и планктический сачок, при применении которых можно получить полную картину смены биотопов и биоценозов на всём протяжении ручья.
При полном обследовании всего ручья или более или менее длинного его отрезка было бы желательно не упускать следующие вопросы:
1. Характер истока — ключа или другого водоёма.
2. Размывание окружающих пород и влияние этого процесса на русло ручья.
3. Общая длина ручья от истока до конца или длина обследованного участка.
4. Расстояние от начала до конца исследованной части ручья по прямой линии (для представления об его извилистости).
5. Смена участков различного характера (присутствие или отсутствие камней и растительности, состав последней и т. п.) и длина каждого из участков.
6. Ширина, глубина, грунт и скорость течения в различных типичных участках.
7. Характер подступающей к ручью наземной растительности.
8. Характер зарастания водными растениями.
9. Биоценозы типичных участков.
10. Размах температурных колебаний суточных и сезонных (см. соответствующую тему для озера).
Кому придётся прожить некоторое время вблизи ручья, тот может сделать и на одном участке интересную и ценную работу. Ручьевые, а тем более ключевые формы плохо поддаются воспитанию в аквариумах, так как все они в большой мере требуют высокого содержания кислорода и текучей воды; каждый участок ручья может являться естественной лабораторией, где внимательный наблюдатель может многое почерпнуть. Исследование жизни и развития каждого отдельного представителя фа у пы, особенно насекомых с их превращением, представляет задачу интересную и нетрудно выполнимую, а иногда и практически важную, как, например, изучение условий жизни и развития мошек — Melusina, являющихся кровососущими паразитами, которые своими укусами вызывают у домашних животных иногда опухоли и даже судороги.
Здесь может быть разрешён ряд отдельных вопросов.
1. Изменения в организации форм ключевых и ручьевых по сравнению с таковыми же из стоячих вод.
2. Приспособления к жизни на камне, среди мха, на песке.
3. Питание ключевых и ручьевых форм (или одного избранного объекта)1.
4. Дыхание ключевых и ручьевых форм.
5. Представители грунтовых вод в ключе и их особенности.
6. Как противостоят ключевые и ручьевые формы течению.
7. Наблюдения над двумя типами дыхания у Pericoma.
1 Для этого необходим микроскоп. Особенно интересно исследовать под микроскопом добычу ловчей сети Hydropsycbe.
12Г
ГЛАВА ШЕСТАЯ
РЕКА
ФИЗИКО-ГЕОГРАФИЧЕСКАЯ ХАРАКТЕРИСТИКА
Река, как тип текучего водоёма, уже самим фактом своей текучести резко отличается от озера и пруда, как водоёмов со стоячей водой. Этот основной факт вместе с теми последствиями, какие он влечёт за собой, отражается как на физической, так и на биологической стороне жизни водоёма и придаёт ему специфический, своеобразный характер.
Мы уже знаем, что движение воды в форме течения имеет место лишь при различной высоте уровней, причём вода стремится течь от более высокого уровня к более низкому. Скорость этого течения при одной и той же разности высот уровней будет, конечно, обратно пропорциональна горизонтальному расстоянию между уровнями; поэтому здесь могут иметь место все переходы от медленного течения равнинной реки (при очень пологом склоне) через быстрый поток горной реки (при крутом склоне) к бешено низвергающемуся водопаду (при вертикальном или почти вертикальном падении воды).
Всякая река имеет верховье, или исток, ипизовье, которое называется или устьем, если река впадает в какой-нибудь водоём, или слепым концом, если река не достигает никакого водоёма вследствие, например, просачивания в почву, исчезновения через трещины земной коры и т. п. Истоком реки может быть источник, питающийся водой тающих снегов или ледника, как это бывает у горных рек, ключ, озеро или, наконец, болото. Для наших русских равнинных рек типичны именно три последних рода истоков. В частности, исток величайшей реки Европейской части нашего Союза— Волги—начинается ключом, впадающим в торфяное болото. Конечно, одно это болото не могло бы поддерживать такую многоводную реку; из него вытекает только самое начало Волги, которая на дальнейше^м пути питается ручьями, вытекающими из таких же болот и ключей, а также несколькими озёрами. Совокупность этих болот и озёр и представляет собой общее верховье Волги.
Разность высот между верховьем в низовьвгМ реки называется высотой падения её, а прямая, соединяющая обе крайние точки (верховье и низовье), — средним уклоном русла. Этот уклон выражают обычно высотой, приходящейся на каждый километр (1 000 м) длины реки. Так. по прежним исследованиЯхМ (до начала крупного гидростроительства), общая длина Волги равнялась 3 694 клс, а высота её падения — 261,5 м, следовательно, высота падения на 1 км = ^^ = 0,07 м\ эту относительную высоту падения выражают так: 0,07 %0.
Все протяжение более или менее значительных равнинных рек — от истока до устья — делят обычно на три отдела — верхнее течение, или верховье, среднее течение и нижпее течение, или низовье.
Сила, приводящая воду в движение, есть, конечно, сила тяжести. Таким образом, это есть один из случаев падения тела по наклонной плоскости. 128
Поэтому теоретически течение воды должно быть равномерно-ускоренным и в низовье должно по скорости значительно превосходить течение верховья. На самом деле, однако, не только нет такого превосходства, но часто имеет место даже обратное явление: скорость течения у истоков бывает больше, чем у устья.
Основной причиной этого является неодинаковый уклон русла реки: сильнее всего он бывает обычно в верховье, слабее в среднем течении и всего меньше в низовье. Кроме того, здесь играют роль трения, которые испытывает вода при своём движении и совокупность которых сильно замедляет течение. Эти трения бывают различного рода: о дно русла, о воздух и внутреннее взаимное трение частиц воды. Так как все эти виды трения не одинаковы по силе и, следовательно, оказывают различное сопротивление движению воды, то в результате оказывается, что в разных местах поперечного сечения река имеет и различные скорости течения: у д н а и берегов течение всего медленнее и ускоряется по направлению вверх и к середине реки. Но благодаря трению воды о воздух наибольшая скорость течения оказывается не на самой поверхности реки, а несколько глубже. Эти разнообразные формы трения частиц воды служат причиной возникновения кругообразных — так называемых вихревых или турбулентных движений этих частиц.
Скорость течения меняется также в связи с изменением площади поперечного сечения реки; так, в Окей Днепре во время половодья, когда площадь сечения их во много раз увеличивается, скорость течения достигает 2—2,5 м/сек, а в межень на плёсах, на середине реки, снижается у поверхности до 0,4—0,6, у дна до 0,05—0,10, у берегов до 0,12—0,20 м/сек; на перекатах Оки скорости течения в меженное время доходят до 1 м/сек.
Текущая по земной поверхности вода производит, как известно, разрушительное действие на эту поверхность. Такой процесс разрушения грунта от действия на него воды носит название эрозии, или размывания, и состоит в разрыхлении и распадении грунта на куски той или иной величины. Возьмём для примера равнинную реку. Скорость и эффективность эрозионного процесса, а также размеры кусков грунта зависят, с одной стороны, от массы текущей воды и скорости её течения, с другой—от свойств грунта, главным образом от степени его плотности: чем он плотнее, прочнее и твёрже, тем сильнее будет его сопротивление процессу размывания, и наоборот, чем он рыхлее, тем быстрее пойдёт эрозия.
Эродированный (размытый) материал сносится водой вниз по течению, причём скорость и продолжительность этого сноса (следовательно, и расстояние, на которое переносится материал) зависят от отношения между, с одной стороны, водной массой и скоростью течения воды, с другой— размерами и удельным весом переносимых частиц грунта: чем мельче и легче эти частицы (глина, детрит, т. е. совокупность взвешенных в воде остатков погибшего планктона) и чем больше масса воды и сильнее течение, тем дольше частицы будут находиться во взвешенном состоянии и тем на большее расстояние они могут быть снесены; наоборот, чем крупнее и тяжелее кусочки грунта и чем слабее течение, тем быстрее эти кусочки будут погружаться и падать на дно; некоторые из них совсем не могут держаться в воде во взвешенном состоянии и потому будут лишь перекатываться по дну (крупный песок, галька), а наиболее крупные и тяжёлые (камни) останутся совершенно неподвижными и, не поддаваясь действию текущей воды, будут лежать на дне. Процесс отложения, или осаждения, на дне реки размытого и переносимого ею материала носит название накопления, или аккумуляции.
Таким образом, текущая по земной поверхности вода посредством эрозии сама себе прокладывает путь, сама образует и постепенно формирует собственное ложе. Возникновение рек в равнинных областях начинается обычно с образования ручьёв, вытекающих из каких-либо постоянных водоёмов — озёр, болот, а во влажном климате происходящих от частых и продолжительных дождей. Эти ручьи стекают по наиболее низким местам неровной наклон-
0 A. H. Липин 129
ной поверхности и своей эрозионной деятельностью углубляют и расширяют те борозды, ложбинки и рытвины, по которым они текут. При слиянии нескольких ручьёв в один, более мощный, водная масса увеличивается, её размывающая работа усиливается, и ложе ручья ещё более углубляется и расширяется. Эти процессы могут совершаться периодически, прерываясь, например, летними засухами, когда ручьи нацело высыхают, оставляя проделанные ими ложбины и канавы. С появлением вновь воды от дождевых ливней, а особенно от весенних паводков, она опять устремляется по существующим уже руслам, иногда заливая их временно.
Слияние ручьёв ведёт к образованию мелких речек, а последние сливаются в более мощный поток — крупную реку. При каждом таком этапе ложе потока увеличивается в глубину (глубинная эрозия) и в ширину (боковая эрозия). Так на пути этого изменения ложе может превратиться в овраг, дно которого при своёхМ углублении может дойти до уровня грунтовых вод — тогда на дне оврага может появиться постоянная речка.
По мере углубления русла углубляется и сама река, обнажая свои берега, высота которых при этом всё более увеличивается. К этому присоединяется ещё эродирующее действие на эти берега боковых потоков воды, стекающих по поверхности расположенных по сторонам реки водосборных площадей. Эти потоки, происходящие весной от таяния снега, а летом — от ливней, стекая по склонам обнажённых берегов реки, размывают и разрушают их сверху и этим увеличивают поперечное расстояние между ними. В результате такого комбинированного (продольного и поперечного) действия эрозионных процессов образуется широкое, тянущееся вдоль реки по обе стороны углубление суши, никогда полностью не заливаемое водой, — так называемая речная долина.
Самое глубокое место долины занято постоянным руслом реки, которое всегда заполнено водой — даже в меженное время, т. е. в период самого низкого уровня воды в реке, вследствие чего это русло называется меженным. Над ним расположена часть долины, называемая пойменным руслом, или обычно просто поймой, которая заливается водой лишь во время паводков (в половодье), когда вода в реке достигает наивысшего уровня. Наконец, расположенные выше этого уровня, всегда свободные от речной воды боковые склоны долины называются коренными берегами реки. Изгиб на поперечном разрезе между меженным руслом и поймой называется просто берегом реки. Нередко от берега реки по направлению к коренному берегу (склону долины) поднимается один, два или целый ряд уступов, похожих на берег реки. Это так называемые речные террасы, служившие в далёком прошлом последовательно берегами реки. Ширина их бывает очень различна — от очень узких, едва выраженных, до нескольких километров. Линия, соединяющая самые низкие места долины, получила название тальвег.
Реки с долинами эрозионного происхождения свойственны главным образом областям равнинного рельефа, как, например, большинство рек Европейской равнины, в том числе реки Европейской части СССР (Волга с её притоками и др.).
Господствующим (но не единственным) фактором в процессе формирования долин является здесь эрозия. Вспомогательную роль играют некоторые другие факторы, среди которых главное значение имеют процессы выветривания (денудации) под влиянием воздушных течений и колебаний температуры.
Но воздействие разных комбинаций факторов на образование речных долин вызывает и различия в их структуре и размерах, обусловливающие значительное морфологическое разнообразие долин. Молодые долины, находящиеся ещё на ранней стадии своего развития, имеют иногда форму т е с-н и н, т. е. вертикальных щелей в земной коре; «склоны» такой долины представлены двумя отвесными (а иногда даже несколько наклонёнными навстречу друг к другу) стенами, расстояние между которыми не превышает 1—2 м, Ш
а внизу, на большой глубине, непосредственно между этими стенами бурлит речной поток. Такая форма долины обязана своим происхождением г лубин-н о и эрозии. Если представши себе, что стены теснины будут постепенно разрушаться (главным образом под влиянием выветривания) и притом — тем сильнее, чем ближе к поверхности земли, то получится долина с поперечным разрезом в виде латинской буквы V — так называемый каньон, дно которого по всей своей ширине занято рекой; её берегами служат непосредственно склоны каньона, которые в данном случае действительно получают право на такое название (берега).
Тектоническими называют такие долины, направление которых первоначально было определено древними тектоническими процессами — образованием складок, сдвигов, сбросов земной коры. Если возникший в результате этих процессов тектонический рельеф образовывал общую линию углубления земной поверхности, наклонённого в одном направлении, то поэтому углублению устремлялся поток воды, и таким путём возникала река. Однако появившаяся вместе с водой эрозия, комбинируясь с денудацией (выветриванием), в течение протекших с тех пор сотен тысяч лет так видоизменила рельеф, что тектонические долины в современном их состоянии следует считать почти нацело продуктом эрозии.
Коснёмся вкратце ещё вопроса об асимметрии долин. Давно уже подмечено, что у многих русских рек долины имеют асимметричное строение: их правый берег высокий, обрывистый, а левый — низкий, пологий. Примеры: в Европейской части СССР—Волга, в Азиатской — Енисей, Обь, Лена (особенно первый). Для объяснения этого явления акад. Бэр высказал предположение, что главную роль играет здесь вращение земли вокруг своей оси. Как известно, это вращение происходит в направлении с запада на восток. Поэтому в реках северного полушария (например, в Волге) вода, частицы которой связаны между собой гораздо слабее, чем частицы твёрдого тела, и потому обладают большой относительной подвижностью, будет несколько отставать в своём движении от твёрдой суши и прижиматься к правому берегу, который вследствие этого подмывается, разрушается и приобретает крутизну и обрывистость. От левого берега вода при этом отступает и оставляет на нём аллювиальные осадки, сообщая ему характер низкой, полого опускающейся к реке равнины. Этим прижатием речного русла к правому склону долины и размыванием этого склона объясняется и перемещение русла вправо.
Бэр считал, что это правило приложимо только к рекам, текущим в меридиональном или близком к нему направлениях. Однако Б а б и н э доказал, что оно применимо ко всем рекам, независимо от направления их течения. Это правило получило название закона Бэра— Бабин э.
Но дальнейшие наблюдения показали, что в природе этот закон далеко не всегда оправдывается: в ряде случаев не удаётся подметить асимметрии в строении долины, и даже нередки случаи, прямо противоречащие этому закону. Такие случаи, однако, не могут опровергнуть закон Бэра— Бабинэ, потому что они могут быть объяснены вмешательством в процесс отклонения течения рек под влиянием вращения земли каких-либо других факторов, более мощных и имеющих местный характер.
В образовании и формировании речного русла активнейшую роль играет, как мы видели, эрозия. Но не меньшее значение имеют в этом отношении и процессы аккумуляция (накопления), самым тесным образом связанные с размывающей деятельностью реки. Непосредственным результатом аккумуляции является образование на дне реки наносов из принесённого рекой и оседающего на дно материала — так называемого аллювия. Непосредственной причиной возникновения наноса является замедление течения воды, вызывающее оседание на дно некоторой части переносимого ею аллювиального материала, что и является начальным моментом образования наноса. Пока величина водной массы и скорость течения остаются более или менее постоянными^ аллювиальный нанос растёт; оказывая сопротивление
* 131
течению воды и замедляя его, он уже сам по себе является фактором, способствующим процессу накопления и усиливающим его.
Но как только сила течения значительно изменяется, это быстро отражается на характере аккумуляционного процесса: прп ослаблении течения оседание на дно взвешенного материала усиливается за счёт более тонкой части его, т. е. более мелких и лёгких частиц; при усилении же течения, наоборот, процесс эрозии становится более интенсивным, а процесс аккумуляции ослабляется, вследствие чего осевшая ранее (до усиления течения) часть наноса теперь будет подхватываться водой, переноситься далее и заменяться более грубо-зернистой частью. Таким образом, характер строения наноса изменится в первом случае в сторону более нежной и рыхлой структуры, во втором — в сторону грубой и плотной. Так как колебания силы течения имеют более или менее регулярный характер (например, ежегодное наступление и спад весеннего половодья от талых вод и летне-осенних паводков от дождевых ливней), то преобладание эрозии и аккумуляции чередуются во времени, в результате чего нанос приобретает слоистую структуру.
Образование наносов в речном русле играет чрезвычайно важную роль в его формировании. Наносы не представляют собой неизменных, раз навсегда зафиксированных образований; они меняют во времени не только свою форму, но и своё место, а в связи с этим изменяет форму и перемещается и самое русло реки. Познакомимся вкратце с некоторыми главными формами наносов, как продуктов комбинированной работы эрозии и аккумуляции, а также с теми изменениями в строении русла, которые эти наносы вызывают.
Естественно, что вследствие замедления течения реки по направлению от истоков к устью размывающая деятельность речной воды больше всего сказывается в верховье. Здесь вода захватывает взвешенный материал и переносит в среднее течение, где он отчасти оседает. Остальная его часть переносится дальше и осаждается в низовье, где скорость течения наименьшая. Таким образом, количество наносов по направлению течения реки увеличивается, и река как бы сама до некоторой степени выравнивает своё ложе, что ещё более замедляет её течение по мере приближения к устью. Следует заметить, что уклон русла при переходе от верхнего течения к среднему уменьшается сильнее, чем при переходе от среднего к нижнему. Поэтому и замедление течения реки, а следовательно, и оседание взвешенного материала на дно в среднем течении выражено резче, чем в нижнем. В связи с этим накопление осаждающегося на дно материала и повышение дна в среднем течении происходит быстрее, чем в нижнем; поэтому дно русла в среднем течении становится несколько выпуклым.
Накопление наносов в устье реки ведёт к образованию здесь так называемых конусов выноса. Чем сильнее и быстрее течение воды и чем крупнее материал, из которого слагаются конусы, что имеет место в горных реках, тем больше крутизна стенок конусов выноса; напротив, слабое и замедленное течение равнинных рек, доходящее в устье до еле заметного движения воды, способствует образованию низких, пологих, но зато расширяющихся на большие площади конусов, в которых невозможно подметить какое-либо возвышение и которые представляют широкую, ровную и плоскую поверхность. В устьях впадающих в море больших рек эта поверхность обычно пронизана сетью рукавов реки, разветвляющейся по направлению от устьевого конца её к морю, отчего вся эта площадь принимает форму треугольника, вершиной упирающегося в конец речного русла, а основанием — в берег моря. Такое устье реки было названо дельтой по сходству его с одной из букв греческого алфавита, имеющей форму такого же треугольника. Чем больше река и чем старте она по возрасту, тем большее число рукавов она образует и, следовательно, тем обширнее занимаемая её дельтой площадь. Какое огромное количество материала сносится крупнейшими реками в устье, показывают размеры образуемых им дельт; так, дельта Волги прежде (до гидростроительства на ней) занимала площадь в 13 с лишним тысяч км2, Аму-Дарьи — свыше 11 тысяч км2. В устьях некоторых больших рек, напри-132 1
мер, наших сибирских Оби, Енисея, образуются особые воронкообразные углубления дна, так называемые эстуарии; они обязаны своим происхождением размывающему комбинированному действию, с одной стороны, речного течения, с другой — приливной волны, противодействующей этому течению, и отливной, усиливающей его. Но эстуарии могут образоваться и другим способом — именно, опусканием дна, в результате чего устьевые пространства затопляются наступающим морем, образующим заливы; таким путём произошли, например, лиманы на северном побережье Чёрного и Азовского морей, на которых нет приливов; лиманы, отделяясь от моря образующимися вследствие наносов полосами суши, превращаются в отдельные, обособленные от моря небольшие водоёмы — лагуны.
Но образование наносов не ограничивается одним лишь устьем реки. Оно имеет место также и по длине русла. Примером могут служить перекаты. Это песчаные валообразные возвышения, идущие по дну русла в поперечном или косом направлении. Чередуясь с соответствующими, вытянутыми в том же направлении впадинами, они придают дну реки волнистую поверхность. Такая поверхность образуется под влиянием непрерывно действующей силы воздушного течения (ветра) на всякое тело, частицы которого более или менее подвижны относительно друг друга (жидкое или твёрдое, но сыпучее), например, поверхность воды, снежной равнины, песчаных дюн. В данном случае под действием постоянно движущейся по дну воды его песчаная поверхность получает характер тоже, так сказать, «подводной дюны», с той разницей, что «волна» имеет здесь ярко выраженное асимметричное строение: сторона её, обращённая против течения воды, широкая и пологая, обращённая по течению — узкая и крутая. Такая форма «волны» объясняется способом её образования. Движущаяся по песчаному дну вода силой своего трения с него передвигает песчинки верхнего слоя, нагромождая их друг на друга по направлению течения реки, отчего получается наклонная поверхность, высота которой увеличивается в направлении к устью реки. Так образуется поперечный по отношению к течению воды песчаный вал, за которым (на стороне его, обращённой вниз по течению) движение воды сразу ослабевает, песчинки прекращают своё перемещение и быстро сваливаются к подножию вала. В результате получается указанная выше ассиметричная форма вала. Она, следовательно, обязана своим происхождением перекатыванию песка по поверхности дна; этот же фактор вызывает и перемещение, пли «перекатывание» самих валов по направлению к устью реки. Такие явления и дали повод к наименованию самих валов перекатами. Естественно, что гребни перекатов, приближаясь к поверхности воды, образуют на реке мели, которые особенно дают себя знать в межень тем, что сильно затрудняют судоходство. Расположенные между этими мелями впадины дна образуют более глубокие места — плёсы.
Продолжающийся в течение многих лет рост переката в вышину приводит к тому, что наиболее высокая часть его гребня появляется наконец над поверхностью воды. Таким путем на реке образуется маленький песчаный островок, медленно передвигающийся вниз по течению. При дальнейшем его увеличении он постепенно покрывается растительностью, которая своими корнями укрепляет его, защищая от размыва, и перемещение его вдоль реки прекращается. Кто проезжал по Волге хотя бы от Горького до Сталинграда, тот имел случай наблюдать целый ряд больших и малых островов.
Мели, острова и другие препятствия постоянно меняют направление русла реки, которая при своём течении всегда выбирает не кратчайший, т. е. прямой путь, а путь наименьшего сопротивления. Таким образом, река никогда не может сохранить прямого направления течения, русло её всегда обладает более или менее многочисленным и извилинами и разнообразными изгибами — так называемыми меандрами, или излучинам и. Достаточно, чтобы вследствие какой-нибудь причины течение реки хотя немного отклонилось в сторону, как в этом месте уже начинается образова-
ние излучины. Допустим, что река повернула влево от основного направления своего течения. В образовавшемся изгибе оба берега реки окажутся в неодинаковом положении по отношению к течению. Поступающая в изгиб вода, стремясь сохранить своё направление и наталкиваясь на изгиб правого (вогнутого) берега, будет размывать последний и сделает его крутым и обрывистым. В то же время этот берег под действием боковой эрозии будет отступать, углубляясь в сторону вогнутости. Но размыву подвергается не только берег, но и прилежащая к нему часть дна, которое в этом месте становится глубже. Понятно, что речной поток при этих условиях должен переместиться к вогнутой части правого берега и стекать с выпуклой части левого, частично обнажая его и оставляя около него на дне часть оседающих здесь переносимых водой продуктов эрозии — наносов. В результате этот берег мелеет и становится низким, плоским и пологим и в то же время ещё более вдаётся в реку. Так создаётся асимметрия берегов в излучинах реки.
После описанного выше первого изгиба река потечёт под некоторым углом к своему первоначальному направлению (до изгиба). Струи воды, ударяющиеся в вогнутый (правый) берег и обладающие наибольшей скоростью течения, отчасти по инерции, отчасти под действием силы тяжести, заставляющей их скатываться по общему уклону долины, будут продолжать прижиматься к правому берегу и несколько подмывать его, хотя и не с такой силой, как в изгибе. Противоположный же берег будет нарастать.
Но под влиянием действующей на воду силы тяжести река при первом же благоприятном случае (когда ей позволят условия рельефа) вернётся вновь к своему первоначальному направлению. Если после этого она сделает поворот направо, то наступит опять тот ряд явлений, который описан выше для изгиба влево, но только эти явления будут протекать в обратном порядке по отношению к берегам реки: теперь будет размываться левый берег, а нарастать правый. Незавпсимо от направления изгибов реки изменениям берегов от действия эрозии и аккумуляции можно дать следующую общую формулировку: в процессе образования меандр размывается всегда вогнутый берег, а нарастает выпуклый, что и создаёт их асимметрию.
Из предыдущего описания видно, что при образовании излучин реки главное размывание её берегов идёт в направлении её основного течения, т. е. течения под влиянием силы тяжести по общему уклону долины. Если основное течение реки направлено с севера на юг, то в меандрах крутыми и обрывистыми будут южные части берегов, а пологими и низкими — северные. Это обстоятельство влечёт ещё одно очень важное следствие, а именно: перемещение меандр и всего русла в целом вниз по течению реки. Так, наблюдения в одном участке Дона показали, что в течение одного только года — с 1927 до 1928 — русло реки передвинулось на довольно значительное расстояние, в месте наибольшего перемещения — на 120 м. Этим явлением объясняются у нас периодические отходы рек от расположенных на их берегах городов и селений и новые их приближения.
Петлеобразные меандры нередко дают начало новым водоёмам — так называемым старицам, получившим своё название оттого, что они пред-ставляютсобойотрезки старых русел реки. Если в наиболее узком месте петли (её «шейке», образующей вход внутрь петли) река прорвётся и проложит себе болеепрямойикороткий путь, соединившись с продолжением основного русла, то главная масса воды устремится по этому новому руслу, минуя петлю, в концах которой течение замедлится, и здесь начнётся образование наносов. Со-временем эти наносы, выйдя на поверхность воды, окончательно отделят петлю от основного русла, превратив её в узкое, длинное озеро, имеющее обычно форму дуги или подковы. Это и есть старица.
Другой способ образования стариц будет указан дальше.
Баланс водыв реке обусловливается теми же факторами, как и в озере; источниками прихода её являются атмосферные осадки и при-Ш
токи (наземные и грунтовые), расход же воды происходит в форме испарения и течения её. Так как атмосферные осадки в испарение в продолжение года сильно меняются, то уровень воды в реке не остаётся постоянным, а испытывает значительные колебания. У наших равнинных рек особенно сильные изменения уровня наблюдаются весной — в сторону повышения, и во второй половине лета — в сторону понижения. Весенние разливы равнинных рек, называемые паводками, или половодьем, и зачастую переходящие в наводнения, обусловливаются таянием снега. Факторами, усиливающими половодье, являются: мощность снежного покрова, дружная весна, вызывающая быстрое таяние снега, и насыщенная влагоёмкость почвы, не воспринимающей в силу этого новых количеств влаги и потому водонепроницаемой, так что текущая по ней талая вода не просачивается, а стекает в реку. У больших рек, как, например, Волга и Кама, разлив достигает иногда громадных размеров; так, в некоторые годы место слияния этих двух рек разливается на десятки квадратных километров. После спада воды многие наши реки в течение лета, особенно жаркого, сухого, сильно мелеют; примером могут служить те же Волга и Кама, которые, будучи столь многоводными весной, в конце лета становятся иногда в некоторых своих частях несудоходными. Однако осенью обычно уровень воды в реке снова поднимается, хотя и далеко не в такой степени, как во время весеннего половодья; этот осенний подъём уровня обусловливается дождевым периодом и носит название осеннего паводка1. К зиме уровень опять несколько падает.
В Оке разница уровней во время паводка и межени достигает 10, а местами и 15 л, в Днепре же она в большинстве случаев не бывает более 5 м\ летний паводок повышает уровень на 1 м и несколько больше.
Основная разница между озером и рекой, именно, текучесть последней, обусловливает целый ряд отличий физ и ко-химического характера. Мы уже видели, что скоростьтечения реки в различных точках её поперечного сечения неодинакова. С первого раза можно подумать — и прежде, действительно, так и принимали, — что это движение с различными скоростями осуществляется путём скольжения разных слоёв воды относительно друг друга. Точные исследования показали, что это не так и что течение реки есть очень сложная форма движения. Здесь возникают в разных местах различно направленные большие и малые водовороты, из которых первые, складываясь с течением реки, в общем результате сообщают воде винтообразное движение. Кроме того, различного рода препятствия, как камни, сваи и т. п., тоже вызывают около себя водоворот и обратное течение. Все эти токи, вместе взятые, обусловливают сильное перемешивание речной воды. Так как основной причиной его является течение реки, т. е. фактор, действующий постоянно и непрерывно, то очевидно, что и перемешивание представляет собой явление столь же постоянное, находящееся в зависимости от скорости течения и, следовательно, имеющее связь с её колебаниями. Этим река сильно отличается от большого озера, в котором такое глубокое перемешивание слоёв воды происходит, как мы знаем, лишь весной и осенью, летом же и зимой в нём устанавливается более или менее спокойное стояние воды по вертикальному направлению. Подобная разница, конечно, не может не наложить резкого отпечатка на весь физико-химический режим реки, что в действительности и было обнаружено соответствующими исследованиями. Рассмотрим несколько примеров.
Изучение распределения температур в некоторых реках, например, в Днепре близ Киева (1892, 1893, 1940), в Волге (1922), а также и в некоторых реках Западной Европы показало, что по вертикали это распределение довольно равномерно, и температура у дна и у поверхности или совершенно одинакова (особенно зимой), или обнаруживает сравнительно небольшие отличия; в горизонтальном же направлении заметные различия наблюдаются
1 Такие же, обусловленные обильными дождями подъёмы уровня воды в реке бывают иногда и летом — летние наводки.
между температурами середины реки и её прибрежных частей: летом температура у берегов несколько выше, чем в середине, осенью наоборот. II на температуре, конечно, скорость течения сказывается вполне заметно: чем она больше, тем сильнее перемешивание п тем равномернее распределяется температура; чем медленнее течение, тем больше река приближается к стоячему водоёму и тем сильнее проявляется характерная для последнего разница в температуре поверхностного и придонного слоёв воды. Кроме того, быстрота течения оказывает сильное влияние — как непосредственно, так и путём того же перемешивания воды — на процесс замерзания реки, сильно замедляя его; охлаждение воды происходит здесь равномерно по всей толщине её, и потому вся вода — от поверхности до дна— достигает 0° почти одновременно; тем не менее образование льда происходит не во всей водной массе, а только с поверхности и у дна (донный лёд) ; это стоит, повидимому, в связи с тем, что эти два слоя — поверхностный и придонный, имея уже температуру в 0°, соприкасаются с ещё более охлаждённой средой: первый с атмосферой, второй с грунтом, температура которого спускается ниже 0°, вследствие отчасти лучеиспускания, отчасти передачи охлаждения по дну от берегов. Окончательный ледостав бывает, разумеется, в разное время не только у разных рек, но и у одной и той же большой реки в разных её участках: так, Волга окончательно замерзает в верхнем своём течении приблизительно в конце ноября, а в нижнем — у Сталинграда и Астрахани — в половине декабря. Точно так же и вскрытие происходит не в одно время: в районе Верхней и Средней Волги —во второй половине апреля, в районе нижнего течения — в первой половине апреля, а у Астрахани даже в половине марта.
Прозрачность воды во всяком водоёме находится в теснейшей связи с количеством взвешенных частиц. Если река и её притоки протекают не по каменистому грунту, а по рыхлым, подвергающимся размыванию породам, то количество взвешенных частиц в их воде будет более или менее значительно. Благодаря перемешивающему действию течения эти частицы в реке держатся во взвешенном состоянии гораздо дольше, чем в озере; из этого уже ясно, что и прозрачность воды в реке будет значительно меньше, чем в озере. Ясно также, что приносимый её притоками в реку взвешенный материал не оседает в ней тем дольше, чем быстрее её течение и чем сильнее, следовательно, перемешивание в ней воды, а это значит, что прозрачность её в этом случае будет меньше. На прозрачность главной реки оказывают значительное влияние её притоки: после впадения в неё рек с мутной водой её прозрачность уменьшается, после впадения притоков со светлой водой — увеличивается; это наблюдается, например, на Волге, прозрачность которой после впадения Оки повышается, после впадения Шексны, Камы и других — понижается. Но количество взвешенных частиц в реке зависит, между прочим, ещё от следующего обстоятельства: если река протекает через озеро или просто вытекает из него, то она выносит из озера сравнительно прозрачную воду, и только на дальнейшем пути эта вода постепенно обогащается взве-шенным материалом, и прозрачность её уменьшается; поэтому реки озёрного происхождения в общем обладают большей прозрачностью, чем неозёрного, например, прозрачность Невы больше, чем Волги. Прозрачность речной воды не остаётся одинаковой в течение года и испытывает сезонные колебания. Наблюдения на Волге около Саратова показали, что наибольшая прозрачность бывает зимой (в декабре), затем она уменьшается и наименьшего значения достигает весной после вскрытия льда (апрель); это и понятно, так как, во-первых, в половодье Волга обогащается талой водой, несущей в неё как непосредственно, так и через притоки громадное количество взвешенного материала, а во-вторых, скорость течения в это время и, следовательно, сила перемешивания увеличиваются; летом прозрачность опять несколько увеличивается, вплоть до осеннего паводка, который её слова понижает, хотя уже не в такой степени, как весенний; после этого в первой половине зимы прозрачность опять возрастает,
К этому ну?кпо добавить ещё влияние взвешенного материала, состоящего из живых организмов (планктон), присутствие которого в воде делает её ещё менее прозрачной; это особенно заметно бывает во второй половине лета в период максимальной вспышки развития планктона.
Химизм речной воды представляет результат сложного влияния многих факторов; он зависит от состава горных пород, на которых расположена речная система, от прихода и расхода воды в реке, от химического состава воды наземных и подземных её притоков, а также озёр, через которые она протекает, и т. д. Если при этом вспомнить, что все эти факторы меняются по сезонам (особенно резко меняется состав речной воды во время весенних паводков), то станет ясно, насколько разнообразен может быть химический состав воды не только в разных реках, но в одной и той же реке в различных её участках и в различное время года.
Отметим один факт, имеющий громадное значение для населения реки,— это вертикальное распределение в ней кислорода. Как и следует ожидать, оно характеризуется, подобно распределению температуры, относительной равномерностью, зависящей от перемешивающего действия течения.
Встречающиеся иногда исключения из этого правила, когда содержание кислорода у дна бывает несколько выше, чем у поверхности, пока ещё не нашли себе общепризнанного объяснения.
Источниками происхождения кислорода и поддержки его содержания в речной воде на довольно высоком уровне служат те же два основных фактора, как и в озёрах: атмосферный воздух и фотосинтез зелёных растений в реке — микро- и макрофлоры. Первый из этих источников является обычно более слабым, он усиливается лишь во время волнений, когда поверхность соприкосновения воды с воздухом сильно увеличивается. Эффективность второго источника — фотосинтеза — зависит от степени развития зелёной растительности — фитопланктона и высших прибрежных растений.
Фитопланктон в равнинных реках с малой и средней скоростью течения обычно хорошо развит и поставляет в воду большое количество кислорода, содержание которого нередко доходит до 100% насыщения. Быстрое течение представляет фактор, неблагоприятный для развития фитопланктона. Что касается макрофлоры, то она в различных реках находит себе различные условия жизни и потому развивается с разной интенсивностью. В равнинных реках, где медленно текущая вода допускает значительное заиление грунта, прибрежные растения имеют возможность хорошо укрепляться на дне, находят в окружающей среде надёжную защиту от механических повреждений и, следовательно, могут успешно произрастать; реки же с быстрым течением, с каменистым, галечным или песчаным дном, естественно, являются неблагоприятным биотопом для высшей растительности, и потому она там или совсем не появляется, или развивается плохо, между тем как фитопланктон может при таких условиях процветать и давать кислорода выше 100% насыщения.
Содержание растворённого в воде кислорода колеблется в течение года в реках (как и в озёрах) в широких пределах: летом оно может быть от 6—7 мг О2 на литр воды (в равнинных реках) до 10—11 мг(ь горных и порожистых речках); осенью и в начале зимы содержание кислородаповышается, а во второй половине зимы вновь падает, доходя иногда до очень низких величия (в Оке, например, перед вскрытием льда наблюдалось количество кислорода 2,35 мг/л). В горных и порожистых реках содержание кислорода в воде в общем выше, чем в равнинных, так как их вода, встречая при своём течении различные неровности дна, разбивается о них, разбрызгивается или растекается тонким слоем (например, по поверхности больших камней) и в это время обогащается атмосферпым кислородом.
Однако не всегда кислородный режим в реках так благоприятен. Существуют исключения, о которых будет сказано ниже.
Количество растворённой в воде свободной углекислоты находится, как известно, в обратном отношении к количеству кислорода. Отсюда ясно, что в реках, как и в озёрах, наибольшее количество СО2 должно 137
быть зимой, наименьшее — летом. Чем богаче река растительными организмами, тем меньше в ней должно быть растворённой СО.а, поглощаемой растениями при фотосинтезе.
Активная реакция (pH) и её колебания в реках также не обнаруживают особо существенных отличий от таковых же в озёрах. Обычно речная вода имеет щелочную реакцию (рН^>7).
Процентное содержание растворённых в речной воде минеральных веществ в течение года сильно колеблется. Меньше всего оно во время паводка, когда громадные количества устремляющейся в реку талой воды во много раз разбавляют концентрацию солевого раствора; после спада полых вод, по мере убывания воды в реке, концентрация возрастает в той или иной степени в зависимости, с одной стороны, от испарения воды и от притока богатых солями грунтовых вод, с другой — от интенсивности развития водной флоры, потребляющей соли на своё питание, и от степени поглощения их дном реки; происходящие на дне и в воде процессы разложения органических веществ также в некоторой степени повышают концентрацию минеральных солей. Зимой и ранней весной (перед паводком) эта концентрация ещё повышается и достигает наибольших значений. С наступлением половодья цикл начинается в основном от притока грунтовых вод снова. Например, в Оке общая жёсткость воды, в известной мере характеризующая общую концентрацию раствора минеральных соединений, при весеннем паводке снижается до 2—3°, летом повышается до 9—12°, а перед ледоходом доходит до 18°.
Наконец, количество растворённых и взвешенных в воде органических веществ, как происходящих в самой реке (автохтонных), так и в приносимых извне (аллохтонных), тоже подвержено сильным колебаниям в одной и той же реке в течение года и может обнаруживать крупные различия при сравнении разных рек. Достаточно вспомнить количественную разницу в развитии водной флоры и фауны летом и зимой, чтобы ясно представить себе пределы тех колебаний, которые испытывает количество органических веществ в реке в течение года. Разница между реками в количестве содержащегося в них органического материала относится в первую очередь за счёт скорости течения в них: чем она больше, тем беднее река живым населением, тем меньше содержит она органических веществ, а чем медленнее течёт река, тем богаче развивается в ней жизнь и тем больше накопляется в ней органического материала. Этим обусловливается крупное различие между горными и равнинными реками.
Количество органических веществ в воде измеряется путём определения так называемой окисляемостиеё, т. е. определения количества О2, идущего на окисление одного литра талой воды. В разных реках это количество колеблется в широких пределах — от нескольких миллиграммов (меньше 10) до 25 мг и выше (в реках, частично питающихся болотными водами).
В тесной связи с накоплением органических веществ в реках стоит явление их загрязнения, наступающего в тех случаях, когда это накопление переходит известный предел. Об этом явлении было уже сказано.
Здесь мы добавим к этому лишь несколько строк. Источником загрязнения рек могут служить органические вещества внутреннего происхождения, т. е. образующиеся в самой реке в результате естественных процессов (главным образом разложения трупов погибших организмов), как это бывает в мелких речках. Для крупных же рек главным источником является приток в реку веществ извне в виде отбросов, отходов и сточных вод из промышленных и сельскохозяйственных предприятий, расположенных на берегах рек. Среди этих веществ могут быть соединения минерального и органического происхождения, оказывающие вредное и губительное действие на организмы. Но и помимо этого непосредственного ядовитого действия сточных вод на организм, эти воды могут приносить большой вред населению реки, если они содержат значительное количество органических веществ, требующих для своей мипе-138
рализации большое количество кислорода и вызывающих дефицит этого газа в воде, доходящий иногда до полного его исчезновения. Такое явление наносит особенно большой вред рыбному хозяйству, вызывая массовую гибель рыбы от недостатка кислорода, т. е. так называемый замор рыбы. Примером речного замора может служить замор на Волге, продолжавшийся в течение четырёх лет — с 1838/39 по 1841/42 г. Причиной этого замора послужило, с одной стороны, загрязнение Волги стоячими водами промышленного и бытового характера, с другой — поступление в реку бедпых кислородом грунтовых вод.
Другой пример массового отхода рыбы в реке представляет Обь. В этой реке происходят периодические (через несколько лет) зимние заморы, при которых гибнет от удушья громадное количество рыбы. Недостаток или даже полное отсутствие растворённого в воде кислорода вызывается здесь, с одной стороны, притоком бедной кислородом болотной воды, с другой — тем, что в определённое время в реке начинает накопляться в огромном количестве закись железа, которая для своего окисления в окись железа поглощает из воды кислород.
Из предыдущего ясно, что в реке господствуют своеобразные физико-химические условия, сильно отличающие её от озера. Это отличие, разумеется, не может не отразиться на живом населении реки. В первую очередь течение воды, затем равномерное распределение в толще воды температуры и кислорода, значительная масса взвешенного в воде материала, небольшая прозрачность воды, характер грунта и т. д. — все эти факторы, различно комбинируясь в разных реках и даже в одной и той же реке, но в различных её пунктах и в различное время года, создают весьма разнообразные комплексы физико-химических условий жизни, разнообразные биотопы, которые в свою очередь обусловливают и соответствующее разнообразие биоценозов.
Поймой реки называется та плошадь дна долины, которая заливается водой весеннего паводка. Если русло реки проходит между обоими склонами долины, на том или ином расстоянии от обоих коренных берегов, то оно, естественно, разделяет дно долины и, следовательно, пойму на две части — правую и левую. Если же основное направление русла прижато к одному из склонов долины, то вся пойма будет расположена по другую сторону реки.
На поперечном разрезе речной долины различают три части поймы, расположенные по одну сторону реки в следующем порядке (по направлению от русла к коренному берегу долины): 1)прирусловая пойма — это песчаный нанос в форме более или менее высокого берегового вала; 2) ц е н т-ральная пойма, представляющая более или менее ровную (с небольшими сравнительно возвышениями и углублениями) поверхность; 3) п р и-террасная пойма — это ближайшая к склону долины полоса, примыкающая или к террасе, если она существует, или непосредственно к склону в случае отсутствия террасы.
Пойма формируется путём аллювиальных наносов, главным образом во время весеннего половодья. В начале паводка вода в русле начинает подниматься вследствие непрерывного пополнения речного потока полыми водами. Чем больше становится водная масса реки, тем быстрее она течёт. Возрастающая сила течения переводит во взвешенное состояние лежащие на дне русла отложения, в том числе и песок, и несёт всю эту взмученную массу вниз по течению. Когда вода начнёт переливаться через берега и разливаться в стороны, сила течения сразу в значительной степени падает, и из воды быстро оседает на берега наиболее тяжёлая фракция взвешенного материала — песок. Этому процессу обязаны своим происхождением береговые валы, образующие прирусловую пойму.
Так как поток воды, разливаясь по пойме, становится всё шире, то скорость течения воды по пойме параллельно реке убывает в направлении от Русла реки к коренному берегу долины. Это замедление вносит различия как в сроки осаждения взвешенного материала на поверхность поймы, так и в структуру этого материала: вблизи прируслового вала (береговой дамбы) 139
раньше всего отлагаются более крупные и тяжёлые частицы, а чем дальше от русла, тем больше замедляется процесс осаждения и мельче и легче выпадающие из воды частицы. В притеррасной части поймы отложения имеют тончайшую пылеватую и иловатую структуру.
Так как этот процесс формирования поймы повторяется ежегодно, то в результате и возникает указанный выше рельеф её, с наиболее высокой прирусловой частью, более низменной, и постепенно понижающейся по направлению к склону долины, центральной и самой низкой притеррасной, имеющей обычно параллельную коренному берегу ложбину.
Понятно, что в период паводка из воды выпадает на дно сравнительно лишь небольшая часть переносимой ею взвеси, основная же масса последней проносится над поймой к устью долины.
При спаде паводка водная масса его и скорость течения постепенно убывают до тех пор, пока вода не войдёт в речные берега и не спустится до своего меженного уровня. При этом прежде всего обнажается прирусловая пойма, затем центральная и позднее всего притеррасная. В последней вода задерживается не только вследствие только что указанной причины, но и благодаря выступающим здесь из уступа террасы или из подножия склона долины грунтовым водам. При обилии этих вод, вытекающих в виде ключей, родников, они стекают в ложбину притеррасной поймы и образуют ток воды (притеррасная речка) с более или менее длительным периодом существования. Если же грунтовых вод мало, то притеррасная пойма превращается в низовое болото или, по крайней мере, в заболоченный участок. После полного спада паводка пойма под действием солнца и ветра высыхает (за исключением притеррасной части, в которой ещё долго может оставаться влага). Раньше всего высыхает, как наиболее высокая часть поймы, песчаный прирусловый вал. При этом связь между песчинками почти совсем исчезает, и песок прирусловой поймы подвергается действию ветров, дующих в речных долинах почти непрерывно и достигающих часто большой силы.
Развеваемый по пойме в разных направлениях песок покрывает её поверхность слоем той или иной толщины. Следующий весенний паводок покрывает этот слой песка вновь топкими пылеватыми и илистыми осадками. Таким путём формируется слоистая структура толщи пойменных отложений.
В образовании поймы участвует ещё один крупнейший фактор — это перемещение речного русла по дну долины. Выше уже было сказано, что русло со всей его меандрической системой в пределах поймы очень медленно передвигается, с одной стороны, по направлению к тому или другому склону долины, обусловливая этим расширение поймы, с другой стороны — вниз по течению реки, оставляя за собой песчаные отложения. Так как этот процесс длится десятками и сотнями лет, то образуется более или менее мощный слой песка, подстилающий пойму.
Кратко описанный выше процесс формирования поймы имеет характер лишь общей схемы. В действительности целый ряд факторов местного значения вносит в него усложнения и детали, обусловливающие большое разнообразие в строении речных пойм.
В водоёмах поймы скорость течения играет роль лишь во время половодья. Эффект воздействия паводка на пойму вообще и на её водоёмы в частности зависит от мощности и продолжительности паводка. При большой массе полой воды и быстром её течении действие паводка на пойменные водоёмы может быть значительным. Полые воды, протекая над ними, в более или менее сильной степени захватывают их водную массу, увлекая её с собой, и таким путём производят в них в той или иной мере смену микрофлоры и микрофауны. С другой стороны, часть взвешенного материалакоторый несёт с собой паводок, оседает на дно этих водоёмов, удобряет его, обогащает питательными веществами, обеспечивая таким образом более богатое развитие в них флоры и фауны, чем в подобных же водоёмах, расположенных вне поймы.
В стоячих водоёмах речной поймы, в противоположность самой реке, распределение температуры и растворённых газов — кислорода и углекислоты — происходит по типу, описанному выше для озера: для них характерна такая же стратификация (слоистость) летом и зимой и циркуляция весной и осенью, как это имеет место в озёрах. Особенно острый характер эта стратификация приобретает в пойменных озёрах и прудах, где летом содержание кислорода у поверхности может дойти до состояния пересыщения (вследствие выделения его водорослями, скопляющимися в верхних слоях воды), а у дна — до очень малых величин благодаря потреблению кислорода гниющими органическими веществами. Зимой кислород может исчезать не только у дна, но и во всей подлёдной толще воды. Углекислота, конечно, претерпевает при этом обратные количественные изменения.
Количество минеральных и органических веществ в пойменных водоёмах также колеблется в довольно широких пределах в течение года — от минимума во время паводка, когда слабо минерализованные воды последнего сильно разбавляют и смывают в значительной степени концентрированный раствор солей, находившийся в пойменных водоёмах перед вскрытием реки, до максимальной концентрации этого раствора, которой он достигает в течение осенне-зимнего периода.
Обычно различают главную, или коренную рекуиеёпри-т о к и, среди которых различают притоки 1-го порядка, впадающие непосредственно в главную реку, и притоки 2-го, 3-го и т. д. порядков, впадающие соответственно в притоки 1-го, 2-го и т. д. порядков. Та площадь, которая занята главной рекой вместе со всеми её притоками, носит название речного бассейна, а совокупность всех рек и ручьёв одного бассейна называется речной системой. Во влажном и умеренном климатах количество воды в главной реке, по мере приближения её к устью, благодаря притокам увеличивается, в жарком же и сухом климате, наоборот, оно вследствие сильного испарения и недостаточного пополнения притоками уменьшается.
Возвышенная местность, с которой в обе стороны стекают воды двух различных речных’систем, носит название водораздела между этими системами. Водораздел представляет обычно более или менее широкую полосу суши. Главный водораздел земли проходит по основным хребтам гор и делит крупнейшие материки (Азия, Америка, Африка) на два главнейших склона, из которых один — атлантико-арктический — направлен к Атлантическому океану и Северному полярному бассейну, а другой — пацифико-индийский — к Тихому и Индийскому океанам. Первый из этих склонов значительно богаче реками (в том числе и мощными), чем второй. На территории СССР по атлантико-арктическому склону стекают такие большие реки, как сибирские — Обь, Енисей и Лена.
БИОЛОГИЧЕСКАЯ ХАРАКТЕРИСТИКА РЕКИ
К сожалению, приходится констатировать, что флора и фауна полоев (пойменных водоёмов) ещё слишком мало обследованы, чтобы можно было дать более или менее цельную картину населения этих водоёмов. Лучше изучены в биологическом отношении коренные реки, причём из рек СССР на первом месте нужно поставить Волгу, на которой была учреждена специальная биологическая станция (в Саратове) для всестороннего изучения жизни Волги, и Оку, для изучения которой много сделала Окская биологическая станция, работавшая сначала в г. Муроме, а затем в г. Горьком. Подобные же станции имеются и на некоторых других реках, например, Киевская, Днепропетровская биологические станции на Днепре и др. И, несмотря на это, всё-таки нужно признаться, что реки даже одной лишь Европейской части СССР исследованы ещё недостаточно, и в этом отношении оставляют желать многого. В настоящее время у нас ведётся интенсивное изучение ряда
Ш
больших рек в связи с крупным гидростроительством на них; в этом направлении наша советская гидробиология сделала уже большой шаг вперёд.
Планктон. Уже в главе о ручьях мы видели, какое влияние на организмы оказывает течение воды. Поэтому, когда мы обращаемся к планктону, невольно возникает вопрос: нельзя ли и здесь подметить это влияние в какой бы то ни было форме? Другими словами, обнаруживает ли планктон реки такие специфические, характерные черты, которые присущи именно этому типу водоёмов (или вообще текучим водам), по сравнению с планктоном стоячих водоёмов? Постараемся осветить этот вопрос, насколько позволяют современные, пока ещё довольно скудные данные.
Первые же исследователи в этой области поставили себе вопрос о существовании особого «речного планктона», или так называемого потамопланктона, в отличие от «озёрного», или лимнопланктона. Сначала этот вопрос решался в положительном смысле, так как в реках находили такие планктические организмы, которые небыли известны для стоячих водоёмов и которые поэтому легко было принять за специально речные, свойственные лишь речному планктону формы. Но потом от этого взгляда пришлось отказаться, потому что эти самые формы стали находить и в стоячих водах. Так, например, описанные сначала в качестве специально речных две водоросли затем были найдены в прудах и озёрах. Да это и вполне понятно: ведь откуда в конце концов река берёт свой планктон? Разумеется, из тех же стоячих водоёмов: вытекая из озёр, прудов, болот, протекая через них или временно соединяясь с ними в период паводка, она сама и её притоки выносят оттуда планктон, который в реке смешивается и на протяжении реки пополняется ещё из стариц, тихих заводей и т. п. Очевидно, стало быть, что в речном планктоне неоткуда взяться таким формам, которые нельзя было бы найти в каком-либо стоячем водоёме. Другой вопрос: какие изменения претерпевает качественный и количественный состав потамопланктона, будет ли он сходен с таковым же озёрного, прудового и т. п. планктона или окажется отличным от него? На этот вопрос нельзя дать, пожалуй, общего ответа, так как здесь всё зависит от свойства самой реки, главным образом, от её протяжения (длины) и скорости течения.
Уже при самом захвате рекой планктона из какого-нибудь стоячего водоёма далеко не все организмы будут в одинаковой степени вовлечены в струю; естественно, что чем большей способностью к активному движению обладает организм, тем меньшему подвергается он риску быть втянутым в струю течения. Поэтому в наибольшей массе в реку попадает растительный планктон, характеризующийся преобладанием совершенно пассивных форм; животные же планктона, которые почти все способны с большей или меньшей силой самостоятельно двигаться в воде, могут рефлекторно избегать вовлечения их в струю, уплывая при первых, ещё слабых проявлениях течения в противоположную ему сторону (реакция на течение воды). Успех этих рефлекторных движений пропорционален той силе, с которой данное животное при своём плавании преодолевает сопротивление воды, и потому наилучшие пловцы — ракообразные, а из них особенно веслоногие — в наименьшем числе попадают в реку; за веслоногими по количеству следуют ветвисто-усые^ачки, а за ними, наконец, и коловратки. Таким образом, уже при самом выходе из озера в реку характерный для первого групповой состав планктона нарушается и приобретает черты, свойственные в общем речному планктону: количественное преобладание фитопланктона над зоопланктоном, а в последнем — коловраток над ракообразными и ветвистоусых рачков над веслоногими.
Но на этом сортировка планктона не останавливается, так как в реке его состав продолжает претерпевать изменения: река, как говорят, «перерабатывает» поступивший в неё планктон. Очутившись в текучей воде, планктические организмы, приспособленные к жизни в стоячем водоёме, подвергаются влиянию непривычных для них условий жизни, более или 142
менее неблагоприятно отражающихся на их основных жизненных отправлениях, главным образом на размножении. И опять-таки степень этого влияния для различных групп планктона будет неодинакова. Растительные организмы, размножающиеся преимущественно делением, не испытывают особенного ущерба от перехода из стоячей воды в текучую и потому продолжают размножаться приблизительно с прежней скоростью; коловратки и ветвистоусые рачки, у которых очень распространён партеногенический способ размножения, хотя и в большей мере, чем фитопланктон, испытывают в этом отношении неблагоприятные влияния течения, но всё же, не имея постоянной надобности во встрече двух полов, могут размножаться более или менее интенсивно; наконец, веслоногие рачки, у которых девственное размножение совершенно отсутствует, в наибольшей степени страдают от течения, затрудняющего встречу самцов и самок. Таким образом, то изменение состава планктона, которое начинается уже при выходе реки из озера, на протяжении её усиливается ещё более. В этом и заключается «переработка» рекой получаемого ею из стоячих водоёмов планктона. Такой переработанный планктон обычно характеризуется следующими чертами:
а) преобладанием фитопланктона над зоопланктоном;
б) преобладанием в фитопланктоне диатомовых водорослей;
в) преобладанием в зоопланктоне коловраток над ракообразными;
г) преобладанием ветвистоусых рачков над веслоногими, взрослые формы которых в летнее время присутствуют в реке в сравнительно небольших количествах, а в массе встречаются только их личинки, так называемые науплиусы (рис. 275); это можно объяснить тем, что науплиусы не являются лучшими пловцами, чем коловратки, и потому вместе с ними в большом количестве выносятся р е к о й из стоячих водоёмов; в добавление к этому играет роль, конечно, и самостоятельное размножение речных веслоногих.
Такая картина планктона характерна для равнинных рек, к каковым относится большинство рек средней части СССР. Таковы, например, Волга и входящие в её бассейн реки — по крайней мере те, которые более или менее исследованы в этом отношении: Ока, Кама, Шошма и др. Но это, разумеется, отнюдь не означает, что все такие реки (даже одного бассейна) должны иметь одинаковый состав планктона. Ведь этот состав зависит прежде всего от тех стоячих водоёмов, из которых река черпает свой планктон, и потому последний может сильно отличаться даже в главной реке и её притоке, как это видно хотя бы на примере Невы и Тосны: в первой—планктон озёрного типа, так как выносится из Ладожского озера, вторая же, вытекая из торфяных болот, несёт планктон совсем другого состава.
Но и сама река, как мы видели, перерабатывая планктон, может сильно изменить его, и вот тут-то и сказывается влияние упомянутых выше факторов — протяжения реки и скорости её течения. Естественно, что чем длиннее река, тем дольше остаётся планктон в ней, и, следовательно, тем более он может быть переработан ею. Отчасти этим объясняется сравнительно мало изменённый озёрный тип планктона Невы: на протяжении 60 км, которые занимает эта река от Ладожского озера до моря, она не успевает настолько переработать выносимый из этого озера планктон, чтобы придать ему специфический речной характер. Напротив, такие реки, как Волга или Ока, в значительной степени изменяют первоначальный состав полученного ими планктона. Само собой понятно, что на протяжении длинной, медленно текущей реки планктон неоднократно может менять, хотя и в слабой степени, свой характер в зависимости от встречающихся на пути водоёмов и от притоков.
Познакомимся вкратце с главнейшими группами планктона Волги. Из фитопланктона на первом месте по количеству стоят диатомовые водоросли (рис. 31), за которыми следуют зелёные (рис. 30): с и н е з е-л ё н ы е (рис. 29) в особенно больших скоплениях (по в небольшом количестве видов) встречаются в нижнем течении; жгутиковые (рис. 32)
и конъюгаты (из нитчатых водорослей) играют .значительную роль. В зоопланктоне главная роль принадлежит коловраткам (рис. 27), которые распространены на всём протяжении реки вплоть до дельты; меньшее значение имеют низшие ракообразные, из которых ветви стоусые рачки (рис. 233) занимают первое, веслоногие (рис. 275) — второе место, причём из последних только науплиусы встречаются в большом количестве; моллюски в планктоне представлены личинками ракушки Dreissena polymorpha, насекомые — личинками Chaoborus (рнс. 434А) и молодыми личинками Tendipes.
Прибрежное и донное население. Главнейшим отличием реки от водоёма со стоячей водой является, конечно, её текучесть, обусловливающая целый комплекс и других особенностей физико-агеографического и физико-химического характера, совокупность которых характеризует реку как определённый, специфический биотоп. Подвижность жизненной среды — не только
самой водпой массы, по и грунта, размываемого и сносимого ею, — создаёт
особые условия существования для водных организмов, в том числе и для
Рис. 62. Ловчая сеть личинки ручейника Neureclipsis bimacu-lata.
донных. В составе грунта дна некоторых из наших крупных рек огромное преобладание имеет песок; например, на дне Волги он составляет приблизительно 95% всего грунта. При значительной скорости течения рек придонный слой воды захватывает поверхностно лежащие песчинки и перекатывает их вниз по течению. Таким образом создаётся неустойчивый субстрат, мало пригодный для поселения на нём многих из организмов, свойственных стоячим водоёмам. Прежде всего это касается укореняющихся в грунте растений. Поэтому берега крупных рек обычно лишены растительности. Наоборот, мелкие речки с медленным течением, а также участки крупных рек часто сильно зарастают.
Что касается животного населения реки, то обитатели стрежня (т. е. линии наибольшей скорости течения) должны обладать специальными приспособлениями для борьбы с опасностью быть сорванными со своего местообитания. Способы эти следующие.
1) Приобретение формы, которая позволила бы остаться на месте, несмотря на течение; наилучшей формой является в данном случае уплощённая, так как через плоское, прижатое к грунту тело вода перека
тывается, не встречая перпендикулярных к своему течению плоскостей, упираясь в которые она могла бы сдвинуть организм (личинки Heptagenia, рис. 3085). 2) Развитие органов прикрепления — постоянного (биссус дрейсены — Dreissena, рис. 211) или временного (нити, которыми временно прикрепляют своё тело личинки мошек — Melu-sina, рис. 437). 3) Способность зарываться в грунт (личинки эфемеры — Ephemera, рис. 302Л, проделывающие в глиняном грунте ходы до 12 см глубины, рис. 65). 4) Устройство прикреплённых домиков, или чехликов (личинки ортокладиин из Tendipedidae) или сетей (сети устраивают, например, личинки ручейника Hydropsyche, Neureclipsis, рис. 62, прячась затем в них, как в тенёта).
Надо заметить, что, помимо основного населения, приспособленного в результате естественного отбора к жизни в том или ином участке, в реке постоянно можно встретить и случайных пришельцев, занесённых сюда течением иногда из очень и очень далёких мест; особенно имеет место подобный
занос весной, когда несущиеся в реку струи талой воды захватывают по пути из стоячих вод массу организмов и их яиц и спор. Какова будет судьба новых пришельцев, зависит от того, насколько окажутся они способными акклиматизироваться в новых условиях; в большинстве случаев им предстоит гибель.
Как и в стоячих водоёмах, в реках можно различить ряд биотопов с характерными для них биоценозами. Главнейшими моментами, обусловливающими характер биотопа, являются в реке скорость течения и грунт. Глубина и освещение имеют первостепенное значение для растений. Что касается прочих факторов, как температура, химизм и пр., то они не могут иметь определяющего значения, так как благодаря течению они постоянно выравниваются на больших протяжениях. Отчасти такое выравнивание течением сказывается и на биоценозах, и потому биоценозы реки обычно не бывают вполне отграничены, а смешиваются и заходят один в другой.
При попытках разгруппировать биоценозы прежде всего должны быть отграничены биоценозы коренной реки с её сильным течением, рукавов — с течением замедленным и затонов — с течением очень слабым или почти без течения.
В каждой из этих областей будет ряд биоценозов, характеризующихся уже грунтом: биоценозы песчаного грунта, илистого, каменистого, глинистого, растительного. В различных реках с большим протяжением разные участки оказываются в различных географических областях, что, конечно, сказывается и на характере биоценозов, находящихся в одинаковых по силе течения и составу грунта условиях.
Попытаемся теперь вкратце охарактеризовать биоценозы коренной реки. При этом заметим, что биоценозам различных речных биотопов присвоены особые названия (на древнегреческом языке), выражающие наиболее существенные и характерные черты тех экологических условий и того субстрата, из которых складывается биотоп каждого биоценоза. Само собой разумеется, что все биоценозы коренной реки должны быть обозначены термином реофильные, т. е. течелюбивые (живущие в текучей воде). К этому термину прибавляются приставки, характеризующие уже субстрат, на котором обитает биоценоз. Так, биоценоз песков будет псаммореофиль-ный, биоценоз камней — литореофильный и т‘. д. Все эти термины в дальнейшем нашем изложении заключены в скобки.
Биоценозы песков (псаммореофильные) в отношении макрофауны бедны количественно и качественно. Лишь микроскопическая фауна этого грунта богата. Из типичных представителей бентоса можно указать на ресничных червей — Turbellaria, круглых червей — Nematodes, малощетинковых червей — Potamodrilus, Aeolosoma, Propappus volki, Chae-togaster; из моллюсков упомянем зарывающихся в песок ракушек-горошинок — Pisidium (рис. 216) и др.; из ракообразных — Ostracoda, Cladocera (Rhynchotalona), Metamysis и бокоплава Pontogammarus sarsi; из насекомых — личинки некоторых Tendipedidae, как Cryptochironomus, Ро-lypedilum, Paracladopelma и др.
Биоценозы камней (литореофильные) богаче в количественном отношении, чем биоценозы песка; число форм здесь значительно больше. Тут можно встретить: губок Trochospongilla, Ephydatia и др.; олигохет Nais, Tubifex и др.; пиявок Piscicola, Glossiphonia, Helobdella, Herpobdella; мшанок Fredericella, Palpnicella и др.; двустворчатых моллюсков Dreissena и Unio, а из улиток Ancylus, Viviparus и др.; из ракообразных встречаются Corophium, Dikerogammarus и др.; из паукообразных — клещи Hydrachnidae; насекомые представлены личинками подёнок Hepta-genia, Ecdyonurus, Ordella, Baetis, Ephemera, Procloeon и др., личинками ручейников Rhyacophila, Polycentropus, Hydroptila, Brachycentrus, Leptoce-rus, Neuredipsis (рис. 62) и др., личинками двукрылых Simulium, Glyptoten-dipes, Rheotanytarsus, Eutanytarsus, Diamesa, Orthocladiinae и мн. др., а также жуками и их личинками Limnius, Helmis и др. и клопом — Aphelochirus.
Биоценозы глинистых участков (аргиллореофиль-н ы е) имеют обычно немного форм, но в довольно большом количестве особей. В тихих местах их больше, в местах с сильным течением меньше. Глинистый грунт располагается большей частью неширокой полосой вдоль размываемого глинистого берега. Растительности здесь нет, а животные принимают
участие в разрушении берега, проделывая в глубине ходы и углубления, в которые попадают струйки воды. К таким животным относятся: личинки ручейника Hydropsyche, которые образуют на поверхности глины небольшие углубления, придающие грунту решётчатый вид (рис. 63), личинки подёнок — Palingenia, Polymitarcys (рис. 64) и отчасти Ephemera (рис. 64), проделывающие в глине довольно длинные ходы (рис. 65). Этими ходами
Рис. 63. Ходы личинок ручейников Hydropsyche и личинок подёнок в глине.
Рис. 64. Личинки подёнок — Ephemeroptera:
А — Polymitarcys Virgo; Б — Ephemera vulgata.
пользуются некоторые рачки — Gammarus, Dikerogammarus, Corophium (рис. 218). Кроме указанных животных, на глинистых грунтах могут встретиться: Nematodes; Oligochaeta (Limnodrilus, Tubifex, Nais рис. 1656 и др.); пиявки (Piscicola, Helobdella), мшанки (Fredericella, Plumatella, Cristatella, Paludicella); моллюски (Viviparus, Anodonta, Unio, Sphaerium, Pisidium, Dreissena); речной рак Astacus; личинки подёнки Baetis; личинки
ручейников (Brachycentrus, Leptocerus); личинки двукрылых (Tendipes, Glyptotendipes, Cryptochironomus, Stictochironomus, Ablabesmyia, Culi-coides и др.).
Биоценозы илов (п е л о реофил ь и ы е) как по разнообразию, так и по количеству животного населения значительно богаче только что описанных биоценозов. В их состав входят: малощетинковые черви Peloscolex, Limnodrilus, Tubifex, Iliodrilus и др.; некоторые Nematodes, моллюски Valvata, Sphaerium, Pisidium и др.; рачки Gammarus, Dikerogammarus (рис. 219); клещи Hydrachnidae; личинки
Рис. 65. Ходы личинок подёнки
стрекозы Gomphus (рис. 289ZJ), подёнок Ephemera в глинистом грунте.
Baetis, Polymitarcys, Ephemera, С1оёоп и
др.; вислокрылки Sialis; двукрылых Harnischia, Glyptotendipes, Procladius, Tendipes, Protenthes, Sayomyia (Chaoborus) и др.
Биоценозы растительных участков (фиторео-
фил ь н ы е) характеризуются присутствием в них высших водных расте-
ний, количественное развитие которых в реке зависит, как известно, главным образом от двух факторов — от скорости течения и от степени прозрачности воды, и потому может быть очень разнообразным, начиная от полного отсутствия водной макрофлоры в горных и в крупных многоводных равнинных реках и кончая обильными зарослями в мелких речках с медленным течением.
14Ь
В реке, имеющей водную флору, вообще говоря, можно различать две зоны: покрытую растительностью и свободную от растений. Животное население этих двух зон в общем различно, так как присутствие растений создаёт для животных совершенно своеобразный биотоп, в котором растения играют для животных роль субстрата, предоставляют им пищу, защищают от механических повреждений, причиняемых движением воды, улучшают кислородный режим и т. п.
Водные растения в текучих водах могут произрастать лишь при том условии, если скорость течения будет небольшая. Животные биоценозы таких участков водоёмов и получили название фитореофильных (средняя часть этого слова «рео» означает наличие течения воды). К водоёмам с медленным течением относятся не только небольшие речки и рукава крупных рек, но и отдельные участки больших равнинных рек (например, верховьев Волги). Здесь мы можем встретить следующие растения, входящие в состав той или иной растительной ассоциации: Butomus, некоторые виды Potamogeton, более редко Sparganium, Equis£tum, Scirpus, Ceratophyllum, Myriophyllum, Elodea, Chara и др. Компонентами фитореофильного биоценоза могут быть следующие животные: губки Spongilla; кишечнополостные Hydra; круглые черви Dorylaimus; малощетинковые черви Stylaria, Nais, Chaetogaster и др.; пиявки Piscicola, Glossiphonia, Herpobdella, Helobdella; мшанки Plumatella; моллюски — Ancylus, Limnaea, Planorbis, Bithynia, Viviparus и др.; ракообразные — Sida, Asellus, Rivulogammarus; водные клещи Hydrachnidae; насекомые и их личинки: стрекозы Алах, Agrionidae и др.; клопы Gerris; подёнки РгосТоёоп, С1оёоп, Centroptilum, Ordella (Caenis), ВаёНз и др.; ручейники — Ecnomus, Anabolia, Phryganea и др.; личинки комаров — Endochironomus, Cricotopus, Parachironomus, Ablabes-myia, Anopheles и др.
ФИЗИКО-ГЕОГРАФИЧЕСКАЯ ХАРАКТЕРИСТИКА ПРИДАТОЧНОЙ СИСТЕМЫ РЕКИ
Под придаточной системой реки подразумевается совокупность так или иначе связанных с коренной рекой водоёмов, там и сям разбросанных по долине, которую с течением времени прокладывает крупная река с постоянно меняющимся руслом. В меженное (т. е. вне периода паводков) время основное русло реки занимает только часть этой долины, па остальной же её площади расположено то или иное количество водоёмов, которые могут быть разделены на две группы: 1) водоёмы, постоянно связанные с коренной рекой, и 2) водоёмы, связанные с ней лишь во время половодья. К первой группе относятся рукава, или протоки, и затоны, ко второй — пойменные озёра, пруды и болота.
Рукав, или проток, возникает в том случае, если образовавшийся остров расщепляет, так сказать, реку на две части: более широкую — коренное русло — и более узкую, расположенную между островом и берегом, вблизи которого "он находится; вот эта последняя часть и называется протоком.
Затон образуется из протока, когда верхний конец его засыпается и, следовательно, сообщение его с коренной рекой остаётся лишь на нижнем конце. Но если засыпается и нижний конец рукава или, вернее, уже затона, то образуется совершенно замкнутый водоём, который, если он окажется постоянным, может превратиться в озеро. Подобные водоёмы, происшедшие из старого русла реки, называются, как мы видели, старицами.
Замкнутые водоёмы возникают и вне непосредственной связи с руслом реки, именно в тех углублениях, ложбинах, оврагах, канавах и т. п., которые окажутся в захваченном половодьем районе: весенняя полая вода, заливая эти углубления, образует стоячие водоёмы, находящиеся в связи с рекой только во время паводка, в остальное же время эта связь нарушается. Некоторые из таких водоёмов оказываются постоянными, другие временными,
*
пересыхающими, которые в середине или в конце лета исчезают, чтобы на следующую весну вновь народиться. Эти водоёмы расположены на пойме реки.
В придаточных водоёмах реки, подобно коренной реке, физико-химические условия в связи с различиями в скорости течения тоже далеко не одинаковы. Чем медленнее течение, тем ближе водоём подходит по всей совокупности физико-химических факторов к типу стоячего водоёма и тем в большей мере даёт он приют характерной для этого типа флоре и фауне. Замедленное течение (например, в затонах в меженный период), связь с коренной рекой, постоянная (протоки и затоны) или временная во время паводков (пойменные водоёмы), опять создают специфические условия жизни, в которых развивается комбинированное население, заключающее в себе формы, свойственные как текучим, так и стоячим водоёмам, с преобладанием то тех, то других. Но наряду с такими формами, есть, конечно, организмы, обладающие широкой способностью приспособления и потому одинаково хорошо уживающиеся как в текучих, так и стоячих водоёмах.
БИОЛОГИЧЕСКАЯ ХАРАКТЕРИСТИКА ПРИДАТОЧНЫХ ВОДОЁМОВ РЕКИ
Биоценозы протоков (рукавов), где течение медленное и где откладывается в довольно большом количестве ил, отличаются от биоценозов коренной реки; здесь, как и в коренной реке, различают разные биоценозы в связи с грунтом. На песчаных участках биоценозы в общем сходны с таковыми реки, но для них характерно присутствие ещё лпчинок стрекозы дедки — Gomphus (рис. 289В), бокоплавов — Gammarus (рис. 220) и личинки Tendipes из группы Thummi.
Биоценозы затонов. Затоны в полую воду проточны, а по спадении воды закрываются грядой, которая является продуктом самой реки. Гряда замедляет течение в затоне даже и весной, оставаясь под водой, и эта замедленность течения так велика, что ил не сносится, а лишь слегка перемешивается в верхних слоях с песком. Летом температура воды очень сильно повышается сравнительно с коренной рекой: фауна развивается чрезвычайно пышно и напоминает фауну эвтрофных озёр. Часто наблюдается цветение Anabaena и Aphanizomenon (рис. 29В,Т7); ил представляет собой гиттию1, так как количество кислорода у дна невелико, особенно зимой. Наиболее часто встречаются в затонах: лу жанка — Viviparus, перловица — Unio, шаровка — Sphaerium, горошинка — Pisidium, малощетинковый червь — Limnodrilus, пиявки — Piscicola и Glossiphonia, личинка стрекозы дедки — Gomphus, личинки тендипедид Procladius и Tendipes из групп Bathophilus, Plumosus и Reductus; затем встречается обыкновенный рак Astacus и др. Весной к ним присоединяются и формы, живущие в проточной воде. Чем мельче затон, тем больше скопляется в нём ила, тем сильнее прогревается вода и тем более приближается его население к озёрному.
Что касается биологической характеристики водоёмов поймы, то заметим по этому поводу следующее. После спада полых вод пойменные озёра, пруды, лужи развиваются, как стоячие водоёмы, поэтому к ним относится та биологическая характеристика, которая была дана в предыдущих главах озеру, пруду и луже. Поскольку в этих водоёмах по прекращении паводка течение отсутствует, в экологических терминах, обозначающих различные биоценозы, среднее слово <<рео» может быть выброшено, и тогда эти термины приобретают такой вид: пелофильный (вместо пелореофильный), фитофильный (вместо фитореофильный) и т. п. Представление об этих биоценозах можно’ получить из тех глав настоящей книги, в которых дано описание флоры и фауны стоячих водоёмов.
1 Ил, представляющий смесь частиц неорганического (известь, глина) и органического (главным образом испражнения животных) происхождения.
148
ТЕМЫ ДЛЯ РАБОТ
Полное исследование реки является, конечно, делом очень сложным и громоздким, требующим большого оборудования и значительных сил; такие обследования — дело специальных научно-исследовательских учреждений или экспедиций. Но обследование отдельных участков или разработка отдельных вопросов может быть по плечу и одному лицу или небольшой группе, даже при отсутствии солидного инструментария (необходимой, конечно, является лодка).
При обследовании какого-либо отрезка реки желательно остановиться на следующих вопросах:
1. Общая длина изучаемой части реки, измеренная по берегу.
2. Приблизительная ширина её. Ширину небольшой и неглубокой реки можно измерить путём расстановки по поперечной линии шестов (втыкая их в дно) па расстоянии 15—20 м друг от друга; сумма этих расстояний от берега даёт ширину реки.
3. Поперечный профиль дна, насколько позволяет сделать это скорость течения, так как при быстром течении слишком сносит лодку, что затрудняет как правильную установку последней, так и самое измерение глубин.
4. Скорость течения у берегов и на середине; для её определения можно пустить по течению какой-нибудь крупный (чтобы хорошо было видно с берега) поплавок, например, наглухо закупоренную, чтобы не проходила вода, пустую бутылку, и заметить время, в течение которого этот поплавок будет плыть от одного пункта реки до другого, причём расстояние между пунктами может быть измерено по берегу (лучше выбрать возможно более прямой участок реки).
5. Температура — в одно время у поверхности и у дна, посредине реки.
6. Суточные колебания температуры на поверхности (см. соответствующую тему для озера.
7. Прозрачность — до впадения притоков и после.
8. Разные грунты и их биоценозы (работа с драгой).
9. Планктон.
10. Есть ли прибрежная растительность, её характер (см. соответствующую тему для озера).
И. Характер берегов и их растительность.
12. Есть ли поблизости притоки, отметить положение их устьев: с какой стороны реки, выше или ниже обследуемого участка.
13. Есть ли придаточная система реки: рукава, старицы, затоны и т. д.
Как отдельные темы можно выдвинуть следующие:
1. Сравнение скорости течения, температуры и фауны коренной реки и рукава} каков в последнем процент речных форм.
2. То же для коренной реки и затона.
3. Судьба выносимых в реку ручьевых форм (у впадения ручья).
4. Животное население затапливаемых весенней водой растений.
5. Приспособления животных к рытью.
6. Способы прикрепления сидячих животных (или временно прикрепляющихся).
7. Распределение личинок и куколок мошек — Melusina.
МЕТОДИКА
Для работ на реке можно пользоваться тем же инструментарием и теми же способами, какие описаны для озера. Но в связи с текучим состоянием воды здесь необходимо указать некоторые особенности в ходе работы с различными орудиями и инструментами.
1. При работах, требующих неподвижного стояния лодки, как например, измерение глубин и глубинных температур, определение прозрачности, необходимо каким-нибудь способом заякорить лодку.
2. Если скорость течения значительна, то при работах с драгой и планктической сеткой следует ставить лодку параллельно течению, так как в противном случае течение всё время будет поворачивать её и таким образом затруднять управление её движением.
ТАБЛИЦЫ ДЛЯ ОПРЕДЕЛЕНИЯ ПРЕСНО ВОДНЫХ РАСТЕНИЙ И ЖИВОТНЫХ
Предлагаемые таблицы составлены в основном по следующим авторам и источникам:
Листостебельные мхи — по Кацу, Куммеру, 14стоминой, Пидопличке, Ростовцеву и др.;
Высшая водная растительность — по Флоровой и Раменскому, Ростовцеву, Сырейщикову, Федченко и Флерову;
губки — по Резвому;
гидры — по Парамонову;
коловратки — по Неизвестновой-Жадпной, Гарингу, Эйферт-Шёпихсну;
мшанки — составлена Г. Г. Абрикосовым;
малощетинковые черви — составлена И. И. Малевичем;
пиявки — по Иогансону;
моллюски — по Жадину;
ракообразные — по Бнрштейну, Рылову, Смирнову, Кайльгаку;
водные клещи — по Соколову;
вилохвостки — по Римскому-Корсакову, Карнп;
стрекозы (личинки) — по Поповой, Руссо, (имаго) — по Якобсону п Бианки; поденки (личинки) — по Черновой, Руссо, Карни;
веснянки (личинки) — по Лепневой, Руссо, Карни;
клопы (личинки и имаго) — по Кириченко, Руссо, Карни;
жуки (личинки и имаго) — по Рейхардту и Оглоблиной, Рейтеру, Карни; болыиекрылые и сетчатокрылые (личинки) — по Римскому-Корсакову, Руссо; ручейники (личинки) — по Леппевой, Руссо, Ульмеру;
бабочки (гусеницы) — по Римскому-Корсакову, Карни;
мухи и комары (личинки) — по Мончадскому, Карни;
тендипедиды (хирономиды) (личинки) — по Липиной, по Липиной и Чер-новскому;
гелеиды (личинки) — по Липиной.
ТАБЛИЦЫ ДЛЯ ОПРЕДЕЛЕНИЯ ЧАСТО ВСТРЕЧАЮЩИХСЯ ВОДНЫХ РАСТЕНИИ ПО ВЕГЕТАТИВНЫМ ПРИЗНАКАМ
Предлагаемые ниже таблицы дли определения водных растений составлены почти исключительно на основании строения и расположения листьев. Это сделано для того, чтобы растение могло быть определено не только в период своего цветения, но и вне этого периода.
Лист состоит в типичном случае из расширенной части — пластинки и узкой — черешка, или стебелька. Иногда нижняя часть черешка расширена я охватывает стебель — это расширение называется влагалищем (рис. 76). Если влагалище срослось на некотором протяжении в трубку, оно называется закрытым, если жр не срослось — открытым. Если лист совсем не имеет черешка, он называется сидячим. Иногда у основания листа сидят листовидные образования, носящие название прилистников.
По форме пластинок различают листья: перистые (рис. 109, 111), стреловидные (рис. 88), линейные (рис. 83), лировидные и пр.
Характерным является очертание края листа, на основании которого листья делят на це л ь н о к р а й п и е, зубчатые, пильчатые и т.п.
Расположение листьев на стебле может быть попарным (супротивные листья — рис. 93), одиночным (очередные листья — рис. 100, 103) или групповым (мутовчатые листья —рис. 112).
Растения, находящиеся в воде, мы разделим для большей лёгкости определения па четыре группы:
1) м х и;
2) надводные растения, выставляющиеся из воды стеблями или листьями;
3) плавающие, распластывающиеся какими-нибудь вегетативными частями (стеблем, листом) по воде;
4) погружённые, не выходящие на поверхность воды своими вегетативными органами (у некоторых лишь цветок поднимается на воздух).
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ПОДВОДНЫХ ЛИСТОСТЕБЕЛЬНЫХ МХОВ
Растеньица из стеблей и листьев; в период плодоношения — со споровыми коробочками.
Образуют по дну более или менее густые дерновинки, прикреплённые к грунту слабо.
Листья тонкие, зелёные, из одного слоя клеток. По середине листа проходят обычно продольные жилки, одна или две.
Листья удлинённой формы, часто у основания имеют ушки (рис. Ь6).
Стебли или неправильно ветвящиеся, или правильно перистые. Коробочки образуются по бокам стеблей; они или сидячие, или па очень коротенькой н о ж к е, имеют крышечку и колпачок, спадающий ко времени созревания спор.
1. Листья заострённые, удлинённые, до 8 мм длины, с продольной килевой складкой, без жилки. Стебель сильно разветвлённый. Дерновинки крупные, до 50 см длиной, тёмнозелёные (рис. 37А). Fontinalis antipyretica L.
0. Листья разной формы, но с жилкой, хотя бы только у основания листа.2
2. Листья почти всегда с двойной короткой жилкой, яйцевидные, притупленные.
Дерновинки жёсткие......................................................3
0. Листья с простой жилкой. Дерновинки мягкие.......................4
Calliergo- Р и с. 67. Р и с. 68. Рис. 69.
nella cuspi- Scorpidium А — Drepanocladus vernicosus— дрепанокладус зе- Calliergon
data — кал- scorpioi- лёный — лист; Б — Drepanocladlus fluitans Дрена- cordiio-
лиергонел- des—скор- нокладус плавающий — листья. lium калли-
ла — лист. пидиум — ергон—лист,
лист.
3. Листья с ушками (рис. 66). Стебель почти с правильно перистым ветвлением. Дерновинки зелёные или бурые. Calliergonella cuspidata Loeske
(Acrocladium cuspidatum Lindb.).
0. Листья часто серповидно изогнуты, без ушков (рис. 67), длина их 3—3,5 мм, ширина 1,4—2 мм. Стебли с редким ветвлением, с сильно загнутой верхушкой. Коробочки встречаются редко. Дерновинки красноватожёлтые или бурые с жёлтозелёной верхушкой, высота до 30 см. Scorpidium scorpioides (L.) Limpr.
4. Листья серповидно изогнутые, жилки доходят большей частью до середины листа. Ветвление почти или вполне перистое.
р1. Drepanocladus ...............................................б
0. Листья не изогнутые, жилки доходят большей частью почти до верхушки листа. Ветвление неправильное................................................6
5. Листья сильно крючкообразно согнутые, продольноскладчатые, без ясно дифференцированных ушков, длина их 2—3 мм, ширина 0,9—1,2 мм (рис. 6ЯА). Жилка заходит много далее половины листа. Стебли почти правильно перистые, до 15 см.
Drepanocladus vernicosus (Lindb.) Warnst.
0. Листья слабо изогнутые, нескладчатые, с золотистожёлтыми ушками, длиной 2,4—3,5 мм, шириной 0,5—0,6 мм. У верхушки листа есть зубцы, хотя бы небольшие; клетки ушка резко отличаются от прочих клеток листовой пластинки. Жилка иногда доходит только до половины длины листа (рис. 68Б). Стебли перистоветвистые, распростёртые или плавающие, до 20 см длиной.
Drepanocladus fluitans (L.) Warnst.
6. Листья слабо изогнутые, мясистые, линейноланцетовидные, клетки мелкие, округлые.
Cinclidotus minor Lindb. (С. fontinaloides (Hedw.) P. de B.).
0. Листья длиннозаострённые, с ушками, жилка лишь немного не достигает верхушки листа (рис. 69). Клетки листа узкие, очень длинные. _____________ Calliergon.
1 р. перед латинским названием организма означает «род».
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ НАДВОДНЫХ РАСТЕНИЙ
1. Растения с развитыми зелёными листьями на стебле или в прикорневой
розетке.........................................................................4
О, Растения без развитых зелёных листьев ..................................2
Рис. 71. Equisetum pa-lustre — хвощ болотный:
А — общий вид; Б — влагалище.
Рис. 70. Scirpus lacustris — камыш озёрный:
А — общий вид; Б — стебель с чешуевидными листьями; В — часть поперечного разреза стебля.
2. Стебель цилиндрический, полый, ребристый, с узлами (рис. 71 А); нижняя часть каждого междоузлия окружена коротким трубчатым влагалищем (рис. 71Б, налево), образованным из сросшихся листьев, концы которых свободны и поднимаются в виде зубчиков; стебель с системой воздухоносных каналов — центрального и периферических; диаметр его до 1 см, высота до 1,5 м\ на верхушке шишкообразные колоски со спорами.
Корневище гладкое, ползучее.
р. Equisetum — хвощ..............3
0. Стебель без узлов, безлистный (рис. 70А) (только при основании стебля недоразвитые чешуевидные листья с остров верхушкой) (рис. 70Б), тёмнозелёный, слабо блестящий, в разрезе имеет вид сотов, составленных из воздушных камер, разделённых плёнчатыми перегородками (рис. 70В) диаметр стебля до 1,5см, высота до 3 м и более; корневище чернобурое, членистое, ползучее, с многочисленными придаточными корнями. Озёра, пруды, болота, пойменные водоёмы, реки, на берегу или в воде на глубине до 2 м\ повсеместно.
Scirpus lacustris L. — камыш озёрный, чакан.
3. Стебель с глубокими бороздками, шероховатый, с узкой центральной полостью, влагалище широкое, с ложбинками и с 6—7 треугольными, заострёнными зелёными зубцами, имеющими тёмную верхушку и широкую белую кайму; влагалища при основании ветвей чёрные? блестящие, с острыми зубцами (рис. 71); диаметр стебля до 3 мм, высота до 50 см. Болота, болотистые луга, берега озёр, прудов и рек; почти повсеместно.
Е. palustre L. — хв. болотный.
0. Стебель ветвистый илп мало ветвистый, с неглубокими бороздками и неясными рёбрами, гладкий; центральная полость очень широкая; влагалище с 15—20 треугольными или ланцетошиловид-
Р и с. 72. Equisetum limosum (heleocharis)—хвощ иловатый: А — общий вид; Б — влагалище.
ными зубцами, имеющими узкую белую кайму и чернобурые концы; влагалища боковых ветвей зелёные или коричневые; диаметр стебля до 8 мм, высота до 1,5 м (рис. 72),
Е. limosum L. (holeocharie Ehrb.) — х в. иловатый.
4. Листья простые, цельные, большей частью линейные, с 5 или более продольными жилками, из которых средняя обычно не резко отличается от прочих; жилки идут параллельно или дугообразно, сходясь па верхушке. Анастомозы (соединения) между главными жилками идут параллельно друг другу. Листья большей частью с трубчатыми влагалищами ....................................................................5
0. Листья сложные или простые, с перистыми или сетчатыми жилками; всегда ясно выраженные главные жилки; большей частью одна продольная или несколько пальчато-расходящихся, редко несколько продольных, но никогда не сходящихся у вершины .....................................................................20
5. Листья никогда не сбрасываются, расчленены на влагалище и листовую пластинку. Влагалище округлое или четырёхгранное, никогда не трёхгранное. На границе
Рис. 73.
А — Qlycerta flu!tans — манник плавающий; Б — Glyceria spectabills — манник пышный.
Рис. 74. Catabrosa aquatics — поручейница водяная:
А — метёлка; Б — стебель с листьями и корневищем: В — явычок.
с пластинкой влагалище продолжается вперёд в виде «язычкаэ — плёнчатого или состоящего из ряда волосков. Листовая пластинка линейная или линейноланцетная. Стебель полый, с узлами — соломина; нижняя часть междоузлий охвачена влагалищем.
Сем. Gramineae — злаки ............................................6
0. Растения иного строения .......................................11
в. Листья у основания имеют наибольшую ширину. В почке листья сложены вдоль пополам; след этой складки часто остаётся в у вполне развитого листа. Верхушка несколько стянута ......................................................7
0. Листья к основанию суживаются. В почке они не сложены вдоль пополам. Верхушка плоская, иногда немного свёрнутая................................ 9
7. Верхняя поверхность листьев ровная, лишь посередине с сочленовной бороздкой ..............................................................8
0. Верхняя поверхность листьев ясно бороздчатая или многократно ребристая, пижяяя — шероховатая. Ширина листьев 3—10 мм. Язычок до 5 mjh длины, тупой, часто разорванный. Метёлка с трёхгранной осью (рис. 73А). Болота» сырые луга, берега стоячих и текучих водоёмов; повсеместно.
Glyceria fluitaosR. Вг. — манник плавающий.
8. Листовые пластинки шириной 1—2,5 см. лентовидные, тонко заострённые, ярко-велёные, по средней жилке килеватые, сверху гладкие, снизу и по краям шероховатые. Язычок низкий, округлый, сразу резко оттянутый в остриё. Стебель до 1,5—2,э м высоты,
сравнительно толстый, слабо сплюснутый, твердый, гладкий. Метёлка густая, крупная, раскидистая (рис. 73Б). Там же, где fluitans.
G 1 у с о г i a spectabilis Mert. et Koch. (G. a q u a t i c a Wahlb.) — манник пышный.
0. Листовая пластинка уже, 5—9 .и.ч, суживается постепенно, светлосероватозелё-пая, матовая. Язычок довольно высокий, широкий; метёлка редкая, раскидистая (рис. 74). По берегам текучих вод, по ключевым болотам; в воде; повсеместно, но к северу реже.
Catabrosa aquaticaP. В. — поручейница водяная.
9. Язычок — валик с плотным рядом волосков (рис. 75). Метёлка пирамидальная. Стебель гладкий, доверху олиствёпный. Листья серозелёные, i w ”
берегам водоёмов и в воде; по болотам и лугам; повсеместно, не исключая солончаковых лугов в степных районах.
Phragmites communis L. — тростник обыкновенный.
0. Жилки листьев на просвет все или почти все белые. Язычок перепончатый, цельный, разве лишь по краю расщеплённый.......................10
шириной 2,5—3 см. По
Рис. 76. Beckmannia eruci-formis — бекмания гусенице-видная: метёлка и стебель с листьями и корневищем.
Рис. 75. Phragmites communis — тростник обыкновенный!
А — метёлка; Б — стебель; В — язычок.
10. Листья мягкие, несколько желтоватые, тусклые, по краю острошероховатые. По верхней поверхности листа проходят рёбра, разделённые широкими плоскими бороздками. Основание стебля вблизи корня клуоневидно вздуто. Язычок от треугольного до остроклиновидного. Колос из мелких, двурядных, прижатых к цветоножке колосков (рис. 76). По берегам водоёмов, болотам, сырым лугам. Beckmannia eruciformis Host — бекмания гусеницевидная.
0. Листья кожистые, острошероховатые по всей повепхпости, особенно сверху; густо-зелёные, лоснящиеся. На верхней поверхности грубые, солиженные рёбра. Язычок высокий, несколько усечённый, по краю обычно расщеплённый (рис. 77). По берегам водоёмов, болотам; иногда заходит глубоко в воду; не часто.
S с о 1 о с b 1 о a festucacea L. — тростянка (т резубка) овсяничная.
11. Листовая пластинка плотная, линейная, иногда несколько суженная к основанию, с килем, в почке продольноскладчатая; вполне развившийся лист хранит следы этой складчатости; листья по краю и по килю остро шершавые, с трёхгранно заострённой или сплюснутой с боков верхушкой. Влагалище трубчатое, более или менее явственно трёхгранное. Язычок большей частью есть — перепончатый, пизкий. Стебель большей частью трёхгранный (рис. 78). Carex L. — осока.
0. Листья более или менее мясистые, мягкие, по краям не острошершавые. Влагалища никогда не трёхгранные, большей частью не замкнутые. Язычок не цельный, стебель не трёхгранный .....................................................12
12. Основание побега лилово красное. Листья крупные (длина до 1 м и более t ширина до 1,5 см}, зелёные, блестящие, килеватоутолщённые, мечевидные (рис. 79), линейные или 154
ланцетные, без черешка, переходящие книзу во влагалище, образованное как бы расщеплением одного из краёв листа. Влагалища срединных листьев замкнуты в начале цельной, но вскоре разрывающейся, хотя бы на коротком расстоянии, плёнкой. По берегам водоёмов, по пойменным, не слишком сырым болотам, по сырым лугам; повсеместно. А со г u s calamus L. — аир болотный (пахучий).
0. Влагалища срединных листьев открытые или неразвитые .............13
13. Листья мечевидные, линейные или линейнолапцстные, с ясно несимметричной верхушкой, без черешка; снизу переходят во влагалище, образованное как бы расщепле-
нием одного края листа (рис. 80). По берегам водоёмов, травяным болотам, сырым лугам; в Европейской части СССР почти повсеместно.
Iris pseudacorusL. — касатик ложноаирный.
0. Листья не мечевидные; листья п влагалища в разрезе ситовидные от сильно развитых воздушных камер, разделённых плёнчатыми перегородками . 14
Рис. 78. Carex gracilis—осока стройная.
Рис. 77. Scolochloa festucacea — тре-зубка овсяничная;
А — стебель; Б — метёлка; В — явычок.
14. Пластинка’относительно узкая — её длина не менее чем вчетверо превышает ширину. Пластинка плавно переходит во влагалище. Листья двурядные..15
0. Пластинка относительно широкая — её длина пе более чем в три раза превышает ширину. Хотя бы часть листьев на длинных черешках, не прикорневые..18
15. На разрезе листа, в утолщённой части, видно несколько рядов воздушных камер. Листья с килем или трёхгранные (рис. 81) ....................16
0. Листья скучены при основании стебля. На поперечном разрезе листа один ряд камер; листья всегда без киля, длинные, крепкие, тончащие вверх, желобоватые, серо-аелёные; край листа со светлой полупрозрачной каймой, жилкп мало заметны. Влагалища крепкие, толстые, с ушками, внутри слизистые (рис. 84, 85).
р. Typha — рогоз...........................................17
16. Листья все прикорневые, при основании трёхгранные, выше — плоские, линейные, с желобовидно вдавленной верхней стороной, ширина до 10 мм. Жилки незаметны. При разрыве остаются крепкие волокна, стебель безлистный. Соцветие зонтиковидное (рис. 81). По берегам водоёмов, болотам, лугам; повсеместно.
Butomus umbellatus L. — сусак зонтичный.
0. Ширина листьев 7—20 мм. Жилки заметные. При разрыве не обнаруживается волокон. Стебель прямостоящий (рис. 82, 83).
Некоторые виды рода Sparganium L. — ежеголовник.
17. Ширина листа до 18 мм\ в поперечном разрезе 15—17 воздушных камер; верхушка круто заострённая (рис. 84). Длина побега до 25 л<. По берегам водоёмов, болотам (чаше травяным), канавам; повсеместно, к северу чаще.
Typha latifolia L. — рогоз широколистный.
0. Ширина листа 5—10 леи; в поперечном разрезе около 9 воздушных камер. Верхняя сторона листа иногда слабо выпуклая. Верхушка полого заострённая (рис. 85). Длина
Рис. 79. Acorus calamus — аир болотный (пахучий).
Рис. 80. Irispseudacorus— касатик ложноаирный.
Рис. 81. Butomus umbeL latus — сусак зонтичный
Рис. 82. Sparganium га-mosum — ежеголовник ветвистый.
Рис. 83. Sparganium simplex —ежеголовник простой.
Рис. 84. Typha latifolia — рогоз широколистный.
Р и с. 85. Typha angustifolia — рогоз узколистный.
Рис. 86. Calla palustris — белокрыльник болотный.
Рис. 87. Alisma pl anta go частуха подорожник.
побега до 3 м. По берегам водоёмов, илистым, более или менее глубоким болотам; почти исключительно в южных районах Европейской части СССР.
Typha angustifoliaL. — рогоз узколистный.
18» Отдельные листья выходят из подземных или плавающих корневищ; листья густозелёные, лоснящиеся, снизу бледнее, длиной до 10 см и более, у вершины оттянутые п заострённые; от средней жилки отходят многочисленные боковые жилки на разных уровнях и загибаются дугообразно, сливаясь впереди в несколько жилок (рис. 86). Черешок сидит на широком белом перепончатом влагалище; язычок крупный. На болотах, особенно лесных.
С а 1 1 a palustrieL. — белокрыльник болотный.
0. Листья отходят пучком ....................................19
19. Листья двух сортов (рис. 87)! 1) подводные (если есть) — на коротких черешках, линейные; 2) надводные — на длинных черешках, крупные, яйцевидные с заострённой верхушкой, при основании большей частью сердцевидные; 5—9 мощных продольных
Рис. 88. Sagittaria sagittifolia — стрелолист обыкно- Рис. 89. Cicuta virosa—вех венный и его лист. ядовитый.
матовобелых жилок; черешок длинный, в основании расширяется в полуобъемлющее влагалище, без язычкп. Стебель прямой, в верхней половине ветвистый (рис. 87>. По сырым берегам водоёмов, повсеместно.
А 1 i s m a planta^o L. — частуха подорожник (водяной подорожник).
0, Листья, образующие прикорневую розетку, трёх сортов; 1) подводные — сидячие, линейные, прозрачные, с параллельными жилками; 2) плавающие — длинночерешковые, пластинки при основании сужены, закруглены или с выемкой; 3) воздушные — черешковые, стреловидные. Стебель — безлистный цветонос (рис. 88). По берегам водоёмов, болотам (главным образом пойменным), влажным лугам; повсеместно, часто.
Sagittaria sagittifoliaL. — стрелолист обыкновенный
20. Листья сложные, дважды-трижды перистые ..................21
0. Листья простые ‘.............................................24
21. Корневища с поперечными перегородками, видными на поперечном разрезе 22
0. Корневища без поперечных перегородок.........................23
22. Листочки лилейные или узколанцетные, остропильчатые, с хорошо заметной срединной жилкой (рис. 89). Топкие берега водоёмов, болота, канавы,, в воде и у воды; повсеместно.
Cicuta virosa L. — вех ядовитый (бешеница водяная).
0. Листочки поднимающихся над водой листьев яйцевидные, перистонадрезанные на линейные или яйцевидные дольки (рис. 90), с хорошо заметной срединной жилкой. Листочки погружённых листьев нитевидные. По берегам стоячих и текучих водоёмов, болотам в канавам; часто на речных перекатах; в воде или у воды; повсеместно.
Oenanthe aquatics Lam. — омежник водяной (конский укроп).
28. Надводные листья не парноперистые, их доли крупные, продолговатоланцетовидные, у основания не равнобокие, с зубчатыми краями; доли подводных листьев дважды
перистораздельпые с нитевидными вторичными дольками. По берегам водоёмов (главным образом стоячих), канавам; повсеместно.
Sium latifolium L. — поручейник широколистный.
0. Листья дважды-трижды перистораздельные с мелкими линейными листочками, кончики которых красные или белые, заострённые; общее очертание листьев треугольное; нижние листья на длинных черешках, средние на коротких, верхние сидячие. По берегам водоёмов, болотам; довольно часто.
Poucedanum palustre L. — горичник болотный.
24. Нижние листья лировидпые, при основании суженные (рис. 91). В водоёмах, повсеместно. Nasturtium amphibium R. Br. — хрен водяной (жеруха земноводная).
Рис. 90. Oenantbe aqua- Рис. 91. Nasturtium am- Рис. 92. Lysimachia vul-
tica — конский укроп phibium — жеруха земно- garis — вербейник обыкно-
(омежник). водная, хрен водяной. венный.
0. Листья не лировидные, цельнокрайние, на коротких черешках; при рассматривании в лупу видны просвечивающие оранжевые круглые пятнышки.
р. Lysimachia — вербейник . . . . .......................25
25. Листья длинные, ланцетовидные, супротивные или в мутовках, длиной до 15 см, шириной до 5—6 см. Стебель стоячий, плотный, прямой, ребристый или тупо 4-гранный, опушённый нежными волосками (рис. 92). По берегам водоёмов (погружённая форма на глубине 60—80 см), по канавам; повсеместно.
L. vulgaris L. — в. обыкновенный.
0. Листья только супротивные, почти округлые, у основания слегка сердцевидные, в диаметре 2,5 см. Стебель лежачий, довольно тонкий, сплюснуто 4-гранный. По берегам водоёмов, у воды; на сыроватых лугах.
L. nummularia L.—луговой чай (полушечная трава).
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РАСТЕНИЙ С ПЛАВАЮЩИМИ ЛИСТЬЯМИ
1. Растения, не укореняющиеся в грунте .......................... 2
0. Растения укреплённые ............................’.............7
2. Листья внизу густо покрыты волосками, сверху усажены мелкими бородавочками с толстыми и короткими волосками; сидят парами один против другого. В озёрах, старицах, речных заводях, протоках с тихим течением; не часто, но в большом количестве (рис. 93).
Salvinia natans All. — сальвиния плавающая.
0. Плавающие листья или стебли без волосков и бородавочек, сидят одиночно или группами ............................................................ 8
8. Стебель в виде очень мелкой, округлённой овальной пли удлинённой пластиночки — без жилок или только с одной, слабо заметной срединной жилкой.4
0. Стебель длинный, нитевидный, плавающий, несёт в узлах пучки немногочисленных листьев на длинных черешках. Пластинка листа округлая, при основании широко-сердцевидная, 3—5 см. Черешок с 2 прилистниками. От основания листа отходит 1 срединная жилка и 2 пары дугообразно загибающихся к верхушке боковых (рис. 94). В различных стоячих водоёмах (в том числе и пойменных).
Hydrocharis morsus ranae L. — лягушат н и к (водокрас лягушечпый).
4. Стебель — тесьмовидная пластиночка, повторно вилообразно ветвящаяся, без жилок и без корешков. Разрастаясь, образует корочки.
Ricciella fluitana (L.) Вг.
0. Стебли удлинённые или округлые пластиночки, иногда соединённые по нескольку вместе короткими ножками, снизу плоские или слабо выпуклые, с отходящими вниз корешками ...............................................................5
sus гапае — водокрас лягу-шечный. лягушатник.
б. На каждой стеблевой пластинке пучок корней (рис. 95). Пластинка с 3 (иногда до 9) жилками 3—'♦ см длиной, округлая или яйцевидная. В прудах, травяных болотах, в пойменных водоёмах.
Spirodela polyrrhiza Schleid. — многокоренник.
0. На каждой стеблевой пластинке не более одного корня (рис. 96).
р. L е m n a L. — ряска....................................6
Рис. 95. Spirodela po-
lyrrhiza — ряска многоко- Рис. 96. Lemna minor—ряска малая и Lemna trisulca— ренная, многокоренник. ряска трёхдольная (направо).
6. Пластинка плавает по самой поверхности воды, округлая или яйцевидная. 2— 3 мм длиной (рис. 96). В стоячих водах, особенно часто в гумусовых.
Lemna mi nor L. — ряска малая.
0. Пластинка погружена в воду, суженная к одпому концу, прозрачная, ромбоидальная, иногда с выемчатыми краями, 5—10 мм длины, с 3 жилками. Часто по три пластинки соединены вместе (рис. 96). В озёрах, прудах, болотах, заводях рек
Lemna trisulca L.—р. трёхдольнал.
7. Плавающие листья линейные, некоторые виды рода.
Sparganium L. — ежеголовник..................................19
0. Плавающие листья нелинейные................................8
8. Листья двух сортов: плавающие с продолговато-клиновидной или 3-лопастной пластинкой и подводные совсем без пластинки (одни лишь черешки длинные, нитевидные). Стебель обычно плавающий. В болотах и на заболоченных берегах; в воде и у воды, на иловатых и глинистых почвах (рис. 97).
Ranunculus polypnyllus W. К. — лютик ыиоголастпый.
0, Плавающие листья иной формы...................................9
9. Листья округлые, с глубокой сердцевидной выемкой у основания.10
0. Листья овальные или удлинённые, у основания не вырезаны сердцевидно или вырезаны лишь слабо..................................................15
10. Отходящие от корневища стебли доходят до поверхности воды и стелются по ней; плавающие листья 3—10 см в поперечнике, округлые, расположены супротивно, у основания сердцевидные; корневые листья, а также расположенные в очередном порядке на нижнеи (подводной) части стебля листья имеют длинные цилиндрические,зелёные черешки; пластинки листьев тёмнозелёные, лоснящиеся, снизу с буроватыми бугорками;
такие же бугорки на стеблях и черешках. В стоячих водоёмах.
Limnanthomum nymphaeoides Link. — ужовник' (павун) кувшинковидный (нимфейник болотный) из сем. Gentianaceae — горечавковые.
0. От корневища отходят очень длинные (до 5 л), черешки листьев; листья плавающие—овальные или округлоовальные размером 16 х 25 см\ листья подводные — тесьмовидные.
Сем. Nymphaeaceae — кувшинковые ..................................11
11. Нижняя сторона листа большей частью красноватая; черешок с прилистниками, в разрезе круглый (рис. 98).
р. Nymphaea L. — кувшинка . . 12
0. Нижняя стопона листа без красноватой окраски; черешок оез прилистников, в разрезе плоский или трёхгранный (рис. 99).
Рис. 97. Ranunculus polyphyl- р. Nuphar Sm — кубышка..............14
lus — лютик многолистный. 12. Пластинка листа округлоовальная шири-
ной 5—8 см\ лопасти по бокам выреза с длинно заострёнными концами и сильно выгнутыми внутренними краями; длина внутреннего края лопасти равна длине пластинки листа по срединной жилке. В озёрах, медленно текущих реках, пойменных водоёмах. Преимущественно на севере и северо-востоке Европейской части СССР.
Nymphaea tetragons Georgi — кувшинка малая. 0. Листья иного строения ..............#......................13
13. Лопасти по бокам выреза равной или почти равной длины, их выпуклые внутренние края часто налегают один на другой. Главным образом в северных частях СССР (рис. 98).
N. Candida Presl. — к. чисто белая (водяная лилия, нимфея).
0. Лопасти расходящиеся, часто неодинаковой длины; их внутренние края почти прямые. В стоячих и медленно текучих водах. Большей частью на юге и юго-западе Европейской части СССР. N. а 1 b a L. — к. б е л а я.
14. Черешки листьев в разрезе 3-гранные; лопасти по бокам выреза пластинки листа сближены, концы их тупые или притуплённые; размеры пластинки до 16 х 27 см. Есть подводные листья —тонкие, прозрачные, слегка морщинистые, бледнозелёные или светложелтоватозелёные. На быстром течении дают погружённые формы. В стоячих и медленно текущих водах, в пойменных водоёмах. Корневище толстое (рис. 99).
Nuphar luteum Sm. — кубышка жёлтая.
Рис. 98. Nymphaea Candida — кувшинка чисто белая.
Рис. 99. Nuphar luteum—кубышка жёлтая.
Рис. 100. Polygonum amphibium —гречиха земноводная.
Рис. 101. Potamo-geton natane — рдест плавающий.
0. Черешки листьев плоские; лопасти пластинки расходящиеся, концы их слабо заострённые или слабо притуплённые; размер пластинки до 10 х 18 см. Подводных листьев нет. Корневище тонкое.
N. р u mi I и m Sm. — к. м а л а я (к а р л и к о в а я).
15. Листья полукожистые, продол 1юватые, заострённые. От средней жилки боковые расходятся под углом (рис. 100) в поперечном направлении. В стоячих водах, ручьях, реках от границы между водой и берегом до значительной глубины.
Polygonum amphibium L. var. n a t a n s Much. — гречиха земноводная.
0. Листья большей частью довольно топкие, эллиптические или яйцевидные. Главные жилки идут дугообразно, сходясь на вершине.
р. PotamogetonL. — рдест...................................18
16. Черешок листа длиннее листовой пластинки................17
0. Черешок не длиннее листовой пластинки ......................18
17. Черешки длинные, сверху несколько желобчатые. Листья кожистые, эллиптические, яйцевидные или удлинённые, размер их до 2 х 5,5 см, при основании слабосердцевидные, с двумя явственными боковыми складками. Жилки многочисленные, нри-
Рис. 103. Sparganium minimum — ежеголовник малый.
Р и с. 102. Potamugeton rufescens — рдест краснеющий.
чём главные из них выдаются по нижней стороне листа (рис. 101). У речных форм лист суживается к черешку. В озёрах, прудах, пойменных водоёмах, в реках с медленным течением. Р. natansL. — рд. плавающий.
0. Черешки листьев с обеих сторон выпуклые. Плавающие листья плоские, овальные или продолговатые, к основанию суженные или за1«)углённые; подводные листья удлинённоланцетные, прозрачные, черешки их сверху слабо выпуклые; стебель толстый, сильно ветвистый. В озёрах, реках, в пойменных водоёмах.
Р. п о d о а и 8 Poir. (f 1 u i t a n s Rath.) — p д. узловатый (речной, колыхающийся).
18. Черешки равны по длине пластинке или короче. Листья довольно тонкие, эллиптические, при основании почти округлые или короткосуженные, прозрачные, шириной до 8 мм, по краям шероховатые (мелкозубчатые) (рис. 125). В прудах, болотах, речках.
Р. heterophyllus Тг. (gramineus L.) — р д. разнолистный (злачный).
0, Черешки большей частью в 1/3 длины листа, стебель удлинённый. Листья кожистые, обратнояйцевидпые (рис. 102). В озёрах, прудах, болотах; в северной половине Европейской части СССР.
Р. rufescens Schrad. (a I р i n u s) — p д. краснеющий.
19. Листья лентовидные, плоско выпуклые, в разрезе, хотя бы в средней части, имеют несколько рядов воздушной ткани.................................30
0. Плавающие листья в разрезе все с одним рядом камер Стебель до 30 с.и длиной, с несколькими (большей частью 5—7) междоузлиями. Листья короче стебля, большей частью 3—4 ми, иногда до 10 шириной, тонкие, с невыступающей средней жилкой (рис. 103). В болотах, мелких водоёмах, на берегах озёр.
Sparganium minimum Fr. (natans) — ежеголовник малый.
11 А. Н. Лилин
Рис. 104. Sparganium Friesii — ежеюлившк Фриза.
Рис. 105. Sparganium affine —ежеголовник сродный.
20. Стебель плавающий, кверху утолщённый, изогнутый, до 1 м длиной. Листья до 2 м длиной, 1—3 мм шириной, книзу суживающиеся (рис. 104). В чистых озёрах, большей частью несколько отступя от берега.
S р. Friesii Beurl. (1 о ngi folium Mein.) — еж. Фриза.
0. Стебель плавающий, кверху не утолщённый. Листья до 7 мм шириной, чаще 2,5—5 МгИ, плоские или полуцилиядрические (рис. 105). В озерах, преимущественно в северной половине Европейской части СССР.
S р. affine Schnitzl. — еж. сродный.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ПОГРУЖЁННЫХ РАСТЕНИЙ
1. Листья разрезные, перистые или сложные ........................2
0 Листья простые, цельные, цельнокрайние (иногда зубчатые)........7
2. Листья иеристомяогораздельные, с нитевидными долями, усаженными редкими, короткими зубчиками. С пузырьками на листьях. Цветок над водой (рис. 106). Стоячие и текучие воды; преимущественно на юго-востоке Европейской части СССР.
Utricularia vulgaris L. — пузырчатка обыкновенная.
0. Без пузырьков на листьях.......................................&
3. Листья мутовками по стеблю или в прикорневом пучке.............5
0. Лисгья очередные...............................................4
4. Листья на черешках. Дольки листа, на которые рассечена его пластинка, нитевидны, расположены не в одной плоскости; вынутые из воды листья слипаются (рис. 107). В озёрах, прудах, реках, часто в пойменных водоёмах; в воде и на берегу у самой воды. Ranunculus confervoides Fries (е г a d i с a t u s Fr.) — лютик нитчатый.
0. Листья сидячие. Дольки листа расположены в одной плоскости, более твёрдые; поэтому, вынутые из воды, не слипаются (рис. 108). В стоячих и медленно текущих водоёмах, также в пойме.
Ranunculus circinatus Sib th. — лютик жестколистный.
5» Листья в прикорневой розетке гребневидные, с линейными долями. Стебель плавающий или ползучий. Цветонос прямостоячий, цветы в мутовчатых кистях. Листья по стеблю в частых мутовках, а у основания цветоноса сдвинуты почти в розетку (рис. 109). Длина стебля до 60 см\ корней нет. В стоячих и медленно текущих водоёмах; редко в болотах.
Hottonia palustris L. — турча болотная (водяпое перо).
0, Листья мутовками по длине стебля..............................6-
6» Листья вильчаторлздельные, с шиповатыми белыми хрящеватыми зубчиками йрис. 110). Цветы пахучие. Стебель тонкий, к осени ломкий, длиной до 80 см. В озёрах, болотах, пойменных водоёмах.
Ceratopbyllum deniersum L. — роголистник тёмнозелёный.
0. Листья гребневидно-пернстораздсльные, глубоко рассечённые на нитевидные дольки (рис. 111). р. MyriopbyllumL — у р у т ь.
7. Мутовки листьев по всему стеблю или пучок прикорневых листьев....8
О, Листья супротивные или очередные по всему стеблю в одиночку......11
8. Все листья прикорневые...........................................10
0. Мутовки листьев по всему стеблю ..................................9
9. Мутовки из 4 —14 простых линейных листьев; подводные листья мягкие, отогнуты вниз, надводные отходят более или менее горизонтально; мутовки сближенные (рис. 112).
Стебель прямой, цилиндрический, довольно крепкий, длиной до 90 см. Кдрпрвиша ползучие, цветы в пазухах листьев. По берегам озёр, прудов, стариц,рек (в воде, иногда у воды), в болотах, на сырых лугах.
Hippuris vulgaris L.— в о д z
0. всему
Д fl-ная сосенка, х в о ст п и к обыкновенный. Листья расположены мутовками по стеблю, по 3—4 листа* (рис. ИЗ),
Рис. 108. Ranunculus circinatus — лютик жестколистный.
Рис. 106. Utricularia vulgaris — пузырчатка обыкновенная.
Рис. 107. Ranunculus confervoides—лютик нитчатый.
продолговатые, суженные к вершине, не длиннее 12 мм, полупрозрачные, с одной ясной жилкой.
Elodea canadensis Rich. — водяная чума, элодея.
10. Листья длинные, мечевидные, жёсткие, с колючими зубцами по краям (рис. 114), крупные, собраны в виде густых пучков, при основании 3-гранные. Корни шнуровидные. В погружённом состоянии находится только осенью, летом плавает. Stratiotes aloides L. — телорез обыкновенный (сабуровидный).
0. Листья шиловидные, прямые или изогнутые; в основании листьев белые мешочки спорангиев (рис. 115) с гребневидными или волосовидными выростами. Длина листьев до 20 см. В озёрах с чистой водой, большей частью на песке.
I s о ё t е s lacustre L. — полушник (расходник) озёрный.
11. Листья очередные..............................................15
0. Листья супротивные ............................................12
12. Листья на верхушке цельные или с выемкой, или двузубчатые, или трёхзубчатые, с одной или тремя сильно выдающимися жилками, мелкие, светлоаелёные, нейро* зрачные. Стебель тонкий. Длина до 25 см.
Cal litriche — болотник..........................................13
0. Листья на верхушке слегка заострены или закруглены, без выемки.14
13. Листья к основанию расширены, с одной жилкой, розетки не образуют (рис. 116). В стоячих и медленно текущих водах; в прибрежье озёр с прозрачной водой; иногда в старицах и протоках.
С. automnalis — б. осенний.
0. Листья к основанию обычно сужены, овальные или эллиптические, на верхушке цельные, выемчатые или двузубчатые; с тремя жилками, верхние с 1; верхние листья * 163
densis — элодея, видяндя чума.
Рис. 114. Stratiotes aloi-des—телорез сабуровидный.
Рис. 115. 1зоё1ев lacustre — полушник озёрный.
обычно образуют розетку, часто плавающие. В стоячих и текучих водоёмах, в воде или у виды. * С. v е г п а — б. весенний.
14. Листья ланцетные или ланцетояйцевидные, сидячие или стеблеобъемлющие, обычно расширенные у основания и заострённые кверху, с мелкими, редкими зубчиками по краям, реже с ровными краями; нижние листья иногда короткочерешковые; длина листовой пластинки до 6—7 см, ширина до 2,5—3 см. Стебель цилиндрический или почти 4-гранный, прямой или восходящий из лежачего основания, в поперечнике до 1 см. Образует несколько разновидностей. На берегах водоёмов, у воды и в воде, на песчаных отмелях рек, в ключах, канавах, болотах.
Veronica anagallis L. — вероника ключевая.
0. Листья овальные или эллиптические, с ровными или неясно редкогородчатыми 1 краями; длина листа до 4—5 см, ширина до 3 см, длина черешков до 6 мм. Стебель цилиндрический, гладкий, голый, обычно плотный, с многочисленными корнями на узлах (рис. 117). Там же, где предыдущий вид.
Veronica beccabunga L. — вероника - лоточник.
Рис. 117. Veronica beccabunga — вероника-поточник.
Рис. 116. Calli triche autumnalls — болотник осенний.
Рис. 118. Potamogeton рес-tinatus — рдест гребенчатый.
15. Листья выемчатые, шиповатозубчатые, сидячие, без жилок, пластинки отходят от вершины влагалища. В озёрах, реках, заводях. р. Najas — резуха.
0. Листья иногда попарно сближенные, более или менее цельнокрайние, обычно тонкие, перепончатые, прозрачные, отходят почти всегда от основания влагалища; последнее иногда отсутствует.
Й. Potamogeton L. — рдест ................................
истьл узколинейпые, цельнокрайние; влагалища хорошо развиты.
Подрод Coleogeton Raunk......................................П
0. Листья разной формы, без влагалищ.
Подрод Eupotamogeton Raunk .................................IS
17. Листья нитевидные, тонкие, хрупкие; жилка одна; влагалища в нижней часто сросшиеся, с коричневой каймой. В озёрах, речках, ручьях.
Р. f i 1 i f о r m i s Pers. — p Д. нитевидный,
0. Листья шириной 0,5—2,5 мм, более плотные, не ломкие; верхние листья с 1 жилкой, нижние с 3; влагалища не сросшиеся, свёрнутые в трубку, большей частью с белой каймой (рис. 118).
Р. pectinatus L. — р д. гребенчаты и.
18. Листья с мелкоостропильчатым краем (рис. 119). большей частью волинсше. Стебель сплюснутый, 4-гранный. В стоячих и медленно текущих водах.
Р. с г i в р u 9 L. — р д. курчавый.
0. Листья цельнокрайние или слабозубчатые, не волнистые. Стебель цилиндрический или слабосплюснутый.......................................И*.
1 Городчатым называют такой зубчатый край листа, у которого вершинки зубчиков тупые, закруглённые, а промежутки острые.
19. Листья линейные или гпетинковидные, шириной 1—3 мм...................20
0. Листья не линейные и не питипковидные, шириной более 3 мм.............25
20. Листья с закруглённым концом, вытянутым посередине в короткое
остриё.........................................................................21
0. Листья к концу суживаются и заостряются постепенно....................22
21. Продольных жилок много, из них 5 (редко 3) крупные, остальные мелкие; длина листа до 20 см, ширина 2—4 мм. Прилистники светлые, длина их до 4 см, Стебель полый,
ton crispus — рдест курчавый.
Рис. 120. Potamogeton compressus — рдест сплюснутый.
двукрылый, одинаковой ширины с листом (рис. 120). В озёрах, прудах, реже в речках, ручьях. Р. compressus L. (zosterifolius Sebum.) —
рд. сплюснутый (в з м о р н и к о л и с т н ы й).
0. Продольных жилок 3—5 (редко 6); ширина листа 1—2 мм. Прилистники широкие, длиной до 1,5 см. Стебель сжатый или почти цилиндрический, без крыльев (рис. 121). В стоячих и слабо проточных водах.
Р. obtusifolius Mert. et Koch. — рд. туполистпый.
22. Листья ланцетные или линейноланцетные, суженные в черешок, иногда полустебле-объемлющие, с шероховатыми краями. Стебель ветвистый, 1—2 мм толщиной.
Р. heteroph у 1 I u s var. g г a mini f о 1 i и s. Fr. — p д. злачный.
Рис. 121. Potamogeton obtusi-lulius — рдест туполистный.
Рис. 122. Potamogeton pusillus—рдест маленький.
0, Листья иного типа..............................................23
23. Листья очень узкие, шетинковидные; тёмнозелёные, на верхушке длинно заострённые; очень толстая центральная жилка и слабые, едва просвечивающие боковые. В небольших озёрах, прудах.
Р. trichoi des Cham, et Sehl. — рд. волосовидный.
0. Листья линейные, зелёные или светлозелёные; боковые жилки ярпо видны.................................................................24
24. Листья до 1,5 мм шириной, округлые или слабо остроконечные; жилок большей частью 3, иногда 1. Прилистники расщеплённые до конца, свёрнутые (рис. 122). В прудах, болотах, реже в озёрах, в речках. Р. pusillus L. — рд. маленький.
0. Листья торчащие, постепенно заостряющиеся, длиной 4—6 см Прилистники сросшиеся при основании, раструбовидные. В озёрах.
Р. г u t i 1 u s Wojfg. — рд. красноватый,
25. Листья эллиптические или яйцевидные, с округлым или сердцевидным основанием, сидячие, стеблеобъемлющие.......................................26
0. Листья ланцетные, сидячие или черенковые, к основанию обычно суженные ..................................................................27
Рис. 123. Potamogeton рег-foliatus—рдест пронзённо-листный.
Рис 124. Potamogeton praelongus—рдест длиннейший.
26. Листья с глубоко сердцевидным основанием, по краю мелкозубчатые. Прилистники обычно не длиннее 1 см, беловатые, тонкие. Стебель прямой (рис. 123). Главным образом в пойменных водоёмах.
Р. perfoliatus L. — рд. пронзепнолвстпый.
0. Листья с округлённым основанием, цельнокрайние, па верхушке стянутые колпачком. Прилистники 1,5—6 см, соломенножёлтые, жёсткие (рис. 124). В озёрах (иногда на большой глубине); встречается и в реках.
Рис. 126. Potamogeton lucens — рдест блестящий.
Рис. 125. Potamogeton hetero-pbyllus — рдест разнолистный.
27. Листья цельнокрайние, сидячие. В озёрах, прудах, болотах, речках северной половины Европейской части СССР.
Р. г u f е в ое d s Schrad. (a I pi n u 9 Balb.)—рд. краснеющий (альпийский), 0. Листья мелкозубчатые, сидячие или черешковые ............ 28
28. Листья сидячие, не шире 1 см (рис. 125); иногда есть и плавающие листья.
Р. heteropnyllus Schr. — рд. разнолистный.
0. Листья ло 30 см длиной, на коротком черешке. Плавающих никогда нет (рис. 126). Осенняя вариация acuminata с остриями на концах листьев. В озерах, прудах, реках, роже в ручьях. Р, 1 и се п s L. — р д. блестящий.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ГЛАВНЕЙШИХ ГРУПП ВОДНЫХ БЕСПОЗВОНОЧНЫХ ЖИВОТНЫХ
1. Животные с ножками, состоящими из ряда подвижных относительно друг друга члеников; иногда ножки вместе со всем телом прикрыты раковиной.
Тип Arthropoda — членистоногие.
0. Животные без членистых ножек........................................2
2. Животные сидячие, постоянно прикреплённые к подводным предметам .... 8
0. Животные подвижные, свободно перемещающиеся......................6
8. Животные одиночные .................................................4
0. Животные колониальные или облекающие подводные предметы сплошной массой, или разветвлённые......................................................5
Рис. 127. Ресничный червь — Planaria gonocephala.
4. Животные очень мелкие — не более (большей частью менее) 2 мм. На переднем конце тела втяжной мерцательный орган.
Тип Vermes — черви, класс .Rotatoria — коловратки.
0. Животные более крупные — до 2 см. На переднем конце тела отверстие, окружённое венчиком длинных, тонких, нежных щупалец, сильно сократимых, как и все тело (рис. 141,142). В расправленном состоянии тело сильно удлинённое, цилиндрическое.
Тип Coelen te га ta — кишечнополостные, класс Hydrozoa — гидроидные, отряд Hydrida — гидры.
Рис. 128. Малощетинковый червь — Stylaria lacustris.
б» Колонии состоят из отдельных плотных кутикулярных камер, в которых заключены животные; в спокойном состоянии животные выставляют из камер венпы щупалец (рис. 163 Д), так что колония кажется покрытой бархатным налётом. Потревоженные животные очень быстро втягиваются в камеры, и налёт исчезает. Беа специфического запаха. Тип Vermes — черви, класс В г у о z о а — мшанки.
0. Животные не имеют ни кутикулярного остова, ни щупалец и не меняют своего вида в спокойном и потревоженном состоянии. При растирании между пальцами издают резкий специфический запах. Тип Pori fera — губки.
6. Животные одиночные ...........................................7
0. Животные колониальные; колонии обычно шаровидные, отдельные особи расположены внутри их радиально.
Тип Vermes, класс Rotator! а — коловратки.
7. Тело заключено в твёрдую обызвествлённую раковину.
Тип М о 1 1 u s с а — мягкотелые.
Ш
0. Раковины нет ...................................................8
8. Тело удлинённое, плоское, покрыто короткими, тонкими ресничками. Движение скользящее, со слабо заметным изменением формы тела (рис. 127).
Тип Vermes, подтип Plathelminthes — плоские черви, класс Turbellaria — ресничные черви.
0. Тело различной формы, не покрытое ресничками. Движение не скользящее, большей частью связанное с сильным изменением формы тела................9
9. Тело всегда удлинённое, подразделено снаружи на ряд колеи......10
0, Тело некольчатое, различной формы .............................12
10. На переднем конце тела или отдельная хитиновая головная капсула, или хотя бы хитиновые ротовые части, просвечивающие через полупрозрачные покровы тела.
Рис. 129. Малощетинковый червь — Ghaetogaster diaphanus.
Некоторые личинки из класса Insecta — на секомые.
0. Нет ни головной капсулы, ни хитипизированных частей...... 11
11. Кольчатость мелкая. Тело без щетинок. На заднем конце всегда, а на переднем — большей частью имеется присоска (рис. 10). Резко выраженная способность к сокращению и вытягиванию.
Тип Vermes, подтип A n n е 1 i d е s, класс Hi rudi nea — пиявки.
0. Кольчатость более крупная. Присосок нет. Способность к сокращению слабая, но резкая способность к изгибанию. Тело всегда снабжено щетинками (рис. 128, 129).
Подтип Annelides, класс Chaetopoda — щетин к*о н о г ие, отряд Oligochaeta — малощетинковые.
Рис. 130. Круглый червь — Dory lai mus stagnalis.
12» Тело сильно удлинённое. Движение при помощи сильных изгибании всего тела.
Тип Vermes, подтип Nemathelminthes — круглые черви... 18 0. Тело не сильно удлинённое. Движение при помощи колов рашательно го аппарата и «ноги», причём прочие части тела пе претерпевают видимых изменений формы.
Класс Rotator! а — коловратка.
18. Тело очень длинное, волосовидное. Задний конец впльчато разделен.
Класс Nematomorpha— волосатиковые, Gordius aquations DyJ. — волосатик,
0. Тело сильно удлинённое. Задний конец всегда несколько загнут (рис. 130). Класс Nematodes — собственно круглые черви.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ПРЕСНОВОДНЫХ ГУБОК
Колоппи образуют наросты па подвод пых предметах. Молодые губки образуют к о р о ч к и в 2—3 мм толщиной, в дальнейшем болтающиеся до нескольких сантиметров. Иногда колонии образуют ветвистые выросты. Отдельные колонии достигают нескольких килограммов веса. Основная масса колоний слизистая, ноздреватая; на разломе большей частью видны мелкие жёлтые, бурые или коричневые зёрнышки, около 0,5 мм в диаметре — геммулы, представляющие собой зимующие почки (рис. 138); у молодых губок геммулы сконцентрированы в основании, в остатках старых колоний.
Рис. 131. Губка Trochospongilla horrida — общий влд.
Рис. 132. Спикулы Baikalospon-gia bacillifera.
В раздавленных или растёртых кусочках колонии, положенных под микроскопом, при слабом увеличении видны пучки удлинённых, прямых или слегка изогнутых кремне-
вых скелетных игл — так называемые спикулы, или ма кросклеры; кроме того, в толще паренхимы рассеяны ещё более мелкие скелетные образования — микро-склеры, имеющие формы или игл (рис. 134—136), или стержня с сидящими на его концах дисками, иногда надрезанными, —так называемые амфидиски геммул
(рис. 137—139).
1. Вся колония покрыта тонкой, сквозь которую просвечивают тёмные каналов (рис. 131). Поверхность в мелких конических возвышениях (места подхода к плёнке скелетных пучков). Спикулы заострённые (рис. 134 — 140), за единственным исключением. Есть * микросклеры.
легко отделяющейся полупрозрачной плёнкой, пятна полостей и радиально сходящихся к ним
Рис. 134. Спикулы Spongilla arctica: A — ма кросклеры; Б — мимросклеры.
Рис. 133. Спикулы Lu-bomirekia abietina.
В основании колонии или по всей толте большей частью имеются геммулы (рис 138). При растирании тело шероховатое, с острым резким влпахом
Сем. Spongillidae — бадяги ..................................2
0. Плёнка не отделима от колонии. Спикулы или закруглённые, или заострённые (рис. 132, 133). Микросклеритов нет. Геммул нет.
Сем. Lubomi rskiidae — байкальские губки,
2. Микросклеры геммул в виде игл (рис. 134, 135, 136). Форма губки куст исто-пальцевидная. р. Spongilla La mark.
ПО
Рис. Г35. Спикулы Spongilla stanleyi:
А — макросклеры; Б — микросклеры.
Рис. 138. Спикулы и геммула Ephy-datia fluviatilis war. teberdana:
A — макросклеры; Б — амфидиски; В — геммула.
Рис 140. Спикулы Hetero-tneyenia balleyi var. stepanowii л _ мзкросклсры; Б - микроснле-A ри; В -- амфадиеки.
139. Спикулы Ephjdatia miil-
амфидискп;
P и с. 137. Спикулы Tro макросклеры, направо к
0. Микросклеры геммул в виде амфидисков. Губки в форме нароста или корки.......................................................... 8
8. Диски амфидисков цельнокрайние, гладкие или радиально исчерченные (рис. 137). Колония в виде корки или массивного нароста с ровной, шероховатой поверхностью (рис. 131). р. Trochospongilla Vejd., 1 вид Т г. horrida Weltner.
О, Край диска надсечён более или менее глубоко, так что диск имеет вид звезды или зубчатого колеса ............................................... 4
4. Стержни амфидисков все приблизительно равной длины (рис. 138, 139). В виде корковых или массивных наростов, то с ровной поверхностью, то бугристой с невысокими выростами. р. Ephydatia Lam.
0. По длине стержня амфпдискп распадаются на две группы (рис. 140). В виде тонких корок на подводных растениях и древесных остатках.
р. Heteromeyenia Potts, у нас 1 вид Н. baileyi (Bwk.) v а г.
stepan о wi (Dyb.).
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ГИДР
Тело гидры (рис. 141) разделяется на стебелёк, которым она прикрепляется к субстрату, и собственно тело. Конец стебелька, которым он прикрепляется к субстрату (обычно к растениям), носит название подошвы. Тело гидры имеет внутреннюю полость, называемую /астральной. На свободном конце тела расположено ротовое отверстие ведущее в гастральную полость и окружённое щупальцами, число которых варьирует не только у разных видов, по и у особей одного и того же вида, а длина — как аосолютная, так и отнесённая к длине тела со стебельком — является довольно постоянной величиной и потому служит одним из определительных при-
знаков. Общая длина тела со стебельком, но без щупалец у разных видов различна; в спокойном состоянии животных она колеблется в пределах от 0,2 до 2,0 см.
Р и с. 141. Hydra vu Iga ri s — обыкно-
Размножение гидр происходит двояким путём — бесполым, или вегетативным, и половым. Бесполое размножение совершается в форме почкования (рис. 9). Стенка тела материнского организма вследствие бокового разрастания выпячивается в виде бугорка (почка), который при дальнейшем росте принимает форму цилиндрика, прикреплённого одним концом к материнскому телу; затем на свободном его конце начинают вырастать щупальцы, число которых постепенно увеличивается; в это время на свободном конце, внутри кольца щупалец, обра-
венная гиппа ...........-----------------------------
г ’ зуется рот. Через некоторое время выросшая из почки молодая
гидра отделяется от материнского организма, прикрепляется
к субстрату и начинает самостоятельное существование. Почки никогда не развиваются на стебельке; у Pelmatohydra они располагаются по спиральной линии вдоль всего собственно тела, у остальных родов —только в нижней части последнего. Почкование происходит с начала вегетационного периода до наступления половой зрелости.
При половом размножении на теле гидры развиваются в виде набуханий наружного эпителия (эктодермы) гонады — женские (яичники) и мужские (семенники). Есть виды раздельнополые (Pelmatohvdra oligactis, Hydra attenuate) и гермафродитные (H>dra vulgaris, Chlorohydra viridissima). Половозрелость наступает у Pelmatohydra осенью, у других родов летом.
Организм гидр чрезвычайно сократим; при малейшем раздражении гидра вместе со щупальцами сокращается в маленький комочек.
Перемещаются гидры наподобие гусениц-пядениц или пиявок, поочерёдно прикрепляясь к субстрату то ртом, то подошвой.
К зиме гидры отмирают, а весной развиваются из оплодотворённых яиц.
1. Тело резко отграничено от стебелька, значительно толще его. Щупальца значительно длиннее тела со стебельком — в 3—5 раз, большей частью свешиваются вниз. Почки располагаются на теле по винтообразной линии; сначала на почке появляются лишь 2 щупальца, потом их число увеличивается до 6—7, иногда и больше. Раздельнополы; половые продукты развиваются осенью. Окраска бурая, иногда красноватая. Длина до 3 см. Сидят обычно на нижнем поверхности листьев кувшинковых или рдестов, Pelmatohydra oligactis (Pallas) Schulze — гидра
длинностебельчатая.
0. Тело или равно по толщине стебельку, или очепь постепенно суживается в стебелёк. Щупальца короче тела со стебельком, или равной с ними длины, или длиннее их, но не более чем в 21/, раза. Семенники развиваются лишь у свободного конца тела ... 2
2. Тело зелёное, не резко отделено от стебелька, немного толше его. Щупальца равны Vi длины тела со стебельком; число их различно, иногда очень большое. Гермафродиты; половозрелость наступает летом; яичники крупные, расположены ближе к сте-
1 Есть указания на существование ещё другого отверстия, так называемой «аборальной поры», расположенной на подошве стебелька. Функция этого отверстия пока не выяснена.
Рис. 142. Hydra attenuate — тонкая гидра.
л»льку; семенники мелкие. Длина без щупалец „at см. В чистой воде, па ряске, кувшинковых и других водных растениях.
и ‘ Chlorohydra viridissima (Pallas) — гидра зеленая.
0. Тело не зелёное. Есть и раздельнополые и гермафродиты. Длина до 2 см.
И ”p. Hydra............................
8. Тело постепенно суживается в стебелёк (рис. 141). Щупалец 6—12, длина их в 2—21/2 раза больше тела со стебельком. Почки не менее чем с 4 щупальцами. Гермафродиты; половозре-лость наступает летом; семенники появляются раньше яичников. Окраска бурая и серая. На рдестах и других растениях.
Hydra vulgaris (Pallas) — гидра обыкновенная.
0. Тело и стебелёк у голодных особей одинаковой толщины (рис. 142). Щупальца немного длиннее тела со стебельком, образуют конус.
Раздельнополые; яичники появляются летом. Окраска серая и бурая. На листьях рдестов, кувшинковых и других водных растений.'
Hydra attenuate (Pallas) — гидра тонкая.
ТАБЛИЦ! ДЛЯ ОПРЕДЕЛЕНИЯ В О Т А Т О 111 А — КОЛОВРАТОК
Коловратки (Rotatoria) представляют собой довольно обширную группу микроскопических животных, по своей организации ближе всего стоящих к низшим червям. Преобладающая часть их является обитателями пресных вод, некоторые живут в морских и солоноватоводных водоёмах, наконец, третья группа состоит из обитателей влажных мест па суше.
Сильная сократимость и большая подвижность этих животных очень затрудняют их определение, поэтому перед определением коловраток приходится прибегать к некоторой предварительной обработке их.
Коловраток, тело которых одето панцырем, сравнительно нетрудно определять по панцырю, величина, форма и структура которого дают для этого немало точек опоры; вследствие того, что панцырь в громадном большинстве случаев не поддаётся деформации при воздействии на него фиксирующих веществ, панцырных коловраток можно определять при обычном наиболее удобном способе их консервирования путём прибавления к живой пробе продажного 40-процептного формалина в количестве приблизительно Vio объёма пробы. Но зато определение беспанцырных коловраток наталкивается при формалиновой фиксации на очень большие трудности, так как коловратки при этом сильно сокращаются, втягивая внутрь тела те признаки, на которых основывается их определение.
В этих случаях для замедления или полной приостановки движения животных употребляют ряд способов, из которых приводим здесь наиболее простой и доступный. Взятую пипеткой из банки с водой, в которой находятся живые коловратки, каплю помещают на предметное стекло, прибавляют каплю глицерина, осторожно и тщательно перемешивают концом препаровальной иглы, накрывают покровным стеклом — и препарат готов для микроскопического исследования. Количество глицерина для достижения необходимой степени замедления движения коловраток определяется опытным путём при двух-трёхкратпом повторении указанной процедуры. Если есть возможность Достать какое-нибудь наркотизирующее вещество (хлоралгидрат, кокаин, новокаин и т. п.), то вместо глицерпна употребляют 1—2-процентпый раствор такого вещества. При этом коловратки остаются в растянутом и расправленном состоянии, в котором их можно и законсервировать тем же формалином.
Перехожу к краткому описанию тех признаков организации коловраток, которые положены в основу определения этих животных. Считаю долгом предупредить, что мной сознательно опущен в таблицах один из важных определительных признаков — так называемый мастакс (mastax), или жевательный аппарат, во-первых, вследствие того, что на формалиновом материале он почти никогда не виден, а во-вторых, вследствие очень мелких размеров этого органа, требующих применения больших увеличений микроскопа.
Тело коловраток имеет чрезвычайно разнообразную форму. В большинстве случаев оно двусторонне-симметрично, имеет ясно выраженные передний и задний концы, брюшную и спинную стороны. В нём можно различить голову, туловище и ногу (рис. 14 4). На голове имеется особый орган — коловратный аппарат, служащий для передвижения животного и для привлечения мелкой взвешенной в воде пищи кротовому от во р с т и ю, расположенному внутри коловратного органа; при сокращении голова втягивается в т у л о в и ще, которое сзади или закруглено, или — чаще всего — суживается в более или менее ясно выраженную ногу, почти всегда сегментированную (расчленённую) или, в крайнем случае, поперечнокольчатую; она оканчивается
то присоской, то одним или несколькими пальцами, то венчиком коротких ресничек; у некоторых коловраток, кроме пальцев, на ноге могут быть шипы, так называемые «шпоры»; при сокращении нога может целиком или частично втягиваться в туловище.
Нередко на теле имеются того или иного рода придатки, как ресничные у ш-к и, более или менее длинные шипы — игловидные или веслообразные отростки и пр. Наружные покровы тела представляют или хитиновую, мягкую и гибкую кожу, или более или менее твёрдый эластичный панцырь, прикрывающий обычно только туловище и лишь в редких случаях распространяющийся на голову и ногу; он состоит из одного или нескольких кусков (пластинок), поверхность его может быть гладкой, зернистой, разделённой на поля, на ней могут быть полоски, бороздки, мелкие ямки; часто панцырь бывает вооружён шипами. У некоторых коловраток тело окружено студенистой оболочкой., которая иногда становится более твёрдой в результате отложения на пей посторонних твёрдых частиц.
Коловратный аппарат по своему строению может быть чрезвычайно разнообразен. В своей основной и наиболее простой форме он представляет два концентрически параллельных круга, или венчика (внешний и внутренний) ресничек, окаймляющих передний (головной) конец животного, носящий название кроны; наличие или отсутствие, а также различные формы коловратного органа в случае необходимости будут указаны в определительной таблице.
Следует упомянуть ещё о строении полового аппарата коловраток. Главнейшие черты этого аппарата у самок, имеющие большое значение для определения, следующие: яичники, продуцирующие яйца, и желточники, производящие питательный материал для яиц. Уже в самом начале определения коловратки распадаются по количеству этих органов на 2 группы: с 2 яичниками и 2 желточниками (Bdel-loidea или Digononta) и с 1 яичником и 1 желточником (Monogononta) При дальнейшем определении встречаются случаи, r которых имеют значение качественные признаки, как форма и строение гонад и яиц. Яичник значительно меньше желтом ника и обычно трудно различим. Желточник характеризуется присутствием в нём хорошо видных ядер, число которых бывает различно: у Digononta в каждом желточнике 4, у Monogononta — обычно 8 (за немногими исключениями, например, Asplanchna, у которой их гораздо больше). Яичник и желточник тесно прилегают друг к другу и расположены на брюшной стороне тела под кишечником.
Этими краткими сведениями я считаю возможным ограничиться здесь, так как более подробные указания будут даны в соответствующих местах таблицы. Последняя дана по самкам.
1. Два яичника. Головной конец вытянут в сократимый хоботок (только у Adineta на его месте лишь бугорок). Тело червеобразное или веретеновидное, поперечно-кольчатое (ложная сегментация). Голова и нога могут полностью втягиваться в туловище наподобие частей зрительной трубы. Нога или с 1—4 пальцами, или без пальцев.
Отряд Bdelloidea (Digononta).
0. Один яичник. Хоботка нет. Голова и нога не втягиваются указанным выше образом в туловище. Нога или с 1—2 пальцами, или без пальцев.
Отряд Monogonon ta.
Отряд Bdelloidea
1. Тело вытянутое, веретенообразное. Коловратный аппарат состоит из 2 респичных венчиков. На ноге шпоры, на её конце 3—4 пальца или присоска. Обычно передвигаются по способу
пиявок (прикрепляясь поочерёдно передним* и задним концом тела), реже плавают при помощи коловратного аппарата. Сем. Phi lodi niaae.
Сем. Habrotrochidae
0. Коловратный аппарат сильно редуцирован, представляет лишь небольшую ресничную площадку, расположенную на брюшной стороне головы. Нога с 3 короткими пальцами (рис. 143). Передвигаются почти исключительно путём ползания. Живут на водных растениях и между ними.
Сем. Adinetidae. 1 род Adineta Huds.
Рис. 143. Adineta acu-leata.
Отряд Monogononta
1. Взрослые самки или сидячие (прикреплённые), живущие в трубкообразных чехликах и в студенистых оболочках, или свободно живущие одиночно или в виде колоний, заключённых в студенистый шар. Коловратный аппарат или отсутствует, или имеет форму одинарного или двойного венчика ресничек. Нога, если имеется, лишена пальцев и оканчивается ресничным венчиком........................................*2
0. Животные большей частью свободно живущие, не образующие колоний. Нога, если присутствует, обычно прикреплена к телу на брюшной стороне и несёт на конце 1 или 2 пальца, редко венчик ресничек . . . &
2. Коловратный аппарат состоит из 2 незамкнутых, неполных ресничных венчиков — наружного и внутреннего— с перерывом, по бокам которого оба венчика соединяются своими концами, переходя одип в другой . . &
0. Коловратный аппарат иного строения...........................4
3. Перерыв ресничных венчиков расположен на брюшной стороне коловратного аппарата. Одиночные или собранные в шарообразные колонии плавающие животные.
Сем. С о пос h i I i d a e,
0. Перерыв венчиков расположен на спинной стороне. Животные большей частью сидячие, в чехликах. Сем. F I о s с u I а г i i d а е.
4. Коловратный аппарат двулопастпой. Часть сем, TcstudincHidae.
0. Коловратный аппарат или отсутствует, или снабжён несколькими лопастями или выростами. Сем. С о I I о t h е с i d а е.
5. Тело пе одето панцирем ..................................в
0. Тело одето панцырем.........................................11
в. Нога узкая, с 1 пальцем. Коловратный аппарат сердцевидный, наклопёп па брюшную сторону. Длина 170—204 р.. В оолотах и лужах.
Сем. Microcodidae, 1 род и вид Microcodon clavus Ehrb.
0. Ноги пет или, если есть, то с 2 пальцами.....................7
7. Тело более или менее мешкообразное. Заднепроходного отверстия нет. Яичпик непарный, шаровидный или подковообразный. Сем. Asplanchnidae.
0. Тело различной формы. Заднепроходное отверстие есть..........&
8. Ноги нет. Тело цилиндрическое или яйцевидное, с длинными щетинками.
Часть сем. Т е s t u d i n e 1 1 i d a e.
0. Нога есть. Тело без длинных щетинок ........................ft
9. Коловратный аппарат в виде отдельных пучков ресничек, сидящих на возвышениях ........................................................... 10
0. Коловратный аппарат без возвышений и пучков ресничек, у большинства сдвинут на брюшную сторону. (У некоторых родов есть более или менее ясно выраженный панцирь.) Сем. Notommatidae.
10. Головной отдел тела с боковыми ушками, несущими реснички. е
Часть сем. Synchaetidae.
0. Боковых ушков на теле нет.
Часть сем. Bracbionidae.
11. В коловратном аппарате нельзя различить наружного и внутреннего ресничных колец; этот орган преобразован в-воронку, служащую для припятия пищи; внутренние стенки этой воронки покрыты ресничками, а на дне её находится рот.
Часть сем. Brachionidae.
0, Околоротовое поле не в виде воровки.........................12
12. Спипная пластинка панциря выпуклая, брюшная большей частью плоская или немного изогнутая и меньше спинной; передний край панцыря прямой или вогнутый, с более или меЪее ясно выраженными боковыми остриями, задний край округлённый или притупленный. Нога одно- или двучленистая с 1 или 2 пальцами.
Сем. Lecanidae.
0. Панцирь иного строения........................................
13. Панцирь, если имеется, асимметричный, перасщеплённый (из одного куска), в виде прямой или изогнутой трубки; передний край его с шипами. Нога обычно короткая, пальцы щетпяковидные, прямые или изогнутые, большей частью неодинаковой длины — левый (брюшной) длиннее правого. Сем. Т ri chocercidae.
0. Папцырь или отсутствует, или имеет округлые очертания и лишён шипов. Нога,, если имеется, отходит от брюшной стороны тела. Сим. Gastropodidae.
Сем. Philodinidae
1. Нога с 3 пальцами. На хоботке 2 глаза (рис. 144). Длина 230—1 500 р.. В мелких стоячих водоёмах. р. R о t а г i a Scop. (Rot? Г е г Schr).
Виды: R. tardigrade (Ehrbg) (рис. 144) с длинными пальцами и шпорами; R. п о р t и n i a (Ehrbg) с узким, длинным телом и с ногой, в 1,5—2 раза превышающей длину остального тела; и др.
0. Нога с 4 пальцами ...........................м................
2. Кутикула тонкая. Границы между отделами тела (головой, туловищем и ногой) ясно выражены. Шпоры маленькие (рис. 145). Длина 100—200 р. и выше. Широко рас-пространен. р. Р h i 1 о d i n a Ehrbg.
Виды: Ph. roseola Ehrbg (рис. 145) бесцветная или розоватая; Ph. с i t-г i п я Ehrbg жёлтая; и др.
0. Кутикула утолщённая, иногда напоминающая панцирь; на спине складки, иногда шиники. В водоёмах разных типов. Р- Dissotrocha Bryce.
Вид D. aculeata (Ehrbg) с шипами на спине часто встречается.
Рис. 144. Rotaria tardigrade.
Рис. 146. Conochilus unicornis.
Рис. 145. Phi-lodina roseola.
Рис. 148. Testudi-nella patina.
Рис. 147. Lacinularia flosculosa.
Pnc. 150. Pedalia mira.
Сем. Conochilidae
1. Боковых (брюшных) щупалец 2 или 1, образовавшийся путём слияния двух; расположены они внутри кроны. Самки соединяются в плавающие шарообразные колонии диаметром в 0,5—4,0 мм. Длина особи 300—600 р.. В озёрах и мелких водоёмах.
р. Conochilus Ehrbg.
2 вида: С. unicornis Rouss. (рис. 146) с 1 боковым щупальцем, колонии из 2—25 особей; С. hippocrepis Schr. |v о I v о х Ehrbg) с 2 боковыми щупальцами, колонии из 25—100 особей.
0. Два длинных свободных или частично сросшихся боковых (брюшных) щупальца под кроной. Самки плавают одиночно или небольшими колониями. Длина осоои 280— 500 р. В планктоне озёр и прудов. р. С о п о с h i 1 о i d е s Hlava,
2 вида: С. n a t a n s (Sei.) c 2 длинными отдельными боковыми щупальцами, б. я, плавают одиночно; С. dossuariusf Huds.) с частично сросшимися боковыми щупальцами, одиночные или соединённые в неправильные колонии с небольшим числом особей. 176
Сем. Flosculariidae
1. Крупные шарообразные колонии, прикреплённые или свободно плавающие. Коловратный аппарат сердцевидный. Топкая нога в 2 слишком раза длиннее туловища. Длина особи 1,5—2,0 мм. На водных растениях; плавающие колонии в болотистых водах.
р. Lacinularia Schweigg.
Виды: L. flosculosa (Mill!) (рис. 147)— прикреплённая, L. па tans West. — плавающая.
0. Сидячие отдельные особи или ветвистые колонии. Коловратный аппарат из 4 лопастей: большей верхней (брюшной) пары и меньшей нижней (спинной). Длина особи 0,7—2,0 мм. В прудах, мелких водоёмах.
р. Floscularia Cuv. (Meli с е г t a Schr.).
Широко распространённый видР 1. ri n ge n s (L.) с правильным трубкообразным чех-ликом, густо покрытым шаровидными зёрнышками желтоватого цвета.
Сем. Testudi nellidae
1. Тело одето панцырем ...........................................2
0. Панциря пет....................................................3
2. Панцырь плоский, круглый, эллиптический или яйцевидный, совершенно прозрачный. Поперечно-кольчатая нога без пальцев, вместо которых на её конце вепчик ресничек; от тела нога отходит в середине или в последней трети брюшка. Длина 100—200 pi. В литорали озёр, в прудах.
р. Testudinella В. de St. Vine. (P te го di na Ehrbg).
Виды: T. patina (Mull) (рис. 148) с круглым панцырем, чрезвычайно широко распространена; Т. mucronata (Gosse) с овальным панцырем, спинной передний край которого посередине вытянут в остриё (живёт среди водных растений) и др.
0. Панцырь приблизительно круглый или яйцевидный. 2 глаза. Ноги нет. Длина 70—140 р. В озёрах и прудах.
р. Pompholyx Gosse.
Виды: Р. comp 1 a na ta Gosse (рис. 149) с почти круглым, уплощён пым в спинно-брюшном направлении панцырем, спинной передний край которого посредине несколько вытянут вперёд, очень широко распространена; Р. sulcata Huds. с яйцевидным, сзади заострённым, не уплощённым панцырем, разделённым четырьмя продольными бороздами на 4 выпуклых части.
3. На теле 6 толстых прыгательных придатков: 1 спинной, 1 (самый длинный) брюшной, 2 спинно-боковых и 2 брюшно-боковых; на концах всех придатков перистые хитиновые щетинки. Длина без придатков 350—400 р. В планктоне озёр и прудов.
р. Р е d а 1 i a Barr. (Р е d a I i о n Huds.).
2 вида: Р. mi га (Huds.) (рис. 150) в пресных водах иР. о х у иге Zern. в солоноватых.
0. Прыгательные придатки в виде более или менее длинных тонких щетинок ... 4
4. С 2 передними и 1 задней щетинками. Длина без щетинок 95—204 р. В озёрах, прудах, болотах. р. F i 1 i n i a В. de St. Vine. (T г i a r t h г a Ehrbg).
Несколько видов, из которых наиболее распространён F. longiseta (Ehrbg) (рис. 27) с длинными щетинками (передние в 2—3 раза длиннее тела, задние короче передних).
0. С 2 передними и 2 задними щетинками; в передней паре правая щетинка значительно длиннее левой, в задней паре спинная длиннее брюшной. Длина без щетинок 170— 204 р. р. Tetramastix Zach., 1 вид Т. opoliensis Zach.
Сем. Collothecidae
(В этом семействе 5 родов, из которых 2 не найдены в СССР.
Из 3 остальных даём определительную таблицу для двух.)
!•> Края кроны или гладкие, или в виде лопастей или острых зубцов, на которых сидят длинные жёсткие неподвижные щетинки, а в углублениях между ними большей частью короткие тонкие колеблющиеся реснички. Длина 132—635 р. Живут (за исключением одного вида) в студенистых чехликах; в озёрах и прудах.
р. Collotheca Harr. (Fl oscula ria Ok.).
Из многих видов только 3 являются плапктическими. все прочие сидячие. В планктоне С. р е 1 a g i с a (Rouss.) (рис. 151).
0. Края кроны вытянуты в 5 длинных, узких выростов, усаженных по всей длине параллельными рядами ресничек; длинных жёстких щетинок пет. Самки сидячие, в прозрачных чехликах (рис. 152), длина их 1—1,5лш.На водных растениях в реках, старицах, озёрах, прудах, р. Stephanoceros Ehrbg, 1 вид. S. fimbria (us (Gold/.).
Сем. Asplanchnidae
1. Ноги нет. Длина 0,5—1,5 мм. Широко распространены.
р. Asplanchna Gosse.
Широко распространённый вид A. priodonta Gosse (рис. 27).
0. Нога есть, но слабо развита. Длина 0,6—1,0 мм.
р. Asplanchnopus de Guerne.
Сем Notommatidae
1. Нога прыгательпого типа, вместе с пальцами длиннее тела.......
0. Нога не прыгательная, вместе с пальцами короче тела...........
2/ Пальцы короче туловища и ноги по отдельности. Панцырь топкий (рис. 153). Длина тела 360—425 р. В озёрах, прудах, лужах среди растений.
р. Scaridium Ehrbg, 1 вид S. longicaudatum (Mull.).
0. Пальцы длинное туловища вместе с ногой. Панцыря нет. Длина тела 200—250 р. Среди водных растений. р. Monommata Bartsch.
Из нескольких видов наиболее распространён М. о г b i s (МОП.) с очень длинным пальцем ноги.
3. Пальцы длиннее ’/• длины тела ................................4
0. Пальцы значительно короче 1/6 длины тела......................5
се ю
Рис. 151. Со1-lotheca pela-gica.
A
Рис. 153. Scandium longicaudatum.
4. Панцырь, если имеется, не менее чем из 4 тонких пластинок. Есть и беспанцыр-ные формы. Длина 90—600 р. В литорали водоёмов.
р. Cepbalodella (Diaschiza Gosse).
0. Панцырь из 3 пластинок с точечной поверхностью. Длина 180—220 р.
р. Metadi aschiza, 1 вид М. t г i g о n a (Rouss.).
5. Нога ясно отграничена от туловища. Нога около 1/ь длины тела с 2 короткими пальцами (рис. 154). На коловратном аппарате 2 глаза.
р. Enteroplea Ehrbg (Tri р h у 1 u s Ehrbg), 1 вид E. lacustris Ehrbg.
0. Нога неясно отграничена от туловища........................6
6. Ресничные ушки развиты хорошо, но редко находятся в расправленном состоянии. Коловратный аппарат смещён на брюшную сторону. Длина 230—330 р. На дне, в илу или плавают среди растений.
р. Notommata Gosse.
Виды этого рода многочисленны и очень трудны для определения.
0. Ресничные ушки развиты слабо. Короткая нога с коническими или серповидно изогнутыми пальцами. Коловратный аппарат состоит из одного ресничного венчика. Кутикула в крупных поперечных складках. Длина 140—200 р. В литорали озёр у дна,в болотах.
р. Taphrocampa Gosse.
Сем. Synchaetidae
1. Панцыря нет...............................................2
0. Панцырь с сетчатой или ребристой поверхностью. Основание ноги расположено в середине брюшной поверхности. На коловратном аппарате 2 пальцевидных, лишённых 178
респичек отростка. Длина 150 — 500 р. В планктоне и в лито. рали. Р* Р * ое s о щ a Herrick
Виды: Р1. budsoni Imh. /рис. 155) с сетчатой поверхностью панпыря, прочие виды с ребристой поверхностью и со щелью на брюшной поверхности панцыря.
2. Тело обратноконусовидное или в форме^ колокола или бокала. Коловратный аппарат в виде нескольких отдельных бугорков, покрытых ресничками. На головной части 4 очень длинных, тонких, более или менее изогнутых щетинки и 2 боковых больших мускульных, отчасти втяжных придатка, покрытых ресничками,—так называемые «ушки». Длина 100 — 500 р >. В планктоне стоячих водоёмов.
р. Synchaeta Ehrbg.
Рис. 155. Ploesoma hudsoni.
Виды: S. pectinata Ehrbg (рис. 156) с короткой и широкой конической ногой, 8. s t у 1 a t a Wierz. с вытянутой, сравнительно узкой ногой; и др.
0, Тело почти цилиндрическое, по бокам его по 6 мечевидных или листовидных плавательно-прыгательных придатков, концы которых значительно выдаются за задний конец тела. Ноги нет. Длина 120—160 р. В планктоне стоячих водоёмов.
р. Polyarthra Ehrbg.
2 вида: Р. t г i g 1 a Ehrbg (platyptera Ehrbg) с мечевидными придатками (рис. 28); Р. euryptera wierz. с широкими листовидными придатками.
Сем. Brachionidae
Рис. 157. Brachionus angu-laris, налево — типичная форма,направо—var bidens.
Р и с. 158. Euch- Рис. 159. Му-lanis dilatata. tilina spinigera.
1. Нога есть......................2
0. Ноги нет ......................5
2. Нога кольчатая.................3
0, Нога членистая.................4
8. Панцырь широкий, немного длиннее своей ширины; его передний спинной край всегда с шипами (2—6); из впадин между ними срединная наиболее глубокая; задний край закруглен, с шипами или без них. Нога гладкая или кольчатая; пальцы обычно короткие. Длина панцыря вместе с шинами 100—140 р. В озёрах, прудах, лужах.
р. Brachionus Pallas.
Основные виды; Вг. calyci floras Pall, (р а 1 a Ehrbg) с 4 передними шипами, В г. cap-sulifl о г u s Pall, (b а к е г i МОП.) с 6 передними шипами, В г. angularis Gosse (рис. 157) с 2 передними маленькими шипами посредине, В г. цrceus (L.) (urceolaris Miill.) (рис. 27) с 6 передними шипами, без задних.
0, Панцырь значительно длиннее своей ширины; передний спинной край его с 4 шипами — 2 средние маленькие и 2 боковые длинные; 2 задних шипа равные или неравные. Длина панцыря 280— 320 р. Широко распространена.
р. Schizocerca Dad., 1 вид S с h. di versicornis Dad. (рис. 27).
4« Панцырь состоит из спинной (нерасщеплён-пой) и брюшной пластинок, из которых первая равна или больше второй. Длина панцыря до 380 р. Среди водных растений.
р. Euchlanis Ehrbg.
Наиболее распространённый вид Е. dilatata Ehrbg (рис. 158).
0. Панцырь призматический, на спине расщеплён; передний край его всегда,
1 Указанные признаки хорошо различимы лишь на живых животных; при фиксации формалином тело их часто так сильно сокращается, что превращается в шарообразный комочек с гладкой поверхностью.
задний большей частью с шипами. На спине 1 или 2 киля. Длина панцыря 100—260 р, В литорали озёр, среди зарослей. р. М у t i 1 i n a В. de St Vine.
Обычный вид M. s p i n i f e r a (Ehrbg) (рис. 159).
5. Спинная сторона панцыря разграничена па многоугольные поля; передний край его с 6 шипами, задний или без шипов, или с 1—2 шипами. Длина панцыря 100—300 р. В разнообразных водоёмах. р. К е г a t е 1 1 а В. de St Vine. (A n и г а е a Ehrbg).
2 основных вида: К. quadrate (Mull.) (An. aculeata Ehrbg) (рис. 27) с 2 задними шипами и К. cochlearis (Gosse) (рис. 27) с 1 задним шипом.
0. Спинная сторона панцыря с продольными полосками; спереди 6 шипов, сзади большей частью 1 шип разной длины. Длина панцыря 150—700 р. Главным образом в озёрах, в планктоне. р. N о th о 1 с a Gosse.
Обычный вид N. longispina Kellie, (рис. 27) с длинным задним шипом и 1 (из 6) длинным передним.
Сем. Lecanidae
J. нога с 1 пальцем. Длина панцыря 150—170 р. В разных 2 * у \ водоёмах.
_р. Monostyia Ehrbg. // ни W Много видов, из них очень обычен М. 1 u n а г i з Ehrbg, у
/\ которой очертания передних краев спинного и брюшного панцыря I п ляМИкЦя । вместе напоминают серп луны.
Нога с 2 пальцами. Длина панцыря до 220 р. В разных во-I доёма х.
Р- L е с a n е Nitzsch (С a t h у р n е Gosse-f-D i s t у 1 a Bckst.). \д ду Много видов, из них очень обычен L. 1 u п а Мй11 (рис. 160)
уГ гуЗх с передним краем панцыря, как у М. lunaris.
/ Д\ Сем. Tri с h о се г ci dae
\ \) 1. Оба пальца ноги или одинаковой длины, или разной, при-
' чём короткий обычно длиннее 1/з длинного, который меньше V2 длины
Рис. 160. Le- тела. Длина панцыря 100—200 р. Встречаются в планктоне, в при-cane luna. донном слое воды и среди растений.
р. D i и г е 1 1 а В. de St. Vine.
Значительное число видов, из которых наиболее обычен D. s t у 1 a t а Е у for th с 2 тонкими шипами на переднем краю панцыря и пальцами неравной длины.
0. Недоразвитый палец меньше Vs нормального, который превышает 1/i длины тела. Длина панцыря 140—500 р. Есть виды пелагические и прибрежные.
р. Trichocerca Lam. (R a 11 u 1 u s Lam).
Пелагический вид T. cylindrica (Imh.) с 1 передним шипом и с длинным пальцем, приблизительно равным по длине панцырю.
Сем. Gastropodidae
О
Рис. 161. Gastropua etylifer.
1. С панцырем....................................................2
0. Без панцыря (его заменяет крепкая кутикула). 1 большой красный глаз. Коловратный аппарат состоит из одного простого ресничного венчика. Тело мешковидное, по форме напоминает кувшин. Длина 130—200 р. В планктоне.
р. Ascomorpha Perty.
Обычен вид A. ecaudis Perty с пальцевидным отростком на коловратном аппарате.
2. Нога с 1 или 2 пальцами, отходит от середины брюшной поверхности тела. Панцырь окружает всё тело, за исключением двух отверстий — для головы и для ноги. Длина 88—363 р. В планктоне.
р. Gastropus Imh.
Наиболее распространён G. etylifer Imh. с одним пальцем (рис. 161).
0. Ноги нет. Панцырь из 2 пластинок — спинной и брюшной. Внутри ресничного венчика коловратного аппарата 2 или 4 отростка, несущих реснички. Длина 110—200 р. В планктоне; широко распространён.
р. Chromogaster Lauterb. (A n а р u s Berge nd).
2 вида: Ch. о v a I i s Bergend., у которой спинная
пластинка меньше брюшной и на коловратном аппарате 4 отростка, и Ch. testudo (Lauterb.) с одинаковыми пластинками панцыря и с 2 отростками на коловратном аппарате.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ НАИБОЛЕЕ ОБЫЧНЫХ В СССР BKYOZO А —МШАНОК.
Составил Г. Г. Абрикосов
Рис. 162. Cristatella mucedo. Две колонии на листе кувшинки (в натур, вел.).
Мшанки — колониальные животные (рис. 162, 163), в большинстве своём прпкреп лённые. Колонии состоят из отдельных особей (рис. 163Д). У некоторых представителей особи резко отграничены друг от друга и заключены в отдельные ячейки, у других такого разграничения нет, и они помещаются в общей полости колонии.
Отдельная особь (рис. 163Д) состоит из переднего отдела, могущего выпячиваться, и заднего, лежащего в ячейке или в общей полости колонии. На переднем конце особи расположено ротовое отверстие, окружённое венцом щ у п а-л е ц в виде подковы или кольца (так называемый лофофор). Щупальца покрыты рядами ресничек, находящихся в непрерывном колебательном движении, которое возбуждает ток воды по направлению ко рту и таким образом загоняет в него содержащиеся в воде мелкие питательные частицы. При прикосновении к колонии щупальца мгновенно втягиваются, и колония делается как бы неживой.
Мшанки размножаются как половым, так и бесполым путём. При половом размножении у них в процессе развития образуется личинка, которая у пресноводных форм устроена очень просто, свободно плавает в течение нескольких часов, затем опускается на дно и переходит к сидячему образу жизни; личиночный период длится очень короткое время. Бесполое размножение происходит двумя способами — наружным почкованием, благодаря которому колония растёт, и внутренним. В последнем случае внутри колонии образуются особые внутренние почки — статобласты (рис. 164). Статобласты представляют собой покоящиеся почки, заключённые в твёрдую хитиновую капсулу, благодаря которой они могут переносить неблагоприятнее условия (высыхание, промерзание). У ряда форм статобласты имеют особое приспособление для плавания — плавательное кольцо, состоящее из ячеистой, воздухоносной ткани. Такие статобласты носят название плавающих. При разрушении колоний зимой эти статобласты выпадают наружу, разносятся с водой и служат для расселения данного вида >. Для прикрепления к подводным предметам у статобластов некоторых мшанок
имеются хитиновые крючочки (рис. 1641?). Кроме плавающих статобластов, у некоторых видов образуются прикреплённые статобласты, прирастающие к стенкам колоний. Мшанки обитают в водоёмах различного типа, как стоячих, так и текучих, главным образом на водных растениях.
1. Колонии из очень нежных, тонких трубочек. Отдельные особи резко отграничены друг от друга. Венец щупалец округлый. Статобластов нет.
Paludicella articulate (Ehren.).
0. Колонии различные по форме — разветвлённые (рис. 44), массивные (рис. 163,А—Г), червеобразные (рис. 162). Отдельные особи резко друг от друга не отграничены. Венед щупалец в большинстве случаев в форме подковы. Статобласты имеются.....2
2. Колонии червеобразные (рис. 162), не прикреплённые, медленно ползущие по субстрату. Статобласты крупные, круглые с плавательным кольцом и крючочками (рис. 164 Е). Cristatellamucedo Cuv.
0. Колонии прикреплённые, различные по форме, состоящие из кожистых трубочек.
Статобласты без плавательного кольца или с таковым, без крючочков........8
8. Статобласты без плавательного кольца, овальные или бобовидные (рис. 164 Л).
Венец щупалец округлый. Колонии разветвлённые, частично поднимающиеся с субстрата.
p.Fredericella Gervais, наиболее часто встречается вид Fredericelia sultana (Bl).
0. Статобласты с плавательным кольцом, овальные или удлинённо-овальные. Венец щупалец подковообразный. Колонии различной формы.
р. Plumatella Lamarck..............................................4
В этот род входит ряд видов (в СССР известно 9), здесь даются только наиболее обычные.
4. Колонии образуют массивные наросты на подводных предметах, а иногда и на животных (рис. 163 Б, В, Г). Статобласты широко овальные, довольно крупные (рис. 164 Б). Plumatella fungoea (Pall.) — наиболее обычная, наша мшанка.
1 Плавающие статобласты мшанок довольно часто встречаются в планктоне различных водоёмов.
Р и с. 163. Plumatella fungosa:
А — колонии на стеблях водных растений; Б — колония на анодонте; В — на раке; Г — на лужанке в разрезе; чёрные точки на раковине — прикреплённые статобласты; Д — в увеличенном размере.
О, Колонии разветвлённые, стелющиеся по субстрату (рис. 44) или несколько приподнимающиеся от него ..................................................5
5. Трубочки колонии тонкостенные, просвечивающие. Статобласты небольшие, широко овальные (рис. 164 В). Plumatella repens (L.).
Рис. 164. Статобласты различных мшанок
(увел, в 60 раз; Е — в 35 раз):
А — Frederlcella sultana; Б — Plumatella fungosa; В — Plumatella repens; Г — Plumatella emarginata: Д — Plumatelia frutlcosa; E — Crlstatella mucedo.
0, Трубочки колонии плотные, часто инкрустированные песчинками. Статобласты удлиненно-овальные или удлинённые (рис. 164 Г) ........................в
6. Статобласты удлинённо-овальные.
Plumatella emarginate Alim.
0. Статобласты сильно удлинённые (рис. 164 Д). Plumatella frutlcosa Alim.
Ш
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ OLIGO СП ЛЕТ А — МАЛОЩЕТИНКОВЫХ ЧЕРВЕЙ
Составили. И. Малевич.
Олигохеты, или малощетинковые черви образуют один из классов типа кольчатых червей (Annelides). Они характеризуются сегментированным телом, причём количество сегментов у разных видов варьирует от 7—8 до 200 и больше. Нумерация сегментов ведётся от переднего конца. Сегменты отделяются друг от друга неглубокими перетяжками, межсегментными бороздками, которые обозначаются по разделяемым ими сегментам, например, 5/0, 12/1П и т. д. Иногда, особенно в передней части тела, благодаря наличию добавочных кольцевых бороздок, сегменты приобретают вторичную кольчатость и бывают двух- или трёхкольчатыми. Самые мелкие виды пресноводных олигохет имеют длину всего 1—2 мм, самые крупные — до 200—300 мм. Внешний вид их также очень различен (рис. 165).
Рис. 165. Внешний вид различных олигохет:
а— Aeolosoma tenebrarum; б — Nals pseudobtusa; в—Rtpistes paraslta; e — Ch a e togas ter llmnaei; d — Peloscolex velutlnus.
Сегменты тела несут щетинки, располагающиеся обычно четырьмя пучками на каждом сегменте: 2 брюшных и 2 спинных Первый сегмент (околоротовой, peristomium) никогда щетинок не несёт; таким образом, щетинки всегда начинаются со 2-го сегмента.
На передней стороне первого сегмента, с брюшной стороны, располагается рот. Над ним, выступая вперёд в виде придатка первого сегмента, находится предротовая, или головная, ло па с т ь (prostomium). Обычно она простая, округлая, или несколько удлинённая, реже вытянута в хоботок или, наоборот, редуцирована. Иногда имеются глаза (только у некоторых представителей сем. Naididae).
Пищеварительный канал проходит в виде трубы через всё тело, от рта до анального отверстия; обычно имеется мускулистая глотка и сравнительно топкий пищевод, иногда перед средней кишкои обособляются в виде самостоятельных отделов зоб и мускулистый желудок (например, у дождевых червей). Между кишечником и стенкой тела располагается вторичная полость тела ( ц е л 6 м), поделённая тонкими перегородками, диссепиментами пли септами, па отдельные участки, соответствующие наружной сегментации; в ней находится полостная жидкость и форменные элементы (лимфоциты). Выделительная система представлена мета нефридия ми, парными сегментарными органами. Кровен о с-ная система замкнутая; главные её части — пульсирующий спинной сосуд, непульсирующий брюшной и соединяющие их кольцевые сосуды. Дыхание
у большинства только кожное, у некоторых имеются на заднем конце тела особые кожные жабры (роды Aulophorus и Dero из сем. Naididae).
Центральная нервная система состоит из парного надглоточного ганглия (головного мозга) и соединённой с ним окологлоточными нервными тяжами — копнективами брюшной нервной цепочки.
Половая система (рис. 166) играет очень важную роль в систематике олигохет и при точном определении последних с ней всегда приходится считаться, почему мы и вынуждены уделить ей здесь несколько больше места, чем описанию других органов. Все олигохеты — гермафродиты. Мужские и женские половые железы, семенники и яичники, расположены в немногих сегментах передней части тела, между 5-м и 13-м сегментами. Число и расположение их более или менее характерны и постоянны в пределах каждого семейства. Семенников имеется 1 и 2 пары (редко больше, до 4 пар), яичников! или 2 пары, причём всегда семенники располагаются впереди яичников. Образующиеся здесь половые клетки дозревают либо в полости соответствующих сегментов, либо в особых яйцевых и семенных мешках (пузырьках), представляющих собой мешковидные разрастания диссепиментов, направленные иногда вперёд (первая пара), чаще только назад и простирающихся нередко на несколько сегментов. Выведение зрелых половых продуктов совершается с помощью особых протоков, семепроводов и яйцеводов, открывающихся наружу позади сегментов, несущих соответствующие
Рис. 166. Схема строения полового аппарата тубифициды:
1 — оеменянк; £ и з — семенные пузырьки (мешки); / — воронка семе-провода; 6 —• семевьпюснщий канал; 6 — атриум; 7 —простатическая железа; 8 — оемеприёмник, 9 — яичннн; Ю — яйцевой мешок; 11 — воронка яйцевода? 12 и 13 — мужское и женское половые отверстия; 14 — поясок.
железы и по устройству своему напоминающих мета нефридии. Обычно мужские выводные пути бывают устроены довольно сложно: каждый семепровод, начинающийся мерцательной воронкой в сегменте, несущем семенники, продолжается в следующем сегменте в виде тонкого извитого канала, выстланного мерцательным эпителием, и впадает в атриум, более широкий трубчатый, веретеновидный или грушевидный орган с мускулистой стенкой. Атриум либо покрыт снаружи железистыми простатическими клетками, либо снабжён более или менее компактной простатической железой. Атриумы или, если их нет, семепроводы либо открываются непосредственно наружу, либо (в большинстве случаев) их дистальная * 1 часть превращена в особый копулятивный аппарат, состоящий из способного выдвигаться наружу втяжного совокупительного органа (penis), дистальная часть которого заключена у некоторых видов в прочную кутикулярную пени-альную2 трубку. К половой системе относятся, кроме того, семеприёмники (сперматеки), расположенные или в тех же сегментах, что и половые железы, или впереди их. При спаривании двух взаимно оплодотворяющих друг друга червей в семеприёмники поступает либо просто семенная жидкость с нитевидными сперматозоидами, либо особые образования — спермиоцейгмы, представляющие собой скопление сперматозоидов, соединённых белковой массой и имеющих характерную форму; наконец, у некоторых образуются настоящие сперматофоры, г. е. особые капсулы, наполненные спермой и прикрепляющиеся снаружи к телу партнёра при спаривании.
Важную роль в размножении играет поясок (clitellum), развивающийся у половозрелых червей на определённых сегментах передней части тела и представляющий собой железистое утолщение кожного эпизелия. Клетками пояска выделяется кокон, в который откладываются яйца и в котором происходит их оплодотворение. Изучение половой системы представляет значительные трудности. Некоторые детали удаётся рассмотреть на живых или фиксированных червях, осторожно расщипывая их и препарируя тонкими иглами под бинокуляром или сильной лупой, прибегая иногда к помощи про-
1 Дистальный — дальний, удалённый от начала (наружный).
1 От слова penis — совокупительный орган.
светления глицерином, раздавливания и т. п. (так можно рассмотреть пениальные трубки у Limnodrilus и некоторые другие детали внутреннего полового аппарата). Но для полного исследования половой системы обычно приходится готовить серии срезов па микротоме.
Все олигохеты обладают способностью к регенерации: если, например, червя разрезать па 2 части, то каждая из них превратится вскоре в целого червя, регенерируя хвостовые или, соответственно, головные сегменты. У некоторых, например, у Lumbriculus variegatus, наблюдается самопроизвольное распадение на части, регенерирующие затем в целых червей (своеобразная форма размножения). Наконец, в сем. Аео-losornatidae и Naididae преобладающей формой размножения является бесполое, в виде так называемой паратомии: в средней части червя, между двумя соседними сегментами, возникает зона почкования (зона образования новых сегментов). Передняя часть этой зоны образует хвостовые сегменты передней особи, задняя — головные сегменты задней особи. Некоторое время обе особи (зооида) остаются соединенными, образуя цепочку, потом расходятся. Нередко, прежде чем они разделятся окончательно, в каждой из них или только в одной возникает новая зона почкования; таким образом получаются цепочки, состоящие из трёх, четырёх или пяти особей. Характерно, что при регенерации головного конца всегда образуется определённое число сегментов: головная лопасть и 1 лишённый щетинок сегмент у Aelosoinatidae, головная лопасть и 5 передних сегментов у большинства Naididae. У последних, как правило, эти сегменты лишены спинных пучков щетинок.
Большое значение в систематике олигохет имеют щетинки: их расположение, количество в пучках, форма и размеры. Так как щетинки видны достаточно хорошо под микроскопом на живых и фиксированных червях при лёгком придавливании их покровным стеклом и так как они имеются и у неполовозрелых червей, то этот признак следует использовать прежде всего при определении олигохет. Различают следующие основные формы щетинок (рис. 167.) Волосовидные могут быть простыми, гладкими или усаженными с двух сторон двумя рядами тончайших коротких волосков, видимых только при сильном увеличении (перистые), или нести тонкие шипики только по выпуклой стороне (пильчатые). Волосовидные щетинки большей своей частью выдаются над поверхностью тела и бывают различной длины, от сравнительно коротких до относительно очень длинных (свыше 1 мм). Все остальные типы щетинок — более короткие и крепкие, большей своей частью погружённые в толщу стенки тела и лишь незначительно выдающиеся над поверхностью тела своими дистальными концами. Различают: штифтообраз-
ные, или палочковидные, щетинки, почти прямые, лишь со слабо изогнутым проксимальным 1 и притуплённым дистальным концом (характерны для сем. Enchy-traeidae); игловидные — почти прямые или слабо изогнутые, обычно более тонкие с заострённым простым или слабо раздвоённым дистальным концом и, в большинстве случаев, с небольшим утолщением (подул юсом) около середины; крючковидные — слегка S-образно изогнутые крепкие щетипки с нодулюсом около середины, со слабо пли сильно загнутым дистальным концом, который может быть простым, заострённым или вилообразно раздвоённым, оканчиваясь двумя зубцами — двузубчатые (вильчатые), или содержать между двумя крайними зубцами ряд промежуточных более гонких зубчиков или образованную их слиянием перепонку (г ребе нчатые, пли веерные щетинки). В области половых сегментов брюшные щетинки одной или нескольких пар пучков нередко бывают видоизменены в половые щетинки (сперматекаль-ные, пениальные) 2, отличающиеся от обычных своей формой и размерами.
При определении олигохет приходится также принимать во внимание форму тела (цилиндрическое, уплощённое, четырёхгранное; на поперечном разрезе: округлое, четырёхугольное, трапециевидное), форму головной лопасти и её отношение к первому ссгмепту и, наконец, окраску червей, которая бывает коасноватой благодаря просвечиванию крови (Tubificidae, Lumbriculidae, некоторые Naiuidae) или зависит от нали-
Р и с. 167. Основные типы щетинок у олигохет:
/—3 волосовидные щетинки: 1 — простая, 2 — перистая, 3 — пильчатая; 4 — штиф тообразнал щетинка; 5 — простая игловидная щетинка; б—8 крючковидные щетинки: 6— простая, 7—двузубчатая (вильчатая), 8 —веерная.
1 Проксимальный — ближний (к началу, к месту прикрепления; внутренний).
а От слов: «сперматека» (семеприёмник) и «penis» — совокупительный орган.
чия особых пигментов, нередко сконцентрированных только в передней части тела (желтая. до почти оранжевой, окраска переднего конца многих Naididae), или, наконец, бывает обусловлена особыми кожпыми жировыми желёзками — оранжевокрасными, жёлтыми, зелёными пли синеватыми (Aeolosomatidae). В некоторых случаях тело кажется довольно тёмным благодаря просвечиванию кишечника.
Иди навстречу практической потребности, мы старались построить наши таблицы главным образом на внешних признаках, и только там, где это безусловно необходимо, указываем детали строения полового аппарата *. Благодаря этому таблицы, может быть,
проигрывают в точности диагнозов, становятся несколько громоздкими, но зато выигрывают в доступности. Нужно только, пользуясь ими, проверять все признаки, указанные
в каждой тезе, а также прочитать введение н ознакомиться сначала с характеристиками семейств.
Считаем полезным сделать следующие замечания для предварительной ориентировки в семействах. Branchiobdellidae («рачьи пиявки») резко отличаются от других семейств формой тела, отсутствием щетинок и местом обитания (паразиты раков). Aeolosomatidae — очень малыми размерами, скользящим движением, волосовидными щетинками
Рис. 168. Пучки щетинок раз-
личных энхитреид: не только в спинных, но и в брюшных пучках,
, 9 FrirtPHrij». 1 — бесполым размножением («цепочки»), кожными жи-
1 - Pachydrihjs.^z - raoericu, ровыми желёзками (у большинства). Naididae -небольшими размерами и небольшим числом сегментов, бесполым размножением («цепочки»), в большинстве случаев прозрачностью, часто наличием глаз (которых у представителей других семейств никогда не бывает), наличием (у большинства) различных типов щетинок — двузубчатых (вильчатых) в брюшных пучках, игловидных и волосовидных — в спинных, оживлёнными движениями, способностью многих из них плавать, обитанием преимущественно в прибрежной области. Enchytraeidae небольшими размерами полным отсутствием волосовидных щетинок, наличием во всех пучках только простых (не вильчатых), слабо изогнутых или чаще прямых «штифтообразных» щетинок (рис. 168), вялыми движениями, обитанием преимущественно во влажной почве, среди растительных остатков, по берегам водоёмов. Tubificidae — длинным и тонким нитевидным телом, обычно красноватого цвета, состоящим из большого количества сегментов (до 100 и больше), наличием в брюшных“пучках только крючковидных, у большинства двузубчатых (вильчатых), а в спинных, кроме того, иногда и волосовидных щетинок, обитанием преимущественно на дне водоёмов. У всех названных семейств (кроме Branchiobdellidae) количество щетинок в отдельных пучках, как правило, больше двух, иногда до 10 и более. Наоборот, у следующих четырёх семейств количество щетинок в отдельных пучках не превышает двух. Lumbriculidae и Phreo-ryctidae имеют тело длинное и сравнительно тонкое, Glossoscolecidae и Lumbricidae — довольно толстое. Щетинки у Lumbriculidae или двузубчатые, причём верхний зубец всегда меньше нижнего, или простые. У остальных трёх семейств — всегда простые.
Таблицы составлены применительно к более распространённым олигохетам средней полосы Союза. При их составлении не учитывались виды, имеющие ограниченное распространение, встречающиеся только в Байкале, и недостаточно ещё изученная фауна олигохет окраин Советского Союза, в частности Восточной Сибири и Дальнего Востока.
1. На заднем конце тела мускулистая присоска. Щетинки отсутствуют. Глотка с кутикулярными («хитинозными») челюстями. Длина до 12 мм. По внешнему виду напоминают пиявок. Паразитируют на теле пресноводных раков («рачьи пиявки»).
Сем. Branchi obdellidae (Discodriliaae) р. Branchiobdella Odier.
0. Присоски на заднем конце нет. Хвостовой отдел червя простой или видоизменён в жаброносный диск (у немногих видов). Щетинки, как правило, имеются.......2
2» Мелкие и очень мелкие черви от 1—2 до 15—20 мм длиной, редко немного больше. Обычно прозрачные, светлые, реже красноватые. Размножаются преимущественно бесполым путём, образуя «цепочки». У некоторых имеются глаза. В каждом пучке, как правило, больше двух щетинок, иногда до 10 и больше...........................8
0. Черви небольшие, средних размеров или более крупные (от 5—20 до 100—200 мм и больше). «Цепочек» никогда не образуется. Глаза всегда отсутствуют.......4
8. Очень мелкие черви длиной обычно от 1 до 4—5 мм. Цепочки редко достигают 10 мм. Тело состоит не больше чем из 12—13 сегментов, обычно меньше. В брюшных и спинных пучках, начинающихся со 2-го сегмента, имеются волосовидные щетинки (один вид — A. neizwestnowae Last. — совершенно лишён щетинок). Диссепимепты отсутствуют или имеется только передний. Кожа, как правило, содержит небольшие округлые жировые желёзки краснооранжевого (наиболее обычная у нас A. h е m р г i chi. Ehrenb., а также A. quaternarium Ehrenb.), жёлтого, зелёного или голубоватого цвета, иногда бесцветные. На нижней стороне головной лопасти ресничный покров и по бокам головы две мерцательные ямки. Глаз нет. Спинной сосуд развит только в перед-
1 Точные диагнозы семейств и родов, а в значительной части также и видов, основываются на строении внутренних органов, прежде всего половой системы. 186
ней чести тела. Поясок образуется неполный, только на брюшной стороне, па 5—7-м сегментах. Семенники в 5-м, яичники в 6-м, семеприёмники в 3—5-м сегментах.
Сем. Aeolosomatidae, р. А е о I os о m a Ehrbg 1.
0. Мелкие черви, длиной от 1—2 до 15—20 мм. Число сегментов от 8—10 до 40—50, в отдельных случаях несколько больше. В цепочках число сегментов в передней особи (до зоны почкования) обычно не превышает 20. Волосовидные щетинки если есть, только в спинных пучках. В брюшных пучках крючковидные, двузубчатые (вильчатые) щетинки. Спинные пучки у немногих видов отсутствуют вовсе, у большинства видов они начинаются с 6-го сегмента, у некоторых с 5-го, 4-го или 2-го. Диссепименты имеются. Спинной сосуд развит по всей длине тела. Кожных жировых желёзок нет. У многих представителей семейства есть глаза. Семенники в 5-м (у одного рода в 7-м), яичники в 6-м (8-м) сегментах, семеприёмники — в 5-м (7-м) сегменте. Поясок развит у половозрелых червей не только на брюшной, но и на спинной стороне сегментов, несущих половой аппарат.
Сем. Naididae.
4. Небольшие черви, обычно 5—35 мм длиной, в отдельных случаях мельче. Волосовидные щетинки всегда отсутствуют. Как спинные, так и брюшные пучки начинаются со 2-го сегмента и содержат только прямые или слегка изогнутые щетинки без нодулюса, с простым (не вильчатым) заострённым или притуплённым дистальным концом (рис. 168). Только у одного рода — Propappus Mich. — щетинки двузубчатые с более коротким верхним зубцом 2 * * * * *. Семеприёмники — в 5-м сегменте, семенники, как правило, в 11-м, яичники в 12-м. Иногда половые железы сдвинуты на 3—4 сегмента вперёд. Обычно беловатые или слабо желтоватые, реже красноватые черви с довольно плотными покровами. Большинство видов обитает во влажной, богатой перегноем почве. Многие виды, однако, встречаются и в водоёмах, преимущественно болотах, а также в прибрежных зарослях и среди мхов, в озёрах, лужах и т. д.
Сем. Enchytraeidae 8.
О, Черви более крупные, от 10—15 до 200 мм и больше. Волосовидные щетинки имеются или отсутствуют. Семеприёмники, если имеются, расположены не в 5-м сегменте, а дальше кзади............................................................. 5
5. Мужские половые отверстия расположены на 12-м сегменте или ближе кпереди, но всегда в области пояска. Довольно тонкие и длинные, иногда очень длинные черви, чаще красноватого цвета, живущие в илу или в прибрежных зарослях............в
О, Мужские половые отверстия расположены на 15-м сегменте, реже на 13-м; как правило, они хорошо заметны, в виде поперечных щелей, расположенных на небольших железистых бугорках (папиллах). Они находятся всегда впереди пояска. В каждом из четырёх пучков данного сегмента не больше двух простых (не вильчатых) щетинок, имеющих нодулюс и слегка S-образно изогнутых. Пресноводные или (большинство) наземные черви довольно крупные, толстые (2—6 мм толщиной и больше), непрозрачные .... 8
6. ,В каждом пучке, как правило, больше двух щетинок, иногда до 10 и больше. Брюшные пучки содержат только крючковидные щетинки, двузубчатые (вильчатые) или (редко) простые. Спинные пучки, кроме того, могут содержать и волосовидные щетинки. Семенники — в 10-м, яичники в 11-м сегментах. Тонкие нитевидные красноватые черви, небольшие или средних размеров; длина их обычно 20—70 мм, иногда несколько меньше или больше —до 100—150 мм. Живут главным образом в илу на дне водоёмов. Многие строят трубочки из ила (отсюда название «трубочники»). Обычно передним концом погружены в ил, задний поднимают над поверхностью ила и совершают им характерные колебательные (дыхательные) движения. Встречаются также на песчаных и каменистых грунтах рек. *
Сем. Tubificidae.
0. Щетинки расположены попарно в четырёх пучках, т. е. по 8 щетинок на каждом сегменте, или поодиночке, т. е. по 4 на каждом сеИкенте, или, если спинные редуцированы, по 2 на сегмент.............................................................7
7. В каждом пучке по 2 щетипки, т. е. всего по 8 щетинок на сегмент. Щетинки крючковидные или с простым заострённым концом, или вильчатые. В последнем случае верхний зубец меньше нижнего. Головная лопасть простая, округлопродолговатая или вытянута в небольшой хоботок. Мускулистого желудка нет. Мужские половые отверстия на 8-м или 10-м сегменте, у представителей рода Stylodrilusua конце наружных (невтяжных) совокупительных органов. 1 или 2 пары семенников, расположенных в области 7—11-го сегментов, 1 или 2 пары яичников в области 9—12-го сегментов, от 1 до 5 пар семеприёмников. Длина от 20 до 140 мм. Тело обычно более плотное, чем у тубифц-
1 Реже, только в реках встречается у нас второй род этого семейства Potamo-
drilus .Last. Он характеризуется чрезвычайно сближенными спинными и брюшными
пучками щетинок, почти сливающимися с каждой стороны в один пучок, и отсутствием
жировых желёзок в коже, а также ресничных ямок по бокам головы.
а В реках, на песчаных грунтах и на перекатах характерен Р а о р а р р u s volki Mich.,
встречающийся часто и характеризующийся, помимо формы щетинок, вытянутой в не-
большой хоботок головной лопастью. По размерам (5—6 мм) он напоминает наидид.
8 Таблицы для определения родов сем. Enchytraeidae мы не даём, так как вести определение, пользуясь только внешними признаками, невозможно, а изучение внутреннего строения, требующее изготовления срезов на микротоме, является делом для неспециалиста практически недоступным.
пвд. Ползают по дну примерно так же, как дождевые черви по земле. Нередко обвиваются вокруг стеблей мхов и других водяных растений. Иногда образуют па дне большие скопления вместе с тубвфицидами. Некоторые могут плавать, совершая энергичные движения толчками, причём тело извивается наподобие бича.
Сем. Lumbriculidae.
0. В каждом пучке только по 1 щетинке, следовательно, по 4 на сегмент, если же спинные отсутствуют, то по 2. Спинные щетинки, если имеются, значительно меньших размеров, чем брюшные. Все щетинки слабо S-образно изогнуты, с простым (не вильчатым) дистальным концом. Имеется мускулистый желудок в 4—6-м сегментах. Семенники в 10—11-м сегментах, яичники в 12-м и 13-м. Очень длинные и тонкие (до 300 мм) черви, напоминающие по внешнему виду волосатиков. В ручьях, источниках, реках, озёрах. Сем. Phreoryctidae (Haplotaxidae) р. Phreoryctes Hartm., 1 вид Ph. gordioides (Hartm.).
8- Головная лопасть зиголобическая (слита с первым сегментом). Спинных пор нет. Щетинки расположены парами (по 2 в пучке, по 8 на сегмент), простые, не вильчатые, почти прямые, но с сильно изогнутым дистальным концом. Поясок бывает развит только периодически, нерезко отграничен от соседних сегментов, с 16-го по 30—35-й сегмент. Мускулистый желудок слабо развит, рудиментарный. Семеприёмники отсутствуют. Крупные (до 320 лмс), похожие на дождевые черви. У встречающегося изредка у нас в реках и других водоёмах, в том числе и солоноватых Criodrilus 1а-с u u m Hoffm. тело на поперечном сечении трапециевидное, обычно буроватого или оливкового цвета.
Сем. Glossoscolecidae, р. Criodrilus Hoffm.
0, Головная лопасть эпи- или танилобическая (обособлена; со спинной стороны её выступ вдаётся до середины или конца первого сегмента).Спинные поры обычно имеются. Поясок у половозрелых червей имеется постоянно, расположен позади мужских половых отверстий так, что между ними и передним краем пояска имеется во всяком случае больше одного сегмента. От соседних сегментов поясок отграничен, как правило, резко. Щетинки слегка S-образно изогнуты, с простым (не вильчатым) дистальным концом, по 8 на каждом сегменте, иногда сближены попарно, иногда расставлены широко, почти на равном расстоянии друг от друга. Хорошо выраженный мускулистый желудок расположен в 17-м или (чаще) в 17—18-м сегментах. Семеприёмники обычно имеются (2—3 пары в области 8—11-го сегментов), иногда отсутствуют. Средних размеров или крупные черви, до 300 мм длиной и до 8 мм толщиной (настоящие дождевые черви). Держатся преимущественно в почве, вне воды, и только некоторые виды попадаются по берегам водоёмов, среди мхов, частично под водой.
Сем. Lumbricidae1.
Сем. Naididae
1. Спинные пучки отсутствуют на всех сегментах. В брюшных пучках вильчатые крючковидные щетинки. На 3-м,Ч-м и 5-м сегментах отсутствуют и брюшные щетинки. Головная лопасть развита слабо, имеет вид небольшого округлотреугольного выступа первого сегмента или рудиментарна, так что рот занимает почти терминальное (конечное) положение. Глотка развита сильно, пищевод не длиннее глотки. Глаз нет. Тело большинства видов очень прозрачно. Длина от 1—2—4 мм (Ch. 1 a n g i Bretsch, С h. d i a-etrophus Gruith.) до 10—15лсж (Ch. diaphanus Gruith. (рис. 129). Среди прибрежных зарослей рек, озёр, прудов и других водоёмов. Как правило, — хищники, питающиеся мелкими ракообразными, нематодами, коловратками и пр. Один вид живёт на пресноводных моллюсках, питаясь в значительной мере церкариямн (Ch. И m n а е i К. von Baer, рис. 165 г. р. ChaetogasterK. von Baer (7 видов).
0. Спинные пучки щетинок имеются хотя бы на части сегментов.......2
2. Все спинные пучки лишены волосовидных щетинок, содержат только вильчатые или простые, крючковидные, или игловидные щетинки.....................8
0. В спинных пучках, кроме крючковидных или игловидных, имеются ещё и более длинные и тонкие волосовидные щетинки.................................5
8» Третий сегмент удлинённый, значительно длиннее следующих за ним. Головная лопасть округлотреугольная. Глаз нет. Спинные пучки начинаются с 3-го сегмента. Во всех пучках — спинных и брюшных — содержатся однотипные равнозубчатые, слегка изогнутые тонкие крючковидные щетинки, обычно по 3—4 в пучке. В самых передних сегментах количество щетинок меньше. Очень мелкие, 1—1,5 мм длиной, бесцветные, прозрачные, довольно подвижные черви. Преимущественно на песчаном грунте, в реках, реже — в озёрах, встречаются также в прибрежной области морей, до 20°/00 солёности. р. Amphicnaeta Tauber, 1 вид А. I е у d i g i Tauber.
0. Третий сегмент по своей длине не отличается заметно от остальных сегментов. Спинные пучки щетинок начинаются со 2-го, 5-го или 6-го сегментов.....4
1 Так как Lumbricidae являются по преимуществу обитателями почвы, мы не даём таблицы для определения их родов. Около воды и в воде чаще всего встречается Е i в е-niella tetгaёdra (Sav.), f. typica, характеризующаяся средними размерами (длила 30—50 мм, толщина 2—4 мм), буроватой или оливковой окраской, четырёхгранным телом, попарно сближенными щетинками, положением мужских половых отверстий на 13-м сегменте в пояска с 22-го или 23-го по 26-й или 27-й сегмент.
(P. litoralis) преимущест-
I
Рис. 169. Ophidonais serpentina: а — головной конец; б — дистальный конец одной из спинных щетинок.
б
4. Щетинки в спинных и брюшных пучках более или менее одинаковые, вильчатые, крючковидные, слегка изогнутые. Головная лопасть развита хорошо, округлая. Глаза чаще имеются, иногда отсутствуют. Кишечник образует заметное расширение в 8-м сегменте. Спинные пучки начинаются со 2-го (Р. n a i d i n a Bretscn.), 5-го (P. 1 i t о r a-1 i s MUll.) или 6-го (P. u n c i n a t a Orsted) сегмента. Длина 5—18 мм. На илисто-пес-чапых грунтах озёр и рек, среди водяных растений. Один вид ~ воппо в прибрежной области морей, до 2О°/оо солёности, по указан также и для пресных вод.
р. Paranais Gzerniavsky (3 вида).
0. В спинных пучках, начинающихся с 6-го сегмента, только прямые, довольно толстые, игловидные щетинки с еле заметно раздвоённым концом. В брюшных пучках изогнутые крючковидные вильчатые щетинки обычного типа. Головная лопасть округлая. Глаза есть. Окраска буроватая, поверхность тела в значительной мере покрыта оболочкой из инородных частиц, благодаря чему червь почти совершенно непрозрачен. На каждом сегменте 2—3 ряда поперечных, светлых, округлых чувствительных бугорков с волоском на каждом. На 2—5-м сегментах со спинной стороны поперечные тёмнокоричневые перевязки. Кишечник образует расширение в 8-м, 9-м или 10-м сегментах. Длина 7—35 мм. Преимущественно в водоёмах с заиленным дном (рис. 169).
р. Ophidonais Gervais, 1 вид О. serpentina Mull.
5. Спинные пучки начинаются со 2-го сегмента. Каждый спинной пучок содержит простые или тонковильчатые, игловидные щетинки и более длинные, тонкие волосовид-
простые или тонковильчатые, игловидные щетинки и более длинные, тонкие
Haemonais щетинки;
Рис. 171. waldvogeli, а — брюпшан щетинка 2-го сегмента; б — то же 40-го сегмента; в и г — спинные щетинки 30-го сегмента.
Рис. 170. Передние концы тела различных видов р. Pristina:
а — Pristina longiseta; б — Р. aequlseta; в — Р. bllobata; а — Р. ГогеИ.
ные щетинки, в большинстве случаев усаженные с одной стороны очень тонкими, мелкими зубчиками. У одного вида (Р г. longiseta Ehrbg) волосовидные щетинки 3-го сегмента значительно длиннее, чем щетинки всех остальных сегментов (до 0,7 мм). В брюшных пучках только слабо изогнутые, тонкие крючковидные щетинки с двузубчатым концом. Глаза отсутствуют. Головная лопасть либо округлённая, либо более или менее вытянутая в щупальцевидный хоботок. Длина 2—15 мм. Чаще в заболоченных и торфяных водоёмах, среди мхов и водяных растений (рис. 170).
1 р. Pristina Ehrbg. *(8 видов)
1 Род Pristina некоторыми специалистами разделяется на 2 рода: собственно Pristina, характеризующийся более или менее вытянутой в хоботок головной лопастью (Р г. longiseta Ehrbg, Рг. aequiseta Bourne, Р г. f о г е 1 i Piguet), и род Naidium, у представителей которого головная лопасть округлая, не вытянутая в хоботок.
0. Спинные пучки щетинок начинаются дальше кзади............................в
6. Спинные пучки щетинок начинаются только с 12—20-го сегмента или ещё дальше. В каждом спинном пучке содержится обычно по одной игловидной и волосовидной щетинке, редко их по две. Игловидные щетинки сильно изогнутые, довольно толстые, двузубчатые, похожие на брюшные крючковидные. Волосовидные щетинки тонкие, довольно короткие (0,14—0,16 мм). В брюшных пучках по 2—3 двузубчатых (вильчатых) крючковидных щетинки. Головная лопасть удлинённая. Глаз нет. Окраска коричневатокрасная. Кровеносная система в передней части тела имеет сложное строение. Длина 5—12 мм. Число сегментов 50—60 (рис. 171).
р. Haemonais Bretscher, 1 вид Н. waldvogeli Bretsch.
Рис. 172. Aulophorus furcatus, задний конец тела.
Рис. 173. Задние концы тела (жаброносные диски) различных видов рода Dero:
1 — Dero perrterl; 2 — Dero obtusa; 3 — Dero limosa; 4 — Dero incisa; & — Dero digltata; 6 — Dero mlchaelsenl.
0. Спинные пучки щетинок начинаются с 4-го, 5-го или 6-го сегмента...7
7. Задний конец тела расширен и преобразован в особый диск, несущий жабры . . 8 0. Задний конец тела простой, жабер нет..............................9
8. Жаброносный диск, кроме настоящих жабер, несёт на своём нижнем конце пару довольно длинных расходящихся в стороны придатков (пальпы), несущих осязательные волоски. Каждый брюшной пучок содержит по 4—5 двузубчатых крючковидных щетинок. Спинные пучки — с 5-го сегмента, в каждом по одной вильчатой и одной волосовидной щетинке. Длина последних не превышает поперечника тела. Глаз нет. Окраска тела желтоватокрасная. Кишечник образует заметное расширение Хв в 8-м сегменте. Длина 6—12 мм Число сегментов до 35. Живут в иле на
ЗЬй дне водоёмов и отчасти на водяных растениях, строят трубки, из которых
Jg высовывают задний конец с жаброносным диском (рис. 172).
р. Aulophorus Schmarda, 1 вид A. furcatus Ok.
0, Жаброносный диск несёт только жабры, пальцевидных придат-ков (пальп) нет. Число жабер варьирует от двух пар (D. obtusa < JS n Udek.) до четырёх пар (D. d i g i t a t a Mull., D. t u b i с о 1 a Pointner, iCy D. michaelseni Svetl). Головная лопасть обычного вида, округлая. чет Глаз нет. Брюшные пучки со 2-го сегмента, содержат крючковидные дву-и \ зубчатые щетинки, по 2—5 в каждом пучке. Спинные пучки начинаются ’ или с 6“го сегмента (у большинства видов), или с 4-го (у D. t u b i с о 1 а)
и содепжат каждый обычно по одной волосовидной и 1—2 игловидных или < - двузуочатых щетинки. Окраска в большинстве случаев красноватая.
Длина 6—18 мм. В иле на дне водоёмов, отчасти среди водяных растений (рис. Р- D е г о Океп (5 или 6 видов).
< 9. Волосовидные щетинки спинных пучков длинные, более чем в 2
IWk Раза пРевышают поперечник тела. По всей своей длине или только в ди-। стальной части они усажены вдоль выпуклого края мелкими зубчиками.
Сами щетинки крепкие, саблевидно изогнутые, по 4—9 в каждом пучке. Кроме них, в спинных пучках имеются и простые игловидные щетинки. JOpSL Брюшные пучки со 2-го сегмента содержат крючковидные щетинки с про-ксимально расположенным нодулюсом. Брюшные пучки или имеются во всех сегментах у речного*вида (V. i ntermedia Bretsch), или от-сутствуют в 5-м у наиболее широко распространённого, характерного для болот вида (V. с о m a t a Veju.), или в 4-м и 5-м сегментах (V. m а-\ crochaeta Lst). Головная лопасть обычного вида, округлённая. Рис. 174. Глаза имеются или отсутствуют. Прозрачные, бесцветные или слегка Vejdosvkyel- желтоватые мелкие черви длиной 3—8 мм. Обилие длинных и крепких
la comata. саблевидных перистых щетинок в спинных пучках придаёт червю очень
характерный «щетинистый» вид (рис. 174).
р. Vejdovskyella Michlsn (3 вида).
0. Волосовидные щетинки спинных пучков гладкие (без зубчиков).......10
10. Головная лопасть вытянута в щупальцевидный придаток (хоботок) или снабжена длинным нитевидным щупальцем............................................. И
0. Головная лопасть простая, округлая или несколько удлинённая, но не вытянутая в щупальцевидный придаток................................................13
11. Головная лопасть широкая, спереди несколько изогнутая (вырезанная), от середины её переднего края отходит длинное, резко отграниченное подвижное нитевидное 190
Рис. 175. Slavina appendiculate, передний конец.
3—7 в каждом пучке, двузубчатые, крючко-от Rinistes. на 4-м и 5-м сегментах. ”
щупальце. Глаза всегда имеются. Передний конец тела пигментирован, от светложёлтого до жёлтобурого цвета. Остальное тело прозрачное, бесцветное, с хорошо просвечивающим тёмным кишечником, образующим в 8-м (инод да 7-м) сегменте заметное расширение. Брюшные пучки со 2-го сегмента содержат но 4—5 тонких, довольно длинных изогнутых крючковидных щетинок с длинным, сильно изогнутым верхним зубцом и коротким, почти рудиментарным нижним. Спинные пучки с 6-го сегмента, с одной длинной, почти вдвое превышающей поперечник тела волосовидной щетинкой (рядом с которой часто имеются ещё 1—2 более коротких волосовидных) и несколькими простыми игловидными. Характерно расположение длинных волосовидных щетинок почти под прямым углом к продольной оси тела. Длина до 15—18 мм. Подвижные животные, хорошо плавают. Часто в прибрежной области различных водоёмов, среди зарослей и на грунте (рис. 128).
р. Stylaria Lamarck, 1 вид S. lacustris L.
0. Головная лопасть постепенно переходит спереди в довольно тонкий, умеренной длины хоботок.........................................................12
12. Спинные пучки 6-го, 7-го и 8-го сегментов содержат большое количество (от 5 до 15 в каждом) очень длинных, достигающих 1,5 мм длины, волосовидных щетинок. В последующих сегментах волосовидные щетинки короткие — 0,14—0,16 мм длины. Все спинные пучки содержат, кроме того, несколько коротких тонких игловидных щетинок. Брюшные щетинки, начиная со 2-го сегмента, по 3—6 в каждом пучке, кроме 4-го и 5-го сегментов, которые совершенно лишены их. Глаза есть. Передний конец тела, до 8-го или 9-го сегментов, как правило, имеет жёлтую или оранжевую (до красноватой) окраску. Животные обитают обычно в построенных ими слизистых трубках на поверхности подводных частей водных растений, свай, коряг и т. п. Часто, преимущественно в сильно зарастающих водоёмах. Могут плавать, иногда попадаются в планктоне. Длина до 7 мм (рис. 165 в).
р. R i р i s t е s Dujardin (2 или 3 вида).
0. Спинные пучки 6-го, 7-го и 8-го сегментов несут такие же волосовидные щетинки (не увеличенные), как и пучки последующих сегментов, длиной 0,2— 0,39 мм, по 10—12 щетинок в пучке. В спинных пучках имеются также очень тонкие и короткие (до 0,06 мм), лишь с трудом различимые игловидные щетинки, до 12 в каждом пучке. Брюшные щетинки: начиная со 2-го сегмента, по видные. Они имеются также, в отличие
8 мм. Редко, преимущественно в реках, в прибрежной области, встречается и в озёрах.
р. Arcteonais Piguet, 1 вид А. 1 о ш о n d i Martin.
13. Волосовидные щетинки 6-го сегмента значительно длиннее, чем в последующих сегментах, в 3—4 раза превышают поперечник тела (до 0,80 jwjw), по 1—3 в пучке. Кроме волосовидных, во всех спинных пучках по 1—2 простых (не вильчатых) игловидных щетинки; в каждом брюшном пучке по 1—5 двузубчатых крючковидных щетинок. Головная лопасть короткая, округлая. Глаза обычно имеются. Окраска светлокоричневая. Тело почти непрозрачное благодаря оболочке из мелких инородных (иловых) частиц. Кожа с осязательными папиллами (сосочками), расположенными кольцами на каждом сегменте. До 20 мм длиной*. Движения обычно вялые. Встречается довольно часто, преимущественно в зарастающих водоёмах с илистым грунтом, особенно в болотах, канавах и т. п. (рис. 175).
р. Slavina Vejdovsky, 1 вид S. appendiculate Udek.
0. Волосовидные щетинки 6-го сегмента не отличаются заметно по своей длине от щетинок последующих сегментов; они вообще тонкие, гладкие (без зубчиков); длина их меньше, равна или немного (в редких случаях в 2 с небольшим раза) превышает поперечник тела. Кроме того, в спинных пучках имеются игловидные щетинки с простым заострённым концом (N. о b t u s a Gervais, N. pseudobtusa Piguet), или с двумя очень маленькими, иногда плохо заметными даже при сильном увеличении зубчиками на конце (большинство видов), или, наконец, вместо игловидных имеются двузубчатые крючковидные щетинки, такого же типа, как и набрюшных пучках (N. b 1 a n ci Piguet). Брюшные пучки содержат двузубчатые крючковидные щетинки, в передних четырёх сегментах обычно более тонкие, длинные и слабее изогнутые, чем в последующих; верхний зубец этих щетинок у некоторых видов (особенно yNais bebningi Michlsn) длиннее и сильнее изогнут, чем нижний. Головная лопасть простая, округлая. Глаза у большинства имеются. Передний конец тела у многих видов окрашен в желтоватый, буроватый или красноватый цвет. Кишечник в 7-м или 8-м сегменте расширяется либо постепенно (N. communis Piguet, N. е 1 i n g u i s MiilL), либо резко, сразу (N. v a r i a b i 1 i s Piguet, N. par-da 1 i s Piguet). Наиболее богатый видами род (у нас И или 12 видов), представители которого часто встречаются в прибрежной области различных водоёмов, среди зарослей, отчасти в грунте. Многие хорошо плавают. Некоторые виды свойственны преимущественно рекам (N. bebningi Michlsn, N. bretscheri Michlsn, N. b I a n c i
4
Рис. 176. Щетинки различных видов рода Naie.
1. Nals bland: а— спинная; б— брюшная щетинки 6-го сегмента. 2. Nais bretscheri: а — спинная щетинка 6-го сегмента; б — брюшная 2-го; в — брюшпая 6-го; г — брюшная 8-го сегментов. 3. Nals behningl, две щетинки из брюшного пучка 2-го сегмента. /. Nals pardalis: а — спинная щетинна 6-го сегмента; б — брюшная 2-го; в — брюшная 6-го сегмента. 5. Nals elinculs: а — спинная щетинка 6-го сегмента; б — брюшная 2-го; е— брюшная 6-го сегмента. 6. Nais communis, обозначения те же. 7. Nals varlablls, обозначения те же. S. Nals simplex, обозначения те же 9. Nais pseudobtusa, обозначения те же.
10. Nais obtusa, обозначения те же.
Piguet), другие — ключам (N. е 1 i n g ui s Mull.), остальные присущи озёрам, прудам или канавам и низинным болотам (N. с о m m u n i s Pig.) (рис. 176).
p. Nais Mull.
Сем. Tubificidae
1. В брюшных пучках много (до 10—13) довольно тонких, крючковидных двузубчатых шетинок, верхний зубец которых заметно меньше и много тоньше нижнего; в спинных пучках либо только двузубчатые шетинки такого же типа, как в брюшных, но с несколько меньшей разницей в величине зубцов (А. 1 i ш п о b i u s Bretscner), либо наряду с ними имеются и довольно тонкие, короткие волосовидные щетинки до 8 в каждом пучке (A.pluriseta Piguet). Черви бледяокрасноватого или желтоватого цвета. Длина 10— 20 мм, редко больше. 3 пары семенников в 4—6-м сегментах, мужские половые отверстия на 7-м сегменте. Не часто, преимущественно в прибрежной зоне рек и озёр.
р. A u 1 о d г i 1 u s Bretscher.
0. В брюшных пучках сравнительно немного щетинок (обычно 3—5, редко больше, до 8—10). Брюшные двузубчатые щетинки, как правило, или с равными зубцами, или с более длинным верхним зубцом; если же, как это имеет место у немногих видов, верхние зубцы щетинок короче нижних, то сами черви относительно крупные и более толстые, чем Aulodrilus, или число щетинок в каждоу пучке меньше. 1 пара семенников в 10-м сегменте, мужские половые отверстия на 11-м сегменте. Длина 15—80 мм и больше (обычно 30— 50 мм).....................................................................2
2. Волосовидные щетинки в спинных пучках отсутствуют; спинные и брюшные пучки содержат только одинаковые двузубчатые крючковидные щетинки, зубцы которых более или менее равпой величины (L. h о f f m е * ticus Piguet), или верхний длиннее нижнего (L. L. claparcdeanus Ratzel), или верхний немного короче нижнего (L. parvus South.), или оба зубца укорочены, причём верхний короче нижнего (L. п е-w а е n si 8 Michlsn, L. m i с h а е 1 s е n i Last.). Penis почти у всех видов снабжён особой, довольно крупной, прозрачной кутикулярной трубкой, хорошо заметной при раздавливании живых половозрелых чсовей или при просветлении глицерином фиксированных. Отношение длины этой трубки к её наибольшей ширине (у проксимального конца) является хорошим видовым признаком (3—4 :1 у L. udekemianus, 5— 6:1 у L. h el-vet i с u s, И—12 : 1 у L. h о f f m е i 8 t e r i, 23—30 : 1 у L. cl a p a rede a n us). Длина до 130 мм. Встречаются часто (рис. 177).
р. Limnodrilus Clap. (8 видов).
0. Волосовидные щетинки имеются (за одним исключением); спинные двузубчатые щетинки в передних сегментах заменены гребенчатыми или веерными щетинками .... 3
3. Поверхность тела, начиная со второго сегмента, за исключением пояска, густо покрыта кутикулярными папиллами, благодаря чему тело кажется почти непрозрачным. Кроме того, на каждом сегменте имеются расположенные кольцевыми рядами осязательные папиллы (сосочки). Голова и первый сегмент могут сильно втягиваться. В брюшных пучках либо крючковидные двузубчатые (вильчатые) щетинки (у Р. f е г о х Eisen), либо простые или неясповильчатые (у более редкого Р. velutinus Grube). В спинных пучках двузубчатые щетипки (очень мелкие, почти незаметные у Р. velutinus), причём в передних сегментах они имеют
isteri Clap., L. helve-udekemianus Clap.,
а б
Рис. 177. Пениальные трубочки некоторых видов рода Limnodrilus: а — L. udekcmlanus; б — L. helveticus; в — L. hotfmcisteri, г — L. claparcdeanus.
характер веерных; кроме того, до семи довольно крепких волосовидных щетинок в каждом пучке. Длина 25—40 мм. Нередко (рис. 165 Д).
0, Поверхность тела более или менее гладкая, без кутикулярных и осязательных папилл ................................. 4
4. Семепроводы много короче атриумов, почти рудиментарны. Атриумы в виде длинной, довольно широкой, изогнутой коленом (рис. 178) трубки. Простатическая железа маленькая или отсутствует. Втяжной penis лишён кутикулярной трубки. В брюшных пучках крючковидные двузубчатые щетинки обычного типа; в спинных пучках передних сегментов крючковидные щетинки гребенчатого типа, в средней и задней части
р. Peloscolex Leidy.
Рис. 178. Мужские выводные протоки llyodrilus hammonie-nsis:
а — воронка ссмепровода; б — простата; в — атриум; г — penis; д — мужское половое отверстие;
е — семеп ровод.
kJ Р
Рис. 179. Щетипки Jlyodrilus hammoniensis: а и б — спинная и брюшная щетинки 3-го сегмента; в — спинная щетлнка 5-го сегмента; г — сперматекальная щетинка.
тела — обычного типа. Кроме того, имеются волосовидные щетипки (они отсутствуют у редкого речного вида I. m ol da vie n si s Vejd. et Mrazek). У нас в различных водоёмах, преимущественпо на илистых грунтах, чрезвычайно широко распространён 1. hammoniensis Mich., выдерживающий значительное загрязнение (мезосапроб) и часто встречающийся вместе с Tubifex tubifex Mull., от которого он почти неотличим в неполовозрелом состоянии; половозрелые черви легко различаются по наличию у I. hammoniensis в брюшных пучках 10-го сегмента более крупных
13 А. Н. Липин 193
ложкообразных половых (сперматекальных) щетинок, отсутствующих у Т u b i f е х t и b i f е х. Длина 20—50 мм (рис. 178, 179). В озёрах встречается и па значительной глубине.
р. Ilyodrilus Eisen.
0. Семепроводы длинные, значительно длиннее атриумов. Простатическая железа массивная. Penis втяжной, снабжённый небольшой кутикулярной («хитинозпой») трубкой. В брюшных пучках крючковидные двузубчатые щетинки обычного типа; в спинных пучках имеются волосовидные щетинкп и крючковидные вильчатые; последние в передних сегментах представлены гребенчатыми пли веерными щетинками (рис. 180). Длина 20—100 мм и больше. Преимущественно на илистых грунтах как в прибрежной, так и в открытой части различных водоёмов. Один из принадлежащих сюда видов Т. t u-bi fe х (Mull.) является чрезвычайно широко распространённым, выдерживает значительное загрязнение и встречается нередко массами, сплошь и рядом вместе cLimno-drilus noffmeisteri и Ilyodrilus hammoniensis. G последним очень сходен по внешним признакам (см. выше). р. Т u b i f е х Lamarck (6 видов).
Рис. 181. Sty-lodrilus herin-gianus: передний конец тела и спинная щетинка 6-го сегмента.
Рис. 180. Щетинки Tubifex albicola:
2С зсбс — спинные гребенчатые щетинки соответствующих сегментов; 2$ — брюшные щетинки соответ-
ствующих сегментов.
Сем. Lumbriculidae
1. Головная лопасть вытянута в более или менее длинный щупальцевидный хоботок. Число сегментов до 200. Тело полупрозрачное, розоватокрасного цвета с фиолетовым оттенком; на поперечном разрезе в передней части круглое, в средней и задней — трапециевидное. Задний конец несколько уплощён. Щетинки крючковидные, простые (не вильчатые), с несколько притуплённым концом, по 2 в каждом пучке. Мужские половые отверстия на 10-м сегменте. Встречающиеся у пас в средней полосе вид R. 1 i m о s е 1 1 а Hoffm. является степоту>мно-холодолюбивым, попадается преимущественно в родниках, ключах, ручьях и т. п. Половое размножение преимущественно зимой. Длина 80—140 мм, толщина 2-3 мм. Встречается не часто. р. Rhynchelmis Hoffm.
Головная лопасть округлая, не вытянута в хоботок...............2
2. Мужские половые отверстия открываются на вершине наружных (не втяжных) совокупительных органов (penis), расположенных на 10-м сегменте и представляющих собой прозрачные трубчатые образования, постепенно утончающиеся к дистальному концу (рис. 181). Небольшие, довольно подвижные желтоватые или слегка красноватые черви. Во всех пучках по 2 щетинки, простых или неясно раздвоённых, с маленьким рудиментарным верхним зубцом. Длина 25—40 мм. Не часто. Среди корней растений, в иле. Преимущественно в ручьях, ключах, реках.
р. S t у 1 о d г i 1 u s Clap., у нас более или менее обычен 1 вид S. heringi anus Clap.
6. Наружных (невтяжных) совокупительных органов пет. Мужские половые отверстия на 8-м сегменте. Во всех пучках по 2 крючковидных щетинки, верхний зубец которых меньше нижнего. Черви очень подвижные и ломкие; часто даже небольшого раздражения бывает достаточно, чтобы они распались на части (автотомия), из которых каждая восстанавливается в целого червя. Эта особенность приобрела у данного вида значение основного способа размножения: половозрелые особи встречаются крайне редко. Черви способны плавать, быстро изгибаясь. Окраска довольно тёмная, краснобурая, кольчатость резко заметна, благодаря концентрации зеленоватого пигмента в диссепиментах. Длина 40— 194
80 л<.и, толщина 1 —V/g.u-w. Число сегментов до 200. Встречается часто, преимущественно в прибрежной зоне различных водоёмов, особенно небольших — лужи, канавы, пруды, болота и т. п., как в иле на дне, так и среди водяных растений, особенно мхов.
р. Lumbriculus Grube, у пас 1 вид L. variegatus Mull. L
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ HIRUDINEA — ПИЯВОК
~ Тело пиявок подразделено поперечными бороздками па узкие кольца; эти кольца не соответствуют внутренней сегментации, так как каждый сегмент подразделён па несколько колец. На обоих концах тела (редко на одном только заднем) имеются присоски; задняя присоска всегда крупнее передней.
Рот расположен на брюшной стороне, на переднем конце тела. Стенка ротовой полости образует ротовой диск.
Глот к а иногда образует способный к выпячиванию хоботок — хоботные пиявки (рис. 183) иногда передний отдел глотки бывает снабжён 3 зазубренными челюстями (рис. 184 А) — челюстные п и я в к и. Иногда глотка без челюстей, длинная и очень мощная, с трёхграппым просветом, ограниченным тремя мощ-
ними мускульными тяжами в стенке глотки — глоточные пиявки 2.
Желудок у некоторых пиявок расширяется за счёт его сегментально расположенных боковых выпячиваний (хоботные и челюст-пые пиявки — рис. 190).
la peledina.
Рис. 185. НегроЬ-della—расположение глаз.
Рис. 183. Rhyncho-bdella — хоботок.
Рис. 184. Hirudo:
А — разрез глотки; Б — расположение глаз.
Рис. 186. Glossiphonia heteroclita: а — в цельном виде; б — расположение глаз.
Анальное отверстие располагается близ заднего конца на спинной стороне и окружено обычно задней присоской. По бокам задней части тела иногда имеются ряды наполненных кровью пульсирующих пузырьков (рис. 187, 188). Тело способно всегда к очень сильному сжатию и сокращению. Приводимые ниже в определительной таблице размеры относятся к пиявкам, находящимся в покое.
1. Передней присоски нет; на 2—6-м сегментах по две пары щетинок. Глаз 3 пары; глаза 1-й пары сближены, 2-й и 3-й — раздвинуты к краям тела. Тело из 30 сегментов. Пиявки мелкие (рис. 182).
Подотряд Acanthobdellae, сем. Acanthobdellidae, р. А с antbob d е 1 1 а Grube.
0. Передняя присоска (рис. 182) имеется, щетинок нет, тело более чем из 30 сегментов ...............................................................2
1 Мы не включаем в таблицу рода Lain prod ril us, представленного целым рядом видов в озере Байкал, по в средней полосе Европейской части Союза до последнего времени не отмечавшегося. Один из видов этого рода недавно был указан для озёр я рек севера Европейской части Союза.
9 Чтобы видеть строение глотки, надо разрезать её, введя кончик тонких ножниц в ротовое отверстие, и развернуть.
2* Есть хоботок (рис. 183). Глаз 1—4 пары, расположенные едва параллельных ряда (редко ряды позади расходятся). Желудок с парными сегментально расположенными придатками. Тело или плоское и широкое, или очень узкое — вытягивается почти в нить.
Подотряд Rhynchobdellae — хоботные пиявки.................в
0. Хоботка нет. Тело слабо уплощённое. Глаз не менее 4 пар...8
8. Глотка без челюстей, с 3 продольными крупными мускульными валиками. Как ' правило, 4 пары глаз, расположенных в 2 изогнутых поперечных ряда (рис. 185), но иногда число их увеличено или уменьшено.
Подотряд Pharyngobdellae — глоточные пиявки...............5
0. Глотка с челюстями, без продольных валиков, 5 пар очень мелких глаз; между 3-й и 4-й парами промежуток в одно кольцо (рис. 184Б).
Подотряд Gnatholbdellae — челюстные пиявки, сем. Н i г п-dinidae ............................................... 4
4. Челюсти маленькие. Около 20 тупых зубов в два неправильных ряда. Спина без продольных полос. Питаются кровью червей, личинок насекомых и др. (в стоячих и текучих водах, особенно в прудах). р. Haemopis Sav.
0. Челюсти хорошо развиты; около 100 острых зубов в один ряд (рис. 184). Вдоль спины красножёлтые полосы. Питаются кровью водпых позвоночных, особенно млекопитающих (предпочтительно в торфяных болотах). р. H i г u d о L.
Рис. 187. Piscicola geometra:
А — в цельном воде; Б — передняя присоска отдельно; В — часть тела с боковыми пузырьками.
Р и с. 188. Cystobranchus respirans:
А — общий вид; Б — часть тела с пульсирующими пузырьками.
5. Сегменты из 6 колец; 4-е из них уменьшенное. Иногда одно или два последних кольца раздвоены, так что число колец в сегменте доходит до 8. Тело одноцветное, только иногда вдоль спины тёмная полоска. Размер (80—140) X (6—И) мм. Питаются кровью" червей и личинок насекомых (в болотах и речках). Trochaeta Dutr
0. Сегменты из 5 колец или равных, или 5-е крупнее прочих. Тело или одноцветное, или с рисунками в виде полосок или пятен. Тело значительно короче (не более 60 мм) и относительно толще. Питаются кровью червей и других животных (в болотистых водах, мелких канавах). Л
' р. Herpobaella Blain.
6» Тело узкое (в сокращённом состоянии длина его по крайней мере в 5 раз превышает ширину), почти равноширокое, утолщённое; только передний отдел (*/4 или 1/л) несколько уже. Передняя присоска хорошо развита и резко отграничена. Покровы тела гладкие, без бородавок. По бокам тела 11 пар пульсирующих пузырьков. Одна или две пары глаз на ротовом диске. Паразитируют на рыбах и ракообразных; паразиты постоянные, покидают хозяина только перед кладкой яиц.
Сем. Ichthyobdellidae — рыбьи пиявки......................7
0. Тело широкое и плоское, имеет форму овала (рис. 186), при сокращении длина его не более чем в три раза превышает ширину; передняя присоска не резко отграничена. Покровы тела или гладкие, или с бородавками. Пульсирующих пузырьков пет. Яйца и молодые пиявки прикрепляются к брюшной стороне матери. Глаз от 1 до 4 пар. Под камнями и на растениях; питаются кровью разных животных (водных беспозвоночных и позвоночных); ползают, но никогда не плавают.
Сем. Glossiphoniidae — плоские пиявки ....................8
7. Пульсирующие пузырьки мелкие, часто едва заметные. Тело цилиндрическое, суживается постепенно (рпс. 187). Длина более чем в 15 раз превышает ширину. Сегменты 196
из 14 колец. 2 пары глаз па ротовом диске. Задняя присоска яйцевидная, с 14 тёмными лучами и тёмными крупными точками, напоминающими глаза. Паразитируют на щуке, окупе, налиме, сиге и др.; хорошо ползают и плавают.
р. Р i s с i с о 1 a Blain (Ichthyobdella Blain.)
0. Пульсирующие пузырьки крупные, ясно заметные (рис. 188). На задней присоске нет резких точек. Тело уплощённое. Передний отдел довольно резко отличается по ширине. На передней присоске 1 или 2 пары глаз. Сегменты из 7 колец. Тело сплющенное, длина его лишь в 5—6 раз превосходит ширину, ширина вдвое превосходит толщину (рис. 188). Паразитируют на форели, хариусе, палимо и др.
р. Cystobranchii9 Dies.
Рис. 190. Hemiclepsis margin ata:
Л — нид co спины; Б — с просвечивающим кишечником; В—расположение глав.
Рис. 192. Batrachobdella paludosa: расположение глаз.
8. Через 10 колец кзади от глаз на спине тёмная пластинка. Спина беловатая, иногда усеянная мелкими коричневыми точками, п тогда коричневатая или сероватая. Желудок с 6 парами боковых выступов, без добавочных карманов. Глаз 1 пара (рис. 189). Форма мелкая: 5 х (10 — 4) лме. Питаются кровью червей и моллюсков.
р. Н е 1 о b d е 1 I a Blanch.
0. Без тёмной пластинки на спине. Окраска различная. Желудок большей частью более чем с 6 парами боковых придатков, причём придатки с добавочными карманами. Глаз 2, 3 или 4 пары..........................................................9
9. Ротовой диск отграничен, хотя и не резко. Спипа зеленовато- или красповатоко-ричповая с жёлтыми пятнами и с 4 продольными рядами бородавок. 2 пары глаз, вторая из них значительно крупнее (рис. 190 В). Размеры: (25 — 30) X (4 — 5) мм. Питаются преимущественно кровью рыб и тритонов. Hernielen^is Void
0. Ротовой диск не отграничен...........................................(0
10. 4 пары глаз (рис. 191). Тело очень мягкое, или с 4 продольными рядами мелких бородавочек на фоне жёлтых пятен, или без бородавочек, с неправильными жёлтыми
Рис. 194. Наешсп teria (Placobdella) — расположение глаз.
Рис. 193. Glossiphonia complanata:
А — в цельном впде; Б — с просвечивающим кшиеч-ником; В — расположение глаз.
Рис. 195. ВогеоЬ-della verrucata — положение сосочков.
пятнами. Тело у взрослых плоское п широкое. Питаются кровью водо. плавающих птиц (в реках и озёрах)
р. Protoclepsis Livan
0. Менее 4 пар глаз . . . . п
11. Тело мягкое, почти гладкое, только лишь с чуть заметными бородавочками. 2 пары глаз, более или менее слитых (рис. 192). Питаются кровью амфибий (в болотах).
р. Batrachobdella Vig.
0. Тело всегда с ясно заметными бородавочками .......... jg
12. Бородавочки в 4 продольных ряда; глаз 1 или 3 пары ... 13
0. Бородавочки в 6—8 или много продольных рядов. Желудок с 6 парами боковых придатков без карманов. Глаз 3 пары; у одних они расположены в два параллельных ряда, у других 1-я пара сближена почти до соприкосновения (рис. 193). Питается как Helobdella.
р. Glossiphonia Joh.
(С 1 е р s i n е Sav.).
13. Тело плотное. 4 продольных ряда крупных бородавок и
1 пара крупных сближенных глаз (рис. 194). р. Н а е m е п
2 ряда мелких по краям тела. В реках.
teria Filip. (Placobdella Fil.).
0. Тело очень мягкое. По обе стороны от срединной линии тела по 4 продольных ряда бородавок, расположенных следующим образом: бородавки 1-го, 3-го, и 4-го рядов (считая от середины тела к краям) на одном кольце, бородавки 2-го ряда — на следующем (рис. 195). 3 пары глаз в два параллельных ряда. Питаются кровью моллюсков (на берегах озер и прудов, на камнях и растениях). pBoreobdella Job.
в
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ MOLLUSCA — МЯГКОТЕЛЫХ
Таблица эта составлена на основании строения раковины. Раковина состоит то из о д н о г о к у с к а (у л и т к и — рис. 196), то из д в у х створок (ракушки — рис. 216). Входное отверстие, или устье, раковины улиток бывает или свобод н о (у дышащих атмосферным воздухом), или прикрыто крышечкой (у дышащих растворённым в воде воздухом). Если держать раковину верхушкой кверху и устьем к себе, то последнее может оказаться или направо, или налево от продольной оси раковины; соответственно и раковина будет называться завитой вправо (рис. 199—203) или з а-витой влево (рис. 197, 198). Внутри раковины но её оси проходит образованный примыкающими друг к другу частями внутренних стопок раковины столбик (веретено), пронизанный каналом; вход в него снаружи, называемый пупком, далеко не у всех улиток ясно выражен.
Створки раковины ракушек соединены на спинном крае посредством эластической связки, стремящейся своим постоянным натяжением держать их всё время раскрытыми; этому препятствуют находящиеся вблизи узких краёв раковины два м у с к у л'а- з а-мыкателя, с помощью которых раковина может закрываться наглухо. У некоторых ракушек по краю раковины внутри — там, где створки соединяются, — имеются выступы (зубцы) и углубления—так называемый замок (рис. 214 Б). От выступающего на каждой створке бугра, называемого верхушкой, на наружной поверхности раковины идут параллельные друг другу и краю раковины полосы.
Короткие зубцы в передней1 части замка называются кардинальными; удлинённые складкообразпые зубцы, идущие от кардинальных зубцов кзади, называются латеральными. Складка кожи, выстилающая внутреннюю поверхность створок ра-
ковины, называется мантией.
1. Раковина цельная, не створчатая. Класс Gastropoda — брюхоногие......................... 2
0. Раковина из двух симметричных створок. Класс Bi valvia — двустворчатые.......................18
2. Раковина без крышечки. Отряд Pulmonata — лёгочные.............................8
0. Раковина с крышечкой. Отряд Prosobranchi а — переднежаберные................10
1 Передним называют более широкий, тупо закруглённый конец раковины.
3. Раковина в форме шапочки или щита, без завитка, с заострённой верхушкой, загнутой несколько назад и наклонённой набок (рис. 196). В стоячих и текучих водах Европейской части СССР и Кавказа.
Сем. Ancylidae, р. А п с у 1 из Mull..— чашечка.
0. Раковина завитая...........................................б
4. Раковина с приподнимающимся завитком....................4
0. Раковина завита в одной плоскости, так что имеет форму диска; только ипогда средняя часть слегка приподнята. Устье никогда не бывает круглым.
Сем. Planorbidae............................................8
5. Раковина завита влево, без пупка, тонкостенная, прозрачная. Завиток более или менее острый. Устье удлинённо-овальное. Формы довольно мелкие: высота не более 17 мм.
Сем. Physidae...............................................в
0. Раковина завита вправо, различной формы. Сом. Limnaeidae.............................................7
6. Завиток низкий, раковина вздутояйцевидная (рис. 197). В стоячих водах, а также в небольших речках и ручьях бассейна Днепра, Дона, на Кавказе и в Средней Азии.
р. Physa Drapar. — пузырчатая улитка.
0. Завиток высокий, раковина веретенообразная (рис. 198). В болотах, лужах и за-росших ручьях,- по всему СССР. р А р ] е х а Пет.
7. Раковина пузыревидная, с 3—Ь оборотами; первые обороты образуют маленький, слабо приподнятый завиток, последний оборот очень крупный. Устье широкое, округлояйцевидное. Раковина очень нежная, топкая, просвечивающая, светлояптарная. Мантия заворачивается на раковину и обволакивает её снаружи почти нацело, так что раковина живого животного
слизистая, скольз-
Рис. 196.
A — Ancylus lacustris; fluviatilis.
Б — Ап.
Р и с. 197. Physa fontiualis.
Рис. 198. Ар-lexa hypnorum.
Р и с. 199. Ат-phi peplea gluti-nosa.
кая. Высота раковины до 15 мм, устье—до 13 мм (рис. 199). В озерах, прудах и лужах всей Европейской части Союза и в бассейне р. Оби.
р. A m р hi peplea Nilss. — плащеносная улитка.
0. Раковина различной формы, большей частью башпевидная или уховидная. Завиток различной высоты. Устье большое, округлое или овальное. Раковина тонкостенная, но не очень прозрачная. Мантия не загибается на раковину (рис. 200). В стоячих водах разных типов, по всему Союзу. р L j rn n а е а Lain. —прудовик.
8. Раковина завита в одной плоскости, завиток не приподнят (рис. 201). В стоячих водах, почти по всей территории СССР.
Подсем. Planorbinae, р. Planorbis Mull. — катушка.
0. Завиток приподнят ....................................... 9
9. Если держать раковину верхушкой кверху, устье будет справа; устье относительно большое, различной формы. Диаметр до 11 мм. Верхушка тупая. Формы байкальские.
Подсем. Choanomphalinae, р. Choanomphalus Gorst.
0. Если держать раковину вершиной кверху, устье будет слева. Верхушка острая. Устье продолговатое, почти эллиптическое, высота его более высоты завитка. Диаметр 4 мм, В бассейне р. Амура. Подсем. Bulininae, р. Glyptophysa Cross, 10. Раковина низкая, с очень маленьким завитком, с наклонностью к двусторонней симметрии, с резким рисунком, большей частью пёстро окрашенная (рис. 202). Кры-
Р и с. 201.
Л, Б — Planorbis planorbis; В, Г — Pl. vortex; Д — Pl. septemgyratus; Е, Ж — PI. conturlus; 3, И — Pl. corneus; К. Л — PI. albus; М, Н, О — Pl. complanatus; 77, Р, С — nitidus; Т — Pl. crista; У — Pl. carinatus.
шечка известковая, спиральная, большей частью полукруглой формы, с отростком. Наибольший диаметр до 13 мм. В реках бассейна Чёрного моря.
Сем. Neritidae, р. Theodoxus Mont. — лунка.
0. Раковина резко не двусторонне симметричная, редко с пёстрым рисунком. Крышечка никогда не полукруглой формы.......................................11
11. Раковина кубаревидная, иногда дисковидная. Устье или округлое, или угловатое сверху. Крышечка всегда роговая, концентрическая . . 12
Рис. 203.
А— Viviparus contectus; Б— Viv. vlviparus.
Рис. 202. Theo-doxus fluviatilis.
0. Раковина яйцевидная, яйцевидноконическая, — конусовидная или башневидная; крышечка спиральная, роговая, за исключением одного рода Bithynia, где опа концентрическая, но известковая. Устье сверху угловатое......................13
12. Раковина кубаревидная. Устье и крышечка сверху угловатые, крышечка коп-, центрическая. Высота до 40 мм (рис. 203). В мелких стоячих водах, в реках и пойменных водоемах. Сем. Viviparidae, р. Viviparus Mont. — живородка.
0. Раковина кубаревидная или дисковидная, завиток расположен в одной плоскости.
Устье и крышечка круглые, крышечка вдавленная, ядро её лежит центрально (рис. 204). В разнообразных стоячих и текучих водоёмах по всему Союзу.
Сем. Valvatidae, р. V а 1 v ata Muller — затворка.
18. Раковина башвевидная, удлинёнпоконическая........................14
0. Раковина яйцевидная или яйцевиднокоиичоская............................16
14. Раковина удлипённо-овальноконическая, с низким завитком. Нижний край 5стья с лункой. Большей частью чёрная (рис. 205). В бассейне Днепра, Южного Буга и Днестра, па Кавказе, в ручьях Черноморского и Каспийского бассейнов, в Средней Азии. р Melanopsis Fer. — чернушка.
0. Раковина башневидная. Устье без лунки..................................15
15. Раковина твёрдостенпая, с рёбрами; крупная — до 45 мм (рис. 206). В бассейне р. Амура. р. Melania Lam.
0. Раковина тонкая, просвечивающая, бледножёлтая, очень мелкая—до 1,6 мм (рис. 207). В Аджаристане. р. Bythinella М. Tand.
Рис. 204.
А — Valvata pisclnalls; Б, В, Г — V. pulchella; Д — V. crlstata.
Рис. 205.
А — Melanopsis espert; Б — М. aclcularls.
16. Крышечка концентрическая, известковая. Высота до 15 мм (рис. 208). В стоячих и текучих водоёмах. р. Bithynia Leach.
0. Крышечка спиральная, роговая, с малым количеством оборотов. Высота до 9 мм..........................................................................17
17. Раковина высокая, яйцевидноконусовидная, с острой верхушкой. Устье яйцевидное (рис. 209). Высота до 5 В пресных и солоноватых водах. р. Hydrobia Hartm.
p. Hydrobia Hartm.
p и с. 206, Melania amuren-sis.
Рис. 207. Bythinella adscha-rica.
Рис. 209. Hydrobia Steini.
Рис. 208.
A — Blthynia tentacula-ta; Б — P. leachl.
0. Раковина округлая, с низким завитком. Устье косое, круппое, грушевидноокруглое (рис. 210). В бассейнах южных рек Европейской части СССР.
р. Lithoglypbus Hartm.
18. Раковина трёхгранпая. Замок из одно1ю слабого кардинального зуба па одной створке и соответствующей ему ямки на другой. Прикрепляется к подводным предметам посредством пучка роговых нитей — б и с с у с а (рис. 211). В реках и некоторых озёрах Европейской части Союза, в Каспийском и Аральском морях.
р. Dpeissena v. Bened.
0. Раковина не трехгранпая. Замок или совершенно без зубов, или с зубами двоякого рода — кардинальными и латеральными (боковыми); только у одного рода имеются одни лишь кардинальные зубы в число 3, крупные........................19
19. Раковина крупная, до 95 мм. Верхушка приближена к более широкому концу.
Сем. Unionidae............................................20
0. Раковина мелкая, не крупнее 20 мм. Верхушка или на середине, или ближе к бо-
лее заострённому концу.
Сем. Sphaeriidae...............................................24
20. Раковина без зубов на замке...................................21
0. Замок с зубами.................................................22
21. Замка нет совсем (рис. 212). В стоячих водоёмах.
р. Anodonta Cuv. — беззубка.
0. Рудиментарный замок имеется.
р. Pseud anodonta Bourg. — ложнобеззубка.
22. Имеются только кардинальные зубы, боковых же пет (рис. 213). В бассейне Амура в реках Карелии, Кольского полуострова, Архангельской области.
р. М а г g а г i tana Schumach. — жемчужница.
0. Замок из кардинальных и боковых зубов..................23
Рис. 210. Lithoglyphus naticoides. polymorpha.
23. Раковина очень узкая и длинная, приблизительно в 5 раз длиннее ширины. Верхушка на !/з длины. Йа Дальнем Востоке в бассейне Амура и в Приморье.
р. Lan ceolaria Gonrad.
0. Раковина более короткая, овальной или клиновидной формы (рис. 214). В реках Европейской части СССР, на Кавказе, в Крыму, в Закавказье, на Дальнем Востоке.
р. U n i о Retz. — перловица.
24. Длина раковины большей частью больше 10 мм. Верхушка в середине раковины, выдаётся мало (рис. 215). В водоёмах разных типов по всему СССР.
р. Sphaerium Scop. — шаровка.
0. Длина раковины мепее 10 мм. Верхушка большей частью смещена из центра (рис.
216). В реках в стоячих водоёмах (болотах, пересыхающих лужах).
р. Pisidium С. Pfeiffer — горошинка.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ГЛАВНЕЙШИХ ГРУПП ВОДНЫХ ARTHROPOD А — ЧЛЕНИСТОНОГИХ
Три большие группы водных животных — насекомые (Insecta), паукообразные (Arachnoidea) иракообразные (Crustacea) — принадлежат к типу членистоногих животных — A rthropoda. Как показывает самое название, этот тип характеризуется присутствием у его представителей ра счле нённых, т. е. состоящих из члеников, ножек. Тело у громадного большинства их тоже явственно или скрыто расчленено, причем число члеников, или сегментов, из которых оно состоит, у разных групп членистоногих очень разнообразно. Обычно в теле этих животных можно различить два или три главных отдела. Так, у насекомых тело ясно разделено па три части — голову, грудь и брюшко, или аб доме я; у пауков, относящихся к паукообразным, голова и грудь так тесно сливаются между собой, что по отдельности их различить невозможно, и потому получается только два отдела тела — головогрудь и б р ю ш к о; v другой группы паукообразных — клещей — слиты не только голова и грудь, но и брюшко, так что тело состоит лишь из одного отдела; наконец, у большинства ракообразных —так же как у пауков — наблюдаются две части тела — головогрудь и брюшко. Каждый из отделов тела состоит из нескольких сегментов, которые или явственно различимы, или, напротив, совсем неразличимы. Так, у насекомых па голове совершенно невозможно усмотреть разделение на сегменты, па груди же и, в особенности, на брюшке — довольно легко; особенно ясно это разделение можно видеть у многих ракообразных как на головогруди, так и — ещё лучше — на абдомене. Однако и в тех случаях, когда никаких члеников различить нельзя (как, например, в голове), разделение па сегменты в скрытом виде всё же существует. Одной из существенных сторон организации членистоногих является то, что каждый сегмент тела несёт на себе не более одной пары членистых конечностей; поэтому, если последние сохраняются, то, даже в случае слияния сегментов тела, по числу пар конечностей можно судить пе только вообще о разделении на сегменты, но и оо их числе. В частности, по отношению к голове нужно заметить, что у членистоногих все членистые придатки головы — усики, или антенны, и ротовые органы — считаются, на основании истории развития, видоизменёнными конечностями, а потому по ним можно судить о разделении головы на сегменты.
Рис. 215.
Рис. 216.
Л Б - Plsldlum pusilium; 8 — Р. millum; Г — ₽• д;.
3 — Р henslowauum; И— Р. casertaoum, К «
J г. neusiuYvauu • н _ р subtruncatum; О — р. pulcheliuin.
Е — Р. suplQuni; — р. ten ui line a turn;
1. ясно на
0.
2.
0.
е
Рис. 217. Речной рак — Astacus astacus:
а —глав; б —первая пара усиков; в—усик второй пары; г клешня; д — 2 — 5-я грудные ножнм; е — 5 брюшных ножек; ж— 6-я брюшная ножка, видоизменённая d хвостовые лопасти; а — 7-й сегмент брюшка.
Ножек 3 пары (у некоторых личинок ног нет); тело взрослого животного расчле-три отдела: голову, грудь и брюшко.
Класс I nsecta — насекомые.
Ножек более 3 пар ........................................2
Ножек 4 пары . . Класс Arachnoidea — паукообразные . . .8 Ножек обычно более 4 пар; в теле можно различить или три, или два отдела;
грудной или брюшпой (абдомен) отделы тела, пли и тот и другой, явственно расчленены на сегменты. Класс Crustacea — ракообразные.
3. Тело расчленено па два отдела — головогрудь и брюшко.
Отряд Araneina — пауки. 0. Тело совсем не расчленено.
Отряд Acarina — клещи.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ CRUSTACEA - РАКООБРАЗНЫХ
Чтобы познакомить читателя с принципом организации ракообразных и с терминологией частей и органов их тела, знание чего необходимо для пользования определительными таблицами, мы разберём наружное строение тела обычного речного рака, являющегося крупнейшим представителем пресноводных беспозвоночных. Благодаря именно своей большой величине он больше всех других ракообразных пресных вод подходит для указанной цели.
Речной рак — Astacus L. (рис. 217). Хотя этот рак, если не всем, то громадному большинству, достаточно известен, мы всё же дадим краткую его характеристику, тем более что на нём можно познакомиться с некоторыми особенностями организации ракообразных вообще. Рак — самый крупный представитель из всех вообще пресноводных ракообразных, достигающий в длину 12 и более сантиметров. Всё тело его одето жёстким панцырем тёмнокоричневого цвета, иногда с примесью оливковозелёных тонов (при варке панцырь становится красным). Если смотреть со спины, то легко можно различить два отдела тела — головогрудь и уплощенное сверху вниз брюшко (абдомен), которое в общежитии называют то «хвостом», то «шейкой». Головогрудь как со спины, так и с боков покрыта одним сплошным, не расчленённым куском панцыря — щ и т о м, между тем как па абдомене панцырь расчленён соответственно разделению самого брюшка на с е гм е н т ы, причём части панцыря, которыми одеты отдельные членики абдомена, сочленены между собой так, что брюшко легко может подгибаться книзу и вновь распрямляться. Сосчитав число члеников абдомена, увидим, что их шесть: пять сходных между собой, последний же отличается от них и состоит из 5 веерообразно расположенных в ряд пластинок; на самом же деле брюшко состоит не из 6, а из 7 сегментов. 5 сходных члеников представляют сегменты от 1 до 5 включительно, а последний является сложным образованием: средняя из 5 пластинок есть 7-й (последний, хвостовой) сегмент, или тельсон, расположенные же по сторонам её 2 боковые пары пластинок представляют видоизменённые копечности 6-го сегмента.
Рассмотрим конечности речного рака. Для конечностей ракообразных характерен тип так называемой расщеплённой ножки; такая ножка состоит из основной части, прикреплённой одним концом к телу, и двух сидящих на другом её конце ветвей, из которых одна, расположенная ближе к средней линии тела, называется внутренней, а другая — дальше от оси тела — наружной, или внешней. Таким
з
внешней. Таким образом, расщеплённая конечность ракообразных является двуветвистой. Этот тип конечностей особенно распространён среди низших ракообразных, но встречается и у высших раков. Передние усики речного рака (а н т е н н у л ы) являются двуветвистыми, так как состоят из основного членика и двух сидящих на нём ветвей; однако эта двуветвистость имеет не первичное, а вторичное происхождение, и потому аптеннулы не могут считаться истинно расщеплёнными конечностями. Задние антеппы, или вторая пара усиков, — то, что называют обыкновенно усами у рака, — построены по типу настоящей расщеплённой конечности: длинная нить, из которой построен такой ус, представляет собой внутреннюю ветвь, наружная же ветвь здесь очень мала и сводится лишь к треугольной пластинке при основании 2-й аптепны. За двумя парами усиков следуют окружающие рот 204
3 пары ротовых конечностей, назначение которых заключается в том, чтобы размельчать поступающую в рот пищу; ввиду малых размеров этих конечностей мы задерживать своё внимание на них не будем. Эти 5 пар конечностей — 2 пары усиков и 3 пары ротовых органов — относятся к голове. Далее идут 8 пар, принадлежащих уже г р уд-пом у о т д е л у. Из них первые 3 пары — челюстные ножки — служат для проталкивания пищи в рот; в этих конечностях, особенно в двух последних парах, тип расщепленной ножки выражен довольно ясно. Следующие 5 пар грудных конечностей представляют собой так называемые ходильные ножки, отличающиеся тем, что в них уже нельзя распознать характер расщеплённой пожни благодаря полному исчезновению внешней ветви; первая из этих 5 пар ножек — всем известные клешни — резко отличается (главным образом, величиной) от следующих 4 пар. Переходим, наконец, к конечностям последнего отдела тела — брюшка , или абдомена; их 6 пар, из которых 1-я недоразвита (у самки — еле заметна) и не обнаруживает характера двуветвистых ножек; следующие 5 пар сохранили тип расщеплённых ножек и помогают раку при плавании; G-я (последняя) пара брюшных ножек, как мы уже раньше видели, сильно отличается от предыдущих конечностей, так как в пей внутренняя и внешняя ветви разрослись в широкие пластинки и вместе с 7-м сегментом (тельсоном) образовали хвостовую лопасть, служащую раку прекрасным органом плавания. У самки 5 первых пар брюшных конечностей служат также для прикрепления икры.
Попробуем теперь подсчитать число сегментов, из которых состоит тело речного рака. Количество конечностей у него будет следующее: 5 пар головных + 8 пар грудных -|- 6 пар абдоминальных (брюшных) = 19 пар. Если прибавим сюда последний (хвостовой) членик, лишённый всяких конечностей, то получим общее число сегментов тела у речпого рака 19 +1 =20. Однако, как мы убедились на примере даже такого крупного рака, как речной, сразу различить и подсчитать число члеников тела у ракообразных — дело не столь уж лёгкое.
В тех случаях, когда непосредственно различить границы между сегментами на каком-либо участке тела трудно или совсем нельзя, об их числе можно судить по количеству пар конечностей на этом участке, так как каждой паре конечностей соответствует один сегмент.
Класс ракообразных — Crustacea — делится па два подкласса: в ы с ш и е— Malacostraca и низшие — Entomostraca. Основные признаки различия между ними приведены в следующей определительной таблице.
1. Тело разделено всегда на 20 сегментов (члеников), из которых 5 приходится на голову, 8 — на грудь и 7 — на брюшко (абдомен). Все грудные сегменты и 6 из брюшных несут по паре конечностей (последний — 7-й сегмент — лишён их); большей частью крупные животные. Подкласс Malacostraca — высшие ракообразные.
0. Число сегментов тела не постоянно, может быть больше и меньше 20. Только грудные сегменты несут по паре конечностей, брюшные не имеют их. Большей частью микроскопические животные.
Подкласс Entomostraca — низшие ракообразные.
MALACOSTRACA — ВЫСШИЕ РАКООБРАЗНЫЕ.
1. Без головогрудного щита. Только 1-й сегмент груди срастается с головой.
Группа Arthrostraca — членистогрудые раки ...........2
0. С головогрудным щитом, срастающимся с рядом грудных сегментов. Все 8 грудных сегментов срастаются с головой в один отдел — головогрудь.
Группа Thoracostraca — щитогрудые раки .........;....3
2. Тело сплющено с боков и изогнуто в дугу, выпуклую сторону которой представляет спинная поверхность тела, а вогнутую — орюшная. Передняя и задняя пары антенн (усиков) почти равны между собой или первая несколько длиннее. Грудных конечностей 8 пар: 1-я пара ногочелюсти, 2-я и 3-я служат для схватывания пищи, остальные 5 пар — ходильные ножки; 2—7-я пары снабжены листовидными придатками — жабра м и. 6 пар абдоминальных (брюшных) двуветвистых конечностей образуют 2 группы: 3 передние пары представляют собой плавательные ножки (обе ветви густо усажены длинными щетинками), 3 задние пары превращены в палочковидные прыгателъные ножки.
Отряд Amphipoda — двояконогне раки.
0. Тело сплющено в спинно-брюшном направлении, прямое. Передние антенны короче задних. У 5-й пары абдоминальных конечностей обе ветви —- внутренняя и наружная — превращены в широкие нежные пластинки, которые, налегая друг на друга под орюшком, прикрывают его нижнюю поверхность. Отряд Isopoda — равноногие раки.
8. Головогрудь, за исключением 1—2-го сегментов, покрыта панцырем. Все грудные конечности двуветвпсты, сходного строения, из них 1-я и 2-я пара видоизменены и приспособлены для целей питания, остальные 7 пар служат ходильными ножками.
Отряд Mysidacea (Schizopod а) — расщеп л ённон огне раки.
0. Грудные конечности не все одинакового строения: 3 передние пары — двуветвистые — превращены в ротовые органы (ногочелюсти), 5 задних пар — одноветвистые — служат ходильными ножками. По числу последних этот отряд ракообразных и получил своё название. Отряд Decapoda — десятиногие раки.
Даём таблицы для определения лишь некоторых представителей высших раков, наиболее распространённых и часто встречающихся в пределах СССР.
AMPHIPOD Л — ДВОЯКОНОГИЕ РАКИ
1. Тело сжато с боков. Первая аитепна с добавочным жгутиком, состоящим более чем из двух члеников......................................................2
0. Тело сжато в спинно-брюшном направлении. Первая антепна без добавочного жгутика. Предпоследний членик 2-й грудной конечности не расширен, без выроста (рис. 218). р. Gorophium.
2. Второй членик последней грудпой конечности не расширен в лопасть. Жабры не имеют пальцевидных выростов (рис. 219). р QammarU8 Fabr.
О, Второй членик последней грудпой конечности расширен в направленную назад лопасть. На спинной стороне 4-го и 5-го брюшных сегментов по одному коническому бу-горку с шипиками на их вершинах (рис. 220). р Di kerogam шаги а.
Отряд Amphipoda по количеству видов является самым богатым из всех выс-п!пх раков, населяющих пресные воды. Громадное большинство относящихся сюда видов принадлежат к сем. Qammaridne.B котором первое место по распространению в пресных водах нашего Союза занимает род Ga m m a rus Fabr. (подр. R i v u 1 о g a Dini а г u s). Особенно широко распространены виды: О. (R.) pule х Col. (рис. 219), встречающийся почти во всей Европейской части СССР, и О. (R.) lacustris G. О. Sara, населяющий почти всю Азиатскую часть Союза. Длина G. (R.) р u 1 е х 15—2O.w,w. Из трёх видов f). Dikerogammarus наибольшее распространение имеет D. h а е m о b а р h е з Eichw.), встречающийся в Волге и её притоках и в южных реках (Урал, Дон, Дунай, Днепр, Терок, Кубань и др.). Из 4 пресноводных видов р. С о г о р h i u rn самым распространённым является С. curvispinumG. О. Sars, обитающий в Волге с её притоками, в Урале, Допу, Днепре и других реках.
Отряд I s о р о d а включает 4 рода, из которых самым обычным и широко распространённым является принадлежащий к сем. A s е 1 1 i d а е, р. A s е 1 I u s: его вид A. aquaticusL. (рис. 221) можно часто встретить в мелких водоёмах и в прибрежной области более крупных. Длина его 10—15 мм.
Из отряда Mysidacea в пресных водах СССР водятся представители только одного семейства — М у s i d а е. Из 10 относящихся сюда родов наибольшее распространение на территории нашего Союза имеют: М у s i s, вид которого М. оси la ta г е I i с t a (Loven) встречен в устьях северных и западных рек и в озёрах северных районов; М eso my si s, виды которого М. Kowalcvskyi Czern. и М. in-to г me d i a’ Czern, живут главным образом в бассейнах южных рек; М е t a m у si з, именно M.strauchi G. О. Sars, живущая в Волге и реках её бассейна, а также в Урале, Дону, Днепре. Приводим таблицу для определения этих трёх родов.
НЕКОТОРЫЕ РОДЫ СЕМ. MYS1DAE
1. Тело и панцырь довольно узкие. Передний и задний спинные края панцыря пред ставляют изогнутые по направлению к голове линии с изломами посередине (по продольной оси тела), образующими тупые углы, направленные вершинами вперёд. Тельсоп (по следний членик брюшка) узкий — его длина в 2х/2 раза больше ширины (рис. 222).
р. М у з i s.
Рис. 223. Metamysis strau- Рис. 224. Mesomysis ko-chi. walevskyi.
0. Тело и панцырь несколько расширены. Задний край панцыря округловогпутый. Отношение длины тельсона к его ширине менее ..............................
2. Передний край панцыря почти прямой (рис. 223). Metamysis.
0. Передний край панцыря плавновыпуклый, без излома (рис. 224).
р. М е s о ш у s i з.
В отряде Decapoda имеются обитатели как морских, так и пресных вод. Из последних наибольшее распространение в СССР имеет род Ast а сиз — речной рак. Он
характеризуется:
1) присутствием клешней па трёх первых парах грудных ножек, 2) отсутствием отростка на 3-м членике 2-й и 3-й грудных ножек и
3) наличием шипов позади глаз. Этот род заключает в себе 6 видов, из которых лишь один морской (A. pachypus), остальные пресноводные. Приводим определительную таблицу для этих видов.
РЕЧНОЙ РАК — ASTACUS
1. Неподвижный отросток клешни на стороне, обращённой к подвижному, снабжён выемкой................................................................2
0. Такой выемки нет...............................................3
2. Боковые еторбны спинного панцыря брюшных сегментов, особенно 3-го, несимметричны. В Белоруссии, в областях Смоленской, Ленинградской, отчасти на Украине (рис. 217). А nstacu8 (L.)
0. Эти стороны симметричны и закруглены. В бассейнах рек Риона, Чороха и
A. colchicus Kessler.
8. Расстояние между боковыми уголками рострума 1 превышает длину его отрезка от боковых уголков до конца рострума. В мелких речках Азербайджана.
А. р у 1 z о w i Skor.
0. Указанное расстояние не больше копцевой части рострума.........? 4
4. Это расстояние равно концевой части рострума. В Средней Азии.
A. kessleri Schimk.
0. Это расстояние мепьше концевой части рострума. В Европейской части СССР, за исключением районов, в которых распространён A. astacus, а также в северо-западной части Азии. A. leptodactylus Esch.
ENTOMOSTRACA — НИЗШИЕ РАКООБРАЗНЫЕ
Из трёх относящихся сюда отрядов, имеющих своих представителей в пресных водах, именно листоногих — Phyllopod а, веслоногих — Copepoda и ра к у ш новых — Ostracoda, последняя группа представляет, на наш взгляд, для новичка столь неодолимые трудности в смысле определения принадлежащих к этому отряду форм, что давать специальную определительную таблицу для ракушковых рачков нам кажется излишним. Каждый из двух других отрядов — листоногие и веслоногие — распадается, в свою очередь, на группы: первый на 2 подотряда — истиннолист о-ногих — Euphyllopoda — и ветвистоусых — С 1 a d о с е г а, второй — на две группы, из которых первая — истинновеслоногие — Еисореро-d а — заключает в себе всех свободно живущих веслоногих рачков, а вторая состоит исключительно из паразитических форм и потому выходит из поля нашего внимания. Поэтому таблицы для определения, до рода (или вида) включительно, даны лишь для трёх групп низших ракообразных: истиннолистоногих, ветвистоусых и свободно живущих веслоногих.
1. Всё тело целиком заключено в двустворчатую не расчленённую в поперечном направлении раковину — мягкую или жёсткую, полупрозрачную или непрозрачную, которая может раскрываться и посредством "особого мускула-замыкателя плотно закрываться; в раскрытом состоянии из неё выставляются, самое большее, щетинки ножек; длина раковины не более 2 мм. ™ .. л ~
1 Отряд Ostracoda — ракушковые.
0. Тело или без раковины, или в раковине, которая или в виде спинпого щита прикрывает тело животного сверху, или, так же как у ракушковых, с боков, в виде двух соединённых на спине створок. В* последнем случае опа может быть двоякого рода: 1) раковина отличается от таковой же Ostracoda только лишь более крупными размерами (длина не менее 5 мм), 2) раковина всегда мягкая, в большинстве случаев прозрачная, не может по произволу животного закрываться и позволяет различить две части, из которых одна прикрывает голову, а другая — остальное тело................2
2. Тело состоит из многочисленных, ясно различимых сегментов и в этом случае имеет сравнительно большую длину (большинство не менее 8—10 мм) и несёт большое число — не менее 10 пар — листовидных ножек * * * * * 8 или, напротив, укорочено, неясно расчленено, заключено в двустворчатую раковину и несёт 4—6 пар листовидных ножек 8.
Отряд Phyllopoda — листоногие.................................. . .8
0. Тело без раковинки; состоит из немногих (9 или 10) ясно различимых сегментов, имеет в длину не более 5 мм и несёт 5 нар грудных двуветвистых ножек, из которых 5-я
1 Рострум — вырост срединной части переднего края головогрудного панцыря,
суживающийся и заострённый к переднему концу.
1 Листовидная ножка есть особая форма расщеплённой ножки, в которой внутренняя
и внешняя ветви уплощены, расширены и превращены в прозрачные, более или менее ши-
рокие пластинки, усаженные по краям щетинками и служащие для дыхания.
8 Исключение представляет Leptodora, тело которой при длине около 1 см состоит из немногих сегментов и несёт 6 кар грудных не листовидных, а простых палочковидных ножек.
пара более или менее недоразвита, так что а большинстве случаев можно различить лишь 4 пары. Отряд Eucopepoda — свободно живущие веслоногие.
8. Тело состоит из многих ясно различимых сегментов; не менее 10 пар плаватель-пых ножек; длина тела (за исключением одного рода) около 1 см и более.
Подотряд Euphyllopoda — истинполистоногие.
0. Тело (за очень редкими исключениями) заключено в двустворчатую раковинку с обособленной головной частью и состоит из немногих, большей частью трудно или совсем не различимых сегментов; 4—6 пар ножек; длина тела (за исключением одного рода) пе превышает 2—3 мм\ у громадного большинства размеры микроскопические.
Подотряд Cladocera — ветвистоусые.
EUPHYLLOPODA — ИСТИННОЛПСТОНОГИЕ РАЧКИ
Тело более или менее удлинённое, покрытое или непокрытое раковинкой и состоящее из большого числа сегментов, многие из которых (не менее 10) несут листовидные ножки; характерным признаком таких ножек является присутствие на них прозрачных пластинок, служащих для дыхания. Эти рачки часто населяют вешние пересыхающие л/жи. Так как нет гарантии в том, что на одном и том же месте будет каждый год возникать лужа, то обитающие в таких временных водоёмах организмы — в том числе и истиннолистоногие рачки — подвергаются риску прекратить своё активное существование иногда на целый ряд лет. Так оно и бывает в действительности. Случается, что в данной местности несколько лет не бывает ни одного рачка из Euphyllopoda, а потом вдруг они появляются в изобилии. Это объясняется тем, что по способу своего размножения они прекрасно приспособлены к таким неустойчивым условиям существования: их яйца не только способны переносить высыхание, но даже нуждаются в нём для своего развития, так что, не претерпевши в высохшем состоянии известного минимального периода покоя, они не могут развиваться. В таком виде они могут вынести очень продолжительную засуху; известны случаи, когда яйца Euphyllopoda, пролежавшие в высохшем иле 14 или 15 лет, при помещении этого ила в воду развивались в рачков.
1. Тело вытянутое, раковины нет. Голова свободная, с двумя стебельчатыми сложными глазами и одним непарным науплисовым глазом. 1-я антенна простая нитевидная; 2-я, превращённая у самцов в хватательный орган, очень большая и крючкообразная; у самок она тоже сильно развита. И пар плавательных ножек, все с хорошо развитыми жаберными придатками. Объединённые яйцеводы образуют мешковидное расширение — выводковую камеру. Брюшко (абдомен) состоит из 8—9 сегментов, лишённых ножек; к концу оно сужено. На конце его имеется вилка (фурка), Состоящая из 2 длинных, плоских, покрытых щетинками придатков — фуркальных пластинок.
Сем. Branchi podidae — жаброногие.
0. Тело более или менее укороченное, снабжённое раковиной..........2
2. Раковинка в виде широкого и плоского мягкого кожистого щита прикрывает со спины голову и тело, за исключением его заднего конца. Количество сегментов тела велико, из них И, принадлежащих переднему — грудному — отделу, несут каждый по одной паре плавательных ножек, между тем как из 14—23 абдоминальных сегментов передние снабжены несколькими парами ножек каждый, а задние совсем лишены их. Каждая ножка состоит из основной части и сидящих па пей 2 наружных и 6 внутренних придатков, из которых 5 (от 2-го по 6-й) носят название эндитов. Хвостовая вилка с длинными щетинками. Длина щита 1—3 см, ширина немного меньше.
Сем. Apodidae (Triopsidae) — щитни.
0. Тело покрыто двустворчатой, сжатой с боков раковинкой...........3
3. Раковинка сильно сжата с боков, с концентрическими полосами. Голова и клюв средних размеров. Сем. Limnadiidae.
0. Раковинка более выпуклая с боков, без полос. Голова большая с очень длинным хоботком. Сем. L у п с е i d а е (L i m n е t i d а е).
1 род и вид Lynceus brachyurus О. F Muller (Limnetis brachy-u г а) распространён почти во всех водоёмах весной.
Сем. Branchipodidae
1. Фуркальные пластипки короткие, самое большее — вдвое длиннее своей ширины (рис. 225). Длина 8—И мм. В солёных водах, широко распространена в южных водоёмах СССР. рв А г t е m i a Leach, 1 вид. A. salina (L.).
О, Длина фуркальных пластинок не менее длины трёх последних сегментов брюшка...............................................................2
2. Для самок: длина выводковой сумки равна её высоте; она очень мало вытянута назад (рис. 226 Л). Для самцов: лобные выросты нитевидные (рис 226 Б). Длина 11,5 мм. В мелких лужах. р в г a n с b i р u s — ж а б р о п о г; впд В г. stagnalis (L.) (schafferi Fiscber) широко распространён в СССР, доходя на восток до западных районов Казахстана.
14 А. Н. Липин
Рис. 227. Streptocephalus torvicornis: A — абдомев самки сбоку; Б — голова самца сбоку.
Рис. 228. Chirocephalus grubii:
0. Для самок: длина выводковой сумки больше её высоты; её брюшпая часть вытянута назад. Для самцов: лобные выросты складчатые и зубчатые, лопастные . 3
3. Для самок: основание удлинённой выводковой сумки толще, чем брюшко. Для самцов: лобные выросты свободны до основания.
р. Chi rocephalus Prev.
0. Д л я самок: основание длинной выводковой сумки значительно тоньше брюшка (рис. 227 А). Д л я самцов: лобные выросты сидят на общем основании (рис. 227 Б), складчатые и зазубренные.
р. Streptocephalus Baird, 1 вид S. torvicornis (Waga).
Род Chi г о с е р h а 1 u 8 Prevost
1. Дл я самок: направленное назад удлинепие выводковой сумки гораздо короче её основания (рис. 228 Л). Для самцов: между лобными выростами и нижней губой срединный рог; лобные выросты большие с маленькими зубовидными придатками на обоих краях (рис. 228Б). В заболоченных ямах и прудах с прозрачной водой и на залитых лугах.
Ch. grubii Dybowsky.
0. Для самок: заднее удлинение выводковой сумки длиннее её основания. Для самцов: рога нет..................................................2
2. Для самок: основание выводковой сумки занимает лишь один 1-й брюшной сегмент (рис. 228 а —Л). Для самцов: лобные выросты очень большие, 5-пальча-тые (рис. 228 а —Б), Длина около 20 мм, Ch. diaphanus Prevost.
0. Для самок: основание выводковой сумки занимает более одного брюшного сегмента (рис. 229 Л). Для самцов: лобные выросты маленькие, на внутреннем крае с зубцами (рис. 229 Б). Длина 14 мм. Ch. (Р г i s t i с ep halus) josephinae Grube; обычен в средних широтах Европейской части СССР в весенних водоёмах.
Сем. Apodi d а е — щ и т я и
1. Последний (задний) сегмент тела не вытянут в пластинку (рис. 230). Предпоследние эндиты (внутренние придатки) первой пары ножек очень длинные, нитевидные, далеко выдаются за боковые края папцыря. Спцннои щит овальный, снабжён килем. Тело состоит
Рис. 230. Apus cancrifor-mis с брюшной стороны.
Рис. 231. Lepidurus producing со спинной стороны.
из 34 сегментов, несущих 60 пар плавательных ножек. Длина шита 1—3 см, ширина 0,9— 2,8 см. В мелких ямах и лужах.
р. Apus. широко распространённый вид A. cancriformis Schaff.
0. Последний сегмент тела вытянут в пластинку, снабжённую килем (рис. 231). Предпоследние эндиты первой пары ножек лишь немного выдаются за края спинного щита. Последний имеет киль только в задней своей части. Тело состоит из 29 сегментов с 41 парой плавательных ножек. Длина щита 1,2—3,0 см. ширина 1.2—2,9 см. В заболоченных ямах с прозрачной водой и на заливных лугах (в поименных водоёмах), в весенних лужах и т. п. Lepidurus productus Возе.
Сем. Limnadiidae
1. Раковинка на вершине с ясно заметным бугорком (рис. 232), жёсткая и мало прозрачная; её конпентрическая шт^иховатость распространена по всей её поверхности, распространяясь и на вершинный бугорок, и состоит из 20—26 полос. Голова вытянута в довольно длинный и острый клювик, на конце которого нет шяппка. Па теле 21—24 пары плавательных ножек. Длина раковины до 12 ле.и, ширина (высота) 7,5 лме, толщина
Р-
0.
горком вытянута которого
4,5 мм. Cyzicus Audoin (Е s t b е г i a Rupp), 1 вид С. te Ira се ru 9 (Kryn.).
~ с бу-
Голова конце шипик р. L е р-виды этого рода
Раковинка также штриховатая, на вершине, по прозрачная.
1 в заострённый клювик, на имеется подвижной tos'theria G. О. Sars;
встречаются главным образом на юге, в степных водоёмах.
Рис. 232. Cyzicus tetracerus.
CLAD ОСЕВ А — ВЕТВИСТОУСЫЕ РАЧКИ
Ветвистоусые рачки характеризуются укороченным телом, заключённым в двустворчатую раковину (рис. 241); голова (г) тоже покрыта нан-цырем или составляющим продолжение раковины, или, напротив, отделённым от нее более или менее ясной бороздой на спине. В шлове всегда имеется г л а з (в), а у многих
форм, кроме того, ешс добавочный глаз, или глазок. На голове прикреплены две пары усиков, или антенн, — передние усики (а), обычно очень маленькие, простые, палочковидные, задние же (е), напротив, сильно развиты и превращены в органы плавания; они устроены по типу расщеплённой ножки и состоят из основной части, одним конном которой усик прикрепляется к телу (между головой и туловищем), и двух сидящих па пей ветвей; обе ветви членистые и более или менее густо*усажены перистыми щетинками, чем достигается необходимое им как плавательным органам увеличение поверхности. Туловище несёт 4—6 пар скрытых под раковинкой плавательных листовидных ножек (п). Следук щий за туловищем задний отдел тела, так называемый постабдомеп (л), загнут на брюшную сторону под некоторым углом к туловищу; его спинной край несёт различного рода придатки — спинные отростки (и), хвостовые щетинки (к), ряды ш и п и к о в, расположенные по бокам заднепроходного отверстия (м), на самом конце постабдомеп снабжён парой концевых, или хвостовых, коготков (о), на вогнутой стороне которых иногда бывают более мелкие шипики (н).
Все перечисленные части тела являются основными систематическими и определительными признаками.
Все ветвистоусые рачки, за исключением разве лишь Leptodora и Bytho-tre р h es, являющихся хищниками, питаются растительной пищей (фитопланктоном) или же детритом. По способу принятия ппщи эти рачки являются фильтрат о р а м и: быстрые колебательные движения их ножек, густо усаженных щетинками, возбуждают ток воды, направленный к ротовому отверстию; приносимые этим током мелкие питательные частички оседают па щетинках и, собираясь в комочки, направляются в рот.
Размножение ветвистоусых рачков происходит двумя способами: так называемым партеногенетическим, т.е. девственным (без участия самцов), и о б о е п о-л ы м (с участием сампов). Обе эти формы определённым образом чередуются между собой, причём обычно через несколько партеногенетических поколений, состоящих, как это само собой попятно, исключительно из самок, появляются самцы, т. е. наступает обоеполое размножение; у одних видов это бывает только один раз в году — осенью, у других, кроме осеннего поколения самцов, бывают ещё летние. Число поколений самцов зависит от тех условий, в которых живёт данный вид, и подчиняется следующему обшему правилу: чем более неблагоприятны условия жизни, тем большее количество поколений самцов появляется в течение года. Соответственно этому, чем мельче водоём и, следовательно, чем большему риску он подвергается в смысле высыхания (например, лужи), тем чаше у обитающих в этом водоёме ветвистоусых рачков нарождаются поколения самцов, чередующиеся с партеногенетическими генерациями; число этих поколений бывает различно даже у одного и того же вида в зависимости от того, в каких условиях он живёт. Среди обитателей больших глубоких озёр известны даже такие виды, у которых до сих пор не найдено самцов и которые, повидимому, всё время размножаются партеногенетически.
Яйца, из которых развиваются партеногенетические самки, т. е. яйца не оплодотворяющиеся, часто называемые «летними» яйцами, характеризуются тем, что они начинают развиваться сейчас же после их откладки, без всякой предварительной стадии покоя; такие яйца вынашиваются самками в особых помещениях — так называемых выводковых камерах, или выводковых сумках (рис. 241 ж); это — полость, расположенная со спинной стороны тела, но внутри раковины, и ограниченная снизу спинной поверхностью туловища, а сверху и с боков — складками эпителия, выстилающего внутреннюю поверхность стенок раковинки *, Сюда откладываются летние яйца в количестве различном у разных видов, и здесь же они развиваются до стадии маленького рачка, уже способного вести самостоятельную жизнь; достигши этой стадии, такой рачок выходит наружу и начинает плавать.
Оплодотворённые самцами яйца носят название «покоящихся», потому что перед началом своего развития они должны выдержать тот или иной период покоя (часто их называют, хотя и не совсем правильно, «зимними» яйцами). Эти яйца откладываются в ту же выводковую сумку, но последняя в этом случае претерпевает особые изменения, в результате которых она превращается в довольно жёсткую и прочную хитиновую капсулу — так называемое «сёдлышко», или «эфиппий» (рис. 16); пока самка жива, она, конечно, носит это сёдлышко на себе, но после её смерти оно или падает на дно, или, наоборот, поднимается к поверхности и здесь плавает. По миновании периода покоя из заложенных в эфип-пии яиц развиваются самки первого партеногенетического поколения. Число покоящихся яиц бывает обычно мало — одно или два, редко больше. Эфиппии разносятся ветром или на ногах птиц по соседним водоёмам и служат средством распространения ветвистоусых.
Довольно распространённое среди Cladocera явление представляют сезонные вариации. Так, среди многочисленных форм, принадлежащих к виду Daphnia longi spina, встречаются такие, у который головной шит вырастает вперёд, заостряется и приобретает вид шлема (рис. 241); этот шлем меняется в зависимости от времени года — летом он удлиняется, зимой укорачивается.
Среди ветвистоусых рачков встречаются настолько мелкие формы, что их определение приходится вести под микроскопом.
1. Тело вместе с ножками покрыто двустворчатой, снизу открытой раковиной . . 3
1 У некоторых родов, например, у Leptodora, раковина недоразвита настолько, что представляет собой лишь одну выводковую сумку и никакой защитной роли для самого животного играть не может. 212
0. Голова, туловище и пожни свободны от раковинки, которая сильно редуцирована и выполняет лишь роль выводковой сумки, наполняющейся во время размножения яйцами ............................................................. 2
2. Тело длинное, голова тоже. Глаз большой. Антенны 2-й пары очень длинные, обе ветви их 4-членистые. 6 пар грудных ножек, из которых 1-я пара самая длинная, 5-я, 6-я — самые короткие. На конце тела два крупных, направленных назад коготка. Животные чрезвычайно прозрачные. Длина до 10 мм. В планктоне крупных озёр.
Сем. Leptodoridae, р. Lep todora Lili jeborg, 1 вид L. k i n d ti Focke; широко распространена в нашем Союзе.
0. Тело короткое, голова тоже. Глаз чрезвычайно большой (рис. 234, 235). Антенны второй пары не очень большие; одна из ветвей состоит из четырёх члеников, другая из трёх. 4 пары ножек. Сем. Polyphemidae.
8. Плавательные усики (вторая пара антепн) у самки одповетвистые с тремя расположенными на их концах длинными щетинками, у самца двуветвистые — одна ветвь с тремя, другая с двумя концевыми щетинками. Всё тело одето толстой студенистой, чрезвычайно нежной, совершенно прозрачной оболочкой (рис. 233). В планктоне озёр с мягкой водой. Сем. Holopedidae,p. Holopedium Zaddach., у пас (и вообще в Европе) 1 вид— Н. gibberum Zad.
0. Плавательные усики у обоих полов двуветвистые.................4
4. Каждая из ветвей плавательных усиков состоит не более чем из трёх члеников . 5
0. Одна из ветвей плавательных усиков состоит из трёх, другая из четырёх члеников (за исключением рода Bosminopsis из сем. Bosminidae).................6
5. Одна из ветвей плавательных усиков состоит из двух, другая из трёх члеников. Голова ясно отграничена от туловища перехватом на спинной стороне.
Сем. S i d i d a e.
0. Обе ветви плавательных усиков состоят из трёх члеников; между головой и туловищем перехвата нет. Сем. Cbydoridae.
6. Передние антенны самки маленькие, мало подвижные (за исключением рода Moina). Сем. Daphnidae.
0. Передние антенны самки длинные................................7
7. Передние антенны самки прикреплены неподвижно, более или менее изогнуты назад. Глазка нет. Конец постабдомена обрезан прямо (перпендикулярно); концевые коготки сидят на длинном выросте постабдомена, на вогнутой стороне имеют добавочные шипики. Сем. В о s m i n i d а е.
0. Передние антенны подвижно сочленены с головой. Глазок есть. Концевые коготки только с базальными шипиками, без добавочных шипиков на вогнутой стороне.
Сем. Macrothricidae.
Сем. Polyphemidae
1. Грудные ножки убывают в длине от первой пары к последней более или менее равномерно. Глаз очень большой (рис. 234). Постабдомен узкий, удлинённый, но короче тела с 2 длинными хвостовыми щетинками па конце. Длина 1,4—1,6 мм. Часто в мелких водоёмах и в прибрежной области больших озёр.
р. Polyphemus О. F. Muller, в пресных водах 1 вид Р. pediculus (L.); широко распространён преимущественно в северных водоёмах СССР.
0. 1-я пара грудных ножек длинная, 2-я и 3-я гораздо короче, 4-я совсем малепькал. Глаз большой. Конец постабдомена вытянут в виде очень длинной прямой иглы (длиннее тела) с 2 очень мелкими хвостовыми щетинками. Длина без хвостовой иглы 2—3 мм. В планктопе больших, глубоких озёр. р. Bythotrephes Leydig.
Род Bythotrephes Leydig.
1. Игла совершенно прямая, без всяких изгибов (рис. 235). Длипа без иглы 2—3 В планктоне больших озёр северной половины СССР, Rinnr»;mnn.ia г pvdio-D• 1 О 11 IT I Ill u II ц □ LlcyLllK.
0. Игла с небольшим изгибом. Длина самки без иглы 2—4,5 ле.к. В мелких водоёмах и озёрах, главным образом на севере Союза, по по Волге доходит до Астрахани.
В. cederstroemi Schoedl.
Сем. S i d i d a e
1. Передние антенны самки короткие. Верхняя спинная ветвь плавательных усиков трёхчленистая, пижпяя двучленистая ................................. 2
0. Передние антенны самки более длинные. Верхняя ветвь плавательных антенн двучленистая, пижпяя трёхчленистая...................................3
2. Голова округлая, па спинной стороне её затылочная присоска (рис. 236). Концевые коготки с мелкими щетинками и более крупными шипиками по вогнутому краю коготков. Длина 3—4 мм. Встречается почти исключительно в прибрежной области, где часто сидит, присосавшись затылочным органом, па растениях (например, на нижней поверхности листьев кувшинки).
р. S i d a Straus, у нас 1 вид — S. crystalline (О. F. МйНег).
Рис. 234. Polyphemus pediculus: A — самка, Б — самец.
Рис. 235. Bythotrephes longimanus.
Рис. 236. Sida crystalline. Рис. 237. Diaphanosoma brachyurum.
О, Голова в виде треугольника, в нижнем углу которого помещается глаз. По вогнутому краю концевых коготков сплошной гребневидный ряд длинных щетинок без шипи-ков. В пелагиали озёр северной полосы СССР.
Li m п о s i d а, вид L. frontosa Sars.
3. Оба членика верхней ветви длинные, один несколько длиннее другого (рис. 237). Длина тела 1 мм. Очень часто в пелагической и прибрежной зонах больших озёр, также в прудах, р, Diaphanosoma Fisch., у нас 1 вид — D. brachyurum Lievin.
0. Первый членик верхней ветви короткий, но несёт длинный боковой отросток, усаженный длинными щетинками; второй членик длинный. 2-я пара антенн, орюшнои и задний края раковинки густо усажены длинными перистыми щетинками, из которых самые длинные сидят на брюшном углу заднего края раковины и вблизи спинного угла (2 щетинки). Длина самки 2—3 мм. В иле прибрежной области и в более глубоких частях озёр, на севере СССР.
р. La ton a Straus, у нас 1 вид — L. setifera (О. F. Muller).
Сем. Dapbnidae
1. Нижняя часть головы самки вытянута в клювик, направленный вниз. Между головой и туловищем на раковине нет перетяжки или она мало заметна...2
0. Голова без клювика. Между нею и туловищем на спинной стороне раковины перехват ............................................................ 4
2. Задпий конец раковины вытянут в более или мепее длинный шип....3
0. Задний копец раковины без шипа. Голова маленькая, туловище большое (рис. 238). р. S i m о с е р h а I и 8 Schodler.
8. Шип отходит от более или мепее закруглённого заднего края раковипы приблизительно в середине его или выше (рис. 241). р Daphnia Miiller.
0. Шип отходит от вертикально срезаппого заднего края раковины и составляет непосредственное продолжение прямого нижнего (брюшпого) края раковины (рис. 246 А).
р. Scapholeberis Schodler.
4. Передние антенны (1-я пара) очень короткие. Ливия, соединяющая центр глаза с местом прикрепления плавательных антенн (2-я пара), круто наклонена, иногда почти отвесна. Есть глазок (придаточный глаз) (рис. 249). р. Ceriodaphnia Dana.
0. Передние антенны длинные, сочленены с головой подвижно. Лилия между глазом и основанием плавательных усиков наклонена слабо, иногда почти горизонтальна. Глазка пет (рис. 251 A). Moina Baird.
Род Simocephalus Schodler
1. Лобная часть головы под глазом в виде заострённого клюва, па копце которого имеются очень мелкие шипики (рис. 238 Б]. В мелких заболоченных водоёмах.
S. serrulatus (Koch.)
О, Эта часть закруглённая, гладкая (без шипиков)..................2
2. Глазок веретенообразно вытянут (рис. 238 Л). Постабдомен одинаковой длины и ширины, с каждой стороны несёт по ряду шипиков (9—10); концевые коготки длинные,
Рис. 238. Simocephalus:
А — S. vetulus; Б — голова S. serrulatus; В — концевой коготок (на заднем конце постабдомена) S. exsplnosus.
слабо изогнутые, внутри и снаружи по всей длине тонко заштрихованные. Длина самки 2—3 мм. В прибрежных зарослях озёр и мелких водоёмах. Наиболее распространённый в СССР вид Simocephalus. S. v е t u 1 u s (О. F. Muller).
0. Глазок маленький, большей частью ромбовидный, иногда круглый. Постабдомен очень широкий, у крупных особей с 18—20 шипиками с каждой стороны; концевые коготки (рис. 238 В) при основании с 9—12 крупными зубцами с наружной стороны и с тонкой штриховатостью по всей длине. Длина самки около 4 мм.
S. exspinosus (Koch).
Род Daphnia Miiller
1. Задний (спинной) край постабдомепа с довольно глубокой выемкой (рис. 239); с каждой его стороны сидит приблизительно по 20 шипиков, расположенных в два ряда; концевые коготки с 2 базальными рядами шипиков и с тонкой штриховатостью. Длина самки 5—6 мм. Часто в прудах, ямах — вообще в мелких водоёмах.
D. m a g n a Straus.
0. Спинной край постабдомена ровный, без выемки...................2
2. Свод (fornix) сзади снабжён шипом (рис. 240)1, от переднего (глазного) конца свода через туловищный отдел раковипы тянется острый киль. Постабдомсн к концу суживается, по его спинному краю проходят два ряда шипиков по 9—10 в каждом ряду; кроме того, на боковых сторонах постабдомепа есть тонкие шипики; концевые коготки с двумя базальными рядами шипиков: в первом ряду по 9—10, во втором ряду 13 14 топких шипиков. Шип раковины отходит выше середины её заднего края. Длина самки 2,7 мм> встречается не часто. .
D. psittacea (Baird.).
1 В самом начале 2-х усиков.
Рис. 239. Daphnia magna — постабдомен.
Рис. 240. Daphnia psit-tacea — голова самки.
Рис. 241. Daphnia pulex (самка):
a — усик (антенна) 1-й пары; б — клювик; в — глаз; г — голова; d — кишка; е— усик (антенна) 2-й пары; ж—выводковая камера с яйц ми; з— 3-й шип на заднем конце раковинки; и — спинные придатки; к — хвостовые щетинки; л — постабдомен; м — шипини на постабдомене; н— шипики на внутренней стороне кинцевых коготков; о — концевые коготки постабдомена;
п — пять ножек.
Рис. 242. Daphnia longispina var. longispina:
A — вид сбоку; Б — голова.
Рис. 243. Daphnia longispina var. hyalina:
справа — вид сбоку, слева — голова со спинной стороны.
Рис. 244. Daphnia longispina var. cu-cullata.
0. Шипа па заднем конце свода нет..............................................8
3. Концевые коготки с 2 базальными рядами шипиков: внутренний ряд состоит большей частью из 5—6 очень тонких, наружный из 6—8 более грубых шипиков. Постабдомен по спинному краю с 12—18 шипиками с каждой стороны (рис. 241). Длина самки 3—4 мм. Широко распространена в мелких водоёмах.
D. pulex (De Geer.).
• 0. Концевые коготки без базальных шипиков. Шип на заднем конце раковины всегда
есть, по большей части длинный.
D. longispina Muller............................................4
4. Глазок есть, хотя иногда очень мал и трудно заметен..........5
0. Глазка почти всегда нет. Голова у летних форм с высоким острым шлемом . . в б. Голова без киля (рис. 242). Живёт в мелких водоёмах.
D. longispina v а г. longispina Mull.
0. Голова с килем (рис. 243), который ясно выделяется при рассматривании со спинной стороны. Живёт в пелагиали озёр.
D. longispina var. hyalina Leydig.
6. Клюв тупой, обонятельные палочки на конце передних антенн выдаются за конец клюва (рис. 244). В пелагиали озёр.
D. 1 о n g i s р i n a var. cucullata Sars (H у a 1 о d a ph n i a cucullata).
О, Клюв острый, обонятельные палочки не достигают до его конца (рис. 245). У летних форм шлем высокий, направлен вперёд по продольной линии тела. В пелагиали озёр.
D. longispina var. cristata Sars. Каждая из этих четырёх вариаций распадается ещё на ряд форм.
Род Scapholeberis Schodlcr
1. Клюв самки короткий и тупой. Брюшной край раковины прямой, сзади удлиняется в шип (рис. 246 Л). Глазок ясно виден, расположен очень близко к концу клювика. Передние антенны очень маленькие, немного выдаются за конец клювика. ПоЙтабдомен
Рис. 246. Scapholeberis:
А — S. mucronata — раковинка с сёдлышком (с)\ Б — S. aurita — голова.
короткий и широкий, с 5—6 шипиками с каждой стороны; концевые коготки короткие, со слабой штриховатостыо. Длина самки 1 мм. Очень часто, в малых водоёмах и в прибрежье крупных. g mucronata (Muller).
0. Клюв самки длинный и острый. Глазок удлинённый, значительно удалён от конца клюва (рис. 246 Б). Передние антенны не достигают до него. Спинной край постабдомена с 8—9 шипиками с каждой стороны; концевые коготки сильно изогнуты, снаружи с грубой штриховатостыо. Длина самки 1,5 мм. В мелких водоёмах, не часто.
S. aurita (Fischer) (о b t u s a Schodler).
Род Ceriodaphnia Dana
1. При основании концевых коготков 2—7 коротких грубых шипиков (рис. 247). Раковина отчётливо разделена на многоугольные поля. Постабдомен короткий, его выпуклый спинной край с каждой стороны песёт по 7—10 шипиков. Длина самки 0,8—1,5 мм. Широко распространён, часто в маленьких водоёмах и в прибрежье больших. С. reticulata (Jurine).
0. Коготки без грубых шипиков при основании..............................2
2. Постабдомен на конце имеет выемку (рис. 248), на которой с каждой стороны сидят по 7—9 шипиков, постепенно увеличивающихся к концу постабдомена; концевые коготки с топкой штриховатостыо на внутренней стороне. Длина самки 1—1,3 мм. В маленьких водоёмах. с m е g о р s Sars.
0. Постабдомен на конце закруглён, без выемки............................8
8. Постабдомен в середине не расширен....................................4
0. Постабдомеп в середине сильно расширен................................7
4. Раковина без шипиков..................................................5
0. Раковина с шипиками па поверхности. Передние антенны довольно большие, выдаются за нпжний край головы; чувствительная щетинка каждой антенны отходит приблизительно от середины её. Постабдомен в основании широк, к концу сильно суживается,
Рис. 247. Ceriodaphnia reticulata — ковцепой коготок по-стабдомеяа.
по спинному краю с каждой стороны по 7—ю мелки шипиков, из которых средние — наибольшие. дЛИп’ самки 0,7—0,8 мм. В заболоченных лужах, в богатых растительностью местах. q s е t о s a Matile
5 Голова между глазом п передними антеннами имеет вздутие, большей частью снабжённое по краю очень мелкими шппиками (рис. 249В). Постабдомеп копоткий и широкий, песет с каждой стороны по две группы шипиков; ближе к концу постабдомена 7— 9 грубых п непосредственно рядом с ними (по направлению к началу постабдомена) 3—4 более топких и ипаче направленных шипика (шипикп первом группы к концу постабдомепа увеличиваются — рис. 249 Г). Длина самки 0,7—0,8 мм. Широко распространена и часто встречается, особеппо в прибрежье озёр, иногда в планктоне. с. р u 1 с h е 1 1 a Sara.
0. Голова не имеет указанного вздутия .... в в. Концевые коготки снабжены в основании длинным рядом тонких шетинок (видны только при сильном увеличении — рис. 249 Л). Постабдомен (рис. 249 Б) не широкий, к концу суживается, с 10—12 шипиками
каждой стороны, из которых средние - наибольшие. Длина самки 1—1,4 мм. прибрежье озёр. С. a f f i n i s Lilljeb.
О Конпевые коготки без шетинок. Постабдомен узкий, на выпуклой части его конца с каждой стороны ГТшипиков. Длина самки 0,6-0,8 мм. В мелких водоемах и в при-брежье крупных.
с
В
C. quadrangula (Muller).
Рве. 249. Ceriodaphnia:
А — С. afflnls; Б — её постабдомен; В — С. pulcbella, голова; Г — её же постабдомен; Д — С. rotunda; Е — её постабдомен.
7. Лобная часть головы более или мепее заострена и покрыта мелкими шипиками (рис. 249 Д). Снабжены шипиками также верхний и задпий края раковины и её шип. Постабдомен (рис. 249 Е} длинный, с 7—9 шипиками с каждой стороны. Длина самки 1 мм В заболоченных местах прибрежья крупных (иногда и более мелких) озёр.
С. rotunda Sara.
0. Лобная часть закруглена, без шипиков. Передние антенны короткие, едва доходят до нижнего края головы. Постабдомен (рис. 250) перед концом сильно расширен, с 9—10 шипиками с каждой стороны. Длина самки 1 мм. Пре имуществен по в прудах и л^жах. С. laticaudata Moller.
Род Moina Baird
Рис. 250. Ceriodaphnia laticaupata — постабдомен.
1. Концевые коготки постабдомепа без добавочных щетинок на вогнутой стороне Голова равномерно закруглённая, без вдавления над глазом (рис. 251 Г). Постабдомен (рис. 251 Д) с 7—И перистыми щетинками с каждой стороны. Длина самки около 1 мм. В прудах и мелких водоёмах.
М. m а с г о с о р a Straus (flagellate Hudendorff).
0. Концевые коготки с добавочными щетинками на вогнутой стороне ..................................... 2
*2. Нижний (брюшпой) край раковины усажен щетинками по всей своей длине. Лобная часть головы закруглённая, над глазом углубление (рис. 251 А). Постабдомен (рис. 251 Б) довольно широкий, к концу сильно суживается, с каждой стороны несёт по 9—14 перистых шипиков, из которых последний (у самого начала концевого коготка) длиннее и шире остальных и па конце расщеплён; концевые коготки с 9—14 щетинками па вогнутой стороне. Длина самки 1—1,6 мм. Всюду распространена, преимущественно илистым дпом и мутной водой. м recti rostris Leydig.
0. Нижыий край раковины несёт щетинки только в передней своей части. На лобной части головы глубокая выемка (рис. 251 В). Постабдомен с 8—10 перистыми зубцами с ка-
в мелких водоёмах с
Р и с. 251. Moina:
А — М. rectirostris; Б — её постабдомен; В — М. brachiata; Г — М. macrocopa; Д — её пост абдомен.
ждой стороны; концевые коготки с 8—9 длинными щетинками на вогнутой стороне. Длина самки 1,2—1,6 мм. В загрязнённых лужах. м brachiata (Jurine).
Сем. Bosmi nidae
1. Обе ветви плавательных антенн трёхчленистые. Голова ясно отчленена от туловища. Длина самки до 0,5 мм (рис. 252), Типичен для планктона рек (Волга, Днепр и др.), но живёт и в стоячих водоёмах речных систем, как старицы и т. п.
р. Bosminopsis, вид В. deitersi z е г п о w i (Linke).
0. Верхняя (спинная) ветвь плавательных антенн четырёхчленистая. Голова не отчленена от туловища.
P-
Baird
n a Baird.
Род В о s m i п 1. Концевые коготки длинных щетинок
Рис. 252. Bosminopsis daitersi ze- выроста постабдомена) и, кроме того, с рядом rnovi. мелких шипиков почти по всему вогнутому краю
рядом при основании
тонких n (на конце
В
a
c
о s m I
коготка (рис. 253 Г). Лоб щетинка отходит приблиз? тельно в середине расстояни между глазом п концом кп. вика (рис. 253 Я). Шип pQI'^ вины развит обычно хорошо перед его началом на пижпе°’ (брюшном) крае раковипы при’ креплена маленькая Щетинка Концевые коготки изогнуты не’ равномерно, образуя два слабо выраженных коленообразных изгиба — у самого основания коготка и вблизи его конца (рис. 253 Г). Длина самки 0,4-, 0,6 мм. Широко распространена, особенно в открытых местах прибрежной зопы крупных озёр, а также в планктоне.
В. longirostrig (Muller). 0. Короткий ряд из 5—7 шипиков расположен лишь у основания концевых, довольно равномерно изогнутых коготков (рис. 253 Б). Лобная щетинка ближе к концу клювика, чем к глазу (рис. 253 Л). Передние антенны большей частью очень длинные, иногда длиннее тела. Щетинка перед задне-нижним углом раковины может отсутствовать, шип раковины тоже. В планктоне крупных озёр.
В. coregoni Baird. Эти два вида распадаются на ряд форм, определительные таблицы которых мы не даём.
Сем. Macrothricidae
1. Концевые коготки постабдомена длинные, слабо изогнутые, с 2 базальными (расположенными у основания коготка) шипиками. Задний и нижний края раковины, постепенно закругляясь, переходят один в другой, оба усажены непрерывным рядом перистых щетинок. Клювик короткий и тупой. Передние антенны 2-членистые, 1-й членик маленький; антенны второй пары с 8 плавательными щетинками. Постабдомен очень большой, по спинному краю сильно вооружён многочисленными шипами (рис. 257—259).
р. Iliocryptus Sars.
Рис. 253.
A— Bosmina coregoni; Б — ее концевой коготок; В — В. iongirostris; Г — её концевой коготок.
Рис. 254. Lathonura rectiro- Рис. 255. Drepanothrix den-stris. tata.
0. Концевые коготки короче половины длины передних антенн; их базальный шипик короткий или совсем отсутствует............................................2
2. Передние антенны узкие, немного изогнутые, па конце суженные (рис. 254). Нижний край раковины с рядом придатков, имеющих форму листочков. Обе ветви плавательных усиков несут по 5 членистых и перистых щетинок. Постабдомен очень короткий, спинной край его пильчатый со щетинками; концевые коготки тоже короткие, без базальных шипиков. Длина самки 1 мм. На дне небольших болот и торфяных ям, в мелких водоёмах, иногда также в литорали озёр.
р. Lathonura Lili job., 1 вид L. rectirostris (Miiller).
0. Передние антенны на конце пе сужены................................3
3. На спине зубец, обращённый назад (рис. 255). Голова и туловище разделены спинной перетяжкой. Наружная (4-членистая) ветвь второй пары антенн с 3 плавательными шетинками, внутренняя (3-членистая) с 5. Постабдомеп короткий и широкий, на спинном краю зазуорен и густо усажен шипиками; концевые коготки короткие, изогнутые, 220
с маленьким базальным шипиком каждый. Длина самки 0,6—0,8мм. Среди водной растительности прибрежья озёр, в иле, также в мелких водоёмах; на севере СССР.
р. Drepanothrix Sars, 1 вид D. dentata (Ehren).
0. Без зубца на спине .......................................4
4. Передние антенны загнуты вперёд (рис. 256). Нижний и задний края раковины со щетинками. Задние антенны с 9 плавательными щетинками: четыре на 4-членистой ветви и пять па 3-членистой. Постабдомен короткий, суживающийся к копцу, спинной край посредине между концом и хвостовыми щетинками вдавлен и усажен шипиками; конце-вые коготки маленькие с очень мелкими доба- С
вочными шипиками. Длина самки 0,48—0,56 мм.
В мелких торфяных и заболоченных ямах, в прибрежной растительности озёр. /f
р. Streblocerus Sars, 1 вид / /
S. serricaudatus (Fischer). / I
0. Передние антенны не загнуты вперёд . . 5 I I (£>
б. Верхний край раковины со спинным ки- *
лем. 4-членистая ветвь задних антенн с 4 ще-тикками. Постабдомен большой, с шипами на * и с. 256. Sterblocerus serricau-спинном краю; концевые коготки мелкие. datus.
р. Macrothrix Baird.
0. Верхний край раковины без киля. 4-членистая ветвь задних антенн с 3 щетинками. Передние антенны несколько изогнуты назад и на конце слабо расширены. Щетинки заднего края раковины сверху вниз увеличиваются в длине и у нижне-заднего края достигают очень большой длины. Постабдомен небольшой, по спинному краю усажен многими рядами шипиков; концевые коготки в основании широкие, с 2 крепкими базальными шипиками каждый. Длина самки 1—1,7 мм. В мелких заболоченных водоёмах; особенно широко распространён на севере СССР.
р. Acantholeberis Lilljeb., 1 вид A. curvirostris (Muller).
Род I 1 i о с г у р t u.s Sars
1. Постабдомен на середине спинной стороны имеет небольшое углубление, разделяющее её на 2 выпуклые части................................................2
0. Постабдомен без такого углубления на спинной стороне; его равномерно загибающийся, выпуклый спинной край несёт шипы различной формы и величины (рис. 257):
Рис. 257. Iliocryptus acutif- Рис. 258. Iliocryptus sordi-rons — постабдомен. dus — общий вид.
1) рядом с хвостовыми щетинками 2 загнутых вперёд крепких зуба; 2) 1 непарный ряд, состоящий из 7 коротких, прямых, убывающих в величине к заднему концу постабдомена шипиков; 3) по одному ряду с каждой стороны, состоящему из 15 крупных, увеличивающихся в длине к концу постабдомена шипов. Концевые коготки на брюшном (выпуклом) краю у самого конца зазубрены. Длина самки 0,7 мм. Встречается редко.
1. acutifrons Sars.
2. Углубление на спинном краю постабдомена делит его на 2 приблизительно равные части, из которых передняя (около хвостовых щетинок) несёт И—14 непарных, загнутых назад и уменьшающихся по направлению к концу постабдомена шипиков, а задняя усажена двумя рядами шипиков с каждой стороны: внутренним рядом мелких шипиков и наружным, состоящим из 8—10 крупных, увеличивающихся к концу постабдо-___________ мена шипов. Задний и нижний края раковины уса-ь ( А *7 жены по всей длине крупными, перистыми шетин-
\ ... ками; верхний край раковины со слабым спин-
х. Р aj* ним килем (рис. 258). Длина самки 0,9—1,0 .км.
Большей частью на илистом дне озёр, чаше на ^^**ч*г 1 Xv глубине нескольких метров; наиболее обычный
вид в СССР. L so г d i d us (Lievin).
Рис. 259. llyocryptus agilis — no- 0. Углубление делит спинной край постаб-стабдомеп. домена на две неравные части (рис. 259)
меньшую переднюю, и большую заднюю, из которых первая несёт 8—9 непарных крепких прямых шипиков, вторая же усажена восемью маленькими шипиками, а сзади — двумя рядами их: внутренними маленькими и 7—8 длинными шипами. Длина самки 0,9—1 мм, Встречается редко; в илистых местах прибрежья крупных озёр. a gi 1 i s К
Сем. Chydoridae
1. Постабдомеп короткий и очень широкий, слабо выпуклый, спинной край его густо усажен многочисленными мелкими шипиками (рис. 260); между задним концом этого края и основанием концевых коготков глубокая выемка, на спинной стороне которой перед коготками ряд мелких шппиков; концевые коготки с двумя базальными шипиками. Спинной край раковипы сильно выпуклый, высокий, с килем, брюшной слабо вогнутый с густым рядом коротких щетинок. Длина самки до трёх с лишним миллиметров. Часто в мелких заросших водоёмах и в растительной зоне больших озёр. ™ „ „ „ „ Л „ „ „ e я -
r р. hurycercus Baird, 1 вид Е. lamellatus (Muller).
0. Постабдомеп другого строения. Рачки меньшей величины....................................2
Рис 260 Енгусеreus laniel- 2. Голова с высоким килем. Постабдомен очень ]atus. длинный и узкий, по бокам спинного края песёт по
ряду крепких шипов. На задне-нижнем углу раковины большей частью мелкие шипики, нижний край со щетинками. р Camptocercus Baird.
0. Эти признаки не встречаются вместе .... 8
8. Концевые коготки в середине вогнутого края с 1 или 2 шипами............................4
0. Этих шипов нет. Голова без киля......6
4. Постабдомеп средней длины, без шипиков, на боках с рядами маленьких пучков тонких щетинок (рис. 261); концевые коготки с большим базальным Рис. 261. Acroperus harpae. шипиком и рядом тонких шетинок на вогнутой стороне коготка, увеличивающихся от основания коготка к середине его, где прикреплена самая крупная конечная щетинка. Голова и спина большей частью с килем. На нижне-заднем углу раковинок обычно несколько зубцов. Длина самки 0,9—1 мм. Очень часто в прибрежной зоне больших озёр, а также в прудах и мелких водоёмах. р. Acroperus Baird.,
у нас широко распространённый вид A. harpae Baird.
0. Кроме рядов пучков щетинок, на постабдомепе есть ешё ряд крепких шипиков, идущий по его спинному краю. Голова без киля............................5
5. Клювик короткий и несколько притуплённый. Постабдомен приблизительно одинаковой ширины по всей его длине; на каждой его стороне ряд мелких и тонких шипиков и ряд пучков топких шетинок; концевые коготки с базальным шипом, а па вогнутой стороне с 1 или 2 большими и несколькими меньшими шипиками. р А 1 о п о р s i з Sars.
0. Клювик длинный и острый (рис. 262). Постабдомеп узкий, к концу суживается, по бокам спинного края несёт 10—12 шипиков и ряд пучков топких щетинок. Длина самки 0,6 мм. Редко, преимущественпо в мелких водоёмах.
р. К u г z i a Dyb. и Groch. (Pseudalona Sars), 1 вид К. 1 a t i s s i m a (Kurz).
6. Задний край расходящихся створок раковипы лишь немного пиже её наибольшей высоты................7
0. Задний край раковины значительно ниже её наибольшей высоты..................................10
7. Головной щит при виде сверху (со спины) шире раковины, при виде сбоку немного загнут вперёд (рис. 263). Раковина со спинным килем; нижний край её почти прямой, покрыт щетинками, впереди длинными и наиболее перистыми; на пижпе-заднем углу 1—3 зубца. Постабдомен в основании широкий, к концу суживается; спинной край его на каждой стороне с рядом отдельно стоящих мелких шипиков и рядом пучков (в числе 6—8) длинных топких щетинок; концевые коготки короткие с мелким базальным шипиком и со щетинками на наружном крае. Длина самки 0,50—0,68 мм. Часто, широко распространён, особенно в литорали крупных озёр, но также и в мелких водоёмах.
р. Graptoleberis Sars, 1 вид G. testudinaria (Fischer).
Рис.
262. Kurzia lalis-sima.
Рис.
263. Graptoleberis testudinaria.
0. Головной щит при виде сверху пе шире раковины, при виде сбоку пе загнут вперёд......................*................................................8
8. Постабдомеп очепь большой, сильно расширенный; па его спинном краю мелкие шипики, па боках группы щетинок, которые к концу постабдомепа становятся крепкими и далеко выдаются за ого край; концевые коготки длинные, базальный шип маленький пли его пет. р. Leydigia Kurz.
0. Постабдомеп не слишком расширен, вооружение его спинного края слабее . . 9
9. Клювик вытянут в более или менее длинное остриё. На последнем членике внутренней ветви задних антепп три плавательных щетинки. На нижне-заднем углу раковипы
1 или несколько зубцов. Конец постабдомепа закруглён; концевые коготки с маленьким базальным шитиком.
р. Rhynchotalona Norman (Leptorhynchus).
0. Клювик не вытянут в длинное остриё, короткий и не очень острый. На последнем членике внутренней ветви задних антенн 4 или 5 плавательных щетинок. На обеих сторонах постабдомепа шипы, расположенные рядами или группами, и большей частью пучки щетинок; концевые коготки с базальным шипом, но без срединного шипа на вогнутой стороне.
р. А 1 о n a Baird.
10. Тело при виде сбоку более или менее шарообразное..............11
0. Тело не шарообразное .... 18 край раковины с довольно глубокой выемкой, позади которой оп усажен щетинками (рис. 264). Клюв длинный, острый, конец его несколько загнут назад. Постабдомеп на конце узкий, его верхне-задний угол сильно выдаётся за основание концевых коготков, па конце края постабдомена два ряда — в каждом по 10—И мелких шппиков; концевые коготки очень длинные, с двумя очень маленькими базальными шипиками и рядом щетинок на коготке. Длина самки около 0,5 мм. Нередко, па дне или среди растительности озёр, на гидрах.
р. Anchistropus Sars., 1 вид Л. е m a rgi natus Sars.
0. Брюшной край раковины без выемки...............................12
12. Есть глаз и глазок. Нижний край раковины в задней половине складчатый и густо усажен щетинками. Спинной край постабдомена с пятью рядами шипиков. Концевые коготки с 2 базальными шипами.
р. Ghydorus Leach.
0. Есть только глазок, глаза нет. Во время линьки раковина пе сбрасывается, а
Рис. 264. Anchist-ropus emarginatus.
’ll. Брюшной
A
Рис. 266. Peracantha truncata.
— Camptocercus recti rostrls; Б — его постабдомеп.
накладывается па старую, в результате чего общая раковина животного оказывается состоящей из песколысих черепицеобразно покрывающих одна другую раковинок с концентрическими краями, что придаёт ей некоторое сходство с раковиной двустворчатых моллюсков (рис. 265). Из-под последней (верхней, наименьшей) раковинки почти вертикально свешивается узкая голова с заострённым клювом и довольно крупным глазком. Постабдомеп короткий и широкий, к концу суживается; с каждой его стороны сидят 5—7 шипиков, а над ними пучки очень топких щетинок; концевые коготки с одним базальным шипом и с несколькими тонкими щетинками. Длина самки 0,5 мм. Нередко на илистом или песчаном грунте литорали озёр.
р. Monospi 1 и з Sars, 1 вид М. d i s р а г Sars.
13. Клювик короткий и тупой. На пижпе-задпем углу раковины зубцы или неровности, ппжний край её со щербинками; раковина со структурой в виде полей или полосок. Постабдомеп без боковых пучков щетинок, только с рядом шипиков на каждой стороне спинного края; концевые коготки с 1 или 2 базальными шипами.
р. А 1 о n е 1 1 a Sars.
0. Клювик длинный и острый..........................................14
14. Задний край раковины зазубрен по всей длине (рис. 266), нижний край усажен пористыми щетинками; раковина продольно исчерчена. Постабдомен небольшой, к концу суживается; на спиппом краю его два ряда мелких шипиков по 12—20 в каждом ряду; концевые коготки с 2 базальными шипиками. Длина до 0,7 мм. В мелких водоёмах и в прибрежье озёр. р. Peracantha Baird, 1 вид Р. truncate (Muller).
0. Задний край раковины не зазубрен.................................15
15. Постабдомен вооружён рядом отдельных или расположенных мелкими группами шипиков; концевые коготки с 2 базальными шипами. На пижне-заднем углу раковипы большей частью зубцы. р. Pleuroxus Baird.
0. Постабдомен с густыми поперечными рядами тонких шипиков; концевые коготки с одним базальным шипом. На нижне-заднем углу раковины 1 крупный, притуплённый, направленный назад зубец. Длина самки 0,4—0,5 мм. В мелких водоёмах и в литорали озёр; распространён на юге СССР.
р. Dunheveaia King (С г е р i d о с е г с u s Birge), 1 вид D. crassa King (С. s е t i g е г Birge).
Род Camptocercus Baird
Рис. 268. Alonopsis elongata.
1. Постабдомен самки узкий, с 15—17 короткими крепкими шипиками с каждой стороны, увеличивающимися к его концу. Кроме того, на его боках по ряду пучков очень тонких шетинок; концевые коготки длинные с крепким базальным шипом и с рядом тонких щетинок на вогнутой стороне, увеличивающихся к середине коготка. На задне-нижнем углу раковины 3—5 зубцов; нижний край волнистый, со щетинками. Голова направлена вперёд, клювик острый. Длина самки до 1,4 мм. Встречается часто, преимущественно в прибрежной зоне крупных водоёмов и в мелких заросших водоёмах; плавает тяжело, обычно ползает в иле. С. rectirostris (Schodler).
0. Постабдомен самки очень длинный и узкий, с 20—23 шипиками с каждой стороны спинпого края; концевые коготки, как у предыдущего вида. Нижний край раковины впереди со щетинками, на задне-нижнем углу её 1—4 зубца. Кончик клюва срезан. Длипа самки 0,9—1,1 мм. Встречаемость и образ жизни, как у предыдущего вида.
С. lilljeborgi Schodler.
Род Alonopsis Sars
1. Постабдомен с 16—17 шипиками, увеличивающимися к его концу (рис. 268); концевые коготки с большим перистым базальным шипом, с рядом тонких щетинок и с отдельно стоящим у заднего конца коготка крепким шипом. Раковина ясно исчерчена в косом направлении вниз; её задне-нижний край обычно с мелким зубчиком. Длина самки до 1 мм. Довольно часто в зарослях прибрежья озёр.
A. elongata Sars.
0. Постабдомен с шипиками по спинному краю и с 18—20 пучками па боках, в каждом пучке по 3— 4 тонких шипика; концевые коготки, как у предыдущего вида, за исключением отсутствующего здесь последнего шипа на вогнутой стороне коготка. На нижне-заднем углу раковины зубцов нет.
A. ambigua Lilljeborg. Рис. 269. Rhynchotalona rostrata.
Род Leydigia Kurz
1. Концевые коготки постабдомена с маленьким базальным шипом; передние пучки на постабдомене состоят из нескольких тонких щетинок. Раковина гладкая. Длина самки 0,8—0,96 мм. На дне илистых водоёмов, в озёрах, также на глубоких местах.
L. leydigi (Schodler).
0. Концевые коготки без базального шипа; передние пучки на постабдомене состоят лишь из двух довольно крепких шипиков. Раковина большей частью ясно исчерчена. Длина самки 1—1,1 мм. Там же, где и предыдущий вид.
L. acanthocercoides (Fischer).
Рол Rhynchotalona Norman.
1. Клюв вытянут в длинный, тонкий, более или менее изогнутый вырост. 11а задне-нижнем* углу раковины один зубец, нижний край со щетинками; раковина большей частью с ясно продольной исчерченностью. На обеих ветвях задних антенн одна иа трёх конечных щетинок меньше двух остальных, а шины на конечных члениках этих антенн очень длинные. Постабдомен довольно толстый, короткий, на копне его спинного Жая с каждой стороны 4 крепких шипика, а на боках по ряду длинных тонких щетинок, тина самки 0,5 мм. В песчаных местах прибрежной зоны озёр.
R. f а 1 с a t a Sara.
0. Клюв несколько удлинённый, но без указанного выше выроста (рис. 269). Раковина косо исчерчена, на её задне-нижнем углу 1—3 зубца. Нижний к^ай несколько волнистый, с короткими щетинками. Из трёх конечных щетинок внутренней ветви задних антенн одна короче двух прочих. Постабдомен на спинном крае несёт с каждой стороны по 10—13 простых шипиков. Длина самки 0,5 мм. На песчаных местах прибрежья больших водоёмов в СССР, особенно часто в реках.
R. rostrata (Koch).
Род А 1 о о a Baird
1. Постабдомен к концу суживается, пе закруглён..........................2
0. Постабдомеп к концу не суживается, конец закруглён...................4
2. Удлинённый, не широкий постабдомен на спинном крае усажен с каждой стороны 10—20 шипиками, из которых 3—5 последних (самых задних) гораздо больше осталь
ных; вторичное вооружение 1 слабое; концевые коготки с S-образно изогнутым базальным шипом. Длина самки 0,6 мм. Преимущественно в болотистых водах.
A. tenuicaudia Sars.
0. Последние шипы постабдомена лишь немного больше прочих........3
3. Постабдомен короткий и довольно широкий, его спиннои край вооружён двумя рядами — по 10—13 в каждом — коротких шипов, постепенно увеличивающихся к концу постабдомена (рис. 270 В}\ параллельно с этим рядом на боках вторичное вооружение
1 Ряд мелких шипиков или щетинок, идущий параллельно спинному краю постабдомена на некотором расстоянии от этого края.
15 А. Н. Линии * 225
в виде двух рядов (по 1 с каждой стороны) пучков очень мелких и тонких щетинок. Нижний край раковины по всей длине густо усажеп короткими щетинками; штрпховатость раковины ясно выражена. Длина самки 0,5 мм. Часто в прудах, а особенно в мелких зарос-in их водоёмах и в прибрежных зарослях озёр; этот вид и три следующих имеют наибольшее распространение в СССР.
A. costata Sars.
0. Постабдомен с 7—10 шипиками с каждой стороны спинного края, без вторичного вооружения (рис. 270 Г). Длина самки 0,4 мм. В тех же местах, где и предыдущий вид.
A. guttata Sars.
4. Постабдомен большой, широкий, с расширенным концом, с 16—18 шипиками с каждой стороны спинного края и с вторичным вооружением из пучков топких щетинок (рис. 270 В). Длина самки 0,65—0,70 мм (рис. 270 А). Очень часто, особенно в прибрежной зоне крупных водоёмов.
A. quadrangular) s (Muller).
0. Постабдомен короткий, умеренно широкий, на конце закруглённый; с каждой стороны спинного края по 7—9 групп шипиков, в каждой группе 1—3 шипика (рис. 270 Д); вторичного вооружения нет. Длина самки 0,25—0,50 мм. В прибрежной зоне больших водоёмов, но также и в мелких водоёмах.
A. rectangula Sars.
Род Chydorus Leach
1. Постабдомен длинный и узкий (рис. 271 А); конец спинного края выдаётся за основание концевых коготков; по спинному краю два ряда очень мелких щетинок; концевые коготки очень длинные, с 2 базальными шипами и рядом тонких щетинок на вогнутой стороне. Клюв длинный и острый. Длина самки 0,8—0,9 мм. В литорали озёр и в мелких водоёмах, плавает между растениями.
Ch. globosus Baird.
0. Постабдомеп короткий и широкий...................................2
2. Спинной край постабдомена с рядам и не более чем из 10 шипиков (рис. 271 В). Форма тела то шарообразная,то овальная. Длина самки меньше0,5лелс. Наиболее распространённая и часто встречающаяся форма из всех Cladocera всяких водоёмов.
Ch. sphaericus Muller.
0. Спинной край постабдомена с рядами из 10—15 мелких шипиков. Длина больше 0,5 мм ... 8
3. Спинной край постабдоменов с 2 рядами —
Рис. 271. Из 12—15 каждый —мелких
А — Chydorus globosus; Б — Ch. sphaericus^ шипиков; брюшной край с
5—6 мелкими надсечками. На переднем крае антен 1-й пары в середине длинная щетинка, ближе к концу антенны 2-я шетинка; 2-я пара антенн с 8 плавательными щетинками. Нижнии край раковины и нижняя часть заднего края со щетинками, наиболее длинными в задней половине нижнего края. Длина самки 0,6 мм. В мелких водоёмах; форма пе частая.
С h. ovalis Kurz.
0. Спинной край постабдомепа с 2 рядами — из 10—12 каждый — мелких шипиков; брюшной край гладкий. Передний край антенн 1-йпары лишь с одной щетинкой (в середине); 2-я пара антенн с 7 плавательными щетинками, только задняя половина нижнего края раковины несет щетинки. Длина самки 0,6 мм. Довольно редкая форма.
Ch. 1 a t u s Sars.
Род А 1 о n е 1 1 a Sars
1. Спинной край постабдомена с 6 шипиками с каждой стороны; концевые коготки с одним базальным шипом. На закруглённом задне-нижнем углу раковины один зубец. Продольная исчерченность раковины резко выражена и направлена спереди и спизу назад и вверх; полоски нижней половины раковины впереди загибаются вниз. Длина самки 0,26 мм. В литоральной зоне озёр и в мелких водоёмах.
А. папа (Baird).
0. Спинной край постабдомена с 9—10 шипиками с каждой стороны. Задне-нижний угол раковины заострённый. Штрпховатость раковины направлена спереди и сверху назад и вниз.................................................................. *
2. Задний край раковипы около задне-нижнего угла волнистый пли тупо зазубренный; несколько вогнутый нижний край покрыт перистыми щетинками (рис. 272). Концевые коготки с 2 базальными шипами — большим и очень маленьким. Длина самки 0,4 -и.м.
Встречается в общем там же, где предыдущий вид.
0. Задне-нижний угол раковипы с довольно крупным зубцом, обращённым назад; кроме того, 1 или 2 зубца над ним и! впереди его (па нижнем краю). Постабдомеп с 10 шипами с каждой стороны. Длина самки 0,35 мм. Встречаемость, как у предыдущих видов.
А. е х i g u a (Lilljeb.).
А. о х ci ч а (F'ischer).
Рис. 273. Pleuroxus striatus.
Рис. 272. Alonella cxcisa и её постабдомеп.
Род Pleuroxus Baird
УТ У Г"
1. Постабдомен длинный, спинной край более или мепее вогнутый (рис. 273), на нём с каждой стороны по 20—21 мелких шипиков. Раковина удлинённая с продольной штриховатостью; на задне-нижнем углу её 1 чрезвычайно мелкий зубчик. Концевые коготки с 2 базальными шипиками — коротким и длинным — и рядом тонких щетинок за ними. Длина до И мм. Р. з t г i a t u s Schodl.
0. Постабдомеп короткий, спинной край прямой илп выпуклый . . 2
2. На спинном краю постабдомепа 2 ряда очень тонких щетинок, расположенных группами; внутренний базальный шип концевых коготков очень мелкий. На задне-нижнем углу раковины 1 — 3 маленьких зубчика. Длина самки 0,66 мм. Часто в прибрежной зоне озёр и в мелких водоёмах.
Р. aduncus (Jurine).
0. На спинном краю постабдомепа 2 ряда отдельно стоящих крепких шипов...........3
3. Концевые коготки большие, равномерно изогнуты, маленький базальный шип не более */2 большого. На задпе-пижнем углу раковипы 1 — 3 мелких зубчика. Клюв длинный и острый, его конец направлен вниз и назад. Длина самки 0,6 мм. Встречаемость, как у предыдущего вида.
Р. trigonellds (Muller).
Б
Рис. 274. Pleuroxus uncinatus:
А — клювик; Б — постабдомеп.
0. Концевые коготки волнистые (с вогнутостью посредине), маленький базальный шип более х/г большого (рис. 274). На задне-нижнем углу раковины большей частью 2— 4 зубца. Раковина в большинстве случаев покрыта посторонними частицами. Конец клюва чаще всего несколько загнут вперёд, иногда лишь направлен прямо вниз. Длина до 0,6 мм\ значительно распространён, хотя и не часто встречается.
Р. unci natus Baird
EUCOPEPODA — СВОВОДНОЖИВУЩИЕ ВЕСЛОНОГИЕ РАЧКИ
Eucopepoda (рис. 275) имеют удлинённое, ясно расчленённое и лишённое раковины тело с более или мепее заметным подразделением на два отдела: головогрудь и брюшко (а б д о ме и), оканчивающееся вилкой, ветви которой носят название хвостовых ветвей и снабжены тем или иным количеством щетинок. На переднем конце головогруди прикреплены две пары усиков (антен н), из которых первая всегда длинное второй. Головогрудь песет, кроме того, целый ряд конечностей: за 2-ми антеннами следуют 4 пары ротовых конечностей — жвалы (мандибул ы), пе редкие челюсти (м а кс и л л ы), задние челюсти и ч е л юст н ы е (м а к с и л-лярные) ножки, а затем идут плавательные ножки в количестве 5 пар, но 5-я пара обычно сильно недоразвита. У веслоногих рачков, в отлично от ветвистоусых, плавательные ножки пе листовидные, так как не обладают никакими пластинками, а устроены по типу двуветвистых конечностей.
Партеногенетический способ размножения у свободно живущих веслоногих почти не встречается х. Оплодотворённые яйца самка откладывает и вынашивает в особых я й ц е-
1 За исключением двух видов подотряда Harpacticoida. *
вых мешках, прикрепляемых к абдомену в количестве одного (рис. 275 Б) или двух (рис. 275Л). Здесь яйца проходят первое своё развитие, пока из них не вылупятся личинки науплиусы (рис. 275 В}, несущие три пары конечностей, из которых две первые пары представляют зачатки двух пар усиков взрослого рачка, а третья — зачатки жвал. Выйдя наружу, эти личинки начинают вести самостоятельную жизнь, во время которой постепенно развиваются во взрослых рачков.
Пресноводные Eucopepoda делятся па 3 подотряда — Calanoida, Cyclopoida и Награ-cticoiaa.
1. Формы мелкие — меньше 1 лм<. Тело удлинённое, узкое. Головогрудь незаметно переходит в абдомен. Передние антенны очень короткие, состоят из 5—8 члеников (у самца обе антенны превращены в хватательные органы). У самки яйцевой мешок один.
Подотряд Harpactieoida.
0. Формы более крупные — в большинстве случаев более 1 мм. Головогрудь ясно отграничена от абдомена. Передние антенны более или менее длинные,состоят из 8—25члеников ................................................................ 2
Рис. 275. Веслоногие рачки — Copepoda: Л—Cyclops; Diaptomus; В — лмчшша науплиус; Г — Oanthocamptus.
2. Передние антенны состоят из 8—17 члеников,длина их пе превышает длину голове-груди; у самца обе антенны превращены в хватательные органы. Яйцевых мешков два.
Подотряд Cyclopoida.
. 0. Передние антенны состоят из 22—25 члеников, длина их превосходит длинуголово-3>уди; у самца только одна из них, обычно правая, превращена в хватательный орган, йцевой мешок один.
Подотряд Calanoida.
Подотряд Calanoida заключает в себе 4 семейства — Centropagi d а е, Di aptomidae, Temoridae и Pseudodi a ptomi dae. Наибольшее число видов, часто встречающихся в пресных водах, содержит сем. Diaptomidae, а из него— р. Di а р t о ш u s Westvv., который на территории Союза представлен более чем 60 видами; из них наиболее распространены в СССР 2 вида! D. g г а с i loides Lili, и D. g г а с i 1 i s Sars, населяющие пелагиаль озёр северной и средней части СССР; первый вид живёт и в прудах.
Подитряд С у с 1 о р о i d а представлен в СССР лишь одним семейством, содержащим обитателей пресных вод — сем. Cyclopidae. Не так давно к этому семейству относили всего лишь один род Cyclops. В настоящее время из него выделен ещё целый десяток новых родов, названия которых произошли путём прибавления к слову Cyclops (впереди его) различных приставок; Eucyclops, Macrocyclops, Ectocyclops, Mesocyclops и т. д. 228
Подотряд Harpacticoida (рис. 275 Г) включает в большинстве очень мелкие прибрежные и придонные формы. Разбираться в них неопытному человеку настолько трудно, что мы считаем излишним останавливаться на них. Желающих ознакомиться с ними более подробно отсылаем к работе Боруцкого (см. список определителей).
Определение семейств и родов Eucopepoda основано главным образом па различиях в строении конечностей, из которых наиболее важное систематическое значение имеет 5-я пара грудных ножек, в большинстве случаев редуцированных в той или иной степени, а также на числе и строении члеников антенн. Таким образом, определение веслоногих обязательно сопряжено с их препаровкой, состоящей в отделении копечностей от тела, абдомена от головогруди и т. п. Для новичка это представляет большие трудности, особенно в отношении мелких форм, каковыми являются очень многие Cyclopoida; поэтому мы не приводим для веслоногих ракообразных таблицу определения, а ограничиваемся лишь описанием представителей двух главных подотрядов — Calanoida и Cyclopoida, именно — Diaptomus и Cyclops (виды которых наиболее распространены в паших пресных водах), чтобы дать общее представление об организации этих животных.
Если кто-нибудь пожелает познакомиться с методами определения свобод ножи пущих веслоногих, то рекомендуем обратиться к определителям Рылова (см. список в конце книги).
Род Diaptomus Westwood (рис. 275 Б)
Головогрудь явственно отграничена от брюшка и разделена на 6 сегментов, из которых два последних вследствие взаимного слияния не ясны или совсем не отличимы друг от друга. Брюшко самки состоит из трёх сегментов, самца — из пяти. Длина ветвей хвостовой вилки не более чем в три раза превышает их ширину; на конце каждой ветви сидят 6 веерообразно расходящихся щетинок, из которых одна — крайняя внутренняя — маленькая и простая, пять же остальных — более длинные и перистые. Усики первой пары 25-членистые, очень длинные — не короче, чем до абдомена, иногда же —до конца хвостовых ветвей или даже (редко) ещё несколько длиннее. В период размножения самка вынашивает яйца в одном мешке. Длина наиболее распространённых видов — 1—2,5 мм. но некоторые достигают даже 5 мм. Встречается преимущественно в пелагической области озёр, но некоторые виды живут в прудах и даже в пересыхающих лужах.
Род Cyclops Mull. (рис. 275 А)
Головогрудь ясно отличима от брюшка и состоит из 5 сегментов. Брюшко разделено у самки на 4, у самца — на 5 сегментов. На конце каждой из хвостовых ветвей, иногда очень длинных, по 4 щетинки, из которых 2 средние всегда значительно длиннее двух крайних. Усики первой пары, состоящие не более чем из 17 члеников, никогда не длиннее головогруди. В период размножения самка вынашивает яйца в двух мешках. Длина относящихся к этому роду многочисленных видов колеблется в очень широких пределах, приблизительно 0,5—5 мм, но большинство — около 1—2 мм. Немногие лишь виды встречаются в пелагической области озёр, большинство является обитателями прибрежной области; кроме озёр, живут и в водоёмах других типов — в прудах, канавах, лужах и пр.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ НЕКОТОРЫХ СЕМЕЙСТВ ГРУППЫ HYDRACHNELLAE - ПРЕСНОВОДНЫХ КЛЕЩЕЙ
Мелкие животные, не достигающие длины 1 см, шарообразные, яйцевидные или сжатые в спинно-брюшном направлении, большей частью ярко окрашенные. Нет расчленения ни на отделы тела, ни на сегменты. На переднем конце тела расположены 4 г да з а — по паре на каждой стороне (рис. 277). Конечностей 6 пар: 2 первых пары — ротовые, 4 следующих — плавательные ножки, все на брюшной стороне. 1-я пара — 5-членистые щупики, или п а л ь п ы, прикреплённые к так называемому м а-ксиллярному органу, у многих форм вытянутому вперёд в виде более или менее длинного, обычно изогнутого хоботка(рострума) с ротовым отверстием на конце; 2-я пара ротовых конечностей — ж в а л ы, или хелицеры, помещаются внутри максиллярного органа. 4 пары 6-членистых плавательных ножек прикреплены соответственно к 4 парам расположенных на брюшке крупных хитоновых пластинок — так называемых э п и м е р, то распределённых в группы, то сросшихся в один общий щит. Позади эпимер помещается половой орган, наружно представленный половым отверстием в виде продольной щели и расположенными по бока^м её двумя мягкими кожистыми полукруглыми или полуовальными пластинками — половыми заслонками, часто покрытыми мелкими присосками.
По местообитанию пресноводных клещей можно разбить па 2 главных группы: 1) обитателей стоячих водоёмов — озёр (в которых они населяют главным образом прибрежную зарослевую зону) прудов, луж и т. п. и 2) обитателей текучих вод — преимущественно ключей и ручьёв. Отсутствие или наличие движения воды (течения) накладывает отпечаток не только на образ жизни, по и на морфологию водных клещей. Так. обитатели стоячих вод, к которым принадлежит громадное большинство этих животных, характеризуются относительно крупным выпуклым (вальковатым) телом, длинными ножками с плавательными волосками, большим числом мелких яиц и пр., между тем как для обитателей текучих вод характерны малая и часто сплюснутая в спинно-брюшном направлении
форма тела, короткие, л пшённые полоскоп ножки с крупными коготками па копне, малое число крупных яиц и т. д.
Из 26 семейств группы Н у d г а с h n е I 1 а е в приводимую определительную таблицу включены только 8 семейств, содержащих наиболее обычные и широко распространённые роды.
Представители всех приведённых в таблице семейств характеризуются 5-члепистыми пальцами и 6-члепистымп плавательными ножками.
1. Глаза сближены и расположены па общей хитиновой пластинке пли в капсулах, соединённых между собой хитиновым мостиком. Половой орган без присосок, которые разбросаны по коже. Тело красное.............................................2
0. Глаза в капсулах, раздвинутых к бокам тела п не соединённых мостиком. Половой орган с присосками, расположенными около полового отверстия. Тело разной окраски .................................................................... 3
2. 2 пары глаз па боковых выступах переднего конца узкой продольной хитиновой пластинки. Пальпы маленькие (рис. 27Ь). Ножки без длинных волосков. Тело мягкое, изменчивой формы. Длина до 4 мм. Живёт в иле мелких водоёмов.
Сем. Limnocbaridae, 1 вид Limnochares aquaticaL.
0. 2 пары глаз па концах поперечно расположенного хитинового мостика (рпс. 277 £>, В). Пальпы длинные, выходящие за пределы переднею конца тела. Только задняя пара ножек без длинных волосков (рис. 277 Л). Длина до 7—8 мм. В стоячих водоёмах — прудах, лужах, канавах, в прибрежных зарослях озёр.
Рис. 278. Hydrachnai
А — с брюшной стороны; Б — рисунок па спине.
3. Глаза в сильно хитинизировапных капсулах............... 4
0. Глаза без капсул, на каждой стороне тела в виде двух раздельных глаз или в виде одного двойного глаза ............................................. 6
4. Максиллярный орган с узким, длинным, загнутым в виде клюва хоботком. Ноги в густых волосках (рис. 278 Л). Тело шарообразное, красное, иногда с симметричным рисунком из частично слившихся чёрных пятен (рис. 278 Б). Крупные формы до 8—9 лги длиной (Н. geographic а). В мелких стоячих водоёмах.
Сем. Hydrachnidae, у нас 1 род Hydrachna О. F. Muller.
9
мелкие сосочки. Длина
Рис. 279. Piona сагпеа — конец ноги.
0. Максиллярный орган без клювообразного хоботка. 2-й членик пальп с бугорком, несущим щетинки. Длина до 2 мм (большей частью меньше 1 мм). В текучих водах — ключах в ручьях.
Сем. Sperchonidae
5. 4-й членик пальп с 2 зубчиками на брюшной стороне.
Sperchon Kramer.
0. Этот членик без зубчиков, но с 1 бугорком.
р. Sperchonopsis Piersig, у нас 1 вид S. verrucosa (Protzi).
6. Глаза на каждой стороне тела раздельные...................7
0. Глаза на каждой стороне тела соединены в 1 двойной глаз...8
7. На конце каждой пальпы клешня, образуемая удлинением конца 4-го членика и 5-м члеником ; на 2-м членике пальп нет придатков. На конце задних ножек коготки. Эпимеры образуют 4 группы. Половые заслонки густо покрыты сосочками. Тело обычно оранжевокрасное, иногда с чёрным рисунком; на поверхности до 2 мм. Очень часто, во всевозможных стоячих водоёмах.
Сем. Diplodontidae, у нас 1 вид Diplodontus despi-с i е n s (Muller).
0. Клешней на пальпах нет; на 2-м членике бугорок или зубчик. На задних ножках, вместо коготков, шипик. Эпимеры расположены в 4 группы. Половые заслонки с 3 сосочками каждая. Длина до 2 мм (очень изменчива). Широко распространена в стоячих водоёмах.
Сем. Limnesiidae, у нас 1 род L i m n е s i a Koch.
8. Спина не покрыта панцирем, кожистая, иногда лишь с хитиновыми пластинками. Тело обычно не уплощённое. Эпимеры расположены в 4 группы. Ножки с плавательными волосками; их коготки с ногтевой пластинкой и добавочным коготком (рис. 279). До 3 мм, чаще меньше. Широко распространена в стоячих водоёмах.
Сем. Pi о n i d а е.
Это семейство состоит из 4 подсемейств, содержащих 9 родов с многочисленными гидами.
0. Спина покрыта панцырем. Тело сжато в спинно-брюшном направлении. Эпимеры срослись в одну группу; их задние края, соединяясь, образуют общую глубокую выемку, внутри которой помещается половой орган (рис. 280). 4-и членик пальп тонкий, по длине превосходит 2-й и 3-й, вместе взятые. Длина до 0,8 мм. В стоячих, реже в медленно текущих водах.
Сем. М i d е i d а е, у нас 1 вид Mideaorbiculata (MOIL),
Рис. 280. Midea orbt-culata — брюшная сторона.
INSECT! — НАСЕКОМЫЕ
Свободноживущие водные насекомые и водные личинки насекомых
Мы видели, что у ракообразных три отдела тела — голова, грудь и брюшко, или абдомен, — если не целиком, то более или менее сливаются между собой, образуя в большинстве случаев лишь два отдела — головогрудь и брюшко. Не то мы находим у насекомых; для них чрезвычайно характерно постоянное разделение тела на все три упомянутых отдела, причём грудной отдел всегда состоит из 3 сегментов, а брюшной — то из 9, то из 10. Что касается числа конечностей, то по сравнению с ракообразными оно значительно меньше. Прежде всего у насекомых имеется не две, как у раков, а только одна пара членистых йнтенн, или у с и к о в, число члеников которых часто является определительным признаком, а затем ротовых конечностей у взрослых насекомых бывает разное количество, во не более 3 пар, причём они могут быть видоизменены до неузнаваемости их основного первоначального типа; наконец, ходильных ножек у насекомых всегда 3 пары — по одной паре на каждом из трёх члеников груди, на спинной стороне которой в большинстве случаев прикреплены две (редко одна) пары крыльев; па брюшке никогда не бывает конечностей, лишь па своём заднем кон^е оно иногда несёт различного рода придатки, вроде, например, хвостовых нитей у веснянок и подёнок, дыхательной трубки у водяного скорпиона и т. п.
Все водные насекомые развиваются с превращением (метаморфоз). У крылатых насекомых существует два главных вида превращения: полное (сетчатокрылые, двукрылые, перепончатокрылые, жуки, бабочки) и неполное (клопы, стрекозы, подёнки, веснянки). При полном метаморфозе насекомое, по вылуплении из яйца, проходит три стадии развития; личинку, куколку и взрослую стадию—так называемую имаго; характерной чертой полного превращения является стадия куколки, которая в большинстве случаев бывает неподвижной и вносит очень резкие различия между личинкой и имаго (взрослым насекомым) как по строению, так я по образу жизни. При н е п о л-
ном метаморфозе личинка более или менее (за исключением крыльев) с х о а н а со взрослым насекомым и превращается в него постепенно, посредством целого ряда линек; пока у личинки не появились зачатки крыльев, она сохраняет своё название личинки, личинку же с зачатками крыльев часто называют нимфой. У некоторых насекомых (клопы) различия между нимфой и имаго очень незначительны и сводятся почти исключительно к длине крыльев. Различают ещё один вид превращения — так называемый анаморфоз, имеющий очень ограниченное распространение среди водных насекомых и встречающийся лишь у низшей группы насекомых — бескрылых. При анаморфозе из яйца выходит насекомое, незначительно отличающееся от взрослой формы, в которую превращается путём линек.
В подавляющем большинстве случаев различия между видами распространяются как на стадии личинки и куколки, так и на стадии имаго (исключение составляют лишь некоторые тендипедиды, стрекозы). С другой стороны, личинки водных насекомых (в широком смысле этого слова) как по количеству, так и по своему влиянию на общую экономику водоёма занимают отнюдь не меньшее, а скорее, большее место, чем сами эти насекомые во взрослом состоянии, так что знание личинок и умение их определять не менее важно, чем то же по отношению к имаго. Поэтому приводимый ниже определитель включает как личинок, так и имаго, причём центр тяжести перенесён скорее на первых.
Остановимся кратко на тех органах и частях тела, которые имеют коренное систематическое значение и положены в основу определения.
Крылья: имеет значение их количество (одна или две пары), длина, строение-(одинаковы обе пары или разные) и жилкование (число и расположение жилок на крыле и образуемых ими ячеек).
Три пары грудных членистых ножек: их присутствие или отсутствие имеет значение лишь при определении личинок.
Ложные ножки: это мягкие членистые выступы, или бугорки, расположенные большей частью на нескольких задних брюшных сегментах личинок некоторых насекомых — по одной паре на каждом сегменте (характерны главным образом для гусениц бабочек).
Ротовые органы, как и у ракообразных, — это видоизменённые конечности (в числе трёх пар, редко менее), причём основное, типичное их строение (жевательные органы, приспособленные для раздробления пищи) сохраняется лишь у некоторых групп насекомых (например, у прямокрылых, у жуков), у других же эти органы претерпевают то или иное изменение, приспособляясь к способу питания насекомых; так, у клопов они образуют острый хоботок для прокалывания внешних покровов и для высасывания внутренних жидкостей из организмов (сок растений или кровь животных); у бабочек ротовые органы превращаются тоже в хоботок, но другого устройства, чем у клопов, приспособленный к высасыванию нектара из цветов, и т. п. В связи с ротовыми органами следует упомянуть и о таком органе, как «маска», по которой легко и безошибочно можно отличить личинок стрекоз от личинок всех других насекомых (описание маски даётся дальше).
Хвостовые (концевые) придатки брюшка, их строение, форма, величина, количество и расположение характеризуют некоторые группы водных насекомых как личинок, так и имаго, например, хвостовые нити подёнок и веснянок, хвостовые жабры личинок некоторых стрекоз и т. д.
Боковые придатки брюшка — это так называемые трахейные жабры различной формы (листовидные, нитевидные, кистевидные) у личинок подёнок и некоторых ручейников.
Ч е х л и к и, которые многие личинки строят себе для защиты, как личинки ручейников, играют тоже некоторую роль для систематики, а следовательно, и для определения этих личинок; точно так же имеют значение в этом отношении те крючки на заднем конце тела, посредством которых указанные личинки цепляются за чехлик с внутренней стороны и удерживают его на себе.
Краткое описание более мелких признаков различия приводится в каждом случае отдельно, т. е. предпосылается соответствующим определительным таблицам.
Ещё одно замечание к определению насекомых. Рамки книги не позволяют нам дать таблицы для определения (хотя бы до рода) всех насекомых, которые по циклу своей жизни и развития имеют то или иное отношение к воде. Поэтому мы доводим (по возможности) до рода определения лишь настоящих водных обитателей, т. е. водных личинок и тех взрослых насекомых, которые и в состоянии имаго проводят*свою жизнь исключительно или преимущественно в воде. По отношению же к тем имаго, которые являются настоящими воздушными обитателями и которые только в стадии личинки живут в воде, мы ограничиваемся лишь определением до крупных групп — отрядов и семейств, отсылая интересующихся более детальным определением их к специальным книгам, примерный перечень которых приводится в конце этой книги; исключение мы делаем лишь для стрекоз, детализируя их определительную таблицу до той степени, как и для водных обитателей; это исключение объясняется той популярностью и известностью, которой пользуются стрекозы даже среди людей, мало интересующихся природой, благодаря отчасти сравнительно крупной величине, отчасти красоте некоторых представителей этой группы насекомых. Кроме того, исключаются те насекомые, которые ведут паразитический образ жизни.
1. Без членистых ножек. Иногда без ясно развитой головы (рис. 444, 445). Беа
зачатков крыльев.................................................................2
0. Есть членистые ножки. Всегда ясно обособленная голова....................3
2. Тело не всегда членистое. OpianoB передвижения (ножек, валиков) нет. Личинки паразитируют на водных насекомых.
отр. Hymenoptera — перепончатокрылые1.
0. Тело всегда ясно членистое. Есть органы передвижения — ложные ножки или ползательные валики. Личинки не паразитируют на водных насекомых.
отр. Di р t е г а — двукрылые.
8. Брюшко или без ясной сегментации, или не более чем из 6 сегментов. На первом брюшном сегменте — выпячивающаяся брюшная трубка; па конце тела — «прыгательная вилка». Мелкие формы. Живут на поверхности воды (рис. 281).
отр Collembola — вилохвостки.
0. Брюшко всегда ясно сегментированное, более чем изб сегментов, большей частью из 10—И. Выпячивающейся брюшной трубки и прыгательной вилки нет.. 4
4. Взрослые насекомые (имаго). Всегда есть две пары вполне развитых крыльев: 1-я пара — более твёрдые, прикрывающие всё или почти всё брюшко, 2-я — мягкие, перепончатые .................................................... 5
0. Личинки (larvae). Крыльев нет совсем или есть только недоразвитые.8
5. Крылья 1-й (верхней) пары твёрдые, роговые, лишённые жилок — «надкрылья». Ротовые части грызущие.
отр. Coleoptera imago — жуки взрослые.
0. Крылья 1-й пары, налегающие одно на другое, неоднородные, состоят из более твёрдого кожистого основного участка и перепончатого концевого, причём оба участка с жилками; иногда эти крылья редуцированы или даже совсем отсутствуют. Ротовые части сосущие, видоизменены в колющий хоботок, подогнутый в спокойном состоянии под голову.
отр. Н е mi ptera (Rh у n ch о t a) imago — клопы взрослые.
6. Есть только зачатки крыльев, не доходящие до брюшка и лежащие вдоль тела со спинной стороны................................................7
0. Нет зачатков крыльев..................................... 10
7. Есть хоботок, подогнутый под голову. Личинки очень похожи на взрослых насекомых, отличаются только недоразвитием крыльев.
отр Hemiptera larvae — личинки клопов.
0. Нет хоботка ...............................................8
8. Голова с нижней стороны имеет так называемую «маску» (рис. 285 В)—орган, состоящий из двух колен: одно отходит кзади, другое снова возвращается кпереди и прикрывает нижнюю поверхность головы своим плоским или ложковидно вогнутым концом; маску легко отвести булавкой, и тогда она торчит в виде длинного придатка.
отр. Odonata larvae — личинки стрекоз.
0. Без маски..................................................9
9. Ноги с двумя коготками. Челюстной щупик из 5 члеников. На заднем конце тела 2 длинные многочленистые нити. Брюшко по бокам без листообразных или кустистых выростов (рис. 263).
отр. Р I е с о р te г a larvae — личинки веснянок.
0. Челюстной щупик из 2—3 члеников. На заднем конце тела 3 длинные многочленистые нити (очень редко 2). По бокам брюшка всегда листообразные или кустистые выросты — трахейные жабры, иногда загнутые на спину. На ногах по одному коготку (рис. 299).
отр. Ephemeroptera larvae — личинки подёнок.
10. На брюшных сегментах по бокам длинные, довольно массивные членистые трахейные жабры (рис. 380).
отр Megaloptera larvae — личинки вислокрыло к.
0. Трахейных жабер нет, или они нечленистые, нитевидные, иногда кустистые..........................................................11
11. На заднем конце тела пара сильных крючков на более или менее развитых ложных ножках (рис. 387 В).
отр. Trichoptera larvae — личинки ручейников.
0. Без пары крючков на заднем конце..........................13
12. На брюшке 5 пар хорошо развитых ложных ножек с венчиком крючков.
отр. Lepidoptera larvae — гусеницы бабочек.
0, Брюшных ножек большей частью нет, если же есть, то слабо развитые, без венчика крючков................................................... 13
1 Церепончатокрылые живут в водоёмах только в состоянии личинки и куколки и то. собственно, не в самой воде, а внутри тела других насекомых (большей частью в яйцах), в состоянии же имаго (взрослого насекомого) они появляются в водоёмах редко и только на короткое время.
18. Очень длинные жвалы, выдающиеся далеко вперёд, много длиннее головы <рис. 382).
отр. N е и го р t е га larvae — личинки сетчатокрылых. U. Жвалы короче головы (рис. 373 и далее).
С о 1 е о р t е г a larvae — личинки жуков.
COLLEMBOLA — ВИЛОХВОСТКИ (НОГОХВОСТКИ)
Мелкие насекомые, бескрылые. Усики большей частью 4-членистые. Кожный покров волосистый, иногда с чешуйками. Дыхание в большинстве случаев кожное. Б р ю ш к о не более чем из 6 сегментов, иногда частично слитых. Своеобразными органами являются: прыгательная вилка, в спокойном состоянии подогнутая под брюшко (рис. 281 Б), и брюшная трубка (рис. 281). Прыгательная вилка расположена на брюшной стороне 4-го или 5-го сегментов и состоит из непарной основной части — рукоятки — и пары зубцов с придатками на концах, так называемыми м у к р о. Брюшная трубка находится на брюшной стороне 1-го брюшного сегмента и представляет собой непарный выступ, на свободном конце которого расположены 2 клейких, наполняющихся кровью и выпячивающихся пузырька; этот орган служит для временного прикрепления. Для определения видов р. Sminthurides имеет большое значение так называемый т и-биальпый орган; это — чувствительный орган, расположенный вблизи дистального конца голени (tibia) и состоящий из 2 коротких зубчиков и чувствительной щетинки, на строении которой и основано различие видов р. Sminthurides.
1. Тело приблизительно шаровидное или яйцевидное, короткое; сегменты груди разграничены слабо, брюшные частично слиты. Конечные отделы прыгательной вилки (mucro), расширены в пластинки.
Подотряд Sympbypleona — цельнобрюхие............2
О, Тело удлинённое; сегменты груди и 4 первых сегмента 6-членистого брюшка ясно различимы. Брюшная трубка с короткими выпячивающимися мешочками.
Подотряд ArthropleoD® — членистобрюхие ............5
2. Грудь с небольшими, соответствующими сегментам бороздками. Длина до 1,5 мм. Па поверхности воды в заросших стоячих водоёмах.
Род Sminthurides Borner............................3
0. Грудь без бороздок. Длина до 1,5 мм. На поверхности воды в заросших стоячих водоёмах.
Род Deuterosminthurus, у нас 1 вид D. novemli neatus Tullb.
3. Чувствительная щетинка тибиального органа листовидной формы с расщеплённым надвое концом. Длина 1 мм. На воде, покрытой ряской и плавающими листьями других растений.
S. penicillifer Scbaff.
0. Эта щетинка не расщеплена на конце............................4
4. Чувствительная щетинка тибиального органа своим концом далеко выдаётся за конец лапки (tarsus). Концевые придатки прыгательной вилки (мукро) на спинной стороне пластинки несут до 12 зубчиков.
S. aquaticus Borner (Bourlet).
0. Эта щетинка лишь приблизительно доходит до конца лапки. Длина до 1 мм.
8. in а 1 m g г е n i Tullb.
5. Тело массивное. Все 3 грудных сегмента хорошо видны со спины (рис. 281 Л). Поверхность тела неровная, бугристая, хитиновыи покров мелкозернистый. Усики 4-членистые, чуть длиннее головы. Зубцы прыгательной вилки тонкие, к концу заостряющиеся (рис. 281 Б). Длина до 1,2 мм. В стоячих водоёмах, на поверхности воды около берегов, также на траве у самой воды.
Сем. Poduridae, р. Podura (L.) Tullb., 1 вид Р. aquatica Nic.
0. Тело более тонкое, цилиндрическое или слабо сплюснутое; со спины видны только 2-й и 3-й грудные сегменты. Поверхность тела гладкая, не бугристая, хитиновый покров не зернистый.........................................................6
в. Зубцы вилки довольно тонкие, ясно суживающиеся к концу (рис. 283, 284) . . . 7
0: Зубцы вилки довольно массивные, к концу не суживающиеся. Длина до 1,2 мм. На воде и у воды (рис. 282).
Сем. Isotomidae, р. Proisotoma, у нас 1 вид Р. ripicola Linnan.
234
7. На концах вилки по 3 зубца (рис. 283 Б). Брюш-ко с короткими волосками. Усики но длинные. Длина до 3,7 мм. На воде и в сырых местах (рис. 283 Л).
Сем. Isotomidae, р. I s о to m a, I. v i-ri d i s (Burl.) Schott.
0. На концах вилки по 4 зубца (рис. 284 Б). Усики довольно длинные. Брюшко с длинными волосками. Длина до 4,5 лш. В сырых мостах и на воде (рис. 284 А).
Сем. Entomobry-i d а е, р. I s о t о m u-r u s, 1 вид, I. p a lust r i s (Muller) Borner.
A — самка; Б — конец её пилки.
ODONATA LARVAE — ЛИЧИНКИ СТРЕКОЗ
Личинки стрекоз бывают двух форм: одни обладают длинным тонким телом и тремя хвостовыми листовидными придатками на конце брюшка с ясно просвечивающими сквозь тонкую стенку разветвлениями трахей — Zygoptera— равнокрылые; у других личинок брюшко не имеет листовидных придатков на конце — A ni soptera — разнокрылые.
У последних личинок тело толще, чем у первых, иногда заострено, иногда тупо срезано; на конце анальная пирамида из 5 хитиновых лопастей.
Рис.’ 285.
А — лич. стрелки — Coenagrlon sp.; Б — лич. красотки — Agri on splendens; В — схема строения личинки Anlsoplera (со спинкой и с брюшной стороны).
На спинпой стороне груди личинок стрекоз находятся 2 пары зачатков крыльев, лежащих вдоль брюшка. Голова большей частью крупная с большими глазами и отходящими вблизи них усиками. На нижней стороне головы личинок стрекоз находится так называемая маска (видоизмененная нижняя губа — рис. 285 В, 286 и далее), представляющая собой длинное образование из 2 колен: первое отходит снизу от середины головы и направляется к заднему концу тела вдоль груди; второе колено причле-няется к первому подвижным суставом и направляется в обратную сторону— вперёд, доходя снова до головы и прикрывая её снизу в виде плоского (у Zygoptera) или ложкообразного (у Anisoptrae) расширения; от наружных углов этого расширения отходят две подвижные боковые лопасти, снабжённые шипами и подвижным зубцом; лопасти эти, складываясь, сходятся по срединной линии и часто снабжены по этой линии зубцами (рис. 286). Маска служит личинкам для схватывания добычи.
1. Тело топкое, удлинённое, стремя листовидными жаберными придатками на заднем конце (рис. 285 А, Б). Zygoptera........................2
О, Тело массивное, без листовидных жабер. На конце тела анальная пирамида (рис. 285 В, Г}, Anisoptera...................... 9
2. Первый членик усика более, чем вдвое длиннее совокупности остальных. Маска в середине с ромбическим окошечком (рис. 286 Г). Боковые жаберные листочки толстые, треугольные, срединный короче, уплощённый (рис. 287 Л, Б).
Сем. Agrionidae (Calopterygidae), р. Agrion L. (Calopteryx Внггп.) — красотка.
0. Первый членик усика короче суммы остальных. Маска без окошечка (рис. 286
А, Б, В, Д). Сем. Coenagrionidae ...................8
8. Срединная часть маски посредине слегка надрезана; боковые лопасти трёхраздельные, на их подвижных зубцах длинные щетинки (рис. 286 В, Д). Жаберные листочки
Рис. 286. Маски личинок стрекоз:
А — Enallagma cyathlgera; Б — Platycnemis pennipes; В — Sym-pycna fusca; Г— Agrion splendens; Д — Lestes.
с крупной срединной трахеей, от которой отходят почти под прямым углом боковые слабые веточки (рис. 287 В).
Подсем. Lestinae — лютки.............4
0. Срединная часть маски без надреза; боковые лопасти двухраздельные (рис. 286 Л, Б). Боковые веточки трахей в жаберных листочках отходят от главного ствола под косым углом (рис. 287 Г—М).
Подсем. Coenagrioninae — стрелки. .5
4. Маска (рис. 286 Д) очень удлинённая, заходит за основания задних ног, ложкообразная. Длина личинки 22—25 мм. В мелких стоячих пересыхающих летом водоёмах, заливах, заводях.
р. Lestes Leach (А п а р е t е s).
0. Маска (рис. 286 В) значительно короче, доходит только до основания средних аог, квадратная. В мелких пересыхающих водоёмах среди водной растительности.
р. Sympycna.
5. На боковых лопастях маски по 3—4 щетинки (рис. 286 Б). Задние углы VII—1 X сегментов брюшка вытянуты в зубцы. Жаберные листочки вытянуты на конце в узкий, 1линный придаток (рис. 287 Г). Ползают на дне проточных водоёмов.
р. Platycnemis Charp. — плосконожка.
0. На боковых лопастях маски по 5—7 щетинок (очень редко 4, рис. 286 Л). Задние углы VII—IX сегментов без шипов. Жаберные листочки без вытянутого придатка (рис. 287 Д—М)................................................................в
6. Жаберные листочки непрозрачные, в чёрных пятнах (рис. 287 Л/), по краю не опушены волосками, вытянуты на конце. На дне стоячих или медленно текущих водоёмов, среди отмерших листьев.
р. Р у г г h о 8 о m a Charp.
0. Жаберпые листочки (рис. 287 И, К), подразделены на два отдела, отличающиеся по вооружению края: дистальный отдел большей частью с волосками, базальный — с шипами; листочки прозрачны, разветвления.трахей хорошо видны.............. . 7
7. Граница между отделами жаберных листочков проходит перпендикулярно к длинной оси листочков, в середине длины листочка или дистальнее; часто граница отмечена поперечной тёмной полосой (рис. 287 3). . ............. ................9
0. Граница между отделами проходит вкось; тёмной полосы на границе нет* (рис. 287 Я)............................................................9
8. Жаберные листочки широкие, закруглённые, просвечивающие зеленоватым (рис. 287 Л). Среди зарослей различных водоёмов.
н р. Erythromma Charp.
0. Жаберные листочки или заострены, или закруглены — в последнем случае они узкие, коричневатые (рис. 287 Ж, 3). В стоячих водоёмах среди растений; редко в реках. р. Coenagrion Kir
9. Жаберные листочки (рис. 287 Е) совершенно прозрачные. Тело светлозелёное или светложёлтое, с нежным рисунком. В стоячих водоёмах среди растений.
Е n a I I a g m a Sei.
Жаберные листочки личинок Zygoptera:
А — Agrion । Д — Ischnura pulchelhim;
Рис. 287. Жаберные листочки личинок Zygoptera:
splendens; Б — Agrion Virgo; В — Lestes sp.; Г — Platyc^emlsr^°nJ^Sn;
I (Mtcronympba) elegans; fe - Enallagma cyathlgera; Ж -
3 —Coenagrion puella; И — Agrion mercuriale; К — Agrion arma turn.
Л — Erythromma najas; M — Pyrrhoaoma nymphula.
0. Листочки (рис. 287 Д) мало прозрачные. Тело серозелёное или желтоватое, ярко и пёстро окрашенное. В стоячих и текучих водах, в прибрежных зарослях.
Ischnura Charp (Micronympna ruroy).
10. Маска плоская,закрывающая снизу лишь заднюю половину головы; без щетинок на срединной части и на боковых лопастях (рис. 288 Л) ц
Сем. Aeschnidae.......................................г • * ’ * ’ * >о
0. Маска выпуклая, совершенно закрывающая всю голову с нижнеи стороны, щетинками на срединной части и на боковых лопастях (рис. 288 Б) ... . • ♦ • • • •
11. Усики 4-членистые; 4-й членик рудиментарный. Лапки передних и средних н 2-членистые, задних — 3-членистые (рис. 289).
Подсем. Gomphinae — дедки.....................................
0. Усики 7-членистые. Все лапки 3-члениртые. Подсем. Aeschninae............................• • •/ ’ ’ * *
12. Без бугорков на спинке (рис. 289 Б). Внутренние зубцы боковых лопастей маски заострены и изогнуты (рис. 290 Б). В реках, зарываются в 9 Leach.
0. Задние края IV—IX брюшных сегментов по срединной линии со спины вытянуты бугорки (рис. 289 А). Внутренние зубцы боковых лопастей маски закруглены (рис. 290 А, В) ..........................................................13
18. Задние углы VII—IX сегментов вытянуты в шип. Бугорки па спинке крупные, укрупняющиеся от V к IX. В текучих водоёмах, зарываются в грунт.
р. О ph i ogomphus Sei.
Рис. 288. Схема строения маски личинок Anisoptera: А—Acschuidae; Б — Llbeilulidae.
Рис. 289. Личинки подсемейства Gomphinae:
А — Onychogomphus uncatus; Б — Gomphus vulgatissimus.
0. В шипы вытянуты только углы VI—IX или VIII—IX сегментов. Спинные бугорки мелкие (рис. 289 Л).
р. Onychogomphus Sei.
14. Задние углы головы вытянуты, образуют острые зубцы. Задние углы сегментов V—IX вытянуты в шины (рис. 291 Г), а на краях сегментов II—IX, кроме того, ещё бугорки.
р. В о у е г i a Me Lachl.
0. Задние углы годовы закруглены. Боковые шипы только на VI—IX или VII—IX сегментах; боковых бугорков нет (рис. 291 А—В) .....................15
15. Внутренние края боковых лопастей маски или закруглены, или прямо срезаны и в последнем случае не оттянуты в зубец. Боковые шипы хорошо развиты только на VII—IX сегментах, а на VI сегменте шипы очень мелкие.................16
Рис. 290. Маски личинок подсемейства Gomphinaei А — Onychogomphus uncatus; Б — Gomphus vulgatissimus; В — Ophlogompbus serpentlnus.
0. Хорошо выраженные боковые шипы на VII—IX сегментах (рис. 291 В). Анальная пирамида длиннее суммы IX + X сегментов. Внутренние края боковых лопастей маски прямо срезаны, внутренний их угол оттянут в зубец (рис. 292 В). В стоячих и текучих водоёмах.
р. А □ а х Leach. — дозорщик.
16. Внутренний край боковых лопастей маски округло изогнут, постепенно суживается и заостряется в шип (рис. 292 Г). Глаза почти сферические, выдающиеся. Анальная пирамида чуть длиннее X сегмента (рис. 291 В). В стоячих водоёмах и текущих с умеренной скоростью течения. р в г а с h у t г о n Ev.
0.4.Внутренний край боковых лопастей маски прямо срезан, без шипа (рис. 292 А). Глаза полусферические, уплощённые (рис. 291 А). Анальная пирамида равна сумме IX + X сегментов. В стоячих, реже в медленно текущих водоёмах, среди растений.
р. А е в с h n a Fabr. — коромысло.
17» Срединная часть маски посредине с раздвоённым выступом; внутренний край боковых лопастей крупно и неправильно зазубрен (рис. 294). Брюшко удлинённое, так что концы распрямленных задних ног не достигают его конца (рис. 293). В иле рек и ручьёв. ,
Подсем. Cordulegastrinae, р. Cordulegaster Leach.
0. Срединная часть маски посредине без выступа (рис. 295). Брюшко короче, так
что концы распрямлённых задних ног заходят за его конец (рис. 296, 297). <
Сем. Libelluiidae — стрекозы...........................18
A — Aeschna Juncea; Б — Anax imperator; В — Brachytron ha f Qi ease; Г — Boyeria Irene.
Рис. 292. Маски личинок подсемейства Aeschninae:
А — Aeschna juncea; Б — Anax imperator; B?— Boyeria Irene; Г — Brachytron halnlense.
18. Боковые лопасти маски по внутреннему краю с ясно выраженными зубцами; высота зубцов достигает приблизительно половины их ширины у основания (рис. 295).
Подсем. Corduliinae — бабки............................... 10
0. Боковые лопасти маски по внутреннему краю с очень плоскими зубцами, высота
которых значительно менее половины их ширины у основания. Подсем. Libellulinae................................................22
19. Конец бедра задних ног не заходит за передний край VII сегмента
(рис. 297 Б. В).....................................................20
Конец бедра задних ног доходит до VIII или IX сегмента (рис. 297 Л) ... 21
20. На срединной части маски, вблизи её сочленения с боковыми лопастями несколько коротких сильных шипиков; вблизи краёв боковых лопастей, приблизительно
посредине, группа мелких, довольно толстых волосков (рис, 295 £). В больших озёрах с чистой водой и песчаным дном и в
мелких
0.
Рао. 293. Личинка Cordule-gaster annulatus. загрязнённых водоёмах (озерки,
Рио. 294. Маска личинки
Gordulegaater.
пруды, болота) с гниющим илом на дне. р. Sornatoohlora Sdl.
На срединной части маски нет ни шипиков вблизи сочленения с боковыми лопа-
стями, ни групп коротких волосков у боковых краёв (рис. 295 Б).
р. Oxygastra 8ё1
21. На III—IX сегментах брюшка, по срединной линии спины, мелкие бугорки; самые крупные из них — на VI—VII сегментах — достигают приблизительно */« высоты несущего их сегмента (рис. 296 Б). Задние углы VIII—IX сегментов вытянуты в корот-кии шип (рис. 297 Л). В оаёрах, прудах, болотах. р. С о г d u 1 i a Leacb.
0. Но срединной части маски, по бокам, группа коротких волосков (рис. 295 Г). Бугорки по срединной линии спины на Ш—IX сегментах крупные и сплюснутые с боков; самые крупные — на сегментах VI—VIII —по длине равны высоте несущего их сегмента или лишь чуть-чуть короче (рис. 296 А). Задние углы VIII — IX сегментов вытянуты в длинный, слегка изогнутый шип; шипы IX сегмента значительно длиннее высоты
Рис. 295. Маски личинок подсемейства Corduliinae:
А — Cordulla аеоеа; Б — Oxygastra curtisl; В—Somatochlora me tallica; Г—Epitheca blmaculata.
несущего их сегмента и почти достигают конца анальной пирамиды (рис. 297 Г). Главным образом в стоячйх водоёмах; в реках только в прибрежье. Epitheca Gharp
22. Усики отходят ясно кпереди от линии, соединяющей передние края сетчатых глаз; глаза мелкие. Тело в длинных зазубренных волосках. Ноги короткие и сильные (рис. 298 Г, Д) .........................................................28
Рис. 296. Брюшко (сбоку) личинок подсемейства Gordu-liinaei
0. Тело гладкое или лишь в коротких гладких волосках. Ноги длинные и тонкие. Усики отходят едва заметно кпереди от линии, соединяющей передние края глаз; глаза крупные (рис. 298 А—В).......23
23. На VII —VIII сегментах по срединной линии спины имеются бугорки (рис. 298 Д). В стоячих и медленно текущих водах.
р. Libellula L. (L е р t е t г u m Kirby).
0. Без бугорков на VII — VIII сегментах спины (рис. 298 Г). Маска короткая; доходит только до 1-й пары ног. Встречаются в стоячих водоёмах, но предпочитают проточную, иногда с довольно быстрым
течением.
р. Orthetrum Newm.
А—Epitheca blmaculata; Б — Cordulla аепеа; В — Somatochlora me-tallica.
24. Задние углы VIII — IX сегментов вытянуты в шипы, равные по длине высоте X сегмента. Бугорков по срединной линии спинки нет (рис. 298 В), р. Grocothemis Brauer.
0. Задние углы VIII — IX сегментов вытянуты в шипы, много превышающие по длине высоту X сегмента 24
25. Голова крупная, кзади слабо суженная, с выпуклым боковым контуром. Глаза мало выпуклые. Бугорки по срединной линии спины мелкие, не достигают 1/8 следующего сегмента (рис. 298 А). В мелких пересыхающих водоёмах, среди растений.
р. Sympetrum Newm (Di р I е х Gharp.).
0. Голова меньше, чем у Sympetrum, резко суживающаяся кзади, с прямыми боковыми контурами. Глаза сильно выпуклые. Бугорков по срединной линии спины или совсем нет (рис. 298 Б), или, напротив, они крупные и достигают середины следующего сегмента. В стоячих водоёмах, среди растений, реже в медленно текущих водах.
р. Leucorrhinia Britt.
EPHEMEROPTERA LA RVAE — ЛИЧИНКИ ПОДЁНОК
Личинки подёнок (рис. 299) имеют большей частью удлинённое тело с тремя (редко двумя) длинными хвостовыми нитями на конце. Голова то эллиптическая, с глазами по бокам (рис. 310, 313, 316), то уплощённая — с глазами на спинной стороне (рис. 306, 308). Для определения большое значение имеют ротовые части (рис. 300), особенно строение губных и челюстных щупиков и строение антенны. Кроме сетчатых глаз, часто очень крупных, имеются ешё 3 личиночных глазка, расположенных треугольником между сетчатыми и иногда сидяших на стебельках. От груди отходят зачатки двух пар крыльев, лежащих вдоль спины, и три пары ног. Боковые края брюшных сегментов на заднем конце иногда вытянуты в зубцы (рис. 318). Дыхание происходит посредством нежных парных выростов на брюшке — ж а б е р. Жабры или 240
Рис. 298. Личинки подсемейства Libellulinae:
А - Sympetrum depresslusculum; Б- Leucorrhiniarublcunda; В — Ciwoiheinls erytrea; p — Orthetrum canccllatum; Д — Libellula quadrimaculata.
Рас. 299. Схема строения личинок подёнок Epheme-roptera.
Рис. 300. Схема строения ротовых частей личинок подёнок Ephemeroptera:
а—нижняя губа; б — нижние челюсти в — верхние челюсти (жвалы, мандибулы); ринке; д — верхняя губа.
(максиллы); г — гипофа-
Р и с. 301. Личинки:
А — Polymitarcya Virgo; Б — Ephemera vulgata; В — PotamanthUB ер.
Рис. 302. Жабры личинок:
а — Ephemera vulgata, 1-я пара; Б — её 2-я пара: В — Polymitarcys virgo, жаберный листочек.
ложатся на спинную сторону (рис. 301 А, В) или отходят вбок (рис. 308, 310); они обычно находятся в быстром движении. Жабры бывают из 1 или из 2 ветвей; форма их различна, причём нередко различны между собой две ветви одной жабры (рис. 322). Кроме брюшных жаоер, изредка встречаются ешё добавочные пучки жаберных нитей на челюстях (рис. 307). Ноги устроены несколько различно соответственно с образом жизни: мощные, приспособленные для рытья, — у форм, живущих в грунте (рис. 301), сильно уплощённые — у форм, живущих на камнях в области прибоя (рис. 309). Коготок на ногах почти всегда один, иногда с зубчиками. Хвостовые нити
всегда снабжены щетинками или располагающимися равномерно по всей окружности (рис. 310, 313, 316), или более длинными и густыми по одной продольной линии, в виде бахромы (рис. 318, 321).
1. Голова с массивными жвалами, выступающими за её передний край или значительной своей частью (рис. 301 Л, Б), или хотя бы одним зубцом (рис. 301 В). Ноги роющие, очень массивные (рис. 303). Брюшные жабры из двух перистых ветвей (рис. 302). 2
0. Жвалы не выступают за передний край головы. Ноги не роющие, более тонкие.
Жабры не перистые.........................................................5
Рис. 305. Личинка Proso pi stoma: Рис. 306. Личинка Oligo-
А — сверху, Б-снизу. . neuriella rhenana.
2. Жвалы широкие, но не длинные (рис. 304 F), за край головы выступает только один зубец. Жабры направлены в стороны (рис. 301 Я), все перистые. В реках, речках, ручьях, под камнями и в зарослях.
Сем. Potamanthidae, р. Potam antbus
Pictet, у нас 1 вид Р. 1 u t е и s L.
0. Жвалы очень длинные, далеко выступают за передний край головы (рис. 304 .1). Жабры загибаются на спинную сторону (рис. 301 Л, Б)............. 8
8. Антенны в очень длинных ресничках (рис. 301 Б). Жвалы изогнуты кнаружи, без зубцов. На передних углах головы направленные вперёд выступы, благодаря чему передний край оказывается вогнутым (рис. 301 Б)> Жабры перистые, кроме I пары (рис. 302 Л). В глинистом грунте озёр и рек.
Сем. Е р h е гп е г i а а е, 1 род Ephemera L. Наиболее обычный вид Ephemera vulgataL.
0. Антенны или с более короткими ресничками, или голые. Жвалы изогнуты внутрь t с зубцами по выпуклому краю (рис. 304 Л). Передний край головы закруглён, пе обра-эует выступов (рис. 301 Л) ...................................................... 4
Рис. 307. Личинка ОН gone uri ell а rhenana:
А — передний конец тела снизу; Б — подчелюстная жабра; В — i-я пара жабер.
4. По выпуклому краю жвалы 6—8 зубцов. Тело довольно короткое и широкое. Голени передних ног широкие, с длинными, крепкими шипами на наружном краю. Первая пара жабер редуцирована. В глинистом грунте.
Сем. Palingeniidae, р. Palingenia Вигщ.
0. Зубцов на жвале много. Тело длинное, довольно узкое. Хвостовые нити почти одной длины, приблизительно равны длине брюшка (рис. 301 Л). Жабры все одинако-
Р и с. 308. Личинки:
А—Rhltrogena aurant!аса; Б—Epeorus torrent! шп; В — Hepta-genia со ecu la ns; Г — Ecdyonurus flumlnum.
вые, перистые (рис. 302 В). Делают горизонтальные ходы в глинистом грунте прибрежья рек.
Сем. Epboronidae, р. Polymitarcys Etn., 1 вид Р. virgo Oliv.
5. Брюшные жабры прикрыты направленной кзади щитовидно расширенной спинкой среднего грудного сегмента (рис. 305). В быстро текущих реках, под камнями.
Сем. Prosopistomatidae, р. Prosopi stoma Latr.
0. Жабры всегда видны...........................в
в. Голова округлая, ширина её больше длины. Глаза на спинной сторону, (рис. 306). Тело сильно сплюснуто. Жабры листовидные, на нижней стороне, при основании с пучками нитей.............................................................................7
г
0. Глаза по бокам головы. Тело цилиндрическое или слабо уплощённое.....12
7. Есть подчелюстные жабры в виде пучков длинных нитей (рис. 307). Передние
ноги с длинными волосками.
Сем. Oligoneuriidae
Рис. 309. Жабры личинок:
rentlumhl пер?дни О конец тела с бРюшн°й стороны; Б —Epeorus tor-
Я °к₽а жоабеР; в — ее же 2-я пара жабер; Г — Heptagenta coerularu 1-я папа абер, Д ее же 3-я пара жабер; Е — Ecdyonurus fluminum, 1-я пара жабер- Ж — её же 2-я пара жабер; 3 — её же 7-я пара жабер.
ногах^* П°Дчелюстнь1Х жабер нет. Передние ноги с короткими волосками, как на прочих
Сем. Heptageniidae (Ecdyonuridae)............................9
8. Брюшные жабры сильно редуцированы (рис. 306), имеют вид маленьких чешуек с зубчатым краем и пучком коротких ветвистых нитей. Тело широкое, короткое. Найдена в Волге, Днепре, Сев. Двине.
р. 01 i g о n е u г i е 11 а, 1 вид. О. rehnana Imh.
Жабры развиты хорошо. Бёдра средних и задних ног в виде широких пластинок. Тело вытянутое. Найдена в Днепре.
р. О 1 i go n е u ri s с a, 1 вид О. borysthenica Tschern.
9. Жаберные листочки первой пары очень длинные, заходят на брюшную сторону до взаимного соприкосновения (рис. 308 А, 309 А).
Подсем. Rhitrogeninae, р. Rhi trogena Etn.
0. Первая пара жабер не длиннее прочих.
Подсем. Heptageniinae(Ecdyonurinae).........................10
10. Брюшные жабры с пучком коротких нитей у основания (рис. 309 Б, В). Средняя хвостовая нить редуцирована (рис. 308 Б). Под камнями, на быстром течении.
р. Е ре о газ Etn.
0. Брюшные жабры с пучком длинных нитей (рис. 309 Г — Ж). Все три хвостовые нити вполне развиты (рис. 308 В, Г)...............................11
11. Переднеспинка всегда с прямым задним краем. Нити в пучках на брюшных жабрах не короче несущего листочка (рис. 309 Г, Д). В озёрах и реках, под камнями.
р. Н ер t age n i a Walsh.
0. Задние углы переднеспинки большей частью оттянуты кзади; образующиеся выступы охватывают верхнюю часть среднеспинки (рис. 308 Г). Нити жаберных пучков короче несущего их листочка (рис. 309 Е, Ж, 3).
F J н р. Ecdyonurus (Ecdyurus) Etn.
12. Боковые хвостовые нити с короткими волосками или шипиками вокруг (рис. 310
313, 316). Голова направлена вперёд...............................13
0. Боковые хвостовые нити с длинными волосками по внутреннему краю (рпс. 31.8, 321). Голова направлена вниз....................... • • ’ -ши’
18. Семь пар брюшных жабер — или все нитевидно рассеченные (рис. oiuj, или первая пара в виде узкой полоски, все же прочие — листовидные.
Сем. ........................................................
г 243
Рис. 310. Личинки;
А — НаЬгорЫеЫа fusca; Б — Habroleptoides modesta; В — Leptophlebla vespertine: Г—Choroterpes plcteti
0, Пять или шесть пар брюшных жабер, причём часть из них прикрыта другими и снаружи не видна (рис. 313, 317).................18
14. Все семь пар брюшных жабер развиты одинаково, узковетвистые (рис. 310 Л, Б, 311 А-Г)................................ 15
0. Первая пара рудиментарна, шесть прочих двойные, листовидные (рис. 311 Д—ж). . . . 17
15. Брюшные жабры из двух узких ветвей (рис. 311 А). Гипофаринкс (подглоточник) по наружному краю закруглён (рис. 312 4). Хвостовые нити вдвое длиннее тела. В ручьях и пойменных болотах центральной и северной зон Европейской части СССР.
р. Paraleptophlebia Lest.
0. Брюшные жабры из одной ветви— раздвоённой (рис. 310 В} или раз-\ ветвлённой (рис. 310 4, 311 Б). Гипо-
фаринкс по наружному краю клювовидно вытянут (рис. 312 Б). Длина хвостовых \ нитей не больше длины тела (рис. 310 4 4, Б)........................16
16. Жабры разветвлённые (рис. 310 4, 311 Б). В мелких речках и ручьях севера и центра СССР.
р. Habrophlebia Ent.
0. Жабры раздвоённые (рис. 310 Б).
р. Habroleptoides.
17. Все брюшные жабры двойные: первая пара в виде очень узких полосок, все прочие расширены листовидно и к концу нитевидно утоньшены (рис. 310 Б, 311 Г). В прудах, ручьях, речках.
р. Leptophlebia (Etn.) Lest.
0. Первая пара брюшных жабер из одного простого, очень узкого листочка, все прочие из двух сросшихся асимметричных листочков, имеющих глубокие надрезы (рис. 310 Г, 311 Д, Е. Ж).
р. Choroterpes Etu.
246
A.
Рис. 311. Жабры личинок:
А — Paraleptophlebla ctncta; В — HabrophlebU; В, Г — Leptophlebla ар., 1-я пара жабер; Д — Choroterpea plcteti, i-я пара жабер; Е — её 2-я пара жабер; Ж — Choroterpea plcteti, жабра.
Рис. 312. Гипофаринкс личинок:
А — Paraleptophlebla submarglnata; Б — Habroleptotdea modesta.
Рио. 313. Личинки:
А — Ordella ba Iter ata; Б- TrtcorythuB (Ordella maxima).
Рис. 314. Жабры личинок:
А — Ordella balterata, 3-я пара жабер; В — Tricory-thus, t-я пара жабер; В — её же 3-я пара жабер.
18. Шесть пар брюшных жабер на I—VI сегментах; первая пара редуцирована (рис. 314 £>), вторая (рис. 313) изменена в крупные четырёхугольные пластинки, распростёртые по спине и вполне прикрывающие четыре прочие пары длинпобахромчатых жабер (рис. 314 В).
Сем. Brach у се г-cidae (Caenidae) ................. 19
О, Пять пар брюшных жабер на III—VII сегментах (рис. 316), причём последняя, а иногда и предпоследняя, совершенно прикрыты предыдущей парой.
Сем. Ephemerelli-
d а е............21
Рис. 316. Личинки:
halterata, голова.
А — Chitonopbora krteghofti; Б — Torleya belgica.
19. Вторая пара жабер с и пучком нитей снизу (рис. 313
короткой бахромой по F. 314 В).
краю, прочие с длинной бахромой р. Т г i с о г у t h u s Etn
Рис. 317. Жабры личинок:
А — Torleya belgica» 1-я пара, Ь—её же 2-я пара; В — её же 5-я пара; Г — Chitonopbora sp., 1-я (налево) и 2-я пары; Д — 4-я пара; Е — 5-я пара- Ж — Ch. k.rlegbo(f|; з — Ephe-тлегеИа Ignita.
0. Вторая пара жабер с длинной бахромой, прочие без пучка нитей (рис. 314 4) . 20
20. Глаза неволосистые, сидят непосредственно на голове. Второй членик антенны чуть длиннее третьего (рис. 315). Ноги короткие и сильные (рис. 313 4) с крупными, у основания зазубренными коготками. Губные и челюггные щупики трёхчленистые. В реках и озёрах (среди зарослей), на глинистых и каменистых грунтах.
р. О г d е I I a Camp. (С а е n 1 a Steph.).
0. Глаза волосистые, сидят на выступах. Второй членик антенны втрое длиннее третьего. Губные и челюстные щупики двухчленистые. Поги довольно гонкие, с прямыми коготками.
р. В га с b у се reus Curt. (Eurycaenia Bgtn.).
21. Тело короткое й широкое, густо одетое волосками. Брюшные жабры (рис. 317 4, В) слегка сдвинуты на спину. Четвёртая и пятая пары брюшных жабер 246
Ене прикрыты третьей, совершенно не видны снаружи. Ноги длинные и сильные, езвычайно расширенными бёдрами (рис. 316 Б). р Т о г I е у <i Lest
0. Тело удлиненное, голое или лишь слабо волосистое. Брюшные жабры совершенно нуты на спину; четвёртая пара хорошо видна снаружи. Бёдра обычной формы (рис. 316 А).............................................................22
22. Пятая пара брюшных жабер с ясно отшнурованными придатками у основания (рис. 317 Е). Боковые шины на брюшке слабо развиты (рис. 316 А). В быстро текущих реках, па каменистом, иногда песчаном грунте, на севере Европейской части СССР и на Алтае. р Chi tonophora Bgtn
0. Пятая пара жабер без придатков (рис. 317 3). Боковые шипы на брюшке развиты хорошо. В горных реках Алтая; в реках и озёрах севера и центра Европейской части СССР.
р. Epheioerella Walsh.
Рис. 318. Личинки!
А - Lsonycbia tgnotus; Б — Slpbonurus lacustris; В — Slphlurella Hnneana.
28. Тело цилиндрическое (рис. 321). Задние углы брюшных сегментов без шилов. Сем. Baetididae................................................ 28
О, Тело слегка приплюснуто. Задние углы брюшных сегментов вытянуты в плоские заострённые зубцы (рис. 318).
Сем. Siphlonuridae............................................24
24. Жаберные листочки все яйцевидные, одинарные (рис. 318 А, 319 А. Б. 321 А, Б)............................................................25
0. Некоторые жаберные листочки двойные (рис. 318 Б, Б, 319 В)... 27
25. Передние ноги в очень длинных, густо сидящих волосках (рис. 318 А, 320 А); голени на конце вытянуты в длинный, заострённый придаток. Кроме брюшных жабер, есть ещё две пары пучков: одна пара на челюстях (рис. 320 Б), вторая — у основания передних ног. В центре Европейской части СССР и в Закавказье.
р. Isonychia Albarda (Chi rotonetes Walk.).
0. Передние ноги с более короткими волосками. Жабры только брюшные ... 26
26. Жаберные листочки заострены, без зубчиков по наружному краю. Задние углы брюшных сегментов вытянуты в слабо выраженные шипы. Щупик нужней губы с характерным выступом на 2-м членике. В текучих водах.
р. Parameletus Bgtn.
0. Жаберные листочки закруглены, с зубчатым или волнистым краем (рис 319 А,Б). В задних углах брюшных сегментов сильные шипы. В текучих водах.
р. А ш е i е t u a Etn.
Ряс. 321. Личинки:
А — ВаёЦв rhodani; Б — С1оёоп diptenun; В — Centroptilum diaphanum*
27. Все брюшные жабры двойные: у последней пары верхний листочек редуцирован (рис. 318 В). р, Siphlurella Bgtn.
0. Только первая и вторая пары жабер двойные (рис. 318 Б, 319 В). Наиболее распространён в Европейской части СССР вид 8. И о n е a n u a Eat. характеризуется присутствием двойных жаберных листков; в стоячих и медленно текущих водах.
р. Sipblonurua Etn.
28. Брюшные жабры сильно асимметричны; шесть первых пар — двойные, с более крупными верхними листочками, седьмая пара из одинарных листочков (рис. 322 А—Д).
Подсем. Cloeoninae....................................... £9
Рис. 322. Жабры личинок:
А — Cloeon simile, 1-я пара; Б — 2-я пара; В — 7-я пара; Г — ₽гос!оёоп sp., 3-я пара; Д — 7-я пара; Е— ВаёНв rhodani; Ж — Oentroptllum luteolum
0. Брюшные жабры в виде простых симметричных листочков (рис. 322, £—ЯС).
Подсем. Ba§tidinae........................................80
29. Большие жаберные листочки III — VI пар более или менее закруглены. 3-й членик губного щупика на конце заострён (рис. 323 А). Самый обычный вид С. d i-pterum (L.) характеризуется тем, что маленький листок 1-й пары жабер вдвое уже большего листка, а по длине или равен ему, или превосходит его. В прудах, болотах-я медленно текущих водах среди зарослей.
р. Cloeon (L.) Bgtn.
Рис. 323. Нижняя губа личинок:
А — Cloeon simile; Б — Ргос1оёоп зр.; В—ВаёИз rhodani; Г—Gentroptllum dlaphanum.
0. Большие листочки треугольные (рис. 322 Р, Д). 3-й членик губного щупика ня конце закруглён (рис. 323 Б). р. Pro cloeon.
80. Две хвостовые нити. Жаберные листочки обратнояицевидные.
р. Acentrella Bgtn.
0. Три хвостовые нити (рис. 321 Л—В).............................81
81. Жаберные листочки яйцевидные, закруглённые (рис. 321 Л, 322 Е). Два последних членика губного щупика слились (рис. 323 В), закруглённые.
р. В а ё t i s Leach.
0. Жаберные листочки удлинённояйцевидные или ланцетные, заострённые (рис. 321 Bt 382 Ж). Последние членики губного щупика широко притуплены (рис. 323 Г).
р. Centroptilum Etn.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ PLECOPTERA LARVAE - ЛИЧИНОК ВЕСНЯНОК
Личинки веснянок несколько сходны по наружному виду с личинками подёнок. Они имеют удлинённое, уплощённое тело с 2 хвостовыми нитями на конце брюшка (рис. ЗЙ). Последние два-три сегмента брюшка устроены весьма различно у разных родов и даже у разных полов одного вида. Хвостовые нити или голые, или одеты волосками или шипиками (рис. 326, 329). На спине имеются зачатки крыльев, которые могут быть или направлены прямо кзади, почти параллельно друг к другу (рис. 326 В), или раздвинуты концами врозь (рис. 326 Б). Большей частью имеются наружные жабры. Они бывают четырёх родов: 1) плевральные — по бокам сегментов груди, представляющие собой пучки разветвлённых нитей (рис. 326 Л); 2) анальные — на конце брюшка, между хвостовыми нитями и анальным отверстием (рис. 326 А); 3)простернальные — на переднегруди с брюшной стороны, состоящие из пальцевидных выступов (рис. 332 А) или пучков нитей (рис. 338 Б); 4) к о ксаль н ы е — на основном членике ног в виде мешковидных выростов (рис. 336 Л),
Рис. 325. Схема ротовых частей лич. Perla: а — верхняя губа; б — жвалы; в — челюсти; а — нижняя губа.
Рис. 324. Схема строения личинки веснянки Perla.
Определение личинок веснянок основано, в первую очередь, на строении ротовых частей (рис. 325) — нижней губы, жвал и челюстей.^ Нижняя г у б а (г) состоит из двух пар губных лопастей (наружной и внутренней) и пары 3-членистых губных щупиков. На челюсти^ важно строение 5-членистого челюстного щупика и относительная длина и толщина отдельных его члеников. На жвалах (6) — количество и строение зубцов.
Живут личинки веснянок в текучей воде, под камнями.
1. Личинки большей частью крупные (больше 20 мм). Внутренняя лопасть нижней губы рудиментарная, много короче наружной (рис. 327, 333, Л, Б). Два первых членика лапки очень коротки, 3-й значительно длиннее двух первых вместе (рис. 326, Г). Иногда есть наружные жабры, помешаюшиеся на груди или на конце брюшка — плевральные или анальные (рис. 328). Челюстные щупики шиловидные....................2
0. Личинки мелкие или средней величины (меньше 20 мм). Нижняя губа приблизительно четырёхугольная с почти равными внутренней и внешней лопастями, достигающими одного уровня и отделёнными друг от друга неглубокой и неширокой бороздой (рис. 332 Б, Г). Членики лапок (рис. 335) приблизительно равные или увеличиваются постепенно; иногда 2-й членик недоразвит или даже не заметен; 3-й членик иногда не короче двух первых вместе. Жабры иногда есть — простерпальпые (рис. 335) или коксаль-ные (рис. 336). Челюстные щупики коротконитевидные......................7
2. Всегда есть три пары наружных жабер на груди, у некоторых видов ещё одна пара на конце брюшка, частично прикрытая последним сегментом. Ноги с густой и длинной бахромой щетинок (рис. 326 А,). Личинки больше 20 мм. В текучих водах преимущественно горных районов.
Сем. Perlidae, р. Perla Klpk.
0. Никогда нет наружных грудных или брюшных жабер..................3
8. Личинки мелкие или средние. 3-й членик челюстного щупика короче, чем 4-й; 5-й заострён.
Сем. Chloroperlidae...............4
Рис. 326. Личинки:
А — Perla abdomina-11s; Б — Chioroperla sp.; В — lsopteryx sp.; Г — Perlodes dis-раг (передний нонен тела).
Крупные личинки — 18—20 мм длиной, 3-й членик челюстного щупика не короче 4-го; 5-й не заострён. В текучих водах разного типа.
Сем. Perlodidae.........5
4. Длина тела 10—И мм, массивные личинки с голым телом (рис. 326 Б). Членики губного щупика утончаются постепенно; последний, сравнительно с предыдущим, не особенно утончён (рис. 327 Б]. Чехлы крыльев сильно расходятся в стороны. Хвостовые нити с мутовками коротких волосков (рис. 329 Л). Покровы спины с длинными, но не густо сидящими волосками.
р. Chioroperla Newm.
Рис. 327. Нижняя губа личинок:
А — Perla abdominalls; Б — Chioroperla sp.; В — lsopteryx sp.; Г — Perlodes dispar.
0. Длина до 5 мм. Тело узкое, с редко сидящими волосками (рис. 326 В). Членики губного щупика гораздо более массивны; последний гораздо уже предыдущего (рис. 327 В). Чехлы крыльев закруглены и сходятся по срединной линии. На хвостовых нитях кроме коротких волосков имеются более длинные щетинки (рис. 330).
р. lsopteryx Pict.
5. Переднеспинка поперечновытянутая, эллипсоидная, ясно закруглённая на боках, несколько уже головы (рис. 326 Г). Наружные лопасти нижней губы сильно развиты, на вершине не вытянуты в отросток (рис. 327 Г].
р. Perlodes Banks.
0. Переднеспинка прямоугольная, с почти прямыми передним и задним краями, лишь чуть-чуть закруглёнными (рис. 331 А). Наружные лопасти нижней губы менее выступают (рис. 332 Б) и оканчиваются маленьким отростком...........6
6. Голова приблизительно равна по ширине переднеслияке (рис. 331 А). Верхняя губа по внутреннему краю с ресничками, посредине выдаётся углом. Жвалы с 5 зубцами в две группы (рис. 333 А). Длина тела до 20 лсз<. р jSOgenus Newm.
0. Верхняя губа со слабо волнистым передним краем (рис. 334). На жвалах 6 несимметричных зубцов (рис. 333 Б). Длина до 17 мм.
r р. Dictyogenus Klpk
Рис. 328. Трахейная система лигч. Perla adbomi-nalis.
jnrnnrmw
Рис. 329. Конец брюшка личинок:
А — Chloroperla sp., Б Leuctra sp; В — Oapnia nigra.
Рис. 330. Конец брюшка лич. Iso-pteryx sp.
Рис. 331. Личинки:
а — laogenus nubecula (передний конец зела); Б — Taeniopteryx sp.; В — Nemura margin ata; Г — Leuctra sp.; Д — Capnia nigra.
в Рис. 332. Нижняя губа:
А — Dictyogenus Imhoffi; Б — Isogenus nubecula;
В — Leuctra sp.
7. Довольно крупные личинки, до 14 мм. Членики лапки постепенно удлиняются; третий почти той же длины, как два первых вместе (рис. 335 А). Хвостовые нити голые, по длине почти равны телу (рис. 331 Б). Жабры, если есть, расположены у основания ног и состоят из сократимого трёхчленистого выступа (рис. 336 А).
Сем. Taeniopterygidae ..........8
Рис. 333. Жвалы (мандибулы) личинок:
А — Isogenus nubecula; Б — Dictyogenus ер.; В — Leuctra sp.
Рис. 334. Верхняя губа лич. Dictyogenus sp.
Рис. 335. Ножка личинок:
А — Nephelopteryx sp.; Б — Leuctra sp.
Рис. 336. Передний конец тела личинок:
А — Nephelopteryx sp.; Б — Protonemura altlda.
Рис. 337. Хвостовая нить:
л — Protonemura humeralls; Б — Ampbinemura cl-аегеа.
0. Более мелкие личинки, не более 10 льи, с узким телом; 2-й членик лапки очень мал, иногда почти не виден, иногда отсутствует (рис. 331 В, Г, Д). Хвостовые нити обычно смутовчато расположенными волосками (рис. 337)..................9
8. Есть втяжные трёхчленистые жабры у основания ног (рис. 336 А). На брюш ных сегментах I—VII у заднего края есть бугорок. X сегмент с брюшной стороны сильно укорочен.
р. Nephelopteryx Klpk
0. Нет жабер, нет спинных бугорков (рис. 331 Б). X сегмент с брюшной стороны вытянут в параболическую пластинку.
р. Taeniopteryx Pict
9. Тело довольно массивное, обычно сплюснутое (рис. 331 В). Три последних членика челюстного щупика почти равной величины. Чехлы крыльев идут вкось. Жабры большей частью есть: они отходят с брюшной стироны переднегруди в виде слепых трубок или пучков нитей (рис. 338). Если жабер нет, то 2-й членик лапки виден, хотя и мал; и 3-й членик лишь немного длиннее, чем два первых вместе. Хвостовые нити довольно длинные, с явственно мутовчатым расположением волосков (рис. 337). Ноги в коротких шипиках, чередующихся с более длинными волосками. Длина 8—10 мм. В стоячих и текучих водах.
Сем. Nomuridae............10
0. Тело очень узкое, вытянутое. 5-й членик челюстного щупика длиннее 4-го (рис. 339). Чехлы крыльев направлены параллельно оси тела (рис. 331 Г, Д). Никогда нет наружных жабер...............................................18
10. Есть наружные жабры, трубчатые, нитчатые, сидящие на груди (рис. 338). . 11 0. Наруж’ных жабер нет......................................12
11. Две группы жабер, каждая из трёх трубчатых выростов. Бока грудных сегментов почти прямые (рпс. 336 Б). Шипики хвостовых нитей вдвое короче несущего их сегмента (рис. 337 А).
р. Protonemura Pict.
Рис. 338. Жабры 1-го грудного сегмента:
А — Protonemura humeral is; Б — Amphinemura cinerea.
Рис. 339. Челюсть:
А — Leuctra sp.; Б — Cap-nia nigra.
0. Две пары жабер, каждая из двух групп: одна — из 5—6, вторая — из 8 нитчатых выростов (рис. 338 Б). Бока грудных сегментов закруглены. Хвостовые нити из узлообраяных сегментов, снабжены шипиками длиной в а/з несущего их членика (рис. 337 Б). р. Amphinemura Ris.
12. Первый членик лапки 3-й пары ног приблизительно равен последнему. Шипики хвостовых нитей длиной по меньшей мере равны несущему их сегменту.
р. Nemurelia Klpk.
0. 1-й членик лапки задней пары ног не более чем в половину длины 3-го. Шипики хвостовых нитей короче несущего их сегмента (рис. 331 В). р. N е m u г a Lair.
13. 2-й членик лапки очень короткий или его нет. 3-й членик лапки передней пары ног по крайней мере в 5 раз длиннее двух первых вместе (рис. 335 Б). Лопасти нижней 1убы равны в ширину и длину и отграничены одна от другой неглубокой бороздой (рис. 332 В). На теле и на ногах вертикально стоящие редкие щетинки. X сегмент с внутренней стороны надрезанный (рис. 329 Б). Тело очень узкое, ноги довольно короткие (рис. 331 Г). Длина 5—7 мм. В текучих водах среди камней.
Сем. Leuctridae, 1 род Leuctra Steph.
0. Два первых членика лапки почти равны, 3-й в два раза длиннее двух первых вместе. Лопасти нижней губы удлинённые и отграничены глубокой бороздой. Ноги с длинными и густыми волосками (рис. 331 Д). X сегмент со спинной стороны продолжается (у самца) в тупотреугольный или (у самки) в параболический выступ (рис. 329 В). Длина 8—9 мм. Сем. С а р n i i d а е, род С а р n i a Pict.
HEMIPTERA (RHYNCHOTA) — КЛОПЫ И ИХ ЛИЧИНКИ
Форма тела водных клопов разнообразна. Голова, грудь и брюшко ясно разделены. Хоботок, отходящий снизу от головы и обращённый кзади, иногда прикрыт пластинкой и выступает из-под неё только своим кончиком. Усики на голове то ясно видны, то скрыты, сверху не видны. Передние ножки в большинстве случаев приспособлены для захватывания и умерщвления добычи (рис. 349 Б); лишь у сем. Cori-xtdae — единственной группы растительноядных клопов — эти ложки имеют «лопаточку* (видоизменённая лапка) как приспособление для соскабливания водорослей с подводных предметов.
На конце пожек, на последнем членике лапки, имеются коготки (рис. 342 Л, Б), иногда отходящие пе от самого конца, а несколько выше, сбоку (рис. 342 Я, Г). Л и-чинки отличаются от взрослых лишь отсутствием или меньшей степенью развития крыльев. Однако нужно заметить, что и взрослые стадии (имаго) некоторых клопов или имеют недоразвитые крылья, или совершенно лишены их.
Питание: все клопы, за исключением сем. Corixidae, — хищники, сосущие кровь различных животных.
1. Тело снизу покрыто очень густым, несмачиваемым бархатистым серебристым пушком. Усики далеко выдаются вперёд, много длиннее головы. Все ножки приблизительно одного строения, ходильные, без каёмки волосков. Форма тела более или менее близкая к линейной. Живут на поверхности воды (рис. 340) и плавающих на ней предметов или у воды.
Gymnocerata — явноусые ...2
и
Рис. 341. Личинки Gerris sp. разных стадий.
Рис.
А — Hydrometra sp.; В — Vella currens; Г ___________
Д — Hebrus pusillus (длиннокрылая форма)
340. Клопы — Rhynchota:
Б — Meso veil a furcata; — Mlcrovellla pygmaea;
: . );
E — Hebrus ruficeps (короткокрылая форма).
0. Усики незаметны сверху: они прилегают к голове с брюшной стороны. Передние ножки приспособлены для хватания, задние — для плавания (рис. 343—348). Форма тела уплощённая или торпедная. Живут в толще воды и только для дыхания поднимаются к поверхпости (за исключением Aphelochirus).
Cryptocerata — скрытпоусые................7
2. Усики из 5 члеников; два первые более массивны, без щетинок. Коготки на самом конце последнего членика ножек. Тело короткое, лишь раза в два с половиной длиннее своей ширины. Очень мелкие, не длиннее 2 мм (рис. 340 Д, Е). Хищники. На мокрых берегах рек и стоячих водоёмов, во мху торфяных болот.
Сем. Hebridae, у нас 1 род Hebrus Curt. (Naeogeus Kirk.).
0. Усики из 4 члеников (рис. 341). Коготки или па самом конце последнего членика, или немного выше, сбоку (рис. 342). Тело большей частью длинное. Величина различная. 3
8. Коготки на самом конце последнего членика (рис. 342 Л, Б).................4
0. Коготки пе на самом конце последнего членика ножек, немного выше — сбоку (рис. 342 В, Г) ..................................................................5
4. Тело очень длинное, приблизительно раз в десять длиннее своей ширины. Голова кпереди от глаз вытянута далеко вперёд, расширяясь кпереди. Длина до 12 мм (рис. 340 Л). Передвигаются медленно. По мокрым берегам водоёмов, на надводных листьях или скользят по поверхности воды.
Сем. Hydrometridae — палочковидные водомеры, 1 род Hydrometra Latr. — водяной бегун.
0. Тело овальное, раза в три длиннее своей ширины. Основания каждой пары ног сближены па брюшной стороне по срединной линии. Первый членик усиков длинный, изогнутый, с одной или двумя щетинками на середине своей длины. Личинки мелкие, не более 3,5 мм (рис. 340 Б). На поверхности воды и на широких плавающих листьях; в стоячих и медленно текущих водоёмах.
Сем. Mesoveliidae, 1 род М е з о v е 1 i a Muis.
5. Все сегменты груди приблизительно одной длины. На поверхности воды ... 6
0. Средний сегмент груди много длиннее I и III (рис. 341). Основа иля задней нары ног расставлены далеко. В разнообразных стоячих и текучих водоёмах, скользят по поверхности воды.
Сем. Gerridae — водомерки, р. Gerris F. (Limnotrechus).
6. Очень мелкие формы, не более 2,5 мм. Первый членик усика короче головы, почти прямой (рис. 340 Г). В стоячих водоёмах, заросших ряской.
Сем. Veliidae, р. Microvelia Westw.
0. Крупные формы, до 8 мм. Первый членик усика длиннее головы, дугообразно изогнутый (рис. 340 В). В быстро текущих водах.
Сем. Veliidae, р. Velia Latr.
7. На конце тела 2 длинные дыхательные трубки (рис. 343). Задние ноги ходиль-
ные. Живут в толще воды.
Сем. Nepidae. ...... 8
0. Тело без дыхательных трубок.............................9
8. Тело плоское, листовидное (рис. 343). Голова значительно уже переднеспинки. В стоячих водоёмах, иногда и в реках; повсюду, кроме Крайнего Севера.
р. N е р a L. — водяной скорпион, широко распространён вид N е р а ci погеа L. (с длинными дыхательными трубками на конце тела).
Рис. 343. Личинка водяного скорпиона—Nepa cinerea.
Рис. 342. Коготки на лапках Ger-rididae:
А — Hydrometra sp.; Б — Mesovelia sp.; В — Velia sp.; Г — Gerris sp.
0. Тело очень длинное, узкое, цилиндрическое. Голова шире переднеспинки.
р. RanatraF.,1 европейский вид R. linearis L.,‘обычен на юге, но редок на севере.
9. Лапки передних ног одночленистые, расширены в лопаточку. Хоботок прикрыт полукруглым выступом верхней губы.
Сем. Corixidae............10
0. Лапки передних ног пе расширены. Хоботок не прикрыт...........14
10. Усики 4-членистые. Щиток закрыт выростом переднегруди, сверху не заметен,
Подсем. Corixinae — гребняки .............11
0. Усики 3-членистые (рис. 345). Щиток виден. Длина 2—4,5 мм (рис. 344 Б). В литорали рек и стоячих водоёмов.
Подсем. Micronectinae, 1 род Micronecta Kirk.
И. Лопаточка длинная, почти цилиндрическая, с длинным когтем на вершине (рис. 346 В). Переднеспинка одноцветная, без поперечных полосок. Длина до 6 мм. Широко распространены.
р. С у m a t i a Flor.
0. Лопаточка 3- или 4-угольпая. Переднеспинка поперечнополосатая..12
12. Глаза сильновыпуклые. Лоб густо волосистый. Встречается в Арктике, в водоёмах на граните, в таёжной зоне как ледниковый реликт.
р. Glaenocorisa, 1 вид G. cavifrons Thoms.
0. Глаза слабовыпуклые. Лоб почти голый..........................13
18. Переднеспинка и надкрылья, кроме поперечных полосок, с тонкими, продольными, параллельными, короткими бороздками. Распространение очень широкое.
р. S i g а г a Leach (Arctocorisa Wall, С а 1 1 i с о г i х a White).
0. На переднеспинке и надкрыльях нет продольных бороздок (рис. 344 Л, 346 Л,В).
р. С о г i х a Latr.
14. Тело сплюснуто с боков......................................16
A — Corlxa ~ -
necta minutissima.
Рис. 345, Усик Micronecta minutissima.
Рис. 347.
Plea leachi.
P и с. 348. Гладыш — Notonecta glauca.
8р.; Б — Micro-
Рис. 346. Corixa dentipes:
A — передняя; Б — средняя ножка; В — ножка Cymatia coleoptrata.
A — плавт — Naucoris clmtcoldcs; Б — его клешня; В — Aphelochirus montandoni.
0. Тело сплюснуто сверху вниз...............................16
15. Ножки с 2 коготками. Формы мелкие, не более 5 мм (рис. 347). Большей частью среди растений в стоячих водах — пресных и солоноватых.
Сем. Pleidae, р. Plea Leach.
0. Ножки без коготков. Формы более крупные (рис. 348).
Сем. Notonectidae, р. Notonecta L. — гладыш.
16. Тело овальное, немного длиннее своей ширины. Голова между глазами не выступает треугольником, срезана почти прямо. Хоооток короткий, доходит лишь до основания первой пары ножек (рис. 349 А, Б). Длина И—16 мм.
Сем. Naucoridae, р. Naucoris F. — плавт.
0. Тело приблизительно треугольное, не длиннее или чуть длиннее своей ширины (рис. 349 Б). Голова между глазами выступает треугольником. Хоботок длинный, заходит за основание последней пары ножек. На дне рек, редко озёр; типичный реофил; ведёт ночной образ жизни. Сем. Aphelochiridae, р. Aphelochirus Westw.
COLEOPTERA IMAGO — ВЗРОСЛЫЕ ЖУКИ
Тело жуков (рис. 359) ясно расчленено на голову, грудь из 3 сегментов и б р ю ш к о из 6 сегментов, каждый, из которых прикрыт снаружи хитиновым кольцом; спинная часть его носит название тер гит а, брюшная стернита. Голова снабжена усиками, большей частью из 11 члеников; они отходят вблизи глаз, направлены в большинстве случаев вперёд и хорошо заметны, во иногда направлены назад я прижаты,
так что сверху не видны. Часто на голове бросаются в глаза тонкие придатки челюстей — челюстные ш у п и к и; с первого взгляда их можно принять за успки, но они ясно отличаются от усиков по числу члеников, всегда небольшому. Все три грудныхсег-мента ясно видны только снизу, где от каждого из них отходит пара ножек; сверху вядпа лишь спинка первого сегмента — так называемый грудной шит, или переднеспинка — и часть спинки второго в виде маленького треугольника — ш ит к а; ко второму же сегменту прикрепляются жёсткие надкрылья, закрывающие второй и третий сегменты и всё брюшко. Образуемая сомкнутыми внутренними краями надкрылий продольная срединная линия называется швом. Иногда эти края смыкаются вплотную, так что шитка не видно. Наружные края надкрылий перегибаются иногда на нижнюю сторону, образуя узкие боковые полосы — эпиплевры. Ножки состоят из нескольких отделов; самый последний из них, носящий название лапки, состоит из нескольких члеников; конечный членик лапки большей частью снабжён коготками. Предпоследний отдел ножки, носящий название голени, снабжён обычно двумя шпорами (рис. 356).
1. Лапки 5-членистые или кажущиеся 4-членистыми, но в таком случае задние ножки плавательные, густо усажены длинными волосками................2
0. Лапки 4-членистые, расширенные, их 3-й членик имеет 2 лопасти, между которыми расположено
2. Последний
Рис. 350. Hygrobia tarda.
начало последнего членика; все ножки ходильные....8
членик лапки небольшой, коготки тоже..............3
0. Последний членик лапки длиннее совокупности всех остальных; ножки тонкие.
Сем. Dryopidae — коготники.
3. Усики щетинковидные или нитевидные иногда пильчатые 4
0. Усики короткие, булавовидные или неправильные, с очень крупным 2-м члеником..................... 7
4. Задние ножки плавательные............б
0. Задние ножки не плавательные. Жуки окружены пу-
зырьком воздуха. Сем. Helodidae — тряс инни ки.
5. Усики кажутся 10-членистыми. Передняя часть брюшка (2—3 первых сегмента) прикрыта расширенными кзади в виде пластинок основными (первыми) члениками задних ножек (рис. 351). Сем. н а 1 i plidae — плавунчики.
0. Усики И-членистые. Основные членики задних ножек расширены не кзади, а кпереди, прикрывая заднегрудь (рис. 359)......................................6
в. Между основными члениками задних ножек на заднегруди короткий отросток, отделённый от остальной частя заднегруди поперечным швом. Длина 8,5—10 мм (рис. 350). В илистых прудах и лужах (встречен только в Житомирской области).
Сем. Н ygrobiiaae — водожилы.
р. Hygrobia,у нас 1 вид Н. t а г d a Hbst.
0. Между основными члениками задних ножек на заднегруди клиновидный заострённый отросток, не отделённый поперечным швом.
Сем. Dytiscidae — плавунцы.
7. Каждый глаз разделён вдоль на 2 участка (рис. 368). Передние ножки вдвое длиннее средних и задних, которые сильно расширены я уплощены.
Сем. Gyrinidae^— вертячки.
0. Глаза цельные. Передние ножки не длиннее прочих. Усики короткие, булавовидные, направлены назад и прижаты к голове, так что сверху не видны *. Спинная сторона тела сильно выпуклая, брюшная плоская или несколько вогнутая.
Сем. Hydrophi lidae — водолюбы.
8. Передняя часть головы нормальная, не вытянута. Усики не коленчатые и не булавовидные.
Сем. Chryaomelidae — листоеды.
0. Передняя часть головы вытянута в трубку. Усики коленчатые, булавовидные. Сем. Curculionidae — долгоносики.
Сем. Haliplidae — плавунчики.
1. Боковые края переднеспинки (грудного шита) почти параллельны между собой, с 2 глубокими бороздками почти во всю её длину. В текучих водах, среди нитчагок и урути.
В г у с h i us Thoms. — килевик.
0. Переднеспинка сильно сужена кпереди........................2
2. В задней половине надкрылий около самого их шва параллельная ему тонкая, гладкая бороздка (рис. 351 В). На Украине, в Поволжье, Закавказье, Средней Азия.
Peltodytes Reg. (Cnemidotus llljg.), у нас 1 вид Р. сае s us Daft.
1 При отсутствии навыка в процессе определения за усики могут быть приняты челюстные щупики, более длинные, чем усики, я направленные вперёд.
О' Укрзаппой бороздки нет. Длина до 5 мм, В стоячих и медленно текущих водах, часто в пойме (рис. 351 Л, Б].
Н а 1 i р 1 u s Latr. — плавунчик.
Рис. 351. Haliplua lineatus:
А — снизу; Б — сверху; В — Peltodytes caesus.
Сем. Dytiscidae — плавунцы
1. Лапки 1-й и 2-й пары ножек кажутся четырёх член истыми; три первых членика всегда несколько расширены, снизу уплощены и снабжены губчатой поверхностью. Щиток обычно не виден. Величина до 3 мм. Подсем. Hydropori nil.
2
0. Лапки 1-й и 2-й пар ножек пятичленистые, расширены лишь у самцов. Величина различна.................................•'.............................£
Рис. 352. Note-rus crassicornis.
Рис. 353. Laccophi-lus obacurus.
Рис. 354. Глаза жуков:
А — Colymbetini; Б — Dy-tiscinl.
2. Щиток не виден (рис. 352). Длина 3,5—4,5 мм ................... 3
0. Щиток виден (рис. 355). Длина не менее 7 мм...4
3. Переднеспинка по переднему и боковым краям с узким бортиком (рис. 352). В лужах и прудах; часто зарываются в ил. р. Noterus Clairv. — толстоуо.
0. Без бортика (рис. 353). В стоячих в медленно текущих водах.
р. Laccophilua Leach — л у ж н и к.
4. Глаза с выемкой близ осповапия усиков (рис. 354 Л). Длина 7—19 мм. ... 5
0. Глаза без выемки (рис. 354 Б). Передние лапки с круглой присоской, образованной из 3 сильно расширенных первых члеников...........................10
5. Надкрылья с поперечной штриховкой (рис. 355 Л). Длина 17—19 мм. В прудах и озёрах. р Colymbetes Clairv. — прудовик.
0. Надкрылья без поперечной штриховки................................6
в. Коготки задних лапок разной величины .............................7
0. Koi’otkh одинаковой величины .....................................8
1 Относящиеся к этому подсемейству поды Hydroporua Clairv. — нырялка, Hyphydrua III. — пузанчик, Hygrotus Steph. — пеструшка, Сое-la m b u з Thoma. — подводник в некоторые другие настолько мелки, что разбираться в их систематических признаках с непривычки очень трудно, и потому мы ограничиваемся определением их лишь до подсемейства.
7. Жук бронзовочёрпый. Задние бёдра снизу у конца с косой ямкой, усаженной щетинками (рис. 356 Л). Длина 9—14 мм. Быстро плавают в стоячих и медленно текущих водах (рис. 355 Z?, В). р I 1 у b i u s Ег. — т и н н и к.
0. Задний край переднеспинки почти прямой. Бёдра без ямки (рис. 356 Б). Жук охряножёлтый. Длина 9—13 мм. В стоячих водоёмах (рис. 355 Г).
р. Rhantus Lac. — и л ь н и к.
8. Надкрылья, сложенные вместе, образуют своими концами остриё (рис. 355 Д), Длина 7—8 мм. р Copelatus Ег., у нас 1 вид С. г u f i с о 1 1 i s Schaff.
0. Надкрылья образуют концами закругления.......................9
9. Эпиплевры одинаковой ширины почти по всей своей длине. Длина 7—8 мм. В проточной воде и в прибрежной области крупных озёр.
р. Plata m bus Thoms. — пёстрый гребец.
А — Colymbetes fuscus; Б — llybius fuligfnosus; В — Ily-
bius angustlor; Г — R ha nt us punctatus; Д — Copelatus ru- A — нога llybius sp.; Б — бедро ficollis; E — Agabus undulatus, Ж — Agabus nebulosus. Rhantus.
0. Эпиплевры резко суживаются немного далее половины своей длины (рис.355 Е, Ж). Длина 6—15,5 мм В лужах, прудах, часто в холодноводных ручьях.
р. A g а Ъ u s Leach — гребец.
10. Крупные жуки, не менее 22 мм. Членики задней лапки без кольцевой бахромы щетинок (рис. 357 Б) ................................................ 11
0. Жуки мельче, не более 18 мм. Членики задней лапки о кольцевой бахромой щетинок (рис. 357 А) .................................................12
11. Ппедпоследняя (перед лапкой) часть ножки — голень — длинная, с тонкими шпорами, задние лапки с 2 коготками (рис. 357 Б, 359). Длина 22—44 мм. В прудах, лужах, торфяных ямах ит. п. р, DytiscusL. — плавунец.
0. Голень широкая и короткая, внешняя шпора тоже сильно расширена. Задние лапки с одним коготком. Длина 30—35 мм (рис. 358 А). В крупных озёрах и в реках с медленным течением.
р. Cybieter Curt. — скоморох.
12. Тело овальное, выпуклое (рис. 358 В, 360 А). Более длинная шпора задней голени много длиннее первого членика лапки (рис. 357 4). Длина И—16 мм. В мелких стоячих водоёмах.
р. Hydaticus Leach — водяник, болотник.
0. Тело уплощённое, наибольшая ширина его ближе к заднему концу. Более длинная шпора задней голени чуть длиннее первого членика лапки. Одна или обе шпоры с выемкой у вершины...................................................18
13. Надкрылья или гладкие, или зернистые (иногда у самки). Длила 13—15 мм (рис. 358 Г). В стоячих водоёмах, затенённых лужах и т. п.
р. G га р h ode res Thoms. — поводень.
0. Надкрылья точечные или (у самки) с глубокими бороздками, покрытыми волос-~ - * и,—jg мм £ стоячих водоёмах с глубиной пе менее 1 м.
А с i 1 i u s
ками (рис. 358 Д, 360 Б). Длипа
Р-
Leach — полоскун (п л а в у я ч
и к).
'4.
Б
Рис. 357. Лапки:
А — Hydatlcus sp.;
Б — Dytlscus margl-nails.
Рис. 359. Dytis-cus latissimus с брюшной стороны.
Рис. 358. Жуки:
А — Cybister laterimarginalls; Б — Dytiscis latissimus; В — Hydatlcus sp.; Г — Graphoderes cine re us; Д — Aclllus suicatus (слева — самец, справа — самка).
Сем. Hydrophili dae — водолюбы
1. Спинная поверхность грудного шита с продольными бороздками или с борозд* ками и вдавлениями.....................................................2
0. Спинная поверхность грудного щита без продольных бороздок.......8
2. На грудном щите 5 продольных бороздок (рис. 361). Последний членик задней лапки короче суммы прочих. Длина 2,2—9 (большинство 2,5—4) мм. В заросших прудах, во всевозможных лужах —торфяных, лесных, пойменных и т. д.
р. Helophorus F. — мор шинник.
0. Переднеспинка с другой скульптурой. Последний членик задней лапки не короче совокупности остальных её члеников.....................................4
3. Основание надкрылий (т. е. их верхние края) в виде поперечного киля. Усики из 7 члеников, из которых 3 мохнатых. В стоячих водоёмах, среди подводной растительности (рис. 362 Б). р. Hydrochus Leach — в л а г о л ю б.
0. Основание надкрылий с косой тонкой линией. Усики из 8—9 члеников, из них 5 последних мохнатые ................................................. 5
4. Последний членик челюстных щупиков много короче предпоследнего. Голова с несколькими ямками. В прибрежной области стоячих и текучих водоёмов (рис. 362 В). р. Ochthebius Leach — прибрежии к.
0. Указанный членик не короче предпоследнего; челюстные щупики сильно выдаются своими концами за передние углы переднеспинки (рис. 362 Л). Надкрылья с рядами точек или бороздками. Длина до 3 мм. В стоячих и текучих водоёмах, под камнями или мхом, а также на водных растениях.
р. Н yd гае о a Kug. — водобродка.
v 5. Конечный членик задней лапки много длиннее суммы всех остальных. Передний край головы с вырезкой (рис. 363). Эпиплевры равной ширины на всём протяжении. Длина ^>5 7 мм. р s ре г с he us Kug.
0. Конечный членик задней лапки короче суммы всех остальных. Передний край головы без вырезки. Эпиплевры к концу сужены...................................в
Рис. 360. Передние ножки самцов жуков:
А — Hydatlcus sp.; Б — Cyblster laterimargl-nalis; В — Acillus sulcatus.
P и c. 361. He-lophorus sp.
6. Надкрылья не закрывают один или два последних сегмента (рис. 364 А). Усики из 9 члеников, из которых 5 последних мелкобугристые. Длина 1—2,5 мм. В стоячих и медленно текущих водах. р. Limnebius Leach — прудожил.
0. Надкрылья закрывают всё брюшко целиком. Усики из 7—9 члеников; бугорками покрыты лишь 3 последних ................................................7
7. Щиток длинный и узкий, заострённый. Между основаниями надкрыльев и грудным щитом перехват. Лапки и голени двух последних пар ножек покрыты волосками. Усики семичленистые. Длина 3,5—5,5 мм (рис. 364 Б). В стоячих водоёмах, большей частью в солоноватых. рв Berosus Leach — пискун.
Рис. 363. Sper-cheus emargi-natus.
Рис. 362. Жуки:
А — Hydrae па sp.; Б — Hydrocbus elongatus; В — Ocbthebius sp.
0. Щиток не длиннее ширины. Тело без перехвата. Ножки без волосков или с волосками лишь на лапках.................................................8
8. 11 и Ш сегменты груди образуют с брюшной стороны по срединной линии выдающийся киль, который заканчивается назади острым зубцом..............9
0. Киля нет....................................................10
9. Надкрылья с ясными сближенными попарно полосками в задней части. Киль заходит за основание задней пары ножек. Длина 32—47 мм (рис. 364 В]. Широко распространён в больших и малых водоёмах.
р. Hydrous Dahl. — водолюб большой.
0. Бороздки на надкрыльях неясные. Киль короче, не заходит за основание задних ножек. Длина 13—18 мм (рис. 364 Г). В небольших водоёмах.
р. HydrophilusDe Geer — водолюб малый.
10» Надкрылья в задней половине с относительно глубоко врезанными точечными бороздками, промежутки между которыми усеяны беспорядочно разбросанными более крупными точками. Киль на брюшной стороне среднего грудного сегмента низкий, кзади Ш
заострённый, кпереди косо уплощённый. В разных водоёмах, особенно в стоячих с загрязненной водой (рис. 364 Е).
р. Hydrobius Leach — в о д о ж у к, 1 вид Н. f u з с i р е s L.
0. Надкрылья без углублённых бороздок, разве только около самого шва..И
11. Надкрылья в задней половине с углублённой бороздкой около шва....12
О, Надкрылья без такой бороздки......................................14
12. Последний членик челюстного щупика длиннее предпоследнего. Надкрылья в беспорядочно рассеянных точках. 1-й членик задних лапок очень короткий. В стоячих водоёмах, особенно — в заросших ряской.
р. Anacaena Thoms. — ш и п о и о ж к а.
Рис. 364. Жуки:
А — ЫтпеЫиз ер.;
Б — Berusus sp.:
В— Hydrous piceiis;
Г — Hydropbllus sp.;
Д — PhHhydrus ше!а-nocepiidius;
E — Hydrobius fusclpes.
0. Последний членик челюстного щупика не длиннее предпоследнего или немного длиннее................................................................18
13. Челюстные щупики длиннее головы; их последний членик короче предпоследнего. Длина до 7 мм. В стоячих водоёмах (рис. 365).
р. Heiochares Muis. —омутник.
0. Челюстные щупики короче головы; их последний членик немного длиннее предпоследнего. На надкрыльях ряды густо расположенных точек. Длина до 4 мм. В стоячих, реже в текучих водоёмах (рис. 366). р. Laccobi us Ег. — бол от о л ю б.
Рис. 365. Helo- Рис. 366. Lacco- Рис. 367. Gyrinus лова жука Аи-chares. bins. natutor. lonogyrus.
Сем. Gyrinidae — вертячки
1. Спинная поверхность густо волосистая. Надкрылья в беспорядочно рассеянных мелких точках. Длина до 6,5 мм. В быстротекущих водах с чистой, прозрачной водой; днём прячутся; на поверхности появляются перед восходом солнца и на закате.
р. Orectochilus Lac. — сумеречная вертячка.
0. Спинная поверхность гладкая, без волосков. Надкрылья с рядами точек или с матовыми бороздками. На поверхности держатся днём...............2
2. Переднеспинка с неполными поперечными бороздками. Надкрылья с И рядами точек. Длина до 4,5 мм. В стоячих водоёмах и речных затонах (рис. 367).
р. GyrinusL. — вертячка.
0. Переднеспинка без поперечных бороздок. Надкрылья с 10 матовыми бороздками. Длина до 7 мм. В стоячих водоёмах, в оросительных каналах.
р. Aulonogyrus Reg., у нас 1 вид А. со n ci n n us Klug.
Рис. 369. Helmis.
Сем. Dryopida — коготники (прицепыши)
1. Усики короткие, 9-членистые, с крупным 2-м члеником. Спинная поверхность тела густо волосистая. Надкрылья в сложенном состоянии образуют на заднем конце слегка заострённый угол.................................................2
0. Усики нитевидные. Спинная поверхность тела голая или в мелких волосках . . 3
2. На боковых сторонах переднеспинки по 1 резкой продольной бороздке. Глаза в густых волосках. В стоячих и текучих водоёмах, на подводных растениях.
р. D г у о р s Oliv. — жук водолаз.
0. Продольных бороздок на переднеспинке нет. Глаза почти голые. Длина до 5,5 мм. У берегов рек и ручьёв, под камнями.
р. Н е 1 i с h u s Ег.
3. На переднеспинке резкие продольные бороздки, соединённые поперечной бороздкой. Длина до 2,5 мм. В быстротечных ручьях, под камнями (рис. 369).
р. Helmis Latr.
0. Поперечной соединительной бороздки нет. Щиток округлый. Задний край (основание) переднеспинки образует перед щитком выемку. Длина до 1,7 мм. В текучих водах, под камнями и во мху.
р. L i m n i u s Mull.
Сем. Helodidae — трясинники
1. Надкрылья с широкими бороздками. 4-й членик лапки не двулопастной. На водных и болотистых растениях.
Е u b г i a Latr. — волнушка, 1 вид Е. £ а 1 u s 1 г i s Germ.
0. Надкрылья без глубоких бороздок. 4-й членик двулопастной......2
2. Задние ножки прыгательные; голени с длинными шипами.
На болотных растениях. р. Scirtes — лжеблошка.
0. Задние ножки не прыгательпые; голени с короткими VJj
шпорами.............................................3
3. Переднеспинка в 2 с лишним раза шире своей длины, с Д у незагнутым вверх передним краем (рис. 370). П\ Vl
р. Су phon Payk — ярмоноска. jtl
0, Переднеспинка не более чем вдвое шире своей длины с заг- ч J путым вверх передним краем...........................4 И
4. Основание переднеспинки с 2 вдавленными ямками. 3-й [I \ /п
членик усика короче 2-го. На болотистых лугах. U \ /к
р. Н е 1 о d е s Latr. — трясинник. £ \
0. Основание переднеспинки без ямок. 3-й членик усика длиннее 2-го. На болотных растениях. Рис. 370. Cyphon.
р. М i с го с а га Thoms. — малоголов.
Сем. Chrysomelidae — листоеды
1. Надкрылья с продольными точечными рядами; их основание значительно шире переднеспинки, бока которой не окаймлены.............................2
0. Надкрылья с рассеянными точками. Бока переднеспинки окаймлены. Усики отходят от головы между глазами. Спина с мелкими волосками. На водных растениях.
р. Galerucelfa Crotch.
2. Первый членик усиков длиннее своей толщины. Глаза цельные. На брюшной стороне тела густой покров из серебристых или золотистых волосков.•. . . . 8
0. Первый членик усиков короткий, не длиннее своей толщины. Глаза с выемкой. Коготки при основании сросшиеся. На водных злаках, например, на рисе.
р. L е m a Lac. — пьявица, 1 вид L. suvorovi Jacobs.
8. Последний членик лапки короче совокупности остальных члеников. Голени прямые, тонкие. На водных растениях. р. Donacia Fabr. — радужница.
0. Последний членик лапки длиннее совокупности прочих члеников. На заднем конце надкрылий вырезка, наружный угол их с шипом. На подводных частях урути, рдеста и др. Н а е m о n i a Latr.
COLEOPTERA LARVAE—ЛИЧИНКИ ЖУКОВ
одного рода имеются на о рюшке л о ж-
Р и с. 371. Личинка Hygrobia tarda.
Тело личинок удлинённое, с резким расчленением на отдельные сегменты и с ясно обособленной головой, причём голова по отношению к грудным сегментам склонена вниз, или загнута кверху, или же ось ее совпадает по направлению с осью тела; покровы головы обычно хитинизированы, причём переднюю часть её составляет верхняя губа, прикреплённая своим задним краем к поперечной хитиновой пластинке — так называемому головному щиту (эпистом). Из рот о-в ы х частей яснее всего выделяются массивные, темнее окрашенные, сильно хити-низировапные верхние челюсти (мандибулы, жвал ы), иногда с добавочными зубчиками. Грудные сегменты несут всегда по паре в о ж е к — иногда обычного для наземных жуков строения, иногда более или мепее видоизменённых для целей плавания: расширенных и снабжённых волосками. На конце ножки всегда снабжены одним или двумя сильными коготками. Брюшко иногда спабжено дыхательными придатками (рис. 372), ные ножки — нечленистые, снабжённые крючками выступы, подобные ложным ножкам гусеницы. На конце брюшка часто бывают палочковидные придатки (рис. 373) — церки.
Личинки большинства жуков живут там же, где и взрослые формы.
1. Ножки 6-членистые, в большинство случаев длинные; лапки с 2 коготками. Церки (придатки на конце брюшка) почти всегда есть.................2
0. Ножки обычно короткие, иногда отсутствуют; лапки с 1 коготком.....4
2. Брюшко из 10 ясно различимых сегментов, из которых последний несёт снизу 4 сильно изогнутых крючка. На всех брюшных сегментах по 1 паре нитевидных, покрытых волосками жабер (рис. 372).
Сем. Gyri nidae — вертячки.
0, Брюшко из 8 сегментов, 4 крючков на конпе тела нет........; . . . 8
3. Голова наклонена вниз. На брюшной стороне грудных и брюшпых сегментов — парные жаберные отростки. В заиленных лужах и прудах (рис. 371).
Сем. Hygrobiidae — водожилы, 1 род Н у g г о b i a Latr. у нас 1 вид Н. tarda Hrbat.
0. Голова направлена вперёд по оси тела. На брюшной стороне грудных сегментов нет жаберных отростков. Сем. Dyti scidae — плавунцы.
4. Ножки 6-членистые; лапка состоит из членика и подвижно соединённого с ним коготка. Брюшко из 9—10 ясных сегментов.
Сем. Н a I i р 1 i d а е — плавунчики.
0. Ножки 5-членистые (членик лапки и коготок слиты) или редуцированы .... 5 5. Церки имеются, хотя иногда трудно различимы. На боках головы по 5—6 глазков. Верхние челюсти с острыми зубцами на внутреннем краю.
Сом. Hydropbiiidae — водолюбы.
0. Церки отсутствуют..............................................6
6. Усики длиннее головы и передпеспинки, с крупными 1-м и 2-м члеником и многочисленными, очень мелкими остальными члениками. На боках головы по 2 глазка. На конце брюшка анальные втяжные трубки — простые или ветвистые. Ножки значительной длины. В воде. Сем. Helodidae — трясин ник и.
0. Усики короче головы, число члепиков в них пе превышает 4.................7
7. Ножки есть, хотя иногда и короткие.......................................8
0. Ножек нет. Внутри водных растений.
Сем. Сиге и li ooidae — долгоносики.
8. На заднем конце тела 3 пучка втяжных анальных жабер. На внутреннем краю жвал узкий, длинный волосистый придаток. Личинки присасываются к подводным камням. Сем. D г у о pi d а е - к о г от и и к и.
0. На заднем конце тела жаберных пучков нет. Внутренний край жвал без указанного придатка. Личинки па водных растениях..
Сем. Cbrysoraeli d а е — листоеды.
Сем. Gyrinidae — вертячки
1 Передний край головного щита без зубцов.
Orectochilus Lac. — сумеречная вертячка.
0, Передний край головного щита с зубцами посредине...........*
2, Передний край головного щита с 2 зубцами. Жаберные отростки очень длинные.
Gyri ous L. — вертячка.
267
Рис. 372. Личинка Aulono gyrus.
0. Передний край головного щита с 4 зубцами. Жаберные отростки короткие (рис. 372).
Aulonogyrus Reg., у нас 1 вид А. с on ci о п из Klug.
Сем. Dytiscidae—плавунцы
1. Голова без шейки. Посредине внутреннего края жвал зубец. Ножкн копательные. В прудах, зарываются в ил.
р. Noterus Clairv. — толстоус.
0. Голова без шейки. Жвалы без зубца. Ножки плавательные . ,2
2. Голова вытянута вперёд в длинный выступ или округлотреугольная. Нижнечелюстные щуппки 3-членистые.........g
0. Голова без выступа. Нижнечелюстпые щуппки 4- (или более) членистые ........................................g
8. Нижняя губа очень узкая и длинная. Головной выступ длинный, узкий. Голова значительно уже переднегруди, суживается в выступ сразу, резко. Усики такой же длины, как голова. Ножки покрыты волосками. В прудах и лужах (рис. 373 Л).
р. Hyphydrus Illig — пузанчик, у нас широко распространённый вид Н. ferrugineus L.
0. Нижняя губа короткая, поперечная или трапециевидная . . 4
4. Тело очень широкое; брюшко немного длиннее своей ширины. Церки короткие, подлипе почти равны VIII брюшному сегменту. Головной выступ узкий, приблизительно втрое короче головы. В стоячих водоёмах.
р. Hydrovatus Motsch.; 1 из видов — Н. cuspidatus Kunze — найден в Средней Азии и Азербайджане.
0. Тело удлинённое; брюшко, по меньшей мере, вдвое длиннее своей ширины. Церки длиннее VIII брюшного сегмента. Головной выступ широкий, округлотреуголь-ныи, перед вершиной на боках с выемками...............................5
* 5. 1-й членик церок короче VIII брюшного сегмента.
р. Hygrotus Steph. — пеструшка.
0. 1-й членик церок длиннее VIII брюшного сегмента.
С о е 1 a m b u s Thoms.— подводник и р. Hydroporus Clairv. — ныряли а.
в. VIII брюшной сегмент на боках без плавательных волосков........7
0. VIII брюшной сегмент с плавательными волосками по бокам.......13
7. На внутреннем краю жвал мелкие зубчики. Ножки без плавательных волосков.
р. Со pel a tus Erichs., у нас 1 вид С. ruficollis Schall.
0. Внутренний край жвал без зубчиков..........................8
8. На дистальном конце 3-го членика усиков чувствительный придаток. Ножки с плавательными волосками (рис. 373 Ж).
р. Laccophilus Leach — л у ж н и к.
0. Чувствительного придатка па 3-м членике усика нет. Ножки без плавательных волосков, за очень редкими исключениями (у некоторых видов Agabus)........9
9. Последний членик усика в 2 с лишним раза короче предпоследнего. На церках большей частью 2 пучка шетипок...........................................10
0. Последний членик усика по длине почти равен предпоследнему. На церках много шетинок, расположенных по их внешнему и внутреннему краям................12
10. Конец брюшка тупо округлён. Жвалы в 2 с лишним раза длиннее своей ширины (рис. 373 Е). р. Platambus Thoms. — пёстрый гребец.
0. Конец брюшка несколько вытянут в виде острого или тупого конуса; церки прикреплены несколько отступя от самого конца брюшка.....................11
11. Жвалы менее чем втрое длиннее своей ширины (рис. 374).
р. A g a b и 8 Leach — гребец.
0. Жвалы более чем втрое длиннее своей ширины.
р. 1 1 у b i u s Erichs. — т и н н и к.
12. Длина жвал при измерении по внутреннему краю более чем в 2,5 раза превышает их наибольшую ширину. р. Khantus Lac. — и л ь н и к.
0. Длина жвал менее чем в 2,5 раза превышает их наибольшую ширину (рис. 375).
р. Coiymbetes Clairv. — прудовик.
18. На переднем краю головного шита нет зубцов. Церки хорошо развиты ... 14
0. На переднем краю головного щита глубокие вырезы и 3 крупных срединных зубца.
Церки рудиментарные (рис. 373 В).
р. Cybister Curt. — скоморох.
14. Голова широкая (рис. 377 Б). Церки в половину длины последнего брюшного сегмента, несут волоски (рис. 373 Д). р. DytiscusL. — плавунец.
0. Голова узкая. Церки короче половины длины последнего сегмента, голые, лишены волосков (рис. 373 Г).
р. А с i 1 i u a Leach — полоскун.
Рис. 374. Личинка Agabus sp.
Рис. 375. Личинка Colymbetes.
Сем. Н а 1 i р 1 i d а е — плавунчики
1. На груди и брюшке есть стигмы. Брюшных жаберных отростков нет. X сегмент брюшка на конце расщеплён, от IX отделён швом.......................2
0. На груди и брюшке стигм пет. На сегментах тела по 2 пары длинных, тонких жаберных отростков, только на VIII сегменте 1 пара. IX и X сегменты брюшка слиты (рис. 376 Б). Последний сегмент с 2 очень длинными перками.
р. Pel t od у tesfCne m i dot us Ill.), у нас 1 вид P. c a e s u s Duft.
2. Предпоследний членик усиков не длиннее каждого из остальных. Посредине внутреннего края жвал короткие щетинки.
р. В г у с h i u s Thoms. — килевик.
О, Предпоследний членик усиков в 2—3 раза длиннее каждого из прочих. На внутреннем краю жвал нет щетинок. На спинной стороне каждого сегмента тела по 4 шиповидных, направленных назад отростка. Последний сегмент брюшка очень длинный, на конце раздвоённый (рис. 376 Л).
р. II а 1 i р 1 u s Latr. — плавунчик.
Рис. 377. Голова личинок:
А — Hydrophilus sp.; Б — Dytlscus sp.
Рис. 376. Личинки:
А—Hallplus sp.; Б—Pel tody tes
caesus.
Hydrophili dae — водолюбы различимых сегментов; X сегмент очень короткий. Церки
Сем.
1. Брюшко из 10 ясно
хорошо развиты. Тело довольно сильно склеротизировано
0. Брюшко из 8 сегментов; IX и X сильно редуцированы. Церки укороченные.
Тело склеротизировано слабо (за исключением Hydrobius)...................5
2. На боках головы по 6 глазков. Жвалы 3-зубчатые, без подвижной лопасти на внутреннем краю. Церки длинные, 3-членистые (рис. 378 Ж).
р. Helophorus Норе — моршинник.
0. На боках головы по 5 глазков. Жвалы с подвижной узкой лопастью на внутреннем краю. Церки короткие, 2-членистые....................................3
Рис. 378. Личинки водолюбов:
д - Berosus sp.; Б, В — Philydrus sp. сверху и сбоку, Г - Hydrobius (uscipes, Д - Hydrophilus sp.;
Е— Spercheus sp.; Ж ~ Helophorus sp. * F вр..
8. Церки, при основании сближенные, к концам расходятся. Предпоследний члепик усиков пе имеет чувствительного придатка. Живут в прибрежпои области стоячих и текучих водоёмов, не избегают и солоноватых.
р. Ochthebius Leach — прибрежии к.
0. Церки, при основании широко раздвинутые, идут параллельно друг другу. Предпоследний членик усиков несёт 1—2 чувствительных придатка.......4
4. Предпоследний членик усиков с 1 придатком; последний членик без придатков.
р. Hydr-aena Kug. — водобродка.
0. Предпоследний членик усиков с 2 придатками, последний с одним.
р. Limnebius Leach — прудожил.
5. Голова слабо наклонена вниз. Усиковые ямки, в которых первые члепики усиков прикреплены к голове, расположены ближе к боковым краям головы, чем к жвалам. 6
0. Голова загнута вверх. Усиковые ямки расположены ближе к жвалам, чем к боковым краям головы..................................................7
6. По бокам головы по 5 глазков. Жвалы на конце раздвоены. На брюшных сегментах с каждой стороны по 1 коническому короткому отростку. Тело широкое (рис. 378 Е).
р. S percheus Kug., у нас 1 вид S. emarginatus Schall.
0. По бокам головы по 6 глазков. Жвалы па конце обрублены. Брюшко без боковых отростков. Тело узкое. р. Н yd го cn us Leach — влаголюб.
7. На боках брюшных сегментов I — VII очень длинные жаберпые отростки; VIII сегмент очень мал, но ясно виден; IX и X незаметны (рис. 378 Л).
В е г о s u s Leach — пискун.
0. Боковые отростки брюшка короткие или отсутствуют; IX и X сегменты слиты и образуют стигмальную полость........................................8
8. На бёдрах ножек густые плавательные волоски..............9
0. Бёдра без волосков........................................ 10
9. Боковые жаберные отростки на брюшке есть. Левая жвала с 3 крупными зубцами (рис. 378 Д). р. Hydrophilus De Geer — водолюб малый.
0. Брюшко без боковых отростков. Левая жвала с 1 маленьким зубчиком посредине .
р. Hydrous Dahl. — водолюб большой.
10. Ножки хорошо развиты, заметны сверху. Глазковое поле овальное; глазки ясно разграничены. Левая половина переднего края головного шита значительно крупнее правой. . Р- Laccobius Erich. — болотолюб.
0, Передний край головного щита слабо асимметричен............11
И. Чувствительный придаток предпоследнего членика усиков пе короче их 1-го членика. Ножки короткие. р. Anacaena Thoms. — шипоножка.
0. Чувствительный придаток на усиках короткий. Ножки длинные, видны сверху. 12
12. Жвалы асимметричны: на левой 1 зубец, на правой 2. На брюшных сегментах 1П —VII парные двигательные бугорки (ложные ножки).
р. Enochrus Thoms. — т и н о л ю б.
0. Жвалы симметричные. Брюшко без двигательных бугорков........13
18. Внутренний край жвал с 3 зубцами (рис. 378 Г).
Hydrobius Leach — водожук, 1 вид Н. fuscipesL.
О, Внутренний край жвал с 2 зубцами............................14
14. Передний край головного щита с 6 зубчиками.
р. Helochares Muis. — омутник.
На переднем краю головного щита более 6 зубчиков.
Cymbiodyta Bedel., 1 вид С. marginella L.
0.
1. ленный 0.
Сем. Dryopidae — к о г отн п к и (прицепыши)
Тело голое, блестящее. Ножки короткие, сверху пе видны. Конец брюшка округ-
..................................................................2
Тело с густозсрНистым покровом, часто несущим волосистые бугорки. Тергиты 1 брюшка с продольными килями посредине. Ножки довольно длинные, заметные сверху. 8 2. Тергит VIII брюшного сегмента с 1 зубцом, IXc2. р. Helichus Erich. 0. Тергиты VIII и IX брюшных сегментов без зубцов.
D г у о р s Oliv. — жук водолаз.
3. Брюшные и грудные сегменты с боковыми лопастями, несущими на краях короткие толстые щетинки. р. Н е I m i s Latr.
0. Брюшные сегменты не имеют лопастей. Тело удлинённое, впереди слегка расширение. Задние края тергитов усажены длинными волосками. р. L i m n i u s MOIL
J Спинные хитиновые пластинки брюшных сегментов тела.
Сем. Crhysomelidae — листоеды
1, Стигмы VIII брюшного сегмента в виде длинных, сближенных, направленных назад хитиновых шипов. На подводных частях растений (стеблях или корнях) .... 2 0. Стигмы VIII брюшного сегмента нормальные. На //j надводных частях растений или внутри стеблей.3
\ I / 2. На наружном краю жвал 1 шетипка (рис. 379).
/V I г... Z f \ р. Donacia ТЬг. — радужница.
0. На наружном краю жвал 2 щетинки.
Р’ Hecmonia Latr.
8. На боках головы по 5—6 глазков.
р qtq п п Анальное отверстие находится на спиппой стороне X
гис. 379. Личинка Do- брюшного сегмента. В живом состоянии личинки покрыты паС1а- слизью и экскрементами. Личинки выгрызают листья риса.
Lerna suvorovi Jacobs. — пиявица.
0. На боках головы по 1 глазку. Личинки выгрызают верхнюю сторону листьев Nymphaea alba и Nuphar lute urn.
Galerueella nymphaeae L.
MEGALOPTEBA LARVAE —ЛИЧИНКИ БОЛЫПЕЕРЫЛЫХ, СЕМ. SIALIDIDAE - ВИСЛОКРЫЛКИ.
Личинки крупные, достигают 2,5 см. Голова крупная, глаза маленькие, усики короткие, четырёхчленистые. Массивные жвалы с острыми зубцами (рис. 381). На семи первых сегментах брюшка по бокам отходят сегментированные дыхательные придатки с бахромой из длинных волосков (рис. 380). В СССР известен один род S i а 1 i s Latr. — в и с докрыл к а.
NEUROPTERA LARVAE — ЛИЧИНКИ НАСТОЯЩИХ СЕТЧАТОКРЫЛЫХ
Из этого отряда насекомых имеют отношение к воде представители двух семейств — О я m у I i d а е nSisyridae — и то лишь в личиночной стадии. Тело личинок веретенообразное, сплюснутое в спинно-брюшном направлении. Голова маленькая, с выставляющимися вперёд за её пределы чётковидными усиками и сосущими ротовыми частями. На поверхности тела со спинной стороны — а иногда и с брюшной — многочисленные бугорки со шетинками.
Рис. 380. Личинка Si alia 1 atari а — вислокрылки.
Рис. 381. Личинка Stalls 1 atari а —голова с брюшной стороны.
Рис. 382. Личинка Osmylus chrysops.
1* Тело тёмное, удлинённое, веретенообразное, покрытое короткими щетинками, сидящими на бугорках (рис. 382). Из апального отверстия выставляются две расходящиеся, выворачивающиеся наподобие пальцев перчатки трубки, поверхность которых покрыта крючочками, служащими для удерживания личинок на субстрате (рис. 383). Усики короткие. Жабер нет. Длина до 20 мм. В сыром мху, чуть покрытом водой.
Сем. О s in у И а а е, р.0 sm у I us Latr., наиболее распространённый вид О. fulvicephalus Scop.
2. Тело удлинёпно-яйцевидпое, слегка сплюснутое, покрытое бугорками со щетинками. Усики d длины тела. На брюшке 1 пар трахейных жабер в виде членистых, подогнутых под брюшко трубочек (рпс. 384). Длина до 6 льи. На поверхности тела и внутри канальцев губок, бадяг, иногда в мшанках.
Сем. Sisyridae, р. Sisyra Burm.
TRICROPTERA LARVAE — ЛИЧИНКИ РУЧЕЙНИКОВ
Средп личинок ручейников различают два типа. Одни имеют тело сплюснутое, ось головы является у них непосредственным продолжением оси тела, так что ротовое отверстие обращено вперёд; эти личинки, за немногими исключениями, живут совершенно свободно; их называют камподеоидными личинками за пх сходство с бескрылыми чинками кампод У других личинок
'ЯИР1
Рис. 383. Личинка Osmylus — задний конец.
I ли-е а (рис. 385 Б), тело цилиндрическое, и ось головы перпендикулярна к оси тела, так что ротовое отверстие обращено вниз; за сходство с гусеницами они получили название г у-се ницеобраз-п ы х (рис. 385Л); эти личинки всегда живут в чехли-ках, построенных из самых разнообразных материалов (песчинок, обрезков листьев и т. п.), скреплённых особыми вы-олове (рис. 386)
Рис. 384. Личинка Sisyra fuscata: А — вид сверху; Б — вид снизу.
определения в некоторых случаях не-> и нижнюю губы (для этого нужна цельной, нерасчленённой пластинки непосредственно у переднего края головы, а нижняя лежит первой снизу; чтобы рассмотреть её на цельной голове, надо положить голову нижней стороной кверху. Все три сегмента груди ясно разграничены; со спинной стороны они то мягкие, то более или менее хитинизированные. На первом брюшном сегменте у гусеницеобразных личинок бывают иногда выступы, служащие для задерживания тела в чехлике. Для той же цели у этих форм служит пара выступов на заднем конце тела, несущая крючки — прицеп-к и; свободно живущим формам эти прицепки позволяют зацепляться за неровности твёрдого грунта или за какие-
нибудь подводные предметы. У гусеницеобразных форм вдоль всего брюшка с боков тянется полоска волосков — боковая линия. Иногда личинки бывают снабжены нитевидными жабрами па брюшке.
1. Личинки камподеоидные; голова направлена вперёд по продольной оси тела (рис. 385 Б). Тело сжато в спинно-брюшном направлении; сегменты его разграничены ясно. Живут большей частью свободно, редко в чехликах; некоторые плетут ловчую сеть.
Подотр. Annulipalpia — кольчат о-щ упиковые.................2
0. Личинки почти или вполне гусеницеобразные; голова наклонена вниз под углом к оси тела (рис. 385 А ). Тело почти цилиндрическое или лишь слегка уплощённое в спинно-брюшном направлении. Перетяжки между сегментами неглубокие. Живут всегда в чехликах.
Подотр. Integripalpia — цельнощупиковые
18 А. Н. Липин
делениями. На
обходимо рассмотреть верхнюю лупа;)
верхняя выступает в
виде
Tri-
Р и с.
385. Типы личинок choptera:
А — гусеницеобразный; Б — кампо-деоидный.
ДЛЯ
Рис. 386. Схема строения головы личинки Tri с ho -
ptera:
А — сверху; Б — снизу.
. 20 273
2, Грудь вся склеротпзирована (твёрдая)...........................8
0. Склеротизирована только переднеспинка (I сегмент груди)........6
3. На брюшке жабры ветвистые или в виде пучков. Прицепки длинные, 2-членистые;
на 1-м членике длинные щетинки. Чехлик неподвижный с ловчей сетью......4
0. Жабер нет. Прицепки иначе устроены ............................5
4. Жабры ветвистые (рис. 387). Сем. Hydropsychidae.
0. Жаберные нити отходят от общего стволика. В прохладных текучих водах, на камнях, поросших водорослями или мхом.
Сем. Arctopsychidae, р. Arctopsyche.
5. Формы очень мелкие. Прицепки короткие. Чехлик из выделений или из песчинок.
Сем. Hydroptilidae.
Рис. 387. Личинка Hydropsyche pellucidula:
А — общий вид личинки сбоку; Б — передняя нога; В — прицепка.
0. Формы сравнительно крупные. Прицепки длинные. Чехлика нет. Длина до 8 мм. В озёрах и медленно текущих речках.
Сем. Psychomyidae, подсем. Ecnominae, р. Ecnomus Me Lachl.
1 вид Е. tenellus Ramb.
6. Жабры ветвистые у основания 2-й и 3-й пар ног и на 8 первых члениках брюшка» Переносных чехликов нет. В текучих прохладных водах (рис. 388 А, В, Г; 389 Л — В).
Сем. Rhyacophilidae, подсем. Rhyacophilinae, р. R h у а с о-р h i 1 a Pict., группа A (R. n u b i 1 a Zett., R. septentrionis Me Lachl. и др.).
0. Жабер нет ................................................ 7
7. Прицепки короткие, частично сросшиеся с последним члеником брюшка. Есть переносный чехлик ................................................8
0. Прицепки длинные (рис. 392 Б). Переносного чехлика нет.....3
8. На среднеспинке (II грудной сегмент) и заднеспинке (III сегмент) по паре небольших щитиков. На коготках ножек нет бугорка у основания базального шипа (рис. 388 Б, 389 Г, 390). Длина до 7 мм. В реках и ручьях.
Подсем. Agapetinae, 1 род Agapetus Curt.
0. На среднеспинке и заднеспинке щитиков нет. На коготках ножек у основания базального шипа есть бугорок. Чехлик из крупных песчинок, нижняя (брюшная) сторона плоская, верхняя выпуклая. Подсем. Glossosomatinae Ч
9. Верхняя губа мягкая, белая, втягивающаяся под головной щит (рис. 392 А).
Сем. Philopotamidae...........10*
0. Верхняя губа склеротизирована ............................12
10. Голова и переднеспинка красноватожёлтобурые. Без чехлика (рис. 391). Дли па до 22 мм. В горных ручьях с более или менее быстрым течением, под камнями или между ними. р. Philopotamus Leach.
0. Голова и переднеспинка светлые ...........................11
11. Голова и переднеспинка жёлтые, с неясным рисунком. Длина до 7 мм. В горных ручьях с быстрым течением. р. Wormaldia Me Lachl.
1 У нас 3 рода — Glossosoma, Diploglossa HMystrophora, различающиеся очень мелкими признаками.
Рис, 388. Личинки сем. Rhyacophilidae: А — Rhyacophlla septeu-trlonis; Б — Agapetus comatus; В — Rhyacophl-la sp.; Г — Rhyacophlla nublla.
Рис. 389. Ноги личинок Rhyacophlla:
A — передняя, Б — задняя; прицепки: В — Rhyacophila trlstls, Г — Agapetus comatus.
Рис. 390. Чех-лики куколок Agapetus comatus.
Рис. 391. Личинка Philopotamus montanus сбоку.
Рис.392. ЛичинкаPhilopotamus montanus: А—верхняя губа; Б — прицепка.
0. Голова и переднеспинка почти белые, без рисунка. Личинки крупные. В Азиатской части СССР. р. Dolophilodes.
12. От нижней губы (с нижней стороны головы) отходит очень длинный узкий вырост, выдающийся за передний конец головы (рис. 393).
Сем. Psychomyidae, подсем. Psychomyinae ..........13
Рис. 393. Жвалы Psychomyia pusilia: А — правая; Б — левая
Рис. 394.
А — коготок ножки Psychomyia pusllla; Б — то же Tinodes rostocki; В — приценки Ecnomus tenellus.
О, Без длинного отростка на нижней губе...............................15
13. Окраска головы жёлтая или коричневая без ясных точек. Жвалы прямые (рис. 393), правая с большим зубцом. Переднеспинка жёлтая, ближе к заднему краю коричневатая, без ясных точек. Коготки ножек короткие (рис. 393 Л). Длина до 4,5 мм. В озёрах и реках на подводных предметах. р. Psychomyia Latr.
0. Рисунок на голове резкий (рис. 395). Коготки ножек длинные (рис. 394 Б) . . 14
Рис. 395. Голова личинки Tinodes woeneri.
Рис. 396. Правая жвала Tinodes.
Рис. 397. Личинка Rhyacophlla si-birica.
Рис. 398. Жвала Neu-reclipsis bi-maculata.
f la vidus.
14. На внутреннем краю правой жвалы ясные зубцы. Длина до 8 мм. В стоячих водоёмах.
р. Lype Me Lachl.
0. На внутреннем краю правой жвалы неясные зубцы (рис. 396). Длина до И мм. В стоячих и текучих водах.
р. Tinodes Leach.
15. Прицепки короткие, из 2 твёрдых (флеротизированных члеников, не густо покрытые волосками. Коготки ножек короткие, сильно изогнутые. Мелкие виды, до 10 мм (рис. 397).
Сем. Rhyacophilidae, подсем. Rhyacophillnae, р. Rhyacophlla Pict., гр. В 4-С (R. в i b 1 г i с a Me Lachl. и др.)« 276
0. Прицепки длинные, из трёх члеников, из которых лишь третий хитинизировап, густо покрытые ресничками, довольно длинные, слабо изогнутые (рис. 401 В). Ноги в длинных волосках; голени большей частью с двумя длинными черноватыми шпорами. Коготки ножек длинные и почти прямые.
Сем. Polycentropidae.............16
16. Только последний членик прицепки с волосками; два первых членика короткие (рис. 401В); коготки припепок без крючочков как на спинной,так и на брюшной стороне; на брюшной (вогнутой) стороне лишь ряд мелких шипиков. Длина до 22 мм. В быстро (реже медленно) текущих водах; делают ловчие сети (рис. 62).
р. Neureclipsis Me Lachl, 1 вид N. blmaculata L.
0. Все 3 членика прицепки в волосках........................17
Рис. 400. Личинки:
А — Polycentropus flavomaculatus; Б — Plectrocnemia сопзрегза; В — Cyrnus flavidus.
17. Крючок прицепки с брюшной стороны с 4 маленькими зубчиками (рис. 399). Длина до 15 мм (рис. 400 В). В стоячих водоёмах, на глубине в несколько метров.
р. Cyrnus Steph,
0. Крючок прицепки или без зубчиков, или с 1 зубчиком на брюшной стороне . . 18
18. Голова широкая. Крючок прицепки изогнут под косым углом (рис. 401 4). Длина до 22 мм (рис. 400 В). Главным образом в горных ручьях.
р. Plectrocnemia Steph.
0. Голова удлинённая. Крючок прицепки изогнут под прямым углом (рис. 401 В)..................................................... 19
Рио. 401. Прицепки:
А — Plectrocnemia conspersa; Б — Polycentropus flavomacuiatus; В — Neureclipsis blmaculata.
19. На крючке прицепки со спинной стороны не более 1 шипа. Длина до 12 мм (рис. 400 Л). В быстрых речках и ручьях с каменистым дном.
р. Polycentropus Curt.
0. На крючке прицепки по меньшей мере 2 шипа. Длина до 16 лм». В озёрах и прудах.
р. Holocentropus Me Lachl.
20. Личинки почти гусеницевидные. Тело слегка уплощено....................21
0. Личинки гусеницевидные. Тело цилиндрическое............................23
21. Среднеспинка мягкая, иногда лишь с небольшим твёрдым шитиком посредине. На брюшной стороне переднегруди заострённый рог. Задняя жаберная нить всегда с волосками. Чехлик из спирально расположенных растительных кусочков или заменён трубкой тростника. Сем. Phryganeidae.
0. Среднеспинка твёрдая, разделена на 2 половинки. На брюшной стороне переднегруди рога нет. Чехлик из песчаной трубки с крыловидными также песчаными выростами по бокам.
22. Коготки задних лапок очень короткие (рис. 402 Л). Жаберные пучки из 3—4 нитей (рис. 403). Длина 17 мм. Чехлик с широкими боковыми крыльями (рис. 404); длина Curt.
Сем. Molannidae
22
Чехлик с широкими боковыми крыльями (рис. 404);
его до 26 мм. В озёрах. р. Molanna
Рис. 403. Личинка Molanna angustata.
Рис. 404. Чехлик личинки Molanna angustata.
В
Г
Рис. 402. Коготок задней ножки личинок:
А—Molanna angustata; Б — Mol anno des zelleri.
личинок: Б — Seri-
Рис. 405. Чехлики
А — Notidobla clliaris; _____
costoma pedemontanum; В — Goera pilosa; Г — Beraeodes mlnuta.
0. Коготки задних лапок очень длинные, щетинковидные (рис. 402 Б). Жаберные пучки из 2 нитей. Длина до 12 л<лс. Чехлик с узкими крыльями; длина его до 20 мм. От мелких дистрофных водоёмов до горных озёр. р. Molannodes Me Lachl.
23. На брюшной стороне I грудного сегмента есть рог........................24
0. На брюшной стороне I грудного сегмента рога нет.........................28
24. Среднеспинка груди сплошь роговая, заднеспинка с 3 парами хитиновых шитиков. I брюшной сегмент с 3 выростами — 2 боковых и 1 спинной. 1-я пара ножек короче и массивнее прочих, 2-я пара — длиннее и толще 3-й.
Сем. Limnophilidae.
0. Среднеспинка груди с отдельными хитиновыми шитиками, заднеспинка большей частью мягкая. 2-я пара ножек не длинней 3-й
Сем. Sericostomatidae..................25
25. Жабры простые нитевидные, толстые, длинные, на II—VII брюшных сегментах. На передних голенях по 2 шипа (рис. 407Л), на средних и задних по 1. На I брюшном сегменте 2 боковых выступа. Прицепки короткие. Чехлик — 4-гранная (у зрелых личинок) или коническая (у молодых) трубка из растительного материала. В текучих водах.
Подсем. Lepidostomatinae.........27
0. Голова сильно втянута в переднегрудь, передний край которой сильно вогнут. Среднеспинка груди с 3 парами щитиков, заднеспинка мягкая с 3 или 4 парами хитиновых шитиков. Жабры маленькими пучками по 3—4 нити, сидящими на переднем краю сегментов брюшка. Чехлик из мелких песчинок в виде трубки с крыльями из крупных песчинок или камешков (рис. 405 В). В прибойных частях берегов озёр и в текучих водах.
Подсем. Goerinae........26
26. На среднеспинке груди 2, на заднеспинке 3 пары щитиков. Боковая линия от середины III до конца VIII брюшного сегмента. Чехлик сильно расширенный (рис. 405 В). Длина личинки до 14 мм. В ручьях и в прибойной зоне озёр.
р. Goera Leach.
Рис. 406. Личинки сем. Sericostomatidae:
А — Brachycentrus subnubilus; Б — Lepidostoma hlrtum: В — Beraea maurus; Г — Notidobta cllLarts; Д — Sertcostoma personatum.
0. На среднеспинке 3, на заднеспинке 4 пары щитиков. Боковая линия от конца III до начала VIII брюшного сегмента. Чехлик крыловидно расширен (слабее, чем у Goera), с камешками по краям (рис. 57—58 Б). Длина личинки до 10 мм. В ручьях.
р. S i 1 о Curt., 1 вид S. р а 11 i р е s Fbr.
27. Голова тёмнобурая со светлыми штрихами или точками.
светлее головы, кзади постепенно темнеет до черноты, вся со светлыми штрихами и точками. Среднеспинка вся склеротизирована. Заднеспинка вся мягкая, с 3 парами мелких бурых щитиков со светлыми штрихами и точками. Жабры на II—VI (иногда и VII) брюшных сегментах, нитевидные, довольно длинные (рис. 406 Б). Длина до 11 мм. Чехлик у зрелых личинок из растительных частиц, 4-гранный, длиной до 17 мм. В равнинных, медленно текущих, сильно заросших ручьях и небольших прохладных речках.
р. Lepidostoma Ramb.
0. Голова жёлтобурая, красноватая, со светлыми точками. Переднеспинка с такой же окраской, кпереди иногда несколько темнее. На заднеспинке с каждой стороны по группе щетинок. Длина до 7 мм. Чехлик, как у Lepidostoma, длина его до9лм*. В горных ключах и на скалах, поросших сырым мхом.
р. Crunoecia Me Lachl.
28. Выступов на I брюшном сегменте нет (рис. 406 Л).
Сем. Sericostomatidae, подсем. В г а с h у-
Переднеспинка спереди
Р и с. 407. Передние ножки личинок:
А — Lepidostoma birtum; Б — Brachycentrus subnubi-
1US.
centrinae .................29
0. Выступы на I брюшном сегменте есть.................80
29. Средние и задние голени на дистальном конце вытянуты в длинный конический отросток. На внутренней стороне средних и задних бёдер гребешки. Длина до 12—13* мм. ‘ Чехлик 4-гранный (по крайней мере у молодых личинок). В крупных реках, большей частью на дне, среди растительности, особенно в бассейне Волги (рис. 406 Л).
Рис. 408. Личинка Odontocerum albi-сотое.
р. Brachycentrus, у нас вид В.
широко распространён subnubilus Curt.
0. Отростков па средних и задних голенях пет. Длина до 10 мм. В быстро текущих водах, на камнях, среди водорослей и мхов. р. Micrasema McLachl.
80. Спинка III сегмента груди с 2 поперечными, лежащими один под другим, хитиновыми щитиками и с 2 меньшими у начала ножек. I и II сегменты груди хитиновые.
Жаберные пучки крупные (рис. 408). Чехлики конусовидные, изогнутые, гладкие, из мелких песчинок. В текучих водах.
Сем. Odontoceridae, р. Odontocerum
Leach, 1 вид О. albicorne Scop.
0. Спинка III сегмента груди без шитиков......................81
81. II грудной сегмент или целиком мягкий, или с отдельными хитиновыми пятнами и роговой полоской по переднему краю, или только с отдельными пятнами. Личинки массивные. Тело цилиндрическое. Жабры хорошо развиты. Выросты I брюшного сегмента мало выдаются. Голени 1-й пары ног с двумя шпорами. Прицепки частично слиты с последним сегментом, крючки их крупные, с 2—3 крючочками на снинной стороне. Чехлики из
песчинок (рис. 405 А, Б).
Сем. Sericostomatidae, подсем. Sericostomatinae ...82
0. II грудной сегмент склеротизирован........................83
82. Среднеспинка груди мягкая, разве лишь с несколькими хитиновыми пятнами на боках (рис. 406 Д). Длина до 12 .иле. Чехлик из песчинок, конический, изогнутый (рис. 405 Б). В горных и равнинных ручьях.
р. Se ri cos tom a Latr., у нас 1 вид S. personatum Spen.
0. Среднеспинка спереди хитинизирована, с многочисленными тёмными пятнышками, образующими по бокам большие тёмные пятна (рис. 406 Г). Длина до 17 мм. Чехлик конический, слабо изогнутый (рис. 405 Л).
р. Notidobia Steph., 1 вид N. ciliaris L.
38. Жвалы с крупными зубами и щётками волосков на внутреннем краю. Брюшко кзади постепенно суживается (рис. 406 В). Вместо боковой линии ряд маленьких твёрдых точек с волосками (на III—VIII брюшных сегментах). Чехлик из песчинок конический, сильно изогнутый, гладкий (рис. 405 Г). В стоячих и текучих водах.
Сем. Beraeidae.........34
0. Жвалы долотовидные, иногда (редко) плоские, ножевидные. Брюшко кзади не суживается. Боковая линия на III—VII брюшных сегментах очень тонкая, с короткими волосками. Чехлик из песчинок или растительных частиц, большей частью конических. Сем. Leptoceridae.
84. На наружном краю жвал, ближе к вершине, щёточка из волосков. Жабры в пучках. Щетинок на ножках немного. Длина до 9 мм (рис. 405 Г).
р. Beraeodes Eaton, у нас 1 вид В. m i n u t a L.
0. На наружном краю жвал щёточки нет. Жабер нет. Щетинок на ножках много. Длина до 6 мм (рис. 406 В). р. В е г а е a Steph., у нас 1 вид В. р u 1 1 a t a Curt.
Сем. Limnophilidae
1. Жвалы без зубов. Заднеспинка без срединных хитиновых пластинок. Голова и переднегрудь тёмные, со светлыми пятнами. Жабер по бокам тела нет, есть только на спине и на брюшке, нитевидные (рис. 409 Е). Шип у основания коготка 2-й и 3-й пар ног длинный, не короче коготка. Чехлики конусовидные, изогнутые, на брюшной стороне слегка уплощённые, из песчинок, с более крупными зёрнами по бокам. Длина личинки 7—8 мм. В литорали озёр, в реках и ручьях на камнях, гальках, обломках дерева.
Подсем. Apataniinae, р. Apatania Koi. и р. Apatelia.
0. Жвалы с зубами. На заднеспинке две пары срединных хитиновых шитиков (рис. 409 А—Д). Голова (рис. 410) и грудь или бледно окрашены с тёмными точками, или тёмные без светлых точек. Боковые жабры есть. Шип у основания коготка на всех ногах короче самого коготка (рис. 412). Чехлики различные (рис. 411). Длина личинки до
32 мм. Подсем. Limnophilinae..........................................2
2. Жабры в виде пучков из 2—4 нитей ..,....................3
0. Жабры из изолированных нитей ..........................8
3. У зрелых личинок в каждом пучке по 4 нити. Голова тёмнобурая со светлыми точками и штрихами.Чехлик конический, у молодых личинок из детрита, у зрелых передняя часть из песчинок, задняя из детрита.
р. Dicosmoecus, 1 вид D. р а 1 a t u s Me Lachl.
0. У зрелых личинок в каждом пучке по 2—3 нити...................4
4. Голова жёлтая с тёмной, изогнутой в виде петли полосой и тёмной срединной полосой внутри этой петли (рис. 409 Г, 410 В), или с тёмным затылочным шитиком. Чехлик или почти цилиндрический, прямой, составленный из неправильно расположенных растительных частичек, или в форме цилиндрической трубки, на которую налегают с двух сторон крупные куски мёртвых листьев, так что получается плоский чехлик (рис. 411 Л, Af, Я). Длина личинки до 32 мм. В прудах и лужах, дно которых покрыто опавшей листвой. р. Glyphotaelius Steph.
0. Голова без тёмной петли. Чехлики никогда не сплюснутые и не покрытые крупными кусками листьев, трубковидные или иногда треугольные в разрезе; иногда отягощённые камешками......................................................5
5. Затылочный щитик жёлтый с чёрным рисунком и виде гриба, по бокам сплошные поля из точек и пятен (рис. 409 Л, 410 А, Б). Чехлик из песчинок и крупных растительных палочек, большей частью расположенных с одного бока (рис. 411 Р). Длина личинки до 280
Рис. 409. Личинки сем. Limnophilidae:
А — Grammotaullus a tom a rl us ; Б — Llmnophllus ро-!ПН8у В Anabolla nervosa; Г — Glyphotaelius pcl-lucldus; Д — Stenophylax latipennis; Е — Apatanla (imbriata.
Рис. 410. Голова личинок:
А — Anobolla nervosa; Б — Anoboll a laevls; В — Glyphotaelius pimctatoltneatus.
М мм. В стоячих и медленно текущих водах; на открытых песчаных участках берегов озёр и рек; в ручьях.
р. A n а b о 1 i a Steph.
0. Нет рисунка в форме гриба ....................................в
6. Голова одноцветная, тёмная, только вокруг глаз светлая кайма. Дистальная шиповидная щетинка переднего бедра вдвое длиннее проксимального шипа; дистальная шиповидная щетинка заднего бедра в 2—3 раза длиннее проксимальной. Чехлик трёхгранный, равной по всей длине ширины, из плоских растительных частичек, главным образом листьев. Длина его 20 мм, ширина 6 мм (рис. 411 3). Длина личинки до 12 мм. В мелких стоячих водоёмах с опавшими листьями на дне.
р. Phacopteryx Kolenoti, 1 вид Ph. b г е v i р е n n j s Curt.
0. Голова иначе окрашена. Чехлик или цилиндрический, или трёхгранный в последнем случае расширенный кпереди ................................• • ?
1. Голова коричневая или красноватокоричневая. В задней части головы и к бокам тёмные пятна; по затылочному шитику ряд тёмных точек (рис. 409 А). Коготки передних ног короче лапки. Чехлик узкий и длинный (длина 40—80 лм<, ширина 6 мм], прямой цилиндрический, лишь слегка суженный кзади, из черепичато налегающих крупных растительных частиц (рис. 411 П). Личинки крупные, до 28 мм. В заросших водоёмах.
р. Grammotaullus Kolen., у нас обычен Q. a to m a ri us rbr.
A
b
Рис. 411. Чехлики личинок сем. Limnophilidae:
А — Limnophllus stigma; Б—L. declpiens; В — L. politus; Г—L. flavicornis; Д — Stenophylax rotund! pennis; E — Halesus auricollis (ч. куколки); Ж — H. teasel a tus; 3 — Phacopteryx brevlpennls; И, К — Chaetopteryx villosa; Л, M, H — Glyphotaelius puncta toll neat us; О — (поперечное сечение) Limnophllus sp.; П — Grammotaulius nitldus; P — Anaholia nervosa; C — Micropterna sequox.
0. Голова иной окраски. Коготки передних ног равны длине лапкщили даже длиннее (рис. 409 F). Чехлики не более 30 мм в длину при ширине основной трубки 3,5 мм\ большей частью прямые, иногда трёхгранные, суживающиеся кзади, иногда конические, изогнутые, иногда с отягощением из растительных или минеральных частичек или раковинок; часто составные частицы располагаются в поперечном направлении или вкось (рис. 411 Л, Б, В, Г). Длина личинки до 28 мм. В разнообразных водоёмах, преимуще-в мелких заросших водоёмах, часто в весенних р. Liranophilus Leach.
крупные тёмные точки ...................9
мелкие тёмные точки ....................10
ственно в стоячих; лужах (рис. 412). 8. На голове 0. На голове 9. Два передних щитика на заднеспинке слиты в один. Чех-
лик из детрита, неровный (рис. 411 Д). Длина личинки до 25 мм. В ручьях и прибойной зоне озёр.
р. Stenophylax Colen. (часть видов).
0. Два первых щитика на заднеспинке раздельны. Чехлик из крупных частиц детрита с палочками на концах (рис. 411 ЯГ). Длина личинки до 26 мм. В ручьях и небольших речках.
Рис 412 Задняя Р- " а е 8 u 8 Steph,
ножка Limnonhi- Спинные жабры есть на брюшных сегментах II—VI, а
lus ffriseus Р ' брюшные, кроме того, и па VII. Срединные щитики заднеспинки ® * ясные, почти слившиеся посредине. Чехлик (рис. 411 Я, К)
почти прямой, спереди из песчинок, сзади из растительных кусочков (чехлик молодых личинок целиком из детрита, зрелых почти весь из песка). Длина личинки до 15 мм. В ручьях, речках, в прибойной зоне озёр.
р. Chaetopteryx Steph.
0. Спинные жабры есть на брюшных сегментах 11—VII, а брюшные на II—VIII. (рис. 409 Д). Чехлик (рис. 411 Д) немного изогнутый, из песчинок или мелких камешков, гладкий (у одного вида из растительных частиц).
р. Stenophylax Colen. (другая часть видов).
Сем. Hydropsychi d а е
1. На коготках средних и задних ножек по 1 шипу; у конца 1-го членика прицепок пучок щетинок. Лопая жвала со щёткой волосков на внутреннем краю.
Подсем. Hydropsychina е..........2
0. На коготках средних и задних ножек по 2 и более шипов; у конца 1-го членика прицепок отдельные длинные волоски и щетинки. Левая жвала без щётки на внутреннем краю. В больших реках Азиатской части СССР.
Подсем. Macronematinae...........4
2. Брюшные сегменты 1—IV постепенно расширяются, потом снова суживаются. 5 анальных жабер. Длина до 10 мм. В быстро текущих ручьях.
р. Diplectrona Westw.
0. IV брюшной сегмент не шире предыдущих. 4 анальных жабры.....3
8. На жвалах наибольший зубец проксимальный, несуший щёточку волосков; такая же щёточка у вершины жвалы. Жабры на VII брюшном сегменте не перистые. Длина до 11 мм. В реках. р. Cheumatopsyche.
0. На жвалах дистальные зубы больше проксимальных. Жабры VII брюшного сегмента перистые (рис. 387). Длина до 20 мм. В текучих водах, главным образом в реках.
р. Hydropsyche Pict.
4. Боковые щитики I грудного сегмента с коротким конусовидным отростком. Голова плоская с подковообразным валиком .................................5
0. Боковые щитики 1 грудного сегмента с очень длинным копьевидным отростком. Голова не плоская, без валика. Жвалы удлинённые. Базальный край 1-го членика ножек без отростка. р. Aethaloptera, 1 вид А. г os si с a Mart.
б. Голова широкая, без рисунка; валик высокий с волосками. Базальный край бёдер (3-й членик ножек) с крючковидным отростком. Личинки крупные.
р. М a crone m а, 1 вид М. radiatum McLachl.
0. Голова слегка расширена спереди, с тёмным рисунком. Валик не высокий, без волосков. Личинки не крупные.
р. Amphi psyche, 1 вид А. р г о 1 u t a McLachl.
Сем. Hydroptilidae
1. На спинной поверхности брюшных сегментов небольшие щитики. Чехлик из очень мелких песчинок или створок диатомовых водорослей, склеенных выделениями; иногда в форме бочонка с косыми отверстиями спереди и сзади. Длина личинки до 3 мм. В быстро текущих горных ручьях, часто на скалах, покрытых влажным мхом.
р. Stactobia McLachl.
0. Брюшные сегменты без спинных шитиков........................2
2. На брюшных сегментах III—VIII спинные и брюшные выступы (рис. 413). Чехлик в виде тыквенного семечка с круглым передним и щелевидным задним отверстием. Длина личинки до 3 мм. В медленно текуших водах между растениями, иногда на камнях.
р. Ithytrichia Eat , 1 вид I. I a me 1 laris Eat.
0. На брюшке нет выступов .....................................8
3. Ножки короткие, почти одинаковой длины (рис. 414 Л). Чехлик из сравнительно крупных песчинок, сплюснутый с боков, бобовидный или почковидный (рис. 415 Л). Длина личинки до 4 мм. Как в стоячих, так и в текучих водах преимущественно в реках с умеренной скоростью течения. р. Hydroptila Dalm.
0: Чехлик из выделений, с водорослями или без них..............4
4. Чехлик (рис. 416) тёмный, в виде тминового зёрнышка, с глубокими бороздками на поверхности. Длина личинки до 4 мм. В стоячих, сильно заросших водоёмах.
р. Orthotrichia Eat., наиболее распространённый вид О. t е t е n s i i Kolbe.
0. Чехлик светлый, жёлтый или зеленоватый. Задние ножки в 2 с лишним раза длиннее передних ......................................................&
б. Чехлик (рис. 415 Л) в форме бутылочки; задний конец (со щелевидным отверстием) гораздо шире переднего (с круглым отверстием). Средние и задние ножки одинаковой длины с телом. Длина личинки до 4 мм. В зарослях стоячих вод.
р. О х у е t h i г a Eat.
0. Чехлик (рис. 415 Б) неправильно эллиптический, посредине более широкий, чем на концах. Средние и задние пожки значительно короче тела. Длина личинки до 6 мм. В озёрах и медленно текущих реках, на растениях и камнях. р. Agraylea Curt.
Сем. Leptoceridae
1. Жабры в пучках (рис. 417 Л). Чехлик (рис. 418 Л) или из выделений, лишь иногда с примесью посторонних частиц, или, наоборот, полностью из посторонних частичек — песчинок или растительных кусочков. Длина личинки до И мм. В стоячих и медленно текущих водах; в прибрежной области среди растений, в глубинном на дне.
р. Leptocerus Leach.
0. Или жабер совсем нет, или они не в пучках, одинарные........8
283
Рис. 413. Личинка Ithytrichia la-mellaris.
Рис. 416. Чехлик Orthotrichia teteiisii.
Рис. 414. Личинки сем.
Hydroptilidae:
А — Ну drop til a MacLachlani; Б — Ptilocolepus granulatus.
Рис. 415. Чехлики личинок сем. Hydroptilidae:
А — Oxyethira tristella; Б — Ag ray lea pallidula; В — Ну drop til a MacLachlani.
Рис. 417. Личинка сем. Leptoceridae:
A — Leptocerud aterrimus; Б — Oecetls (urva; В — Trlaeuodes bleolor; Г — Mystacides azurea.
2. Жабры всегда есть, одиночные. Жвалы удлинённые, узкие. Челюстные щупики длиннее верхней губы. Голова и переднеспинка свотложёлтые с тёмным рисунком (рис. 417 Б). Ноги 3-й пары без плавательной бахромы. Чехлики слабо изогнутые, из песчинок, перемешанных с растительными кусочками (рис. 418 Е). Длина личинок до 14 мм. В стоячих водоёмах — на дне, часто среди растений; иногда в реках, р. Oecetis Me Lachl.
Рис. 418. Чехлики личинок:
А — Leptocerus a terrim us; Б, В, Г — чехлик куколки Trlaenodes bicolor; Д — Му-stacides longicornis; Е — Oecetis furva.
0. Жабры имеются не всегда. Жвалы широкие. Челюстпые тупики короткие ... 8
8« Задняя голень цельная. Чехлик то крупный (до 15 мм длиной) конический, очень узкий, гладкий, состоящий целиком из выделений, то мелкий (до 6 мм длиной), изогнутый, шероховатый, построенный из песчинок. Длина личинки до 8 мм. В зарослях стоячих и медленно текущих вод. р. Setodes Ramb.
Рис. 419 Голова лич. Му-etacidee nigra.
0. Задняя голень двураздельная.....................4
4. Задние ноги в длинных, густых плавательных волосках. Жабры имеются (рис. 417 В). Чехлики прямые, гладкие, конусовидные, из спирально расположенных растительных кусочков (рис. 418 Б, В, Г). Длина личинки до 13 мм. В стоячих и очень медленно текущих водах. р. Triaenodes Me Lachl.
0. Задние ноги без плавательных волосков. Жабер большей частью нет — в этом случае на голове рисунок из тёмных точек; или жабры развиты хорошо — и тогда на голове рисунок в виде буквы Н (рис. 417 Г, 419). Чехлики различные (рис. 418 Д). Длина личинки до 12 мм. В стоячих и медленно текущих водах.
р. Mystacides Latr.
Сем. Pbryganeidae
1. У переднего края среднеспинки есть 1—2 хитиновых щитика.......2
0. У переднего края среднеспинки нет хитиновых щитиков ..........3
2, У переднего края среднеспинки 1 крупный щитик с 2 продольными тёмными полосками. Длина личинки до 45 мм. В стоячих и медленно текущих водах.
р. Holostomis.
0л У переднего края среднеспинки 2 небольших щитика
Nour onia reticulata L.
8. На спинной поверхности головы есть продольная срединная тёмная полоска (рис. 420, 421 Л) ....................................................4
0. Такой полоски на голове нет (рис. 421 В). Длина до 30 мм. Чехлик (рис. 422 В) — слегка коническая трубка из различных растительных кусочков, расположенных спирально. В стоячих и медленно текущих заросших водах. р. Neuronia Leach.
4. На брюшной поверхности головы тёмные полоски или многочисленные тёмные точки. Длина до 26 мм. В качестве чехлика часто пользуется куском трубки тростника (рис. 422 Г). Главным образом в прудах и лужах, среди растений.
р. Agrypnia Curt.
0. На брюшной стороне головы или совсем нет тёмных точек, или они немногочисленны и неясны. Длина до 44 мм. Чехлик (рис. 422 4, В) со спиральным расположением частиц строительного материала (только у вида Phryganea minor) с расположением этих частиц вдоль чехлика. В стоячих водоёмах с большим количеством растительности.
р. Phryganea L.
яял
Рис. 421. Личинки сем. Phryganeidae:
Рис, 420. Рисунок головы личинки Phryganea sp.
Рис. 422. Чехлики личинок
сем. Phryganeidae:
д — Phryganea gran dis; Б — Neuronla ruflcrus.
Рис. 423. Гусеница Paraponyx stratiotata.
а, Б — Neuro nla
Phryganea grandls; В — ruficrus; Г — Agrypnia pagetana.
Рис. 424. Acentropus niveus:
“S?0®1 Б ~ лист, повреждённый гусеницей; В — куколка; Г — гусе-иица; м кучка яиц и только что вылупившаяся гупряипл« е — бескрылая самка (имаго).
Б Рис. 425. Cata-clysta lemnata: A — гусеница в чех-лике ив отревна листа; Б — чехлик куколки.
LEPIDOPTFRA LARVAE — ГУСЕНИЦЫ БАБОЧЕК
Взрослые бабочки — почти без исключения—типично воздушные насекомые, по некоторые их гусеницы питаются водпыми растениями и живут па них п внутри их и, следовательно, имеют отношение к воде. Их нельзя назвать настоящими водпыми животными, так как в самой воде они не живут. Эти гусеницы принадлежат к нескольким семействам, из которых среди «водных» 1усепиц богаче всего представлены семейства Pyralididae и Noctuidae. Мы приводим таблицу для определения лишь 4 родов из сем. Pyralididae, относящихся к той группе, представители которой не минируют обитаемые ими растения, а используют их только как материал для построения чехликов. Эти 4 рода являются наиболее обычными среди гусениц, живущих на водных растениях.
1. С каждой стороны тела по 3 продольных ряда кустистых сероватобелых нитевидных трахейных жабер (рис. 423). Голова очень маленькая, коричневая. Чехлик — из 2 кусочков водного растения телореза (Stratiotes). Длина до 26 мм.
Рагаропух stratiotata — огнёвка телорезная.
О, Трахейных жабер нет .............................................2
2. На теле продольные ряды мелких бородавок, несущих волоски; окраска желтоватая или зеленоватая. Голова светлокоричневая с чёрной каёмкой. Личинка строит чехлик иэ частей растений — урути, рдеста и т. д. (рис. 424).
Acentropus niveus Oliv.
0. Нет ни рядов бородавок, ни чёрной каймы на голове...............3
8. Тело гладкое, чёрное или оливковозелёное. Голова спереди и щиток с жёлтым. Чехлик обычно из ряски, в виде мешочка, внутренняя поверхность которого покрыта белой паутиной (рис. 425).
У нас 1 вид Cataclysta lemnata L.
0. Тело оливковое (окраска очень изменчива). Голова светлобурая или коричневая. Чехлик из 2 кусочков листа, соединённых между собой паутинными нитями (рис. 426).
р. N у m р h u 1 a Schr. (Hydrocampa Gart.)
DIPTERA LARVAE —ЛИЧИНКИ ДВУКРЫЛЫХ
Организованы личинки крайне разнообразно. У одних тело состоит из трёх явных отделов — голова, грудь и брюшко, у других головы нет или она мало развита и явного различия между грудными и брюшными сегментами нет.
Голова иногда обладает способностью втягиваться в тело; у многих личинок она развита вполне, но у многих представителей претерпевает редукцию, иногда доходящую до того, что остаются всего лишь пара или две крючков.
На теле иногда имеются разные выросты, то в виде ложных ножек различной формы — валиков или бугорков, то в виде боковых выступов, то в виде «лопаточек». Настоящие членистые ножки отсутствуют у личинок всех двукрылых.
Личинки, ведущие прикреплённый образ жизни, имеют специальные органы прикрепления: венчик крючков на заднем конце тела или ряд присосок на брюшной стороне. Иногда имеется вторичная сегментация.
По числу и расположению стигм, т. е. отверстий, которыми дыхательная (трахейная) система личинок может сообщаться с атмосферным воздухом, можно различить несколько типов. 1-й тип: 1 пара стигм на переднегруди и несколько (до 8) пар на брюшке (сем. Itonidae); 2-й тип (наиболее распространённый) : 1 пара на переднегруди и 1 на VIII сегменте брюшка (сем. Tipulidae); 3-й тип: 1 пара па VIII сегменте брюшка (сем. Culici-dae, за исключением подсем. Chaoborinae); 4-й тип: отсутствие стигм (подсем. Ghaoborinae, сем. Tendipedidae и др.). Стигмы открываются или на конце дыхательной труб-к.и, или непосредственно на поверхности кожи — на так называемой стигмальной пластинке, часто несущей различные образования, назначение которых — закрывать или открывать стигмы, смотря по надобности. Дыхательная трубка то обладает неизменной длиной (у форм, всплывающих для дыхания на поверхность воды), то может удлиняться или укорачиваться (у форм, живущих на дне или на погружённых предметах и вытягиваюших трубку до поверхности).
1. Головная капсула развита хорошо, склеротизирована (отвердевшая) вся целиком или только на спинной стороне.
Подотр. Nematocera — длинноусые.................................2
О, Головная капсула отсутствует или редуцирована, имеется лишь ротоглоточный аппарат; жвалы двигаются в вертикальпай плоскости.
Подотр. ВгасЬусега — короткоусые ..............................14
2. На брюшной стороне среднегруди «лопаточка» для прыганья (рис. 427, 428). Головная капсула втяжная, только её передний отдел на спинной стороне склеротизи-Вован. Антенны 2-членистые. Тело из 13 сегментов, часто ярко окрашено. Длина 6—15 мм, {ивут в тканях водных растений, вызывая их разрастания — галлы.
Сем. I t о n i d а е (= Cecidomyidae) — г а л л и цы.
0. Личинки без «лопаточки». Голова невтяжная, склеротизирована вся целиком. 3 8. Голова, грудь и I сегмент брюшка слиты между собой, 3 последних сегмента тоже. Тело широкое, плоское; на брюшной стороне по средней линии 6 сильных присосок, посредством которых личинки прикрепляются к субстрату и передвигаются (только в боковом направлении); бока брюшных сегментов резко выдаются в виде конических
выростов (рис. 430). Длина до 12 мм. В быстро текущих горных реках и ручьях, при-асываются к камням, водорослям и т. п. Сем. Blepharoceridae.
0. Голова, грудь и I сегмент брюшка раздельны; сегменты груди или раздельны, !ли слиты между собой........................................4
4. На брюшке 7 пар крупных боковых нечленистых выростов — ложных ножек, юоружённых па концах концентрическими рядами мелких коготков. Антенны длинные, (вуветвистые (рис. 429). В быстро текущих горных ручьях.
Сей. Deuterophlebiidae, 1 вид Deuterophlebia mirabilis Edw.
Рис. 426. Nymphula nymphaeata?
Л — лист, повреждённый гусеницей; Б — гусеница в чехлике; В — гусеница сбоку.
Рис. 430. Личинка В1е-pharocera sp.: А — о брюшной стороны; Б — сбоку.
А
А Б
Рис. 428. Прыгатель-ная лопаточка личинок сем. Itonidae:
А — Thurauia sp.; Б — rLasioptea sp.
Рис. 427. Личинки Itonidae:
А — Retinodiplosis зр.; Б — Thuraula sp.
Рис. 429. Личинка Deuterophlebia mirabilis:
а — со спинной; Б — с брюшной стороны.
0. Без длинных ложных ножек, а если они есть, то их меньше 7 пар. Антенны не двуветвистые ............................................................б
б. Голова втянута в грудь; брюшная поверхность её слабо склеротизирована; задний край выемчатый.......................................................6
0. Голова не втянута, с цельным задним краем........z...............8
6. Тело с длинными, упругими нитевидными или листовидными выростами. На некоторых высших водных растениях, чаще на мхах. Сем. Cylindrotomidae.
0. Тело без выростов ...............................................7
7. Голова хорошо хитинизирована, на спинной стороне выпуклая, с двумя продольными щелями, на брюшной — с одной. Нижняя губа заострённая, никогда не расщеп-ЯМ
лённая вдоль посредине; передний её край зубчатый. Усики длиннее челюстного щупика. Тело вальковатое, толстое, с правильно расположенными щетинками (рис. 431). На заднем конце площадка, окружённая 6 звездообразно расположенными лопастями, на которой открываются сильно хитинизированпые стигмы. Среди прибрежной растительности или в иле. Сем. Ti pulidae — долгоножки.
0. Голова хитинизирована слабо. Усики не длиннее челюстного щупика. Тело мягкое, вальковатое (рис. 432 Л) или уплощённое; часто в густых волосках, роже — с правильно расположенными щетинками. Передний грудной сегмент (а иногда п брюшные)
со втяжными ложными ножками или ползательными валиками. На заднем конце тела дыхательная трубка или площадка со стигмами (рис. 432 Б). Нижняя губа или глубоко растеплённая продольно, и тогда на заднем конце тела 6 отростков, или не расщеплённая, а тогда па заднем конце не более 5 отростков, расположенных не звездообразно.
Сем. Li mooildae.
8. Без ложных ножек и заменяющих их валиков или подушечек...............9
0. С ложными ножками .....’...............................................11
9. Грудные и несколько первых брюшных сегментов подразделены па 2 кольца,
прочие брюшные — на 3. На спинке вторичных сегментов узкие поперечные хитиновые
пластинки. Задние стигмы иногда на короткой дыхательной трубке. Иногда на конце тела пара придатков, покрытых длинными волосками. Тело в многочисленных щетинках (рис. 433). Во влажных местах берегов, во мхах, в иле и т. д.
Сем. Psychodidae — бабочницы.
0. Вторичной сегментации нет. Поперечные пластинки, если имеются, слабо развиты только на передних брюшных сегментах...........................................10
Рис. 432. Личинка Limnobia sp.: А — общий вид; Б — задний конец.
10. Грудные сегменты слиты в один отдел, который шире брюшка, состоящего И8 9 сегментов; дыхательная система или замкнутая, характеризующаяся присутствием внутри тела просвечивающих через кожу трахейных пузырьков (рис. 434 Л), пли открытая, сообщающаяся с внешней средой посредством стигм на спинной стороне VIII и частью IX брюшного сегмента, расположенных то непосредственно на их поверхности, то на конце отходящей от сегмента дыхательной трубки (рис. 434 Б, В). Или: грудные сегменты свободны и не шире брюшка, состоящего из 10 сегментов; на VIII брюшном сегменте имеется пара стигм, расположенных на стигмальной пластинке, частично заходящей и на IX сегмент. Ложные ножки или отсутствуют (подсем. Culicinae в Ghaoborinae) или имеются по паре на 1—II грудных и на V—VI (иногда и VII) брюшных (подсем. Dixinae). В воде стоячих и текучих водоёмов разной глубины.
Сем. С u 1 i cidae — комары, подсем Culicinae.
0. Грудные сегменты разве чуть шире брюшка. Трахейных пузырьков никогда нот. Тело-червеобразно удлинённое, беловатое; на заднем конце несколько креиких щетинок. Голова хитинизированцая. удлинённая. Стигм нет (рис. 435). В иле, в детрите, среди ра
стительных остатков.
Сем. Н е 1 е i d а е (сем. Tendi pedidae, подсем. Culicoidinae).
11. Ложные ножки на трёх первых сегментах брюшка в виде отростков или валиков. Задние сегменты вытянуты в длинную трубку, могут телескопически (одна в другую) складываться; у основания трубки пара нитевидных придатков; на конце трубки поме-
MS
тается последняя пара стигм (рис. 436). Тело из 12 сегментов. Длина до 7—8 мм. В слабо проточных или в стоячих, сильно загрязнённых водоёмах.
Сем. Liriopidae (Ptychopteridae), 1 род L i г i о р е Mgn. (Ptycboptera Mgn).
0. Ложные ножки на концах тела — или на обоих, или на каком-нибудь одном. Стигм нет ...............................................................12
Рис. 433. Личинки сем. Psycbodi-dae:
Maruina sp. A — c брюшной стороны; Б — co спинной стороны, В, Г — Perlcoma sp.; Д — Psy-choda sp.
12. Одна непарная ложная ножка на переднем конце тела, непосредственно за головой; 3—4 последних сегмента раздуты; на последнем из них прикрепительный диск с кольцевыми концентрическими рядами шипиков, при помощи которых личинка держится за погружённые предметы (рис. 437). Исключительно в проточной воде.
Сем. Melusinidae (Simuliidae) — мошки.
Рис. 434. Личинки сем. Culi-cidae:
А— Chaoborus sp.; Б — Culex эр.; В — Anopheles sp.
0. Ложные ножки на переднегруди имеются всегда; на заднем койце тела —или пожки, или специальный аппарат из крючков............................ 13
18. Стигм нет. Одна пара ложных ножек на переднегруди и одна пара на последнем сегменте (отталкиватели — рис. 438).
Сем. Tendi pedidae (С bi го п о mi d ае) — звонцы.
0. Одна пара коротких ложных ножек на переднегруди. Стигмы на боковых выростах переднегруди и на выростах VI11 сегмента орюшка. Последний сегмент несколько
Рис. 437. Личинка Melusina sp. (из сем. Melusinidae).
Рис. 438. Личинка Tendipes (Chirono-mus, из сем. Tendi-pedidae):
А—схема общего строения; Б — голова.
Рис. 436. Личинки Li-trope (Ptychoptera) sp. из сем. Ptychopteridae.
Р и с. 439. Личинка Thmimalea (Orphne-phila) sp. (из сем. Thaumaleidae).
Рис. 440. Личинки сем. Stratiomyidae:
А — Stratlomylu sp.: Б — Neraotelus sp.; В — Pacbygasier sp., задний конец; Г — Eupachygaster sp., аадний конец (часть пустой шкурки).
291
загнут к брюшной стороне; на конце его, вокруг анального отверстия, венчик из загнутых крючков разной величины (рис. 439), выполняющий функцию отталкивателя. Длина до 14 мм. В быстро текущих ручьях и родниках, на камнях.
Сем. Thaumaleidae (Orphnephilidae), 1 род Thaumalea (О г р h n е р h i 1 а).
14. Головная капсула на спинной стороне большей частью хорошо развита; только часть головы втягивается в грудь...........................................16
О, Головной капсулы нет; голова может втягиваться в грудь целиком.....18
15. Задние стигмы находятся или непосредственно на конце тела, пли на дыхательной трубке; они погружены в полость, образуемую выпячиванием кожи и открывающуюся наружу посредством щели ................................................ .16
0. Задние стигмы или есть, или отсутствуют; в первом случае они взаимно удалены и, если задний конец личинки не втянут, то расположены непосредственно на поверхности (пе погружены) ............................................................17
16. Личинки заметно уплощены, причём спинная поверхность выпуклее брюшной. Голова со щетинками, не втянута. Задний конец тела или округлённый, или вытянут в дыхательную трубку; щель, ведущая в стигмальную полость, расположена горизон
Р и с. 441.
Личинки сем. Tabani dae.
Рис. 442. Личинка Atherix sp. (из сем. Rhagionidae— Leptidae).
Рис. 443. Личинка Hemerodro-mia sp. (из сем. Empididae).
Рис. 444. Личинка Erista-lis sp. из сем. Syrphidae: А — общий вид; Б — ложная
ножка.
А
тально, с перистыми волосками вокруг. Нет ни ложных ножек, ни двигательных валиков (рис. 440 Л). Длина до 5 см. В стоячих водоёмах, среди растений, иногда на дне.
Сем. Stratiomyiidae — л ь в и в к и.
0. Личинки вальковатые. Голова без шетинок, втянута. Щель стигмалыюй полости расположена вертикально. Есть втяжные ложные ножки или двигательные валики (рис. 441). Длина до 4,5 см. В стоячих водоёмах — болотах, лужах и пр. (на илистой почве берегов и дна), реже в проточных (ручьях, речках).
Сем. Tabanidae — слепни.
17. Головная капсула па спинной стороне длинная, достигает среднегруди. Стигм пет. Брюшные сегменты имеют по паре втяжных крупных ложных ножек с когтевидными шипами на концах. 6 первых сегментов брюшка несут по 2 пары (спиппых и брюшнобоковых) загнутых назад подвижных шипов. На последнем брюшном сегменте 2 длинных, покрытых крупными волосками выроста, выполняющих, возможно, дыхательную функцию (рис. 442). В проточной воде, на различных погружённых предметах.
Сем. Rhagionidae (Leptidae), 1 род Atherix.
0. Головная капсула па спинной стороне округлая, короткая. Задние стигмы могут быть расположены на стигмальной пластинке с лопастями. На брюшке утолщения в виде валиков или слабо развитых ложных ножек с шипами на концах. Конец последнего сегмента с 5—6 радиально расходящимися волосками (рис. 443). В проточных водах (ручьях, реках), в иле или во мху. Сем. Empididae — толкунчики.
18. Задние стигмы или на конце одной тонкой, сильно вытягивающейся дыхательной трубки, или на концах двух коротких хитинизированных, взаимно параллельных трубочек. Тело то гладкое, то с мелкими шипиками на поверхности; иногда на нём имеются различные отростки, часто втяжные ложные пожки или валики (рис. 444). В стоячих, сильно загрязнённых водоёмах.
Сем. Syrphi dae — журчалки; паиболее известны личинки рода Eristalis — крыски.
0. Задние стигмы на двух самостоятельных, ярко окрашенных пластинках или на двух расходящихся трубочках......................................19
19. Задние стигмы внутри общего поля, снабжённого по краям лопастями (рис. 445). Длина 8—15 мм. В стоячих и слабо проточных водоёмах у поверхности воды среди растении. Сем. Sciomyzidae(Tetanoceridae).
0. Общее стигмальное поле без наружных лопастей.............20
20. Личинки на водных растениях, минирующие.............21
0. Личинки, свободно живущие в воде ........................22
21. Личинки с короткими, снизу пе зазубренными ротовыми крючками. Длина до 13 мм. На листьях жёлтых кувшинок.
Сем. Cordyluridae (Scatophagidae).
0. Личинки с зазубренными снизу ротовыми крючками. Длина до 5 мм. На разных водных растениях, растущих у берега.
Часть сем. Ephydridae — береговушки.
22. Ротовые крючки, заострённые, пе зазубренные снизу. Тело из 11 сегментов, из которых 2 первых сегмента груди могут втягиваться внутрь остального тела, а па последнем (заднем) сегменте имеются 2 спинных выроста с расположенными на их концах задними стигмами; на брюшной стороне тоже есть пара коротких втяжных выростов, покрытых шипиками.
Сем. Muscidae — мухи настоящие.
0. Ротовые крючки зазубрены снизу или оканчиваются пластинкой с зубцами или выростами. Задние стигмы на двух длинных дыхательных отростках, на заднем конце тела или на двух коротких трубках (рис. 446). Образ жизни чрезвычайно разнообразен — свободно живущие (в стоячих водоёмах среди растений), сапрофаги и паразиты.
Часть сем. Ephydridae — береговушки.
Сем. Culicidae larvae —личинки комаров
Тело состоит у большинства личинок из головы, обособленной груди, состоящей из более или менее слившихся и раздутых сегментов (нет слияния только у рода Di ха) и ясно сегментированного брюшка из 9—10 сегментов.
Голова большей частью сильно хитинизирована. Усики одпочлениковые, часто с пучком длинных чувствительных волосков; у личинок Ghao-borinae усики видоизменены в хватательный орган, причём волоски утолщены и изогнуты. Личиночных глазков одна пара, мелкие, простые. У личинок последней (иногда и предпоследней) стадии имеются очень крупные зачатки сетчатых глаз имаго, видимо, уже функционирующих. Спинная сторона головы ограничена главным образом крупным фронтальным склеритом, или лобным щитком; часто в передней части склерит этот сужен за счёт сближения основания усиков.
Грудь всегда с большим количеством волосков; из них в систематическом отношении важны группы плевральных волосков, сидящих на брюшной стороне вблизи боковых краёв, группы эти состоят из одного длинного перистого волоска и пучка более коротких, отходящих от общего цоколя с сильным коп'ем.
Брюшко у личинок подсем. Dixinae состоит из 10 хорошо разграниченных сегментов, у прочих из 9 сегментов за счёт слияния VIII и IX сегментов в один; таким образом, последний сегмент здесь является, в сущности, X. На VIII сегменте (за исключением подсем. Ghaoborinae) располагаются стигмы. Стигмы открываются или на конце дыхательной трубки — сифона, или непосредственно па поверхности сегмента и окружены клапанным аппаратом — стигм альной пластинкой. Дыхательная трубка — крупный, твёрдый, почти всегда тёмноокращенный вырост на спинной стороне VIII брюшного сегмента, несущий часто пару боковых гребней из зубцов, шипов или волосков. Под основанием сифона иногда имеется так называемая щетка — одиц или несколько рядов своеобразных шипиков. Урода Mansonia сифон превращён в буравящий аппарат.
Стигмальпая пластинка — сложная система из нескольких лопастей и поддерживающего их скелетного образования располагается она на предпоследнем сиг-293
мепто (у личинок с девятью сегментами брюшка) или па двух предпоследних (у личинок с десятью сегментами) со спинной стороны; у различных родов стигмальпая пластинка устроена различно, но всегда сразу бросается в глаза благодаря тёмной окраске и ряду сильных шипов, обращённых кзади. От стигм отходят два крупных трахейных ствола, которые идут до последнего отдела грудп. У некоторых форм эти стволы (или их боковые ветви) образуют пузыревидные расширения; в живых личинках пузырьки кажутся серебряными и хорошо просвечивают через покровы тела. Последний сегмент брюшка обычно уже прочих; кончается он анальным отверстием, вокруг которого располагаются две пары жабер; поверхность сегмента илп окружена сплошным хитиновым кольцом, или несёт хитиновую спинную пластинку — седло, иногда с сидящим на теле трубкообразным выростом. На брюшной стороне по срединной линии проходит полоса из сильно ветвящихся волосков, расходящихся веерообразно — анальный плавник.
1. Грудные сегменты не слиты. Брюшко из 10 сегментов, пе уже груди. Тело густо покрыто шипиками и редкими волосками. На I и 1П сегментах по паре небольших ложных ножек с крючками. На V и VI, а иногда п на VII сегментах ползательные валики с шиппками и щетинками. На VIII и IX сегментах по паре лопастей стигмальной пластинки. На боках последнего сегмента по 1 хитиновой пластинке, несущей 2—3 щетинки: па спинной стороне его хитиновое «седло», переходящее назад в длинную узкую трубку — хвостовой отросток — с 3 парами длинных щетинок на конце.
Подсем. Dixinae .........2
0. Грудные сегменты слиты. Брюшко из 9 сегментов, уже грудп.3
2. Усики с группой волосков по внутреннему краю (рис. 447 Б). Хвостовой отросток покрыт шипиками (рис. 448 Б). На боковых пластинках по 3 шетинкн. На спинной стороне брюшных сегментов нет розетки из щетинок. В различных стоячих водоёмах.
р. Paradixa Топп.
0. Усики без группы волосков по внутреннему краю. Хвостовой отросток без ши-пиков (рис. 448 А). На боковых пластинках по 2 щетинки. На спинной стороне брюшных сегментов розетка из коротких, но крепких шетинок (рис. 449). У берегов быстротечных речек и ручьёв. р. Di ха Meig.
3, Усики видоизменены в хватательные органы. В VII или VI—VII сегментах брюшка имеются трахейные пузырьки (рис. 450).
Подсем. Chaoborinae.
0. Усики не видоизменены в хватательные органы. Трахейных пузырьков нет.
Подсем. Culicinae.........6
4. Основания усиков раздвинуты на далёкое расстояние (рис. 451 Б). Дыхательная трубка длинная. Две пары воздушных пузырьков: в груди и в VII брюшном сегменте (рис. 450). В лужах и небольших ямах лесных водоёмов.
р. Mochionyx Loew. (С о г е t b г a Meig.).
0. Основания усиков сближены. Дыхательная трубка рудиментарна или отсутствует ............................................................ 5
5. Между усиками вклинивается суженный передний отдел лобного щитка. Голова на уровне глаз сильно расширена. Глаза крупные (рис. 451 В), Три пары трахейных пузырьков: в груди, в VI и VII сегментах (рис. 450 Б). Есть рудиментарная дыхательная трубка. В лесных дистрофиых водоёмах.
р. Cryophila Edw., 1 вид С. lapponica Mart.
0. Основания усиков сильно сближены (рис. 451 Л). Лобный щиток редуцирован. Голова на уровне глаз расширена слабо. Дыхательной трубки пет. Две пары трахейных пузырьков: в груди и в VII орюшном сегменте (рис. 434 Л). В разнообразных стоячих водоёмах — от крупных озёр до болот.
р. Chaoborus Licht.
6. Стигмы помещаются непосредственно на спинной стороне заднего края VIII брюшного сегмента, окружены стигмальной пластинкой (рис. 452, 453 Л, 454). Личинки держатся горизонтально под поверхностной плёнкой воды в очень разнообразных стоячих водоёмах. р. Anopheles Meig.
0. Стигмы на конце дыхательной трубки, длина которой, по меньшей мере, равна её ширине у основания .................................................7
7. Дыхательная трубка и клапаны стигмальной пластинки видоизменены в буравящий аппарат (рис. 455 Б], посредством которого они пробуравливают ткани подводных растений и получают воздух для дыхания из воздухоносных полостей растений.
р. Manson la Blanch.
0. Буравящего аппарата нет ...........................................8
8. Дыхательная трубка с несколькими парами пучков щетинок (рис. 434 Б), В загрязнённых водоёмах. р. С п 1 е х L.
0. Дыхательная трубка с одной парой пучков щетинок....................9
9. Пучки у основания дыхательной трубки (рис. 455 Л, 458 Л).
р. Theobaldia Nev — Lem.
0. Пучки около середины дыхательной трубки или ближе к её вершине.10
10. По бокам VIII сегмента брюшка хитиновые пластинки, от заднего края которых отходят шипы щётки. Зубцы гребня расшеплены на конце (рис. 456). Личинки дер-254
Рис. 447. Личинка Paradixa sp.: А общий вид личинки; Б — голова.
Рис. 449. Личинка Di ха sp. — брюшной сегмент со спинной стороны.
Рис. 448.
Dixiuae,
Рис. 450. Личинки подсем. Chao-borinae;
Личинки подсем. задний конец:
А — Dlxa sp.; Б — Paradixa sp.
А — Mochlunyx sp.; Б — Cryopiiiia sp.
1
Рис. 452. Задний конец личинки Anopheles sp.
Рис. 453. Голова личинок подсем. Culicinae (схематически):
А — Anopheles: Б — Aedes.
Рис. 454. Личинка Anopheles sp.: А — грудные щетинки; Б — основание грудной шетинки.
Рис. 455. Сифоны личинок: А — Theobald!a sp.; Ь — Manson!a sp.
жатся горизонтально иод поверхностью воды на расстоянии длины дыхательной трубки от неё. р. Uranotaenia Arrib.
0. По бокам VIII сегмента только щётки из чешуек, в один ряд или пятнами............................................................ И
11. На груди и брюшке, кроме длинных боковых пучков, есть ещё очень длинные одиночные волоски. На спинной стороне VL—VIII сегментов хитиновые пластинки, ширина которых увеличивается по направлению к заднему концу тела. Гребень на сифоне отсутствует (рис. 458 В). р. Orthopodomyia Theob.
0. Отдельные волоски не длиннее пучков. Пластинок на VI—VIII сегментах нет. Гребень на дыхательной трубке есть (рис. 453 Б, 457, 458 Б). Во всяких стоячих водах.
р. А ё d е s Meig.
Сем. Heleidae
1. На последнем сегменте группа из нескольких крючков (редуцированная задняя ножка). Передние сегменты имеют приблизительно одинаковую длину и ширину (рис. 435, 460 Б, 461). Жвалы с 3 зубцами. В пресных и солоноватых водах, среди водорослей.
Триба Intermedia. 1 род — D а 8 у be I е a Meig.
0. Задних ножек нет. Длина сегментов больше их ширины. Жвалы с одним зубцом (рис. 459). В стоячих и медленно текущих водах, в иле или среди растений; особенно часто в плаваюших подушках водорослей.
Триба Vermiformes............2
2. Анальный сегмент на конце закруглён и несёт или бледные, слабые щетинки на некотором расстоянии от заднепроходного отверстия, или пару очень длинных и пару более коротких у самого заднепроходного отверстия (рис. 460 Б). В мелких озёрах — в иле, в крупных — на глубинах.
р. Culicoides Latr.
0. Анальное отверстие окружено 8 очень длинными щетинками, стоящими парами, в 4 короткими (рис. 460 Л, 462).
Прочие роды трибы Vermiformes: Be i ii о Kieff; P го be z z i a Kief!., P a 1 p о m у i a Megerle и др.
296
Нис. 457. Личинка Аёйеь ер.
Рис. 459. Схема строения головы личинки Vermi formes.
Рис. 460. Задний конец личинок:
А— Bezzla ер.; Б — Cullcoides ер.; В—Daeybolea ер.
Рис. 461. Глоточное кольпо Dasyhelea sp.
Рис. 462. Жвала личинки Bezzia sp.
Рис. 463. Схема строения головы личинок:
А — Chlronomariae; Б — Tan у tarsari ае; В — Ortbociadilnae; Г — Tanypodinae.
Р и с. 464. Схема строения головы личинок Tanypodinae:
А — с вытянутыми усиками; Б — со втянутыми усиками; В — голова лич. Chirinomariae сбоку.
Сем. Tendipedidae (Chironomidae)
Длина взрослых личинок колеблется от 2 до 30 мм.
Основание головы (рис. 463, 464) срезано несколько вкось, так что спинная сторона длиннее брюшной; по этому признаку легко отличить спинную и брюшную стороны на оторванной голове, повернув ее в профиль (рис. 464 В).
С каждой стороны, ближе к спинной поверхности, расположены личиночные глазки но два с каждой стороны, но иногда два сливаются в один, или с каждой стороны бывает по 3 глазка; глазки представляют скопление пигмента иногда со светопреломляющими линзами; вследствие более быстрого роста переднего копна головы расстояние между ним и глазками с ростом личипки постепенно увеличивается. Кпереди от глазков отходят антенны или усики, состоящие из нескольких — большей частью 5 — члеников; у некоторых форм они могут втягиваться внутрь головной капсулы, у других неподвижно прикреплены к ней — или непосредственно, или посредством хитинового цилиндрического выступа — цоколя. Первый членик усика носит название основного, совокупность прочих — жгутика, или бича. На основном членике имеются один или несколько кольцевых органов, каждый из которых представляет круглый утончённый участок хитинового покрова, окружённый кольцевым хитиновым валиком. Между кольцевым органом и концом основного членика имеется щетинка, 298
ипогда незаметная. На конце основного членика еще 1 относительно крупная глет инка, у многих личинок двуветвистая, с неровными ветвями.
На 2-м членике усика (иногда и на 3-м) располагаются своеобразные органы в виде луковицы — на ножке или сидячие — л а у те р бо р н о в ы органы. Они состоят из центральной палочки, несущей па конце тонкий волосок, и целого ряда нежных, хп-типовых, листовидных пластиночек, охватывающих волосок наподобие чешуек луковицы.
Отношение длины основного членика усика к длине жгутика называется индексом усика; индекс с возрастом личинки увеличивается.
Рот (рассматривать в лупу или микроскоп) находится на брюшной сторопе; снизу он ограничен нижней губой, представляющей собой или плоскую дугу из зубцов (тёмных или с более светлой срединной частью), или мягкую, бесцветную треугольную пластинку с примыкающими к пей зубчатыми паралабиальными гребнями. Если губа зубчатая, то иногда по бокам от неё, более поверхностно, слегка налегая па губу своими концами, лежат две пара лабиальные пластинки — тонкие хитиновые образования, напоминающие по форме или развёрнутый веер о радиальпо расходящимися тонкими лучами, или сильно удлинённый прямоугольник с закруглёнными углами и поперечной штриховкой. Различение их без навыка представляет некоторые трудности. Лучше всего их отыскивать следующим образом: повернув голову брюшной стороной кверху, наставить на фокус нижнюю губу, а затем слегка приподнять лупу или тубус микроскопа. На препаратах хорошо видна штриховка, на живых объектах большей частью лучше заметен общий контур.
На дне ротовой полости, у входа в глотку располагается сложное хитиновое образование — п о д г л о т о ч н и к, или гипофаринкс; ипогда па нём имеется хитиновый, большей частью тёмноокрашенный, состоящий из 4—6 зубцов язычок.
По бокам от нижней губы расположены челюсти. Они слабо хитиппзированы, представляют собой невысокое образование с небольшими придатками на конце; один из придатков — челюстной щупик, большей частью недлинный, одночлепистый — выдаётся иногда довольно сильно. Ипогда на нём имеется кольцевой орган, подобный таковому же усика. Несколько выше челюстей расположена жвалы, или верхние челюсти — мандибулы, сильно хитипизированиые, сразу бросающиеся в глаза зубчатые органы. Жвалы представляют собой 3-гранную пирамиду, обращённую основанием назад (к телу личинки), а вершиной вперёд. Жвалы изогнуты вершинами навстречу друг к другу (к оси тела). Вогнутый край жвалы надрезан по длине на две части — внутренний (верхний, спинной) край и внешний (нижний, брюшной), сходящиеся на вершине жвалы. По внешнему краю расположен большей частью ряд из 3—5 зубцов, сильнее хити-ппзированных и темнее окрашенных, чем сама жвала, — внешние зубцы; концевой, или вершинный зубец этого ряда обычно крупнее остальных (у хищных личинок имеется только концевой зубец); последний в ряду внешних зубцов, считая от вершины жвалы, своим задним краем часто сливается с основной частью жвалы и выделяется лишь своей тёмной окраской (ложный зубец). На внешнем краю жвалы, непосредственно позади ряда зубцов, сидит плоская щетинка различной формы и длины, иногда выдающаяся своим концом за край жвалы. По внутреннему краю жвалы сидят 1—2 светло окрашенных зубца — внутренние зубцы. Выпуклый край жвалы обычно гладкий, но ипогда имеет надсечки. Часто на нём сидят 2 длинные наружные шетинки.
Со спинной стороны рот ограничен верхней губой. Верхняя губа представляет собой непарную лопасть, непосредственно сливающуюся с головной капсулой; с внешней стороны па ней сидит несколько пар щетинок или пузырьков. С внутренней стороны к верхпей губе примыкает нэдглоточник (эпифаринкс) — весьма сложное образование со щетинками, выростами и хитиновыми гребнями и палочками; особеппо выдаются на нём две крупные подвижные хитивовые жевательные лопасти, напоминающие челюсти, — премандибулы.
Тело личинки состоит из 13 сегментов (3 грудных 4- 10 брюшных) приблизительно одинаковой удлинённой формы, но кажется состоящим из 12, вследствие неясной границы между двумя последними сегментами. У некоторых личинок слиты также II и III грудные сегменты. Ко времени окукливания грудные сегменты сильно вздуваются, становясь приблизительно вдвое толще. Сегменты I—VII у большинства личинок несут в задних углах щетинки, простые или перистые. Первый сегмент имеет с брюшной стороны пару небольших бугорков, сидящих на коротких толстых основаниях (ипогда па общем) и усаженных втяжными хитиновыми крючками, зазубренными на вогнутой стороне, — передние ложные ножки, которые служат для передвижения и для плетения сети.
XII сегмент имеет па заднем конце со спинной стороны два стоящих почти рядом бугорка, на которых сидят тонкие длинные жёсткие преанальные щетинки. Несколько кзади от них, на спинной стороне по срединной линии, около анального отверстия, находятся 4 (редко 6) удлинённых тонкостенных мешочка, всегда сильно напряжённых, — анальные жабры (папиллы).
Заканчивается тело двумя удлинёнными нечленистыми придатками, служащими при передвижении по субстрату для подталкивания; эти задние ножки (н од-тал к и в а те л и) несут на конце вепчик из втяжных хитиновых крючков.
Живая личинка быстро передвигается при помощи передних и задних ножек; у консервированных передние ножки часто втянуты, так что видны только бугорки, а у задних втягиваются крючки.
Ш
В трёх первых сегментах (а иногда также и в тфёх последних) просвечивают разветвления трахей (лучше заметны на живых объектах или уже на готовых препаратах). У взрослых личинок видны зачатки сетчатых глаз взрослого насекомого, располагающиеся то в голове, то в I сегменте.
Покровы тела большей частью вполне прозрачны и бесцветны, и окраска тела зависит от окраски внутренних органов и полостной жидкости, а также от цвета принятой пищи.
Брюшко большей частью вальковатое, сжатое в спинно-брюшном направлении. Сегменты IV—X (брюшные I—VII) у многих личинок имеют продольные боковыекалики,
Рис. 465. Личинка Tanypodinae:
А — усик; Б — конец усика при большом увеличении; В, Г — передние ножки, расправленные и втянутые; Д — нижняя губа.
которые на заднем конце X сегмента вытянуты в пальцевидные, направленные назад вдоль тела выросты; обычно у молодых личинок эти выросты короткие, но с каждой линькой удлиняются.
"Определение желательно вести на зрелых личинках, так как у Tendipedidae между разными стадиями развития иногда имеются довольно крупные различия.
1. Усики (рис. 465 Л, Б) могут втягиваться внутрь головы (рис. 464 Л, Б); на консервированном материале они почти всегда втянуты вполне, но ход их легко может быть прослежен, для чего необходим микроскоп (рассматривать нужно со спинной стороны головы, слегка придавив голову покровным стеклом); живые личинки непрерывно то втягивают, то выдвигают свои усики: жгутик усика очень мал по сравнению с основным члеником. На подглоточнике (гипофаринксе) / хитиновый, большей частью тёмноокрашенный
и-* язычок (глосса) с 4—6 зубцами на дистальном
1 копце. Нижняя губа бледная, без зубцов, иногда
I с зубчатыми паралабиальными гребнями. Перед-
J / ние ножки втяжные; венчик крючков на задних
—л ножках может сжиматься и втягиваться нацело;
\ У. отдельные крючки часто окрашены различно —
от светложёлтого до тёмнокоричневого и даже до чёрного; окраска крючков с возрастом стано-'s/у/ вится интенсивнее. Преанальные щетинки (пучки
' щетинок на XII сегменте) всегда сидят на удли-
нённой хитиновой подставке, которая, по мень-шей мере, в 3 раза длиннее своей ширины
Рис 466 Зачяий конеп тела (Рис- 466)’ Никогда =е СТРОЯТ чехликов и ни-с. . задний конец тела когда не минируют. Встречаются в разнообраз-anypoainae. ных водоёмах. Хищники.
Подсем. Pelopiinae (Tanypodinae).
0. Усики прикрепляются к головной капсуле и не могут быть втянуты внутрь. С каждой стороны головы большей частью 2 глазка, редко 1. Язычка большей частью нет, а если есть, то без зубцов. Преанальные щетинки большей частью на невысоких бугорках. Личинки часто в чехликах .............................................2
2. По бокам нижней губы есть паралабиальные пластинки с веерообразно расходящейся штриховкой, или удлинённые, с поперечной штриховкой. Нижняя губа бывает очень часто прикрыта спущенными вниз жвалами — чтобы их поднять, нужно надавить на покровное стекло. Внешние контуры паралабиальной пластинки иногда трудно различить, но лучистая или поперечная штриховка почти всегда ясно видна. По два личиночных глазка с каждой стороны, расположенных рядом в поперечном направлении, ближе кзади; иногда бывает по 3 глазка с каждой стороны. Зачатки глаз взрослого насекомого помещаются в I сегменте груди. Личинки различной величины.
Подсем. Tendipedinae (Chironominae).
0. Паралабиальных пластинок большей частью нет. По два личиночных глазка, лежащих рядом в продольном направлении; большей частью глазки соприкасаются или 300
вполне разделены, иногда слиты в один. Зачатки сетчатых глаз в голове. Личинки большей частью мелкие. Подсем. Orthocladiinae.
Подсем. Tendipedinae
1. Усик 5-членистый, всегда на цоколе, в большинстве случаев высоком; длина основного членика большей частью вдвое превышает длину жгутика, редко равна ей; лаутер-борповы органы хорошо развиты, обычнопротивостоящие, редко чередующиеся (рис. 467 Б). Паралабиальные пластинки большей частью узкие, длинные, сходящиеся внутренними
Рис. 467. Усики личинок*
А — Chlronomariae; Б — Tanytarsariae; В — Agrayloides; Г — Abbreviates.
Рис. 468. Нижняя губа:
А — Chlronomariae; Б — Tanytarsariae; В — Mi его tendipes; Г — Zavreltella; Д — Lauterborniella.
Рис. 470. Щетинки брюшных сегментов: А — Tanytarsariae; Б — Agrayloides.
Рис. 469. Жвалы:
А — Chlronomariae; Б — Tanytarsariae; В — Abbreviate.
концами (рис. 468 Б), реже укороченные и несходящиеся. Щетинка рядом с зубцами, жвалы длиннее (рис. 469 Б) или лишь немного короче ряда зубцов. Срединный зубец нижней губы обычно тройной (рис. 468 Б), редко простои. Щетинки на брюшных сегментах двуветвистые, перистые (рис. 470 Л). Трахеи, кроме груди, видны ещё в трёх последних сегментах брюшка. Крючки задних ножек расположены подковообразно (рис. 471 Б). В большинстве личинки очень мелкие — до 5—6 мм, очень редко свыше 10 мм. В литорали озёр среди обрастаний прибрежных растении, камней и свай; живут среди растений в чехликах (большей частью прикреплённых) или минируют растения; очень чувствительны к загрязнению среды. Триба Tanytarsi ni.
кольцеобразно
Б
472. Задний
Рве. 471 Расположение крючков задних ножек:
А — Chironomariae; Б — Tan у tarsari ае.
Рис. конец тела:
А — Lauterbomiella ag-rayloides; Б — Zavrelfeb* la clavatlcrus.
0. Усик 5- или 6-членистый, обычно без цоколя; основной членик большей частью короче жгутика или равен ему. но бывает и длиннее; лаутерборновы органы рудиментарные (часто совершенно не видны), противостоящие (рис. 467 Л), за исключением групп Agra у I oides и Abbreviatus. Паралабпальные пластинки в большинстве случаев не сходятся внутренними концами (рис. 468 А). Щетинка рядом с зубцами жвалы много короче ряда зубцов (рис. А69 А). Срединный зубец пижнен губы парный или непарный (рис. 468 ,4). Щетинки в задне-боковых углах брюшных сегментов простые или кисточкой (рис. 470 Б). Крючки задних ножек расположены (рис. 471 А). Длина до 30 ми и выше. Обычно строят чехликн; в стоячих и текучих водах; питаются большей частью детритом, немногие — хищники; большинство мало чувствительны к загрязнению среды. Триба Tendipedini.
Триба Tendi pedi ni
1. Усики 5-членлстые; основной членик большей частью короче жгутика или равен «ему, но иногда бывает и длиннее.^ Лаутерборновы органы руди-" ментарпые, противостоящие (рис. 467 Л). Щетинки в заднебоковых углах брюшных сегментов простые.
Собственно Tendi pedi ni. 0. Усики 6-члепистые; основной членик не короче жгутика. Лаутерборновы органы хорошо развиты, чередующиеся. Щетинки в задне-боковых углах брюшных сегментов в виде кисточек.......2
2. Осповной членик усика по длине равен жтутику. Нижняя губа с парным срединным зубцом, который всегда выше первых боковых (рис. 468 Г, Д), и одного цвета с боковыми (светлые). На спинной стороне XI сегмента выросты, направленные вперёд или назад (рис. 472).
Группа Agrayloides. . . .3 0. Основной членик усика всегда длиннее жгутика (рис. 467 Г). Зубцы средней части нижней губы {1—4) светлее прочих (рис. 468 В). На спинной стороне XI сегмента выростов нет. Группа Abbreviatus...........4
8. Передний край паралабиальных пластинок изогнутый (рис. 468 Г). На спинной стороне XI сегмента обращённый вперёд выступ. На боках X сегмента ближе к брюшной стороне цилиндрические придатки с очень короткой щетинкой. Первая пара анальных жабер своеобразной изогнутой формы (рис. 472 Б). Длина до 4 мм.
р. Z a v г е И е 1 1 a Kieff.
0. Передний край паралабиальных пластинок прямой (рис. 468 Д). Выступ на XI сегменте обратен назад. Придатки на X cei-менте заострённые, с очень крупной щетинкой. Анальные жабры обычной формы (рис. 472 Л). Длина до 3 мм. По берегам озёр и луж, среди мха. р. Lauterborniella Thien.
4. Зубцы средней части нижней губы светлые.................... 5
Зубцы средней части нижней губы не светлее боковых...........в
В средней части нижней губы светлых зубцов 2 или 3 (рис. 468 В). Длина до В литорали эвтрофных озёр, в прудах, в реках.
р. Microtendipes Kieff.
В средней части нижней губы светлых зубцов 4. Длина до 8 мм. В литорали озёр на песчано-илистых грунтах; в реках на песке (на течении)
р. Paratendipes Kieff.
6. Нижняя губа со срединным парным зубцом и 6 парами боковых, из которых 2-й мельче 1-го» и 3-го. Паралабиальные пластинки довольно широкие, трапециевидные. Длина до 13 мм. На глубоких местах мезотрофных (средней питательности) озёр.
р. Stictochironomus Kieff.
0. Нижняя губа с непарным срединным зубцом и 5 парами боковых, из которых 5-е зубцы с выпуклостью на внешней стороне Паралабиальные пластинки в форме приблизительно равностороннею треугольника. В прибрежной области озёр и рек.
Cbiro nomi пае genu i пае №6 Lip. (g. larva 4e^novi Lip.).
Собственно Tendipedini
1 Щетинка усика в осповной своей части срастается с основанием 2-го членика: на конце 2-ю членика расположен лаутерборпов орган. Край нижней губы вогнутый, без 302
0.
ъ.
15 лмс.
0.
Рис. 473. Нижняя губа личинки Cbironominae ge-nuinpe Яг И Lip.
зубцов (рис. 473). Сегменты тела удлинённые, задние вдвое длиннее своей ширины. Анальные жабры узкие, оадние ножки тонкие, длинные. Длина до 9 мм. В реках, на течении, на чистом песке.
Cbironominae genuinae № 11 Lip. (р. Cry pt ос hi ron omus г о 11 i Kirpitsch.)‘. 0. Нижняя губа зубчатая .........................2
2. Нижняя губа состоит из широкого цельного, более светлого среднего куска и боковых зубчатых частей; ширина среднего куска не менее ширины суммы 4 боковых зубцов. Длина челюстного щупика большей частью не менее чем в 3 раза превышает его ширину, так что резко бросается в глаза на цельной голове (па первый взгляд
кажется, что усиков две пары — рис, 477, 478 Б). Пара-
лабиальпые пластинки большей частью длинные. Щетипка усика (рис. 475 Л, Б) почти всегда частично срастается со 2-м или с 3-м члеником.........................Я
0. Нижняя губа вся сплошь зубчатая. Челюстной щупик (рис. 478 А) почти всегда короткий, длина его не более чем в два раза превышает ширину. Паралабиальные пластинки (за единственным исключением) не достигают края головы. Щетинки усика (рис. 475 Г, Д, Е) отходят от конца основного членика........................8
Рис. 474. Нижняя губа личинок:
А — Cbironominae genulnae К? 12 Lip.; Б — Cbironominae genuinae № 10 Lip.; В — Cryp-tochironomus sp. Kieft.; Г — Ilamischla fu-scimanus Kieff.;4 — Paracladopelma Harn.; E — Lauterborniella (brachylabis? Zavrel)-Cbi-ronomlnae connectens № 4 Lip.
Рис. 475. Усики личинок:
A — Cryptochironomus Kleff; Б — Harnlschia Kleff.; В —- Chir. gen. M 12 Lip.; Г — Cbin gen. № 6 (larva zernovl Lip.); Д — Chir. gen. № 7 Lip.; E — Cblr. gen. № 8 Lip.
1 Теперь род Cryptochironomus принимается в более широком объёме, чем раньше, в него включён ряд форм, выделенных ранее в самостоятельные роды. В настоящей книге в определительной таблице оставлены прежние роды, но рядом с ними приводятся их синонимы по Cryptochironomus.
303
8. Челюстной щупик короткий. Щетинка усика отходит от конца основного членика. Внешних зубцов жвалы три — длинные, закруглённые. Паралабиальные пластинки со слабой штриховкой. Срединная часть нижней губы полукруглая, боковые зубцы очень узкие, на конце закруглённые (рис. 474 Е}. В иле сублиторали озёр; в реках.
р. Lauterborniella (brachylabis? Zavrel) (С h i го о о mi пае connectens № 4 Lip.).
0. Челюстной щупик длинный (рис. 477). Щетинка усика отщепляется от 2-го или 3-го членика (рис. 475 Л, Б). Внешних зубцов жвалы не более двух, не считая концевого (рис. 476Б). Паралабиальные пластинки с резкой штриховкой. Средняя часть нижней губы или закруглённая, или почти прямая ...........................4
4. Средняя часть нижней губы почти полукруглая (рис. 474 А, В, В) .5
0. Средняя часть нижней губы почти прямая (рис. 474 Г, Д}..........7
5. Концы зубцов нижней губы и верхний край паралабиальных пластинок образуют сильно изогнутую линию (рис. 474 В}, срединный непарный зубец широкий, округлый, боковые зубцы острые, резко чёрные, сплошь окрашенные. Щетинка усики отщепляется от 2-го членика (рис. 475 Л). Жвала светлая, с резко чёрными, крупными внешними зубцами (рис. 476 Б). В литорали озёр и рек, преимущественно на песке
р. Cryptochironomus, гр. defectus Kieff.
0. Копцы зубцов нижней губы и верхний край паралабиальных пластинок обпазуют почти прямую или слабо изогнутую линию (рис. 474 Л, Б), Жвала вся светлая. Щетинка усика отщепляется от 3-го членика (рис. 475 В}..............................6
Рис. 476. Жвалы личинок:
А — Hamiscbia fuscimanus, Б — Cryptochi ronomus delectus.
Рис. 477. Челюстной щупик Cryptochironomus defectus.
Рис. 478. Челюсть личинок:
А — Cladopelma virescens; Б — Paracladopelma camptoiabis.
6. Пять широких боковых зубцов нижней губы1; срединная часть уже ряда боковых зубцов одной стороны (рис. 474 Л). Chironominae genuinae№12 Lip.
0. Семь узких зубцов нижней губы; срединная часть приблизительно равна ряду боковых зубцов одной стороны (рис. 474 Б}. В сублиторали озёр и рек, на песке, на быстром течении.
Chironominae genu i пае №10 Lip. (Cryptochironomus vulneratus Zett.).
7. 1-й и 2-й членики усика хорошо видны, прочие очень мелки, трудно различимы; кольцевой орган у основания 1-го членика; шетинка усика на конце 1-го членика; между основаниями усиков пара плоских, двураздельных щетинок (рис. 478). Парный срединный зубец нижней губы низкий, широкий, светложёлтый; 1-й боковой также светложёлтый; прочие светлокоричневые (рис. 474 Д}. Паралабиальные пластинки с крупной штриховкой. Тело сзади суживается. Длина до 7 мм. В крупных эвтрофных озёрах, в иле сублиторали и профундали; чаще в реках.
р. Paracladopelma Harn. (Cryptochironomus, гр. camptoiabis Kieff.)
0. 2-й и 3-й членики усиков (рис. 475 Б) приблизительно равной длины: 4-й и 5-й едва заметны. Бугорка рядом с челюстным щупиком нет. Жвалы (рис. 476 А) с едва заметными выступами, вместо зубцов. Типична для рек, иногда в озерах, большей частью на песке.
р. Harnischia Kieff. (Cryptochi ronomus, гр. fuscimanus — см. прим, на стр. 304).
8. Нижняя губа с парным срединным зубцом..................8
0. Нижняя губа с непарным срединным зубцом...............26
1 Число боковых зубцов везде указывается лишь для одной стороны нижней губы; если в середине её 1 зубец, то он называется «срединным непарным», если 2, то «срединным парным»; в последнем случае стоящий рядом с ним боковой зубец считается «первым боковым».
9. Место отхождения щетинки усика расположено на 1/ъ от конца 2-го членика. Жвала с 3 чёрными зубцами. Нижняя губа с 4 боковыми зубцами (на каждой стороне), с вогнутым передним краем. Паралабиальные пластинки с 2—4 крупными неясными штрихами. I и II грудные сегменты значительно шире всех остальных сегментов тела. Пучок щетинок на лПсегменте без подставки. Задние ножки короче несущего их сегмента, с мелкими крючками. Анальные жабры прямые, пальцевидные. Длина до 16 мм. В прибрежье рек и в зарослях озёр и прудов;
минируют упавшие в воду растения и листья осок р. Stenochironomus Kief!.
(Cryptochironomus sp. Zavr. Chironomidae g? Lip. =C h i-ronominae incongruens Tschern.).
0. Нижняя губа с 6 или 7 боковыми зубцами, выпуклая. Паралабиальные пластинки со штриховкой........10
10. Нижняя губа с 6 боковыми зубцами .......................11
0. Нижняя губа с 7 боковыми зубцами ;.......‘..............18
11. Все зубцы нижней губы светлые, все приблизительно одной величины, разделены неглубоко, широкие, низкие . . 12
0. Все зубцы нижней 1убы тёмные; образуют две резко различные по величине зубцов группы: срединные и 1-е и 2-е боковые — крупные, боковые с 3-го по 6-й — мелкие (рис. 479 В). Заросли озёр (ил), реже рек.
Cbironominae genuinae
№ 3 Lip. (Polypedilum «genuinae № 3 Lip.» Tschern.).
Рис. 479. Нижняя губа личинок:
А — Chir. gen. № 3 Pancr.; Б — Chir. gen. № 5 Pancr.; В — Chir. gen. № 3 Lip.
12. Боковые зубцы нижней губы одинаковы по величине; срединный равен по ширине сумме 4 боковых одной стороны (рис. 479 А).
Cbironominae genuinae №3 Pancr. (из Аму-Дарьи).
0. Боковые зубцы убывают по величине; срединный равен по ширине сумме всех боковых одной стороны (рис. 479 Б).
Cbironominae genuinae №5 Pancr. (из Аму-Дарьи).
18. Различие между зубцами по величине не особенно резкое...............14
0. Различие в величине зубцов резкое ...................................17
Рис, 480. Нижняя губа личинок:
А — Endochironomus, гр. nymphotdes; Б — Pentapedllum ар.; В — Ohtr. gen. № 7 Lip.; Г — Chir. gen. M 8 Lip.
14. Все зубцы нижней губы сходны между собой, светлые, острые, длинные, приблизительно одинаковой длины, глубоко надрезанные; срединный немного крупнее. Длина личинки 8 мм...................................................15
0. Зубцы нижней губы недлинные, резко суживающиеся, жёлтые или коричневые, закруглённые или заострённые ........................................16
15. Первый боковой зубец значительно мельче срединного и 2-го бокового. Паралабиальные пластинки округлённотреугольные, слегка волнистые ио верхнему краю (рис. 480 В). Cbironominae genuinae №7 Lip.
(Cryptochironomus «genuinae № 7 L i p.» Tschern.).
0. Первый ^боковой зубец одинаковой длины с соседними; все боковые зубцы стоят косо (наклонены к середине). Паралабиальные пластинки веерообразные, слегка скло-305
нёнпые, по верхнему краю гладкие (рис. 480 Г). Длина до 8 мм. В зарослях рек, много среди обростов. Chironominae genuine е. №8 Lip.
16. Зубцы нижней губы тёмные (рис. 480 Б). Длина до 9 мм. В зарослях озёр и прудов свободно или в инкрустациях, или в покинутых ходах более крупных личинок, р. Pentapedilum Kieff.
0. Зубцы нижней губы светложёлтые или светлокоричневые (рис. 480 Л). В иле, среди растений, по берегам озёр; в старых стеблях камыша.
р. Endocnironomus Kieff., гр. nympboides.
17. Срединный и 1-пи 2-й боковые зубцы нижней губы образуют группу из убывающих мелких зубцов;3-й боковой самый крупный,4-й меньше, остальные мелкие (рис. 481Л). В реках на песке, на течении.
Chironominae connectens № 3 Lip. (Paratendipes «connect e n s № 3 Lip.» Tschern).
Рис. 481. Нижняя губа личинок:
А — Chlr. conn. № 3 Lip.; Б — Chlr. gen. № 1 Lip.
0. Нижняя губа иного строения ................................... 18
18. Срединный и 1-й боковой зубцы нижней губы одинаковы; тот и другой (иногда также и 2-и боковой) — мелкие, прочие боковые — крупные; из них наиболее крупным является 2-й (или 3-й) боковой; таким образом, в середине губы образуется группа из 4 или 6 мелких зубцов (рис. 481 Б). Паралабиальные пластинки в 4 раза длиннее (перпендикулярно к оси тела) их ширины (параллельно оси тела), внутренними концами почти сходятся на оси тела. Усики расставлены широко. На XII сегменте в задне-боковых углах по 1 короткому придатку. Длина до 16 мм. В реках и крупных озёрах, на песке.
Chironominae genuinaeNslLip.
0. Срединный и 1-й боковой зубцы не одинаковы.....................19
19. 1-й боковой зубец ниже срединного и 2-го бокового.........20
0. 1-й боковой зубец ниже срединного, но приблизительно равен 2-му боковому 21 20. Разница между зубцами ясная (рис. 482 Л, Б). Жвала с 3 внешними зубцами, внутренний зубец крупный, коричневый, далеко выступает над концевым зубцом. Длина до 13 мм. В иле среди растений. р. Polypedilum Kieff.
Рис. 482. Нижняя губа личинок!
А, Б — Polypedilum sp.; В — Microchi-ronomus laccophi lus.
0. Разница между зубцами менее ясная (рис. 480 Б). Внешних зубцов на жвале более трёх; внутренний зубец не длиннее концевого. Длина до 9 мм. (Обитание см. выше, п. 16.) р. Pentapedilum Kieff.
21. Срединные зубцы светлее боковых; боковые спускаются довольно круто; разделение между срединными зубцами слабое; боковые зубцы с 1-го по 3-й приблизительно равны; 4-й очень мал, иногда только в виде придатка на 3-м; 5—7-й отстоят далеко отдельной группкой, резко отличаются от прочих боковых тёмной окраской (рис. 482 В). Длина до 7 мм. В литорали и сублиторали эвтрофных озёр, на песчано-глинистом грунте.
Microchironomus laccophi! us Kieff. (Cryptochi ronomus, rp. v i ri d u 1 u s Fabr. — см. прим, на стр. 304).
0. Срединные зубцы не выше боковых; все зубцы окрашены одинаково и расположены в 1 непрерывный ряд, последние боковые не отделены от остальных; дуга зубцов ноло1ая ................................................... 22
22. Кольцевой орган на х/ь—1/в длины основного членика от начала усиков. Различие между зуоцами нижней губы (рис. 480 А) нерезкое. Голова светлая. Длина личинки до 15 мм, (Обитание см. выше, п. 16—0.)
р. Endochironomus Kieff., гр. nymphoides.
0, Кольцевой орган на Vs—’Л длины основного членика. Различие между зубцами резкое. Голова тёмная .....................................23
Рис. 484. Усик личинки Sergentia.
Рис. 483. Нижняя губа личинок:
А, Б — Stictochironomus sp.; В — Aiiochlronomus sp.; Г — Sergentia.
23. Кольцевой орган на V< длины основного членика. Дуга, соединяющая основания зубцов нижней гуоы, под третьими боковыми зубцами значительно вдавлена (рис.4835) Жвала с 4 истинными внешними зубцами. Длина личинки И лос В зарослях озёр и прудов.
р. Allochironomus Kieff.
0, Кольцевой орган на х/3 длины основного членика (рис. 484). Дуга оснований зубцов почти без вдавлений под третьими боковыми (рис. 483 Г). Жвала с 5 истинными
Рис. 485. Личинка Chir. gen. № 14 Lip.:
А — верхняя губа и усик; Б — нижняя губа.
Рис. 486. Нижняя губа личинок:
А — Chir. gen. № I Pancr.; Б — Chir. gen К? 6 Lip.; В — Chir. gen. № 13 Lip.
зубцами. Длина личинки 14 мм. На глубинах озёр, в сублиторали (в иле) слабо эвтрофных озёр п в литорали олиготрофных озёр; иногда в реках. р. Sergentia Kieff.
24. Нижняя губа почти прямоугольной формы; ближайшие к срединному боковые зубцы расположены на одной горизонтальной прямой с ним, прочие боковые расположены по бокам нижней губы в направлении, параллельном её продольной оси..........-5
0. Нижняя губа иной формы; все зубцы расположены по дуге или по прямой линии ...................................................................... ,
26. На передней части головы мощные пучки очень длинных щетинок (рис. 484 А) 26
0. Беа таких пучков щетинок. В стоячих водоёмах, живёт в губках. Demejerea rufipee L.
£6. Крупные зубцы переднего края нижней губы разделены широкими промежутками с мелкими зубчиками в них (рис. 485).
Cbironominae «genuinae №14 Lip.».
0. Срединный непарный зубец с неглубокой 3-угольной выемкой на вершине; 1-й я 3-й боковые очень малы. Длина до 9 мм. В реках и озёрах, минирует губки.
Xenochironomus xenolabis Kieff.
27. Боковых зубцов нижней губы менее 6 .................28
0. Боковых зубцов 6 или 7 ..............................30
28. Боковых зубцов 4; срединный зубец крупнее боковых (рис. 486 4); все зубцы резко чёрные, расположены по прямой линии. Щетинка усика отщепляется от 2-го членика. Нет передних ножек (рис. 487).
Cbironominae genuinae №1 Panor. (иэ Аму-Дарьи).
0. Боковых зубцов 5................................... 29
29. Зубцы нижней губы расположены по дуге; срединный зубец в основании шире боковых, довольно резко суживается, слегка эаоотрён; на продолжении последнего зубца сбоку округлый выступ. Паралэ-биальные пластинки узкие, треугольные (рис. 486 В). Челюотной тупик обычной формы, длина его чуть больше ширины. Щетинка усика отходит от начала основного членика. Жвалы с двумя внутренними зубцами (рис. 488). Cbironominae ge-n u i n а е № 6 Li р. (larva z e r n о v i Lip.).
Рис. 489. Личинка Chir. gen. № 13 Lip.:
A — усик; Б — челюстной щуп ин
Рис. 488. Жвала
Chir. gen. № 6 Lip.
Рис. 487. Передний конец личинки Chir. gen. № 1 Pancr.
0. Зубцы нижней губы расположены по прямой линии; срединный зубец узкий, длинный, заострённый, сходный по форме с боковыми (рис. 486 В). Основной членик челюстного щупика жёлтый, сильно хитинизированный, поперечно разделённый на две части; длина его превышает ширину в три раза; на конце его бледный широкий листовидный вырост с пальцевидными выступами и длинный 3-членистый придаток (рис. 489 В). Щетинка усика отшепляется от конца 2-го членика (рис. 489 4).
Cbironominae genuinae №13 Lip.
30. Боковых зубцов 6..........................................31
0. Боковых зубцов 7...........................................38
81. Срединный зубец много выше и шире боковых............... 32
0. Срединный зубец если и больше первых боковых, то лишь немного, чаще же меньше первых боковых или им равен................................34
82. Срединный зубец очень широк — почти равен по ширине совокупности боковых одной стороны; боковые узкие и длинные, слабые. Пара лабиальные пластинки округлые (рис. 490 4). Длина до 5 мм. Б больших озёрах и реках, на песчано-илистых грунтах.
Cbironominae genuinae № 9 Lip. (Cryptochi ronomus «genuinae № 9 L i p.» — см. прим, на стр. 304).
0. Срединный зубеп не так широк, боковые—массивные; паралабиальные плаотинки обычные ........................................................ 83
88. Срединный зубец светлее боковых; 4-й боковой очень мал, часто трудно заметен, все зубцы закруглённые (рис. 490 Г). Длина личинки 6 мм. В стоячих водоёмах при хороших кислородных условиях. р. Prochi ronomus Kieff.
0. Срединный в боковые зубцы одинаково тёмные; боковые приблизительно равны по величине (рис. 490 Д), последний мал, трудно различим. Длина личинки 5 мм, В литорали и сублит орали эвтрофных озёр.
р. Microchironomus conjungens Zavr, (р. Cryptochironomus, гр. conjungens Kieff.).
84. Срединный и 1-й боковой зубцы при не расплющенной губе кажутся одной длины, прочие неразличимы. При расплющивании 1-е боковые становятся несколько ниже срединного и отделяются от него довольно глубокой выемкой; 2-й боковой зубец ниже 1-го и слегка с ним сливается. Длина до 10 мм..........................86
0. Все зубцы видны и при нерасплющеиной губе, 1-й и 2-й боковые разделены довольно ясно (рис. 491 В).......................................Зв
* 85. На XI сегменте ближе к брюшку длинные выросты (рис. 493 И). Длина личинки до 12 мм. В иле сублиторали эвтрофных озёр.
р. Е i n i e 1 d i а, гр. с а г b о n a r i a Meig. (Chironominae genuinae larva kruglovicola Lip.).
0. Выростов па XI сегмент© нет. Длина до 10 мм.
р. Paracladopelma Kieff. (Cryptochironomus camptolabis Kieff.) и p. Limnochironomus Kieff.
86. Щетинка жвалы довольно широкая, листовидная. Длина до 17 мм, личинки довольно массивные.
р. Glyptotendipes Kieff..........37
Рис. 490. Нижняя губа личинок:
А — Chir. gen. № 9 Up.; Б, В — Elnfeldla g, гр. carbonarla; Г—Prochironomus ер.; Л—Mlcrochlronomib sp. (conjungens).
0. Щетинка жвалы узкая. Длина
87. XI сегмент с отростками.
до 12 лиг.
р. Phytochi г о п о m и з Kieff.
Gl. polytomus Kieff.
0. XI сегмент без отростков. Длина до 15 мм. В литорали и сублиторали эвтрофных озёр, в иле и в старых стеблях камыша.
G1., гр. gripekoveni Kieff.
88. Все зубцы нижней губы светлые; срединный зубец в виде острого треугольника отходит не в общем ряду, а значительно ниже, снаружи, и налегает сверху на 1-е боковые зубцы (рис. 492 Л). Длина до 5 мм.
Chironominae genuinae №2 Pancr. (из Аму-Дарьи).
Рис. 491. Нижняя губа личинок: д — Cladopelma эр.; Б — Glyptotendipes эр.
0. Нижняя губа иного строения ................................. 88
89. 1-е боковые зубцы меньше срединного и двух боковых; 2-е боковые выше срединного .............................................................40
0. 1-е боковые зубцы не меньше прочих боковых...................41
40. Первые боковые зубцы равны по длине срединному (рис. 492 Д). На песчаноилистых хунтах рек. Chironominae genuinae Л» 2 Lip.
0. Жвала с 4 внешними чёрнокоричневыми зубцами, постепенно убывающими по величине от вершины к основанию жвалы. Срединный зубец нижней гуоы имеет иногда зарубки на боках, и тогда боковых зубцов 6 нар; все зуоцы нижней губы чёрнокоричневые; срединный зубец вместе с первыми боковыми ^же общей ширины прочих боковых
У)9
одной стороны (рис. 492 F). Зубцов гребешка надглоточника около 15. Анальные жабры грушевидные, большей частью очень крупные, часто посередине перетянутые, в солоноватых водах редуцированные до бугорков. На X и XI сегментах или на одном из них поЧти всегда имеются выросты (рис. 493 Л). Живут в иле.
р. Tendi pes Meig. (Chironomus Meig.)......42
41. Боковые зубцы почти равны по длине; срединный выше и шире; все боковые заостренные, не очень тёмные, иногда жёлтые (рис. 492 В). Длина личинки 9 мм. В стоячих водах и в реках, преимущественно на растениях.
Parachironomus Lenz (Cryplochironomus, гр. р а г а г о s t г a t u s).
Рис. 492. Нижняя губа личинок:
А — Chir. gen. №2 Pancr.; Б — Tendipes sp.; В — Parachi гоп om us sp.; Г — Endo-chironomus, гр. signaticornis; Д — Chlr. gen. № 2 Lip.
0. Первые боковые зубцы пмтпе срединного и вторых боковых, Зубцы тёмные, закруглённые (рис. 492 Г). Длина личинки 15 мм. Среди растении, некоторые минируют.
р. Endochironomus, гр. signaticornis Kieff. (Tendipes signaticornis Kieff.).
42. С каждой стороны головы но 3 глаза. Жвала с крупным концевым и тремя мелкими коричневыми внешними зубцами. На X сегменте отростков нет, на XI одна пара крупных. Длина до 12 мм. В небольших эвтрофных водоёмах на грубом детрите.
Т. larva appendiculate Krugl.
0. С каждой стороны головы по 2 глаза. Жвала с 4 внешними зубцами.43
Рис. 493. Задний конец телаз
А — Tendipes (Chironomus) plumusus; Б — Elnfeldla, гр. carbonarla.
43. Выростов пет ни на X, ни на XI сегментах. Длина до 15 мм. На глубинах олиготрофных озёр; в солоноватых водоёмах морского побережья
Т., гр. з а И n а г i u з Thien.
0. Выросты есть ................................................44
44. Выросты на X сегменте есть...............................45
0. На X сегменте выростов нет...................................48
46. На XI сегменте выросты есть.................................40
0. На XI сегменте выростов нет. Длина до 18 мм. В равнинных реках, в песке, на течении. Т. гр. reductin Lip.
46. Выростов на XI сегменте 1 пара. Длина до 15 мм. В дельтах ранни иных рек, на течении. Т., гр. plumosus-reductus Lip.
0. Выростов на XI сегменте 2 пары. В иле озёр и прудов; выдерживают длительное существование без кислорода .........................................47
47. На XI сегменте выросты длинные. Длина личинки до 20 мм и более.
Т., гр. plumosua Thien.
0. На XI сегменте выросты короткие, не длиннее задних ножек. Длина личинки до 25 мм. Тгр. semi reductus Thien.
48. Выростов па XI сегменте 1 пара на месте задней пары; они короче задних ножек, на концах заострены. Длина до 12 мм. В глубоких озёрах и прудах.
Т., гр. р a g a n u s (Е i о f е 1 d i a larva р a g a n a Meig.).
0. Выростов па XI сегменте 2 пары....................................49
49. Выросты XI сегмента короче задних пожек, с тупыми концами. Длина до 18 мм. Главным образом в олиготрофных озёрах, в иле, на глубинах.
Т., гр. bathophilus Kieff.
0. Выросты XI сегмента длиннее задних ножек, с заострёнными концами (в реках выросты бывают и короче задних ножек). Длина до 20 мм. В реках и заросших загрязнённых прудах, в иле. Т., гр. th u m mi Kieff.
Триба Tanytarsini
1. Голова коричневая. 2-й членик усика в 4—4,5 раза короче 1-го. Жвала с одним концевым и одним широким внутренним зубцом. Нижняя губа с 3 жёлтыми зубцами (рис. 494). Чехлики — иловые трубки. Длина до 7 мм. В мелких озёрах.
Рис. 494. Нижняя губа личинки Tanytarsus pedicelliferusg?(larva uralicola).
Tanytarsus pedicelliferus Bir. (секция Eutanyta rsi, поде. U r a 1 i a, g? larva uralicola Lip.)
0. Жвала c 4 внешними зубцами. Нижняя губа с б парами боковых зубцов (рис. 496)......................................................2
2. Усики 5- или 6-членистые.....................................3
0. Усики 5-членистые; основной членик приблизительно вдвое длиннее жгутика. Нижняя губа с 5 боковыми зубцами; паралабиальные пластинки внутренними концами сходятся на оси головы. Чехлики-трубки одинакового по всей длине диаметра, свободные или прилегающие к субстрату.........................................4
Рис. 495. Усики личинок:
А — Paratanytaraus la u terb oral; Б— Atany tarsus bausel; В — Rheotan у tarsus raptortus; Г — Mlcropsectra longimaaus; Д — StempeUina bausel; E — Zavrella.
8. Усики 6-членистые; 2-й членик несёт 2 крупных лаутерборновых органа -—один у основания и один у конца членика; цоколь с крупным^ шипом (рис. 495 Е). Жвала с 5 коричневыми внешними зубцами, из которых первый (считая от основания жвалы) очепь мал. Чехлики (рис. 497 Г) переносные, из мелких песчинок. Длина до З.н.и. В лужах и прибрежье озёр и прудов. р. Zavrelia Kieff.
0. Усики 5-членистые; на вершине 2-го членика пара крупных противостоящих лаутерборновых органов на недлинных стебельках. Чехлики (рис. 497 Д) — песчаные, конические трубки. Длина до 4 мм. В песчаных участках литорали озер, в текучих водах только в мелких. . . ... .. .
р. Stempellina, Thien. (Tanytarsus bausel Kieff.).
Я^вала с 4 внешними зубцами. Боковых зубцов
4. Лаутерборновых органов неТ*сретлее боковых; 2-й боковой значительно мельче нижней губы 6; срединный зубец не иле, в эвтрофных и слабо дистрофных озёрах; 1-го и 3-го (рис. 496 В), Длина до 7 на глубинах.
большей частью в литорали, изредка nomj п. Pseudochi ronomus Staeg.
Секция Р s е u d о с h i г0 " к 6
.........................................о
0. Лаутерборновы органы есть противостоящие на коротких стебельках или
5. Лаутерборновы органы крупны Paratanytarsi................в
сидячие. Боковых зубцов нижнеи
л „ е на стебельках ...........................7
О, Лаутерборновы органы м яка почти на середине основного членика усика.
в. Цоколь усика невысокий, $ Все боковые зубцы нижней губы прибли-Лаутерборновы органы сидячие (рис. ' j к
Р и с. 496. Нижняя губа личинок:
А — Tanytarsus; Б — Ata-nytarsus; В — Pseud ос hl топот us; Г — Stcmpellina bau-sei ; Д — Zavrella bausel.
зительно равны между собой; срединный зубец простой. Жвала с 3 внешними зубцами. Длина до 8 мм. р. Р а г a t а п у t а г s и э.
0. Цоколь высокий; щетинка на основном членике усика на длины этого членика от его основания. Лаутерборновы органы на коротких стебельках (]эис. 495 Б). 2-й боковой зубец нижней губы мельче 1-го и 3-го; срединный зубец тройной (рис. 4962э). Жвала с 4 внешними зубцами. Кладка и молодые личинки в литорали озёр, по мере роста, спускаются на глубины; в реках только на медленном течении.
Секция Atanytarsi, р. Atanytarsus Zavr. (Tanytarsus «Attersee» В a use, Tanytarsus, rp. mancus. v. d. Wulp.).
Рис. 497. Чехлики личинок;
A — Rhcotanytareus raptorius; Б — Rh. pentapoda; В — Rb. sp.; Г — Zavrella pentatoma, Д — Stempellina bausel. 7 8
7. Стебельки лаутерборновых органов короткие, по длине равны 3-му членику (волжские формы) или совокупности 3 последних члеников (обычные формы — рис. 495 В) усика. Чехлики конические, с килевыми нитями ~ (рис. 497 А — В). Длина до 15 мм. Преимущественно в быстро текущих водах; большей частью на каменистых грунтах или в зарослях; в озёрах только в зоне сильного прибоя.
Секция Rheotanytarsi, р. Rheotanytarsus Thien.
0. Стебельки длинные, не менее чем в 2,5 раза длиннее 3 последних члеников усика (рис. 495 Г). Чехлики—длинные иловые трубки. Длина до 15 мм. Большей частью в стоячих водоёмах; в текучих только на заиленных участках при медленном течении, среди растений; некоторые виды на глубинах олиготрофных озёр.
Секция Eutanytarsi, подсекция Eutanytarsi.............k.8
8. Цоколь с шипом ........................................9
0. Цоколь без шипа
р. Tanytarsus v. d. Wulp., гр. gregari us Bause.
9. Цоколь с очень мелким, часто незаметным шипом на внутреннем краю, дистально. Стебельки в 2,5 раза длиннее совокупности 3 последних члеников. 2-й членик усика в 4 раза короче основного. Чувствительная щетинка ня основном членике сдвинута к дистальному концу до а/8 или 8/4 длины членика. Анальные жабры крупные: спинные крупнее, вздутые, брюшные тоньше в короче. Длина до 10 мм. В озёрах в сублиторали, в олиготрофных озёрах спускается до больших глубин; требовательна к кислороду.
р. Lauterborniella Kieff.
0. Цоколь с шипом (рис. 495 Г). Чувствительная щетинка приблизительно в середине основного членика усика. Анальные жабры: мешочки, слабо изогнутые вентрально (в брюшном направлении), короткие, равной длины.
р. Micropsectra Kieff., гр. i п е г mi pes Ba use.
10. Шип цоколя не длиннее V» высоты цоколя. Длина до 15 мм. В ручьях и олиготрофных озёрах. р. Micropsectra, гр. ргаесох Meig.
0. Шип цоколя равен 1/3 его высоты. В луговых канавах и в прудах.
р. Micropsectra, гр. trivialis Kieff.
Подсем. Orthocladii пае
1, Нижняя губа из 6 слабо выпуклых зубцов, лежащих по прямой линии (рис. 498 Б). Всё тело в довольно массивных, коротких коричневых волосках, особенно густо сидящих на задне-боковых частях последних сегментов. Анальные жабры в числе одной пары. Крючки задних ножек расположены подковообразно; из них пять почти чёрного цвета. Длина до 4 мм. На теле подёнки Ephemera vulgata, питается детритом, застревающим в волосках и на жабрах.
р. Epoicocladius Zavr. (Smittia ephemerae Kieff.)
Рис. 498. Личинка Epoicocladius:
A — верхняя губа; Б — нпяшня губа.
Рис. 499. Голова личинок Corynoneurariae:
А — Согупопеига; Б — Thlenemannlella
0. Зубцы нижней губы лежат по выпуклой линии. Покровы >ела или гладкие, или лишь с немногочисленными волосками. Анальных жабер 2 пары или их совсем пет. Крючки на задних ножках расположены по кругу.............................; -2
2. Голова узкая, удлинённо-овальная, в наиболее широком месте ширина её вдвое меньше длины. Длина усиков не меньше половины длины головы; 2-й членик усика коленчато изогнут; 2-й, а иногда и 3-й членик темнее прочих (рис. 499). Передние ножки сидят на общем основании. У основания задних ножек шип. Формы всегда мелкие, не длиннее 5 мм..................................................................3
Рис. 500. Щетинки преанального сегмента личинок: А — Paracricotopus; Б — P&ectrocladius psi lopterus; В — Syndlameaa lepnevi.
0. Голова относительно короче. Длина усиков большей частью меньше половины длины головы; 2-й членик не изогнут и не темнее прочих.............4
8. Усики в половину длины головы; осповной членик их короче жгутика; 2-й и 3-й членики тёмные (рис. 499 Б). В обростах и среди мхов, преимущественно в текучих водах. р. Thienemanniella Kieff. (Microlenzia Kieff.).
0. Усики длиннее головы; основной членик одинаковой длины со жгутиком; 2-й членик тёмный (рис. 499 Л). Длина до 5 мм, В озёрах, прудах, среди растений.
р. Corynoneura Winn.
4. В задне-боковых углах I—VI сегментов пучки щетинок.........5
0. В задне-боковых углах I—VI сегментов нет пучков щетинок, большей частью только по одной простой щетинке ...................................7
5. По одному пучку щетинок с каждой стороны. Длина до 6 мм.
р. Е и с Н cotopus Thien. (С г i с о t о р u s v. d. Wulp.).G
0. По два пучка щетинок.
р. Synorthocladius Thien. (Orthocladius — D a c t у 1 о с 1 a-d i u s, rp. semivirens).
6. Голова конусообразная. Усики редуцированы. Тело сильно сжато с боков. Минирует листья Potamogeton natans.
р. Eucricotopus, гр. brevipalpis Thien.
0. Голова овальная. Усики обычные. Тело не сплюснуто с боков.
р. Eucricotopus, гр. Silvestris Thien.
Рис. 502. Лобное и ротовое поле Psectrocladius psi-lopterus.
Рис. 603. Усик лич. Euphaeno-cladius.
Рис. 501. Нижняя губа личинок:
А — Psectrocladius psi lop terus; Б — Euphaenocladius rhitrogenae.
7. Пучок щетинок XII сегмента на подставках, несущих у основания сильный выступ с сильно склеротизирова иными шипиками (рис. 500 Б). По бокам нижней губы всегда складки, иногда с рядом тонких щетинок — бородкой (рис. 501 Л). Жвала всегда с длинным концевым и маленькими внешними зубцами. На лобном поле большей частью дланевидные щетинки (рис. 502). Длина до 8 мм. В зарослях.
р. Psectrocladius Kieff.
0. Подставки без выступа у основания (иногда с шипом) или совсем отсутствуют...................................................м- ... 8
8. Подставки с более тёмным хитиновым полем, на котором крепкий, острый, тёмный шип (рис. 500 Л).
р. Paracricotopus Zavr. н некоторые виды Rheocri cotopus.
0. Подставки, если они есть, без шипа ......................9
9. Очень короткие, почти рудиментарные усики. Пучки преанальных щетинок XII сегмента на очень высокой подставке. Длина 6,5 мм. В родниках и лужах среди мха.
р. Metriocnemus v. d. Wulp.
0. Усики обычные...........................................10
10. Преанальных подставок с пучками щетинок нет. На их месте разве лишь несколько слабых щетинок. Анальных жабер тоже нет (или они втяжные). Основной членик усиков короткий; 2-й членик наиболее длинный (рис. 503).
р. Euphaenocladius Kieff.
0. Пучки преанальных щетинок есть. 2-й членик усиков не длиннее основного . . 11 11. На лобном поле плоские, по дистальному краю зазубренные щетинки, лопато-видные или расширенные на конце (рис. 504 Б).
р. Dyscamptocladius Thien.........12
0. Нет дистально зазубренных щетинок.......................18
12. Основной членик усика вдвое длиннее жгутика. Анальные жабры короткие, закруглённые. р. Dyscamptocladius, гр. v i t е 1 1 i n u s Thien.
0. Основной членик усика не длиннее или даже короче жгутика.
р. Dyscamptocladius, гр. га i n u t us,
3U
13. По бокам нижней губы есть крупные пластинки, по положению и контурам напоминающие паралабиальные пластинки Tendipedini, но без штриховки; нижняя губа с парными срединными зубцами (рис. 505)............................ 14
0. Нрт ясно выраженных пластинок................................% . 16
14. Паралабиальные пластинки с волосками (рис. 505 Б,. В). Усики с лаутерборно-выми органами. Премаялибулы раздвоены на конце (рис. 506)............... 15
Рис. 504. Лобное поле личинок:
А — Euphaenocladius; Б — Dyscamptocladtus.
0. Паралабиальные пластинки без волосков (рис. 505 А). Усики без лаутербор-новых органов (рис. 507 А). Премандибулы одновершинные. Длина личинки 7—8 мм.
Prodiamesa (Monodiamesa) bathyphila Kieff.
15. Нижняя губа с широкой, бледной, незубчатой частью и 5 мелкими боковыми зубцами (рис. 505 Б). Усики из 5 члеников, основной коленчато изогнут (рис. 507 Б). Зубчатая часть жвалы относительно мала; на внешней стороне три пучка щетинок; внутренняя щетинка представлена длинным рядом веерообразно расходящихся щетинок (рис. 508 A). Prodiamesa (Monodiamesa) f 1 a b е 1 1 a t a Kieff.
0. Нижняя губа вся зубчатая, с мелким парным срединным зубцом (рис. 505 В). Усики четырёхчлепистые, основной прямой. На внешней стороне жвалы простые щетинки; внутренняя щетинка перисто рассечена с одной стороны (рис. 508 Б). Длина до 15 мм. В родниках и холодных ручьях, среди грубого детрита.
Prodiamesa praecox Kieff. (Р. о I i та се a Meig).
Рис. 506. Премандибула личинки Monodiamesa flabellate.
Рис. 505. Нижняя губа личинок:
А — Monodiamesa batbypbila; Б — Monodiamesa Habellata; В — Prodiamesa praecox
Рис. 507. Усики личинок:
А — Monodiamesa batby pbi la; Б — Monodiamesa flabellata.
16. Нижняя губа с широкой, бледной, незубчатой, разве иногда слабо надсечённой средней частью (рис. 509)............................................... 17
0. Вся губа зубчатая, более или менее тёмная....................... . 20
17. Голова светлая, округлённо-квадратная. По одному почковидному глазу с каждой стороны. По голове рассеяны волоски (рис. 511). Жвалы с очень длинным, узким концевым зубцом и 5 (редко 6) мелкими узкими внешними (рис. 512). Премандибулы дистально слабо надсечены на четыре частя. Средняя часть нижней гуоы светлая, с мелкими зубчиками, боковые части с чёрными, острыми, наклонёнными к середине зубцами (рис. 509 Б). Подставки нреанальных щетинок высокие (рис. 510) Не строит чехликов; очень подвижна. Длина личинки 15 мм. В олиготрофных озёрах.
Protanypus morio Zett. (Diaiamesa miriforceps Kieff.).
Рис 509. Нижняя губа личинок.
„пп^тряная' Ь — Didi amesa пип-”«S“- 6г““'а'"1“' ”
Рис. 510. Задний коней личинки Didiamesa mi-riforceps.
Рис. 511. Передняя часть головы личинки Didiamesa miriforceps с брюшной стороны.
Рис. 508. Жвалы личинок:
А — Monodlamesa Г1а-bellata; Б — Prodi amesa ргаесох
Рис. 512. Жвала личинки Di di amesa mi ri forceps.
Рис. 513. Усик личинки Ortbocla-diinae g? (larva о bi cola).
Рис. 514. Лобное и ротовое поле Orthocladiinae g? (larva obi cola).
0. На голове только редкие волоски. Жвалы с более коротким концевым зубцом и 4 боковыми. Средняя честь нижней губы не шире */а всей губы......18
18. Нижняя губа с прозрачной бледной плёнкой, соединяющей концы первых боковых зубцов (рис. 509 В). Подставки преанальяых шетинок мелкие, высота их не менее ширины. Trichocladius inequalis Kieff.
0. Нижняя губа без прозрачной плёнки. Ширина подставок преанальных щетинок больше их высоты; подставки с тупым клювовидным выступом в дистальной части ... 19
19. Голова широкая. Усики четырёхчленистые; длина основного членика в 8,5 раз больше его ширины; щетинка неуплощённая, несколько длиннее жгутика (рис. 513). Боковые зубцы жвалы довольно широкие и заострённые. Премандибулы надсечены на 3 ветви (рис. 514). Средняя часть нижней губы при расплющивании шириной в Vs всей губы; боковых зубцов 6 (рис. 509 Г). Длина личинки до 13 мм.
Orthocladiinae g? larva obi cola Lip.
0. Голова удлинённая. Усики пятичленистые; длина основного членика в 2 раза больше его ширины; шетинка уплощённая, короче жгутика. Боковые зубцы жвалы до-316
вольно узкие, закруглённые. Премандибулы рассечены на 4 ветви. Средняя часть нижней губы при расплющивании не более 7в всей губы; боковых зубцов 7 (рис. 509 Л). Длина личинки до 12 мм. Di а те за bathyphila Lip.
20. Нижняя губа с парным срединным зубцом (рис. 515) ................21
0. Нижняя губа с непарным срединным зубцом (рис. 516)...............22
21. Основной членик усика слабо изогнут .............................28
Рис. 515. Нижняя губа личинок:
А — Euktefferiella; Б — Orthocladilпае g? (larva altalcola).
0. Основной членик усика прямой; отношение его к жгутику равно 2:1. Срединные зубцы нижней губы разделены не глубоко; боковых зубцов 5, из них при нерасплю-щенной губе видны 3 (рис. 515 Л). Тело в длинных волосках.
Eukiefferiella (Tricbocladi ns)coerulescens Kieff,
22. Отношение основного членика к жгутику по длине равно 1,5 : 1. 2-й членик усика не разделён. Срединные зубцы нижней губы разделены глубоко; боковых зубцов 6 пар; 1-й боковой ниже срединного; от 2-го бокового идёт складка, прикрывающая все боковые зубцы при не расплющенном состоянии губы (рис. 515 Б).
Orthocladiinae g? larva altalcola Lip.
Рис. 516. Нижняя губа личинок:
А — Lapporthocladlus; Б — Rheorthocladlns; В — Paraortho cladlus; Г — Orthocladiinae g? (larva aibirtcola).
0. 2-й членик усика разделён на 2 части — короткую проксимальную и длинную дистальную, на конце которой мелкие лаутерборновы органы; щетинка усика длинная, заходит за его конец. Нижняя губа тёмнокоричневая, боковых зубцов 5 пар; 1-й боковой значительно выше срединного. Длина личинки до 6 мм. В реках, ручьях, в открытой литорали озёр. р. В г i 1 1 i a Kieff.
28. Преанальные щетинки на подставке с выступом на дистальном конце. В родниках и холодных ручьях р. S у в d i a m е s а . . . 24
0. Подставка без выступов или её совсем нет.......26
24. Челюсти и гипофаринкс с волосками.............24
0. Челюсти и гипофаринкс без волосков. Длина личинки до 12 мм.
Syndiamesa branicki Thien.
25. В ротовом поле 7 шлемовидных чешуек (рис. 517 Л). Премандибулы рассечены дистально на 9—10 ветвей (рис. 517 В). Нижняя губа с 7 боковыми зубцами (рис. 517 Б). Длина подставок преанальных щетинок вдвое больше их ширины (рис. 500 В). Длина личинки 20 мм.
Syndiamesa lepnevi Lip.
Рио. 517. Личинка Syndiameaa lepnevi:
А — лобное и ротовое поле; Б—нижняя губа; В — конец арен анд иб у лы.
0. В ротовом поле 5 шлемовидных чешуек. Премандибулы рассечены на 7 ветвей. Нижняя губа с 9 боковыми зубцами. Ширина подставок преанальных щетинок больше их высоты* Длина личинки до 11 мм. Syndiamesa hygropetrica Kieff.
26. Щетинки преанального сегмента сидят непосредственно на сегменте, подставки
нет....................................................27
0. Щетинки на обычной подставке, без выступов, р. Orthocladius v. d. Wulp..........................28
27. Усики пятичленистые; 3-й членик пекольчатый. На жвале, на месте внутренней щетинки, придаток, усаженный щетинками (рис. 518 Л). 1-й боковой зубец нижнеи губы равен срединному. Длина личинки 5,5—7 мм.
В г а с by d i a in е s a (Dianiesa steinbocki Goetgh).
Рис. 518. Личинка Brachydiamesa steinbocki:
A — жвала; Б — лобное и ротовое поле.
Рис. 519. Усик личинки Di а mesa.
0. Усики четырёхчленистые; 3-й членик большей частью кольчатый (рис. 519). Внутренняя щетинка жвалы из многих простых ветвей. В олиготрофных озёрах на глубинах, в родниках, в холодных ручьях. р. Di a mesa Meig.
28. На лобном поле вентрально пара дву ветвистых щетинок (рис. 520 Л) ... 29
0. Щетинки на лобном поле простые ......................................84
29. Задние ножки большей частью очень длинные, равны по длине совокупности XI и XI1 сегментов. Основной членик усика длиннее жгутика в 2 с лишним раза (рис. 521)...................................................................30
Рис. 521. Усики личинок:
А — лобное поле личинки Rheorthocladius; Б — ротовое поле личинки Lapporthocladlus.
A— Lapporthocladlus; Б — Rheorthocladius.
0. Ножки не длиннее XI сегмента. Основной членик усика приблизительно в 2 раза длиннее жгутика.
р. Trichocladius Kieff.....38
30. Ветви двуветвистой щетинки длинные ....................31
0. Ветвя двуветвистой щетинки короткие и тонкие. Ротовое поле с 5 длинными листовидными чешуйками (рис. 520 В).
Подр. Lapporthocladius Thien.
31. Голова тёмная; лаутерборновы органы широкие, хорошо заметные, по длине равны 3-му членику усика. Нижняя губа с 9 или с 6 боковыми зубцами. Ротовое поле или с расходящимися в стороны, как усики, пучками изогнутых узких лопастипок, или с 5 закруглёнными на концах узкими чешуйками.
Подр. Euorthocladi us Thien. (Orthocladius, гр. thienemanni Thien).
0. Голова более светлая; лаутерборновы органы незаметны. Нижняя губа с 6 боковыми зубцами. Ротовое поле иного строения....................82
32. Ротовое поле с треугольной, тупо заоенгённой, короткой срединной чешуйкой и с несколькими более длинными и узкими на боках.
Подр. Rheorthocladius Thien. (Orthocladius, гр. saxicola Thien).
О, Ротовое поле с 3 закруглёнными на концах чешуйками.
р. Eudactylocladius Thien. (О г f h о с 1 a d i u s, гр. о 1 i v о c e u s Р о t t h.).
88. Усики массивные, основной членик не более чем в 3 раза длиннее своей ширины. Анальные жабры сильно редуцированы или отсутствуют. Длина до б мм. В солоноватых водоёмах; у морских берегов.
(Cricotopus, гр. vitripennis Mg., р. Trichocladius Kieff., гр. h а 1 о р b i 1 u s).
0. Основной членик усика в 4—5 раз длиннее своей ширины. Анальные жабры обычные. р. Trichocladius (Cricotopus), гр. algarum Kieff.
84. Внутренний край жвалы гладкий. По бокам нижней губы есть бородка.
Orthocladius, гр. nudipennis Kieff.
0. Внутренний край жвалы зазубрен. Бородки нет. В родниках, ручьях, реках, в литорали крупных озёр.
р. Eukiefferiella, гр. 1 о п g i с а 1 с а г Kieff.
Подсем. Pelopiinae (Tanypodinae)
1. Тело сжато в спинно-брюшном направлении. Сегменты тела широкие и короткие. По бокам тела каёмка волосков (рис. 522). Анальные жабры плоские, широкие, короткие, треугольные, в количестве 2 или 3 пар; 2-я и 3-я пары сдвинуты на ножки (рис. 522) . . 2
Рис. 522. Личинка Tanypi.
0. Тело вальковатое. Сегменты тела узкие и длинные, без боковой каёмки волосков, иногда лишь с рассеянными отдельными волосками. Индекс головы менее 0,7. Длина усика равна 1/г или 7з длины головы. Язычок мягкий, треугольный, с пузыревидными выступами и с 5 чёрнокоричневыми зубцами. Анальные жабры узкие, веретеновидные, в количестве 2 пар; обе пары у самого анального отверстия (рис. 523 Л).
Триба Micropelopiae, р. Ablabesmyia Joh. (Micropelopia
Zavr.)........................................
2. Голова узкая, длинная, индекс 0,7, спереди сильно сужена. Усики в 8/4 длины головы; кольцевой орган у конца 1-го членика. Язычок из 5—6 жёлтых или жёлтокоричневых зубцов, убывающих к середине по величине. Параглоссы одновершинные (рис. 542 В). Жвалы сильно загнуты в виде крючка (рис. 524 А), жёлтокоричневые с чёрным кончиком. Личинки красные. Длина до 15 мм. В литорали стоячих, слабо текущих вод.
Триба Clinotanypi, р. Clinotanypus Kieff.
Рис. 523. Задний конец личинки Ablabesmyia:
А — со спинной стороны; Б — сбоку.
Рис. 524. Личинка СИ -notanypus:
Л — жвала; Б — язычок и параглоссы.
0. Голова приблизительно сердцевидная, усики в */< или 7э длины головы. Нижняя губа с тёмножёлтыми паралабиальными гребнями. Язычок из 4—5 жёлтых зубцов или из 5 чёрных с наименьшим средним зубцом. Параглоссы почти всегда с 2 вершинами (рис. 529). Жвалы серповидные.
Триба Tanypi..........13
3
Ряс. 525. Личинки Ablabesmyia:
А — нижняя губа и язычок гр. fulva, Б — язычок гр. talcigera.
3. Анальные жабры очень короткие, чуть-чуть больше в длину, чем в ширину, приблизительно раз в семь короче задних ножек, изогнуты в виде оога. Глаз простои или двойной из неравных по величине частей. Окраска зеленоватая с белыми пятнами. Длина до 10 мм. А. гр. g u t t i р е n n i s Lenz L.
0. Анальные жабры узкие, веретеновидные. Глаз простой................4
4. Основной членик усика в два-три раза длиннее жгутика..........5
0. Основной членик усика в четыре-пять раз длиннее жгутика. Индекс головы около 0,6. Средний зубец язычка наименьший.....................................12
5. Индекс головы 0,66. Усики длиной в головы, вдвое длиннее жвал. Некоторые крючки проксимального ряда на задних ножках с добавочным зубчиком на вогнутой стороне. Средний зубец язычка наименьший (рис. 525 Л). Окраска красная или красноватая. Длина до 9 мм.
Ablabesmyia. гр. fulva Lenz (Zavr. — Thien.),
9. Усики равны половине длины головы и втрое длиннее жвал. Крючки задних ножек иногда все простые, иногда некоторые с добавочным зубчиком. Зубцы язычка или равной длины, или с наименьшим средним зубцом.................................. в
6. Усики одноцветные, жёлтые. Все крючки задних ножек одноцветные, жёлтые. Язычок тёмнокоричневый................7
0. Второй членик усика тёмнокоричневый, остальные жёлтые. На задних ножках по одному коричневому зубцу..........11
7. Кольцевой орган усика на УБ длины основного членика от его основания. На вогну
той стороне жвалы 2 зубца: ближний к основанию жвалы крупный и тупой, а расположенный между ним и конпевым — мелкий и острый. Средний зубец язычка наименьший. Все крючки задних ножек простые, жёлтые. Личинки белые. Длина до 6 мм. В ключах.
А., гр. bi о о ta t a Lenz ........................................8
0. Кольцевой орган усика посередине основного членика или несколько сдвинут по направлению к его концу. 2 зубца жвалы широкие, треугольные, почти равной длины. Все зубцы язычка почти равные или средний наименьший. Крючки задних ножек все простые, жёлтые или один из них с сильным добавочным зубцом. Личинка коричневатая. Длина до 8 мм. В ключах и лужах.
А., гр. tetrasticta Lenz (Nigropunctata Thien.)....................9
8. Усик вдвое длиннее жвалы; основной членик втрое длиннее жгутика; кольцевой орган на 1/в от конца основного членика до 6 мм.
А., подгр. bi п о t a t a Lenz (mi Dime Zavr. — Thien.).
0. Усик втрое длиннее жвалы; основной членик вдвое длиннее жгутика; кольцевой орган на */s длины основного членика от его конца. Длина личинки до 7 мм. В ключах.
А., подгр. pecteni pbora Lenz (Ablabesmyia pecteniphora Goetgh).
9. Зубцы язычка почти равны между собой. Один крючок задних ножек с крупным добавочным зубном. Личинка красноватокоричневая с коричневыми пятнами. Длина до 8 мм. В ключах и лужах. А., подгр. tetrasti eta Lenz.
0. Средний зубец язычка наименьший. Все крючки задних ножек простые ... 10
10. Передний край язычка слабо вогнут. Основной членик усика в 3,5 раза длиннее жгутика. Личинка жёлтая. Длина до 7 мм. В ключах. А., подгр. f 1 a vi da Lenz.
0. Передний край язычка сильно вогнут. Основной членик усика в 3,5—4 раза длиннее жгутика. Личинка беловатая. Длина до 5 мм. В ручьях, лужах. А., подгр. curti calcar Lenz.
11. Зубцы язычка равной длины (рис. 525 Б). Крючки задних ножек, расположенные олиже к основанию последних: 3 гребневидных, изогнутых, жёлтых и 2 (один жёлтый, другой коричневый) с 3 добавочными зубчиками. Личинка коричневая. Длина до 8 мм. В ключах, лужах, прудах. А., гр. f а 1 ci ge г a Zavr. (Lenz)
0. Средний яубеп язычка немного короче прочих. Большей частью крючки задних ножек, ближние к их основанию, с 3—6 добавочными зубчиками; 4 крючка жёлтых и 1 коричневый, имеющий иногда только 1 зубчик (рис. 526). Личинка желтая или зелёная. Длина до 5—6 мм. В стоячих водоёмах, в ключах.
А., гр. t е n u i с а I с а г Zavr. (Lenz).
Рис. 526. Крючки задних ножек личинки Ablabesmyia, гр. temilcalcar.
12. Кольцевой орган усика на г/3—’/< длины усика от его основании. Сегменты тела с длинными, крепкими разбросанными волосками. Все крючки задних ножек простые, жёлтые. Личинка белан, желтоватая или зеленоватая. Длина до И мм. В самых разно* образных водоёмах. А., гр. 1 е n t i g i n о s a Lenz (cos ta I i s iavr).
0. Кольцевой орган усика на середине основного членика. 1-й членик челюстного щупика подразделён па 2 части, между которыми расположен кольцевой орган. Все крючки задних ножек гладкие, 2 наиболее сильных чёрнокоричневые, остальные жёлтые.
Рис. 528. Жвала Psectrota-nypus, гр. varius.
Личинка желтоватая, с коричневым мраморным рисунком. Длина до 9 мм. В стоячих о текучих водоёмах,’среди растений. А., гр. monilis Zavr. (Lenz).
13. Голова бледная. Усик втрое короче головы; кольцевой орган в последней 1/< основного членика усика. Жвала с маленьким концевым зубцом и с мелкими зарубками у его основания. Паралабиальные гребни вогнутые. Язычок узкий и длинный с 5 (редко с 4) жёлтыми зубцами равной длины. Анальных жабер большей частью 3, иногда 2 пары (рис. 527). Длина до 12 мм. р. Pelopia Meig. (Protenthes Job.).
0. 2 пары анальных жабер...............................................14
14. Кольцевой орган на 1/9 от конца основного членика усика. Язычок с 4 жёлтыми зубцами равной длины. Жвалы по внутреннему краю пильчато зазубрены (рис. 528).
Рис. 530. Язычок личинки: Рис. 531. Нижняя губа
А — Anatopynia plumipes; Б — Psectrotanypus, гр. trifa- Psectrotanypus, гр. trifasci-
scipennls. pennis.
Паралабиалъные гребни почти прямые, зубцов более 6. Личинка зеленоватая, красноватая или жёлтокоричневая. Длина до И мм. В лужах и загрязнённых прудах.
р. Psectrotanypus Zett., гр. varius Lenz (brevicalcar Zavr.).
0. Язычок с 5 зубцами, из которых средний наименьший, иногда очень мал. Жвалы не зазубрены...................................................15
15. Язычок с чёрными зубцами, средний очень мал; параглоссы листовидные, иногда с зубчатыми краями, с одним концевым выступом (рис. 529 В). Длина до И мм. В стоячих водах от прибрежья до больших глубин; встречаются и в текучих водах.
р. Procladius. Skuze (Trichotanypus Kieff.) и p. Psilotanypus Kieff.
0. Язычок иногда с жёлтыми, чаще с коричневыми зубцами. Параглоссы удлинённые, 2- или 3-раздельные на вершине, не зубчатые или только со слабыми зубчиками.
о. Anatopynia Job.........16
16. Язычок широкий, жёлтый, с очень крупными крайними зубцами. Параглоссы короткие с 2—3 неравными короткими концевыми выступами (рис. 530 А). Окраска тёмная, оливковокоричневая. Длина до 19 мм. В мелких прудах.
А. р 1 u m i р е s Fries.
0. Язычок большей частью с красножёлтыми, редкое тёмно коричневыми зубцами; крайние зубцы не очень резко отличаются по величине. Параглоссы длинные с 2 выступами неравной длины (рис. 530 Б) ....................................17
17. Голова тёмнокоричневая. Кольцевой орган на 1/в основного членика от его конца. Длина челюстного щупика вдвое больше его ширины при основании. Паралабиальные гребни с 6 зубцами (рис. 531). Личинка тёмнокоричневая с жёлтым мраморным рисунком. Длина до 11 мм. Выдерживают сильное загрязнение.
р. Anatopynia (Psectrotanypus Kieff.) trifascipennis Zett.
0. Голова красножёлтая. Кольцовой орган усика на 1/< основного членика от его конца. Длина челюстного щупика в 4 раза больше его ширины при основании. Паралабиальные гребни с 7 зубцами. Личинка кровавокраспая. Длина до 14 мм. В лужах, прудах, в реках с замедленным течением.
р. Anatopynia, подр. Macropelopia Thien.
ВОЗДУШНЫЕ НАСЕКОМЫЕ, ЛИЧИНКИ КОТОРЫХ ЖИВУТ В ВОДЕ
1. Крыльев 2, только передние; всегда перепончатые, с сетью жилок.2
0. Крыльев 4 .....................................................3
2. Крылья с небольшим количеством древовидно разветвлённых жилок. На месте задних крыльев мелкие отростки в виде щетинок с расширением на конце (щетинки большей частью прикрыты чешуйками). На конце брюшка нет длинных нитей (рйс. 532). Ротовые части развиты — колющие пли сосущие.
Отряд Diptera — двукрылые (комары, мухи «и др.).
Рис. 532. Двукрылые имаго:
А — Eristalis; Б — Tendipes, самец.
Рис. 533. Крыло Ordella dimidiata.
0. Крылья с большим количеством жилок (рис. 533). На месте задних крыльев никаких отростков нет. На конце брюшка 2—3 хвостовые нити. Ротовые части мягкие, неразвитые.
Сем. Caenidae и часть сем. В а ё t i d а е из отр. Ephemeroptera (Agnatha) — подёнки.
3. Переднее и заднее крылья устроены одинаково ...............5
0. Переднее и заднее крылья различного устройства. Переднее более или менее твёрдое, роговое (целиком или частично), заднее — мягкое, перепончатое, прозрачное, в покое сложенное под переднее ...................................4
4. Рот с хоботком, направленным кзади и подогнутым под брюшную поверхность тела в состоянии покоя. Передние крылья не однородны: более твёрдая — основная часть и более нежная, перепончатая — конечная (рис. 534 Б, В).
Отряд Hemiptera (Heteroptera) — клопы.
0. Рот без хоботка, ротовые части грызущие. Передние крылья сплошь жёсткие, без жилок (рис. 534 А). Отряд Coleoptera — жуки.
5. Крылья покрыты чешуйками или волосками ....................6
0. Крылья прозрачные; перепонка без чешуек и волосков; по жилкам бывают волоски ......................................................... 7
6. Обе пары крыльев одинаковой плотности; передние больше задних; в покое или поднимаются, прикладываясь над спинкой друг к другу, или складываются над телом кровлеобразно. Ни передние, ни задние крылья никогда не складываются в продольные складки. Есть хоботок, большей частью закрученный спирально.
Отряд Lepidoptera — бабочки.
Рис. 534. Имаго:
А — Coleoptera (Dytiscus latissimus); Б и В — Н emi р tera (Б — Ran at га linearis, В — Nepa cinerea).
Рис. 536. Имаго Agriotypus (из перепончатокрылых).
Рис. 537. Имаго Perla bipunctata.
Рис. 538. Крылья:
А — Ephemera vulgata; Б Osmylus tulvi-cephalus.
0. Крылья и тело в волосках. Передние крылья уже задних и большей частью несколько твёрже. Задние иногда складываются продольными складками. Хоботка нет (рис. 535). Отряд Triohoptera — ручейники.
7. Крылья с большим количеством жилок, идущих в продольном и поперечном направлениях и образующих большое количество (не менее 20) ячей . . . . ..8
0. Крылья с небольшим количеством жилок, образующих не более 20 ячей (рис. 536).
Отряд Hymenoptera — перепончатокрылые.
8. На конце тела 2—3 длинные нити. Ротовые части мягкие, недоразвитые ... 9
0. Длинных нитей на конце тела нет. Ротовые части вполне развитые..10
Ш
9. Передние и задние крылья равны по длине; задние часто шире передних и тогда складываются продольными складками (рис. 537). В покое крылья складываются плоско вдоль спины. Нитей на конце брюшка две.
Отряд Plecoptera — веснянки.
Рис. 539. Имаго Sialis flavilatera (] atari а).
Рис. 540. Нижняя губа Cordulegaster.
0. Задние крылья много меньше передних (рис. 538 Л). В покое крылья приподняты кверху. Нитей на конце брюшка 2 или 3.
Отряд Ephemeroptera (Agnatha) — подёнки.
10. Крылья большие, буроватые; заднее немного меньше переднего. Жилки не очень многочисленны. В покое крылья сложены на спине под углом, кровлеобразно (рис. 539).
Отряд Megaloptera — вис л окрыли и.
0. Передние и задние крылья одной длины. Жилки очень многочисленны. В покое крылья распластаны горизонтально или подняты кверху. На конце брюшка очень короткие придатки.
Отряд Odonata — стрекозы.
ODONATA IMAGO - ВЗРОСЛЫЕ СТРЕКОЗЫ
Тело стрекоз удлинённое, с ясным расчленением на голову, грудь и б рюш-к о. На голове всегда очень резко выступают глаза, иногда сходящиеся на верхней стороне головы до полного соприкосновения (рис. 541). Крылья большей частью имеют у переднего края, ближе к концу, окрашенные пятнышки — глазки. Крылья в покое у одних стрекоз распластаны горизонтально, у других приподняты кверху. Н о ж-к и всегда слабые, иногда с расширенными голенями.
1. Крылья одинаковы по форме. Сложные глаза разделены, расстояние между ними больше диаметра глаза. Брюшко очень длинное, тонкое (рис. 547—549).
Zygoptera...........6
0. Крылья не одинаковы по форме: заднее при основании значительно шире переднего. Сложные глаза или соприкасаются, или раздвинуты на расстояние, меньшее диаметра глаза. Тело более толстое, брюшко к концу суживается (рис. 541—546).
Anisoptera ...........2
2. Крыловые треугольники 1 переднего и заднего крыла одинаковой формы и вытянуты по длине крыла (рис. 541, 543, 545)....................... 3
0. Крыловые треугольники переднего и заднего крыла неодинаковы по форме; кроме того, на переднем крыле треугольник вытянут в поперечном направлении, а на заднем — в продольном (рис. 542, 546). Сем. Libellulidae — стрекозы..б
3. Сложные глаза не соприкасаются (рис. 545).
Сем. Gomphidae — дедки.
0. Глава соприкасаются .......................................4
4. Глаза соприкасаются только в одной точке (рис. 543). Средняя лопасть нижней губы с продольным надрезом (рис. 540).
Сем. Cordulegastridae.
0. Глаза соприкасаются на некотором расстоянии (рис. 541, 544). Средняя лопасть вижвей губы без надреза.
Сем. Aeechnidae — коромысло.
б. Задний край глаз посередине образует полукруглый выступ. Тело зелёное с металлическим блеском или бронзового цвета (исключая Epitheca) (рис. 542).
Сем. Corduliinae.
0, Задний край глаз посередине без выступа (рис. 546). Тело зелёное, без металлического блеска.
Подсем. Libellulinae.
6. Крылья довольно широкие, к основанию суживаются очень постепенно (рис. 547), в покое приподняты кверху. Крылья у самца всегда металлически блестящие, синие или с буросиней перевязью.
р. Agrion L. (Calopteryx Burm.) — красотка.
1 Крыловыми треугольниками называются образуемые жилками крыла треугольные ячейки, расположенные приблизительно в середине трети крыла, обращённой к телу. 324
<
Рис. 542. Cordulia aenea, самец.
Рис. 541. Aesch-na sp.
Ряс. 543. Gordalega-ster annulatus, самка.
Рис. 544. Aeschna суапеа, самец.
Рис. 545. Gomphus vu]gatissimus, самец.
и
Рис. 547. Agrion splendens, самец.
Рис. 546. Libellula quadri maculate, самец.
0. Крылья узкие, невдалеке от основания сразу довольно сильно суживаются вследствие изгиба заднего контура (рис. 548, 549) .......................7
7. Передние и средние голени сплюснуты и сильно расширены. Тело беловатое, на груди чёрные линии. Глазок на крыльях равной длины и ширины, р. Platycnemis Sharp.
0. Голени не расширены..........................................8
8. Глазок почти одинаков в длину и ширину (рис. 548). Крылья в покое приподняты кверху. Тело не бронзовозелёпое.....................................9
Рис. 549. Lestes eponsa, самец.
0. Глазок вытянутый (рис. 549). Крылья в покое большей частью распластаны. Тело бронзовозелёное.
р. Lestes Leach. — лют к а.
9. Грудь и брюшко со спинной стороны чёрнозелёные, с слабым металлическим блеском.
р. Ischnura Sharp.
0- Грудь и брюшко со спинной стороны с бронзовотёмным фоном..
10. Рисунок по чёрному фону голубой.
р. Coenagrion Kieff. (Agrion Fabr.) —стрелка.
О. Рисунок голубой или зелёный.
р Erythromma Sharp.
СПИСОК ОПРЕДЕЛИТЕЛЕЙ ПРЕСНОВОДНЫХ ОРГАНИЗМОВ НА РУССКОМ ЯЗЫКЕ
Общие
«Жизнь пресных вод СССР» (под ред. В. И. Жадина, т. 1, 1940; т. II. 1949. Академия наук СССР, Ленинград.
Ламперт К., Жизнь пресных вод, 1900.
Липин А. Н., Пресные воды и их жизнь, изд. 1, 1926; изд. 2, 1941, Учпедгиз Москва.
Водные растения
Ануфриев Т. И.. Таблицы для определения важнейших болотных и лесных мхов Европейской части СССР. 1932.
Аржанов С. П., Среди вод и болот, 1926.
Еленкин А. А., Определители растений. Мхи и лишайники, 1930.
Кузнецова Н. А. и Милованов В. К., Определитель осок, 1927.
Маевский П. Ф., Флора средней полосы Европейской части СССР, изд. 7,1940.
Нейштадт М. И., Наши растения, 1935.
Его же, Определитель растений, Учпедгиз, 1940.
Ростовце в С. И., Морфология и систематика печёночных мхов, 1913.
«Растительность СССР», изд. Академии наук СССР, 1938.
Р ы чи в Ю. В., Флора гигрофитов, 1948.
Рычин Ю. В. иСергееваП. В., Водная прибрежная флора, 1939.
Сырейшиков П. Д., Иллюстрированная флора Московской губернии, ч. I — IV, 1906—1914.
Его же, Определитель растений Московской губернии, 1927.
Т а л и е в В. И., Определитель высших растений Европейской части СССР, 1940.
Федченко Б. А.. Водяные растения. Библ. журн. «В мастерской природы», 1928.
Ф е д ч е н к о Б. А. и Ф ле ров А. Ф., Водяная флора Европейской России, 1913.
Их же. Водяные растения Средней России, 1900.
Их же, Флора Европейской России, 1908—1911.
«Флора СССР», Акад, наук СССР, 1934—1937.
Флорова В. М. и Раменский Л. Г., Определение растений в нецветущем состоянии для средней части СССР, 1937.
Водные животные (общее)
Бродский А. Л.\ Жизнь в пресной воде, 1925.
Б р о н ш т е й н 3. С., Водные животные, 1935.
Гентшель Е., Жизнь пресных вод, 1914.
Павловский Е. Н. и Ле п н е в а С. Г., Очерки из жизни пресноводных животных, 1948.
Р ы л о в В. М., Жизнь пресных вод.Планктон, 1924.
«Фауна СССР», изд. ЗИН АН СССР.
Губки
Резвой П. Д., Пресноводные губки, Академия наук СССР, 1936.
Коловратки
Неизвестноеа-Жадина Е. С., Коловратки — Rotatoria. Жизнь пресных вод, т. 11, Академия наук СССР, 1949.
Мшанки
К л ю г е Г. А., Мшанки (Bryozoa). Жизнь пресных вод, т. И, Академия наук СССР, 1949.
Черви малощетинковые
Ласточкин Д. А., Кольчатые щетинковые черви (Char topodal. Жизнь пресных вод, т. II, Академия паук СССР, 1949.
Малевич И И., Материалы по фауне Oligochaeta рек СССР. Бюллетень Московского общества испытателей природы, 49, 1940.
Пиявки
Иогансон Л., Определитель пиявок, изд. ЦУЕГМС, 1935.
ИогансонЛ. и Васильев Е., Определитель пиявок. Труды отдела гидрологии Ленинградского областного гидрометеорологического управления, 1 при лож., 1935.
Щеголев Г. Г., Пиявки. (Hirundinea). Жизнь пресных вод, т. II, Академия наук СССР. 1949.
Мягкотелые (моллюски)
Владимирский Н. Д., Наши пресноводные моллюски. 1927.
Жадин В. И., Наши пресноводные моллюски, 1926.
Его же, Пресноводные моллюски СССР, Ленснабтехиздат, 1933.
Его же, Промысловые моллюски пресных вод СССР. НК местной промышленности, 1937.
JI и н д г о л ь м В. А., Мягкотелые, или моллюски. Животный мир СССР, 1938.
Членистоногие (общее)
Л и п и н ы А. Н. и Н. Н., Таблицы для определения членистоногих (водных). Главное управление по заповедникам, зоопаркам и зоосадам, 1941.
Ракообразные
Бирштейн Я. А., Высшие раки. Жизнь пресных вод, т. 1, Академия паук СССР, 1940.
БоруцкийЕ. В., Пресноводные и солоноватоводные Harpaticidae СССР. Определители организмов пресных вод, вып. 3. Изд. Научно-исследовательского ихтиологического ин-та, 1931.
Его же, Harpacticoida. ВАСХНИЛ, 1936.
Б р о п ш т е й н 3. С., Ракушковые раки (Ostracoda). Жизнь пресных вод, т. 1, Академия наук СССР, 1940.
Л и н к о А., К познанию фауны Phyllopoda Европейской России. Труды СПБ общества естествоиспытателей, 31, 1901.
Рылов В. М., Свободноживущие веслоногие. Пресноводная фауна Европейской России, вып. 1. Гл. упр. по рыболовству и рыбным промыслам, 1922.
Его же, Пресноводные Calanoida СССР. ВАСХНИЛ, 1930.
Его же, Ветвистоусые ракообразные (Cladocera). Жизнь пресных вод, т. 1, Академия наук СССР, 1940.
Смирнове. С., Листоногие раки (Phyllopoda). Жизнь пресных вод, т. 1, Академия наук СССР, 1940.
Паукообразные
Мусселиус А. А.,К познанию фауны Hydracarina России. Труды Гидробиолог, ст. оз. Глубок., V, в. 1, 1913.
Соколов И.И., Hydracarina водяные клеши, ч. 1, Hydrachnellae. Фауна СССР, Паукообразные, V, 2, 1940.
Его же, Водяные клещи (Hydracarina). Жизнь пресных вод, т. 1, Академия наук СССР, 1940.
Харитонов Д. Е., Пауки (Aranelna). Жизнь пресных вод, т. 1, Академия наук СССР, 1940.
Насекомые (общее)
Б о г д а н о в-К а т ь к о в Н. Н., Практическая энтомология, в. 1 (определитель насекомых до отрядов), 1923.
Мирам Э. Ф., Определение отрядов взрослых насекомых и их личинок, Академия наук СССР, 1933.
УльмерТ., Пресноводные насекомые. Изд. «Задруга», 1918.
Ногохвостки (вилохвостки)
Римски й-К орсаков М. Н., Вилохвостки, или ногохвостки (Collembola). Жизнь пресных вод, т. 1, Академия наук СССР, 1940.
Стрекозы
Бартенев А. Н., Насекомые ложносетчатокрылые. Фауна России и сопредельных стран, т. 1, в. 1—2, 1915—1919.
ДьяконовА. М., Наши стрекозы. Определитель стрекоз и их личинок, 1926.
Попова А. Н., Стрекозы (Odonata). Жизнь пресных вод, т. 1, Академия наук СССР, 19'*0.
Якобсон Г. Г. и Б и а н к и В. Л., Прямокрылые и ложносетчатокрылые, гл. VII — стрекозы, 1905.
Подёнки
Чернова Л. А., Подёнки (Ephemeroptera). Жизнь пресных вод, т. 1, Академия наук СССР, 1940.
Веснянки
Л е п н е в а С. Г., Веснянки (Plecoptera). Жизнь пресных вод, т. 1, Академия наук СССР, 1940.
Клопы
Бианки В. Л. и Кириченко А. Н., Насекомые полужесткокрылые. Практическая энтомология, в. IV, 1923.
Кириченко А. Н., Водные полужесткокрылые. Раб. Северокавказской гидробиолог. ст., Ill, 1930.
Его же, Настоящие полужесткокрылые (клопы) (Hemiptera). Жизнь пресных вод, т. 1, Академия наук СССР, 1940.
Жуки
Б а у А., Определитель жуков Средней Европы. СПБ 1914.
Рейхардт А. Н., Наши водяные жуки, 1926.
Р-е пхардтА. Н. и Оглоблин Д. А., Жуки (Coleoptera). Жизнь пресных вод, т. 1, Академия наук СССР, 1949.
Якобсон Г. Г., Жуки России и Западной Европы, 1905—1915.
Его же, Определитель жуков. Практическая энтомология, в. VII, 1931.
Большекрылые
Римски й-К орсаков М. Н., Большекрылые (Megaloptera). Жизнь пресных вод, т. 1, Академия наук СССР, 1940.
Сетчатокрылые
Римски й-К орсаков М. Н., Сетчатокрылые (Neuroptera planipennia). Жизнь пресных вод, т. 1, Академия наук СССР, 1940.
Ручейники.
Л е п не в а С. Г., Ручейники (Trichoptera). Жизнь пресных вод, т. 1, Академия наук СССР, 1940.
Чешуекрылые (бабочки)
Римски й-К орсаков М. Н., Чешуекрылые, или бабочки (Lepidoptera). Жизнь пресных вод, т. 1, Академия наук СССР, 1940.
Двукрылые
Липина Н. Н., Личинки и куколки хирономид, 1928.
Б ё ж е, Семейство Heleidae. Жизнь пресных вод, т. 1, Академия наук СССР, 1940.
Липина Н. Н. и ЧерновскийА. А., Семейство Tendipedidae. Там же.
Мончадский А. С., Личинки комаров (сем. Culicidae) СССР и сопредельных стран. Определители по фауне СССР. Академия наук СССР, 24, 1936.
Его же. Двукрылые (Di ptera). Жизнь пресных вод,т. 1, Академия наук СССР, 1940.
Рубцов И. А., Мошки (Simuliidae). Фауна СССР, т. VI, в. 6, 1940.
Черновский А. А., Определитель личинок комаров семейства Tendipedidae. Определители по фауне СССР, изд. ЗИН Академии наук СССР, 1949.
УКАЗАТЕЛЬ РУССКИХ НАЗВАНИИ ОРГАНИЗМОВ
(* — означает
А.ир болотный 155, 156 ♦ — пахучий, 155, 156 ♦
Бабка 85 * 239 240 ♦
241 ♦
Бабочки 82, 85, 87 233 286, 287, 288, 322 ’
Бабочпицы 289, 290 ♦ Бадяга 170, 170 ♦ Бактерии 75, 107, 112 — гнилостные 112 — денитрифицирующие 52 — железные 50 — нитрифицирующие 52 Бегун водяной 257 Беззубка 88, 91, 202 Бекмания 154, 154 * Белокрыльник 117; б. болотный 156 ♦, 157
Битинелла 201, 201 * Битиния 84 ♦, 91,201,201 ♦ Блоха водяная (дафния) 37, 215, 216 ♦, 217 *
Бокоплав 91, 125, 126, 148, 206, 206 ♦
Болотник (жук) 262, 263 ♦, 264 ♦
Болотник (раст.) 163, 165 Болотолюб 265, 265 ♦, 271 Брюхоногие 198
Вербейник обыкновенный 158, 158 ♦
Вероника 165, 165 ♦,
Вертячка 82, 260, 265, 265 ♦, 266, 267, 268
Вех ядовитый 157, 157*
Вилохвостки 233, 234,
234 ♦, 235, 235 ♦
Вислокрылка 81, 82 ♦, 87, 88, 91, 272, 272 *
Влаголюб 271
Водобродка 271
Водожил 260, 260 •, 267, 267 ♦
Водожук 265, 265 *, 270, 270 *
Водокрас (лягушатник) 157,
Водолаз (жук) 266, 271
Водолюбы 55 *,83, 260, 267, 270, 270 ♦, 271
присутствие рисунка на данной странице)
Водомерка 37, 82, 83 ♦ 86 ♦, 257, 257 ♦, 258* 258 * ’
Водоросли 40, 50. 57 75 89, 142
— диатомовые 40, 66 67 ♦ 89, 143
— жгутиковые 65. 67, 68 ♦ 69, 143
— зелёные 44, 45, 66 ♦, 67 89, 143
— конъюгаты 143
— нитчатые 84, 143
— синезелёные 40, 65 ♦, 67 68, 69, 111, 143
Водяник, см. болотник Волосатик 169
Галлицы, 287
Гидра 26, 26 ♦, 84, 87, 168, 172, 172 *, 173, 173 *
Гидроидные 168
Гладыш 54 ♦, 259, 259 ♦
Горичник болотный 158
Горошинка 91, 148, 202
Гребец 268, 269 ♦
Гребняк 86 ♦, 258
Гречиха земноводная 78, 88, 160 ♦, 161.
Губка 30, 50, 87, 145, 168, 170, 170 ♦, 171 *, 172
Дафния (блоха водяная) 69, 70, 111, 215, 216 ♦
Дедка 148, 237, 238 ♦ Долгоножка (карамора), см. комары
Долгоносик 260
Дрейссена 91, 144, 202 ♦
Ежеголовник 155; виды 156 ♦, 161, 161 *, 162, 162 ♦
Жаброног 122, 209, 210 »
Жемчужница 202, 203
Жеруха земноводная (хрен водяной) 158, 158 *
Живородка (лужанка) 2UU, 200 ♦
Журчалки 292
Затворка 91, 200, 201 ♦
Иловой мухи личинка, см. крыска
Ильник 262, 262 ♦, 268
Камыш 57, 79, 86, 88. 152 152 ♦ ’
Касатик ложноаирный 155,
Катушка 84 ♦, 199
Килевик 270
Кишечнополостные 168
Клещи 63, 87, 229; кладка 86 •
Колодезный рачок 125 Комары
— дергуны 74, 82, 90, 122
— долгоножки (карамора) 289, 289 ♦
— звонцы, см. тендипедиды — толкунчики 292
Коромысло 55, 57, 58 ♦, 238 239 *
Красотка 54 *,84, 224, 235, 236, 236 ♦, 237, 240
Крыска 123, 292, 292 •
Кубышка 84, 85, 160, 160 ♦, 161
Кувшинка 30, 53, 57, 78, 84, 84 ♦, 88, 89,160, 160 •
Кукушкин лён 118, 118 ♦, 119, 120
Лилия водяная — см. кувшинка
Листоеды 260, 266, 267, 272, 272 ♦
Ложнобеззубка 202
Ложноконская пиявка, см.
Пиявки
Лужанка (живородка), см. Живородка
Лужник 261, 261 *, 268, 269 ♦
Лунка 200, 200 *
Лучица 50, 78, 79 *, 85 *,
Львинка 54 ♦, 83, 291*,292
Лютик 78 w ф
— жестколистный 162,1 w — многолнстный 159, 160
Лютик нитчатый 162, 163 * Лютка 85 ♦, 236, 236 ♦, 237 *, 327, 327 *
Лягушатник (водокрас) 80 •, 84, 88, 89, 158, 159 ♦
Манник обыкновенный (плавающий) 153, 153 *
— пышный 153 ♦, 154 Многокоренник 159, 159 ♦ Морщинник 270, 270 ♦ Мотыль (лич. Tendipes) 37, 74, 74 ♦, 103, 310, 310 ♦
Мох 78, 117, 120
Мох водяной 78
Мох торфяной 117
Мошка 290, 291 *
Мухи 89, 89 ♦, 125, 233 Мхи листостебельные 152, 152 ♦
Науплиус 143, 144, 228, 228 •
Ногохвостики, см. вилохвостки
Нырялка 261, 268
Огнёвка 84, 87, 286 ♦, 287, 288 ♦
Омежник водяной (конский укроп) 157, 158 ♦
Ослик водяной 54 *, 58 ♦, 88, 147, 206 ♦, 207
Ослик слепой 125
Осока 117, 118, 154, 155 ♦, 157
Остракоды, см. ракообразные
Павун 89, 160
Пароводяной (серебрянка)
Пауки 125, 204
Паукообразные 202, 204
Перепончатокрылые 233 Перикома 126, 290 ♦ Перловица 127, 148, 202 Пеструшка 261, 268 Пискун 271
Пиявки 31, 37, 84, 87, 90, 145, 148, 169, 195, 195 ♦, 196, 196 ♦, 197, 197 ♦, 198 198 ♦
Плавт 86 ♦, 88, 259, 259 ♦ Плавунцы 54, 55, 55 ♦, 57, 58 ♦, 83, 86 ♦, 114, 260* 261, 262, 267, 268, 269 ♦’ 270 ♦
Плавунчик 261, 267, 270 270 ♦
Планария 54 ♦, 86, 168 ♦
Плосконожка 236, 236 ♦ 237 ♦
Подводник 261
Подорожник водяной (ча-стуха — подорожник) 156 ♦, 157
Подура 83
Полоскун 268, 269 ♦
Полушник (расходник) 79 *, 163, 164 *
Поручейница 153, 154
Почки зимующие 37
Прибрежник 271
Прицепыши (коготники) 266, 271
Прудовик (жук) 261, 262 , 268, 269 •
Прудовик (моллюск) 55 55 *, 83, 84, 84 *, 199, 199 *
Пузанчик 261, 268
Пузырчатка 36 ♦, 57, 78, 162, 163 ♦
Пушица 120
Радужница 84, 87, 266, 272, 272 *
Ракообразные 37, 63, 75, 111, 142, 202, 204, 205 — веслоногие 64, 70, 121, 142, 143, 144, 227, 228, 228 ♦
— ветвистоусые 64, 111, 121, 142, 144, 208, 211
— высшие 205
— двояконогие 205
— десятиногие 205
— жаброногие 122, 209
— истинновеслоногие ‘(свободно живущие) 208
— истиннолистоногие 208
— листоногие 208
— низшие 144, 205
— остракоды (ракушковые рачки) 122,125, 127,208
— равноногие 205
— расщепленноногие 205
— членистогрудые 205
— щитогрудые 20^
Рак речной (обыкновенный) 204, 204 ♦, 207, 208
Расходник, см. полушник Рачок колодезный 125 Рдест 36, 50, 85, 89; виды 160 *, 161, 165, 165 *, 166, 166 ♦, 167, 167 ♦
Резуха 165
Рогоз 88, 155, 156 ♦
— узколистный 156 ♦, 157
— широколистный 155, 156 ♦
Роголистник 78, 89, 162; р. подводный (тёмнозелён-ный) 162, 164 ♦
Росянка 117, 118
Ручейники 57, 81, 83, 84, 85*, 87, 88, 90, 91, 111, 125, 125 ♦, 126, 127, 233, 273, 273 ♦
Ряска 78, 84, 89, 159; виды 159, 159 ♦
Сабельник 117
Сальвивия плавающая (водяной папоротник) 158,
Серебрянка, см. паук водяной
Сида 84, 89, 213, 214 ♦
Скоморох 262, 263 ♦ 268
269 ♦
Скорпион водяной 54 ♦ 57 87, 258, 258 •
Скрытноусые 257
Слепни 292, 292 •
Сосенка водяная (хвостник) 80*, 163, 164 •
Стрелолист 117, 157, 157 ♦ Сусак зонтичный 155, 156 • Сфагнум (мох торфяной) 118, 118 ♦, 119, 120
Телорез сабуровидный (обыкновенный) 36*, 50, 57 • 163, 164 ♦
Тендипедиды 53, 56, 74, 75 84, 85, 87, 89, 91, Юз’ 108, 114, 121, 127, 298 ’
Тинник 262, 262 ♦, 268
Тинолюб 271
Толкунчики 292, 292 ♦
Трезубка (тростянка) овся-ничная 88, 154, 155 ♦, 157 ♦
Трифоль 117
Тростник 30, 79, 88, 154, 154 •
Трубочник 53, 54 ♦, 74 *
Трясинник 260, 266, 267
Турча 78; т. болотная 162, 164 ♦
Укроп конский (омежник водяной) 157, 158 ♦
Улитка пузырчатая 57, 125, 199, 199 ♦
Улитка гребенчатая (затворка) 54 *, 91, 200, 201 ♦
УрутьЗб ♦, 78, 89, 162,164*
Физа 84
Хара (лучица) 50, 79 ♦
Хвощ 79, 88, 117, 152, 152 ♦, 153
Хирономиды, см. тендипедиды
Циклоп 64, 68, 121, 122, 123, 228, 228 ♦, 229
Цикута, см. вех ядовитый
Чашечка 84, 90, 198, 199 •
Черви 84, 168, 168 *, 169, 169 •
Чернушка 201
Членистоногие 168, 202
Чума водяная 50, 78, 80 ♦, 85 ♦, 163, 164 *
Шаровка 148, 202
Шипоножка 265, 271
Щитень 122, 209, 211, 211 *
Элодея, см. чума водяная Эфемера 242, 243, 243 *
Ярмоноска 266
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ ОРГАНИЗМОВ
Abbreviates 301 ♦, 302
Ablabesmyia 146, 147, 319, 319 *, 320, 320 *; группы 320, 320 ♦
Acanthobdella 195; A. pele-dina 195, 195 ♦
Acanthobdellae 195
Acanthobdellidae 195
Acantholeberis curvirostris 221
Acarina 204
Acentrella 251
Acentropus niveus 286 ♦, 287
Acilius 268, 269 ♦; A. sul-catus 264 ♦
Acorus calamus 155, 156 ♦
Acrocladium cuspidatum 151
Acroperus 222; A. harpae 222, 222 *
Adineta 174; A. aculeata 174 ♦
Adinetidae 174
Aedes 296, 296 *, 297 ♦
Aelosoma 145, 187; виды 183 ♦, 186
Aeolosomatidae 185, 186, 187
Aeschna 58 ♦, 86, 238, 325 ♦; виды 239 ♦, 325 ♦
Aeschnidae 82 ♦, 237, 324
Aeschninae 237, 239 ♦
Aethaloptera rossica 283 Agabus 268, 269 ♦
Agapetus 274; A. comatus 275 ♦
Agapetinae 274
Agnatha — cm. Ephemeroptera
Agraylea 283; A. pallidula 284 ♦
Agrayloides 301 *, 302
Agrionidae 147, 236
Agrion 54 *,236, 326 *; виды 235 ♦, 236 ♦, 237 ♦
Agriotypus 323 •
Agrypnia 285; A. pagetana, чехлик 286 ♦
Akiefferiella coerulescens 317, 317 ♦ .
algarum, Trichocladius 31. Alisma plantago 156 ♦, 157
Allochironomus 307, 307
Alona 223, 225; виды 225 225 ♦
Alonella 224, 226; виды 226 227, 227 ♦
Alonopsis 222, 224; виды 224, 224 ♦
Altaicola 317 *
Ameletus 249, 250 ♦
Amphichaeta leydigi 188
Amphinemura 256; A. cine-rea 256 ♦
Amphipeplea 199; A. gluti-nosa 199 ♦
Amphipoda 205, 206
Amphipsyche proluta 283
Anabaena 65 ♦, 67, 68, 148
Anabolia 147, 281; виды 281, 281 ♦; чехлик 282 ♦
Anacaena 265, 271
Anapetes, см. Lestes Anapus, cm. Chromogaster . Anatopynia 321; A. plumi-pes 321 ♦, 322
Anax 147, 238; виды 239, 239 ♦
Anchistropus emarginatus 223, 223 ♦
Ancylidae 198
Ancylus 84, 90, 145, 147, 198; виды 199 *
Anisoptera 235, 236, 238 *, 324, 325 *
Anne li des 169
Annulipalpia 273
Anodonta 88, 91, 146, 202;
A. cygnea 203 ♦
Anopheles83, 87, 147; 290* 294, 296, ♦
Anuraea, см. Keratella
Apatania 125, 280; A. fim-briata 281 *
Apataniinae 280
Apatelia 280
Aphanisomenon 65 ♦, 67, 68, 111, 148
Aphelochirus 145, 259; A. montandoni 259 ♦
Aplexa 199; A. hypnorum 199 ♦
Apodidae 209, 211
Apus 211; A. cancriformis 211, 211 ♦
Arachnoidea 204
Araneina 204
Arcteonais lomondi 191
Arctocorisa 258
Arctopsyche 274
Arctopsychidae 274
Argulus foliaceus 114
Argironeta aquatica 55*,
Artemia salina 209, 2-10 •
Arthropleona 235
Arthropoda 168 Arthrostraca 205
Asellus 54 *, 58 ♦, 88, 147, 207; виды 125, 206 ♦, 207
Asplanchna 64 ♦, 69, 174, 177; A. priodonta 178
Asplanchnidae 175, 177
Asplanchnopus 178
Astacus 146, 148, 207, 208;
виды 204 ♦, 208
Asterionella 67, 67 ♦
Atanytarsi 312
A tanytarsus 312, 312 ♦; A. bausei 311 ♦
Atherix 292, 292 ♦
Aulodrilus 192; виды 192
Aulonogyrus 265 *, 266
Aulophorus 184, 190 ♦
Baetididae 249, 322, 322 *
I^tidinae 251
Babtis 83 ♦, 146, 147, 251;
В. rhodani 250 *, 251 *
Baicalospongia bacillifera 170 ♦
batophilus, Tendipes 148, 311
Batrachobdella 198; B. pa-ludosa 197 *
Bdellbidea 174
Beckmannia eruciformis 154, 154 *
Beggiatoa 51
Beraea 280; виды 279 ♦, 280
Beraeidae 280
Beraeodes 280; B. minuta, чехлик 278 *, 280
Berosus 264, 265 ♦, 270
Bezzia 296, 297 *
binotata, Ablabesmyia 320
Bithynia 91, 147, 200, 201;
виды 121, 201 *; кладка 84 *
Bi ttacomorpha 291 * Bivalvia 198
Blepharocera 288 ♦ Blepharoceridae 126 *, 127, 288
Boreobdella 198; B. verru-cata 198 ♦
Bosmina 64, 69, 219; виды 220, 220 •
Bosminidae 219 Bosmi nopsi s 213, 24 9 * Boyeria 238; B. irene 239 ♦ Brachionidae 175, 179 Brachionus 69; виды 64 *, 179, 179 ♦ Brachycentrinae 279 Brachycentrus 146, 279; B. subnubilus 279, 279 • Brachycera 287 Brachycercus 248 Brachydiamesa steinbocki 318, 318 ♦
Brachytron 238; B. haf-niense 239 *
Branchiobdellidae 186 Branchipodidae 209 Branchipus 122, 123, 209;
B. stagnalis 209, 210 * brevipalpis, Eucricotopus, 314
Brillia 317
Bryozoa 168, 181 Bulininae 199 Butomus umbellatus 155, 156 ♦
В у thine Ila 201; B. adscha-rica 201 ♦
Bythotrephes 212, 213; виды 213, 214 ♦
Caenidae 322, 322 ♦ Caenis, cm. Ordella Calanoida 228, 229
Calla 117, C. palustris 156 * 157
Callicorisa 258
Calliergon 78, 89, 151; C. carai folium 151 •
Calliei^onella cuspidate 151,
Calliophrys 293 *
Callitnche 163 ;C. autumna-
lis 163, 165
Calopteryx, см. Agrion
Camptocercus 222, 224; ви-
ды 223 ♦, 224
Candona 123, 127
Canthocamptus 228 • Campnia 256; C. nigra 256 * Capnidae 256
Carex 117, 155 •
154; C. gracilis
Catabrosa aquatica 153 ♦. 154
Cataclysta lemnata 287;
чехлик 286 *
Cecidomyiidae 287
Centropagidae 228
Centroptilum 147, 251 ; ви-
ды 250 *, 251*
Cephalodella 178 Ceratium 40, 65, 68 *, 69;
C. hirundinella 69
Ceratophyllum 78, 89, 147, C. aemersum 162, 164
Ceriodaphnia 64, 215, 217; виды 217, 218, 218 , 219 •
Chae toga ster 188; виды 147, 183, 188
Chae topoda 169
Chaetopteryx 282; Ch. vil-losa 283 ♦
Chaoborinae 287, 289, 293, -294, 295 ♦
Chaoborus 74, 75, 106, 107, 108 144, 146, 290 *, 294
Chara 50, 78, 79 ♦, 89, 147
Cheumatopsyche 283 Chirocephalus 210, виды 210, 210 •
Chironomariae 298 *, 301 ♦, 302 ♦
Chironomidae, см. Tendipe-didae
Chironomidae g?, cm. Steno-chironomus
Chironominae, cm. Tendipe-dinae
— connectens Lip. № 3, 306, 306 *
----- 4, 303 ♦, 304
— genuinae Lip. № 1, 306, 306 ♦
----- 2 309, 310 ♦ ----- 3 305, 305 ♦ ----- 6 303 *, 307 *, 308, 308 *
----- 7 303 ♦, 305, 305 ♦ ----- 8 303 ♦, 305 *, 306 ----- 9 308, 309 * ----- 10 303 ♦, 304 ----- 11 303, 303 * ----- 12 303 *, 304 ----- 13 308, 308 * ----- 14 307 *, 308 -----larva kruglovicola
309, 309 ♦
-----larva sernovi, cm. gen. № 6
Chironominae genuinae Pancr. № 1, 308, 308 * ----- 2 309, 310 ♦ ----- 3 305, 305 ♦ ----- 5 305, 305 * Chironomus, cm. Tendipes Chirotonetes, cm. Isonychia
Cbitonophora 248 *, 249; Ch. krieshoffi $48 *
Chlorohydra 172; Ch. viri-dissima 173
Chioroperla 253, 253*, 254*
Chloroperlidae 252 Chlorophyceae 66 * Choanomphalinae 199 Choanomphalus 199 Choroterpes 246; C. picteti 246 *, 247 ♦
Chromogaster 180; Ch. ova-lie 180*; Ch. testudo 180 ♦
Chrysomelidae 266, 267
Chydoridae 213, 222
Chydorus 64, 111, 121, 223; виды 226, 226 *
Cicuta virosa 157, 157 ♦
Cinclidotus minor 151
Cladocera 25, 62, 64, 111, 121, 208, 209, 212, 226
Cladopelma 309 •; C. vires-cens 304 *
Cladophora 66 ♦, 89
Clathrocystis 65 ♦
Clepsine, cm. Glossiphonla
Clinotanypi 319
Clinotanypus 319, 319 ♦
С1оёоп 146, 251; виды 83 ♦, 147, 251, 251 ♦
С1оёотпае 250
Closterium 67 •
Cnemidotus, см. Peltodites
Coelambus 261
Coelenterata 168
Coenagrion 85 ♦, 235 ♦, 237, 326 ♦, 328; виды 237 *
Coenagrionidae 236
Coleogeton 165
Coleoptera 233, 322
Collembola 233
Collotheca pelagica 177, 178 ♦
Collo thecidae 175, 177
Colymbetes 261, 268, 269 *;
C. fuscus 262 *
Colymbetini 261 ♦
Comarum 117
Conochilidae 175, 176
Conochiloides 176; виды 176*
Conochilus 176; виды 176, 176 ♦
Copelatus 268; C. ruficollis 268
Copepoda 25, 64, 70, 121, 208, 228 *
Cordulegaster 239 *, 324 •;
C. annulatus 239 *, 325 *
Cordulegastridae 324
Cordulegastrinae 238
Cordulia 88, 239; виды 325 ♦;
кладка 85 ♦
Corduliinae 239, 324
Corethra, см. Chaoborus, Mochionyx
Corixa 258; C. dentipes 259 *; кладка 86, 86 *
Gorixidae 258
Corophium 145, 146, 206, 206 ♦
Corynoneura 89, 314
Corynoneurariae 313 ♦
Cosmarium 66 * costalis, Ablabesmyia 321 Crenothrix 50
Cri co topus, cm. Eucricotopus
Criodrilus 188; C. lacuum 188
Crista tella mucedo 181, 181 ♦, 182 *
Crocothemis 240; C. ery-thraea 241 *
Crunaecia 126, 279
Crustacea 204, 205
Cryophila 294; C. lapponica 294
Cryptocerata 257
Cryptochironomus 145 303 ♦, 305; группы 304* 304 *, 306, 308, 310; ви^ ды 303, 304
Culcx 83, 123, 290 ♦, 294
Culicidae 287, 289 ’ 290 ♦ 293
Culicoides 75, 146 296
297 ♦ ’
Curcuiionidae 267 curticalcar, Ablabesmyia
Cyanophyceae 65 ♦
Cybister 268; C. laterali-marginalis 264 ♦
Cyclopidae 228
Cyclopoida 228, 229
Cyclops 121, 122, 123, 228, 228 ♦, 229; C. strenuus 68
Cylindrotomidae 288
Cymatia 258
Cymbiodita marginalia 271
Cyphon 266, 266 ♦
Cyrnus 277; C. flavidus 277 *
Cystobranchus 197; C. respirans 196 ♦
Cyzicus 211; C. tetracerus 211, 211 ♦
Daphnia 37 ♦, 64, 111, 215;
D. longispina 212; var. cristata, cucullata, hya-lina, longispina 216 ♦, 217 Daphnia magna 111, 215, 216 ♦
Daphnidae 213, 214
Dasyhelea 291 ♦, 296, 297 ♦
Decapods 205, 207, 208
Demejerea rufipes 308
Dero 184, 190 ♦; виды 190, 190 *
Desmidium 66 ♦
Deuterophlebia mirabilis 288, 288 •
Deuterophlebiidae 288 Deuterosminthurus 234 Diamesa 145, 318, 318 ♦; D. bathyphila 316 *, 317
Diamesini 108
Di apha nosoma brachyurum
214, 214 *
Diaptomus 122, 123, 228, 228 ♦, 229; виды 228
Diaschiza 178
Diatomeae 40, 50, 67 *, 69, 87, 89, 143
Dicosmaecus palatus 280
Dictyogenus 253; D. im-hofi 255 *
Didiamesa miriforceps 315, 316 *
Digononta 174
Dikerogammarus 145, 146, 206 ♦; D. haemobafes
207
Dinobryon 65, 68 *, 69
Diplectrona 283
Dip lex, см. Sympetrum
Diplodontidae 231
Diplodontus despiciens 231
D.
Diploglossa 274
Diptera 233, 287, 322 322 ♦
Discodnlidae, cm. Bran-chiobdellidae
Dissotrocha aculeata 175 Distyla, cm. Lecane Diurella stylata 180 Dixa 126 ♦, 294, 295 maculata 125
Dixinae 294, 295 ♦ Dolophilodes 276
Donacia 84, 87, 266, 272-яйца 87
Dorylaimus 147; D. stagna-lis 169 ♦
Dreissena 91, 144, 145, 146, 202P°lymOrPha
Drepanocladus 151; виды 151, 151 •
Drepanothrix dentata 220 *, 221
Drose i*a 117
Dryopidae 266, 267, 271
Dryops 266, 271
Dyscamptocladius 314, 315 ♦; группы 314
Dytiscidae 267, 268
Dytiscus 55 ♦, 83, 268, 269 », 270 ♦; D. latissimus 323 ♦; кладка 86 •
Ecdyonuridae, cm. Hepta-geniidae
Ecdyonurinae, см. Hepta-geniinae
Ecdyonurus 90, 145, 245;
E. fluminum 244, 245 ♦
Ecnominae 274
Ecnomus 147, 274; E. tenel-lus 274, 276 ♦
Ectocyclops 228
Einfeldia, rp. carbonaria 309
Einfeldia, larva paganus, cm. Tendipes rp. paganus
Eiseniella tetraedra 188
Elodea 50, 78, 80, 89, 147;
E. canadensis 163, 164 *
Elosa worallii 121 Empididae 292, 292 ♦ Enallagma 237; E. cyathi-gera 236 ♦, 237 ♦
Enchytraeidae 186, 187 Enchytraeus 186 ♦ Endochironomus; группы 305 ♦, 306, 307, 310, 310 ♦
Enochrus 271
Enteroplea lacustris 178
Enthomobryidae 234 Entomostraca 205, 208 Epeorus 245; E. torrentium 244 ♦, 245 *
Ephemera 144, 146,
E vulgata 146 ♦, 323
Ephemerella 249; E. ignita 83 ♦, 248 *
.Ephemercllidae 248 Ephemeroptera 83 , 146 ,
H233, 322, 322 •
Ephydatia 145, 172; виды 171 ♦
Ephydra 293
Ephydridae 293, 293 * Epytheca 88, 240; E. bima-
KUala85?°*-241-:^-
Epoicocladius 313, 313 ♦
Equisetum 88, 117, 147
152; виды 152, 152», 153
Eriophorum 120
Eristalis 123, 292 292 ♦
322, 322 ♦
Eristalomyia, cm. Eristalis Erythromma 86, 237 327-
E. najas 237 *
Es the ria, cm. Cyzicus
Eubria palustris 266
Euchlanis dilatata 179
179 •
Eucopepoda 208, 209 227 229
Eucricotopus 314, группы 314 ™
Eucyclops 228
Eukiefferiella, rp. longi-calcar 319
Euorthocladius 318
Eupachygaster 291 ♦
Euphaenocladis 314, 314 ♦,
315 ♦; E. rhitrogenae 314
Euplyllopoda 208, 209
Eupotamogeton 165
Eurycaenis 248, см. Brachy-cerus
Eurycercus 222; E. lamella tus 222, 222 ♦
Eutanytarsi 311, 312
Eutanytarsus 145
Eylais 230; виды 230 *
falcigera, Ablabesmyia 320
Filinia 64, 177; F. longiseta 64 ♦, 177
Flagella ta 68 •
flavida, Ablabesmyia 320
Floscularia ringens 177
Fontinalis 78, 79 •, 89, 90;
F. antipyretica 151
Fragilaria 67, 67 *, 68
Fredericella 146; F. sultana 181, 182 ♦
Fridericia 186 ♦
fulva, Ablabesmyia 320
Galerucella 84, 266, 272 Galionella 50 Gammaridae 207 Gammarus 91, 127, 146, 1*8, 206, 206 ♦; виды 125, 206, 207
Gastropoda 198
Gastropodidae 175, 180
Ga stropus sty lifer 180, 180
Gerrididae 86
Gerris 83 *, 147: !«ладна 86
Glaenocorisa cavifrons 258
Gloeocapsa 65 ♦ Gloeotrichia 65 Glossiphonia 27 ; 145^14 .
198; виды 195 iv» Glossiphoniidac 196 Glossoscolecide 186, 1»» Glossosoma 274 Glossosomatinae 27ч
Glyceria flultans 153, 153 *;
CL spectabilis 153, 153 ♦ Glyphotaelius 280; виды 281 ♦; чехлик 282 ♦ Glyptophysa 199 Glyptotendipes 145, 146, 309, 309 ♦; G. poiytomus 106, 108, 309 Gnathobdellae 196 Goera 90. 278; G. pilosa 278 ♦
Goerinae 278
Gomphinae 237, 238 ♦, 324 Gom phone ma 67 * Gomphus85 *, 91, 146, 148, 237; G. vulgatissimus 238 ♦, 326 *; яйца 85 ♦ Gonium 66, 66 * Gordius aquaticus 169 Gramineae 153 Grammotaulius 281; виды 281, 281 ♦, 282 *
Grap hod ere s 263; G. cine-reus 263 ♦
Graptoleberis 222; G. testu-dinaria 222, 222 * gregarius, Tanytarsus 312 guttipennis. Ablabesmyia 320
Gymnocerata 257 Gyrinidae 265, 267 Gyrinus 82, 266, 267; G.
natator 265 ♦
Habroleptoides 246; H. mo-desta 246 ♦. 247 ♦ Habrophlebia 246, 247 ♦;
H. fusca 146 ♦ Habrotrochidae 174 Haementeria 198, 198 * Haemonais waldvogeli 190 Haemonia 272 Haemopis 27, 27 ♦, 196;
H. sanguisuga 196 Haleeus282; виды и чехлик 282 ♦
Haliplidae 267, 270
Haliplus 270 ♦; H. lineatus 261 ♦
ha lophilus, см. Tri choc la-di us
Haplotaxidae 188 Harnischia 146, 303 *, 304;
H. fuscimanus 303 * 304 ♦
Harpacticoida 228, 229 Него us 257; виды 257 ♦ Heleidae 289, 296 Helicbus 266, 271 Helmis 127, 266, 266 * 271 Helobdella 145, 146 ’147 197, 197 ♦ ’
Helochares 265, 265 *, 271 Helophorus 263, 264 * ’ 270 270 ♦ *
Hemiclepsis 197; H. marginate 197 *
Hemiptera 83 •, 86 ♦ 233 322, 323 ♦
Heptagenia 90, 144, 145, 245; H. coerulans 245 ♦ Heptageniidae 245
Heptageniinae 245 Hermione 126
Herpobdella 27 •, 90, 145.
147, 195, 195 ♦ Heteromeyenia 171 ♦, 172;
H. bayleyi v. stepanowi 172
Heteroptera 322, 323 *
Hippurus vulgaris 80 *, 163, 164 ♦
Hirudinea 169, 195 Hirudinidae 196 Hirudo 195 ♦, 196 Holocentropus 125 *, 277 Holopedium 213; H. gib-berum 213, 214 *
Hottonia 78; H. palustris 162, 164 ♦
Hydaticus 264 *
Hydra 84, 172; виды 172, 173, 173 ♦
Hydracarina 86 Hydrachna 230, 230 ♦ Hydrachnellae 229, 230 Hydrachnidae 145, 146, 147, 230
Hydraena 127, 263, 264 ♦, 271
Hydride 168
Hvdrobia 201; H. steini '201 ♦
Hydrobius fuscipes 265, 265 ». 270, 270 *, 271
Hydrocampa, cm. Numphula Hydrocharis morsus ranae
♦. 84. 88, 89, 158, 159 * Hydrochus 271; H. elongates 264 ♦
Hydrometra 257 Hydrometridae 82 Hydromysa livens 89, 89 ♦ Hydrophilidae 263, 270 Hydrophilus 55, 83, 264,
265 * 270 *, 271; H. fuscipes 265
Hydroporini 261 Hydroporus 268, 269 ♦ Hydropsyche 127, 144, 146
146 ♦, 283; H. pelluci-dula 274 ♦; чехлик 125 ♦ Hydropsychidae 274, 283 Hydropsychinae 283 Hydroptila 145, 284 ♦; чех-
лик 125 ♦; H. Mac Lach-lani 284
Hydroptilidae 274, 284 Hydrous 264, 271; H. piceus
265 ♦
Hydrovatus 268; H. cuspi-datus 268
Hydrozoa 168
Hydryphantes 86 *
Hygrobia 267; H. tarda 260 ♦, 267 *
Hygrobiidae 260, 267 Hygrotus 261, 268 Hymenoptera 233, 323 Hyphydrus 268, 269 ♦ Hypnum 112, 117, 118,
Icbtyobdella 197 Ichtiobdellidae 196
Hiocryptus 220; виды 221 221 ♦
Ilybius 268; виды 262 * Ilyodrilus 146,194; виды 75
193, 193 ♦ inermipes 313 Insecta 169 Integripalpia 274 Intermedia 296 Iris pseudacorus 155, 156 ♦ Ischnura 237, 327; I. ele-gans 237 *
Isoetes 79*; I. lacustris 163 Isogenus 253; I. nubecula
254 ♦, 255 ♦ Isonychia 249 Isopoda 205, 207 Isopteryx 254, 254 ♦ Isotoma 234; I. viridis 234, 235 ♦
Isotomidae 234
Isotomurus 234; I. palustris 234, 235 ♦
Ithytrichia lamellaris 283, 284 *
Itonidae 287, 288 *
Ke rate Ila 64, 68, 69, 180; виды 25, 64, 64 ♦, 68, 180 kruglovicola (larva) 309, 309 ♦, 310 ♦
Kurzia 222; K. latissima 222, 222 •
Laccobius 265, 265 ♦, 271 Laccophilus 261. 268; виды 261 ♦
Lacinularia 177; L. floscu-losa 177; L. natans 177
Lamprodrilus 195 Lanceolaria 202 Lapporthocladius 317 ♦, 318 Lartetia 125 Lasioptera 288 ♦
Lathonura rec tiros tris 220, 220 ♦
Latona setifera 214 Lauterborniella 302, 313;
L. agrayloides 302 • — (brachylabis?) 303 ♦, 304 Lecane 180; L. luna 180, 180 ♦ Lecanidae 175, 180 Lemna 78, 84, 89, 159; виды 159, 159 *
len tiginosa, Abblabesmyia
321
Lepidoptera 233, 287, 322 Lepidostoma 279; L. hirtum
279 ♦
Lepidostoma tinae 278 Lepidurus 121, 122; L. pro-auctus 211, 211 ♦
Leptestheria, cm. Cyzicus. Leptetrum 240 Leptidae 292, 292 ♦ Leptoceridae 283, 284 * Leptocerus 145, 146, 283;
L. aterrimus, чехлик 285 Leptodora 62, 64, 111, 212, 213; L. kindti 213 Leptodoridae 213
Leptophlebia 246 ;L. vesnei*-tina 246 ♦ p
Leptophlebiidae 245
Lestes 86, 236, 236 ♦, 327-
L sponsa 327 ♦; кладка 85 ♦, 86
Lestinae 236
Leucorrhinia 240; L. rubi-cunda 241 *
Leuctra 254 ♦, 255 ♦ 256 256 ♦ ’
Leuctridae 256
Leydigia 223, 224; виды 224
Libellula 88; L. quadrimacula ta 326 ♦; кладка 85 * 86
Libellulidae 238, 238 *
Libellulinae 239, 324
Limnadiidae 209, 211
Limnaea 55 ♦, 57, 57 ♦, 83, 199; виды 199 ♦; кладка 84 *
Limnaeidae 199
Limnanthemum 89; L. nym-phaeoides 160
Limnebius 264, 265 *, 271
Limnesia 231
Limnesiidae 231
Limnetidae 209
Limnetis brachyura 209
Limnius 145, 266, 271
Limnobia 289 ♦
Limnobiidae, cM.Limoniidae
Limnochares aquatica 230t 230 ♦; кладка 86 ♦
Limnochironomus 309
Limnodrilus 91, 146, 148, 193, 193 ♦; виды 193 ♦, 194
Limnophilidae 278, 280,
281 *, 282 ♦
Limnophilinae 280
Limnophilus 282; виды 282 ♦; кладка 85 ♦
Limnosida frontosa 214
Limoniidae 289
Liriope 291 *
Liriopidae 290, 291 ♦
Lithoglyphus 201; L. nati-coides 202 ♦
Lubomirskiidae 170, 170 *
Lumbricidae 186, 188
Lumbriculidac 185, 188, 194
Lumbriculus 195; L. vario-gatus 185, 195
Lynceidae 209
Lynceus 122
Lynceus brachyurus 209
Lype 276
Lysimochia vulgaris 158, 158 *
Macrocyclops 228
Macronema radiatum 283
Macronematinae 283
Macropelopia 322
Macro thricidae 213, 220
Macrothrix 221
Malacostraca 205
Mansonia 294, 296 ♦
Margaritana 202; M. marga-ritifera 203 ♦
Marionina 121
Maruina 290 *
Megaloptera 233, 272
Melania 201; M. amurensis
Melanopsis 201; виды 201 ♦
Melosira 66, 67 •
Melusina 127, 144,149 291 *
Melusinidae 209
Menianthes 117
Mesocyclops 228
Mesomysis 207, 207 * Mesovelia 257*’, 258, 258 ♦ Mehdiaschiza trigona 178 Metamysis 207, 207 ♦
Metriocnemus 314
Micrasterias 66 *
Microchironomus conjun-gens 308, 309 ♦; M. lac-cophilus 306, 306 •
Microdon clavus 175
Microlenzia 314
Micronecta 258; M. minu-tissima 259 ♦
Micronympha, cm. Ischnura
Micropelopia, cm. Ablabesmyia
Micropelopiae 319
Micropsectra; виды 311 ♦, 313
Micropterna sequox 282 ♦
Microtendipes 301 ♦; 302
Microvelia 258; M. pygmaea 257 ♦
Midea orbiculata 231 minima, Ablabesmyia 320 Mochionyx 294 ; 295 ♦ Moina 215, 219; виды 219, 219 ♦
Molanna 91, 278; M. angu-stata 278 ♦; чехлик 278 ♦
Molannidae 278
Molannodes 278; M. zelleri 278 ♦
Mollusca 77, 88, 146, 169, 198; кладка 84 ♦
monilis, Ablabesmyia 321
Monodiamesa ba thyphila 315, 315 ♦; M. flabellata 315, 315 ♦, 316 *
Monogononta 174
Monommata orbis 178
Monospilus dispar 223, 223 ♦
Monostyla lunaris 180
Moraria 121
Muscidae 293
Myriophyllum 36 *, 78, 89, 147, 162; виды 164 ♦
Mysidacea 205, 207
Mysis oculata relic ta 206 , 207
Mystacides 285, виды 284 ♦, 285 ♦; чехлик 285 ♦
Mystrophora 274
Mytilina spinifera 179 ♦, 180
Naididae 183, 184, 185, 186, 187, 188
Nais 89, 91, 145, 146, 147;
виды 183 *, 191, 191
Najas 165
Nasturtium amphibium 158, 1 Do *
Naucoridae 259
Naucoris 86, 86 *, 88 259-N. cimicoides 259 ♦; кладка 86, 86 *
Nemathelminthes 169
Nematocera 288
Nematodes 145, 146, 169
Nematomorpha 169
Nemotelus 291 ♦
Nemura 256; N. marginata 254 ♦
Nemurella 256
Nemuridae 256
Nepa 54 ♦, 86, 87, 258; N. cinerea 258 ♦, 322, 322 ♦; кладка 86
Nephelis 86
Nephelopterix 255, 255 ♦
Nepidae 258
Neritidae 200
Neureclipsis 144, 145; N. bimaculata 144 ♦, 276 ♦, 277, 277 ♦, ловчая сеть 58 ♦, 145 ♦
Neuronia 285; виды 285, 286 ♦; чехлик 286 •
Ne гор to га 233, 272 nigropunctata, Ablabesmyia 320
Niphargus puteanus 125
Nitella 78, 79 ♦, 89
Nitrobacter 52
Nitrosomonas 52
Nostoc 65 ♦, 89
Noterus 261; N. crassicornis 261 ♦
Notholca 65, 180; N. longispina 25, 64 ♦, 180
Notibodia ciliaris, чехлик 278 ♦, 279 ♦, 280
Notommatidae 175, 178
Notonecta 54 *, 86, 259;
N. glauca 259 ♦ Notonectidae 259 nudipcnnis, Orthocladius
319
Nuphar 84, 88, 160; виды 160, 160 ♦, 161
Nymphaea 84, 88, 160; виды
160, 160 ♦
Nymphaeacea 160 nymphoides, Endochirono-mus 307
Nvmphula 84, 85, 86, 87, 287; N. nymphaea ta 288 •; кладка 85 •
obi cola larva 316, 316 *
Ochthebius 264 *, 271
Odonata 233, 235; кладка 85 *
Odontoceridae 280
Odontocerum 280; O. albi-corne 279 *,280
Oecetis 285; O. furva 284 *, чехлик 285 ♦
Oenanthe aquatica 157, 158 *
Oligochaeta 37, 75, 87, 121, 146, 169, 183
Oligoneuriella 245; О. rhena-na 245, 245 •
Oligoneuriidae 244
Ollgoneurisca 245
Onychogomphus 238; O. un-catus 238 ♦
Ophidonais serpentina 189, 189 *
Ophiogomphus 238; O. ser-pentinus 238 ♦
Ordella 145, 147, 247 ♦, 248, 248 *, 322 ♦
Orphnephila, cm. Thauma-lea
Orphnephilidae, cm. Thau-maleidae
Orthetrum 240; O. cancel-latum 241 ♦
Orthocladiinae 84, 144, 145, 298 ♦, 301, 313; O. g? larva altaicola 317, 317 ♦;
O. g? larva obicola 316, 316 ♦; O. g? larva sibi-ricola 317 ♦
Orthopodomyia 296, 297 *
Orthotrichia tetcnsii 283, 284 ♦
Oscillatoria 65 ♦
Osmylidae 272
Osmylus 272, 273 *; виды 272 ♦, 323 ♦
Ostracoda 122, 127, 208
Oxycera 126
Oxyetbira 283; чехлик 284 ♦
Oxygastra 239; 0. curtisi 241 *
Pachydrilus 186 ♦
Pachygaster 291 ♦ paganus, Tendipes 311 Palingenia 146, 244 Palingeniidae 244 Palpomyia 296
Paludicella 145, 146; P. articulata 181
Pandorina 66 *
Parachi ronomus 147, 310 310 ♦
Paracladopelma 145, 309; P. camptolabis 303 ♦, 304. 304 ♦
Paracricotopus 313 ♦ Paradixa 294, 295 ♦ Paraleptophlebia 246, виды 83 ♦, 247 *
Paramaecium 27 ♦ Parameletus 249 Paranais 91; виды 190 Paraponyx stratiotata 286 ♦.
287
Paratanytarsi 312
Paratanytarsus 312; P. lau-terbornii 311 •
Paratendipes 302, 306 pecteniphora, Ablabesmyia
Pedalia (Pedalion) 177; P. mira 176 ♦, 177; P. oxyura 177
Pediastrum 66 ♦, 67
Pedizia 126
Pelopia 321, 321 ♦; P.
kraatzi 321 *
Pelopiinac 319
Peloscolex velutinus 183 , 193; P. ferox 75, 193
Peltodytes 270, 270 * Pentapedilium 305 ♦. 306 Peracantha 224; P. truncata
223 ♦, 224
Pericoma 126,127,128,290 * Perla 55 ♦, 252, 252 ♦; виды
253 ♦, 254 *
Perlidae 252
Perlodes 253; P. dispar 253 ♦
Perlodidae 253
Peucedanum palustre 158
Phacoptcryx brevipennis 281; чехлик 282 ♦
Phalacrocera 289 ♦ Pharyngobdellae 196 Philhydrus 270 ♦; P. mela-nocephalus 265 ♦
Philodina 175; виды 175, 176 ♦
Philodinidae 174, 175
f Philopotamidae 274
Philopotamus 274; P. mon-tanus 275 ♦
Phragmites 88; Ph. communis 154, 154 *
Phreoryctidae 186, 188 Phryganea 147, 285, 286 ♦;
P. grandis 286 ♦; кладка 85 *
Phryganeidae 278, 285, 286; чехлик 286 ♦
Phyllopoda 208
Physa 84, 199; P. fontina* lis 199**
Physidae 199
Phytochironomus 309 Pinnularia viridis 67 ♦ Piona, кладка 86 ♦; P. car-nea 231 *
Pionidae 231
Piscicola 27 *,145, 147,148, 197; P. geometra 196 ♦
Pisidium 91, 145, 146, 148, 202; виды 203 ♦
Placobdella, cm. Haementeria
Planaria 54 ♦; P. gonoce-phala 168
Planorbinae 199
Planobris 147, 199; виды 200 ♦; кладки 84 ♦
Platambus 268, 269 * Plathelminthes 169 Platicnemis 236; P. penni-pes 236 ♦, 237 *
Plea 259
Plea Leachi 259 ♦ Plecqptera 233, 324 Plectrocnemia 125, 277; P.
conspersa 277 ♦ Pleurosigma 67 ♦ Pleosoma hudsoni 179, 179* Pleuroxus 224, 227; виды
227, 227 *
Plumatella 146, 147, 181, 182; виды 84, 84 *, 87, 182, 182 ♦
plumosus, Tendipes 74, 74 *, 310, 310 ♦
plumosus—reductus, Tendipes 110
Podura 83, 234; P. aquatica 234 *
Poduridae 234
Polyarthra trilga 64, 64*, 179
Polycentropidae 277 Polycentropus 145, 277; P. fla vomacula tus 277, 277 ♦; ловчая сеть 58
Polygonum 88; P. amphi-bium 160 ♦, 161
Polymitarcys virgo 146, 146 ♦, 242 ♦, 243 *, 244
Polypedilum 145,306,306 • Polyphemidae 213 Polyphemus pediculus 213, 214 ♦
Poly trichum 118, 118 * Pompholyx 176 *, 177 Porifera 168 Potamantidae 243 Potamantus 242 ♦, 243, 243 ♦; P. luteus 243
Potamodrilus 145 Potamogeton 36 ♦, 50, 78, 147, 161, 165; виды 89, 160 *, 161, 161 *, 165, 165 *, 166, 166 *, 167, 167 ♦
Prionocyphon 123
Pristicepnalus josefinae 210 ♦, 211
Pristina 121, 189; виды 189, 189 ♦
Probezzia 296 Prochironomus 308, 309 * Procladius 146, 148, 321;
P. serratus 321 ♦
Procloeon 145, 147, 251, 251 ♦
Prodiamesa praecox 315, 315 ♦
Proisotoma 234, 234 * Propappus 187; P. volki 145 Prosobranchia 198 Prosopistoma 243 *, 244 Prosopi stoma tidae 244 Protanypus morio, cm. Di-diamesa miriforceps
Protenthes 146, 321, 321 * Protoclepsis 198; P. maculosa 197 ♦
Protonemura 256; виды 255 ♦, 256 *
Psammoryctes 91
Psectrocladius 314; P. psi-lopterus 313 *, 314 ♦
Psectrotanypus 321, 321 ♦;
P. rp. varius 322 Pseudalona, см. Kurzia Pseudanodonta 202 Pseudochi ronomus 312, 312 *
Pseudodiaptomidae 228 Psilotanypus 321 Psychoda 290 ♦ Psychodidae 289, 290 ♦ Psychomyia 276, виды 276 *
Psychomyidae 274 276 Psycbomyinae 276 ’ Pterodina, cm. Testudinella Ptilocolepus 125 * 127 Ptychoptera cm. Liriope Pt^e°Pteridae’ CM> Liri°Pi-Pulmonata 198 Pyrrhosoma 236; P nvm.
phula 237 * У
Ranatra 87; R. linearis uZu *
Ranunculus 78; R. circin-natus 162, 163 *; R. Con-rervoides 162, 163 ♦; R. polyphyllus 159, 160 ♦
Rattulus, cm. Trichocerca reductus, Tendipes 310
Retinodiplosis 288 ♦ Rhagionidae 292, 292 * Rhantus 262, 268; R. punc-tatus 262 ♦
Rheorthocladius 317 ♦, 318 Rheotanytarsi 312 Rhe о tanytarsus 127, 145, 312; R. pentapoda; чехлик 312*; R. raptorius 311 ♦, 312 ♦
Rhitrogena 245; R. auran-tiaca 244 ♦, 245 ♦
Rhyacophila 127, 145, 274, 276; виды 274, 275 ♦, 276^ 276 *; чехлик 125 ♦ Rhyacophilidae 274, 275 ♦ 276
Rhyacophilinae 274, 276 Rhynchelmis 194; R. limo-sella 194
Rhynchobdella 195 ♦ Rhynchobdellae 196 Rhynchota 83 ♦, 233; кладка 86 ♦
RhynChotalona 145, 224; виды 224 ♦, 225, 225 ♦
Riciella fluitans 159
Ripistes 191; R. parasita 183 ♦
Rotaria 175
Rotatoria 64, 168, 173
Sagitta ria sagitti folia 157, 157 ♦
Salvinia natans 158, 159 ♦ Sayomyia, cm. Chaoborus Scapholeberis 121, 215, 217, 217 *; виды 217, 217 ♦
Scaridium longi cauda tum
178, 178 ♦
Scenedesmus 66 ♦
Schizocerca diversicornis64, 111, 179
Schizopoda 205 Sciomyzidae 293, 293 ♦ Scirpus 88, 147; S. lacustris 152, 152 ♦
Scolochloa 88; S. festucacea 154, 155 *
Scorpidium scorpioides 151, 151 •
Sepedon 293 Sorgentia 307, 307 *
279,
S,g?'.toS'. , num ’ 280’ S' Pedemonta-num, чехлик 278 ♦
Sericostoma tidae 278,
280
Sericostomatinae 280
Setodes 285
Sialidae 272
Sialis 81, 82 •, 87, 91 146-виды 82 ♦, 272 ♦, 324 ♦ ’
Side89>147, 213; S. crystalline 84, 213, 214 ♦
Sididae 213
Sigara 258
Silo 90, 127; чехлик 125*
Simocephalus 215, виды 215,
Simuliidae 290, 291 ♦
Simulium 90
Siphlonuridae 249
Siphlurella 250; S. linneana 249 ♦
Siphlonurus 250; S. lacustris 249 ♦
Sisyridae 273
Sisyra 273; S. fuscata 273
Sium la tifolium 158
Slavina appendiculata 191 191 ♦
Sminthurides 234; виды 234
Smittia ephemerae,cm. Epoi-cocladius
Somatochlora 239; S. metab lica 240 ♦
Sparganium 147, 155, 159; виды 156 ♦, 161, 161 ♦, 162, 162 ♦
Spercheus 264, 270 ♦, 271;
S. emarginatus 264 ♦, 271
Sperchon 231
Sperchonopsis 231
Sphaeriidae 202
Sphaerium 88, 146, 148, 202; виды 203 ♦
Sphagnum 118,*118 ♦, 119
Spirodela 84; S. polyrrhiza 159, 159 ♦
Spongilla 147, 170; виды 170 ♦, 171 ♦
Spongillidae 170
Stactobia 283
Staurastrum 66, 66 *
Stempellina bausei 311, 311 ♦, 312 ♦; чехлик
312 ♦
Stenochironomus 305
Stenophylax 282; виды 281 ♦, 282 ♦; чехлик 282 ♦
Stictochironomus 302, 307 * Stratiomyia 54 ♦, 83, 291 ♦ Stratiomyiidae 291 ♦, 292 Stratiotes 36 ♦, 50, 57 ♦; S. aloides 36 *, 163, 164
Streblocerus serricandatus 221, 221 *
Streptocephalus 210; виды 210, 210 ♦ •
Stvlodrilus 187, 194; S. he-ringianus 194, 194 *
Surirella 67 *
Sympetrum 240, 241 » кладка 85 ♦
Symnycna fusca 236, 236 ♦ ^ta и её вп«ьг 179,
Syndiamesa 317; виды 313 • о 317, 317 *, 318 ’
Synorthocladius 314 Syrphidae 292, 292 ♦
Tabanidae 292, 292 * Taeniopterygidae 255 Taeniaptoryx 254 • 255 Tanypi 319, 319 ♦ Tanypodinae 298 ♦ 300 300 ♦, 319 ’ ’
Tanypus 74
Tanytarsini (Tanytarsariae) 75, 107, 298 ♦, 301, 301 • 302 ♦, 311
Tanytarsus 74, 106, 312 ♦; группы 311, 311 *, 312
Tendipedidae 53, 57 ♦ 82 85, 86, 87, 89, 90, 91 ,’10з’ 108, 114, 121, 122, 127* 144, 145, 287, 290, 291 *
Tendipedinae Tendipedini, tus 302
Tendipes 37, 144, 146,
310 ♦;
300, 301 rp. Abbrevia-
74, 106, 123, 148, 291 ♦, группы 74 ♦, 75,’ 106, 108, 310, 311; виды 123, 148, 310
Testudinella 176 ♦, 177
Tetanocera 293 ♦
Tetramastix 177
Thaumalea 125, 291 ♦, 292
Theobaldia 294, 296 ♦, 297 ♦
Theodoxus 200; T. fluvia ti-lis 200 ♦
Thienemaniella 126, 127, 313 ♦, 314
Thoracostraca 205
Thurauia 288
Tinodes 276, 276 ♦
Tipula 289 ♦
Tipulidae 126, 287, 289, 289 ♦
Torleya 249; T. belgica 248 ♦
Triaenodes 285; T. bicolor, чехлик 285 ♦; кладка 85
Triarthra, см. Filinia
Trichocerca cylindrica 180
Trichoceridae 175, 180
Trichocladius coerulescens 317; T. inequalis 316, 316 ♦; группы 319
Trichoptera 88, 91, 125 ♦, 126, 233, 273, 273 ♦, 322, 322 ♦; кладка 85 ♦
Trichotanypus 321
Tricorhytus 247 ♦, 248
Triogma 289 ♦
Trochaeta 196
Trochospongilla 145; T. horrida 170 ♦, 171 *, 172
Tubifex 53, 54 ♦, 74, 74 % 145, 146, 193; виды 193, 194, 194 ♦
Tubificidae 184, 184 ♦, 185, 186, 187, 192
Turbellaria 84, 145, 169
Typha 88, 155: виды 155, 156 ♦, 157
Unio 127, 145, 146, 148, 202: виды 203 *
Unionidae 202
Uralia 311 uralicola 311, 311 ♦ Uranotaenia 296, 297 * Utricularia 36 *, 78: U. vulgaris 162, 163 ♦
Valvata 54 *, 91, 146, 200; виды 201 ♦
Valvatidae 200
Vejdowskiella 121, 190; виды 190, 190 ♦
Velia 258; V. currens 257 *
Vermes 168
Vormi formes 296, 297 ♦
Veronica anagalla 165; V. beccabunga 165, 165 ♦
vi tellinus, Dyscamptocla-dius 314
vitripennis, Trichocladius, 319
Viviparidae 200
Viviparus 145, 146, 147, 200; виды 200 ♦
Volvox 65, 66 *
Wormaldia 274
Xanthidium 66 ♦
Xenochironomus xenolabis 308
Zavrelia 311, 311 ♦; воды 312 ♦; чехлик 312 ♦
Zavreliella 302; Z. clava-ticrus 302 ♦
Zygoptcra 235, 236, 237 *, 324.
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
Азот 46, 51, 52
Азота кругооборот 52
Аквариум 97, 98 Аккумуляция 130, 132 Активная реакция среды
57, 59, 60, 108, 120, 138 Активное движение водных организмов 25
Алкалит^офные озёра 106
Аллювии 131
Аммиак 51
Аммиачные соли 51
Амфидиски 170, 171 ♦
Анаморфоз 232
Анаэробные условия 46 Ассимиляция 27, 50, 52 Атмосферные осадки 28
Базисная линия 16
Баланс водный в озере 28
--- реке 134
Барьер зарослей 88
Бассейн озёрный 61
— речной 142
Белки 51
Бентос 74, 76
Берег 130
Береговая линия 21
--- длина 21
--- развитие 21 — мель И
— область И
Береговой откос 10
Биогенные вещества 105, 108, 110
Биогенная декальцинация 50
Биологическая продуктивность 100, 101
Биологическое очищение 109
Биомасса 101, 102, 103, 104
Биотоп 61, 106, 126
Биоценоз 61
— болот 121
— временных водоёмов 122,
123 с
— глинистых участков 145
— зарослей 127, 146
— затонов 148
— — надводных 88
— — погружённых 89
— — с плавающими листьями 88
--- рдестов 89
— илов 146
- камней 90, 126, 145 ключей 124, 125
— коренной реки 145 — песков 145
— прибойной зоны 90 рек 141—147
— ручьёв 126, 127
— сублиторали 91
Биссус 144, 201
Болото 116—121
— атмосферного питания 118, 119
— лесное 120
— луговое (низовое, низинное) 116, 119, 120, 121 — моховое (верховое, возвышенное) 117, 119, 120
— осоковое 117
— сфагновое — тростниковое 120 — торфяное, см. моховое — травяное, см. луговое Брожение 47
Буссоль 15, 15 ♦
Вариации сезонные 69, 212
Величина озёр 9 .Верховье 128 Водная масса 22 Водоёмы 5
Водоёмы временные 122, 123
— грунтовые 5
— стоячие 5
— текучие 5
Водопад 5, 129
Водораздел 141
Водород 57, 58
Водохранилище 8
Воды и суши взаимодействие 5
Водяной пар 42
Воздушный колокол паука 55 *
Возрастные периоды озера
Волнение 29, 30, 89, 90
Волнообразное движение Волны 29, 30
Вскрытие рек 136
Выводковая сумка (камера)
Высота падения реки 128
Высыхание водоема, 1ч,
— организмов 12, 122
Вычерчивание плана водоёма 16, 17
Выщелачивание горных пород 8
Вязкость воды 23, 62
Газы 43
Газовый баланс 43
— режим озера 43, 69, 70
Газообмен 54
Галька 10
Гелокрен 124
Геммула 170, 171 ♦
Гемоглобин 74
Гербарий 93
Гидроксил 57, 58
Гидрохимический режим ПО
Гиполимнион 34
Гипофаринкс 299
Гиттия 73, 74, 106, 148 Глоточное кольцо 298 ♦
Глубина озера 10, 61 ----- максимальная 10 ----- средняя 10 Глубины измерение 19 Глубоководная (пелаги-
ческая) область 11
Гниение 47, 52, 73
Годовые колебания температуры — см. Температура — колебания.
Гравий Ю
Грунт 61, 75, 77, 78, 90, 123, 146
Грунтовые (подземные) воды 119, 125, 135
Гуминовые вещества 52 Гумусовые кислоты 52. 73, ‘74, 120
Гусеницеобразные личинки ‘273
Давление атмосферное 42, 43 77
Движение в воде 20—./ — ВОДЫ 90 — волнообразное 29 — животных активное 2о
27, 63, 64 •
----- пассивное 62 -----реактивное 26 Двууглекислая закись же-
леза 50
344
— известь (кальций) 48, 89
Декальцинация биогенная 50
Дельта 132
Детрит 12, 39, 40, 44,46, 52,
56, 87, 106, 108, 139
Диоптры 15
Диск Секки 41
Диссоциация 57, 58
Дистрофные озёра 105, 106
Диффузия газов 44
Длина озера 9
Днепр 8, 135, 141
Дно 61, 73
Дночерпатель 98
Дождевые потоки 47, 123
Долина речная 131
Донная фауна — см. Бентос
Донные отложения 73, 74
Драга 76
Драгирование 76
Дыхальца 54
Дыхание водных животных 53, 56
---растений 53
— подо льдом 5, 55, 55 ♦
Дыхания водный тип 53, 56
— воздушный тип 54
Дыхательная полость 54
— трубка 288, 294
Дю (дью) 73, 74, 106 •
Жабры 53, 54, 56
Жабры анальные 252, 299
— коксальные 252
— плевральные 252
— простернальные 252
— трахейные 55, 56
Железа соединения 50
Жёсткая вода 48, 49
Жиры 51
Заболачивание 118, 119
Заводь 126, 127
Загрязнение водоёма 52, 107, 138,139
Закон минимума 49
— совокупного действия факторов 49
Закраина 77
Замерзание озёр 35
— рек 136
Замор рыбы 46, 139
Запруда 8
Зарастание озера 108, 116
Заселение новообраэовав-шегося водоёма 12
Засуха 130
Затон 147
Зимняя спячка 74
Зимовка животных 1, 54 77, 89
— растений 36, 37, 89
Зимовка рыбы 114
Зимующие почки 36 * 37
122, 170, 171 * ’
Зообентос 100
Зоопланктон 63, 64, 65 69 70, 143, 144
Известковые соли, см. соли кальция
Ш
Известняк 48
Известь, см. Кальция окись и Кальций углекислый
Излучение тепла 32
Изобаты 20
Икра 55 ♦
Ил И, 71, 74, 75, 108
Ила органические остатки 73
Индикаторы 107, 109
Инкрустации 50, 89
Ионы 58
Испарение 28
Исток реки 128
Источник 5
Кальций двууглекислый 48
— углекислый 4 9
Кальция окись 48
— соли 49
Камни 90
Камподеоидные личинки 273
Кислород 46, 78, 81, 88, 89, 108, 111, 124, 137
Кислорода вертикальное распределение 44, 44 ♦
Кислородная слоистость 105
Кислородный баланс 61 — режим 45, 126 — скачок 44, 44 ♦, 45
Кислоты гумусовые 73
Кладки 84 *—86 *
Классификация озёр 104
Ключ 124—126
Кокон 87, 286 ♦
Количественный учёт водных организмов 98, 99, 100
Коловратный аппарат 173, 174
Колориметрия 59
Компас 15, 127
Конвекционные токи 32, 33
Конечная продукция 99
Консервирование 97
Консументы 60, 81
Конус накопления (наносный) 132
Кремнёвая кислота 50
Кремнезём 50
Круговорот веществ 53
Круговые волны 29
Лаутерборновы органы 299,
Лёд 35, 89
Лёд донный 136
Ледник 8, 128
Ледниковые озёра 8
Ледостав 136
Ледоход 35
Лес, влияние на водоём 118
Лимнокрен 124
Лимнопланктон 142
Линь 19
Литоральная область 11 61, 77—91
Ловчая сеть (ручейников)
Лопаточка прыгательная 287, 288 *
Лофофор 181
Лужа 5, 122
Лужи весенние 122, 123
— дождевые 123
— на пнях 122 — пойменные 148 Лучеиспускание 32
Макросклеры, см. Спикулы
Малярия 92
Маска стрекоз 232, 235
Мастакс 173
Меандры 133, 134
Межень 130, 135
Мели 133
Мель береговая И Мельничный газ 71 Металимнион 34, 50 Метаморфоз 231 Метан (оолотный гав) 47 Миграции вертикальные и горизонтальные планктона 69, 70
Микросклеры, см. Спикулы
Минеральные вещества 111, 138, 141
Минирующие животные 57
Моренные озёра 8
Морены боковые 8 — донные 8 — конечные 8 Мутта 117, 117 • Мягкая вода 48
Надглоточник 299
Накопление, см. аккумуляция
Нанос 132
Наросты 87
Насыщение воды газами 43
Нектон 63
Неорганические вещества— см. Минеральные вещества
Низовье рек 128
Нимфа 232
Нитраты 52
Нитриты 52
Ножка листовидная 209 — расщеплённая 205
Обмеление озера 13, 14 — реки 135
Обмен газов — см. Газообмен
Обросты 87
Обь 8, 131, 139, 141
Объём водной массы 21
Озеро алкалитрофное 106 — аргиллотрофное 106 — ацидотрофное 106 — балтийского типа 44, 59, 105, 106
— бентос 74
— бессточное 28
— величина 9—10
— водная масса 22
— горное 44
— гумусовое 46, 59 Ю5 106 ’ ’
— дистрофного типа 105 106, 108
— замерзание 35 запрудное (плотинное) 8
— искусственное 8
— испарение 28
— классификация 7—9 104, 105
колебание температурное 3 2
— конечное 28
круговорот веществ биологический 53
^ледниковое (моренное) 8,
— методика 15—22
— олиготрофного типа 105, 106, 108
— остаточное 8
— плотинное 7
— плотность воды 22
— пойменное (полойное) 148
— прибой 30
— провальное 8
— происхождение 7—9
— проточное 28
— равнинное 44
— реликтовое (остаточное) 8
— солёное 8
— сточное 28
— субальпийского типа 44,
Озёрная руда 6
Озёрное ложе 7, 13
Озёрный бассейн 61
Окисляемость 138
Окраска воды 40
Олигосапробы 107
Олиготрофные водоёмы 105, 106
Определение положения пункта па водоёме 18
— прозрачности 41
Оптимум температурный 36
Оптические свойства воды 39—41
Организмы: мезосапробные 107
— олигоса пробные 107
— полисапробные 107
— стеноионные 60
— эвриионные 60
Органические вещества 51, 108, 111, 138, 141
Органы передвижения водных животных 26—29
Органы плавания 62
Остаточный вес 24
Откос береговой 10
— подводный И
Отложепия донные в озёрах 73
— — в реках 73, 74, 129
Паводки 135, 137, 138, 139, 140
Палетка 21
Пальпы 229
Паралабиальные пластин-ки 299
Партеногенетический спо-соб размножения 212
Парциальное давление 43 Пассивное движение 62 F/В коэфициент 104 Пелагическая область (пе-лагиаль) И, 61, 62
Пена 30
Перегной 80
Передвижение животных 64 Перекаты 133 Переработка планктона в реке 142
Перемещение русла реки
Пересыпь 8
Пересыщение воды кислородом 43, 44
Песок 10, 90
Питание водных животных 56, 57
Пища 99
Пищевые цепи 100 Плавание 26, 62 Плавательные пузырьки 25 — усики 62
Плавательный пузырь рыб 26 План водоёма 15—18 — окружающей водоём местности 18
Планиметр 21
Планктическая сетка 71 71 ♦, 98
Планктический сачок 95, 127
Планктон 40, 52, 63—72, 96, 142
— вертикальное распределение 69
— горизонтальное распределение 69
— разложение 45
— речной 142
— сезонные изменения 68 Плотность воды 32, 33 — льда 35 Площадь водоёма 21 — зарослей 93
— озера 9, 10 Поверхностная плёнка 54, 82, 83
Поглощение света водой 39 — цветов спектра 40 Подводный откос И Подталкиватель 299 Пойма 130, 139, 140, 141 Пойменные водоёмы 140 Полисапробы 107 Половодье 132, 135 Постабдомен 212 Потамопланктон 142 Потребители органического вещества 60
Почки зимующие 37, см. Зимующие почки
Превращение озера в пруд 13, 14
Прибой 30
Прибрежная область см. Литоральная область
•Прибрежная раститель-ность 78—91
— фауна 81—91
Придаточная система реки
Придаточные водоёмы 148
Прикрепление организмов в воде 30
Притоки 47
Приход воды в озере 28
— воды в реке 134
— солен 47, 48
— тепла в озере 31
Прицепки ручейников274 — 277
Продуктивность 99—101 104
— биологическая 100, 101
Продукция 101—104
~ конечная 99
— первичная 100
— промежуточная 100
Продуценты 60, 81, 104
Продуцирование J01
Прозрачность воды 39, 41
108, 136
— измерение 41
Производители органического вещества — см. Продуценты
Происхождение озёр 7
Промерзание 77
Проток 147
Профиль дпа 20
Профундальная область (профундаль) 11, 61, 75
Пруд ПО—115
— вредители рыб 114
— дикий 110
— естественный ПО—ИЗ
— загрязнение 112
— искусственный 110, ИЗ
—114
— колхозный 111, 112, 115
— мельничный 112
— образование 13
— превращение в болото 14
— рыбоводный 113, 115
— фабрично-заводской 112
Прыгательная лопаточка 287, 288
Прямолинейные волны 29 pH (активная реакция) 57— 60
Пупок в раковине моллюсков 198
Равнинные озёра 44
Развитие береговой линии 21 w
Разлив весенний 135
Разложение 51
— гнилостное 52
Размножение 102
— вегетативное растении 79
Размывание берега 10, 134
— русла 129
Ракушник 91
Распределение водной растительности в озере 13, 40
— водных животных в зарослях 81
Растворение воздуха в воде 43 — твёрдых веществ 41 Растворимость газов 41—43 Растения водные 30—31, 78—81, 150—167 — надводные 79, 152 — подводные (погружённые) 78, 89, 162 — с плавающими листьями 78, 79, 158
Расход воды в озере 28 — — в реке 135 — тепла в озере 32 Реактивное движение 56 Регенерация 185 Река 128—149 — асимметрия берегов 134 — высота падения 128 — главная (коренная) 141 — горная 128, 138, 139 — равнинная 128, 129,143 Реликты Б Рельеф дна 19 Реокрены 124
Речная система 141
Речной бассейн 141
Родник 5 Рострум 208 Руда озёрная 6 Рукав 147 Руление 24 Рулетка 15 Русло реки 130, 131 — уклон 128, 129 Ручей 126
Рыбоводный пруд ИЗ, 115
Самоочищение водоёма 109, 112
Сапробность 107 Сапропелит 117 * Сапропель 6, 73, 74, 75, 81, 106
Сачок 94
— воздушный 96 — планктический 95 Сбор животных 95, 96 — растений 93, 94 Свет 39, 40 — поглощение 39 Седло 294
Сёдлышко, см. Эфиппий Сезонные вариации 69, 212 — изменения бентоса 37 — — планктона 37 — — среды 68 — — прибрежной флоры и фауны 93
Сера 50, 51
Серные источники 51 Сероводород 47, 51 Сила сцепления воды 23 Сифон^личинок двукрылых Скачок кислородный 44.
44 ♦. 45, 45 * — температурный 34, 35 ♦ 44, 45, 49
Скорость течения в реке 129, 135, 140, 143
Скребок 94, 95, 127
Слоистость кислородная 105 — тепловая обратная 32, 33, 33 ♦
--- прямая 32
Снежный покров 36, 40
Солевой режим озера 47
Солёные озера 8
Сопротивление воды движению 25
— трения 24
— формы 24
Спикулы губок 170, 170 *, 171 ♦
Сплавина 14, 117, 117 *
Споры 12, 37
Способы передвижения водных животных 63, 64
Спячка зимняя 74
Средний уклон русла 128
Стагнация 32, 33, 50
Старица 134, 147
Статобласты 181, 182 ♦
Стеноионные организмы 60
Стигмы 287
Сток 5, 28
Стоячие воды или водоёмы 125
Стержень 144
Сублиторальная область (сублитораль) И, 61, 91, 92
Суточные колебания температуры см. Температура, суточные колебания
Съёмка плана водоёма 15, 18
Тальвег 130
Тектонические долины 131
Текучие воды 5
Тельсон 205
Температура воды 42—47, 44 *, 45 ♦
— вертикальное распределение 43, 44 ♦, 45 ♦, 135 — влияние на организмы
35, 36, 69
— измерение 38, 149
— колебания периодические 32
— — сезонные 68, 69, 77
— — суточные 70, 77
— максимальная 36, 68
— оптимальная 36, 68
Температурные пределы существования организмов 36
Температурный режим 77 — скачок, см. Скачок температурный
Тепловая слоистость, см. слоистость тепловая
Тепловой баланс 32
Тепловые свойства воды 31
Теплоёмкость воды 31
Теплопроводность воды 31 — льда 35
Теплопрозрачность воды 37
Тёрка улитки 57, 57 *
Термометр 38
Террасы речные 130
Теснина 130
Течения 29
Тина 84
Типы озёр 105, 106
Тление 80
Токи конвекционные 32, 33, 49
Торф 81, 119
— сапропелитовый 117 ♦
Трахеи 54, 55
Трофические процессы 105
Турионы 36 ♦
Углеводы 54
Углекислая известь — см.
Кальций углекислый Углекислота 137
Углекислый газ 46, 48, 55, 56, 137
— — распределение вертикальное 46 ♦
Углекислые соли 48
Углерод 51
Угольник 17, 17 ♦
Удельная поверхность 25
Умирание озера 14
Уровень воды 10, 135 ♦ Устье 128
Устьица 56
Фабричные стоки 51
Фауны количественный учет 99
Физико-химические свойства озера 22
Фиксирование 97
Фильтраторы 56, 212
Фитопланктон 63, 65, 70, 137, 143
Фосфор 51 Фотосинтез 137 Фронтальный склерит 293
Хелицеры 229
Химизм озёрной воды 41 — ключевой 124
— речной воды 137
X имические свойства воды 41
Хоботок 256
Хозяйственное значение
внутренних водоёмов 6
Цвет воды 40
Цветение воды 40, 45, 68, 69, 111
Цикл жизни озера 10
Цикломорфоз 69
Циркуляция воды 29, 33, 33 •, 44
— — весенняя 33 — — осенняя 33 — — полная 29 — — частичная 29 Циста 122
Чехлики (домики) ниц бабочек 286 ’ 288 *
.— ручейников 275, 282 ♦, 284 ♦, 285 «
— хирономид 312 ♦
Ширина озера 9
Шлем (дафний) 69
гусе- Щёлочность 58 ‘, 287,
278 ♦, Эвриионные организмы
, 286 ♦ 60
Эвтрофный тип озёр 105, 108
Эпилимнион 34, 50
Эпимеры 229
Эпиплевры 260
Эфиппий 37, 37 ♦, 212 Эфир 96
Яйца 12, 31, 37, 45 •, 55 ♦, 66 •, 81, 82, 82 ♦, 84 ♦, 85 ♦, 86, 86 ♦, 92, 96, 100, 122, 145, 229, 229 *, 286 ♦ — зимние 212 — летние 212 — покоящиеся 122, 212 Яйцевые мешки 228, 228 *
ОГЛАВЛЕНИЕ
Предисловие................................................................ 3
Введение................................................................... 6
Глава первая.
ОЗЕРО.
I. Физико-географическая характеристика.
Озёрное ложе ........................................................ 7
Что мы называем озером............................................. 7
Классификация озёр по происхождению................................ 7
Величина озёр...................................................... 9
Цикл жизни и возрастные периоды озера.............................. Ю
Темы для работ................................................. 14
Водная масса озера, главнейшие физико-хймические свойства озера.... 22
Свойства воды как жидкого тела.................................... 22
Тепловые свойства ................................................ 31
Темы для работ................................................. 38
Оптические свойства воды.......................................... 39
Темы для работ............................................... 41
Химические свойства воды......................................... 41
П. Качественная биологическая характеристика. Биотопы и биоценозы.
Пелагическая область ................................................. 62
Водная масса как среда............................................. 62
Планктон .......................................................... 63
Темы для работ.................................................. 70
Условия жизни на дне................................................ 73
Бентос............................................................. 74
Темы для работ.................................................. 75
Литоральная область. Общая характеристика............................. 77
Участки литорали, покрытые растительностью......................... 78
Сублитораль........................................................... 91
Темы для работ.................................................. 91
III. Значение количественного учёта населения озера.
Понятие о продуктивности. Классификация озёр..................
Глава вторая.
ПРУД.
Естественные пруды............................................. И®
Искусственные пруды............................................ И*
Темы для работ............................................ И®
Глава третья.
БОЛОТО.
Темы для работ ... ...................................... 121
Глава четвёртая.
ВРЕМЕННЫЕ ВОДОЕМЫ.
Характеристика................................................ 122
Темы для работ.......................................... 123
Глава пятая.
КЛЮЧ И РУЧЕЙ
Характеристика ключа............................................... 124
Характеристика ручья............................................... 126
Темы для работ............................................... 127
Глава шестая.
РЕКА.
Физико-географическая характеристика реки ......................... 128
Биологическая характеристика реки.................................. 141
Физико-географическая характеристика придаточной системы реки...... 147
Биологическая характеристика придаточных водоёмов реки............. 148
Темы для работ............................................... 149
Таблицы для определения пресноводных растений и животных.
Водные растения.................................................... 150
Подводные и листостебельные мхи................................. 151
Надводные растения.............................................. 152
Растения с плавающими листьями.................................. 158
Погружённые растения............................................ 162
Главнейшие группы водных беспозвоночных животных................... 168
Пресноводные губки — Spongillidae............................... 170
Гидры — Hydri dae.............................................. 172
Коловратки*— Rotatoria......................................... 173
Мшанки — Bryozoa. . ............................................ 181
Малощетинковые черви — Oligohaeta............................... 183
Пиявки — Hirundinea ............................................ 195
Мягкотелые — МоПизса............................................ 198
Членистоногие — Arthropoda...................................... 202
Ракообразные — Crustacea........................................ 204
Высшие ракообразные — Malacostraca ............................. 205
Низшие ракообразные — Entomostraca.............................. 208
Пресноводные клещи — Hydrachnellae ............................. 229
Насекомые — Insecta............................................. 231
Вилохвостки (ногохвостки) — Collembola........................ 234
Личинки стрекоз — Odonata..................................... 235
Личинки подёнок — Ephemeroptera larvae........................ 240
Личинки веснянок — Plecoptera larvae............................ 252
Клопы и их личинки — Hemiptera (Rhynchota)...................... 256
Жуки взрослые —Coleoptera imago................................. 259
Личинки* жуков — Coleoptera larvae................................
Личинки большекрылых — Megaloptera larvae.........................
Личинки настоящих сетчатокрылых — Neuroptera larvae...............
Личинки ручейников — Trichoptera larvae...........................
Гусеницы бабочек — Lepidoptera larvae.............................
Личинки двукрылых — Diptera larvae................................
Воздушные насекомые, личинки которых живут в воде.................
Взрослые стрекозы —- Odonata imago................................
Список определителей пресноводных организмов на русском языке............
Указатель русских названий организмов....................................
Указатель латинских названий организмов..................................
Предметный указатель, ...................................................
328
331
333
341
Редактор И. А. Пал амар чу к.
Техн, редакторы М.И. Натапов и Н. Н. Махова.
Подписано к печати 16/V 1950 г. A-0I56I. Булата 70х10в*/хв. Бумажн. л. 10,88. Печати, листов 29,80. лчётно-изд. листов 35,07.
Заказ Лв 469. Цена без переплета 10 р. 30 к. Переплет 75 к.
Отпечатано в типографии Т-13 с матриц 2-й типографии •Печатный Двор* им. А. М. Горького Главполиграфиздата при Совете Министров СССР. Ленинград.