/




































































































































































































































































































































































































Автор: Ливанов А.Н.
Теги: животный мир зоология морфология многоклеточные животные пути эволюции
Год: 1955




Текст
Л.Л.ЛиВаноВ
ПУТИ
эволюции
! ' животного
! МИРА
Н. А. Л ИВ АНО В
ПУТИ ЭВОЛЮЦИИ
ЖИВОТНОГО МИРА
АНАЛИЗ ОРГАНИЗАЦИИ
ГЛАВНЕЙШИХ ТИПОВ МНОГО
КЛЕТОЧНЫХ ЖИВОТНЫХ
ГОСУДАРСТВЕННОЕ ИЗДАТЕЛЬСТВО
«СОВЕТСКАЯ НАУКА»
Москва — 1955
ПРЕДИСЛОВИЕ
Ч. Дарвин говорил в «Происхождении видов», что наука о
строении животных (морфология) представляет собой один из
самых интересных отделов естествознания. Но сам творец дарви-
низма в эту область мало углублялся. В морфологию вдохнули
жизнь исследования замечательных русских ученых 60—70-х годов
прошлого века, впитавших в себя передовые философские идеи ре-
волюционеров-демократов того времени. Эмбриологические работы
А. О. Ковалевского и И. И. Мечникова, палеонтологические —
В. О. Ковалевского легли в основу эволюционной морфологии. Сам
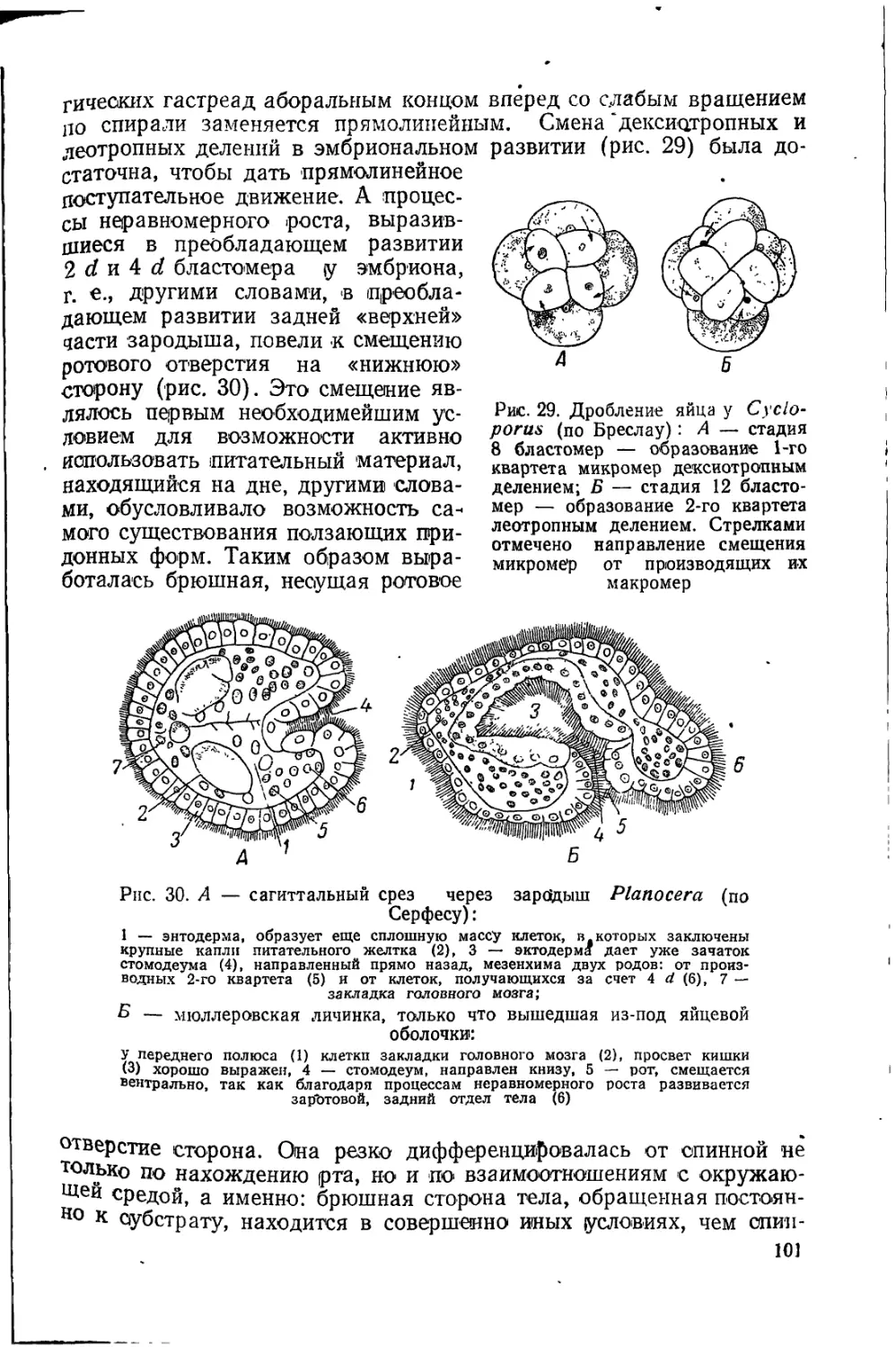
Ч. Дарвин высоко ценил эти работы, и они сделались образцом для
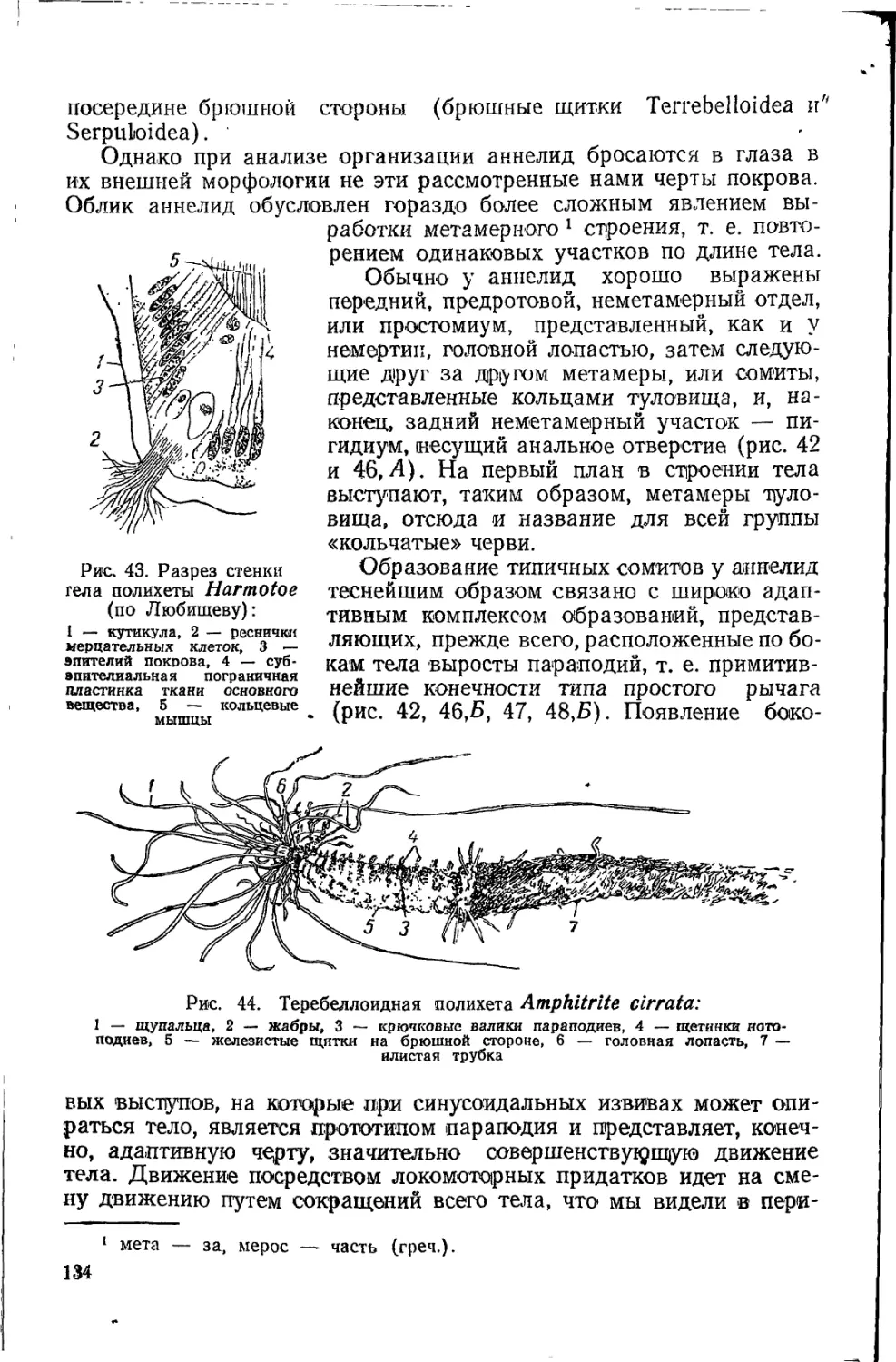
большинства зоологов 60-х и 70-х годов. Застывшие формы Кювье
перестали быть предметом научной морфологии.
Появившаяся в 1866 г. «Общая морфология» Э. Геккеля, разви-
вавшая материалистическое ядро дарвинизма, но носившая отпеча-
ток немецкой натурфилософии первой половины XIX века, сначала
оказала сравнительно мало влияния на зоологов; лишь позднее
формальные и схематические построения в духе Геккеля распростра-
нились среди морфологов. Однако в России эволюционная морфо-
логия сохраняла традиции своих основателей. Наряду с критикой
Э. Геккеля она развивала свои методологические положения и
углубляла фактический материал. Наши известные дарвинисты
К. А. Тимирязев, М. А. Мензбир, В. М. Шимкевич ставили требо-
вание детального исследования и безупречности фактов в проти-
вовес уйвлЙчению схемами геккелианства.
Еще в 60-х годах И. Сеченов выдвинул положение 9 един-
стве организма и среды.4 В 80-х годах мой учитель Э. А. Мейер
блестяще показал в своих работах необходимость для зоолога
изучать организацию животных в неразрывной связи с условиями
существования. Его исследования по седентарным аннелидам были
Широко известны морфологам беспозвоночных. Методологически
они были прогрессивны.
Мы не будем здесь останавливаться на ряде морфологических
работ, появившихся у нас в начале XX века и продолжавших преж-
ние пути. Тон науке в то время стали давать искания уже в других
3
областях биологии, морфология отошла на второй план. Нарастав-
ший кризис буржуазного естествознания в эпоху империализма от-
ражался в шатании научной мысли и в разноголосом хаосе гипотез,
теорий, мнений. Под давлением реакции биология часто теряла
твердую материалистическую основу и скатывалась к идеализму.
Лишь некоторые работы, относящиеся уже к советскому периоду,
дали возможность внести необходимую ясность в накопленные био-
логией факты и позволили правильно поставить широкие проблемы
благодаря тому, что в нашей стране постепенно, в борьбе с бур-
жуазными течениями вырабатывался необходимый единый фило-
софский подход к вопросам теории и практики, подход'с единствен-
но- научных позиций диалектического_материализма.
Вместе с этим снова на первый~план выдвигается эволюционная
морфология — входящие в ее состав анатомия, эмбриология, ги-
стология.
Крутой поворот был сделан в области гистологии. Она разви-
валась в конце XIX и в начале XX веков в направлении детальней-
шего изучения тонкостей структур. Эволюционный подход к их по-
ниманию почти отсутствовал, преобладали тенденции отказа от
возможности уложить эволюцию тканей в рамки общей эволюции
животных организмов. В 20-х годах формулируются идеалистиче-
ские положения А. А. Заварзина (1925) о параллелизме структур и
их "расщеплении, как особых основных закономерностях тканевой
эволюции. Только путем длительных перестроек своих положений
А. А. Заварзин в дальнейшем более или менее близко подошел к
материалистической эволюционной гистологии.
А. Н. Северцову принадлежит бесспорная заслуга продвижения
вперед другой области морфологии, а именно эмбриологии. Его
работы и работы его сотрудников дали богатейший фактический
материал по эмбриологии и сравнительной анатомии позвоночных,
что позволило А. Н. Северцову (1927, 1931, 1939) развить свою тео-
рию филэмбриогенеза и вступить на новый путь изучения самого
хода эволюции и его закономерностей. Он дает своеобразное мор-
фо-биологическое истолкование хода эволюционного процесса, хотя
и далеко не удовлетворительное в философско-теоретическом отно-
шении.
Однако и в области эмбриологии беспозвоночных у нас накапли-
вается материал, позволяющий подойти к материалистическому
решению важнейших вопросов индивидуального развития
(П. П. Иванов, А. А. Захваткин, Г. А. Шмидт).
В 1944^г. появляется книга В. Н. Беклемишева «Основы сравни-
тельной анатомии беспозвоночных». Она посвящена преимуществен-
но проблеме «планов строения» — архитектонике животных орга-
низмов, совершенно особому отделу эволюционной морфологии.
«Без такого архитектонического раздела из нашего рассмотрения
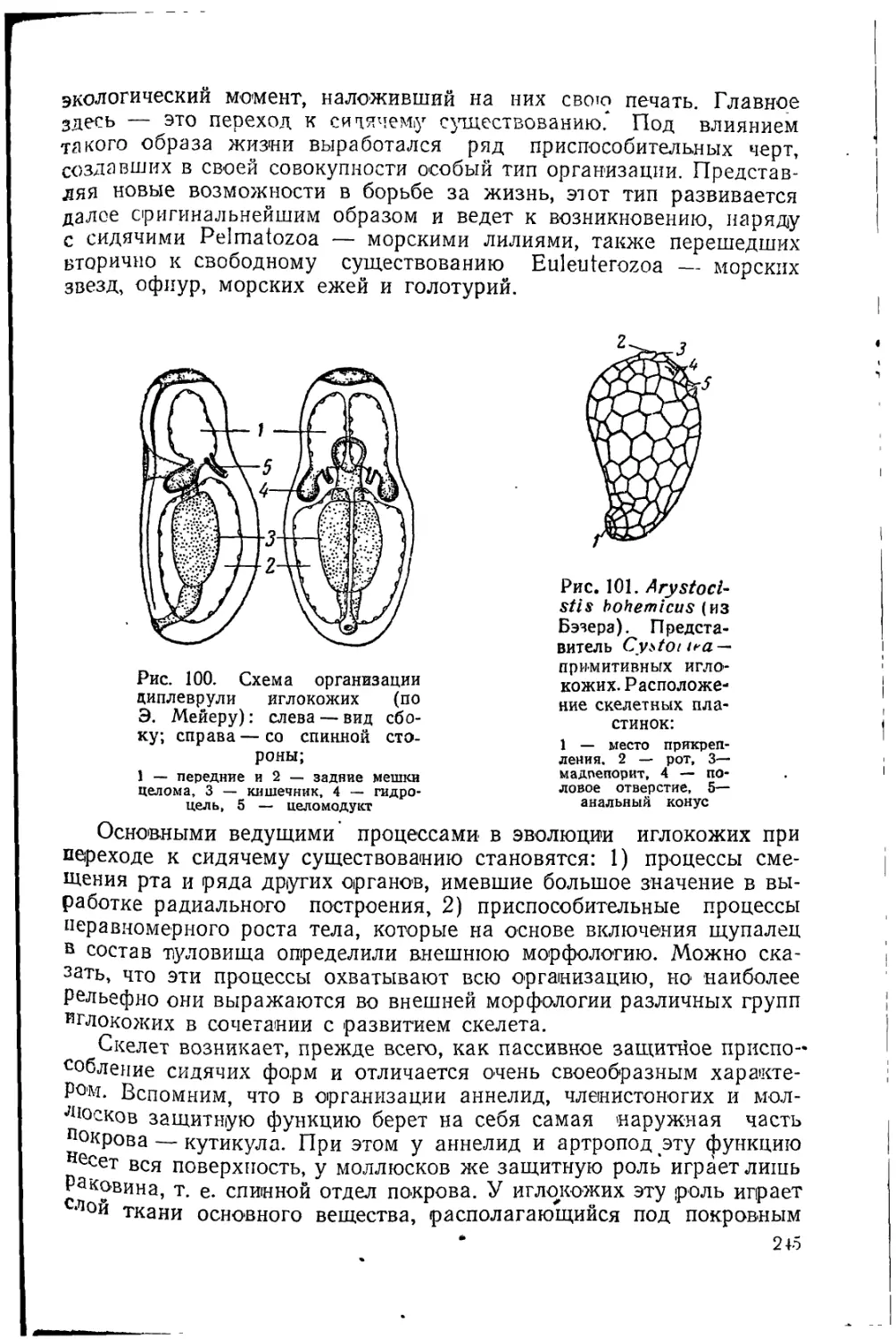
выпадает организм как целое и вся его эволюция», говорит
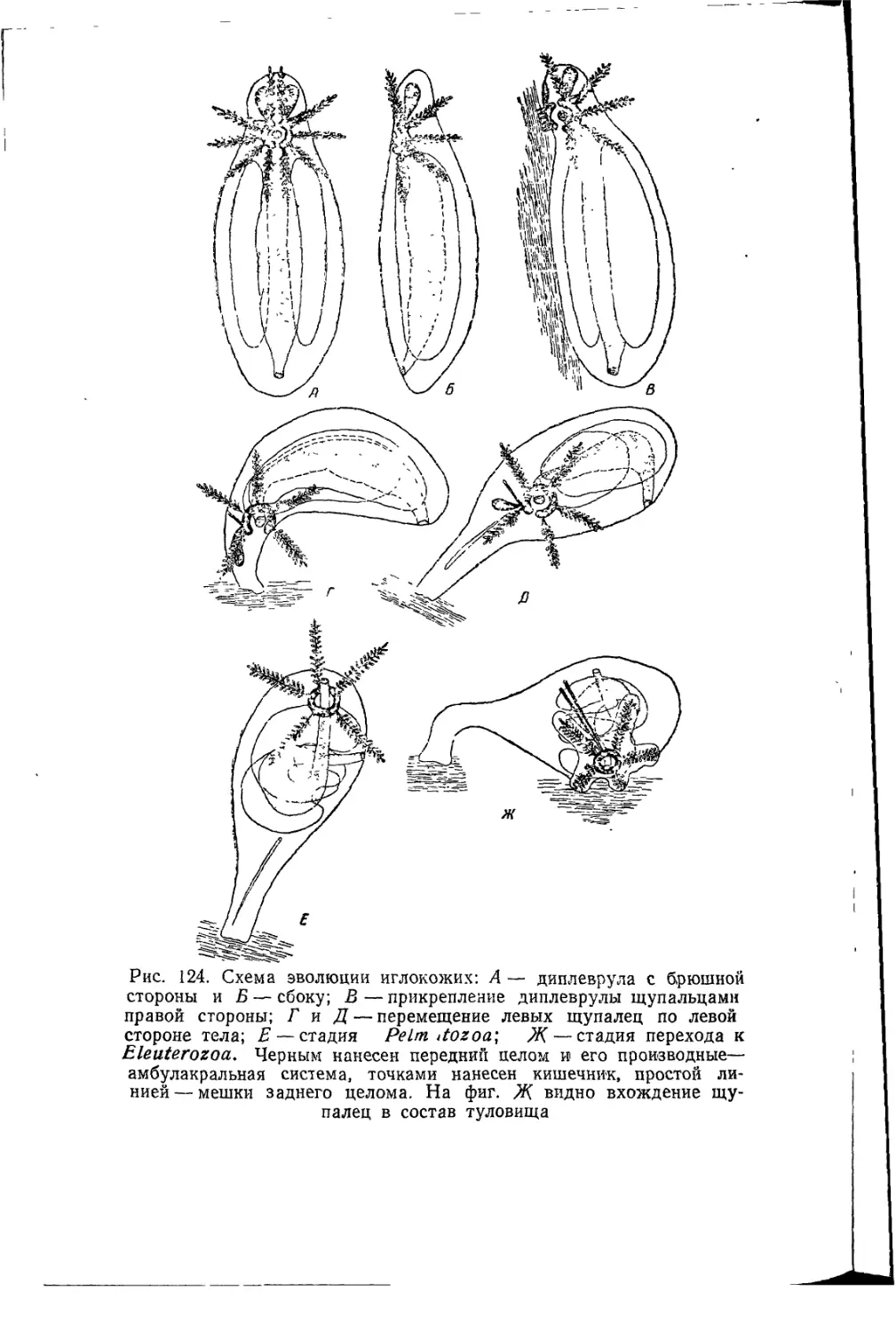
В. Н. Беклемишев, формулируя свои устремления (1944 и 1952,
стр. 3). В книге В. Н. Беклемишева основной предмет общей мор-
4
фологии — «целостный организм» — вполне правильно выдвигает'
ся на первый план.
Этому же пониманию целостной организации больших групп
животного царства посвящена была моя книга «Пути эволюции жи-
вотного мира», появившаяся в 1946 г. Однако подход к решению
поставленных задач был совершенно другой, чем.у В. Н. Бекле-
мишева. Давался анализ всех сторон организации данной группы,
выявлялись их связи друг с другом и, наконец, их ведущие комп-
лексы, которые легли в основу возникновения того1 или иного
тина, — через анализ к синтезу целостной организации, но всегда
на базе единства организма и среды, решающей роли условий су-
ществования в развитии организмов. Особое внимание уделялось
мною при этом биоценотическим отношениям, выявлялось их веду-
щее значение в развитии данного типа животного царства. Я исхо-
диА’при этом из положений, одинаковых с положениями И. П. Пав-
лова, сформулированными еще в 1904 г., о том, что животный
организги представляет собой очень сложную систему, состоящую из
почти бесконечного ряда частей, связанных как друг с другом, так
и с окружающей природой.
Такой подход естественно должен был возникнуть в нашей
науке. В 1913 г. С. А. Зернов дал картину биоценозов Черного
моря у Севастополя и тем самым привлек общее внимание к подоб-
ным исследованиям. В 20-х годах изучение биоценозов становится
основным методом работы наших экологов. Вместе с этим разра-
батывались теоретические положения биоценологии, и обойти это
мощное течение нашей научной мысли эволюционная морфология
не могла; напротив, она черпала в нем глубокий импульс для
своего дальнейшего1 развития. На этой базе мы и строили наш ана-
лиз организации типов — он исходит из биоценотических установок
о единстве организма и среды. Биоцентическим исследованиям
в науке принадлежит большое будущее.
Огромное, решающее значение для эволюционно-морфологиче-
ских исследований и построений имеет в наше время советский
творческий дарвинизм. Он позволяет, наконец, построить эволю-
ционную морфологию на основе точного, последовательно материа-
листического понимания «природы» организма.
Обобщая, мы можем сказать — эволюционная морфология ста-
ла наукой, в прогрессе которой русская мысль всегда идет впереди.
В борьбе двух идеологий — прогрессивной социалистической й
реакционной буржуазной — эволюционной морфологии принадле-
жит большое место. Она должна дать научное понимание организа-
ции животных и знание закономерностей эволюционного процесса,
что необходимо при выработке цельного материалистического ми-
ровоззрения; это необходимо и для преобразования природы в ин-
тересах общества.
Дух истинной науки — постоянное движение вперед. Остановка,
оцепенение научной мысли подобно смерти. «Общепризнано, —
говорит И. В. Сталин, — что никакая наука не может развиваться
а
и преуспевать без борьбы мнений, без свободы критики»1. На осно-
ве единственно правильной философии диалектического материа-
лизма должны подвергаться принципиальной научной критике и
имеющиеся в области морфологии «общепринятые» догмы, за фа-
садом которых в наших старых научных течениях скрывается много
нерешенных вопросов первостепенной важности. Уже назрела необ-
ходимость и в творческом научном обсуждении вопросов эволюцион-
ной морфологии в связи с воззрениями А. Н. Северцова, А. А. За-
варзина и др. Теперь приходится встречаться с неоправданным
усложнением в решении задач эволюционной морфологии, как мы
это видим у С. Г. Крыжатовского. Подобные явления подчеркивают
необходимость свободного обмена мнениями по вопросам морфоло-
гии, что мы и попытаемся сделать в меру наших возможностей.
Предлагаемая вниманию читателей книга предполагает знаком-
ство их с курсом зоологии, но содержит в себе и элементарные
сведения, которые необходимы для уяснения и развития идей авто-
ра. Она не претендует на сколько-нибудь исчерпывающий охват
фактического и литературного материала. Ее задача — дать по воз-
можности экологическое понимание организации главнейших типов
животных.
Для выпуклости, резкости картины различные группы проанали-
зированы неравномерно: то лишь в общих, грубых штрихах, то
более детально-. Несистематично, на первый взгляд, используется и
материал: то приводятся в качестве иллюстрирующих примеров
обширные систематические группы, то, напротив, мелкие конкретные
единицы системы. Однако- всем этим преследуется одна цель —
сделать картину определенной организации насколько возможно
дающей целостное впечатление, конкретной, хотя в основе и лежит
ее анализ, ее расчленение.
Естественным результатом и завершением изучения последова-
тельного ряда типовых групп является, в конце концов, картина
эволюции животного мира. Вместе с тем исправляется и уточняется
формулировка тех закономерностей, которые в свое время пытался
дать А. Н. Северцов.
Ряд зоологов, принадлежащих к иным научным направлениям,
чем я и мои учителя — проф. М. М. Усов и Э. А. Мейер, уже ука-
зывал на несогласие с некоторыми моими положениями. Я пред-
вижу немало возражений такого рода и после выхода книги в
свет. Хотелосьоднако, предупредить, что констатация есте-
ственных при современном уровне науки разногласий между раз-
личными морфологическими школами не может выдаваться за кри-
тику. В таких случаях происходит борьба мнений, критика же долж-
на указывать на прямые недостатки исследований и мышления.
Так, я не сочту за критику, если мне скажут, что спиральное дроб-
ление не связано с выработкой прямолинейного движения, или что
не согласны с «теорией жидкой ткани» в вопросе о происхождении
1 И. В. Сталин, Марксизм и вопросы языкознания, 1952, стр. 31.
6
кровеносной системы, с пониманием целома как «миоцеля» и т. д.
Возможно иное понимание олигомерности целома у иглокожих, иное
толкование происхождения и эволюции хордовых и т. п. Но не сле-
дует каждый раз при расхождении взглядов упрекать автора, что
он стоит на других позициях, а тем более обвинять в незнании или
игнорировании фактов, литературы, взглядов других исследовате-
лей, тем более не следует, что, несмотря на все достижения, наша
морфология не находится еще на таком уровне, чтобы какая-либо
школа могла претендовать на абсолютную роль, вычеркнув факты,
теории, гипотезы других направлений морфологической мысли.
Следует сказать, что я вполне приму упреки в тяжелом стиле
языка во многих местах моей книги — за перегрузку фраз, за
обилие терминов, наконец за повторения. Однако* в этом приходит-
ся винить не только автора, но и громоздкость терминологии, обще-
принятой в морфологических работах, а отчасти и сложность вопро-
сов, затрагиваемых в книге. Насколько мог, я стремился упростить
язык изложения.
Я и впредь буду признателен за всякую деловую критику, ибо
без нее, как и без самокритики, невозможно движение вперед в
строительстве нашей науки и нашей жизни. В этом отношении со-
ветским морфологам предстоит сделать немало дружных творче-
ских усилий. Хотелось бы особо подчеркнуть, что существование
различных научных направлений и разногласий между ними не
только законно, не только не причинит вреда нашим целям, но спо-
собно обогатить наши искания и ускорить процесс создания единой
общепризнанной системы взглядов.
С благодарностью отмечаю, что указания и советы моих това-
рищей и учеников (проф. Б. С. Матвеева, проф. В. В. Изосимова,
проф. М. В. Маркова, проф. С. В. Жданова и др.) много помогли
мне в оформлении моих взглядов, а при подготовке книги в печать
в высокой степени ценны были замечания моих рецензентов проф.
Л. А. Зенкевича, проф. Д. М. Федотова, В. М. Каганова и взявше-
го на себя редактирование книги А. А. Передельского,
ВВЕДЕНИЕ
• Неорганическая природа, ступени ее эволюции. Живое вещество
в эволюции материи, его закономерности. Одноклеточные и много-
клеточные организмы, их характерные особенности. Колонии
и популяции, биоценозы.
В вечном круговороте, в котором движется материя, говорит
Энгельс^ «каждая конечная форма существования материи — без-
различно, солнце или туманность, отдельное животное или жи-
вотный вид... — одинаково п?реходяща и... ничто не вечно, кроме
вечно изменяющейся, вечно движущейся материи и законов ее дви-
жения и изменения» К Наука изучает различные формы существова-
ния, движения материи, ряд различных ступеней ее развития. «Но-
вая атомистика, — отмечает Энгельс, — отличается от всех прежних
тем, что она... не утверждает, будто материя только дискретна, а
признает, что дискретные части различных ступеней (атомы эфира,
химические атомы, массы, небесные тела) являются различными
узло_выми точками, которые обусловливают различные качественные
формы существования всеобщей материи...»1 2.
«Элементарными частицами» материи, насколько смогли в на-
стоящее время проникнуть вглубь ее строения физика и химия,
являются отрицательно заряженные электроны, положительно за-
ряженные протоны, беззарядные нейтроны, мезоны и т. д. Йх массы
ничтожно малы, а их энергии, относительно, чрезвычайно велики.
Они составляют космическую среду.
При некоторых условиях движение элементарных частиц ведет
к образованию неопределенных скоплений, потоков их или дает
комплексы, в которых они закономерно связаны, т. е. системы эле-
ментарных частиц. В последнем случае получается дальнейшая
ступень эволюции материи — атом. Довольно полно зная свойства
элементарных частиц, мы не в состоянии непосредственно из них
вывести качества атомов. Системы элементарных частиц обнаружи-
вают новые, специфические черты, присущие атому того или иного
элемента — водорода, гелия, кислорода и т. д., но совершенно от-
1 Ф. Энгельс, Диалектика природы, 1946. стр. 20.
2 Там же, стр. 238.
8
сутствующие у элементарных частиц. Так количественные изменения
с образованием системы закономерно приводят ж образованию но-
вой ступени организации, к коренным качественным изменениям,
которые наступают скачкообразно.
Напротив, проявления нового качества при скоплении тех же
электронов, при образовании их потоков, выявляются и накопляют-
ся постепенно, нарастая от минимальных проявлений к таким, где
новое качество действительно становится полноценным. Поток
электронов дает электрический ток; полностью проявляются электро-
магнитные свойства поля.
Эволюция материи при образовании атома выражается и в зна-
чительном увеличении числа качественно отличных друг от друга
систем элементарных частиц, т. е. атомов различных элементов (по
сравнению с количеством качественно различных элементарных
частиц), достигая порядка сотни.
Вместе с появлением атомов начинает существовать новая
среда — среда атомов, которую мы находим в космических туман-
ностях и в раскаленных звездах, являющихся юными солнцами все-
ленной. Те закономерности, которые выявляются в мире элементар-
ных частиц, сохраняют для последних все свое значение и в атом-
ном мире, но опосредствуются: ведущими будут уже новые, спе-
цифические закономерности, которым подчиняются атомы. Законы
атомного мира будут свои, а законы мира элементарных частиц —
свои, но они всегда взаимосвязаны. Само существование атома, как
качественно определенной единицы, обусловливается этими ©посред-
ственными закономерностями. Согласно им, выбивается из системы
один из электронов — и атом становится уже качественно другим,
прежняя система изменяется, заменяется новой или разрушается.
Система существует постольку, поскольку сохраняются ее составные
части и их всесторонние отношения. Это — условие существования
всякой системы.
Атомная среда дифференцируется в связи с различием свойств
разных атомов (их массы и т. д.) pi в связи с различием энергети-
ческих состояний атомной среды — отсюда солнечная атмосфера,
внутренняя часть солнца и т. п.
При дальнейших изменениях в некоторых условиях среды эволю-
ция материи ведет к появлению закономерных комплексов атомов,
систем из атомов. Получается новая, еще более высокая ступень
в организации материи — молекула !.
Зная прекрасно^ все качества атомов водорода и атомов кисло-
рода, мы, однако, не можем непосредственно из них вывести те
свойства, которые присущи относительно простой системе из двух
атомов водорода и одного атома кислорода, т. е. воде, имеющей
огромнейшее значение в жизни нашей планеты. Опять количествен-
ные изменения при образовании системы молекулы приводят к ка-
11 Мы сознательно, для резкости картины, опускаем здесь некоторые субмо-
лекулярные стпуктуры, вроде атомных решеток и т. и., которые не меняют об-
Щего положения, но в рамки молекулярной структуры не укладываются.
9
чественным изменениям, к возникновению нового качества, причем
происходит увеличение и числа качественно отличных друг от
друга молекул по сравнению с числом качественно различных ато-'
мов — увеличение огромное.
Закономерности мира атомов сохраняют для последних все свое
значение и в мире молекул, но характеризуется этот мир уже свои-
ми особыми, новыми законами, они являются ведущими, закономер-
ности атомные ими опосредствуются.
Среда молекул свойственна стареющим звездам и в соответ-
ствии с различными свойствами молекул и различными энергети-
ческими условиями в этой среде происходит дифференцировка мо-
лекулярных масс, их распределение в различных участках среды.
Дальнейшим шагом в эволюции материи являются гомогенные
системы, состоящие из молекул, — многие кристаллы, мицеллы
и т. д. с их совершенно- новыми свойствами, выражающимися, на-
пример, в правильном ограничении кристаллов определенным обра-
зом расположенными плоскостями. При этом- выявляются свои
особые закономерности, изучаемые наукой о строении кристаллизо-
ванного вещества.
Вместе с тем мы доходим в эволюции материи до появления
гетерогенных систем характера коллоидно-дисперсных и им подоб-
ных. Большие поверхности раздела их фаз характеризуются воз-
никновением особых проявлений энергии, так называемой энергии
поверхности. Ранее имевшиеся дисперсные системы, большинство
молекулярных и все атомные, характера «истинных» растворов, не
обладали еще этим свойством. Получаются системы с огромным
разнообразием, с особыми закономерностями, раскрываемыми физи-
ческой химией и являющимися ведущими при опосредствованном
состоянии закономерностей молекул, атомов и элементарных частиц
микромира. Полностью сохраняя свое значение для соответствую-
щих систем, эти последние закономерности становятся подчинен-
ными, неспецифичными для рассматриваемой нами ступени орга-
низации в ходе эволюции материи.
Дифференцировка в среде коллоидно-дисперсных систем ведет к
тому разнообразию, которое мы видим, например, в атмосфере,
водной оболочке, породах земной коры и т. д., ведет к тем процес-
сам, которые разыгрываются, изменяя лик и недра Земли.
Итак, в эволюции материи от ничтожного количества качествен-
но различных элементарных частиц, почти беспредельно устойчивых,
путь шел к атомам, далее к молекулам, затем к кристаллам и кол-
лоидно-дисперсным системам. Последние чрезвычайно мало устой-
чивы, но представлены огромнейшим количеством качественно раз-
личных форм. Этот космический процесс завершается в следующей
ступени организации — комплексных коллоидно-дисперсных систе-
мах, переходя в бесконечное количество качественно различных
систем.
Как предполагает А. И. Опарин, они представляют собой коацер-
ватные системы, образующиеся из соединения простых коллоидных
10
растворов, резко обособляющиеся, индивидуализирующиеся от
окружающей среды, бесконечно разнообразные по своему составу и
соответственно в каждом отдельном случае индивидуально различ-
ные в качественном отношении. Такая индивидуальная различность
комплексных коллоидно-дисперсных систем взаимосвязана с их пол-
ной лабильностью, неустойчивостью, так как при всяком изменении
среды изменяется и динамическое состояние системы, а вместе с
ним и ее состав. Конечно, неустойчивые системы должны были
появляться и появлялись на любой ступени организации. Мы знаем
их еще в мире атомов, как радиоактивные элементы, в мире моле-
кул — как непредельные соединения и т. д., Но, появляясь, все
такие системы не развивались далее, а вследствие своей неустойчи-
вости постоянно, более или менее скоро или более или менее мед-
ленно, кончали свое эфемерное существование.
Совершенно особое место среди лабильных коллоидно-дисперс-
ных систем занимают белковые гидрофильные системы, отличаю-
щиеся чрезвычайной сложностью своих молекул. Появление этой
единственной в своем роде категории белковых систем является в
эволюции материи отправным пунктом для возможности нового
цикла все большего и большего усложнения. Первый цикл от эле-
ментарных частиц до комплексных коллоидно-дисперсных систем —
это цикл неорганической, мертвой природы, начинающийся немно-
гими качественно различными элементарными частицами и закан-
чивающийся бесконечностью качественно различных комплексных
коллоидных систем. Второй цикл, начинающийся относительно не-
многочисленными качественно различными гидрофильными белко-
выми системами, есть цикл развития живой природы.
«Жизнь есть способ существования белковых тел, и этот способ,
существования состоит по своему существу в постоянном самооб-
новлении химических составных частей этих тел» !. Другими слова-
ми, «жизнь — это способ существования белковых тел, существен-
ным моментом которото является постоянный обмен веществ с окру-
жающей их внешней природой, причем с прекращением этого обме-
на веществ прекращается и жизнь»2. Так совершенно точно
охарактеризовал жизнь Энгельс. Она начинается индивидуализацией
комплексной коллоидно-дисперсной системы гидрофильных белков,
причем появляется резкое отграничение особи и возникает целая
серия сорбционных явлений на образовавшейся отделяющей поверх-
ности, которые ведут к проникновению веществ извне внутрь
системы и обратно. Благодаря очень большой сложности своих
компонентов, и прежде всего белковых молекул с ферментативными
свойствами, такая система, частично' изменяясь в одних частях,
в то же время частично восстанавливается в других — в ответ
на изменения внешней цреды. Таким образом возникает сложный
ток веществ, позволяющий неопределенно долгое время существо-
з п Э н г е л ь с, Анти-Дюринг, 1948, стр. 77.
ин же, Диалектика природы, стр. 246.
11
вать системе, которая иначе должна была бы разрушиться, по-
гибнуть.
При некоторых условиях в такой комплексной белковой коллоид-
ной системе могут преобладать процессы распада, и вся система
будет постепенно деградировать. При других условиях, а именно —
при наличии в системе соединений, богатых запасами энергии, на-
против, будут преобладать процессы накопления вещества, и си-
стемы будут продолжать существование, увеличивать до известных
пределов свою массу — расти и затем, в силу диспропорции по-
верхности и массы, делиться. Возникающие таким образом про-
цессы ассимиляции и диссимиляции и процессы, связанные с ними,
являются характерной чертой жизни. Вместе с этим выявляется и
определенная ориентировка вещества, образуется некоторая струк-
турность системы.
Дальнейшая судьба живой системы зависит как от условий
внешней среды, в которой она существует, так и от внутренней
организации, т. е. от известной структуры и процессов, в ней про-
текающих, причем определяющую роль играют условия внешней
среды, становящиеся условиями существования системы. При ла-
бильности системы и при способности ее к размножению, это ведет
в ряде поколений к возможности ее постепенных изменений под
воздействием внешней среды в сторону освоения условий этой
среды, соответственно в сторону все большего соответствия усло-
виям существования и в сторону дальнейшей внутренней диффе-
ренцировки. Системы, не подходящие к условиям среды, разруша-
ются; продолжают свой жизненный путь только соответственно
изменяющиеся. В них определенная организация продолжается,
наследуется и в то же время закономерно изменяется, так как свой-
ства, признаки, приобретаемые системой в течение индивидуальной
жизни под влиянием окружающих условий существования, также
являются наследственными. Таким образом, выявляется противоре-
чивое единство наследственности и изменчивости.
Возникает качественно новое состояние вещества, которое, начи-
наясь немногими подобными системами, различно эволюционирует
дальше в направлении образования многочисленнейших новых,
более сложных систем живой природы, в которых проявляется
сложнейшая эволюция материи от протестов до организма человека
включительно.
Характерный для живой материи исторический процесс развития
по существу своему является необратимым. Если даже получается
при определенных условиях состояние системы, сходное с одним из
пройденных этапов, все равно оно получается в результате изме-
нений, непрерывно идущих все дальше и дальше от отправного
пункта. Попятного движения времени нет и быть не может. Этот
поступательный характер эволюции и выявляется так называемым за-
коном Долло — законом необратимости эволюции. Получающийся
при определенных условиях результат, в некоторых случаях как
будто сходный с одним из пройденных этапов, есть лишь конвер-
12
гентное сходство, совпадение, обусловленное сходством условий, в
которых оказывается в тот или иной момент существования данная
система, но- ее исторический путь, ее эволюционное развитие, ее
«природа» будут уже иными, а потому и сходство обычно является
лишь внешним совпадением.
Необратимый процесс эволюции гидрофильных белковых кол-
лоидных комплексов приводит к доклеточному состоянию живого
вещества., а затем к появлению тёх низших форм живой материй,
в которых мы теперь ее знаем как самостоятельно живущие орга-
низмы — к бактериям, к клеткам. При этом путь эволюции отмечен
бесконечной массой погибших на нем лабильных систем, просу-
ществовавших тот или иной промежуток времени, но в конце кон-
цов разрушившихся под влиянием воздействия изменяющейся
среды. Сохраняются лишь системы, наиболее соответствующие
условиям среды, осваивающие их, превращающие их в необходи-
мые условия существования, системы все более сложные и гибкие,
ставшие монерой, бактерией и, наконец, типичной клеткой.
Возникла клеточная ступень организации материи с ее особыми
качественными чертами. Ей присущи не только индивидуализация,
процессы ассимиляции, диссимиляции и в связи с ними рост и
размножение. Но в клетке на основе обмена веществ и вытекаю-
щих отсюда энергетических состояний живого вещества развива-
ются раздражимость, возбудимость и движение протоплазмы, на
основе своеобразных энзимных процессов — пищеварение, секре-
ция и т. д., а также, что особенно важно, процесс, регулирую-
щий поддержание и в то же время перестройку определенной орга-
низации и способствующий повышению жизненности, при котором
две клетки взаимно ассимилируют друг друга — половой процесс.
Связанная со всем этим дифференцировка в пределах клеточной
ступени организации ведет к появлению того, что хорошо известно
как типичная клетка с ее протоплазмой, ядром, органоидами, сокра-
тительными и тому подобными волокнами, вакуумом, различного
рода оболочками и т. д.
Вместе с появлением живых организмов, точнее — первоначаль-
ных доклеточных форм жизни, появляется образуемая ими биоти-
ческая среда; она далее в большей и большей мере усложняется с
развитием клеток, а затем и многоклеточных организмов.
В конце концов исторически слагающиеся в биотической среде
отношения между организмами, животными и растительными, ве-
дут к дифференцировке их специфических группировок — ценозов.
Биоценоз мы определяем как исторически развившийся закономер-
нътй комплекс, т. е. систему животных и растительных форм, свя-
занных общей средой обитания, общим биотопом.
Такие системы эволюционируют вместе с прогрессирующим
освоением различных сторон биотической и абиотической среды и
соответственно все более и более усложняющейся организацией
входящих в них форм.
Закономерности, являющиеся ведущими для рассматриваемой
13
нами ступени организации и для организма вообще, определяются
теми противоречивыми отношениями, во власти которых находится
организм. Это, с одной стороны, противоречивое единство организма
и окружающей среды в системе биоценоза, с другой — единство
частей в системе целостного организма.
Первая закономерность была дана Ч. Дарвином в принципе
естественного отбора, т. е. «выживания наиболее приспособленного
в борьбе за существование».
Однако Дарвин употребляет понятие «борьбы за существование»
в чрезмерно широком смысле взаимоотношений со всеми факторами
внешнего мира, т. е. и с неорганической природой, и с биотической
средой.
Расчленение понятия борьбы за существование позволяет глуб-
же его проанализировать, заглянуть в его сущность. Подобные
попытки делались рядом ученых, и такой анализ, показывая каче-
ственное различие разных категорий «борьбы за существование»,
позволяет глубже понять и оценить значение качественно различ-
ных форм естественного отбора.
Как мы видели, в неорганической природе полную силу имеет то
положение, что, во-первых, существование любой молекулы, любой
дисперсной системы и т. п. возможно' лишь при определенных физи-
ко-химических условиях, сохраняющих соотносительное постоянство
ее компонентов: при определенных температурах, определенном дав-
лении, определенном химизме среды и т. д. Вне их данная молекула
или дисперсная система претерпевает различные изменения, веду-
щие к ее разрушению как самостоятельной системы, или переходит
в качественно иную систему. Во-вторых, для всякой такой системы
ее существование возможно только при условии существования и
сохранения систем низшего порядка, входящих в нее компонентами.
Для живых организмов, т. е. в данном случае для клетки,
физико-химические условия среды играют такую же роль. При
температуре выше 40° протоплазма обычно свертывается, и клетка
гибнет. При воздействии различных химических веществ, например
алкоголя, сулемы и т. д. получается тот же эффект. Не будем
стремиться перечислять все эти условия, скажем в общей форме:
жизнь возможна только в свойственных ей определенных рамках
физико-химических взаимодействий с окружающей неорганической
природой. Далее, для клетки как и для живого вещества, не имею-
щего клеточной структуры, это связано с сохранением ее клоллоид-
но-дисперсных, молекулярных и, наконец, атомных систем. Специ-
фична относительная узость границ возможности существования
вследствие того, что клетка — это система, главным образом опре-
деляемая лабильными гидрофильными белковыми телами, их свой-
ствами, возникшая в определенных условиях среды, соответствовав-
шая им и эволюционировавшая в ограниченных условиях своего
дальнейшего существования.
Но в эволюции живого единство организма и неорганической
среды выражается специфически в том, что организм сохраняется,
14
выживает во взаимодействии с меняющимися факторами среды,
осваивай'эти факторы. Они являются уже необходимыми для раз-
вития и существования организма и таким образом из внешних
факторов среды превращаются во внутренние условия, без которых
невозможно развитие и существование организма. Возьмем яркий
пример из области высших животных форм — пример выработки
постоянной температуры тела у птиц и млекопитающих.
Таким образом выявляются качественно особые отношения орга-
низма и неорганической среды, выражающиеся в приспособленно-
сти к условиям последней и их освоении и становящиеся на место
отношений при существовании тел неживой природы, не имеющих
свойства приспособляться к изменяющимся условиям среды.
И. В. Мичурин показал все огромное значение в эволюции орга-
низмов момента освоения внешних факторов среды. В организме
ничего не может происходить вне связи с условиями внешней
среды.
Конечно, естественный отбор получает при этом свои особенные
черты — он ведет к большей выживаемости тех форм, которые
адэкватно соответствуют изменениям окружающей неорганической
среды и все больше овладевают ее разными сторонами.
Обращаясь к взаимосвязям организма в биоценотической среде,
легко видеть, что здесь действительно можно говорить о борьбе за
существование в собственном смысле этого слова, т. е. конкурент-
ных и антагонистических отношениях между различными организма-
ми. Таковы противоречия хищника и его жертвы или растительно-
ядных форм и их пищи, характерные для взаимоотношений между
различными видами животных и растений в биоценозе. Они могут
найти свое выражение и в обоюдной защите или симбиотических
отношениях, но это в любом случае борьба за жизнь.
Иначатобстоит дело в пределах одного и того же вида, т. е.
в популяции данного вида, где взаимоотношения особей определя-
ются не столько борьбой за жизнь между отдельными особями,
сколько моментом сохранения и продолжения всего вида, т. е. яв-
ляются качественно иными. Все идет на пользу вида. На первый
план выступает совместное обеспечение выживаемости потомства.
Это ведет, в конце концов, к образованию семей, стад, стай и т. п.
Но каковы бы ни были факторы внешнего мира, с которыми
взаимосвязан организм, при всех условиях происходит естественный
отбор в качественно различных его формах, причем ему принад-
лежит ведущая роль в эволюции организмов, так как длящаяся
изменчивость идет в направлении отбора, поддерживается отбором.
Он, можно сказать, творит новое и в то же время отметает все
неподходящее и старое.
Путем естественного отбора вырабатывается чрезвычайно харак-
терное свойство организации всяких живых существ — их приспо-
собленность к определенным условиям того или иного биотопа, соот-
ветственно биоценоза. Это проявляется как в широких масштабах,
когда получается общее повышение организации, значительно рас-
15
ширяющее возможности организма в борьбе за жизнь, так и в
узких рамках тонкого приспособления к различным деталям опре-
деленного биотопа и биоценоза. В некоторых случаях возможно,
наконец, достижение чрезмерной, как говорят, эксцессивной одно-
сторонности приспособлений.
Хотя в начальных биоценозах сложность не могла быть особен-
но велика, однако она была достаточна, чтобы выработать целый
ряд адаптивных черт, при разнообразной эволюции которых развил-
ся своеобразный и обильный мир протистов: свободно плавающие
жгутиковые; перешедшие к использованию донного пищевого мате-
риала ползающие амебоидные формы; вторично приспособившиеся
к использованию пищевых ресурсов пелагиали фораминиферы и
радиолярии; выработавшие служащий для движения и захватыва-
ния пищи универсальный аппарат ресничек инфузории и т. д. и т. д.
Другая ведущая закономерность в эволюции живой материи
отображает целостность организма и его частей. Это — закономер-
ность дифференцировки функций и структур, получившая название
«принципа разделения физиологического труда», или, лучше, физио-
логических отправлений. В результате получается специализация
частей организма.
Закономерность разделения физиологических отправлений, осно-
ванная на выявлении и развитии противоречивых возможностей,
присущих каждому жизненному процессе, ведет к дифференцировке
функций и структур со все большей и большей детализацией про-
тиворечивых сторон процессов до самых крайних их ступеней. Этим
обусловливается возможность совершенствования функций и все-
стороннего' выявления их качеств. В конечном же счете дело1 сво-
дится к обмену веществ, к процессам ассимиляции и диссимиляции
в организме, а отсюда и к ведущему моменту — взаимодействию
между организмом и средой.
В одноклеточном организме дифференцировка проявляется
прежде всего1 в появлении и обособлении центров химической дея-
тельности протоплазмы (органоидов), кинетических центров (кле-
точного' центра) и т. п., в дифференцировке компонентов ядра и,
наконец, в образовании экзоплазматических клеточных оболочек. Но
далее она проявляется не менее ярко в таких дивергентно эволю-
ционирующих деталях, как органеллы движения — жгутики, реснич-
ки и т. п., как сократительные волоконца, экскреторные вакуоли
и т. д. Не будем вдаваться в детали, но в общем можно сказать,
что в пределах клетки дифференцировка и соответствующая специа-
лизация, в силу внутренних противоречивых моментов выявления
функциональной деятельности и строения, порожденных воздей-
ствием среды, могут идти очень далеко, и клетка инфузории пред-
ставляет тому прекрасный пример.
Но, конечно, неизмеримо дальше заходит процесс специализации
в организмах многоклеточных.
К ним мы и обратимся.
Многоклеточные системы не раз возникали на основе образова-
16
нйя разнообразных колоний простейших, но из всех таких колоний,
в конце концов, далее эволюционировали лишь -немногие колонии
жгутиковых, в которых отдельные особи объединялись студенистым
«основным» веществом экзоплазмы.
Многоклеточный организм представляет закономерный комплекс
тканей и органов, состоящих из клеток и из основного вещества,
не оформленного в клетки, но участвующего в построении организ-
ма. Это — та ступень эволюции материи, которая будет являться
специальным объектом нашего внимания, раскрытию которой будет
посвящена вся дальнейшая наша работа. Она качественно отлична
от клеточной ступени эволюции материи.
Это особенно подчеркивается наличием объединяющего компо-
нента системы в виде основного вещества — «экзоплазмы». Послед-
няя происходит за счет клеток, но не имеет клеточной структуры и
в противоположность клеткам неиндивидуализирована. Однако, она
является наравне с клетками обязательной и далее сложно эволю-
ционирующей составной частью многоклеточного животного.
Далее, в многоклеточном организме разделение функций приоб-
ретает чрезвычайно широкий размах по сравнению с одноклеточ-
ным. Происходит обособление целых вторичных систем в единой
целостной системе многоклеточного животного организма, что дает
на основе дифференцировки клеток и участвующего в построении
тела основного вещества и его производных сначала первоорганы,
а далее как системы тканей, так и сложные системы органов.
Наконец, как мы видели, уже у одноклеточных организмов
выработался половой процесс. Он обусловливает наследственную
передачу характерных черт организации и в то же время некоторое
изменение этих черт, а также повышение жизненности, что необхо-
димо для существования вида в изменяющихся условиях среды.
Для многоклеточного животного развитие каждой новой особи на-
чинается поэтому обычно также одноклеточным состоянием, в ко-
тором процесс взаимной ассимиляции половых клеток единственно
возможен. Таким образом, половой процесс и размножение объеди-
няются — вырабатывается характерное для многоклеточных поло-
вое размножение.
В связи с этим многоклеточный организм обладает периодом
эмбрионального и постэмбрионального развития, которые в основ-
ном отражают длительный исторический процесс эволюции данного
вида, но в то же время двигают его дальше и изменяют.
Дав общую характеристику своеобразия организации многокле-
точных животных, возвратимся к закономерностям живой природы.
В неорганической, мертвой природе не может быть речи о раз-
делении функций. Дифференцировка идет просто' на основе разницы
систем той или иной среды по их физико-химическому характеру
и энергетическим особенностям. Аналогично этому и приспособле-
ние заменяется в мертвой природе простой возможностью сохране-
ния системы в определенных рамках физико-химических условий.
И та, и другая закономерности в живых организмах, в силу спе-
2 Пути эволюции животного мира 17
цифических свойств живых систем, перешли на новый, более
высокий уровень, стали качественно иными, чем в мертвой природе.
При этом сильнее выявляется и взаимообусловленность этих
закономерностей. Эволюция частей в системе целостного организма,
выражающаяся их специализацией на основе разделения функций,
направляется освоением условий окружающей среды, превраще-
нием их в необходимые для организма (условия существования
и развития. Другими словами, дифференцировка частей подпадает
под действие ведущей закономерности системы организма — сре-
ды, под действие естественного отбора. Однако в свою очередь и
эволюция целого организма определяется теми возможностями,
которые дает дифференцировка частей, обусловленная его природой.
У многоклеточных бросается в глаза еще одна закономерность,
которая теснейшим образом связана с обеими рассмотренными
выше закономерностями естественного отбора и разделения физио-
логических отправлений. Закономерность эта касается выявления
определенных направлений взаимодействия организма и среды,
определенных направлений дифференцировки частей организма. В
основе лежит то, что в процессе приспособления и специализации
одинаковые по своему происхождению формы, равно как и гомоло-
гичные образования, изменяются по-разному в своем потомстве, в
дальнейших поколениях, — происходит дивергентная эволюция.
Но весь путь целостного эволюционного процесса определяется
не одной дивергенцией. Наряду с ней имеет место и конвергенция.
Конвергентное, сходящееся, развитие выражается в явлениях
аналогии, т. е. морфологического сходства тканей и органов в связи
с осуществлением одинаковой или подобной функциональной дея-
тельности при различном исходном материале. Оно выражается
также в явлениях далеко1 не редкого сходства в организации между
представителями разных групп животного царства при одинаковых
или близких условиях их жизни.
В последнем случае получаются «жизненные формы» экологов.
Например, у ведущих сидячий образ жизни животных имеется
своеобразный характерный^ облик, приближение к радиальному
строению и другие черты, вырабатывающиеся под действием есте-
ственного отбора и показывающие, что налицо некоторые общие
условия в жизни сидячего организма, на которые он реагирует
развитием сходных адаптивных черт как в одной* так и в другой
п в третьей группе животных. Неограниченного1 разнообразия в
этих общих ответах нет — они в известной мере определены харак-
терными комплексами условий среды. В результате — конвергент-
ное развитие, т. е. «общее сходство» при большом разнообразии в
деталях организации. Получаются аналогичные варианты органи-
зации в ответ на ту или иную крупную категорию условий суще-
ствования, однако возникающие из разного исходного материала.
Такова характерная организация животных при своеобразных усло-
виях жизни в грунте, при особых условиях существования эндопа-
разитов и т. д.
18
Сходство главных направлений осуществления и разделения
физиологических отправлений в организме вызывает конвергенцию
в эволюций и тканей, и органов, и целых организмов в разных
группах животных. Получается иногда поразительное сходство, как,
например, глаз высших головоногих моллюсков и глаз позвоночных,
но всегда тут же и характерные различия в деталях: у первых
глаза не инвертированные, у вторых инвертированные, что обуслов-
лено разницей исходного материала, из которого развились эти
органы в обеих группах. К этому присоединяется и возможность
разнообразия вариантов хода эволюции, причем она может пойти
и по совершенно новым путям, не повторяющим тот или иной из
осуществляемых в ряде других случаев, но по путям все же немно-
гочисленным. Наряду с глазом позвоночного' или каракатицы, выра-
ботались сложные глаза ракообразных или насекомых и т. п. Общее
сходство, конвергентное сходство и частное несходство, разница в
деталях — вот что постоянно взаимосвязано в этой категории
явлений, потому что органы, не сходные у предков, становятся
сходными у потомков.
В целом весь путь развития животных форм — это единый про-
цесс, рассматриваемый лишь с двух различных сторон, ~ расхож-
дения внутри данной группы животных и схождения между разны-
ми группами.
Общие для всех живых систем закономерности естественного
отбора и разделения физиологических отправлений дают у много-
клеточных организмов качественно различные результаты по сравне-
нию с клеткой. Мир одноклеточных не имеет, например, дифферен-
цировки нервных образований, соответственно нервной деятельности,
характерной для мира многоклеточных животных и завершающейся
выработкой высшей нервной деятельности. Поэтому хотя конвер-
гентные явления могут иметь место в эволюции протестов и много-
клеточных организмов, они дают лишь самое общее сходство. Про-
стейшие и многоклеточные сравнимы только в ограниченной мере,
как принадлежащие к двум разным ступеням организации с их
особыми качественными различиями.
Следующей за многоклеточным организмом ступенью являются
колонии многоклеточных, которые представляют, собственно, один
из частных случаев в образовании населения того или иного вида
многоклеточных животных. Обычно популяции не образуют резко
отграниченного, индивидуализированного целого, представляя не-
определенные скопления отдельных индивидов, стада и т. д.
Однако в случае выработки бесполого размножения, выгодного
для вида в борьбе за существование, спорадически создаются в
различных группах животного царства колонии особей. Особи в ко-
лонии органически связаны друг с другом, благодаря чему возни-
кает возможность образования индивидуальности высшего порядка
сРавнению с одиночной многоклеточной особью. Таковы колонии
асцидийНЬсаль°ЛИПОВ> СИФОНОФ°Р> коралловых полипов, мшанок,
19
Обладая органической связью друг с другом, особи в этих ко-
лониях подчиняются закономерности разделения физиологических
отправлений, что создает крайне своеобразную дифференцировку
отдельных из них для выполнения той или иной функции. Напри-
мер, в колонии сифонофор создается дифференцировка особей: под-
держивающих колонию во взвешенном состоянии в воде — так
называемых пневматофоров; двигающих колонию — плавательных
колоколов; защищающих части колонии пассивно — кроющих брак-
тей, или гидрофиллий; защищающих активно — дактилозоидов; пи-
тающих — гастрозоидов; половых — гонозоидов и т. п. Такая диф-
ференцировка ведет к крайне своеобразной организации, в которой
грань между органом и индивидуумом стирается, особь в известных
отношениях низводится до состояния органа. Понимание зоологами
колоний сифонофор может служить примером того, насколько труд-
но расшифровать такую организацию, выявить, что в ней от органа
и что от особи.В общем получается организация, по сложности и
оригинальности, конечно, значительно превосходящая то, что можно
найти в индивидуумах гидроидного полипа или медузы, с которыми
сифонофоры теснейшим образом связаны по своему происхождению.
Конечно, в борьбе за существование естественный отбор может
вести такую колонию к чрезвычайно сложным приспособлениям,
значительно превышающим размах адаптаций в отдельной много-
клеточной особи.
Сложность организации многоклеточного индивидуума в высших
типах животного царства не давала возможности, особенно при
свободном образе жизни, к развитию бесполого размножения, ве-
дущего к образованию колоний. Этим делалось невозможным суще-
ствование у них живых индивидуализированных систем высшего
порядка.
Этого не произошло и потому, что для многоклеточных харак-
терно половое размножение, при котором новые особи органически
не связаны друг с другом. В результате получается лишь популя-
ция из отдельных индивидуумов, относящихся к данному виду, не-
которое видовое единство, состояние качественно различное по
сравнению с отдельно существующим многоклеточным организмом.
Эволюция живых индивидуализированных систем таким образом не
пошла дальше образования в некоторых случаях колоний много-
клеточных.
Однако эволюция живой материи, как мы видели, выражалась и
в других формах, а именно в организованности самой биотической
среды — в образовании крайне подвижных, не ограниченных инди-
видуально, но закономерных комплексов животных и растений, объ-
единяемых общностью условий существования, — в образовании
уже упомянутых нами выше биоценозов. Они представляют гетеро-
генные системы, в состав которых входят различные животные и
растительные особи, соответственно — популяции каждого вида,
свойственного данному биоценозу.
Таким образом, развитие живой природы идет в двух напра-
20
влениях, тесно связанных друг с другом, оказывающих влияние
чруг на" друга. Развитие биоценотических систем‘является мощным
фактором эволюций индивидуализированных систем, и в то же
время эволюция этих последних обусловливает эволюцию ценозов.
Конечно, с появлением живых организмов началась и выработка
соответствующих биоценотических систем.
Сначала эти ценозы были обусловлены только наличием тех или
иных доклеточных форм в определенных условиях абиотической
среды. Особое значение приобретало наличие близкородственных
форм, между которыми создавались отношения, способствовавшие
сохранению их среди других. Создавались внутривидовые отноше-
ния, отличные от отношений с несходными, более или менее значи-
тельно разнящимися формами. Первые при взаимодействии их,
ассимилируя друг друга, только повышали свою жизненность. На-
против, во втором случае одна из форм разрушала, уничтожала
другую, используя ее для построения своего тела. У последних
вырабатывались антагонистические отношения хищника и жертвы,
в особенности если принять гетеротрофное питание за присущее уже
исходным доклеточным формам жизни, как это допускает А. И. Опа-
рин. Однако выработка автотрофного питания вела одинаково к
развитию биотических отношений с противоречивыми тенденциями
между различными видами.
Вначале это были ценозы доклеточных форм, затем сюда присое-
динились одноклеточные организмы, а далее, с появлением много-
клеточных, и эти последние. Из популяций разных видов создались
особые системы гетерогенного характера, в которых каждая эколо-
гическая возможность использована своим представителем, и каж-
дый представитель в своей организации приспособлен, «пригнан»
к условиям соответствующего биоценотического аспекта. И чем
дальше идет процесс освоения различных сторон абиотической и
биотической среды данндго биоценоза развивающимися видами, тем
более выявляются противоречивые тенденции компонентов биоце-
ноза и последний становится сложнее, а входящие в его состав
виды постояннее.
В пределах определенного биоценоза популяции представляют,
как мы видели, известное внутреннее единство, в котором возмож-
но дальнейшее развитие отношений между входящими в них осо-
бями, начиная с постоянно имеющегося подразделения полов. И
можно в различнейших группах животного царства наблюдать
возникновение временных или постоянных обособленных единств
характера семей и более обширных объединений в виде стай, стад
и т. п. «Общины» термитов, пчел, муравьев, с одной стороны, и
семьи у млекопитающих, с другой, представляют примеры таких
далеко ушедших популяций.
Глава I
ТКАНИ И ОРГАНЫ
Дифференцировка вторичных систем — тканей и огранов в единой
системе организма. Происхождение многоклеточных животных. Тка-
ни и органы у губок. Дифференцировка тканей и органов у низших
кишечнополостных. Два пути дифференцировки и их характеристика.
Примордиальные ткани, первоначальное единство тканей и органов,
дальнейшее расхождение обеих систем. Определение органа и ткани.
Специализация тканевых элементов и дефинитивные ткани. Зако-
номерности дифференцировки тканей и органов.
В начале книги мы очертили ту область, которая будет предме-
том нашего изучения. В ее пределах задача не столько описать,
сколько, на основе установленных закономерностей, дать понимание
организации главнейших типов животных, используя обильный фак-
тический материал, накопленный массой исследователей.
В связи с этим сначала проанализируем организм как цело-
стную систему, в границах которой дифференцируются вторичные
системы — органы и ткани. И те и другие не имеют свойства са-
мостоятельного существования. Для понимания их мы должны
обратиться к вопросу о происхождении многоклеточных животных.
Свою последнюю зоологическую работу И. М. Мечников закон-
чил так (1886, стр. 159): «Насколько возможно большее выяснение
вопроса о первичных состояниях многоклеточных животных необ-
ходимо-, чтобы приобрести базис для сравнительной морфологии».
По смыслу этих знаменательных слов, нисколько не утративших
своего значения и в настоящее время, мы прежде всего постараемся
наметить основные черты организации первичных многоклеточных
животных.
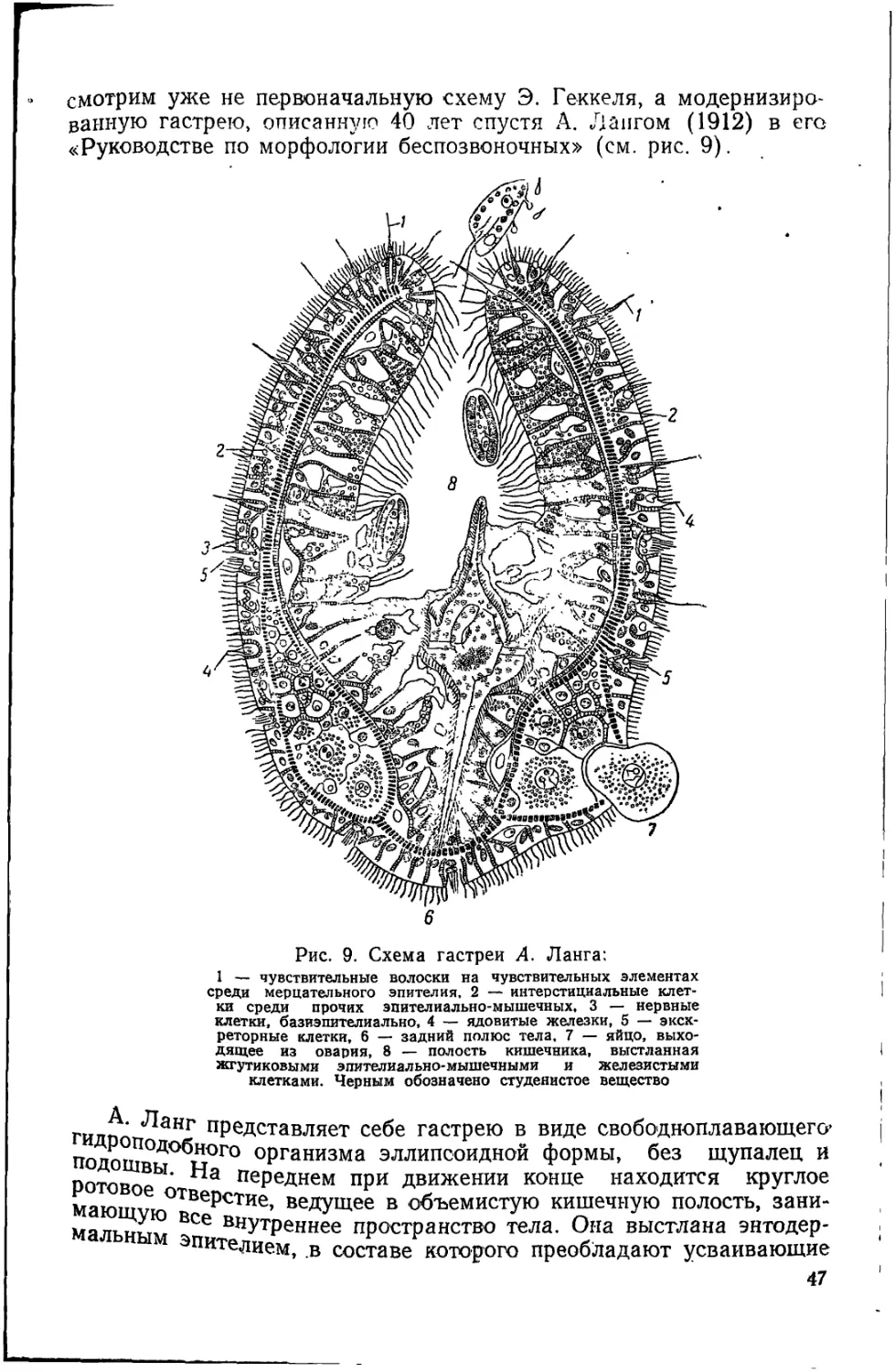
Для губок (рис. 1) характерно чрезвычайно слабое развитие
эпителиев тела по сравнению с мощным и довольно сложным раз-
витием среднего слоя, образованного основным веществом. Покров
у них представлен тонкой пластинкой из одинаковых, часто совер-
шенно плоских элементов, а пищеварительный аппарат имеет вы-
стилку, состоящую только из воротничковых жгутиковых клеток.
Слабость, примитивность эпителизации такова, что можно просто
говорить о кроющих клетках — «ппнакоцитах» и воротничковых
22
жгутиковых элементах, выстилающих пищевз1рителыныи аппа-
рат, — «хоаноцитах». В связи с этим невольно "вспоминается, что
Рис. 1. Разрез через Leucosolenia (по Пренану) (X) и через пресноводную
губку (по Гиман) (5) (Пояснения см. на рис. 2)
Рис. 2. Разрез через стенку тела Clathrina coriacea: А — в спокойном со-
стоянии; Б — то же в слегка сокращенном состоянии; В — то же в состоя-
нии полного сокращения (по Минчину);
- ^амебоциты, 2 — археоцит, з — хоаноциты, 4 — покровный эпителий (пинакоциты),
основное вещество соединительной ткани, 6 — пороцит, 7 — спикулы, 8 — произ-
водящие их клетки
при длительных раздражениях, как было показано опытами, ворот-
ничковые клетки втягивают свои жгуты и воротнички, теряют эпи-
телиальное расположение, образуя кучки клеток неправильной
Ф рмы (рис. 2). По окончании раздражений они снова располага-
23
ются в один слой, ограничивая образующуюся внутреннюю полость,
и восстанавливают жгуты и воротнички.
Такого рода факты давали повод называть губок «Zoa anhistia»—
«нетканевыми», или, по крайней мере, обладающими факультатив-
ной дифференцировкой тканей.
Конечно, для подобных взглядов есть некоторые основания,
однако эпителизация, соответственно обособление срединной массы
основного вещества, представляющей внутреннюю среду организма,
у губок уже совершились. При этом, наряду с примитивностью их
эпителиев, мы видим значительные осложнения в строении ткани
основного вещества. Эволюция губок пошла под знаком дифферен-
цировки именно последней, составляющей главную массу тела
губки и всегда содержащей различного рода клетки — недифферен-
цированные археоциты, затем производящие само основное веще-
ство амебоциты, далее дающие защитный и опорный скелет склеро-
бласты, наконец, сократительные мышечные клетки и половые
элементы.
Список различных клеток нами не исчерпан, но вывод ясен:
губки с самого начала их эволюции отличались присутствием кле-
точных элементов в основном студенистом веществе, а в дальней-
шем развитие этой ветви животного мира шло по пути специализа-
ции указанных клеток при разделении функций между ними. В
основном веществе, кроме производящих его элементов, дифферен-
цировались защитные клетки, дающие спикулы, а затем и клетки
другого характера.
К.ЦПнейдер удачно подчеркнул эту ведущую черту организации,
выделив Fyffol как «Pleromata», т. е. как организмы, характеризую-
щиеся прежде всего развитием заполняющей ткани.
Слабость, примитивность эпителизации выявляется как типичная
черта губок, а в связи с этим и отсутствием дифференцировки эпи-
телогенных образований — нервных и мышечных.
Итак, губки отличаются в гистологическом отношении, во-пер-
вых, некоторой неустойчивостью тканевой дифференцировки, во-
вторых, примитивностью эпителиев, в-третьих, отсутствием нервной
ткани и эпителогенных мышц и, наконец, в-четвертых, наличием
клеток в ткани основного вещества и их сложной дифференциров-
кой до выработки мезенхимных мышечных элементов включительно.
Губки показывают нам чудесную организацию, которую Л. Ги-
ман (1940) не без основания, но односторонне, называет «клеточ-
ной ступенью организации метазоа». Для нас в ней крайне ценно
видеть первые шаги эволюции эпителия. У губок, таким образом,
выработалась, во-первых, система, отграничивающая собственную
среду организмами1 внешней среды — орган покрова и тот постоян-
ный способ, та структура, которыми это осуществляется, — ткань
эпителия в его примитивнейшем виде, т. е. в виде слоя одинаковых
клеток. Во-вторых, выработалась система, захватывающая и пере-
варивающая *пйщу и отдающая питательный материал в массу
основного вещества — система пищеварения с тем же способом
24
структурного осуществления, а именно эпителизациёй одинаковых
зоротничковых жгутиковых клеток Питающая система у губок со-
хранила характер слоя из примитивных воротничковых жгутиковых
элементов, тогда как покров образован вторично измененными эле-
ментами, утратившими жгуты и воротнички.
Примитивное состояние образований внутренней среды мы на-
ходим в другом типе животного царства, а именно—у низших пред-
ставителей кишечнополостных, например у гидры. Здесь эволюция
пошла в совершенно ином направлении,
чем у губок, и картина получается абсо-
лютно другого рода. Отношения многими
зоологами расцениваются так, что если
губки выделяются в особую ветвь живот-
ного царства — «паразоа», то кишечнопо-
лостные, соответственно гидроидные поли-
пы, ставятся в начале другой ветви жи-
вотного царства — «метазоа», берущих на-
чало от иных форм простейших, чем губ-
ки. И насколько- у губок бросается в гла-
за слабая выраженность эпителизации и
значительное развитие основного веще-
ства, настолько для гидры характерны
сложность эпителия и примитивность об-
разований основного вещества.
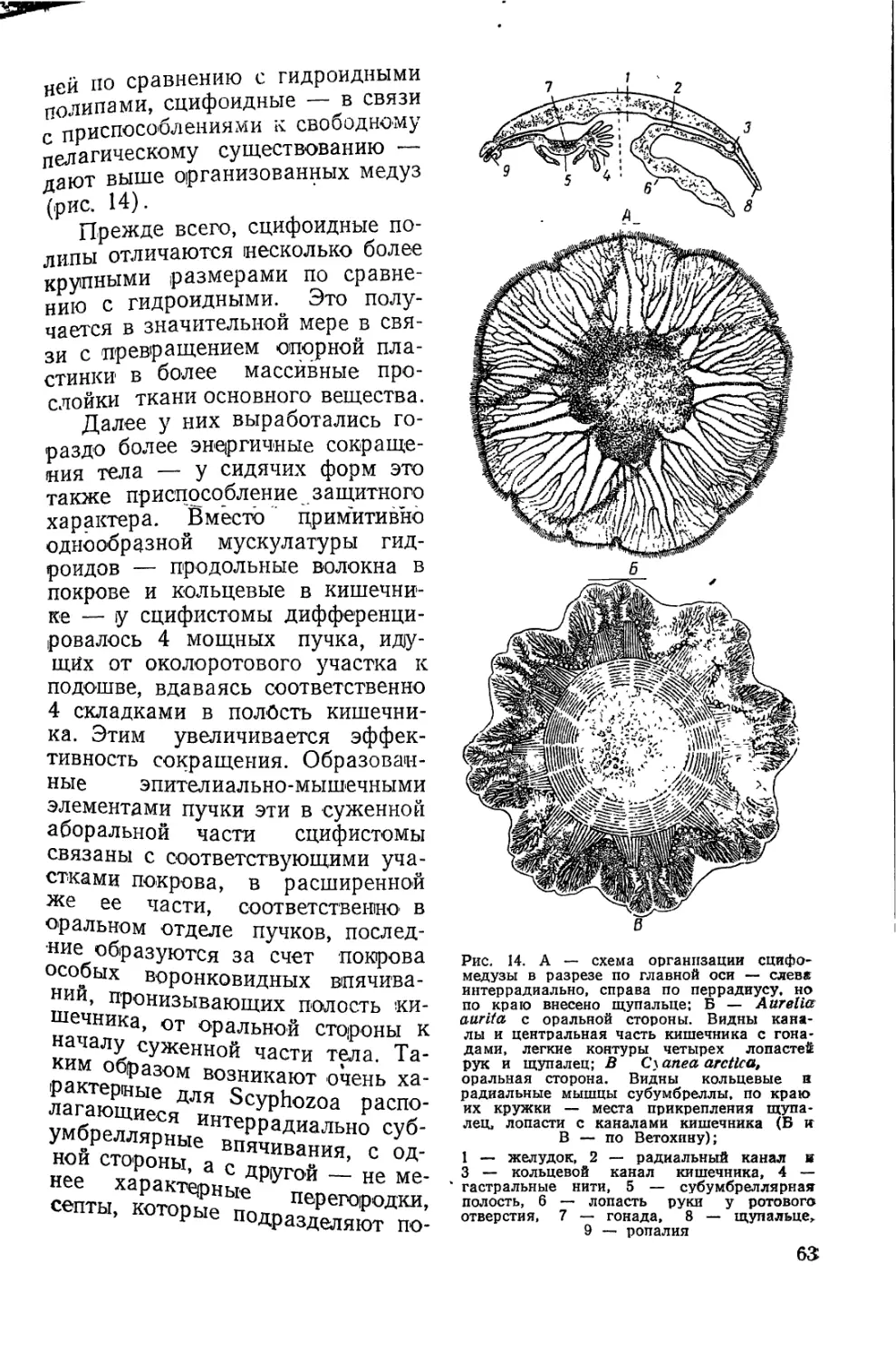
Покров гидры представлен эпителием,
состоящим из эпителиально-мышечных
клеток, расположенных в один слой
(рис. 3). Наружная поверхность эпителия
покрыта защитным зернистым ободком.
Мерцательный слой ресничек при переходе
к сидячему образу жизни у полипов
исчез, а у морских форм выработалась
настоящая плотная защитная кутикула.
Базально каждая клетка имеет неболь-
Рис. 3. Поперечный разрез
через стенку тела гидры в
средней его части (по
Гиман):
1 — кольцевые мышечные во-
локна, 2 — кишечный эпителий,
3 — покровный эпителий, 4 и
5 — железистые клетки, 6 —
опорная пластинка, 7 — интер-
стициальные клетки и книдо-
бласты, 8 — продольные мышеч-
ные волокна. Нервных и чув-
ствительных клеток на срезе не
видно
шие протоплазматические выросты, содер-
жащие внутри по сократительному волокну. Превращение у .гидры
всех кроющих элементов в эпителиально-мышечные —• очень харак-
терная черта ее покрова.
Таким образом, у кишечнополостных эволюция кроющих эпите-
лиев пошла по пути выработки сократимости, с одной стороны, и
защитной кутикулы, с другой. Норма — отсутствие обособленных
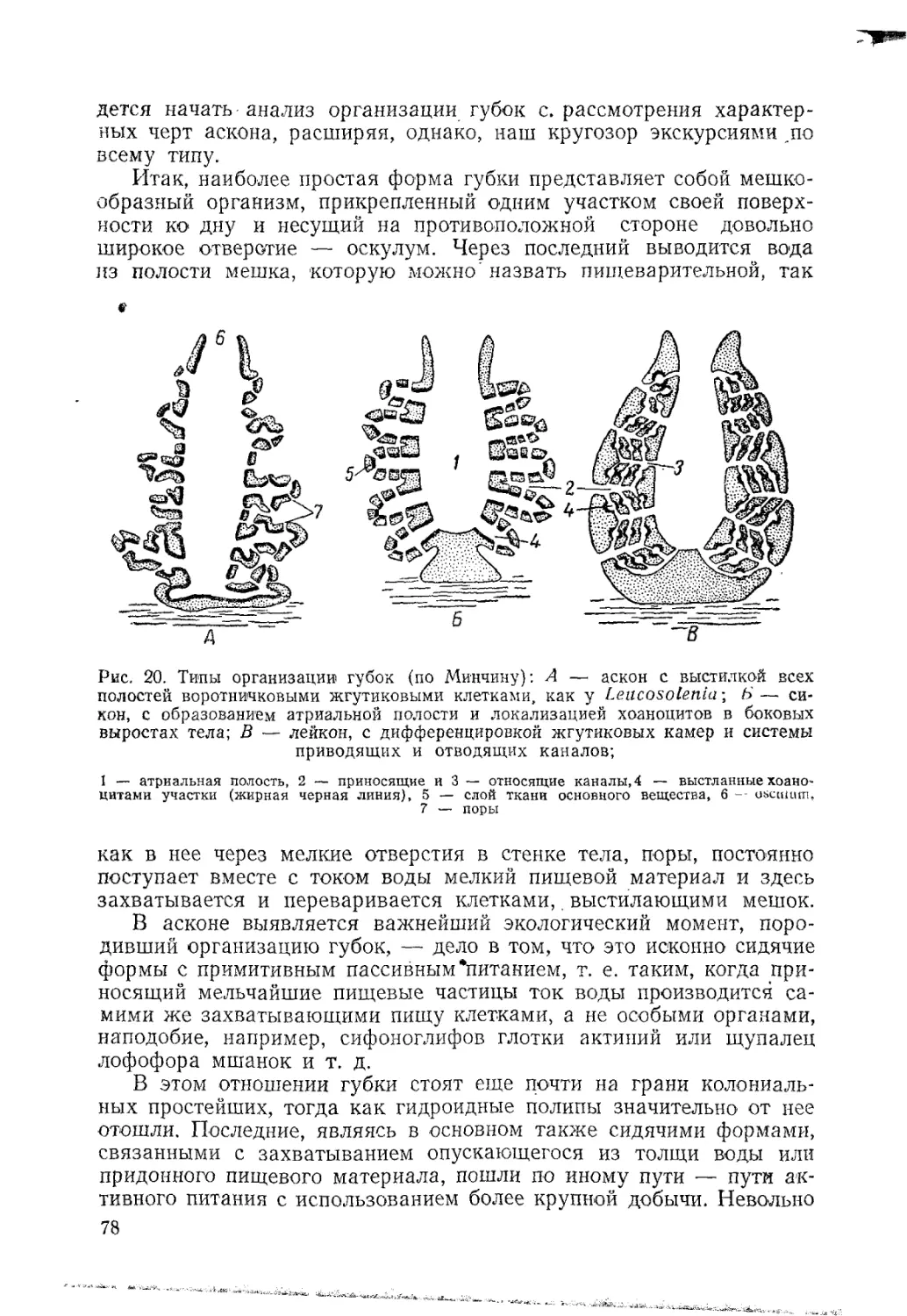
мышечных элементов, хотя в ряде случаев, как, например, на щу-
пальцах актиний (рис. 4), вырабатывается обособленная мускула-
тура путем образования базальных складок в эпителии и дальнеп-
его их отшнуровывания и погружения в слой основного вещества.
добЛУЧаеТСЯ настояЩая эпителогенная мускулатура. Чего-либо по-
как °ГО мезеихимогенной мускулатуре губок у кишечнополостных.
' правило, нет. Перед нами два разных пути развития.
25
Постоянством эпителизации и сложностью самой эпителиальной
клетки дело не ограничивается. У гидры имеется дальнейшее услож-
нение, выражающееся в дифференцировке на подошве некоторых
эпителиально-мышечных клеток в железисто-мышечные и в при-
сутствии между эпителиальными элементами клеток, сохраняющих
свой индифферентный характер, так называемых промежуточных,
или интерстициальных. За их счет может идти в дальнейшем диф-
Рис. 4. Поперечный
разрез через щупаль-
це актинии (по бр.
Гертвиг):
1 — стрекательные кап-
сулы, 2 — эпителий ки-
шечника. 3 — покров-
ный эпителий, 4 — во-
локнистая ткань основно-
го вещества, 5 — обо-
собившиеся мышечные
пучки, 6„— нервное спле-
тение
ференцировка чувствительных и нервных кле-
ток, с одной стороны, и книдобластов, выраба-
тывающих стрекательные капсулы, с другой, а
также образование половых элементов. Под-
черкнем, что все эти разнообразные элементы
располагаются всегда интраэпителиально.
Как видно, эпителий покрова кишечнополо-
стных достигает очень большой сложности, да-
же большей, чем в других, вышестоящих, ти-
пах животного царства. Это особого рода ком-
плексный эпителий.
Все сказанное вполне приложимо и к дру-
гому эпителию тела гидры, а именно — обра-
зующему выстилку кишечника, только здесь
клетки более обнаруживают свои типовые чер-
ты. Во-первых, они сохраняют жгуты, обуслов-
ливающие в известной мере движение пищевых
частиц, и, во-вторых, сохраняют способность к
внутриклеточному перевариванию этих частиц,
предварительно до некоторой степени уже об-
работанных секретами железистых клеток ки-
шечника. Последние располагаются в нем глав-
ным образом непосредственно у ротового
отверстия. Далее, в кишечном эпителии слабее
развиты чувствительные и нервные элементы,
а также в малом количестве встречаются ин-
терстициальные клетки. Наконец, у гидры в
эпителии кишечника не дифференцируются по-
ловые клетки, однако у вышестоящих сцифоид-
ных и коралловых полипов имеет место как
раз обратное: отсутствие половых элементов в :
эпителии покрова и постоянное их образова-
ние интраэпителиально в кишечнике.
В силу всего этого кишечнополостные заслуживают названия по |
преимуществу «эпителиальных» животных. В полном соответствии ‘
с этим, или вопреки всему этому, стоит примитивность и простота
образований внутренней среды, представленных у гидрополипов
только бесструктурной и не содержащей клеток прослойкой ткани
основного вещества, так называемой опорной пластинкой. Про-
исходя путем дифференцировки экзоплазмы за счет обоих клеточ-
ных слоев стенки тела, она в то же время представляет собой
6
единое образование, так как производные различных клеток сли-
ваются в сплошную массу. В противоположность мультифункцио-
нальности клеточных слоев, опорная пластинка несет только две
функции — функцию механической и химической связи и функцию
опоры. Последняя^становится особенно характерной для нее, так
как ни тот, ни другой эпителий, будучи образованы пластичной
массой протоплазмы, не могут служить в качестве опорного обра-
зования, возвращающего' тело после деформации в прежнее состоя-
ние. Это происходит благодаря эластичности основного вещества
экзоплазмы.
Таким образом, прослойка основного' вещества у гидры является
особым неклеточным органом связи и опоры наравне’с клеточными
органами покрова и кишечника и, добавим, с нервной системой,
представленной расположенными внутриэпителиально чувствитель-
ными и нервными элементами, а иногда даже примитивными чув-
ствительно-нервными клетками.
Сложность отношений у гидры выявляется в присутствии, вместе
с рассмотренными первичными органами, образований иного харак-
тера — щупалец и подошвы (см. рис. 8, Д), представляющих уже
типичные органы. Щупальца несут функции аппарата защиты и на-
падения и в то же время захватывания добычи. Подошва представ-
ляет орган прикрепления и движения. И те и другие являются
результатом дифференцировки всех трех слоев тела. Щупальца
образованы покровным эпителием, особенно богатым стрекательны-
ми элементами (стрекательные батареи), кишечным эпителием,
превращающимся у гидроидных полипов в осевой опорный тяж
вакуолизированных клеток, и прослойкой опорной пластинки. По-
дошва содержит слой железисто-мышечных клеток покровного
эпителия, прослойку основного вещества и внутренний слой кишеч-
ного эпителия. Чувствительные и нервные элементы присущи обоим
органам, но книдобласты в подошве отсутствуют.
Все эти факты показывают своеобразие организации кишечно-
полостных ПО' сравнению с организацией губок, выявляют ориги-
нальность эволюционного пути, ведущего к кишечнополостным и
прочим многоклеточным, и совершенно особого пути, ведущего к
губкам. Оба вместе они дают возможность воссоздать интересней-
шую картину зарождения и первых шагов существования много-
клеточных организмов.
Они показывают, что у исходных форм многоклеточных живот-
ных, наряду с клеточными образованиями, роль непременного ком-
понента организации играет основное вещество экзоплазмы. Клетка
является первичной единицей в построении многоклеточного орга-
низма, но, происходя от клеток и являясь деградированной прото-
плазмой, студенистое основное вещество экзоплазмы становится
особой составной частью многоклеточного организма, постоянной и
существенной, как и клетки. Рассмотренные факты далее показы-
вают, что происхождение многоклеточных животных теснейшим об-
разом связано с одновременно протекавшими процессами: с эпите-
27
лизацией, соответственно образованием внутренней среды организ-
ма в виде его основного вещества, т. е. тканеобразованием, и с диф-
ференцировкой первичных органов — кишечника и покрова, а так-
же опорной пластинки, т. е. с органообразованием. При это-м в це-
лостном организме система ткани и система органа тесно взаимо-
связаны.
Уже первая дифференцировка в многоклеточном организме ведет
к образованию клеточных и неклеточных органов — питающего
кишечника, изолирующего и защищающего покрова и связующей
опорной пластинки. Одновременно создаются и две исходные ткане-
вые дифференцировки: клеточная — кроющий и усваивающий эпи-
телии и неклеточная — ткань основного вещества. При этом в при-
митивных состояниях орган может быть образован из одинаковых,
т. е. сходно дифференцированных и сходно связанных, элементов
или из однообразной массы основного вещества. Другими словами,,
орган может состоять из одной ткани, как, например, покров губок
из пинакоцитов, жгутиковые камеры из хоаноцитов, нервная систе-
ма гидры из сплетения нервных клеток, опорная пластинка из гомо-
генного слоя ткани основного1 вещества, не содержащей клеток,
Таким образом, ткань и орган в приведенных примерах до некото-
рой ,степени едины, но дальше они расходятся, эволюционируя,
как два особых направления дифференцировки вторичных систем
в целостном организме.
Этим поставлены некоторые вехи для решения ряда интересую-
щих нас вопросов. Можно видеть, что тканевая дифференцировка и
дифференцировка органов представляют, как отмечено, два пути
возникновения вторичных систем в единой целостной системе мно-
гоклеточного организма. Из них первая идет как в направлении
дифференцировок в пределах клетки, существующих уже у про-
стейших, так и в направлении выработки взаимосвязей .элементов
и дифференцировок экзоплазмы, возможных и неизбежных в много •
клеточных организмах.
Органы всегда образуются из тканей, обеспечивающих как спе-
циальную функцию, так и общие жизненные отправления органа,
и, в свою очередь, обусловливают соответствующую дифференци-
ровку тканей, как, например, орган покрова и его главная ткань —
кроющий эпителий — с ее тонофибриллярными, кутикулярными и
тому подобными образованиями.
Примордиальная клеточная ткань, эпителий, представляет си-
стему клеток, обладающих полярной дифференцировкой и связан-
ных непосредственно друг с другом в единый слой. И то и другое
является выражением отношений в многоклеточном организме. При
этом полярность обусловлена тем, что один участок клетки стоит в
непосредственном соприкосновении с внутренней средой организма,
а другой или с 'Внешней средой, окружающей организм, или со
средой внутренних полостей организма. Одним словом, две части
эпителиальной клетки, базальная и дистальная, оказываются в раз-
ных условиях в смысле протекания их жизненных процессов. Пол-
28
или студня колокола
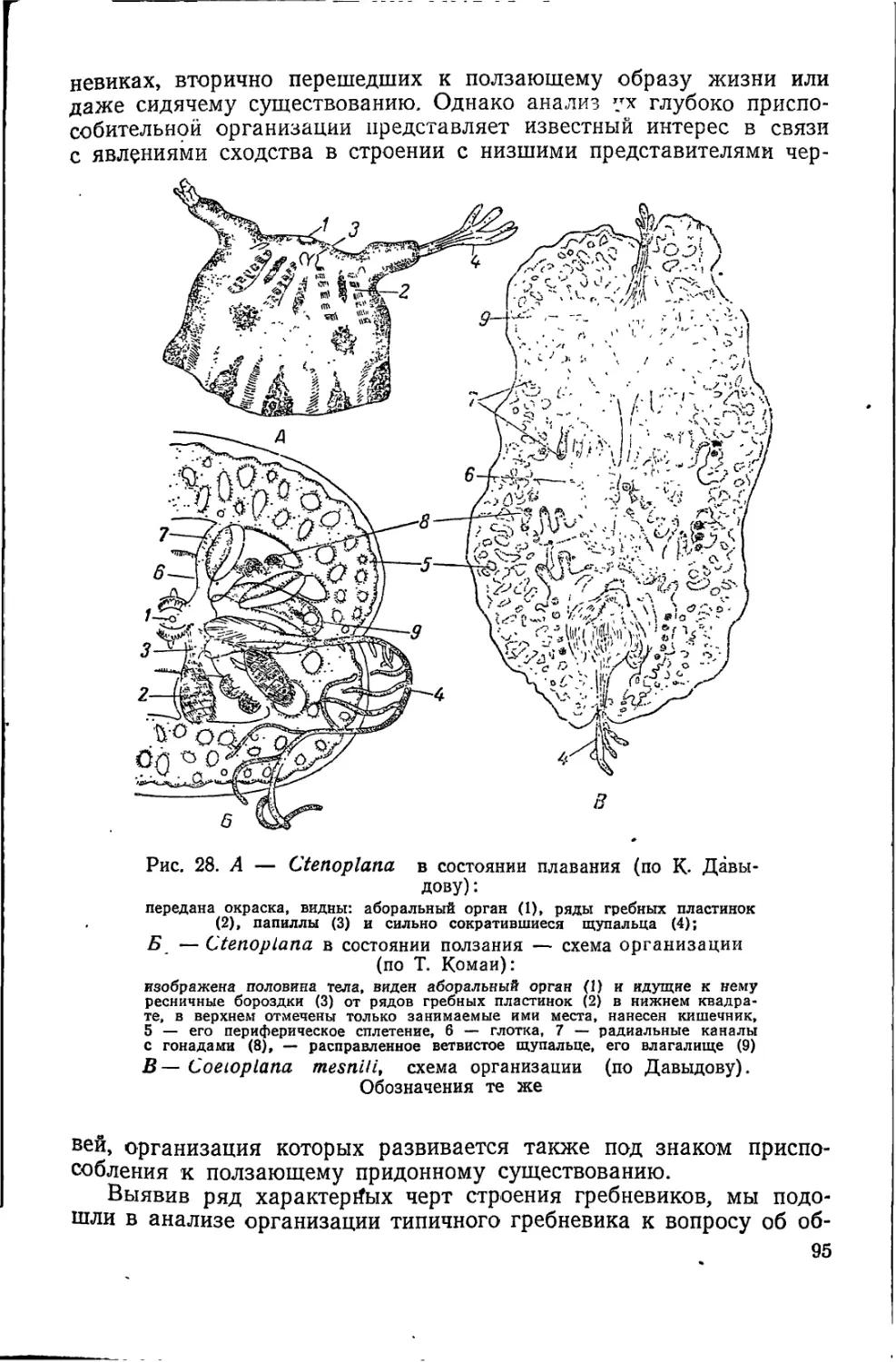
Рис. 5. Колония Prote-
rospongia (по Кенту)
-i-jcTbio это выражено уже в бластуле (см. рис. 8, Л),1. е. на пер-
вых шагах онтогенетического развития многою л ^очного организма.
Ее клетки полярно дифференцированы, но, конечно, подобная диф-
ференцировка ничуть не может помешать дальнейшей специализа
пни клеток в различнейших направлениях. Клетки, собственно, со-
храняют еще полностью свой индифферентный характер, и даль-
нейшая эволюция идет за счет специализации эпителиальных эле-
ментов.
Другой примордиальной тканевой дифференцировкой является
гкань основного вещества, происходящая за счет протоплазмы кле-
ток эпителиев покрова и кишечника, но становящаяся самостоя-
тельной наподобие опорной пластинки гидры
гидроидной медузы. Оба последних обра-
зования имеют совершенно индифферент-
ный характер и еще не обнаруживают ка-
кой-либо специализации структурных ча-
стей в своей однородной массе.
Вдаваясь несколько глубже в вопросы
эволюции многоклеточных животных,
можно утверждать, что на пути паразоа,
при выработке организации губок, первым
этапом являлась дифференцировка в не-
подвижной колонии наподобие проблема-
тичной Proterospongia (рис. 5), изолирую-
щих, кроющих пинакоцитов, с сохране-
нием в более защищенных участках
неизменных питающих воротничковых
жгутиковых клеток. Последние сближа-
ются в один слой до соприкосновения друг с другом. При этом в
основном веществе мезенхимы локализуются клеточные элементы,
полностью сохраняющие свой индифферентный характер. В дальней-
шем они могут заменять утраченные или поврежденные хоано- и
пинакоциты, а после полового процесса — давать начало новой
многоклеточной особи; или, наконец, они могут специализироваться
в качестве клеток, вырабатывающих основное вещество мезенхимы,
образующих защитные, а в дальнейшем и опорные отложения в
виде спикул, клеток с сократительными волоконцами и т. п.
Однако дифференцировка пинакоцитов и других элементов у
губок еще крайне примитивна, специализация их настолько слаба,
что легко может изменяться, и пинакоцит превращается в пороцит,
а последний при определенных условиях, например при постоянных
Раздражениях, может переходить в индифферентное состояние и
обратно.
По пути эволюции, ведущему как к кишечноплостным (полипы
и медузы), так и к гребневикам и к ресничным червям, отношения
развивались иначе. Свободноподвижное существование их исходных
Форм обусловило начальную дифференцировку в направлении вы-
29
работки элементов, несших функцию изоляции внутренней среды
организма от окружающей наружной водной стихии и в то Ж(
время функцию передвижения животного. Дифференцируется, мер-
цательный покров, значительно более устойчиво специализирован-
ный, чем пинакоциты у губок. В то же время неизменные питающие
жгутиковые клетки обособляются, скопляясь в одном участке тела,
а именно — в заднем. Эпителизируясь, они образуют первоорган
пищеварения — первичный кишечник. При относительно слабом
развитии основного вещества в теле исходных форм кишечнополо-
стных индифферентные (интерстициальные) элементы первоначаль-
но рассеяны всюду среди эпителиальных клеток, придавая эпителию
значение особого рода «эпителиальной среды», в которой они сво-
бодно двигаются и подвергаются дальнейшей дифференцировке и
специализации. За их счет идет замена и восстановление элементов
покрова и кишечника, происходит выработка иногда очень сложно-
оформленных секретов в виде стрекательных капсул и т. п., диф-
ференцируются специальные чувствительные и нервные клетки, и,
наконец, формируются половые клетки. Специализация элементов
эпителия у кишечнополостных заходит очень далеко не только в
связи с образованием такой «эпителиальной среды», играющей ве-
дущую роль в их организации, но и в связи с превращением клеток
эпителия в эпителиально-мышечные путем дифференцировки сокра-
тительных мышечных волоконец в протоплазме. Внутренняя же
среда организма у низших представителей типа кишечнополостных,
у гидроидных полипов, соответственно представлена только тонкой
прослойкой ткани основного вещества.
По пути, ведущему к гребневикам и червям, эволюция шла в
направлении увеличения массы и значения основного вещества, как
внутренней среды организма.
Дальше углубляться в эти вопросы здесь нет необходимости,
так как речь о них будет еще впереди. Обратимся к завершающим
положениям о дифференцировке тканей и органов.
По мере усложнения организации многоклеточных животных
тождество тканей и органов очень быстро пропадает. Но у низших
паразоа и метазоа, организацию которых мы могли проанализиро-
вать, единство ткани и органа в той или иной мере у тех или иных
органов еще сохраняется. Система тканей только что начинает от-
ходить от системы органов.
Далее, в процессе эволюции организмов, в образовании огром-
ного большинства органов уже участвуют различные ткани, вторич-
но присоединяющиеся к главной, первоначальной и специальной
для данного органа. Они обеспечивают полноту жизненных отправ-
лений главной ткани и всего органа: Так, например, соединительная
ткань объединяется с эпителием, как это имеет место в покрове и
кишечнике. Или вместе с главной тканью вторично дифференци-
руется побочная; так, например, в нервной системе нейроглийная
ткань развивается из общего зачатка с нервной за счет покровного
эпителия.
30
Таким образом, орган — это дифференцированная, в связи с
рыполнением той или иной специальной функции, часть тела, пол-
нота жизненных отправлений которой обеспечивается входящими в
состав тканями. Тка^же или представляют локальные диффе-
ренцировки в соответствии’ с необходимостью осуществления орга-
ном какой-либо специальной функции, или образуют диффузные
системы, охватывая весь организм, для выполнения общих функций.
К последним относятся обмен веществ, возбудимость, сократимость,
механическая функция, присущие каждому многоклеточному орга-
низму в любом участке его тела.
Для обеспечения различных сторон жизнедеятельности в построе-
ние органов втягиваются и части других органов и их систем,
например скелет, сосуды, нервы и т. п. Наконец, сложные органы
образуются путем дифференцировки целых комплексов, так, напри-
мер, образование примитивных конечностей — параподий анне-
ЛИд —. происходит за счет всей боковой стенки тела каждой мета-
меры. Соответственно сходно образование жабцр у них же и т. д.
В подобных случаях разнообразие комбинаций,, в которых сочета-
ются и далее дифференцируются тканевые элементы, бывает крайне
пестро и велико — от самых простых, как, например, в щупальцах
гидры, до самых сложных, как в глазу птицы.
Итак, ткань и орган отражают единство процессов дифференци- |
ровки, ведущей к обособлению, и процессов интеграции, ведущей *
ко все более совершенному объединению частей в едином, целост-
ном, организме. Обычно, в то время как общие тканевые диффере!
цировки охватывают организм диффузно и непрерывно связаннс
дифференцировка специальных тканей, соответственно органо?
протекает локализованно и обособленно. Только единством, взаимс
проникновением обоих процессов объясняются их взаимоотношени
и кажущиеся трудности их понимания.
Общим выражением осложнения отношений в тканях является
все дальше и дальше идущая специализация тканевых элементов.
Примитивные эпителий и бесструктурная масса основного вещества,
равно как и скопления недифференцированных клеток в виде интер-
стициальных элементов в эпителиях или недифференцированных
клеток мезенхимы, специализируясь, дают дефинитивные ткани.
Ряд такого рода дифференцировок носит характер постепенных
переходов. Но в различных группах животного царства они могуг
идти в разных направлениях и останавливаются на разных ступенях.
Получается своеобразная тканевая эволюция с ее дивергентными и
конвергентными проявлениями. Создаются большие тканевые систе-
мы со все дальше и дальше идущей специализацией элементов,
возникают в конце концов комплексы дефинитивных тканей, уклады- •
кающиеся в рамки двух широких категорий: 1) эпителий и эпитело-
генные ткани и 2) ткань основного вещества и мезенхимогенные
ткани.
Центр тяжести тканевой эволюции при образовании дефинитив-
ых тканей переносится, с одной стороны, на дифференцировку
31
(главным образом) в пределах клетки, т. е. на клеточную ступень
организации, а с другой стороны, на дифференцировку в массе
основного вещества.
Специфика тканевых систем заключается в том, что они обус-
ловлены проявлением процессов и структур, свойственных уже одно-
клеточным, но развивающихся у многоклеточных организмов неиз-
меримо дальше на основе разделения физиологических отправлений
в новых условиях, переходя, так сказать, на высшую ступень. Так
вырабатывается мышечная, нервная и другие клетки, независимая
жизнь которых вне организма становится невозможной. Даже в са-
мом организме их существование обусловлено постоянным положе-
нием и постоянными взаимоотношениями с другими элементами,
как, например, нервной клетки с глийной. Клетка в организме
теряет свойства самостоятельно живущего одноклеточного индиви-
дуума.
Однако имеются все переходы, все ступени специализации,
начиная от таких индифферентных элементов, как интерстициальные
клетки гидры, способных образовывать в дальнейшем любые специа-
лизированные клетки, и кончая специфическими гранулоцитами
позвоночных, сохраняющими амебоидный характер, но специализи-
ровавшимися на того или иного рода секреции, или эритроцитами,
уже совершенно утрачивающими характерные черты клетки. При
этом стрекательный 'элемент гидроидного полипа, обладающий
чрезвычайно высоко специализированным оформленным секретом в
виде стрекательной капсулы, может еще независимо продолжать
свою жизнь и функционировать, даже попадая в других животных—
некоторых турбеллярий, голожаберных моллюсков. Напротив, выра-
ботка, например, поперечнополосатых мышц идет настолько под
знаком потери клеткой своей индивидуальности, подчинения нуждам
многоклеточного организма, что о клетке, собственно, говорить не
приходится, — перед нами живое вещество протоплазмы, ядерные
образования, органоиды и над всем — доминирующая сократитель-
ная структура большой сложности. Сердечная мышца млекопитаю-
щего — яркий образец того, как тканевая система переходит на
новый, высший, уровень, где вторично стирается грань между клет-
кой, тканью и органом. ;
Этот ряд «клетка — неклетка», как мы видим, является впол-
не естественным и понятным.
Выяснив сущность дифференцировки вторичных систем — тканей
и органов — в единой целостной системе многоклеточного организ-
ма, уделим внимание тем закономерностям, которым они подчинены.
Прежде всего, как в тканевой эволюции, так и в эволюции орга-
нов, проявляется общая закономерность соотношений части и целого
в форме специфичной для рассматриваемой нами ступени органи-
зации. Это, как мы уже выше отметили, — принцип разделения
физиологических отправлений, закономерность выявления и обособ-
ления противоречивых сторон жизненного процесса, противоречивых
тенденций обмена веществ в животном организме, выражающаяся
32
«1К В дифференцировке функций и структур, так и в интеграции их,
и приводящая к специализации частей в организме. Ч Дарвин в
«Происхождении видов» (по русск. изд. 1896, стр. 63 и 75) говорит:
«Конечно ни один натуралист не сомневается в полезности принци-
па так называемого «физиологического разделения труда» — пред-
мет этот превосходно выяснен Мильн-Эдвардсом». У последнего
МЫ читаем (по русск. изд. 1859, стр. 44): «Природа совершенствует
организм преимущественно посредством разделения труда», которое
« во всех направлениях и во всех частях животного идет параллель-
но совершенствованию отправлений».
Разделение физиологических отправлений основывается на том,
чт0 Igg-gb или °В~1 в соответствии с обеспечением необходимого
уровняжизнедеятельности каждой части целостного организма
бывают мультифункциональны. В эволюции происходит усиление
той или иной ' из функций/ соответственно — структур. В то же
время из прочих структур и функций одни сохраняются, коррелятив-
но развиваясь как вспомогательные, а другие отходят на задний
план, ослабляются и могут совсем исчезнуть. Так, в электрическом
органе ската необыкновенно сильно развиваются нервные оконча-
ния, и совершенно подавляется развитие мышечной ткани. Однако
исходным состоянием для этого органа являлся обычный мышечный
пучок с хорошо выраженными сократительными волокнами и нерв-
ными окончаниями на них. Такой пучок обнаруживает лишь слабые
изменения электрического потенциала в нервных окончаниях, но в
электрическом органе эта функциональная черта усиливается. Со-
ответственно увеличивается и развитие нервной ткани. Напротив,
мышечные волокна становятся слабее выраженными, и функция
сокращения утрачивает свое. значение. Таким образохМ выявляются
противоречивые тенденции, имеющие место в мышечном пучке: с
одной стороны — сократимость, а с другой — возбудимость. Пред-
ставить себе, что и та и другая функции могут одновременно
развиваться в одном и том же образовании, конечно, до известной
степени возможно, однако обычно процесс протекает иначе, и одна
из функций преобладает.
^Обратимся опять к классическому объекту многих исследова-
нии — пресноводной гидре. Покров ее образован, как мы видели,
эпителиально-мышечными клетками, открытыми в 1872 г. Н.Клейнен-
бергом. Последний, однако, неправильно понимал их значение. Он
считал, что настоящий покровный эпителий у гидры в онтогенезе
с расывается, и снаружи оказываются располагавшиеся ниже эле-
менты, которые он принял за нервно-мышечные, т. е. клетки,
воспринимающие падающие извне раздражения, перерабатывающие
и соответственно проводящие импульс к мышечным волоконцам,
^одержащимся
в их базальной части. Таким образом, весь рефлек-
нео Ь1^ аппаРат У гидры, по Н. Клейненбергу, представлен одной
Фео Н°'МЬ1Шечн°й клеткой, нервная и мышечная ткань еще не диф;
та^НЦ1ИР°ваны. В дальнейшем происходит поперечное деление
1 го Двойственно специализированного элемента, что ведет к
эволюции животного мира 33
появлению особых клеток нервных и особых мышечных, — появ-
ляются нервная и мышечная ткани.
По. пошли Гегенбаур, Геккель и др., раз-
вив ш^^^^е^но^мшп' еч ну ю » теорию, а среди'советских "гистоло-
гов А. Заварзин (1925, 1934, 1936, 1941, 1947).
Однако сначала А^Ко£отнев (£876), а затем О. и Р. Гертвиги
(1880) дали более др авил ьнде"тепр^Д^т а в лени я о строении покрова
гидры. Они пбказалиГчт^ клеток базально
располагаются особые, настоящие многоотростковые нервные клет-
ки, образующие нервное спДёпщи^^ ы элементах,
таким образом, говорильне приходится. Описанные Н. Клейненбер-
гом клетки представляю^только’ эпителиально-мышечные элементы,
так как кроющий эпителий в онтогенезе не утрачивается. Впослед-
ствии среди эпителиальных клеток были найдены единичные чув-
ствительные элементы, базальные отростки которых, ветвясь, стоят
в связи с нервным сплетением покрова.
Таким образом, О. и Р. Гертвиги выдвинули положение о дивер-
гентной эволюции чувствительных, нервных и мышечных элементов
из^рбщего^ недифференцированного зачатка. Они утверждали, что
гистологическая дифференцировка происходит не таким путем, что
одна клетка особенно развивает одновременно две функции, а затем
соответственно двум ставшим различными частям распадаются на
два функционально различных клеточных индивидуума; скорее это
постоянно уже особые, первоначально одинаковые клетки, которые
делят между собой работу для той или иной функции и особым
образом развиваются дальше. Процессы гистологической дифферен-
цировки покоятся це _на обособлении и дальнейшей самостоя-
тельности различно дифференцированных частей клетки, а на раз-
личной дифференцировке отдельных первонач альпо одинакова
клеток. ~ ~
Эти взгляды приняты большинством морфологов. В пользу их
говорят и более новые исследования относительно1 происхождения
нервных и чувствительных элементов при почковании гидры. Те и
другие образуются за счет недифференцированных, так называемых
промежуточных клеток, разбросанных обычно в базальной части
покровного эпителия. Вклиниваясь между эпителиально-мышечными
элементами, они либо дают начало веретеновидным чувствительным
кледвдм, образующим дистально чувствительный волосок, а прокси-
мально вытягивающимся в тонкие отростки к нервному сплетению,
либо, оставаясь базиэпителиальными, превращаются в двух- И
многоотростковые нервные клетки. Последние своими отростками
объединяются в нервное сплетение, но иногда, в виде исключения,
тот или иной отросток направляется к поверхности эпителия,
образуя перцепирующий аппарат. Таким образом, несомненна пер-
вичная связь нервных и чувствительных клеток и их дивергентное
развитие от одной исходной формы, тогда как сократительные .эле-
менты у гидры уже всегда обособлены от нервно-чувствительных-
Вышеупомянутые промежуточные клетки могут развиваться и я
34
овеошенно ином направлении, а именно: в протоплазме некоторы <
С них дифференцируются овальные капли секрета, дающего затем
начало так называемым стрекательным капсулам (рис. 23, Б).
_______ своеобразный оформленный секрет защитного типа, харак-
терный для всех кишечнополостных, объединяемых по этому приз-
наку также под общим названием «Cnidaria» — стрекающих». Да
пее промежуточные клетки, содержащие стрекательные капсулы,
передвигаются наподобие амебоцитов к оральному участку тела п
помещаются здесь среди эпителиальных элементов поодиночке или
целыми группами, образуя оригинальный аппарат защиты и наш -
деНИЯ__стрекательные батареи. При этом клетки достигают наруж-
ной поверхности эпителия и здесь образуют небольшой чувствитель-
ный вырост — книдоциль, при прикосновении к которому происхо-
дит выворачивание*нйти из стрекательной капсулы. Такой аппарат
защиты и нападения развивается у кишечнополостных очень разно-
образно — описано ДО' 20 форм стрекательных капсул. Они лока-
лизуются особенно обильно на -щупальцах. Последние становятся
очень эффективным специальным органом «активной» защиты, на-
падения и ловли добычи.
Все приведенные выше факты и выводы из них о дивергентной
эволюции тканей и органов в связи с разделением физиологических
отправлений, т. е. с дифференцировкой функций и структур, в силу
противоречивых тенденций, обусловленных различными сторонами
процессов обмена веществ и энергии в живом организме, — все эти
факты и выводы стоят в резком противоречии с положениями
А. А. Заварзина о закономерностях эволюции тканей, широко
пропагандировавшимися в последнее время, на которых поэтому
нельзя не остановиться. В частности, А. А. Заварзин утверждал
(1941), что гораздо глубже вопрос о происхождении нервной систе-
мы был в свое время поставлен Клейненбергом (1872), чем рабо-
тавшими после него и исправившими его наблюдения О. и Р. Герт-
вигами (1878, 1880). В соответствии с этим Заварзин отрицал
дивергентную эволюцию тканей и закономерность «раздё,пения
физиологического труда». По его словам, эта закономерность недо-
статочна, и прежде всего потому, что придает исключительное значе-
ние функции. На долю же дифференцирующейся системы остав-
ляется лишь роль инертной пластической массы, из которой внеш-
ние условия делают все. При этом не принимаются в расчет специ-
фические свойства самой дифференцирующейся системы, т. е.
специфические свойства живого вещества.
В этой связи должно иметь в виду, что, когда мы говорим о
разделении физиологических отправлений, то, в силу единства фор-
-ЛРлнкции, мы говорим о процессе изменения всей организации,
гим И стР°ения и отправлений дифференцирующейся системы. Дру-
ФоомСЛОВаМИ’ здесь Ч£ может быть и речи об отрыве функции от
цов Ы’ Э nPotle’cc охватывается во всей его полноте, и, в конце кон-
„ ’ всл-’1Цес значение внешних условий для развития процесса,
конечно, непререкаемо.
3* . _
Вместо общего закона для развития вторичных систем, т. е. тка-
ней и органов, закона разделения физиологических отправлений,
А. А. Заварзин формулировал закон эволюционного расщепления
гистологических структур и закон параллельных рядов. При этом, по
А. А. Заварзину, в расщеплении мы имеем дело с той закономер’
костью, которая лежит в основе параллельных рядов, так как парал-
лелизм рядов обусловлен эволюционным расщеплением. И то и дру-
гое являются лишь двумя сторонами одного и того же элементар-
ного эволюционного процесса, а термин «расщепление» выбран
вместо более индифферентного «дифференцировка». Следовательно,
иа.„место закономерности разделения физиологических отправлений
/Г. А. Заварзиным ставится лишь морфологическая дифференциров-
ка структур, т. е. он удовлетворяется ничего не объясняющей кон-
статацией факта вместо выявления физиологической его значимо-
сти, причем функция отрывается от формы.
В 1925 г., когда А. А. Заварзин формулировал свои «законы»,
они были в полном соответствии со стремлением найти особые пути
з явлениях конвергентного развития в противовес прежним дарви-
нистическим построениям, выдвинуть конвергентное развитие на
первое место и определить закономерности его путей как ведущие.
Это была эпоха «номогенеза» и «параллельных рядов наследствен-
ной изменчивости», и А. А. Заварзин формулирует свои законы
«параллелизма структур» и их «расщепления» как специальные
закономерности только эволюции тканей, как совершенно особые,
являющиеся закономерностями элементарного порядка, покоящиеся
на основных качествах живого вещества, связанные с обязательным
наличием специфических качеств у живых белков.
А. А. Заварзин в дальнейшем перерабатывает свои основные
положения и в 1941 г. он уже пишет, что эволюция органического
мира шла путем все большего и большего приспособления органи-
ческих форм к внешним условиям среды путем естественного отбо-
ра. Следовательно, одни и те же факторы эволюции должны были
привести и к появлению многообразия видов и к сохранению одно-
образия в строении тканей. Это произошло, по А. А. Заварзину,
потому, что ткани представляют собой такие биологические системы
организма, которые обусловлены его общим взаимодействием со
средой. А отсюда следует вывод: одинаковые ткани у всех живот-
ных, даже самых различных по своему происхождению, проделы-
вают эволюцию параллельными рядами по закону конвергентной
эволюции, и механизмом филогенетического усложнения структуры
является «эволюционное расщепление» ее, причем более универ-
сальные, менее точно работающие и менее многочисленные звенья
расщепляются на более многочисленные, менее универсальные,
но более точно и разнообразно работающие. В системе увеличи-
вается число элементов и уменьшается универсальность каждого из
них. Однако, по А. А. Заварзину, термином «расщепление» обозна-
чается самый факт усложнения и его историческая обусловленность,
совершенно никак не предрешая вопроса о механизме этого явления,
36
ирповательно, теряется всякая определенность. Больше того,
С\ А Заварзин говорит, что самый процесс расщепления остается
явлением мало понятным
Таким образом, у А. А. Заварзина, в конце концов, выпала^
лбшая для тканей и органов закономерность разделения физиоло-
^"чёских отправлений* и дифференцировки структур. Для тканей ои
нёШчнб "стремится заменить эту закономерность констатированием
«мало понятного» явления расщепления структур.
Собственно, от первоначалных построении 1925 г. у А. А. Завар-
зина в 40-х годах осталось очень мало, а именно многочисленные
факты конвергентного развития, которые он, обычно искусственно,
Рис. 6. Строение кутикулы дождевого чер-
вя Elsenia rosea (по К. Шнейдеру).
Плоскостной препарат:
1 — волокна кутикулы, образующие две взаимно
перекрещивающиеся системы, 2 — железистые
протоки между ними
стремится втиснуть в рамки
параллелизмов, как, напри-
мер, при сравнении нервной
системы членистоногих и по-
звоночных. Иногда это стрем-
ление Заварзина проявляет-
ся и рискованно, как, напри-
мер, при толковании брюш-
ной цепочки полихеты Ne-
phthys в параллель к кругло-
ротым. Однако, несмотря на
это, исследования А. А. За-
варзина дали много нового и
оригинального и проникнуты
эволюционным духом, что де-
лает их весьма ценными для
развития нашей гистологщ.
Еыше 'уже было указано,
что ткани являются состав-
ными частями организма, ко-
торые характеризуются определенными структурами. Последние
обусловлены в своем появлении теми разнообразными сторонами
обмена веществ и энергии, соответственно теми мицеллярными
структурами, которые присущи живой материи вообще. Это- имеет
силу в многоклеточном организме также для основного вещества
соединительной ткани и для кутикулы, т. е. для живых образова-
ний наболее простого типа, в которых индивидуализации нет,
ферментативные процессы, повидимому, не играют такой роли, как
в клетке, сократимость и возбудимость не выявлены, и о вполне
самостоятельном продолжении организации которых, т. е. о само-
стоятельной жизни, едва ли можно говорить.
Существенные элементы гистоструктур, представленные волок-
нистыми образованиями экзоплазмы, появляются в виде преколла-
генных, коллагенных, эластических и т. п. волокон основного ве-
щества или в виде волокон кутикулы. По отношению к ним действи-
льно можно говорить об «элементарных» закономерностях их
явления в многоклеточном организме. Натяжения определенного
37
I
характера в слое кутикулы земляного червя (рис. 6) или в субэпи-
телиальном слое ткани основного вещества у полигордиуса (рис. 7)
обусловливают дифференцировку чрезвычайно правильно проходя-
щих диагональных кутикулярных, соответственно преколлагенных
зол окон. Это — процессы, связанные с мицелярным строением
основного вещества или вещества кутикулы, и их закономерности —
элементарные закономерности появления гистологических структур.
Что касается клеточных элементов в основном веществе соедини-
;ельной ткани, то их тканевая дифференцировка связана с разделе-
Рис. 7. Фронтальный
срез через стенку тела
hoLjgot ,ць:
1 — кутикула, 2 — крою-
щий эпителий, 3 — погра-
ничная перепонка из ткани
основного вещества, содер-
жащая две взаимно пере-
крещивающиеся системы пре-
коллагенных волокон, 4 —
продольная мускулатура под
ней
нием физиологических отправлений: одни
накапливают экскреторные гранулы, другие
содержат резервные вещества, третьи стано-
вятся отлагающими кальциевые соли осте-
областами или дающими вещества, харак-
терные для образования хряща и т. д. и т. д.
Все эти дифференцировки развиваются jg
базе разнообразных сторон обмена веществ
в клетке.
Элементарные закономерности выра-
жаются прежде всего- в «специфичности
структур», т. е. постоянном соответствии оп-
ределенных процессов с определенной струк-
турой. Вакуома никогда не сможет служить
проведению возбуждения, а миофибрилла
для переваривания пищевых частиц. Где
причина этого специфичного соответствия,
этого выражения двух сторон одного и того
же процесса? Вероятно, она должна заклю-
чаться в известном единообразии воздейст-
вия механических, физических и химических
агентов на организацию живого вещества,
как сложной дисперсно-коллоидной системы
гидрофильно-белкового характера в прото-
плазме или образованиях экзоплазмы. Но- в
настоящее время мы еще не в состоянии
точно формулировать закономерности, результатом которых являет-
ся «специфичность гистоструктур». Мы рассматриваем их только
как две нераздельные стороны жизненного процесса, взаимообуслов-
ленные форму и функцию. Однако в работах советских биохимиков
(Энгельгардт, Бреслер и др.) уже выявляется ряд важнейших фак-
тов, вскрывающих в известной мере значение этой связи.
В отличие от дифференцировок основного вещества и кутикуляр-
ных, присущих только многоклеточным организмам, автоплазмати-
ческие клеточные структуры ведут свое происхождение от структур,
имеющихся уже в одноклеточных организмах, следовательно, они
характерны и для нижестоящей, по сравнению с многоклеточными,
ступени организации. У протистов, появившись на основе элемен-
тарных закономерностей, они эволюционируют, как органеллы.
38
Для многоклеточных миофаны, опорные волоконца и т. д. яв-
яются как бы этапом в эволюции мышечных элементов, приводя-
щим к поперечнополосатой мускулатуре членистоногих и позвоноч-
ных, к тонфибриллам и т. п. При этом выявление элементарных'кле-
точных закономерностей переходит на высшую ступень, получает
гораздо больший размах, становится дающим новое в условиях
развития тканей. Такова, например, эволюция глийной и нер-вной
тканей в мякотных нервных волокнах позвоночных или дифферен-
цировка клеток хряща и кости, действующих на окружающее основ-
ное вещество.
Если А. А. Заварзин перешел к чисто эктогенетическим пред-
ставлениям и почти совершенно отбросил свои первоначальные авто-
генетические концепции, оставив «расщепление» лишь как «малопо-
нятный процесс», то А. Н. Северцов (1939), также стоя на чисто
онтогенетических позициях, отдал все в организме во власть естест-
венного отбора, во власть закономерности системы организм — сре-
да. Система целостного организма и его частей с ее закономер-
ностью разделения физиологических отправлений и дифференциров-
ки структур у А. Н. Северцова выпала. Однако теснейшая взаимо-
связь обеих закономерностей привела А. Н, Северцова к тому, что
для него результаты процесса дифференцировки функций и структур
становятся только различными способами выражения естественного
отбора, или, как он их называет, «типами», или «принципами», фи-
логенетических изменений. Последние, по его словам, рассматри-
ваются с биологической точки зрения, т. е. центр тяжести переносит-
ся с анатомических изменений органов на их физиологическую сущ-
ность. При этом каждое морфологическое изменение органа оцени-
вается как способ развития,активной функции или пассивного при-
способления в полезном для всего организма направлении. Другими
словами, всякое морфологическое изменение оценивнается лишь по-
стольку, поскольку оно обусловливает собой выживание организма
в борьбе за существование и биологическое процветание вида. Во-
прос о движущих силах процесса дифференцировки функций и
структур совершенно отпадает, их место занимает детерминирующая
закономерность естественного отбора, и дифференциация, по
А. Н. Северцову, является необходимым общим последствием типов
филогенетических изменений.
Таким образом, для А. Н. Северцова задача заключается не в
1РМ, чтобы свести все известные ему филогенетические изменения
к общим основным принципам (интенсификация функций и мульти-
ФУНкциональность органов), ро в том, чтобы проследить во всех
Деталях и объяснить весь путь филогенетических изменений органов.
Д^Довательно, это — чисто описательный путь, ведущий А. Н. Се-
п2ЦОВа к Установлению, в конце концов, двенадцати «принципов»,
вернее —- -
органов,
и Функций»^'"
~ способов прогрессивных филогенетических изменений
и среди них последнего - «принципа разделения органов
39
Но все эти «двенадцать» способов представляют, собственно,
лишь различные формы и варианты проявления закономерности
«разделения физиологических отправлений», т. е. выявления проти-
воречивых тенденций обмена веществ и энергии в системе единого
целостного организма при образовании вторичных систем органов
и тканей.
И сам А. Н. Северцов, в конце концов, отмечает, что все типы
прогрессивного филогенетического изменения органов животных
покоятся на принципе интенсификации функций и па мультифунк-
циональности органов — т. е. вплотную подходит к закономерности
разделения физиологических отправлений, но не решается объеди-
нить как будто бы очень различные явления, охватываемые его
двенадцатью принципами. Это можно и должно сделать только на
основе признания того, что противоречивые тенденции, выражаю-
щиеся, прежде всего, в обмене веществ и энергии в организме, яв-
4 ляются результатом взаимодействия его с окружающей средой. В
единстве организма и среды те стороны последней, которые осваи-
ваются организмом, и будут ведущими в процессе его дифферен-
цировки. «Процесс органического развития как отдельного индиви-
да, так и видов путем дифференцирования является убедительней-
шим подтверждением рациональной диалектики»1, — сказал в свое
время Ф. Энгельс.
J Ф. Энгельс, Диалектика природы, стр. 162.
Глава II
ТИП КИШЕЧНОПОЛОСТНЫЕ — COELENTERATA
Анализ организации типов — адаптация и специализация. Значе-
ние филогенетического подхода к анализу организации. Теория
гастреи Геккеля. Характеристика типа кишечнополостных как имею-
щих только полость кишечника. Организация гидры. Гастрея по
Лангу. Происхождение гастреи. Паренхимелла Мечникова и бластея
Ланга. Происхождение многоклеточных. Выработка радиальной
симметрии и организация гидрополипа. Регенерация, почкование и
колониальность гидроидных полипов. Полиморфизм и диморфизм,
смена поколений. Гидромедуза. Организация сцифополипа. Сцифо-
медуза. Эволюция сцифополипа и сцифомедузы. Организация
Anthozoa. Итоги анализа организации кишечнополостных.
Проанализировав многоклеточный организм в отношении диф-
ференцировки в нем вторичных систем — фканей* и органов, мы
можем пойти в его понимании дальше по иному пути, а именно п<у
пути выявления главнейших компонентов организации и их ведущих
комплексов у различных типов животных, по пути выявления каче-
ственного своеобразия типов.
В основу кладется тот факт, что каждая черта организации пред-
ставляет, с одной стороны, результат взаимоотношений организма
и биоценотической среды в их противоречивом единстве, т. е. яв-
ляется так или иначе приспособительной. С другой стороны, она в
то же время представляет выражение специализации частей орга-
низма на основе дифференцировки функций и структур, на основе
разделения физиологических отправлений в системе целостного’
организма. При этом оба процесса взаимосвязаны и протекают в
единстве с неорганической средой, условия которой так или иначе,
в той или другой степени осваиваются организмом.
На этом мы строим паш анализ организации типов многоклеточ-
ных животных.
пые^ дальне™ем мы рассмотрим лишь те главнейшие этапы, кото-
терныаМаГИЛИСЬ В Рез'Ультате образования качественно новых харак-
Угл из прежде существовавших.
новых 6 ЛеНН°е п°пимание организации последовательного ряда ти-
ства И °РМ позволит нам уточнить пути эволюции животного цар-
филогене ° И теснейшим способом взаимосвязано' — без
ического подхода мы не можем правильно вести анализ'
41
организации, а этот последний в свою очередь дает нам ключ к
пониманию филогении.
Так, совершенно различны будут попытки анализа организации
губок в том случае, если мы видим в них представителей много-
клеточных животных, близких к типу целентерат, или если мы пред-
ставляем их в виде самостоятельной ветви животного царства, т. е.
как паразоа. В последнем случае естественна расшифровка особен-
ностей тканевой дифференцировки губок, дававшая основание ви-
деть в них «нетканевых животных» — «Zoa anhistia». Точно так же
понимание развития зародышевых пластов у губок возможно с иных
позиций, чем у прочных многоклеточных, откуда характерное их
название «Enantiozoa» — животные с извращенными пластами,
животные «шиворот-навыворот». И, наконец, выявляется особое
представление об индивидуальности у губок, откуда определение
«Zoa impersonalia» — животные, лишенные индивидуальности.
Итак, переходим к анализу типовых групп животного мира.
80 лет тому назад в «Монографии известковых губок» (1872),
а затем в «Этюдах по теории гастреи» (1873—1876) Э. Геккель
попытался набросать облик гипотетической исходной формы для
всех многоклеточных животных. И, приступая к анализу организа-
ции кишечнополостных, как низших многоклеточных, вполне есте-
ственно обратиться к его взглядам, получившим широкую изве-
стность и возбудившим в свое время много горячих споров.
Ядром целого ряда обобщений, которые Э. Геккель объединяет
под названием «теории^гастреи», является мысль, что, базируясь на
«основном биогенетическом законе», по которому история развития
особи повторяет историю развития вида, необходимо признать фи-
логенетическое значение за всегда имеющейся в онтогенезе стадией
гаструлы. Так как гаструла в различных типах животных представ-
лена или чисто палингенетической, т. е. первоначальной архига-
струлой, или сводимыми на нее ценогенетическими формами, следует
далее заключить, что, все метазоа происходят от единственной об-
щей предковой формы, сходной в существенных чертах организации
с архигаструлой (рис. 8, Б), Свободно двигавшаяся посредством
жгутов или ресничек покрова, она отличалась от теперешних архи-
гаструл только дифференцировкой половых клеток, что делало ее
взрослым половозрелым животным. Эту первичную, давно вымер-
шую форму, которая жила в докембрийский («лаврентийский»)
период существования Земли и, вероятно, была представлена в
свое время различными видами, Э. Геккель назвал «гастреей» и
соответственно всех вымерших самых древних метазоа, ближайших
к гастрее, объединил в группу гастреад (Gastreadae).
Непосредственным предком гастреи, по Э. Геккелю, была «пла-
чея» или «бластея», повторяемая в онтогенезе студией планулы или
42
^1астулы (,рис, 8, Л). При разделении физиологических отправле-
ний между однородными еще элементами бластеи‘дифференцирова-
лись двигательные клетки покрова и клетки, служащие для захва-
тывания и переваривания пищи. Последние образовывали впячи-
зание, где пища могла дольше задерживаться и перевариваться.
дто — зачаток первичного кишечника, в противоположность дви-
Рис. 8. Схема бластеи (Л) и гастреи (Я) по Геккелю;
В, Г.— прикрепление и выработка организации гидро-
идного полипа; л — схема организации гидры:
1 — стрекательные батареи на щупальцах, 2 — выстилка
кишечника, 3 — покров, 4 — опорная пластинка (черная)
соот-
экто-
гательному покрову. Так возникли два первооргаиа гастреи,
ветственно в онтогенезе первичные зародышевые пласты —
дерма и энтодерма гаструлы.
В дальнейшем потомки гастреи эволюционировали по двум на-
правлениям в зависимости от приспособления к различному образу
Жизни- Одни оставляют свободноподвижное существование, прикре-
ляются аборальным полюсом и вырабатывают «тем самым», как
ч °Р'ИТ Э. Геккель, «радиальный план» построения (кишечнопо-
заюНЫе^ (рИС‘ 8,В> другие переходят от плавания к пол-
посто МУ °браз^ жизни на Дне МОРЯ и Дают «билатеральный план»
иттЛтТ;г°ения (чеРви и четыре высших типа — моллюски, иглокожие,
Так°Н°ГИе’ гоночные).
ляет по*** Обра'ЗОМ те°Рия гастРеи> по словам Э. Геккеля, представ-
ПЫТКУ на основе биогенетического закона и учения о заро-
43
дышевых пластах «причинно» обосновать «естественную систему жи-
вотного царства, понимание развития его типов и естественного
родства его главных групп».
Хотя подобные взгляды высказывались и раньше, однако цело-
стная концепция Э. Геккеля в 70-х годах прошлое столетия была
принята большинством зоологов как блестящее обобщение, откры-
вавшее широкие перспективы, и до> настоящего времени геккелев-
ская гастрея пользуется большим вниманием. Но уже в 80-х годах
Н. Клейненберг (1886) указывал на чрезвычайную схематичность
построений Э. Геккеля. По его остроумному выражению, гастерея—
только «тощий призрак животного Лаврентийских ночей».
И. Мечников (1882, 1886) считал теорию гастреи в общем стояв-
шей ниже уровня эмбриологии 80-х годов, которая обладала уже
большим запасом фактов, не учтенных Геккелем, и стремился пока-
зать, «что гастрея не представляет первичную форму многоклеточ-
ных животных, но является одним из более поздних состояний». Он
выдвинул свою теорию происхождения метазоа, теорию «паренхи-
меллы».
Чтобы иметь правильное представление об этих вопросах, уже
отчасти затронутых в первой главе, необходимо обратиться к фак-
там. Просмотрим в этой связи организацию типа Coelenterata —
кишечнополостных.
Самим названием типа должна подчеркиваться характернейшая
черта организации всех относящихся сюда форм — наличие у них
единственной полости в теле, полости кишечника. Однако* Р. Лей-
карт (1849), выделивший этот тип, понимал отношения иначе, счи-
тая, что в полости кишечнополостных объединены и система ки-
шечника и система собственно полости тела — целом и энтерон. В
настоящее время такое толкование приемлемо только для
приверженцев теории происхождения вторичной полости тела путем
отшнуровывания от кишечника, — так называемой «энтероцельной
теории». В дальнейшем мы оценим эту категорию гипотетических
построений, в данный же момент просто выдвинем положение, что
настоящие полости тела, и первичная, как у ротаторий, и вторичная,
как у аннелид, ничего общего с кишечником Coelenterata не имеют.
Следовательно, для последних характерно, как уже подчеркивал
Э. Геккель, именно отсутствие какой бы то ни было системы поло-
сти тела и наличие единственно системы кишечника, в которой да-
же потенциально не заложено ничего подобного целому, а тем более
так называемой «первичной полости тела».
Эта черта организации подчеркивает примитивность строения
всего типа и выражена в Геккелевской схеме гастреи; однако вне
связи с прочими системами органов нельзя полностью оценить эту
черту. Поэтому обратимся опять к классическому объекту многих
замечательных исследований, к одному из низших представителей
типа кишечнополостных — пресноводной гидре (рис. 8, Д).
Прежде всего, как мы уже видели, гидоа имеет вполне выра-
женную дифференцировку трех первичнЙхорганов: двух клеточ-
44
покрова, 'Кишечника и опорной
органы адаптивного характера:
конца полые выросты стенки
нападения, защиты и захватыва-
— подошву, служащую для при-
ых __ покрова и кишечника и одного пеклеточного — опорной
пластинки. Далее, гидра обладает постоянными тканевыми диффе-
ренцировками (рис. 3), по чрезвычайно своеобразного характера.
Именно эпителий покрова и кишечника является комплексным, и в
частности сократительным, так как образован эпителиально-мышеч-
ными клетками, т. е. мышечная ткань еще не выделена в самостоя-
тельную систему; вполне дифференцированная нервная ткань
располагается в том же эпетцлии базально; ткань основного веще"
ства не имеет особого камбия и представлена обособленной некле-
точной студенистой прослойкой, производимой обоими эпителиями.
Наконец, гидра является вполне оформленной индивидуальностью,
свойственной многоклеточным животным. Все компоненты тела
гидры объединяются в единое целое не только механически и хими-
чески через основное вещество опорной пластинки, как это мы
видим у губок^нр и с помощью специальной клеточной дифферен-
цировкиГЪредставляющей простейшую нервную систему.
Следует оговорить, что, кроме —— ~ ------л
пластинки, гидра имеет и другие
расположенные вокруг орального
тела •— щупальца, служащие для
ния пищи, а на аборальном конце
крепления животного к субстрату и для движения. Однако у бли-
жайшего родича гидры, хорошо изученной Protohydra, нет ни того,
ни другого, и она, можно сказать, идеально представляет наиболее
простую форму гидроидного полипа.
Покров гидры нами уже был рассмотрен. Отметим, что мульти-
функциональное состояние покрова гидры очень характерно для
исходных форм многоклеточных. Покров гидры отграничивает орга-
низм от. внешней среды, пассивно и активно защищает его, воспри-
нимает падающие извне раздражения, производит сокращения тела,
ведает газовым обменом, получая кислород из внешней среды и
отдавая туда углекислоту, и, наконец, выполняет экскреторную
Функцию, выделяя наружу жидкие продукты распада, получаю-
щиеся при жизнедеятельности его клеток. Кроме того, в покрове же
заключено диффузное сплетение нервных клеток, представляющее
примитивнейшую нервную систему, и локализованы скопления поло-
вых клеток: — примитивнейшие гонады.
Равным образом мультифункционален и другой клеточный
первоорган гидры — кишечник. Он также образован эпителиально-
мышечными элементами. Однако, сохраняя более примитивный ха-
рактер, чем покровные, последние несут на свободной поверхности
один-два жгута. Пищевая частица прикасается к протоплазме таких
гослрК И обволакивается ею. Так происходит принятие пищи с
куолеА^ЮШ'ИМ Свариванием и усвоением в пищеварительной ва-
от У простейших. Это — примитивнейшее, унаследованное
УсложИШИХ внутриклеточное пищеварение.
ДифференпиНИе выРажено> однако, в том, что в кишечном эпителии
Р ОДрованы железистые клетки, вырабатывающие секрет,
45
действующий на пищу еще перед захватыванием ее внутрь клеток.
Таким образом, пищеварение является в известной мере полостным,
хотя анатомически кишечник представляет только мешок, происхо-
дящий за счет эктодермального слоя зародыша, с ротовым отвер-
стием, служащим как для. введения пищи, так и для выбрасывания
непереваримых остатков. Это — примитивнейший, так называемый
«целентерический» аппарат.
Вполне естественно, что у кишечника отсутствуют присушке
покрову функции — изоляционная, защиты и нападения. Но прочие,
в дополнение к главной, пищеварительной, функции, сохраняются
Широкопетлистым бдзиэпителиальным нервным сплетением пред
ставлена нервная система кишечника. Что касается подрвых эле-
ментов, то у ближайших родичей гидры — морских гидроидных
полипов, они временно мигрируют из покрова в кишечный эпите-
лий, где растут за счет обильного притока пищевого материала,
после чего снова уходят в эктодермальный эпителий.
Лишь неклеточный орган опорной пластинки является более
ограниченным функционально. Но и в нем объединены две, на пер-
вый взгляд взаимоисключающие, функции: 1) опоры и механиче-
ской связи, что требует известной прочности й плотности образова-
ний, передачи питательного материала от моста его выработ-
ки — кишечника — к покрову, что у высших типов животных в
значительной мере связано с жидкой тканью крови и лимфы.
^по^на^пластинка выполняет функции, характеризующие преж-
де всего именно многоклеточный организм, — механическую и хи-
мическую связь клеточных первоорганов. Однако представлена она
лишь бесструктурной прослойкой студенистого вещества, сжимаю-
щегося при сокращении мускулатуры покрова и кишечника и по
окончании сокращения восстанавливающего благодаря своей упру-
гости и эластичности прежнюю свою “форму. Создается постоянство
формы тела, чего не могут дать клеточные первоорганы, образован-
ные пластичной массой протоплазмы. Опорная пластинка, происхо-
дящая за счет обоих клеточных слоев тела, функционально и
морфологически представляет очень важную составную часть тела.
И если орган определять как обособленную часть организма, с ту-
жащую для выполнения специальной функции, то опорная пластин-
ка представляет особый неклеточный орган, так как функции связи
и опоры не присуши ни покрову, ни кишечнику. Это — неклетлч-
иый первоорган связи и опоры.
Господствовавшая многие годы в науке узкоортодоксалгная
форма клеточной теории, которая ничего в организме, кроме клеток,
не хотела видеть, не могла оценить значения этого неклеточного
первооргана — опорной пластинки, не уясняла себе филогенетиче-
ской важности подобных образований. Однако с эволюционной
точки зрения перед нами тот компонент многоклеточного организма,
появление которого обусловило самую возможность развития много-
клеточных животных из одноклеточных.
Этим мы естественно подошли снова к jpggTpee, но здесь рас-
46
смотрим уже не первоначальную схему Э. Геккеля, а модернизиро-
ванную гастрею, описанную 40 лет спустя А, Лангом (1912) в его
«Руководстве по морфологии беспозвоночных» (см. рис. 9).
Рис. 9. Схема гастреи А. Ланга:
1 — чувствительные волоски на чувствительных элементах
среди мерцательного эпителия, 2 — интерстициальные клет-
ки среди прочих эпителиально-мышечных. 3 — нервные
клетки, базиэпителиально, 4 — ядовитые железки, 5 — экск-
реторные клетки, 6 — задний полюс тела. 7 — яйцо, выхо-
дящее из овария, 8 — полость кишечника, выстланная
жгутиковыми эпителиально-мышечными и железистыми
клетками. Черным обозначено студенистое вещество
гидпп ^анг представляет себе гастрею в виде свободноплавающего’
подо1пвДОбм°ГО °Рганизма эллипсоидной формы, без щупалец и
ротовое**1 пеРеднем ПРИ движении конце находится круглое
маюпгую°ТВеРСТИе’ веДУВДее в объемистую кишечную полость, зани-
мальным Г внУтРеннее пространство тела. Она выстлана энтодер-
питедием, .в составе которого преобладают усваивающие
47
жлетки эпителиально-мышечного типа, но также содержащие ре-
зервные вещества, или, ближе к ротовому отверстию, железистые
клетки. Базально среди этих клеток располагаются нервные эле-
менты и интерстициальные клетки. Последние служат для регенера-
ции поврежденных участков стенки кишечника. Таково строение
этого первооргана гастреи, соответствующего архентерону гаструлы,
лишенному какой-либо дифференцировки.
Другой первоорган образован покрывающим наружную поверх-
ность гастреи эктодермальным эпителием. В нем преобладают
мерцательные эпителиально-мышечные клетки. Но среди них рас-
полагаются чувствительные, железистые с ядовитым защитным сек-
ретом и, наконец, мерцательные экскреторные элементы. Базально
в эпителии находятся нервные клетки и, наконец, интерстициаль-
ные, служащие для регенерации покровного эпителия.
Э. Геккель считал гастрею двухслойным организмом. А. Ланг
идет дальше. Он полагает, что половые клетки были у гастреи
обособлены от первоорганов покрова и кишечника и дифференци-
рованы в виде третьего первооргана — гонады, располагавшейся
между эктодермальным и эктодермальным слоями и состоявшей из
клеток как эволютивного, так и абортивного характера. Таким обра-
зом в гастрее имелась, по А. Лангу, «средняя, или мезодермальная,
масса половых клеток». Следовательно, гастрея была трехслойным
организмом, если не принимать во внимание студенистую погранич-
ную перепонку, которая охватывала гонады и на которой распола-
гались кроющий и кишечный эпителии. Она имела достаточную
для поддержания определенной формы тела консистенцию и доста-
точную эластичность для возвращения тела к нормальному состоя-
нию после той или иной деформации. Кроме этих кратких указа-
ний, А. Ланг не уделяет сколько-нибудь внимания опорной пла-
стинке и представляет ее пронизанной протоплазматическими
мостиками, соединяющими все клетки организма между собой.
Конечно, в гастрее А. Ланга недооценено значение опорной
пластинки как неклеточного первооргана связи и опоры и переоце-
нено значение гонад как среднего клеточного слоя, так как у кишеч-
нополостных еще нет подобной дифференцировки, и половые клетки
располагаются в составе того или другого эпителия. Наконец, в
гастрею А. Ланга фантастически много внесено от организации
гидроидного полипа, например ядовитые железы или мышечные
образования в виде эпителиально-мышечных элементов. А. А. За-
хваткин (1949) в значительной мере прав, когда говорит, что пора-
зительные гипотетические организмы, составленные таким способом
Лангом, могут служить образцом беспочвенных филогенетических
реконструкций. Поэтому вернее держаться ближе к геккелевской
гастрее, так как организация этой исходной формы все же остается
для науки гипотетической в деталях, будучи несомненной в общих
очертаниях.
Тот факт, подчеркнутый А. Лангом, что гастрея по своей орга-
низации является, собственно, одинаковой с гидроидным полипом
48
вроде Protohydra, т. е. взрослой формой кишечнополостных, а также
факт нахождения гаструлы в онтогенезе этого типа только у более
сложно построенных форм (некоторые Scyphozoa и Anthozoa), тогда
как у всех низших (Hydrozoa) она отсутствует, заставлял многих
исследователей, начиная с И. И. Мечникова (1882), с полным осно-
ванием сомневаться и отрицать правильность предположения, что
гастрея должна считаться исходным, первичным многоклеточным
организмом. Филогенетическое значение гастреи общепризнано, но
она является лишь п^сд^щщм звеном в той цепи форм, которая
вела от колониальных простейших к целентерическим животным,
таким,"собственно, и должна считаться гастрея. Поскольку кишечно-
полостные оказываются низшими из ныне существующих многокле-
точных, постольку мы должны признать и значение гастреи в
филогении.
Но имеется ли основание полагать, что гастрея получилась в, ре-
зультате крайне простого процесса впячивания одной части шаровой
поверхности под другую у бластеи, стоявшей на границе Протозоа
и Метазоа? — это вопрос. Э. Геккель слишком упрощенно пред-
ставлял себе ход вещей. В данном случае И. Й. Мечников был
совершенно прав, указывая, что процесс впячивания совершенно- не
характерен для низших кишечнополостных. В соответствии с этим
разные авторы ставили на первое место при образовании энтодермы
в онтогенезе то деламинацию, то иммиграцию, придавая при этом
соответствующее толкование и филогении.
Образование гаструлы путем деламинации представляет доволь-
но редкий процесс, сводимый на иммиграцию, поэтому не будем
останавливаться па связанных с деламинацией интересных, но мало
приемлемых теоретических соображениях. Сконцентрируем внима-
ние на явлениях иммиграции.
Наиболее близок к инвагинации, как показал И. И. Мечников,
широко распространенный у свободно развивающихся представите-
лей Hydrozoa процесс унипоуш^^ и, несоменно, раз-
витие гаструлы можно считать лишь дальнейшим шагом в этом
направлении. На филогенетическую важность процесса униполяр-
ной иммиграции указывает присутствие его как раз у свободножи-
вущих зародышей, т. е. у проводящих период своего эмбриональ-
ного развития в тех приблизительно условиях, в которых суще-
ствовали взрослые формы предков. У зародышей, развивающихся
под защитой кутикулярных оболочек, имеет место1 мультиполярная
иммиграция.
Биологическое значение этого факта вполне убедительно выяс-
няется наблюдениями над личинками кишечнополостных. Э. Кор-
Kofi1^) И Фейдер (1910) приводят подобные наблюдения над губ-
ппи 3уСаГе^а' ^ичинка движется всегда вокруг своей главной оси;
пути МеРВДтельное движение ее клеток гонит попадающиеся на
потюсуеЛКИе ЧасТИЦЫ по П0Л0Г|°и спирали к заднему при движении
время в Те^а’ где и собираются, задерживаясь на некоторое
о разующемся здесь мертвом пространстве. Таким образом
4 Пути эволюции животного мира 49
создается место, наиболее удобное для захватывания пищевых
частиц располагающимися здесь клетками — «оральный» полюс,,
тогда как на остальном протяжении тела захватывание почти не-
возможно вследствие движения частиц. Этот показательный биоло-
гический момент представляется очень важным. Он должен был
обусловливать у исходных форм — колониальных простейших —
такое разделение физиологических отправлений между одинаковыми
сначала клетками, что клетки, располагавшиеся у заднего полюса,
становились элементами, захватывающими пищу и затем, мигрируя
внутрь, переваривающими ее для нужд организма. Прочие же клет-
ки играли роль главным образом двигающих и чувствительных эле-
ментов. Так создавалась предпосылка возникновения,' углубления и
дальнейшей дифференцировки кишечника.
Как видно, представления А. Ланга о движении гастреи ротовым-
отверстием вперед неприемлемы, так как отдаленная аналогия с
инфузориями (например Stentor), имеющими при мерцательном
движении рот у переднего конца тела, мало соответствует рассмо-
тренным фактам.
Неудачны и более новые представления А. Нефа (1931) ж
Н. Г. Хлопина (1946), которые допускают, что шарообразная исход-
ная форма, двигаясь по богатому пищей дну наподобие очень мел-
ких ресничных червей, при уплощении нижней стороны тела могла
на месте захватывать пищевые частицы всей поверхностью. Снача-
ла временное, а потом постоянное впячивание последней и дало
кишечник гастреи. Подобные схемы совершенно неприемлемы пото-
му, что не стоят в соответствии с фактами, наблюдающимися при
движении бластулы и гаструлы и им соответствующих личинок.
Набросав общими штрихами вероятный путь к гастрее. мы по-
дошли к принимаемому обычно за отправную точку в развитии мно-
гоклеточных бластулоподобному организму, бластее Э. Геккеля или
паренхимелле И. Мечникова. Такой организм уже мыслится лишен-
ным дифференцировки первоорганов. Это — шарообразный или
овальной формы организм, все клетки которого, располагающиеся
по поверхности в виде эпителиального слоя, одинаковы. Внутри же, в
студенистой, бесструктурной массе, находятся амебоидные элементы.
По И. Мечникову, последние и служили для переваривания пищи.
По этому воззрению, первичное состояние энтодермы есть стадия
паренхимная, т. е. простого скопления клеток, расположенных без
особого порядка. Отсюда название «паренхимеллы» для гипотетиче-
ской исходной формы многоклеточных, которые предлагает И. Меч-
ников. Далее он его меняет, вводя вместо чисто морфологического
понятия, связанного с паренхимным строением, широко биологиче-
ское понятие «фагоцителла», связанное с представлением о деятель-
ности клеток — захватывании пищевых частиц и дальнейшей ми-
грации. Такая форма всего ближе может быть выведена, по И. Меч-
никову, из колоний жгутиковых простейших вроде Proterospongia
(см. рис. 5). Последняя представляет неправильной формы комочек
студенистого вещества, в котором у поверхности расположены от-
50
дельные особи воротничковых жгутиконосцев; из них образуются
амебоидные особи, уходящие внутрь студня для переваривания за-
хваченной пищи и затем для размножения.
В настоящее время некоторыми исследователями выдвигается
предположение, что Proterospongia в действительности представляет
1
Рис. 10. Схема бластеи А. Ланга. Сзади на теле дефект, воспол-
няемый регенерационными клетками. Черным представлен сту-
день колонии, пронизываемый межклеточными протоплазматиче-
скими мостиками. Внутри половые элементы
не колониальную форму воротничковых жгутиковых, а лишь кусочек
тела одной из известковых губок. Однако едва ли можно считать
вопрос решенным, и теоретическая сторона взглядов Мечникова от
этого не меняется.
А. Ланг и большинство других исследователей рисуют гипотети-
ческую бластею иначе наподобие Volvox (рис. 10). Клетки, обра-
зующие ее, расположены, по Лангу, эпителиально, в студенистом
4+ 51
основном веществе у поверхности. Л внутри студня находятся бли-
же к эпителию регенерационные, резервные, элементы, глубже ~
половые. Все клетки соединяются друг с другом протоплазматиче-
скими мостиками, проходящими через основное студенистое веще-
ство. Движение тела происходит постоянно одним полюсом впе-
ред. Поверхностные клетки более или менее одинаковы между со-
бой. У клеток имеются хорошо развитые жгутики. Служа главным
образом для движения, они посредине на теле и сзади становятся
многочисленнее и напоминают реснички. Каждая клетка захваты-
вает пищу или посредством клеточного рта или, наподобие амеб,
псевдоподиями и переваривает ее. Каждая клетка обладает сокра-
тительной экскреторной вакуолью и капельками ядовитого секрета,
выделяемого при защите и нападении. Некоторые из жгутиков кле-
ток передней части тела дифференцируются для восприятия раздра-
жений.
Такая бластея значительно отличается от паренхимеллы И. Меч-
никова, в основном и прежде всего, значением внутренней массы,
воплощающей в себе собственно половой зачаток, гонаду гастреи
А. Ланга. Как было выше отмечено, И. Мечников учитывает факты
онтогенеза низших кишечнополостных, принимая миграцию внутрь
студня для клеток, переваривающих пищу. И его пареихимелла
гораздо больше соответствует уже рассмотренному нами возмож-
ному ходу эволюции гастреи многоклеточных.
Итак, бластея Э. Геккеля является буквально перенесением са-
мой ранней стадии эмбрионального развития многоклеточных жи-
вотных в начальный этап филогенеза, представляя простую схему
архибластулы. Однако в ряду предков такая форма едва ли могла
существовать, так как процесс эпителизации, соответственно обо-
собления внутренней среды организма, должен был совершаться
одновременно- с дифференцировкой клеток в покровные и питаю-
щие. Другими словами, переход от колоний простейших вел уже
прямо к прогастрее с униполярной иммиграцией питающих клеток
у заднего полюса, а затем и с инвагинацией, что подчеркивает
И. И. Мечников. Недифференцированные элементы располагались
всюду в эпителии базально, в виде интерстициальных клеток, как
мы это находим еще у гидры и гидроидных полипов. Постоянно обо-
собленных гонад не было, а интерстициальные клетки при опреде-
ленных условиях переходили в состояние половых элементов, при
других же условиях, например при регенерации, превращались в
другие элементы тела. Таким образом, единый эпителий был еще
самым примитивным тканевым образованием — «архибластом» с
определенной дифференцировкой клеток, в противоположность не-
клеточному внутреннему студенистому основному веществу эксо-
плазмы — «парабласту».
В последнее время А. А. Захваткин (1949), развивая идеи И. И.
Мечникова, значительно углубил наши представления о начальных
этапах онтогенеза метазоа. Используя богатый фактический мате-
риал, накопленный протистологами, он показал, что предками мно-
52
гоклеточных могли быть только представители жгутиковых, при-
надлежавшие к группе Protomonadina. Цикл их жизни слагался из
тех же 4 -периодов, как у протомонад; 1) прогамного (гаметоге-
нез), дающего яйцевую клетку и спермин, 2) сингамного — поло-
вого, 3) метагамного, представленного дроблением оплодотворен-
ного яйца и заканчивающегося образованием бластулы, соответст-
венно — синзооспоры у простейших, 4) вегетативного, включающего
весь остальной отрезок развития и формирования половозрелой осо-
би, соответственно' — ряд делений у простейших.. Поэтому сингам-
пый характер жизненного цикла многоклеточных и его общая кон-
струкция не могут считаться следствием их многоклеточности,
результатом ее возникновения. Напротив, оформление дифференци-
рованного цикла у простейших явилось предпосылкой возникнове-
ния многоклеточности. Противоположная трактовка цикла Metazoa,
как выработанного ими новообразования, по мнению А. А. Захват-
кина, не встречает никакой поддержки в относящихся сюда фактах.
Но А. А. Захваткин не может среди простейших указать формы, ко-
торые бы осуществили полностью метазойный цикл, хотя все его от-
дельные периоды у них — несомненный факт и принятие его в
целом вполне оправдано.
Дальнейшей важной предпосылкой для возникновения организа-
ции многоклеточных животных, по А. А. Захваткину, явилась вы-
работка у жгутиковых палинтомищ т. е. повторения быстро1 следую-
щих друг за другом делений без периода питания и роста клетки
между ними. Это связано с гипертрофическим состоянием исходной
материнской клетки после полового1 процесса и последовательным
уменьшением размеров у ее потомков. Таким образом, создается
прочная предпосылка процесса дробления у многоклеточных. А
вместе с тем, добавим мы, создается и его экологическое обоснова-
ние: а именно постоянная палинтомическая колониальность выра-
боталась у простейших как защитное приспособление, предохра-
няющее делящихся особей от нападения мелких хищников.
А. А. Захваткин приравнивает, наконец, получающуюся в ре-
зультате дробления бластулу синзооспоре, т. е. расселительной ста-
дии сидячих простейших, и только дальнейшие состояния будут
соответствовать вегетативной стадии одноклеточных с обычным де-
лением и войдут в онтогенез многоклеточных как первые его шаги
после бластулы у сидячих исходных форм. Таким образом, бластула
в их развитии является, по- А. А. Захваткину, с самого начала пер-
вичной свободной личинкой, тогда как дальнейшие состояния свя-
заны с образованием сидячих взрослых животных.
Мы не будем углубляться в детальный разбор этих последних
с°ображений А. А. Захваткина. Отметим только, что вполне допу-
стимы и другие возможности развития многоклеточных организ-
мов, чем рассмотренные им для вольвоксовых, губок и книдарий. А
именно, для гребневиков- и червей, как будет показано далее, при-
водится допустить существование свободноживущих, несидячих
исходных форм, и потому пет необходимости вести путь развития
53
многоклеточных обязательно через сидячие колонии простейших и
соответственно допускать некоторые вторичные явления в качестве
необходимых для всех, например позднее образование просвета ки-
шечника и рта у планулы и т. п.
Мы выше уже видели, что возможно через планулу—паренхи-
меллу ?Лечникова с униполярной иммиграцией перейти к инвагина-
ционной гаструле и на прочной основе характера движения и пита-
ния зародышей перейти без обязательной бластеи к гастрее. Но от
этого общая ценность уточнения вопроса о происхождении много-
клеточных организмов в том направлении, как мы находим у
А. А. Захваткина, ничуть не уменьшается и дает значительное уг-
лубление наших представлений о ходе процесса, как он был нами
изложен выше.
Выяснив вопрос об исходных формах многоклеточных животных,
возвратимся к типу кишечнополостных.
Уже Э. Геккель отметил, что гастреады эволюционировали, при-
способляясь к различному образу жизни, по двум путям — одни,
сохраняя свободное существование, становятся ползающими по дну
формами, другие прикрепляются аборальным участком тела к суб-
страту и делаются сидячими организмами. Очевидно, оставался и
третий путь — пелагическое существование в свободновзвешенном
состоянии, как жили их предки. В этих условиях, как мы видели,
уже у гастреи имелось движение постоянно одним полюсом вперед.
Соответственно имелась одна особая ось тела, проходившая через
передний и задний полюсы. Создалось так называемое монаксонное
(одноосное) гетерополюсное состояние. Биологическое значение его
ясно. Это широко адаптивная черта организации, позволявшая жи-
вотному использовать наиболее продуктивно, в смысле питания и
защиты, свое жизненное пространство — свободную водную стихию
с взвешенными в ней животными и растительными организмами.
Возможность использования придонных пищевых ресурсов, особен-
но обильных на дне моря, ведет к донному существованию: к пол-
занию по дну или прикреплению на субстрате. При этом на основе
монаксонного гетерополюсного строения развертываются адаптив-
ные явления иного порядка, вырабатываются особые планы построе-
ния. Сидячие формы приспособляются к питанию за счет организ-
мов, опускающихся из верхних слоев водной толщи; происходит
прикрепление аборальным концом, оральный подставляется под
обильно падающие пищевые частицы. Однако уже у гидроидов вы-
работались приспособления для использования в пищевом отноше-
нии более широкого пространства и более крупной и активной до-
бычи, а именно образовались выросты стенки тела около ротового
конца — щупальца, служащие для активного захватывания пищи.
В одно и то же время они служат в качестве органов и защиты, и
нападения, обладая массой стрекательных батарей. Эти адаптивные
образования присущи всем кишечнополостным и, как норма, рас-
полагаются по радиусам вокруг ротового отверстия, причем центром
является проекция главной оси монаксонного тела. Такое радиаль-
54
ное расположение необходимо следует из простого биологического
факта; прикрепленное животное в силу отсутствия возможности
перемещения тела должно равномерно по окружности располагать
свои органы нападения и защиты, так как шансы нападения врага
или захвата добычи одинаковы вокруг орального конца. Вообще же
образование щупалец наиболее важно именно у орального отдела.
Так у сидячих, полипоидных форм кишечнополостных путем есте-
ственного подбора создался радиальный план расположения орга-
нов и прежде всего органов взаимодействия с внешней средой, а
затем и прочих. У гидры он выражен очень примитивно, у гидроид-
ных полипов часто гораздо более резко с их большим количеством
шупалец, но наиболее полно он представлен в более сложной орга-
низации Scyphozoa и Anthozoa, о чем речь будет при рассмотрении
этих групп.
Другой адаптивной чертой гидроидных полипов при условии,
что сидячее животное никуда не может уйти от врага, являемся вы-
работка пассивных защитных образований покрова в виде толстого
кутикулярного слоя, делающего животное в той или иной мере
неуязвимым для нападающего. Приспособившись к жизни в прес-
ных водах, гидра почти не имеет здесь серьезных врагов, способ-
ных противостоять ее батареям стрекательных элементов, и у нее
кутикулярный покров рудиментарен, имеется только зернистый обо-
док в клетках, утративших уже у давно* прикрепившихся предков
характер мерцательного эпителия. У морских форм, живущих в
условиях гораздо' более острой борьбы за существование, защит-
ная кутикула развивается настолько сильно, что у Thecophora, об-
разуя чашечку, прикрывает и несущий щупальца участок тела.
Первоначально, же он был лишен ее, защита велась аппаратом
щупалец. Конечно, развитие чашечки вносит своеобразные измене-
ния и в самую жизнедеятельность обладающего ею полипа. Мы не
будем здесь вдаваться в детали эволюции кутикулы, чашечки и
т. п., как и щупалец и стрекательных элементов. Направляясь из
одного исходного пункта по различным путям, все они представ-
ляют очень большое разнообразие узких приспособлений к разным
условиям существования. Выражаясь Дарвиновским термином, на-
лицо многообразное «дивергентное» развитие. Они не затрагивают,
не меняют организации по существу, а пригоняют ее к определен-
ным узким рамкам того- или иного биоценоза и биотопа.
Другим широким приспособлением гидроидных полипов, обус-
ловленным в значительной мере простотой организации и делаю-
щим животное способным противостоять многим неблагоприятным
воздействиям, является чрезвычайная способность к регенерации,
которая со времени исследований еще XVIII века прославила гид-
РУ- Выражаясь в способности восстанавливать поврежденные и ут-
раченные части, она достигает своего кульминационного пункта в
разрастании какого-либо- участка стенки тела, которое ведет к диф-
ференцировке
Щупалец дает
почки; последняя при образовании рта и венчика
новую особь. Таким образом возникает бесполое раз-
множение — процесс значительно повышающий способность сохра-
нения вида, теряющего массы особей в борьбе за существование.
При наличии плотной защитной кутикулы почкование, во-пер-
вых, ограничивается определенными отделами тела, где такая ку-
тикула отсутствует, во-вторых, ведет к объединению почкующихся
особей в своеобразные колонии, отличающиеся тем или иным типом
роста в зависимости от характера почкования. Таким образом диф-
ференцируется ветвящийся столон, иногда образующий сложные
сплетения и как-бы ползущий по субстрату. Иногда колонии под-
Рис. 11. Колония Hydractlnia (по Гн-
ман):
1 — гастрозоиды, 2 — гонозоиды, почкующие
медузоидов женских (3) и мужских (4), 5 —
шипы, 6 — спиралозоиды
нимаются в виде кустика или
деревца с характерным разви-
тием ветвей, на которых рас-
полагаются отдельные особи
полипов. В разнообразии форм
колоний, частью являющихся
прямым ответом на условия су-
ществования в том или другом
биотопе, например в условиях
сильного прибоя, гидроидные
полипы обнаруживают богатую
приспособительную диверген-
цию.
Пресноводная гидра в связи
с крайним ослаблением кутику-
лы, о чем говорилось выше,
перестает давать постоянные
колонии. При унаследованной
способности почкования у нее
образуются лишь временные
колонии, так как почкующиеся
особи, возможно потому, что
не удерживаются кутикулой,
рано или поздно отделяются от
материнского организма и ведут далее самостоятельное существо-
вание. Выгодная для вида приспособительная черта сохранилась,
но результат уже иной, чем в первоначальных условиях/ у типич-
ных исходных морских гидроидных полипов.
Наконец, крайне важным приспособительным процессом, возник-
шим в результате развития колониальности, является обусловлен-
ная разделением функций дифференцировка первоначально одина-
ковых особей колонии в различных направлениях. Возникает ди-
морфизм или полиморфизм особей в колонии.
Одним из ярких примеров далеко пошедшего полиморфизма яв-
ляются колонии Hydractlnia (рис. 11).
Мы не будем здесь касаться организации чрезвычайно полиморф-
ных колоний сифонофор. Норма для гидроидных полипов — димор-
физм. Среди обычных полипоидных дифференщфуются медузоидные
особи, т. е. такие, в которых сосредоточиваются половые продукты.
56
Организация медуз, на первый взгляд резко разнящаяся от орга-
низации полипоидных особей, является лишь ярким примером при-
способления последней к свободному планктонному существованию
(рис. 12). При этом вид получает возможность широкого распро-
странения благодаря отделению от колоний тех особей, которые»
содержат половые продукты и могут свободно плавать.
Приспособительной является прежде всего форма, которую при-
нимает полип; тело его становится широким и несет по краю щу-
пальца, образуется зонтик медузы, причем происходит очень боль-
Рис. 12. Выработка организации гидромедуз; на всех фигу-
рах слева — разрез по интеррадиусу, справа — разрез по
перрадиусу;
I — кольцевой канал, 2 — ‘эктодермальная пластинка и 3 — централь-
ный отдел кишечника, 4 — покров менубриума, 5 — мезоглея эксумб-
релли, 6 — радиальный канал, 7 — опорная пластинка, в которую
мезоглея переходит на субумбрелле, 8 — парус, черные точки в нем--
мышечные волокна
шое увеличение поверхности тела между щупальцами и ротовым
отверстием, развивается субумбрелла. Тело обычно является чаше-
видным или настоящим зонтиковидным,- т. е. таким, при котором
животное может держаться, несясь течениями, парить в воде бла-
годаря увеличению трения с водной массой при погружении.^ Далее,
его удельный вес приближается к удельному весу морской воды,
а положение в водной массе стабилизируется, благодаря тому, что
опорная пластинка эксумбреллы, т. е. защупальцевой части тела,
чрезвычайно увеличивается в объеме, превращаясь в мезоглею,
студень колокола медузы. Вместе с тем неклеточная масса мезо-
! леи является и хорошим защитным образованием для прикрывае-
мого ею участка тела, так как ее повреждения не приносят особого
вреда для животного. Наконец, при исчезновении мышечных воло-
кон в покрове эксумбреллы, развивается мускулистая складка по
краю колокола, непосредственно примыкающая к щупальцам, склад-
ка паруса. Залегающие в ней мышечные волокна образуют кольце-
57
вую мышцу, при сокращении которой вода выталкивается из
субумбреллярной полости и животное движется обратным толчком,
что вполне гарантирует от погружения и используется далее для
активного продвижения аборальным полюсом вперед.
Исходным, ведущим моментом в этой серии взаимосвязанных
явлений было, конечно, образование студня медузы за счет абораль-
ной части опорной пластинки полипа. Это уменьшало удельный вес
тела, стабилизировало тело определенным образом в водной массе
и, наконец, защищало важные для жизни органы. Что касается
развития паруса, то наличие подобной складки у полипов Bonne-
viellidae еще вовсе не доказывает, что она имелась и у исходных
форм гидроидных полипов вообще. Напротив, ее отсутствие у огром-
ного большинства их заставляет видеть в ней как раз характерную
черту гидроидных медуз, выработавшуюся у их начальных форм
вместе с выработкой зонтика, как очень важное вспомогательное
приспособление для удерживания тела во взвешенном состоянии в
воде, а затем для движения.
Вторично происходят характерные изменения и в других орга-
нах. Так, вытягивается несущий на конце ротовое отверстие прида-
ток «manubrium» — приспособление для лучшего захватывания
пищи ртом, первоначально оказавшимся в глубине субумбредлы.
Далее, единая полость кишечника полипа при развитии зонтика
дифференцируется в сложную гастроваскулярную систему медузы,
а именно: ее центральный участок обособляется в виде более или
менее объемистого желудка, тогда как далее к периферии зонтика
эксумбреллярная и субумбреллярная стенки кишечника сближаются
и превращаются в единую эктодермальную пластинку, в которой
лишь ПО' самой периферии сохраняется просвет кишечной полости
в виде кольцевого1 канала, и по четырем радиусам в виде каналов,
соединяющих последний с желудком. Таким образом получается
характерная система каналов, приносящих пищевой материал от
желудка к периферии, откуда и само название — гастроваскуляр-
ная, т. е. кишечно-сосудистая, система. Соответственно, половые
клетки, располагающиеся в покровном слое, скопляются в виде го-
над или на манубрии, как это присуще Athecata—Antomedusae, или
на пути радиальных каналов, как у Thecophora—Leptomedusae.
одним словом, в защищенном положении на субумбрелле и близ
источников питания.
Независимо от выработки этих характерных черт организации
идет сосредоточение стрекательных элементов по самому наружному
краю колокола медузы — защитное приспособление активной обо-
роны от врага. Пассивная оборона обеспечивается развитием массы
студня эксумбреллы. Далее, по краю же сосредоточиваются чувстви-
тельные клетки, образующие более или менее значительный коль-
цевой пояс. Здесь же у основания щупалец, являющихся как орга- ;
нами защиты и нападения, так и осязательными придатками, раз-
виваются органы чувств, ориентирующие свободноподвижный орга-
низм при его движении в воде — это примитивные глазки анто-
58
мрдуз. Они представляют скопления светочувствительных элемен-
тов, с одной стороны охватываемых клетками, содержащими пиг-
мент. благодаря чему животное ориентируется в интенсивности- и
направлении световых лучей и может двигаться к ним или от них.
Это примитивные «глаза направления света».
У другой ветви гидроидных медуз — лептомедуз и трахимедуз
развиваются органы другого рода, а именно1 ориентирующие жи-
вотное в положении его тела в водной среде. Это органы равнове-
сия. В наиболее простых случаях они представляют собою ради-
ально расположенные углубления покрова субумбрелли у основания
паруса. В них возле обычных чувствительных клеток, снабженных
чувствительными волосками, дифференцируются клетки с известко-
выми отложениями, которые давят на чувствительные при измене-
нии положения тела. В других случаях (Trachilina) в органы рав-
новесия превращаются щупальца, находящиеся по определенным
радиусам тела. Щупальца, укорачиваясь, становятся как бы була-
вообразными придатками, надавливающими при изменении положе-
ния тела на те или иные участки расположенных вокруг чувстви-
тельных клеток. Получаются своеобразно специализированные ме-
ханорецепторы, отмечающие наклонение тела в ту или иную
сторону. Органы равновесия того или другого рода могут образо-
вываться и в более защищенном положении в особых нишах по
краю колокола на субумбрелле. При совершенном же отшнуровы-
вании ниш от покрова развиваются хорошо защищенные пузырьки,
выдающиеся к эксумбрелле.
Так выявляются два пути, по которым шла эволюция Hydrozoa.
Путь Athecata — антомедуз, не имеющих защитных чашечек у по-
липов, снабженных глазками у медуз и с гонадами на манубрии.
Другой путь Tecophora — лептомедуз и Trachilina с выработкой
защитных чашечек у полипов, органов равновесия у медуз и с
гонадами на радиальных каналах. У обеих групп в нервном спле-
тении покрова одинаково развивается при основании паруса, соот-
ветственно кольцевой мышце последнего, скопление нервных клеток
и волокон в виде кольца двигательного характера. Параллельно
первому кольцу образуется другое кольцо чувствительного характе-
ра соответственно расположению органов чувств по краю колокола
в эксумбрелле. Как говорят, нервная система дифференцируется
«функционально», т. е. в теснейшей взаимосвязи с двигательными
и чувствительными образованиями. Из диффузного сплетения поли-
а с АИФФузным расположением мускулатуры и чувствительных
кольтГ Развивак>тся кольцевые нервные образования медуз при
Расположении мускулатуры и органов чувств по краю
ная^и3 РассмотРенных фактов ясно, что раз выработавшись, гидроид-
совепщм^ЗОИдная формы с их различным образом жизни могут
правлениях независимо эволюционировать дальше в разных на-
частности по отношению к медузам соотношения организма и
59
^реды могут быть таковы, что для вида становится выгоднее
выбрасывать медузу, а вместе с ней и потомство, на волю воль а
ветров, а сохранить в колонии под надежной защитой, увеличив тем|
продукцию потомства, которое имеет, хотя бы в личиночной стадии!
иланулы, возможность расселения. Борьба этих противоположных!
моментов ведет в различных случаях к тому, что медуза не отде-1
ляется от материнской колонии, становится так называемым!
медузоидом, сохраняя все типичные черты организации свободножи!
вущей формы. Однако в дальнейшем начинается процесс регрессив^]
ной эволюции органов, ненужных при прикрепленном образе жизни/
Редуцируются органы чувств, редуцируется субумбреллярная
полость и парус, редуцируются каналы кишечника, и развивается
лишь гонада. Из медузоида получается наполненный половыми про-
дуктами мешок гоносака, сидящий на полом выросте кишечника
колонии,
У пресноводной гидры, в условиях жизни в пресной воде, меду-
зоидные особи также полностью отсутствуют. Но чем вызвано это
явление — примитивной ли невыработанностью диморфизма особей
или вторичной редукцией медузоидов — это пока открытый вопрос,
Можно допустить, что естественный отбор позволил проникнуть в
пресные воды тем формам, у которых имелась хотя и слабая спо-
собность к передвижению полипов, и у которых отсутствовали сво-
бодные медузоидные особи, не способные противостоять течению,
постоянно1 сносившему их к морю. Полная редукция таких особей
могла быть лишь вопросом времени.
Диаметрально противоположный результат получается у тех
форм, медузы которых, достигнув значительной степени преуспева-
ния в борьбе за существование, стали, можно- сказать, ведущими в
жизни вида, тогда как значение полипоидной формы отошло на
второй или задний план. Естественно, развитие колоний становится
все слабее и слабее, и, наконец, полипоидное состояние вполне вы-
падает, что и имеет место у гипогенетических медуз — трахи-
и наркомедуз. Таким образом, ход эволюции радиального сидячего
полипа завершился в свободной хищной медузе герионии, сохраня-
ющей, однако, радиальное строение прикрепленного полипоидного
предка, не утрачивающее своего значения и легко используемое
также в условиях пелагической жизни. В этих условиях студень
зонтика прикрывает и пассивно защищает весь комплекс прочих
органов, которые эксумбрелла песет на себе, причем органы актив-
ной защиты и нападения — щупальца — и органы ориентировки
во внешней среде располагаются правильно по радиусам, по краю
колокола, где могут реагировать на внешние условия равномерно
во все стороны, полностью обслуживая пассивно носящуюся
медузу.
Брукс и в последнее время Л. Гиман (1936) держатся совер-
шенно' иного взгляда -на отношения полипа и медузы. Полипоидные
формы, по их мнению, представляют собой только личинок, наподсн
бие актинулы, вторично выработавших сложную организацию и бсс-
60
полое размножение. «Гидроидная колония является таким образом
постоянной личиночной общиной», а первичными в эволюции кишеч-
ноПолостных были медузоидные формы наподобие Trachymedusae.
1 Этот взгляд нельзя без особых натяжек поставить в соответствие
< ,с фактами. Во-первых, радиальный план построения экологически
связан с сидячим существованием исходных форм, а не мог выра-
I фотаться в условиях свободного существования. Во-вторых, доказа-
I но, ЧТО половые клетки, обособляясь в‘ различных участках покров-
i ного эпителия колонии, далее мигрируют для питания в эпителий
1 кишечника и, наконец, оттуда собираются в определенных пунктах
тела образующихся медузоидных особей, давая гонады. Следова-
тельно, половой является вся гидроидная колония, а вовсе не ме-
дузоидная особь только. Предположить, для спасения положения,
раннюю дифференцировку половых клеток вообще возможно, но
тогда нужно допустить еще и особую природу этих клеток, скопляю-
щихся только в медузах, причем прочие особи колонии уже кончают
свое существование, не переходя в конечную медузоидную стадию
развития. Такого рода «неотения» с половым размножением для
некоторых лишь избранных особей, конечно мало приемлема, и все
это построено крайне искусственно.
В связи с изучением циклов жизни простейших вопрос о смене
поколений при метагенезе у Hydrozoa в совершенно повоут свете
ставится А. А. Захваткиным (1949). Он считает, что преобразова-
ние характерного для простейших многочленного жизненного цикла
(см. стр. 52—53) в «нормальный» и единый онтогенез, типичный для
высших Metazoa, проходит через ряд последовательных ступеней.
На первой ступени стоят гидроидные полипы, у которых настоящим
индивидуальным развитием будет начальный этап — от яйцеклетки
до оформления полипа — основателя колонии и этап, заключаю-
щийся в образовании поколения медуз, большая же часть онтогенеза
осуществляется неопределенно многочисленными, но плохо разгра-
ниченными поколениями гидрантов, еще не выкристаллизовавшихся
в самостоятельные особи. В этой части цикла понятие об индиви-
дуальности одинаково приложимо и к колонии в целом, и к каждо-
му из составляющих его гидрантов, однако многочленность этой
части цикла все же несомненна.
Таким образом, по мнению А. А. Захваткина, почкование и обра-
зование колоний у гидрополипов представляет своеобразное выра-
жение множественности вегетативных поколений, присущей простей-
шим. Другими словами, выработка эмбрионального развития у гидро-
идов еще не закончилась формированием единого целостного эмбрио-
еза> что имеет место уже у выше стоящих кишечнополостных.
как ^Нако мы рассматривали процесс почкования у гидрополипов
к пегеСП°ЛСе Размножение> выработавшееся на основе способности
ДимотЬ^иЦН1*’ а в че’РеА°вании поколений видели лишь явления
биолошчЗМа ос°беч колонии, развившегося на основе разделения
специа тиеСКИХ отпРавлен™ между последними. Медузоидные особи
зированы для расселения вида, так как разносят половые
продукты, а полипоидные обеспечивают жизнь всей колонии. Это —.
процесс биологически сравнимый с дифференцировкой колоний у
ряда сидячих же форм из высших типов, как, например, у мшанок
или
ное
В
13. Схема строения
сцифополипа: А — про-
дольный разрез слева по
интеррадиусу, справа по
пер радиусу; Б — попе-
речный разрез на уров-
не 1, В — то же на
уровне 2:
3 — кольцевой канал, 4 —
радиальные камеры кишеч-
ника, 5 — субумбреллярное
впячивание септ
Рис.
оболочников. У Doliolum он получает еще более парадоксаль-
выражение, когда отдельные особи колонии начинают свое
существование в виде свободных индивидуу-
мов, вторично затем объединяющихся в по-
лиморфную колонию. Однако никому не
придет в голову считать развитие таких вы-
сокоорганизованных форм за недостаточно
выработанный эмбриогенез, а в образовании
полиморфных колоний видеть «плохо разгра-
ниченные поколения» организмов, еще не
выкристаллизовавшихся в самостоятельные
особи. Поэтому и колониальность гидроид-
ных полипов с выработкой ди- и полимор-
физма не имеет ничего общего с завершен-
ностью или незавершенностью их онтогенеза.
ПолиморфизхМ находит свое биологиче-
ское объяснение во взаимоотношениях орга-
низма и среды, являясь полезным для сохра-
нения и биологического прогресса вида.
«Незавершенность» же онтогенеза представ-
ляет только пустую аналогию бесполого раз-
множения гидроидов с вегетативным раз-
множением простейших.
Как мы видели, выработка индивидуаль-
ности у гидроидов настолько завершена — в
особенности, если прнять во внимание отно-
шения у губок, о чем речь еще будет впере-
ди, — что теряется всякая возможность рас-
сматривать бесполое размножение как явле-
ние, связанное с невыработанностыо эмбрио-
генеза, другими словами — со слабой сте-
пенью целостности индивидуума.
Наконец, известно, что гидромедузы
иногда почкуются. Следовательно и послед-
ний этап «настоящего оформленного инди-
видуального развития» подрывает построение А. А. Захваткина.
Не будем дольше останавливаться на подобных взглядах, а
перейдем к организации выше стоящих групп кишечнополостных,
к сцифоидным и коралловым полипам, которые одним своим суще-
ствованием говорят о первичности полипоидной формы в эволюции
кишечнополостных.
Сцифоидный полип (рис. 13) представляет, по сравнению с гид-
роидным, значительное усложнение организации, дающее характер-
ный облик Scyphozoa. Начинаясь формами, организованными слож-
62
ней по сравнению с гидроидными
полипами, сцифоидные — в связи
приспособлениями к свободному
пелагическому существованию —
дают выше организованных медуз
(рис. 14).
Прежде всего, сцифоидные по-
липы отличаются несколько более
крупными размерами по сравне-
нию с гидроидными.^ Это полу-
чается в значительной мере в свя-
зи с превращением опорной пла-
стинки в более массивные про-
слойки ткани основного вещества.
Далее у них выработались го-
раздо более энергичные сокраще-
ния тела — у сидячих форм это
также приспособление защитного
характера. Вместо цримитивно
однообразной мускулатуры гид-
роидов — продольные волокна в
покрове и кольцевые в кишечни-
ке — у сцифистомы дифференци-
ровалось 4 мощных пучка, иду-
щих от околоротового участка к
подошве, вдаваясь соответственно
4 складками в полбсть кишечни-
ка. Этим увеличивается эффек-
тивность сокращения. Образован-
ные эпителиально-мышечными
элементами пучки эти в суженной
аборальной части сцифистомы
связаны с соответствующими уча-
стками покрова, в расширенной
же ее части, соответственно в
оральном отделе пучков, послед-
ние образуются за счет покрова
особых воронковидных впячива-
TTiLu’tr п'РОНизЬ1вающих полость ки-
ияиа?Ка’ от оральной стороны к
ким гЗ^СУЖенн°й части тела. Та-
рактеп1нЛ^°М возникают очень ха-
лагаю₽щиесяДЛинт8СУрЬ°20а распо’
умбреллярные 1пРраДИаЛЬН0 Суб’
ной стороны а о апИВа»ИЯ’ С °Д-
нее характерные1^1'011 ~ не ме’
септы, которое п0ЛпяПереГОрОДКИ’
р подразделяют по-
Рис. 14. А — схема организации сцифо-
медузы в разрезе по главной оси — слева
интеррадиально, справа по перрадиусу, но
по краю внесено щупальце; Б — Aurelia
aurita с оральной стороны. Видны кана-
лы и центральная часть кишечника с гона-
дами, легкие контуры четырех лопастей
рук и щупалец; В Су anea arctlca,
оральная сторона. Видны кольцевые и
радиальные мышцы субумбреллы, по краю
их кружки — места прикрепления щупа-
лец, лопасти с каналами кишечника (Б и
В — по Ветохину);
1 — желудок, 2 — радиальный канал ж
3 — кольцевой канал кишечника, 4 —
гастральные нити, 5 — субумбреллярная
полость, 6 — лопасть руки у ротового
отверстия, 7 — гонада, 8 -- щупальце,.
9 — ропалия
63
лость кишечника на центральный отдел и 4 боковых кишечных кар-
мана. Последние соединяются друг с другом периферически, за суб-
умбреллярными впячиваниями, так называемым кольцевым кана-
лом.
Однако в кишечнике сцифополипа имеет место и другого1 рода
усложнение, связанное уже с захватыванием и перевариваниехм бо-
лее крупной добычи. На септах дифференцируются продольные ряды
щупальцеобразных выростов, гастральных нитей, на которых скон-
центрированы стрекательные элементы и железистые клетки, по
преимуществу вырабатывающие пищеварительные ферменты.
Все эти адаптивные черты сцифополипа достигают полного
своего выражения при переходе в свободпоподвижное состояние
медузы (рис. 14). Как и у гидроидных полипов половое размноже-
ние у Scyphozoa соединено обычно с расселением вида, т. е. с
отделением особи от субстрата, в связи с чем получаются конвер-
гентные с Hydrozoa изменения всей организации. Животное прини-
мает зонтикообразную или колоколообразную форму медузы, спо-
собствующую парению в воде. Развивается сильная мускулатура,
но не в виде мускулистой складки паруса гидромедуз, а как коль-
цевые и идущие по радиусам дельтовидные и радиальные пучки
мышц субумбрелли. При сокращении их нижняя сторона зонтика,
обычно снабженная лопастями по краю, ударяет о воду, что доста-
точно для предохранения животного от погружения. Этому же спо-
собствует мощное развитие студня мезоглеи в колоколе, причем
процесс распространяется, как в эксумбрелле, так и в субумбрелле
на все части опорной пластинки, за исключением щупалец, а не
ограничивается лишь эксумбреллей, как это имеет место у гидроид-
ных медуз.
Конвергенция, сходство в общем и несходство в деталях между
организацией гидро- и сцифомедузы, находит свое яркое выражение
в строении самой мезоглеи. Она, во-первых, всегда содержит во-
локнистые структуры в гомогенном основном веществе, что значи-
тельно увеличивает ее плотность, резистентность. Во-вторых, у
дискомедуз она содержит клеточные элементы, превращаясь в до-
вольно сложную соединительную ткань. Как в образовании самого
студня, так и клеточных его элементов главную роль играет энто-
дерма, хотя и эктодерма не теряет еще своего значения.
Таким образом, морфологически выявляются те общебиологиче-
ские моменты, которые были разобраны выше у гидромедуз, но
выявляются на иной основе организации сцифополипов. Конвер-
гентно происходят вторичные изменения и в других системах орга-
нов. Относительно небольшой манубриум низших сцифомедуз эво-
люционирует у крупных хищных форм, какими являются
Semaeostomeae и Rhizostomeae, в 4 мощных радиальных лопдсти
рук, которыми Aurelia просто захватывает свою добычу и которые,
срастаясь своими гофрированными краями, превращаются в слож-
ную систему как бы сосущих каналов Cassiopea, или Rhizosloma.
Далее, в кишечнике у Semaeostomeae к исходному центральному
64
отделу и 4 боковым карманам присоединяется система отходящих
ст них периферических радиальных каналов, лр ветвящихся или
более или менее значительно' ветвящихся и иногда развивающих
кольцевой канал. В такой гастроваскулярной системе устанавли-
вается вполне определенный ток содержимого. У Rhizostomeae ради-
альные каналы в зонтике объединены или кольцевым каналом с
значительной сетью анастомозирующих канальцев, или только
сплошным сплетением этих последних. Такое развитие гастрова-
-кулярной системы стоит в теснейшей связи с выработкой зонтика
сцифоидной медузы путем вторичного разрастания сократительного,
подвижного края и особенно расположенных адрадиально участков
первоначально 8-лопастной эфиры (рис. 16, Б). При этом 4 перво-
начальные мускулистые септы с их субумбреллярными впячивания-
ми, утрачивая свое сократительное значе-
ние в организме медузы, редуцируются, а
гастральные нити получают более диф-
фузное расположение.
В строении гастроваскулярной систе-
мы труднее уловить приспособительные
моменты, которые так ясны в развитии
аппарата оральных рук. Скорее прихо-
дится говорить о коадаптивных явлениях
и дифференцировке системы на основе
разделения функций, но в том и другом
случае сцифомедузы обнаруживают бога-
тую дивергенцию, вдаваться в подробно-
сти которой не наша задача.
Значительная адаптивная дивергенция
Рис. 15. Разрез через
край колокола и аппарат
ропалии:
1 — чувствительные ямки,
2 — кроющая пластинка,
3 — глаз. 4 — статолит
выявляется также в развитии по краю зон-
тика сцифомедуз щупалец, разнообразных
по числу и характеру, равно как и в выработке располагающихся
тут же органов чувств. Последние представлены ропалиями — слож-
ными образованиями, являющимися прежде всего органами равнове-
сия, а затем у многих форм несущими в том или ином количестве
примитивные глазки и ямки хеморецепторного характера. Развива-
ются они при основании щупалец сцифистомы (рис. 15) как була-
вовидные выпячивания стенки тела, в эктодермальном эпителии
конца которых образуются отложения сернокислого кальция. Соот-
ветственно базально в покровном эпителии получаются сгущения
нервного сплетения покрова и скопления нервных клеток, дающие
ганглии ропалий. Чего-нибудь подобного нервному кольцу гидроме-
дуз здесь нет. Это подчеркивает коррелятивность развития нервной
системы с развитием органов чувств и мускулатуры и вносит резкий
штрих своеобразия в поразительно полную картину общего сходства
сцифо- и гидромедуз. Организация их —как бы два варианта на
один экологический мотив при разном исходном материале.
Обладая большей сложностью, этот исходный материал для
сцифомедуз давал возможность эволюции крупных хищников пела-
5
v Пути эволюции животного мира
65
гиали. В результате центр тяжести в жизненной борьбе переносился
на медузоидных особей, полипоидные теряли свое значение для
существования вида, отходили на второй план, как это мы видели
уже и у Trahilina среди Hydrozoa. Сцифополипы выпочковывают
себе подобных сходно с обычным для гидрополипов процессом поч-
кования с образованием столона, но колонии обыкновенно незначи-
тельны и не могут идти в сравнение с полиморфными колониями
гидроидов.
Однако в колонии сцифополипов не приходится говорить не
только о полиморфизме, но даже диморфизм, ставший нормой для
Рис. 16. Л — схема
полидисковой строби-
лы; Б — эфирула
стка тела полипа в
гидроидов, здесь отсутствует, так как в конце
концов каждый сцифополип переходит в поло-
возрелое состояние сцифомедузы. Таким обра-
зом, сцифомедуза представляет завершение
развития сцифополипа, который становится
как бы только личинкой в онтогенезе Scypho-
zoa. Однако этим ничуть не умаляется значе-
ние полипоидной стадии, так как она соответ-
ствует зрелой форме предков, выработавшей
радиальную симметрию, а не представляет
ново.образования в онтогенезе' в связи с при-
способлением'развивавшегося зародыша к осо-
бым условиям самостоятельного существова-
ния, что имеет место при выработке настоящих
личиночных форм.
Совершенно иным по сравнению с почкова-
нием Scyphozoa и Hydrozoa является и самый
процесс образования сцифомедузы из сцифо-
полипа, так называемая стробиляция (рис. 16).
При ней происходит отделение половой особи
от субстрата, переход в свободноподвижное
состояние, сопровождаемый регенерацией ос-
тающегося небольшого прикрепленного уча-
новую полипоидную особь. В монодисковой
стробиле Cassiopea такой процесс как раз имеет место.
Обычно же стробиляция бывает полидисковой, т. е. процесс
образования медузы и процесс регенерации полипа чрезвычайно
сближены для, целого ряда медуз, которые последовательно разви-
ваются из регенерирующей особи сцифополипа.
Живущие в условиях прибрежной, мелководной полосы, Lucer-
nariidae утратили период свободного существования, и стробиляция
исчезла. Биологически это связано с тем, что взрослая медузоидная
форма не отделяется от субстрата, не бросается на волю волн,
гибельную в прибрежье.
Последним штрихом, завершающим нашу картину организации
Scyphozoa, будет то, что половые клетки у сцифополипов не обна-
руживают такого раннего появления и таких миграций, какие
имеются у гидроидных полипов. Гонады дифференцируются только
66
в сцифомедузе, причем все время связаны с эктодермальной стенкой
кишечника, откуда половые элементы попадают" прямо в полость
кишечника. Другими словами, временное пребывание половых кле-
ток гидроидной колонии в энтодерме, т. е. у источников питания,
у Scyphozoa заменилось постоянной локализацией гонад в стенке
кишечника, а именно в хорошо защищенном положении на септах.
Это — в высокой степени характерная, широко адаптивная черта
рассматриваемого класса, присущая и последней группе кишечно-
полостных, наиболее сложно построенным их формам — Anthozoa.
Anthozoa, или коралловые полипы, представляют дальнейший
и последний шаг в эволюции сидячих форм кишечнополостных,
причем значительная сложность организации, достигнутая ими, за-
вершается половозрелым полипоидным состоянием. Биологический
момент распространения вида, ведущий к образованию свободно-
подвижной формы, что характерно для низших, дающих медуз
групп кишечнополостных, у Anthozoa не выявился, и чнет абсолютно
никаких данных, которые могли бы указывать на вторичное выпаде-
ние у них подобного состояния. В этом приходится видеть решаю-
щий факт для признания полипоидной формы за исходную, первич-
ную в эволюции всех Cnidaria, как мы освещали ее и раньше при
нашем анализе организации рассмотренных классов кишечнопо-
лостных.
Среди них не раз вздымалась могучая творческая волна. Снача-
ла на гребне ее выросли богатые разнообразием приспособлений
сплошные заросли гидроидных полипов, достигающие своего куль-
минационного пункта в мощных рифах Hydrocorallia Индийского
океана. Далее развернулись крупные сцифоидные медузы, огром-
ные стада которых не только носятся в течениях теплых морей, но
и в наших морях Aurelia и Суапеа собираются несметными массами.
Наконец, последним могучим взлетом поднялись громады коралло-
вых рифов Hexacorallia в тропических морях и глубокие барьеры
Octocorallia в норвежских фиордах.
Не раз отдельные всплески этих волн жизни или, замирая, пре-
рывались, или широким прибоем рассыпались в жизненной борьбе.
Такими были среди Hydrozoa всплески гипогенетических медуз и
сифонофор, среди Scyphozoa — Cubomedusae и Peromedusae, среди
Anthozoa — немногочисленные Ceriantharia, Anthipatharia Zoantharia
и т. п., с одной стороны, и обильные сады актиний, с другой.
Если организация гидроидного' полипа развивалась под знаком
активного использования придонных пищевых ресурсов сидячими
животными, а сцифомедузы •— под знаком использования свободно-
подвижным хищником ресурсов пелагиали, то у коралловых поли-
пов перед нами организация, теснейшим образом связанная с вы-
работкой пассивного питания сидячих форм, которые черпают пищу
из придонных и поверхностных течений, несущих массы мелких
Пелагических организмов.
Коралловые полипы по размерам в среднем немногим превосхо-
дят сцифополипов, но в противоположность последним они путем
67
почкования на столонах у Octocorallia или путем деления у Неха-
corallia дают обычно большие колонии со многими особями, так
что получаются не мелкие, немногочисленные полипчики, а значи-
тельные массы. Это дает известную защиту от врага и некоторое
обеспечение питанием. Напротив, у одиночных форм Anthozoa,
какими являются актинии и Ceriantharia, мы встречаемся с укруп-
нением особей, достигающих у некоторых видов значительных, ги-
гантских по сравнению с коралловым полипчиком, размеров —
явления одинакового порядка в том смысле, что также ведут к
лучшей защищенности и обеспечиванию питанием, но не путем об-
разования колоний, а путем увеличения размеров отдельной особи.
Колонии развиваются в теснейшей взаимосвязи с развитием
поддерживающих и защитных образований, причем у одиночных
актиний и цериантусов собственно нет скелета. У Cerianthus обра-
зуется защитная трубка из посторонних частиц, а у актиний стенка
тела получает значительную плотность благодаря превращению
сравнительно тонкого слоя гомогенной мезоглеи цериантусов в
основательную прослойку соединительной ткани, содержащей клетки
и состоящей из слоев правильно пробегающих преколлагенных воло-
кон (рис. 4). Такая стенка тела хорошо защищает внутренности,
особенно при сокращении животного, и в то- же время служит проч-
ной поддерживающей прослойкой, располагаясь между покровным
и кишечным эпителиями. Таким образом эволюция мезоглеи идет
здесь в том же направлении, как и у сцифополипов. Один путь
выработался у Octocorallia, где в наиболее простых случаях
(Cornularia) столон защищен плотной ороговевающей кутикулой,
но обычно видим иное, а именно: в клетках, мигрирующих из по-
крова в мезоглею, развиваются характерные неправильно' палочко-
видные отложения извести, получается внутренний скелет. Он об-
разован изолированными спикулами в стенке тела полипов, отсут-
ствуя почти совсем в щупальцах, а в цененхиме, т. е. в ткани,
заполняющей пространства между полипами, превращается в ком-
пактную массу, которая в колонии, например, благородного коралла,
дает плотный осевой ствол (рис. 17).
Другой путь развития скелета представляют НехасогаШа —
рифообразующие кораллы. У них отложение извести клетками по-
кровного эпителия происходит не во внутренней мозоглее, а обыз-
вествляются наружные клетки покрова. Получается известковая
подошва полипа, валик вокруг его основания, радиально идущие
пластинки склеросепт, образующиеся за счет соответственных впячи-
ваний стенки тела внутрь, и т. д., и т. п. Скелет достигает большой
мощности и дает неисчерпаемое разнообразие вариантов развития
его частей.
Можно ли здесь говорить об адаптивной дивергенции, придавая
приспособительное значение развитию каждой детали скелета!
Скорее происходит дифференцировка по различным возможным ва-
риантам связанных коадаптивно скелетных образований, лишь в
общем дающая животному преимущества в борьбе за жизнь.
68
Последним звеном в цепи защитных приспособлений, которые
вСе теснейшим образом взаимосвязаны, является способность
Anthozoa к сильнейшему сокращению тела при раздражении. Акти-
Рис. 17. Л — часть колонии Coralllum rubrum (по Пфуртшеллеру):
полипы в разных степенях сокращения и сифонозоиды (1), внизу вскрыт ценосарк (6> и
Два полипа, видна скелетная ось (2), гастродермальные каналы, (3), рот, глотка (4),
септы (5) — в полипе;
Б— поперечный разрез через полипа на уровне глотки (по Гиксону):
1 периферические камеры кишечника, отделенные друг от друга восемью септамп (5) с
характерными мышцами ретракторами по одной стороне (3), посредине глотка (4), сложен-
ная в складки, снизу в ней сифоноглиф (2), соответственно «брюшная» направляющая
камера
ния превращается в плотную подушечку, внутри которой упакованы
Все органы, становящиеся недоступными для врага, а коралловый
полип частично или полностью втягивается при этом под защиту
69
скелета. Это обусловлено главным образом развитием мышечных
пучков, идущих от орального диска к подошве, и кольцевой мышцы,
смыкающей край орального диска над щупальцами. Но и другие
более слабые мышечные пучки, например в щупальцах, играют в
этом известную роль (рис. 18).
В противоположность Hydrozoa и Scyphozoa, имеющим диффе-
ренцировку мощных мышечных пучков преимущественно за счет
эпителиально-мышечных элементов покрова, например, в парусе и
субумбрелле медуз или в ретракторах сцифополипа, коралловые
Рис. 18. А — организация актинии и Б — схема расположе-
ния септ на уровне глотки (слева) и ниже глотки (справа)
(по Паркеру);
1 — три круга щупалец на ротовом диске, 2 — рот. 3 — глотка, 4 —
главные септы, 5 — септы второстепенные, 6 — гастральные мезенте-
рии, в центре аконтии (7), 8 — мышцы ретракторы, 9 — гонады, 10 —
отверстия в стенке тела
полипы развивают мощные ретракторы септ, сфинктеры диска у
актиний и т. д. за счет эктодермальных эпителиально-мышечных
клеток. Их сократительные волоконца располагаются елочковид-
ными или еще более сложными складками, чем чрезвычайно сильно
увеличивается их количество в каждой мышце. Однако на щупаль-
цах актиний дифференцируется и настоящая эпителогенная мышеч-
ная ткань, в результате того, что складки покровного эпителия,
погружаясь в мезоглею, совершенно обособляются, отшнуровывают-
ся от производящего эпителия (рис. 4). В меньшем масштабе
подобная дифференцировка имеет уже место и в нижестоящих
группах, у Hydrozoa и Scyphozoa.
И если развитие септ в кишечнике сцифополипа приходится
поставить в связь с развитием ретракторов за счет утолщений по-
крова в ножке и субумбреллярных впячиваниях орально, то, несом-
ненно, складки эктодермальной выстилки кишечника, в которых
дифференцировались пучки мышц ретракторов, обусловливали
70
развитие септ у Anthozoa. При этом развитие мышечных пучков в
септах, вдающихся в кишечник, могло итти гораздо дальше, дать
гораздо более мощную мускулатуру, чем в подобных складках
покрова, самим своим положением несколько ограниченных в этом
отношении.
Естественно, что развитие мускулатуры, соответственно септ,
должно было происходить в известном соотношении с щупальцами,
являющимися у Anthozoa полыми выростами стенки тела. Ретрак-
торы могли орально прикрепляться только между вдавлений щупа-
лец. Отсюда норма, что каждой камере кишечника, расположенной
между септами, соответствует обычно одно щупальце, и возникно-
вение новых щупалец должно было быть взаимосвязано с возник-
новением новых пар септ. Это ,
закономерность и появления, и
числа как щупалец, так и септ
у коралловых полипов, причем,
возникая сначала в небольшом
числе и стабилизируясь в раз-
ных группах в количестве 4—
6—8, они достигают у актиний,
Ceriantharia, Zoantharia очень
значительных количеств с уве-
личением размеров тела (рис.
19).
Характерны два пути эво-
люции Anthozoa, отчетливо вы-
разившиеся как в числе септ и
Рис. 19. Схема расположения и роста
септ: А — у Zo ant harin', Б — у
Ceriantharia (по Деляжу). Цифрами
обозначен ход их последовательного
появления
щупалец, так и в форме послед-
них; Octocorallia с 8 ветвистыми щупальцами и НехасогаШа с 6
неветвистыми щупальцами или кратными шести (рис. 17, 18).
Характерно далее для каждого из этих путей и вполне стабильное
в различных их группах расположение пучков ретракторов с той
или иной стороны соответствующих септ. В строении и расположе-
нии щупалец соответственно септ проявляется богатейшее разно-
образие Anthozoa. Правда, определить в каждом частном случае,
какие приспособительные моменты обусловливают развитие того
или иного числа и характерного строения или расположения, нет
возможности, так как один и тот же эффект сохранения в борьбе
за Жизнь может получаться различнейшими путями.
Значение развития септ, однако, не ограничивается рассмотрен-
ными выше сторонами их организации, оно значительно шире, так
как септы теснейшим образом связаны и с деятельностью кишеч-
фДа' О™ подразделяют последний на центральный отдел и пери-
полуЧеСКИе камеРы- В соответствии с этим аксиальная часть септ
стре ЭеТ СВОИ ос<э^ые чеРты — она несет скопления железистых и
мезен ТеЛьных элементов, образуя утолщение так называемых
ний мГериальных нитей, которые близ подошвы, например у акти-
огут продолжаться в свободно свисающие в кишечник нити
71
аконтиев. Таким образом, попадающая в кишечник пища прежде
всего соприкасается с образованиями, по преимуществу выраба-
тывающими пищеварительные соки, чтобы уже далее подвергнуться
характерному для кишечнополостных внутриклеточному пищеваре-
нию.
Выше было отмечено, что питание коралловых полипов является
пассивным, характерным для большинства сидячих форм и выра-
батывающимся здесь впервые среди кишечнополостных. Однако у
крупных актиний мы видим опять активное хищничество- за счет
более крупных животных.
Пассивное питание коралловых полипов связано с появлением
токов воды, несущих пищевые частицы по оральной стороне щупа-
лец и далее по- оральному диску к ротовому отверстию. Сохранение
на этих участках примитивного мерцательного покрова, а тем более
увеличение его действенной площади ветвлением’ щупалец, как у
Octocorallia, вполне достаточно1 для продвижения мелких планктон-
ных организмов по длине щупалец к ротовому отверстию. Однако
этого мало1, попадание пищи по назначению в центральный отдел
кишечника гарантируется дальнейшим приспособлением, а именно:
развивается впячивание покровного мерцательного эпителия, кото-
рое, наподобие рукава, свисает в полость кишечника. Впервые в
животном царстве образуется глотка. Первичный рот гидроидного
полипа оказывается на внутреннем ее конце там, где эктодермаль-
ная выстилка глотки переходит в эктодермальный эпителий кишеч-
ника, и рот Anthozoa будет уже вторичным ртом.
Наконец, в связи с током воды, идущим через глотку в эктодер-
мальный отдел кишечника, вырабатываются приспособительные
черты для регулирования этого тока, а именно1: в одном отделе
глотки, который принимает форму более или менее глубокой склад-
ки желоба, идущего от рта до центрального кишечника, особенно
сильно развиты мерцательные реснички, получается так называемый
сифоноглиф. Соответствующая сифоноглифу сторона тела полипа
получила условное название «брюшной», и соответственно радиаль-
ная симметрия расположения органов в известной мере наруши-
лась, перешла из вполне радиальной в бирадиальную. Часто в пло-
скости сифоноглифа глотка сплющивается или, даже у актиний,
образуется другой сифоноглиф — «спинной». С этой «спинной» сто-
роны мерцательный эктодермальный эпителий глотки разрастается
па пару прилежащих септ и идет по их мезентериальному утолще-
нию глубоко в эктодермальный кишечник до самого основания септ.
Таким образом создается мерцательный аппарат, регулирующий
ток воды в гастроваскулярнойГсистеме кораллового полипа или
актинйй через сифоноглиф снаружи в кишечник, соответственно
по эктодермальным мерцательным участкам мезентериев из послед-
него в глотку и наружу. У вышестоящих представителей сидячих
животных с пассивным питанием, например у прозопигий, иглоко-
жих и т. д., подобных приспособлений не вырабатывается, так как
трубчатый кишечник уже сам по себе обусловливает движение со-
держимого к анальному отверстию. У кишечнополостных пассивное
питание стало возможным только при выработке регулярного тока
воды в гастроваскулярной системе, и как раз у низших представи-
телей Cnidaria оно еще отсутствует. Как мы видели с ним связана
прежде всего бирадиальная дифференцировка глотки, однако далее
им обусловлено и расположение септ, прикрепляющихся аборально
к подошве, а с оральной стороны — к околоротовому диску и све-
шивающейся глотке. Только вторично появляющиеся, например
у актиний, дальнейшие пары септ, так называемые • септы второго,
третьего и т. д. порядков, могут не доходить до глотки, а свободным
краем идут до орального диска. Таким образом и в расположении
септ по бокам глотки, вытянутой щелеобразно соответственно пло-
скости соленоглифа, получается бирадиальное расположение. Свое
завершение оно находит в соответственно двухстороннем располо-
жении мышц ретракторов на септах и в последовательности появле-
ния септ парами, причем камеры кишечника, соответствующие
сифоноглифам, получили наименование «направляющих». Плоско-
стью бирадиально-й симметрии каждая из них делится пополам, с
симметричным расположением мускулатуры их септ.
Особенно ярко выражается бирадиальность строения тела у
несколько аберрантных групп Ceriantharia и Zoantharia в процессах
неравномерного роста их тела, а соответственно и появления септ
и щупалец (рис. 19).
В то время как у какой-нибудь актинии при росте новые пары
септ и щупальца вставляются равномерно по- всей окружности тела
между имеющимися уже септами, и рост идет, можно сказать, рав-
номерно во всех дочках окружности поперечного сечения, у Zoantha-
ria новые септы появляются только в промежутках справа и слева
от «брюшной» камеры кишечника. Следовательно, рост тела идет
неравномерно, усиленно растут части тела, располагающиеся справа
и слева «вентрально». У Ceriantharia новые перегородки септ обра-
зуются у «брюшной» стороны, постоянно- отодвигая ранее образо-
вавшиеся септы к «спинной» стороне, т. е. рост идет только посре-
дине «вентральной» стороны в «брюшной» направляющей камере.
Эти факты говорят о чрезвычайно резко выраженной бирадиаль-
пости организации некоторых Anthozoa и выявляют значение про-
цессов неравномерного роста, начинающихся с простого- образова-
ния бирадиальности в глоточном отделе.
При относительной сложности организации Anthozoa их нервная
система несет очень примитивный характер сплетения нервных кле-
ток, более обильных только- на ротовом диске и в глотке. Отсут-
ствие дифференцировки органов чувств, особенно дистантных ре-
цепторов, обусловленное сидячим существованием и пассивным
питанием, не дает эффекта образования ганглиев, что мы видели,
например, у сцифоидных медуз.
Примитивен и половой аппарат, представленный скоплениями
половых клеток в эктодермальном эпителии на септах возле мезен-
териальных валиков. Расселение вида связано только со свобод-
ноплавающей стадией планулы, но, как норма, чрезвычайно мощно
разыгрывается бесполое размножение путем почкования, а затем- и
деления, ведущее к образованию гигантских коралловых рифов. Зна-
чение его уже было выяснено в начале анализа организации
Anthozoa, как один из ведущих моментов образования последней.
Организация кишечнополостных нами рассмотрена. Подводя
итоги, прежде всего можно наметить типовые черты той группы
гастреад, которая являлась исходной в эволюции кишечно-полост-
ных, другими словами, можно выявить примитивные черты органи-
защ1£^^следних. Сюда относятся: I) мбнаксонное гетерополюсное
строение овального тела с оральным концом, обращенным при дви-
жении назад; 2) выработка постоянных тканей — арх'ибластических,
представленных эпителием, который состоит из эпителиально-мы-
шечных и чувствительно-нервных клеток, и парабластических —
в виде бесструктурной прослойки ткани основного вещества, не
содержащей клеток, причем в эпителии сохраняются неспециализи-
рованные интерестициальные элементы, а также половые клетки;
3) дифференцировка двух клеточных первоорганов (мешковидного
кишечника и покрова) и одного неклеточного (опорной пластинки),
причем первые отличаются мультифункциональностью, включитель-
но до того, что содержат в себе in statu nascendi дифференцировку
нервной системы и мускулатуры, равно как и полового зачатка;
опорная же пластинка несет только функцию .связи: и механиче-
ской, и химической, и опоры; 4) выработка примитивного рефлек-
торного аппарата из двух компонентов, чувствительно-нервной клет-
ки и эпителиально-мышечной; 5) выработка индивидуальности мно-
гоклеточных не только в механическом и химическом отношении, а
также морфологически — через единую нервную систему, однако
без всякой централизации последней; и, наконец, 6) очень большая
способность к регенерации.
Все перечисленные характерные черты организации исходных
гастреад, как было' детально разобрано при рассмотрении гастреи
и низших целентерат, являются результатом широких приспособле-
ний в системе организма — биоценотическая среда, с одной сторо-
ны, и разделения функций со специализацией частей в единой цело-
стной системе организма, с другой.
На основе этого комплекса черт предков вырабатываются далее
характерные черты типа кишечнополостных, а именно; 1) адаптив-
ный комплекс, связанный с прикреплением особи для использования
придонных пищевых ресурсов, 2) комплекс, обеспечивающий раз-
множение вида и 3) комплекс, обеспечивающий расселение, распро-
странение вида.
К первому относятся: выработка радиального плана строения;
развитие пассивнозащитной кутикулы или скелета; дифференци-
ровка стрекательных капсул, равно как и щупалец для активной
защиты и нападения; развитие полостного пищеварения, наряду с
примитивным внутриклеточным, для использования более крупной
добычи.
74
Ко второму комплексу принадлежит развитие, способности раз-
множения путем почкования или деления в связи с очень большой
способностью к регенерации, с одной стороны, и с другой—появле-
ние постоянной колониальное™ в связи с развитием кутикулярных
и скелетных образований.
К третьему относится образование свободноподвижных личинок
и появление свободноподвижных пелагических половых форм.
Ведущими в выработке организации низшей группы кишечнопо-
лостных — класса Hydrozoa — явились два последних комплекса
в крайне своеобразном взаимодействии. А именно: на основе разде-
ления труда между особями колонии развивается ди- и полимор-
физм ее особей; эта особенность приспособительного характера очень
многосторонняя в своем выявлении, но прежде всего выразившаяся
в приспособлении к расселению вида, а именно — в дифференци-
ровке свободно живущих пелагических особей — медуз, в которых
сосредоточиваются половые продукты.
В организации медуз приспособления к свободному пелагиче-
скому существованию выражены чразвычайно ярко обширным
комплексом адаптивных черт; 1) в форме тела в виде зонтика,
2) в образовании массы мезоглеи за счет эксумбреллярной ткани
основного вещества, 3) в развитии складки паруса, 4) в располо-
жении щупалец и образовании стрекательного кольца по1 краю ко-
локола, 5) в развитии тут же чувствительного пояса и в образова-
нии глаз или органов равновесия, а в связи с этим, 6) в образовав
нии двойного кольца нервной системы, 7) в образовании манубрия,
8) в развитии радиальных и кольцевых каналов кишечника, 9) в
сосредоточии гонад на манубрии или радиальных каналах.
У Scyphozoa тот же момент расселения вида выразился вне вся-
кой связи с диморфизмом колоний простым превращением полипа
в сцифомедузу с последующей регенерацией полипоидной особи.
Таким образом из разного исходного материала получаются сход-
ные, в общем, формы гидро- и сцифомедуз — явление далеко иду-
щей конвергенции, но с глубокой разницей в деталях: 1) мезоглея
сцифомедуз развивается за счет всех частей опорной пластинки по-
липа, 2) вместо паруса развивается субумбреллярная мускулатура,
3) вместо простых глазков и органов равновесия — сложные ропа-
лии, 4) вместо нервного кольца паруса — ганглий ропалий, 5) вме-
сто простого манубрия — оральные руки или сосущий аппарат кор-
неротов, 6) образование сложной гастроваскулярной системы,
?) расположение гонад в эктодермальном эпителии септ.
Последним приспособительным штрихом дифференцировки ме-
Дузоидной особи будет увеличение ее значения, по сравнению с
полипоидной, в жизни вида, как имеющей большие преимущества
в борьбе за существование. Это ведет к подавлению или выпадению
полипоидного состояния у Trachilina и у сцифоидных медуз.
Однако ведущая роль в эволюции всего типа принадлежит дру-
гим адаптивным чертам, выявляющимся в организации Scyphozoa
и Anthozoa, а именно — защитным приспособлениям, не обусловлен-
75
ным обязательно сидячим образом жизни: 1) увеличению размеров
особей, связанному прежде всего' с развитием клеточной формы
ткани основного вещества, 2) выработке мощного аппарата сокра-
щения тела, выражающейся в дифференцировке мышц ретракторов
у сцифополипов в эктодермальных впячиваниях, вызывающих обра-
зование септ в кишечнике, а у коралловых полипов уже в эктодер-
мальных септах кишечника, соответственно в дифференцировке цен-
трального отдела и боковых камер гастроваскулярной системы с
правильным движением ее содержимого.
Завершение эволюционного пути кишечнополостных в Anthozoa
связано с переходом к пассивному питанию и с полным отсутствием
приспособлений к разнесению половых продуктов, соответственно
особо мощно развивается прикрепленная полипоидная форма.
Комплекс приспособлений к пассивному питанию выражается в
следующих чертах организации: 1) мерцательный покров сохра-
няется в околоротовой части и соответственно на щупальцах; от-
сюда возникновение токов воды, несущих пищевые частицы, 2) об-
разование глотки, 3) дифференцировка в ней сифоноглифов, разра-
стание эктодермальных ресничных полос на мезентериальные нити,
4) появление бирадиальности, проявление ее в аппарате септ и щу-
палец и максимальное развитие в процессах неравномерного роста
Ceriantharia и Zoantharia. При всем этом у Anthozoa сохраняется
примитивность нервной системы и гонад.
Итак, в типе кишечнополостных на основе широко' .адаптивных
черт организации гастреад имеет место богатое развитие приспо-
соблений более узкого характера, ведущее к богатейшему их разно-
образию, но не к возникновению таких черт организации, которые
могли бы, накопляясь, дать новый этап в эволюции Coelenterata,
открывали бы новые для нее возможности. Такие широкие адапта-
ции у несложно организованных форм мало совместимы с сидячим
существованием, при котором развиваются прежде всего приспо-
собления для использования богатого пищевого материала и при-
способления защитного характера, а не расширяются и не услож-
няются взаимоотношения с живой и мертвой внешней средой в си-
стеме биоценоза, что мы находим, напротив, особенно выраженным
у свободноживущих животных.
Организацию кишечнополостных мы сможем еще глубже понять,
а также еще глубже осветить вопрос о происхождении многокле-
точных животных, проанализировав другую примитивнейшую груп-
пу последних — тип губок.
Г л а в a III
ТИП ГУБКИ — SPONGIA, S. PORIFERA
Три этапа осложнения организации губок. Аскон. Ткани у губок.
Органы губок. Индивидуальность у губок. Происхождение губок.
Губки как Parazoa.
Появившаяся в 1872 г. монография об известковых губках
Э. Геккеля, а затем исследования Ф. Э. Шульце (1876) на долгие
годы направили по неверному пути представление об организации
этой группы. Совершенно правильные наблюдения И. И., Мечни-
кова над их развитием были под гипнозом предвзятых идей отбро-
шены как якобы незаслуживавшие внимания, и лишь в 90-х годах
они получили подтверждение в исследованиях И. Деляжа, Мааса
и Минчина. Этим было положено начало перелому во взглядах на
организацию губок, нашедшему свое полное выражение в отличной
сводке Минчина (1900). Подобную трактовку организации губок мы
находим и у А. Резвого (1938).
В организации губок обычно' различают как бы три этапа посте-
пенного осложнения: наиболее простой, соответствующий строению,
например Leucosolenia, этап «ascon»; более сложный «sycon», или
«rhagon», и самый распространенный и в то же время самый ос-
ложненный .«leucon» (рис. 20). Эти три этапа отлично характери-
зуют организацию губок и потому нашли широкое применение у
зоологов, но совершенно иначе обстоит дело с пониманием исход-
ного состояния «аскон».
Как показывает само название (ascos — мешок), дело идет о
мешкообразной форме или во взрослом состоянии, или в разви-
вающемся организме, так называемой «Olynthus-стадии» Геккеля
(рис. 22, В), Этот исходный этап имеет уже все характерные чер-
ты губок, которые далее эволюционируют в различных направле-
ниях, а потому является особенно- важным как для понимания орга-
низации всей группы, так и для выяснения ее происхождения и для
сравнения с другими формами животного царства.
Геккель считал возможным видеть в асконе, или стадии «Олин-
туса», типичный целентерический организм наподобие гидры, —
отсюда его дальнейшие выводы о ближайших родственных отноше-
ниях Spongia и Coelenterata. Вполне естественно', что и нам при-
77
дется начать анализ организации губок с. рассмотрения характер-
ных черт аскона, расширяя, однако, наш кругозор экскурсиями .по
всему типу.
Итак, наиболее простая форма губки представляет собой мешко-
образный организм, прикрепленный одним участком своей поверх-
ности ко дну и несущий на противоположной стороне довольно
широкое отверстие — оскулум. Через последний выводится вода
из полости мешка, которую можно' назвать пищеварительной, так
Рис. 20. Типы организации губок (по Минчину): А — аскон с выстилкой всех
полостей воротничковыми жгутиковыми клетками, как у Leucosolenia\ Ь — си-
кон, с образованием атриальной полости и локализацией хоаноцитов в боковых
выростах тела; В — лейкон, с дифференцировкой жгутиковых камер и системы
приводящих и отводящих каналов;
1 — атриальная полость, 2 — приносящие и 3 — относящие каналы, 4 — выстланные хоано-
цитами участки (жирная черная линия), 5 — слой ткани основного вещества, 6 -- uscuium,
7 — поры
как в нее через мелкие отверстия в стенке тела, поры, постоянно
поступает вместе с током воды мелкий пищевой материал и здесь
захватывается и переваривается клетками,. выстилающими мешок.
В асконе выявляется важнейший экологический момент, поро-
дивший организацию губок, — дело в том, что это исконно сидячие
формы с примитивным пассивным *питанием, т. е. таким, когда при-
носящий мельчайшие пищевые частицы ток воды производится са-
мими же захватывающими пищу клетками, а не особыми органами,
наподобие, например, сифоноглифов глотки актиний или щупалец
лофофора мшанок и т. д.
В этом отношении губки стоят еще почти на грани колониаль-
ных простейших, тогда как гидроидные полипы значительно от нее
отошли. Последние, являясь в основном также сидячими формами,
связанными с захватыванием опускающегося из толщи воды или
придонного пищевого материала, пошли по иному пути — пути ак-
тивного питания с использованием более крупной добычи. Невольно
78
рождается вопрос, возможно ли сравнить мешкообразный организм
аскона с мешкообразной же формой гидроидного полипа?
Однако к этому общеморфологическому вопросу мы вернемся
несколько позже, с более богатым фактическим материалом. • Сна-
чала же проанализируем гистологическую сторону организации гу-
бок.
Для губок, как мы отметили в первой главе, очень характерно
весьма сильное, часто мощное развитие среднего- слоя тела, состоя-
щего из ткани основного вещества. Это бросается в глаза особенно
при сравнении с покровом, который образован тонкой пластинкой
часто совершенно плоского эпителия, или при сравнении с выстил-
кой пищеварительных полостей, которые покрыты сплошь очень
примитивным воротничковым жгутиковым эпителием (см. рис. 1,2).
Слабость, примитивность эпителизации в организме губок действи-
тельно заслуживает особого внимания — и тот и другой эпителии
состоят только из клеток одного сорта. По сравнению со сложно^-
стью дифференцировки эпителиев у гидры картина, наблюдаемая
у губок, поражает своей простотой. Но этого мало — как мы ви-
дели, сама устойчивость эпителиальной дифференцировки у губок
оказывается очень своеобразной. Известны опыты с длительным ме-
ханическим раздражением губки, причем воротничковые элементы
втягивают свои жгуты, сглаживают воротнички, теряют эпителиаль-
ное расположение, образуя кучки клеток неправильной формы. По
окончании раздражений они снова начинают располагаться в один
слой, каждая образует воротничок и жгут и в конце-концов дают
воротничковый эпителий, выстилающий внутреннюю полость ас-
кона.
Такого рода факты приводили исследователей к мысли, что губ-
ки, собственно, являются «Zoa anhistia» —• бестканевыми или, по
крайней мере, с «факультативной дифференцировкой» тканей, в
противоположность всем прочим многоклеточным организмам. Эту
идею можно подкрепить и фактом отсутствия у губок эпителоген-
ных тканей, а именно мышечной и нервной. Однако- мышечные эле-
менты мезенхимного типа у губок встречаютсС^асГТбЙ^аясь^ПР^
да форм под эпителием вокруг osculum и на некоторых других уча-
стках системы каналов; здесь они образуют своего рода сфинктеры,
регулирующие ток воды через организм губки.
В связи с этим мы может констатировать, что эволюция губок
идет по пути дифференцировки ткани основного вещества — она
образует главную массу тела, в ней происходит дальнейшая диффе-
ренцировка клеточных элементов. Как у гидры гомогенная опорная
пластинка не представляет развития каких-либо структур, прису-
щих основному веществу, так и у губок однородность основного
вещества — обычное явление. Но, в противоположность гидре, для
губок характерно постоянное присутствие в нем клеток. Здесь на-
годятся индифферентные элементы — археоциты, амебоциты раз-
ного характера, резервные клетки, производящие скелет, — склеро-
бласты, мышечные элементы, отмеченные выше, и, наконец, поло-
79
вне клетки. Производимые клетками скелетные образования, раз-
ного рода спикулы и аморфные отложения, в конце-концов также
оказываются свободно лежащими в массе геля основного вещества.
Такого этапа в развитии основного вещества, когда мы имели бы
только бесструктурную, не содержащую клеток гомогенную массу,
у губок нет. И, очевидно, в животном царстве есть два пути эволю-
ции; один пуА связан с образованием настоящих эпителиев,
клетки которых берут на себя и образование основного соединяю-
щего вещества, как у гидры; другой путь ведет к чрезвычайно при-
митивным эпителиям, но с самого начала отличается присутствием
клеточных образований в основной массе соединительного студени-
стого вещества. Имеют ли оба эти пути общее начало — вопрос, к
которому мы вернемся в дальнейшем.
Итак, губки отличаются в гистологическом отношении, во-пер-
вых, неустойчивостью тканевой дифференцировки, во-вторых, отсут-
ствием нервной ткани и эпителогенных мышц, в-третьих, примитив-
ностью эпителиев и, наконец, в-четвертых, первичной клеточностыо
ткани основного вещества, проделавшей у губок своеобразную эво-
люцию на основе разделения физиологических отправлений между
различными категориями клеток. Здесь мы подходим к анализу ор-
ганов и их систем у губок.
У олинтуса и аскона легко можно, как и у гидры, различить
покров, кишечник и средний слой тела, функционирующий в каче-
стве органа опоры и связи, но в строении этих органов у тех и дру-
гих имеется огромная разница. Примитивность покрова губок пора-
зительна — это' только тонкий слой протоплазмы, изолирующий
внутреннюю среду организма, массу студня основного вещества, от
наружной водной среды. Таким образом создается первая возмож-
ность передачи питательных веществ от одних клеток организма к
другим, и, стало быть, к чисто механической функции связи через
посредство студия, что дало колонию простейших, присоединяется
новый вид связи — связь химизма, что на основе колонии создает
уже новый этап развития животного мира — многоклеточный ин-
дивидуум.
Осложнение покрова выражается у губок только в усилении по-
граничной функции путем превращения эктоплазматического слоя
в ороговевающую пленку. Это дает возможность дальнейшей диф-
ференцировки в различных направлениях: ведет, с одной стороны,
к появлению защитной функции в покрове, а с другой — функциям
прикрепления и скелетной. •
Пищеварительный аппарат у аскона (рис. 20, Л) на первый
взгляд представлен типичным целентерическим кишечником, напо-
добие имеющегося у гидры, как это- и принимал Геккель, а за ним
далее большинство зоологов. Однако гидра, как мы выше видели,
имеет настоящий кишечник с объединенной работой его элементов,
благодаря чему возможно использование крупной добычи. Напро-
тив, у губок это чрезвычайно примитивный аппарат, выстланный
слоем одинаковых воротничковых жгутиковых клеток, из которых
S0
каждая работает совершенно независимо, захватывая мелкие пище-
вые частицы. В этом процессе могут участвовать й амебоидные эле-
менты среднего слоя, вставляющиеся местами среди жгутиковых
клеток. Какой-либо дифференцировки клеток пищеварительной по-
лости и объединенное™ их работы совершенно не видно. Таким об-
разом перед нами исключительно' примитивный аппарат по сравне-
нию с имеющимся у гидры, притом же построенный совершенно по
иному плану. А именно: у гидры это мешкообразное вдавление,
где сконцентрированы железистые и пищеварительные клетки и в
которое пища вводится активно через ротовое отверстие, через него
же затем вторично удаляются и непереваримые остатки. У губки
типа аскон это полость внутри тела, куда через мельчайшие по-
ры в стенке поступает вода с пищевыми частицами, удаляющаяся
далее через более крупное, отточное отверстие — оскулум, распо-
ложенный против места прикрепления. Можно' сказать, что между
кишечником гидры и пищеварительным аппаратом губки существует
глубокая принципиальная разница, усугубляющаяся в дальнейшей
эволюции последнего.
У сикона (рис. 20, Б) внутренняя полость выстлана таким же
плоским кроющим эпителием, как и остальная поверхность тела;
воротничковые жгутиковые клетки располагаются в боковых вы-
ступах этой полости, получающих название жгутиковых камер, при-
чем сама полость становится атриальной. Таким образом создается
тип пищеварительного аппарата с дифференцировкой собственно’
переваривающего участка и системы приводящих и отводящих ка-
налов, из которых первые у сикона представлены лишь в зачаточном
виде пространствами между боковыми выростами, а вторые — ат-
риальной полостью. У типа лейкон (рис. 20, В) и та и другая части
системы уже выражены настоящими, сильно ветвящимися приводя-
щими и отводящими каналами, а иногда и еще особыми полостями
в начальном и конечном отделах системы, причем ток воды может
регулироваться благодаря появлению мышечных элементов, обра-
зующих сфинктеры на каналах.
В нашу задачу не входит дать описание различных вариантов
эволюции пищеварительного аппарата губок, мы можем ограни-
читься лишь установлением общего ее хода. Он выражается в ис-
чезновении единого резко отграниченного усваивающего отдела
аскона, в распадении его на большое число отдельных жгутиковых
камер с одновременной выработкой все более и более сложной си-
стемы приводящих и отводящих каналов, т. е. вспомогательной
части аппарата. Едва ли правильно будет характеризовать процесс
(в терминах, обычных для многоклеточных организмов вообще и
для гидры в частности) как переход единого кишечника в состоя-
ние диффузное, правильнее будет видеть в нем просто значительное
расширение и увеличение очагов пищеварения — важнейший мо-
мент в борьбе за жизнь.
1аким образом самый принцип построения пищеварительного
парата у губок совсем иной, чем у кишечнополостных.
$ Пути эволюции животного мира 81
У губок не выработалось устойчивой тканевой дифференциров-
ки, примитивен эпителий, имеются только мезенхимогенные мышцы
и совершенно не развилась нервная ткань. И в отношении орга-
нов: наряду с примитивнейшим покровом у них можно говорить
не столько об едином кишечнике, сколько о крайне экстенсивной
системе пищеварительного аппарата, несущего в некоторых своих
частях весьма примитивные* черты, а в других обнаруживающего
значительное осложнение.
Такое же своеобразие мы находим и в не рассмотренной еще
нами системе, образованной тканью основного вещества, •— это си-
стема механической и химической связи и опоры. Важнейший эко
логический момент — сидячий придонный образ жизни — выдви-
нул на первый план еще и другую функцию, связанную с тем же
образованием, функцию защиты. Исконно сидячие формы могли
сохраниться в борьбе за существование лишь при условии выра-
ботки таких защитных приспособлений, которые могли бы предо-
хранить от гибели животное, лишенное возможности уйти, скрыться
от врага. У губок клетки, лежащие в основном веществе, взяли на
себя роль таких защитных элементов в связи с выработкой в их
протоплазме известковых или кремниевых игл в виде макро- и мик-
росклер (рис. 22, 5).
Внутри склеробласта на органической основе происходит до-
вольно сложный процесс развития известковых (у небольшой ветви
губок) или кремниевых (у большинства) спикул, достигающих иног-
да очень значительных размеров, своеобразной формы и большой
сложности. Во всяком случае это те пассивно защитные образова-
ния, которые делают губку не только несъедобной, но и опасной
для нападающего — гарантируют жизнь форме, беспомощной по
другим чертам организации.
Первоначально защитные спикулы далее используются как ске-
летные образования, что ведет к возможности заселения разнооб-
разных биотопов^ особенно разнящихся по силе движения воды, и
к увеличению размеров животного. В конце-концов скелет разви-
вается крайне многообразно, и сами кремневые спикулы, редуци-
руясь, могут заменяться роговым скелетом, первоначально служив-
шим лишь в качестве защитного слоя, а затем и материала, связы-
вающего иглы в единое целое. Завершающим звеном в.этом ряде
эволюции скелета губок явились, с одной стороны, формы, совер-
шенно утратившие в связи с особенностями их существования ске-
летные образования, с другой же, — формы, у которых при обра-
зовании роговых отложений поверхностно расположенными клетка-
ми захватываются в растущий скелет посторонние частицы напо-
добие песчинок и т. п. при редукции настоящих спикул.
В общем и целом адаптивное явление образования защитных
спикул предками губок стало в эволюции этой группы ведущей
чертой, создавшей почти все многообразие их форм на путях узкого,
иногда чрезмерного («каменные» губки) приспособления к разно-
образным условиям существования в водной среде. В теснейшую
взаимосвязь с эволюцией скелета следует поставить и развитие в
высокой степени характерного для губок пищеварительного аппа-
рата в его типах'аскона, сикона и лейкона, так как то или иное на-
правление в развитии скелета обусловливало и соответствующее
расположение жгутиковых камер и образование системы их кана-
лов.
Последний штрих в рассмотрении организации’ губок дается их
крайне примитивным половым аппаратом. Здесь собственно нет еще
определенной (пространственно и морфологически) дифференциров-
ки гонад — половые продукты образуют скопления половых клеток
в любом почти пункте массы основного вещества, где далее про-
исходит и оплодотворение, и начальные этапы развития зародыша.
Отношения более простые, чем у гидры.
Рассмотрев все системы органов, мы должны подчеркнуть пол-
ное отсутствие у губок как органов чувств, так и нервной системы,
наряду с присутствием крайне примитивной мускулатуры. Послед-
няя работает в ответ на общее раздражение протоплазмы мышеч-
ной клетки или прилежащих элементов. Этот факт выявляет еще
одну оригинальнейшую черту губок — у них нет системы, морфо-
логически объединяющей в единое целое деятельность всех частей
тела. Индивидуальность губок выражена менее полно, чем у всех
прочих многоклеточных животных, завершивших ее оформление
выработкой нервной системы. Если губок мы не можем выделить
как «Zoa impersonalia», т. е. не имеющих индивидуальности, хотя
бы потому, что в развитии их имеется хорошо выраженная личин-
ка, то во всяком случае они еще остаются на более низком этапе
эволюции индивидуальности, чем прочие типы животного царства.
Их индивидуальность обусловлена, как было выше отмечено, во-
первых, механической связью клеток через посредство студня ос-
новного вещества, во-вторых, общим химизмом на этой же основе.
Если первый момент присущ уже колониальным простейшим, то
второй характеризует многоклеточную особь, однако без достаточ-
ной морфологической определенности. Последняя дается только с
развитием нервной системы. Поэтому, собственно, едва ли правиль-
но говорят в широком смысле о почковании у губок, так как поч-
кование представляет процесс образования бесполым путем новых
особей, что мы и видим в отдельных частных случаях, например об-
разование геммул у пресноводных губок или зародышей у Thetia. В
типичном экземпляре Euspongia (рис. 21) перед нами не образо-
вавшаяся почкованием колония, число особей в которой определяет-
ся количеством особых систем oscula или жгутиковых камер, а
только своеобразно растущая и дифференцирующая особь иного,
изшего, качества, чем особь гидры и других многоклеточных.
губоРкеЗЮМИРУЯ все сказанное> видим, что несложная организация
пле с ХаРактеРизУется ведущим защитно-опорным адаптивным ком-
стемы°жЧерТ’ С КОТОРЬ1М теснейшим образом связано развитие си-
гутиковых камер, а также приносящих и относящих кана-
6*
83
лов пищеварительного аппарата. Прочие черты организации носят
характер чрезвычайной примитивности — слабость эпителизации,
отсутствие эпителогенных*мышц и нервной ткани, местное развитие
мезенхимогенной мускулатуры, простота дифференцировки основ-
ного вещества, а в соответствии с этим — простота покрова, отсут-
ствие органов чувств и нервной системы, простота пищеварения,
диффузность-образования половых продуктов и в завершение все-
го — низшее качество индивидуальности по сравнению со всеми
прочими многоклеточными животными. Всего этого вполне доста-
точно, чтобы поставить и
Рис. 21. Euspongia officinalis (По Пфурт-
шеллеру). Справа и слева вырезки, справа
удален наружный слой, видны системы кана-
лов, стрелками отмечены направления тока
воды
решить вопрос об органи-
зации губок в иной пло-
скости, чем это было сде-
лано в свое время Гекке-
лем.
Губок нельзя сближать
с кишечнополостными, да-
же с наиболее простыми
представителями послед-
них, вроде гидры и гидро-
полипов. Губки являются
гораздо более примитив-
ною, чем кишечнопо-лост-
ные, ветвью животного
царства, пошедшей по
своему совершенно осо-
бенному пути, не обнару-
живающей чего-либо об-
щего с прочими много-
клеточными животными. Это ярко выражается, между прочим,
в невыработанности у губок радиального плана строения. Хотя
губки и являются искони сидячими формами, но симметрия у них
почти отсутствует, а радиальное строение их прими^Йб, не имеет
тех черт законченности, которые присущи сидячим потомкам
гастреад.
Решительным свидетельством в пользу этого взгляда может слу-
жить и эмбриональное развитие губок. Геккель находил у них ти-
пичную эктодерму и энтодерму, но уже Мечников утверждал иное,
а в дальнейшем пришлось признать у губок «извращение пластов».
Это — животные «шиворот-навыворот», Enantiozoa Деляжа. У них,
по А. А. Захваткину, имеется табличное дробление с образованием
изгиба и затем двуслойности; первоначально впячивающаяся часть
далее выворачивается и становится кроющим слоем, а первона-
чально поверхностные клетки становятся элементами, выстилающи-
ми жгутиковые камеры (рис. 22, А, Б). Таким образом, зародыше-
вые пласты губок оказываются несоответствующими этим образо-
ваниям прочих метазоа.
Естественный вывод из этих фактов, что нельзя крайне прими-
84
тивную в одном, и крайне адаптивную в другом, организацию гу-
бок укладывать в привычные рамки организации кишечнополост-
ных-
Решение вопроса об исходных предковых формах губок давно
VJKe намечено рядом исследователей благодаря одной замечатель-
ной особенности организации губок. Только в этой группе и нигде
более у многоклеточных животных постоянно присутствуют ворот-
Рис. 22. Развитие известковых губок: А к Б — Sycon (по Тайме-
ру), амфибластула и прикрепление; В — стадия олинтуса у
Sycandra (по Маасу):
1—поры, 2—спикулы
ничковые жгутиковые элементы. Это кладет резкую грань между
губками и прочими многоклеточными, но это же является указанием
на некоторые связи в другом направлении. Только среди однокле-
точных организмов, у Choanoflagellata, мы встречаем совершенно
такое же, и притом в высокой степени своеобразное строение. Одна
из колониальных их форм уж^ давно была названа «первогубкой»—
Proterospongia (см. рис. iy;
• .> Pterospongia представляет собой неправильной формы комочек
студенистого вещества, в котором периферически располагаются на
близком расстоянии друг от друга воротничковые жгутиковые осо-
би. Они движением жгута, каждая вне зависимости от других, вы-
зывают движение воды и улавливают при этом взвешенные части-
могущие служить питательным материалом.
Внутри студня располагаются клетки амебоидного характера,
здесь делящиеся и проходящие половой процесс. Происходят они
а счет жгутиковых элементов, удаляющихся с периферии вглубь
85
студня. Правда, в настоящее время природа Proterospongia под-
вергается сомнениям, и есть основания предполагать, что она пред-
ставляет только «реституционные» тельца губок.
В свое время И. Й. Мечников (1886) от подобных форм пытал-
ся произвести все многоклеточные животные организмы.
Думать, что организация Choanoflagellata и воротничковых жгу-
тиковых клеток губок развились путем простой конвергенции до-
вольно трудно, ввиду очень большого их своеобразия. Представить
же себе самый процесс эволюции типичной губки за счет колониаль-
ных Choanoflagellata чрезвычайно просто. Первым шагом в этом
направлении являлось расположение питающих элементов колонии
в более защищенном положении, на нижней поверхности, в углуб-
лениях студенистого вещества. Таким образом, должна была возник-
нуть система воротничковых жгутиковых полостей и связанных с
ними каналов. Этот процесс мы наблюдаем и ныне в развитии гу-
бок, когда личинка садится на дно поверхностью, несущей жгутико-
вые клетки (рис. 22, Б). Вместе с этим происходила и изоляция
внутренней среды губки. Путем расположения амебоидных клеток
на поверхности студня образовалась примитивная плазматическая
пленка покровного эпителия, колония стала многоклеточным орга-
низмом. Дальнейшим шагом, шагом, крайне характерным для гу-
бок, была выработка клетками защитных отложений характера
спикул внутри студня (рис. 22, В), что сразу значительно повыша-
ло возможность успешной борьбы за существование для этой
группы.
Таким образом, если губок можно характеризовать как живот-
ных, у которых тканевое строение крайне примитивно, анатомиче-
ская дифференцировка идет особым образом, развитие индивидуума
происходит совершенно своеобразно, а выработка индивидуально-
сти не завершена, то обобщая все это, мы должны сказать: перед
нами совершенно иной путь эволюции по сравнению с гастреадами!
Это заставляет выделить губок в независимую от прочих многокле-
точных животных группу Parazoa, берущую начало от иных коло-
ниальных простейших.
В нашем анализе мы попытались показать всю оригинальность
эволюционного пути губок по сравнению с путем гастреад. Вместе
они дают поразительную картину рождения и первых шагов много-
клеточной ступени организации, имеющей свои специфические чер-
ты, которые совершенно отсутствуют, даже потенциально, в орга-
низме одноклеточных животных и делают тех и других несравни-
мыми друг с другом.
Переходим к истокам высших типов животного мира.
Глава /V
ТИП ГРЕБНЕВИКИ — CTENOPHORA
Клейкие клетки, щупальца. Дифференцировка мерцательного покро-
ва. Происхождение гребневиков. Кишечник гребневиков Общий
план построения тела гребневиков. Мускулатура и нервная система.
Предки гребневиков. Итоги анализа организации.
Лишь в последнее время получает более широкое распростране-
ние взгляд Б. Гатчека (1889) на гребневиков как на представите-
лей особого типа — Ctenophora, наравне с типом Coelenterata.
Большинство же поныне верно1 традиционным представлениям, по
которым гребневики целиком и полностью включаются в тип ки-
шечнополостных.
Несомненно, что гребневики обладают кишечником целентериче-
ского строения, т. е. в виде мешка с одним отверстием, служащим
как для введения пищи, так и для выведения непереваримых остат-
ков. Однако если руководствоваться только этим, хотя и очень
важным, признаком, было бы последовательным в тип кишечнопо-
лостных включить также низших представителей червей — плоских
червей. Но разрывать эволюционный ряд типа Vermes, это значит •>
культивировать искусственную систему. Поэтому более рационально
выделить три независимые ветви целентерических животных, про-
исходящих от гастреадных предков, в три независимых типа: тип
Coelenterata, тип Ctenophora и отчасти тип Vermes, что мы и
сделаем.
Уже традиционным приемом стало характеризовать эти три фи-
логенетических ветви адаптивным признаком, бросающимся в глаза
по его оригинальности, а именно: в качестве аппарата защиты и
нападения у них функционируют оформленные секреты покровного
эпителия. В этом выявляется особенно далеко идущая дифферен-
цировка в пределах одной клетки, что у выше стоящих организмов
обычно заменяется дифференцировками целых клеточных комплек-
сов. В силу этого А. Ланг включает в характеристику своей гастреи,
как мы видели выше, клетки покрова с ядовитым секретом.
У кишечноплостных они представлены книдобластами, содержа-
щими в высокой степени специализированные стрекательные кап-
улы различных сортов, у гребневиков также сложно устроенными
87
Б
23. А — клейкий ап-
Рис.
парат Coeloplana (по Т. Ко-
маи); Б — стрекательная
капсула гидры типа стено-
тели с вывернутой нитью
(по Л. Тиман);
1 — клейкие зерна, 2 —
превращенное в длинную
мую нить, 3—протоплазма ____
кой клетки, 4 — спирально за-
витая нить тонофибриллярного
характера
клейкими элементами (рис. 23). у турбелляоий— характерными клет-
ками, содержащими рабдиты. Отсюда довольно употребительные
названия подтипов кишечнополостных, принимаемых большинством
зоологов: Cnidaria — «стрекающие» и Collaria — «клейкие».
Однако если действительно стрекательные капсулы кишечнопо-
лостных и рабдиты турбеллярий являют-
ся только оформленными секретами, став-
шими как бы своеобразными органеллами
клеток, этого нельзя сказать про клейкие
элементы. В них капельки клейкого се-
крета покрывают свободную поверхность
протоплазмы, а особая дифференцировка
происходит не в секрете, а в клетке—яд-
ро превращается в прямую нить, вокруг
которой в протоплазме дифференцируется
спиральное волокно характера тонофиб-
рилл, фибрилл сопротивления. Получается
аппарат, благодаря которому приклеив-
шийся предмет крепко, как на аркане,
удерживается на щупальце гребневика,
покрытом такими элементами.
Покров прочих участков тела обильно
содержит обычные железистые элемен-
ты — слизистые и белковые, но диффе-
ренцировки, подобной клейким клеткам, в
нем уже нет, и, конечно, последние мож-
но считать доведенным до крайних степе-
ней приспособлением для захватывания
щупальцами добычи. В этом смысле мож-
но и следует противопоставить широкий
ряд форм, обладающих стрекательными
капсулами —> тип кишечнополостных, как
единое филогенетическое направление
другому ряду форм, предки которых не
обладали подобными образованиями, а
выработали совершенно иной аппарат
клейких клеток на щупальцах — тип гре-
бневиков.
Сами щупальца последних резко отли-
чаются от подобных образований кишеч-
нополостных, Являясь, как и щупальца
кишечнополостных, приспособлениями
для защиты, а также для захватывания добычи и передачи ее в рот,
они представляют собой только длинные ветвящиеся выросты по-
кровного слоя, т. е. эпителия, и прилежащей мезоглеи с мощным
пучком или несколькими пучками мышц внутри. Этим обусловлена
очень большая способность сокращения щупалец, могущих полно-
стью втягиваться в особые углубления покрова — щупальцевые
88
ядро
пря-
клей-
влагалища (рис. 27). Таким образом перед нами по существу со-
вершенно иного характера образования, чем щупальца кишечнопо-
лостных, представляющие выпячивания всей стенки тела и обязатель-
но содержащие по оси эктодермальные элементы. Скорее их исходное
состояние можно видеть в небольших простых или даже ветвящихся
выростах покровного слоя, встречающихся среди гребневиков же
аборально у Ctenoplana и Coeloplana, Вегое и др. Закладка щупа-
лец в онтогенезе идет из эктодермы также близ' аборального по-
люса эмбриона, т. е. совершенно иным образом, чем в оральных
щупальцах кишечнополостных
Рис. 24. Цидиппоидная
стадия в развитии
Coeloplana go'ioctena
(по Кремпфу):
1 — аборальный орган
чувств, 2 — закладки греб-
зых пластинок, сближенные
попарно, 3 — глотка, 4 —
закладка щупалец и щупаль-
цевых влагалищ
(рис. 24). Очевидно, выработку
щупалец гребневиков следует
представлять себе, как образова-
ние у аборального полюса пары
клейких придатков, служащих в
дальнейшем при их удлинении для
захватывания добычи при движе-
нии этим полюсом вперед. В соот-
ветствии с этим ротовая полость
и глотка, которой захватываются
Рис. 25. Строение гребных пласти-
нок (по Р. Гертвигу):
1 — клетки, дающие начало гребным
пластинкам, 2 — покровный эпителий
пойманные планктонные животные, сплющиваются, образуя в попе-
речном сечении овал, широким краем обращенный к соответствую-
щему щупальцу. Таким образом, как приспособление, гарантирую-
щее поступление пойманной добычи в кишечник, создается опре-
деленное положение плоскости, в которой лежит пара щупалец, к
плоскости, в которой сплющена глотка, а именно, под углом в 90°.
У Вегоё, где сама глотка становится необычайно широкой, совер-
шенно исчезает и аппарат щупалец.
В покрове гребневиков кроме присутствия клейких клеток
имеется другая характерная черта — сохраняется примитивное
ресничное его состояние, но здесь оно связано х крайним приспо-
соблением — реснички по большей части склеиваются в тонкие
гребневидные пластинки (рис. 25). Отсюда и название типа —
гребневики.
Образуя движущий аппарат, значительно более сильный, чем
простые реснички, гребные пластинки располагаются по округлен-
ному грушевидному телу, как норма в виде восьми простых мери-
диональных рядов, которые, однако, при формировании особи пред-
89
ставлены сначала четырьмя парными рядами — очевидно более при-
митивное, исходное состояние.
Близ аборального полюса ряды гребных пластинок продолжают-
ся просто в мерцательные бороздки, достигающие, соединяясь по-
парно, аборального органа (рис. 26). Здесь ресничный аппарат
претерпевает опять очень характерное изменение, становясь суще-
ственнейшим компонентом своеобразного механорецептора, функ-
ционирующего как аппарат
Аборальный орган состоит
из склеенных в виде купола рес-
ничек, прикрывающих и защи-
щающих расположенный под
ним собственно орган равнове-
сия. Последний состоит из шаро-
образного комочка клеток, про-
исходящих из эктодермального
эпителия, содержащих каж-
дая отложение извести в своей
протоплазме и вместе давящих
на поддерживающие их как
пружинки 4 пучка склеенных
ресничек. При наклоне тела
гребневика в ту или иную сто-
рону меняется давление на
пружинки, в связи с чем изме-
няются темп движения соответ-
ствующих рядов гребных плас-
тинок, и тело гребневика вы-
равнивается. *
Однако своеобразный сам
по себе орган осложняется в
аборальном аппарате тем, что
Рис. 26. А — схема строения" абораль-
ного органа; Б — комплекс мерцатель-
ных образований, связанных с абораль-
ным органом;
1 — аборальное ресничное поле, 2 — мерца-
тельные бороздки, 3 — купол органа, 4 — с ООКОВ К Нему ПрИМЫКаЮТ ДВН
мышечные пучки, 5 — пружинки 6 — стато- J
лит ресничных поля,- реснички кото-
рых, также обычно склеиваясь,
образуют небольшие пластинки, вызывающие ток воды по направ-
лению к куполу аборального органа. Этот ток проникает под купол
через пару отверстий в нем, соответствующих мерцательным по-
лям, и выходит наружу через мерцательные бороздки, идущие к
рядам гребиых пластинок. Оба аборальных ресничных поля утол-
щены по краям, а у некоторых форм (Вегоё, Platyctenea) эти утол-
щения превращаются в соответствующие ряды уже упомянутых
щупальцеобраэных придатков, повидимому, с чувствительной
функцией.
Таким образом получается сложный аборальный аппарат, ука-
зывающий на значительную приспособленность гребневиков к сво-
бодному существованию, при котором только возможно и необхо-
димо развитие подобных образований. Напротив, аборальный полюс
90
у кишечнополостных, как пункт; служащий для прикрепления поли-
поидных форм, никогда не дает каких-либо рецегпорных аппаратов, ,
а затем далее и у медуз уже является филетически бесплодным,
бесперспективным участком в смысле развития новых образова-
ний. Если у Trachilina и имеются некоторые дифференцировки
на эксумбрелле, то они идут сюда от края зонтика.
Наличие у гребневиков аборального органа равновесия говорит
за то, что их нельзя связать с кишечнополостными, нельзя, исходя
й3 организации полипа или медузы, представить гребневиков как
дальнейший шаг в эволюции этих форм. Наконец, естественно, от-
сюда следует, что' Ctenophora представляют искони свободноживу-
ших обитателей моря, для предков которых придонное существова-
ние сидячих или ползающих форм было совершенно чуждо. Это
животные, сохранившие свободный пелагический образ жизни искон-
ных гастреад, причем их аборальный комплекс органов должен был
развиться на основе имеющихся хотя бы у планулы кишечнополо-
стных чувствительных образований аборального конца тела, как
дальнейшая дифференцировка этих образований. Подобный про-
цесс мы увидим далее и в другой ветви свободноподвижных целен-
терических животных, у турбеллярий.
Некоторое сходство с этими последними имеется у гребневиков
также в наличии двигательного ресничного покрова, пошедшего,
однако, по совершенно иному пути в своей эволюции. Образование
первоначально четырех парных рядов гребных пластинок, перехо-
дящих у взрослых в восемь ординарных, является, во-первых, вы-
ражением более сложной формы мерцательного двигательного по-
крова, а во-вторых, не может быть отделено' от дифференцировки
аборального аппарата равновесия и должно было развиваться в
тесной взаимосвязи с последним. Это единый комплекс образова-
ний, что опять-таки говорит за примитивный характер свободно-
пелагического существования гребневиков и за отсутствие момента
прикрепления в их эволюции. Она шла по пути выработки более
энергичного движения и перехода его из вращательного гастреад
в прямолинейно-поступательное при использовании для питания
более крупной добычи и при развитии пары длинных щупалец. При
этом четыре гребных ряда представляют наиболее простой случай
возможности равновесия тела при дифференцировке ресничного '
движения.
Как яркая приспособительная черта к пелагическому образу
ныхНИ’ напод°бие выработки организации медуз у кишечнополост-
небопПРедетавляется и весь габитус гребневиков (рис. 27). Обычно
тело ЬШое’ но иногда, как у Mertensia, Cestum, довольно крупное
homv*!^ пр03Рачно и У исходных форм приближается к шарообраз-
зрачнос И ГрушевиДному. Это происходит в связи с развитием про-
Р ти тканей и образованием обильной мезоглеи. Последняя
она до^ТсихКапопТСЯ пР°блематичной Hydroctena К. Давыдова, то, к сожалению,
Р ае нашла признания и подтверждения других исследователей.
91
представляет значительную массу основного вещества, несколько
уплотненную под покровным эпителием и содержащую волокна,
свободные клетки и мезенхимные мышечные элементы. Таким обра-
зом, тело по удельному весу приближается к морской воде. Его фор-
ма менее подходяща для парения в воде, чем зонтик медузы, и ни-
как не может быть сведена на последний, однако сохранение мер-
цательного покрова по восьми меридианам гарантирует животному
взвешенное состояние и поступательное движение в морской воде.
Рис. 27. Схема организации гребневиков (по Паркеру):
А — разрез в плоскости щупалец в правой половине, в
плоскости прилежащего ряда гребных пластинок слева;
Б — разраз в экваториальной плоскости, видна система
каналов кишечника;
1 — аборальный орган, 2 — гребные пластинки, 3 — центральный
отдел, 4 —воронка, 5 — меридиональные, 6 — окологлоточные,
7 — щупальцевые каналы, 8 — гонады, 9 — мезоглея, 10 — мы-
шечные волокна, 11 — глотка с железистыми валиками
Вероятно, при равномерном развитии студня за счет первоначально
незначительной прослойки основного вещества и сама шарообраз-
ная или грушевидная форма обусловлена сохранением мерцатель-
ного покрова по меридианам гетерополюсного тела в направлении
его движения. Отклонения в сторону выработки большей приспо-
собленности формы тела к парению у гребневиков часты и связаны
с процессом неравномерного роста. Укажем на лопасти у Lobata,
удлиненное в виде пояса тело Cestum, наконец, на глубокий ко-
локол Вегоё. Все это явления адаптивных изменений на основе ис-
ходной шарообразной или грушевидной формы, постоянно сохра-
няющейся в онтогенезе ктенофор, как цидиппоидная стадия.
Здесь же следует отметить характерное для гребневиков при-
сутствие двух разветвленных щупалец, исчезающих лишь у вторич-
но измененных Вегоё. Они также могут играть некоторую роль при
парении в пелагиали. Однако главное значение щупалец, конечно,
в активном нападении и защите, что определяется присутствием на
них клейких клеток. Щупальца вносят некоторую диспропорцию в
92
равномерное развитие мезоглеи ктенофор, так как их основания
располагаются глубоко в теле животного, что связано, с одной сто-
роны, с более прочным их укреплением, а с другой — с образованием
защитных влагалищ. Диспропорция выражается в слабом развитии
мезоглеи под основанием щупальца, тогда как кругом она превра-
щается в мощную массу ткани основного вещества.
В этой массе кишечник приобретает своеобразное развитие в
виде системы каналов, обусловливающих и функцию переваривания,
я функцию движения пищевого материала. Усвоение последнего
происходит, как у всех форм с целентерическим кишечником, в
эктодермальном отделе путем внутриклеточного пищеварения. Осо-
бенно интенсивно функционирующие части располагаются под во-
семью меридианами гребных пластинок, где, как и у Scyphozoa и
Anthozoa, постоянно залегают базально в эктодермальном эпителии,
у источников своего питания, половые клетки. Последние, распола-
гаясь в виде меридиональных тяжей, образуют гонады, по семен-
нику и яичнику соответственно с той и другой стороны меридио-
нального канала кишечника. Последние в онтогенезе первоначаль-
но закладываются в виде четырех широких карманов, что вызывает-
ся развитием пары щупальцев в одной меридиональной плоскости и
эктодермального' участка кишечника в другой, лежащей под углом
в 90° к первой. Эти карманы, далее, при дифференцировке восьми
рядов гребных пластинок и массы мезоглеи превращались в восемь
меридиональных каналов и в каналы, соединяющие их с централь-
ным отделом. Последний принимает вытянутую в поперечнике фор-
му, проходя от одного щупальца до другого, а по направлению к
аборальному полюсу опять-таки суживается в канал, доходящий до
аборального органа. Получилась крайне характерная картина ра-
диального расположения частей энтодермального кишечника, в ко-
торый, однако, чего-нибудь подобного развитию септ Scyphozoa и
Anthozoa совершенно нет.
Вторично, с разрастанием оральных участков тела у Lobata или
у Cestum происходит проникновение в них оральных отделов соот-
ветствующих меридиональных каналов и соединения их друг с дру-
гом. У ползающих Platyctenea в уплощенных краях лепешкообраз-
ного тела развивается настоящее сплетение гастро-васкулярных ка-
налов, связанное с меридиональными и щупальцевыми каналами
средней утолщенной части тела. В известной мере сходные отно-
шения вырабатываются и у Beroidea, но здесь они связаны осо-
бенно тесно с развитием глоточного эктодермального отдела кишеч-
ника гребневиков.
Характерная картина пищеварительной системы последних за-
вершается выработкой мощного эктодермального впячивания, полу-
чившего очень большую роль в переваривании особенно более
крупной добычи. Железы в проксимальном отделе его стенки выра-
батывают пищеварительные секреты, что ведет к интенсивному по-*
лестному пищеварению. Крайне своеобразный путь дифференциров-
ки! Пища здесь подвергается такой переработке, что этот отдел,
93
соответствующий глотке кораллового полипа или актинии, совер-
шенно правильно может получить название желудка в части, при-
лежащей к энтодермально-му отделу. Конвергентное развитие в
филетических ветвях гребневиков и стрекающих сказывается в об-
разовании глоточного отдела и системы гастро-васкулярных кана-
лов, но их эволюция у тех и других идет иными путями — эго лишь
общее сходство на основе приспособления к пелагическому суще-
ствованию и питанию у гребневиков и медуз и несходное развитие
в деталях.
Как уже было выше отмечено, глоточный отдел ктенофор в по-
перечном сечении представляет не округлые очертания, а, приспо-
собляясь к захватыванию краями рта добычи со щупалец, имеет
форму овала, сплющенного в плоскости, перпедикулярной к пло-
скости щупалец. Соответственно в этой же плоскости от централь-
ного эктодермального отдела отходит пара каналов или карманов,
располагающихся параллельно стенке глотки и образовавшихся при
вдавливании глотки в пространство, занятое эктодермальным меш-
ком кишечника.
Развитие глотки адаптивно может достигать колоссальных раз-
меров. Такая чрезмерно развитая, соответственно дифференцирован-
ная глотка образует всю зияющую полость глубокого колокола Вегоё,
на которой сверху располагается небольшой центральный отдел
эктодермального кишечника. ЛАеридиональные каналы, соответст-
вующие рядам гребных пластинок, разрастаются здесь далеко к
оральному краю глотки, где соединяются друг с другом и с около-
глоточным каналом посредством кольцевого канала. На всем своем
протяжении они дают массу ветвящихся выростов в стенке коло-
колообразного тела. Можно сказать, что в организации Beroidea
глотка стала ведущей. В известной мере подобное значение приоб-
ретает глотка и в организации Platyctenea — ползающих Ctenoplana
и Coeloplana (рис. 28) и сидячей Tjalfiella. У первых двух боль-
шая, дистальная часть глотки, покрытая мерцательным эпителием,
выворачивается, образуя обращенную при ползании к субстрату
«брюшную» поверхность лепешкообразного тела. Таким образом
Cteno- и Coeloplana ползут, собственно, на вывернутой глотке, в
связи с чем гребные пластинки утрачивают свое значение. У Ctenop-
lana они уже незначительно развиты/а у Coeloplana совершенно
исчезают. Однако и та и другая в онтогенезе имеют типичную ци-
диппоидную стадию, что подчеркивает чрезмерное развитие глотки,
ставшей ведущей в своеобразном процессе приспособления к пол-
зающему образу жизни с питанием придонными организмами.
Tjalfiella после цидиппоидной стадии переходит к сидячему су-
ществованию, теряя гребные пластинки и прикрепляясь оральным
краем глотки, часть которой однако разрастается около щупалец,
образуя вокруг последних два трубовидных рукава. Через них пища
поступает в центральный отдел, а в стенке их вторично ветвятся
обильные выросты гастро-васкулярных каналов.
Не будем больше распространяться об этих аберрантных греб-
94
невиках, вторично перешедших к ползающему образу жизни или
даже сидячему существованию. Однако анализ ух глубоко приспо-
собительной организации представляет известный интерес в связи
с явлениями сходства в строении с низшими представителями чер-
Рис. 28. А — Ctenoplana в состоянии плавания (по К- Давы-
дову) :
передана окраска, видны: аборальный орган (1), ряды гребных пластинок
(2), папиллы (3) и сильно сократившиеся щупальца (4);
Б — Ctenoplana в состоянии ползания — схема организации
(по Т. Комаи):
изображена половина тела, виден аборальный орган (1) и идущие к нему
ресничные бороздки (3) от рядов гребных пластинок (2) в нижнем квадра-
те, в верхнем отмечены только занимаемые ими места, нанесен кишечник,
5 — его периферическое сплетение, 6 — глотка, 7 — радиальные каналы
с гонадами (8), — расправленное ветвистое щупальце, его влагалище (9)
В — Coeioplana mesnill, схема организации (по Давыдову).
Обозначения те же
вей, организация которых развивается также под знаком приспо-
собления к ползающему придонному существованию.
Выявив ряд характерных черт строения гребневиков, мы подо-
шли в анализе организации типичного гребневика к вопросу об об-
95
щем плане построения его тела. Вопрос, представляющий немалый
интерес, так как выше нам уже пришлось раскрыть адаптивное
значение радиального плана построения кишечнополостных в связи
с прикреплением аборальным концом тела и с сидячим образом
жизни полипа. Для гребневиков обычно также говорят о радиаль-
ном плане построения, который выражается в расположении мери-
диональных рядов гребных пластинок, в ходе соответствующих кана-
лов в гастро-васкулярной системе, в расположении гонад. Действи-
тельно, известное радиальное расположение всех этих образований
по отношению к орально-аборальной оси — несомненный факт, и
если принимать, что гребневики происходят от кишечнополостных
(будет ли это Ctenaria Геккеля, Hydroctena Давыдова или еще ка-
кая-либо медузоидная форма), то их радиальное построение есть
лишь унаследованная от предков, выработавшаяся средй ниже
стоящих кишечнополостных черта организации.
Наш анализ показал, что указанные черты радиального строе-
ния у гребневиков обусловлены развитием комплекса двигательного
ресничного покрова и аборального аппарата, тогда как у сидячего
полипа радиальность строения связана с определенными взаимоот-
ношениями прикрепленного организма с окружающей биоценоти-
ческой средой. Как раз этот последний момент у пелагического
гребневика находит совершенно иное выражение —. развивается
пара аборальных щупалец, располагающихся в плоскости, переводя-
щей радиальную симметрию в бирадиальную наподобие имеющейся
у коралловых полипов при развитии сплющенной глотки с сифоно-
глифами и т. д. Однако этим дело не ограничивается, в теснейшей
взаимосвязи с развитием пары щупалец стоит выработка щелеоб-
разного рта и сплющенной глотки, причем их плоскость симметрии
пересекает плоскость симметрии щупалец под углом в 90°. Таким
образом через монаксонное гетерополюсное тело гребневика можно
провести две взаимно перпендикулярные плоскости — плоскость
щупалец и плоскость глотки, — делящие тело на симметричные по-
ловины, плоскости же, проведенные через меридиональные ряды
гребных пластинок, уже не дадут симметричных частей, хотя, если
откинуть щупальца и глотку, получается радиальная симметрия.
Итак, дважды бирадиальная симметрия гребневиков совершенно
иного характера и иного происхождения, чем простая бирадиальная
симметрия Anthozoa. Перед нами два совершенно различных плана
построения: у гребневиков под маской радиальной симметрии —
тетрамерное строение, у кишечнополостных под маской бирадиаль-
ности —• настоящее радиальное строение. Разные адаптивные мо-
менты породили и разные отношения в расположении органов.
Последним, очень характерным штрихом организации гребневи-
ков является совершенно иной по сравнению с кишечнополостными
путь дифференцировки у них мышечных, сократительных образова-
ний. В то время как у кишечнополостных мы'находим образование
эпителогенной мускулатуры или в виде эпителиально-мышечных
элементов, или настоящих, отшнуровывающихся у актиний от покро-
96
ва мышечных пучков, у гребневиков совершенно нет подобной мус-
кулатуры. У них эволюция последней пошла по пути дифференци-
ровки мезенхимных мышц, по мнению одних исследователей разви-
вающихся за счет клеток, уходящих из эктодермы, по мненщр дру-
гих — из особого эмбрионального зачатка мезодермы, но никогда
не имеющих характера и не проходящих стадию эпителиально-мы-
шечных элементов. В этом отношении мускулатура гребневиков при-
ближается к мускулатуре плоских червей. В области глотки разви-
ваются особенно характерные многочисленные мышечные пучки,
разветвленные концы которых с одной стороны заканчиваются, под-
ходя к эпителию глотки, а с другой — под эпителием орального уча-
стка покрова. «Адаптивное значение этой мускулатуры ясно — она
обусловливает у гребневиков сократимость и подвижность краев
рта и глотки, играющих такую важную роль при захватывании
добычи. О специальных мощных мышечных пучках по оси щупалец
уже было в свое время упомянуто.
В противоположность мускулатуре, дифференцировавшейся пол-
ностью, нервная система сохраняет свое примитивное базиэпите-
лиалыюе положение и представлена диффузным нервным сплете-
нием покрова, в котором рассеяны чувствительные клетки. Вероят-
но в связи с развитием мускулатуры глоточного отдела тела можно
подметить некоторое сгущение нервного сплетения в виде кольца,
охватывающего оральную часть тела. Однако факты иннервации
мускулатуры у гребневиков еще совершенно не выяснены. Также
под некоторым вопросом стоит и принимаемое некоторыми исследо-
вателями образование ганглиозного скопления под аборальным ор-
ганом чувств, хотя теоретически следовало1 бы ожидать его появле-
ния, поскольку правильно общее положение, что нервная система
развивается функционально, т. е. коррелятивно с одной стороны с
органами чувств, а с другой — с мускулатурой. У гребневиков, ус-
тановлен в настоящее время и еще один интересный факт, а именно:
параллельно рядам гребных пластинок проходят, в виде меридио-
нальных сгущений нервного1 плексуса, как бы зачаточные нервные
стволики. Однако едва ли можно на основании этого факта делать
какие-либо далеко идущие выводы кроме обычного утверждения,
что нервная система «развивается функционально» и у гребневиков
несет еще черты большой примитивности.
Мы закончили анализ организации гребневиков, оставив в сто-
роне некоторые мелкие ее черты и еще некоторые, недостаточно
выясненные. Подведем итоги.
Во-первых, несомненна основная разница в организации гребне-
виков и кишечнополостных, которая ни в коем случае не позволяет
°бъ,единять их в один тип — традиционных Coelenterata. Это два
Разных пути эволюции примитивных гастреад, причем у гребневи-
ков сохраняется исконный пелагический образ жизни свободнопод-
вижных хищников. Те гастреады,* которые дали начало гребневи-
кам, должны были разниться от рассмотренных нами ранее исход-
ных для целентерат. Имея такое же овальное монаксонное гетеро-
7 Пути эволюции животного мира 97
полюсное тело с ротовым отверстием, обращенным при движение
назад, они отличались от последних характером дифференцировки
тканей, а именно: клетки эпителиев не являлись эпителиально-мы-
шечными элементами, а представляли простой мерцательный эпи-
телий, так как мышечные, сократительные ‘волокна дифференциро-
вались здесь не в эпителии, а в мезенхимных элементах, т. е. утра-
тивших эпителиальную связь и ушедших в основное вещество. Со-
ответственно и ткань основного вещества, имея в себе клеточные
элементы, получала характер мезенхимы. Таким образом, уже среди
гастреад должны были наметиться два различные пути дифферен-
цировки мышечной ткани и, соответственно, развития ткани основ-
ного вещества. Далее, у предков гребневиков чувствительно-нервные
элементы, повидимому, были хорошо развиты в эктодермальном
эпителии, их присутствие в эктодермальном является минимальным,
или, может быть, здесь они совершенно не дифференцировались.
Этот вопрос пока неясен. Наконец, половые клетки локализирова-
лись в эктодермальном эпителии, т. е. у источников питания.
В связи с такой дифференцировкой тканей гастреады, давшие
начало гребневикам, отличались и в отношении органов и их си-
стем: если нервная система была присуща покрову и, может быть,
в малой степени кишечнику, то мускулатура безусловно расположи-
лась в прослойке ткани основного вещества, а гонады локализиро-
вались в стенке кишечника.
Сравнительно слабо была развита способность к регенерации.
Всего это достаточно для того, чтобы представить себе прими-
тивные черты организации предков гребневиков. На их основе, в
известной мере, как их дальнейшее узко- и даже крайне адаптив-
ное развитие и осложнение возникает организация гребневиков.
Ведущими в ней являются два комплекса черт: один комплекс,
связанный с свободным пелагическим существованием, другой —
обусловленный питанием.
К первому относятся следующие черты организации: 1) прозрач-
ность тканей как защитное приспособление, 2) шарообразная или
грушевидная форма тела вместе с развитием ткани основного ве-
щества в виде массы мезоглеи как приспособления для уменьшения
удельного веса животного, 3) выработка, за счет ресничного покро-
ва, аппарата восьми рядов гребных пластинок для поддержания во
взвешенном состоянии и для более энергичного передвижения; и в
теснейшей взаимосвязи с этим 4) образование аборального аппа-
рата с органом равновесия, 5) неравномерный рост орального от-
дела у Lobata в виде сократительных лопастей, расположенных в
плоскости глотки, у Cestum образование лентовидного тела, у
Вегоё — глубокого колокола как 'приспособления к парению в воде
и к движению.
Последние черты связывают комплекс приспособлений к пелагй*
ческой жизни со вторым комплексом узко приспособительного же
характера к захватыванию добычи и питанию, в который входят:
1) развитие пары ветвящихся щупалец у аборального полюса, слу-
98
жащих для облавливания более значительного пространства окру-
жающей водной среды, 2) выработка на щупальцах клейких кле-
ток как приспособления для удержания более крупной добычи,
3) развитие эктодермальной глотки, чрезвычайно подвижной в свя-
зи с выработкой мышц в оральном отделе тела, -как приспособле-
ния для заглатывания пойманной добычи; при этом глотка сплю-
щена в плоскости, перпендикулярной к плоскости расположения
щупалец, что является приспособлением, гарантирующим попа-
дание пищи в глотку, 4) выработка меридиональных рядов греб-
ных пластинок, в связи с переходом к прямолинейному движению
вместо движения по спирали, 5) превращение проксимальной части
глотки в желудок, в связи с развитием здесь пищеварительных же-
лез — приспособление к питанию более крупной добычей, а отсюда
и 6) развитие эктодермального кишечника, усваивающего' перерабо-
танную пищу путем внутриклеточного пищеварения, в виде системы
каналов с определенным током содержимого и расположением их
соответственно другим образованиям тела для питания последних—
меридиональные каналы с половыми железами, каналы щупалец,
канал к аборальному органу, каналы около глотки.
С тем и другим комплексом связаны своеобразные отношения
симметрии в строении гребневиков, а именно тетрамерность под
маской радиального построения. Также с обоими ведущими ком-
плексами коррелятивно связано, наконец, развитие примитивной
нервной системы в виде диффузного сплетения в покрове, где рас-
сеяны чувствительные клетки, с образованием сгущения сплетения
у гребных рядов и вокруг подвижной глотки. Однако моторный от-
дел ее остается неизученным, а сведения об .иннервации мускула-
туры собственно отсутствуют.
В заключение остается отметить, что особенно резко в органи-
зации гребневиков разнообразие приспособлений выразилось в
приспособлениях для использования пищевых ресурсов, причем ряд
форм перешел в придонные биотопы и биоценозы, обычно совер-
шенно не свойственные гребневикам. Такими крайними филогенети-
ческими ветвями, развивающимися, однако, всегда через исходную
цидиппоидную стадию, являются прикрепляющаяся ко дну Tjal-
fiella и ползающие по субстрату Platyctenea (рис. 28).
От форм гастреад, очень близких к предкам гребневиков, берет
начало третья ветвь целентерических животных, представители ко-
торой являются исходными для всех высших типов животного цар-
ства — это плоские черви в типе червей.
7*
Глава V
ПОДТИП ПЛОСКИЕ ЧЕРВИ — PLATHELMINTES
Билатерально-симметричное построение. Нестабилизированность
рта. Покров турбеллярий и выработка кожно-мышечного мешка.
Опорная паренхима. Дифференцировка нервной системы. Органы
чувств. Разнообразие полового аппарата. Примитивность прото-
нефридиев. Кишечник Acoela. Итоги анализа организации турбелля-
рий. Происхождение турбеллярий.
k В последний раз мы встречаемся с настоящими целентерически-
ми организмами уже в типе червей. Это будут плоские черви, и
прежде всего их свободноживущие формы — турбеллярий или пла-
нарии, наиболее сложно построенные непосредственные потомки
примитивных гастреад. Название планарии, собственно, принято
лишь для части ресничных червей, но мы будем им пользоваться,
как характеризующим наиболее типичные формы.
Планарий по их целентерическому кишечнику можно было бы,
как указывалось выше, поставить среди наиболее высоко организо-
ванных кишечнополостных, но мы видели, во-первых, насколько ис-
кусственен в таком широком понимании этот тип. Во-вторых, пла-
нарии одновременно являются низшими представителями иного ти-
па — типа червей. Этот этап в эволюции животного мира чрезвы-
чайно важен, как начало пути всех вышестоящих форм, как этап
выработки билатерально-симметридного плана строения, легшего
в основу построения всех остальных типов.
Если экологический момент использования обильного пищевого
материала, оседающего в море из верхних слоев воды, повел, как
мы видели, у очень примитивных гастреад к прикреплению або-
ральным концом ко дну и создал, в конце-концов, радиальный план
строения кишечнополостных, то переход к использованию находя-
щегося на дне пищевого материала у других, более сложно по-
строенных гастреад, уже обладавших захватывающей пищу глоткой,
повел к придонному существованию с ползанием по дну и; отсюда,
к созданию билатерально-симметричного построения червей, что
Э. Геккель считал понятным «само собой». Расшифруем эти
отношения.
У планарий могли дифференцироваться брюшная и спинная сто-
роны в связи с тем, что поступательное движение исходных пела-
100
гических гастреад аборальным концом вперед со слабым вращением
по спирали заменяется прямолинейным. Смена ‘дексицтропных и
деотропных делений в эмбриональном развитии (рис. 29) была до-
статочна, чтобы дать прямолинейное
поступательное движение. А процес-
сы неравномерного роста, выразив-
шиеся в преобладающем развитии
2 d и 4 d бластомера у эмбриона,
г. е., Другими словами, <в преобла-
дающем развитии задней «верхней»
части зародыша, повели -к смещению
ротового отверстия на «нижнюю»
сторону (рис. 30). Это смещение яв-
лялось первым необходимейшим ус-
ловием для возможности активно
использовать питательный материал,
находящийся на дне, другими слова-
ми, обусловливало возможность са-
мого существования ползающих при-
донных форм. Таким образом выра-
боталась брюшная, несущая ротовое
Рис. 29. Дробление яйца у Cyclo-
porus (по Бреслау): А — стадия
8 бластомер — образование 1-го
квартета микромер дексиотропным
делением; Б — стадия 12 бласто-
мер — образование 2-го квартета
леотропным делением. Стрелками
отмечено направление смещения
микромер от производящих их
макромер
Рис. 30. А — сагиттальный срез через зародыш Planocera (по
Серфесу) :
1 — энтодерма, образует еще сплошную массу клеток, в.которых заключены
крупные капли питательного желтка (2), 3 — эктодерма дает уже зачаток
стомодеума (4), направленный прямо назад, мезенхима двух родов: от произ-
водных 2-го квартета (5) и от клеток, получающихся за счет 4 d (6), 7 —
закладка головного мозга;
Б — мюллеровская личинка, только что вышедшая из-под яйцевой
оболочки:
У переднего полюса (1) клеткп закладки головного мозга (2), просвет кишки
(3) хорошо выражен, 4 — стомодеум, направлен книзу, 5 — рот, смещается
вентрально, так как благодаря процессам неравномерного роста развивается
зарЪтовой, задний отдел тела (6)
отверстие сторона. Она резко дифференцировалась от спинной не
только по нахождению рта, но и по взаимоотношениям с окружаю-
щей средой, а именно: брюшная сторона тела, обращенная постоян-
но к субстрату, находится в совершенно иных условиях, чем спин-
10]
пая, обращенная к свободной водной стихии. С брюшной стороны,
со стороны грунта, возможность нападения и необходимость ориен-
тировки и защиты были значительно менее остры, чем со стороны
спинной, из открытой водной стихии.
Однако и по-длине тела экологические отношения не везде оди-
наковы, а именно — аборальный конец'типичных гастреад, при дви-
жении являющийся передним, попадает в новые, неизведанные уча-
стки среды, тогда как задний проходит уже по местам, в которых
животное ориентировалось и которые сравнительно мало таят не-
ожиданного. В результате на переднем конце, а у ползающих при-
донных организмов преимущественно со спинной стороны, сосредо-
точиваются образования, служащие для ориентировки в окружаю-
щей среде —> механо-, хемо- и, особенно, фоторецепторы. Получает-
ся головной комплекс органов чувств, и впервые дифференцируется
несущий его1 головной отдел тела.^По продольной оси тела турбел-
лярий, являющейся осью движения животного, располагается и ча-
сто встречающийся у них орган равновесия, как довольно обычный
компонент головного комплекса. И лишь справа и слева от продоль-
ной оси, вернее от сагиттальной плоскости, взаимодействия орга-
низма с окружающей средой оказываются одинаковыми, в связи с
чем симметрично справа и слева оказывается построенным и все
тело червя, расположены все его органы (рис. 31—33).
Так вырабатывается в особых условиях ползания по дну била-
терально-симметричный план построения. Каждая его деталь вы-
деляется и усиливается естественным отбором в условиях придон-
ных биоценозов моря. Перед нами такого рода явление, которое
следует отнести к широким адаптациям, т. е. приспособлениям, зна-
чительно повышающим возможности организма в борьбе за суще-
ствование. Можно назвать это ароморфозом в смысле А. Н. Север-
цева.
Выяснив значение экологического момента в образовании била-
терально-симметричного строения, мы можем далее сделать и еще
один вывод — придонные свободноползающие формы стали источ-
ником эволюции всех вышестоящих типов животного мира. Ни ра-
диально-симметричные сидячие кишечнополостные, ни даже тетра-
мерные свободные пелагические гребневики не дали ничего
подобного. Однако к этому вопросу мы должны будем вернуться
в дальнейшем при разборе различных современных взглядов на
происхождение турбеллярий. Сначала нам необходимо проанализи-
ровать организацию последних.
После выработки билатеральной симметрии с образованием го-
ловного комплекса органов чувств и со смещением рта на брюш-
ную сторону, характернейшей чертой организации турбеллярий
является нестабилизированность ротового отверстия (рис/31—34).
Обычно расположение его от задней трети тела до средины брюш-
ной стороны. Однако нередко оно сдвинуто далеко вперед, иногда
даже находится на переднем конце тела; изредка, напротив, у
самого заднего конца. Эти факты представляют большой интерес.
102
Они вскрывают самую сущность процесса перехода к использова-
нию пищевого материала, находящегося на дне, показывая, что
с выработкой глотки, активно захватывающей пищу, ротовое от-
верстие изменяло свое положение. Во-первых, как было уже отме-
чено, оно сместилось на сторону, обращенную ко дну, в результате
Рис. 32. Окема экскре-
торного аппарата прес-
новодной триклады (по
Вильгельми). Внесена
глотка, чтобы продемон-
стрировать отношения у
форм с неразвитыми
периферическими частя-
ми полового аппарата:
18 — экскреторные поры, 19-
трубка протонефридия
Рис. 3.1. Схема органи-
зации пресноводных
гриклад (по Бреслау) :
1 — аденодактиль, 2 — поло-
вой атриум, 3 — мозг, 4 —
ложный семенной пузырек,
5 — кишечник, 6 — брюшной
нервный ствол с отходящими
нервами, 7 — рот, 8 — глаз,
9 яйцевод, 10 — оварий,
11 — пенис, 12 — половое
отверстие, 13 — глотка, 14 —
семяприемник. 15—семенники,
16 — семевод, 17 — желточ-
никп
усиленного роста задней части спинной стороны тела, во-вторых,
аа брюшной стороне оно сдвигается постепенно к головному концу,
то полезно для все более и более рационального его использова-
дви П^И захватывании пищевого материала, находящегося по пути
рий^ния зкивотного- Этот процесс в различных группах турбелля-
Ное 3аШел то менее, то более далеко. Несомненно одно, что обрат-
еРемещение рта по направлению к заднему концу, т. е. назад
103
по пути движения животного в обычных условиях экологически
немыслимо, отбор не мог сохранить такого рода отклонения как
полезные. Это обстоятельство крайне существенно, так как сейчас
же заставляет отбросить возможность предположения о происхож-
дении плоских червей от примитивных гастреад (Неф и др.) или
форм -вроде Platyctenea (Ланг и др.)
путем превращения оральной поверхно-
сти в брюшную при переходе к донно-
му ползающему существованию. Такие
формы должны бы были иметь ротовое
отверстие или посередине брюшной сто-
роны, пли последняя полностью пред-
Рис. 34. DlopistoporiiS'
представитель Acoela с
расположением рта (7)
на заднем конце тела и
значительно развитой,
глоткой (13) (по Вест-
бладу):
Рис. 33. Схема организации
Rhabdocoela (по Граффу).
Обозначения, как в рис. 31
20 — покровный эпителий,
21 — скопление железистых
клеток, 22 — ядра мускула-
туры, 23 — пищеварительная
паренхима, 24 — статоцист;
прочее, как в рпс. 31
ставляла бы своеобразный расширенный рот и часть глотки (Plaiy-
ctenea). Если подобные формы среди турбеллярий действительно
имеются, по предположению, например, А. Ланга — Poliycladida, то
все же вопрос решается иначе — от них не могли произойти пла-
нарии с ртом, расположенным у заднего конца тела, и поликлады
на самом деле представляются только одним из этапов в процессе
передвижения ротового отверстия по брюшной стороне тела от зад-
него конца к головному.
Рассмотрение внешней морфологии турбеллярий показало нам
ряд широких приспособлений, создавших тип червей, но, наряду
с этим, они обладают и характерными примитивными чертами,
сохранившимися от исходных форм гастреад. Само название «тур-
104
беллярии» или «ресничные» черви подчеркивает одну такую черту—
наличие ресничного покрова (рис. 35), который сохраняет и свою
первичную функцию передвижения животного. Вторично реснички
в тех или иных группах плаиарий могут исчезать, покров становится
кутикулярным. Это — защитное приспособление, например при пе-
Рис. 35. Поперечный разрез стенки тела Glanduloderma (по
Иегерстену):
1 —• клетки эпителия, с рабдитами (2), 3 — зерна секрета, образую-
щиеся в клетках, лежащих глубже, 4 — субэпителиальная перепонка
ткани основного вещества, 5 — кольцевые и 6 — продольные мышцы,
7 — ядра мышечных клеток
реходе к наземному существованию, но двигательные участки с
ресничным покровом остаются в виде ползательных валиков, и
норма для турбеллярий.— однообразный ресничный покров наподо-
бие имеющегося у планул кишечнополостных. Сохранение при-
митивного ресничного покрова говорит о том, что едва ли пред-
ками турбеллярий могли быть формы, имевшее сложные реснич-
ные образования вроде, например, гребных пластинок гребневи-
ков, а тем более кишечнополостные, утратившие ресничный по-
кров.
Наряду с этим покров турбеллярий имеет специфичные особен-
ности в смысле его защитной функции, которые приходится считать
ужо адаптивными чертами рассматриваемой группы червей,
т. е. такими, которые, не открывая для них широких возможностей
дальнейшей эволюции, тем не менее весьма существенны для
сохранения в борьбе за существование. Такими являются различ-
ного рода рабдиты в клетках покровного эпителия, представляю-
щие оформленные секреты сходно со стрекательными капсула-
ми кишечнополостных. Перед нами, очевидно, один из примитив-
ных способов активной защиты первичных форм ядовитыми
секретами.
К узко-адаптивным чертам относится в покрове турбеллярий
также развитие железистых участков, секрет которых имеет зна-
чение для удерживания, закрепления животного на субстрате дна—
105
таковы кантовые железы (рис. 36) и железистые поля, вентрально
расположенные на переднем конце тела. Благодаря им планария
прочно держится на субстрате и не смывается, не уносится бес-
помощно волной. Опять-таки у кишечнополостных мы находим раз-
витие железистых участков с подобной функцией, как, напри-
мер, в подошве гидры или актинии. Связь с примитивными гаст-
реадами!
К защитному комплексу у планарий относится еще одна чрез-
вычайно существенная черта их организации — способность произ-
Рис. 36. Схема поперечного разреза через планарию на уровне глотки
(по Лейкарту):
1 — покровный эпителий, 2 — субэпителиальная перепонка. 3 — кантовые железы,
4 — средняя кишка, 5 — кольцевые и 6 — продольные мышцы, 7 — паренхима пла-
нарий, 8 —• брюшной нервный ствол. 9 — яйцевод, 10 — глотка, 11 — семенники,
12 — семевод. 13 — желточнпкп
водить быстрые и очень сильные сокращения и изменения формы
тела. Она обусловлена дифференцировкой в стенке тела, сходно с
гребневиками, мезенхиматозной мускулатуры в виде главным обра-
зом значительных кольцевых и продольных мышечных слоев. Эпи-
телиально-мышечного состояния покрова, что характерно для га-
стреад из ряда, ведущего к Coelenterata, здесь обычно не встре-
чается. Мускулатура стенки тела, совершенно обособленная от по-
кровного эпителия, связана с ним лишь через посредство тонкой
прослойки из примитивной неклеточной ткани основного вещества,
образующей базальную перепонку.’
Таким образом создается характерный для червей кожномы-
шечный мешок, образующий стенку тела (рис. 36). В дальнейшем
он получает мощное развитие, причем мускулатура его превра-
щается в активный орган движения животного. *
Как дифференцировку мускулатуры стенки тела планарий, так
и образование кожно-мышечного мешка необходимо поставить в
100
число широких приспособлений, вырабатывающихся в группе тур-
беллярий и создающих особый тип организаций, тип червей.
В теснейшей взаимосвязи с развитием мускулатуры стоит раз-
витие опорной системы и нервной системы турбеллярий. Первая из
них выражена чрезвычайно несложно в виде пузырчатой или звезд-
чатой паренхимы, расположенной внутрь от мускулатуры (рис. 37,
А и Б). Она представляет массу мезенхимных клеток, между кото-
рыми основное вещество не развивается и в протоплазме которых
накопляется в вакуолях жидкость. Это тот же принцип внутренней
опоры на несжимаемую массу жидкости, какой, мы видим, осуще-
Рис. 37. Строение паренхимы турбел-
лярий (по Забусову): <4 — разрез
через Rimacephalus pttlvinar, ясно
видны границы клеток; Б — разрез
через Sorocells alba, переход к ти-
пичной паренхиме
ствляется, например, в экто-
дермальном осевом клеточном
тяже щупалец гидроидных по-
липов или в хорде ланцетника.
Однако выражен он крайне
примитивно, что выявляется в
диффузности распространения
и различных вариантах офор-
мления паренхимы. Во всяком
случае дальнейшей эволюции
этих опорных образований не
произошло — они представля-
ют специфическую особенность
плоских червей, возникающую
с развитием мускулатуры кож-
но-мышечного мешка.
Нервная система турбелля-
рий, развиваясь как всегда в
теснейшей корреляции с разви-
тием мускулатуры, с одной стороны, и с развитием чувствительных
образований, в особенности головного комплекса органов чувств,
с другой, сама несет черты типовой дифференцировки, легшей в
основу эволюции нервной системы у всех вышестоящих типов жи-
вотного царства. Во-первых, в ней при выработке мускулатуры
кожно-мышечного мешка соответственно обособляется моторная
периферия от сензорной, остающейся в связи с покровом, где
находятся чувствительные образования. Во-вторых, в соответствии
с развитием головного комплекса органов чувств дифференцируется
ганглиозное скопление головного мозга, являющееся прежде всего
сенсорным центром. Его наличие характеризует головной отдел тела
червей в равной степени с обособлением головного комплекса орга-
нов чувств. Далее, по всей длине тела дифференцируются двига-
тельные продольные нервные стволы, располагающиеся билатераль-
но-симметрично в количестве от 4 до 1 пары и соединяющиеся
поперечными комиссурами, иногда проходящими крайне непра-
вильно, как примитивное сплетение, иногда же дающими харак-
терную «ортогональную» систему с ясно выраженной мета мерностью
кольцевых комиссур.
107
Наряду с дифференциацией нервной системы на центральную
и периферическую с их чувствительным и двигательным отделами,
протекает и процесс их интеграции в его первых этапах, а именно:
головной мозг объединяется с туловищными продольными стволами,
располагаясь на их переднем отделе и принимая особо ассоциатив-
ное значение. Адаптивные процессы выражаются в нервной системе
турбеллярий удалением ее из покрова в глубинное положение, под
мускулатуру — узко' приспособительная защитная черта.
Нельзя здесь не упомянуть, что органы чувств головного ком-
плекса турбеллярий представляют расположенные справа и слева
па переднем конце тела участки очень примитивного чувствитель-
ного покрова, дифференцированные как осязательные поверхности,
и более обособленные участки хеморецепторного характера в виде
обонятельных ямок. Комплекс завершается выработкой примитив-
ных глаз и органов равновесия, находящихся на спинной стороне
в области около головного мозга, но уже утративших первоначаль-
ное эпителиальное положение, располагаясь более или менее глу-
боко в паренхиме.
, Глаз планарий представляет группу ретинальных клеток. Часть
их поверхности, несущая ободок, воспринимающий световые раз-
дражения, изолирована от световых лучей пигментным кубком, со-
стоящим из одной или нескольких пигментных клеток. Получается
очень простой инвертированный глаз, ориентирующий животное в
интенсивности света и в направлении световых лучей. Ориентиров-
ка совершенствуется с увеличением количества таких глазков и
расположением их на значительной части поверхности головного от-
дела тела. Но здесь едва ли еще можно говорить о картинном
зрении.
Так же просты и статорецепторы, нередко имеющиеся у турбел-
лярий. Они представлены лишь небольшими эпителиальными пу-
зырьками, часто заложенными в самом мозгу и содержащими вну-
три статолит (рис. 34). Как видно, и глаз, и орган равновесия раз-
виты у плоских червей даже более примитивно, чем у некоторых ки-
шечнополостных и гребневиков. Во всяком случае их дифференци-
ровка просто выводится из скопления чувствительных клеток на
апикальном полюсе гастреад, близких с описанными нами, как ис-
ходные .для типа гребневиков.
Мы уже попутно коснулись одной существенной^ черты органи-
зации плоских червей, характерной для вышестоящих форм вооб-
ще. В противоположность кишечнополостным у них пространство
между покровным эпителием и эктодермальной стенкой кишечника
заполняется разного рода образованиями, частью уходящими вглубь
тела в более защищенное положение, частью развивающимися здесь,
на мес4е. Неклеточная по строению прослойка из ткани основного
вещества низших кишечнополостных, утолщаясь, заселяется, дсак
мы видели, различного рода клетками. У гребневиков здесь уже,
сверх того, располагается и мускулатура, создается некоторый ком-
плекс, который, конечно, нельзя назвать единым средним пластом
108
?дли внутренней средой организма. Это — комплекс различных об-
разований, уходящих из эпителия при их дифференцировке и рас-
полагающихся в ткани основного вещества, заполняющей простран-
ство между эктодермальным покровом и эктодермальным кишеч-
ником. У турбеллярий (рис. 36) здесь периферически, под субэпи-
телиальной пограничной перепонкой, дифференцированы слои мус-
кулатуры стенки тела, а глубже расположена диффузно опорная
паренхима. Сюда же, на внутреннюю поверхность мышц, ушла цен-
тральная нервная система, занимая более защищенное положение.
Как подобного же рода приспособление, мы склонны толковать и
нахождение гонад у турбеллярий в этом комплексе, тогда как у
прочих целентерических организмов они всегда еще располагают-
ся в том или другом эпителии. Эта характерная черта, выработав-
шаяся у турбеллярий, далее присуща всем вышестоящим типам.
Однако она в половом аппарате турбеллярий отходит на второй
план, на первое место здесь выступает чрезвычайно сложная и так
далеко’пошедшая его дифференцировка, что по сравнению с други-
ми системами органов здесь можно говорить об эксцессивном, чрез-
мерном развитии (рис. 31, 33, 34). Это определение правильно вы-
ражает отношения — половая система эволюционировала у тур-
беллярий чрезмерно по сравнению с эволюцией других органов и
их систем. Она в конце-концов сделалась ведущей чертой строения
плоских червей, она сдвинула их с пути общего повышения органи-
зации на путь самого сложного* разнообразия узких приспособле-
ний, в частности сложнейшей диференцировки и приспособлений,
обеспечивающих оплодотворение, а далее питание и защиту заро-
дыша, ставящих гарантию продолжения рода на чрезвычайную вы-
соту. Благодаря им группа плоских червей, оставаясь в остальном
на низком уровне организации, смогла в борьбе за жизнь вы-
держать конкуренцию с вышестоящими формами, дожить из отда-
леннейших эпох прокембрия до наших дней. ,
Во-первых, у турбеллярий, как и у гребневиков, выработался
гермафродитизм, другими словами, часть гонад становится семен-
никами, другая яичниками. Во-вторых, большая часть яичников у
ряда групп турбеллярий превращается в желточники, т. е. дает лишь
абортивные яйца, служащие для питания развивающихся эмбрио-
нов. В-третьих, развивается сложный выводящий аппарат в виде
семеводов, яйцеводов, вителлодуктов. В-четвертых, периферический
отдел выводящего аппарата, принимая на себя функцию оплодо-
творения, дает сложнейший копуляционный аппарат со спермато-
форным оплодотворением. Таким образом, дифференцируется муж-
®Ои а^РиУм> содержащий разнообразно построенный пенис, в кото-
мУскулатура служит для его движения, а сложная
ЗИСТЗД Дифференцировка служит для образования и заполнения
мускулТ°Ф°Ра’ У Ряда Ф°РМ образуются, кроме того, железисто-
лении ИСТЬ1е °Рганы — аденодактили, играющие роль при закреп-
постооеи В^еМя копуляции. В половом атриуме развиваются сложно
тр ные семеприемники. Наконец, в-пятых, с женским атриумом
109
связаны бывают сложные железистые образования, служащие для
формирования и заполнения кокона, содержащего яйцо и материал
для питания эмбриона в дальнейшем. Иногда дело осложняется
еще и тем, что дифференцируются участки атриума, в которых по-
мещаются коконы с развивающимися в них эмбрионами, таким
образом все развитие может протекать под защитой материнского
организма.
Здесь намечен только общий план выработки полового аппара-
та турбеллярий. В деталях он неисчерпаемо разнообразен в своей
дифференцировке и специализации, иногда парадоксален в своих
чрезмерно адаптивных проявлениях. В итоге можно сказать, что
систематика турбеллярий строится, прежде всего, на строении поло-
вого аппарата, и часто определение вида становится совершенно
невозможным, если мы имеем дело с неполовозрелыми экземпля-
рами. Среди планарий хорошо известны факты, когда формы как
будто тождественные по всем чертам организации оказывались
при ознакомлении с их половым аппаратом относящимися к-разным
родам и видам. Таковы европейская Crenobia alpina и Penecurva
sibirica Прибайкалья. Не различая их, Арндт считал Cr. alpina
транспалеарктом. Это только иллюстрация к сказанному выше по
поводу дифференцировки и эволюции полового аппарата турбел-
лярий.
Итак, пространство между покровом и стенкой кишечника у пло-
ских червей оказывается заполненным целой серией органов — мус-
кулатура, опорная паренхима, нервная система и половой аппарат.
В теснейшей связи с этим стоит появление системы экскреторных
органов, которые выводят из организма как продукты азотистого
распада, накопляющиеся при жизнедеятельности всех перечислен-
ных образований, так и поддерживают определенное осмотическое
давление внутри организма (рис. 32). У кишечнополостных подобная
дренажная система отсутствовала, так как немногочисленные обра-
зования, располагавшиеся между покровом и кишечником, давали
при их жизнедеятельности относительно мало отравляющих орга-
низм азотистых продуктов распада, и в соответствии с этим эпите-
лии покрова и кишечника были вполне достаточны, чтобы обеспе-
чить удаление экскретов из организма. У турбеллярий впервые диф-
ференцируется система протонефридиев, т. е. идущих к покрову
трубок, стенки которых выбирают экскреты из прилежащих тканей,
а концевые внутренние клетки — «клетки с мерцательным пламе-
нем» являются осморегуляторами; они фильтруют через свою про-
топлазму воду из организма, и с ней выводятся наружу азотистые
продукты распада.
Дифференцировка протонефридиев может быть поставлена в
ряд с теми широкими приспособлениями, которые мы уже выше
отметили при анализе организации турбеллярий; она завершает
появление нового своеобразного типа в эволюции гастреад, типа
строения плоских червей. Несомненно, что дифференцировку прото-
нефридиев А. Н. Северцов должен был бы считать «ароморфо-
110
зом» — энергия жизнедеятельности должна повышаться с появле-
нием системы, удаляющей продукты распада, отравляющие орга-
низм. Тем не менее, повторяю, трудно оценить этот критерий «по-
вышения энергии жизнедеятельности», гораздо легче подойти с
другой точки зрения — у вышестоящих групп экскреторная4 система
протонефридиев эволюционирует, все более усложняясь. Следова-
тельно, выработавшие ее организмы получали в борьбе за суще-
ствование более широкие возможности сохраняться и развиваться,
а отсюда и вывод о широко адаптивном значении этой черты орга-
низации турбеллярий, наряду с ее значением в разделении физио-
логических отправлений в едином целом организме.
Последним штрихом в нашем анализе можно поставить рас-
смотрение кишечника турбеллярий. Сохраняя всю примитивность
целентерическбго^аппарата с внутриклеточным пищеварением, обла- .
дая в локализации рта на брюшной стороне широко адаптивной
чертой, он представляет далее как в строении глотки, так и в строе-
нии усваивающего энтодерм а л ьно го отдела огромное разнообразие'
специфических черт строения. В этом отношении ею можно поста-
вить до некоторой степени в параллель к тому, что мы могли кон-
статировать в половом аппарате турбеллярий, однако в значитель-
но меньшей мере.
Глоточный отдел дает, в связи с дифференцировкой в нем муску-
латуры и желез, разнообразные узкие приспособления к различней-
шему пищевому материалу при его захватывании и переварива-
нии — pharynx simplex, ph. plicatus, ph. bulbosus, ph. tubulosus
и т. д. Имеется или простая трубка глотки, ведущая из ротового
отверстия в среднюю кишку (рис. 34, 33), или за счет дистального
отдела глотки дифференцируется особое влагалище для мускулисто-
го отдела (рис. 31, 32). Получается мускулистый аппарат в виде
рукава, охватывающего добычу, способный сильно изменять свою-
форму, как у поликлад, или функционирующий, наподобие хобота,
мускулистый цилиндр глотки триклад и т. д., и т. п. в
С другой стороны, эктодермальный усваивающий отдел пред-
ставляет массу характерных изменений, главным образом в его вет-
влении, связанных с обеспечением различных систем органов выра-
батываемым питательным материалом. Это- обстоятельство нередко
придает некоторым частям кишечника характер ветвящихся кана-
лов, проходящих в периферические отделы тела, причем филогене-
тически рано наметились несколько путей, по которым пошла диф-
ференцировка: путь Rhabdocelida — мелких прямокишечных турбел-
лярий и пути Tricladida и Polycladida — более крупных, трех- и
многоветвистых турбеллярий.
Ьольшое внимание зоологов привлекает к себе аберрантная по
строению кишечника группа Acoela (рис. 34). У ее представителей
просвет кишечника отсутствует, откуда и само название группы.
Гулетки эктодермального отдела образуют сплошную массу, зани-
мающую всю центральную часть тела. Через ротовое отверстие,
расположенное обычно близ средины брюшной стороны или ближе*
ш
•к головному концу, клеточная масса кишечника выдается наружу,
и эктодермальные клетки, наподобие амеб, захватывают каждаядо
отдельности мелкие пищевые частицы, перемещаясь затем вглубь
для их переваривания; более крупная добыча захватывается целым
синцитием. Таким образом получается чисто внутриклеточное пи-
щеварение в противоположность прочим турбелляриям, у которых
комбинируется и энзиматозное, и внутриклеточное пищеварение.
Очень многие исследователи видят в таком процессе вторичного
приспособления к питанию мелкими донными организмами первич-
ное состояние, исходное для всех плоских червей. Однако с этим
едва ли можно согласиться, так как 1) и у кишечнополостных, и у
гребневиков нигде нет подобного примитивного состояния кишеч-
ника, но всегда уже имеется в той или иной мере энзиматозное
пищеварение и хорошо выраженная полость кишечника; предпола-
гать сохранение примитивной простоты в обоих отношениях у тур-
беллярий рискованно — этот процесс мы отнесли в предгастрейное
состояние; 2) против такой примитивности говорит обычное, за
редкими исключениями, перемещение рта у Acoela от заднего кон-
ца тела на брюшную поверхность к ее середине; 3) при таком пони-
мании отношений нельзя представить возможности образования
глотки, которая как раз хорошо выражена у форм, имеющих рот
на заднем конце тела, тогда как у Acoela она обычно или отсут-
ствует, или представляет как бы отрывки различных состояний
усложнения. Все факты укладываются без всяких натяжек в посте-
пенный ряд упрощения организации в связи с переходом к более
Простому способу питания путем захвата находящихся на субстрате
мелких пищевых частиц непосредственно эктодермальными клетка-
ми. Ясно, что поэтому терялась необходимость в железах глотки, ее
мускулатуре и, наконец, в просвете кишечника для пищеварения. В
связи с этим развивалась амебоидность эктодермальных элементов,
что вело вторично к использованию более крупной добычи целым
синцитием, с одной стороны, и к исчезновению отграничения кишеч-
ника от прилежащих тканей, к диффузному распространению его
клеток внутри тела, с другой.
Подводя итоги рассмотрению организации турбеллярий, мы мо-
жем сказать, что она представляет высший этап в эволюции форм
о целентерическим кишечником, связанный со свободным существо-
ванием в сложных условиях придонной жизни. Получается общее
повышение организации, чего не дает прикрепленное существование
кишечнополостных, ведущее к узким приспособлениям защитного
характера, чего не может дать и пелагическая жизнь, более простая
по условиям и представляющая менее сложные требования к орга-
низации гребневиков. У турбеллярий вырабатывается характерный
комплекс черт организации, значительно повышающий возможности
организма в борьбе за жизнь, получается особый план построения.
Этот план типа червей эволюционирует далее чрезвычайно широко,
112
давая все высшие типы животного мира. С такой точки зрения ор-
ганизация турбеллярий представляет в то же время первый шаг,
низший этап в развитии червей.
В соответствии с этим к ряду примитивных черт, сохраняющихся
от исходных форм, — целентерическое состояние кишечника, нали-
чие в значительной мере внутриклеточного пищеварения, реснич-
ный покров, простота механо- и хеморецепторов, примитивность
фото- и статорецепторов, наличие не содержащих клеток погранич-
ных перепонок ткани основного вещества — присоединяется два
комплекса прогрессивно эволюционирующих, широко приспособи-
тельных черт организации.
Первый из них связан с использованием возможностей придон-
ного существования. Сюда относятся: 1) при выработке прямоли-
нейного движения смещение ротового отверстия на сторону, на-
правленую ко дну, с перемещением этого отверстия в той или иной
степени вперед; 2) при ползающем движении развитие билатераль-
но-симметричного построения, 3) дифференцировка головного ком-
плекса органов чувств, а в теснейшем соотношении с этим и обра-
зование сензорного центра нервной системы — головного мозга.
Второй комплекс широко адаптивных черт является в основе
связанным с защитой животного, для чего служит 1) дифференци-
ровка мезенхимной мускулатуры в стенке тела, 2) удаление вглубь
тела нервной системы и гонад. На этой почве получается 3) обо-
собление двигательной периферии и дифференцировка продольных
стволов, имеющих моторный характер в нервной системе и 4) появ-
ление системы протонефридиев, очищающих глубокие части организ-
ма от продуктов азотистого распада, образующихся во всех пере-
численных образованиях.
Объединяя оба эти комплекса, развивается, наконец, единая
центральная нервная система с выработкой особого ассоциативного
значения головного мозга, вступающего в связь с брюшными ство-
лами. Эта черта широко адаптивного характера завершает этап ор-
ганизации плоских червей.
Дальнейшее его выявление у турбеллярий связано1 уже со специ-
фичными для них узкоприспособительными чертами, имеющими
очень мало отношения друг к другу, однако легко объединяемыми
В характерный «турбеллярный» комплекс узких адаптаций, перехо-
дящих иногда в чрезмерные: 1) защитного характера — выработка
активно защитных оформленных секретов в виде рабдитов, диффе-
ренцировка кантовых желез, защищающих от захвата движением
воды, образование диффузного, крайне примитивного опорного аппа-
рата для мускулатуры стенки тела в виде пузырчатой или звездча-
той паренхимы, 2) связанные с питанием — дифференцировка гло-
точного отдела кишечника для захватывания пищи и ветвление
эктодермального, усваивающего его отдела, для подачи питатель-
ных веществ к органам, 3) сложнейшие детали строения полового
аппарата, стоящие в связи с выведением половых продуктов, опло-
дотворением яиц, питанием и защитой развивающегося зароды-
S Пути эволюции животного мира 113
ша. Последняя категория узко или крайне адаптивных черт орга-
низации является особенно характерной для турбеллярий и по своей
сложности далеко превосходит имеющееся в других системах орга-
нов, является ведущей чертой группы в разнообразии ее приспосо-
бительной эволюции.
Какие же выводы мы можем сделать на основаннии нашего ана-
лиза о формах, давших начало типу червей? Вопрос, который уже
не раз ставился, и попытки решения которого делили зоологов на
два главных лагеря. Ярким примером этого могут служить наши
учебные руководства — студенческий учебник зоологии беспозво-
ночных В. А. Догеля (1947), в котором проводится взгляд А. Ланга
на происхождение турбеллярий от ползающих ктенофор (Cteno-
plana и Coelopland), и руководство по зоологии (1937, т. 1), в кото-
ром обработка турбеллярий принадлежит В. Н. Беклемишеву. Он
рассматривает организацию последних согласно с представлениями
Л. Граффа о происхождении плоских червей от Acoela, т. е. тур-
беллярий, лишенных просвета кишечника. Наличие этих взглядов
представляет яркое выражение значения филогении для понимания
организации, но в то же время может показать и значение анализа
организации для правильного решения вопросов филогении — их
взаимообусловленность.
В 80-х годах прошлого столетия открытие А. О. Ковалевским в
Красное море Coeloplana metchnikovii и А. Коротаевым в морях
Малайского архипелага Ctenoplana kovalevksii произвело большое
впечатление, как находка настоящих переходных форм от гребне-
виков к червям. А. Ланг изображал процесс таким образом. При4
переходе гребневиков к ползанию по дну, оральной стороной книзу,
рот располагался посредине последней. Соответственно главная ось
тела проходила через рот. Но ее аборальный конец смешался из
первоначального положения посредине спинной стороны к передне-
му концу животного, выработавшего постоянное направление дви-
жения. Главная ось как бы сгибалась, так как аборальный ком-
плекс органов соответственно перемещался из срединного положе-
ния на участок тела, передний при движении. Ctenoplana и Coelo-
plana имеют аборальный комплекс еще типично посредине спинной
стороны. Они еще не выработали продольной оси при движении,
но представляют уже плоские червеобразные формы. От них со
стабилизацией направления движения и с переходом аборального
комплекса на головной конец произошли Polycladida.
Эта на первый взгляд очень простая и вероятная картина про-
исхождения ресничных червей от гребневиков, однако, тускнеет и
теряет свою определенность при 6ojfee глубоком ознакомлении с
организацией Plathyctenea, и в настоящее время наукой собственно
уже отброшена, как пережитый этап. Период увлечения переходны-
ми формами, семидесятые и восьмидесятые годы прошлого столе-
тия, сменился более глубоким изучением и критическим подходом
114
к фактам. Так, с одной стороны, Coeloplana и Ctenoplana являются
типичными, хотя и сильно измененными гребневиками, приспособив-
шимися вторично к придонной жизни и ползанью по субстрату,- с ко-
торого они собирают свою пищу. Их предками были пелагические
гребневики, о чем вполне определенно говорит онтогенез, в кото-
ром цидиппоидная форма постепенно переходит в плоскую, лепеш-
кообразную форму взрослого организма. При этом нижняя сторона
тела образуется за счет разрастающейся глотки. Ничего подобного
у турбеллярий мы не найдем. С другой стороны, в онтогенезе тур-
беллярий имеется типичное спиральное дробление, значение кото-
рого было выше отмечено. Далее, характерны процессы неравно-
мерного роста задней части спинной стороны тела и, наконец, у
поликлад имеется пелагическая мюллеровская личинка, которая с
цидиппоидной стадией ничего общего не имеет. Приведенного впол-
не достаточно, чтобы признать, что сходство Platyctenea с поликла-
дами — явление вторичное, конвергентное. Это — экологически
обусловленное общее сходство очень своеобразных гребневиков с
плоскими червями при коренной разнице в деталях организации и
развития.
Иначе обстоит дело с представлениями о происхождении турбел-
лярий от исходных форм, подобных Acoela. К этому взгляду примы-
кают в настоящее время очень многие, основываясь на тщательном
изучении этой группы. В. Н. Беклемишев, например, рисует такую
картину (1925): «У большинства представителей, Acoela имеется
хорошо обособленный наружный эпителий, но у некоторых... эпите-
лий этот не более обособлен по сравнению с остальной паренхи-
мой, чем слой пинакоцитов в теле губок, и представляет не более,
как пограничный слой паренхимы...». Последняя «представляет сме-
шение или, точнее, соединение компонентов опорных, мышечных,
железистых, пищеварительных, нервных, половых. В этой массе веб
эти типы компонентов расположены, в сущности, строго правильно,
а именно — по аппаратам... Имеется весьма совершенно развитой
мышечный аппарат, кое-какой нервный аппарат, половой аппарат
и т. д., но эти аппараты построены не из тканей, а непосредственно
из клеток и волокон. Однотипные компоненты структуры вовсе не
собраны вместе (мышечный аппарат), или собраны мелкими груп-
пами по 5—10 клеток, не заслуживающими названия тканей
(сперматогонии), или, если однотипные клетки и собраны вместе в
довольно значительную массу, она вся пронизана посторонними
элементами, как мышцы и железы пронизывают по всем направ-
лениям главное скопление нервных элементов — мозг...».
Весьма соблазнительно принять, что при общей простоте органи-
зации^ Acoela отсутствие полости кишечника у них является прими-
тивной чертой и что далее эволюция ведет к типичным формам рес-
ничных червей.
«У большинства турбеллярий — по В. Н. Беклемишеву, —
прежде всего резко обособляется от остальной паренхимы на-
ружный кроющий слой, принимающий вид кожного эпителия. Ба-
8* 115
зальные концы кроющих и части железистых клеток подравнивают-
ся, между ними и остальной толщей тела выделяется пограничная
пластинка. Этот кожный эпителий образует обособленную ткань, со-
стоящую из нескольких определенных родов клеток: кроющих, чув-
ствительных и железистых. Они обособились от всех остальных кле-
ток и образуют ткань, которая как бы выкристаллизовалась из ос-
тальной паренхимы... Прогресс структуры в пределах отряда Acoela
заключается в последовательном выделении из первоначальной
смешанной паренхимы отдельных тканей, одной за другой, т. е. в
переходе структуры из паренхимной в тканевую,., разного типа
клетки группируются в отдельные ткани», причем «каждая оформ-
ленная тканевая масса уже не просто ткань, но известная конст-
руктивная единица, элементарный орган, построенный из данной
ткани», тогда как «в понятии ткани не мыслится никакой опреде-
ленной формы».
Такой путь эволюции от более простого к более сложному ка-
жется очень естественным.
Однако подобного рода построения В. Н. Беклемишева, являясь
чисто формальными, конструктивными схемами, не могут удовле-
творить требованиям современной эволюционной морфологии в «их
обосновании». Дифференцировка тканей и органов обусловлена
теми или иными взаимоотношениями организма с внешней средой,
что мы и видели на примерах возникновения первоорганов покрова
и кишечника, обособления опорной пластинки гидры и дальнейшей
дифференцировки таких образований у вышестоящих форм. С этой
точки зрения совершенно' необъяснимы нахождение в паренхиме
Acoela рассеянных железистых, мышечных, нервных и т. д. элемен-
тов, их первичная дифференцировка и дальнейшее объединение в
органы — ткани. Создается впечатление об их спонтанном возник-
новении в однородной паренхиме, что совершенно несовместимо с
основными установками научной материалистической мысли. На-
против, все факты получают вполне четкое объяснение, если в его
основу будет положен факт развития амебоидности у клеток ки-
шечника, связанной с особенностями питания исходных форм Acoela.
Выше уже был отмечен тот экологический момент, который тре-
бует принять втрричное упрощение Acoela. А именно •— у них имел
место переход от питания активного хищника, какими обычно яв-
ляются турбеллярий, к питанию малоподвижными мелкими орга-
низмами, в массах встречающимися на дне. Это достигалось очень
своеобразным способом — путем выпячивания эктодермального
эпителия через рот и захватывания его клетками всяких пищевых
частиц с субстрата, как бывает у амеб или плазмодиев. Такое пи-
тание приближается к пассивному, но не путем захватывания всех
мелких взвешенных в воде и несущихся с током воды организмов,
а путем собирания всех попадающихся по пути на субстрате и мел-
ких и более крупных форм. При этом глоточный отдел теряет свое
значение и подвергается редукции, развивается амебоидность кле-
ток кишечника, просвет последнего исчезает, отграничение его от
116
прилежащих тканей теряет свою определенность, его амебоидные
элементы распространяются всюду в теле, получается строение по
аппаратам, но их компоненты дезинтегрированы, тканевая целост-
ность нарушена. При подобном питании, конечно, взаимоотношения
с внешним миром упрощаются. В общем, организация должна уп-
рощаться, и на явления регрессивной эволюции указывают остатки
сложных аппаратов, которые имеются у Acoela. В. частности, у не-
которых из них наблюдается мускулистая глотка, развитие которой
при отсутствии просвета кишечника и всецело внутриклеточном пи-
щеварении объяснимо только как сохранение некоторого рудимента,
у прочих уже совершенно исчезнувшего. Также и в строении поло-
вого аппарата встречается то, у одних форм, рудимент сложно уст-
роенного пениса, то, у других форм, семеприемника и т. д., прогрес-
сивное развитие которых в сложный комплекс полового аппарата
однако отсутствует. Наконец, и в эмбриональном развитии ярко
выражены редукционные явления —• при спиральном дроблении
вместо типичных квартетов образуются дуэты — это едва ли может
найти свое объяснение в признании за таким дроблением прими-
тивной, исходной формы процесса.
Не будем вдаваться в дальнейшие детали. Сказанного достаточ-
но, чтобы видеть, насколько экологический момент упрощения в спо-
собе питания может быть принят ведущим в выработке организа-
ции Acoela, как вторично упрощенных форм.
Наконец, из признания Acoela исходными формами в эволюции
турбеллярий необходимо следует, что предками их были уже не
гастреады, а организмы, лишенные первооргана кишечника, имев-
шие только обособление массы питающих клеток от покровных дви-
гающих ресничных. Согласно изложенному выше относительно про-
исхождения многоклеточных, это были начальные шаги по пути к
гастрее. Таким образом теряется связь с потомками настоящих
гастреад — типом кишечнополостных и типом гребневиков. Выра-
ботка кишечника становится процессом конвергентного развития, с
одной стороны, у гастреад, с другой — у Acoela среди плоских
червей, если только не допустить, что кишечнополостные и гребне-
вики также происходят от Acoela. Считая нереальным такой взгляд,
так как факты упрямо свидетельствуют о единстве всех организмов
с целентерическим кишечником и в смысле выработки тканевых
комплексов и в смысле дифференцировки систем органов, перейдем
к тому, как же конкретно следует представить предковые формы
турбеллярий. *
Этот вопрос собственно уже был решен, когда, проанализировав
организацию кишечнополостных и гребневиков, пришлось установить
эволюцию двух ветвей гастреад, одну с преобладанием в организме
эпителиев над тканью основного вещества и с эпителогенной мус-
кулатурой и другую с сильным развитием мезенхимы и с мезенхи-
могенными мышцами. Первые дали начало типу кишечнополост-
Ных> вторые гребневиков. Несомненно, что общий предок рабдоце-
ЛиД> поликлад и Alloiocoela, т. е. трех дивергентно развивавшихся
117
ветвей турбеллярий, обладающих кишечной полостью, также при-
надлежал к категории гастреад с развитой мезенхимой и с мезен-
химогенной мускулатурой, т. е. истоки как типа гребневиков, так и
червей являются общими в противоположность типу кишечнопо-
лостных.
Такое сближение уже допускалось рядом авторов, но оно не
имеет ничего общего1 с рассмотренными выше взглядами А. Ланга
на происхождение ресничных червей, в частности поликлад, от пол-
зающих гребневиков — Ctenoplana и Coeloplana.
Итак, общими предками обоих типов были гастреады, которые
в дальнейшей эволюции дали, с одной стороны, сохранив пелаги-
ческий образ жизни, очень специализированных гребневиков цидип-
поидного характера, а с другой, перейдя к придонному, ползающе-
му существованию, — турбеллярий с типичным целентерическим
кишечником.
Однако можно отметить детальнее некоторые черты, характери-
зовавшие гастреадных предков плоских червей. Покров их был об-
разован типичным мерцательным эпителием, содержавшим также
железистые и чувствительные клетки, с нервным сплетением бази-
эпителцально, со скоплениями чувствительных клеток и, быть мо-
жет, примитивным статоцистом на аборальном отделе тела, т. е. пе-
реднем при движении животного, и соответственно со скоплением
здесь ганглиозных элементов. Далее, располагавшаяся под покро-
вом, в прослойке ткани основного вещества, мускулатура стенки
тела состояла из кольцевых и продольных слоев мезенхимных
мышц. Здесь же в мезенхиме, под мускулатурой, были заложены
гонады. Наконец, целентерический кишечник, обладавший муску-
листо-железистой глоткой, открывался на заднем конце тела рото-
вым отверстием, направленным назад и ко дну, что' связано с вы-
работкой поступательного прямолинейного движения. В онтогенезе
таких гастреад соответственно должен был вырабатываться спи-
ральный тип дробления с преобладающим развитием d-квадранта,
т. е. неравномерным ростом спинной стороны тела.
Подобные гастреады отличались от пелагических гастреад, дав-
ших начало типу гребневиков, выработкой прямолинейного движе-
ния в связи с переходом к придонному существованию, более зна-
чительным развитием мускулатуры и относительно плотной мезен-
химой, наконец, перемещением полового зачатка в более защищен-
ное положение, в глубь тела.
К появлению на этой основе организации червей, вообще, и пло-
ских червей, в частности, вели: 1) выработка билатерально-сим-
метричного строения; 2) смещение рта на брюшную сторону, 3) об-
разование головного комплекса органов чувств и соответственно
головного мозга; 4) дифференцировка кожно-мышечного мешка;
5) обособление чувствительной и двигательной периферии и цен-
трального отдела в нервной системе; 6) дифференцировка протоне-
фридиев и, наконец, 7) выработка мешетчатых гонад (саккогонад)
с половыми протоками.
118
Это — довольно сложный комплекс черт организации, давший в
непрерывном процессе эволюции новый этап по сравнению с гаст-
реадами — этап червей с его характерными особенностями. И со-
временные турбеллярий представляют ветвь наиболее просто пост-
роенных червей, дошедших до нашего времени благодаря выработке
в покрове защитных образований — рабдитов и кантовых желез,
опорной паренхимы, сильно дифференцированных глотки и эктодер-
мальной кишки и, особенно, разнообразнейших частей полового ап-
парата, обеспечивающих оплодотворение, а также развитие и за-
щиту зародыша. Acoela среди плоских червей являются группой,
деградировавшей в связи с упрощением питания, но отделившейся
очень рано, а потому сохранившей и некоторые примитивные черты
строения.
Дальнейший путь эволюции типа червей обозначился яркими
вехами подтипа немертин и подтипа аннелид, к рассмотрению ор-
ганизации которых и переходим.
Глава VI
О
ПОДТИП НЕМЕРТИНЫ — NEMERT1NI
Внешняя морфология немертин, развитие кожно-мышечного мешка,
стабилизация рта, выработка головной лопасти, анального отверстия,
полостного пищеварения. Примитивность мезенхимы тела и сакко-
0 гонад. Органы чувств немертин, развитие нервной системы. Кро-
веносная система и теории ее происхождения. Нефридии. Итоги
анализа организации. Хобот. Взгляды на происхождение немертин.
Следующим за турбелляриями этапом эволюции типа червей
являются немертины. По сравнению с плоскими червями они пред-
ставляют значительный шаг вперед в осложнении организации и ее
повышении. Однако попытки объединить их в одну группу с выс-
шими представителями червей — аннелидами имеют в настоящее
время лишь исторический интерес. Большинство- зоологов видит в
них или вместе с О. Бюргером (1897) особый класс паренхиматоз-
ных червей, наряду с турбелляриями, трематодами и цестодами,
или выделяет их как группу с неопределенным систематическим по-
ложением, что мы находим, например, в учебниках зоологии Герт-
вига или В. Догеля. Наша точка зрения по этому вопросу была
формулирована в «Руководстве по зоологии» (1937, стр. 703) при-
мерно следующим образом: немертины занимают среднее место
между плоскими червями и аннелидами и должны вести свое про-
исхождение от примитивных плоских червей, как в свою очередь
от примитивных немертинообразных форм должны были в свое
время развиться аннелиды. Анализ организации немертин позво-
лит нам* в дальнейшем получить ясную ориентировку в этом во-
просе.
Что касается осложнения и общего повышения организации, то
оно связано с важнейшим экологическим фактом — немертины яв-
ляются не только свободноживущими придонными формами, но это
очень подвижные, энергичные хищники. Такой образ жизни требует
постоянного совершенствования различных сторон организации для
все более и более тонкой ориентировки в окружающей среде, что
необходимо как для нахождения добычи и нападения на нее, так
и для защиты от врага. Он требует при этом все большей и боль-
шей подвижности, а то и другЪе ведет соответственно к усложне-
но
'нию нервной системы, а затем и других систем органов. Таким об-
разом, с условиями существования свободноподвижного хищника
связан прежде всего всесторонний общий прогресс организации,
чего в борьбе за существование не дают условия какого-либо друго-
го образа жизни. Уже вторично сюда присоединяются узкие, ино-
гда крайне адаптивные черты вроде выработки, например, своеоб-
разнейших орудий нападения и т. п.
У немертин в соответствии с этими общи-
ми положениями движение посредством мер-
цательного покрова, свойственное турбелля-
риям, заменяется гораздо более энергичными
его формами — перистальтическими сокра-
щениями всего тела или синусоидным дви-
жением путем образования извивов тела, в
связи с чем развивается мощная мускула-
тура стенки тела на смену слабым мышеч-
ным слоям, имевшимся у турбеллярий. Од-
новременно в тесной взаимосвязи происходит
выработка удлиненно-червеобразной формы
тела и увеличение его размеров (рис. 38).
Таким образом создается внешний облик
немертины, значительно разнящийся с об-
ликом плоских червей, — тело длинное, не-
сколько сплющенное дорзо-вентрально, в по-
перечном разрезе овальное (рис. 39, Л) или
приближающееся к круглому. Хотя покров
сохраняет свою примитивность в том смыс-
ле, что образован мерцательным эпителием,
но развитие тонофибрилл в его 'клетках при-
дает ему большую по сравнению с турбелля-
риями плотность, а богатство железистыми
клетками делает его гораздо более мощным.
Эта черта усиливается значительным развитием
го вещества, располагающегося под эпителием и переходящего в про-
слойки соединительной ткани между мышечными пучками стенки
тела. При сравнении с турбелляриями поражают своей мощностью
слои кольцевых и под ними продольных, мышц, а у гетеронемертин
еще и слои наружной продольной мускулатуры. Главным компонен-
том в построении тела становится у немертин кожно-мышечный ме-
шок. •
Рассмотренный комплекс широко приспособительных черт в ор-
ганизации немертин тесно связан СО' стабилизацией рта близ пе-
реднего конца тела (рис. 40). Если у придонных турбеллярий вы-
работалось вентральное положение рта, то у немертин при длинном
червеобразном теле закреплено уже его наиболее выгодное для хищ-
ника положение, а именно, он располагается типично близ перед-
него конца, богато снабженного чувствительными образованиями.
Таким образом, впервые в животном царстве, у немертин вырабаты-
121
вается голова, ограниченная сзади ротовым отверстием и представ-
ленная так называемой «головной лопастью».
При стабилизации рта у переднего конца и постепенном удли-
нении тела немертинооб-
Рис. 39. А — схема поперечного разреза
через немертину Tubulanus polymor-
pbus в участке тела за хоботом:
1 — покровный эпителий, 2 — субэпителиаль-
ный слой ткани основного вещества, 3 — кро-
веносный сосуд, 4 — средняя кишка, 5 — коль-
цевые и 6 — продольные мышцы, 7 — внут-
ренняя мезенхима, 8 — кольцевые мышцы
кишечника, 9 — семенники. 10 — боковой
нервный ствол
Б — мезенхима нс
мертин (по К. Шней-
деру) , видны 2 клет-
ки в основном веще-
стве:
1 — кольцевые и 2 — про-
дольные мышечные клетки и
волокна стенки кровеносного
сосуда
немыслимым целентерическое состояние кишечника, когда непере-
варимые остатки пищи должны были бы снова пройти весь длинный
кишечник, чтобы быть выкинутыми через ротовое отверстие. В тес-
нейшей взаимосвязи с обеими указанными чертами и вырабатывает-
ся трубкообразный кишечник, образующийся в результате прибли-
жения заднего участка усваивающего эктодермального отдела киш-
ки к покрову на заднем конце тела и образования здесь анального
отверстия путем простого расхождения клеток обоих эпителиев.
У личинки немертин пилидия кишечник еще типичный целен-
терический, и только при формировании молоденькой немертины,
т. е. сравнительно поздно, образуется анальное отверстие, кишеч-
ник становится трубкообразным (рис. 41).
Таким образом получается возможность более полного по срав-
нению с кишечнополостными использования пищевого материала
при его продвижении по всей длине кишечника. Эта черта органи-
зации, присущая, начиная с немертин, далее всем вышестоящим
группам животного царства, обусловливает возможность полного
перехода от внутриклеточного пищеварения к полостному, энзима-
тозному, что также крайне характерно как широко адаптивное
явление, вырабатывающееся в этой группе.
122
-11
Рис. 40. Схема организации немертины Cerebratulus (по Бенге-
му) : А — виден кишечник:
1 — глотка, 2 — средняя кишка, 3 — анальное отверстие, далее нервная
система, 4 — дорзальные и 5 — вентральные ганглии головного мозга,
6 — церебральный орган, 7 — головные щели, 8 — боковой ствол, 9 —
аппарат хобота, 10 — хоботковое влагалище, И — ринходеум;
А — кровеносная система:
1 — спинной сосуд, 2 — боковые сосуды, 3 — ’ ротовое отверстие, 4 —
черифарингеальные сосуды и синусы, 5 — апикальная анастомоза и отно-
шения сосудов в головном конце
—• удалена спинная сторона и хобот, видны отношения голов-
ного мозга (1) и церберальных органов (2) к кровеносным си-
нусам головы, нефридиев (3) к боковым сосудам (4), располо-
жение гонад (5). кишечника и кровеносных сосудов, сзади
в поперечном разрезе
Обычно средняя, усваивающая кишка сохраняет у немертин
форму простой трубки, иногда лишь снабженной боковыми расши-
рениями (рис. 40), увеличивающими ее усваивающую поверхность.
Осложнение идет в переднем, глоточном, отделе, который может
достигать и порядочной длины и значительной дифференцировки
мускулистого и железистых участков в результате узких, а иногда и
крайних (Amphiporus, Malacobdella) приспособлений.
Наряду с прогрессивно эволюционировавшими кожно-мышечным
мешком и кишечником пространство между ними занято у немертин
примитивной тканью ОС-
Рис. 41. Молоденькая немертина после
метаморфоза (по Лейкарту):
1 — головной мозг, 2 — церебральные органы,
3 — кишечник, еще без анального отверстия, 4 —
рот, 5 — глаз, 6 — хобот
вания, получившие у турбеллярий в их
новного вещества харак-
тера мезенхимы (рис. 39,
е Б); последняя заключает
в себе также крайне при-
митивные гонады в виде
мешков, которые лишь при
созревании половых 'Про-
дуктов дают трубчатые
выросты, открывающиеся
наружу — примитивные
яйцеводы и семепроводы.
Как раз все эти образо-
опорной клеточной паренхиме
и в их сложнейшем половом аппарате особо сильное и своеобраз-
ное развитие, сохраняют, таким образом, у немертин черты значи-
тельной примитивности, показывая исходное их состояние в эволю-
ции всего типа червей. *
Выработавшийся у турбеллярий головной комплекс органов
чувств, сохраняя все свое значение, у немертин концентрируется на
головной лопасти (рис. 40, Л). Она становится прежде всего от-
делом тела, ориентирующим животное в условиях окружающей сре-
ды, что мы находим далее, как норму, у вышестоящих форм. При
этом тактильная чувствительность диффузна; можно сказать, вся
головная лопасть представляет мощный механорецептор. Глаза со-
храняют приблизительно то же примитивное состояние, которое при-
суще уурбелляриям. Сильно дифференцируются хеморецепторы в
виде расположенного апикально фронтального органа и особенно
характерных церебральных, затылочных органов. Последние, раз-
виваясь на основе образований, подобных аврикулярным органам
турбеллярий, эволюционируют в сложно построенные хеморецепто-
ры, играющие ведущую роль в жизни немертин. Так создается ряд
черт широко адаптивного характера, связанных с совершенствова-
нием ориентировки хищного червя в окружающей среде.
С этим связано, наконец, и развитие в головной лопасти у не-
мертин головного мозга, а именно, прежде всего, значительных сен-
зорных центров в виде парных спинных ганглиев, иногда в заднем
отделе непосредственно' объединяющихся с церебральными органа-
ми (рис. 40, 41) и содержащих дифференцировку особых ассоциа-
124
тивных элементов. Это значительный шаг по пути эволюции мозга ♦
от турбеллярий к высшим формам червей.
Однако в строении головного мозга сказался и другой момент.
Со стабилизацией ротового отверстия, а вместе с тем и глоточного
отдела кишки, у немертин в головном мозгу дифференцировались
скопления ганглиозного характера, так называемые брюшные ган-
глии мозга, являющиеся центрами стоматогастр.ического отдела
нервной системы. Таких ганглиев и постоянных связей с той или
иной частью нервной системы у турбеллярий при отсутствии стаби-
лизации положения рта, а следовательно, и глотки, не могло раз-
виться. Напротив, для вышестоящих групп подобные отношения
уже обычно имеют место.
В неменьшей мере эволюция мускулатуры немертин отразилась
в развитии нервной системы туловища. Наметившиеся уже у тур-
беллярий два продольных ствола дифференцированы у немертин
всегда в виде мощных боковых продольных стволов, имеющих
прежде всего характер двигательных. Располагаются они или в ос-
новании эпителия, или непосредственно под покровом (рис. 39, А),
или, наконец, уходят в мускулатуру, занимая более защищенное
положение. На смену диффузному расположению клеток и волокон,
что1 имеет место в продольных стволах у турбеллярий, выступает
резкая дифференцировка центральной волокнистой массы и распо-
ложенных на ней вентрально и дорзалыю скоплений униполярных
ганглиозных клеток, что связано с лучшим питанием последних.
Таким образом рассмотренные нами широко приспособительные
черты организации немертин отражаются в строении их нервной
системы, но последняя, в свою очередь, обусловливает появление,
вернее дифференцировку, образований, прежде всего служащих для
питания головного мозга. А* именно — при стабилизации рта и обра-
зовании трубчатого кишечника обособившаяся головная лопасть с
ее чрезвычайно важными для животного органами чувств и особен-
но головным мозгом становится в мало благоприятные условия в
отношении получения питательного материала от кишечника, усваи-
вающий эктодермальный .отдел которого1 начинается лишь на неко-
тором расстоянии позади рта. У турбеллярий мы видели сильное
ветвление эктодермального отдела кишечника, подходящего всюду
к нуждающимся в пищевом материале органам и их частям. У не-
мертин эволюция пошла другим путем. Чрезвычайно повысилась
возможность ткани основного вещества передавать пищевые веще-
ства от места их усвоения к местам потребления благодаря выра-
ботке подвижности основного вещества мезенхимы, а именно, ее
гель переходит в состояние золя. Таким образом получается жьщ-^
основного вещества — кровь, вместе с тем создается и
кровеноснатГ“сйстсма, как система лакун и каналов, наполненных
жидким содержимым — золем крови в неизмененном геле основно-
го вещества мезенхимы (рис. 40, Б). Важнейшая роль возникающей
кровеносной системы, как мы видим, трофическая.
Кровеносная система представлена у примитивных форм (па-
125
леонемертины, например Tubulanus) прежде всего двумя боковыми
сосудами (рис. 39, Л), подающими кровь из средней части тела к
головной лопасти, и большими лакунарными пространствами, рас-
положенными в головной лопасти и как бы являющимися резервуа-
рами питательных веществ, из которых органы головы, и прежде
всего головной мозг и церебральные органы, черпают необходимый
им питательный материал (рис. 40, Б и В). Вероятно' и глоточный
отдел кишечника получает питание из того же источника, но несом-
ненно, что цефализация (процесс образования головы) явилась ве-
дущим моментом в появлении кровеносной системы.
Дальнейшая эволюция последней у немертин шла по пути диф-
ференцировки правильного кровообращения, чем значительно совер-
шенствовалась ее основная трофическая функция и что морфоло-
гически выразилось в развитии спинного и комиссуральных сосудов
наряду с боковыми (рис. 40, Б).
! Отношение кровеносной системы к мезенхиме прекрасно харак-
теризуется классическим сравнением, что они относятся друг к дру-
! гу как неразрывно связанные, как «гора и долина». Представление
ЭТО' вполне согласуется с нашим об образовании сосудов, как жид-
ких участков, золя, в студне основного вещества мезенхимы немер-
лин. Поэтому характерным для сосуда .является це строение стенки,
а его содержимое — ткань ^жидким основным веществом — кровь.
У немертин стенки сосудов местами образованы лишь интимой из
уплотненной пластинки основного вещества окружающей их мезен-
химы, местами на интиме могут располагаться клетки или разроз-
ненно или наподобие эпителия.
Первые попытки поставить вопрос о происхождении кровеносной
системы относятся к 70-м и 80-м годам прошлого столетия. Не бу-
дем останавливаться на отброшенных уже взглядах Э. Геккеля. До-
вольно долго господствовал взгляд, разработанный О. Бючли
(1883), по которому кровеносная система представляет остаток по-
лости бластулы — «бластоцель-теория». Однако у немертин, на-
пример, никакой связи между бластоцелем и кровеносными сосуда-
ми нет. По точным наблюдениям Бюргера и Нусбаума и Окснера
они образуются путем разжижения студня основного вещества у
личинки.
Уже* несколько раньше О. и Р. Гертвиги (1881) в своей работе
о целоме выдвигают представление о кровеносной системе, как о
ряде щелей, образующихся между различными тканями и частями
органов — «схизоцель-теория». Однако приведенные для немертин
факты решительно заставляют отказаться как от подобных пред-
ставлений о происхождении кровеносной системы, так и от выска-
занных позже Ф. Вейдовским (1907) и А. Лангом (1903) их ва-
риантов, которые основаны на изучении отношений, наблюдаемых
у аннелид.
В частности, Ф. Вейдовский в своей «теории гемоцеля» 1 считает,
1 Гемоцель —; полость кровеносной системы.
126
что кровеносные сосуды первично связаны в своем происхождении
с эктодермальным эпителием усваивающей кишки; в основании эпи-
телия здесь образуются скопления трофической жидкости, которые
оформляются в сосуды благодаря тому, что находящиеся в основа-
нии эпителия клетки располагаются в виде тонкой пластинки «вазо-
телия»; все прочие сосуды развиваются затем путем разрастания
первично образовавшихся в стенке кишки. Факты, наблюдаемые у
немертин, не дают никаких подтверждений построениям Ф. Вейдов-
ского и, прежде всего, нет основного — постулируемой связи раз-
вития кровеносных сосудов с клетками стенки средней кишки.
Также неприложима к кровеносной системе немертин «трофо-
цель-теория» А. Ланга, по- которой у животных, обладающих вторич-
ной полостью тела, при развитии мешков целома первоначально на-
копляющаяся по бокам средней кишки жидкость трофического ха-
рактера далее покрывается стенкой, происходящей за счет базаль-
ной перепонки охватывающего ее снаружи целотелия; отсюда же
берет начало и мускулатура обособляющихся сосудов. Таким обра-
зом, единственным компонентом стенки последних является собст-
венно- стенка целома. Сам А. Ланг отмечает, что важнейшим де-
фектом его теоретических построений является неприложимость их
к немертинам, не имеющим целома, но обладающим кровеносной
ситемой. Однако факты не только- делают трофоцель-теорию А. Лан-
га неприложимой к немертинам, но заставляют вообще ее отверг-
нуть, так как выдвигают на ее место- другие взгляды, как развитые
нами выше, так и формулированные почти одновременно учеником
А. Ланга — Фернандесом' (1905).
По Фернандесу, в кровеносной системе необходимо различать, с
одной стороны, мезенхимный, первичный, «проводящий аппарат»,
образованный соединительнотканным слоем стенки сосуда, клетки
которого могут давать в конце концов у ряда форм и эндотелиаль-
ную выстилку, а с другой стороны, вторичный, главным образом
продвигающий «пропульзаторный аппарат», который происходит за
счет прилегающего снаружи мышечного слоя. Совершенно правиль-
но расценивая факты, констатированные при изучении строения
стенки кровеносных сосудов, Фернандес видит, однако, в проводя-
щем аппарате лишь схизоцель, т. е. вторичные щели и полости в
соединительной ткани, и далее не дает объяснения этим фактам. Мы
видим, что кровеносная система должна быть рассматриваема преж-
де всего с точки зрения на кровь как на жидкую ткань основного
вещества, как на золь среди гелеобразных участков последнего —
«теория жидкой ткани» — а не как на жидкость, диффундировав-
шую в щели, образующиеся вторично в основном веществе.
Развитие кровеносной системы у немертин оказало своеобразное
воздействие на их экскреторные органы, которые, сохраняя свой
примитивный характер, а именно — будучи представлены как и у тур-
беллярий протонефридиями, в то же время обнаруживают ясную
взаимосвязь с кровеносной системой. Во-первых, экскреторные ка-
налы вступают в теснейшие отношения с боковыми кровеносными
127
1
сосудами, соответственно лакунами, благодаря тому, что их участки,
, заканчивающиеся клетками с мерцательным пламенем, оказывают-
ся лежащими в разжиженном основном веществе, вдаются в про-
свет сосуда (рис. 40, В). Таким образом, как извлечение экскретов,
так и осморегуляторная функция могут активизироваться; поступ-
ление жидких продуктов распада из прилегающих тканей в жидкую
среду постоянно подвижной крови должно способствовать большей
энергии их извлечения нефридиями.
Во-вторых, в связи с этим обычно нефридии распространены
уже не диффузно по всему телу, а находятся в виде одной пары
в участке, расположенном непосредственно за головной лопастью,
т. е. у лакунарных пространств кровеносной системы, и в то же вре-
мя там, где в круг их действия попадают прежде всего органы го-
ловного отдела тела, особо важные в жизнедеятельности организма.
Вторично, в особых условиях существования, например у пресновод-
ной Prostoma graecense, протонефридии опять распространяются по
всему телу. Здесь ясно сказывается влияние изменения осмотиче-
ских явлений с переходом в пресную воду.
Подводя итоги нашему анализу организации нимертин, мы мо-
жем прежде всего отметить ее типовые черты, т. е. такие, которые
унаследованы немертинами от их исходных форм, очевидно от плос-
ких червей, предшественников современных турбеллярий. Сюда от-
носятся примитивный мерцательный покров, значительно развитая
ткань основного вещества, главным образом в виде внутренней ме-
зенхимы тела, экскреторный аппарат в форме протонефридиев и,
наконец, простые мешкообразные гонады с примитивными вывод-
ными протоками, образующимися при созревании половых продук-
тов. На этом фоне прогрессивно и характерно эволюционирует не-
сколько комплексов черт, связанных с хищным образом жизни. Во-
первых, обусловливающие активную подвижность—выработка удли-
ненно-червеобразного тела немертин и мощной мускулатуры кож-
но-мышечного мешка. Во-вторых, связанные с захватыванием добы-
чи и питанием — стабилизация рта у перднего конца тела и обра-
зование головной лопасти, а далее и выработка трубчатого кишеч-
ника с образованием анального отверстия и полостным пищеваре-
нием. В-третьих, ряд черт, связанный с более совершенной ориенти-
ровкой в окружающей среде — прогрессивное развитие органов
чувств, особенно церебральных органов. В-четвертых, соответствую-
щее рассмотренным .комплексам развитие нервной системы с диф-
ференцировкой сензорных спинных ганглиев головного мозга, обла-
дающих ассоциативными центрами, с обособлением вентральных
ганглиев мозга, имеющих значение стоматогастрических отделов
центральной нервной системы, и с дифференцировкой пары боковых
моторных нервных стволов в туловище. В-пятых, при стабилизации
рта и трубчатом кишечнике образование кровеносной системы в свя-
зи с обеспечением питания важнейших органов передней части тела,
особенно мозга и церебральных органов, а затем и передней кишки.
Все эти черты дают значительное повышение организации по
128
сравнению с плоскими червями и переходят далее, как прочное
приобретение в вышестоящие группы животного' царства. Вместе
л<е взятые они создают своеобразный план построения, своеобраз-
ный этап, эволюций, резко отличающийся от присущего плоским чер-
вям, и турбелляриям в частности, — этап немертин. Этот план
организации в пределах группы крайне стойко1 сохраняется и варьи-
рует лишь в деталях в связи с приспособлением к тем или иным
частностям условий существования в различных биоценозах.
Завершается организация немертин выработкой характернейшего
органа нападения и защиты — хобота, присущего всем представи-
телям этой группы, но не передающегося далее ни в одну из вы-
шестоящих групп (рис. 40 А, 41). Он построен как трубкообраз-
ное впячивание стенки тела на переднем конце головной лопасти,
выстланное мощным железистым эпителием. Как было в свое вре-
мя нами указано, подобные апикальные скопления железистых кле-
ток нередко встречаются у турбеллярий. Иногда (Calyptorhynchia)
они используются в связи с развитием сответстйующих мышечных
пучков для нападения и защиты. Но все это только первые шаги в
в том направлении, которое завершилось у немертин образованием
особой полости — ринхоцеля, или хоботкового влагалища, вокруг
впячивающегося участка стенки тела, дающего собственно хобот.
Благодаря образованию ринхоцеля трубка хобота получила воз-
можность выворачиваться как палец перчатки, могущий вворачи-
ваться обратно благодаря особому мышечному пучку, как бы про-
должающему слепой конец трубки. Хобот в связи с развитием мы-
шечных слоев его стенки может иметь значительную твердость, а
выстилающий железистый эпителий вырабатывает ядовитый секрет.
Так, благодаря образованию ринхоцеля создался эффективный
орган нападения и защиты,' который в силу своей эффективности
быстро эволюционировал в группе немертин, сделавшись мощным
образованием, во- многих случаях даже крайне адаптивным по ха-
рактеру, как, например, сложнейший хобот вооруженных немертин.
Он является настолько специфичным, что, эволюционируя в
различнейших направлениях, сделался в руках зоологов одним из
необходимейших и надежнейших признаков в систематике немертин
как для выделения основных групп — «невооруженные» и «воору-
женные» иемертины, — определяющих главные линии эволюции,
так и для дробного и точного разделения отдельных видов, что свя-
зано с его далеко идущим, разнообразием в деталях строения.
Главное же—быстрая и доминирюущая эволюция хобота вышла
за рамки общей прогрессивной эволюции немертин, сдвинула их с
широкого пути повышения организации, переведя на рельсы частных
приспособлений все более и более односторонних. Этим был завер-
шен этап развития немертинообразных форм, создались собственно
иемертины, представители современного подтипа в типе червей.
В резком противоречии со всем изложенным стоят взгляды
В. Заленского и К. Давыдова, по которым немертины являются
лШць деградированными потомками аппелид, и в этом ряду регрес-
Пути эволюции животного мира 129
сивной эволюции турбелляриям принадлежит дальнейшее и послед-
нее место. Основывается этот взгляд на данных эмбриологии, а
именно: всем им свойственно спиральное дробление и присутствие
телобластической закладки мезодермы за счет бластомера 4 d.
О значении спирального дробления в эволюции червей было сказа-
но при рассмотрении турбеллярий. Мы вйдели в выработке этого
процесса явление адаптивного порядка — приспособление к пря-
молинейному движению при переходе к придонному существованию.
Что касается мезодермального зачатка, то у аннелид он дает
типичные метамерные сомиты, в которых появляется вторичная
полость тела — целом, ограниченный сомато- и спланхноплеврой.
У немертин, по В. Заленскому, метамерия отсутствует, но соответ-
ствующий зачаток также дает начало сомато- и спланхноплевре, и
лишь в дальнейшем происходит путем расхождения их клеток обра-
зование мезенхимных элементов. Наконец, .у турбеллярий процесс
не доходит до образования целома, так как клетки зачатка рано
начинают расходиться, давая типичные мезенхимные элементы.
В. Заленский видит в этих фактах процесс постепенной редукции и
утраты целома, т. е. допускает регрессивную эволюцию в ряде
аннелиды—немертины—турбеллярий. Однако выводы В. Заленского
стоят в противоречии с тем общим положением, что у свободно-
подвижных хищников организация развивается прогрессивно, и у
немертин нет никаких оснований допускать регрессивную эволюцию.
Кроме того, данные В. Заленского стоят в противоречии с наблю-
дениями других исследователей, которые не находят у немертин
настоящего целома соответственно сомато- и спланхноплевры, а
лишь участки с разжиженным гелем основного вещества.
Мы рассматриваем появление у немертин зачатка, соответствую-
щего закладке целомической мезодермы у аннелид, как раннюю
дифференцировку закладки мощной мускулатуры туловища. Первые
шаги в этом направлении мы видим уже у турбеллярий (прликла-
диды), а завершается процесс у аннелид образованием полостей с
транссудативной жидкостью—целома в мышечном зачатке туловища.
Также ряд прогрессивной эволюции, в связи с приспособлением
к возможностям питания развивающегося зародыша, представляют
личиночные стадии в ряду турбеллярий — немертины — аннелиды.
Появившись у турбеллярий в виде примитивной Мюллеровской
личинки как приспособление зародыша к жизни и питанию в мор-
ском планктоне, у немертин личинки достигают большей сложности.
В организации пилидиума гетеронемертин это ведет к некробиоти-
ческому метаморфозу, когда большая часть тела личинки, как
крайнее приспособление к пелагическому образу жизни отбрасы-
вается при переходе к донному существованию взрослых. Кишечник
у личинки является при этом целентерическим. У аннелид процесс
заканчивается в образовании личинки трохофоры, обладающей ужо
трубчатым кишечником.
Мы подходим вплотную к подтипу аннелид..
Гл а в а VII
ПОДТИП КОЛЬЧАТЫЕ ЧЕРВИ — ANNELIDES
Кутикулярный покров. Сохранение мерцательного покрова. Образо-
вание трубок. Метамерия и образование комплекса параподий.
Мускулатура стенки тела. Система целома. Теории происхождения
целома. Система пограничных образований и ее значение. Крове-
носная система. Развитие жабер. Половой аппарат и эволюция
нефридиев. Головной комплекс органов чувств и мозг. Брюшной
мозг, ди- и тетранейрия. Кишечник. Итоги анализа организации
аннелид. Архитектоника анмелид.
Аннелиды представляют высший этап в эволюции типа червей.
Это гигантский шаг осложнения и совершенствования организации
по- сравнению с немертинами. Если у плоских червей вырабатывает-
ся билатеральное строение и появляются впервые экскреторные
органы, а у немертин — трубчатый кишечник и кровеносная систе-
ма, то у аннелид налицо уже все системы органов, присущие выс-
шим типам животного царства: органы движения, органы дыхания,
вторичная полость тела и появляется метамерия тела.
Строение аннелид выявляет прежде всего замечательное общее
повышение организации на основе того, что было достигнуто немер-
тинообразными формами. И, как это было показано для последних,
повышение организации здесь также теснейшим образом связано
с подчеркнутым нами выше важнейшим экологическим моментом —
свободноподвижным существованием хищников. При таком образе
жизни естественный отбор обусловливает наиболее интенсивную
прогрессивную эволюцию всех черт строения. И лишь вторично, в
пределах создавшейся организации ' аннелид, чрезвычайно богато
вырабатываются формы, приспосабливающиеся к всевозможным
условиям существования — растительноядные, обитатели грунта,
живущие в трубках сидячие формы, паразиты и т. п.
В системе аннелид мы видим чрезвычайно яркое выражение
'/того момента. Наиболее сохранившими типовые черты формами,
на которых мы и сосредоточим наше внимание, являются, как это
в настоящее время общепризнано, полихеты. Они богато представ-
лены во всевозможных биоценозах мирового океана и * лишь в
отдельных случаях перешли в пресные воды. Их крайне измененны-
и потомками являются также обитатели моря эхиуриды и сипун-
9*
131
кулиды. Напротив, приспособившись к жизни в грунте пресновод-
пых водоемов, коренными обитателями пресных вод и далее назем-
ными формами являются происходящие от полихет олигохеты.
Наконец, своеобразное хищничество, соединенное с паразитизмом,
дало на основе олигохет оригинальнейшую группу пиявок.
Конечно, в связи с этим, облик аннелид крайне разнообразен,
но у них у всех выступают в то же время некоторые типовые
черты, ярче всего выраженные у свободных придонных полихет
(Nereidae, Eunicidae и т. дО (рис. 42).
Прежде всего, примитивный ресничный покров, имевшийся как
у турбеллярий, так и у немертин, заменяется кутикулярным, что
представляет ярко выраженное приспособление защитного характе-
ра. Однако кутикула аннелид настолько эластична, что она не пре-
пятствует изменению формы тела животных и не сковывает его
неподвижным футляром. Покров сохраняет возможность всюду
иметь чувствительные образования в виде, например, чувствитель-
ных волосков, и выработка кутикулы сочетается, наконец, с сохра-
нением железистости покрова, так как выводные протоки железистых
клеток всюду могут ее пронизывать. Это — обычное состояние для
олигохет и пиявок, У полихет мерцательные участки покрова неред-
ко еще сохраняются в разных семействах, но уже как узкие при-
способления специального характера. Так, например, они имеются
на хеморецепторах, обусловливая движение воды, в химизме кото-
рой ориентируется животное (затылочные органы, боковые органы
и т. п.). Они развиты на жабрах, где движение воды способствует
газообмену (Nephthydidae и т. д.). Они образуют в ряде семейств
мерцательные полукольца на спинной стороне, определяющие ток
воды по телу, к жабрам у Spionidae, под элитрами у Aphroditidae
(рис. 43), к пластинкам параподиальных усиков Phyllodocidae, или
образуют брюшной и спинной продольные желобки, обусловливаю-
щие выкидывание экскрементов из трубки Serpulidae.
Гораздо сильнее узкоадаптивные черты проявляются в разви-
тии железистых образований покрова аннелид. Для групп, пере-
шедших к жизни в грунте, общее богатство покрова железками
преимущественно слизистого типа облегчает движение и предохра-
няет от поранений. Но своего крайнего развития железистость по-
крова достигает в образовании временных (Nereidae, Eunicidae)
или постоянных (Eunicidae и Sedentaria) защитных трубок (рис. 44)
за счет затвердевающего секрета. Это ведет в конце концов у ряда
полихет к своеобразному сидячему образу жизни. Животное, соб-
ственно, свободноподвижно в узких пределах его трубки, но сама
трубка обычно прикреплена к подводным предметам. В связи с
этим появляется целый комплекс изменений организации, являю-
щихся приспособлениями как к жизни в трубке, так и к сидячему
существованию.
Сами железки покрова при этом дифференцируются в грандиоз-
ные железистые поля, располагающиеся или по бокам тела, — так
называемые параподиальные железы (Nereidae — рис. 46, Б), или
132
Рис. 42. А — внешняя морфология типичной полихеты Eunice,
передняя часть тела:
1 — антенны, 2 — жабры, 3 и 4 — спинные и брюшные усики параподий,
5 — затылочные органы, 6 — глаза, 7 — хетоподий, 8 — пальпы, 9 — голов-
ная лопасть, 10 — первый, 11 — второй, 12 — третий сомиты;
Б — головной конец тела с брюшной стороны:
13 — ротовое отверстие;
В — то же, рот раскрыт и виден аппарат челюстей (14); Г —зад-
ний конец тела:
15 — анальное отверстие, 16 — лигидиум с его усиками (17), 18 — зона обра-
зования новых сомитов;
Д — схема внутреннего строения Eunice. Поперечный разрез, сле-
ва на уровне ^иссепимента, справа посредине сомита:
1 — нотоподий, 2 — невроподий, 3 —- целом, 4 — железистые скопления,
* — кишечник, 6 — кольцевая и 7 -- продольная мускулатура, 8 — косые
мышцы, 9 — мезентерий, 10 — метанефридий, П — брюшная нервная цепочка,
12 — средний нерв сомита, 13 — оварий, 14 — подиальный ганглий, 15 --
комиссуральные, 16 — спинной и 17 — брюшной сосуды, 18 —• сплетение со-
судов в стенке кишки, 19 — жабра, 20 — спинной и 21 — брюшной
усики.
посередине брюшной стороны (брюшные щитки Terrebelloidea и"
Serpuloidea).
Однако при анализе организации аннелид бросаются в глаза в
их внешней морфологии не эти рассмотренные нами черты покрова.
Облик аннелид обусловлен гораздо более сложным явлением вы-
работки метамерного 1 строения, т. е. повто-
рением одинаковых участков по длине тела.
Обычно у аннелид хорошо выражены
передний, предротовой, неметамерный отдел,
или простомиум, представленный, как и у
немертин, головной лопастью, затем следую-
щие друг за другом метамеры, или сомиты,
представленные кольцами туловища, и, на-
конец, задний неметамерный участок — пи-
гидиум, несущий анальное отверстие (рис. 42
и 46,4). На первый план в строении тела
выступают, таким образом, метамеры туло-
вища, отсюда и название для всей группы
Рис. 43. Разрез стенки
гела полихеты Harmotoe
(по Любищеву):
1 — кутикула, 2 — реснички
мерцательных клеток, 3 —
эпителий покрова, 4 — суб-
эпителиальная пограничная
пластинка ткани основного
вещества, 5 — кольцевые
мышцы
«кольчатые» черви.
Образование типичных сомитов у аннелид
теснейшим образом связано с широко адап-
тивным комплексом образований, представ-
ляющих, прежде всего, расположенные по бо-
кам тела выросты параподий, т. е. примитив-
нейшие конечности типа простого рычага
(рис. 42, 46,5, 47, 48,5). Появление боко-
Рис. 44. Теребеллоидная полихета Amphitrite cirrata:
1 — щупальца, 2 — жабры, 3 — крючковые валики параподиев, 4 — щетинки ното-
подиев, 5 — железистые щитки на брюшной стороне, 6 — головная лопасть, 7 —
илистая трубка
вых выступов, на которые при синусоидальных извивах может опи-
раться тело, является прототипом параподия и представляет, конеч-
но, адаптивную черту, значительно совершенствующую движение
тела. Движение посредством локомоторных придатков идет на сме-
ну движению путем сокращений всего тела, что мы видели в пери-
1 мета — за, мерос — часть (греч.).
134
стальтическом и синусоидном движении у немертин и что, в свою
очередь, заменило более примитивное ресничное'- движение тур бел -
дярий.
Расположение параподий в строгом соответствии с кольчатостью
аннелид невольно заставляет связывать появление метамерии с
двигательным моментом, и в свое время (1892) Э. А. Мейером была
выдвинута теория локомоторного происхождения метамерии. Хотя
впоследствии она была оставлена автором, но, как отмечает
А. Ланг (1903, стр. 67), локомоторный момент, несомненно, мог
играть некоторую роль в расчленении тела через посредство соот-
ветствующей дифференцировки мышечных групп, относящихся к
каждому параподию, и через посредство дифференцировки более
подвижных участков покровной кутикулы, т. е. путем образования
поперечных бороздок и соответственно следующих друг за другом
колец. Можно добавить к этому и дифференцировку опорных обра-
зований, однако здесь мы переходим уже к другому комплексу —
полости тела и прочим внутренним органам, которые, несомненно,
играли важнейшую роль в выработке метамерий. Таким образом,
следует полагать, что метамерия представляет явление сложное,
которое, в известной мере, «во внешней» метамерии может
быть связано с движением животного и с выработкой локомоторных
придатков, параподий, а во всей полноте будет раскрыто далее при
анализе «внутренней» метамерии тела.
Наличие параподиальных выростов, как подвижных и наиболее
выступающих пунктов тела в каждом метамере, используется далее
для более совершенной ориентировки во внешней среде, что связы-
вается с концентрацией здесь чувствительных образований —
развиваются чувствительные придатки — спинной и брюшной уси-
ки (рис. 42, 45,5, 47). Таким образом, из выроста параподия и
усиков создается основа так называемого подиального комплекса
полихет, или, короче, «подия».
Подиальный комплекс прежде всего получает узкоприспособи-
тельный характер к условиям движения аннелиды, а далее и к
Другим условиям существования. В нем обычно прежде всего и
ярче всего отражается процесс видообразования, и в систематике
полихет он играет важнейшую роль. К его узкоадаптивным чертам
относится выработка аппарата кутикулярных щетинок (рис. 45),
располагающихся в особых апикальных разрастаниях эпителия
параподий, щетинковых мешках. Образуются так называемые
«хетоподии». Ряд исследователей ставит их в качестве обязательного
компонента подиального комплекса вместе с выростом параподия
и усиками. Наличие аппарата выставляющихся наружу тонких ще-
тинок значительно уточняет и утончает функционирование парапо-
дий, как простых боковых выростов стенки тела; при разнообразии
поверхности субстрата, по которому должно' двигаться животное,
°ни прежде всего приходят в соприкосновение с этим субстратом и
используются как тонкий аппарат механической опоры, необходи-
мый для действия простого рычага параподия.
135
По существу, каждая щетинка представляет пучок слитых вн
одно целое кутикулярных волокон, который образуется одной эпи-
телиальной клеткой покрова, занимающей погруженное положенйе.
Совокупность таких клеток и дает вдающийся внутрь параподия
щетинковый мешок. Узкоадаптивный характер выражается прежде
всего в чрезвычайном разнообразии форм щетинок, начиная с тон-
ких простых нитевидных и кончая сложными щетинками; выраба-
тываются длинные плавательные щетинки пелагиче'
Рис. 45.
Щетинки
Eunice, за
исключе-
иием крю-
чочка из не-
вроподия
теребеллиды
(О
ских Alciopidae или гетеронереидных форм, крепкие
крючковидные щетинки живущих в грунте Drilo-
morpha, цепляющиеся за трубку крючки Terebellidae
(рис. 45,7) и Serpulidae, защитные палеи Amphicte-
nidae, крепкие шипы Aphroditidae, и т. д. и т. п.
Происходит дифференцировка различных форм
щетинок и внутри одного и того же хетоподия —
прежде всего дифференцировка особо мощных опор-
ных aciculae у Errantia, которая создает внутренний
скелет хетоподия, служащий опорой для движения
всех прочих щетинок (рис. 47).
Наконец, в результате разницы в деятельности
щетинок брюшной и спинной сторон параподия вы-
рабатывается дифференцировка всего подия на две
резко выраженные части, иногда совершенно отде-
ленные друг от друга, невроподий и нотоподий
(рис. 46). Это, собственно, обычное состояние для
современных полихет, и те случаи, когда параподий
является как бы примитивным, одноветвистым, обыч-
но связаны с вторичной редукцией нотоподия, как,
например, у Eunicidae (рис. 42,Д, 47). В связи с этим
Б. Гатчак (1893) характеризует исходную форму
параподия — «полный» параподий, как двуветви-
стый с брюшным и спинным усиками и с дорзаль-
ной жаброй. О последнем образовании мы еще бу-
дем говорить в дальнейшем.
Крайней адаптацией является «разделенный», по
терминологии Б. Гатчека, параподий Terebelloidea
(рис. 47) и Serpuloidea, у которых в связи с жиз-
нью в трубке невроподий или нотоподий пре-
вращается в длинный валик, располагающийся в поперечном на-
правлении и несущий ряды крючковых щетинок («tori uncinigeri»
систематиков). Таким образом, червь может чрезвычайно прочно
закрепляться в своей трубке. *
Однако узкоадаптивные процессы захватывают и прочие части
подиального комплекса, обусловливая то их необычайное разви-
тие, как, например, длинные лопастные параподий пелагического
Tomopteris, или элитры Aphroditidae и пластинки Phyllodocidae,
развивающиеся за счет спинных усиков, то, напротив, их редукцию.
Так, спинной и брюшной усики, а также и жабры редуцируются и
136
исчезают у многих живущих в грунте форм, например Lumbricone-
reina из Eunicidae или различных Drilomorpha, или, наконец, у
Oligochaeta. У последних и самый выступ параподия совершенно
сглаживается, сохраняются лишь брюшной и спинной хетоподии.
часто с единичными щетинка-
ми.
Таким образом, широко
адаптивная черта прогрессив-
ной эволюции червей, связан-
ная с большим повышением их
возможностей в борьбе за
жизнь — выработка конечно-
стей и затем подиального ком-
плекса у аннелид, переходит с
пути общего повышения орга-
низации на путь узких и край-
них приспособлений, обуслов-
ливая чрезвычайное разнообра-
зие их формы и биоценотиче-
ских отношений.
Во внутреннем строении
аннелид выработка подиально-
го комплекса не могла, как
уже мы указывали, не отра-
зиться. В связи с появлением
параподиев стоит дифференци-
ровка мускулатуры кожно-мы-
шечного мешка (рис. 42, Д,
46,Б, 47).
В стенке тела имеются ти-
повые отношения — эпителий
Nereis
Рис. 46. А —
(по
головной конец
Мак Интошу):
1 — головная лопасть, 2 — передние .. . ..
3 — пальпы, 4 — антеннальные усики, 5 —
антенны, 6 — выдвинутая глотка с аппаратом
челюстей (7) и крючков;
Б — схема строения типичного сомита
Nereis (по Мак Интошу). Полный па-
раподий:
1 — нотоподий, 2 — невроподий, 3 — пара-
подиальные железы, 4 — спинной и 5 —
брюшной усики, 6 — нефридий с простой во-
ронкой (8), 7 — продольная мускулатура,
9 — брюшная цепочка, 10 — комиссуральный
сосуд, 11 — кишка, 12 — целом, 13 — брюш-
ной и 14 — спинной сосуды
метамеры,
подостлан слоем ткани основу
него вещества в виде тонкой
субэпителиальной попраничной
пластинки с сильно развитыми
преколлагенными волокнами и
обычно лишенной клеток (рис.
43). За ней следует слой коль-
цевых мышц, подразделяемый
на большие или меньшие пучки
такими же пограничными пла-
стинками, и, наконец, слой про-
дольной мускулатуры, подсти-
лаемый снаружи тонкой суб-
мускулярной пограничной перепонкой (рис. 47). Пограничные обра-
зования стенки тела полихет представляют такое же неразрывное
Целое, как и более мощные прослойки ткани основного вещества,
в которые заключены пучки мышц стенки тела у немертин. Здесь
137
гчмеет место только конденсация этой ткани, заполняющая и свя-
зующая функция которой сохраняется, однако, в полной мере. .
Образование боковых выростов параподия вызывает изменения
кольцевой мускулатуры на
соответствующих участках стенки тела
рис. 42,Д, 46,Б, 47). От кольцевых
мышц обособляются, во-первых, пучки,
двигающие самый вырост параподия.
Во-вторых, дифференцируются пучки,
прикрепляющиеся на щетинковом меш-
ке с одной стороны и на стенке пара-
подия с другой. И, в-третьих, пучки,
соединяющие одни части щетинко-
вых мешков с другими, как, например,
пучкр, идущие от внутреннего конца
опорных щетинок — ацикул к прочим
частям щетинкового аппарата. При со-
кращении этих последних пучков аци-
кулы используются как скелет хэтопо-
дия, благодаря которому становятся
возможными разнообразные движения
прочих щетинок параподия.
Если кольцевая мускулатура в свя-
зи с развитием конечностей аннелид
дифференцируется по бокам сомитов в
Рис. 47. А — схема располо-
жение пограничных образова-
ний у Eunice, нанесенных жир-
ной черной линией:
1 — пограничная перепонка спин-
ного мезентерия, содержащая спин-
ной сосуд, 2 — субэпителиальная
пограничная перепонка покрова,
3 — такая же перепонка кишечни-
ка, содержащая сплетение кровенос-
ных сосудов, 4 — суб мускул ярная
перепонка, дающ выросты в про-
дольную мускулатуру, 5 — нотопо-
дий с жаброй, спинным усиком и
редуцированным пучком щетинок,
Ф — невроподий с брюшным усиком
и пучком щетинок с акцикулой по-
средине;
К — схема жаберных капилля-
ров Eunice:
1 — целотелнй, 2 — кутикула, 3 —
покровный эпителий, 4 — субэпи-
телиальная перепонка, 5 — перепон-
ки, отходящие от субэпителиальной,
проникающие между клеток эпите-
лия и содержащие капилляры в
свободном крае (6)
специальные мышцы параподиального
комплекса, то и продольная мускула-
тура по всей длине тела претерпевает
характерные изменения, резко подраз-
деляясь параподиями на спинные ц
брюшные мышечные ленты. Их разви-
тие тесно связано в то же время с дру-
гим чрезвычайно важным моментом —
появлением у аннелид вторичной поло-
сти тела, или, по принятой теперь
всеми Геккелевской терминологии,
«целома». Последний в виде парных
метамерных мешков занимает прост-
ранство, где у намертин находится
масса внутренней мезенхимы тела. Он
вдается и в выросты подиев (рис. 48,
Б, 42,Д, 46,5, 47, 49).
Целом представляет систему поло-
стей, располагающихся между стенкой
тела и стенкой кишечника и имеющих
жидкое содержимое. В силу своего положения и тесного соотноше-
ния с различными другими внутренними образованиями, а также, в
связи с этим, в силу возможности значительных изменений состава
содержимого целом может нести и действительно несет очень
338
разнообразные функции. Создается комплекс целома, который, вме-
сте со связанными с ним образованиями, по многообразию отноше-
ний и по значению в организации аннелид становится чрезвычайно
важным, можно сказать ведущим. Утверждение, что комплекс це-
Рис. 48. Схема организации полихет (по Э. Мейеру): А — нервная
система и нефридии с брюшной стороны:
1 — головной мозг, 2 — коннективы и 3 — ганглии брюшной цепочки, 4, 5—
передний и задний кольцевые, моторные нервы сомита, б — средний, чувстви-
тельный нерв, 7 — нефридий, 8 — нефридий половых сомитов, 9 — рот;
Б — вид со спинной стороны-
10 — целом, 12 — диссепименты, И — кишка, 13 — мезентерии, 14 — пищевод,
15 — ротовая полость, 16 — глотка с ее мышцами — 17, 18 и 19 — кольцевая
п продольная мускулатура, 20 — затылочные органы, 21 — глаза, 22 — яйчннки;
В — нервная система, кишечник и кровеносная система, вид сбоку:
23 и 24 — спинной и брюшной сосуды, 25 — сплетение сосудов на кишке и
26 — сосуды, соединяющие его со спинным и брюшным, 27 — сосуды глоточ-
ного отдела, 28 — комиссуральные, 29 — палыгы
-лома создал организацию аннелид, не будет парадоксальным, тем
более, что образованный в каждой метамере парой мешков — пра-
вым и левым, он обусловливает внутреннюю метамерию тела поли-
хет. Во внешней морфологии с ней, как мы видели, объединяются
Развитие параподиев и выработка кольчатости тела. Таким образом
139
создается сложное явление метамерии аннелид с обособлением неме-
тамерной головной лопасти и пигидия.
Правый и левый мешки целома в каждом сомите, образуя зна-
чительные полости между стенкой тела и кишечником и вдаваясь
в параподий, сходятся друг с другом по спинной и брюшной средин-
ной линиям. Получаются спинной и брюшной мезентерии, как бы
подвешивающие кишечник в полости тела. Точно так же задние
стенки одной пары мешков в поперечном направлении сходятся
с передними стенками следующей пары, образуя поперечные пере-
городки — диссепименты, отделяющие один сомит от другого.
Однако, как видно' из происхождения диссепимента, он представляет
не единую поперечную перегородку, а пару образований — правое
и левое, которые могут находиться иногда и на некотором расстоя-
нии вперед или назад по отношению друг к другу. В головной ло-
пасти собственного целома не образуется, туда заходят лишь вто-
рично выросты целомических мешков первой метамеры тела, а
пигидиум вообще целома не имеет.
Чтобы закончить характеристику типичного целома, необходимо
отметить, что мешки целома имеют собственную стенку, образован-
ную обычно плоскими клетками целотелия (перитонеального эпи-
телия), который располагается непосредственно на проксимальной
поверхности продольных мышечных лент стенки тела и на мышеч-
ном слое кишечника, образуя с ними единое целое. В результате
получается чрезвычайно характерная черта строения продольной
мускулатуры стенки тела полихет, а именно: ее спинной и брюшной
отделы, соответственно правому и левому мешкам целома, образо-
ваны каждый двумя, правой и левой, обычно мощно развитыми,
мышечными лентами.
В ряде случаев единство целотелия и продольной мускулатуры
особенно ясно выявлено наличием в целотелии целотелиально-мы-
шечных элементов или невозможностью отделить мышечные клетки
от целотелиальных. В других частях сомита, например в парапо-
диях, диссепиментах, мезентериях, целотелий располагается на
пограничных образованиях, представляющих внутренний слой стен-
ки параподия, щетинкового мешка и т. п. и уже в комплекс целома
не входящих.
Такие отношения объясняются происхождением продольного
мышечного слоя стенки тела и кишечника. Стенка целомического
мешка в эмбриональном развитии происходит из сначала сплошного
зачатка сомита, отшнуровывающегося от мезодермальной телобла-
стической полоски (рис. 49). Внутри этого зачатка путем расхож-
дения клеток получается полость — зачаток целома, а образую-
щаяся клеточная стенка дифференцируется в тонкий слой целотелия
и скопление клеток, за счет которых далее получается мускулатура
париетальная и висцеральная. Жидкое содержимое целома пред-
ставляет транссудат из окружающих тканей и по солевому составу
изотонично морской воде, следовательно, не сходно с жидкостью
крови, представляющей, собственно, как мы видели у немертин,
140
основное вещество в состоянии золя, тогда как в прилежащей ме-
зенхиме оно является в виде геля. На этом основании необходимо
отграничивать друг от друга жидкость целома и кровь, как совер-
шенно разнородные образования, а соответственно и проводить рез-
кую границу между полостью целома и полостью кровеносной
системы. Первая имеет целотелиальную стенку, вторая обычно вы-
стлана интимой из несколько уплотненного слоя основного вещества,
в образование здесь клеточной выстилки, эндотелия, может происх'о-
дить только вторично за счет мезенхимных
В функциональном отношении система
целома очень многообразна в связи с мно-
гообразием органов, которые к нему при-
лежат и могут так или иначе влиять на
жидкое содержимое целома как в отноше-
нии его химизма, так и в отношении
его форменных элементов. У полихет
в целом вдаются, а иногда глубоко про-
никают щетинковые мешки и железы па-
раподий и отдельные мышечные пучки,
гонады, нефридии, лимфоидные органы;
в целоме пробегают кровеносные сосуды,
части нервной системы; в его жидкости
элементов.
вие. 49. Мезодермальный за-
чаток Polygordius (по
Э. Мейеру):
б — мезодермальный телобласт,
6 — клетки мезенхимы, одна из
которых (7) в стадии деления,
8 — стенка кишки, 1, 2, 3, 4 —
мезодермальные сомиты, из ко-
торых второй содержит уже
полость (9)
находятся различные амебоидные эле-
менты и половые клетки. Однако основ-
ная функция целома, присущая всем его
участкам — опорная. А именно, являясь
системой заполненных жидким содержи-
мым полостей, он во время сокращения
окружающих его мышечных слоев стано-
вится для них, при несжимаемости жидко-
стей, настоящим опорным образова-
нием, «пассивным органом движения» по формулировке Нир-
штраса (1923). Мешки целома с жидким содержимым при-
дают определенную плотность телу; распространяясь в парапо-
дии, придают плотность этим последним и, наконец, вторично про-
никая в головную лопасть, также и ей. Особенно рельефно опорная
функция целома выступает в некоторых частных случаях узких
приспособлений: при выработке перистальтических движений тела
для передвижения в ходах грунта и в трубках у некоторых полихет
(Arenicolidae, Terebellidae). При этом происходит вторичная утрата
диссепиментов и мезентериев, целом превращается в единую по-
лость в большей части тела, благодаря чему волны сокращений и
расширений могут пробегать почти по всему телу червя. Или другой
случай узкой адаптации при образовании содержащих целом при-
датков вроде щупалец теребеллид или жабер некоторых полихет,
когда эти придатки расправляются и удлиняются при вхождении в
иих целомической жидкости и спадаются при оттоке последней. Это
граничит уже с выработкой двигательного целома.
141
Другие функции далеко не так постоянны. Из них особенное,
внимание обращалось на питательную и половую. Трофическая
функция обусловлена тем, что в участках тела, содержащих усваи-
вающий отдел кишечника, питательные вещества могут диффунди-
ровать из стенки кишки в жидкость целома, представляющую собой
собственно межтканевую среду, по солевому составу изотоничную
морской воде. Отсюда питательные вещества могут поступать в
прилежащие ткани и органы, а также и в форменные элементы
самой целомической жидкости. Однако в сомитах, содержащих
переднюю кишку, трофическая функция целома не может иметь ме-
ста и, очевидно, всегда отсутствовала. Кроме того, и содержание
питательных веществ в целоме разных полихет очень различно.
Так, у Aphroditidae жидкость целома дает лишь очень слабый оса-
док при свертывании. По М. Кено, в ней в среднем имеется 3%
растворенных альбуминов, но ряд исследований дал отрицательные
результаты на содержание белков. Напротив, у Arenicola целомиче-
ская жидкость очень быстро свертывается, давая сетку фибрина.
Несомненно, развитие этой функции является только вторичным ре-
зультатом скопления больших количеств жидкости возле органов,
вырабатывающих питательные вещества, и диффузии этих послед-
них в омывающую жидкость. Дальнейшим связанным с этим,
вторичным, явлением будет отложение питательных материалов
(белков и жира) в клетках стенки или в элементах целома, что
достигает особо сильного’ проявления у Terebellidae в элеоцитах,
служащих для питания половых клеток. ।
Половая функция связана с тем, что гонады при их росте и |
созревании половых элементов начинают вторично вдаваться в при- 1
лежащий целом, а в дальнейшем, с нарушением целостности цело- 1
телия, половые клетки, отделяясь от гонад, попадают в жидкость |
целома, где и проделывают часть своего жизненного цикла. При на-
личии в целомической жидкости питательных веществ вполне по-
нятны такого рода отношения, которые гарантируют образование
зрелых половых клеток, и в то же время полости целома могут
служить резервуарами для их сохранения. Эта функция присуща
только тем сомитам, где образуются гонады, следовательно, являет-
ся далеко не общей и не основной, но она бросается в глаза и ха-
рактерна для аннелид.
Кроме того, в жидкость целома могут поступать и поступаю’;’
путем диффузии из прилежащих тканей в различных количествах
жидкие продукты азотистого распада, с чем связана его экскретор-
ная функция; затем в целом проникают амебоидные элементы; об-
разуя лимфоидные и фагоцитарные скопления — фагоцитарная
функция целома. Последняя функция очень непостоянна, но еще
более спорадично встречается развитие у целома функции газового
обмена, когда жидкость целома через тонкие участки стенки тела
может участвовать в удалении из организма углекислоты путем
диффузии в окружающую воду при разнице парциальных давлений,
и в получении таким же образом из окружающей воды кислорода.
142 1
Углекислота при этом поступает из прилежащих тканей и находя -
щихся в целоме клеток, а кислород потребляется-ими. В конце кон-
цов это, как узкоадаптивное явление, ведет к появлению гемоглоби-
на в целоме, образованию настоящих целомических эритроцитов
(Glyceridae, Capitellidae) и даже к возникновению целомических
жабер, т. е. содержащих целом и служащих для дыхания ветви-
стых выростов стенки тела.
Итак для аннелид характерен метамерный, и притом полимер-
ный, т. е. состоящий из большого количества метамер, целом, обра-
зующий единый комплекс с так называемой целомической мускула-
турой и в то же время имеющий ближайшие отношения с целым
рядом важнейших систем органов в теле. Естественно поставить
вопрос о происхождении системы целома, отсутствующей еще у
плоских червей и немертин, которых поэтому иногда и объединяют
как червей паренхиматозных, противополагая аинелидам — червям
целомическим.
Уже в 1881 г. братья О. и Р. Гертвиги уточняют и разрешают
вопрос о целоме на основе изучения его развития и строения, при-
чем они решительно высказываются за его происхождение путем
дифференцировки от кишечника — так называемая «энтероцель-
теория». 15 годами позже выдвигается другая попытка объяснения
происхождения целома—«гоноцель-теория» Р. Берга и Э. Мейера, не-
которой полость целома соответствует своеобразно дифференцирую-
щейся полости гонад низших червей. Эти два взгляда делят зооло-
гов до настоящего времени на два лагеря. Для одних особенно
важным является факт развития целомических зачатков в некото-
рых случаях (типы иглокожих, хордовых и др.) путем отшнуровы-
вания боковых выростов от энтодермальной закладки, а отсюда за-
манчивые представления о происхождении целома за счет боковых
камер кишечника кишечнополостных путем обособления от цен-
трального отдела. Для других вопрос решается соблазнительным
сходством между гонадами низших червей и содержащими половые
продукты мешками целома по их положению относительно других
органов и по их половой функции. А именно: полости гонад разра-
стаются, превращаясь в мешки целома, создаются мезентерии и дис-
сепименты и развивается целомическая мускулатура, первоначально
служащая для опорожнения полости гонады.
Циглером, а затем Фаусеком была выдвинута и еще одна
теория происхождения целома — «нефроцель-теория», по которой
полость целома представляет часть экскреторного аппарата и пер-
воначально связана с накоплением в ней жидких продуктов распада.
Однако этот парадоксальный взгляд имел за собой так мало- фактов,
что, не получив хоть сколько-нибудь широкого распространения,
постепенно сошел со сцены и лишь в последнее время снова выдви-
гается Сцодграсом.
Как видно, многообразие функций целома в известной мере
нашло свое отражение в этих взглядах. Что же, собственно, по су-
ществу представляет целом, что это за орган в основном?
ИЗ
Гоноцель-теория видит в целоме своеобразно измененную полость
гонады и те метамеры целома, в которых отсутствует образование
половых клеток, считает вторично их утратившими, сделавшимися
стерильными. Эта вторичная стерильность крайне проблематична, но
окончательно1 губит гоноцель-теорию тот факт, что часто в онтоге-
незе половой .зачаток бывает совершенно независим от зачатка
целома’ и лишь вторично гонада сближается с целомическим меш-
ком, начинает прилегать к его стенке, наконец, входить в ее состав.
Таким образом, очень красивая схема, на которой гонады паренхи-
матозных червей по положению ило содержанию как-будто вполне
соответствуют парным мешкам целома аннелид, оказывается ио
существу совершенно неправильно выражающей истинные отноше-
ния целома и гонад. Мы выше рассматривали половую функцию
целома, как вторичную.
Вторичной является также и его трофическая функция, которую
энтероцель-теория кладет в основу понимания целома. Факт отшну-
ровывания, в эмбриональном развитии некоторых типов (Echino -
dermata, Chordata и т. д.) метамерных зачатков целома от эктодер-
мальной закладки говорит собственно только о пространственном
сближении двух, может быть, и совершенно независимых образова-
ний, или, выражаясь словами П. П. Иванова, мезодермальный за-
чаток закладывается, может быть, и независимо от энтодермы, цо
всегда в тесном контакте с нею.
Этот факт вполне естественно включается в понимание целома,
как отшнуровавшихся отделов кишечника, при предположении
Седжвика, а в последнее время Ламера, о происхождении аннелид
от актиниеподобных предков. Фантастическое представление о пе-
реходе актинии к свободному ползанию на оральной поверхности
(причем происходит срастание краев щелеобраэмого рта, за счет
сифоноглифов которого получаются рот и анус аннелиды, а из сред-
ней части брюшная цепочка), далее превращение щупалец актинии
в параподии полихет, наконец отшнуровывание периферических
камер кишечника для образования целома — все это одна в высо-
кой степени недоказуемая концепция периода увлечения гипотети-
ческими построениями в филогении. Она совершенно несовместима
с представлениями об эволюции Vermes от турбелляриеобразных
форм через немертинообразные к аннелидам. В свое время Циглер
(1898) уже указывал на это.
В.Заленский (1907) логически завершил взгляды Седживика, при-
нявши, как было указано выше, что немертины произошли от анне-
лид путем некоторой деградации их организации, а турбеллярий пред-
ставляют дальнейший и конечный этап в этом ряду регрессивной эво-
люции. Абсурдность подобного рода филогенеза была нами показана.
Если же допустить, что предками аннелид были паренхиматозные
черви, то ни у турбеллярий, ни у немертин нет ничего подобного
отшнуровыванию боковых выростов кишечника и нет биологических
предпосылок для такого процесса. Геккель, Гертвиги, Рей-Ланке-
стер, В. Заленский и др., наконец, Неф (1929) видели такую пред-
144
юсылку в том, что стенка кишечника у актиний^ содержит половые
продукты, и при отшнуровывании боковых его камер могли полу-
чаться трофические полости, защищенные от внешних воздействий
и используемые как вместилища для половых клеток. Но у турбел-
лярий и немертин гонады уже совершенно обособлены и в то же
время не обнаруживают каких-либо отношений к ветвям кишечника,
следовательно, нет никаких данных в этом направлении для образо-
вания энтероцеля.
Едва ли стоит пытаться представить себе происхождение целома,
как это делает нефроцель-теория, из разрастания концевой части
протонефридиев в виде экскреторной полости значительных разме-
ров и, конечно, также бесплодны будут попытки подойти к целому
как к органу газового обмена или фагоцитарному, хотя и та и
другая функция имеются в числе вторичных, побочных его функций.
Совершенно иначе приходится поставить вопрос о происхожде-
нии целома, если обратиться к той функции, которую мы назвали
основной — к его опорной функции. Хотя А. Ланг, например, и
упоминает о ней, тем не менее в попытках понять происхождение
целома она до недавнего времени полностью выпадала, ее засло-
няли вторичные функции, так или иначе особенно бросавшиеся в
глаза. Между тем опорная роль целома неразрывно связана с чрез-
вычайно важным фактом эмбрионального развития — целомиче-
ская мезодерма дает не только полость целома, но всегда также и
ту или иную часть прилежащей мускулатуры. Во взрослом состоя-
нии, как мы видели, приходится говорить о целомическом мешке и
прилежащих мышечных образованиях, как неразрывно связанных
друг с другом. Этот факт первостепенной важности обычно не учи-
тывался в должной мере при попытках разрешить вопрос о проис-
хождении целома. Слабым намеком звучит; «Мы, пожалуй, с боль-
шим правом могли бы предположить теорию мускульного про-
исхождения целомов... Мышцы составляют основную ткань целоми-
ческих мешков, судя по тому, что даже у тех целомических
животных, у которых целом как таковой почему-либо не появляется
(личинки асцидии), мышечная ткань все-таки существует»
(П. П. Иванов, 1937, стр. 20).
В недавнее время Нирштрас (1923) обратил внимание на зна-
чение оперной функции целома, в частности доходя до своеобраз-
ного представления о том, что следующие друг за другом мешки
Целома .аннелид являются расчлененным осевым скелетом, обуслов-
ливающим возможность независимого движения отдельных уча-
стков тела.
В 1933 г. я выступил с «миоцель-теорией» происхождения вторич-
ной полости тела, по которой целом представляет лишь полость в
Мускульной закладке, развившуюся у немертинообразных предков
Целомических животных; она заполнена жидким содержимым, по со-
левому составу изотоничным морской воде, а поэтому может играть
роль опорного образования для окружающей ее мускулатуры и
придавать твердость телу аннелиды. Таким образом, единство це-
10 Пути эволюции животного мира 145
лома и прилежащей мускулатуры является исходным моментом в
понимании его- происхождения — в мышечный зачаток диффунди-
рует жидкость из окружающих тканей, в конечном счете вода из
окружающей животное морской среды. Все вторичные функции
обусловлены или изменением химического состава целомической
жидкости, в связи с деятельностью прилежащих органов, а именно:,
трофическая функция в связи с деятельностью усваивающего отде-
ла кишечника и кровеносных сосудов/экскреторная в связи с жиз-
недеятельностью всех прилежащих ткйней, газового обмена в связи
с покровом. Или вторичные функции обусловлены проникновением
в целом клеточных образований — нефро-фагоцитарная функция
при попадании в него и дифференцировке амебоцитов разного
рода, половая — при попадании половых клеток, трофическая —
при дифференцировке элеоцитов и т. д., т. е. опять-таки целом здесь
играет роль своеобразного посредника, «медиатора» для прилежа-
щих клеточных образований.
Зачаток целома у аннелид представляет завершение эволюции,
идущей от турбеллярий через немертин. У тех и других уже имеется
закладка, происходящая за счет того же 4 d микромера, которая
дает и целомический комплекс аннелид, но эта закладка у паренхи-
матозных червей производит мускулатуру, не образуя чего-либо
подобного мешкам целома. Таким образом, можно считать основ-
ной функцией 4 d микромера в онтогенезе червей образование
мускулатуры. У поликлад, повидимому, за счет этого зачатка оора-
зуется также паренхима, а у немертин — своеобразная мезенхима,
некоторыми исследователями принимаемая за целенхиму. Однако
принимать за целом содержащие основное вещество пространства
немертин, конечно1, нет достаточных оснований. И у поликлад, и
у немертин перед нами, собственно, типичная, дающая мышцы мезен-
хима, совершенно такая же в этом отношении, как мезенхима, про-
исходящая в онтогенезе Vermes за счет 2-го и 3-го квартетов микро-
мер. Сериально все эти закладки совершенно одинаковы, и лишь у
аннелид 4 d дает еще целом.
Постулируя единство мезенхиматозных зачатков 2-го, 3-го и 4-го
квартетов, как дающих начало прежде всего мускулатуре тела,
естественно поставить вопрос, чем обусловлена дифференцировка
этого зачатка по разным квартетам. И можно видеть, что мезенхима
2-го квартета дает мышцы стенки тела, а именно1 — кольцевую му-
скулатуру аннелид, 3-го — глоточную мускулатуру — это эктомезен-
хима, то-есть происходящая за счет эктодермы и тесно связанная
с ее дериватами. Естественно, что 4-й квартет находится в ближай-
шей связи с образованием кишки. Об этом говорит и происхождение
энтеробластов из того же 4 d зачатка и развитие средней части ки-
шечника из макромеров 4-го квартета. Это — энтомезенхима, т. е.
связанная по происхождению с энтодермой. В дальнейшем рас-
сматриваемый зачаток дифференцируется в сравнительно слабо
развитую висцеральную и значительную продольную париеталь-
ную мускулатуру аннелид. Однако ставить формальную границу
146
между экто- и энтомезенхимой по происхождению от эктодермы или
энтодермы, как мы видим у Г. А. Шмидта (1951), не соответство-
вало бы существу образования мезенхимы вообще как скоплений
клеток, дающих начало мышечным и соединительнотканным эле-
ментам.
Так раскрывается эволюция целомической мезодермы аннелид
из первоначально чисто мезенхимной закладки уполиклад. Целом
же раскрывается как дифференцировка в прилежащей *к кишечнику
мускулатуре опорного образования в виде скопления жидкости, по-
ступающей путем диффузии из окружающих тканей, причем при-
лежащие клетки образуют выстилку целомического мешка. Пред-
полагать, что этому должно было предшествовать наличие уже ка-
кой-то полости с выработанной целотелиальной стенкой, нет ника-
ких оснований. Такая стенка должна была образоваться одновре-
менно и в теснейшей связи с образованием скопления жидкости в
мышечном зачатке. При этом первоначально должны были появить-
ся правый и левый мешки целома, функционировавшие как анта-
гонисты во время сокращений мускулатуры стенки тела связанных
с синусоидными движениями червя.
Последний вопрос, который связан неразрывно- с вопросом о
происхождении целома, касается факта метамерности целома анне-
лид. Почему целом образован парами следующих друг за другом
мешков? Ответ дают факты, констатированные у Palaeonemertini,
а именно: у Tubulanus (syn. Carinelld) имеются в мезенхиме захо-
ботковой части туловища поперечные септы, которые образованы
двойными пучками тонких преколлагенных волокон, идущими от
слоя ткани основного вещества под покровным эпителием через
слои мышц и внутреннюю мезенхиму до кишечника. Септы эти по-
разительно сходны с теми пограничными образованиями, которые
характерны для диссепиментов полихет. Эта дифференцировка, по-
видимому, связана у Tubulanus с возможностью обрывов длинного
тела червя и имеет место на несколько неопределенных расстояниях
по длине туловища. Подобную дифференцировку, производящую
впечатление настоящей метамерности, в свое время (1901) конста-
тировал у Carinina из Севастопольской бухты Т. Е. Тимофеев.
У аннелид она могла подвергнуться некоторому упорядочению в
связи с развитием параподий, т. е. в связи с особенностями их дви-
жения, и далее выразиться в подразделении развивавшихся цело-
мических полостей.
С этой точки зрения интересны взгляды П. П. Иванова (1912).
Он наблюдал, что первоначальная закладка целомической мезодер-
мы дифференцируется в связи с развитием передних пар параподий
на определенное число сомитов, щелом в которых появляется одно-
временно и полости которых не резко отграничены друг от друга.
Эти сомиты, образующиеся характерно для разных групп полихет
в количестве от 3 до 7, П. П. Иванов называет личиночными —
«ларвальными», так как они развиваются в личиночной стадии
Метатрохофоры. Дальнейшие сомиты образуются несколько позже,
10* 147
почему и названы им «постларвальными». Они закладываются в
' концевой зоне роста червя строго обособленно пара за парой .вне
зависимости от образования параподий. '
В одних случаях постларвальные сомиты, по П. П. Иванову, раз-
виваются за счет тех же телобластов, которые дают и личиночную
мезодерму (Aricia, Arenicold). В других случаях начало им дают
особо дифференцирующиеся зачатки, образующиеся при концевом
росте тела позади телобластических полосок (Spionidae, Serpuli-
dae), наконец, у Polygordius имеется только «постларвальная» те-
лобластическая закладка, а личиночные сомиты отсутствуют.
Такого рода факты дают повод П. П. Иванову к расчленению
единого зачатка целомической мезодермы на «личиночную» и «има-
гинальную».
Однако с нашей точки зрения на целомическую мезодерму как
лишь на особо дифференцированный в ряду прочих мезенхимный
зачаток, резкая грань между ларвальной и постларвальной мезо-
дермой, соответственно между различными способами развития ме-
зодермы, имеющимися у полихет, стирается. А именно — у Drilo-
inorpha вся мезодерма сосредоточена в телобластах, связанных с
4 d микромером, у Polygordius ларвальная мезодерма при этом
редуцируется. У Spiomorpha в 4 d микромере концентрированы толь-
ко начальные целомические зачатки — «ларвальные», тогда как
дальнейшее развитие «постларвальных» сомитов идет за счет ме-
зенхимных закладок терминальной зоны роста, у Lopadorhynchus,
повидимому, вся мезодерма происходит за счет этих последних.
Припомним, наконец, и тот факт, что у Nereis и некоторых других
форм роль 4 d играет 3 d микромер.
Таким образом, у полихет процесс образования мезодермы до
некоторой степени еще не стабилизирован и может идти разными
путями. В некоторых случаях (Spiomorpha) резко обособляется
зародышевая закладка «ларвальных» сомитов с дифференцировкой
особого мезодермального их зачатка. В других случаях (Drilo-
morpha) такого особого зачатка не выделяется, те и другие про-
исходят от общего зачатка — «ларвальные» сомиты обособлены от |
«постларвальных» только тем, что их дифференцировка происходит I
после некоторого перерыва. В третьих, как у Lopadorhynchus, I
сглажен и такой перерыв, вся целомическая мезодерма развивается
из мезенхимного зачатка.
В известной мере это связано с условиями жизни животного, в
частности, с переходом лрчинки от пелагической жизни к донному
существованию, характерному для взрослых полихет и типовому
вообще для червей. Пелагическая личиночная стадия является вто-
ричной вставкой, обеспечивающей питание и развитие зародыша до
достижения окончательного состояния донного червя.- Целомический
же комплекс аннелид мог выработаться только у донной формы с
ее синусоидными движениями по грунту и с дальнейшей опорой на
параподиальные выросты. Лишь вторично появляется ранняя диф-
ференцировка хетоподий и зачатков мезодермы и в связи с этим
148
развитие «ларвальных» сомитов уже у нектохетьт Это ведет в конце
концов к обособлению личиночной и имагинальной мезодермы с
дифференцировкой соответствующих зачатков и с обособлением
«ларвальных» и «постларвальных» сомитов у ряда форм полихет. У
Lopadorhynchus, ведущего пелагический образ жизни, эта разница
стирается, исчезает.
В дальнейшем мы еще вернемся к вопросу о происхождении
сомитов целома при анализе организации других типовых групп
животного мира. Здесь подчеркнем еще раз, что для полихет целом
в силу многообразия его
функций и взаимоотноше-
ний с другими системами
органов стал ведущей
чертой организации всей
группы.
Прежде всего, как на-
стоящая антитеза разви-
вающемуся целому диф-
ференцируется в значи-
тельной ее части система
пограничных образований
аннелид (рис. 47, 50), ко-
торая представляет типо-
Рис. 50. Схемы взаимоотношений мезенхи-
мы, целома и кровеносной системы у ис-
ходных форм аннелид (по Э. Мейеру):
Л — фронтальный и Б — поперечный раз-
рез ьв;
1 — мешки целома, 2 — слой мезенхимы, 3 —
стенка кишки, 4 — кожно-мышечный мешок, 5 —
кровеносные образования на кишке, 6 — комис-
суральные сосуды
вую систему ткани основ-
ного вещества, заполняю-
щую и связывающую раз-
личные компоненты тела
механически в единое не-
разрывное целое. Ее пери-
ферический отдел, зало-
женный в стенке тела,
субэпителиальная и по-
граничные перепонки кольцевых мышц уже были рассмотрены выше;
центральная часть, соответствующая внутренней мезенхиме немер-
тин, вгоняется в определенные границы с появлением парных меш-
ков целома. Эти мешки вместе с прилежащей мускулатурой окру-
жены всюду субмускулярной пограничной перепонкой, получившей
свое название от того, что целомическая мускулатура всегда ею
подстилается снаружи. На местах диссепиментов и мезентериев
субмускулярная перепонка переходит в пограничные перепонки
этих последних, переходящие, в свою очередь, в перепонки мышеч-
ных пучков кишечника. Эта единая система завершается субэпите-
лиальной перепонкой кишечника.
Обычным приходится считать отсутствие пограничной перепонки
под целотелием там, где за ним следует целомическая мускулатура,
т- е. на продольных мышечных лентах стенки тела и на кишке. Од-
нако вторично пограничные образования в той или иной мере про-
никают в комплекс целома, т. е. в продольные мышечные ленты
149
от субмускулярной перепонки, и от участка диссепиментальных пе-
репонок, пронизывающих продольную мускулатуру. Таким обра-
зом, в конце концов, эта последняя может быть, как например у
Capitellidae, разбита полностью на отдельные пучки, из которых
каждый заключен в чехол из пограничной перепонки, и подобная
перепонка оказывается подстилающей даже целотелий. Конечно, это
совершенно иные отношения, чем вторичное усиление продольной
мускулатуры Serpulidae или Lumbricidae, где из первоначально тон-
кого мышечного слоя, благодаря развитию простых или ветвящих-
ся складок этого' слоя, получается мощная мускулатура. Соответ-
ственно развивается и сложная система пограничных пластинок,
проходящих по оси этих складок и образующихся за счет типичной
субмускулярной перепонки.
Итак, система пограничных образований полихет представляет в
значительной мере измененную появлением целома и целомической
мускулатуры заполняющую ткань основного вещества паренхима-
тозных червей, и ее отношения с Другими органами с этой точки
зрения ясны и закономерны.
В свою очередь, как мы видели у немертин, с тканью основного
вещества теснейшим образом связано п о я в лен и е к р ове н о с но и си сте -
мы, и развитие пограничных образований у полихет также опрёде-
ТТяет, ставит в узкие рамки ход и взаимоотношения кровеносных со-
судов (рис. 50). Вторично в' различных группах полихет в связи с
усложнением кровеносной системы изменяются отношения сосудов
к целомическому комплексу — с сосудами или, вернее, под видом
сосудов, пограничные образования могут глубоко проникать в це-
лом. В основном же через посредство пограничных образований
ведущему комплексу целома у аннелид подчиняется кровеносная
система, вырабатывается ее аннелидный тип (рис. 48,В; 42,Д; 47).
Последний отличается одной примитивной чертой строения со-
судов: их стенка, как и у немертин, не имеет других постоян-
ных компонентов, кроме интимы, образуемой пограничной пере-
понкой ткани основного вещества. Для сосуда, поэтому, являет-
ся существенным не столько стенка, сколько содержимое — жидкая
ткань основного вещества — кровь, в противоположность жидко-
сти целома, представляющей транссудат из окружающих тка-
ней.
Дальнейшим, местным осложнением стенки сосуда, стоящим уже
в связи с выработкой сократительных отделов кровеносной системы,
является образование на ней мышечных волокон за счет приле-
жащих клеток целомического комплекса. Таким образом, к первич-
ному аппарату, который, суживая его значение, М. Фернандес
(1907) называет проводящим, прибавляется со стороны целома вто-
ричный, «пропульзаторный» аппарат совершенно иного про-
исхождения.
Анатомически аннелидный тип кровеносной системы опреде-
ляется следующими характерными чертами. Центральным отделом
является, в связи с основной трофической функцией кровеносной
150
системы, сплетение сосудов в субэпителиальной перепонке усваи-
вающей кишки, в некоторых случаях переходящее даже в сплошной
кровеносный кишечный синус. Здесь кровь получает питательные
вещества от клеток усваивающего эпителия. Этот отдел отсутствует
у немертин, так как у них еще нет непосредственного поступления,
аккумуляции питательных веществ из стенки кишечника кровенос-
ной системой. При сохранении массы внутренней мезенхимы,
прилежащей к кишке, основное вещество здесь является еще по-
средником при проведении питательных веществ от кишки к со-
судам.
Однако и собственно проводящую часть кровеносной системы —
продольные сосуды, снабжающие все тело, и комиссуральные сосу-
ды, снабжающие отдельные метамеры у аннелид едва ли можно
непосредственно сравнить с одноименными сосудами немертин. Как
показывает развитие продольных сосудов, и спинной и брюшной со-
суды представляют обособляющиеся части кишечного сплетения,
проникающие в спинной и брюшной мезентерии и здесь развиваю-
щиеся как главные коллекторы и регуляторы движения крови.
Последняя функция особенно сильно проявляется на спинном сосу-
де, часто развивающем продвигающий мышечный аппарат за счет
элементов правого и левого целомических мешков. Однако дру-
гие сосуды также могут дифференцироваться в этом направ-
лении. Повидимому, и вообще развитие кольцевой мускулатуры на
кишечнике в значительной мере служит для продвижения крови,
так как сокращения этой мускулатуры идут антиперистальтически;
вероятно, здесь перед нами первичный пропульзаторный аппарат
полихет.
На первоначальное, неметамерное строение кровеносной системы
налагается ангиомерия, обусловленная метамерностью целомических
и параподиальных зачатков. Первые расположением диссепиментов
определяют ход комиссуральных сосудов, необходимых для подачи
крови к нуждающейся в питании стенке тела и, позже, к сосудам,
врастающим в целом и в прилежащие органы. Вторые влияют раз-
витием специальных периферических сосудов параподиального
комплекса, связанных с теми же комиссуральными сосудами. Си-
стема всех периферических сосудов, развиваясь во> взаимосвязи
с развитием различнейших органов, является крайне разнооб-
разно построенным отделом с собственно трофической функ-
цией.
У ряда семейств полихет (Phyllodocemorpha) только эта первич-
ная функция обусловливает строение кровеносной системы. Однако
в связи с развитием периферического отдела ее в параподиях, она
У многих форм используется функции газового обмена.
В жидкости крови появляются дыхательные пигменты (гемоглобин
и др.), первоначально (у Phyllodocemorpha и немертин) располагав-
шиеся в нервной системе и обслуживавшие только нервную ткань.
Ааким образом усиливаются окислительные процессы во всех систе-
мах органов. Далее разделение физиологического труда ведет к
151
дифференцировке определенных участков в виде выростов жабер,
дорзально около параподий, там, где условия особо благоприятны
по движению воды для диффузии газов через покров. Обычно жаб-
ры, как выше уже было упомянуто, наравне с параподиальным вы-
ростом, хэтоподиями и усиками считаются входящими в состав
подиального комплекса, настолько они характерны для полихет
(рис. 42, Д; 47; 42,А).
Так впервые среди червей происходит дифференцировка органов
дыхания, основанная на усилении функции газового обмена, что
можно рассматривать как широко адаптивную черту организации,
значительно повышающую возможности организма в борьбе за
существование. Это один из редких, но типичных примеров саромор-
фозов» по А. Н. Северцову. Для нас это частный случай широких
адаптаций.
Обычны для полихет гребневидные жабры, располагающиеся на
сомитах туловища при основании нотоподия дорзально (рис. 42, А).
Они представляют выросты стенки тела, в которые заходит цен-
трально целом с дистальным участком петли комиссурального сосу-
да. Нисходящая и восходящая ветви этой петли в большинстве
случаев соединяются дугами капилляров, уже проникающих с по-
граничными пластинками в самый эпителий жабры и располагаю-
щихся здесь периферически близ покровной кутикулы, т. е. в усло-
виях, наиболее благоприятных для газового обмена с окружающей
водной средой (рис. -47, Б).
Жабры полихет обнаруживают значительное разнообразие даль-
нейших узкоадаптивных изменений в связи с приспособлением к
различнейшим биоценотическим отношениям. Они очень часто под-
вергаются большей или меньшей степени редукции при переходе к
жизни в грунте, где чисто механически оказываются мало приемле-
мыми. При жизни в трубках сохраняются и сильно развиваются
лишь жабры, расположенные на самых передних сомитах, где они
могут свободно функционировать, выставляясь из трубки в окру-
жающую воду (рис. 44). У сидячих форм, например у Serpnloidea,
они могут подвергаться редукции в связи с тем, что другие придат-
ки головы, вероятно пальпы, обусловливающие приток воды с пище-
выми частицами к ротовому отверстию, чрезмерно развиваются и
принимают на себя функцию газового обмена — откуда и само
название для этих придатков серпулид «жабры». Наконец, редукция
жабр у полихет может быть связана в некоторых случаях, например
у Capitellidae, с расширением функций целома вообще и с перехо-
дом функции газового обмена к целому в частности. Его сильное
развитие и вторичное проникновение далеко в стенку тела отра-
жается на кровеносной системе, обусловливая ее болею слабое раз-
витие и редукцию, а соответственно и исчезновение жабер. С другой
стороны, у некоторых форм (Dasybranchas) в связи с этим происхо-
дит и новообразование целомических жабер, т. ё. образование выро-
стов стенки тела у параподия, заключающих целом и специализи-
рующихся для газового обмена.
152
Это явление нельзя смешивать с развитием целомических жабер
у некоторых полихет (например, Nephtys), которые в своей эволю-
ции не дошли до выработки типичных жабер, а пошли по совершен-
но иному пути образования выростов на параподиях, заключающих
в своем целоме обильные петли кровеносных сосудов. Невидимому,
у Clyceridae, при слабом развитии кровеносной системы вообще,
произошла также самостоятельно выработка целомических жабер,
лишенных кровеносных сосудов.
Если в развитий жабер несомненно видно очень, значительное
влияние узких приспособлений, то относительно развития кровенос-
ной системы в различных группах аннелид приходится'отметить ее
бескод^цнре, разнообразие в деталях. Но здесь на первый план
выступает уже момент ее чрезвычайно тесной связи с различнейши-
ми., системами органов, включая васкуляризацию покрова, нервной
системы, целома и даже стенок самих крупных сосудов. Однако к
своеобразнейшим проявлениям приходится отнести и обратное явле-
ние уже упомянутой редукции кровеносной системы, находящее свое
завершение во вторичном ее исчезновении, например у Polycirrus
из теребеллид.
Наконец, в соответствии с эволюцией целома и кровеносной си-
стемы идет развитие полового аппарата полихет (рис. 46, Б; 42, Д).
Как было выше указано, гонады, располагающиеся во внутренней
мезенхиме тела немертин, с вытеснением этой последней целомом,
оказываются приходящими в соприкосновение с целомическими
мешками и обычно у полихет вдаются в них в виде выростов, прикры-
тых целотелием или вторично лишенных этого покрывающего слоя.
Таким образом, на всем протяжении тела, где находится усваиваю-
щая часть кишечника, начинает развиваться половая функция цело-
ма, которая осложняется дальнейшим выпаданием половых клеток
в полость целома. Здесь они находят подходящую среду в отноше-
нии их питания и сохранения. Первоначально функция питания
гонад тесно связана с кровеносной системой; располагаясь в целоме,
они развиваются на тех или иных сосудах, но в дальнейшей жизни
половых элементов эту роль полностью берет на себя жидкость це-
лома. Своего наивысшего выражения функция питания половых
элементов достигает в образовании массы элеоцитов, периодически
развивающихся за счет амебоцитов целома и отдающих затем свои
питательные запасы созревающим яйцевым клеткам.
Своеобразное разделение физиологического труда обусловли-
вает, наконец, дифференцировку сомитов туловища аннелид, сна-
чала равномерно снабженных гонадами, на сомиты с гонадами и
на „лишенные гонад, т. е. вторично их утратившие. Здесь на пер-
Вь1й план выступают соотношения половой и экскреторной функ-
ций целома.
Примитивный типовой экскреторный аппарат в виде протонефри-
диев, в количестве одной или двух пар, сначала появляется в раз-
витии полихет у личинки, трохофоры, вне какой бы то ни было
связи с целомическим зачатком (рис. 51). Но затемв связи с образо-
153
ванием целома, во-первых, развиваются новые пары нефридиев
соответственно развитию каждой метамеры, т. е. экскреторный
аппарат получает метамерное строение (рис. 48, А), а личиночные
экскреторные органы редуцируются; во-вторых, нефридии начинают
вдаваться в целом и образуют в нем извитые трубки, покрытые
целотелием (рис. 46, Б). Концевые отделы, соответствующие клет-
кам с мерцательным пламенам,
Рис. 51. Личинка Psygmobranchus
сбоку (полусхема по Э. Мейеру):
1 — зачаток головного мозга, 2 — глаз,
3 — железистый зачаток, 4 — кишечник,
5 — мезенхимные мышцы, 6 — закладка
брюшной цепочки, 7 — протонефридий, 8—
глотка, 9 — мезодермальный телобласт,
10 — мезодермальные сомиты, 11 — за-
кладка щетинковых мешков, 12—протрох,
13 — аданальная железка
у ряда семейств полихет, неви-
димому вторично, прямо' омы-
ваются целомической жидко-
стью, наподобие того как у не-
мертин они вдаются в просвет
кровеносных сосудов. Наблю-
дается лишь характерная спе-
циализация клеток с мерца-
тельным пламенем в длинные
трубки «соленоцитов», специ-
Рис. 52. Схема нефромиксия Phyllo-
ci о се:
1 — целомическая воронка, 2 — протоне-
фридий с соленоцитами (3)
фичных для полихет (рис. 52). Следовательно, примитивный экскре-
торный аппарат типа протонефридиев вступает в теснейшие отноше-
ния с ведущей системой аннелид — с целомом, и дальнейшая его
эволюция всецело связана с ним же.
Это взаимообусловленное развитие идет в разных направле-
ниях — и по линии осморегуляции, и по линии половой функции.
Так, у Phyllodocidae, например, мерцательные воронки целома,
выработавшиеся для выведения половых продуктов, открываются в
проток протонефридия, образуя так называемые нефромиксии
(рис. 52). Но обычно происходит редукция осморегулирующего
аппарата соленоцитов. Проток нефридия открывается прямо в по-
лость целома и получается открытый нефридий (рис. 46, Б), могу-
щий непосредственно выводить жидкость из целома наружу. У неко-
торых форм, например Protodrilus, в проксимальном участке нефри-
дия клетки просто расходятся, и получается отверстие, прикрытое
154
одной клеткой. У Nereis этот участок расширяется в виде неболь-
шой воронки. У большинства полихет проксимальный конец откры-
того нефридия переходит в образованную целомическим эпителием
мерцательную воронку, особенно мощно развитую в сомитах, содер-
жащих половые продукты, причем длина самого нефридиального
протока здесь может значительно уменьшаться (рис. 42, Д). Таким
образом получаются типичные для полихет метанефридии с глав-
ной функцией выведения половых продуктов из целома. При даль-
нейшей дифференцировке, как было выше отмечено, ведущей у не-
которых семейств, например у Terebellidae, к выработке стериль-
ного переднего, «предиафрагмального» целома с экскреторной
функцией и «постдиафрагмального» с половой, тип метанефридия
сохраняется. Лишь мерцательные воронки в переднем отделе тела
являются небольшими по размерам, а экскреторный отдел нефри-
диев развивается чрезмерно.
Итак, эволюция экскреторного аппарата полихет проходит
всецело под знаком приспособления к разнообразным функциям це-
лома. Особым направлением ее, наконец, будет сравнительно редко
наблюдающаяся тесная связь с цилио-фагоцитарными образования-
ми целома. Последние развиваются за счет мерцательных участков
в целотели, причем трудно сказать, представляют ли мерцатель-
ные'участки новообразование, или перед нами сохранение прими-
тивных отношений, вроде имевших место в выводных протоках го-
над немертинообразных предков. Мерцательные участки целома,
как мы видели, превращаются в широкие воронки для выведения
половых продуктов. Но у ряда форм они взяли на себя другую
функцию, а именно функцию движения содержимого целома, что
достигает своего завершения в правильной циркуляции целоми-
ческой жидкости у Polynoidae или Nephthydidae. В других случаях
это движение специализировано для накопления в определенных
отделах целома фагоцитирующих клеток, что ведет затем к обособ-
лению мешков пилио-фагоцитарных органов, как имеется, напри-
мер, у Glycera. Эти образования становятся в ближайшие отноше-
ния к нефридиям, чем облегчается удаление из организма получаю-
щихся при фагоцитозе продуктов. Вероятность такого своеобразного
пути эволюции на почве использования мерцательного эпителия
подтверждается отсутствием у дох семейств полихет, где развиваются
Цилиофагоцитарные органы, каких-либо выводных протоков гонад
и выведением половых продуктов при этом наружу прямо через
Разрыв стенки тела.
Закончив рассмотрение широко адаптивного, ведущего в органи-
зации аннелид, комплекса целома и связанных с ним систем орга-
нов, переходим к третьему комплексу, связанному^ более совершен-
ной ориентировкой полихет в окружающей среде, что для свободно-
подвижных животных, особенно хищников, имеет, конечно, большое
значение в борьбе за существование. В соответствии с этим у
n-rrantia прогрессивно развиваются органы чувств. В головном их
наборе (рис. 42, 46, 48), унаследованном от немертинообразных
155
предков, механорецепторы с развитием длинных антенн обычно в
количестве от 5 до 1, превращаются в известной мере из ближних
в дальние, «дистантные», органы осязания. Далее, перед ротовым
отверстием дифференцируется пара пальп как специальных хемо-
рецепторов, достигающих у Terebelioidea и Serpuloidea в их аппара-
тах «щупалец» и «жабер» кранного развития. Затылочные органы
полихет едва ли можно считать далеко ушедшими от состояния,
свойственного немертинам, хотя сложно построенные карункулы
Arriphynomidae и представляют далеко зашедшую крайнюю диф-
ференцировку. Зато глаза у свободноподвижных полихет, особенно
у пелагических, как например у Alciopidae, развиваются с образова-
нием диоптрического аппарата в органы картинного зрения,
у Eunicidae это еще довольно простые ямочные глаза, но у Alciope
вырабатываются уже хрусталик и стекловидное тело, а ретина
дифференцируется на главную и придаточную части. Однако с пере-
ходом к жизни в трубках и особенно в грунте происходит редук-
ция фоторецепторов при ненужности картинного зрения в этих
условиях существования. И в то же время имеет место распростра-
нение очень простых глазков не только по голове и ее придаткам,
но и по туловищу и даже (Fabricia sabella) на пигидии, чем
обусловливается широкая общая чувствительность к изменениям
освещения, важная при сидячей жизни как защитная реакция.
В соответствии с этим головной мозг у Errantia дифференци-
руется в три хорошо выраженных отдела (рис. 48): передний —>
пальпарпый, средний — антеннально-глазной и задний — затылоч-
ный. Из них в первом и последнем имеются особые участки нервных
контактов, так называемые гломерулы, впервые здесь дифференци-
рующиеся, а пальпарный отдел дает начало мощным ассоциатив-
ным центрам с мелкими ганглиозными элементами, так называемые
глобулы: Подобные образования наблюдались уже и у немертин.
Таким образом головкой мозг на типовой основе мощного сензор-
кого центра пргрессивно эволюционирует у полихет как в связи с
развитием более сложных органов чувств в головном комплексе, так
и по пути интеграции деятельности организма с развитием глобул.
Приспособительные процессы у трубкоживущих полихет, резко
выражающиеся в редукции и своеобразном развитии органов
чувств, также сказывается и в дифференцировке головного мозга
значительным его упрощение^м — исчезает выраженность трех отде-
лов, редуцируются глобулы.
Также «функционально» развивается у полихет и центральная
нервная система метамерного туловища (рис. 48, А и В; 42, Д). На
основе пары продольных моторных стволов немертин создается
брюшная нервная цепочка аннелид. В_связи с развитием боковых
выростов параподий боковые стволы умещаются вентрально и всту-
пают в ближайшую связь друг с другом через систему поперечных
комиссур, причем интеграция их выражается в сближении обоих
стволов. Дифференцировка центральной волокнистой массы и распо-
ложение ганглиозных клеток на ней в две группы — епинную и
156
брюшную — сохраняются такими же, как у немертинообразных пред-
ков. Часто также встречается и примитивное расположение брюш-
ной цепочки в покровном эпителии, как это имеет место у Errantia;
однако имеются все переходы к нахождению ее глубоко в целоме,
как, например, у Capitellidae, в результате адаптивного' процесса
перемещения в более защищенное положение.
Дифференцировка в брюшной цепочке идет в связи с дифферен-
цировкой мощного среднего, чувствительного нерва сомита
(рис. 42, который развивается за счет нервных, волокон от
чувствительных почек, расположенных в покрове посредине кольца
сомита и за счет чувствительных спинного и брюшного усиков каж-
дого параподия. Эти усики, входящие в качестве непременной со-
ставной части в состав параподиального комплекса, обладают как
осязательный функцией, так и функцией восприятия химических
раздражений и развиваются аналогично антеннам головы в своеоб-
разные дистантные рецепторы. При основании параподия на чув-
ствительном нерве сомита, в связи с поступлением в нерв волокон
из различных частей параподия, дифференцируется обычно значи-
тельный сензорный, так называемый пар апо ди альный ганглий. У ме-
ста вхождения среднего, чувствительного нерва в брюшную цепочку
концентрируются ганглиозные элементы последней. Соответственно
дифференцируются ганглии и коннективы брюшной цепочки
(рис. 48, А. и В). Отходящие от нее к мускулатуре стенки тела
двигательные нервы пробегают в виде двух кольцевых нервов,
располагающихся у передней и задней границ сомита, т. е. там, где
оставляет им место развившийся с боков тела параподиальный
комплекс. Так, в теснейшей связи, с одной стороны, с органами
чувств, а с другой, — с развитием мускулатуры, создается аннелид-
ный тип центральной нервной системы — брюшная цепочка и типич-
ный певросомит аннелид.
В нервной системе полихет пути дифференцировки и специализа-
ции тестю объединяются с путями интеграции. Своеобразным край-
ним выражением последних является описанное О. Шторхом
(1912) явление тетранейрии у Amphinomidae. Здесь параподиальные
ганглии соединяются друг с другом мощными продольными пучками
нервных волокон, образующих как бы подобные стволам брюшной
Цепочки боковые нервные стволы. Однако нельзя принять их вместе
с О. Шторхом за такую же часть центральной нервной системы,
как и стволы брюшной цепочки с ее коннективами и ганглиями.
Нельзя принять потому, что, во-первых, боковые нервные тяжи рас-
полагаются на значительной части их хода в основаниях параподий,
как. своеобразные местные нервы, и, во-вторых, как сензорные обра-
зования они имеют иной характер, чем моторные стволы брюшной
Цепочки. Против утверждения Шторха говорит также то, что он
искусственно разрывает имеющийся у Amphinomidae единый типич-
ный средний, чувствительный нерв сомита па две части, а именно:
па собственно чувствительный нерв параподия, вступающий в па-
раподиальный ганглий, и на комиссуру между этим последним
157
и брюшной цепочкой, считая ее лишь за часть центральной нервной
системы. Прямым выводом из тетранейрии Amphinomidae является,
по Шторху, особая примитивность организации этого семейства —
что признать однако нет веских оснований. Совершенно определен-
ные филогенетические связи, которые нами были отмечены при ана-
лизе организации турбеллярий, немертин и аннелид, заставляют
считать предковыми формами аннелид немертинообразные организ-
мы г у которых в центральной нервной системе туловища уже стойко
выработалась всего одна пара продольных моторных стволов. А
поэтому и встречающаяся среди аннелид только у Amphinomidae
тетранейрия представляет исключительно свойственное им выраже-
ние эволюционного пути наравне с тетранейрией моллюсков, о ко-
торой нам еще придется говорить в дальнейшем.
В заключение обратимся к нерассмотренной еще нами пище-
варительной системе аннелид (рис. 48). Она имеет типовые черты
трубчатого кишечника с хорошо развитым стомодеальным отделом
и незначительным — задним проктодеальным и относительно мало
затронута общим повышением организации аннелид.
В соответствии с выработкой метамерии у Errantia на усваиваю-
щем, энтодермальном, отделе кишечника появляются метамерные
расширения в бока, достигающие у Aphroditidae своего крайнего
развития в виде длинных метамерных придатков. Однако, с другой
стороны, при вторичной редукции диссепиментов эти расширения
сглаживаются.
Гораздо сильнее выражены явления узкой, а иногда и чрезмер-
ной адаптации в глоточном отделе кишечника, в связи с использо-
ванием различнейшего пищевого материала. В основном тут намети-
лось два пути — один путь ведет к выработке мускулистой трубча-
той глотки, имеющей вид хобота, выдвигающейся наружу и снаб-
женной обычно кутикулярными зубцами челюстей (рис. 48 и 46, А).
Это путь большинства Errantia, путь хищников, приводящий к
чрезмерному развитию. Например у Glycera, глотка, почти равня-
ясь длине тела животного, выворачивается наподобие хобота немер-
тин. Другой путь ведет к образованию глоточного мешка, открываю-
щегося в глотку вентрально. Среди Errantia это присуще только
Eunicidae, у которых имеется мускулистый и иногда очень длинный
мешок, снабженный мощным аппаратом кутикулярных челюстей
(рис. 42, Б и В), что связано с хищным образом жизни. Однако у
большинства сидячих полихет глоточный мешок оказывается годным
для захватывания пищи иного рода, представляя из себя подобие
ковша для забирания мелкого пищевого материала или, у роющих
форм, даже для заглатывания ила. У трубкоживущих происходит
переход к пассивному питанию. Животное, приспосабляясь к исполь-
зованию в качестве пищевого материала мельчайших организмов,
вырабатывает за счет пальп придатки, снабженные мерцательным
эпителием. С током воды, вызываемым этим мерцательным аппара-
том, пища несется к ротовому отверстию. Крайним приспособлением
такого рода являются многочисленные длинные, тянущиеся во все
158
стороны от трубки по грунту щупальца теребеллид (рис. 44), имею-
щие вентрально мерцательную борозду, по которой и идет ток воды
к ротовому отверстию. Глоточный мешок, теряя аппарат челюстей,
сохраняется у них уже в редуцированном виде. У Serpuloidea раз-
вивается за счет пальп аппарат ветвистых «жабер», который также
несет пищевые частицы с током воды ко рту. Глоточный мешок
подвергается при этом полной редукции.
Не будем дальше вдаваться в частности. Разнообразие приспо-
соблений в глоточном аппарате полихет выражено чрезвычайно
сильно и может заходить чрезвычайно далеко,
Подводя итоги рассмотрению организации аннелид, ее можно
характеризовать многогранностью, многообразием тех путей, по ко-
торым идет узкая или даже крайне узкая приспособленность. Здесь
нет одного, чрезмерно развитого образования наподобие хобота не-
мертин, у последних определившего почти все разнообразие приспо-
соблений. У полихет различнейшие черты их организации эволюцио-
нируют по своим путям, и в связи с этим для всей группы остаются
открытыми различнейшие возможности повышения организации. Это
находит свое яркое выражение и в многочисленности происходящих
от полихет групп аннелид и в происхождении от них же высшего
по организации типа членистоногих. Полихеты становятся одной из
важнейших узловых групп животного царства.
Анализ организации полихет позволил отметить в ней несколько
комплексов прогрессивно эволюционирующих, широко адаптивных
черт, связанных с образом жизни хищного животного, с его защи-
той и нападением.
Первый комплекс связан с усложнением движения тела. Он
выражается в появлении метамерно расположенных по бокам те-
ла примитивных конечностей типа простого рычага — движение
посредством локомоторных придатков идет на смену движению
исключительно путем сокращения всего тела. Однако в параподиях
полихет всегда налицо и узко приспособительная черта — наличие
аппарата кутикулярных щетинок, значительно совершенствующих
действие параподий, но в конце концов доводящих их до состояния
крайней узости приспособлений, особенно при использовании щети-
нок в качестве защитных приспособлений (Aphroditidae, Amphicte-
nidae).
В связи с появлением параподий стоит отделение спинных
продольных мышц от брюшных и соответственная дифференцировка
мускулатуры конечностей, выражающаяся в образовании парапо-
Диальных и щетинковых мышечных пучков за счет, главным обра-
зом. кольцевой мускулатуры стенки тела.
Наличие параподиальных выростов, как подвижных, наиболее
выдающихся пунктов тела, используется для ориентировки во
внешней среде — развиваются спинной щ брюшной усики, и для
базового обмена появляются ветвистые выросты жабер. Таким
образом создается сложный комплекс подия, в котором развитие
спинных усиков, как образований, используемых вторично для за щи-
159
ты животного (пластинки Phyllodocidae, элитры Aphroditidae),
может достигнуть крайних степеней адаптивного развития.
С рассматриваемым комплексом связано образование опорной
системы полихет. Это система целома. Она далее широко исполь-
зуется с различнейших сторон и таит в себе массу возможностей.
Комплекс целома и связанных с ним образований является вто-
рым в нашем рассмотрении, но по свому значению он оказывается
в организме полихет ведущим, можно сказать, создавшим организа-
цию аннелид.
Целом, образованный в соответствии с параподиями рядом пар-
ных мешков, обусловливает внутреннюю метамерию тела полихет.
Во внешней морфологии с ним объединяются развитие параподий
и выработка межсомитных борозд, связанная с кутикуляризацией
покрова. Таким образом получается внешнее расчленение тела на
метамерные сомиты и дифференцировка неметамерных головной
лопасти и пигидия.
Во внутренней организации образование целома ведет к своеоб-
разной дифференцировке целомической мускулатуры, т. е. прежде
всего к образованию пары спинных и пары брюшных продольных
мышечных лент. Далее оно ведет к развитию системы пограничных
образований, которая получается в результате вытеснения целомом
и целомической мускулатурой ткани основного вещества, сводящей-
ся на пластинки, тяжи и волокна. Целом же обусловливает выра-
ботку особого типа кровеносной системы, которая, как система
полостей в ткани основного вещества, вгоняется в рамки распределе-
ния пограничных образований, но, вместе с тем, осложняется с
выработкой функции газового обмена. Целом вызывает метамерное
развитие экскреторной системы протонефридиального типа и в
дальнейшем замену ее открытыми нефридиями. Гонады также всту-
пают в теснейшую связь с целомом, который делается вместилищем
половых продуктов, а с развитием элеоцитов используется для ро-
ста и созревания яиц. В конечном итоге это ведет к использованию
нефридиев для выведения половых продуктов путем присоединения
к нефридию половой воронки, т. е. путем образования нефромиксиев
и метанефридиев. Наконец, с целомом связана выработка цилио-
фагоцитарных образований.
Однако все эти системы, вступающие в ближайшие взаимоотно-
шения с целомом, сохраняют во многих отношениях примитивные
черты. Пограничные образования обычно во взрослом состоянии не
содержат клеточных элементов, кровеносные .сосуды имеют основ-
ным слоем интиму из пограничной пластинки основного вещества,
гонады представляются примитивными скоплениями половых кле-
ток. Более дифференцирована система нефридиев в связи с ослож-
нением ее функций. К описываемому комплексу следует отнести и
еще одну дифференцировку — а именно развитие в сомите системы
двух двигательных нервов — переднего и заднего кольцевого.
Третий комплекс адаптивного характера развивается в связи с
более совершенной ориентировкой полихет в окружающей среде и
160
их подвижностью, выражается он прежде всего в прогрессивном
развитии органов чувств: дифференцируются хеморёцепторы—паль-
пы, механорецепторы —• антенны. Осложняются глаза с развитием
сложного диоптрического аппарата или, не усложняясь, а иногда
даже упрощаясь, распространяются широко по головной лопа-
сти, туловищу и пигидию. Чрезмерное развитие пальп у Seden-
.taria в связи с присоединением функции улавливания пищи заменяет
их сначала широкоадаптивное развитие. Соответственно усложняет-
ся строение головного мозга, особенно с развитием мощных скопле-
ний ассоциативных элементов и выработкой сложных проводящих
путей. В метамерах к этому комплексу относится уже выше рассмо-
тренная дифференцировка органов чувств параподиального комплек-
са — спинных и брюшных усиков. В связи с этим развивается си-
стема чувствительного среднего нерва сомита. В результате, как мы
видели, получается характерный тип невросомита полихет, разви-
вающийся функционально на основе нервных сплетений и двух
продольных стволов нервной системы немертинообразных червей.
Четвертый комплекс адаптаций выражается в приспособлениях
защитного характера: а именно, в выработке кутикулярного покро-
ва на смену ресничному покрову низших червей и в дифференциров-
ке железистых участков покрова, служащих для образования защит-
ных трубок. Последнее при дальнейшей эволюции прикрепленного
существования переходит уже в крайнюю адаптацию сидячих форм
Наконец, остается указать, что мало затронут общим повышени-
ем организации кишечник, дающий лишь иногда в энтодермальном
отделе метамерные выросты и представляющий ряд узких и крайне
узких приспособлений в глоточном своем отделе в связи с использо-
ванием различного пищевого материала.
В заключение нельзя не. сказать несколько слов об общем плане
построения аннелид, а вместе с тем и всего типа червей.
Мы видели в турбелляриях, немертинах и аннелидах генетиче-
ский ряд последовательных этапов эволюции придонньдх ползающих
форм, и анализ организации был в полной гармонии с этим взгля-
дом. Характерный аппарат параподий у аннелид мог развиться,
конечно, только как приспособление к донным условиям существо-
вания, к движению по грунту ползающего животного с синусоидны-
ми извивами тела. Он не мог бы развиться у пелагического взрос-
лого организма вроде трохофоры — у трохозоона — проблемати-
ческого предка высших червей по Гатчеку. И мы принимали, что
личиночные формы выработались как «вставочные» стадии в онтоге-
??езе донных организмов, обеспечивавшие питание зародыша в более
благоприятных для него пелагических условиях существования.
Исходными формами для первичных червей были гастрееподоб-
НЬ1е> свободно жившие предки, которые с выработкой спирального
Дробления, по нашему мнению, приобрели прямолинейное поступа-
тельное движение по главной оси, ставшей продольной осью тела,
месте с тем в придонных условиях существования стабилизируется
°стоянное движение той стороной к грунту, на которую смещается
Уги эволюции животного мира 161
первоначально терминальное ротовое отверстие. Дифференцируются
брюшная и спинная стороны, и вырабатывается билатеральная сим-
метрия тела. Смещение рта связано' с процессами неравномерного
роста, а именно- с преобладающим ростом заднего участка спинной
поверхности. Таким образом гетерополярная форма с одинаковыми
побочными осями, какой мыслится предок многоклеточных живот-
ных, превращается в билатериально-симметричиую, причем главная
ось не просто становится продольной, а, проходя через задний конец
тела, она в известной мере делает изгиб на брюшную сторону.
Однако только в редких случаях эта ось, сохраняя примитивные от-
ношения, проходит через ротовое отверстие. Обычно- далее рот са-
мостоятельно передвигается по брюшной стороне в той или иной
мере кпереди. Экологическое значение всех этих процессов нами
уже было в свое время выяснено. Таким образом, задний конец
оси характеризуется уже не отношениями ко рту, а только- к зад-
нему концу тела.
В этой связи приходится несколько исправить положение
В. Н. Беклемишева (1944), что смещение рта турбеллярий кпереди
не отражается у них на остальных системах органов и является по-
этому перемещением отверстия, а не искривлением оси тела и еще
менее — развитием особой оральной стороны. Нам приходится
утверждать, что первоначально имело место искривление оси — рот
стал смотреть книзу, но затем, как это предполагает и В. Н. Бекле-
мишев, он только смещался независимо от продольной оси, характе-
ризуя однако брюшную сторону.
Подобные отношения имеют место дрлее у немертин, но о прими-
тивном положении рта на заднем конце здесь говорить уже не при-
ходится. Закладка эктодермального глоточного отдела связана с
типичным бластопором гаструлы, но открывается в архентерон спе-
реди, т. е. ротовое отверстие всегда уже стабилизировано близ пе-
реднего конца тела. Процесс передвижения рта здесь завершен и
стоит вне зависимости от каких-либо смещений продольной оси тела.
Задний ее конец проходит, как и у турбеллярий, через задний конец
тела, особо характеризующийся при образовании трубчатого кишеч-
ника прорывом анального отверстия, появляющегося у немертин
впервые и притом совершенно независимо от бластопора. Что здесь
имеются отношения осей такие же, как у планарий, можно судить
по очень своеобразным чертам развития пилидия. Во-первых, при
образовании головной лопасти червя все части преорального отдела
пилидия во время некротического метаморфоза заново перестраи-
ваются. Эта коренная перестройка происходит благодаря наличию
пары имагинальных дисков головы, расположенных впереди рта и
связанных с передней лопастью пилидия. Поэтому вопрос о перед-
нем конце главной оси и ее смещения отпадает. Во-вторых, раз-
витие туловища у немертин связано, так же каку планарий, с процес-
сом неравномерного роста спинной стороны, выражающемся в диф-
ференцировке особого спинного имагинального диска. Итак, взрос-
лые формы немертин и турбеллярий построены по одному плану.
162
У аннелид, как и у немертин, эктодермальная глотка заклады-
вается у головного конца тела и открывается в передний участок
бластопора, замыкающегося в направлении от заднего его отдела к
переднему. При этом .рот, стабилизированный уже у немертин, не
обнаруживает какой-либо связи с продольной осью тела. Процесс
неравномерного роста спинной стороны, находящий выражение в
дифференцировке 2d и 4й? микромеров в виде соматобластов, обус-
ловливает такое же развитие туловища, как у немертин. При этом
задний конец тела, как и у последних, характеризуется расположе-
нием анального отверстия, которое здесь образуется, независимо1 от
Рис. 53. Схема архитектоники трубеллярий и аннелид (по Беклемишеву): А—
общий радиальный предок; Б — тербеллярия, вид сбоку, рот сместился с ораль-
ного полюса, чем определилось положение брюшной стороны; изображены две
стадии сдвигания рта вдоль брюшной стороны; В — протрохула — первичный
рот щелевидно вытянулся, на концах его сифоноглифы, резкое усиление двулу-
чевой симметрии; Г— первая билатерисальная форма—сифоноглифы дифферен-
цировались на рот и анус, оральная половина животного начинает вытяги-
ваться кзади; Д — дальнейшее развитие того же процесса; Е— верхнее и ниж-
нее полушария поворачиваются по отношению друг к другу и ложатся вдоль
одной прямой (регуляция осей у аннелиды);
1 — аборальный, 2 — оральный полюса и оральная сторона, 3 — передний, 4 — задний концы
тела, 5 дефинитивный рот и 6 — анус, между ними бластопоральная поверхность аннели-
ды, 7 — передняя, или брюшная, 8 — задняя, или спинная, стороны тела
бластопора, хотя и на месте его заднего края, за счет производ-
ных 2d, Таким образом, отношения осей тела приходится считать
тождественными с имеющимися у немертин, а далее и у турбел-
лярий.
Совсем иначе рисует архитектонику аннелид В. Н. Беклемишев
(рис. 53). По его словам, в наиболее примитивных случаях аннели-
представляет собой спинной вырост трохофоры, образовавшийся
П°Д прямым углом к ее главной оси. Таким образом, продольная ось
тела аннелиды проходит под прямым углом к продольной оси тела
трохофоры, и только позже происходит регуляция этих осей, т. е.
продольная ось взрослого червя приближается по положению к про-
дольной оси личинки, однако не совпадает с ней полностью.
В основу такого взгляда положены некоторые своеобразно истол-
кованные факты развития полихет. По В. Н. Беклемишеву, у тро-
хофоры (рис. 51) спинная сторона идет от переднего конца тела до
анального отверстия, брюшная же доходит от переднего конца
только до рта. Все пространство между ртом и анусом представля-
ет особую «бластопоральную поверхность личинки». А именно,
вслед за рядом других авторов В. Н. Беклемишев принимает, что
бластопор у наиболее примитивных в этом отношении трохофор
вытягивается в длинную щель и, замыкаясь посередине, дает нача-
ло дефинитивному рту и анусу. При этом середина замкнувшегося
бластопора может оставаться приблизительно против аборального
полюса (личинка Lopadorhynchus, Mitraria и пр.) или же сдвигает-
ся в сторону квадранта В, как у личинки Polygordius и многих дру-
гих. Таким образом, у трохофоры возникает «бластопоральная по-
верхность», которую можно сравнить с оральной поверхностью
гребневиков или актиний, и брюшная сторона взрослого червя пред-
ставляет не что иное, как разросшуюся в длину «бластопоральную»
сторону трохофоры. Рот и анус, лежащие на обоих концах бласто-
пор альной поверхности трохофоры, оказываются расположенными
на обоих концах туловища аннелиды. Брюшная сторона плоских
червей, образованная за счет В-квадранта, у аннелид всецело вытес-
нена бластопоральной стороной тела, совершенно отсутствующей у
плоских червей. Задний конец аннелиды, с расположенным на нем
анусом, соответствует не оральному полюсу зародыша, как задний
конец турбеллярий, а представляет собой как бы задний край ораль-
ной поверхности, на стыке ее со спинной поверхностью.
В основу этих соображений В. Н. Беклемишевым кладётся судь-
ба бластопора в развитии аннелид. По широко распространенному
взгляду, ставшему почти общепризнанной догмой и принятому
В. Н. Беклемишевым, бластопор, как мы указали выше, вытяги-
вается в длинную щель, которая, замыкаясь посредине, дает начал^
дефинитивному рту и анусу. Однако сам В. Н. Беклемишев дела;
оговорку, что у большинства личинок полихет бластопор замыкаете
сзади наперед и анус-впоследствии образуется наново, однако мест
его образования соответствует все же бывшему заднему кра
бластопора. А в своей наиболее примитивной форме, например
личинки Polygordius, процесс, по В. Н. Беклемишеву же, заиль
чается в том, что задняя часть бластопора через некоторое врел
закрывается, но диферентивный задний проход возникает вслед :
тем на том же самом месте.*
Таким образом, для категорического утверждения, что рот и за -
ний проход трохофоры Polygordius образуются за счет бластопора
путем его разделения, собственно нет оснований. Можно лишь
утверждать, что бластопор закрывается, а анальное отверстие
образуется после, хотя и на том же месте. Следовательно, скорее
можно утверждать, что бластопор у Polygordius не дает ануса,
164
последний развивается после закрытия бластопора, и в большинстве
случаев у аннелид бластопор вообще не остается- открытым в зад-
нем участке, а постепенно замыкается сзади наперед, и анус
впоследствии образуется наново. Образование заднего прохода за
счет бластопора можно надумать, но факты этого не говорят, а го-
ворят лишь то, что анус наново образуется именно там, где обра-
зуется задний конец тела червя, при этом обычно вне всяких непо-
средственных преемственных отношений к бластопору. В эволюции
у немертин такие отношения бросаются в глаза, так как анальное
отверстие появляется лишь у молодого червя. У аннелид эти процес-
сы протекают в эмбриональном и личиночном состоянии, причем
замыкание бластопора непосредственно связано с образованием рта
у переднего полушария трохофоры, а развитие ануса идет за счет
производных 2d, первичного соматобласта, т. е. к бластопору бли-
жайшего отношения не имеет.
Следовательно, бластопоралы-гой поверхности, в смысле В. Н. Бек-
лемишева, у аннелид нет. И заменять представление о единой
брюшной поверхности червей представлением о бластопоральной
поверхности у аннелид нет достаточных оснований. Вместе с тем
отпадает парадоксальный вывод В. Н. Беклемишева, что истори-
чески вряд*'ли трохофорные животные происходят от плоских чер-
вещ и что непосредственное сравнение между взрослыми животными
обеих групп невозможно, так как их планы строения радикально
различны. Высшие черви, по В. Н. Беклемишеву, могут сравниваться
с низшими червями только через своих личинок, и планы строения
гребневика и трохофоры ближе между собой, чем планы строения
гребневика и турбеллярий. В конце концов, по В. Н. Беклемишеву,
аннелиды произошли от ктенофорообразных примитивных свободно-
плавающих предков при переходе их к ползающему образу жизни,
причем ползательной подошвой становится оральная сторона. Пло-
ские же черви делаются особой ветвью, развившейся совершенно
независимо от аннелид.
Все это напоминает построения Седжвика о происхождении
аннелид от актиний со всеми дальнейшими сравнениями и носит
«конструктивный» характер без учета экологических отношений.
Дальнейшим положением В. Н. Беклемишева, с которым трудно
согласиться, но которое играет важную роль в его построениях,
является утверждение, что тело аннелиды представляет разрастание
задней губы бластопора, или, другими словами, спинной вырост
нижнего полушария трохофоры. Продольная ось червя при этом
проходит под углом в 90° к продольной оси трохофоры.* Только
впоследствии происходит «регуляция» осей. Основывается это поло-
жение на фактах, наблюдаемых в развитии трохофоры Lopador-
hynchus, которая, по В'. Н. Беклемишеву, обладает примитивным
Расположением осей, свойственным самой молодой трохофоре
Polygordius, причем у* большинства других аннелид эти примитив-
ные отношения бывают затушеваны, и лишь сравнение способов
развития различных аннелид позволяет показать, что все они вы-
165
водятся из наиболее примитивного способа, свойственного Lopador-
-hynhcus.
Однако едва ли правильно исходить из фактов, наблюдаемых у
Lopadorhynchus. Этот «выродок» среди полихет безусловно является
формой вторично сильно измененной в связи с его планктонным
существованием, тогда как обычно полихеты ведут придонный образ
жизни, и им обусловлена выработка всей их организации.
Судя по строению трохофоры Lopadorhynchus, она, как и взрос-
лый червь, должна быть вторично измененной, на что указывал еще
такой знаток развития полихет, как Э. А. Мейер (1897). Он предо-
стерегал против искушения принять ее черты за первичные. А имен-
но, для развития полихет характерны процессы усиленного роста
Спинной стороны, выражающиеся в дифференцировке 2d и 4d микро-
мер, как соматобластов, что имеет место и в развитии низших чер-
вей. У Lopadorhynchus, напротив, развитие спинной стороны перво-
начально подавлено — отношения явно атипичные. Приведем здесь
другой пример — образование так называемой эндолярвы Polygor-
dius. И у Lopadorhynchus, и у Polygordius нижняя часть трохофоры,
хотя и разными путями, сохраняет на некоторое время упрощенную
форму в связи с длительным пелагическим существованием. Таким
образом у Lopadorhynchus получается изгиб продольной оси, и толь-
ко при дальнейшем росте спинной стороны трохофоры происходит
регулирование, т. е. восстанавливаются отношения продольной оси,
типичные для большинства полихет. «Предварительной» регуляции
осей, которая допускается В. Н. Беклемишевым и якобы является
основным отличительным признаком развития большинства полихет,
на самом деле нет. На самом деле первичная ось зародыша у поли-
хет совпадает с продольной осью взрослого, так как проходит через
аборальный комплекс «розетки» и «креста» спереди и через аналь-
ное поле сзади без каких бы то ни было отношений ко рту аннели-
ды, т. е. так же, как и у низших червей.
Наконец, развивая идеи ленинградской школы морфологов о
«ларвальном» и «постларвальном» теле аннелид, В. Н. Беклемишев
облекает свои взгляды на понимание организации аннелид пример-
но в следующую фррму: у громадного большинства аннелид туло-
вище состоит из двух отделов, из них ларвальный отдел образуется
на протяжении нижнего полушария трохофоры, постларвальный
образуется как заднебластопоральный вырост трохофоры, несущий
на своем конце анальное отверстие и состоящий из длинного ряда
постларвальных сегментов.
Теорйя двойственного характера метамерии аннелид П. П. Ива-
нова, по словам В. Н. Беклемишева, в настоящее время является
одной из основ морфологии Protostomia. Эта теория, по П. П. Ива-
нову (1944), ведет к представлению о первичной гетерономности
ларвальных и постларвальных отделов тела, как об общей «новой
закономерности в возникновении метамерии». У В. Н. Беклемишева
она получает свое архитектоническое утверждение. *
166
В свое время П. П. Иванов выдвигал положение, что у аннелид
и членистоногих исходной и в филогенетическом,-и в онтогенети-
ческом отношении формой метамерных животных является олиго-
мерная водная личинка аннелид, метатрохофора или нектохета,
представляющая «ларвальное тело». Однако мы уже показали, что
взгляд на личиночные формы как на предковые (в этих условиях
среды организация взрослых донных форм ни в коем случае не мог-
ла выработаться) должен быть отброшен, а вместе с ним должны
быть отброшены и различные варианты «трохофорных теорий»,
начиная с Гатчековского трохозоона. С этой точки зрения метатро-
хофора — лишь личиночная форма с закладками некоторого коли-'
чества метамер, свойственных взрослой аннелиде. Число их, по
В. Н. Беклемишеву, у различных видов различно: 3 (Nereis, Serpu-
lidae), 7 (Polynoe'), 9 (Capitellidae), 9—13 (Spionidae) и т. д., хотя
сам П. П. Иванов указывает только 3—7 сомитов.
Что касается образования ларвальных и постларвальных соми-
тов, то В. Н. Беклемишев отмечает, что у тех аннелид, которые
имеют короткое ларвальное тело (например, Serpulidae), путем
терминального роста образуется только задний отдел, т. е. постлар-
вальные сегменты. Наоборот, у форм, имеющих длинное ларвальное
тело (например у Spionidae), путем терминального роста обра-
зуются не только постларвальные, но и часть ларвальных; при этом
вслед за образованием последних ларвальных сомитов без перерыва
начинается образование постларвальных. В -таком случае нам ка-
жется, что «ларвальное» и «постларвальное» тело нельзя уже при-
знать «первично гетерономными» отделами. Разница между обоими
отделами, «двойственный характер» их метамерии стирается, вме-
сте с тем стирается и различие в архитектонике.
Пытаясь удержать позиций, В. Н. Беклемишев утверждает, что,
однако, и здесь между теми и другими остается существенное раз-
личие и, прежде всего, лишь постларвальные сомиты являются поло-
выми. Таким образом способ образования — путем зависимой диф-
ференцировки ларвальных сомитов или путем терминального роста
с независимой дифференцировкой постларвальных — является ме-
нее существенным различием между ларвальными и постларвальны-
ми сомитами полихет, чем морфологические особенности, выражаю-
щиеся в присутствии или отсутствии гонад. И П. П. Иванов также
отмечал, что при анализе происхождения метамерии аннелид преж-
де всего мы должны различать у них ларвальный отдел, представ-
ляющий передний отдел, не содержащий половой аппарат.
Конечно, присутствие или отсутствие гонад характеризует опреде-
ленные отделы тела у аннелид, но связать с этим определенные со-
миты как ларвальные й постларвальные едва ли правильно. Гонады
У аннелид располагаются в ряду сомитов очень разнообразно, что
ВЬ1Ра-жается уже в приводимых В. Н. Беклемишевым числах лар-
вальных сомитов от 3 до ‘13, тогда как П. П. Иванов дает от 1 до
• Кроме того, и в пределах постларвальных сомитов наблюдается
0 Же разнообразие отношений, что особенно резко выступает у оли-
167
гохет и пиявок. Последние обладают всего двумя парами гонад,
принадлежащими 10-му и 12-му сомитам.
Далее, половые зачатки у аннелид во многих случаях совершен-
но не связаны первоначально с закладками тех или иных целоми-
ческих мешков, а вступают с ними в ближайшие отношения лишь
вторично, в связи с чем и половая функция целома является вторич-
ной. Отсюда и положение о «первичной гетерономности» ларваль-
ных и постларвальных сомитов по половому аппарату становится
мало приемлемым.
Этим мы здесь ограничимся, в дальнейшем нам еще придется
коснуться вопроса о происхождении гетерономности целомических
закладок при анализе организации иглокожих и хордовых.
Глава VIII
КЛАСС ПИЯВКИ —HIRUD1NEA
Образование задней присоски и внешняя морфология пиявок. Ста-
билизация числа сомитов, выработка областей тела, вторичная*
кольчатость сомита. Во внутренней морфологии — выработка диа-
гональной мускулатуры, редукция целома, развитие внутренней
мезенхимы. Кишечник, кровеносная система и целом. Сенсиллы и
глаза пиявок, нервная система. Нефридии и половой аппарат.
Покажем на конкретном примере небольшой систематической
группы пиявок, как типовая организация аннелид может, ориги-
нальнейшим образом изменяясь в пределах одного класса, давать
новую качественно форму. Крайне сжато мы приведем тот пример,
который был нами разработан 20 лет тому назад в качестве первой
конкретной попытки анализа организации. Он интересен в качестве
образца, прямо вводящего в сущность эволюционно-морфологиче-
ского понимания животных форм на основе понимания единства
организма и среды.
В организации пиявок ведущим является то, что одновременно
с переходом их предков к своеобразному хищничеству за счет круп-
ных животных у них выработалось особого рода состояние времен-
ного наружного паразитизма. Этим экологическим моментом обус-
ловлены почти полностью характерные черты пиявок.
Прежде всего привлекает внимание ряд характерных особенно-
стей внешней морфологии, тесно связанных с прикреплением на
Добыче. Прикрепление, как и в ноге моллюсков, происходит вслед-
ствие плотного приставания к тому или иному предмету посред-
ством слизистой поверхности расширенных задних сомитов. При
этом имеет место выработка куполообразной присоски из четырех-
семи последних сомитов, входящих друг в друга наподобие частей
подзорной трубы (рис. 54, 55). Подобную присоску, но всего из
Двух сомитов, мы находим уже у бранхиобделлид среди олигохет.
С образованием присоски был положен предел новообразова-
нию сомитов, которое-у аннелид при их терминальном росте проис-
ходит как раз в заднем конце тела. Таким образом, число сомитов
стабилизировалось, а именно стало равным 30—33.
В связи со стабилизацией числа сомитов, выработалось извест-
ное ^постоянство в положении различных органов по отношению к
169
12 П
13 ! ' 'J
Рис. 54. Схема ор-
ганизации Hirudo
medicinalis (no
Ливанову). Пунк-
тиром справа отме-
чены границы об-
ластей тела, сле-
ва — сомиты. На
спинной стороне
нанесены метамер-
но расположенные
чувствительные
почки и глаза, с
брюшной стороны
d — мужское и
$ — женское по-
ловые отверстия,
остальные обозна-
чения см. рис. 55.
170
определенным метамерам, что повело к диффе-
ренцировке характерных гетерономных областей
тела. Здесь определяющими оказались два об-
стоятельства: 1) выработавшееся уже у олигохет
и связанное теснейшим образом с дифференци-
ровкой полового аппарата у пресноводных форм
обособление 10-го, 11-го и 12-го сомитов в об-
ласть пояска и 2) обособление 7 задних сомитов
(лишь у акантобделлид, 4) в область задней при-
соски.
Далее и передние 6 сомитов, служившие сво-
ими щетинками у акантобделлид для фиксации
Рис. 55. Схема организации Acanthobdella peledina (по
Ливанову). Пунктиром нанесены границы сомитов и гра-
ницы областей тела, светлые цифры означают номера со-
митов.
I — анальное отверстие, 2 — задняя присоска, 3 — щетинки, 4 —
рот, 5 — брюшная цепочка, 6 — средняя кишка, 7 — средняя усва-
ивающая кишка, 8 — нефридий, 9 — семенник, 10 — яйцевой ме-
шок, 11 — задняя полость проводящего аппарата, 12 — сензиллы,
13 — глаз, 14 — головной мозг, 15 — глотка'
рта при насасывании пищи, обособились в головную область, при-
чем за счет передних 3—4 сомитов этой области компенсаторно
образовалась передняя присоска (рис. 56 и 57), а именно: она заме-
нила первоначальный щетинковый аппарат 2—б-го сомитов аканто-
бделлид, а затем стала использоваться и при передвижении живот-
ного. Наконец, 13—20-я метамеры образовали область средней
кишки, куда насасывается и где сохраняется запас пищи, обеспе-
чивающий паразита на более или менее продолжительный срок,
а 21—26-й сомиты образовали примыкающую к задней присоске
область усваивающей кишки, где запасы пищи постепенно перера-
Рис. 57. Пе-
редняя при-
соска Hlrudo
medicinalis с
брюшной сто-
роны (по Ли-
ванову)
Рис. 56. Схема головного
конца Protoclensls mac lo-
sa (по Ливанову). Пункти-
ром отмечены границы со-
митов, римскими цифрами
даны номера сомитов.
6 — задняя губа присоски, 7 —
ротовое отверстие, 8 — глаз
батываются и используются. С выработкой задней присоски аналь-
ное отверстие смещалось по спинной стороне в последние метамеры
этой области. В теле пиявки лишь 7-й, 8-й и 9-й сомиты остались
неспециализированными, образуя таким образом как бы особый,
преклителлярный отдел тела.
Отметим, что рассматриваемый ряд своеобразных черт органи-
зации стоит в связи с исчезновением характерного для олигохет
аппарата параподиальных щетинок, утрачивающих свое значение
Для передвижения при особом образе жизни пиявок и выработке
ими иного рода движения, в котором главную роль играют при-
соски.
Стабилизация числа сомитов повлекла за собой также и рост
тела не за счет увеличения числа, а только за счет увеличения
размеров сомита и, прежде всего, его длины. Это обусловило появ-
ление намечающейся уже у олигохет и характерной для пиявок вто-
ричной кольчатости. Закономерно у клепсин сомит состоит из 3 ко-
лец, у акантобделлид — из 4, а у челюстных пиявок — из 5 колец.
Вторично число колец либо уменьшается до 2 (Thorix), либо, чаще,
увеличивается (до 14 у Piscicold). Но как в переднем, так и в зад-
Нем отделах тела, в связи с объединением сомитов при образовании
присосок, число колец в соответствующих сомитах вторично не уве-
171
Рис. 58. Тангентальный разрез покрова Hirudo
medlcinalis под эпителием (по Гахлову):
1 — целомические каналы, 2 — волокна мезенхимы, 3 —
железистые клетки, 4 — кольцевые мышечные волокна,
5 — пигментные клетки
личивается, но по мере приближения к концам тела даже умень-
шается. Происходит постепенная редукция межкольцевых бороздок
в головной области и в области усваивающей кишки, а в области
задней присоски исчезают и границы сомитов, сливающихся полно-
стью в единый орган прикрепления — присоску.
Таким образом вся внешняя морфология пиявок развивается под
знаком приспособительного образования задней присоски. Однако
это обстоятельство от-
ражается и во внутрен-
ней организации. Так, в
типовом кожно-мышеч-
ном мешке аннелид
между кольцевой и про-
дольной мускулатурой у
пиявок дифференциру-
ется диагональная мус-
кулатура, невидимому,
за счет первой из них.
Диагональная мускула-
тура имеет особое зна-
чение. Она обусловли-
вает разнообразие дви-
жений животного путем
извивов тела при фик-
сированном заднем кон-
це. С этим связана да-
лее редукция целома,
который был опорой
для мускулатуры стен-
ки тела при движе-
нии аннелид посред-
ством синусоидных из-
гибов и перистальтиче-
ских сокращений, осо-
бенно когда имеются параподий. При иных способах движения,
как в данном случае при извивах у пиявок, целом утрачивает свое
опорное значение. Зато, в связи с развитием диагональной мускула-
туры, мощно развивается мезенхима тела, как упаковывающая мас-
са. Она часто содержит особую систему трубчатых волокон, несу-
щих механическую функцию. Волокна правильно расположены под
эпителием покрова, с одной стороны, в слое кольцевой и диагональ-
ной мускулатуры (рис. 58) и, с другой, — во внутренней мезенхи-
ме под продольной мускулатурой.
Первый комплекс черт организации, связанный с развитием зад-
ней присоски, вступает в ближайшие отношения с другим харак-
терным для пиявок комплексом узко приспособительных черт, свя-
занных с питанием паразитарного хищника (рис. 54, 55, 60). Гло-
точный отдел кишечника дифференцируется для добывания пищи
172
з мускулистый хобот или в глотку с челюстями, причем их железы
начинают вырабатывать секрет, препятствуотнк свертыванию кро-
ви. Средний отдел кишечника на протяжении 8 сомитов обычно
дифференцируется, образуя боковые выросты, задняя
пых особенно объемиста и занимает почти весь задний
рых особенно объемиста и занимает почти
Таким образом получается резервуар
для значительного запаса пищи, обес-
печивающий существование пиявки на
продолжительное время, до нового бла-
гоприятного случая попадания на хо-
зяина.
С этим связана редукция кровенос-
ного синуса на средней кишке, сохра-
няющегося только на небольшом про-
тяжении ее заднего участка в 21—24
сомитах, где происходит постепенное
усваивание пищи (рис. 60). В резуль-
тате, при слабом вообще развитии кро-
веносных сосудов у олигохетных пред-
ков, значение их у пиявок уменьшается,
и у хоботных развиваются лишь сосу-
пара кото- •
отдел тела.
крове-
Рпс. 60. Схема
поеной системы Hemi-
clepsis т irglnata
(по Ока):
1 — боковые выросты сред-
ней кишки, 2 — кровеносный
синус на усваивающей киш-
ке, 3 — спинной сосуд, 4 —
брюшной сосуд, 5, б, 7, 8 —
вторая, третья, четвертая и
пятая пары комиссуральных
сосудов передней части тела,
9 — сосудистая петля в хо-
боте, 10 — семь пар комиссу-
ральных сосудов задней при-
соски
10
9
8
3
Z
3
1
Рис. 59. Схема целома qlossosvphonia в
поперечном разрезе (по А. Ковалевскому).
1 — спинной канал целома, 2 — брюшной канал
Целома с кровеносными сосудами внутри (3), и с
брюшной цепочкой (4), 5—гиподермальный коль-
цевой канал, 6 — промежуточные каналы це-
лома, 7 — боковой канал, 8 — кишечник, 9 —
мезенхима тела, 10 — цилпофагоцитарный орган
4
ды головных сомитов и задней присоски. У челюстных же пиявок
происходит полное исчезновение кровеносной системы.
• Коррелятивно имеет место усиление трофической функции цело-
ма, принимающего форму каналов с правильным движением содер-
жимого и по всему телу, и по отдельным метамерам, входящим в его
состав (рис. 59). Получается система, состоящая из продольных
173
сов .целома в мускулистый
Рис. 61. Чувствительная почка
Placobdelia со st at а (по
А. Ковалевскому):
1 — кроющий эпителий, 2 — сое-
динительная ткань, 3 — нерв, 4 —
ретинальная клетка, 5 — чувстви-
тельные клетки
синусов — спинного, брюшного и боковых •— с сетью соединяющих
каналов в каждой метамере тела. Имеет также место дальнейшее
их разрастание в периферической части мезенхимы, проникновение
в стенку тела ш здесь, сверх того, при выработке и усилении дыха-
тельной функции образование богатейшего сплетения целомических
капилляров в покрове. Последним шагом в этом 'направлении у
челюстных пиявок является присутствие гемоглобина в целомиче-
ской жидкости и дифференцировка двух продольных боковых сину-
пропульзаторный аппарат, а у рыбьих
пиявок развитие боковых сократитель-
ных ампул и даже кожных целомиче-
ских жабер (Branchelliori).
В эволюции целома первый веду-
щий приспособительный комплекс ха-
рактерных черт организации пиявок
теснейшим образом связывается со вто-
рым. Вместе оба они настолько значи-
тельно и многосторонне охватывают
различные стороны организации, что
действительно создается особый этап в
эволюции ветви аннелид — особый
план организации пиявок.
Переход к хищничеству временно
эктопаразитарного характера обусло-
вил выработку третьего комплекса, до-
полняющего два рассмотренных. Если свободноподвижное сущест-
вование типичного хищника ведет к многостороннему повышению
организации — усложнению органов движения и -мускулатуры,
органов чувств и центральной нервной системы, то переход к па-
разитизму обусловливает значительную односторонность приспо-
соблений. Это мы видели в рассмотренных комплексах. На первый
план выдвигаются свойственные паразитам приспособления для
прикрепления на хозяине и для извлечения и сохранения макси-
мального пищевого запаса. Однако при временном паразитизме
периоды свободого существования обусловливают приспособления,
поддерживающие на известном уровне как аппарат органов чувств,
так и органы активного и пассивного движения и нервную систему,
но ведущая роль принадлежит в организации уже не им,
Соответственно третий комплекс черт организации пиявок выра-
жается в развитии на выдающемся участке сомита, его среднем
' кольце, пояса чувствительных почек в виде сензилл, содержащих
одиночные ретинальные клетки (рис. 61). В головном отделе рети-
нальные клетки сензилл развиваются возле чувствительной почки в
большом количестве и дифференцируются, покрываясь пигментным
кубком, в характерные, единственные в своем роде глаза пиявок
(рис. 62). Соответственно сильно развивается средний чувствитель-
ный нерв, образуя мощные брюшной и спинной нервы в типичном
невросомите (рис. 63), а в головной области развиваются большие
174
передние нервы — производные спинного
нерва. В продольной мускулатуре проис-
ходит дифференцировка системы особых
мышечных волокон, связанных с диффе-
ренцировкой аппарата проприоцептив-
ных клеток. Это значительно осложняет
типовые пары кольцевых двигательных
нервов аннелид, располагающиеся у гра-
ниц сомита. Наконец, в центральной
нервной системе имеется резкое обособ-
ление коннективов и ганглиев брюшной
цепочки. Однако образование задней и
передней присосок вызывает смещение
соответствующих ганглиев в ближайшие
к ним сомиты (рис. 55), сближение йх и
образование надглоточной ганглиозной
массы из двух, подглоточной из четырех
и анальной из семи слитых ганглиев.
В связи с крайне ограниченным количе-
ством элементов глийной ткани при их
чрезвычайном укрупнении, что нередко
встречается у паразитов, в каждом из
ганглиев пиявок далее имеет место диф-
ференцировка шести пакетов ганглиозных
клеток соответственно числу шести глин-
ных клеток.
Совершенно своеобразно и самостоя-
тельно идет эволюция нефридиев и поло-
вого аппарата пиявок, представляя как
Рис. 62. Медианный разрез
через глаз Hi' ” п >nedi-
cinalis (по Гессе)’:
1 — кроющий эпителий, 2 —
нерв, 3 — глазной нерв, 4 —
пигментный кубок глаза, 5 —
ретинальные клетки, 6 чув-
ствительная почка
Рис. 63. Схема невросомита Protoclepsis tessellata (по
Ливанову):
1, 2, 3, — первое, второе и третье кольца сомита, 4 — коннектив и
5 — ганглий брюшной цепочки, 6, 7 — передний и задний двигатель-
ные кольцевые нервы, 8 — задняя ветвь заднего кольцевого нерва,
9 — чувствительный нерв сомита, 10 — спинной чувствительный нерв,
11 — клетка мышечной чувствительности, 12 — чувствительная почка,
13 — сензилла
175
бы особый комплекс прогрессивно типовых черт, унаследованный
от олигохетообразных предков, но пошедший значительно дальше,
развившийся в некоторых отношениях, можно сказать, чрезмерно.
Открытый нефридий олигохет, регулировавший содержание жид-
кости в целоме, замыкается теперь со стороны целома, который
принимает здесь трофическую функцию, заменяя кровеносную си-
стему (рис. 64). Это замыкание происходит на основе особо силь-
ного^ развития внутриклеточных экскреторных капилляров в прокси-
мальном отделе нефридия, что можно видеть уже у бранхиобдел-
лид среди олигохет. С этим связана и редукция ресничек, играв-
ших большую роль при движении содержимого в открытом нефри-
дии олигохет. Нефростомы или редуцируются, или, специализиру-
ясь, обособляются в виде очень сильно развитых цилиофагоцитар-
ных органов (рис. 59).
Рис. 64. Схема нефридия Acanfhobdella peledina
(по Ливанову):
1 — отдел нефридия с ветвящимся каналом, 2 — то же
с неветвящимся каналом, 3 — впячивание покрова, в
которое открывается нефридий
Еще дальше ушла эволюция полового аппарата пиявок (рис. 65,
66). Это выразилось не только в стабилизации половых • сомитов,
10-го как мужского и 12-го как женского, и не только в полном
отделении полового целома от прочих его участков, что выработа-
лось уже у олигохет, но и в сильнейшем развитии присущих олиго-
хетам семенных, соответственно яйцевых мешков.’ Эти мешки вда-
ются из 10-го, соответственно 12-го, сомита на все протяжение обла-
сти средней кишки (рис. 54 и 55). При этом, что характерно для
пиявок, мужская гонада переходит в диффузное состояние, распо-
лагаясь в семенном мешке, который подразделяется на метамерные
участки, получившие название «семенников», а суженные его уча-
стки дают «семяводы».
Так создается крайне характерный половой аппарат пиявок, на-
ходящий свое завершение в выработке непарного дистального
отдела у первоначально парных выводных половых протоков. Обра-
зованием особого непарного «проводящего» спермин участка за
счет сперматек олигохет и прилежащих частей стенки тела, так на-
зываемой «проводящей» ткани, совершенствуется сперматофорное
оплодотворение. Развитие в этом направлении достигает, наконец,
крайней степени при гиподермальной импрегнации, т. е. при проник-
новении спермиев внутрь на любом участке тела, где прикреплен
176
сперматофор. У Hirudinidae, напротив, развивается. типичная копуля-
ция с превращением дистальных частей выводных протоков в копу-
ляционные органы, но тем не менее с сохранением при этом типич-
ных сперматофоров. Эволюция не обратима.
Таким образом завершается выработка организации пиявок с ее
характерными чертами. Это особый этап в эволюции аннелид, на-
Рис. 66. Половой аппарат Pisctcola geo-
metra(по Бренту): А — анатомия; Б —
наружный вид, <? — мужское, § —
женское половые отверстия;
I — копулятивная зона, 2 — семеизвергатель-
ный канал, 3 — брюшная цепочка, 4 — яйце-
вой мешок, 5 — prostata, 6 — семенник, 7 —
проводящая ткань, 8 — vas deferens, 9 — vas
efferens, 10 — vesicula seminalls
Рис. 65. Схема полового ап-
парата AcantlwbileUa pele-
dina (по Ливанову):
X, XI, XII — клителлярные со-
миты, 1 — атриум, 2 — сооб-
щение с целомом у семейных
мешков, 3, 4 — семеизвергатель-
ный канал, 5 — половая ворон-
ку, 6 — яичник, 7 — спермате-
ка, 8 — проводящий аппарат,
у — uterus, 1U -vesicula seminaHs,
$ 9 мужское и женское
половые отверстия
павших свое развитие с морских форм — класс полихет, продол-
жавших его в условиях узкоадаптивной
форм — класс олигохет, и закончивших
ках> какими являются пиявки.
эволюции пресноводных
в эктопаразитах—хищни-
организация пиявок после предельно сжатого анализа ее глав-
нейших компонентов раскрыта для нас и в целом, т. е. в больших
1(ли меньших комплексах взаимосвязанных черт, и в частностях —
3 отдельных органах и их частях. При этом выявлены ведущие,
определяющие черты организации. Она раскрыта как выражение
‘ риспособительных процессов, происходящих под ведущим влияли-
среды на основе типовых черт наследия предков, и как выраже-
спр процессов Дифференцировки, происходящих при освоении
Р ДЫ на основе специализации взаимно связанных образований.
^2 Пути эволюции животного мира
Глаза IX
ТИП МОЛЛЮСКИ — MOLLUSCA '
Эмбриональное развитие. Трохофора и ее значение. Выоаботка
раковины, ноги, мантии и мантийного комплекса как ведущих мо-
ментов в организации моллюсков у гастеропод, у Scaphopoda и плач
стинчатожаберных, у головоногих. Amphineura. Покров, внутренняя
мезенхима, целом и мускулатура у моллюсков. Кровеносная систе-
ма. Экскреторный аппарат. Кишечник. Органы чувств и нервная
система. Итоги анализа организации моллюсков. Филогения
моллюсков.
Чрезвычайная характерность организации моллюсков вскрыва-
ется их эмбриональным и постэмбриональным развитием, показы-
вающим необычайную близость к развитию полихет. А именно: на-
чиная с первых делений оплодотворенного яйца и кончая стадией
личинки-трохофоры, процессы протекают одинаково.
В дроблении имеет место спиральный его тип, т. е. выработав-
шийся уже у турбеллярий (рис. 29) процесс образования четырех
квартетов микромер и их производных за счет четырех макромег
последовательно чередующимися дексио- и леотрогтными делениям !.
Приспособительное значение такого процесса мы пытались показать
при рассмотрении организации турбеллярий, конечно гипотетично
в переходе от поступательно-вращательного движения гастреад г
прямолинейному движению червей.
Дальнейшее образование различных зачатков за счет потомков
четырех квартетов микромер протекает у полихет и моллюсков оди-
наково, за исключением специфического отличия. Крест моллюсков
представлен промежуточными клетками полихет, а промежуточные
клетки моллюсков соответствуют кресту полихет (рис. 67). И это
еще более подчеркивает близость обеих дивергентно развивавшихся
групп между собою.
Характерна для обеих групп также ранняя закладка .и значи-
тельная дифференцировка зачатков. Так, во-первых, обособлены за-
кладки головного отдела (розетка, крест, промежуточные клетки),
сосредоточенные в производных первого квартета, и закладки ту-
ловища в производных прочих квартетов. Во-вторых, в туловищном
отделе, с одной стороны, происходит дифференцировка второго
квартета для образования почти всей эктодермы, третьего — для об-
разования мезенхимных зачатков и четвертого — для целомиче-
178
ской мезодермы (рис. 53) и вместе с микромерами для энтодермы.
С ДРУг°й стороны, преобладающее значение в связи с процессами
неравномерного роста получают спинные закладки: 2 d—первичный
соматобласт и 4 d — вторичный соматобласт, или целомезобласт.
Они обусловливают 1) смещение бластопора на брюшную сторону
и загладку стомодеума впе-
реди вентрально, т. е., иначе
говоря, перемещение перво-
начально терминального рта
на брюшную сторону и
2) дифференцировку сперва
однообразных мышечных за-
кладок.
Наконец, и для полихет и
для моллюсков обычен ран-
ний переход к свободному
существованию в виде пела-
гической личинки «трохофо-
ры». При этом резко выяв-
ляется свойственная обеим
группам строгая детермини-
рован1ность развития, веду-
щего к трохо-форе. В конце
концов за счет клеток розет-
ки образуется темянной ор-
ган личинки; клетками кре-
ста, а у моллюсков промежу-
точными, образуется голов-
ной ганглий; клетками про-
межуточными, а у моллю-
сков клетками креста-, —
вся эктодерма верхнего
полушария трохофоры до
предротового мерцательного
обруча. Этот последний об-
разуется за счет части кле-
Рис. 67. А—дробление у Are nicola, ста-
дия 64 бластомеров (по Чайльду), вид с
анимального полюса:
1 — клетки розетки, 2 — клетки креста. 3 — про-
межуточные клетки, 4 — первичные трохобласты;
производные II квартета (редкий пунктир),
5 — вторичные трохобласты и 2 d — первичный
соматобласт;
Б—схема дальнейшего развития сбоку (по
Коршельту и Тендеру). Обозначения те же
и so п — первичный соматобласт, еп
эктодермальный зачаток; производные 3-го
квартета отмечены косой штриховкой (6);
В, Г, Д — последовательные стадии разви-
тия зародыша полюег сбоку (по Давы-
дову) :
som — производные первичного соматобласта,
еп — энтодерма, 7 — прототрох, 8 — теменная
пластинка
первого квартета, так
называемых трохобластов, и
части второго квартета.
пижнее полушарие личинки
ооразует свою эктодерму за
счет 2 cl соматобласта. Лишь
вентрально у ротового отверстия принимают участие в ее образова-
ли и отчасти в образовании мезенхимы производные прочих мик-
л меР второго квартета. Третий квартет дает стомодеальный отдел
ЧитИНКИ ’И ее мезенхимные элементы, дифференцирующиеся в зна-
Проельн°й мере в личиночные мышцы. У заднего конца личинки
Р взводные 2 d соматобласта дают зачаток задней кишки. Вся же
179
ее средняя часть развивается за счет энтодермы, происходящей
главным образом за счет макромер. Наконец, характерный для тро-
хофоры зачаток, отсутствующий у личинок немертин и турбеллярий,
это — помещающиеся в заднем конце тела трохофоры между киш-
кой и покровом пара (4 d) телобластов, дающих сначала несколько
энтеробластов, а затем образующих справа и слева по мезодер-
мальной полоске, зачаток будущего целомического комплекса взрос-
лой полихеты и моллюска.
Конечно, о конвергентном развитии в обеих группах цри таком
тождестве процессов говорить не приходится, речь может идти
лишь о ближайшем родстве, об общности эволюции до трохофоры
включительно. При этом сама трохофора представляет лишь вста-
вочный личиночный этап, выработавшийся уже у низших червей и
далее, в известной мере независимо от взрослой формы, эволюцио-
нировавший в своих приспособительных и в своих Чиповых чертах.
Очевидно, для достижения взрослого состояния уже у исходных
форм червей было необходимо дополнительное получение питатель-
ного материала. Таким образом, адаптивно создается Мюллеровская
личинка Polycladida, наряду с выработкой прямого развития у дру-
гих турбеллярий. Последние вторично бывают богато снабжены
пищевым запасом от материнского организма в виде желточных
элементов, т. е. массы абортивных яйцевых клеток. У немертин
развитие соответственно ведет к пелагическому пилидию, а у анне-
лид и. моллюсков к трохофоре, которая представляет последний этап
на пути приспособительной эволюции личинки червей. При этом, не-
сомненно, трохофора по организации стоит выше пилидия немер-
тин, что выражается в наличии трубчатого кишечника, тогда как у
пилидия сохраняется еще целентерическое состояние последнего, и
анальное отверстие появляется поздно, а именно — у молодой не-
мертинки.
Узкоадаптивные черты, обусловленные приспособлением трохо-
форы к свободному пелагическому существованию, следующие:
1) форма тела яйцевидная или дискообразно вздутая благодаря
развитию студенистой массы основного вещества, что связано с бо-
лее легким поддерживанием животного во взвешенном состоянии в
воде при пелагическом образе жизни, 2) образование мерцатель-
ных шнуров, особенно преорально го и адорального, обусловливаю-
щих движение и питание личинки, 3) развитие апикальной темен-
ной пластинки — своеобразного органа чувств.
С иной точки зрения ^рассматривали трохофору эмбриологи
прошлого столетия, видевшие вообще в личиночных формах пред-
ковые состояния той или иной группы животного царства. И как ь
свое время А. Дорн (1871) заселил моря палеозоя навплиусами,
принимая их за предков ракообразных животных* так Б. Гатчек
(1878, 1888) превратил трохофору в «трохозоона» — предковую
форму аннелид, членистоногих, моллюсков, сохранившуюся и
до наших времен в ротаториях. По Гатчеку, трохофора есть пов-
торение трохозоона, т. е. общей исходной формы трохофорных чер-
180
вей — Trochhelmintes, другими словами,-— червей, стоящих выше
плоских. Предок последних «протрохозоон» отличался от трохозо-
она целентерическим состоянием кишечника и был общим предком
всех червей, представляя лишь дальнейший этап эволюции после
гастреи.
Идеи Б. Гатчека о филогенетическом значении трохофоры широ-
ко распространились в науке, и К. Гейдер в 1914 г. проводит их в
своей «Филогении беспозвоночных». Однако те факты, что трохо-
фора представляет пелагическую форму, тогда как взрослые явля-
ются донными организмами, и что вся организация трохофоры под-
вергается сильнейшей перестройке, а иногда и катастрофическому
метаморфозу при переходе во взрослое состояние, вполне достаточ-
ны, чтобы усомниться в правильности упрощенного’ филогенетиче-
ского подхода эмбриологов прошлого столетия к эволюции донных
животных, какими являются по всей их организации турбеллярий,
немертины и аннелиды.
Итак, предками как аннелид, так и моллюсков были дошще
организмы, у которых в развитии имелась давно выработавшаяся
свободная пелагическая стадия, давшая трохофору. Она обладает
типовыми чертами высших червей, обнаруживая в то же время ряд
приспособительных особенностей, и является дальнейшим шагом в
эволюции личинок, представленных у паренхиматозных червей
Мюллеровской личинкой турбеллярий и пилидием немертин.
Переходим к организации взрослых моллюсков. Она отличается
от широко приспособительной организации червей. Отдельные чер-
ты строения моллюсков являются узко приспособительными, но в
своей совокупности они создают особый новый тип организации,
дающий широкие возможности в борьбе за жизнь и богато развер-
тывающийся при овладении различнейшими биотопами моря, прес-
ных вод и суши. Ведущим моментом организации моллюсков явля-
ется выработка защитных'известковых отложений в кутикулярном
покрове на спинной стороне удлиненного туловища, образование
раковины, что вполне соответствует потребностям в защите для
Донных ползающих червеобразных форм (рис. 68). Далее, в тесней-
шей связи с развитием раковины стоит выработка за счет вен-
тральной стенки тела мускулистого органа движения — ноги. Это
происходит там, где сохраняется возможность функционирования
мускулатуры при условии сравнительно медленного передвижения
нехищных животных. Лишь передний отдел тела, голова с ее ком-
плексом органов чувств, остается у большинства моллюсков мало
измененным по сравнению с червями.
Таким образом создается характерный внешний вид моллюс-
ков, который обычно осложняется еще с дальнейшим увеличением
защитного значения раковины благодаря образованию по бокам
>ела складки покрова, так называемой мантии, увеличивающей по-
верхность известковых отложений раковины (рис. 69 и 70). Эта
кладка прикрывает возникающую при этом мантийную полость и
- вляется особенно развитой в задней части тела. Здесь в мантий-
181
Рис. 68. Схема организации моллюсков, вид сбоку (по Э. Мейеру).
Черным выделена нервная система:
1 — анальное отверстие, 2 — спинной кровеносный сосуд, 3 — жабра, 4 —
сердце, 5 — раковина, б — целом (перикард), 7 — плевровисцеральный кон-
йЬктгтв, 8 — гонада, 9 — церебральный. 10 — педальный. 11 — париетальный,
12 — плевральный и 13 — висцеральный ганглии, 15 — печень. 17 — мантий-
ная полость, 18 — нефридий, 19 — рот, 20 — глаз, 21 — осфоадий. 23 — перед-
няя кишка, 25 — радуля, 26 — слюнная железа, 27 — статоцист, 28 — сто-
матогастрическая система, 29 — щупальце, 30 — желудок (сравни с рис. 69, 70)
Рис. 69. А — схема организации моллюсков, вид ()о спинной сто-
роны; Б—схема нервной системы моллюсков, образованной нерв-
ными стволами без дифференцировки соответствующих ганглиев и
коннективов;
22 — педальные стволы, 24 — плевро-висцеральный нервный ствол (остальные
обозначения см. на рис. 68, сравни с рис. 70)
ную полость открывается медианно анальное отверстие, а далее с
боков пара нефридиальных и половых пор и, наконец, располага-
ется пара перистых жабер, 1ак называемые ктенидии, с прилежа-
щими к ним осфрадиями, т. е. хеморецепторами, ориентирующими
животное в качествах воды, омывающей жабры Расположение этого
так называемого мантийного комплекса органов в заднем, особенно
глубоком, участке мантийной полости, конечно, представляет при-
способительную черту организации моллюсков, связанную с лучшей
Рис. 70. Схема поперечного разреза через моллюска: А — в обла-
сти желудка и печени; Б — в области сердца, внесена часть ман-
тийного комплекса.
14 — кишка, 16 — мышечные пучки (остальные обозначения см. на рис. 68 и 69)
защитой и функционированием всех образований, входящих в со-
став мантийного комплекса.
Нарисованная нами картина внешней морфологии моллюсков
характерна для всего типа, и ее обычно относят к некоторой гипо-
тетической исходной форме, так называемому «первичному моллюс-
ку». В различных группах этого типа она претерпевает значитель-
ные изменения в зависимости от приспособления к различным част-
ным условиям жизни в тех или иных биотопах и биоценозах. Полу-
чается ряд узко, а иногда и крайне узко адаптивных черт органи-
зации моллюсков.
Однако проникновение моллюсков в новые биотопы (между
камней, в грунт или в пелагические условия существования) часто
связано со вторичной редукцией защитной раковины, можно ска-
зать, с возвратом в этом отношении к примитивному состоянию без
раковины, но с характерными чертами уже выработавшейся орга-
низации моллюсков. Особенно это сказывается в эволюции высшей
их группы — головоногих, перешедших к образу жизни подвижных
хищников и выработавших сложно построенные органы чувств и
Движения и сложную центральную нервную систему. Утеряв рако-
вину, головоногие одинаково приспособлены к придонному и пела-
гическому существованию и представляют по организации един-
ственную группу беспозвоночных животных, успешно конкурирую-
183
щую в настоящее время в море с представителями водных позво-
ночных, или, говоря другими словами, по А. Нефу, выдерживаю-
щую активную борьбу за существование с преуспевающими позво-
ночными.
Наиболее сохранившей типовые черты является раковина брю-
хоногих (рис. 71). Для нее характерно увеличение защитного зна-
чения путем разрастания в виде конуса из плоской, кроющей пер-
воначальной формы. Благодаря этому, во-первых, часть внутренних
Рис. 71. Схема организации гастропод
' (по Нефу):
I — анальное отверстие, 2 — передняя аорта, 3 —
задняя аорта, 4 — жабра с приносящим и относящим
сосудами, 5 — сердце, б — раковина. 7 — крышечка,
8 — гонада, 9 — церебральный, 10 — педальный,
И — париетальный, 12 — плевральный, 13 — висце-
ральный ганглии, 14 — печень, 15 — кишка, 1G —
мантийная полость, 17 — нефридий, 18 — рот, 19 —
глаз, 20 — перикард, 21 — глотка, 22 — радуля, 23 —
статоцист, 24 — желудок
Рис. 72. Pleurotomaria
(по Нефу):
1 — передняя аорта, 2 —
жабра, 3 — сердце, 4 —
щель раковины, 5 — кишка
органов вдается в образующуюся полость раковины, развивается
так называемый внутренностный мешок; во-вторых, под защиту ра-
ковины при раздражениях может втягиваться и головной отдел, и
нога, причем на заднем спинном участке последней иногда разви-
ваются известковые отложения крышечки, наглухо замыкающей
доступ к телу втянувшегося моллюска. В связи с образованием
внутренностного мешка происходит обособление головы и ноги, в
виде особой области тела. Образуется втягивающая, так называе-
мая колумеллярная мышца, с одной стороны прикрепляющаяся на
раковине, с другой — разделяющаяся на пучки, которые входят в
голову и йогу. •
Последним моментом в развитии раковины брюхоногих было
образование спирального завитка раковины с перемещением вперед
всего мантийного комплекса из его типового* положения сзади
(рис. 71 и 72). Эти два процесса, неразрывно связанные друг с
184
другом, обусловлены, как это экологически наиболее реально пред-
ставлено А. Лангом, а затем Нефом, приспособлением к ползающей
донной жизни. При ней, по Лангу, нарастающий внутренностный
мешок и принимавшая коническую форму раковина должны были
во время движения животного отклоняться назад и несколько
вбок, сохраняя возможность функционирования мантийного ком-
плекса. В силу этого должны были происходить процессы неравно-
мерного роста раковины. При этом более интенсивно растущая
передняя и верхняя ее поверхность должна вызывать завертывание
Рис. 73. Схема ор-
ганизации S си pho-
poda (по Нефу).
Обозначения см
на рис. 74
конуса в спираль, вызывая в то же время смешение входа в ман-
тийную полость, а соответственно и всего мантий-
ного комплекса по правой стороне все более и
более вперед из первоначального положения пря-
мо назад. Процесс заканчивается образованием
спиралыюкопической раковины, обращенной сво-
им входом и мантийным комплексом прямо впе-
ред, как это мы и видим у переднежаберных. В
связи с этил! произошли характерные смешения в
ряде внутренних органов, как, например, явления
перекреста плевровисцеральных коннективов
нервной системы, о которых речь будет еще впе-
реди. Завершается процесс редукцией первона-
чально левой половины мантийного комплекса у
Monotocardia.
Не наша задача рассматривать адаптивные
процессы в группах Opistobranchiata и Pulmo-
nata, которые повели к обратному смещению
мантийного комплекса, редукции и исчезновению
раковины и дали крайне своеобразную организа-
цию пелагических Pteropoda или донных Nudi-
branchiata, с одной стороны, и легочных моллю-
сков — с другой.
Отметим одно приспособление брюхоногих
моллюсков, которое выразилось в существовании
медианной щели, идущей от края раковины над
мантийным комплексом (рис. 72), или ряда отвер-
стий у некоторых Prosobranchia (Emarginula,
Pleurotomaria, Polutremaria, Haliotis, Fissurella).
Оно служит для функционирования органов
мантийного комплекса при любом состоянии
гела моллюска, когда в то же время через узкую щель они не
подвергаются опасности повреждений.
Это приспособление защитного порядка характерно проявляется
Далее в раковине Scaphopoda, с одной стороны, и Lamellibranchi-
ata> с другой.
В классе Scaphopoda переход к жизни в илистом или песчаном
Рунте стал возможен в связи с тем, что выработалась раковина с
Руглым отверстием (апикальной щелью) на конце (рис. 73). Бла-
185
годаря .этому приспособлению животное могло углубляться в грунт,
сохраняя при этом возможность функционирования мантийного
комплекса; далее в грунте отпадала необходимость поддерживать
раковину в равновесии, вызывающая развитие завитка раковины
брюхоногих. Напротив, примитивная, удлиненная конусовидная
форма даже лучше защищала животное, позволяя ему глубже ухо-
дить в грунт гфи условии все же сохранения свободным апикаль-
ного конца раковины, несущего отверстие мантийной полости.
В этом направлении раковина Dentalium крайне адаптирована, и
соответственно развиты внутренностный мешок, голова с длинными
нитевидными щупальцами и якоревидная нога.
Совершенно иначе происходит развитие раковины у Lamelli-
branchiata (рис. 74). У них, во первых, повышение ее защитной
Рис. 74. Схема организации Ntcula (по Пель-
зенеру и Нефу):
1 — рот, 2 — щупальце, 3 — нога, ее ползательная
поверхность, 4, 5 — биссусовая железа. 6 — край ман-
тии, 7 — мантийная полость, 8 — жабра, 9 — анальное
отверстие, 10 — парус, 11 — мышцы аддукторы, 12 —
кишка, 13 — желудок, 14 — печень, 15 — гонада, 16 —
целом, 17 — сердце, 18 — связка, 19 — головной, 20 —
плевральный, 21 — педальный, 22 — париетальный и
23 — висцеральный ганглии
функции пошло не по
пути увеличения ее апи->
калькой части с образо-
ванием внутренностно-
го мешка, а по другому
возможному пути —
пути увеличения ман-
тийной складки, следо-
вательно, увеличения
периферического отде-
ла раковины, т. е. более
полного прикрытия те-
ла моллюска. Во-вто-
рых, соответственно с
увеличением задней ме-
дианной щели над ман-
тийным комплексом и с
образованием такой же
щели спереди, эволю-
ция раковины идет од-
новременно по пути
расчленения на правую
и левую половины, сое-
диненные лишь на
спинной стороне так называемой «связкой». Таким образом соз-
далась полностью охватывающая животное с боков и замыкаю-
щаяся спереди, сзади и снизу раковина «двухстворчатых» моллюс-
ков — идеальное защитное приспособление при переходе к жизни
на мягких грунтах и при переходе одновременно к пассивному пи-
танию. При этом вырабатываются мощные передняя и задняя
мышцы аддукторы, прикрепляющиеся к правой и левой створке
каждая и при своем сокращении наглухо притягивающие их друг к
Другу. Нога превращается в мускулистый килевидный вырост, вы-
ставляющийся между створками раковины и своими сокращениями
обусловливающий продвижение сплющенной с боков раковины по
186
^лу или песку; лишь у Protobranchia, невидимому, сохранилась
еще типовая ползательная подошва ноги, могущая, однако, склады-
ваться по длине в киль. В ногу в той или иной степени вдаются
внутренние органы — антитеза внутренностному # мешку брюхо-
ногих.
Другим важным моментом в развитии двустворчатых моллюс-
ков был переход к пассивному питанию. Развитие, прикрывающих
все тело створок являлось возможным для животного при условии
утраты необходимости в активных движениях при получении пищи.
Это в конце концов в крайнем своем выражении привело к полной
утрате головного отдела — «безголовые» моллюски, и стало воз-
можным благодаря использованию мерцательного покрова мантии,
для установления тока воды, несущего пищевые частицы. Послед-
ний идет от нижней части задней щели раковины, омывая жабры
вперед к ротовому отверстию и оттуда по верхнему участку мантий-
ной полости назад к отверстиям мантийного комплекса и, наконец,
наружу через верхнюю часть задней щели раковины. В результате
произошло образование мощных мерцательных складок справа и
слева у рта, так называемый «парус», сильное развитие нитей жабер
или превращение их в мощные пластинчатые образования, и, нако-
нец, дифференцировка на заднем крае мантии вводного и выводно-
го отделов для тока воды, превращающихся у ряда форм в длин-
ные трубки сифонов. Погребенное, иногда на 20—30 см, в грунте
животное благодаря сифонам обеспечено постоянным притоком во-
ды с необходимыми для питания мельчайшими организмами и с
кислородом для нормального газового обмена, равно как и возмож-
ностью оттока экскрементов, экскретов и углекислоты, но ценою по-
тери головы.
Вторично двустворчатые моллюски распространяются ' и на
твердые грунты, переходя от малоподвижного к настоящему сидя-
чему образу жизни, что вполне сочетается с пассивным питанием и
килевидной ногой, уже выработавшимися ранее. При этом в высо-
кой степени своеобразен и способ прикрепления, а именно вен-
тральной поверхностью ноги, на которой развивается так называе-
мая биссусовая железа, дающая секрет для прикрепления в виде
нитей биссуса. Устрицы или мидии, хорошо защищенные своей ра-
ковиной, прикрепляющиеся к скале, образуя банки, являются
образцами форм моллюсков, преуспевающих в борьбе за жизнь.
Развитие раковины Cephalopoda сходно в некоторых отношениях
о описанным для Gastropoda. Увеличивая свое защитное значение,
раковина у них также становится конической и завитой в спираль
(рис. 75). Но у головоногих ее образование связано с переходом к
плаванию благодаря тому, что между эпителием покрова и проду-
цируемой им раковиной апикально скопляется газ. Раковина ста-
новится как бы поплавком, который поддерживает в воде соб-
ственно тело моллюска, занимающее базальную ее часть и лишь в
Вйде тонкого тяжа доходящее до ее вершины. При этом своеобраз-
ное приспособление выражается далее в периодически повторяю-
187
Рис. 75. С^ема организации Nautilus (по
Нефу). Обозначения см. на рис. 76
щемся образовании известковых поперечных перегородок, по мере
роста животного и раковины. За каждой перегородкой происходит
снова накопление газа; так что раковина становится образованной
рядом следующих друг за другом «газовых» камер, пронизываехмых
посредине тонким «сифонохм», т. е. продолжением туловища.
Нарастающая раковина образует спираль, завиток которой на-
висает над голодным отделом тела Nautilus в связи с тем, что ман-
тийная полость, содержащая типичный комплекс органов, разрас-
тается и получает особое значение для движения животного.
В задней стенке мантийной полости развиваются сильные мышеч-
ные пучки, так называемая ’«мышечная» мантия; при сокращении
вода с силой. выбрасы-
вается из мантийной по-
лости через разраста-
ющуюся в виде двух до-
пастей заднюю часть но-
ги, так называемую во-
ронку. Таким образом жи-
вотное обратным .толчком
движется задним концом
тела вперед — приспособ-
ление опять-таки защит-
ного характера, позволяю-
щее уходить от врага, от
раздражений вредных для
животного, делающее его
в конце концов отличным
пловцом.
В то же время перед-
няя часть ноги развивае!
щупальцеобразные выро-
сты, которые могут слу-
жить для захватывания
добычи, а рот смещается
так, что оказывается со
всех сторон окруженным
ими. Процесс образования особого отдела тела из головы и ноги,
резко выраженный уже и у брюхрногих, здесь проявляется в пол-
ной мере, что нашло свое отражение и в названии класса Cephalo-
poda — головоногие.
Описанная дифференцировка развивалась в теснейшей связи с
переходом к хищному существованию. Современные спруты и кара-*
катицы (рис. 76) завершили эволюцию группы в этом направлении,
имея 8 или 10 мощно развитых щупалец, снабженных по всей дли-
не присосками и служащих как чувствительными образованиями,
так и орудиями нападения и защиты и органами передвижения.
Воронка сохранила свое значение при плавании. Но и то и другое
могло развиваться наряду с редукцией раковины. Подвижность,
188
ловкость и сила хищника превращали раковину из защитного обра-
зования в препятствие движению, и она подвергается рудимен-
тации, а у осьминога — и полному исчезновению. Будучи узко
адаптивной чертой, опа, как было указано выше, преодолевается,
лто позволяет головоногим при выработке сложно построенных
органов чувств и движения и высоко дифференцированной и инте-
грированной центральной нервной системы
достичь па основе типовой организации мол-
люсков высшей степени сложности организа-
ции, имеющейся в этом типе.
В противоположность всем рассмотрен-
ным классам моллюсков раковина Amphi-
neura не представляет единого образования,
а у хитонов (рис. 77) состоит из восьми
пластинок, располагающихся друг за другом
цорзально по длине тела. Такое состояние
едва ли можно производить от единой ра-
ковины всех Conchifera. Скорее у Placopho-
га перед нами особый приспособительный
путь, начавшийся раньше выработки типово-
го строения раковины моллюсков. Для
последнего характерно, в конце-концов, при-
сутствие трех слоев: наружного — periostra-
cum, являющегося неизмененной кутикулой,
среднего — призматического слоя, или
ostracum, образованного известковыми приз-
мами из аррагонита, и внутреннего — пер-
ламутрового слоя hypostracum, состоящего
из тонких известковых пластинок. Послед-
ний выделяется всей поверхностью мантий, а
призматический слой образуется за счет кле-
ток постоянно нарастающего края мантии.
Таким образом структура характерной ра-
ковины моллюсков теснейшим образом свя-
зана с образованием края мантии и, прежде
всего, с отложением в ее кутикуле извести
8 виде призм, поднимающихся над соответ-
ствующими клетками. Образования перла-
мутрового слоя в ряде случаев не происхо-
Развитие раковины у хитонов также тес-
та связано с мантией, которая у них, одна-
та, своеобразно развита в мясистый край
тела — «зону», защищающий бока и распро-
страняющийся не только на заднюю часть
гела; но и прикрывающий голову. Его за-
щитная, функция значительно повышена отложением в кутикуле со
' 'Интай и брюшной сторон как одиночных известковых игол, про-
Рис. 76. Схема организа-
ции Sepia (по Нефу).
I — газовые камеры, 2 —
жабра, 3 — сердце, 4 —
раковина, 5 — целом, 6 —
гонада, 7 — церебральный.
8 — педальный, 9 — висце-
ральный ганглии, 10 — киш-
ка, И — челюсти, 12 —
мантийная полость, 13 —
передняя кишка, 14 — не-
фридий, 15 — радула, 16 —
сифон, 17 — перегородки
раковины, 18. 19 — щупаль-
ца
го
внутреннего и наружно-
кругов, 20 — воронка,
21 — желудок
189
аудируемых каждая одной отдельной клеткой, так и известковых
чешуй различного размера, производимых уже группой однородных
клеток. Восемь пластинок раковины доходят только до «зоны», по
существу представляя сплошные участки отложения извести, в кото-
рых, однако, на местах расположения в .эпителии групп чувстви-
тельных клеток не происходит обызвествления кутикулы. Эти так
называемые мегаль- и микрэстеты как бы пронизывают раковину.
Перед нами, повидимому, отношения более примитивные, чем у про-
чих моллюсков, что вообще характерно для Amphineura. Ostracum
и hypostracum построены у них также иначе.
Своеобразие хитонов выразилось, наконец, в том, что защитное
значение известковых отложений на спинной стороне связано сс
Рис- 77. Схема организации хитонов:
Г— анальное отверстие, 2 — спинной кровеносный сосуд, 3 — жабры, 4 — сердце,
5 — куски раковины. 6 — целом поповой, 7 — .целом перикарда, 8 — гонада. 9 —
церебральный ганглий, 10 — педальный, 11—боковой нервные стволы, 12 — кишка,
13 — печень, 14 — мантийная полость, 15 — нефридий, 16 — рот, 17 — глотка, 18 —
• радула, 19 — зона
свертыванием животного при нападении врага в кольцо по брюш-
ной стороне, наподобие гусениц некоторых бабочек. Такого рода
приспособление подставляет врагу со всех сторон хорошо защи-
щенную спинную поверхность и могло выработаться только у жи-
вотных, еще не имевших сплошной единой плотной раковины, а
лишь более подвижные известковые отложения, но уже дифферен-
цировавших в связи с этим ногу и своеобразный мантийный ком-
плекс. Однако эволюция самой мантии у Placophora пошла, как мы
видели, по иному пути, чем у прочих моллюсков. В этом отноше-
нии характерно и сильное распространение раковины вперед по
телу, почти на голову. У соленогастров вторично, в связи с жизнью
за счет колоний гидроидных полипов, по которым моллюск ползает,
происходит редукция раковины — имеются только отдельные отло-
жения извести в кутикуле наподобие игол. Нога при этом представ-
лена длинным килем, проходящим медиовентрально и имеющим с'
боков узкие бороздки мантии. Большая часть поверхности червеоб-
разного тела образована разрастанием его спинной части. Мантий-
ный комплекс сохраняется лишь в небольшом углублении на зад-
нем конце тела.
В заключение еще раз подчеркнем, что мы проследили разнооб-
разные пути приспособлений ведущего комплекса в организации
190
моллюсков (раковина — нога — мантийные органы) только в
главнейших группах, т. е. в главнейших направлениях эволюции
типа. Но этого достаточно, чтобы видеть, насколько эти направле-
ния разнообразны в соответствии с разнообразием биотопов и био-
ценотических отношений внутри их. И мы можем заключить, что'
хорошо подогнанный, хотя и узкоадаптивный комплекс черт орга-
низации моллюсков дает, в конце концов, массу разнообразнейших
форм, во многих случаях обнаруживающих крайний характер при-
способлений. Только лишь в группе головоногих, с переходом к хищ-
ному образу жизни и с исчезновением раковины, организация шаг-
нула значительно выше.
В связи с рассмотренными приспособительными явлениями, об-
ратимся к внутренней организации моллюсков.
Стенка тела всегда несет однослойный покровный эпителий,
большей частью кутикулярный. Его продуктом, как мы видели,
является и раковина моллюсков. Он сохраняет на многих участках
характер примитивного мерцательного покрова. Особенно сильно
развит мерцательный эпителий у Bivalvia, у которых он покрывает
ногу, мантию, жабры и парус, в связи с поддержанием постоян-
ного тока воды в мантийной полости этих моллюсков, перешедших
к пассивному питанию. Таким образом, перед нами отношения,
близкие с выработавшимися у аннелид, но несколько более при-
митивные.
Это сказывается и в другом компоненте стенки тела, а именно:
покровный эпителий подостлан тканью основного вещества в виде
мезенхимы, близкой к мезенхиме немертин. Мезенхима, как запол-
няющая, но не опорная ткань занимает далее и все простоанство
между различными внутренними органами (рис. 70). Перед нами
как бы антитеза целому аннелид.
Однако в развитии моллюсков, у трохофоры, как мы видели,
всегда имеется закладка правой и левой мезодермальных полосок.
Моллюски, как и аннелиды, принадлежат к настоящим целомиче-
ским животным, но судьба мезодермальных закладок у них не-
сколько иная. Во-первых, от передней части каждой мезодермаль-
ной полоски отделяется часть элементов в виде разрозненных клеток
мезенхимы. Эти клетки дают начало мышечным образованиям кож-
но-мышечного мешка, из которых лишь брюшные сохраняются до
некоторой степени типично в виде кольцевых и продольных мышц
ноги. Спинные же участки особо специализированы в связи с разви-
тием раковины дорзально. К ним мы еще вернемся. Во-вторых, в
Дальнейшем отделе мезодермальной полоски клетки располагаются
Л эпителиальный слой, ограничивающий полость целома, но целом
не обнаруживает ясно выраженной метамерности. Таким образом,
Развивающийся у аннелид терминальный рост тела и целома, с об-
разованием все новых и новых пар мешков, у моллюсков не имеет
места. Целом моллюсков в противоположность полимерному состоя-
нию целома аннелид остается олигомерным в соответствии с огра-
191
ничением терминального роста благодаря образованию раковины
на спинной стороне тела.
В этом отношении интересны исследования П. П. Иванова, стре-
мившегося, как мы видели, показать, что у аннелид первые мета-
меры целома, «личиночные^ сомиты, закладываются несколько ина-
че, чем дальнейшие, «постларвальные» сомиты, а именно: сначала в
мезодермальном зачатке образуется несколько неправильная по-
лость, за счет которой впоследствии дифференцируется от трех до
семи метамер. У моллюсков с этой точки зрения мы могли бы гово-
рить о закладке, соответствующей «личиночным» сомитам мезодер-
мы, и об отсутствии дальнейшей дифференцировки, что ведет в кон-
це-концов к олигомерному целому взрослых Mollusca и ставит их в
резко определенные отношения к близкородственной группе анне-
лид. Эволюция последних пошла по пути выработки червеобразного
тела с терминальным ростом и образованием все новых и новых*'
метамер. Это путь хищных червей, для которых целом являлся необ-
ходимым опорным образованием для мускулатуры стенки тела и
параподий, «пассивным органом движения», по парадоксальному
выражению Нирштраса (1923).
Эволюция моллюсков, как мы видели, определилась совершенно
иными моментами — развитием защитной раковины, ноги и ман-
тийного комплекса органов, т. е. узко адаптивными чертами, вовсе
не связанными с выработкой длинного подвижного тела и с обра-
зом жизни хищника, отсюда и судьба целома оказывается совер-
шенно иной.
Целом представлен среди моллюсков, повидимому, в близком к
примитивному состоянии (рис. 69, 7Q) у Amphineura, причем у со-
леногастров в виде тянущейся дорзально над кишечником по длине
тела неширокой полости, передний отдел которой — парный у
Neomenia и непарный у Chaetoderma — является половым цело-
мом, а задний, короткий — перикардиальным. У Placophora (рис.
77) целом резко и полностью подразделяется на половой и пери-
кардиальный. У Gastropoda (рис. 71) и Bivalvia в некоторых случа-
ях соединение полового отдела с перикардиальным наблюдается, но
целом в общем значительно редуцирован, занимая небольшое про-
странство дорзально. Однако у голово-ногих (рис. 73) снова наблю-
дается более значительное развитие целома, входящего тонким ка-
налом в «сифон». Он охватывает в некоторой мере извивы кишки,
причем в конце-концов происходит дифференцировка широких от-
делов, содержащих гонаду, сердце и экстреторный аппарат, и уз-
ких каналообразных соединительных участков.
В наши задачи не входит прослеживать в подробностях лутй
дифференцировки в связи со специализацией целома. Достаточно ,z
ясно, что целом моллюсков не имеет резко выраженной метамер-
ности и несет черты значительной редукции, располагаясь над ки-
шечником дорзально. Исходным будет иногда наблюдающееся
состояние, когда правый и левый мешки целома охватывают с бо-
ков кишку, но когда дело не доходит до образования брюшного
192
мезентерия, или когда он только намечается, спинной же мезен-
терий хорошо развит, содержа спинной кровеносный сосуд. Кроме
того, целом подразделяется на перикардиальный и половой отделы,
так как гонада, располагающаяся у стенки целома в передней его
части, вторично вдается в него, а половые продукты, подобно тому,
как и у аннелид, падают в его полость.
Эти главные черты целома моллюсков получают полное объяс-
нение в свете тех положений, которые были высказаны выше отно-
сительно целома и мускулатуры аннелид и протоцеломат. Основная
функция целома опорная для окружающей мускулатуры. С эволю-
цией защитной раковины у моллюсков функцйя опоры постепенно
переходит к этому твердому образованию. У Placophora, с их вось-
мичленистой раковиной и удлиненным телом, еще сохраняется типо-
вое наличие мышечных пучков на спинной стороне. Однако они
дифференцируются в специальные мышцы, прикрепляющиеся опре-
деленным образом на участках раковины. Последняя приобретает
вполне естественно значение опорного образования и для мышц,
идущих в «зону», и для мускулатуры ноги, представленной пучка-
ми, которые, проникая в ногу с боков, заканчиваются на различных
участках подошвы. Соленогастры, утрачивая раковину, вместе с
выработкой длинного червеобразного тела вторично восстанавлива-
ют и кольцевую и продольную мускулатуру кожно-мышечного меш-
ка для своих крайне медленных и немногообразных движений.
У брюхоногих при выработке единой спирально завитой раковины
мускулатура спинной стороны тела превращается во втягивающую,
колумеллярную мышцу, которая одним концом прикреплена на ра-
ковине, а дистально подразделяется на пучки, проходящие в боль-
шом количестве в ногу и в меньшем в голову. Первоначально эта
мышца парная (у ряда Diotocardia), но обычно, в связи с общей
асимметрией тела брюхоногих сохраняется и мощно развита лишь
левая мышца, прикрепляющаяся на столбике раковины. Bivalvia
эволюционировали в направлении не только выработки мышц, обус-
ловливающих высовывание, втягивание, подымание и опускание
ноги и гомологичных колумеллярным мышцам брюхоногих, но и в
направлении развития пары мощных аддукторов, идущих от одной
створки к другой, биологически чрезвычайно важных и характерных
Для двустворчатых моллюсков (рис. 74). Кроме того, мелкие мы-
шечные пучки вторично развиваются у них по краю мантии, обу-
словливая его сократимость, и находят свое особо характерное,
иногда крайнее выражение в мускулатуре очень сократимых сифо-
нов. Этими фактами исчерпываются главнейшие пути развития
мускулатуры у Conchifera, и совершенно ясно, что ведущую роль
здесь играло появление и эволюция раковины, как твердого опор-
ного образования.
Несколько иначе обстоит дело у головоногих, но и у них целом
не имеет никакого опорного значения. Как выше было отме-
нено, вырабатывается движение по принципу ракеты, связанное
использованием тока воды из мантийной полости, причем самый
13 Пути эволюции животного мира 1S3
ток производится благодаря образованию мышечной стенки,
гак называемой «мускульной мантии», которая дополняет обычную
мантийную складку, производящую край раковины. Регулируется
же ток образованием воронки за счет задней части ноги (рис. 75)
и дифференцировкой особых мышц в передней части мускульной
мантии, так называемой «затылочной» мышцы. Особенного развития
и сложности достигает этот аппарат при утрате раковины современ-
ными Dibranchiata (рис. 76), становящимися прекрасными пловца-
ми «Tintenfische» — «чернильные рыбы», как их называют немцы.
Но, кроме плавания, головоногие выработали и другого рода
движение — своеобразное ползание на выростах переднего отдела
ноги. Последние являются крайне подвижными и сократительными
придатками благодаря дифференцировке ножной мускулатуры; при-
крепляющейся справа и слева на раковине, а затем и на хрящевой
коробке, которая прикрывает, как защитное образование, централь-
ную нервную систему — мозг головоногих.
Итак, кратко рассмотрев мускулатуру моллюсков, мы видим, что
она развивается всецело в связи с развитием защитной раковины,
принимающей также роль опорного образования характера наруж-
ного скелета. Утрата единства мускулатуры кожно-мышечного ме-
шка при дифференцировке последней в связи с развитием раковины
и ноги, лишает целом основного опорного значения. На первый
план выступают вторичные функции целома. Они уже и обусловли-
вают как значение, так и морфологическое и физиологическое выяв-
ление целома в организации моллюсков.
Мы видели большое многообразие побочных функций целома у
аннелид; его значение у моллюсков резко определенное — он под-
разделяется на отделы половой и перикардиальный, в известной
мере являющийся также экскреторным.
Половая функция присуща целому аннелид в такой же мере,
как и целому моллюсков. В обеих группах гонады закладываются
независимо от целома, и лишь вторично, вместе с разрастанием це-
ломических мешков, они покрываются целотелием, а далее начина-
ют вдаваться в полость тела. У моллюсков они представлены неме-
тамерными правой и левой'гонадами и совершенно не затрагивают
заднего участка целома, тогда как у аннелид такой дифференци-
ровки обычно не наблюдается, и гонады могут быть равномерно
распределены по телу. Кроме того, гонады у моллюсков располага-
ются дорзально, так как брюшная сторона занята образованиями
ноги, а выше кишечником. Это соответствует и лучшей защите го-
над раковиной. Bivalvia представляют в этом отношении исключе-
ние. Гонады, при полной защищенности створками раковины и при
отсутствии внутренностного мешка, проникают в верхнюю часть
ноги. Соответственно расположены и вмещающие гонады парные
мешки целома, обычно называемые здесь яичниками или семенни-
ками.
У брюхоногих наблюдается другая особенность в развитии поло-
вого целома. Парные первоначально мешки уже у Diotocardia раз-*
194
виты различно, а именно — правый значительно-слабее левого, что
является одним из выражений ассиметричного развития всего ман-
тийного комплекса и связанных с ним внутренних органов. У боль-
шинства брюхоногих сохраняется только одна левая гонада и соот-^
ветственно левый мешок полового целома, заключающий ее. У хи-
тонов и головоногих получается также единая гонада, .но другим
путем — путем слияния двух первоначально симметричных зачат-
ков. В результате и вмещающий ее целом, сохраняя симметричное
расположение, становится, однако, непарным, причем парность вы-
водных протоков обычно сохраняется.
Крайне важен тот факт, что и у моллюсков и у аннелид целом
используется для сохранения половых продуктов, падающих в пего
при разрыве целотелиальной выстилки. Это подчеркивает далеко
идущие общйе пути обеих групп.
В том же направлении чрезвычайно интересные отношения вы-
являет дифференцировка перикардиального целома моллюсков.
Задний отдел целома охватывает здесь участок спинного сосуда на
месте впадения в него двух (Placophora — рис. 77 — и Nautilus), а
обычно (рис. 71, 74) одной пары комиссуральных сосудов, по кото-
рым кровь движется от двух, соответственно одной пары жабер
мантийного комплекса. При этом совершенно ясно, что комиссу-
ральные сосуды проходят в самом заднем участке целома, т. е. в
непосредственной близости к расположению жабер.
Только здесь обнаруживаются характерные черты целомического
комплекса аннелид, т. е. ближайшая взаимосвязь целома и целоми-
ческой мускулатуры, совершенно утраченная в других участках при
потере целомом опорной функции и при специализации мышечных
пучков. Даже в развитии моллюсков мы видели, что мускулатура
образуется за счет расхождения части клеток мезодермальных по-
лосок, дающих «вторичную» мезенхиму. Рассматриваемый задний
отдел целома сохраняется у моллюсков в связи с такой его функ-
цией, которая у аннелид совершенно не привлекает к себе внима-
ния, а именно—с образованием за счет спинного мезентерия целома
пропульзаторпого аппарата кровеносной системы, т. е. мускулатуры
спинного сосуда и прилежащих частей комиссуральных сосудов.
Этот факт очень знаменателен потому, что мы с ним столкнемся в
Другом типе животного царства и при совершенно иных условиях,
а именно — у членистоногих.
Последний штрих, завершающий наш анализ целома моллюсков,
касается вопроса об его олигомерности. Наличие комиссуральных
сосудов — это показывает организация аннелид — обычно связано
с присутствием пластинки ткани основного вещества, находящейся
в Диссепименте и предопределяющей возможность образования кро-
ВеноснОго сосуда. В дальнейшем развитие у аннелид жабры на
соответствующей петле комиссурального сосуда определяет в ряде
случаев и самый сосуд. Основываясь на этом, мы можем, по присут-
ствию одной или двух пар жабер и соответствующих комиссураль-
Ь1Х сосудов, заключать о наличии у моллюсков в заднем отделе
195
тела одного' или двух диссепиментов, что соответствует двум или
трем парам мешков целома. Как и личиночные сомиты аннелид, эти
целомические сомиты моллюсков были, позидимому, нерезко выра-
жены и слабо дифференцированы. Лишь при терминальном росте
аннелид произошла полная дифференцировка дальнейших пар
целомических мешков. Это — шаг, не пройденный моллюсками при
выработке их организации, остановившейся на трех или двух слабо
дифференцированных сомитах.
Нет оснований принимать вместе с Пельзенером и Нефом, что
у моллюсков произошла -вторичная редукция полимерного целома,
т. е., другими словами, что предками моллюсков были типичные
аннелиды, близкие к Eunicidae из полихет, как это утверждает
первый из них. Редукции полимерного целома не наблюдается в
эмбриональном развитии. Нельзя в этом направлении истолковы-
вать и данные сравнительной анатомии, а именно: увеличение
количества пар жабер у хитонов представляет собой местное вто-
ричное явление, не сопровождающееся увеличением соответственных
пар комиссуральных сосудов (рис. 77). Наконец, и это не менее
важно, моллюски не обнаруживают никаких данных, которые эко-
логически могли бы объяснить вторичную редукцию метамерного
строения аннелидных форм.
Уже у аннелид нами было отмечено, что ткань основного веще-
ства можно назвать антитезой целома. И вполне естественно, что
целом при потере им функции опоры не может взять на себя функ-
ции заполняющей ткани. Происходит редукция целома. Последний
вытесняется соединительной тканью с ее студенистым основным ве-
ществом, с ее волокнистыми структурами, с ее клеточными образо-
ваниями, с кровеносными сосудами, являющимися в ней участками
с жидким основным веществом. Этот процесс можно наблюдать у
аннелид и пиявок, с ним мы встретимся и у вышестоящих типов.
Итак, в известной мере коррелятивно с целомом развивается >
соединительная ткань, заполняющая все пространства между раз-
личными органами в теле моллюсков (рис. 70) и обусловливающая !
возможности развития кровеносной системы. Здесь выявляется зна- ;
чимость кровеносной системы предков моллюсков в двух отноше-
ниях. Во-первых, трофическая ее функция была, как и уряда анне-
лид, доминирующей нгд такой же функцией целома. Благодаря
этому целом мог утратить или очень ослабить ближайшие отноше-
ния к усваивающей кишке, откуда он получал питательный мате-
риал. Во-вторых, уже у предков моллюсков кровеносная система
имела не только трофическую, ио и функцию газового обмена я
связи с образованием типичных жабер в виде ктенидиев.
Благодаря этим двум типовым чертам кровеносной системы со-
хранился перикардиальный целом моллюсков со вторичной фун^'
цией пропульзаторного аппарата. Но эти же черты указывают
возможность редукции тех частей целома, которые прилежали *
усваивающей кишке и образовывали брюшной мезентерий. Крове'
ноская система в связи с этим в известной мере теряет резкост
1S6
своей дифференцировки, какая наблюдалась у аннелид. При диф-
фуз пости ткани основного вещества, сосуды на большой части сво-
его хода имеют характер лакунарных и ян юобЛ'-ных прост-
ранств. В КОНЦе-КОНЦОВ собираюишхся В " "ЛГИ:? синус,
дающий полую вену. Эти образования х? < гравя я пъ с брюш-
ным сосудом аннелид. От них берет начали одна или две пары со-
судов к соответственным жабрам. Тип самих жабер близок к ха-
рактерным жабрам полихет. Части комиссуральных сосудов, отхо-
дящие от жабер, являются наиболее хорошо выраженными наравне
с частью спинного сосуда, в который они впадают — сердце мол-
люсков. Весь комплекс предсердий и сердце располагается, как мы
видели, в соответствии с положением жабер мантийного комплекса,
т. е. занимая самое заднее положение, что обусловливает, как было
отмечено, дифференцировку заднего отдела целома в виде перикар-
диального целома.
Часть спинного сосуда, обычно идущая от сердца вперед, выра-
жена довольно хорошо (рис. 68, 71). Но ее ведви уже теряют свою
определенность, переходя в систему лакунарных пол- тей. Задний
конёгГспинного сосуда — отходящая от сердца зад”.ы аорта —
выражен слабо, так как и снабжаемый ею отдел отель невелик.
В связи с развитием мантии компенсаторно бывают выражены ман-
тийные сосуды. В некоторых случаях они развиваются очень значи-
тельно, принимая на себя по своему положению и взаимоотноше-
ниям с внешней средой функцию газового обмена. У Pulmonata
это ведет в конце-концов к развитию за счет капилляров мантий-
ной полости так называемого легкого моллюсков, заменяющего
жабру при переходе к наземному существованию.
Другая особенность брюхоногих, стоящая в связи с уже отме-
ченной выше общей асимметрией органов мантийного комплекса,
выявляется в редукции у Monotocardia правой жабры и комиссу-
рального сосуда. Это ведет к рудиментации правого предсердия, а
затем и к полному его исчезновению. У Nudibranchiata с исчезнове-
нием раковины развиваются ничего общего не имеющие с ктениди-
ями вторичные жабры в виде выростов спинной стороны тела, чем
сильнейшим образом изменяется и ход соответствующих сосудов.
Единственный в своем роде случай представляют Scaphopoda с
полной редукцией жабер и кровеносной системы, причем в виде
Рудимента сохраняете^ мешочек перикардиального целома.
Напротив, особенно сильного развития с хорошей выражен-
ностью всех сосудов достигает кровеносная система головоногих,
То вполне соответствует развитию и дифференцировке мускулату-
Р1 У подвижных хищных каракатиц, спрутов и т. д. Отметим здесь
СтКЖе.Поязление у них образований, экскреторного характера в
енке целома. Они располагаются на предсердиях и венозных при-
-ах, Кеся ФУНК1Щю извлечения жидких продуктов распада из
Р°3и, что в сильной степени специфично для головоногих.
Кой аконец» своеобразные взаимоотношения сердца с задней киш-
Развиваются у Diotocardia (рис. 71) и Bivalvia (рис. 74). Зави-
197
сит это от положения жабер, которые фиксированы в мантийном
комплексе, кишка же постепенно смещается дорзально в соответ-
ствии с постепенным смещением анального отверстия. Получается
сердце «пронзенное» кишкой, причем налицо все переходы от типо-
вого его положения над кишкой.
Итак, вместе со значительной редукцией целома у моллюсков
их кровеносная система выпадает из-под его ведущего влияния.
В связи с развитием целома у моллюсков развиваются, наконец, и
экскреторные органы (рис. 68, 69). Представленные первоначально
парой таких же протонефридиев в заротовом отделе трохофоры,
как и у трохофоры аннелид, они далее подвергаются у взрослой
формы подной редукции. В качестве экскреторных органов функ-
ционируют здесь одна или две (Nautilus, Chiton cojetanus) пары
метанефридиев, которые определяются в своем положении тем, что
их парные отверстия входят в состав мантийного комплекса. Эти
одна-две пары метанефридиев соответствуют одной-двум парам жа-
бер и принадлежат, следовательно, двум-трем парам дифференци-
рующихся мешков целома.
Здесь мы не будем касаться вопроса о происхождении метане-
фридиев, он был рассмотрен при анализе организации аннелид.
Затруднение появляется в том, гомологичны ли метанефридии в
обеих группах или перед нами случай конвергентного развития, так
как, повидимому, у свсбодноживущих полихет можно проследить
весь процесс эволюции от типичного протонефридия или через
нефромиксин или через открытый нефридий к метанефридию.
Однако не исключена возможность образования метанефридия, ха-
рактерного для Eunicidae, и у предковых форм, общих обеим груп-
пам и рано отошедших от протоцеломат.
У хитонов имеется резко выраженная дифференцировка перед-
ней и задней пар метанефридиев. В то время как первая из них
функционирует в качестве половых протоков, вторая достигает очень
значительного развития в качестве экскреторного органа (рис. 77).
и перикардиальный целом у них никогда не содержит половых
продуктов. Напротив, у соленогастров сохраняются только задние
нефридии, но они несут и половую и экскреторную функции, а пе-
рикардиальный целом не дифференцирован от полового — через
него половые продукты выводятся далее в нефридии. Подобные отно-
шения имеются также у Gastropoda и Bivalvia. У них развивается
одна пара нефридиев. Но разница в том, что половой целом совер-
шенно обособлен от перикардиального, и реноперикардиальное от-
верстие ведет в нефридий с одной стороны, а с другой — туда л<е
открывается половой целом (рис. 71). У Bivalvia сохраняется еще
связь обоих целомов в виде канала, ведущего из перикардиального
целома в участок, где открывается в нефридии половой целом. По-
следний шаг дифференцировки выражается в полном подразделе-
нии полового и экскреторного выводных отверстий.
Не вдаваясь в дальнейшие подробности, можно констатировать,
что у Mollusca в эволюции экскреторного аппарата видны
198
два пути: один — с сохранением и с дифференцировкой двух пар
метанефридиев, другой — с редукцией, передней пары и с пере-
ходом обеих функций (и половой и экскреторной) к задней
паре.
Таким образом, комплекс целома и связанных с ним систем ор-
ганов у моллюсков продолжает играть важную роль в организации,
хотя по сравнению с целомом аннелид он отошел- на задний план
по своему объему и связям с другими органами. В этом аспекте
кишечник моллюсков представляет очень своеобразные отношения
(рис. 68, 70). Обычно только задний отдел средней кишки прохо-
дит под перикардиальным целомом. У некоторых Diotocardia и
Bivalvia он уже вдается в него. У головоногих ряд извивов кишеч-
ника расположен, наконец, вторично в целоме. Однако нет ничего
подобного охватыванию целомом кишечника на всем его протяже-
нии, что имеет место у аннелид.
Возможным объяснением этой независимости кишечника от це-
лома является прежде всего увеличение железистой и усваивающей
поверхности кишечника путем образования присущих всем моллю-
скам правого и левого мешков печени (рис. 68—70). Последние до-
стигают очень большой мощности и, становясь крайне характер-
ными для всей передней части эндодермальной кишки, в высокой
степени ограничивают возможность развития целома, отбрасывают
его, так сказать, в самый задний участок тела. Соответственно
развивается и другая особенность, а именно: у кишечника или во-
круг него появляется полость лакунарного характера, у брюхоногих
часто довольно сильно выраженная. Она входит уже в состав кро-
веносной системы.
В общем на кишечнике моллюсков лежит отпечаток приспособ-
ления исходных форм к особым условиям питания. На это указы-
вает рассмотренное выше увеличение усваивающей поверхности
образованием выростов печени, а также и другая характерная при-
способительная черта кишечника моллюсков — развитие в глоточ-
ном отделе с брюшной стороны мешковидного впячивания, несущего
на мускулистом валике особый аппарат кутикулярных зубцов, так
называемая радула или терка (рис. 68, 71). Она служит для пере-
тирания пищи. '
Радула, встречающаяся с огромным постоянством у некоторых
гРупп моллюсков, представляет узкоприспособительную черту ор-
ганизации, обнаруживающую большое разнообразие и присущую
Уже первичным их формам. Пельзенер считает возможным на этом
основании говорить даже о происхождении моллюсков от полихет,
й и частности от семейства Eunicidae, которое обладает действи-
тельно очень сходным с радулой вентральным глоточным мешком,
несущим кутикулярные зубцы «челюстей». Однако этого признака
ЗДя решения вопроса, конечно, недостаточно, хотя сходство край-
не соблазнительно. - *
Переход к пассивному питанию у Bivalvia вторично влечет за
обою исчезновение радулы, но вызывает сильное развитие пару-
199
•са, представляющего как бы оттянутые в бока углы рта с боль-
шой площадью мерцательного покрова (рис. 74). С другой сто-
роны переход к образу жизни хищников у головоногих вызывает
дифференцировку, кроме радулы, еще и мощного аппарата челю-
стей, напоминающих клюв попугая (рис. 75, 76).
Особняком стоит крайне оригинальное защитное приспособле-
ние, связанное у каракатицы и близких форм с задней кишкой и
являющееся почти единственным в своем роде в животном царст-
ве — это чернильная железа.
Независимо от комплекса целома развиваются органы чувств
и нервная система моллюсков (рис. 69, 68).
У брюхоногих наиболее сохранились типовые отношения го-
ловного комплекса органов чувств, как мы их видели уже у раз-
личных подтипов червей (рис. 71). Хорошо развитый головной
отдел тела несет пару щупалец — механорецепторов, и пару
илаз, причем у Stylommatophora из Pulmonata они располагаются
на щупальцеобразных выростах головы. Но хеморецепторы как бы
вынесены в другое место и специализированы для другой цели,
а именно — в мантийном комплексе органов развиваются так назы-
ваемые осфрадии, которые помещаются при основании каждой
жабры и служат для ориентировки в химизме воды, омывающей
последние.
У хитонов (рис. 77) бороздки чувствительного эпителия име-
ются по бокам ноги в мантийной складке, ближе к голове. Но го-
лова у них вообще оказывается, как мы видели, слабо развитой г
связи с тем, что мантийная складка, а соответственно и раковине
получают своеобразное развитие, прикрывая головной отдел тела.
В результате головной комплекс органов чувств редуцируется,
зато развиваются органы чувств, расположенные в покрове спин-
ной стороны тела в области раковины — микро- и мегалэстеты
несущие функцию механо- и фоторецепторов. Случай единственный
в своем роде у моллюсков, тесно связанный и с особенностями ус-
тройства 8-членистой раковины хитонов.
По сходному пути редукции головного комплекса органов
чувств пошли также Bivalvia (рис. 74). У них, как мы видели,
полное исчезновение головного отдела связано с развитием двух
створок раковины, полностью покрывающих тело моллюска. Необ-
ходимые механо-, хемо- и фоторецепторы компенсаторно развива-
ются совершенно независимо от головного комплекса органов
чувств там, где сосредоточиваются взаимоотношения с внешней
средой, т. е. по краю мантии и на сифонах. Являясь приспособи-
тельными новообразованиями, возникающими путем дифференци-
ровки чувствительного эпителия края мантии, они очень различно
выражены у разных двустворчатых моллюсков вплоть до довольно
сложных щупалец и глаз Pecten включительно.
Перешедшие к хищничеству головоногие обычно имеют в вы-
сокой степени сложно построенные глаза (рис. 79) и в качестве
хемо- и механорецепторов очень многочисленные присоски восьми,
200
соответственно десяти, рук каракатицы или спрута (рис. 76).
У Nautilus отаошшшя бил ее примитивны (рис. 75, 78). Глаза пред-
ставлены еще парой простых ямочных глаз, далее имеются щу-
пальца, соответствующие щупальцам или эпиподиальным выро-
стам по бокам ноги у Diotocardia из брюхоногих и, наконец, мно-
гочисленные руки развиты гораздо слабее, чем у прочих голово-
ногих. Мы не будем касаться более подробно всех этих образо-
ваний. Отметим лишь, что пара глаз головоногих представляет
Рис. 78. Ямочный глаз
Nautilus (из Ланга по Ген-
зену):
I — глазная ямка, 2 — ганг-
лиозные клетки. 3 — зрительный
нерв, 4 — пигментный слой, 5—
светочувствительный слой, 6 —
ретинальные клетки
Рис. 79. Глаз Sepia (из Ланга по Ген-
зену) :
1 — передняя камера глаза, 2 — цилиарная
мышца, 3 — роговица, 4 — хрящевая капсу-
ла, 5 — ретина, 6 — оптический ганглий, 7 —
радужина, 8 — хрусталик, 9 — зрительный
нерв
высшую ступень дифференцировки относительно простых фоторе-
цепторов, имеющихся в головном комплексе, например каких-либо
брюхоногих и очень близких по строению к глазам некоторых
Eunicidae. Глаза осьминога по своей сложности не уступают гла-
зам позвоночных, с которыми имеют и очень большое сходство
строения в результате конвергентного развития, но именно общее
сходство при разнице в деталях строения каждой отдельной черты.
Нервная система моллюсков развивается, как говорят, функ-
ционально, т. е. в теснейшей взаимосвязи с органами чувств и
мУскулатурой. Поэтому, наряду с типовым развитием головного
мозга, близким к состоянию мозга хотя бы Eunicidae, она в своем
туловищном отделе имеет совершенно своеобразные отношения,
развита здесь в соответствии с характерной дифференциров-
кой ноги и мантийного комплекса.
От головного мозга у хитонов, солеяогастров и ряда Diotocar-
ша отходят две пары нервных стволов (рис. 69,Б, 70, 77). Из
201
них одна пара — педальные стволы соленогастров, хитонов и не-
которых прозобранхов — заложена в соединительной ткани среди
мускулатуры ноги и является двигательным отделом центральной
нервной системы моллюсков. Другая пара — боковые стволы со-
леногастров и хитонов — располагается выше, возле мантийной
складки, и имеет в значительной мере характер сензорного отдела,
связанного с образованиями края мантии и мантийным комплек-
сом органов.
У большинства моллюсков имеется дальнейшая дифференци-
ровка, идущая, как и у аннелид, в направлении обособления ган-
глиозных центров и коннективов, т. е. усложнения связей между
ганглиозными элементами (рис. 68, 71, 73, 74). Вместо боковых
стволов образуются ганглии: плевральные близ мозга, париеталь-
ные близ осфрадиев и висцеральные у задней кишки с связываю-
щими их плевро-висцеральными коннективами, а вместо педаль-
ных стволов — педальные ганглии.
Казалось бы, наличие двух пар продольных стволов у более
примитивных форм моллюсков вполне естественно производить от
типовых отношений, присущих нервной системе турбеллярий сих
несколькими парами продольных стволов. Так это и делают Тиле,
Шторх, Ганстрем и др. Однако тесная филогенетическая связь
моллюсков с аннелидами и протоцеломатами не позволяет при-
нять такое понимание отношений у моллюсков. Скорее приходится
признать, что у них из состояния, подобного немертинам — анне-
лидам, т. е. из двух продольных нервных стволов брюшной цепоч-
ки, вторично получались характерные две пары.
Неф (1923) поэтому принимает, что пара боковых, плевровис-
церальных стволов моллюсков гомологична стволам брюшной це-
почки аннелид, а педальные стволы представляют новообразова-
ния, связанные с дифференцировкой передней части брюшной
поверхности тела в мускулистую ногу. Этому представлению реши-
тельно противоречит тот факт, что брюшная цепочка аннелид со-
держит главным образом двигательные нейроны, чего как раз
нет v плевровисцеральных стволов моллюсков.
Шторх (1913) правильнее оценивал факты, принимая, что
брюшная цепочка аннелид соответствует педальным стволам
моллюсков. Плевровисцеральные стволы он приравнивает боко-
вым стволам, имеющимся среди аннелид только в семействе
Amphinomidae, и создает свое представление о тетранервных фор-
мах. К последним он относит Mollusca и Amphinomidae, происхо-
дящих, по его мнению, непосредственно от турбелляриеподобных
предков. Мы уже отметили, при рассмотрении организации анне-
лид, неправильность обобщений Шторха относительно боковых
стволов Amphinomidae и показали, что это лишь местная диффе-
ренцировка прежде всего в подвальной, т. е. главным образом
сензорной части нервной системы полихет. . Аналогично и у мол-
люсков боковые нервные тяжи связаны прежде» всего с чувстви-
тельными образованиями по краю мантии и с мантийным ком-
202
плексом органов. Конвергентно на этой основе, совершенно неза-
висимо в обеих группах, развиваются боковые нервные стволы,
не имеющие ничего общего с продольными нервными стволами
двигательного по преимуществу характера у турбеллярий, а у ан-
нелид дифференцировавшиеся из сензорного кожного сплетения
только в одном семействе Amphinomidae.
Едва ли можно допустить на основании наблюдающегося у
Chaetoderma слияния обоих стволов соответственной стороны в
задней части их хода, что из состояния подобного немертинам-
апнелидам, т. е. из двух продольных стволов, вторично дифферен-
цировались характерные для моллюсков две пары. Прежде всего,
соленогастры являются вторично крайне измененными формами из
Amphineura, и слияние двух стволов наблюдается только у Chaeto-
derma. Затем против этого говорит двигательный по преимуществу
характер продольных стволов немертин-аннелид.
Рассматривая нервную систему моллюсков, нельзя не упомя-
нуть о крайне оригинальной «хиастонейрии» брюхоногих, особенно
ярко выраженной у прозобранхов (рис. 71). Это — явление пере-
креста плевро-висцеральных коннективов, так что левый из них
проходит под кишкой и направляется к париетальному ганглию,
расположенному справа, а правый коннектив идет над кишечни-
ком к такому же левому ганглию. Мы берем типичный случай—
Diotocardia. Еще Шпенгель (1880) связал перекрест с явлением
перемещения мантийного комплекса органов у прозобранхов по
правой стороне тела из первоначального положения в такое,
когда все образования мантийного комплекса обращены вперед,
смещены, следовательно, на 180° по сравнению с типовым располо-
жением их у моллюсков и.у Monotocardia в то же время еще по-
вернуты на 90° по отношению к продольной оси. Адаптивное зна-
чение этого явления, в связи с развитием завитка конической ра-
ковины, выше было выяснено. Здесь для нас важно, что переме-
щаются и париетальные ганглии, а в результате происходит сме-
щение связанных с ними частей плевровисцеральных коннехтивов
при остающихся в прежнем положении плевральных ганглиях. Эти
факты еще раз подчеркивают значение мантийного комплекса
органов для развития боковых стволов центральной нервной
системы. Педальные стволы сохраняют свой ход в ноге Proso-
branchia неизменным.
Итак, у моллюсков имеет место сохранение типовой пары про-
дольных педальных стволов, обусловленных прежде всего диффе-
ренцировкой мускулатуры ноги, и появление ч боковых стволов,
связанных по преимуществу с дифференцировкой чувствительных
образований мантии, с одной стороны, и мантийного комплекса
(оссЬоадии) с другой.
Развитие педальных стволов нервной системы у головоногих
Моллюсков дает дальнейшие интересные факты, выявляющие неко-
торые стороны ее эволюции.
Главная часть ноги превращается у большинства из них в 8 или
203
10 щупалец — рук, служащих как чувствительные и двигательные
придатки, так и являющихся органами нападения и защиты.
В соответствии с этим па брюшной поверхности рук осьминога
или каракатицы располагается большое количество присосок
(рис. 76), играющих в то же время роль очень чувствительных
хемо- и механорецепторов. А мускулатура руки дифференцируется
в пучки, обусловливающие общую ее подвижность, и в специаль-
ные мышцы, связанные с функционированием присосок. В резуль-
тате в каждой руке проходит мощный нерв — n. brachialis, име-
ющий характер ганглиозного тяжа, который развивается в зависи-
мости от развития чувствительных образований присосок. Ган-
глиозные клетки первоначально расположены равномерно по всей
его длине, но у ряда форм происходит дифференцировка в направ-
лений развития на нем особых ганглиев соответственно с располо-
жением присосок.
Далее, в руке развиваются и небольшие двигательные ганглии,
стоящие в связи с ганглиями n. brachialis, а именно — субацета-
булярные и интермускулярные ганглии. Таким образом рука по-
лучает самостоятельные рефлекторные пути и с выработкой ган-
глиозных центров приобретает значительную автономность в сво-
их движениях.
В параподиях полихет мы видели образование мелких чувстви-
тельных ганглиев по ходу среднего чувствительного нерва соми-
та. Видели далее образование параподиального ганглия — мощ-
ного скопления чувствительного характера, однако до выработки
самостоятельных рефлекторных аппаратов дело не доходило. У го-
ловоногих с многообразием и важностью функций рук для хищ-
ника происходит значительное осложнение и дифференцировка их
сензориума и моториума. В результате разделения физиологиче-
ской деятельности соответственно появляются чувствительные и
двигательные ганглии, ведущие к выработке в руке особых рефлек-
торных путей. Конечным этапом в эволюции рук головоногих
являются гектокотилизированпые руки Argonautcg достигающие
такой степени автономности, что отрываясь от тела моллюска,
совершенно самостоятельно плавают и производят оплодотворение
самки.
Наряду с этим процессом, не имеющим себе подобного в живот-
ном царстве и обусловленным далеко пошедшим разделением
функций под ведущим действием естественного отбора, мы видим
у головоногих и процесс интеграции, выражающийся в объединении
всех п. п. brachiales в одной большой массе — ganglion brachiale
у основания рук (рис. 80). Последний непосредственно связан уже
с педальным ганглием. Далее происходит концентрация и типичных
педальных и висцеральных ганглиев в одну большую массу мозга,
охватывающую пищевод и сливающуюся с церебральным ган-
глием. Таким образом создается чрезвычайно централизованная
неовная система, получается возможность развития сложнейших
межнейрональных связей с соответственном развитием сложной
204
нервной деятельности. При этом мэ°г головоногих защищен хряще-
вой капсулой наподобие того, как это имеет место у позвоночных.
Головной мозг моллюсков, представленный церебральным ган-
глием (рис. 69, 68) обнаруживает первоначально, у исходных форм
вроде прозобранхов, или брюхоногих вообще, типовые черты, кото-
рые уже были нами выявлены для немертин и аннелид. А именно,
наличие у этих моллюсков щупалец, как механо- и хеморецепторов,
глаз, а в ноге статорецепторов обусловливает соответственно суще-
ствование нервных скоплений церебрального ганглия, имевшихся
уже у червеобразных предков. Они
располагаются под покровом в соеди-
нительной ткани головы. Сюда же
присоединился центр стоматогастриче-
ской системы.
Не повторяя уже сказанного по
этому поводу для червей, обратимся к
отношениям, имеющим место у голово-
ногих. Выше было описано своеобраз-
нейшее развитие рук с их хемо- и ме-
ханорецепторами, играющими в жизни
хищника — головоногого — огромную
роль. На долю головы остались фото-
рецепторы. Эволюционируя из состоя-
ния простого ямочного глаза Nautilus,
они достигают в глазах высших пред-
ставителей класса чрезвычайной слож-
ности и больших размеров \ играя ве-
дущую роль в жизни каракатицы или
спрута (рис. 79).
Неинвентированный глаз, обладаю-
щий очень совершенным диоптрическим
аппаратом, защищенный хрящевой кап-
сулой и приводимый в движение рядом
специальных мышц, сходен в общих
чертах и не уступает по сложности строения инвертированному
глазу позвоночных, но совершенно своеобразен в деталях сзоего
конвергентного развития. Соответственно и в нервных образованиях
глаза за светочувствительным слоем ретинальных клеток распола-
гается толща связанных с ними ганглиозных элементов, состоящая
из двух слоев нейронов «глубокой ретины» (retina profunda), и
Далее, «конгломерата» нейронов оптического ганглия (ganglion
opheum). Таким образом развивается большой сложный ганглиоз-
ный центр зрительной доли (lobus opticus), которая отдает мощ-
ные пучки нервных волокон в головной мозг.
Рис. 80. Центральная нервная
система Argonaut i ar jo (по
Пфеферкорну из Ганстрема):
1 — буккальный ганглий с губны-
ми нервами, 2 — брахиальный ган-
глий с нервами рук — 3, 1 —- не-
ребральный га»ггт1й с верхней лоб-
ной (5\ и теменной (6) долями, 7 —
педальный ганглий, — висцераль-
ный ганглий, 9 — зрительный нерв.
1 Головоногие имеют самые большие в животном царстве глаза по их абсо-
' ютным размерам и по их относительной величине, по сравнению с размерами
и весом тела.
205
Однако получающийся в результате осложнения и детализации
функций сложнейший чувствительный ганглиозный комплекс не
ограничивается периферией. В церебральном ганглии дифференци-
руется вторичный зрительный центр в виде ganglion pedunculare,
стоящего в связи с различными частями головного мозга.
Как видно, вместе со сложным развитием глаза гигантским ша-
гом идет дифференцировка ганглиозных центров. К этому остается
добавить, что в голов-ном мозге у головоногих развиваются и особые
высшие ассоциативные центры в виде передней и задней фронталь-
ных, передней и задней базальных и вертикальных долей (рис. 8Э).
Подведем итоги рассмотрению организации моллюсков.
Если у аннелид мы должны были говорить о многообразии пу-
тей, по которым протекает их приспособительная эволюция, и в
связи с этим могли видеть широкие возможности повышения их ор-
ганизации, что делало их важнейшей узловой группой животного
царства, то по отношению к их ближайшим сородичам — моллюс-
кам картина представляется совершенно иной. Вся организация их
идет под знаком выработки узкоадаптивного защитного образова-
ния — раковины, хотя и свободноживущими, но нехищными фор-
мами.
Во внешней морфологии развитием раковины обусловлено обра-
зование ноги, мантийной складки и мантийного комплекса. Во в ут-
реннем строении в теснейшей взаимосвязи с раковиной стоит диф-
ференцировка мускулатуры, редукция целома и развитие соедини-
тельной ткани. Но далее, в связи с целомом находится кровенос-
ная система с органами дыхания, происходит развитие экскретор-
ного аппарата и половых органов. Так же, вероятно, благодаря
образованию раковины возникает и олигомерность целома. Наконец,
развитие раковины налагает свой отпечаток на дифференцировку
органов чувств, а в тесной взаимосвязи с ними и с мускулатурой
стоит своеобразие нервной системы моллюсков. Только кишечник
проходит в известной мере особо свою приспособительную эволю-
цию. Новые пути для узко-адаптивной организации моллюсков,
хорошо подогнанной к довольно различным условиям жизни, а по-
тому дающей обильнейшее разнообразие приспособлений, становят-
ся возможным и при вторичной утрате раковины, что нередко и про-
исходит в разных группах моллюсков. Но значительный прогресс и
Повышение организация имеют место только в классе головоногих,
становящихся мощными подвижными хищниками. Таким образом
пластичность организации не утрачивается, но узлового положения
в смысле развития новых типовых форм моллюски уже не имеют.
Какие же выводы относительно филогении моллюсков дает наш
анализ их организации?
Выше не раз указывалось, что, основываясь на специальном изу-
чении архаических форм моллюсков, известный знаток этого типа
Пельзенер считал возможным производить их непосредственно от
полихет, в частности от семейства Eunicidae. Также и Неф (1924,
1931), базируясь на исследованиях, с одной стороны, над Lepidop-
206
leurus cajetanus, а с другой — над цефалоподами, приходит к за-
ключению о ближайших связях моллюсков с подахетами. Он гово-
рит, что мягкотелые произошли от древнейших аннелид и что пере-
ходными формами были ползающие, небольшие черви с уменьшен-
ным числом сегментов (однако, по крайней мере, тремя), похожие ч
по внешнему виду на Dinophilidae или Myzostomidae. Напротив,
Тиле поддерживает свои прежние предположения о непосредствен-
ной связи соленогастров с турбелляриями.
Анализ организации Mollusca показывает, что с турбелляриями
их сближать, конечно, нельзя. Трубчатый кишечник' с ротовым от-
верстием у переднего конца тела и хорошо развитая кровеносная
система определенно указывают на то, что предками моллюсков
были червеобразнйе формы, стоявшие по организации не ниже не-
мертин. Точнее определяются филогенетические связи развитием у
моллюсков целомического зачатка, дающего половой и перикар-
диальный отделы вторичной полости тела. Следовательно, перед
нами дальнейший этап по сравнению с немертинообразными чер-
вями — этап целомических червей. К нему, мы видели, относятся
и аннелиды. Но в отличие от аннелид, обладающих полимерным
целомом и характеризующихся терминальным ростом тела с ново-
образованием сомитов, моллюски имеют олигомерный целом и не
обнаруживают терминального роста, как не обнаруживают и обра-
зования новых сомитов; более того — они не обнаруживают и
чего-либо подобного процессу вторичной редукции полимерной за-
кладки целома.
Таким образом, можно прийти к выводу, что моллюски произо-
шли от олигомерных исходных форм целомических червей—Proto-
или Archicoelomata, являвшихся потомками немертинообразчых
форм, еще лишенных целома. Это были придонные, свободноживу-
щие, но сравнительно малоподвижные, нехищные, весьма возможно
растительноядные, червеобразные организмы с относительно ко-
ротким телом, в противоположность предкам полихет, являвшихся
хищниками с длинным подвижным телом и терминальным энер-
гичным ростом.
Однако и Mollusca и Polychaeta прошли значительный этап
своей эволюции совместно, как это видно из эмбрионального и
постэмбрионального развития. Включая трохофору, это был общий
^Уть. Следовательно, отношения между обеими группами самые
тесные. Лишь с дифференцировкой целомического зачатка начи-
нается их расхождение — в то время как у хищных предков поли-
хет вырабатывается терминальный рост, а в «вязи с ним полимер-
ный целом и параподиальные комплексы, у нехищных предков
Моллюсков нет ни того, ни другого, а развивается защитная рако-
вина, мантийный комплекс, нога. Целом сохраняет свои примитив-
ные отношения в смысле развития только таких участков, которые
соответствуют личиночным сомитам полихет с их нерезко выра-
снным разграничением. Однако с утратой своей главной, основной
Функции опоры для мускулатуры кожно-мышечного мешка, с пре-
207
вращением части последней в специальные мышцы, использующие в
.качестве опоры раковину, целом Mollusca подвергается вторичной
редукции. Он сохраняется лишь постольку, поскольку сохраняли
значение его вторичные функции — половая, экскреторная и «мы-
шечная», или, правильнее, пропульзаторная, для спинного сосуда в
участке его, взявшем на себя роль сердца и связанном с мантий-
ным комплексом. Целом редуцируется также постольку, поскольку,
теряя свою трофическую функцию, вытесняется и оттесняется назад
парой боковых разрастаний кишечника, образующих так называе-
мую печень моллюсков.
Итак, предками моллюсков были те Protocoelomata, которые
пошли в своей эволюции по линии аннелид, в противоположность
Другой их ветви, пошедшей в направлении хордовых и иглокожих,
о чем речь будет при анализе организации этих типов. Здесь мы не
будем предвосхищать дальнейших выводов в этом отношении. Кон-
кретизируем лишь некоторые черты организации общих предков
аннелид и моллюсков. У этих протоцеломат выработался уже спи-
ральный тип дробления, детерминированные закладки (2d — пер-
вичный соматобласт и 4d — целомезобласт), личинки трохофора,
а у взрослого: олигомерный целом, жабры типа ктенидиев, глоточ-
ный мешок, наконец, — метанефридии. И как раз три последние
черты организации заставляют задуматься над их значением, так
как у полихет мы могли проследить их возникновение среди группы
Polychaeta errantia, и, в частности, выявить их эволюцию до пол-
ного выражения в семействе Eunicidae. Пельзенер, как было указа-
но, шел в этом направлении до конца, производя моллюсков от
Eunicidae и не придавая решающего значения другой категории
фактов, а именнб—образованию олигомерного целома и отсутствию
терминального роста у Mollusca. Однако легче допустить даже
конвергентное развитие ктенидия и перистой жабры Eunice, радулы
и глоточного мешка с челюстями у Eunicidae, наконец, образование
у тех и других метанефридиев. На этом мы и остановимся, предо-
ставляя окончательный ответ на вопрос будущим исследователям.
Глава X
ТИП ЧЛЕНИСТОНОГИЕ — ARTHROPODA
Хитииизация покрова как ведущая черта организации. Расчленение
кутикулы, периодичность роста, линьки. Дифференцировка мускула-
туры и наружный скелет. Членистые ноги и их дифференцировка —
антенны, ротовые части. Гетерономное расчленение. .Полость тела
членистоногих — целом, нефридии и половой аппарат. Миксоцель
и кровеносная система. Пограничные образования. Экскреторный
аппарат. Трахейная система. Легкие пауков. Кишечник. Органы
чувств и нервная система. Итоги анализа организации артропэд.
В своих истоках аннелиды чрезвычайно тесно связаны с моллю-
сками, организация которых вырабатывается под знаком образова-
ния защитного покрова в виде характерной раковины, прикрываю-
щей тело дорзально. Прямым продолжением эволюции аннелид яв-
ляются членистоногие — многочисленнейшая группа населения вод
и суши, организация которой характеризуется прежде всего той же
адаптивной чертой — выработкой защитного покрова. Этот веду-
щий признак развивается, однако, совсем в ином направлении, чем
у моллюсков, а именно: кутикула на всем теле членистоногих, хи-
тинизируясь, становится значительно белее явочной, чем кутикула
аннелид. Выработка хитина делается одной из характернейших черт
организации всего типа. С другой стороны, с этим связано полное
исчезновение мерцательных эпителиев.
Хитин, азотсодержащее производное углеводов, ацетилглюкоза-
мин, не является присущим исключительно членистоногим. Хотя
прежние указания на нахождение хитиновых образований у многих
животных являлись выражением чисто внешнего сходства производ-
ных кутикулы, однако более точные исследования химизма у ряда
представителей животного мира показали, что перидерм... тидроидов,
Ш1етиннн и челюсти аннелид, радула моллюсков, в известной мере
раковина головоногих и т. п. в той или иной мере содержат хитин.
Становясь значительно более прочной благодаря хитинизации,
кугикула членистоногих сохраняет в общем обычное строение, при-
сУЩее кутикуле аннелид. Однако вместе с тем она получает воз-
можность развивать различные, иногда довольно сложные, образе-
Вапия в виде волосков, шипов, чешуй и т, п., что значительно ус-
Л Пути эволюции животного мира 209
ложняет строение покрова и его придатков (рис. 81)..Кроме того
у ракообразных и Diplopoda твердость кутикулы увеличивается
отложениями в ней известковых солей. Таким образом создается
оболочка, эффективно защищающая все тело животного, — приспо-
собление, очень широко используемое в борьбе за жизнь.
Лишь у перипатуса (рис. 82) кутикула еще однообразно гибка
почти по всей поверхности тела. У прочих групп — обычно образова-
ние своего рода панцыря, покрывающего и тело и его придатки.
В связи с этим стоит понижение возможности движения и сокраще-
ний частей покрова, одетых такой сильно выраженной защитной
оболочкой. И как уже у некоторых полихет,
например у Eunicidae, можно наблюдать
появление более тонких межкольцевых бо-
роздок в обычной кутикуле, что увеличивает
подвижность тела, так у членистоногих
всегда имеется характерная дифференциров-
ка отделов толстой защитной хитиновой ку-
тикулы и соединяющих их утонченных, со-
членовных участков. Основными такими со-
членовными участками являются метамер-
ные бороздки, близкие к межкольцевым, но
обычно дело идет значительно дальше, и
хитиновая кутикула, как собственно тела,
так и его придатков — антенн, ног и т. п.,
дифференцируется на многочисленные от-
дельности — склериты. На теле членистоно-
гого получается сильно расчлененный пан-
цырь.
В связи с этим идет дифференцировка
покровного эпителия. Функция образования
хитинизированной кутикулы выступает резко
на первый план, и большинство клеток эпи-
телия связано с ней, тогда как железистые
и чувствительные клетки резко локализиру-
ются в покрове. Получается гораздо меньшее, чем у аннелид, разно-
образие и богатство клетками в покровном эпителии, — это прежде
всего слой сравнительно простых элементов, производящих кути-
кулу.
Другая характерная особенность, обусловленная выработкой
твердого хитинового панцыря по всей поверхности тела связана с
невозможностью постепенного увеличения размеров животного при
росте. При благоприятных условиях происходит накопление массы
живого вещества, но твердая, не поддающаяся растяжению и мало
способная к росту путем интусусцепции хитинизированная кутикула
является препятствием для постепенного роста всего тела. В резуль-
тате, образование кутикулы у членистоногих происходит периодиче-
ски. Клетки покровного эпителия время от времени под старой за-
твердевшей кутикулой дают новую. Старая кутикула отделяется от
210
Рис. 81. Покров насеко-
мого (по Веберу из Рим-
ского-Корсакова) :
1 — наружная кожица кути-
кулы, 2 — наружный и 3 —
внутренний слой кутикулы,
4 — чувствительный воло-
сок, 5 — шипы кутикулы,
6 — кроющий эпителий, 7 —
каналы в кутикуле, 8 —
субэпителиальная погранич-
ная перепонка, 9 — трихо-
генная клетка
тела и сбрасывается — происходит так называемая линька. Новая
кутикула быстро растягивается, давая возможность увеличения
размеров тела, но вскоре, затвердевая, теряет эту способность.
Таким образом наступает опять некоторый период накопления,.
увеличения массы живого вещества без увеличения роста живот-
ного, до новой линьки, когда произойдет опять и соответствующее
увеличение размеров тела. Очевидно, процесс развития твердого
защитного хитинового покрова у членистоногих шел в теснейшей
взаимосвязи с выработкой периодичности в образовании кутикулы
и росте, связанной с повторяющимися линьками.
Период линьки является критическим в жизни животного, когда
оно почти беззащитно против врагов и неблагоприятных условий
биотопа. И если у ракообразных обычно1 число линек не ограничено,
и они происходят в течение всей жизни животного, т. е. сопровож-
дают рост организма и в половозрелом состоянии, то у трахеат про-
исходит ограничение количества линек в эволюции различных
групп. Речной рак, например, сначала несколько, раз в год, а по-
том один раз ежегодно переживает этот критический период, скры-
ваясь в норах, пока не окрепнет новый панцирь. У насекомых же,
имеющих крылья, линьки в половозрелом состоянии у подавляю-
щего числа видов не бывает, вероятно, в известной мере в связи с
трудностями смены кутикулы трахей и крыльев.
В конце концов сохраняется одна лишь линька во взрослом
состоянии — переход из куколки в стадию имаго, т. е. половозре-
лого животного. Уменьшение числа линек идет и в дополовозрелом
периоде, и если американская Cicada septendecim проделывает
почти два десятка линек, — у Aphididae имеется всего две линьки.
Закон необратимости эволюции резко выявляется в случаях про- -
грессирующего уменьшения числа линек. Выработавшееся у пред-
ков членистоногих явление роста с линьками сохраняется и в тех
случаях, когда число линек ничтожно, и, казалось бы, организм
мог вернуться к исходному безлиночному состоянию, равно как
и в случаях, когда хитинизированная кутикула становится на-
столько тонкой, что, как, например, у комаров, не препятствует
росту между линьками. И все-таки линьки сохраняются, чему спо-
собствует развитие довольно сложной системы эндокринных отно-
шений L
В то же время линьки ограничивают размеры артропод — соз-
дается масса малых форм, приспособляющихся в связи с этим к
использованию самых ничтожных жизненных пространств, а соответ-
ственно крайне приспособляющихся и к ограниченнейшим жизнен-
ном возможностям таких «пустот» в природе, которые они запол-
Таким образом выработка хитинизированного покрова представ-
ляет яркий пример приспособления, котороё переходит в конце-
v 1 См. А. А. Передельский, Эндокринология беспозвоночных. —
,УСп- совр. бион.», т. X, вып. 1, 1939.
концов в существеннейшее препят-
ствие к дальнейшему повышению
организации, переводит ее на
рельсы узких приспособлений, хо-
тя и крайне разнообразных, —
одних насекомых насчитывают
более миллиона видов.
Возвратимся к дальнейшему
рассмотрению хитшшзированного
покрова у членистоногих. Как уже
было указано, приобретая значи-
тельную прочность и твердость, он
в то же время состоит из легко
гнущихся участков, так как меж-
ду не поддающимися сгибанию
частями происходит дифференци-
ровка сочленений. Этот процесс
получает свой более глубокий
смысл во взаимосвязи с характер-
ным развитием мускулатуры ар-
тропод. Типовой кожно-мышечный
Рис. 82. Поперечный разрез через
стенку тела Pennants
(по Шнейдеру):
1 — амебоциты, 2 — кроющий эпителий,
3 — соединитетьвотканный слой с всиок-
па'Ч1. 4 — миксоцель. 5 — кочьцевые, 6 —
диагональные п 7 — продольные мышеч-
ные волокна
Рис. 83. Поперечный разрез туловища Peripatus
tnohoni (по Федорову):
1 — спинной кровеносный сосуд, 2 — нервная комиссура,
3 — диафрагма, 4 — покровный эпителий. 5 — соедини-
тельнотканный фиброзный слой, 6 — кишечник, 7 —
кольцевые, 8 — диагональные, 9 — продольные мышцы,
10 — боковой нервный ствол, 11 — задний кольцевой
нерв сомита и его ветви, 12 — перикард, 13 — слюнные
железы, 14 — слизистые железы
мешок сохраняется
лишь у перипатуса при
его гибкой кутикуле
(рис. 82, 83). Однако и
здесь фушшия спопы
ч чя ко чьцевых, диаго-
нальных и продольных
мышц, равномерно ппо-
ходяших по1 в^ей л пине
тела, уже теряется у це-
лома и связана с по-
p’V'bom. Кроме хитпни-
'ЩП0ВЯННОЙ кутикулы, у
:: пшятуса вырабаты-
вается мощный слой
плотной фиброзной тка-
ни поп покровным эпи-
телием, и таким обоя-
том покров получает
значение опорного об-
разования.
У прочих артропод
мускулатура под твер-
дыми участками панцы-
ря теряет свой смысл #
соответственно редуцй'
212
руется. Она сохраняется только у подвижных соитеновиых участков
g" связи с этим единая мускулатура кожно-мышечного- мешка рас
членяется на отдельные специальные мышцы (рис. 84). Целим
имеющий у червей опорную функцию для мускулатуры кожно-мы
шейного мешка, уже лишается
своего основного значения. Твер-
дый покров может служить и дей-
ствительно служит для прикрепле-
ния концов отдельных мышечных
пучков (рис. 85). Защитная хити-
низированпая кутикула становит-
ся, таким образом, наружным ске-
летом членистоногих.
Соответственно этому покров-
ный эпителий в местах прикрепле-
ния мышечных пучков приобре-
тает особенную прочность благо-
даря развитию дтощного аппарата
нередко хитшшзированных тоно-
фибрилл в протоплазме эпптели-
< льных клеток. Таким образом,
мыпша при своем сокращении
может использовать в качестве
Рис. 84. Поперечный разрез через
чУ'Чочище (из Еернара):
4 ~ серлце- 2 — диафрагма, 3 — гонада,
половые протоки, 5 — кишка, 6 —
ирзрвентРальные и 7 — продольные мыш-
ку’ мь,и,ечиые пучки, идущие в нож-
У (9). ю — лакуны миксоцеля. 11 —
нервная цепочка, 12 — перикард
Рис. 85. Срез через место при-
крепления к покрову жевательных
мышц st<ciis(uo Шнейдеру):
1 — наружная кожппа кутикулы (2),
3 — покровный эпителии с тонофибрнл-
лами в протоплазме клеток, 4 — ткань
основного вещества с волокнами, иду-
щими от эпителия до поперечнополоса-
тых мышечных пучков (5), одетых
саркоплазмой (6)
пункта опоры хитиновую-кутикулу, прикрепляясь к эпителию ба-
зально посредством прослоек ткани основного вещества. В других
естах эта последняя сохраняется лишь в виде незначительных
ластинок, тяжей, нитей и т. п.
213
При дальнейшем развитии отдельных мышц, например жева-
тельных в голове насекомых, вырабатывается и внутренний скелет
в виде апофизов, аподемов, фрагм и т. п. Но он представляет все
тот же наружный хитиновый панцырь, лишь образующий вдаю-
щиеся внутрь и даже от!ннуровывающиеся складки соответственно
мускулатура кожно-мышечного
складкам всего покрова.
Первоначальная типовая
мешка, состоящая из
кольцевых наружных и
продольных внутренних
слоев мышц, уже у
полихет осложняется диф-
ференцировкой мышц па-
р аподи ев, равно' как и
обр а зов ани ем мускул ату-
ры диссепиментов и ме-
зентериев в виде дорзо-
вентральных, косых и по-
перечных пучков. У чле-
нистоногих (рис. 84, 86)
все эти категории мышц
сохраняются, но обособ-
ляются, во-первых, в пуч-
ки, двигающие отдельные
соседние или более от-
даленные по отношению
друг к другу метамеры,—
это главным образом про-
изводные продольной му-
скулатуры; во-вторых,
происходит обособление
пучков, обусловливающих
Рис. 86. А — мускулатура таракана со спин- движение частей ОДНОЙ И
ной стороны (по Майоллю и Денни):
1 — аорта, 2 — сердце, 3 — абдоми-
нальные мышцы, 4 — крыловидные
мышцы, 5 — грудные мышцы;
Б — мускулатура ножки Lepisma (из Эше-
риха):
I — соха, 2 — femur, 3 — trochanter, 4 — tibia, 5 — tarsia, г т л
6 — praetarsus, 7 — мышечные пучки НаКОНвЦ, В-ТрвТЬИХ, ПрО'
исходит обособление пуч-
ков, обусловливающих движение конечностей — производ-
ные косой и параподиальной мускулатуры. При этом дифференди-
тои же метамеры, т. е. ее
склеритов,
имуществу
кольцевой
тральной
— это по пре-
производные
и дорзо-вен-
ровка отдельных мышц и отдельных склеритов тесно взаимосвяза-
ны. Оба процесса эволюционируют совместно и обычно в направле-
нии концентрации в одну резко выраженную мышцу первоначаль-
но обособленных, но сходных по функции пучков. Таким образом,
не только в пределах классов членистоногих, но и в пределах раз-
личных групп одного класса, например насекомых, вырабатываю1"
214
ся свои определенные характерные комплексы мускулатуры. Вяд
точных описаний анатомии мускульных образований, например
Astacus, Gryllus, Dytiscus, гусениц бабочек и т. д., показали очень
большую сложность подразделения и скелета, и соответственно
мускулатуры, причем число резко дифференцированных мышц
достигает большого количества — нескольких сотен.
Сверх того, эволюция мускулатуры членистоногих завершается
характернейшей чертой их гистологии — вся мускулатура, за нич-
тожным исключением некоторых мышечных образований внутрен-
ностей, является поперечнополосатой (рис. 85). Однако, с одной
стороны, у перипатуса вся мускулатура еще сохраняется гладкой,
с другой, уже у некоторых полихет, например у Nephtys, имеются
пучки поперечнополосатых мышц, образующих строго определенные
системы среди остальной мускулатуры, типично гладкой.
Выработка поперечной полосатости обычно связывается с более
энергичными сокращениями таких мышц в противоположность
медленным тоническим сокращениям гладкой мускулатуры. И, ко-
нечно, раз появившись, поперечная полосатость закрепляется естест-
венным отбором, как полезная для вида в борьбе за жизнь, но
самый момент образования поперечной исчерченности связан со
строением сократимых структур вообще, и в частности с мицелляр-
но-фибриллярными образованиями клетки, с процессом обмена ве-
ществ в них.
Очень большое значение в выработке организации артропод
имеет образование членистых конечностей с соответствующей спе-
циализированной мускулатурой (рис. 86, Б). Это настолько харак-
терно для рассматриваемого типа, что дало и само название
Arthropoda—«членистоногие».. На место «подия» полихет, построен-
ного по типу простого рычага, появляется конечность типа слож-
ного рычага, создаются возможности гораздо более сложных и со-
вершенных приспособлений при дифференцировке частей ноги и
при разделении физиологического труда между различными парами
ног, что далее глубоко отражается на всей организации.
Уже у Peripatus примитивная конусообразная нога, напоминаю-
щая по внешности параподий какой-нибудь полихеты, например
uiopatra, имеет два-три дистальных пленника, заканчивающихся
парным хитиновым коготком (рис. 87). Последний заменил пучки
Щетинок, уточняющих действие параподия полихет, а присущая
ацикулам полихет опорная функция осуществляется здесь покровом,
прочих членистоногих конечности гораздо совершеннее.
В ряду наземных членистоногих произошла переделка ноги,
одобной ножке Peripatus, в длинную, многочленистую «унисери-
ьную», ходильную конечность (рис. 86, В), которая при разнооб-
** Усл°вий существования дает бесконечное разнообразие при-
ныхС°бЛеНИЙ:г Достигающих крайнего своего выражения в бегатель-
’ ирьтгательных, плавательных, хватательных и т. д. ножках.
Нй *Се ЭТ° конечности унисериального типа. У водного ряда чле-
погих трилобиты и ракообразные сохраняют большое сходство
215
с полихетами в дифференцировке бисеральных ножек, т. е. облада-
ющих внутренней и наружной ветвями (эндо- и эксоподитом) -и не-
сущих со спинной стороны жабру (эпиподит) (рис. 88). Это соот-
ветствует типичному мультифункциональному комплексу подия по-
лихет с невролоднем, нотоподием и жаброй.
Уже у разных семейств полихет наблюдается своеобразный про-
цесс использования параподиев, а именно—их усиков, па первом и
втором сомитах тела в качестве чувствительных придатков голов-
ного отдела дополнительно к антеннам головной лопасти (рис. 43).
I
3-)-
&
3
V -
о
/г::.- с».
нтеннам головной лопасти (рис. 43).
Таковы cirri tentaculares нереид и
филлодоцид. Факты эти легко по-
нятны с той точки зрения, что дви-
гательная функция параподиев
в первых сомитах тела у взрослых
аннелид имеет мало значения. На-
против, чувствительные их при-
датки в высокой степени выгодны
для ориентировки животного,
проникающего в новые условия
среды прежде всего передним кон-
цом тела. У членистоногих только
лишь эти придатки двух первых
сомитов развиваются па переднем
конце тела в виде антенн или \си-
ков, которыми животное пользует-
ся как важнейшими механо- и
хеморецепторами.
Уже у перипатуса первая пара
конечностей является смещенной
на передний конец тела и обра-
зует пару членистых антенн. Свя-
занный с ним ганглий брюшчсш
цехлочки смещается соответственно
дорзально, непосредственно при-
мыкая к ганглиозным образова-
ниям головной лопасти в виде
deuterocerebrum (рис. 90). Таким
образом создается сложный го-
ловной мозг членистоногих. Рако-
образные представляют иные отношения, имея две пары антенн (рис-
88). У многоножек и насекомых сохраняется только первая пара, тог-
да как вторая подвергается полной редукции (рис. 92). Однако зна-
чение их как важнейших хемо- и механорецепторов ничуть не
уменьшается. Напротив, пара антенн, например, у насекомых, раз-
вивается в чрезвычайно мощные органы чувств, являющиеся ваЖ'
нейшими в ориентировке и поведении животного. Происходит замена
нескольких сходно функционирующих образований одним, раз-
вивающим соответственную функцию особенно интенсивно. У паук0'
216
Рис. 87. Ножка перила туса.
Полу-
схематический разрез (по Федорозу):
1 •- мешок целома, 2 — воронка нефри-
дия, * — мышцы, входящие в ножку, 4 —
продольные мышцы, 5 — брюшной нервный
сгвол, б— нефропор, 7 — коготок ножки,
8 — чувствительные почки. 9 — чувстви-
тельный нерв, 10 — расширение канала
нефрпдия
образных процесс пошел иначе — редуцировались обе пары. При
всем разнообразии в развитии антенн членист<ллйнх мозг одинако-
о состоит из прото-, дейтеро- и тритоцеребрума, о чем речь будет
еще впереди (рис. 98, 99).
Переходим к дальнейшей чрезвычайно важной черте членисто-
ногих, выражающейся в том, что для них характерно изменение
нескольких следующих за антеннальными пар конечностей в рото-
Рис. 88. Artwli salvia (по Вегштедту):
I—XI — грудные сомиты, 1—8 — брюшные сомиты, 2 — первая ангенна, 3 — вторая антен-
на, 4 — копулятивный аппарат, 5 — спинной сосуд с остиями (6), 7 — печеночный прида-
ток, 9 — жвалы, 10 — макспллярная железа, 11 — иауплпусовый глаз, 12 — фасеточный
глаз, 13 — перикард, 14 — ножки, 15 — семенник, 16 — кишечник
Рис. 89. Peripatoides novae Zeahmdiae
вне части. Этот процесс у трилобитов и Limulus выражается
наиболее примитивно. Ротовая щель, как бы врезаясь в сомиты,
следующие за антеннальными, оказывается ограниченной с боков
смещенными к середине брюшной поверхности тела типичными
конечностями. Однако базальные членики этих конечностей своеоб-
разно изменены в связи с использованием их в качестве органов,
Удерживающих, а затем и /механически измельчающих захватывае-
мую добычу. У трилобитов эту функцию несут 3 пары, а у
Limulus даже б паРр ножек.
Совершенно особняком стоит Peripaius (рис. 89, 90), у которого
тодько одна пара конечностей превращена в ротовые части, пред-
ъявляя пару хитиновых зубцов — челюстей. Последующие же ко-
ечности ко рту никаких отношений не обнаруживают — состояние
3Условно крайне примитивное и совершенно особое.
217
У прочих членистоногих процесс пошел дальше иным образогл.
Первая пара ротовых конечностей превращается в специфический
аппарат разжевывания пищи — мандибулы, утрачивающие всякое
сходство с ногами, но закладывающиеся в онтогенезе, как типичные
конечности. Вторая и третья пары ротовых конечностей образуют
первую и вторую пары максилл, сохраняющих еще ряд черт типич-
ных конечностей -членистоногих, но являющихся функционально
только ротовыми частями (рис. 93). У ракообразных и следующие
пары ног в большем или меньшем количестве (максимально
Рис. 90. А — голова Peripatoidps с брюш-
ной стороны (из Снодграса):
2 — антенны, 5 — челюсти, 6 — ротовые папил-
лы; 9 — околоротовой валик;
7 пар у Stomatopoda) участ-
вуют в обслуживании рта,
но изменения их не так зна--
чительны, что характеризу-
ется самим названиехм «ного-
челюсти». Процесс протека-
ет различно в различных
группах ракообразных, да-
вая обильную адаптивную
дивергенцию.
Напротив, у многоножек
и насекомых имела место
резкая дифференцировка од-
ной пары мандибул и двух
пар максилл, причем послед-
няя из них превращается в
нижнюю губу, замыкая ро-
Б — схема метамерии переднего кон-
ца тела перипатуса (по Федорову):
1 — акрон, 2 — усики, 3 — protocerebrum, 4 —
deuterocerebrum, 5 — челюсти, 6 — ротовые папил-
лы, 7 — брюшной нервный ствол, 8 — ножки,
I, II, III, IV, V, VI — первая и т. д. метамеры
тела
товую щель сзади, последу-
ющие пары конечностей ко
рту уже никакого отношения
не имеют (рис. 91). Это не-
сколько иной путь эволюции
по сравнению с ракообраз-
ным. Приспособление к различнейшим родам пищи дает бесконеч-
ное разнообразие ротового аппарата насекомых, очень часто
ведущее к крайним формам адаптивного их развития, например
хоботок бабочек или колющий аппарат комаров.
В теснейшей взаимосвязи с выработкой ротовых частей — челю-
стей и жвал — стоит образование головы Arthropoda. Ее хитиновый
покров уплотнен и объединен для создания надежной опоры для
мускулатуры, движущей ротовые части (рис. 88, 89, 90, 91). Этим
кладется начало характерной черте организации — гетерономному
расчленению тела членистоногих, создающемуся на базе гомоном-
ной членистости аннелид.
У перипатуса мы видим обособление головы, образующейся за
счет неметамерной головной лопасти — «акрона» (простомиума)
аннелид плюс антеннальный, постантеннальный и мандибулярный
сомиты (рис. 90). У прочих членистоногих в состав головы уже
почти всегда входят акрон, два антеннальных, один мандибулярный
218
п два максиллярных сомита, т. е. неметамерная головная лопасть
плюс пять передних метамер тела (рис. 92). Как у Crustacea, так и
v Tracheata это норма. Однако особенно резко голова выделяется
v насекомых, так как здесь она адаптивно приобретает большую
подвижность, соединяясь с грудным отделом тела посредством су-
женного участка — шеи. Peripatus, Chilopoda и Diplopoda не выра-
ботали дальнейшей дифференцировки гетерономных областей тела,
вероятно в связи со значительным числом сомитов в туловище,
Рис. 91. А—голова черного таракана (по Богданову-
Катькову); Б — челюсть; В — максилла; Г—нижняя
губа черного таракана (по Гертвигу);
1 — усики, 2 — челюсти, 3 — глаз, 4 — palpus maxillaris, 5 —
palpus labialis, 6 — carao, 7 — glossae, 8, 9 — lobus externus u 1.
interims, 10 — подбородок, 11 — submentum, 12 — stipes, 13 — palpus
labialis, 14 — palpus maxillaris
что, в свою очередь, связано со своеобразным движением этих форм.
Ножки у них сохранились на всех сомитах и не обнаруживают
каких-либо региональных дифференцировок. Иное имеет место
У ракообразных, паукообразных и насекомых.
Crustacea имеют в различной степени выраженную приспособи-
тельную дифференцировку некоторого числа задних сомитов, на-
правляющих движение плавающего животного — вырабатывается
Длинный брюшной отдел. Конечности этого отдела часто редуци-
рованы, как, например, у Branchipus (рис. 88), Cyclops и т. д., или
ноеобразно изменены для выполнения иных специальных функций,
219
I
1
Рис . 92. Схемы эмбриональной метамерии на-
секомого (по Эшериху): А — ранняя стадия;
Б — поздняя стадия; в первой арабские циф-
ры обозначают сомиты по порядку, во вто-
рой— отдельно метамеры головы (1—5), ме-
тамеры груди (1—3), метамеры брюшка
(/-И);
20 — акрон, 21 — анальное отверстие, 22 — усики,
23 — ceivi. 24 — надглоточный ганглий, 25 — под-
глоточный ганглий. 26 — in . । (н> нас, 27 — max . а •,
28 — брюшная нервная цепочка, 29 — рот. 30 — нож-
ки, 31 — ьунсегеишш, 32 — тельсон
и их развитие в связи с особенностями
как, например, для прикрепления яиц у Масгига, для закрепления
в раковине у Anomura и т. д. Сюда же мы должны отнести и обра-
зование заднего плавника речного рака за счет тельсона и послед-
ней пары конечностей.
В результате получается более или менее резкая дифференци-
ровка хвостового отдела тела ракообразных. При этом внутренние
органы сосредоточиваются перед хвостовым отделом, а некоторой
границей, определяющей
его начало, является зад-
ний участок полового ап-
парата. Выводные отвер-
стия последнего у Maia-
costraca стабилизирова-
ны на 11—13-ом сомитах,
а число сомитов хвост?
доходит до 6, тогда как у
Phyllopoda оно примитив-
но велико и пестабилизи-
ровано.
В соответствии с диф-
ференцировкой хвостового
отдела для направления
движения (что может пе-
реходить и в особые виды
того же движения, напри-
мер плавание задним кон-
цом тела вперед у речно-
го рака) конечности сред-
него отдела тела — гру-
ди — сохраняют свою ос-
новную функцию. Они мо-
гут быть плавательными
(у Phyllopoda) или хо-
дильными и бегательными
(у большинства Malacos-
traca). У Decapoda число
их стабилизировано до
пяти пар, тогда как у
Entomostraca оно крайне
варьирует, причем крайне
разнообразна и их форма
условий существования и
организации. Так у Cladocera, а особенно у Ostracoda, с развитием
защитных складок кутикулярного покрова по бокам тела, образую-
щих раковину или панцирь, конечности груди подвергаются значи-
тельной редукции. Подобный процесс разыгрывается и у сидячих
Cirripedia. Дивергентная эволюция приспособлений здесь очень
многообразна.
220
Совершенно другим является путь эволюции у насекомых
<рнс. 92, 93). При стабилизации у них числа сомитов стабилизи-
руется и число пар конечностей, служащих для передвижения по-
средством хождения, бегания, плавания и т. д., и т. п. Именно
сохраняются и адаптивйо развиваются три пары конечностей на
следующих за головой сомитах, т. е. на 6-м, 7-м и 8-м. Однако дело
этим не ограничивается. На 7-м и 8-м сомитах, кроме того, разви-
заются характерные дня насекомых органы передвижения в воз-
душной среде — крылья, представляющие собственно боковые
Рис. 93. Внутренности- таракана сбоку (по Холодковскому):
1 — аорта, 2 — анальное отверстие, 3 — пилорические придатки, 4 —• сердце, 5 —
семеизьепгатечьный канат, 6 — надглоточный у?ел, 7 подглоточный узел нервной
ci стемы. 8 — чоб. 9 — средняя кишка, 10 — задняя кишка, 11 — меза Кения, 12 —
мальпигиевы сосуды, 13 — эг аденин, 14 — брюшная нервная цепочка, 15 — роговое
отвопстие, 16 — стоматогасгрический нерв, 17 — 18 — ректальная кишка, 19 —
слюнная железа, 20 — резервуар слюнной железы, 21 — семенник, 22 — мышечный
желудок, 23 — семейровод
екладки стенки тела, внутрь которых проходят сосуды, трахеи, нер-
вы и проникают разного рода амебоциты. Мускулатура, необходи-
мая для движения крыльев, заходит в основание крыла лишь у стре-
коз. У прочих насекомых она туда даже не проникает, а крылья
движутся пассивно в результате сокращения чрезвычайно развитых
мышц, проходящих от брюшных склеритов кутикулярного наружно-
го скелета к спинным в 6—8-м сомитах. В результате для их при-
крепления развивается уплотненный участок наружного скелета, ох-
ватывающий три сомита, а именно 6-й, 7-й и 8-й, которые образуют
рез^о обп-пбдепрнй двигательный отдел тела — thorax — грудь.
Разнообразнейшее развитие крыльев становится буквально неис-
тощимым источником в приспособительном разнообразии миллиона
видов насекомых. Однако далее мы не будем вдаваться в частности
этой, хотя и в высшей степени интересной стороны морфологии на-
секомых. Отметим лишь, что задний отдел тела — брюшко утрачи-
вает у них значение для продвижения животного при полете, в
связи с чем редуцируются соответственные конечности при сохра-
нении их возле половых отверстий, находящихся на 8-м и 9-м соми-
тах заднего отдела тела. Сменяя свои функции, эти конечности пре-
вращаются в части копуляционного аппарата, а также приспособля-
ется для .откладки яиц самкой. Конечности 10—11-го сомитов
221
становятся задними придатками с чувствительной функцией. Весь
отдел, заключающий по общему счету 9—19-й сомиты, становится,
прежде всего, вместилищем всех внутренностей, превращается в
характерное брюшко — abdomen насекомых, тогда как грудь за-
полнена мощной мускулатурой.
Последним шагом во внешней морфологии членистоногих
является дифференцировка областей тела у пауков. Будучи назем-
ными животными, пауки параллельно насекомым при стабилизации
числа метамер до 18—19 образуют за счет 9—10 задних сомитов
типичное брюшко, вмещающее внутренности, т. е. главнейшие отде-
лы пищеварительного аппарата, сердце, половые органы. Голова же
и следующие за ней четыре сомита, несущие четыре пары ходиль-
ных ног, превращаются в единое целое. Здесь мускулатура ротовы.
частей и мускулатура следующих пар конечностей объединены
опорном отношении. Образуется единый хитиновый панцырь голово-
груди. Ограничиваясь этим, мы не будем входить в дальнейшие по-
дробности прочих вариантов приспособительных явлений подобного
рода у паукообразных.
Итак, вся внешняя морфология членистоногих определяется
выработкой защитной хитиновой кутикулы, превращающейся в пан-
цырь наружного скелета. Отсюда и периодичность роста и линьки,
отсюда — членистость тела и его придатков, отсюда — дифферен-
цировка конечностей, отсюда — гетерономное подразделение тела,
но отсюда же характерное строение покровного эпителия и диффе-
ренцировка мускулатуры.
Однако сказанным дело не ограничивается. Изменяется не
только стенка тела, но в связи с этим и глубже лежащая система
целома претерпевает глубокие изменения вместе со всеми тесней-
шим образом связанными с ней органами. Такой эффект является
прямым следствием отмеченной нами закономерности, что основная
функция целома есть функция опоры для мускулатуры стенки тела.
У членистоногих с развитием хитиновой кутикулы функция
опоры переходит на наружный скелет, причем одновременно идет и
дифференцировка мышц стенки тела, а целом постепенно лишается
своей опорной роли. В результате происходит редукция целома,
сохраняющегося постольку, поскольку имеют значение его вторич-
ные функции, как это мы установили и для моллюсков — половая,
экскреторная. Они теперь выступают на первый план.
У перипатуса мы находим в этом отношении поучительную кар-
тину (рис. 94). Во всех метамерах туловища целом сохраняется в
виде небольших мешков, заложенных в основании ножек. Иначе
говоря, имеется типовое состояние парных мешков целома в каЖ'
дом сомите. Целомические мешки теснейшим образом связаны
здесь с экскреторным аппаратом, представляющим типичные мета-
нефридии, открывающиеся в целом хорошо развитой мерцательно11
воронкой, а наружу небольшим отверстием, расположенным при ос'
ковании соответствующей ножки (рис. 87). Этот комплекс целоМИ'
ческого мешка и соответствующего метанефрйдия, характерный ДлЯ
222
каждой метамеры, осложняется лишь в предпоследнем сомите тем,
что его целом объединяется с гонадой и таким образом получает
значение части полового аппарата. Соответствующий метанефри-
дий становится сильно развитым выводным половым протоком —
яйцеводом у самки, семяпроводом у самца.
Перед нами другая характерная черта членистоногих — сведе-
ние полимерных гонад аннелидообразных предков до небольшого
их количества, всего до одной пары, с выработкой более сложного
состояния прежде всего выводных протоков полового аппарата
Рис. 94. Схема организации перипатуса (по Лейкарту):
I — головной мозг, 2 — глаз, 3 — глотка, 4 — нервный ствол, 5 — слюнная железа, 6 —
слизистая железа, 7 — нефридий, 8 — целом, 9 — ножка, 10 — средняя кишка, 11 — кок-
сальная железа, 12 — гонада, 13 — целом тестикула, 14 — семепровод, 15 — семеизверга-
1 ельнып к хичл
(рис. 94, 88, 93, 96). Обычно парные образования, они, соединяясь,
переходят в непарный отдел, проходящий медианно. Во взаимосвязи
с этим развивается популяционный аппарат, и обычным является
внутреннее оплодотворение. При огромном различии в экологии раз-
ных rpvnn членистоногих оплодотворение происходит иногда крайне
оригинально, как, например, у стрекоз среди насекомых или у книж-
ных лжескорпионов среди паукообразных. Соответственно крайне
разнообразны и приспособления для оплодотворения в различных
случаях. В подробности вдаваться здесь нет возможности, но как
°бщее, отметим на эктодермальном конечном отделе выводных про-
токов полового аппарата образование различного характера желез
и железистых полостей (эктадении), а затем популяционных при-
датков— гонапофизов у самцов, у самок же придатков, связанных
с откладыванием яиц. Здесь приспособительные явления чрезвы-
чайно разнообразны. Для ряда групп, например у бабочек, перепон-
чатокрылых и т. д., -определение видов ведется систематиками
рожде всего по периферическим частям полового аппарата, и их
Развитие дает совершенно своеобразный, самостоятельный, но, ко-
плек°’ связанны® с другими чертами организации адаптивный ком-
л возвращаемся нецелому. У перипатуса уже наметился путь эво-
Ции целома членистоногих — общая редукция и частичное со-
223
хранение в связи с экскреторным и половым его значением,
'В эмбриональном развитии у него закладываются еще большие,
охватывающие кишку целомические полости, которые, однако, не
сохраняют своих отношений к другим образованиям зародыша,
являясь впоследствии лишь в виде небольших зачатков.
Подобный процесс имел место и в больших, связанных с трило-
битами группах — Crustacea, Xyphosura и Arachnoidea. Среди по-
следних у скорпионов в развитии закладывается много пар целоми-
ческих мешков, из которых мешки, принадлежащие 5-му сомиту,
развивают выводные протоки и становятся коксальными железами,
открывающимися у 3-й пары ног. Мешки, относящиеся к 8-му со-
миту, делаются выводными протоками полового аппарата, а прочие
дезинтегрируются.
У Limutus пара коксальных желез, состоящих каждая из четы-
рех железистых долей и одного длинного выводного протока, откры-
ваются также у основания 3-й пары ног и развиваются за счет
двух пар целомических мешков.
Подобные сложные коксальные железы имеются и у пауков,
сольпуг и т. д. Не будем вдаваться в частности их формы, числа
и расположения, отметим лишь, что процесс постепенного уменьше-
ния их количества до одной пары легко можно проследить.
У ракообразных этот процесс находит свое завершение в сохра-
нении .одной лишь пары целом*
ями в максиллярном сомите —
traca (рис. 88), или в антенне
traca. Однако у Nebalia и ря
состоянии обе пары желез от
У Tracheata уже нет цело?
ческих мешков с их метанефриди-
«скорлуповая» железа у Entemos*
ом — «зеленая» железа у Malacos-
зругих форм имеются во взрослом
пеменно.
Имеющиеся в эмбриональном раз-
витии парные зачатки мезодермальных сомитов, в конце-концов,
дезинтегрируются, распадаясь на мышечные зачатки и скопления
элементов иного характера, подобных различным целомоцитам, как,
например, клетки жирового тела.
Таким образом создается совершенно иная картина внутреннего
строения, чем у исходных аннелидных предков членистоногих сих
сильно развитой, полимерной вторичной полостью тела, окружаю-
щей центрально расположенный кишечник. Между кишечнико?л и
стенкой тела членистоногие имеют пространство, пронизываемое
мышечными пучками и пластинками и тяжами ткани основного
вещества и содержащее придаточные образования кишечника, экс-
креторные каналы и жировое тело, а у трахеат еще и трубочки
трахей и т. д. (рис. 83, 84, 93). Получается система лакунарных или
щелеобразных полостей, лишенная собственной клеточной выстилки.
Местами, как, например, под покровом, эта система отделена от
кроющего эпителия пограничными образованиями и заполнена
гемолимфой с разнообразными форменными элементами характера
амебоцитов, жировых клеток и т. п.. Обычно этой системе дается
название «миксоцель», которое должно говорить о двояком ее ха-
рактере — это «смешение» первичной полости тела и целома. Так
224
трактуются факты происходящего в эмбриональном развитии распы-
пеиия, дезинтеграции* целомических зачатков — полость целома
теряет свое отграничение, свои стенки в результате расхождения их
клеток и таким образом как бы вступает в соединение с ретроцело-
мическими пространствами.
Однако помириться с толкованием эмбриологических фактов, как Ц
«хаоса», морфология не может. И уже в конце прошлого столетия *'
рей-Лэнкестер выдвинул гипотезу «флебэдезиса», позволяющую
иначе осветить наблюдаемые явления. Миксоцель,. по Рей-Ленке-
стеру, есть прежде всего гемоцель, т. е. «расширенная и неправиль-
но вздутая сосудистая система, эквивалент кровеносной системы
аннелид». Решающим „в ютом вопросе будет строение кровеносной
системы
^ Наиболее постоянной и характерной ее частью является ее ти- ,
повой компонент — спинной сосуд, пропульзаторный отдел кото- ।
рого, снабженный мускулатурой, дифференцируется как сердце.^
Иных хорошо выраженных сосудов может и совсем не быть, как,
например, у Peripatus или Phyllopoda (рис. 83, 88). В других слу-/.
чаях, кроме спинного, имеется брюшной сосуд, как, например, у \
многоножек, затем ряд сосудов, отходящих от сердца, и ряд сосу- I
дов, связанных с жаберным аппаратом, как у ракообразных и пау- |
кообразных.
Мы не будем входить в детали анатомии кровеносной системы,
которые представляют большое разнообразие, особенно в связи с
развитием жаберного аппарата и областей тела. Возьмем лишь два
конкретных примера. Из филлопод Artemia (рис. 88) имеет только
один резко дифференцированный сосуд, а именно — сиицпой. ко-
торый на всем протяжении от головы до тельзона построен почти
одинаково, продолжаясь вперед в очень короткую аорту. В каж-
дом метамере тела он имеет по паре остий (всего 18 пар) и зад-
ний непарный остиум, через которые кровь поступает в сосуд из
окружающей его перикардиальной полости. Последняя представ-
ляет обособленный отдел миксоцеля, расположенный дорзально
ОТ кишечника и ограниченный снизу тонкой перепонкой — спин-
ной диафрагмой, состоящей из пластинки ткани основного веще-
ства. Диафрагма прикрепляется с боков к стенке тела под силь-
ными продольными мышечными пучками, оставляя, однако, здесь
отверстия, через которые гемолимфа протекает в перикард из ни-
жележащих отделов миксоцеля и, прежде всего, из лежащих по
бокам тела синусов, связанных с лакунарными пространствами
жаберных придатков ног. При сокращении спинного сосуда кровь
поступает отчасти в передний отдел перикардия, в значительной
же мере в лакуны головного отдела, откуда направляется далее
^азад в брюшной синус туловища. Из него она продвигается в
боковые синусы, идя далее или к жабрам, а оттуда в перикардий, -
или, в заднем отделе тела, непосредственно в перикардий.
^Совершенно иная и очень сложная картина строения кровенос-
ной системы у речного рака. Сердце, представляющее мускулистый
1 С п ' . ~
ю пути эволюции животного мира ’ "225
мешок с тремя парами остий, располагается в задней части тело-
вогруди и со всех сторон охвачено перикардием. Полость послед-
него замкнута, кровь в нее поступает только с боков из трех пар
жаберных сосудов. Диафрагма, образующая брюшную стенку по-
лости, представляет пластинку ткани основного вещества, в которой
заключены идущие в бока мышечные волокна. От сердца кровь
направляется в хорошо развитые сосуды — медиапно проходящую
переднюю и заднюю аорты, пару идущих вперед и вбок печеночных
артерий и непарную нисходящую артерию, продолжающуюся в суб-
невральную артерию. По ве точкам этих сосудов кровь поступает
в лакуны самой неопределенной формы и собирается в брюшной
синус, откуда в жаберном отделе тела проходит в жабры, а из них
в жаберно-перикардиальные каналы.
Как видно из этих списаний, кровь из спинного сосуда или
сложно развитой системы его разветвлений в виде разного рода—
от самых крупных до самых мелких — артерии всегда далее по-
ступает в систему лакун. Здесь ее ток регулируется наличием
соединительнотканных перепонок — перегородок. Особенно это
хорошо выражено в жабрах. И, наконец, кровь собирается в свое-
образном синусе, в перикардии, охватывающем сердце. Особых
сосудов, которые приносили бы кровь в спинной сосуд, нет. Име-
ются лишь остии, т. е. отверстия для вхождения крови из пе-
рикардия в сердце. Такой тип кровеносной системы свойствен
только членистоногим. Ни у немертин, ни у аннелид или моллюс-
ков нет подобного развития артерий, несущих кровь от спинного
сосуда, как нет и приносящей системы остий и синуса пери-
кардия.
Если далее обратиться к строению стенки спинного сосуда и его
ветвей у членистоногих, то картина оказывается очень типичной.
Внутри располагается тонкая интима в виде пластинки ткани
основного вещества. Далее идет слой кольцевых мышц и снаружи
располагается соединительнотканная adventitia. Таким образом,
отношения такие же, как, напри?тер, в спинном сосуде некоторых
аннелид. В лакунарных пространствах картина иная. Тут полного
отграничения лакун интимой из ткани основного вещества уже
нет. Имеется только неправильная и незначительно развитая
система тяжей и пластинок ткани основного вещества. Как будто
бы еще хорошо выраженная в сосудах взаимосвязь последних и
тка-ни основного вещества исчезает в лакунах.
Во всех нижестоящих группах, обладающих кровеносной систе-
мой,— у немертин, аннелид, моллюсков — хоц сосудов был обус-
ловлен прежде всего присутствием и расположением ткани основ-
ного вещества, которая являлась единственным непременным ком-
понентом стенки сосудов, так как и кровеносная система, по су-
ществу, представляла лишь систему полостей в этой ткани с
содержимым, состоящим из той же ткани, но с основным вещест-
вом, перешедшим из состояния геля в состояние золя. Таким
образом, к отмеченным выше двум характерным чертам кровснос-
226
ной системы членистоногих прибавляется еще третья — дезинте- L
грация стенки большей части сосудов за исключением спинного. ▼
Понимание всех этих особенностей кровеносной системы члени-
стоногих приходится связать с судьбами их целома. Уже у анне-
лид развитие целома вытеснило «заполняющую» ткань основного
вещества, которая дала систему пограничных образований. Это
определило ход кровеносных сосудов, неразрывно связанных с
тканью основного вещества. Создался тип кровеносной системы
аннелид. У моллюсков этот тип в основном выявлен, хотя значи-
тельно упрощен в связи со значительной редукцией целома.
Взаимосвязанность обеих систем выразилась крайне оригинально.
Сохранился тот участок целома, который необходим для образо-
вания' мускулатуры пролульзаторного отдела — перикардиальный
целом. А из сосудов сохранились те сосуды, которые прежде всего
направляют ток крови, т. е. спинной сосуд в виде сердца с одной
или двумя парами приносящих участков комиссуральных сосу-
дов — предсердиями.
’ Аналогичное явление наблюдается и у Arthropoda. При редук-
ции целома сохраняется также пропульзаторный отдел кровенос-
ной системы — спинной сосуд. Однако перикардиального целома,
за счет стенки которого у моллюсков, сходно с аннелидами, обра-
зуются мышечные элементы спинного сосуда, у членистоногих нет.
Пропульзаторная функция целома выражается здесь в ранней
дифференцировке из мезодермального зачатка кардиобластов,
дающих мускулатуру сердца, а образование вторичной полости
тела уже утрачено. Масса ^мезодермальных клеток дифференци-
руется непосредственно в мышцы и в амебоцитов. Этим ограничи-
вается процесс в примитивных формах кровеносной системы у
членистоногих, как у Phyllopoda, Peripatus, или во вторично упро-
щенных, как у насекомых. Однако кровеносная, система услож-
няется^ Crustacea, Arachnoidea, Myriapoda. Образуются ответвле- j
ния спинного сосуда, как, например, артерии речного рака. Совер- |
шенно отсутствуя у аннелид, у членистоногих эти ответвления вто- |
рично дифференцируются, определяя ток крови от сердца. , *
Иная участь постигает комиссуральные сосуды аннелид, прино-
сящие кровь к спинному сосуду. Они у членистоногих полностью
утратили свою целостность и их последними следами являются
остии, через которые кровь поступает в сердце. С редукцией це-
лома исчезло отграничение сплетений кровеносных сосудов на
средней кишке от кровеносной сети стенки тела. Дезинтеграция же
Целома в эмбриональном развитии не могла не отразиться на ука-
занных сосудах, придавши им своеобразный характер лакун и
вызвав также утрату целостности их интимы.
Совершенно особым процессом явилось образование перикар-
диальной полости и спинной диафрагмы. Рей-Ленкестер считал,
что произошел процесс «флебэдезиса», т. е. расширение и слияние
между собой сосудов, приносящих кровь к сердцу, последним сле-
дом которых и являются остии. Понимание остий как устий при-
U* 227
. носящих сосудов/ соответствующих комиссуральным сосудам анне-
лид, есть констатирование несомненного факта. Но расширение и
слияние сосудов в перикардиальный синус является еще просто
предположением. Возможно, что с исчезновением целомических
Полостей ясно выраженные комиссуральные сосуды переходят в
состояние лакун, из последних же вторично, с развитием мышеч-
ных пучков и регулирующей ход гемолимфы пластинки основного
вещества постепенно выработался перикардиальный синус со
спинной диафрагмой.
I Итак> несомненно, что миксоцель Arthropoda представляет со-
I бой "Wch'b кровеносной системы, развитой в теснейшей связи с
^развитием у членистоногих ткани основного вещества в виде по-
граничных образований, унаследованных от аннелидообразных
предков. У Argulus, например, эти образования также имеют ха-
рактер непрерывной системы, причем находящаяся под покровным
эпителием перепонка переходит в нити, тяжи, пластинки в миксо-
целе, а эти последние переходят далее в субэпителиальную пере-
понку внутренностей. Такие оболочки мышц и нервной системы
стоят в связи с пограничными мембранами покрова или внутрен-
них образований. Таким образом в отличие от полихет здесь от-
сутствует субмускулярная перепонка, что является результатом
редукции целомических мешков. Однако здесь при редукции
целома не происходит восстановления сплошного слоя ткани
основного вещества наподобие имевшегося у немертин. Эволюция
необратима. И у членистоногих, на основе пограничных образо-
ваний аннелид, располагающихся ретроцелотелиально в виде тя-
жей и пластинок, получается подобная же неправильная сеть,
среди которой разбросаны дифференцирующиеся из мезодермы
закладки мышц, амебоидных элементов жирового тела и нефро-
фагоцитарных клеток. Более того, наступающая редукция целома
обусловила и дезинтеграцию кровеносных сосудов: кровь из спин-
ного сосуда поступает не в пространства, стенка которых образо-
вана целостной интимой, а в щели между различными органами и
их частями, где пробегают и пограничные образования, т. е. посту-
пает в миксоцель, содержащий уже не кровь, а гемолимфу. Итак,
j миксоцель Arthropoda представляет собой часть кровеносной си-
I стемы вместе с лимфатическими щелями в тканях и между раз-
I личными органами?
Редукция целома, сопровождаемая резкой дифференцировкой
различных мезодермальных зачатков и, далее, дезинтеграцией его
стенки, ведет не только к образованию характерного миксоцеля,
НО' и к появлению в миксоцеле скоплений элементов целотелиаль-
ного происхождения, имеющихся уже и у аннелид —> нефрофаго-
цитов и элеоцитов. Последние особенно богато представлены,
образуя скопления клеток характерного для членистоногих жиро-
вого тела, располагающиеся в брюшке между внутренностями 11
имеющими адаптивное значение, как запасы питательных веществ,
особенно при метаморфозе.
228
Роль нефрофагоцитов также возрастает у членистоногих с ре-
дукцией экскреторного аппарата. В различных -.участках миксо-
целя, например на диафрагме перикарда насекомых и т. д., распо-
лагаются группы нефрофагоцитов. Однако главную роль в выве-
дении из организма жидких продуктов распада принимают на
себя придатки тех или иных отделов кишечника. Таким образом,
коррелятивно развиваются пара, а далее и большое число харак-
терных трубчатых мальпигиевых сосудов, располагающихся в
брюшке и относящихся у многоножек и насекомых к началу зад-
ней эктодермальной кишки (рис. 93, 96), а у паукообразных к
средней, эктодермальной кишке. Развитие мальпигиевых сосудов с
большой долей вероятности можно поставить в связь с утратой
Рис. 95. Трахеи перипатуса (по Шнейдеру):
1 — кутикула, 2 — кроющий эпителий, 3 — трубочки
трахей
нефридиями осморегуляторной функции уже у ракообразных,
а особенно у наземных членистоногих. Последние в высокой сте-
пени экономят запасы воды в организме, извлекая ее даже из со-
держимого самого заднего отдела кишечника — из ректальной
кишки, где дифференцируются для этого ректальные железы у
насекомых.
Если у наземных членистоногих не резко выражено адаптивное
значение мальпигиевых сосудов, то у них приспособительный мо-
мент ясен для органов газообмена. В замену типовых органов вод-
ного дыхания, представленных у Crustacea и Xyphosura как и у
аннелид аппаратом жабер, связанным с конечностями, развивается
чрезвычайно характерная система трахей. Это — система выстлан-
ных тончайшей кутикулой трубок, по которым кислород воздуха
приносится ко всем внутренним органам, непосредственно обеспечи-
вая их газообмен.
Уже у Peripatus по всей поверхности тела разбросаны неболь-
шие отверстия стигм, ведущие в очень короткие впячивания по-
крова, от которых отходят пучки тончайших трубок—трахей (рис. 95).
У многоножек и насекомых стигмы располагаются совершенно пра-
вильно — по одной паре в каждой метамере с боков. Правда, име-
ется они уже не во всех метамерах, и обычно голова лишена их.
Стигмы ведут в крупные трубки, стенки которых образованы одно-
229
слоимым эпителием и выстланы кутикулой с особым спирально про-
ходящим утолщением, являющемся приспособлением, гарантирую-
щим от спадания стенок, В более простых случаях трубки образуют
отдельные метамерные пучки или, в большинстве случаев, дают
сложную систему ветвящихся, анастомозирующих друг с другом
Рис. 96. Внутреннее строение таракана со спин-
ной стороны (по Холодковскому):
1 — надглоточный ганглий, 2 — зрительная лопасть. 3 —
глаз, 4 — слюнный резервуар, 5 — слюнная железа,
6 — зоб. 7 — мышечный желудок, 8 — пилорические
придатки, 9 — средняя кишка, 10 — задняя кишка, 11 —
мальпигиевы сосуды, 12 — ректум, 13 — трахеи, 14 —
стигмы, 15 — брюшная нервная цепочка. 16 — семенник,
17 — семепровод, 18 — мезадении, 19 — семеизверга-
тельный канал.
трубочек и мешков, по-
степенно утрачиваю-
щих свое первоначаль-
ное 1метамерное распо-
ложение (рис. 96). От
этих трубочек в глуби-
ну тела отходят ли-
шенные спирального
утолщения тончайшие
в ну тр икл еточн ые тр а -
хейные капилляры, оку-
тывающие внутреннос-
ти и проникающие во
все органы и даже
внутрь их клеток.
В тесной связи с
развитием системы тра-
хей подвергается упро-
щению часть кровенос-
ной системы, которая
была связана у водных
форм с дыханием жаб-
рами. Это повело в
конце-концов к вторич-
ному упрощению состо-
яния кровеносной си-
стемы насекомых, име-
ющих один только на-
стоящий сосуд — спин-
ной сосуд. В этом ас-
пекте интересно выяс-
нить происхождение
трахейной системы.
Она представляет
одну из важнейших со-
ставных частей того
комплекса черт органи-
зации, который являет-
ся приспособительным к особым условиям наземной жизни члени-
стоногих. Ранее мы уже коснулись относящихся к этому комплексу
образования унисериальной конечности, развития крыльев, диффе-
ренцировки мальпигиевых сосудов. Конечно, исходным состоянием
трахеальной системы были .метамерные углубления покрова, в ко-
230
юрые открывались пучки трахей. Такие трахеи, как мы видели,
еще имеются у перипатуса, но их разбросанное расположение по
всей поверхности тела в сопоставлении с постоянство-м метамер-
ного их расположения у многоножек и насекомых кажется вторич-
ным изменением. С другой стороны, чрезвычайно многообразной
приспособительной чертой у насекомых можно считать развитие
снабженных спиральной нитью трахейных трубок и образование
сложной системы их анастомоз.
Таким образом, первоначально метамерные образования превра-
щаются в единый аппарат дыхания, сохраняющий иногда только
одну пару стигм, как, например, в условиях вторичного приспособ-
ления к водной жизни у личинок комаров. У гусеницы Рагаропух,
живущей в воде, единая система трахей становится замкнутой и
газообмен через ее посредство происходит благодаря образованию
выростов тела, в которые заходят трахеи, создаются так называе-
мые трахеальные жабры. Через тонкую стенку таких жабер раство-
ренный в воде кислород диффундирует в трахеи, и далее по ним
разносится по всему телу, ко всем внутренним органам. Это —
крайний этап адаптивной эволюции, можно сказать, превращающий
свободный приток атмосферного воздуха к внутренним органам в
свою прямую противоположность. Получается замкнутая система,
обеспечивающая газообмен наподобие кровеносной, но только не с
жидкой средой, а с газовой.
Для понимания происхождения трахеальной системы решающим
является ее примитивное, метамерное состояние с расположением
стигм по бокам тела соответственно закладкам конечностей. Трудно
было бы для высоко дифференцированной системы у высоко стоя-
щей группы допустить, что образующиеся здесь впячивания могут
быть новообразованиями, которые не имеют перед собой предшест-
венников иного физиологического значения. Морфологи не решали
так вопроса. Кеннель считал возможным видеть в трахеях пучки
запустевших кожных железок, принявших на себя вторично функ-
цию газообмена. Однако у аннелид мы не знаем подобных желез
с кутикулярной выстилкой на всем их протяжении, а тем более рас-
положенных правильно метамерно по бокам тела. Подобное распо-
ложение имеется зато у нефридиев аннелид, и Ланг (1903) склонен
пидеть в протонефридиях червей образования гомологичные трахеям
членистоногих. Но против такого предположения говорит факт
присутствия метамерных метанефридиев во всех сомитах туловища
"eripaius наравне с пучками трахей. Таким образом нефридии сами
По себе, а трахеи сами по себе — никакого сходства в строении и
никакой зависимости или замещения одних другими.
Наконец, Бернар (1892) развивает мысль о возможном проис-
хождении трахей насекомых от пучков нотоподиальных щетинок,
как он называет, от «щетинковых желез» полихет.> За это гово-
рит, одной стороны, метамерное и сходное их расположение, а с
другой, — сплошная кутикулярная выстилка. И действительно, при
3Ученни параподий полихет нередко приходится наблюдать запу-
* 231
стевшие участки щетинковых мешков, в которых щетинки выпали,
но сохранились их оболочки в виде пучков тонких кутикулярных
трубок, очень напоминающих трубочки трахей.
Значительно осложняется вопрос наличием системы трахей в
другой ветви типа членистоногих, а именно у паукообразных. Сход-
ные по строению с трахеями насекомых пучки трахей у различных
групп Arachnoidea открываются своими стигмами не на боках, а
вентрально на брюшке, у сольпуг же еще и на груди. В соответ-
ствии с этим Бернар принимает, что здесь аппарат трахей разви-
вается не за счет нотоподиальных щетинковых мешков, а сходным
образом дифференцируются в трахеи щетинковые мешки невропо-
диев.
Однако дело крайне осложняется присутствием у скорпионов,
Pedipalpi и пауков других органов дыхания — легких. Они образо-
ваны рядами тонких пластинчатых выростов" стенки тела, которые
вдаются в небольшую полость. Располагаясь метамерно попарно',
такие полости открываются наружу щелями на передних сомитах
вентральной поверхности брюшка. Эти оригинальные органы газооб-
мена закладываются в эмбриональном развитии на зачатках брюш-
ных конечностей и постепенно углубляются в прилежащий отдел
стенки тела. В соответствии с этим в настоящее время почти обще-
принятым является взгляд на происхождение легких Arachnoidea от
жабер, располагающихся на основных члениках абдоминальных но-
жек, наподобие имеющихся у Xiphqsura. Предполагается, что при пе-
реходе к наземному существованию жаброносные членики конечно-
стей как бы замкнулись в углублении брюшного покрова, и таким
образом жабры, будучи в известной мере предохранены хитиновой
кутикулой от иссушающего влияния воздуха, приняли на себя роль
органов воздушного дыхания.
У скорпионов имеются только эти своеобразные легкие в коли-
честве четырех пар на передних сомитах брюшка. У пауков в груп-
пе Tetrapneumones наблюдаются сходные отношения, но у них лег-
ких две пары, у Dipneumones вместо задней пары легких имеется
пара типичных трахей. У прочих Arachnoidea легких уже нет, раз-
виваются лишь трахеи. Эти соотношения заставили ряд зоологов
признать, что первоначально' имевшиеся легкие, унаследованные от
ксифозурообразных предков и связанные с особыми частями крове-
носной системы, в последующем, путем своеобразного изменения,
дали начало по существу совершенно иному аппарату — трахеям,
несущим кислород непосредственно к нуждающимся в нем частям
организма без опосредствующей системы кровеносных сосудов.
Однако вывести типичное строение трахей из строения легкого чрез-
вычайно затруднительно. Еще труднее представить конвергентное
развитие системы трахей у Arachnoidea и Antennata.
Поэтому ряд зоологов становится на совершенно противополож-
ную точку зрения, выводя легкие пауков из своеобразно развитых
пластинчатых трахей. Однако против подобного1 допущения говорит
то, что в развитии легких и трахей нельзя найти необходимого сход-
232
ства. Ка*к видно, вопрос о происхождении трахей у членистоногих >
ждет^пщ решедя.
Итак, 1-й комплекс черт организации, связанный с образованием |
защитной хитиновой кутикулы, создал особый тип строения I
Arthropoda. Он характеризуется специализацией покрова, выработ- 1
кой наружного скелета, соответствующей дифференцировкой муску- *
натуры, дифференцировкой конечностей, гетерономным расчленени-
ем тела, редукцией целома, с сохранением лишь его полового отде-
ла, и редукцией нефридиев, ранней дифференцировкой мезодермы и
ее дезинтеграцией, выработкой миксоцеля и редукцией пограничных
образований и кровеносной системы, образованием жирового тела,
периодичностью роста с линьками, резко выраженными личиночны-
ми стадиями, часто развитием с метаморфозом.
2-й комплекс черт организации, приспособительных к особым ус- \
ловиям наземного существования, дает уже характерные особенно- I
сти наземных Arthropoda, возникающие на основе водных типовых I
форм. Этот комплекс выражается выработкой конечностей наземно-
го' типа — унисериальных, а у насекомых и образованием крыльев,
развитием органов воздушного дыхания — трахейной системы и у
Arachnoidea, кроме того, развитием легких при редукции жаберно-
го- аппарата и соответственно упрощении кровеносной системы и,
наконец, развитие мальпигиевых сосудов при редукции нефридиев.
Переходим к кишечнику членистоногих, в своем развитии очень
мало связанному с рассмотренными двумя комплексами черт орга-
низации. Его специфическая приспособленность обусловлена* тем
или иным способом использования различного' пищевого материала,
и чем разнообразнее этот материал, чем многообразнее он исполь-
зуется, тем богаче адаптивное разнообразие группы. В этом отноше-
нии членистоногие представляют неисчерпаемое богатство форм
морских, пресноводных и наземных, использующих всяческие воз-
можности питания, начиная от типичного хищничества, переходя к
употреблению в пищу всевозможных растительных ресурсов, остат-
ков животного и растительного происхождения, гниющих веществ и
кончая настолько далеко заходящим паразитизмом, что кишечник
У них подвергается редукции.
При редукции вторичной полости тела кишечник членистоногих
утратил отношения к целому, присущие аннёлидам. Он на всем про-
тяжении погружен в миксоцель и не обнаруживает каких-либо приз-
наков метамерности. В связи с этим во всех группах членистоногих
он эволюционирует ОТ ИСХОДНОГО ТИПОВОГО1 состояния трубки с не-
большими стомодеальным и проктодеальным отделами, как, напри-
мер, у Peripatus или Branchipus (рис. 94, 98) в направлении все-
большего и большего увеличения этих отделов с расширением их
Функций, а также сокращения средней, эктодермальной кишки при'
Условии все далее идущей дифференцировки частей во всех трех
отделах (рис. 93, 96). .
Первой характернейшей чертой пищевого тракта Arthropoda, от-
личающей их от прочих типов животного царства, является уже
233
рассмотренное выше превращение (конечностей передних метамер
юла в ротовые части, служащие для удерживания, а далее и для
механической переработки пищевого материала. Повторять уже ска-
занное при рассмотрении дифференцировки ног мы здесь, конечно,
не будем.
В связи с выработкой ротового аппарата произошло в переднем,
стомодеальном отделе кишечника исчезновение примитивных кути-
кулярных челюстей, присущих полихетообразным предкам
Arthropoda. Однако в ряде случаев дополнительно к ротовому ап-
парату дифференцируется особый отдел для механической обработ-
ки пищи в передней кишке. Таков, например снабженный
хитиновыми кутикулярными зубцами жевательный желудок
Malacostraca или мышечный желудок насекомых. В первом, кроме
того, вырабатывается фильтрующий аппарат ворсинок, не пропу-
скающий дальше недостаточно измельченные частицы пищи. Неред-
ко стомодеальный отдел приспособляется и к хранению запаса по-
лучаемой в изобилии пищи. В крайнем выражении мы это- находим
в зобе бабочек. У большинства наземных членистоногих в глоточ-
ный отдел открываются высокодифференцированные железы, на-
пример различные слюнные железы насекомых. Все это придает пе-
редней кишке характер особой сложности.
Параллельно с увеличением и дифференцировкой стомодеально-
го отдела идет тот же процесс и в заднем, проктодеальном участке
кишечника членистоногих. Так, у речного рака этот участок в виде
тонкой трубки проходит через все брюшко. Здесь очевидна связь
проктодеального отдела с развитием очень короткого среднего, эк-
тодермального, усваивающего отдела. Однако укорочение средней
кишки связано1 с сохранением и даже увеличением ее сецернирую-
щей и всасывающей поверхности. У всех высших раков (Malacostra-
ca) компенсаторно развиваются боковые выросты этого отдела
кишечника, достигающие огромной мощности, как, например, в пече-
ни речного рака. Если наличие подобного hepatopancreas характер-
но в той или иной форме для Malacostraca, то в известной мере
сходную роль, хотя часто с меньшим развитием, несут метамерные
печеночные выросты Arachnoidea и «пилорические придатки» в ки-
шечнике некоторых насекомых. У наземных форм часто1 имеет место
своеобразное развитие длинной задней кишки, дифференцирующей-
ся на два отдела в связи с новой функцией, важной при наземном
существовании — функцией всасывания тех небольших количеств
воды, которые имеются в выводимых из кишечника каловых мас-
сах. Как было уже отмечено, эта экономия воды имеет связь с рф
дукцией типичного аппарата метанефридиев и их осморегуляторной
функции. Последняя заменена выведением экскретов через клетки
кишки или посредством специальных трубчатых' выростов мальпи-
гиевых сосудов, развивающихся у Arachnoidea за счет заднего уча-
стка эктодермальной кишки, a Antennata за счет передней части
задней кишки. Следовательно у тех -и других получаются два со-
вершенно независимых ряда образований. Часто это сильно ветвя-
234
jjjiecn трубочки, расположенные всюду во внутренностях и убираю-
щие из миксоцеля продукты распада у насекомьГх обычно в виде
твердых отложений мочевой кислота и уратов, и гуанина у пауко-
образных. Соответственно, у насекомых дифференцирован в задней
кишке отдел, куда впадают мальпигиевы сосуды, и отдел, где фор-
мируются экскременты. Последний содержит в стенке всасывающие
воду элементы. Часто процесс ведет к образованию ректальных
желез, специально при разделении физиологического труда диффе-
ренцированных для функции всасывания воды, сохранения ее для
нужд организма. У водных форм, например у ракообразных, подоб-
ных приспособлений, естественно, нет.
Итак, кишечник Arthropoda развивается приспособительно к
особым условиям использования разнообразнейших пищевых воз-
можностей малого масштаба. Он развивается из типового состоя-
ния аннелид в значительной мере независимо от двух ранее рас-
смотренных главных комплексов черт строения и обнаруживает вы-
работку характерных, хотя и немногочисленных особенностей. Го-
раздо сложнее у членистоногих отношения в органах чувств и нерв-
ной системе, рассмотрением которых мы и закончим выявление всей
сложной организации этой группы.
Богато развитый у полихет головной комплекс органов чувств
сохраняет всю свою значимость для свободноподвижных их потом-
ков, т. е. для исходных форм членистоногих, эволюционировавших,
однако, прежде всего под знаком развития защитного хитинового
покрова. Так, у Peripatus (рис. 89, 94) имеется пара глаз, по строе-
нию совершенно сходных с глазами полихет, в частности Phyllodo-
ddae — единственный в своем роде случай у членистоногих, и при-
том чрезвычайно важный для понимания их родственных отноше-
ний. Однако у того же перипатуса, как выше было указано, уже
завершился процесс наметившейся еще у полихет субституции чув-
ствительных придатков иеметамерной головной лопасти — антенн
и пальп — метамерными образованиями, относящимися к парапо-
дням первого сомита (рис. 90, 98). Таким образом создались чрез-
вычайно характерные членистые антенны Arthropoda, расположен-
ные на передней поверхности головы. У ракообразных (рис. 88) за
ними следует и вторая пара антенн, принадлежащих второму соми-
ту» вошедшему в состав головного отдела, как это имеет место уже
У полихет, где cirri tentaculares развиваются не только на первом,
но и на следующих сомитах. Однако у многоножек и насекомых
°Дна пара антенн редуцировалась в связи с интенсификацией функ-
ций первой пары (рис. 93, 96, 91). Невидимому, .нечто подобное
имело место и у Peripatus, Очевидно это — процесс, общий для на-
земных членистоногих. Свое завершение редукция антенн находит
У паукообразных, где обе пары совершенно' исчезли, но и водные
Aiphosura уже лишены антенн, еще хорошо развитых у трилобитов
а Gigantostraca.
Антенны членистоногих внешне сходны с придатками головной
л°пасти полихет, особенно с антеннами последних, но функциональ-
235
но они значительно сложнее, так как играют не только роль механо-
рецепторов, но и хеморецепторов. В этом отношении эволюция ,их
особенно далеко зашла у'насекомых, где антенны стали настолько
мощными органами хеморецепции, что в жизни большинства насе-
комых химические раздражения стали ведущими. Как пример при-
веду хотя бы майското жука (Melolontha hippocastani) , у которого
на пластинчатых усиках самок имеется около 8 000 органов хими-
ческого чувства, а у самцов около 25 000. Тонкость настройки этих
хеморецепторов такова, что самцы некоторых бабочек (например,
Saturnla pavonia) летят за несколько километров к клетке, в кото-
рой заключена самка. Важность их в жизни насекомых видна хотя
бы на примере муравьев.
Таким образом хеморецепторы головной лопасти полихет —
пальпы и затылочные органы — утратили свое значение у предков
Arthropoda и редуцировались в связи с развитием у последних
антенн. Возможными их следами являются только1 отделы головного
мозга членистоногих, представленные globuli с glomeruli и цен-
тральным телом, Oi чем речь будет дальше. Таким образом у пери-
патуса головной комплекс органов чувств уже иного рода, чем
у аннелид.
Для понимания рассмотренных отношений следует подчеркнуть,
что ведущее или во всяком случае немалое значение здесь имел
процесс смещения рта назад между конечностями 3-го, 4-го1 и т. д.
сомитов с превращением этих конечностей в ротовые части, для
мускулатуры которых и дифференцировался консолидированный
хитиновый панцырь головы. Этот процесс мог вести к смещению
вперед конечностей 1-го и 2-го сомитов и превращению их в мощ-
ные антенны, с редукцией, в то же время, головной лопасти и ее
чувствительных придатков. Взаимосвязанность всех этих процессов
самая тесная. В них первый, ведущий комплекс черт организации
Arthropoda сближается с адаптациями кишечника и с головным
сосредоточением органов чувств.
Как было указано, только перипатусы сохраняют ямочный глаз
взрослых полихетообразных предков, прочие же членистоногие те-
ряют его. У ракообразных компенсаторно сохраняются личиночные
глазки трохофоры, построенные по типу инвертированных глазков
немертин и турбеллярий; они развиты у личинки раков — науплиу-
совый глаз, далее переходят к Иоёа и, наконец, проникают и во
взрослую форму как «медианные» глаза Entomostraca, а изредка и
высших раков. Обычно при этом соединение трех или четырех
глазков в одно сложное образование, но число ретинальных клеток
в каждом остается всегда небольшим. •
Для членистоногих обычно развитие глаз совершенно1 иного
рода — неинвертированных, обладающих своеобразным диоптри-
ческим аппаратом кутикулярной линзы и кристаллического конуса
и характерными образованиями, воспринимающими световые ра3"
дражения — рабдомами (рис. 97). Такие глаза в виде простых
глаз имеются 'То "всех группах Arthropoda за исключением
236
protracheata. Их специфическую особенность — Развитие корнеаль-
ной линзы —• можно в известной мере поставить в связь с характер-
ным развитием кутикулы членистоногих. Она при своей мощности
мало подходяща для образования ямочных глаз, но, линзообразно
утолщаясь, используется как диоптрический аппарат. Другая харак-
терная черта — это особый путь повышения чу ветви гелыности
глаза у членистоногих. Опа выражается в специфическом объеди-
нении нескольких воспринимающих исчерченных ободков у опре-
деленного количества ретинальных клеток в одно палочковидное
тело, рабдом.
Простые глаза, очевидно, развились совершенно независимо от
медианного науплиусового глаза личиночного типа и от взрослого
Рис. 97. Схема фасеточного глаза бабочки Sphingidae (по Бюньону и
Попову):
2 — клетки покровного эпителия, производящие корнеальную линзу, 2, 3 — кристаллический
конус, 4 — пограничная перепонка, 5, б — пигментные клетки, 7 — рабдом, 8 — ретиналь-
ные клетки, 9 — чувствительные нервные волокна
долихетного глаза типа Eunicidae из ретинальных элементов, при-
сутствующих у аннелид в покровном эпителии и под ним на различ-
ных участках тела. Это небольшие фоторецепторы, рассеянные в
головном отделе тела первичных Arthropoda. Они наиболее значи-
тельно развиваются в своеобразных глазах Arachnoidea и Myria-
poda, стабилизируясь в небольшом количестве.
У ракообразных и насекомых, независимо и конвергентно в обеих
группах, за счет объединения значительного количества простых
глаз развиваются сложные, или фасеточные, глаза. Опи чрезвычай-
но характерны для членистоногих и имеются обычно в количестве
одной пары боковых глаз, причем у ракообразных одновременно
присутствуют и медианные глазки, а у насекомых в личиночном
состоянии развиты только простые глаза, тогда как во взрослом,
наряду с простыми, существуют фасеточные.
Можно легко проследить, например, у насекомых все этапы
объединения отдельных простых глаз в сложный, начиная от кучек
Довольно рыхло расположенных простых глаз и кончая единым
мощным органом картинного зрения — фасеточным глазом. За
счет расположения отдельных глазков в виде мозаики получается
картинное зрение, при котором небольшие участки окружающей
°Реды, воспринимаемые каждым омматидием, т. е. глазком, входя-
щим в состав фасеточного сложного глаза, слагаются сротвётствен-
в большую мозаичную картину. И начиная со сравнительно
„ 737
небольших .комплексов, например у рабочих особей Ponera panda-
tissima из 1—5 омматидиев, Solenopsis fugax 6—9, Asellus
aquatlcus 4—8, Cladocera — 11, Porcellio scaber — 20, Limnetis —
50 омматидий, дело доходит до очень значительных комплексов,
например у Limnadia — 5б0, Branchipus — 1000, шмелей — 4000*
пчел — 5000, крапивницы — 4500, комнатной мухи — 4900, Aeschna
grahdis — 10 000, бабочки мертвой головы — 12 400 и, наконец,
у Necrophorus germanicus — 30 000 омматидиев. Сложные глаза
членистоногих по числу омматидиев, их расположению, развитию и
дифференцировке представляют большое разнообразие, которое еще
увеличивается тем, что развитие фасеточных глаз может сочетаться
с развитием простых глаз. В этом отношении знаменательно
характерное отсутствие в глазах членистоногих возможностей
аккомодации. Неизменная форма диоптрических аппаратов кор-
неальной линзы и кристаллического тела и постоянство расстояния
между ними и ретинальным слоем делают глаза членистоногих
установленными для зрения на постоянном расстоянии. Поэтому
развиваются различные глаза, одни для зрения на близком расстоя-
нии, другие — вдаль. То и другое сразу возможно при различной
дифференцировке различных частей одного сложного глаза, как,
например, у Bythotrephes или Simuliuni, С1оё. или при дифферен-
цировке у одной и той же особи одних глаз на дальнее зрение, дру-
гих на ближнее, как у многих насекомых, и т. д. Приспособительная
эволюция завершается, наконец, огромным разнообразием катоптри-
ческих приспособлений, обусловленных развитием и распределением
пигмента в простых и, особенно, в сложных глазах, и рефлектирую-
щего аппарата — tapetum. Получается возможность использования
световых раздражений от чрезвычайно слабых их источников, что
особенно важно для ночных животных и для обитателей глубин.
Мы наметили основные пути богатейшего адаптивного дивергент-
ного развития органов зрения членистоногих. Вдаваться в дальней-
шие подробности здесь не место.
В теснейшей взаимосвязи с головным комплексом органов чувств
находится развитие головного отдела нервной системы Arthropoda.
Это азбучная истина. Но факты приобретают здесь, однако, совер-
шенно- иное выявление, чем у аннелидообразных предков членисто-
ногих, у которых голова была представлена только их неметамерчой
головной лопастью. Как выше было выяснено, голова членистоногих
вторично дифференцируется за счет как неметамерног© отдела, го-
ловной лопасти аннелид, получившего здесь название acron, так и
присоединяющихся к нему метамер. Из них 1-я и 2-я —это ан-
теннальные сомиты, а несколько следующих дают ротовые част?
(см. рис. 92). Повидимому, приходится отнести и глаза членистоно'
гих, за исключением глаз перипатуса и науплиусовых медианных
глаз раков, к производным уже объединившихся вместе акрона *
первых двух метамер, а не считать их образованиями головной ло-
пасти, как глаза полихет и вышеуказанных форм. В результате
нервная система головы резко обособлена в два участка: предр0'
238
товой — собственно головной мозг аргропод ь кротовой — ПОД-
ГЛОТОЧНЫЙ ганглий. П^слсдшш представляет объединившиеся ганг-
лий брюшной цепочки трех ротовых сомитов, развитые главным об-
разом в соответствии с мощней мускулатурой ротовых частей, кзх
двигательные центры.
Совершенно иной характер представляет головной мозг члени-
стоногих. Его можно' назвать «сложным» головным мозгом —
syncerebnim потому, что, кроме иеметамерного отдела, принадлежав-
шего головной лопасти — archicerebrum, или protocerebrum, в сто
состав вошли переместившиеся
1-й и 2-й метамеры — deute-
ro и tritocerebrum. Этот про-
цесс является уже завершив-
шимся еще у Peripatus (рис.
98), т. е. характерен для всех
Arthropoda.
Смешение центров антен-
нальных сомитов в предротовое
положение резко противопола-
гается сохранению заротового
положения ганглиями мандибу-
лярного и следующих сомитов,
что особенно сказывается в
постоянстве заротового положе-
ния комиссуральных пучков,
связывающих * коннективные
стволы соответственно каждо-
му ганглию. Переход комиссу-
ральных образований антен-
нальных ганглиев из заротово-
дорзально по' коннективам ганглии
Рис. 98. Схема мозга перипатуса
(по Федорову)
1 — антеннальный нерв с его ветвями
(2, 3, 4); 5 —гигантская нервная
клетка, 6 — окологлоточный коннектив,
7 — дорзальный нерв, 8 — глобули, 9 —
гломерули, 10 —глаз, 11 — зрительный
перв, 12— п. recurrens, 13 — трабе-
кулы, 14 — центральное тело
го в предротовое состояние, ко-
нечно, нельзя представлять как
простое передвижение вперед
по окологлоточным конвектив-
ным стволам вместе с соответ-
ствующим передвижением ан-
тенн. Очевидно, при превращении подиальных образований антен-
нальных сомитов в мощные чувствительные придатки головы соот-
ветствующие нервные центры все теснее и теснее связывались с
всецело чувствительными и ассоциативными очагами головного
мозга. Возникли новые связи через этот последний. Развивался
вторичный предротовой комиссуральный аппарат, тогда как пер-
вичный заротовой, моторный, редуцировался. Подобный процесс
не мог иметь места в ганглиях ротовых частей, так как самое зна-
чение их по существу было иное, а именно — двигательных центров,
связанных всегда в известной мере с перекрестом отростков мотор-
нейронов, для мощной мускулатуры челюстей и жвал.
Неметамерный archicerebrum, при перемещении антенн* вперед.
239
у Peripatus как бы зажат между атеннальными нервами, прохо-
дящими отчасти с боков, отчасти дорзально от него, так что глаза
оказались сдвинутыми этими нервами в стороны и вниз. У прочих
Arthropoda (рис. 99) глаза, простые или фасеточные, развиты го-
раздо сильнее. Соответственно и антеннальные нервы проходят в
антенны позади и не задевая мощно развитых зрительных центров.
В общем protocerebrum у Arthropoda, обнаруживая в основном
характерные типовые черты полихетного головного мозга, эволю-
ционировал, с одной стороны, в направлении большей его интегра-
ции, т. е. по линии выработки более сложных ассоциативных цен-
тров, и с другой — в направлении мощного развития зрительного
отдела при редукции пальп и антенн аннелидного типа. Последний
процесс шел в теснейшей взаимосвязи с нарастающим значением
антенн артроподного типа. Происходила субституция ими первич-
Рис. 99. Схема мозга насекомого (по
Заварзину):
1 — протоцеребрум, 2 — дейтоцеребрум.
3 — тритоцеребрум, 4 — подглоточный
ганглий, 5 — брюшная цепочка, б — около-
глоточные коннективы, 7 — антеннальный
нерв, 8 — оптический ганглий, 9 — зри-
тельная лопасть, 10 — глобули, 11 — цент-
ральное тело, 12 — фронтальный ганглий
пых хемо- и механорецепторов
головы. Возникшие же в связи с
пальпами глобули, а в связи с
затылочными органами corpora
striata, ставшие ассоциативными
образованиями, развиваются чрез-
вычайно сильно. Своим крайним
развитием они, может быть, отча-
сти угнетают развитие соответ-
ствующих органов чувств. Таким
образом protocerebrum членисто-
ногих прежде всего содержит
впереди мощную систему globuli,
грибоводные или стебельчатые те-
ла, и сзади — систему централь-
ного тела или corpus striatum с
гломерулярными образованиями.
С боков в этом отделе мозга- рас-
положены зрительные центры, ко-
торые через tractus opticus пере-
ходят в зрительные доли, содер-
жащие один-три зрительных ганглия и уже не принадлежащие
мозгу. Последние представляют последовательно осложняющуюся
дифференцировку ганглиозного скопления ретины в более и более
сложные экранные центры зрительного типа.
Так создается характерный головной мозг Arthropoda, строение
которого в различных группах довольно сильно варьирует, в зави-
симости, с одной стороны, от развития глаз и антенн, с другой, -у
от высоты интеграции, т. е. сложности ассоциативных образовании,
достигая наибольшего ее выражения в мозгу «общественных» на-
секомых.
Набросанная нами схема понимания морфологии головного М°3'
га членистоногих в некоторых отношениях отличается от принятых
другими авторами. Так, ряд исследователей (Гольмгрен, ГанстреМ)
240
считает возможным производить антенны артропод прямо от пальп
аннелид и в соответствии с этим признает, что головной мозг анне-
лид представлен прото- и дейтероцеребрумом членистоногих. Трито-
церебрум при этом получает значение главным образом стомато-
гастрического центра. По более старым представлениям антенналь-
ный отдел мозга — дейтероцеребрум представляет первый присоеди-
нившийся к голове сомит, вторым таким сомитом является уже ман-
дибулярный. Не вдаваясь глубже во- все разнообразие высказанных
различными исследователями взглядов, отметим только, что, во-пер-
вых, теперь можно считать доказанной иннервацию мандибул от
участка, лежащего позади тритоцеребрума. Следовательно, соот-
ветствующая поправка должна быть внесена в более ранние взгля-
ды. Во-вторых, достаточных оснований гомологизировать пальпы
полихет и антенны членистоногих в настоящее время не имеется.
Поэтому взгляды, проводимые Гольмгреном, Ганстремом, Снодгра-
сом, не приемлемы.
Что касается заротового отдела нервной системы, то лишь у
Peripetias (см. рис. 83, 87, 90, Б) он представлен двумя стоволами,
покрытыми более или менее равномерно латерально и вентрально
ганглиозными клетками. Обычна для артропод типовая брюшная
цепочка с хорошей дифференцировкой ганглиев и коннективов,
расположенная в миксоцеле медиовентрально (см. рис. 84, 92, 93,
96). И опять-таки, лишь перипатус имеет два ствола отодвинутыми
далеко в бока, к основанию выростов ног, что можно поставить в
связь с мощным развитием у него медиовентральных продольных
мышц, которые у полихет обычно бывают представлены лишь не-
большими пучками, лежащими на брюшной цепочке проксимально.
Однако брюшная цепочка членистоногих далеко ушла в своей
эволюции, во-первых, по дифференцировке, специализации ее ганг-
лиозных клеток и по редукции глийных элементов, во-вторых, по
слиянию метамерных ганглиев в сложные образования вплоть до
концентрации их всех в один мощный центр, как это встречается
у крабов или у некоторых насекомых. Подобная тенденция часто
наблюдается в различных группах животного царства, ведя к по-
вышению ассоциативных отношений и интеграции деятельности
нервной системы.
При этом, конечно, подвергается коренной перестройке тип
невросомита полихет. Теряются характерные двигательные кольце-
вые нервы, дифференцирующиеся вместе с дифференцировкой спе-
циальных мышц на отдельные специальные пучки. Правда, у
Peripatus можно еще, в связи с сохранением кожно-мышечного
Мешка, не только установить присутствие моторных — переднего и
заднего кольцевого — нервов сомита, но и появление между ними
еЩе четырех пар подобных нервов, сопровождаемых хорошо выра-
женными чувствительными ветвями в дополнение к паре мощных
срединных чувствительных ножных нервов. Но эти изменения свя-
8аны с удлинением сомита, с одной стороны, и с резкой дифферен-
цировкой ног, с другой.
16 Пути эволюции животного мира * 241
У прочих Arthropoda удается еще иногда подметить следы быв-
ших кольцевых нервов. Но в норме — полная утрата характерного
невросомита аннелидных предков при постоянном сохранении ме-
та мерности ганглиев и отходящих нервов. Обычно наблюдающееся
перемещение задних ганглиев брюшной цепочки вперед и слияние
их друг с другом идет совершенно независимыми и разными путя-
ми в разных группах, больших и малых, и по-разному в различных
возрастных состояниях. В этом отношении членистоногие представ-
ляют огромное разнообразие, которое можно поставить в параллель
ко многим другим приспособительным явлениям в уже рассмотрен-
ных выше комплексах черт организации.
Подведем.„итоги нашему анализу организации членистоногих.
Характерная для" аннелид возможность эволюции в различных на-
правлениях* благодаря многообразию и пластичности их организа-
ции, находит свое завершение в типе членистоногих, но завершение
оригинально однобокое. Оно идет под знаком выработки защитной
хитиновой кутикулы покрова и выражается в конце концов в край-
них проявлениях этой характерной черты — в образовании наруж-
ного хитинового скелета, расчленении тела и конечностей, в далеко
идущей специализации последних, в росте с линьками и в незначи-
тельных размерах тела. В связи с этим перерабатываются взаимо-
отношения почти всех систем органов, гармонично’ развивавшихся
у червей. Особенно затрагивается целомический комплекс аннелид
с его многочисленными отправлениями. Это находит свое выраже-
ние в расчленении мускулатуры, редукции целома и связанных с
ним нефридиев, а также в развитии кровеносной системы и образо-
вании миксоцеля.
Организация аннелид коренным образом перестраивается — со-
здается особый тип организации членистоногих.
Обладая уже весьма сложным строением, этот тип обнаруживает
значительное повышение организации в связи с разнообразием
условий существования, которые ими осваиваются. Он дает огромный
размах приспособлений особенно с переходом к наземной жизни и
с проникновением в воздушную среду. Этому прежде всего способ-
ствовало развитие хитиновой кутикулы. Многообразие аспектов
наземного существования находит свое выражение в эволюции орга-
нов газообмена, экскреторного’ аппарата, кишечного тракта, органов
чувств и органов движения, полового аппарата, наконец у насеко-
мых в органах полета. В конце концов получается неисчерпаемое
богатство* форм членистоногих, которое определяется также и отно-
сительно малыми их размерами, соответственно относительно не-
значительным количеством клеточного материала, образующего
различные системы органов. Это ведет к далеко идущей дифферен-
цировке клеток последних, что особенно выражается в чувствитель-
ных аппаратах и нервной системе. Относительно незначительное
количество клеток в нервной системе обусловливает и своеобразие
нервной деятельности членистоногих, особенно' же насекомых с пре-
обладанием у них цепных рефлексов в виде сложных инстинктов.
242
Биологически судьбы Arthropoda определяются тем же ведущим
моментом развития хитиновой кутикулы, обусловливающим неболь-
шие размеры их тела. При возможностях огромного разнообразия
приспособлений это превращает членистоногих в организмы, могу-
щие заполнить малейшую нишу в биоценозе. Вместе с этим и в
результате этого Arthropoda становятся самой многочисленной груп-
пой животного населения Земли, далеко оставляющей позади все про-
чие типы животного царства как по количеству видов, так часто и по
числу особей. Однако их эволюция в силу некоторой односторонно-
сти организации не ведет к сложным выражениям высшей ступени
развития материи, к социальным явлениям. Дальше «социального
зачатка», как называет Ф. Энгельс общины пчел, муравьев и т. п.,
они не пошли.
16*
Глава XI
ТИП ИГЛОКОЖИЕ — ECHINODERMATA
Симметрия иглокожих. Прикрепление как ведущий момент в выра-
ботке'их организации. Смещение рта и- ряда других органов, вклю-
чение щупалец в состав туловища и процессы неравномерного
роста. Скелет и покров, оральный и аборальный отделы тела.
Мускулатура. Диплеврула. Пищеварительный аппарат, питание
иглокожих. Целом и его расчленение. Амбулакральная система. Пе-
ригемальная и кровеносная системы. Развитие зачатков левой и
правой сторон тела и процесс прикрепления. Осевой комплекс
органов. Камерный орган морских лилий. Органы чувств. Экто- и
гипоневральная системы и аборальная нервная еистема и их про-
исхождение. Эволюция мезодермальных зачатков. Итоги анализа
организации иглокожих.
Понимание организации иглокожих стало возможным только
после самого тщательного изучения их строения и онтогенетического
развития, но еще и теперь для зоолога это одна из увлекатель-
ных задач, решение которой дается глубоким анализом организации.
Было время, когда по чисто внешнему признаку — пятилучевой
симметрии тела — иглокожих ставили рядом с кишечнополостными
(см. рис. 111, 112, 113). Однако строение их оказалось неизмеримо
сложнее. Это типичные целомические животные, и уже в 1869 г.
И. И. Мечников, а затем в 1888 г. Земон и в 1892 г. Бючли вводят
е общий обиход положение, что иглокожие происходят от свободно
живших двустороннесимметричных прародителей. Эта гипотетиче-
ская форма получила название «диплеврулы» — двубокой (рис.
100). С переходом к сидячему существованию она дает уже ради-
ально построенную, непосредственную исходную форму всех игло-
кожих — «пентактею».
В настоящее время можно считать принятой большинством
зоологов такую формулировку: Echinodermata, обладая радиальной
симметрией, на самом деле являются собственно асимметричными
животными, происходящими от билатерально построенных предков.
Среди зоологов существуют и совершенно иные взгляды на игло-
кожих, но эти взгляды недостаточно обоснованы. Мы не станем на
них останавливаться.
При анализе организации иглокожих выявляется важнейший
244
.••• .... • .....
экологический момент, наложивший на них свою печать. Главное
здесь — это переход к синичему существованию.* Под влиянием
такого образа жизни выработался ряд приспособительных черт,
создавших в своей совокупности особый тип организации. Представ-
ляя новые возможности в борьбе за жизнь, этот тип развивается
далее оригинальнейшим образом и ведет к возникновению, наряду
с сидячими Pelmatozoa — морскими лилиями, также перешедших
вторично к свободному существованию Euleuterozoa — морских
звезд, офиур, морских ежей и голотурий.
Рис. 101. Ary stati-
st is bohemicus (из
Бэзера). Предста-
витель Cystoi
примитивных игло-
кожих. Расположе-
ние скелетных пла-
стинок:
Рис. 100. Схема организации
диплеврули иглокожих (по
Э. Мейеру): слева — вид сбо-
ку; справа — со спинной сто-
роны;
1 — передние и 2 - задние мешки
целома, 3 — кишечник, 4 — гидро-
цель, 5 — целомодукт
1 — место прикреп-
ления. 2 — рот, 3—
мадрепорит, 4 — по-
ловое отверстие, 5—
анальный конус
Основными ведущими’ процессами в эволюции иглокожих при
переходе к сидячему существованию становятся: 1) процессы сме-
щения рта и ряда других органов, имевшие большое значение в вы-
работке радиального построения, 2) приспособительные процессы
неравномерного роста тела, которые на основе включения щупалец
в состав туловища определили внешнюю морфологию. Можно ска-
зать, что эти процессы охватывают всю организацию, но1 наиболее
Рельефно они выражаются во внешней морфологии различных групп
иглокожих в сочетании с развитием скелета.
Скелет возникает, прежде всего, как пассивное защитное приспо-
собление сидячих форм и отличается очень своеобразным характе-
ром. Вспомним, что в организации аннелид, членистоногих и мол-
люсков защитную функцию берет на себя самая наружная часть
п°крова — кутикула. При этом у аннелид и артропод эту функцию
Несет вся поверхность, у моллюсков же защитную роль играет лишь
Раковина, т. е. спинной отдел покрова. У иглокожих эту роль играет
слой ткани основного вещества, располагающийся под покровным
* 245
эпителием и содержащий отложения извести (рис. 103). Таким об-
разом у иглокожих покровный слой тела остается собственно неза-
щищенным. Защищены лишь глубже лежащие, существеннейшие
части организма.
Отложение известковых солей происходит чрезвычайно характер-
но. В основном веществе рыхлой соеди-
нительной ткани, содержащей коллагец-
пые волокна, получается сетчатый или
губчатый известковый остов, в петлях ко-
торого находится основное вещество, а в
нем отдельные клетки и клеточные ско-
пления (рис. 104). Картина получается
совершенно своеобразная — нигде боль-
ше такого скелета нет. Достаточно' одного
взгляда на шлиф из кусочка скелета, что-
бы безошибочно сказать, имеем ли мы де-
ло с представителем иглокожих или нет.
Так создается тангентальный скелет в
виде пластинок, облекающих тело плот-
ным панцырем, т. е. как бы броня, пас-
сивно защищающая внутренности и при-
сущая, конечно, прежде всего туловищу,
как это хорошо видно у ряда примитив-
Рис. 102. Морская
ЛИЛИЯ Ну О' пи us из
Карского моря.
Внешний вид
2
~ оо
°ГО Q
о О О о
<^оУо О о о 0
О о о О
о О оУд
о о
Рис. 103. Разрез через стенку тела голотурии:
1 — покровный эпителий, 2 — соединительнотканный
слой кожи, 3 — пластинки скелета. 4 — мускулатура,
5 — целомический эпителий
ных Cystoidea (см. рис. 101). Однако у большинства форм <к тан-
гентальному скелету присоединяется скелет, представленный игла-
ми, образующимися в соответствующих выростах покрова. Выстоя-
щий скелет может играть уже роль активной защиты в дополнение
к пассивно защитному панцырю. Этот в известной мере второстепен-
ный признак организации получает такое развитие и так бросается
246
Рис. 104. Разрез через пластинку ске-
лета Astropecten aur mtiacus
(по К. Шнейдеру):
I — отложения извести. 2 — соедини-
тельнотканные тяжи с клетками (3)
з глаза у ряда свободноживущихформ (см. рис. Ill, 112) —морских
звезд, офиур, морских ежей, что весь тип характеризуется назва-
нием ’«иглокожие». Однако ведущей чертой организации иглокожих
является выработка пластинок тангентального скелета, а не игл.
В связи с развитием скелета находится то или иное состояние
покрова иглокожих. У морских звезд и ежей последний сохраняется
пОд защитой игл в виде мерцательного эпителия. Но на чашечке мор-
ских лилий, равно как и у офиур, он становится своеобразным кути-
кулярным и как бы теряет свою обособленность от подстилающего
соединительнотканного слоя. Эта черта выступает также у голотурий,
у которых тангентальный ске-
дет в известной мере редуциро-
ван в связи с выработкой чер-
веобразного тела. Ведущий,
хотя и узко адаптивный, приз-
нак организации был ими
преодолен — скелет редуциро-
ван. Но, тем не менее, покров-
ный эпителий оказался слабо
развитым. И защищает орга-
низм не своеобразная кутикула
покровного эпителия, а, как и у
прочих Euleuterozoa, сильно вы-
раженный слой ткани основно-
го вещества с мощным разви-
тием волокнистых структур cu-
tis, — эволюция необратима.
Несомненно, примитивной
чертой покрова является при-
сутствие мерцательного эпите-
лия, заменяющегося далее у
ряда форм своеобразным кути-
кулярным эпителием.
Очень важным в развитии ио-крова, как мы видели, представ-
ляется момент защиты, выраженный в развитии тангентального ске-
лета. Так, у Pelmatozoa тангентальный скелет образован хорошо
Развитыми пластинками, покрывающими все тело1 плотным панцы-
Р*ем. Панцырь этот представлен либо в виде мешка, прикрепленного
евоим суженным основанием к подводным предметам (см. рис. 101),
либо в виде чашечки, сидящей на более или менее длинной и гиб-
кой ножке или стебельке (см. рис. 102). У современных морских
лилий такая чашечка обладает значительной мощностью известко-
вьтх образований и вполне закономерным 5-радиальным расгГоложе-
йием рядов своих пластинок (рис. 105). Пластинки образуют либо
°Дин ряд — моноциклические лилии, либо два ряда из радиальных
и интеррадиальных пластинок, окружающих центральную, — ди-
^вкличеокие лилии. *
Морские лилии — это последние этапы длинного пути эволюции,
247
начинающегося такими формами, как
Cystoidea, с неопределенным коли-
чеством пластинок, своим располо-
жением образующих до некоторой
степени подвижный еще папцырь
(см. рис. 101).
Во всех случаях на полюсе, про-
Рис. 105. Схемы моноциклической
(А) и дициклической (Б) чаше-
чек морских лилий (по Кирх-
иеру):
1 — базальные, 2 — радиальные, 3 —
брахиальные, 4 — пнфрабазальные
пластинки чашечки. 5 — пластинка
стебелька
тивоположном месту прикрепления
животного, располагается ротовое
отверстие, а прилежащая к нему
«оральная» часть тела не имеет та-
ких мощных скелетных отложений,
как чашечка, и покрыта хорошо раз-
витым мерцательным эпителием,
Рис. 106. Продольный разрез через чашечку мор-
ской лилии Rhizocrinus lofotensis (по Деляжу
из Людвига):
1 — камерный орган с его нервным центром (2) и отхо-
дящим нервом (3), 4 — нерв, проходящий в брахиаль-
ных пластинках скелета рук, 5 — скелет основания ча-
шечки, б и 7 — радиальные пластинки. 8 — рот, 9 —
кишечник, 10 — целом чашечки, 11 — осевой орган, 12 —
амбулакральная система, 13 — мышечные пучки между
радиальными пластинками скелета, 14 — целом, входя-
щий в руки
в противоположность
«аборальной», несущей
деградирующийся кути-
кулярный эпителий.
Как показывает ис-
тория развития, между
брюшной и спинной сто-
ронами диплеврулы иг-
локожих (см. рис. 100),
с одной стороны, и меж-
ду оральной и абораль-
ной частями тела взрос-
лого животного, с дру-
гой, — нет ничего об-
щего. Поэтому обозна-
чения «оральная» в
«аборальная» части те-
ла подчеркивают ха-
рактерные черты орга-
низации иглокожих, вы-
работавшиеся именно в
эволюции этого типа.
Пр испособительное
значение оральной по-
верхности совершенно
определенно. Это та по-
верхность, которая сво-
им мерцательным по-
кровом обусловливает
возможность пассивно-
го питания сидячих
форм. Создается т°к
воды, несущий пищевые
248
Рис. 107. Прикрепленная стадия морской лилии Antedon rosacea
(по Карпентеру). Видно образование рук путем объединения щу-
палец и выростов чашечки. Пластинки скелета:
1 — оральные, 2 — базальные, 3 — радиальные, 4 — дальнейшие радиаль-
ные пластинки в основании рук, 5 — стебелек, 6 — руки
частицы к ротовому отверстию. И у цистидей, как, например, у Aris-
tocystis (см. рис. 101), это лишь оральная поверхность окружав-
ших рот щупалец, в противоположность всей остальной поверхно-
сти мешковидного тела.
У современных криноидей, имеющих чрезвычайно мощный або-
ральный скелет в виде прикрывающей внутренности массивной и
правильно построенной 5-радиалыюй чашечки (рис. 106) к базаль-
ным и радиальным ее пластинкам присоединяются далее косталь-
ные, а также брахиальные, располагающиеся уже в ветвящихся
руках. Они образуют аборальный скелет рук. В соответствии с этим
руки собственно являются выро-
стами туловиша, прикрывающими
и включающими в себя, «инкор-
порирующими», первоначальную
оральную поверхность, т. е. около-
ротовые щупальца, (рис. 107 и
108).
Можно сказать, что у кринои-
дей процессы неравномерного ро-
ста выразились в чрезвычайно
сильном развитии аборальной ча-
сти тела, тогда как оральная раз-
вивается слабее, в виде мерца-
тельных поверхностей рук. В
организации этих форм момент
защиты при сидячем существова-
нии выявляется особенно сильно
в развитии мощного аборального
скелета.
По иному пути пошли Euleu-
terozoa. У них на выростах пяти
неветвящихся рук чувствительные
придатки, имеющиеся на щупаль-
цах криноидов, превращаются в
сократимые «амбулакральные но-
жки», снабженные на конце при-
соской (рис. 109).
В дальнейшем при использовании щупальцевых придатков для
передвижения всего тела животного происходит переход к свобод-
ному существованию, и пассивное питание сидячих форм заме-
няется активным захватыванием пищи. Создаются двигательные
придатки, совершенно непохожие ни на параподий аннелид, ни на
ножки членистоногих. Вместе с тем на первый план выступает
скелет оральной поверхности, вырабатываются амбулакральные и
адамбулакральные пластинки, прилежащие к мерцательной борозде
рук между амбулакральными ножками. Это, в свою очередь, стоит
в теснейшей связи со значительным увеличением размеров ораль-
ного отдела тела — он у морских звезд становится равным або-
250
Рис. 108. Схема поперечного разреза
через руку морской лилии (из Лан-
га):
I — брахиальная пластинка скелета, 2 —
чувствительный придаток руки с амбула-
кральным каналом внутри, 3 — радиаль-
ный амбулакральный канал, 4 — перйге-
мальный канал, 5 — эктоневральный и 6—
гипоисвральный отделы нервной системы,
7 — аборальный нерв, 8 — спинной и 9 —
брюшной отделы целома. 10 — половой
синус целома с половым тяжем
16
15
Рис. 109. Поперечный разрез через руку морской звезды (по
Лангу):
1 — гипоневральная и 4 — эктоневральная нервные системы, 2 — радиальный
канал амбулакральной системы и канал в ножку (6) 3 — кровеносный сосуд,
5 — перигемальная система, 7 — ампула амбулакральной ножки, 8 — мыш-
цы, соединяющие амбулакральные пластинки скелета (9), 10 —адамбула-
кральные пластинки, И — иглы, 12 — гонады, 13 — печеночные выросты
кишечника, 14 — целом, 15 — палулы для газообмена, 16 — аборальный
нервно-мышечный тяж
Рис. НО. Поперечный разрез через руку офиуры (по Лангу). Обо-
значения, как на рис. 109, кроме того:
9 — позвонок, образованный амбулакральными пластинками, 17 — эпине-
вральный канал, 18—эпиневральная пластинка скелета, 19—прикрепления мышц
ральному (рис. 111). Таким образом выявляется несколько иное
направление процессов неравномерного роста, чем это имеет место
у морских лилий. Проводя разрез по маргинальной линии лучей
морской звезды, мы делим ее тело на две равные половины —
нижнюю, «оральную», и верхнюю, «аборальную».
Кроме того, неравномерный рост выражается в развитии пяти
лучей морской звезды соответственно пяти первоначальным около-
Рнс. 111. Организация морской звезды Astropecten aurantiaciis (по Пфурт-
шеллеру). Правая сторона вскрыта и видны внутренние органы; правый луч
• отпрепарирован, чтобы показать скелет:
1 — мадрепоровая пластинка, 2 — «позвонки», 3 — амбулакральные ножки, 4 — ампулы
амбулакральной системы, 5 — полиевы пузыри, 6 — иглы, 7 — желудок, 8 — печеночные
придатки, 9 — гонады
ротовым щупальцам. У них в терминальных пластинках рук нахо-
дятся концы первоначальных радиальных щупалец.
Процессы неравномерного- роста по пяти радиусам достигают
своего крайнего выражения у офиур. Их длинные руки сами ста-
новятся членистыми придатками для передвижения тела и резко
обособлены от диска, а амбулакральные ножки теряют свое зна-
чение придатков, передвигающих тело (рис. 110). Но интеррадиаль-
ный рост оральной половины у офиур, можно сказать, отсутствует.
Процессы неравномерного роста, выражающиеся в преобладаю-
щем развитии орального отдела, достигают почти полного заверше-
252
яия у морских ежей (рис. 112), у которых от ^орального отдела
20 слабо развитым скелетом сохраняется лишь небольшой участок
2 виде «перипрокта» — околоанального поля (рис. 113). Следова-
Рис. 112. Организация морского ежа (по Пфуртшеллер}) :
1 — мадрепоровая пластинка, 2 — амбулакры, 3 — интерамбулакры, 4 — амбула-
кральные ножки, 5 — иглы, 6 — начальная часть и 7 — извивы кишки, 8 — аналь-
ное отверстие, 9 — радиальный канал и 10 — околоротовое кольцо амбулакральной
системы, 11 — полиевы пузыри, 12 — каменистый канал, 13 — ампулы, 14 —аристо-
телев фонарь, 15 — гонады
тельно, вся скорлупа ежа представляет, собственно, чрезвычайно
развитый оральный отдел. Радиальные ряды амбулакральных пла-
стинок у морских ежей заканчиваются около перипрокта глазными
пластинками, т. е. заключающими, как и
terminalia звезд, концы щупалец. Кроме то-
го, развиты интеррадиальные ряды, как ин-
терамбулякры. И здесь, в отличие от офиур,
нельзя говорить об отсутствии роста интер-
радиально: мадрепоровая пластинка, распо-
ложенная у офиур возле рта, у морских
ежей находится около перипрокта.
Таким образом, морские ежи являются по
характеру процессов неравномерного роста
прямой противоположностью по отношению
к криноидеям. У первых максимум развития
орального отдела, у криноидей аборального.
Голотурии обладают теми же отношения-
ми процессов неравномерного роста, как и
еЖи, но перипрокт уже совсем отсутствует.
Рис. 113. Перипрокт и
прилегающая часть скор-
лупы морского ежа (по
Пфуртшеллеру). Обозна-
чения см. на рис. 112,
кроме того:
16 — генитальные и 17 — глаз-
ные плас!инки скелета
253
Голотурии, в силу своеобразных условий существования, перешли к
ползанию посредством трех радиальных
Рис. 114. Организация Hofothnria tubulosa (по
Мильн-Эдвардсу):
I — околоротовые щупальца, 2 — окологлоточное кольцо
амбулакральной системы, 3 — полнев пузырь, 4 — гона-
да, 5 —- кольцевая и 7 — продольная мускулатура, 6 —
радиальный амбулакральный канал, 8 — аначьное отвер-
стие. 9 — водное легкое. 10 — кишечник, 11 — крове-
носный сосуд, 12 — чудесное сплетение на легком
рядов амбулакральных
ножек (trivium), т. е. к
ползанию на боку. Про-
тивоположные два ряда
ножек (biviumj для пе-
редвижения не служат,
направлены в окружаю-
щую водную среду и
несут, в соответствии с
этим, роль шупальцеоб-
разных придатков.
Подобные отноше-
ния стали возможными
вместе с выработкой
более длинного, черве-
образного тела и с
утратой сплошного пан-
цыря, заменяющегося
прочной соединительной
тканью кожи. Она со-
держит относительно
мелкие известковые от-
ложения. Таким обра-
зом, как уже было от-
мечено, крайность адап-
тивного развития- скеле-
та преодолевается голо-
туриями. Они стано-
вятся донными черве-
образными формами,
способными проникать
в узкие пространства
между камнями и т. п.
С этим связана и
выработка характерно-
го' для голотурий аппа-
рата для захватывания
пищевых частиц. Аппа-
рат этот развивается
за счет второй пары
амбулакральных ножек
каждого из пяти р&~
диусов. Вместе они об-
разуют вокруг рта вен-
чик из 10 Ш|УпалеП
• (рис. 114), чего совершенно нет у других Eleuterozoa, где для за-
хватывания пищи служат так называемые зубы, развившиеся за
254
счет скелетных частей, прилегающих к ротовому отверстию (см.
□ис. 112). Таким образом, аппарат щупалец голотурий не имеет
ничего общего с руками морских лилий.
Итак, процессы перестройки организации иглокожих связаны с
усилением возможностей питания — развивается оральный отдел
тела. Но в то время как у сидячих морских лилий одновременно и
. преобладающе происходит также еще и развитие аборального за-
щитного скелета, у Eleuterozoa, с эволюцией амбулакральных но-
жек и игл, гигантски развивается оральный отдел с. амбулакраль-
ным скелетом. При этом вместе с включением щупалец в состав
туловища редуцируется или полностью исчезает аборальный
скелет.
Во внутренней организации в связи с развитием скелета сто-
ит развитие мускулатуры стенки тела. У иглокожих, как и в типах
членистоногих или моллюсков, образование скелета повело к ре-
организации единой мускулатуры кожно-мышечного мешка двусто-
роннесимметричных предков. Мускулатура дифференцировалась
в специальные пучки, идущие от одного куска скелета к дру-
гому.
Таковы парные мышечные пучки криноидей между соседними
костальными (см. рис. 106), между костальными и дистихальными
и между дальнейшими брахиальными пластинками, пучки, произво-
дящие в общем сгибание рук морских лилий к оральной стороне.
Таковы мышечные пучки между «позвонками» у офиур (см.
рис. 110), относящиеся уже к оральному скелету и дающие значи-
тельные возможности движения рук, особенно в соединении с ра-
диальными мышечными пучками, проходящими у морских звезд
(см. рис. 109) по аборальному отделу руки медианно до терминаль-
ных пластинок.
У морских ежей при сплошной скорлупе имеет место исчезнове-
ние мускулатуры тангентального скелета (рис. 1161, но развиваются
специфические мышечные пучки при основании игл, служащие для
движения последних. Напротив, у голотурий при редукции скелета
образуется мускулатура, сходная с мускулатурой типа червей, коль-
цевые и продольные мышцы (рис. 114). Но она возникает на другой
основе. Вдоль по длинному телу, соответственно’ каждому радиусу
и рядам амбулакральных ножек, проходят от переднего конца до
заднего по паре рядом лежащих мышечных лент, т. е. всего 10
мощных продольных тяжей. Это продольная мускулатура. А между
Денгами двух соседних радиусов, от одной до другой ленты, пре-
Рьгваясь под ними, идут пучки, в совокупности дающие кольцевую
мускулатуру стенки тела (см. рис. 117).
Таким образом, вместо непрерывных слоев мышц кожно-мышеч-
ного мешка червеобразных форм, оказывается резко дифференциро-
ванная мускулатура голотурий, развивающаяся из радиальных
пУьков, сходных с межпозвоночными пучками морских зврзд, и из
иДуЩих под углом в 90° к ним интеррадиальных пучко-в. Пои этом
Те и другие происходят, как типичная мезодермальная мускулатура,
255
за счет париетального листка. Все говорит о вторичном и очень
характерном развитии мускулатуры голотурий за счет уже выра,
ботавшихся мышечных пучков, специфичных для иглокожих и обу-
словленных дифференцировкой скелета.
Переходим к рассмотрению других комплексов характерных черт
внутреннего строения иглокожих, прежде всего к системе кишечни-
ка и целома, понимание которых становится возможным при допу-
щении смещения некоторых органов в связи с особыми условиями
сидячей жизни. Вместе с этим мы подходим и к пониманию воз-
никновения пятилучевой симметрии у иглокожих.
В прошлом, одновременно с выяснением вопроса о значении
прикрепления у иглокожих, шло выявление зоологами основных
черт строения их двусторонне-симметричных исходных форм.
Здесь, как подчеркнул еще И. И. Мечников (1869), решение
вопроса зависело от понимания строения кишечника и от факта
наличия целома и его развития. Р. Земон (1888), О. Бючли (1892),
а затем Э. А. Мейер (1904) представляли себе гипотетическую
диплеврулу (см. рис. 100) следующим образом: эллипсоидной
формы, несколько уплощенная вентрально, с ротовым отверстием в
передней части брюшной стороны и анальным близ заднего конца
последней, с парой задних мешков целома справа и слева от трубки
кишечника и с парой передних мешков по бокам рта (гидроцели),
имевших по пяти выростов, входивших в соответствующие щупаль-
ца, причем левый гидроцель, кроме того, имел канал, начинавший-
ся между вторым и третьим щупальцами и открывавшийся наружу
па спинной стороне.
Такая диплеврула в основном близка к личинке иглокожих, как
бы дополненной щупальцами, которые появляются в развитии особи
лишь позже и только с левой стороны.
Конечно, как уже было выше отмечено, при настоящем состоя-
нии науки в большинстве случаев мысль, что личинки представляют
анцестральные формы, давшие начало тем или иным группам жи-
вотного мира, является упрощением.
Эпоха увлечения подобными теоретическими построениями —-
это уже давно пройденный этап, и в настоящее время гипотетиче-
ская диплеврула должна мыслиться как взрослый организм, черты
которого' находят выражение и в личиночных формах иглокожих*
Так, он имел двусторонне-симметричное строение тела, но по су-
ществу это был представитель первичных целомических животных,
т. е. принадлежал к олигомерным целомическим червям, ведшим
придонное существование и обладавшим, как и все черви, пелаги-
ческими личинками.
Подобные формы мы уже постулировали, исходя из анализа
организации аннелид и моллюсков.
Представить себе, что протоцеломаты, давшие начало иглоко-
жим, могли быть какими-нибудь иными формами, чем уже выве-
денные нами немертинообразные предки моллюсков и аннелид, это
значит признать две независимые ветви целомических животных,
*256
конвергентно развившиеся из разных исходных групп. Такое допу-
0НИе делают в настоящее время большинство зоологов, следуя по
пути, проложенному Гатчеком и Гроббеном.
Но имеются ли достаточные основания идти по этому пути? На
этот вопрос мы ответим позже и ответим отрицательно.
Обратимся к пищеварительному аппарату иглокожих. Он в очень
малой степени несет в себе черты радиального построения, а в’го-
раздо большей степени говорит о смещении органов в процессах
неравномерного роста.
Б онтогенезе всех групп иглокожих-всегда имеется’прямой труб-
чатый кишечник, одетый правым и левым мешками целома. Иное
мы обычно находим у взрослых. Кишечник образует за вертикально
идущим глоточным отделом горизонтально' лежащую петлю, которая
продолжается в вертикальный задний участок. Целом является
всегда эпи- и гипогастрическим, т. е. правый его мешок помещается
над петлей кишечника, а левый, находящийся у рта, под ней. При
этом вертикальные мезентерии становятся горизонтальными. Теряя
свою целостность, мезентерии распадаются далее на отдельные тяжи
или редуцируются. Получается общая полость тела, окружающая
кишечник (см. рис. 112).
Свое завершение процесс находит, с одной стороны, у крино-
идей, которые развиваются в направлении усиления пассивного пи-
тания сидячих форм. При этом разнообразно' и мощно дифферен-
цируется мерцательный аппарат рук, обусловливающий ток воды
к ротовому отверстию. Рот вместе с окружающими его щупальцами
находится на полюсе, противоположном пункту прикрепления жи-
вотного, а кишечник имеет типичную форму петли (рис. 106). Как
показывает онтогенез, такие отношения получаются в результате
передвижения ротового* отверстия по левой стороне тела из перво-
начального положения у места прикрепления-па.противоположный
конец (рис. 115).
При мощном развитии в стенке тела скелета целом, теряющий
свое первичное значение опорного образования, заполняется в цен-
тральной своей части рыхлыми тяжами ткани целотелиального
происхождения.
С другой стороны, у Eleuterozoa — свободных потомков сидячих
форм — пути приспособительного1 развития кишечника значительно
разнообразнее. Это понятно в связи с большими возможностями
использования пищевого материала свободно живущими животными.
*ак, эволюция, пошедшая по пути хищничества, проявляется у
Морских звезд и офиур (см. рис. 111). Кишечник в проксимальном
отделе — так называемый желудок — становится крайне растяжи-
мым, способным в некоторых случаях буквально выворачиваться
через рот, как мешок, и охватывать добычу, например двустворча-
тых моллюсков. При этом теряется значение дальнейшего участка
кишки и анального отверстия. И если последнего нет уже у неко-
торых морских звезд, то для офиур это обычное явление. Конечно,
такое «целентерическое» состояние кишечника — явление вторич-
17 ПутИ эволюции животного мира 257
вое, не имеющее ничего общего с первичным целентерическим
аппаратом кишечнополостных или турбеллярий.
Эволюция хорошо развитых лучей у морских звезд связана и с
другой характерной особенностью их кишечника, именно с появле-
нием некоторой вторичной радиальности строения, которая выра-
жается в развитии на аборальной части желудка печеночных
Рис. 115. Личинки морской лилии Antedon rosacea: А — только
что вышедшая; Б— после прикрепления (по Зейлигеру):
1 — vestibulum 2 — место прикрепления. 3 — апикальный пучок ресничек,
4 — ресничные кольца, 5 — передний левый целом — гидроцель. 6 — задний
левый и 7 — задний правый целомы, 8 — передний правый целом, 9 — ки-
шечник, 10 — пластинки стебелька, прочие пластинки относятся к чашечке
придатков. Они располагаются попарно в руках и служат для уве-
личения поверхности кишечника, усваивающей питательный мате-
риал, поступающий из желудка.
Приспособление к использованию в качестве пищи покрываю-
щих дно порослей мшанок, гидроидов и т. п., породило кишечник
морских ежей совершенно иного характера (см. рис. 112). Он
образует типичную петлю, как и у криноидей, но отличается зна-
чительным удлинением с образованием извивов, обособлением си-
фональной кишки, хорошо выраженными подвесками. Как уже было
отмечено, для кишечника ежей характерна в связи с их питанием
выработка жевательного аппарата аристотелева фонаря за счет
околоротового отдела скелета. Отсюда приспособительное развитие
ведет к заглатыванию грунта, что имеет место у спатангид и
•клипеастрид.
^58
В полной мере сохраняется петля кишечника и у голотурий,
идущих в известной мере по пути, сходному с ежами в эволюции
иглокожих (рис. 114).
При удлиненном теле голуторий вытягивается в длину и петля
кишечника, крепко подвешиваемая к стенке тела па трех своеоб- -
разных отделах мезентерия. При этом, как мы видели, вторая пара
амбулакральных ножек привлекается для захватывания пищевого
материала, т. е. мельчайших организмов, слизываемых с каменистых
н песчаных грунтов. Вторично, при приспособлении к жизни на
илу, происходит просто захватывание грунта вместе с пищевыми
частицами в кишечник. Развиваются формы наподобие синаптид —
постоянные обитатели грунта, соответственно вторично упростившие
свою организацию, что некоторыми исследователями неправильно
принималось за первичную примитивность. ’
Итак, кишечник в виде петли характерен для иглокожих, причем
петля располагается в горизонтальной плоскости и получается в
результате смещения ротового отверстия от места прикрепления у
криноидей на пункт, противоположный последнему, а у Eleuterozoa
не доходит до него, смещаясь вентрально. При этом анальное от-
верстие занимает место на аборальном полюсе.
Своеобразные отношения эпи- и гипогастрического' целома ста-
новятся естественным следствием перемещения ротового отверстия,
так как соответствующий отдел кишечника при движении рта по
левой стороне тела оттесняет левый целомический мешок. Последний
располагается под петлей кишки у рта, тогда как правый занимает
пространство над петлей аборально.
Что касается функциональных отношений, то, вместе с* разви-
тием скелета при утере целомом опорной функции, на первый план
выступает его' трофическая функция. Однако сохраняется и экскре-
торная функция целома, имеющая особое значение при отсутствии
системы нефридиев, заменяемых нефрофагоцитами. Спорадически
особо развивается функция газообмена через посредство тонких
участков стенки тела: через папулы у морских звезд (см. рис. 109),
через околсротовые жабры — у морских ежей.
Всего сложнее отношения выявляются у голотурий (рис. 114),
которые при редукции скелета вновь используют целом как опору
для мускулатуры стенки тела. Кроме того, они вырабатывают для
экскреции цилиофагоцитарные органы типа урн, и, наконец, за счет
заднего отдела кишки образуют водные легкие. Последние, в соеди-
нении с чрезвычайным развитием кровеносных сосудов в мезенте-
риях кишечника, через целом, как промежуточное звено, обусловли-
вают газообмен организма.
Из сказанного уже видно разнообразие функций целома иглоко-
жих, что указывает на возможности гораздо больших и дальше
идущих дифференцировок целома, какие в действительности и име-
ются у иглокожих.
Как мы видели, общая полость тела развивается за счет задней
пэры целомических мешков, прилежащей к усваивающему кишеч-
17* 259
нику. Первоначальная парность, обусловленная у протоцеломат ан-
тагонизмом при деятельности правой и левой половин кожно-мы-
шечного мешка, теряет свое значение при утрате целомом опорной
функции. Более того, целом, приобретая преимущественно трофиче-
ское значение, утрачивает свою парность и с образованием петель
кишечника сохраняет мезентерии лишь для фиксации па стенке тела
определенных участков кишки. Развитие вдобавок к этому газооб-
мена у звезд и морских ежей также связано- с образованием в конце
концов единой общей полости тела.
В соответствии с этим стоит проникновение общей полости тела
в околородовые участки, собственный целом которых, изолирован-
ный от усваивающего отдела кишечника, не имеет уже трофическо-
го значения.
У морских лилий в мощно развитые пять рук входят выросты
гипо- и эпигастрического целома в виде каналов, тянущихся до са-
мых концевых разветвлений рук (см. рис. 108). Один из каналов
«дорзальный», берущий начало от эпигастрического целома и про-
ходящий непосредственно под брахиальными члениками скелета, и
пара каналов, лежащих под ним, «вентральных» от гипогастриче-
ского целома. Таким образом, создается система, питающая руки
морских лилий, и, вместе с тем, первичные щупальца охватываются
аборальным отделом тела и входят в его состав.
У морских звезд и офиур этот процесс получает очень яркое и
несколько' иное выражение в связи с тем, что оральные придатки
резче обособлены (см. рис. 109 и 110) благодаря развитию амбу-
лакральных и адамбулакральных пластинок скелета, и включение
их в состав туловища происходит в онтогенезе в некоторой адере
раздельно. Над пятью закладками оральной части лучей морской
звезды появляются соответственно аборальные выросты стенки те-
ла, содержащие целом. Эти' выросты увеличиваются и сливаются с
оральными закладками. Образуется единый луч морской звезды. Но
процесс осложняется тем, что каждая аборальная закладка при ро-
сте смещается и сливается не прямо с нижележащей оральной, а с
соседней по возникновению. Это является у морских звезд своеоб-
разным выражением общего смещения околоротового участка к
брюшной стороне диплеврулы, не доходя до> апикального положе-
ния. Последнее, напротив, наблюдается у сидячих форм.
У морских ежей аборальный участок тела крайне редуцирован,
а потому не происходит, с одной стороны, соответствующих разра-
станий целома и стенки тела аборально, а с другой стороны, и за-
кладки амбулакральных пластинок скелета. Процесс идет так, что
оральная часть просто располагается на общей полости тела, т. е.
полностью включена в туловище, чем полностью обеспечено и ее
питание (рис. 116). У голотурий имеют место подобные же отно-
шения (рис. 117).
Все это' приходится поставить в теснейшую связь с развитием
образований амбулакральной борозды, т. е. своеобразнейшего ком-
плекса внутр еййих органов, свойственных первоначальным оральным
260
Рис. 116. Поперечный разрез через стенку тела мор-
ского ежа по радиусу (из Ланга). Обозначения см.
рис. 117
Рис. 117. То же у голотурии (из Ланга):
1—гипоневральнаЯ и 4 — эктоневральная нервные системы, 2— радиальный канал
амбулакральной системы и канал в ножку (6), 3 — кровеносный сосуд, 5 — периге-
мальная система, 7 — ампула амбулакральной ножкп, 8 — продольные и 9 — коль-
цевые мышцы, 10 — соединительнотканный слой кожи, 11 — покровный эпителий,
12 — целотелий, 13 — целом, 14 — двигательный и 15 — чувствительный нервы,
16 — пластинка скелета, 17 — эпиневральный канал
Рис. 118. А, Б, В, Г — схема развития целома у
иглокожих (из Гейдера); схема развития целома
у офиуры Ophiothrix; Д — общая закладка целома;
Е— подразделение на правый и левый целомы;
Ж — увеличение левого целома; 3 — подразделение
его на передний и задний; И — такое же подраз-
деление правого целома; К и Л — дальнейшее раз-
витие (по Мак Брайду);
1 — передний и 2 — задний целимы, 3 — гидроцель,
4 — поровый канал, 5 — рот, 6 — анальное отверстие, 7 —
мезенхима, 8 — кишечник
щупальцам, включе-
ние которых в состав
туловища, как мы
видели, играло веду-
щую роль во ^внеш-
ней морфологии иг-
локожих. Среди ком-
плекса амбулакраль-
ной борозды особен-
ное значение имеют
целомические систе-
мы — амбулакраль-
ная и перигемальная,
к анализу которых
мы и переходим.
Эволюция перед-
них целомических за-
кладок, дающих на-
чало амбулакраль-
ной системе, идет со-
вершенно’ иначе, чем
задних мешков це-
лома (см. рис. 100,
118). Во-первых,
обычно развивается
левый зачаток, а
правый или соврем
отсутствует, или яв-
ляется рудиментар-
ным. Во-вторых,. их
функции теснейшим
образом .связаны у
сидячих форм с ап-
паратом оральных
шу п а л ец пр и пас-
сивном питании, а
у свободноживуших
они служат для ак-
тивного захватыва-
ния пищи и для пе-
редвижения всего те-
ла. Простая плот-
ность щупальцеоб-
разных придатков
морских лилий, обус-
ловленная жидким
содержимым входя-
щих в них выростов
262
переднего целома, переходит в настоящую двигательную функцию
амбулакральных ножек, специфичных для иглокожих.
Закладывается амбулакральная система в виде дугообразно изо-
гнутого мешочка, расположенного слева от глоточного отдела ки-
шечника личинки и несущего на выпуклой стороне дуги пять выро-
стов, соответственно щупальцам. В дальнейшем дуга принимает
форму подковы и, наконец, замыкается в кольцо вокруг глотки
(рис. 119). Другими словами глотка, смещаясь на пункт, противо-
положный месту прикрепления животного, вдавливается в левый
мешок переднего целома. При этом глотка увлекает в своем дви-
жении этот последний, и его отходящие в пять щупалец каналы
постепенно располагаются вокруг глотки правильно по пяти радиу-
сам, определяя пятилучевое строение взрослого животного. Значе-
ние для сидячих форм такого плана построения, при котором жи-
вотное ориентируется и равномерно реагирует по всем радиусам
вокруг пункта прикрепления, мы видели уже у кишечнополост-
ных.
Зачаток переднего целома правой стороны у свободноживущих
иглокожих или отсутствует совсем или сохраняется в виде рудимен-
та — небольшого пузырька, расположенного в стенке тела, близ
аборального отдела и обычно утрачивающего функциональное зна-
чение.
У морской лилии Antedon сначала закладывается один зачаток
переднего целома, который, однако, затем подразделяется на две
части. При прикреплении личинки одна из них смещается на про-
тивоположный месту прикрепления полюс, т. е. к будущему ораль-
ному отделу чашечки, и замыкается в кольцо, от которого отходят
5 радиальных каналов. Следовательно, этот зачаток ведет себя ха-
рактерно для закладки амбулакральной системы (см. рис. 115).
Напротив, другой зачаток (см. рис. 122, Л) сохраняет теснейшую
связь с головным участком личинки, образующим при прикреплении
к субстрату аборальный отдел чашечки и стебелек. Он проходит от
основания чашечки по оси стебелька.
Однако у Eleuterozoa и зачаток левого переднего целома, дающий
начало амбулакральной системе, не утрачивает связей с передним
концом тела личинки при своем перемещении вместе с глоткой на
оральный полюс прикрепившегося животного. Тонкий канал, откры-
вающийся у личинки наружу слева дорзально и являющийся цело-
модуктом переднего левого целома, сохраняется с частью прилежа-
щего апикального участка последнего (рис. 118). Другая часть
дает так называемый «каменистый» канал и сохраняет свои перво-
начальные положение и отношения, с одной стороны, с околорото-
вым кольцом амбулакральной системы, а с другой, — с ампулой
переднего целома близ места бывшего прикрепления. Ампула при-
нимается рядом исследователей за особый передний целом, так
называемый аксоцель, или «первый целом», причем «вторым» —-
будет уже зачаток амбулакральной системы — гидроцель. Однако
разделять эти два отдела, открывающиеся наружу единым целомо-
263
г
дуктом, едва ли имеются основания. Перед нами, как показал
Э. А. Мейер, единый целом аппарата щупалец, обусловливающий
их плотность.
У взрослой морской звезды (рис. 119) или морского ежа амбу-
лакральная система представлена околоротовым кольцом и •вторич-
ными резервуарами (полиевы пузыри), расположенными на нем ин-
террадиально. От кольца отходят радиальные каналы, которые от-
дают в бока многочисленные канальцы к амбулакральным ножкам.
Последние представляют щупальцеобразные придатки, превращен-
ные в органы движения (см. рис. 109). Каждый радиальный канал
заканчивается на конце луча у морской звезды в непарном щу-
Рис. 119. Схема вертикального разреза морской звезды (из
Пфуртшеллера):
1 — рот, 2 — кольцо амбулакральной системы с полиевым пузырем
(3), 4 — каменистый канал, 5 — мадрепорит, 6 — анальное отверстие,
7 — печеночный придаток кишечника, 8 — радиальный канал и 9 —
амбулакральная ножка с ампулой (10), И — непарное щупальце с
глазком, 12 — «позвонки» скелета
пальце, а у морского ежа — в его рудименте, у перипрокта — в
гак называемых «глазных» пластинках, завершающих амбулакраль-
ные ряды. Таким образом, создается система целомических каналов,
обладающая запасными резервуарами для жидкого содержимого.
Эта система всецело принадлежит аппарату околоротовых щупа-
лец, т. е. только оральной области, и располагается под покровом
оральной стороны. Через интеррадиальный каменистый канал, т. е.
канал с неспадающимися обызвествленными стенками, она откры-
вается наружу посредством ампулы и отверстия мадрепоровой пла-
стинки вблизи аборального поля перипрокта.
Создается система с функцией механически поддерживать в
плотном состоянии оральный щупальцевый аппарат, а затем и с
активной функцией передвижения всего тела при преобразовании
щупальцевых придатков в присасывательные ножки. Жидкое содер-
жимое амбулакральной системы служит, прежде всего, этой меха-
нической функции. Вместе с этим для регулирования наполнения
различных отделов системы вырабатываются запасные резервуары,
связанные с каналами, а также аппарат каменистого канала с мад-
репоритом, обусловливающий возможность поступления воды из ок-
ружающей среды внутрь амбулакральнойтистемы, что превращает
ее действительно в «водно-сосудистую» систему.
264
У голотурий (см. рис. 114) с редукцией перипрокта амбула-
кральная система охватывает все червеобразное тело, причем осо-
бенно ярко выступают вторичные изменения. Так, каменистый канал
не проходит к аборальному полюсу через все длинное червеобраз-
ное тело, а в виде короткого протока открывается наружу медиан-
но, недалеко' от орального конца или, чаще, не достигая стенки тела,
заканчивается мадрепоритом свободно в целоме. Таким образом, со-
держимое амбулакральной системы становится в связь уже не с
наружной водной средой, а с жидкостью целома, что для механи-
ческого значения амбулакральной системы не представляет разницы.
Далее, при ползании на боку амбулакральные ножки развиваются
по трем радиальным каналам тривиума, образующего как бы
брюшную сторону, тогда как на двух каналах бивиума они имеют
характер небольших щупальцеобразных придатков. Наконец, вто-
рая пара амбулакральных ножек по- всем радиусам превращается
в 10 ветвистых щупалец, окружающих рот, и служит для захваты-
вания пищевых частиц, часто вместе с грунтом. Переход к пита-
нию организмами и органическими остатками, находящимися в са-
мом грунте, ведет к заглатыванию и пропусканию через кишечник
массы грунта и к жизни в нем. При этом теряется значение для
передвижения тела и самих амбулакральных ножек, которые под-
вергаются редукции. И у Apoda редуцируется вообще вся система
радиальных амбулакральных каналов с их придатками — типичней-
шая система иглокожих.
У морских лилий механическая функция амбулакральной систе-
мы представлена наиболее просто в связи с сохранением примитив-
ных отношений оральных щупалец при сохранении примитивного
пассивного питания. В соответствии с ветвлением рук радиальный
канал амбулакральной системы дает боковые ветви в располагаю-
щиеся по бокам медианной мерцательной борозды чувствительные
придатки щупалец, обусловливая их плотность (рис. 108). Каких-
либо придаточных резервуаров здесь нет, соответственно и около-
ротовое кольцо лишено их, а канал гидропора у взрослой морской
лилии редуцируется, причем амбулакральное кольцо вторично об-
разует воронки, открывающиеся в общий целом. Таким образом на
примитивном общем фоне получаются свои особенности, которые
не были присущи первоначальным Pelmatozoa, характеризовавшим-
ся относительно небольшим аппаратом околоротовых щупалец и
простым околородовым кольцом переднего целома, который давал
пять выростов в щупальцы и обладал гидропором для регулирова-
ния содержания жидкости во- всей этой небольшой системе.
В уже охарактеризованном нами комплексе образований амбу-
лакральной борозды иглокожих, вместе с увеличением значения
орального отдела тела, сложно развивается не только амбулакраль-
ный целом, берущий на себя механическую функцию, но и другого
РсДа образования, обеспечивающие трофику — кровеносная и так
Назьшаемая перигемальная системы.
Как показывает само название, перигемальная система, т. е. си-
265
стема, располагающаяся вокруг кровеносной (см. рис. 123, А), тесно
связана с кровеносной, а именно—с той ее частью, которая принад-
лежит оральным щупальцам и представлена окологлоточным коль-
цом и отходящими от него пятью радиальными сосудами с их вет-
влением. Радиальные кровеносные сосуды располагаются под ра-
диальным каналом амбулакральной системы и являются питающи-
ми для всего комплекса образований амбулакральной борозды, т. е.
для образований, находящихся кнаружи от амбулакральных и адам-
булакральных скелетных пластинок морских звезд (см. рис. 109).
Радиальный сосуд заканчивается в концевом щупальце луча
морской звезды или в его рудименте, находящемся в глазной пла-
стинке у перипрокта ежа. Таким образом, кровь подается во все
образования оральной части, но это только колебательные движе-
ния — о настоящем кровообращении говорить не приходится.
Дальнейшие связи оральной кровеносной системы идут в двух
направлениях. С одной стороны, к^ источникам питания, т. е. к ка-
пиллярам и лакунам кровеносной системы в стенке усваивающей
кишки — они отдают кровь в окологлоточное кольцо. С другой, то
же окологлоточное кольцо переходит в интеррадиусе каменистого
канала в осевой орган. Последний представляет, обычно, мощный
тяж лимфатического характера, аборально переходящий в лимфа-
тическое кольцо, которое окружает перипрокт и образует интерра-
диально небольшие выросты, вступающие в расположенные здесь
гонады (рис. 120).
У голотурий при типичном развитии оральной части кровеносной
системы отсутствуют как осевой орган, так и аборальное кольцо.
Таким образом, с редукцией перипрокта редуцирован и аборальный
отдел кровеносной системы с осевым органом. На первый план вы-
ступает туловищный отдел кровеносной системы, связанный с ки-
шечником. Этот отдел представлен главным образом хорошо выра-
женными «спинным» и «брюшным» сосудами с сетью капилляров на
передней петле кишечника, а у обладающих водными легкими еще
и с «чудесными сплетениями» на левом из них. Так, особенно выяв-
ляется в этой части кровеносной системы функция газообмена в
придачу к основной трофической функции (см. рис. 114).
Сходные отношения в туловищном отделе кровеносной системы
имеются также у морских ежей, обладающих богатым сплетением
капилляров на передней части петли кишки и хорошо выражен-
ным продольным сосудом на сифональной кишке, тогда как задняя
часть петли ни того, ни другого не имеет. Если принять, что сифо-
нальная кишка с ее током воды может служить и для газообмена,
то перед нами появление дыхательной функции в этом отделе крове-
носной системы, происходящее у ежей совершенно независимо й
другим путем, чем у голотурий.
У морских звезд, в связи с выработкой мешковидного желудка,
способного выворачиваться для захватывания пищи, туловищная
часть кровеносной системы представлена только сплетением сосу-
дов в стенке кишечника, без дальнейшей их дифференцировки.
266
Лакунарный, неопределенный характер сплетений имеет крове-
носная система другой пинии эволюции иглокожих — у морских
лилий, особенно в туловищном отделе. Как уже было выше отмече-
но, общая полость тела лилий типично представлена в руках хо-
Рис. 120. Схема строения осевого комплекса морской звезды. Вертикаль-
ный разрез через интеррадиус (по Стрелкову):
1 — мадрепорит, 2 — ампула, 3 — левый передний целом с оральным кольцом (4),
5 — оральная часть осевого органа, 6 — его аборальная часть, 7 — правый передний
целом, 8 — гипоневральное и 9 — эктоневральное кольца нервной системы, 10 —
околоротовое кольцо кровеносной системы, 11 — кольцо перигемальной и 12 — амбула-
кральной системы, 13 — каменистый канала, 14 — стенка кишечника. 15 — стенка
тела, 16 — аборальный кровеносный сосуд с лимфоидными образованиями на нем
(17) в половом целоме (18)
рошо выраженными каналами (парой «брюшных» и непарным
«спинным»), пробегающими в центре каждой руки под аборальным
скелетом «брахиальных» пластинок (см. рис. 108). В чашечке по-
лость тела вторично заполняется рыхлыми целенхимными образо-
ваниями (см. рис. 106). В них проходят мезенхимные тяжи и про-
слойки с кровеносными сосудами и лакунами. Оральный отдел по-
следних образован кольцевым лакунарным сплетением вокруг глот-
ки, но хоршо -выраженных радиальных сосудов в руках нет. Ораль-
267
ное кольцо-посредством неопределенно пробегающих сосудов и ла-
кун соединяется с чрезвычайно рыхлым осевым органом, который
проходи? по оси чашечки и, истончаясь, продолжается в ось камер-
ного органа и стебелька. Аборальное кольцо1 отсутствует. В придат-
ки стебелька; невидимому, проникают тонкие веточки от тяжа осе-
вого органа. А в руках, по границе между «спинным» каналом це-
лома и парой «брюшных» каналов, проходит лимфатический ради-
альный тяж, на отростках которого в пиннулах развиваются гона-
ды. Эти пять тяжей при начале рук в чашечке начинаются неопре-
деленными скоплениями лимфоидного характера.
Таким образом, отношения у Pelmatozoa и Eieuterozoa довольно
значительно разнятся. В известной мере это, как уже выше было
указано, зависит от развития оральных и аборальных пластинок
скелета в обеих группах. Кроме того, у Eieuterozoa резко обособ-
ляется от лежащих над ним органов комплекс образований амбула
кральной борозды, располагающийся у звезд и офиур под амбу-
лакральными пластинками. У Pelmatozoa такого обособления нет.
Поэтому образования амбулакральной борозды руки лилии оказы-
ваются в совершенно ином положении в смысле их питания, чем у
Eieuterozoa. Брюшные и спинные каналы целома, являющиеся не-
посредственными выростами центральной части общей полости те-
ла чашечки лилии, обеспечивают комплекс амбулакральной бороз-
ды в отношении питания. В результате редуцируются радиальные
кровеносные сосуды рук, обеспечивающие питание амбулакральной
борозды.
Напротив, у морских звезд, а затем и у других Eieuterozoa эти
радиальные сосуды сохраняют свое значение для питания образова-
ний амбулакральной борозды, так как от трофического целома по-
следние обособлены амбулакральными пластинками скелета. Боль-
ше того, развивается вспомогательная целомическая система, обхва-
тывающая радиальные сосуды и окологлоточное кольцо1 кровеносной
системы, т. е. уже отмеченная нами перигемальная система.
О ее значении можно судить по ее развитию (рис. 121). Зачатки
перигемальной системы появляются в виде пяти интеррадиальных
выростов от оральной части заднего целома, у звезд — четыре от
левого и один от правого его мешка. Относительно последнего Дан-
ные расходятся и требуют уточнения. Но это не меняет сущности
дела. Выросты эти дихотомически ветвятся, и каждая ветвь, приле-
гая к окологлоточному кольцу кровеносной системы, достигает соот-
ветствующего радиального сосуда, около которого далее и проходит
как перигемальпый капал до концевого радиального щупальца. Та-
ким образом, каждый радиальный кровеносный сосуд оказывается
заключенным как бы в мезентерий между двумя прилегающими
правым и левым перигемальнымп каналами. Вторично начальные
участки перигемальных каналов сливаются в одно перигемальное
кольцо, охватывающее снаружи окологлоточное кольцо кровеносной
системы.
Несомненно, что перигемальная система по своему происхожу
268
пию является производным заднего трофического отдела целома. В
соответствии с этим она может нести трофические с функцию для об-
разований амбулакральной борозды, вспомогательную к функции
кровеносных сосудов, не обеспечивающих правильного кровообраще-
ния. Но ей присуща и защитная механическая функция, так как она
отделяет амбулакральную систему с ее изменяющимися давлениями
от радиальных нервных стволов амбулакральной борозды.
За это говорят отношения перигемальной системы у лилий (см.
рис.
108). Она представлена здесь только
ми, идущими в руках под по-
диальными каналами амбула-
кральной системы, 'причем как
в основании рук, так и на кон-
це их, каналы заканчиваются
слепо. Таким образом, их тро-
фическое значение здесь сво-
дится к нулю, но в то же время
несомненно, что «брюшные» п
«спинные» каналы туловищного
целома в руках морокой лилии
обеспечивают трофику образо-
ваний амбулакральных борозд,
не отделенных здесь скелетны-
ми пластинками от указанных
отделов трофического целома.
Перигемальные каналы мор-
ских лилий можно считать на-
стоящими рудиментами образо-
ваний, имевшихся в щупальцах
примитивных исходпы.х форм
иглокожих, у которых аппарат
щупалец еще не вошел в со-
нятью тонкими канала-
Рис. 121. Развитие перигемальной систе-
мы морской звезды (по Деляжу): А —
дихотомически ветвящиеся выросты ле-
вого заднего пелома под зачатком амбу-
лакрального кольца и радиальных кана-
лов; Б, В, Г — образование за счет
выростов заднего целома перигемаль-
ного кольца и зачатков перигемальных
каналов;
став туловища, а орально уже
проникали в них пять интерра-
диальных выростов заднего це-
лома, дихотомически ветвив-
шихся и образовывавших пять
радиальных каналов пориге-
1 — рот, 2 — кишечник, 3 — левый и 4 —
правый задние мешки целома. 5 — амбула-
кральное кольцо и зачатки каналов, 6 —
интеррадиальные выросты заднего целома и их
дихотомическое ветвление (7). для образования
перигемальной системы. 8 — осевой синус це-
лома, 9 — поровой канал, 10 — правый перед-
ний целом
мальной системы. Это усиливало трофику, обеспечивавшуюся по-
средством кровеносных сосудов.
У морских лилий и эти последние подверглись редукции в ре-
зультате их функционального замещения выростами заднего цело-
ма, проникшими, как было отмечено, в руки орально от скелетных
образований стенки тела. При несовершенстве функционирования
сосудоз кровеносной системы, подобная субституция, в связи с все
большим и большим развитием рук криноидов, вполне понятна.
Иное имеет место у Eleuterozoa, именно у морских звезд и офиур,
У которых образования амбулакральной борозды, представляя в из-
269
вестной мере изолированное целое, отделены от проникавшего або-
рально заднего целома рядами амбулакральных скелетных пласти-
нок. В этих условиях сохранились под амбулакральными каналами
и радиальные кровеносные сосуды и подкреплявшие их перигемаль-
ные каналы заднего целома. Однако трофическое значение перигс-
мальной системы теряется в связи с включением оральных образо-
ваний в состав туловища и редукцией аборального отдела при моцц
ном развитии общей полости тела, как мы это видим у морских
ежей и голотурий.
В онтогенезе перигемальные каналы утрачивают первичные от-
ношения к заднему целому (что вторично должно отражаться на
ослаблении их первичного трофического значения) и, вступая в
связь друг с другом, образуют окологлоточное перигемальное коль-
цо, образуют обособленную систему.
Итак, загадку амбулакральной системы иглокожих удалось ре-
шить на основе своеобразных процессов эмбрионального развития,
тесно увязав их с особыми условиями существования. А именно:
амбулакральная система является производным части переднего ле-
вого целома предков иглокожих, которая несла функцию опоры для
аппарата околоротовых щупалец. При прикреплении исходных форм
оральным правым отделом тела ротовое отверстие смещается по
левой стороне и увлекает за собой часть переднего левого целома,
образующего, в конце концов, кольцо вокруг глотки. При этом цело-
мические каналы в пяти переместившихся щупальцах левой стороны
соответственно располагаются по пяти радиусам на образовавшем-
ся кольце.
Рассмотрение кровеносной и перигемальной систем иглокожих
позволяет нам дальше конкретизировать наши представления об
аппарате щупалец у исходных форм. Каждое щупальце по ораль-
ной стороне несло срединную полосу мерцательного покрова,
обусловливавшую ток воды с пищевыми частицами к ротовому от-
верстию. Это — основа пассивного питания сидячих форм. При
ветвлении щупалец образовались располагавшиеся с боков чувстви-
тельные придатки, эволюционировавшие далее в амбулакральные
ножки. В соответствии с этим, по длине каждого из пяти щупалец
между покровным эпителием и амбулакральным каналом проходил
радиальный кровеносный сосуд, служивший для питания образова-
ний амбулакральной борозды и отходивший от окологлотчного
кольца кровеносной системы. В связи с несовершенством кровооб-
ращения при попеременном нагнетании и оттягивании крови в paj
диальных сосудах, для усиления питания указанных образований
присоединяются пять интеррадиальных, далее ветвящихся выростов
левого заднего целома, имеющего, по преимуществу, трофическую
функцию. Образуется перигемальная система.
Таким образом, исходные формы иглокожих облекаются в нашем
представлении в плоть и кровь, но можно пойти и дальше по этому
пути.
Рассматривая судьбы левого переднего целома, мы не нарушили
270
представления о целостности системы вторичной полости тела, так
как при этом выяснили и процессы редукции, захватившие правый
передний целом. Кровеносная система образует не два отдельных
симметрично расположенных образования, подобно правому и ле-
вому целомам, а единый аппарат. Поэтому, чтобы охарактеризовать
ее, необходимо выяснить отношения не только в левой, но и в
правой стороне тела исходных форм.
Уже на том основании, что при редукции правого переднего це-
лома левый образует мощную амбулакральную систему, можно
было утверждать, что прикрепление произошло правой околоротовой
передней областью тела, соответствовавшей положению правого
переднего целома. В этом отношении дальнейшее уточнение карти-
ны, сразу раскрывающее сущность процесса, выявляется в аномаль-
ном развитии у некоторых личинок зачатка правого переднего це-
лома в амбулакральную систему взрослого. При этом левый зача-
ток в таких случаях претерпевает участь, обычно уготованную пра-
вому, т. е. сохраняется в виде рудимента — небольшого мешочка
в аборальной стенке. Отсюда можно заключить, что оба зачатка пе-
реднего целома — правый и левый — у исходных форм вполне го-
мологичны, и как с левым были связаны пять околоротовых щупа-
лец левой стороны, впоследствии давшие пять лучей оральной сто-
роны, так и с правым зачатком стояли в связи пять щупалец правой
стороны, которыми прикреплялись предки иглокожих (рис. 124,
Д 5, В, Г).
Самый факт прикрепления должен был отразиться, прежде все-
го, на аппарате, с которым, была связана подвижность щупалец,
т. е. на опорном для щупалец переднем правом целоме. Произошла
его редукция и рудиментация, тогда как в прогрессивно эволюцио-
нировавших левых щупальцах целом мощно развивается в амбу-
лакральную систему.
В кровеносной системе подвергаются редукции сосуды, питав-
шие щупальца правой стороны тела вместе с превращением послед-
них в образование, служащее для прикрепления животного. Мор-
ские лилии — единственная сохранившаяся до нашего времени
ветвь сидячих иглокожих — дают в этом отношении драгоценные
факты, которые позволяют более точно представить себе аппарат
прикрепления и процессы, разыгрывавшиеся при прикреплении пред-
ков иглокожих.
Однако мы остановимся сначала на других фактах. Развитие
кровеносной системы оральной стороны тела показывает, что кро-
веносные сосуды пяти щупалец отходили от общей кровеносной ла-
куны, которая полукольцом проходила у их основания около гло-
точного отдела кишечника. При передвижении рта к пункту, все
Далее и далее отстоявшему от места прикрепления (что можно хо-
рошо видеть, например, в развитии морской лилий) и с расположе-
нием щупалец соответственно пяти радиусам тела (см. рис. 115),
кровеносная лакуна, подобно амбулакральной системе, замыкается
в окологлоточное кольцо, от которого отходят пять радиальных
271
сосудов. Однако своей связи с правой половиной кровеносная лаку^
на не теряет. Ее медианный участок, по мере удаления левой поло-
вины вместе с ртом и щупальцами дальше и дальше от места при-
крепления, вытягивается в длинный лимфоидный тяж осевого
органа (рис. 120). При этом лакуна вдается в прилежащий голов-
ной участок левого переднего целома вместе с каменистым каналом
гидроцели, также проходящим от оральной части к аборальной. Ко-
нец лимфоидного органа располагается в узком каналообразном
отделе правого переднего целома? Вместе эти два целомических
участка образуют так называемый осевой синус. Так создается
осевой комплекс, столь характерный для иглокожих и когда-то
столь мало понятный для сравнительной анатомии.
Часть кровеносной окологлоточной лакуны, связанная с пятью
редуцирующимися правыми щупальцами, остается у места прикреп-
ления и превращается в аборальное лимфатическое кольцо, в то
время как пятирадиальное построение охватывает все тело.
У морских лилий эволюция аборального отдела пошла иными
путями. Вспомним, что морские ежи перешли от пассивного питания
к активному, используя в пищу организмы, обрастающие каменистое
дно, что морские звезды становятся хищниками, пожирающими
прикрепленных к субстрату моллюсков и т. п., а голотурии облизы-
вают более мелкие камни, гальки и гравий. Морские лилии, будучи
по преимуществу жителями спокойных вод, поднимаются над грун-
том и используют для своего пассивного питания взвешенных в воде
мелких животных и растительные организмы; у лилий аппарат при-
крепления вторично выносит их ввысь и становится длинным сте-
бельком, тогда как у прочих иглокожих он теряет свое значение и
атрофируется. Соответственно, осевой орган кровеносной системы у
морских лилий продолжается абсрально в лимфоидный тяж, прохо*-
дящий центрально по оси стебелька, не образуя чего-либо подоб-
ного аборальному кольцу.
Но зато у лилий, как свидетель процессов, имевших место при
прикреплении, развивается замечательный аппарат целомических
полостей (рис. 106), которые в виде пяти каналов пробегают по
всей длине стебелька, охватывая со всех сторон центральный тяж
кровеносной системы. Начальные участки этих каналов в основании
чашечки, расширяясь, образуют камерный орган. Происходит вся
система камерного органа путем образования пяти выростов на
передней части заднего правого целомического мешка (рис. 122)
совершенно сходно с тем, как образуются на левом мешке выросты
зачатков перигемальной системы Eleuterozoa. Конечно, эти пять ка-
налов, представляя выросты трофического отдела целома, должны
нести функцию снабжения стебелька питательными веществами, что
не выполняет уже сильно измененная кровеносная система и что
необходимо при значительном развитии стебелька. Гомология пери-
гемальной системы и системы полостей камерного органа, как об-
разований, происходящих от левого и правого задних мешков це-
лома, несомненна. И в этом мы видим еще указание на первояа-
272
чальный характер прикрепления предков пятые околоротовыми
щупальцами правой стороны тела. Это указываем также и на суще-
ственную черту строения щупалец предков иглокожих, а именно на
проникновение в щупальца для усиления питания, кроме кровенос-
ной системы, также выростов трофического заднего целома.
Рис. 122. Целомические образования личинки морской лилии Antedon rosacea
(по Зейлигеру): А — сагиттальный разрез только что выведшейся
личинки; Б — то же перед прикреплением;
1 — задний левый и 2 — задний правый целомы, 3 — передний целом, 4 —
зачаток амбулакральной системы, 5 — зачаток камерного органа, 6 — кишечник,
7 _ vestlbulum, 8 — место прикрепления
В связи с этим у морских лилий замечательным образом выяс-
няются особые черты еще нерассмотренного нами, очень своеобраз-
ного компонента в комплексе образований амбулакральной борозды
иглокожих — их нервной системы и органов чувств.
Сидячий образ жизни наложил свою неизгладимую печать на
развитие органов чувств иглокожих. Эти органы чрезвычайно обед-
нены, а важнейший в организации их червеобразных предков голов-
ной комплекс органов чувств подвергся редукции при прикреплении
Диплеврулы передним правым участком тела.
18 Пути эволюции животного мира 273
Чувствительные образования развиваются на оральной стороне
рук морских лилий, главным образом в виде чувствительных почек
на щупальцевых придатках, располагающихся справа и слева у
амбулакральной борозды (см. рис. 108). Таким образом ориенти-
ровка в окружающей среде связана здесь, прежде всего, с ориен-
тировкой в отношении питания. Для защиты сидячего животного,
обладающего мощным скелетом в стенке тела, острота ориентиров-
ки не имеет уже такого значения, и чувствительные клетки по оди-
ночке разбросаны везде в покрове рук морской лилии.
Также и у Eieuterozoa главную роль в качестве чувствительных
образований играют развивающиеся за счет щупальцевых придат-
ков амбулакральные ножки, ряды которых у морских звезд распро-
страняются по всей оральной половине тела, а у морских ежей
проходят по всему телу до перипрокта. У голотурий два ряда
амбулакральных ножек бивиума представляют только чувствитель-
ные придатки, тогда как обращенные к грунту три ряда тривиума
служат в качестве типичных амбулакральных ножек. Однако у
Eieuterozoa увеличивается и общая чувствительность покрова и
развиваются некоторые специальные образования. Так, у морских
звезд и ежей непарное концевое щупальце несет своеобразные
глаза. В наиболее сложной форме они представляют ямочные
глаза, покрытые общей прозрачной кутикулой, образующей хоро-
шо выраженную кутикулярную линзу. У морских ежей с характер-
ным развитием аппарата игол связана выработка за их счет сфе-
ридиев — органов равновесия.
Не вдаваясь в частности морфологии органов чувств, обратимся
к нервной системе иглокожих, которая, как и всегда, в своем раз-
витии обусловлена, с одной стороны, развитием органов чувств, а
с другой — мускулатуры, но, конечно, на той основе, которая была
присуща предкам данной группы животных. Добраться до этой
основы у иглокожих очень нелегко, настолько она затемнена вто-
ричными изменениями и новообразованиями, обусловленными при
переходе к сидячему образу жизни процессами неравномерного
роста и передвижения органов. Но она, безусловно, находит свое
отражение в комплексе образований амбулакральной борозды.
Так, в нервной системе иглокожих определенно выделяется сен-
сорная часть, связанная теснейшим образом с чувствительными
образованиями амбулакральной борозды — прежде всего с щупаль-
цевыми придатками, соответственно с амбулакральными ножками.
Она представлена расположенными в покровном эпителии борозды
ганглиозными клетками и нервными волокнами (рис. 123), так что
получается пять радиально пробегающих в покровном эпителии
нервных тяжей. Это — «эктоневральная» нервная система.
Наиболее примитивно выражена она у морских лилий (см.
рис. 108), где образована сравнительно слабыми тяжами, иннерви-
рующими части рук только у соответственной амбулакральной
борозды, главным образом щупальцевые придатки, тогда как рас-
положенные далее в бока участки покрова отдают свои нервные
274
пучки уже глубже лежащим парным радиальным стволом так
называемой «гипоневральной» системы. Последние у глотки объ-
единяются околоротовьш кольцом, располагающимся в соедини-
тельнотканном слое покрова. Подобного кольца в эпителии, которое
Рис. 123. Образования амбулакральной борозды у морской звезды Astropecten
aurantiacus: А—весь комплекс в поперечном разрезе (по Людвигу); Б— по-
перечный разрез нервной системы (по К. Шнейдеру);
I — каналы перигемальной системы, 2 — радиальные кровеносные сосуды, 3 — гипэневраль-
ная система, ее клетки н волокна, 4 — волокна эктоневральной системы, 5 — клетки эпите-
лия с их волокнами, 6 — кутикула, 7 — нервные клетки эктоневральной системы, 8 —
пограничная перепонка
Должно бы объединять пять радиальных стволов эктоневральной
системы, у морских лилий не найдено, но оно хорошо развито у
Eleuterozoa, и поэтому вполне возможно, что будет найдено1 и у
Морских лилий, хотя бы в слабо дифференцированном состоянии.
Во всяком случае, у морских звезд эктоневральная нервная
система образована околоротовым нервным кольцом (см. рис. 120)
!8* 275
и отходящими от него пятью радиальными стволами (см. рис. 109 и
123), принимающими нервы от всей оральной поверхности рук
кончая термиальным щупальцем, несущим глаз.
У офиур,'.морских ежей и голотурий (см. рис. 1.10, 116, 117) ота
эктоневральная система располагается также в покровном эпителии
амбулакральной борозды, но вместе с разрастанием оральной части
тела у ежей доходит до перипрокта, а у голотурий прямо до зад-
него конца тела, причем теряет свое поверхностное положение бла-
годаря образованию краями амбулакральной борозды двух складок,
срастающихся далее друг с другом. Получается своеобразная си-
стема полостей, представляющая не что иное, как обособившиеся
участки внешней среды, замкнувшиеся в до некоторой степени
особый аппарат эпиневральных каналов. Она является защитным
приспособлением для первоначально поверхностно расположенной
эктоневральной нервной системы. Конечно, эпиневральные каналы
мы должны рассматривать вместе с эктоневральной нервной систе-
мой, как ее вспомогательное образование, с целомом они ничего
общего не имеют.
Совершенно обособленные от эктоневральных, гипоневральные
нервы содержат у морских лилий, наряду с чувствительными, также
и двигательные пучки (см. рис. 108). У Eleuterozoa (см. рис. 109,
НО, 116, 117, 123) это — хорошо дифференцированная двигатель-
ная система, два тяжа которой располагаются под целотелием
перигемальных каналов справа и слева на радиальных эктоневраль-
ных стволах, но всегда резко отделены от них субэпителиальной
пластинкой ткани основного вещества. Они посылают нервы к мы-
шечным пучкам прилежащего скелета позвонков морских звезд и
офиур, а у'глотурий к мускулатуре стенки тела. У морских ежей
с редукцией таких мышечных образований редуцируются соответ-
ствующие части гипоневральной нервной системы. Что касается ее
околоротового кольца, то оно или отсутствует, как это имеет место
у голотурий, или представлено пятью ганглиозными скоплениями в
соответствии с развитием мускулатуры жевательного аппарата у
прочих Eleuterozoa.
Сам собой напрашивается вывод, что в рассмотренном комплексе
образований амбулакральной борозды эктоневральная система игло-
кожих представляет хорошо развитую сенсорную систему оральной
части тела. Эта система является не чем иным, как дифференциров-
кой и дальнейшим усложнением чувствительных нервов, проходив-
ших в покровном эпителии пяти первоначальных щупалец двусто-
ронне-симметричных предков иглокожих. Ввиду того, что' гипонев-
ральная система настолько резко обособлена от эктоневральной, что
объединить их в единое образование нельзя, приходится признать
в гипоневральной системе двигательные нервы, располагавшиеся
под эпителием тех же щупалец. Но и чувствительный и двигатель-
ный компоненты у иглокожих пошли в своей эволюции самостоя-
тельно и дали две особые системы — эктоневральную и гипонев-
276
рзльную, с их особыми центрами, независимыми друг от друга, хотя
и тесно связанными.
Однако, наряду с экто- и гипоневральной системами оральной
части тела, которые только и имеются у голотурий, лишенных або-
рального отдела тела, у прочих иглокожих развита в той или иной
мере также отдельная аборальная нервная система. Особое положе-
ние занимают, конечно, морские лилии, обладающие сильно разви-
тии аборальным отделом тела со стебельком и чашечкой. У
Eieuterozoa, с редукцией в той или иной степени аборальной части
т?ла, также соответственно слабо развивается и аборальная нерв-
ная система. Так, у морских ежей под целотелием аборального
кольцевого синуса около лимфатического кольца проходит «апи-
кальное» нервное кольцо, дающее нервы к располагающимся возле
него частям тела. У морских звезд имеется такое же кольцо, от
которого отходят пять слабо развитых радиальных нервов, прохо-
дящих под целотелием аборальной стороны лучей соответственно
имеющемуся здесь мышечному пучку.
Но совершенно особое впечатление производит апикальная
нервная система морских лилий (см. рис. 106). У них в мощной
скелетной массе основания чашечки под целотелием вокруг пяти
расширений целомических каналов камерного органа, заложен круп-
ный нервный центр, образованный скоплением ганглиозных клеток
и нервных волокон. Этот центр посылает нервные пучки в стебелек
и усики, а также—значительные интеррадиальные нервы. Последние
сейчас же дихотомически ветвятся, а затем попарно соединяясь, про-
бегают уже радиально в толще аборального скелета рук (см. рис.
108). Эти нервы связаны главным образом с чувствительными обра-
зованиями аборальной поверхности рук, но содержат также и дви-
гательные волокна к мышечным пучкам скелета последних.
Трудно сказать, соответствует ли апикальная система морских
лилий аборальной системе Eieuterozoa. Разницу можно1 видеть в
том, что у морских лилий, в противоположность морским звездам
и ежам, мощно развиты аборальные образования. Вполне есте-
ственно, что редукция аборальных образований у Eieuterozoa повела
и к редукции части апикальной системы. Так, у морских звезд еще
видны остатки радиальных стволов, у ежей сохранилось лишь
аборальное кольцо.
Но как бы то ни было, присутствие мощной массы нервных
клеток и волокон в толще скелета в камерном органе морских
лилий кажется зоологическим парадоксом. И единственное объяс-
нение нахождения здесь нервного центра заключается в предполо-
жении, что он сохранился тут от предков, которые обладали иными
чертами организации и соответственно находились в других отно-
шениях с внешней средой. Приходится обратиться к' двухсторонне-
симметричным исходным формам иглокожих, к диплевруле.
Попытаемся представить себе весь процесс формирования орга-
низации иглокожих в целом (рис. 124).
277
Рис. 124. Схема эволюции иглокожих: А — диплеврула с брюшной
стороны и Б — сбоку; В — прикрепление диплеврулы щупальцами
правой стороны; Г и Д — перемещение левых щупалец по левой
стороне тела; £ —стадия Pelm <tozoa\ Ж— стадия перехода к
Eleuterozoa. Черным нанесен передний целом и его производные—
амбулакральная система, точками нанесен кишечник, простой ли-
нией— мешки заднего целома. На фиг. Ж видно вхождение щу-
палец в состав туловища
II Как уже было указано, у предков иглокожих с переходом к
|идячему существованию, в связи с прикреплением правым перед-
ним участком тела и смещением ротового отверстия По левой сто-
роне тела, произошло- смещение и особое развитие- пяти левых око-
поротовых щупалец, которые, обеспечивая пассивное питание, в
конце концов располагаются вокруг рта по радиусам и образуют
оральную часть тела. Часть переднего левого целома в своем от-
деле, связанном со щупальцами и несшем функцию опоры для них,
принимает при этом сначала форму подковы, а затем замыкается
в кольцо. От кольца отходят пять радиальных каналов первичных
оральных щупалец — образуется амбулакральная система с опорной
функцией для придатков этих последних, а далее и для амбула-
кральных ножек. Отдел переднего левого целома, заходивший перед
щупальцами в головную часть диплеврулы, сохраняет свое преж-
нее положение и отношения, так как сама головная лопасть не
смещается вместе со ртом, а остается у места прикрепления. При
этом целом вытягивается в виде длинного канала, идущего от
орального отдела в аборальный. Таковы «париетальный» канал
морских лилий, подвергающийся во взрослом состоянии редукции,
и, по данным Д. М. Федотова, левый осевой целомический синус
осевого комплекса у Eleuterozoa.
У сидячих иглокожих при перемещении рта и образовании пяти-
лучевой оральной системы ветвящихся щупалец в связи с наличием
у двусторонне-симметричных их предков в каждом из оральных
щупалец питающего кровеносного сосуда, отходившего от крове-
носной лакунарной дуги головной лопасти, развивается околоротовое
кровеносное кольцо и пять радиальных сосудов, питающих амбула-
кральные борозды и их придатки.
Однако при дальнейших изменениях кровоснабжения для усиле-
ния питания важнейших образований амбулакральной борозды
развиваются идущие сюда же интеррадиальные выросты заднего
. левого целома, имеющего трофическое значение. Таким образом,
получается особая перигемальная система. Вторично утрачивая
затем связь с задним левым целомом, она сохраняет лишь механи-
ческое значение.
Наконец, присущие пяти первоначальным щупальцам чувстви-
тельные нервы, проходившие под амбулакральной бороздой от кон-
ца до основания щупальца образуют эктоневральную нервную
систему в виде пяти радиальных стволов, объединяющихся около-
ротовым кольцом и расположенных у иглокожих в эпителии амбу-
лакральной борозды. В то же время парные двигательные нервы
каждого щупальца, располагавшиеся глубже под целотелием пери-
гемальных каналов, образуют пять радиальных двойных тяжей
гипоневральной системы, проходящих под стволами эктоневральной.
системы, от которой они, однако, резко обособлены.
Так протекают процессы в левой стороне тела формирующегося
иглокожего, в которых ведущее значение приобретают пять ораль-
279
ных щупалец с их чувствительными и мышечными, нервными, цело
мичсскими и сосудистыми, а затем и защйтноскелетпыми образова-
ниями. Этот оральный комплекс приобретает все большее и боль-
шее значение, входя постепенно в состав туловища.
Что касается правой стороны тела, то она, так же как и левая
сторона, несла около рта щупальца в числе пяти. Но', в связи с при-
креплением исходных форм, щупальца правой стороны при форми-
ровании тела иглокожего должны были подвергнуться совершенно
иным приспособительным процессам, чем пять левых. На правой
стороне они превращаются в аппарат прикрепления (рис. 124,
В, Г, Д, Е). Правый передний целом, утрачивая значение опорного
аппарата для щупалец, сохраняется при этом лишь в виде неболь-
шого пузырька, примыкающего аборально к осевому органу и
иногда пульсирующего. Таким образом этот целом, гомологичный
амбулакральному целому левой стороны, становится столь руди-
ментарным, что с выпадением функций щупалец не сохраняется
следов присутствия пяти его выростов, связанных со щупальцами.
Трудно сказать что-либо определенное относительно отдела пра-
вого целома, связанного с головной лопастью.
Кровеносная система правой стороны представлена продолже-
нием осевого органа аборально, а именно той его частью, которая
вдается в остаток правого переднего целома. Так что и здесь
нельзя найти более определенных следов прежних отношений. Мож-
но только сказать, что осевой орган, соединяющийся далее с або-
ральным лимфатическим кольцом кровеносной системы Eleuterozoa
проходит и в аппарат прикрепления, в частности и в стебелек мор-
ских лилий, но здесь он рудиментарен, так как трофическая функция
осевым органом утрачивается. Он превращен в орган лимфоидно-
экскреторного типа. Таким образом, как мы видим, в едином кро-
веносном аппарате головного отдела тела предков иглокожих левая
половина образует околоротовое кольцо и пять радиальных сосудов
щупалец, соответственно рук иглокожих, тогда как правая часть
в значительной мере рудиментирована и изменена функцио-
нально.
Иные отношения выявляются в трофическом целоме стебелька
морских лилий, гомологичном перигемальной системе левой стороны,
и в их нервной системе. Это вполне понятно, так как, с изменением
у иглокожих функции пяти правых щупалец в функцию аппарата
прикрепления и с ослаблением значения трофической функции кро-
веносной системы, в щупальцах ничуть не уменьшается, напротив,
даже повышается трофическая роль заднего правого целома и его
выростов. И мы видим, что как левый задний целом дает пять
выростов — зачаток перигемальной системы, так и правый задний
целом образует пять выростов, входящих в аппарат прикрепления
и образующих в centro-dorsale камерный орган. Конечно, можно
считать, что число выростов случайно совпадает в том и в другом
случае, но гораздо более вероятно предположить иное. Как справа,
так и слева от рта двусторонне-симметричного предка иглокожих
280
располагалось по пяти щупалец орального аппарата, и пятью пра-
выми такая форма прикрепилась, используя пять свободных левых
щупалец при пассивном питании.
Условия, в которых такое прикрепление может быть мыслимо,
легко представить на сравнительно круто спускающихся скалистых
побережьях. Использовать эти склоны, богатые скатывающимся
сверху пищевым материалом, было, конечно, непросто, но в высокой
степени выгодно, и это приспособление создало организацию иг-
локожих, прикрепившихся правой половиной, т. е. правыми
пятью оральными щупальцами, и прекрасно использовавших ле-
вую половину для тока воды, пассивно несущего пищевые части-
цы ко рту.
Последним штрихом, завершающим картину, будет выяснение,
при нашем понимании организации иглокожих, значения загадоч-
ного нервного центра в камерном органе морских лилий. У
Eieuterozoa с редукцией аборальной‘части тела такой центр реду-
цируется, функционально теряя свое значение, но в организации
морских лилий, обладающих хорошо развитым стебельком и або-
ральными образованиями, он сохранен. Он располагается в основа-
нии предполагаемых пяти правых щупалец и охватывает продол-
жение осевого органа. Последний же, как мы видели, представляет
рудимент кровеносной системы головы, а целомический канал, охва-
тывающий осевой орган, является рудиментом головного целома.
Все это заставляет допустить, что при образовании аппарата при-
крепления головная лопасть исходной двусторонне-симметричной
формы подверглась редукции и вошла в состав этого аппарата, но
не перемещалась вместе со ртом и пятью левыми оральными щу-
пальцами дальше и дальше от места прикрепления по левой
стороне тела. Соответственно, важнейший нервный центр головной
лопасти — головной мозг у морских лилий мог сохраниться у осно-
вания щупалец, служивших для прикрепления вместе с рудиментом
головной лопасти. У Eieuterozoa головной мозг, как было указано,
становится вне функции, а соответственно нервы щупалец приобре-
тают полную самостоятельность, превращаясь в сенсорную экто-
невральную нервную систему и двигательную — гипоневральную.
У морских лилий эти системы развиты более примитивно, чем у
свободноживущих иглокожих.
Мы сделали только общий набросок эволюции исходной двусто-
ронне-симметричной формы — диплеврулы до1 образования общего
предка всех иглокожих, не вдаваясь в частности. Дальнейшая их
эволюция пошла разными путями в связи с приспособлением к раз-
ным биотопам. Морские лилии, как типичные сидячие потомки при-
митивных иглокожих, усиливают свой аппарат пассивного питания,
развивая ветвистые руки и аборальный скелет, рот перемещается
на пункт, противоположный месту прикрепления, а тело поднимает-
ся на стебельке. Благодаря этому, животные осваивают обширные
площади с песчаными и илистыми грунтами.
281
Иначе пошла эволюция Eleuterozoa (рис. 124, Ж). Оставаясь на
скалистых грунтах, предки звезд и ежей используют более крупную
добычу, находившуюся, можно сказать, под боком. Они используют
околоротовой скелет в качестве ротовых частей, а придатки щупа-
лец для активного захватывания пищи и переходят от пассивного
питания к активному. Рот при этом не перемещается до места, про-
тивоположного пункту прикрепления, а располагаясь на некотором
расстоянии на левой стороне и вентрально, приходит в соприкосно-
вение с такими источниками пищи, как обрастающие скалы ракуш-
ки и т. п. животные (пища морских звезд и офиур) или мшанки и
растения (пища ежей). Наконец, организация голотурий связана с
крупно-песчаными участками между скал, где своими ближайшими
ко рту парами амбулакральных придатков голотурии собирают
мелкие и мельчайшие организмы, располагающиеся на частицах
гравия и песка.
Переход Eleuterozoa к активному питанию ведет и к переходу
от сидячего существования к свободноподвижному образу жизни,
а далее и к выявлению характерной организации морских звезд,
морских ежей, голотурий. Впоследствии она позволяет Eleuterozoa
заселить другие биотопы и стать одним из наиболее широко распро-
страненных типов, обитающих в море.
Подводя итоги нашему анализу организации иглокожих, мы
можем видеть, какое огромное значение в ее выработке играет ап-
парат щупалец, тогда как головная лопасть предков отходит на
задний план и редуцируется. Но для туловищного отдела — это в
то же время своеобразное объединение со щупальцевым аппара-
том,— это включение последнего в состав туловища. Таким обра-
зом, перед нами процесс совершенно особого рода, показывающий
все возрастающее значение щупалец для питания прикрепленного
животного, выработавшего надежный защитный аппарат танген-
тального скелета.
В этой организации на первый взгляд все необычно. Можно
сказать, что она представляет собой целостный аппарат щупалец,
осложненный до невероятной степени. В то же время это1 относи-
тельно простое туловище с несложным целомом, с кишечником,
обладающим кровеносным сплетением, с примитивными (мы о них
до сих пор даже не упоминали) гонадами типа мешетчатых гонад,
т. е. имеющими свои особые выводные протоки, открывающиеся
наружу и не обнаруживающие никаких связей с целомом. Наконец,
туловище лишено экскреторных органов типа нефридиев, и отсут-
ствуют какие-либо следы туловищного мозга.
Исходная форма — диплеврула — по всем признакам могла
быть только билатерально-симметричным червеобразным организ-
мом, обладавшим ресничным покровом, трубчатым кишечником,
кровеносной системой, олигомерным целомом из двух пар мешков
(передняя пара — это головной и щупальцевый целомы, вторая
большой туловищный целом) и имевшим мешотчатые гонады вне
282
всякой связи с целомом. Все эти признаки говорят о протоцеломатах,
близких к немертинообразным червям, затылочныегО|рганы которых,
создающие мощный ток воды, стали использоваться в соответствую-
щих условиях донного существования для притока мелких организ-
мов к располагавшемуся возле ротовому отверстию. Другими
словами, затылочные органы дали возможность выработки пассив-
ного питания для форм, живших в грунте или в узинах каменистых
грунтов (рис. 124, А, Б),
В настоящее время нет достаточных оснований видеть в диплев-
руле червеобразный организм, развившийся на базе более сложной
организации аннелид, а именно—сидячих полихет, как это делал
Э. А. Мейер (1904). Полихеты имеют совершенно иной тип эмбрио-
нального развития, терминальный рост и полимерную метамерию,
органы движения в виде типичных параподий, экскреторный аппарат
характера нефридиев, постоянную связь гонад с целомом и т. п.
Мы видели, как шаг за шагом в ряду многоклеточных животных
вырабатывались и сохранялись и трубчатый кишечник, и кровенос-
ная система, и вторичная полость тела. Нашему анализу противо-
речило бы представление, что все эти важнейшие- приобретения
организации могли с легкостью возникать неоднократно в разных
группах как конвергентные новообразования. Поэтому приходится
признать, что «первичные» целомические организмы не.могли раз-
виться непосредственно от очень примитивных форм, вроде кишеч-
нополостных или гребневиков.
На этом основании следует сказать, что попытки рассматри-
вать гребневиков как непосредственных предков иглокожих пред-
ставляются мало убедительными. В последнее время подобные пред-
ставления развивались Д. М. Федотовым в В. Н. Беклемишевым.
В. Н. Беклемишев (1952, стр. 287—288) следующим образом
рисует картину происхождения диплеврулы. «Сравнивая гребневика
и молодую диплеврулу на стадии, когда у этой последней уже обра-
зовались целомические карманы, но еще нет рта, мы находим у
обоих животных монаксонно-гетерополярное строение с теменным
органом чувств на апимальном полюсе и ртом или бластопором на
вегетативном... У молодой диплеврулы в одной и той же плоскости
ее тела (будущей фронтальной) отходят три яруса целомических
мешочков, которые, подобно каналам гастроваскулярной системы,
представляют выросты кишечника. Отходящий к анимальному по-
люсу передний целом парный или подобно акрогастеру (ктенофор)
непарный, так же как и этот последний, открывается наружу двумя
поровыми каналами. Целомы вторые у всех вторичноротых,.имею-
щих щупальцевый аппарат, всегда бывают с ним связаны, подобно
Щупальцевым каналам гребневиков... Целомы третьи диплеврулы,
'гак же как и глоточные каналы гребневиков, никаких особых при-
мет не имеют, и о гомологии их друг другу приходится заключить
лишь по одинаковому положению, которое те и другие занимают в
составе гастроваскулярного или гастроцеломического аппарата
283
сравниваемых животных... В оральной полости вытягивается рот у
гребневиков... В этой же плоскости образуется и рот диплеврулы.
Образование его происходит очень просто: на будущей брюшной
стороне личинки возникает небольшое впячивание эктодермы —
глотка или'стомодеум. К этому впячиванию прикладывается перед-
ний конец кишечника, эпителии их спаиваются и в месте спайки
возникает отверстие; это и есть рот...». В 1944 г. В. Н. Беклемишев
указывал (стр. 252): «Такое новообразование рта напоминало бы
независимое от бластопора возникновение анального отверстия у
немертин». «Но представить себе филогенетически замену первона-
чального рта новым очень трудно... (стр. 254). У De'uterostomia
первичный рот превращается в анальное отверстие, а где-то в той
же оралытой плоскости независимым путем возникает вторичный
рот». В 1952 г. В. Н. Беклемишев ограничивается указанием
(стр. 288), что «гомология возникшего' таким образом рта представ-
ляет труднейшую проблему в морфологии Deuterostomia. Все по-
пытки ее разрешения кончались до сих пер неудачей».
Так рисуется В. Н. Беклемишему возникновение на основе орга-
низации ктенофор организации диплеврулы с ее трубчатым кишеч-
ником и целомическими мешками. Однако если мы стоим на пози-
циях миоцель-теории, другими словами, если принимаем, что цело-
мическая мезодерма представляет собой дифференцировавшийся из
обшей мезенхимы бластопоральный ее участок, в котором в каче-
стве опорного приспособления для развивающейся мускулатуры
образуется целомическая полость с ее жидким содержимым, то все
построение В. Н. Беклемишева, а ранее — Мак Боайда (1915),
теряет почву под ногами, так как целентерический аппарат ктенофор
не может иметь каких-либо отношений к образованию целома. С
этой точки зрения параллель в образовании трех пар выростов
гастро-васкулярной полости ктенофор и трех пар мешков целома
диплеврулы представляет чисто поверхностное сходство. К этому
можно- добавить и другие соображения. Во-первых, у иглокожих
имеется, собственно, две пары мешков целома, так как «первый^
и «второй» целомы не обособлены политотою друг от друга. Во-
вторых, бифуркация акрогастера гребне: >в имеет совершенно
иной характер и располагается в иной скости, чем «первые»
целомы диплеврулы иглокожих. В-треты щупальцы ктенофор
являются массивными придатками тела, а каналы пелентерического
аппарата не имеют для них опорного значения, как это, нацпотив,
всегда характерно для «вторых» целомов иглокожих и вызвало их
дифференцировку. И, в-четвертых, «третьи» целомические мешки у
иглокожих охватывают усваивающий эктодермальный кишечник,
тогда как у гребневиков соответствующие каналы расположены по
бокам эктодермальной глотки.
В этой связи следует подчеркнуть, что вся концепция целомиче-
ской мезодермы подвергается решающему испытанию при анализе
огранизации иглокожих.
284
В рассмотренных типах аннелид, моллюсков, членистоногих за-
даток целомической мезодермы располагался всегда в непосред-
ственной близости к энтодермальной закладке по краю бластомера.
I даже единичные клетки эюго зачатка оказывались включенными
, стенку энтодермального участка кишечцика. В дальнейшем <мезо-
шасты уходили из эпителиального расположения вглубь и давали
начало мезодермальным полоскам, уже совершенно независимым о г
энтодермы. Однако далее у полихет при образовании терминально
новых и новых сомитов мезодерма последних вторично могла полу-
чаться за счет клеток, располагавшихся в эпителии препигидиальной
зоны и несущих характерные черты мезенхимы.
В связи с этим П. П. Иванов различал, как мы видели, лар-
}алыную и постларвальную закладки сомитов. Мы считаем возмож-
ным видеть в мезодермальной закладке особо дифференцированную
мезенхимную закладку, стоящую в ряду прочих мезенхимных за-
кладов и первоначально особенно тесно1 связанную с эктодермаль-
ным кишечником.
У иглокожих мы встречаем, на первый взгляд, совершенно иную
картину. Мезодермальные мешки передней и задней пар образуются
путем отшнуровывания от кишечного впячивания, т. е. из
эктодермальной закладки. Здесь как будто нет ничего общего с
аннелидами, например, но есть сходство с кишечником целентерат
и ктенофор. Однако точные исследования И. И. Мечникова, а затем
и многих других показали, что первоначально от образующегося
впячивания отделяется значительное количество мезенхимных
клеток, уходящих в прилежащий бластоцель. А далее происходит
здесь же отшнуровывание мезодермальных мешков. И можно
сказать, что у иглокожих, как и у аннелид, это — образования
одного1 и того же ряда, т. е. мезодерма представляет лишь особо
дифференцированные мезенхимные закладки. Нет в отличие от анне-
лид и те и другие сохраняют свою теснейшую связь с эктодермаль-
ным впячиванием, образуя как бы часть последнего. То, что у анне-
лид было минимальным «клеточным воспоминанием», у иглокожих
представляет основную часть мезодермы и вкраплено в эктодермаль-
ный зачаток. Но в дальнейшем, как и у аннелид, мезодерма
выделяется из этого зачатка наподобие образования мезодермаль-
ных полосок и дает те же самые производные — мышцы и целом.
Таким образом, намечаются два пути эволюции одного и того
же первичного мезодермального аппарата, стоявшего по его поло-
жению в теснейшей связи с эктодермальным зачатком. В том ,и
другом случае это первоначальный миоцель. Лишь у иглокожих в
связи с выработкой скелета мышечная часть подавляется, а осо-
бенно^ сильно и сложно дифференцируется целом, на первый план
выступают вторичные его функции, соответственно развиваются
особые целомические системы и целенхима. При рассмотрении
организации хордовых мы найдем дальнейшие подтверждения
этому взгляду.
285
Возвращаясь к приведенным нами выше представлениям
В. Н. Беклемишева относительно диплеврулы, нельзя обойти молча-
нием вопрос об образовании у нее трубчатого кишечника, другими
словами, — проблему рта и ануса.
Если считать предками иглокожих протоцеломат, близких к
немертинообразным червям, — что мы должны были сделать, как
необходимый вывод из анализа организации, — то- ни о- каком
развитии нового вторичного рта и превращении первичного рта в
анальное отверстие не может быть и речи. Рот иглокожих соответ-
ствует рту червеобразных предков. Следовательно, он является пер <
вичным ртом, сместившимся из своего первоначального положения.
Будучи у низших червей на брюшной стороне, рот у немертин и
аннелид смещен к задней границе головной лопасти. Для дипле-
врулы характерна закладка стомодеального- впячивания уже непо-
средственно у головной лопасти. Благодаря этому получается впе-
чатление, что рот закладывается заново у переднего конца, а бла-
стопор закрывается у заднего конца тела. Так как образование
анального отверстия всегда, как мы точно могли установить у
немертин, связано с задним же концом тела, то у диплеврулы в
развитии образование ануса происходит также у заднего конца,
т. е. там же, где ранее закрывался бластопср. Получается впечат-
ление, что- бластопор становится анальным отверстием. Более того,
сокращение процессов может привести к фактическому переходу
замыкающегося бластопора в отверстие ануса. Но- ни в коем случае
нельзя перенести эти вторичные изменения отношений бла стопор а
и ануса в филогению и признать, что первоначальный рот стал
функционировать как анус, а в связи с этим возник, как говорит
В. Н. Беклемишев, «где-то независимым путем» новый рот. Это- —
экологически ничем не оправдываемое допущение, а сравнение
процесса образования заново рта у диплеврулы с процессом обра-
зования анального отверстия у немертин физиологически несостоя-
тельно.
Диплеврулы, по В. Н. Беклемишеву (1952, стр. 312—313), как
представлял и Мак Брайд, прикрепляются головной лопастью, это
«всегда ведет к особенно глубоким изменениям организации... Важ-
нейшим последствием прикрепления является миграция рта на
задний конец диплеврулы, сказавшейся верхним полюсом прикреп-
ленного животного. Уступая ему место, анальное отверстие сдви-
гается в сторону, кишечник вследствие этого образует спирально^
изогнутую петлю. Диссимметрическое смещение анального отверстия
является, вероятно, одной из главных причин возникновения дис-
симметрии всех внутренних органов. Из двух гонад прототипа
остается одна, половое отверстие располагается асимметрично
между ртом и заднепроходным отверстием... Мадрепоровое отвер-
стие оказывается рядом с половым, тогда* как передний конец ле-
вого переднего целома... остался близ аборального полюса... В
силу этого левый передний целом протягивается в виде узкого
286
канала — «аксоцеля» — вдоль всего тела животного. От... правого
переднего целома сохраняется лишь небольшой ‘участок... Левый
средний целом (гидроцель) остается вблизи рта и передвигается
вместе с ним, причем он обвивает пищевод, образуя вокруг послед-
него сначала подкову, а затем и кольцо... Правый средний целом
исчезает... Левый задний целом... оказывается захваченным петлею
кишечника и в силу этого сдвигается в оральном направлении,
тогда как правый задний целом располагается по отношению к нему
аборально».
Весь этот круг идей в настоящее время является в основных
чертах общепринятым. У В. Н. Беклемишева он разработан не-
сколько своеобразно, но здесь не будем вдаваться в детали. Мы же
стремились показать процессы в связи с условиями существования
современных иглокожих и вероятными условиями жизни их предков,
поскольку об этом можно судить по данным анализа организации.
Главное отличие наших положений состоит в том, что мы, в
соответствии с Э. А. Мейером, допускаем прикрепление не
головной лопастью, а посредством части околоротовых щупалец.
При этом принимаем, что уже у диплеврул имелся аппарат около-
ротовых щупалец, обладавших мерцательными поверхностями, цело-
мическими полостями, кровеносными сосудами, соответствующими
нервными образованиями, т. е. обладавших комплексом образований
амбулакральной борозды. Такое допущение вполне соответствует
необходимости видеть в диплевруле не чрезвычайно упрощенного
хтенофорообразного представителя животного мира, а гораздо более
сложный организм, обладавший и трубчатым кишечником, и крове-
носной системой, и целомом. Одним словом диплеврулу следует
считать взрослой формой протоцеломат.
В этом случае радиальная симметрия иглокожих представляется
результатом эволюции аппарата части оральных щупалец при сме-
щении рта на полюс, противоположный месту прикрепления у пред-
ков иглокожих, перешедших к сидячему существованию. Не нужны
будут и те туманные построения, которые В. Н. Беклемишевым
(1952, стр. 315) формулируются следующим образом. «Радиальная
симметрия появляется у иглокожих благодаря возникновению у них
радиально построенного амбулакрального набора органов, в кото-
рый постепенно включаются один за другим органы различных-
систем: первыми его членами являются приводящие борозды, затем,
включается эктоневральная система, амбулакральная система, за-
тем, вероятно, кровеносная и перигенальпая системы и во всяком
случае скелет».
Можно надеяться, что все это сложнейшее и довольно пробле-1
матичное изображение развития радиального строения иглокожих
становится совершенно излишним.
Однако нельзя не согласиться с В. Н. Беклемишевым (1952,
стр. 335) в общей оценке им эволюции иглокожих: «Оглядываясь
На эволюцию иглокожих в целом, мы видим неисчерпаемые преоб-
287
разования, непрерывную перестройку, громадное разнообразие на-
правлений развития и почти полное отсутствие прогресса».
Действительно, ведущие в организации иглокожих* системы'
целом и скелет — несут в себе известного рода односторонность.
Особенно это касается целома. Хотя иглокожие по развитию этой
именно системы несомненно могут быть поставлены выше всех дру-
гих типов. Но’для целома основной характерной чертой является
его жидкое содержимое. Оно не открывает широких возможностей
выхода иглокожих в наземные условия существования, где имеют
место гораздо более сложные отношения, чем в водной среде. Но
даже и в водной среде целом ограничивает иглокожих возможно-
стью использовать лишь воды мирового океана, вне которых они не
встречаются.
Всестороннее развитие организации, обусловленное единством
организма и среды, мы находим в типе хордовых, к которому и пе-
реходим.
Глава XII
ТИП ХОРДОВЫЕ — CHORDATA
Пассивное питание' исходных форм, строение и развитие переднего
отдела кишечника. Аппарат жаберных щелей и аппарат щупалец.
Дифференцировка печеночного отдела. Целом и его дифференци-
ровка. Миохордальный комплекс. Гонады. Экскреторная система.
Кровеносная система и сердце хордовых. Органы чувств и нервная
система. Головной мозг хордовых. Итоги анализа организации
хордовых. Подтипы хордовых.
Обычно подчеркивают, что для хордовых типовыми чертами
организации являются: 1) ходда, 2) аппарат жаберных щелей и
3) дорзально расположенная трубка спинного и головного мозга.
Однако это лишь слабый абрис сложной картины, нанесенный в
одном плане. Развернем его.
В организации хордовых, как и в других типах, одним из веду-
щих является комплекс черт, связанный с питанием. Эта категория
отношений со> средой выявлена у них наиболее характерно, тогда
как связанные с защитой черты организации хордовых более раз-
нообразны и являются ведущими уже в эволюции подтипов. Таковы,
например, развитие туники — органа пассивной защиты Tunicata,
или развитие боковой мускулатуры, служащей для активной защиты
быстрым движением у Acrania. У позвоночных эта мускулатура
используется и^для движений при активном нападении на добычу.
Таким образом, у них отношения, связанные с питанием, объединя-
ются с чертами защиты, обусловливая богатейшую прогрессивную
эволюцию подтипа.
Характерно для исходных форм хордовых пассивное питание,
при котором мерцательное движение на оральном отделе тела вызьы
вает ток воды, влекущий мелкие взвешенные в воде пищевые части-
цы ко рту. В наиболее простом виде мы находим это у сидячих
Phabdopleura и Cephalodiscus (см. стр. 306 и далее, рис. 125, 126,
127, 128).
Ток воды направляется у Rhabdopleura по ветвящимся щупаль-
цам справа и слева прямо в эктодермальный кишечник, так как
эктодермальный глоточный отдел в кишечнике отсутствует. Его ре-
дукция при пассивном питании, когда механическое захватывание и
19 Пути эволюции животного мира 289
переработка мельчайших пищевых частиц це требует особых при-
способлений вроде мускулистой глотки, челюстей и т. п., вполне
понятна. И мы должны допустить редукцию сзюмодеума у предков
хордовых, перешедших к пассивному питанию и сидячему или
близкому к тому образу жизни. Это могли быть лишь червеобраз-
ные организмы из Protocoelomata, для которых, как и для всех
свободноживущих червей, типично было наличие эктодермальной
глотки и присутствие мерцательных борозд с боков за головой напо-
добие затылочных или аврикулярных органов. Таким образом,
раскрывается одна из очень характерных черт организации хордо-
вых — пища приносится с током воды, вызываемым околоротовым
мерцательным аппаратом, и в развитии кишечника отсутствует
; эктодермальный глоточный отдел, а за ротовым отверстием непо-
средственно идет эктодермальный отдел.
У Cephalodiscus намечается и другая характернейшая черта
хордовых, а именно — по бокам передней части эктодермального
кишечника образуется по одному отверстию, отводящему наружу
избыток воды, пригоняемый в кишечник мерцательным движением.
Это — первая закладка жаберных щелей.
У прочих подтипов хордовых на подобной основе вырабатывает-
ся чрезвычайно характерный сложный аппарат жаберных щелей,
производящий своим мерцательным эпителием постоянный ток воды
при пассивном питании, причем околоротовые щупальца теряют
значение и редуцируются (см. рис. 132, 133, 134), и в то же время
это аппарат, служащий (см. рис. 135 и 145) для задерживания
мельчайших пищевых частиц и для проведения их в усваивающий
отдел кишечника. Тот способ, которым осуществляются эти своеоб-
разные, противоречивые задачи, является едиственным в своем роде.
На пути тока частиц у баланоглосса находится слизистый секрет'
железок головной лопасти или у ланцетника и оболочников ставит-
ся завеса из слизистых нитей, вырабатываемых железистыми клет-
ками расположенной медиовентрально железы эндостиля. Вся эта
слизистая сеть с прилипшими и охваченными слизью пищевыми
частицами гонится мерцательным движением сначала по брюшной
стороне глотки вперед, затем у входа в жаберный 'отдел передви-
гается по бокам на спинную сторону и по ней назад уже, в усваи-
вающий отдел кишечника.
Небольшой первоначально эндостиль разрастается по глотке
назад, параллельно с чем идет увеличение количества жаберных
щелей. У ланцетника развивается вместе с тем комплекс опорных
образований в виде хрящевых пластинок и их синаптикул в стенках
жаберных щелей. А у баланоглосса к этой системе присоединяется
дорзальный щитовидный хрящ и боковые его выросты, охватываю-
щие начало глотки. Оба аппарата поддерживают в зияющем состоя-
! пии глотку и жаберные щели. На основе их получается сложнейший
жаберный отдел, который с сильным развитием в его стенке капил-
лярных сосудов обеспечивает также функцию газообмена. Он ха-
рактерен для Acrania, Tunicata и Enteropneusta, и лишь с переходом
290
к хищничеству у Vertebrata в известной мере редуцируется, сохра-
няя преимущественно функцию газообмена.
Существование подобного жаберного аппарата вполне понятно
у Tunicata при сидячем образе жизни их асцидиеобразных исходных
форм. То же можно сказать про полусидячего в песке ланцетника.
Но этого' нельзя принять для баланоглосса, живущего в фунте и
пропускающего его через свой кишечник. Несомненно, баланоглосс
сохранил выработавшийся в других условиях аппарат при переходе
к питанию заключенными в поверхностных слоях грунта органиче-
скими частицами. Это тоже пассивное питание фильтраторов, но
только в этом случае охваченные слизью пищевые частицы попа-
дают в кишечник вместе с частицами твердого грунта. Здесь вполне
возможно предположить в эволюции переход в песчаные и .заилен-
ные грунты, но невозможно' представить, чтобы имела место перво-
начальная выработка совершенно сходных приспособлений у бала-
ноглосса и ланцетника в условиях роющего образа жизни. Несом-
ненно, что это наследие от предков, живших в трубках или в ходах,
проделанных в грунте.
Весь комплекс сложных превращений, давший на место экто- .
дермальной глотки червей эктодермальную глотку хордовых, ото- V
бразился еще в одной характерной черте организации последних.
С редукцией стомодеального отдела у взрослых форм лесно связа-
но и исчезновение его закладки в онтогенезе. Бластопор — первич-
ный рот — постепенно закрывается. Прилежащий к нему участок
энтодермы, в силу процессов неравномерного роста, особенно невро-
хордальной закладки, идущей от заднего края бластопора, смещает-
ся па 180° к будущему переднему концу тела с его чувствитель-
ными (ольфакторными) и железистыми (гипофизарными) заклад- ;
ками. Здесь прорывается, как у немертин или аннелид, дефипи- j
тивный рот, на границе которого эктодерма покрова переходит ‘
в прехордальную энтодерму кишечника. Исчезла преемственность
с бластопором, в связи с исчезновением стомодеума. Но рот от- '
«врывается в тот же самый передний участок энтодермальной киш-
ки, и, конечно, это тот же самый рот, что и у типичных Proto-
stomia.
Мы должны пополнить- анализ комплекса характерных черт хор-
довых, связанного с питанием, указанием на развитие у низших
форм своеобразной дифференцировки особого отдела тела, распо-
ложенного между головной лопастью и передней частью туловища,
несущей жаберные щели. Речь идет о том, что вырабатывается «во-
ротничок». Он охватывает у Pterobranchia (рис. 129 и 130) с боков
и дорзально ротовое отверстие и имеет по бокам полые выросты
стенки тела в виде щупалец или жабер. По ним ток воды при пас-
сивном питании несется к ротовому отверстию. В соответствии с
этим от парного целома туловища обособляется пара целомиче-
ских мешков с опорной функцией для щупалец — целом воротнич-
ка. У баланоглосса (рис. 132 и 133) воротнмчковый отдел тела со-
храняется, но уже с иной функцией. Он становится железистым
19* 291
утолщенным участком, гарантирующим благодаря массовому вы-
делению слизи возможность образования, ходов в грунте и движе-
ния в них без повреждений нежного червеобразного тела. Щупаль-
ца при этом редуцировались, но воротничковый целом, обусловли-
вающий их плотность, сохранился. В дальнейшем мы еще вернем-
ся к нему. У ланцетника воротник, как таковой, отсутствует, но его
целом сохраняется и, проникая в складки метаплевр, распростра-
няется от переднего конца назад. Складки метаплевр прикрывают
жаберный аппарат и гонады.
Отметим в заключение, что у хордовых в ближайших отношениях
к питанию, а также и к экскреции, вырабатывается общая для них,
очень характерная черта кишечника — дифференцируется пе^ць.
Она уже выражена у баланоглосса, но железистые клетки в 'ё^^ю-
щем за 'жаберным печеночном отделе кишки располагаются более
или менее равномерно по всему ее просвету. У ланцетника (см.
рис. 145, 147) они локализованы в вентральном, слепом печеноч-
ном выросте. У позвоночных — это резко обособленная массивная
железа, приобретающая огромную роль в химизме организма. Од-
нако у Pterobranchia печень отсутствует, а у туникат она еще да-
леко не стабилизирована. Таков путь дифференцировки крайне важ-
ного и своеобразного органа, каким является печень в типе хордо-
вых. Конечно, наряду с особенностями развития кишечника и ори-
гинальнейшей дифференцировкой глоточно-жаберного аппарата, об-
разование печени трудно принять за выражение конвергентной эво-
люции в различных подтипах хордовых.
От рассмотрения ведущего комплекса, связанного с пассивным
питанием, переходим к другому чрезвычайно важному комплексу,
охватывающему различнейшие стороны жизнедеятельности хордо-
вых и соответственно различные экологические аспекты. Это — ком-
плекс целома. Как мы уже видели, такое значение целома, помимо
его"'первичной связи с мускулатурой, обусловлено многообразием
его функций как внутреней среды в организме/ Наряду с главной
функцией опоры, он несет у хордовых, как и в других типах, ряд
побочных функций — трофическую, экскреторную и половую.
Обратимся к Pterobranchia (см. рис. 127, 128, 129, 130). Голова
их превращена в своеобразный щит, который секретом своих желез
образует трубку, защищающую животное. Необходимая при пост-
ройке трубки плотность хорошо развитой головной лопасти обуслов-
лена дифференцировкой особого головного целома с его жидким со-
держимым. Дальнейший отдел — воротничковый целом, что мы уже
отмечали и теперь еще раз особо подчеркнем, несет также функцию
опоры, обусловливая плотность воротничка и его выростов —- щу-
па л ец-жабер.
У баланоглосса (см. рис. 132, 133, 134) сходные отношения зна-
чительно осложняются. Во-первых, в связи с переходом к роющему
образу жизни редуцируются наружные околоротовые выросты щу-
палец, но правый и левый мешки воротничкового целома сохра-
няются, хотя и подвергаясь некоторой редукции и в известной мере
292
заполняясь целенхимой. Во-вторых, головная лопасть превращается
в мускулистый овальный придаток, благодаря сокращениям кото-
рого животное при опоре на головной целом и далее на хорду про-
бивает ход в грунте. В-третьих, тело значительно удлиняется в
связи с терминальным ростом туловища, причем движение живот-
ного обусловлено сокращениями типичной для червей мускулатуры
стенки тела—слабой кольцевой, более значительной продольной.
Опорным образованием здесь является целом туловища в виде
двух — правого и левого — мешков, идущих от воротничка до зад-
него конца. Чего-либо подобного придаткам конечностей, вроде па-
раподий аннелид, здесь не выработалось, мускулатура стенки тела
равномерно, без перерывов идет по всей периферии, как у немер-
тин. Перед нами отношения, очевидно, близкие к характерным для
Protocoelomata и сходные с имевшимися у немертанообразных
предков. Дифференцировка воротничкового целома, связывающаяся
с выработкой околоротовых плотных боковых придатков, наподо-
бие жабер Pterobranchia, говорит об основной дифференциров-
ке, вызвавшей появление олигомерное™ целома, и о своеобраз-
ных путях эволюции олигомерных форм целомат — иглокожих
и хордовых.
Обращаясь к другому опорному образованию баланоглосса — к
.хорде, мы видим, что она представлена у хордовых как бы в двух
вариантах — с одной стороны, в комплексе образований головной
лопасти у рассмотренных Pterobranchia и Enteropneusta, с другой—
в миохордальном комплексе туловища ланцетника и позвоночных,
В первом случае это опорное образование для укрепления хорошо
развитой головной лопасти. Во втором, при редукции последней —
опора для мускулатуры туловища и хвоста. Бесчерепные в этом от-
ношении представляют интересную особенность, так как туловищная
хорда у них вторично проникает до головного конца, принимающего
функцию лопатки для раздвигания песчаного грунта, в котором
скрывается ланцетник. У личинок туникат и у позвоночных обычно
хорда доходит спереди лишь до средины головного мозга.
Обе формы хорды одинаково представлены эпителогенной пузьгр-
чатой тканью. Заключенное во внутриклеточных вакуолях жидкое
содержимое хорды, окруженной плотной общей наружной оболоч-
кой, дает хорошую опору, в одном случае, основанию головной ло-
пасти, в другом — для соответствующей продольной мускулатуры
стенки тела в туловище. Головная хорда не теряет связи с передней
частью глотки, .что может только способствовать прочности опоры
головы. Напротив, спинная хорда совершенно обособлена от киш-
ки, служа опорой для мощных продольных мышц и располагаясь
среди них (см. рис. 147, 148). Однако тем более бросается в глаза
сходство происхождения и строения этих оригинальнейших органов <
опоры. Их гомологичность говорит и за близость их в филогенети-
ческом отношении.
Балапоглосс обладает, однако, и еще одной категорией опорных
образований, совершенно иных, чем укрепляющая основание хобота
293
хорда или целом, придающий ему плотность при сокращении. Это—
соединительнотканный хрящевой скелет начала глотки, поддержи-
вающий ее в иеспадающемся состоянии. По существу это третья
категория опорных образований с особой, по сравнению с двумя
вышеописанными, функцией. Невольно бросается в глаза функцио-
нальное сходство этой третьей категории с имеющимися у ланцет-
ника околоротовыми хрящами, и с ротовым кольцевым хрящем круг-
лоротых среди позвоночных.
У бесчерепных, позвоночных и туникат дифференцировка хорды
и целомических мышц представляет важнейший комплекс в их ор-
ганизации. Вместо относительно- слабой мускулатуры стенки тела
червей, какую мы видим у Entenopneusta (рис. 134), за счет спин-
ного отдела ее продольных мышечных пучков развиваются мощные
«боковые» мышцы рыбообразных ланцетника (см. рис. 145, 147,
148) и низших позвоночных. Их образование связано с дифферен-
цировкой нового- осевого скелета — хорды на спинной стороне тела.
То и другое взаимно обусловлено. Вместе с тем теряют значение и
редуцируются боковые и брюшные продольные и также кольцевые
мышцы стенки тела. Так, на место относительно слабых перисталь-
тических сокращений туловища червя с опорой па целом, возни-
кают энергичные сгибания тела с опорой на хорду, достигающие
своего полного выражения в быстрых движениях ланцетника и
позвоночных.
Дифференцировка миомеров в онтогенезе ланцетника и позво-
ночных является механическим выражением упорядоченного распо-
ложения продольных мышечных волокон при развитии синусои-
дальных извивов тела, обусловливающих поступательное движение
при плавании. При этом границы миомеров справа смещены на
полсомита по сравнению с миомерами левой стороны тела.
Свободное плавание у ланцетников обычно выражается в быст-
рых бросках, за которыми следует моментальное проникновение в
грунт и полный покой в нем. В этом можно видеть крайне ориги-
нальное освоение глубоко различных жизненных аспектов — биото-
пов песчаного грунта и придонной толщи воды.
У личинок туникат и у позвоночных отношения несколько иные.
Это — свободноплавающие организмы бентоса с равномерным по-
ступательным движением, обусловленным правильными синусоид-
ными извивами удлиненного тела, а при значительном развитии
хвостового, заанального его отдела — ударами хвостового плавни-
ка. Каких-либо локомоторных придатков здесь, как .и у баланоглос-
са, совершенно не имеется. Это указывает, что исходные червеоб-
разные и рыбообразные формы хордовых не обладали ими даже в
наиболее простом виде, наподобие, например, параподий аннелид.
Метаплевры ланцетника в этом отношении, невидимому, не имеют
значения, а должны быть поставлены в связь с развитием ворот-
ника низших хордовых. Основное значение при выработке плавания
у высших хордовых переходит к усиленному росту заднего отдела
тела.
29 £
Крайнего выражения подобный процесс достигает у аппендику-
лярий (см. рис. 143), у которых за жаберным и половым отделами
•гела идет хвостовой придаток, смещенный на 180°, т. е. направлен-
ный прямо вперед. Он содержит хорду, мускулатуру и нервную си-
стему, но лишен всяких следов кишки и представляет собой разра-
стание заанальной спинной части тела. Будучи резко обособлен от
остального туловища, он играет важнейшую роль- органа, который
обусловливает своим движением в «домике» ток воды, несущий пи-
щевые частицы при пассивном питании карликовым планктоном.
Это — своеобразнейшая узкоприопособительная черта организации
аппендикулярий.
Обычно происходит образование хвостовой почки за задним от-
делом кишечника, причем иногда развивается даже небольшая
петля постанальной кишки. Таким образом, получается хвостовой
вырост, содержащий хорду в центре, мощную продольную мускула-
туру с боков от нее и дорзалыно типичный для хордовых спинной
мозг. Этот процесс неравномерного роста заднеспинной части ту-
ловища, в котором взаимосвязаны дифференцировка хорды, миото-
мов и нервной трубки, являлся ведущим в выработке из сидячих
или полусидячих червеобразных предков организации свободнопо-
движных, а затем и хищных форм, а именно Acrania, а затем и поз-
воночных. При эволюции хордомышечного комплекса хвоста соз-
дается характерная черта усваивающего отдела кишечника рыбо-
образных хордовых — диспропорция его длины с общей длиной
тела. Заанальный хвостовой отдел не содержит или почти не содер-
жит кишки. Кишка является в виде короткой прямой трубки.
И лишь дифференцировка железистого переднего отдела последней
в виде печени, невидимому, в известной мере компенсирует корот-
кость усваивающей кишки. У ланцетника эти отношения выступают
особенно рельефно. У него печеночный вырост кишечника, выпячи-
вая брюшную стенку тела, вызывает образование длинного при-
датка, лежащего на значительном протяжении параллельно жабер-
ному отделу тела. Такой монстр не мог бы существовать, если бы
вентрально и с боков на протяжении жаберной области уже небыли
развиты защитные складки метаплевр, вторично давшие для пече-
ночного выроста атриальную полость и новую стенку тела.
Развитие хордомышечного комплекса происходило во всем туло-
вище. Оно шло беспрепятственно в хвостовом отделе. Но впереди
развитие хордо-мышечного комплекса выражалось своеобразно в
силу того, что при переходе к сидячему образу жизни туникат или
к своеобразному существованию предков бесчерепных и позвоноч-
ных редуцировалась головная лопасть исходных форм, представлен-
ная у Pterobranchia в виде щитка, а у Enteropneusta в виде хобо-
та. Возможно предположить, что, в силу далеко ушедшей специа-
лизации, такое узко адаптивное образование, как годовная
лопасть низших хордовых при изменении условий существования
Утрачивало свое значение, не принимая каких-либо иных функ-
ций.
295
Как бы то ни было, факт редукции головной лопасти налицо
В голове Acrania и Vertebrata миомеры идут до самого передней
конца, и в онтогенезе не сохранилось передней неметамерной за-
кладки.
С образованием хордо-мышечного комплекса опорная функция
целома в туловище утрачивается. Но побочные функции целома —
экскреторная и трофическая — сохраняются. К ним присоединяется
лишь “у ланцетника и позвоночных и половая функция. В этом отно-
шении чрезвычайно демонстративно, что половой зачаток у туни-
кат (см. рис. 135) совершенно независим от целома, у птеробрап-
хов и баланоглосса гонады вдаются в целом, а у ланцетника поло-
вой зачаток вторично становится в ближайшие взаимоотношения с
целомом. Так, мешотчатые гонады баланоглосса (см. рис. 134) рас-
полагаются дорзально в отделе туловища, непосредственно следую-
щем за жаберным и содержащем уже усваивающую кишку. Гонады
эти очень сходны с мешотчатыми гонадами немертин. Полость их
при созревании половых продуктов также открывается наружу ко-
ротким протоком яйцевода, соответственно семевода. Но внутрь
гонады глубоко вдаются в целом в виде простых или ветвистых
мешков, и только в этом выражаются взаимоотношения гонад с це-
ломом. Таким образом, здесь полости гонады и целома существуют,
можно сказать, бок о бок и в то же время совершенно независимо
друг от друга. Для теории происхождения целома за счет полости
гонад (гоноцель-теория) это едва ли преодолимое препятствие.
У ланцетника (рис. 145, 148) гонады, расположенные также в
зажаберном отделе тела, связаны уже с особым отделом целома —
гонотомом, в который они вдаются. Гонотом лежит под мышечным
зачатком — миотомом, а дальнейший, вентральный отдел диффе-
ренцируется как околокишечный целом. В последнем на спинном
мезентерии подвешен кишечник и отсутствует метамерия, характер-
ная для мезодермальных сомитов. Этот факт говорит за в^орщшое
происхождение полимерности целома в миотомах на почве выработ-
ки синусоидного движения с опорой на хорду в туловищном и хво-
стовом отделах тела у Tunicata, Acrania и Vertebrata.
Гонады ланцетника с заключающими их мешками целома при
созревании половых продуктов вдаются в стенку тела при начале
метаплевральных складок, и эти последние прикрывают и жабры и
гонады. При дифференцировке стенки гонад и стенки целома над
ними в виде примитивного протока половые продукты попадают в
атриальную полость и выводятся наружу через атриальную пору
при сокращении кольцевых мышц метаплевр. Таким образом, с
развитием метаплевральных складок, создается защищающий аппа-
рат не только для жабер от забивания их грунтом и для печеноч-
ного выроста брюшной стенки тела, но также и для гонад и поло-
вых продуктов. Это последнее обстоятельство вызывает продолже-
ние атриальной полости на значительное расстояние позади жабер-
ного отдела, создает из нее сложное в функциональном отношении
приспособительное образование.
296
У туникат, точнее у их сидячих асцидиеобразных форм (см. рис.
.35), главную массу тела образует чрезвычайно-сильно развитый
жаберный отдел, доводящий фильтрацию воды, а следовательно, й
ток пищевых частиц при пассивном питании сидячих животных до
максимальных пределов. При прикреплении личинки асцидий (см.
рис. 138) мускулатура типа ланцетника дегенерирует и утрачива-
ются все отделы, целома, за исключением перикардиального, о чем
речь будет впереди. Их гонады, расположенные в петле кишки,
представлены одним мешком яичника и парой семенников по бокам,,
причем каждый бывает снабжен хорошо развитым выводным про-
током. Последние открываются в клоакальный сифон, куда откры-
вается и анус. Особенно рельефно отношения гонад и целома
выступают у аппендикулярий (рис. 143), у которых нет клоакаль-
ного сифона, а снабженная хордой и метамерной мускулатурой
хвостовая часть тела смещена на 180° сильно развитым отделом,
содержащим гонады. Впереди последнего расположен жаберный
отдел и петля кишки, тогда как обычно у Tunicata, у которых гона-
ды лежат в петле кишки, половой отдел не дифференцирован.
Своего полного выражения взаимоотношения гонад и целома
достигают у позвоночных. У большинства из них яичники и семен-
ники лежат, глубоко вдаваясь в туловищный целом, причем яйце-
вые и семенные клетки попадают в него через разрыв стенки гона-
ды и из него выводятся уже наружу через посредство определенных
частей экскреторного аппарата. Создается мочеполовая система. Так,
мешетчатые гонады, являющиеся типовой примитивной чертой орга-
низации, вступают у высших хордовых в связь с целомом. Отноше-
ния, как видим, аналогичные имеющимся у аннелид и членистоно-
гих, развивающиеся в известной мере конвергентно.
Замечательные отношения целома и экскреторной системы су-
ществуют у ланцетника. Примитивные протонефридии, развиваю-
щиеся в области жаберного отдела тела в количестве, соответству-
щем количеству первичных жаберных щелей, лежат под миотомами.
Пучки их концевых клеток, чрезвычайно сходных с соленцитами
полихет, вдаются в целом, а выводные протоки открываются в дор-
зальную часть жаберной щели. У Cephalodiscus имеется одна пара
таких же протонефридиев возле пары жаберных щелей. Больше ни
У кого из хордовых ничего подобного не наблюдается. Как будто
эволюция нечаянно обронила свой след в организации птеробран-
хов и ланцетника, сохранив протонефридии в глоточной области^
как у немертинообразных червей или личиночных форм аннелид и
моллюсков.
У червеобразного баланоглосса нефридии отсутствуют, и эскре-
ты удаляются из организма нефро-фагоцитарными клетками, обра-
зующими большие скопления на переднем конце спинного сосуда.
При этом амебоидные клетки извлекают жидкие продукты распада,
содержащиеся в крови, и отлагают их в своей протоплазме в виде
окрашенных зернистостей. Далее амебоциты падают в целом голов-
ой лопасти, откуда, в конце концов разрушаясь, выводятся через
297
«хоботковую» пору, т. е. через парные, или обычно непарные отвер-
стия при основании хобота. Таким образом, целом одукты головного
целома баланоглосса, наряду с функцией регулирования плотности
хобота, что присуще иногда нефридиям целомических червей, ока-
зываются своеобразно вовлеченными и в экскреторный процесс,
который в своих начальных этапах, однако, сосредоточивается в
кровеносной системе.
В воротничковом целоме баланоглосса, в связи с наличием пар-
ных целомодуктов очень простого характера, сохранена функция
регулирования плотности тела. В туловищном же целоме уже нет
ни регулирующих, ни экскреторных образований.
То же приходится подчеркнуть и относительно зажаберного от-
дела туловищного целома ланцетника. У ланцетника подъем нефро-
фагоцитарной деятельности ярко выявляется в больших скоплениях
нефро-фагоцитов в стенке зажаберного отдела атриальной полости.
Подобные отношения, наконец, характерны и для Tunicata, которые
при сидячем существовании их исходных асцидиеобразных форм
утрачивают, как мы видели, типично выраженные целомические
полости. Экскреторная деятельность у них всецело выполняется
нефро-фагоцитами, сосредотачивающимися, в конце концов, на стен-
ках клоакального сифона, как и в атриальной полости ланцетника.
У всех рассмотренных форм при их сидячем или приближаю-
щемся к тому образу жизни и при пассивном питании энергия про-
цессов жизнедеятельности в общем невелика, процессы обмена
веществ протекают на низком уровне. Иное мы видим у позвоноч-
ных, перешедших к свободному образу жизни и активному пита-
нию. С повышением интенсивности жизнедеятельности и соответ-
ственно обмена веществ нефро-фагоцитарная функция отходит на
задний план, в туловищном отделе тела развиваются характерные
для позвоночных экскреторные органы в виде открытых трубочек
сначала пронефроса, затем мезо- и метанефроса. Они проникают
все далее и далее назад в туловище. Часть их протоков исполь-
зуется и для выведения половых продуктов. Прилежащий к почеч-
ным канальцам отдел целома, располагающийся ниже миотома,
дифференцируется в виде нефротома. В дальнейшее изложение
фактов мы не будем вдаваться, хотя эволюция мочеполового аппа-
рата позвоночных в высокой степени интересна как пример разви-
тия конвергентного с имеющимся у аннелид и некоторых других
групп беспозвоночных.
Для завершения анализа целома в типе хордовых обратимся
к трофической его функции. У червеобразного баланоглосса она ни-
чем не отличается от отношений, присущих аннелидам. В развитии
ланцетника она выражается, повидимому, в проникновении целома
между хордой и мускулатурой и в образовании здесь узких целоми-
ческих пространств и некоторых полостей, связанных с ними (см.
рис. 148). Однако у взрослого животного эти части целома редуци-
руются, так как за счет их мезодермальных элементов образуется
вторичная мезенхима или, «целенхима». Аналогичные явления на-
29S
блюдаются также в других группах целомических животных. Так,
например, целомические полости в слое кольцевых мышц стенки
тела полихет из Eunicidae часто в значительной мере заполняются
.деленхимой. У позвоночных в развитии также дифференцируются
по бокам хорды участки, где происходит образование вторичной
мезенхимы. Это так называемые «склеротомы», располагающиеся
между миотомами и нефротомами. Здесь, кроме того, и париеталь-
ный слой мезодермальных сомитов становится также источником
вторичной мезенхимы-
Таким образом, трофическая функция сравнительно слабо выра-
жена в целоме хордовых, и это должно быть поставлено в прямую
связь с развитием кррвдаоРДоД системы. Последняя у высших
Chordata сильно развита и усиливает как свою функцию газообме-
на, так и трофическую функцию. Кроме того, у позвоночных резко
выявляется функция эндокринной химической регуляции жизнедея-
тельности всего организма через посредство крови, а кровеносная
система в известной мере специализируется как система, поддер-
живающая единство особи в отношении химизма.
Целом становится в своеобразные взаимоотношения с кровенос-
ной системой, обусловливая пропульсаторную деятельность сердца.
Аналогичное явление было уже описано нами при анализе органи-
зации моллюсков, где парный перикардиальный целом образует •
мускулатуру сердца, и этим обусловлено сохранение соответствую-
щего отдела целома в этом типе. У хордовых обычно дифференци-
руется непарный мешок целома в качестве перикардиального и
обусловливающего своей мускулатурой сокращения сердца. Но даль-
нейшие соотношения в разных подтипах очень своеобразны и во
всяком случае совершенно отличны от моллюсков. Перед нами при-
мер того, как эволюция различных органов может вести совершенно
различными путями к сходным физиологическим результатам.
Сердце баланоглосса (см. рис. 133) и Pterobranchia (см. рис.
128, 129) теснейшим образом связано с головной хордой этих под-
типов. Оно представляет собой передний участок спинного сосуда, с
одной стороны, непосредственно прилежащий к хорде, а с другой—
охватываемый полукольцами мышечных волокон, которые разви-
ваются в прилежащей стенке перикардиального мешка целома. .
Функционирование подобного аппарата возможно только при на-
личии хорды. Можно сказать, что наличие «головной» хорды обу- I
словливает и наличие «головного» сердца, тогда как у прочих под-Лл
типов хордовых с редукцией головной лопасти редуцируется соот- Л#
ветствующий отдел кровеносной системы и вырабатывается «брюш- р
н°о сердце», расположенное на брюшном сосуде. При этом меняется
и направление тока крови в сосудах. Вспомним, что у Pterobranchia
и Enteropneusta наблюдаются отношения, характерные для Vermes,
Mollusca и Arthropoda, когда по спинному сосуду кровь движется
вперед, а по брюшному — назад. Но у ланцетника (см. рис. 149) и
позвоночных кровь по брюшному сосуду направляется вперед к рас-
пложенным здесь жабрам, а по спинному сосуду и его ветвям
299
артериальная кровь гонится главным образом назад, ко всем орга-
нам и, прежде всего, к развивающейся здесь мощной продольной
мускулатуре.
Вместе с этим вырабатывается и большая самостоятельность
кровеносной, cngffgMU в смысле образования сплошной эндотелиаль-
ншР^ютилки во всех сосудах и в смысле преобладающего значе-
ния клеточных элементов в построении сосудов. Если у баланоглосса
стенка Сосудов образована только пограничной пластинкой основ-
ного вещества, а у асцидий в сердце имеется уже .настоящий эндо-
телий, то у ланцетника и позвоночных все сосуды, включая и
капилляры, обычно имеют собственную эндотелиальную выстилку.
Таким образом, кровеносная система гораздо сильнее и резче диф-
ференцируется, и в эмбриональном развитии позвоночных появляет-
ся клеточный «ангиобласт» в виде кровяных островков и т. п.
В анатомии кровеносной системы уже у ланцетника (см.
рис. 149) мы находим существенную особенность. В соответствии с
мощным развитием боковой мускулатуры тела развиваются два
мощных боковых сосуда, по которым венозная кровь уносится от
мускулатуры стенки тела вновь к жаберному аппарату. Образуются
передняя и задняя боковые вены на правой и левой стороне, соеди-
няющиеся в правый и левый кювьеровы протоки, т. е. сосуды, при-
носящие кровь в брюшной продольный сосуд и впадающие в него
непосредственно позади жабер. Таким образом, с развитием туло-
вищной хорды и связанной с ней мощной целомической мускулату-
рой, с одной стороны, а с другой — с образованием жаберных
щелей в переднем отделе кишечника создается типичная кровенос-
ная система позвоночных. В ней, кроме того, за счет брюшного
сосуда и сосудистого сплетения кишечника дифференцируется еще
особая система капилляров печени — воротная система печени,
располагающаяся позади сердца.
У червеобразного баланоглосса жаберная часть кровеносной
системы уже выражена, но характерная для червей мускулатура
стенки тела баланоглосса еще не имеет каких-либо особенностей
в кровеносной системе, как нет и обособления воротной системы
печени. Развиты спинной и брюшной сосуды (см. рис. 134), связан-
ные с системами капилляров стенки тела и стенки кишки, т. е. ти-
пичные для червей отношения.
У Tunicata (см. рис. 135), у асцидиеобразных исходных сидячих
форм чрезвычайно сильно развивается жаберный аппарат, обеспе-
чивающий и обильное питание и достаточный газообмен под плот-
ной защитной туникой. Хорда же и головная лопасть при прикреп-
лении животного вентральным отделом переднего конца тела реду-
цируются и исчезают. Соответственно сердце развивается не впере-
ди жаберного мешка, а за ним. Но сердце построено подобно тому,
как у птеробранхов и баланоглосса, т. е. непарный мешок целома
с полукольцевыми мышечными волокнами во внутренней стенке
охватывает наподобие муфты брюшной кровеносный сосуд и обус-
ловливает продвижение крови. Однако масса капилляров, которая
300
развивается в. жаберном отделе кишечника, оказывается непреодо-
лимым препятствием для работы сердца. Протолкнув кровь до
некоторого предела вперед, сердце прекращает сокращаться, а за-
тем сокращения мускулатуры перикарда начинают проталкивать
кровь в обратном направлении, назад к кишке. Так происходит до
новой остановки через несколько минут и новой перемены тока
крови. Перед нами единстве1нный в своем роде, оригинальнейший
случай в животном царстве' — кровообращение собственно отсут-
ствует, заменяясь колебательными движениями нагнетания и оття-
гивания массы крови, с одной стороны, в жаберный отдел, а с дру-
гой — в кишечное сплетение пищеварительного аппарата.
У ланцетника (см. рис. 149) при редукции головного сердца, при
мощном' развитии жабер и мускулатуры и при сильном развитии
сосудистого сплетения в печеночном отделе кишечника процессы
кровеснабжения происходят иначе. На каждом сосуде, приносящем
кровь в каждую первичную жаб^явуф развивается мускули-
стый участок, так называемый bulbillus, сокращений которого дос-
таточно, чтобы продвинуть кровь через капилляры соответствую-
щей жабшрно^ щели. В итоге, вместо обычного единого сердца у
ланцетника оказывается 90 сердец слева и 90 сердец справа.
У позвоночных мы видим продолжение пути выработки единого
сердца, осуществленного у туникат. Но жабры у позвоночных не
достигают того крайнего развития, которое присуще асцидиям. На-
против, мускулатура стенки тела и капиллярная система печени и
кишок развиты мощно. В связи с этим позвоночные вырабатывают
такой тип единого сердца, который обеспечивает возможность пра-
вильного и сложного кровообращения путем дифференцировки в
сердце венозного синуса, предсердия, желудочка и артериального •
конуса. Кровь под нарастающим в них давлением прогоняется к
жаберным капиллярам, проталкивается через последние и цирку-
лирует по телу.'
Из приведенных фактов можно видеть, что головное сердце с Vх
редукцией головной лопасти заменяется в типе хордовых туловищ-
ным. Далее можно видеть, что ведущую роль в этой замене играет
выработка, аппарата жаберных щелей?‘и мощной боковой мускула-
туры, а также своеобразной печени. Наконец, в тесной связи с этим
происходит образование особого клеточного зачатка ангиобласта.
Дифференцировка перикардиального целома хордовых невольно
заставляет ставить вопрос, не могло ли произойти смещение голов-
ного перикарда птеробранхов и баланоглосса в туловище у хордо-
вых, обладающих туловищной хордой? Однако отсутствие единого
сердца у взрослого ланцетника при полном отсутствии даже соответ-
ствующих закладок в развитии говорит скорее за то, что появление
брюшного сердца связано с редукцией головной лопасти и ее
перикардиального целома. Но такое сердце налицо у туникат и
Позвоночных, обладающих первоначально малым числом жабер-
пых^ щелей. Это последнее обстоятельство позволило выработать
иной ток крови, чем первоначально имевшийся у низших червеоб-
301
разных форм. Tunicata являют собой живой образец возможности
периодических изменений тока и во взрослом состоянии.
В итоге -кровеносная система хордовых становится в своей эво-
люции высоко самостоятельным компонентом их организации.
В силу Сложности своих связей и функций она стоит в организации
позвоночных наряду с комплексом целома.
На зтом мы заканчиваем выявление того значения, которое
имеет развитие целома и кровеносной системы для создания типо-
вых черт хордовых. Многообразие функций целома, а в связи с
этим разнообразие эволюции ряда других тесно связанных с ним
систем органов находят свое отображение уже на ранних стадиях
развития хордовых в дифференцировке закладки целомической
мезодермы на миотомы, склеротомы, нефротомы, гонотомы и, нако-
нец, общую полость тела. В выработке сложной внутренней орга-
низации комплекс целома и связанных с ним мускулатуры, экскре-
торных и половых органов, как мы это видели, играет ведущую-
роль и у аннелид, моллюсков, членистоногих, и у иглокожих. Что
же касается кровеносной системы, то она с обособлением особой
своей закладки становится мало зависимой от целома, наряду с
ним ведущей в организации высших хордовых.
В анализе организации Chordata нам остается выявить еще один
характерный комплекс, развившийся в эволюции всего типа в зна-
чительной мере своим самостоятельным путем. Это — комплекс
органов чувств и нервной системы. Представленный у исходных
форм в высокой степени примитивно, он развивается далее чрезвы-
чайно оригинально и сложно, и у высших представителей позвоноч-
ных становится определяющим всю их организацию, а вместе с тем
их победу в борьбе за жизнь. Ему обязан «Homo sapiens» своим
именем. С этим комплексом связана возможность развития высшего
цикла в эволюции материи — цикла социальной жизни с прогрес-
сом человеческого общества, развитием человеческого мышления,
достижениями науки и техники и овладением природой.
У живущих в трубках Rhabdopleura и Cephalodisens, как и
вообще у сидячих форм с пассивным питанием, нет сложно разви-
тых органов взаимоотношения с окружающим миром, то же прихо-
дится сказать о живущем в грунте баланоглоссе. Однако пора-
жает мощное развитие нервного волокнистого слоя базально в по-
кровном эпителии головной лопасти у баланоглосса, что, конечно,
связано с высокой чувствительностью последней. Особенно сильно
развит этот слой, переходящий далее на воротник, в основании
хобота.
В туловищном отделе тела отношения меняются. У Pterobran-
chia медиодорзально в покрове воротника находится ганглий (см<
рис. 131), в который поступают нервные пучки спереди пз голов-
ного щита, с боков из щупалец и сзади из спинного покрова туло-
вища, а кроме того, пара глоточных коннективов проходит от ган-
глия к брюшной стороне, где они соединяются в продольный ствож
У баланоглосса (см. рис. 133) вся срединная спинная полоса по-
302
крова воротника с его эпителиальными, чувствительными и нерв-
ными элементами погружается под кроющий эпителий, образуя бо-
розду, отшнуровывающуюся в виде трубки. Остальной покров .во-
ротника превращается в мощное железистое образование. Таким
образом, создается спинной мозг хордовых, очень сильно отличаю-
щийся от брюшного мозга Vermes. Последний содержит только
нервные и глийные клетки, дифференцирующиеся в базальной части
покровного эпителия и затем от него обособляющиеся, тогда как у
хордовых целая полоса эпителия со всеми его элементами отде-
ляется в качестве зачатка мозга. Следовательно, уже у низших хор-
довых появляется, происходя от кроющего эпителия, нервный
центр с большим запасом нервных элементов и, сверх того, с чув-
ствительными и кроющими клетками. Это открывает широкие пер-
спективы дальнейшей эволюции, по сравнению с нервными центра-
ми большинства беспозвоночных.
Трудно решить, имеет ли нервная трубка воротника баланоглос-
са свое особое происхождение или связана с какими-либо образо-
ваниями червеобразных предков из Protocoelomata. Вполне естест-
венно было бы ожидать найти в головной лопасти Enteropneusta и
Pterobranchia нервный центр сензорно-ассоциативного характера,
как это имеет место в головном мозгу, присущем головной лопасти
немертин или аннелид. Однако ни в хоботе баланоглосса, ни в го-
ловном щите Rhabdopleura или Cephalodiscus не имеется ничего
подобного. Это понятно, так как комплекс фото-, хемо- и механо-
рецепторов головной лопасти червей у низших хордовых не разви-
вается, редуцируясь в связи с их образом жизни в трубках или
ходах грунта. Вместе с исчезновением эктодермальной глотки реду-
цируются также и центры стоматогастрической нервной системы.
Таким образом, здесь отсутствуют органы, порождающие надгло-
точный ганглий немертин и аннелид. Соответственно происходит
редукция этого центра или же он теряет свою обособленность, пре-
вращаясь в мощный нервный слой в эпителии основания хобота
баланоглосса. При таком понимании отношений следует думать, что
у высших хордовых, утративших головную лопасть, не должно сох-
раняться уже никаких следов головного мозга червей. Их голодной
мозг должен был возникнуть и эволюционировать далее на основе
спинной нервной закладки переднего отдела туловища, т. е. на ос-
нове ганглия Pterobranchia и затем нервной трубки в воротнике
баланоглосса.
Такое решение вопроса является наиболее приемлемым при на-
стоящем состоянии наших знаний относительно нервной системы
низших хордовых. Однако при своеобразных приспособительных
процессах, которые превращают головную лопасть червей или в
головной шит Pterobranchia, производящий защитную трубку, или в
хобот Enteropneusta, пробивающий ход в грунте, возможно предпо-
ложить, что произошло некоторое смещение назад, в воротник за-
датка головного мозга из специализированного органа, каким явля-
ется у них головная лопасть. В таком случае спинная нервная
303
трубка воротника баланоглосса представляла бы из себя участок
покрова с чувствительными и кроющими образованиями и с задат-
ком головного мозга червеобразных Protocoelomata, погрузив-
шийся в более защищенное положение.
Как бы ни представлять себе судьбы головного мозга червеоб-
разных протоцеломат, несомненным являются значительно большие
возможности развития переднего отдела спинного мозга, соответ-
ственно воротничкового мозга, у высших хордовых по сравнению с
головным мозгом червей или даже членистоногих.
Образование жаберных щелей ставит в определенные рамки рас-
положение продольных нервных стволов туловища. Становится воз-
можным только медиодорзальное и медиовентральное их положе-
ние, что мы действительно находим в туловище баланоглосса. Но
уже у ланцетника с развитием мускулатуры из спинных зачатков
миотомов редуцируется брюшной ствол, двигательный нервный
центр туловища, вместе с редукцией соответствующей мускулатуры
типичного кожно-мышечного мешка. Напротив, сохраняется и раз-
вивается спинной ствол, входящий в состав образующейся нервной
трубки спинного мозга, которая продолжается уже до заднего
конца тела. Тем самым увеличиваются эволюционные возможности
и этого отдела нервной системы.
Так, у ланцетника нервная трубка спинного мозга не только
является двигательной частью центральной нервной системы, но
содержит в себе почти по всей длине тела также фоторецепторы
чрезвычайно простого характера — так называемые «глазки Гес-
се», состоящие из одной ретинальной и одной прикрывающей ее
пигментной клетки. Подобные отношения вполне понятны, если
твердо усвоить, что спинной мозг хордовых включает в себя часть
спинного покрова с его чувствительными и прочими элементами,
а не только нервные образования.
Передний конец нервной трубки ланцетника (см. рис. 150) обра-
зует расширение, и этот отдел, несомненно, представляет первый
этап в развитии головного мозга хордовых. В нем, как мы видели,
возможно включен и сензорный центр головного мозга червеообраз-
ных предков. Во всяком случае он является исходным в развитии
сложнейшего головного мозга позвоночных. У ланцетника в рас-
ширении мозга находится и рудимент глаза в виде скопления пиг-
мента в передней стенке, статорецептор, в заднем вентральном
участке, и, наконец, хеморецептор в виде передней обонятельной
ямки.
У свободноплавающих личинок асцидий, в отличие от живущего
в грунте ланцетника, головной мозг (см. рис. 137), построенный в
общем сходно, содержит, однако, и хорошо развитый глаз и своеоб-
разно дифференцированный орган равновесия. Несомненно, что эта
часть нервной системы представляет из себя сложный сенсорный
центр. Однако при переходе во взрослое состояние сидячей асци-
дии, органы чувств ее головного мозга подвергаются гораздо боль-
шей редукции, чем у ланцетника. У последнего глаз в головном
304
мозгу рудиментарен, но зато по всей длине спинного мозга разви-
ваются «глазки Гессе», и функционирование фоторецепторов стано-
вится крайне оригинальным. А именно, для ланцетника, прикры-
того слоем песка, световые раздражения, попадая на туловище,
служат «сигналом бедствия», нарушения спокойного существова-
ния. И животное реагирует на них энергичными сокращениями
мускулатуры, проделывает молниеносные броски в воде и момен-
тально снова скрывается в грунте. Глазки существуют, чтобы не
видеть* Настоящий же глаз, располагающийся в том участке тела,
который постоянно направляет ток придонной воды в жаберный
отдел кишечника и естественно подвергается постоянному воз-
действию световых лучей, редуцируется, так как теряет свое
ориентирующее значение.
В противоположность сидячим асцидиеобразным исходным фор-
мам туникат или полусидячему живущему в песке ланцетнику,
позвоночные, как свободноживущие и активно питающиеся живот-
ные, вырабатывают сложнейший головной мозг со сложными орга-
нами чувств. Они возникают на основе головного комплекса чув-
ствительных образований и сензорного нервного центра, сходного
с имеющимся у Acrania и личинок асцидий. Уже у низших рыбооб-
разных позвоночных мозговой пузырь, соответственно трем важ-
нейшим рецепторам головы — органу обоняния, глазу и органу
равновесия, подразделяется на три отдела — прозенцефалон, мезен-
цефалон и метенцефалон. Очень сложный и совершенный глаз, в
соответствии с имеющимся у низших хордовых, закладывается в
своих основных частях за счет стенки среднего мозга. Орган обо-
няния и орган восприятия равновесия и колебательных движений
окружающей среды развиваются за счет впячиваний прилежащего
чувствительного эктодермального покрова.
Однако, по сравнению с головным мозгом исходных Protocoelo-
mata, строение головного мозга позвоночных осложняется тем, что
он представляет не только сенсорный центр, но в его состав вхо-
дят и двигательные компоненты, по крайней мере соответствующие
передним 5—6 миомерам. Перед нами процесс до известной степе-
ни сходный с образованием «сложного» мозга членистоногих.
За счет сензорных и двигательных компонентов головного мозга
позвоночных идет далее выработка сложнейших ассоциативных цен-
тров — мозжечка, среднего мозга ц полушарий переднего мозга.
При этом огромную- роль играет большое богатство клеточного
материала, вовлекающегося в построение мозга при образовании
нервной трубки. Напротив, у беспозвоночных осложнение идет уже
не за счет нервной закладки мозга, а в значительной мере со сто-
роны независимо закладывающихся органов чувств, как это имеет
место, например, у членистоногих в «lobi optici», в их экранных
Центрах, связанных с глазами.
Строение спинного мозга позвоночных, как двигательного отдела
нервной системы, подвергается значительной дифференцировке. Но
особенно осложнение его связано с образованием симпатической и
20 Пути эволюции животного мира 305
парасимпатической систем, вырабатывающихся за счет боковых
частей закладки спинного мозга и служащих для иннервации внут-
ренностей. Не будем вдаваться в дальнейшие подробности — в об-
щем картина ясна.
Итак, если* комплекс образований, связанный с пассивным пи-
танием, в значительной мере определил основные черты организа-
ции хордовых, то другой комплекс — комплекс целома, многообраз-
ный в своих функциональных и экологических связях и чрезвычайно
разнообразный в своих морфологических взаимоотношениях, вместе
с комплексом кровеносной системы создал сложную организацию
типа. Однако в конце концов, с переходом к активному питанию эта
организация получила свое завершение у позвоночных в развитии
ведущего комплекса органов чувств и нервной системы.
Сложная картина организации хордовых была кратко набро-
сана нами но не голой схемой, а в возможной полноте и связях ее
компонентов. Как можно видеть из всего приведенного, труднейшей
задачей морфологии хордовых является сведение организации
рыбообразных форм на организацию червеобразных. Взгляды раз-
личных исследователей в этом отношении сильно расходятся. Пра-
вильное решение, возможное лишь на основе выяснения взаимосвя-
зей организма и среды, требует анализа целостной организации
различных групп, в которых она характерно представлена. В этой
связи мы должны перейти к анализу тех этапов, через которые
шла эволюция хордовых. Звенья ее сохранились в подтипах
Pterobranchia, Enteropneusta, Acrania, Tunicata и Vertebrata. В из-
вестной мере их организация уже была раскрыта, когда создава-
лось общее представление о всем типе. Теперь наша задача —
дать целостную картину организации каждой из этих столь своеоб-
разных групп, конечно, сделав это в общих чертах и с неизбежным
повторением, но в другом освещении уже приведенного фактиче-
ского материала.
ПТЕРОБРАНХИ — PTEROBRANCHIA
Приспособления к защите определили всю внешнюю морфоло-
гию этого подтипа и, прежде всего, выработку защитных трубок
или домиков, в которых Cephalodisctis (рис. 125 и 126) в известной
мере свободен, а особи Rhabdopleura (рис. 127) связаны столона-
ми. Выставляясь своими жабрами-щупальцами из трубок в свобод-
ную воду, эти животные втягиваются под защиту трубок при
опасности извне. Для образования трубок служит головная ло-
пасть, которая приобретает своеобразную форму щита (рис. J28,
129). Брюшная ее поверхность представляет обширное железистое
поле, секрет которого при соответствующих движениях головной
лопасти и используется для построения трубки. Движения эти
становятся возможными в связи с образованием суженного отдела
головной лопасти, на котором как бы сидит щитовидный ее отдел.
В суженном отделе находится головная хорда, укрепляющая
головную лопасть на туловище, и мышечные пучки, двигающие
306
головной щиток. Необходимая для работы щитки/-плотность обес-
печивается дифференцировкой головного отдела целома. Таким
образом, оригинальное строение головной лопасти Pterobranchia
всецело определяется функцией образования трубки, и головной
Рис. 125. Cer'ha1o'ifscus dodecalophus вид сбоку (по Щепо-
тьеву). Обозначения см. на рис. 126
Щиток представляет оригинальнейший в своем роде строительный
орган.
Несомненно, выработка защитной трубки у птеробранхов связа-
на с пассивным питанием, и это нашло свое выражение в диффе-
ренцировке участка, следующего за головным щитком, в виде ворот-
ника, т. е. отдела, окаймляющего ротовое отверстие и несущего
выросты стенки тела — жабры. По ним, благодаря присутствию
мерцательного эпителия, пищевые частицы с током воды несутся в
кишечник. Во внутренней организации с этим связана дифференци-
ровка правого и левого мешков воротничкового целома, дающего
выросты в жабры, являющиеся для последних специальными опор-
ными образованиями (рис. 130).
21* 307
Если у Protocoelomata наличие головной лопасти легко позво-
ляет представить себе использование ее как железистого образова-
ния для построения трубки защитного характера, то не менее
естественно, как мы уже указывали, мерцательные дифференци-
ровки, имеющиеся в заднем отделе головы на границе с тулови-
щем — аврикулярные и затылочные органы турбеллярий, немертин,
Рис. 126. Часть трубки: А — вид снару-
жи; Б — поперечный разрез через трубку:
В — тонкое строение стенки (по Щепотье-
ву):
1 — животное в полости трубки (2), 3 — пере-
городка, 4 — отверстие, ведущее внутрь трубки,
5 — головной щиток, б — его загнутая предрото-
вая часть, 7 — воротник, 8— щупальца-жабры
и их ветвтение (9), 10 — экскреторная пора,
11 —жаберная щель, 12 — стебелек, 13 — присо-
ска, 14 — две почкп
Рис. 127. Phabdopleura
пог muni, вид сбоку
(по Щепотьеву): •
1 — щупальца-жабры, 2 — голов-
ной щиток, 3 — воротник, 4 — сте-
белек, 5 — туловище, 6 — трубка
Внизу вид колонии
аннелид — принять за отправной пункт в развитии мерцательного
аппарата, несущего пищевые частицы с током воды ко рту при
образовании трубки.
За воротничковым отделом у Pterobranchia следует мешковид-
ное туловище, продолжающееся вентрально в ножку или стебелек.
Сокращение последнего обусловливает втягивание животного в за-
щитную трубку. Понимание значения этого отдела теснейшим обра-
зом связано с фактом перемещения заднепроходного отверстия к
308
переднему концу тела. Экологически смысл подобного явления
сводится к тому, что трубка, в которой живет животное, не загряз-
няется экскрементами. Они выбрасываются из кишечника непосред-
ственно в окружающую воду, вода в трубке свободна от них. У сер-
пулид, из трубкоживущих полихет, для этой цели служит мерца-
Рис. 128. Схема строения Cephalodis-
€its (по Щепотьеву). Обозначения см
на рис. 129
Рис. 129. Передняя часть
тела Rhabdopteura (по Ще-
потьеву) :
1 — головной щиток, 2 — рот,
3 — воротник и его целом, 4—
жаберная щель, 5 — глотка,
о — кишка, 7 — яичник, 8 —
анальное отверстие, 9 — щу-
пальца-жабры. 10 — ганглий
воротника, 11 — перикард, 12-
хорда, 13 — стебелек, 14 — поч-
ки, 15 — желудок, 16 — экскре-
торный канал, 17 — головная
пора
тельная борозда, идущая медиовентрально от ануса вперед и здесь
переходящая на спинную сторону, таким образом, экскременты
уносятся из трубки. У Phoronis этот эффект достигается так же,
как у Pterobranchia. А. Ланг в свое'время и объединил по Этому
признаку Phoronoidea с мшанками и брахиоподами в тип Piosopy-
gia.
Здесь мы не будем вдаваться в детали, укажем лишь, что в
развитии Phoronis с чрезвычайной ясностью выявляется основной
момент происхождения прозопигии (проз—вперед, лиги — зад). Он
заключается в том, что процессы неравномерного роста во время
постэмбрионального развития обусловливают чрезвычайное разра-
стание брюшной стороны тела по сравнению со спинной. Благодаря
309
этому анальное отверстие оказывается очень близко к ротовому а
кишечник образует полную петлю, расположенную в червеобразно'
туловище. Последнее представляет в известной мере не что иное
как своеобразно разросшуюся брюшную сторону тела. У Pleroh'
ranchia неравномерный рост брюшной стороны выражается в дна.
ференцировке стебелька, т. е. обособляется сократительный участок
туловища, в который не заходит уже петля кишки, а который
слу-
жит для прикрепления к стенке трубки
и для втягивания тела под защиту по-
следней.
У Cephalodiscus намечается жабер-
ный отдел, следующий за во-
3
8
4
?
Рис. 131. Схема нервной си-
стемы Rh2b <о (по Ще-
потьеву):
I — головной щиток, 2 — рот, 3 —
целом головы, 4 — целом воротни-
ка, 5 — целом тутовища, 6 —
хорда, 7 мозговой ганглий, 8 —
передний нерв из головного щитка,
9 — нерв щупалец-жабер, 10 —
заднпй спинной неов, 11 — около-
глоточный коннектив, 12 — брюш-
ной нерв, 13 — глотка
Рис. 130. Целом воротника Rhabdopleura.
Схема (по Щепогьеву):
1 — целом в щупальцах, 2 — целом воротника.
3 — мезентерий, отделяющий правый мешок от
левого, 4 — целомодукт, 5 — рот
ротником и имеющий единственную пару жаберных щелей спе-
циально для оттока из кишечника воды, несущей пищевые частицы.
Далее выявляется и отдел, содержащий гонады, выводные протоки
которых открываются по бокам дорзально. У Rhabdopleura жабер-
ных щелей нет, жаберный отдел не дифференцирован. Это, конеч-
но, более примитивное состояние.
Во внутреннем строении птеробранхов уже имеется вторичное
упрощение строения кишечника, выражающееся в редукции стомо-
деального отдела, а также редукция органов чувствительного ком-
плекса головы (рис. 131). Наличие двух мешков целома в туловище
и пары протонефридиев, снабженных соленоцитами, в области жа-
берных отверстий Cephalodiscus, сохранение спинного и брюшного
сосудов в кровеносной системе, наличие пары мешетчатых гонад-—
все это приходится отнести к примитивным типовым чертам Proto-
coelomata. Совершенно своеобразное развитие хорды и сердца с
310
перикардом в головном отделе и спинного ганглия в воротничковом
деле являются характерными чертами низших хо'рдовых, облада-
ющих головной хордой, как мы это уже видели при общем рас-
смотрении типа.
Краткий анализ организации Pterobranchia позволяет видеть,
насколько она является уже приспособительной как в отношении
зашиты — сидячий образ жизни в трубках, так и в отношении
питания—пассивное питание/ Трудно на основании этих данных сде-
лать определенные выводы об организации исходных форм хордо-
вых, поскольку нет критерия для определения, где начинаются спе-
цифические черты Pterobranchia и где кончаются типовые черты
хордовых. Эти вопросы можно решить лишь путем анализа органи-
зации других групп низших Chordata.
КИШЕЧНОДЫШАЩИЕ — ENTEROPNEUSTA
Защита баланоглосса обеспечивается его жизнью в грунте.
С этим же связано и его питание. В качестве пищевого материала
используются органические частицы и
мелкие организмы верхних слоев грунта,
захватываемые вместе с частицами ила
и песка. Как уже было отмечено выше,
перед нами своеобразное пассивное пита-
ние, при котором ток воды, вызываемый
мерцательным эпителием головной лопа-
сти, связан с пассивным захватом пище-
вых частиц. Активный выбор пищевого
материала отсутствует. Конечно, и про-
никновение в грунт и использование его
ресурсов при питании развивались одно-
временно и в теснейшей связи.
Этим всецело обусловлена внешняя
морфология подтипа (рис. 132 и 133).
Головная лопасть становится мускули-
стым придатком, пробивающим ход в грун-
те. Ее кроющий эпителий, являясь мер-
цательным, обладает в то же время зна-
чительной плотностью, очень богат сли-
зистыми железками и содержит базально
хорошо выраженный слой нервно-волок-
нистого характера. Плотность головной
лопасти обусловлена мощным развитием
мускулатуры — кольцевой и, особенно,
продольной, для которой в качестве опоры служит центрально и
базально расположенный целом. Прочность прикрепления всего
«хобота» связана с образованием головной хорды, являющейся
как бы передним придатком начала кишки. Таким образом, ряд
своеобразных приспособлений к продвижению в грунте превраща-
311
Рис. 132. Balanoglossus
kovjiei'skii (по Бэтсону):
1 — хобот, 2 — воротник, 3 —
жаберные щели, 4 — область
гонад, 5 — рот, 6 — анальное
отверстие
ют головную лопасть в образование крайне узко приопособител
ного характера. К этому следует прибавить еще редукцию голов
ного комплекса органов
чувств на хоботе, теряю,
щих свое значение в по-
добных условиях. А с
этим связана и редукция
сенсорного центра — го,
ловного мозга червей, в
хоботе становится веду-
щей первоначально мало
значащая механическая
функция, а основная —
сенсорная — отходит на
задний план, сохраняясь
только в виде общей чув-
ствительности «хоботка».
В связи с проникнове-
нием в грунт, не менее
своеобразно изменяется и
передний отдел тулови-
ща — воротник. Образо-
вавшись
как отдел,
пальца, по
ток воды
пассивном
ротник у
10
17i
щу-
шел
при
во-
первоначально
несший
которым
ко рту
питании,
живущего в
грунте баланоглосса теря-
ет в этих условиях щу-
пальца. Однако правый и
левый мешки целома во-
ротника, служившие для
поддерживания щупалец,
сохраняются, а покров-
ный эпителий над ними
превращается в желези-
стый орган массового вы-
деления слизи, защищаю-
щей нежное туловище от
?,1еханических
ний грубыми частицами
грунта и способствующей
образованию хода в пос-
леднем.
Одновременно протекает и другой процесс. В воротнике проис-
ходит погружение под покров, т. е. в более защищенное положение,
срединного спинного участка покровного эпителия, содержащего
^20 21 22
Рис. 133. Схема строения передней части тела
баланоглосса (по Деляжу и Эруару):
1 — хобот, 2 — продольные и 3 — кольцевые его
мышечные слои, 4 — головной целом. 5 — перикар-
диальный его отдел, 6 — спинной кровеносный синус
в сердце, 7 — головная хорда, 8 — скелетный щи-
ток в основании хобота с выростами, охватывающи-
ми глотку, 9 — рот, 10 —целом воротника (11), 12 —ту-
ловище и 13 — его целом, 14 — глоточный отдел, 15 —
нервная спинная трубка, 16 — лежащий вбок от нее
вырост туловищного целома, 17 — целомодукт ворот-
ничкового целома, 18 — жаберная перегородка, 19 —
жаберная пора, 20 — жаберный мешок, 21 — язычки
и 22 — синаптикулы жаберных щелей
поврежде-
312
х^увствительные клетки и особенно богатого нервными элементами.
Так возникает характерная черта хордовых --зачаток спинной
нервной трубки.
Непосредственно .под нервной трубкой, прилегая с боков к про-
ходящему здесь спинному сосуду, в воротник проникают два выро-
ста правого и левого мешков туловищного целома. Просвет этих
выростов почти исчезает с образованием за их счет пары мощных
продольных мышечных пучков. Спереди они заканчиваются у осно-
вания головной хорды и, очевидно, функционально с ней связаны.
Итак: хорда в голов-
ной лопасти и мышеч-
ные пучки, идущие к
ней, справа и слева в
воротнике. Таким об-
разом, уже здесь,-прав-
да, еще в совершенно
своеобразной форме,
возникают элементы
Аыохордального ком-
плекса, характерного
для высших хордовых.
Узким приспособлени-
ем кажется и выработ-
ка в основании хобота
щитовидного хряща с
двумя боковыми отро-
стками, проходящими в
воротнике и служащи-
ми для поддерживания
в зияющем состоянии
глотки, что необходимо
при питании балано-
глосса.
Следующий за во-
ротником отдел тела,
Рис. 134. Поперечный разрез через бал аноглосса:
/ — в области гонад, II—в жаберной области
Схематизировано (по Лейкарту);
1 — покров, 2 — слой продольных мышц, 3 — полость
целома, 4 — кишка, 5 — жаберная щель, G — ее язы-
чок, 7 — жаберный мешок, 8 — жаберная пора, 9 —
брюшной и 10 — спинной нервные стволы. И — брюшной
и 12 — спинной кровеносный сосуд, 13 — гонада, 11 —
ее выводной проток
не обнаруживая никакой метамерности внешней и внутренней, пред-
ставляет собой типичное туловище червя наподобие немертинооб-
разных форм. Чего-либо подобного хотя бы самым примитивным
конечностям здесь нет. В стенке тела (рис. 134) слои продольных и
кольцевых мышц расположены по всей периферии непрерывно, а
задний целом идет через все туловище в виде двух мешков, разде-
ленных только спинным и брюшным мезентериями и в середине
прямой трубкой кишечника. Движение тела обусловлено перисталь-
тическими сокращениями стенки тела с опорой на жидкость це-
лома.
Примитивность отношений выражается и в развитии ткани ос-
новного вещества, что, в свою очередь, определяет развитие крове-
носной системы. Образование двух мешков целома ставит в опре-
313
деленные рамки развитие ткани основного вещества, пограничные
пластинки которой располагаются под покровным эпителием и под
эпителием кишечника. Здесь образуются сосудистые сплетения
соединяемые продольными сосудами коллекторами — брюшным и
спинным. Эти сосуды проходят в соответствующих мезентериях, где
под целотелием имеются пластинки ткани основного вещества
соединяющие друг с другом субэпителиальные ее перепонки. В ка-
честве интимы везде в сосудах оказывается только уплотненный
слой того же основного вещества.
Таким образом, примитивное состояние кровеносной системы
немертинообразных форм, не обнаруживающее ближайшей -связи со
стенкой кишечника, с развитием целома переходит у всех целомат
в более дифференцированное состояние с обособлением кишечного
сплетения от системы, питающей стенку тела. Новые коллекторы
вырабатываются там, где одна система переходит в другую — брю-
шной и спинной сосуды в соответствующих мезентериях.
Однако уже у низших хордовых отношения значительно ослож-
няются. Так создаются, в связи с проникновением целома в голов-
ную лопасть и с укреплением ее посредством развивающейся хорды,
крайне оригинальные взаимоотношения между кровеносной лаку-
ной, заканчивающей спинной сосуд, твердым тяжем хорды с брюш-
ной стороны от нее и мускулистой стенкой прилежащего мешка
целома со спинной стороны. Создается пропульзаторный аппарат
сердца, крайне характерный для низших хордовых — Pterobran-
chia и Enteropneusta — «головное сердце».
Завершая рассмотрение внешней морфологии баланоглосса,
остается сказать, что дальнейшая дифференцировка туловища этого
животного, как и у птеробранхов, связана с обособлением средней
части, содержащей примитивные мешетчатые гонады. Но еще в
большей степени дифференцировка туловища стоит в зависимости
от характерных для хордовых дифференцировок кишечника, не вы-
раженных или только намеченных у Rhabdopleura и Cephalodiscus.
А именно: за воротником обособляется область со многими парами
жаберных щелей, развитых очень типично для хордовых, а не так
примитивно в виде простых отверстий, как у Cephalodiscus, Далее
идет половой отдел, но одновременно это — отдел с дифференци-
ровкой печеночных складок в стенке кишки. И та и другая черта
организации, в особенности первая, позволяют считать баланоглосса
типичным представителем Chordata.
Однако дифференцировка печеночного отдела крайне примитив-
на. Он характеризуется только скоплением специфических желези-
стых клеток на небольшом протяжении хода кишечной трубки. Зато
жаберные щели закладываются в большом количестве, каждая
подразделяется на вторичные щели язычком, идущим со спинной
стороны, и благодаря развитию поперечных перемычек оинаптикул
весь аппарат становится более сложным. Для поддержки его и для
сохранения в зияющем состоянии вырабатывается своеобразная
«аркадная система» — система утолщенных, хрящеобразных участ-
314
ков в субэпителиальной перепонке ткани основного вещества
кишечной стенки. Это коренным образом отличается от «жабер-
ных» щелей Cephalodiscus, но, как уже было указано, чрезвычайно
сходно с наблюдающимся у ланцетника. При сложности дифферен-
цировки системы не вызывают сомнений связи родства между
баланоглоссом и ланцетником. И установление этих связей чрезвы-
чайно важно для понимания отношений среди типа хордовых.
Жаберные щели баланоглосса представляют уже типичную систему
газового обмена, обладающую массой кровеносных капилляров. Это
не просто система оттока воды при пассивном питании, и не прихо-
дится сомневаться в ее широко приспособительном значении. Но не-
сомненно и то, что подобная система не могла развиться в усло-
виях роющего образа жизни баланоглосса, а является наследием
предков, фильтровавших воду при пассивном питании, ибо только '
•в этом случае возможно превращение ее в систему газового обмена.
В заключение нашей характеристики баланоглосса скажем еще
несколько слов о чрезвычайно простом характере нервной системы
туловища. Ее центральный отдел образован двумя нервными ство-
лами — спинным и брюшным, представляющими лишь утолщение
базиэпителиального нервного слоя покрова, характерного для
Enteropneusta. Трудно сравнивать ее с брюшной цепочкой кольча-
тых червей или с боковыми стволами немертин. Скорее это местная
дифференцировка нервного сплетения покрова, служащая также
для иннервации равномерно развитой мускулатуры кожно-мышеч-
ного мешка туловища малоподвижного животного. Однако разви-
тие спинного ствола несомненно связано с обособлением дорзально
нервной трубки воротника. Перед нами совершенно особый путь V
эволюции нервной системы из состояния ее у немертинообразных
протоцелломат.
Теперь мы уже в известной мере можем сделать выводы об
исходных формах Chordata и попытаться из сопоставления Ptero-
branchia и Enteropneusta вывести первичные черты организации
хордовых.
Конечно, ни Pterobranchia, с их очень далеко ушедшим приспо-
соблением к жизни в трубках, что повело даже к выработке беспо-
лого размножения путем почкования, ни Enteropneusta, с их
жизнью в грунте и своеобразным питанием, не могут быть постав-
лены в ряд предков хордовых. Все это очень узко приспособленные
формы, являющиеся боковыми ветвями тех или иных этапов в раз-
витии типа хордовых.
Однако может быть поставлен вопрос, чем, собственно, доказы-
вается, что рассмотренные два подтипа находятся в родственных от-
ношениях друг к другу? Первое и очень существенное это то, что и
не и другие представляют олигомерные формы с тремя парами
Целомических мешков (головной, воротничковый и туловищный це-
ломы). При этом передние две пары обладают целомодуктами,
н задняя пара лишена последних. За счет одного из головных
мешков образуется перикард. Второе это то, что оба подтипа могут
31S
быть объединены, как обладающие «головной» хордой. Но этого
мало. На основе хорды создается сложный комплекс: хорда, крове-
носный сосуд'и перикард, т. е. аппарат сердца, неповторимый в жи-
вотном царстве. Третье — кишечник у обоих является чисто экто-
дермальным, закладка стомодеума отсутствует. Четвертое — в обо-
их подтипах имеются сходно расположенные мешетчатые гонады,
вдающиеся в целом. Пятое — в воротничке дорзально дифференци-
руется нервный центр, тогда как головная лопасть не имеет ничего
подобного сенсорному центру головного мозга червей. Шестое — в
начале туловищного отдела кишечника образуются отверстия жа-
берных^ щелей, ведущие из него наружу. Последняя черта органи-
зации может возбудить некоторые сомнения относительно гомоло-
гии рассматриваемых образований. С одной стороны, у Rhabdo-
pleura видим отсутствие подобных отверстий, za у Cephalodiscus их
только одна пара, но без функции газообмена, с другой — наблю-
даем большую сложность жаберного аппарата баланоглосса. Во
всяком случае даже аналогия их дает возможность понять, как мог
возникнуть столь характерный для хордовых аппарат жаберных
шел ей.
Однако всех прочих сходных черт вполне достаточно, чтобы при-
знать ближайшие филогенетические отношения Pterobranchia и
hnteropneusta. В этом смысле особенно показателен сложный ком-
плекс образований, дающий сердце. Но трудно поддается понима-
нию прозопигия птеробранхов при полном ее отсутствии у балано-
глосса. Приходится допустить вторичное перемещение у них аналь-
ного отверстия при преобразовании заднего отдела туловища в виде
стебелька.
На этой основе исходную форму можно представить себе как
червеобразного представителя Protocoelomata, обладавшего уже
олигомерным целомом. У него, в соответствии с подразделением
тела на головную лопасть, воротничок и туловище, имелись три
пары мешков целома. Трехмерность целома была обусловлена
обособлением воротничкового целома, как аппарата, поддерживаю-
щего придатки около рта, вызывавшие мерцательным движением
своего эпителия ток воды, несший пищевые частицы в кишечник.
Такое пассивное питание отразилось и на кишечнике, обусловив
утрату эктодермальной глотки. Стенка тела была представлена ти-
пичным кожно-мышечным мешком с равномерным развитием коль-
цевой и продольной мускулатуры. Каких-либо зачатков конечностей
совершенно не существовало, и движение тела было ограничено
его перистальтическими сокращениями. Головной и воротничковый
целомы обусловливали, прежде всего, плотность соответствующих
отделов тела, и для регуляции ее служили соответственно две
пары целомодуктов. Задний целом, охватывавший усваивающий
отдел кишечника, имел и трофическую функцию, в связи с чем не
обладал целомодуктами. В него вдавались гонады типа мешетча-
тых гонад, но половой функции целом не получил. Половые про-
дукты скоплялись в полости гонад, откуда выводились через прими-
316
тив'Ные протоки яйцеводов и семеводов. В средине туловища диф-
ференцировался половой отдел тела, содержавший гонады. Выра-
ботка целома и целомической мускулатуры была связана с образо-
ванием системы пограничных образований в виде пластинок ткани
основного вещества, что обусловило появление особого типа крове-
носной системы со сплетениями сосудов в стенке кишки и в стенке
тела и объединяющими их коллекторами — спинным и брюшным
сосудами в мезентериях. Мерцательный покров был богат желези-
стыми и чувствительными клетками, но высоко развитые органы
чувств отсутствовали. Однако имелась общая высокая чувствитель-
ность головной лопасти и поверхности щупалец в области воротника.
В связи с этим здесь дорзально дифференцировался сенсорный нерв-
ны!! центр. В головной лопасти такая дифференцировка отсутство-
вала, по всей вероятности, в результате редукции головного мозга
червей при редукции органов чувств их головного комплекса. Туло-
вищная нервная система дифференцировалась на месте в виде при-
митивных брюшного и спинного продольных нервных стволов.
Наша картина была бы не полна, если бы в нее не был внесен
еще один штрих, хотя и очень специального характера. Для укре-
пления основания головной лопасти выработался плотный придаток
кишечника — «головная хорда», а затем за счет ее и прилежащего
к ней спинного кровеносного сосуда и в известной мере охватываю-
щего последний дорзально участка головного целома развилось
«головное сердце». Такой комплекс образований, чрезвычайн©
характерный для птеробранхов и баланогласса («Hemichorda») и в
то же время сложный, конечно, должен был развиться еще у общей
исходной формы хордовых. Его дальнейшие судьбы мы освети?л
впоследствии.
Таким образом, выявляется существенный факт особой специа-
лизации головной лопасти низших хордовых, чего у немертинообраз-
ных Profocoelomata не было. Эта специализация не могла не быть
связанной с важнейшей категорией условий существования — пас-
сивным питанием первичных Chordata. Оно вызвало образование
околоротовых придатков воротника и дифференцировку воротнич-
кового целома. Головная лопасть оказалась при пассивном питании
утратившей свою основную роль носителя головного комплекса
органов чувств. На первый план выступила ее механическая функ-
ция при движении у баланоглосса и железистая и механическая
при построении защитной трубки у Pterobranchia. При этом, в свя-
зи с механической функцией существенное значение приобретает
укрепление головной лопасти, для которого и дифференцируется
головная хорда.
Жаберные щели еще отсутствовали у первичных Chordata, не
процесс редукции стомодеума уже подготовил условия для тока
воды, приведшие к образованию пары боковых отверстий у Cepha-
lodiscus. Однако лишь на дальнейшем этапе образование жаберных
щелей, как сложного аппарата для движения воды при пассивном
питании, а равно и для газообмена, стало совершившимся фактом.
317
В этом отношении показательным является сложное строение жа-
берного аппарата баланоглосса, может быть, и возникшее совер-
шенно независимо от пар боковых отверстий птеробранхов.
Таким образом, первым этапОлМ в эволюции хордовых было
состояние птеробранхиоподобных форм, но современные Rhabdo-
pleura и Cephalodiscus очень далеко ушли от него, ставши настоя-
щими, живущими в трубках, сидячими формами, обладающими
прозопигией. Мы видели, как главнейшие категории условий суще-
ствования — защита и питание —• определили всю их внешнюю
морфологию и внутреннее строение, уменьшение размеров и харак-
терное бесполое размножение путем почкования. На дальнейшем
этапе эволюции хордовых, который характеризовался уже вырабо-
танным жаберным аппаратом, отделились Enteropneusta, современ-
ные представители которых далеко ушли по пути приспособления
к жизни в грунте.
Мы разобрали, как этот биотип, можно сказать, создал ориги-
нальнейшую внешнюю морфологию и в значительной мере внут-
реннее строение баланоглосса, а также и его способ питания с по-
путным заглатыванием грунта.
Как же мы должны теперь представлять гипотетических пред-
ков хордовых, давших начало Pterobranchia, а затем и Entero-
pneusta? ~
Безусловно, это были первичные целомические червеобразные
формы, по строению и образу жизни приближавшиеся к немерти-
нообразным червям, как нами было выявлено выше и для предков
иглокожих. Но головная лопасть их уже специализирована как при-
даток, позволявший продвигаться в грунте, образуя в известной
мере постоянные ходы, благодаря выделению слизистого секрета.
Небольшой червеобразный представитель Prochordata — так назо-
вем этих примитивных хордовых — жил, таким образом, в постоян-
ном ходе, проделанном им в грунте. Это было сопряжено с выра-
боткой пассивного питания благодаря развитию мерцательного ап-
парата по краю рта у основания головной лопасти. Дальнейшее
его осложнение выражалось в образовании щупалец и обособлен-
ного воротника, в связи с обособлением отдела целома, опорного
для щупалец. Жизнь в грунте и пассивное питание вели при этом
к редукции сложных хемо- и фоторецепторов, а затем и сенсорных
центров головной лопасти.
Вместе с образованием защитной трубки стал возможен пере-
ход в другие биотопы. Так возникли первоначальные Pterobran-
chia с их головным щитком, щупальцами воротника, а впоследствии
с прозопигией и дифференцировкой стебелька. Прямая линия эво-
люции прохордат, с развитием аппарата жаберных щелей, пошла,
однако, дальше по пути большего совершенствования пассивного
питания. Enteropneusta являются представителями этого направле-
ния в эволюции, но у них и защита и питание всецело обусловлены
существованием червя в песчаном и илистом грунте. Самый грунт в
известной мере может использоваться при-пропускании через кишеч-
318
ник для пассивного захвата пищевых частиц, а _головная лопасть
превращается в таран для пробивания хода в Грунте. При разви-
тии мерцательного эпителия в аппарате жаберных щелей, заменяю-
щего околоротовые мерцательные поверхности щупалец, последние
редуцируются. Напротив, мощно развиваются жаберные щели. Та-
ким образом, баланоглосс превратился в исключительного обитателя
грунта, и его организация всецело отражает это. .
Однако возможности иного использования выработанных пер-
спективных черт этим не были исчерпаны, а повели к возникнове-
нию более сложных форм Chordata, обладающих епинной хордой.
ОБОЛОЧНИКИ — TUNICATA
Оболочники представляют среди хордовых группу, исключитель-
ную по разнообразию и необычности входящих в нее форм. Наря-
ду с сидячими, донными асцидиями имеются и носящиеся во власти
морских течений планктонные колонии сальп и Doliolum и, наконец,
подвижные аппендикулярны в -их маленьких домиках.
Анализ организации туникат вполне естественно раскрывает
картину их строения только при правильном филогенетическом под-
ходе, т. е. при правильном выборе той группы, в условиях суще-
ствования которой развились характерные черты подтипа. Поэтому
обратимся к организации асцидиеобразных форм, из которой уже
достаточно просто выводятся особые, связанные с упрощением и
узкой приспособленностью, черты сальп и Doliolum.
Ведущим моментом в развитии организации асцидий (рис. 135)
является прикрепленный, сидячий образ жизни взрослого животного.
Прежде всего, с таким образом жизни связано образование ха-
рактерной для всего подтипа мощной защитной оболочки — туники,
гарантирующей сохранение животного, не имеющего возможности
активно избежать врага. Туника представляет собой особого рода
кутикулу, образующуюся на однослойном покровном эпителии и
содержащую — единственный случай в животном царстве — клет-
чатку, «туницин». Эта кутикула отличается чрезвычайной сложно-
стью, так как из нижележащих эпителиального и соединительно-
тканного слоев в нее продвигаются разнообразные эпителиоидные и
амебоидные элементы, в ней дифференцируются волокнистые струк-
туры, иногда отлагаются известковые соли и зерна пигмента, в нее
нередко проникают кровеносные сосуды, а на поверхности обра-
зуются выросты в виде шипов, щетинок и т. п. Здесь может даже
за счет амебоидных клеток вторично получаться подобие слоя крою-
щего эпителия. Одним словом, кутикула превращается как бы в мощ-
ный слой плотной соединительной ткани сложного строения
(рис. 136).
С -выработкой туники возможность надежной защиты обеспе-
чивается так основательно, что при общей пластичности организа-
ции для туникат становится нетрудным проникновение и освоение
иных биотопов и биоценозов, чем исходные. При этом, на основе
319
уже выработанного жаберного аппарата предков, имевших голов-
ную хорду, происходит использование в новых условиях возможно-
стей пассивного питания. Жаберный аппарат достигает у асцидий
крайней сложности, занимая большую часть тела животного, тогда
как все прочие органы оттесняются в небольшой задний отдел,
Рис. 135. А —внешний вид асцидии Ciona intestine 'is; Б — схема ее внут-
реннего строения (из Лейкарта);
I — ротовой сифон, 2 — клоакальный сифон, 3 — просвечивающие через тунику мышечные
пучки, 4 — нервный ганглий, 5 — жаберный мешок, 6 — эндостиль, 7 — клоакальная
полость, 8 — половые выводные протоки, 9 — анальное отверстие, 10 — задняя кишка, 11 —
пищевод, 12 — желудок, 13 — кишка, 14 — перикард, 15 — брюшной сосуд, 16 — половые
железы
образуя так называемый nucleus. Таким образом важнейшее в
жизни — защита и питание — надежно обеспечены. Животное за-
щищено своей туникой так, что никакие нападения не страшны и в
то же время постоянный приток пищи вполне достаточен для пол-
ного обеспечения всех жизненных процессов. В борьбе за жизнь
асцидии завоевали себе прочное положение. .В море нередко можно
встретить большие их скопления на небольших площадях.
320
Конечно, их организация узко приспособите^:.-.а, широких гори-
зонтов она не открывает, но поразительно, сколько возможностей
нашел в ней естественный отбор для дальнейшей эволюции.
Сидячее существование, вызвав вме-
сто мерцательного покрова прохордат
образование туники, одновременно обус-
ловило и всю внешнюю морфологию ас-
Рис. 136. А — поперечный разрез че-
рез тунику асцидии Ascidiella asper-
se, при малом увеличении (по К.
Сент-Илеру). В основном веществе
туцики видны мелкие амебоидные
клетки (/) и капсулы, содержащие
клетки другого характера (2)
Б — клетки туники
цидий, приспособительный характер кото-
рой особенно хорбшо виден в их онтогене-
зе. Свободноживущая личинка асцидий
(рис. 137) прикрепляется к подводным
предметам своим приротовым брюшным
участком, несущим сосочки и занимаю-
щим соответственно апикальное положе-
ние. В связи с этим у личинки рот переме-
щен на спинную сторону, причем головной
лопасти не остается места. Она без еле-
21 Пути эволюции животного мира
Рис. 137. Строение личинки
асцидии в момент прикреп-
ления (по А. Ковалевскому):
1 — папиллы, служащие для
прикрепления, 2 — эндостиль в
переднем отделе эктодермальной
глотки, 3 — рот, 4 — переднее
расширение нервной трубки (12),
5 — статорецептор, 6 — глаз,
7 — закладка околожаберной
полости, 8 — перикард, 9 —
вторичный вырост глотки, даю-
щий кишку, 10 — мезодермаль-
ная закладка, 11—эктодермаль-
ный тяж в хвосте, 12 — нерв-
ная трубка, 13 — хорда
321
да исчезает. Далее начинается радикальная перестройка организа-
ции (рис. 138). Подвергается гистолизу хвостовой отдел личинки,
и в то же время ротовое отверстие, вследствие процессов неравно’
мерного роста участка между ртом и местом прикрепления, выно
Д
Рис. 138. А, Б, В — схема
чинки
последовательных изменений прикрепившейся ли-
асцидии (из Деляжа и Эруара);
I — папиллы, служащие для прикрепления, 2 — эндостиль в жаберном отделе кишечника,
3 — рот, 4 — переднее расширение нервной трубки со ста то рецептор ом (5) и глазом (6),
7 — околожаберная полость, 8 — перикард, 9 — желудок, 10 — жаберные щели, 11 — руди-
мент кишки, 12 — нервная трубка, 13 — хорда, 14 — дупликатура покрова, 15 — аналь-
ное отверстие, 16 — рудимент хвоста
сится на пункт, противоположный месту прикрепления, т. е. в по-
ложение, наиболее выгодное для тока воды через рот при пассив-
ном питании сидячего животного. При этом участок, несущий рот,
при отсутствии головной лопасти вытягивается в трубку ротового
сифона. Возле жаберного отдела тела личинки со спинной стороны
образуется впячивание, разрастающееся далее в виде клоакального
322
выводного сифона. Асцидия принимает вид мешка, прикрепленного
одним концом, а на противоположном — несущего два сифона.
Только в утонченных краях сифонов развиваются мышечные волок-
на, только здесь возможен эффект от Сокращения последних в
связи с тем, что здесь туника является значительно более тонкой,
чем на прочих участках мешковидного тела, где она плотна и не
поддается деформации. При сокращении мышц сокращаются сифо-
ны, и внутренние образования совершенно изолируются от воздей-
ствий внешней среды — предельная степень защиты для сидячего
животного. Мешок асцидии делается неуязвимым.
В этих условиях мускулатура стенки тела, при плотности туники
становясь вне функции, обычно в той или иной степени подвер-
гается редукции.
Комплекс черт организации асцидий^ связанный с их сидячим
образом жизни и выработкой туники, естественно- пополняется
редукцией органов чувств, имевшихся у свободноживущей личинки.
Сохраняются лишь чувствительные клетки по краю ротового сифона
и на усиках и ямка хеморецептора, расположенная в сифоне на
спинной стороне. Соответственно здесь, между сифонами, находится
нервный ганглий, представляющий единственный нервный центр
асцидий и являющийся остатком переднего отдела спинной нервной
трубки личинки.
Другой ведущий комплекс черт организации асцидий связан с их
пассивным питанием. Об этом уже говорилось выше, но в другой
связи, а потому считаем желательным повторить сказанное. Жабер-
ный отдел кишечника, вследствие разрастания и вторичного увели-
чения числа жаберных щелей, имевшихся первоначально в неболь-
шом количестве,. становится настолько объемистым, что занимает
большую часть тела. При этом строение его чрезвычайно ослож-
няется. Благодаря появлению в жаберных щелях перемычек и
выростов стенки, которые срастаются друг с другом в сложные
сети, образуется своеобразнейшее жаберное сито. Большая слож-
ность и разнообразие получающихся структур дают в руки система-
тика много вариантов, на которых и строится главдым образом
систематика асцидий. Перед нами ярко выраженный процесс раз-
вития многообразия очень узких приспособлений.
Благодаря наличию в стенках жаберного сита массы кровенос-
ных капилляров, передний отдел кишечника может являться мощ-
ным органом газообмена. Он вызывает удивление у физиолога тем
более, что у асцидий отмечается слабая интенсивность жизненных
процессов. Однако мощное развйтие жаберного мешка первона-
чально имеет значение, прежде всего, для подъема его фильтрующей
функции при лучшем обеспечении организма питанием. В этом же
направлении идет дифференцировка медиовентрально в жаберном
мешке железисто-мерцательного аппарата энлостиля. А именно —
образуется глубокая борозда, доходящая до половины или больше
длины жаберного мешка и переходящая па переднем его крае р
чисто мерцательные правую и левую дуги. Последние продолжаются
2i* 323
медиодорзально в эпибранхиальную складку, доходящую у дна
жаберного мешка до пищевода. Таким образом, создается аппарат,
продвигающий выделяемую железистыми клетками эндостиля слизь
от заднего конца эндостиля вперед к мерцательным полукольцам
при начале жаберного мешка, далее по ним и, наконец, от них по
спинной стороне назад, в пищевод. Нитями слизи обволакиваются
взвешенные в воде пищевые частицы и передаются вместе со слизью
в усваивающий отдел кишечника через пищевод, вода же филь-
труется через жаберную сеть в околожаберную полость. Последняя
представляет парную полость, образованную врастанием справа и
слева со спинной стороны двух мешков покрова к жаберным ще-
лям. Это — приспособление, защищающее нежную выстилку жабер-
ного отдела от каких-либо непосредственных повреждающих влия-
ний извне, что для сидячего животного является существенно
важным. Первоначально это простые вдавливания стенки тела на
спинной стороне. С разрастанием жаберных щелей вдавливания
растут до срединной брюшной линии и превращаются в околожа-
берные мешки, которые далее объединяются своими спинны-
ми частями через отшнуровывающийся участок спинной поверх-
ности тела. Туда же в зажаберном отделе открываются половые
протоки и анальное отверстие. Образуется выводной клоакальный
сифон.
Так создается сложный аппарат, обеспечивающий защиту жабер
и отток из них воды, и одновременно осуществляется возможность
безопасного функционирования кишечника и протоков половых
желез у сидячего животного, покрытого со всех сторон туникой.
При этом, в связи с редукцией хвостового отдела кишечника ли-
чинки, передняя, расширенная его часть вторично дает вырост в
виде петли, и вновь образующийся на ее конце анус открывается
в дорзально расположенный клоакальный сифон. В петле кишки
помещается яичник и семянники. Все эти внутренности распола-
гаются чрезвычайно компактно, так как целом здесь редуцируется.
Он теряет свою основную функцию опоры для мускулистой стенки
тела или для его выростов. Теряет и вторичные функции — тро-
фическую при хорошем развитии у туникат кровеносной системы и
экскреторную при переходе последней к нефрофагоцитам.
Остается лишь один отдел целом, охватывающий в виде муфты
брюшной сосуд позади жаберного мешка. Это — перикард, мышеч--’
ные волокна внутренней стенки которого используются для продви-
жения крови. Используются они крайне оригинально, а именно:
сокращения кольцевых мышечных волокон прогоняют кровь в
брюшном сосуде так, что она или нагнетается в систему капилляров
жаберного мешка, или в капиллярные сосуды усваивающей части
кишечника. В последнем случае кровь по стенке кишечника посту-
пает далее уже со спинной стороны в спинной сосуд, отдающий
кровь в каппиляры жаберного мешка. После нескольких минут
такой работы, когда капилляры переполняются, йо двигающей силы
оказывается недостаточно, чтобы прогнать кровь через них, сокра-
'324
щения останавливаются, а затем возобновляются уже в обратном
направлении. Кровь начинает нагнетаться в жабры через брюшной
сосуд, от которого идут жаберные капилляры с брюшной стороны.
В это же время кровь оттягивается от кишки. Таким образом, при
чрезмерном развитии системы жаберных капилляров, через которую
сердце должно прогонять кровь, вырабатывается вместо обычной ее
циркуляции единственное в своем роде колебательное движение.
Она нагнетается то в одном направлении, то в другом, причем про-
исходит периодическое изменение пульсации сердца то в одну сто-
рону, то обратно. Таков результат мощного развития жаберного
мешка асцидий!
Заканчивая анализ организации взрослой асцидии, нельзя не
обратить внимания еще раз на тот факт, что здесь нет никаких
следов образований, подобных головной лопасти и воротнику
Pterobranchia и Enteropneusta. Приходится признать фактом, что
при переходе прохордат, обладавших головной лопастью, к жизни
с прикреплением на твердом грунте брюшным участком заротового
отдела, как это имеет место у асцидий, головная лопасть теряла
значение, редуцировалась и исчезала. Усиленные 'же процессы
неравномерного роста заротового брюшного отдела, переносившие
рот на пункт, противоположный месту прикрепления, совершенно
стирали ее даже в онтогенезе. Точно так же исчезает воротник с
его придатками, ресничный аппарат которых в качестве аппарата,
производящего ток воды, заменяется мерцательным эпителием
самого жаберного отдела кишечника. На месте воротника сохра-
нялись лишь железистые элементы, расположенные вентрально и
служившие для прикрепления предка, покрывавшегося мощной
кутикулой.
Регрессивная эволюция асцидий, обусловленная переходом к
сидячему существованию, обнаруживает развитие приспособлений
к расселению вида, которые связаны с наличием свободноживущей
стадии в онтогенезе в виде личинки, обладающей большой подвиж-
ностью. Как можно представить подобный процесс?
У червеобразного предка асцидий, имевшего туловище, развитое,
как у Enteropneusta, произошла характерная дифференцировка
мио-хордального комплекса, дававшего возможности иного рода
движений, чем первоначальные перистальтические сокращения
кожно-мышечного мешка баланоглосса. Это было обусловлено
распространением клеток хордального типа по всей длине кишеч-
ника. Процесс, первоначально локализованный в переднем спинном
участке энтодермальной -кишки и давший головную хорду, распро-
странился назад в связи с особым развитием прилежащих продоль-
ных мышечных пучков целомической мускулатуры. Появление
возможности энергичных движений используется далее для передви-
жения тела при свободном плавании. Особенно резко это выражено
у асцидий, переходящих во взрослом состоянии к сидячему суще-
ствованию. Из состояния свободножившего предка происходит
выработка оригинальной личинки, гарантирующей в топ или иной
мере возможность расселения прикрепленных форм. Это — два
взаимообусловленных процесса.
Крайность переделки предковой организации в личиночную
здесь выразилась в том, что зажаберный отдел тела всецело
связывается с функцией движения. В соответствии с этим редуци-
руется пищеварительная функция у большой части кишечника и
выведение неперевариваемых остатков происходит уже через вто-
рично образующийся и идущий к спинной стороне вырост средней
кишки. От дальнейшего отдела кишки сохраняется лишь рудимент
в виде тяжа клеток, проходящего по брюшной стороне до заднего
конца тела. Центральное положение занимает хорда, представляю-
щая, таким образом, главный зачаток, входивший в комплекс экто-
дермальной закладки кишечника. Над ней располагается нервная
трубка, с боков — продольные мышечные волокна. Зачатки мезо-
дермальной мускулатуры уже обособлены от кишечной закладки, и
существеннейший компонент мезодермы — целом — полностью
редуцируется, теряя все свои функции. Основная функция опоры
переходит к хорде, а вторичные' — трофическая, экскреторная и
половая — отпадают в связи с дифференцировкой почти всего зажа-
берного отдела тела в орган движения — хвост личинки.
Таким образом, анализ организации личинки асцидий показывает
превращение большей части туловища червеобразных предков в
хвост, используемый в качестве полезного для расселения вида
органа движения, что обусловливает само появление этой личиноч-
ной формьп Последнюю приходится считать вторичной, новой, по
сравнению с первичными личинками типа трохофоры или дипле-
врулы, которая у баланоглосса еще представлена торнарией. Этот
анализ позволяет уточнить и понимание организации взрослой асци-
дии, как развивающейся всецело за счет переднего отдела тела ли-
чинки, включая сюда, главным образом, содержащий жабры уча-
сток кишечника. Разделение на туловище и хвост у личинки выра-
жено чрезвычайно резко и усугубляется полным гистолизом всех
частей хвоста личинки при переходе к прикрепленному образу
жизни взрослой асцидии и соответственно при перестройке органи-
зации переднего отдела тела. Это находит свое особое выражение
в процессах неравномерного роста брюшной стороны между ртом и
местом прикрепления, что ведет, как было уже выше указано, к
расположению ротового сифона на полюсе, противоположном месту
прикрепления, т. е. к положению рта, наиболее выгодному для осу-
ществления тока воды, несущего пищевой материал сидячему орга-
низму.
Так, под знаком выработки высоко совершенных защитных при-
способлений при максимальном использовании возможностей пас-
сивного питания создалась крайне своеобразная организация сидя-
чих асцидий.
Раз выработавшись, она широко используется в различных на-
правлениях. Она оказалась достаточно пригодной и для перехода
в пелагические условия существования, где также могут быть
использованы большие количества мелкого питательного материала
для пассивного питания.
Это мы находим у сальп и Doliolum. Переход их асцидиеобраз-
ных предков к свободному существованию отразился прежде всего
на утрате значения подвижной личиночной стадии, важной для рас-
селения сидячих асцидий и становящейся излишней для свободно-
плавающих животных. Однако у Doliolum она еще сохранилась,
хотя и в редуцированном виде (рис. 139) — хвостовой отдел ли-
чинки резко отделён образующимся вторично пузыреобразным
зздутием от жаберного, переднего участка тела. Личинка как бы
Рис. 139. Личинка Doliolwn (по Ульянину):
I — яйцевая оболочка, 2 — эктодерхмальный покров, 3 — нервный зачаток, 4 — ро-
товая закладка, 5 — мезодермальный зачаток, 6 — хорда, 7 — срединная питатель-
ная часть личинки
разделена на две части, из которых передняя передается в органы-
зацию. взрослого Doliolum, а задняя, содержащая хорду и мышцы,
подвергается гистолизу. Такая личинка производит впечатление
уродливого результата резко выраженного процесса деградации, ко-
торому подвергается бывшая когда-то важной для жизни стадия.
У сальп уже не осталось и следа чего-либо подобного личиночным
формам Doliolum и асцидий.
Организация прозрачных сальп (рис. 140), или Doliolum
(рис. 141), пассивно несущихся морскими течениями, поражает
упрощенностью отношений с окружающей средой. Клоакальный
сифон, закладывающийся на спинной стороне, у сальп смещается
значительно кзади. У Doliolum он обращен своим отверстием прямо
назад, причем животное получает боченкообразную форму благо-
даря сильному развитию как жаберной, так и клоакальной поло-
стей. Nucleus внутренностей становится крайне незначительным по
сравнению с размерами всего тела. У сальп он сохраняет еще свое
типичное положение вентрально у заднего конца, но у Doliolum,
будучи смещен широким клоакальным сифоном, глядящим прямо
назад, nucleus уже образует только небольшое утолщение посре-
дине брюшной стороны. Мускулатура как клоакального, так и ро-
тового сифонов, имевшаяся у асцидий, развивается в виде значи-
тельных кольцевых пучков. В конце концов у Doliolum они охваты-
вают наподобие обручей на бочонке все раздутое тело. Они обу-
словливают при своем сокращении ток воды через жаберную
полость, которая через немногочисленные жаберные щели широко
327
сообщается с клоакальной полостью. У сальп жаберные щели пред-
ставлены даже всего одной парой зияющих отверстий. Упрощение
жаберного аппарата вызвало и упрощение кровеносной системы,
представленной широкими лакунарными пространствами между
покровом и внутренними стенками сифонов. Но сердце асцидий, с
Рис. 140. А — общий вид и мускулатура сальп; 5 —схема их
внутреннего строения (по Деляжу и Эруару);
1 — рот, 2 — мускулатура губ, 3 — кольцевые мышечные ленты, 4 — муску-
латура клоакального сифона, 5 — кили стенки тела, 6 — столон, 7 — глаз,
8 — нервный ганглий, 9 — эндостиль, 10 — жаберная полость, И — жабры,
12 — клоакальная полость, 13 — перикард, 14 — задняя кишка, 15 — желу-
док, 16 — пищевод, 17 — клоакальный сифон
его попеременными сокращениями, сохраняется как аппарат, регу-
лирующий ток крови.
Мы не будем вдаваться в детали приспособлений, превращаю- •
щих организацию сидячих асцидий в раздутые, прозрачные кожи-
стые тела Doliolum и сальп, взвешенные в морской воде. Пассив-
ность этих животных в отношениях с окружающей средой ведет
эволюцию группы в другую сторону, в сторону развития нового
приспособления в борьбе за жизнь, а именно — бесполого размно-
жения, принимающего здесь исключительные формы.
Уже в развитии асцидий происходит новообразование петли
кишки за счет заднего участка жаберного отдела, когда при появле-
нии хвоста личинки почти весь туловищный отдел кишечника реду-
цируется. В то же время имеет место и дифференцировка особого
328
мезодермального зачатка у переднего концу .корды. Создается
активный бластогенный очаг, который особенно ‘резко выделяется
у личинки Doliolum, как передний отдел мезодермы, оторванный
пузыревидным вздутием от хвостового ее отдела.
У сложных асцидий на основе этого бластогенного очага выра-
батывается бесполое размножение, путем почкования, что у сидячих
форм представляется вообще широко распространенным явлением
Рис. 141. А — общий вид и мускулатура Doliolum: Б — схема его
внутреннего строения (по Деляжу и Эруару);
1 — рот, 2 — кольцевые мышечные ленты, 3 — спинной вырост тела, 4 — почки,
двигающиеся по поверхности тела от столона (6), 5 — перикард, 7 — эндостиль, 8—
ганглий, -9 — статоцист, 10 — жаберные щели, 11 — пищевод, 12 — желудок, 13 —
задняя кишка, 14 — анальное отверстие
и развивается в разных группах совершенно независимо. У свобод-
ноживущих животных это — гораздо более редкое явление по са-
мому существу дела, так как ставит почкующуюся особь, а затем
и колонию, в менее благоприятные условия. Но раз выработавшись
у первоначально сидячих колониальных форм, оно сохраняется при
переходе к свободному существованию и эволюционирует. Это про-
исходит особенно в связи с развивающимся полиморфизмом особей
на основе разделения между ними жизненных отправлений, как мы
видели, например, у сифонофор.
Общим для туникат, начиная со сложных асцидий, является об-
разование столона, находящегося вентрально у бластогенного очага
и заключающего в себе бласто-гонные зачатки эктодермального
329
кишечника — так называемые эпикардиальные выросты, затем за-
чатки перикарда и, наконец, недифференцированные мезодермаль-
ные клеточные образования.
У сальп развившаяся из яйца одиночная особь путем подразде-
ления растущего столона на последовательно расположенные друг
аа другом участки образует цепочку половых особей. Таким образом,
развивается смена поколений, а именно — размножающаяся беспо-
Рис. 142. Почкование Do’io’um (по Деляжу и Эруару):
I — спинной вырост тела, 2 — гастрозоиды, образующие два боковых ряда, 3 —
форозоиды, образующие два средних ряда, 4 — половые особи, располагающиеся
на ножке форозоидов. Внешний вид и внутреннее строение
дым путем одиночная форма дает цепную форму, размножающую-
ся уже половым путем. И цепочки прозрачных сальп в течениях
теплых морей несутся массами.
Однако крайней оригинальности достигает развитие колоний у
Doliolum (рис. 142). Небольшие почки, отделяющиеся от столона,
одна за другой перемещаются здесь с брюшной стороны на задний
спинной выступ тела. Для этого служат особые амебоидные «слетки,
как бы несущие на себе почку. Почки располагаются на растущем
заднем выросте тела дорзально и, в конце концов, образуют два
ряда особей, принимающих на себя функцию питания колонии —
гастрозоиды. В связи с этим кишечник материнской особи постепен-
но дегенерирует, а у гастрозоидов значительно сильнее развивается
жаберный аппарат. Дальнейшие порции почек образуют на спинном
ззо
дьгросте еще два ряда особей, располагающихся м<жду предыдущи-
ми рядами ближе к спинной срединной линии. Уже на ножках
этих особей — форозоидов — располагаются почки, дающие неболь-
ших половых индивидуумов Dolioltun. Из их яиц развиваются
личинки, дающие (родоначальников новых колоний.
Таким образом, возникла вся сложная и необычайная смена по-
колений свободноплавающего Doliolum, являющаяся завершением
Рис. 143. Схема строения аппендикулярий Falta ethiopica
(из Н. Бобринского):
] —• рот, 2 — головной нервный ганглий, 3 — жаберный отдел, 4 —
ресничные его образования, 5 — желудок, 6 — семенник, 7 — яичник,
8 — эндостиль, 9 — жаберная щель, 10 — анальное отверстие, 11 —
ганглий хвоста, 12 — хорда, 13 — зона ойкопластов
начавшегося у сидячих асцидий процесса бесполого размножения.
Он выгоден для сохранения вида и в конце концов значительно
осложнился дифференцировкой различных особей колонии на основе
разделения физиологического труда между ними.
Чрезвычайной оригинальностью выделяется группа ашщндаку-
лярий (рис. 143), которую обычно также относят к Tunicata. Одна-
ко самый вопрос о принадлежности их к этому подтипу может быть
поставлен под сомнение, настолько они своеобразны. Их организа-
цию некоторые рассматривают как сочетание черт строения взрос-
лой формы туникат с нарастающим выявлением личиночных осо-
бенностей или как настоящую •неотению, т. е. как сохранение орга-
низации личинки, достигающей преждевременно половой зрелости.
Личинка становится способной к размножению в этом состоянии, и
дальнейшие этапы органогенеза выпадают. Однако ряд исследова-
телей видит в аппендикуляриях прямых потомков примитивных
хордовых, а не вторично упрощенных представителей туникат.
Ведущим моментом в создании организации аппендикулярий
(рис. 143) приходится считать переход к использованию карлико-
вого планктона — наннопланктона — в качестве нормального
источника питания. Такое узкое приспособление связано с выработ-
кой на основе организации близкой к личиночным формам асцидий,
во-первых, общих малых размеров тела и, во-вторых, характерных
331
образований, служащих для использования карликового планктона
при пассивном питании. В частности околоротовой покров личинко-
подобной формы продуцирует кутикулу, которая отделяется от про-
изводящих ее эпителиальных клеток — ойкопластов и образует
подобие капюшона. К нему подвешена ртом аппендикулярна
(рис. 144). При движении заднего отдела туловища аппендикуля-
рии, превращенного в хвостообразный придаток, получается сильный
ток воды. Кутикулярный капюшон при этом раздувается, и через
Рис. 144. Домик аппендикулярий: г! — Olcopleura; Б — KovaUvskla
(по Фолю);
1 — фильтр, стрелкой обозначен ток воды, 2 — сито для лова наннопланктона, 3 —- вывод-
ное отверстие» 5 — отверстие выхода аппендикулярий из домика, 4 — хвост и 7 — туловище
аппендикулярии в домике, 6 — рот
него фильтруется вода, наннопланктон которой скопляется у рта и
далее заглатывается аппендикулярней.
Это — наиболее простое состояние аппарата. У большинства
форм он становится гораздо сложнее. Продуцируемая ойкопластами
кутикула, охватывая все тело, превращается в домик, не только
фильтрующий наннопланктон, но и защищающий помещающуюся
в центре аппендикулярию. При нападении врага она выскакивает
из своего кутикулярного домика и образует новый, что связано с
непрекращающейся деятельностью ойкопластов. После отделения
первого домика ойкопласты продолжают продуцировать новые пор-
ции кутикулы. Животное, даже ничем не тревожимое, пробыв не-
которое время в старом домике, должно покидать его, так как новая
кутикула отделяется от эпителия и дает начало новому домику.
Таким образом, для аппендикулярий характерна периодическая
смена кутикулы, как бы линьки, дающие все новые и новые аппа-
раты фильтрации по мере засорения старых, соответственно новые
и новые домики. • '
Не вдаваясь в детали, отметим, что собственно фильтрующие
части домика образованы тончайшими волоконцами кутикулы. Со-
здается своего рода сито, которое задерживает более крупные ча-
стицы, а карликовый планктон беспрепятственно доходит до рта.
Этим определяется и другая сторона процесса. Различная структура
332
т мощность кутикулярного слоя в разных местах домика обуслов-
ливают образование в нем полостей, которые определяют направле-
ние тока воды с пищевым материалом через фильтрующие сита по
направлению к ротово’му отверстию. Движущей же силой, как было
отмечено выше, являются волнообразные движения хвостового от-
дела, вызывающие появление токов воды. Перед нами совершенно
своеобразное использование движений животного вообще и его
специального органа движения — хвоста — в частности. Как видно,
домик аппендикулярий не имеет собственно ничего общего с туникой
асцидий. Это только кутикулярная обкладка приротового участка
тела, принявшая на себя функцию отбора поступающего пищевого
материала и требующая постоянной смены по мере ее загрязнения.
И химически она не обнаруживает ничего общего с туникой, так как
не содержит чего-либо подобного «туницину» — животной клетчат-
ке, характерной для Tunicata. Наконец, морфологически в ней нет
клеточных элементов и других структур, превращающих тунику
асцидий в особого рода ткань, покрывающую тело снаружи. Сле-
довательно, домик аппендикулярий не дает оснований для сближе-
ния их со взрослыми формами типичных Tunicata.
Но, наряду с выработкой защитного покрова, характернейшей
чертой организации последних является чрезвычайно сильное раз-
витие жаберного мешка. Аппендикулярий, напротив, отличаются
слабым развитием жаберного отдела и имеют всего одну пару про-
стых жаберных щелей, причем нет и следов чего-нибудь подобного
околожаберной полости. Ток воды при пассивном питании нанно-
планктоном поддерживается, как было указано, в значительной
мере движением заднего отдела туловища, который резко обособ-
ляется в виде хвостика и смещается на 90° по отношению к про-
дольной оси тела.
Хвостик, как придаток, осуществляющий движение воды, дости-
гает при этом большей специализации, чем у личинок асцидий. В
нем совершенно исчезает остаток кишечника, представленный у
асцидий тяжем клеток на брюшной стороне и так же закладываю-
щийся в развитии аппендикулярий. Этот участок туловища превра-
щается в тонкую пластинку, содержащую посредине хорду, надпей
тонкий нервный тяж, а по бокам правильно следующие друг за
Другом мышечные элементы. Таким образом, бывшее туловище ста-
новится прежде всего двигающим воду органом аппендикулярий, и
вторичный характер этих изменений несомненен.
Последнее обстоятельство приходится поставить в связь с тем,
что отдел тела аппендикулярий, расположенный непосредственно за
жаберным, значительно разрастается в результате развития нахо-
дящихся здесь гонад — одного или, вторично, пары семенников и
расположенного между ними яичника. Их выводные протоки реду-
цированы, и половые продукты выбрасываются непосредственно
наружу. Хвост, место прикрепления которого соответствует началу
хорды, в жаберном отделе на 180° смещается к брюшной стороне
развивающимся половым отделом, т. е. направляется не назад, а
333
вперед, параллельно переднему участку тела, поворачиваясь при
этом на 90е, т. е. широкой своей стороной к брюшной стороне тела
. Такова живущая в свободно плавающем домике аппендикулярна
как будто бы не имеющая на первый взгляд ничего общего с асци-
диями.
Видеть в ней примитивные черты исходных форм хордовых хотя
и заманчиво, но не соответствует действительным отношениям
целостной организации. Так, прежде всего, приходится расценивать
факт полной редукции головной лопасти, которая несомненно была
присуща исходным червеобразным формам и имеется у Pterobran-
chia и Enteropneusta. Вместе с этим полное развитие миохордаль-
ного комплекса' при простоте жаберного аппарата резко изменяет
те отношения, которые представлены уже у червеобразного бала-
ноглосса (мощный жаберный отдел и первые шаги в развитии
миохордального комплекса), или у Cephalodiscus (с одной парой
жаберных щелей и примитивными зачатками хорды и пр.).
Совсем иначе представляется картина, если подойти к организа-
ции аппендикулярий, как к вторично упрощенной’ в связи с особыми
условиями существования, особенно в связи с питанием карликовым
планктоном. Отсюда, во-первых, малые размеры тела и характерное
постоянство числа клеточных элементов для определенных органов
и даже видов, а вместе с тем минимальное развитие мезенхимных
образований, вторичная простота кровеносной системы, рудимента-
ция перикарда; отсюда, во-вторых, рудиментация эндостиля и упро-
щение жаберного аппарата до одной пары жаберных щелей; отсюда,
в-третьих, незначительность развития нервной системы. Но отсюда
же, с другой стороны, такие узкие приспособления, как аппарат
кутикулярного домика для сбора наннопланктона, а затем и для
защиты, и дифференцировка хвоста, вместе с полной редукцией в
нем кишечника и целома и с поворотом на 90э для использования
при пассивном питании. При этом происходит необычайно сильное
развитие области примитивнейших гонад.
Все это говорит о вторичном упрощении и в то же время о край-
ней специфичности организации аппендикулярий. Как она могла
создаться, показывает анализ организации туникат. Прежде всего
анализ дает ответ на основную характерную особенность аппенда-
кулярий, выражающуюся в отсутствии головной лопасти. Прикреп-
ление предков асцидий передним участком брюшной стороны —
вот единственное экологическое объяснение редукции головной ло-
пасти у туникат. Момент прикрепления мы должны принять и для
других групп хордовых, утративших головную лопасть, так как иных
возможностей полного ее исчезновения экология не дает, особенно
если принять во внимание исходное пассивное питание хордовых:
Эволюция последнего отчасти направлялась тем же моментом при-
крепления, как мы видим у асцидий в чрезмерном развитии жабер-
ного мешка. Но перенесение ведущей роли при пассивном питании
с мерцательного покрова околоротовых образований на мерцатель-
ные образования жаберных щелей произошло уже у низших под-
типов хордовых, обладавших еще головной лопастью, что наглядно*
показывает организация баланоглосса.
Поэтому нельзя преуменьшать значение бросающегося в. глаза
сходства аппендикулярий с личинками асцидий. Развитие тех и дру-
гих идет одинаково. У тех и других полная редукция головной
допасти — совершившийся факт, и у асцидий он лишь прямое след-
ствие прикрепления взрослой формы. У тех и других обособлен
жаберный отдел с одной парой жаберных щелей у аппендикулярий,
с одной — двумя у личинок асцидий, которые при прикреплении
образуют, однако, сложнейшее жаберное сито. У тех и других вто-
рично образующаяся петля кишечника и вторично образующийся
анус. У тех и других одинаково построенный хвостовой отдел. Но
в то время как личинки асцидий — лишь кратковременный этап
жизни, служащий для расселения и лишенный самостоятельного
питания, аппендикулярий приспособляются к самостоятельному пи-
танию мельчайшим, карликовым планктоном.
С этим связаны как общие черты упрощения организации, так к
специфические — малые размеры, выработка домика и дифферен-
цировка хвоста, которые уже были выше рассмотрены и которые
говорят, что туника асцидий действительно отсутствует у аппенди-
кулярий с самого начала, а образование хвоста, как движущего
момента в пассивном питании наннопланктоном привело- к сохране-
нию личиночного состояния закладки жаберного мешка с одной па-
рой жаберных щелей.и с отсутствием дополнительно развивающихся
околожаберных полостей и клоакального сифона.
Итак, аппендикулярий, всего вероятнее, представляют собой
своеобразные неотенические формы Tunicata, выработавшие свою
особую специфическую организацию.
БЕСЧЕРЕПНЫЕ — ACRANIA
Если у туникат, при сидячем образе жизни исходных асцидие-
образных форм, распространение мышечно-хордального зачатка на
протяжении всего туловища привело к образованию личиночного
органа движения — хвоста, то у свободноподвижных ланцетника
и позвоночных мио-хордальный комплекс становится ведущим в
формировании всего рыбообразного тела. Это обусловлено возмож-
ностью использовать развивающееся энергичное движение тела в
качестве широкого приспособления для активной защиты. Ланцетник
быстрыми движениями прекрасно ускользает от врага. У позвоноч-
ных движение тела используется не только для активной защиты,
но и для активного добывания пищи, чем значение движения еще
более усиливается.
Ланцетник полностью сохраняет пассивный тип питания исход-
ных форм хордовых, с которым крайне оригинально сочетает воз-
можность свободного движения, вырабатывая своеобразнейший об-
раз жизни. Ланцетник живет в грубом песчаном грунте, зарываясь
в поверхностные слои песка всем телом за исключением переднего
335
Рис. 145. .4 — общий вид ланцетника Branchiosto>na lanceolatum (по Клейну;) Б — схема внутреннего строения
ланцетника (по Краузе);
1 — лопаточка, 2 — обонятельная ямка, 3 — головной мозг, 4 — околоротовые усики, 5 — жаберный отдел кишечника с жаберными
щелями, 6 — спинной непарный плавник, 7 — миомеры, 8 — ротовая воронка, 9 — спинной мозг, 10—хорда, 11— усики гч.пп (12), 13—средняя кишка
(желудок), 14 — задняя кишка, 15 — печень. 16 — хвостовой плавник, 17 — анальное отверстие, 18 — атриальная пора, 19 — го-
нады, 20 — метаплевральная складка, 21 — атриальная полость
конца, несущего ротовую воронку. Она открыта в-придонную воду.
Таким образом, животное, живущее в грунте и защищенное этим
от врагов, использует в то же время неистощимый источник пита-
ния в виде мелких животных и растительных организмов, взвешен-
ных в воде (рис. 145).
В соответствии с этим сочетанием двух различных аспектов
жизни вырабатывается весь наружный облик ланцетника. Образует-
ся кутикулярный покров, предохраняющий наружный эпителий от
повреждения грубым'грунтом. Форма тела становится ланцето-
видной, хорошо проникающей в грунт и в то же время при сокра-
щениях боковой мускулатуры способствующей быстрому продви-
жению тела синусоидными движениями в водной среде. Этому по-
могает и развитие складки спинного, хвостового и заанальнрго непар-
ного плавников. Наконец, передний конец тела приобретает форму
лопаточки, благодаря чему животное легко1 проникает в песчаный
грунт. В связи с этим хорда как опорное образование разрастается
до самого конца лопаточки и покрыта здесь с боков мышцами
двух передних сомитов. От головной лопасти червей — первичной
головы птеробранхов и баланоглосса — не остается и следов, как
мы это видели уже у асцидий.
Указания на то, что прикрепление передним концом тела, обу-
словившее редукцию головной лопасти у асцидий, имело1 место и у
предков ланцетника, дают факты, связанные с происхождением у
него рта (рис. 146). В развитии совершенно нет закладки первич-
ного рта хордовых, который мы находим у баланоглосса или асци-
дий, а дефинитивный рот ланцетника представляет из себя настоя-
щую вторую жаберную щель левой стороны тела. Эта щель разра-
стается, образуется ведущее к ней спереди углубление ротовой
воронки, в которое втягивается закладка первой жаберной щели,
остающейся в виде рудиментарной «гатчековской» ямки. При этом
медианная складка покрова лопаточки распространяется от перед-
него конца тела по правой стороне ротовой воронки, охватывает ее
сзади и продолжается на левую сторону, отделяя от следующих
жаберных щелей.
Таким образом несомненно, что функцию рта у ланцетника вы-
полняет вторая жаберная щель девой стороны. И это можно объяс-
нить прикреплением его предков передним концом тела подобно
асцидиям, причем и рот и ближайшая жаберная щель становятся
в невыгодное положение для поддержания тока воды в жаберном
отделе. Рот совершенно редуцируется и уже не закладывается, а
первая жаберная щель не прорывается. Что же касается дальней-
ших жаберных щелей, они стали при прикреплении не в одинако-
вых условиях с первой и второй, и функция рта была постепенно
взята второй левой щелью. При этом произошло значительное ес
расширение и отрыв от следующих щелей, перемещающихся на
брюшную сторону, в связи с чем жаберные щели правой стороны
начинают закладываться значительно позднее. Лишь при последую-
щем переходе от прикрепленного образа жизни к свободному суще-
Пути эволюции животного мира 337
ствованию в песчаном грунте произошло смещение всего ротового
комплекса на брюшную сторону и, в соответствии с этим, полно»
обрастание вторичного рта складкой покрова, шедшей с правой
стороны.
Однако ряд компетентных зоологов держится иного взгляда
относительно значения рта у ланцетника и относительно тех уело-
Рис. 146. Схема мезодермальных закладок в головной части ланцетника: А —
первая закладка рта; Б — рот и прилежащие образования вполне выражены (по
Легро); В, Г, Д — последовательные стадии закладок жаберных щелей (по
Деляжу и Эруару);
I, II, Ш, IV — передние миотомы, 1 — лопаточка, 2 — передний целом (ростральная
полость), 3 — закладка преоральной ямки, 4 — боковые пластинки целома, 5 — закладка
рта и личиночный рот. 6 — гатчековский нефридии, 7 — мышечные пучки закладок прео-
ральной ямки и ротовой, 8 — зачаток зндостиля, 9 — закладки жаберных щелей левой сто-
роны, 10 — то же правой стороны, 11 — складки метзплевр, образующие атриальную
полость, 12 — ротовая воронка
вий существования, в которых могла выработаться подобная орга-
низация. Так А. Н. Северцев (1939) принимает, что первичный рот
у ланцетника в развитии сохраняется и дает рот взрослой формы.
Асимметрия положения рта и развития жаберных щелей объясняет-
ся своеобразным приспособлением к донному существованию, когда
рыбообразный организм лежит наподобие камбал на дне правой
стороной тела, а левая обращена в придонные слои воды и сюда
перемещаются все жаберные щели для свободного функционирова-
ния. Только при дальнейшем проникновении ланцетника в грунт
вторично восстанавливается правильное двусторонне-симметричное
строение тела.
В этой связи следует отметить, что головная лопасть у ланцет-
338
ника несомненно редуцирована, и этот факт большой важности не
находит объяснения в построениях А. Н. Северцева. Совершенно
игнорируется далее гомология рта и жаберных щелей, о чем ’ еще
будет речь дальше.
Обращаемся к дальнейшему анализу организации ланцетника.
К защитному комплексу черт при жизни в грунте у ланцетника
приходится отнести образование метаплевральных. складок стенки
тела, прикрывающих весь отдел туловища, несущий очень сильно
развитые жаберные щели, и далее сле-
дующий отдел, содержащий гонады и
вырост печени (рис. 147). Начинаясь
над уровнем спинных концов жаберных
щелей, метаплевральные складки идут
к брюшной стороне и срастаются по
брюшной срединной линии. Таким об-
разом создается околожаберная или
атриальная полость, защищающая жа-
берный аппарат живущего в песке жи-
вотного от засорения' грунтом и выз-
вавшая появление выроста тела, содер-
жащего печень. Развитие подобного
придатка было бы невозможно для
свободноживущего ланцетника, если
бы уже не имелось соответствующей,
защищающей его околожаберной по-
лости. В основания складок правой и
левой метаплевры вдается целом и
7 /7
вместе с ним гонады, но дальше в
брюшной стенке атриальной полости
идут только кольцевые мышцы. По-
следние образуют два независимых,
правый -и левый, дугообразных пучка,
как след развития за счет срастания
двух независимых правой и левой скла-
док. Лишь в заднем конце, где мета-
плевры не срастаются, остается отвер-
стие — атриальная пора. Атриальная
полость через нее свободно сообщается
с внешней средой, представляя, таким
образом, часть той же среды, но как бы
включенную в организм животного на-
Рис. 147. Схема поперечно-
го разреза взрослого лан-
цетника в жаберной обла-
сти тела (по Деляжу и
Эруару):
1 — хорда, 2 — спинной мозг
с отходящим слева пучком
двигательных волокон, 3 —
спинная аорта. 4 — продольные
мышцы, 5 — целом, 6 — атри-
альная полость. 7 — эндостиль,
8 — жаберный отдел кишечни-
ка, 9 — жаберная щель, 10 —
жаберные септы, 11 — нефри-
дий, 12 — печеночный выроет,
13 — гонада. 14 — мстаплев-
ральные складки. 15 — непар-
ный плавник
подобие клоакального сифона асцидий. Функции их сходны, нс
гомологии здесь нет. Только тут у ланцетника имеются остатки
кольцевых мышц, обусловливающие возможность сужения атриаль-
ной полости и выбрасывания ее содержимого. В других частях тела
они отсутствуют, так как лишь продольные спинные мышечные пуч-
ки из кожно-мышечного мешка червеобразных предков сохраняют
все свое значение с выработкой туловищной хорды.
22*
339
Образование метаплевр и атриальной полости сглаживает типич-
ное подразделение туловища на следующие друг за другом жабер-
ный, половой и печеночный отделы тела, присущие червеобразному
баланоглоссу или оболочникам. Но все же у ланцетника сохраняет-
ся общий план расположения этих органов по длине тела.
Функции атриальной полости, как мы видели, сложны — защита
жаберных щелей, гонад и печеночного выроста. Повидимому, в из-
вестной мере и увеличение количества жаберных щелей с первона-
чального состояния 10—12, которые некоторое время держится в
постэмбриональном развитии ланцентника, а затем быстро начинает
нарастать до 90—100 у взрослой формы, связано с образованием
надежной защиты их метаплеврами при переходе к жизни в грунте.
Тогда же начинается и образование печеночного выроста немысли-
мого без атриальной полости. Увеличение числа жаберных щелей
гарантирует достаточное питание ланцетника и достаточный газо-
вый обмен, что в особых условиях его существования представляет
почти все необходимое для благополучия животного. Оно ушло с
широкой арены борьбы за существование в однообразную жизнь в
грунте. На этой арене борются, гибнут или преуспевают его роди-
чи — свободноживущие водные позвоночные, а ланцетник, при-
способившись к своему песку, проходит, мало изменяясь, через тол-
щу веков.
Активная защита ланцетника обусловлена всецело использо-
ванием придонных слоев воды. Когда что-либо тревожит ланцетни-
ка, он вырывается из песка, делает быстрые броски в воде и момен-
тально снова уходит в грунт. Развитие мио-хордального комплекса
ио всей длине тела, что первоначально служило для защиты самим
। фактом сокращений, дает возможность подобных движений. При
J этом особенно развиваются которые из сравнительно
| небольших зачатков превращаются в мощные боковые поперечно-
J полосатые мышцы и сегментируются. Без сегментирования, при ко-
; тором миосепты фиксируются на оболочке хорды, были бы невоз-
можны подобные энергичные сокращения боковой мышцы с опорой
на хорду. Слабые поперечнополосатые мышечные пучки в хвосте
личинки асцидий или у аппендикулярий уже показывают правильно
метамерное расположение их клеточных элементов, хотя септ здесь
нет. Поэтому-в литературе уже высказывалась мысль, что метамерия
целома в области миотомов возникла у ланцетника и, соответствен-
но, у позвоночных втдщгчно, в связи с чисто механическими усло-
виями мощных сокращений боковой мышцы. Исходным было неме-
тамерное состояние целома туловища птеробранхов и баланоглосса,
т. е. олигомерный целом протоцело-мат. К этому вопросу нам еще
придется вернуться в дальнейшем. 1
Огромное значение целома мы уже могли оценить при анализе
организации аннелид, а затем моллюсков, членистоногих и, осо-
бенно, иглокожих. И у ланцетника многосторонние связи целома
. являются ведущими в выработке внутренней организации.
Дифференцировка целома у него идет очень своеобразно — по
340
пути, общему с позвоночными и характерному ддя обеих групп, но
на основе, уже выясненной нами для других типов животного цар-
ства. Главный момент; обусловивший само появление цел о мл у
Protocoelomata, был связан с выработкой опорной массы жидкости
в мышечном зачатке (миоцель-теория). Полным подтверждением
этого взгляда являются отношения, наблюдающиеся у ланцетника
(рис. 148), а именно—закладка целома у него прежде всего связана
с образованием мускулатуры стейки тела. Но если у баланоглосса
эта мускулатура является сплошным слоем продольных мышц
кожно-мышечного мешка, имеющих в качестве опоры жидкое со-
держимое целома, то у ланцетника такой опорой становится прохо-
дящий медио-дорзально тяж хорды, дающий большие возможности
рыбообразного движения ’’’вместю^перистальтических сокращений
тела баланоглосса. При опоре на хорду из целомической мезодер-
мы развиваются в виде мощных мышечных образований только
прилежащие к хорде медио-дорзальные мышечные пучки — правая
и левая боковые мышцы ланцетника. Таким образом, создается мио-
хордальный комплекс, а париетальный слой мезодермального соми-
та остается лишь в виде тонкого' целотелиального листка, прилега-
ющего к субэпителиальному слою ткани основного вещества стенки
тела. Сама целомическая полость, утратив здесь свое опорное зна-
чение, редуцируется, а париетальный целотелий начинает непосред-
ственно прилегать к продольным мышечным волокнам, которые
своего целотелия не имеют. Таким образом здесь хорошо видна ।
равнозначность мезодермальной мускулатуры и целотелия, лежа- *
щая в основе миоцель-теории целома.
Образование мешков целома вытесняет и вводит в определенные
рамки заполняющую ткань основного вещества. У ланцетника, как
и у баланоглосса, имеются не содержащие клеток слои этой ткани
под эпителием покрова и кишечника, и особенно сильно выражен-
ные в наружной стенке метаплевральных складок. При отсутствии
в стенке тела мезенхиматозных кольцевых мышц отсутствует ди-
ференцировка отдельной субмускулярной перепонки ткани основного
вещества, мешки целома непосредственно' прилежат к субэпите-
лиальным пластинкам. Зато особое развитие получают пограничные
образования спинного мезентерия, в котором заложены и хорда и
спинной мозг. Оболочки их усиливаются слоем, образующимся уже
за счет вторичной мезенхимы склеротома, т. е. из целенхимы. Осо-
бенно характерно наличие метамерных септ, отделяющих пучки
продольных волокон одной миомеры от другой, идущих от субэпи-
телиальной перепонки покрова к спинному мезентерию и представ-
ляющих неклеточные пластинки пограничной ткани основного' веще-
ства. Таким образом, каждая миомера с прилежащим к ней снару-
жи париетальным листком целотелия оказывается заключенной в
резко выраженную коробку из пограничных образований. Брюшной
край миомеры внутри коробки, разрастаясь, загибается кнутри и
поднимается кверху до хорды и нервной трубки, плотно прилегая к
остальной массе продольных мышечных волокон миомеры. Чрезвы-
341
чайно показательно полное отсутствие пограничных образований
внутри миомер, чем резко подчеркивается различный характер раз*
вития системы ткани основного вещества и системы целома.
Крайне оригинально исходная опорная функция целома частич-
но сохраняется у ланцетника; именно в личиночном состоянии це-
лом дает многочисленные слепые выросты в медианные складки
спинного и брюшного непарных плавников, чем повышается плот-
ность последних. В дальнейшем эти выросты отделяются от дав-
ших им начало спинного и брюшного отделов целомических соми-
тов, образуя в лопаточке и хвостовом плавнике целую обособлен-
ную систему каналов. На остальном протяжении тела они дают
метамерные камеры, которые покрывают плавниковые лучи, пред-
ставляющие врастания ткани основного вещества в основания це-
ломических камер.
Как в самых передних двух-трех миомерах, так и в десяти зад-
них заанальных и хвостовых не происходит дальнейшей дифферен-
цировки в пределах каждого мезодермального сомита, но в части
тела, содержащей кишечник, имеет место своеобразнейшее их рас-
членение в соответствии со вторичными функциями целома — тро-
фической, половой и экскреторной. Оно характерно далее и для по-
звоночных. В основе этого расчленения лежит дифференцировка
спинного отдела, берущего на себя выработку мощной мускулату-
ры миохордального комплекса — мнотом. Брюшной отдел, охва-
тывающий кишечник, вторично утрачивает метамерное строение и
брюшной мезентерии. Спланхногомы отдельных сомитов сливают-
ся в единую полость тела, трофическая функция которой обуслов-
лена теснейшей близостью с усваивающими частями кишечника.
Более ограничены отношения целома к экскреторному и поло-
вому аппаратам, но у ланцетника они чрезвычайно ясно показы-
вают сложность целомического комплекса. На месте перехода мио-
томов в спланхногомы, медиально на протяжении жаберного отдела
кишечника вдаются в целом, соответственно каждой первичной
жаберной щели, экскреторные органы типа протонефридиев, снаб-
женных соленоцитами.
Открытие их в начале нынешнего столетия показало сохранение
примитивнейших черт в организации ланцетника, наряду с вполне
выраженными особенностями строения, свойственными высшим хор-
довым. Подобного рода нефридии имеются у некоторых полихет и
ряда личиночных форм, но они были совершенной неожиданностью
для хордовых. Очевидно, что для немертинообразных протоцело-
мат, предков хордовых, это — типовое состояние, сохраняющееся и
у современных немертин, имеющих протонефридии в передней ча-
сти туловища. Таким образом, «нефритом». т. е. отдел целому,
имеющий ближайшую связь с экскреторным аппаратам, уже у лан-
цетника обладает дифференцировкой, характерной далее для поз-
воночных.
Одновременно происходит обособление дальнейшего участка це-
лома, как гонотома в связи с тем, что гонады, располагающиеся в
342
задней части жаберного отдела и позади него, вдаются под мышеч-
ными закладками в целом. Далее, участки цело\т;е гонадами внут-
ри обособляются, располагаясь в боковой стенке атриальной поло-
сти. Зрелые половые продукты выводятся в последнюю через обра-
зующиеся за счет цело-
мической стенки вре-
мецньге половые прото-
ки. Примитивные гона-
ды баланоглосса с их
собственными вывод-
ными протоками заме-
няются, таким образом,
более сложным состоя-
нием, свойственным да-
лее и позвоночным,
когда устанавливается
более тесная связь с
целомом. Однако гоно-
дукты у ланцетника
еще настолько прими-
тивны, что долгое вре-
мя в науке господство-
вало мнение о выхож-
дении половых продук-
тов из гонад в атриаль-
ную полость просто
через разрыв стенки те-
ла.
Последним шагом в
дифференцировке цело-
ма у ланцетника явля-
ется образование скле-
ротома. В постэмбрио-
нальном развитии (рис.
148) целом дает снача-
ла полые выросты,
Рис. 148. Схема поперечного разреза молодого
ланцетника в жаберной области (по Бовери):
1 — хорда, 2 — спинной мозг, 3 — спинная аорта, 4 —
боковая продольная мышца, 5 — целом околокишечный,
6 — атриальная полость, 7 — эндостиль. 8 — жаберный
отдел кишечника, 9 — брюшной сосуд, 10 — протонефри-
проникающие между
мускулатурой и хордой
от брюшного края мио-
тома до спинного. За-
тем эти выросты пре-
Вращаются в целенхим- дий и И — его проток, 12 — целомические полости
оптгптпг/ по лттлт миотома, 13 — гонады и их целом, 14 — целом мета-
НЫИ зачаТОК, за счет плевральных складок, 15 — целом непарного плавника
которого далее образу-
ется соединительнотканный чехол вокруг хорды и нервной труб-
ки. Этот процесс образования очагов вторичной мезенхимы за счет
Первоначально целотелиальных зачатков у позвоночных не ограни-
чивается склеротомом, а распространяется далее и на париетальную
343
стенку миотома и спланхнотома, так что возникают многочисленные
очаги образования вторичной мезенхимы и последующего развития
соединительной ткани в стенке тела и вокруг хорды и нервной
трубки.
Таким образом, за счет простых мешков олигомерного целома
низших хордовых происходит сложная дифференцировка целома у
ланцетника, а затем и у позвоночных, связанная с выявлением раз-
личных вторичных его функций при утрате первичной опорной. Од-
нако, наряду с этой дорзо-вентральной дифференцировкой, имеет ме-
сто процесс, идущий и по продольной оси тела. У червеобразного
баланоглосса и птеробранхов процесс этот выразился в обособле-
нии переднего опорного целома головной лопасти и среднего, под-
держивающего придатки воротника, дальнейшего расчленения двух
целомических мешков в туловище не было. В связи с выработкой
миохордального комплекса у ланцетника, как мы видели, разви-
лась метамерность туловищного отдела, наложившая глубокую пе-
чать на всю организацию ланцетника и позвоночных. Создались ак-
тивно плавающие формы с синусоидальными движениями тулови-
ща, а вместе с тем и с терминальным ростом длинного тела. Про-
цесс — аналогичный разыгравшемуся у аннелид. Сходные условия
порождают и сходные следствия. Но по существу эти процессы в
двух названных группах глубоко различны. А именно: у ланцетника
и позвоночных выработка осевой хорды вела к расчленению муску-
латуры, располагающейся по бокам; й вместе с тем к подразделе-
нию единого1 мезодермального зачатка на. мпомеры ~ миомерия. У
аннелид, напротив, расчленяется осевой опорный целом при сохра-
нении единой мускулатуры кожно-мышечного мешка, а в результа-
те получается целомерия.
Конечно-, развитие полимерного целома представляло- широко
приспособительную черту, двинувшую организацию^вперед по пути
выработки рыбообразных форм. Но в то же время головной и во-
ротничковый целом у высших хордовых редуцируются. У ланцетни-
ка первый из них теряет свое значение, так как редуцируется сама
головная лопасть, а второй — также не может прогрессивно разви-
ваться при жизни в грунте и утрате придатков у рта. Это тем более
характерно для позвоночных, переходящих к активному пита-
нию.
Однако воротничковый целом, повидимому, все же сохранился,
у ланцетника в виде небольших «головных» полостей, которые да:
ют выросты в складки метаплевр, вторично прикрывшие жаберные
щели. Особенности развитая целомических мешков в головном кон-
це ланцетника показывают их некоторую независимость от поли-
мерной закладки туловищного целома. <
Итак, комплекс цело-ма, дифференцировавшийся в многом ы,
склеротомы, пефротомы, гонотомы и сплапхнотомы, у ланцетника
становится ведущим во внутренней организации,. Им обусловлено
также состояние пограничных образований ткани основного веще-
ства и кровеносной системы (рис. 149).
344
Очень мощное развитие жаберного отделе!
у ланцетника находит свое выражение в уст-
ройстве аппарата, продвигающего кровь через
жабры. Как известно', у низших хордовых эту
роль играет сердце, характеризующееся диф-
ференцировкой перикардиального целома. Лан-
цетник пошел в своей эволюции иным путем.
На сосудах, приносящих кровь из брюшного
сосуда к каждой первичной жаберной щели,
развиваются мускулистые утолщения — bulbilli.
Таким образом, достигается возможность про-
движения крови отдельно' через систему каж-
дой первичной жаберной щели, а 180—200
bulbilli продвигают ее через весь жаберный от-
дел, от чего отказалась, можно сказать, орга-
низация асцидий.
Однако не менее существенно на строении
кровеносной системы ланцетника отразилось
развитие других характерных черт его органи-
зации. Прежде всего отразилась дифференци-
ровка печени, приобретающей особое значение
в обмене веществ. Вклинившийся между жа-
берным отделом и усваивающей кишкой пече-
ночный вырост обусловил распадение брюшно-,
го сосуда на вставочную систему капилляров—
образовалась ворсугная система печени. Она
еще слабо отдифференцирована от кишечного
сосудистого сплетения, но уже вполне выраже-
на. Подобная вставка капилляров имеет место
и на сосудах жаберного отдела вокруг экскре-
торных органов ланцетника.
Последним, но очень важным характерным
штрихом, заканчивающим выработку «позво-
ночного» склада кровеносной системы, будет
образование у ланцетника двух боковых сосу-
дов. Они пробегают под продольными мышца-
ми над гонадами и, с одной стороны, обеспечи-
вают питание гонад, а с другой —> в них соби-
рается кровь, поступающая из боковых отде-
лов тела, главным образом из мощной муску-
латуры. Идущий спереди боковой сосуд —
передняя кардинальная вена — и идущий
сзади — задняя кардинальная вена — соеди-
няются позади последней жаберной щели в
единый сосуд с каждой стороны, в правый и
левый юовьеровы протоки. Последние, направ-
ляясь к брюшной срединной линии, впадают в
Рис. 149. Схема кро-
веносной системы
ланцетника (комбини-
ровано по Гроббену и
Царнику): .
1 — bulbilli, 2 — брю-
шной сосуд, 3 — жа-
берные сосуды, 4 — во-
ротная система печени,
5 — спинная аорта, 6 —
печеночная вена, 7 —
кювьеров проток, 8 —
париетальные сосуды, 9—
подкишечная вена, 10 —
хвостовая вена, 11 —зад-
няя кардиальная вена,
12 — нисходящая аорта,
13 — поперечные вены,
14 — венозный синус,
15 — передняя кардиаль-
ная вена, 16 — гонады
345
брюшной сосуд сейчас же за жаберным отделом. Эта часть брюш-
ного сосуда, так называемый венозный синус, принимает сзади так-
же печеночную вену, несущую кровь из стенки кишки.
Таким образом, создается тип кровеносной системы, находящий
свое наиболее яркое выражение у позвоночных и обусловленный
развитием мощной боковой мускулатуры в стенке тела, а также
развитием жаберного комплекса и дифференцировкой в кишечнике.
Только в брюшной и спинной аорте дифференцировка стенки со-
суда доходит до образования эндотелиальной выстилки, как это
имеет место и в сердце асцидий. В остальных сосудах ланцетника
интима представлена лишь тонким слоем ткани основного веще-
ства, т. е. отношения, близкие к наблюдающимся у немертин. Пе-
ред нами путь, ведущий у позвоночных к эндотелизации всех со-
судов, в противоположность полному отсутствию клеточной выстил-
ки в сосудах у аннелид, моллюсков и членистоногих.
Однако основным представляется тот факт, что везде развитие
кровеносной системы происходит на почве существования тех или
иных образований ткани основного вещества, жидким видоизмене-
нием которой с трофической функцией является кровь. В соответст-
вии с этим развитие системы пограничных образований определя-
ло, как мы видели, ход сосудов кровеносной системы у полихет.
Тот же момент выявляется в полной мере для хордовых у червеоб-
разного баланоглосса, где отношения крайне примитивны, и у рыбо-
образного ланцетника, где они уже значительно осложнены в соот-
ветствии со значительным усложнением организации. Через систему
пограничных образований, .определяемых развитием целома, крове-
носная система ланцетника также ставится в связь с комплексом
целома.
Выработка миохордального комплекса наложила свой неизгла-
димый отпечаток на нервную систему ланцетника (рис. 150). Пер-
воначально сплошной кожно-мышечный меток'стенки тела червей
заменяется осевой мускулатурой ланцетника и позвоночных и в со-
ответствии с этим двигательный отдел центральной нервной систе-
мы, представленный у червей стволами брюшной нервной цепочки,
теряет свое значение и редуцируется. Развививается иннервация
осевой мускулатуры за счет ближайшего участка нервного сплете-
ния покрова первичных хордовых. Срединная часть спинного покрова,
продолжая закладку воротничкового мозга, обособляется в виде
впячивания, дающего нервную трубку спинного мозга. Таким об-
разом, заново создается двигательный туловищный отдел централь-
ной нервной систем^!. Он отличается у ланцетника еще чрезвычай-
но слабой дифференцировкой двигательных нервов, представленных
рыхлыми, короткими пучками волокон, направленных к осевой ча-
сти каждой миомеры.
Характерно для ланцетника совмещение функций центра и пе-
риферии, другими словами, комплексность спинного мозга, имеюще-
го по всей длине, наряду с ганглиозными элементами, фоторецепто-
ры примитивного типа. Это, как мы видели, стоит в теснейшей свя-
346
зи со своеобразием его образа жизни. В комплексности находит вы-
ражение и иной способ зактадки нейтральной ибрвной системы у
высших хордовых по сравнению с прочими типами. Именно — пу-
тем удаления из покрова целого медиодорзального участка покров-
ного эпителия со всем разнообразием и богатством его элементов.
Рис. 150. Нервная система ланцетника: А—головной отдел (по
Купферу); Б— поперечный срез спинного мозга (по Гессе);
1 — переднее расширение нервной трубки, 2 — рудимент глаза, 3 — стато-
реиептор, 4 — центральный канал. 5 — скопление ганглиозпых клеток у щели
переднего расширения, 6 — гигантские нервные клетки, 7 — глазки Гессе,
8 — пучки гигантских волокон, 9 — ганглиозные клетки, 10 — эпендимные
клетки задней борозды, И — нервные волокна
Специфично, наконец, слабое развитие органов чувств в голов-
ном мозгу, представленных рудиментом глаза и простым статоре-
цептором, что обусловлено опять-таки, как было выяснено выше,
оригинальным образом жизни ланцетника. В то же время задний
отдел мозгового пузыря усложняется образованием медиодорзально
щелеобразной складки окруженной скоплением крупных ганглиоз-
ных элементов. Периферические нервы обнаруживают метамерность
до самого переднего конца мозга, где отходит единственный, неви-
димому, неметамерный нерв к хеморецептору келликеровской ямки.
Резюмируем все вышесказанное. Момент защиты многообразно
отразился на организации ланцетника. В качестве пассивной защи-
347
ты он привел к кутикуляризации покрова, далее к ланцетовидной
форме тела со спинным, брюшным и хвостовым плавниками, затем
к образованию складок метаплевр и, соответственно, образованию
атриальной полости, наконец к выработке лопатки переднего конца
тела. В качестве активной защиты выработалась дифференцировка
миохордального комплекса с мощным развитием продольной бо-
ковой мускулатуры, образованной метамерными участками. Весь об-
лик ланцетника обусловлен, следовательно, жизнью в грунте, кото-
рый его защищает, и активными движениями в толще воды, а так-
же проникновением в тот же грунт при бегстве от опасности.
Миохордальный комплекс, первоначально выработавшийся в
онтогенезе сидячих предков как приспособление, дающее самим ак-
том движения возможность защиты, а затем используемое для рас-
селения прикрепленных животных, и форма тела, подобная личин-
кам асцидий, сохраняются у ланцетника. Последующее усиление и
распространение миохордального зачатка до переднего конца тела
происходит уже при переходе к подвижной жизни в грунте и в при-
донных слоях воды. Таков ход выработки рассматриваемого за-
щитного комплекса черт организации ланцетника.
Во внутреннем строении с ним тесно связаны изменения в важ-
нейшей системе, являющейся системой внутренней среды, системе
целома. Дифференцировка мио-хордального зачатка ведет к редук-
ции спинной части целома и к подразделению единой закладки
вторичной полости тела в соответствии с побочными функциями
целома на спланхнотом, охватывающий кишечник, и на боковые от-
делы нефро- и гонотомов. Последняя дифференцировка связана с
сохранением двух примитивных черт организации ланцетника, уна-
следованных от немертинообразных протоцеломат, предков хордо-
вых. Здесь подразумеваются протонефридии с типичными пучками
соленоцитов в жаберном отделе тела и мешковидные гонады в за-
жаберном отделе. Однако гонады уже вступают в некоторую связь
с целомом, за счет стенки которого образуются зачатки выводных
цротоков. Наконец, образование миотомов и мощной боковой мус-
кулатуры влечет за собой дифференцировку приносящих и вынося-
щих кровеносных сосудов, становящихся значительнейшей частью
кровеносной системы. С другой стороны, образование миотомов при
редукции брюшного мозга червеобразных предков обусловливает
образование спинного мозга как двигательной нервной системы ту*-
ловища.
В известной мере, независимо от ведущего защитного комплек-
са черт организации ланцетника стоят характерные, унаследован-
ные от первичных хордовых особенности системы питания, а имен-
но —• пассивного питания сидячих или трубкоживущих предков. У
бесчерепных происходит только крайне значительное увеличение ко-
личества жаберных щелей с редукцией при этом первичного рта и
заменой его второй левой жаберной щелью. В соответствии с этим
в кровеносной системе на приносящих сосудах каждой из 90—100
первичных жаберных щелей происходит выработка сократитель-
348
ных отделов — bulbilli. Единое -сердце отсутствует. С увеличением
числа жаберных щелей происходит вторичное увеличение числа
нефридиев и их кровеносных сплетений, а также удлинение а три-'
альной полости.
Значение рассмотренного комплекса в организации ланцетника
стоит уже на втором плане как своеобразно используемое насле-
дие сидячих предков. В известной мере оно стоит наравне с пече-
нью как с начинающим свою дифференцировку важнейшим образо-
ванием, связанным с кишечником, но принимающим функцию ре-
гулятора внутреннего обмена. Образование печеночного выроста
также ведет к увеличению атриальной полости, ио важнее всего
значение печени выражается* в дифференцировке воротной системы
в и1<ровообр а щен ин.
Как видно, все три комплекса, находя каждый свое выражение
в кровеносной системе, создают характерный для высших хордовых
сложный тип этой системы.
В известной мере независимым, но стоящим в связи с защитным
комплексом, является фоторецепторный аппарат спинного мозга
ланцетника — глазки Гессе. Их развитие одновременно обусловли-
вает рудиментацию глаз в головном -мозгу. Наконец слабое выявле-
ние дифференцировки головного мозга у ланцетника обусловлено
слабым развитием органов чувств, стато- и хемерецепторов у живу-
щего в грунте животного.
На примере ланцетника видно, как приспособительная черта ор-
ганизации высших хордовых — миохордальный комплекс — дости-
гает своего крайнего развития и сопровождается рядом узких при-
способлений при упрощении условий существования, а именно —
при переходе к жизни в малодоступном грубом песчаном грунте с
сохранением пассивного типа питания предков.
В каких же конкретных условиях существования могли проте-
кать подобные процессы? Попробуем на основе анализа организа-
ции ланцетника дать предположительный ответ на этот вопрос.
Выработка организации ближайших родичей Асгапiа — туни-
кат показывает, что их асцидиеподобные предки приобрели все
свои характерные черты в связи с прикреплением к твердому суб-
страту. Они овладели каменистым грунтом в простых биоценозах,
а в сложных поселились на водных растениях, твердых раковинах
и панцырях животных. Лишь немногие'проникли на такие грунты,
как песок и пл, и своеобразно к ним приспособились. Тот же мо-
мент прикрепления приходится принять ответственным и в выра-
ботке организации ланцетника, хотя прикрепленная стадия в его
развитии совершенно' сглажена. Легко можно представить себе
особые условия жизни, к которым приспособились предки ланцет-
ника, если припомнить смешанные биоценозы на неглубоко лежа-
щих каменистых грунтах с наносами песка. Песок подвижен, легю>
перемывается, как это прекрасно описал, например, С. А. Зернов
для Черного моря. Прикреплявшиеся на твердом каменистом суб-
страте животные могли в известной степени покрываться наносными
349
частицами, чем создавалась для них лучшая защита от врагов на- * 5
подобие того, как дело обстоит теперь с камбалами. При этом выра-
батывались защитные приспособления для жаберных щелей в виде
метаплевр и атриальной полости. При проникновении в грубопесча-
ный грунт-предки ланцетника, повидимому, перестали прикреплять-
ся, перешли к свободному существованию, но закономерно сохра-
нили биотоп песка и пассивное питание из придонного слоя воды,
чем создался оригинальнейший образ жизни современного ланцет-
ника. В дальнейшем некоторые формы (Asymmetrdn и Amphioxi-
des) перешли к бонтонному существованию, чем завершается освое-
ние ими биотопов, доступных для свободноподвижных форм. Для й
них вполне допустима и своеобразная эволюция лежавших на |
одном боку животных, как это имеет место у камбал. Но нет ника-
ких достаточных данных принимать такой образ жизни для пони-
мания организации исходных форм предков ланцетника.
ПОЗВОНОЧНЫЕ — VERTEBRATA
Пассивное питание стало для бесчерепных непреодолимым барье-
ром на пути всестороннего повышения организации и вело к узкой
или даже крайней приспособленности подтипа. Этот тормоз был
преодолен предками позвоночных. Они перешли к активному пита-
нию, что в соединении с активной защитой свободноподвижного
животного создало для сложной и богатой широкими приспособле-
ниями организации позвоночных возможности эволюции до самых
высоких ступеней дифференцировки и, в то же время, интеграции
организма. В связи с этим у высших их представителей «основой
всей организации» становится нервная система. По И. П. Павлову,,
влияние нервной системы с развитием больших полушарий мозга
распространяется «на возможно большее количество деятельностей
организма», кора становится органом, выполняющим функцию об-
разования новых связей между организмом и средой, функцию при-
способления животного организма к изменяющимся условиям
среды.
Пути эволюции позвоночных были предметом многочисленней-
ших работ и нашли свое выражение в последнем .обобщающем тру-
де А. Н. Северцова (1939). Поэтому мы ограничимся лишь крат-
ким анализом организаций их представителей, чтобы завершить эту
сторону наших исканий С
У низших позвоночных мы не находим развития какой-нибудь
одной черты, которая наложила бы свою печать на всю их органи-
зацию, стала бы единственно ведущей, как, например, образование
туники у асцидий среди хордовых. Будучи прекрасной пассивной
защитой для сидячего животного, туника превратила асцидию в
недоступный врагу мешок, для которого не нужно ни активного
движения, ни хорошей ориентировки в окружающей среде. Нужен
1 Ввиду краткости очерка и общеизвестности материала по позвоночным
рисунков не дается.
350
только ток воды, пассивно несущей пищу в кишечник, причем сама
пища такого рода, что не требует дальнейшей активности живот-
него при ее захватывании и переработке. Подобного рода образо-
вание представляет и раковина при выработке организации мол-
люсков или хитиновый покров у членйстоногих. Таких черт пассив-
ной защиты, которые стали бы ведущими в организации всех позво-
ночных, в их эволюции нет. Напротив, их защита по преимуще-
ству активна. Она связана, с одной' стороны, "с "подвижностью жи-
вотного, обусловленной развитием миохордального комплекса и его
производных, плавников и парных конечностей. С другой стороны,
защита связана и с ориентировкой животного в окружающей сре-
де, т. е. с поведением, определяющимся развитием комплекса ор-
ганов чувств и нервной системы. Но те же два комплекса разви-
ваются и ради обеспечения питанием активного хищника, какими
явились исходные формы позвоночных. Таким образом, и активная
защита и активное питание объединяются в эволюции подтипа,
обусловливая многообразие и богатство вырабатывающихся форм,
тогда как разница путей приспособления к защите, с одной сторо-
ны, и к питанию, с другой, как, например, у туникат, в известной
мере ограничивает возможности эволюции. У ланцетника пассивное
питание сидячего в грунте животного и активная защита сводобно-
плавающей формы дали специфическую, узкоограниченную в своих
возможностях организацию. Вне песчаного грунта моря мы редко
находим ланцетника, тогда как позвоночные овладели различней-
шими биотопами на всем земном шаре.
Однако в организации позвоночных богато развивается и комп-
лекс, связанный с пассивной защитой в виде разнообразных покров-
ных образований, и комплекс дифференцировок кишечника, связан-
ный с различнейшими формами питания. Наконец, во внутреннем
строении у позвоночных место целомического комплекса, который в
значительной мере отходит на задний план, сохраняя только экскре-
торную и половую функции, занимает кровеносная система. С эндо-
телизацией ее стенок она становится в высокой степени независи-
мой, а вместе с тем вырабатывается сложный комплекс хемо- и
бдарегуляций — эндокринная и фагоцитарная системы.
Совокупность столь многосторонне охватывающих всю жизнь
животного комплексов делает организацию позвоночного способной
приспосабливаться к различнейшим условиям среды от водной жиз-
ни до наземного и даже воздушного существования.
Особенно велико в этом отношении значение воздушной среды,
обладающей большим разнообразием и большим диапазоном ее
факторов по сравнению с водной средой, неизмеримо более однооб-
разной. Освоение наземной среды обусловливает широчайшие мас-
штабы эволюции позвоночных. Создается действительно наиболее
сложная и совершенная организация победителей в жизни, не огра-
ничиваемая какими-либо трудно преодолимыми односторонними
приспособительными чертами вроде хитинового покрова членисто-
ногих или раковины моллюсков, а обусловливаемая необычайным
351
развитием нервной системы, в частности полушарий головного
мозга, их коры.
Для иллюстрации сказанного рассмотрим в самых общих кон-
турах перечисленные комплексы черт организации позвоночных.
Покров, как всегда, прежде всего служит в качестве пассивной
защиты для тела животного, и в этом направлении его эволюция
у позвоночных определяется двумя характерными чертами. Во-пер-
вых, у них выработалась ^косослойность кроющего эпителия, что
при большом количестве входящих в” его состав клеток'обусловли-
вает возможность разнообразных дифференцировок. Во-вторых, под-
стилающий слой ткани основного вещества достигает значительной
мощности и прочности и опять-таки содержит значительное количе-
ство различных элементов, за счет которых могут возникать разно-
образные структуры. При сравнении с ланцетником бросается в
глаза сложность покрова уже у низших представителей позвоноч-
ных. У круглоротых развивается покров с кроющим эпителием, бо-
гатым одноклеточными слизистыми железами, а у рыб, кроме того,
с отложениями извести в соединительнотканном слое. У акуловых
это — плакоидные чешуи, образованные дентином кориума и по-
крытые эмалевым слоем, происходящим уже за счет эпителия. У
прочих рыб на этой основе в соединительнотканном слое разви-
ваются костные чешуи и далее настоящие покровные кости, которые
принимают участие в построении некоторых частей скелета позво-
ночных со специальной функцией, как. например, накладные кости
черепа, защищающие головной мозг.
У амфибий образование многочисленных многоклеточных ^сли-
зистых желез, проникающих в соединительнотканный слой покро-
ва, создало вместе с утратой костных чешуи возможность выхода на
сушу, возможность овладения такими условиями существования, в
которых решающим моментом является способность противостоять
иссушивающему влиянию воздуха. Это приспособление, еще слабо
развитое у амфибий и проявляющееся лишь у взрослых особей —
личинки обычно ведут водное существование — тем не менее откры-
вает широкие возможности эволюции в условиях наземной среды.
Полностью наземные условия существования осваиваются лишь
рептилиями, у которых постоянное увлажнение покрова слизью, ха-
рактерное для амфибий и далеко несовершенно защищающее их от
высыхания и мумификации, заменяется образованием непроницае-
мого для жидкостей ороговевающего слоя покровного эпителия. Вы-
рабатываются роговые чешуи и роговой кожный панцыръ, но реду-
цируется железистый аппарат. Вместе с этим для высших позво-
ночных становится доступным почти все пространство необъятной
суши, и не только суши, а также и водной стихии, куда они и их
потомки вторично проникают.
Эволюция рогового покрова повела к полному освоению назем-
ной среды (в смысле ее сухости), но далее и к освоению свойствен-
ных суше контрастных температурных условий. С выработкой за
счет роговых чешуи и других ороговений кожи своеобразнейшей
352
оболочки из перьев у птиц и из волос у млекопитающих покров в
той или иной степени предохраняет организм от влияния колебаний
температуры наружной среды. Такой изолятор был первым усло-
вием для эволюции животных с постоянной температурой тела, 'так
как создавал независимость от временных повышений и пониже-
ний температур, а вместе с тем и возможность проникновения в об-
ласти постоянно низких температур, овладения всеми частями зем-
ного шара.
У млекопитающих в теплорегуляции участвуют и потовые же-
лезы кожи, способствующие понижению температуры тела при вы-
соких температурах окружающей среды, защищающие от перегре-
вания. Этого нет у птиц, которые, используя покров из перьев для
передвижения при полете в воздухе, освоили прилегающие к поверх-
ности земли слои атмосферы.
Таково значение в организации позвоночных комплекса покрова,
в той или иной мере определяющего’ весь внешний облик этих жи-
вотных и проделывающего сложную и многообразную эволюцию от
сравнительно простого состояния у низших позвоночных с главной
функцией пассивной защиты до покрова птиц и млекопитающих,
сложно дифференцированного и в других направлениях.
Другой комплекс черт организации, к которому мы, естественно,
переходим, это миохордальный комплекс, связанный'прежде всего со
свободной подвижностью высших хордовых, а у позвоночных еще
и обусловливающий! активную защиту от врагов или неблагоприят-
ных условий и обеспечивающий возможность нападения на добычу
при питании. Он играет в известной мере ведущую роль в органи-
зации позвоночных.
У круглоротых полностью выражены отношения, типичные для
низших позвоночных. Имеются хорошо развитая, доходящая до го-
ловного мозга хорда и мощные боковые мышцы, образованные мио-
мерами, еще не подразделенными соединительноткашной прослойкой
на спинной и брюшной отделы. Эта мощная мускулатура длинного
тела круглоротых дает при его синусоидных изгибах прямолинейное
движение. При этом значительная роль выпадает на долю хвостово-
го отдела тела, располагающегося за анальным отверстием. У рыб
па этой основе вырабатывается хвостовой плавник, становящийся
как бы специальным органом, продвигающим тело вперед наподо-
бие винта парохода.
Боковые мышцы на передней части туловища утрачивают свое
значение для продвижения тела. Уже у круглоротых три передние
миомера совершенно потеряли эту функцию, превратившись в
мышцы, двигающие развивающееся здесь значительное по разме-
рам глазное яблоко. Следующие два миомера с развитием слуховой
капсулы редуцируются. Эти отношения характерны и для рыб.
Наконец, у круглоротых появляется еще одна черта, далее при-
сущая рыбам. Речь идет о том, что складки непарного плавника,
имеющегося уже у ланцетника и стабилизирующего прямолинейное
движение, получают мускулатуру, обособляющуюся в виде неболь-
23 Пути эволюции животного мира 353
ших почек от 'миотомов на спинном и брюшном краях, Однако
парных плавников у круглоротых еще нет, благодаря чему их дви-
жения отличаются от движения рыб, у которых появляются парные
плавники. Плавники дифференцируются как боковые складки стенки
тела, в которые проникают метамерные мышечные пучки от каждого
миомера. Таким образом, наряду с единой складкой непарного
плавника, появляются боковые плавники как рули глубин.
Далее в миохордальном комплексе рыб возникают и быстро
развиваются скелетные образования, заменяющие простую прими-
тивную хорду и обусловливающие дальнейшую дифференцировку
мускулатуры. Развитие скелета детерминировано тем, что обычная
система^ пограничных пластинок ткани основного вещества, имев-
шаяся у баланоглосса и ланцетника, превращается у позвоночных
в более или менее значительные прослойки соединительной ткани,
богатой клетками. Богатство клеточными элементами характерно и
для прочих систем органов этого подтипа, и мы уже отметили раз-
витие соединительнотканного слоя кориума в их покрове.
Клеточный материал для соединительнотканных образований
исчерпывается из тех же мезодермальных сомитов, а именно из
склеротомов в осевом комплексе и из париетального листка стенки
пелома. Это так называемая вторичная мезенхима или целенхима.
Обилие клеточных элементов, будучи характерной чертой орга-
низации позвоночных, особенно важно для эволюции подтипа, так
как дифференцировки у них идут в большей степени за счет орга-
нотипического развития, а не цитотипичеокого, чем открываются
большие возможности для осложнения и дальнейшей эволюции. В
этой связи обратимся к скелетным образованиям.
Первые зачатки скелета позвоночных развиваются сходно с хря-
щами жаберного аппарата баланоглосса и ланцетника в соедини-
тельной ткани жаберных щелей, поддерживая их в неспадающемся
состоянии. У круглоротых образуется жаберная коробка, которая
своим задним отделом охватывает сердце, принимая на себя защиту
этого существенно важного органа. У рыб это — жаберные дуги,
хрящевые еще у акуловых, а у прочих уже костные. Во-первых, они
не обнаруживают каких-либо отношений к сердцу. А во-вторых,
этот висцеральный скелет находится в ближайшей связи не с мио-
хордальным комплексом, а со стенкой кишечника.
Ближе к миохордальному комплексу стоят другие скелетные об-
разования, имеющиеся у круглоротых. Таковы зачатки кеврапофи-
зов, дорзально примыкающие к соединительнотканной оболочке
хорды. Но эти зачатки, как и лежащий впереди череп круглоротых,,
являются собственно защитными образованиями для мозга и перво-
начально развиваются в связи с комплексом нервной системы и
органов чувств.
Лишь yj3bi6 возникают тела позвонков, создается позвоночник—
характерйЖшая черта организации позвоночных, — входящий, та-
ким образом, существенной частью в состав миохордального комп-
лекса. Простая хорда заменяется более прочным позвоночником, у
351
низших форм хрящевым, а у высших костным. Первоначально
каждый позвонок закладывается диплоепондиличёски соответствен-
но передней и задней половинам примыкающих миосомитов. Воз-
можность дальнейшей дифференцировки различных участков скелета
стоит в связи с теми хрящевыми и костными образованиями, кото-
рые появляются в различных участках системы соединительноткан-
ных прослоек среди мышц. Таковы, например, ребра и т. и.
У рыб, как мы видели, независимо от позвоночника, но в связи
с миотрмами развиваются парные плавники. Их мускулатура перво-
начально представлена метамерными пучками, идущими ог миото-
?viob/ Соответственно и их опорные образования сначала хрящевые,
а дальше костные также закладываются метамерпо и самостоя-
тельно, как местные дифференцировки соединительной ткани. В
дальнейшем, однако, метамерность сглаживается, и плавники эво-
люционируют в резко обособленные органы, поддерживаемые неза-
висимыми от осевого скедета поясами хрящевого или костного ха-
рактера, происходящими как за счет первоначальных плавниковых
лучей, так, отчасти, и из покровных костей.
Если при переходе к наземным условиям существования у позво-
ночных выработались приспособления к иссушающему влиянию
воздуха и к большому диапазону температурных колебаний воздуш-
ной среды, что нашло свое отражение прежде всего* в эволюции
покрова, то малая плотность воздушной среды по сравнению с вод-
ной отразилась преимущественно на механизме опоры и движения,
т. е. на миохордальном комплексе и плавниках. Синусоидные
изгибы тела и удары хвостового плавника, продвигающие рыбу в
воде, теряют свое значение в воздушной среде при се малой плот-
ности. Сохраняются дающие оперу телу рычаги парных плавников.
Они эволюционируют в характерные, типа сложных рычагов конеч-
ности, поддерживающие тело и способные производить разнообраз-
ные движения, как для перемещения животного, так и служащие
для иного эффекта, например для удара, хватания и т. д. В связи
с этим они развиваются крайне разнообразно, включительно до
образования крыла птицы, служащего для парения и колета, или
руки человека со всем многообразием ее функций.
Необходимая для поддержания и передвижения тела в воздуш-
ной среде прочность прикрепления конечностей обусловливается
плотностью самого костного скелета и прикреплением костей поясов
конечностей на туловище. Вырабатывается непосредственная свяГЗь
заднего пояса с позвоночником, чем обусловлена дифференцировка
крестца. Сначала крестец содержит лишь один-два позвонка, а у
млекопитающих он образован двумя-четырьмя позвонками. У птиц
для образования огромного сложного крестца срастается большое
число позвонков, так как при превращении передних конечностей
в орган полета вся тяжесть тела поддерживается лишь на задних
конечностях. Передний пояс конечностей при выработке грудной
клетки наземных позвоночных связывается с позвоночником при
помощи ребер. Таким образом, миохбрдальный комплекс вступает
в теснейшую связь с конечностями. Это ведет к дифференцировке
соответствующих отделов позвоночника — шейного, грудного, ту-
ловищного, тазового и хвостового. Но особенно сильно выявляются
все эти отношения в дифференцировке первоначально однообразной
мускулатуры, имевшейся у рыб, на специальные мышцы, связанные
с теми или иными костями туловища и 'конечностей. Наиболее свое-
образно развитие летательных мышц и мышц ног у птиц, как ре-
зультат приспособления передних конечностей для полета и задней
пары конечностей для опоры всего тела.
Таким образом ясно, что эволюция миохордалыного комплекса
имеет большое значение в организации позвоночных. Однако' это
не исключает, но скорее подчеркивает своеобразие и значимость
для их организации третьего комплекса — комплекса органов чувств
и нервной системы с их придаточными образованиями. В жизни
животного эти системы играют важнейшую роль в защитен в обеспе-
чении питанием. Они теснейшим образом связаны с рассмотренным
комплексом активного движения. В эволюции все они взаимообу-
словлены, но в то же время развиваются в значительной мере свои-
ми особыми путями.
Основным моментом, характерным для нервной системы позво-
ночных и определяющим ее эволюцию, а вместе с тем и эволюцию
всего подтипа, явилось не только образование спинной нервной
трубки путем обособления вместе с нервными элементами и соот-
ветствующей части покровного эпителия. Это имеет место уже у
аппендикулярий, асцидий и ланцетника и ведет к появлению в цен-
тральной нервной системе чувствительных дифференцировок, ведет
к комплексности мозга. Но, помимо того, чрезвычайно существенно
накопление в закладках мозга позвоночных огромных запасов кле-
точных элементов. В любой системе это обусловливает возможность
дальнейших сложных дифференцировок в эволюции того или иного
комплекса.
В головном мозге аппендикулярий и асцидий имеется, как мы
видели, очень просто построенный глаз, что детерминировано малым
числом входящих в его состав клеток. Имеется также орган равно-
весия, а у ланцетника и предельно простые глазки по всей длине
нервной трубки. Однако при органотипическом развитии позвоноч-
ных то же явление дифференцировки в мозге чувствительных обра-
зований приводит уже у круглоротых к появлению весьма совершен-
ных боковых глаз. В связи с этим развиваются и очень сложные
чувствительные производные прилежащего к мозгу покрова —
орган обоняния и орган равновесия.
Нервная трубка ланцетника по всей длине представляет собой
туловищный спинной мозг. От последнего метамерно отходят с
брюшной стороны двигательные нервные пучки к миомерам, иду-
щим до переднего конца тела. А со спинной стороны в мозг также
метамерно, чередуясь с двигательными, входят из покрова нервы по
преимуществу чувствительного характера. Эта метамерная иннер-
вация имеет место и в переднем расширенном участке нервной
356
трубки ланцетника — его головном мозге, который отличается от
спинного только рудиментом глаза и развитием органа равновесия,
и как бы продолжается вперед в обонятельную ямку.
По существу те же отношения имеются и у круглоротых, но сво-
бодный образ жизни хищника и фрганотипическая дифференциров-
ка до неузнаваемости изменили их выявление. Во-первых, нервная
трубка получает защитную оболочку из хрящевых зачатков невра-
пофизов. На переднем расширенном участке нервной трубки, стано-
вящемся головным мозгом позвоночных, у круглоротых там, где
заканчивается хорда, в соединительнотканной оболочке также раз-
виваются хрящевые образования, защищая мозг снизу, со стороны
ротовой полости, и с боков. Это — первичный череп, открытый
сверху и сзади и не включающий еще обонятельного и отеческого
отделов. Последние входят в .состав черепа только у рыб. У круг-
лоротых они представлены независимыми защитными хрящевыми
капсулами, охватывающими орган обоняния и орган равновесия и
лишь прилежащими к черепу. Затылочный отдел, замыкая череп
сзади, развивается тогда, когда развиваются и тела позвонков, т. е.
лишь у рыб.
Несомненно, что образование сначала хрящевого, а у вышестоя-
щих форм костного черепа имеет важное защитное значение. В си-
стематике это находит свое отражение в названии подтипа, куда
относится ланцетник, «бесчерепные» — «Acrania» — в противопо-
ложность позвоночным — «Craniata». Однако гораздо существеннее
другое. У позвоночных происходит развитие нового головного комп-
лекса органов чувств, не имеющего ничего общего с головным
комплексом червей. Происходит и выработка соответственно повой
дифференцировки головного мозга.
Там, где на переднем конце мозга у ланцетника находится
небольшая обонятельная ямка с ее нервом и непарный рудимент
глаза, там у круглоротых при их свободном образе жизни и хищ-
ничестве дифференцируются важнейшие органы ориентировки во
внешней среде. На переднем конце развивается мощный хеморецеп-
трр в виде обонятельного мешка с отходящей от него первой парой
головных нервов — n. olfactorii. По бокам в мозге дифференци-
руется пара высоко организованных глаз со второй парой нервов —•
и. optici. Субстрат для развития обонятельных мешков был тот же,
самый, что и для обонятельной ямки ланцетника, а именно—чувстви-
тельный хеморецепторный эпителий в области невропора. Но в
жизни свободноподвижиой двусторонне-симметричной рыбообраз-
ной формы обонятельный зачаток подразделяется на правый и левый
отделы и соответственно образует два мощных нервных пучка к
передней части головного расширения мозга — к переднему голов-
ному пузырю (prosencephalon). Соответствующий отдел последнего,
специализируясь как особый сенсорный центр, обособляется в виде
переднего, или обонятельного, мозга (telencephalon).
Также функционально дифференцируется задний участок перед-
него пузыря в виде промежуточного мозга (diencephalon), пред-
357
ставляющего зрительный мозг. В его боковой и спинной стенках,
как и у туникат, локализуются светочувствительные элементы. Раз-
витие глаз в онтогенезе позвоночных наглядно показывает, как про-
текал процесс. Сначала будущие светочувствительные клетки сосре-
доточиваются в дистальных частях двух боковых выступов стенки
мозгового расширения. Затем, подходя к покрову, каждый из этих
выступов благодаря впячиванию слоя, содержащего светочувстви-
тельные клетки, принимает форму бокала. Получается внутренний
ретинальный слой, покрытый наружным изолирующим слоем, содер-
жащим пигмент. За счет покровного эпителия в этом месте обра-
зуется прозрачное линзообразное тело — диоптрический аппарат
глаза. Проксимальный отдел первоначального выпячивания превра-
щается в тонкую ножку. Поэтому дифференцирующееся глазное
яблоко получает возможность некоторого движения при прикрепле-
нии к соединительнотканной оболочке яблока мышечных пучков,
происходящих за счет первого, второго и третьего миотомов. Таким
образом расширяются возможности использования глаза для охвата
большего пространства окружающей среды. Последним шагом в
этом направлении будет выработка подвижности головы у наземных
позвоночных.
Мы видим, что вместо примитивного глазка асцидий и рудимента
глаза ланцетника при обилии соответствующего клеточного материа-
ла в передней части нервной трубки развивается высоко совершен-
ный глаз позвоночных. В связи с этим в качестве зрительного цент-
ра дифференцируется, как было указано, задний зрительный отдел
переднего расширения головного мозга — промежуточный мозг. В
дальнейшем к нему присоединяется и следующий отдел головного
мозга — средний мозговой пузырь (mesencephalon). Уже у кругло-
ротых процесс полностью завершен и их оптические нервы представ-
ляют лишь своеобразно измененные части того же головного мозга.
В переднем мозге мы, следовательно, видим новообразования,
связанные с дифференцировкой двух важнейших органов чувств
головы позвоночных. Создается новый головной комплекс органов
чувств и, в соответствии с этим, возникает и новый головной мозг.
Только в дальнейших отделах мозгового расширения, а именно — в
среднем и заднем мозговых пузырях мы находим нервы, характерные
для спинного мозга и относящиеся к передним пяти-сомитам тела. Из
этих сомитов три передние сохранили свои миотомы в качестве мышц
глазного яблока. И из их нервов два передние — n. oculomotorius
и n. trochlearis — относятся' к среднему мозгу, а задний —
п. abducens — относится уже к заднему мозгу.
Два следующих миотома, как было уже указано, подверглись
редукции в связи с развитием еще одного очень важного для свобод-
ноподвижного животного и характерного для позвоночных головного
органа чувств — органа равновесия. Дальнейшие миотомы дают
уже соматическую мускулатуру.
В мозговом расширении аппендикулярий, асцидий и ланцетника
имеется своеобразный орган равновесия, который расположен в
3 53
брюшной части мозгового» пузыря, У позвоночных мы находим
сходно построенную рейснерову нить, начинающуюся у задней гра-
ницы промежуточного мозга. Однако в новом головном комплексе
органов чувств позвоночных роль совершенного органа равновесия
выполняет иное образование, (развивающееся за счет механорецепто-
ров покрова головы. Развивается внутреннее ухо, не имеющее ни-
чего общего с органом равновесия в мозге низших хордовых.
У низших водных позвоночных для восприятия колебательных
движений водной среды специализировались особые механорецеп-
торы — сейсмосенсорные органы. Они связаны на голове с ветвями
п. facialts, т. е. относятся к сомитам головы, но распространяются
по телу рыбы далеко назад, образуя так называемую «боковую
линию». В этой части их иннервация происходит за счет дальнейших
головных нервов — n. glossopharyngeus и, главным образом,
n. vagus. У наземных позвоночных с выходом из воды сейсмосен-
сорная система теряет свое значение и подвергается редукции.
Однако часть механорецепторов четвертого сомита головы диффе-
ренцируется в несколько ином направлении, чем остальные, и, в
соответствии с этим, из n. facialis выделяется особый нерв —
n. acusticus. Обособляясь, эти механорецепторы располагаются в
мешковидных впячиваниях покрова задней части головы и образуют
у круглоротых слуховой пузырек, защищенный особой хрящевой
капсулой, прилежащей к черепу. Последняя входит в состав черепа
только у рыб. Как уже было указано, основная функция этого об-
разования — статорецепторная. Развитие ее связано с замыканием
слухового пузырька и особенно выражается в дифференцировке на
нем трех полукружных каналов соответственно трем измерениям
пространства. У круглоротых таких каналов дифференцируется
только два. У наземных позвоночных происходит дальнейшая диффе-
ренцировка начальной закладки. Развивается аппарат улитки вну-
треннего уха. С другой стороны за счет покрова образуется мем-
брана для восприятия воздушных колебаний — барабанная пере-
понка и вспомогательные аппараты среднего и наружного уха для
передачи воздушных колебаний на чувствительные образования
среднего уха.
С развитием этого третьего существенного компонента в голов-
ном комплексе органов чувств позвоночных происходит и в головном
мозге обособление особого отдела заднего мозга — мозжечка, содер-
жащего центры координации и точности движения. Он прекрасно
развит и является в известной мере ведущим отделом у акуловых
и костистых рыб, наряду со специфическим развитием у первых из
них обонятельных, а у вторых зрительных центров. Говоря просто —
акулы живут обонянием, а костистые рыбы — зрением.
У низших позвоночных эти образования сравнительно просты, но
у высших — птиц и млекопитающих—эволюция ведет опятькслож-
нопостроенным мозжечку, зрительным и обонятельным отделам.
Особое значение в эволюции высших позвоночных получают
ассоциативные центры, развивающиеся на основе переднего обоня-
359
тельного отдела мозга — создаются полушария головного мозга.
Получаются мощные центры высшей нервной деятельности млекопи-
тающих. У человека кора полушарий большого мозга при огромном
количестве клеток, участвующих в ее построении, становится унивеп-
сальным анализатором, развитие которого определяет весь дальней-
ший ход эволюций Homo sapiens. Вырабатывается вторая сигналь-
ная система, вырабатывается мышление, даюш.ее возможность по-
знавать окружающий мир, использовать и перестраивать его в
интересах человека.
По сравнению со значением последнего из рассмотренных нами
комплексов характерных черт организации хордовых — комплекса
органов чувств и нервной системы, — у высших позвоночных на
задний план отходят все прочие комплексы черт. Однако связанный
с питанием четвертый комплекс играл у позвоночных очень большую
роль. При переходе к активному питанию свободноживущего живот-
ного, этот комплекс радикально изменил направление эволюции
других частей.
Выработка активного движения первоначально служила у при-
крепленного животного лишь для защиты самим актом движения,
а затем и для расселения вида -при отделении от субстрата. Но она
создавала возможность и для активного захватывания пищи. Как
это можно наблюдать у миноги, ее личинка Ammocoetes еще обла-
дает аппаратом эндостиля, характерным для пассивного питания
низших хордовых. Взрослое животное утрачивает эндостиль ш пере-
ходит к питанию активно захватываемой крупной пищей. У прочих
позвоночных стадия, подобная пескоройке, выпала. Соответственно,,
в несущем жаберные щели переднем отделе эктодермального кишеч-
ника, происходят характерные изменения. Во-первых, за счет уча-
стка кишечника, принадлежащего первым трем сомитам, вырабаты-
вается ротовая полость. Но эволюция необратима. У хордовых, как
мы видели, полностью редуцировался стомодеальный отдел кишеч-
ника, происходящий у прочих типов как впячивание эктодермы и
дающий начало ротовому и глоточному отделам. И у позвоночных
ротовая полость образуется из эктодермального переднего участка,
в котором жаберные щели, а именно—первая и вторая пары, не за-
кладываются. Благодаря этому захватываемая пища удерживается
без какой-либо утечки. Механическое изменение пищи идет за счет
покровных образований, вторично присоединяющихся и врастаю-
щих в ротовую полость, — роговых зубов в ротовой воронке у круг-
лоротовых и настоящих зубов, представляющих видоизменения
костных плакоидных чешуи, у прочих -позвоночных.
Во-вторых, не способствуя более приносу пищи, прохождение
воды через жаберные щели может, как раз наоборот, только спо-
собствовать ее утечке, жаберные щели остаются простыми, а число
их не увеличивается вторично, остается небольшим. Обычно это 5
или 4 пары. Но несут они уже исключительно функцию газообмена.
В соответствии с этим у миног произошло обособление нижней
части отдела с жаберными щелями — дыхательной трубки — от
360
верхней, служащей только в качестве пищевода? -При этом служа-
щая для газообмена пове;рхность значительно увеличивается за счет
образования многочисленных складок энтодермального эпителия в
каждой жаберной щели, расширяющейся наподобие мешка. Таким
образом, при малом числе и простом строении жаберных щелей,
дыхание достигает большой интенсивности, необходимой для свобод-
неподвижного животного в противоположность сидячим формам.
У рыб тот же эффект достигается иным путем. Жаберные щели,
будучи простыми и в малом количестве, сближаются, занимая лишь
очень короткий участок глотки, и несут с одной стороны ворсинки,
не пропускающие крупных частиц пищи из кишечника, а с дру-
гой — массу складок жаберных лепестков, происходящих уже за
счет покрова и служащих исключительно для газообмена.
Отношения радикально меняются с переходом к наземному су-
ществованию. Взамен редуцирующегося жаберного аппарата выра-
батываются органы воздушного дыхания — легкие. Первоначально'
это полые выросты в виде придатков энтодермальной кишки за
последней жаберной щелью, содержавшие газ и игравшие у рыб
роль регуляторов водного давления. В дальнейшем содержание на-
ходящегося в них газа стало регулироваться путем газообмена с
кровеносными капиллярами их стенок. При переходе к наземной
жизни роли переменились. Газообмен превратился в дыхательный
процесс, доставляющий кислород в кровеносную систему и удаляю-
щий, из него углекислоту. Интенсификация движения связана у
наземных млекопитающих с большой сложностью и самих легких,
и проводящих дыхательных путей, а у птиц, сверх того, и со слож-
нейшими приспособлениями к дыханию во время полета. Образу-
ются воздушные мешки и развивается пневматичность костей птиц.
В теснейшей взаимосвязи с развитием жаберного аппарата, а за-
тем легких стоит эволюция кровеносной системы позвоночных. Это
наметилось полностью уже у ланцетника, поскольку у него жабер-
ные щели получают кровь от брюшного сосуда и передают ее,
насыщенную питательным материалом из стенки усваивающей
кишки и кислородом из жабер, прежде всего мощной боковой му-
скулатуре, характерной для рыбообразных форм. Отсюда через
боковые вены, а затем Кювьеровы протоки она попадает снова к
жабрам.
У позвоночных, однако, основным и ведущим в кровеносной си-
стеме является ее эндотелизация. У беспозвоночных кровеносный
сосуд представляет собственно участок ткани основного вещества,
где оно, перейдя из состояния геля в состояние золя, стало подвиж-
ным и взяло на себя прежде всего функцию переноса питательных
веществ от усваивающей кишки к другим частям тела. Клеточные
элементы присущи крови постольку, поскольку они присущи вообще
ткани основного вещества. Но в сосудах на стенках, представляю-
щих собой пограничные перепонки, различные клетки особенно
часто оседают или стабильно дифференцируются. Отсюда неверное
утверждение об эндотелии как постоянном и характерном компонен-
361
7'е стенки сосуда немертин и других червей. Эндотелизация, т. е.
специальная клеточная выстилка сосудов с ее особыми свойствами,
вырабатывается лишь у хордовых. У асцидий мы видим первые
шаги в этом направлении. Эндотелий развит только в области
сердца и отходящих больших сосудов.У ланцетника он имеется во
всех крупных сосудах. Наконец, у позвоночных эндотелиальная
выстилка становится непременной составной частью всех сосудов до
капилляров включительно. Лакунарные пространства являются
редким исключением со специальной функцией при кроветворении.
В соответствии с этим в эмбриональном развитии позвоночных за-
кладка кровеносной системы происходит путем обособления спе-
циального клеточного зачатка (ангиобласт некоторых авторов),
дающего начало и эндотелию стенки и элементам крови.
Таким образом, создается известная независимость кровеносной
системы от соединительной ткани. Кровь в некоторой мере изоли-
руется от других жидкостей организма, как своеобразная среда с
трофической функцией. Далее ей присуща функция газообмена и
юна становится также средой, регулирующей химизм организма в
связи с деятельностью органов внутренней секреции. Кровь в то же
время еще больше связывается со всеми другими органами и играет
важнейшую роль в интеграции организма.
Значение кровеносной системы у позвоночных возрастает как
бы в противовес ослаблению значения целома и целомической жид-
кости.
Мы видели, что с образованием миохордального комплекса ше-
лом теряет свою основную функцию опоры. Она переходит к вну-
треннему скелету позвоночных. Побочные его функции — трофиче-
ская, экскреторная и половая — эволюционируют различно. Тро-
фическая функция при мощном развитии кровеносной системы и
при редукции целома в голове позвоночных теряется. Но экскретор-
ная и половая функции, присущие туловищному целому, сохраняют-
ся и за целомом позвоночных. Сохраняются, впрочем, в ограничен-
ной степени постольку, поскольку целомодукты используются для
выведения экскретов и половых продуктов. Основное значение
целома у позвоночных сводится к роли общей полости тела, вме-
щающей кишечник и его придаточные образования, например пе-
чень. Сзади целом простирается до анального отверстия, так как
дальнейший участок туловища не содержит кишки и у рыб пере-
ходит в хвостовой плавник, особенно сильно развитый и служащий
для движения. У взрослых форм позвоночных нет каких-либо при-
знаков вторичной метамерности в целоме, а потому кишечник и пе-
чень свободно в нем подвешены. Для их увеличения и в длину ц в
объеме препятствий нет. Это важно в связи с тем, что первоначаль-
но у низших позвоночных (круглоротых и акуловых рыб) длина
кишечника очень незначительна при наличии длинного заанального
хвостового отдела. Такая дисгармония исправляется у вышестоящих
форм значительным увеличением длины кишечника путем образова-
ния извивов его различных частей. ,
362 /
У круглоротых полость целома едина, хотя за счет его отдела,
примыкающего сзади к жабрам, образуется вокруг брюшного сосуда
перикард. Мышечная стенка перикарда, прилежащая к эндокарду,
т. е. эндотелию сосуда, представляет миокард. Таким образом, у
позвоночных создается единое сердце, которое при малом количе-
стве жаберных щелей оказывается в состоянии продвинуть кровь
через всю систему жаберных капилляров. Это было невозможно у
ланцетника и асцидий с увеличением у них количества жаберных
щелей.
У водных позвоночных благодаря образованию клапанов,ч пре-
пятствующих оттоку крови назад, продвижение ее связано с посте-
пенным увеличением давления от венозного синуса, куда собирается
вся венозная кровь, последовательно в предсердии,'а затем в желу-
дочке сердца.
У рыб и выше стоящих позвоночных перикардиальный отдел
уже дифференцирован* от остального целома — брюшной полости
позвоночных. У наземных их представителей в брюшную полость
спереди вдаются обширные мешки легких, причем у птиц еще и
связанные с легкими воздушные мешки. Лишь у млекопитающих
происходит дальнейшая дифференцировка. Вокруг легких обособ-
ляется участок, образующий полость плевры, в связи с чем про-
цесс дыхания в значительной мере становится независимым от со-
стояния прочих внутренностей брюшной полости.
Таков в общих, конечно, чертах ход эволюции целома у позво-
ночных. Целом все суживается и ограничивается в своих функциях
и строении. Его антипод — кровеносная система осложняется и
дифференцируется. В едином круге кровообращения от лимфати-
ческой системы, неимеющей эритроцитов, обособляется система, со-
держащая красную кровь с эритроцитами и несущая, наряду с
трофической, также функцию газообмена. Уже у амфибий лимфа-
тическая система сложно построена и обладает особыми сократи-
тельными отделами, так называемыми лимфатическими сердцами. У
высших позвоночных она располагает собственными центрами раз-
множения клеточных элементов — лимфатическими железами
и т. п.
Далее, в круге кровообращения, содержащем эритроциты, про-
исходит дифференцировка, возникающая в связи со снабжением
различных органов кровью, в различной степени насыщенной кис-
лородом. Дифференцируется малый, связанный с легкими, и боль-
шой системный круги кровообращения. В соответствии с этим вы-
рабатывается подразделение предсердия на правое и левое —
трехкамерное сердце амфибий, а затем и желудочка на правый и
левый — четырехкамерное сердце птиц и млекопитающих.
Известно, что уже у ланцетника имеется воротная система пе-
чени, а у позвоночных печень становится массивной железой, кото-
рая перерабатывает приносимые кровью жидкие продукты распада
белковых соединений в мочевину, далее удаляемую из крови почка-
ми. У низших позвоночных соответственно развивается воротная
363
система почек, исчезающая, однако, вторично у высших позвоноч-
ных.
Так, кровеносная система получает огромное значение в химиз-
ме организма и, конечно,^ эндотелизация сосудов имеет при этом*
немалое значение. У высших позвоночных к этому присоединяется
дифференцировка желез, обусловливающих содержание в крови
ряда гормонов. Это*главным образом производные аппарата эндо-
стиля и жаберных дуг, а именно—зобная, щитовидная и т. и. желе-
зы, затем органы нейросекреции — надпочечники и т. п. и, наконец,
обусловливающие и регулирующие половую дифференцировку го-
надотропные железы. Создается характерная для высших позвоноч-
ных важнейшая система гормональных дифференцировок1.
Экскреция у позвоночных, как мы видели, связана с массой кро-
веносных капилляров, которые развиваются в связи с метамерными
каналами нефротома. У круглоротых они начинаются сейчас же за
жаберным отделом в области перикарда в виде предпочки (pro-
nephros), но продвигаются дальше назад, образуя первичную поч-
ку (mesonephros), и объединяются единым продольным каналом,
открывающимся у анального отверстия. У акуловых рыб, а также
у наземных позвоночных их.эволюция ведет к установлению по-
стоянных связей с половыми железами. А именно: передние их ка-
нальцы используются для выведения женских половых продуктов,
падающих из гонады в целом, а затем поступающих в экскретор-
ный проток — мюллеров канал предпочки. Мужские половые же-
лезы вступают в прямую связь с канальцами далее идущего отде-
ла почки — мезонефроса. Образуются вольфовы протоки, через
которые спермин и выводятся, уже не попадая в целом. Экскретор-
ную функцию у высших позвоночных несет окончательная почка
(metanephros), представляющая сплетение экскреторных канальцев
тазовых сомитов туловища.
Таким образом, создается относительно несложная мочеполовая
ситема позвоночных, достигающая большой дифференцировки лишь
в дистальных частях с развитием копулятивного аппарата.
Подводя итоги анализу организации позвоночных, мы можем
сказать, то переход к свободному существованию и от пассивного
к активному добыванию пищи разносторонне охватил все системы
органов. Создалось несколько более или менее самостоятельных на-
правлений, по которым эволюционировали те или иные системы ор-
ганов и их комплексы, всегда, однако, находящиеся то в самых
тесных, близких, то более далеких взаимосвязях друг с другом.
Первым комплексом органов, наложившим особый отпечаток на
организацию позвоночных и в этом отношении ведущим, являлся
1 К сожалению, автор обошел молчанием рассмотрение эволюции не менее
важных и характерных эндокринных систем в различных типах беспозвоночных
и у низших хордовых. Ред.
364
миохордальный комплекс. Он выражен в развитии мощной боковой
мускулатуры рыбообразных форм и необходимых-для ее функцио-
нирования скелетных образований хорды (заменяющейся далее хря-
щевым и.костным позвоночником), а также и других частей скеле-
та в системе соединительнотканных прослоек среди и возле мышц.
Особенно разнообразно и богато дифференцируются компоненты
миохордального комплекса при переходе к наземному существова-
нию с выработкой пятипалых конечностей, т. е. при смене движения
водных, рыбообразных форм движением в наземной среде.
Обеспечивая движение животного, необходимое и для защиты и
для нападения, миохордальный комплекс развивается (одновременно
с развитием другого ведущего комплекса характерных черт, пред-
ставленного, органами чувств. Это мощные хеморецепторы, ?лаза и
органы, воспринимающие колебания окружающей среды и обеспе-
чивающие равновесие тела. В связи с переходом к наземному 'су-
ществованию в высокой степени сложно эволюционируют органы
обоняния и особенно органы равновесия, на основе которых разви-
ваются ор?Ьны слуха. В соответствии с выработкой этих органов,
из которых только глаза представляют части закладки самой нерв-
ной системы, а прочие дифференцируются за счет покрова приле-
жащих к ней участков головы, эволюционирует церсдний сенсорный
отдел нервной системы — головной мозг. Он представлен немета-
мерными обонятельным и глазным отделами, тогда как дальнейшие /
его отделы являются уже видоизмененным метамерным туловищ-
ным-мозгом, развитие которого связано с миохордальным комплек-
сом туловища. Как сложные образования выявляются мощные
ассоциативные центры мозжечка и переднего’ мозга, эволюция кото-
рых завершается выработкой в переднем мозге сложнейших струк-
тур коры полушарий большого' мозга. Вместе с этим во всей орга-
низации ведущей становится нервная система.
Сначала покров, по сравнению с двумя приведенными комплек-
сами, занимает второстепенное место в создании организации позво-
ночных. Эволюция покрова, как образования пассивной защиты, на-
ходящая свое особое выражение в выработке многослойное™ эпи-
телия и в дифференцировке кориума, у рыб значительно осложняет-
ся развитием костных чешуй. Однако особое значение эволюция
покрова получает при переходе к условиям наземной среды, при не-
обходимости защиты от иссушающего влияния воздуха и от рез-
ких. колебаний температуры. В конце концов покров из волос у
млекопитающих и из перьев у птиц приобретает и особое механиче-
ское значение и объединяется у последних с активным передвиже-
нием в воздухе. До известной меры комплекс образований покро-
ва сравнивается в своем значении с двумя описанными ведущими
комплексами. * < ' 7
Совершенно своеобразно, но, конечно, в теснейшей органической
взаимосвязи с другими формируется система внутренней среды в
организме — комплекс образований кровеносной системы. Его зна-
чение и функции значительно расширяются благодаря эндотелиза-
365
ции стенок сосудов. Кровеносная система в некоторой мере эманси-
пируется от соединительной ткани, приобретает самостоятельность
и в эмбриональном развитии. С комплексом кровеносной системы
тесно связаны дифференцировки кишечника в области жаберного
отдела и выработка печени. Переход к наземному . суще-
ствованию особенно сильно отразился на дифференцировке жабер-
1 »
ного отдела в развитии органов воздушного дыхания, что вырази-
лось в осложнении кровообращения в направлении широких при-
способлений, связанных с дифференцировкой большого и малого
кругов кровообращения и, соответственно, с образованием из двух-
камерного сердца трех- и четырехкамерного сердца наземных позво-
ночных. Наконец’ сюда же относится дифференцировка лимфати-
ческой и собственно кровеносной систем, первой из которых свой-
ственна трофическая функция при исчезновении функций газово-
го обмена.
Система целома, утрачивая свое былое значение, теряет прежде
всего опорную функцию для мускулатуры стенки тела, а затем и
побочные функции трофическую и экскреторную, переходящие к
скелету л соответственно к кровеносной системе. Однако целомиче-
ские протоки сохраняют свое значение в качестве аппарата пред-
почки, первичной и окончательной почки и выводных протоков по-
ловых желез.
Таким образом, комплекс кровеносной системы в выработке внут-
реннего строения позвоночных приобретает ведущее место, тогда
как у низших хордовых и кольчатых червей эта ведущая роль .при-
надлежит целому.
С огромным влиянием на всю организацию позвоночных разви-
вается кишечник в смысле приспособления его переднего отдела к
переходу на активное питанйё?"Однако морфологически этот ком-
плекс занимает при анализе организации второстепенное место.
Для полноты картины необходимо обратить внимание еше на од-
ну существеннейшую черту организации наземных позвоночных. У
них вырабатываются приспособления для защиты развивающегося
зародыша от влияния воздушной среды. Если у амфибий развитие
протекает еще свободно в условиях водной среды, то у рептилий
и птиц яйцо покрыто плотной оболочкой, под которой имеется и за-
пас необходимого для развития питательного желтка, и зародыш
защищен от иссушающего влияния воздуха, а от механических воз-
действий огражден жидким содержимым развивающихся амниоти-
ческих оболочек. Этим путем совершается освоение наземных ус-
ловий среды зародышем. У млекопитающих отношения развивались
в ином направлении. Яйценесущими среди них остаются только од-
нопроходные, у прочих развивается живсродность. Зародыш форми-
руется в материнском организме, чему служит и образование мат-
ки в половом аппарате самки и развитие питающих приспособле-
ний в виде плаценты. Сюда же следует отнести развитие млечных
желез, служащих для питания молоди. Эти приспособления создают
особый комплекс эмбриональных и постэмбриональных черт органа-
366
зации, имеющих огромное значение для выживаемости млекопитаю-
щих в их борьбе и победе за жизнь.
Мы далеко не исчерпали возможностей более глубоко проник-
нуть в сущность и значение массы различных черт строения в слож-
нейшем организме позвоночных, ограничившись общим наброском
отношений. Но вывод совершенно ясен. Мы видели, что для позво-
ночных характерно- многообразие ведущих комплексов черт,' что
’ каждый комплекс развивался своими особыми путями и что в сво-
Ъх взаимосвязях, особенно под воздействием нервной системы, раз-
витие зтих комплексов определяло широчайшие возможности ус-
ложнений и создания в высокой степени совершенной организации
в противоположность эволюции других высших типов, часто обна-
руживающих однобокое развитие. ‘
Перед нами естественно встает вопрос, как представлять себе?
предков хордовых?
Глава XIII
СИСТЕМА ЖИВОТНОГО ЦАРСТВА И ПУТИ ЭВОЛЮЦИИ
ХОРДОВЫХ
Наш анализ показал, насколько хордовые далеки от членисто-
ногих.
Наш анализ показал также, насколько хордовые разнятся от
аннелид. У ^последних на первый план выступает образование поли-
мерного целома, который ограничен только туловищем. При этом
каждый сомит характеризуется присутствием пары примитивных
конечностей в виде параподий, головная лопасть и пигидиум свое-
го особого целома не имеют, и рост тела идет терминально в пре-
пигидиальной зоне, где дифференцируются один за другим новые
сомиты с их целомическими мешками.
Иное мы находим у хордовых. Целом их в основе олигомерен,
дифференцировка его связана не с выростами метамерных пара-
подий, а с обособлением от головного и туловищного участков осо-
бого воротничкового отдела, несущего плотные придатки щупалец.
Лишь вторично, вместе с исчезновением головной лопасти, происхо-
дит утрата головного целома и редукция воротника, в то время как
туловищный отдел с образованием спинной хорды подразделяется
на многочисленные мета мерные участки. Иными словами, разви-
вается полимерный целом, не связанный с образованием выступов
тела, но с расчленением мускулатуры туловища.
'Отличие основ организации хордовых и аннелид иллюстрирует-
ся и в другом. Таково- отсутствие у хордовых типичных для аннелид
нефридиев, хотя в свое время Земпер (1875) именно их ошибочно
клал в основу сближения позвоночных с кольчатыми червями; это
относится и к отсутствию связи мешотчатых гонад с целомом, что
наблюдается у низших хордовых, тогда как у аннелид они всегда
теснейшим образом объединены. Наконец, у хордовых имеются
спинной мозг и головной мозг, не имеющие ничего общего с брюш-
ТТой^цепочкой и надглоточным ганглием аннелид.
Приходите5т поэтому просто откинуть так называемую «аннелид-
ную теорию» происхождения хордовых, представляющую спекуля-
ции в духе формальных построений Э. Геккеля. Они были даны
одновременна К. Земпером и А. Дорном (1875) и приняты в то
368
время большинством зоологов, да и в настоящее время имеют своих
приверженцев в лице А. Нефа и др. Действительно,. для многих
казалось крайне простым и крайне соблазнительным допустить, что
позвоночные представляют потомков апнелид, перешедших от обыч-
ного ползания на брюшной стороне тела к движению спинной сто-
роной книзу и брюшной кверху. В таком случае обычное располо-
жение органов в теле апнелид — вентрально брюшная цепочка, над
ней брюшной кровеносный сосуд, а далее кишка и спинной крове-
носный сосуд — должны были бы принять расположение, харак-
терное для хордовых — брюшной сосуд, над ним кишка, а далее
спинной сосуд и спинной мозг. В общем тот же, но только перевер-
нутый план построения.
Однако экологическое обоснование процесса «переворота» до сих
пор остается в области фантазии, а простая схема превращения
при этом кольчатого червя в позвоночное животное оказывается не-
состоятельной при более глубоком сравнении. Прежде всего ясно,
что развитие и строение спинного мозга хордовых и брюшной
цепочки аннелйд совершенно различны. Но особенно большие
трудности возникают при сопоставлении отношений головного
мозга и рта у апнелид с теми отношениями, что имеются у хор-
довых.
Таким образом вместе с аннелидной теорией происхождения хор-
довых рухнули попытки поставить кольчатых червей в центре эво
люции высших типов. Эти попытки находили свое выражение или
в сравнительно скромных схемах, по которым от аннелйд произо-
шли два важнейших типа — тип членистоногих и тип хордовых,
или в более широких, тогда от аннелйд производили также тип мол-
люсков (Пельзенер), или, наконец, в наиболее грандиозных схемах,
когда потомками аннелйд считали и тип иглокожих и тип прозопи-
гий (Э. А. Мейер). Следовательно, согласно последней схеме все
высшие типы животного царства нужно считать потомками аннелйд!
Мы видели, что в конечном счете от всех этих построений со-
временная эволюционная морфология должна отказаться, и можно
считать твердо установленным, что только тип членистоногих яв-
ляется завершением эволюции аннелйд, все же прочие произошли
от преаннелидных предков.
Однако недавно у нас была сделана попытка возродить анпе-
лидную теорию в несколько необычной форме. Е. К» Сепп (1936,
1949) находит возможным смотреть (1949, стр. 40—42) на ли-
чинку хордовых «как на дупликатуру аннелиды», как «на сложен-
ного пополам многочленистого червя». При этом происходит «сги-
бание оси тела червя» «вплоть до срастания соприкасающихся по-
верхностей». «Благодаря срастанию эпителия червя, жившего в
трубке в сложенном пополам виде», развивается хорда, причем
«перистальтика невозможна и змеевидное движение становится
единственной формой локомоции». Пытаясь обосновать свои взгля-
ды экологически. Е. К. Сепп пишет: «Мы знаем, что у животных,
укрывающихся в тех или других оболочках, перестраиваются внут-
24 Пути эволюции животного мира 369
ренние органы и прежде всего пищеварительный тракт таким обра-
зом, что к отверстию их оболочек сходятся все органы, по своей
функции общающиеся с внешней средой: ротовое отверстие, аналь-
ное отверстие, дыхательные органы, органы активного воздействия
на внешнюю среду и рецепторы... Нужно предположить, что усло-
вия изменились, и вместо того, чтобы получить достаточное пита-
ние, почти не сходя с места, оказалось необходимым совершать в
поисках пищи далекие перемещения. Таким образом, борьба за су-
ществование выражается на данном этапе в выработке более мощ-
ной локомоторной способности на новой основе — развилась хор-
да, прежний нервно-мышечный аппарат... заменился новым с цент-
ральной нервной системой и с поперечнополосатой мускулатурой...»
Так, по словам Е. К. Сеппа, «могут быть устранены основные воз-
ражения против аннелидной теории». Они «отпадают при анализе
эмбриологических данных, приводящих к представлению об эмбрио-
нальном метаморфозе, соответствующем филогенетическому процес-
су превращения полимерного червя в хордовое животное».
Е. К- Сепп считает при этом, что нервная трубка личинки лан-
цетника представляет собой просто эпителиальную трубку кишеч-
ника, к которой только вторично присоединяется сначала с боков
вентрально, а затем все дальше по периферии нервный слой, проис-
ходящий за счет брюшной цепочки червя. Е. К. Сепп считает так-
же, что1 с боков к хорде примыкают миотомы, а к кишке спланхно-
томы, представляющие два ряда отдельных целомических мешков
соответственно верхней и нижней половинам петли, образуемой те-
лом червя. Таким образом, создается некое первичное высшее хор-
довое.
Построения Е. К. Сеппа с экологической стороны фантастичны
и произвольны, а с морфологической не вяжутся с фактами образо-
вания хорды, нервной системы и простых мешков целома в разви-
тии хордовых. Поэтому мы не будем на них серьезно останавли-
ваться.
Своеобразным выражением подобных же представлений можно
считать попытки свести как организацию хордовых, так и аннелид к
трохофорным формам, т. е. к формам, имеющим личинку в виде
трохофоры и происходящим от трохофорообразных предков. Подоб-
ная попытка в новое вермя была сделана Дельсманом (1913, 1922)
и, конечно, представляет немалый интерес, потому что взаимоотно-
шения аннелид, моллюсков и членистоногих, имеющих трохофор-
мую личинку, довольно хорошо определены, их положение в систе-
ме ясно', и установление связи хордовых с аннелидами через трохо-
фору дало бы возможность точно ориентироваться в положении са-
мих хордовых.
По Дельсману, трохофора в результате процессов неравномерно-
го роста дала-начало трем характерным направлениям развития.
Одно определялось усиленным ростом заротового отдела тела тро-
хофоры, так что получалось длинное метамерное туловище аннели-
ды со ртом у переднего конца и анальным отверстием на заднем.
370
Другое было связано с ростом спинной стороньетрохофоры и вело к
образованию типичного моллюска. Третье, наконец, выразилось в
росте и метамеризации предротового отдела, причем первоначаль-
ный рот отодвигался к заднему концу тела, образуя неврэнтериче-
ский канал, а к нему вторично подходила спереди эктодермальная
глотка, тянувшаяся до переднего конца тела. Эта трубка глотки в
дальнейшем превращается в нервную трубку хордовых и теряет
связь с кишечником, который вторично получает новое ротовое от-
верстие у переднего края тела. Превращение глотки в нервную труб-
ку обусловлено тем, что в стенке глотки с самого начала имелись
чувствительные клетки, а в дальнейшем в ее стенки врастают нерв-
ные элементы двух стволов брюшной цепочки, расположенных спра-
ва и слева возле глотки. Хорда развивается, как пузырчатая ткань
мезодермального происхождения наподобие имеющейся у некото-
рых моллюсков.
Вся эта Дельсмановская фантастика, получившая у него назва-
ние «новой теории» происхождения хордовых, была основана на пе-
реходе нервной трубки зародыша хордовых через canalis neurente-
ricus в кишечную трубку. Однако представление о подобном обра-
зовании трубки-глотки и превращении ее в нервную трубку не на-
ходит себе подтверждения в фактах.
В свое время Б. Гатчек (1888) пришел к выводу, что трохофор-
ные типы (черви, моллюски, членистоногие и прозопигии) резко раз-
нятся от иглокожих и хордовых, настолько резко, что он считает
нужным отказаться от монофилетических представлений о проис-
хождении всех целомических животных. По его мнению, трохофор-
ные типы берут начало от ктенофорообразных предков, а иглокожие
и хордовые — от актиниеподобных. Лишь в низших формах кишеч-
нополостных объединяются два главных направления эволюции мно-
гоклеточных животных — один путь Ecterocoelia, т. е. обладающие
целомом эктодермального происхождения, другой — Enterocoelia с
целомом эктодермального происхождения.
К. Шнейдер (1902) еще более углубил разрыв, рассекая много-
клеточных животных с самого начала на две ветви, а именно на
Pleromata, куда относятся губки, гребневики, черви, членистоногие,
моллюски и на Coelenteria с Cnidaria, иглокожими, прозопигиями и
хордовыми.
В этой связи отметим, что анализ происхождения губок показал,
насколько они далеки от всех прочих Metazoa, в частности от греб-
невиков, и насколько правильно видеть в них особую, имеющую
свои собственные корни среди простейших и пошедшую по своему
собственному пути группу Parazoa. Что касается гребневиков; то об
их близости к типам иглокожих и хордовых для нас не может быть
речи. Поэтому разрыв всех настоящих Metazoa на два независимых
ряда теряет почву под ногами, и нет оснований идти в этом отно-
шении за К- Шнейдером. Зато очень многие зоологи склонны при-
нять систему Б. Гатчека, взявши, однако, ее в варианте, данном
К. Гроббеном (1910) с изменением названий Ecterocoelia и Entero-
24* 371
coelia на Protostomia — первичноротые и Deuterostomia — вторич-
норотые.
О новой характеристике, основанной на происхождении рта,
Б. Гатчек говорил (1911, стр. 28): «Насколько мало сможет это
новое обоснование (вводимых групп) воздействовать на критически
мыслящего зоолога, который знает трудность проблемы первичного
рта, настолько же привлекающе и убедительно оно сможет действо-
вать на неискушенного юнца в науке».
В полном соответствии с этим В. Н. Беклемишев в 1944 г. пи-
шет (стр.‘252): «Гомология... рта представляет труднейшую, еще не
разрешенную проблему в морфологии Deuterostomia... представить
себе филогенетически замену первоначального рта новым очень
трудно». Однако В. Н. Беклемишев заканчивает формальным ут-
верждением (стр. 254): «у Deuterostomia первичный рот превра-
щается в анальное отверстие, а где-то в той же оральной плоскости
независимым путем возникает вторичный рот».
Когда же мы читаем в учебнике зоологии беспозвоночных
В. А. Догеля (1947, стр. 71), что «у Protostomia бластопор гаструлы
переходит в дефинитивный рот..., а мезодерма образуется телобласти-
чесКим способом..., дробление в наиболее типичных случаях носит
детерминативный характер..., у Deuterostomia бластопор переходит
в заднепроходное отверстие..., мезодерма формируется выпячива-
нием боко-вых стенок кишечника, т. е. энтероцельно», то в конце
концов из всего этого создается впечатление полной определенности
установления двух групп типов — Proto- и Deuterostomia — на ос-
нове происхождения у них рта.
Более или менее сходные формулировки мы находим и у других
авторов, например Горст (1939) подчеркивает, что бластопор га-
струлы у Protostomia, закрываясь сзади наперед, делается ртом, и
шов находится за ним, а у Deuterostomia бластопор закрывается
спереди назад и дает анальное отверстие, так что шов находится
перед ним, и рот является новообразованием. В соответствии с этим
нервный зачаток окружает рот Protostomia и не имеет никакого
отношения к вторичному рту Deuterostomia.
Гораздо осторожнее говорит А. Н. Северцов (1939, стр. 324): у
первичноротых «ротовое отверстие развивается непосредственно из
бластопора или первичного рта зародыша», у вторичноротых «ро-
товое отверстие взрослого животного является новообразованием,
развивающимся независимо от бластопора». И только.
Выше мы пришли к выводу, что ротовое отверстие у хордовых и
иглокожих в эмбриональном развитии закладывается вне связи с
бластопором, но представляет собою тот же первичный рот, лишь
вторично перемещенный у сидячих предков на участок тела, проти-
воположный месту прикрепления.
У компетентнейшего нашего эмбриолога П. П. Иванова мы чи-
таем (1937, стр. 719): «Бластопор хордовых, хотя и не превращает-
ся в ротовое отверстие, но не превращается также и в анальное и
становится эмбриональным образованием», а «энтероцельное воз-
372
нжновение мезодермы приходится считать лишь-.кажущимся явле-
нием, обусловленным временным соединением на более ранних ста-
диях развития различных по происхождению зачатков мезодермы и
энтодермы».
Если к этому добавить абсурдность с экологической точки зре-
ния превращение рта в анус, то невольно' приходится вспомнить ци-
тированные выше слова Б. Гатчека о неудачном творчестве терми-
нов Proto- и Deuterostomia. Однако факт остается фактом. Две
группы типов — аннелиды, членистоногие и моллюски, с одной сто-
роны, а с другой — иглокожие и хордовые резко разнятся, пред-
ставляя два пути эволюции.
Но гораздо правильнее было бы взять в качестве их характери-
стики не рот, а другие черты организации, не вызывающие сомне-
ний в их понимании, как, например, непререкаемые факты морфо-
логии нервной системы. Все черви, членистоногие и моллюски обла-
дают первичными головным и туловищным брюшным мозгом и со-
ответственно головным комплексом органов чувств, тогда как игло-
кожие и хордовые утратили их, выработавши вторичные формы
нервной системы. У иглокожих это экто- и гипоневральная систе-
мы, у хордовых — головной и спинной мозг. Невольно напраши-
ваются названия — Protencephala вместо Protostomia и Dcuten-
cephala вместо Deuterostomia, т. е. «первично- и вторичномозговые».
Д. М. Федотов (J932, 1938) и В. Н. Беклемишев (1944, 1952)
вместе с Гатчеком раскалывают целомических животных на два
независимых ряда в отношении происхождения вторичной полости
тела. Но в то время как Б. Гатчек производит червей от ктенофоро-
образных предков, а иглокожих и хордовых, подобно Седжвику и
Дамеру, производит от актиниеподобных форм, наши отечественные
зоологи производят их от ктенофорообразных или общих с ктено-
форами предков. «Очевидно», делает вывод Л. А. Зенкевич (1937,
стр. 46), «все разногласия сводятся к тому, от какого места ствола
беспозвоночных отводить ветвь вторичноротых».
В своей работе, посвященной специально происхождению хор-
довых, Д. К. Третьяков (1929, стр. 580) следующим образом харак-
теризует предполагаемых предков этого типа (рис. 151, А, Б, В).
«Первичной формой прехордат была... бептонная ктенофора, кото-
рая на спинной стороне имела медуллярную пластинку... Перед ней
находился орган обоняния», происходивший из двух полярных полей
ктенофоры. Она «имела экскреторные органы в виде протонефри-
диев, которые образовались из погрузившихся вглубь гребных пла-
стинок и поэтому уже обнаруживали метамерное расположение.
Эктодермальная глотка была редуцирована и втянута в область зад-
него концевого отдела медулярной пластинки. Перед органом обо-
няния образовался новый рот... С его образованием старый рот мог
временно функционировать как анус. При превращении медулляр-
ной пластинки в трубку передняя половина старого рта сделалась
неврентерическим каналом; ее эктодермальные и чувствительные
клетки уже ранее присоединились к медуллярной пластинке. Зад-
373
няя половина старого рта сохранила значение анального отверстия...
С отделением частей кишечника для образования целома и с мета-
целомических сегментов также и мерца-
мерным расположением
Рис. 151. Схемы эволюции хордовых, данные
Третьяковым: А — ктенофорообразный предок
прехордат; Б — типичный представитель прехор-
дат; В — первичные бесчерепные;
1 — аборальный орган, 2 — модулярная пластинка, 3 —
аборальное поле, дающее орган обоняния, 4 — радиаль-
ные нервы, 5 — боковые органы, 6 — новый рот, 7 —
первичный рот, переходящий в анальное отверстие, 8 —
кишечник с боковыми выростами, дающими три пары
целомических мешков (9), 10 — щупальце ктенофор, 11—
головная лопасть, 12 — усики околоротовые 13 — жабер-
ный аппарат, 14 — атриальная пора, 15 — зачаток
хорды
тельные розетки полу-
чили метамерное распо-
ложение. Они сдела-
лись в то же время
целомодуктами. Заднее
щупальце должно было
при образовании ме-
дуллярной пластинки
исчезнуть? переднее
вступило в соотноше-
ние с новым ротовым
отверстием... и приоб-
рело... еще большую
функциональную цен-
ность, так как пойман-
ная добыча благодаря
сокращению щупальца
непосредственно притя-
гивалась в рот».
Все это построение
Д. К. Третьякова явля-
ется типичным приме-
ром изображения пред-
ковых форм в духе
старых филогенетиче-
ских фантазий, почти
не учитывающих веду-
щего момента влияния
среды на эволюцию ор-
ганизмов. Я привел его
как образчик очень де-
тальной разработки ма-
териала, относящейся
к последнему времени,
однако разбирать его
критически не имеет
смысла.
Связи прехордат с
другими группами идут,
по Д. К. Третьякову
(стр. 581), через ктено-
фор: «Исходная форма прехордат произошла от ктенофор при изме-
нении организации в том же направлении, которое характеризует
также и происхождение турбеллярий, их пути эволюции разошлись,
однако, очень рано... Также, может быть, немертины стоят близко
374
к источнику происхождения прехордат... Они выработали, как и
прехордаты, кровеносную систему. Наконец, также и аннелиды род-
ственны первичной форме прехордат, ибо свою метамерию, целоми-
ческие сегменты, про- и метанефридии они получили из тех же ис-
точников (ктенофоры), как и прехордаты...» Таким образом, для
Д. К. Третьякова гребневики оказываются исходным звеном в эво-
люции обоих рядов типов, обладающих целомом, но этот последний
развился из общего источника — выростов кишечника гребневи-
ков—в том и другом’ряду независимо. Следовательно, перед нами
своеобразный вариант дифилетических представлений относительно
происхождения высших типов животного царства, имеющих вторич-
ную полость тела.
Итак, Protostomia и Deuterostomia! Новейшие дифилетические
представления о происхождении целомических животных против мо-
нофилитических представлений старых зоологов-эволюционистов!
Конечно, о губках — йастоящих паразоа — здесь нет .речи, так как
дело идет только о типах, обладающих целомом.
Для большинства зоологов прошлого столетия, начиная с Гекке-
ля, важнейшей двигающей силой было стремление раскрыть в эво-
люции животных непрерывный ряд постепенных изменений, ведших
от низших форм к самым сложно организованным. В соответствии
с этим жило стремление найти переходные формы между различ-
ными группами и эмбриологически и сравнительноанатомически
проследить судьбы гомологичных образований. Монофилетические
представления являлись естественным выражением таких умона-
строений. Гребневиков связывали с кишечноплостными, происхо-
дившими от исходной гастреи; далее считалось, что гребневики, в
свою очередь, через Ctenoplana и Coeloplana дали начало червям.
Гипотетический трохозооп, представленный в онтогенезе трохофор-
ной личинкой, был сделан исходной формой высших червей и мол-
люсков. Кольчатые черви через перипатуса связывались с членисто-
ногими. Позвоночных произвели от анцелид, перевернув их со спин-
ной стороны на брюшную.
Работая в этом направлении, морфологи собрали богатейший
фактический материал, но в то же время должны были убедиться,
насколько1 трудно' решить поставленные задачи, и насколько шатки
первоначальные построения. Нарастал кризис. Центр научных инте-
ресов переместился в другие отрасли биологии, а вместе с наступ-
•лением эпохи империализма изменились и умонастроения. В 20-х
годах и у нас на первый план выступают измышления несущест-
вующих закономерностей в аналогичных, конвергентных явлениях,
формулируются положения о «параллелизме структур», о «захожде-
нии» или «трансгрессии признаков», их «индукции», «уподоблении»,
«инерции» и т. д. и т. п. У В. А. Догеля выставляется тезис, что в
области беспозвоночных ведущими представляются явления конвер-
генции, а не гомологии.
Необходимы были новые методологические, философские основы
для решения поставленных еще в XIX веке вопросов системы жи-
375
вотыопо царства, так как в рамки плоской эволюции они не укла-
дывались, и преобладание анализа перед синтезом, конвергенций
перед гомологиями суживало возможности правильного решения.
| Лишь методологические установки диалектического материализма
| дают нашей биологии надежную опору и возможности справиться
I с поставленными вопросами на новом уровне советской эволюцион-
* ной морфологии.
Обратимся к Нашим филогенетическим положениям. Трудно до-
пустить, чтобы переход от внутриклеточного пищеварения кишечно-
полостных, гребневиков и плоских червей, имеющих целентериче-
ский кишечник, к полостному пищеварению и трубчатому кишечни-
ку вышестоящих форм осуществлялся конвергентно в разных груп-
nax^Prot- и Deutencephala (соответственно Proto- и Deutero-
stomia). Не менее трудно предполагать, что кровеносная система
происходила несколько раз в животном мире независимо в разных
группах. То же самое приходится сказать относительно возникнове-
ния целома и соматической мускулатуры, протонефридиев и т. п.
Все это в конечном счете приспособления, открывающие широкие
возможности эволюции. Но их появление возможно только в опре-
деленных условиях окружающей среды и соответствующей органи-
зации. Раз появившись, приспособления такого рода обеспечивают,
можно сказать, волну образования новых форм и прочно закрепля-
ются в эволюции потомков как важнейшие приобретения их орга-
низации.
Все это заставляет с крайней осторожностью подходить к вопро-
су о ди- или полифилетическом происхождении кровеносной систе-
мы, целома и т. д. Допустить повторение сочетания определенных
условий во взаимоотношении с определенной организацией, при не-
обратимости эволюции возможно лишь, когда факты неодолимо
требуют этого. С таких позиций мы будем подходить к вопросу о
построении системы животного царства. Добавлю, что при построе-
нии системы ведущая роль среды ранее обычно не находила доста-
точного выражения.
Как было указано выше, идея Б. Гатчека о дифилетическом
происхождении двух больших групп типов метазоа — Protostomia
от гребневиков, и Deuterostomia от актиниеобразных предков — не
нашла поддержки. Фантазиями, не имеющими достаточных факти-
ческих оснований, явились старые представления Седжвика, разде-
ляемые в настоящее время Ламером, о переходе актиниеобразных
форм к ползанию на щупальцах, о срастании при этом средних ча-
стей сплющенной глотки, так что образуются рот и анус, об отшну-
ровывании боковых камер целентерического кишечника для обра-
зования метамерных мешков целома.
Как у Д. М. Федотова, так и у Д. К. Третьякова мы видим иное.
Обе группы типов развиваются от ктенофорных предков, а в своей
дальнейшей эволюции идут разными путями. В известной мере это
сходно со взглядами А. Ланга, которые мы находим еще и теперь,
например в учебнике В. А. Догеля, и которые мы уже рассмотрели.
376
Но Д. К. Третьяков (1929) связывает турбеллярий, немертин ан-
нелид и хордовых не с Ctenoplana и Coeloplana, а с типичными
гребневиками, перешедшими к движению параллельно дну на одной
из боковых поверхностей тела.
В. Н. Беклемишев (1952, стр. 102) следующим образом пред-
ставляет отношения ктенофор к турбелляриям: «Наиболее правдо-
подобным остается мнение Граффа о происхождении турбеллярий
от планулообразных предков кишечноплостных. Или, что сводится
к тому же, наиболее вероятными предками и несомненным прото-
типом турбеллярий можно считать первично планктических кишеч-
нополостных, снабженных аборальными органами чувств, но еще
лишенных сложной симметрии; прямыми, но односторонне специа-
лизированными потомками этих форм являются' гребневики... И
гребневики и турбеллярий составляют две самостоятельные группы,
которые развились независимо друг от друга».
Ко взглядам В. Н. Беклемишева очень близки те выводы, к ко-
торым пришли мы при анализе организации кишечнополостных,
гребневиков и турбеллярий, а именно: все эти группы произошли
от первичных гастреад, ведших свободноподвижной образ жизни во
взвешенном состоянии в водах мирового океана, но дальнейшие
пути их эволюции радикально различны, в связи с использованием
различных пищевых ресурсов и освоением разных условий суще-
ствования. Кишечнополостные перешли к использованию придонно-
го пищевого материала, прикрепившись своим аборальным полюсом.
Гребневики не изменили образ жизни предков, сохранивши преж-
нее пелагическое существование и питание пелагическими организ-
мами. Турбеллярий же используют богатые запасы пищи, имеющие-
ся на дне, и перешли к ползающему донному образу жизни, при-
чем связи их с предками гребневиков более тесные, чем с формами
гастреад, перешедших к сидячему существованию.
Однако предки всех трех типов были уже целентерическими
организмами, а не личиночными, как думает Графф, планулообраз-
ными формами, лишенными полости кишечника. На присутствие у
них этой полости указывает постоянный целентеризм и гребневиков
и кишечнополостных, по организации более простых, чем низшие
черви.
Не будем повторять сказанное при анализе организации тур-
беллярий о положении их в системе, а также об их филогенезе как
низших, еще целентерических представителей типа червей. Обратим-
ся к вышестоящим формам. Мы видели, как с организацией ктено-
фор расправился Д. К. Третьяков (1929), чтобы превратить их в
предполагаемых предков прехордат. Произвольность его построения
не требует особых комментариев. Но, по Д. К. Третьякову (стр. 581),
и турбеллярий происходят от общего с прехордатами предка, и не-
мертины стоят близко к ним («они выработали, как и прехордаты,
кровеносную систему»), и, наконец, аннелиды по метамерии, целому
и нефридиям родственны прехордатам. Очевидно, что при такой
точке зрения нельзя ожидать решения вопроса, но кажется можно
377
полагать, что автор склоняется к монофилетическим представле-
ниям.
К монофилетическим взглядам заставляет придти и проведенный
нами анализ организации главнейших типов животного царства.
Прежде всего приходится установить, что действительно «дистанция
огромного размера» отделяет всех выше стоящих червей от целен-
терических турбеллярий, а. тем более от гребневиков и кишечнопо-
лостных. Однако все вышестоящие черви сохраняют ряд характер-
нейших черт, присущих турбелляриям. Во-первых, им свойственно
билатерально-симметричное строение с преобладающим ростом зад-
него отдела спинной стороны и расположением рта вентрально, вы-
работка головного комплекса органов чувств, дифференцировка
кожно-мышечного мешка и образование периферической и централь-
ной нервной системы с сенсорным головными моторным туловищ-
ным мозгом. Во-вторых, для них характерно наличие мезенхимной
мускулатуры и внутренней мезенхимы с располагающимися в ней
гонадами и экскреторными органами типа нефридиев. Этот слож-
ный комплекс черт становится типовым для немертин, аннелйд и,
в известной мере, для моллюсков и червеобразного баланоглосса.
Но никаких этапов выработки этого комплекса мы не находим у
гребневиков, или кишечнополостных. Породившие его условия су-
ществования придонных ползающих хищников дали эффект, на-
столько своеобразный и сложный, что, вопреки мнению, например,
В. Н. Беклемишева, представляется мало вероятным конвергентное
его развитие у разных высших групп на основе организации, с одной
стороны, планулоэбразных форм для турбеллярий, с другой —
трохофорных форм для аннелйд и моллюсков, с третьей — ктено-
форообразных форм для баланоглосса.
Поэтому турбеллярные формы следует считать низшими, исход-
ными в эволюции всех червей, причем характернейшей чертой их
потомков — современных турбеллярий оказывается сложнейшее
развитие полового аппарата, обеспечивающее сохранение последних
в борьбе за жизнь. Развиваемые здесь взгляды представляют лишь
современный вариант построений, господствовавших до Б. Гатчека
и принимаемых многими и в настоящее время. В них на первый
план как естественный результат анализа организации выдвига-
ются те комплексы черт, которые являются ведущими, создающими
определенный тип строения. На задний же план отодвигаются от-
дельные, сходные с другими формами, черты, которые сами по себе,
вне комплекса, не могут иметь решающего значения.
Дальнейшим шагом в эволюции червей, как было нами в своем
месте отмечено, является этап немертин. Немертины резко разнятся
от турбеллярий важнейшим комплексом черт, далее присущим уже
всем вышестоящим типам. Действительно, со стабилизацией рото-
вого отверстия близ переднего конца тела и тем самым с диффе-
ренцировкой головы у активно подвижных хищников, обладавших
длинным мускулистым телом, создается трубчатый кишечник с
полостным пищеварением. Это дает возможность полного исполь-
378
зования пищевого материала по мере его продвижения через кишеч-
ник. В этом огромная разница по сравнению с целентерическим
аппаратом турбеллярий. Это. же ведет, с другой -.стороны, к выра-
ботке кровеносной системы, передающей питательные вещества от
места усвоения их, т. е. средней эктодермальной кишки к такому
жизненно важному пункту потребления, как голова с ее органами
чувств и головным мозгом.
Таков сложный комплекс черт, который превращает турбеллярие-
образного предка в немертинообразный организм. И современные
немертины отличаются лишь выработкой чрезвычайно разнообразно
развивающегося органа нападения и защиты — хобота, котовый
гарантирует сохранение немертин в борьбе за жизнь до наших дней,
но который еще не был развит у начальных форм. Конечно, слож-
ность комплекса трубчатого кишечника и кровеносной системы со
своеобразными деталями их развития делает маловероятным воз-
можность появления той или иной из его составных частей незави-
симо в разных группах животного царства, например хотя бы у
аннелид, минуя немертинообразных предков. Однако подобные
гипотезы проскальзывали и проскальзывают иногда при недооценке
значения и положения в системе этого подтипа червей. Значимость
данного комплекса настолько велика, что опа заставляет резко от-
делить немертин от низших целентеричеоких червей, т. е. от тур-
беллярий.
Однако связи обеих групп резко подчеркиваются рядом фактов,
и прежде всего тем, что1 целентерическое состояние кишечника в
течение продолжительного времени сохраняется в> развитии немер-
тин. Лишь при переходе к донному существованию взрослой немер-
тины образуется анальное отверстие, и притом прорывается оно на
растущем заднем конце тела независимо от закладки рта, но как
новоприобретение немертинообразных предков. Уже одним этим
фактом определяется положение немертин в системе червеобразных
форм как резко выраженного этапа эволюции последних.
Совершенно своеобразные отношения вырисовываются далее в
развитии немертин. Они привлекали внимание эмбриологов уже со
времени И. И. Мечникова (1869), но только значительно позже они
были в достаточной мере выяснены. Так, у немертин были выявле-
ны черты спирального типа дробления. И можно сказать, что здесь
в эволюции мы наблюдаем как бы грань спирального1 и радиаль-
ного типа дробления. Далее, в развитии червеобразного зародыша,
как у турбеллярий, так особенно у аннелид, ясно видно значение
процесса неравномерного роста туловищного отдела спинной сторо-
ны тела. Здесь у пилидия существенную роль играет непарный
спинной зародышевый диск, дающий весь покров спины и боков
тела. Наконец, присутствие ярко выраженного мезодермального
зачатка характерно для развития немертин. Но он не дает начала
целому, а стоит в ряду типичных мезенхимных зачатков, помещаясь,
как и у аннелид, сначала по краям впячивания энтодермы и давая
клеточный материал отчасти в эту последнюю, а главным образом
379
для образования мускулатуры кожно-мышечного мешка за счет
получающейся целенхимы. Некоторые авторы, например В. В. За-
ленский (1912), описывают даже расположение клеток целенхимы в
виде правильных париетального и висцерального листков, но без
образования настоящего целома.
Эти факты мы расцениваем с точки зрения миоцель-теории
происхождения вторичной полости тела. Именно у немертинообраз-
пых червей резко дифференцированные зачатки мезодермы, как за-
чатки прежде'всего мышечные, должны были внутри себя дать
начало опорным скоплениям жидкого транссудата, с выработкой за
счет их клеточного материала целотелиальных стенок, другими сло-
вами, дать начало целому. Каких-нибудь особых мешковидных за-
чатков, являвшихся предшественниками целома, не могло быть,
так как единство мышечной и целомической закладки является
фактом, не требующим дальнейших пояснений, и оформление кле-
точной стенки целома для жидкого содержимого шло одновременно
с дифференцировкой целомической мускулатуры. Так развились
первичные немертинообразные целомические черви — Archi- или
Protocoelomata.
Как же можно себе представить дальнейшее развитие аннелид
и других целомат из немертинообразных предков?
Эволюция двух групп типов животного царства Proto- и Deu-
terostomia, или Prot- и Deutencephala, как будто бы требует при-
знать и два пути эволюции целома, но их начало может быть об-
щим. Так мы и представляем себе отношения.
^Нелом появляется у немертинообразных предков в виде двух
независимых полостей — правой и левой, простирающихся от пе-
реднего конца тела до заднего, в_ качестве опоры для мускулатуры
кожно-мышечного мешка. Но в силу антагонистических отношений
мышц правой и левой сторон при синусоидном движении целом
появляется обязательно как независимая правая и левая полости,
и в таком виде может играть уже свою немалую роль в жизни
свободноподвижного хищника.
Дальнейшая его эволюция у кольчатых червей связана уже с
появлением боковых выростов тела — параподий, являющихся пер-
вичными конечностями, значительно совершенствующими синусоид-
ное движение свободноподвижных полихет. Целом становится опо-
рой и для параподий, обусловливая их плотность. В связи с этим,
соответственно независимой деятельности каждой параподий при
движении тела, возникает подразделение первоначально единого
зачатка на ряд отдельных мешков. Как это совершенно правильно
представляет П. П. Иванов (1945), метамерия целома аннелид
индуцируется внешним моментом появления параподий. И это на-
ходит свое отражение в первоначальной закладке личиночных со-
митов у полихет точно по этой схеме. Но в дальнейшем процесс
эмбрионизируется, и постларвальные метамеры закладываются уже
строго правильно без выявления их взаимозависимости с образова-
нием параподий. Более того, ларвальные сомиты могут совершенно
380
выпасть, как мы это видим у Polygordius с его высоко специализи-
рованной трохофорой. Теория ларвальных сомитов П. П. Иванова
дает нам ключ к пониманию отношений целома у Protencephala на
основе возникновения его как мио цел я в виде двух, правого и ле-
вого, мешков у немертинообразных Archicoelomata. При этом уже
рано в развитии из зачатка, связанного и по положению и отчасти
функционально с образованием эктодермального кишечника, диффе-
ренцируются два телобласта, уходящие из состава эпителия вглубь
или сохраняющие в той или иной мере свое первоначальное поло-
жение. Это происходит вместе с нарастающей детерминированностью
развития Protencephala.
Иное бросается в глаза у Deutencephala. Но новейшие исследо-
вания заставляют иначе и глубже оценивать факты, чем делалось
это раньше. Противополагать энтероцельное образование мезодермы
у Deutencephala телобластическому у Protencephala приходится с
большой осторожностью. Вспомним здесь мысль П. П. Иванова
(1937, стр. 719), что' энтероцельное возникновение мезодермы при-
ходится считать лишь кажущимся явлением, обусловленным вре-
менным соединением на более ранних стадиях развития различных
по происхождению зачатков мезодермы и энтодермы. Действитель-
но, тщательное изучение этих процессов у ланцетника показало, что
мезодермальные правая и левая полоски происходят за счет клеток,
лежащих при гаструляции на границе энтодермы и эктодермы в
задней части гаструлы, как и у Protencephala. В дальнейшем,
не выделяясь из эпителиального положения, они врастают вмес-
те с энтодермой по бокам во впячивание архентерона, продви-
гаются по длине энтодермального зачатка и обособляются, нако-
нец, от закладки кишечника в виде двух боковых складок
мезодермы.
При этом у низших хордовых дифференцируются три мешка
целома: передний — головной, средний — воротничковый и зад-
ний — туловищный. Движущий момент этой дифференцировки, как
мы видели, ясен. При переходе к жизни в ходах-трубках или при
прикреплении у них развивается пассивное питание мелкими, взве-
шенными в воде организмами. За счет мерцательного покрова
юколоротовых частей тела — таких, как, например, мощные заты-
лочные органы немертин — образуются участки, производящие ток
воды к ротовому отверстию. Эти участки затем при содействии
естественного отбора превращаются в придатки щупалец. С этим
связано обособление части целома, образующей воротничковый
целом, который поддерживает щупальца в твердом состоянии. В
результате мы имеем трехраздельный целом хордовых, у низших
форм которых головная лопасть сохраняет передний участок целома
обособленным для специальных функций головного отдела, напри-
мер, в виде целома хобота баланоглосса.
У иглокожих головная лопасть утрачивает свое значение и в
значительной мере или совсем {редуцируется. Соответственно этому
передний и воротничковый отделы целома не дифференцируются как
381
Prgtomonadtna
Рис. 152. Схема филогенетических отношений главнейших типов многоклеточных,
животных (по Б. С. Матвееву, но со значительными изменениями)
два независимых образования, а остаются соединенными. Создает-
ся двухраздельный целом.
Заключая наше рассмотрение путей развития хордовых, мы мо-
жем сказать, что эволюция целомат, начавшись общим состоянием
двух мешков (правого и левого) целома немертинообразных Archi-
coelomata, пошла в дальнейшем по двум путям Proto- и Deutero-
stomia, или, лучше Protencephala и Deutencephala. Пути эти опре-
делились прежде всего условиями существования: у Protencephala—
свободных форм с активным питанием, а у Deutencephala — трубко-
живущих и прикрепленных форм с пассивным питанием. Соответ-
ственно характеризуется и положение хордовых в системе. Вместе
с иглокожими они представляют два независимых типа, входящие
в достав Deutencephala. Взаимоотношения внутри типа хордовых
мы уже рассмотрели при анализе организации входящих в него
главнейших групп. Хотя следует оказать, что здесь наш анализ
натыкается на самые большие трудности. Так, не изучена в доста-
точной мере организация Pogonophora, которых некоторые иссле-
дователи ставят в ближайшее родство с хордовыми. Темны эколо-
гические моменты появления организации бесчерепных и рыбооб-
разных хордовых и т. д.
Представления о системе животного царства и путях эволюции
хордовых, которые вытекают из нашего анализа организации типов,
иллюстрируем заимствованной у Б. С. Матвеева (1935) и несколько
видоизмененной схемби (рис. 152).
Глава XIV
ЗАКОНОМЕРНОСТИ ПУТЕЙ ЭВОЛЮЦИОННОГО ПРОЦЕССА
В рамки нашего анализа не вошли многие группы, представляю-
щие немалый интерес в различных отношениях, как, например,
круглые черви, ротатории и т. д., но наша задача заключалась не в
том, чтобы дать полную «систематическую филогению» животного
царства, а в том, чтобы в достаточной мере осветить организацию
на главных этапах эволюции, чтобы выявить общую картину эво-
люционного процесса. Думаю, что наш материал достаточен для
поставленной цели.
В заключение бросим общий взгляд на ту картину, которая вы-
является из рассмотрения организации главнейших типов многокле-
точных животных.
Прежде всего внимание исследователя останавливает своеобраз-
ный ход эволюции в низших типах — кишечнополостных, гребне-
виков, червей. Мы видим смелый взлет организации от гидры до
аннелид, «прямая большая дорога» эволюции, ведущая от двух кле-
точных первоорганов (покрова и кишечника) и одного неклеточного
(опорной пластинки) к выявлению всех систем органов, присущих
далее высшим типам моллюсков, членистоногих, иглокожих, хордо-
вых. На этом отрезке хода эволюции произошла дифференцировка
особых аппаратов для всех главных категорий функциональной
деятельности организма. Это — стремительный ряд приспособле-
ний, открывающих широкие перспективы. Их бурный поток, по
сравнению с которым выработка, например, трех-, а далее четырех-
камерного сердца у позвоночных — берем пример ароморфозов
А. Н. Северцова — лишь мелкая деталь.
Первым широким приспособлением, собственно создавшим мно-
гоклеточный организм, была дифференцировка кишечника и покро-
ва, и все богатство форм кишечнополостных представляет размен
этотО' основного достижения на мелкую монету частных, узких при-
способлений к условиям существования. Далее, у турбеллярий мы
находим целый ряд других широких приспособлений. Таковы диф-
ференцировка кожно-мышечного мешка, образование головного
комплекса' органов чувств, дифференцировка "центральной нервной
384
системы, а в ней головного и туловищного мозга, появление экскре:
торного аппарата в виде протонефридиев. Немертины обладают
уже головной лопастью, у них вырабатывается трубчатый кишечник
с полостным пищеварением, возникает кровеносная система. У
аннелид, наконец, налицо система целома со всем многообразием
ее'функций, дифференцируются органы движения в виде параподий,
возникают органы газообмена — жабры, создаются ассоциативные
центры в головном мозге — центры высшей нервной деятельности.
Иной дорогой пошли в своей эволюции высшие типы животного
царства. На базе достигнутого выявления и дифференцировки всех
основных сторон жизнедеятельности организма происходит только
применение и специализация их в направлении приспособления к
тем или иным большим категориям условий внешней среды, главным
образом по линиям питания и защиты. Под ведущим влиянием
этих двух важнейших категорий условий существования возникает
организация типов, подтипов и т. д. — членистоногих, моллюсков,
прозопигий, иглокожих, хордовых — организация как бы особо,
идогда односторонне и даже уродливо перерабатывающая достиже-
ния кишечнополостных, гребневиков и червей.
Так, для членистоногих ведущим моментом в создании типа "
явилась, как мы видели, выработка защитной хитиновой кутикулы,
поведшая к редукции и почти полному исчезновению системы цело-
ма и к своеобразному развитию кровеносной системы. Но в то же
время хитиновая кутикула обусловила появление наружного скелета
и разнообразных приспособлений конечностей, обладающих харак-г
тером сложного рычага. Образование такой кутикулы вызвало диф-
ференцировку на базе кожно-мышечного мешка поперечнополоса-
той специальной мускулатуры и вызвало развитие приспособлений,
связанных с овладением наземной средой жизни. К ним относятся:
система газового обмена в виде аппарата трахей, экскреторная си-
стема мальпигиевых сосудов, а у насекомых дифференцировка ти-
пичнейших органов полета — крыльев. Одним словом, вырабаты-
вается организация хорошо защищенных, очень подвижных хищных
животных, которые в условиях наземной среды развиваются иск-
лючительно разнообразно, переходя к использованию различнейших
возможностей наземного существования.
У моллюсков ведущими чертами организации являются образо-
вание “на спинной стороне'защитной известковой раковины за счет
кутикулы покрова и превращение брюшной части тела в орган
движения — ногу. Это, как и у членистоногих, обусловливает
частичную редукцию кожно-мышечного мешка и значительную ре-
дукцию целома. Создается огранизация хорошо защищенных, но
слабо подвижных нехищных животных.
Тип иглокожих определяется выработкой в соединительнотканном
слое покрова защитного известкового панциря, что при сидячем
образе жизни и пассивном питании, связанными с упрощением ряда
систем органов, ведет к развитию организации, поражающей своей
оригинальностью. По существу организация иглокожих представ-
25 Пути эволюции животного мира 385
ляется усложнением целома. При редукции мускулатуры стенки
тела, наступающей в связи с обызвествлением последней, целом
дифференцируется в ряд систем на основе исходного многообразия
его функций. За его счет образуется амбулакральная система с дви-
гательной функцией, перигемальная система с трофической функ-
цией и система общей полости тела. Создается организация, позво-
ляющая использовать другие источники питания, чем при первона-
чальном пассивном питании, и способная дать при переходе к сво-
бодному существованию крайне оригинальные формы морских
звезд, ежей, голотурий. В них, однако, нет чего-либо существенно
нового, но выявляются лишь своеобразные приспособления уже
выработавшегося ранее комплекса организации. Иглокожие не
смогли выйти из водной среды, так как ведущий признак — целом—
теснейшим образом с ней связан. Суша осталась им недоступной.
В противоположность иглокожим, пассивно питавшиеся и вед-
шие близкое к сидячему существование предки хордовых пошли в
отношении защиты разными путями. Одни ведутЗкйзнь в трубках,
другие переходят к жизни в грунте, оболочники вырабатывают
мощную тунику. Характерная организация типа создается в конце
концов на основе разных зачатков. Сюда вошли, с одной стороны,
аппарат жаберных щелей, обусловленный пассивным питанием,
с другой — миохордальный комплекс, обусловленный активным
движением, с чём связано и образование спинной нервной трубки.
При'переходе к свободному существованию хищников на этом фоне
различных компонентов развивается организация позвоночных, даю-
щая многообразие широких приспособлений. При переходе же в
условия наземной, крайне разнообразной среды, эта организация в
более позднее время как бы завершает большую дорогу эволюции
кишечнополостных, гребневиков и червей. Мезозойская эра стано-
вится эрой рептилий, кайнозойская — эрой птиц и млекопитающих,
а современная — веком человека. Чего не могли достичь в богат-
стве наземных условий существования членистоногие с их односто-
ронней доминантой хитинового покрова, то смогли выполнить позво-
ночные с их многообразным типовым комплексом организации
хордовых.J
В свете фактов, полученных при рассмотрении эволюции глав-
нейших типов животного царства, подойдем к вопросу о выработке*
реальной организации животных, соответствующей понятию типа.
Типы — это узловые пункты в развитии животного мира, кото-
рые характеризуются образованием совокупности черт организации,
связывающихся в единый новый комплекс, новую систему с особы-
ми новыми качествами. Входящие в нее компоненты могут быть
очень разнообразны и неравноценны: наряду с открывающими
широкие перспективы для эволюции есть и такие, которые выраба-
тываются как узкие приспособления к особым условиям возникно-
вения того или иного типа или даже носят характер редукции, деге-
нерации. Все связано с осваиваемыми условиями среды — они яв-
ляются ведущими. При этом решающая роль принадлежит эколо-
386
гическим соотношениям защиты и питания, их активным и пассив-
ным формам, их сочетаниям.
С этой точки зрения еще раз и более углубленно рассмотрим
организацию высших типов.
Так, организация иглокожих возникла в условиях перехода к
прикрепленному существованию. В этих (условиях в соединительно-
тканном слое покрова образуются известковые отложения, создаю-
щие надежную пассивную защиту, а вместе с тем происходит упро-
щение кроющего эпителия и редукция мускулатуры кожно-мышеч-
ного мешка и соответствующих частей нервной системы. Питание
также приобретает пассивный характер — пища поступала в рот
с током воды, вызывавшимся мерцательным движением эпителия на
придатках-щупальцах, окружающих рот.
Выработка типовой организации иглокожих идет в направлении
усиления возможностей пассивного питания. Происходит перемеще-
ние рта и аппарата щупалец на место, противоположное пункту
прикрепления, увеличивается оральная поверхность, соответственно
усиливается ток воды, приносящий пищу, и щупальца входят в со-
став туловища, а вместе с тем перестраивается система вторичной
полости тела. При этом в качестве особой, амбулакральной системы
дифференцируется участок целома, двигающий щупальцевые при-
датки, а трофическая для щупальцевых образований часть целома
обособляется в качестве перегимальной системы. В соответствии с
этими процессами перестраивается и нервная система. Можно ска-
зать, что у иглокожих при прикреплении происходит упрощение
отношений со средой, упрощается ряд систем органов, но при од-
новременно прогрессирующем пассивном питании усложняется ряд
других систем. Это накладывает отпечаток -на всю организацию
иглокожих, особенно в связи с их характерным защитным панцырем.
Такие животные переходят к использованию возможностей в на-
правлении развития активного питания, т. е. прежде всего к иному
использованию своего щупальцевого аппарата и его придатко-в,
связанных с амбулакральной системой и т. д. Создается этап сво-
бодноживущих иглокожих, вырабатывающих и активное движение
посредством амбулакральных ножек, а вместе с тем через процессы
неравномерного роста перестраивающих все тело в направлении
преобладающего развития оральной части, связанной с амбула-
кральной системой. В конце концов иглокожие освоили для своей
жизни все участки морского дна, некоторые из них — правда, еди-
ницы — перешли даже в пелагические условия существования.
Пример иглокожих ярко подтверждает наше положение, что
выработка типовой организации это — комплекс различного харак-
тера приспособлений, в котором выявляется качественно новый тип
организации. Новые качества используются как исходный пункт
для дальнейшей эволюции в других ноцых для типа условиях суще-
ствования, причем эти условия как были ведущими в выработке
организации типа, так и дальше обусловливают ее изменение в том
или ином направлении.
25* . 387
Комплекс характерных черт типа многообразен. На примере
иглокожих можно отчетливо видеть, что в него входят как сравни-
тельно узкие приспособления, не поднимающие организацию вы-
ше, например такие, как пассивный защитный скелет или придатки
для пассивного питания, так и представляющие некоторый рег-
ресс — редукция мускулатуры стенки тела, редукция глоточного
отдела кишечника и т. д. Но в эволюции целома имеется значи-
тельный прогресс, широкие приспособления, открывающие перспек-
тивы — дифференцируются амбулакральная система и перигемаль-
ная система. Их можно назвать широкими приспособлениями, но
правильнее будет сказать, что широко приспособительным является
весь комплекс организации типа — он открывает новые пути и воз-
можности, поднимает организацию на новую ступень эволюции, И
расцвет типа иглокожих определен далее возможностями перехода,
во-первых, от пассивной защиты к активной, вместе с тем от сидя-
чего существования к свободноподвижному, во-вторых, от пассив-
ного питания к активному и в конце концов к хищничеству, требую-
щему всестороннего прогресса органов движения, органов чувств,
нервной системы, насколько это возможно в данном типе.
Организация моллюсков создалась иными условиями существо-
вания. Она характерна для свободноподвижных придонных форм,
для которых защита была связана с обызвествлением кутикулы
спинной стороны тела — образованием раковины. Наряду с этой
пассивной защитой активное питание (но не питание хищника)
обеспечено возможностью передвижения благодаря дифференциров-
ке брюшной стенки тела в мускулистую ногу. Защитное значение
раковины усиливается развитием мантийной складки, соответствен-
но образованием мантийного комплекса органов. Одновременно с
этим происходит редукция мускулатуры спинной стороны тела и
исчезновение целома за исключением его специализированных —
полового и эскреторно-перикардиального — отделов, расположен-
ных под раковиной на спинной стороне. Получается комплекс типо-
вых черт не хищного, но подвижного моллюска. Он может быть
использован и при переходе к пассивному питанию с крайне вы-
раженным развитием раковины у двустворчатых, часто1 сидячих
форм и связан здесь с полной редукцией головы. Напротив, у голо-
воногих происходит переход к самому активному хищничеству,
связанный с превращением единой ноги в серию рук — щупалец,
служащих для захватывания добычи, а затем и для движения.
Кроме того, имеет место дифференцировка задней части ноги в
орган плавания—воронку. При этом происходит редукция и полное
исчезновение пассивного органа защиты — раковины, затрудняю-
щей быстрое движение хищника. Соответственно сложно развивает-
ся мускулатура и органы чувств, а также центральная нервная
система.
Я не решусь назвать широкими приспособлениями, поднимающи-
ми энергию жизнедеятельности, выработку раковины, ноги или
мантии. В комплексе организации моллюсков налицо и редукция
388
мускулатуры, редукция целома и специализации его полового и
перикардиального отделов. Но в общем и целом происходит образо-
вание нового типа организации с такими новыми качествами, что
моллюски далее используют их, распространяясь в различнейших
частях мирового океана, переходя в пресную воду и выходя даже
на сушу. Главными моментами были пассивная защита и активное
питание — они создали типовые черты организации и, можно ска-
зать, весь их комплекс является широко приспособительным. Создал-
ся тип моллюсков на иной почве, чем тип иглокожих, и результат
получился иной. Моллюски — это в основном малоподвижные, но
хорошо защищенные донные формы. Но в их дальнейшей эволюции
мы уже находим резко различных представителей. С одной стороны
возникают двустворчатые моллюски, совсем теряющие активное
питание и в значительной мере активное движение. С другой сторо-
ны, головоногие переходят к активному питанию хищников и к
энергичному движению при редукции раковины. И это верх прогрес-
са в типе моллюсков.
На совершенно иной основе формируется тип членистоногих.
Они продолжают путь эволюции апнелид и представляют прежде
всего свободноподвижных хищников. Ведущим в создании органи-
зации членистоногих явилось развитие прекрасного средства пассив-
ной защиты в виде хитинизированной кутикулы, покрывающей все
тело и превращающейся в конце концов в наружный скелет. Ведя
к периодичности роста и ограничивая размеры тела и количество
образующих его клеток и вызывая редукцию целома, как опорного
образования, а также обусловливая своеобразное развитие крове-
носной системы, хитинизация кутикулы создает в то же время из
простой параподии аннелйд• конечность характера сложного рычага.
Это ведет и к чрезвычайно разнообразной дифференцировке ножек
членистоногих в связи с их приспособлением к разным функциям
при разных условиях существования, и к сильнейшей дифференци-
ровке мускулатуры и к выработке гетерономных отделов тела.
Кроме того, образование хитинового покрова, предохраняя от ис-
сушающего влияния воздуха, позволило членистоногим выйти на
сушу и стать в лице многоножек, насекомых и паукообразных самой
значительной, в известной ме^е главной группой наземного' живот-
ного населения. При этом в новых условиях членистоногие выраба-
тывают новые характерные для них системы. Так, на смену жабер
и нефридиев водных родичей, у наземных членистоногих появились
системы трахеального дыхания и мальпигиевых сосудов. Членисто-
ногие используют всяческие условия среды, используют различней-
шие минимальные возможности существования в биоценозах, а в
ряде случаев переходят к своеобразным зачаточным формам обще-
ственной жизни.
Однако сама односторонность ведущего момента в создании ти-
па членистоногих, а именно—выработка пассивной защиты посред-
ством хитиновой кутикулы, накладывает неизгладимый отпечаток
на всю эволюцию членистоногих. Казалось бы, что их активное пи-
339
тание и средства' активной защиты могут' послужить основой для
всяческого прогресса. Однако1 преобладание пассивной защиты —
хитиновый скелет — ведет к ограничению размеров, соответственно
(к ограничению количества клеток, входящих в состав тела, а в
связи с этим к крайней специализации, что становится препятствием
для дальнейших дифференцировок систем у членистоногих и для
перехода к новым широким приспособлениям.
Для низших хордовых характерны пассивное питание полусидя-
чих животных и, в связи с этим, выработка аппарата жаберных
щелей и полная редукция эктодермальной глотки, а также образо-
вание плотных окблоротовых придатков и трехмерного целома. У
туникат — типичных сидячих животных - - к этому присоединяется
мощная и сложно построенная кутикула в виде туники, обешечи-
вающей пассивную защиту. И мы видим, что пассивный характер
этих двух экологических моментов ведет к значительной деградации
всей организации взрослого животного. Редуцируется мускулатура,
органы чувств, нервная система, целом, но весьма сильно разви-
вается туника, жаберный аппарат, атриальная полость и крайне
своеобразна кровеносная система.
Совсем иначе выражается момент защиты у высших представи-
телей типа. У ланцетника пассивная защита, создаваемая пребыва-
нием в грунте, усложняется при пассивном питании из придонных
слоев воды тем, что возникает особый способ активной защиты
посредством энергичного плавания с последующим зарыванием в
грунт. Далее происходит полная редукция головной лопасти, и ве-
дущим становится развитие миохордального комплекса и спинной
нервной трубки. Две последние черты организации выработались у
низших хордовых как приспособления для укрепления головной ло-
пасти при ее активном функционировании и для иннервации перед-
ней части тела. У ланцетника они превращаются в мощное приспо-
собление, заменяя мускулатуру кожно-мышечного мешка и ее опор-
ный целом комплексом хорды и прилежащих продольных мышц,
получающих метамерное расположение. Соответственно спинная
нервная трубка распространяется на всю длину туловища и диффе-
ренцируется на головной и спинной мозг.
Всякие следы пассивного питания и защиты, свойственные низ-
шим хордовым, стираются в организации позвоночных благодаря
переходу к активному питанию и активной защите хищника. Этим
же порождается ряд широких приспособлений, дающих исключи-
тельный эффект подъема организации. Это особенно стоит в связи
с многообразием условий существования в наземной среде, куда в
середине палеозоя проникают позвоночные и где проделывают даль-
нейшую, чрезвычайно сложную и далеко идущую эволюцию. Богат-
ство клеточного и неклеточного материала и, соответственно1, разно-
образие приспособлений, возникающих под ведущим воздействием
условий существования, вызывают мощный подъем организации.
Происходит дифференцировка покрова и его придатков (чешуй,
волос, перьев), внутреннего скелета и мускулатуры, образование
390
конечностей характера сложного рычага и их специализация, про-
исходит дифференцировка областей тела, развитие аппарата легоч-
ного дыхания и усложнение кровеносной системы, происходит диф-
ференцировка очень совершенных -органов чувств. Наконец, появ-
ляется своеобразная сложность нервной системы и особенно голов-
ного мозга, становящегося мощным комплексом ассоциативных
центров, который обусловил возможность развития сложнейшей
нервной деятельности до психики человека включительно. Здесь
уместно вспомнить меткие слова Ф. Энгельса, что у позвоночных
нервная система — «основа всей организации», она «завладевает
всем телом и организует его сообразно своим потребностям»1.
Таков результат эволюции позвоночных, достигнутый благодаря
объединению сложных форм активной защиты и активного питания
в условиях сложной среды. Здесь еще раз подчеркнем, что и до
позвоночных те же эко-логические моменты активного питания и
активной защиты при освоении разных условий среды создавали
основные этапы эволюции по пути морфо-физиологического про-
гресса, а именно — организацию плоских червей, организацию не-
мертин и организацию целомических червей.
Не будем повторять частностей. Из анализа типов видно, что
типовые комплексы черт организации являются прежде всего обу-
словливающими биологический прогресс организмов, т. е. увеличе-
ние числа особей, расширение ареалов обитания и распадение на
подчиненные систематические группы.
Какие же общие выводы из всего рассмотренного можно сделать
относительно хода эволюции?
В общем процесс эволюции всегда является результатом взаи-
модействия между «природой» организмов и меняющимися условия-
ми их существования. Значение природы организма, т. е. его наслед-
ственной организации, связанной с определенными условиями среды,
мы особенно рельефно видели в эволюции членистоногих. Однако
ведущую, определяющую роль в выработке типов играют условия
существования. Высшие типы дали нам яркие примеры того, как
на основе организации первичных целомических форм сложились
типовые комплексы черт членистоногих, моллюсков, иглокожих и
хордовых, из которых каждый порожден особой комбинацией био-
ценотических отношений в питании и защите, а при переходе чле-
нистоногих, моллюсков и хордовых к наземному существованию
особо ясно выступает значение абиотических условий наземной
среды по сравнению с абиотическими условиями водной.
Возможные взаимные сочетания черт активной и пассивной за-
щиты и активного и пассивного питания в рассмотренных типах,
можно сказать, исчерпаны, однако варианты, в которых они выяв-
ляются в природе, неисчерпаемы. Легко видеть, как на такой основе
создались все типы в качестве узловых групп эволюции. Но в то же
время они показывают, что создавшиеся типовые комплексы могли
1 Ф. Энгельс. Диалектика природы, 1952, стр. 250.
391
эволюционировать дальше опять-таки на ведущей основе измене-
ния и перехода пассивных черт защиты и питания в активные и
наоборот, т. е. опять-таки под ведущим влиянием условий суще-
ствования. Так, например, создается организация Eleuterozoa среди
иглокожих на основе организации Pelmatozoa, организация дву-
створчатых моллюсков и головоногих на основе первичных Con-
chifora, организация оболочников и позвоночных на основе низших
хордовых.
Как основное положение приходится признать, что эволюция
не ограничена. Критериями ее хода являются, во-первых, ее необра-
тимость, как процесса, протекающего во времени, и, во-вторых, на-
правляющее действие условий существования. Сидячие или близкие
к тому формы низших хордовых дали грандиозную картину эволю-
ции позвоночных на основе овладения всемозможными аспектами
условий среды. Сидячие цистидеи через освоение разнообразных
условий водной морской среды повели к богатой эволюции свобод-
ноживущих морских звезд, офиур, морских ежей, голотурий.
Эти примеры показывают, что формы, выработавшиеся при
условии сужения взаимоотношений с окружающей средой, как это
имеет место у прикрепленных животных при их пассивной защите
и пассивном питании, вовсе не обречены на прозябание,' застой и
гибель — обычное утверждение плоских дарвинистов, а имеют
шансы на дальнейший расцвет, если их организация становится
качественно новым комплексом черт, новой системой, новым звеном
в эволюции. Вступая во взаимосвязи с условиями среды, уклоняю-
щимися от первоначальных, новые комплексы организации осваи-
вают новые условия и под их ведущим влиянием сами изменяются.
При этом возможности дальнейшего развития или расширяются
или остаются на прежнем уровне, или суживаются и тогда происхо-
дит дегенеративное упрощение. Результат один — многообразие
условий порождает и многообразную эволюцию нового типа орга-
низации, ограничиваемую только природой организма и тем же
многообразием условий существования, в которых он развивается.
В этом отношении, конечно, организация, например, кишечнополо-
стных по своей природе естественно более ограничена, не может ид-
ти в сравнение хотя бы с организацией сидячих же иглокожих.
Однако у колониальных сифонофор она делает скачок, поражая осо-
бенной, только им свойственной сложностью.
В высокой степени поучительны в рассматриваемом аспекте
паразитные группы. Так, выработавшийся на основе олигохетных
форм класс пиявок пошел по пути эктопаразитизма. Его оригиналь-
ная приспособительная организация, рассмотренная нами выше в
основных ее чертах, показывает пример появления качественно
новой группы. Она овладевает широкой ареной жизни. Пиявки из
пресных вод проникают в море, а в тропиках «Старого Света» пе-
реходят к наземному существованию, что мы видим у Haemadlpslna..
С другой стороны, они вступают и на путь настоящего хищничества
герпобделлид, теряя даже типичнейшие присоски, что мы видим у
лиманной и каспийской Archaeobdella.
392
Однако крайнее приспособление эндопаразита* например ленточ-
ных глист, к совершенно исключительным условиям существования
внутри другого организма делает организацию, представляющуюся
в .целом качественно новой, уже неспособной по своей природе
осваивать условия свободной жизни в водной или наземной среде.
С утратой кишечника, например, солитер теряет возможность за-
хватывать и переваривать пищу, доступную свободноживущим пло-
ским червям. Благодаря исключительности, обособленности условий
среды и крайностям организации подобных форм эволюция при-
водит их к границам возможностей существования. Часто бывает,
что совершенное полезное приспособление становится родовым при
всяких существенных изменениях условий среды. Такое приспособ-
ление чрезмерно и, доходя до своего завершения, переходит в свою
противоположность, становится гибельным.
। Не будем здесь останавливаться на многочисленных построениях
|и теоретических обобщениях относительно' хода эволюции, идущих
со столоны зарубежных палеонтологов. Они в достаточной мере
освещены и оценены в нашей литературе, и лишь недавно (1941,
1948), появилась большая сводка «История эволюционной палеонто-
логии от Дарвина до наших дней» Л. III. Давиташвили, в которой
ярко показана несостоятельность и реакционность многих построе-
ний.
Прямым завершением всего нашего анализа организации ти-
пов является сопоставление наших выводов с теми заключениями
относительно хода эволюционного процесса и его закономерностей,
к которым пришел А. Н. Северцов (1931, 1939). От этого сопостав-
ления мы не должны и не можем уклониться.
А. Н. Северцов отмечает' (1939), что биологически прогрессив-
ная эволюция приводит к образованию крупных систематических
групп, состоящих из большого числа подчиненных, причем (стр. 309)
ароморфозы «становятся характерными признаками высших систе-
матических групп: семейств, отрядов, классов», и (стр. 308) «перио-
ды ароморфоза представляют собой как бы узловые точки эволю-
ционного процесса».
Общую картину эволюционного процесса А. Н. Северцов ри-
сует так (стр. 328): «Многие крупные группы животных, например
все типы, развивались в течение первых стадий своего филогенеза
путем ароморфоза, после чего для них обычно наступал период
идиодаптический или, в некоторых случаях, период общего регресса
(дегенерация)», причем ароморфозы и идиоадаптация (стр. 332)
«свободно переходят друг в друга. Вместе с тем идиоадаптивный
путь эволюции способен перейти, во-первых, в тип регрессивной
эволюции, во-вторых, что бывает чаще, в тип специализации. Мы
находим много примеров такого перехода. Наоборот, — говорит
А. Н. Северцов, — я не нашел примеров того, чтобы группа спе-
циализированных животных или группа животных, вступивших на
путь общего регресса или общей дегенерации, пошли по пути про-
грессивной эволюции». Здесь не наблюдается обратимости... Эволю-
ция с пути идиоадаптаций может перейти на путь специализаций,
/деградации, но обратный переход, повидимому, не имеет места.
«Во всяком случае — добавляет, несколько противореча себе,
А. Н. Северцов, — такие переходы очень редки и представляются
чем-то исключительным... Специализация, а тем более дегенерация,
служат препятствием к появлению прогрессивных изменений у жи-
вотных и вступлению их на путь ароморфоза». При этом общая де-
генерация, наблюдающаяся у сидячих форм и у эктопаразитов, ве-
Рис. 153. Схема эволюционного процесса по А. Н. Северцову. Смены пе-
риодо® ароморфозов (а1—я2, а3—а4) периодами идиодаптаций (Ь), или
периодами регрессивной эволюции (г1—г2). В частности: А — предки выс-
ших хордовых, г1—г2 — Tunicata, а1—а2 — Асгаша, а3—сА—Craniata
дет к редукции органов, несущих активную функцию, а прогрес-
сивно развиваются пассивные.
Чтобы завершить изложение этих идей А. Н. Северцева, приво-
дим его чрезвычайно наглядную схему (рис. 153) хода эволюцион-
ного процесса.
Свои выводы и обобщения А. Н. Северцов делает, однако, с су-
щественной оговоркой (стр. 341): «Мы, конечно, никоим образом не
можем утверждать, что изменение эволюционных направлений
всегда происходит именно так, как мы это только что описали, но
сказанного, думается нам, совершенно достаточно для того, чтобы
показать, что здесь мы имеем дело с необычайно сложной пробле-
мой... Мы лично имели в виду лишь указать читателю на то, что
здесь перед исследователем открывается огромное поле интересней-
ших и еще почти никем не затронутых проблем и что проблемы
далеко не так неразрешимы, как это кажется на первый взгляд».
Необходимость этой оговорки А. Н. Северцова ярко подтверж-
дается всем нашим материалом по анализу организации типовых
групп животного царства, будь это основные типы, принимаемые
зоологами — губки, иглокожие, хордовые, или такие небольшие
группы, как пиявки.
-394
Мы должны констатировать, что путь смены гаромарфозов пе_
риодами идиоадаптации, приводимый А. Н. Северцовым как" норма
хода процесса эволюции, представляет в действительности только
определенный частный случай эволюции, связанный с морфо-физио-
логическим прогрессом и присущий позвоночным, т. е. той группе жи-
вотных, которая была объектом работ этого ученого. Однако тот же
путь, как мы это показали в начале заключительной главы, имел
меЪго в более широком масштабе и при выработке типовых’ групп
кишечнополостных, плоских червей, немертин, аннелйд (правильнее
первичных целомат) — одним словом, путь этот вел по «прямой
большой дороге» эволюции, завершающейся позвоночными. Попыт-
ки А. Н. Северцова распространить его шире, например на игло-
кожих, не соответствуют фактам. Можно сказать, что А. Н. Север-
цов, будучи глубоким знатоком позвоночных, имел очень общие
представления о беспозвоночных. Но слабость его в этом отношении
стала источником его силы в другом, направив его мысль на раз-
решение вопросов о закономерностях хода эволюционного процесса,
которые были по уровню знаний уже назревшими для позвоноч-
ных, но еще почти не затрагивались у беспозвоночных. Только те-
перь, после долгого периода работы над происхождением различ-
ных систем органов, над филогенией отдельных групп и т. д., изу-
чение беспозвоночных подошло вплотную к морфологии целостного
организма, к анализу организации типовых групп.
На этой основе мы неизбежно должны прийти к выводу, что
высшие типы животного царства — членистоногие, моллюски, игло-
кожие, хордовые — шли в своей эволюции от первичных целомиче-
ских форм иными путями, чем данная А. Н. Северцовым норма
смены периода ароморфозов периодом идиоадаптаций, а далее или
дегенераций, или снова ароморфозом. В действительности на осно-
ве организации первичных протоцеломат вырабатывались новые
качественно характерные комплексы организации, которые, каждый
своим особым образом, были приспособлены к определенным глав- >
ним категориям условий существования. Так, аннёлиды и членисто-
ногие приспособлены к свободноподвижному существованию хищ-
ников, моллюски ™ к свободному существованию нехищных живот-
ных, иглокожие и хордовые — к сидячему образу жизни пассивно
питавшихся форм. Наконец, пиявки — к эктопаразитическим усло-
виям существования (рис. 154).
Освоение и использование главнейших категорий условий суще-
ствования, другими словами, ведущая роль среды, — вот что обус-
ловливает ход эволюционного процесса и завершается возникнове-
нием качественно новых типовых групп. Конечно, сужения активно-
сти организма, происходящие при приспособлении в пассивной за-
щите у животных, переходящих к сидячему существованию, или
при переходе к пассивному питанию никак нельзя назвать аромор-
фозом и отнести к периоду ароморфных изменений. Но сам факт
возникновения качественно новой организации открывает .широкие
возможности для освоения и разнообразного использования иных
395
условий существования, чем те, в которых данный тип выработал-
ся, а следовательно, и к дальнейшим изменениям как идиоадаптив-
ного характера, так и к ароморфным, и к дегенеративным. Таким
образом теряет свое универсальное, по мнению А. Н. Северцова,
значение его утверждение об ограничении возможностей эволюции,
т. е. об ограничении возможности перехода к ароморфозам, раз ор-
ганизм вступил на путь дегенерации или специализации. В этой свя-
Рис. 154. Общая схема эволюционного процесса в дополнение к схеме А. Н. Се-
верцева. Смены периодов выработки типовых групп (а*—&2, cfy—а4, г1—г2, г3—г4)
периодами их приспособительной эволюции (6). В частности: А — Protoccelo-
mata, О — кольчатые (полимерные) черви, 7? — членистоногие, Р — олиго-
мерные черви типа диплеврулы, Q — иглокожие ****
зи мы видим, что А. Н. Северцов недостаточно ясно представлял
себе появление качественно новой организации узловых групп и
поэтому недостаточно правильно расценивал значение различных
форм взаимоотношений организма и среды в выработке типов.
Можно сделать более общий вывод. Эволюция неограниченна.
Но в каждом конкретном случае ее направление, ее формы опре-
деляются, с одной стороны, той организацией, той природой орга-
низма, которые выработались в длительном процессе эволюции и
являются результатом всей филогенетической истории, а с дру-
гой — условиями биотической и абиотической среды и возможно-
стью их освоения организмом.
Из актиний не могла развиться аннелида, минуя низших червей,
что допускал Седжвик, равно как из аннелиды не могли произойти
позвоночные животные, как предполагали Земпер и Дорн. Анало-
гично этому высшая нервная деятельность насекомого выливается
в выработку сложнейших инстинктов, тогда как у позвоночных она
идет в сторону сложнейших условных рефлексов. Наконец, органи-
зация губок пошла совсем иными путями, чем весь прочий живот-
ный мир. Эти примеры доказывают значение организации, природы
организмов, но весь наш анализ типов дает ярчайшие доказатель-
ства ведущего значения условий существования в выработке типо-
вых групп и в дальнейшей их эволюции.
список
важнейшей из приводимой литературы
Беклемишев В. Н., 1925, Морфологическая проблема животных структур.
Извест. биол. н.-и. инет. (Пермь), т. 3, пр. 1.
Беклемишев В. Н., 1951, К построению системы животных. Успехи совр.
биол., т. 32.
Беклемишев В. Н., 1944, Основы сравнительной анатомии беспозвоночных.
Москва.
Беклемишев В. Н., 1952, то же, изд. 2.
Бер нар1 X., 1892 (Bernard Н. М.), An Endeavour to show that the tracheae
of the Artropoda arose from setiparous sacs. Zool. Jahrb-, Anat., v. V.
Бючли О., 1892 (Butschli О.), Versuch der Ableitung des Echinoderms, Zeit-
schrift wiss. Zoologie, V. 53, Suppl.
ВейдовскийФ., 1905 (Vejdovsky F.), Zur Hamocoltheorie. Zeitschr. wiss.
Zoologie, V. 82.
Ган стрем Б., 1928 (Hanstrom В.), Vergleichende Anatomie des Nervsystems
der wirbellosen Tiere. Berlin.
Гатчек Б., 1878 (Hatschek B.), Studien fiber Entwicklungsgeschichte der An-
neliden. Arb. Zool. Inst., Wien, V. I.
Гатчек Б., 1888, Lehrbuch der Zoologie, eine morphologische Obersicht Tier-
reichs. Jena.
Г атчек Б., 1911, Das neue zoologische System. Leipzig.
Гей дер К. (Heider К.), 1913, Entwicklungsgeschichte und Morphologic der
Wirbellosen. Kultur der Gegenwart, 2, Zoologie.
Геккель Э., 1866 (Haeckel E.), Generelle Morphologic. Berlin.
Геккель Э.,* 1872, Die Kalkschwamme. Berlin.
Геккель Э, 1873—1876, Biologische Studien zur Gastraea-Theorie. Jena,
Zeitschr., V. 8, 9, II.
Гертвиг О. и P., 1878 (Hertwig О. und R.). Die Actinien. Jena.
Г e p т в и г О. и Р.» 1880, Die Coelomtheorie. Jena.
Гиман Л., 1940 (Hyman L. Н.), The invertebrates: Protozoa through Ctenopho-
ra. New York.
Гольмгрен H., 1916 (Holmgren N.), Zur vergleichenden Anatomie des
Gehirns. Svensk. Vetenskap. Ak. Handl, V. 50.
Гроббен К.. 1910 (Grobben К.), Lehrbuch der Zoologie Claus-Grobben. Mar-
burg.
Давиташвили Л. Ш., 1948, История эволюционной палеонтологии от Дар-
вина до наших дней. Ленинград.
Давыдов К., 1903, Hydroctena salenskii. Mem. Ac. Sc. Petersbourg, Ser. 8,
Cl. phys.-mat, V. 14, № 9.
Давыдов K-, 1928, Traite d’embryologie comparee des invertebres. Paris.
Дарвин 4.» 1859, Происхождение видов.
Деляж И. и Эру ар Е„ 1897—1903 (Delage Y. et Herouard Е.), Traite de
zoologie concrete. Spongiaires, Vermidiens, Piocorde. Echinodermes. Paris.
397
Дельсман X., 1913 (Delsman Н. С.). Der Ursprung der Vertebraten. Mitt
d. Zool Stat. Neapel, V. 20.
Дельсман X., 1922, The ancestry of vertebrates. Amersfoort (Holland).
— Догель В. A, 1940, Сравнительная анатомия беспозвоночных, ч. 2. Ленинград,
Д or е л ь В А., 1947, Зоология беспозвоночных, 4 изд. Москва.
Догель В. А., 1954, Олигомеризация гомологичных органов. Ленинград.
Заварзин А. А., 1925, Der, Parallelismus der Strukturen als ein Grundprincip
der Morphologic. Zeitschr. wiss. Zoologie, V. 124.
Завапзин A. A., 1934, Об эволюционной динамике тканей. Архив биол. наук,
т. 34, А, в. 1.
Заварзин А. А., 1941, Очерки по эволюционной гистологии нервной системы,
Ленинград.
Заварзин А. А., 1947, Очерки эволюционной гистологии крови и соедини’
тельной ткани. Москва.
Заленский В. В,-1907, Morphogenetische Studien an Wurmern. Mem. Acad.
Sc. Petersbourg, Cl. phys-mat, V. 19.
3 a x в а т к и н A. A., 1949, Сравнительная эмбриология низших беспозвоночных.
' Москва.
Зе мон Р., 1888 (Semon R.), Die Entwicklung der Synapta digitata und die
Stammesgeschichte der Echinodermen Jena.
3 ем пер 1875 (Semper C.), Die Stammesverwandschaft der Wirbeltiere und
Wirbellosen Arb. Zool. Inst., Wurzburg, V. 2.
Зенкевич Л. A., 1937, История системы беспозвоночных. Руководство по
зоологии, т. I.
Зернов С. А., 1913, К вопросу об изучении жизни Черного моря. Зап. Ак.
наук, Петербург, сер. 8; т. 32, в. 1.
Иванов А. В. 19г5. Основные черты организации погонофор. Докл. Ак. наук
СССР, т. 100, № 1.
И в а н о в А. В, 1955, О принадлежности класса погонофор к особому типу
вторичноротых. Докл. Ак. наук СССР, т. 100, № 3.
Иванов П. П., 1937, Общчя и сравнитетьная эмбриология. Ленинград.
Иванов П. П., 1944, Первичная и вторичная метамерия тела. Журн. общ.
биол., т. 5.
К е г ь Ж 1^91 (Kennel J.), Die Verwandschaftsverhaltnisse der Arthropoden.
Schrift Nat. Gesell Dorpat, V. 6.
Клейненберг H., 1872 (Kleinenberg N.), Hydra. Leipzig.
Клейненберг H.3 1886. Die Entstehung des Annelids aus der Larve von
Lopadorhynchus. Zeitschr wiss. Zoologie, V. 44.
Ковалевский A. O.. 1866, r "vO r-iuncrSgeschichte der einfachen Ascidien.
Mem. Ac. Sc.; Petersbourg, 7 Ser., V. 10.
К о в а л e в с к и й А. О., 1807, Entwicklungsgeschichte des Amnhioxus lance-
olatus, ibid. V. 11.
Коротнев A. A, 1885, Ctenoplana Kowalevskii. Zeitschr. wiss. Zoologie, v. 43.
* Корш ел ьт К- и Гей дер К-, 1890—1910 (Korschelt К- und Heidcr К.).
Lehrbuch der vergleichenden Entwicklungsgeschichte der wirbellosen Tiere.
Jena.
Ла мер A., 1929 (Lameere A.), Precis de Zoologie, v. I. Bruxelles.
Ланг A. 1903 (Lang A.), Beitrage zu einer Trophccoeltheorie. Jena, Zeitschr.,.
V. 38. 1
Ланг A, 1912, Handbuch der Morphologic der wirbellosen Tiere, Jena.
Лейка nT ppj 1849 (Leukart R.), Uber die Morphologic und Verwandschaft-
verhaltnisse der wirbellosen ТЩге Braunschweig.
Ливанов H. A., 1931, Die Organisation der Hirudineen. Ergebn. und
Fortschr. der ZooMgie, V. 7, H. 4
Ливанов H. A., 1946, Hvth эволюции животного мира, ч. 1. Учен, записки
Казанск универе, т. 105, кн. 3.
Лысенко Т. Д, 1948, Агробиология. Москва.
Мак Боайд Е. В., 1914 (Mac-Bride Е. W.). Text-Воок of embryology, v. С
London.
v ЧП .
Мечников И. И., 1882, Vergleichend-embryologische Studien Zeitschr. wiss.
Zoologie, V. 36. 37.
Мечников И. И., 1886, Embryologische Studien an Mcdusen. Wien.
Мейер Э. A., 1887—1888, Studien fiber den Korperbau der Anneliden. Mitt.
A Zoolog Stat. Neanel, V. 7, 8.
Мейер Э. A., 1897, Исследования над развитием кольчатых червей. Труды
Казанск. общ. естественен., т. 31, вып. 4.
Мейер Э. А., 1904, Theoretische Betrachtungen fiber die ersten Anfange
ambulacralen Wassergefassystems der Echinodermen. Zool. Jahrb , Anat., V. 21.
Мильн Эдвардс A., 1859 (Pharis, 1853), Введение в общую зоологию.
Москва.
Минчин Е. А., 1900 (Minchin Е. A.), Sponges (Lankester, A Treatise
on zoology. P. 2).
Неф A, 1911—1924 (Naef A.), Studien zur generellen Morphologic der
Mollusken. Ergebn. und Fortschr. Zool., V. 3, 6.
Неф A., 1931, Phylogenie der Tiere. Handbuch d. Vererbungswissenschaft, V. 3S
Leif I, Berlin.
Нирштрас Г., 1923 (Nierstrass H. F.), Die Verwandtschaftverhaltnisse
zwischen Mollusken und Anneliden. Bidr. Dierk, V. 22.
Опарин А. И., 1945, Происхождение жизни на земле. Москва.
Пельзенер П., 1895 (Pelseneer Р.), Recherches morphologiques et ohyloge-
netiques sur les mollusques archaiques. Mem cour. Ac. Sc. Belg., v. 57.
Резвой П. Д., 1937, Тип губок Руководство по зоологии, т. 1.
Рей Л анкестер Е., 1900 (Ray Lankester Е.) The enterocoela and coelomo- <
coela. A traetise on zoology, p. 2, London.
Севеоцов A. H., 1931. Morphologishe Gesetzmassigkeiten der Evolution.
Jena.
Северцов A. H., 1934, Главные направления эволюционного процесса»
Москва.
Северцов А. Н., 1939, Морфологические закономерности эволюции. Москва.
Сед ж вик А., 1884 (Sedgwick A.), On the origin of metameric segmentation.
Quart. Journ. micr. Sc., v. 24.
Сепп E. K., 1936, Развитие нервной системы. Проблемы теорет. и практ. мс- -
дацины, Москва.
С е п п Е К., 1949, История развития нервной системы позвоночных Москва
Третьяков Д. К., 1929, Ursprung der Chordaten. Zeitschr. wiss. Zoologie
V.434.
Фа усек В., 1897, Исследования над историей развития головоногих. Труды
Петерб. общ. ест., т. 28.
Федотов Д. М., 1935, Очерк эволюции животного мира. Тр. Пал. инет. Акад.
наук, т, 4.
Федотов Д. М., 1924, К морфологии осевого комплекса иглокожих. Zeitschr
wiss. Zoologie. V. 123.
Федотов Д. М., 1951, Тип иглокожих. Руководство по зоолог,, т. 3
Фернандес М., 1905 (Fernandez М), Zur mikroskopischen Anatomie de&
Blutgef assy stems der Tuntcaten. Jena Zeitschr. V. 39.
X л о п и н H. Г., 1946, Общебиологические и экспериментальные основы г ист о-
логии. Москва.
Циглё'р X., 18.98 (Ziegler Н. Е ), Uber den derzeitigen Stand der Colomfrage.
Verh. Deutsch. Zool. Gesell, 8 Vers.
Шмидт Г. A., 1951, Эмбпиология животных, ч. I Москва.
Шнейдер К-, 1902 (Schneider К. С.). Lehrbuch der vergleichenden Histologic
der Tiere. Jena
Шпенгель И., 1893 (Spengel J. W.), Die Enterouneusten. Fauna u. Mora
Golf. Neapel, V. 18.
Шторх О.. 1912 (Storch О.), Vergleichend-anatondsche Polychatenstudien
Sitzber. Ak. Wien, Math.-natur. KI.. V. 122.
ОГЛА ВЛ ЕН И Е
Предисловие...................................................... £
Введение . ....................................8
Глава I. Ткани и органы.......................................22
Глава II. Тип кишечнополостные — Coelenterata ' 41
Глава III. Тип губки — Spongia, s. Porifera . . . ' . . 77
Г лава IV. Тип гребневики — Ctenophora............................87
Глава V. Подтип плоские черви — Plathelmintes . ... 101
Глава VI. Подтип немертины — Nemertini...........................120
Глава VII. Подтип кольчатые черви — Annelides . . . .131
Глава VIII. Класс пиявки — Hirudinea . . .... 169
Глава IX. Тип моллюски — Mollusca..........................178
Глава X. Тип членистоногие — Arthropods......................209
Глава XI. Тип иглокожие — Echinodermata.....................244
Глава XII. Тип хордовые — Chordata ...... 289
Птеробранхии — Pterobranchia.................................306
Кишечнодышащие — Enteropneusta . . . . . .311
Оболочники — Tunicata........................................319
Бесчерепные — Acrania........................................335
Позвоночные — Vertebrata.................................... 350
Глава XIII. Система животного царства и пути эволюции хордовых 368
Глава XIV. Закономерности путей эволюционного процесса . . 384
Список важнейшей из приводимой литературы .... 397
Н. А. Ливанов
ПУТИ эволюции животного МИРА
Редактор А. Передельскай
Техн, редактор Л. И. Королёва
Сдано в набор 5/1 1955 г. Подписано к печати 22/VII—55 г.
Бумага 60X92/16=25,0 печ. л., 28,8 уч.-изд. л. Тираж 5.000 экз. Т—05925-
Издательство «Советская наука». Заказ 1. Цена 15 руб. 40 коп.
Типография изд-ва «Советская наука», Неглинная, 29/14. Зак. 9
ЗАМЕЧЕННЫЕ ОПЕЧАТКИ
Стра- ‘ ница ’ Строка Напечатано Следует читать
3 11 снизу 1 * увеличению увлечению
8 9 сверху переходяща преходяща
13 17 снизу ; вакуумом вакуомом
24 19 » ! отсутствием отсутствие
30 5 сверху неизменные неизмененные
71 15 снизу соответственно септ , соответственно септ,
74 15 » организма организм
79 2 » скелет,— диференцировки и скелет
109 24 » дифференцировки
194 13 сверху
329 1 снизу i бластогонные зачатки зачатки
376 13 сверху пах. пах
396 подпись к 1 рис. 154 (сР~а5, а2—а4, (а1—а2, а3—а4,
Н. Л Ливанов, Пути эволюции животного мира.