/

















































































Автор: Левина Р.Е.
Теги: морфология растений экология растений этология растение и окружающая среда биология ботаника
Год: 1987




Текст
Р. Е. Левина
Морфология
и экология
плодов
АКАДЕМИЯ НАУК СССР
ВСЕСОЮЗНОЕ БОТАНИЧЕСКОЕ ОБЩЕСТВО
Р. Е. Левина
Морфология
и экология
плодов
Ответственный редактор
Н. Н. ЦВЕЛЕВ
е
ЛЕНИНГРАД
ИЗДАТЕЛЬСТВО «НАУК А»
ЛЕНИНГРАДСКОЕ ОТДЕЛЕНИЕ
198 7
УДК 581.47:581.522.6
|jl е в и н а Р. Е.| Морфология и экология и лодок. ' ЛНнукп, 1987. — 160 с.
В книге на фактическом материале мировой флоры рпссмнтрипается широкий
круг вопросов и проблем современной карпологии: типология и номенклатура
плодов, их классификация, карпоэкология, гетерокарпия, типология соплодий.
Содержание и изложение материала отражают эволюционный подход. Являясь
монографическим' исследованием, книга вместе с тем служит и справочным посо-
бием по карпологии.
Для специалистов-ботаников, преподавателей вузов, аспирантов и студентов,
а также специалистов в области прикладной ботаники. Библиогр. 148 назв.
Ил. 60. Табл. 9.
Рецензенты:
М. Э. КИРПИЧНИКОВ, В. Н. ТИХОМИРОВ
2004000000-603
Л 042(02)-87 22°-87~Z
©Издательство «Наука», 1987 г.
ОТ АВТОРА
Книга автора «Плоды», опубликованная в 1967 г., представ-
ляла собой первый опыт обобщающей работы по вопросам карпо-
логии в нашей литературе. Настоящая работа первоначально была
задумана как 2-е издание «Плодов», но изучение новой литературы
и переосмысление многих фактов привели к тому, что книга факти-
чески написана заново. От первого варианта сохранились лишь
наименование глав и их последовательность.
В названии книги отражено содержание основных по объему
II и V глав; в IV главе также дается морфолого-экологическая
характеристика плодов при гетерокарпии.
Не вводя новых принципов классификации, типологии и номен-
клатуры плодов, автор предлагает вместе с тем естественную
морфологическую классификацию плодов с четкой иерархией
классификационных категорий. Основной такой категорией, свя-
занной с номенклатурой плодов, служит «вид плода». Для поли-
морфных видов плода вводится категория ниже видового ранга —
вариация. Предложенная классификация представлена в виде
таблиц. Описание всех видов плодов, названных в таблицах,
составляет содержание II, морфологической, главы. В ней (как
и в V главе) помимо плодов растений флоры СССР много места
уделено плодам тропических и субтропических растений. Можно
ожидать, что возражения вызовет III глава книги, посвященная
соплодиям — наименее разработанному разделу карпологии. Я ду-
маю, однако, что традиционное понимание соплодия как группы
сросшихся плодов слишком узко и характеризует лишь небольшую
часть соплодий. В предложенном более широком определении
отмечен и тот факт, что не всякое соцветие превращается в сопло-
дие. Помимо определения понятия мной намечены и первона-
чальные принципы типологии соплодий, отражающие степень их
специализации. Надеюсь, что предложенная схема классификации
соплодий может служить отправной для обсуждения и усовершен-
ствования. В главе, посвященной гетерокарпии, использованы
новые материалы последних лет, отражающие не только класси-
ческий — морфолого-экологический — но и другие аспекты изуче-
ния этого явления: физиологический, онтогенетический и отчасти
филогенетический. V, экологическая, глава дополнена и факти-
ческим материалом, и некоторыми теоретическими/положениями;
существенно иное и построение главы.
з
Обновлен и иначе представлен иллюстративный материал.
Оригинальные рисунки выполнены О. В. Фиминой.
Латинские названия растений флоры СССР даны по С. К. Чере-
панову (1981), остальные — по А. Энглеру (Engler, 1964) и по
«Жизни растений» (1980—1982).
В соответствии с очень детальными, весьма критическими
(хотя и более чем доброжелательными) рецензиями В. Н. Тихоми-
рова и М. Э. Кирпичникова в первоначальный вариант рукописи
внесено много исправлений, уточнений и дополнений. Заключи-
тельный анализ рукописи выполнил ответственный редактор книги
Н. Н. Цвелев. Всем названным коллегам — моя самая искренняя
глубокая благодарность.
Приношу свою признательность А. Л. Тахтаджяну. Его под-
держка и интерес к работе очень помогали в процессе написания
книги.
Особую благодарность выражаю Президиуму ВБО за предо-
ставленную возможность публикации работы.
4
Глава I
ОБЩИЕ ВОПРОСЫ
ЗНАЧЕНИЕ ПЛОДОВ.
ОПРЕДЕЛЕНИЕ ПОНЯТИЯ «ПЛОД»
Плод, свойственный только покрытосемянным, относительно
других органов высших растений представляет собой морфологи-
ческое новообразование. Вместе с тем вследствие быстрой экологи-
ческой эволюции плоды достигли почти такого же разнообразия,
как и виды цветковых растений.
Функцией плодов являются защита и рассеивание семян (дис-
семинация), при этом защитная функция может ограничиваться
лишь периодом созревания семян (сухие раскрывающиеся плоды)
или продолжаться вплоть до их прорастания (односемянные и рас-
падающиеся сухие плоды, костянки).
Значение плодов, однако, не ограничивается только'их ролью
в жизни растений. Для многих групп животных плоды составляют
основу пищи (плодо- и зерноядные птицы, обезьяны, рукокрылые,
некоторые грызуны и др.). Экология и ряд морфофизиологических
особенностей этих животных, как и признаки поедаемых ими пло-
дов, являются результатом длительной сопряженной эволюции.
Л. ван дер Пейл (Pijl van der, 1969) устанавливает «синдромы»
признаков плодов, по которым легко можно узнать агентов их рас-
пространения — рептилий, птиц, летучих мышей, приматов или
муравьев. И в настоящее время плоды растений определяют в зна-
чительной степени характер трофических связей в биоценозе.
Наконец, невозможно переоценить значение плодов в жизни
человека. Этому вопросу посвящается небольшая глава нашей
книги. Здесь же только отметим, что изучение плодов имеет еще
и важное научно-прикладное значение. Карпологические (от греч.
«карпос» — плод) признаки отдельных видов, родов, семейств
достаточно устойчивы, поэтому по плодам оказывается возможным
определение тех или иных таксонов цветковых растений. Из исто-
рии ботаники известно, что система растений, созданная в конце
XVI в. Андреа Чезальпино, была основана на строении плодов.
В настоящее время эта система имеет лишь чисто исторический
интерес. Однако и современные систематики при разработке эво-
люционных систем цветковых растений используют наряду с дру-
гими карпологические признаки.
Значение морфологии плодов и семян, умение их распознавания
совершенно необходимы в контрольно-семенном деле, в частности
5
для карантинной службы, при анализе растительного сырья,
в особенности лекарственного, иными словами, для специалистов
по фармакогнозии, судебной медицине, товароведов различного
профиля, а также для лесоводов, агрономов и т. п.
Очень трудно дать лаконичное и вместе с тем полное определе-
ние плода. Не случайно в учебниках по морфологии растений такого
определения вообще не дается или плодом называют видоизме-
ненную завязь, что справедливо лишь для немногих «настоящих»
плодов. А большинство плодов, образованных многими завязями
или с участием других частей цветка или даже соцветия (кроющих
листьев, оси и т. п.), при таком узком определении оказываются
либо «сборными», либо «ложными». Неправомерность этих поня-
тий убедительно показана Н. Н. Каденом (1947).
Объем книги не позволяет обсуждать определения плодов,
предложенные разными авторами. Существующие виды плодов
более полно, как нам кажется, охватывает следующее определение
(Левина, 1967а): плод — это видоизмененный вследствие оплодо-
творения (или апомиксиса) гинецей одного цветка с прирастаю-
щими или сохраняющимися при гинецее другими частями цветка
и соцветия. Для простоты можно принять менее точное, но лако-
ничное определение: «Плод — это зрелый цветок» (Имс, 1964).
МОРФОЛОГИЧЕСКОЕ РАЗНООБРАЗИЕ ПЛОДОВ
ТИПЫ ГИНЕЦЕЯ
Гинецей, т. е. совокупность плодолистиков цветка, обычно
составляет структурную основу плода. Чтобы получить представ-
ление о морфологическом разнообразии плодов, познакомимся
сначала с типами и вариантами гинецея.
Плодолистики, как известно, несут на себе семязачатки. Зона
их возникновения получила название плаценты, а способ разме-
щения — плацентации. Характер плацентации определяется
отчасти способом смыкания краев плодолистика и числом семяза-
чатков, а также обособленностью или, напротив, типом срастания
плодолистиков.
Гинецей из свободных, т. е. не сросшихся между собой, плодо-
листиков называют апокарпным (греч. «апо» — приставка,
обозначающая отделение от чего-либо, и «карпос» — плод). Число
плодолистиков апокарпного гинецея может варьировать от многих
до одного. Поэтому приходится различать 2 варианта апокарпного
гинецея: полимерный (из многих плодолистиков) и мономерный.
Для полимерного гинецея характерно спиральное расположение
плодолистиков. С уменьшением их числа укорачивается ось
цветка и расположение плодолистиков становится циклическим
(мутовчатым).
Характер плацентации апокарпного гинецея зависит от количе-
ства семязачатков в каждом отдельном пестике. Если семязачатки
6
очень многочисленны, плаценты располагаются почти по всей
внутренней поверхности плодолистика, т. е. на его пластинке.
Такая ламинальная (пластинчатая) плацентация свойственна
некоторым представителям порядка Magnoliales\ из растений на-
шей флоры встречается у нимфейных, сусаковых.
Уменьшение числа семязачатков приводит к тому, что они
располагаются двумя рядами вдоль краев плодолистика. Такую
плацентацию называют различно: сутуральной (шовной), марги-
нальной (краевой) или субмаргинальной (почти краевой). Наибо-
лее точен последний термин, так как семязачатки располагаются
не по шву или краю плодолистика, а вблизи края. Субмаргиналь-
ная — это наиболее распространенный тип плацентации. Различ-
ные ее варианты встречаются в апокарпном и во всех других типах
гинецея.
Процесс сокращения числа семязачатков приводит к формиро-
ванию плодолистика с одним-единственным семязачатком, чаще
всего расположенным у самого основания плодолистика. Поэтому
такую плаценту иногда называют базальной (лютик, гравилат,
земляника и др.). Эволюционный путь развития от многих к одному
семязачатку легко прослеживается благодаря наличию недоразви-
тых семязачатков (например, Hydrastis canadensis, Potentilla
recta), но главным образом по постепенному сокращению и упро-
щению всей проводящей системы плодолистика (различные виды
лютика и др.).
Вследствие срастания плодолистиков друг с другом возникает
ценокарпный гинецей (от греч. «койнос» — общий, «карпос» —
плод). В зависимости от способа срастания и характера плацен-
тации различают 3 варианта ценокарпного гинецея: синкарпный
(от греч. «син» — вместе), паракарпный (от греч. «пара» —
возле), лизикарпный (от греч. «лизис» — растворение, уничтоже-
ние) .
Синкарпный гинецей образуется при срастании замкнутых
плодолистиков боковыми сторонами. Естественно, что завязь
в этом случае оказывается многогнездной, а сближенные в центре
плаценты отдельных плодолистиков образуют так называемую
центрально-угловую плаценту1 (очень распространенный вариант
субмаргинальной плаценты).
Чаще всего синкарпия возникает при циклическом расположе-
нии плодолистиков. Но у представителей некоторых семейств
происходит частичное срастание плодолистиков спирального гине-
цея (магнолиевые). В иных случаях гинецей остается еще спираль-
ным апокарпным, но плод из него развивается синкарпный, т. е.
срастание происходит в процессе онтогенеза (Аппопасеае и неко-
торые другие). У барбариса спиральность плодолистиков опреде-
ляется лишь анатомически.
1 Субмаргинальная плацента в замкнутом плодолистике апокарпного гинецея
тоже является угловой. Поэтому для синкарпной завязи целесообразно применение
термина «центрально-угловая».
7
Известно немало таких случаев, когда плодолистики сраста-
ются лишь у основания, а в верхней части остаются свободными,
образуя гемисинкарпный гинецей (греч. «геми» — полу; водосбор,
спирея, шейхцерия, сусак). Гораздо реже слияние захватывает
лишь область рылец, а на остальном протяжении плодолистики
остаются свободными. Из такого гинецея образуется по существу
апокарпный плод, так как завязи совершенно свободны (ласто-
вневые — Asclepiadaceae, кутровые — Аросупасеае).
Паракарпный гинецей характеризуется одногнездной завязью
с постенным расположением семязачатков вдоль ее швов. Такую
плацентацию называют париетальной (лат. parietalis — постен-
ный) . Каждому плодолистику соответствуют 2 плаценты, таким
образом, по числу семяносцев легко определить, из скольких
плодолистиков образована завязь. Париетальные плаценты
нередко сильно разрастаются (например, у тыквенных) и несут
очень большое число семязачатков. Иногда плаценты, разраста-
ясь радиально к центру завязи, сталкиваются и образуют много-
камерную, как бы вторично- или ложносинкарпную завязь [неко-
торые колокольчиковые, зверобой, лобелия (Тахтаджян, 1964),
горечавковые].
Лизикарпный гинецей имеет также одногнездную завязь, но
со свободной центральной (колончатой) плацентой. Морфологи-
ческая природа такой плаценты, по-видимому, может быть различ-
ной или даже двойственной. В некоторых случаях характер пла-
центарных пучков показывает, что колончатая плацента образу-
ется от срастания субмаргинальных плацент всех плодолистиков
(пучки инвертированы так же, как и в центрально-угловой пла-
центе) . Одногнездность завязи возникает вследствие разрушения
перегородок, т. е. боковых участков плодолистиков (отсюда
и наименование гинецея «лизикарпный»). Гистогенетическими
исследованиями установлено также, что центральная плацента
ряда семейств имеет осевое происхождение и никаких связей
с плодолистиками не обнаруживает (Pankow, 1962). Для очень
многих гинецеев доказано участие оси в образовании центральной
колонки (Волгин, Тихомиров, 1980).
В нашей литературе широко принято различать названные
здесь три типа ценокарпного гинецея, выделяемые в работах
А. Л. Тахтаджяна (1942, 1948, 1964). Но у зарубежных авторов
синкарпный гинецей соответствует ценокарпному в нашем понима-
нии, а паракарпный, следуя Троллю, называют одногнездный
ценокарпный гинецей, независимо от характера плацентации
(Eames, 1961; Kaussmann, 1963; Эзау, 1980).
Как будет показано далее, разграничение трех вариантов
ценокарпного гинецея в значительной степени условно.
Число плодолистиков, образующих ценокарпный гинецей,
может быть различным, но обычно оно невелико и часто сокращено
до двух. Число семязачатков варьирует от очень многих (иногда
нескольких тысяч — орхидные, заразиховые, колокольчик) до
одного, при этом плацентация обычно становится базальной.
8
Рис. 1. Продольный разрез нижней завязи опунции
(Opuntia dillenii).
Пунктирная линия — флоэмная часть пучка (из Эзау
по Тиаги).
Когда сохраняется лишь один семяза-
чаток, все плодолистики,, кроме одного,
сильно редуцируются и в зрелых плодах
их следы распознаются с большим трудом.
Такой гинецей получил название псевдо-
мономерного, так как внешне он выглядит
как одиночный плодолистик. Псевдомоно-
мерный вариант ценокарпного гинецея
распространен очень широко: крапивные, тутовые, маревые,
ильмовые, валериановые и ряд других. Эккардт, детально изучив-
ший этот тип гинецея, обнаружил его почти у 40 семейств (Eckardt,
1937, 1938).
При полной редукции рудиментов стерильных плодолистиков
возникает гинецей, который фактически является вторичномоно-
мерным. По Эккардту, таков гинецей у рогоза и ежеголовника.
Общеизвестно, что завязь в цветке может быть свободной, т. е.
не сросшейся с другими элементами цветка (верхняя), или при-
росшей к соседним частям цветка. Второго типа завязь принято
называть нижней, так как геометрически (но не морфологически!)
она располагается ниже остальных органов цветка.
При апокарпном гинецее завязи всегда остаются свободными.
У разных типов ценокарпного гинецея нижняя завязь возникала
многократно и независимо в различных эволюционных рядах.
Очень большая литература посвящена вопросу о происхож-
дении нижней завязи. Классики морфологии растений (Гёте, Де-
кандоль, ван Тигем, Варминг и др.) полагали, что нижняя завязь
образуется вследствие срастания гинецея с основанием около-
цветника и андроцея; ось цветка не принимает участия в форми-
ровании нижней завязи. Эту гипотезу обычно связывают с именем
Декандоля и называют «теорией срастания».
Немецкой школой ботаников была выдвинута «осевая теория»
происхождения нижней завязи, согласно которой она формируется
главным образом из тканей оси цветка и представляет собой
вогнутое цветоложе, окружающее гинецей. Справедливость этой
точки зрения пытались обосновать ходом морфогенеза цветков
с нижней завязью. Позднее утвердился более достоверный васку-
лярно-анатомический метод изучения цветка, в частности гинецея.
Если цветоложе образует чашу, охватывающую гинецей, то пучки,
снабжающие завязь, должны круто изгибаться вниз и при этом
инвертироваться (рис. 1).
Васкулярный метод исследования приводит к заключению, что
в подавляющем большинстве случаев нижняя завязь образуется
вследствие срастания гинецея с цветочной трубкой. Вместе с тем
установлены факты и осевого происхождения нижней завязи.
9
Наиболее четко это прослеживается у ряда видов кактусовых
и близкого к ним сем. аизооновых (Aizoaceae) (Buxbaum, 1944;
Замятнин, 1951; Тахтаджян, 1964; Волгин, 1982). Б. Кауссман
(Kaussmann, 1963) отмечает, что у некоторых видов бромелиевых
(Pitcairnia xanthocalyx), камнеломковых (Saxifraga aizoides) и др.
нижняя завязь — следствие конгенитального срастания плодо-
листиков с осью цветка. При осевом формировании нижней
завязи доля участия плодолистиков в образовании перикарпия
может быть весьма незначительной, как это наблюдается у жимо-
лостных (Артюшенко, 1951).
Хотя гинецей чаще всего составляет морфологическую основу
плода, только в простейших случаях зрелые плоды отличаются
от гинецея лишь размерами (лютиковые, некоторые бобовые, лап-
чатка). У подавляющего большинства видов имеет место различ-
ной степени специализация плодов, связанная с выполнением ими
функций защиты и рассеивания семян. В результате такой специ-
ализации плоды приобретают настолько своеобразные признаки,
что черты гинецея распознаются в них с трудом.
Морфологическое разнообразие плодов помимо гинецея опре-
деляется тремя группами приспособлений: а) строением около-
плодника; б) способом вскрывания или распадения (сухие много-
семянные плоды); в) характером придатков и других особенностей,
обеспечивающих диссеминацию.
СТРОЕНИЕ ОКОЛОПЛОДНИКА
Околоплодник (перикарпий, от греч. «пери» — вокруг) пред-
ставляет собой разросшуюся и нередко сильно видоизмененную
стенку завязи (включая и ткани прирастающих к ней других частей
цветка).
В околоплоднике различают 3 зоны: наружную — внеплодник
(экзокарпий), среднюю — межплодник (мезокарпий) и внутрен-
нюю — внутриплодник (эндокарпий). Но во многих случаях эти
зоны очень трудно разграничиваются не только морфологически,
но и гистологически.
У незрелых плодов околоплодник состоит из более или менее
однородных живых паренхиматических клеток (за исключением
проводящих пучков), частично хлорофиллоносных. Характер видо-
изменения и дифференциации тканей околоплодника в процессе
его созревания определяется способом диссеминации. Чаще всего
исчезает зеленая окраска. Плоды либо высыхают и становятся
бурыми («бесцветными»), либо, наоборот, делаются сочными
и приобретают очень темную или яркую окраску благодаря образо-
ванию каротиноидов, антоциана и т. п. В редких случаях зрелые
плоды бывают чисто-белыми — омела (Viscurn album), снежно-
ягодник (Symphoricarpus) из-за полного отражения света обиль-
ными межклетниками. Наконец, известны и яркоокрашенные сухие
плоды, например крылатки клена татарского.
10
Рис. 2. Анатомическое строение околоплодников.
а — косточка тёрна (Prunus spinosa), шлиф; б — перикарпий крыжовника (Grossularia
reclinata)-, в — орешек шиповника (Rosa majalis), шлиф; г — перикарпий подсолнечника
(Melianthus annuus), д — лещины (Corylus avellana) (а, в — по Зажурило; б, г, д —
по Кауссманну).
Наиболее четко все 3 зоны околоплодника различаются в плодах
типа костянки. Экзокарпий представлен тонким кожистым слоем;
мезокарпий обычно сильно разросшийся, мясистый или очень
сочный и, наконец, эндокарпий — твердый, из нескольких слоев
сильно одревесневших клеток, образующих стенку косточки
(рис. 2). Достаточно ясная дифференциация наблюдается также
в околоплоднике яблока, но в типичных случаях эндокарпий
яблока хрящеватый, волокнистый. У типичных ягод весь около-
11
плодник сочный (кроме наружной кожицы) и границы между экзо-,
мезо- и эндокарпием очень нечетки (рис. 2, б).
Дифференциация околоплодника сухих многосемянных плодов
связана с формированием вскрывающего механизма. Так, напри-
мер, у бобов со скручивающимися створками в мезокарпии образу-
ется несколько продольных полос механических волокон, располо-
женных под углом 30—40° к продольной оси плода. Скручивание
створок происходит вследствие неравномерного сокращения воло-
кон при высыхании (подробнее см.: Эзау, 1980). Кроме того,
в околоплодниках вскрывающихся или распадающихся плодов
дифференцируются особые участки «отделительных» тканей. В этих
участках и происходит вскрывание или распадение плодолистиков.
В перикарпиях сухих односемянных плодов в одних случаях
обнаруживаются хорошо заметные слои различно дифференциро-
ванных клеток (шиповник, подсолнечник; рис. 2, в, г), а в других —
весь околоплодник состоит из однородной ткани (орех лещины;
рис. 2, <5).
СПОСОБЫ ВСКРЫВАНИЯ И РАСПАДЕНИЯ ПЛОДОВ
При всей своей элементарности понятие «вскрывание плодов»
нуждается в уточнении. Некоторые авторы полагают, что невскры-
вающиеся плоды — это биологическая бессмыслица, так как
прорастание семян всегда предполагает разрыв околоплодника.
Однако под вскрыванием плодов обычно понимают такой процесс,
который способствует освобождению семян до их прорастания.
Анализируя различные способы вскрывания плодов, Н. Н. Каден
(1962, 19646) выделяет как особый тип неправильное вскрывание,
которое характерно для очень тонких пленчатых перикарпиев (не-
которые виды клевера, маревых и др.). Нов этих случаях, по суще-
ству говоря, никакого вскрывания плодов не происходит. В около-
плодниках нет ни анатомических, ни физиологических механизмов
вскрывания: они просто разрушаются от механического пов-
реждения, воздействия микроорганизмов и т. п. Действительное
вскрывание плодов происходит лишь при условии формирования
в строго фиксированных участках перикарпия специальных раз-
делительных тканей.
Таким образом, под вскрыванием плодов мы понимаем гисто-
логически обусловленные разрывы околоплодника, способствую-
щие освобождению семян. В подавляющем большинстве случаев
вскрывание свойственно сухим многосемянным плодам. Эта группа
плодов очень обширная и совершенно разнородная по своей
морфологической природе. Поэтому и способы вскрывания таких
плодов исключительно разнообразны.
Еще в конце прошлого столетия делались попытки характеризо-
вать и классифицировать способы вскрывания плодов не чисто
описательно (по внешней картине), а морфологически. Из анало-
гичных работ середины нашего века надо указать работы Штоппа
(Stopp, 1950) и Н. Н. Кадена (1962, 1964). В классификации
12
способов вскрывания, которую дает Н. Н. Каден, действительно
выдерживается единая морфологическая терминология. При этом
автор пытается охватить все единичные и даже теоретически воз-
можные варианты вскрывания плодов. В результате получился
слишком громоздкий, излишне детализированный перечень слож-
ных наименований, которым трудно пользоваться. Мы предлагаем
упрощенную схему Кадена, несколько изменив его терминологию.
Основным типом вскрывания является вскрывание продоль-
ными щелями. Топографически, т. е. по месту образования разры-
вов, вскрывание может быть: 1) сутуральное1 (шовное).— по шву,
т. е. по линии срастания краев плодолистика; 2) дорсальное (спин-
ное) — по спинной жилке; 3) ламинальное1 2 — по пластинке плодо-
листика.
Одногнездные плоды вскрываются либо одним из этих трех
способов, либо путем различного сочетания двух из них. В синкарп-
ных плодах кроме этих трех типов происходит еще нередко и раз-
рыв по перегородкам — септицидно, т. е. в плоскости срастания
боковых сторон плодолистиков. Септицидное вскрывание соче-
тается с одним или двумя из трех основных.
Назовем наиболее распространенные конкретные типы
продольного вскрывания плодов (рис. 3).
Апокарпные'плоды: сутуральное — типичные листовки (люти-
ковые и др.); дорсальное — листовки типа магнолии; сутурально-
дорсальное — все типичные бобы.
Ценокарпные многогнездные: септицидно-сутуральное (некото-
рые лилейные); септицидно-дорсально-ламинальное (дурман,
наперстянка); дорсальное (грушанка, кислица, вероника, конский
каштан); сутурально-дорсальное (многие лилейные, ирисовые
и др.).
Ценокарпные одногнездные: сутуральное (многие гвоздичные,
первоцветные, горечавковые); дорсальное (фиалка, ивовые, неко-
торые первоцветные); ламинальное (орхидные); сутурально-
дорсальное (гвоздика, первоцвет и др.).
В природе встречаются и другие варианты, но все они в конеч-
ном счете сводятся к различным комбинациям трех основных типов
вскрывания.
Продольное вскрывание плодов может быть полным — это
вскрывание створками (дурман, ива, лилейные, вероника и мн.др.),
или неполным, т. е. зубцами (гвоздичные, первоцветные) или
щелями (орхидные, грушанка).
Сутуральные и дорсальные разрывы околоплодника всегда
бывают продольно-линейными, а ламинальные помимо продольно-
линейного могут давать и другие типы вскрывания: а) продольно-
кольцевое, б) поперечно-кольцевое, в) подковообразное.
1 Это название точно определяет место вскрывания и применимо к любому типу
гинецея, поэтому оно предлагается вместо обычно принятого «вентрального».
2 Предлагается вместо «латерального» (бокового), так как линия разрыва
может проходить и вдоль спинной жилки, и в углу завязи близ плаценты, т. е. совсем
не сбоку.
13
Рис. 3. Типы продольного вскрывания плодов (схемы поперечных срезов).
а—в — апокарпии: а — сутуральное, б — дорсальное, в — сутурально-дорсальное; г—ж —
ценокарпии многогнездные: г — септицидно-сутуральное, д — септицидно-дорсально-
ламинальное, е — дорсальное, ж — сутурально-дорсальное; з—л — ценокарпии одногнезд-
ные: з — дорсальное, и — ламинальное, к — дорсальное, л — сутурально-дорсальное
(по Кадену с изменениями).
Такие варианты ламинального вскрывания свойственны раз-
личным типам ценокарпных плодов и гораздо реже встречаются
у апокарпиев.
При продольно-кольцевом вскрывании линия разрыва пред-
ставляет собой замкнутую кривую, вытянутую продольно, т. е.
по длине плодолистика. Такое вскрывание характерно для типич-
ных стручков и стручочков крестоцветных, стручковидной коро-
бочки чистотела. В этих случаях линия разрыва проходит по плас-
тинке одного плодолистика вдоль плацент и сутуральных пучков.
Иногда она может захватывать участки двух соседних плодо-
листиков (Catalpa). Плод катальпы двугнездный, у нее кольце-
вое вскрывание сочетается с ламинальным линейным разрывом
по перегородке (Stopp, 1950).
В отдельных случаях продольно-кольцевое вскрывание может
быть неполным. Так, например, у некоторых крестоцветных
стручки вскрываются только снизу (Tri.chochi.ton inconspicuum)
или, наоборот, только сверху (Strigosella).
Поперечно-кольцевое вскрывание приводит к образованию
крышечки. Оно известно у белены, разных видов подорожника,
некоторых первоцветных (Anagallis). ।
При подковообразном вскрывании участок плодолистика,
отгибаясь в виде клапана, открывает отверстие, через которое
14
и рассеиваются семена. Число отверстий соответствует числу пло-
долистиков (мак, колокольчик, льнянка, львиный зев),?
Особую, более специализированную группу многосемянных
плодов представляют собой распадающиеся плоды. От вскрыва-
ющихся они отличаются тем, что семена их не освобождаются
из околоплодника, а, наоборот, каждое семя опадает, окруженное
участком перикарпия. Таким образом, защитная функция плода
в этом случае сохраняется вплоть до прорастания семени.
Существуют два основных способа распадения плодов: про-
дольное — в плоскости срастания стенок соседних плодолистиков,
и поперечное — в плоскости, перпендикулярной к продольной оси
плодолистика. Первая группа плодов носит название дробных
и образуется из двух- или многогнездной завязи. Вторая группа —
членистые плоды — известны среди апокарпных (членистые бобы)
и ценокарпных (членистые стручки).
К распадающимся плодам могут применяться те же термины,
что и к вскрывающимся. Так, дробные плоды распадаются септи-
цидно. При этом образуются односемянные замкнутые фрагменты
плодов, соответствующие одному плодолистику и называемые мери-
карпиями — от греч. «мерис» — часть, доля (большинство видов
зонтичных, клены, мареновые и др.). Септицидное распадение
плода может сочетаться с ламинальным продольно-кольцевым
разрывом плодолистика близ плаценты. В этом случае мерикарпий
оказывается открытым с брюшной стороны (молочай, многие
мальвовые).
У членистых плодов околоплодник разрывается ламинально
в поперечно-кольцевом направлении, но членики оказываются
замкнутыми благодаря формированию поперечных ложных перего-
родок плода, в плоскости которых и происходит отделение члени-
ков. В более редких случаях образование члеников происходит
вследствие ламинальных продольно-кольцевых разрывов около-
плодника, а перегородки и пучки плодолистиков остаются в виде
рамки, из которой выпали членики. Поэтому такие плоды называют
рамчатыми, или окончато-членистыми, что точнее. Наконец, осо-
бого типа распадающиеся плоды свойственны губоцветным, бурач-
никовым, вербеновым. Более детальное описание различных распа-
дающихся плодов дается в соответствующих разделах следующей
главы. (О дробных плодах см. также Каден, 1964а).
ОСТАЮЩИЕСЯ ПРИ ПЛОДАХ. ЧАСТИ ЦВЕТКА И СОЦВЕТИЯ
СПЕЦИАЛЬНЫЕ ПРИДАТКИ
Мы уже говорили о том, что в формировании плода нередко
принимают участие кроме гинецея и другие части цветка или даже
соцветия. Это относится не только к случаям образования нижней
завязи, когда имеет место срастание гинецея с околоцветником
или цветоложем. При зрелых плодах, развивающихся и из верхней
завязи, часто сохраняются, не срастаясь с перикарпием, или около-
15
цветник, в разной степени видоизмененный, или особые образова-
ния различной морфологической природы. Так, например, у боль-
шинства видов клевера околоцветник засыхает, но сохраняется
при плодах и опадает вместе с ними, создавая дополнительную
защиту для семян. Есть указания на то, что околоцветник в данном
случае благодаря гигроскопичности способствует прорастанию
семян. Околоцветник сохраняется и при плодах многих гречишных.
У щавелей внутренние листочки околоцветника заметно разрас-
таются после цветения и окружают плод. В иных, более редких
случаях околоцветник при плодах становится сочным и ярко-
окрашенным [некоторыетропические семейства; из нашей флоры —
жминда (Chenopodium foliosum)]. Но чаще при плодах сохраня-
ется чашечка, выполняющая также защитную функцию (гвоздич-
ные, белена и др.) или, кроме того, играющая роль в диссеминации
[губоцветные, бурачниковые, первоцвет весенний (Primula veris), по-
гремок большой (Rhinanthus alectrolophus), мытник (Pedicula-
ris), лапчатка и др.]. В некоторых случаях чашечки не только со-
храняются, но и разрастаются к моменту созревания плодов, как
например у клевера земляничного (Trifolium fragiferum). Хорошо
известны оранжево-красные чашечки физалиса (Physalis), во
много раз превышающие плод. Именно из-за чашечек некоторые
виды физалиса разводят как декоративные растения.
Для ряда розоцветных (Rosaceae— Rosoideae) характерны
плоды, окруженные особым образованием, которое получило наз-
вание гипантия. Чаще всего он представляет собой базальную
часть околоцветника, сросшуюся в трубку. При плодах гипантий
иногда частично одревесневает (Alchemilla, Agrimonia, Poterium,
Sanguisorba). Мясистая яркоокрашенная ткань плода шиповника,
которую можно принять за околоплодник, в действительности
представляет собой гипантий.
Не менее редко сохранение и видоизменение при плодах вне-
цветковых органов, обычно прицветников или кроющих листьев.
Так, у большинства злаков зерновки опадают окруженные цветко-
выми чешуями, а иногда и колосковыми вместе с осью колоска
(так называемый антеций). Плоды осок заключены в замкнутые
мешочки из кроющего листа. Сросшиеся или сомкнутые при-
цветнички, окружающие плод, известны у ряда маревых. Своеоб-
разная плюска при плодах лещины (Corylus) также образована
прицветными листьями. В формировании плюски буковых помимо
брактей участвуют и осевые части редуцированного соцветия (Имс,
1964; Меницкий, 1984).
Разнообразие плодов определяется также видоизменениями
перикарпия и образованием разного типа придатков — крыловид-
ных, цепких, железистых и т. п. Все они служат приспособлениями
к распространению плодов.
Заметим в заключение, что внешнеморфологическое много-
образие плодов обусловлено также грандиозным размахом измен-
чивости таких признаков, как размеры, форма, окраска, характер
поверхности и т. п.
16
О ПРИНЦИПАХ КЛАССИФИКАЦИИ И НОМЕНКЛАТУРЕ
ПЛОДОВ
В любой области науки вопрос о принципах классификации
объектов исследования, как правило, является «вечно юным», т. е.
постоянно дискутируемым. В области ботаники к числу «вечно
юных» относится и вопрос о принципах классификации и номен-
клатуре плодов. В нашей литературе эти вопросы оживленно
обсуждались в работах 60-х годов (Каден, 1961, 1964а, 1965;
Левина, 1961, 1967а, 1968; Тахтаджян, 1964; Каден, Кирпичников,
1966; Грудзинская, 1968).
Эти дискуссии не устранили всех разногласий во взглядах
названных авторов, но выявились и некоторые более или менее
общепринятые положения. Сущность их сводилась к тому, что
современные классификации плодов должны отражать пути их
морфогенеза, а как основа классификации принималась схема
эволюции гинецея от апокарпного к пара- и лизикарпному через
синкарпный. Эти положения отражены, в частности, в графической
классификации автора (Левина, 1961), но в ней выделены и ана-
логичные экологические группы плодов, занимающие разное
положение в системе. В то же время и в названной, как и в других
работах, посвященных общим вопросам карпологии, отмечалось,
что разработка полной последовательной морфогенетической си-
стемы плодов — задача отдаленного будущего.
Детальный анализ многих фактов заставляет признать, что
сейчас мы еще более далеки от решения такой задачи, чем это
казалось два десятилетия тому назад. Впрочем, еще Л. вандер Пейл
(Pijl van der, 1969) справедливо отметил, что многие восточно-
европейские системы плодов (в числе их авторов Гоби, Тахтаджян,
Каден, Левина) конструируются на двойной основе — морфологи-
ческой и экологической. И хотя их морфологическая основа имену-
ется «генетической», по существу она остается лишь онтогенети-
ческой.
Однако и с онтогенетической точки зрения жесткое разграниче-
ние трех типов ценокарпного гинецея далеко не всегда оправдано.
У многих видов базальный участок гинецея является синкарпным,
а вышележащие — пара-, лизи- или даже апокарпным (в случае
свободных стилодиев). Так возникают плоды гемипаракарпные
(некоторые вьюнковые), гемилизикарпные (часть гвоздичных),
вторичносинкарпные (колокольчик). Как указывает Л. И. Ива-
нина (1967, с. 10), в сем. геснериевых наиболее распространены
гемипаракарпный и паракарпный гинецеи, реже встречаются син-
карпный и гемилизикарпный.
Рассматривая вертикально-зональную структуру ценокарпных
гинецеев в свете концепции Ляйнфельнера, С. А. Волгин и В. Н. Ти-
хомиров (1980) приходят к заключению, что гинецеи с паракарп-
ным и синкарпным участками нельзя трактовать как переходные
между двумя типами структуры, так как они составляют едва ли
не подавляющее большинство. А. Л. Тахтаджян также отмечает
2 Р. Е. Левина
17
(1980, с. 93) : «Однако далеко не всегда можно отличить паракарп-
ный гинецей от синкарпного». Надо еще добавить, что весьма
распространенный тип псевдомономерного гинецея с базальной
плацентой чаще всего невозможно отнести к какому-либо из трех
вариантов ценокарпии.
Типы гинецея как основа морфогенетической классификации
плодов не оправдывают себя и с другой точки зрения.
В схеме эволюции типов гинецея, предложенной А. Л. Тах-
таджяном в 1964 и 1980 гг., в отличие от предыдущих (Тахтаджян,
1942, 1948) паракарпный гинецей оказывается конвергентной
структурой, возникающей либо из апокарпного, либо из синкарп-
ного гинецея. О двойственной природе паракарпии пишет также
Имс (Eames, 1961), а Б. М. Козо-Полянский (1922) считал, что
паракарпия — это один из путей возникновения покрытосемян-
ности, наравне с апокарпией. Имс утверждает даже, что «сраста-
ние плодолистиков друг с другом, так же как их прирастание
к другим органам явно предшествовали смыканию краев плодо-
листика», а «сомкнутый плодолистик не был присущ наиболее
ранним покрытосеменным» (Имс, 1964, с. 195, 196).
Возникновение синкарпного (многогнездного) гинецея из апо-
карпного тоже, вероятно, не единственно возможный путь разви-
тия. Если учесть, что замыкание плодолистиков возможно путем
прирастания к цветоложу, то синкарпия оказывается независимой
в своем происхождении от апокарпии.
Выше уже отмечалось, что свободная колончатая плацента
может имёть разную (либо фолиарную, либо осевую) или двой-
ственную морфологическую природу. Следовательно, и лизикарп-
ный гинецей в морфогенетическом смысле — структура гетероген-
ная.
Напрашивается вывод, что построение единой морфогенети-
ческой системы плодов для всех покрытосемянных в целом вообще
невозможно. Реально лишь создание частных карпологических
классификаций для таксонов невысокого ранга (рода, семейства).
Классификация, которая охватила бы все существующее мно-
гообразие плодов, по-видимому, может быть лишь морфоло-
гической.
В ботанической литературе широко принято любую неэво-
люционную классификацию снабжать эпитетом искусственной,
независимо от объектов, задач и методов исследования. Эта оши-
бочная точка зрения берет начало в систематике, где постепенно
стерлись различия, с одной стороны, между искусственной и естест-
венной системами, а с другой — между естественной и филогене-
тической. В результате любая неэволюционная система стала име-
новаться искусственной (Левина, 1971). Это представление некри-
тически воспринято морфологами и так прочно укоренилось, что,
например, специалисты по жизненным формам, очевидно, боясь
прослыть антиэволюционистами, называют искусственной габи-
туальную классификацию жизненных форм. Но ведь габитус
18
является наиболее существенной их чертой, следовательно, габи-
туальная — это и есть сущностная, т. е. самая естественная
классификация жизненных форм.
Морфологическая классификация плодов может быть и искус-
ственной, если ее строить по какому-либо одному несущественному
признаку (размеры, форма и т. п.). Но классификация, учитыва-
ющая совокупность морфологических признаков и охватывающая
все многообразие плодов, неизбежно будет отражать и модусы их
морфологического преобразования, и характер экологических
приспособлений. Иначе говоря, такая классификация будет сущно-
стной, т. е. естественной.
Морфогенетическая и морфологическая классификации плодов
не могут и не должны заменять одна другую: у них разные задачи,
принципы и сферы приложения. Первая решает чисто теоретиче-
скую задачу карпологии и эволюционной морфологии и может быть
использована в филогении конкретных таксонов невысокого ранга.
В морфологической классификации нуждаются специалисты раз-
ного профиля — палеоботаники, зоологи, лесоводы, агрономы,
семеноводы, товароведы и др., все, кто связан с распознаванием
плодов.
Естественные классификации, учитывающие совокупность при-
знаков объекта, обычно строятся по иерархическому принципу,
благодаря которому диагностические признаки приобретают раз-
ное рангово-таксономическое значение.
В существующих классификациях плодов фактически тоже
присутствует иерархический принцип, но неупорядоченно, непос-
ледовательно. Классификационные категории не установлены
и не обозначены, поэтому под типом плода в одних случаях пони-
мают апокарпии или синкарпии, а в других — желудь или яблоко.
В работах, в которых обсуждаются вопросы классификации плодов,
можно встретить выражения: «мелкие подтипы», «карпологические
виды» (кавычки авторов) и т. п.
Опыт иерархической морфологической классификации плодов
представлен в табл. 1 и 2. Из них видно, какие выделяются класси-
фикационные категории и каковы диагностические признаки
морфотаксонов разного ранга.
Поскольку плод — это в первую очередь преобразован-
ный гинецей, по его характеру устанавливаются типы плодов —
апокарпии и ценокарпии. По мотивам, изложенным выше, син-,
пара- и лизикарпные плоды не выделяются как единицы класси-
фикации.
В основу классовых различий положена много- и односемян-
ность, потому что с этим признаком коррелируют глубокие морфо-
логические различия плодов и существенно разное соотношение
их защитной и диссеминационной функций. Поли- и мономерные
апокарпии выделяются как подклассы. Среди ценокарпиев под-
классы составляют верхние и нижние плоды.
Консистенция околоплодника — наименее устойчивый признак;
он определяет лишь групповые различия. Подгруппы выделяются
2*
19
только в группах сухих многосемянных плодов, так как типичные
сочные и односемянные плоды не освобождают семян.
Центральной классификационной категорией является «вид
плода». С ней связана далеко не решенная проблема номенклатуры
плодов.
Из таблиц видно, что, за небольшими исключениями, видовые
названия плодов сохраняются те же, что давно вошли в нашу
научную и учебную литературу. Уточнение, дополнение и усовер-
шенствование номенклатуры — процесс постепенный и медленный.
Только при этом условии новые наименования будут жизненны
и устойчивы, так как всякий неологизм проходит через «лингвис-
тический отбор».
Новым в таблице является наименование плода «финик»,
предложенное Г. С. Ландсбергом (1981). По положению его
в системе ясно, что это апокарпный мономерный односемянный
сочный плод. Введено и старое название «калачик» для дробных
плодов мальвовых, так как у них образуются особые незамкнутые
мерикарпии.
Из таблиц видно, что кроме костянки названия апокарпных
плодов не повторяются у ценокарпиев. Но апокарпные костянки
имеют приставку «много» или «одно», а ценокарпные — эпитет
«верхняя» или «нижняя»; кроме того, многосемянные костянки
имеют и второй эпитет — «многокосточковая». Таким образом,
апо- и ценокарпные костянки четко различаются по видовым
названиям. Костянка — в типе сочный плод, поэтому эпитет «соч-
ная» лишний; сухая костянка — это уже особый вид плода.
Другое исключение — «ценокарпная многолистовка» — подчерки-
вает только сохранение структуры этого апокарпия при ценокарпии.
Только к апокарпиям применяется термин «плодик» в обще-
принятом понимании. Плодик — часть полимерного апокарпного
плода, образованная одним пестиком. Плодиком может быть лис-
товка, орешек или костяночка.
Трудности наименования многих видов плодов связаны с недо-
статочной их изученностью, а также с отсутствием общепринятых
критериев для разграничения близких карпологических видов,
таких, например, как семянка и орех. Для одного и того же плода
у разных авторов приводятся разные названия. Так, плод конопли
называют и сухой костянкой (Каден, 1965), и орехом с неопадаю-
щим околоцветником (Яценко-Хмелевский, 1980).
Иногда восстанавливают явно неудачные названия только
потому, что они были предложены ранее других и ни к чему не обя-
зывают, так как не отражают морфологии плода. Таким восстанов-
ленным названием является «мешочек» для плода маревых (Каден,
Смирнова, 1968), с чем трудно согласиться, поскольку это назва-
ние связывается обычно с плодами осок. Во всех «Флорах» и опре-
делителях «мешочком» называют оболочку плода осок, образован-
ную кроющим листом; при этом признакам мешочка придается
диагностическое значение. Кроме того, в старых работах, перевод-
ных с немецкого, мешочком им-енуют листовку.
СО
X
X
Ч
\о
СО
Н
X
X
X
СО
X
X
е
X
20
21
Таблица 2
Классификация ценокарпных плодов
Тип • Класс Подклас^ Группа Подгруппа Вид плода Вариации
Ценокарпии Многосемян- ные Верхние Сухие Вскрывающиеся Ценокарпная многоли- стовка Коробочка Стручок Сочная По типу плаценты и спо- собу вскрывания Стручочек
Распадающиеся Дробная коробочка Калачик Двукрылатка Дробный ценокарпий Ценобий Членистый стручок Трех-четырехкрылатка
Сочные Ягода Костянка многокосточ- ковая Померанец •
Нижние и полу- нижние Сухие Вскрывающиеся Коробочка Вскрывающаяся ко- стянка Потипу плаценты и спо- собу вскрывания
Распадающиеся Дробный ценокарпий Двусемянка Вислоплодник Дробная костянка
Сочные Ягода Костянка многокосточ- ковая Яблоко Тыквина Гранатина Костянковидное Вскрывающаяся
Односемянные Верхние Сухие Односемянная коробочка Семянка Орех Ореховидный стручочек Сухая костянка Крылатка
Сочные Костянка Односемянная ягода
Нижние Сухие Семянка Орех Желудь Сухая костянка Крылатка
Сочные Костянка Односемянная ягода
Для плодов, не имеющих общепринятого названия, мы даем
самую краткую морфологическую характеристику, например, плод
якорцев — верхний дробный ценокарпий.
Легко предвидеть упрек в том, что видовые названия плодов
очень неравноценны по степени своей мономорфности, такие,
например, как ценобий и коробочка. Но, видимо, это неизбежно;
виды в систематике тоже очень различны по объему, но это прини-
мается как должное. Коробочка относится к числу самых распро-
страненных ценокарпиев, возникавших многократно и независимо
в ходе эволюции; естественно поэтому, что в деталях строения
коробочки очень разнородны. Но видовые названия плода не могут
быть такими, как: «коробочка верхняя, лизикарпная, пятичленная,
с покрывалом, вскрывающаяся несовершенно дизъюнктивно-дор-
сально» (Каден, 1965, с. 778). Такое название-описание может
быть уместно в диагнозе нового таксона или в специальной класси-
фикации коробочек, т. е. в узкокарпологическом исследовании.
Полиморфность многих видов плодов заставляет ввести катего-
рию ниже вида. Она названа вариацией. В тех случаях, когда вари-
аций немного, основные из них названы в таблице, например для
многоорешка или боба. Но перечислить все существующие вари-
ации коробочек просто невозможно в таком кратком обзоре, как
таблица. Для коробочек указаны только признаки, по которым
различаются их вариации.
24
Глава II
МОРФОЛОГИЯ ПЛОДОВ
В настоящей главе дается описание всех видов плодов, назван-
ных в классификационных таблицах.
В силу господствующего параллелизма в изменчивости морфо-
логии плодов в нашем изложении, как и в таблицах, неизбежно
повторяются одноименные классы в разных типах и одинаковые
группы и подгруппы плодов в разных классах.
АПОКАРПИИ
МНОГОСЕМЯННЫЕ ПОЛИМЕРНЫЕ
Таким видом плода является многолистовка (multifolliculus).
Листовкой (folliculus) называют сухой многосемянный плод,
образованный одним плодолистиком и вскрывающийся по шву
(сутурально). Семена в листовке обычно располагаются субмар-
гиально в 2 ряда, но встречаются листовки и с ламинальной пла-
центацией. Как мы увидим ниже, в природе нередки различные
видоизменения типичной листовки. Листовка чаще представляет
собой не целый плод, а только плодик, т. е. является частью много-
листовки.
Многолистовка — обычно сухой плод со спиральным располо-
жением плодиков. В нашей флоре она известна у представителей
лютиковых (Trollius europaeus, Caltha palustris) (рис. 4, a). У рас-
тений тропиков и субтропиков многолистовка встречается в семей-
ствах цефалотовых (Cephalotaceae), диллениевых (Dilleniaceae),
триурисовых (Гriuridaceae).
Своеобразна многолистовка магнолии. Ее многочисленные
плодики сидят тесной спиралью на удлиненной оси. Зрелый плод
может достигать в длину 10—12 см. Однако в строении отдельной
листовки наблюдаются существенные отклонения от типичной.
Во-первых, плодики магнолии вскрываются не сутурально, а дор-
сально (по медианной жилке). Это связано с тем, что своей брюш-
ной частью (где находится шов) листовки плотно прижаты к оси
плода. Во-вторых, околоплодник у них не тонкий, кожистый, как
у типичных листовок, а значительно утолщенный и плотный,
с заметно склерифицированным эндокарпием. Наконец, в листовке
развивается лишь несколько семян (2—6) с сочными ярко-крас-
25
Рис. 4. Типы многолистовок.
а, б — сухие спиральные (калужница — Caltha ,palustris, магнолия — Magnolia virgini-
апа); в — сухая циклическая (иллициум — IlUcium anisatum); г — трехлистовка (жи-
вокость— Delphinium elatum); д — сочная спиральная (аннона — Annona reticulata)
[б— п0: Деревья и кустарники СССР, в — по Кернеру,, г — по Троллю, д — по: Жизнь
растений).
ними оболочками. Из раскрытых плодиков семена свисают
на длинных тонких семяножках (рис. 4, б).
С уменьшением числа плодиков образуется олигомерная цикли-
ческая многолистовка. Число листовок у таких плодов варьирует
от 10—12 {Illicium; рис. 4, в; Limnocharis) до 3—5, как у видов
борца (Aconitum), живокости {Delphinium; рис. 4, г), различных
видов толстянковых (Crassulaceae), спиреи, рябинника (Sorbaria)
и пузыреплодника \{Physocarpus) ;из ^розоцветных, сусака зонтич-
ного (Butomus umbellatus). Для листовок сусака характерны
многочисленные семена, расположенные по всей внутренней по-
верхности плодолистика. .Мнополистовки _с небольшим числом
плодиков известны также у пиона, шейхцерии болотной (трехлис-
товка), тропического рода Brunellia (Brunelliaceae), реликтового
эндемика о-ва Робинзон Крузо (группа Хуан-Фернандес) Lacto-
ris. fernandeziana {Lactoridaceae). В циклических многолистовках
может происходить частичное срастание плодолистиков друг с дру-
гом в их базальной части. Такие плоды иногда называют гемисин-
карпными (сусак, водосбор, толстянковые, некоторые виды спи-
реи).
К числу олигомерных многолистовок следует отнести плоды
некоторых мимозовых — бразильского рода Affonsea и новогви-
нейского Archidendron. Н. М. Дудик (1981) предлагает называть
эти плоды «многобоб» и рассматривает их как специализирован-
26
ные многолистовки. В чем выражается специализация, автор
не указывает. Плодики этих растений раскрываются сутурально,
таким образом, морфологически они ничем не отличаются от лис-
товок. Надо согласиться с X. Я. Гоби (1921), который называет
плоды аффонсеи и архидендрона многолистовками.
Специальной оговорки заслуживают двулистовки, свойствен-
ные кутровым (Аросупасеае) и ластовневым (Asclepiadaceae).
В цветках этих семейств плодолистики срастаются лишь в области
стилодиев, образуя один столбик, или только в области рылец.
К моменту созревания плодики оказываются совершенно свобод-
ными.
Особую группу многосемянных полимерных апокарпиев состав-
ляют сочные многолистовки. Они свойственны большинству видов
субтропического сем. анноновых (Аппопасеае; рис. 4, д), некото-
рым видам сем. Lardizabalaceae, обоим родам сем. схизандровых —
лимоннику (Schisandra) и кадсуре (Kadsura). У лимонника
к моменту созревания плода ось цветка сильно удлиняется (до
8 см), благодаря чему листовки сидят на некотором расстоянии
друг от друга. Плод кадсуры, напротив, представляет собой
головку нескольких сантиметров в диаметре, образованную скучен-
ными плодиками, сидящими на расширенной мясистой оси. Сочная
многолистовка известна также у золотой печати (Hydrastis cana-
densis) — лекарственного растения сем. Hydrastidaceae, близкого
к лютиковым. Сочные листовки обычно не раскрываются, но у неко-
торых видов они вскрываются сутурально, как и типичная сухая
листовка (Decaisnea из сем. Lardizabalaceae-, Гоби, 1921).
МНОГОСЕМЯННЫЕ МОНОМЕРНЫЕ
К этому подклассу апокарпиев относятся однолистовка и боб.
Однолистовка (unifolliculus) хорошо известна у видов рода кон-
солида (Consolida regalis) и др. (рис. 5, а). Это вполне типичная
сухая многосемянная листовка с субмаргинальной плацентой
и сутуральным способом вскрывания. Из лютиковых с таким же
видом плода назовем еще цимицифугу (Cimicifuga simplex), а из
других семейств — багрянник (Cercidiphyllum japonicum), энде-
мичный для Японии, но довольно широко культивируемый в СССР
как парковое дерево. Однолистовка известна и у некоторых Prote-
асеае.
Небогата видами группа сочных однолистовок. Она свойст-
венна монотилному тропическому сем. дегенериевых (Degeneria
vitiensis). Плод дегенерии не вскрывается, плацентация ламиналь-
ная, хотя семена сидят в„2 ряда. Такие же невскрывающиеся сочные
однолистовки свойственны в нашей флоре различным видам во-
ронца (Actaea) из лютиковых. Благодаря окраске (черной или
красной) и сочной консистенции эти листовки напоминают ягоду, но
п@. продольному желобку на их поверхности легко угадывается шов
единственного плодолистика. Сочная ткань околоплодника раз-
27
Рис. 5. Однолистовки.
а — сухая (консолида — Consolida re galls), б — сочная (воронец — Actaea spicata) {а —
по Левиной, 1957; б — по Левиной, 1967а).
Рис. 6. Многосемянные бобы.
а — чина (Lathyrus vernus), б — люцерна посевная (Medicago saiiva}, в — л. щитко-
видная (М. scutellata), г — л, полевая (М, agrestis), д, е — пузырник (Colutea arbores-
cens) (а, в, г — по Кернеру, б — по Хюбнеру, д, е — по Троллю).
28
вита слабо. Основную массу плода составляют семена, лежащие
двумя плотными рядами и заполняющие всю его полость (рис. 5, б).
Боб (legumen) отличается от листовки сутурально-дорсальным
вскрыванием с образованием двух створок. Полимерные плоды
с таким способом вскрывания неизвестны, таким образом, боб
является не плодиком, а мономерным плодом. Для него характерна
только субмаргинальная плацента.
О механизме, вскрывающем боб, уже говорилось в I главе.
Здесь только напомним, что у типичного боба при вскрывании про-
исходит внезапное скручивание створок и разбрасывание семян.
Типичные, т. е. сухие, многосемянные, вскрывающиеся двумя
створками бобы широко представлены у мотыльковых: дрока
(Genista), караганника (Caragana), гороха (Pisum), фасоли
(Phaseolus), лядвенца (Lotus), горошка (Vicia), чины (Lathyrus)
и многих других (рис. 6, а). Плоды этих растений сравнительно
мало варьируют по форме. Они удлиненные, цилиндрические или
более или менее сплюснутые и имеют в длину от 2—3 до нескольких
сантиметров. Вообще же бобы достигают такого изумительного
разнообразия, что по форме их совершенно невозможно отнести
к одному виду плода. Даже в пределах одного рода иногда наблю-
дается крайнее морфологическое разнообразие плодов, например
у астрагалов или различных видов люцерны.
Чтобы представить, насколько бобы изменчивы по размерам,
достаточно сравнить гигантские плоды тропической лианы Entada
pursaetha из мимозовых, достигающие 15 см шир. и 1.5 м дл.,
с односемянными плодами клевера в 2—3 мм дл.
У ряда видов клевера бобы содержат несколько семян и вскры-
ваются по шву (например, клевер ползучий, к. гибридный). Такие
бобы по существу ничем не отличаются от листовки, но в данном
случае сутуральное вскрывание вместо сутурально-дорсального
вторично. Как известно, плоды клевера опадают полностью окру-
женные околоцветником, поэтому вскрывание является для них
функцией рудиментарной. Во всяком случае оно не способствует
рассеиванию семян, но, возможно, способствует их прорастанию.
Весьма своеобразны бобы многих видов люцерны: для них
характерно спиральное скручивание. Такие плоды хорошо известны
у люцерны посевной (рис. 6, б). Нередко благодаря многочислен-
ным виткам спирали плоды становятся шаровидными. Особенно
причудливый вид таким плодам придают обильные шипики, густо
сидящие по краю боба (рис. 6, г). Виды люцерны с подобными
плодами (Medicago agrestis, М. arabica, М. orbicularis, М. trunca-
tula и др.) — это обитатели нашего юга и юго-востока (Крым,
Кавказ, Средняя Азия) и Средиземноморья.
Известны также бобы сильно вздутые, у которых семена зани-
мают лишь незначительное место в большой воздушной полости
плода. Такие бобы имеют овально-округлую обтекаемую форму
и очень низкую плотность. Так, у растения пустынь Средней Азии
смирновии (Smirnovia turkestana) бобы достигают 25—30 см3
в объеме при массе 0.17 г. Сильно вздуты плоды у пузырника
29
Рис. 7. Плоды астрагалов.
а — Astragalus skorniakovii, б — A. schelkovnlkovli, в — A. commixtus (по: Флора СССР).
(Colutea arborescens\ рис. 6, д, е) — растения Южного Крыма,
нередко разводимого; у безлистного кустарничка песков Прикас-
пия и Казахстана эремоспартона (Eremosparton aphyllum).
Сильно вздутые плоды встречаются у многих астрагалов (рис. 7, а).
Существенно отклоняются от типичных ложнодвугнездные
бобы. Настоящими принято называть гнезда завязи (и плода),
возникающие в результате срастания боковых сторон соседних
замкнутых плодолистиков. Ложнодвугнездные бобы образуются
вследствие загибания в полость завязи краев плодолистика или
впячивания его стенки со спинной стороны; в зависимости от сте-
пени разрастания плодолистика в полости завязи перегородка
может быть неполной как по длине, так и по ширине плода или
полной, т. е. образующей два замкнутых гнезда с одной плацентой
в каждом. Ложнодвугнездные бобы свойственны большинству
видов астрагала (Astragalus) и остролодочника (Oxytropis).
Плоды астрагалов крайне разнообразны по форме, размерам
и способу вскрывания (рис. 7). У ряда видов вскрывание плодов
типичное, т. е. сутурально-дорсальное (Astragalus albicaulis,
A. arenarius и др.), у других — сутуральное (Л. dasyanthus) или
дорсальное (A. testiculatus). Возможно также неполное сутураль-
но-дорсальное вскрывание лишь верхушки прямостоячих бобов.
В отдельных случаях плоды расщепляются по перегородке и полу-
бобы опадают замкнутыми. Наконец, известны и невскрывающиеся
плоды, как например у довольно обычных наших видов — астра-
гала солодколистного (A. glycyphyllos) и серповидного (A. fal-
catus).
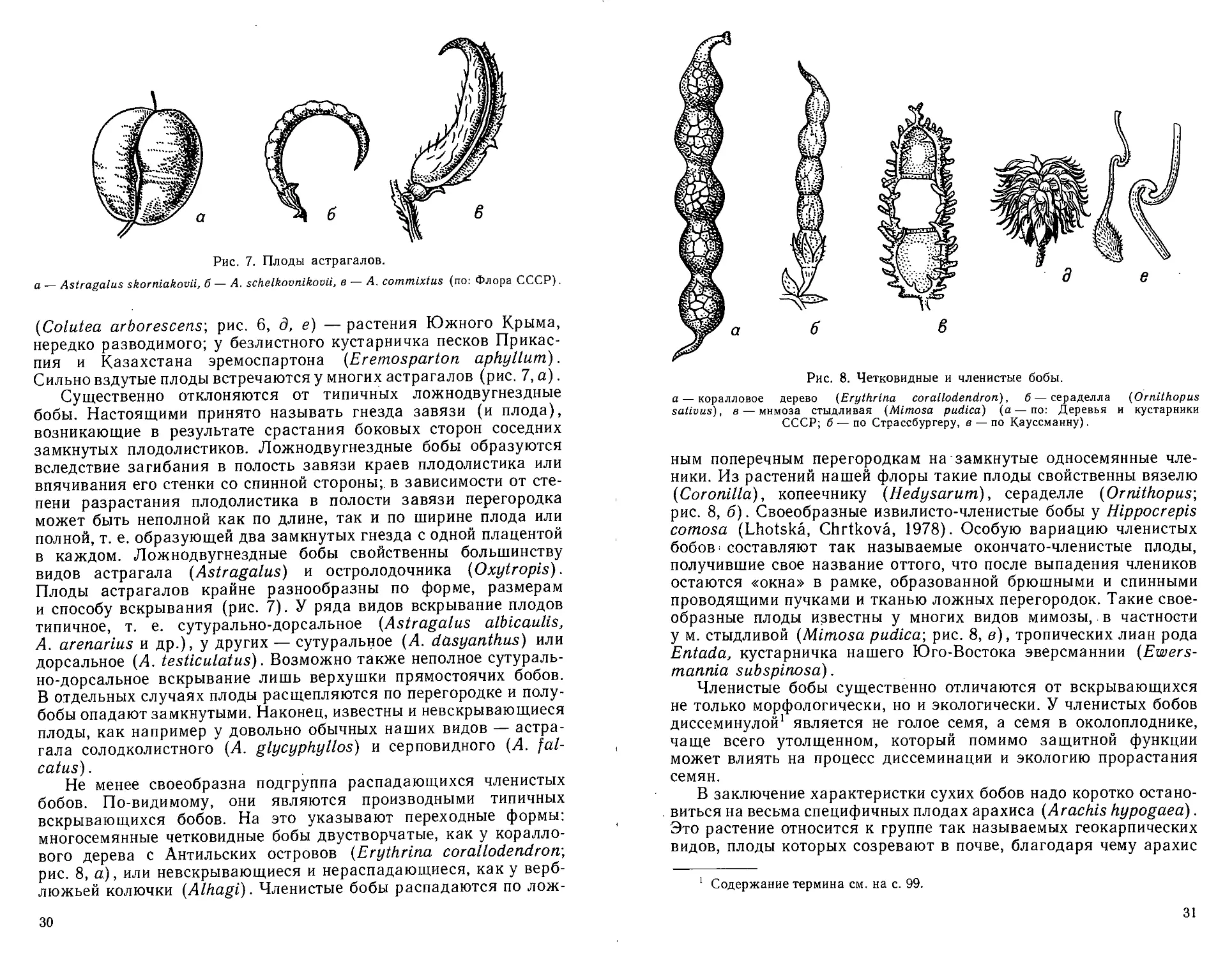
Не менее своеобразна подгруппа распадающихся членистых
бобов. По-видимому, они являются производными типичных
вскрывающихся бобов. На это указывают переходные формы:
многосемянные четковидные бобы двустворчатые, как у коралло-
вого дерева с Антильских островов (Erythrina corallodendrorr,
рис. 8, а), или невскрывающиеся и нераспадающиеся, как у верб-
люжьей колючки (Alhagi). Членистые бобы распадаются по лож-
30
Рис. 8. Четковидные и членистые бобы.
а — коралловое дерево (Erythrina corallodendron), б — сераделла (Ornithopus
salivas), в — мимоза стыдливая (Mimosa pudica) (а—по: Деревья и кустарники
СССР; б — по Страссбургеру, в — по Кауссманну).
ным поперечным перегородкам на замкнутые односемянные чле-
ники. Из растений нашей флоры такие плоды свойственны вязелю
(Coronilla), копеечнику (Hedysarum), сераделле (Ornithopus-,
рис. 8, б). Своеобразные извилисто-членистые бобы у Hlppocrepis
comosa (Lhotska, Chrtkova, 1978). Особую вариацию членистых
бобов составляют так называемые окончато-членистые плоды,
получившие свое название оттого, что после выпадения члеников
остаются «окна» в рамке, образованной брюшными и спинными
проводящими пучками и тканью ложных перегородок. Такие свое-
образные плоды известны у многих видов мимозы, в частности
у м. стыдливой (Mimosa pudica-, рис. 8, в), тропических лиан рода
Entada, кустарничка нашего Юго-Востока эверсманнии (Ewers-
mannia subspinosa).
Членистые бобы существенно отличаются от вскрывающихся
не только морфологически, но и экологически. У членистых бобов
диссеминулой1 является не голое семя, а семя в околоплоднике,
чаще всего утолщенном, который помимо защитной функции
может влиять на процесс диссеминации и экологию прорастания
семян.
В заключение характеристки сухих бобов надо коротко остано-
виться на весьма специфичных плодах арахиса (Arachis hypogaea).
Это растение относится к группе так называемых геокарпических
видов, плоды которых созревают в почве, благодаря чему арахис
1 Содержание термина см. на с. 99.
31
и называют «земляным орехом». Бобы его невскрывающиеся,
могут содержать до 5 семян, но чаще одно-двусемянные, овальные
или неправильно-цилиндрические, с перетяжками, слегка вздутые.
Околоплодник грязновато-палевый, с очень своеобразной ямчато-
сетчатой поверхностью, кожистый, плотный, но хрупкий.
Группа сочных бобов отличается от типичных сочных плодов
и формой, и консистенцией. По форме сочные бобы сходны с сухими
и характеризуются слабым развитием сочных тканей. Так, напри-
мер, у гледичии обыкновенной (Gleditschia triacanthos) в ее очень
крупных (20—45 см дл. и до 3 см шир.) плоских плодах лишь эндо-
карпий становится слегка мясистым и сладковатым, экзокарпий,
напротив, остается плотным и кожистым. Бобы гледичии невскры-
вающиеся. В природных условиях (леса центральной части Север-
ной Америки) они, по-видимому, поедаются животными, а в зеле-
ных насаждениях долго еще остаются на дереве и постепенно
опадают. Высыхая, они становятся хрупкими, легко разламыва-
ются и освобождают семена. Плоды рожкового дерева (Cer atonia
siliqua), так называемые «царьградские рожки», также представ-
ляют собой невскрывающиеся крупные бобы (до 20 см дл., 2.5 см
шир. и до 1 см толщ.) с очень плотным кожистым экзокарпием
и мясистой сахаристой тканью мезоэндокарпия, образующей пере-
городки между семенами. Мясистый мезокарпий характерен также
для перегородчатых бобов тамаринда (Tamarindus indicus),
употребляемых в незрелом виде как овощ. Очень своеобразны
четковидные, мясистые, слегка морщинистые бобы софоры япон-
ской (Styphnolobium. japonicum). Отдельные односемянные уча-
стки боба по внешнему виду и консистенции напоминают изюм.
У софоры становится мясистым весь околоплодник. При хранении
подсохшие плоды софоры распадаются на членики.
К числу сочных бобов следует отнести и плоды бундука (Gym-
nocladus dioic us). Этот североамериканский вид давно культивиру-
ется у нас и рекомендуется как мелиоративное, декоративное
и источник ценной древесины. Бобы бундука широкие, плоские
(до 4 см шир., 10—15 см дл. и до 1.5 см толщ.), с прочным острым
носиком на верхушке; экзокарпий плотный, кожистый, мякоть
сладковатая, клейкая.
ОДНОСЕМЯННЫЕ ПОЛИМЕРНЫЕ
Плоды этого подкласса состоят из многих односемянных плоди-
ков. Строго говоря, такой плод является многосемянным, так как
в сумме дает много семян. Однако ни морфологически, ни эколо-
гически он не равноценен таким плодам, как многолистовка, боб
или коробочка. У односемянных апокарпиев число семяпочек
в завязи редуцируется до одной, а диссеминулой является не семя,
а плодик или весь плод.
К односемянным полимерным плодам относятся многоорешек
и многокостянка.
32
Рис. 9. Многоорешки и их плодики.
а, б — лютик едкий (Ranunculus acris), в, г — ломонос вииоградиолистный (Clematis
vitalbd), д, е — гравилат речной (Geum rivale) (а, б — по Троллю, в, г — по Веберу,
д, е — по Ульбриху).
Многоорешек (multinucula) легко представить как производ-
ное многолистовки, у которой число семяпочек в завязи редуциро-
валось до одной и утратился вскрывающий механизм. Замеча-
тельно, что у одного из садовых сортов землянки плодики вскрыва-
ются.
Поскольку орешек функционирует как диссеминула, защитная
роль околоплодника сохраняется до прорастания семени. Морфо-
логически это проявляется в более или менее выраженном одревес-
нении эндо- и мезокарпия.
Многоорешек является одним из наиболее распространенных
видов апокарпных плодов. Он характерен для большинства родов
лютиковых, очень многих розоцветных подсемейства Rosoideae,
Пастуховых, некоторых магнолиевых и др. Среди других много-
орешков выделяется плод мышехвостника (Myosurus); его мелкие
очень многочисленные орешки густой спиралью сидят на плодо-
ложе настолько вытянутом, что плод нередко превышает осталь-
ную часть генеративного стебля. Относительно удлиненное плодо-
ложе свойственно и адонису (Adonis). У лютиков и лапчаток плод
спиральный, но плодоложе обычно выпуклое (рис. 9, а). Форма
и характер поверхности орешков у обоих названных родов заметно
варьируют, что служит диагностическим признаком для распозна-
вания видов. Плодики ветрениц могут быть либо коротко опушен-
ными (Anemonoides ranunculoides), либо покрыты очень длинными
густыми волосками (Anemone sylvestris).
Подобно многолистовкам, у многоорешков также наблюдается
уменьшение числа плодиков. Так, малочислены орешки у васи-
листника (Thalictrum), лабазника (Fillpendula), частухи (Alisma).
Плоды платана (Platanus orientalis), состоящие из нескольких
(5—9) орешков, собраны в очень компактное шаровидное сопло-
3 Р. Е. Левина
33
Рис. 10. Вариации многоорешков.
а — погруженный многоорешек лотоса (Nelumbo nucifera), б — земляничина в продольном
разрезе (Fragaria vesca), в — цинародий каликаитуса (Calycanthus sp.) в продольном
разрезе (а — по Веттштейну, б — по Троллю, в — по Кернеру).
дне. Трехорешки айланта (Ailanthus altissima) также сближены
в кистевидное соплодие до 10—12 см дл. Плоды рдестов (Potamo-
geton) чаще всего четырехорешки, а цирцеастеру полевому
(Circaeaster agrestis) свойствен двуорешек.
В связи с функцией диссеминации орешки часто бывают снаб-
жены анемохорными или эпизоохорными придатками. Это либо
сильно разросшиеся перистые стилодии, как у прострела (Pulsa-
tilla), княжика (Atragene), ломоноса (Clematis-, рис. 9, в, г), либо
крыловидные выросты околоплодника (Anemonastrum narcissiflo-
rum, Thalictrum aquilegifolium.). Очень своеобразен спиральный
многоорешек тюльпанного дерева (Liriodendron tulipifera, Magno-
liaceae): сильно удлиненное плодоложе несет многочисленные
орешки, снабженные крылом, во много раз превышающим размеры
плодика.
Цепкие плодики известны у цирцеастера и многих других.
Прицепками также служат стилодии, одревесневшие и заострен-
ные, как например у рогоглавника (Ceratocephala) из лютиковых.
.Жесткие, слегка загнутые стилодии сохраняются также на ореш-
ках платана. Кроме того, его плодики у основания несут пучок
волосков. Более своеобразны стилодии на орешках гравилата
(Geum). Они очень тонкие и удлиненные и примерно на середине
имеют крючковидный изгиб с отделительной тканью. При созрева-
нии плодов стилодий обламывается в месте перегиба, а нижняя его
часть, крючковидно загнутая и очень острая на верхушке, сохраня-
ется на орешке (рис. 9, д, е) как эпизоохорный придаток.
В тех случаях, когда в формировании плода существенную роль
играет цветоложе, типичный многоорешек сильно видоизменяется
и превращается в специализированный плод. Такими вариациями
многоорешка являются плоды индийского лотоса (Nelumbo nuci-
fera) , земляники, шиповника. У лотоса сильно разрастается в диа-
метре ось цветка, и каждый плодик оказывается погруженным
в особое углубление в ткани плодоложа (погруженный много-
орешек, по: Тахтаджян, 1948) (рис. 10, а).
34
Плод земляники — земляничина (fragum) — характеризуется,
как известно, также сильно разросшимся (особенно у культурных
форм!) сочным и яркоокрашенным цветоложем, на поверхности
которого редкой спиралью расположены мелкие сухие орешки
(рис. 10, б). Такого же типа плод, но несъедобный, встречается
у рода Duchesnea из розоцветных.
У шиповника орешки расположены на внутренней поверхности
чашевидного или кувшинчатого гипантия, который к моменту со-
зревания становится мясистым и яркоокрашенным. Такой плод,
получивший название цинародия (cynarrhodium, от гр. «цинарро-
дон» — «собачья роза»), мы именуем шиповником. Кроме рода
Rosa он свойствен семействам Monimiaceae и Cyclanthaceae
(рис. 10, в).
Многокостянка (multidrupa) составляет подгруппу сочных
односемянных полимерных апокарпиев. Она широко известна
в роде Rubus. Как показывает название, такой плод состоит из
многих отдельных костяночек, сидящих на общем плодоложе. Все
многокостянки достаточно однотипны; они варьируют лишь
по числу и размерам плодиков и степени развитости плодоложа.
Так, например, у малины (/?. idaeus) на удлиненном коническом
плодоложе размещается несколько десятков мелких (около 2 мм
в диам.) костяночек, а плод костяники (/?. saxatilis) состоит
из немногих (3—5—6) плодиков, достигающих 5 мм в диам.
и сидящих на почти плоском плодоложе. Промежуточными фор-
мами можно считать плоды ежевики (/?. caesius) и морошки
(/?. chamaemorus). Кроме рода Rubus в сем. розоцветных много-
костянка встречается у двух близких монотипных родов, эндемич-
ных для Китая и Японии, — керрии (Kerria japonica) и розовика
(Rhodotypos scandens). Оба кустарника культивируются в СССР
в парках как декоративные. Плоды их состоят из нескольких
темно-коричневых довольно крупных костяночек (4—6—7 мм
в диам.); плодики розовика с мучнистой мякотью.
Из других семейств, в которых известны многокостянки, можно
назвать Menispermaceae, Amborellaceae (Amborella trichopoda —
вид монотипного рода из Новой Каледонии; плод амбореллы
из 3—6 костяночек) и близкое к рдестовым сем. Ruppiaceae.
Многокостянка руппии — обитателя внетропических солоноватых
водоемов — очень своеобразна: ее плодики сидят на длинных
тонких ножках, их экзо-мезокарпий мясистый, богатый крахма-
лом. Плодики распространяются эндозоохорно птицами и рыбами
(Цвелев, 1982а).
ОДНОСЕМЯННЫЕ МОНОМЕРНЫЕ
Среди апокарпиев плоды только этого подкласса являются
односемянными в строгом смысле слова. Они возникли как есте-
ственное следствие процесса олигомеризации, наблюдаемого у всех
видов полимерных плодов — многолистовок, многоорешков, мно-
3*
35
Рис. 11. Одноорешки.
а — рогоз широколистный (Typha latifolia), б— роголистник темно-зеленый (Ceratophyl-
lutn demersum), в — репешок обыкновенный (Agrimonia eupatoria), продольный разрез
орешка в гипантии (а — по Маевскому, б — по Кадену, в — по: Жизнь растений).
гокостянок. Примеры всех видов олигомерных плодов были на-
званы выше.
Олигомеризация охватывает и семязачатки. Так, для сем.
Cabombaceae характерны невскрывающиеся сухие плодики с 1 —
3 семенами. Известны и аналогичные бобы, например у арахиса.
Типичными, можно сказать, хрестоматийными односемянными
мономерными плодами являются одноорешек, односемянный боб
и однокостянка.
Одноорешек (uninucula) нельзя назвать широко распростра-
ненным видом плода, однако он встречается у некоторых розоцвет-
ных, а также в семействах Ceratophyllaceae, Eupteleaceae, Naja-
daceae, Nyctaginaceae, Proteaceae, Typhaceae и некоторых других.
Одноорешек, как и плодики многоорешков, часто бывают снаб-
жены анемохорными или зоохорными придатками различной мор-
фологической природы. Очень маленький овально-веретеновидный
одноорешек рогоза (Typha) сидит на длинной нитевидной ножке,
а на верхушке несет такого же типа стилодий. Кроме того, от осно-
вания ножки отходит пучок направленных вверх волосков (видо-
измененный околоцветник) (рис. 11, а).
Одноорешки, снабженные хохолком из волосков или перистыми
выростами, известны у некоторых протейных. Крылатый однооре-
шек — у эуптелеи. Для плодов роголистника (Ceratophyllum)
характерны шиповатые или более или менее уплощенные выросты,
нередко превышающие размеры орешка. Верхушечный вырост —
это видоизмененный стилодий (рис. 11, б).
У представителей розоцветных орешки нередко заключены
в гипантий. Число орешков плода может колебаться от одного
до двух. Орешки в гипантиях свойственны манжетке (Alchemilla),
кровохлебке (Sanguisorba), но особенно широко известны одре-
36
весневшие гипантии репешка (Agrimonia), усаженные крючковато
загнутыми шипиками (рис. 11, в). По происхождению эти прицепки
представляют собой видоизмененные листочки подчашия. К вариа-
ции «орешек в гипантии» следует отнести и мономерный плод
ld.iosperm.um из сем. каликантовых, цинародий которого содержит
лишь один орешек. Сюда же можно включить и плоды некоторых
видов никтагиновых, например Mirabilis nyctaginea, у которой
орешек окружен антокарпием,1 и виды Pisonia с плодом в железисто
опушенной чашечке.
Разноречиво наименование плода лоховых (Elaeagnaceae).
Нередко его называют ложной, или псевдомономерной, костянкой,
сфалерокарпием1 2 (Каден, 1965). Поскольку гинецей лоховых моно-
мерный, а плод односемянный, сухой, заключенный в мясистую
трубку чашечки (Engler, 1964), его также можно рассматривать
как орешек в гипантии.
Односемянный боб (tegumen monospermum) — гораздо более
распространенный вид плода, чем одноорешек, что связано
с исключительным разнообразием видов бобовых. В разных таксо-
нах этого обширного порядка происходила редукция числа семяза-
чатков в завязи до одного. Следы такого преобразования легко
обнаруживаются в одно-двусемянности невскрывающихся бобов
и, напротив,во вскрывании односемянных; в наличии много-
и односемянных плодов в пределах политипных родов и в других
особенностях.
Как начальный этап формирования односемянного боба можно
рассматривать вариацию так называемых крылатковидных бобов.
Своеобразие их заключается в том, что при наличии лишь одного
семени стерильная часть околоплодника не редуцируется, а пре-
вращается в крыловидный придаток. При этом семя может зани-
мать самое разное положение — базальное, как у центролоби-
ума (Centrolobium robustunv, рис. 12, а), верхушечное (Spatho-
lobus roxburgii, Hardwickia binata, рис. 12, б) или центральное.
В последнем случае плод может оставаться удлиненным, как бы
двукрылым, как у дальбергии (Dalbergia stipulacea), или округ-
лым, как у видов птерокарпуса (Pterocarpus) (рис. 12, в). Ориги-
нальны крылатые бобы песчаной акации (Ammodendron conollyi),
достигающие 30 мм дл. и 7 мм шир. Семя помещается в центре во
вздутой и винтообразно перекрученной части боба, на остальном
протяжении его боковые части сплюснуты. Весь плод напоминает
пропеллер (рис. 12, г).
Односемянные бобы наряду с многосемянными известны у раз-
ных видов дрока, клевера и некоторых других. У клевера виды под-
рода Trifoliastrum имеют плоды с 2—6 семенами, а подрода
Lagopus — односемянные, реже о двух семенах (Бобров, 1945).
Из видов среднерусской флоры односемянные бобы свойственны
клеверу луговому, мясокрасному (Trifolium incarnatum), среднему,
1 Околоцветник, окружающий зрелый плод и срастающийся с ним у основания.
2 Восстановленное название.
37
a—г — крылатковидные: а — центролобиум {Centrolobium robustum), б — хардвикия
{Hardwickia binatd), в — красное дерево (Pterocarpus indicus), г — песчаная акация
(Ammodendron conollyi)-, д — клевер луговой {Trifolium pratense}', е — эспарцет равно-
зубчатый (Onobrychis aequidentata) (а, е — по Кернеру, б — по Русанову, в — по Ветт-
штейну, г — по Коровину, д — по Хюбнеру).
горному, красноватому (Т. rubens), пашенному, альпийскому,
днепровскому (Т. borysthenicum). У клевера лугового, альпийского,
мясокрасного и некоторых других боб вскрывается ламинально
поперечно-кольцевой щелью, т. е. с образованием крышечки
(рис. 12, д). У клевера среднего сохранилось сутуральное вскры-
вание. Виды дрока с односемянными бобами: Genista hispanica,
G. monosperma, G. radiata, G. sylvestris и др.
В большинстве случаев с уменьшением числа семян редуциру-
ется и околоплодник; односемянный боб становится невскрыва-
ющимся, сходным с орешком. Такие орешковидные бобы известны
у люцерны хмелевой (Medicago tupulina), донника (Melilotus),
эспарцета (Qnobrychis) и др. Плоды ряда видов эспарцета снаб-
жены шиповатыми выростами, способствующими эпизоохории
(рис. 12, е). Цепкие орешковидные бобы характерны и для солодки
иглистой (Glycyrrhiza echinata). Замкнутые односемянные бобы
язвенника (Anthyllis macrocephala) заключены во вздутую ча-
шечку, что повышает парусность плода. Односемянные бобы изве-
стны также у некоторых наших декоративных кустарников
(Amorpha fruticosa, Carmichaelia australis, Lespedeza).
Однокостянка (unidrupa) свойственна всем представителям
подсем. сливовых (Prunoideae) сем. розоцветных. Этот вид плода
широко известен благодаря многим культурным формам, у кото-
рых в результате искусственного отбора оказался особенно сильно
развитым сочный мезокарпий (персик, абрикос, слива). Выше уже
отмечалось, что у костянок особенно четко проявляется дифферен-
38
Рис. 13. Косточка черемухи (Padus avium) (по Левиной, 1967а).
--------------------------------------------------
циация всех слоев околоплодника, при этом сложного
анатомического строения достигает главным образом
эндокарпий (Зажурило, 1931; рис. 2, а). Костянки В
в целом достаточно однотипны, различаясь лишь
размерами, окраской, отчасти очертаниями. Значи---|
тельно разнообразнее строение косточек, вклю-
чая скульптуру их поверхности. Хорошо известны, например,
совершенно гладкие косточки абрикоса и косточки персика с ха-
рактерными глубокими, причудливо изгибающимися бороздами.
Очень своеобразный скульптурный рисунок наблюдается и на
поверхности косточек черемухи {Padus avium.-, рис. 13). Морфоло-
гия косточек может служить надежным диагностическим призна-
ком для распознавания родов и видов. Это относится и к много-
костянкам, и ценокарпным костянкам.
Среди сливовых у миндаля известна сухая костянка, у которой
мезокарпий становится относительно тонким, кожистым и довольно
плотным; ко времени созревания плодов экзомезокарпий растрес-
кивается. Так называемые «орехи миндаля» представляют собой
косточки.
Кроме розоцветных однокостянка встречается у некоторых
протейных, луносемянниковых, у дидимелеса (Didymelaceae) —
узкоэндемичного реликта флоры Мадагаскара. К однокостянкам
следует отнести и плоды некоторых пальм (главным образом
из подсем. Coryphoideae}, у которых из трехчленного апокарпного
гинецея развивается одна костянка, а 2 плодолистика дегенери-
руют. Сухая однокостянка с волокнистым мезокарпием и дере-
вянисто-твердым эндокарпием характерна для нипы {Nypa fruti-
cans, подсем. Nypoideae).
В заключение обзора апокарпиев рассмотрим некоторые одно-
семянные мономерные плоды неясной типологии. Среди других
к ним относятся плоды пальм рода Phoenix, наиболее известные
по плодам культурной финиковой пальмы {Ph. dactylifera). Ее плод
именуют костянкой, ягодой или сочным орешком (Левина, 1974).
Однако исследование анатомического строения околоплодника
ряда видов Phoenix (Ландсберг, 1981) показало своеобразие этого
вида плода. В его перикарпии четко различаются экзо-, мезо- и эн-
докарпий; последний не содержит одревесневших клеток и вполне
обособлен от семени. Характерной чертой экзокарпия является
слой каменистых клеток, залегающий под гиподермой. Г. С. Ланд-
сберг предлагает называть такой плод «фиником». В нашей клас-
сификации (см. табл. 1) финик помещен рядом с однокостянкой,
т. е. в группе сочных односемянных мономерных апокарпиев.
Очень своеобразный мономерный плод тропического сем. мус-
катниковых {Myristicaceae) вообще не имеет наименования.
Только в старых работах его называют односемянной мясистой
коробочкой (Варминг, 1893;. Веттштейн, 1912), а в современных
(Engler, 1964; Тахтаджян, 1966; Трифонова, 1980) дается лишь
39
описание плода. Нетипичность его состоит в том, что это односе-
мянный плод с хорошо развитым мясистым околоплодником и
вместе с тем вскрывающийся сутурально-дорсально; экзокарпий
кожистый. По совокупности признаков плод мускатниковых можно
назвать односемянной сочной листовкой.
Плод наиболее известного «мускатного дерева» (Myristica
fragrans) в раскрытом виде обладает целым набором средств
привлечения агентов диссеминации: оранжево-желтая мякоть
плода и на ее фоне темное душистое семя, окруженное рассеченным
ярко-алым сочным ариллусом с сильным запахом. Пряность миро-
вого рынка — «мускатный орех» — представляет собой семя.
До сих пор остается спорной морфологическая природа зер-
новки злаков (Цвелев, 19826). Чаще ее трактуют как ценокарпный
(паракарпный) тримерный плод (Тахтаджян, 1966) или как псев-
домономерный на тримерной основе (Eckardt, 1938). Ряд авторов
считают зерновку мономерным плодом (Каден, 1958а, 19586, 1959;
Potztal, 1964). В работах Н. Н. Кадена показано, что в гинецее
злаков нет никаких следов недоразвитых плодолистиков, на осно-
вании чего автор причисляет зерновку к апокарпиям.
Злаки несомненно относятся к наиболее специализированным
семействам цветковых растений, занимающим высокое положение
в системе. У таких таксонов можно ожидать полной редукции
нефункционирующих структур. С этой точки зрения гинецей злаков
можно рассматривать как вторичномономерный, в отличие от псев-
домономерных гинецеев, у которых сохраняются рудименты
стерильных плодолистиков. Исходя из этих соображений, в нашей
морфологической классификации зерновка помещена среди одно-
семянных мономерных апокарпиев.
Зерновка (caryopsis) — односемянный сухой плод с тонким
перикарпием, плотно прижатым к семени и неотделяющимся
от него. Наименование «ягодовидная зерновка», применяемое
к сочным плодам бамбуков, едва ли правильно. Так, например,
плод Melocanna bambusoid.es по ряду признаков не имеет ничего
общего с зерновкой. В его зрелых плодах семена без эндосперма.
У большинства видов злаков зерновка опадает вместе с окру-
жающими ее цветковыми, а иногда и колосковыми чешуями и при-
легающим участком оси колоска. Как отмечалось ранее, такую
диаспору называют антецием. Цветковые чешуи, полностью скры-
вающие зерновку, хорошо известны, например у овса, проса,
ковыля, костра, овсяницы и многих-многих других. Остающиеся
при плодах чешуи несомненно создают дополнительную защиту
семени, поэтому у таких зерновок перикарпий тонкий, слаборазви-
тый.
Чешуи, одевающие зерновку, играют не только защитную роль.
Они в значительной степени способствуют и распространению пло-
дов. Чешуи даже сами по себе, без всяких придатков, заметно уве-
личивают парусность мелких зерновок (например, Alopecurus,
Beckmannia, Briza, Phalaris) и служат, таким образом, летатель-
ным или плавательным приспособлением. Кроме того, на цветковых
чешуях часто развиваются различного рода придатки: перистые,
40
цепкие или спирально закрученные гигроскопичные, которые
несомненно облегчают распространение плодов ветром, живот-
ными или зарывание их в почву. Общеизвестны, например, пери-
стые и вместе с тем гигроскопичные ости ковылей; своеобразны
трехраздельные перистые ости Aristida; хорошим парашютом
служит хохолок волосков на каллусе вейника наземного (Calamag-
rostis epigeios).
ЦЕНОКАРПИИ
К ценокарпиям отнесены все виды плодов, образованные двумя
или несколькими сросшимися друг с другом плодолистиками.
Классификация плодов в пределах типа ценокарпиев представлена
в табл. 2. Чтобы избежать повторений, аналогичные виды верхних
и нижних плодов описываются параллельно.
МНОГОСЕМЯННЫЕ ВЕРХНИЕ И НИЖНИЕ
СУХИЕ ВСКРЫВАЮЩИЕСЯ
Представителями этой подгруппы плодов являются цено-
карпная многолистовка, коробочка и стручок.
Ценокарпная многолистовка (multifolliculus coenocarpus)
от аналогичного апокарпного плода отличается полным сраста-
нием плодолистиков боковыми стенками, а от типичной коро-
бочки — характером вскрывания. Она вскрывается неполно суту-
рально лишь в области верхних свободных участков плодолистиков.
Плоды этого вида известны в семействах Tetracentraceae, Trocho-
dendraceae, Winteraceae. Многолистовка экзохорды (Exochorda)
из розоцветных отличается от типичной односемянными листов-
ками.
Из растений нашей флоры ценокарпная многолистовка встре-
чается у чернушки (Nigella), канатника (Abutilon), ясенца (Dlcta-
mnus), некоторых видов спиреи. Очень своеобразную сочную вари-
ацию этого вида плода с перикарпием из губчатой паренхимы мы
находим у нимфейных (Снигиревская, 1980).
Коробочка (capsula) — наиболее распространенный вид плода
среди многосемянных ценокарпиев. Она свойственна самым
разным представителям нашей среднерусской флоры и очень мно-
гим семействам тропиков и субтропиков (табл. 3 и 4).
Вполне очевидно, что коробочки возникали независимо в раз-
личных филогенетических рядах, поэтому и существует такое
множество их вариаций. В табл. 3 отражено варьирование по со-
четанию только трех основных признаков: способу вскрывания,
типу плацентации и характеру завязи (верхней или нижней),
а в табл. 4 — по двум последним признакам.
Анализ таблиц позволяет сделать некоторые обобщения. Во-
первых, верхних коробочек значительно больше, чем нижних.
41
Таблица 3
Вариации коробочек у растений среднерусской флоры
Тип плацентации и характер завязи
Способ вскрывания ' 1 -------------------------—
(полного или неполного)_________центрально-угловая париетальная
£
S
X
X
X
X
По видимому, параллельно с форми-
рованием нижней завязи возникали
и более специализированные, чем
коробочка, виды плода. Во-вторых,
для коробочек более типична цент-
рально-угловая плацента; иначе го-
воря, дву- или многогнездные коро-
бочки встречаются чаще, чем одно-
гнездные. В-третьих, наблюдается
явная корреляция между типом
плацентации и способом вскрывания
коробочек: при центрально-угловой
плаценте эти способы наиболее раз-
нообразны и чаще всего комбиниро-
ванные. Париетальная плацентация
сочетается преимущественно с дор-
сальным вскрыванием. При этом
число створок раскрытой коробочки
соответствует числу плодолистиков,
но каждая створка образована смеж-
ными половинками двух соседних
плодолистиков. Наконец, при колон-
чатой плаценте более обычно не-
полное сутуральное вскрывание (т. е.
зубцами, число которых соответ-
ствует числу плодолистиков).
Разнообразие коробочек обус-
ловливается не только типом пла-
центации и способом их вскрывания,
но и целым рядом других морфологи-
ческих особенностей: размерами и
формой, консистенцией околоплод-
ника к моменту созревания, различ-
ного типа выростами и придатками,
сохраняющимися частями цветка и
т. п. Достаточно сравнить плод
конского каштана и какой-либо веро-
ники, чтобы представить, насколько
могут различаться коробочки даже
с одинаковым способом вскрывания.
У конского каштана (Aesculus hippo-
castanum) формируются крупные
(4—5 см в диам.) более или менее
шаровидные трехгнездные коробочки,
покрытые сильно развитыми шипова-
тыми выростами. В каждом гнезде
закладываются по 2 семязачатка, но
развивается не более одного семени
(рис. 14, а, б). Характерной особен-
42
43
Таблица 4
Вариации коробочек в некоторых семействах тропической и субтропической
флоры 1
Тип плацентации и характер завязи
центрально-угловая париетальная
верхняя НИЖНЯЯ верхняя нижняя
Acanthaceae Amarylidaceae Bixaceae Begoniaceae
Аросупасеае Саппасеае Capparaceae Corsiaceae
(Allemanda) Caprifoliaceae (Cleome, Isomeris) Datiscaceae
Bignoniaceae (Diervilla, Weigela) Dioncophyllaceae Loasaceae
Bombacaceae Costaceae Escalloniaceae Melastomataceae
Bonnetiaceae Euphorblaceae Flacourtiaceae Taccaceae
Bromeliaceae (Securinega) Frankeniaceae
Buddlejaceae Hamamelidaceae Gesneriaceae
Byblidaceae Heliconiaceae Passifloraceae
Clethraceae Lowlaceae Pittosporaceae
Commelinaceae Diapensiaceae Dioscoreaceae Elaeocarpaceae Epacridaceae Erlocaulaceae Guttiferae Loganiaceae Medusagynaceae Meliaceae Oleaceae (Forsythia) Sapindaceae ( Koelreuteria) Sarcolaenaceae Theaceae Vochysiaceae Str elit ziaceae Zingiberaceae Turneraceae
1 В литературе нет указаний на способы их вскрывания.
ностью плодов конского каштана является их вскрывание в зеле-
ном, внешне недозрелом состоянии. Мясистые стенки коробочки
растрескиваются дорсально и освобождают крупные сплюснуто-
шаровидные темно-коричневые семена с округлым серым рубчиком,
достигающим 15 мм и более в диам. Иногда во всей коробочке
развивается всего два или даже одно семя, особенно крупное.
Из вероник с дорсально вскрывающимися коробочками можно
назвать в. простертую (Veronica prostrata). Плоды ее настолько
сильно сплюснуты с боков, что становятся почти плоскими. Они
обратносердцевидной формы, 3—4 мм дл. и около 3 мм шир. Зре-
лая сухая коробочка вскрывается двумя продольными щелями по
краю сверху вниз. Семена начинают осыпаться сразу же по мере
образования щели.
Нет возможности и необходимости описывать все вариации
коробочек. Остановимся лишь коротко на некоторых из них с наи-
более оригинальным способом ламинального вскрывания.
Коробочка недотроги из сем. бальзаминовых (Impatiens noli-
tangere) вскрывается будучи еще зеленой и сочной. Вследствие
44
Рис. 14. Плод конского каштана (Aesculus hippocastanum).
а — раскрытая коробочка, б — коробочка в поперечном разрезе, схема (а — по: Деревья
и кустарники СССР, б — по Троллю).
образования неполных продольно-кольцевых щелей в стенках
плодолистиков вскрывание происходит 5 створками от основания
к вершине. В коробочке развиваются всего 2 семени, расположен-
ные у расширенного основания плода, где стенки его оказываются
наиболее тонкими. Стенки коробочки в верхнем ее конце утолща-
ются, и в них развивается особый слой клеток с очень высоким
осмотическим давлением (до 25 атм, или более 2.5- 106 Па).
В результате большого напряжения, которое создается в стенках
плода, при малейшем прикосновении створки разверзаются,
закручиваются улиткообразно и с силой выбрасывают семена
(рис, 15, а). Аналогичный способ вскрывания наблюдается у сухих
коробочек герани. Для них характерно образование так называе-
мой колонки (из сильно разросшихся в длину брюшных участков
плодолистиков), которая остается после вскрывания плода
(рис. 15, б).
Очень своеобразны длинные цилиндрические коробочки ка-
тальпы — декоративных деревьев из сем. бигнониевых. Так, у Са-
talpa bignonioides коробочки достигают 15—45 см дл. при 6—
8 мм в диам. Они вскрываются двумя опадающими «створками»,
каждая из которых образована спинными участками двух соседних
плодолистиков. Помимо продольно-кольцевой линии разрыва,
захватывающей оба плодолистика, образуются еще два линейных
продольных разрыва по перегородке. После опадения створок
многочисленные семена остаются открыто сидящими на плаценте
и постепенно рассеиваются.
При ламинальном продольно-кольцевом вскрывании одногнез-
дных коробочек возникают оригинальные «рамчатые коробочки»,
В этом случае линия разрыва проходит по пластинке плодолистика
вдоль швов, а сутуральные участки плодолистиков с плацентами
образуют рамки (рис. 16, а). В коробочке из двух плодолистиков
всего одна рамка (рис. 16, б). Такую коробочку называют струч-
ковидной; от типичного стручка она отличается только отсутствием
перегородки. Стручковидные коробочки известны у хохлатки,
некоторых каперсовых (Саррагасеае), у чистотела. В сем. маковых
встречается и типичный стручок, (мачок — Glaucium).
В заключение обзора коробочек отметим, что у некоторых
весьма специализированных сорно-полевых видов типичные по
45
'Рис. 15. Раскрытые коробочки.
а — недотрога (Impatiens noli-tangere), б—герань (Geranium palustre) (по Кернеру).
Рис. 16. Рамчатые коробочки маковых.
а — Argemone mexicana, б — чистотел (Chelldonium majus) (по Троллю).
Рис. 17. Плод лунника однолетнего
(Lunaria annua) после опадения ство-
рок (по Страссбургеру).
Рис. 18. Стручок горчицы белой (Sinapis
alba) (по Веттштейну).
46
внешнему виду коробочки утрачивают способность вскрывания,
как это произошло у засорителей льна торицы льняной и крупной
(Spergula arvensis subsp. linicola, S. arvensis subsp. maxima).
Стручок (siliqua), широко известный в сем. крестоцветных,
представляет собой своеобразную верхнюю коробочку. Для стручка
характерно наличие тонкой пленчатой перегородки, вырастающей
между плацентами от краев плодолистиков внутрь гнезда завязи.
Вскрывание стручка происходит так же, как у стручковидных
коробочек, т. е. ламинально по замкнутой продольно-кольцевой
линии. После опадения «створок» остается рамка с перегородкой
и сидящими на плаценте семенами (рис. 17).
Стручок представляет собой двучленный плод, хотя в очень
редких случаях у некоторых крестоцветных {Nasturtium, Draba)
образуются четырехстворчатые плоды из четырех плодолистиков.
В. Е. Подколзиной (1974) описан как наследуемый признак трех-
створчатость части стручков у некоторых, преимущественно гиб-
ридных сортов горчицы.
Плоды крестоцветных, однотипные по происхождению, крайне
разнообразны морфологически, поэтому карпологические признаки
широко используются для диагностики родов и видов внутри
семейства.
Типичный стручок — вскрывающийся, многосемянный, удли-
ненно-цилиндрический или более или менее четырехгранный, одно-
родный на всем своем протяжении; длина его во много раз превы-
шает диаметр. Такой стручок, свойственный гулявнику {Sisymb-
rium), сурепице {Barbarea), левкою {Matthiola), вяжечке (Turri-
tis) и многим другим, рассматривается как исходный для всех
других модификаций стручка (Zohary, 1948).
Укорачивание стручка и соответственно уменьшение числа
семян привели к образованию стручочка, длина которого более
или менее равна ширине. По очертанию стручочки довольно раз-
нообразны: округлые, овальные, грушевидные, треугольные,
но среди них различают два основных варианта — широкоперего-
родчатые, т. е. сжатые со спинки {Alyssum, Berteroa, Draba, Luna-
ria, Schivereckla и др.), и узкоперегородчатые — сжатые с боков,
т. е. со стороны швов {Capsella, Lepidium, Subularia, Thlaspi и др.).
Другое направление эволюции стручка выразилось в его диф-
ференциации по длине, в результате чего возникли стручки с так
называемым носиком, или клювиком, на верхушке, как например
у горчицы (Sinapis; рис. 18, а) или редьки (Raphanus). Носик
может достигать различной длины относительно остальной части
стручка и оставаться бесплодным или содержать одно—несколько
семян. Эволюция стручка с носиком шла двумя путями: в одних
случаях плодущей оставалась основная, базальная часть стручка,
а носик оказывался бесплодным; в других — наоборот: совершенно
редуцировалась базальная часть, а носик развивался и превра-
щался в основную плодущую часть стручка, нередко со многими
семенами. Такого типа плод у дикой редьки {Raphanus raphani-
strum\ Rytz, 1936; Zohary, 1937; Stopp, 1950).
47
Невскрывающиеся модификации стручков и стручочков мы
рассмотрим в других разделах этой главы.
Помимо крестоцветных стручки встречаются редко. Уже упоми-
нался мачок из маковых; можно назвать еще виды Cleome и Pola-
nisia из каперсовых — семейства, вообще близкого к крестоцвет-
ным.
К подгруппе вскрывающихся ценокарпиев надо отнести очень
редкий вид плода — сухую вскрывающуюся костянку. Такие плоды
встречаются у некоторых крушиновых: полунижние Colletia hor-
rida, Colubrina asiatica, Ceanothus azureus или нижние: Hellnus
ovatus. У всех названных видов плоды очень мелкие (4—7 мм
в диам.) и содержат по 3 косточки, окруженные тонким мезокар-
пием. Косточки, активно раскрываясь, разрывают наружные слои
околоплодника и разбрасывают семена (Вихирева, 1952).
РАСПАДАЮЩИЕСЯ
Эта подгруппа ценокарпиев очень разнородна по своей морфо-
логии, но подавляющее большинство ее видов — дробные плоды,
т. е. распадающиеся продольно в плоскости срастания плодолисти-
ков (см. главу I). Число образующихся мерикарпиев (частей
плода) соответствует числу плодолистиков.
Распадающиеся ценокарпные плоды возникают как из верхней,
так и из нижней завязи. Рассмотрим вначале верхние дробные
плоды.
Переходной структурой между вскрывающимися и распадаю-
щимися плодами можно считать дробные коробочки. Распадаясь,
они освобождают голые семена, тогда как у типичных распада-
ющихся плодов диссеминулой служит семя, окруженное «своей»
частью околоплодника.
Дробные коробочки известны у молочайных — молочаев
(Euphorbia), клещевины (Ricinus communis) и др. Их коробочки
распадаются септицидно на 2—3 доли (по числу гнезд), кроме
того, в результате продольно-кольцевого разрыва по перегородке
гнездо отрывается от плаценты. Частично происходит разрыв и
дорсально. Вследствие неравномерного сокращения различных
слоев околоплодника при высыхании коробочки молочайных раз-
рываются внезапно и разбрасывают семена (рис. 19, а). Много-
гнездная коробочка эукрифии сердцелистной (Eucryphia cordifolia,,
Eucryphiaceae) расщепляется по перегородкам, а гнезда отрыва-
ются от оси плода и раскрываются вдоль шва, обнажая одно или
несколько семян (рис. 19, б). Коробочка вильгельмсии пузырчатой
(Wilhelmsia physodes, Caryophyllaceae) распадается на три взду-
тых почти замкнутых пленчато-кожистых гнёзда. Они разносятся
ветром и рассеивают семена.
Дробные плоды, распадающиеся на незамкнутые односемянные
части, наблюдаются у многих мальвовых (Mala a, Althaea, Lava-
tera). Они распадаются септицидно с образованием, дополни-
48
тельно, продольно-кольцевого разрыва близ плаценты. Мерикар-
пии оказываются открытыми с брюшной стороны. Такой плод на-
зывают калачиком (dieresilis).
У некоторых родов мальвовых плоды распадаются на вполне
замкнутые мерикарпии, иногда несущие цепкие шипы (Pavonia
spinifex; рис. 19, в). Такие плоды, не имеющие общепринятого
наименования, мы называем дробным ценокарпием. Верхние дроб-
ные ценокарпий свойственны также триостренникам {Triglochin)
из сем. ситниковидных (7uncaginaceae), якорцам (Тribulus terre-
stris) из парнолистниковых (Zygophyllaceae). Плоды триостренни-
ков очень мелкие: до 10 мм дл. и 1 мм шир. (Triglochin palustre)
или еще короче — около 4 мм дл. и до 2.5 мм шир. (Т. maritimum).
У триостренника болотного плод распадается на 3 части снизу
вверх, причем каждая из них книзу заметно суживается и заканчи-
вается короткой очень острой остью. У другого распространенного
у нас вида — триостренника морского — образуется 6 частей
плода, и каждая из них притупленная у основания, но суженная
и крючкообразно загнутая на верхушке.
Очень своеобразны плоды якорцев, распадающиеся на 5 тре-
угольно-клиновидных частей; каждая из них до 7 мм дл., 4—5 мм
шир. и 2—3 мм толщ. Многочисленные короткие и длинные шипы
на спинной стороне мерикарпиев придают им причудливую форму
(рис. 19, г). Каждый мерикарпий содержит 3—'5 семян, лежащих
в отдельных линейно расположенных полостях.
Верхние дробные плоды встречаются в сем. рутовых — у неко-
торых видов цельнолистника (Haplophyllum acutifolium, Н. latifo-
lium, Н. perforatum), у киркии (Kirkia, Simaroubaceae).
Широко известна дробная двукрылатка (disamara) клена.
После ее растрескивания мерикарпии-крылатки опадают не сразу,
а некоторое время висят еще на обособившихся проводящих
пучках (рис. 20, а). Плоды разных видов клена вполне однотипны,
хотя и различаются размерами и деталями морфологии. В этом
семействе встречаются, однако, плоды совсем иного типа, как на-
пример двукрылатка диптеронии китайской (Dipteronia sinensis),
мерикарпии которой более сходны с плодами ильмовых (рис. 20, б).
В близком к кленовым сем. сапиндовых также известны дву- и
трехкрылатки (Diatenopteryx, Thinouia) «кленового» типа, но от-
личающиеся тем, что у клена прямым и утолщенным является
абаксиальный край крылатки, а у сапиндовых — адаксиальный
(рис. 20, в, г). Верхние дву- и трехкрылатки свойственны пантро-
пическим лианам сем. мальпигиевых. Оригинальна по форме дву-
крылатка Tetrapterys citrifolia (рис. 20, д). В сем. рутовых у севе-
роамериканской гелиетты (Helietta parvifolia) плоды очень сходны
с плодами клена, но они представляют собой не дву-, а четырех-
крылатки.
Нижними дробными ценокарпиями являются плод урути
(Myriophyllum), распадающийся на 4 мерикарпия, и двусемянки
многих мареновых. Плоды подмаренника (Galium) и ясменника
(Asperula) распадаются на 2 плоско-выпуклые, слегка почковид-
4 Р. Е. Левина
49
Рис. 19. Дробные плоды.
а — коробочка клещевины (Ricinus communis), б — коробочка эукрифии (Eucryphia
cordifolia), в — мерикарпий плода павонии (Pavonia spinifex), г — мерикарпий плода
якорцев (Tribulus ierrestris) (а — по Кернеру, б — по: Жизнь растений, в — по Ветт-
штейну, г — по Броуверу).
Рис. 20. Дробные крылатки.
а — клеи (Acer pseudoplatanus), б — диптероиия (Dipteronia sinensis), в — диатеиопте-
рикс (Dlatenopteryx sorbifolia), г — трехкрылатка тинуйи (Thinouia scandens), д — дву-
крылатка тетраптериса (Tetrapterys citrifolia) (а — по Страссбургеру, остальные —
по: Жизнь растений).
50
Рис. 21. Нижние дробные плоды.
а — двусемянка подмаренника (Galium aparine), б — вислоплодник тмина (Carum carvi),
в — его поперечный срез, г — плод борщевика (Heracleutn sphondylium), д — орлайи
(Orlaya grandiflora), е, ж — дробные костянки крушиновых: е — трималиума (Trymalium
helianthoides), ж — гуанин (Gouania blanchetiana) (а — по Троллю, б, в — по Страссбур-
геру, г, д — по Веттштейну, е, ж — по Вяхиревой).
ные или почти шаровидные части от 1.5—2 до 3—4 мм дл. (или
в диам.). Поверхность их то гладкая блестящая, то матовая ямча-
тая, иногда с частыми прицепками (рис. 21, а). Нижний дробный
ценокарпий, распадающийся на 3 мерикарпия, свойствен сем.
Heliconiaceae, близкому к банановым.
Наиболее специализированным дробным плодом является вис-
лоплодник (cremocarpium) зонтичных, который, по заключению
ряда исследователей (Н. В. Первухина, В. Н.Тихомиров и др.),
развивается из полунижней завязи. Большинству зонтичных свой-
ственны плоды, распадающиеся в плоскости срастания плодоли-
стиков на 2 мерикарпия. Некоторое время они остаются висеть
на цельном или двураздельном плодоносце (карпофоре), или ко-
лонке, откуда и происходит наименование плода (рис. 21, б).
Колонка представляет собой сутуральные участки двух соседних
плодолистиков (Тахтаджян, 1948). В типе строение мерикарпия
у самых различных видов зонтичных довольно однородно. Он
снабжается 5 проводящими пучками, которым соответствуют вы-
ступающие на поверхности 5 главных ребер. В ложбинках между
ними в ткани околоплодника проходят секреторные канальцы
(рис. 21, в). Между главными ребрышками могут развиваться
еще дополнительные; число канальцев также может возрастать.
При однотипности общей структуры плоды зонтичных очень разно-
4*
51
образны по деталям анатомического строения и внешней морфо-
логии. На этом основано широкое использование карпологических
признаков в диагностике родов и видов этого обширного семейства.
Морфологические различия этих плодов связаны со способами
их распространения. Наиболее четко выражены приспособления
к разносу ветром и животными. Анемохорные плоды настолько
сжаты со спинки, что становятся плоскими; кроме того, на их
краевых ребрах развиваются относительно широкие крыловидные
выросты. Такого типа плоды известны у борщевика (рис. 21, г),
ферулы, пастернака, дудника (Angelica) и др. Не менее распрост-
ранены цепкие плоды, усаженные по ребрышкам щетинками или
шипами [морковь, торилис, тургения, подлесник (Sanicula) и др.;
рис. 21, д].
Помимо зонтичных вислоплодник встречается у рода Myodo-
carpus из аралиевых (эндемичный род для о-ва Новая Каледония).
Исследование его плодов привело к заключению, что вислоплод-
ник является производным нижней сухой костянки (Baumann,
1946). Этот вывод находит подтверждение в морфологии своеоб-
разных дробных костянок некоторых крушиновых. Сухие полуниж-
ние костянки Pomaderris apetala и Trymalium helianthoides очень
мелкие (2 мм в диам.), шаровидные, с тонким пленчатым мезокар-
пием и тремя косточками. Распадение плодов происходит в пло-
скости срастания косточек на 3 части. У помадерриса при
растрескивании плода мезокарпий отделяется от косточек и оста-
ется висеть на плодоножке, опадают только косточки. Мерикарпии
трималиума представляют собой косточку с приросшими к ней
остальными слоями околоплодника. Опавшие мерикарпии раскры-
ваются (Вихирева, 1952; рис. 21, е). Более своеобразны плоды
гуании Бланше (Gouania blanchetiana). Это нижние сухие трехко-
сточковые костянки 3—4 мм в диам., снабженные тремя продоль-
ными крыловидными выростами, равными по ширине диаметру
плода. Каждое крыло образовано тканями двух соседних плодо-
листиков. Плод распадается также на 3 части в плоскости сраста-
ния косточек, при этом крылья расщепляются продольно, вслед-
ствие чего каждый мерикарпий несет по 2 крыла. Подобно
крылаткам клена, части плода гуании также повисают на вычле-
нившихся краевых пучках плодолистиков (Вихирева, 1952;
рис. 21, ж).
В отличие от дробных членистые ценокарпий очень однотипны:
к ним относятся лишь членистые стручки. Подобно членистым бо-
бам они распадаются по поперечным ложным перегородкам, вслед-
ствие чего образуются замкнутые односемянные членики. Типич-
ные членистые стручки, достигающие нескольких сантиметров
длины, известны у дикой редьки (рис. 22, а). Такого же типа
членистые стручки свойственны роду Sterigmostemum. У средне-
азиатского вида Diptychocarpus strictus нижние стручки также
являются членистыми. Более своеобразны плоды хориспоры
(Chorispora). Ее стручок можно назвать рамчатым. Поперечными
ложными перегородками он делится на двусемянные участки,
52
Рис. 22. Членистые стручки.
а — редька (Raphanus raphanistrum), б — репник (Rapistrum rugosum), в — гипекоум
(Нуресоит albescens) (а — по Вармингу, б — по: Сорные растения СССР, в — по: Жизнь
растений).
которые в свою очередь расщепляются продольно. Таким образом
по всей длине стручка формируются односемянные замкнутые по-
лучленики. После их опадения остается рамка, образованная
швами плодолистиков и поперечными перегородками стручка.
У некоторых крестоцветных плоды представляют собой члени-
стые стручочки, заключающие всего по 2—3 семени и распада-
ющиеся на соответствующее число члеников. Нечетко выражена
членистость стручочков у видов гольдбахии {Goldbachia)-, плод
репника морщинистого {Rapistrum rugosum) состоит из двух резко
различающихся односемянных члеников (рис. 22, б).
Кроме крестоцветных членистый стручок встречается у гипеко-
ума беловатого {Нуресоит albescens, Papaveraceae s. 1.; рис. 22, в)
и других видов этого рода.
По данным биохимических исследований, семейства маковых
и крестоцветных не имеют ничего общего; в современных системах
они занимают очень разное положение. Тем замечательнее кон-
вергенция в морфогенезе плодов, наблюдаемая в этих семействах:
и у маковых, и у крестоцветных мы находим стручковидную коро-
бочку, стручок типичный и членистый.
Самый своеобразный вид распадающегося плода — ценобий
(coenobium, «колония»), развивающийся из ценокарпного верх-
него двучленного гинецея. Он характеризуется сочетанием про-
дольно-кольцевых разрывов в ткани плодолистиков с поперечно-
кольцевыми у их основания. Вследствие таких разрывов от зрелого
плода отделяются 4 замкнутых односемянных фрагмента, получив-
ших название эремов (eremus, «келья»; рис. 23, а, б).
Таким образом, морфогенетически эрем отличается и от мери-
карпия, и от членика; тем более он несравним с плодиком
53
Рис. 23. Ценобии и эремы.
а — полный ценобий воробейника полевого (Buglossoides arvensis), б — тот же плод
с неопадающим эремом, в — эрем ляллеманции (Lallemantia iberica), г — эрем криво-
цвета (Lycopsls arvensis) (а, б — по Любину, в — по: Сорные растения СССР, г —
по: Флора Юго-Востока).
апокарпного многочленного плода, поскольку плодик — это произ-
водное целого плодолистика.
В редких случаях образуется диэремный ценобий, у которого
опадают 2 двусемянных эрема (Cerinthe, Boraginaceae).
Ценобий представляет собой высокоспециализированный вид
плода и, как правило, характеризует крупные таксоны. Широко'
известен такой плод в семействах губоцветных и бурачниковых;
он свойствен также вербеновым, близким к губоцветным, и неболь-
шому сем. болотниковых (Callitrichaceae). Ценобии мало варьи-
руют по размерам, отдельные эремы измеряются миллиметрами,
но они могут существенно различаться по форме и характеру
поверхности (рис. 23, в, а). У всех губоцветных при плодах
сохраняется чашечка, играющая большую роль в диссеминации
(Левина, 1957); то же наблюдается и у многих видов бурачнико-
вых, но в этом семействе известны виды и с цепкими плодами:
хорошо знакомые всем плоды липучки (Lappula), чернокорня (Су-
noglossum) и др.
При внешнеморфологической однотипности ценобии в разных
семействах существенно различаются по анатомии околоплод-
ника (Федосеева, 1935; Артюшенко, Коновалов, 1951).
СОЧНЫЕ
Среди сочных многосемянных ценокарпных плодов известны
и конвергентные виды, свойственные самым разным таксонам, и
весьма специализированные плоды, характерные лишь для одного
семейства.
Самым распространенным видом плода рассматриваемой
группы является ягода — верхняя (uva) и нижняя (Ьасса). Реже
встречаются многокосточковые костянки. Известны и однокосточ-
ковые костянки, заключающие несколько семян, например у неко-
торых пальм (Orbignya barbosiana, Phytelephas), у вербеновых
54
Таблица 5
Ягоды и многокосточковые (многосемянные) костянки у растений флоры СССР
Ягоды Костянки
верхние нижние верхние нижние
Актинидия Белокрыльник Вороний глаз Дереза (Lycium) Зверобой красиль- ный Купена Ландыш Майник Паслен Спаржа Хурма Культурны Виноград Картофель Перец овощной Томат Брусника Голубика Жимолость Клюква Омела Переступень Плющ Черника е растения Крыжовник Смородина Фейхоа (Feijoa sel- lowiana) Бархат амурский Водяника Жестер Крушина Сажереция чайная Толокнянка Аралиевые Бузина Снежноягодник
Таблица 6
Ягоды и многокосточковые (многосемянные) костянки в некоторых семействах
тропической и субтропической флоры
Ягоды Костянки
верхние нижние верхние нижние
Агасеае Buddlejaceae Caricaceae Ebenaceae Flacourtiaceae Gesneriaceae Loganiaceae Meliaceae Musaceae Oleaceae (Jastninium) Philesiaceae Resedaceae (Ochradenus) Smilacaceae Theophrastaceae Trilliaceae Araliaceae Bromellaceae Cactaceae Campanulaceae ( Canarlna) Dioscoreaceae (Tamas) Lobeliaceae Loranthaceae Melastomataceae Taccaceae Aquifoliaceae Arecaceae Davidiaceae Lennoaceae Myoporaceae Nitrariaceae Pandanaceae Rhamnaceae (Scutia indica) Tiliaceae (Grewia biloba) Araliaceae Haloragaceae Helwingiaceae Myrsinaceae (Maesa) Rubiaceae Symplocaceae
(Vitex agnus-castus), в сем. Symplocaceae. Конвергентный харак-
тер ягод и костянок явно отражается в табл. 5 и 6: видно, сколь
различным таксонам свойственны эти виды плодов.
Благодаря тщательным анатомо-карпологическим исследова-
ниям К. К. Зажурило (1935, 1936) широко принято считать
ягоды производными коробочек. Им выявлены рудиментарные
55
структуры вскрывающих механизмов в околоплодниках зверобоя
красильного {Hypericum androsaemum), волдырника, спаржи
и др. Вскрывающиеся косточки в плодах амурского бархата, по
Зажурило (1939), также следы происхождения этой костянки из
коробочки. Вскрывающиеся косточки встречаются и в плодах не-
которых крушиновых {Sageretia thea, Rhamnus alaternus, Alphito-
nia excelsa, Colletia horrida и др.). Несомненный интерес представ-
ляют вскрывающиеся ягоды. Так, у некоторых филезиевых
{Eustrephus и др.) сочная ягода после длительного полного высы-
хания вскрывается тремя створками. К верхним ягодам относят
обычно плоды каперсов. Однако плод каперса травянистого {Cap-
paris herbacea), наблюдавшийся мной в Крыму, скорее является
сочной коробочкой. Плоды этого стелющегося растения, еще бу-
дучи зелеными, раскрываются неправильной продольной трещиной
(иногда с поперечными разрывами). Относительно мясистый око-
лоплодник оказывается распластанным, почти прижатым к почве.
На фоне зеленовато-белой внутренней поверхности плода резко
выделяется тонкий слой сочной ярко-малиновой мякоти с темными,
довольно крупными (около 3 мм), почковидными семенами.
Разное происхождение ягод и сопряженная эволюция с аген-
тами диссеминации привели к исключительному внешнеморфоло-
гическому разнообразию этого вида плода. Так, ягоды майника
двулистного имеют лишь 3—4 мм в диам., а у многих видов
страстоцвета — от 3—5 до 12—22 см и являются съедобными.
Плоды дикорастущих видов дынного дерева {Carica) 4—Юсмдл.,
тогда как овально-удлиненная или грушевидная ягода культиви-
руемого в тропиках С. papaya достигает 30 см дл. и 7—15 см
в диам.
Плод банана {Musa) одноименного сем. банановых по внеш-
нему виду мало похож на ягоду привычного типа. Он удлиненно-
цилиндрический или слегка гранистый, изогнутый у основания.
Плоды культурных форм достигают более 15 см дл. и 3—4 см
в диам. Экзокарпий их кожистый и относительно толстый
(рис. 24, а). Мезэндокарпий образует очень нежную кремовую
мякоть. Выведены бессемянные сорта банана, но дикорастущие
формы имеют типичную трехгнездную ягоду с двумя рядами
семязачатков в каждом гнезде (рис. 24, б).
Как известно, у типичной ягоды сочная ткань образуется из
мезо- и эндокарпия, а у костянки — из мезокарпия. Но нередки
случаи, когда сочная ткань плодов имеет иную морфологическую
природу. Так, у ряда видов смородины {Ribes), особенно у с. чер-
ной, мякоть плода образована в значительной степени ариллусами
семян. Сочность плодов крыжовника {Grossularia) также опреде-
ляется некоторым участием строфиолей семян (Комар, 1965).
В сочных плодах пасленовых (картофель, томат) основу их мякоти
составляют разросшиеся плаценты (рис. 24, в). Попутно отметим,
что ягоды пасленовых являются двугнездными. Многокамерность
плодов большинства культурных сортов томата возникает вслед-
ствие увеличения числа плодолистиков в результате срастания
56
Рис. 24. Ягоды банана и томата.
а — плод банана (Musa sp.), на верхушке удален экзокарпий, б — его поперечный срез,
в — поперечный срез плода томата (Lycopersicon esculentum) (по Троллю).
Рис. 25. Косточки.
а — крушина ломкая (Frangula alnus),
б — жестер (Rhamnus cathartica) (по
Левиной, 1967а).
Рис. 26. Специализированные сочные плоды.
а — плод лимона на поперечном срезе, б — отпрепарированный плод яблони, в — отдель-
ный плодолистик с семенем, г — плод граната (Punica granatum) в продольном разрезе
(а—в — по Троллю, г — по Веттштейну).
57
цветков (Данилова, 1952). Таким образом, многокамерный плод
томата, строго говоря, является соплодием.
У нижних ягод и костянок в случае осевого происхождения
нижней завязи мякоть плода также может развиваться из тканей
оси цветка. На основании исследования нижней завязи у жимоло-
стных 3. Т. Артюшенко (1951) называет костянки этого семейства
(бузина, снежноягодник) аксиальными. Такое понятие, вероятно,
приложимо к ягодам и костянкам некоторых видов кактусовых.
Независимо от происхождения сочной ткани костянки разных
видов могут быть внешне очень сходными, но, как и апокарпные
костянки, они четко различаются по косточкам (рис. 25).
К специализированным видам рассматриваемой группы плодов
относятся померанец, яблоко, тыквина и гранатина.
Верхний сочный плод цитрусовых, хорошо известный у лимона,
апельсина, мандарина и др., так называемый померанец, или геспе-
ридий (hesperidium), весьма своеобразен. Для него характерен
плотный кожистый экзокарпий с огромным количеством желёзок,
заполненных эфирным маслом. Мезокарпий более или менее губча-
тый, слаборазвитый. Эти слои околоплодника различаются не
только по консистенции, но и по окраске. Экзокарпий, окрашен-
ный каротиноидами в желтый или оранжевый.цвет, получил назва-
ние «флаведо», в отличие от белого мезокарпия — «альбедо».
Тонкая кожица, непосредственно окружающая сочные «дольки»
плода, по-видимому, представляет собой эндокарпий. Съедобная
мякоть плодов цитрусовых образована сильно разросшимися и
заполненными жидким соком волосками внутренней эпидермы пло-
долистиков. По мере созревания плодов эти волоски плотно запол-
няют всю полость завязи (рис. 26, а).
Все остальные названные специализированные плоды являются
нижними. Морфологическая природа их сочной съедобной ткани
совершенно различна.
Яблоко (pomum) представляет собой многолистовку, оброс-
шую мясистой тканью цветочной трубки (гипантия). При этом
дифференцируется и околоплодник: эндокарпий оказывается жест-
ким, хрящеватым и выстилает гнезда плода; наружные ткани
плодолистика становятся мясистыми и совершенно сливаются
с тканями цветочной трубки (рис. 26, б, в). В учебной и научной
литературе нередко указывается, что сочная ткань яблока осевого
происхождения, т. е. представляет собой вогнутое цветоложе. Но
сравнительно-морфологические исследования приводят к заключе-
нию, что основная масса плода у яблока образована цветочной
трубкой (Кузнецов, 195'1; Имс, 1964). Яблоко характерно для под-
сем. яблоневых (Maloldeae) сем. Rosaceae s. 1.
Типичное яблоко известно у яблони, груши, рябины, айвы
(Cydonia), эриоботрии (Eriobotrya) и др. Но в некоторых случаях
плодолистики образуют твердую одревесневшую косточку, заклю-
чающую семя. Такой плод можно назвать костянковидным ябло-
ком (Левина, 1961); он встречается у боярышника (Crataegus),
кизильника (Cotoneaster), мушмулы (Mespilus germanica).
58
Не менее своеобразен плод тыквенных, получивший название
тыквины (реро). Отличительным признаком ее является твердый,
иногда очень прочный экзокарпий и сильно развитый мясистый
мезокарпий. Полость плода заполнена разросшимися (чаще всего
тремя) плацентами, обычно очень сочными. Съедобная мякоть
плода арбуза представляет собой плаценты. В значительной сте-
пени это относится и к плодам огурца.
Плоды бутылочной тыквы — горлянки (Lagenaria) отлича-
ются не только формой (колбовидной или грушевидной), но глав-
ным образом очень твердым водонепроницаемым экзокарпием. На
этом основано использование сухих плодов в качестве сосудов
для воды. Мезокарпий удлиненно-цилиндрических плодов люффы
(Luffa) снабжен мощно развитым сосудистым скелетом, ради
которого и культивируются некоторые виды люффы (L. cylindrica,
L. acutangula), дающие хорошо известную мочалку. Зрелые плоды
люффы вскрываются на верхушке колпачком, т. е. ламинально
поперечно-кольцевым способом.
У некоторых тыквенных развиваются крайне своеобразные
сочные вскрывающиеся плоды. К моменту их созревания в клетках
внутренних слоев околоплодника возникает очень высокое осмоти-
ческое давление, благодаря которому семена выбрасываются с си-
лой на довольно значительные расстояния (Ecballium, Cyclan-
thera, Momordica).
Плод монотипного сем. гранатовых (Punicaceae) настолько
специфичен, что получил название по роду растения — гранатина
(balausta). Он развивается из нижней завязи,.имеет сухой, кожи-
стый, очень плотный околоплодник, раскрывающийся неправиль-
ными трещинами. Гнезда плода плотно заполнены крупными семе-
нами с ярко-красной очень сочной кожурой, которая и составляет
съедобную часть плода. Весьма своеобразно протекает его разви-
тие у культивируемого вида Punica granatum-, вследствие-разра-
стания цветоложа плодолистики наружного круга смещаются
вверх и оказываются над плодолистиками внутреннего круга.
Гинецей, таким образом, становится двухъярусным, причем в ниж-
нем его круге плацентация центрально-угловая, а в верхнем она
переходит в париетальную. Плод граната в продольном разрезе
двухъярусно-четырехгнездный (рис. 26, г).
ОДНОСЕМЯННЫЕ
Виды односемянных ценокарпных плодов многочисленны и
крайне разнородны по^ своей морфологической природе и систе-
матической принадлежности. НО' все они, по-видимому, являются
производными многосемянных плодов. На это указывают и одно-
семянные вскрывающиеся плоды в некоторых семействах, и, напро-
тив, невскрывающиеся 2—3-семянные, которые встречаются на-
ряду с односемянными плодами у одного и того же вида или
59
рода. Еще чаще в завязи односемянного плода закладываются
2—3—несколько семязачатков, из которых только один развива-
ется в семя, а остальные дегенерируют. Таким образом, только
один плодолистик оказывается фертильным. Стерильные же плодо-
листики редуцируются в разной степени, и их участие в формирова-
нии плода может оказаться совсем незначительным. Так возни-
кают плоды, получившие название псевдомономерных (Eckardt,
1937, 1938). Псевдомономерность гинецея и плода — явление
широко распространенное: Эккардт называет около 40 семейств
с таким гинецеем.
Полное исчезновение рудиментов стерильных плодолистиков
приводит к образованию гинецея, который мы назвали вторично-
мономерным; соответствующие им плоды (рогозовых, злаков)
мы отнесли к типу апокарпных. Все псевдомономерные плоды
рассматриваются как ценокарпии. Сочные односемянные плоды
этого типа достаточно однообразны: чаще это однокосточковая
костянка (верхняя или нижняя), реже односемянная ягода. Сухие
плоды, напротив, крайне разнородны, в то же время их типология
и номенклатура разработаны совершенно недостаточно. Вот по-
чему один и тот же плод липы — это и односемянный орешек
(Веттштейн, 1912), и сухая костянка (Артюшенко, Коновалов,
1951), и орех (Strasburger et al., 1962; Васильев, 1981), и карце-
рула (Каден, 1965), и, наконец, просто невскрывающийся плод
(Engler, 1964; Тахтаджян, 1966). Аналогичный перечень названий
можно привести для плодов маревых, осоковых и многих других.
В цитированной работе 3. Т. Артюшенко и И. Н. Коновалов
предлагают типологические критерии для сухих односемянных
плодов. Авторы относят к орехам все плоды, у которых склери-
фицированы наружные слои перикарпия (или хотя бы наружная
эпидерма); к сухим костянкам — плоды со склерифицированной
внутренней эпидермой или вообще внутренними слоями перикар-
пия. Семянкой авторы именуют только такой плод, у которого
весь перикарпий состоит из клеток с целлюлозными оболочками.
Однако авторы не всегда придерживаются своих критериев. Они
называют орехом и плод гречишных с одревесневшим экзокар-
пием, и плод осоковых, у которого наружная эпидерма с цел-
люлозными оболочками. Кроме того, семянка в понимании авто-
ров — очень редкий вариант плода. У общепризнанных семянок
сложноцветных склерификация слоев или отдельных участков пе-
рикарпия наблюдается очень часто (Александров, Савченко, 1951;
Kaussmann, 1963; Бойко, 1972; Меликян, Мурадян, 1975; Ханд-
жян, 1983).
В цитированных работах показано, насколько различны се-
мянки по анатомическому строению околоплодника. Как уже отме-
чалось ранее, разнятся анатомически и такие морфологически
специализированные плоды, как вислоплодник и ценобий. По-
видимому, детали анатомического строения перикарпия должны
служить основанием для выделения вариаций в пределах таких
конвергентных морфологических видов плода, как орех, семянка
60
или костянка, а разграничение этих карпологических видов целе-
сообразно основывать главным образом на внешнеморфологиче-
ских критериях.
В отношении костянки важен и такой признак, как эколого-
функциональная дифференциация зон перикарпия. У костянок
функцию защиты семени выполняет эндо- или мезэндокарпий,
образующий косточку, а остальные слои околоплодника специали-
зируются в связи со способом диссеминации — эндозоохорией
(сочные), синзоохорией, гидро- или анемохорией (сухие). Поэтому
к костянкам мы будем относить только такие плоды, в которых
эндокарпий образует косточку, четко отграниченную от остальных
слоев околоплодника.
У ореха и семянки много общих признаков: это плоды цено-
карпные, односемянные, сухие, невскрывающиеся, с околоплод-
ником, легко отделяющимся от семени. Различаются они степенью
склерификации околоплодника: у ореха он сильно склерифици-
рован, очень твердый, ломается, распадаясь на куски. У семянки
околоплодник кожистый, относительно тонкий, сгибается не ло-
маясь.
Названные здесь признаки различий костянки, ореха и семянки
будут приняты в дальнейшем изложении.
СУХИЕ ВЕРХНИЕ И НИЖНИЕ
Своеобразным видом плода этой группы является односемян-
ная коробочка, развивающаяся из верхней завязи. Такой плод
довольно обычен в семействах плюмбаговых (Plumb aglnaceae s. 1.)
и амарантовых (Amaranthaceae). У видов Plumbago и Cerato-
stigma коробочка вскрывается пятью створками, а у Acantholi-
топ — и створками, и поперечно-кольцевой щелью (крышечкой).
Большинству видов щирицы (Amaranthus) также свойственны
односемянные коробочки, вскрывающиеся крышечкой (рис. 27, а).
Односемянные коробочки можно встретить и в сем. маревых
(свекла, габлиция — Hablitzia tamnoides). Своеобразен псевдомо-
номерный односемянный плод казуарины (Casuarina): его кожи-
стый околоплодник образует крыловидный вырост, но зрелый плод
вскрывается двумя створками. Удлиненная коробочка с одним
крупным семенем характерна для палеотропического рода эгице-
рас (Aegiceras), выделяемого в особое подсемейство сем. мирси-
новых (Myrslnaceae).
Вскрывание односемянных плодов можно рассматривать как
рудиментарную функцию, теряющую свое биологическое значение.
Обычно при односемянности диаспорой служит не семя, а весь
плод, опадающий нередко еще с дополнительным покровом из
околоцветника, прицветников и т. п.
Наиболее распространенным видом односемянного сухого
ценокарпия является семянка или сходный с ней семянковидный
плод. По признакам, названным ранее, к верхним семянкам и
61
Рис. 27. Односемянные ценокарпий.
а — коробочка ширицы (Amaranthus retroflexus); б—е—плоды гречишных: б — горца
(Potygonum linicola), в — щавеля (Rumex crispus), г — ревеня (Rheum turkestanicum),
д — джузгуна безлистного (Calligonum aphyllum), е — д. голова медузы (С. caput-medu-
sae) (а — по Маевскому, б, в — по: Сорные растения СССР, г, е—по Коровину, д —
по Бушу).
семянковидным плодам мы относим плоды гречишных (Polygo-
пасеае), маревых (Chenopodiaceae), ряда осоковых (Сурегасеае),
невскрывающиеся плоды амарантовых, плюмбаговых и др.
Гинецей гречишных образован чаще всего тремя, иногда двумя
и реже четырьмя плодолистиками. Их числом определяется форма
семянки: трех-, двух- или четырехгранная. У богатого видами рода
Polygonum при плодах сохраняется засыхающий околоцветник,
который полностью или лишь частично закрывает семянку
Трис. 27, б). Плоды курчавки (Atraphaxis) и щавеля (Rumex) ок-
ружены сильно разросшимися листочками внутреннего круга око-
лоцветника, которые служат анемо- и гидрохорным приспособле-
нием (рис. 27', в). Особенно хорошо плавают плоды щавеля, у ко-
торых на листочках околоцветника развиваются особые вздутия
(«желвачки») из губчатой паренхимы. У древесных растений нео-
тропического рода Coccoloba околоцветник при зрелых плодах
становится мясистым и является, по-видимому, приманкой для
птиц или других животных.
Более своеобразны голые плоды ревеня (Rheum) и особенно
джузгуна (Calligonum), снабженные крыловидными или иного
типа выростами околоплодника. Трехгранные семянки ревеня не-
сут на каждом ребре по широкому крылу (рис. 27, г). У джузгуна
безлистного (С. aphyllum) и некоторых других видов на каждом из
четырех ребер плода развивается двойное крыло, так что весь
плод снабжен восемью меридиональными относительно широкими
62
Рис. 28. Соплодие осоки вздутой (Carex physodes) (по:
Флора СССР).
крыльями (рис. 27, д). Для плодов джузгуна
характерны частые, радиально расположенные,
упругие, тонкие выросты, разветвленные на I
концах (рис. 27, е). Семянки, покрытые такими .
выростами, представляют собой геометрически II |
правильный шарик, очень легкий и упругий. Л S
Они с большой скоростью переносятся ветром \ /
по поверхности песка — обычного субстрата Ц
местообитаний этих видов. Многие виды джуз- \ Ш
гуна разводят как декоративные кустарники: их \
ажурные шаровидные плоды, достигающие 3— Mil
5 см в диам., бывают ярко окрашены (красные, WII
розовые, зеленые) и долго сохраняются на вет- Ж
ВЯХ.
К верхним семянкам мы относим и плоды
осок с тонким кожистым околоплодником. Плод осоки (Carex)
заключен в замкнутый мешочек, образованный кроющим листом.
Размеры, форма и детали строения мешочков служат диагности-
ческими признаками для распознавания видов осоки. Нередко
мешочки по объему заметно превышают размеры самого плода,
и в этом случае они служат хорошим плавательным приспособле-
нием (С. atherodes, С. pseudocuperus, С. vesicaria и др.). Особенно
сильно вздуты мешочки при плодах осоки вздутой (С. physodes) —
характернейшего растения песчаных пустынь Средней Азии. Они
овальные, достигающие в длину 2 см (рис. 28), очень легкие,
благодаря чему даже слабым ветром далеко перекатываются
по поверхности песка.
У некоторых лесных видов осок (С. digitata, С. montana, С. orni-
thopoda) базальная часть мешочков становится сочной и функцио-
нирует как элайосом, привлекающий муравьев — агентов расселе-
ния. Оранжево-красные и более или менее мясистые мешочки
С. baccans способствуют орнитохорному распространению
этого вида.
Своеобразные плоды некоторых тропических осоковых будут
описаны среди других видов плодов.
Наименование плодов маревых — самое неопределенное. Ранее
уже было показано, что название «мешочек» ничем не оправ-
дано. Едва ли можно считать правильным и наименование «орех»
(Eckardt, 1964; Тахтаджян, 1966, и др.). Скорее можно согла-
ситься с 3. Т. Артюшенко и И. Н. Коноваловым (1951), опреде-
ляющими плод маревых как семянку. Однако от типичной семянки
он отличается пленчатым околоплодником (вместо кожистого)
и плотной частично склерифицированной семенной кожурой.
Поэтому будем называть плод маревых семянковидным верхним
ценокарпием.
Редукция околоплодника у маревых, очевидно, связана с тем,
63
Рис. 29. Плоды маревых.
а — кохия (Kochia laniflora), б — климакоптера (Сlimacoptera lanata), в — сведа (Suaeda
baccifera), г — бассня (Bassia hyssopifolia), д — лебеда (Halimione pedunculata), е —
терескен (Ceratoides pungens) (a — по: Сорные растения СССР, остальные — по: Флора
Юго-Востока).
что их плоды бывают окружены покровом из прицветников или
листочков околоцветника. Морфологическое разнообразие плодов
маревых и определяется главным образом характером их вторич-
ного покрова (рис. 29).
Для большинства родов маревых характерны плоды в около-
цветнике. Иногда он сохраняется таким же, как при цветении
(почти все виды Chenopodium), но чаще всего заметно видоизме-
няется, образуя широкие крыловидные выросты, как у кохии
(Kochia), солянки (Salsola), некоторых видов сведы (Suaeda),
саксаула (Haloxylon) и ряда других. Реже околоцветник при
плодах становится сочным и даже яркоокрашенным (Chenopo-
dium foliosum, Suaeda baccifera) или образует колючие, иногда
цепкие выросты (Bassia, Horaninovia, Spinacia).
Плоды лебеды (Atriplex), рогача (Ceratocarpus), терескена
(Ceratoides) заключены в прицветники, как правило, разраста-
ющиеся к моменту созревания плодов. В одних случаях они
срастаются лишь у самого основания, но нередко образуют совер-
шенно замкнутое вместилище плода своеобразной формы. За-
метно отличаются от других маревых плоды свеклы (Beta):
полунижние, с хрящеватым околоплодником, вскрывающиеся (при
набухании) крышечкой. У борщовии (Borszczowia) плод оказы-
вается нижним вследствие полного срастания околоцветника
с перикарпием; при этом у зрелых плодов околоцветник стано-
вится мясистым.
Образование соплодий у маревых и весьма характерное для
них явление разноплодия будут рассмотрены в следующих
главах.
Семянковидные плоды некоторых гвоздичных и плюмбаговых
часто окружены сохраняющимся околоцветником или внецветко-
64
Рис. 30. Односемянные плоды, окруженные дополнительными покровами.
а — дивала (Scleranthus annuus), б — армерия (Armeria maritima), в — плюмбаго (Plum-
bago capensis) (а — по: Сорные растения СССР, б, в — по Броуверу).
выми элементами, которые нередко сильно разрастаются и видо-
изменяются к моменту созревания плодов. Так, плод дивалы опа-
дает вместе с плотно охватывающим его вогнутым отвердевшим
цветоложем и сидящей на нем чашечкой (рис. 30, а). Плоды мно-
гих родов плюмбаговых (Armeria, Limonium, Plumbago, Acantho-
litnon) заключены в чашечку с разрастающимися зубцами, пере-
пончатой воронковидной каймой или усаженную железистыми
волосками (рис. 30, б, в).
К верхним семянкам или семянковидным плодам относятся
невскрывающиеся плоды амарантовых.
Среди нижних семянок первое место по праву принадлежит
плодам сложноцветных. При однотипности их морфологической
природы они крайне разнообразны внешне (размеры, форма,
скульптура поверхности, характер придатков) и по анатомии пери-
карпия и семенной кожуры.
Как показано в цитированных ранее и других работах, семянки
сложноцветных различаются и расположением механической
ткани в различных зонах перикарпия, и соотношением толщины его
и семенной кожуры. Обычно функция защиты зародыша выполня-
ется преимущественно одним из этих покровов, а другой реду-
цируется. Так, у сушеницы топяной (Gnaphalium uluginosum)
слои околоплодника сильно смяты, весь он очень тонкий в отличие
от хорошо развитой семенной кожуры. У подсолнечника, особенно
культурного, напротив, облитерирована семенная кожура, а пери-
карпий толстый, многослойный, почти весь склерифицированный.
Поэтому плод культурного подсолнечника правильнее было бы
называть орехом. Реже, например у козлобородника (Tragopo-
gori), при сильно развитом, значительно склерифицированном око-
лоплоднике полностью сохраняется и многослойная семенная
кожура.
Внешнеморфологическое разнообразие плодов сложноцветных
в значительной степени определяется характером тех или иных
придатков на плоде. Наиболее распространенным типом придатка
является хохолок волосков, сидящий либо непосредственно на
5 Р. Е. Левина
65
Рис. 31. Семянки сложноцветных.
а — одуванчик (Taraxacum erythrospermum), б — василек (Centaurea nigra), в — суше-
ница (Gnaphalium sylvaticum), г — бодяк (Cirsium arvense) (а—в — по Броуверу, г —
по: Сорные растения СССР).
верхушке семянки, либо приподнятый на так называемом носике,
иногда сильно вытянутом (рис. 31, а). Степень развития хохолка
очень различна: от едва заметных коротких щетинок до пышного
султана волосков, превышающих во много раз семянку (рис. 31,
б—г).
О морфологической природе хохолка сложноцветных сущест-
вует большая литература, и мы не будем останавливаться на
этом вопросе. Отметим только, что у некоторых сложноцветных
верхушка плода несет хорошо развитые листочки, которые можно
принять за чашечку (рис. 32, а). Но чаще сохраняется коронка
из мелких зубчиков или кольцевое утолщение в виде гладкого
валика; наконец, семянки могут быть полностью лишены каких-
либо придатков (рис. 32, б—г).
Хохолок из волосков служит весьма совершенным анемохорным
приспособлением. Но известны семянки с цепкими придатками:
различные виды череды (Bidens; рис. 33, а) или кельпинии (Koelpi-
nia linearis-, рис. 33, б). В последнем примере прицепки несомненно
представляют i собой выросты околоплодника. Их не следует
смешивать с колючими, крючковидно загнутыми листочками
обертки дурнишника (Xanthium).
Нижней семянкой является также плод ворсянковых (Dipsaca-
сеае). От семянки сложноцветных он существенно отличается
тем, что бывает окружен (обернут) сросшимися прицветными
листьями, образующими на верхушке плода перепончатую ворон-
ковидную окраину. Обычно ее называют наружной чашечкой.
Над нею сохраняется настоящая (так называемая «внутренняя»)
чашечка из 5 сросшихся у основания щетинковидных листочков
(рис. 34).
66
Рис. 32. Редукция чашечки на семянках сложноцветных.
а — урсиння (Ursinia pulchra), б—цикорий (Cichorium intybus), в — ромашка (Chamo-
milla matricarioides), г — полынок (Artemisia austriaca) (а — по Пидоттн, б — по Броу-
веру, в, г — по: Сорные растения СССР).
Рис. 33. Цепкие семянки сложно-
цветных. '
а — череда (Bidens tripartita), б —
кельпиння (Koelpinia linearis) (по:
Сорные растения СССР).
Рис. 34. Плод скабиозы
(Scabiosa maritima) (по
Броуверу).
5*
67
Рис. 35. Крылатки.
а — птелея (Ptelea trifoliata), б — ясень (Fraxinus pennsylvanica), в — береза (Betula
sp.) (а — по: Жизнь растений, б — по: Деревья и кустарники СССР).
К нижним семянкам следует относить и плоды валериановых
(Valerianaceae). Их называют орехом или сухой костянкой, но оба
эти названия неоправданны. В околоплоднике валерианы склери-
фицирован лишь внутренний эпидермис, не образующий обособ-
ленной косточки.1
Из семейств тропической флоры с плодом нижней семянкой
можно назвать небольшое южноамериканское сем. калицеровых
(Calyceraceae). Они близки к сложноцветным, что отражается и
в морфологии плодов, несущих на верхушке лопасти чашечки, или
снабженных шиповатыми выростами. У некоторых калицеровых,
как и у сложноцветных, наблюдается резкая гетероморфность пло-
дов в пределах корзинки.
Семянки с крыловидными выростами околоплодника широко
принято называть крылаткой. Выделение крылатки в особый вид
плода неоправданно; ведь семянки с хохолками или прицепками
не имеют специальных названий. Но поскольку «крылатка» по-
стоянно встречается в научной, учебной и популярной литературе,
это название сохраняется в нашей классификации как вариация
семянки.
Верхние крылатки с широким крылом, окружающим гнездо
завязи, хорошо известны у разных видов вяза {UImus').
Внешне очень сходна с плодами вяза крылатка птелеи (Ptelea
trifoliata, Rutaceae), но их сходство поверхностное. Эти плоды
различаются не только характером васкулярного скелета (Зажу-
рило, 1945), но и более существенно: плод вяза односемянный,
псевдомономерный (Eckardt, 1937; Черник, 1981), а у птелеи —
двугнездный и нередко двусемянный (рис. 35, а). У эвкоммии
ильмовидной (Eucommia ulmoides, Eucommiaceae) крылатка одно-
семянная, хотя завязь двугнездная с двумя семязачатками. Зре-
1 Н. Н. Каден (1965) восстанавливает для плода валериановых название
«аггедула».
68
лый плод удлиненный (3—4 см дл. и 0.6—1.2 см шир.), слегка
асимметричный, с относительно узкими крыльями.
Верхние крылатки с удлиненным однобоким крылом свойст-
венны видам ясеня (Fraxinus-, рис. 35, б), фонтанезии (Fontanesia,
Oleaceae).
Нижняя крылатка с двумя симметричными крыльями —
хорошо известный плод березы (рис. 35, в). Менее развиты крылья
у плодов ольхи.
Орехи и ореховидные плоды, как и семянки, встречаются
в самых различных семействах покрытосемянных, следовательно,
также могут возникать конвергентно. Вместе с тем в одном и
том же семействе могут быть представлены и семянки, и орехи,
как например в сем. ильмовых. Так, очень мелкие орехи несколь-
ких миллиметров в диаметре свойственны видам дзельквы (Zel-
kova). Своеобразный крылатый орех развивается у филлостилона
бразильского (Phyllostylon brasiliense) из того же семейства
(рис. 36, а). В близких к ильмовым семействах крапивных
(Urticaceae) и коноплевых (Cannabaceae) также формируются
очень мелкие ореховидные плоды.
К верхним плодам этой же группы относятся довольно широко
распространенные ореховидные стручочки крестоцветных. Это не-
вскрывающиеся, обычно одногнездные и односемянные плоды
с очень твердым склерифицированным околоплодником. Они из-
вестны у свербиги (Вunias), таушерии (Тauscheria), миагрума
(Myagrum), неслии (Neslia) и др. В плодах неслии и свербиги
нередко развиваются 2 семени, а невскрывающиеся плоды Eucli-
dium всегда являются двугнездными с одним семенем в каждом
гнезде. Размеры ореховидных стручочков невелики, измеряются
миллиметрами, а форма и характер поверхности очень различны
(рис. 36, б—г). Несколько своеобразен плод катрана (Crambe).
Он представляет собой стручочек, состоящий из двух члеников.
Верхний членик — односемянный, невскрывающийся, относи-
тельно крупный, нижний — очень мелкий, рудиментарный, бесплод-
ный (рис. 36, д). В некоторых случаях односемянные стручочки бы-
вают снабжены хорошо развитыми крыловидными выростами, как
например у различных видов вайды (Isatis; рис. 36, е). Помимо
крестоцветных верхний ореховидный плод свойствен также дымян-
кам (Fumaria).
Как уже отмечалось ранее, очень разноречиво наименование
плода липы. Морфологически он ближе всего к верхним орехо-
видным плодам. От типичного ореха его отличают неполная
склерификация околоплодника и хорошо развитая семенная ко-
жура.
Своеобразные, мелкие, яркоокрашенные орехи свойственны не-
которым тропическим и субтропическим видам осоковых из родов
Gahnia и Scleria. По своей экологии это эндоорнитохорные плоды,
хотя гания не предлагает своим агентам никаких съедобных
тканей. У основания плодов склерии разрастается яркоокрашен-
ный мясистый гинофор.
69
Рис. 36. Ореховидные плоды.
а — филлостнлон (Phyllostylon brasilien.se), б—е—ореховидные стручочки крестоцвет-
ных: б — миагрума (Myagrum perfoliatum), в — неслни (Neslia paniculata), г — эуклиди-
ума (Euclidium syriacum), д — катрана (Crambe maritima), е — вайды (Jsatis tinctoria)
(а — по: Жизнь растений, б—г—по: Сорные растения СССР, д, г — по Вармингу).
Рис. 37. Орехи и желуди.
а, б— плоды лещины обыкновенной и л. маньчжурской (Corylus avellana, С. mandshu.-
rica), в — граба (Carpinus betulus), г — желудь дуба грузинского (Quercus iberica), д —
соплодие каштана настоящего (Castanea sativa) (а, в, д — по Троллю, б, г — по: Деревья
и кустарники СССР).
70
Наиболее типичным орехом является нижний плод лещины
(Corylus). Он характеризуется полной склерификацией всех слоев
околоплодника (см. рис. 2, д). Плюска, окружающая орех лещины
обыкновенной (С. avellana), образована прицветниками, которые
сохраняют свою листовую природу (рис. 37, а). У других видов
(С. cornuta, С. mandshurica, С. tubulosa) плюска кувшинчатой
формы, полностью скрывающая орех (рис. 37, б). Наконец,
в более редких случаях плюска густо усажена острыми длинными
колючками, как у л. устрашающей (С. ferox). Несколько иного
типа плод граба (Carpinus). Его обратногрушевидный продольно-
ребристый орех достигает нескольких миллиметров в диаметре
и сидит у основания открытой, листовидной, обычно трехлопастной
плюски (рис. 37, в)'. У хмелеграба (Ostrya carpinifolia) мелкие
(несколько мм дл.) орехи заключены в совершенно замкнутую
пленчатую плюску.
Среди нижних ореховидных плодов наиболее специализирован-
ный вид плода — желудь (glans), характерный для сем. буковых
(Fagaceae). От типичного ореха он отличается более тонким око-
лоплодником и сложным происхождением плюски, возникающей
от сращения укороченных осей редуцированного соцветия с много-
численными чешуйчатыми прицветниками. Но желудь, как и орех,
развивается из многогнездного гинецея, у которого фертильны
только одно гнездо и один семязачаток. Плоды разных родов
буковых очень различны по своей морфологии. Желуди дуба
варьируют по форме и размерам не только у разных видов
(Меницкий, 1984). Даже в одной ценопопуляции дуба обыкновен-
ного (Quercus robur) наряду с типичными овальными плодами
встречаются более или менее шаровидные или, напротив, удлинен-
ные заостренно-цилиндрические. Более устойчивым видовым при-
знаком является характер плюски. Чаще она мелкочешуйчатая,
шероховатая с наружной поверхности и охватывает плод лишь
у основания (Q. robur, Q. ilex и мн. др.). Но у иных видов
плюска отличается многочисленными длинными ланцетными че-
шуями и окружает до половины шаровидный плод (Q. dentata,
Q. iberica; рис. 37, г).
У бука (Fagus) и каштана (Castanea) в одной плюске разви-
ваются по 2—3 желудя. Таким образом, плюска с заключенными
в ней плодами представляет собой соплодие. Желуди бука, трех-
гранные, с острыми ребрами, и плоды каштана, гладкие, плоско-
выпуклые или сжато-шаровидные, полностью скрыты в плюске,
но при созревании она растрескивается и освобождает плоды.
Плюска каштана густо покрыта длинными очень жесткими
острыми колючками (рис. 37, д).
Нижние ореховидные плоды свойственны и травянистым расте-
ниям, например в сем. санталовых (Santalaceae). Представленные
в нашей флоре виды ленца (Thesium) образуют очень мелкие
(около 3 мм) овальные плоды с твердым склерифицированным
околоплодником.
Обзор односемянных сухих ценокарпиев закончим описанием
71
Рис. 38. Плод кокосовой пальмы (Cocos nucifera) в продольном разрезе (а), ко-
сточка— вид снизу (б) и в поперечном разрезе (в) (по: Жизнь растений).
сухих костянок. Хбтя не всегда легко разграничить сухую и сочную
костянки, первая нередко встречается в самых различных семей-
ствах и ее следует считать особым видом плода.
Под сухой костянкой мы понимаем такой плод, околоплодник
которого в зрелом состоянии не содержит сочных тканей, а эндо-
карпий образует четко обособленную косточку. Сухие костянки
чаще всего являются гидрохорными или синзоохорными диаспо-
рами.
Верхние псевдомономерные сухие костянки свойственны мно-
гим видам пальм. Среди них хорошо известен плод кокосовой
пальмы (Cocos nucifera), называемый обычно кокосовым орехом.
Он отличается очень крупными размерами (до 30 см дл. и 20 см
в диам.), четкой дифференциацией слоев околоплодника и особым
характером эндосперма единственного семени. Завязь кокосовой
пальмы образована тремя плодолистиками. Первоначально она
трехгнездная с одним семязачатком в каждом гнезде. Но 2 гнезда
облитерируются и обнаруживаются лишь в виде небольших щелей
в ткани эндокарпия (Имс, 1964). Зрелый плод одногнездный
и односемянный. Его экзокарпий относительно тонкий, кожистый,
но очень плотный и гладкий. Мезокарпий мощно развитый, сухой,
волокнистый, достигает в толщину нескольких сантиметров. Эндо-
карпий состоит из очень твердых каменистых клеток. На нем выда-
ются 3 меридиональных гребня — следы срастания плодолистиков,
а у основания имеются 3 зародышевых отверстия, 2 из них зара-
стают, а третье служит местом выхода зародышевого корня.
Эндосперм семени очень богат маслом, первоначально он жидкий,
но к моменту созревания часть эндосперма твердеет, образуя слой
до 2 см толщ., прилегающий к эндокарпию (рис. 38).
Сходное строение имеет гигантский плод знаменитой сейшель-
ской пальмы (Lodoicea maldivica), достигающий массы 13—18 кг.
От кокосового «ореха» он отличается менее развитым мезокар-
пием (2—2.5 см толщ.) и двулопастными косточкой и семенем.
В некоторых плодах развиваются 2—3 семени. Сухие костянки
с волокнистым мезокарпием известны и у других видов культиви-
72
руемых пальм: бетелевой (Areca catechu) и пальмиры (Borassus
flabellifer), волокно плодов которой имеет техническое применение.
Как будет показано в V главе, названные здесь плоды пальм по
своей морфологии являются отличными «мореплавателями». То же
можно сказать и о костянках некоторых хризобалановых (Chry-
sobalanaceae). У прибрежных и водных травянистых растений
также известны сухие костянки, обладающие плавучестью. Так,
в сем. осоковых у тропических родов Марата и Scirpodendron
развиваются костянки с губчатым мезокарпием. Из видов нашей
флоры детально изучены аналогичные плоды ежеголовника (Spar-
ganium). Они характеризуются относительно мощным каменистым
эндокарпием; мезокарпий — из рыхло расположенных паренхим-
ных клеток; экзокарпий плотный, с механическим слоем склерифи-
цированной гиподермы (Зубкова, Шабес, 1983).
Своеобразная сухая крылатая костянка встречается у каркасо-
вых (Celtidiaceae)—птероцельтиса (PteroceUis); крылья явля-
ются выростами эндокарпия.
Наконец, отметим верхние сухие костянки в сем. анакардиевых
(Anacardiaceae). Фисташка настоящая (Pistacia vera) образует
удлиненно-овальные плоды 8—24 мм дл. и 6—14 мм шир. и дает
съедобные семена со светло-зелеными семядолями. Косточки этих
плодов часто раскрываются по шву. У других видов фисташки
плоды шаровидные, около 5—6 мм в диам. Такие же мелкие
костянки у скумпии (Cotinus) и сумаха (Rhus). Плоды послед-
него — со смолистым мезокарпием, опушенные железистыми во-
лосками.
Нижние сухие костянки характерны для сем. ореховых (Juglan-
daceae). Наиболее известны среди них плоды разных видов ореха
(Juglans), в особенности грецкого (/. regia). Их женские цветки
окружены плюской из двух прицветников, которые срастаются
с завязью и участвуют в формировании наружных слоев около-
плодника. Экзомезокарпий представляет собой относительно
мясистую ткань толщиной в несколько миллиметров, которая рас-
трескивается неправильными трещинами и быстро сгнивает или
целиком присыхает к эндокарпию. Эндокарпий сильно склерифи-
цированный, очень твердый, образует более или менее бугристую
косточку с двумя—несколькими сильно выступающими ребрами.
Косточку с заключенным в ней семенем и называют орехом. Семя
такого «ореха» имеет причудливую форму: двумя неполными пере-
городками завязи оно разделено на 4 более или менее обособлен-
ных участка, а складчатость семядолей обусловливает их крайне
неровную бугристо-бороздчатую поверхность. Косточки («орехи»)
различных видов Juglans варьируют по размерам, очертанию,
характеру поверхности «скорлупы» и по числу ребер, которые
соответствуют числу плодолистиков.
Плоды карии (Сагуа) сходны с плодами Juglans. Они отлича-
ются лишь тем, что наружная часть околоплодника к моменту
созревания древеснеет и растрескивается четырьмя створками.
У других представителей семейства развиваются сухие крылатые
73
Рис. 39. Сухие крылатые костянки.
а — лапина (Pterocarya fraxinifolia), б — энгельхардия (Engelhardia wallichiana)
(по:. Жизнь растений).
костянки, иногда очень мелкие. Так, плоды платикарии (Platy-
сагуа) достигают всего 5 мм и собраны в шишковидное соплодие
до 40 мм дл. Виды лапины (Pterocarya) образуют двукрылые
костянки около 3—4 см шир. (рис. 39, а) или плоды, очень
похожие на костянки держи-дерева благодаря крыловйдному
диску, развивающемуся вокруг косточки (Р. paliurus). Свое-
образны плоды энгельгардии. У Engelhardia wallichiana мелкая
костянка (5—10 мм) снабжена большим трехлопастным крылом
из прицветника. Главная лопасть крыла достигает 7.5 см дл. и до
2 см шир. (рис. 39', б),.
В других семействах нижние сухие костянки встречаются на-
ряду с сочными плодами. В сем. крушиновых, например у держи-
дерева (Paliurus spina-christi), обитателя каменистых склонов
Крыма и Кавказа, развивается сухая костянка с широкой крыло-
видной каймой и губчатым мезокарпием, окружающим шаровид-
ную очень твердую косточку. Нередко она заключает 2—3 семени.
Сухие костянки известны и в сем. волчниковых (Thymelaeaceae)
у видов Thymelaea и Edgeworthia. Кольквиция прелестная (Kol-
kwitzia amabilis) образует небольшие шаровидные костянки (5—
7 мм в диам.), несущие на верхушке чашечку, поднятую на узкой
трубке, и густо опушенные щетинистыми волосками. Псевдомоно-
мерная нижняя сухая костянка свойственна водяной сосенке
(Hippuris vulgaris).
СОЧНЫЕ ВЕРХНИЕ И НИЖНИЕ
Ценокарпные плоды этой группы чаще представлены одноко-
сточковой костянкой, реже односемянной ягодой.
Верхние однокосточковые костянки можно встретить в сем.
пальм. Наиболее известен из них плод масличной пальмы (Elaeis
74
Рис. 40. Косточка чилима (Trapa natans)
(по: Жизнь растений).
guinensis) — важнейшей культуры
тропической Африки и Азии. Ко-
стянки масличной пальмы в оранже-
вой мякоти мезокарпия содержат
ценное техническое масло; пищевое
масло получают из семян. Масляни-
стый мезокарпий характерен также для съедобных плодов маури-
ции извилистой (Mauritia flexuosa) — величественной пальмы
бассейна Амазонки. Сочные костянки свойственны и подсем. ка-
риотовых.
Верхние костянки нередки в сем. маслиновых (Oleaceae). Ши-
роко известные плоды оливкового дерева (Olea еигораеа), дости-
гающие у некоторых сортов до 3 см дл., содержат в мезокарпии
пищевое масло. У дикорастущих видов, например О. chrysophylla,
плоды мелкие (4—7 мм в диам.), со слаборазвитым мезокарпием.
Такие же мелкие удлиненно-овальные костянки бирючины (Ligu-
strum vulgare, L. ovalifolium) отличаются мучнистой зеленоватой
горькой мякотью мезокарпия. У декоративного кустарника форе-
стиеры новомексиканской (Forestiera neo-mexicana) костянки соч-
ные, кислые, сине-черные, как и у бирючины.
Верхние однокосточковые костянки, чаще псевдомономерные,
свойственны многим представителям тропической и субтропиче-
ской флоры: хризобалановым (Parinari curatellifolia), дафнифил-
ловым (Daphniphyllum macropodum), баланитовым. У Balanites
aegyptiaca плоды съедобны. Они отличаются маслянистым мезо-
карпием; из семян добывают масло. Знаменитый кокаиновый куст
(Erythroxylum coca, Erythroxylaceae) образует небольшие продол-
говатые красные костянки. Среди тропических семейств, виды ко-
торых имеют верхние костянки, назовем еще мирсиновых (Ardisia
japonica, Embelia ribes).
Очень мелкие псевдомономерные костянки (3—4 мм в диам.)
образуют виды кондалии из крушиновых (Condalia obovata,
С. spathulata). Несколько крупнее (до 1 см) плод рейнозии
(Reynosia laxifolia) того же семейства.
В отличие от сухих плодов ильмовых каркас (Celtis), выделя-
емый по ряду признаков в особое подсемейство или семейство,
характеризуется сочной костянкой. Красновато-желтые шаровид-
ные плоды С. caucasica съедобны. Мясистые яркоокрашенные или
темные костянки свойственны другому роду каркасовых — треме
(Trema).
Нижние костянки, как и верхние, встречаются в самых раз-
личных семействах. Типичная нижняя однокосточковая костянка —
хорошо известный плод калины (Viburnum opulus). Его легко
отличить по своеобразной плоской сердцевидной косточке. Одно-
косточковые костянки встречаются у некоторых видов боярыш-
ника, например однопестичного (Crataegus monogyna).
75
Таблица 7
Виды плодов и соответствующие им таксоны
Виды и вариации плодов Таксоны
Апокарпии Многолистовка сухая: полимерная Диллениевые, триурисовые, цефалотовые; калуж-
олигомерная ница, купальница, магнолия Толстянковые; архидендрон, афонсея, борец, бру-
М. сочная неллия, живокость, иллициум, лимнохарис, пион, пузыреплодник, рябинник, спирея, сусак, шейхце- рия; лакторис фернандесовый Анноновые, лардизабаловые; кадсура, лимонник;
Однолистовка сухая гидрастис канадский Протейные (часть видов); багрянник, консолида,
О. сочная: многосемянная цимицифуга Воронец, дегенерия
односемянная Мускатниковые
Боб: типичный Горох, горошек, дрок, караганник, лядвенец, фа-
спирально свернутый соль, чина и мн. др. «Люцерна (ряд видов)
ложнодвугнездный Астрагал, остролодочник
невскрывающийся Астрагал солодколистный, а. серповидный
Б. членистый Вязель, копеечник, сераделла, эверсманния, энтада;
Б. сочный мимоза стыдливая Бундук, гледичия, рожковое дерево, софора япон-
Многоорешек: полимерный ская (стифнолобиум), тамаринд Адонис, ветреница, гравилат, лапчатка, лютик, про-
олигомерный стрел, тюльпанное дерево Айлант, василистник, лабазник, платан, рдест, цир-
погруженный цеастер, частуха Лотос
земляничина Дюшенея, земляника
шиповник Каликантовые, монимиевые; роза
Многокостянка:
полимерная Рубус (большинство видов)
олигомерная Луносемянниковые (часть видов); амборелла, ко-
Одноорешек: типичный стяника, керрия, руппия; розовик лазящий Никтагиновые, протейные, эвптелейные; наяда, ро-
о. в гипантии голистник, рогоз Лоховые; идиоспермум, кровохлебка, манжетка, пи-
Односемянный боб: типичный зония, репешок; мирабилис ночецветный Аморфа, донник, дрок (ряд видов), кармихелия, кле-
крылатковидный вер (подрод Lagopus), леспедеца, эспарцет, язвен- ник; люцерна хмелевая, солодка иглистая Птерокарпус, центролобиум; аммодендрон Конноли,
Однокостянка сухая дальбергия прилистниковая, спатолобус Роксбурга, хардвикия двойная Миндаль; иипа кустистая
Зерновка Злаки
Однокостянка сочная Луносемянниковые (часть видов), протейные (часть
Финик видов), розоцветные, подсем. сливовые; дидимелес, пальмы (главным образом подсем. корифовых) Пальмы подсем. фениксовых
76
Таблица 7 (продолжение)
Виды и вариации плодов Таксоны
Ценокарпии Ценокарпная многолистовка: сухая Винтеровые, тетрацентровые, троходендровые; ка-
натник, спирея (часть видов), чернушка, эксо-
сочная хорда, ясенец Нимфейные
Коробочка См. табл. 3 и 4
Стручок Каперсовые (клеоме, поланисия), крестоцветные (большинство родов), маковые (мачок) Бурачок, икотник, крупка, лунник, пастушья сумка,
стручочек
Вскрывающаяся костянка шиверекия, шильница, ярутка Крушиновые (коллеция колючая, колубрина азиат-
Дробная коробочка ская, хелинус яйцевидный, цеанотус голубой) Клещевина, молочай; вильгельмсия пузырчатая,
Калачик эукрифия сердцелистная Мальвовые
Двукрылатка: Клен; диатеноптерис рябинолистный, диптерония
трех-четыре^скрылатка китайская, тетраптерис цитрусолистный Гелиетта мелколистная, тинуйя лазящая
Ценобий Болотниковые, бурачниковые, вербеновые, губо-
Членистый стручок цветные Крестоцветные (гольдбахия, двоякоплодник, реп-
Дробный ценокарпии: верхний ник, стеригмостемум, хориспора; редька дикая); ги- пекоум беловатый Киркия, павония, триостренник, цельнолистник,
нижний якорцы Геликониевые; уруть
Двусемянка Подмаренник, ясменник
Вислоплодник Зонтичные (большинство видов); миодокарпус
Дробная костянка Крушиновые (гуания Бланше, помадерис безлепест-
Ягода ный, трималиум подсолнечниковидный) См. табл. 5 и 6
Костянка многокосточковая То же
Померанец Цитрусовые
Яблоко: типичное Айва, груша, рябина, эриоботрия, яблоня
костянковидное Боярышник, кизильник, мушмула
Тыквина: типичная Тыквенные (большинство родов)
вскрывающаяся Тыквенные (бешеный огурец, люффа, момордика,
Гранатина циклантера) Гранат
Односемянная коробочка Амарантовые, маревые (габлиция, свекла), плюмба-
Семянка: верхняя: типичная говые (часть видов); казуарина, эгицерас Гречишные, осоковые
крылатка Вяз, птелея, фонтанезия, ясень; эвкомия ильмовид-
иижняя: типичная ная Валериановые, ворсянковые, калицеровые, сложно-
крылатка цветные Береза, ольха
Семянковидный Амарантовые (часть видов), маревые, плюмбаговые
цеиокарпий (акантолимон, армерия, кермек, плюмбаго); дивала
77
Таблица 7 (продолжение)
Виды и вариации плодов Таксоны
Орех:
верхний Ильмовые (дзельква, филостилон бразильский), осоковые (гания, склерия)
нижний Граб, лещина, хмелеграб
Ореховидный стручочек Вайда, миагрум, неслия, самерариа, свербига, тау-
Ореховидный ценокарпий: шерия, эвклидиум
верхний Коноплевые, крапивные; дымянка, липа
нижний Санталовые
Желудь Сухая костянка: Буковые
верхняя Анакардиевые (скумпия, сумах, фисташка), осоко- вые (мапания,сцирподендрон), пальмы (кокосовая,
сейшельская, бетелевая, пальмира) .хризобалановые; ежеголовник, птероцельтис
нижняя Волчниковые (тимелея, эджевортия), ореховые; во- дяная сосенка, держи-дерево; кольквиция прелест-
ная
Сочная костянка (одноко- сточковая) :
верхняя Баланитовые, дафнифилловые, каркасовые (каркас, трема), маслиновые (бирючина, маслина, форести- ера), мирсиновые (ардисия японская, эмбелия ри- бес); крушиновые (кондалия обратнояйцевидная, к. лопатчатая, рейнозия широколистная), пальмы (масличная, мауриция извилистая, ряд видов под- сем. кариотовых), хризобалановые, эритроксиловые (кокаиновый куст)
нижняя Кизиловые, крушиновые (зизифус мавританский, унаби); волчник (ряд видов), калина, чилим; боя- рышник однопестичный
Ягода односемянная:
верхняя Омела; мирсина африканская, ремнецветник евро- пейский
нижняя Аукуба японская, волчник алтайский, в. обыкновен- ный
Полунижние костянки, сочные, яркоокрашённые, свойственны
многим видам волчника {Daphne, Thymelaeaceae). Псевдомоно-
мерная полунижняя костянка, довольцо крупная (до 2 см дл.),
овальная, с мясистым мезокарпием и крупной заостренной косточ-
кой — плод унаби (Zizyphus jujuba) из сем. крушиновых. Этот
вид давно окультурен и насчитывает до 400 сортов. Съедобны
плоды и зизифуса мавританского.
Очень своеобразна полунижняя костянка чилима, или водяного
ореха (Trapa natans). Слаборазвитый наружный мясистый слой
околоплодника быстро разрушается, и остается каменистая твер-
дая косточка, которую обычно и называют плодом водяного
ореха. Другое название растения — рогульник — больше соответ-
ствует морфологическим особенностям плода, несущего 2—4 очень
прочных острых шипа (рис. 40).
Нижние однокосточковые костянки характерны для сем. кизи-
78
ловых (Cornaceae). В подсем. Mastixioideae косточка всегда одно-
семянная, у кизила может быть двугнездной и двусемянной.
Костянки разных видов кизиловых очень различны по окраске —
от оранжево-красных до черных или белых. В плодах свиды
кроваво-красной (Swida sanguinea) в мезокарпии и семенах до
45 % невысыхающего (технического) масла. Кизил мужской (Сог-
nus mas) дает ценные съедобные плоды, широко используемые
в свежем и переработанном виде.
Односемянная ягода — довольно редкий вид плода. Верхняя
такая ягода известна у ремнецветниковых (Loranthaceae): ремне-
цветника европейского (Loranthus europaeus) и видов омелы
(Viscum). В сем. мирсиновых наряду с костянками можно встре-
тить и односемянную ягоду (Myrsine africana).
Нижняя односемянная ягода характерна для декоративного ку-
старника аукубы японской (Aucuba japonica) сем. кизиловых.
Как показали исследования К- К. Зажурило (1935), плоды волч-
ника алтайского (Daphne altaica) следует считать.односемянной
ягодой, а не костянкой. В мезокарпии ее автор обнаружил руди-
менты вскрывающего механизма — следы происхождения этой
ягоды из коробочки. Такой же ягодой является и плод в. обык-
новенного (D. mezereum).
Все названные в настоящей главе виды и вариации плодов
представлены в обзорной табл. 7.
79
Глава III
СОПЛОДИЯ
«Соплодие», как и многие широкоупотребительные понятия,
не имеет общепринятого определения. Тем более не существует
разработанной классификации и номенклатуры соплодий.
Часто под соплодием понимают группу тесно сближенных
и сросшихся плодов. Но такое понимание несомненно слишком
узко. Ему противопоставляется широкое понимание соплодия как
всякого соцветия в фазе плодоношения. Казалось бы, такое
широкое определение вполне правомерно и логично. Если плод —
это зрелый цветок, то соплодие — это зрелое соцветие. Таким об-
разом, как будто снимается вопрос о наименовании и клас-
сификации соплодий, так как к ним применимы все те понятия,
которые приняты для соцветий. Однако в работах последних лет,
вслед за Троллем (Troll, 1964, 1969), классификация и номенкла-
тура соцветий настолько усложнены и детализированы (Федоров,
Артюшенко, 1979), что практически пользоваться ими невозможно.
Следует учесть и то обстоятельство, что не всякое соцветие
превращается в соплодие. Если период цветения сильно растянут,
а плоды опадают (или раскрываются) по мере созревания, то
в этом случае соплодие не образуется. Так, например, у эспарцета
песчаного (Onobrychis arenaria) у основания соцветия могут на-
блюдаться зрелые опадающие плоды, а на верхушке его — цветки
и бутоны. Аналогичная картина характерна для ряда крестоцвет-
ных и многих других растений. Соплодие образуется только тогда,
когда плоды всего соцветия созревают до начала диссеминации.
Под соплодием мы понимаем совокупность зрелых плодов
одного соцветия, четко обособленную от вегетативной части по-
бега. Такое понимание соплодия оправдано и его биологическим
значением. Подобно тому как сближение цветков в соцветия
способствует более успешному опылению, соплодия обеспечивают
более эффективную диссеминацию. Так, соплодия яркоокрашен-
ных плодов сильно контрастируют с зеленой листвой
деревьев и кустарников, благодаря чему даже издали хорошо
заметны для плодоядных птиц. Сухие соплодия, такие как у зон-
тичных или сирени, создают гораздо большее сопротивление воз-
душному потоку, чем одиночные плоды. Далее будет показано,
что соплодие сложноцветных является функционально целостным
морфологическим образованием.
Разнообразие соплодий в нашем понимании исключительно
80
велико, но объем книги не позволяет дать хоть сколько-нибудь
полное их описание. Мы ограничимся отдельными примерами, на
которых можно проследить, как изменяются характер и степень
интегрированности плодов в процессе специализации соплодий.
С этой точки зрения мы выделим следующие группы соплодий.
А. Соплодия из свободных плодов
1. Соплодия, сохраняющие структуру соцветия
а) рыхлые соплодия: плоды на плодоножках
б) плотные соплодия: плоды сидячие
2. Соплодия с видоизмененной структурой соцветия
Б. Соплодия из сросшихся плодов
1. Соплодия из сросшихся голых1 плодов (или завязей)
2. Соплодия из сросшихся цветков и других элементов со-
цветия
В подавляющем большинстве соплодий плоды остаются свобод-
ными и морфология соцветия повторяется в соплодии. Следова-
тельно, наша группа А. 1 — самая многочисленная. Но даже в пре-
делах одного семейства специализация соплодия может достигать
разного уровня, поэтому представители того или иного семейства
будут попадать в разные группы.
A.l.a. Соплодия этой самой распространенной группы хорошо
известны у видов с сочными плодами — винограда, рябины, бу-
зины, калины, черемухи, боярышника и многих других, и сухими —
зонтичных, орхидных, сирени, валерианы, видов клена, ясеня,
айланта и т. п.
А. 1.6. В этой группе также встречаются соплодия и сочные,
и сухие. Из сочных назовем шаровидное соплодие плюща золо-
тистоплодного (Hedera chrysocarpa); початковидное — бело-
крыльника болотного (Calla palustrls). Сочные соплодия широко
представлены у растений тропической флоры (в семействах панда-
новых, банановых и др.). Так, у фрейсинетии (Freycinetia, Panda-
пасеае) образуется массивное початковидное соплодие из ягод,
сидящих на утолщенной оси. Костянки пандануса простого (Рап-
danus simplex) также собраны в виде початка до 60 см дл.,
20 см толщ, и массой до 25 кг.
Исключительной обильностью отличаются соплодия банановых
(Musaceae). Так, у мадагаскарского вида энсеты Перрье (Enseta
perrieri) соплодия содержат до 200 плодов с общей массой 25—
30 кг. Культурные сорта банана образуют до 300 плодов в соплодии
с массой 50—60 кг.
Широко представлены и сухие соплодия этой группы. Из расте-
ний нашей флоры они хорошо известны у сложноцветных, ворсян-
ковых, рогоза, ежеголовника. В соплодиях сложноцветных ли-
сточки обертки выполняют не только защитную функцию в период
созревания плодов, но играют и очень существенную роль в про-
цессе диссеминации. Благодаря гигроскопичности листочков
обертка попеременно открывается и закрывается в зависимости
1 Плоды, образованные только из завязи.
6 Р. Е. Левина
81
Рис. 41. Соплодие банксии (Banksia
verticillate.) (по Веттштейну).
Рис. 42. Соплодие ежеголовника (Spar-
ganium neglectum) (по Комарницкому).
от влажности атмосферы. При этом одним видам свойственна
ксерохазия, т. е. раскрывание корзинок в сухую погоду (Senecio,
Crepis и др.), другим — гигрохазия [раскрывание при увлаж-
нении (Asteriscus, Cichorium, Evax; Zohary (1950)]. Такое попере-
менное изменение положения обертки способствует выталкиванию
семянок, которое облегчается различными выростами ложа кор-
зинки (волосовидными, пленчатыми и т. п.). Значение обертки
для распространения плодов и целостность соплодия еще нагляд-
нее можно проследить у видов с цепкими обертками, таких как
виды лопуха (Arctium), василька (Cetitaurea aggregata, С. calci-
trapa, С. squarrosa, С. virgata), кузинии (Cousinia) и др. Цепля-
ясь за шерсть проходящих мимо животных, такие соплодия отры-
ваются целиком, а затем постепенно из них рассеиваются семянки.
Изящные цилиндрические соплодия рогоза (Typha angustifolia,
Т. Latif olid), состоящие из огромного числа мелких орешков,
отличаются от соцветий лишь своей темно-коричневой окраской.
Некоторые початковидные или головчатые соплодия можно
принять за полимерный апокарпный плод. Так, соплодие Banksia
verticillata (Proteaceae; Австралия), образованное множеством
листовок, внешне напоминает многолистовку магнолии (рис. 41).
Еще более сходно с многоорешком соплодие ежеголовника (Spar-
ganiurrr, рис. 42). А шаровидное соплодие платана восточного
(Platanus orientalis), 25—30 мм в диам., может быть названо
мультиплицированным многоорешком, поскольку оно состоит из
олигомерных многоорешков отдельных цветков. В процессе дис-
семинации соплодие платана распадается на отдельные орешки,
которые и являются диаспорами.
82
Рис. 43. Соплодие (а) и отдельный плод (б) фителефаса крупноплодного (Phytele-
ph.as macrocarpa) (по: Жизнь растений).
Из этой группы рассмотрим еще оригинальное соплодие сухих
костянок неотропической пальмы фителефаса крупноплодного
(Phytelephas macrocarpa). Шаровидные (до 25 см в диам.)
соплодия этой пальмы кажутся состоящими из очень многих
плодов (рис. 43, а). В действительности их немногим более 10,
но каждый плод по периферии соплодия имеет бугристую поверх-
ность с конически заостренными выростами (рис. 43, б).
А.2. К этой группе отнесены соплодия, отличающиеся от со-
цветий видоизмененными при плодах осевыми или иными их
элементами. Очень своеобразны соплодия скумпии {Cotinus coggy-
gria), несущие несколько мелких сухих костянок (около 5 мм дл.
и до 2 мм в диам.). У основания соплодия сильно вытягиваются
в длину и становятся перистыми цветоножки недоразвитых цвет-
ков, которые придают всему соплодию вид пышного султана.
Иногда волоски приобретают пурпурную окраску, что делает со-
плодие особенно эффектным. Зрелые соплодия отламываются
целиком и уносятся ветром.
У многих видов из самых разных семейств морфологические
особенности соплодий определяются видоизменением прицвет-
ников.
Довольно широко известны соломенно-желтые легкие овальные
соплодия хмеля {Humulus lupulus), состоящие из сильно разрос-
шихся, черепитчато налегающих друг на друга прицветников,
в пазухах которых сидят плоды (рис. 44). Совершенно такого же
типа и той же морфологической природы соплодия хмелеграба
(Ostrya carpinifolia) и разных видов граба (Carpinus betulus,
С. cordata, С. japonica, С. orientalls и др.). У лещины прицветники
образуют плюску, более или менее полно окружающую плод.
У нашей лещины обыкновенной {Corylus avellana) нередко наблю-
даются соплодия из нескольких плодов, плюски которых частично
срастаются.
Существенную роль в формировании соплодий прицветники
и прицветнички играют в сем. ворсянковых (Dipsacaceae). Харак-
терная для их цветков наружная чашечка (epicalyx) из двух
в*
83
Рис. 44. Соплодие хмеля (Humulus
lupulus) (по Кернеру).
Рис. 45. Соплодие скабиозы (Scabiosa
graminifolia) (по Веттштейну).
Рис. 46. (Соплодие инжира (Ficus carica), внешний вид и продольный разрез
(по: Деревья и кустарники СССР).
Рис. 47. Соплодие циноксилона (Су-
noxylon capitata) (по: Деревья
и кустарники СССР).
Рис. 48. Соплодие пандануса (Pandanus
tectorius) и отдельная фаланга (по: Жизнь
растений).
84
сросшихся прицветничков при плодах обычно разрастается, опре-
деляя облик всего соплодия (рис. 45).
«Ворсовальная шишка», издавна и широко известная в тек-
стильной промышленности, представляет собой удлиненно-голов-
чатое соплодие культурного вида ворсянки посевной (Dipsacus
sativus). Ось соплодия несет густую спираль удлиненных упругих
крепких и эластичных прицветников, заостренных и отогнутых
книзу в виде зацепов. Эти особенности прицветников и определяют
применение соплодий ворсянки для ворсования тканей. Селекцион-
ные сорта ворсянки отличаются цилиндрическими, до 15 см дл.,
соплодиями и прицветниками с высокими техническими свойст-
вами. До сих пор нет полноценной замены ворсовальной шишки,
и она не утратила своего значения для изготовления высокоценных
ворсованных тканей.
Благодаря разросшимся и одревесневшим прицветничкам, по
2 при каждом цветке, соплодие казуарины прибрежной (Casuarina
litorea-, syn. С. equisetifolia) оказывается сходным с шишкой
хвойных. Из растений нашей флоры аналогичная картина наблю-
дается у ольхи, у которой одревесневают ось соплодия и прира-
стающие к ней кроющие чешуи соцветий. После опадения плодов
изящные черные «шишечки» ольхи еще долго висят на ветвях.
Видоизменения обертки при плодах у некоторых сложноцвет-
ных приводят к образованию весьма специализированных со-
плодий, функционирующих как один плод. Широко известны
корзинки — соплодия разных видов дурнишника (Xanthium)
с одревесневшими замкнутыми обертками, покрытыми крепкими
цепкими шипами. Корзинки дурнишника — двугнездные, с одной
семянкой в каждом гнезде — отрываются целиком и распростра-
няются эпизоохорно. У амброзии (Ambrosia) очень мелкие (2—
5 мм) корзинки, также с замкнутой прочной оберткой, заключают
единственную семянку.
Таким образом, преобразования, которые произошли в ходе
эволюции в соплодиях сложноцветных, аналогичны процессу пре-
вращения коробочки в односемянный невскрывающийся плод.
Среди соплодий со свободными плодами наиболее специфично
соплодие инжира, или фигового дерева, из сем. тутовых (Ficus
carica, Moraceae). Оно развивается из соцветия, у которого
срастаются многочисленные прямые мясистые веточки, образуя
полость, открытую на верхнем конце.1 К моменту созревания
плодов соцветие превращается в сочное грушевидное образование
до 5 см и более дл. зеленовато-желтой, красноватой или фиолето-
вой окраски. Всю его полость изнутри покрывают мелкие сухие
односемянные плоды. Основную пищевую ценность так называе-
мых плодов инжира представляют мясистые стенки соплодия
(рис. 46).
1 Соцветия (и соплодия) тутовых, в частности фикуса, нередко называли
сиконом (sycon). По современной терминологии, соцветие тутовых — ценосома,
а у фикуса — ценосома гипантиевидиая (Федоров, Артюшенко, 1979).
85
Рис. 49. Соплодие горанино^ии (Но-
raninovia ulicina) (по: Сорные ра-
стения СССР).
Б. 1. Целостность сопло-
дия выражена более полно
при срастании плодов, но сте-
пень специализации таких со-
плодий тоже может быть раз-
личной. В простейших слу-
чаях срастаются тесно сбли-
женные завязи или даже го-
лые плоды, чаще сочные. Та-
кого типа соплодия встречаются у некоторых кизиловых (Супоху-
lon capitata, С. japonica; рис. 47), у панданусов. Своеобразное со-
плодие развивается у пандануса кровельного (Pandanus tecto-
rius). Его голые женские цветки сидят очень тесными группами,
срастаясь в так называемые фаланги. Зрелая фаланга состоит
из 8—9 сросшихся костянок; эти фаланги собраны на утолщенной
оси, образуя головчатое соплодие второго порядка (рис. 48).
Могут срастаться и сухие плоды. Очень оригинальные шаровид-
ные соплодия известны у ликвидамбара (Liquidambar, Hamemeli-
daceae). У него срастаются своими стенками одревесневшие дву-
створчатые коробочки. Свободной остается лишь верхушка коро-
бочки, увенчанная одревесневшим столбиком. Даже после вскры-
вания коробочек и осыпания семян такое соплодие сохраняет
свою структуру: оно представляет собой многокамерное шаровид-
ное тело 25—40 мм в диам. с многочисленными, слегка отогнутыми
радиальными отростками.
Б.2. В формировании соплодий этой группы в большей или
меньшей степени участвуют элементы соцветия; это в равной мере
относится и к сочным соплодиям, и к сухим. Последние нередки
в сем. маревых, парциальные соцветия которых — «клубочки» —
состоят из нескольких тесно сближенных цветков. У некоторых
родов — свеклы (Beta), шпината (Spinacia), гораниновии (Нога-
ninovia) — околоцветники при плодах разрастаются, одревесне-
вают, срастаются с околоплодником и друг с другом. Так, клубочек
превращается в соплодие, иногда снабженное крепкими шипова-
тыми выростами (Spinacia tetrandra, Horaninovia ulicina', рис. 49).
У мари многолистной (Chenopodium foliosum) околоцветники
при плодах становятся сочными, яркоокрашенными (красными),
превращая клубочек в типичное сочное соплодие. Совершенно
такой же морфологической природы и сочные съедобные соплодия
шелковицы (Morus) сем. тутовых. Но в этом семействе известны
соплодия, более сложные по происхождению.
У палеотропического рода артокарпус — знаменитого хлебного
дерева (Artocarpus altilis) и джекфрута (Л. heterophyllus) —
образуются огромные соплодия вследствие срастания плодов
с околоцветниками, брактеями и осью соцветия. В процессе
86
созревания ткани соплодия сильно разрастаются и превращаются
в однородную мучнистую мякоть. Соплодия хлебного дерева —
до 30 см в диам. и 12 кг массы; удлиненные соплодия джекфрута —
до 1 м дл. и массой в 16—20 кг. Оба вида являются важнейшими
пищевыми растениями тропиков и широко культивируются.
Оригинальные соплодия свойственны и другому представителю
трибы артокарпусовых — маклюре. Американский вид Maclura
pomifera образует многочисленные сухие односемянные удлинен-
ные плоды, погруженные в сильно разрастающуюся мясистую
ось соцветия. Формируется шаровидное соплодие до 10—15 см
в поперечнике, по окраске и внешнему виду напоминающее апель-
син, но несъедобное.
Широко известны соплодия ананаса (Ananas sativus) из нео-
тропического сем. бромелиевых (Bromeliaceae). Формирование
такого соплодия очень сходно с тем, что наблюдается у арто-
карпусов. С сильно развитой осью соцветия срастаются завязи
многочисленных цветков и основания кроющих листьев. Все эти
элементы соплодия образуют мясистую сочную ткань, в которой
очень трудно обнаружить следы срастания. Только с поверхности
соплодие покрыто довольно жестким панцирем, образованным
верхушками кроющих листьев. Особенностью соплодий ананаса
является пролификация его оси, вследствие чего на верхушке
соплодия всегда имеется укороченный побег с пучком зеленых
листьев. Культурные формы ананаса чаще всего образуют бес-
семянные плоды (партенокарпия), но характеризуются очень
крупными соплодиями с массой от 2 до 15 кг (в зависимости
от сорта).
Названные в этой главе соплодия не только отражают очень
разную степень специализации, многие из них обнаруживают
явный параллелизм путей морфогенеза и в относительно близких
семействах одного подкласса (казуариновые и березовые, березо-
вые и коноплевые), и в столь отдаленных, как маревые и тутовые,
тутовые и бромелиевые.
87
Глава IV
ГЕТЕРОКАРПИЯ
Морфология плода является, как известно, устойчивым видо-
вым признаком и широко используется в систематике покрыто-
семянных. В то же время давно было установлено, что у некоторых
видов на одной и той же особи формируются плоды морфологи-
чески столь различные, что их свободно можно отнести к разным
родам. Это явление назвали гетерокарпией.
В конце прошлого и в начале нашего столетия были опубли-
кованы обширные списки гетерокарпных видов и описаны разные
формы гетерокарпии.1 В настоящее время известно около 30 се-
мейств, в которых установлены гетерокарпные виды. Среди них
такие крупные и широко распространенные, как сложноцветные,
маревые, крестоцветные, зонтичные, бобовые, и сравнительно
небольшие семейства валериановых, дымянковых, кисличных,
фиалковых, калицеровых и др.
Морфологические различия плодов при гетерокарпии могут
проявляться в разной степени и касаться их формы и размеров,
анатомии околоплодника, наличия или отсутствия тех или иных
выростов перикарпия, характера вторичных покровов плода, если
они имеются. Разнотипные плоды могут различаться и числом
семян; наряду с односемянными невскрывающимися плодами
образуются и многосемянные вскрывающиеся (неэквивалентная
гетерокарпия по Зохари). У распадающихся плодов гетерокарпных
видов названные здесь различия относятся к частям плода —
членикам, мерикарпиям или эремам. Членики также могут разли-
чаться числом семян, т. е. быть эквивалентными или неэквива-
лентными.
Различные формы гетерокарпии можно сгруппировать и обо-
значить следующим образом:
А. Гетероголокарпия — разнотипны целые плоды
а) эквивалентная
б) неэквивалентная.
Б. Гетерофрагмокарпия — разнотипны части плода
1. Гетероартрокарпия — членики
а) эквивалентная
б) неэквивалентная.
1 Эти работы здесь не перечисляются, так как они многократно цитировались
ранее (Левина, 1957, 1967а; Войтенко, 1968, 1974).
88
2. Гетеромерикарпия — мерикарпии
3. Гетероэремокарпия 1 —эремы.
Последняя форма (Б.З) свойственна только ценобиям, форма
А — всем нераспадающимся плодам. Остальные термины понятны
из сказанного ранее.
Неэквивалентные формы гетерокарпии проявляются в том, что
на особи формируются диаспоры разной морфологической при-
роды: и голые семена, и односемянные плоды (форма А.б) или
членики (форма Б. 1.6), т. е. семена, окруженные околоплодником.
Наиболее характерной чертой гетерокарпии являются сущест-
венные различия семян из разнотипных плодов или их частей.
Семена различаются и морфологически (размеры и масса, форма
и скульптура поверхности, положение зародыша), и главным
образом физиологически (по глубине покоя, жизнеспособности,
характеру прорастания, наконец, по фенологии развивающихся
из них растений).
Наличие или отсутствие тех или иных придатков у гетероморф-
ных диаспор также имеет несомненное биологическое значение;
благодаря такому различию потомство гетерокарпной особи и
завоевывает относительно широкое пространство, и закрепляется
на месте произрастания материнского растения.
К общей характеристике гетерокарпии необходимо еще доба-
вить, что разнотипные плоды занимают разное и всегда определен-
ное положение на цветоносе или в компактном соцветии (точнее
соплодии). Расположение гетероморфных частей плода при гетеро-
фрагмокарпии также всегда строго фиксировано.
При всем своеобразии гетерокарпии ее не всегда легко отгра-
ничить от других форм неоднородности плодов и семян, свойст-
венных растительной особи. Наиболее сходна с гетерокарпией
топографическая неоднородность семян (Левина, 1981); она
обусловлена онтогенетической изменчивостью метамеров, отра-
жающей ярусные физиологические различия цветоносов и гене-
ративных побегов разного порядка (и возраста).
Как показали наши исследования, в пределах даже такого
укороченного соцветия, как у клевера, наблюдаются различия
в интенсивности созревания семян нижних- и верхних цветков
головки (Левина, 1965). Тем более будут различаться плоды
и семена в пределах открытых сильно разветвленных соцветий
при растянутом периоде цветения.
Но топографическая неоднородность плодов и семян обычно
проявляется в их количественных признаках и не приводит к су-
щественным изменениям их структуры. Такие глубокие морфологи-
ческие различия плодов и их семян на одной особи, какие наблю-
даются при гетерокарпии, — это уже особый наследственный
признак вида.
Таким образом, гетерокарпия — это генетически обусловленное
' Термин предложен С. Н. Опариной (Войтенко, Опарина, 1985). Остальные
термины уже использовались в литературе с указанием их авторов.
89
свойство вида формировать на одной особи морфологически разно-
типные диаспоры, семена которых также различаются своей
морфологией и физиологией.
Хотя отличительная черта гетерокарпии — структурная разно-
типность диаспор, проблема в целом выходит далеко за рамки
морфологической карпологии. Аспекты изучения гетерокарпии и ее
биологическое значение уже обсуждались в ряде работ (Левина,
1957, 1967а, 19676, 1980, 1981; Войтенко, 1969а; Левина, Вой-
тенко, 1975), поэтому здесь мы не будем останавливаться на этих
вопросах. Перейдем к рассмотрению конкретных гетерокарпных
видов в отдельных семействах.
Вполне очевидно, что форма гетерокарпии определяется морфо-
логией плода, поэтому можно говорить о характере проявления
гетерокарпии в том или ином семействе в целом. Так, форма А.а —
эквивалентная гетероголокарпия — свойственна семействам с од-
носемянными плодами: сложноцветным, маревым, гречишным;
форма Б.2 — зонтичным; Б.З — бурачниковым и т. д.
Очень широко и своеобразно гетерокарпии представлена в сем.
маревых. Как мы уже знаем (глава II), их сухие односемянные
плоды характеризуются тонким пленчатым перикарпием, который
не определяет морфологических особенностей плода. Поэтому
и гетерокарпии в этом семействе связана с признаками вторичных
плодовых оболочек (чаще прицветников) и самих семян. В зависи-
мости от относительной роли этих признаков у маревых можно
выделить 4 варианта разноплодия: 1) существенно различаются
покрывальца из прицветников, семена более или менее одинаковы;
2) сильно различаются вторичные плодовые оболочки и семена;
3) разнотипны голые плоды; 4) неоднородны только семена.
К первому варианту относится разноплодие рогача песчаного
(Ceratocarpus arenarius), плоды которого заключены в замкнутые
покрывальца из прицветников. Различия базальных плодов и вер-
хушечных видны на рис. 50, но между ними — постепенный ряд
переходных форм. Покрывальца базальных плодов сильно опу-
шены, но по анатомическому строению они более мезоморфны,
чем верхушечные. Покрывальца, подобные базальным, появля-
ются и на самой верхушке растения в углах разветвлений (Тах-
таджян, 1934).
Базальные плоды, семена которых отличаются глубоким по-
коем, всегда остаются близ материнского растения. Остальные
«рогатые» плоды с дружно прорастающими семенами разносятся
либо животными, либо ветром на оторвавшихся растениях (пере-
катишоле).
Наиболее распространена гетерокарпии второго варианта. Она
встречается почти у всех видов лебеды (Atrip lex), видов сведы
(Suaeda), бинерции (Bienertia cycloptera) (Ильин, 1930).
В одной группе гетерокарпных видов лебеды все женские
цветки не имеют околоцветника и заключены в 2 почти свободных
прицветника. Семена у них развиваются двух типов; очень мелкие
(1 —1.5 мм), выпуклые, черные, с плотной кожурой и вдвое круп-
90
Рис. 50. Разнотипные плоды рогача (Ceratocarpus arenarius) (по Тахтаджяну).
нее, плоские, желтовато-зеленые, в тонкой пленчатой кожуре
(Atriplex micrantha, A. patens, A. patula). Для первого вида
характерен также диморфизм прицветничков: очень мелкие —
при плодах с черными семенами и значительно крупнее при плодах
второго типа. У лебеды блестящей (Л. nitens) и садовой (Л. horten-
sis) из женских цветков в прицветничках развиваются 2 таких же
типа семян, как описаны выше. Кроме того, у них имеются еще
женские цветки с околоцветником (не скрытые в прицветничках),
которые приносят горизонтальные плоды с мелкими черными
выпуклыми семенами. Плоские светлые семена прорастают без
периода покоя, тогда как черные семена, даже скарифицирован-
ные, не прорастают без длительного покоя (по-видимому, не
меньше года).
У видов сведы из различных цветков также образуются 2 типа
семян, подобных семенам лебеды. Так, у сведы рожконосной
(Suaeda corniculata) мелкие черные вертикальные семена с твер-
дой кожурой развиваются из верхних цветков дихазия («клу-
бочка»), а боковые цветки дают плоские желтоватые семена
в тонкой пленчатой кожуре; у с. льнолистной (S. linifolia) семена
первого типа образуются из более ранних, преимущественно
обоеполых цветков, а второго типа — из позднеосенних женских
цветков. Характер околоцветника при разнотипных плодах не-
сколько варьирует.
Разнотипные голые плоды свойственны аксирису (Axyris ата-
ranthoides). Часть плодов — удлиненно-овальные, сжатые с боков,
красновато-бурые, крапчатые, с хорошо развитым раздвоенным
крыловидным выростом на верхушке. Семена их легко прорастают
без периода покоя. Плоды другого типа — более короткие, об-
ратнояйцевидные, однотонные, без крыловидного придатка
(рис. 51). Семена их прорастают лишь после длительного покоя.
Плоды обоих типов встречаются на протяжении всего генератив-
ного стебля, но на главном цветоносе, особенно , в нижних его
ярусах, и у основания веточек преобладают плоды второго типа.
Наконец, неоднородность семян (гетероспермия) характерна
для различных видов мари. Она хорошо изучена у мари белой
(Chenopodium album), образующей семена трех типов: а) крупные
91
Рис. 51. Крылатый и бескрылый плоды аксириса (Axyris amaranthoid.es) (по:
Сорные растения СССР).
(до 2 мм в диам.), плоские, с заостренным краем, светло-коричне-
вые, матовые; б) более мелкие, менее сплюснутые, черные и
в) очень мелкие, почти шаровидные, черные, блестящие. Семенная
кожура черных семян значительно более плотная, почти в 4 раза
толще, чем у коричневых семян. Семена первого типа прорастают
быстро и дружно в год созревания, второго типа — на второй год
и третьего типа — лишь на третий год после созревания.
Очень широко и в различных формах гетерокарпия представ-
лена в сем. сложноцветных, при этом разнотипные плоды на-
ходятся в одной корзинке, занимая в ней разное положение.
Гетерокарпия свойственна представителям обоих подсемейств
сложноцветных, и гетероморфные плоды могут развиваться как из
однотипных (язычковых) цветков, так и из разнотипных (ложно-
язычковых и трубчатых). В первом случае между крайними
морфотипами плодов обычно имеются переходные формы; во
втором диморфизм плодов проявляется резко, без промежуточных
форм.
Степень выраженности и характер признаков, определяющих
гетерокарпию у сложноцветных, весьма различны, что позволяет
выделить несколько типов (Zohary, 1937). Мы остановимся только
на некоторых вариантах, более или менее типичных.
В политипном роде Tragopogon много гетерокарпных видов,
хотя полиморфизм плодов у них выражен нерезко. Цветки в кор-
зинках все однородные — обоеполые, фертильные, язычковые. Су-
щественно различаются краевые семянки и центральные, а между
ними постепенный ряд переходных форм. Все семянки с хорошо
развитым носиком и хохолком, но краевые значительно массивнее,
темноокрашенные, слегка изогнутые, 5-гранные, груборебристые,
с чешуйчатыми выростами по ребрам. Центральные семянки
прямые, округлые в поперечном сечении, светлые и почти гладкие.
Детальное карпологическое изучение козлобородника сомнитель-
ного (Т. dubius) обнаружило заметные различия краевых и цент-
ральных семянок по анатомии околоплодника, характеру прора-
стания, дальности диссеминации (Войтенко, 1981). Аналогичным
образом гетерокарпия проявляется у ряда других видов козло-
бородника (Т. capitatus, Т. gratninifoUus, Т. kultiassovii, Т. para-
doxus, Т. heterospermus, Т. tuberosus и др.).
92
Другим вариантом можно считать гетерокарпию крестовника
Якова (Senecio jacobaea). У этого вида краевые семянки, развива-
ющиеся из ложноязычковых цветков, в зрелом состоянии лишены
хохолков, а центральные — производные трубчатых цветков —
снабжены хорошо развитым хохолком. Кроме того, центральные
семянки обильно опушены, груборебристые, с четкой скульптурой
поверхности. У краевых семянок эти признаки слабо выражены,
однако они (семянки) отличаются утолщенным околоплодником
за счет более развитых механических и паренхимных тканей.
В связи с этим масса 1000 краевых семянок — 295 мг против
175 мг для центральных. Числовое соотношение краевых и цент-
ральных семянок в корзинке в среднем 1 : 4.8.
Вполне очевидно, что разнотипные семянки в данном случае
различаются по способу и дальности диссеминации, что подтверж-
дено наблюдениями. Вместе с тем центральные семянки характери-
зуются более высокими показателями общей всхожести, друж-
ности и скорости прорастания.
Гетерокарпия типа крестовника, с некоторыми вариациями,
свойственна многим родам и видам сложноцветных. Она касается
не только хохолков (как у Carthamnus, Heteropappus, Heterotheca,
Crepis pulchra, Hedypnois cretica), но и другого рода придатков —
крыловидных (Ximenesia encelioides), цепких (Heterospermum),
Третий вариант характеризуется резким поли- или диморфиз-
мом плодов в пределах корзинки. Классическим многократно
описанным примером такого типа гетерокарпии являются разные
виды ноготков (Calendula'), у которых различают не менее трех
видов семянок, связанных переходными формами. Семянки, разви-
вающиеся от периферии к центру корзинки, называют соответ-
ственно когтевидными, ладьевидными и кольцевидными. Все они
образуются из обоеполых язычковых цветков; трубчатые цветки
остаются бесплодными.
и, Еще более разительный диморфизм плодов без всяких пере-
ходов обнаруживают виды диморфотеки (Dlmorphotheca) — оли-
готипного южноафриканского рода из трибы Calenduleae. В ее
корзинках фертильны и ложноязычковые (периферические)
цветки, и трубчатые. Из ложноязычковых цветков образуются
клиновидные, слегка изогнутые семянки с выраженными ребрами
и скульптурой поверхности; трубчатые цветки производят упло-
щенные гладкие более светлые семянки, окруженные широкой
крыловидной каймой, утолщенной по краю (рис. 52). В зрелых
корзинках листочки обертки (по числу равные краевым цветкам)
отгибаются книзу и краевые семянки постепенно опадают; цент-
ральные плоды разносятся ветром.
Ярко выраженный полиморфизм плодов наблюдается у колюче-
головника Бентама (Acanthocephalus benthamianus) из трибы
Lactuceae. Помимо внешней формы (рис. 53) семянки существенно
различаются по анатомии околоплодника. Кроме того, сжатые
со спинки краевые семянки опадают вместе с прирастающим
листочком обертки.
93
Рис. 52. Крылатая и клиновидная семянки диморфотеки (Dimorphotheca pluvi-
alis).
Гетеромерикарпия в нашем узком ее понимании проявляется
лишь у дробных плодов. В сем. зонтичных, например у лизеи
разноплодной (Lisaea heterocarpa), некоторых видов Torilis, Dau-
cus и др., абаксиальные мерикарпии несут грубые цепкие выросты,
а другой мерикарпий того же плода — лишь небольшие бугорки.
В семействах, которым свойствен плод ценобий, гетерокарпия
может проявляться в форме гетероморфное™ эремов одного
плода. Эта форма гетерокарпии, для которой мы приняли название
«гетероэремокарпия», остается почти неизученной. Но специальное
карпологическое исследование некоторых родов и видов бурачни-
ковых выявило несколько вариантов гетероэремокарпии в этом
семействе (Войтенко, Опарина, 1985). Общий признак всех гетеро-
карпных видов — один (реже 2) неопадающий эрем, прочно
связанный с карпобазисом.1 Этот эрем всегда занимает адаксиаль-
ное положение в цветке, крупнее остальных эремов плода и отли-
чается от них морфологически. Если все 3 остальные (опадающие)
эремы однотипны, имеет место диморфный вариант гетероэремо-
карпии (Buglossoides arvensis, Lappula barbata, L. microcarpa,
L. drobovii, Paracaryum intermedium.), но наблюдается и триморф-
ный вариант, когда среди опадающих эремов можно выделить
2 морфотипа (Lappula semialata, L. sessiliflora). Наконец, в наи-
более специализированном тетраморфном варианте все 4 эрема
различаются по форме, характеру придатков и другим признакам
(Heterocaryum subsessile, Н. szovitsianum). Остается добавить,
что все гетероморфные эремы различаются по анатомии около-
плодника и занимают в плоде строго фиксированное положение.
В семействах, которым свойственны разнотипные плоды —
многосемянные и односемянные, вскрывающиеся и распадаю-
щиеся, и гетерокарпия может проявляться в разных формах.
Примером таких семейств могут служить бобовые. Среди них
известны амфикарпические виды или разновидности: Vida amphi-
carpa, V. angustifolia var. amphicarpa, Lathyrus sativus var.
' Часть ценобия, остающаяся на плодоножке после опадения всех эремов.
94
Рис. 53. Краевая, центральная и переходная семянки колючеголовника {Acantho-
cephalus benthamianus).
amphicarpus, L. setifolius, L. saxatilis, Pisum fulvutn var. amphi-
carpum и др. Амфикарпия у всех этих видов сопряжена с неэквива-
лентной гетероголокарпией: их надземные бобы линейные, 4—5-се-
мянные, а подземные — односемянные, округлые, с плотнокожи-
стым околоплодником, невскрывающиеся.
Неэквивалентная гетероголокарпия, но не связанная с амфи-
карпией, свойственна цейлонскому виду десмодиума (Desmodium
heterocarpum): из нижних цветков соцветия развиваются одно-
семянные бобы, из верхних — членистые из 5—7 односемянных
члеников.
У видов с членистыми бобами может наблюдаться гетероартро-
карпия, как например у копеечника Гмелина (Hedysarum gmelinii)
с резко различающимися члениками (рис. 54).
Разнотипность плодов, аналогичная бобовым, характерна и для
крестоцветных. Разносторонние исследования гетерокарпии в этом
семействе обнаружили многообразие ее форм, различия в диссеми-
нации и характере прорастания гетероморфных диаспор и неко-
торые другие закономерности. Результаты этих исследований
опубликованы (Войтенко, 1968, 1969а, 19696, 1970, 1974; Марасов,
1974а, 19746; Левина, Марасов, 1976), и здесь мы ограничимся
по возможности кратким изложением основного фактического
материала.
С эквивалентной гетероголокарпией крестоцветных ознако-
мимся на двух типичных примерах, однолетниках флоры Средней
Азии: серпоносике (Spirorhynchus sabulosus) и самерарии Буассье
(Satneraria boissieriana). Насколько различны гетероморфные
плоды серпоносика, видно на рис. 55. Оба типа плода односемян-
ные; прямые, веретеновидные, более массивные плоды развива-
ются у основания цветоносных побегов. Опадая, они остаются
близ материнских растений и внедряются в песок. Плоды другого
типа занимают верхушки цветоносов; они легко срываются ветром
и катятся по поверхности песка на несколько метров (их коэф-
фициент парусности 75—90).
У самерарии также образуются 2 типа односемянных плодов;
основания цветоносов — главного и боковых — занимают массив-
95
Рис. 54. Гетероартрокарпия у копееч- Рис. 55. Разнотипные плоды серпоносика
ника (Hedysarum gmelinil) (по: Сор- (Spirorhynchus sabulosus) (по Вой-
ные растения СССР). тенко).
ные овальные плосковыпуклые ореховидные стручочки, глубоко
поперечно-морщинистые, с очень узкой прижатой крыловидной
закраиной, переходящей на верхушке плода в более широкий
слегка изогнутый вырост. Эти плоды сменяются сплюснутыми
с боков, ширококрылатыми, яйцевидного очертания, почти глад-
кими плодами (рис. 56). Средняя масса бескрылого плода 45.9 мг,
крылатого — 19.2 мг; коэффициент парусности соответственно
10—15 и 80—85. Массивность бескрылых плодов обусловлена
сильным развитием мезокарпия. К моменту созревания около-
плодник становится деревянистым, очень грубым.
Вполне очевидно, что бескрылые плоды барохорного типа,
а крылатые — анемохорного. Первые остаются близ материнской
особи (более 80 % в радиусе до 0.7 м); вторые улетают до 8 м,
но основная масса их оседает в радиусе до 3 м.
Эквивалентная гетерокарпия с многосемянными разнотипными
стручками встречается у Matthiola, Torularia, Strigosella circinata,
Diptychocarpus strictus. У последнего вида базальные плоды
цветоносов членистые, а верхние — вскрывающиеся.
Неэквивалентная гетероголокарпия наиболее ярко выражена
уэтионемы (Aethionema heterocarpum) и кампилоптеры (Campylo-
ptera carnea). У последней формируются 2 типа резко различаю-
щихся плодов: раскрывающиеся двугнездные многосемянные стру-
чочки до 8 мм дл. и невскрывающиеся одногнездные односемянные,
вдвое меньших размеров. Существенно различны и семена этих
плодов по размерам, форме, характеру поверхности, положению
зародыша. У основания цветоносов разного порядка сначала
образуются многосемянные плоды, их сменяют односемянные,
и такие смены повторяются 3—5 раз, а в условиях культуры —
до 7 раз. Таким образом, для кампилоптеры характерны 2 типа
диаспор: голые семена и односемянные плоды. На их количествен-
ном соотношении и особенностях прорастания мы остановимся
позже.
96
Рис. 56. Бескрылый и крылатый плоды самера-
рии (Sameraria boissieriana}.
Подобно бобовым, амфикарпия
у крестоцветных также сопряжена
с неэквивалентной гетерокарпией.
Так, бразильский вид Cardamine che-
nopodiifolia и обитающий на о-вах
Хуан-Фернандес Heterocarpus fer-
nandezianus образуют надземные
многосемянные вскрывающиеся ли-
нейные стручки, а из подземных цвет-
ков — односемянные невскрывающи-
еся стручочки.
В трибе Brassiceae широко пред-
ставлена гетероартрокарпия. Плоды
у видов этой трибы состоят из двух
частей — базальной (вальварной), развивающейся из завязи, и
апикальной (стилярной, так называемый носик), образующейся
из столбика. Эти части плода могут различаться числом семян и
другими существенными признаками.
Установлены различные варианты гетероартрокарпии у кресто-
цветных (Zohary, 1948; Войтенко, 1968). Наиболее типичный
вариант неэквивалентной ее формы свойствен всем видам Sinapis
и Hirschfeldia, части видов Brassica, Diplotaxis и др. Вальварная
часть их стручка многосемянная, вскрывающаяся, а короткий
носик — односемянный, невскрывающийся. Противоположный ва-
риант — плод дикой редьки (Raphanus): ее членистый стручок —
стилярная часть плода, а вальварный членик сильно редуцирован,
1—2-семянный, невскрывающийся. Такой же плод у всех видов
Enarthrocarpus.
Проращивание семян гетерокарпных крестоцветных выявило
закономерности, характеризующие гетерокарпию в целом: разно-
типные диаспоры существенно различаются по глубине покоя
и интенсивности прорастания семян. Глубокий покой или понижен-
ная всхожесть свойственны барохорным диаспорам; противо-
положные показатели — аллохорным 1 (Войтенко, 19696). Так,
после 9 мес сухого хранения интактные диаспоры, проращиваемые
в почве, обнаружили следующие показатели всхожести (%; первая
цифра — аллохорные диаспоры, вторая — барохорные):
Hirschfeldia incana 98—О
Campyloptera carnea 96—О
Sameraria boissieriana 100—1
Sinapis arvensis 88—4
Diptychocarpus strictus 97—12
Brassica sisymbrioides 100—16
Онтогенетические факторы, регулирующие размещение и
числовое соотношение гетероморфных плодов на особи, очень
1 Аллохорные диаспоры — это диаспоры, разносимые различными агентами;
барохорные — опадающие под действием силы тяжести.
7 Р. Е. Левина
97
мало изучены. Однако известные факты позволяют сделать
некоторые обобщения.
При гетерофрагмокарпии любые разнотипные части плода
(мерикарпии, членики, эремы) всегда занимают в плоде строго
фиксированное положение. Для гетероголокарпии однозначного
ответа дать нельзя. В этом случае решающую роль играет струк-
тура соцветия.
В специализированных соцветиях типа сложноцветных, т. е.
ограниченных в росте, с плотным размещением цветков и плодов,
действуют морфогенетические корреляции; в силу этого форма
плода определяется его положением в соплодии. Так, у ноготков
когтевидные семянки всегда являются краевыми, а кольцевид-
ные — центральными. Строго фиксировано периферическое по-
ложение барохорных семянок у крестовника, диморфотеки и др.
Числовое соотношение разнотипных плодов в корзинке также
зависит от структуры соцветия и его общей плодовитости. У ногот-
ков очень высокая положительная корреляция ( + 0.798) между
плодовитостью корзинки и числом кольцевидных плодов (Левина,
Марасов, 1976). То же наблюдается у диморфотеки в отношении
крылатых семянок.
В соцветиях типа крестоцветных, с неограниченным ростом
и осями ветвления разного порядка (и возраста), в роли регули-
рующего фактора выступает физиологическое состояние всей
особи. У самерарии и кампилоптеры в течение трех лет на всех
модельных особях мы наблюдали синхронную (в один и тот же
день) смену типа плода .на цветоносах всех порядков ветвления.
При удалении части цветков или цветоносов плоды обоих типов
смещались топографически, но последовательность их формирова-
ния, как и числовое соотношение, сохранялись- (Левина, Марасов,
1976) / Таким образом, у самерарии, например, бескрылые и крыла-
тые плоды правильнее характеризовать не топографически (ниж-
ние и верхние), а хронологически (ранние и поздние).
Независимо от типа соцветия у гетероголокарпных видов
в оптимальных условиях развития наблюдается преобладание
плодов аллохорных с легко прорастающими семенами (60—80 %).
Можно полагать, что преобладание этого типа диаспор повышает
генетическое разнообразие и устойчивость популяции (Левина,
1980).
98
Глава V
ЭКОЛОГИЯ РАСПРОСТРАНЕНИЯ
ПЛОДОВ И СЕМЯН
ОСНОВНЫЕ понятия и ТЕРМИНЫ
Одна из важнейших функций плода — распространение семян
(диссеминация). Именно с этой функцией связано морфологи-
ческое разнообразие плодов. Однако следует учитывать, что те
или иные морфологические особенности плодов возникали в силу
морфогенетических корреляций или параллелизма морфогенеза,
а механизм диссеминации может быть заключен в морфологии
вегетативных органов плодоносящего растения.
Таким образом, по., морфологии плода, изолированного от
растения, далеко не всегда можно судить о способе диссеминации.
Вместе с тем морфология плода определяет природу образований,
выполняющих функццю размножения и расселения. Ими могут
быть голые семена, части плода (плодики, мерикарпии, членики,
эремы, косточки), односемянные плоды или даже соплодия. Все эти
образования в русской литературе обозначались термином «зача-
ток» (по Хитрово), Но этот же термин чаще употребляется в дру-
гом смысле — как начальная стадия развития какого-либо органа.
Поэтому в сфере экологии расселения растений лучше пользо-
ваться международными терминами «диссеминула», или «диа-
спора», равнозначными по смыслу и этимологии. Оба термина
давно приняты в литературе,- но более употребителен «диаспора»
(Sernander, 1906), очевидно, как более короткий и удобный
в произношении.
Для обозначения процесса рассеивания диаспор термин «дис-
семинация» предпочтительнее «дисперсии» (Pijlv, d., 1969, 1982).
Дисперсия — слишком широкое понятие, принятое в математике,
физике, химии, тогда как диссеминация — понятие чисто биологи-
ческое.
Отрасль науки, изучающая процесс диссеминации, не имеет
общепринятого названия;: Чаще всего ее именуют, по Ульбриху
(Ulbrich, 1928), карпобиологией. Но это название явно неудовле-
творительно. Во-первых, диаспора даже у покрытосемянных расте-
ний.— не только плод, не говоря уже о спорах или, вегетативных
диаспорах. Во-вторых, диссеминация — это сложный экологи-
ческий процесс биоценотического уровня, который нельзя сводить
к биологии плода.
Надо признать удачным термин «диаспорология», предложен-
7:
99
ный Мюллером и Лотцкой (Miiller-Schneider, Lhotska, 1971).
В это понятие можно включить и экологическую морфологию
диаспор, и характер их экспонирования на растении, и разные
стороны процесса диссеминации (агенты, механизмы, эффектив-
ность). В цитированной работе авторы предлагают и классифика-
цию растений по способам диссеминации. Но в ней ряд положений
дискуссионен, кроме того, она слишком детальна для наших целей
и изобилует многими непривычными терминами.
В настоящей главе освещаются основные способы диссемина-
ции, наименование которых широко известно: зоохория, анемо-
хория, баллистохория, гидрохория, автохория и антропохория.
Под способом здесь понимается и агент, и механизм диссеминации,
т. е. характер функционирования агента в зависимости от типа
адаптаций диаспор или всего растения. Перечисленные выше спо-
собы диссеминации названы лишь по агентам. Дальнейшая клас-
сификация каждого из способов по механизмам диссеминации
будет дана в соответствующих разделах главы. Особой оговорки
заслуживает «баллистохория». Этот термин предлагается впервые,
хотя понятие «баллисты» используется часто. Баллистохория
в отличие от других способов названа не по агенту, а по механизму
диссеминации (лат. ballista — метательная машина). Как будет
показано далее, этот механизм не сопряжен с каким-либо одним
агентом.
Принятая нами последовательность изложения материала в из-
вестной мере отражает эволюционный уровень специализации
того или иного способа и степень его распространенности в при-
роде.
ЗООХОРИЯ
Связи между животными и плодами (семенами) растений
относятся к числу древнейших, многообразных и устойчивых
консортивных связей, сложившихся в биоценозах. Об этом свиде-
тельствуют, с одной стороны, экологическая морфология многих
видов плодов и семян, с другой — наличие разных групп живот-
ных, преимущественно и даже исключительно плодоядных.
Эти отношения «животные—плоды» можно определить как тро-
фофорический мутуализм: «платой» за пищу является распростра-
нение семян этими агентами, или иначе зоохория. Существенно
различаются 2 формы зоохории, связанные с поеданием плодов
или их частей: 1) эндозоохория, при которой диаспоры проходят
через пищеварительный тракт животного, а затем семена выбра-
сываются с погадкой или пометом; 2) синзоохория — активное
растаскивание диаспор в гнезда для поедания или в «кладовые»
запаса кормов. При этом часть диаспор теряется, а некоторые
запасы остаются неиспользованными. Третья форма зоохории
осуществляется животными «бескорыстно»: они пассивно перено-
сят на поверхности тела прицепившиеся или прилипшие диаспоры.
100
Таблица 8
Сопряженность агентов и форм зоохории
Основные агенты Формы диссеминации
эндозоохория синзоохория эпизоохория
Муравьи Рептилии + +
Птицы +
Грызуны Крыланы + 4-
Летучие мыши +
Приматы +
Травоядные + +
Хищники" + (?) +
Эта форма диссеминации — эпизоохория — может иметь разную
степень специализации.
Агентами разных форм зоохории являются животные, принад-
лежащие к различным систематическим группам, что показано
в табл. 8.
Перейдем к более детальной характеристике различных форм
зоохории.
ЭНДОЗООХОРИЯ
Регулярная адаптивная эндозоохория обусловлена соответ-
ствующей специализацией диаспор. Морфологически они крайне
разнообразны: семена с саркотестой, костянки и ягоды, плоды
особого вида — яблоко, гесперидий; очень разные сочные соплодия
от таких мелких, как у шелковицы, до гигантских хлебного дерева.
Но все эти диаспоры экологически сходны двумя чертами: нали-
чием съедобной ткани, которая своей окраской, запахом, вкусом
служит специальной приманкой для агентов диссеминации, и проч-
ной защитой зародыша семени (склеротеста, эндокарпий, семен-
ная кожура). Пока семена не созреют и не разовьются их защит-
ные ткани, плоды остаются зелеными, неприметными на фоне
листьев, жесткими и несъедобными на вкус.
Крайняя разнородность морфологической природы сочных диа-
спор, их принадлежность к семействам, занимающим самое разное
положение в филеме, систематический состав потребителей сочных
диаспор от рептилий до приматов — все это говорит о том, что
эндозоохория возникала многократно и независимо в ходе сопря-
женной эволюции животных и растений. Но есть основание по-
лагать, что именно этот способ диссеминации был самым древним
у семенных растений. Об этом свидетельствует характер семян
таких архаичных голосемянных, как гинкговые и саговниковые,
с их хорошо развитой саркотестой и прочной склеротестой. Потре-
бителями этих семян были мезозойские рептилии, что подтвержда-
ется некоторыми палеонтологическими находками.
101
Современные рептилии не играют заметной роли в зоохории,
и в нашей таблице они названы в числе основных агентов с учетом
их значения в прошлом. Семена современных саговниковых по-
требляют самые различные животные: обезьяны, грызуны, летучие
лисицы и др.
Сопоставляя различия сочных плодов с поведением и физио-
логическими особенностями их потребителей, Пейл (Pijl v. d.,
1969) устанавливает «синдромы» (сочетания) признаков диаспор,
поедаемых рептилиями, птицами, рукокрылыми. Но эти категории
крайне условны, так как ряд признаков разных синдромов сов-
падает, а одни и те же плоды, в особенности тропических видов,
поедаются самыми различными животными.
В биоценозах внетропической зоны основными агентами
эндозоохории являются птицы. И хотя среди них нельзя назвать
виды исключительно плодоядные, практически все сочные плоды
наших лесов, болот и тундры потребляются птицами. Так,
по данным орнитологов, дрозды поедают более 30 видов плодов,
рябчик — более 20, тетерев—15, куропатки — 9, зарянка — 8
и т. д. С другой стороны, плодами брусники и черники питаются
23 вида птиц, водяники — 20, шиповников — 13, голубики — 10,
рябины— 14 видов и т. д. (Левина, 1957).
Однако поедание плодов — это лишь начальный этап эндо-
зоохории. Для реального расселения вида необходимо, чтобы
семена, прошедшие через пищеварительный тракт, сохраняли
свою жизнеспособность. С этой точки зрения, как отмечает
А. Н. Формозов, наиболее полезны мелкие насекомоядные птицы
с относительно слабым мускулистым желудком: дрозды, зарянки,
свиристели, славки и др. Не случайно сочные плоды обычно
созревают к осени, когда значительно убывает количество насеко-
мых и птицы переходят на растительную пищу. Некоторые виды
плодов не опадают всю зиму и поедаются постепенно, вплоть
до весны (рябина, кизильник, бирючина и др.).
Орнитохорные плоды характеризуются яркой или контрастной
окраской и лишены запаха, что соответствует острому зрению
птиц и слаборазвитому обонянию.
Специальные исследования показали, что семена, прошедшие
через кишечник дроздов и других видов воробьиных, сохраняют
всхожесть на 80—88 % (Дементьев, 1940). Крупные семена
и косточки отрыгиваются из зоба с погадкой и полностью
сохраняют свою всхожесть.
Кроме птиц сочные плоды в наших лесах поедаются и млеко-
питающими, главным образом грызунами и хищниками. Из копыт-
ных кабан, олень европейский и косуля едят плоды диких яблони
и груши, косуля сибирская — плоды актинидии и амурского
винограда.
Из мышевидных грызунов потребителями многих видов сочных
плодов являются рыжая полевка и желтогорлая мышь. В их кор-
мах нередки плоды боярышника, брусники, бузины, калины,
костяники, рябины, черники, шиповника и др.
102
Сочные плоды входят в пищевой рацион многих типичных
хищников — шакала, песца, барсука, соболя, куницы, лисицы,
а при недостатке животной пищи — россомахи и горностая.
Лисица и горностай поедают плоды брусники, водяники, голубики,
черники, а лисица — еще и ягоды ландыша. В очень больших
количествах сочные плоды поедают медведи: маньчжурский —
плоды актинидии, винограда амурского, черемухи Маака; бурый
медведь — плоды яблони и груши, малины, рябины, черемухи,
водяники, черники, клюквы.
Роль названных здесь групп млекопитающих в распростране-
нии семян остается неизученной. Имеются лишь указания на то,
что на помете медведя появляются обильные всходы рябины или
водяники.
В тропических лесах птицы также относятся к числу основных
агентов эндозоохории. Среди них много видов крупных плодояд-
ных птиц (попугаи, голуби, птицы-носороги, фазаны и др.), способ-
ных заглатывать целиком крупные диаспоры, такие как костянки
пальм или семена какао. Птиц особенно привлекают ярко и контра-
стно окрашенные плоды или семена. Так, темно-голубые костянки
с оранжевой купулой и плодоножкой сассафраса беловатого
(Sassafras albidum, Lauraceae) поедаются 18 видами птиц. Охотно
потребляются плоды и других видов лавровых, ягоды логаниевых,
яркоокрашенные костянки пальм, плоды анноновых, многих
сапотовых и ряда других (Имханицкая, 1980а, 19806, 1985).
К числу «избранных» орнитохорных диаспор относятся ярко-
окрашенные семена (виды бобовых), черные семена с ярким
ариллусом (Polygala, Myristica, Sterculia) или погруженные
в яркую мякоть раскрытых плодов (Theobrotna). Некоторые
птицы (вороны, сороки, дикие голуби, грифы) поедают плоды
масличной пальмы; чаще масличные плоды употребляют хищники.
Для тропической фауны характерны особые группы плодояд-
ных млекопитающих — рукокрылые и обезьяны, которые также
заслуживают «ранга» основных агентов эндозоохории.
Из рукокрылых исключительно плодоядными видами представ-
лен подотряд крыланов — крупных рукокрылых: летучие собаки,
летучие лисицы и др. Подотряд летучих мышей — преимущест-
венно насекомоядные, но есть среди них и плодоядные виды.
У рукокрылых хорошо развит зубной аппарат, острое обоняние
и слабо развито зрение, особенно у видов, ведущих ночной образ
жизни. Этим особенностям соответствуют и признаки плодов,
поедаемых преимущественно рукокрылыми: крупные, блеклоокра-
шенные, часто с прочным экзокарпием и сильным запахом (обычно
неприятным). Характерно также и положение этих плодов,
свисающих на длинных плодоножках или сидящих на стволах.
Излюбленную пищу рукокрылых составляют плоды кауликарпи-
ческих пальм, многих тутовых, включая легендарный анчар
(Antiaris toxic aria} и виды Ficus, плоды семейств анакардиевых,
анноновых, сапотовых, хризобалановых. Очень охотно поедаются
плоды банановых, сочные плоды и соплодия мареновых и др.
юз
Приматы (обезьяны) обладают более развитым, чем у других
млекопитающих, цветовым зрением, хорошим обонянием, креп-
кими зубами, хватательными передними конечностями. Кроме
того, среди обезьян имеются виды, живущие на деревьях и ходя-
щие по земле. Поэтому им доступны самые разные плоды, поеда-
емые и птицами, и рукокрылыми. Они легко справляются с круп-
ными «панцирными» плодами с прочным кожистым или твердым
экзокарпием. Так, любой другой пище орангутанг предпочитает
плоды дурьяна (Durio zibethinus) и мангустана (Garcinia tnango-
stana), а шимпанзе — фикуса и дынного дерева (Carica papaya).
Плоды дикорастущих гарциний поедаются мартышками (Pij 1 v. d.,
1982). «Хлебом обезьян» называют плоды широко известного
африканского баобаба (Adansonia digitata), крупные толстостен-
ные коробочки, заполненные белой кисловатой мякотью с множе-
ством погруженных в нее семян. А бразильский «обезьяний
банан» — плод порцелии (Porcelia goyazensis, Annonaceae).
Охотно и в больших количествах поедаются обезьянами плоды
и других анноновых, многих пальм, цитрусовых, настоящего
банана, мареновых (Gardenia), логаниевых (Strychnos), маранто-
вых, стеркулиевых и ряда других. В плодах какао они поедают
только мякоть, выплевывая семена.
Потребителями плодов тропических растений являются
и другие млекопитающие: слоны, жвачные и другие копытные,
отчасти грызуны и хищники. Африканский слон поедает плоды
пальм — гифены, пальмиры (Borasus flabellifer) и особенно
охотно крупные (до 20 см в диам.) костянки пальмы делеб
(В. aethiopum). В состав его пищи входят и плоды баобаба,
многих бобовых, некоторых сапотовых. Вид этого семейства
Tieghemella heckelii — источник ценной древесины — распростра-
няется вдоль дорог слонов. Индийский слон потребляет плоды
дурьяна и манго (Mangifera).
Жвачные, в частности антилопы, играют большую роль в рас-
пространении тропических бобовых (Fabaceae s. 1.) с мясистыми
плодами (виды Acacia, Cassia, Tamarindus, Ceratonia и др.).
Их чрезвычайно твердые семена не разрушаются жевательными
зубами, но вполне вероятно, что происходит скарификация семян,
что способствует их прорастанию. Замечено, что пионерами
древесных видов на травянистых пространствах являются
Acacia arabica и A. horrida, по-видимому заносимые травоядными
(Pijl V. d., 1982).
Опавшие плоды пальм поедают тапиры, олени, лани, козы,
крупный рогатый скот. Травоядных и хищников привлекают
масличные плоды пальм. На Яве плодами кариоты питаются
шакалы, пальмовая куница, виверры. Жирафы, антилопы, буйволы
потребляют сочные плоды мареновых (Имханицкая, 1985).
В заключение обзора адаптивной эндозоохории очень коротко
о рептилиях. Современные рептилии в подавляющем большинстве
питаются животной пищей. Немногие растительноядные виды
поедают плоды обычно вместе с зеленым кормом. Так, в желудке
104
коробчатой черепахи с остатками растений обнаружены зрелые
«семена» Fragaria, Rubus, Vitis, Morus, Diospyros, Polygonum.
Но известны факты потребления рептилиями сочных плодов.
На Галапагосских островах черепахи поедают плоды кактусов
и томатов, при этом стимулируется прорастание семян. На Борнео
черепах привлекают плоды базикарпических видов дурьяна.
Вараны и черепахи также охотно поедают плоды салакки
съедобной (Salacca edulis) — бесстебельной пальмы Малайского
архипелага. Лазящие игуаны едят костянки «своего» каркаса
игуанового (Celtis .iguanaea) — лианы неотропических лесов.
В рацион рептилий входят также плоды анноновых: асимины
карликовой (Asimina pygmaea) — пища наземных черепах, ан-
ноны голой (Annona glabra) — «яблоко» аллигатора, но им «уго-
щаются» и игуаны (Pijl v. d., 1969; Имханицкая, 19806).
Инадаптивной эндозоохорией можно считать распространение
сухи-х диаспор, поедаемых попутно с зеленым кормом. Для ряда
видов грызунов и зерноядных птиц сухие плоды и семена являются
не случайной, а постоянной пищей, но здесь имеет место лишь
истребление семян. Грызуны так «обрабатывают» резцами и пе-
режевывают семенной корм, что в нем уже не остается живых
семян. Такому же разрушению подвергаются семена, и в сильных
мускулистых желудках зерноядных птиц.
Таким образом, эндозоохорное распространение сухих диаспор
может осуществляться лишь травоядными. Во многих регионах
практически агентами такой диссеминации являются виды домаш-
него скота, поскольку степи повсеместно распаханы, а фауна
диких травоядных крайне оскудела.
Нередко для оценки роли домашнего скота в распространении
семян используют данные о содержании всхожих семян в навозе,
вносимом в пахотные земли. Но этот показатель абсолютно не пра-
вомерен. Во-первых, в навоз попадает большое количество
интактных семян из подстилки и остатков корма. Во-вторых,
собирание, вывозка и разбрасывание огромного количества
навоза — это целенаправленная деятельность человека, а не зоо-
хория. О действительной роли травоядных в эндозоохории можно
судить лишь по видовому составу и количеству всхожих семян
в помете этих животных. Некоторые данные по результатам таких
исследований (своих и других авторов) приводит Мюллер
(Muller-Schneider, 1977). Более 15 видов всхожих семян обнару-
жено в помете северного и европейского оленей; в их числе мятлик
луговой, тимофеевка альпийская, некоторые осоки, щавель кислый,
лютик ползучий, марь белая, клевер и др. Значительный процент
семян сохраняет всхожесть, пройдя через пищеварительный
тракт домашних животных (овцы, крупный рогатый скот). Это
преимущественно виды с мелкими хорошо защищенными семенами
(плодами): марь белая, лебеда садовая, подорожники большой
и ланцетолистный, горец щавелелистный, щавель кислый и щаве-
лек, крапива двудомная, виды клевера и др. Всхожесть таких
семян — от 25—30 до 100 % (клевера луговой и ползучий).
105
Для обогащения пастбищ клевером ползучим в некоторых странах
используют овец, выпасая их предварительно на участках, где
этот вид обильно плодоносит.
Количество и видовой состав семян в помете животных могут
быть очень различны в зависимости от сезона, типа пастбища,
вида скота. По данным С. А. Котта (1947), на пастбищах
Подмосковья в 3 кг свежего коровьего помета содержится не-
поврежденных семян: в июле — 294, августе — 1537, сентябре —
7610. Если принять среднюю всхожесть этих семян в 25 %,
то масштабы такой формы эндозоохории становятся очевидны.
СИНЗООХОРИЯ
Адаптивная синзоохория также связана с поеданием диаспор
или их частей. Но характер этих диаспор, как и адаптаций агентов,
совершенно иные. С этой точки зрения явно различаются 2 типа
диаспор и соответствующих им агентов: 1) крупные ореховидные
диаспоры, распространяемые птицами и грызунами; 2) мелкие
сухие «муравьиные» диаспоры, снабженные специальными соч-
ными придатками.
Диаспоры первого типа.богаты концентрированным (масло,
крахмал, белки) запасом пищи для зародыша; этот запас привле-
кает и агентов диссеминации. Птицы и грызуны, поедая такие
диаспоры, полностью их разрушают. Но крупные с прочным
одревесневшим покровом диаспоры не потребляются на месте
произрастания. Они уносятся в гнезда или специальные «кла-
довые» запаса кормов. Успешная диссеминации осуществляется
за счет диаспор, утерянных при переносе или неиспользованных
из запасов. Наиболее специализированные агенты синзоохории
отличаются не только инстинктом собирания и запасания корма,
но и морфологическими особенностями: мешки защечные (бурун-
дук, хомяки, суслики) или подъязычные (кедровка), особое
строение лап, клюва, пищевода (сойка) и др.
Типичные синзоохорные диаспоры — это семена и плоды
деревьев, реже кустарников: семена всех видов «кедровых» сосен
(Pinus sibirica, Р. koraiensis, Р. pumila, Р. cembra, Р. pined),
желуди дуба, бука и каштана, семена конского каштана, орехи
лещины, плоды грецкого ореха и других ореховых, плоды липы.
Но в запасах мышевидных грызунов встречаются и сочные плоды
(см. выше), и анемохорные (клена, ясеня, ильма), что можно
рассматривать как факультативную синзоохорию. Таким же
необязательным способом диссеминации является растаскивание
белками и мышевидными грызунами плодов некоторых пальм
или мелких сухих диаспор травянистых растений.
Обратимся к реальным примерам синзоохории. Среди ее аген-
тов — птиц первое место принадлежит кедровкам и сойке, некото-
рую рол играют клесты, дятлы, ворон.
Кедровка, или ореховка (Nucifraga caryocatactes) в разных
106
ее разновидностях распространена очень широко в лесной зоне
всей Евразии. Ее можно назвать всеядной птицей, но основным
кормом для нее служат ореховидные плоды и семена: орехи
лещины, желуди, семена «кедровых» сосен. Существование сибир-
ской разновидности кедровки тесно связано с урожаями семян
сибирской, корейской и карликовой сосен. Кедровки уничтожают
до половины всего урожая семян, но благодаря резко выражен-
ному инстинкту запасания корма они играют основную роль
в расселении этих видов (Бибиков, 1948).
В особом подъязычном мешке у кедровки помещается до 40—
80 штук «кедровых орехов», а иногда и больше. Кедровки устраи-
вают свои кладовые как в лесу, так и в горной тундре выше
границы леса, очень часто на гарях и вырубках, способствуя
возобновлению сосны. В кладовые они иногда зарывают целые
шишки по 5—6 штук, но чаще всего запасы состоят из семян,
в среднем по 6—12 штук (от 2 до 22 и более). Количество кладовых
в зависимости от обилия семенных деревьев может достигать
нескольких сотен и даже 2—3 тыс. на гектар. Хотя часть этих
запасов потребляется «хозяевами» и очень много уничтожается
грызунами, кедровки несомненно способствуют семенному возоб-
новлению «кедровых» сосен. Расстояния, на которые кедровки
разносят семена, измеряются сотнями метров или немногими
километрами (по единичным указаниям, до 7—9 км). Радиус
возобновления кедра на гарях обычно составляет 0.5—1 км
от семенников (Бибиков, 1948; Новиков, 1953).
В Подмосковье и Ярославской обл., по наблюдениям А. Н. Фор-
мозова, в ельниках с подлеском из лещины ее орехи в массе запа-
саются кедровками. В западноевропейских лесах в числе других
кормов они питаются семенами сосны европейской (Pinus cembra).
Расселение этого вида в Альпах выше границы леса осуществля-
ется также за счет запасов кедровок (Muller-Schneider, 1977).
Сойка (Garrulus glandarius) также относится к числу широко
распространенных всеядных птиц, но большое место в ее питании
занимают желуди, на что указывают ее видовое название, строение
клюва, лап и пищевода. Благодаря растяжимости пищевода
сойка может заглатывать до 7 желудей среднего размера, что
облегчает устройство значительных зимних запасов. В расселении
дуба сойка играет очень существенную, если не основную роль.
Семенной подрост дуба отмечается в радиусе 100—150 м от семен-
ников, а в единичных случаях — в нескольких километрах от дуб-
рав. На юге сойки распространяют плоды бука, каштана и грецкого
ореха.жДо 200 м от дерева уносят его плоды и вороны (Новиков,
1948; Холодный, 1949).
Из числа грызунов как агенты синзоохории наибольшее зна-
чение имеют белки и бурундуки, гораздо меньшее — мышевидные.
Основу пищи белки обыкновенной составляют семена хвойных
(в тайге), желуди, орехи лещины, а белки закавказской, кроме того,
плоды бука и каштана. Зимние запасы белок способствуют рас-
селению этих видов. Обитатель тайги — бурундук — к осени
107
собирает в своих норах большие запасы плодов и семян. В этих
кладовых особенно много «кедровых орехов».
Из отряда мышевидных зимние запасы плодов и семян соби-
рают курганчиковая и лесная мыши, рыжая полевка, серый хо-
мячок, туркестанская крыса и др. Большие (по 10—15 кг) запасы
названных здесь видов мышей мало способствуют расселению
растений. Эффективнее маленькие (5—10 диаспор) запасы рыжей
полевки, спрятанные в случайных местах. Серый хомячок собирает
свои запасы в радиусе 10—50 и даже 100—200 м от норы. В отдель-
ных случаях он уносит в своих защечных мешках семена кустар-
ников и трав даже на 500 м. Туркестанская крыса — эндемик
Средней Азии — является агентом расселения грецкого ореха.
Мирмекохория — распространение диаспор муравьями (гр.
myrmex — муравей) — является, по-видимому, наиболее прогрес-
сивной формой синзоохории и зоохории в целом. В пользу такого
вывода говорит ряд фактов. Во-первых, муравьи — молодая
ветвь в эволюции насекомых, а муравьи-собиратели обладают
сложным инстинктом запасания и обработки корма. Весьма
специализированы и мирмекохорные диаспоры (семена, односе-
мянные сухие плоды или плодики, мерикарпии, эремы): они
защищены от повреждения прочным покровом и снабжены специ-
альной приманкой для муравьев. Ею служит особый сочный
придаток—элайосом, богатый маслом и другими питательными
веществами. Установлено (Bresinsky, 1963), что чрезвычайно
быстрая реакция муравьев на элайосомы вызывается присутствием
в них особых летучих аттрактантов.
Морфологическая природа элайосома крайне различна: у се-
мян это ариллусы или разного рода ариллоиды, у плодов — сочные
базальные участки перикарпия, основания столбика и т. п.
(рис. 57; Левина, 1957; Pijl v. d., 1969). Специализация растений-
мирмекохоров выражается не только в характере их диаспор,
но и в особенностях плодоношения. Они рано цветут, и осыпание
диаспор происходит уже в конце весны—начале лета, т. е. в период,
когда муравьи выкармливают личинок и особенно активны в соби-
рании пищи. Кроме того, у многих плодоносящих мирмекохоров
стебли слабые, поникающие или даже полегающие, что облегчает
растаскивание диаспор муравьями.
По нашим наблюдениям и данным других авторов, муравьи
собирают только свежевыпавшие диаспоры, пока не засыхают
их элайосомы. У мирмекохоров тенистых лесов уносится более
80—90 % опавших диаспор. Добыча утаскивается в муравейники,
где потребляются только придатки, а сами диаспоры, вполне
жизнеспособные, периодически выбрасываются. Некоторая часть
диаспор теряется и по пути к муравейнику.
Муравьи широко распространены во всех наземных биоцено-
зах, за исключением тундр. Особенно богата муравьями фауна
тропиков обоих полушарий. В СССР и Западной Европе больше
видов обитает в широколиственных лесах.
Мюллер (Miiller-Schneider, 1977) называет более 30 видов
108
Рис. 57. Мирмекохорные диаспоры.
а, б — семена фиалок волосистой и полевой (Viola hirta, V. arvensis), в — семя чистотела
(Chelidonium majus), г — плодик перелески (Hepatica nobilis), д — эрем незабудки (Муо-
sotis sp.), е — плод осоки горной (Carex montana) (а—в — по Левиной, 1967а; г—е —
по Ульбриху).
растений, распространяемых муравьями в гариге близ Монпелье;
среди них есть злаки и бобовые (включая виды люцерны и кле-
вера) , губоцветные, сложноцветные и др. О мирмекохории в тро-
пиках дают представление работы Пейла (Pij'l v. d., 1955, 1969).
Здесь назовем только два-оригинальных мирмекохора: древовид-
ная Sebastiana из молочайных (Аргентина), плоды которой,
взрываясь, разбрасывают семена с карункулой — элайосомом,
и Dendromecon (Papaveraceae) — ксерофитный кустарник из ча-
парали Калифорнии. Много сведений о мирмекохорах австралий-
ской флоры и их диссеминации можно найти в работе Р. Берга
(Berg, 1975).
Подавляющее большинство мирмекохоров в любых ценозах —
это травянистые растения. В нашей флоре — преимущественно
лесные виды: фиалки, марьянники, хохлатки, кислица, копытень,
пролесник (Mercurialis), мерингия (Moehringia trinervia), меду-
ницы, Петров крест, перловники, осоки (Carex digitata, С. mon-
tana, С. sylvatica), пролеска (Scilla sibirica), звездчатка (Stel-
laria holostea).
Степные группировки беднее мирмекохорами, чем леса, но в се-
верных степях и на сухих склонах их встречается немало. Среди
них адонис весенний, касатик безлистный, молочай, истод (Poly-
gala'), ленец (Thesium), птицемлечник (Ornithogalum), гусиный
лук, живучка (Ajuga), осоки (Carex ericetorum, С. ornithopoda,
С. ргаесох) и др. Мирмекохорные виды можно назвать и среди
луговых и рудеральных растений: окопник лекарственный, молочай
болотный, бодяк съедобный, чистотел, яснотка, нонея и др.
109
Благодаря широкому ареалу муравьев, обилию среди них
растительноядных видов, исключительно высокой численности
особей и их большому «трудолюбию», т. е. активности в собирании
корма, мирмекохория является одним из важнейших способов
массовой диссеминации, но на малые расстояния. Обычный радиус
разноса диаспор муравьями — в пределах 10—12 м. Только в еди-
ничных случаях отмечался занос на 15, 27, 45 и 70 м.
ЭПИЗООХОРИЯ
Эта форма зоохории носит характер комменсализма, поскольку
животное лишь пассивно переносит на теле диаспоры, не извлекая
для себя никакой пользы. Адаптивная эпизоохория связана со
специализацией диаспор, которые могут быть цепкими, липкими
или ослизняющимися. Не меньшее значение имеет инадаптивная
(факультативная) эпизоохория, при которой любые мелкие диа-
споры разносятся вместе с прилипшей сырой почвой или илом.
Прицепками иногда служат видоизмененные стилодии орешков
(рогоглавник, гравилат), но чаще всего это выросты околоплод-
ника или вторичных покровов плода (гипантия, прицветников)
и даже соплодия (обертки сложноцветных). По морфологической
природе и систематической принадлежности цепкие диаспоры
крайне разнородны. Это орешки голые или в гипантии (репешок),
бобы (виды люцерны, эспарцета, солодки), семянки (череда,
кельпиния); мерикарпии зонтичных (морковь, тургения, торилис)
или мареновых (подмаренник цепкий); эремы бурачниковых
(липучка, чернокорень). В этом перечне, как и в природе, отсут-
ствуют цепкие семена.
Цепкие соплодия известны у сложноцветных (лопух, дурниш-
ник, некоторые васильки), реже у маревых (гораниновия). Цепкие
плоды и соплодия не опадают самостоятельно, а срываются
животными, к телу которых цепляются. У эпизоохоров сухих
и пустынных пастбищ плоды опадают или развиваются на лежа-
чих стеблях. Эти плоды с прочным деревянистым околоплодником,
снабженные острыми и твердыми шипами, переносятся на ногах
животных, вонзаясь в копыта и мягкие ткани пальцев и ступни.
Такого типа плоды встречаются в самых различных семействах,
особенно разнообразны они у кунжутовых и парнолистниковых;
в нашей флоре — у якорцев.1
На поверхности липких плодов (или их вторичных покровов)
развиваются клейкие желёзки, а в плодоножках — слой отдели-
тельной ткани. Приклеившись к телу проходящего животного,
плоды очень легко отрываются по отделительному слою (шалфей
клейкий, линнея, виды плюмбаговых).
Ослизнение диаспор, чаще всего семян, в зависимости от усло-
вий обитания может иметь разное биологическое значение. В ряде
случаев оно способствует и эпизоохории.
1 Рисунки различных эпизоохорных диаспор см. в главе II.
НО
Агентами распространения цепких и липких диаспор являются
крупные млекопитающие — хищники и главным образом травояд-
ные, а в современных условиях — это в основном домашний
скот. Поэтому эпизоохоры — чаще всего сорно-рудеральные
и пастбищные или прибрежные растения. Последние и распростра-
няются хищниками и лесными копытными, постоянно посещаю-
щими водоемы для питья.
Домашний скот является массовым агентом и факультативной
эпизоохории. Вместе с сырой почвой животные разносят на ногах
множество мелких плодов и семян луговых и сорно-рудеральных
видов, расселяя их вдоль дорог, на выгонах и т. п.
Особую роль в факультативной эпизоохории играют водо-
плавающие и болотные перелетные птицы. Они переносят на ог-
ромные расстояния диаспоры водных и болотных растений, при-
липшие к ногам и оперению вместе с илом и слизью. Такой способ
расселения Дарвин считал одним из факторов.космополитности
ряда гидрофитов.
ЗНАЧЕНИЕ И ЭФФЕКТИВНОСТЬ ЗООХОРИИ
С момента появления семенных растений зоохория стала
существенным фактором их расселения. В ходе эволюции с увели-
чением многообразия фауны и флоры Земли специализировались
и формы зоохории. В настоящее время она несомненно является
важнейшим способом расселения цветковых растений почти
во всех наземных биоценозах.
В ходе географической, экологической, видовой и структурной
дифференциации фитоценозов возникла более или менее тесная
приуроченность форм и агентов зоохории не только к типам расти-
тельных группировок, но и к их ярусам. Так, во внетропических
широтах основные агенты эндозоохории — птицы, а в тропических
лесах не меньшую роль играют особые группы млекопитающих
(крыланы и обезьяны). В то же время во внетропических лесах
эндоорнитохория свойственна деревьям и кустарникам второго
и третьего ярусов, а мирмекохория — видам травяного покрова.
Инадаптивная эндозоохория и эпизоохория, осуществляемые
травоядными, характерны для сорно-рудеральных и пастбищных
растений. Наконец, эпиорнитохория играет большую роль в рас-
селении водно-болотных видов.
Оценивая эффективность зоохории, как и любого способа
диссеминации, необходимо различать массовость и дальность
регулярного распространения диаспор. Формами наиболее мас-
совой диссеминации можно считать эндозоохорию травоядными
и мирмекохорию. Но и у некоторых крыланов (калонг) наблюда-
ются скопления в сотни и тысячи особей, поедающих огромные
количества плодов. «Кладовые» кедровок иногда насчитываются
тысячами на 1 га, и сойки могут переносить за сезон тысячи
желудей.
111
По дальности диссеминации выделяются 2 формы: самая
ближняя — мирмекохория, измеряемая единицами, реже десят-
ками метров, и самая дальняя — эпиорнитохория; благодаря ей
в некоторых случаях возможна миграция за пределы ареала.
Остальные формы зоохории при участии птиц и млекопитающих
более или менее равноценны по дальности расселения: в основном
оно осуществляется в радиусе сотен метров или единичных кило-
метров.
Итак, мы можем дать следующую оценку зоохории в целом:
она охватывает все без исключения наземные биоценозы, гранди-
озна по своим масштабам, свойственна множеству древесно-
кустарниковых и травянистых растений, обеспечивает постепенное
и регулярное расселение вида в пределах или по периферии его
ареала, а в виде исключения может способствовать очень далеким
миграциям.
АНЕМОХОРИЯ
Для наземных растений наряду с животными к основным
агентам диссеминации относится ветер. В сравнении с животными
он представляется вполне однородным по характеру воздействия
на диаспоры. В действительности ветер крайне изменчив по напра-
влению и скорости; помимо горизонтальных постоянно возникают
вихревые движения, обусловливающие порывистость ветра, и вер-
тикальные восходящие токи воздуха. Также хорошо известно,
что скорость ветра резко меняется с высотой, а в приземном слое
воздуха — в зависимости от характера напочвенного покрова.
На скорости ветра в сильнейшей степени сказываются и особен-
ности растительного покрова: плотность крон и ярусность леса,
высота и плотность травостоя в степях и т. п.
ФОРМЫ АНЕМОХОРИИ И ТИПЫ ДИАСПОР
С выходом растений на сушу они, естественно, стали анемохо-
рами, так как споры при их ничтожной массе легко переносятся
ветром. Диаспоры семенных растений для разноса ветром нуж-
даются в специальных аэродинамических структурах, т. е. таких,
которые помогали бы им держаться в воздухе, замедляли их паде-
ние и оказывали бы сопротивление воздушному потоку.
В ходе эволюции у семенных растений действительно возникли
крайне разнообразные анемохорные диаспоры разной степени
специализации. В зависимости от того, каким образом ветер
перемещает эти диаспоры, различают несколько форм анемохории:
1) анемоаэрохория 1 — перенос диаспор по воздуху; 2) анемогео-
хория — перекатывание по поверхности земли; 3) анемогидро-
1 «Метеорохория» зарубежных авторов.
112
хория — перенос диаспор ветром по поверхности стоячей воды;
4) анемохионохория — перенос ветром по снежному насту.
Самая распространенная форма — аэрохория,1 которой и соот-
ветствуют самые разнообразные типы диаспор. Хионохория,
напротив, свойственна немногим видам, у которых опадение
диаспор, хотя бы частично, происходит зимой. Геохория также
характеризуется особой структурой диаспор, а по воде и насту
переносятся плоды и семена аэрохорного типа.
Аэрохорные диаспоры при всем их морфологическом разно-
образии можно разбить на 2 группы по степени их аэродинамиче-
ской специализации: эванемохорные — диаспоры, способные
к полету, особенности которых понятны из их названий (см. ниже),
и гемианемохорные — низкого аэродинамического качества
(неспособные к полету), у которых ветер лишь удлиняет траекто-
рию падения.
В дальнейшем мы рассмотрим следующие типы диаспор.
А. Аэрохорные
1. Эванемохорные
а) парящие
ai) очень мелкие с низкой абсолютной массой
аг) с парашютом
б) планирующие
б]) без вращения
бг) авторотирующие
2. Гемианемохорные
Б. Геохорные
а) типа воздушного шара1 2
б) типа воздушной турбины
АЭРОХОРИЯ. АЭРОХОРНЫЕ ДИАСПОРЫ
Парящие диаспоры нашей первой группы — это так
называемые пылеватые семена ряда семейств, отличающихся
чрезвычайно высокой плодовитостью: грушанковых, орхидных,
близких к ним бурманниевых (Burmanniaceae) и корсиевых
(Korsiaceae), непентовых, заразиховых, некоторых геснериевых
и др. Размеры таких семян почти микроскопические, а масса
одного семени — сотые или тысячные доли миллиграмма. Кроме
того, у иных видов семя имеет пузыревидно вздутую оболочку
(рис. 58, а). Такие идеальные парители поднимаются даже очень
слабыми восходящими токами воздуха и длительное время не осе-
дают, попадая в зоны различных воздушных потоков. Виды
с пылеватыми семенами — это чаще всего либо эпифиты тропи-
ческих лесов, либо наземные низкорослые растения густых тени-
1 В дальнейшем для краткости, но без ущерба для смысла, приставка «анемо»
опускается.
2 Pneumatocarpia некоторых авторов (Roth, 1977).
8 Р. Е. Левина ] (3
стых лесов, глубоких ущелий, расселин скал, т. е. таких место-
обитаний, где всегда царит безветрие.
Диаспоры с парашютом, напротив, свойственны обитателям
открытых пространств. Парашютным приспособлением чаще всего
служит пучок волосков — хохолок. Волосовидные придатки раз-
ного типа встречаются на семенах, орешках, семянках, зерновках
в самых различных семействах. Независимо от морфологической
природы волоски гигроскопичны. Только в сухом воздухе хохолки
расправляются и могут служить летательным аппаратом. Поэтому
в сухую жаркую погоду диаспоры уносятся порывами ветра
из своих вместилищ (корзинок, коробочек и т. п.), а затем,
попадая в восходящие потоки воздуха, поднимаются на значитель-
ную высоту.
Волоски могут покрывать равномерно всю поверхность плода
или семени, находиться у их основания (семена ив и тополей,
орешки рогозов; см. рис. 11, а) и т. п. Но более эффективны
и гораздо чаще встречаются хохолки на верхушке семени
(рис. 58, б, в) или плода. Такие хохолки типичны для сложноцвет-
ных (см. рис. 31). Положение хохолка выше центра тяжести
придает диаспоре устойчивое положение в воздухе, особенно
если хохолки приподняты на особых выростах («носиках»), как
у одуванчиков, козлобородников и др. (рис. 31, а, 58, г).
Хохолки на верхушке семян и плодов, как это видно на рисун-
ках, обычно имеют форму обращенного конического парашюта,
что также повышает поперечную устойчивость диаспор. Но основ-
ное значение такой формы парашюта заключается в том, что
он образует положительный угол атаки относительно горизонталь-
ных потоков воздуха и, следовательно, повышает динамическую
подъемную силу, действующую на диаспору. Важное аэродинами-
ческое значение имеет также лабильность формы хохолков: при
горизонтальных порывах ветра они сплющиваются, т. е. угол
атаки возрастает до 90° и парашют превращается в парус,
оказывающий максимальное сопротивление воздушному потоку,
что способствует дальнему заносу плодов и семян. Когда ветер
стихает, форма парашюта восстанавливается и эти диаспоры
снова могут парить до новых порывов ветра. Наиболее совершенны
перистые хохолки, свойственные многим сложноцветным (рис. 31,
г, 58, г).
Перистые очень длинные и коленчато изогнутые ости на зернов-
ках ковылей и аналогичные им выросты (см. рис. 9, г) при горизон-
тальных порывах ветра служат парусом, а в турбулентных потоках
действуют по принципу геликоптера, т. е. способствуют подъему
диаспоры.
Планирующие плоды и семена принципиально отлича-
ются от парящих и своей морфологией, и аэродинамическими
свойствами, и, следовательно, характером полета.
Под планированием понимают полет со снижением по наклон-
ной траектории под действием силы тяжести. Траектория такого
полета будет тем длиннее, чем медленнее происходит снижение,
114
Рис. 58. Анемоаэрохорные диаспоры.
а — семя заразихи (Orobanche с и mana), б — семя кипрея (Epllobium montanum), в —
семя ваточника (Asclepias syriaca), г — плод козлобородника (Tragopogon dubius),
д, е — семена бигноний (Bignonia sp., В. fraxinifolia), ж— семя диоскореи (Dloscorea
dodecaneura) (а—в, д — по Ульбриху, г, е, ж— по Веттштейну).
что достигается наличием аэродинамических поверхностей, таких,
например, как крылья планера. Планирующие плоды и семена
также снабжены крыловидными придатками самого различного
происхождения, очень разных размеров, очертаний и других
параметров.
Из аэродинамики известно, что при планировании дальность
полета по горизонтали определяется высотой взлета или, для
наших объектов, высотой, с которой падает диаспора. С каждым
метром высоты взлета (или падения) дальность перемещения
по горизонтали возрастает на 10—30 м, а при ветре еще больше.
Вот почему крылатые плоды и семена свойственны преимущест-
венно высокоствольным деревьям или лианам.
Выделенные нами 2 группы планирующих диаспор существенно
различаются аэродинамически. У первой группы центр тяжести
совпадает с центром давления,1 в силу чего они снижаются без
вращения. Вторую группу составляют диаспоры, у которых эти
центры не совпадают, что и вызывает вращение планера при
падении.
У плодов и семян первой группы семя занимает центральное
положение, а крылья или двусторонне-симметричные, или равно-
1 Центром давления называют точку приложения полной аэродинамической
силы, т. е. результирующей подъемной силы и лобового сопротивления.
8»
115
Рис. 59. Авторотирующие диаспоры.
а, б — крылатки клена и ясеня, в — семя сосны, г — орешек айланта; сплошной стрелкой
показаны оси вращения, штриховой — направление вращения (по Левиной, 1967а).
мерно окружают семя. К этой группе относятся семена многих
бирнониевых, диоскорейных, некоторых логаниевых (Norrisia),
мареновых (Cinchona), фиалковых (древесные лианы Anchietea,
Agamea), а также крылатковидные бобы птерокарпуса, крылатки
березы, вяза, птелеи, птерокарии и др. (рис. 12, в, 35, а, в, 58, д—
ж).
Группу авторотирующих диаспор составляют крылатые семена
хвойных, орешки тюльпанного дерева, однокрылый боб центро-
лобиума (см. рис. 12, а), крылатки ясеня, всех видов клена и близ-
ких к ним сапиндовых (см. рис. 20, в, г), плодики айланта. У всех
названных здесь семян и плодов, кроме айланта, единственное
крыло резко асимметрично относительно центра тяжести. Поэ-
тому при падении диаспоры продольная ось ее наклонена. Оси
вращения некоторых планеров показаны на рис. 59. Своеобразная
морфблогия плодика айланта обусловливает и иной характер его
вращения (рис. 59, г). Авторотация диаспор создает силу тяги,
замедляющую падение, а наклонное положение крыла обусловли-
вает горизонтальный полет под действием ветра.
Гемианемохорные диаспоры — группа очень сборная. Общая
их черта — низкое аэродинамическое качество: они либо вовсе
не имеют никаких анемохорных придатков, либо эти придатки
малоэффективны для несущих их массивных диаспор.
К первой подгруппе мы относим мелкие семянки полыней
и зерновки многих злаков (Agrostis, Eragrostis, Koeleria, Phleum,
Роа и т. п.). Вторая подгруппа более пестрая. В нее включаются
семена с крыловидной каймой некоторых лилейных, льнянки,
погремка; зерновки в опушенных чешуях (Alopecurus, Еппеаро-
gon, Melica); односемянные бобы во вздутых чашечках (Anthyl-
lis macrocephala, Trifolium aureum). В этой же категории диаспор
следует назвать относительно массивные семянки наголоваток
116
(Jurinea) и некоторых васильков с короткими щетинистыми
хохолками; семянки ворсянковых с воронковидной закраиной
наружной чашечки и т. п.
Отметим, что все названные здесь виды — травянистые, часто
низкорослые растения. Их крайне разнородные плоды и семена
не парят и не планируют в воздухе. Ветер лишь относит их на
некоторое расстояние от материнского растения.
АНЕМОГЕОХОРИЯ И ДРУГИЕ ФОРМЫ АНЕМОХОРИИ
Перекатывание плодов по поверхности почвы возможно лишь
в местообитаниях с разреженным травяным покровом, поэтому
анемогеохорные плоды особенно характерны для песчаных пу-
стынь. Благодаря своей легкости и подвижности они передвига-
ются быстрее песка и «спасаются бегством» от погребения.
Анемогеохорные плоды имеют обтекаемую форму (что умень-
шает их трение о почву) и высокое профильное сопротивление
воздушному потоку при низкой плотности. Плоды типа воздушного
шара — это чаще всего сильновздутые бобы, например у некото-
рых астрагалов, пузырника (см. рис. 7, а, 6, д, е), эремоспартона,
смирновии. У осоки вздутой плод заключен в эллипсоидальный
сильновздутый замкнутый мешочек (см. рис. 28). Полые почти
шаровидные плоды леонтики сомнительной (Leontice incerta,
Berberidaceae) достигают 45 мм в диам.
Плоды типа воздушной турбины, шаровидные или эллип-
соидальные по общему очертанию, напротив, не имеют гладкой
поверхности, а несут несколько меридиональных крыльев, как
у ревеня, многих видов джузгуна (см. рис. 27, г, д'), некоторых
стираксовых (Halesia, Pterostyrax). Сила трения о почву у таких
плодов ничтожная при очень большой аэродинамической поверх-
ности. В любом положении плода воздушный поток ударяется
в лопасть крыла, образующего угол атаки. Таким образом очень
полно используется энергия порывов ветра. К этому же типу
плодов можно отнести и плоды с многочисленными радиальными
выростами (см. рис. 27, е), у которых высокое профильное сопро-
тивление создается за счет трения воздушного потока.
Особую форму анемогеохории представляет собой диссемина-
ция растений, именуемых «перекати-поле».1 Однако такая жизнен-
ная форма не всегда связана с анемохорией. У некоторых много-
летников надземная часть растения отрывается уже после плодо-
ношения, что является способом самоочистки (Алехин, 1939).
Но чаще всего у многолетних видов перекати-поле, как например
качима метельчатого (Gypsophyla paniculata), синеголовника рав-
нинного (Eryngiutn campestre), отрыв и перекатывание растения
служат средством и самоочистки, и диссеминации. Лишь у одно-
летников — рогача песчаного, клоповника сорного, василька рас-
1 М. Э. Кирпичников предложил называть их первольвеитами (Левина, 1967а).
117
кидистого, солянки южной (Salsola australis) — форма перекати-
поле связана только с диссеминацией.
Характер рассеивания диаспор у видов перекати-поле принци-
пиально отличается от всех других форм анемохории, так как
сами диаспоры обычно не анемохорного типа.
Анемогидрохория, а тем более анемохионохория, не могут рас-
сматриваться как регулярные способы диссеминации. Уже было
отмечено, что они не связаны с особыми типами диаспор, поскольку
служат лишь дополнением к анемоаэрохории у тех же видов.
По глади водоемов чаще всего переносятся опушенные плоды
и семена прибрежных растений (волоски не смачиваются во-
дой!) — ивовых, рогозов, пушицы, посконника (Eupatorium сап-
nabinum), а также крылатки ольхи и березы.
По снежному насту переносятся крылатые семена хвойных,
плоды ясеня и других древесных растений, у которых обсеменение
продолжается и зимой.
ЭФФЕКТИВНОСТЬ И ЗНАЧЕНИЕ АНЕМОХОРИИ
Напомним, что эффективность того или иного способа диссеми-
нации необходимо оценивать не только по дальности, но и по
массовости заноса диаспор. Единичные семена, унесенные штор-
мовым ветром на многие километры от материнского растения,
не могут обеспечить реального расселения вида. Поэтому важна
не дальность заноса в километрах, а дальность эффективной
диссеминации, т. е. расстояние действительного «шага рассе-
ления».
По массовости распространения диаспор анемохория является
бесспорно самым эффективным способом диссеминации, что опре-
деляется тремя обстоятельствами: число анемохорных видов
чрезвычайно велико; они отличаются очень высокой плодови-
тостью; все полноценные диаспоры, производимые особью, рано
или поздно полностью разносятся ветром в любой сезон года.
В силу того, что ветер — постоянно действующий фактор
и часто достигает большой силы, укоренилось представление,
что он может разносить семена и плоды на расстояния порядка
многих километров. Конкретных данных о дальности анемохории
крайне мало из-за трудности прямых наблюдений. Но те реальные
цифры, которые имеются в литературе, не подтверждают бытую-
щих представлений.
Решающую роль в дальности анемохорного разноса диаспор
играют высота растения и характер его окружения. По нашим
’наблюдениям, мелкие орешки ветреницы лесной, снабженные
длинными волосками, оседают на расстоянии 0.15—0.30 м от
материнской особи, так как этот вид находится в 3—4-м ярусе
густого травостоя. Семена иван-чая, обитающего на вырубках
и полянах и достигающего 1—2 м высоты, разносятся обильно
на расстояние 3—15 м и единичные семена — на 45—60 м. Наши
118
наблюдения в степи над другими видами анемохоров с прекрасно
развитыми хохолками на плодах или семенах (ластовень, черто-
полох, крестовник) показали: значительное число диаспор рассеи-
вается на 0.5—4 м и единичные — на 7.5—9 м.
Для древесных растений приводятся иные показатели даль-
ности разноса семян и плодов: для осины, березы, сосны — 1 —
2 км, для кленов — 4—5 км (Нестеров, 1950; Miiller-Schneider,
1977). Но едва ли это расстояния массового заноса диаспор.
Семенной подрост осины, березы, ели, сосны наблюдался в 300—
450 м от ближайших семенников (Молчанов, Шиманюк, 1949).
Эти цифры, по-видимому, отражают реальную картину расселения
древесных анемохоров. Лесоводы отмечают, что воздушные тече-
ния, пройдя через лес в открытое поле, начинают снижаться
и достигают почвы на расстоянии, равном примерно десятикратной
высоте насаждения.
Многолетняя лесоводственная практика подсказывает опти-
мальную ширину лесосек, рассчитанных на естественное Семенное
возобновление, в 100 м. Считается, таким образом, что надежный
массовый самосев осуществляется на расстоянии 50 м от стены
леса.
Итак, анемохория свойственна огромному числу видов семен-
ных растений самой различной систематической принадлеж-
ности — от хвойных до сложноцветных и орхидных. Естественно,
что и анемохорные диаспоры чрезвычайно разнородны по проис-
хождению, морфологии, степени аэродинамической специали-
зации.
Анемохория играет большую роль во всех, без исключения,
наземных фитоценозах от таежных лесов до жарких пустынь;
при этом различные формы анемохории приурочены к тем или иным
растительным группировкам и их ярусам.
Все эти черты анемохории позволяют заключить, что она
наряду с зоохорией явилась одним из факторов морфологической
эволюции диаспор и экологической дифференциации раститель-
ного покрова. Некоторой иллюстрацией к сказанному могут слу-
жить следующие цифры. Среди 153 видов — эдификаторов расти-
тельного покрова СССР анемохоры (всех групп) составляют 68 %.
В числе эдификаторов отдельных ботанико-географических об-
ластей анемохоры составляют: в Евразиатской таежной области —
16 видов из 17, Евразиатской степной — 18 из 35, Азиатской пу-
стынной— 42 из 49 видов (Левина, 1958).
Различные фитоценозы довольно четко характеризуются сво-
ими типами анемохорных диаспор: 1) все древесные анемохоры,
за исключением ивовых, имеют крылатые плоды или семена;
это относится также к деревьям и кустарникам пустынь; 2) для
травяного покрова хвойных лесов, сфагновых болот и эпифитов
тропических лесов характерны виды с пылеватыми семенами;
3) плоды и семена с перистыми парашютами свойственны расте-
ниям низинных болот, степным и сорным видам; 4) в песчаных
пустынях широко распространены анемогеохорные плоды.
119
Анемохория не способствует скачкообразно дальним мигра-
циям видов, она надежно обеспечивает заселение всех благоприят-
ных территорий в пределах ареала или по его периферии.
БАЛЛИСТОХОРИЯ
Под баллистохорией мы понимаем разбрасывание диаспор
метанием. Этот способ диссеминации отличается от всех других
двумя особенностями: 1) морфологический механизм диссемина-
ции заключен в структуре не плода или семени, а вегетативных
и флоральных органов растения в период обсеменения; 2) агент,
обеспечивающий диссеминацию, не специфичен; «метателем»
может оказаться любой фактор, раскачивающий плодоносящее
растение. В природе такими факторами обычно являются ветер
или животные. На основании этого некоторые авторы относят
баллистов и к анемохорам, и к зоохорам (Miiller-Schneider, Lhot-
ska, 1971), а иногда выделяют еще и гидробаллистов (влияние
дождя). Такое размещение баллистов по разным группам не
оправдано, так как анемобаллисты будут успешно раскачиваться
животными, а еще успешнее сельскохозяйственными машинами
(лобогрейка, комбайн). В то же время баллисты четко характери-
зуются общими морфологическими чертами. Их прямостоячий
стебель, цветоносы и цветоножки к периоду созревания становятся
упругими, пружинящими. Все они обладают приспособлениями,
препятствующими самопроизвольному осыпанию диаспор.
Целому ряду баллистов свойственны анемохорные плоды или
семена; эти виды составляют особую категорию баллистов-анемо-
хоров.
По характеру препятствий, задерживающих свободное опаде-
ние диаспор, можно выделить 4 группы баллистов.
1. Тип гвоздичных. Это самая многочисленная и разно-
родная по систематическому составу группа. У всех ее представи-
телей многосемянные вскрывающиеся плоды и диаспорой служит
семя. Коробочки, листовки или бобы занимают вертикальное
положение и вскрываются вверху одним или несколькими неболь-
шими отверстиями. Такое положение коробочек типично для гвоз-
дичных, норичниковых, лилейных, колокольчиков, мака и многих
других. К баллистам этого типа относятся лютиковые с полимер-
ными или олигомерными многолистовками, а из бобовых —
многие астрагалы и остролодочники.
Для многих баллистов типа гвоздичных характерны карпо-
тропические изгибы цветоножек. Сущность этого явления понятна
из рис. 60. В тех редких случаях, когда зрелые коробочки остаются
поникшими, они открываются в базальной части, т. е. геометри-
чески вверху (некоторые колокольчики, ортилия однобокая —
Ortilia secunda).
К баллистам-анемохорам типа гвоздичных относятся орхидные,
грушанковые, заразиховые, колокольчики и др. Семена с крыло-
120
Рис. 60. Карпотропические изгибы цветоножек у баллистов.
а, б — положение цветков и плодов у чемерицы (Veratr ит nigrum), в, г — то же у зимо-
любки (Chimaphilla umbellata) (по Левиной, 1957).
видной каймой известны у некоторых лилейных, погремка, льнянки
обыкновенной.
2. Тип губоцветных. К нему относятся преимущественно
виды этого семейства. Диаспорами являются эремы, характер
рассеивания которых определяется морфологией чашечки. Имеют
значение также упругие сильно пружинящие плодоножки (виды
шалфея, змееголовника, Черноголовки): достаточно толкнуть ча-
шечку сверху, чтобы эремы из нее выбросились. Такие толчки
могут производить, по-видимому, крупные прыгающие насекомые.
К баллистам типа губоцветных относятся и некоторые бурач-
никовые (виды синяка, оносмы).
3. Тип сложноцветных. Кроме видов этого семейства
он включает некоторых ворсянковых. У сложноцветных-баллистов
в отличие от анемохоров листочки обертки при плодах не отгиба-
ются книзу, а поднимаются, препятствуя свободному опадению
семянок. Высота обертки различна у разных видов, но она коррели-
рует с размером плодов. При горизонтальном положении корзинок
и таких мелких семянках, как у видов Anthemis, Leucanthemum,
Pyrethrum, Tanacetum, обертка в 2 мм выс. служит достаточным
барьером. Кроме названных к баллистам относятся виды наголо-
ватки, василька, цикория, тысячелистника.
У ворсянковых обертка не играет заметной роли при обсемене-
нии; свободному опадению семянок препятствуют обильные вы-
росты цветоложа (щетинки, пленки).
4. Тип зонтичных включает виды только этого семейства.
Они не имеют никаких внешнеморфологических приспособлений,
препятствующих опадению плодов, но даже совершенно зрелые
мерикарпии довольно прочно сидят на карпофоре и рассеиваются
121
только при раскачивании стебля или соплодия. У огромного
большинства видов зонтичных стебли высокие, прямостоячие
и полые, что придает им упругость. В период плодоношения
сухими и упругими становятся также лучи зонтика и плодоножки.
Среди зонтичных встречаются и баллисты-анемохоры с плоскими
крылатыми мерикарпиями (Heracleum, Ferula, Pastlnaca и др.).
Баллистохория свойственна очень многим видам травянистых
растений из самых различных семейств. Вместе с тем можно
назвать ряд семейств, у которых все или почти все виды являются
баллистами: гвоздичные, грушанковые, губоцветные, заразиховые,
зонтичные, колокольчиковые.
Баллисты встречаются в различных ценозах, но преобладают
в степях, чаще всего в первом и втором ярусах. Среди характерных
видов разных вариантов степей баллисты составляют 24—40 %.
По эффективности баллистохория относится к способам массо-
вой диссеминации на близкие расстояния. Наблюдения показали,
что у баллистов-неанемохоров основная масса диаспор рассеива-
ется в радиусе, равном высоте растения, и только единичные из них
преодолевают расстояние вдвое большее.
ГИДРОХОРИЯ
Вода, конечно, не является таким универсальным агентом
массовой диссеминации наземных растений, как ветер или живот-
ные. Но с точки зрения топографии расселения гидрохория носит
более постоянный характер, чем анемо- или зоохория, так как
границы морей и океанов, направление морских течений, русла рек,
направления стока и гидрографическая сеть определенной терри-
тории остаются более или менее постоянными в течение очень
длительного времени.
В распространении диаспор водой четко выделяются адаптив-
ная регулярная гидрохория, связанная со специализацией диа-
спор, и факультативная, или случайная. Первая кроме гидрофитов
свойственна растениям морских побережий и речных долин. Вто-
рая осуществляется потоками дождевых и талых вод или искус-
ственных оросительных каналов. Все эти временные потоки пере-
носят самые разные диаспоры сухопутных растений, не приспо-
собленные к гидрохории. Масштабы и эффективность этой случай-
ной гидрохории совершенно не изучены. Имеются лишь некоторые
данные о переносе диаспор сорных видов водой оросительных
каналов, что можно назвать антропогидрохорией (Левина, 1967а).
Но в последнее время основным способом орошения становится
дождевание, при котором поливные воды фильтруются и, следо-
вательно, антропогидрохория уже не имеет места.
При всем морфологическом разнообразии гидрохорных диа-
спор характер их приспособлений однотипен. Они сводятся к не-
смачиваемости и низкой плотности диаспоры и к надежной защите
зародыша от воды, особенно морской. Несмачиваемость достига-
ется восковым налетом, наличием масла в оболочке плода или
122
семени, густым коротким опушением, ямчатой поверхностью и т. п.
Низкая плотность обусловлена различного рода воздухоносными
тканями или полостями.
К распространению морскими течениями отлично приспособ-
лены плоды ряда пальм. Во II главе подробно описано строение
сухой костянки кокосовой пальмы с ее прочным гладким несмачи-
ваемым экзокарпием, сухим рыхловолокнистым мезокарпием
и маслянистым эндоспермом. Аналогичное строение имеют плоды
и других пальм — сейшельской, бетелевой, пальмиры, нипы,
а также некоторых хризобалановых. Далеко не у всех пальм плоды
таких гигантских размеров, как у сейшельской или кокосовой;
у некоторых они измеряются миллиметрами. Вместе с тем гидро-
хория играет большую роль в расселении очень многих пальм,
обитающих на морских побережьях, по краю мангровых зарослей,
по заболоченным лесам и берегам рек, особенно обильно Амазонки,
Ориноко и их притоков. Широко разносятся реками и паводковой
водой плоды мауриции извилистой, рафии, метроксилона и др.
(Имханицкая, 1985).
В числе гидрохоров тропических широт назовем еще эритьеру
прибрежную (Heritiera littoralis, Sterculiaceae), килеватые плоды
которой по нескольку недель могут плавать в морской воде;
Donax cannaeformis (Marantaceae) с морских побережий Малай-
ского архипелага — его плоды с пробковым эндокарпием раз-
носятся морскими течениями; церберу (Cerbera manghas} из
кутровых, широко распространенную по морским побережьям
от Мадагаскара до Новой Гвинеи.
Плоды пресноводных гидрохоров обычно очень мелкие, но и они
снабжены разнообразными плавательными приспособлениями; их
можно объединить, в 3 основные группы.
1. Плавательный пояс из крупных воздухоносных опробковев-
ших клеток залегает под эпидермисом. Семя защищено от намока-
ния плотным эндокарпием или семенной кожурой (частуха, стрело-
лист, ольха клейкая, лютик, многие виды зонтичных, кирказон).
2. Плотный водонепроницаемый слой покрывает диаспору
снаружи, а- плавательная ткань, залегает под ним (ежеголовник,
меч-трава, сабельник).
3. Плавучесть обеспечивается воздушными пузырями (плоды
осок со вздутыми мешочками; семена: белозора и касатика аиро-
видного с воздушными камерами в семенной кожуре; семена
кувшинки с воздухоносным ариллусом и кубышки в пенистой
слизи). Напомним, что водой могут успешно разноситься анемо-
хорные плоды и семена, снабженные хохолком.
По дальности заноса диаспор морские течения несомненно
эффективнее рек, но дальних гидрохорных миграций видов практи-
чески; не наблюдается, главным образом из-за потери всхожести
семян. Известно, например, что плоды сейшельской пальмы
обильно заносятся на Мальдивские острова, но этот вид так и
остается эндемиком Сейшельских островов. Легкие весьма плаву-
чие костянки сакоглоттиса {.Sacoglottis amazonica, Humiriaceae)
123
заносятся от устья Амазонки к Азорским островам и берегам
Западной Европы, но семена их теряют всхожесть.
Морские течения, конечно, способствуют расселению гидро-
хоров, но главным образом вдоль побережий и на ближних остро-
вах. Так, в 1963 г. в 20 милях южнее Исландии возник вулкани-
ческий остров. В 1965 г. на нем появился Cakile maritima —
обитатель морских побережий.
Реками расселяются растения вниз по течению, о чем свиде-
тельствуют ареалы некоторых видов, вытянутые вдоль речных
долин (белокопытник, девясил германский). В пределах поймы
наблюдается массовое распространение гидрохорных диаспор
полой водой.
АВТОХОРИЯ
В отличие от всех остальных аллохорных (гр. alios — другой)
способов диссеминации при автохории распространение диаспор
осуществляется без посредников (если не имеет места дипло-
хория). Существенно различаются 2 формы автохории: 1) актив-
ное разбрасывание семян благодаря функционированию особых
структурных механизмов—автомеханохория (Левина, 1957);
2) самопроизвольное опадение диаспор под влиянием силы тя-
жести — барохория.
Автомеханохория свойственна и сухим, и сочным плодам.
В деталях морфология разбрасывающих механизмов очень раз-
лична, но принцип их действия однотипен: у сухих плодов напряже-
ние создается неравномерным сокращением различных слоев
околоплодника при высыхании, в сочных плодах, напротив, на-
буханием отдельных слоев клеток и резким повышением в них
осмотического давления. Иногда механизм разбрасывания заклю-
чен в самих семенах.
В I и II главах коротко описаны механизмы вскрывания бобов,
коробочек герани и молочая, костянок крушиновых, при которых
происходит разбрасывание семян (рис. 6, а, 15, 19, а). Такого же
типа механизмы в стручках сердечника (Cardamine hirsuta,
С. impatiens), в коробочках акантовых и альстремерии (Alstro-
emeriaceae, Liliales). У пилеи (Pilea, Urticaceae) плод выбрасы-
вается сидящими у его основания и внезапно раскручивающимися
стаминодиями. Одногнездные коробочки фиалок вскрываются
дорсально тремя створками, а затем семена выталкиваются сжа-
тием створок в зоне плацент. У анаксогореи (Anaxogorea javanica)
семена также выбрасываются щелчком при вскрытии листовок.
Семенной механизм разбрасывания известен у кислицы. Под
давлением набухающего нижележащего слоя клеток наружный
слой семенной кожуры внезапно разрывается и сообщает толчок
семени. У эшольции (Eschscholtzia) семена, снабженные пружин-
ками, с силой отскакивают от плацент.
Во II главе были названы и сочные плоды, разбрасывающие
семена (недотрога, некоторые тыквенные). Упомянем еще дорсте-
124
нию (Dorstenia contra.]erva, Moraceae), у которой зрелые плоды
«катапультируются» благодаря разбуханию ткани цветоложа.
Дальность разбрасывания семян при автомеханохории прямо
коррелирует с высотой растения, силой толчка и массой семени.
Но во всех случаях радиус разброса измеряется единичными
метрами (1—4.5 — у фиалок, 2.5—6 — у гераней, 10—11—у ка-
раганы) или десятками метров (акантовые, пилея).
Автомеханохория нередко сочетается с мирмекохорией: у фиа-
лок, молочайных, кислицы семена с элайосомом.
Категория барохоров в литературе — совершенно неопределен-
ная. Так, например, Мюллер (Miiller-Schneider, 1977) включает
в нее и мангровые, и виды с ореховидными плодами, и мирмеко-
хоров с полегающими коробочками.
Барохория в нашем понимании — это специализированный
способ диссеминации, направленный на сохранение диаспор на
месте произрастания. В противоположность баллистам у баро-
хоров ничто не препятствует свободному опадению плодов и даже
имеются механизмы, способствующие' этому процессу: ломкий
колос у некоторых злаков (эгилопс, дикая рожь), отделительная
ткань на плодоножке или плодовом рубчике (овсюг, горец шеро-
ховатый, конопля сорная) и т. п. Кроме того, барохорные диаспоры
лишены придатков или приманок в виде пищевых тканей.
Барохория свойственна видам весьма специфических местооби-
таний и при этом сопровождается дополнительными особенностями
процесса диссеминации. Так, тяжелые плоды мангровых растений
опадают уже проросшими с гигантскими корешками, заякори-
вающими в иле эти необычные диаспоры.
В сухих и жарких пустынях барохория сочетается с ослизне-
нием диаспор, что также закрепляет их на месте опадения.
Наконец, барохория широко распространена у сорно-полевых
растений, для которых характерны высокая плодовитость, значи-
тельная долговечность семян и растянутый период прорастания.
Как было показано в IV главе, дифференциация диаспор одной
особи при гетерокарпии приводит к утрате анемо- или зоохорных
придатков у части плодов. Так возникают барохорные диаспоры,
имеющие явно вторичный характер.
АНТРОПОХОРИЯ
По силе и характеру воздействия на природу человек занимает
особое место среди всех экологических факторов, что общеиз-
вестно. Как агент диссеминации человек также принципиально
отличается от всех природных агентов. Поэтому под антропохорией
мы понимаем только такое распространение диаспор, которое
связано с трудовой деятельностью человека. Антропохория может
быть преднамеренная (перевозка, высев, заделка семян культур-
ных растений) и сопутствующая ей непреднамеренная, которая
относится к дикорастущим и сорным видам. Преднамеренная
антропохория выходит за рамки диаспорологии. Мы рассмотрим
125
только непреднамеренную антропохорию, как наиболее вторичный
и действенный способ диссеминации и расселения растений.
По приуроченности к тем или иным группам растений, по эф-
фективности и степени специализации диаспор четко различаются
3 формы антропохории: 1) агестохория (гр. agesthaj — везти
с собой) — распространение диаспор средствами транспорта;
2) эргазиохория (гр. ergasia — обрабатывать землю) — распро-
странение диаспор сельскохозяйственными орудиями; 3) спейро-
хория 1 (гр. speiro — сеять) — распространение диаспор сорных
видов путем высева.
АГЕСТОХОРИЯ
Эта форма антропохории не связана со специализацией диа-
спор или строго определенной группой растений. Наиболее харак-
терной чертой агестохории является дальность распространения
и быстрота роста создаваемых ею искусственных ареалов. Транс-
океанические миграции многих видов растений происходят чаще
всего благодаря межконтинентальной торговле, главным образом
зерном, хлопком и шерстью. Так, были завезены из Европы в Аме-
рику бодяк, овсюг и «русская чума» — Salsola australis-, из руде-
ральных видов — подорожник большой. Американскими пришель-
цами в Европе и в СССР оказались крупносемянные повилики,
агрессивные карантинные сорняки — амброзия (Ambrosia artemi-
siifolia) и циклахена (Cyclachaena xanthii(olia), не считая многих
общеизвестных рудеральных видов.
В расселении заносных растений внутри материка большую
роль-играет железнодорожный транспорт. Откосы железнодорож-
ных насыпей и территории небольших станций — самые обычные
местонахождения адвентивных видов. Об этом свидетельствует
большая литература прошлых лет и наблюдения последнего вре-
мени (Гусев, 1977; Цвелев, 1977; Малышева, 1980; Ильминских
с соавт., 1981; Игнатов, Игнатова, 1982; Босек, 1983; Пузырев,
1984; Мотекайтите, 1985). В иных случаях возможен и повторный
занос чужеземцев (Lhotska, Slavik, 1969; Недолужко, 1984).
Несомненно имеет значение и автомобильный транспорт. В цити-
рованной работе Игнатовы отмечают, что некоторые заносные
виды встречаются у бензоколонок.
Агестохория больше всего обогащает рудеральную флору.
Но широкому и массовому распространению адвентивных руде-
ральных видов способствует их очень высокая плодовитость и при-
родные агенты диссеминации — анемохория (мелколепестник, га-
линсога), факультативная эпизоохория (амарантусы, ромашка паху-
чая) и т. п. Легко натурализуются заносные сорно-полевые виды,
так как условия обрабатываемых полей в разных странах со сход-
1 Эти термины были предложены давно (Левина, 1944) и довольно широко
используются в нашей специальной и справочной (БСЭ, 1970, 2) литературе.
Первый и. третий термины принимают и зарубежные авторы (Muller-Schneider,
Lhotska, 1971).
126
ным климатом достаточно однородны. Гораздо реже и лишь
единичные чужеземные виды входят в состав природных фитоце-
нозов в новой для них стране.
ЭРГАЗИОХОРИЯ
Эта форма антропохории, в противоположность агестохории,
во-первых, свойственна определенным группам растений: обита-
телям сенокосов и засорителям посевов; во-вторых, способствует
распространению диаспор в пределах обрабатываемых угодий.
Многие виды сенокосных лугов отличаются ранними сроками
плодоношения, а у остальных семена быстро дозревают в скошен-
ном травостое. Вот почему в процессах сенокошения (в особен-
ности лобогрейками), ворошения, копнения, стогования и пере-
возки сена человек рассеивает огромные количества диаспор.
На полях действуют и уборочные машины, и орудия обработки
почвы, поэтому эргазиохории подвержены и те диаспоры, которые
опадают до уборки.
При раздельной комбайновой уборке хлебов обсеменению
сорняков сначала помогает срезание травостоя (особенно дейст-
вие мотовила), а затем в валках и при их подборке происходит
дозревание и осыпание плодов и семян сорных видов. Прямое
комбайнирование приводит к иному распределению сорных диа-
спор. По нашим наблюдениям, на засоренных полях под дейст-
вием режущего агрегата осыпается около 17 % диаспор сорных
видов, более 65 % уходит в полову и около 13 %—в зерно.
Отмеченная ранее барохория многих сорно-полевых растений
сочетается с эргазиохорией. При высокой плодовитости барохоров
и долговечности их семян постоянная обработка почвы приводит
к тому, что пахотный горизонт наших полей оказывается равно-
мерно насыщенным миллионами жизнеспособных семян.
СПЕЙРОХОРИЯ
Эта форма диссеминации человеком характеризуется высокой
степенью специализации диаспор и является следствием засорен-
ности посевного материала. Поэтому для осуществления спейро-
хории решающее значение имеет трудность отделения плодов
или семян засорителей от семян (в агрономическом смысле)
высеваемой культуры. Так, например, зерновки щетинника сизого
очень легко отсортировать от семян ржи или пшеницы, но очень
трудно — от проса, с которым обычно и высевается этот сорняк.
Специализация спейрохоров выражается в том, что и период
их плодоношения, и характер диаспор совпадают с соответствую-
щими признаками засоряемой культуры. Так возникла группа
специальных засорителей: овсюг, засоряющий овес; гречиха
татарская и фаллопия (горец) вьюнковая — гречиху; конопля
сорная — коноплю посевную; щетинник сизый и ежовник — просо,
и др. Плоды названных здесь видов, как и многих других, частично
127
осыпаются в поле, другая часть засоряет семена культур. Эту
группу сорных видов можно рассматривать как факультативные
спейрохоры. Дальнейшая специализация засорителей привела
к редукции анемохорных (погремок бескрылый) или зоохорных
(прицепник) придатков и, наконец, к утрате способности обсемене-
ния в поле. Эта черта характерна, в частности, для специальных
засорителей льна — торицы льняной с нераскрывающимися коро-
бочками, очень сходными с коробочками льна; горца льняного,
утратившего сочленение в плодоножке. Не высыпаются из раскры-
тых коробочек и семена куколей (Agrostema githago, A. linicola,
A. macrosperma-, Левина, 1957).
Такая степень специализации привела к возникновению группы
облигатных спейрохоров, для которых этот способ диссеминации
стал единственно возможным. С усовершенствованием методов
очистки посевного материала облигатные спейрохоры постепенно
исчезнут. Исчезающим растением стал уже куколь обыкновен-
ный — злостный засоритель зерна хлебов в прошлом.
Даже такая, самая краткая, характеристика антропохории
позволяет судить о своеобразии и эффективности этого способа
диссеминации в сравнении с природными агентами. Только вслед-
ствие антропохории (агестохории) возникают трансокеанические
дизъюнкции и космополитные ареалы; независимо от экологи-
ческих условий обеспечивается постоянная и равномерная дис-
семинация луговых и сорных видов в свойственных им ценозах
(эргазио- и спейрохория). Диссеминации приводит к возникнове-
нию видов или форм специальных засорителей (спейрохория).
СОЧЕТАНИЕ СПОСОБОВ ДИССЕМИНАЦИИ
В природе очень часто сочетаются 2 или даже более способов
диссеминации, что обозначают как дипло- или полихорию. В раз-
ных разделах настоящей главы и в IV главе уже говорилось
о сочетаниях тех или иных способов диссеминации.
Диплохория — явление крайне разнородное. Она может быть
регулярной и эпизодической; обусловливаться только морфологи-
чески, т. е. наличием двойного механизма диссеминации, или мор-
фолого-экологически. Во втором случае условия обитания опреде-
ляют характер одного из способов диссеминации или, наконец,
бифункциональность одного и того же механизма.
Эпизодической диплохорией можно назвать, например, рас-
таскивание грызунами типично анемохорных плодов клена, ясеня,
вяза, по-видимому, в условиях бескормицы.
Необходимо различать диплохорию диаспор, когда одни и те же
диаспоры распространяются разными способами, и диплохорию
вида, когда разными способами распространяются диаспоры одной
особи (при гетерокарпии) или однородные диаспоры разных
особей и даже популяций (при агестохории).
Ниже впервые предлагается предварительная классификация
форм регулярной дицлохории с некоторыми пояснениями.
128
I. Диплохория диаспор
А. Морфологически обусловленная
1. Двойной механизм диссеминации
а) оба механизма вне диаспоры (бешеный огурец —
автомехано- + эпизоохория)
б) один механизм вне диаспоры, второй в диаспоре
(фиалки, молочайные — автомехано- -Т мирмеко-
хория; грушанковые, орхидные и др. — баллисты-
анемохоры)
в) оба механизма в диаспоре (кислица — авто-
механо- + мирмекохория; аннона голая и др. —
эндозоо- + гидрохория; вишня — эндозоо- + син-
зоохория; турретия — эпизо- + анемохория)
Б. Морфолого-экологически обусловленная
1. Бифункциональность механизма
а) побережья рек, озер, болот (ивовые, рогоз, бело-
копытник и др. — анемо- + гидрохория)
2. Действие адаптивного и инадаптивного способов
а) посевы (марь белая, овсюг, щетинник, ежовник
и мн. др. — баро- + эргазиохория, баро- + спейро-
хория)
б) рудеральные местообитания (большинство руде-
ральных видов — баро- или баллистохория + фа-
культативная эпизоохория)
II. Диплохория вида
А. Морфологически обусловленная
1. Разнотипность диаспор одной особи — гетерохория
(Левина, 1957) (анемо- и барохория; эпизоо- и баро-
хория — см. главу IV)
2. Бифункциональность механизма (ячмень гри-
вастый — эпизоо- или анемогеохория; ковыли — ане-
мо- или эпизоохория)
Б. Морфолого-экологически обусловленная
1. Действие адаптивного и инадаптивного способов
(мелколепестник, галинсога — агесто- и анемохория).
Многие примеры, приведенные в классификации, уже описаны
ранее. Поясним только некоторые. Аннона голая, александрий-
ский лавр (Calophyllum inophyllum, Clusiaceae) и др. — обитатели
побережий. Их сочные плоды поедаются разными животными,
а семена гидрохорного типа долго плавают в воде. У вишни
синзоохорно распространяются косточки. Турретия (Turretia,
Bignoniaceae) образует коробочки, покрытые острыми прицеп-
ками, с мелкими анемохорными семенами. Коробочки срываются
и уносятся на теле животных, а семена постепенно разлетаются.
У ячменя гривастого (Hordeum jubatum) отдельные колоски разно-
сятся эпизоохорно, а клубки соплодий перекатываются ветром.
Наша классификация диплохории в значительной мере условна
и не отражает всех вариантов и комбинаций, происходящих
в природе. Но и такой предварительный обзор заставляет при-
9 Р. Е. Левина
129
знать, что процесс диссеминации — понятие более сложное, чем
способ. Процесс диссеминации вида может реализоваться раз-
лично в зависимости от условий обитания и складываться из
разных этапов: и способов диссеминации.
ЗАКЛЮЧЕНИЕ
Материал настоящей главы позволяет утверждать, что мор-
фологическое многообразие плодов в значительной степени свя-
зано со способами диссеминации. Ине только морфология плодов,
но часто и многие черты всего растения обеспечивают этот процесс
(мирмекохоры, баллисты, перекати-поле).
Каково же биологическое значение процесса диссеминации,
определившегово многом морфологическую эволюцию плодов
и семян? Диссеминация для прикрепленных растений — обяза-
тельное условие их расселения, благодаря которому достигается:
а) расширение площади, занимаемой популяцией или даже видом;
б) равномерное распределение особей в популяции, что снижает
конкуренцию; в) более высокий эффект перекрестного опыления
(Дарвин) благодаря «растеканию генов» (Pijl v. d., 1969);
г) заселение освободившихся участков суши или вновь возникших
островов; д) «уход» от неблагоприятных факторов (очагов болез-
ней и вредителей, изменившихся эдафических условий и т. п.).
Кроме того, расселение растений — это не только функция видовой
жизни. Оно лежит в основе флорогенеза и смены фитоценозов.
Можно выделить- З^ категории способов диссеминации по их
эффективности: мирмекохория, баллистохория, барохория и эрга-
зиохория — способы массовой ближней диссеминации; остальные
формы зоохории; анемон и гидрохория обеспечивают относительно
дальнюю диссеминацию в пределах десятков или сотен метров
(до единичных километров). Совершенно особое место по быстроте
расселения и дальности диссеминации занимает агестохория,
создающая трансокеанические дизъюнкции. Из природных агентов
наиболее дальний занос осуществляют морские течения, но для
действительного расселения рекордно далекий «заплыв» отдель-
ных диаспор не эффективен: они либо теряют всхожесть,, либо
не находят необходимых для развития условий.
Но если разные способы диссеминации более или менее равно-
ценны по своей эффективности, то почему они столь.разнообразны
и еще комбинируются в разных сочетаниях? Во-первых, на протя-
жении эволюции покрытосемянных экологические адаптации диа-
спор возникали многократно на разной морфологической основе;
во-вторых, многообразие диаспор и способов диссеминации под-
держивается отбором; оно позволяет максимально использовать
самые разные агенты расселения; куда не занесет ветер, занесет
белка; то, что выбросит птица, утащит мышь, и т. п. Только много-
образие способов диссеминации позволяет растениям занимать
самые разные экологические ниши — от стволов в тропическом
лесу (эпифиты) до низин в жарких песчаных пустынях.
130
Глава VP
ПРАКТИЧЕСКОЕ ЗНАЧЕНИЕ ПЛОДОВ
Плоды растений сыграли выдающуюся роль в истории чело-
вечества. Начало земледелия (а вслед за ним оседлости и государ-
ственности) связано с использованием плодов. Но их значение
не умалилось и в наши дни: ради плодов возделываются важней-
шие культуры мирового земледелия — пшеница, рис, кукуруза,
хлопчатник, виноград, кофе, яблоня, цитрусовые..
Плоды доставляют к нашему столу хлеб и,крупу, макароны
и другие мучные изделия, отличные растительные масла — коко-
совое, оливковое, кунжутное, подсолнечное; - растительные белки
и такие привычные овощи, как томаты, огурцы. Натуральные
или переработанные плоды — это всевозможные фрукты и ягоды,
варенье, джемы и компоты, кофе и какао, шоколад и халва и. . .
всего не перечислишь.1
Плоды «заботятся» и о нашем здоровье: они дают такие сильно-
действующие средства, как опий й кофеин, строфантин и стрихнин,
касторовое и облепиховое масла, вату и бинты; многие плоды —
настоящие кладовые витаминов; самые различные виды плодов
успешно применяются в народной и официальной медицине.
Плоды являются источником; ценнейших эфирных и техни-
ческих жирных масел; нередко масло одного и того же вида имеет
пищевое, техническое и медицинское значение. Плоды дают пре-
восходное прядильное (хлопчатник) и набивочное (виды Ceiba)
волокна и множество: других необходимых материалов и веществ.
В натуральном виде употребляют лишь немногие виды плодов.
Значительное большинство их подвергается различной пере-
работке; при этом используются либо плод полностью, либо
отдельные части околоплодника или семени. Но известны и плоды-
«комбинаты», удовлетворяющие самые разные потребности. Так,
плоды хлопчатника кроме прядильного волокна дают жирное
масло (семена), вату (хлопковый пух) и целлюлозу (хлопковый
пух и стенки коробочек). Из плодов кокосовой пальмы получают
питательный освежающий напиток — кокосовое молоко (незрелый
эндосперм), пищевое масло (из зрелого эндосперма), ценный
поделочный материал (эндокарпий)У
Перечисление и описание всех культурных и дикорастущих
1 Описание многих плодов пищевых растений можно найти в книге «Плоды
земли» (Франке с соавт., .1979).
9*
131
Таблица 9
Характер использования плодов
Категории и названия растений 1 Используемые части плодов
в натуральном виде в переработан- ном виде в виде экстрагиро- ванных веществ
ПИЩЕВЫЕ
Крахмалоносные (хлебные и крупяные)
Пшеница, рис, рожь, кукуруза, ячмень, овес, просо, могар (Se- taria), сорго (Sorghum) Гречиха Хлебное дерево, джекфрут Весь плод Семя Соплодие
Белковые
Горох, фасоль, чечевица, соя, нут (Cicer), бобы (Faba), чина Семена
Жирномасличные
Подсолнечник, арахис, мак Кунжут (Sesamum), соя Горчица, рыжик, сафлор (Carthamnus), конопля, хлоп- чатник Кукуруза Маслина Кокосовая пальма Масличная пальма Семена Жидкий эндо- сперм » » Семена » » Зародыш семени Мезокарпий Твердый эндо- сперм (копра) Семя
Плодовые Сочноплодные
Актинидия, земляника, ма- лина, крыжовник, смородина Авокадо (Persea) Виноград, финик, хурма, дын- ное дерево, мангустан Яблоня, груша, вишня, слива, абрикос, персик, манго, унаби Айва (Cydonia) Банан Фейхоа Арбуз Дыня Апельсин, мандарин, грейп- фрут (Citrus paradisi) Лимон Гранат Дуриан цибетиновый Ананас, инжир, шелковица Весь плод Весь около- плодник То же Экзо- и мезо- карпий Мезэндокар- пий То же Плацента Мезокарпий Выросты эндо- карпия Весь плод Семенная ко- жура Ариллусы Соплодие Весь плод Весь около- плодник Экзо- и мезо- карпий То же Мезэндокар- пий Околоплодник Мезокарпий Весь плод Соплодие
132
Таблица 9 (продолжение)
Категории и названия Используемые части плодов
растений 1 в натуральном в переработан- в виде экстрагиро-
виде ном виде ванных веществ
Орехоплодные
Грецкий орех, каштан настоя- щий, лещина, миндаль, фи- Семя Семя
сташка
Овощные
Томат Весь плод Весь плод
Огурец Весь плод (не- Весь плод (не-
зрелый) зрелый)
Баклажан (Solanum rnelon- gena), перец овощной (Capsi- cum) Весь плод
Кабачки, тыква Мезокарпий
Маслина Экзо- и мезо- карпий
Пряные
Ваниль (Vanilla planifolia), перец черный (Piper nigrum), п. душистый (Pimenta officina- Незрелый плод
Us) Перец красный (Capsicum) Мускатное дерево Семя Зрелый плод
Горчица Тмин, анис Весь плод Семена
Кардамон (Elettaria cardamo- тит) Весь плод
Напиточные
Виноград Околоплодник
Какао, кофе Семена
КОРМОВЫЕ Зерновые
Овес, ячмень, просо, могар Весь плод
Кукуруза Весь плод, не- зрелое сопло- дие
Бобы, вика, люпин, соя, нут, Семена
чина
Сочноплодные
Арбуз, тыква Весь плод
лекарственные
Безвременник (Colchicum), клещевина, чилибуха (Strych- Семена
nos пих vomica), строфант
(Strophanthus), термопсис (Thermopsis), облепиха, кофе
Кола (Cola) Семядоли
133
Таблица 9 {продолжение)
Категории и названия растений 1 Используемые части плодов
в натуральном виде в переработан- ном виде в виде экстрагиро- ванных веществ
Миндаль Лимонник Ажгон (Caruin ajowan), анис, тмин, фенхель, гранат Мак Боярышник, жестер слабитель- ный, кассия (Cassia), колоцинт (Citrulus colocynthus), со- лянка Рихтера Малина, черника, черемуха Бузина черная Тыква Шиповник Весь плод Весь плод Семена (без кожуры) Семя Весь плод Семя Плод, семена Околоплодник Незрелый около- плодник Весь плод » » Гипантий
ХИМИКО-ТЕХНИЧЕСКИЕ
Эфирномасличные
Ажгон, анис, борщевик, кори- андр, тмин, фенхель Цитрусовые Миндаль Околоплодник Экзокарпий Семя
Жирномасличные
Маслина, масличная пальма Клещевина, лен, мак, рапс, сафлор, тунг (Aleurites), пе- рилла (Perilla), ляллеманция (Lallemantia), хлопчатник Мезокарпий Семена
ВОЛОКНИСТЫЕ
Хлопчатник Шерстяное дерево (Ceiba) Выросты се- менной кожуры Выросты эндо- карпия
ПОДЕЛОЧНЫЕ
Кокосовая пальма Люффа Бутылочная тыква Васкулярный скелет около- плодника Экзокарпий Эндокарпий
1 Латинские названия даются только для малоизвестных растений, не упоминав-
шихся в других главах.
плодов, так или иначе используемых человеком, потребовало» бы
слишком много места. Приходится ограничиться конспективным
изложением. В табл. 9 представлен характер использования пло-
дов наиболее важных полезных растений, в которых выделены
лишь основные категории, без детальной их классификации.;
134
в частности, это относится к лекарственным растениям. В пре-
делах каждой категории в одну группу объединены растения
с одинаковым характером использования их плодов.
В заключение настоящей главы мы коснемся еще одного
вопроса, связанного с использованием плодов,— ботанической
характеристики посевного материала. В агрономической литера-
туре все, что высевается в почву, называют «семенами», хотя
хорошо известно, что с морфологической точки зрения посевным
материалом служат различные генеративные органы или их части.
Ниже приводится ботаническая характеристика посевного ма-
териала наших основных полевых и огородных культур.
Морфологическая природа «семян» Культуры
Семена Все бобовые, в том числе и кормовые травы; бах- чевые (тыквенные); крестоцветные (капуста, редис, редька, горчица, рыжик, рапс); все пасленовые, кроме картофеля (томат, баклажан, перцы, табак); хлопчатник; лен; кунжут; мак; клещевина
Плоды (голые или с вторич- ными покровами) Мерикарпии Все злаки; гречиха; подсолнечник; сафлор; ко- нопля Все зонтичные (морковь, петрушка, укроп, тмин, ажгон, анис, кориандр, фенхель)
Эремы Все губоцветные (мята, мелисса, лаванда, шал- фей мускатный, перилла, ляллеманция и др.)
Соплодия Свекла
Мы не даем экономической оценки плодов, потому что это
означало бы дать экономическую характеристику всех важнейших
культур мирового земледелия. Естественно, что такая задача
по своему содержанию и объему выходит далеко за пределы
настоящей книги.
135
ЛИТЕРАТУРА
Александров В. Г., Савченко М. И. Об особенностях истории развития
плода и семени в семействе сложноцветных//Тр. Ботан. ин-та АН СССР.
Сер. 7. 1951. Вып. 2. С. 5—98.
Алехин В. В. Некоторые соображения относительно жизненной формы
«перекати-поле» // Юбил. сб. в честь 70-летия акад. В. Л. Комарова. Л., 1939.
С. 97—107.
Артюшенко 3. Т. Развитие цветка и плода у жимолостных//Тр. Ботан.
ин-та АН СССР. Сер. 7. 1951. Вып. 2. С. 131—168.
Артюшенко 3. Т„ Коновалов И. И. Морфология плодов типа орех и орешек //
Тр. Ботан. ин-та АН СССР. Сер. 7. 1951. Вып. 2. С. 170—192.
Бибиков Д. И. К экологии кедровки // Тр. Печоро-Илыч. заповед. 1948.
Т. 4, вып. 2.
Бобров Е. Г. Клевер (Trifolium) // Флора СССР. 1945. Т. 11. С. 189—261.
Бойко Э. В. Анатомическое строение околоплодника семянок некоторых
видов рода Senecio L. // Вести. ЛГУ. Биология. 1972. № 15. С. 42—47
Босек П. 3. Новые материалы по адвентивной флоре Брянской области //
Ботан. жури. 1983. Т. 68, № 5. С. 672—674.
Буш И. А. Систематика высших растений. М., 1959. 3-е изд. 535 с.
Васильев И. В. Семейство липовые (ТШасеае) //Жизнь растений / Под
ред. А. Л. Тахтаджяна. М., 1981. Т. 5 (2). С. 117—120.
Вихирева В. В. Морфолого-анатомическое исследование плодов крушино-
вых //Тр. Ботан. ин-та АН СССР. Сер. 7. 1952. Вып. 3. С. 241—292.
Войтенко В. Ф. Формы гетерокарпии в семействе Brassicaceae Burn, и их эво-
люционная оценка // Ботан. жури. 1968. Т. 53, № 10^ С. 1428—1439.
Войтенко В. Ф. Опыт изучения гетерокарпии в семействе крестоцветных
(Brassicaceae): Автореф. дис. . . . канд. биол. наук. Л., 1969а. 23 с.
Войтенко В. Ф. Особенности прорастания семян некоторых гетерокарпных
крестоцветных // Науч. докл. высш, школы. Биол. науки. 19696. № 3. С. 72—77.
Войтенко В. Ф. Эколого-географический и филогенетический анализ гетеро-
карпии в пределах семейства крестоцветных // Бюл. МОИП. Отд. биол. 1970.
Т. 75, № 3. С. 77—84.
Войтенко В. Ф. Основные черты дисперсии гетерокарпных крестоцветных //
Вопросы биологии семенного размножения. Ульяновск, 1974. С. 67—90.
Войтенко В. Ф. Гетерокарпия у растений флоры СССР. 1. Tragopogon
dubius Scop. (сем. Asteraceae) // Вопросы биологии семенного размножения.
Ульяновск, 1981. С. 44—64.
Войтенко В. Ф., Опарина С. Н. Гетерокарпия в семействе Boraginaceae //
Ботан. журн. 1985. Т. 70, № 7. С. 865—875.
Волгин С. А. Система и филогения подтрибы Rhipsalinae семейства Cactaceae:
Автореф. дис. . . . канд. биол. наук. М., 1982. 17 с.
Волгин С. А., Тихомиров В. Н. О структурных типах моноциклического синкарп-
ного гинецея покрытосеменных // Бюл. МОИП. Отд. биол. 1980. Т. 85, № 6.
С. 63—74.
Гоби X. Я. Генетическая классификация плодов семенных растений // Зап.
Лабор. по семеновед, при Гл. ботан. саде РСФСР. Пг., 1921. Т. 4, вып. 4. 30 с.
Грудзинская И. А. Системы классификаций и номенклатуры плодов в свете
их применения в ботанических руководствах // Бюл. МОИП. Отд. биол. 1968. Т. 73,
№ 3. С. 78—85.
136
Гусев Ю. Д. Проникновение новых адвентивных растений в Марийскую АССР
по железной дороге // Ботан. журн., 1977. Т. 62, № 3. С. 429—431.
Данилова М. Ф. О природе многокамерности плодов у томатов // Тр. Ботан.
ин-та АН СССР. Сер. 7. 1952. Вып. 3. С. 87—146.
Дементьев Г. П. Птицы // Руководство по зоологии. М.; Л., 1940. Т. 6. 856 с.
Деревья и кустарники СССР. М.; Л., 1951—1962. Т. 2—6.
Дудик И. М. Морфология плодов бобовоцветных (Fabales Nakai) в связи
с эволюцией: Автореф. дис. . . . докт. биол. наук. Новосибирск, 1981. 35 с.
Жизнь растений // Под ред. А. Л. Тахтаджяна. М., 1980— 1982. Т. 5—6.
Зажурило К. Д. Классификация орнитохорных плодов и семян // Журн. Рус.
ботан. о-ва. 1931. Т. 16, № 2—3. С. 169—187.
Зажурило К. К. Современные проблемы анатомии в карпологии // Тр.
Воронеж, ун-та. 1935. Т. 7. С. 21—42.
Зажурило Д. Д. Следы эволюции плодов в их анатомическом строении // Тр.
Воронеж, ун-та. 1936. Т. 9,1. С. 5—27.
Зажурило Д. Д. Функциональные рудименты в плодах амурского бархата
Phellodendron amurense Rupr.//Тр. Воронеж, ун-та. 1939. Т. 11, 2. С. 7—12.
Зажурило Д. Д. Сравнительная анатомия вяза и птелеи // Тр. Воронеж,
ун-та. 1945. Т. 13: Науч, сообщ. и автореф., 1. С. 45.
Замятнин Б. Н. Основы морфологической интерпретации нижней завязи:
Автореф. дис. . . . канд. биол. наук. Л., 1951. 21 с.
Зубкова И. Г., Шабес Л. Д. Анатомическое строение околоплодника видов
Sparganium (Sparganiaceae) // Ботан. журн., 1983. Т. 68, № 3. С. 381—385.
Иванина Л. И. Семейство геснериевых (карпологический обзор). Л., 1967.
126 с.
Игнатов М. С., Игнатова Е. А. Новости адвентивной флоры Барнаула и его
окрестностей (Алтайский край) // Ботан. журн. 1982. Т. 67, № 10. С. 1421 —1424.
Ильин М. М. Chenopodiaceae — Маревые // Флора Юго-Востока, вып. 4 // Тр.
Гл. ботан. сада, Л. 1930. Т. 36, вып. 1. С. 123—239.
Ильминских И. Г., Димитриев А. В., Мильчаков Л. В. О некоторых редких
и новых адвентивных растениях во флоре Волжско-Камского края//Ботан.
журн. 1981. Т. 66, № 8. С. 1221 — 1224.
Имханицкая И. И. Семейство лавровые (Lauraceae) // Жизнь растений /
Под ред. А. Л. Тахтаджяна. М., 1980а. Т. 5 (1). С. 158—166.
Имханицкая И. И. Семейство анноновые (Аппопасеае) // Жизнь растений /
Под ред. А. Л. Тахтаджяна. М., 19806. Т. Т. 5 (1). С. 132—139.
Имханицкая И. И. Пальмы. Л., 1985. 244 с.
Даден И. И. К вопросу о ложных плодах // Вести. МГУ. Сер. 16: Биология.
1947. № 12. С. 21—30.
Даден И. И. Плоды и семена среднерусских роголистниковых // Бюл. МОИП.
Отд. биол. 1953. Т. 58, № 3. С. 86—89.
Даден И. И. Апокарпия гинецея и плода злаков по данным сравнительной
морфологии //Науч. докл. высш, школы. Биол. науки. 1958а. № 3. С. 113—123.
Даден И. И. Апокарпия гинецея и плода злаков по данным тератологии //
Науч. докл. высш, школы. Биол. науки. 19586. № 4. С. 111 —117.
Даден И. И. Апокарпия гинецея и плода злаков по данным васкулярной
анатомии и онтогении // Науч. докл. высш, школы. Биол. науки. 1959. № 3. С. 147—
159.
Даден И. И. О некоторых основных вопросах классификации, типологии
и номенклатуры плодов // Ботан. журн. 1961. Т. 46, № 4. С. 498—504.
Даден И. Н. Типы продольного вскрывания плодов // Ботан. журн. 1962.
Т. 47, № 4. С. 495—505.
Даден И. И. Основы эволюционной морфологии плодов: Автореф. дис... докт.
биол. наук. М., 1964а. 18 с.
Даден И. И. К вопросу о дробных плодах // Ботан. журн. 19646. Т. 49, № 7.
С. 966—973.
Даден И. И. Типы плодов растений средней полосы европейской части
СССР // Ботан. журн. 1965. Т. 50, № 6. С. 775—787.
Даден И. И., Дирпичников М. Э. Проект новой системы и номенклатуры
плодов // Ботан. журн. 1966. Т. 51, № 4. С. 473—483.
137
Каден Н. Н„ Смирнова С. А. Типы плодов и карпологические признаки
родов сорных маревых СССР // Морфология высших растений. М., 1968. С. 70—84.
Козо-Полянский Б. М. Введение в филогенетическую систематику высших
растений. Воронеж, 1922. 167 с.
Комар Г. А. О структуре ариллуса некоторых представителей семейства
крыжовниковых (Grossulariaceae) // Морфология цветка и репродуктивный
процесс у покрытосеменных растений. Л., 1965. С. 114—130.
Комарницкий И. А., Кудряшов Л. В., Уранов А. А. Ботаника: Систематика
растений. М., 1975. 7-е изд. 608 с.
Коровин Е. П. Растительность Средней Азии. М.; Ташкент, 1934. 480 с.
Котт С. А. Биологические особенности сорных растений и борьба с засорен-
ностью почвы. М., 1947. 240 с.
Кузнецов М. Д. К вопросу о происхождении яблока: Рефер. докл. Моск,
с.-х. акад. 1951. Т. 13.
Ландсберг Г. С. Анатомо-морфологическое строение перикарпа Phoenix
(Агесасеае) // Ботан. журн. 1981. Т. 66, № 3. С. 388—391.
Левина Р. Е. К вопросу об антропохории // Сов. ботаника. 1944. № 3. С. 42—46.
Левина Р. Е. Способы распространения плодов и семян. М., 1957. 358 с.
Левина Р. Е. Способы распространения плодов и семян эдификаторов
растительного покрова СССР//Учен. зап. Ульянов, пед. ин-та. 1958. Т. 11,
вып. 1. С. 23—35.
Левина Р. Е. О классификации и номенклатуре плодов // Ботан. журн.
1961. Т. 46, № 4. С. 488—495.
Левина Р. Е. О ритме плодоношения травянистых многолетников (на примере
Trifolium montanum L. и Т. alpesire L.) 11 Ботан. журн. 1965. T. 50, № 7. С. 917—
933.
Левина Р. Е. Плоды (морфология, экология, практическое значение).
Саратов. 1967а. 215 с.
Левина Р. Е. Аспекты изучения гетерокарпии // Ботан. журн. 19676. Т. 52,
№ 1. С. 3—12.
Левина Р. Е. О задачах морфологического изучения плодов и принципах
их классификации // Реф. докл. Всесоюз. межвузов, конф, по морфологии расте-
ний. М„ 1968. С. 171 — 173.
Левина Р. Е. Очерки по систематике растений. Ульяновск, 1971. 192 с.
Левина Р. Е. Морфология и типы плодов. Ульяновск, 1974. 32 с.
Левина Р. Е. Новый аспект в трактовке гетерокарпии // Журн. общ. биологии.
1980. Т. 41, № 5. С. 680—684.
Левина Р. Е. Репродуктивная биология семенных растений (обзор проблемы).
М., 1981. 96 с.
Левина Р. Е., Войтенко В. Ф. Гетерокарпия, или разноплодие // Природа.
1975. № 5. С. 87—95.
Левина Р. Е., Марасов А. Н. Закономерности проявления гетерокарпии
в онтогенезе // Ботан. журн. 1976. Т. 61, № 12. С. 1686—1695.
Любин Ф. П. Гетерокарпия некоторых растений, ее биологическое и практи-
ческое значение // Сборник-материалов по флоре и растительности Юго-Востока.
Саратов, 1950. С. 3—35.
Малышева В. Г. Адвентивная флора Калининской области: Автореф. дис. . . .
канд. биол. наук. Л., 1980. 17 с.
Марасов А. Н. Последовательность образования гетероморфных плодов
у вайды Буассье (Isatis boissieriana Rchb.) и этионемы мясокрасной [Aethionema
carneum (Soland). Fedtsch.] //Вопросы биологии семенного размножения.
Ульяновск, 1974а. С. 101 —112.
Марасов А. Н. О соотношении типов плодов у вайды Буассье (Isatis boissieriana
Rchb.) и этиоиемы мясокрасной [Aethionema carneum (Soland.) Fedtsch.] // Воп-
росы биологии семенного размножения. Ульяновск, 19746. С. 113—124.
Меликян А. П., Мурадян Л. Г. Основные направления эволюции перикарпия
и спермодермы в подтрибе Chrysantheminae (Asteraceae) // Ботан. журн. 1975.
Т. 60, № 8. С. 1123—1133.
Меницкий Ю. Л. Дубы Азии. Л., 1984. 316 с.
Молчанов А. А., Шиманюк А. П. Восстановительные процессы на концентри-
рованных лесосеках. М.; Л., 1949. 127 с.
138
Мотекайтите В. П. О новых и редких рудеральных видах Литовской ССР //
Ботан. журн. 1985. Т. 70, № 10. С. 1410—1414.
Недолужко В. А. Распространение Ambrosia artemisiifolia (Asteraceae)
в Приморском крае // Ботан. журн. 1984. Т. 69, № 4. С. 527—529.
Нестеров В. Г. Главные рубки в лесах СССР. М.; Л., 1950. 136 с.
Новиков Г. А. О распространении дуба сойкой // Природа. 1948. № 3.
С. 69—70.
Новиков Г. А. Роль позвоночных животных в жизни леса // Животный мир
СССР. М.; Л., 1953. Т. 4. С. 599—630.
Пидотти О. А. Определитель семян декоративных растений. М.; Л; 1952. 116 с.
Подколзина В. Е. Явление многостворчатости плода у видов Brassica // Ботан.
журн. 1974. Т. 59, № 5. С. 688—691.
Пузырев А. Н. Новые адвентивные виды сем. Brassicaceae в Удмуртии//
Ботан. журн. 1984. Т. 69. № 9. С. 1269—1270.
Русанов Ф. Н. Особо интересные плоды и семена растений Индии. Ташкент,
1969. 49 с.
Снигиревская Н. С. Семейство нимфейные, или кувшинковые (Nymphaea-
сеае) //Жизнь растений / Под ред. А. Л. Тахтаджяна. М., 1980. Т. 5 (1).
С. 182—188.
Сорные растения СССР. Л., 1934—1936. Т. 1—4.
Тахтаджян А. Л. К экологии Ceratocarpus arenarius L. // Сов. ботаника.
1934. № 4. С. 22—28.
Тахтаджян А. Л. Структурные типы гинецея и плацентация семязачатков //
Изв. Арм. фил. АН СССР. 1942. № 3/4. С. 91—112.
Тахтаджян А. Л. Морфологическая эволюция покрытосеменных. М., 1948.
301 с.
Тахтаджян А. Л. Основы эволюционной морфологии покрытосеменных.
М.; Л., 1964. 236 с.
Тахтаджян А. Л. Система и филогения цветковых растений. М.; Л., 1966.
611 с.
Тахтаджян А. Л. Плод // Жизнь растений / Под ред. А. Л. Тахтаджяна.
М„ 1980. Т. 5 (1). С. 91—96.
Трифонова В. И. Семейство мускатниковые (Myristicaceae) // Жизнь расте-
ний / Под ред. А. Л. Тахтаджяна. М., 1980. Т. 5(1). С. 141 —143.
Федоров Ал. А., Артюшенко 3. Т. Атлас по описательной морфологии растений:
Соцветие. Л., 1979. 295 с.
Федосеева А. И. К анатомо-карпологической характеристике сем. Boragina-
сеае // Тр. Воронеж, ун-та. Ботан. отдел. 1935. Т. 7. С. 43—67.
Флора Юго-Востока европейской части СССР. Л., 1927—1936. Вып. 1—6.
Ханджян Н. С. Анатомия семянок некоторых видов рода Achillea (Astera-
ceae) //Ботан. журн. 1983. Т. 68, № 3. С. 346—351.
Холодный Н. Г. Как расселяется дуб в естественных условиях? // Среди
природы и в лаборатории. М., 1949. Вып. 1. С. 31—43.
Цвелев Н. Н. О некоторых адвентивных растениях Ленинградской области //
Новости систематики высших растений. Л., 1947. Т. 14. С. 244—255.
Цвелев Н. Н. Семейство руппиевых (Р up place ае) // Жизнь растений / Под
ред. А. Л. Тахтаджяна. М., 1982а. Т. 6. С. 34—35.
Цвелев Н. Н. Семейство злаки (Роасеае) // Жизнь растений / Под ред.
А. Л. Тахтаджяна. М., 19826. Т. 6. С. 341—378.
Черепанов С. К. Сосудистые растения СССР. Л., 1981. 509 с.
Черник В. В. Псевдомономерия гинецея представителей ильмовых и кар-
касовых // Ботан. журн. 1981. Т. 66, № 7. С. 958—967.
Яценко-Хмелевский А. А. Семейство коноплевые (Cannabaceae) // Жизнь
растений /Под ред. А. Л. Тахтаджяна. М., 1980. Т. 5 (1). С. 279—282.
Baumann М. С. Myodocarpus und die Phylogenie der Umbelliferen-Frucht.
Umbellifloren-Studien. 1//Ber. Schweiz, bot. Ges. 1946. Bd 36. S. 13—111.
Berg R. Myrmecochorous plants in Australia and their dispersal by ants//
Austral. J. Bot. 1975. Vol. 23. P. 475—508.
Bresinsky A. Bau, Entwicklungsgeschichte und Inhaltsstoffe der Elaiosomen //
Bibliot. Botan., Stuttgart. 1963. N 126.
139
Brouwer W., Stahlin A. Handbuch der Samenkunde. Frankfurt am Main,
1955. 655 S.
Buxbaum F. Untersuchungen zur Morphologie der Kakteenbliite. I: Das Gynae-
ceum // Bot. Archiv. 1944. 45. S. 190—247.
Eames A. J. Morphology of the Angiosperms. New York etc., 1961.
(Eames A. J.) Имс А. Морфология цветковых растений. Пер с англ. М., 1964.
497 с.
Eckardt Th. Untersuchungen fiber Morphologie, Entwicklungsgeschichte
und systematische Bedeutung des pseudomonomeren Gynoeceum // Nova acta
leopold. N. F„ Halle. 1937. Bd 5, N 26. S. 1 — 112.
Eckardt Th. Das pseudomonomeren Gynoeceum // Chron. Bot. 1938. № 4.
S. 206—208.
Eckardt Th. Reihe Centrospermae // A. Engler. Syllabus der Pflanzenfamilien
(Angiospermen). Berlin, 1964. 12. Aufl. C. 79—101.
Engler A. Syllabus der Pflanzenfamilien (Angiospermen). Berlin, 1964. 12.
Aufl. Hrsg. v. H. Melchior. 666 S.
(Esau К.) Эзау К- Анатомия семенных растений. Пер. с англ. М., 1980.
Кн. 1 и 2. 558 с.
(Franke G. et al.) Франке Г. и др. Плоды земли. Пер с нем. М., 1979. 270 с.
НйЬпег R. Der Same in der Landwirtschaft. Radebeul, 1955. 224 S.
Kaussmann B. Pflanzenanatomie. Jena, 1963. 624 S.
Lhotska M., Chrtkova A. Carpologie a diasporologie cescoslovenkych zastupcufie-
ledi Fabaceae. Praha, 1978. 296 s.
Lhotska M., Slavik B. Zur Karpologie, Karpobiologie und Verbreitung der
Art Iva xanthiifolia Nutt, in der Tschechoslowakei // Folia geobot. et phytotaxon,
Praha. 1969. № 4. S. 415—434.
Muller-Schneider P. Verbreitungsbiologie (Diasporologie) der Bliitenpflanzen.
Zurich, 1977. 2. Aufl. 226 S.
Muller-Schneider P., Lhotska M. Zur Terminologie der Verbreitungsbiologie
der Bliitenpflanzen //Folia geobot. et phytotaxon, Praha. 1971. № 6. S. 407—417.
Pankow H. Histogenetische Studien an den Bliiten einiger Phanerogamen //
Botan. Stud. hrsg. v. Troll u. Guttenberg. Jena, 1962. 106 S.
Pijl L. van der. Some remarks on myrmecophytes // Phytomorphology. 1955.
Vol. 5, N 2—3. P. 190—200.
Pijl L. van der. Principles of dispersal in higher plants. Berlin etc., 1969, 1st ed.,
154 p.; 1982, 3rd ed.
Potztal E. Reihe Graminales // A. Engler. Syllabus der Pflanzenfamilien
(Angiospermen). Berlin, 1964. 12. Aufl. S. 561—579.
Roth I. Fruits of Angiosperms // Berlin; Stuttgart, 1977. Vol. 10, pt 1. 675 p.
Rytz H7. Systematische, okologische und geographische Problem bei den
Brassiceen // Ber. Schweizar. Ges., Bern. 1936. Bd 46. S. 517—544.
Sernander R. Entwurf einer Monographic der europaischen Myrmekochoren.
Upsala, 1906. 410 S.
Stopp K. Karpologische Studien (I, II) // Abhandl. d. Mathemat.-Naturwissen.
Klasse Akad. d. Wissen. und Liter, in Mainz. 1950. N 7. S. 3—56 (165—218).
Strasburger E. et al. Lehrbuch der Botanik. Jena, 1962. 28. Aufl. 732 S.
Troll W. Praktische Einftihrung in die Pflanzenmorphologie. 2. T: Die bliihende
Pflanze Jena 1957 420 S
Troll W. Die Infloreszenzen. Jena, 1964. Bd 1. 615 S.; 1969. Bd 2. 630 S.
Ulbrich E. Biologie der Friichte und Samen (Karpobiologie). Berlin, 1928. 230 S.
(Warming E.) Варминг E. Систематика растений. Пер. с дат. М., 1893. 624 с.
Weber R. Friichte und Samen. Wittenberg. 1955. 84 S.
(Wettstein R.) Веттштейн P. Руководство по систематике растений. Пер. с нем.
М., 1912. Т. 2, ч. 2. 501 с.
Zohary М. Die verbreitungsokologischen Verhaltnisse der Pflanzen Palastinas.
1. Die antitelechorischen Erscheinungen//Beih. z. Bot. Centralbl. 1937. Bd 56,
Abt. A, H. 1. S. 1—155.
Zohary M. Carpological studies in Cruciferae 11 Palestine J. Bot. 1948. Vol. 14,
N 3. P. 158—165.
Zohary M. Evolutionary trends in the fruiting head of Compositae // Evolution
(New York). 1950. Vol. 4, N 2. P. 103—109.
140
УКАЗАТЕЛИ ТЕРМИНОВ
Автомеханохория 124
Автохория 124
Аггедула 69
Агестохория 126
Амфикарпия 95
Анемоаэрохория 112
Анемогеохория 112
Анемогидрохория 112, ИЗ
Анемохионохория ИЗ
Антеций 16
Антокарпий 37
Антропогидрохория 122
Антропохория 125, 126
Апокарпий 21, 25
Баллистохория 120
Баллисты-анемохоры 120
Барохория 124
Боб 29, 30
— крылатковидный 37
— ложнодвугнездный 30
— односемянный 37
— орешковидный 38
— сочный 32
— членистый 15
— неэквивалентная 88
— эквивалентная 88
Гетероголокарпия 88
— неэквивалентная 88
— эквивалентная 88
Гетерокарпия 88—90
— неэквивалентная 88
Гетеромерикарпия 89
Гетероспермия 91
Гетерофрагмокарпия 88
Гетерохория 129
Гетероэремокарпия 89
Гигрохазия 82
Гинецей 6
— апокарпный 6
---- мономерный 6
---- полимерный 6
— вторичномономерный 9
— гемисинкарпный 8
— лизикарпный 8
— паракарпный 8
— псевдомономерный 9
— синкарпный 7
— ценокарпный 7
Гипантий 16, 58
Гранатина 59
Вислоплодник 51
Внеплодник 10
Внутриплодник 10
Вскрывание плода 12
---- вентральное 13
---- дорсальное 13
----ламинальное 13
----латеральное 13
---- подковообразное 14
---- поперечно-кольцевое 14
---- продольно-кольцевое 14
---- септицидное 13
----септицидно-дорсально-ламинальное
13
----септицидно-сутуральное 13
----сутуральное 13
Двукрылатка 49
— дробная 49
Двуорешек 34
Двусемянка 49
Диаспора 99
Диаспорология 99, 100
Диаспоры авторотирующие 113
— аллохорные 97
— барохорные 97
— гемианемохорные 113
— эванемохорные 113
Диплохория 128, 129
Желудь 71
Гесперидий 58
Гетероартрокарпия 88
Земляничина 35
Зерновка 40
141
Калачик 20, 49 Карпобазис 94 Карпотропические изгибы 120 Карпофор 51 Карцерула 60 Коробочка 42, 44 — дробная 48 — односемянная 61 — рамчатая 45 — стручковидная 53 Костянка 55, 61 — дробная 22 — многокосточковая 23 — сухая 23 Крылатка 68 Ксерохазия 82 — центрально-угловая 7 Плод 6 — гемипаракарпный 17 — гемисинкарпный 26 — дробный 15 — окончато-членистый 15 — псевдомономерный 60 — распадающийся 15 — членистый 15 Плодик 20 Плюска 16 Полихория 128 Померанец 58 Семянка 60, 61 Сикон 86
Листовка 25 — односемянная сочная 40 Синзоохория 100 Соплодие 80 Спейрохория 126 Спейрохоры облигатные 128
Межплодник 10 Мезокарпий 10, 11 Мерикарпий 15 Мирмекохория 108 Многобоб 26 Многокостянка 35 Многолистовка 25 — олигомерная циклическая 26 — полимерная 25 — факультативные 128 Стручок 47 — членистый 52 Стручочек 47 — ореховидный 69 Сфалерокарпий 37 Трехкрылатка 49 Трехлистовка 26
— сочная 27 — ценокарпная 41 Тыквина 59
Многоорешек 33 — погруженный 34 Финик 39
Однокостянка 38 — сухая 39 Однолистовка 27 — сочная 27, 29 Одноорешек 36 Околоплодник 10 Орех 60 — кокосовый 72 — мускатный 40 Орешек в гипантии 36, 37 Ценобий 53 — диэремный 54 Ценокарпий 41 — дробный 49 Ценосома 86 — гипантиевидная 86 Цинародий 35 Четырехкрылатка 49 Четырехорешек 34
Партенокарпия 87 Первольвенты 118 Перикарпий 10 Плацента 6 — колончатая 8 Плацентация 6 — базальная 7 — ламинальная 7 — маргинальная (краевая) 7 — париетальная 8 — субмаргинальная 8 — сутуральная (шовная) 7 Экзокарпий 10, 11 Эндозоохория 100 — инадаптивная 105 Эндокарпий 10, 11 Эпизоохория 101 — факультативная 111 Эргазиохория 126 Эрем 53 Шиповник 35
142
Яблоко 58
— костянковидное 58
Ягода 54
— односемянная 74, 79
Васса 54
Balausta 59
Capsula 41
Caryopsis 40
Coenobium 53
Cremocarpium 51
Cynarrhodium 35
Dieresilis 49
Disamara 49
Epicalyx 83
Eremus 53
Folliculus 25
Fragum 35
Glans 71
Hesperidium 58
Legumen 29
Multidrupa 35
Multifolliculus 25
Pepo 59
Pneumatocarpia 113
Pomum 58
Siliqua 47
Sycon 86
Unidrupa 38
Uninucula 36
Unifolliculus 27
Uva 54
УКАЗАТЕЛЬ
РУССКИХ НАЗВАНИЙ РАСТЕНИЙ 1
Абрикос 38, 133
Авокадо 133
Авран 43
Адонис 76
— весенний 109
Ажгон 135, 136
Аизооновые 10
Аистник 42
Айва 58, 77, 133
Айлант 34, 76, 81, 116*
Акантовые 124, 125
Аканталимон 77
Аксирис 91*
Актинидия 55, 102, 133
Александрийский лавр 129
Альстремерия 124
Амарантовые 61, 62, 65, 77
Амарантус 126
Амборелла 76
Амброзия 85, 126
Аммодендрон Конноли 76
Аморфа 76
Анакардиевые 73, 78, 103
Анаксогорея 124
Ананас 87, 133
Анис 134—136
Аннона 26*
— голая 105, 129
Анноновые 27, 76, 103—105
Анчар 103
Апельсин 58, 133
Аралиевые 52, 55
Арахис 31, 36, 133
Арбуз 59, 133, 134
Ардисия японская 78
Армерия 65*
Артокарпус 86
Архидендрон 27, 76
Асимина карликовая 105
Астрагал 29, 30*, 76, 117, 120
— серповидный 76
— солодколистный 30, 76
Аукуба японская 78, 79
Аффонсея 27, 76
Багрянник 27, 76
Багульник 43
Баклажан 134, 136
Баланитовые 75, 78
Бальзаминовые 44
Бамбук 40
Банан 56, 57*, 81, 103, 104, 133
Банановые 51, 81
Банксия 82*
Баобаб 104
Барбарис 7
Бархат амурский 55, 56
Бассия 64*
Безвременник 43, 134
Белена 16, 42
Белозор 42, 123
Белокопытник 124, 129
Белокрыльник 55
— болотный 81
Береза 68*, 77, 116, 118, 119
Березовые 87
Бересклет 42
Звездочкой отмечены страницы с рисунками соответствующих таксонов.
143
Бешеный огурец 77, 129
Бигнониевые 45, 116
Бигнония 115*
Бинерция 90
Бирючина 75, 78, 102
Бобовые 10, 88, 94, 95, 103, 104, 109, 136
Бобы 133, 134
Бодяк 66*, 126
— съедобный 109
Болотниковые 54, 77
Болотный мирт 43
Борец 26, 76
Борщевик 51*, 52, 135
Борщовия 64
Боярышник 58, 77, 81, 102, 135
— однопестичный 75, 78
Брандушка 43
Бромелиевые 10, 87
Брунеллия 76
Брусника 55, 102
Бузина 55, 81, 102
— черная 135
Бук 71, 106, 107
Буковые 16, 71, 78
Бундук 32, 76
Бурачниковые 15, 16, 54, 77, 90, 94, 121
Бурачок 77
Бурманниевые 113
Вайда 69, 78
Валериана 81
Валериановые 9, 68, 77, 88
Ваниль 134
Василек 66*, 82, ПО, 117, 121
— раскидистый 117, 118
Василистник 33, 76
Ваточник 115*
Вахта 42
Вейник наземный 41
Венечник 42
Вербейник 42
Вербеновые 15, 54, 77
Верблюжья колючка 30
Вереск 43
Вероника 13, 42, 43
— простертая 44
Ветреница 33, 76
— лесная 118
Вика 134
Вильгельмсия пузырчатая 48, 77
Виноград 55, 81, 132—134
— амурский 103
Винтеровые 77
Вишня 129, 133
Водосбор 8, 26
Водяная сосенка 74, 78
Водяника 102, 103
Водяной орех 75*, 78
Волдырник 56
Волчник 78
— алтайский 78, 79
— обыкновенный 78, 79
Волчниковые 74
Воробейник полевой 54*
Воронец 27, 28*
Вороний глаз 55
Ворсянка посевная 85
Ворсянковые 66, 77, 81, 117, 121
Вьюнковые 17
Вьюнок 42
Вяжечка 47
Вяз 68, 77, 116, 128
Вязель 31
Габлиция 61, 77
Галинсога 126, 129
Гания 78
Гардвикия 38*
Гарциния 104
Гвоздика 13
Гвоздичные 13, 16, 17, 42, 64, 77, 120, 122
Гелиетта 49
— мелколистная 77
Геликониевые 77
Герань 42, 46*, 124, 125
Геснериевые 17, 113
Гибискус 42
Гидрастис канадский 76
Гипекоум беловатый 53*, 77
Гифена 104
Гледичия 76
— обыкновенная 32
Голубика 55, 102, 103
Гольдбахия 53, 77
Гораниновия 86*
Горец 62*
— льняной 128
— шероховатый 125
— щавелелистный 105
Горечавковые 8, 13
Горлянка 59
Горох 29, 76, 133
Горошек 29
Горчица 47, 133, 134, 136
— белая 46*
Граб 70*, 71, 78, 83
Гравилат 7, 34, 76, ПО
— речной 33*
Гранат 57*, 77, 133, 135
Гранатовые 59
Гребенщиковые 42
Грейпфрут 133
Гречиха 127, 133, 136
— татарская 127
Гречишные 16, 60, 62*, 77, 90
Груша 58, 77, 102, 103, 133
Грушанка 13, 42
Грушанковые 113, 120, 122, 129
Гуания Бланше 51*, 52
Губоцветные 15, 16, 54, 77, 19, 122, 136
144
Гулявник 47
Гусиный лук 42, 109
Дальбергия 37
— прилистниковая 76
Дафнифилловые 75, 78
Двоякоплодник 77
Девясил германский 124
Дегенериевые 27
Дегенерия 76
Дербенник 43
Дереза 55
Держи-дерево 74, 78
Десмодиум 95
Джекфрут 86, 133
Джузгун 62, 117
— безлистный 62*
— голова медузы 62*
Дзельква 69, 78
Диатеноптерикс 50*
— рябинолистный 77
Дивала 65*, 77
Диди мелес 39, 76
Диллениевые 26, 76
Диморфотека 93, 94*, 98
Диоскорейные 116
Диптерония китайская 49, 50*
Донник 38, 76
Дрок 29, 38, 76
Дуб 71, 106, 107
— грузинский 70*
— обыкновенный 71
Дудник 52
Дуриан 104, 105
— цибетиновый 133
Дурман 13, 43
Дурнишник 66, 85, 110
Дымянка 69, 78
Дымянковые 88
Дынное дерево 56, 104, 133
Дыня 133
Дюшенея 76
Ежевика 35
Ежеголовник 9, 78, 81, 82, 123
— забытый 82*
Ежовник 127, 129
Ель 119
Жестер 55, 57*
— слабительный 135
Живокость 26*, 76
Живучка 109
Жимолостные 10, 58
Жимолость 55
Жирянка 42
Жминда 16
Заразиха 115*
Заразиховые 8, 42, 113, 120, 122
Звездчатка 109
Зверобой 8
— красильный 55, 56
Земляника 7, 35, 76, 133
Земляной орех 32
Зизифус мавританский 78
Зимолюбка 121*
Злаки 16, 40, 60, 76, 109, 116, 136
Золотая печать 27
Зонтичные 15, 51*, 77, 80, 81, 88, 90, 94,
НО, 121 — 123, 136
Ива 13
Иван-чай 118
Ивовые 13, 42, 114, 118, 119, 129
Идиоспермум 76
Икотник 77
Иллициум 26*, 76
Ильм 106
Ильмовые 9, 75, 78
Инжир 84*, 85, 133
Ирисовые 13
Истод 43, 109
Кабачки 134
Кадсура 27, 76
Казуарина 61, 77
— прибрежная 85
Казуариновые 87
Какао 103, 104, 134
Кактус 105
Кактусовые 10, 58
Каликантовые 37, 76
Каликантус 34*
Калина 75, 78, 81, 102
Калицеровые 68, 77, 88
Калужница 26*, 76
Кампилоптера 96, 98
Канатник 41, 77
Каперсовые 45, 48, 56, 77
Каперс травянистый 56
Капуста 136
Караганник 29, 76, 125
Кардамон 134
Кариота 104
Кариотовые 75, 78
Кария 73
Каркас 75, 78
— игуановый 105
Каркасовые 73, 75, 78
Кармихелия 76
Картофель 55, 56
Касатик 42
— аировидный 123
— безлистный 109
Кассия 135
Катальпа 14, 45
10 Р. Е. Левина
145
Катран 69, 70*
Качим метельчатый 117
Каштан 71, 106, 107
— настоящий 70*, 134
Кельпиния НО
— линейная 67*
Кермек 77
Керрия 35, 76
Кизил 79
— мужской 79
Кизиловые 78, 79, 86
Кизильник 58, 77, 102
Кипрей 115*
Кипрейные 43
Кирказон 43, 123
Киркия 49
Кислица 13, 42, 124, 125, 129
Кисличные 88
Клевер 12, 16, 29, 37, 76, 105, 109
— альпийский 38
— гибридный 29
— горный 38
— днепровский 38
— земляничный 16
— красноватый 38
— луговой 37, 38*, 105
— мясокрасный 37
— пашенный 38
— ползучий 29, 105, 106
— средний 38
Клен 15, 50*, 77, 81, 106, 116*, 128
— татарский 10
Клеоме 77
Клещевина 48, 50*, 77, 134—136
Климакоптера 64*
Клоповник сорный 117
Клюква 55, 103
Княжик 34
Ковыль 40, 41, 114, 129
Козлобородник 65, 114, 115*
Кокаиновый куст 75, 78
Кола 134
Колокольчик 8, 15, 120
Коллеция колючая 91
Колокольчиковые 8, 42, 122
Колоцинт 135
Колубрина азиатская 77
Кольквиция прелестная 74, 78
Колючеголовник Бентама 93
Кондалия 75
— лопатчатая 78
— яйцевидная 78
Коноплевые 69, 78, 102
Конопля 20, 133, 136
— посевная 127
— сорная 125, 127'
Конский каштан 13, 42, 43, 45*, 106
Консолида 27, 76
Копеечник 31, 76
— Гмелина 95, 96*
Коралловое дерево 30, 31*
Кориандр 135, 136
Корифовые 76
Коровяк 43
Корсиевые 113
Костер 40
Костяника 35, 102
Кофе 132, 134
Кохия 64*
Крапива двудомная 105
Крапивные 9, 69, 78
Красное дерево 38*
Крестовник 93, 98, 119
Крестоцветные 14, 47, 53, 69
88, 95, 97, 136
Кривоцвет 54*
Кровохлебка 36, 76
Крупка 77
Крушина 55
— ломкая 57*
Крушиновые 48, 56, 74, 75, 77
Крыжовник 11*, 54, 55, 133
Кубышка 123
Кувшинка 123
Кузиния 82
Куколь 128
— обыкновенный 128
Кукуруза 132—134
Кунжут 133, 136
Кунжутовые ПО
Купальница 76
Купена 55
Курчавка 62
Кутровые 8, 27, 123
Лабазник 33, 76
Лаванда 136
Лавровые 103
Лакторис фернандесовый 76
Ландыш 55, 103
Лапина 74*
Лапчатка 10, 16, 33, 76
Лардизабаловые 76
Ластовень 119
Ластовневые 8, 27
Лебеда 64*, 90, 91
— блестящая 91
— садовая 91, 105
Левкой 47
Лен 43, 47, 128, 135, 136
Ленец 71, 109
Леонтика сомнительная 117
Леспедеца 76
Лещина 11*, 16, 71, 78, 83, 106,
— маньчжурская 70*
— обыкновенная 70*, 71, 83
— устрашающая 71
Лизея разноплодная 94
Ликвидамбар 86
Лилейные 13, 116, 120, 121
Лимон 57*, 58, 133
146
Лимнохарис 76
Лимонник 27, 76, 135
Линнея НО
Липа 60, 69, 78, 106
Липучка 54, НО
Лобелия 8, 42
Логаниевые 103, 104, 116
Ломонос 34
— винограднолистный 33*
Лопух 82, 110
Лотос 34*, 76
— индийский 34
Лоховые 37, 76
Лук 42
Лунник 77
— однолетний 46*
Луносемянниковые 39, 76
Львиный зев 15, 42
Льнянка 15, 42, 116
— обыкновенная 121
Лютик 7, 33, 76, 123
— едкий 33*
— ползучий 105
Лютиковые 10, 13, 25, 27, 33, 120
Люффа 59, 77, 135
Люцерна 28*, 29, 76, 109, ПО
— полевая 28*
— посевная 28*, 29
— хмелевая 38, 76
— щитковидная 28*
Лядвенец 29, 76
Ляллеманция 54*, 135, 136
Медуница 109
Мелисса 136
Мелколепестник 126, 129
Мерингия 109
Метроксилон 123
Меч-трава 123
Миагрум 69, 70*, 78
Мимоза 31
— стыдливая 31*, 76
Мимозовые 26, 29
Миндаль 76, 134, 135
Миодокарпус 77
Мирабилис ночецветная 76
Мирсина африканская 78
Мирсиновые 61, 75, 77, 79
Могар 133, 134
Молл ю го 42
Молочай 15, 48, 77, 109, 124
— болотный 109
Молочайные 48, 109, 125, 129
Мамордика 77
Монимиевые 76
Монция 42
Морковь 52, НО, 136
Морошка 35
Мотыльковые 29
Мускатниковые 39, 40, 76
Мускатное дерево 40, 134
Мушмула 58, 77
Мытник 16, 42
Мышехвостник 33
Мята 136
Мятлик луговой: 105
Магнолиевые 7, 33
Магнолия 13, 25, 26*, 76
Майник 55
— двулистный 56
Мак 15, 42, 120, 133, 135, 136
Маклюра 87
Маковые 45, 48, 53, 77
Малина 35, 103, 133, 135
Мальвовые 15, 48, 77
Мальпигиевые 49
Манго 104, 133
Мангустан 104, 133
Мандарин 58, 133
Манжетка 36, 76
Макания 78
Марантовые 104
Маревые 9, 12, 16, 60, 62, 64, 77, 86—88,
90
Мареновые 15, 49, 103, 104, НО, 116
Марь 91
— белая 91, 105, 129
— многолистная 86
Марьянник 42, 109
Маслина 78, 133—135
Маслиновые 75, 78
Мауриция извилистая 75, 78, 123
Мачок 45, 48, 77
Наголоватка 116, 121
Наперстянка 13, 43
Наяда 76
Недотрога 42, 44, 46*, 124
Незабудка 109*
Непентовые 113
Неслия 70*, 78
Никтагиновые 37, 76
Нимфейные 7, 41, 77
Нипа 39, 123
— кустистая 76
Ноготки 93, 98
Нонеа 109
Норичниковые 120
Нут 133, 134
Облепиха 134
Овес 40, 127, 133, 134
Овсюг 125, 126, 127, 129
Овсяница 40
Огурец 59, 134
Одуванчик 66*, 114
Ожика 42
Окопник лекарственный 109
Оливковое дерево .75
10‘
147
Ольха 69, 77, 85, 118
— клейкая 123
Омела 10, 55, 78, 79
Оносма 121
Опунция 9*
Орех грецкий 106—108, 134
Ореховые 73, 106
Орлайя 51*
Ортилия однобокая 120
Орхидные 8, 13, 42, 113, 119, 120, 129
Осина 119
Осока 16, 63, 105, 109
— вздутая 63*, 117
— горная 109*
Осоковые 60, 62, 69, 73, 77, 78
Остролодочник 30, 120
Очанка 42
Павония 50*, 77
Паланисия 77
Пальма 39, 54, 72, 83, 104
— бетелевая 73, 78, 123
—делеб 104
— кокосовая 72*, 78, 123, 132, 133, 135
— масличная 74, 78, 103, 133, 135
— сейшельская 72, 78, 123
Пальмира 73, 78, 104, 123
Пандановые 81, 84
Панданус кровельный 84*
— простой 81
Парнолистниковые 49, 110
Паслен 55
Пасленовые 56, 136
Пастернак 52
Пастушья сумка 77
Первоцвет 13, 42
— весенний 16
Первоцветные 13, 14
Перелеска 109*
Переступень 52
Перец душистый 134
— красный 134
— овощной 55, 134
— черный 134
Перилла 135, 136
Перловник 109
Персик 38, 133
Песчаная акация 37, 38*
Петров крест 42, 109
Петрушка 136
Пизония 76
Пи лея 124, 125
Пион 26, 76
Платан 33, 76, 82
— восточный 82
Платикария 74
Плюмбаго 65*, 77
Плюмбаговые 61, 62, 64, 65, 77, ПО
Плющ 55
— золотистоплодный 81
148
Повилика 42, 126
Повойничек 43
Погремок 42, 116, 121
— бескрылый 128
— большой 16
Подбел 43
Подлесник 52
Подмаренник 49, 51*, 77
— цепкий 110
Подорожник 42
— большой 105, 126
— ланцетолистный 105
Подсолнечник 11*, 12, 65, 133, 136
Подъельник 43
Полынок 67*
Помадеррис 52
— безлепесный 77
Портулак 42
Порцелия 104
Посконник 118
Пролесник 109
Проломник 42
Просо 40, 127, 133, 134
Прострел 34, 76
Протейные 36, 39, 76
Птелея 68*, 77, 116
Птерокария 116
Птерокарпус 37, 38*, 116
Птероцельтис 73, 78
Птицемлечник 109
Пузыреплодник 26, 76
Пузырник 28*, 29, 117
Пузырчатка 42
Пушица 118
Пшеница 127, 132, 133
Рапс 135, 136
Рафия 123
Рдест 34, 76
Ревень 62*, 117
Редька 47, 53*, 136
— дикая 47, 52, 53*, 77, 97
Резеда 42
Рейнозия 75
Ремнецветник европейский 78, 79
Ремнецветниковые 79
Репешок 37, 76, ПО
— обыкновенный 36*
Репник 53, 77
— морщинистый 53*
Рис 132, 133
Рогач 64, 91
— песчаный 90, 91*, 117
Рогоглавник 34, ПО
Рогоз 9, 36, 76, 81, 118, 129
— широколистный 36*
Рогозовые 61
Роголистник 36, 76
— темно-зеленый 36*
Рогульник 78
Рожковое дерево 32, 76
Рожь 127, 133
— дикая 125
Роза 76
Розовик 35
— лазящий 76
Розоцветные 16, 26, 33, 35, 36, 38, 39, 41,
76
Ромашка 67*
— пахучая 126
Росянковые 42
Рубус 76
Руппия 35, 76
Рутовые 49
Рыжик 133, 136
Рябина 58, 77, 81, 102
Рябинник 26, 76
Сабельник 123
Сажереция чайная 55
Сакоглоттис 123
Саксаул 64
Салакка съедобная 105
Самерария 78, 98
— Буассье 97*
Санталовые 71, 78
Сапиндовые 49, 116
Сапотовые 103, 104
Сассафрас беловатый 103
Сафлор 133, 135, 136
Сведа 64*, 90
—льнолистная 91
— рожконосная 91
Свекла 61, 64, 77, 86, 136
Свербига 69, 78
Свида кроваво-красная 79
Седмичник 42
Селезеночник 42
Сераделла 31*, 76
Сердечник 124
Серпоносик 95, 96*
Синеголовник равнинный 117
Синюха 43
Синяк 121
Сирень 42, 81
Ситник 42
Ситниковидные 49
Скабиоза 84*
Склерия 78
Скумпия 73, 78, 83
Слива 38, 39, 133
Сливовые 38, 76
Сложноцветные 60, 65, 66, 77, 80, 81,
85, 88, 90, 92, 98, 109, НО, 114, 119,
121
Смирновия 29, 117
Смолевка 42
Смородина 55, 56, 133
Снежноягодник 10, 58
Солнцецвет 42
Солодка НО
— иглистая 38, 76
Солянка 64
— Рихтера 135
— южная 118
Сорго 133
Сосна 116*, 119
Софора японская 32, 76
Соя 133, 134
Спаржа 55, 56
Спатолобус Роксбурга 76
Спирея 8, 26, 41, 76, 77
Стеригмостемум 77
Стеркулиевые 104
Стираксовые 117
Стифнолобиум 76
Страстоцвет 56
Стрелолист 123
Строфант 134
Сумах 73, 78
Сурепица 47
Сусак 8, 26, 76
— зонтичный 26
Сусаковые 7
Сушеница 66*
— топяная 65
Схизандровые 27
Сцирподендрон 78
Табак 136
Тамаринд 32, 76
Таушерия 69, 78
Терескен 64*
Терн 11*
Термопсис 134
Тетраптерис 50*
— цитрусолистный 77
Тетрацентровые 77
Тимелея 78
Тимофеевка альпийская 105
Тинуйя 50*
— лазящая 77
Тмин 51*. 134, 135
Толокнянка 55
Толстянковые 26, 76
Томат 57*, 58, 105, 134, 136
Торилис 52, ПО
Торица крупная 47
— льняная 47, 128
Торичник 43
Тофиельдия 43
Трема 75
Трималиум 51*
— подсолнечниковидный 77
Триостренник 49, 77
Триурисовые 25, 76
Троходендровые 77
Тунг 135
Тургения 52, НО
Турретия 129
149
Турча 42
Тутовые 9, 85, 86, 87, 103
Тыква 134, 135
— бутылочная 59, 135
Тыквенные 8, 59, 77, 124, 136
Тысячеголов 43
Тысячелистник 121
Тюльпан 42
Тюльпанное дерево 34, 76
Циклантера 77
Циклахена 126
Цикорий 67*, 121
Цимицифуга 27, 76
Циноксилон 84*
Цирцеастер 34
— полевой 34, 76
Цитрусовые 58, 77, 104, 132, 135
Унаби 78, 133
Урсиния 67*
Уруть 49, 77
Фасоль 29, 76, 133
Фейхоа 55, 133
Фениксовые 76
Фенхель 135, 136
Ферула 52
Фиалка 13, 42, 109, 124, 125, 129
— волосистая 109*
— полевая 109*
Фиалковые 88, 116
Фиговое дерево 85
Физалис 16
Фикус 86, 104
Филезиевые 56
Филлостилон бразильский 70*
Финик 133
Фисташка 73, 78, 134
— настоящая 73
Фителефас крупноплодный 83*
Фонтанезия 69, 77
Форестиера 78
— новомексиканская 75
Франкения 42
Фрейсинетия 81
Частуха 33, 76, 123
Частуховые 33
Чемерица 121*
Череда 67*, 110
Черемуха 39*, 81, 103, 135
— Маака 103
Черника 55, 102, 103, 135
Чернокорень 54, НО
Чернушка 41, 77
Чертополох 119
Чечевица 133
Чилибуха 134
Чилим 75*, 78
Чина 28*, 29, 76, 133, 134
Чистотел 14, 42, 45, 46*, 109*
Чубушник 43
Шалфей клейкий ПО
— мускатный 136
, Шейхцерия 8, 76
— болотная 26
Шелковица 86, 133
Шерстяное дерево 135
Шиверекия 77
Шильница 77
Шиповник 11*, 12, 16, 34, 102, 135-
Шпинат 86
Хардвикия 38*
— двойная 76
Хвойные 116, 118, 119
Хелинус яйцевидный 77
Хлебное дерево 86, 133
Хлопчатник 132, 133, 135, 136
Хмелеграб 71, 78, 83
Хмель 83, 84*
Хориспора 52, 77
Хохлатка 42, 45
Хризобалановые 73, 75, 78, 103, 123
Хурма 55, 133
Царьградские рожки 32
Цеа нотус голубой 77
Цельнолистник 49, 77
Центролобиум 37, 38*, 116
Цербера 123
Цефалотовые 25, 76
Щавелек 105
Щавель 16, 62*
— кислый 105
Щетинник 129
— сизый 127
Щирица 61, 62*
Эверсманния 31, 76
Эвклидиум 78
Эвкомия ильмовидная 68, 77
Эвптелейные 76
Эгилопс 125
Эгицерас 61, 77
Эджевортия 78
Экзохорда 41, 77
Эмбелия рибес 78
Энгельхардия 74*
Энсета Перрье 81
Энтада 76
Эремоспартон 30, 117
150
Эриоботрия 58, 77
Эритроксиловые 78
Эритьера прибрежная 123
Эспарцет 38, 76, 110
— песчаный 80
— равнозубчатый 38*
Этионема 96
Эуклидиум 70*
Эукрифия сердцелистная 48, 50*, 77
Эуптелея 36
Эшольция 124
Яблоневые 58
Яблоня 57*, 77, 102, 103, 132, 133
Язвенник 38, 76
Якорцы 49, 50*, 77, 110
Ярутка 77
Ясенец 41, 77
Ясень 68*, 69, 77, 81, 106, 116, 118, 128
Яснотка 109
Ясменник 49, 77
Ячмень 133, 134
— гривастый 129
УКАЗАТЕЛЬ
ЛАТИНСКИХ НАЗВАНИЙ РАСТЕНИЙ
Abutilon 41
Acacia 104
— arabica 104
— horrida 104
Acanthaceae 44
Acanthocephalus benthamianus 93, 95*
Acantholimon 61, 65
Acer pseudoplatanus 50*
Aconitum 26
Actaea 27
— spicata 28*
Adansonia digitata 104
Aegiceras 61
Aesculus hippocastanum 43, 45*
Aethionema heterocarpum 96
Affonsea 26
Agamea 116
Agrimonia 16, 37
— eupatoria 36*
Agrostema githago 128
— linicola 128
— macrosperma 128
Agrostis 116
Ailanthus altissima 34
Aizoaceae 10
Alchemilla 16, 36
Aleurites 135
Alhagi 30
Alisma 33
Allemanda 44
Alopecurus 40, 116
Alphitonia exelsa 56
Alstremeriaceae 124
Althea 48
Alyssum 47
Amaranthaceae 61
Amaranthus 61
— retroflexus 62*
Amarylidaceae 44
Amborella trichopoda 35
Amborellaceae 35
Ambrosia 85
— artemisiifolia 126
Ammodendron conollyi 37, 38*
Amorpha fruticosa 38
Anacardiaceae 73
Anagallis 14
Ananas sativus 87
Anaxogorea javanica 124
Anchietea 116
Anemonastrum narcissiflorum 34
Anemone sylvestris 33
Anemonoides ranunculoides 33
Angelica 52
Annona glabra 105
— reticulata 26*
Annonaceae 7, 27
Anthemis 121
Anthyllis macrocephala 38, 116
Antiaris toxicaria 103
Apocynaceae 8, 27, 44
Aquifoliaceae 54
Araceae 54
Arachis hypogeae 31
Araliaceae 54
Archidendron 26
Arctium 82
Ardisia japonica 75
Areca catechu 73
Arecaceae 44
Argemone mexicana 46*
Aristida 41
Armeria 65
— maritime 65*
Artemisia austriaca 67*
Artocarpus altilis 86
— heterophyllus 86
Asclepiadaceae 8, 27
Asclepias syriaca 115*
Asimina pygmaea 105
Asperula 49
Asteriscus 82
Astragalus albicaulis 30
— arenarius 30
— commixtus 30*
— dasyanthus 30
151
— falcatus 30
— glycyphyllos 30
— schelkovnikovii 30*
— skorniakovii 30*
— testiculatus 30
Atragene 34
Atraphaxis 62
Atriplex 64
— hortensis 91
— micrantha 91
— nitens 91
— patens 91
— patula 91
Aucuba japonica 79
Axyris amaranthoides 91, 92*
Balanites aegyptiaca 75
Banksia verticilata 82*
Barbarea 47
Bassia hyssopifolia 64*
Beckmannia 40
Begoniaceae 44
Berberidaceae 117
Berteroa 47
Beta 64, 86
Betula 68*
Bidens 66
B. tripartita 67*
Bienertia cycloptera 90
Bignonia sp. 115*
— fraxinifolia 115*
Bignoniaceae 44, 129
Bixaceae 44
Bombacaceae 44
Bonnetiaceae 44
Boraginaceae 54
Borassus aethiopum 104
— flabellifer 73, 104
Borszczowia 64
Brassica 97
— sisymbrioides 97
Brassiceae 97
Briza 40
Bromeliaceae 44, 54, 87
Brunellia 26
Brunelliaceae 26
Buddlejaceae 44, 54
Buglossoides arvensis 54*, 94
Bunias 69
Burmanniaceae 113
Butomus umbellatus 26
Byblidaceae 44
Cabombaceae 36
Cactaceae 54
Cakile maritima 124
Calamagrostis epigeios 41
Calendula 93
Calenduleae 93
Calla palustris 81
Calligonum 62
— aphyllum 62*
— caput-medusae 62*
Callitrichaceae 54
Calophyllum inophyllum 129
Caltha palustris 25, 26*
Calycanthus 34*
Calyceraceae 68
Campanulaceae 54
Campyloptera carnea 96, 97
Canarina 54
Cannabaceae 69
Cannaceae 44
Capparaceae 44, 45
Capparis herbacea 56
Caprifoliaceae 44
Capsella 47
Capsicum 134
Caragana 29
Cardamine chenopodiifolia 97
— hirsuta 124
— impatiens 124
Carex 63
— atherodes 63
— digitata 63, 109
— erice torum 109
— montana 63, 109*
— ornithopoda 63, 109
— physodes 63*
— praecox 109
— pseudocyperus 63
— sylvatica 109
— vesica ria 63
Carica 56
— papaya 56, 104
Caricaceae 54
Carmichelia australis 38
Carpinus 71
— betulus 70*, 83
— cordata 83
— japonica 83
— orientalis 83
Carthamnus 93, 133
Carum ajowan 135
— carvi 51*
Carya 73
Caryophyllaceae 48
Cassia 104, 135
Castanea 71
— sativa 70*
Casuarina 61
— equisetifolia 85
— litorea 85
Catalpa 14
— bignonioides 45
Ceanotus azureus 48
Ceiba 132, 135
Celtidiaceae 73
Celtis 75
— caucasica 75
152
— iguanaea 105
Centaurea aggregata 82
— calcitrapa 82
— nigra 66*
— squarrosa 82
— virgata 82
Centrolobium robustum 37, 38*
Cephalotaceae 25
Ceratocarpus 64
— arenarius 90, 91*
Ceratocephala 34
Ceratoides 64
— pungens 64*
Ceratonia 104
— siliqua 32
Ceratophyllaceae 36
Ceratophyllum 36
— demersum 36*
Ceratostigma 61
Cerbera manghas 123
Cercidiphyllum japonicum 27
Chamomilla matricarioides 67*
Chelidonium majus 46*, 109*
Chenopodiaceae 62
Chenopodium 64
— album 91
— foliosum 16, 64, 86
Chimaphilla umbellata 121*
Chorispora 52
Chrysobalanaceae 73
Cicer 133
Cichorium 82
— intybus 67*
Cimicifuga simplex 27
Circeaster agrestis 34
Cirsium arvense 66*
Citrulus colocynthus 135
Citrus paradisi 133
Clematis 34
— vitalba 33*
Cleome 44, 48
Clethraceae 44
Climacoptera lanata 64*
Clusiaceae 129
Coccoloba 61
Cocos nucifera 72*
Cola 134
Colletia horrida 48, 56
Colubrina asiatica 48
Colutea arborescens 28*, 30
Commelinaceae 44
Condalia obovata 75
— spathulata 75
Consolida regalis 27, 28*
Cornaceae 79
Cornus mas 79
Coronilla 31
Corsiaceae 44
Corylus 16, 71
— avellana 11*, 70*, 71, 83
— cornuta 71
— ferox 71
— mandshurica 70*, 71
— tubulosa 71
Coryphoideae 39
Costaceae 44
Cotinus 73
— coggygria 83
Cotoneaster 58
Cousinia 82
Crambe maritima 70*
Crassulaceae 26
Crataegus 58
— monogyna 75
Crepis 82
— pulchra 93
Cyclachaena xanthiifolia 126
Cyclanthaceae 35
Cyclanthera 59
Cydonia 133
Cynoglossum 54
Cynoxylon capitata 84*, 86
— japonica 86
Cyperaceae 62
Dalbergia stipulacea 37
Daphne 78
— altaica 79
— mezereum 79
Daphniphyllum macropodum 75
Datiscaceae 44
Daucus 94
Davidiaceae 54
Degeneria vitiensis 27
Delphinium 26
— elatum 26*
Dendromecon 109
Desmodium heterocarpum 95
Diapensiaceae 44
Diatenopteryx 49
— sorbifolia 50*
Dictamnus 41
Didymelaceae 39
Diervilla 44
Dilleniaceae 25
Dimorphotheca 93.
— pluvialis 94*
Dioncophyllaceae 44
Dioscoreaceae 44, 54
Diospyros 105
Diplotaxis 97
Dipsacaceae 66, 83
Dipsacus sativus 85
Dipteronia sinensis 49*
Diptychocarpus strictus 52, 96, 97
Donax cannaeformis 123
Dorstenia contrajerva 125
Draba 47
Duchesnea 35
Durio zibethinus 104
153
Ebenaceae 54
Ecballium 59
Edgeworthia 74
Elaeagnaceae 37
Elaeis guinensis 74, 75
Elaeocarpaceae 44
Elettaria cardamomum 134
Embelia ribes 75
Engelhardia wallichiana 74*
Enneapogon 116
Enseta perrieri 81
Entada 31
— pursaetha 29
Epacridaceae 44
Epilobium motanum 115*
Eragrostis 116
Eremosparton aphyllum 30
— briobotrya 58
Eriocaulaceae 44
Eryngium campestre 117
Erythrina corallodendron 30, 31*
Erythroxylaceae 75
Erythroxylum coca 75
Escalloniaceae 44
Euclidium syriacum 70*
Eucommia ulmoides 68
Eucommiaceae 68
Eucryphiaceae 48
Eucryphia cordifolia 48, 50*
Eupatorium cannabinum 118
Euphorbiaceae 44
Eupteleaceae 36
Eustrephus 56
Evax 82
Ewersmannia subspinosa 31
Exochorda 41
Faba 133
Fabaceae 104
Fagaceae 71
Fagus 71
Feijoa sellowiana 55
Ferula 122
Ficus 103
— carica 84*, 85
Filipendula 33
Flacourtiaceae 44, 54
Fontanesia 69
Forestiera neo-mexicana 75
Forsyth!a 44
Fragaria 105
— vesca 34*
Frangula alnus 57*
Frankeniaceae 44
Fraxinus 69
— Pennsylvania 68*
Freycinetia 81
Fumaria 69
Gahnia 69
Galium aparine 51*
Garcinia mangostana 104
Gardenia 104
Genista 29
— hispanica 38
— monosperm a 38
— radiata 38
— sylvestris 38
Geranium palustre 46*
Gesneriaceae 44, 54
Geum rivale 33*
Glaucium 45
Gleditschia triacanthos 32
Glycyrriza echinata 38
Gnaphalium sylvaticum 66*
— uliginosum 65
Goldbachia 53
Gouania blanchetiana 51*, 52
Grewia biloba 54
Grossularia 56
— reclinata 11*
Guttiferae 44
Gymnocladus dioicus 32
Gypsophyla paniculata 117
Hablitzia tamnoides 61
Halesia 117
Halimione pedunculata 64*
Haloragaceae 54
Hdloxylon 64
Hamamelidaceae 44, 86
Haplophyllum acutifolium 49
— latifolium 49
— perforatum 49
Hardwickia binata 37, 38*
Hedera chrysocarpa 81
Hedypnois cretica 93
'Hedysarum 31
— gmelinii 95, 96*
Helianthus annuus 11*
Heliconiaceae 44, 51
Helietta parvifolia 49
Helinus ovatus 48
Helwingiaceae 54
Hepatica nobilis 109*
Heracleum sphondylium 51*
Heritiera littoralis 123
Heterocarpus fernandezianus 97
Heterocaryum .subsessile 94
— szovitsianum 94
Heteropappus 93
Heterospermum 93
Heterotheca 93
Hippocrepis comosa 31
Hippuris vulgaris 74
Hirschfeldia 97
— incana 97
Horaninovia 64, 86
— ulicina 86
154
Hordeum jubatum 129
Humiriaceae 123
Humulus lupulus 83, 84*
Hydrastidaceae 27
Hydrastis canadensis 7, 27
Hypecoum albescens 53*
Hypericum androsaemum 55, 56
Idiospermum 37
Illicium 26
— anisatum 26*
Impatiens noli-tangere 46*
Isatis 69
— tinctoria 70*
Isomeris 44
Jasminium 54
Juglandaceae 73
Juglans 73
— regia 73
Juncaginaceae 49
Jurinea 117
Lespedeza 38
Leucanthemum 121
Ligustrum ovalifolium 75
— vulgare 75
Liliales 124
Limnocharis 26
Limonium 65
Liquidambar 86
Liriodendron tulipifera 34
Lisaea heterocarpa 94
Loasaceae 44
Lobeliaceae 54
Lodoicea maldivica 72
Loganiaceae 44, 54
Loranthaceae 54, 79
Loranthus europaeus 79
Lotus 29
Lowiaceae 44
Luffa 59
— acutangula 59
— cylindrica 59
Lunaria 47
— annua 46*
Lycium 55
Lycopersicon esculentum 57*
Lycopsis arvensis 54*
Kadsura 27
Kerria japonica 35
Kirkia 49
Kochi a 64
— lanifolia 64^
Koeleria 116
Koelpinia linearis 66, 67*
Koelreuteria 44
Kolkwitzia amabilis 74
Korsiaceae lil3
1 Lactoris fernandeziana 26
Lactoridaceae 26
Lagenaria 59
* Lagopus 37, 76
Lallemantia 135
— iberica 54*
Lappula 54
— barbata 94
— drobovii 94
— microcarpa: 94:
' — semialata 94
— sessiliflora 94
Lardizabalaceae 27
Lathyrus 29
— sativus var. amphicarpus 94, 95
— saxatilis 95
— setifolius 95
— vernus 28*
Lauraceae 103
Lavatera 48
Lennoaceae 54
Leontice incerta 117
Lepidium 47
Maesa 54
Magnolia virginiana 26*
Magnoliaceae 34
Magnoliales 7
Malva 48
Mangifera 104
Mapania 73
Maranthaceae 123
Mastixioideae 79
Mattiola 47, 96
Mauritia flexuosa 75
Medicago agrestis 28*, 29
— arabica 29
— lupulina 38
— orbicularis 29
— sativa 28*
— scutellata 28*
— truncatula 29
Medusaginaceae 44
Melastomataceae 44, 54
Meliaceae 44, 54
Melica 116
Melilotus 38
Melocana bambusoides 40
Menispermaceae 35
Mespilus germanica 58
Mimosa pudica 31*
Mirabilis nyctaginea 37
Moehringia trinervia 109
Momordica 59
Monimiaceae 35
Moraceae 125
Mor us 86, 105
155;
Musa 56, 57*
Musaceae 54, 81
Myagrum 69
— perfoliatum 70*
Myoporaceae 54
Myosotis 109*
Myosurus 33
Myriophyllum 49
Myristica 103
— fragrans 40
Myristicaceae 39
Myrsinaceae 54, 61
Myrsine africana 79
Najadaceae 36
Nasturtium 47
Nelumbo nucifera 34*
Neslia 69
— paniculata 70*
Nigella 41
Nitrariaceae 54
Norrisia 116
Nyctaginaceae 36
Nypa fruticans 39
Nypoideae 39
Ochradenus 54
Olea chrysophylla 75
— europaeae 75
Oleaceae 44, 54, 69, 75
Onobrychis 38
— aequidentata 38*
— arenaria 80
Opuntia dillenii 9*
Orbignya barbosiana 54
Orlaya grandiflora 51*
Ornithogalum 109
Ornithopus 31
— sativus 31*
Orobanche cumana 115*
Ortilia secunda 120
Ostrya carpinifolia 71, 83
Oxytropis 30
Padus avium 39*
Paliurus spina-christi 74
Pandanaceae 54, 81
Pandanus simplex 81
— tectorius 84*, 86
Papaveraceae 53, 109
Paracaryum intermedium 94
Parinari curatellifolia 75
Passifloraceae 44
Pastinaca 122
Pavonia spinifex 49, 50*
Pedicularis 16
Perilla 135
Persea 133
Phalaris 40
Phaseolus 29
Philesiaceae 54
Phleum 116
Phoenix 39
— dactylifera 39
Phyllostylon brasiliense 69, 70*
Physalis 16
Phytelephas 54
— macrocarpa 83*
Pilea 124
Pimenta officinalis 134
Pinus cembra 106, 107
— koraiensis 106
— pinea 106
— pumila 106
— sibirica 106
Piper nigrum 134
Pisonia 37
Pistacia vera 73
Pisum 29
— fulvum var. amphicarpum 95
Pitcairnia xanthocalyx 10
Pittosporaceae 44
Platanus orientalis 33, 82
Platycarya 74
Plumbaginaceae 61
Plumbago 65
— capensis 65*
Poa 116
Polanisia 48
Polygala 103, 109
Polygonaceae 62
Polygonum 62, 105
— linicola 62*
Pomaderris apetala 52
Porcelia goyazensis 104
Potamogeton 34
Potentilla recta 7
Poterium 16
Primula veris 16
Proteaceae 27, 36, 82
Prunoideae 38
Prunus spinosa 11*
Ptelea trifoliata 68*
Pterocarpus 37
— indicus 38*
Pterocarya 74
— fraxinifolia 74*
— paliurus 74
Pteroceltis 73
Pterostyrax 117
Pulsatilla 34
Punica granatum 57*, 59
Punicaceae 59
Pyrethrum 121
Quercus dendata 71
— iberica 70*, 71
— ilex 71
— robur 71
156
Ranunculus acris 33*
Raphanus 47, 97
— raphanistrum 47, 53*
Rapistrum rugosum 53*
Resedaceae 54
Reynosia latifolia 75
Rhamnaceae 54
Rhamnus alaternus 56
— cathartica 57*
Rheum 62
— turkestanicum 62*
Rhinanthus alectrolophus 16
Rhodotypos scandens 35
Rhus 73
Ribes 56
Ricinus communis 48, 50*
Rosa 35
— majalis 11*
Rosaceae 16, 58
Rosoideae 16, 33
Rubiaceae 54
Rubus 35, 105
— caesius 35
— chamaemorus 35
— saxatilis 35
Rumex 62
— crispus 62*
Ruppiaceae 35
Rutaceae 68
Sacoglottis amazonica 123
Sageretia thea 56
Salacca edulis 105
Salsola 64
— australis 118, 126
Sameraria boissieriana 95, 97*
Sanguisorba 16, 36
Sanicula 52
Santalaceae 71
Sapindaceae 44
Sarcolaenaceae 44
Sassafras albidum 103
Saxifraga aizoides 10
Scabiosa graminifolia 84*
— maritima 67*
Schivereckia 47
Scilla sibirica 109
Scirpodendron 73
Scleranthus annuus 65*
Scleria 69
Scutia indica 54
Sebastiana 109
Securinega 44
Seneci о 82
— jacobeae 93
Sesamum 133
Setaria 133
Simaroubaceae 49
Sinapis 47, 49
— alba 46*
— arvensis 97
Sisymbrium 47
Smilacaceae 54
Smirnovia turkestana 29
Solanum melongena 134
Sorbaria 26
Sorghum 133
Sparganium 73, 82
— neglectum 82*
Spatholobius roxburgii 37
Spergula arvensis subsp. linicola 47
-------maxima 47
Spinacia 64, 86
— tetrandra 86
Spirorhynchus sabulosus 95, 96*
Stellaria holostea 109
Sterculia 103
Sterculiaceae 123
Sterigmostemum 52
Strelitziaceae 44
Strigosella 14
— circinata 96
Strophanthus 134
Strychnos 104
— nux vomica 134
Styphnolobium japonicum 32
Suaeda 64, 90
— baccifera 64*
— corniculata 91
— linifolia 91
Subularia 47
Swida sanguinea 79
Symphoricarpus 10
Symplocaceae 54, 55
Taccaceae 44, 54
Tamarindus 104
— indicus 32
Tamus 54
Tanacetum 121
Taraxacum erythrospermum 66*
Tauscheria 69
Tetracentraceae 41
Tetrapteris citrifolia 49, 50*
Thalictrum 33
— aquilegifolium 34
Theaceae 44
Theobroma 103
Theophrastaceae 54
Thermopsis 134
Thesium 71, 109
Thinouia 49
— scandens 50*
Thlaspi 47
Thymelaea 74
Thymelaeaceae 74
Tieghemella heckelii 104
Tiliaceae 54
Torilis 94
Torularia 96
157
Tragopogon 65, 92
— dubius 92
— capitatus 92
— graminifolius 92
— heterospermum 92
— kultiassovii 92
— paradoxus 92
— tuberosus 92
Trapa natans 75*, 78
Trema 75
Tribulus terristris 49, 50*
Trichochiton inconspicum 14
Trifoliastrum 37
Trifolium aureum 116
— borysthenicum 38
— fragiferum 16
— incarnatum 37
— pratense 38*
— rubens 38
Triglochin 49
— maritimum 49
— palustre 49
Trilliaceae 54
Triuridaceae 25
Trochodendraceae 41
Trollius europaeus 25
Trymalium helianthoides 51*, 52
Turneraceae 44
Turretia 129
Turritis 47
Typha 36
— angustifolia 82
— latifolia 36*, 82
Typhaceae 36
Ulmus 68
Ursinia pulchra 67*
Urticaceae 69, 124
Valerianaceae 67
Vanilla planifolia 134
Veratrum nigrum 121*
Veronica prostrata 44
Viburnum opulus 75
Vici a 29
— amphicarpa 94
— angustifolia var. amphicarpa 94
Viola arvensis 109*
— hirta 109*
Viscum 79
— album 10
Vitex agnus castus 55
Vitis 105
Vochysiaceae 44
Weigela 44
Wilhelmsia physodes 48
Winteraceae 41
Xanthium 66
Ximenesia encelioides 93
Zelkova 69
Zingiberaceae 44
Ziziphus jujuba 78
Zygophyllaceae 49
158
ОГЛАВЛЕНИЕ
Стр.
От автора
Глава I. Общие вопросы .............................................. 5
Значение плодов. Определение понятия «плод» ....................... 5
Морфологическое разнообразие плодов................................ 6
Типы гинецея.........................................: . . . 6
Строение околоплодника ...................................... 10
Способы вскрывания и распадения плодов......................... 12
Остающиеся при плодах части цветка и соцветия. Специальные при-
датки ......................................................... 15
О принципах классификации и номенклатуре плодов . ................ 17
Глава II. Морфология плодов ................ 25
а
Апокарпии......................... . ................. 25
Многосемянные полимерные............................... 25
Многосемянные мономерные .............................. 27
Односемянные полимерные................................ 32
Односемянные мономерные................................ 35
= Ценокарпии........................ '................... 41
Многосемянные верхние и нижние........................ 41
Сухие вскрывающиеся................................. 41
Распадающиеся...................................... 48
Сочные.............................................. 54
Односемянные.......................................... 59
Сухие верхние и нижние............................. 61
Сочные верхние и нижние............................. 74
Глава Ш. Соплодия........................................... 80
Глава IV. Гетерокарпия................................................ 88
Глава V. Экология распространения плодов и семян..................
Основные понятия и термины.....................................
Зоохория ......................................................
Эндозоохория ...............................................
Синзоохория ................................................
Эпизоохория ................................................
Значение и эффективность зоохории . .
Анемохория ............................
Формы анемохории и типы диаспор . .
Аэрохория. Аэрохорные диаспоры . . .
99
99
100
101
106
ПО
111
112
112
113
3S
159
Анемогеохория и другие формы анемохории . . . ............... 117
Эффективность и значение анемохории....................... . 118
Баллистохория................................................... 120
Гидрохория...................................................... 122
Автохория.........................................................124
Антропохория....................................................125
Агестохория............................................... . 126
Эргазиохория ................................................ 127
Спейрохория...................................................127
Сочетание способов диссеминации................................ 128
Заключение ......................................................130
I
*
Глава VI. Практическое значение плодов.......................... . 132
Литература..........................................................136
Указатели терминов................................................ .141
Указатель русских названий растений.................................143
Указатель латинских названий растений...............................151
Роза Ефимовна Левина
МОРФОЛОГИЯ и экология плодов
Утверждено к печати
Всесоюзным ботаническим обществом
Академии наук СССР
Редактор издательства Е. А. Чекулаева
Художник А. И. Слепушкин
Технический редактор Г. А. Смирнова
Корректор И. А. Корзинина
ИБ № 33027
Сдано в набор 2.12.86. Подписано к печати 15.04.87.
М-17136. Формат 60X90l/i6. Бумага офсетная № 1.
Фотонабор. Гарнитура литературная. Печать офсетная.
Усл. печ. л. 10. Усл. кр.-отт. 10.25. Уч.-изд. л. 11.70.
Тираж 2150. Тип. зак. 1044. Цена 1 р. 20 к.
Ордена Трудового Красного Знамени
издательство «Наука». Ленинградское отделение
199034, Ленинград, В-34, Менделеевская линия, 1.
Ордена Трудового Красного Знамени
Первая типография издательства «Наука».
199034, Ленинград, В-34, 9 линия, 12.