/

































































































Текст
К. ФРИШ
ПЧЕЛЫ,
ИХ ЗРЕНИЕ, ОБОНЯНИЕ,
ВКУС И ЯЗЫК
Перевод с английского
проф. В. В. АЛПАТОВА
Под редакцией и с прьдцдлови&м,
акад. Е. Н. ПАВЛОВСКОГО
И ж Л
ИЗДАТЕЛЬСТВО
ИНОСТРАННОЙ ЛИТЕРАТУРЫ
Москва — 19 5 5
BEES
THEIR VISION, CHEMICAL
SENSES, AND LANGUAGE
by
K. FRISCH
19 6 0
Scan AAW
ПРЕДИСЛОВИЕ
К РУССКОМУ ИЗДАНИЮ
Имя К. Фриша широко известно среди биологов и
пчеловодов. Ему принадлежат многочисленные работы по
изучению органов чувств и поведения медоносной пчелы.
Предлагаемая вниманию советского читателя книга К.
Фриша содержит описание чрезвычайно интересных опытов
автора по изучению поведения медоносной пчелы. Особый
интерес своей новизной представляют данные по
исследованию способности пчел ориентироваться по поляризации
света и не только сообщать другим пчелам о наличии
взятка в природе, но и указывать направление, в котором
следует лететь за ним, и расстояние его от улья.
Эти работы Фриша, выполненные им в послевоенные
годы, по началу вызывают удивление и даже недоверие,
что повлекло за собой естественное желание их повторить.
В настоящее время можно указать по крайней мере три
места, где эти работы были проверены с положительными
результатами, если не считать некоторых деталей,
требующих доработки. В Кембридже их повторил известный
энтомолог Тори, который пишет: «Мои наблюдения
затронули все основные стороны работы, и я мог сам оценить
эффективность указаний пчел о расстоянии до источника
корма и направлении к нему... Этот незабываемый опыт
позволил мне разрешить некоторые сомнения и трудности,
которые возникают при первом чтении работ Фриша, и
убедил меня в обоснованности его выводов в целом».
На кафедре зоологии Корнеллского университета (США)
©пыты Фриша проверил Гриффин, также получивший
положительные результаты.
На биологическом факультете Московского
государственного университета Н. П. Смарагдова с помощью
двух студентов-физиологов провела проверку основных
опытов Фриша и также подтвердила правильность его
выводов.
о
Заслуживает внимания не только чрезвычайно
интересный материал, приводимый автором, но и способ его
изложения. Книга написана живо и увлекательно, простым
и доступным языком, правда, иногда с примесью
некоторого антропоморфизма. Фриш описывает свои опыты в
строгой логической последовательности, ясно показывая
читателю, как разрешение одной проблемы порождало новую
и как постепенно, шаг за шагом, ему удалось проникнуть
в сущность явлений, кажущихся на первый взгляд
необъяснимыми.
Мы считаем нужным отметить, что метод,
применявшийся Фришем для изучения объективного восприятия пчелами
внешнего мира и их поведения, представляет собой
прекрасную иллюстрацию плодотворности учения И. П.
Павлова об условных рефлексах в применении к физиологии
органов чувств и высшей нервной деятельности у
представителей столь далекой от млекопитающих группы
животных, как насекомые, хотя сам Фриш нигде и не
упоминает об этом.
Помимо теоретического интереса работы Фриша имеют
серьезное практическое значение, так как понимание
поведения пчелы крайне важно для пчеловодства. В СССР
практическое использование результатов работ Фриша
началось на 6 лет раньше, чем в других странах. Это
признает и сам Фриш. В предисловии к своей книге
«Пчелы, направленные по запаху, на службе сельского
хозяйства и пчеловодства» (Вена, 1947) Фриш пишет:
«Русские ученые-пчеловоды разработали способ повышения
посещаемости пчелами обширных красыоклевериых полей
с целью увеличить опыляемость и семенную продукцию...
Русские исследователи исходили из возможности
дрессировать пчел на ароматическое вещество, и им удалось,
скармливая пчелам в улье ароматизированный сироп,
направлять их на посещение определенных растений».
Книга Фриша безусловно заинтересует не только
работников в области научного и практического пчеловодства,
физиологов, биологов и психологов, но и широкие круги
читателей не-специалистов.
Акад. Е. Павловский
Глава I
ЦВЕТОВОЕ ЗРЕНИЕ ПЧЕЛ
Медоносная пчела, обитающая в ульях,—насекомое
общественное. В обычных ульях живет около шестидесяти
тысяч пчел, но среди них есть только одна вполне развитая
самка. Это пчелиная матка (рис. 1, А)—единственная особь в
семье, откладывающая яйца. Самцы, или трутни (рис. 1, В),
Рис. 1. Различные особи медоносной пчелы.
А—матка; Б—рабочая пчела; В—трутень. 1—голова;
2—грудь; 3—брюшко; 4—антенна; 5—глаз.
по размерам крупнее, но более неуклюжи, несколько
бестолковы и ленивы. Все остальные обитатели улья—это
рабочие пчелы (рис. 1, Б). В обычных условиях рабочие
пчелы неспособны откладывать яйца, так как их яичники малы
и недоразвиты; однако в остальном их следует считать
самками, так как они обладают инстинктами, свойственными
самкам. Рабочие пчелы кормят личинок, строят соты,
убирают мусор и вылетают из улья для сбора меда и
пыльцы, которыми кормится семья.
Рабочие пчелы собирают корм, посещая цветы. Некот
торые из них берут с цветков капли нектара, богатого
о
сахаром; другие собирают пыльцу, доставляя растущим
личинкам белковую пищу. Используя цветки в качестве
источника пищи, пчелы, в свою очередь, приносят
растениям пользу, производя их опыление: перелетая от цветка
Рис. 2. Разные типы цветков.
А —цжзток злака, опыляемый ветром; Б—цветок,
опыляемый насекомыми.
к цветку, они переносят пыльцу, приставшую к
поверхности их тела, с одного цветка на другой.
Как известно, цветки высших растений бывают двух
типов. Многие растения обладают мелкими зелеными
цветками, не имеющими запаха (рис. 2, А); пыльца таких
растений разносится ветром и в порядке чистой случайности
может попасть на другие цветки того же самого вида.
Цветки других растений имеют привлекающую внимание
яркую окраску или же резкий запах, а иногда и окраску
и запах одновременно—эти-то цветки мы обычно и
называем «цветами» (рис. 2, Б). Лишь цветки последнего типа
выделяют нектар, и^поэтому их посещают насекомые,
осуществляющие опыление при перелете с одного цветка
на другой (рис. 3). Биологи давно пришли к убеждению,
6
что яркая окраска и запах цветков служат для
привлечения насекомых. Эти качества облегчают насекомым
нахождение цветков, а следовательно, и добычу корма,
обеспечивая вместе с тем и осуществление опыления.
Однако эта точка зрения разделялась не всеми
биологами. Примерно в 1910 г. известный офтальмолог Гесс
(1913) произвел многочисленные
опыты с рыбами, насекомыми и
другими низшими животными. Он
обнаружил, что животные,
проявляющие положительный
фототаксис (т. е. активно движущиеся по
направлению к самому яркому из
имевшихся в наличии источнику
света), всегда собираются в
зеленой или желто-зеленой зоне
спектра, которая для глаз человека с
цветовой слепотой представляется
наиболее яркой. На основании
этого Гесс утверждал, что рыбы
if беспозвоночные, в частности
пчелы, отличаются полной
цветовой слепотой. В таком случае
окраска цветков теряла всякий
биологический смысл. Я никак не
мог согласиться с этим, что и
послужило толчком к моим работам
над пчелами, начатым примерно
40 лет назад. Я поставил своей
задачей выяснить, обладают ли пчелы способностью
различать цвета.
Поместив на опытные столики небольшие количества
меда, можно заставить пчел, привлекаемых запахом меда,
посещать эти столики. Можно, например, кормить их на
кусках синего картона; они забирают мед, переносят его
в улей, а затем возвращаются вновь и вновь на
обнаруженный ими богатый источник пищи.
После того как пчелы летали на корм в течение
некоторого времени, кусок синего картона, приобретший запах
меда, убирают, и по обе стороны от того места, где он лежал,
кладут два куска чистого картона—налево синего, а
направо красного цвета. Если пчелы запомнили, что они
получали корм на синем фоне и если они в состоянии отличить
Рис. 3. Насекомое,
сосущее нектар.
Нектар выделяется на дне
цветка; поэтому, высасывая
нектар, насекомое
приходит в соприкосновение с
пыльцой.
7
красный цвет от синего, то они доляшы прилетать на синий
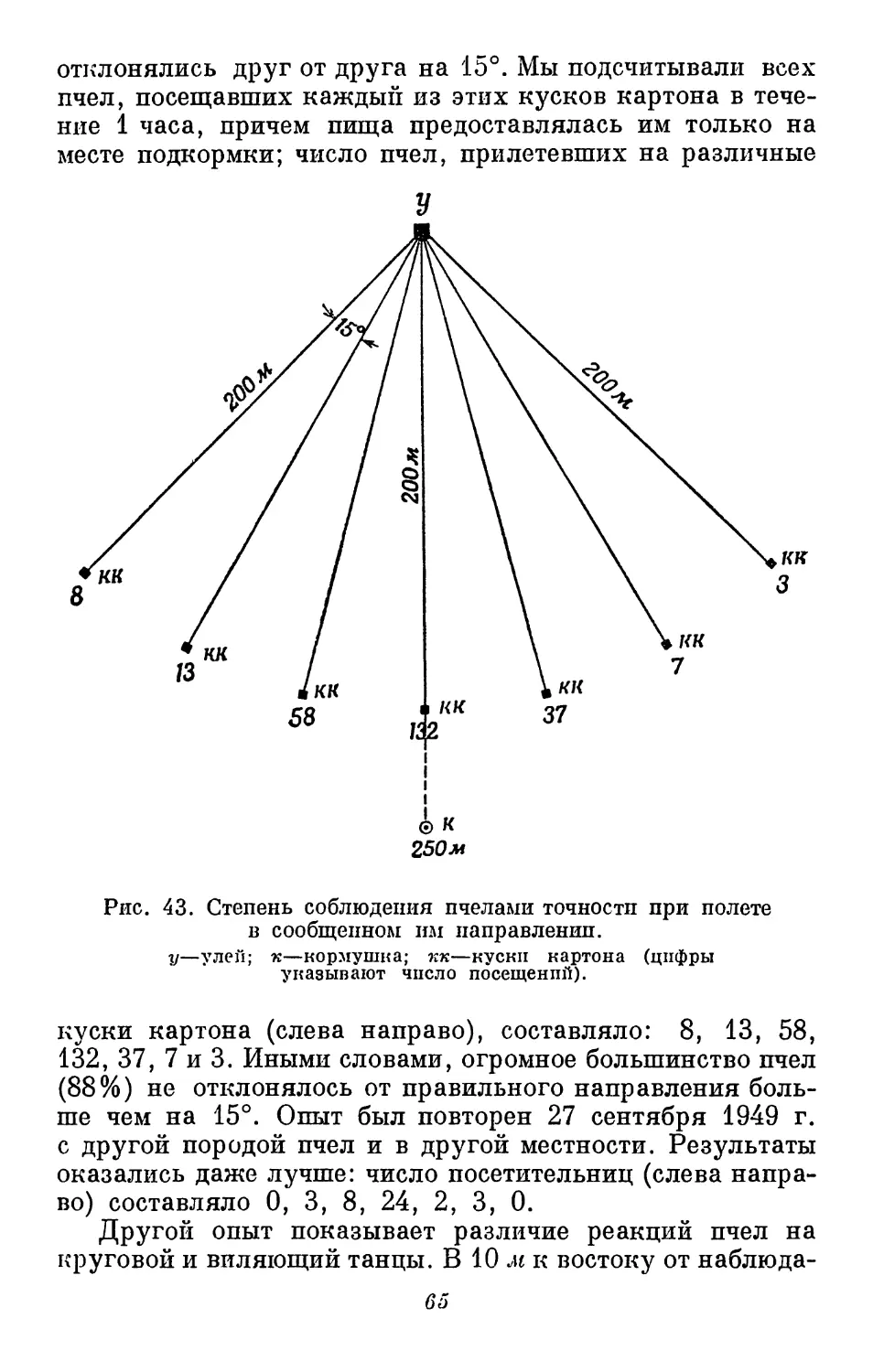
картон, Именно так и происходит (рис. 4).
Этот опыт был сделан давно. Он показывает, что пчелы
способны различать цвета; однако это еще не значит, что
они обладают цветовым чувством, или способностью
к восприятию цветов, так как это не всегда одно и то же.
Встречаются, хотя и очень редко, люди с абсолютной
цветовой слепотой. Для них все предметы выглядят так, как
Рис. 4. Опыт по приучению пчел к цвету.
Пчелы, предварительно получавшие корм на синем
картоне в середине столика (помечено звездочкой),
прилетают на синий картон, не содержащий корма
(слева), отличая его от красного картона (справа).
они выглядели бы для нас на обычном (не цветном)
фотографическом снимке. Эти люди, однако, способны отличать
красный цвет от синего, так как красный цвет кажется
им очень темным, а синий—более светлым. Таким
образом, только что описанный опыт с пчелами не дает
возможности установить, отличали ли пчелы красный
картон от синего по окраске или по яркости подобно тому, как
это делает человек с цветовой слепотой. Человеку
с абсолютной цветовой слепотой все цвета кажутся серыми,
отличаясь лишь по яркости. Мы не знаем, как
воспринимается яркость наших кусочков цветного картона
насекомым с цветовой слепотой. Поэтому мы поставили
следующий опыт.
На стол кладут кусок синего картона и окружают его
кусками серого картона разных оттенков—от белого до
черного—и на каждый кусок картона помещают небольшое
часовое стекло; в стекло, лежащее на синем картоне,
наливают корм (сахарный сироп). Таким путем пчел приучают
прилетать на синий цвет. Так как пчелы обладают очень
s
хорошей памятью на место, то относительное
расположение кусков картона часто меняют. Сироп же остается на
синем картоне, так что синий цвет все время служит
указанием на наличие корма. По прошествии нескольких
часов ставят решающий опыт. Куски картона и стекла,
обсиженные пчелами, удаляют. На столе располагают
куски чистого серого картона различного оттенка с пустыми
часовыми стеклами на них, и среди них помещают кусок
чистого синего картона, на котором также стоит пустое
часовое стекло, Оказывается, что пчелы запоминают синий
цвет, так как они прилетают только на синий картон,
отличая его от серого картона различных оттенков. Это
означает, что пчелы обладают истинным цветовым зрением.
Достоверность результатов этого эксперимента
оспаривали, ссыдаясь на то, что синий картон, возможно,
обладал специфическим запахом, по которому пчелы его
и узнавали. То обстоятельство, что человек не
воспринимает этого запаха, еще не доказывает, что картон лишен
запаха и для пчел. Поэтому следует учитывать
возможность того, что пчелы узнавали синий картон не по цвету,
а по запаху. Эта возможность, однако, исключается. Мы
повторили опыт, покрыв все куски картона стеклом, так
что если бы они и имели запах, он не мог бы проникнуть
через стекло (рис. 5), При этом результаты опыта
оказались совершенно такими же, как и без стекла.
Приучение пчел прилетать на корм, помещенный на
картоне оранжевого, желтого, зеленого, фиолетового
и пурпурного цветов, дает те же результаты. Если же
приучать пчел находить корм на яркокрасном картоне, то они
прилетают не только на красный, но также на все куски
темносерого картона. Таким образом, красный и черный
цвета воспринимаются глазами пчел одинаково, иными
словами, пчелы слепы на красный цвет. Из этих опытов
вытекает, что пчелы обладают цветовым зрением, но оно
у них не совсем такое, как у человека с нормальным
зрением.
Для того чтобы лучше разобраться в способности пчел
различать цвета, мы ставили несколько иной опыт. Мы
приучали пчел находить корм на синем фоне, а затем
выкладывали на стол все куски цветного картона, за
исключением серых. Пчелы искали синий картон, но мы с
изумлением обнаружили, что они не способны сделать это
безошибочно. Они путали синий картон с фиолетовым и
9
пурпурным. Далее оказалось, что пчелы, приученные к
желтому цвету, садятся не только на желтый картон, но также
на оранжевый и зеленый, приученные же к оранжевому
цвету предпочитают садиться на желтый картон; то же
самое получается, если приучить пчел к зеленому цвету.
Создается впечатление, что пчелиному глазу оранжевый,
желтый и зеленый цвета представляются одинаковыми
Рис. 5. Пчелы, приученные к синему
цвету, прилетают на чистый синий картон без
корма, покрытый стеклом, отличая его от
других кусков картона, окрашенных в
различные оттенки серого цвета.
и наш желтый картон кажется пчелам более насыщенным
по цвету и потому более заметным и привлекательным,
чем оранжевый и зеленый. Для пчел, приученных к синему,
пурпурному и фиолетовому цветам, синий и пурпурный,
повидимому, более привлекательны, чем фиолетовый
(Фриш, 1915).
В 1927 г. Кюн повторил наши опыты, но вместо
цветного картона он пользовался цветовым спектром.
Результаты опытов Кюна подтвердили наши выводы: пчелы
слепы на красный цвет, оцк могут отличать другие
цвета от всех оттенков серого цвета, но не отличают
желтый цвет от оранжевого и зеленого или синий от фио-
10
летового. Но, кроме того, Кюн обнаружил два совершенно
новых факта. Прежде всего оказалось, что пчелы
воспринимают лучи, расположенные в узкой сине-зеленой зоне
(480—500 т(х), как самостоятельный цвет. Пчелы,
приученные к сине-зеленому цвету, отличают его от синего
и желтого. Мы не заметили этого обстоятельства, так
как у нас не было картона такого сине-зеленого цвета.
Далее он обнаружил, что если в течение некоторого
времени кормить пчел на опытном столике, подвергая их
действию ультрафиолетовых лучей, то они будут прилетать
в любое место, куда падают ультрафиолетовые лучи. Для
нас этот свет невидим, пчелы же отличают его от всех
оттенков белого или серого цвета. Они воспринимают
ультрафиолетовые лучи как совершенно особый цвет.
При сравнении цветового восприятия пчел и людей
оказывается, что для пчел видимая часть спектра
укорочена в области красных лучей и удлинена в области
ультрафиолетовых. Таким образом, видимая часть спектра
для пчел просто сдвинута в сторону коротких волн.
Гораздо более важное отличие состоит в том, что глаз
человека различает около шестидесяти отдельных цветов
видимого спектра, тогда как пчела, повидимому,
различает лишь четыре цвета: желтый, сине-зеленый, синий
и «ультрафиолетовый» (рис. 6).
Рассматривая окраску цветков в связи с восприятием
цветов пчелами, мы обнаружим много интересного. Нам
сразу становится понятной редкость в Европе яркокрас-
ных цветков, поскольку насекомые, посещающие цветки,
слепы на красный цвет. Вместе с тем многие растения
Америки и Африки имеют яркокрасные цветки, однако еще
задолго до изучения восприятия цветов насекомыми было
известно, что яркокрасная окраска типична для цветков,
посещаемых и опыляемых птицами. Из опытов Хониг-
мана (1921) известно, что глаза птиц особенно
чувствительны к красному цвету. Цветки многих европейских растений,
которые мы называем красными, на самом деле имеют
пурпурную окраску, а пчелам кажутся синими.
Некоторые виды европейских растений имеют цветки более
нежного красного цвета, например Adenostyles alpina
Bluff et Fingerh., многие виды рода Dianthus, Daphne
striata Tratt, Erigeron uniflorus L., Erigeron alpinush.,
Silene acaulis Jacq. и Viscaria alpina G. Don.
Биологам давно было известно, что эти растения посе-
11
8
Е
!
Яэ 3
з
щаются и опыляются
бабочками. Ильзе (1928) изучала
способность бабочек
различать цвета; она установила,
что бабочки являются
единственными известными нам
насекомыми, способными вос-
§ принимать красный цвет.
| Другое кажущееся
исключение из правила —это мак
(Papaver), цветки которого,
я
ев
ё и- несмотря на их яркокрас-
§ S ную окраску, часто
посещаем | ют пчелы. Однако цветки
~ | мака отражают ультрафиоле-
о § I товые лучи, и путем соответ-
qj | | ствующих экспериментов мо-
5 rt a жно показать, что пче-
?г § лы, приученные к цветкам
о сз | ? мака, в действительности
f 5 i2 ° § различают отраженные уль-
о | Jg 8 трафиолетовые лучи(Лотмар,
?8 1933).
4 м Мы неспособны воспри-
Я g нимать эти лучи и видим
? g лишь красную окраску; пче-
g 1 лы же, напротив, не видят
g, § красной окраски, а воспри-
ф ? нимают только «ультрафио-
| я летовую». Иными словами,
g^l* пчелы, будучи слепыми на
g w красный цвет, тем не
мерз нее воспринимают мак как
^ окрашенный цветок, так как
для них он имеет «ультра-
н фиолетовую» окраску.
Известно, что пчелы по-
| сещают некоторые растения,
<§ имеющие неприметные
цветки, как, например, черника
(Vaccinium myrtillus L.) и
дикий виноград {Ampelopsis
quinfuefolia Michaux). Можно
12
было бы предположить, что цветки этих растений обладают
резким запахом и пчелы обнаруживают их при помощи
обоняния. Однако человек не слышит запаха эуих цветков, и
можно показать, что они лишены запаха и для пчел. В опытах,
проведенных много лет назад, мы обнаружили, что
восприятие пчелами окраски зеленых листьев черники сильно
меняется, если устранить при помощи светового фильтра,
помещенного над цветками, ультрафиолетовые лучи. Хотя
нам до сих пор не удалось исследовать листья черники
при помощи спектрографа, тем не менее мы предполагаем,
что для пчел эти листья, вероятно, имеют
«ультрафиолетовую» окраску.
Реакция пчел на белые цветки, повидимому, несколько
сложнее. Очень легко приучить пчел к «настоящим»
(окрашенным, или хроматическим) цветам; что же касается
белого цвета, то иногда пчел легко удается приучить к
белой бумаге или картону, иногда же это оказывается очень
трудным. Это своеобразное обстоятельство было
объяснено в работах Герц (1937а, Ь, с, (1939). Она исследовала
различные образцы белой бумаги и обнаружила, что
некоторые из них поглощают ультрафиолетовые лучи. К такой
белой бумаге пчелы приучаются легко. К бумаге же,
также белой, но отражающей ультрафиолетовые лучи
подобно другим лучам, видимым для человека, пчел приучить
трудно; они не запоминают ее цвета и не приучаются
уверенно находить ее среди бумажек других оттенков белого
и серого цвета. Однако глаз человека не в состоянии
различать эти два «сорта» белого цвета. Чтобы понять это,
необходимо разобраться в сущности «белого» цвета.
Свет, излучаемый солнцем, представляется человеку
белым; если же пропустить его сквозь призму, то свет
разлагается, в итоге чего получается цветовой спектр.
Если при помощи линзы вновь собрать цветные лучи, то
мы получим опять белый свет. Таким образом, свет,
содержащий цветные лучи в таком же соотношении, как и
солнечный свет, представляется глазу человека белым. Если
удалить из него какие-либо лучи, то оставшиеся лучи
в сумме уже не дадут белый свет. Если удалить желтый
луч, то свет будет иметь синюю окраску, так как желтый
цвет является дополнительным к синему (рис. 7).
Перейдем теперь к глазу пчел. Белый свет,
воспринимаемый глазом пчел, должен содержать в себе лучи
всех длин волн, видимые для пчел; как указывалось Ьыше,
13
Рис. 7. Разложение света.
А—пучок белого света, проходя через призму, разлагается на составные
цвета; В—проходя через линзу, разложенный в призме свет вновь
становится белым; В—без синих лучей свет имеет желтую окраску.
мы обнаружили, что глаз пчел чувствителен также и
к ультрафиолетовым лучам. Если удалить
ультрафиолетовые лучи, то свет воспринимается пчелами
не как белый, а как дополнительный к
ультрафиолетовому, возможно как сине-зеленый. Создается впечатление,
что белый свет, включающий в себя все видимые для пчел
лучи, не является для них приметным; повидимому, для
привлечения пчел свет должен быть цветным. В связи
с этим интересно отметить, что почти все белые цветки
действительно поглощают ультрафиолетовые лучи, тогда
14
как желтые и синие цветки часто их отражают. Вследствие
этого цветки, кажущиеся нам белыми, пчелам, возможно,
представляются сине-зелеными.
Таким образом, становится ясным, что окраска
цветков возникла в качестве приспособления к зрительным
восприятиям их посетителей. Совершенно очевидно, что
она не предназначена для того, чтобы радовать
человеческий глаз; это, однако, не мешает нам наслаждаться
красотой цветов.
В Австрии, как и в Германии, пчеловоды ставят ульи
друг на друга, устраивая павильон. В большом павильоне
возвращающимся домой пчелам трудновато отыскивать
свой улей среди множества других; и в самом деле, часто
приходится наблюдать залет пчел в чужие летки. В
большинстве случаев это обстоятельство не столь важно,
потому что пчел, приносящих нектар, всюду встречают
радушно. Тем не менее иногда разгораются драки и
чужаков зажаливают насмерть. Особенной опасности
подвергается матка, возвращающаяся после ориентировочного
или брачного вылета; попади она в чужой улей, ей
грозит смерть. Поэтому часто можно видеть ульи,
окрашенные в разные цвета, чтобы помочь пчелам узнавать свой
улей; однако не все пчеловоды считают это
целесообразным.
Для того чтобы разобраться в этом вопросе, мы
поместили рой пчел в один из стоящих в ряд пустых белых
З'льев и покрыли его переднюю стенку листом жести,
окрашенным в синий цвет. Справа от него стоял такой же
белый улей, покрытый желтой жестью, а слева—просто
белый улей. Спустя несколько дней мы решили изменить
цвета. Если бы мы ограничились только тем, что поменяли
бы местами окрашенные листы жести, после чего пчелы
принялись бы летать не в свой улей, мы не могли бы
решить, привлек ли пчел синий цвет или же запах пчел,
прилетавших раньше на синюю жесть. Чтобы избежать
этого, оборотная сторона синей жести была окрашена
в желтый цвет, а оборотная сторона желтой—в синий,
что давало возможность менять цвета, просто
переворачивая листы жести, а не перенося их с одного улья на другой.
Перевернув оба листа, мы убедились, что многие пчелы
стали летать к пустому улью, покрытому синей краской.
Другие пчелы проявляли признаки нерешительности
и лишь спустя некоторое время начинали залетать в свой
15
Рис. 8. Значение окраски для нахождения пчелами
своего улья.
Семья пчел была помещена в синий улей (4, верхний ряд), по бокам
которого стояли белый (of, верхний ряд) и желтый (5, верхний ряд) ульи. После
того как синий щиток перевернули желтой стороной наружу (4У нижний
ряд), а желтый—синей стороной наружу (5, нижний ряд), многие пчелы
стали входить не в свой улей, руководствуясь окраской, к которой они
привыкли.
улей, не обращая внимания на желтый цвет его передней
стенки (рис. 8). Тогда у нас возникла мысль, что, быть
может, пчелы обращали внимание не только на свой улей,
но и на соседние. В этом случае мой опыт привел бы к
неверному заключению. Пчелы были приучены прилетать к
синему улью, справа от которого стоял желтый улей, а слева—
белый. Теперь же пчелы видели синий улей, слева от
которого стоял желтый, а справа—белый, т. е.
расположение было иным; этим, быть может, и объяснялось, почему
пчелы колебались в своем выборе и в конце концов
находили свой улей при помощи обоняния. Затем этот опыт
16
был повторен, причем синяя шесть была перевернута
желтой стороной вперед, а желтая—синей стороной вперед
и перемещена справа налево. Относительное
расположение цветов при этом оставалось неизменным: пчелы видели
синий улей с желтым справа от него и белым—слева
(поскольку все остальные ульи в ряду, за исключением
ульев, снабжегных листами окрашенной жести, были
белыми). При этом все пчелы без исключения прилетали
не к своему улыо и даже входили в него, хотя он был
совершенно пустой (рис. 9). Эти опыты показывают, что окраска
улья—хороший опознавательный признак, помогающий
пчелам узнавать свой улей и отличать его от соседних
(Фриш, 1915).
Чем же объясняются разногласия пчеловодов по
поводу действенности окраски ульев? Просто тем, что
пчеловоды не принимают во внимание природу цветового
зрения пчел. Часто приходится видеть ульи, поставленные
в несколько этажей, причем соседние ульи окрашены в
красный и черный цвета. Эти цвета являются
контрастными для глаза человека, но воспринимаются совершенно
одинаково глазом пчел. Это касается и тех случаев, когда
зеленый улей ставят под желтым. Во всех этих случаях
окраска ульев не дает никакого эффекта, так как не
создается никаких цветовых контрастов для глаза пчел.
Для окраски ульев пчеловоды должны использовать
исключительно синюю, желтую, черную и белую краски (рис. 10).
В тех случаях, когда приходится пользоваться одним и тем
же цветом для окраски нескольких ульев, следует
учитывать комбинации окрасок, образуемые с соседними
ульями, так как пчелы обращают внимание на окраску не
только своего улья, но и соседних. Более того, при
окраске ульев в белый цвет важно правильно выбрать краску.
Цинковые белила, повидимому, пригодны для этой цели,
так как они поглощают ультрафиолетовые лучи, а потому
кажутся пчелам сине-зелеными. Свинцовые же белила
отражают ультрафиолетовые лучи, а потому кажутся
пчелам действительно белыми и менее приметными, чем
«настоящие» (т. е. хроматические) цвета. Среди
пчеловодов США павильоны не приняты; ульи обычно
располагают на значительном расстоянии друг от друга. Иногда
на лугу располагают множество ульев без каких-либо
опознавательных предметов вокруг них. Нам не известно,
изучался ли этот вопрос эшшеряментально, однако весьма
вероятно, что и на
таких пасеках пчелы часто
не могут найти свой
улей среди множества
других. Нам
представляется, что окраска
ульев по описанным
выше правилам окажется
целесообразной и при
расположении ульев,
принятом в США.
Вернемся к окраске
цветков. Каково ее
биологическое значение?
Биологи давно пришли
к мысли о том, что
окраска цветков играет
важную роль в привлечении
к ним пчел и других
насекомых. Это,
несомненно, справедливо в
отношении пчел,
летающих в поисках новых
цветков для сбора
пищи . Оттинген-Шпиль-
берг (1949) поместила
в закрытом дворике,
затянутом
мелкоячеистой сеткой, семью пчел,
никогда до того не
покидавших
наблюдательный дворик и не
посещавших цветков. В
разных местах дворика
были размещены
окрашенные бумажки, а в
других его точках были
расставлены издающие
запах цветы, которые
были прикрыты, так что
пчелы не могли их
видеть. Среди пчел было
\М/>*/Ш№Ш№/№///ШШУ»М»ЯЩ
Белый
«nvw/WW/W Ф
Рис. 9. Значение окраски соседних ульев для нахождения пчелами своего улья.
Нижний ряд показывает, что после изменения окраски синего улья на желтую (4) и снятия желтого щитка,
прикрывавшего улей 5, и изменения окраски улья 3 на синюю все возвращающиеся домой пчелы вместо своего улья (4)
залетают в пустой улей (3), имеющий теперь синюю окраску. В верхнем ряду показано исходное расположение ульев,
среди которых улей 4 занят пчелами, а ульи 2, з и 5 пустые.
несколько пчел-разведчиц, и их одинаково привлекали
как цветные бумажки, так и пахучие цветки. Из этого
следует, что пчел, отыскивающих новые места добычи
корма, в равной мере привлекают как запах, так и
окраска.
Мы также обнаружили, что лишь очень немногие
пчелы—только пчелы-разведчицы—обращали внимание на
Ряс. 10. Пример расположения окрашенных ульев,
облегчающего пчелам отыскивание своего улья.
Использованы 4 цвета: белый, синий, желтый и черный.
цветки или окрашенные предметы и запахи,
предлагаемые их вниманию. Оттинген-Шпильберг (1949) провела
40 опытов общей продолжительностью 45 час. В опытах
участвовали сотни пчел и 15 различных видов цветущих
растений; однако каждую группу растений посещали
лишь 1—2 пчелы в 1 час (в среднем 1,27). Максимум
посещений составлял 6 пчел в 1 час. Огромное большинство
рабочих пчел вовсе не искало пищу, даже если в улье
ее было очень мало. Вообще большинство особей
предпочитает ждать, пока немногочисленные разведчицы
дадут им указания об обнаруженном ими источнике пищи
(см. гл. III).
Однако окраска может быть полезна пчелам и в другом
отношении. Очень часто приходится видеть цветки, у
которых вход в трубочку, содержащую нектар, окрашен иначе,
нежели другие части; он бывает либо темнее, либо
20
светлее, а часто просто другого цвета. Замечательно, что
почти во всех случаях это различие в окрасках создает
резкие контрасты для глаза пчел. По-немецки такие
окрашенные места называются нектарными пятнами (Saftmale).
Вероятно, их следует называть
указателями нектара (рис. 11).
Более 150 лет назад Шпренгель
(1793) высказал мысль, что такие
окрашенные пятна представляют
собой опознавательные знаки,
помогающие насекомым-опылителям
отыскивать нектар. Позднее
биологи частенько высказывали
сомнения поэтому поводу. Однако мы
уверены в правоте Шпренгеля,
потому что ее можно доказать при
помощи простого опыта. Если
положить на стол большой кусок
синего картона с небольшим
желтым пятном, то оно привлекает
пчел, и в поисках пищи они садятся
преимущественно на это пятно
(Келлермайер, неопубликованные
данные). Вероятно, аналогичным
образом пчел привлекают
«указатели нектара» на таких цветках,
каких они раньше никогда не
посещали.
Окраска цветков полезна и еще в одном отношении. Если
пчелы начали посещать то или иное растение с цветками
определенного типа, они остаются верными цветкам этого типа в
течение многих дней и даже недель; данная пчела во время
фуражировок всегда посещает растения определенного вида.
Это выгодно пчелам, так как все цветки растений данного
вида имеют одинаковое строение и благодаря знакомству
с ним пчелы экономят время. Растениям же это выгодно
потому, что для осуществления опыления необходимо,
чтобы пчела прилетала с других цветков растений одного
с ними вида. Так как пчелы специализируются к посещению
определенных цветков, они должны уметь отличать друг
от друга различные виды растений, встречающиеся на их
пастбище. Окраска, конечно, очень помогает пчелам в
распознавании определенного вида цветков, но ее нельзя
Рис. 11. Синие цветки
незабудки (Myoso t is),
снабженные желтым
кольцом (нектарное
пятно) вокруг входа к месту
выделения нектара.
21
считать безупречным указателем. В отличие от биологов
прежних лет нам теперь известно, что пчелы неспособны
распознавать столь большое разнообразие окрасок, как
это можем делать мы, а различают лишь четыре цвета,
тогда как человек различает примерно шестьдесят цветов.
Поэтому в распознавании цветка определенного вида
растений пчелы не могут руководствоваться только его окраской,
Рис. 12. Примеры фигур, легко различаемых пчелами.
поскольку весьма вероятно, что рядом с ним находятся
другие цветки, окраска которых кажется пчелам сходной.
Отсюда следует, что пчелы руководствуются какими-то
другими признаками, помогающими им с уверенностью
находить цветки растений определенного вида.
Можно предполагать, что отличительным признаком
для распознавания разных видов растений служит форма
цветков. Для проверки этого предположения мы приучали
пчел узнавать цветные бумажки определенной формы,
наклеенные вокруг входа в картонные коробки с
сахарным сиропом; к другим коробкам, не содержавшим сиропа,
прикреплялись бумажные фигуры того же цвета, но иной
формы. Пчел удалось за короткое время приучить к
коробкам, помеченным такими фигурами (рис. 12). Так,
например, в одном опыте корм в течение некоторого времени
находился в коробке, украшенной фигурой с лучами
(рис. 12, Б). Затем эту коробку убрали и поставили на
столик для опытов несколько чистых, не обсиженных
пчелами коробок, украшенных такими же фигурами из
цветной бумаги. Коробку, украшеную фигурой с лучами,
посещало много пчел, хотя на этот раз корма не было ни
в одной мг коробок; в коробку же, украшенную дру-
22
гой фигурой (рис. 12, А), забредали лишь случайные
пчелы.
Попытки приучить пчел различать геометрические
фигуры оказались безуспешными. Мы отнесли это за счет
того, что пчелы способны различать только те фигуры,
которые характерны для формы цветков, и что они не
могут приучиться узнавать треугольные или квадратные
• ¦^ /
XO"»Y
Рис. 13. Пчел нельзя научить отличать друг от
друга фигуры в пределах ряда А или ряда Б.
Однако они четко отличают любую фигуру ряда А
от любой фигуры ряда Б.
фигуры потому, что никогда не встречали их в природе
(Фриш, 1915). Однако несколько лет спустя, продолжая
эти опыты, Герц (1929) установила, что пчелы легко
приучаются отличать фигуры, изображенные в ряду А на
рис. 13, от фигур, изображенных на том же рисунке в
ряду Б. Но при этом оказалось, что пчелы неспособны
приучиться отличать друг от друга фигуры в пределах
ряда А или ряда Б. На основании этих и других
наблюдений Герц пришла к заключению, что восприятие формы
у пчел и человека обусловлено различными моментами.
Для глаза пчелы наиболее важным моментом, по Герц,
является степень «расчлененности» формы. Человек легко
отличает друг от друга все фигуры ряда А\ для пчел же
они неотличимы, потому что степень «расчлененности»
формы у всех у них примерно одинакова. Протяженность
границ, или линий контраста, между формами и фоном
у всех этих фигур незначительна, и все они представляют
собой компактные пятна. У фигур же ряда Б протяженность
23
границ значительна—это фигуры растянутые. Пчелыг
повидимому, улавливают, расчленена ли фигура или
компактна; однако они не в состоянии уловить иные отличия
этих фигур друг от друга. Таким образом, восприятие
формы пчелами основано на совершенно других
критериях, чем восприятие формы человеком; это, видимо,
связано с оптическими особенностями сложного глаза
насекомого.
Вполне возможно, однако, что главная причина этого
различия в восприятии формы состоит в том, что пчела
наблюдает формы во время полета. Так как глаз пчелы
неподвижно укреплен на ее голове, то возможно, что
расчлененность формы создает у пролетающей пчелы
мелькающее зрительное восприятие. Недавние опыты Аутрума
(1948, 1949) подтвердили это предположение. В итоге
становится понятным, почему расчлененность фигуры может
оказаться более существенной для восприятия формы, чем
ее общий контур. Поскольку степень расчлененности
формы цветков различных растений, общие контуры которых
кажутся нам явно различными, часто бывает сходной,
форма цветков, так же как их окраска, не может служить
для пчел достаточным опознавательным признаком.
Повидимому, пчелы используют какие-то более надежные
признаки для различения растений определенного вида.
Возможно, что для этого им служит запах, столь
характерный для каждого растения. Подобные соображения
заставили нас заняться изучением обоняния пчел, чему и
посвящена следующая глава.
Глава II
ЧУВСТВО ВКУСА И ОБОНЯНИЕ ПЧЕЛ
В гл. I уже указывалось, что обычно каждая пчела
посещает цветки растений какого-либо одного вида. Это
обстоятельство весьма выгодно самой пчеле, так как она
каждый раз встречается с одним и тем же, хорошо
знакомым ей строением цветка. Важно это также и для
растений, поскольку при этом приносимая пчелой пыльца
принадлежит цветкам растений одного с ними вида.
Каким же образом пчелы узнают цветки растений
определенного вида среди цветков растений других видов,
растущих рядом с ними?
Цветки бывают самых различных цветов и оттенков,
но глаз пчелы различает лишь четыре цвета: желтый, синий,
сине-зеленый и «ультрафиолетовый». Как показано в
предыдущей главе, форма цветков также не может служить
критерием, позволяющим пчеле безошибочно отличать одни
виды растений от других. Остается предположить, что
пчелы выбирают цветки по их специфическому аромату.
Цветки почти каждого вида характеризуются своим
особым запахом, по которому человек отличает их от цветков
других видов. Однако, начав изучать эту проблему, мы
не знали, относится ли это и к пчелам; мы не были уверены
даже в том, что пчелы вообще способны воспринимать запах
цветков.
При изучении этих вопросов мы применяли для
приучения пчел к запаху те же методы, которые
использовали для приучения их к фигурам различной формы.
На опытный столик помещают ряд картонных коробок,
открывающихся сверху и снабженных входом для пчел
на передней стороне (рис. 14). В одну из коробок
помещают блюдечки с сиропом, пахучий цветок или несколько
капель какого-нибудь эфирного масла. Местоположение
коробки многократно меняют, чтобы исключить
возможность цриучения пчел к определенному месту, поскольку
25
нам нужно, чтобы в поисках пищи они руководствовались
только запахом. Спустя несколько часов все коробки
убирают и выставляют новый набор чистых коробок, в одну
из которых наливают капельку пахучего вещества,
использовавшегося для приучения пчел; ни в одну из
коробок никакого корма не ставят. Пчелы летают от коробки
к коробке, обнюхивая входные отверстия, однако они
заходят только в ту коробку, которая содержит пахучее
Рис. 14. Картонная коробка, используемая
для приучения пчел к запаху.
А—общий вид; В—открытая коробка; приучая пчел к запаху,
следует помещать капельку пахучего вещества на картонную
пластинку, расположенную над входом в коробку.
вещество. Отсюда следует, что пчелы ощущают этот запах
и воспринимают егокакуказательналичия источника корма.
Затем было решено выяснить, различают ли пчелы
столь же большое разнообразие запахов, как и человек.
Для этого их приучали к эфирному маслу из
итальянских апельсинов. После приучения в течение одного дня
на стол выставили 24 коробки, из которых только одна
содержала пахучее вещество, уже известное пчелам,
тогда как остальные 23 коробки содержали различные
другие эфирные масла (рис. 15). Затем этот опыт был
повторен с использованием того же знакомого запаха и 23 новых,
которые никогда до этого нф использовались при даче
корма. В обоих опытах мы подсчитывали число пчел,
посетивших коробки в течение пятиминутных наблюдений. В
коробку, содержавшую пахучее вещество, к которому пчелы
26
были приучены, в первом опыте зашло 205 пчел, а во
втором—120. Из остальных 46 коробок внимание пчел
привлекли лишь 3 коробки, содержавшие лимонную
эссенцию (148 посещений), бергамотное масло (93
посещения) и эфирное масло из корки испанских апельсинов
(60 посещений). В другие коробки заходили лишь
единичные пчелы, а в некоторые коробки вообще не заходила
ни одна пчела. Все три названные эфирные масла, запах
ддддддддддю,
1 LI, l>" 'HiwiwOb 'ттж^УЛ 'ггнгаУ!ч\ 'iwiuiBW 'тТшкЛ *тттттт^\ Ут*г* Tnq*W Iihili' liirtn' ft'irw j |
Рис. 15. Расположение коробок в опыте для обнаружения
способности пчел различать запахи.
которых привлек пчел, так же как и масло, к запаху
которого предварительно приучали пчел, были добыты из
плодов рода Citrus. Человеку запах этих масел кажется весьма
сходным и отличным от запаха всех других
использовавшихся в опыте веществ. В результате ряда других опытов
удалось установить, что пчелы предпочитали запах
эфирного масла из корки итальянских апельсинов, на который
шло приучение, запаху масла из корки испанских
апельсинов, лимонной эссенции и бергамотного масла, однако
разница в числе пчел, привлекавшихся этими четырьмя
запахами, была невелика.
И а основании этих и других опытов мы можем сделать
вывод, что пчелы способны различать разные запахи,
подобно людям с хорошо развитым обонянием. Кроме
того, запахи, которые человеку кажутся похожими,
пчелами, повидимому, также воспринимаются как запахи
сходные.
В других опьиах обнаружилось, что вещества,
лишенные запаха для человека, лишены запаха и для пчел.
Таким образом, несмотря на совершенно различное
строение органов обоняния пчел и человека, их обонятельные
реакции почти одинаковы (Фриш, 1919).
Где же расположены у пчел органы обоняния?
Энтомологам было давно известно, что насекомые перестают
реагировать на запахи, если у них отрезать антенны, или
27
«усики». Поэтому антенны считали органами обоняния
насекомых. Однако другие энтомологи возражали против
такого толкования этих опытов; по их мнению, перерезка
антенн, в которых проходит относительно крупный нерв,
вызывает у насекомых шоковое состояние* и отсутствие
обонятельных реакций объясняется именно этим шоком,
а не потерей чувства обоняния. В течение ряда лет
взгляды на этот вопрос были противоречивы. Между тем решить
его можно следующим образом.
Пчел приучают к определенному запаху, например
к запаху мятного масла. Для выполнения описываемых
Рис. 16. Приучение пчел к запаху.
А—стеклянное блюдце, содержащее корм и окруженное несколькими
каплями мятного масла; Б—Г—пустые блюдца, окруженные
каплями другого эфирного масла.
опытов картонные коробки неудобны, так как в процессе
опыта приходится ловить пчел на кормушках и
производить с ними различные операции. Поэтому была
использована установка для приучения пчел к окраскам,
описанная в предыдущей главе. На кусок картона, пропитанный
пахучим веществом, ставили часовые стекла с
сахарным сиропом. Рядом с ним располагались куски картона,
пропитанные различными другими пахучими веществами,
но без всякого корма (рис. 16). После нескольких часов
тренировки пчелы начали отличать запах, к которому их
приучали, и безошибочно отыскивали картон с кормом.
Затем приученных пчел, возвращающихся из улья на
кормушку, вылавливали и ртрезали им антенны, после
чего их снова выпускали. Выпущенные на волю пчелы
продолжали разыскивать корм, но уже не могли найти
его. Они обнаруживали корм лишь чисто случайно;
28
у таких пчел было не больше шансов прилететь на картон
с запахом, к которому их приучили, чем у любых других
пчел. В поведении таких пчел не наблюдалось никаких
признаков того, что они страдают от шока; они упорно
Ряс. 17. У одной из пчел, приученных к запаху
мятного масла, были обрезаны антенны; пчела
ищет запах мятного масла, но не может его
обнаружить.
Рис. 18. Приученная к синему цвету пчела,
несмотря на удаление антенн, может отличать
синий картон (А) от желт ого (Б—Г) и прилетает в
поисках корма на пустое блюдце.
продолжали поиски корма (рис. 17). Для подкрепления
этих результатов мы приучали других пчел находить корм
на куске синего картона, после чего удаляли им антенны.
Эти пчелы без всяких колебаний находили синий картон
и ни разу не путали его с желтым (рис. 18). Это доказывает,
что операция сама по себе не вызывает серьезного шока,
так как пчелы сохраняли в памяти то, к чему они были
приучены, но не были в состоянии отличить запах. Иными
29
словами, можно считать, что органы обоняния
действительно расположены на антеннах.
Рассмотрение антенн пчелы под микроскопом
показывает, что они густо покрыты чувствующими органами;
часть из них представляет собой органы осязания, а
другая часть—органы обоняния (рис. 19 и 20). Строение
последних отличается тем, что кутикула, прикрывающая
б-Цгб
А Б
Рис. 20. Органы чувств пчел.
А—членик антенны с органами чувств
при большом увеличении; Б—часть
членика (отмеченная на рис. А черной
рамкой) при большом увеличении: 1—орган
осязания; 2—пористые пластинки
(органы обоняния); 3—конусовидьые органы
обоняния.
нервные окончания, чрезвычайно тонка, вследствие чего
она проницаема для пахучих веществ (рис. 21). На
длинном основном членике антенн, или стебельке, эти органы
отсутствуют; нет их также и на первых трех сегментах
гибкой дистальной части антенны, носящей название
жгутика. Они приурочены к восьми дистальным членикам
жгутика. После удаления восьми дистальных члеников
одной антенны (рис. 19,6—б) и семи члеников другой антенны
пчела была еще в состоянии отличать запах, к которому
ее приучали, от других; ее даже можно было приучить
к новому запаху. Если же удаляли восьмой членик
второй антенны, то чувство обоняния полностью исчезало.
Этот опыт также показывает, что последствия удаления
Рис. 19. Антенна рабочей
пчелы.
После перерезки по линии
а—а сохраняется лишь один
членик с органами
обоняния. После же перерезки по
линии б—б на антенне не
остается органов обоняния.
30
антенн нельзя объяснять только шоком; эти последствия
становятся понятными лишь в том случае, если
допустить, что органы обоняния расположены на антеннах
(Фриш, 1921).
У большинства других насекомых органы обоняния
также расположены на антеннах. Однако у некоторых
Рис. 21. Схема строения органов чувств пчелы.
А—разрез через орган осязания. Нервное окончание расположено в
основании волоса и раздражается при движении последнего; Б—разрез
через конусовидный орган обоняния; В—разрез через пористую пластинку.
Нервные окончания у очень тонких хитиновых пластинок возбуждаются
проникающими сквозь них пахучими веществами. Черным цветом
обозначена хитиновая кутикула.
насекомых органы обоняния находятся также на других
головных придатках, например на щупиках (бабочки,
жук плавунец).
Мы пытались также определить степень
чувствительности органов обоняния у пчел, сначала приучая их
к определенному запаху, а затем ослабляя его до тех пор,
пока пчелы не переставали отличать «пахучие» коробки
от «непахучих». Когда запах становился столь слабым,
31
что человек его уже не воспринимал, мы обнаружили,
что и пчелы потеряли способность его воспринимать. Таким
образом, чувствительность обоняния пчел примерно
соответствует таковой человека, откуда следует, что аромат
большинства цветков недостаточен для привлечения пчел
с большого расстояния. Роль обоняния прекрасно
выявляется в следующем опыте.
Цвет и запал
Цвет Запах
Рис. 22. Роль цвета и запаха в привлечении пчел к корму.
Мы приучили пчел отличать «пахучую» коробку,
передняя стенкакоторой окрашена в синий цвет,от
других,неокрашенных и «непахучих» коробок. Затем мы изменили
условия, поставив справа неокрашенную «пахучую» коробку
(рис. 22, Е), а слева—«непахучую» коробку с окрашенной
синей стенкой (рис. 22, Г). Пчелы решительно устремились
к окрашенной коробке, но на расстоянии примерно 3 см
они проявляли признаки колебания, явно стараясь
уловить запах, к которому они были приучены. После этого
они обследовали другие коробки и, приблизившись к
«пахучей», садились и входили внутрь. Очевидно окраска
привлекала пчел с большего расстояния, однако
ощущение запаха оказалось более убедительным. Аналогичным
образом окраска цветков привлекает внимание пчел на
большом расстоянии, запах же, будучи специфичен для
32
каждого вида растений, позволяет пчелам сделать
окончательный выбор на близком расстоянии (Фриш, 1919).
В одном лишь отношении обоняние пчел превосходит
обоняние человека. Орган обоняния человека расположен
внутри носовой полости, и частицы пахучих веществ
попадают в него с током воздуха, используемого для дыхания.
Поскольку этот воздушный поток по пути в нос хорошо
перемешивается, у человека не возникает ассоциации
между формой пахучего предмета и ощущением
исходящего от него запаха. Между тем у пчел органы обоняния
расположены на подвижных антеннах, покрытых
органами осязания, а потому ощущение запаха связано у пчел
с осязательными ощущениями. Круглый пахучий предмет
может вызвать у пчелы совершенно иное ощущение по
сравнению с угловатым предметом. Форель (1910) много лет
назад высказал мысль, что вследствие тесной связи между
расположенными на антеннах органами осязания и
обоняния пчелы могут воспринимать форму предметов «по
запаху». Ассоциации между осязательными и
обонятельными восприятиями у пчел аналогичны постоянной, с
раннего детства, ассоциации зрительных и осязательных
восприятий у человека. Такая ассоциация между
осязанием и обонянием очень полезна для пчел при посещении
цветков. Нередко они приближают свои антенны к
цветкам почти вплотную, возможно, улавливая при этом даже
очень слабые запахи. Более того, если различные части
цветка обладают неодинаковым запахом, пчела может
различать и определять расположение этих частей весьма
точно. В связи с этим мы решили выяснить, нет ли в
цветках «указателей нектара», воспринимаемых не только при
помощи зрения, но и при помощи обоняния. Оказалось,
что у многих цветков они имеются. У нарцисса, например,
желтый указатель нектара (рис. 23) отличается от белого
венчика не только по окраске, но и по запаху. Отделив
желтые части цветка от белых и приучив пчел к запаху
тех или других частей, можно показать, что они различают
эти запахи. Человек также способен различать эти запахи,
но только в том случае, если он нюхает разные части цветка
порознь; нюхая же целый цветок, мы не улавливаем двух
запахов, так как они попадают в наш орган обоняния,
будучи смешанными друг с другом. Пчелы же, благодаря
расположенным на поверхности их антенн органам
обоняния, легко улавливают эти различия и направляются
33
Рис. 23. Белый цветок нарцисса с желтым
указателем нектара (показан точками).
к местоположению нектара, руководствуясь этими
пахучими указателями.
Такие же пахучие указатели нектара были
обнаружены и у других растений. Как показали опыты Губер и
Леке (не опубликовано), у цветков многих растений запах,
издаваемый разными частями, качественно одинаков,
однако у входа в более глубоко расположенные части
цветка, где помещается нектар, его интенсивность
возрастает.
Человек обладает двумя химическими чувствами—
обонянием и вкусом. Органы обоняния расположены в
носовой полости, а органы вкуса—на языке; они иннерви-
руются различными нервами. При помощи обоняния мы
можем различать столь разнообразные по качеству запахи,
что у нас даже нехватает слов для их обозначения. По
вкусу же (если на помощь не приходит запах пищи)
человек различает лишь четыре качества: сладкое, горькое,
кислое, соленое. Надо отметить, кроме того, что обоняние
гораздо более чувствительно, чем вкус. Следовательно,
можно думать, что биологическая роль этих чувств весьма
различна. Очень многие животные с хорошим обонянием
могут обнаруживать пищу или врагов на значительном
расстоянии, так как переносимые с током воздуха частицы
34
вызывают раздражение обонятельных рецепторов при
очень низких концентрациях. Однако для того, чтобы
химические вещества вызвали раздражение органов вкуса,
концентрация их должна быть гораздо выше, и поскольку
орган вкуса расположен во рту, он способен оценить пищу
только после того, как животное уже пришло в контакт
с пищей, а именно—непосредственно перед трапезой или
во время ее.
Сходное разделение химического восприятия
наблюдается также у пчел и у других насекомых. Насколько
нам известно, органы чувств, расположенные на антеннах,
могут обнаруживать столько же качественно различных
запахов, сколько и нос человека. Будучи весьма
чувствительными, они могут обнаружить присутствие пищи и
других предметов на некотором расстоянии. Однако у пчел
имеются, кроме того, органы вкуса, расположенные на
ротовых частях и предназначенные для исследования пищи
в момент ее потребления. Пчелы могут обнаружить сладкий
сироп лишь при контакте с ним, однако они очень
разборчивы в отношении его концентрации. Если давать
пчелам сироп, содержащий 20% сахара (примерно 2/3 моль/л),
то они обычно его берут. Если же сироп содержит только
10% сахара, то можно наблюдать, как это бывает у людей,
индивидуальные особенности вкуса: одни пчелы сосут,
другие проявляют признаки нерешительности, третьи же
совсем отказываются от сиропа. Если сироп содержит
только 5% сахара, пчелы его пробуют, но отказываются
сосать. Здесь лежит их порог приемлемости, причем он
варьирует в зависимости от условий кормления. Если
в природе цветет много растений, этот порог очень высок,
подчас около 40%. Осенью же, когда цветущих растений
становится мало, порог падает до 5%. Подобно людям,
пчелы в более благоприятных условиях становятся более
разборчивыми.
Кроме того, существует другой предел, а именно—
порог восприятия, т. е. наиболее низкая концентрация,
способная вызвать раздражение органа вкуса. Этот порог
постоянен и не зависит от изменения условий. Для
определения этого порога необходимо дать пчелам поголодать
в течение нескольких часов. После этого они готовы брать
любой сироп, лишь бы он давал ощущение сладости.
Порог восприятия голодных пчел всегда соответствует
концентрации 1—2% (Фриш, 1934).
35
Примечательно, что пчелы сосут сироп с низкой
концентрацией сахара, однако никогда не используют его для
создания запасов в улье. Дело в том, что такой сироп,
если его отложить в сотах, не сохранится до наступления
зимы. Одно из важных химических отличий любого меда
от нектара, из которого он получается, состоит в том, что
в нем в результате удаления воды повышается
концентрация сахара. Поэтому для превращения сиропов с низким
содержанием сахара в мед требуется затрата
дополнительного времени или труда со стороны пчел. Цветки растений
приспособились к этому требованию, выделяя нектар с
высоким содержанием сахара—в среднем около 40% (Р. Бейт-
лер, 1930). Химикам известно много различных Сахаров,
большинство которых обладает для человека сладким
вкусом. Из 34 испытанных нами Сахаров и сахароподоб-
ных веществ сладкими для пчел оказались лишь 9:
сахароза, мальтоза, виноградный сахар, фруктоза, трегалоза,
мелецитоза, фукоза, а-метилглюкозид и инозит.
Большинство Сахаров, воспринимаемых нами как сладкие, для
пчел оказались безвкусными; независимо от того,
предлагать ли их в чистом виде или в смеси с тростниковым
сахаром, пчелы реагируют на них так же, как на чистую
воду. К таким сахарам относятся: лактоза, мелибиоза,
целлобиоза, тетрагликозан, галактоза, манноза, сорбоза,
рамноза, ксилоза, /- и d-арабиноза, триметилглюкоза,
а-метилманнозид. эритрит, кверцит, маннит, сорбит и
дульцит; несколько отталкивающим действием обладают
генциобиоза, раффиноза, тетралеван, гликоген, Р-метил-
глюкозид, (3-метилфруктозид и Р-метилгалактозид.
Выявить общие признаки, определяющие сладкий вкус этих
веществ, для всех насекомых не удалось, так как
вещества, сладкие для одного насекомого, лишены всякого
вкуса для другого. Так, например, трисахарид
раффиноза для пчел безвкусен, однако для муравьев его надо
считать самым сладким из всех испытанных нами Сахаров.
Медоносные пчелы различают не только сладкое, но
также соленое, кислое и горькое. Если добавить к
сахарному сиропу соли, то пчелы отказываются от него. Мы
испытали ряд методов оценки чувствительности пчел к
соли. Нам удалось установить количество соли, добавление
которого к сахарному сиропу делает его неприемлемым
для пчел, или, точнее, заставляет пчел проявлять
признаки нерешительности, прежде чем начать сосать такой
36
сироп. Лучший метод для выяснения этого вопроса
основан на том, что обычно пчелы забирают большие объемы
сиропа, если концентрация в нем сахара высока. Чем
слаще сироп на вкус, тем большее количество его набирает
пчела в свой зобик. Так, например, опыт, проведенный
9 сентября 1929 г., дал следующие результаты:
Средний объем
хтстптгп Сахар жидкости взятой
JH5" в сиропе, 8а одно посеще-
пчел % ние, мм*
70 17 (0,5 М) 42
55 34 (1 М) 55
49 68 (2 М) 61
Для определения пороговой концентрации соли,
которую пчелы едва выдерживают, мы предлагали им два
блюдца с сиропом; оба сиропа содержали одинаковое
количество сахара, но к одному из них было подмешано немного
соли. Количество соли было столь незначительным, что
пчелы без колебаний сосали подсоленный сироп, но брали
его в меньших количествах, чем чистый. По всей
вероятности, чистый сироп казался им вкуснее. Эти опыты
позволили нам установить нижний порог концентрации соли,
при котором пчелы еще замечали изменение вкуса.
В другом опыте, выполненном 17 сентября 1929 г.,
пчелы получали 0,5 М сахарный сироп, которого они
набирали в зобик в среднем по 42 мм* за одно посещение.
Немедленно после этого им предложили такой же сироп с
добавлением 0,03 моль/л поваренной соли, что соответствует
0,017-процентному солевому раствору. Этого смешанного
сиропа они брали лишь по 37 мм3 за одно посещение. При
повторении этого опыта через год результаты составляли 44
и 37 мм? соответственно. Если 0,5 М раствор сахарозы
содержит 0,015 молъ/л поваренной соли, то пчелы берут
его меньше, чем чистого сахарного сиропа. Разница
исчезает лишь тогда, когда поваренная соль добавляется
в количестве 0,0075 молъ/л.
На основании этих опытов мы пришли к заключению,
что медоносная пчела несколько более чувствительна
к соли, чем человек; она чувствительнее также и к
веществам, имеющим кислый вкус. К горьким же веществам
пчела гораздо менее чувствительна, чем человек. Пчеле,
повидимому, нравится такая смесь хинина с сахаром,
37
которую любой человек, взяв в рот, выплюнул бы
немедленно (Фриш, 1934).
Установив все эти факты, мы решили использовать их
практически. Во многих странах пчеловодам дают
возможность приобретать сахар по удешевленным ценам для
подкормки пчел зимой. Это содействует росту численности
пчелиных семей, повышает выход меда и улучшает
опыление цветов и некоторых сельскохозяйственных культур.
Предполагается, что дешевый сахар, предоставляемый
пчеловодам, поступает только в пчелиные ульи,- а не на
стол человеку. Чтобы этот сахар действительно
доставался пчелам, желательно подмешивать к нему
вещество, делающее этот сахар непригодным для
потребления человеком. Однако найти подходящее вещество
для этого было нелегко. Оно оказывалось либо
недостаточно неприятного вкуса для человека, либо делало сахар
неприятным и для пчел. Мы предложили смешивать сахар
с небольшим количеством октоацетилсахарозы, горькой
для человека, но совершенно безвкусной для пчел.
Вначале применение ее осложнялось тем, что этого вещества
было мало и оно стоило дорого. Однако химическая
промышленность освоила производство этого вещества, сделала
его дешевым и стала выпускать под названием октозан.
Это вещество оказалось особенно подходящим, так как
оно представляет собой соединение тростникового сахара,
или сахарозы, и уксусной кислоты и совершенно
безвредно как для пчел, так и для людей. Более того, в процессе
созревания меда в улье оно разлагается в течение
нескольких недель, благодаря чему его горький вкус не
передается меду (Валь, 1937).
Октозан с успехом применяется на протяжении ряда
лет в Чехословакии, Польше, Румынии, Болгарии,
Голландии и Бельгии. В других же странах пчеловоды не
согласились кормить своих любимцев столь неприятным
на вкус сахаром. Возможно также, что это в значительной
степени объясняется именно непригодностью такого сахара
для потребления человеком.
Можно показать, что пчелы, так же как и человек,
различают соленое, кислое и горькое. Методы для
установления этого несколько сложны, так как невозможно
приучить пчел находить корм по вкусу, подобно тому
как мы приучали их обнаруживать корм по цвету и запаху.
Мы приведем лишь один пример для того, чтобы
показа
зать, как нам удалось установить, что пчелы отличают
горькое от соленого. К сахарному сиропу определенной
концентрации мы добавляли ровно столько соли, чтобы
пчелы отказывались брать его. Затем мы приготовляли
смесь сиропа с хинином, тоже настолько горькую, что
пчелы отказывались от нее. Если бы вкус этих двух веществ
был для пчел совершенно одинаков, наши два раствора
вызывали бы у них совершенно одинаковые вкусовые
ощущения. Однако нам удалось показать, что это не так.
В самом деле, после того как пчелы поголодают, они берут
горькую смесь, содержащую хинина в 8 раз больше, чем
раньше. Что же касается смеси сахара с солью, то у
голодных пчел не наблюдается никакого смещения порога
чувствительности, и они продолжают отказываться от нее.
Таким образом, голодные пчелы берут более горькую
смесь, но отказываются от более высоких концентраций
соли. Отсюда следует, что горькое и соленое
воспринимаются пчелами как различные вкусовые качества.
Подобным же методом можно доказать, что кислое
воспринимается пчелой как особое вкусовое качество (Фриш, 1934).
Способность насекомых различать четыре вкусовых
качества (сладкое, горькое, соленое и кислое) может быть
непосредственно показана в опытах с водяными жуками
{Hydrous piceus, Ditiscus latissimus и Cybister laterimargi-
nalis). Эти насекомые при поисках пищи в воде благодаря
диффузии могут ощущать вкус на некотором расстоянии.
Поэтому их можно приучить находить пищу,
руководствуясь вкусовыми восприятиями. Водяные жуки,
приученные обнаруживать пищу по приданному ей соленому,
кислому, сладкому или горькому вкусу, отличают каждое
из этих качеств от трех других (Риттер, 1936; Бауэр, 1939).
У некоторых животных органы вкуса расположены
не только во рту или около него, но и на других частях
тела. Миннич (1922а) открыл у бабочек способность
определять вкус пищи кончиками ног. Как только бабочка
наступает на сладкое вещество, она мгновенно вытягивает
свой хоботок и нередко начинает сосать пищу. В этом
случае восприятие вкуса приспособлено не только для
отыскивания пищи, но и для регулировки ее принятия,
и поэтому нижний порог чувствительности имеет большое
значение, так как наличие даже небольшого количества
сахара служит указателем источника пищи. И в самом деле,
вкусовые рецепторы на ногах бабочек можно считать
39
наиболее чувствительными из всех доныне известных.
У мух и у других насекомых мы также находим хорошо
развитое восприятие вкуса кончиками ног (Миннич, 1922b;
Гаслингер, 1935).
Из позвоночных лишь рыбы обладают очень низким
порогом вкусовой чувствительности. У них это также связано
с особенностями биологии питания, поскольку
растворенные в воде вещества могут привлекать рыбу к корму даже
с довольно больших расстояний. Если пища находится на
некотором расстоянии от рыбы, то в результате диффузии
в воде частицы ее доходят до рыбы в сильном разведении.
Рис. 24. Морской петух (Trigla).
Пальцеобразные лучи грудных плавников снабжены большим
количеством органов вкуса.
Молекулы пищи могут попасть на любую часть тела
рыбы, и поэтому органы вкуса имеются у нее не только
в области рта, как у высших позвоночных животных, но
широко рассеяны по всей поверхности тела. Морской петух
(Trigla)—рыба донная, и у нее, подобно бабочке, имеется
специальное приспособление для распознавания вкуса
предметов, лежащих на дне моря; часть лучей грудных
плавников превращена у нее в пальцеобразные придатки
(рис. 24), на которых размещены многочисленные органы
вкуса (Шаррер, 1935).
Рисунок 25 наглядно иллюстрирует большие различия
в степени чувствительности органов вкуса у разных
животных.
В заключение мы остановимся на некоторых
практических приложениях наших знаний в этой области. Сельские
хозяева часто заинтересованы в увеличении посещаемости
40
пчелами цветков некоторых растений для повышения
их опыляемости. Особенно плохо обстоит дело с опылением
красного клевера, если посевы его занимают большие
площади. Обычно цветки клевера посещают и опыляют
в основном шмели, обладающие достаточно длинным
хоботком для того, чтобы использовать весь нектар из
трубочек венчиков в цветках красного клевера. Медоносная
же пчела часто предпочитает посещать другие растения,
Рис. 25. Диаграмма, показывающая различный
порог ощущения сладости для разных
животных и человека.
Белые кучки показывают количество сахара, которое
нужно растворить в 1 л воды, чтобы создать едва
ощутимую сладость: А—для пчелы (0,083 М); Б—для человека
(0,0125 М); В—для рыбы (0,0002 М); Г—для ног бабочки
адмирала (0,000078 М).
цветущие одновременно с клевером, но более удобные для
сбора пищи. Между тем шмелей слишком мало, чтобы
обеспечить опыление всех растений на больших полях
красного клевера. Примерно в 1930 г. русские исследователи,
используя способность пчел узнавать определенные запахи,
разработали новый прием для улучшения опыления
красного клевера. Ульи с пчелами подвозили к клеверным
полям и приучали пчел к запаху клевера. Приучение шло
с вечера путем скармливания пчелам в ульях сиропа,
принявшего запах цветков красного клевера (для
получения такого ароматизированного сиропа цветки красного
клевера погружали в сахарный сироп, и после того, как
41
сироп принимал аромат клевера, цветки удаляли).
Значительное число подкормленных таким образом пчел на
следующий день начинало летать на красный клевер.
Этот прием дал возможность в значительной мере повысить
урожай семян красного клевера.
За несколько лет до этого нами был предложен
сходный метод. Для того чтобы описать его, нам придется
несколько забежать вперед и коснуться материала
следующей главы. Дело в том, что как только пчелы
обнаружат хорошее место для кормежки, они сообщают об
этом в улей при помощи особых танцев, выполняемых ими
на сотах. Пчелы, находящиеся в улье, не только узнают
о наличии корма, но и получают указание о том, в каких
цветках его следует искать. Они узнают об этом благодаря
запаху цветков, приставшему к телу пчел-танцовщиц.
Если кормить пчел на искусственной кормушке,
пахнущей цветками тех растений, на которые мы хотим
направить пчел, то пчелы-сборщицы начнут танцевать в улье,
побуждая других пчел к поискам этого запаха, что
вызовет посещение цветков того же самого вида. На основании
этих данных мы считали возможным искусственно
направлять пчел на поля, требующие повышенного опыления.
Ввиду катастрофической нехватки продовольственных
продуктов в Европе в последние годы эта проблема приобрела
серьезное значение и разрабатывалась нами совместно
с пчеловодами и сельскохозяйственными опытными
станциями. Из ряда методов, давших хорошие результаты,
я опишу лишь один весьма простой метод.
Когда начиналось массовое цветение красного клевера,
мы подвозили несколько семей пчел к полю и каждое утро
кормили их небольшой порцией сахарного сиропа в
кормушке, расположенной у передней стенки улья, так что
пчелы, входя в улей и выходя из него, должны были
проходить через нее (рис. 26). Кормушка состояла из трех
отделений: в среднее помещали цветки красного клевера,
а в боковые—сироп (рис. 27). Для того чтобы добраться
до сиропа, пчелы проходят через проволочную решетку
в среднее отделение кормушки и пробираются между
цветками; можно с уверенностью сказать, что при этом их тело
приобретает сильный аромат. После этого пчелы
принимались танцевать в улье. Другие обитательницы улья,
чувствуя запах клевера от тела танцовщиц, вылетали из
улья в поисках красного клевера. Для оценки эффектив-
42
Рис. 26. Улей с кормушкой, крышка которой открыта
чтобы показать три отделения (см. рис. 27).
\ЧЧ\\ЧЧЧЧЧ\\Ч\ЧЧЧЧЧЧЧЧЧЧ\ЧЧ\Ч\\ЧЧ^\\\Ч\\Ч\\\ЧЧЧЧЧ^^
• •••••оэ<
в
Рис. 27. Устройство кормушки.
А—вид сверху; видна проволочная решетка, приходящаяся над
средним отделением, содержащим цветы; Б—вертикальный разрез.
ности этого метода мы провели опыт с двумя клеверными
полями, находящимися на таком расстоянии друг от
друга, что пчелы с одного поля не могли залетать на другое.
Характер почвы, размеры площади, семена,
удобрения и другие факторы были по возможности уравнены.
У каждого поля было поставлено одинаковое число ульев
с пчелами. Каждое утро у одного поля пчелы получали
43
сахарный сироп, пахнувший клевером, а у другого—точно
такое же количество сиропа, но без аромата клевера; второе
поле служило контролем. Этот опыт был повторен 12 раз
в разных местностях. Во всех случаях на тех полях, где
пчел приучали к запаху клевера, посещаемость была в
3—4 раза выше, чем на контрольных полях. В 9 из 12
опытов был учтен урожай, и оказалось, что на полях, где
применялся ароматизированный сироп, урожай был на
40% (по весу) выше.
Обнадеживающие результаты были получены также
в опытах с другими сельскохозяйственными растениями:
гибридным клевером (Trifoliam hybridum L.), брюквой
(Brassica napus L.), репой (Brassica rapa L.), гречихой
(Fagopyrum esculentum Moench). Однако данные опытов по
этим растениям недостаточны, чтобы можно было сделать
определенное заключение об эффективности этого метода;
вопрос этот, несомненно, заслуживает более подробного
изучения.
Подобные мероприятия могут принести пользу как
пчеловодству, так и сельскому хозяйству в целом. В наших
опытах мы неоднократно наблюдали, что если пчел
направляли таким образом на медоносные растения, служащие
богатым источником нектара, то они давали на 50%
больше меда. Это происходило потому, что пчелы, получавшие
ароматизированный сироп, с большим упорством
разыскивают цветки, с запахом которых их познакомили в
кормушке, и собирают нектар более прилежно, чем пчелы,
получившие обычный сахарный сироп (Фриш, 1947).
Применяя эти приемы на практике, следует
учитывать многие обстоятельства, которые мы не могли
разобрать здесь в деталях. Касаясь прикладного значения этих
опытов, мы только хотели лишний раз иллюстрировать
хорошо известное положение, не всегда признаваемое
государственными деятелями: исследования, проведенные
из чисто научного интереса, часто приносят большую
пользу практике в таких областях, где этого меньше всего
можно было ожидать.
Глава III
«ЯЗЫК» ПЧЕЛ
Когда нам нужно привлечь пчел для опытов по
приучению, мы обычно выкладываем на стол несколько листов
бумаги, смазанных медом. После этого приходится ждать
часто часами, а иногда и днями, пока какая-нибудь одна
пчела обнаружит место подкормки. Но как только хоть
одна пчела обнаружит мед, в самом непродолжительном
времени на него прилетает множество других пчел. Все
они прилетают из того же улья, что и первая
пчела-сборщица, и поэтому совершенно очевидно, что она каким-то
образом сигнализировала в улье о своем открытии.
Нам было интересно выяснить, каким образом пчела
сообщает другим пчелам о наличии пищи в новом, не
известном им месте. Мы не можем наблюдать за поведением
пчел в обычном улье, где они ползают между сот. Поэтому
мы сконструировали наблюдательный улей, в котором все
соты расположены в одной плоскости, так что их
поверхность можно свободно обозревать через стеклянные окна
(рис. 28). Необходимо было также пометить каждую
подлежащую изучению пчелу, с тем чтобы ее можно было
узнавать среди массы других пчел на сотах. С этой целью пчел
помечали небольшими метками пяти различных цветов:
белое пятно на переднегруди означало цифру 1, красное—2,
синее—3, желтое—4, зеленое—5, белое пятно на задне-
груди означало цифру 6, красное—7, синее—8, желтое—9
и зеленое—0. Используя комбинации этих пятен, можно
выразить любое двузначное число, тогда как пятна на
брюшке обозначали сотни (рис. 29). Эта система позволяла
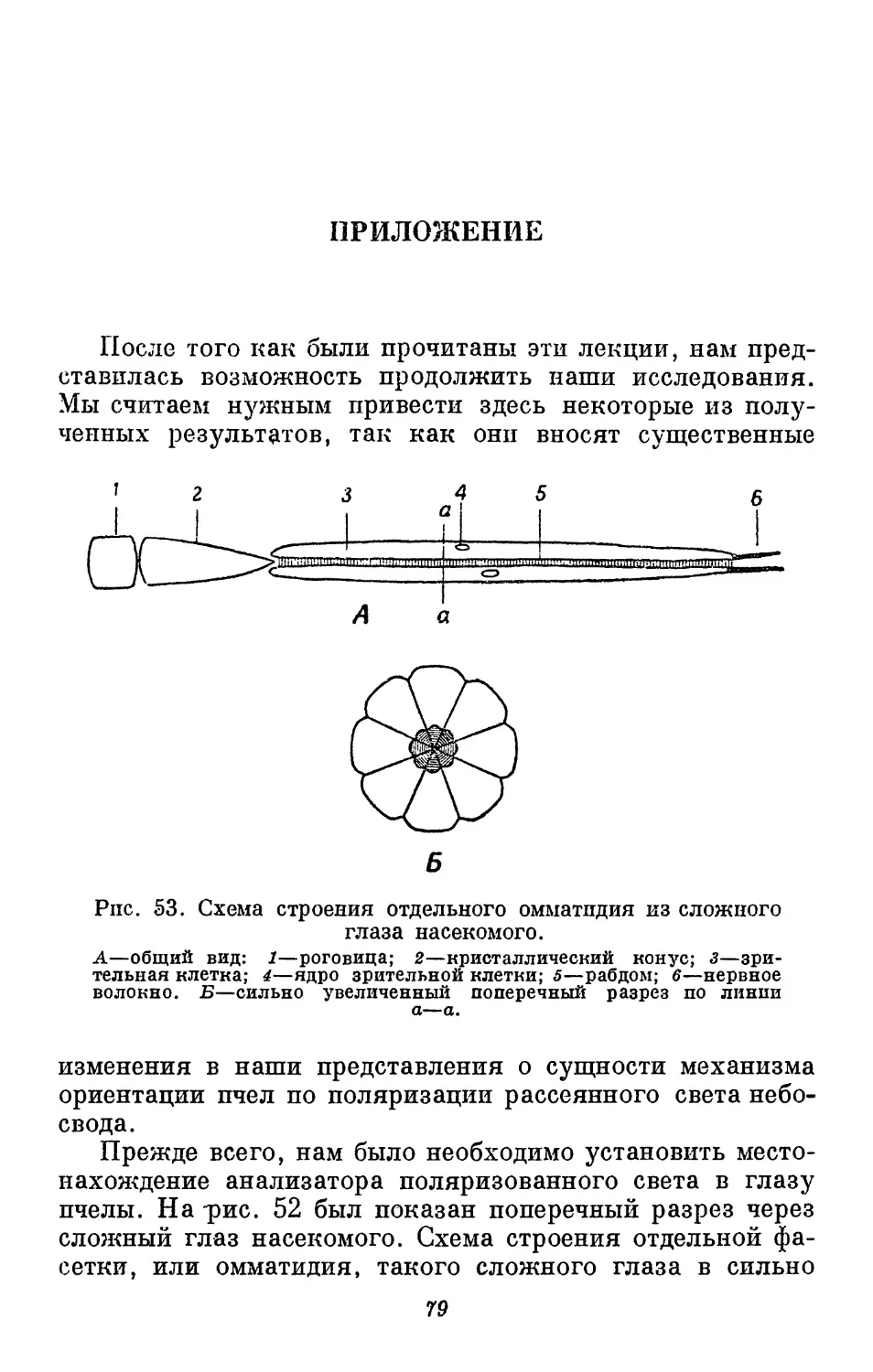
пометить одновременно 599 пчел. Краска приготовлялась
из сухого цветного порошка, замешанного на спиртовом
растворе шеллака. Такая краска быстро высыхает и
хорошо пристает к телу пчелы.
Для изучения поведения пчел, только что
обнаруживших богатый источник корма, вблизи от наблюдательного
45
Рис. 28. Наблюдательный улей.
Леток для пчел расположен справа.
К 28 f3i 599
Рис. 29. Способ метки пчел.
Для метки применялось пять разных цветов, показанных на
рисунке следующим образом: белый кружек—белое пятно; белый кружок
с горизонтальной чертой—красное пятно; белый кружок с крестом—
синее пятно; белый кружок с точкой—желтое пятно; черный
кружок—зеленое пятно.
пункта выставляют тарелку с сахарным сиропом. Как
только пчела-сборщица появится на месте подкормки и начнет
сосать сироп, ее метят, с тем чтобы в дальнейшем можно
было следить за ее поведением в улье. По возвращении
в улей меченая пчела прежде всего отдает почти весь
46
сироп другим пчелам. Затем оиа начинает делать
движения, которые я назвал круговым танцем. Она кружится
на одном месте, сначала направо, а затем налево,
неоднократно повторяя эти круговые движения с большой
энергией. Нередко она продолжает этот танец полминуты и
дольше, не сходя с одного места. Часто танцовщица
переходит на другое место сотов и повторяет круговой танец,
после чего она обычно возвращается на место подкормки
за новой порцией сиропа.
Во время танца ближайшие к танцовщице пчелы
приходят в сильное возбуждение; они толпой следуют за ней,
и их антенны почти касаются ее тела. Внезапно одна из
них поворачивается и покидает улей; другие
устремляются за ней, и вскоре некоторые из этих пчел появляются
на месте подкормки. По возвращении в улей они также
начинают танцевать, и чем больше танцующих пчел, тем
больше пчел оказывается на кормушке. Очевидно, что
танец в улье указывает на наличие корма. Однако неясно,
каким образом пчелы, возбужденные танцем, находят
место подкормки.
Для того чтобы выяснить, дает ли круговой танец
указания о том, в каком направлении следует искать корм,
мы кормили несколько меченых пчел из наблюдательного
улья на кормушке, расположенной в 10 ж к западу от
улья. В четырех точках лужайки, где стоял улей, к
северу, югу, востоку и западу от него, мы поместили на земле
по стеклянному блюдцу с сахарным сиропом, к которому
для запаха было подмешано немного меда. Через
несколько минут после начала кругового танца в улье на всех
блюдцах, независимо от их расположения по отношению
к улью, внезапно появились новые пчелы. Весть,
принесенная в улей и выраженная в танце, повидимому,
означала: «Вылетай и ищи по соседству с ульем».
Нельзя, однако, считать естественным для пчел сбор
корма со стеклянных блюдец. Если кормить меченых пчел
на тех же местах подкормки сахарным сиропом,
нанесенным на свежие цветы, например цикламен (рис. 30), л если
по возвращении пчел-сборщиц в улей они исполняют
круговой танец, из улья, как и прежде, вылетают новые пчелы,
но на этот раз они ищут что-то определенное. Поблизости
от улья мы устроили наблюдательный пункт, поместив
на земле две большие тарелки: в одной из них стояли
цикламены, а в другой—флоксы. Новые пчелы, вылетевшие
47
Рис. 30. Кормление меченых пчел
на цветках цикламена.
Рис. 31. Пчелы, мобилизованные танцовщицами, прилетев-
пиши с цикламена, в поисках корма прилетают только на
цветки цикламена (А), не обращая внимания на цветки флокса (Б).
из улья, интересовались только цикламенами, не замечая
флоксов (рис. 31). Затем мы заменяли цветы яа месте
подкормки, нанося сироп на цветки флокса. После замены
меченые пчелы, раньше бравшие сироп на цикламенах,
48
Рис. 32. Кормление меченых пчел на
цветках флокса.
начинали собирать его на флоксах (рис. 32). Через
несколько минут положение на наблюдательном пункте в корне
изменялось: новые пчелы переставали интересоваться
цикламеном; они прилетали только на флоксы, обследуя их
так, как будто были убеждены, что там есть корм.
Мы всегда получали сходные результаты в таких
опытах, если только пользовались цветками, издающими хотя
бы слабый аромат. Однако эти опыты не имели успеха,
если брались цветки без всякого запаха. При кормлении
меченых пчел на цветках черники (Vaccinium myrtillush.),
не имеющих запаха, новые пчелы, вырывавшиеся роем
из улья, деятельно искали по соседству корм, о котором
они получили извещение; однако тарелка с цветками
черники, поставленная на лужайке, привлекала их
внимание не больше, чем окружающая трава или другие
непахучие предметы. Те же результаты получались с
лишенными аромата Holcus lanatus L., лилией Tritonia crocos
maeflora Voss. и диким виноградом Ampelopsis quinque-
folia Michaux.
Совершенно сходный результат можно получить и без
цветов. Мы кормили меченых пчел сахарным сиропом из
стеклянных блюдечек, поставленных на куски картона,
пропитанные мятным маслом. По соседству с ними мы
располагали ряд сходных кусков картона, часть которых
49
была пропитана мятным маслом, а часть—другими
эфирными маслами. Новых пчел-сборщиц привлекает только
запах мятного масла; они садятся на любые предметы со
следами этого масла. Повидимому, эти пчелы знакомятся
с запахом цветков, только что посещавшихся пчелами-
танцовщицами, и, вылетев в поисках этого запаха,
попадают на цветки того же вида. В этом состоит одна из
биологических функций запаха цветков, не известная ранее.
Новые сборщицы прекрасно запоминают запах, с
которым их ознакомили пчелы-танцовщицы, и безошибочно
отыскивают его. Мы наблюдали однажды изумительное
проявление этой способности пчел. В Мюнхене в середине
июля на одном из участков ботанического сада
находилось в цвету одновременно семьсот видов растений.
Одним из этих растений был Helichrysum lanatum DC,
занимавший одну маленькую грядку; цветки этого
растения посещались одним видом одиночной пчелы и никогда
не привлекали к себе медоносных пчел. Согласно данным
биологической литературы, медоносные пчелы вообще
никогда не берут нектар с цветков растений этого вида.
Мы произвели следующий опыт: некоторое число меченых
пчел из наблюдательного улья, стоявшего у этого участка
ботанического сада, кормили сахарным сиропом из
стеклянной чашки, окруженной цветками Н. lanatum. Пчелы-
танцовщицы, возвращаясь с этой кормушки в улей,
уносили на своем теле аромат Н. lanatum, и в течение
последующих часов много пчел принялось посещать грядку
с Н. lanatum и садиться на цветки в поисках корма.
Таким образом, пчелы отыскали специфический запах этого
растения среди запахов 700 других видов растений,
цветущих по соседству с ними (Фриш, 1923b).
Пчелы, толпящиеся позади возвратившейся в улей
танцующей пчелы-сборщицы, воспринимают запах
цветков двояким образом. Во-первых, вытягивая свои
антенны по направлению к танцовщице, они воспринимают запах,
приставший к телу танцовщицы в результате ее
соприкосновения с цветком; оказывается, спинная поверхность тела
пчелы обладает способностью на долгое время удерживать
запахи (Штейнгофф, 1948). Во-вторых, в перерывы между
танцами танцующая пчела кормит сопровождающих ее
пчел, отрыгивая из зобика капельку нектара, собранного
е цветка и насыщенного свойственным цветку запахом.
Таким образом пчела-танцовщица приносит в улей запах
50
посещавшихся ею цветков как на своем теле, так и с
собранным ею нектаром.
Читатель вправе задать вопрос, каким образом удалось
установить все эти подробности. На самом деле их совсем
не трудно выявить при помощи следующего опыта. Мы
кормили меченых пчел, прилетающих на цветки
цикламена, лишенным аромата сахарным сиропом. Для
предотвращения пропитывания сиропа запахом цветков
цикламена мы помещали его в небольшую сферическую склянку,
из которой пчелы могут брать сироп только через узкую
щель (рис. 33, А). Эти пчелы переносят запах цветков
Рис. 33. Постановка опытов по перенесению пчелой-сборщицей
запаха цветков на поверхности своего тела и в зобике.
А—перенос запаха цветков поверхностью тела; Б—перенос вапаха
цветков в вобике (с нектаром). 1—сосуд с водой; 2—стеклянная трубка;
3—узкая щель, из которой можно сосать сироп.
цикламена лишь на поверхности своих телец, и другие
пчелы, мобилизуемые ими в улье, разыскивают этот
аромат. В другом опыте мы кормили других пчел-сборщиц,
прилетающих на лишенный запаха картон, сахарным
сиропом, предварительно насыщавшимся в течение часа
запахом цветков цикламена (рис. 33, Б). В этом случае
пчелы-сборщицы переносили запах только с нектаром,
находящимся в их зобике; однако пчелы, побужденные
к вылету танцовщицами, точно так же разыскивали
цикламены.
Интересно сравнить относительную эффективность этих
двух путей переноса запаха цветков пчелам,
находящимся в улье. Этот вопрос можно решить
экспериментально, подкармливая пчел, прилетающих на цветки
цикламена, сахарным сиропом, предварительно насыщенным
запахом цветков флокса. При такой постановке опыта
запах, переносимый телом танцующих пчел, будет отли-
51
Рис. 34.
Пищеварительный тракт пчелы.
чаться от запаха нектара, находящегося в их зобиках.
Если место подкормки расположено вблизи от улья, то
пчелы, возбужденные танцовщицей, будут прилетать на
цветки цикламена почти так же часто, как и на цветки
флокса. Если же место подкормки расположено на
расстоянии примерно 800 м от улья, то новые сборщицы
интересуются только цветками флокса, запах которых
переносился с нектаром, находившимся в зобиках
пчел-танцовщиц. Это означает, что за время дальнего обратного пути
в улей тело пчелы утрачивает приставший к нему запах
цикламена. Следовательно, когда пчелы собирают нектар
на далеком расстоянии от своего улья, запах нектара,
переносимого в закрытом зобике (рис. 34), имеет большое
здачение (Фриш, 1946а).
Круговой танец наблюдается лишь в том случае, если
пчелы обнаружили богатый источник корма (рис. 35), так
что, очевидно, он выражает следующее сообщение:
«Имеется много корма и сладости». Заменив чашку, полную
сиропа, на которой кормились пчелы, чашкой с
фильтровальной бумагой, смоченной снизу небольшим количеством
сиропа, мы создаем нехватку корма, сохраняя его состав
(рис. 36). До тех пор, пока объем сиропа будет выше
определенного критического уровня, пчелы продолжают его
52
брать., старательно высасывая его из пор фильтровальной
бумаги. Однако, возвращаясь после такой подкормки
в улей, они уже не танцуют, и поэтому другие рабочие
пчелы не прилетают на место подкормки. То же самое
происходит и в том случае, если понизить концентрацию
сахарного сиропа до известного уровня, даже не
уменьшая при этом объем жидкости в кормушке. Чем слаще
сироп, тем энергичнее проходят танцы (Фриш, 1923b, 1934).
Рис. 35. Рис. 36. Недостаток корма.
Обилие корма. Фильтровальная бумага смачивается
снизу сахарным сиропом при помощи шприца.
Такое же поведение пчел можно наблюдать и в
естественных условиях, при посещении ими цветков; оно имеет
чрезвычайно важное значение, так как очень часто
цветение наступает одновременно у многих видов растений.
При этом разные пчелы-сборщицы из одного и того же улья
могут обнаружить разные медоносные растения, однако
благодаря тому, что пчелы, обнаружившие самый
богатый источник корма, танцуют наиболее энергично,
наибольшее число новых сборщиц устремляется именно на
растения, наиболее богатые нектаром. В результате
опыление таких растений обеспечивается наилучшим образом,
а пчелЬг приносят в свой улей наилучший и
наисладчайший нектар.
После того как пчелы столь успешно собрали
нектар из богатого источника корма, что количество его
у данного вида растений уменьшается, пчелы перестают
танцевать и мобилизация рабочих пчел на данный вид
растений прекращается. Таким образом, активность всей
группы пчел-сборщиц приспосабливается к относительному
обилию нектара у различных видов растений.
53
Описанные наблюдения ясно показывают, что пчела-
танцовщица направляет других пчел к обнаруженным ею
цветкам и использует запах цветков как указатель
местонахождения богатого источника корма. Для этой же цели,
помимо запаха цветков, используется и другой запах,
который выделяет сама рабочая пчела. На брюшке пчел в
кожном кармане, выстланном железами, имеется особый
пахучий орган. Обычно карман закрыт и никакого запаха
Рис. 37. Три пчелы у блюдечка с большим
количеством корма (сахарный сироп).
А—пчела с открытым пахучим органом; Б и В—пчелы,
уже втянувшие свои пахучие органы.
не выделяется. Однако, возвращаясь к богатому
источнику корма, пчелы открывают этот пахучий орган
в тот момент, когда они приближаются к месту кормления
и садятся на него (рис. 37). Этим самым они, видимо,
сообщают источнику корма запах, весьма привлекательный
для других пчел, который как бы означает: «Иди сюда,
этой дорогой». Значение выделения этого запаха при
посещении пчелами цветков, лишенных запаха,
обнаруживается следующим опытом. Мы кормили примерно по дюжине
пчел на двух разных кормушках сиропом, не имевшим
никакого запаха. На одной из этих кормушек каждой
прилетающей пчеле карман, в котором помещается
пахучая железа, замазывали шеллаком. Все пчелы,
возвращаясь в улей, энергично танцевали, как и раньше,
однако пчелы с замазанным карманом, прилетая на место
подкормки, не могли распространять свой запах. В итоге
число пчел, привлеченных танцовщицей к этой тарелке
с сиропом, оказалось в 10 раз меньше, чем число пчел,
привлеченных к контрольной тарелке, где пчелы поль-
54
зовались своим пахучим органом нормальным путем
(Фриш, 1923b и 1934).
Проводя подобные опыты в течение ряда лет, мы всегда
помещали корм поблизости от улья, так как это было
удобнее и давало возмояшость наблюдать пчел одновременно
и на месте подкормки и в наблюдательном улье. Однако
случайные наблюдения подсказали нам, что, повидимому,
пчелы могут сообщать кое-что другим пчелам и о
расстоянии места подкормки от улья. Поскольку пчелам очень
Рис. 38. Деревянный столик, служащий местом
подкормки пчел, со стоящим на нем стеклянным
блюдцем с сиропом.
часто приходится собирать корм на расстоянии 1,5—
2 км от улья, им было бы очень выгодно, если бы
сборщицы обладали способностью сообщать другим пчелам
не только о запахе, но и о местонахождении пищи.
Для решения этого интересного вопроса мы поставили
в августе 1944 г; следующий опыт. В 10 и в 300 м от
наблюдательного улья были устроены две кормушки,
посещавшиеся мечеными пчелами из этого улья. Для того чтобы
установить, куда летят вновь мобилизованные пчелы из
улья, мы использовали лавандовое масло. На одном из
мест подкормки стеклянный сосуд с сахарным сиропом
был поставлен на кусок картона, пропитанный
лавандовым маслом (рис. 38). В разных точках лужайки мы
расставили куски картона без корма, поместив на них
маленькие пробирки с лавандовым маслом (рис. 39,6). Пчелы из
улья, мобилизованные танцовщицами, возвращавшимися
55
с кормушки, отыскивали этот запах и, приблизившись
к кускам картона, часто садились на них (рис. 39,Л).
После этих подготовительных процедур мы начали
самый опыт. В кормушку, находившуюся в 10 ж от улья,
поместили обильный корм, в результате чего пчелы,
возвращаясь с нее в улей, принимались танцевать. Некоторое
время на этой кормушке и соседних кусках картона
появилось много новых пчел, тогда как около кормушки,
отстоящей от улья на 300 м, были обнаружены лишь единичные
Рис. 39. А—пчелы, мобилизованные
танцовщицами, обнюхивают три маленьких сосудика с тем же
запахом, что и запах на кормушке, и садятся на
картон; Б—схема расположения сосудиков с
пахучим веществом. 1—картон; 2—сосудик;
3—пахучее вещество.
пчелы. Так, в одном опыте (отнюдь не представлявшем
исключения) мы насчитали в течение одного и того же
промежутка времени 12 пчел около дальней (в 300 м)
кормушки и 174 пчелы—на ближней (в 10 м). Изменив
условия опыта, т. е. давая обильный корм на дальней
кормушке и не давая его на ближней, мы обнаружили,
что в улье танцевали только те пчелы, которые вернулись
с дальней кормушки. Теперь вновь мобилизованные
сборщицы быстро появились на дальней кормушке, тогда как
на ближней мы наблюдали только единичных пчел. В
одном из таких опытов в течение часа мы насчитали 61 пчелу
около дальней кормушки и лишь 8—около ближней.
Иногда разница была еще более разительной. На
основании этих данных становится очевидным, что
возвращающиеся с места подкормки пчелы-сборщицы сообщают в улье
56
о расстоянии, на котором ойи обнаружили богатый
источник корма.
Каким же образом пчелы сообщают об этом? Если
обильно снабдить кормом обе кормушки и пометить пчел,
собирающих корм на каждой из них, то, заглянув в
наблюдательный улей, мы видим изумительную картину:
пчелы, вернувшись в улей с ближней кормушки, исполняют
описанный выше круговой танец (рис. 40, А); пчелы же,
прибывающие в улей с дальней кормушки, исполняют
/
У
Рис. 40. Танцы пчел.
Л—круговой танец; Б—виляющий танец (а—прямолинейная часть).
совершенно особый танец, названный нами «виляющим
танцем». Они пробегают немного по прямой линии,
быстро виляя брюшком из стороны в сторону, затем делают
полный (на 360°) поворот налево, снова бегут по прямой
и делают полный поворот направо (рис. 40, В); весь этот
цикл пчелы многократно повторяют. Виляющий танец
мы наблюдали много лет назад, но мы всегда считали его
типичным танцем пчел, приносящих в улей пыльцу
(обножку). Теперь же оказалось, что этот танец
выполняется весьма энергично пчелами, приносящими сахарный
сироп с экспериментальной кормушки, находящейся
в 300 м от улья.
Скоро стало ясно, что наши первоначальные
представления о значении этого танца были неверны. Ошибка
возникла в рез}'льтате того, что мы всегда подкармливали
пчел сиропом на близком расстоянии от улья, в
основном в целях удобства наблюдений. Пчелы же,
приносящие в улей обножку, прилетали с естественных пастбищ,
67
находящихся на большем расстоянии от улья (пчел,
приносящих обножку, легко узнать по плотно набитым
пыльцевым корзиночкам на задних лапках). Поэтому мы могли
наблюдать круговые танцы только у пчел, приносящих
нектар или сахарный сироп, а виляющие танцы—только
у пчел с обножкой. Однако как только мы обнаружили,
что эти два типа танцев связаны с различным расстоянием
до источника корма, а не с его качеством, было нетрудно
при помощи соответственных опытов показать, что танцы
сборщиц пыльцы и сборщиц нектара, возвращающихся
в улей с одного и того же расстояния, ничем не отличаются.
К счастью, нам самим удалось обнаружить эту ошибку.
Пусть этот пример послужит предостережением молодым
исследователям, с нетерпением стремящимся опубликовать
первые же результаты своих исследований. Пусть они
дважды и трижды проверят свои наблюдения, прежде
чем считать свои выводы правильными. Ибо очень часто
природа идет иными путями, неожиданными для человека.
Мы изменили описанные выше опыты, постепенно
отодвигая ближнюю кормушку с 10 м на все большее и
большее расстояние; при этом эту кормушку продолжала
посещать все та же группа меченых пчел. Однако при
расстояниях 50—100 м от улья круговой танец этих пчел
сменился виляющим. Одновременно мы перемещали дальнюю
кормушку все ближе и ближе к улью, и ее также
продолжала посещать прежняя группа меченых пчел. Однако
в зоне между 100 и 50 м от улья виляющий танец этой
группы пчел сменился круговым танцем. Таким образом,
стало ясно, что круговой и виляющий танцы означают на
языке пчел два различных понятия: первый из них
указывает на наличие источника корма вблизи от улья, а
второй—на источник, удаленный от улья на 100 м и более
(Фриш, 1946а,Ь).
Обитатели ульев, повидимому, понимают смысл этих
танцев, что вытекает из следующего опыта. Одну группу
меченых пчел приучили брать корм из кормушки,
расположенной в 10 ж от улья, а другую—на кормушке,
расположенной в 200 м от него. В обоих пунктах сосуды с кормом
были поставлены на куски картона, лишенные какого-либо
запаха. После этого мы прекратили подачу сахарного
сиропа в обоих пунктах, и оба сосуда в течение 1—2 час.
оставались пустыми. К концу этого срока пчелы обеих
групп оставались в улье, не вылетая на волю; лишь время
58
от времени одна из них появлялась на кормушке, чтобы
выяснить, нет ли там корма. Если затем мы наполняли
сиропом дальнюю кормушку, то виляющий танец
первых же сборщиц, вернувшихся в улей с наполненными
зобиками, возбуждал главным образом пчел той самой
группы, которая и раньше посещала дальнюю кормушку.
Когда же мы наполняли сиропом ближнюю кормушку, то
пчелы выполняли круговой танец, опять-таки возбуждая
главным образом тех пчел, которые и прежде кормились
на этой кормушке.
При хорошем медосборе пчелы часто улетают за 1,5—
2 км от улья. При таких расстояниях сигнал о том, что
корм находится ближе, чем за 100 м, или дальше, едва ли
может иметь большой биологический смысл. На самом же
деле виляющий танец не только сигнализирует о том, что
богатый источник пищи расположен на большом
расстоянии, но также и о том, сколь велико это расстояние.
Указание о дальности расстояния выражается довольно точно
числом поворотов пчелы, исполняющей виляющий танец,
в единицу времени. Постепенно перемещая кормушку на
все большее и большее расстояние от улья, мы, после
довольно напряженных и увлекательных опытов, удалили
ее от улья на 6 км. Мы подсчитывали число поворотов
в единицу времени (15 сек.) у танцовщиц, возвращающихся
с кормушек, удаленных от улья на различные расстояния
от 100 м до 6 км. Итоги 3885 наблюдений этого рода
изображены графически на рис. 41. При расстоянии 100 м
пчела совершала 9—10 полных циклов (см. стр. 57)
за 15 сек., при расстоянии 200 м—7 циклов, при 1000 м—
4,5 цикла, а при 6000 м—только 2. Установив эту
зависимость между числом поворотов и расстоянием до
кормушки, мы могли при помощи секундомера определять, как
далеко летала танцующая пчела. Пчелы в улье также
понимают значение частоты поворотов пчелы-танцовщицы
и таким образом узнают, на какое расстояние надо лететь,
чтобы добраться до корма.
Не все отдельные точки на рис. 41 (каждая из них
представляет собой среднее многих наблюдений) точно
укладываются на кривую, выражающую зависимость между
числом поворотов и расстоянием до корма. Причины таких
отклонений в основном сводятся, повидимому, к
следующим. Во-первых, в этом играют роль
индивидуальные особенности пчел, хотя в обычных условиях они
59
удивительно мало различаются в этом отношении;
во-вторых, каждая семья характеризуется определенной частотой
поворотов для данного расстояния, иными словами,
кривые зависимости между частотой поворотов в танце и
расстоянием до корма варьируют у разных сеъюй пчел; наконец,
200 6001 7000 1500 2000 2500 3000 3500 4000 4500 5000 5500 6000
400 800
Расстояние от кормушки ,м
Рис. 41. Зависимость между числом поворотов при виляющем
танце в единицу времени и расстоянием до кормушки.
в-третьих, частота поворотов варьирует в зависимости от
направления ветра. Встречный ветер по пути к месту
подкормки обладает тем же действием, что и удлинение
пути,—он замедляет темп танца; попутный же ветер
обладает противоположным действием. Повидимому, оценка
пчелой удаленности корма основана на времени или
усилиях, необходимых для того, чтобы до него добраться
(Фриш, 1948Ь).
Если кормушка стоит неподалеку от улья, пчелы,
возбужденные круговым танцем, разлетаются в разных
направлениях и ищут корм в непосредственной близости;
если же кормушка стоит дальше, то благодаря
виляющему танцу они узнают и о расстоянии, на котором
обнаружен корм. Однако, помимо этого, они получают
GO
сведения о направлении, в котором им надлежит лететь.
Этот поистине изумительный факт выявляется при помощи
следующего опыта. Мы кормили известное число меченых
пчел на пахучем куске картона, расположенном в 200 м
к югу от улья. Вокруг улья, на расстоянии также 200 м
от него, мы расположили куски картона с тем же запахом,
но без корма. Спустя несколько минут на кормушке и на
кусках картона, прилегающих к ней, появляется много
новых пчел. На кусках же картона, расположенных в
других направлениях от улья, пчел либо не видно совсем,
либо их только единицы. Стоит переместить пищу,
поставив ее на расстоянии тех же 200 м, но на восток от улья,
как мы обнаружим, что через некоторое время новые
сборщицы будут лететь на восток. Язык пчел поистине
совершенен, и их способ указания направления, в котором
расположен источник кор.ма, можно считать одной из самых
замечательных тайн их сложного общественного
устройства.
Когда мы наблюдаем пчел-танцовщиц, вернувшихся
с кормушки, местоположение которой нам известно, мы
с удивлением обнаруживаем, что все они выполняют один
и тот же танец; в частности, направление прямолинейной
части виляющего танца (см. рис. 40) у всех танцующих пчел
совершенно одинаково. В типическом случае пчелы,
вернувшиеся с кормушки, расположенной на 200 м к югу от
улья, танцевали на соте таким образом, что при
прямолинейной части танца все они двигались влево. Другие
же пчелы, получавшие в это же самое время сироп в
кормушке, поставленной на 200 м к северу от улья, в
прямолинейной части своего танца направлялись вправо.
Иными словами, оказывается, что направленность
прямолинейной части виляющего танца каким-то образом связана
с направлением, в котором находится источник корма.
Наблюдая за танцами на протяжении ряда часов
и не меняя при этом места подкормки, мы обнаружили,
что направление прямолинейной части пути не остается
неизменным, а постепенно изменяется так, что после
полудня оно оказывается иным, нежели утром. Более
тщательные наблюдения показали, что это направление всегда
изменялось примерно на такой же угол, на какой
повернулась за это время Земля или переместилось солнце на
небе. Это заключение не было неожиданным, так как
опыты, выполненные за несколько лет до этого Вольфом (1927),
61
Санчи (1911,1923) и другими, показали, что пчелы и
муравьи пользуются солнцем как компасом при передвижении
по гладкой поверхности без каких-либо особенностей,
могущих служить вехами. Эти наблюдения касались пчел
и муравьев, выходящих из гнезда в поисках пищи и
возвращающихся назад раньше, чем положение солнца
успевало существенно измениться. Если рабочая пчела летела
от улья по направлению к солнцу, ей оставалось только
лететь в противоположную от солнца сторону, чтобы найти
дорогу домой; если по пути из улья солнце находилось
справа от пчелы, то во время обратного полета она должна
иметь его слева от себя. Поэтому, обнаружив, что
направление прямолинейной части виляющего танца сдвигается
по мере изменения положения солнца, мы были убеждены,
что танцы указывают направление, в котором
находится источник корма по отношению к солнцу.
Ключ для понимания этих сигналов весьма любопытен.
Прежде всего необходимо вспомнить, что в нормальных
условиях пчелы танцуют в улье на вертикально
расположенных сотах, причем в улье царит темнота. В обычном
улье пчела не может ощущать положение солнца, а
вместо этого полагается, повидимому, на направление силы
тяжести. Направление прямолинейной части их танца
отклонено от направления силы тяжести под углом,
равным углу направления их полета от улья к источнику
корма по отношению к солнцу (рис. 42).
Если прямолинейная часть танца направлена прямо
вверх, то это, очевидно, означает: «Источник корма
находится в том же направлении, что и солнце». Если же она
направлена вниз, то это, вероятно, означает: «Лети в
противоположную от солнца сторону, чтобы найти корм».
Если прямолинейная часть танца направлена под углом
в 60° влево от направления силы тяжести, то это означает,
что место кормежки расположено на 60° влево от солнца;
сходным образом, если она направлена на 120° вправо, то
это указывает, что место кормежки расположено на 120°
вправо от солнца (Фриш, 1946а, b; 1948b).
Весьма примечательно, что полету пчелы по направлению
к солнцу соответствует движение вверх во время исполнения
прямолинейной части виляющего танца. Невозможно
представить себе, чтобы такое поведение возникло у пчел сразу.
Мы совершенно уверены в том, что это столь
знаменательное соответствие возникло, подобно другим особенностям
62
У
т
V
"/рч?
¦г 7—-«Л
V
^
К
к х- 1 <
V 120°\
\
/рГс
4F
\
\
\
Ж«
/77
Рис. 42. Ориентированность танцев пчел относительно положения
солдца.
у—улей; к—кормушка; с—солнце; т—схема танца.
поведения, постепенно, в процессе исторического
развития вида. Было бы чрезвычайно интересно изучить
более примитивных общественных насекомых и
попытаться обнаружить у них более простые формы «языка»,
которые позволили бы нам представить себе пути
возникновения чрезвычайно сложного поведения медоносной
пчелы.
63
Даже при облачном небе пчелы продолжают при
помощи виляющего танца указывать правильное
направление к месту подкормки по отношению к положению солнца.
Следовательно, пчелы, невидимому, имеют представление
о положении солнца на небосклоне, даже если оно закрыто
облаками. Быть может, пчелы по опыту знают, где должно
находиться солнце в то или иное время суток? Пчелы
обладают великолепной памятью на время. Если
несколько дней подряд кормить пчел в определенном месте между
10 и 12 час. утра, они в течение ближайших дней будут
продолжать прилетать на это место по утрам, от 10 до 12
час, даже если кормушка будет пустой. Таким образом,
вполне возможно, что на хорошо известной пчелам
территории они могут быть осведомлены о положении солнца
в каждый час суток. Для проверки этого предположения
мы перенесли наблюдательный улей подальше, в
местность, совершенно не знакомую пчелам, преднамеренно
выбрав для этого пасмурное утро, когда небо было плотно
закрыто облаками. В этой новой местности мы установили
кормушку, а затем постепенно приблизили ее на
расстояние 100 м от улья. Даже при этих обстоятельствах
виляющие танцы пчел правильно указывали направление пути
к кормушке по отношению к положению солнца, несмотря
на то, что в этой незнакомой им местности пчелы не могли
знать из опыта положение солнца в данное время. По всей
вероятности, пчелы обладают способностью
непосредственно ощущать положение солнца, несмотря на пелену
облаков. Однако мы пока еще не знаем, каким образом они это
делают.
Пчелы, возбужденные танцами на сотах, узнают угол
танца по отношению к силе тяжести; они запоминают его
и., вылетая из улья, направляют свой полет под таким же
углом к положению солнца. Для выяснения того,
насколько точно мобилизованные пчелы выдерживают
направление, сообщенное им пчелами-танцовщицами, мы
поставили ряд опытов; результаты одного из них графически
представлены на рис. 43. Кормушка стояла в 250 м к югу
от улья; несколько меченых пчел получили здесь корм на
кусках картона, пропитанных лавандовым маслом. На
расстоянии 200 м от улья на землю было положено 7
других кусков картона, также пахучих, но без корма; один
из них лежал на прямой линии между ульем и местом
подкормки, 3—влево от нее, а 3—вправо, причем все они
64
отклонялись друг от друга на 15°. Мы подсчитывали всех
пчел, посещавших каждый из этих кусков картона в
течение 1 часа, причем пища предоставлялась им только на
месте подкормки; число пчел, прилетевших на различные
© к
250м
Рис. 43. Степень соблюдения пчелами точности при полете
в сообщенном им направлении.
у—улей; к—кормушка; кк—куски картона (цифры
указывают число посещений).
куски картона (слева направо), составляло: 8, 13, 58,
132, 37, 7 и 3. Иными словами, огромное большинство пчел
(88%) не отклонялось от правильного направления
больше чем на 15°. Опыт был повторен 27 сентября 1949 г.
с другой породой пчел и в другой местности. Результаты
оказались даже лучше: число посетительниц (слева
направо) составляло 0, 3, 8, 24, 2, 3, 0.
Другой опыт показывает различие реакций пчел на
круговой и виляющий танцы. В 10 м к востоку от
наблюдала
тельного улья мы поставили кусок картона с
ароматизированным сиропом, а в 25 ж к северу, югу, востоку и
западу от него мы разложили куски картона с тем же запахом,
но без корма (рис. 44). Меченые пчелы, возвращаясь
в улей с этой кормушки, как обычно, выполняли
круговые танцы. В течение 1 часа после этого мы насчитали на
С
0 к к
1 Юм Л
¦ш * о в
кн
И кк
Ю
Рис. 44. Постановка опыта для определения
различия реакций пчел на круговой и виляющий танцы.
V—улей; к—кормушка; кк—куски картона (см. рис. 45).
пахучих кусках картона, лежавших вокруг улья на
расстоянии 25 м от него, следующее количество пчел: на
картоне, лежавшем к востоку от улья,—27 пчел, а на других
кусках—37, 20 и 19 пчел (рис. 45,-4). Эти результаты
показывают, что круговые танцы не дают никаких указаний
о направлении, в котором следует искать корм, и пчелы
разыскивают его по всем сторонам вокруг улья. Затем ту
же кормушку мы поместили на 100 м к северу от улья.
а пахучие куски картона без корма также на 100 м к
северу, югу, востоку и западу и повторили опыт. В течение
66
I часа мы насчитали 278 пчел, посещавших пахучие
куски картона, лежавшие к северу от улья, а на трех
других кусках картона—лишь 6, 2 и 1 пчелу (рис. 45, Б).
100 м
10.
ш
Ш
27 37 20 19
278
Рис. 45. Различие реакций пчел на круговой и
виляющий танцы.
А—пчелы, собирающие корм на расстоянии 10 м от улья,
выполняют круговые танцы и вызывают прилет примерно
одинакового числа пчел на все четыре куска картона,
расположенные в разных направлениях; Б—пчелы, собирающие
корм на расстоянии 100 м от улья, выполняют виляющие
танцы и вызывают полет других пчел в определенном
направлении.
Высота столбцов наглядно показывает относительное число
пчел на 4 кусках картона (см. рис. 46), а цифры под
столбцами—абсолютное число пчел. Черные столбцы относятся к
картону, расположенному в одном направлении с
кормушкой; заштрихованные столбцы—к картону, лежащему в
противоположном направлении, а столбцы, покрытые точками,—к
двум кускам картона, лежащим под прямым углом к
направлению на кормушку.
Пчелы, возвращавшиеся с кормушки, находившейся в
100 м от улья, выполняли виляющий танец, дававший
четкие сигналы о направлении, в котором следует искать корм.
Горный ландшафт моей родины дал возможность
поставить опыт, при котором место подкормки
располагалось за гребнем так, чтобы пчелам пришлось лететь за
67
кормом в обход (рис. 46). Было чрезвычайно интересно
выяснить, указывают ли пчелы-танцовщицы направление
первой или второй части пути. В действительности
оказалось, что пчелы не указывали ни того, ни другого,
а указывали направление от улья к корму по прямой.
Одна местность в горах была особенно удобна для
проведения опытов, в которых пчелы были вынуждены
добираться до корма, делая большой крюк.
Наблюдательный улей стоял по одну сторону крутого гребня, а место
подкормки—по другую (рис. 47). Мы постепенно
передвигали кормушку с сахарным сиропом вокруг гребня, в то
Рис. 46. Расположение кормушки («) за холмом;
при этом пчелам, летящим из улья (у),
приходилось огибать этот холм.
время как пчелы продолжали брать с нее корм. Однако
результаты этого опыта разочаровали нас, так как пчелы,
к нашему удивлению, летели вверх и перелетали через
гребень. Повидимому, они быстро обнаружили, что этот
путь был короче, чем путь в обход, вокруг оконечности
гребня. Они сохраняли правильное направление, хотя
фактически их перелет был длиннее, чем прямой путь
от улья к месту подкормки, если бы он проходил сквозь
гребень. Определяя продолжительность танцев в
наблюдательном улье, можно было установить, что пчелы
указывали длину фактического перелета через гребень, а не
расстояние по прямой сквозь него. Преимущество такого
указания состоит в том, что вновь мобилизованные пчелы
получают сведения не только об абсолютном направлении,
в котором следует искать корм, но и о фактическом
расстоянии до него по наиболее подходящему пути. Однако,
несмотря на то, что мы видели все это своими глазами, мы
не в состоянии понять эт^у способность пчел (Фриш, 1948b).
25 лет назад нам казалось, что мы разобрались в языке
пчел. Между тем дальнейшие опыты принесли много
68
неожиданностей, одна из которых заключалась в том, что
круговой и виляющий танцы пчел сигнализируют о
расстоянии до источника корма, а не о его характере. Кроме
того, неожиданным было открытие, что виляющий танец
может дать указания о направлении пути к источнику
Рис. 47. Еще одии опыт, в котором
пчелам приходилось делать крюк по
пути за кормом.
Место подкормки расположено в левой
стороне снимка (белый крест), а улей—
примерно на том же уровне по другую
сторону гребня.
корма по отношению к положению солнца. Однако самое
удивительное открытие мы сделали совсем недавно.
Было время, когда результаты наших опытов казались
столь фантастичными, что им трудно было верить. Нам
не раз приходило в голову, что, быть может, пчелы из
наблюдательного улья превратились в каких-то «ученых»
пчел. Поэтому мы решили выяснить, происходят ли такие
танцы и в обычном улье. Однако наши опасения оказались
напрасными, так как, вынимая соты из обычного улья
и держа их в руках, можно было видеть, что, несмотря
69
на причиненное им беспокойство, пчелы продолжали свои
танцы. Однажды, держа в руках соты из обычного улья,
покрытые пчелами, мы решили посмотреть, что
произойдет, если держать соты не вертикально, а горизонтально;
пчелы продолжали танцевать, но при таком положении
сотов они не мбгли ориентировать свои танцы
применительно к направлению силы тяжести, так как в
горизонтальной плоскости не может быть направления вверх или
вниз. Однако при этих условиях прямолинейная часть
танца пчел была направлена прямо в сторону места
подкормки. При вращении сотов на месте, наподобие
вертящегося столика, пчелы продолжали прямо указывать
направление к месту подкормки, как бы уподобляясь игле
компаса. Для более детального исследования этого
явления мы вновь воспользовались нашим наблюдательным
ульем вместо отдельного сота, взятого из обычного улья,
но при этом мы положили наблюдательный улей набок.
Эта перемена, видимо, не мешала пчелам, даже если улей
продолжал оставаться в горизонтальном положении в
течение нескольких дней. В горизонтально лежащем улье
направление танцев не менялось при изменении
положения солнца; они всегда указывали направление прямо
к месту расположения кормушки.
Проще всего было бы объяснить это поведение,
допустив, что танцовщицы продолжают указывать направление
к месту подкормки относительно положения солнца, но
в последнем случае непосредственно, а не применительно
к направлению силы тяжести. Если прямолинейную часть
виляющего танца пчела ориентирует под тем же углом
к солнечным лучам, под каким она летела из улья к месту
подкормки, то при горизонтальном положении улья она
будет направлять свои танцы прямо в сторону корма
(рис. 48). Это объяснение предполагает, что
пчела-танцовщица может видеть положение солнца на небосводе.
Однако в наших опытах наблюдательный улей был всегда
затенен крышей. Повидимому, дело обстояло гораздо
сложнее.
Прежде всего надо было выяснить, нужен ли вообще
солнечный свет для ориентировки танцев на горизонтально
расположенных сотах. Для проведения такого опыта
мы построили переносный кожух, которым можно было
накрывать улей вместе с наблюдателем (рис. 49). Под этим
кожухом можно было вести наблюдения над танцами либо
70
Рис. 48. Ориентация танцев пчел по отношению к положению
солнца.
А—направление от улья (у) к кормушке (к) на 40° правее
положения солнца; Б—улей (показана лишь некоторая его часть)
находится в горизонтальном положении; если пчелы ориентируют
прямолинейную часть виляющего танца под таким же углом к
солнечным лучам, как путь к кормушке, то они указывают путь прямо
к кормушке.
Рис. 49. Наблюдательный улей,
заключенный в непрозрачный кожух.
Леток помещается на скамье.
при красном свете, при котором пчелы ничего не видят,
либо при рассеянном белом свете. В обоих случаях танцы
столь же энергичны, как обычно, но лишены какой-либо
ориентации. Их направление все время менялось, и вновь
мобилизованные пчелы, совершенно сбитые с толку,
искали пищу во всех направлениях. Из этого следует,
71
что при горизонтальном положении сотов ни в темноте,
ни на рассеянном свету пчелы не могут дать указания
о направлении, в котором следует искать пищу. В обычном
же улье танцы происходят в темноте, но на вертикальной
поверхности и ориентированы относительно направления
силы тяжести. Смысл танцев на горизонтальной
поверхности можно хорошо понять, если обратиться к поведению
пчелиной семьи в нормальных условиях. В теплую погоду
на горизонтальной прилетной доске среди пчел-сборщиц,
останавливающихся перед летком и передающих нектар
пчелам-приемщицам, часто можно видеть танцующих пчел.
В этом случае танцы происходят при дневном свете и пчелы
прямо указывают направление к кормушке. Внутри же
темного улья в такой ориентировке, не связанной с
направлением силы тяжести, нет никакой необходимости,
так как обычно в улье нет горизонтальных поверхностей,
подходящих для выполнения танцев.
Физиолог может задать вопрос: как же могут пчелы
указывать направление к кормушке, танцуя на
горизонтальной поверхности, если они не видят при этом
солнца?
Как мы уже упоминали, при заключении
наблюдательного улья в непрозрачный кожух пчелы теряли
способность указывать направление на горизонтальной
поверхности. Наш следующий опыт состоял в удалении одной
из стенок кожуха с тем, чтобы пчелы могли видеть кусок
неба, расположенный на некотором расстоянии от солнца.
Как только пчелы получали возможность видеть голубое
небо, танцы мгновенно приходили в порядок и пчелы
вновь начинали указывать направление к кормушке;
для этого достаточно было сделать в стенке щель шириной
лишь в 10 см. Более того, пчелы начинали правильно
указывать направление, даже если им давали возможность
видеть небо через трубку длиной 40 см и диаметром 15 см
(рис. 50). Как только мы закрывали трубку, танцы
становились беспорядочными, стоило ее открыть—они вновь
приобретали определенную ориентацию.
В этих опытах трубка была направлена вверх, в
сторону северной части небосвода, причем прямолинейная
часть танцев была направлена к месту подкормки,
находившемуся на запад от улЬя. Затем мы поместили у самого
отверстия трубки зеркало, так что пчелы попрежнему
видели синее пятно, но теперь это было отражение южной
Р
части неоосвода; в результате направление танцев
изменилось и они указывали не на запад, а на восток.
Результаты этого опыта ясно показали, что пчелы улавливают
в небе какое-то явление, зависящее от положения солнца,
даже в том случае, если они не имеют возможности видеть
солнце непосредственно. Как только пчелы установили
положение солнца на небе, они получают возможность
указывать путь к месту подкормки, ориентируя свой танец
<п
Рис. 50. Вертикальный разрез через улей,
находящийся в кожухе,
1—улен в горизонтальном положении; 2—нормальное
(вертикальное) положение улья; 3—трубка, позволяющая пчелам
видеть во время танцев кусок голубого неба.
под таким же углом к солнцу, какой им пришлось бы
сохранять вне улья при полете к источнику корма.
Иногда кусок неба, видимый пчелами через трубку,
закрывадо облако; при этом танцы становились
беспорядочными и пчелы теряли способность указывать
направление к месту подкормки. Этот опыт показал, что явление,
помогавшее пчелам ориентировать танцы, сильно
нарушалось, если небо затягивалось облаками. Нам известно
явление, обнаруживаемое при чистом небе и связанное
с положением солнца на небосводе, интенсивность
которого значительно снижается при облачности,—это
поляризация света (Фриш, 1948Ь).
Солнечный свет состоит из колебаний, происходящих
во всех возможных направлениях, перпендикулярных
к направлению солнечного луча. Однако, прежде чем
попасть на землю, солнечный свет рассеивается в
атмосфере. Рассеянный cbqt, падающий с неба, частично
73
поляризован, т.е. его колебания совершаются преимущест
венно в некотором определенном! направлении. Призма
Николя или пластинка поляроида пропускают свет,
состоящий из колебаний, происходящих в одной определенной
плоскости; поэтому их иногда называют анализаторами
поляризованного света. Если держать поляроид перед глазами
и смотреть сквозь него на небо, то при вращении прибора
небо будет казаться то темнее, то светлее. Рассматривая
сквозь поляроид различные участки неба, легко
обнаружить, что степень поляризации изменяется по мере
удаления от солнца. Наибольшие изменения яркости
наблюдаются в том случае, если, вращая поляроид,
смотреть сквозь него на участок неба, расположенный на
расстоянии 90° от солнца; при этом свет поляризован на 70%.
Однако в участках неба, расположенных ближе к солнцу
и дальше от него, степень поляризации уменьшается,
а непосредственно подле солнца и вокруг прямо
противоположной ему точки небосвода поляризация очень слаба
или же вообще отсутствует.
Существует также другая зависимость между
положением солнца и степенью поляризации рассеянного
солнечного света. Плоскость поляризации света, идущего
от любой точки небосвода, всегда перпендикулярна к
другой плоскости, определяемой тремя точками: глаз
наблюдателя, точка на небе, на которую смотрит наблюдатель,
я солнце. Это правило нарушается лишь в зоне,
непосредственно прилегающей к солнцу, и в точке небосвода,
противоположной солнцу. (Эти геометрические
соотношения зависят от характера рассеяния света в атмосфере,
детальное рассмотрение вопроса можно найти в любом
курсе оптики.)
Таким образом, теоретически возможно определить
положение солнца путем осмотра любого участка синего
неба, если только имеется какой-либо анализатор для
определения направления поляризации света.
Эти теоретические соображения оставались, однако,
чисто спекулятивными, пока мы не установили, что
направленность танцев пчел меняется под влиянием изменения
направления поляризации падающего на них света. Эти
опыты проводились при помощи пластинок поляроида,
сходных со щитками, которые иногда прикрепляют к
ветровому стеклу автомашин для защиты от света фар
встречных машин. Мц пользовались пластинками размером
74
15x30 см, любая часть которых действовала в качестве
анализатора. При прохождении через такой поляроид
свет поляризуется в одном направлении. Мы поместили
поляроид над стеклянным окном наблюдательного улья,
и это тотчас же сказалось на танцах пчел. Для упрощения
условий эксперимента улей с трех сторон был закрыт
непрозрачным экраном; крышу также затенили, чтобы
исключить свет, падающий сверху. Таким образом,
пчелы-танцовщицы могли видеть небо лишь через поляроид. Мы
наблюдали sa танцами, меняя положение поляроида,
лежащего непосредственно на застекленном окне. В результате
62 опытов этого типа (Фриш, 1949а) были выявлены
следующие закономерности в реакциях пчел. При таком
положении пластинки поляроида, когда плоскость колебаний
светового луча при прохождении сквозь нее существенно
не изменялась, танцующие пчелы вполне правильно
указывали направление к месту подкормки (рис. 51, Аг).
Если мы поворачивали поляроид, подобно турникету,
так что плоскость колебаний поляризованного света
смещалась, танцы пчел отклонялись в том же направлении
(рис. 51, Б2); если мы поворачивали поляроид в
противоположном направлении, то танцы опять-таки
отклонялись в соответствии с поворотом. Если мы поворачивали
поляроид на угол, равный 55° и больше, то танцы пчел
становились беспорядочными, как в темноте (рис. 51, Вг).
Однако если плоскость поляризации рассеянного
солнечного света действительно является фактором,
определяющим ориентацию танца пчел, то мы должны получить
одинаковые результаты как при вращении поляроида
описанным выше способом, так и при изменении
направления, в котором пчелы видят небосвод. Для проверки
этого мы изменили условия опыта, смещая не поляроид,
а непрозрачный экран, покрывавший улей. В предыдущих
опытах пчелы, находясь в улье, могли видеть северную
часть небосвода, а на этот раз, повернув темный экран
на 90°, мы давали им возможность видеть лишь западную
часть неба. В результате мы обнаружили, что танцы
сместились в соответствии с различием плоскости колебаний
света в двух областях небосвода.
Направление танцев изменялось и в том случае, если
мы, не изменяя условий опыта, производили наблюдения
в разные часы суток. Несмотря на то, что положение
поляроида и непрозрачного экрана оставалось неизменным,
75
А
34°^
**^
-•?
/
\
в<
Рис. 51. Изменение направления танцев пчел при различных
положениях поляроида.
?™?™П?222^?И? поляРоиДа в каждом опыте; сплошные линии
указывают плоскость колебаний поляризованного света паитт?рп по
свеДтПаЮГпоРпУрЧаС^^Неба' а прерЙвистые-плоскость поляризации
S5SJ-» л прохождения через поляроид: Лх—Вг—сплошныр
линии означают направление к месту подкормки a Sm
стае—направление, указываемое танцовщицами, когда ул?й пшт"
крыт пластинкой поляроида (среднее ив прим™ ЛиЖ
воздействие на нчел поляроида в утренние часы отличалось
от воздействия его после полудня. Это также соответствует
зависимости между плоскостью поляризации рассеянного
солнечного света и положением солнца на небе. Следует
76
все же отметить, что в тех случаях, когда утром мы
показывали пчелам восточную часть небосвода, в полдень—
южную, а после полудня—западную, наблюдались
некоторые отклонения от этих закономерностей; однако эти
отклонения, повидимому, объясняются слабостью и
изменчивостью поляризации в участках неба,
расположенных в непосредственной близости к солнцу. Необходимо
Рис.. 52. Микрофотография среза
через глаз стрекозы.
Видно множество отдельных глазков, расположение
которых позволяет охватить взглядом весь небосвод.
поставить опыты с более совершенной аппаратурой,
чтобы получить точные количественные данные (см.
Приложение). Однако уже сейчас можно считать твердо
установленным, что поляризация рассеянного солнечного света
играет крайне важную роль в способности пчел к
ориентации (Фриш, 1949а).
Мы уверены в том, что поляризованный свет имеет
значение не только для пчел. При определенных условиях
муравьи также используют солнце как своего рода компас.
Много лет назад Санчи (1911) установил, что муравьи
сохраняют способность к ориентировке и определенному
направлению движения даже в том случае, если они
накрыты экраном, позволяющим им видеть лишь один
участок неба. Он высказал предположение, что муравьи,
77
вероятно, способны видеть звезды даже при дневном свете.
Нет сомнений, что при повторении опытов Санчи с
применением поляроидов его теория потерпит крушение и ее
придется заменить другой, не менее романтичной. Летом
1949 г. Шифферер (неопубликованные данные) показала,
что муравьи Lasius niger ориентируются по поляризации
рассеянного солнечного света.
Сложный глаз насекомых (рис. 52), помещающийся
на голове, повидимому, особенно пригоден для
установления поляризации всего небосвода, если допустить, что
он может действовать в качестве анализатора
поляризованного света, подобно призме Никол я или пластинке
поляроида. Однако это еще требует доказательств.
Таким образом, мы пришли к выводу, что до конца
пути еще далеко и мы стоим на пороге новых проблем,
требующих разрешения.
ПРИЛОЖЕНИЕ
После того как были прочитаны эти лекции, нам
представилась возможность продолжить наши исследования.
Мы считаем нужным привести здесь некоторые из
полученных результатов, так как они вносят существенные
J 7
j—г
I'l'll'll.illllliyM'dllHHMIIIIlll
Рис. 53. Схема строения отдельного омматидия из сложного
глаза насекомого.
А—общий вид: 1—роговица; 2—кристаллический конус;
3—зрительная клетка; 4—ядро зрительной клетки; 5—рабдом; 6—нервное
волокно. Б—сильно увеличенный поперечный разрез по линии
а—а.
изменения в наши представления о сущности механизма
ориентации пчел по поляризации рассеянного света
небосвода.
Прежде всего, нам было необходимо установить
местонахождение анализатора поляризованного света в глазу
пчелы. На -рис. 52 был показан поперечный разрез через
сложный глаз насекомого. Схема строения отдельной
фасетки, или омматидия, такого сложного глаза в сильно
79
увеличенном виде представлена на рис. 53, А. Каждая
фасетка пропускает лишь узкий столбик света, падающий
примерно вдоль ее оси. Оси соседних омматидиев несколько
расходятся и обращены до некоторой степени в разные
стороны. В итоге 8—10 тысяч омматидиев двух сложных
глаз воспринимают такое же число световых точек, из
которых наподобие мозаики складывается все поле
зрения. Кристаллический конус служит для проведения к
светочувствительной клетке пучка лучей, идущих вдоль оси
< >
Рис. 54. Способ вырезания треугольников из
листа поляроида для изготовления модели ом-
матидия насекомых.
Двойная стрелка указывает плоскость колебаний света,
проходящего через поляроид.
омматидия. Первоначальное предположение о том, что
кристаллический конус и есть анализатор, не
подтвердилось нашими исследованиями. В омматидии пчелы к
внутреннему концу кристаллического конуса примыкает 8
зрительных клеток, расположенных радиально (рис. 53, Б).
Их внутренние части (заштриховано) образуют
зрительную палочку, или рабдом, в котором свет вызывает
нервное возбуждение. Каждая зрительная клетка снабжена
нервным волокном. Аутрум в письме к нам высказал
предположение, что эти радиально расположенные зрительные
клетки, в зависимости от своего положения, могут
поляризовать свет в разных плоскостях. Если это допущение
справедливо, то можно считать, что теоретически даже
отдельный омматидии может определять направление
колебаний поляризованного (?вета, падающего с любой части
неба. Как указывалось выше, для того чтобы человек
мог определить направление этих колебаний, он должен
80
смотреть на небо через пластинку поляроида, вращая ее.
Если радиально расположенные зрительные клетки
действительно функционируют как поляроиды, то пчелы
должны одновременно получать все те впечатления, которые
человек получает одно за другим, вращая анализатор
перед своими глазами.
Следующим шагом было изготовление сильно
увеличенной модели омматидия пчелы, которая
воспроизводила бы его гипотетические свойства как поляроида.
Рис. 55. Модель омматидия пчелиного
глаза, построенного из 8
треугольников поляр оида.
Двойная стрелка в каждом треугольнике
показывает направление колебаний
проходящего света.
Из пластинки поляроида мы вырезали 8 равнобедренных
треугольников (рис. 54). Направление колебаний
проходящего света (показанное на рисунке двойной стрелкой)
в каждом треугольнике было параллельно его основанию.
Расположив эти треугольники радиально (рис. 55),
подобно расположению зрительных клеток в омматидии, мы
прикрепили их к стеклянной пластинке. Затем была
построена установка, в которой стекло со звездообразно
расположенными поляроидными треугольниками было
смонтировано таким образом, что оно могло легко
вращаться в любом направлении или наклоняться под
любым углом (рис. 56).
81
Если смотреть сквозь этот прибор на синее небо, от
яркость составляющих его треугольников оказывается
различной (рис. 57). Причину этих различий легко понять.
Свет того участка неба, на который направлен инструмент,
содержит колебания преимущественно в направлении,
отмеченном двойной стрелкой; это соответствует
направлению колебаний, которые могут проходить через
Рис. 56. Модель одшатидия,
смонтированная для производства наблюдений.
Круглый диск на столе представляет собой
лист поляроида, который можно положить на
обращенную к небосводу сторону
искусственного глаза и вращать в любом направлении.
треугольники, обозначенные буквой А, и поэтому они
кажутся светлыми. Треугольники, обозначенные буквой Б,
расположены перпендикулярно к треугольникам А\ они
препятствуют прохождению поляризованного луча света
и поэтому кажутся темными. Чем сильнее поляризован
падающий луч света, тем резче контраст между
треугольниками. Треугольники В ж Г имеют промежуточную
яркость, так как они пропускают лишь часть
поляризованного света.
Если смотреть сквозь этот «искусственный пчелиный
глаз» на различные участки неба, то картина все время
меняется, так как направление колебаний
поляризованного света в этих участках различно. Типичный пример
82
этого показан на рис. 57. На нем представлены фотография
неба, сделанные через этот прибор в 10 час. утра 25
сентября 1949 г. Небо фотографировалось под углом 45°
к горизонту в четырех направлениях: на запад, северо-
запад, север и северо-восток. Каждое направление дает
типичное соотношение яркостей, не наблюдаемое в эти
Рис. 57. Фотографии синего неба через
искусственный глаз.
Прибор направлен: 1—на запад; 2—на
северо-запад; 3—на север; 4—на северо-восток.
часы суток ни при каком другом направлении.
Естественно, что в другие часы суток это соотношение смещается,
так как оно зависит от положения солнца.
Если теперь допустить, что такая же картина возникает
и в глазу пчелы и каким-то образом ею воспринимается,
то, следовательно, насекомые способны использовать
любой кусочек синего неба в качестве солнечного компаса.
(Нет необходимости допускать, что пчела видит несколько
тысяч таких маленьких звездообразных фигур.
Индивидуальные возбуждения могут интегрироваться в единое
восприятие целого, подобно тому как картины,
возникающие в двух глазах человека, сливаются в единое
восприятие. Конечно, мы не можем знать, каковы
действительные зрительные ощущения пчелы.)
83
Пчела должна сохранять лишь определенное
положение тела, при котором картина яркостей в каждом омма-
тидии остается неизменной. В разбираемом нами
частном случае полета из улья к месту подкормки каждый
омматидий видит характерную картину в той небольшой
зоне небосвода, к которой он направлен. Во время танца
на горизонтальной плоскости пчелы видят кусочек синего
неба и ориентируют прямолинйную часть своего танца
таким образом, чтобы картина, воспринимаемая одной
зоной их поля зрения, совпадала с картиной,
запечатлевшейся в их памяти во время полетов к месту подкормки.
Это и позволяет пчелам правильно указать направление,
в котором следует искать корм.
Для проверки соответствия этой гипотезы
наблюдаемым фактам мы провели параллельные опыты с
танцующими пчелами и с искусственным глазом. Рассмотрим
сначала группу пчел, танцующих в горизонтально
расположенном наблюдательном улье и имеющих возможность
ясно видеть ограниченный участок неба. Покроем
стеклянное окно улья пластинкой поляроида, ориентированной
таким образом, чтобы рассеянный свет неба проходил
через поляроид без заметного изменения направления
колебаний. При этих обстоятельствах пчелы будут продолжать
правильно указывать направление к месту подкормки.
Это было обнаружено в 1948 г. и подтверждено в 14
дополнительных опытах в 1949 г. Если при проведении таких
опытов посмотреть через искусственный глаз на видимый
для пчел участок небосвода, можно увидеть типичную
картину соотношения яркостей. Поместив перед этим
прибором пластинку поляроида, ориентированную так же,
как и поляроид, покрывающий окно улья (т. е. так, что
направление колебаний поляризованного света неба не
меняется), мы не замечаем каких-либо изменений в
соотношении яркостей составляющих прибор треугольников.
Мы видим лишь более контрастную картину, так как
проходящий через поляроид свет поляризуется полностью,
тогда как исходный свет неба поляризован лишь частично.
В подобных условиях, согласно принятой нами гипотезе,
настоящий сложный глаз пчелы будет видеть одну и ту
же картину как с поляроидом, так и без него. Поэтому
направление танцев не изменится.
В опытах, проведенных 5 сентября 1949 г., мы давали
возможность пчелам, находившимся в наблюдательном
84
улье, видеть небольшой участок синего неба на зацад
от улья. Место подкормки лежало в том же направлении,
в 200 м от улья. Как и в первом опыте, если мы клали на
улей пластинку поляроида так, что направление колебаний
света неба не менялось, то пчелы сохраняли правильную
ориентировку и танцы их продолжали указывать на
запад.
Когда же мы повернули пластинку на 30°, вращая ее
против часовой стрелки, то направление прямолинейной
части танцев сместилось на 35° к юго-западу (среднее из
10 наблюдений). Мы направили искусственный глаз в
сторону западного сектора неба, видного танцующим
А Б В
Рис. 58. Фотографии синего неба,
сделанные через искусственный глаз между 9 час.
40 мин. и 10 час. утра 5 сентября 1949 г.
А—вид западного сектора неба через прибор;
Б—то же при помещении над инструментом
поляроида в таком же положении, как и
поляроид над наблюдательным ульем; В—картина,
сходная с Б, которую можно видеть сквозь
инструмент без поляроида, повернув его с запада
к северу на 34°.
ачелам, и увидели при этом картину яркостей,
воспроизведенную на рис. 58,-4. После этого мы поместили перед
прибором пластинку поляроида, ориентировав ее таким
же образом, как и при накрывании ею наблюдательного
улья. Картина яркостей изменилась (рис. 58,/>). Затем,
удалив поляроид, мы поворачивали искусственный глаз
в других направлениях и при повороте на 34° к северо-
западу получили ту же картину яркостей (рис. 58,В),
но эта картина не наблюдалась ни при каком другом
направлении.
Эти результаты позволяют понять, почему пчелы
поворачивали к югу на несколько больший угол, чем угол 30р,
на который был повернут поляроид. При полете от улья
к месту подкормки они видели прямо перед собой и в за-
Я5
падном направлении картину, соответствующую рис 58 А
Однако на 34° правее они видели картину,
соответствующую рис. 58, Б я В. Поэтому при повороте пластинки
Рис. 59. Изменение ориентации танцев пчел при помощи
поляроида.
поляроида перед окошком наблюдательного улья на 30°
картина, соответствующая рис. 58, Б, предстала пчелам
как единственная видимая зона поляризованного света
Все это схематически изображено на рис. 59. На этом
рисунке исходная картина, которую пчелы видели прямо
перед собой в западном направлении, обозначена цифрой 1
тогда как картина, видимая ими правее, на 34° к северо-
86
западу, обозначена цифрой 2. В наших опытах мы сначала
показывали картину 1 и пчелы указывали направление
прямо к ней. Покрыв же окно поляроидом, мы в том же
окне показали картину 2\ эта последняя была совершенно
сходна с картиной, которую пчелы, вылетая из улья к
месту подкормки, видели на 34° вправо от себя, и в
соответствии с этим они направляли свой танец на 35° левее
того положения, в котором их застала эта картина.
Таким путем ониг указывали направление, отклонявшееся
на 35° от линии полета к месту подкормки, но лишь на 1°
от того направления, которое мы предсказывали, исходя
из нашей гипотезы.
В 1948 г. в сходных опытах мы установили, что при
вращении пластинки поляроида танцы смещались в том же
цаправлении. Однако в то время мы не могли объяснить,
почему иногда направление танцев смещалось на такой же
угол, на какой мы повернули поляроид, иногда же угол
смещения был совершенно иной. Контрольные опыты
с искусственным глазом показали, что направление
танцев всегда смещается на такой же угол, на какой смещается
картина яркостей неба в конкретных условиях опыта.
В 48 опытах подобного рода разница между
наблюдаемым направлением и направлением, предсказанным по
этой теории, составляла в среднем 8° при максимуме 20°.
Это можно считать хорошим совпадением, принимая во
внимание источники ошибок в подобных опытах.
В 16 опытах из 83, выполненных летом 1949 г., пчелы
не указывали никакого определенного направления,
если поляроид был на месте, а танцы их носили
совершенно беспорядочный характер. Одновременные
наблюдения при помощи искусственного глаза показали, что
именно в этих случаях—и только в них—картину, получаемую
после помещения на прибор поляроида, нельзя было
обнаружить в момент опыта ни в каком участке неба,
наблюдая за ним через искусственный глаз, не покрытый
пластинкой поляроида.
В 4 из этих 83 опытов было обнаружено
примечательное явление—пчелы указывали два различных
направления, причем одно из них соответствовало зеркальному
отображению другого. Нередко пчелы смещали
направление на 180° по нескольку раз в течение одного танца.
Именно в этих случаях—и только в них—картина, видимая
в искусственный глаз после помещения на него поляроида,
87
возникала на небе в двух точках, лежащих друг против
друга (Фриш, 1950).
Подробное описание этих новых опытов увлекло бы нас
слишком далеко от основного содержания книги. Тем не
менее нам хотелось в этой краткой заключительной части
показать, что способность пчел ориентироваться по
поляризации рассеянного солнечного света нашла себе
подтверждение и что мы можем получить представление
о механизме осуществления этой ориентировки. Следует,
однако, отметить, что наши опыты не дают
неопровержимых доказательств полной идентичцости механизма
пчелиного глаза и нашей модели. Возможно, что в сложном
глазу анализ поляризованного света осуществляется
каким-либо иным путем. Однако недавние исследования
Аутрума (1950), применявшего совершенно другие методы,
свидетельствуют о том, что наши представления, по всей
вероятности, правильны и что каждый омматидий
функционирует как анализатор поляризованного света.
ЛИТЕРАТУРА
А у т р у м (Autrum H.), 1948. Об энергетических и временных
границах ощущений, Die Naturwissenschaften, 35, 361—369.
— 1949. Новые опыты по зрительному восприятию у летающих
насекомых, Experientia, 5, 271—277.
— иШтумпф (Stumpf H.), 1950. Глаз пчелы как анализатор
поляризованного света, Zeitschr. f. Naturforsch., 5b, 116—122.
Бауэр (Bauer H.), 1939. Исследования по физиологии вкуса у
водных жуков, Zeitschr. f. vergl. Physiol., 26, 107—120.
Б е й т л е р (Beutler R.), 1930. Биохимические исследования
нектара медоносных растений, Zeitschr. f. vergl. Physiol., 12, 72---176.
Б е л и н г (Beling G.), 1929. О памяти на время у пчел> Zeitschr. /.
vergl. Physiol., 9, 259—338.
Валь (Wahl О.), 1932. Новые исследования над памятью на время
у пчел, Zeitschr. f. vergl. Physiol., 16, 529—589.
— 1933. К познанию биологического значения памяти на время
у пчел, Zeitschr. f. vergl. Physiol., 18, 709—717.
— 1937. Изыскания наилучшего денатурирующего вещества для
сахара, раздаваемого для подкормки пчел, Zeitschr. f. vergl.
Physiol., 24, 116—142.
Вольф (Wolf L.), 1927. О способности пчел находить дорогу
домой, Zeitschr. /. vergl. Physiol., 6, 221—254.
Гаслингер (Haslinger F.), 1935. Восприятие вкуса у С alii-
phora erythrocephala и использование этой мухой Сахаров и
спиртов, являющихся продуктами их восстановления, Zeitschr. /.
vergl. Physiol., 22, 614—640.
Герц (Hertz M.), 1929 и 1931. Строение оптического поля у пчелы,
I, II и HI, Zeitschr. /. vergl. Physiol., 8r 693—748 (1929); И,
107—145 (1929); 14, 629—674 (1931).
— 1933. Оптическое восприятие пчелами фигур, отличающихся
по четкости и характеру, Biol. ZenlralbL, 53, 10—40.
— 1935. Исследования восприятия формы у медоносной пчелы,
Die Naturwissenschaften, 23, 618—624.
— 1937а. К вопросу о познании восприятия окраски и формы у
пчелы, Zeitschr. /. vergl. Physiol, 24, 413—421.
— 1937b. Опыты по цветовому зрению пчел, Die
Naturwissenschaften, 25, 492—493.
— 1937с. К вопросу о технике и методике опытов с пчелами с
применением цветной бумаги и стеклянных фильтров, Zeitschr. /.
vergl. Physiol., 25, 239—250.
— 1939. Новые опыты над цветовым зрением у пчел, Journ. of ex*
per. Biology, 16, 1—8.
89
Р с с с (Hess С), 1913. Зрение, см. Wintersteins Handbuch der ver-
gleichenden Physiologie, vol. 4, 555—840, Jena, Fischer.
11 л ь з е (Use D.), 1928. Цветовое зрение у дневных бабочек,
Zeitschr. f. vergl. Physiol., 8, 658—692.
Клебер (Kleber E.), 1935. Имеет ли память на время у пчел
какое-либо биологическое значение?, Zeitschr. f. vergl. Physiol.,
22, 221—262.
К н о л л (КЙ6Ч1 F.), 1921. Насекомые и цветы, Abhandl. der Zool.-
Botan. Gesellsch. in Wien, 12, 1—377.
К ю н (Kiihn A.), 1927. Цветовое зрение пчел, Zeitschr. /. vergl.
Physiol., 5, 762—800.
Л о т м a p (Lotmar R.), 1933. Новые исследования по цветовому
зрению пчел, в частности по восприятию «ультрафиолетового»
цвета, Zeitschr. f. vergl. Physiol., 19, 673—723.
M и н н и ч (Minnich D.), 1922а. Химическая чувствительность
лапок бабочки адмирала Pyrameis atalanta L., Journ. of exper.
Zool., 35, 57—81.
— 1922b. Количественные исследования по чувствительности лапок
бабочки адмирала Pyrameis atalanta L. к растворам сахарозы,
Journ. of exper. Zool., 36, 445—457.
— 1929. Химическая чувствительность ножек мясной мухи СаШ-
fora vomitoria L. к различным сахарам, Zeitschr. f. vergl. Physiol.,
И, 1-55.
— 1932. Контактные хеморецепторы медоносной пчелы, Apis melli-
fera L., Journ. of exper. Zool., 61, 375—393.
Оттинген-Шпильберг (Oettingen-Spielberg Т.), 1949.
Об основных свойствах пчел-разведчиц, Zeitschr. /. vergl.
Physiol., 31, 454—489.
Парк (Park О. W.), 1928. Исследования содержания сахара в
нектаре различных растений, Report of the Iowa State Apiarist, 1928,
80—89.
П о р ш (Porsch 0.), 1931. Яркокрасный цвет в окраске цветков,
опыляемых птицами, Biologia generalis, 7, 647—674.
Р и т т е р (Ritter E.), 1936. Исследования химического восприятия
жука водолюба Hydrous piceus, Zeitschr. f. vergl. Physiol., 23,
543—568.
С а н ч и (Santscbi F.), 1911. Наблюдения и критические замечания
по поводу механизма ориентации у муравьев, Revue Suisse de
Zool., 19, 303—338.
— 1923. Ориентировка по звездам у муравьев и соображения по
поводу их различных возможностей ориентировки, Memoires de
la Soc. Vaudoise de Sc. Nat., 1, 137—176.
T о р п (Thorpe W.), 1949. Ориентировка и способы установления
связей у медоносной пчелы и ее восприимчивость к
поляризованному свету, Nature, 164 (4157), И—14.
Фогель (Vogel В.), 1931. Зависимость между сладостью и
питательной ценностью Сахаров и спиртов, являющихся продуктами
их восстановления, на примере медоносной пчелы, Zeitschr. f.
vergl. Physiol., 14, 273—347.
Форель (Forel A.), 1910. Чувства у насекомых, Munchen, Rein-
hardt.
Фриш (Frisch К.), 1915. Восприятие цвета и формы у пчел, Zool.
Jahrb., Abt. f. allg. Zool. und Physiol., 35, 1—182.
90
Фриш (Frisch К.), 1919. Обоняние у пчел и его значение в
биологии цветка всех растений, Zool. Jahrb., Abt. f. alls. Zool. und
Physiol., 37, 1—238.
— 1921. О местонахождении органа восприятия запаха у
насекомых, Zool. Jahrb., Abt. f. alls. Zool. und Physiol., 38, 1—68.
— 1923a. Проблема восприятия цветов у животных, Die Naturwis-
senschaften, 11, 470—476.
— 1923b. О «языке» пчел, Zool. Jahrb., Abt. f. allg. Zool. und
Physiol., 40, 1—186.
— 1927. Об одном предложении кочевым пчеловодам, Bienenzucht
und Bienenforschung in Bayern (Wacholtz-Verlag, Neumunster),
S. 1—4.
— 1934. О восприятии вкуса пчелами, Zeitschr. f. vergl. Physiol..
21, 1—156.
— 1943. Теория цветка Х. Шпренгеля 150 лет назад и теперь,
Die Naturwissenschaften, 31, 223—229.
— 1946а. «Язык» пчел и его практическое применение в сельском
хозяйстве, Experientia, 2, 397—404.
— 1946b. Танцы пчел, Osterreichische zool. Zeitschr., 1, 1—48.
— 1947. Пчелы, приученные к определенному запаху, на службе
сельского хозяйства и пчеловодства, Wien, Springer Verlag.
— 1948а. Из жизни пчел, 4-е изд., Wien, Springer Verlag.
— 1948b. Решенные и нерешенные загадки «языка» пчел, Die
Naturwissenschaften, 35, 38—43.
— 1949а, Поляризация рассеянного света неба как ориентирующий
фактор в танцах пчел, Experientia, 5, 142—148.
— 1949b, Ты и жизнь, Berlin, Verlag Druckhaus Tempelhof.
— 1950. Солнце как компас в жизни пчел, Experientia, 6, 210—221.
Хонигман (Honigmann П.), 1921. Исследования по восприятию
света и адаптированию глаз птиц, Pfliigers Archiv f. die gesamte
Physiol., 189, 1—72.
Ш а л л е р (Schaller A.), 1926. Исследование восприятия и
психологии водных жуков и рыб, Zeitschr. f. vergl. Physiol., 4, 370—464.
Ш a p p e p (Scharrer E.), 1935. Восприимчивость свободных
плавниковых лучей морского петуха (Trigla) к химическим
раздражителям, Zeitschr. f. vergl. Physiol., 22, 145—154.
Шпренгель (Sprengel С.), 1793. Открытие тайны строения
и оплодотворения цветков, Berlin. (Переиздана в 1894 г. в
оствальдекой серии Klassiker der exakten Wissenschaften, Nr.
48—51, Leipzig, Engelmann.)
Штейнгоф (Steinhoff H.), 1948. Исследование способности*
пахучих веществ удерживаться на теле пчелы, Zeitschr. /.„
vergl. Physiol., 31, 38—57.
ОГЛАВЛЕНИЕ
Предисловие к русскому изданию . . . 3
Глава I. Цветовое зрение пчел , , 5
Глава II. Чувство вкуса и обоняние пчел 25
Глава III. «Язык» пчел • , * 45
Приложение 79
Литература . • . . , 89
к. Фриш
ПЧЕЛЫ, ИХ ЗРЕНИЕ, ОБОНЯНИЕ, ВКУС И ЯЗЫК
Редактор Я. О. ЮЗБАШЕВА. Обложка художника А, Я. Завьяловой.
Технический редактор Б. М. Ильин. Корректор Е. А. Яковлева.
Сдано в производство 22/XII 1954 г. Подписано к печати 17/11 1955 г.
Т-00788. Бумага 84хЮ81/з2=1»4, 6yiK. л., 4,7 печ. л. Уч.-издат. л. 4,7.
Изд. № 4/2430. Цена 3 р. 30 к. Зак. № 658
ИЗДАТЕЛЬСТВО ИНОСТРАННОЙ ЛИТЕРАТУРЫ
Москва, Ново-Алексеевская, 52
16-я типография Главполи-'рафпрома Министерства культуры СССР
Москва, Трехпрудный пер., д. 9.
ОПЕЧАТКИ
Страница
56
67
S2
Строка
7—8 св.
Подпись
к рис. 45
1 св.
Напечатано
появилось
(см. рис. 46)
от
Следует читать
появлялось
(см. рис. 44)
то
Чак. 658