/

































































































Текст
АКАДЕМИЯ НАУК УКРАИНСКОЙ ССР
ИНСТИТУТ ФИЗИОЛОГИИ ИМ. А. А. БОГОМОЛЬЦА
А. Л. ЧИЖЕВСКИЙ
ЭЛЕКТРИЧЕСКИЕ И МАГНИТНЫЕ СВОЙСТВА ЭРИТРОЦИТОВ
«НАУКОВА ДУМКА
КИЕВ-1973
57.04
Ч—59
УДК 612.111; 612.117 7
Рассматриваются электролиты плазмы и электрический заряд эритроцита. Приведены приближенные модели электростатического и магнитного взаимодействия эритроцитов в кровотоке, а также приближенная модель гироскопической устойчивости в кровотоке вращающегося эритроцита.
Книга рассчитана на медиков и биологов, интересующихся применением физико-математических методов к физиологии кровообращения.
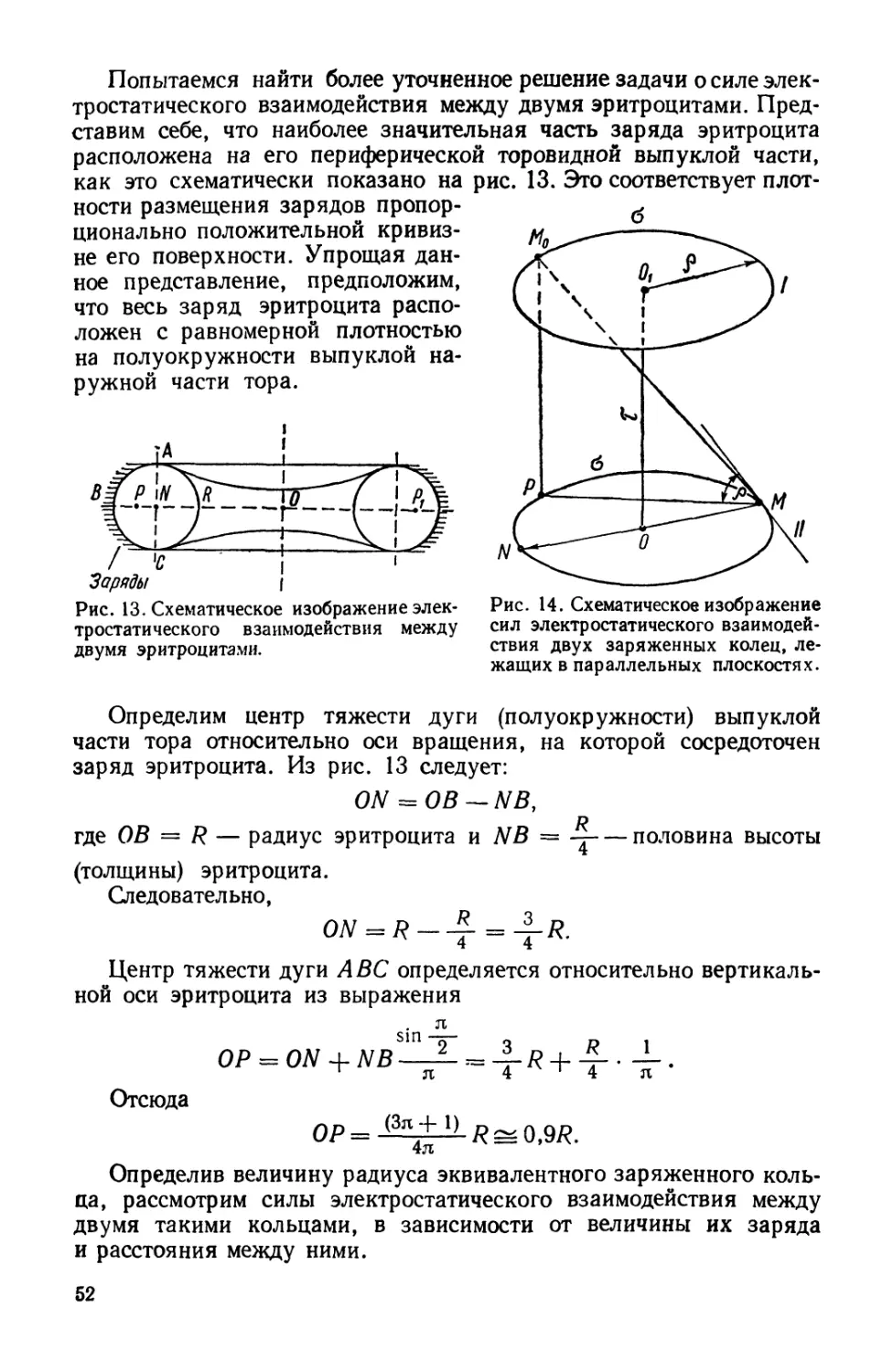
Ответственные редакторы:
доктор медицинских наук М. И Гуревич, доктор физико-математических наук В. Т. Черепин
Редакция физико-математической литературы
Зав. редакцией И. В. Евсеенко-Мисюренко
0236-1М
М22Ц04)—73
С Издательство «Наукова думка>, 1973 г.
ПРЕДИСЛОВИЕ
Профессор А. Л. Чижевский (1897—1964) первым в мире в конце 40-х годов обратил внимание на структуру крови при ее движении по кровеносным сосудам. Его монография «Структурный анализ движущейся крови», изданная в 1959 г. издательством Академии наук СССР, получила известность в научных кругах как у нас в стране, так и за рубежом. Этой работой А. Л. Чижевский впервые ставил на обсуждение чрезвычайно важную проблему, основанную на том, что патофизиология и медицина часто вплотную соприкасаются с явлениями, лежащими в области нарушений пространственной структуры крови. «По-видимому, нет ни одного заболевания, хотя бы самого легкого, которое не нашло бы немедленного отражения в пространственной структуре крови, в этом «зеркале организма». И, наоборот, уклонения в распределении частиц крови от нормы отражаются на состоянии всего организма»,— писал А. Л. Чижевский. Ученый открыл пространственно-структурные ансамбли эритроцитов в движущейся по сосудам крови, исследовал статико-геометрические модели этих систем, а также их кинематику и динамику, наметил пути и области дальнейших теоретических и экспериментальных поисков, поставил перед физиологами и биофизиками ряд новых задач. Открытие А. Л. Чижевским в кажущемся хаосе гуморальных явлений стройной и взаимозависимой во всех частях и функциях динамической микросистемы, прочно связанной с внешней средой, с макросистемами организма, внесло определенную ломку в старые воззрения на кровяное русло.
В предлагаемой читателю монографии А. Л. Чижевский развивает теорию динамического строения красной крови: исследует электрические и магнитные явления в крови, рассматривая ее как электрическую систему, балансирующую около точки равновесия. К силам, ответственным за устойчивость радиально-кольцевых структур, он относит гидродинамические, электрические и магнитные силы. Для сугубо предварительного выяснения электростатического взаимодействия между эритроцитами А. Л. Чижевский пользуется^аконрт^ Калона.
В монографии автор неоднократно делает оговорки насчет того, что он не претендует на исчерпывающее рассмотрение вопроса, а лишь ставит его на обсуждение специалистов разных отраслей науки. Однако при знакомстве с работой нетрудно убедиться, сколь смела и плодотворна эта постановка вопроса. Подходя к работе с критериями современного естествознания, нетрудно убедиться, насколько ученый опередил свое время, дав фундаментальные предпосылки для изучения движущейся крови как целостной системы взаимосвязанных и органически соподчиненных процессов, регламентированных в пространстве и во времени физиологическими, биохимическими и биофизическими факторами.
3
Мы старались по возможности сохранить оригинальный стиль изложения и язык монографии, внося лишь самые необходимые коррективы. Безусловно, наши усилия в этом направлении не могли заменить непосредственного участия автора в окончательном редактировании рукописи. Связанные с этим обстоятельством неизбежные погрешности, как нам кажется, не смогут умалить значения публикации этой работы для дальнейшего углубления представлений о природе, характере и взаимодействии между форменными элементами в потоке крови.
С этой точки зрения издание монографии профессора А. Л. Чижевского явится важным вкладом в развитие науки о крови.
М. И. Гуревич
В. Т, Черепиц
ВВЕДЕНИЕ
Современная физиология занимается изучением явлений динамического электричества — токов действия и токов покоя. Электростатические явления в организме долгое время не привлекали внимания исследователей. Но за последние десятилетия наука обогатилась рядом новых воззрений на физиологическую роль электростатических зарядов, несомых клетками, внутриклеточными органоидами и белковыми частицами протоплазмы. Было также выяснено, что протоплазма имеет определенную стерическую структуру: дифференциальные показатели двулучепреломления были найдены для многих ее видов. Это говорит о наличии ориентированных в пространстве белковых макромолекул, а ориентировка макромолекул — об электростатических полях между ними. Можно количественно описывать электрические свойства протоплазмы, исходя из соотношения между ее сопротивлением, реактивным сопротивлением, фазовыми углами и импедансами. В связи с развитием этих воззрений возникли новые отрасли электрофизиологии: биоэлектростатика, электрогистология, учение о воздушно-гуморальном и гуморально-тканевом электрообмене; была открыта электростатическая функция легких.
Микрометоды экспериментального изучения числа и полярности электрических зарядов в органических структурах и субстратах получили широкое распространение. Созданы микроманипуляторы, позволяющие вводить электроды внутрь микроскопических структур, чувствительнейшие электрометры и гальванометры для регистрации ничтожных количеств электричества, счетчики Гейгера, усилители и катодные осциллографы для записи быстрых электрических колебаний, стал широко применяться рентгенографический анализ. В лабораторной практике используются методы изучения окислительно-восстановительных процессов, методы, связанные с изучением концентрации водородных ионов, методы микроэлектрофореза, метод меченых атомов. К изучению строения ультрамикроструктур, в том числе структуры эритроцитов, были применены наиболее тонкие методы физико-химического исследования: с использованием явлений интерференции и дифракции света, фотоэлектрические методы, ультрацентрифугирование, микрохирургические операции, электронная микроскопия. Было уделено внимание
5
как динамическим, так и статическим свойствам микроструктур в витальном и постмортальном состояниях. Применяются физические и химические стимуляторы, раздражители, индикаторы. Широко используются физико-химические анализы.
Благодаря успехам клеточной электрофизиологии можно с достаточной достоверностью утверждать, что большинство клеток несут на своей поверхности и на поверхности своих органоидов электрический заряд. С помощью электрофореза можно измерить потенциал частиц и клеток любого размера. Это достигается наложением постоянного электрического поля на взвесь или суспензии клеток, а при микроэлектрофорезе — на отдельные клетки или даже на внутренние части клеток. Клетки или их дифференциаты при наложении электрического поля начинают двигаться в нем к аноду или к катоду в зависимости от знака присущего им заряда, по скорости миграции частиц на единицу напряжения вычисляется величина их электрического потенциала, а по направлению движения судят о полярности последнего.
Поскольку поверхность клеток заряжена отрицательно по отношению к вмещающей их среде, при электрофорезе клетки движутся к аноду. Морфологические элементы крови обладают тем же свойством. Органоиды клетки могут обладать различной полярностью. Так, ядро яйца, ядрышко и хромосомы тритона при электрофорезе движутся к аноду. Ядро клетки слюнной железы личинки Sciara движется к катоду, а масса хромосом — к аноду. Цитоплазматические гранулы яйца тритона, кристаллы амебы и хлоропласты Е1о-dea мигрируют к катоду.
Исследователям удалось приближенно выяснить количество и знак электрических зарядов в некоторых тканевых и органных клетках, например в клетках мозга, печени, почках, селезенке и т. д., и тем самым приблизиться к пониманию роли, которую играют электрические заряды при выполнении этими органами тех или иных функций как в норме, так и при патологии. В свете этих работ каждая живая клетка может быть грубо схематично рассматриваема как специализированная электростатическая система, каждый орган как совокупность таких систем, а весь организм как комбинат этих совокупностей, взаимосвязанных и непрерывно взаимодействующих одна с другой.
Данная схема становится понятной, если вспомнить о том, что химические соединения возникают вследствие электростатического притяжения атомов, несущих заряды противоположной полярности, т. е. начало химических реакций в организме обусловлено электрическими зарядами или, иначе говоря, ионизацией реагирующих веществ. Отрицательный заряд энергично проявляет себя при всякой химической реакции в клетке. Периферические орбиты, электронные слои атомов участвуют во всех каталитических реакциях в клетке. Каталитические реакции возникают при условии глубокого изменения электронной структуры реагирующих между собой атомов. Механизмы энергии активации тканевых структур
6
состоят в сдвигах электронной ковалентной связи, в дипольной индукции, в деформации слоев периферических электронов и т. д. Роль периферических электронов, число которых обусловливает отрицательный или положительный заряд данной молекулы, становится ясной в свете положений физической и электростатической химии: прежде чем атомы или молекулы реагирующих между собой веществ вступят в химическую реакцию, встречаются периферические электроны сблизившихся атомов, в результате чего и происходит та или иная химическая реакция.
Электрические заряды, расположенные на поверхности или вокруг морфологических элементов крови, были использованы для объяснения ряда существенных явлений, развивающихся в крови, в основном для объяснения феномена стабилизации эритроцитов. Впервые Джевонс (1870) указал на то, что отталкивание одноименно заряженных взвешенных в жидкости частиц является силой, препятствующей сближению этих частиц, их слипанию и выпадению из раствора. С тех пор эта идея получила широкое распространение и легла в основу ряда представлений коллоидной химии. Она была экспериментально проверена и теоретически обоснована в трудах многочисленных исследователей. Идеи о роли электрических зарядов в коллоидных системах проникли в биологическую физико-химию. Живой организм стали рассматривать как сложную коллоидную систему. В настоящее время принято считать, что наличие электрических зарядов, их количество и полярность обусловливают устойчивость дисперсной системы белковых тел в живой клетке.
Роль электрического заряда эритроцитов была изучена, главным образом, с клинической точки зрения. Скорость потери электрических зарядов эритроцитами при оседании красной крови была поставлена в связь с нормальным или патологическим состоянием организма, что позволило вскрыть ряд клинически важных закономерностей.
Фарэусв 1917 г. одним из первых сделал попытку определить заряд эритроцитов в оседающей с различной быстротой крови. Он нашел, что в быстро оседающих пробах крови заряд эритроцитов меньше, чем заряд в пробах крови, оседающих медленно. Это наблюдение позволило ему сделать предположение о том, что скорость оседания эритроцитов зависит от их склеивания — агглютинации, а агглютинация — от величины электрического заряда эритроцитов. В том же году Герцфельд и Клингер предложили теорию, согласно которой такие грубо дисперсные белковые частицы крови, как глобулины и фибриноген, являющиеся продуктами клеточного распада, в том числе распада лейкоцитов, легко адсорбируют частицы более высокой степени дисперсности, какими являются альбумины, обнаруживающие при электрофорезе отрицательную полярность и достаточно высокий электрокинетический потенциал.
В следующем году Старлингер сделал попытку объяснить быструю реакцию оседания эритроцитов усиленным образованием в крови грубо дисперсных частиц клеточного распада, которые для того,
7
чтобы удержаться в крови во взвешенном состоянии, адсорбируют частицы более высокой дисперсности или даже снимают их с поверхности эритроцитов. Вследствие этого эритроциты теряют свою стабильность, начинают склеиваться один с другим и быстро осаждаются.
Соломон также пришел к убеждению, что основным фактором, обусловливающим изменение скорости оседания эритроцитов, является взаимоотношение между глобулинами и альбуминами. Галль, Гейзер, Кауфман, Пилибрам, Клейн и другие признают, что лабильность плазмы и белковая формула имеют определенное значение для хода процесса оседания эритроцитов.
Особое значение в этом вопросе может иметь тот факт, что некоторые слабо отрицательные белковые частицы способны иногда к перезарядке, т. е. к изменению своей электрической полярности с минуса на плюс. Копачевский показал, что при некоторых заболеваниях электрический заряд глобулинов сыворотки не только резко уменьшается, но, более того, перейдя через нуль, становится положительным. Волиш допускает, что фибриноген, выпадая на поверхности эритроцитов, влияет на скорость их оседания вследствие разницы в величине электрического заряда и полярности у эритроцитов и частиц фибриногена.
В 1922 г. Гёбер и Монд, работая над изучением изоэлектрической точки различных белковых тел, пришли к выводу, что присутствие в плазме значительного числа глобулинов влечет падение электрического заряда эритроцитов. По мнению упомянутых авторов, это явление аналогично тому, которое происходит после прибавления к коллоиду так называемого коагулятора или разряжающего электролита. Коагулятор, несущий на своих частицах электрический заряд иной полярности, чем заряд частиц коллоида, способствует снижению электрического заряда частиц, силы взаимного отталкивания между частицами падают, коллоидные частицы постепенно агрегируются, склеиваются и седиментируют под влиянием силы тяжести. Наоборот, увеличение количества альбуминов, несущих достаточно высокий электроотрицательный потенциал, содействует сохранению заряда на поверхности эритроцитов.
В 1925—1926 гг. появился ряд работ по изучению электрических свойств эритроцитов. Пондер показал, что если разность потенциалов между плазмой и эритроцитами становится ниже некоторого уровня, наступает быстрая агглютинация эритроцитов. На исключительное значение электроотрицательного заряда эритроцитов при оседании красной крови указывают работы Колло. Электрический заряд эритроцитов ставится этим автором во главу угла при изучении реакции оседания.
Поппер и Краиндлер, Папаниколау и Вейлер и другие рассматривают систему эритроциты — плазма как коллоидную. Первые два автора сравнивают кровь с коллоидным раствором, в котором седиментация представляется аналогичной седиментации или флоккуляции коллоидных суспензий.
8
Доньон (1931) считает, что кровь со взвешенными в ней форменными элементами и белковыми телами должна быть уподоблена коллоиду: форменные тела и белки крови несут электрические заряды и благодаря взаимному отталкиванию находятся в состоянии электростатического равновесия. Достаточно это равновесие нарушить, как вся электрическая система крови разрушается и наступает осаждение ее морфологических элементов и белковых мицелл. Это и случается тогда, когда в плазму поступает более значительное, чем обычно, количество грубо дисперсных частиц глобулинов, электрический заряд которых гораздо меньше, чем заряд альбуминов. Аналогичные высказывания и наблюдения были сделаны в отношении электрического заряда лейкоцитов и тромбоцитов.
Таким образом, морфологические и корпускулярные элементы крови представляют собой электрические системы. У поверхности этих тел сосредоточен двойной слой электрических зарядов. Внутри морфологических элементов крови, на внутренней поверхности мембраны и на органоидах также распределены электрические заряды. Система морфологических и корпускулярных элементов крови окутана силовыми линиями электрического поля, а между отдельными элементами действуют силы электростатического распора, предотвращающие полное их сближение, соприкосновение и слипание.
Сложнейшая электрическая система крови находится в непрерывном и многообразном движении по кровеносным сосудам разного диаметра, а следовательно, и разного режима движения. Расстояние между клеточными и коллоидными элементами крови постоянно в ограниченной мере изменяется, пружины электростатического распора находятся в непрерывной работе. Электростатическая система крови все время балансирует около точки своего равновесия. В кровотоке морфологические компоненты крови стремятся сохранить свои основные физические векторы при всех возможных динамических перемещениях данных компонентов в пространстве плазмы. Электростатический вектор все время удерживает частицы крови на определенном расстоянии одну от другой, обеспечивая их относительную эквидистантность и определенную ориентировку в пространстве. Как бы гидродинамические силы при известных условиях турбулентного режима ни нарушали эту систему, электростатические поля, т. е. силы электростатического взаимодействия и распора в каждый данный момент, имеют тенденцию выравнивать нарушенное положение элементов системы и частично или полностью восстанавливать их пространственную ориентацию, в известной степени специализированную для данного калибра кровеносного сосуда. Так как при прохождении одного эритроцита или их ассоциации мимо другого эритроцита или их ассоциации, нахо-цящихся в соседнем слое, происходит электростатическое взаимодействие в форме отталкивания, то, естественно, в каждый данный момент эритроциты имеют тенденцию, несмотря на их движение, вращение и смещение эритроцитных слоев друг относительно друга. сохранять известное расстояние один относительно другого и
9
быть обращенными один к другому соответствующими сторонами или находиться под соответствующим углом согласно условиям гидродинамического и электростатического равновесия.
Электростатические заряды, несомые поверхностью морфологических тел, при ротационном или вихревом движении последних должны возбуждать конвекционные токи. Трансляционные движения этих тел и их систем, обладающих электрическими полями, приводят к возникновению электродинамических явлений. Система движущихся эритроцитов может быть представлена как система электродинамически взаимодействующих частиц *. Радиально-кольцевая модель эритроцитов, построенная нами для ламинарного и отчасти турбулентного режимов движения крови, позволяет наметить качественную картину электродинамических взаимодействий, которая в общих чертах может быть представлена так:
1) взаимодействие магнитных полей двух токов (прямолинейного и кольцевого);
2) поступательное движение системы в целом в совокупном поле круговых колец тока — соленоидов, обусловливающее возникновение электрических токов от периферии к центру;
3) взаимодействие прямолинейного тока и соленоидального поля, приводящее к пондеромоторному движению от центра к периферии;
4) относительное движение двух соседних кольцевых цилиндров, т. е. наложение двух пар электрических и магнитных полей разной частоты.
Из перечисленных факторов следует путем анализа выделить основные, определяющие электромагнитные процессы в кровотоке и, возможно, связанные с этим электромагнитные излучения в окружающее пространство.
Не исключена возможность существования и того явления, что после бифуркации сосудов, например после бифуркации брюшной аорты на две подвздошных артерии — правую и левую, текущая с большой скоростью кровь начинает совершать винтообразное движение, причем оба потока крови могут вращаться в разных направлениях. А так как подвздошные артерии переходят в бедренные без больших изгибов, то направление винтообразного движения крови может распространяться на обе нижние конечности. Опыты на моделях показали, что возможность такого рода винтообразного хода жидкости при изгибе сосудов не исключена. Гиндеркс доказал, что для трубки, округленно изгибающейся под углом 90°, это явление объясняется следующим образом: центральные части жидкости, движущейся быстрее чем периферические при изгибе трубы, под действием центробежной силы, оттесняются к стенкам, где они вытесняют в свою очередь части жидкости, движущиеся медленнее. Это приводит к возникновению результирующего винтового течения.
* А. Л. Чижевский. Структурный анализ движущейся крови. Изд-во АН СССР, М., 1959.
10
Если такого рода явление имеет место в крови, то можно говорить об электрической и магнитной асимметрии правой и левой нижних конечностей человеческого организма.
Устойчивость и, следовательно, физиологические функции радиальнокольцевых систем эритроцитов зависят не только от гемодинамических факторов, но и от количества электричества, несомого каждым отдельным эритроцитом, ибо устойчивость эритроцита в кровотоке обусловлена электрическим распором между эритроцитами, а последний является функцией числа зарядов, несомых эритроцитом, которые в свою очередь обусловливают величину конвекционных электрических токов, а эти токи вызывают появление магнитных полей между вращающимися эритроцитами.
Следовательно, для организма не безразлична величина электрического заряда эритроцитов, более того — эта величина является одним из важнейших фа кторов транспортно-обменной работы всего кровяного русла. При достаточном числе зарядов, несомых эритроцитами, достаточен и электрораспор между ними, т. е. величина пути, по которому плазма осуществляет функцию подвоза к поверхности эритроцита питательных материалов, которые эритроцит относит далее, к месту их отдачи. Величина электрораспора обусловливает процесс и обратного направления — увоз отработанных материалов. Это способствует нормальному обмену веществ. При недостатке зарядов на эритроците распор также недостаточен, пути прохождения плазмы между эритроцитами ограничены, в связи с чем транспортно-обменная функция падает, обмен веществ становится недостаточным, а отсюда следует возникновение ряда патологических явлений. Электрический заряд эритроцитов играет, по-видимому, очень существенную роль в предотвращении образования эритроцитных тромбов. Таким образом, в области электростатики и электродинамики крови появляется широкое поле интереснейших и многообещающих исследований.
Электрические свойства эритроцитов обычно принято выражать не в виде числа элементарных электрических зарядов, несомых поверхностью эритроцита, а в форме дзета-потенциала, т. е. электрического потенциала на границе двух фаз — поверхности эритроцита и плазмы. Сумма электрических зарядов на поверхности эритроцитов и дзета-потенциал не идентичны. Тем не менее в литературе иногда встречается отождествление этих понятий. Некоторые авторы, говоря о дзета-потенциале, употребляют термин «электрический заряд». Это, конечно, неверно, ибо значение электрического заряда выражается в элементарных электростатических единицах, а дзета-потенциала — в милливольтах. Можно считать, что дзета-потенциал дает лишь некоторое весьма ограниченное и относительное представление об электрическом заряде эритроцитов. Он вычисляется из величины их электрофоретической подвижности. Истинные абсолютные значения потенциала, несмотря на все поиски, вот уже десятки лет ускользают от исследователей, истинных значений дзета-потенциала реальных поверхностей неправильной формы
11
мы также пока вычислять не умеем. Тем не менее мы при прочих равных условиях можем следить за изменениями значений дзета-потенциала. Плотность электрического заряда на поверхности эритроцитов может быть весьма приближенно получена путем постепенного добавления во взвеси эритроцитов лантана — электроположительного элемента — до тех пор, пока эритроциты станут электрически нейтральными. Тогда взвесь эритроцитов при электрофорезе останавливается, определяется изоэлектрическая точка эритроцитов и по количеству прибавленного для нейтрализации лантана вычисляется степень отрицательного заряда эритроцитов. Однако такой метод определения величины заряда эритроцитов нельзя считать удовлетворительным.
Точное определение величины электрического заряда взвешенных частиц даже в коллоидной или физической химии считается до сих пор задачей не вполне разрешенной, а полученные рядом авторов результаты не вполне достоверными. Различные методы измерения заряда дают иногда большие расхождения, ибо явления электрофореза осложняются электрическими силами между дисперсной частицей и дисперсионной средой и другими факторами. Из многочисленных и разнообразных наблюдений в этом направлении авторы обычно делают лишь один основной вывод, свидетельствующий о том, что частица несет тем больше элементарных зарядов, чем больше ее диаметр.
В таком крайне неудовлетворительном состоянии находится вопрос об истинной величине электрического заряда взвешенных частиц. И тем не менее мы должны приблизиться к точному определению электрического заряда эритроцитов, ибо этого требует открытие нами радиально-кольцевой системы красной крови. Точное представление об электрических параметрах эритроцитов будет содействовать прогрессу новой области науки — физиологии кровотока. Однако для нахождения интересующих нас электрических параметров придется не только преодолеть некоторые теоретические трудности, но и провести впоследствии большую экспериментальную работу по разработке новых методик, которые в общей форме уже могут быть намечены в настоящее время.
Глава I
ЭЛЕКТРОЛИТЫ ПЛАЗМЫ
И ЭЛЕКТРИЧЕСКИЙ ЗАРЯД ЭРИТРОЦИТА
§ 1. Водородные и гидроксильные ионы крови
Изучение электролитного состава плазмы имеет кардинальное значение для понимания основных физических процессов, протекающих на поверхности и вокруг морфологических и корпускулярных элементов крови и обусловливающих в значительной мере те явления, которые мы уже отчасти рассмотрели во введении и к которым вынуждены еще вернуться.
Согласно современным воззрениям, силы, способствующие стабилизации, т. е. препятствующие коагуляции коллоидных частиц и взвесей, а значит, и частиц крови коллоидной и микроскопической дисперсности, есть в основном силы электрические. Униполярные электрические заряды, находящиеся на поверхности или сосредоточенные около частиц, отталкивают последние одну от другой и тем самым препятствуют стягиванию частиц друг к другу, или ат-траксии, чем предупреждают их столкновение, или контакцию, объединение, или агрегацию, слипание, или коагуляцию, и, наконец, осаждение, или седиментацию.
Механизм процесса электрозарядки поверхности частиц и образования двойного слоя вокруг частиц в растворах электролитов еще не вполне выяснен. Несомненно, что появление электрических зарядов на поверхности и вокруг частиц следует в основном приписать ионам электролитов, находящимся в растворе. Известно, что факторы, уменьшающие величину электрического заряда частиц, например ионы противоположного знака или коллоиды с зарядом противоположной полярности, способствуют коагуляции частиц.
Электролитный состав крови несет функцию стабилизатора корпускулярных компонентов крови. Электролиты крови являются тем основным источником, из которого эритроциты и вообще морфологические элементы, а также различные дифференциаты крови черпают электрические заряды (ионы) для образования поверхностного заряда или двойного электрического, либо диффузного слоя. Для устойчивости дисперсных систем и взвесей необходимо, помимо дисперсной фазы и дисперсионной среды, наличие стабилизатора, который создает систему коллоида, объединяет дисперсную фазу с дисперсионной средой и обеспечивает системе стабильность и эквидистантность пространственного распределения дисперсных
13-
частиц, а также динамическую пластичность всей системы при ее движении и возможных деформациях в целом (кровоток).
Рассмотрим вопрос о количестве Н+ и ОН~ ионов в 1 мм3 крови. Если pH = 0, то:
[Н+] = 10“° = 1 г/л = 10-6 г/мм3.
Для определения количества ионов Н+ в 1 мм3, полученное количество граммов Н следует разделить на вес 1 иона Н+, т. е. на 1,669 • 10-24 г. Получим
Ю“6 п =---------- = 6,061 • 1017.
1,669 • 10“24
При возрастании pH на одно деление шкалы количество ионов в 1 мм3 убывает в 10 раз. Следовательно, по одному значению для количества Н+ ионов, соответствующему значению pH = 6, можно построить таблицу количества Н+ ионов для всей шкалы pH (табл. 1).
Таблица 1
Таблица 2
Количество ионов и соответствующее ему количество ионов ОН" при разных значениях pH в 1 лслс3 крови Значения числа положительных и отрицательных зарядов для любого значения pH и рОН Г* TnUU/V'TLin ПЛ П 1 onuuuilLI
pH Количество н+ ионов в 1 мм9, деленное на число Авогад-РО рОН Количество ОН~ ионов в 1 мм9, деленное на число Аво-гадро шкалы Vfl w^riiin
pH рОН Количество ионов в 1 мм9 крови • 10“11
0 1017 14 103 0,1 1,0 6,061
1 10™ 13 104 0,1 0,9 4,759
2 10™ 12 10е 0,2 0,8 3,781
3 1014 11 10е 0,3 0,7 3,003
4 10™ 10 Ю7 0,4 0,6 2,385
5 1012 9 108 0,5 0,5 1,895
6 10й 8 109 0,6 0,4 1,504
7 10™ 7 10™ 0,7 0,3 1,195
8 109 6 1011 0,8 0,2 0,949
9 108 5 10™ 0,9 0,1 0,754
10 Ю7 4 10™ 1,0 0,0 0,606
11 10е 3 1014
12 10б 2 10™ । 1
13 104 1 10™
14 Ю3 0 1017 Проведем также расче-
ты и составим таблицу для
нахождения десятых долей одного деления логарифмической шкалы pH и рОН (табл. 2). Принцип построения этой таблицы аналогичен принципу построения табл. 1. Последняя таблица носит универсальный характер. В ней данные приведены с точностью до 0,1 одного деления шкалы pH или шкалы рОН.
Если pH + рОН = 14, то и (п + 0,4) + (13 — п + 0,6) - 14, где п — целое число по шкале pH и рОН.
Пользуясь этими таблицами, можно быстро получить точное значение Н+ и ОН“ ионов в крови.
14
Многочисленными измерениями установлено, что концентрация водородных ионов в крови величина весьма стабильная. Она составляет 0,3—0,7 • 10”7, или в величинах pH: 7,3—7,4 (в среднем 7,35). В венозной крови pH на несколько сотых ниже, чем в артериальной, за счет повышенного содержания СО2 на 5—10%.
Значения по шкале pH указывают на преобладание щелочности^ т. е. числа ОН~ ионов над Н+ ионами, а именно: на 0,35 одной единицы шкалы.
Поскольку pH = 7,35, то
[Н+] = 10”7’35 —= 10”8 • 10+°’65 г/л - 4,467 • 10”8 г/л = л
= 4,467 . 10”14 г/мм3.
Но так как масса одного иона Н+, т. е. масса одного протонаУ равна 1,669 • 10~24 г, то, чтобы узнать количество ионов Н+ в 1 мм3, следует общую массу ионов в 1 мм3 разделить на массу одного, иона Н+. Получим
п = 4Л 67 = 2 67б . юю.
1,669 • 10~24
Поскольку 1Н+] ЮН”] = const и для нейтральной реакции [Н+] ЮН”] = (6,061 • 1010)2 = 36,736 • 1020, то для количества ОН” в 1 мм3 крови получим
_ 36,736 • 1020 _ «Q 1Q1O
2,676 • 1010 “ 1и •
Таким образом, в крови число ОН” ионов превышает число Н+ 13,733 - 1016
И0Н0В В 2,676~ ~10™ = 5,13 ра3а*
Величина pH = 7,35 носит название активной реакции крови и играет огромную физиологическую роль. Всякий сдвиг pH в сторону щелочности крови (алкалоз) или в сторону кислотности (ацидоз) вызывает ряд серьезных осложнений в организме. Поэтому организм обладает буферной системой, которая в случае внезапного поступления в кровь излишнего количества кислых продуктов,, например после тяжелой мышечной работы, нейтрализует их вредное действие, приводя в состояние активности «щелочной резерв». В крови имеется целый ряд так называемых буферов, которые обеспечивают ей постоянство слабо щелочной реакции, единственно пригодной для всех физиологических реакций организма, а потому организм обладает большей емкостью буферного действия на кислые, чем на щелочные растворы. Главными буферами являются карбонаты, бикарбонаты, фосфаты и отчасти протеины.
Отклонения от нормального состояния ткани сопровождаются изменением pH. При воспалениях наблюдается гипертония: в
15
центральном гнойном очаге pH находится в пределах 6,1—5,4; в поясе воспалительного инфильтрата pH возрастает до 6,3—6,1; в зоне периферического отека pH приближается к норме 7,2—6,9; далее следует переход к нормальному состоянию pH = 7,3—7,4. Следовательно, при воспалительных явлениях число Н+ ионов в очаге воспаления и в смежных точках увеличивается.
§ 2. Изоэлектрическая точка и провизорный электрический заряд эритроцитов и некоторых белковых компонентов крови
Величина электрического заряда различных компонентов крови различна. При нормальном состоянии организма величина заряда находится в зависимости от величины разности А между активной реакцией крови pH и точкой нейтрализации электрического заряда или изоэлектрической точкой pH, данного компонента крови, т. е. А = pH—pH,.
Чем значительнее эта разность А — «сдвиг», «зона» или «1 ’e’cart» (Влес), тем больше электрический заряд данного компонента крови. Это правило имеет исключение лишь в том случае, когда pH = pH,.
Во всех других случаях получим либо pH > pH,, и тогда заряд компонента крови имеет отрицательную полярность, либо pH < < pH,, тогда заряд компонента крови имеет положительную полярность.
Разные компоненты крови характеризуются различными изоэлектрическими точками: pH, альбуминов равна 4,6 шкалы водородного показателя; глобулинов — 5,4; эритроцитов — от 5,6 до 6,5, т. е. в среднем 6,0; гемоглобина — 6,7; фибриногена — около 8. Изоэлектрические точки эритроцитов млекопитающих, согласно измерениям ряда авторов, составляют 3,5—4,7. Наиболее часто приводится значение изоэлектрической точки эритроцитов около 4,7. Влес считает изоэлектрическую точку эритроцитов в среднем равной 6,0. В сыворотке человеческой крови обнаруживается, по данным Влеса и Кулона, наличие трех изоэлектрических точек: а, pH, = 5,5; (J, pH, = 10,0—12,0 (в среднем 11,0); у, pH, = 5,0.
Из них точки р и у изменчивы; а постоянна в крови здоровых людей и сдвигается только при острых инфекциях. Можно отметить, что величина изоэлектрической точки целого эритроцита есть величина среднего значения всех изоэлектрических точек различных структурных элементов эритроцита.
В то время как активная реакция крови благодаря ее совершенным буферным механизмам чрезвычайно устойчива и постоянна по своей величине, изоэлектрические точки отдельных компонентов крови лабильны и могут изменяться в заметных пределах под воздействием содержащихся в крови электролитов — катионов К, Na, Са, Mg и неэлектролитов — органических соединений, а также под воздействием ряда других эндогенных и экзогенных причин.
Лб
Основной чертой, характеризующей величину электрического заряда корпускулярных компонентов крови, является, таким образом, смещение их изоэлектрических точек в ту или иную сторону по отношению к точке активной реакции крови, т. е. величина «сдвига», или «зоны», между pH, и pH.
Схема, представленная на рис. 1, дает представление об условиях увеличения или уменьшения электрического заряда компонентов крови со смещениями pH ч pH,. Из этой схемы следует, что
увеличение электрического заряда данного компонента крови может происходить при таких обстоятельствах:
а) сдвиге pH, в кислую сторону и при постоянстве активной реакции pH крови;
б) при постоянстве pH, и сдвиге pH в щелочную сторону;
в) при наличии обоих указанных сдвигов одновременно.
Из рис. 1 следует также, что увеличение расстояния по шкале или расширение зоны между pH и pH, означает увеличение электрического заряда того или иного компонента крови.
При уменьшении электрического заряда компонента крови реакция сдвига протекает таким образом:
17
aj pH, сдвигается в щелочную сторону, pH постоянна;
6J pH сдвигается в кислую сторону при постоянстве pHf;
в/) при обоих указанных сдвигах.
Обычно pH крови не дает сколько-нибудь заметных сдвигов. Но отдельные компоненты крови в зависимости от различных условий обнаруживают заметное смещение своих изоэлектрических точек в ту или другую сторону. Но и в норме А = pH—pHt- компонентов крови обнаруживает достаточно отчетливую величину рас-
рОН’
Рис. 2. Изоэлектрические точки основных компонентов крови.
хождения, которая позволяет иметь суждение об электрическом заряде этих компонентов, их полярности и даже подойти к провизорному определению порядка величины заряда.
Построим диаграмму (рис. 2) расположения изоэлектрических точек ряда компонентов крови. По оси абсцисс отложим четырнадцать делений шкалы pH, по оси ординат — четырнадцать делений шкалы рОН, крайние точки соединим прямой линией. На эту прямую должны лечь изоэлектрические точки компонентов крови в порядке их последовательных значений: альбуминов, глобулинов, эритроцитов, гемоглобина, цельной крови и фибриногена. Биссектриса угла, выходящая из точки начала координат, разделит пополам указанную прямую в точке нейтральной реакции pH. Если опустить перпендикуляры из каждой полученной точки на
18
оси х и у. то длина этих перпендикуляров будет означать сравнительное количество в данной точке ионов Н+ и ОН-.
Из этой диаграммы следует, что в изоэлектрической точке альбуминов по сравнению с другими компонентами крови есть наибольшее число Н+ ионов и, следовательно, среда изоэлектрической точки, где разряжаются альбумины, имеет наибольший по сравнению с изоэлектрическими точками других компонентов крови электрический заряд положительной полярности. Отсюда вытекает следующее: чтобы альбумины могли только в одной среде найти свою изоэлектрическую точку и, разрядившись в данной среде, стать электрически нейтральными, их частицы должны нести соответственно равный электрический заряд отрицательной полярности. Иначе говоря, заряд частицы эквивалентен величине заряда в данной точке шкалы pH, но противоположен по знаку полярности. Действительно, альбумины обладают наибольшим электрическим зарядом отрицательного знака из всех основных компонентов крови.
Собственно говоря, такого рода определение поверхностного заряда частиц является сугубо провизорным. С одной стороны, ввиду одновременного действия сдвига pH на все компоненты крови величина абсолютного заряда, определенная на основании этих соображений, будет заведомо преувеличенной, хотя качественная картина сравнительной величины плотности поверхностных зарядов остается верной. С другой стороны, наличие диффузного слоя противоположно заряженных ионов вокруг каждой частицы обусловливает противоположный эффект.
Далее по значению своего электрического заряда идут глобулины. Их электроотрицательный заряд, судя по положению изоэлектрической точки, по своей абсолютной величине меньше заряда альбуминов, но сравнительно еще достаточно велик. За глобулинами следуют эритроциты, обладающие зарядом отрицательной полярности меньшим, чем глобулины, но также достаточным, чтобы нейтрализоваться в кислой области pH. Изоэлектрическая точка гемоглобина лежит еще в кислой области шкалы pH. Но изоэлектрическая точка фибриногена уже находится по другую сторону нейтральной точки шкалы, т. е. лежит уже в щелочной стороне pH и, следовательно, фибриноген несет не отрицательный, а положительный заряд.
Зная величину pH, указанных компонентов крови, можно вычислить количество ионов Н+ и ОН~ или число элементарных электрических зарядов е+ и которые содержатся в 1 мм3 раствора или крови, в точках шкалы, где происходит нейтрализация этих компонентов, т. е. в их изоэлектрических точках. Имея эти данные, вычислим с помощью табл. 2 разность между pH, и pH и разность между рОН и рОН, в значениях заряда электрона:
для положительных зарядов
^^КОМП 2£Кр0ВИ
19
Количество водородных и гидроксильных ионов в изоэлектрических точках корпус- i
Корпускулярные и морфологические компоненты крови pH; Количество водородных ионов в 1 мм3
комп 2е+ крови
Альбумины 4,6 1,504-1013 2,676-1010
Глобулины 5,4 2,385-1012 2,676-1010
Эритроциты 6,0 6,061-10й 2,676-1010
Гемоглобин 6,7 1,195-10й 2,676-1010
Фибриноген 8,0 6,061 -109 2,676-1010
и соответствующий коэффициент
Ур+
_ КОМП
скрови
для отрицательных зарядов
= 2^омп - Se-оаи и соответствующий коэффициент
TS комп
Ке~ = скрови
Результаты вычислений представлены в табл. 3, которая дает возможность сравнить соотношение сумм положительных и отрицательных зарядов в 1 мм3 крови для изоэлектрических точек рассмотренных выше компонентов крови и, таким образом, определить, на сколько и во сколько раз число положительных или отрицательных зарядов в изоэлектрической точке данного компонента больше или меньше числа положительных или отрицательных зарядов в 1 мм3 pH цельной крови. Следовательно, можно сравнить, на сколько и во сколько раз электрический заряд крови — среды, в которой помещаются те или иные ее компоненты, меньше или больше (для фибриногена) электрических зарядов самих компонентов.
Предварительно необходимо вспомнить о величине площади поверхности отдельных фракций белка. В 1 мм3 крови содержится 0,0007 г белка (Негели, Абдергальден), которые имеют, согласно нашим расчетам, суммарную поверхность 838,52 см2 в 1 мм3 крови. Табл. 4 дает представление о распределении указанной величины площади поверхности между основными фракциями белка.
Из левой части табл. 3 видно, что альбумины разряжаются в изоэлектрической точке, где положительный заряд равен 15,04 • 1012е+. Эта цифра может дать известное представление о по-
20
Таблица 3
кулярных и морфологических элементов крови
крови Количество гидроксильных ионов в 1 мм3 крови
. комп крови pOHz Хеками Хе~ крови 2{?комп k 2скрови
568: 1 9,4 2,385-108 13,733-1010 1 :568
90:1 8,6 1,504-10» 13,733-1010 I : 90
23:1 8,0 6,061-10» 13,733 10’° 1 :23
4,5: 1 7,3 3,003-1010 13,733-1010 1 :4,5
1 : 4,4 6,0 6,061-10й 13,733-1010 4,4: 1
рядке величины отрицательного заряда, несомого суммарной поверхностью всех частиц альбумина в 1 мм3 крови. Этот отрицательный заряд должен быть равным 15,04 • 1012е-. Таким образом, открывается возможность рассчитать число отрицательных зарядов, находящихся на 1 мк2 условной поверхности частиц альбумина.
Очевидно, это количество выразится так: - П8 = 259е .
□OZ, 1У • 1U
Глобулины несут значительно меньший заряд отрицательной полярности, чем альбумины. Число положительных зарядов в изо-
электрической точке глобули- ~ . r о not mis т-г Таблица 4 нов равно 2,385 • 1012е. Поэто- D г Величина поверхности отдельных му можно считать, что число фраКцИЙ белка в 1 мм* крови
\J X JtX ХДСХ 1 V-v 1 DAIU*Zx <JCX /Х^Ддуи 9 ходящихся на 1 мк2 условной поверхности глобулинов, СО- Фракции 2,385 • 1012 1п, _ ставит „ „ ft8 = 104е . Относительное количе ство фракции, % Поверхность, см3
Наибольший интерес, сточ- Альбумины ки зрения теоретического ВЫ- Глобулины яснения величины отрицатель- Фибриногон ного заряда, представляют эритроциты. Концентрация положительных зарядов в их 66,7 28,9 4,4 582,19 230,30 26,03
2 = 100,0 S = 838,52
изоэлектрической точке сос-
тавляет 6,061 • 10пе в 1 лш3. Чтобы эритроциты, число которых в 1 мм3 крови равно 5 • 106, могли разрядиться и стать нейтральными, среда должна нести эквивалентный электрический заряд. Рассмотрим, какое число электрических зарядов приходится на 1 мк2 поверхности эритроцитов: в’06^ = 1105е“, где 109 — число
квадратных микронов, составляющих поверхность одного нормоцита. Произведя обратное действие, получаем величину заряда нормоцита: 1105 . 109 = 120 445г-.
По-видимому, плотность данного заряда достаточно велика, чтобы
21
стабилизировать нормоцит в кровяной плазме. Допустимое число зарядов, приходящихся на 1 мк2 поверхности эритроцита, больше числа зарядов, приходящихся на 1 мк2 альбуминов и глобулинов.
Сравнительно недавно Абрамсон показал, что целая и ничем не поврежденная поверхность эритроцита человека имеет pH, = 1,7 при условии измерения в первые 0,5 мин после ее внесения в буферный раствор. Более длительные измерения искажают указанное значение pH, вследствие адсорбирования на поверхности эритроцита белков и ее механического повреждения. Необходимо все-же отметить, что столь низкое положение изоэлектрической точки эритроцита не соответствует pH, ни одному из его компонентов: гемогло-
Т а б л и ц а 5
Сравнительные значения заряда нормоцита при различных изоэлектрических точках
pH, Заряд нормоцита
в 1 мм* крови на поверхности нормоцита на 1 мк2 нормоцита
1,7 1,195-10'» 2,39-10’ 2,19-Ю7
3,5 1,895-1014 3,80-107 3,49-105
4,7 1,195-1013 2,39-10е 2,19-104
бин имеет pH, = 6,8; отмытые белки стромы — pH, = 4,7; эмульгированные липоиды эритроцита — pH, = 2,6. Тем не менее наблюдения Абрамсона интересны в том отношении, что они как будто проливают свет на зависимость между величиной заряда и количеством времени от начала наблюдений. По-видимому, поверхность
эритроцитов после экстравазирования и помещения в кислую среду быстро изменяет свои свойства и теряет большую часть присущего ей отрицательного заряда. В табл. 5 приведено количество
электрических зарядов, несомых эритроцитом при других значениях pH,.
Гемоглобин, содержащийся в эритроцитах, также несет отрицательный электрический заряд, ибо разряжается в кислой стороне шкалы pH. Изоэлектрическая точка гемоглобина лежит сравнительно близко от нейтральной точки pH, а потому отрицательный заряд
гемоглобина меньше, чем заряд эритроцита. Вычислим число е , приходящихся на 1 мк2 условной поверхности гемоглобина in toto.
В 1 лш3 крови содержится 1,5 • 10 4 г гемоглобина. Число корпускул гемоглобина в 1 мм3 крови составляет = Х
X Ю15, где 1,096 • 10 19 г — абсолютный вес одной молекулы гемоглобина.
Вычислим поверхность корпускулы гемоглобина, имеющей офор-му эллипсоида вращения, полуоси которого равны а = 16А и b = 77,5А (по Нейрату). Поверхность всех молекул гемоглоби-на в 1 мм3 крови (1,37 • 1015) • (12,46 • 10~5) = 17,07 • 1010 мк2.
22
Отсюда число электрических зарядов, приходящихся на 1 мк* услов-нои поверхности гемоглобина in toto, выразится так: =
= 0,7е“.
Ниже будет показано, что количество корпускул гемоглобина, содержащихся в одном эритроците, равно 2,7 • 108. Следовательно, приходящийся на эти части заряд равен (2,7 • 108) • 0,7 = 1,89х — 2 39 • 109
X 108е , т. е. ровно в = 12,65 раз меньше, чем число
электрических зарядов на поверхности эритроцита, полученное из данных Абрамсона при рН£ = 1,7 ив { 8Q 1Q5- = 1572 раза больше, чем число зарядов, полученных исходя из значений pHz = 6.
Изоэлектрическая точка фибриногена лежит в щелочной стороне pH. Это значит, что частицы фибриногена несут преимущественно положительный заряд и для нейтрализации фибриноген должен быть помещен в среду с равным числом отрицательных ионов. Зная количество отрицательных ионоз в 1 лм? изоэлектрической точки фибриногена и общую поверхность всех мицелл фибриногена в 1 крови, можно вычислить количество положительных заря-дов на 1 мк* условной поверхности фибриногена: -^8 = 233^ .
Приведенные расчеты, касающиеся абсолютной величины электрического заряда тех или иных компонентов крови, следует рассматривать только как ориентировочные, дающие возможность лишь приблизиться к определению полярности и порядка величины электрического заряда того или иного компонента крови. Выяснение электрического заряда основных компонентов крови представляется чрезвычайно важным, ибо все эти составные части крови функционально связаны друг с другом и изменение заряда одного компонента вызывает соответствующие изменения во всех других.
Это особенно касается вопроса об изменении заряда морфологических и дисперсных составляющих крови, непрерывно электростатически взаимодействующих друг с другом. Как только электрический заряд под влиянием тех или иных причин изменяется, например уменьшается, на одном из компонентов крови распор между одноименными элементами крови меняется соответствующим образом и возникают явления аттраксии, агрегации и седиментации.
Принято считать, что когда отрицательный заряд альбуминов или глобулинов понижается либо уменьшается заряд эритроцита, электростатический распор между ними падает и частицы указанных белковых фракций оседают на поверхности эритроцита. Это, по мнению ряда авторов, приводит к «отяжелению» эритроцита, слипанию с другими эритроцитами и их оседанию. Вопрос об отяже-лении эритроцита мы ставим под сомнение на основании некоторых расчетов, приведенных в нашей монографии *, посвященной реакции
* А. Л. Чижевский. Структурный анализ движущейся крови. Изд-во АН СССР, ?А., 1959.
23
оседания. Но прилипание глобулинов с пониженным отрицательным зарядом или фибриногена с его положительным зарядом на поверхности эритроцита может вызвать частичную разрядку поверхности эритроцита. Копачевский показал, что при сифилисе заряд глобулинов не только претерпевает сильное уменьшение, но даже извращается, т. е. присущий глобулинам в норме отрицательный электрический заряд доходит до нуля и сменяется зарядом положительной полярности. Фибриногену также приписывается способность оседать на поверхности эритроцита и содействовать уменьшению его электрического заряда. Аналогичным разряжающим действием обладают частицы, поступающие в кровь при патологических процессах в организме, при распаде тканей и т. д.
§ 3. Ионы основных элементов и химических соединений крови, их число, радиусы и проекции на плоскость
Рассмотрим, какие же вещества являются электролитами плазмы или сыворотки крови? Ионы Н+ и ОН~" не вступают во взаимодействие с ионами сильных кислот и сильных оснований, какими являются ионы крови: Na+, Cl~, Mg++, HPO" и т. д., поэтому мы выясним специальную роль этих последних ионов отдельно. Участие Н+ и ОН“ в качестве стабилизаторов морфологических и корпускулярных элементов крови, по-видимому, очень невелико. Ниже мы увидим, что общее число ионов Н+ и ОН-" в 47 000 раз меньше числа всех диссоциированных ионов крови. Очевидно, что интересующие нас электролиты образуются в основном из солей, давая в результате диссоциации ионы Na+, К+, Са++, Mg++ и т. д., и кислотных остатков, например СП, НСОГ и НРО7~".
Кровь содержит около 0,95% солей. Главное место среди солей отведено поваренной соли NaCl. Поваренная соль определяет осмотическое давление крови, постоянство которого управляется почками. Другие ионы крови, например К+, Са++ и Mg++, содержатся в крови в незначительном количестве.
По силе своего воздействия на коллоидное состояние плазмы эти соли располагаются по рядам Гофмейстера. При слабо щелочной реакции крови наибольшее значение имеют катионы, между которыми в организме выдерживается определенное соотношение Na : К : Са, и оно при помощи особых физиологических механизмов поддерживается постоянным.
Из неорганических электролитов крови NaCl и другие соли полностью диссоциированы. Ранее предполагали, что кальций на 33,0% находится в ионизированной форме. Теперь считается, что кальций диссоциирован в количестве около 50%. Число ионов фосфатов и бикарбонатов в крови колеблется в зависимости от изменений кислотно-щелочного равновесия. По мнению Штрауба, электролиты сыворотки диссоциированы на 80,0%. Поскольку в крови имею
24
тся сильные электролиты, надо принять, что они диссоциированы полностью. Это вполне согласуется с исследованиями Дебая, Бьер-рума, Мильнера и др. Приняв это наиболее достоверное допущение, произведем расчеты соединений в 1 мм3 крови и затем вычислим проекцию их на площадь.
Количество Q ионов основных элементов крови в 1 г!мм3. В основу расчета положены следующие соотношения:
катионы: 135Na + 5,6К + 5,3 (1/2Са) + 1,6 (^Mg) мэкв!л; анионы: 100С1 + 26НСО3 + 5 (1/2SO4) + 2 С/2НРО4) мэкв!л. Получим:
QNa = 135 • 23 • 10“3 • КГ6 = 31,05 • 10~7,
где 10—3 — пересчет миллиэквивалента в граммы; 10-6 — количество кубических миллиметров в 1 л; 23 — вес 1 г • экв Na;
Qk+ = 4,6 • 39,1 • Ю-3 • 10-6 = 1,799 • 10-7; вес. г эквК = 39,1;
QCa++ = 5,3 • 20 • 10-3 -’IO-6 = 1,06 • 10-7; вес. г эквСа = 20;
QMg++ = 1,6 • 12,16 • 10~3 • 10-6 = 0,194 • 10~7;
вес. г • экв Mg = 12,16;
Qcl_ = 100 • 35,46 • 10~3 • 10-6 = 35,46 • 10-7;
вес. г • же Cl = 35,46;
О_______= 26 • 61 • 10~3 • 10-6 = 15,86 • IO-7;
HCO3
вес. г • экв HCO3 = 61,0;
(J____= 5 • 48 • 10-3 10~6 = 2,4 • 10“7; вес. г экв SO4 = 48,0;
SO 4
QHDn_ _ = 2 • 48,5 • 10“3 • 10-6 = 0,96 • 10-7;
вес. г • экв HPO4 = 48,5.
Для сравнения укажем, что в лимоннокислом натрии
QNa+ = 23 • 7,05 • 101в • 1,66 • 10-24 = 26,91 • 10~7;
Qr„n_______= 189 • 2,35 • 1016 • 1,66 • 10-24 = 73,73 • 10~7.
свн
Число ионов в 1 мм3 крови. Расчет проводим счедующим обрезэм.. Если 1 г-ат элемента (Мг-ат) содержит 6,06 • 1и23 ионов (N), то какое количество ионов электролитов (/э) будет содержаться в уже подсчитанном нами количестве ионов в 1 г!мм3 крови (Л»? Запишем пропорцию:
^г-ат N,
Ki-l3,
25
-откуда
r _ N Kt
3 Ms.am ‘
Следовательно:
/Na+ = J^JQ^.J.,05 IQ-? = 8>18 iQle;
/к+= 6,06-10^8.JO-7 ^ozg.ioxe.
/ca++= 6,06 -l-02-34b:i’°-6'10~7 = °’16 •101e;
. _ 6,06 • 1023 • 0,194 • 10-7 = 0 048 • IO16,
fMg++~ 24,3
, 6,06 • 102s • 35,46 • 10~7 c nc lnl«
/г,_ =-------А-ё-.д----------= 6,06 • 101в;
ci 35,46
, = 6,06-1023-15,86 • 10~7 = I(57 . I016;
HCO3 I
^SO7-“
^HPO--
6.06.10-^^'.-’=0|15. 10„.
6.06-102».70,96.1(Г7 = 00604 . 1QM
При экспериментах с кровью приходится прибегать к лимоннокислому натрию как дефибринирующему средству. Диссоциация лимоннокислого натрия в водном растворе происходит по следующей схеме:
Лимоннокис- Ион кислот- Натрий лый натрий но го остатка
СН2—COONa СН2—СОО-
| .ОН I /ОН
С( -» С( + 3Na+
|xCOONa рСОО-
СН2—COONa СН2—COO-
В наших расчетах принята схема полной диссоциации (третья ступень) лимонной кислоты: во-первых, используется раствор незначительной концентрации, во-вторых, константа диссоциации для лимонной кислоты в нашем случае /< = 15,1 • 10“7, для углекислоты К = 3 • 10“7 и для фосфорной кислоты К — 5,5 • 10“8, т. е. лимонная кислота в водном растворе распадается примерно так, как углекислота и фосфорная кислота.
Подсчитаем, сколько анионов лимонной кислоты и катионов натрия находится в 1 мм3 дефибринированной крови, если для де-фибринирования крови принимается соотношение: одна часть
26
5%-ного раствора лимоннокислого натрия и четыре части цельной крови.
Молекулярный вес цитрата натрия C6H6O7Na3 равен 258 г; 258 г цитрата натрия содержит 6,06 • 1023 молекул; 5 г цитрата натрия содержит 6,06 of/ ~ молекул; 1 см3 5%-ного раствора цитрата натрия содержит 6,06 2gg—— = 10“2 г-мол\ \мм3 5%-ного раствора
5%-ного раствора цитрата натрия с кровью в отношении 1 : 4 со-
6,06 . 1023 • 5 • Ю“5
держит
= 2,35 • 1016 молекул.
258 • 5
Таким образом, в 1 мм3
указанной смеси содержится ионов:
/Na+ = 2,35 • 1016 • 3 = 7,05 • 1016;
/Свн5о7 = 2,35 • 10lG • 1 = 2,35 • 1016.
Радиусы, ионов основных химических соединений крови. Пользуясь данными Гольдшмидта и Пайлинга, найдем радиусы ионов основных химических соединений крови.
Рассмотрим радиус иона НСО3 (R
Этот ион схематически
нсо~)
можно принять сложенным из центрального иона С++++ и трех ионов О++. Ион Н+ в расчет не принят ввиду его весьма малых размеров. Площадь проекции рассматриваемого иона на рис. 3 изображена в форме круга, описанного вокруг трех ионов кислорода,
27
соприкасающихся друг с другом, причем ион углерода находится между ионами кислорода. Из этой схемы видно, что радиус описанного круга равен = г + G» н0 ri — • Тогда
(' Уз \
1 + 2 —— I, где г — радиус иона кислорода 7?0-- = .= 1,32А. Подставив значение 7?0— в формулу, имеем А’нсо, = = 1,32 (1 + 2-|-1 = 2,84А.
Рис. 4. Проекция на плоскость иона Н2РО4
Радиусы ионов SO?-(PSO--) и НРО4 (/?Нро.) вычисляются оди-наковыми способами; для первого случая мы приводим описание, для второго — рис. 4.
Для вычисления Rso____принимаем конфигурацию этого иона в
форме круга, описанного около четырех взаимосоприкасающихся кругов (ионов О ), в центре которых находится ион серы.
Имеем R________= г + G. Так как гг = г]/2 то R________=
= г (1 + 1^2). Подставив значение г = /?о—в последнюю формулу, находим ______= 1,32 (1 4- 1,41) = 3,18А.
оО^
Величина R_____получена при допущении, что ион серы на-
зо4
столько мал, что дает возможность для взаимного соприкосновения четырех ионов кислорода. Для проверки этого допущения сделаем расчет /?so_, исходя из того, что R$Q_= /?s + 2/?0, так как Rs =
28
= 0,34; 7?о = 1,32, следовательно, /?so_= 0,34 + 2 • 1,32 —
= 2,98 А.
Сопоставив значение радиусов ионов SO7 - в первом и во втором случаях, т. е. /?so.— = 3,18А и A’so.— =2,98А, видим, что в случае взаимосоприкасающихся шаров (ионов) кислорода ион S свободно располагается между ними, причем образуется даже некоторый зазор, величина которого определяется как н q разность 7?so.--#so«—, т, е. 3,18А — 2,98А = I II
= 0,2Д. Н-С-С-ДО
Радиус иона НРОГ-(ЯНро—) примем равным I
Янро-- = г<1 +2) = 1,32 (1 + 1,41) = 3,18А. Н0“ |~С*о
Необходимо отметить, что принятые нами рас- Н - С - С — ОН положения ионов в одной плоскости не соответству- I II
ют действительности, а потому и значения ионных н ®
проекций на плоскость несколько преувеличены, Рис- 5- Струк-хотя и пригодны для наших ориентировочных рас- ^лекул^ли^-четов. НОЙ КИСЛОТЫ.
Проекции на плоскость ионов основных химичес-
ких элементов и соединений крови. Проекции ионов основных химических элементов и некоторых соединений крови на площадь подсчитываем по формуле площади круга, т. е. S = яг2, так как выше мы условно приняли конфигурацию ионов в форме шара:
SNa+ = л (T?Na+)2 = 3,14- 0,982 = 3,01 А2;
Sk+ = л (/?к+)2 = 3,14 • 1,332 = 5,56А2;
SCa+ + = л (/?Са+ +)2 = 3,14 • 1,062 = 3,53А2;
SMg++ = л(*Mg++)2 = 3>14 • О-732 = 1.91 А2;
SCI_ = л (Яс1_)2 = 3,14 • 1,812 = 10.29А2;
SHCO_ = л (Янго-)2 = 3,14 - 2,842 = 25,31 А2;
___= Л(Р______)2 = 3,14 • 3,182 = 31,74 А2;
S„POT- = "<«„РО4-J’ - 3.14 3,18- - 31.74А".
Значение проекции иона лимонной кислоты на площадь получаем, исходя из нашего исследования параметров этого иона.
Структурная формула молекулы лимонной кислоты может быть изображена в виде, представленном на рис. 5. Расчет площади большого иона не может быть проведен в результате расположения всех
29
компонентов иона в одной плоскости и подсчета расстояний между центрами компонентов как сумм соответствующих радиусов составляющих его ионов, ибо при таком рассмотрении получается намного преувеличенный результат. В самом деле: 1) элементарные слагающие ионы располагаются не в плоскости, а в пространстве, что сокращает величину проекции молекулы; 2) длина ионной связи меньше сумм длин радиусов ионов, так как при образовании
Рис. 6. Пространственная структура молекулы лимонной кислоты.
связи ионы, так сказать, входят один в другой.
Для расчета проекции нужно построить, хотя бы в первом приближении, пространственную струк-
А
Рис. 7. Схема для вычисления параметров тетраэдра.
туру формулы лимонной кислоты. Известно, что для четырех связей углерода, расходящихся в разные стороны, может быте принята тетраэдрическая конструкция, т. е. атом углерода может быть представлен в центре тетраэдра, а четыре атома, от него отходящих,— в вершинах тетраэдра. Пространственную структуру карбоксильной группы можно построить по аналогии со структурой сходной группы:
С1\
\=О и Z.C1CC1 = 117°.
С1/
Поэтому без большой погрешности предположим, что группа расположена в одной плоскости и углы все равны между собою: 7 ССО = 7 ОСО = 7 ОСС = 120°. Направление гидроксильной связи ОН подвижно, а потому среднее значение проекции гидрок-
2
сильной связи можно принять равным —(О — Н).
Таким образом, пространственная структура молекулы изобразится в виде трех вдвинутых один в другой тетраэдров (рис. 6) с отходящими от трех вершин тетраэдров карбоксильными группа
30
ми. Вычислим угол, под которым отходят друг от друга отрезки, соединяющие центр тетраэдра с его вершинами. Пусть О — центр тетраэдра ADEB (рис. 7). Опустим из вершины А перпендикуляр на основание DEB, который пройдет через О и пересечет основание центра С равностороннего треугольника DEB.
Опустим в плоскости ААСВ перпендикуляр ОК. из точки О на АВ. Тогда АК = КВ. Обозначая сторону тетраэдра DB = а, по-
Рис. 8. Длина проекции цитратного иона.
лучим: _
АВ = а; СВ=-^-; СА = ^-. О О
Обозначим / АОВ = <р. Тогда
-|- = ХАОК= А АВС =
= arcsin -4^- = arcs in [ а-~~-: а I = 54° 30',.
отсюда <р = 109°.
Рис. 9. Ширина проекции цитратного иона.
Длины необходимых
нам связей можно определить так:
Связь
С —Н С —С с = о с —о (C-N) О —н
Длина связи, А
1,09
1,55
1,25
1,33
0,97
Длина проекции иона лимонной кислоты равна длине проекции ломаной НСССН, т. е.одлине прямой НН (рис. 8).
Длина СН = 1,09А; длина СС = 1,55А. т. е. МС = 0,46А, z иск = 54°.
Из этих условий получим:
НМ = 2 • 2,09 • arccos 54°30' = 2,18 • 1,581 = 1.27А.
MN = 2 • 0,46 • 0,581 = 0,54А.
Отсюда
НН = 2,54 + 0,54 = З.ОвА.
81
Для определения ширины иона рассмотрим рис. 9. Расстояние ОС с небольшой мажорацией может быть рассчитано так:
ОС = 2 • 1,55-0,582 = 1,80А.
Проекция ОС на направление ширины, т. е.
OG = СО • cos 30° = 1,25 - 0,866 = 1,08 А.
Длина СО перпендикулярна направлению ширины, а потому €е можно не учитывать. Длина ОН = 0,97А, но ввиду неопределенности ориентации ОН усредненная длина проекции ОН на направление ширины может быть принята равной
— • 0,97 А = 0,64 • 0,97 = 0,62А. л
Итак, окончательная проекция ширины будет равна
0,62+ 1,08+ 1,80 = 3,50А.
Для площади проекции иона лимонной кислоты получим
•^citr = 3,08 • 3,50 = 10,8А2.
Для определения длин связей мы можем воспользоваться также ковалентными радиусами по Паулингу. Приведем часть его таблицы, необходимую для расчетов (табл. 6).
Таблица 6
Ковалентные радиусы по Паулингу
Связь Ковалентный радиус атомов, А
Н В с 1 °
Единичная 0,37 (0,37) 0,88 0,771 0,66 (0,74)
Двойная — 0,76 0,665 0,55
Для интересующих нас длин связей получим:
Связь Длина связи, А
С—Н 0,37 +0,771 = 1,141
С —С 0,771+0,771 = 1,542
С = О 0,665+0,55 =1,215
С —О 0,771+0,74 =1,511
О —Н 0,74 +0,37 =1,11
Как видим, длины связей, вычисленные вторым способом, мало отличаются от длин связей, вычисленных первым. В таблицах ковалентных радиусов в скобках указаны значения, принятые в последнее время Шомекером и Стивенсоном.
32
Подставляя в рис. 8 другие данные для длин связей, для длин проекции по длине, получим следующее выражение: НН = = 4,141 • 0,581 + 2 • 0,401 • 0,581 = 2,65 + 0,466 = 3,116А.
Длину проекции по ширине получим, подставляя эти же данные в рис. 9: 2 • 1,542 • 0,581 + 1,215 • 0,866 + 0,64 • 1,11 = 1,80 + + 1,05 + 0,71 = 3,56А.
Отсюда находим площадь проекции иона лимонной кислоты
Scitr =3,116 • 3,56 = 11,1А2.
Таким образом, двумя способами получено для площади проекции почти одно и то же число. Принимая для расчетов среднее арифметическое значение площади проекции из двух величин, полученных разными путями, будем иметь
Scitr==JM+lbL=11>0A2.
Сумма проекций ионов на плоскость. Эта сумма проекций подсчитывается как произведение проекций ионов основных химических элементов крови на число их в 1 л<л<3 крови, т. е.
2S3 = S373 [A2];
2SNa+ = SNa+ • ZNa+ = 3,01 • 8,18 • 10le = 26,62 • 10leA2;
SSk+ = 5,56 • 0,28 • IO16 = 1,557 • 10wA2;
2SCa++ = 3,53 • 0,158 • 10le = 0,558 • 1016Aa;
2SMg++ = 1,91 0,048 • 10le = 0,0917 10ieA2;
2SC1_ = 10,29 • 6,06 • 10le = 62,357 • 10leA2;
2S„„rt_ = 25,31 • 1,57 • 101" = 39,74 • 10“A2;
HCO3
2S______= 31,74 • 0,15 • 10м = 4,761 • 10leA2;
so4 ’
2SHpo____= 31,74 • 0,0604 • 10й = 1,917 • 10leA2.
Укажем для сравнения, что для лимоннокислого натрия:
2SNj+ = 3,01 • 7,05 • 101в = 21,22 • 101вА2;
2Sc,hso, = 11,0 • 2,35 • 10le = 26.50А2.
Все приведенные вычисления сведены в табл. 7. Видно, что сумма положительных и отрицательных зарядов в 1 л«ж3 крови (кроме ионов Н+ и ОН-) составляет
2е+ + 2е- = 16,4 • 10м.
33
Ионы основных элементов и химических соединений в 1 мм* крови и их проекции на плоскость
Таблица 7
Ионы Полярность Концентрация ионов, мэкв/л Вес ионов • Ю7, г Число ионов Радиус ионов Яэ, 1 Проекция на площадь 5Э, X Сумма проекций ионов на плоскость ZS3.10»e, А»
Na + 135 31,05 8,18 0,96 3,01 24,62
К + 4,6 1,80 0,28 1,33 5,56 1,557
Са ++ 5,3 * 1,06 0,158 1,06 3,53 0,558
Mg ++ 1,6* 0,194 0,048 0,78 1,91 0,0917
= 8>5бб = 26,827
Cl — 100 35,46 6,06 1,81 10,29 62,357
нсо3 — 26 15,86 1,57 2,84 25,31 39,74
so4 — 5* 2,40 0,15 3,18 31,74 4,761
НРО4 1— — 2 * 0,97 0,0604 3,18 31,74 1,917
Se- = 7,84 SSS” = 108,781
Na + ** 26,91 7,05 0,98 3,01 21,22
с.н5о7 ** 73,73 2,35 ** 11,0 26,50
2е+ = 15,616 S2S+= 48,047
2е- = 10,19 = 135,281
• С учетом заряда.
•* Данных нет.
В 1 лш3 цитратной крови число положительных и отрицательных ионов возрастает до 25,8 • 1016. Число ионов Н+ и ОН~" в 1 мм? представляет собою сравнительно очень небольшую величину» а именно
1,67 • 10В * * * 12 * * * + 2,71 • 1012 = 4,38 • 1012,
или 0,000438 • 1016, т. е. в 47 тысяч раз меньше, чем число ионов, основных химических элементов и соединений крови.
Сумма проекций на плоскость всех положительных ионов основных химических элементов и соединений в 1 мм3 крови относится к сумме проекций всех отрицательных ионов, как
SSS+ __ 26,827 • 10’6 __ 1
~ 108,781 • IO’® ~ 4,04 *
В цитратной крови это отношение составляет
48,047 __ 1
135,281 ~ 2,83 *
Зависимость электропроводности крови от ионного состава. Нами сделана попытка выяснить вопрос о количестве основных ионов в плазме. Экспериментальное определение количества активных ионов отдельных веществ в плазме, кроме ионов Н+, представ-
84
ляет большие затруднения. Выяснение же общей суммы ионизированных молекул в плазме или в сыворотке может быть произведено с достаточной точностью путем определения электропроводности плазмы или сыворотки. Электропроводность плазмы и сыворотки одинакова. Электропроводность цельной крови значительно меньше, чем плазмы или сыворотки, ибо способность морфологических элементов проводить ток весьма мала. Электропроводность плазмы и сыворотки является достаточно постоянной величиной и изменяется только в патологических случаях.
Величина электропроводности плазмы и сыворотки в основном зависит от содержания NaCl, который обусловливает большую часть электропроводности. Оставшаяся часть электропроводности приходится главным образом на долю бикарбонатов. Не отражается на величине электропроводности плазмы и сыворотки содержание в крови СО2 вследствие процесса обмена ионами между эритроцитами и плазмой при изменении содержания СО2 в крови. Величина электропроводности плазмы и сыворотки не зависит от превалирующего содержания в ней ионов минеральных электролитов или ионов белковых частиц. Но так как взвешенные в крови форменные элементы и частицы коллоидов препятствуют прохождению тока через кровь, то, естественно, величину электропроводностей гетерогенных плазмы или сыворотки нельзя сравнивать с величиной электропроводности гомогенного раствора. На этом основании Бу-дарским и Тангльем введено понятие коррегированной электропроводности. Эта величина вычисляется по формуле
_ 100 +2,5Р
Лсогт - 100 ,
где Л — величина, полученная при непосредственном измерении электропроводности сыворотки; Р — процентное содержание белков.
Упомянутые авторы установили, что 1 г белка, растворенный в 100 см3 жидкости, уменьшает электропроводность на 2,5%. По данным тех же авторов, температура влияет на электропроводность, ибо с ее повышением увеличивается диссоциация электролитов. Так, повышение температуры на Г С вызывает увеличение электропроводности на 2,2%.
Многие другие факторы также влияют на величину электропроводности плазмы и сыворотки, а именно: время дня, состояние здоровья, рацион, покой или мышечная работа (Фаркас, Гурьев и др.). Хотя прием пищи и не влияет на электропроводность, но предел электропроводности при этом повышается, свидетельствует об изменении в плазме или сыворотке количества ионов (Виола). При ряде заболеваний разной этиологии и патофизиологии размах колебаний электропроводности сыворотки или плазмы значительно больше, чем в норме (Виола). Наблюдаются случаи как резкого понижения, так и резкого повышения электропроводности. Но вопрос о том, какие заболевания и почему обусловливают эти отклонения,
35
остается в известной мере открытым до сих пор. Есть мнение, что состояние беременности не влияет на величину электропроводности.
Известно, что нарастающее растворение электролита увеличивает его диссоциацию. Поэтому разжижение сыворотки влечет за собой уменьшение снижения концентрации ее ионов и, следовательно, ее электропроводности. Это дало повод Окер-Блому ввести определение «физиологической электропроводности», которая выражается цифрой специфической электропроводности, умноженной на степень разведения (А^, Л2, Л3, ... и т. д.). Разводя сыворотку, можно дойти до такого момента, когда все те ионы, которые находятся в недиссоциированном состоянии, т. е. потенциальные ионы будут диссоциированы. Наибольшая физиологическая электропроводность носит название предела электропроводности и обозначается Хоо.
Чтобы вычислить приближенную степень ионизации сыворотки, пользуются формулой
По данным Виола, электропроводность сыворотки у разных людей при t = 25° С и Л2 колеблется в пределах X = (106 4- 119) х X 1(Г4 ом~‘. При t= 18° С Х= (90 4- 102) • 10“4 ом.-1. По данным Энгельмана, при /=18° С X = (101 4- 107) • 10-4 ол_|.
Величина Лоо у разных людей колеблется в узких пределах. Так, у восьми человек обнаружено = (149,33 -4- 164,00) х X 10-4 олС1, несмотря на то, что эти величины были получены при разных степенях разведения сыворотки (от -gj Д0^)-
При заболеваниях предел колебания электропроводности больше. По Виола этот предел при t = 25° С составляет Л = = (98,29 4- 142,01) • 10~4 ом~{.
Для сравнения приведем электропроводность плазмы, цельной крови и кашицы из форменных элементов у лошади и собаки (Л = = п • 10’4 сш""1):
Плазма Кровь Кашица из форменных элементов
Лошадь 105,3 63,4 1,63
103,7 62,8 1,67
Собака 112,9 36,9 1,70
107,4 43,3 2,17
Видно, что наибольшей способностью проводить ток обладает плазма, вдвое меньшей способностью обладает цельная кровь и очень малою способностью обладает кашица из форменных элементов. Укажем, что Бетмен с сотрудниками (1937) получили следующую величину электропроводности цельной дефибринированной крови: 82,5 • 10“4 ом~{.
36
Гебер показал, что электропроводность эндоглобулярного содержания эритроцитов соответствует содержанию в растворе NaCl в количестве 0,2%, т. е. в 4,5 раза меньше, чем содержание NaCl в цельной крови, равное 0,9%. Переводя данные Гебера на число ионов Na+ и С1“, получаем для 1 мм3 эндоглобулярного содержания эритроцитов 2,07 • 1016 ионов Na+ и столько же ионов С1“. Отсюда находим число положительных и отрицательных ионов в эндогло-булярном содержании эритроцитов:
1 мм3 0,2% NaCl содержит 2,07 • 1016 ионов положительных и от-. о 2,07 • 101в 3 2,07 . 1016 • 63
рицательных, 1 мк? содержит—— ионов, --------------------=
= 1,2 • 109 ионов Na+ и столько же С1~. Здесь 63 мк3 — приблизительный объем нормоцита.
Таблица 8
Электропроводность отдельных химических компонентов крови
Ион Концентрация ионов в крови Подвижность ионов, см 1 сек • F Удельная электропроводимость 10*, ом~1
мэкв/л экв! л • 10*
Na+ 135,0 1,350 43,5 58,72
к+ 4,6 0,046 64,6 2,97
Са++ 5,3 0,053 * *
Mg++ 1,6 0,016 * *
сг 100,0 1,00 65,2 65,20
НСО^ 26,0 0,260 60,0 15,60
SO" 5,0 0,050 * *
НРО^- 2,0 0,020 S = 142,49
* Этих данных автор получить не мог.
На основании данных о электропроводности можно сделать грубо ориентировочный расчет числа ионов в плазме, ибо в неконцентрированных растворах электропроводность прямо пропорциональна числу ионов. Если 1,8 • 10“4 ом~1 соответствует наличию в плазме 2,07 • 1016 положительных и отрицательных ионов в 1 мм3, то ПО • 10“4 ом~1 соответствует 63 • 1016 положительных и отрицательных ионов в 1 мм3.
Возвращаясь к табл. 7, где приводится число ионов основных элементов и химических соединений в 1 мм3 плазмы крови, мы видим, что последнее число оказывается в четыре раза большим.
Рассмотрим значения электропроводности отдельных химических составляющих крови, изученных вне крови. Как известно, удельная электропроводность ионов при электролизе вычисляется по формуле
X = Favp,
37
где F — константа Фарадея; а — степень диссоциации электролита; т] — концентрация электролита; v — скорость движения иона.
Вместо абсолютных скоростей вводим величину пропорциональности
Fv = Vt
где V — подвижность иона.
Удельная электропроводность крови состоит из отдельных электропроводностей слагающих ионов. Например, для Na имеем
1,35 • 10-4 • 43,5 = 58,72 • 10~4 ом~1.
Прочие данные приводятся в табл. 8, из которой видно, что сумма электропроводностей основных ионов крови совпадает со значением электропроводности крови, полученным экспериментально.
§ 4. Ориентировочные вычисления электрического заряда эритроцита
Дзета-потенциал эритроцитов человека и других млекопитающих хорошо изучен с помощью электрофореза. Скорость переноса в электрическом поле эритроцитов разных животных в одинаковой среде оказалась различной. Подвижность эритроцитов человека равна 1,31 мк/сек • в • смГх. Некоторые авторы отмечают, что величина электрофоретической подвижности эритроцитов характеризуется большим постоянством: она остается в некоторых случаях на одном уровне сутки и более. Изменение среды, естественно, влияет на величину подвижности. В изотоническом растворе глюкозы с небольшим количеством фосфатного буфера эритроциты человека приобретают подвижность 2,33 мк/сек • в • см~х. В изотоническом растворе сахарозы дзета-потенциал эритроцитов равен 25—26 в/см (Складовский и Шредер). Роль пола, расы или кровяной группы не оказывает никакого влияния на величину подвижности эритроцитов человека. Беременность также заметно не влияет на эту величину. Еще Фарэус показал, что при беременности скорость оседания эритроцитов резко возрастает и одновременно уменьшается необходимая для перезарядки эритроцитов концентрация ионов лантана. Сопоставление этих двух фактов говорит о том, что значения дзета-потенциала не находятся в прямом соответствии с необходимой для перезарядки концентрацией многовалентного катиона, иначе говоря, величина дзета-потенциала не отражает количества электростатических зарядов на поверхности эритроцита и может иметь только условное значение. Впрочем, это явствует и из самой теории образования дзета-потенциала.
Можно считать, что скорость электрофоретического переноса почти одинакова для частиц самых разнообразных размеров. Независимость величины дзета-потенциала от размеров частицы может быть объяснена практически одинаковой поверхностной плотностью электрического заряда у частиц всех размеров, т. е. одинаковым
38
количеством элементарных зарядов, приходящихся на единицу поверхности коллоидных или взвешенных частиц. Но одинаковая плотность электрического заряда наблюдается лишь в том случае, когда частицы находятся в чистой дисперсионной среде. В присутствии электролита происходят отступления от данного правила.
Было показано, что скорость электрофореза коллоидных частиц лишь немногим меньше, чем скорость движения ионов в электролите. Скорость движения иона водорода равна 33 мк/сек при градиенте поля 1 в/см, скорость иона хлора 6,8 мк/сек, а скорость
Таблица 9
Зависимость электрического заряда нормоцита от радиуса и поверхности
Частица Радиус частицы, ммк Поверхность частицы, ммк2 Электрический заряд, е
Коллоид серебра 25 7852 62
Эритроцит 2940 108 618 588 370 396
коллоидных частиц 2—4 мк/сек. Это говорит о том, что электрические заряды высокодисперсных коллоидных частиц достаточно велики, иначе их электрофоретические скорости были бы чрезвычайно малыми. Зигмонди подсчитал, что заряд коллоидных частиц серебра радиусом 25 ммк равен 297 • 1О“10 абс. эл. ед., или 297 х х 1О“10 • 0,21 • 1010 = 62е. Отсюда получаем площадь поверхности частицы, на которой располагается один заряд:
S лб/2 7850 юг? с 2
-V— = “v— = —= 126,5 ммк2, Se Ye 62
Упомянутый факт, что коллоидные частицы и некоторые ионы переносятся в электрическом поле одинакового градиента с почти одинаковой скоростью, привел к мысли о том, что число электрических зарядов на поверхности частицы находится в известном соотношении с величиной развития поверхности, т. е. с радиусом частицы. К тому же порядку величин мы приходим, исходя из теоретических подсчетов Зигмонди величины заряда на коллоидной частице серебра. Допустив, что нормоцит имеет форму шара с радиусом 2,94 мк, находим значение его электрического заряда (табл. 9).
Приведенная величина заряда эритроцита хорошо совпадает со значением, теоретически найденным Льюисом для заряда шарика эмульсии (840 ОООе).
Абрамсон показал, что некоторые бактерии, поверхность которых значительно меньше поверхности эритроцита, несут до 300 000 зарядов. Виленд и Франенбургер применили теорию коагуляции к образованию дождей и теоретически получили размеры капель
39
воды в тумане и ихо заряд. По их расчетам, эти размеры колеблются от 3000 до 30 000 А, а заряд — от 300 до 3 000г.
Герат, Гунн, Чэлмерс, Пасквиль, Гшвэнд и другие приводят значения электрического заряда дождевых капель от 0,1 до 0,001 ед. CGSE, т. е. 2 • 108 4- 2 • 105 элементарных электростатических зарядов на одну каплю.
Таким образом, видно, что между величиной частицы и ее электрическим зарядом как будто нет никакой зависимости. Само собой разумеется, что химическая природа частицы и вмещающей ее среды играет роль в отношении величины заряда, однако основной причиной несоответствия между величиной заряда и размерами частиц следует считать недостаточность знаний в этой области.
Если рассматривать коллоидную частицу, окруженную двойным слоем электрических зарядов как шаровой конденсатор, а расстояние между обкладками принять равным толщине двойного слоя, то дзета-потенциал такой частицы выразится так:
r _ g ('1 — И “ Drtr ’
где гг — радиус частицы с внешней поверхностью двойного электрического слоя; г — радиус частицы; D — диэлектрическая постоянная.
Отсюда электрический заряд частицы
Р ’ где р = г2 — г.
Гевеей, пользуясь этой формулой, теоретически получил заряд коллоидных частиц при возрастающем радиусе. Приняв величину элементарного заряда е = 4,80 • 1О“10 ед. CGSE, дзета-потенциал равным 0,076 = 0,07 : 300 = 0,00023 ед. CGSE, D == 81 (воды) и толщину двойного электрического слоя р = 5 • 10“7 см, находим
0,00023.81 • г (г + 5 . 10“7) r PrQr.
е=----------ГЪЬ------------ |ед. CGS£]
ИЛИ
__ 0,00023 -81 • г (г + 5 • 10“7) I элементарных 5 • 10“7 • 4,80 • 1О“10 [ зарядов
Пользуясь приведенными ниже рядами Гевеей, сделаем попытку определить электрический заряд сферулированного по поверхности нормоцита, имеющего г = 2,94 мк = 2940 ммк. Из рассмотрения рядов третьей колонки табл. 10 легко заметить, что заряд частиц приблизительно равен квадрату радиуса. Следовательно, можно искать зависимость параболического типа второй степени
е = Ах2 4- Вх.
В результате исследования находим уравнение
е = 0,8х2 + 4х.
40
Подставляя в это уравнение вместо х значения радиусов, получим четвертую колонку той же таблицы.
Льюис для теоретического определения заряда частиц масляной эмульсии и коллоидных частиц серебра и платины пользовался той же формулой, что и Гевеей, приняв в числителе гг = г:
VDr2 е =------,
Р где V = С — разность потенциалов между частицей и жидкостью. Для заряда коллоидных частиц им было найдено значение е — = 168 000.
Таблица 10
К расчету электрического заряда нормоцита и коллоидных частиц
Частица Радиус частицы, мм к Заряд частицы по Гевеей, е Вычисления по формуле е = 0,8х2 4Х
Коллоидная 1,2 6,0 5,952
» 2,4 14,3 14,208
» 12,0 164,0 163,200
» 24,0 550,0 556,800
» 240,0 46 600,0 47 040,000
Нормоцит 2 940,0 — 6 926 640,000
Из последней формулы следует, что заряд частицы обратно пропорционален толщине двойного слоя. Имея значения дзета-потенциала эритроцитов (в милливольтах), диэлектрическую постоянную цельной крови D = 85,5 (Фюрт) и размеры частиц, можно отыскать более или менее достоверные величины для толщины двойного* электрического слоя р вокруг эритроцита.
Чтобы найти значения р, необходимо остановиться на рассмотрении вопроса о том, ионы каких элементов или химических соединений принимают главное участие в образовании двойного слоя. Вопрос этот до сих пор нельзя считать разрешенным.
Электропроводность крови в основном зависит от ионов Na и С1, т. е. от хлорида натрия NaCl и бикарбоната натрия NaHCO3. В самом деле, принимая в расчет только ионы Na и ионы С1, получаем значение электропроводности крови
X = 58 72 • 10~4 + 65,20 • 10~4 = 123,92 • 10-4 ом~\
хорошо совпадающее со значениями электропроводности, полученными Виола, Энгельманом и другими авторами. Это обстоятельство дает нам основание с известной достоверностью считать в наших расчетах ионы Na и С1 активными, участвующими в образовании двойного электрического слоя вокруг эритроцитов, и, таким образом, получить приближенное значение толщины р двойного слоя.
Бригг показал, что расстояние между электрическими зарядами двойного слоя может быть переменным. Гуи получил значение толщины р двойного слоя, в зависимости от концентрации раствора,
4L
•в котором помещается частица с двойным слоем: для 0,1-нормального раствора NaCl 0,96 ммк, для 0,001-нормального раствора NaCl 9,6 ммк, для чистой воды 10,1 ммк
Мак Клендон нашел такие значения толщины двойного слоя: для 0,1-нормального раствора NaCl 1,94 ммк, для 0,001-нормального раствора NaCl 3,25 ммк.
Из этих данных следует, что с понижением концентрации раствора толщина двойного слоя возрастает.
Как известно, нормальным раствором называется раствор, содержащий 1 г • экв соответствующих ионов, т. е. такое количество ионов, которое соответствует в химической реакции 1 г водорода или 8 г кислорода на 1000 мл воды. 1 г • экв соли NaCl состоит из 1 г • экв натрия, равного 23 г, и 1 г • экв хлора, равного 35,46 г. Это значит, что 5,846 г NaCl, разведенных в 1 л воды, дают 1-нормальный раствор. В соответствии с этим 5,846 г NaCl дают в 1 л воды 0,1-нормальный раствор, а 0,5846 — 0,01-нормальный раствор. Переводя в проценты, получаем:
58,46 г NaCl, растворенных в 1 л воды, дают 5,846%-ный раствор, 5,846 г NaCl, растворенных в 1 л воды, дают 0,5846%-ный раствор, равный 0,1-нормальному раствору.
Отсюда следует, что если 0,5846% раствора NaCl — 0,1-нор-мальный раствор, то 0,95% раствора NaCl (изотонического) дадут
0,95 0,1 л 1рОг жт
0 5845 ' 0*1625 нормального раствора NaCl,
где 0,95— содержание NaCl в крови, вес.%.
Допустим, что содержание NaCl в основном обусловливает образование двойного электрического слоя. Приняв это допущение, определим толщину двойного слоя р на частицах крови, в согласии с данными Мак Клендона. Получаем следующую пропорцию: р : 1,94 = 0,1 : 0,162.
Отсюда
1,94 -0,1 . о
р — 0,162 — 1»2лшК-
Полученное непрямым путем значение для толщины двойного слоя р = 1,2 ммк = 12А точно совпадает с новейшими данными о параметрах двойного слоя при различных концентрациях разных типов электролитов. Принимая, что Na и С1 составляют в крови 100 ммоль/л, находим для одновалентных Na и С1 значения 6 = = 10А (Булл).
Зная основные значения физических параметров эритроцита, входящие в формулу Гевеей, рассчитаем эффективный заряд нормоцита
£ _ _ мад ,»-•)• as _ 5 3 ! №S
б 300 . 1,2 Ю“7
где £ — электрокинетический потенциал, в (для эритроцита 0,025 в), г — радиус сферулированного по поверхности эритроцита (2,94 мк},
42
D — диэлектрическая постоянная крови (85,5); 6 — толщина двойного слоя (1,2 ммк).
Значения электрического заряда нормоцита в зависимости от толщины слоя и разности потенциалов между нормоцитом и средой приведены в табл. 11.
Невольно возникает вопрос: каков же действительный заряд эритроцита? Ответить на этот вопрос в настоящий момент еще нельзя. Надо полагать, что с помощью достаточно современного эксперимента такая задача будет разрешена.
Таблица 11
Теоретические значения электрического заряда нормоцита в электростатических единицах и элементарных зарядах в зависимости от толщины слоя и разности потенциалов между нормоцитом и средой
V, ед. CGSE Заряд нормоцита
6 « 0,04 см 0,03 0,02 0,01 0,005
0.5.10"“7 19,72-10“3 4,1412.107 14,8-10“3 3,1059-10’ 9,86-10“3 2,0706-10’ 4,93-10“» 1,0353-10’ 2,46 КГ3 0,5176-107
1,0-10“7 9,86-10“3 2,0706-107 7,4-10“3 1,554-10’ 4,93-10“3 1,0353-10’ 2,465-10“3 0,5176-10’ 1,23-Ю-з 0,2588-1О7
2,0-10—7 4,93-10“3 1,0353-107 3,7-10-3 0,777-107 2,465-Ю-з 0,5176-107 1,2325-10“3 0,1294-10’ 0,308-Ю-з 0,0647-1О7
30-Ю"7 3,29-10-3 0,691-10’ 2,47-10“3 0,519-10’ 1,64-10“3 0,344-10’ 0,822-10“3 0,1726 10’ 0,205-IO-3 0,0430-107
4,0-10“7 2,47.10-з 0,519-107 1,85 IO”3 0,389-107 1,233-10“3 0,259-10’ 0,308-10“3 0,647-10’ 0,154-Ю-з 0,08235-1О7
5,0-10“7 1,972-10—3 0,414-10’ 1,47-10“3 0,311 • 10’ 0,986-10“ 3 0,207-10’ 0,493-10“3 0,104-10’ 0,123-10“3 0,0518-10’
Если определять величину заряда эритроцита, исходя из значения его изоэлектрической точки, то получим, как уже было показано, величину заряда эритроцита, которая достаточно хорошо совпадает с результатами расчета по методу Льюиса — Гевеей. Впрочем, в этом нет ничего удивительного, так как в основе формулы Льюиса — Гевеей лежит величина дзета-потенциала, в соответствии с которой определяется и изоэлектрическая точка. Если же считать, в согласии с Абрамсоном, что истинная pHt- эритроцита равна 1,7, тогда заряд эритроцита чрезвычайно возрастает, достигая величины 2,4 • 109е.
При сопоставлении величины суммарного заряда, несомого всеми отрицательно заряженными частицами 1 мм3 крови (эритроциты, альбумины и глобулины), величина которого составляет для эритроцитов 6,061 • 10пе, для альбуминов 1,504 • 1019е, для
43
глобулинов 2,385 • 1012е (S = 180,31 • 10пе), с общим количеством отрица1ельных ионов 1 мм3 крови, которое приближенно равно 8,0 • 101ве, легко увидеть, что лишь 0,023% всего количества отрицательных ионов плазмы принимает участие в образовании электрического заряда основных частиц крови.
Итак, теоретически полученные нами, по примеру Зигмонди, Гевеей, Льюиса и других исследователей, значения абсолютного электрического заряда эритроцита выражаются в сотнях тысяч и миллионах элементарных зарядов и варьируют в зависимости от метода вычислений в больших пределах. Эффективный заряд эритроцита должен быть значительно меньшим. Ориентировочным определениям эффективного заряда эритроцита посвящена следующая глава.
Глава II
ПРИБЛИЖЕННЫЕ МОДЕЛИ ЭЛЕКТРОСТАТИЧЕСКОГО ВЗАИМОДЕЙСТВИЯ ЭРИТРОЦИТОВ В КРОВОТОКЕ
§ 1. Кулоновское взаимодействие
Левин и независимо от него Ленгмюр изучили роль электрических сил в устойчивости коллоидных систем и пришли к следующим заключениям. При сближении частиц гидрофобного золя между ними появляется электростатическое притяжение даже в том случае, если частицы несут заряд одной и той же полярности. Данное притяжение возникает в результате взаимодействия между первой частицей и двойным слоем второй частицы, а также между второй частицей и двойным слоем первой частицы. Когда частицы несут одинаковый заряд, они отталкивают одна другую. В случае приближения частиц друг к другу сила отталкивания быстро возрастает. Следовательно, между коллоидными частицами существуют электростатические силы притяжения и отталкивания, взаимодействие этих сил обусловливает поведение двух частиц. При очень тесном сближении приходят в действие значительные вандерваальсовские силы притяжения и частицы соединяются.
Дерягин и Ландау, исходя из уравнения Дебая — Гюккеля, показали, что взаимодействие одноименно заряженных поверхностей всегда сводится к отталкиванию (в противоположность расчетам Левина и др.). Принимая во внимание действие вандервааль-совских сил, упомянутые авторы нашли, что кривая результирующего отталкивания может иметь максимум на близких расстояниях и минимум — на удаленных.
Для сугубо предварительного выяснения электростатического взаимодействия между эритроцитами, которые не являются частицами коллоидной дисперсности, можно применить закон Кулона. Правда, следует ожидать, что применение закона Кулона к изучению интересующих нас явлений встретит резкие возражения. Но еще Лоренц в своих лекциях по термодинамике указал, что вследствие невозможности дать полное исследование того, что происходит в жидкости, окружающей взвешенную частицу, приходится ограничиваться рассмотрением весьма упрощенных задач.
Коль скоро мы станем на ту точку зрения, что в кровяном русле белковые частицы, эритроциты и другие взвеси электростатически взаимодействуют одна с другой, а против такой точки зрения вряд ли кто-либо найдет достаточно аргументов, мы будем вправе в качестве первого приближения обратиться к работе кулоновых сил.
45
Как известно, закон Кулона гласит: одноименно заряженные тела отталкиваются в вакууме с силой, прямо пропорциональной их электрическому заряду и обратно пропорциональной квадрату расстояния между центрами взаимодействующих тел, т. е. так, как если бы весь электрический заряд, находящийся на поверхности этих тел, был бы сосредоточен в их центрах:
__р ___ 61е2 (Гг + г^ ’
где ек и е2 — число электрических зарядов на поверхности одного и другого взаимодействующих тел; гг и г2 — радиусы этих тел; г± + г2 — расстояние между их центрами. Знак минус перед F указывает, что речь идет об отталкивании тел. В дальнейшем он опускается.
Закон Кулона позволяет установить единицу электрического заряда: за абсолютную единицу принимается такой заряд, который отталкивает равный ему заряд, расположенный на расстоянии 1 см в воздухе, с силой 1 дин.
Приведенная формула пригодна для вычисления силы отталкивания между двумя одноименно заряженными телами в вакууме. Но, как известно, взаимодействие двух наэлектризованных тел зависит от того, находятся ли вблизи них другие тела или нет: наэлектризованные тела индуцируют на них электрические заряды и эти заряды вступают во взаимодействие с первыми. Вопрос сильно осложняется в том случае, когда постороннее тело заполняет собою все пространство между заряженными телами. Таким промежуточным телом может быть только диэлектрик, но не проводник. Кровь и представляет собою диэлектрик. При погружении в диэлектрик сила электростатического взаимодействия между наэлектризованными телами уменьшается соответственно электрическим особенностям диэлектрика. Вокруг заряженного тела из диэлектрика собираются положительные заряды, которые соответственно уменьшают силы взаимодействия между наэлектризованными телами.
При внесении тел в среду с известной диэлектрической постоянной D сила электростатического взаимодействия соответственно уменьшается:
Д’ 1 ^1^2 ^1^2
= * (Г1 + r2y D (гх + *
Здесь D — диэлектрическая постоянная плазмы крови, характеризующая препятствие, оказываемое двум телам при приближении их один к другому. Согласно Фюрту, D = 85,5. Таким образом, взаимодействие двух заряженных тел, находящихся в крови, в 85,5 раз слабее, чем взаимодействие таких же тел, также заряженных и также расположенных, но находящихся в вакууме.
Если F выражена в динах, а е — в электростатических единицах, то D имеет размерность отвлеченного числа, причем для пустоты D » 1, а для хороших проводников приближается к бесконечности. В диэлектриках D = 1 4- 100.
46
Допуская, что на поверхности обоих взаимодействующих эритроцитов находится равное число зарядов, т. е. ег = е2 = е, и что* радиусы эритроцитов равны, т. е. г± = г2 или гг + г2 = /?, предыдущее выражение можно переписать так:
F = - е —
DR2 *
Формулу Кулона можно представить в следующем виде, удобном для получения числа е:
с е2 (4,8 • 10~ 10е)2 _ 23,49 • 10~15 • е2
D(10“4 R)2 • 981 “ DR2
DR2
Отсюда
о__ D 1/^Ю16 е R V 23,49 ’
где е выряжено в электронах, R — в микронах, F — в граммах.
Закон Кулона рассматривает электростатическое взаимодействие между центрами заряженных тел сферической формы. Хотя характер ньютоновских и кулоновых сил различен, но формулировка законов идентична: в законе Ньютона идет речь о том, чго два однородных тела шаровой формы притягиваются друг к другу с силой, прямо пропорциональной произведению их масс и обратно пропорциональной квадрату расстояния между центрами этих масс. В случае тел нешаровой формы этот закон приобретает другую формулировку и аналитический вид выражения сил взаимного притяжения в этом случае приобретает форму пертурбационной функции.
При условии электростатического взаимодействия между телами нешаровой формы закон Кулона также значительно усложняется. При взаимодействии между двумя одноименно наэлектризованными пластинками или дисками, подвешенными на шелковых нитях, максимально устойчивое положение этих дисков будет наблюдаться только тогда, когда эти плоскости дисков расположатся параллельно. При иных взаиморасположениях, например при косом положении одного диска относительно другого, состояние косостоящего диска вследствие электрических сил отталкивания будет неустойчивым, в результате чего возникнет вращающий момент, который будет стремиться привести эти диски к взаимно параллельному положению.
Это явление легко проверить на опыте, подвесив на тонких шелковых нитях два бумажных кружка диаметром 0,5 см. Пусть один кружок стоит под углом к другому, не касаясь последнего. Достаточно поднести снизу к этим кружкам натертую эбонитовую палочку, как оба кружка в силу электростатической индукции однозначно зарядятся и будут стремиться стать во взаимно параллельное положение своими плоскими сторонами. В этом простом опыте легко подметить наличие вращающего момента, особенно у кружка, стоящего косо, сдвига и параллельного выравнивания обоих кружков
47
относительно друг друга, с одновременным их отталкиванием друг от друга.
Из рис. 10 видно, что взаимодействие между электрическими зарядами, находящимися на поверхности двух пластинок, стоящих на рисунке в профиль, будет неодинаковым: > г2 > г3, а следовательно, силы электростатического отталкивания будут неодинаковы, а именно fх < f2 <z f3. Это обстоятельство должно повлечь за собой появление вращающего момента и соответственно сдвинуть пластинки, как показано на рис. 10. Можно составить такую схему:
Неустойчивое равновесие------
I *
I Отталкивание
Вращающий момент------------
I *
। Отталкивание
Выравнивание
Ф
Устойчивое равновесие
Этой схеме подчиняются все возможные взаимные расположения
бумажных кружков, висящих на шелковых нитях, кроме случая, когда кружки расположены в одной плос-
Рис. 10. Схема электростатического взаимодействия двух пластин.
кости и обращены друг к другу своими ребрами. При поднесении к ним натертой эбонитовой палочки кружки расходятся, отталкиваясь друг от друга на соответствующее величине заряда расстояние, не отклоняясь от заданного им до электризации положения, т. е. их оси, проходящие через центр симметрии, остаются параллельными друг другу. Описанные выше опыты имеют значение для теоретической интерпретации вопроса о взаиморасположении эритроцитов в плазме крови.
Электростатические силы, действующие между эритроцитами, образуют свое-
образное силовое поле. Качественная сто-
рона этого силового поля может быть выявлена на основании тех предпосылок, которые были положены в основу наших представлений об эритроците, егоформе и плотности распределения его заряда как функции положительной кривизны его поверхности. Вне зависимости от ко-
личественной стороны заряда эритроцита принятая гипотеза о плотности его распределения позволяет нам создать приближенную картину электростатического силового поля эритроцитов. Для составления картины силового поля эритроцитов, показанной на рис. 11, были приняты следующие положения.
1. Плотность распределения зарядов на поверхности эритроцита является функцией положительной кривизны его поверхности.
48
2. Силовые векторные линии направлены в ту сторону, куда круче всего падает потенциал; величина этих векторов будет равна падению потенциала на единицу длины нормали к эквипотенциаль-
Рис. 11. Схема электростатического распора между эритроцитами: -----------силовые линии; —-----------эквипотенциальные поверхности.
ной поверхности в данной точке. Но поскольку каждая линия дает только направление, а не величину вектора, то размещение линий векторов на рисунке гуще там, где градиент потенциала больше и реже, т. е. дальше друг от друга там, где градиент потенциала меньше.
49
3. Все векторные линии градиента потенциала во всех точках встречи с эквипотенциальными поверхностями должны быть к ним перпендикулярны.
4. Густота линий зависит от расположения эквипотенциальных поверхностей. Большая густота векторных линий наблюдается там, где эквипотенциальные поверхности подходят ближе друг к другу и наоборот.
Таблица 12
Зависимость сил электростатического взаимодействия между двумя эритроцитами от величины заряда и расстояния между центрами эритроцитов
е R, мк F‘ = 10“ [Л"<1
50 1,885 0,748 0,420 0,270 0,187 0,137 0,105
100 6,72 2,99 1,68 1,08 0,748 0,549 0,42
200 27,0 12,0 6,73 4,31 2,99 2,2 1,68
300 60,7 26,95 15,2 9,70 6,74 4,95 3,79
400 108,0 48,1 27,0 17,5 12,0 8,8 6,75
500 188,5 74,8 42,0 27,0 18,7 13,7 10,5
750 379 168 94,7 60,6 42,0 30,9 23,7
1000 672 299 168,0 107,6 74,8 54,9 42,0
1500 1520 672 378 243 169 124 94,9
2000 2700 1200 673 431 299 220 168,0
2500 4210 1870 1050 674 468 344 263,0
3000 6070 2695 1520 970 674 495 379,0
4000 10 800 4810 2700 1750 1200 800 675,0
5000 18 850 7480 4200 2700 1870 1370 1050,0
6000 23 600 10 800 6060 3880 2700 1980 1515,0
7000 33 100 14 700 8250 5300 3670 2700 2067,0
8000 43 200 19 200 10 800 6900 4800 3520 2700,0
9000 54 600 24 300 13 650 8740 6060 4450 3410,0
10 000 67 200 29 900 16 800 10 760 7480 5490 4200,0
Силсвэе поле эритроцитов, показанное на рисунке, изображено в соответствии с приведенными выше положениями. Эта схематическая картина силового поля дает представление о том электростатическом «распоре», который имеет место в крови между отдельными эритроцитами.
Теорию электростатического взаимодействия между эритроцитами можно рассмотреть по частям и начать ее изложение с простейшего случая. Применяя формулу Кулона и взяв различные значения для расстояний между центрами сферулированных эритроцитов /?, находим соответствующие значения силы электростатического взаимодействия Fe между эритроцитами
л2
= 85,5 • /?2 [дгш],
где е — число электрических зарядов, k\ D = 85,5 — диэлектрическая постоянная крови; R — расстояние между центрами эритроцитов, см, т. е. все значения представлены в системе CGSE. Но так как в дальнейшем заряд исчисляется в электронах, a R —
50
в микронах, вводим переводные множители: I электрон = = 4,8025 • 10~'° ед. CGSE; 1 мк 10-4 см.
Если на поверхности каждого из двух взаимодействующих эритроцитов имеется один электрический заряд, то, следовательно, сила электростатического взаимодействия между эритроцитами будет F __ е2 _ (4,8025 • 1О~10)2 23,04 • 10“2 0,2696 • 10~12
^—85,5^— 85,5 (1 О'”4/?)2 ~ 85,5 • ЦТ8/?2 “ *2 [диН]>
ИЛИ
г 0,27 . 10“15 г .
Fe —-----&----1г].
Подставляя в это выражение значения R от 1 до 8 мк, получаем соответствующие значения Fe. То же выполняем и для значений е
Рис. 12. Сила электростатического взаимодействия между эритроцитами при разных значениях £, вычисленная по формуле Кулона.
от 500 до 10 000 элементарных зарядов. Результаты приведены в табл. 12 и на рис. 12, где показана зависимость Fe = f (R).
§ 2. Электростатическое взаимодействие между двумя эритроцитами, расположенными в параллельных плоскостях
Определение электростатической силы взаимодействия между эритроцитами по закону Кулона при предположении о точечной локализации зарядов не является точным. Чем крупнее взаимодействующие частицы по сравнению с расстоянием между этими частицами и чем более форма поверхности частиц отличается в действительности от шарообразной, тем более величина сил кулоновского взаимодействия зависит от формы поверхности и взаимного относительного расположения частиц.
51
Попытаемся найти более уточненное решение задачи о силе электростатического взаимодействия между двумя эритроцитами. Представим себе, что наиболее значительная часть заряда эритроцита расположена на его периферической торовидной выпуклой части, как это схематически показано на рис. 13. Это соответствует плот-
ности размещения зарядов пропорционально положительной кривизне его поверхности. Упрощая данное представление, предположим, что весь заряд эритроцита расположен с равномерной плотностью на полуокружности выпуклой наружной части тора.
Рис. 14. Схематическое изображение сил электростатического взаимодействия двух заряженных колец, лежащих в параллельных плоскостях.
Рис. 13. Схематическое изображение электростатического взаимодействия между двумя эритроцитами.
Определим центр тяжести дуги (полуокружности) выпуклой части тора относительно оси вращения, на которой сосредоточен заряд эритроцита. Из рис. 13 следует:
ON = OB — NB,
D
где OB = R — радиус эритроцита и NB = -^------половина высоты
(толщи ны) эр итр оцита.
Следовательно,
ON = R--$- = ±R.
Центр тяжести дуги АВС определяется относительно вертикальной оси эритроцита из выражения
л
OP = ON + NB------+ * 1.
1 л 4 1 4 л
Отсюда
4л
Определив величину радиуса эквивалентного заряженного кольца, рассмотрим силы электростатического взаимодействия между двумя такими кольцами, в зависимости от величины их заряда и расстояния между ними.
52
Рассмотрим силы взаимодействия двух таких заряженных колец (рис. 14), которые лежат в параллельных плоскостях и центры которых находятся на одном перпендикуляре к этим плоскостям. На каждом кольце расположен заряд Е — 2лро, где о — плотность заряда на единицу длины кольца.
Возьмем на первой окружности произвольную точку Мо с плотностью заряда а. Действие точки Мо на точку М, находящуюся на второй окружности, выразится так:
F __ c2dS г 9
а на всю вторую окружность — г с c2dS
где (At0Af)2 = г2 + 4р2 sin2 ср, так как (Л!^)2 = (At0Р)2 + (Л4Р)2. Но MqP = г, ибо плоскости окружностей параллельны, a MGP — перпендикуляр (расстояние между окружностями); МР == 2р sin <р — катет прямоугольного треугольника MPN; ср — угол между касательной и проекцией М0М на вторую окружность МР.
Отсюда
dS = У(МР)2 + (MP")2dq.
Так как хорда МР представляет собой радиус-вектор, являющийся функцией угла ср, то МР = 2р sin <р.
Производная МР' = 2р cos <р. Следовательно,
dS = 4р2 sin2 <р 4- 4р2 cos2 cprfcp = 2pd(p.
Подставляя в выражение для Fe(Mj значения dS и MGM, получаем л л
р (М \ -= С 2P°2d(P = 2оа2 ——d(p =
е ' Q' J г1 4- 4р2 4- sin2 <р “ J г1 4- 4р2 4- sin2 (р о о
л
_________dtp_________
г2 4- 4р2 4- sin2 ф
о
Ввиду того, что подынтегральная функция четная, то пределы интегрирования можно заменить: вместо — от 0 до л принять от О до и удвоить все интегральное выражение. В результате получим
Л/2
Fe(M) = 2-2ро2 (
е ’ r J г2 4- 4р- 4- sin2 ф о
Производим замену tg ср = и, учтя, что sin <р =----------------------,
(14-1^ф)^
dip
53
получим
F,(M.) = 4ро! С --------%—---------,
«к,+">,-«г+эг](,+“,)
причем пределы интегрирования для и, равного 0 и оо: оо
е (Ч) = 4pO2 j f2 + (f2 4р2) •
О
Вынося г2 как общий множитель, получаем:
du
r* + 4Р2 2 ’
------2---- “ г2
. /г2 + 4р2 Т arctg------у н и I.
о
г2 \ о
Fe(M0)~ +
Интегрируя, находим F (Л1)= 2яРр
*' °' rVr»-i-4pt
Действие силы F по всей окружности параллельных колец будет выражено так:
2^(4)= у ;2,Тг-г у г2 4рг
Упрощая это выражение, получаем в окончательном виде
F — —Е [дим]. е /- 1<г24- 4р2
Пользуясь выведенной формулой и подставляя численные значения для Е, г и р, а также вводя диэлектрическую постоянную D, определяем силу электростатического отталкивания между двумя эритроцитами как функцию заряда Е и расстояния между ними г.
Пример:
р =___________(1,44 10~7)2______ =
е 3 • 10~4 ]/(3 • 10-4)2 + 4 (3,37 • 10~4)2 D _________2,074 • 10~14____2,074 • 10~14 =*
З Ю"41^9 • 108 + 45,44 • 10“8D 3 • Ю"4 V54,44 • lO^D
______________________________=2&^=11. Ю-10 дим, 3 • 10~4 • 7,38 • 10~4 • 85,5_1890 • 10“8
где Е = 300 ед. CGSE ~ 300 • 4,8 • 1О~10 = 1,44 • Ю-7 электростатических единиц; г = 3 • 10-4 см — расстояние между эритроцитами; р = 3,37 • 10“4 см — эквивалентный радиус эритроцита; D = 85,5 — диэлектрическая постоянная крови.
54
Вычисленные по выведенной формуле силы электростатического взаимодействия между двумя эритроцитами в зависимости от величины заряда и расстояния между центрами эритроцитов приведены в табл. 13.
При сопоставлении величин, приведенных в данной таблице, с соответствующими им величинами, рассчитанными при допущении точечной локализации, видно, что в случае малых расстояний между
Таблица 13
Зависимость сил электростатического взаимодействия между двумя эритроцитами, расположенными параллельно, от величины заряда и расстояния между эритроцитами
Е Fe«1010, дин
г=2 • 10—4 см з-ю—4 4-ю-4 5-Ю-4 6-10—4 7-10-4 8-10—4
50 0,478 0,304 0,215 0,158 0,124 0,10 0,08
100 1,91 1,22 0,86 0,62 0,50 0,40 0,32
200 7,64 4,87 3,44 2,52 1,98 1,59 1,30
300 17,2 11,0 7,72 5,63 4,48 3,56 2,91
400 30,5 19,5 13,7 10,05 7.96 6,35 5,19
500 47,8 30,4 21,5 15,75 12,4 22,3 18,2
750 108,0 68,4 48,2 35,3 28,0 22,3 18,2
1000 191 122 85,7 62,4 49,6 39,5 32,3
1500 430 274 193 141 112 77,5 72,5
2000 764 487 344 252 198 159 130
2500 1190 761 537 393 318 248 202
3000 1720 1100 772 563 448 356 291
4000 3050 1950 1370 1005 796 635 519
5000 4780 3040 2150 1575 1240 990 807
6000 6750 4880 3440 2520 2040 1585 1295
7000 9360 5970 4200 3080 2440 1950 1590
8000 10 760 7800 5500 4020 3180 2540 2070
9000 15 450 9860 6950 5240 4030 3200 2620
10 000 19 100 12 200 8570 6240 4960 3950 13 230
эритроцитами (например, 2 мк) при этом предположении значения силы более чем в три раза превышают аналогичные им значения, полученные из рассмотрения двух параллельных колец с зарядами. Это следует считать вполне понятным, если учесть то обстоятельство, что диаметр эритроцита (d = 7,5 • 10~4 см) в этом случае намного больше расстояния между двумя эритроцитами, что исключает возможность получения правильного результата при изучении сил взаимодействия в предположении сосредоточения зарядов в центре. Чем больше увеличивается расстояние между эритроцитами, тем ближе значения F, определяемые по обычной формуле Кулона, к значениям Fe, определяемым по формуле, выведенной нами для двух параллельных колец. Например, при г = = 8 мк разница между теми и другими значениями составляет уже только 30%.
55
Рассматривая формулу, выведенную для случая двух заряженных колец, убеждаемся, что подкоренное выражение (г2 + 4р2) всегда будет несколько больше г2, в связи с чем силы взаимодействия, определяемые по этой формуле, всегда будут несколько меньше кулоновых. Но при величине г, намного превышающей величину р» разница между результатами,
Рис. 15. Электростатическое взаимодействие между двумя эритроцитами, вычисленное по формуле Кулона (/) и по формуле взаимодействия двух заряженных колец (2).
получаемыми по формуле Кулона и по формуле двух колец, будет становиться все меньше и меньше, что и соответствует физической стороне явления. Для иллюстрации этого положения на рис. 15 приведены две кривые F (г) и Fe (г) (при Е = = 10 000), значения для которых получены обоими методами.
Необходимо оговориться, что значения силы Fe, приведенные в табл. 13, верны лишь в том случае, когда рассматриваются силы взаимодействия двух изолированных эритроцитов. В реальных условиях, в русле кровотока мы имеем дело со множеством эритроцитов, сгруппированных в кольца, причем каждый отдельный эритроцит находится под воздействием сил окружающих его соседних эритроцитов. Таким образом, в действительности, каждый отдельный эритроцит взаимодействует не с одним эритроцитом, а с двумя, находящимися в параллельных (или расположенных под небольшим углом) с ним плоскостях, в связи с чем силы взаимодействия должны быть при дальнейших расчетах уменьшены в четыре раза по сравнению с цифрами, показанными в таблице. Уменьшение силы взаимодействия в четыре раза связано с тем, что при расчете силы Fe величина заряда фигурирует в числителе последней формулы во второй степени, а при уменьшении действующей величины заряда вдвое сила Fe уменьшается в четыре раза.
§ 3. Электростатическое взаимодействие между двумя эритроцитами, расположенными в одной экваториальной плоскости
При рассмотрении радиально-кольцевой модели эритроцитов, движущихся в русле кровотока, необходимо допустить, что отдельные кольца эритроцитов, следуя вперед одно за другим, удерживаются
56
ана-
подойти к
V
Рис. 16. К расчету потенциала точки Мо.
на определенных расстояниях друг от друга силами электростатического отталкивания. Эти же силы в значительной степени определяют расстояния между отдельными концентрически расположенными кольцами вращающихся эритро лизу этих сил, попытаемся в порядке первого приближения рассмотреть силы взаимодействия между двумя эритроцитами с общей экваториальной плоскостью.
Для решения этой задачи представим себе заряд эритроцита в виде кольца с равномерной плотностью зарядов подобно тому, как это было сделано в предыдущем параграфе. Радиус р такого эквивалентного кольца был определен как 0,9 радиуса эритроцита, т. е. 3,37 • 10-4 см.
Найдем потенциал точки Л40, расположенной в пространстве, вне плоскости заряженного кольца с радиусом р и плотностью заряда о (рис. 16). По-
тенциал точки Мо, находящейся в силовом поле заряженного кольца:
V(г, 0) = \-^г.
Рассматривая на рис. 16 треугольники OMoN и 0NM, можно записать
ЛШо = ОМг + О№ — 20 М ON cos q> + NM20,
где ON = r sin 0 и NMQ = r cos 0.
Подставляя значения ON и NM0 в последнее выражение, получаем
ММо = р2 + г2 — 2pr sin 0 cos <р.
Рассмотрим случай, когда р < г, т. е. тот случай, который соответствует двум несоприкасающимся эритроцитам:
1__________________1_____________
Л1Л10 Г р \2 р
г 1 / 1 + I— ) — 2 — sin 0 cos <р
Разложим это выражение в ряд по полиномам Лежандра:
OQ
------ ' 77 ' = S (-Т-)" рп <sin ° cos ф)-
г Л/ 1 + 1—j +2 — sin 0 cos ф п=0
57
Выражение для потенциала точки Мо примет вид 2л JC
у (r’е) = f —/
о r I / 1 + ( — I — 2 — sin 0 cos ф
2л ар р
dtp________________
я Р | — 2 — sin 0 cos tp
так как dS = рб/ф. Пользуясь разложением по полиномам Лежандра, получим
V(r, 0)=-^-
2П - QQ
\ У (—) Рп (sin 9 cos ф) t/ф, О L п=о \ / J
или
2л
V (г, 0) = -у- S (-7-)" У Рп <cos ч’sin е) d(₽-' ' о
Ввиду того, что а Рп<1> обрывая раложение на
п == 4, находим Г 2л 2л
V (г, 0) = Ройф + — Pi (cos ф sin 9) </ф 4-г о г о
2л 2л
' ° ' Г ' :os ф sin 0) йф + ( —) [ Р3 (cos ф sin 0) dtp 4-' ' о
2л
+ (-“У I jP4 (cos ф sin 0) б/ф . \ / о
о
При этом
Ро = 1; Pr = cos ф sin 0; Р2 = (3 sin2 0 cos ф —
Р3 = Ц- (5 sin3 0 cos3 ф — 3 sin 0 cos ф);
Р4 = -у- sin4 0 cos4 ф-----sin2 0 cos2 ф 4- .
Взяв от каждого полинома интеграл по б/ф, получаем:
2л 2л 2л
| б/ф = 2л; | sin 0 cos фб/ф = sin 0 • 0 = 0;
b ob
2л 2л
P2dtp = Т- (3 sin2 9 cos q> — 1) t/ф = (3 sin2 0 — 2);
о 6
58
i Ptdq> = | f-y- sin4 9 cos4 <p-sin2 9 cos2 <p + dq> =
ob' '
3,35л . 4A 15 . 2n 3л
“32“sln 9-----T-nsin29-----4-
Интеграл P3 = 0 из-за нечетности степеней тригонометрических функций и получающихся при этом синусов в нечетной степени. Принимая во внимание, что пределами интеграции являются нуль и 2л, значение синусов окажется равным нулю.
Подставляя после интегрирования значения отдельных членов в выражение потенциала, получаем:
V(Г, 9) = [2л + 4-(sin29-2) +
+1 (-г)4 (лг3 sin‘9 —гsin2 9 + 4) ] •
£
По условию заряд Е = 2лра, откуда о = .
Подставляя вместо о его значение и отбрасывая третий член выражения потенциала, поскольку он представляет собой
Рис. 17. К расчету силы взаимодействия между двумя заряженными кольцами, расположенными в общей экваториальной плоскости.
дробь, меньшую единицы (дробь (jyj меньше единицы, а /35 15 з \
13 “8"sin40 — -g-sin20 +“g"l меньше или равно единице), находим для потенциала в точке Л1о следующее выражение:
V (г, 9) = -у- [ 1 + —п24-~2
59
Составляющие напряженности поля выразятся так:
dv _ av dr dv ае
дх dr дх ' дв дх ’
ст _ dy dr аг ао
ду дг ду **" ав ду ’
av av ar av ао
dz ~ dr ’ дг ' d0 дг *
Для нашего случая:
аг £х г 1 . 3 р2 /о . 2 п п\1 • е £ • д д ао
-г— =------1 + -г- -Цг- + (3 sin2 0 — 2) + 6 sin 0 cos 0 -г—
дх г3 | 1 4 г2 ’ v 71 г3 dx
aV Еу I « . 3 p2 /Q . 2 n c E . n д dO
-г— =------f- 1 + (3sin20 — 2) 4- 6—r sin0cos 0-7—;
dy r3 i 1 4 r2 v 71 ' r2 dy ’
dV Ez Г i । 3 p2 /Q . 2 д 1 । a E • д д ^0
-Д— =------x- 1 + -r- -V(3sm20 — 2) + 6-^sin0cos0-7-.
dz r3 [ 4 r2 v 7 1 r2 dz
Таблица 14
Зависимость сил электростатического взаимодействия между двумя эритроцитами, лежащими в общей экваториальной плоскости, от величины заряда и расстояния между центрами
Е Ffi-10*0, дин
а = 8-10* см 8,5-104 9-104 9,5-10* 10,5 ю4
b = 0,5 • 10* см 1,0 10* 1,5-10* 2,0-10* 3,0-10*
50 0,115 0,085 0,065 0,055 0,04
100 0,43 0,34 0,27 0,23 0,16
200 1,74 1,36 1,08 0,98 0,63
300 3,85 3,08 2,43 2,08 1,4
400 6,85 5,45 4,33 3,68 2,48
500 10,7 8,5 6,7 5,75 3,9
750 24,2 19,2 15,2 13,0 8,8
1000 42,8 33,8 27,0 23,0 15,5
1500 96,3 76,3 60,8 51,5 34,8
2000 171,5 136 107,5 92 62
2500 268 213 169 144 87
3000 385 307 243 207 140
4000 685 544 432 367 247
5000 1070 850 673 574 386
6000 1538 1223 970 825 558
7000 2100 1650 1320 1125 760
8000 2925 2175 1725 1470 994
9000 3475 2750 2185 1860 1258
10 000 4275 3388 2700 2295 1545
60
Для случая, когда точка Мо лежит в плоскости заряженного кольца, т. е. когда 0 = -у, можем записать:
dV Ex л з р2\
dx г2 \1 + 4 г2 у;
дУ ______Еу_[} , _3_ PLV
ду “ г2 + 4 ’ г2 р
2К_ = _ _Ё£/1 4-А АЛ
dz г2 Р + 4 ’ г2 /•
Напряженность поля по радиусу вектора г будет
Рис. 18. Зависимость электростатического взаимодействия между двумя эритроцитами, лежащими в общей экваториальной плоскости, от величины заряда и расстояния между их
Н ., / / dV\2 ldV \2 if dV \2
H=±v м--(*)+w=
Отсюда сила, действующая на точку М0‘.
где D — диэлектрическая постоянная среды.
Перейдем к определению силы взаимодействия между двумя заряженными кольцами, расположенными в общей экваториальной плоскости (рис. 17).
Сила взаимодействия между двумя заряженными кольцами I и II выразится так:
Fei(a) = $F(r)dS, или
FUa) = f-^-(1 + -r- Я45,
центрами.
где г — функция дуги S.
Разбивая первый интеграл на два интервала, взятых отдельно для каждой полуокружности f(NM и KM.L, получим
Ftl(a) — ( F(r)dS+ f F(r)dS. KNM NLK
61
Применим теорему о среднем значении:
FC1 (а) = F (r0) J dS + F (r0) J dS, или Fei (а) = F (г0) 2лр, KNM MLK
где г0— среднее значение переменной г функции F (г).
Подставляя значения из выражения для этой функции и интегрируя, имеем
р (а\ — 2лрст£ (J _1_____. Р2 \
е'Щ) D(a2 + p2) 4 а2 + Р2 /
Заменяя 2лро == Е в последнем выражении, получаем окончательную формулу для случая двух заряженных колец в одной экваториальной плоскости, где в основу расчета берется электростатическое воздействие всей длины одного кольца на всю длину другого кольца:
Fe. (а) = н (1 + -4- • 2 ? 2 ) [дин],
1' 7 D (а2 + Р2) \ 1 4 а2 -4- р2 j L п
где D — диэлектрическая постоянная среды.
Если при рассмотрении сил взаимодействия двух заряженных колец сделать допущение, что взаимодействие вызывается только зарядами, расположенными на обращенных друг к другу полуокружностях (PRS и NKL), последнее выражение примет вид pin---fw-—
Это выражение следует считать наиболее приемлемым для определения сил электростатического взаимодействия между двумя эритроцитами в русле кровотока, если учесть, что эритроциты следуют один за другим, причем одна половина их заряда обращена в сторону идущего впереди эритроцита, а вторая половина заряда взаимодействует с эритроцитом, идущим следом за ним.
Результаты вычислений сил взаимодействия Fe по окончательной формуле в зависимости от заряда Е и расстояния между центрами эритроцитов (а) представлены в табл. 14 и на рис. 18.
Глава III
ПРИБЛИЖЕННАЯ МОДЕЛЬ МАГНИТНОГО ВЗАИМОДЕЙСТВИЯ ЭРИТРОЦИТОВ В КРОВОТОКЕ
§ 1. Магнитное взаимодействие двух вращающихся эритроцитов, находящихся в параллельных плоскостях
Опытами Эйхенвальда, Роуланда и Хистеда, проведенными в разное время, было доказано, что вращающийся электрический заряд-возбуждает электрический ток и магнитное поле.
Согласно общепринятому определению, электрический ток — это количество электричества, проходящее за 1 сек через поперечное сечение проводника. Отсюда следует, что можно получить электрический ток механически, зарядив изолированный проводник некоторым количеством электричества и двигая его вместе с зарядом.
Приклеим к стеклянному кружку концентрическое станиолевое кольцо, зарядим его электричеством и приведем стеклянный кружок в быстрое вращение. Заряд кольца будет действовать как электрический ток и будет вызывать магнитные явления.
Несмотря на все трудности экспериментальной проверки этой идеи (получающееся магнитное поле слабее земного магнитного поля в 100 ООО раз), опытами Эйхенвальда было доказано, что движущиеся электрические заряды производят магнитные действия. Оказалось также, что на результат опыта не влияет способ вращения кольца: вращается ли оно как целое или будет разрезано по радиусам на изолированные одна от другой части. Следовательно, можно сказать, что всякое движущееся заряженное тело эквивалентно некоторому электрическому току. Ток, вызванный электростатическими зарядами, перемещающимися вместе с материей, получил название конвекционного или кругового тока.
Поступательно-вращательное движение эритроцитов, сгруппированных в кольца в русле кровотока, и наличие на эритроцитах зарядов обусловливают возникновения конвекционных токов. Эти токи, в свою очередь, вызывают появление магнитных полей, в результате действия которых вращающиеся эритроциты уподобляются торовидным магнитам, взаимно притягиваются, а тем самым обеспечивают устойчивость радиально-кольцевых систем в кровеносных путях.
Величина конвекционных токов определяется произведением числа оборотов эритроцита в секунду п на величину его заряда Е, т. е. i = пЕ.
Чтобы получить величину кругового тока в амперах, необходимо величину заряда перевести в абсолютные электростатические
63
единицы, т. е. умножить на 4,8 • 10 7, а величину кругового тока — на „ • Например, при заряде эритроцита Е = 1000 эле-
ментарных зарядов или 4,8 • 10“7 абсолютных электростатических единиц и числе его оборотов в секунду п = 50 величина кругового тока ia составит 4,8 • 10~7 • 50 • 3 = 80 • 1Г16 а.
В табл. 15 приводятся значения круговых токов в зависимости от заряда Е и числа оборотов п.
Таблица 15
Зависимость величины конвекционных токов, возникающих при вращении эритроцита, от его заряда Е и числа оборотов п в секунду
М01в, а
Е п = =10 об[сек 20 30 40 50 70 90
50 0,8 1,6 2,4 3,2 4,0 5,6 7,2
100 1,6 3,2 4,8 6,4 8,0 11,2 14,4
200 3,2 6,4 9,6 12,8 16,0 22,4 28,8
300 4,8 9,6 14,4 19,2 24,0 33,6 43,2
400 6,4 12,8 19,2 25,6 32,0 44,8 57,6
500 8 16 24 32 40 56 72
750 12 24 36 48 60 84 108
1000 16 32 48 64 80 112 144
1500 24 48 72 96 120 168 216
2000 32 64 96 128 160 224 288
3000 48 96 144 192 240 336 432
4000 64 128 192 256 320 448 576
5000 80 160 240 320 400 560 720
6000 96 192 288 384 480 672 864
7000 112 224 336 448 560 784 1008
8000 128 256 384 512 640 896 1152
9000 144 288 432 576 720 1008 1296
10 000 160 320 480 640 800 1120 1440
10 000 000 160-103 32103 48103 64 -103 80 103 1 112-103 1 144 103
Магнитное поле эритроцита, возникающее в результате круговых токов, имеет вид, показанный на рис. 19. Эритроциты, расположенные в параллельных (или под небольшим углом) плоскостях, обращены друг к другу разноименными полюсами и поэтому взаимно притягиваются.
Силы магнитного взаимодействия, т. е. взаимного притяжения, могут быть приближенно определены по законам Ампера и Био-Савара — Лапласа.
Представим себе эритроцит с его круговым током как один виток кругового тока. Радиус этого витка р примем равным 0,9 радиуса эритроцита, т. е. р — 0,9 • 3,75 • 10“4 = 3,75 • 10~4. Это соответствует такому распределению заряда на поверхности эритроцита, при котором предполагается, что весь заряд сосредоточен на
64
его торовидной выпуклой части пропорционально положительной кривизне его поверхности.
Пондеромоторное взаимодействие двух параллельных круговых витков тока в дифференциальной форме определяется следующей формулой (в векторном виде):
г12
Рис. 19. Магнитное поле эритроцита, возникающее в результате действия конвекционных токов при вращении оси S — N.
Значения входящих в формулу величин явствуют из рис. 20.
Интегрируя это выражение, получаем:
Рассматривая подынтегральную функцию как тройное векторное произведение, можем записать:
[б//2 • G2)] = ^1012 * ^2) 12 0^1 ’ ^2)’
Разложим вектор г12 на два взаимно ортогональных вектора а и гх (вектор г\ равен вектору M2M2)t т. е.
'12 = CL + Гр
Вектор а перпендикулярен плоскости круговых витков и по величине равен расстоянию между ними, а вектор rY лежит в плоскости кругового витка II.
Следовательно, тройное векторное произведение можно представить в следующем виде:
\dl2 (dlY • r12) ] = dlY (г12 • d/2) (я g) (Zx • d/2).
65
Заменяя в выражении для Fm векторное произведение последним выражением, получаем
Рис. 20. Взаимодействие двух параллельных круговых витков тока.
Рассмотрим второй интеграл и докажем, что он равен нулю:
Вынесем постоянные множители за знак интеграла:
Первый интеграл
так как подынтегральное выражение есть скалярное произведение и интегралы берутся по замкнутым контурам.
Второй интеграл
66
так как вектор г19 вращаясь вокруг точки М'2, может изменять свой аргумент от 0 до л, причем его значения симметричны относительно его максимального значения при .
Скалярное произведение
(d^ • dl2) = d/x • dl2 cos (dlly dl2),
но угол dllt dl2 = 2ф, следователь-л
но, при аргументе, равном у, скалярное произведение меняет знак на обратный, значит, и второй интеграл также равен нулю.
Отсюда
Рис. 21. К расчету г1а.
Подставляя скалярное произ-
ведение (гп • d/2) в виде — r18 dl2 cos (rla, dl2) в последнее выражение, находим
Т Т * ^12 ’ ^2 C^S (^12» ^2)
Я
</ и » 12
Вектор dlx = pd (2фх) = 2pd<px. Вектор d<px параллелен вектору а, т. е.
^Ф1 — [ а | ^Ф1> следовательно, векторы
= 2р б/фх,
dl2 — 2рб/ф2.
Вводим значения дифференциалов под знак интегрального выражения FM и получаем
Fm = /,/2
р dtp, • г12 • 2pd<p2cos (r12> dl2)
12
Вынося постоянные множители за интеграл, находим
FM = Wp8 Л- cos (гх,, у dq^ .
I а I J J /jg
Определяем cos (r12, dl2). Из рис. 21 следует:
Г12
67
Далее,
но
следовательно,
(MJ)2 = (ЛШ)2 + (М2Т)2,
М2Т = р ctg ф2, (MJ)2-a2 + p2ctg2<p2.
Из косоугольного треугольника МгТМ2 величина (Л^Т)2 может быть выражена через cos (r12, dl^ так:
(М,Т)2 = rf2 + (М2Т)2 - 2г12 (Л42Т) cos (r12,"d/2).
Сторона треугольника М2Т как касательная к окружности в точке М2 равна р ctg <р2. Из последних двух выражений получаем
а2 + р2 ctg3 <р., = а2 + 4р2 sin2 ф2 +
+ р2 ctg2 ф2 — 2р ctg ф2 Vа2 + 4р2 sin2 ф2 cos (r12, dZ2).
Далее,
cos (r12, dl2) =
2р sin ф2 cos ф2 Vа- + 4р2 sin2 ср2
Подставляя значение cos (r12, dl2) в интегральное выражение для FM, находим
FM = / / 4р2 -Д- 6$ Р Sin ф2 C°S
I а I J J (a2 + 4p2 sin2 (p2)
Подставляем пределы интегрирования:
FM
2л л
= Wp2-[7j-J d(Pl J
0 0
или
Далее,
2л
гЛ, = J аФ1
6
p sin ф2 cos ф2^ф2 (a2 + 4p2 sin2 ф2)*/в
р sin ф2 cos ф2^ф2 (а2 + 4р2 sin2 ф2)*/е
Л
2
1
Fm — —/х/24лр
а
! а2 + 4р2 sin2 ф2 I а I }
Подставляя пределы, получаем
^1'
или
1Л„| = 4Яр/Л^—jT—r)-
68
После перевода размерностей в систему CGS
F„ - 0,044,7,7, (4 - |<М.
Здесь ток / выражен в амперах. Магнитная проницаемость р, в формулу не -водится, так как предполагается, что для плазмы р = 1.
В качестве примера рассчитаем величину силы магнитного взаимодействия FM между двумя вращающимися эритроцитами, имеющими заряд Е =-- 1000, при п = 10 об/сек и при расстоянии между эритро
Рис. 22. Зависимость магнитного взаимодействия двух эритроцитов от заряда, расстояния и числа оборотов в секунду (для сравнения приведена зависимость Fe — f (г)).
цитами а = 2 • 10 4 см. Величина кругового тока /х = /2 для данного
Рис. 23. Зависимость магнитного взаимодействия двух эритроцитов от п (в точке FM = Fe силы электростатического отталкивания уравновешиваются силами магнитного притяжения).
случая составляет 16 • 10 16 а
X Ю-16 • 16 • Ю-16 • 9- 1О20
и
Ли = 0,04 • 3,14-3,37 • 10-16 х
1________________1__________=
2 : 10~4 4 - 10-8 + 4 • 11,4 - 10-8
= 975,2 - 10“16 • 0,36 • 10* =351 • 10“12 - 3,51 • 10“'° дин.
Вычисленные значения сил магнитного взаимодействия (притяжения) FM в зависимости от величины заряда £, расстояния между эритроцитами а и числа оборотов эритроцитов в секунду п приведены в табл. 16 и на рис. 22, 23.
69
Таблица 16
Зависимость величины силы магнитного взаимодействия между двумя эритроцитами от заряда Е, расстояния а и количества оборотов п_____________________________________________________________________________________________
Е • Ю10» дин
п = 10 об/сек п = 20 об/сек
a = 2-10“4 см з-ю~4 4 • io-4 5 -10—4 2-10-4 3-10~4 4-Ю-4 5 • 10“4
50 0,009 0,005 0,003 0,002 0,035 0,019 0,012 0,08
1СЭ 0,035 0,019 0,012 0,008 0,14 0,08 0,05 0,03
200 0,141 0,007 0,048 0,032 0,56 0,31 0,19 0,13
300 0,315 0,174 0,107 0,071 1,26 0,69 0,43 0,23
500 0,88 0,483 0,298 0,198 3,52 1,93 1,20 0,79
700 1,72 0,946 0,585 0,388 6,89 3,78 2,34 1,55
1000 3,51 1,93 1,195 0,79 14,05 7,7 4,78 3,16
2000 14,05 7,72 4,78 3,16 56,2 30,8 19,1 12,7
3000 31,5 17,4 10,74 7,10 126,4 69,2 43,0 28,4
5000 87,9 48,3 29,8 19,75 352 193 119,5 79,1
7000 172 94,6 58,5 38,8 689 378,5 234 155
10 000 351 193 119,5 79 1405 772,5 478 316
10 000 000 0,0351 10~10 0,0193 Ю-10 0,0119Х Х10-10 0,0079 X X Ю-1» 0,1405х Х10-» 0,773-10—10 0,0478X Х1(Г“ О,О315Х X10“w
Е п = 30 об/сек п = 50 об/сек
50 0,08 0,044 0,027 0,018 0,22 0,12 0,075 0,049
100 0,32 0,17 0,11 0,07 0,88 0,48 0,30 0,20
200 1,23 0,70 0,43 0,29 3,52 1,93 1,19 0,78
300 2,76 1,57 0,97 0,64 7,9 4,35 2,69 1,77
500 7,9 4,36 2,69 1,78
700 15 8,55 5,28 3,50
1000 31,6 17,4 10,76 7,12
2000 123 69,8 43,0 28,6
3000 276 157 37 64,1
5000 790 436 269 178
7000 1500 855 528 350
10 000 3160 1740 1076 712
10000 000 0,316-ю-10 0,174-10—10 0,108-Ю-10 0,071 1О-“
Е п = 70 об!сек.
50 0,43 0,24 0,147 0,097
100 1,72 0,95 0,59 0,39
200 6,89 3,79 2,24 1,55
300 15,5 8,5 5,26 3,5
500 43 23,7 14,7 9,7
700 84,2 46,4 28,6 19,0
1000 172 94,6 58,5 38,8
2000 689 379 224 155
3000 1550 851 526 350
5000 4300 2370 1470 970
7000 8420 4640 2860 1900
10000 17 200 9460 5850 3880
10000 000 1,72-10““ 0,946- 1(Г“ 0,585.10-“ 0,388.10““
1 22 12,1 4,9
43 23,7 14,7 9,65
88 48,3 29,9 19,7
352 193 119 78
790 435 269 177
2200 1210 748 493
4300 2370 1465 965
8800 4830 2990 1970
0,88-10“10 0,483 10~“ 0,299-10“10 0,197.10““*
n — 90 об /сек
0,71 0,39 0,24 0,16
2,84 1,56 0,97 0,64
11,4 6,24 3,87 2,56
25,6 14,0 87 57,7
71,0 39 24,2 16
139,5 76,4 47,4 31,4
284 155,6 96,7 64,1
1140 624 387 256
2560 1400 870 577
7100 3900 2420 1600
13 950 7640 1740 3140
28 400 15 600 9670 6410
2,84 10“ 1,56 10““ 0,967 10-“ 0,641.10““
Сопоставляя вычисленные значения сил магнитного взаимодействия Fm между эритроцитами с аналогичными значениями ранее вычисленных сил электростатического взаимодействия Fe, можно убедиться в том, что величина этих сил примерно одного порядка. Силы электростатического отталкивания уравновешиваются в значительной мере силами магнитного притяжения, которые становятся равными отталкивающим силам при п = 70—80 об/сек. Точка, при которой Fm = Fe, показана на рис. 23. Как видно, это соответствует п = 75 об!сек.
Для сравнения сил электростатического отталкивания с силами магнитного притяжения на рис. 22 приведена зависимость электростатического отталкивания Fe = f (г), которая иллюстрирует наше предположение о примерном равенстве этих сил, что должно обеспечивать необходимую стабильность эритроцитов в радиально-кольцевой системе при их движении по сосудам.
При рассмотрении системы, состоящей из нескольких (более двух) эритроцитов, необходимо иметь в виду, что величина силы магнитного взаимодействия FM должна быть уменьшена в четыре раза по сравнению с вычисленной, так как каждый эритроцит взаимодействует с двумя другими эритроцитами, из которых один расположен по одну, а другой — по другую сторону от него. Следует предположить, что в таком случае с соседним эритроцитом взаимодействует лишь одна половина заряда, принимающего участие в создании магнитного поля. Ввиду того, что в формулу взаимодействия величина круговых токов (являющихся производными от заряда) входит во второй степени, сила взаимодействия Fm должна уменьшиться в четыре раза.
§ 2. Приближенная модель равновесия электрических, магнитных и гидродинамических сил, действующих на эритроцит в потоке плазмы
Выше было показано, что при движении эритроцитов в кровеносных сосудах наблюдается эффект Бернулли, в результате чего скорость движения плазмы является функцией расстояния от геометрической оси сосуда. Изменение скорости движения плазмы по ортогональному сечению сосуда имеет параболический характер с максимумом у геометрической его оси. Разность скоростей движения отдельных слоев плазмы ориентирует эритроциты ортогонально к поперечному сечению сосуда и радиально к его стенкам, создавая значительную разность гидродинамических усилий, приложенных к диаметрально противоположным точкам эритроцита, центр которого находится между геометрической осью и стенкой сосуда. Для определения величины этих гидродинамических усилий в гл. III приведен расчет для случая расположения эритроцита на середине радиуса данного сосуда.
Гидродинамические усилия, испытываемые эритроцитом при его движении в плазме, стремятся переместить его по радиусу сосуда
72
в направлении к его геометрической оси и тем самым сблизить его* с эритроцитами, находящимися на более близких к оси концентрических окружностях. Этим усилиям противодействуют силы электростатического отталкивания, действующие между отдельными эритроцитами, расположенными в разных концентрических кольцах, экваториальная плоскость которых совпадает. Силы электростатического отталкивания, возрастая с квадратом уменьшения расстояния между эритроцитами, создают надежный электростатический распор между отдельными концентрически расположенными кольцами эритроцитов и стабилизируют их расположение при движении в плазме, предохраняя тем самым отдельные эритроциты от взаимного соприкосновения, что имеет кардинальное физиологическое значение.
Гидродинамические усилия, стремясь передвинуть отдельные эритроциты в направлении к оси сосуда, сжимают кольца эритроцитов, чем могут уменьшить диаметр этих колец. Уменьшение диаметра колец влечет за собой уменьшение расстояний между соседними эритроцитами данного кольца, что, в свою очередь, вызывает быстрое возрастание электростатических сил отталкивания, противодействующих гидродинамическим силам, стремящимся сжать кольца.
Силы магнитного притяжения, действующие между отдельными эритроцитами внутри колец, также возрастают, но в меньшей мере, чем электростатические. Величина данных магнитных сил несколько меньше электростатических. Они примерно уравниваются лишь при значительном числе оборотов эритроцитов. Действие гидродинамических сил на движущиеся в потоке плазмы эритроциты, а также противодействующие им силы электростатического отталкивания схематически показаны на рис. 24.
Изложенные соображения о взаимодействии сил, действующих на эритроцит, подтверждаются расчетами, в которых определяются те заряды эритроцитов, при которых силы электростатического взаимодействия эквивалентны гидродинамическим силам.
Выше были приведены расчетные данные о величине гидродинамических усилий, возникающих в сосудах различного диаметра. Для приближенного определения величины электростатического-заряда эритроцита, соответствующего приложенным к нему гидродинамическим усилиям, воспользуемся ранее выведенной формулой (см. гл. II, § 3) для определения сил электростатического взаимодействия между двумя эритроцитами, находящимися в одной экваториальной плоскости:
ЕЕ 2 [ 1 + 0,75 —-
= 4D (а —(р)8~ 1даН^
Е ~ (а — ей2 1/ w
у [(а_р)2 + о,75р21 9
7.Т
где Е — величина электростатического заряда эритроцита, ед. CGSE, а — расстояние между центрами эритроцитов, см; р = = 0,9/? = 3,37 • IO”"4 * * * см — эквивалентный радиус эритроцита; D = 85,5— диэлектрическая постоянная крови, F (а) — сила электростатического взаимодействия между двумя эритроцитами в одной экваториальной плоскости, дин.
Стенка сосуда
Стенка сосуда
Рис. 24. Схема равновесия электрических, магнитных и гидродинамических сил, действующих на эритроцит в кровотоке (магнитные силы перпендикулярны плоскости чертежа).
Подставляя цифровые значения в приведенную выше формулу и приравнивая величину гидродинамической силы FH к величине электростатической силы F(fl), определим величину заряда Е, эквивалентную силе FH.
Полученные значения Е представлены в табл. 17. Таблица показывает, что величина гидродинамической силы FH, действующей на
эритроцит, эквивалентна силам электростатического взаимодействия,
имеющим место при величине «действующего» заряда порядка 25—50 тысяч элементарных зарядов. Приведенные в табл. 17 величины силы FH следует считать несколько преувеличенными вследствие намеренно допущенного при их вычислении мажорирования.
В качестве примера приведем более точный расчет. Рассмотрим случай, соответствующий сосуду с радиусом R = 0,407 см. Как видно из табл. 17, в этом случае расстояние между центрами эритроцитов, расположенными в разных кольцах, но в одной экваториальной плоскости составляет 9,1 • 10“4 см, а величина гидродинамической силы FH = 376,2 • 10“8 дин. Этот случай соответствует
74
Таблица 17
Величина электрического заряда, соответствующая приложенной к эритроциту гидродинамической силе
Радиус сосуда Я, см Величина гидродинамической силы • 10е, дин Расстояния между центрами эритроцитов а-10*. см Величина электростатического заряда Е
ед. CGSE элементарные зарядо!
1,4 105,7 9,1 966 23 200
0,61 210,5 9,1 1360 18 600
0,407 376,2 9,1 1825 38 200
0,245 463,13 9,0 1980 41 600
0,138 778,45 9,0 2570 53 800
0,072 839,36 9,0 2670 55 800
0,0356 885,95 8,9 2680 56 200
0,0076 953,13 8,8 2720 57 000
0,0033 971,05 8,6 1620 54 900
0,0014 988,07 8,5 2580 54 000
средней скорости вращения эритроцита и = 40 об/сек. Угол между двумя эритроцитами в кольце можно считать равным 17°.
Гидродинамическая сила FHi действуя на эритроцит, может быть разложена на два вектора (рис. 25): вектор а, действующий в ради
альном направлении, прижимающий эритроциты одного кольца к эритроцитам другого концентриче
ского кольца, и вектор d, который может рассматриваться как равнодействующая векторов бис, стремящихся сжать эритроцитное кольцо и уменьшить его диаметр, т. е. уменьшить расстояние между отдельными эритроцитами кольца.
Действие вектора а уравновешивается силой электростатического отталкивания F (а), действующей между эритроцитами разных колец, находящимися в одной экваториальной плоскости, а действие векторов
b и с уравновешивается разностью сил между электростатическим отталкиванием и магнитным притяжением, имеющим место между эритроцитами, расположенными в од
ном кольце, т. е. разностью сил Fe — FM.
Из рис. 25
FH = а, + d,
Рис. 25. Схема гидродинамических сил, действующих на эритроцит.
75
а также
d= b + c9 причем b = с, следовательно, d2 = b2 + c2 — 2bc cos a. Отсюда определяется величина
d = У 2b2(l —cos a).
(a \
2 sin2 — L получаем
d=
Угол a между эритроцитами в рассматриваемом случае равен 17°, следовательно, sin = sin 8°30'; sin 8°30' = 0,148.
Можно записать
d = 2b sin 8° 30' = 2b . 0,148 = 0,2966.
Учитывая, что вектор b или равный ему вектор с представляет собой разность Fe — FM, находим
d = 0,296 (Fe — FM).
Таким образом, сила FH может быть выражена через F (a), Fe, FH так:
FH = F(a)+0,296(Fe-FH).
Решение этого уравнения даст искомую величину эквивалентного этим силам заряда F.
Для решения воспользуемся ранее выведенными формулами для F (а) (гл. II, § 3), Fe (гл. II, § 2) и FM (§ 1 настоящей главы). Подставим их:
Fh =
_______(a Р)а 1 4- 0 2°6 I______Е2____
4D(a-p)2 4-U,z.O| —_2
— 0,04np/2/-
л V2
V а- + 4р2 )
Используем численные данные для рассматриваемого случая, а именно: FM = 376,2 • 10~8 дин, D = 85,5; d = 9,1 • 10”4 см, г = 2 • 10~4 см\ р = 3,37 • 10~4 см, п = 40 об/сек, с2 = 9 . 1020 — переводной коэффициент (скорость света в квадрате) и аг = 2 X X 10~4 см. Приведем в формуле магнитного взаимодействия 76
необходимое преобразование — замену I равным ему произведением заряда на число оборотов эритроцита:
376,2 - 10-8 = £2
. Л„е 11,4-Ю-8 1 + 0,75----------
_________32,8 • 10~8 342 • 32,8 • 10-8
0,296
85,5 • 2 • 10-4 К4 • IO”8 + 45,6 • 10-8
0,296 • 0,04 • 3,14 • 3,37 • 10-4 • Ю2 (-------------—!-----------=Ь • Ю20
\2 • 10-4 4 • 10~8 + 45,6 • 10~8/
9 • 1018
После вычислений получаем
376,2 - 10-8 = £2 (---1’-2-6 -|----0,296 -7--7220),
\ И 260 • 10~8 1192-Ю-8 /
ИЛИ
376,2 • 10“® = 32080, откуда __________
£ = У 37632 0М~’ = '-Об 10-S ед. CQSE,
либо, переведя величину Е в элементарные заряды:
Е = 1,06 • 10“5 • 0,21 • Ю10 = 22 260 элементарных зарядов.
В ламинарном кровотоке число оборотов п меньше, поэтому величина Е будет несколько большей.
Приведенный расчет хорошо подтверждает очень важное обстоятельство, а именно: действующие между эритроцитами гидродинамические, электростатические и магнитные силы относятся к величинам одного и того же порядка.
Иллюстрацией характера упругих взаимодействий гидродинамических, электростатических и магнитных сил, которые имеют место в радиально-кольцевой системе эритроцитов, может служить рис. 26, на котором изображен ряд упругих кольцевых пружин, соединенных, в свою очередь, между собою также упругими пружинами. Концентрически расположенные кольцевые пружины соответствуют кольцам эритроцитов в потоке плазмы; пружинные соединения между отдельными кольцами внутри любого концентра, а также между кольцами разных концентров соответствуют рассмотренному выше упругому взаимодействию гидродинамических, электростатических и магнитных сил, при котором действие любой упругой силы может быть немедленно эластично воспринято всей упругой радиально-кол ьцевэй системой эр итроцитов.
Следует сказать еще несколько слов об электростатической амортизации эритроцитов при их возможных соударениях в момент замедления движения потока крови при фуркациях сосудов, при ответвлении сосудов под большим углом и т. д. Это замедление может
77
Рис. 26. Схема упругих взаимодействий гидродинамических, электростатических и магнитных сил радиально-кольцевой системы эритроцитов.
произойти при появлении местных сопротивлений в системе кровеносных сосудов, при внезапном изменении направления кровотока, когда могут появиться инерционные силы, вызывающие отклонения эритроцитов от линии потока и частичную потерю ими живой силы. В таких случаях электростатический распор должен сыграть роль амортизатора, предохраняющего эритроциты от соударения и повреждения. Расчеты, которые мы здесь опускаем, говорят в пользу такового допущения даже при максимальных скоростях движения крови.
Наконец необходимо указать, что все расчеты в этой главе сделаны без учета вязкости плазмы, действующей на эритроциты при их небольших колебательных движениях относительно друг друга, возникающих при движении эрит-роцитных систем в кровотоке. Ориентировочные расчеты показали, что незначительная вязкость плазмы хотя и играет определенную роль в изучаемых явлениях, но ею в этих прелиминарных исследованиях для простоты можно временно пренебречь. В дальнейшем, при углублении и уточнении формул, описывающих процессы взаимодействия между эритроци
тами, необходимо будет ввести постоянный коэффициент вязкости плазмы при нормальной температуре тела.
В расчетах гидродинамических, электростатических и электромагнитных взаимодействий, приведенных в этой главе, не учтены некоторые существенные факторы. Учет влияния электролита лишь путем внесения диэлектрической постоянной в выражение закона Кулона не учитывает подвижности ионов электролита (плазмы), образующих вокруг заряженных частиц двойные и диффузные слои, которые вносят значительные изменения в характер взаимодействия между заряженными частицами. Дерягин и Ландау справедливо замечают в одной из своих работ, что всякие расчеты, которые пытаются найти силы электростатического взаимодействия без выяснения вопроса об изменениях в локализации ионов, наступающих
78
в результате сближения двух мицелл, рискуют привести, даже в качественном отношении, к неверным результатам, а формулы, при этом выводимые, в лучшем случае имеют характер эмпирических.
Однако взаимодействие эритроцитов в крови в отличие от взаимодействия заряженных мицелл в растворах обычных электролитов имеет свою специфику. Эта специфика вытекает из следующих обстоятельств: 1) эритроциты находятся в крови в сравнительно очень большой концентрации, занимая до 36% общего объема крови; 2) эритроциты по размерам значительно превосходят объемы заряженных частиц и имеют специфическую форму, отличную от шарообразной; 3) эритроциты, по сравнению с их размерами, расположены очень близко относительно друг друга на расстояниях, составляющих всего примерно 15% их собственных размеров; 4) эритроциты при своем движении по сосудам располагаются в виде упорядоченной радиально-кольцевой модели, т. е. представляют в движущейся крови упорядоченную структуру.
Как следствие получаем: 1) гидродинамические силы во взаимодействии эритроцитов выдвигаются на первый план; 2) появляются электромагнитные силы взаимодействия; 3) вандерваальсовы силы или вовсе не играют, или почти не играют никакой роли; 4) при расчете сил электростатического взаимодействия форма и взаимное расположение эритроцитов начинают играть значительную роль, что возможно лишь в том случае, если относительно меньшую роль начинает играть взаимодействие ионных диффузных слоев, при котором форма и взаимное расположение частиц, ввиду их относительной малости, почти не играет никакой роли. Собственно говоря, уже нельзя приписать каждому эритроциту в крови независимого существования со своим отдельным диффузным слоем. Каждый эритроцит есть лишь составная часть единой эритроцитной динамической структуры, движущейся также в единой ионной среде с периодически закономерно сменяющейся локальной концентрацией ионов в зависимости от периодически меняющихся взаимных расположений движущихся друг относительно друга эритроцитных. колец.
Глава IV
ГИРОСКОПИЧЕСКИЕ ЭФФЕКТЫ ПРИ ДВИЖЕНИИ ЭРИТРОЦИТОВ
§ 1. Приближенная модель гироскопической устойчивости в кровотоке вращающегося эритроцита
При микроскопическом наблюдении ротации эритроцитов в движущейся плазме нами была обнаружена стабилизация проекции движения эритроцита, принимающей при достаточных скоростях вид прямой линии*. Данная стабилизация является результатом гироскопического эффекта, возникающего при поступательно-вращательном движении эритроцитов. В кровеносных сосудах и особенно в крупных ротационные скорости эритроцитов достигают большого количества оборотов в секунду. При этом гироскопический эффект должен обусловить стабилизацию направления движения эритроцитов, чтобы противостоять действующему на них моменту электростатических сил, стремящемуся повернуть эритроцит в его экваториальной плоскости и тем самым изменить величину угла между эритроцитами, находящимися в одном кольце радиально-кольцевой системы. Этот момент сил возникает вследствие того, что экваториальные плоскости эритроцитов в преде
лах одного кольца непараллельны, а составляют некоторый угол а, величина которого определяется частным от деления 2л на количество эритроцитов в данном кольце.
Найдем порядок сил, стремящихся изменить положение эритроцита в кольце. Для этого вычислим момент сил Ме, который возникает между двумя эритроцитами в пределах одного кольца, в зависимости от величины заряда £ и угла а между их экваториальными плоскостями.
Для упрощения расчетов смажорируем действие электростатического момента сил, представив себе весь электростатический за
* А. Л. Чижевский. Структурный анализ движущейся крови. Изд-во АН СССР, М., 1959.
80
ряд эритроцита сконцентрированным в двух диаметрально противоположных точках так, как это показано на рис. 27. Далее представим себе, что изображенные на рисунке эритроциты закреплены так: один в точке М, а другой в точке 7ИХ, вращением вокруг которых можно расположить эритроциты под любым углом друг к другу.
Так как расстояние гх фиксировано, г2 будет зависеть от угла а. На расстоянии гх будет действовать сила электростатического от-
Таблица 18
Зависимость момента сил от расстояния между зарядами и угла между экваториальными плоскостями
QVds *Ъ rt -10* (при rt = 2,2), см FflO10, дин дин Afe«1010, эрг rt • 10* (при г = 2,3), см Ft *1010, дин о кГ Afe«1010, эрг
1 2,321 17 300 16 400 0,152 2,421 16 600 15 600 0,168
3 2,552 17 300 14 700 0,438 2,652 16 600 14 200 0,405
5 2,794 17 300 13 300 0,674 1,894 16 600 12 800 0,640
10 3,374 17 300 10 700 1.11 3,474 16 600 10 300 1,07
20 4,56 17 300 7300 1,69 4,66 16 600 7000 1,62
30 5,70 17 300 5200 2,04 5,80 16 600 5100 1,94
45 7,88 17 300 3300 2,36 7,98 16 600 3200 2,26
50 8,96 17 300 3000 2,41 9,06 16 600 2900 2,28
талкивания Flt а на расстоянии rf — сила F2. Момент сил Ме представляет собой половину разности сил Fx — F2, умноженную на плечо /?, т. е.
где R = 3,37 • 10-4 см — эквивалентный радиус эритроцита.
Принимая в качестве примера число зарядов эритроцита Е = = 10 000 элементарных зарядов, получаем таблицу значений Ме (табл. 18), где показано, в каких пределах изменяется величина момента сил Ме в зависимости от угла а. Чем больше угол а между экваториальными плоскостями эритроцитов, тем больше момент сил Ме, абсолютная величина которого при заряде Е — 10 000 и расстоянии г между эритроцитами (2,2 -4- 9) 10-4 см составляет (0,152 4- 2,41) 1О“10 эрг.
Можно отметить, что наименьший угол а между экваториальными плоскостями эритроцитов наблюдается в наиболее удаленных от геометрической оси сосуда периферических кольцах. Следова тельно, и момент сил Ме в периферических кольцах значительно меньше, чем в кольцах, расположенных вблизи геометрической оси сосуда.
Моменту сил Л4е, стремящемуся изменить положение экваториальной плоскости эритроцита, противодействует гироскопический
81
Рис. 28. К расчету моментов инерции эритроцита.
момент Л1Г, стабилизирующий эритроцит в его ротационном движении. Гироскопический момент Мг может быть определен так:
Л4Г = Лоф, где J — момент инерции эритроцита относительно его оси вращения; <0 — угловая скорость; ф—угловая скорость отклонения оси вращения эритроцита под действием момента сил Ме.
Найдем моменты инерции эритроцита. Для их приближенного расчета воспользуемся формулой, выведенной ранее для торовидного кольца (рис. 28):
Jy = m2nRr2|/?2 + (2г)а ];
Л = т2л/?г2[4- + 4"<2г)2]-
Подставляя соответствующие
численные параметры эритроцита, т. е. массу нормоцита т — 68,7 х х 10-12, R = 3,75 • 10-4 см, г = -5- 0,94 • 10-4 см, получаем:
Jy = 68,7 • 10“12 -3,14-2- 3,75 • 1(Г4 (0,94 • 1(Г4) х
X [(3,75 • 10“4)2 + -^-(1,875 • 10~4)2| = 264 • Ю-30 г • см2;
Jx = 68,7 • 10~12 • 3,14 • 3,75 • 10-4 (0,94 • 10-4) х
X [ (3’75 910~4)2 + -4- (1.875 • 10-4)2] = 54 • 1О“30 г • см.
I <5^ J
Величину угловой скорости отклонения оси вращения ф находим, исходя из равенства производной момента количества движения системы относительно данной оси моменту внешних сил относительно этой оси, т. е.
dM г ;
-2Г = Ч = 44’,
где М — момент количества движения.
Отсюда
Определяя угловую скорость ф путем интегрирования, получаем
4>= \J^-dt = -4-\Medt.
J JX JX J
0 О
Применяя теорему о среднем значении интеграла к Ме, находим
* X
82
Подставляя в формулу гироскопического момента полученное выражение для ф, имеем
Л4Г = j/(O2gL = -^®ZA4f.
Эритроцит в своем вращательном движении может считаться устойчивым при условии, что гироскопический момент Мг Ме. Устойчивость эритроцита начинается с того момента, когда гироско-
пический момент Мг начинает становиться равным моменту электростатических сил Мв. В этом случае Мг = М„ и последнее выражение примет вид -^-®/= 1. J X Подставляя ранее вычисленные значения Jy и Jx, получаем 264 • ИГ30 . . = 1, 54 • 10-3° или 4,9®/ = 1, откуда ®/ = 0,204. Придавая величине t различ- Зависимость эритроцита Таблица 19 времени стабилизации от числа оборотов
Л сек Угловая скорость со Число оборотов п эритроцитов в 1 сек
1 0,1 0,01 0,005 0,003 0,002 0,001 0,0005 0,204 2,04 20,4 40,8 68,0 102 204 408 0,0325 0,325 3,25 6,50 10,8 16,2 32,5 65,0
ные значения в пределах от 1 до 0,0005 сек. получаем соответствующие им значения со, а также число оборотов эритроцита в секун-
(0
ду П= ЪГ
Вычисленные значения приведены в табл. 19, которая дает воз-
можность судить о том, в течение какого интервала времени гироскопический момент успевает стабилизировать эритроцит в его поступательно-вращательном движении в кровотоке. Время, необходимое для стабилизации, зависит от числа оборотов эритроцита в секунду. Например, при п = 65 об/сек (что соответствует угловой скорости со = 408) стабилизация эритроцита, под действием гироскопических сил, наступает в течение времени t = 0,0005 сек. Этот результат вытекает, как мы установили выше, из предположения о равенстве гироскопического момента моменту электростатических сил. Не исключены случаи, когда гироскопический момент будет больше момента сил, тогда время стабилизации t будет соответственно уменьшаться, а стабилизация эритроцита возрастать.
Эритроциты периферических колец, обладающие наименьшей поступательно-вращательной скоростью, имеют сравнительно небольшой гироскопический момент. Но это не значит, что они менее устойчивы чем эритроциты, находящиеся в близких к геометрической оси кольцах и обладающие значительно большими скоростями ротаций: момент сил у периферических эритроцитов вследствие того, что они находятся в почти параллельных экваториальных
83
плоскостях, очень мал, и поэтому даже ничтожный гироскопический момент достаточен для их стабилизации.
Необходимо заметить, что расчеты в этом параграфе проведены без учета вязкости, что, однако, не меняет качественной стороны рассматриваемого явления.
§ 2. Возможная частота электромагнитных колебаний в кровеносных сосудах при движении радиально-кольцевых систем эритроцитов
Ввиду неравномерности распределения скоростей по радиусу сосуда, кольца, движущиеся в цилиндрических слоях, отстоящих на разных расстояниях от своей оси, имеют различные скорости поступательного движения.
Рис. 29. Схема изменения расположения экваториальных плоскостей колец соседних слоев при движении радиально-кольцевых систем эритроцитов.
Следовательно, для двух соседних колец (цилиндрических слоев) имеется относительная скорость перемещения одного слоя относительно другого, а потому имеется известная периодичность, или частотность, совпадения экваториальных плоскостей колец соседних слоев.
Пусть в момент а (рис. 29) экваториальные плоскости колец двух соседних слоев совпадают. Тогда в момент б благодаря тому, что ско
84
рость их слоя, ближе расположенного к оси, больше, чем скорость и2, произойдет относительное смещение слоев, и кольца внутреннего слоя уже будут расположены не строго под кольцами наружного слоя, а окажутся смещенными одно относительно другого. Положения а и б будут периодически сменять друг друга, и можно говорить об известной частоте процесса. Для выведения формулы частоты воспользуемся формулой параболического распределения скоростей по сечению
2 + а д / п» Vp = —-—k(R — р).
Тогда для скорости поступательного движения внутреннего слоя получим выражение
где х — расстояние центров эритроцитов внутреннего слоя от оси.
Для скорости наружного слоя получим
vP, = + d + 6)“],
где d — диаметр эритроцита, 6 — распор между соседними кольцами, величина которого зависит от радиуса сечения сосуда. Относительная скорость перемещения внутреннего слоя относительно наружного будет
vo = уР1 — VP, = -- ц а- kl(x + d + 8)а — ха].
Расстояние между центрами эритроцитов соседних колец в одном слое равно d + 6, а потому для величины периода встречи получим
т =_________(d+6)a_________
(2 + a)ft[(x + d+6)“-x“]
Следовательно, частота встреч
m-JL= (2 + «)*Цх-М+д)а-ха] т (d+6)a
Коэффициенты k и а должны меняться от сосуда к сосуду. Принимая в качестве первого приближения универсальную формулу для средней скорости по всему артериальному кровотоку в виде
Кер = kF,
получим приближенные значения a = 0,776; k — 36,4.
Поскольку эритроциты несут на своей поверхности электрический заряд, постольку периодические изменения относительных положений кольцевых слоев будут обусловливать периодические изменения соответствующих магнитных и электрических полей с той же частотой v, т. е. с длиной волны Л = £•.
85
Не следует забывать, что картина рассматриваемых явлений фактически значительно сложнее, ибо в настоящем предварительном
Таблица 20
Приближенные значения электромагнитных колебаний в сосуде с R = 100 мк
X V, сек~^ X, м
0,25 28,6 10 500
0,50 24,9 12 050
0,75 20,9 14 380
значения X будут еще большими.
расчете мы принимаем во внимание лишь относительное поступательное перемещение эрит-роцитных колец, не учитывая вращательного движения, в результате которого возникает еще относительное взаимодействие циклоидальных токов.
В качестве примера приведем расчет частот и длин волн для сосуда с /?= 100 мк (табл. 20).
Для ламинарного кровотока Ввиду пропорциональности для
ламинарного кровотока ир— иР4-б отношению и приходим к выводу о возрастании частоты от центра сосуда к периферии (в противоположность данным табл. 20), и о возрастании частоты от крупных артерий к капиллярам.
§3. Провизорная величина суммарного электрического тока вращающихся эритроцитов и некоторые другие приближенные значения
Как известно, в научной литературе приняты «наглядные» сравнения. Были сделаны многократные попытки образно представить количество эритроцитов, содержащихся в человеческой крови, путем различных сравнений. Физик Цингер пишет, что если бы все
Общая кинетическая энергия эритроцитов при вращательном движении в сосудах
Фуркации Радиус сосуда. см р. см Число оборотов
1 1,40 0,915 32 287 0,0003= 12
2 0,61 0,400 32 287 0,0004= 13
3 0,41 0,266 32 287 0,00044= 14
4 0,245 0,160 32 287 0,0005= 16
5 0,138 0,090 32 287 0,00052= 17
6 0,072 0,047 32 287 0,00059= 19
7 0,0356 0.023 32 287 0,00069=22
8 0,0169 0,011 32 287 0,00081=26
9 0,0076 0,005 32 287 0,00096=31
10 0,0033 0,002 32 267 0,00116= 37
И 0,0014 0,0009 32 287 0,00136=44
12 0,0005 0,0003 32 287 0,00163=53
86
эритроциты, заключающиеся в пяти литрах крови человека, сложить в стопку, то высота этой стопки будет равна 41 000 км. а если их положить в ряд так, чтобы они касались друг друга, то этот ряд будет иметь длину 210 000 км. Гистолог Заварзин для соответствующих величин дает такие цифры: 62 000 и 187 000 км. Внося соответствующее изменение в значение высоты эритроцита, получим соответственно 45 000 и 187 000 км. Прибегая к аналогичному методу, коснемся некоторых других вопросов.
1. Для определения примерной величины суммарного электрического тока, возникающего в человеческом организме при вращении эритроцитов, можно произвести следующий расчет.
В нашей книге «Структурный анализ движущейся крови» (ч. III, гл. 2, § 5, стр. 322) приведены результаты расчетов скоростей поступательного движения плазмы в точках нормоцита, центр которого находится в середине радиуса сосуда. В табл. 21 приводятся данные о количестве эритроцитов в литре крови большого круга с разбивкой их на группы определенных скоростей вращения.
Скорость вращения эритроцитов, выраженная числом оборотов в секунду, показана во второй графе табл. 21. Соответствующие данным скоростям количества эритроцитов в одном литре крови показаны в пятой графе табл. 22. Умножая величину конвекционного тока одного эритроцита при данной скорости вращения на количество эритроцитов, вращающихся с этой скоростью, получаем величину конвекционных токов для эритроцитов данной группы. Суммируя величину токов, получаем суммарную величину их для одного литра крови большого круга, которая выражается в 194,2 а. Ввиду значительно меньшего числа оборотов в ламинарном кровотоке суммарная величина круговых токов будет в несколько раз меньше.
При наличии в человеческом организме 5 л крови, из которых
Таблица 21 артериальной части большого круга кровообращения
Угловая скорость Кинетическая энергия • 1012, эрг Количество эритроцитов • 10* Общая кинетическая энергия в сосуде, эрг
6,28.12=75,36 0,028 196 278 0,0055
6,28.13=81,64 0,033 404 616 0,0134
6,28-14=87,92 0,038 449 625 0,0171
6,28-15=100,48 0,051 514 085 0,0262
6,28-17=106,76 0,057 522 122 0,0298
6,28-19=119,32 0,071 514 842 0,0366
6,28-22=138,16 0,095 492 946 0,0468
6,28-26=163,28 0,133 467 849 0,0622
6,28-31=194.68 0,190 420 206 0,0798
6,28-37=232,36 0,270 369 308 0,0996
6,28-44=276,32 0,382 346 076 0,1322
6,28-53=332,84 0,554 301 865 0,1673
2 = 0,7165
87
в циркуляции находятся 2,5 л, общая величина конвекционных токов в циркулирующей крови достигает 194,2 • 2,5 = 485 4- 500 а. Естественно, что это только наглядное представление о порядке суммарной величины конвекционных токов.
2. Если пересечь кровеносный сосуд М — N ортогонально к к линиям потока крови, то через живое сечение ее С — С (рис. 30) в единицу времени будет протекать некоторый объем крови с мор-м фологическими и дисперсными
> —1 элементами. Зная скорость про-
с ____ текания крови через площадь
— ^Ft числ0 частии и их по-верхность, легко вычислить вели-- чину суммарной их поверхности,
проходящей через поперечное се-
'"/V чение сосуда в единицу времени:
Рис. 30. К расчету электростатических SS = SSft-----V,
зарядов и конвекционных токов эритро- 4
цитов, проходящих в 1 сек через по-перечное сечение сосуда. где суммарная поверх-
ность частиц крови в 1 мм3.
Зная, что суммарная поверхность всех основных частиц в 1 мм3
крови составляет 85 000 мм2 и взяв 1% поверхности при допущении, что на 1 мк2 расположено 100 элементарных зарядов (что вполне возможно) получаем табл. 23. Таким образом, через поперечное сечение аорты проходит в 1 сек 3 • 1016 элементарных электрических зарядов, непрерывно «работающих» в кровотоке.
Таблица 22
Расчет суммарного тока для одного литра крови большого круга кровообращения
Число оборотов эритроцита в секунду п t’lO3 одного эритроцита Количество эритроцитов в одном литре крови-10 10 а
ед CGSE а
12 57,6 19,2 19,628 3,76
13 62,4 20,8 40,462 8,42
14 67,1 22,4 44,962 10,1
16 76,7 25,6 51,408 13,2
17 81,5 27,2 52,212 14,2
19 91,1 30,4 51,484 15,6
22 105,6 35,2 49,295 117,4
26 125,0 41,6 46,785 19,5
31 148,0 49,6 42,021 20,8
37 178,0 59,2 36,931 21,26
44 211,5 70,4 34,608 24,4
53 254,5 84,8 30,187 25,56
2 = 499,633 2= 194,2а
88
Таблица 23
Количество элементарных электрических зарядов, занимающих 1% поверхности морфологических и корпускулярных элементов крови, проходящей в 1 сек через поперечное сечение сосудов разных диаметров
Диаметр сосуда d, мм Объем крови v, проходящей через площадь сечения сосуда в 1 сек, мм3 Суммарная поверхность форменных и корпускулярных элементов крови в объеме V, м2 1% поверхности форменных и корпускуляр-нь х элемен- тов, л2 Количество элементарных зарядов, занимающих 1% поверхности
0,01 78,54-10-е 6,63-10“® 6,63-10-8 6,63-10е
0,1 0,047124 0,00398 3,98-10-6 3,98-109
1 28,27 2,385 0,02385 0,02385-1014
5 2434,74 205,5 1,055 2,055-1014
10 16729,02 1413,0 14,13 14,13-1014
15 51600,78 4350 43,5 43,5-10й
20 112154,76 9460 94,6 94,6-1014
25 213039,32 17 980 179,8 179,8-1014
30 253429,0 29 750 297,5 297,5-1014
Таблица 24
Величина электростатических зарядов и конвекционных токов эритроцитов, проходящих в 1 сек через поперечное сечение сосудов разных диаметров
Диаметр сосуда d, мм Площадь сечения сосуда nd* —5 , ММ* 4 Средняя скорость кровотока, vcp, мм/сек Объем крови, проходящей через площадь сечения сосуда в 1 сек па2 ~4 ГсР’ мм3 Суммарный заряд отрицательной полярности, проходящей через поперечное сечение сосуда, ед. CGSE Величина конвекционного тока 1, а
0,01 0,00007854 1 78,54-10“® 37,6-10“4 1,257-10“12
0,1 0.007854 6 0,047124 226 0,752-10“9
1,0 0,7854 36 28,27 1345 45,3- 10“9
5,0 19,635 124 2434,74 116 800 38,9-10“®
10,0 78,54 213 16729,02 803 000 267-10“®
15,0 176,71 292 51600,78 2 475 000 825-10“®
20,0 314,16 357 112154,76 5 390 000 18-10“*
25,0 490,87 434 213039,32 10 230 000 34-10”3
30,0 706,86 500 353429,0 16 950 000 56,6-10“3
Таблица 25
Электростатическое взаимодействие между двумя суммарными зарядами, помещенными на расстоянии 1 см один от другого
d, мм 1 Суммарный заряд отрицательной полярности, проходящий через сечение сосуда в J сек, ед CGSE Сила электростатического взаимодействия F
дин кг т
0,01 (капилляр) 1,0 (сосуд) 10 (сосуд) 30 (аорта) 37,6-10“4 1345 16729,02 16 950 000 14,14-10“* 1,809-10* 279-10® 287,3-1012 14,14-10—12 1,809 279 187,3-10» 14,14-Ю-15 0,001809 0,279 287 300
89
Аналогичным образом составим табл. 24. Число эритроцитов в 1 мм3 крови принято равным 5- 10е, заряд эритроцита — 20 000 элементарных зарядов.
Можно привести еще один пример. Возьмем из табл. 24 значение суммарного электрического заряда отрицательной полярности, проходящего через ортогональное сечение аорты в 1 сек. Он равен 16 950 000 ед. CGSE. Поместим его в одной точке, а на расстоянии 1 см от него расположим заряд такой же величины и того же знака. Между этими зарядами по закону Кулона возникнет взаимодействие в виде отталкивания одного заряда от другого, которое будет равно F = = 1695 • 101 • - 104 = 287,3 • 1012 дин = 287,3 • 10е кг=
= 287 • 300 т.
В табл. 25 даны соответствующие расчеты для капилляра и сосудов, диаметры которых равны 1 и 10 мм.
Приведенные примеры являются не более как воображаемыми образами некоторых электрических величин и их взаимодействий применительно к явлениям, происходящим в кровеносных сосудах. И поверхности частиц крови, и электрические заряды, связанные так или иначе с этими поверхностями, диспергированы в кровеносных путях, чрезвычайно раздроблены и как бы незаметны, но интегральное их значение колоссально. Оно велико не только в отношении абсолютных цифр, но и в отношении их физиологических функций. Эти поверхности и эти заряды развивают интенсивные физико-химические явления, вмещающиеся в ничтожных обьемах материи.
ЛИТЕРАТУРА
Аринкин М. И. Клиника болезней крови и кроветворных органов. «Практическая медицина», Л., 1928.
Балаховский С. Д. Реакция оседания эритроцитов. Госиздат, М.— Л., 1928.
Баркрофт Д. Основные черты архитектуры физиологических функций. Биомедгиз, М.— Л., 1937.
Б у л л Г. Б. Физическая биохимия. ИЛ, М., 1949.
Васильев Б. Д. Атмосферный воздух, жизнь и кровь.— В кн.: Земля во вселенной. «Мысль», М., 1964, 443.
Вишневский Г. Б. К вопросу о колебаниях величины диаметра эритроцитов у различных народностей.— Русский журнал тропической медицины, 1928, 3.
Вишневский Г. Б. Метод измерения диаметра эритроцита.— Лабораторная практика, 1929, 2.
Владос X. X. Клиническая гематология. Биомедгиз, М., 1937.
Г а т ч е к Э. Вязкость жидкостей, ОНТИ, М.— Л., 1935.
ГиршфельдГ. Учебник болезней крови. Госиздат, М.— Л., 1930.
Гортнер Р. Основы биохимии. Ч. 1. Коллоиды. Снабтехиздат, М., 1933.
Думанский А. В. Дисперсность и коллоидное состояние вещества. ОНТВУ, Коксохимия, X.— К., 1932.
Ж‘у к о в Е. Дыхательные функции крови. Изд. ЛГУ, Л., 1937.
Земля во вселенной. Сб., «Мысль», М., 1964.
К о р ж у ев П. А. Эволюция дыхательной функции крови. Изд-во АН СССР, М.— Л., 1949.
Коржуев П. А. Гемоглобин. «Наука», М., 1964.
К р о г А. Анатомия и физиология капилляров. Изд. Мосздравотдела, М., 1927.
Кулакова И. Н. К вопросу об электрореакции оседания эритроцитов. — Клиническая медицина, 1958, 8.
Лоренц Г. А. Статистические теории в термодинамике. Л.— М., ОНТИ, 1935.
Лукьянов С. М. Основания общей патологии сосудистой системы. Варшава, 1893.
МлодзиевскийА. Б. Молекулярная физика. Гостехиздат, Л., 1941.
О т т А. Гидравлика. Изд. Ленинградского заочного ун-та, Л., 1937.
П и н е й А. Последние достижения гематологии. Госиздат, М.— Л., 1947.
Скульский Н. А. Капилляроскопия и капилляротонометрия. Госиздат, М.—Л., 1930.
Солнце, электричество, жизнь. Сб., Изд. Московского общества испытателей природы, М., 1969.
Сретенский Л. Н. Теория волновых движений жидкостей. ОНТИ, М.— Л., 1936.
Уинттекер А. Аналитическая динамика. М., 1934.
Чижевский А. Л. О методике измерения электрического заряда крови. Проблемы ионификации. Труды ЦНИЛИ, 3. Воронеж, 1934, 504.
Чижевский А. Л. Электрические характеристики крови, М., 1941.
Чижевский А. Л. Структурный анализ движущейся по сосудам крови. Вестник АН Каз.ССР, Алма-Ата, 1951, 12 (81), 58.
91
Чижевский А. Л. Структурные образования из эритроцитов в движущейся по сосудам крови. — Бюллетень экспериментальной биологии и медицины, 1951, 12, 443.
Чижевский А. Л. Ориентация и кинематика эритроцитов в кровотоке. Изв. АН СССР, Сер. биолог., 1953, 5, 72.
Чижевский А. Л. Электрореакция оседания красных кровяных телец.— Клиническая медицина, 1953, 31, 60.
Чижевский А. Л. Об истинной величине диаметра нормоцитов крови человека. ДАН СССР, 1954, 94, 3, 565.
Чижевский А. Л., Трофимов Г. К. Образуются ли эритроцитные монетные столбики вне организма. Бюллетень экспериментальной биологии и медицины, 1955, И, 70.
Чижевский А Л. Структурный анализ движущейся крови. Изд-во АН СССР, М., 1959.
Щелкунов С. Н. Капилляры. БМЭ, изд. 2-е, 1959, 12, 191—197.
Biggs R., Macfarlane R. G. Human Blood. Coagulation and its Disorders. Oxford, 1957.
BremyP. Die Gewcbsmastzellen im menschlichen Knochenmark. Stuttgardt, 1950.
Carter R., Buseh E., StargV. The effect of vitamin B12 on the leukopenia induced by radiation. Blood, 1950, 5, 8.
Deutseh E. Blutsgrinnungs factoren. Wien, 1955.
D о w m у H. Handbook of hematology, III. V., 1938.
F e г r a t a A., S t о г i E. Le malatu del sangue. Milano, 1948.
JiirgensR., GraupnerH. Darstellung eines Entwieklungssystems der Thrombocyten. Folia heamat, 1937, 57, 263—282.
К о m i у a E. Zentralnervise Regulation des Blutbildes. Stuttgardt, 1956.
L a n d s t e i n er K., W i e n e r A. An agglutinable factor in human blood recognized by immune sera for rhesus blood. Proc. Soc. exp. biol. and med., 1940, 43, 223.
Maupin B. Indications de la serie plaquettaire. Le Sang., 1956.
M a n d e m a F. et al. Proc, of the VII Intern. Congress of the Intern. Society of Haemet. Roma, 1960.
M о e s e h 1 i n S. et al. Experimental investigations of relationship between plasma cells and antibody formation (phase contrast microscope). Acta hae-matol, 1951.
Prankerd H. The red cell, Oxford, 1961.
S о u 1 i e r J. P. Liste des produits du sang (gamma-globulines excepte’es). Le sang, 1956.
Withby L., Britton C. Disorders of the blood. London, 1953.
Wright C. Survey of the 1953 hematology literature. Acch. int. med., 1954.
ОГЛАВЛЕНИЕ
Предисловие ............................................................ 3
Введение ............................................................... 5
Г лава /. Электролиты плазмы и электрический заряд эритроцита
§ 1. Водородные и гидроксильные ионы крови ...................... 13
§ 2. Изоэлектрическая точка и провизорный электрический заряд эритроцитов и некоторых белковых компонентов крови .... 16
§ 3. Ионы основных элементов и химических соединений крови, их число, радиусы и проекции на плоскость ......................... 24
§ 4. Ориентировочные вычисления электрического заряда эритроцита 38
Глава II. Приближенные модели электростатического взаимодействия эритроцитов в кровотоке
§ 1. Кулоновское взаимодействие ................................. 45
§ 2. Электростатическое взаимодействие между двумя эритроцитами, расположенными в параллельных плоскостях........................ 51
§ 3. Электростатическое взаимодействие между двумя эритроцитами, расположенными в одной экваториальной плоскости................. 56
Г лава III. Приближенная модель магнитного взаимодействия эритроцитов в кровотоке
§ 1. Магнитное взаимодействие двух вращающихся эритроцитов, находящихся в параллельных плоскостях ............................ 63
§ 2. Приближенная модель равновесия электрических, магнитных и гидродинамических сил, действующих на эритроцит в потоке плазмы.......................................................... 72
Глава IV. Гироскопические эффекты при движении эритроцитов
§ 1. Приближенная модель гироскопической устойчивости в кровотоке вращающегося эритроцита . . ’.............................. 80
§ 2. Возможная частота электромагнитных колебаний в кровеносных сосудах при движении радиально-кольцевых систем эритроцитов 84
§ 3. Провизорная величина суммарного электрического тока вращающихся эритроцитов и некоторые другие приближенные значения ....................................................... 86
АЛЕКСАНДР ЛЕОНИДОВИЧ ЧИЖЕВСКИЙ
ЭЛЕКТРИЧЕСКИЕ И МАГНИТНЫЕ СВОЙСТВА ЭРИТРОЦИТОВ
Печатается по постановлению ученого совета Института физиологии им. А А. Богомольца АН УССР
Редактор Н. И. Воронцова
Художественный редактор И. П. Антонюк
Оформление художника К. Г. Лычаковского
Технический редактор Г. Р. Боднер
Корректор И. В. Столярчук
Сдано в набор 9.Х 1972 г. Подписано к печати 26.11 1973 г. БФ 01140. Зак. №2—1997. Изд. №42. Тираж 1450. Бумага № 1, бОхЭО1/^. Усл. печ листов 6,0. Учетно-изд. листов 5,75 Цена 58 коп.
Издательство «Наукова думка», Киев, Репина, 3.
Отпечатано с матриц Киевского полиграфического комбината в Несте-ровской городской типографии Львовского областного управления по делам издательств, полиграфии и книжной торговли, г. Нестеров, ул. Горького. 8, зак. 2499