/
























































































































































































































































































































































































































































































































































































































































Текст
ПОЗДНЕПАЛЕОЗОЙСКОЕ ВРЕМЯ (D - девон, С - карбон, Р — пермь)
РАННЕПАЛЕОЗОЙСКОЕ ВРЕМЯ (С - кембрий, О - ордовик, S - силур)
ПОЗДНЕПРОТЕРОЗОЙСКОЕ ВРЕМЯ (V - венд или эдиакарий)
КАЙНОЗОЙСКОЕ ВРЕМЯ (А - антропоген)
КАЙНОЗОЙСКОЕ ВРЕМЯ (₽ - палеоген, N - неоген)
МЕЗОЗОЙСКОЕ ВРЕМЯ (Т - триас, J - юра, К - мел)
'Орский растительноядный динозавр Diplodocus длиной 26.5 м имел стройную шею и длинный
vboct. Реконструкция прижизненного облика рептилии основана на находках скелетов в
северо-восточной части Юта
THE
FOSSIL BOOK
A Record of Prehistoric Life
CARROLL LANE FENTON
and MILDRED ADAMS FENTON
Revised and expanded by
Patricia Vickers Rich, Thomas Hewitt Rich,
and Mildred Adams Fenton
DOUBLEDAY
NEW YORK LONDON TORONTO SYDNEY AUCKLAND
НАУЧНЫЙ ЦЕНТР МОНАШ
ПАЛЕОНТОЛОГИЧЕСКИЙ ИНСТИТУТ РАН
МОСКОВСКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ
ФОНД ХАНСА РАУСИНГА
КАМЕННАЯ
КНИГА
Летопись доисторической жизни
Кэрролл Лейн Фентон
и Милдред Адамс Фентон
Переработали и дополнили
Патриция Викерс Рич, Томас Хьювитт Рич
и Милдред Адамс Фентон
Перевели и переработали
О.Б. Бондаренко (зам. ответственного редактора ),
В.Н.Голубев, Ю.М. Губин, Д.Н. Есин, Т.В. Кузнецова,
Е.Н. Курочкин, ИА. Михайлова, С.В. Наугольных,
А.Ю. Розанов (ответственный редактор )
Москва
Международная академическая
издательская компания “Наука”
1997
ББК 28.1
Р56
УДК 56
Патриция Викерс Рич, Томас Хыовитт Рич, Милдред Адамс Фентон
Каменная книга. Летопись доисторической жизни / Пер. с англ, с
дополнениями и изменениями. — М.: МАИК “Наука”, 1997. - 623 с.,
433 ил.
ISBN 5-7846-0009-5
Вниманию читателя предлагается увлекательная научно-популярная книга по па-
леонтологии, в которой изложены разнообразные сведения об органическом мире
близкого и далекого геологического прошлого. Книга снабжена богатым иллюстратив-
ным материалом.
Для школьников, студентов, преподавателей, научных сотрудников и всех любите-
лей палеонтологии.
ISBN 5-7846-0009-5 © 1958 by Carroll Lane Fenton and Mildred Adams Fenton
© 1989, 1996 by Mildred Adams Fenton, Patricia Vickers Rich, Tho-
mas Hewitt Rich
© 1997, перевод на русский язык с дополнениями и изменениями
О.Б. Бондаренко, В.Н. Голубев, Ю.М. Губин, Д.Н. Есин,
Т.В. Кузнецова, Е.Н. Курочкин, И.А. Михайлова, С.В. Науголь-
ных, А.Ю. Розанов
© 1997, Патриция Викерс Рич, Томас Хьювитт Рич
Предисловие
к англоязычному изданию книги
1989 года
Книга представляет собой обзор ископаемых организмов от пер-
вых следов докембрийской жизни до зверей и птиц, которые жили
еще несколько столетий назад. О человеке в книге приведены только
краткие сведения. Это связано с тем, что литература о происхожде-
нии и развитии человека столь обширна (а нередко и противоречи-
ва), что лучше обратиться к специальным источникам.
Обзор дан в общих чертах, так как затронутые темы выходят дале-
ко за пределы собственного опыта авторов, чьи исследования ограни-
чены водорослями, ранними беспозвоночными, птицами и млекопи-
тающими. Мы поэтому широко использовали сведения, опублико-
ванные в специальных статьях и монографиях, а также в учебниках,
руководствах, сводках и справочниках. У нас также была возмож-
ность ознакомиться с рукописью книги РА. Кэрролла (R. A. Carroll),
представляющей собой современный аналог книги А.Ш. Ромера “Па-
леонтология позвоночных”.
Новые данные исключительно важны для глав, где описаны рыбы
и низшие четвероногие. Мы также очень благодарны за помощь в
подготовке второго издания книги многим палеонтологам, кто прове-
рил главы по своей специальности: Н. Арчболд (N. Archbold, брахио-
поды), Р. Бэрд (R. Baird, птицы), Д. Блэйк (D. Blake, мшанки),
Ф.М. Карпентер (F.M. Carpenter, насекомые), Х.В. Дэли (H.V. Daly,
насекомые), ТА Дэррак (Т.А. Darragh, двустворки), Дж.В. Дарэм и
П.Джелл (J.W. Durham и Р. Jell, иглокожие), Т.Ф. Фланнери
(T.F. Flannery, млекопитающие), Д. Холловей (D. Holloway, трилоби-
ты), С.РА. Келли (S.R.A. Kelly, двустворки), Дж. Лонг (J. Long, ры-
бы), Дж.Р. Макдональд (R. Macdonald, несколько тем), К. Маллетт
(С. Mallett, фораминиферы), Дж. МакЭвен-Мэзон (J. McEwen-Mason,
растения), Р.Э. Мольнар (R.E. Molnar, амфибии и рептилии), Дж. Пи-
кетт (J. Pickett, губки), П.А. Рэйвн (Р.А. Raven, растения), А. Ричч
(A. Ritchie, рыбы), Я. Стъюорт (I. Stewart, граптолиты), К. Тесселл
(С. Tassell, гастроподы и др. моллюски), Т. Тальборн (Т. Thulborn,
рептилии), Г.Ф. ван Terne (G.F. van Tets, птицы), Л. Верделин (L. Wer-
delin, млекопитающие), Б. Вагштафф (В. Wagstaff, растения),
А. Уоррен (A. Warren, амфибии), Дж. Уоррен (J. Warren, докембрий-
ские организмы) и М. Уайтло (М. Whitelaw, растения).
Рукопись книги первого издания прочли и внесли изменения:
Э.Дорф (Е. Dorf, растения), К.Э. Кастер (К.Е. Caster, все беспозво-
6
КАМЕННАЯ КНИГА
ночные), Б. Шэффер (В. Schaeffer, рыбы), Э.Х. Кольберт (Е.Н. Col-
bert, амфибии и рептилии) и Г.Л. Джепсен (G.L. Jepsen, птицы и
млекопитающие). Миссис Рейчел Николъс (Rachel Nichols), которая
впоследствие присоединилась к редколлегии справочного издания
“Библиография ископаемых позвоночных”, возглавляемого
К.Л. Кэмп (C.L. Camp), познакомила К.Л. и МА Фентонов с многи-
ми важными книгами и рукописями библиотеки Осборна, находя-
щейся в Американском музее Естественной Истории.
Большинство фотографий ископаемых беспозвочных и некоторых
позвоночных были сделаны в Национальном музее Естественной Ис-
тории (Washington, D.C.), где нам помогали Р.С. Бэсслер
(R.S. Bassler), ГЛ Купер (G.A. Cooper), Г.Л. Газин (G.L. Gazin),
Д.Данкл (D. Dunkle), Дж.Б. Рисайр (J.B. Reeside) и Р.В. Браун
(R.W. Brown). Для первого издания книги Е. Йохелъсон (Е. Yochelson)
представил прекрасную пермскую улитку с сохранившейся окраской,
а АР. Палъмэр (A.R. Palmar) передал фотографии и информацию об
исключительно сохранившихся насекомых пустыни Мохаве. Образ-
цы для фотографирования были представлены С. Веллее (S. Welles),
РА. Стиртон (R.A. Stirton) и Р.В. Чэйни (R.W. Chaney) из Калифор-
нийского университета Беркли (Berkeley), а также Э.Дорф (Е. Dorf) и
Б.Ф. Хоуэлл (B.F. Howell) из Принстонского университета. Негативы
срезов средне-позднекаменноугольных растений представил ХА. Энд-
рюс (Н.А. Andrews, Вашингтонский университет). Ф.М. Карпентер
(F.M. Carpenter, Гарвардский университет) дал негативы с изображе-
ниями ископаемых насекомых и прислал сведения об этих экземпля-
рах. Цветные фотографии диорам Демонстрационного музея универ-
ситета Мичигана были представлены И.Г. Рейманном (I.G. Reimann,
директор музея во время первой публикации книги в 1958 году).
Для второго издания многие исследователи щедро делились цвет-
ными слайдами и черно-белыми фотографиями образцов или место-
нахождений, с которыми они работали: Д. Бэрд (D. Baird, Принстон-
ский университет), Ф.М. Карпентер (F.M. Carpenter, Гарвардский
университет), М. Чжоу (М. Chow, Институт палеонтологии позвоноч-
ных и палеоантропологии, Пекин), М. Доусон (М. Dawson, музей Кар-
неги), Й. Хасагава и К. Оно (Y. Hasagawa and К. Ono, Национальный
научный музей Японии), К.Б. Хэнди (Q.B. Hendey, Южно-Африкан-
ский музей), Дж А. Дженсен (J. A. Jensen, Brigham Young University),
3. Келан Яворовска (Z. Kielan-Jaworowska, Польская Академия на-
ук, Институт палеобиологии), М. Лики (М. Leakey, Национальный
музей Кении), М.К. МакКеннэ (М.С. МсКеппе, Американский музей
Естественной Истории), К. Море (С. Mourer, университет Клода Бер-
нара, Лион), Г.М. Филип (G.M. Philip, университет Сиднея),
Н.С. Пледж (N.S. Pledge, Южно-Австралийский музей), Д.Е. Саваж
(D.E. Savage, Калифорнийский университет, Беркли), Б. Шелтон
(В. Shelton, Крэнборн, Виктория), Б. Сижё (В. Sige, университет Мон-
пеллье), Я. Стъюорт иДж.А. Уоррен (I. Stewart and J. A. Warren, уни-
верситет Монаш) и Г. Вел (G. Viohl, Юрский музей, Эйхштатт).
Ф. Коффа (F. Coff а, музей Виктории) и С. Мортон (S. Morton, универ-
ситет Монаш) предоставили превосходные новые фотоматериалы для
второго издания. Б. Вильсон (В. Wilson), Д. Андерсон (D. Anderson) и
Э. МакКлеллан (Е. McClellan) оказали исключительную помощь,
предоставив образцы и архивные материалы музея Виктории.
Предисловие к англоязычному изданию книги 1983 года
7
Для второго издания книги многие другие исследователи снабди-
ли нас литературой и иллюстрациями, которые оказались важными
при ревизии глав. Особенно следует отметить РЛ. Кэрролла
(R.A. Carroll), М. Чиба (М. Chiba), Э.Х. Кольберта (Е.Н. Colbert) и
ЕМ. Томсона (Е.М. Thomson).
Рисунки фоссилий были сделаны непосредственно с самих эк-
земпляров или с фотографий и опубликованных иллюстраций. Боль-
шинство рисунков в обоих изданиях являются оригинальными, но
некоторые заимствованы из других изданий. Среди последних:
брахиоподы из тома VIII “Палеонтология Нью-Йорка”, иглокожие
из монографии “Мезозойские и кайнозойские иглокожие Соеди-
ненных Штатов” В.Б. Кларка (W.B. Clark) и М.В. Твитчелла
(M.W. Twitchell), опубликованной как 54-я монография Геологичес-
кой службы Соединенных Штатов Америки.
Реконструкции по беспозвоночным основаны преимущественно
на исследованиях К.Л. и МЛ. Фентонов (C.L. and М.А. Fenton). Ре-
конструкции позвоночных в первом издании включали рисунки
Найта (Knight), Хорсфолла (Horsfall), Прайса (Price), Германна
(Germann) и других художников, а также иллюстрации таких пале-
онтологов, как Осборн (Osborn), Мэттью (Matthew), Скотта
(Scott), Кейза (Case), Виллистона (Williston), Мерриам (Merriam),
Стока (Stock), Симпсона (Simpson) и Ромера (Romer). Реконструк-
циям растений мы обязаны Съюорду (Seward), Вейланду (Wieland),
Голдрингу (Goldring) и другим авторитетным специалистам.
Новые иллюстрации для второго издания были подготовлены пре-
имущественно Д.Джелтом (D. Gelt, университет Монаш), а реконст-
рукции и рисунки были сделаны П. Траслером (Р. Trusler, Бэкбарн,
Виктория) и Ф. Найтом (F. Knight, Канберра, А. С. Т.). За возмож-
ность использования Ф. Найтом реконструкций из работы “Kadi-
makara: вымершие позвоночные Австралии” мы благодарим
Д. Стоуна (D. Stone) и Пайонир Дизайн Студию (Pioneer Design Stu-
dio, Лилидэль, Виктория), а также музей Виктории.
Большую техническую помощь в оформлении текста, переводе и
проверке названий и ссылок на источники оказали сотрудники уни-
верситета Монаш М. Лейчестер (М. Leicester), П. Германсен
(Р. Hermansen), П. Камаровер (Р. Komarower), С. Сабатье (S. Sabati-
er), Р. Шихэн (R. Sheehan), П. Котс (Р. Coates), Т. МакКонелл
(Т. McConell) и М. Уайтло (М. Whitelaw). Микрокомпьютер Осборна
несомненно помог оформить рукопись легче и быстрее.
Особенно мы чрезвычайно благодарны профессору Минчену Чжоу
(Minchen Chow, Институт палеонтологии позвоночных и палеоант-
ропологии, Пекин), который познакомил Милдред Фентон с Томом
и Пэт Рич, будущими соавторами этой книги. Во время встречи воз-
никла и расцвела (укрепилась) идея о втором издании “Fossil Book”.
Университет Монаш и музей Виктории помогали нам на всем протя-
жении осуществления этого проекта, за что мы им чрезвычайно благо-
дарны. А также мы благодарим свою юную дочку, Лиэллин (Leaellyn),
за ее терпение в течение всех трех лет, когда готовилась эта книга.
Патриция Викерс Рич,
Томас Хъювитт Рич
Предисловие
к русскоязычному изданию
Научно-популярная книга “The Fossil Book. A Record of Prehis-
toric Life” 1989 года издания была подготовлена к печати амери-
канкой Милдред Фентон и австралийской семейной парой Патри-
цией Викерс Рич и Томасом Рич (М. Fenton, Р. Vickers Rich and
Т. Rich). Все они известные и авторитетные палеонтологи высоко-
го профессионального уровня. Идея перевода книги на русский
язык возникла у А.Ю. Розанова и П. Викерс Рич в 1995 году при
посещении Научного центра университета Монаш в Австралии. На
книгу “Fossil Book...” также обратили внимание и другие русские
палеонтологи. Книга богато иллюстрирована, содержит характе-
ристику огромного множества ископаемых, начиная от бактерий и
до млекопитающих, включая человека. В книге отражены основ-
ные успехи палеонтологии и геологии. Актуальность перевода
продиктована и отсутствием научно-популярных книг такого объ-
ема и изложения на русском языке, хотя просто популярные и по
современному красочно иллюстрированные серии по палеонтоло-
гии в России издаются. Тем не менее с повестки дня не снимается
необходимость создания отечественной научно-популярной книги
по палеонтологии.
В начале 1996 года к переводу “Fossil Book...” приступил автор-
ский коллектив в следующем составе: из МГУ - Московского госу-
дарственного университета кафедры палеонтологии - О.Б. Бонда-
ренко, Т.В. Кузнецова, ИА. Михайлова; из Палеонтологического
института РАН - В.К. Голубев, М.Ю. Губин, Е.Н. Курочкин, А.Ю. Ро-
занов; из Геологического института РАН - С.В. Наугольных; из му-
зея Вернадского РАН - Д.Н. Есин. Главы были распределены следу-
ющим образом (нумерация глав дана по русскому изданию): I-V,
VII, VIII, Х1-Х1П, XX, XXI - Бондаренко; VI, X - Бондаренко, Ро-
занов; IX, XIV-XIX - Михайлова; XXII - Есин; XXIII, XXXII - На-
угольных; XXIV-XXVI, XXXI - Голубев; XXVII-XXIX - Губин;
XXX - Курочкин; XXXIII, XXXIV - Кузнецова.
Научным редактором (А.Ю. Розанов) были сформулированы три
основные задачи:
- учесть достижения палеонтологии и геологии, появившиеся
в период с 1989 года (время выхода “Fossil Book...”) по конец
1996 года (время завершения подготовки русскоязычной рукописи
к печати);
Предисловие к русскоязычному изданию
9
- по возможности включить, а в необходимых случаях частично
заменить текст и рисунки оригинала сведениями по России и ближ-
нему зарубежью;
- объем книги на русском языке сократить на треть по сравне-
нию с оригиналом, учитывая возможности издания и покупатель-
ский спрос.
Последняя задача решалась легко. К выполнению первой и вто-
рой задачи русские палеонтологи-переводчики подошли по-разно-
му. Одни не меняли и не дополняли ничего, считая, что оригинал
тем и интересен, что отражает стиль и знания англоязычных иссле-
дователей на 1989 год. Другие сопроводили перевод примечаниями
и сносками, содержащими новые данные или другой взгляд на про-
блему. Третьи снабдили главы новыми рисунками, отсутствующи-
ми в оригинале, и включили небольшие фрагменты нового текста в
текст оригинала, отметив его в некоторых случаях соответствую-
щим знаком. Некоторые написали новые варианты глав. Почти все
авторы переводов сократили число фотографий, так как не пред-
ставлялось возможным добиться их качественного изображения
при перепечатке. Сокращение объема “Fossil Book...” на треть в ос-
новном было достигнуто за счет изъятия двух глав, связанных с
тектоникой плит, а также за счет глав и четырех приложений, со-
держащих описания полевых и лабораторных работ, а также спра-
вочные данные. На наш взгляд, сокращение объема оригинала
“Fossil Book...” (1989 г.) не нарушило основную информативность
книги.
В русском издании книги для иллюстраций использованы и фор-
зацы. На первых двух даны изображения наиболее характерных
форм для разных отрезков времени. На последних двух приведены
общие стратиграфические шкалы фанерозоя и докембрия, приня-
тые в России и СНГ (Стратиграфический кодекс, 1992). Цифры ука-
зывают, на сколько миллионов лет отстоят от современности те или
иные рубежи систем (периодов) и т.д.
На обложке дано изображение знаменитой находки - скелета
археоптерикса (Archaeopteryx) из верхнеюрских известняков Гер-
мании, выбранной в свое время символом для англоязычного из-
дания.
При подготовке русскоязычной рукописи к печати палеонтоло-
ги-переводчики консультировались с научными сотрудниками
МГУ и РАН. Материал по диатомовым водорослям, фораминифе-
рам и радиоляриям был обсужден с Л.Г. Пирумовой, Т.Н. Горбачик
тлД.И. Витухиным; по кораллам - с Л.М. Улитиной; по мшанкам -
с ЛА. Висковой и Р.В. Горюновой-, по членистоногим - с А.С. Алексе-
евым., В.В. Жерихиным, Е.М. Тесаковой-, по граптолитам — сА.Р. Ор-
ловой; по конодонтам - с Л.И. Кононовой иА.Н. Реймерсом; по ры-
бам - с ОА. Лебедевым и С.В. Николаевой; по динозаврам - с
С.М. Курзановым; по млекопитающим - с В.И. Жегалло иА.К.Ага
джаняном. Всем им авторы приносят свою искреннюю благодар-
ность.
Большая работа по оформлению текста большинства глав была
проделана Е.К. Миклашевской. Она также составила сводный ука-
затель латинских и русских названий видов, родов и т.д., встреча-
ющихся в тексте русского издания книги. В подготовке книги к пе-
10 КАМЕННАЯ КНИГА
чати большую помощь оказала Е.А. Жегалло. Без их помощи не
удалось бы так быстро подготовить книгу к печати, за что автор-
ский коллектив им признателен и благодарен.
Выражаем свою благодарность и художнице Е.П. Загозиной, на-
рисовавшей к книге новые иллюстрации и переоформившей неко-
торые из прежних.
Перевод семи глав (IX, XIV-XIX) осуществлен при непо-
средственном участии И.С. Сурковой. При подготовке рукописи в
очень жесткие сроки постоянную помощь оказывали А.В. Долицкий
и М.В. Кнорина. Мы им искренне признательны.
Коллектив авторов русского издания книги
Глава I
О ЧЕМ РАССКАЗЫВАЮТ ИСКОПАЕМЫЕ
• Северные области Евразии и Северной Америки освободились от ма-
терикового льда 20 тысяч лет тому назад, а где-то это произошло
только 12 тысяч лет назад. Холмы и долины этих территорий приоб-
рели современный вид еще позже под влиянием дождей, образующих ручей-
ки, ручьи и реки. Хотя холмы и долины образовались недавно, породы, их
слагающие, в основном являются древними: палеозойскими и мезозойски-
ми. Они когда-то представляли собой илы и пески, которые отлагались в мо-
рях сотни и десятки миллионов лет тому назад. Моря были заселены водорос-
лями, кораллами и другими организмами. Маленькие двустворки вспахива-
ли дно; улитки-гастроподы ползали по нему; морские лилии колыхались над
ним. Там были орды брахиопод, или раковинок-фонариков; некоторые из
них свободно лежали на дне, другие прикреплялись к дну или к другим рако-
винам. Далекие предки наутилуса плавали или прятались рядом; морские
ежи пожирали пленки водорослей и морские отбросы; а животные, назван-
ные мшанками, строили колонии, которые выглядели как кораллы. Рыбы
лениво проплывали рядом (фиг. 1.1.).
Долгая история жизни на планете - это история смен, которая вела мир от
бактериальных образований до огромных дубов и красноствольных деревь-
ев; от очень простых существ к гастроподам, слонам и динозаврам. Такие из-
менения привели к появлению миллиона или около того современных видов,
как и к более чем миллионам видов, которые еще не открыты или вымерли.
Некоторые из живших ранее типов исчезли полностью. То, что они когда-то
существовали, устанавливается по другим звеньям в истории жизни Земли.
Другие организмы погибли, но превратились в ископаемые - фоссилии. Они
и помогают частично восстановить историю жизни далекого прошлого.
ЧТО ТАКОЕ ИСКОПАЕМЫЕ, ИЛИ ФОССИЛИИ
Слово “фоссилии” в дословном переводе обозначает то, что выкапывают. Но
в геологии и палеонтологии фоссилиями называют, как правило, остатки до-
исторических животных и растений, которые были минерализованы, то есть
“превращены в камень”. На карикатурах и в комических сериалах некоторым
из них восстанавливают внешний облик, превращая растительноядных дино-
завров в коней первобытных людей, преследующих на них шерстистых ма-
монтов или убегающих на них от хищного динозавра Tyrannosaurus rex.
© 1997. О.Б. Бондаренко, перевод на русский язык и изменения.
12
КАМЕННАЯ КНИГА
Фиг. 1.1. Реконструкция сообщества жизни позднедевонского моря
Эти комические сериалы, хотя и забавны, имеют мало общего с фактами.
По крайней мере 40 млн. лет отделяют “тиранистых” динозавров от длинно-
шеих растительноядных, а последний динозавр вымер за 60 млн. лет до рас-
селения мамонта из Европы в Америку, то есть до того как человек научился
создавать каменные изделия. Эти различные существа оказываются вместе
только тогда, когда их кости помещают в музеи Естественной Истории. Но
даже там они обычно находятся в разных залах.
О ЧЕМ РАССКАЗЫВАЮТ ИСКОПАЕМЫЕ
13
Мнение, что фоссилии являются минерализованными остатками доисто-
рических организмов, тоже неоднозначно. Часть фоссилий представляет
следы передвижения, обугленные деревья, пропитанные битумом кости и за-
мороженные или высохшие тела. Эти остатки значительно варьируют по воз-
расту от примерно 3500 млн. лет для очень древних фоссилий и до несколь-
ких столетий для гигантской птицы моа Новой Зеландии. Обычное понятие
о фоссилиях поэтому слишком расплывчато. Следующее определение явля-
ется более точным: фоссилии — это остатки или следы организмов, живших
в прошлые геологические времена и погребенных в отложениях, которые на-
капливались в внешнем чехле Земли, то есть в земной коре.
Это определение заполняет брешь, если иметь в виду два уточнения. Пер-
вое: термин “прошлые геологические времена” включает всю историю Земли
от самых первых стадий, заключенных в породах, и до эпохи, наступившей
сразу после последних оледенений. Второе: слово “порода” не соответствует и
часто не означает, что это “камень”. Для геолога породой могут быть любые
отложения, которые слагают внешний чехол Земли, так называемую кору.
В этом смысле гравий и глина являются породами; также и почвенные льды,
то есть вечная мерзлота Сибири и Аляски; парафиновые и озокеритовые пла-
сты Польши и асфальт, который проникает на поверхность в Калифорнии.
Остатки, найденные в таких отложениях, являются фоссилиями, такими же
очевидными, как любые другие, заключенные в “твердых камнях”.
КАК ОБРАЗУЮТСЯ ФОССИЛИИ
Мы сказали, что фоссилии представлены от следов передвижения до за-
мерзших тел, от минерализованных костей и раковин до стволов и высушен-
ной кожи. Ниже мы перечислим способы захоронения.
Замораживание. Идеальными фоссилиями являются те, которые оказа-
лись замороженными после смерти с минимальными нарушениями. Такие
идеальные остатки редки, и они никогда не являются очень древними. Среди
животных они представлены мамонтами и носорогами последнего великого
оледенения, которые проваливались во впадины или расщелины в вечной
мерзлоте и оставались там в течение тысяч лет. Такие остатки сохраняют ко-
сти, мышцы, кожу и волосы так же хорошо, как и внутренние органы. Со-
храняются высохшие кровеносные сосуды артерий и вен и не до конца пере-
варенная пища в желудке. Знаменитые замерзшие мамонты Сибири и Аляс-
ки являются примерами такого способа захоронения. Один из мамонтов
Аляски, замороженный до сих пор, был помещен в американский музей. Си-
бирские мамонты, из которых были изготовлены чучела или смонтированы
скелеты, хранятся в различных музеях России и зарубежья, в том числе в
Санкт-Петербурге и в Москве. Широко известен найденный 23 июня 1977 го-
да в районе Магадана мамонтенок “Дима”, мумия которого хранится в Зоо-
логическом музее Санкт-Петербурга. Мамонтенок получил свое имя от на-
звания ручейка, текущего рядом с местом его находки.
Высушивание. Следующими по качеству после замороженных фоссилий
являются те, которые были полностью высушены. Такие остатки верблюдов,
наземных ленивцев, птиц моа, сумчатых волков и некоторых других живот-
ных были найдены в пещерах полупустынь юго-запада США, Южной Аме-
рики, Новой Зеландии и Австралии. Ленивцы одной патогонской пещеры
были, очевидно, заключены туда древними индейцами, которые часто так
содержали животных для еды, перед тем как убить их.
14
КАМЕННАЯ КНИГА
Битумизация. Естественный парафин, асфальт и озокерит такие же хоро-
шие консерванты, как и лед. В 1907 году из озокерита в восточной Польше
были извлечены голова, конечности и значительная часть кожи шерстистого
носорога. Асфальт обычно сохраняет только твердые части, такие как кости,
зубы и покровы насекомых. Огромное число их было обнаружено в асфальто-
вых образованиялх Калифорнии. В Хэнкокском парке в Лос-Анджелесе по-
сетители могут и сейчас познакомиться с ископаемыми костями, все еще ле-
жащими в асфальтовых ямах. Другое подобное местонахождение, включаю-
щее огромное количество плейстоценовых позвоночных, находится около
Тал ара в Перу.
Простое погребение. Остатки растений и известковых раковин часто ле-
жат долгое время без больших изменений. Например, послеледниковые тор-
фяники содержат почти неизмененные шишки, стволы, стебли и пыльцевые
зерна, которые накопились в болотах. Широко известны ископаемые леса,
представленные стволами, пнями и корневидными образованиями (Англия,
Россия, Монголия и другие страны). Стволы, погребенные в германских лиг-
нитах около 40 млн. лет тому назад, обесцвечены и слегка разрушены, но тек-
стура и строение их древесины почти из изменились. Морские ежи и ракови-
ны моллюсков, чей возраст колеблется от нескольких тысяч до 75 млн. лет,
обычно утрачивают свою окраску, но их твердое известковое вещество испы-
тывает незначительные изменения.
Обугливание. Это процесс неполного разрушения, при котором исчезают
нестойкие компоненты, но не затрагивается углистая составляющая. Остат-
ки растений в торфяниках несут следы ранней стадии обугливания; еще
сильнее обугливание проявляется в древесных завалах многих лигнитовых
образований. В исключительных случаях обугливание приводит к замеще-
нию первичного состава растений и животных блестящими черными пленка-
ми, которые тоньше, чем папиросная бумага. При этом процессе сохраняется
строение листьев, так же как и частей тела древних водных рептилий, и стро-
ение внутренних органов некоторых морских беспозвоночных и рыб.
Окаменение (петрификация) и замещение. Эти процессы, приводящие к
“каменистым” фоссилиям, протекают двумя сходными путями. Простей-
ший из них, названный перминерализацией, заключается в том, что органи-
ка разрушается, а вода, содержащая минеральные компоненты, заполняет
все полости и поры известковых структур. Там и отлагаются минералы, со-
здавая каменистые фоссилии, которые сохраняют большую часть своего пер-
вичного “твердого” материала. Большинство фоссилий захоронились имен-
но таким способом.
Процесс замещения заключается в том, что вода растворяет первичные
твердые части и замещает их другими минеральными компонентами. Это
может происходить так медленно, что новая минеральная составляющая
полностью дублирует микроструктуру раковины, коралла, кости или дере-
ва. Замещение может происходить и таким образом, что не остается и следа
первичной структуры. Разница хорошо видна при сравнении массивной хал-
цедоновой бесструктурной древесины с растительными остатками из так на-
зываемых угольных почек (колболлы), которые сохраняют детали каждой
клеточки (фиг. 1.2). Многие замещенные кости и раковины также сохраняют
все детали, но другие теряют все следы своей первичной структуры. Замеще-
ние является первичным, если оно происходит, когда органические остатки
сначала окаменевают. Вторичное замещение уничтожает микроструктуру,
которая была в начале фоссилизации. Обычно это случается, когда известко-
вые скелеты растворяются и замещаются кремнеземом. С другой стороны,
О ЧЕМ РАССКАЗЫВАЮТ ИСКОПАЕМЫЕ
15
Фиг. 1.2. Примеры прекрасно сохранившихся растений и грибов, обнаружен-
ных в угольных почках (фото Henry Andrews)
а - клеточное строение стебля Lyginopteris в поперечном разрезе, сильно увел,
(птеридоспермы, отдел Голосеменные); б — споры в стробиле (шишка) Lepldocar-
роп, сильно увел, (отдел Плауновидные); в — грибница (мицелий), проросшая
сквозь стебель папоротника, х210 (царство Грибы)
многие окаменелости, сформированные благодаря кремнезему, имеют наи-
лучшую сохранность среди известных фоссилий.
Химические фоссилии. Эти фоссилии являются органическими составля-
ющими, сформированными организмами при жизни. Обычные остатки этих
организмов, как правило, отсутствуют. Примером служит обогащение угле-
родом-12 в отложениях группы Онвервахт докембрийского возраста, о чем
смотреть дальше в главе IV.
Полости, слепки, ядра. Раковины, пни деревьев и другие остатки часто по-
гружаются в осадок, пока он не уплотнился. Позже умершие объекты сгни-
вают или растворяются, оставляя полости. В таком виде полости можно со-
бирать и изучать, но еще лучше заполнить их алебастром, гипсом, воском
или каким-нибудь другим веществом. Такие образования, которые повторя-
ют форму и поверхность ископаемого, называют слепками.
Ядра образуются, когда ил, песок или другие тонкие частицы заполняют
полости. Ядра (иногда их ошибочно называют слепками) внутренних полос-
тей двустворчатых моллюсков и гастропод встречаются очень часто. Ядра
других скелетов, а также деревьев тоже довольно обычны, особенно в песча-
никах. Наилучшие из всех полостей, тем не менее, принадлежат ископае-
мым насекомым в европейских янтарях. Янтарь сначала в виде смолы выте-
кал из древних деревьев, и смола, как капкан, нередко захватывала насеко-
мых. С тех пор смола затвердела, а насекомые высохли почти нацело. В связи
с этим они сохранились только как пустоты, отражающие специфику строе-
ния головы, антенн, тела и конечностей. С помощью микроскопа можно изу-
чить строение и расположение чешуек на крыльях или сосчитать щетинки
на этом уже несуществующем теле.
Другим интересным, но редким примером полости является полость от но-
сорога, вероятно Diceratherium, обнаруженная в базальтовой лаве около Го-
лубого озера в штате Вашингтон. Очевидно, когда-то, в позднем олигоцене
или раннем миоцене, носорог погиб и его раздувшееся тело оказалось в не-
глубоком озере. Лава была достаточно пластичной, она облекла тело носоро-
га, но, охлажденная водой озера, быстро остыла и затвердела. Таким обра-
зом, внешний облик животного был закреплен, а некоторые из его костей со-
16
КАМЕННАЯ КНИГА
Фиг. 1.3. Последовательность реконструкций ископаемого носорога Dicera-
therium (поздний олнгоцен или ранний миоцен, США)
а — слепок полости, оставшейся от носорога; б — реконструкция вздувшегося те-
ла носорога до погребения в базальтах; в — реконструкция живого носорога и его
образа жизни (составил D.E. Savage, департамент палеонтологии Калифорний-
ского университета, Беркли)
хранились внутри полости после того, как мягкие ткани разрушились. То,
что данная полость принадлежала погибшему носорогу, догадались не сразу.
Сначала в полости нашли несколько костей носорога. Впоследствии стали
О ЧЕМ РАССКАЗЫВАЮТ ИСКОПАЕМЫЕ
17
Фиг. 1.4. Кожные покровы ископаемых рептилий
а - отпечаток в песчаниках кожного покрова грудной клетки утконосого
динозавра Anatosaurus (фото из Американского музея Естественной Истории);
б - окаменевшие чешуйки кожи олигоценовой ящерицы Placosaurus (“Glypto-
saurus”)
подозревать, что сама полость может представлять собой слепок тела умер-
шего носорога. Чтобы убедиться в этом, полость обклеили марлей и мешко-
виной, пропитанной гипсом. Полученный “саван” длиной 2.4 м действитель-
но совпал по форме и строению с телом носорога (фиг. 1.3а), после чего были
осуществлены последующие реконструкции (фиг. 1.36, Зе).
Отпечатки. Они напоминают внешние полости, слепки и ядра, но только
тонких объектов, таких как, например, листья и кожные покровы (фиг. 1.4).
Их находят в песчаниках, сланцах и туфах. Значительное число раститель-
ных отпечатков обнаружено в угольных пластах карбона и юры.
Следы, ходы и норы. Эти образования даже более, чем отпечатки, полости
и ядра, объясняют, почему мы говорим, что фоссилии являются не только ос-
татками организмов, но и следами их жизнедеятельности. Некоторые из са-
мых знаменитых ископаемых следов оставлены динозаврами, которые шага-
ли по влажной илистой отмели или по мягкой топи; следы затем отвердева-
ли. Были обнаружены следы и других рептилий, а также отпечатки ног
амфибий, птиц, слонов, наземных ленивцев и даже людей каменного века.
Огромное разнообразие следов оставили также ползающие и зарывающиеся
беспозвоночные. Многие губки, актинии (морские анемоны), гастроподы и
2. Каменная книга
18
КАМЕННАЯ КНИГА
Фиг. 1.5. Ископаемые ходы и норы, названные Daemonelix (а) и реконструкция (б)
создавшего их грызуна (миоцен, штат Небраска, США)
крабы известны по своим следам зарывания или сверления, а не только по
скелетам и панцирям. Образования, названные Cruziana, по-видимому,
представляют собой следы трилобитов. То, что названо Daemonelix (“штопор
дьявола”), очевидно, представляет собой подземные ходы и норы грызуна,
чьи кости описаны под названием Ceratogaulus. Это животное закапывалось
вертикально вниз по винтовой спирали на глубину 1.8-2.4 м, а затем проде-
лывало туннель, идущий полого вверх и слабо расширяющийся на конце
(фиг. 1.5). Паводки иногда заполняли эти необычайно странные ходы пес-
ком, благодаря чему они и сохранились в ископаемом состоянии.
Конечные продукты пищеварения и копролиты. Продукты пищеварения
(следы проедания) являются остатками непереваренной пищи, проглочен-
ной роющими беспозвоночными. Черви, например, поглощают песок вместе
с мелкими организмами. После того как пищевые частицы будут извлечены
в процессе пищеварения, осадок извергается в виде искривленных валиков.
Вымершие черви несомненно действовали сходным образом, и их ископае-
мые следы проедания осадка довольно обычны в некоторых морских форма-
циях. Другие следы проедания относятся к роющим голотуриям (“морские
огурцы”), которые в настоящее время питаются так же, как черви.
Термин “копролиты” (“навоз-камень”) относится к фекалиям, сохранив-
шихся как “окаменелости” либо как ядра или слепки и следы. Некоторые из
них образованы в результате прохождения переваренной пищевой массы че-
рез спиральный клапан кишечника акулы. Некоторые, вероятно, являются
отвердевшим пометом амфибий, а другие остались от рептилий и маммалий,
в то время как остальные относятся к неизвестным позвоночным.
Гастролиты, или желудочные камни. Каждый, кто разделывал птицу для
готовки, знает, что они проглатывают камушки. Камушки застревают в му-
скулистом мешке в одном из отделов желудка, где они дробят пищу на кусоч-
ки, пока она там перемещается.
Многие вымершие рептилии и другие позвоночные используют этот спо-
соб дроблёния пищи с помощью желудочных камешков. Эти камешки, на-
званные гастролитами, распознаются по их округлым краям и гладким, поч-
О ЧЕМ РАССКАЗЫВАЮТ ИСКОПАЕМЫЕ
19
ти полированным поверхностям при условии, что они найдены вместе с ос-
татками позвоночных. Эти выделенные слова очень важны, потому что
камушки могут округляться и становиться гладкими благодаря движению
воды и работе ветра. Надежным доказательством, несомненно, являются на-
ходки гастролитов среди ископаемых костей в месте, где когда-то был желу-
док. На худой конец, такие гальки должны лежать около костей, чтобы не
было сомнений, что их отшлифовали не вода или ветер. Гладкие гальки, ко-
торые просто находятся в слоях с костями рептилий или птиц, нельзя счи-
тать гастролитами.
Гастролиты, по-видимому, не связаны с какой-то определенной диетой
(определенным типом еды). Плезиозавры, питающиеся рыбами, имеют их;
но и растительноядные формы, подобные гигантским новозеландским пти-
цам моа, так же обладают ими, как и современные цыплята.
КАК МНОГО РАССКАЗЫВАЮТ ИСКОПАЕМЫЕ?
Теперь мы подходим к вопросу: как много эти различные типы ископае-
мых рассказывают о жизни в прошлом.
Ответы зависят от конкретных ископаемых, найденных в определенных
местах, и от задач, которые ставит исследователь. Если ископаемые пред-
ставлены только следами ползания и зарывания в таких породах, как песча-
ники, то они могут рассказать, что делали древние существа, но почти ничего
не скажут о них самих. Например, следы хождения указывают, что диноза-
вры ходили, бегали и отдыхали на илистом побережье, но только имеющиеся
там несколько окаменевших костей могут сказать, на что были похожи эти
вымершие рептилии. Некоторые из древнейших извивающихся следов яв-
ляются просто колеями, оставшимися от ползания в иле, поэтому, как вы-
глядят эти извивающиеся существа и к кому они относятся, установить труд-
но. Тем не менее можно оценить разнообразие следов и, кроме того, уровень
сложности поведенческих реакций. Так сравнительный анализ вендских,
раннекембрийских и более поздних следов показывает, что поведение дон-
ных животных неуклонно усложнялось (фиг. 1.6).
Даже наиболее привлекательные ископаемые вызывают досаду у людей,
которые хотели бы узнать о вымерших фаунах (животные) и флорах (расте-
ния) столько же, сколько они могут узнать (но редко это делают) о современ-
ных. Ископаемые являются остатками или следами, которые захоронились;
но большинство вымерших организмов были уничтожены; они были съеде-
ны или быстро разрушены после смерти. Кости были раздроблены и обглода-
ны хищниками и грызунами; стволы и мертвые листья сгнили; раковины
разрушили волны до мельчайших кусочков. Следы хождения и ползания
были размыты; слои песка, содержащие подземные ходы и норы, обруши-
лись и были переработаны прежде, чем они окончательно окаменели. Только
малая часть организмов, которая фактически существовала, избежала этих
разнообразных сил разрушений и в конце концов превратилась в ископае-
мые. Некоторые критически настроенные исследователи говорят, что вос-
станавливать прошлую жизнь по ископаемым все равно, что пытаться рекон-
струировать город по нескольким остаткам с его кладбищ.
Такое сравнение содержит долю правды, но сильно преувеличено. Хотя
ископаемые и не отражают весь мир, который существовал в прошлом, они
все же представлены внушительным количеством организмов, многие из ко-
торых хорошо сохранились и содержат сведения об их условиях жизни. Та-
2*
20
КАМЕННАЯ КНИГА
Фиг. 1.6. Схема строения следов грунтоедов (Федонкин, 1978)
Слева - изменение следов грунтоедов от венда к кембрию, справа — сложно из-
гибающиеся следы грунтоедов в фанерозое
кие ископаемые рассказывают не только об общей истории, они содержат
также огромное количество поражающих деталей, характеризующих мно-
гие эпохи во многих разных местах.
Потрясающую сохранность имеют ископаемые из миоценовых конкреций
пустыни Мохаве (фиг. 1.7). Экземпляры были извлечены после растворения
конкреций, которые широко известны под названием “пуговичные слои”.
Находки включают окремнелых пауков, насекомых и ракообразных, кото-
рые когда-то жили в озере или около него, похожего на Мыльное озеро в рай-
оне современного Вашингтона или на одно из эфемерных соленых озер цент-
ральной Австралии. Выводковая сумка самки ракообразных содержит яич-
ки. Другие насекомые до сих пор лежат в своих коконах, другие погибли при
выходе из него. На некоторых экземплярах видны антенны и шипики на ко-
нечностях. Особенно хорошие ископаемые, заключенные в карбонатные
осадки, сохранили мускулы и воздухоносные трубочки, которые пронизыва-
ли тело.
Таких ископаемых по сохранности превосходят только редкие насекомые
и пауки, чье строение мягких тканей, так же как строение конечностей, со-
хранились в янтаре. У одного янтарного паука сохранились брюшные (абдо-
минальные) органы и мускулы конечностей, у другого - структура клеток
брюшной стенки. У одного вида можно проследить, как идут нити паучьего
шелка к отдельным прядильным трубочкам.
О ЧЕМ РАССКАЗЫВАЮТ ИСКОПАЕМЫЕ
21
Фиг. 1.7. Миоценовые насекомые пустыни Мохаве (юг Северной Америки)
а - куколка двукрылого насекомого из семейства мокрецов, х43; б — личинка
стрекозы, х4; в - личинка жука-плавунца, х9.5; (фото N.W. Shupe)
Многочисленные виды, сохранившиеся хуже вышеописанных, все же до-
статочно информативны, что часто не учитывают критики, которые настаи-
вают на “неполноте палеонтологической летописи”. Так многие окаменев-
шие деревья сохраняют клеточное строение стволов, корней и шишек, най-
денных в углях или известковистых угольных почках. Пальмоподобные
цикадовые до сих пор сохранили структуры, являющиеся прообразом цвет-
ков, а растения, которые напоминают папоротники, содержат семена в
“стручках” на верхушках своих листьев. Ископаемые споры весьма много-
численны в кеннельском угле, а некоторые из них содержат ядра, которые
находились в центрах их желатинообразных клеток. Ядра даже сохраняют
иногда хромосомы, те сложные комплексы, которые называются генами, и
хотя они микроскопические, но их структуры контролируют наследствен-
ность. Шишки, корни и даже семена являются обычными ископаемыми, и
даже цветки не так редки, как принято думать. Один из ископаемых какту-
сов, живший 50 млн. лет назад, имел как цветы, так и плод. Обугленные
морские беспозвоночные гораздо более древнего возраста сохраняют внут-
ренние органы так же хорошо, как щетинки, чешуйки и другие внешние об-
разования.
Ископаемых позвоночных часто относят к категории почти превосходных
ископаемых, если они содержат все основные кости, а их челюсти сохранили
большинство зубов. Остатки рыб нередко представлены “обугленными” тка-
нями и плавниками, такую же сохранность имеют некоторые водные репти-
лии. От динозавров, найденных в тонкозернистых песчаниках, сохранились
отпечатки кожи вокруг их костей. Мамонты из вечной мерзлоты демонстри-
руют почти все детали строения, что и слоны, застреленные охотниками в
Африке, но редко ими изучаемые.
Мышечная ткань (мускулы), головной мозг и поведение. Даже голые кос-
ти могут много рассказать об анатомии мягкого тела. Основой тела любого
позвоночного являются мускулы, так как они формируют облик животного,
22
КАМЕННАЯ КНИГА
а в совокупности с костями определяют его движения. Мускулы, прикрепля-
ясь к костям, часто оставляют следы, указывающие на их размеры, форму и
функции.
Головной мозг и нервная система определяют интеллект и образ жизни,
их принципиальное строение можно определить по пустотам и каналам че-
репной коробки. Благодаря им мы знаем способ поведения и органы чувств
многих позвоночных от бесчелюстных рыбообразных до млекопитающих.
Так, летающие рептилии и птицы видят превосходно, но запахи различают
плохо. Они охотятся с помощью зрения, на что указывают огромные разме-
ры “оптических” долей головного мозга, отвечающих за зрительные импуль-
сы (сигналы). Ранние млекопитающие находили свою пищу по запаху. Са-
мые громадные динозавры имели такую маленькую черепную коробку, что
большинством реакций, по-видимому, управляли расширения (ганглии)
спинного мозга. Некоторые мелкие двуногие тероподы тем не менее имели
увеличенный головной мозг (относительно размеров тела), напоминающий
мозг птиц. Полости черепной коробки динозавров и некоторых ископаемых
млекопитающих иногда сохраняют мельчайшие детали строения мозга. От-
печатки, сохранившиеся на внутренней поверхности черепов, могут помочь
разгадать поведение животных, как, например, стремление собираться в
стаи у собаковидных хищников.
Рост, увечья и болезни. Современные организмы изменяются по мере рос-
та и с возрастом, и это правило также выдерживается среди организмов про-
шлого. Ископаемые беспозвоночные, жившие от 500 до 300 млн. лет тому на-
зад, называемые трилобитами, развиваются из малюсенькой овальной ли-
чинки во взрослое членистоногое длиной до 76 см. Многие моллюски и
брахиоподы меняют в течение жизни форму и скульптуру. Косточки и скеле-
тики неродившейся молоди динозавров, сохранившиеся в яйцах, очень сход-
ны между собой, и однозначно определить их видовую принадлежность чрез-
вычайно трудно, в то время как взрослые особи этих видов четко отличают-
ся. Другой пример изменений во время роста иллюстрируют ископаемые
млекопитающие, у которых многие кости имеют на обоих концах расшире-
ния из губчатого костного вещества (эпифизы), отделенные от кости хряще-
видной пластинкой роста. Когда млекопитающие достигают зрелости, эпи-
физы сливаются с главным стволом кости, образуя одно целое. Эпифизы раз-
ных животных срастаются в процессе роста индивидумов в различное время.
Археологи используют эти закономерности для определения возраста жи-
вотных, включая человека, найденных при раскопках.
Некоторые окаменевшие стволы несут рубцы, полученные от пожара при
жизни этих деревьев. Другие ископаемые растения имеют патологические
узелки, грибковые пятна и вздутия (галлы) от внедрившихся насекомых.
Некоторые скелеты искривлены, по-видимому, от сокращения и спазм
мышц, связанных со смертью. Другие ископаемые кости несут следы перело-
мов, полученные при жизни животного, как, например, экспонат крокоди-
ла, хранящийся в Москве в Палеонтологическом музее.
Форма и функция. Можно много узнать об образе жизни организмов исхо-
дя из строения скелета, будь то беспозвоночные или позвоночные. Ныне жи-
вущие двустворки, обитающие на мягком нестабильном илистом дне, обыч-
но имеют низкий удельный вес раковины с тенденцией к мелким размерам;
их поверхность обычно покрыта шипиками. Все эти особенности помогают
моллюскам не проваливаться в ил. Интересно, что у бегающих птиц, таких
как додо о-ва Маврикий, кости крыльев сильно редуцированы, а грудная
клетка не имеет киля. У птиц, таких как пингвин, которые используют свои
О ЧЕМ РАССКАЗЫВАЮТ ИСКОПАЕМЫЕ
23
крылья как весла, кости крыльев утолщены и не имеют воздухоносных по-
лостей, но вместо этого они довольно плотные и большого удельного веса. Эти
приспособления (адаптации) создают обтекаемую структуру достаточно
мощную, чтобы позволить птице передвигаться в водной толще под поверх-
ностью океана. Существует бесчисленное число разнообразных особенностей
для растительноядного и хищного стиля жизни у передвигающихся (бегаю-
щих), парящих (летающих) и зарывающихся (копающих) форм. Растения
имеют специальные адаптации для жизни в районах с обильными ливнями,
такие как гребни на листьях, помогающие быстро избавиться от воды. Такие
специализации, конечно, оставляют свои характерные метки у организмов,
и наблюдательный палеонтолог может узнать очень много о прежнем образе
жизни этих ископаемых, изучая такие особенности строения.
Деятельность и инстинкты. Увеличенные доли мозга говорят нам, что не-
которые животные охотятся, используя преимущественно зрение, а другие
полагаются на запахи. Большие размеры и скелетная конструкция ископае-
мых ленивцев указывают на то, что они были наземными животными, в то
время как их современные потомки обитают на деревьях. Краб, найденный в
древней норе червя, жил там так же, как это делают его современные пред-
ставители, забирающиеся в норки и ходы, сделанные сходными червями.
Клыки так называемых “гигантских свиней” (энтелодонов) имеют глубокие
зазубрины - признак, который стал понятным, когда догадались, что зазуб-
рины связаны с корнями растений, которые животные выкапывали для еды.
Образ жизни беспозвоночных иногда понятен по их внешней форме, ино-
гда - по взаимоотношениям, а иногда по находкам в прижизненном положе-
нии. Древние устрицы, например, были прикреплены к другим раковинам
или к камням; белемниты плавали как кальмары, которых они напоминают;
некоторые улитки (гастроподы) высверливают отверстия в других ракови-
нах и поедают их содержимое. Другие улитки приползают к вершинам чаше-
чек морских лилий (криноидеи) и там питаются остатками пищи, что оста-
ются от хозяек. Такие улитки почти не передвигаются и присасываются
столь плотно, что их раковины в процессе роста начинают отражать строение
морской лилии (Platyceras). Некоторые морские звезды также приспособле-
ны к подобному образу жизни, при этом они плотно обхватывают хозяина,
соединяя вместе свои лучи.
Некоторые современные улитки просверливают отверстия в других мол-
люсках, внедряя плотное языковидное образование, содержащее рашпиль-
ные зубы (радулу), и поглощая мягкое содержимое, находящееся внутри ра-
ковин их жертв. Такой способ питания существовал и в древнее время, по
крайней мере уже с ордовика. Многие рыбы и рептилии проглатывали свои
жертвы целиком. Ископаемые морские рептилии, найденные в отложениях
Германии, содержат непереваренные остатки таких трапез; такое же явле-
ние наблюдается у некоторых американских рыб. Одна такая хищная рыба,
вероятно, рассталась с жизнью внезапно, о чем свидетельствует положение
жертвы, заглоченной только частично. Другая хищная рыба - 4.2-метровый
“бульдожий тарпон” (Portheus molossus) проглотила 1.7-метровую жертву
целиком, но вскоре после этого погибла. Смертельные схватки и борьба
жертвы за жизнь, вероятно, вызывает фатальные внутренние повреждения
и у хищника.
Некоторые морские рептилии были быстрыми сильными пловцами, про-
плывающими большие расстояния. Отдельные находки гастролитов показы-
вают дальность путешествий, но не скорость передвижения. Один мозазавр
проглатывал куски специфического розового кварцита, распространенного в
24
КАМЕННАЯ КНИГА
западной части штата Айовы и в штате Миннесота, но погиб он в Канзасе, бо-
лее чем в 650 км вдали от площади распространения этих кварцитов. Следо-
вательно, мозазавр мог проплывать расстояния более чем 650 км.
Еще более удивительными являются ходы грызуна Ceratogaulus, полу-
чившие родовое имя Daemonelix, но более известные под названием “штопор
дьявола”. Ходы сначала идут винтообразно круто вниз, а затем переходят в
прямые туннели, воздымающиеся полого вверх (см. фиг. 1.5). Такие ходы
требуют интенсивного и точного забуривания; это означает, что такой слож-
ный тип зарывания является унаследованной чертой, одной из тех, которая
заставляет щегла или малиновку строить свои характерные гнезда. Если мы
считаем, что построение гнезда является инстинктом, то этот термин можно
также использовать для деятельности этого древнего грызуна. Образно гово-
ря, под названием Daemonelix скрывается окаменевший инстинкт, сущест-
вовавший около 15 млн. лет тому назад.
Тафономия. Кости и раковины фиксируют, что происходит с организмами
с того момента, когда они умерли, до того времени, когда они погребаются, а
затем их извлекает “охотник за фоссилиями”. Изучением этого процесса за-
нимается тафономия. Большое количество длинных тонких костей в косос-
лоистых песчаниках может означать, что это ископаемое сообщество форми-
ровалось в потоке, который отсортировал кости определенного размера и
формы. Это не означает, что только животные с длинными тонкими коцтями
существовали в окрестностях этого потока. Много факторов определяет веро-
ятность перехода живого организма в ископаемое: его размеры, имеет ли он
твердые части, число твердых частей, молодой он или старый, образует коло-
нии или является одиночным, где живет и какой образ жизни ведет.
Палеонтологи, изучающие тафономический аспект ископаемых, должны
тщательно документировать полевую ситуацию, брать как можно более
крупный монолит, внимательно изучать ископаемых, их форму, размеры и
число, а также внешнюю текстуру и степень повреждений, прежде чем пере-
ходить к обобщениям. Такие исследования очень ценны для понимания, как
формировались ископаемые сообщества. Они помогают установить, сущест-
вовали ископаемые вместе при жизни (биоценоз) или они объединены друг с
другом только после смерти (танатоценоз, тафоценоз, ориктоценоз).
Очень важно, чтобы коллекционеры тщательно документировали поле-
вые наблюдения и были уверены, что они исследовали каждый клочок, что
обнажается. Если такую информацию не собирают с самого начала изуче-
ния, то теряется навсегда огромное число сведений об ископаемых.
УСЛОВИЯ ОБИТАНИЯ И ТЕМПЕРАТУРА
Ископаемые также много рассказывают о своем окружении и условиях, в
которых они жили. Деревья несомненно произрастали на суше, а морские во-
доросли существовали в соленой воде. Мы можем утверждать, что кораллы,
устрицы и кальмароподобные существа жили как и водоросли, исходя из то-
го, что все их ныне живущие родственники обитают сегодня в морях.
Более подробную информацию можно узнать и о таких факторах сущест-
вования, как степень освещенности, характер субстрата, на котором обитает
животное или растение, степень загрязнения воды и насколько соленой она
была (фиг. 1.8, 9).
Рифостроящие кораллы, например, содержат водоросли зооксантеллы и
зоохлореллы; поэтому они встречаются где много света, преимущественно не
О ЧЕМ РАССКАЗЫВАЮТ ИСКОПАЕМЫЕ
25
2Ий.
СОЛОНОВАТОВОДНЫЕ
ПЕРЕХОДНЫЕ
—^АГГЛЮТИНИРОВАННЫЕ ФЮРАМЙНИФЁРЬ1<;
НОРМАЛЬНО
МОРСКИЕ
30
—<Екрасныё вбд6робли?>
<±: ^зеленые водоросли
ДИАТОМОВЫЕ водоросли:
<СЕКРЁЦЙбННО-ИЗВЕСТКОВЫЕ ФОРАМИНИФЕРЫ
:-брахиоподьг$>
гастроподы
-остракоды;
ПЕРЕХОДНЫЕ
ГИПЕРСОЛЕНЫЕ
40
цианобионты:
бфГУБКИЁ:
кораллы:
бИЗВЁСТ. ТРУБКИ червей:
МШАНКИ?
<ЕжЛОКОЖЙЁ?>
УСОНОГИЕ РАЧКИ'
прочие ракообразные:
<<хитоны:-:>
рСАФОПОДЫ;
--------<ЦЕФАЛОПОДЫ^
<:двуствоРКЙу«-:-:«-у777:
Фиг. 1.8. Распространение циаиобионтов, растений и животных в бассейнах
различной солености (R.L. Anstey, T.L. Chase, 1974. Environments Through
Time. Burgess Publishing Co., Minneapolis; с изменениями)
глубже 30 метров. Географическое распространение рифостроящих корал-
лов обычно ограничено широтой ±30 от экватора. Кораллам необходима от-
носительно чистая прозрачная вода, они требуют также определенных мор-
ских условий, где соленость колеблется между 30 и 40 промиллями.
26
КАМЕННАЯ КНИГА
ПРОЗРАЧНАЯ ВОДА
УВЕЛИЧЕНИЕ ТУРБУЛЕНТНОСТИ
ИНТЕНСИВНОЕ
ОСАДКОНАКОПЛЕНИЕ
СЕСТОНОФАГИ
--- ДЕТРИТОФАГИ---
ХИЩНИКИ, ПАДАЛЕЕДЫ
КРАСНЬIE ВОДОРОСЛИ.
ЗЕЛЕНЫЕ ВОДОРОСЛИ:
:ЦИАНОБИОнты::
фораминиферы>:::
ПРОЧИЕ БРАХИОПОДЫ
ГУБКИ:
КОРАЛЛЫ.:;:
МШАНКИ:
КРИНОИДЕИ-
МОРСКИЕ ЗВЕЗДЫ
МОРСКИЕ ЕЖИ
хитоны:
СКАФОПОДЫ;
ГАСТРОПОДЫ;
ОСТРАКОДЫ
УСОНОГИЕ РАЧКИ:
ПРОЧИЕ ЧЛЕНИСТОНОГИЕ
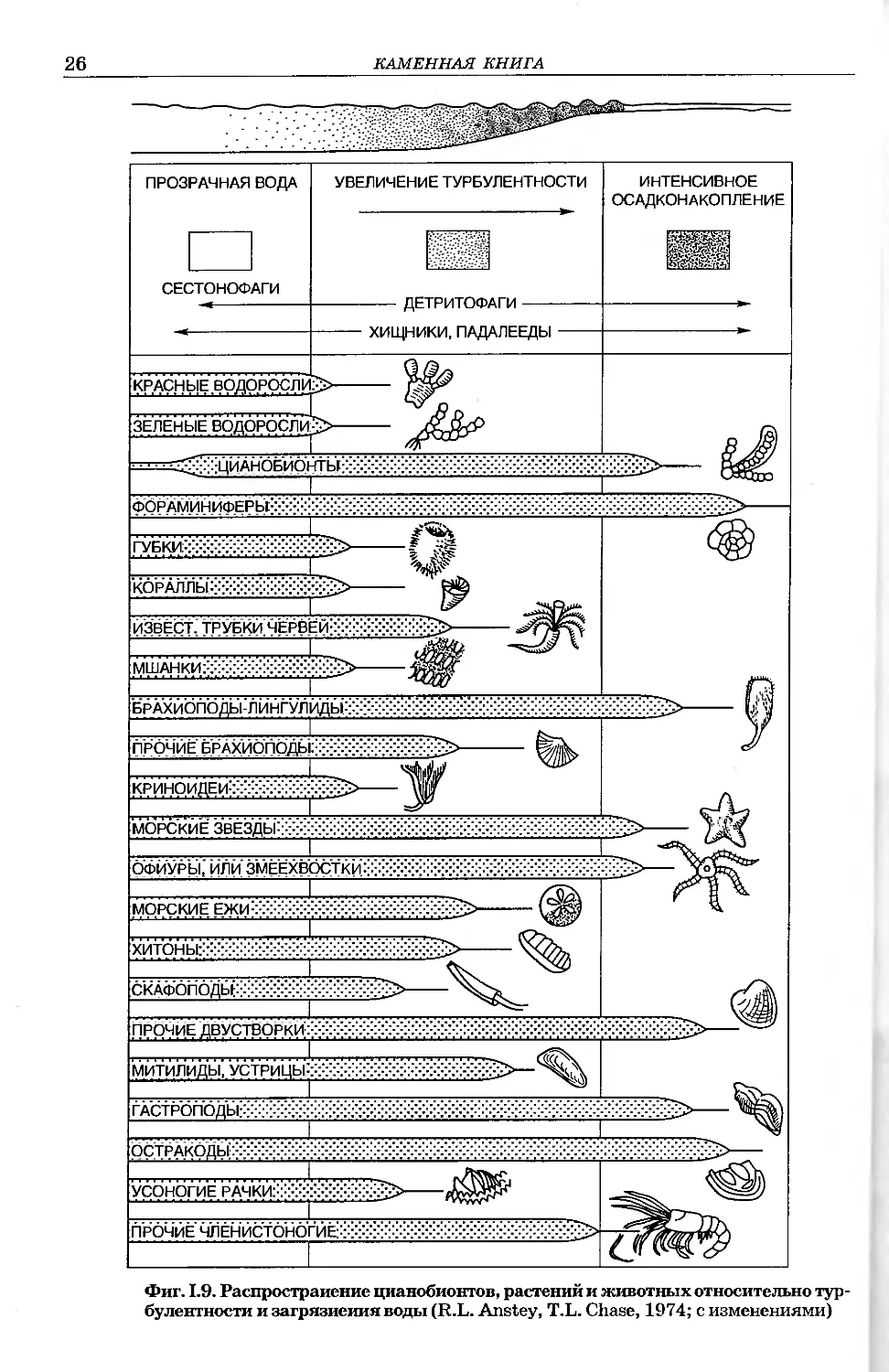
Фиг. 1.9. Распространение цианобионтов, растений и животных относительно тур-
булентности и загрязнения воды (R.L. Anstey, T.L. Chase, 1974; с изменениями)
ИЗВЁСТ- ТРУБКИ ЧЕРВЕЙ
БРАХИОПОДЫ-ЛИНГУЛИДЫ,:;:
ОФЙУРЫ, ИЛИ ЗМЕЕХВОСТКИ
ПРОЧИЕ двустворки;?:
МИТИЛИДЫ, УСТРИЦЫ
О ЧЕМ РАССКАЗЫВАЮТ ИСКОПАЕМЫЕ
27
Субстраты, с которыми связана жизнь организмов, тоже можно восстано-
вить. Обычно наблюдается уменьшение числа видов, если двигаться от пес-
чаных субстратов к илистым. Улитки, двустворки и рыбы, принадлежащие
к группам, обитающим в настоящее время только в пресноводных водоемах,
могут указывать на сходные условия в прошлом. Животные, такие как ло-
шади и верблюды, несомненно существовали в степных и пустынных услови-
ях, в то время как животные, похожие на гиппопотамов, обитали на заболо-
ченных пространствах.
Протисты, растения и животные также свидетельствуют о древних кли-
матах. Пальмы, например, живут в теплых районах, хотя некоторые из них
встречаются в настоящее время и на юге Новой Зеландии. Ели преуспевают
там, где климат холодный. Папоротники нуждаются в огромном количестве
влаги, а травы и кактусы обходятся гораздо меньшим ее количеством. Север-
ный олень указывает на холодный климат, так же как шерстистые мамонты,
чьи остатки находят в вечной мерзлоте.
Факты, такие как эти, тщательно уточняют, чтобы восстановленные кар-
тины древней суши, морей и климата были максимально точными. До
1950 года, тем не менее, точную температуру по ископаемым установить бы-
ло нельзя. В тот год Урей (Н.С. Urey) исследовал юрского белемнита, живше-
го около 150 млн. лет тому назад (см. главу XVII). Благодаря изотопам кис-
лорода Урей установил, что белемнит погиб летом, а прожил почти четыре
года. В течение жизни белемнита летняя температура колебалась между
20-21°С, а зимняя была около 15-17.7°С.
Многие палеонтологи использовали метод Урей при изучении ископае-
мых разного геологического возраста и получили соответствующие резуль-
таты. Одна из серий температур показывает, как она изменялась в конце ме-
зозоя. Эти изменения хорошо сопоставимы с расцветом и вымиранием мело-
вых динозавров. Сводные данные температурных измерений можно найти в
работах Д.П. Найдина, С.А. Ясаманова и у других русских и зарубежных
авторов.
Глава II
ФОССИЛИИ, ПОРОДЫ И ВОЗРАСТ
Фоссилии, или ископаемые, являются остатками или следами ког-
да-то живших существ, погребенных в породах земной коры. Поче-
му одни породы содержат фоссилии, а другие нет и как определяют
их геологический возраст - читайте ниже.
ПОРОДЫ И ИХ ФОССИЛИИ
Породы, сильно отличающиеся по строению и плотности, отличаются и
происхождением. Происхождение в свою очередь определяет наши шансы
найти или не найти в них фоссилии.
Изверженные породы. Термин “изверженные” означает огненный, пла-
менный, хотя породы этого типа на самом деле не горели. Изверженные по-
роды представлены лавами (которые попали на поверхность Земли во время
извержений) и близкими им породами, остывшими и затвердевшими под по-
верхностью Земли. Лавы появлялись из трещин или вулканов в виде пото-
ков, а далее они разливались по поверхности. Подземные массы были гораз-
до более вязкими; их называют магмой, от греческого слова “magma” - меси-
во. Это общий термин для того состояния, пока масса не остыла. По мере
остывания из магмы образуются граниты и другие породы, не содержащие
следов течения и газовых пузырьков, что так характерно для лав.
Изверженные породы, которые отвердевают под поверхностью Земли, не
содержат ископаемых, так как ничто не может жить при высокой температуре
глубоко под поверхностью. Фоссилии очень редки в лавах, которые, как пра-
вило, уничтожают растения и животных, попадающих на их пути. Исключе-
ние представляет собой носорог, описанный в первой главе, и стволы деревьев,
оставляющих полости в постепенно остывающих лавах. Исключение состав-
ляют и ископаемые из палеозойских гранитов Урала, сохранившиеся благода-
ря постепенному замещению (метасоматоз) карбонатных пород гранитами.
Лавы при извержении могут выбрасываться в виде “брызг” в воздух, где
они быстро застывают. Падая на землю, они образуют породы, называемые
“аггломератами”, если размер частиц крупный, и “туфами”, если они очень
мелкие, пылевидные. Обе эти породы, частицы которых упали на земную по-
верхность, образуют своеобразный мостик между изверженными и осадоч-
ными породами.
© 1997. О.Б. Бондаренко, перевод на русский язык и изменения.
ФОССИЛИИ, ПОРОДЫ И ВОЗРАСТ
29
Аггломераты засыпали многие ископаемые деревья, некоторые из кото-
рых уже были мертвыми ко времени погребения. Наиболее знаменитые ис-
копаемые деревья находятся в Йеллоустонском Национальном парке, где из-
вестно по крайней мере восемнадцать последовательных уровней погребен-
ных лесов, состоящих из секвой, сосен, платановидных сикамор и дубов,
засыпанных вулканическими образованиями. Стволы секвой, великолепно
сохранившиеся, найдены в светлоокрашенных туфах в несколько милях от
Калистога в Калифорнии. Пепел, смешанный с илом, отложился в озерах
около Флориссанта в Колорадо, образовав светло-серые сланцы. Они содер-
жат ошеломляющее количество листьев и около 1100 видов бабочек, сверч-
ков, кузнечиков, мух, жуков и других насекомых. Бабочки до сих пор сохра-
нили свой характерный рисунок в виде полос и пятен.
В пепловых отложениях, то есть в туфах, иногда можно обнаружить следы
передвижения. Знаменитым примером является местонахождение Л это ли в
Восточной Африке, образовавшееся 3.5 млн. лет тому назад. Тысячи следов
антилоп, львов и даже предков современных людей сохранили все детали.
Осадочные породы. Хотя фоссилии являются исключением в извержен-
ных породах, в осадочных отложениях - они правило. Слово “осадочный”
очень удачное, так как означает, что “нечто осело”. Осадочные породы содер-
жат пыль, песок, ил и другие компоненты, которые осаждаются под водой
или на суше. Они образуют толщи, пласты, пачки, слои, прослои; название
зависит от мощности и соотношения пород друг с другом. Некоторые осадоч-
ные породы формировались в горячих источниках. Уголь является широко
известной осадочной породой, состоящей почти целиком из растений.
Тем не менее не все осадочные породы содержат фоссилии. В конгломера-
тах, например, их может не быть, так как скелеты организмов, связанных с
грубыми песчаными и галечными субстратами, вскоре после смерти истира-
ются и ломаются галькой. Тонкозернистые известняки могут быть пустыми
без фоссилий в том случае, если они хемогенные или бактериальные. Рако-
вины и кораллы редки в большинстве морских песчаников - кораллы пото-
му, что они обычно не живут на песчаных субстратах, а раковины потому,
что они были разрушены растворами, циркулирующими в песке, а затем в
порах отвердевших пород. Даже когда фоссилии сохраняются, они часто не
более чем пустоты или сильно разрушенные остатки.
В контрасте с этими почти пустыми отложениями существуют и такие, где
фоссилии многочисленны или даже образуют почти всю породу. Нередко от-
ложения содержат так много остатков, что исследователь, не пренебрегаю-
щий ни мелкими, ни крупными фоссилиями, может собрать около десяти
тысяч экземпляров в день. На верхних поверхностях плитчатых известня-
ков карбона Подмосковья наблюдается множество остатков морских лилий,
а девонские и каменноугольные песчаники и известняки Воронежской обла-
сти содержат многочисленные раковины брахиопод. Меловые породы юга
России образованы скоплениями крошечных раковинок фораминифер. Ри-
фы содержат хорошо сохранившиеся почти не измененные известковые во-
доросли (см. главу IV). Даже грубозернистые песчаники могут заключать ко-
сти крупных динозавров, в то время как тонкозернистые слои, например
слюдистоглинистые, часто переполнены следами передвижения и зарыва-
ния. Другие тонкозернистые песчаники, которые образуются на суше, бога-
ты ископаемыми млекопитающими. В знаменитом местонахождении Небра-
ски, известном под названием “Среднекайнозойский агатовый костеносный
слой”, тонкозернистый песчаник заполняет только пустоты между тесно рас-
положенными черепами и другими костями (см. главу ХХХШ).
30
КАМЕННАЯ КНИГА
Метаморфические породы. Среди них известны настоящий мрамор и кри-
сталлические породы, часто называемые гранитами, хотя правильнее их на-
зывать гнейсами и сланцами. Некоторые из метаморфических пород вначале
были осадочными, другие - изверженными. Все они были изменены за счет
жара и испарений от магмы и(или) за счет давления, которое подняло поро-
ды вверх, превратив их со временем в горы. Этот процесс иногда заходил
столь далеко, что мы не можем уверенно установить первичное происхожде-
ние породы: магма, лава или осадочные отложения.
Обычно метаморфические породы не содержат фоссилий, а если они и бы-
ли, то, как правило, полностью уничтожены. Есть и другие примеры: стро-
матолиты весьма многочисленны в слабо измененном мраморе, а разнообраз-
ные раковины весьма обычны в слабо метаморфизованных сланцах, но там
они часто расплющены и разломаны. Тем не менее раковины можно опреде-
лить, хоть их облик может сильно отличаться от того, когда эти существа бы-
ли живыми. Интересно, что искажение облика иногда помогает геологам по-
нять направление и интенсивность тех сил, которые деформировали первич-
ные породы.
ГДЕ НАХОДЯТ ФОССИЛИИ?
Теперь можно ответить на вопрос, где и как искать фоссилии. Первый
шаг, конечно, связан с выяснением, не является ли порода изверженной или
метаморфической, и тогда почти наверняка фоссилий в ней нет. Затем надо
исключить формации и слои, где остатки редкие или плохие, кроме тех слу-
чаев, когда исследователь изучает докембрийские породы и заинтересован в
любых ископаемых. И тогда перед нами предстает огромное разнообразие
формаций, в которых фоссилии многочисленные, а по качеству хорошие и
даже великолепные.
Многие породы поэтому не вызывают сомнений, а перспективность дру-
гих устанавливают по составу и облику, сходному с известными “фоссило-
носными” отложениями, или благодаря тому, что похожие образцы демонст-
рируют на выставках и в музеях. Следуя этим правилам, исследователь тща-
тельно изучает каждое обнаруженное обнажение, пока не соберет
внушительный ряд образцов или докажет, что их невозможно найти.
Сборы ископаемых беспозвоночных осуществлять легче, чем сборы иско-
паемых позвоночных. Хотя некоторые слои переполнены зубами и костями,
все же ископаемые позвоночные встречаются гораздо реже, чем раковины
или кораллы, и гораздо хуже поддаются идентификации. Многие гастропо-
ды, если не все, узнаваемы как гастроподы, но череп или даже скелет может
выглядеть в породе не более как светло- или темноокрашенная шишка или
конкреция. В руках неопытного исследователя конкреция может быть легко
разрушена и уничтожена, в то время как многих беспозвоночных (если не
всех!) легко извлечь из пород, в которых они заключены.
ФОССИЛИИ И ФОРМАЦИИ
Каждый исследователь вскоре замечает, что определенные ископаемые
связаны с определенными типами пород. Болотные растения, например,
наиболее многочисленны в сланцах и песчаниках между угольными пласта-
ми. Граптолиты обычно приурочены к темным тонкозернистым сланцам, ко-
ФОССИЛИИ. ПОРОДЫ И ВОЗРАСТ
31
/91 02 04 ^5 06 07 (З8 й9 0^10 011 012 Q13 ©14 <s>15 ^16 017
<0)18 <919 020 (gb21 4 22 4,23 ZX24 X3)25 £)26 =>27 ^28 ©29 t^30 ^31 <032 ^33
Фиг. II. 1. Биофациальный профиль позднемелового моря южной части Восточ-
но-Европейской платформы (время В. mucronata—B. langei)
I — песчаные осадки; II - карбонатно-алевритовые осадки; III - карбонатно-гли-
нистые осадки; IV - микрозернистые карбонатные илы. 1-20 - двустворчатые
моллюски (в том числе: 1 - Nucula; 3 - Area; 4 — Inoceramus; 3 - Chlamys; 18 -
Astarte, 19 - Tellina), 21-24 — брюхоногие моллюски; 25 - лопатоногие моллю-
ски; 26 - аммониты; 27 — белемниты; 28 — губки; 29 - шестилучевые кораллы;
30 — черви; 31 - мшанки; 32 — брахиоподы; 33 - морские ежи. А - приконтинен-
тальная часть моря; Б - открытая часть моря (Собецкий, 1978)
торые расслаиваются на тонкие пластиночки. Следы хождения рептилий и
следы ползания беспозвоночных характерны для тонкозернистых песчани-
ков, особенно если они переслаиваются с сланцами. Кораллы приурочены к
известковистым сланцам и массивным известнякам, многие из которых яв-
ляются фациями древних рифов.
Свидетельства изменений. Установлено, что фоссилии могут не изменять-
ся на протяжении нескольких слоев, хотя в других случаях они отличаются.
Эти отличия и сходство показывают, какие слои надо группировать вместе, а
какие должны быть отделены.
Надо иметь в виду, что разные фоссилии могут быть одновозрастными,
просто они обитали в разных условиях, что отражается в составе и текстуре
отложений. Вот один из примеров (фиг. II. 1). На биофациальном профиле
позднемелового моря юго-западной части Восточно-Европейской платформы
показаны отложения и фауна четырех батиметрических зон. Первая зона (I)
представлена песчаными осадками с довольно однообразными двустворка-
ми; она отвечает прибрежному мелководью, самой верхней части сублитора-
ли. Вторая зона (II) представлена карбонатно-алевритовыми осадками с мно-
гочисленными двустворками, гастроподами, лопатоногими моллюсками,
аммонитами, кораллами, морскими ежами и червями; вторая зона соответ-
ствует морскому мелководью, то есть верхней и началу средней суб литорали.
Третья зона (III) представлена карбонатно-глинистыми осадками с богатой
32
КАМЕННАЯ КНИГА
ПЕСЧАНИКИ
FOX HILLS
Вулкани-
ческие
пеплы
и туфы
О lmS
5^3 ФОРМАЦИЯ
HELL CREEK
ПЕСЧАНИКИ
FOX HILLS
ФОРМАЦИЯ
NIOBRARA
(морской мел)
ПЕСЧАНИКИ EAGLE
ФОРМАЦИЯ (сопоноватые- морские)
ПЕСЧАНИКИ ТЕ^гс5РН
V1RGELLE
^сланцы:
Z-ZPIERRE-Z
z-( морские);
I
<:±=
Осадочная
свита
СЛАНЦЫ
-CLAGGETT
ФОРМАЦИЯ JUDITH RIVER- -
(пресноводные.
-континентальные
—солоноватые) к —
ФОРМАЦИЯ HELL CREEK-7-
Футы Метры
ЮОО-иЗОО
ЗЕ
(пресно-
водные
континен-
тальные) Ш| °
СЗСЛАНЦЫ BEARPAW;
* :-(морские)з--_
ЛЕСЧАНИКИ HORSETHIEF=Z-
- “-(у морского 6epera)-z=z=2=
z'cnAHUb!‘z-z-z-z-z-z-z-z:
^EARpAW-z=z-z-z-z-z-z-z
-(морские)'------------ —Z
Z-Z---Z-Z-Z-Z-OOPMAUHRZ-
z-z=z-z-z-z-zJUDnH river:
ФОРМАЦИЯ JUDITH RIVER-
JcpnoHOBaTbie^McpcKMej^
-j- Z-СЛАНЦЫ--
2J zGLAGGETT
=сланцьс-:
GAMMON-
-200
500-
100
0 50 ЮО км
Фиг. П.2. Соотношение верхнемеловых формаций по траверсу от северной ча-
сти Монтаны (слева) до Дакоты (справа)
Местонахождения всех пяти (1-5) колонок показаны вдоль траверса на левой
карте. На правой карте серым закрашено пространство, которое занимали позд-
немеловые моря на западе и юге Северной Америки
разнообразной фауной, похожей на таковую из предыдущей зоны, но здесь
уже появляются представители более глубоких зон моря, такие как белемни-
ты, стеклянные губки, брахиоподы; третья зона более глубоководная, чем
вторая, и соответствует средней и нижней сублиторали. И наконец, четвер-
тая зона (IV), самая глубокая в позднемеловом бассейне, представлена мик-
розернистыми карбонатными илами с очень бедной фауной морских глубин:
стеклянные губки, белемниты, мшанки и однообразные двустворки. Еще бо-
лее сложные соотношения осадков (а значит и систематического состава фос-
силий) проиллюстрированы на фиг. II.2.
Руководящие ископаемые. Мы не всегда можем проследить, как одни от-
ложения переходят в другие, или найти отложения достаточной мощности,
которые перекрывают друг друга, как кровля на крыше. В этом случае необ-
ходимо обратиться к руководящим ископаемым.
По настоящему хорошее руководящее ископаемое должно соответство-
вать трем основным критериям. Первый критерий - четкость опознавания,
это означает, что его нельзя перепутать с фоссилиями, которые жили совсем
в другое время. Второй критерий - руководящее ископаемое должно быстро
трансформироваться в другой вид или вымереть, но при этом успеть широко
распространиться, становясь обычным ископаемым для маленького отрезка
времени почти по всей планете. И наконец, третий критерий - данный вид
должен хорошо сохраняться в ископаемом состоянии. Редко встречающиеся
виды иногда имеют ценность руководящих (если мы их находим), но польза
от них намного меньше, чем от тех, кого много.
ФОССИЛИИ, ПОРОДЫ И ВОЗРАСТ
33
Фиг. П.З. Корреляция обнажений методом руководящих ископаемых (слева
изображена сводная колонка)
1—5 - номера обнажений (объяснения см. в тексте)
Корреляция. Чтобы разобраться, как используют руководящие виды, да-
вайте рассмотрим следующий пример. Предположим, мы изучили пять об-
нажений, или разрезов (фиг. П.З). Все они содержат слои, сменяющие друг
друга в определенном порядке, но сразу сопоставить их между собой не уда-
ется, а только постепенно, не более двух-трех разрезов за одну операцию. Оз-
начает ли это, что разные отложения накопились в разных местах, или эти
слои образуют одну непрерывную серию с древними породами внизу и моло-
дыми наверху?
В данном примере на эти вопросы невозможно ответить, прослеживая
слои по простиранию, так как они скрыты или отсутствуют между нашими
участками и холмами. Вместо прослеживания мы соберем фоссилии из каж-
дого места, а затем скоррелируем, то есть сопоставим их. На вершине холма
обнажения 1 мы находим морских ежей, идентичных с теми, что были собра-
ны в основании другого холма в обнажении 2. Здесь выше пород с морскими
ежами находятся известняки с устрицеподобными раковинами, называемы-
ми Gryphaea, которые также встречаются на участке обнажения 3. Спираль-
но-конические раковины гастропод позволяют сопоставить этот участок с об-
нажением 4 другого холма, а двустворки - митилиды показывают, что слои
у вершины холма встречаются в основании обнажения 5 еще одного участка.
Сопоставляя слои, которые содержат эти руководящие ископаемые, мы вы-
страиваем наши обнажения в одну непрерывную серию, то есть в единый
сводный разрез - колонку (см. фиг. П.З, слева).
Проблемы корреляции могут возникать, когда ископаемые приурочены
только к одному типу пород (фациальные проблемы). Другими словами, ког-
да эти организмы были живыми, они могли существовать в узком диапазоне
условий, или захоронялись только при специфических условиях. Например,
граптолиты часто приурочены к черным сланцам, в то время как брахиопо-
ды нормально сохраняются в известняках и песчаниках. Итак, когда мы пы-
таемся скоррелировать сланцы и известняки, которые отлагались в одно вре-
мя, но в разных местах, нам необходимо найти таких ископаемых, которые
3. Каменная книга
34
КАМЕННАЯ КНИГА
встречаются и там и тут. Пыльца часто отвечает такому требованию; она осе-
дает из воздушного пространства на все типы осадков и в результате образует
“нить времени”, соединяющую их всех.
ПОДРАЗДЕЛЕНИЯ ГЕОЛОГИЧЕСКОЙ ИСТОРИИ
Метод корреляции пород по ископаемым впервые применил в течение
1790-х годов английский инженер Вильям Смит и два французских палеон-
толога - Жорж Кювье и Александр Броньяр. Они также использовали разли-
чия между ископаемыми, чтобы разделить мощные толщи на группы, фор-
мации и слои. Оба предложенных приема (корреляция и расчленение) при-
зывали геологов отказаться от догмы, которая гласила, что все породы,
содержащие фоссилии, возникли благодаря Божьему творению. В течение
сорока лет после того, как В. Смит обнародовал свои идеи, его последователи
создали классификацию формаций и крупных подразделений, начиная с
“Раннего Кембрия” и до “Молодого Плиоцена”. Хотя термины использовали
самые разные и много деталей было упущено, эта последовательность вклю-
чает отложения трех последних эр истории Земли, в которых встречаются
разнообразные и хорошо сохранившиеся ископаемые.
И, как результат “Смитовской организации пород”, была построена гео-
хронологическая шкала. Следует отметить, что шкала не всегда сконструи-
рована по одному и тому же критерию. Некоторые подразделения были пер-
воначально установлены по физическим характеристикам пород, а другие
сразу по ископаемым. Юрский период был первоначально установлен по по-
родам, которые слагают Юрские горы восточной Франции и западной Швей-
царии. Эоценовая эпоха была выявлена благодаря тому, что ее породы в Па-
рижскому бассейне Франции содержали множество морских моллюсков, из
которых 3.5% живут до сих пор. Новые исследования позволили в XIX веке
и в начале XX века расширить и стандартизировать шкалу геологического
времени, включив туда много подразделений, отсутствующих первоначаль-
но (фиг. II.4-8; табл. II. 1).
Геохронологическая шкала основана на трех типах геологических подраз-
делений: литологических (литостратиграфические), стратиграфических
(биостратиграфические) и хронологических (хроностратиграфические). Ли-
тологические подразделения - это те, которые указывают на физические ха-
рактеристики пород; например, красноцветные слои, речная формация или
фосфоритовый горизонт. Хронологические подразделения (time units) отно-
сятся только к интервалам времени; таким образом, кайнозойская эра в це-
лом соответствует времени между рубежом, который был 65 млн. лет назад и
по настоящее время. Стратиграфические подразделения (time-rock) включа-
Таблица П.1. Подразделения третичного периода сэра Чарльза Ляйеля
Формация и эпоха Смысл названий Процентное содержание современных видов, %
Молодой Плиоцен Обновление нового (почти современность) 90-95
Древний Плиоцен Усиление нового 35-50
Миоцен Начало нового 17
Эоцен Пробуждение нового 3.5
ФОССИЛИИ. ПОРОДЫ И ВОЗРАСТ
35
ют одновременно время и последовательность пород, а точнее, ископаемых в
породах. Стратиграфические подразделения геологи используют чаще всего,
когда они выстраивают осадочные породы мира в определенном порядке.
Кайнозойская эратема, таким образом, представляет собой породы, которые
отложились в течение кайнозойской эры, и именно ископаемые определяют
их границы.
Хронологические подразделения Стратиграфические подразделения
Эра Эратема
Период Система
Эпоха Серия (отдел)
Век Этаж (ярус)
Фаза (время) Зона
Итак, кайнозойской эре отвечает продолжительность времени, которое
началось 65 млн. лет назад и продолжается по настоящее время. Кайнозой-
ская эратема не столь непрерывна. Она соответствует всем породам и ископа-
емым, отложившимся на протяжении 65 млн. лет, но в этой последователь-
ности какие-то породы исчезли в результате эрозии, или они вообще не на-
капливались или их невозможно установить, так как характерные
ископаемые отсутствуют.
Мы должны быть осторожны, когда думаем и говорим об этих трех геоло-
гических подразделениях, чтобы не путать хроно-, лито- и биостратигра-
фию. Эти три типа подразделений отражают разные концепции.
От ископаемых к развитию Земли. Подразделения Ляйеля (табл. II. 1) “ра-
ботали” достаточно хорошо, пока геологи знали об ископаемых больше, чем
о породах и Земле. Со временем, конечно, они обнаружили, что такой метод
разделения истории Земли приводит к выяснению причин изменений. Мело-
вой период начался не потому, что какие-то моллюски и рептилии перестали
существовать, и окончился этот период не благодаря тому, что они вымерли.
Это было время низкого стояния суши, когда моря широко распространи-
лись по планете. Пермский период был не только временем папоротниковид-
ных и примитивных деревьев; это было время, когда площадь морей умень-
шилась и огромные пространства превратились в болота, где накапливались
растительные массы будущих угольных месторождений. Плейстоцен, или
Ледниковое время, с другой стороны, характеризуется высоким стоянием
суши, изменившим климат, и материковые ледники снова и снова возника-
ли в Северном полушарии. Эти события и отражают фоссилии, которые при-
спосабливались к тому миру, в котором они оказывались.
Число хроностратиграфических подразделений росло по мере того, как
знания об истории Земли увеличивались. Когда Чарльз Ляйель опубликовал
свои “Основы геологии” в 1838 году, он описал 16 эпох и формаций, относя-
щихся к трем крупным подразделениям: Первичному (Первоначальному
или Переходному), Вторичному и Третичному. В настоящее время мы разли-
чаем по крайней мере девять эр, около двадцати периодов и огромное число
эпох. Все они перечислены на фиг. II.5-8, где дана история Земли, указаны
события и все данные скоррелированы с выдающимися изменениями в исто-
рии органической жизни.
В то время как число периодов и эпох увеличивается, число вариантов
формаций и групп пород растет все меньше и меньше. “Меловая группа”
Ляйеля в настоящее время является системой; его “формация Лейас” стала
отделом. Обычные формации по существу соответствуют слоям, которые об-
з*
36
КАМЕННАЯ КНИГА
в
Продолжительность эр
Фиг. П.4. Соотношение различных геохронологических подразделений
а — соотношение эр; б — соотношение периодов в кайнозойской эре; в - соотно-
шение подразделений в четвертичное время (а - Салоп, 1970; Долицкий, 1980;
б, в - Михайлова, Бондаренко, Обручева, 1989)
разевались в течение короткого отрезка Земной истории практически в одно-
родных условиях и в одном ограниченном регионе. Многие формации поэто-
му имеют один доминирующий тип пород и получают названия по местнос-
ти, в которой они хорошо развиты. Другие формации содержат два и более
вариантов пород, но так связанных между собой постепенными переходами,
что в целом представляют одно целое.
ВОЗРАСТ ФОССИЛИЙ И ПОРОД
Итак, мы выяснили, как использовать фоссилии, чтобы установить после-
довательность пород. Динозавры, как мы знаем, появились после того, как
вымерли трилобиты, и до того, как на “сцене” появились шерстистые мамон-
ты и люди. Но фоссилии сами по себе не могут дать возраст в летоисчисле-
нии, в годах. По фоссилиям была создана относительная временная шкала,
явившаяся основой для современной геохронологической шкалы, опираю-
щаяся на последовательность появления и исчезновения разнообразных
ФОССИЛИИ, ПОРОДЫ И ВОЗРАСТ
37
Четвертичное
200 -I
300 -
400 -I
500 -4
900 -I
1300 Ч
1400 Ч
Cz
&
Формация Биттер-Спрингс. Австралия
Разнообразие прокариот
Гренвилский
Эльзевитский
Шунгиты ^водорослевые угли),
1600
8
1700 -
8.
Гудзонский
1800 -
1900 -
2000 -
2100 -
2200 -
2300 -
2400 -
Сланцы Нонсач.
Мичиган, США
ПП обычных
красноцветов
ПЯ полосчатых
кварцитов
*
Стертское
Микробиота Аделаидской
геосинклинали (примитивные
водоросли, прокариоты,
эукариоты)
*.
Гуронское
*
Лаксфордский цикл
(Великобритания)
ОТНОСИТЕЛЬНЫЕ ИЗМЕНЕНИЯ
УРОВНЯ МОРЯ
_ Современный
100 -I
600 -I
700 Ч
600 Ч
1000 Ч
1100 Ч
1200 Ч
1500 Ч
Альпийский
орогенез
&
би
2300-
2400-
ГЕОЛОГИЧЕСКИЕ СОБЫТИЯ
Гуронское
*
I Гондванское
*
&
, Поздне-
Ж ордовикское
Варисций-
скии
орогенез
2500
Великая Дайка,
Зимбабве
Кеноранский
1ОН-
орогенез
г IАссинтский
I [орогенез
Позднесинийское
Эдиакорская мегаэойная микробиота
db .Варангское
Авалонский
Киларнейский
I
Кислородная
атмосфера
Бескислородная
атмосфера
I
Железорудная формация Ганфлит;
фотосинтезирующая биота (эукариоты).
(?) Грибы: колониальные бактерии;
цианобионты. (?) Строматолиты в
кремнях Ганфлинт, США и Канада
Моранский
Диабазовые дайки Ниписсинг, Канада
Блезардский
Система Витватерсранд, Южная
Африка (бактерии, кокки, бациллы,
водорослевые нити)
5
ф
5
ъ
!
&
£
2600-
2700-
2600-
2900-
3000 -
Скурийский цикл (Великобритания)
ПГТ красноцветов w
Витватерсранд
Булавайские строматолиты. Лаврентийский
Зимбабае, Африка
Зеленокаменные пояса Провинции
озера Верхнего, Канада
Ванипиговский
Wanipigowan
3100 - 3200 - 3300 - »ЗЕ S о О) Ал, ПП микроорганизмов (бактерии + _ цианобионты) группы Фиг-Три. Трансвааль, Южная Африка Деформация гнейсов, район Барбертон, Свазиленд _ Формация Онвервахт группы Свазиленд, Южная Африка (строматолиты)
3400 - X С’2 увеличено) Юивакский ”1П появление микроорганизмов Пильбара, Австралия)
3500 - ПП строматолитов (первый фотосинтез)
3600 -
3700 -
3800 -
3900 -
4000
4100
4200
4300
4400
4500 -
ЛУНА - вторичная дифференциация,
образование базальтовой магмы богатой
железом, образование морей
Гнейсы Амитсок, Западная Гренландия
Кремни группы Исуа, Зал. Гренландия
(древнейшие осадочные породы)
Деформация Родезийско-
каапваапьского кратона
(гнейсы фундамента)
ЛУНА - образование светлых равнин
ЛУНА - активное кратерообразование;
формирование морских бассейнов
первые датированные породы Земли (Австралия)
ЛУНА вероятно, основная метеорная
бомбардировка
ЛУНА - дифференциация за счет частичного
плавления, образование коры (в настоящее
время сохранилась на возвышенностях)
4600 - Рг Аккреция Земли и Луны
Фиг. II.5. Глобальная геохронологическая шкала в едином масштабе, показы-
вающая ужасающую длительность докембрия по сравнению с фанерозоем, с
указанием основных геологических и биологических событий (Harland и др.,
1982, с изменениями). Снежинки указывают время основных оледенений
38
КАМЕННАЯ КНИГА
25
30
35 4
40-
45 —
50-
60
ЮЛЯРНОСТЬ
ЛАГНИТНОГО
ПОЛЯ
нормальна?
обратная
смешанная
Td
20 4
КЬ
1004
2004
Поздняя
\Пог-
Бозды- Груже
мание \ ние_
О (4
Четв.
“СТ
Плвйстоген
ид"
55 4
65-
70 -
Плейстоце!
Tie п
Плиоцен "Рн
Pli П
Миоцен С
Рн
Mio
п
Олиго-
цен
Рн
Oli
п
Эоцен с
Еос Рн
Палео- п
цен —
Рн
Pai
Поздняя
Ранняя
Средняя
/скифская
Scy
РАССЧИТАННЫЕ
ЭВСТАТИЧЕСКИЕ
ИЗМЕНЕНИЯ
УРОВНЯ МОРЯ
Метры выше и
ниже современ-
ного уровня моря
+400 -30(
ГЕОЛОГИЧЕСКИЕ И
БИОЛОГИЧЕСКИЕ СОБЫТИЯ
ПП - первое появление
ПЯ - последнее появление
Континентальное оледенение
в Северном полушарии
Осушение Средиземного моря
ПП гиппариона в Палео-
Средиземноморской области
ПП гоминид
Раскрытие Красного моря
Разделение Австралии и
Антарктиды
ПП трав цветковых растений
ПП Equidae, ПП грызунов
Диварсификация млекопитающих
ПЯ глоботрунканид, рудистов.
аммонитов и динозавров
Раскрытие Тасманова моря
ПП сумчатых
ПП диатомовых
ПП цветковых растений
ПП приматов и др. плацентарных
Начало раскрытия моря Лабредор
Раскрытие Южной Атлантики
Разделение Индии, Мадагаскара,
Африки, Антарктиды
ПП Archaeopteryx
ПП рудистов
ПП планктонных фораминифер
Разделение Гондваны и Лавразии
Раскрытие Центральной
Атлантики
ПЯ конодонтов
ПП птиц, ПП кокколитофорид
Быстрая диверсификация
зверообразных рептилий
ПП гексакораллов.
ПП динозавров
Фиг. II.6. Глобальная геохронологическая шкала с увеличенным масштабом
для фанерозоя, показывающая основные биологические и геологические собы-
тия, уровни моря, полярность магнитного поля и время основных оледенений
(Harland и др., 1982, с изменениями). П - поздняя; Рн - ранняя; С - средняя.
Снежинки указывают время основных оледенений
ФОССИЛИИ. ПОРОДЫ И ВОЗРАСТ
39
смешанная
350 —
4004
450-
500-
550 —
590
600 Ч
10004
20004
10ЛЯРН0СП
МАГНИТНОГО
ПОЛЯ
нормальна»
обратная
Пермскии
Реп
€-О
Эдиакарская
Var
3000Н
5000
Относительные
изменения
к суше
Бурзянский
В
РАССЧИТАННЫЕ
ЭВСТАТИЧЕСКИЕ
ИЗМЕНЕНИЯ
УРОВНЯ МОРЯ
Метры выше и
ниже совр. уровня
+ моря-*- -
Возды- Погру-
мание женив
250 >
300-
Рн
Гжельская
Касимовская
Московская
Башкирская
Серпуховска?
Визейская
Mis
Турнейская
РР
ГЕОЛОГИЧЕСКИЕ И
БИОЛОГИЧЕСКИЕ СОБЫТИЯ
ПП - первое появление
ПЯ - последнее появление
ПЯ трилобитов, табулятоидей, ругоз,
брахиопод-ортид rfTts
Окончательное закрытие г
Герцинского Аппалачского океан;
ПП крылатых насекомых
ПП пеликозавров
ПП котилозавров
ПЯ граптолитов
Девонский
D
Силурий-
ский g
П
С
Рн
. /Пржидол- х
Лудловская
Венлокская
Лландовер.
Ашгиллская
Карадокская
1ландейлска?
1ланвирнска?
Аренигская
Гремадокска?
Вендский
________V
Стертскиир
Юрматин-
ский v
Гуронский
Н
Рэндский
Ran
Исуанскии!
Хэдский
Hde
Мерионвт
Сент-Дэвид
Керфай
Edi
Варангская
D-M
O-S
ПП амфибий (лабиринтодонты)
ПП голосемянных
ПП бескрылых насекомых
ПП аммоноидей
ПП челюстноротых (рыбы)
ПП наземных высших растений
ПП мшанок, ругоз, морских ежей
ПП бесчелюстных позвоночных
ПП хордовых
ПП граптолитов
ПП большинства беспозвоночных
ПП минеральных скелетов
ПП многоклеточных
ПП эукариот
ПП обычных красноцветов
ПЯ полосчатых железистых кварцитов
ПП строматолитов, первые прокариоты
Древнейшие осадочные породы
Древнейшие датированные породы
Фиг. II.6. Окончание
ископаемых в породах. В XIX веке эта относительная временная шкала име-
ла уже почти современное содержание.
Вопрос о геохронологическом возрасте ископаемых долго оставался откры-
тым. Вскоре после открытия радиоактивности в 1896 году Анри Беккерелем
40
КАМЕННАЯ КНИГА
ЭОН ЭРА/ ЭРАТЕМА СУБ- ЭРА
ПЕРИОД/
СИСТЕМА
ЭПОХА/
ОТДЕЛ
ЕВРОПЕЙ-
СКИЙ
СТАНДАРТ
ГЕОХРОНОЛОГИЧЕСКИЕ (СТРАТИГРАФИЧЕСКИЕ^
СХЕМЫ ПО МОРСКИМ ОТЛОЖЕНИЯМ
СЕВЕРНАЯ
АМЕРИКА
ВЕК/ЯРУС
АВСТРАЛИЯ
ВЕК/ЯРУС
НОВАЯ ЗЕЛАНДИЯ
ЭПОХА/ОТДЕЛ
ВЕК/ЯРУС
QUAT
— Q
Pleistogene
5
Ruscinian
Messinian
Several
Villafranchian
"_2 Pleistocene
Ptg- Pie | l~
Pliocene e
~Pli
M -ь о сл о о о о ослослослослЗслосло ! 1 1 1 1 1 1 1 1 1 1 1 1 1 ‘ 1 1 1 1 1 1 1 1 1 1 1-1 1 l_t_l_l_L-r.il 1 1 1 1 1 1 !< 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 Г 1 1 1 I ! 1 1 1 < 1 1 1 Phanerozoic n Cenozoic N -j Tertiary Neogene s MiocB Mio L Tortonian (Helvetian)
M
Langian _(Vindobon ian Burdigalian
le E
Aquitanian
Paleogene Oligo- свпе Oli L Chattian
E Rupelian (Stampian)
Eocene Eoc L Bartonian
M Lutetian
E Ypresian
Paleo- cene Pal L Thanetian
E Mon tian Danian
Mesozoic Cretaceous Late Senonian Maastrichtian
Campanian
Santonian
— Coniacian
\ Turanian
Early \ Cenomanian
Albian
Aptian
c Barremian
1 Neocomi Hauterivian
Vaianginian
Berriasian
Jurassic c_ Late E co 5 Tithonian
Kimmeridgian
Oxfordian
Middle Dogger Callovian
Bathonian
Bajocian
Aalenian
Early Lias Toarcian
Pliensbachian
Sinemurian
Hettang ian
Late
Rhaetian
Norian
Carnian
(Л
СЛ
со
Middle
248 -
Ph
Mz
Landian
Anisian
/----Scythian^
Haitian
Wheelerian
Venturian
Delmonlian
Mohnian
Luisian
Rulisian
Saucesian
Zemorrian
Refugian
Narizian
Ulatisian
Bulitian
Ynezian
Gulfian
Comanchean
/ Spathian
Smithian
Dlenerian
Griesbachian
Mitchellian
Batesfordian
Longfordian
Janjunkian
Aldtngian
Johannian
Wangerripian
Barnsdalian
Balcombian
Werrikooian
— Yatalan
Kalimnan
^XCheltenhamian
\ Hawera Castlechffian
Wanganui \ Nukumarian
X Mangapantan
Taranaki x\ Waiptptan
\\ Opotian
\ kapitean
x^Tongaporutuan
Southland
X Waiauan
Pareora \ Lillbumian
\ Clifdinian
Altonian
Otaian
Landon Waitakian
Duntroonian
Whaingaroan
Arnold Runangan
Kaiatan
Bortonian
Dannevirke Poarangian
Heretaungan
Mangaorapan Waipawan
Teurian
Haumurian
Piripuan
Raukumara Teratan
Mangaotanian Arowhanian
Clarence Ngatenan Motuan Urutawan
Taitai Korangan Puaroan
Oteke
Kawhia Ohauan
I Hetenan t 1— ? —J Temaikan
Herangi Ururoan
Aratauran
Balfour Otapirian
Warepan
Otamitan
Oretian
Gore
Kaihikuan
Etalian
Malakovian
Фиг. II.7. Корреляционная таблица основных геохронологических (стратигра-
фических) подразделений морских отложений Европы, Северной Америки,
Австралии и Новой Зеландии (Harland и др., 1982)
ФОССИЛИИ, ПОРОДЫ И ВОЗРАСТ
41
ВРЕМЯ, МЛН. ЛЕТ эон < ПЕРИОД/ СИСТЕМА ЭПОХА/ ОТДЕЛ i i - i ГЕОХРОНОЛОГИЧЕСКИЕ (СТРАТИГРАФИЧЕСКИЕ)
ЭРА/ ЭРАТЕМ СУБ- ЭРА ЕВРОПЕЙ- СКИЙ СТАНДАРТ СХЕМЫ ПО M СЕВЕРНАЯ АМЕРИКА BEK/ЯРУС ОРСКИМ отло> АВСТРАЛИЯ BEK/ЯРУС КЕНИЯМ НОВАЯ ЗЕЛАНДИЯ ЭПОХА'ОТДЕЛ BEK/ЯРУС
250 _ 300- 350- 400- 450- 500- 550- 590 - g Phanerozoic Paleozoic г? Permian Р Late Tartarian < Kazanian C\ Ufimian Ochoan Guadalupian Leonardian \ D'Urville / Ара ima ч Malakovian . \ Makarewan W Waiitian V \ Puruhauan \ ’X Braxtonian
Early
\ Kiingurian \ Artioskian \\ Sakmarian
Carboniferous о >. с w « if Pen Gzelian _/ Wolfcampian \
Kasimovian - \ / Atokan \ \ \Mangapirian
Moskovian : 14 named Missourian Desmoinesian Morrowan \ Telfordran
Bashkirian : ages
<Z Missis- es sippian Serpukhoviar Springerian Chesterian Meramacian Osagean Kinderhookian
Visean - 11 named ; ages ;
Tournaisian
Devonian D Late Famennian Chautauquan
Frasnian Senecan
Middle Givetian s, Erian
Lite han
Early Emsian Ulsterian
Siegeriiari
J PridolianV- Geofoflian . .
Silurian S Ludlovian 9 named E Cayugan Melbournian
Wenlockian ages = Niagran Eildonian
Llandoverian Keilorian
Ordovician О Ashgillian Caradocian 16 named ages Medinan Cincinnatian Bolmdian ' Eastonian 4
Llandeilian Champlainian Darriwilian
Lianvirnian
Arenigian Yapeenian
Tremadocian Canadian Castlemanian
\\ Chewtoman
Cambrian 6 Merioneth Dolgellian Croixian \\ Bendigonian
Maentwrogian \,\ Lancefieldian
Albertan
Menevian
St. David’s Sohan \\ Idamean
Z Laman
Caerfai Atdabanian Tommottan Waucoban \\ Templetonian
\ Ordian
Фиг. П.7. Окончание
некоторые геологи догадались, что радиоактивность можно использовать
для определения возраста пород. Основанием для такого вывода служило то,
что радиоактивные процессы идут с постоянной скоростью, несмотря на ко-
лебания температуры, давления и химических условий на поверхности Зем-
ли. В XX столетии было разработано много технических методов для измере-
ния радиоактивности. Каждый метод имеет свои преимущества, что дает
возможность установить возраст тех или иных типов образцов, который
другими методами восстановить нельзя. Хорошо известный метод по углеро-
ду-14 обычно используют для датировки древесного угля и растительного
материала из позднечетвертичных отложений, имеющих возраст не древнее,
чем сорок тысяч лет. Калий-аргоновый метод эффективен для датировки об-
разцов не моложе, чем 1 млн. лет. Широкое применение этого метода при да-
тировке лунных пород позволило установить, что они сформировались около
4 млрд, лет тому назад.
Хотя порядок цифр, получаемый этими методами, отличается, но они оба
основаны на одном и том же фундаментальном принципе: после известного
периода времени, названного полураспадом, количество первичного радио-
активного материала, оставшегося в образце породы, уменьшается наполо-
вину. После второго периода полураспада останется половина от прежней по-
ловины, то есть одна четвертая от первичной радиоактивности материала.
Таким образом, измеряя количество оставшегося радиоактивного материала
и определяя, сколько было вначале, можно определить, как долго продол-
жался этот процесс с тех пор, как он начался в данном образце породы.
42
КАМЕННАЯ КНИГА
ГЕОХРОНОЛОГИЧЕСКИЕ (СТРАТИГРАФИЧЕСКИЕ) ВРЕМЯ, МЛН. ЛЕТ
ЕВРОПЕЙСКАЯ СХЕМА Схемы по континент СЕВЕРОАМЕРИКАНСКАЯ СХЕМА АЛЬНЫМ ОТЛОЖЕНИЯМ ЮЖНОАМЕРИКАНСКАЯ СХЕМА АФРИКАНСКАЯ СХЕМА
Oldenburg ian Biharian _ Viliafranchian Rancholabrean Irvingtonian Blancan Lujanian . Ensenadan/Uguian Chapadmalaian Montehermosan 5 10 - 15 т 20 ~ 25 ” 30 “ 35 " 40 - 45 “ 50 - 55 60 - 65 - 70 - 100 _
। Rodolfian
Lothagamian
Ruscian уЯ' Hemphillian Huayquarian
Ngororan
Turolian
Clarendonian Chasicoan
"" Vallesian Tarnanian
Astaracian Barstovian Friasian
Rusingan
Orleanian Hamingfordian Santacrucian
Agenian Monte Leon
Arikarean Colhuehuaptan Fayumian
Arvernian
Whitnayan
Orellan
Suevian Chadronian
Deseadan
Headonian Duchesnaan
Divisaderan
Uintan
Mustenan
Bridgertan
Wasatchian
Casamayoran
Ciarkforkian
Tiff an ian Riochican
Torrejonian
Salamancan (marine)
” \ Puercan /
Фиг. II.8. Корреляционная таблица основных геохронологических (стратигра-
фических) подразделений континентальных отложений Европы, Северной
Америки, Южной Америки и Африки (Harland и др., 1982)
Чтобы лучше понять этот процесс и разницу между двумя методами, пред-
ставим себе, что у нас имеется ведро воды, но оно дырявое и теряет воду с оп-
ределенной постоянной скоростью. По объему оставшейся воды можно вы-
числить, сколько времени тому назад ведро было полным. Если ведро сильно
протекает, то оно быстро становится пустым, и определить время, когда оно
было полным, можно только для небольшого отрезка времени. Слабо проте-
кающее ведро станет пустым очень нескоро, но при этом возрастают ошибки
в измерениях.
На примере с дырявым ведром ясно, что по углероду-14 с полураспадом,
равным 5 730 лет, ошибка в измерении будет равна только нескольким годам.
Но количество радиоактивного углерода-14 столь незначительно, что уже
спустя миллион лет не остается ни одного его атома. За этот период количест-
во радиоактивного вещества уменьшалось вдвое 174.52 раз. Или, другими
словами, по прошествии 1 млн. лет остаток радиоактивного вещества состав-
ляет от первоначального лишь величину 3434, стоящую после 49 нулей.
Вероятность, что хотя бы один атом при этом сохранится, равен одной сотой.
ФОССИЛИИ. ПОРОДЫ И ВОЗРАСТ
43
Фиг. IL9- Основные климатические события на Земле за последний 1 млрд, лет
с выделением гляциоэр и термоэр, объединяющих ледниковые и неледниковые
периоды времени (Чумаков, 1995)
По этой причине углерод-14 эффективно работает только для датировки со-
бытий последних 40 тысяч лет. По геологическим меркам это очень близкое
прошлое. После этого относительно короткого времени в образце остается
1/125 часть первичного радиоактивного вещества.
С другой стороны, древнейшие породы на Земле и Луне, получившие да-
тировки по калий-аргоновому методу, равные 4 млрд, лет и более, еще имеют
1 /8 изотопа калия-40, сохранившегося благодаря тому, что его период полу-
распада равен 1.4 млрд. лет.
Цифра 14 для углерода-14 относится к общему числу частиц (протоны + ней-
троны), которые образуют ядро атома. Углерод-14, или С14, один из изотопов
углерода, который является радиоактивным. Углерод-12, или С12, являю-
щийся другим изотопом, имеет 12 частиц в ядре и он не радиоактивен. Почти
99% углерода на Земле относится к С12, а большая часть оставшегося пред-
ставлена другим стабильным изотопом - С13.
С14 образуется, когда космические лучи из открытого пространства
бомбардируют азот-14 в верхних слоях атмосферы. Таким способом в земной
атмосфере образуется примерно 70 тонн С14 ежегодно. С14 поглощается
растениями, а затем теми животными, которые поедают растения. Когда
44
КАМЕННАЯ КНИГА
Фиг. II.1O. Геохронологическое распространение организмов, часто встречаю-
щихся в ископаемом состоянии. Черным отмечено время, для которого группа
имеет важное стратиграфическое значение (M.S. Petersen, J.К. Rigby. Голо-
вная лаборатория исторической геологии, W.C. Brown Со., Dubuque, Айова,
1969; с изменениями)
ФОССИЛИИ. ПОРОДЫ И ВОЗРАСТ
45
животные и растения умирают и больше не поглощают углерод, то каждые
5 730 лет количество изотопа С14 уменьшается наполовину. Геологический
возраст устанавливают по соотношению двух изотопов С12 и С14, которое все
время меняется, так как количество С14 уменьшается, а С12 остается посто-
янным.
Ежегодный прирост С14 в земной атмосфере в количестве 70 тонн является
средней величиной за последние несколько тысяч лет. Эта средняя величина
является одной из причин возможных ошибок при определении возраста.
Поэтому мы и даем геологический возраст с определенным допущением (±);
например, возраст копролитов одного из южноамериканских ленивцев, со-
держащих растительные остатки, датирован как 10 832 ± 400 лет. Другими
словами, ленивец жил не раньше, чем 11 232 года тому назад, но и не позже,
чем 10 432 лет тому назад.
Кроме летоисчисления, построенного по С14 и калий-аргоновому методам,
существует и много других, основанных на других радиоактивных изотопах.
Один из них уран-ториевый метод. В основе всех перечисленных методов ле-
жит один и тот же принцип: постоянная скорость процесса распада изотопов.
Некоторые химические процессы в специфических условиях идут при по-
стоянной скорости. Аминокислоты, являющиеся строительными кирпичи-
ками для протеинов, присутствуют во всех живых клетках. Во всех живых
организмах структуры аминокислот являются левосимметричными. Когда
организм умирает, аминокислоты из-за изменения температуры становятся
правосимметричными, или зеркальносимметричными. В конце концов об-
разуется смесь, где лево- и правосимметричные аминокислоты достигают со-
отношения 50 на 50, и далее эти пропорции уже не меняются. С помощью
аминокислот удается индексировать образцы возрастом до одного миллиона
лет, если температура в прошлом оставалась постоянной, как, например, на
дне глубокого моря или внутри пещеры.
Методы, основанные на радиоактивной бомбардировке образцов, разрабо-
таны тоже хорошо. Один из них связан с термолюминисценцией. Следы (тре-
ки) излучений радиоактивных изотопов наблюдаются во всех образцах по
мере их трансформации. Если образец нагреть до определенной температу-
ры, то начинается свечение. Чем больше поток электронов, тем ярче свет. Ко-
личество света становится тем инструментом, с помощью которого определя-
ется возраст образца. Этот метод эффективен для датировки образцов не
древнее одного миллиона лет и не моложе двенадцати лет.
Годовые кольца деревьев можно использовать довольно точно для дати-
ровки последней тысячи лет. Главная проблема с годовыми кольцами заклю-
чается в том, что в некоторые годы они не образуются. Относительная толщи-
на колец отражает климатические условия, а последовательность изменения
ширины колец позволяет сравнивать поперечные сечения разных деревьев.
Например, сначала идут пять широких полос, а затем четырнадцать узких,
за ними еще семь широких, и такая последовательность является специфи-
ческой только для определенного отрезка геологического времени. Если та-
кая последовательность обнаружена у одного дерева в внешней части ствола,
а у другого во внутренней части, то это указывает, что первое дерево является
более зрелым и старым, чем второе. Суммируя последовательность толщины
колец обоих деревьев, можно построить общую серию изменений, характер-
ную для более длительного отрезка времени. Обычно определенные последо-
вательности годовых колец характерны для какой-то части суши, но не для
всей Земли, так как климатические изменения могут отличаться в зависимо-
сти от места обитания. Лучше всего последовательность годовых колец, или
46
КАМЕННАЯ КНИГА
Фиг. 11.11. Уменьшение числа дней в году в течение фанерозоя
а — ископаемый колониальный коралл с линиями роста на эпитеке; Закавказье,
поздняя пермь; б - график изменения числа дней в году, построенный по лини-
ям роста различных кораллов (Wells, 1963; Николов, 1986; с изменениями)
“дендрохронология”, разработана для сосен с колючковидными шишками
Сьерры-Невады в Калифорнии, охватывающая последние девять тысяч лет.
Для определения возраста используют и последовательность палеомаг-
нитных данных аналогично методу “дендрохронологии”. Время от времени
магнитное поле Земли меняет полярность. Это значит, что при смене поляр-
ности стрелка компаса, указывающая, например, сейчас на север, будет по-
казывать на юг. По мере накопления осадков частицы вещества, богатые ни-
келем и железом, ориентируются согласно магнитному полю Земли. Когда
отложения “окаменевают”, превращаясь в породу, ориентация частиц “за-
мораживается”, что дает возможность определить положение магнитных по-
лей в прошлом. В лавах после остывания такие частицы также сохраняют
ориентировку магнитных полей Земли.
Еще один метод определения возраста связан с чередованием темных и
светлых глинистых слоев, которые отлагались в озерах умеренного пояса и
прилегающих к ним районах. Каждая пара, состоящая из темного и светлого
слоя, соответствует году. Чтобы узнать время, за которое образовались дан-
ные отложения, подсчитывают число всех слоев, а после деления на два мы и
ФОССИЛИИ, ПОРОДЫ И ВОЗРАСТ
47
получаем количество лет. Этим методом нашли, что прошло 13 500 лет с тех
пор, как растаял материковый ледник Швеции.
Следует остановиться на одном из временных аспектов палеонтологичес-
кого метода, связанном с ростом организмов. У современных и ископаемых
животных и высших растений наблюдаются струйки, линии, морщины и
кольца роста. Одни из них соответствуют суткам или ночному-дневному вре-
мени суток (струйки, тонкие линии), другие - месяцам (морщины), третьи -
годам (кольца). У ископаемых животных линии и кольца роста особенно хо-
рошо фиксируются на внешнем морщинистом слое (эпитеке) кораллов, на
раковинах двустворчатых и головоногих моллюсков. По линиям роста эпи-
теки ископаемых кораллов было подсчитано число дней в году в прошлые
геологические периоды (Wells, 1963). Оказалось, что в кембрии год состоял
из 420-425 дней, а в настоящее время -из 365.25 дней (фиг. 11.11). Уменьше-
ние числа дней в году в течение фанерозоя указывает на замедление враще-
ния Земли вокруг своей оси при условии, что длина орбиты вокруг Солнца ос-
талась прежней. Правомерно и другое объяснение: Земля вращается вокруг
своей оси с той же скоростью, но сокращается длина орбиты, т.е. Земля при-
ближается к Солнцу. Возможно, что действуют оба процесса. Так, палеонто-
логический метод позволил посчитать число суток в году для разных перио-
дов геологического прошлого, что оказалось важным аргументом для плане-
тологов, утверждающих, что скорость вращения Земли вокруг своей оси
замедляется.
Глава III
ГРУППЫ ОРГАНИЗМОВ, НАЗВАНИЯ
И ОТНОШЕНИЯ МЕЖДУ НИМИ
• В большинстве книг современные организмы разделены на два цар-
ства — растения и животные. Первые объединяют тех, кто имеет зе-
леный цвет, не двигаются и создают “еду” из двух групп неживого
материала - углекислого газа и воды. Типичные животные, наоборот, не зе-
леные или у них зеленая только внешняя поверхность, они обычно свободно
передвигаются, а едой для них служат другие животные и растения.
Такое деление уже существует 2.5 тыс. лет. Втечение этого времени состав
и объем царств менялся. Еще в 345 году до новой эры греческий философ
Аристотель отнес к растениям грибы и водоросли, а неподвижных кораллов
и губок перенес в царство животных. С тех пор объем царства растений вырос
ужасающе, а его состав ставит ученых в тупик. В царстве оказались организ-
мы от миниатюрных и простых до больших и сложных; они могут быть зеле-
ными, красными или других цветов; они живут и растут, прикрепленные к
одному месту, или свободно “парят” в воде всю жизнь, они сами создают соб-
ственную “еду” или пожирают живые и мертвые организмы. Такая же пест-
рая картина сложилась и в царстве животных. И многие формы можно отне-
сти как к одному, так и другому царству.
ОТ ДВУХ ЦАРСТВ К ПЯТИ (А МОЖЕТ, ЕЩЕ БОЛЬШЕ?)
Число биологов, считающих, что выделение только двух царств не отвеча-
ет реальности, все более увеличивается. Держаться за два царства — это озна-
чает, “что одни и те же группы надо то впихивать, то выталкивать”, объеди-
нять непохожее и разъединять родственное. Результат получается хуже чем
сомнительный, так как он искажает структуру жизни, существующую на
планете.
Тем не менее оказалось, что для многих естествоиспытателей легче счи-
тать старые царства абсолютно правильными, чем выделять и описывать но-
вые. Почти каждый из нас выучил, что есть растения и животные и все жи-
вущее (за исключением непонятных групп) находит среди них место. Неко-
торые ученые склонны выделять самостоятельное царство бактерий; другие
делают то же самое для свободноплавающих “растений”; некоторые выделя-
ют и третье царство для грибов и близких к ним групп. Один специалист раз-
делил водоросли на шесть групп, каждую из которых рассматривает как цар-
© 1997. О.Б. Бондаренко, перевод на русский язык и изменения.
ГРУППЫ ОРГАНИЗМОВ, НАЗВАНИЯ И ОТНОШЕНИЯ МЕЖДУ НИМИ
49
Фиг. III.1. Пять основных царств жизни (House, 1979; с изменениями)
ство. Затем, “что и следовало ожидать”, он эти шесть групп плюс еще не-
сколько объединил в категорию “Доминион растения”. Другие утверждают,
что даже среди организмов, имеющих ядро, можно выделить по крайней ме-
ре 18 царств. Подобные и другие высказывания существуют в огромном ко-
личестве.
У палеонтологов нет острой необходимости совершать такие радикальные
перемены. Ископаемые легко укладываются в пять царств: монеры, протес-
ты, грибы, растения и животные (фиг. III. 1). Последние два царства являют-
ся “стандартными”, но переосмысленными и дополненными новыми харак-
теристиками. Первые два, монеры и протесты, были выделены и названы
так немецким биологом Эрнстом Геккелем в 1866 году. Хотя Геккель считал
монер частью царства протистов, последующие авторы их разделили, уточ-
нив и прояснив характеристики обеих групп. Определения, приведенные в
этой книге, были опубликованы Р.Х. Уиттэкером (Whittaker) в 1969 году.
Царства Уиттэкера в большинстве случаев не происходят от одного общего
предка, исключая, возможно, только монер. В результате его группы оказы-
ваются не монофилетическими, а полифилетическими или парафилетичес-
кими, то есть происходящими от нескольких предков. Таким образом, его
царства отражают уровни структурной организации, а не ветви (кладии),
растущие от общего “родителя”. Уровни структурной организации трех наи-
более сложных групп основаны на способе питания. Растения, например, со-
здают свои собственные “пищевые продукты” с помощью фотосинтеза (о чем
будет ниже), и они - продуценты. Грибы питаются путем абсорбции, то есть
поглощают всей своей поверхностью и объемом обычно неживое органичес-
кое вещество, разрушая и вовлекая его вновь в круговорот живого; грибы по-
этому можно относить и к деструкторам (“разрушителям”) и к редуцентам
(“восстановителям”). Животные являются в основном консументами (“по-
4. Каменная книга
50
КАМЕННАЯ КНИГА
требители”), они поглощают живое (реже мертвое) органическое вещество,
заглатывая его.
Монеры — наипростейшее царство. Хотя часть монер Геккеля - это ре-
зультат неверных наблюдений, царство в целом реально; оно объединяет две
группы очень простых существ: цианобионтов (бывшее название “синезеле-
ные водоросли или цианофиты”) и бактерий. Их сходство в очень маленьких
размерах и в том, что они одноклеточные. Хотя монеры довольно разнообраз-
ны, основной план клеточного строения остается простым. Клетка монер в
отличие от клеток растений и животных не подразделена на две части: цито-
плазму и ядро. От того, что у них отсутствует оформленное ядро, их называ-
ют прокариотами, то есть доядерные. Структуры, соответствующие со-
держимому ядра у монер рассеяны в клетке, поэтому кажется, что материн-
ская клетка просто расщепляется на две дочерние клетки, когда наступает
время репродукции. Этот тип деления отражен в термине “шизофиты” (“рас-
щепленные растения”), который употребляют во многих англоязычных кни-
гах, где придерживаются деления только на два царства и относят монер к
растениям.
Синезеленые “водоросли”, а точнее Cyanobionta (от греч. kyanos — лазур-
ный), как указывает их название, имеют одновременно синюю и зеленую
субстанцию, последняя соответствует хлорофиллу. Как и для чего служит
синяя субстанция, неясно, но хлорофилл “подслащивает еду”, комбинируя
воду с углекислым газом, которого в настоящее время предостаточно в возду-
хе, морях, озерах и реках. Этот процесс (фотосинтез) использует энергию
солнечного света, часть этой энергии накапливается в виде конечного про-
дукта, являющегося простой формой сахара.
Большинство современных организмов базируются на продуктах, со-
зданных хлорофиллом. Этот факт плюс простая структура цианобионтов
привела к выводу, что на планете Земля они были первыми организмами,
создавшими продукты, которыми потом стали питаться “дегенеративные”
бактерии. В настоящее время эта последовательность рассматривается пря-
мо наоборот. Похоже, что первыми появились бактерии, питающиеся не-
живым и почти живым материалом, который накопился до их возникнове-
ния. Со временем некоторые из первичных бесцветных бактерий преврати-
лись в слабоокрашенных в пурпур потомков, чья окраска возникла из
смеси красной и зеленой субстанций, создававшими конечные продукты
почти тем же способом, что хлорофилл. Цианобионты возникли тогда, ког-
да эта окрашенная субстанция превратилась в хлорофилл и к нему еще до-
бавилось загадочное синее вещество; параллельно у цианобионтов исчез
жгутик для плавания.
Эта последовательность была реконструирована по современным организ-
мам, а не по ископаемым. Если вывод правильный, то цианобионты появи-
лись позже бактерий, хотя они и очень древние. Цианобионты несомненно
участвовали в построении строматолитов, так как их клетки обнаружены в
докембрийских органогенных постройках. Обе группы, цианобионты и бак-
терии, были обнаружены в нижнепротерозойских породах (формация Гурон
и ее аналоги). Бактерии также найдены и в верхнепротерозойских породах;
они часто встречаются в фанерозойских углях, окаменевших растениях и ко-
пролитах. Наиболее обычная форма бактерий - это сферы и нити, которые
могут соединяться с образованием цепочек.
Протесты — сложное царство. Протистов обычно называют Protozoa —
Простейшими, или “первыми животными”, и помещают их в царство живот-
ных, хотя некоторых из них относят к зеленым водорослям. Такое двойное
ГРУППЫ ОРГАНИЗМОВ, НАЗВАНИЯ И ОТНОШЕНИЯ МЕЖДУ НИМИ
51
Фиг. Ш.2. Строение монер (а, б), протистов (в) и многоклеточных животных (г, д')
а — две современные пурпурные бактерии, тип строения которых мог быть ис-
ходным для растений; б — цепочка колониальных цианобионтов рода Nostoc
(справа) и их студнеобразная оболочка (слева); в - современные протисты
Chrysamoeba, выпускающие ложноножки во время захвата пищи (справа); г —
Hydra, современное пресноводное животное с двумя слоями разнообразных
клеток, принадлежащее книдариям; д — продольный разрез через гаструлу —
двуслойную стадию развития, характерную для животных
толкование вызвано тем, что некоторые протисты содержат хлорофилл. Так,
хризамеба (Chrysamoeba) то ведет себя как растение, то как животное. В те-
чение какого-то времени хризамеба создает питательные вещества через фо-
тосинтез, форма ее при этом яйцевидная и двигается она с помощью жгутика
(фиг. III.2). В течение другого времени хризамеба, потеряв жгутик, выпуска-
ет ложноножки (псевдоподии) и питается как животное, “заглатывая” пищу.
Царство протистов трудно диагностировать потому, что его представите-
ли, принадлежащие разнообразным группам, чрезвычайно отличаются друг
4*
52
КАМЕННАЯ КНИГА
Macroporella
Зеленая водоросль
Зеленые'водоросли
Cylindroporella
Фиг. Ш.З. Ископаемые водоросли (увел.)
а — Oldhamia, отпечаток, вероятно, красной водоросли из кембрия штата Нью-
Йорк; б - Primocorallina, реконструкция зеленой водоросли из ордовика штата
Нью-Йорк; в, г - Macroporella, зеленая водоросль из перми в поперечном и ско-
шенном продольном сечениях; д, е — харофиты из девона-карбона (д) и юры-ме-
ла (е); ж, з - Cylindroporella, зеленая водоросль из мела с внешней поверхности
и в продольном срезе
от друга. Тем не менее все они одноклеточные, все имеют ядро, многие строят
раковинки или другие твердые покровы, которые легко сохраняются в иско-
паемом состоянии. Протисты входят в состав группы эукариот, имеющих
четкое и сложно построенное ядро. Остатки некоторых протистов образуют
известняки, а другие — кремневые породы. Они довольно многочисленны в
кварцитах и в диатомовых почвах.
Растения и грибы — еще два сложных царства. Растения могли возник-
нуть как потомки зеленых одноклеточных протистов, которые плавали с по-
мощью двух жгутов. Они создавали из углекислого газа и воды сахаристые
ГРУППЫ ОРГАНИЗМОВ, НАЗВАНИЯ И ОТНОШЕНИЯ МЕЖДУ НИМИ
53
вещества, используя его частично и для создания клеточных оболочек, назы-
ваемых целлюлозой. Целлюлоза помогала пережить потерю хлорофилла,
что случалось несколько раз в сложных путях развития грибов (шляпочные
грибы, плесень, ржа и т.д.). Многие современные грибы являются паразита-
ми или живут на мертвых растениях; оба способа существования являются
очень древними. Трубочки и споры грибов находили в растениях, произрас-
тавших 300 млн. лет назад. Шаровидные грибы (ball fungi) встречаются на
ископаемых каменноугольных листьях. Другие паразитические варианты
грибов были обнаружены и в посткаменноугольных отложениях. Некоторые
из них напоминают современные грибки, вызывающие ржу, приносящую
огромный вред зерну и фруктам.
Наземные растения появились с середины силура, но стали многочислен-
ными к концу девона. Большинство ископаемых растений (кроме известко-
вых водорослей) происходят из девонских и более молодых формаций.
Зеленые водоросли, или хлорофиты, иногда помещают среди растений, но
многие палеоботаники теперь относят их к протистам. Три семейства хлоро-
фитов являются важными ископаемыми. Одно из этих семейств образует ма-
ленькие известковые трубочки, которые периодически разветвляются
(фиг. III.3). Современные.дазикладиевые состоят из коротких стволиков с
пучками листьеподобных структур; ископаемые — представлены короткими
трубочками или “шариками”, принимающими участие в образовании рифо-
вых известняков. Харовые, или харофиты, выглядят как кустики высотой до
61 см, растущие в настоящее время в пресноводных бассейнах; они выделяют
снаружи известковые корочки. Ископаемые харовые представлены изгибаю-
щимися столбиками, корневидными структурами и шаровидными образова-
ниями, которые когда-то содержали споры. Многие ископаемые харовые яв-
ляются морскими формами.
Животные — единое унифицированное царство. Тело животных хотя и до-
стигает большей сложности, чем “тело” растений, в целом они образуют еди-
ную группу, представители которой тесно связаны друг с другом. Первые
животные, так же как растения, произошли от жгутиконосных протистов,
но их предки потеряли хлорофилл и превратились в многоклеточные шари-
ки с полостью внутри, действовавшие как единая скоординированная коло-
ния. У животных эти шарики сплющивались и удлинялись так, что каждая
колония становилась единым организмом, состоящим из двух слоев клеток.
Все животные до сих пор проходят эту двуслойную стадию (см. фиг. III.Зд).
Кроме того, процесс репродукции у животных стал начинаться от вздутий
или “почек” внутри тел своих родителей. У многих животных возник и тре-
тий зародышевый слой клеток - мезодерма, находящийся между двумя пре-
дыдущими, эктодермой и энтодермой.
Губки — животные или протесты? Утверждение, что царство животных
унифицированное, верно, если исключить губковых. Их обычно считают
животными, хотя их ранние стадии развития очень непохожи на те, что ха-
рактерны для царства животных в целом. Более того, воротничково-жгути-
ковые клетки, выстилающие полость губок, по-видимому, унаследованы от
протистов, имеющих воротничково-подобную структуру вокруг своего жгу-
тика. Многоклеточность в этом случае возникла от многих протистов, соеди-
нившихся в одной колонии. Некоторые клетки специализировались для пе-
реваривания, другие для движения. В конце концов колония стала скоорди-
нированным целым. Губки, скорее всего, являются свободной федерацией
клеточных субъектов в царстве протистов. Губки представляют в эволюции
тупиковую ветвь, не давшую потомков.
54
КАМЕННАЯ КНИГА
ПУТИ И СПОСОБЫ КЛАССИФИКАЦИИ
Классификация позволяет одни организмы объединить, а другие разде-
лить. Такая практика не является чем-то новым, ее уже использовал первый
человек, отмечавший свое сходство с другими людьми или использовавший
для разных оленей одно общее название, потому что все они имеют ветвистые
рога. Уже с тех пор люди пытались объединять разные существа в группы оп-
ределенного порядка. Так они систематизировали свои знания об окружаю-
щем мире.
Линнеевская система. Имеется много систем классификаций, от люби-
тельских до научных. Система, которую используют в науке, берет начало с
работ XVIII века, она создана ученым, доктором, профессором Карлом фон
Линнеем (Каролус Линнеус). Он построил классификацию, опираясь на ви-
ды, которые, как он считал, являются легко узнаваемыми категориями. Ви-
ды, похожие друг на друга по структуре или облику, были сгруппированы в
роды. Сходные роды были объединены в семейства; семейства - в отряды; а
отряды - в классы. Линней установил шесть классов для животных и не-
сколько больше классов для растений.
Линней считал, что виды произошли по Божьему замыслу, и описывал их,
не задумываясь, как и почему они появились. Впоследствии натуралисты
стали сомневаться в таком происхождении видов, особенно когда скептики
установили, что виды могут изменяться или они могут превращаться в другие
виды. Впоследствии установили, что роды, семейства, отряды и классы так-
же непостоянны. Духовенство, чтобы сокрушить эту ересь, выдвинуло догму
божественного порядка, в чем им помогали некоторые натурфилософы, объ-
яснявшие, что порядок отражает “силу, мудрость и доброту Бога, отражен-
ную в Его Божьих созданиях”. Другие предполагали, что серия повторяю-
щихся катастроф уничтожала ископаемые растения и животные. После чего
новый акт творения, благодаря Божественной мудрости, приводил к появле-
нию все более прогрессивных организмов. Кульминацией этого процесса яв-
ляется создание человека, он вершина улучшенного плана, который уже пы-
тались осуществить человекообразные обезьяны, но безуспешно. Была указа-
на и дата создания человека - начало ноября 4004 года до рождества
Христова.
Такие рассуждения преобладали до 1859 года, пока Чарльз Дарвин и Аль-
фред Рассел Уоллес не доказали, что виды не только могут, но и на самом де-
ле сменяют друг друга, о чем свидетельствуют ископаемые организмы. Дар-
виновская теория, хотя и вызвала озлобленную критику, но переориентиро-
вала биологов и придала новый смысл классификации. Ученые начали
изучать изменчивость и развитие ископаемых, чтобы выяснить, как они эво-
люционируют по мере того, как “век сменяет век”. Классификация, кроме
распознавания и названия видов и других категорий, объясняет, почему ви-
ды относятся к определенным родам, семействам, отрядам и т.д. Проиллюс-
трируем эту последовательность на примере атлантической устрицы, начи-
ная от царства.
Regnum (Царство) Animalia (все животные в узком понимании или вклю-
чая также протистов).
Phylum (Тип) Mollusca (моллюски - большинство имеют известковые ра-
ковины).
Classis (Класс) Pelecypoda или Bivalvia (двустворки - ползающие и зары-
вающиеся, а также устрицы, мидии и др., чьи раковины имеют две створки).
Subclassis (Подкласс Pteriomorpha (птериоморфы).
ГРУППЫ ОРГАНИЗМОВ, НАЗВАНИЯ И ОТНОШЕНИЯ МЕЖДУ НИМИ
55
Order (Отряд) Dysodonta (беззубые или беззамковые двустворки, ракови-
ны которых не имеют под макушкой системы выступов и ямок, осуществля-
ющих жесткое крепление створок, как в замке; аномииды, морские гребеш-
ки, устрицы, мидии и т.д.).
Family (Семейство) Ostreidae (настоящие устрицы и близкие к ним роды).
Genus (Род) Ostrea (настоящие устрицы).
Species (Вид) Ostrea virginica (устрица виргинская; обычная устрица вос-
точного побережья Америки, современные и ископаемые формы).
Царство, тип и т.д. являются основными категориями, используемыми
при классификации всех организмов. Ученые также применяют и дополни-
тельные категории, такие как надтип, надкласс, надотряд, подрод, подвид,
раса, варьетет и т.д.
Что означают названия. Каждая группа организмов нуждается в собст-
венном имени, которое выбирают согласно правилам, установленным Лин-
неем, и принятыми последующими учеными. За редким исключением, Лин-
ней давал для категорий выше вида одно название, например, Ostrea — для
всех настоящих устриц, Aves — для всего класса птиц и т.д. Название вида со-
стоит из двух слов: первое указывает род, а второе вид. Таким образом, чер-
но-синяя съедобная мидия была названа Mytilus edulis (от лат. edulis — при-
годный в пищу). Родовое название всегда идет первым, как и фамилия чело-
века (сначала Кузнецов или Смит, а потом имя — Иван или Джон). Родовое
имя дают в сокращении по первой букве в том случае, если оно сначала было
указано полностью, а затем многократно повторяется в тексте.
К правилам Линнея добавили еще несколько. Одно из них касается выде-
ления новых подродовых и подвидовых категорий. Если род подразделяют
на несколько подродов, то их называния пишут в скобках после названия ро-
да, но один из подродов должен быть номинативным, то есть повторять на-
звание первоначального рода; например, Ostrea (Ostrea) lutraria. Когда под-
разделяют вид, то подвидовое название прибавляют к видовому; например,
Homo sapiens sapiens. Палеонтологи стремятся избежать тройных названий,
переводя подвиды в ранг видов. Такое укорачивание названий вызывает воз-
ражения, так как родственные отношения становятся неясными, что умень-
шает ценность классификации. Но следует иметь в виду, что те маленькие
отличия, по которым различают современные подвиды, у большинства иско-
паемых не сохраняются.
Появление другого правила вызвано тем, что люди иногда независимо
друг от друга выделяют одни и те же роды или виды и называют их по-разно-
му или наоборот, дают одинаковые названия группам, отличающимся друг
от друга. Фамилию автора прибавляют к названию вида, чтобы указать, кто
и что назвал, например, Mytilus Linnaeus (род выделил Линней) или Ното
sapiens Linnaeus (вид Человек разумный выделил Линней). Если вид был
вначале отнесен к одному роду, а затем к другому, фамилию автора заключа-
ют в скобки. Так, например, вид Atrypa reticularis (Linnaeus) вначале нахо-
дился в составе рода Anomia, но впоследствии был отнесен к роду Atrypa. Па-
леоботаники пошли еще дальше, они не только заключают фамилию перво-
начального автора в скобки, но и прибавляют фамилию того автора, который
перенес данный вид в другой род. Таким образом, вид Metasequoia occidenta-
lis (Newberry) Chaney, 1951 был описан Ньюберри как Taxodium occidentale,
но в 1951 году был переведен Чани в состав Metasequoia.
Древние языки — основа для названий систематических категорий. Лин-
ней писал свои труды на латинском языке и выбирал свои названия из этого
языка или использовал латинизированные греческие слова. С тех пор уче-
56
КАМЕННАЯ КНИГА
ные так и поступают. Кроме того, они латинизируют слова современных язы-
ков, когда классические слова становятся неадекватными, или комбиниру-
ют слова различного происхождения по своему авторскому замыслу. Резуль-
таты обычно получаются хорошими, даже полезными, но существуют и
невнятные и даже загадочные названия. Практика использования древних
языков вызывает нескончаемые жалобы людей, которые хотят, чтобы объек-
ты имели знакомые названия. Так зачем все же использовать слова и буквы
древних языков?
Самая важная причина, чтобы так поступать, связана с тем, что латин-
ский язык был международным языком общения ученых Европы еще до но-
вой эры. Большинство видов, родов и других систематических групп не были
известны и не имели названий, пока ученые их не описали. Англичане, на-
пример, вначале не имели своих названий для ископаемых в таких огром-
ных группах, как млекопитающие, земноводные, позвоночные или моллюс-
ки. Но повседневные названия для некоторых растений и животных сущест-
вовали у разных народов издавна; например, Valeriana officinalis (Валериана
лекарственная) была известна людям под названием “кошачья трава” или
“мяун”. Научные названия необходимо было придумать, а латынь и гречес-
кий, языки таких ученых, как Аристотель, Платон и другие, подходили луч-
ше всего. Но как сделать близкими и по-настоящему русскими или англий-
скими словами такие названия, как рептилии, протисты или Astylospongia
praemorsal И почему считать эти названия менее родными, чем телефон, ре-
волюция, фонограф и т.д. В России обычно научным латинским названиям
находили русские (чаще старославянские) эквиваленты; например, репти-
лии = пресмыкающиеся, протисты = простейшие, маммалии = млекопитаю-
щие, амфибии = земноводные, брахиоподы = плеченогие и т.д. Поиски и да-
же создание новых славянских эквивалентов особенно увенчались успехом в
XIX веке, когда старались максимально приблизить международные знания
к пониманию соотечественников. Особенно в этом преуспели преподаватели
и энциклопедисты. Не случайно в Биологическом энциклопедическом сло-
варе 1989 года названия “рептилии”, “амфибии” и т.д. даны как синонимы
русских терминов - “пресмыкающиеся”, “земноводные” и др., а не как само-
стоятельные термины. Такая же картина наблюдается во многих русских
учебниках средней и высшей школы.
Важным аргументом в защиту использования латинских слов в научной
литературе является их международная узнаваемость благодаря принятым
правилам. А “народные” названия каждодневного общения меняются от
страны к стране. Так, морские беспозвоночные, известные в научной литера-
туре как Echinoidea (от греч.-лат. echinos — еж), в России носят название
морские ежи, в англоязычных странах - sea urchins (реже poundstone), во
Франции - oursins, в Германии - Seeigeln, в испаноязычных странах - erizos
de maro.
Если использовать эти названия для обозначения рода и вида, то они, по-
нятные для одной языковой группы, ничего не означают для другой. Исполь-
зование латинских и латинизированных названий, как обязательное между-
народное правило, не снимая “чужеродности”, имеет то преимущество, что
буквы и звуки латинского языка простые, понятные и общедоступные, а по- I
этому универсально пригодные для всего мира. Невозможно представить се-
бе, например, что русскоязычные и англоязычные исследователи способны
легко овладеть звуками и иероглифами китайского и японского языков для
обозначения морских ежей и других многочисленных современных и иско-
паемых организмов!
ГРУППЫ ОРГАНИЗМОВ, НАЗВАНИЯ И ОТНОШЕНИЯ МЕЖДУ НИМИ
57
Требовать “знакомых” слов бесполезно; большинство из них просто не су-
ществует или не будет “работать”, если мы даже попытаемся их использо-
вать. Самое большее, что можно сделать, это дать перевод или транскрипцию
на своем родном языке. Например, вместо Hyalospongia можно использовать
название “Стеклянные губки”, а вместо Reptilia, Amphibia, Mollusca — репти-
лии, амфибии, моллюски. Некоторые научные названия вошли в русский
язык как новые, а затем стали обычными словами: например, гидра, мамонт,
инд рикотерий.
Почему изменяют названия. Правила Линнея позволяют однозначно ис-
пользовать названия по всему миру, но они также и не мешают изменить их
в случае необходимости, что предусмотрено Международным кодексом бота-
нической номенклатуры (1980, 1993), Международным кодексом зоологи-
ческой номенклатуры (1988) и Международным кодексом номенклатуры
бактерий (1978).
Некоторые названия систематических категорий изменены потому, что
ученые выяснили, что характеристики прежних групп понимали столь ши-
роко, что в них оказались далекие и совсем не родственные формы. В 1758 го-
ду, например, Линней описал одну из ископаемых раковин, которую в насто-
ящее время называют Atrypa reticularis. В течение последующих 160 лет па-
леонтологи отнесли к A. reticularis огромное количество разнообразных
раковин из силурийских и девонских отложений разных регионов мира.
Другие названия для этих раковин были проигнорированы или забыты; в ре-
зультате объем A. reticularis стал непонятным и захламленным. Видовое на-
звание A. reticularis означало только то, что раковины определенного плана
строения жили в течение двух периодов, длившихся 90 млн. лет.
Так продолжалось до 1920 года, когда палеонтологи провели ревизию это-
го геологически долгоживущего вида. Внимательно изучая все раковины и
следуя современным стандартам классификации, палеонтологи восстанови-
ли забытые виды и описали несколько новых. Таким образом, ископаемые,
определенные когда-то как A. reticularis, получили названия A. nuntia, A. de-
voniana, A. nevada и др. Другая ревизия превратила раковины, определенные
как Platystrophia lynx Eichwald, в P. ponderosa, P. moritura, P. biforata и т.д.
Состав родов после ревизий тоже оказывается отвечающим нескольким
самостоятельным родам, а не одному. Хорошим примером является род
Spiritег, в состав которого включали большое число разнообразных видов из
силура, девона и карбона. Ревизия позволила род Spirifer разделить на под-
роды, а также выделить из его состава роды Choristites, Eospirifer, Delthyris,
Syrinogothyris и т.д. Более двадцати из них отвечают всем критериям руко-
водящих ископаемых.
Другие названия ископаемых были изменены благодаря правилу, которое
гласит, что первое наименование, данное роду или виду, сохраняется навсег-
да (старший синоним), несмотря на то что последующие названия (младшие
синонимы) могут быть более подходящими по смыслу или лучше известны-
ми. В 1839 году, например, кости и зубы ископаемого кита назвали Zeugl-
odon (“зубцы ярма”). Это название использовали много лет, пока специалист
по китам не обнаружил, что идентичные кости были описаны в 1835 году как
Basilosaurus. Хотя в переводе слово Basilosaurus означает “король репти-
лий”, оно является научным именем этих китов, а название zeiglodon (без за-
главной Z) оставлено для свободного пользования.
Название рода Orthoceras иллюстрирует другое правило, которое гласит,
что если одно и то же название было независимо дано более чем одному роду
и виду, оно принадлежит тому, кто первый его получил. На протяжении бо-
58
КАМЕННАЯ КНИГА
Фиг. Ш.4. Прямые раковины палеозойских головоногих, ранее относимые к
одному роду Orthoceras
лее чем ста лет всех палеозойских и триасовых головоногих с прямой ракови-
ной называли Orthoceras. Затем в 1836 году два исследователя установили,
что это название использовали для двустворчатых моллюсков раньше, чем
для головоногих. Кроме того, головоногие, названные Orthoceras, оказались
столь различными, что пришлось многим из них дать самостоятельные на-
звания Michelinoceras, Spiroceras т.д. (фиг. III.4).
Названия крупных систематических категорий, от отрядов до царств, не
подчиняются этим правилам; их названия варьируют от привычек и точек
зрения авторов. Таким образом, царство Protista в данной книге соответству-
ет (с некоторыми оговорками) подцарству или типу Protozoa большинства
старых трудов. Кораллы входят в состав класса, называемого Anthozoa или
Actinozoa, а для граптолитов используют названия Graptolithina, Grapto-
loidea или Graptozoa. Дублирующие наименования известны и у групп, дале-
ко отстоящих друг от друга. Название Cyclostomata (Круглоротые), напри-
мер, используют для одного из классов позвоночных, куда входят миноги и
миксины; также называли один из отрядов мшанок. Когда это обнаружили,
то по правилу приоритета название Cyclostomata оставили за миногами и
миксинами, а отряд мшанок переименовали в Tubuliporida.
РОДОСЛОВНОЕ ДРЕВО ТИПОВ И КЛАССОВ
В прошлом биологи обычно располагали организмы по принципу “восхо-
дящей лестницы”. Это означает, что наипростейшую группу помещали на
нижнюю “ступеньку”, а те группы, которые выглядели более сложными, по-
мещали выше и по такому принципу распределяли все известные организ-
мы. Каждый согласится, что водоросли (бактерии тогда были фактически не-
известны) должны были находиться у основания, а животные, имеющие по-
звоночник, на самом верху. Между этими крайними членами располагались
остальные организмы, но с некоторыми вариациями, потому что исследова-
тели не всегда однозначно оценивали степень усложнения.
ГРУППЫ ОРГАНИЗМОВ, НАЗВАНИЯ И ОТНОШЕНИЯ МЕЖДУ НИМИ
59
Затем пришел Дарвин, и классификации стали отражать родственные от-
ношения. Биологи стали рисовать “древа родословных”. Такие “древа” фик-
сируют предков и потомков, как и родословные династий людей.
Если в этой книге мы хотим только описать типы и классы по плану род-
ственных отношений, то надо начать с монер, затем перейти к протистам и
далее продолжать описывать растения, грибы и животных. Ниже даны ха-
рактеристики типов и других групп внутри трех царств - растений, грибов и
животных, что позволяет построить так называемое родословное древо
(фиг. III.5).
Царство растений начинается с зеленых водорослей, чьи предки, очевид-
но, были зелеными жгутиконосными протистами. Некоторые современные
зеленые водоросли представлены только одноклеточными формами, другие,
как морской салат или морская капуста, являются многоклеточными. Неко-
торые из наиболее примечательных зеленых водорослей покрыты известью в
виде табличек и пластиночек или сплошных покровов и внешне напоминают
кораллы. “Коралловые водоросли” часто являются каркасостроителями ри-
фов вместе с кораллами или даже замещают их.
Растения более сложные, чем водоросли, образуют восемь или девять ха-
рактерных групп, которые разные авторы выделяют в четыре или девять ти-
пов. Наиболее простыми из них и, вероятно, древнейшими являются настоя-
щие и печеночные мхи. Они растут на сырых почвах и камнях, а также на
стволах деревьев. Некоторые исследователи думают, что мхи произрастали
на влажных полузатопленных равнинах с очень давних времен, в то время
как лишайники (симбиоз грибов и цианобионтов) покрывали преимущест-
венно сухие скалы, как и в настоящее время.
Высшие растения имеют корни, стебли, листья и сосуды, по которым осу-
ществляется перенос питательных веществ от одних частей растения к другим.
Эти сосуды впервые появляются у водных или прибрежных растений, кото-
рые, очевидно, произошли от зеленых водорослей. Эти первые высшие расте-
ния, вероятно, сначала существовали около устий рек; некоторые из них до-
стигали значительных размеров и образовывали мощные заросли, особенно в
течение среднекаменноугольного времени, более чем 280 млн. лет тому назад.
Семенные папоротники достигли своего расцвета в позднекаменноуголь-
ное время, и они, по-видимому, дали начало предковым группам всех после-
дующих растений, включающих цикадовые, хвойные и близкие к ним груп-
пы, а также цветковые растения (ангиоспермы), развивающихся в течение
последних 200 млн. лет.
Родословное древо животных начинается от предка, чье тело имело, веро-
ятно, чашеобразную форму и два слоя клеток. Этот организм дал начало
двум ветвям потомков: гребневикам и книдариям (кишечнополостным). По-
следние распались на несколько классов, но прежде дали начало еще двум
группам потомков, обладавших третьим зародышевым слоем - мезодермой,
находившимся в пространстве между эктодермой и эндодермой. В одной из
групп мезодерма возникла за счет деления клеток, находящихся в кольце во-
круг “ротового” отверстия чашеобразного тела (шизоцёльный или телоблас-
тический способ закладки мезодермы). В результате мезодерма стала треть-
им слоем, окружающим полость тела. В другой группе мезодерма возникла
за счет впячивания и отшнуровки эндодермального слоя в пространство
между эктодермой и эндодермой (энтероцёльный способ закладки мезодер-
мы). Впячивания (“карманы”) и отшнурованные участки (“мешочки”) пре-
вратились в мезодермальные органы, а полости внутри них получили науч-
ное название - целом.
PLANTAE
Глоссоптериевые
Glossopteridales
Покрытосеменные
A ngiosperm opsida
Пресмыкающиеся
> Reptilia
ANIMALIA
актовые
PROTISTA
MONERA
рилобить
Thlobita
Археоциаты
ocyatb
Губки
Хвойные
Coniferales
Насекомые
Insecta
Унирамии
Jniramia
золотистые водоросли
Chrysophyta
Динофла-
гелляты
Цикадовые
Cycadales
Cycadeoidales
Аневрофитовые
Aneurophytales
есчелюстные
"рыбы" Плакодермы
. Agnatha Placodermi
Земноводные
Amphibia
апортники и др. Хвощевидные
Fihcales, etc. Equisetopsida,
Sphenopsida, etc.
Археоптериевые
Archaeopteridales
Ракообразные
Crustacea
Риниопсиды
Rhyniopsida
Плауновидные
Lycopodiopsida
FUNGI
Хелицеровые
ri Chelicerata
Плоские черви
Platyhelminthes
Красные
водоросли Y
Rhodophyta Зеленые
водоросли
Chlorophyta
Бактерии
Bacteria
Двоякодышащие
Dipnoi
Иглокожие
Echinodermata
Кольчатые
черви (
Annelida I
Книдарии
Cnidaria'(
Ресничные
Ciliophora
Фнг. III.5. Родословное древо органического мира
Птицы
Aves
Млекопитающие
Mammalia
Cl
о
КАМЕННАЯ КНИГА
ГРУППЫ ОРГАНИЗМОВ, НАЗВАНИЯ И ОТНОШЕНИЯ МЕЖДУ НИМИ
61
Энтероцёльный вариант закладки мезодермы обычно считают более высо-
ким и продвинутым, так как им обладают позвоночные животные. Энтеро-
цёльный план закладки мезодермы характерен для небольшого числа типов:
иглокожих (морские звезды, морские лилии и др.), полухордовых, хордовых
(оболочники, бесчерепные, позвоночные), брахиопод и некоторых других.
Шизоцёльный план закладки мезодермы считают более низким, чем энтеро-
цёльный, но он характерен для большого числа типов, хотя ни один из них не
приближается к позвоночным ни по размерам, ни по умственной деятельно-
сти или сложности строения. Шизоцёльная группа превосходит энтероцёль-
ную по разнообразию и многочисленности; так, например, насекомые пред-
ставлены значительно большим числом особей и видов, чем все остальные
группы животных вместе взятые. Так кто и как должен определять “успех”
и “неуспех” этих групп? Несомненно, что “успех” является категорией весь-
ма спорной.
Русский ученый А.Н. Северцов считал (1912,1914,1925,1939), что следу-
ет различать биологический и морфофизиологический прогресс. Биологиче-
ский прогресс определяют по трем критериям: 1) большая численность осо-
бей, 2) огромный ареал распространения (как географический, так и эколо-
гический), 3) дифференциация групп на новые систематические категории
(популяции —» подвиды —» виды). Морфофизиологический прогресс характе-
ризуется усложнением структуры и поведения организмов. Обычно биологи-
ческий прогресс совпадает с морфофизиологическим, но в некоторых случа-
ях прямо наоборот.
РОДОСЛОВНОЕ ДРЕВО И КЛАССИФИКАЦИЯ
Одна из задач изучения животных и растений - это понять их отношения
между собой. Как только приходит понимание, можно строить классифика-
цию. Как было показано в этой главе, эти классификации со временем меня-
ются. Дополнительная информация по определенной группе животных или
растений, усиливающая наше понимание, меняет акценты и положение их в
классификации.
Другая причина изменения позиций в классификации связана с различ-
ными правилами, используемыми учеными. В течение последних двух деся-
тилетий идет активная дискуссия, что надо предпринять для родословного
древа. Существуют по крайней мере три школы. Первая школа является фо-
нетической; она пересчитывает все характеристики, которые являются об-
щими для тех или иных групп животных и растений. Те группы, которые об-
ладают наибольшим числом общих признаков, считают наиболее близкими
между собой. Родословные древа строят из степени сходства, которое высчи-
тывают с помощью компьютера, перебирающего сотни характеристик, что-
бы выявить особенности родственных отношений.
Другая школа исследователей, строящая классификации и родословное
древо, основывает свои выводы, используя только те характеристики, кото-
рые являются специфическими. Чем больше комплекс содержит общих
черт, как, например, перья у птиц, тем больше шансов, что они свидетельст-
вуют о структуре, которая развивалась лишь однажды. Таким образом, груп-
па будет монофилетической, возникшей из одной предковой ветви. Иногда
смущает, что у неродственных животных и растений развиваются сходный
облик и структуры, потому что у них сходный способ питания или похожий
тип передвижения. Такое явление называют конвергентной эволюцией, в
62
КАМЕННАЯ КНИГА
результате которой возникают внешне сходные организмы, генетически со-
вершенно не связанные друг с другом. Так, слоны имеют хобот, а также и та-
пиры, хотя у последних он короче. Но эти две группы не являются близкими
родственниками. Аналогичная ситуация с нелетающими птицами, такими
как додо и эму, с редуцированными крыльями, свидетельствующими не об
общности происхождения, а о сходных путях развития, приведших к такому
образу жизни.
Среди второй школы ученых, использующих специфические признаки,
существуют по крайней мере две группировки, каждая из которых придер-
живается разных правил. Кладисты утверждают, что если однажды живот-
ное или растение приобрело определенный специфический признак, то его
надо объединить в одну группу со всеми остальными, которые также облада-
ют этим признаком. Используя классификацию, созданную кладистами,
можно легко реконструировать родословное древо. Другая группа ученых
при построении того же родословного дерева создает свою классификацию
несколько другим способом. Они учитывают не только специфические чер-
ты, но также имеют в виду и общее сходство тех или иных групп друг с дру-
гом. При этом организмы, которые только что произошли от предковой груп-
пы, благодаря развитию нового специфического признака нередко объеди-
няют с предковой группой, так как у них много общих черт. И не так просто
нарисовать родословное древо, исходя из такой классификации.
Итак, часто не существует только одного ответа, куда надо поместить жи-
вотное или растение в классификационной схеме и родословном древе. Чита-
тель любых статей и книг по таксономии (классификации) или филогении
(родословное древо) должен это учитывать и относиться внимательно к фило-
софии и своду правил, которыми руководствуется каждый автор. Тогда ста-
нет яснее, почему классификации и родословные древа несколько отличают-
ся, хотя они связаны с одними и теми же организмами.
Глава IV
ОСТАТКИ ДРЕВНЕЙШИХ ОРГАНИЗМОВ
• Основными компонентами первичной атмосферы 4 млрд, лет тому
назад, кроме водной составляющей, были водород, аммиак и метан;
или азот, углекислый газ и моноксид углерода. Какая группа из
трех составляющих (плюс вода) и ни была бы, другие компоненты присутст-
вовали в ничтожных количествах. Не важно, какие составляющие были до-
минантными, ясно, что свободный кислород отсутствовал. Не было и озоно-
вого слоя в верхах атмосферы, выполняющего в настоящее время роль щита
от злостной радиации.
Начиная с 2 млрд, лет назад широкое распространение получили полосча-
тые железистые кварциты и сланцы, а также химически не окисленные ми-
нералы, такие как пирит и уранит. Многие считают, что если бы в атмосфере
существовал кислород даже в количестве 0.1% (от современного), то такие
отложения не смогли сформироваться.
Большинством исследователей принимается, что атмосфера оставалась
почти такой же, как описано выше, до рубежа 1.5-2 млрд, лет назад. В это
переходное время кислород, который, по-видимому, возник в результате
биологической активности, вошел в состав атмосферы и океанов. Впервые в
геологической летописи появляются красноцветные отложения, указываю-
щие, что кислорода стало достаточно, чтобы окислить в осадках ионы желе-
за. После этого времени уже не наблюдается широкого распространения не-
окисленных осадков. Высокое содержание кислорода позволило появиться
намного более разнообразным формам жизни, чем прежде, и это началось до
рубежа 1.5 млрд, лет назад. Возможно, первое свидетельство о появлении
жизни содержится в породах группы Варравон Западной Австралии, сфор-
мировавшихся почти 3.5 млрд, лет назад (см. ниже).
НАЧАЛО ЖИЗНИ
Ученые, чтобы понять, с чего начинается жизнь, используют два пути.
Один из них заключается в тщательном изучении ископаемых в докембрий-
ских породах. Второй путь связан с химиками-органиками, которые пыта-
ются в лабораторных экспериментах создать базисные блоки жизни из про-
стых компонентов, которые, вероятно, составляли часть первичной атмосфе-
ры и гидросферы Земли более чем 3.5 млрд, лет назад. Один из знаменитых
© 1997. О.Б. Бондаренко, перевод на русский язык и изменения.
64
КАМЕННАЯ КНИГА
экспериментов был выполнен аспирантом Стенли Л. Миллером (Stanley L.
Miller) в Чикагском университете в 1953 году. Он соорудил установку, через
которую циркулировала смесь паров метана, аммиака и водорода. Через эту
смесь он пропускал мощные электрические разряды, имитирующие мол-
нии. Затем Миллер конденсировал пар, прежде чем снова запустить его в ус-
тановку. К концу недели вода покраснела и стала клубиться. Он проанали-
зировал смесь и нашел, что она содержит какое-то количество аминокислот,
являющихся основными строительными кирпичиками протеинов. Продол-
жая эксперименты, Миллер перешел от простых составляющих к сложным
молекулам, которые прямо связаны с живыми системами. Спустя годы по-
сле работ Миллера, в других экспериментах, в которых использовались раз-
нообразные “атмосферные” компоненты, жесткая солнечная радиация, ура-
ганные звуковые волны, высушивание и(или) замораживание, получили
разнообразные сложные молекулы, включающие и наиболее важные нукле-
оиды, структурные комплексы, которые формируют ДНК. Но до сих пор в
лабораторных экспериментах не удалось создать молекулу ДНК, способную
к репродукции. Экспериментальная работа убедительно показала, как мог-
ла возникнуть сложная молекула. Впереди еще долгий путь исследований,
прежде чем мы поймем, как сложные молекулы превратились в клетки. Из-
вестно только, что под влиянием особых условий такие молекулы, находя-
щиеся в воде во взвешенном состоянии, собираются в маленькие шарики,
отделяясь от воды мембраной. Как и почему это происходит, пока не
понятно.
“КАНАДСКАЯ ЗАРЯ ЖИЗНИ”
Там, где когда-то была жизнь, возможно, сохранились ископаемые; про-
блема в том, как найти их. А обнаружив, надо дать их интерпретацию. Что и
кто они такие и как существовали? Что они говорят о развитии жизни в то
весьма древнее время?
Первая серьезная попытка дать ответ на эти вопросы была предпринята в
1859 году. В тот год сэр Вильям Логэн (William Logan), директор Геологиче-
ской службы Канады, представил экземпляр “вероятно органического про-
исхождения” из пород несколько моложе, чем так называемая первичная ко-
ра. Логэн отдал найденных ископаемых своему другу, палеонтологу
Дж. Вилльяму Досону (J. William Dawson). Он определил их как колониаль-
ных фораминифер, покрывших себя известковыми корочками (фиг. IV. 1).
Досон назвал вид Eozoon canadense (от греч. eos — заря, zoon — животное, са-
nadense - из Канады, канадская).
Досон высказал такое суждение в 1864 году с одобрения одного из автори-
тетов по ныне живущим фораминиферам. Некоторые ученые встретили со-
общение Досона с энтузиазмом, другие были более сдержанными, а часть от-
неслась к этому враждебно. В 1866 году два ирландских минералога усомни-
лись в “заре живого”, посчитав, что это просто смесь различных минералов,
упорядоченных в результате диагенеза, превратившего известняк в мрамор.
Другой критик сравнил эти предполагаемые фоссилии с глыбами известня-
ка, считая, что они подобны глыбам, выброшенным древним вулканом.
Теперь рассмотрим аргументы и факты. С одной стороны имеется Досон и
его сторонники, которые пытаются доказать, что Eozoon и подобные ему яв-
ляются настоящими ископаемыми. А с другой стороны имеется оппозиция,
включающая геологов и минералогов, которая утверждает, что все эти на-
ОСТАТКИ ДРЕВНЕЙШИХ ОРГАНИЗМОВ
65
Фиг. IV.l. Eozoon canadense из протерозоя Квебека, Канада (гренвилльский
мрамор)
ходки имеют неорганическое происхождение. Большинство критиков осно-
вывают свои суждения, изучая только шлифы, игнорируя утверждение До-
сона, что Eozoon надо видеть и изучать в целом, то есть в породе и полевых ус-
ловиях. Один немецкий специалист даже пересек Атлантику, чтобы изучить
досоновское животное в обнажениях обрывов реки Оттава, на той стороне,
где находится Квебек. Его вердикт совпал с выводами его друзей: Eozoon -
это просто структуры и текстуры пород, обусловленные диагенезом, и поэто-
му он - никакой не ископаемый организм.
ЗАГАДОЧНЫЕ СТРОМАТОЛИТЫ
Если бы Досон не нашел еще и других ископаемых в породах округа Хэс-
тинкг в Онтарио, напоминающих Eozoon, то победили бы его противники.
Мало того, что эти эозооны практически не были изменены диагенезом, они
еще удивительно напоминали то, что было описано под названием “строма-
толиты”, что в переводе означает “слоистые камни”.
Это сходство, тем не менее, не привело к немедленной реабилитации
эозоонов в статусе настоящих фоссилий. Строматолиты были известны уче-
ным на протяжении многих лет, и некоторые из них были описаны как изве-
стковые Protozoa. Такое мнение держалось недолго, и когда интерес к стро-
матолитам благодаря ископаемым находкам снова возрос, они были включе-
ны в царство растений как водоросли. Но некоторые геологи настойчиво
продолжали называть их неорганическими конкрециями, а некоторые авто-
ритетные палеоботаники укрепляли такое мнение, утверждая, что только
ископаемые, сохранившие клеточное строение, можно отнести к водорос-
лям. Другие критики, не столь категоричные, считали, что все дискуссии о
строматолитах являются “никчемными” и “неудачными”.
Неудачно или наоборот, но беспокойство оставалось. В настоящее время
доказано, что строматолиты являются продуктами жизнедеятельности циа-
нобионтов, хотя в образовании их также могут принимать участие бактерии,
5. Каменная книга
66
КАМЕННАЯ КНИГА
Фиг. IV.2. Шаровидные и головчатые формы роста современных строматоли-
тов в Акуловом заливе Западной Австралии в условиях отсутствия преоблада-
ющих течений
зеленые водоросли, фораминиферы и процессы осадконакопления. Циано-
бионты долгое время рассматривали как группу синезеленых водорослей
или цианофитов на основании того, что у них, так же как у водорослей и дру-
гих растений, кроме специфического синего компонента имеется и зеленый
хлорофилл. Но в отличие от водорослей у цианобионтов клетки не имеют
оформленного ядра, что заставляет их рассматривать вместе с бактериями в
группе прокариот, то есть доядерных организмов. Растения, грибы и живот-
ные, в отличие от бактерий и цианобионтов, имеют оформленное ядро,
ограниченное оболочкой; и они входят в группу эукариот, то есть ядерных
организмов.
Строматолиты преимущественно состоят из углекислого кальция и маг-
ния, отлагающихся цианобионтами в виде слоистых корочек. Цианобион-
ты обитают в настоящее время в озерах, морях и горячих источниках. В За-
падной Австралии в Акуловом заливе (Shark Вау) современные стромато-
литы были изучены Брайоном Логаном (Brian Logan). Там они
представлены слоистыми продуктами жизнедеятельности цианобионтов и
зеленых водорослей - хлорофитов, образующих разветвления и покровы -
маты, покрытые слизью. Цианобионты и хлорофиты, извлекая двуокись
кальция (carbon dioxide) из воды, создают с его помощью питательные ве-
щества, отлагая известковый материал слоями поверх слизи. Таким обра-
зом, организмы, пока они живы, строят твердые “каменные” опоры для
своих колоний, вовлекая в свои постройки также раковинки разных мик-
роорганизмов. Таким способом и возникают слоистые осадочные структу-
ОСТАТКИ ДРЕВНЕЙШИХ ОРГАНИЗМОВ
67
Фиг. IV.3. Вытянутые формы роста современных строматолитов в Акуловом
заливе Западной Австралии, отражающие направления преобладающих тече-
ний. Зная эти соотношения, можно восстановить направления течений в местах
скопления древних строматолитов
ры - строматолиты, которые обычно выступают над поверхностью осадка на
несколько сантиметров или даже многих метров в виде шаровидных и хол-
мистых тел.
Современные строматолиты встречаются в литорали и в начале верхней
сублиторали, то есть в мелководье, примерно до 5-метровой глубины.
Строматолиты также обнаружены в минеральных источниках. Характер
роста и типы построек строматолитов могут рассказать об окружающей
среде, в которой они обитали (фиг. IV.2, 3). Строматолиты, находящиеся в
зоне волн, имеют головчатую форму, а растущие в спокойной воде образу-
ют пластинчатые структуры. Современные колонии строматолитов в зоне
течений образуют линейно вытянутые валики по направлению этих тече-
ний. Учитывая эту закономерность, можно восстановить направления те-
чений в древних строматолитовых толщах. Внутренняя структура строма-
толитов также меняется в зависимости от глубины, количества дождей,
эрозии, степени осушения во время отлива и субстрата, на котором они
растут, а также от систематического состава видов, формирующих маты.
Строматолиты служат местом обитания и прикрепления для микроскопи-
ческих инкрустированных фораминифер, миниатюрных моллюсков и из-
вестковых трубочек червей. По мере роста строматолитов они оказыва-
ются замурованными и благодаря этому сохраняются в ископаемом состо-
янии вместе с ними.
Современные формы цианобионтов, тем не менее, возможно, не отража-
ют все возможности разнообразия. В девонских отложениях Австралии
(Canning Basin) гигантские строматолитовые рифы почти в 1.5 км шири-
5*
68
КАМЕННАЯ КНИГА
Фиг. IV.4. Hadrophycus immanis, одно из древнейших и мощнейших местона-
хождений строматолитов, отдельные постройки которого достигают в попереч-
нике более 5 м (докембрийские породы горной гряды Medicine Bow, Вайоминг)
ной были сформированы на глубинах от 3.7 до 9 м. Вероятно, диапазон глу-
бин, где формируются современные строматолиты, меньше, чем был в
прошлом.
Вернемся теперь к Eozoon и другим древнейшим ископаемым. Eozoon был
найден в обычных и мраморизованных известняках, названных сериями
Гренвилл (Grenville Series), которые, возможно, образовались более чем
800 млн. лет назад. Ископаемые Eozoon имеют выпуклую или столбчатую
форму. Они представлены изогнутыми или выпуклыми слоями, состоящими
из образований, напоминающих слившиеся трубочки и прутики.
Строматолиты Hadrophycus представлены колониями разнообразной
формы: лепешковидной, пластинчатой, головчатой и караваеобразной, их
структура меняется в зависимости от сохранности и окружающей породы
(фиг. IV.4). В породах, где доломиты чередуются с грубозернистыми окрем-
нелыми известняками (chert), ископаемые представлены только изгибаю-
щимися пластинами. Породы, состоящие из тонкозернистых окремнелых
известняков, содержат как пластинчатые, так и неправильно-столбчатые
строматолиты. Они лучше всего видны на сильно выветрелой поверхности.
Строматолиты, состоящие из дуговидно изогнутых слоев, выполаживаю-
щихся к центру, получили название Collenia (фиг. IV.5). Они образуют плас-
тинчатые или столбчатые колонии, причем последние могут объединяться в
одну общую большую колонию.
ОСТАТКИ ДРЕВНЕЙШИХ ОРГАНИЗМОВ
69
Фиг. IV.5. Collenia frequens (слева) и Collenia versiformis (справа)
Два типа строматолитов из верхней части докембрийских пород Глетчерного
национального парка, Монтана
Наиболее красивые ископаемые строматолиты обнаружены в темно-крас-
ных железистых кварцитах, называемых джеспелитами. Из них можно из-
готовлять облицовочные плиты, а из наиболее красивых - брошки и камеи.
Внимательное изучение современных строматолитов Австралии и Пер-
сидского залива показало, что родовые и видовые названия, используемые
для тех или иных форм, нереальны, так как они отражают экологические
модификации, а не систематическую принадлежность. Лучше классифици-
ровать строматолиты по морфотипам, связанным с определенными условия-
ми обитания. Обозначение LLH-морфотип (Laterally Linked Hemespheroids -
латерально связанные друг с другом полусферы) означает, что данные стро-
матолиты образуют маты из био- и литокластического материала в морских
илистых отложениях мелководья, преимущественно в защищенных участ-
ках, таких как пространства за барьерным рифом. Дополнительно к таким
обозначениям морфотипа строматолитов можно давать родовые и видовые
названия остаткам собственно цианобионтов и водорослей, найденных сов-
местно с этими образованиями.
Строматолиты имеют важное стратиграфическое значение. Их используют
для расчленения и корреляции верхнепротерозойских пород России, Север-
ной Америки, Африки, Индии и Австралии, учитывая преимущественно их
микроструктурные характеристики. Особенности микроструктурного строе-
ния строматолитов, по-видимому, в основном связаны с видовой спецификой
цианобионтов. Строматолиты более многочисленны и разнообразны в позд-
нем докембрии, чем в другое время. Это, вероятно, связано с широким рас-
пространением мелководных шельфовых морей в протерозое (в архее они бы-
ли развиты незначительно), а также с отсутствием сверлильщиков и тех, кто
использовал цианобионты и строматолиты в качестве пищи. Возможно, что
массовое появление в кембрии карбонатных раковинных организмов привело
к понижению кальция в морях и его стало недостаточно для цианобионтов.
70
КАМЕННАЯ КНИГА
Фиг. IV.6. Поверхность строматолитов Cryptozoon proliferum
Поздний кембрий, “Окаменевшие морские сады” около источников Saratoga,
Нью-Йорк
НЕКОТОРЫЕ СТРОМАТОЛИТЫ И ВОДОРОСЛИ ФАНЕРОЗОЙСКОГО ВРЕМЕНИ
Распространение строматолитов не ограничивается докембрием, они бы-
ли распространены в кембрии и в последующее время, включая и современ-
ность. Некоторых из них можно называть Collenia, но других надо помещать
в другие роды.
Род Cryptozoon (от греч. cryptos — скрытый, тайный; zoon - животное) на-
зван бал так, когда его и Eozoon относили к фораминиферам. Большинство
видов Cryptozoon растет в виде расползающихся лепешек или караваев, чья
верхняя поверхность напоминает цветную капусту (фиг. IV.6). Вид Cryptozo-
on proliferum является наиболее эффектным. Он участвовал в образовании
барьерного рифа, который сейчас слагает горы Адирондак в штате Нью-Йорк
(Adirondack Mountains). Поверхность этого рифа, сглаженного ледниками,
можно увидеть в “Окаменевших морских садах” около Саратога-Спрингс.
Род Solenopora относится, вероятно, к красным водорослям. Его пластин-
чатые и головчатые колонии образованы миниатюрными известковыми тру-
бочками, которые формировались вокруг нитей (филаментов) водорослей.
Род Girvanella также представлен трубочками, но его маленькие колонии
имеют шаровидную и яйцевидную форму. Колонии Ottonosia выглядят как
маленькие, стелющиеся образования, обволакивающие раковину, гальку
или другие твердые образования.
Водоросли пресноводных озер. Вдоль побережий кайнозойских пресно-
водных озер около 50 млн. лет назад были развиты заросли Chlorellopsis, фор-
ОСТАТКИ ДРЕВНЕЙШИХ ОРГАНИЗМОВ 71
мы, вероятно, относящейся к цианобионтам. Клетки Chlorellopsis были по-
крыты маленькими известковыми пластиночками, они аккумулировались в
слои, а те в лепешки и пластины, напоминающие Cryptozoon. Цианобионты
рода Chlorellopsis также обрастали стволы или поверхности пней, которые
затем замещались халцедоном. Некоторые из них были ошибочно описаны
как трубки и конусы гейзеров.
ПРОЯВЛЕНИЯ РАННЕЙ ЖИЗНИ: ДРУГИЕ ДОКЕМБРИЙСКИЕ ИСКОПАЕМЫЕ
Существуют две группы доказательств присутствия ранней жизни в до-
кембрийских породах. Одна группа доказательств - это непосредственно ос-
татки организмов; другая группа доказательств представлена физическими
и биохимическими проявлениями жизнедеятельности организмов. Строма-
толиты являются продуктами жизнедеятельности цианобионтов; иногда они
сохраняют и оболочки когда-то существовавших цианобионтов.
Древнейшим доказательством жизни служат строматолиты, образовав-
шиеся около 3.8 млрд, лет назад, хотя органическое происхождение этих
слоистых структур из Западной Австралии до сих пор под сомнением.
Другие доказательства, что жизнь существовала, дают сложные карбонат-
ные соединения, обнаруженные в породах группы Онвервахт (Onverwacht) в
Южной Африке. Состав углеродов этих пород от подошвы до кровли был про-
анализирован Д.З. Элер (D.Z. Oehler). Она обнаружила, что в нижней и средней
частях пород Онвервахт было непропорционально высокое количество углеро-
да С13 относительно С12, в то время как в верхних частях наблюдалось заметное
падение количества С13. Оба углерода, С13 и С12, являются естественными изо-
топами, где С13 имеет в ядре на один нейтрон больше. В атмосфере присутствует
определенное соотношение С13 к С12, но когда зеленые растения поглощают уг-
лекислый газ — СО2 из воздуха, они избирательно концентрируют более легкий
углерод С12 в карбогидраты (carbohydrates), которые создаются во время фото-
синтеза. Эти карбогидраты поглощаются другими организмами и вовлекаются
в их жизненный цикл. В результате соединения углерода в живых системах
обогащены С12. Таким образом, исследования Элер пород Онвервахт подтверж-
дают, что фотосинтез начался в то давнее время, когда накапливались породы
верхней части Онвервахт, и этот процесс зафиксирован в соотношении изото-
пов углерода (химические фоссилии или хемофоссилии).
Остатки первых клеток. На планете существует несколько местонахож-
дений, где в докембрийских породах обнаружены одноклеточные организ-
мы. Древнейшие из них несомненно органического происхождения; они
найдены в кварцитах, сланцах и песчаниках Трансвааля Южной Африки.
Эти породы образовались около 3.2 млрд, лет назад. В группе Фиг Три
(Fig Tree) микроскопические ископаемые сохранились в окремнелом изве-
стняке. Их лучше всего исследовать в тонких срезах пород - шлифах, при
очень большом увеличении в оптическом или электронном микроскопах.
Если окремнелый известняк обработать плавиковой кислотой, то можно
извлечь остатки ископаемых, обычно сильно деформированные. По край-
ней мере, извлекается два типа остатков: тоненькие прутикообразные фор-
мы (напоминающие оболочки современных бактерий по структуре стенок,
размерам и облику) и сферические формы (напоминающие цианобионтов).
Большинство из них размером не более 20 микрон! Все эти клеточные
структуры, скорее всего, относятся к прокариотам, не имеющим оформлен-
ного ядра.
72
КАМЕННАЯ КНИГА
Фиг. IV.7. Скопления клеток из протерозойских кремнистых известняков Бит-
тер Спрингс Австралии (возраст 900 млн. лет назад)
Диаметр клеток 10 микрон (1/1000 мм). В центре клеток кристаллики пирита
или другие включения, вероятно, не связанные с ядром (с разрешения
I.W. Warren и I. Stewart, университет Монаш)
Окремнелые известняки (Ганфлинт) (Gunflint Chert) в северной Миннесо-
те и соседнем Онтарио возрастом около 1.9 млрд, лет и формация Биттер
Спрингс Центральной Австралии (Bitter Springs Formation) возрастом
0.9 млрд, лет содержат намного более разнообразных прокариот, чем породы
Фиг Три возрастом 3.2 млрд, лет назад. В породах Ганфлинта, содержащих
также строматолиты, присутствуют: 1 — тонкие нитевидные образования,
разделенные на участки стенкоподобными структурами, очень напоминаю-
щими современных нитевидных бактерий и цианобионтов; 2 - разнооб-
разные сферические, звездчатые, зонтиковидные тельца неизвестного про-
исхождения. Более молодые породы Биттер Спрингс содержат еще более
разнообразные формы, большинство которых, видимо, относятся к цианоби-
онтам. Некоторые исследователи утверждают, что разнообразие жизни было
еще выше, так как часть “внешне сходных” видов могли возникнуть различ-
ными путями. Дальнейшие совместные исследования современных и иско-
паемых форм помогут решить эту запутанную проблему.
Являются ли какие-нибудь из этих ископаемых эукариотами, то есть ор-
ганизмами, имеющими оформленное ядро, до сих пор неясно. Некоторые
клетки выглядят так, как будто они имеют ядро (фиг. IV. 7); но при внима-
тельном изучении такое утверждение кажется сомнительным, так как “сгу-
стки” в центре клеток оказываются кристаллами пирита или других неорга-
нических включений, либо трещинами растрескивания. Первое достоверное
появление эукариот произошло 700 млн. лет назад, когда появились первые
многоклеточные организмы.
ОСТАТКИ ДРЕВНЕЙШИХ ОРГАНИЗМОВ
73
Возраст
(млн. л.)
ФАНЕРОЗОЙ
500
О
С9
О
Массовое
вымирание
водорослей
ДИВЕРГЕНЦИЯ
МИКРОБИОТ
И ШИРОКОЕ
РАСПРОСТРАНЕНИЕ
СТРОМАТОЛИТОВ
ДРЕВНЕЙШИЕ
ОСАДОЧНЫЕ
ПОРОДЫ
(Гренландия)
ДРЕВНЕЙШИЕ
ДАТИРУЕМЫЕ
ПОРОДЫ ЗЕМЛИ
(Австралия)
ФОРМИРОВАНИЕ
ЗЕМЛИ
^СИЛЬНОЕ <
ОЛЕДЕНЕНИЕ
‘ 1000 ' -
1500
2000
ОЛЕДЕНЕНИЕ
' 2500 ' -
3000
3500
4000
| 4500
ЭДИАКАРСКАЯ ФАУНА
(глобальное распространение)
Первые эукариоты, первые
многоклеточные животные
БИОТА БИТТЕР СПРИНГС
(Австралия)
BECK SPRING
(США) ? Эукариоты
БИОТА КРЕМНИСТЫХ
ИЗВЕСТНЯКОВ
(США/Канада)
ПЕРВОЕ ШИРОКОЕ
РАПРОСТРАНЕНИЕ
КРАСНОЦВЕТОВ
(процессы окисления)
МИКРОБИОТА ФИГ-ТРИ
(Ю. Африка)
Первые прокариоты
ОБОГАЩЕНИЕ С12
(Ю Африка)
Первый фотосинтез
МИКРОБИОТА WARRAWOONA
(Австралия)
? Первые строматолиты
Резкий подъем
температуры
до 10000° F
Фиг. IV.8. Последовательность выдающихся событий во время возникновения
жизни и ее ранней эволюции (с разрешения I.W. Warren)
74
КАМЕННАЯ КНИГА
Фиг. IV.9. Эдиакарская фауна (реконструкция Robert Allen и Mary Wade, с раз-
решения Alan Bartholomai, музей Квисленда, Австралия)
Многоклеточные организмы. Каким образом организмы, состоящие из
многочисленных клеток, произошли от одноклеточных форм, до сих пор не-
понятно. Одна из теорий утверждает, что оболочки одноклеточных организ-
мов, оказавшись внутри другого одноклеточного организма, оформились в
ядро, митохондрии и хлоропласты. Митохондрии и хлоропласты осуществ-
ляют энергетические и окислительные процессы. У прокариот все эти функ-
ции выполняют клеточные оболочки. Другая теория утверждает, что опреде-
ленные прокариоты внедрились в другие прокариоты (эндосимбиоз) и воз-
никшее “содружество” стало функционировать как единое целое.
Каким бы способом это ни случилось, но уже около 700 млн. лет назад по-
явились первые многоклеточные организмы. Они обнаружены в докембрий-
ских породах почти всех континентов, причем их находки сконцентрирова-
ны в регионах, находящихся в настоящее время в зоне низких широт; но,
по-видимому, наилучшей сохранностью обладают те, которые были обнару-
ОСТАТКИ ДРЕВНЕЙШИХ ОРГАНИЗМОВ
75
Фиг. IV. 10. Эдиакарская фауна (реконструкция Frank Knight)
Преобладают животные, сходные с червями и медузами: 1,2 — животные неяс-
ного систематического положения Tribrachidium (1) и Parvancornia (2); 3 - губ-
ки; 4, 5 - червеподобные организмы Dickinsonia (4) и Tomopterla (5); 6 - Pennat-
ula, животные, напоминающие кораллы из группы морских перьев; 7 - черве-
подобное животное в песчаной норке; 8 — Ediacaria, медузоподобное животное
жены в Эдиакаре Южной Австралии в 1947 году. Ископаемые эдиакарского
местонахождения послужили основанием для выделения временного отрез-
ка под названием эдиакарского периода, охватывающего интервал от 670 до
550 млн. лет назад, который предшествовал кембрию. В России и странах
СНГ этот временной отрезок соответствует вендскому периоду. В конце
вендского периода и в самом начале кембрия произошло сильное похолода-
ние и оледенение (фиг. IV. 8). В течение вендского периода появились почти
все основные типы и большинство классов беспозвоночных.
В породах Эдиакары обнаружены беспозвоночные животные, которые
можно отнести к кораллам (Charniodiscus), медузам (Ediacaria), червям
(Dickinsonia), членистоногим (Praecambridium), а также к другим группам
животных неясного или дискуссионного систематического положения, как,
например, Tribrachidium (фиг. IV.9-11). Все обнаруженные животные были
мягкотелыми, они не имели минеральных скелетов и поэтому сохранились
только в виде отпечатков, реже ядер. Они были несомненно более высокоор-
ганизованными, чем их одноклеточные предшественники, хотя и сравни-
тельно проще, чем большинство многоклеточных животных, сменивших их
76
КАМЕННАЯ КНИГА
Charniodiscus longus
Parvancornia minchami
Фиг. IV. 11. Некоторые бесскелетные животные эдиакарской фауны (поздний
докембрий, Флиндерс Рэндж, Южная Австралия)
Наиболее древние многоклеточные животные, имеющие уже оформленное ядро
в своих клетках (фото N.S. Pledge, репродукции сделаны с разрешения музея
Южной Австралии): а — Mawsonites spriggi, медузоподобное животное; б - Sprig
gina floundersi длиной около 4 см, напоминающая червеобразное сегментиро-
ванное животное (? тип Annelida, ? тип Arthropoda); в — Charniodiscus longus
шириной около 5 см, сходен с кораллами из группы морских перьев
(Petalonamae); г - Dickinsonia costata длиной около 6.5 см, червеобразное сег
ментированное животное; д - Parvancornia minchami длиной около 2 см, систе-
матическое положение неясное (? тип Arthropoda); е — Tribrachidium heraldicum
около 2.5 см в диаметре, систематическое положение неясное (? тип Cnidaria,
? тип Echinodermata)
ОСТАТКИ ДРЕВНЕЙШИХ ОРГАНИЗМОВ
77
в кембрии. Некоторые эдиакарские животные, например часть червеобраз-
ных форм, были, очевидно, очень тонкотелыми. Так думают некоторые пале-
онтологии, считая, что дыхательная и кровеносная системы этих животных
были слабо развиты, а необходимый кислород они получали через поверх-
ность тела. Они поэтому не могли быть толстыми, так как кислород не смог
бы проникать далеко внутрь тела от его внешней поверхности.
Эдиакарские животные, по-видимому, могли размножаться половым пу-
тем. Такой путь репродукции значительно увеличивал число комбинаций и
разнообразие организмов. В свою очередь интенсивное формообразование
служило тем исходным материалом, над которым “работал” естественный
отбор, что приводило к дивергенции и формированию разнообразных жиз-
ненных форм, которые характеризуют последующее фанерозойское время,
то есть следующие 550 млн. лет.
Долгое время эдиакарских животных объединяли с ныне живущими
группами, такими как кольчатые черви и медузы. Другая точка зрения сво-
дится к тому, что эдиакарские животные могли принадлежать к группам
животных, не оставивших потомков. Тогда эдиакарские животные свиде-
тельствуют об одной из попыток усложнения организмов, которая потерпела
неудачу и окончилась вымиранием.
Глава V
РАЗНООБРАЗИЕ ПРОТИСТОВ
• Предполагают, что протисты возникли благодаря тому, что два или
более одноклеточных организма, не содержащие оформленного яд-
ра (то есть прокариоты), слились и создали симбиотические отноше-
ния (обоюдно полезное сосуществование) в единой общей оболочке. Это мог-
ло случиться благодаря “фаготрофности”, то есть при поглощении - “поеда-
нии” клеток друг другом. Возможен другой путь, когда клетка или клетки
проникали сквозь оболочку внутрь другого организма. Такие прокариоты,
вероятно, стали строительными кирпичиками, создавшими первую эукари-
отную клетку, содержащую следующие структуры: отчетливое ядро, ограни-
ченное собственной мембраной; клеточные органеллы, ответственные за про-
изводство энергии и поддержание жизни; а в некоторых случаях маленькие
кнутоподобные жгутики, используемые для движения. Современная амеба
Pelomyxa напоминает таких ранних эукариот, так как она лишена многих
специализированных структур, имеющихся у большинства современных
клеток, содержащих ядро.
Вышеизложенное является только одной из теорий, но есть и другие тео-
рии, объясняющие пути развития ранних организмов. Несомненно необ-
ходима дополнительная информация, чтобы окончательно понять, что же
произошло на самом деле. Например, могут ли организмы, создающие собст-
венную пищу (автотрофы, такие как фотосинтезирующие растения), разви-
ваться после тех, кто потребляет еду из живого окружения (гетеротрофы)?
Геологические данные свидетельствуют, что первые организмы были ана-
эробными и они не могли существовать в кислородной среде. Только позднее
появились организмы, способные существовать в условиях, богатых кисло-
родом (аэробные формы), что характерно для большинства организмов, жи-
вущих в современном мире.
ПРОТИСТЫ
Вопрос о том, кто принадлежит царству протистов, не обходится без про-
блем. Протисты, являющиеся элементарнейшими из эукариот, сложнее про-
кариотных организмов (которые не содержат оформленного ядра) и проще,
чем наземные растения и многоклеточные животные. Учитывая это, в состав
протистов можно включить простейших (Protozoa), одноклеточные водорос-
© 1997. О.Б. Бондаренко, перевод на русский язык и изменения.
РАЗНООБРАЗИЕ ПРОТИСТОВ
79
ли и, может быть, низшие грибы. Протисты включают как организмы, кото-
рые сами создают собственную пищу (по варианту растений), так и организ-
мы, которые потребляют другие организмы (по варианту животных). Следу-
ет иметь в виду, что в дальнейшем может оказаться, что протисты являются
искусственной группой организмов, которые содержат формы, более близ-
кие другим царствам. Однако в настоящее время категория Protista - хорошо
работающая модель и ею можно пользоваться, пока не появится что-нибудь
лучшее. Царство протистов было выделено Эрнстом Геккелем в 1866 году.
Многие протисты широко распространены в ископаемом состоянии; они
входят в группу микрофоссилий. Микрофоссилии часто имеют огромное
практическое значение, так как их крошечные размеры позволяют сохра-
няться тысячам и даже миллионам скелетиков в небольшом образце породы.
Особенно это справедливо по отношению к морским отложениям, где скеле-
тики протистов составляют основную часть “седиментационного дождя”, па-
дающего на поверхность дна океанических глубин и образующего глубоко-
водные илы. По скелетикам протистов удается определить возраст пород
керна, извлекаемом при бурении из глубин Земли. По ним также можно вос-
становить историю изменения среды обитания, включая и температурный
режим морских бассейнов.
ЖГУТИКОНОСНЫЕ КЛЕТКИ
Динофлагелляты названы так за присутствие двух жгутикоподобных об-
разований, осуществляющих движение. В переводе название динофлагелля-
ты означает “ужасные жгутики”. Динофлагелляты имеют размеры от 5 мик-
рон до 2 миллиметров, по типу питания среди них имеются как “раститель-
ные”, так и “животные” формы (фототрофы и гетеротрофы). Некоторые
современные динофлагелляты на суше вызывают “красный дождь” и “крас-
ный снег”, а в морях и океанах — “красный прилив”. Другие представители
динофлагеллят принимают основное участие в жутком зеленоватом свече-
нии, вызывающем ночное сияние морской воды. Динофлагелляты в основ-
ном являются обитателями морей, но некоторые существуют в пресной воде,
редко в песчаных почвах, а небольшое число даже являются паразитами.
Динофлагелляты проходят две фазы в своем жизненном цикле: подвиж-
ную и неподвижную. Во время подвижной фазы они свободно плавают, ис-
пользуя свои жгутики; во время неподвижной фазы они донные неподвиж-
ные формы (стадия цист). На стадии цист идет основное формообразование
оболочек динофлагеллят, которые сохраняются в ископаемом состоянии и
имеют огромное значение для геологии. Оболочки цист состоят из прочного
органического материала, очень похожего на спорополленин, который обра-
зует внешние стенки спор и пыльцы наземных растений. Каждая циста име-
ет маленькую “предохранительную дырочку”.
Ископаемые динофлагелляты широко распространены в морских отложе-
ниях от перми по настоящее время, но встречаются они и в пресноводных во-
доемах. Наиболее древние находки известны из силура. Динофлагелляты
очень разнообразны по форме, размерам, скульптуре, от гофрированных
складчатых удлиненных конусов и шишечек до коротких разветвленных
шипов (фиг. V.1).
Акритархи, одна из групп микрофоссилий, напоминающая цисты дино-
флагеллят. Акритархи, так же как динофлагелляты, имеют оболочки из
спорополленина. Систематическое положение акритарх неясное. Размеры
80
КАМЕННАЯ КНИГА
Динофлагелляты
Силикофлагелляты
Фиг. V.I. Представители протистов, рассматриваемые во многих работах или
как растения либо как животные (увел, от Х100 до хЗООО)
а, б — динофлагелляты, силур—ныне, наиболее распространены в мезозое: а -
Ctenidodinium, юра; б — Peridinium, мел; в — силикофлагелляты, мел-ныне:
Cannopilus, олигоцен—ныне; г, д - диатомовые, юра-ныне: г - Actinoptychus,
миоцен-ныне, д - Melosira s.l., поздний мел-ныне; е, ж — кокколитофориды,
триас-ныне: в — Cyclococcolithina, современная форма размером 0.017 мм, ж —
Coccolithus, кайнозой; з - Thoracosphaera, сильно увел., входит в известковый
мезо-кайнозойский наннопланктон, родственные связи не ясны (е — рисунок
Frank Knight с изменениями из Brasier,1981)
колеблются от 8-500 микрон до 1 мм. Время распространения от протерозоя
по плейстоцен. Акритархи играли наиболее важную роль в морском планк-
тоне позднего протерозоя и раннего палеозоя. Акритархи обычно представ-
лены сферическими, яйцевидными или треугольными оболочками, от кото-
рых отходят шипы, отростки или поднимаются линейно вытянутые гребни
(фиг. V.2).
Кокколитофориды являются одноклеточными планктонными протиста-
ми с двумя, реже тремя жгутиковидными образованиями. Цитоплазма кок-
колитофорид защищена панцирем из очень маленьких (3—5 микрон) извест-
ковых пластиночек — кокколитов (см. фиг. V. 1). После смерти организма
панцирь распадается и составляющие его кокколиты попадают в глубоко-
РАЗНООБРАЗИЕ ПРОТИСТОВ
81
Фиг. V.2. Раннекембрийские акритархи (сильно увел.)
а - Multiplicisphaeridium, б — Archaeodiscina, в — Tasmanites. Сильно увел.
(Михайлова, Бондаренко, Обручева, 1989)
водные глинистые и известковые илы. Так идет процесс захоронения кокко-
литофорид в современных морях и океанах, так было и в прошлом. Кокколи-
ты обнаружены в породах триасового, юрского и более молодого возраста; с
мелового времени они становятся породообразующими. В современных
морях и океанах кокколитофориды образуют основную часть морского
планктона.
ЕЩЕ О НЕКОТОРЫХ ОРГАНИКОСТЕННЫХ ПРОТИСТАХ
Тасманитиды — еще одна группа микрофоссилий, полностью морская, ко-
торая, вероятно, близка динофлагеллятам. Они обладают стенкой из споро-
полленина, но стенка очень толстая. Обычно тасманитиды имеют сферичес-
кую форму, достигая в диаметре 100-600 микрон. Многочисленные малень-
кие каналы частично или полностью пронизывают толстые стенки. Первое
появление этой группы приурочено к кембрию, в миоцене они практически
вымерли, хотя ныне живущая Pachysphaera несомненно представитель этой
группы.
Хитинозоа. Никто точно не знает, кому принадлежат Chitinozoa — еще од-
на группа органикостенных микрофоссилий, встречающаяся от ордовика по
девон. Точки зрения о систематической принадлежности самые противо-
речивые: одноклеточные животные, одноклеточные водоросли, многокле-
точные животные (близкие граптолитам или гидроидам), капсулы яиц и экс-
крементов червей и гастропод и т.д. Органические оболочки хитинозоа по-
лые внутри, открытые на одном конце и замкнутые на противоположном.
Они радиально-симметричные относительно центральной длинной оси
(фиг. V.3). Размер хитинозоа близок к динофлагеллятам, он колеблется от
30 микрон до 2 миллиметров, но преимущественно около 100 микрон. Хити-
нозоа, скорее всего, планктонные формы. Их используют в биостратиграфии
ордовика, силура и девона.
Многих из вышеперечисленных организмов (динофлагелляты, акритар-
хи, тасманитиды и другие) раньше называли гистрихосферами. В настоящее
время это подразделение считается “мусорной корзинкой”, то есть сборищем
6. Каменная книга
82
КАМЕННАЯ КНИГА
Фиг. V.3. Хитинозоа (поздний кембрий?, ордовик—девон)
а-в — одиночные формы, сильно увел.: а — Lagenochitina estonica, ранний ордо-
вик, аренигский век, б - Angochitina sp., средний девон, живетский век, в-Ап-
cyrochitina sp., поздний силур, пржидольский век; г — колониальная форма
Margochitina margaritana catenaria, сильно увел., ранний девон (Обут, 1973)
форм, которые, возможно, и близки, а может быть и нет, а потому во избежа-
ние недоразумений они разделены на несколько характерных групп, пере-
численных выше.
КРЕМНИСТЫЕ “КАМЕНИСТЫЕ” ЖГУТИКОНОСЦЫ
Эбридии являются жгутиконосными формами, которых можно сопоста-
вить с динофлагеллятами и с силикофлагеллятами. Эбридии обладают внут-
ренним кремнистым скелетом, напоминающим в какой-то степени спикулы
губок или даже скелеты простых радиолярий. Эбридии впервые появляются
в палеоцене, продолжают сохранять разнообразие какое-то время в плиоце-
не, а после этого они резко сокращаются в числе. Эбридии входят в состав
морского микропланктона.
Силикофлагелляты, как указывает их название, используют кремнезем
как основу своего скелета, так же как и эбридии. Они обладают хлыстоподоб-
ным жгутиком и способны создавать самостоятельно для себя еду благодаря
фотосинтезу. Являются ли они “растениями” или “животными” - это пред-
мет дискуссии. Силикофлагелляты составляют незначительную часть мор-
ского планктона начиная с мелового времени, но они приобретают исключи-
тельную важность, когда известковые фоссилии редки или отсутствуют,
как, например, в полярных широтах или в глубоководье, где карбонат каль-
ция растворяется в морской воде. Кремнезем, напротив, не растворяется и
поэтому кремневые скелетики сохраняются.
Скелеты силикофлагеллят состоят из опалового кремнезема, отлагающе-
гося вокруг базального кольца. Обычно более сложные скелетные структуры
возникают только на одной стороне, так что многие виды имеют асимметрич-
ные раковины (см. фиг. V. 1в).
РАЗНООБРАЗИЕ ПРОТИСТОВ
83
Фиг. V.4. Эоценовые тинтинниды Украины
а, б — Savroniella rugosa с аборальной стороны (х200) и сбоку (х165); в, г- Savro-
niella cincinnata с аборальной стороны сверху (хЮО) и изнутри (х100)
(Белокрыс, 1995)
Несмотря на то что силикофлагелляты имеют довольно продолжительную
геологическую историю, изменялись они очень медленно. Вероятно, силико-
флагелляты никогда не станут теми ископаемыми, по которым можно будет
выделять дробные стратиграфические (геохронологические) подразделения.
РЕСНИЧНЫЕ ПРОТИСТЫ
Некоторые протисты имеют не два или три жгутика, а намного больше; их
называют ресничными, или Ciliophora. Только немногие из них являются
важными фоссилиями; в основном это тинтинниды и кальпионеллиды.
Тинтинниды известны с ордовика по настоящее время, биостратиграфи-
ческое значение они приобретают с эоцена (фиг. V.4). Тинтинниды обладают
цилиндрическими, коническими и шаровидными скелетиками, заостренны-
ми или вытянутыми на одном конце в тонкий отросток. Состав скелета орга-
нический, иногда с включениями посторонних частиц. Кальпионеллиды в
этой группе являются известковыми формами, которых используют для вы-
деления зон в мезо-кайнозойских породах. Они преимущественно находятся
в плотных известняках, и поэтому их изучают с помощью тонких срезов -
шлифов данных известняков.
ДИАТОМОВЫЕ
Современные диатомовые особенно многочисленны в холодных морях,
они также встречаются в реках и озерах и даже в почвах. Все диатомовые
нуждаются в освещении, а это означает, что в морской воде они не живут
глубже 250-300 м. Диатомовые являются одиночными или колониальными
формами, лишенными жгутиков; они строят орнаментированные раковин-
ки, или панцири (фрустулы) из кремнезема (см. фиг.У.1г, д). Каждый пан-
цирь состоит из двух половинок, одна из которых частично перекрывает дру-
гую, подобно крышке коробки. Форма и орнамент панцирей весьма разнооб-
разные, но морские формы преимущественно округлые (центрические), в то
время как пресноводные обычно удлиненные (нештатные). Наиболее древние
морские диатомовые найдены в позднемеловых отложениях; пресноводные
виды появляются начиная с палеогена.
Современные диатомовые составляют значительную часть осадков, кото-
рые отлагаются на дне океанов, особенно в Арктических и Антарктических
6*
84
КАМЕННАЯ КНИГА
Фиг. V.5. Современные и ископаемые мезо-кайнозойские радиолярии
а-г - Spumellaria: а - Heliosphaera (палеоген-ныне), б - Centrospira (мезозой-
ныне), в — Hexadoridium (мел-ныне), г — Saturnalidae (мезозой-ныне); д - Nas-
selaria, Pterocorys (эоцен-ныне). Сильно увел., от хЮО до 200
регионах. В течение палеоген-неогенового времени диатомовые были широ-
ко распространены в морях и озерах многих регионов. Морские диатомовые
отложения миоцена широко развиты на юге Украины и России. Озерные ди-
атомовые отложения плиоцена в районе озера Севан Армении достигают
мощности 140 м. Сланцы Monterey Калифорнии миоценового возраста до-
стигают мощности около 60 м. Можно предположить, что многие докайно-
зойские кремнистые породы были первоначально сформированы за счет ди-
атомовых, чьи коробчатые раковинки исчезли, а их кремневая составляю-
щая сохранилась.
КОРНЕНОЖКИ И СТЕКЛЯННЫЕ СКЕЛЕТИКИ РАДИОЛЯРИЙ
Огромное число ископаемых Protozoa принадлежит ризоподам, или Sar-
codina. Название “ризопода” переводится как “корненожки”, что удачно ха-
рактеризует псевдоподии, то есть ложноножки, являющиеся удлиненными
выростами цитоплазмы, которые эти существа выпячивают, когда охотятся,
ползут, плывут или дрейфуют в морской и пресноводной воде. Некоторые
Sarcodina имеют непостоянную форму тела и оно не защищено раковиной,
другие строят красивые и сложные раковины, которые обычно сохраняются
в ископаемом состоянии. Некоторые породы почти нацело состоят из таких
раковинок.
Радиолярии. Если бы ввели конкурс красоты раковинок, то радиолярии
стали бы основными претендентами из ризопод. Типичная радиолярия со-
стоит из сферического и удлиненного тела, заключенного в ажурную рако-
РАЗНООБРАЗИЕ ПРОТИСТОВ
85
винку из стекловидного кремнезема, которая может состоять из нескольких
вложенных друг в друга каркасов, обычно украшенных шипами (фиг. V.5).
Еще одна пористая капсула находится внутри желеподобной цитоплазмы,
делящая ее на две части: внутреннюю и внешнюю. Внешняя часть цитоплаз-
мы выпускает лучевидные псевдоподии, которые улавливают пищу; внут-
ренняя содержит одно или более ядер и отвечает за репродукцию, то есть раз-
множение. В случае необходимости радиолярии могут использовать в пищу
то, что производят крошечные водоросли — зооксантеллы и зоохлореллы,
живущие внутри ее цитоплазмы. Водоросли, в свою очередь, утилизируют
углекислый газ и азотистые продукты, возникающие в процессе жизнедея-
тельности радиолярий.
Этот типичный план строения радиолярий, в свою очередь, имеет много
вариантов. Некоторые радиолярии обладают только внутренней капсулой, а
раковинка (экзоскелет) отсутствует. У других нет шипов, они напоминают
маленькие сосновые шишечки или разделены на радиальные доли. Одна из
групп, ранее относимая к радиоляриям, а теперь рассматриваемая как само-
стоятельный тип Acantharia, строит раковинки из сернокислого стронция
(SrSO4), а не из кремнезема.
Когда-то считали, что радиолярии являются редкими фоссилиями, не
имеющими значения для геологии. В настоящее время установлено их широ-
кое распространение начиная с палеозоя. Многие палеозойские кремнистые
толщи, ранее относимые к докембрию, как, например, в Казахстане, оказа-
лись девонскими, что удалось установить по находкам радиолярий. Особен-
но богаты радиоляриями позднемеловые и кайнозойские отложения Кали-
форнии, Японии, Сахалина и Курильских островов. Скелетики радиолярий
иногда почти нацело составляют породу, которую тогда называют радио-
ляритом.
РАЗНООБРАЗНЫЕ И ПОЛЕЗНЫЕ “ФОРАМЫ”
Многие палеонтологи в течение своей жизни могут так и не увидеть жгу-
тиковых, ресничных или даже радиолярий. Но никому, кто занимается
беспозвоночными, не удалось избежать знакомства с фораминиферами, чье
название часто сокращают до слова “форамы”. Они являются не только наи-
более многочисленными и прекрасно сохранившимися ископаемыми проте-
стами; они также являются важнейшими руководящими формами, то есть
“индекс-фоссилиями”. Трудно представить себе, какое огромное количество
формаций удалось идентифицировать, определив по фораминиферам их воз-
раст и условия формирования, что помогло во многих случаях открыть неф-
тяные месторождения.
Название “foraminifera” означает “несущие отверстия”, то есть поры, ды-
рочки в капсуле или раковине, которую также называют панцирем (test).
Хотя некоторые фораминиферы имеют только плотную органическую обо-
лочку, подавляющее большинство строят раковинки, имеющие одно или
много отверстий. Желеобразная цитоплазма может полностью находиться
внутри раковины, выпуская наружу псевдоподии только через крупное от-
верстие, называемое устьем. Обычно цитоплазма заполняет и обволакивает
раковину, просачиваясь на ее поверхность через множество маленьких пор и
посылая во все стороны псевдоподии. Эти “ложноножки” тонкие и обычно
ветвящиеся, но, соприкасаясь и соединяясь, они образуют сгустки. В том
случае, когда вокруг захваченной пищи возникают сгустки цитоплазмы, пе-
86
КАМЕННАЯ КНИГА
Фиг. V.6. Типы секреционно-известковых и агглютинированных раковин
фораминифер
а — многокамерная спирально-плоскостная раковина; б, в - двукамерные рако-
вины с сферической первой и трубчатой второй камерой, свернутой в плоскую
спираль; г — однокамерная неправильно цилиндрическая раковина; д — много-
камерная спирально-винтовая двухрядная раковина
реваривание происходит в них, а не в основной цитоплазме, находящейся в
раковине (фиг. V.6a).
Клетки и жизненные циклы. Намного раньше, чем установили стратигра-
фическое значение “форамов”, ученые обнаружили, что многие виды пред-
ставлены двумя вариантами раковин: мегасферической и микросферичес-
кой. Мегасферическая раковина начинается с сравнительно большой первой
камеры, называемой “пролокулюс”, но имеет небольшие размеры и малое
число оборотов. Микросферическая раковина начинается с маленького про-
локулюса, но ее размеры и число оборотов намного больше, чем у мегасфери-
ческой формы (см. фиг. V.66, в).
Причина такой разницы стала ясной, когда биологи изучили процессы
размножения у современных фораминифер. Микросферические раковины
возникают за счет полового размножения, но сами являются бесполыми.
При достижении зрелости ядро микросферической особи расщепляется на
множество ядрышек. При этом цитоплазма вытекает из раковины и разделя-
ется на шарики, в каждом из которых находится ядрышко. Эти шарики, ко-
торые являются юными фораминиферами, строят крупные пролокулюсы
мегасферических раковин.
Хотя мегасферические особи возникают за счет бесполого размножения,
сами они размножаются половым способом. При достижении зрелости ядро
с цитоплазмой у мегасферической “особи” разделяется на роящуюся массу
крошечных “частиц” с двумя (реже тремя) жгутиками. Каждая “частица”
РАЗНООБРАЗИЕ ПРОТИСТОВ
87
является самостоятельной клеточкой, похожей на маленькую серую
Chrysamoeba. Многие из этих роящихся клеточек погибают, но некоторые
спариваются и сливаются, образуя новые особи микросферического поколе-
ния. В свою очередь микросферическая особь опять путем деления (а не по-
лового размножения) даст начало мегасферическим особям, и этот цикл аль-
тернативной репродукции будет повторяться вновь и вновь.
Схема вышеописанного цикла имеет варианты. Так, мегасферические
особи могут создавать мегасферические поколения до двух-трех и более раз,
прежде чем настанет очередь микросферического поколения. Пролокулюсы
таких последовательных мегасферических поколений могут отличаться по
размерам, что затрудняет видовые определения.
Различные типы раковин. Размеры раковин некоторых фораминифер
очень маленькие - около 0.02 мм в диаметре, а наиболее крупные достигают
в поперечнике 19.3 см. Обычный диаметр около 0.5 мм.
Хитиновые, то есть органикостенные, раковины являются наиболее
простыми и, вероятно, самыми примитивными. Они округлые, овальные
или колбовидные и состоят из вещества, которое выглядит как прозрачный
пластик либо как жесткий покров насекомых. Это вещество называется тек-
тином; он состоит из протеинов и полисахаридов. Тектиновые раковины ха-
рактерны для аллогромиин. Тектин также развит на ранних стадиях у дру-
гих фораминифер.
Агглютинированные раковины состоят из слипшихся посторон-
них частиц (см. фиг. V.6). Частицы представлены илом, песком или чем-ни-
будь другим; они сцементированы хотиновым, известковым, кремневым
или железистым материалом. Некоторые фораминиферы для построения
агглютинированных раковин используют все, что имеется вокруг; другие -
только определенный материал, либо кварцевые песчинки, либо кошечные
чешуйки слюды или маленькие кусочки раковин. Некоторые, как, напри-
мер, Textularia, если они существуют в холодных водах, используют
кварцевые зерна, а если в теплых водах, то они предпочитают известковые
частицы.
Секреционно-известковые раковины являются наиболее распро-
страненными. У них, в отличие от агглютинированных известковых рако-
вин, кальцит выделяется за счет секреторной жизнедеятельности так же,
как раковины гастропод и других животных. Кальцит может быть зернис-
тым или кристаллическим, а стенки раковин сплошными или пронизанны-
ми множеством пор. Секреционно-известковые раковины характерны для
М Utolina, Fusulinina, Rotaliina и других групп фораминифер.
Стекловатые секреционно-известковые раковины имеют множество пор,
а фарфоровидные лишены их.
Разнообразие форм и строение фораминифер. Агглютинированные рако-
вины являются более древними, они появляются впервые в кембрии, но, воз-
можно, уже существовали в докембрии. Фораминиферы с микрогрануляр-
ными стенками раковин, развивающиеся в течение палеозоя, являются пе-
реходными формами между агглютинированными и секреционными
фораминиферами. Микрогранулярные раковины состоят из маленьких
кальцитовых частиц, соединенных кальцитовым цементом, придающим ра-
ковине “сахаристую” текстуру.
Фораминиферы имеют разнообразное строение и форму. Они могут состо-
ять из одной или многих камер (фиг. V.6, 7). Обычно первая камера сфериче-
ская или яйцевидная, последующие камеры могут быть такими же или труб-
чатыми. Камеры нарастают разнообразными способами. Способ нарастания
88
КАМЕННАЯ КНИГА
Фиг. V.7. Типы секреционно-известковых многокамерных раковин форами-
нифер
а — камеры расположены по винтовой спирали в два ряда; б - камеры располо-
жены бисериально в плоскую или коническую спираль; в — удлиненные камеры
расположены по конической спирали, обычно по четыре в каждом обороте;
г, д- раковины спирально-плоскостные, вытянутые по оси навивания, инво-
лютные, так как последний оборот перекрывает все предыдущие, перегородки
между камерами внизу складчатые; е-и - раковины спирально-плоскостные,
сжатые по оси навивания, инволютные; к—л — раковины спирально-конические
сбоку (к) и сверху (л); м - раковина диморфная: в начале - спирально-плоскост-
ная, а затем — прямая однорядная
РАЗНООБРАЗИЕ ПРОТИСТОВ
89
характеризует крупные систематические категории. У однорядных (унисе-
риальных) фораминифер камеры располагаются линейно в один ряд, у дву-
рядных (бисериальных) в два ряда и т.д. Если камеры нарастают по спирали
в одной плоскости, они называются плоскоспиральными (планиспираль-
ные), если более чем в одной плоскости, то коническими (трохоспиральные).
Строение устий - апертур, наличие или отсутствие пор, внешняя скульптура
раковины и т.д. - все эти признаки учитывают при определении видов и дру-
гих систематических категорий.
Классификация фораминифер. Фораминифер разделяют на несколько
больших групп: англоязычная школа считает их подотрядами, а русско-
язычная - надотрядами и отрядами. Самая примитивная группа, вероятно,
Allogromiina, представлена однокамерными раковинами, состоящими почти
полностью из органического материала. Они редки в ископаемом состоянии,
впервые появляются в позднем кембрии и живут по настоящее время.
Textulariina имеют агглютинированные раковины, построенные из орга-
нического или минерального материала, склеенного известковым и органи-
ческим цементом или даже окислами железа. Текстуляриины встречаются
от девона (кембрия?) по настоящее время.
Fusulinina имели известковые микрогранулярные стенки раковин. Они
были в основном распространены в палеозое, начиная с девона, но некоторые
дожили до мела (эндотириды). Фузулинины, вероятно, дали начало ныне
живущим роталиинам. Как указывает название фузулинин, они были вере-
теновидными. Фузулинины существовали в мелководье тропических бассей-
нов, расцвет приурочен к позднему палеозою (карбон-пермь).
Miliolina имели плотные непористые известковые раковины с фарфоро-
видной внешней поверхностью. Они впервые появились в карбоне, испытали
расцвет в мезозое и существуют поныне.
Rotaliina обладают стекловатыми известковыми пористыми раковинами с
несколькими отчетливыми слоями. Группа впервые появляется в раннем ме-
зозое; испытывает расцвет в мелу, успешно обитая как в мелководье конти-
нентальных морей, так и в открытой Атлантике.
Определение времени с помощью фораминифер. Фораминифер использу-
ют для дробного расчленения отложений и корреляции их друг с другом, да-
же когда они географически удалены друг от друга на огромные расстояния.
Практика использования ископаемых для этих целей составляет содер-
жание биостратиграфии. Фораминиферы имеют большое значение для гео-
логии, особенно эффективно их используют в биостратиграфии позднего
палеозоя, позднего мела и кайнозоя.
Для позднего палеозоя наиболее важными являются фузулинины. Изучая
внутреннюю структуру их стенок в шлифах, удалось выделить дробные под-
разделения во многих частях света, особенно в древнем Средиземноморье,
называемом Тетисом, протягивающемся через Европу, Северную Африку и
Центральную Азию. В России фузулиниды широко распространены в карбо-
не Московской синеклизы, а также в карбоне и перми Предуралья. Каменно-
угольные фузулиновые известняки использовали в России как строитель-
ный камень, например, при постройке зданий в древней Москве и Владими-
ре. Фузулинины приурочены к тропическим морям позднего палеозоя, они
неизвестны в местах холодного климата, таких как Австралия и Антаркти-
ка, которые в то время находились в зоне оледенения. Фузулинины были
крупными фораминиферами, обладавшими раковинами, напоминающими
по форме и размерам пшеничные или рисовые зерна. Раковины имели очень
сложную структуру стенки.
90
КАМЕННАЯ КНИГА
В среднем и позднем мезозое начали развиваться настоящие планктонные
фораминиферы. И снова фораминиферы стали важными биостратиграфиче-
скими фоссилиями. Планктонные фораминиферы имели широкое географи-
ческое распространение, а геохронологическое время существования видов
было сравнительно коротким, охватывающим несколько миллионов лет и
даже меньше, чем 1 млн. лет.
Дробные временные отрезки в кайнозое во многом выделены благодаря
планктонным фораминиферам, таким как Globigerinidae с шарообразными
камерами и Globorotaliidae с уплощенными килеватыми камерами. Оба се-
мейства из роталиин, хотя в последнее время их выделяют в самостоятель-
ную категорию, равную роталиинам. Используя появление и исчезновение
этих протистов, можно выделить временные отрезки продолжительностью
около 200 лет. Планктонные формы широко распространились в кайнозое
(особенно в “теплом” эоцене и миоцене), где стали играть решающую роль
при выделении дробных зон в морских породах всего мира.
Другой знаменитой кайнозойской группой фораминифер были нуммули-
тиды (Nummulites и др.). Это крупные дисковидные формы, которые появи-
лись на границе мела/палеогена, но продолжали существовать и в течение
кайнозоя; их представители до сих пор существуют в морях Индо-Пацифи-
ки. Нуммулиты были чрезвычайно многочисленными в морской акватории
Тетиса. Известняка, состоящие в основном из раковин нуммулитид, исполь-
зовали в Египте как строительный камень для постройки пирамид. Гречес-
кий историк Геродот принимал раковины нуммулитид за окаменевшие чече-
вицы, считая их остатками еды, которой египтяне кормили своих рабов.
Орбитоиды были также крупными дисковидными фораминиферами, су-
ществовавшими от позднего мела по миоцен. Одна из хорошо изученных
форм - это Lepidocyclina, которая обитала только в тропических водах от
эоцена по средний миоцен.
Образ жизни и условия обитания. Хотя большинство современных фора-
минифер являются морскими формами, некоторые из них живут в опреснен-
ных солоноватых водах. Фораминиферы обнаружены в пресных озерах Вен-
грии и в колодцах пустынь Азии и Северной Африки. Подавляющее боль-
шинство палеозойских и раннемезозойских видов свободно лежало или
передвигалось по дну мелководных морей. Такой же способ существования
вели нуммулиты и орбитоиды. Мелководные фораминиферы были наиболее
многочисленными в прибрежных водах; такие условия донные животные
предпочитают и сейчас.
В течение второй половины мезозойской эры разнообразные маленькие
планктонные фораминиферы освоили жизнь в открытых пространствах мо-
рей и океанов. Там они дрейфовали в верхней толще воды, где течения разно-
сили их по всему Земному шару. Некоторые планктонные фораминиферы
эволюционировали довольно быстро, со скоростью, сопоставимой с донными
ползающими фузулининами.
Распространение фораминифер лимитировано температурным фактором
и особенностями морского дна. Например, вдоль современного Атлантичес-
кого побережья Северной Америки четыре разные группы протистов образу-
ют четыре зоны обитания, отличающиеся по глубинам и температурам. Ни-
жнемеловая формация в Северном Техасе содержит в темных тонкослоистых
сланцах один комплекс бентосных видов, в серых сланцах и мергелях - дру-
гой, а в желтых мергелях и известняках - третий.
О чем рассказывают фораминиферы. Вышеперечисленные факты пока-
зывают, что ископаемых можно использовать для реконструкции характе-
РАЗНООБРАЗИЕ ПРОТИСТОВ
91
MILIOLINA
Г иперсоленые топи
Нормально-соленые топи
Шельфовые моря
ROTALIINA
Г ипосоленые топи
Нормально-соленые
лагуны
Г ипосоленые
лагуны __
TEXTULARIINA W
Фиг. V.8. Треугольное поле, где по соотношениям видов подотрядов Miliolina,
Rotaliina и Textulariina установлена соленость н зоны моря
ристик давно исчезнувших морей. Обилие донных свободноживущих видов
указывает на небольшие глубины, а если они становятся более разнообраз-
ными по мере продвижения в заданном направлении, то мы приближаемся к
побережью (и наоборот).
Существует методический прием, позволяющий определить, какой тип
донных условий преобладает, если вычислить процентные соотношения ви-
дов в комплексе по трем систематическим категориям: Miliolina, Rotaliina и
Textulariina. Процентное содержание наносится на поле треугольника.
Зная, какие существуют комплексы современных фораминифер в разных ус-
ловиях, и нанося их процентное содержание на подобное треугольное поле,
можно определить условия обитания: открытое море, солоноватоводная ла-
гуна и т.д. (фиг. V.8).
Разные группы донных фораминифер характеризуют типы дна, а разнооб-
разные планктонные формы отражают температурные изменения. Если фос-
силии близкородственны современным видам, можно сказать, какой ком-
плекс указывает на теплую воду, а какой, что вода была охлажденной или
холодной.
До сих пор самым большим практическим значением обладают планк-
тонные, то есть дрейфующие, виды; они являются превосходными руково-
дящими ископаемыми. Значение их еще более возрастает благодаря тому,
что размеры большинства планктонных фораминифер достаточно малень-
кие. Благодаря этому они сохраняются полностью и во множестве экземпля-
ров в небольшом кусочке породы, в том числе и в керне, добытом при буре-
нии. Таким образом, их можно использовать для идентификации форма-
ций, залегающих под поверхностью Земли, на различных глубинах, что
важно при поисках нефтегазоносных месторождений. Так, одна из форма-
92
КАМЕННАЯ КНИГА
ций в нефтесодержащих сериях была прослежена по фузулининам от Не-
браски до Нью-Мексико. Особенно больших успехов в поисках нефти с
помощью фораминифер достигли русские и американские палеонтологи-
биостратиграфы. Следует отметить, что практическое использование фузу-
линид для расчленения, корреляции и прослеживания толщ было убеди-
тельно продемонстрировано русскими палеонтологами-биостратиграфами
(Д.М. Раузер-Черноусова, А.В. Фурсенко и др.) на примере нефтяных мес-
торождений Предуралья и других регионов. Для дробного расчленения
позднепалеозойских толщ Предуралья были использованы микроструктур-
ные изменения стенок раковин фузулинин, а также строение раковин и
перегородок.
Не только секреционно-известковые фузулинины, но и агглютинирован-
ные раковины позволяют проследить толщи на огромных расстояниях. По-
зднемеловые формации, например по Vulvulina advena, были прослежены от
Кубы до Алабамы и восточной Мексики. Таким образом, практическая важ-
ность фораминифер приводит к тому, что нефтяные компании каждой стра-
ны имеют лаборатории экспертов для их определения, чтобы указывать гео-
логам и буровикам направления поисков.
Планктонные фораминиферы благодаря своему дрейфующему образу
жизни обычно быстро распространяются по всему Мировому океану. Их ске-
летики можно использовать для межрегиональной корреляции с большой
точностью. Тем не менее, вследствие зависимости планктонных форамини-
фер от температуры, приходится строить отдельные зональные схемы для
тропических, умеренных и высокоширотных морей.
Глава VI
ГУБКОВЫЕ:
НАСТОЯЩИЕ И ПРЕДПОЛАГАЕМЫЕ
• Основу типа Губковых (Spongiata) или Пороносцев (Porifera) состав-
ляет класс Губки, или Spongia. Кроме них, в состав типа Spongiata
включают класс Склероспонгиа (Sclerospongiae) и с разной степенью
убедительности и достоверности такие группы ископаемых, как сфинктозоа,
археоциаты, рецептакулиты, соаниты, афросальпингидеи, строматопораты,
хететиды и т.д. Подавляющее большинство является фильтраторами, проце-
живающими воду и улавливающими таким способом пищевые частицы.
НАСТОЯЩИЕ ГУБКОВЫЕ: ГУБКИ И СКЛЕРОСПОНГИИ
Класс Губки, или Spongia
Губки — примитивные многоклеточные царства животных или колони-
альные одноклеточные царства протистов? Когда говорят о типе губковых,
то прежде всего имеют в виду современных представителей класса губок, из-
вестных человечеству с незапамятных времен. Тем не менее дискуссии о по-
ложении и таксономическом ранге губок в системе органического мира бу-
дут продолжаться еще долго. Сначала губок относили к царству растений,
потом к сводному царству животные—растения (Zoophyta), затем к прими-
тивным многоклеточным царства животных. В настоящее время все больше
склоняются к мысли, что губки относятся к царству протистов, являясь од-
ним из вариантов их колониального существования, наподобие знаменитого
Вольвокса (Volvox), но без хлоропластов.
У губок много характерных черт строения, но укажем только следующие:
фильтрационный способ добычи пищи через систему, состоящую из множе-
ства пор, каналов, камер и центральной полости, оканчивающейся круп-
ным отверстием - устьем округлой или звездообразной формы (специфиче-
ский признак); наличие воротничково-жгутиковых клеток (общий признак
с воротничковыми жгутиконосцами из протистов); пищеварение только
внутриклеточное и пристеночное (общий признак с протистами); минераль-
ный скелет представлен иголочками - спикулами, а органический скелет -
спонгиновыми волокнами (специфический признак); зародыш проходит
стадию “инверсии”, то есть воротничково-жгутиковые клетки наружного
слоя погружаются внутрь, в результате клетки внутреннего слоя оказыва-
© 1997. О.Б. Бондаренко и А.Ю. Розанов, новый вариант главы.
94
КАМЕННАЯ КНИГА
а Спикулы
Protospongia
в Спикулы
Haptonia
Фиг. VI. 1. Кембрийские губки с кремневыми спикулами
а, б — отряд Triaxonida-. Protospongia erixo, крестовидные спикулы с различно
развитыми лучами, ранний кембрий, Якутия (Основы палеонтологии, том 2,
1962); б — Diagoniella hindei, реконструкция скелетного каркаса с ромбовидны-
ми ячеями, средний кембрий, Канада, Британская Колумбия (Walcott, 1920);
в — отряд Tetraxonida: Hamptonia bowerbanki, одноосные спикулы, средний
кембрий, Канада, Британская Колумбия (Walcott, 1920)
Фиг. VI.2. Девонские кремневые губки отряда Triaxonida Северной Америки
а - Dictyospongia sceptrum; б - Prismodictya prismatica; в - Bothryodictya гото-
ва-, г - Hydnoceras tuberosum; все четыре губки из девона США (Hall, Clarke,
1898; Основы палеонтологии, том 2, 1962)
ГУБКОВЫЕ: НАСТОЯЩИЕ И ПРЕДПОЛАГАЕМЫЕ
95
ются снаружи (специфический признак); клетки не стабильны в своей диф-
ференциации, легко трансформируются одни в другие, а отсюда отсутст-
вуют ткани (специфический признак). Все эти особенности губок резко от-
личают их от настоящих многоклеточных царства животных, которые не
содержат воротничково-жгутиковые клетки; не проходят стадию инверсии,
но имеют стадию гаструлы, неизвестную у губок; клетки стабильны, ткани
имеются и т.д. Более того, у всех настоящих многоклеточных животных
развито внутриполостное (“автоклавное”) пищеварение. Таким образом,
особенности строения губок указывают на их большую близость к протис-
там, чем к настоящим многоклеточным животным. Недарок губок называ-
ют еще и Parazoa, то есть “идущими параллельно животным”, противопос-
тавляя их таким образом Eumetazoa, то есть настоящим многоклеточным
животным.
Ископаемые губки. Губки существуют на протяжении 600-700 млн. лет,
начиная, по-видимому, с рифейского времени (достоверно с кембрийского
периода) и по настоящее время. В подавляющем большинстве ископаемые
губки представлены мелководными формами морских бассейнов, в меньшей
степени глубоководными (начиная с юры) и пресноводными (с юры).
Из вышеперечисленных особенностей строения губок в ископаемом состо-
янии остаются минеральные каркасы из спикул, сохраняющие форму тела
губки и систему пор, каналов, камер и полостей. Состав спикул: кремневый
(SiO2 • Н2О) или известковый (СаСО3, арагонит, кальцит). Если спикулы не
связаны в жесткий каркас, то они, как правило, находятся в породе в разроз-
ненном состоянии. От губок могут сохраняться и спонгиновые волокна, со-
став которых близок к коллагену.
Достоверные ископаемые губки с кремневым и спонгиновым скелетом
встречаются с раннего кембрия, а с известковым скелетом - с девона
(фиг. VI. 1-5). Теоретически губки должны были существовать уже в венд-
Фиг. VI.3. Кремневая литистидная губка Siderospongia sirensis из карбона
Подмосковья (отряд Tetraxonida)
а - общий вид внешней поверхности, б - фрагмент при увеличении (Traut-
schold, 1869)
96
КАМЕННАЯ КНИГА
Elasmojerea
Peronidella
г Eudea
Stellispongia
Фиг. VI.4. Известковые губки мезозоя Европы (Calcispongiae)
а - Epitheles hemisphaerica, xl.5, юра, Германия; б — Elasmojerea sequana, х2,
нижний мел, Крым, Симферополь; в - Peronidella sp., нижний мел, Крым; г -
Eudea cribraria, х2, нижний мел, Крым, Симферополь; д - Stellispongia glomer-
ata, xl.5, верхняя юра, Германия (Основыпалеонтологии, том 2, 1962)
ское время. Действительно, образования, напоминающие спикулы и спон-
гин, обнаружены в докембрийских породах, но их губковая природа одно-
значно не доказана. В кембрии встречаются достоверные губки с кремневы-
ми трехосными, четырехосными и одноосными спикулами, неспаянными в
жесткий каркас (см. фиг. VI. 1).
В ордовике широко распространены кремневые губки с четырехосными
спикулами, спаянными в литистидную решетку (Tetraxonida). Они образу-
ют губковые горизонты в ордовике северо-запада России, Эстонии, Герма-
нии, Англии и Северной Америки. В девоне наблюдается расцвет кремневых
губок с трехосными (шести- и пятилучевыми) спикулами, особенно в Север-
ной Америке (Triaxonida, фиг. VI.2). В девоне впервые появляются и досто-
верные известковые губки (Calcispongiae). Начиная с карбона кремневые и
известковые губки становятся все разнообразнее, но по-прежнему встреча-
ются только в морских бассейнах (фиг. VI.3). В мезозое, в юрское время,
впервые появляются пресноводные формы, представленные кремнероговы-
ми губками. В это же время морские губки начинают осваивать глубоково-
дье. Наибольшего разнообразия и распространения губки достигают в позд-
ней юре и в мелу. В раннемеловое время известковые губки участвовали в об-
разовании малых рифовых тел, биостромов и биогермов (фиг. VI.4). В
позднем мелу наиболее известны кремневые трехосные шестилучевые губ-
ки, образующие разнообразные заросли в мелководье сантонского века и в
глубоководье кампанского века России и других стран (фиг. VI.5; см.
фиг. II. 1).
Современные губки встречаются в морских, реже в пресноводных бассей-
нах всех широт от тропиков до Арктики и Антарктики и на различных глу-
бинах, от зоны прилива-отлива (литораль) и до океанических желобов (уль-
траабиссаль). Губки обитают также и в абиссальных оазисах, образуя внеш-
нее кольцо вокруг курильщиков.
ГУБРЮВЫЕ. НАСТОЯЩИЕ И ПРЕДПОЛАГАЕМЫЕ
97
Фиг. VI.5. Разнообразные кремневые губки из сантонских отложений верхнего
мела Поволжья (отряд Triaxonida)
а — Craticularia cylindrical б, в — Guettardiscyphia stellata, внешний вид со сторо-
ны трех лопастей (б) и схема поперечного сечения, где видны все четыре лопасти
(в); г - Polyscyphiapseudocoeloptychium; д - Sporadoscinia sp.; е - Zittelispongia al
cyonoideai ж — Ventriculites pedester. Все губки почти в натуральную величину
Класс Коралловые губки, или Sclerospongiae
Коралловые губки были впервые обнаружены в конце XIX века в Кариб-
ском море у Кубы, однако систематическое положение находок было непо-
нятно. Начиная с 1970 года В.Д. Хартман и Т.Ф. Горо (Hartman, Goreau,
1970, 19) приступают к тщательным исследованиям прежних и новых нахо-
док. Оказалось, что склероспонгии - формы колониальные, обладающие
смешанным составом скелета, состоящим из известковых микрокристаллов
и зерен (арагонит или кальцит), кремневых одноосных спикул и органичес-
ких волокон. Известковая составляющая образует сплошной массивный или
мелкотрубчатый скелет, который пронизан кремневыми игольчатыми спи-
кулами. На поверхности массивных колоний наблюдаются звездчатые кана-
лы, сходные с устьями губок и астроризами строматопорат (фиг. VI.6). Кро-
ме того, у найденных форм обнаружили воротничково-жгутиковые клетки,
которые выстилали внутренние полости, что позволило включить склеро-
спонгий вместе с губками в тип Spongiata. Вместе с тем специфика строения
найденных форм заставила Хартмана и Горо (1970) выделить их в самостоя-
тельный класс Sclerospongiae. Склероспонгий также называют коралло-
7. Каменная книга
98
КАМЕННАЯ КНИГА
а ис кс
Склероспонгии- Sclerospangiae
Фиг. VI.6. Склероспингии и строматопораты
а-в - класс Sclerospongiaet а — Ceratoporella nicholsoni, х75, известковые стенки
трубочек пронизаны кремневыми спикулами, современная форма; б — Stromato-
spongia, увел., внешняя поверхность с порами и звездчатыми каналами, совре-
менная форма (а, б-Hartman, Goreau, 1969,1972; Журавлева, Мягкова, 1987);
в — реконструкция фрагмента колонии склероспонгий (Bergquist, 1978; Журав-
лева, Мягкова, 1987); г, д - класс Stromatoporata: г — продольное сечение при
большом увеличении, хорошо видно слоистое нарастание, д -Aculatostroma ver-
rucosum, х5, звездчатые (астроризальные) каналы в поперечном сечении, ран-
ний девон, Салаир (Халфин, 1971); вж - камеры с воротничково-жгутиковыми
клетками, вп - входящие поры, зс - звездчатые каналы (выводящая система),
ис — известковые стенки трубочек, кс — кремневые игольчатые спикулы
выми губками, так как они обитают в гротах, пещерах и каналах коралло-
вых рифов. Некоторые считают их потомками палеозойских археоциат,
строматопорат и хететид.
Ископаемые склероспонгии известны с триаса, если не принимать во вни-
мание вышеназванные палеозойские группы.
ПРЕДПОЛАГАЕМЫЕ ГУБКОВЫЕ: СФИНКТОЗОА, АРХЕОЦИАТЫ,
РЕЦЕПТАКУЛИТЫ. СТРОМАТОПОРАТЫ
Сфинктозоа, или Sphinctozoa
Сфинктозоа указывают с среднего кембрия по палеоген, возможно, они
встречаются и в современных морях. Сфиктозоа представлены одиночными
ГУБКОВЫЕ: НАСТОЯЩИЕ И ПРЕДПОЛАГАЕМЫЕ
99
Amblysiphonella Guadalupia Vaceletia
Фиг. VI.7. Сфинктозоа
а - Amblysiphonella barroisi, xl, продольное сечение, карбон, Испания; б-Guad-
alupia cylindrica, х10, поперечное сечение, пермь, Сицилия; в — Waagenella cf.
W. elongata, x2, продольное сечение, поздний триас, Молуккские острова; д, г -
Thalatnopora cribrosa; г — вид сверху, х2, д - вид сбоку, xl.5, мел, Германия;
е, ж - Vaceletia cripta: е — продольное сечение, сильно увел., ж - внешний вид,
современная форма, акватория о. Новая Каледония (Основы палеонтологии,
том 2,1962; е, ж - Vacelet, 1977)
и колониальными формами, ведущими прикрепленный донный образ жиз-
ни. Скелет состоит из известковых шаровидных камер, нарастающих друг
над другом вверх. Стенки камер пронизаны многочисленными порами. Осе-
вая часть верхней стороны камер обычно вогнута внутрь, в результате чего
образуется центральный канал (фиг. VI.7). Камерное нарастание создает ха-
рактерную систему пережимов с внешней стороны сфинктозоа, с чем связано
их название (греч. sphinkter от sphingo — сжимаю). Известковые стенки ка-
мер состоят из сросшихся кальцитовых или арагонитовых микрокристаллов
и зерен; присутствие известковых спикул подвергается сомнению.
Наличие системы из многочисленных пор и центрального канала застав-
ляет относить сфинктозоа к типу губковых, а присутствие известковых мик-
рокристаллов и зерен, после изучения склероспонгий, такому выводу не про-
тиворечит. С сфинктозоа некоторые объединяют кембрийских археоциат,
особенно камерные формы (см. фиг. VI.8, 9).
Сходное с сфинктозоа, но более сложное, строение скелета обнаружено у
современного вида Vaceletia cripta, имеющего воротничково-жгутиковые ка-
меры. Большинство исследователей не сомневаются в истинной губковой
природе Vaceletia и сфинктозоа и даже относят их к Calcispongiae, реже к
Sclerospongiae.
Геологическую историю сфинктозоа одни начинают с кембрия или ордо-
вика и девона, другие только с позднего карбона. Такое разночтение связано
7*
100
КАМЕННАЯ КНИГА
с наличием кремневого скелета (первичный?, вторичный?) у некоторых до-
карбоновых находок. Расцвет истинно известковых сфинктозоа начинается
с перми и продолжается в триасе. В этом временном промежутке класс
сфинктозоа — один из каркасостроителей рифовых сооружений Азии (Па-
мир, Кавказ, Индия и др.), Европы (Альпы), Северной и Южной Америки.
Археоциаты, или Archaeocyathi
Археоциаты - вымершая группа, чрезвычайно важная для стратиграфии
отложений нижнего кембрия Сибири, Монголии, Китая, Северной Африки,
Америки, Австралии и др. регионов. По ним устанавливают не только яру-
сы, но и зоны. Кроме того, по археоциатам и сопутствующей фауне в основа-
нии кембрия был выделен томмотский ярус. Археоциаты вместе с цианоби-
онтами и водорослями являются основными каркасостроителями рифовых
образований раннего кембрия. Более чем 130-летнее изучение археоциат
позволило установить большое разнообразие строения этих организмов
(фиг. VI.8-10). Некоторые формы, особенно камерного строения, сходны с
Фиг. VI.8. Одностенные и двустенные правильные археоциаты атдабанского и
ботомского времени (ранний кембрий)
Прорисовки шлифов: а — Yadjaicyathus astashkini, х10, косопоперечное сечение
кубка, атдабанский век. Сибирь, р. Лена; б — Pretiosocyathus subtilis, хб, попе-
речное сечение кубка, атдабанский век, батеневский кряж; в — Capsulocyathus
subcallosus, х15, продольное сечение кубка, ботомский век, Западный Саян; г —
Plicocyathus rosanovi, хб, продольное сечение кубка, ботомский век, Канада, го-
ры Маккензи; д — Carinacyathus sp., х27, продольное сечение кубка, атдабан-
ский век (?), Сибирь, р. Лена (Ф. Дебренн, А.Ю. Журавлев, А.Ю. Розанов,
1989); 1 - стадия одностенного кубка, кзн — каналы с козырьками, тб — тумулы-
бугорки, тс — толстая внутренняя стенка
ГУБКОВЫЕ: НАСТОЯЩИЕ И ПРЕДПОЛАГАЕМЫЕ 101
сфинктозоа. Отсюда и дискуссии о месте археоциат среди губкоподобных
форм, их систематике и геохронологическом интервале существования:
только ранний кембрий или ранний, средний и поздний кембрий, либо не
только палеозой, но и мезозой.
Археоциаты - морские, преимущественно одиночные организмы, пред-
ставленные известковыми коническими, цилиндрическими, реже грибооб-
разными и блюдцеобразными формами, называемыми кубками. Кубки при-
Фиг. VI.9. Поперечные сечения некоторых неправильных археоциат и рекон-
струкция трех кубков двустенных правильных археоциат раннего кембрия
а — Archaeocyathus regularis, еланский век, Сибирь (ориг. В.Д. Фонина); б-Pseu-
dosyringocnema eleganta, ботомский век, Саяны (ориг. В.Д. Фонина); в - Ajaci-
cyathus nevadensis, Невада (Р. Vickers Rich, Т. Rich, М. Fenton, 1989); г — дву-
стенный кубок с каблучком прирастания, септами и гребенчатыми днищами;
д - двустенный кубок Coscinocyathus dianthus, состоящий из субсферических
камер (г, д - Ф. Дебренн, А.Ю. Журавлев, Ф.Ю. Розанов, 1989)
5-10 м
Фиг. VI. 10. Археоциатовый биогерм, содержащий многочисленных археоци-
ат, редких циаиобионтов и водоросли
Ранний кембрий, Сибирская платформа, р. Лена (Журавлева, Зеленов, 1955)
102
КАМЕННАЯ КНИГА
растают к субстрату или свободно лежат на нем. Размеры археоциат от 3 мм
до 70 см в поперечнике и до 1.5 м в высоту. Археоциаты состояли из одной
или двух стенок, в пространстве между которыми (в интерваллюме) развиты
разнообразные скелетные элементы; некоторые археоциаты имели камерное
строение. Стенки археоциат пронизаны многочисленными порами так же,
как и большинство скелетных элементов; имеется центральная полость. Ми-
кроструктура стенок гранулярная, но не спикульная. Система пористости,
подобная губкам, с самого начала изучения археоциат заставляла относить
их вместе с настоящими губками к одной группе организмов, сначала внутри
типа Porifera (Spongiata), затем в качестве самостоятельного типа Archaeo-
cyathi и даже подцарства Archaeata отдельно от подцарства Porifera, но все
же в одном царстве, но каком?
В России раннекембрийские археоциаты широко распространены на Си-
бирской платформе, Саяно-Алтайской области, Туве, Дальнем Востоке (Яку-
тия и другие регионы), где они нередко были рифообразующими (фиг. VI. 10).
В европейской части России археоциаты отсутствуют (? кроме Урала).
Рецептакулиты, или Receptaculita
К рецептакулитам (Receptaculita) относятся вымершие палеозойские оди-
ночные, реже колониальные организмы, представленные кубками с цент-
ральным отверстием и с характерной “чешуевидной” внешней поверхнос-
Ischadites
Receptaculites
Hexabactron
Фиг. VI.11. Рецептакулиты
а — Ischadites iowensis, х0.25-0.5, средний ордовик, центральные районы Север-
ной Америки (Р. Vickers Rich, Т. Rich, М. Fenton, 1989); б, в - Ischadites
murchisoni, х0.25—0.5; б — внешний вид широким концом вверх, в - продольное
сечение того же кубка широким концом внйз, средний ордовик, Эстония; г - Re-
ceptacuiites, хЗ, поперечное сечение через наружную стенку с чешуями и ради-
алями, ордовик, Северо-Восток России; д - Hexabactron, увел., схема строения
стенок кубка, силур, Австралия (Campbell et al., 1974, Журавлева, Мягкова,
1987). п - поры внутренней стенки; р — радиаль; тб - таблички-“чешуи” с неров-
ной внешней поверхностью; щ - щель между табличками
ГУБКОВЫЕ: НАСТОЯЩИЕ И ПРЕДПОЛАГАЕМЫЕ
103
тью. Отсюда их другое название - Squamiferida, или Чешуеносцы. Форма
кубков от конической, грушевидной и шаровидной до блюдцеобразной и уп-
лощенно-меандрической. Максимальная высота кубков до 20 см, попереч-
ник до 80 см. “Чешуи” наружной стенки кубков представлены известковыми
табличками, имеющими очертания ромба, прямоугольника или шести-
угольника (фиг. VI. 11). Между табличками имеются щели, выполняющие,
вероятно, функцию пор. Таблички расположены спиральными. На внутрен-
ней стороне табличек находятся лучевидные элементы, расположенные, как
правило, крестообразно. От центра пересечения “креста” внутрь кубка отхо-
дят различной толщины и формы столбики, называемые радиалями. Радиа-
ли оканчиваются крестовидными структурами, подобными предыдущим
или расширениями, которые срастаются друг с другом и образуют решетку
внутренней стенки. Вдоль оси кубка располагается центральная полость, от-
крывающаяся отверстием. Все скелетные элементы состоят из известковых
микрогранул.
Систематическое положение рецептакулит весьма дискуссионно: Protis-
ta, Spongiata, водоросли (? Dasycladaceae), высшие растения, самостоятель-
ный тип или царство. Даже ориентировка кубков широким концом с отвер-
стием вверх подвергается сомнению (а может быть, вниз!). Несомненно, ре-
цептакулиты были донными прикрепленными или свободнолежащими
организмами мелководья (? фильтраторы).
Рецептакулиты встречаются со среднего ордовика по пермь почти повсе-
местно; в ордовике они участвовали в рифостроении. В России рецептакули-
ты обнаружены на Урале, Сибири, Алтае, Дальнем Востоке, Чукотке.
Строматопораты, или Stromatoporata
Вымершие строматопораты (средний ордовик - палеоген) представлены
слоистыми известковыми образованиями (см. фиг. VI.6) различной внешней
формы: от пластинчатой, полусферической и шаровидной до грушевидной,
неправильно желваковидной с пальцевидными выростами и цилиндричес-
кой. Под микроскопом видно, что известковые слои, называемые ламинами,
обычно соединены или пронизаны вертикальными элементами - столбика-
ми. Строение ламин разнообразное: пузырчатое, петельчатое, сплошное, губ-
чатое и т.д. Также разнообразны и вертикальные элементы: зубчики, шипи-
ки, короткие и длинные столбики, пластины и т.д. Ламины и столбики могут
утолщаться с образованием губчатой ткани. Скелет строматопорат пронизан
разнообразными пустотами. Самыми примечательными из них являются ас-
троризальные каналы, имеющие на верхней поверхности ламин вид звездча-
тых пустот, обычно с длинными и дихотомически ветвящимися горизон-
тальными каналами. Они сходны с звездчатыми каналами склероспонгий и
некоторых губок (см. фиг. VI.6), а также напоминают циклосистемы гидро-
кораллов. Отсюда дискуссионность их систематического положения: тип
Spongiata (класс Sclerospongiae) или тип Cnidaria (класс Hydrozoa).
Известны находки строматопорат, у которых известковый скелет прони-
зан кремневыми спикулами. Такие находки еще больше убеждают, что стро-
матопораты относятся к губковым. В любом случае строматопораты являют-
ся колониальными организмами мелководья. Они, как правило, участвова-
ли в рифостроении уже начиная с ордовика и далее, вплоть до мезозоя. Но в
каменноугольное и пермское время они крайне редки, что заставляет некото-
рых исследователей считать, что мезозойские строматопораты (сферактино-
иды) не являются потомками палеозойских.
104
КАМЕННАЯ КНИГА
Фиг. VI.12. Разнолучевые
а, б - скелетные структуры - археастеры у Chancelloria, ранний кембрий, Си-
бирская платформа (Журавлева, Кордэ, 1955; Розанов, 1986); в — археастераАг
chaeospongium, ордовик-карбон (Hinde, 1877 1912); г — Eiffelia globosa, рекон-
струкция “тела” губки, археастеры с шестью лучами в одной плоскости и од-
ним, перпендикулярным к ним, средний кембрий Британской Колумбии,
Канада (Walcott, 1920, Основы палеонтологии..., том 2, 1962)
Другие ископаемые губкоподобные организмы
В палеозое встречено много губкоподобных и археоциатообразных иско-
паемых, кроме вышеописанных: соаниты, радиоциаты, афросальпингидеи,
о которых подробно можно прочесть в книге И.Т. Журавлевой и Е.И. Мягко-
вой “Низшие многоклеточные фанерозоя” (1987).
Кратко опишем только представителей отряда Многолучевых, или Разно-
лучевых (Heteractinida) из класса Radiocyatha, распространенных на протя-
жении всего палеозоя по всему миру, но особенно многочисленных в кемб-
рии. От них чаще всего сохраняются звездчатые многолучевые структуры, у
которых центральный луч ориентирован вертикально, а остальные в числе
6-13 расположены радиально от него, почти в единой горизонтальной плос-
кости (фиг. VI.12). Эти звездчатые структуры, называемые в прежней науч-
ной литературе многолучевыми спикулами, а в новой - незастерами и архе-
астерами, слагали внешние стенки кубков. Центральный луч в последних
работах рассматривают как аналог радиали рецептакулит. Максимальный
размер кубков - 120 мм, обычный - 20-30 мм, высота кубков до 50 мм. Раз-
меры незастер и археастер в диаметре 1-12 мм.
Самым известным среди Heteractinida был род Chancelloria, археастеры
которого были полыми внутри, а по составу - известковыми или фосфатно-
известковыми.
Глава VII
ПРОСТЫЕ КИШЕЧНОПОЛОСТНЫЕ:
ТИП CNIDARIA, ИЛИ СТРЕКАЮЩИЕ
• Книдарии, куда входят кораллы, являются наиболее простыми
многоклеточными, за исключением губковых и археоциат. В отли-
чие от современных губок у книдарий клетки организованы в на-
стоящие ткани, имеется нервная система, внутриполостное пищеварение и
т.д. Кораллы, тем не менее, не рассматривают как самых простых книда-
рий. Эта привилегия, но не только она одна, принадлежит современной гид-
ре, которая обитает в пресноводных протоках, спокойных реках, озерах и
прудах.
Хотя “гидра” названа именем мифического водяного змея, имеющего
множество голов, сама она представляет собой малюсенькое существо, чье
тело в виде стройной трубочки содержит два слоя клеток, разделенных желе-
образной упругой пластинкой. Основание этой трубочки свободно прикреп-
лено к прутикам или гальке; на верхнем конце находится рот (см.
фиг. III. 2г). Полые щупальца вытянуты для захвата пищи, которая усваива-
ется в центральной полости, соответствующей желудку, кишкам и некото-
рым другим органам высших животных. Клетки в обоих слоях тела разнооб-
разные: одни сокращаются как мускулы и посылают сигналы через нервную
сетку, другие ощущают вкус или жалят добычу и обвивают ее и т.д. Некото-
рые клетки имеют ложноножки-псевдоподии или жгутики, но последние не
окружены воротничком, как у губок. Это одно из убедительных доказа-
тельств, что гидра не является близким родственником губок.
План строения тела гидры простой, но он имеет три больших достоинства.
Первое из них — это высокая эффективность, которая дает возможность жи-
вотному жить и процветать неопределенно долгое время. Второе - план стро-
ения может меняться, создавая существа от тоненьких маленьких зооидов у
колониальных форм до крупных одиночных полипов и свободно плавающих
медуз, образно называемых “желе-рыбы” (jellyfish). Третье достоинство -
ранние предки гидры оказались способны к созданию третьего зародышево-
го слоя клеток и, таким образом, дали начало всем другим животным, более
сложным, чем тип Cnidaria. Книдарии, или стрекающие - группа живот-
ных, характеризующаяся: наличием “крапивных” жалящих капсул (нема-
тоцисты), бесполым размножением, что свойственно полипам; половым раз-
множением, что свойственно медузам; и обычно наличием скелета (внутрен-
ним или внешним) известкового (кальцит, арагонит), хитинового или
хитиново-фосфатного состава.
© 1997. О.Б. Бондаренко, перевод на русский язык и изменения.
106
КАМЕННАЯ КНИГА
КОЛОНИАЛЬНЫЕ ГИДРОИДНЫЕ (HYDROZOA)
Класс Hydrozoa представлен полипами и медузами. Полипы преимущест-
венно колониальные, ведущие обычно бентосный прикрепленный образ
жизни, реже планктонный (сифонофоры). Медузы - одиночные формы, оби-
тающие в толще воды. Медузы гидроидного плана строения обнаружены в
докембрийских породах; достоверные гидроидные полипы с хитиноидными
скелетами найдены в кембрии, ордовике и более молодых формациях Евро-
пы, Азии, Африки, Австралии и Северной Америки.
Многие биологи и палеонтологи считают, что Hydrozoa были самой древ-
ней и исходной группой для всех остальных книдарий.
Миллепоры и стиластерины (гидрокораллы). Миллепоры и стиластерины
являются колониальными гидроидными, строящими известковые скелеты.
Внешняя конструкция скелетов сходна с настоящими кораллами класса Ап-
thozoa. Еще одно сходство заключается в том, что те и другие могут быть ри-
фостроящими. Миллепоры и стиластерины поэтому и носят общее название
гидрокораллы, которое подчеркивает, что они хотя и похожи на кораллы, но
относятся совершенно к другому классу, чем настоящие кораллы.
Каждя колония гидрокораллов содержит три группы зооидов (фиг. VII. 1).
Зооиды первой группы имеют вид коротких полипов с щупальцами; они ло-
вят пищу, заглатывают и переваривают ее (гастрозооиды). Зооиды второй
группы длинные, как пальцы (дактилозооиды), без рта, ветвящиеся, содер-
жащие на концах гроздья стрекательных капсул. Дактилозооиды захваты-
вают пищу, но заглатывать ее не приспособлены, поэтому питание они полу-
чают от гастрозооидов по внутренним каналам. Третья группа зооидов нахо-
дится в округлых полостях (ампулы), где они формируют молодь в форме
крошечных медуз. Через мелкие отверстия в верхней части ампулы медузки
“выходят” наружу и попадают в окружающее водное пространство. Зооиды
ампул так же, как и дактилозооиды, получают питание от гастрозооидов по
внутренним каналам.
Дактилозооид
Фиг. VII1. Гидрокораллы "класс Hydrozoa)
а - реконструкция фрагмента колонии с тремя типами зооидов; б — Congre-
gopora, фрагмент взточки при увеличении с вскрытыми полостями-ампулами;
в — внешний вид ветвистой колонии Congrepora nasiformis, xl, поздний мел,
Германия
ПРОСТЫЕ КИШЕЧНОПОЛОСТНЫЕ: ТИП CNIDARIA
107
Гидрокораллы впервые появляются в палеогене и продолжают встречать-
ся в современных морях. Некоторые из них имеют вид уплощенных покро-
вов — матов или пластин, но большинство форм ветвистые (см. фиг. VII. 1).
Внешняя поверхность известковых скелетов гидрокораллов несет многочис-
ленные отверстия, отсюда и название - Millepora - миллионнодырчатая.
Крупные отверстия — поры соответствуют вместилищам гастрозооидов; в бо-
лее мелких порах находятся дактилозооиды. Округлые скопления еще более
мелких крошечных пор, ведущие в полости ампулы, соответствуют репро-
дуктивным зооидам.
Spongiomorphida. Спогиоморфиды - вымершие колониальные формы,
строящие массивные колонии, где радиальные столбики, разрастаясь в гори-
зонтальном направлении, создают структуры, напоминающие ламины и ас-
троризы строматопорат. Спонгиоморфиды, вероятно, близки губкам, а не
книдариям. В триасовое и юрское время спонгиоморфиды нередко являлись
породообразующими организмами.
ДРЕВНИЕ МЕДУЗЫ
Современные медузы принадлежат двум классам, Hydrozoa и Scyphozoa,
ископаемые - еще и к нескольким другим классам. Медузы и медузоподоб-
ные Cnidaria захоронялись спорадически при определенных условиях суще-
ствования и специфики осадконакопления. Особенно хорошие отпечатки
медуз и медузоподобных форм находят в тонких известково-глинистых
осадках.
Самые ранние убедительные медузоподобные формы происходят из венд-
ских кварцитов Южной Австралии, открытые в 1947 году геологом Сприг-
Фиг. VII. 2. Класс Cyclozoa
а — Cyclomedusa davidi с нижней стороны; б, в — Cyclomedusa plana, схема стро-
ения отпечатка (б) и “объемная” реконструкция с ротовым отверстием в центре
(зачернено); г, д — Spriggia wadea, реконструкция продольного сечения через
центр тела (г) и строение отпечатка (д) (Wade, 1972; Sun, 1986; Федонкин,
1987). d - кольцевые камеры, f - краевой выступ, t - краевые щупальца
108
КАМЕННАЯ КНИГА
1 см
4 см
Rhizostomites
Dickinsonia
ротовой диск Anomalocaris
Фиг. VII. 3. Медузы (а-в) и лжемедузы (г, д)
а, б- Brooksella Cambria (классProtomedusae) с верхней (а) и нижней стороны (б),
xl, ранний кембрий, Алабама; в - Rhizostomites admirandus (класс Scyphozoa
или "IHydrozoa), х0.25, поздняя юра, золенгофенские сланцы, Германия; г — Dick-
insonia (надтип Vermes), xl, докембрий, эдиакарий, Южная Австралия; д - Реу-
toia (тип Arthropoda), ротовой диск среднекембрийского членистоногого Anom-
alocaris (см. также фиг. XI. 17), средний кембрий, сланцы Бёрджесс, Канада,
Британская Колумбия
гом (Sprigg). Некоторые из них относятся к Hydrozoa и, возможно, K<Scypho-
zoa, в то время как другие остаются вне каких-либо известных групп, напри-
мер Cyclomedusa plana, Ediacaria flindersi, Lorenzinites rarus, Mawsonites
spriggi. Медузоидные формы такого типа были также обнаружены в венд-
ском периоде позднего докембрия в других частях мира. В России они найде-
ПРОСТЫЕ КИШЕЧНОПОЛОСТНЫЕ: ТИП CNIDARIA
109
ны в Архангельской области и на Урале; они также известны из вендских по-
род Западной Украины. Особенности строения вендских, а также кембрий-
ских Cnidaria заставили палеонтологов выделить несколько новых классов,
в том числе Protomedusae, Cyclozoa и Inordozoa. Большинство из них, по-ви-
димому, представляют собой ранние эксперименты природы по пути много-
клеточной эволюции животных, которые не привели к возникновению по-
следующих форм.
Среди вендских книдарий самым многочисленным и разнообразным ока-
зался класс Cyclozoa. Реконструкции жизненных форм циклозоа являются
предметом оживленной дискуссии: а - формы медузоидные или полипоид-
ные; б - свободно-плавающие или донные сидячие и т.д. (фиг. VII.2). Неко-
торые медузовидные формы при тщательном изучении оказались не медуза-
ми, а иглокожими, другие червями или членистоногими (фиг. VII.3). Часть
лжемедуз представляет собой отпечатки дождевых капель, газовых пузырь-
ков, трещины усыхания, ядра инволютных аммонитов и т.д.
Несколько групп гидроидных и сцифоидных медуз имеют ископаемых
представителей. Из гидроидных это Trachylina и Chondrophora, из сцифоид-
ных - Coronata, Cubomedusae, Rhizostomida и др. Интервал существования
сцифомедуз отряда Coronata'. юра-современность. Они имеют перетяжку,
разделяющую тело (зонтик) на центральный диск и корону; низ зонтика не-
сет лопасти и щупальца, начинающиеся от выемок между лопастями. Поли-
пы Coronata строят вокруг себя хитиноидную трубочку. Сцифомедузы отря-
да Carybdeida {Cubomedusae) с 4 щупальцами, включают тропические формы
с кубоподобными зонтиками без всяких лопастей. Они обладают чрезвычай-
но сильнодействующими ядовитыми стрекающими капсулами. Cubomedu-
sae существуют от поздней юры по настоящий день. Ископаемые формы,
имеющие низкий тарелкообразный зонтик с фестончатым краем, сопостав-
ляют с медузами семонтэостомидами, известными из поздней юры золенго-
фенских сланцев Германии и в настоящее время. Сейчас они единственные
медузы, часто встречающиеся в умеренных широтах. Сцифомедузы ризосто-
миды очень напоминают эту группу, но не имеют щупалец. В современных
морях ризостомиды в основном приурочены к тропикам, но встречаются они
и в Черном и Азовском морях. Ризостомиды также обнаружены в поздней
юре (золенгофенские сланцы).
Еще одна группа, Lithorhizostomatida, характеризуется караваеобраз-
ным зонтиком и лопастями с 8 гроздьями коротких щупалец. Они известны
только из тонкослоистых юрских литографских известняков Золенгофена,
где так много захоронено медуз и другого типа. Довольно неожиданно, что
такие мягкотелые существа, как медузы, хорошо сохраняются в ископае-
мом состоянии. Но когда они лежат на пляже и высыхают на солнце, они
становятся плотными и таким образом неплохо подготовлены к захо-
ронению.
Конулярии (фиг. VII.4). Эту группу в одних книгах описывают среди чер-
вей или моллюсков, а в других - среди медуз. За морфотип группы принята
Conularia, кембрийско-пермский род, который имеет чертырехгранную пи-
рамидальную хитиновую или хитиново-фосфатную раковину, нередко в раз-
личной степени объизвествленную. Каждая грань несет множество тонких
поперечных морщинок; на стыке граней развиты продольные борозды.
Предполагают, что сначала раковинки могли прикрепляться к морскому дну
или к другим субстратам. Впоследствии раковинки могут обламываться у
своих кончиков, и животные начинают дрейфовать или плавать. На некото-
рых реконструкциях показывают щупальца, растущие от края тела вверх,
110
КАМЕННАЯ КНИГА
Paraconularia
Exoconularia
Archaeoconularia
Фиг. VII. 4. Конулярии (Conulata)
а — Paraconularia, силур-карбон; б - Conularia undulata, средний девон (род 6 -Р)>‘
в - Exoconularia, реконструкция планктонной формы, ордовик; г — Archaeo-
conularia, реконструкция прикрепленной формы, кембрий-силур
когда животное прикреплено к дну, но щупальца свисают вниз, когда живот-
ное становится дрейфующим.
Ископаемые, напоминающие Conularia, были найдены в породах конца
докембрия Австралии. Достоверные конулярии распространены в палеозое
повсеместно. Встречаются они и в мезозое, но редко (юра Европы). Хитино-
идные трубочки современных сцифомедуз Stephanoscyphus из отряда Coro
nata очень напоминают конулярий.
Конулярии - фоссилии небольших размеров от 0.3 до 15 см в длину. Их ча-
сто находят в черных сланцах, и весьма вероятно, что они были преимущест-
венно планктонными животными.
ГИДРОКОНОЗОА, ИЛИ HYDROCONOZOA
Гидроконозоа представлены известковыми кубками конической или поч-
ти цилиндрической формы высотой до 1-2 см (фиг. VII.5). Кубки толстостен-
ные с воронковидным или цилиндрическим углублением в верхней части.
У рода Hydroconus кубок пронизан сложной системой каналов, радиальные
из которых создают рисунок, напоминающий септы. У рода Gastroconus пла-
стинчато-слоистое строение кубка напоминает днищеподобные структуры.
Гидроконозоа, вероятно, образуют самостоятельный класс.
Гидроконозоа широко развиты в раннем кембрии Сибири, Тувы, Монго-
лии, Канады и Австралии. Последние находки гидроконозоа обнаружены в
меловых отложениях.
ПРОСТЫЕ КИШЕЧНОПОЛОСТНЫЕ: ТИП CNIDARIA
111
Фиг. VII.5. Среднекембрийские гидроконозоа Тувы
а, б - Hydroconus в поперечном и продольном сечениях, увел, около х8; в, г -
Gastroconus в поперечном и продольном сечениях, увел, около х8 (Кордэ, 1963;
Бондаренко, Михайлова, 1984)
КОРАЛЛЫ И ИХ СЕМЬЯ: ANTHOZOA
Когда-то кораллы рассматривали как растения или как Zoophyta, то есть
животные-растения. На самом деле они принадлежат типу Cnidaria классу
Anthozoa, существующему только в полипоидном варианте; стадия медузы
отсутствует. Представители Anthozoa имеют трубкообразную глотку, кото-
рая ведет от рта в огромную центральную полость, выполняющую функцию
желудка. Щупальца полые, как у гидры. Одни, как, например, морские ане-
моны, не имеют скелета, другие строят роговые и известковые - “каменис-
тые” скелеты.
Наиболее древняя находка животного, предположительно относящегося
к кораллам, приурочена к нижнекембрийским породам Белых гор Калифор-
нии. Там обнаружены ископаемые Paiutiida, представленные маленькими
(1-5 мм в длину) фосфатными, трубковидными структурами с билатераль-
ным расположением перегородок, напоминающих септы. У них отсутствуют
стенные поры, осевые структуры и диссепименты, но общее строение совпа-
дает с коралловой архитектурой. Тем не менее помещение их в Anthozoa до
сих пор убедительно не доказано.
Другие древние находки происходят из среднего кембрия Алтая (род Bija)
и Австралии (род Cothonion). Первая находка предположительно отнесена к
табулятам аулопоридам, вторая — к ругозам.
Строение ругоз. Роговидные палеозойские кораллы принадлежат руго-
зам, называемым так потому, что их поверхность обычно покрыта морщин-
ками (от лат. rugosus - морщинистый). Первые достоверные ругозы появля-
ются в раннем ордовике, они быстро становятся многочисленными по всему
112
КАМЕННАЯ КНИГА
Фиг. VII. 6. Одиночные ругозы
а-г — Lambeophyllumprofundunr, примитивный коралл с септами, но без днищ:
а — внешний вид, б, в — поперечные сечения на разных стадиях роста, г — про-
дольное сечение, средний ордовик, Северная Америка; д, е — Streptelasma сог-
niculum с септами и пузырчатой тканью, средний ордовик, Северная Америка;
, ж, з — Breviphyllum cliftonense, поперечное и продольное сечения, коралл с сеп-
тами, днищами и зоной пузырей, девон, Теннеси, бс - большие септы, гл -
главная септа, дн — днища, мл - малые септы, пз - пузырчатая ткань
свету; на границе перми и триаса ругозы вымирают. В качестве примитивно-
го варианта строения можно выбрать род Lambeophyllum, который появляет-
ся в среднем ордовике (фиг. VII.6). Это громоздкое название дано для экзо-
скелета маленького одиночного коралла. Род Lambeophyllum представлен
слегка изогнутым кальцитовым конусом, называемом обычно кораллитом.
В верхней части он имеет углубление - чашку, в которой находилось тело по-
липа. Конус покрыт внешним слоем, или эпитекой, несущей линии роста и
продольные борозды; последние соответствуют внутренним вертикальным
перегородкам, называемыми септами. Половина этих септ - короткие, а по-
ловина такие длинные, что некоторые достигают центра чашки.
Во многих книгах Lambeophyllum и других ругоз называют тетракоралла-
ми. Название становится понятным, если рассмотреть как закладываются
септы. Первые две септы начинаются недалеко от основания, они называют-
ся кардинальной (или главной) и противоположной. Затем появляются че-
тыре септы, расположенные как крылья, которые делят конус на неравные
сектора. По мере того, как коралл увеличивается в размере, постепенно по-
являются и другие септы, но только в четырех секторах; одни из них длин-
ные, другие — короткие. Такая закладка и расположение септ, а не поверхно-
стные морщинки, являются теми признаками, которые по существу отлича-
ют ругоз от других кораллов.
Таким образом, один из ранних тетракораллов - Lambeophyllum появился
как очень примитивный конус с септами, но без днищ; по-видимому, он бли-
зок по структурному плану к исходной форме. Его простой план строения,
подобный Hydra, является тем, который в дальнейшем мог усложняться; и
этот процесс начался, вероятно, довольно рано. Так, у среднеордовикской
Streptelasma ниже чашки развиты изогнутые пластинки. Осевые продолже-
ПРОСТЫЕ КИШЕЧНОПОЛОСТНЫЕ: ТИП CNIDARIA
113
Фиг. VII.7. Основные элементы строения одиночных ругоз
а — внешний вид с чашкой вверху, б — внешний вид двухзонной ругозы, час-
тично вскрытой, в - фрагмент кораллита трехзонной ругозы, имеющего кроме
днищ и диссепиментов и осевую колонну
ния септ достигают центральной части кораллита и, видимо, закручиваются
у основания чашки. У силурийских кораллов, похожих на Breviphyllum, до-
бавляется зона пузыревидных пластинок - диссепиментов вокруг днищ (см.
фиг.УП.6 ж, з). У некоторых ругоз около трех первичных септ развиваются
щели-ямки (фоссулы). Эти и дальнейшие усложнения показаны на
фиг. VII. 7.
Размножение и колониальный рост. Современные кораллы размножаются
половым и бесполым способами. Первый способ связан с яйцами и спермой,
продуцируемых железами, находящимися на мягких перегородках - мезен-
териях гастральной полости. Мезентерии, свисая вниз как полотнища, распо-
лагаются между складками основания полипа, формирующими септы. Яйца,
оплодотворенные сперматозоидами, развиваются в личинки. Они дрейфуют
какое-то время, прежде чем опуститься на дно. Дрейф личинок несомненно
дает им хороший шанс для широкого пространственного расселения.
Половое размножение древних кораллов, вероятно, было таким же, хотя
данных об этом не сохранилось. Многочисленные ископаемые, тем не менее,
четко демонстрируют бесполое размножение. При бесполом размножении
возникают новые особи за счет почкования или деления материнской особи,
а затем и за счет себя; в результате возникает колония (фиг. VII.8). У некото-
рых кораллов новые кораллиты развиваются в результате процесса, назван-
ного периферическим краевым размножением, или пристенным почковани-
ем. Периферическая часть материнского кораллита разделяется на секции,
или почки, которые начинают расти вверх и вбок. Родительская особь ино-
гда отмирает; в других случаях она становится меньше, но продолжает суще-
ствовать среди своих отпрысков. При осевом размножении родительский по-
лип разделяется на два и более отпрысков, которые, развиваясь в чашке ро-
дителя, полностью замещают его. При боковом размножении новые
кораллиты прорывают стенки старых, строя ветвистые колонии или увели-
чивая размеры массивных колоний. Бесполое размножение дает начало ко-
лониям, в которых дюжины, сотни или даже тысячи индивидуумов-полипов
формируют компактные массивные, ветвистые или сетчатые массы. Коло-
s. Каменная книга
114
КАМЕННАЯ КНИГА
Фиг. VII.8. Варианты бесполого размножения
а - краевое пристенное, Heliophyllum, средний девон, Нью-Йорк; б — боковое,
Lithostrotion. s.L, ранний карбон, Шотландия; в - осевое, род и вид не определе-
ны, средний девон, Луисвилль, Кентукки
Фиг. VII.9. Периодичность роста у одиночных ругоз (процессы “старения и
омоложения”)
а-в - Blothrophyllum decorticatum, х0.5: а - внешний вид, б, в - поперечное и
продольное сечения, средний девон, Северная Америка; г — Heliophyllum halli,
xl, средний-поздний девон, Северная Америка
нии тетракораллов достигали обычно в высоту 15 см, а в ширину 20-30 см.
(В Музее землеведения Московского универститета есть еще более крупная
колония.) Индивидуумы, или кораллиты, большинства колоний имеют ха-
рактерный облик и такое же внутреннее строение, как и их одиночные род-
ственники.
ПРОСТЫЕ КИШЕЧНОПОЛОСТНЫЕ: ТИП CNIDARIA
115
г
Cystiphylloides D
Фиг. VII. 10. Различные способы закрепления ругоз на субстрате (реконструк-
ции)
а - сохранение вертикального роста при частичном погружении в осадок,
Streptelasma и другие ругозы; б - крышечный коралл Goniophyllum с каблуч-
ком прирастания, силур, венлок; в - корневидные выросты у “Omphyma”, си-
лур; г — расширенное основание-фундамент с выростами у пузырчатого коралла
Cystiphylloides americanus. девон
На ископаемом материале обычно так же хорошо наблюдаются процессы
“омоложения” и “старения”. Все ругозы растут быстрее в одно время, и мед-
леннее - в другое, продуцируя у кораллитов утолщения и морщины разного
масштаба. Кораллиты в какой-то момент могут сильно уменьшиться, поте-
рять часть септ, а затем снова начать интенсивно расти (фиг. VII.9). Это про-
исходит иногда только раз в жизни, но у большинства видов такие измене-
ния циклически повторяются снова и снова в течение всей жизни полипа-ко-
раллита, что, по-видимому, отражает “пир и голод” по отношению к
поступающей пище.
Способы закрепления и защиты. Ругозы цилиндрической и узкоконичес-
кой формы держались прямо, и только их точечные начальные кончики по-
гружались в ил, который также накапливался и вокруг основания кораллов,
удерживая их от опрокидывания (фиг. VII. 10).
Лучший способ прикрепления связан с цементацией к плотным субстра-
там и скелетам животных, в том числе и к другим кораллам. Часть кораллов
формируют устойчивые основания - фундаменты с горизонтальными расши-
рениями. Другие одиночные ругозы, как и губки, имеют корневидные выро-
сты, удерживающие кораллы в вертикальном положении.
Некоторые ругозы решали проблему закрепления, оставаясь маленькими
или прекращая рост вверх. Другие, надстраивая свои кораллиты вверх очень
медленно, росли вширь, как распускающиеся бутоны. Они могли лежать
плашмя на илистом субстрате, не опасаясь быть перевернутыми.
Большинство роговидно изогнутых одиночных кораллов медленно опро-
кидываются по мере того, как растут, но, изгибаясь, держат верхнюю часть
тела вертикально. Те, которые постепенно изгибались в одном направле-
нии, становились роговидно изогнутыми (фиг. VII. 11). Другие, изгибаю-
щиеся то в одну, то в другую сторону, приобретали неправильно искривлен-
8*
116
КАМЕННАЯ КНИГА
Фиг. VII. 11. Свободнолежащие ругозы (реконструкции)
а — крышечный коралл Calceola, ранний-средний девон; б, в - роговидные ко-
раллы на взрослой (б) и ранней (в) стадиях роста; г, д - различно изгибающиеся
кораллы, меняющие направление роста в течение жизни: г — Caninia и другие
ругозы; д — Siphonophrentis gigantea, одна из крупнейших одиночных форм,
в начале роста прикрепление цементацией, затем под собственной тяжестью ко-
ралл опрокидывается, меняет направление роста и снова растет вверх; е, ж —
свободно лежащие ругозы
ные очертания. Один из хорошо изученных родов такого типа - это род
Siphonophrentis, достигающий в длину 18-61 см. В плотных поселениях ба-
нок он сильно изгибается или остается почти прямым, но на открытых про-
странствах он опрокидывается, перекатывается в разных направлениях и
меняет направление роста в каждой новой позиции. Кустистые и массив-
ные колониальные ругозы обычно имеют широкое основание, служащее им
опорой.
Ругозы защищали свое мягкое тело так же, как и современные полипы,
двумя способами: с помощью стрекательных “крапивных” капсул и при со-
кращении тела. Многочисленные “крапивные” капсулы, находящиеся в щу-
пальцах, в момент тревоги выстреливают, как гарпуны. Второй способ защи-
ты заключается в том, что в случае тревоги мускулы сокращаются, тело съе-
живается, втягивая щупальца, и полип погружается в чашку кораллита.
Кроме того, у некоторых ругоз развивались крышечки (одна у Calceola и
Rhizophyllum или четыре у Goniophyllum), подвижно соединенные с верхней
стороной чашки. Когда полипы таких кораллов сокращались, они захлопы-
вали крышки, пряча, таким образом, свое тело.
ПРОСТЫЕ КИШЕЧНОПОЛОСТНЫЕ: ТИП CNIDARIA
117
Cystiphylloides
н
Heliophyllum
Фиг. VII. 12. Некоторые одиночные ругозы
а, б - Siphonophrentis gigantea, внешний вид, х0.5 (а) и поперечное сечение, х0.5
(б), средний девон, Северная Америка; в - Aulacophyllum sulcatum, х2, средний
девон, США; г, д — Homalophyllum ungula, х2, вид сверху (г) и внешний вид сбо-
ку (д), сердний девон. Северная Америка; е-з — Macgeea solitaria, xl, внешний
вид сбоку (е), поперечное (ж) и продольное сечения (з), поздний девон, Северная
Америка; и~л — Cystiphylloides americanus, внешний вид, xl (и), поперечное, х2
(к) и продольное сечения, х5 (л), ранний-средний девон, Северная Америка;
м,н — Heliophyllum obconicum, поперечное, х2 (м) и продольное сечения, х5 (и),
средний девон, Нью-Йорк
118
КАМЕННАЯ КНИГА
Eridophyllum Synaptophyllum
Фиг. VII. 13. Некоторые колониальные ругозы довона и карбона
а-е — кустистые колонии из изолированных кораллитов, соединенных только в
местах почкования и перемычек: а-в - Eridophyllum serials, внешний вид (а),
поперечное (б) и продольное (в) сечения, xl, средний девон, Кентуки; г-е - Syn-
aptophyllum simcoense, поперечное (г), продольное (д) сечения, х4 и внешний
вид (е), х2, средний девон; ж—к — массивные колонии из плотно примыкающих
кораллитов без просветов между ними: ж, з — Hexagonaria percarinata, внеш-
няя поверхность колонии (ж) и поперечное сечение (з), средний девон; u — Phil-
lipsastraea, поздний девон, Айова; к — Petalaxis stylaxis, трехзонный коралл, у
которого, кроме септ, имеются днища, диссепименты и осевой столбик, средний
карбон, московский век, Европа
ПРОСТЫЕ КИШЕЧНОПОЛОСТНЫЕ: ТИП CNIDARIA
119
Срезы-шлифы. У ругоз, или тетракораллов, когда-то изучали только
внешние характеристики, особенно те, что были видны в чашке. В настоя-
щее время большинство видов и родов установлены по внутреннему строе-
нию, которое расшифровывают, делая тонкие поперечные и продольные сре-
зы-шлифы через колонии или одиночные кораллиты. Такие срезы раскрыва-
ют много особенностей, невидимые на внешней поверхности, которые можно
использовать, хотя чашки уничтожены или забиты породой.
Многие направления (тренды) эволюции ругоз, раскрытые после изуче-
ния тонких срезов, оказались связанными с построением сложных и проч-
ных скелетов за счет развития диссипиментов, осевых структур и различных
колониальных конструкций (фиг. VII. 12, 13).
ШЕСТЬ ПЕРВЫХ СЕПТ: СКЛЕРАКТИНИИ
Склерактиний также называют гексакораллами, т.е. шестилучевыми ко-
раллами. Название указывает на то, что число септ и мезентерий основано на
цифре 6, с последующим возрастанием этого числа до 12, 24 и далее. Строе-
ние этих септ имеет важное значение для классификации гексакораллов.
Известно огромное число вариантов строения скелета. Некоторые гексако-
раллы являются одиночными организмами, другие колониальными
(фиг. VII. 14), хотя некоторые колонии могут внешне походить на одиноч-
ных. Как правило, одиночные гексакораллы не имеют роговидного или не-
правильно изогнутого облика, как ругозы. Кораллиты в колониях могут
быть цилиндрическими или призматическими с округлыми и многоуголь-
ными поперечными сечениями, но и здесь много вариантов, вплоть до одно-
временного существования тех и других в одной колонии, как, например, у
современных ветвистых форм. Фоссулы не образуются, и первичные септы
можно узнать только благодаря тому, что они уже есть в основании коралла,
в то время как последующие септы появятся в растущем кораллите позже и
выше первых.
Остатки склерактиний впервые достоверно появляются в отложениях
среднего триаса, и фактически они единственные кораллы в триасовое вре-
мя. Уже к концу триаса они строят мощные рифы в Южной Европе, Юго-Вос-
точной Азии, Калифорнии и Аляске. Другие рифостроящие эпохи приуроче-
ны к средней и поздней юре, раннему и позднему мелу, третичному и после-
дующему времени, включая и современность. Большинство крупных
коралловых рифов, найденных в современных южных морях, существовали
начиная с плейстоцена, то есть они начали формироваться уже 2 млн. лет то-
му назад, а некоторые, возможно, и раньше.
В мезозойских отложениях Северной Америки кораллы обычно редки,
исключая рифы Калифорнии, Аляски и др. Сборы ископаемых склеракти-
ний лучше всего проводить в Западной Европе, в Крыму, на Кавказе и Сред-
ней Азии, где верхнетриасовые, верхнеюрские и меловые породы содержат
большое количество хорошо сохранившихся кораллов, которые жили и
строили огромные рифы на морских путях древнего Тетиса. В США одиноч-
ные кораллы сейчас весьма обычны в Мексиканском заливе, а колониаль-
ные формы (“рифостроители”) были широко распространены в миоцене и
более молодых отложениях. Некоторые из наиболее красивых кораллов
происходят из миоценовых слоев “Silex” у залива Тампа во Флориде. На
внешней поверхности видно, что чашки и септы кораллитов построены из
тонкого белого известняка. Когда кораллы разрушались и пропитывались
120
КАМЕННАЯ КНИГА
Северная Каролина I ермания Англия Archohelia vicksburgensis
Haimesiastraea coriferta Meandrina baretti Stylocoeniella armata
Алабама Ямайка Бикини
₽2 N. О
Фиг. VII. 14. Некоторые одиночные и колониальные склерактинии
кремнеземом, они приобретали полосчатое и концентрическое строение, ха-
рактерное для жеод.
Склерактинии, вероятно, произошли от группы бесскелетных анемонов
или от циатаксонид, небольшой группы одиночных ругоз очень простой ор-
ганизации, известной от ордовика по ранний триас. Некоторые палеонтоло-
ги не склонны рассматривать ругоз предками гексакораллов, потому что
между ними имеется много принципиальных отличий, включая способ при-
крепления личинки, характер заложения септ и их аранжировка на взрос-
лой стадии, состав и форма биокристаллов (кальцитовый у ругоз и арагони-
товый у гексакораллов).
Самые ранние гексакораллы были мелководными формами и преимуще-
ственно строителями рифов; они, вероятно, содержали симбиотические рас-
тительные зооксантеллы из динофлагеллят, которые, нуждаясь в хорошо ос-
вещенных водах для фотосинтеза, ограничивали распространение кораллов
на глубину. За этими герматипными, рифостроящими кораллами появились в
ранней юре агерматипные формы уже без растительных симбионтов, и они,
преимущественно одиночные формы, уже овладели морскими глубинами.
ПРОСТЫЕ КИШЕЧНОПОЛОСТНЫЕ: ТИП CNIDARIA
121
Favosites Alveolites
Фиг. VII. 15. Табулятоидеи: сотовые кораллы фавозитиды
а-д — Favosites, силур: а — призматические кораллиты с порами на стенках, б —
кораллиты с окаменевшими полипами внутри (увел.), ранний силур, Канада,
о-в Антикости (Copper, 1985); в — внешний вид массивной колонии, х0.5; г — по-
перечное сечение кораллитов, стенки прерываются там, где находятся поры; д,
е -Alveolites, д — внешний вид верхней поверхности со стелющейся колонией ау-
лопорид в центре, xl, е - Alveolites suborbicularis, поперечное сечение коралли-
тов при увеличении, поздний девон, Европа
МНОГОЧИСЛЕННЫЕ ТАБУЛЯТЫ
Внушительную группу палеозойских кораллов представляет подкласс,
называемый Tabulata или Tabulatomorpha (табулятоморфы). Табуляты по-
явились в среднем кембрии (раньше ругоз) и вымерли одновременно с ними
на границе перми и триаса. Все табуляты являются колониальными организ-
мами, состоящими из небольших, иногда крошечных кораллитов, которые
могут объединяться в цепочки или кустики, но обычно они так плотно при-
мыкают друг к другу, что образуют массивные колонии, где кораллиты при-
обретают вид призм с четырьмя и более, но обычно пятью или шестью граня-
ми. У них хорошо развиты днища, то есть пластинки, протягивающиеся от
стенки к стенке, как правило, горизонтальные, реже вогнутые воронковид-
ные или пузырчатые. Септы могут отсутствовать, но обычно они есть, но по
сравнению с ругозами малочисленные в виде шипов, реже низких гребней и
пластин. Стенки большинства массивных табулят пронизаны отверстиями,
называемыми стенными порами, которые соединяют внутренние полости
кораллитов. Одно время табулят пытались рассматривать как своеобразных
губок, включая их в класс Sclerospongiae, но после опубликования П. Коппе-
ром (Р. Copper) в 1985 г. о сенсационной находке окаменевших полипов у фа-
возитид от такой трактовки отказались (фиг. VII.15).
Трубчатые, сотовые и цепочечные табуляты. Среди табулят по типу соот-
ношения кораллитов выделяют три основные группы колоний: трубчатые,
122
КАМЕННАЯ КНИГА
Syringopora
Фиг. VII. 16. Табулятоидеи: аулопориды (а—г) и сирингопориды (д-ж)
а — Aulocaulis annectens, стелющаяся колония, использующая в качестве субст-
рата раковину брахиопод, х0.5, поздний девон, Нью-Йорк; б-г - Aulocystis jack
soni, xl, внешний вид кустистой колонии (б), поперечное (в) и продольное (г) се-
чения кораллита, средний девон, Северная Америка; д-ж — Syringopora retifor-
mis, поперечное (б), продольное (в) сечения, хЗ, и внешний вид кустистой
колонии (ж), xl, силур, венлок
массивные и цепочечные. Внешний вид колоний при этом самый разнообраз-
ный: инкрустирующий, стелющийся, розетковидный, кустистый, лепешко-
видный, полусферический, желваковидный, ветвистый и т.д.
У трубчатых табулят кораллиты изолированы друг от друга (фиг. VII. 16).
Они соединены или только в местах почкования (Auloporida), или между
кораллитами имеются также поперечные соединительные трубочки (Syrin
goporida). Аулопориды самые древние достоверные трубчатые табуляты,
они известны с среднего кембрия по пермь. От них произошли многие пале-
озойские кораллы, возможно, включая и ругозы. Род Aulopora представлен •
маленькими коническими или субцилиндрическими трубочками с толсты-
ми морщинистыми стенками, редкими септальными гребнями и несколь-
кими днищами или без них. Новые кораллиты возникают путем почкова-
ния, образуя стелющиеся цепочки, сетки или даже корки, которые высту-
пают над илистым субстратом или они прикреплены к раковинам,
ПРОСТЫЕ КИШЕЧНОПОЛОСТНЫЕ: ТИП CNIDARIA
123
строматопоратам и кораллам. Роды, близкие Aulopora и Aulocaulis, имели
кораллиты различной формы, растущие вверх короткими колоннами или
кустами (см. фиг. VII. 16 б-г).
Массивные колонии состоят из плотно примыкающих кораллитов,
обычно призматических, имеющих облик медовых сот (Lichenariida, Fa-
vositida). К массивным колониям можно отнести и те, у которых пространст-
во между кораллитами занято скелетными структурами в виде пузырей, тру-
бок и стержней (Heliolitoidea). В обоих случаях между кораллитами нет
пустых пространств. Лихенарииды были исходной группой, от которой про-
изошли сотовые кораллы-фавозитиды, цепочечные - хализитиды, “солнеч-
ные кораллы”-гелиолитоидеи и, возможно, тетрадииды и хететоидеи
(фиг. VII.15-18).
Фавозитиды самые известные сотовые кораллы; они появились в среднем
ордовике и просуществовали до конца перми, но особенно широко они были
распространены в силуре и девоне (см. фиг. VII. 15). Колонии Favosites состо-
ят из плотно упакованных призматических кораллитов, имеющих много-
угольные поперечные сечения и тонкие отчетливые стенки, по которым ог-
ромные колонии легко раскалываются. Маленькие поры пронизывают стен-
ки, свидетельствуя, что мягкое тело и внутренние полости соседних полипов
соединялись; днища хорошо развиты, они горизонтальные и протягиваются
от стенки к стенке. Известно много видов Favosites, имеющих разнообразные
колонии от небольших пластинчатых до крупных полусферических кониче-
ских, желваковидных и ветвистых. Некоторые образованы кораллитами од-
ного размера, но другие, как F. argus, имеют дифференцированные коралли-
ты: среди крупных находятся мелкие. Другие роды, похожие на Favosites,
отличаются следующими особенностями. Emmonsia, например, имеет не-
полные днища, которые на продольном срезе выглядят как шипы. Tham-
порога обладает очень толстой стенкой и ветвистой колонией. Кораллиты у
Alveolites имеют полулунное поперечное сечение, отчего поверхность коло-
нии напоминает чешую, и т.д.
Цепочечные, а точнее, трубчато-цепочечные кораллы хализитиды (Нal-
ii sitida) распространены от среднего ордовика по поздний силур, но особенно
они характерны для позднеордовикских и раннесилурийских отложений.
Род Catenipora состоит из маленьких трубочек овального поперечного сече-
ния, которые плотно примыкают друг к другу боковыми сторонами, образуя
ряды, называемые шеренгами или цепочками, особенно четкими в попереч-
ном сечении. Каждая трубочка имеет двенадцать гребневидных септ, чьи
края усеяны шипами. Цепочки покрыты морщинистой эпитекой. Halysites
напоминает Catenipora по общему виду, но отличается наличием маленьких
призматических кораллитов с прямоугольными поперечными сечениями,
расположенных между кораллитами с овальными поперечными сечениями
(см. фиг. VII. 18).
Шизокораллы (от греческого schizo - расщепляю, дроблю). Рост колоний
табулятоморфных кораллов происходит путем продольного деления и
почкования. Первый способ считают более древним и примитивным.
Исходный “материнский” кораллит как бы расщепляется вдоль; возникаю-
щие участки растут вверх и превращаются в новые кораллиты. Такой тип
образования колоний наблюдается у тетрадиид, хететид, некоторых лихена-
риид, фавозитид и частично у гелио литоидей. Наиболее ярко продольное
деление проявляется у тетрадиид, когда четыре длинные септальные плас-
тины, удлиняясь и соединяясь, в центре образуют стенки четырех новых
кораллитов (см. фиг. VII.17а, б).
124
КАМЕННАЯ КНИГА
ТАБУЛЯТОИДЕИ ХЕТЕТОИДЕИ
Tetradium Lichenaria Chaetetes
ГЕЛИОЛИТОИДЕИ
Khangailites Plasmopora
Фиг. VII. 17. Табулятоидеи (а-г), хететоидеи (д, е) и гелиолитоидеи (ж-н)
а, б - Tetradium fibratum, хб, поперечное (а) и продольное (б) сечения, хб, сред-
ний ордовик. Северная Америка; в, г - “Lichenaria” typa, поперечное (в) и про-
дольное (г) сечения, х4, средний ордовик, долина Миссисипи; д, е - Chaetetes
septosus, поперечное (б) и продольное (е) сечения, увеличение до х4, ранний кар-
бон, визе, Франция; ж-н — Khangailites, х0.7, поперечное (ж, и, л) и продольное
(з, л, м) сечения трех видов, последовательно сменяющих друг друга во време-
ни, поздний ордовик, ашгилл, Монголия (ж-м — Атлас фауны ордовика Монго-
лии, 1981); н - Plasmopora, s.l., х5, поперечное сечение, силур, венлок. Север-
ная Америка
Некоторые палеонтологи считали, что табулятоморфные кораллы, обра-
зующие колонии способом продольного деления, составляют единый под-
класс Schizocoralla. Естественность подкласса выглядит сомнительной, учи-
тывая происхождение и пути развития этих кораллов, универсальный ха-
рактер продольного деления и сочетание его с почкованием в одной и той же
колонии, как, например, у гелиолитоидей.
Табулятоморфные кораллы имеют важное стратиграфическое значение
для карбонатных толщ, но для различных интервалов характерны разные
ПРОСТЫЕ КИШЕЧНОПОЛОСТНЫЕ: ТИП CNIDARIA
125
Catenipora
Halysites
Фиг. VII. 18. Табулятоидеи: цепочечные кораллы хализитиды
а — Catenipora micropora, xl, внешний вид, поздний ордовик-ранний силур, Се-
верная Америка; б, в — Halysites amplitubatus, х2, поперечное и продольное се-
чения, силур, Канада, о-в Антикости
группы: для ордовика - тетрадииды, лихенарииды, хализитиды и гелио-
литоидеи; для силура — фавозитиды, хализитиды и гелиолитоидеи; для дево-
на - фавозитиды; для карбона - хететиды, сирингопориды и аулопориды.
В течение перми группа вымирает после долгого существования, измеряю-
щегося более чем 400 млн. лет.
ИСТОРИЯ ПРО РИФЫ И ИХ АРХИТЕКТУРУ
Когда люди думают о рифах, они сразу вспоминают кораллы. Кораллы,
конечно, являются главными, хотя и не единственными строителями тропи-
ческих рифов, таких как, например, Большой Барьерный риф Австралии.
В прошлом кораллы иногда были и не так важны, как сегодня, а в течение не-
которых периодов они даже не являлись частью рифовых комплексов.
Первые рифовые каркасостроители известны из докембрия, они появи-
лись более чем 2000 млн. лет тому назад. Они были представлены в основном
строматолитами, представляющими собой карбонатные постройки цианоби-
онтов, ранее считавшихся синезелеными водорослями. Насколько можно су-
дить, только цианобионты, бактерии и, возможно, водоросли, но не живот-
ные, строили эти массивные известковые бастионы-крепости.
Кембрийские известковые курганы-холмы, или биогермы, по архитекту-
ре мало отличаются от докембрийских рифовых сооружений. Цианобионты-
строматолиты в большинстве случаев по-прежнему доминировали, но их со-
провождали археоциаты и другие губкоподобные беспозвоночные живот-
ные. Перед окончанием кембрия, тем не менее, археоциаты исчезли и оста-
лись одни строматолиты.
В середине ордовика развивается растительно-животный комплекс в
Северной Америке и в Сибири, занимая экологические пространства, заня-
тые прежде археоциатами. Эти рифы созданы красными водорослями,
мшанками, строматопоратами, ругозами и табулятоморфными кораллами.
Это древнейшее известное сообщество коралловых рифов. К концу девона на
границе франского и фаменского веков это сообщество было “сметено” после
130 млн. лет существования.
126
КАМЕННАЯ КНИГА
После катастрофического кризиса среднего палеозоя развились другие
рифовые комплексы из ассоциаций брахиопод, мшанок, известковых губок
и хететид.
Потом в течение перми наступил другой кризис из-за глобального пони-
жения температуры. Огромные массы воды были заморожены в быстро рас-
тущих полярных шапках. Уровень Мирового океана упал, обнажая огром-
ные участки континентального шельфа. Так как рифы находились на конти-
нентальных шельфах, то их распространение в результате кризиса сильно
сократилось; наступило всесветное массовое вымирание.
В течение мезозоя рифы стали содержать огромное количество гексако-
раллов, которые впервые появились в триасе. Они жили, как всегда, бок о
бок с водорослями и строматопоратами - сферактиноидами и спонгиоморфи-
дами, а в мелу со странной группой коралловидных моллюсков - рудистами.
Рудисты, тем не менее, вымерли, как и динозавры, в течение мела.
В конце мела наступил всесветный коллапс многих сообществ. Понадоби-
лось более 10 млн. лет, пока рифовые сообщества оправились и реанимирова-
ли. Фактически коралловые рифы во многих районах мира почти неизвест-
ны в палеоцене. Точная причина этого кризиса однозначно не установлена:
возможно, она связана с сезонными колебаниями температур и количеством
осадков; или с исчезновением мелководных континентальных морей; или
причина — в столкновении Земли с астероидами или кометой.
В течение кайнозоя обилие и разнообразие строения коралловых рифов
усиливалось. Следует отметить, что облик эоценовых и более молодых рифов
поразительно совпадает с современными. Кроме того, многие эоценовые ро-
ды продолжают существовать и сегодня.
Итак, рифы существовали на протяжении более чем миллиарда лет как
специфические биоценозы и рельефообразующие структуры морской среды.
Временами они испытывали расцвет, а в другое время почти исчезали. Со-
вершенно ясно, тем не менее, что систематический состав сообщества орга-
низмов, строящих рифы, постоянно менялся в течение геологического
времени.
Глава VIII
МОХОВИДНЫЕ ЖИВОТНЫЕ:
ТИП BRYOZOA
• Многие мелководные банки среднего и позднего палеозоя перепол-
нены ископаемыми, которые кажутся окаменевшими гидроидами
или кораллами, пронизанными малюсенькими отверстиями. Неко-
торые образуют постройки, похожие на бисквиты или неправильные глыбы.
Другие напоминают веточки, прутики и изгибающиеся листья или образуют
корки на раковинах. Некоторые колонии плоские веерообразные или раз-
лично изгибающиеся вплоть до образования конусов и спирально навиваю-
щихся пластин. Для них характерно сетчатое строение, где крупные поры
представляют собой сквозные отверстия, а мелкие, находящиеся на извест-
ковых прутиках, - вместилища зооидов.
Эти ископаемые не являются кораллами или гидроидами; они даже не
книдарии. Они принадлежат типу, чьи современные представители живут во
многих пресноводных бассейнах, но особенно обильны в морях. Их обычное
название Bryozoa, что переводится как “мхи-животные”, или “моховидные
животные”, что связано с мшистым внешним обликом многих видов. В рус-
скоязычной литературе их зовут мшанками. Британцы называют их Polyzoa
(“многоживотные”), отражая факт их колониального существования.
Как ни кажется неправдоподобным, эти миниатюрные животные родст-
венны брахиоподам, так как оба имеют структуру, называемую “лофофо-
ром”, которую они используют при захвате пищевых частиц. Третья группа,
которая имеет одновременно лофофор и полость тела - целом, представлена
форонидами, червеобразными существами, живущими в трубках. Форони-
ды, возможно, очень близки предкам мшанок и брахиопод.
СТРОЕНИЕ МШАНОК
Несмотря на миниатюрность, индивидуум мшанки, т.е. зооид, организо-
ван более высоко, чем любой представитель книдарий. Их мешкообразное те-
ло содержит три зародышевых листка: те же три листка обнаружены у плос-
ких червей и у всех остальных многоклеточных, более сложных, чем губко-
вые и книдарии. Задний конец тела замкнут; другой незамкнутый (но это не
голова!) несет щупальца (лофофор-щупальценосец), располагающиеся коль-
цом или подковой вокруг рта (фиг. VIII. 1). Реснички на щупальцах колеб-
лются, создавая ток воды, который помогает переносить пищевые частички
© 1997. О.Б. Бондаренко, перевод на русский язык и изменения.
КАМЕННАЯ КНИГА
128
- Щупальца
- - Стенка
Пилорус
3 - Желудок
Рот
Анус
_ .Мускулы
ретракторы
М-й!
- -Зооеций, - -
или автозооеций
Акантозооеции,
или акантостили-
—Аценструла
Диафрагмы.
- - -Апертура,
или устье
Стенка - - -
Фиг. VIII. 1. Строение некоторых современных и ископаемых мшанок
а — схема строения мягкого тела; б — скелет кустистой стелющейся колонии
Stomatopora parvipora с первой ячейкой-анцеструлой; х25, эоцен, Миссисипи;
в — объемная реконструкция массивного скелета Dekayella из трепостомат, х90
ко рту. Пищеварение осуществляется в U-образной или V-образной трубке,
оканчивающейся анусом, находящимся за пределами щупалец несколько
ниже рта. Мшанки, чей анус находится внутри подковы щупалец - Entoproc
ta, — очевидно, образуют отдельный тип.
Хотя зооид мшанки не обладает головой или мозгом, он все же содержит
нервный узел между ртом и анусом. Мускулы, связанные с зооидом, вызыва-
ют его сокращение во время опасности. Другие органы распрямляют его,
когда зооид готов к питанию.
Каждая колония может продуцировать яйца и сперматозоиды, хотя это
может происходить неодновременно. Яйцеклетки после оплодотворения
развиваются в личинки, которые плохо плавают, но часто переносятся тече-
ниями на далекие расстояния. Многих личинок съедают другие беспозвоноч-
ные. Оставшиеся по мере оседания прикрепляются к морским водорослям,
раковинам или другим субстратам и дают начало новым колониям. Зооиды
выделяют различные покровы, мембраны и стенки, которые образуют ячей-
ки, коробочки, мешочки, трубочки и колбочки. Эти покровы могут быть хи-
тиновыми, кожистыми (часто называемыми “роговыми”) или известковыми
из карбоната кальция. Мшанки с известковыми стенками наиболее широко
распространенные ископаемые, их колонии изменяются от стройных ветви-
стых форм до массивных, более 60 см в диаметре. Но последние являются ис-
ключением, так как колонии мшанок более чем 10 см в поперечнике уже счи-
тают необычайно большими.
Стенки зооида формируют коробочки, мешочки, трубочки и т.д., этими
словами описывают их внешний облик. Скелет вокруг одного зооида называ-
ется “зооеций”, а совокупность зооециев образует скелет колонии - зоарий.
Живое существо внутри зооеция — это зооид или полипид. В колониях зоо-
еции наиболее крупных зооидов называют автопорами или автозооециями.
Зооеции меньших размеров называют “мезопорами” или “мезозооециями”.
Еще более мелкие шиповидные структуры получили термин “акантопоры”,
“акантостили” или “акантозооеции” (см. фиг. VIII. 1). В современной палеон-
тологической литературе названия “автопоры”, “мезопоры” и “акантопоры”
МОХОВИДНЫЕ ЖИВОТНЫЕ: ТИП BRYOZOA
129
уже не используют, т.к. они связаны со словом “пора” — отверстие, а не со
скелетом.
Как отличить мшанок от кораллов. Многие известковые мшанки напоми-
нают кораллы. Например, Stomatopora выглядит как миниатюрная Aulopo-
ra, Cyclotrypa напоминает Favosites', Polypora вызывает в памяти современ-
ные морские перья рода Gorgonia. Внешние отличия заключаются в строе-
нии и мелких размерах зооециев (не более 1 мм), а также в строении
колоний. Доказательством разницы между мшанками и кораллами служат
и ранние стадии развития колоний. Коралловые личинки развиваются пря-
мо в полипы, как только они прикрепятся и этот полип в процессе почкова-
ния или деления производит себе подобных. Личинки мшанок становятся
сначала округлой анцеструлой, от которой отпочковывается трубкообраз-
ный зооид. Он в свою очередь почкуется, образуя таким образом колонию.
Анцеструлу наиболее легко наблюдать у прикрепленных стелющихся труб-
чатых форм, таких как Stomatopora (см. фиг. VIII. 1), но при внимательном
исследовании их можно обнаружить и у других мшанок. Итак, округлый
первичный зооид — это то, что окончательно убеждает скептиков, что эти ис-
копаемые не являются кораллами.
Другим отличительным признаком'большинства, но не всех мшанок
является наличие овицелл, или полостей, где находятся яйца, превра-
щающиеся после оплодотворения в личинки. Они могут быть в виде само-
стоятельных увеличенных зооециев или в виде полых вздутий над авто-
зооециями.
КАК КЛАССИФИЦИРУЮТ МШАНОК
Хотя мшанки широко распространены, но в породах древнее среднего ор-
довика найдены единичные формы. Коллекционеры легко собирают мшанок
в глинах и сланцах, но из плотных известняков их извлекают с трудом.
Мнение, что мшанки не оправдывают затраченных усилий, опровергнуто
многими профессиональными палеонтологами. Просмотр старых книг пока-
зывает, что маленькими колониями пренебрегали, в то время как крупные по-
лучали громоздкое имя, как двухэтажный автобус: например, Stenopora fibro-
sa. Описания базировались на таких характеристиках, как форма и внешнее
строение. Впоследствии, особенно в течение XIX века, пришли к открытию,
что ископаемые со сходными внешними данными могут весьма отличаться в
поперечных и продольных тонких срезах - шлифах. Более того, эти шлифы
одинаково хорошо рассказывают нам о мшанках, заключенных как в глинах
и сланцах, так и в известняках. Благодаря тщательному изучению шлифов па-
леонтологи обнаружили более 4 тысяч современных и более чем 15 тысяч ис-
копаемых видов. Изученные виды мшанок принадлежат трем классам.
Класс Phylactolaemata. Филактолематы включают несколько десятков
видов, они известны с мезозоя по настоящее время. Современные формы оби-
тают в мелководье пресноводных бассейнов и в глубоководье некоторых
озер. Филактолематы отличаются от других мшанок тем, что все, кроме од-
ного рода, имеют подковообразный или U-образный лофофор, который
окружает рот и анус. Они также имеют маленький губообразный вырост,
нависающий над ртом. Некоторые колонии (зоарии) имеют вид цепочек или
покровов и наростов, состоящих из кожистых трубочек; другие представ-
лены червеобразными студнеобразными массами, достигающими в длину
20—25 см.
9. Каменная книга
130
КАМЕННАЯ КНИГА
Класс Stenolaemata. Эти мшанки — известковые морские формы, которые
богато представлены ископаемыми от ордовика по настоящее время. Лофо-
форы, несущие щупальца, распрямляются с помощью мускулов, которые,
действуя на мягкую внутреннюю стенку, поджимают тело и толкают его в
сторону фронтальной части зооида, заставляя лофофор распрямиться и рас-
пустить щупальца. Стенолематы обладают известковыми стенками.
Класс Gymnolaemata. Современные представители характеризуются на-
личием замкнутого кольца щупалец. Их лофофоры распускаются с помощью
мускулов, действующих на часть внешней стенки тела. Хотя некоторые ви-
ды живут в солоноватоводных или пресноводных водоемах, подавляющее
число являются морскими. Большинство гимнолематных мшанок строят из-
вестковые стенки, чем и объясняется факт их нахождения, начиная с ордо-
вика и по настоящий день.
ГЛАВНЫЕ ГРУППЫ ИСКОПАЕМЫХ МШАНОК
Наши знания о мшанках начинаются с маленьких ископаемых из нижне-
ордовикских пород России и Северной Америки. Русские находки (Profistu-
lipora) относятся к цистопоридам, в то время как североамериканские, веро-
ятно, относятся к трепостомидам. В среднем ордовике уже известны все от-
ряды стенолемат и гимнолемат.
Расцвет и широкое географическое распространение мшанок наблюдает-
ся в то же время, что и остальных сестонофагов. Некоторые палеобиологи
считают, что это связано с широким распространением благоприятных усло-
вий, которые приводят к эффективному питанию взвешенными пищевыми
частицами. Другие палеонтологи думают, что только глобальный климат,
приводящий к стабильным планктонным пищевым связям, к которым пред-
расположены сестонофаги как дополнительному источнику питания, вызы-
вает широкое распространение мшанок. Собственно говоря, принципиаль-
ной разницы в этих толкованиях нет, в обоих случаях говорится о благопри-
ятных условиях.
Сведения об ископаемых филактолематах скудные и противоречивые,
потому что они лишены известковых скелетов. Противоположная ситуация
наблюдается у стенолемат и гимнолемат. Стенолематы, вероятно, произош-
ли от гимнолемат и довольно рано. В ордовике, когда стенолематы первый
раз появились в геологической истории, они уже были довольно разнообраз-
ными и относились по крайней мере к четырем отрядам: Tubuliporata (рань-
ше называемые Cyclostomata или Cyclostomida), Cystoporata, Trepostomata
и Cryptostomata. Некоторые палеонтологи выделяют пятый отряд Fenestra-
ta, но он еще не всеми принят. Гимнолематы представлены отрядом Ctenos-
tomata, начиная с палеозоя, и еще одним - Chellostomata, впервые появ-
ляющемся в мезозое и чрезвычайно широко распространенном в современ-
ных морях.
Tubuliporata, или трубчатопоровые. Прежнее название этого отряда Cy-
clostomata Международной комиссией по мшанкам упразднено, т.к. оно
оказалось преоккупированным для одного из классов хордовых, куда вхо-
дят миноги и миксины. Мшанки, принадлежащие этому отряду, относи-
тельно крупные, разнообразные и сложные. В типичном случае автозоое-
ции представлены простыми известковыми трубочками с округлыми или
полигональными отверстиями (апертурами), которые не имеют ни гребня,
ни других признаков прикрытия. Стенки простые и тонкие, содержат мно-
МОХОВИДНЫЕ ЖИВОТНЫЕ: ТИП BRYOZOA
131
Фиг.¥П1.2. Отряд Tubuliporata (ордовик—современность)
а - стелющаяся кустистая колония Corynotrypa delicatula, х25, ордовик-мел,
Огайо; б — стелющаяся листовидная колония Flabellotrypa rugulosa, х15, девон,
Теннеси
го крошечных пор. Тубулипораты обладают крупными специализирован-
ными женскими зооидами (гонозооеции), в которых созревают эмбрионы
мшанок.
Тубулипораты появились в ордовике и с умеренным успехом стойко суще-
ствовали на протяжении всего палеозоя. Они оказались единственной группой
известковых мшанок, переживших кризис в конце палеозоя. В юрское время
тубулипориды стали процветать, достигнув своего зенита в меловых морях,
где стали доминантными мшанками, и дожили в сокращенном числе до насто-
ящего времени. Тубулипораты сильно отличаются по размерам и форме: от
простых цепочек из трубчатых или грушевидных зооеций до листовидных,
дисковидных, веерообразных и массивных или многоветвистых колоний, вер-
тикально стоящих на широких основаниях, прикрепленных к субстрату.
Наиболее простые варианты, как Stomatopora и Corynotrypa, состоят из
стелющихся трубочек, цементирующихся к раковинам, кораллам или даже
к другим мшанкам. Трубки разнообразно разветвляются, диафрагмы в них
отсутствуют (фиг. VIII.2; см. также фиг. VIII.16).
Некоторые палеонтологи как тубулипорную мшанку рассматривают Fis-
tulipora и сходные с нею роды. Они имели массивные, обычно желваковид-
ные и полусферические зоарии, довольно крупные, достигающие в попереч-
нике 40 см. Верхняя поверхность зоариев усеяна мелкими дырочками и ок-
руглыми возвышениями; нижняя поверхность покрыта морщинистым
слоем, напоминающим эпитеку кораллов и часто носящим такое же назва-
ние. Крупные трубчатые автозооеции отделены друг от друга пузыревидно
изгибающимися пластиночками, обычно ориентированными колоннами
(цистозооеции). Одна из сторон стенки каждой автозооеции утолщена и рез-
ко изогнута, образуя лунарий, который воспринимают как навес-покрышку
над частью апертуры или как утолщенное основание лофофора (фиг. VIII.3).
Края лунария иногда так резко изгибаются, что на поперечных сечениях на-
поминают септы кораллов.
Некоторые специалисты по мшанкам выделяют Fistulipora и ряд других
родов в отдельный отряд Cystoporata (Пузырчатопоровые), характеризую-
щийся цистифрагмами и лунариями.
Trepostomata, или измененноротые. Этот отряд практически палеозой-
ский, т.к. он появился в раннем ордовике и вымер в триасе. Все колонии тре-
постомат были массивными “каменистыми” и многие - крупными формами.
Благодаря этому они преобладают среди палеозойских мшанок по числу эк-
земпляров, но не видов.
9*
132
КАМЕННАЯ КНИГА
..-И- .
Фиг.УШ.З. Отряд Cystoporata (ордовик—пермь)
а, б — поперечное и продольное сечения Fistulipora s.l., х10, силур-пермь.
Изображен экземпляр из карбона Северной Америки, ав — автозооеций, л — лу-
нарий, ц — цистозооеций с цистифрагмами
Автозооеции трепостомат состоят из длинных призматических или ци-
линдрических трубок. Каждая трубка во внутренней части колонии (эндозо-
на) имеет тонкие стенки и редко расположенные диафрагмы, что свидетель-
ствует о быстром и почти непрерывном росте. Трубки во внешней части коло-
нии (экзозона), однако, имеют толстые стенки и часто расположенные
диафрагмы, которые отражают много остановок для “отдыха”. Кроме того, к
экзозоне автозооеции меняют ориентировку роста от вертикальной к гори-
зонтальной. Изменения, происходящие от эндо- к экзозоне и дали название
отряду.
Характеристики трепостоматных мшанок хорошо иллюстрируют ветвис-
тые колонии рода Dekayella (средний-поздний ордовик). Продольные разре-
зы показывают, что незрелые зоны содержат только крупные трубки, или ав-
тозооеции, обычно с тонкими стенками и редко расположенными диафраг-
мами. При достижении зрелости автозооеции утолщаются и косо
отклоняются наружу. Маленькие трубочки, называемые мезозооециями,
также развиты; их диафрагмы располагаются чаще, чем диафрагмы в авто-
зооециях (фиг. VIII.4; см. также фиг. VIII. 1 в). Поперечное сечение показы-
вает еще более мелкие трубочки (акантозооеции, или акантостили), чьи тол-
стые стенки обычно выступают как короткие шипики. Акантозооеции, воз-
можно, служили опорой для мягких тканей, а мезозооеции, вероятно, были
их вместилищами.
У некоторых видов Monticulipora мезозооеции отсутствовали, а аканто-
зооеции обычно были единичными. У Prasopora, имевшего бутонообразные
и конические колонии, один из видов которого иногда называют “смоляной
каплей”, автозооеции содержали полого изогнутые пластинки (цистифраг-
мы), которые окружали диафрагмы, очень напоминая диссепименты вокруг
днищ у роговидных кораллов - ругоз.
Некоторые трепостоматы, или трепостомы, имеют неполные или перфо-
рированные диафрагмы; другие, как Hallopora, не содержат их. У некото-
рых родов скопления мезозооециев образуют пологие впадины или плоские
участки, нарушающие общий облик поверхности, и поэтому они названы ма-
кулами, то есть пятнами. Привлекательная Constellaria имела вогнутые ма-
кулы, в которых автозооеции и мезозооеции образуют звездоподобный
рисунок. Эти “звездочки” напоминают маленькие цветы (см. фиг. VIII.4).
Фактически почти все виды трепостом были определены по строению, на-
блюдаемому в шлифах. Сейчас, когда эта работа в основном проведена, мно-
МОХОВИДНЫЕ ЖИВОТНЫЕ: ТИП BRYOZOA
133
Фиг.УП1.4. Отряд Trepostomata (ордовик—триас)
а, б— тангенциальное и продольное сечения ветвистой колонии рода Dekayella,
х10, этот экземпляр из среднего ордовика, Миннесоты; в, г — внешний вид и тан-
генциальное сечениеConstellaria constellata, xl.3 и 27, скопления автозооециев
и мезозооециев создают звездчатый рисунок, ордовик, Северная Америка
гие ископаемые мшанки можно определить уже по внешним признакам,
как, например, Constellaria, по звездочкам. Внутренние признаки также
можно узнать по пришлифовкам, слегка протравливая их слабой кислотой.
В результате выступают акантозооеции, мезозооеции, стенки и даже днища,
но, чтобы увидеть их отчетливее, необходимо хотя бы небольшое увеличе-
ние.
Трепостомы были очень разнообразны в ордовике и силуре, но позже ста-
ли угасать, исчезнув в триасе.
Fenestrata, или фенестраты. Многие породы с ордовика по пермь содер-
жат ископаемых, которые выглядят как сетки или миниатюрные копии
современных кораллов из группы морских перьев. Сетки образованы корот-
кими колбовидными автозооециями, растущими в один слой. Отверстия
(устья) автозооеций открываются только в одну сторону колонии; противо-
положная сторона остается “глухой”. Однослойность колоний фенестеллид -
это тот характерный признак, который заставляет рассматривать их как са-
мостоятельный отряд вне отряда Cryptostomata, куда их относили раньше
(у криптостомат колонии двуслойные и устья открываются на обе или на все
стороны колонии).
В основании “горлышка” (вестибюль) колбовидного автозооеция имеется
горизонтальный вырост (гемисепта). Некоторые специалисты думают, что
вестибюль является зрелой зоной зооеция, лежащей над гемисептой. Эти
зрелые участки не содержат диафрагм.
Отряд Fenestrata, как и Cryptostomata, появляется в ордовике, но дости-
гает своего пика в позднем палеозое, то есть в карбоне-перми. Большинство
134
КАМЕННАЯ КНИГА
Fenestella йв
Archimedes
Фиг.VIII.5. Отряды Fenestrata и Cryptostomata (ордовик—пермь)
а, б — Fenestella s.l. сетчатая колония с фронтальной стороны у верхней поверх-
ности (а) и у основания (б), х15, такие экземпляры встречаются в венлокских
породах Онтарио и Нью-Йорка; в — скошенное тангенциальное сечение Fenestel-
la retiformis от внешней поверхности (внизу) до основания (вверху), увеличейо,
ранняя пермь, Русская платформа; г, д — внешний вид сетчатой винтообразной
колонии Archimedes wortheni, х0.5, средний-поздний карбон, Иллийнойс; е -
Lyropora subquadrans, xl, карбон-пермь, долина Миссисипи; ж - Rhombopora,
х8, объемная реконструкция, где крупные отверстия соответствуют автозооеци-
ям, а гребни ромбов — акантозооециям; ав — устья автозооеций на фронтальной
поверхности, ан - полость автозооеций на глубине, к — киль, о - сквозные отвер-
стия между прутьями и перекладинами, ок - основание фронтальной поверхно-
сти, п — прутья, пк — перекладины
МОХОВИДНЫЕ ЖИВОТНЫЕ: ТИП BRYOZOA
135
Фиг.УШ.б. Отряд Ctenostomata (ордовик—современность)
а — Vinella repens, xl, нитчатая бесскелетная мшанка, оставившая пустоты на
раковине брахиоподы, xl, ордовик-мел, Миннесота; б — Ropalonaria venosa, му-
ляж колонии бесскелетной мшанки, полученный после “заливки” пустот, х25,
поздний ордовик; в - Elliasopora siluriensis, “звездчатая” бесскелетная мшанка,
х10, силур, Нью-Йорк
из них требует шлифования для точной идентификации. В позднем ордовике
появляются сетчатые кружевные колонии Fenestella (фиг. VIII.5). Они рас-
тут в виде веерообразных или конических колоний, прикрепленных к рако-
винам и другим субстратам с помощью широких оснований или структур,
напоминающих корни. Их сетчатые колонии представлены прутьями, со-
единенными перемычками. Устья автозооециев находятся только на одной
стороне колонии, называемой фронтальной.
Особенно замечательным был род Archimedes, у которого кружевная сетка
спирально отходила от центральной стержневидной колонны в форме широ-
ких отворотов винта (см. фиг. IX. 5г, д). Вся колония стояла вертикально,
опираясь на широкое основание. Такое строение было эффективным пище-
вым устройством, так как оно совмещало усилия лофофоров с энергией пото-
ка воды, двигающегося сверху вниз по винтовой спирали, обеспечивая таким
образом пищей индивидуумов колонии. Род Archimedes (карбон-нижняя
пермь) многочисленен в отложениях среднего карбона, найден в нескольких
верхнекаменноугольных формациях и последний раз встречен в породах ни-
жней перми.
Ctenostomata, или гребенчаторотые. Название отряда происходит от гре-
бенчатоподобной структуры, которая покрывает каждый зооеций после то-
го, как щупальца втянуты. Ктеностоматы, или ктеностомы, принадлежат к
примитивному, но до сих пор существующему отряду, впервые появившему-
ся в ордовике. Его зооеции почкуются от центральных стержней, или столо-
нов, и образуют пучки, кустики, цепи, сеточки или листовидные пластиноч-
ки. Зооеции студнеобразные или роговые, но столоны могут объизвествлять-
ся. Многие из этих мшанок со столонами и трубками погружены внутрь
раковин и скелетов кораллов. Ископаемые формы были полностью бесске-
летными, от них остались узкие щели и округлые отверстия. При заливке
этих полостей получили объемные формы. Их принадлежность к мшанкам -
дискуссионна, но, тем не менее, в справочниках они отнесены именно к это-
му типу.
У Vinella (ордовик—мел) столоны напоминают нити. Род Elliasopora (ордо-
вик-пермь) представлен сеткой из столонов, которые усеяны пузырями с по-
ристыми стенками, имеющими неясную функцию. Род Elliasopora (силур-
карбон) имеет овальные или грушевидные вздутия, которые образуют пучки
или разбросаны вдоль нитевидных столонов. Перечисленные мшанки, веро-
ятно, “прожигали” субстрат “кислотой”, высверливая таким образом раз-
личные пустоты (фиг. VIII.6).
136
КАМЕННАЯ КНИГА
Фиг.УШ.7. Отряд Cheilostomata (юра—современность)
а — современная мшанка Белого моря Hyppothoa hyalina, внешний вид колонии
при увеличении (рис. Д.В. Гражданкина); б - Steginoporella magnilabris, фраг-
мент современной колонии из Мексиканского залива; в — Floridinella vick.sbu.rgi
са, мел—современность, Алабама
Cheilostomata, или губоротые. Начиная с мезозоя и до современности
встречаются мшанки, чьи автозооеции представлены коробчатыми или бо-
ченковидными ячейками с дистальными устьями - апертурами (фиг. VIII. 7).
Стенки состоят из известкового или рогового материала либо из их комби-
нации. При жизни эти ячейки несут подвижные хитиноидные крышки
(оперкулы), которые действуют в противовес узким “губам”, закрывая
апертуры, когда тело сокращается и щупальца втягиваются. Изящные ко-
лонии представлены пленками, дисками и кустистыми образованиями с
уплощенными или членистыми веточками. Среди автозооециев встреча-
ются как стерильные, так и фертильные формы со вздутиями над аперту-
рой, где шло формирование яйцеклеток. Современные мшанки обладают
авикуляриями, своеобразными индивидумами на ножках, которые в по-
стоянном движении щелкают как клювы птиц, отпугивая хищников и
осаждающихся личинок. Вибракулы представлены жгутиками, которые
как хлысты рассекают воду. Когда их много, то они действуют одновремен-
но, отпугивая, как и авикулярии, хищников и личинок; они также удаля-
ют с поверхности колонии осадок. У одной из групп мшанок вибракулы в
нижней части колонии видоизменены до расширенных подпорок, что со-
здает возможность движения (ползающие мшанки).
Хейлостоматы, или хейлостомы, впервые появились в юре; в течение мела
они стали среди мшанок наиболее многочисленной и разнообразной груп-
пой. Отряд продолжал процветать в кайнозое, а в настоящее время его виды
намного превосходят число всех остальных отрядов. Экспансия хейлостом-
ных мшанок, видимо, связана с угасанием циклостомат. Коллекционер лег-
ко соберет современных хейлостомат после шторма в выбросах водорослей,
раковин и галек, к которым мшанки плотно прикреплены.
МОХОВИДНЫЕ ЖИВОТНЫЕ: ТИП BRYOZOA
137
ЗНАЧЕНИЕ МШАНОК ДЛЯ ГЕОЛОГИЧЕСКОЙ ЛЕТОПИСИ
Палеозойские роды и виды мшанок были приспособлены к определенным
условиям существования, поэтому их находят в определенных типах пород,
преимущественно в известковых сланцах или известняках. В результате их
считают “фациеконтролирующими” ископаемыми. Мшанки в целом не
очень пригодны как руководящие ископаемые. Тем не менее исключения из-
вестны, если палеонтолог изучает комплексы видов внутри одного типа
пород, особенно в хорошо развитой карбонатной последовательности. Рос-
сийские палеонтологи, например, успешно использовали мшанок при выде-
лении дробных биостратиграфических подразделений. Возможно, недоста-
точное использование этой группы в других частях мира отчасти отражает
отсутствие усилий в разработке таких схем. Некоторые меловые и третичные
мшанки имеют ограниченный временной интервал, что может оказаться по-
лезным для определения возраста пород, несмотря на общие тенденции раз-
вития в разных группах.
Несомненно, усиленная работа с мшанками в будущем увеличит их цен-
ность как “часов” и “отрезков времени”, но мшанки, вероятно, никогда не
приобретут такого значения, как планктонные фораминиферы, граптолиты
и другие пелагические группы.
Глава IX
БРАХИОПОДЫ-КРЕПЫШИ
• Среди ископаемых позднего докембрия обнаружено несколько бра-
хиопод. Некоторые ученые анализируют их природу, однако другие
полагают, что допалеозойские брахиоподы весьма проблематичны и
не предоставляют существенную информацию об этом типе царства живот-
ных. В палеозое после медленного старта они заняли место обитателей мел-
ководных морских бассейнов, аналогичное таковому у двустворок. На протя-
жении более чем 300 млн. лет брахиоподы процветали, а затем еще до начала
мезозойской эры начали приходить в упадок. В современных морях обитает
немногим более 200 видов.
В начале этого века брахиопод и мшанок нередко объединяли вместе и от-
носили к типу моллюскоподобных. В дальнейшем эти две группы были раз-
делены, несмотря на то что они располагаются в одной и той же основной ча-
сти родословного древа животных. Места распространения брахиопод как в
прошлом, так и в настоящее время более ограничены по сравнению с
ареалами обитания двустворчатых моллюсков. Им, по-видимому, всегда
требовалась проточная вода, не содержащая большого количества взве-
шенных частиц осадка, так как их щупальцеобразный лофофор, служащий
для улавливания пищи, не может достаточно хорошо функционировать в
мутной воде.
Возможно, тип Brachiopoda имел не одного, а несколько предков, и поэто-
му некоторые палеонтологи считают его полифилетической , а не монофиле-
тической группой. Наиболее близкими родственниками брахиопод являют-
ся обладающие щупальцами черви и форониды. Раковина защищает брахио-
под снаружи, лофофор создает ток воды, доставляющий пищу, а ножка
обеспечивает закрепление на дне.
КТО ТАКИЕ БРАХИОПОДЫ?
Брахиоподы являются исключительно морскими животными, мягкое те-
ло которых заключено в двустворчатую раковину. В этом отношении брахио-
поды похожи на мидий, но в то же время их нетрудно отличить друг от друга.
Створки типичной мидии зеркально отображают друг друга; раковины
равностворчатые, так как могут быть разделены на две одинаковые полови-
ны (створки). Брахиоподы обладают неравными и непохожими створками.
© 1997. И. А. Михайлова, перевод на русский язык и изменения.
БРАХИОПОДЫКРЕПЫШИ
139
Фиг. IX. 1. Положение плоскости симметрии у двустворчатых моллюсков (а, б —
между створками) и брахиопод (в, г — поперек створок)
Чтобы разделить такую раковину на две парные половины, обе створки необ-
ходимо разрезать поперек (фиг. IX. 1).
Мягкое тело брахиопод (фиг. IX.2). Раковина, по существу, является без-
жизненной структурой. Живое тело брахиопод состоит из четырех основных
частей: мантии, лофофора, или рук, внутренних органов и ножки. Мантия
представляет собой широкую полосу, две половины которой покрывают ос-
тавшуюся часть мягкого тела и выделяют раковину. На концах мантии рас-
положена бахрома или волосообразные щетинки, которые у таких родов, как
Lingula, являются длинными и жесткими, а у остальных - короткими и мяг-
кими. Мантия также закрывает полость тела и внутренние органы,
последние располагаются, главным образом, в задней части раковины. Кана-
лы на мантии обеспечивают поток “водянистой” крови, об их направлении
свидетельствуют углубления на внутренней поверхности раковины.
В передней части полости тела под мантией находятся две руки. Руки в со-
вокупности называются лофофор, каждая из них состоит из двух оборотов
спирали, несущих по краям многочисленные щупальца. На них расположе-
ны изгибающиеся в разные стороны реснички, которые направляют ток во-
ды и приносят ко рту пищу в виде липкой слизи. Слизь с мелкими организ-
мами заглатывается ртом, находящимся у основания рук. Существует
мнение, что лофофор обеспечивает дыхание, но, вероятнее всего, эту функ-
цию выполняет мантия.
Фиг. IX.2. Строение внутренних органов брахиопод (я) и схема расположения
мускулов(б)
140
КАМЕННАЯ КНИГА
К внутренним органам относятся мускулы, пищеварительная система,
нервная система и железы нескольких видов; может присутствовать, а мо-
жет и отсутствовать сердце. Нервная система очень простая с неярко выра-
женными органами чувств. Круглый рот ведет в пищевод, далее следует
желудок, затем длинный или короткий U-образный слепой кишечник. Это,
естественно, означает, что отходы выбрасываются через рот, после чего за-
глатывается новая порция слизи.
Мускулы пересекают полость тела и прикрепляются к стенке раковины.
Открывание раковины никогда не бывает очень широким, даже когда бра-
хиоподы питаются, их створки раздвигаются на небольшое расстояние, об-
разуя щель.
Ножка представляет собой кожистое мускулистое образование, выходя-
щее между створками или через отверстие в одной из них. У примитивных
брахиопод ножка длинная и гибкая, ее мускулы обеспечивают сжатие или
даже закручивание. При удлинении ножки мускулы расслабляются и в нее
проходит жидкость из полости тела. У многих форм ножки короткие и негиб-
кие. Они прикрепляют своих владельцев к крупным объектам, так же как и
гибкие ножки незначительной длины. Удлиненные, почти червеобразные
ножки служат для зарывания в песок или мягкий ил.
Название brachiopoda (“рука-нога”) было придумано много лет назад, ког-
да считалось, что ноги моллюсков каким-то образом эволюционировали в ру-
ки брахиопод. Это не так, и данное название больше не имеет прямого значе-
ния, но английский вариант “лампочковидные раковины” - ненамного луч-
ше. Это название подходит только одной створке у таких родов, как Laqueus
и Сгапаепа, которые похожи на античный светильник с носиком для фитиля
(фиг. IX.3). Но данный термин не характеризует противоположную створку
и еще меньше соответствует раковинам клиновидной формы, слегка выпук-
лым, уплощенным, резко и разнообразно ребристым или чашеобразным, по-
крытым иглами. Лучшим решением в данном случае было бы не принимать
во внимание перевод и оставить название брахиоподы.
Фиг. IX.3. Строение раковины брахиопод
а — ламповидная брюшная створка, б — схема светильника с носиком для фити-
ля, в — спинная и г - брюшная створки изнутри, д — раковина со стороны смыч-
ного края, е — брюшная створка снаружи
БРАХИОПОДЫ-КРЕПЫШИ
141
Классификация, основанная на раковине. Брахиоподы можно классифи-
цировать по строению створок и ножки, микроструктуре стенки раковины
или характеру смычного края. Первый способ, вероятно, распределил бы
животных по двум группам, которые были бы близки, но ошибочно походи-
ли друг на друга. Микроструктура раковины позволит выделять подгруппу
раковин с пористыми створками и подгруппу с непористыми створками. Тре-
тья система классификации отражает более очевидное деление на основе на-
личия либо отсутствия структур, расположенных на смычном крае створок.
По такой классификации выделяют два класса: беззамковые - Inarticulata и
замковые — Articulata. И хотя такая система может быть не более значимой,
чем любая другая, большинство ученых используют эти две группы, приня-
тые и в данной книге.
У беззамковых брахиопод на смычном крае отсутствуют зубы. Такие фор-
мы были обнаружены среди древних ископаемых и дожили до сих пор, на-
пример, рода Lingula и Glottidia. У большинства беззамковых раковины тон-
кие, створки не скреплены связкой, как и у обычных съедобных моллюсков,
например устриц, и не имеют зубов на смычном крае. Створки при жизни мо-
гут смещаться из стороны в сторону, а после смерти животного обычно рас-
падаются. Обе створки могут быть очень тонкими, у ископаемых форм почти
плоскими.
У замковых брахиопод створки смыкаются между собой по прямой либо
изогнутой линии. Смычный край имеет два зуба на брюшной створке, им
соответствуют две ямки на спинной, что обеспечивает фиксацию створок.
У обоих классов на створках имеются различные внутренние и наружные
структуры (фиг. IX.4), очень важные для определения. Ниже дано их крат-
кое описание и иллюстрации.
Фиг. IX .4. Строение ручного аппарата брахиопод
а—г — петлевидный ручной аппарат: а, б - Dielasma, в, г - Beachia-, d—u — спи-
ральный ручной аппарат: d-ж - Martinopsis, з — Spirifer, u —Atrypa
142
КАМЕННАЯ КНИГА
Состав и структура раковины. У многих беззамковых брахиопод ракови-
ны хитиново-фосфатные. Это означает, что раковина образована фосфатом
кальция в комбинации с хитиновым материалом. Такие раковины всегда
тонкие и обычно глянцевитые. Последнее свойство является характерной
чертой, сохранившейся и у ископаемых форм, многие из которых имеют чер-
ный цвет. Два основных компонента могут быть смешаны или располагаться
чередующимися слоями, как полоски разноцветной бумаги.
Почти у всех замковых и у некоторых беззамковых брахиопод раковины в
основном состоят из карбоната кальция (кальцита). Такие раковины имеют
тонкий наружный пластинчатый слой и значительно более утолщенный
внутренний слой, образованный наклонными косыми волокнами (фибрами).
У непористых раковин оба слоя сплошные. Пористые раковины имеют поры,
оставленные нитевидными выростами мантии, продолжающимися в плас-
тинчатый слой. Ложнопористые раковины содержат в волокнистом слое та-
леолы кальцита, формирующие на поперечном разрезе пустулы, обманчиво
напоминающие настоящие поры.
Наименование створок. На протяжении многих лет ученые предполага-
ли, что одна створка брахиопод является верхней, или спинной, а другая
нижней, или брюшной. Эти термины используются во многих статьях и
учебниках.
Неправильность приведенных названий можно проиллюстрировать на
примере рода Lingula и других беззамковых брахиопод, зарывающихся в
илистые группы; их створки ориентируются передним концом вверх. Про-
шло значительно большее время, прежде чем был достаточно хорошо изучен
образ жизни замковых брахиопод. Тогда было установлено, что виды с ко-
роткой ножкой располагаются в любом положении и их “брюшная” створка
зачастую оказывается верхней. Прикрепляющиеся формы с длинной нож-
кой ориентируются достаточно разнообразно, они могут быть направлены
вверх, свешиваться вниз, при этом сверху находится любая из створок. Дру-
гие брахиоподы, в особенности ископаемые, могли изменять ориентацию
створок, так как с изменением пропорций тела животные разворачивались
до тех пор, пока педальная створка (с ножкой) не оказывалась в нижнем по-
ложении. Такое положение было также присуще большинству брахиопод,
утративших ножку и лежащих на дне, хотя роды, подобные Strophonella и
Strophomena, вероятно, лежали на брахиальной створке.
Приведенные факты превратили термины “спинная” створка и “брюш-
ная” створка в бесполезные и сфокусировали внимание ученых на других су-
щественных особенностях. В связи с этим створка, через которую у многих
брахиопод выходит ножка, стала называться педальной створкой. Вторая
створка, к которой прикреплялись руки, получила название брахиальной.
Эти термины применяются по отношению к примитивным беззамковым бра-
хиоподам, а также к специализированным родам замковых брахиопод.
ОТРЯДЫ БРАХИОПОД
Мы уже выделили среди брахиопод два класса: Inarticulata - беззамко-
вые, у которых отсутствуют зубы, и Articulata — замковые, у которых зубы
имеются. Первый из них насчитывает по крайней мере четыре отряда, а вто-
рой— восемь.
Класс Inarticulata. Беззамковые брахиоподы включают в себя четыре, а
возможно, и более отрядов, два из которых являются долго живущими: Lin
БРАХИОПОДЫ-КРЕПЫШИ
143
gulida nAcrotretida (фиг. IX. 5). Они представлены ископаемыми и современ-
ными формами. Остальные отрады ограничены ранним палеозоем, и их вза-
имоотношения с другими отрадами и между собой не достаточно ясны. Неко-
торые из них, возможно, могут и не относиться к брахиоподам.
Отряд Lingulida. Типичными представителями этого отрада являются
некоторые древнейшие брахиоподы и современная Lingula. Лингулиды не
имеют четкого отверстия для ножки, на поверхности обеих створок наблюда-
ются желобки, позволяющие ножке проникать между створками, либо нож-
ка может отсутствовать. Примером наиболее распространенной формы явля-
ется род Lingulella, для которого характерны продолговатая заостренная пе-
дальная створка и более короткая тупая брахиальная; смычные края обеих
створок слегка изогнутые. Некоторые роды имели почти круглые, в то время
как другие — овальные створки с субугловатыми макушками.
Формы подобные Lingula - наиболее распространенные современные
представители этого отряда — существуют с раннего кембрия и до настоящего
времени, устанавливая рекорд стойкой продолжительности жизни среди
животных. Они процветают почти 600 млн. лет. В палеозое этот род, видимо,
занимал более широкий географический ареал, обитая и в тропическом, и в
умеренном поясах, тогда как в настоящее время он приурочен только к тро-
пическим морям. Окружающая их среда могла быть значительно более раз-
нообразной, а не ограничивалась хорошим песчаным грунтом, как в настоя-
щее время.
Отряд Acrotretida. Представители этого отряда, существующего с ран-
него кембрия до настоящего времени, имеют либо фосфатные, либо известко-
вые раковины. Контур большинства акротретид круглый, либо почти круг-
лый с различной степенью выпуклости. Ножка присутствует и выходит че-
рез педальную створку, например у Acrotreta, либо полностью отсутствует,
как у рода Crania. Последние имели блюдечкообразную форму и прирастали
к субстрату непосредственно одной из створок. Кранииды являются наибо-
лее необычными беззамковыми брахиоподами, так как для них характерна
комбинация признаков замковых и беззамковых брахиопод, например, со-
став раковины.
Отряды Obolellida, Paterinida и Kutorginida. Эти три отряда беззамко-
вых брахиопод появились в самом начале кембрия и вымерли к концу этого
периода. Оболлелиды имели известковую двояковыпуклую раковину с ок-
руглым либо яйцевидным контуром. Существовала так называемая ложная
внутренняя арея, формировавшая изогнутую поверхность между макушкой
и задней границей педальной створки; ножка могла занимать различное
положение.
Раковины патеринид имели фосфатный состав, круглые либо овальные
створки с ложной внутренней ареей на обеих створках, а также с дельтирием -
выемкой под макушкой педальной створки, в которой располагалась ножка.
У них присутствовал и нототирий, представлявший собой выемку для выхо-
да ножки в брахиальной створке.
Самыми загадочными брахиоподами являются куторгиниды. Палеонто-
логи не имеют четкого представления о том, как их следует классифициро-
вать и, более того, куда их необходимо отнести: к замковым брахиоподам
или к беззамковым. Они имели известковые раковины и были первыми из
брахиопод, у которых присутствовали и дельтирий, и нототирий, но вдоль
смычного края отсутствовали зубы и выемки для них. По многим признакам
куторгиниды напоминают замковых брахиопод, поэтому на сегодняшний
момент они не могут быть помещены в один или в другой класс. Некоторые
144
КАМЕННАЯ КНИГА
Фиг. IX.5. Беззамковые брахиоподы (Hall and Clarke, 1892—1895)
а, б — Lingula anatina, xl, обе створки изнутри, видны мускульные отпечатки,
современная форма, Филиппины; в, г - Lingula elderi, х2, видны отпечатки вну-
тренних структур, средний ордовик, Миннесота, центральная часть США; д -
Dinobolus conradi, xl, педальная створка, ранний силур, центральная часть
США и Европа; е - Trimerella ohioensis, xl, брахиальная створка, ранний силур,
восток Северной Америки; ж - Trematis ottawaensis, xl, педальная створка,
средний ордовик, восток Северной Америки; з-к — Kutorgina cingulata, хЗ, ран-
ний кембрий, Вермонт; л — Lindstroemella aspidium, xl, педальная створка,
средний девон. Северная Америка; м — Schizambon canadensis, х2, педальная
створка, поздний ордовик, Онтарио и Нью-Йорк
БРАХИОПОДЫ КРЕПЫШИ
145
палеонтологи предполагают, что они вообще не являются брахиоподами, а
представляют собой одну из ранних групп уникальных беспозвоночных жи-
вотных, питавшихся с помощью щупалец.
Класс Articulata. Створки замковых брахиопод, в отличие от беззамко-
вых, смыкались жестко, на одной из створок имелись зубы, а на другой - со-
ответствовавшие им ямки. Замковые брахиоподы появились одновременно с
беззамковыми и незначительно изменялись до раннего ордовика, благопо-
лучно пережив период, когда несколько отрядов беззамковых брахиопод вы-
мерли (средний кембрий). К ордовику появилось большинство отрядов зам-
ковых брахиопод, только теребратулиды возникли в позднем силуре. Схема
временных рамок основных групп брахиопод дает некоторое представление
о увеличении и сокращении каждой из этих групп, а также показывает, на-
сколько сильно пермское похолодание воздействовало почти на все группы
замковых брахиопод. В течение палеозоя замковые брахиоподы бурно дивер-
гировали, на сегодняшний день они являются тенью прошлого, поскольку
выжили только три из восьми отрядов замковых брахиопод и два отряда из
беззамковых, но и они значительно сократились в численности.
Кратко охарактеризуем основные группы.
Отряд Orthida. Это достаточно разнообразный отряд, встречающийся с
раннего кембрия до поздней перми (фиг. IX.6). Для ортид характерны широ-
кие и, в основном, двояковыпуклые неравные створки с радиальной скульп-
турой, хорошо развитыми зубами, простым или разделенным замочным от-
ростком и отсутствием ручного аппарата (брахидия). Дельтирий и нототирий
обыкновенно открыты, но иногда могут быть закрыты специальными плас-
тинками. Начав с маленьких субквадратных форм раннего и среднего кемб-
рия, ортиды еще до окончания этого периода достигли значительного разно-
образия и размеров от 3 до 5 см в ширину. Позднекембрийские и раннеордо-
викские роды включали в себя разнообразные раковины с острыми и тупыми
макушками. Створки могли быть гладкими или несущими резкую скульпту-
ру в виде радиальных ребер.
Раковина этих брахиопод, за исключением нескольких представителей,
обычно непористая. По форме некоторые из них напоминают роды других
отрядов. Например, один род ордовикских ортид похож на строфоменид, а
другой развивал такую же форму и скульптуру, как в более позднее время
шипастые продуктиды. Род Platystrophia предвосхитил по форме и характе-
ру обитания отряд спириферид и даже прошел через сходные эволюционные
ряды видов.
Ортиды были наиболее ранними замковыми брахиоподами, и, возможно,
от них произошли и некоторые другие, например, пентамериды, строфоме-
ниды и спирифериды. Ортиды быстро дивергировали в ордовике, а далее в те-
чение оставшейся части палеозоя стали все более и более сокращаться.
Брахиоподы подотряда Dictyonellidina не обладают четко определенными
родственными связями с другими замковыми брахиоподами. Они появились
в среднем ордовике и просуществовали до перми. Род Eichwaldia, имеющий
шаровидную форму и специфическую треугольную пластинку на макушке
педальной створки, является типичным представителем данной группы.
Отряд Strophomenida. Эти брахиоподы из-за их уплощенной формы ча-
ще всего относят к так называемым “окаменевшим бабочкам” (фиг. IX.7).
Раковина строфоменид кажется пористой, но в действительности является
ложнопористой. Это означает, что под внешним слоем раковины располага-
лись кальцитовые столбики (талеолы), а не полые трубочки. Талеолы созда-
вали обманчивое впечатление пористости. Раковина имела широкий смыч-
10. Каменная книга
146
КАМЕННАЯ КНИГА
Фиг. IX.6. Отряд Orthida, все х0.9, кроме а, б (Hall and Clarke, 1892—1895)
а, б - Bllllngsella pepina, обе створки х2.5, поздний кембрий, долина Миссиси-
пи; в, г — Retrorsirostra carleyi, поздний ордовик, долина Огайо; д-з — Hebertella
sinuata, поздний ордовик, Огайо; и, к - Dinorthis pectinella, средний ордовик,
восток Северной Америки; л—п — Tropidoleptus carinatus, средний—поздний де-
вон, восток Соединенных Штатов и Нью-Мексико; р—т — Heterorthis clytie,
средний ордовик, Кентукки; у — Enteletes hemiplicatus, средний-поздний кар-
бон, Небраска и Техас
ный край, тонкую радиальную ребристость и одну створку слабовыпуклую,
а другую вогнутую. Отверстие для ножки было очень маленьким или вообще
отсутствовало, в этом случае отсутствовала ножка, и строфомениды были
свободно лежащими. У большинства отсутствовал ручной аппарат. Появив-
шись в раннем ордовике, они вскоре достигли большого разнообразия и ши-
рокого распространения, достигнув пика в позднем палеозое. После стреми-
тельной экспансии в ранней перми данная группа почти вымерла во время
БРАХИОПОДЫ-КРЕПЫШИ
147
Фиг. IX.7. Отряд Strophomenida, все х0.85 (Hall and Clarke, 1892-1895)
a-в - Strophomena planumbona, поздний ордовик, долина Огайо; г — Schuchertel-
la woolwortharia, ранний девон, запад Северной Америки; д-з — Stropheodonta
demissa, средний девон, Нью-Йорк и Пенсильвания, подобные виды встречают-
ся и в других регионах; и—м — Rafinesquina alternata, поздний ордовик, долина
Огайо; н-р — Leptaena rhomboidalis, силур—девон, Северная Америка
10*
148
КАМЕННАЯ КНИГА
позднепермского глобального кризиса, и только несколько видов продолжа-
ли существовать до ранней юры.
Несмотря на то что мягкое тело строфоменид было очень тонким, о чем
можно судить по маленькой полости раковины, эти формы обыкновенно
строили толстую раковину с выступающими отпечатками мускулов. Такие
раковины были широко распространены в палеозое; хорошо отпрепариро-
ванные педальные створки чрезвычайно привлекательны. У многих видов
имеются линии нарастания, показывающие, как изменялась форма с увели-
чением размеров.
Отряд Productida - группа, существовавшая с девона до поздней перми
(фиг. IX.8). Некоторые палеонтологи рассматривают продуктид в качестве
подотряда строфоменид. Брахиальная створка продуктид обычно плоская,
тогда как педальная створка сильно выпуклая. Продуктиды обычно имели
очень длинные шлейфы, например у Productella и Dictyoclostus, а педальная
створка имела тенденцию развивать длинные окончания или удлинения.
Дельтирий был преимущественно закрыт; ножка обычно отсутствовала.
Продуктиды процветали в значительной степени благодаря характеру оби-
тания, отличавшемуся от всех других брахиопод. Они неглубоко погружа-
лись в осадок - частично являясь “инфауной”. Такой характер обитания,
как и другие факторы окружающей среды, обеспечивал им защиту от хищ-
ников. Шипы, очевидно, играли значительную роль для закрепления в
осадке. К этой необычной группе принадлежал род Gigantoproductus, наи-
более крупная форма из всех брахиопод, достигавшая 21 см в ширину. Пик
расцвета продуктид приходился на поздний палеозой, когда многие группы
брахиопод либо вымерли, либо ареалы их распространения были жестко
ограничены.
Отряд Pentamerida. Эти двояковыпуклые непористые брахиоподы по-
явились в среднекембрийских морях, стали широко распространенными в
ордовике и просуществовали до конца девона (фиг. IX.9). Они имели корот-
кий смычный край, желобок для ножки и, за одним исключением, хорошо
Фиг. IX.8. Продуктиды и хонетиды, все х0.8 (Hall and Clarke, 1892-1895)
а, б- Marginatia burlingtonensis, ранний карбон, Аппалачи и долина Миссиси-
пи; в, г - Whidbornella lachrymosa, поздний девон, Нью-Йорк и Аппалачи;
д-ж - Devonochonetes carinata, средний девон, широко распространен в Север-
ной Америке
БРАХИОПОДЫ-КРЕПЫШИ
149
Фиг. IX.9. Пентамериды, все х0.85 (Hall and Clarke, 1892—1895)
a-в - Stricklandia casteliana, ранний силур, Айова; г - Virgiana decussata, ран-
ний силур, Северная Америка; д-ж - Pentamerus laevis, ранний силур, Север-
ная Америка; з, u - Pentamerella arata, средний девон, Онтарио, Нью-Йорк, Ин-
диана; к, л — Conchidium nysius, ранний силур, Кентукки; м-о-Anastrophia ver-
neuili, ранний девон, Гренландия, Нью-Йорк и Оклахома
развитый на педальной створке спондилий, который, по существу, был мес-
том прикрепления мускулов. На противоположной брахиальной створке ча-
сто располагалась структура, выполнявшая аналогичные функции — крура-
рий. У некоторых пентамерид имелись выступающие радиальные ребра и
складки (синус-седло). Силурийский Pentamerus laevis (часто называемый
Р. oblongus) селился плотными банками, ножка была погружена в ил, а удли-
ненные створки направлены вверх. Такие поселения часто встречаются с
внутренней стороны пластов мелкозернистого песчаника.
150
КАМЕННАЯ КНИГА
Р
п
к
Фиг. IX.10. Ринхонеллиды, все х0.85 (Hall and Clarke, 1892—1895)
a-в — Plethorhyncha speciosa, ранний девон, Квебек, Теннесси и Оклахома; г-е -
Orthorhynchula linneyi, поздний ордовик, Теннесси и долина Огайо; ж-и — Са-
marotoechia congregate, средний девон, Нью-Йорк и Индиана; к-о - Pugnoides
calvini, поздний девон, Айова; п-т - Hypothyridina venustula, средний девон,
Пенсильвания и Нью-Йорк
Отряд Rhynchonellida. Эти клинообразные и обыкновенно маленькие
брахиоподы, наиболее распространенные в некоторых девонских породах,
существуют со среднего ордовика до настоящего времени (фиг. IX. 10). У них
короткий смычный край, выступающие макушки и обыкновенно резкие ра-
диальные ребра. Часто имеется зигзагообразный синус — углубление на пе-
дальной створке, седло — складка на брахиальной створке, и всегда присутст-
вует ножка. Некоторые виды являются отличными руководящими ископае-
мыми (видами-индексами).
Отряд Spiriferida. Эти, как правило, двояковыпуклые брахиоподы,
существовавшие со среднего ордовика до юры, вероятно, произошли от рин-
БРАХИОПОДЫ-КРЕПЫШИ
151
Фиг. IX. 11. Атрипины (Hall and Clarke, 1892—1895)
а~г - Zygospira modesta, x2.5, поздний ордовик, Квебек, долина Огайо, Тен-
несси; d - Spinatrypa hystrix, х0.85, поздний девон, Нью-Йорк и Пенсильвания;
е -Atrypa occidentalis, х0.85, поздний девон, Айова и Иллинойс; ж-u -Atrypa sp.,
х0.85, средний девон, Онтарио и Нью-Йорк
хонеллид. Они объединены вместе спиральной структурой (брахидий), дав-
шей им свое название (фиг. IX. 11). Эта структура — спиральный ручной ап-
парат — поддерживала лофофор. По другим признакам спирифериды в зна-
чительной степени разнообразны. Древнейший подотряд Atrypidina имеет
раковины с коротким смычным краем, невыделяющимися макушками и с
вертикально или горизонтально расположенными спиралями. Скульптура
тонкоребристая или слабо шероховатая; некоторые роды имели широкие
“крылья”. У других форм тонкие слои изгибаются в шипы, которые легко
разрушаются. Дельтирий мог быть как открытым, так и закрытым, поэтому
одни животные закреплялись на морском дне, а другие вели свободнолежа-
щий образ жизни.
К этому подотряду относится Atrypa reticularis, одно время она считалась
обитавшей и в силуре и в девоне. Фактически A. reticularis представляет со-
бой комплекс множества вариаций и видов, ни один из которых не существо-
вал на протяжении целого периода, а многие из них настолько ограничены
по времени обитания, что служат в качестве хороших руководящих иско-
паемых (index fossils).
Раковины подотряда Spiriferina (фиг. IX. 12) в основном имеют характер-
ную спириферидную форму с коротким или длинным прямым смычным кра-
ем, выступающей макушкой и хорошо развитыми синусом и седлом. Спи-
ральный ручной аппарат более ориентирован к боковым сторонам створок,
152
КАМЕННАЯ КНИГА
Фиг. IX.12. Спирнферины, все х0.8 (Hall and Clarke, 1892—1895)
а - Eospirifer radiatus, ранний силур, восток Северной Америки; б — Cyrtospiri-
fer disjunctus, поздний девон, Нью-Йорк, Аппалачи, Юта; в - Neospirifer сате-
ratus, средний карбон, Пенсильвания, Техас; г, д - Cyrtia exporrecta, Ниагара,
Индиана, Кентукки; е, ж - Spinocyrtia granulosa, средний девон, Онтарио, Нью-
Йорк, Пенсильвания
чем внутрь брахиальной створки. Поверхность может быть складчатой, или
ребристой, или украшена более тонким орнаментом.
Некоторые спирифериды внешне напоминали таких ринхонеллид, как
Retziacea, которые известны с позднего силура до перми (фиг. IX. 13). Другие
представляют собой огромное многообразие форм, одни из них имели корот-
кий смычный край (Atrypidina), а другие гладкую раковину (Athyrididina).
В большинстве случаев только изучение внутреннего строения может предо-
ставить необходимую информацию о родственных взаимоотношениях этих
групп. Многие палеонтологи считают, что данные подотряды настолько от-
личаются от остальных спириферид, что их следует выделять в самостоя-
тельные отряды. Различие во мнениях относительно классификации будет
существовать и в дальнейшем.
БРАХИОПОДЫ-КРЕПЫШИ
153
Фиг. IX. 13. Атиридины, все х0.9 (Hall and Clarke, 1892—1895)
а, б - Merlstlna maria, поздний силур, Индиана, Теннесси, Нью-Йорк; в -Athy-
ris spiriferoides, средний девон, Аппалачи, Нью-Йорк; г—е - Ambocoelia umbona-
ta, средний девон, Онтарио, Кентукки; ж, з - Torynifera setigera, ранний кар-
бон, Алабама, Кентукки, Арканзас
Спирифериды были наиболее разнообразны в девоне и сохранили значи-
тельное многообразие до позднего триаса. После этого они быстро пришли в
упадок и вымерли в юре.
Отряд Terebratulida. Эти брахиоподы с пористыми раковинами, вероят-
но, появились в позднем силуре (фиг. IX. 14). Для них характерен короткий
смычный край и петлевидный ручной аппарат, поддерживавший лофофор.
Форма изменялась от почти круглой до удлиненно-овальной, створки глад-
кие либо тонкоребристые. У многих меловых и третичных теребратулид пе-
редний край раковины был крупноскладчатым. Дельтирий частично за-
крыт, и ножка выходит из раковины через круглое отверстие на макушке.
Теребратулиды достаточно широко распространены и в настоящее время,
особенно в Тихом океане. Некоторые виды обитают на скалистых берегах, в
то время как другие встречаются в глубоких, спокойных и холодных водах.
Несомненно, что в современных морях численность видов теребратулид зна-
чительно превосходит таковую у всех остальных отрядов.
Своеобразная группа маленьких замковых брахиопод с толстыми створка-
ми обособлена в подотряд Thecideidina', в прошлом их не раз относили к стро-
фоменидам, спириферидам или теребратулидам. Обычно, в связи с отсутст-
вием ножки, тецидные брахиоподы прикреплялись к какому-либо объекту
и, в отличие от большинства замковых брахиопод, очевидно, при добыче пи-
щи могли очень широко раскрывать свои створки. Ручной аппарат был
крючкообразный, изогнутый назад и направленный параллельно в сторону
рта, он подразделялся на множество долей и назывался птихолоф.
Эта группа, впервые появившаяся в среднем триасе, стала несколько бо-
лее разнообразной в позднем мелу, а к концу этого периода начала сокра-
154
КАМЕННАЯ КНИГА
Фиг. IX. 14. Теребратулиды, все х0.8 (Hall and Clarke, 1892—1895)
а, б - Amphigenia elongata, средний девон, Нью-Йорк, Кентукки; в - Rensse-
laeria ovoides, ранний девон, Нью-Йорк; г, д — Rensselaeria marylandica, ранний
девон, Мэриленд; е, ж - Dielasma bovidens, средний карбон—ранняя пермь, Мис-
сури, Техас
щаться. В незначительном количестве тецидные брахиоподы существуют и
в настоящее время (фиг. IX. 15).
ЕСТЕСТВЕННАЯ ИСТОРИЯ
Как и в случае с другими группами животных и растений, мы изучаем ес-
тественную историю брахиопод, сравнивая древние виды с современными,
исследуя слои, в которых они были обнаружены впервые, и изучая сами ис-
копаемые.
Глубины и места обитания. Многие брахиоподы существуют на глубинах
от 45 до 450 м, а другие процветают на глубинах от 760 до 5500 м. Большин-
ство из них обитало и обитает в настоящее время в мелкой воде или даже в
приливно-отливной зоне. Например, род Terebratalia, живущий на севере
тихоокеанского побережья Северной Америки, часто закрепляется на ска-
лах, обнажаемых отливом, в то время как Lingula и Glottidia зарываются в
грунт на песчаном мелководье. Нередко они вымываются волнами и иногда
БРАХИОПОДЫ-КРЕПЫШИ
155
Фиг. IX.15. Геологическое распространение в фанерозое основных групп бра-
хиопод (изменено D. Gelt no Clarkson, 1979)
выбрасываются в большом количестве на берег, простираясь на мили и на-
считывая миллионы животных. Несмотря на то, что мелководья нередко яв-
ляются солоноватыми и негостеприимными для многих обладающих рако-
винами беспозвоночных, современные лингулиды предпочитают именно их
в качестве места обитания. Они могут переносить значительные колебания
солености. На сегодняшний день лингулиды предпочитают песчаный грунт
и часто встречаются в богатых органикой эстуариях. Ископаемые из песча-
ника, имеющие следы потоков и волн, обнаруживают аналогичные особен-
ности характера обитания начиная с кембрия. Видимо, ранее, до триаса,
лингулиды могли переносить более широкий диапазон колебаний условий
окружающей среды. Они могли обитать в значительно более холодной воде,
например, в Сиднейском бассейне Австралии, недалеко от активно действу-
ющих континентальных ледников. А в настоящее время лингулиды ограни-
чены тропиками.
Закрытые мелководные эпиконтинетальные моря плалеозоя занимали ог-
ромные площади, где глубина редко была более 180 м, а обычно менее, чем
45 м. Здесь брахиоподы и другие беспозвоночные находили освещенную
солнцем спокойную воду и почти невероятные запасы микроскопической пи-
щи. Эти благоприятные условия отразились на разнообразии и многочислен-
ности брахиопод. Разнообразие хорошо иллюстрируется числом обнаружен-
ных родов, видов и подвидов или разновидностей, а многочисленность - ог-
ромным числом образцов и банок, заселенных так же плотно, как у
“голубых” устриц на современном побережье. Хорошим примером этого ти-
па являются банки Pentamerus laevis.
Нет сомнения в том, что многие древние лингулиды, как и их современ-
ные потомки, обитали на песчаном дне. Существует серьезное предположе-
ние, что в аналогичных местах распространения жили и некоторые строфо-
мениды с тонкими раковинами и, возможно, к ним присоединялись спири-
фериды с длинным смычным краем, острые окончания которого служили им
156
КАМЕННАЯ КНИГА
Фиг. IX.16. Ориентация и типы прикрепления брахиопод
а - Paraspirifer, в молодости был прикреплен, но ножка в зрелом возрасте исчез-
ла и раковина свободно лежала на илистом дне; б — Platystrophia также с возра-
стом утрачивала ножку и лежала на мягком илистом грунте, частично погружа-
ясь в него; в — стадии роста (1-5) и прижизненная ориентация раковины At гура-,
г — Tritoechia, ножка выходила через отверстие около макушки, с ее помощью
этот род прикреплялся к другим раковинам; д - Hesperorthis имел толстую нож-
ку, которая могла прикрпляться к раковинам или твердым грунтам; е — Schizo-
phoria использовала ножку на протяжении всей жизни, хотя педальная створка
временами, видимо, лежала на дне; ж — Brachyspirifer имел толстую ножку, с
помощью которой он, вероятно, на протяжении всей жизни прикреплялся к
крупным раковинам, кораллам и др.; з - Composita прикреплялась ножкой, ве-
роятно меняя положение подобно современной теребратуле; и, к - два экземп-
лялра современной Terebratalia, они прикрепляются короткой ножкой, но
педальная и брахиальная створки ориентированы различно; л - Barroisella, ко-
торая при помощи ножки зарывалась в грунт подобно современной лингуле
БРАХИОПОДЫ-КРЕПЫШИ
157
Фиг. IX. 17. Среднедевонские брахиоподы западного Нью-Йорка и Онтарио, по-
казывающие различную ориентацию в течение жизни
а - Spinocyrtia granulosa, б-Mucrospirifermucronatus, в - Meristella barrisi, г -
Athyris spiriferoides, d — Tropidoleptus carinatus, e — Pustulina pustulosa, ж — Рго-
ductella truncata, з — Atrypa reticularis, u - Camarotoechia sappho, к — Rhipi-
domella penellope, л - Stropheodonta demissa
якорем. В этом случае раковины захоронены в том месте, где они жили и
умерли. Многие брахиоподы, обнаруженные в известняках и сланцах, также
захоронены непосредственно вблизи места их обитания, поскольку на них
сохранились хрупкие складки, иглы и поверхностный рельеф, который дол-
жен был разрушиться при переносе в другое место. Некоторые слои содержат
только окатанные раковины либо сдавленные фрагменты. Это свидетельство
того, что волны разбивали мертвые брахиоподы на мелкие кусочки и переме-
шивали их с илом, или сгружали раковины вместе, напоминая современный
ракушняк, например, образованный раковинами современных съедобных
двустворок вдоль побережья Флориды. В других местах течения разъедини-
ли створки, оставляя брахиальные створки с одной стороны потока, а пе-
дальные - с другой. Таким образом, скопления ископаемых брахиопод (тана-
тоценозы) достаточно разнообразны.
Движение и закрепление. Личинки брахиопод пассивно плавают и дрей-
фуют, но расстояние, на которое они могут перемещаться, настолько ограни-
чено, что личинки, как правило, оседают стаями вблизи своих родителей и
сородичей. После этого брахиоподы прекращают передвигаться и всю остав-
шуюся жизнь проводят на одном месте. Наиболее продолжительную личи-
ночную стадию имеют лингулиды. Прежде чем осесть на дно, они около двух
недель обитают в пелагиали, свободно перемещаются и поэтому могут быть
достаточно далеко перенесены океаническими течениями.
Для закрепления на дне различные группы обладают специфическими
приспособлениями (фиг. IX. 16). Примитивные лингулиды имеют очень
длинную, упругую ножку, которая всверливается в песок либо ил. Конец
ножки покрыт вязкой слизью, к которой прилипают песчинки. Поочередно
сокращая и расслабляя ножку, животное перемещает свою раковину вверх и
вниз до тех пор, пока не выкопает маленькую норку (фиг. IX. 17). Во время
сбора пищи раковина примерно наполовину приподнимается из норки. Ма-
лейшее беспокойство приводит к сокращению ножки и втягиванию ракови-
ны внутрь норки.
158
КАМЕННАЯ КНИГА
Мы обнаружили, что штормы часто смывали слои песка, в которые зары-
вались лингулиды. Те, которые были выброшены на берег, погибали, а ос-
тальные зарывались снова. Обычно это осуществляется с помощью ножки, а
поврежденные животные могут использовать свои щетинки. Во втором слу-
чае они перемещаются на короткое расстояние, погружая щетинки в песок и
подтягиваясь боком вперед. Таким же образом животное, собравшееся от-
дохнуть и находящееся в составе банки, может оттолкнуть себя в сторону
свой собственной норки. Древние лингулиды, несомненно, обладали подоб-
ными особенностями обитания.
У некоторых брахиопод ножки были чрезвычайно маленькими. Напри-
мер, у рода Cyrtia, у которого ножка выполняла только функцию прикрепле-
ния, а не поддержки. Многие утратили ножку и были свободно живущими.
Другие, как, например, тецидные брахиоподы, прикреплялись к кораллам
или другим раковинам.
Ножка, по-видимому, присутствовала у ортид, спириферид, у некоторых
пентамерид и строфоменид, а в настоящее время имеется у большинства те-
ребратулид и ринхонеллид. У различных брахиопод ножка отличается по
длине и гибкости. Короткая и негибкая ножка Terebratalia удерживает обе
створки на скалах или раковинах, к которым они могут прикрепляться поч-
ти перпендикулярно. Такие ножки могли быть у ископаемых с толстыми ра-
ковинами. У родов типа Lagueus с тонкими створками ножка более длинная,
тонкая и гибкая, позволяющая раковинам свободно наклоняться. Некото-
рые современные виды прикрепляются к морским водорослям. Возможно,
что такие древние роды, как Oleneothyris, Rensselaeria и Beachia, поступали
аналогичным образом.
У многих палеозойских брахиопод имелись отверстия, соответствующие
очень крупным ножкам. Такие раковины носят следы легкой деформации либо
потертости, напоминающей следы закрепления ножки. Мы делаем вывод, что
эти брахиоподы обладали длинной ножкой, которая становилась упругой или
оставалась настолько сильной, что раковины удерживались достаточно высоко
над грунтом и крупными объектами, к которым прикреплялось животное.
В самом деле, несложно описать спириферов, имеющих резкие синус и седло и
крупное отверстие для ножки, в качестве форм, проявляющих атавистические
признаки. Вероятно, они, подобно роду Lingula, имели крепкую длинную нож-
ку, которая погружалась в ил, но удерживала раковину над дном.
Скорее всего, многие брахиоподы в раннем возрасте закреплялись при по-
мощи ножки, а в период зрелости утрачивали ее. Это позволяло раковинам
ложиться на дно либо частично в него погружаться. Роды Rafinesquina, Stro-
р/ютпепа и другие, имевшие вогнуто-выпуклую форму, вероятно, вели подоб-
ный образ жизни, покоясь на своих выпуклых створках. Род Hypothyridina,
возможно, располагался на своей умеренно изогнутой педальной створке,
как и другие роды сходной формы.
Разнообразные шипы и широкие линии нарастания удерживали взрослые
экземпляры рода At гура в одном положении, хотя не исключено, что некото-
рые из них могли перекатываться на брахиальные створки. Роды Paraspiri
fer и Platy strophia, вероятно, лежали на своих педальных створках, имев-
ших глубокий синус, и все глубже и глубже погружались в ил. На этих рако-
винах иногда поселялись мшанки, кораллы и другие инкрустировавшие
организмы, как бы впечатанные в створки, когда те подвергались атмосфер-
ным явлениям.
Развитые брахиоподы, так же как и примитивные, закреплялись на
твердых грунтах. Многие из них осуществляли это с помощью шипов и при-
БРАХИОПОДЫ-КРЕПЫШИ
159
растания одной из створок. У пермского рода Richthofenia прикрепляющая-
ся створка изменяется настолько, что становится похожей на одиночный ко-
ралл конической формы. Свободная створка уплощена и по форме напомина-
ет крышку.
Наиболее поразительная модификация ножки обнаружена у современно-
го рода Magadina, обитающего вдоль южного побережья Австралии. Эти
формы поселяются в зонах, где осадконакопление идет быстро и осадок часто
может покрывать живых брахиопод. Magadina избегает этой участи, ис-
пользуя свою ножку для быстрого и эффективного удаления от негостепри-
имного места.
Методика изучения. В 1880-х годах два палеонтолога отмыли и просеяли
огромную коллекцию силурийских ископаемых из местонахождения Wal-
dron, штат Индиана. Они обнаружили большое число раковин брахиопод,
размером от очень маленьких до огромных. Четыре вида представлены в пер-
вой части ученых записок Государственного музея Нью-Йорка, опублико-
ванных в 1889 году.
Методы отмывания и просеивания ископаемых значительно улучшились,
тем не менее собрано и описано всего несколько “жизнеописаний” (“историй
жизни”), подобных вышеуказанной из Индианы.
Повреждения на раковинах ископаемых. Многие современные брахиопо-
ды — выносливые животные. Лингулиды живут на омываемом волнами мел-
ководье, переживая штормы, наполняющие воду песком и уменьшающие
соленость. Род Terebratalia терпит обрушивающийся на нее прибой и после-
дующее осушение во время отлива. Этот род также поселяется в мелковод-
ных, освещенных солнцем бассейнах, куда во время шторма долетают толь-
ко брызги, а длительные периоды вода пузырится газом, который выделяют
разлагающиеся водоросли.
Древние брахиоподы, возможно, были столь же выносливыми, как и их
современные сородичи. Ископаемые свидетельствуют о том, что они пережи-
вали повреждения, вызываемые изгибом или даже расколом створок. Осо-
бенно хорошим примером является раковина, створки которой росли, оги-
бая мшанку, оказавшуюся как бы внутри раковины. Несмотря на искривле-
ние и невозможность нормального открывания створок, эта брахиопода
выжила и снова почти обрела свою обычную форму. Такие образцы, как и те,
которые показывают закрепление или изменение ориентации, являются по-
лезным ключом к познанию естественной истории древних брахиопод.
Глава X
ЧЕРВИ, СЛЕДЫ ЖИЗНЕДЕЯТЕЛЬНОСТИ
И ПРОБЛЕМЫ ПАЛЕОИХНОЛОГИИ
• В сборную группу червей, или Vermes, включают двусторонне-сим-
метричных беспозвоночных животных в составе типов: плоские
черви, немертины, круглые (или первичнополостные) черви и коль-
чатые черви. Некоторые исследователи выделяют из их состава самостоя-
тельные типы скребней и приапулид, а другие к червям относят онихофор и
целый ряд довольно подозрительных ископаемых. Для подавляющего числа
червей характерно удлиненное тело уплощенной или цилиндрической фор-
мы, с длиной намного больше ширины, иногда весьма значительно (напри-
мер, у немертин максимальная длина доходит до 30 м, а максимальная ши-
рина только до 9 мм). У большинства червей отсутствуют конечности, а пе-
редний и задний отделы слабо отделены от туловища. Из 44000 видов только
у 9500 видов, принадлежащих кольчатым червям и онихофорам, тело явля-
ется членистым, состоящим из множества сегментов - метамеров, числом от
5 до 600. Длина червей колеблется весьма значительно: от 0.01 мм до 30 м.
Среда обитания свободноживущих червей - моря, океаны, пресноводные
бассейны и почвы. Средой обитания паразитических червей и комменсалов
являются различные полости беспозвоночных и позвоночных животных,
растений и грибов.
Современные свободноживущие черви встречаются от тропиков до Аркти-
ки и Антарктики, преимущественно в морях и океанах, реже в пресновод-
ных бассейнах и еще реже в почвах. Большинство червей ведет донный (бен-
тосный) образ жизни, но некоторые являются пелагическими формами (по-
стоянно или спорадически во время личиночной стадии, реже во время
половой зрелости). Глубины обитания - от литорали до абиссали, включая и
зоны “курильщиков”. Донные черви ведут разнообразный образ жизни: по-
движный - ползающий, зарывающийся и неподвижный, обычно прикреп-
ленный. Все эти экологические группы существовали и в геологическом про-
шлом, о чём свидетельствуют многочисленные фоссилии. От ползающих
червей остаются борозды ползания, валики переваренного осадка после про-
хождения через кишечник, а также отпечатки тела во время отдыха и после
гибели. Борозды и валики получили название биоглифы (от греч. bios -
жизнь; glyphoi — письмена). От зарывающихся червей остаются ходы и норы.
Субстратом для зарывающихся современных форм служат различные грун-
ты: илы, пески, гравий, ракушковый детрит и смешанные осадки. Некото-
рые виды приспособлены к строго определенным грунтам. Более того, часть
© 1997. О.Б. Бондаренко и А.Ю. Розанов, новый вариант главы.
ЧЕРВИ, СЛЕДЫ ЖИЗНЕДЕЯТЕЛЬНОСТИ
161
из них оставляют столь характерные ходы, что по ним можно определить ви-
довую принадлежность. Все это надо иметь в виду при изучении следов зары-
вания геологического прошлого. Многие зарывающиеся черви являются
грунтоедами. Они используют осадок (обычно илистый) как источник еды,
пропуская его через кишечник и извлекая таким образом пищевые частицы.
Переваренный осадок по цвету и составу отличается от остального грунта,
так как благодаря пищеварительной ферментной деятельности кишечника
обогащается железом, магнием, фосфором, калием и кальцием. Процесс из-
менения первичного залегания и состава осадка называют биотурбацией.
Большинство илистых, особенно известковых, осадочных пород прошли
биотурбацию, поэтому их можно рассматривать как биогенные породы.
К особой группе зарывающихся червей относятся роющие формы, кото-
рые образуют норки и ходы в приповерхностном слое субстрата. Норки обыч-
но имеют узкоконическую, цилиндрическую, но обычно U-образную форму,
с двумя наружными отверстиями. Еще более специфической группой зары-
вающихся червей являются черви-сверлильщики, которые благодаря секре-
тирующей деятельности некоторых желез способны растворять известь и по-
селяться в раковинах моллюсков и в известковых образованиях водорослей.
Прикрепленные черви, строящие известковые трубки, известны только у
кольчатых червей, о чем более подробно будет сказано в соответствующем
разделе. Итак, от червей в ископаемом состоянии сохраняются отпечатки те-
ла, щетинки, следы ползания, зарывания и проедания грунта, полости оби-
тания и сверления в раковинах животных и известковых образованиях водо-
рослей, различные хитинизированные чехлики и известковые трубочки, а
также челюсти с хитиновыми зубчиками — сколекодонты. Отпечатки иско-
паемых червей и следы их жизнедеятельности с разной степенью доказатель-
ности известны с венда, сколекодонты - с ордовика.
В ископаемом состоянии плоские черви (тип Platyhelminthes) обнаруже-
ны внутри насекомых (паразиты?) в карбоне, перми, палеогене и неогене.
Круглые черви (тип Nemathelminth.es) были встречены внутри насекомых в
карбоне, мелу, эоцене, олигоцене и плейстоцене, а также у ископаемых чет-
вертичных млекопитающих, включая человека. Кольчатые черви (тип Аппе-
lides) встречаются начиная с венда. Приапулиды (Priapulida) в большом ко-
личестве обнаружены в среднем кембрии сланцев Бёрджесс; но их разнооб-
разный систематический состав указывает на более раннее появление в
геологической истории. Онихофоры (тип Onychophora, или Protracheata) из-
вестны со среднего кембрия. Немертины (тип Nemertini) и скребни (тип
Acanthocephala) в ископаемом состоянии пока не обнаружены.
УДИВИТЕЛЬНЫЙ МИР СЛАНЦЕВ БЁРДЖЕСС (СРЕДНИЙ КЕМБРИЙ)
Сланцы Бёрджесс (Burgess Shales) слагают горные гряды в восточной час-
ти Британской Колумбии (Национальный парк Йохо - Joho недалеко от ту-
ристских центров Банфф - Banff и озера Луиз - Louise). Честь открытия фа-
уны сланцев Бёрджесс принадлежит Ч.Д. Валькотту (Ch.D. Walcott). Нача-
лом открытия считают 1909 год. Тогда при спуске по склону горной гряды
Филд-Вапта к городу Филд (Field) лошадь задела камень, который привлек
внимание Валькотта. В камне оказались странные членистоногие прекрас-
ной сохранности. На следующий, 1910 год Валькотт установил местонахож-
дение коренных пород, откуда скатился камень на тропу. С тех пор сбор и
изучение фауны из сланцев Бёрджесс Валькотт не прекращал почти до самой
11. Каменная книга
162
КАМЕННАЯ КНИГА
Фиг. Х.1. Реконструкция некоторых среднекембрийских видов, обитавших на,
над и внутри илистых осадков (сланцы Бёрджесс Британской Колумбии), на-
капливающихся у подножья рифового уступа, показанного сзади
На рисунке соблюдены относительные размеры животных (Conway Morris,
Whittington, 1985): 1 - приапулида Ottoia; 2 - кольчатый многощетинковый
червь Burgessochaeta-, 3 - приапулида Louisella; 4 - хиолит Hyolithes; 5 - ползу-
ющие Aysheaia по губке Vauxia (22); 6 - членистоногое Leanchoilia; 7 — замковая
брахиопода Nususia; 8 - Opabinia, систематическое положение неясное; 9 - Di
nomischus, систематическое положение неясное; 10 - членистоногое Waptia;
11 - членистоногие Johoia; 12 - членистоногие Canadaspis; 13 — членистоногие
Mollaria-14 - хордовое Pikaia; 15- членистоногие Marrella; 16 - моноплакофо-
ры Scenella; 17 - членистоногое Sydneyia-, 18 - трилобит Ole.noide.s-, 19 - члени-
стоногие Burgessia; 20 - губка Pirania; 21 - иглокожие Echmatocrinus-, 22 - губ-
ка Vauxia-, 23 - Wiwaxia (?моллюски, близкие гастроподам); 24 - Anamalocaris
(?членистоногое); 25 - губка Choia
смерти. Свои удивительные находки прекрасной сохранности Валькотт
обнаружил в небольшой линзе темно-серых сланцев мощностью около 3 м
(8 футов). Здесь им было собрано около 40 000 экземпляров ископаемых жи-
вотных, представленных отпечатками и известковыми корочками. Такую
потрясающую концентрацию ископаемых в сланцах Бёрджесс объясняют
спецификой обитания биоценоза, приуроченного к илистому дну у подножья
рифового уступа, сложенного известковыми водорослями.
С сланцами Бёрджесс связан удивительный мир разнообразных ископае-
мых, представленных червями и червеподобными организмами, членисто-
ногими, сегментированными и несегментированными животными неясного
систематического положения, моллюсками, брахиоподами, хордовыми и
другими животными (фиг. Х.1). По числу экземпляров и биоразнообразию в
сланцах Бёрджесс преобладают черви, червеподобные организмы, членисто-
ногие и различные сегментированные животные, вероятно принадлежащие
новым типам. На первых двух группах остановимся ниже в этой главе.
ЧЕРВИ. СЛЕДЫ ЖИЗНЕДЕЯТЕЛЬНОСТИ
163
КОЛЬЧАТЫЕ ЧЕРВИ, ИЛИ ANNELIDES
Тело кольчатых червей, или кольчецов, состоит из головной лопасти,
длинного туловища и анальной лопасти. Длина кольчатых червей от долей
мм до 3 м. Туловище кольчатых червей, или Annelides (от лат. annelus - ко-
лечко) состоит из множества округлых сегментов — метамеров, за что они и
получили свое название. Число сегментов колеблется от 5—7 до 600. Сегмен-
ты туловища обладают почти одинаковым строением, представляя собой
своеобразные модули, последовательно соединенные друг с другом. У много-
щетинковых кольчатых червей {Polychaeta) каждый сегмент имеет по бокам
пару выростов кожно-мускульного мешка, оканчивающихся щетинками.
Выросты, или параподии, осуществляют двигательную функцию. У малоще-
тинковых кольчатых червей {Oligochaeta) параподии отсутствуют. Разделе-
ние многощетинковых на бродячих и сидячих является искусственным, от-
ражающим скорее образ жизни, чем систематическое положение.
Кольчатые черви являются свободноживущими формами, гораздо реже
комменсалами и паразитами. Они обитают в морях и океанах, реже пресно-
водных бассейнах и в почвах. Кольчатые черви, особенно многощетинковые,
из всех червей представляют наибольший интерес для палеонтологов и био-
логов. Считают, что предки аннелид дали начало онихофорам, членистоно-
гим, моллюскам, а возможно, и некоторым другим группам. Кроме того, сре-
ди ископаемых червей самыми распространенными являются кольчатые.
Ископаемые кольчатые черви. Любые находки ископаемых червей пред-
ставляют интерес, но особенно те, которые сохраняют строение мягкого тела.
К таким находкам относятся отпечатки, встреченные в среднекембрийских
сланцах Бёрджесс Канады и в верхнеюрских золенгофенских сланцах Герма-
нии (фиг. Х.2). Не меньший интерес представляют и ископаемые в виде перга-
ментообразных хитинизированных чехликов и известковых трубок, а также
крышечки трубок и челюсти с зубами-сколекодонтами (см. фиг. Х.4-6). Иско-
паемые следы передвижения грунтоедов, скорее всего, также относятся к
кольчатым червям, для которых характерно змеевидное или сочетание змее-
видного и прямого последовательного движения. Интересной группой коль-
чатых червей являются черви-комменсалы (симбионты), живущие вместе с
губками, строматопоратами, кораллами, моллюсками, иглокожими и други-
ми организмами. В колониях палеозойских кораллов черви-комменсалы жи-
вут между стенками или в стенках кораллитов, образуя различные по конфи-
гурации ходы и трубочки (фиг. Х.З). На Урале разнообразие и сложность та-
ких червей возрастает в эпохи рифообразования (поздний силур-девон).
В вендской (эдиакарской) биоте встречаются многочисленные отпечатки
сегментированных форм, такие как Spriggina, Dickinsonia и др., относимые
одними исследователями к типу кольчатых червей, а другими - к типу чле-
нистоногих (см. фиг. IV. 11). Они широко встречаются в эдиакарии Австра-
лии, венде Подольского Приднестровья, Архангельской области, Предура-
лье, Северной Америке и др. регионах. Из вендских пород преимущественно
методом химического препарирования были извлечены многочисленные хи-
тинизированные чехлики, длиной до 7-10 мм, шириной до 0.6-2 мм, сход-
ные по строению и составу с кольчатыми червями семейства Sabellidae. Кро-
ме того, в вендских отложениях встречаются многочисленные следы пере-
движения, часть из которых несомненно относится к кольчатым червям.
В раннем кембрии, кроме следов жизнедеятельности и хитинизирован-
ных чехликов Sabellidites, обнаружены узкие маленькие, но толстостенные
трубочки, состоящие из аморфного кремнезема {Platysolenites), особенно
11*
164
КАМЕННАЯ КНИГА
в Eunicites
Фиг. Х.2. Ископаемые многощетинковые черви
а, б - Canadia spinosa: а — отпечаток, х2, б - реконструкция, средний кембрий,
сланцы Бёрджесс, Канада (Walcott, 1911; Основы палеонтологии, том 2, 1962;
Gould, 1989); в — Eunicites atavus, х0.75, поздняя юра, Германия (Ehlers, 1869;
Основы палеонтологии, том 2, 1962)
многочисленные в синих глинах Ленинградской области и Эстонии
(фиг. Х.4).
В среднекембрийских сланцах Бёрджесс Канады обнаружены также мно-
гочисленные отпечатки разнообразных сегментированных форм, рассматри-
ваемых то как кольчатые черви, то как членистоногие. Одна из форм Cana-
dia spinosa считается бесспорным представителем многощетинковых коль-
чатых червей (см. фиг. Х.2). У нее туловище состоит из двадцати двух
сегментов, несущих короткие параподии с многочисленными щетинками на
концах. Параллельно параподиям развиты крупные вееровидные жабры.
Такое сочетание, параподии полюс жабры, довольно обычно для современ-
ных многощетинковых червей. Головной отдел среднекембрийской Canadia
несет две пары щупальцевидных отростков, вероятно, с глазами. Задний от-
дел представлен маленькой лопастью с недоразвитыми жабрами, к которой
примыкает туловищный сегмент с такими же маленькими жабрами. Из кем-
брия Австралии был описан отпечаток, сходный с червями семейства Тото-
pteridae, но, возможно, он относится к членистоногим.
Начиная с ордовика и по настоящее время встречаются хитиноидные че-
люсти с зубчиками (сколекодонты) и известковые трубки, достоверно при-
надлежащие кольчатым червям. Сколекодонты характерны для активных
хищных полихет, а известковые трубки для прикрепленных неподвижных
форм. Трубки имеют разнообразную форму и скульптуру, но чаще всего
встречаются неправильно изгибающиеся или спиральносвернутые формы.
Первых обычно относят к роду Serpula, а вторых - к Spirorbis, ведущими си-
дячий неподвижный образ жизни (фиг. Х.5).
ЧЕРВИ, СЛЕДЫ ЖИЗНЕДЕЯТЕЛЬНОСТИ
165
Фиг. Х.З. Палеозойские черви комменсалы в колониях кораллов-фавозитид
(Основы палеонтологии, том 2, 1962)
а, б - Camptosalpinx sibiriensis, х4.5, поперечное и продольное сечения, силур,
венлок, Красноярский край; в —Asterosalpinx asiaticus, х4, поперечное сечение,
ранний девон (?), Новая Земля; г — Streptindytes sp., хб, продольное сечение, си-
лур, поздний лудлов, Средняя Азия, Туркестанский хребет); д, е - Actinosalpinx
uralensis, Х1О, поперечное и продольное сечения, ранний-средний девон, вос-
точный склон Урала
Оба рода были установлены на современном материале. Ископаемые фор-
мы, относимые к ним, скорее всего представляют собой известковые трубки
других кольчатых червей, но установить это без изучения мягкого тела пока
невозможно. Тем не менее серпул указывают с силура, отмечая, что они наи-
более многочисленны в мезозое и начале кайнозоя; а спирорбисов - с ордови-
ка, отмечая их более широкое распространение в палеозое, чем в мезо-кайно-
зое. Известковые трубки Serpula, Spirorbis, Turbinia и других родов семейст-
ва Serpulidae имеют известковые крышечки, закрывающие верхнее
отверстие трубок. Крышечки также могут сохраняться в ископаемом состоя-
нии (фиг. Х.6). Трубки червей семейства серпулид имеют длину от несколь-
ких мм до 10-12 см, максимальный поперечник до 1 см. Они нередко
образуют заросли, банки и малые рифовые тела, формирующие впоследст-
вии органогенную известковую породу, называемую серпулитами.
Некоторые известковые трубки червей Serpulidae внешне можно спутать
с трубками улиток - Vermetidae, но на уровне микроструктуры они отлича-
ются. У большинства серпулид известковые составляющие во внешнем слое
(по отношению к поверхности трубки) располагаются дуговидно, а у верме-
тид - перпендикулярно.
Ископаемые агглютинированные трубочки встречаются с кембрия, но их
принадлежность к червям часто дискуссионна, так как подобные трубочки
166
КАМЕННАЯ КНИГА
Sabellidites
Фиг. Х.4. Трубчатые ископаемые венда и кембрия (Розанов, 1986)
а — Sabellidites, х15, венд, немакит-далдынское время, Сибирь; б - Anabarites,
х25, венд, немакит-далдынское время, Сибирь; в — Platy solenites, х9, ранний
кембрий, Европа
Фиг. Х.5. Известковые трубки кольчатых миогощетинковых червей, ведущих
прикрепленный образ жизни (Р. Vickers Rich, Т. Rich, М. Fenton, 1989)
а, б — Spirorbis: а — современная форма с расправленными щупальцевидными
жабрами, х15, б — ископаемая форма, увел.; в, г - ископаемые формы Serpula и
Hamulus, увел, около х2
могут строить простейшие и членистоногие. В тонкозернистых маастрихт-
ских известняках верхнего мела Поволжья были обнаружены крупные аг-
глютинированные трубки червей семейства Terebellidae, у которых в качест-
ве строительного материала использована чешуя и косточки рыб.
Паразитические многощетинковые черви известны с силура. Их присут-
ствие фиксируют по различным болезненным разрастаниям скелета. Но воз-
можно, эти разрастания принадлежат другим червям. Следы сверления мно-
гощетинковых червей достоверно известны с палеогена.
Малощетинковые кольчатые черви описаны из лагунных отложений ор-
довика-силура США, верхнеюрских морских горючих сланцев Поволжья и
континентальных образований палеогена Северной Америки и Европы.
ЧЕРВИ, СЛЕДЫ ЖИЗНЕДЕЯТЕЛЬНОСТИ
167
и
Turbinia
Фиг. Х.6. Трубки, крышечки и сколекодонты ископаемых кольчатых многоще-
тинковых червей
а, б - Turbinia abbreviata: а, б — поперечное сечение и внешний вид трубки сбо-
ку; в—д - крышечка при увеличении сверху, сбоку и снизу, мел, Западная Евро-
па; е, ж - схема расположения кристаллов в продольном сечении в трубке червя
Ditripula (е) и улитки Vermetus (ж), увеличено в несколько раз, современные
формы; з-н — сколекодонты палеозойских червей: з - Eunicites (D3), и — Arabel
lites (D3), к, л - Ildraites (Sjw), м - Lumbriconereites (SjW), и - Oenonites (Sjw),
увеличено в несколько раз (Основы палеонтологии, том 2,1962; Р. Vickers Rich,
Т. Rich, М. Fenton, 1989).
Наибольшее стратиграфическое значение из перечисленных фоссилий
имеют сколекодонты. Их ископаемые остатки оказались довольно многочис-
ленными и морфологически разнообразными, о чем хорошо свидетельству-
ют, например, фоссилии ордовика Польши и перми центральной части
США. Благодаря миниатюрным размерам количество сколекодонтов в керне
оказывается довольно представительным, а систематическое разнообразие
достаточным, чтобы использовать их как руководящие ископаемые.
ПРИАПУЛИДЫ, ИЛИ PRIAPULIDA
Приапулиды имеют несегментированное тело длиной от 0.2 до 15-20 см
при максимальной ширине до 4 см. В передней части тела имеется хоботок,
а конечная часть несет пучок жабр или они отсутствуют (фиг. Х.7). По осо-
бенностям строения приапулиды ближе всего к низшим червям, а именно к
типу круглых червей, куда некоторые исследователи относят их в ранге
класса. Тело приапулид покрыто рядами хитиновых крючков и шипиков,
168
КАМЕННАЯ КНИГА
Фиг. Х.7. Современные и ископаемые приапулиды
а — Priapulopsis bicaudatus, современная форма; б — схема пентагонального рас-
положения зубчиков в передней части тела Priapulus caudatus, современная
форма; в — Ottoiaprolifica в норке, ископаемая форма, средний кембрий, сланцы
Бёрджесс, Канада, Британская Колумбия (а, б - Conway Morris, 1977; в - Gould,
1989)
число которых кратно пяти - признак, общий с иглокожими. Приапулиды,
скорее всего, являются самостоятельным типом беспозвоночных.
В настоящее время выделяют около пятнадцати видов и семь-десять родов
приапулид, широко распространенных в мелководье морей умеренных по-
ясов (бореальных и нотальных); в меньшей степени они встречаются в глубо-
ководье тропиков и субтропиков. Современные приапулиды - свободно живу-
щие формы, но малоподвижные, преимущественно живущие в норках, кото-
рые они образуют, с силой “забуриваясь” в плотные песчаные или песчано-
глинистые субстраты. Способ внедрения в субстрат гидравлический со скоро-
стью 1 м в час. Большинство приапулид являются хищниками, хватающими
все, что плавает и ползает вокруг; некоторые являются детритофагами.
Интерес палеонтологов к приапулидам сильно возрос после того, как была
изучена среднекембрийская фауна сланцев Бёрджесс Канады (см. фиг. Х.1).
В этой фауне приапулиды более многочисленная группа, чем кольчатые мно-
гощетинковые черви. В кишечниках приапулид сланцев Бёрджесс были об-
наружены хиолиты и мелкие формы других приапулид, что указывает на
хищничество и каннибализм. Резкое уменьшение роли приапулид в совре-
менных морях свидетельствует о вымирании этой группы.
ОНИХОФОРЫ, ИЛИ ONYCHOPHORA
Онихофоры имеют сегментированное удлиненно-цилиндрическое тело
длиной до 15 см, шириной до 2 см. Число сегментов от 13 до 43. Каждый сег-
мент обладает парой членистых конечностей, заканчивающихся хитиновы-
ми коготками. Головной отдел практически сливается с телом, на переднем
его конце имеется два щупальцевидных выроста с глазами у основания. Рот
с хитиновыми челюстями находится на брюшной стороне. По бокам от рта -
два сосочка, выделяющие клейкую слизь.
ЧЕРВИ, СЛЕДЫ ЖИЗНЕДЕЯТЕЛЬНОСТИ
169
Фиг. Х.8. Среднекембрийская Aysheaia — вероятная онихофора (сланцы Бёрд-
жесс; Канада, Британская Колумбия)
а — реконструкция прижизненного положения Aysheaia, обитающих и собира-
ющих пищу на губках (Whittington, 1978), б-одна из реконструкций внешнего
обликаАувйеа/а (Gould, 1989; см. также фиг. XI.26)
Многосегментность, или полимерность, сближает онихофор с типом коль-
чатых червей и с многоножками типа членистоногих. Сторонники объедине-
ния онихофор с червями называют их бархатными червями, а сторонники объ-
единения с трахейными членистоногими называют их первичнотрахейными.
У онихофор, в отличие от кольчатых червей, конечности являются настоящи-
ми выростами тела и притом членистыми, оканчивающимися коготками. Уже
этих признаков, не говоря о других, достаточно, чтобы не считать их червями.
От трахейнодышащих многоножек и насекомых онихофоры отличаются стро-
ением тела и конечностей, иным составом хитиновой оболочки и т.д. Совре-
менные исследователи выделяют онихофор в самостоятельный тип первично-
трахейных — Protracheata, занимающий эволюционный уровень выше коль-
чатых червей, но ниже членистоногих (см. также главы XI и ХШ).
Вновь вспыхнувший интерес палеонтологов к онихофорам, как и в случае
с приапулидами, связан с изучением среднекембрийских сланцев Бёрджесс
Канады, где было найдено онихофороподобное беспозвоночное, названное
Aysheaia. У этого беспозвоночного членистые (или морщинистые?) конечно-
сти; рот занимает терминальное положение и окружен шестью-семью удли-
ненными сосочками (фиг. Х.8). Первая пара конечностей у головного отдела
Aysheaia оканчивается шестью шиповидными отростками. Следующие де-
сять пар конечностей туловища уплощены и несут бахрому крошечных кри-
вых коготков. Кроме того, у основания туловищных ножек развиты крупные
шиповидные выросты, направленные вперед, а у последних двух пар - назад
и т.д. Есть и другая реконструкция Aysheaia (см. фиг. XI.2). В любом случае
отличия между Aysheaia и онихофорами довольно заметные, не позволяю-
щие объединять их однозначно.
Aysheaia — морское существо, в отличие от наземных онихофор. Вместе с
Aysheaia обычно находят спикулы губок, поэтому предполагают, что они
170
КАМЕННАЯ КНИГА
обитали на их поверхности, собирая там пищу. По-видимому, Aysheaia мог-
ли заглатывать и мягкое содержимое губок.
ХИОЛИТЫ И ТЕНТАКУЛИТЫ - ЧЕРВИ?
Среди фоссилий встречается много ископаемых, чье систематическое по-
ложение не удается доказать однозначно, а можно только предположить не-
сколько вариантов интерпретации. Таких ископаемых принято называть
проблематиками. Хиолиты и тентакулиты - одни из них; обычно их относят
к червям или к моллюскам.
Хиолиты (Hyolitha) - морские организмы; они имели известковую уд-
линенно-коническую гладкую раковину длиной от 1 мм до 1.5 см. Раковина
с “брюшной” стороны уплощена, а со “спинной” дуговидно изогнута. Откры-
тый конец раковины закрывался крышечкой. У некоторых хиолитов внутри
раковины была обнаружена сегментация (?перегородки между воздушными
камерами, ?отделы кишечника и т.д.). Варианты интерпретации образа жиз-
ни хиолитов следующие: донные неподвижные свободно лежащие формы;
донные малоподвижные или активно ползающие формы (см. фиг. X, 14);
планктонные, а возможно, и нектонные формы, свободно обитающие в тол-
ще воды. Геохронологический интервал существования хиолитов ранний
кембрий-пермь.
Тентакулиты (Tentaculita), как и хиолиты, были морскими организ-
мами; они обладали известковой узкоконической сильно удлиненной рако-
виной, обычно несущей многочисленные концентрические ребра (Tentacu-
ities S.I.), реже раковина была гладкой (Styliolina s.l.). Длина раковин от 1 мм
до 2 см. Тонкие многочисленные концентрические ребра раковины законо-
мерно чередовались с более толстыми и редкими. Внутренняя полость рако-
вины была подразделена перегородками на камеры, напоминающие камеры
головоногих моллюсков, но без сифона. Тентакулиты, скорее всего, вели
планктонный образ жизни. Они, по-видимому, существовали “стаями”. Мас-
совое захоронение раковинок тентакулитов приводило к образованию тента-
кулитовых известковых прослоев, широко распространенных в девоне мно-
гих стран (Северная Америка, Западная Европа, в России и странах СНГ -
Подольское Приднестровье, Поволжье, Закавказье и др. регионы). Геохро-
нологический интервал существования тентакулитов: ордовик-девон.
СЛЕДЫ ЖИЗНЕДЕЯТЕЛЬНОСТИ И ПРОБЛЕМЫ ПАЛЕОИХНОЛОГИИ
Следы жизнедеятельности червей в современных морях более заметны,
чем следы других беспозвоночных. От подвижных червей-грунтоедов оста-
ются борозды ползания и валики субстрата, пропущенного через кишечник.
Кроме того, могут сохраниться отпечатки тела отдыхающего или умершего
червя. Борозды и валики имеют различную и часто довольно сложную фор-
му. Жизнедеятельность малоподвижных червей, находящихся в дуговидно
изогнутых и в U-образных норках, приводит к образованию на поверхности
субстрата воронковидных углублений и конусовидных холмиков. Углубления
приурочены к переднему концу червя, где находится его рот, а холмики — к
заднему концу, где расположен анус.
Ископаемые образования, сходные с вышеописанными, известны с докем-
брия. Первоначально их считали проявлениями только жизнедеятельности
ЧЕРВИ, СЛЕДЫ ЖИЗНЕДЕЯТЕЛЬНОСТИ
171
Фиг. Х.9. Ассоциации следов жизнедеятельности, обычно встречающиеся сов-
местно в ископаемом состоянии, и их значение для реконструкций условий оби-
тания (из A. Seilacher с изменениями. Trace Fossil Concepts by P.B. Basan et al.,
Soc Econ. Paleo and Mineral Short Course 5, S.E. P.M., 1978)
червей. Позднее, при изучении современных организмов, оказалось, что по-
добные образования оставляют и другие беспозвоночные и даже позвоноч-
ные и растения. Так, например, борозды могут возникнуть при волочении и
соприкосновении водорослей с субстратом, а также при передвижении дву-
створок и улиток. Валики, как конечные продукты деятельности кишечни-
ка, могут остаться от кого угодно. Роющих организмов и сверлильщиков
много и среди других беспозвоночных, а не только у червей. Конусовидные
холмики создают и некоторые рыбы, забрасывая свою икру осадком, кото-
рый они сгребают плавниками по радиусу к центру.
Объектами изучения ихнологии являются следы жизнедеятельности со-
временных, а палеоихнологии - ископаемых животных в основном с точки
зрения морфологии и происхождения (греч. ichnos — след; logos — учение,
знание). Проблемы палеоихнологии проистекают из следующего: принад-
лежность ископаемых следов жизнедеятельности только к какой-нибудь од-
ной группе организмов неоднозначна; одна и та же особь может оставлять
разные следы в зависимости от конкретного поведения в конкретной ситуа-
ции (передвигается быстро или медленно, находится в плотном или редком
поселении и т.д.); кодирование следов, поэтому, через латинские названия
по правилам зоологической номенклатуры, является произволом. Полного
решения этих проблем добиться невозможно, но свести их до минимума
можно. Основным ключом для решения является метод современного анало-
га, или метод актуализма, то есть знание следов жизнедеятельности совре-
менных организмов. Так, следы на поверхности осадка, отражающие посту-
пательное попеременно изгибающееся движение, характерны для червей.
Такое движение недаром называют червеобразным.
Изучение следов жизнедеятельности, особенно тех толщ, где отсутствуют
другие ископаемые, достигло больших успехов, несмотря на все проблемы
палеоихнологии. По морфологии (структуре) следов удается установить, в
результате какой функциональной деятельности (или бездеятельности) ос-
тавлены те или иные следы. По ассоциации следов устанавливают прежние
зоны моря (фиг. Х.9,10), а также тип усложнения поведенческих реакций во
времени, а отсюда и возраст вмещающих отложений (см. фиг. 1.6).
172
КАМЕННАЯ КНИГА
Нижний кембрий
Доломит
Пакистан
Ордовик-девон
Флиш
Португалия
Верхний триас
Песчаники
Германия
Фиг. Х.10. Примеры различных ассоциаций ископаемых следов (Treatise...
Part W., 1979 из A. Seilacher, 1955 с изменениями)
ЧЕРВИ, СЛЕДЫ ЖИЗНЕДЕЯТЕЛЬНОСТИ
173
Фиг. Х.10. Окончание
Глава XI
ЖИВОТНЫЕ ИЗ ТРЕХ ЧАСТЕЙ:
ТРИЛОБИТЫ
• История развития органического мира по ископаемым предстает пе-
ред нами прерывистой и непростой. В мире беспозвоночных многие
типы животных появляются в фанерозое почти одновременно и нео-
жиданно, когда впервые возникает возможность строить твердые минераль-
ные скелеты. Происхождение многих животных поэтому до сих пор непонят-
но. Можно только знакомиться с группой за группой; рассказав историю од-
ной из них, снова необходимо возвращаться к истокам, чтобы начать
повествование о другой. В сложных типах, где классы значительно отлича-
ются друг от друга, приходится так поступать несколько раз.
ЧЛЕНИСТЫЕ И “КОЖИСТО-ПАНЦИРНЫЕ”: АРТРОПОДЫ
Членистость и отвердевшие покровы — черты чрезвычайно характерные
для Arthropoda, типа древнего и разнообразного. Его делят на семь основных
групп, рассматриваемых одними учеными как подтипы, а другими - как
надклассы, причем число классов по мере исследования все более возрастает.
Некоторые из них обособились в кембрийский период или даже раньше, и
только три группы оформились в постсилурйское время. Многие палеонто-
логи утверждают, что тип Arthropoda на самом деле сборище неродственных
животных, отвечающих только определенному уровню организации. Воз-
можно, крабы, насекомые, трилобиты и многие другие артроподы произош-
ли от различных предков и каждая из этих групп образует отдельный тип.
Название “Arthropoda" переводится как “Членистоногие” или “Суставча-
тоногие”. Членистость характерна не только для конечностей, но и для всего
организма в целом. От головы до “хвоста” артроподы состоят из серии сег-
ментов, или сомитов, соединенных подвижно. У многих сегменты способны
сливаться и уменьшаться в числе, и тогда они, когда-то подвижные, образу-
ют неподвижные комплексы. Конечности и другие придатки имеют сходную
членистость.
Полезный экзоскелет, или панцирь. Экзоскелет, или панцирь, обычно бы-
вает столь прочным, что его можно воспринимать как раковину. Зоны сочле-
нения сегментов представляют собой наиболее мягкие части экзоскелета,
что помогает телу изгибаться. Экзоскелет по сути своей представляет собой
корку, выделяемую кожей, причем корка является слоистой и состоит у
© 1997.0.Б. Бондаренко, перевод на русский язык и изменения.
ЖИВОТНЫЕ ИЗ ТРЕХ ЧАСТЕЙ: ТРИЛОБИТЫ
175
Фиг. XI. 1. Продольный срез двух сегментов у членистоногих в местах крепле-
ния мускула
большинства ныне живущих артропод из трех слоев (фиг. XI. 1). Поверхност-
ный слой, представленный воскоподобной пленкой, выполняет водозащит-
ную функцию. Ниже находится средний слой, который придает панцирю
прочность; он развит только между зонами сочленения. Этот слой может
быть хитиновым, но у крабов и их родственников он пропитан микрокрис-
талликами кальцита. Внутренний слой, непосредственно контактирующий
с кожей, представляет собой плотный, но гибкий слой хитина. Он составляет
основную часть покрова у “мягкотелых” насекомых и у только что прошед-
ших линьку “мягко-раковинных” крабов.
Экзоскелет, или панцирь, как известно, содержит внутри мягкое тело и в
основном выполняет ту же роль, что и “доспехи рыцарей”. Панцирь артро-
под, кроме функции защиты, благодаря зонам сочленения выполняет и не-
которые другие функции, присущие костям скелета позвоночных. Многие
мышцы позвоночных, прикрепляясь к костям и протягиваясь от одной к
другой, при сокращении осуществляют движения рук, ног, челюстей и так
далее. Мускулы артропод прикрепляются к коже, которая большей частью
плотно срастается с корковидным панцирем. Когда мускулы, протягиваю-
щиеся от сегмента к сегменту, сокращаются, то они вызывают у артропод та-
кие же быстрые и точные движения, как мускулы позвоночных.
ЛИНЬКА И ПАЛЕОНТОЛОГИЧЕСКИЕ ДАННЫЕ
В двух отношениях панцирь, тем не менее, хуже костей или даже обыч-
ных раковин. Он сковывает движения, и никакая комбинация органов арт-
ропод не может создать то разнообразие движений, которое присуще конеч-
ностям позвоночных (особенно рукам). Панцирь, быстро сформировавшись
и отвердев, становится неподвижным; он не увеличивается в размерах, в то
время как его обладатель растет день за днем. Панцирь начинает препятство-
вать росту тела все больше и больше. В конце концов это приводит к его сбра-
сыванию, а созидатель и носитель панциря выходит из этой “борьбы”, имея
только мягкое тоненькое покрытие. Затем тело снова растет, экзоскелет
утолщается и животное снова оказывается заключенным в панцирь.
176
КАМЕННАЯ КНИГА
Фиг. XI.2. Примитивные артроподы
а, б — Aysheaia pedunculata длиной около 1.5-5 см, вероятно, онихофора: а - от-
печаток, б — одна из реконструкций (см. также фиг. Х.6), средний кембрий,
сланцы Бёрджесс, Канада, Британская Колумбия; в — реконструкция прими-
тивного членистоногого Arthropleura длиной до 1.5 м, средний-поздний кар-
бон, Европа
Этот процесс сбрасывания панциря называется линькой, или экдизисом.
Линька происходит у большинства членистоногих от четырех до десяти раз,
у долгоживущих форм до тридцати раз, как, например, у омаров. На ранних
стадиях линька может также сопровождаться существенными изменениями
в строении и облике животного. Молодой краб, например, начинает жизнь,
как маленькая шипастая планктонная форма, затем проходит стадию, напо-
минающую креветку, прежде чем окончательно обретет свое характерное
строение. После этого линька сопровождается в основном увеличением раз-
меров и незначительными изменениями в облике. Насекомые перестают рас-
ти, когда достигают зрелой стадии.
Древние артроподы также линяли и изменялись. Это значительно увели-
чивает число ископаемых, так как сброшенные плотные оболочки сохраня-
ются так же хорошо, как и панцири мертвых взрослых особей. Таким обра-
зом, одна и та же особь может оставить после себя несколько ископаемых
форм, представленных как фрагментами, так и превосходными панцирями.
Когда сброшенные при линьке панцири удается выстроить в последователь-
ный ряд изменяющихся форм, то по ним можно установить стадии развития
трилобита, принадлежащему одному виду.
ЖИВОТНЫЕ ИЗ ТРЕХ ЧАСТЕЙ: ТРИЛОБИТЫ
177
ПЕРЕХОДНЫЕ ЗВЕНЬЯ МЕЖДУ АРТРОПОДАМИ И ЧЕРВЯМИ
Сведения об ископаемых артроподах начинаются с докембрийских пород
Южной Австралии, откуда происходят ископаемые, описанные в главе IV.
Ископаемое Xenusion из ледникового валуна, найденного в Швеции, по-ви-
димому, тоже происходит из докембрия, оно было морским существом. Тем
не менее его обычно помещают в группу Onychophora, которая сочетает при-
знаки кольчатых червей и членистоногих (см. также главы X и XIII). Так же
морской формой была Aysheaia из среднекебрийских сланцев Бёрджесс, чер-
веподобное существо с кольцами щетинок вокруг тела и с двадцатью морщи-
нистыми ножками, которые оканчивались коготками (фиг. XI.2). Голова
имела одну пару ветвистых антенн и еще два других отростка, которые, веро-
ятно, выделяли ядовитую слизь, когда на это существо нападали. Существу-
ют и другие реконструкции Aysheaia (см. фиг. Х.6). Aysheaia так похожа на
современных онихофор, что последних можно считать живыми ископаемы-
ми, освоившими среду обитания, весьма отличную от той, в которой они ког-
да-то начинали (см. также главу XIII).
ТРЕХДОЛЬНЫЕ ТЕЛА: ТРИЛОБИТЫ
Онихофоры, хоти и освоили наземные условия, все-таки не стали много-
численной и разнообразной группой. Другая группа, называемая трилобита-
ми, осталась в море и там преуспела, о чем свидетельствует ошеломляющее
число семейств, родов (около 2000) и видов (более 10000). На протяжении
около 350 млн. лет ее представители были наиболее многочисленными и пре-
успевающими артроподами. Трилобиты имеют особую ценность для опреде-
ления возраста кембрийских и ордовикских пород.
Название Trilobita, или трилобиты, обозначает “трехдольные”. Можно по-
думать, что такое название отражает план строения - голова, туловище,
хвост. На самом деле название связано с теми частями тела, которые протя-
гиваются вдоль его оси, отделяясь продольными бороздами, а не поперечны-
ми границами сегментов. Эти борозды разделяют каждого трилобита на осе-
вую часть, идущую посередине, и две боковых - плевральных с каждой сто-
роны от нее (фиг. XI.3).
Общепринято считать, что трилобиты возникли от кольчатых червей, у
которых головной отдел увеличился в размерах благодаря слиянию сегмен-
тов, в то время как тело раздалось вширь, развив плевры по обе стороны от
первичных сегментов. С трилобитовым предком, вероятно, совпадает форма,
подобная эдиакарскому роду Spriggina с его огромной уплощенной головой
(см. фиг. IV. 11). Не исключено также, что все известные эдиакарские формы
не связаны ни с какими последующими кембрийскими и другими группами.
Исходя из этого , наиболее примитивной группой трилобитов из известных
являются оленеллиды, включающие род Fallotaspis, который впервые по-
явился в раннем кембрии (атдабанский ярус) Сибири, а затем распростра-
нился по Америке, Европе и Северной Африке (см. фиг. XI.9).
В течение всей истории трилобиты создавали потрясающее разнообразие
форм, сохраняя общий план строения. Остановимся теперь на основном пла-
не строения и опишем некоторые из его наиболее известных вариантов.
Головной щит, или цефалон. Он состоит из нескольких сегментов, кото-
рые слились в единый сплошной щит. Например, у известного североамери-
канского рода Olenellus цефалон выглядит как половинка луны с загнутыми
12. Каменная книга
178
КАМЕННАЯ КНИГА
а
в
Плевральная Осевая
часть I часть
Осевая Плевральная
часть . часть •
Фиг. XI.3. Схема строения трилобитов
а — продолный разрез Ceraurus; б-вид снизу; в, г - вид сверху двух трилобитов
из отряда Phacoptda (в) и отряда Ptychopariida (г), н - неподвижная (фиксиро-
ванная) щека, п - подвижная (свободная) щека
концами, которые оканчиваются двумя щечными шипами. У других родов
цефалоны такие причудливые, что словами трудно описать, и только по изо-
бражениям понятно их строение. Наиболее своеобразный цефалон у шипас-
того Terataspis, хотя округлый цефалон Bumastus также очень специализи-
рован.
Типичный цефалон имеет три продольные доли, как и остальное тело три-
лобита: глабель и щеки по бокам (см. фиг. XI.3). Линии на щеках, разделяю-
180
КАМЕННАЯ КНИГА
окончаниями, направленными назад. Число туловищных сегментов часто
было меньше сорока; некоторые виды Olenellus имели только 18—20 сегмен-
тов. Самое маленькое число было у агностид, чьи туловищные сегменты в
числе двух-трех были очень узкими.
Уменьшение числа туловищных сегментов происходило за счет редукции
или слияния с хвостовым щитом — пигидием. Второй путь иллюстрирует та-
кие роды, как Ogygopsis и Trimerus, чьи пигидии содержали много сегмен-
тов, слившихся в единый щит. У Arctinurus эти сегменты сохранили шипо-
видные окончания своих боковых лопастей.
Некоторые трилобиты, как, например, Trimerus и Bumastus, имели от
восьми до тринадцати сегментов, но сохранили только слабые следы деления
на лопасти.
Антенны и конечности. Конечности плохо сохраняются, поэтому у подав-
ляющего большинства трилобитов они в ископаемом состоянии отсутству-
ют. Тем не менее у некоторых экземпляров они обнаружены. Конечности
(включая и антенны) были описаны у следующих трилобитов, существовав-
ших в разное время в интервале от раннего кембрия по девон: Olenellus, Ole-
noides, Kootenia, Ehmaniella, Westergaardia, Triarthrus, Cryptolithus, Cer-
aurus, Phacops, Asteropyge. Передние конечности начинаются с двух много-
членистых антенн, прикрепленных к нижней поверхнсоти цефалона по
бокам от гипостомы (см. фиг. XI.3). Антенны являются простыми нерас-
щепленными образованиями. При жизни антенны, вероятно, несли малень-
кие сенсорные органы, скорее всего, обоняния и вкуса, как антенны совре-
менных омаров.
Членистые ножки (коксы) прикреплялись парами снизу к каждому сег-
менту цефалона, торакса и пигидия, если последний имел несколько сегмен-
тов. Каждая конечность двуветвистая, разделенная на нижнюю ветвь (эндо-
подит), используемую для хождения, и верхнюю ветвь (экзоподит) с пери-
стыми жабрами, которые поглощали кислород, а у некоторых трилобитов
могли служить как весла при плавании. И все же функции экзоподита, или
жаберной ветви трилобитовой конечности, в отличие от ходильной ветви, до
конца непонятны.
Многие следы передвижения трилобитов по субстрату описывают в разде-
ле ихнофоссилий. Наиболее распространенные трилобитовые следы названы
Cruziana. Они напоминают подошву обуви с спаренными шевронами или по-
хожи на скелет селедки. Доказано, что такие следы оставлены трилобитами,
потому что некоторые из следов вели в норки, названные Rusophycus, в кото-
рых находились трилобиты (фиг. XI.4). Такие норки часто встречаются в
кембрии и ордовике, реже в силурийских и девонских породах и полностью
отсутствуют в постпермских отложениях. Разнообразие, присутствие или
отсутствие таких и подобных следов отражает появление, разнообразие и вы-
мирание трилобитов. Трилобитовые следы свидетельствуют о том, что жи-
вотные передвигались различными способами и не только вперед, но и вбок;
они также неплохо зарывались.
Хвостовой щит, или пигидий. Хвостовой щит состоит из одного или более
слившихся сегментов, находящихся в конце тела трилобита. Под пигидием
вдоль осевой части протягивается кишка, оканчивающаяся анальным отвер-
стием (см. фиг. XI.3). У примитивных родов, таких как Olenellus, размеры
пигидия очень маленькие (микропигиевые трилобиты), но у других ранне-
кембрийских форм он разрастается за счет включения трех или более туло-
вищных сегментов. Формы, у которых пигидий уже не маленький, но все
еще меньше, чем цефалон, называются гетеропигиевыми. Процесс увеличе-
ЖИВОТНЫЕ ИЗ ТРЕХ ЧАСТЕЙ: ТРИЛОБИТЫ
179
щие неподвижные щеки от свободных подвижных, называются лицевыми
швами. Они соответствуют линиям растрескивания панциря, вдоль которых
подвижные щеки отделяются от неподвижных во время линьки, помогая бо-
лее легко сбросить старый панцирь. .
Г лабе ль (или глабелла) Olenellus и других родов представляет собой воз-
вышенную осевую часть, оканчивающуюся у переднего края вздутием, напо-
минающим шарик. Пары поперечных борозд с каждой стороны глабели — это
все, что осталось от сегментации цефалона (см. фиг. XI.9). У большинства по-
следующих родов борозды сгладились и даже исчезли; форма глабели также
менялась.
Нижняя сторона цефалона наблюдается у трилобитов редко. Когда она со-
храняется, то виден рот, три (или ?четыре) пары конечностей и основания от
двух членистых антенн (см. фиг. XI.3). Рот ведет в желудок, представляю-
щий собой огромное расширение под глабелью, переходящее затем в кишку,
идущую к пигидию. Вследствие того, что желудок находится под глабелью и
определяет ее форму, можно по ней определить пищевые пристрастия трило-
бита. У исключительно хорошо сохранившихся экземпляров видно прими-
тивное сердце, расположенное сверху желудка на границе его с кишкой.
Сердце, по-видимому, накачивало кровь в сосуды, идущие вдоль осевой час-
ти тела. Губовидную пластинку (гипостома), находящуюся впереди рта, ино-
гда находят отдельно от панциря трилобита.
Глаза. Трилобитовые глаза являются наиболее древней системой зрения.
Линзы глаз представлены у фоссилий кальцитом (?). Большинство трилоби-
товых глаз сложные, они состоят из множества плотно примыкающих линз
(фасеток), находящихся в капсулах и покрытых единой прозрачной плен-
кой, то есть роговой мембраной. Вначале число линз было небольшим, но у
некоторых родов оно увеличилось до двенадцати-пятнадцати тысяч. У дру-
гих трилобитов известна обратная тенденция, число фасеток уменьшалось до
шестисот, двухсот или даже четырнадцати и двух. Глаза, где все линзы
плотно примыкают друг к другу и покрыты общей оболочкой, называют го-
лохроическими. Такой вариант глаз появился в кембрии и продолжал встре-
чаться, пока не вымерли трилобиты. Другой, более прогрессивный тип глаз,
шизохроический, был свойственен Phacopina (ордовик-девон). Шизохрои-
ческие глаза характеризуются крупными линзами, отделенными друг от
друга толстой кутикулой, называемой склерой. Каждая линза имела отдель-
ную роговую оболочку. Такие глаза, вероятно, позволяли видеть мир в трех-
мерном изображении или использовались трилобитами, ведущими ночной
образ жизни. Одно совершенно ясно: шизохроические глаза были необык-
новенными.
Еще более удивительным является полное отсутствие глаз, или их редук-
ция до двух или нескольких простых линз. Такая тенденция развития на-
блюдается у Paraharpes и у некоторых других представителей, принадлежа-
щих тому же отряду Ptychopariida.
Глаза, кроме того, отличаются по форме и размерам. Простенькие глаза
смотрятся как точки или гранулы. Более изысканные глаза имеют форму по-
лумесяца, линзовидного возвышения или полусферы на концах стебельков.
Несколько родов мелких трилобитов имели огромные выпуклые глаза, кото-
рые были способны смотреть вниз и назад, так же как и вперед, вверх и по
сторонам.
Туловищный отдел, или торакс. Примитивный трилобитовый торакс со-
держал более чем сорок сегментов, из которых четырнадцать или пятнад-
цать несли широкие и хорошо развитые плевральные доли с шиповидными
12*
ЖИВОТНЫЕ ИЗ ТРЕХ ЧАСТЕЙ: ТРИЛОБИТЫ
181
Фиг. XI.4. Ископаемые следы жизнедеятельности трилобитов
а, б — заполнение двух ямок, названных Rusophycus, вырытых, по-видимому,
раннекембрийским Olenellus: а — вид сверху, б — вид сбоку, в — схема выкапыва-
ния ямки. Ямки, вероятно, служили местом отдыха и откладывания яиц
ния пигидия продолжался в течение среднего кембрия и в последующие эпо-
хи. В результате пигидй у некоторых трилобитов стал таким же большим,
как цефалон (изопигиевые формы); они содержали восемнадцать или более
сегментов. Редко пигидии были больше, чем цефалоны, и такие трилобиты
называются макропигиевыми. Нижняя сторона пигидиев обычно сохраняла
конечности всех сегментов, хотя сегменты слились и границы между ними
уже исчезли.
Многие пигидии несли борозды, указывающие на границы уже сливших-
ся сегментов. У некоторых родов борозды оставались глубокими и четкими;
у других становились неотчетливыми и в конце концов исчезали. У других
родов часть борозд исчезала, а часть сохранялась.
Окраска. Окраска панциря трилобитов неизвестна, но некоторые формы
сохранили полосчатый и точечный рисунок. Вид Isotelus maximus из поздне-
го ордовика Северной Америки сохранил полосы вдоль оси тела.
От яйца до совершеннолетия. Трилобиты, по-видимому, откладывали яй-
ца, как большинство современных членистоногих. Самка, скорее всего, ос-
тавляла яйца в слегка присыпанной ямке, а затем уползала, оставляя “гнез-
до” заполняться рыхлым песком. Если это верно, то самец, вероятно, следо-
вал за самкой и ждал около нее, готовясь оплодотворить яйца, как только
она их отложит. Песок, вмываемый в “гнезда”, был достаточно рыхлым, что
давало возможность воде достигать яиц и снабжать их кислородом, а затем
не мешать выбраться из ямки новому юному поколению. В то же время песок
скрывал яйца от хищников и падалеедов, в компанию которых входили дру-
гие трилобиты.
В течение первой стадии развития трилобитов, то есть протаспидной ста-
дии, цефалон и протопигидий (последний состоял из сегментов, которые в
182
КАМЕННАЯ КНИГА
Фиг. XI.5. Стадии онтогенеза
а—д — стадии онтогенеза Shumardiapusilia, х40, на стадии а есть конечности, на
стадии д развиты почти все структуры, хотя по размерам трилобит наполовину
меньше взрослой формы (отряд Ptychopariida); е—з — Paedeumias transitans на
двух ранних - е, ж, х20, и взрослой - з, xl, стадиях (отряд Redlichiida)-, и — О1е-
nelloides armatus, х7, незрелая стадия с шипастым головным щитом (отряд
Redlichiida)
дальнейшем вошли в переднюю часть туловищного отдела) слиты в единый
шит (фиг. XI.5). Невозможно сказать точно, как много сегментов соединены
в этом щите, но цефалон некоторых протаспид включал более чем пять сег-
ментов, а пигидий, вероятно, состоял из других пяти. Начало второй стадии,
то есть мераспидной, знаменуется появлением сегмента между цефалоном и
протопигидием. В течение мераспидной стадии сегменты постепенно влива-
ются в туловищный отдел с фронтальной стороны протопигидия, пока не на-
берется полный комплект туловищных сегментов, указывающий, что голас-
пидная, то есть зрелая, стадия достигнута. Изменения в течение зрелой ста-
дии в основном проявляются в увеличении размеров и в изменении
пропорций.
Сравнение с современными членистоногими не оставляет сомнений, что
трилобиты на личиночных стадиях имели ветвистые шипастые конечности,
используемые при плавании.
Знание стадий онтогенеза, то есть развития индивидуумов от рождения до
смерти, помогает при разработке эволюционных взаимоотношений трилоби-
тов. Сходство между личинками указывает на близкие родственные связи,
хотя зрелые формы могут сильно отличаться.
ЖИВОТНЫЕ ИЗ ТРЕХ ЧАСТЕЙ: ТРИЛОБИТЫ
183
ШВЫ, ЛИНЬКА И КЛАССИФИКАЦИЯ
У подавляющего большинства трилобитов лицевые швы были жизненно
важны для линьки, так как они связаны с ослабленными зонами панциря,
по которым он разламывался. Вероятно, приходилось много раз изгибать-
ся, извиваться, крутиться и выжиматься, прежде чем такое животное, как
Arctinurus, освобождалось от своего панциря (фиг. XI.6). Процесс, возмож-
но, проходил среди кораллов или у оснований водорослей, где “мягко-пан-
цирный” трилобит мог спрятаться, пока построит свой новый жесткий
панцирь.
Выше было сказано, что лицевые швы очень важны для большинства, но
все же не для всех трилобитов, потому что у некоторых трилобитов они отсут-
ствовали. Кроме того, есть роды, такие как Acidaspis, у которых швы почти
не заметны. Вряд ли они имели большое значение при линьке.
Лицевые швы. Лицевые швы в прежних классификациях использовали
для диагностики отрядов трилобитов. Различают три основных типа швов:
пропариевый, описториевый и гипопариевый (фиг. XI. 7). Кроме того, выде-
ляют гонатопариевый и протопариевый варианты. Пропариевый шов харак-
теризуется тем, что задняя ветвь оканчивается на передней стороне щита, не
доходя до щечного угла. Опистопариевый лицевой шов оканчивается на зад-
ней стороне цефалона между его щечным углом и затылочным кольцом. Ги-
попариевый, или краевой маргинальный, шов идет вдоль края цефалона и не
виден на верхней поверхности. У гонатопариевого лицевого шва задняя
ветвь заходит в щечный угол цефалона. Протопариевый похож на гипопари-
евый, он также идет вдоль края цефалона, но по его верхней поверхности.
Фиг. XI.6. Arctinurus, хО.З (отряд Lichida). Слева — ползущий по илистому дну,
справа — во время линьки, частично освободившийся от панциря
Протопариевый
Пропариевый
Опистопариевый
Г ипопариевый
Фиг. XI.7. Типы лицевых швов у трилобитов (показаны толстой черной линией)
184
КАМЕННАЯ КНИГА
Фиг. XI.8. Отряд Agnostida (кембрий—ордовик)
а — Pagetia bootes, х7.5, средний кембрий, Британская Колумбия; б — Hypagnos-
tus parvifrons, х8.2, средний кембрий, род встречается в Северной и Южной
Америке, Евразии и Австралии; в - Eodiscus punctatus, х7.5, средний кембрий,
Ньюфаундленд (Treatise ... Part 0, 1959)
Долгое время типы лицевых швов использовали при классификации как
основной признак. В настоящее время их классификационное значение при
выделении отрядов отрицают, так как выяснили, что одни и те же типы ли-
цевых швов возникали многократно у разных трилобитов. Таким образом,
группы трилобитов, выделенные только по типам швов, содержат смесь
близких и неродственных форм. В настоящее время тип шва - только один из
многих признаков, используемый при определении родственных связей
между трилобитами.
Классификация. Современная классификация трилобитов базируется на
нескольких признаках, рассматриваемых совместно. Класс Trilobita в основ-
ном делят на восемь отрядов, на которых кратко остановимся ниже.
Agnostida (фиг. XI.8). Агностиды — маленькие трилобиты, существовав-
шие начиная с раннекембрийского и кончая позднеордовикским временем.
У большинства агностид глаза отсутствовали; швы краевые (гипопариевые)
или отсутствовали; число туловищных сегментов равно двум (Agnostus,
Hypagnostus) или трем (Eodiscus). Цефалон и пигидий равновеликие (изопи-
гиевые трилобиты).
Redlichiida (фиг. XI.9; см. также фиг. XI.5). Редлихииды существовали в
течение раннего-среднего кембрия. Цефалон в виде крупного полукруга,
обычно с удлиненными щечными шипами. Пигидий крошечный (микропи-
гиевые трилобиты), а туловищные сегменты обычно шиповидно заострены.
В отличие от “слепых” агностид, редлихииды обладали крупными глазами.
Лицевые швы неразличимы или опистопариевые. Роды Fallotaspis, Olenellus
и Paradoxides являются широко известными трилобитами.
Corynexochida (фиг. XI. 10; см. также фиг. XI. 16). Коринексохиды жили
в течение кембрия. Глабель обычно расширялась к передней части или была
почти цилиндрической. Пигидий нередко больше, чем цефалон (макропи-
гиевые трилобиты). Торакс обычно имел семь или восемь сегментов. Лице-
ЖИВОТНЫЕ ИЗ ТРЕХ ЧАСТЕЙ: ТРИЛОБИТЫ
185
Fallotaspis
в
Lermontovia
Фиг. XI.9. Отряд Redlichiida (ранний—средний кембрий)
а - Fallotaspis typica, реконструкция, xl, ранний кембрий, Марокко; б - Olenel
lus thomsoni, хО.6, ранний кембрий, Северная Америка; в - Lermontovia dze-
vanoivskii, xl, ранний кембрий, Сибирь; г - Paradoxides paradoxissimus, х0.5,
средний девон, Швеция (Treatise...Part 0, 1959)
вые швы - опистопариевые. Характерный представитель этой группы - род
Olenoides.
Ptychopariida (фиг. XI.11; см также фиг. XI.5, фиг. XI.16). Птихопарии-
ды существовали с раннего кембрия по конец девона. Они характеризуются
186
КАМЕННАЯ КНИГА
Фиг. XI.10. Отряд Corynexochida (ранний—поздний кембрий)
а, б-Olenoides serratus, реконструкция, xl (а - вид снизу, б - вид сверху), сред-
ний кембрий, Канада, Британская Колумбия; в - Ogygopsis klotzi, х0.7, средний
кембрий, Северная Америка (Treatise... Part 0, 1959)
разнообразными путями развития. Торакс имел от 5 до 42 сегментов. Боль-
шинство птихопариид обладали опистопариевыми лицевыми швами, неко-
торые имели краевые (маргинальные) или пропариевые швы. Глабель обыч-
но суживалась впереди, но здесь имелись варианты. Древние формы облада-
ли большим тораксом и маленьким пигидием, но у последующих форм число
туловищных сегментов уменьшалось, а пигидий увеличивался (например,
Asaphus). Роды Olenus, Elrathia, Cryptolithus, Asaphus и Illaenus являются
типичными представителями птихопариид.
Многие древние Ptychopariida очень сходны с Redlichiida, от которых они,
вероятно, произошли. В свою очередь, птихопарииды, по-видимому, дали
начало большинству посткембрийских трилобитов, исключая агностид, су-
ществовавших с раннего кембрия.
Proetida (фиг. XI. 12). Проетиды были долгоживущей группой, впервые
появившейся в ордовике и вымершей наконец в перми. Как и агностиды,
представители Proetida были изопигиевыми трилобитами, но торакс имел от
ЖИВОТНЫЕ ИЗ ТРЕХ ЧАСТЕЙ: ТРИЛОБИТЫ
187
Фиг. XI.11. Отряд Ptychopariida (ранний кембрий—поздний девон)
а - Dikelocephalus oweni, х0.8, поздний кембрий, Северная Америка; б, в-Сгур
tolithus tesselatus, х27, сверху и сбоку, средний ордовик, Северная Америка; г —
Asaphus expansus, xl.5, со стороны головного щита и панцирь сверху, ранний
ордовик, Швеция
восьми до десяти сегментов. Глабель была крупной и булавовидной, ее стро-
ение указывает на большой и сложный желудок. Имелись щечные шипы.
Глаза крупные голохроические с линзами, плотно примыкающими друг к
другу; лицевые швы опистопариевые. Роды Proetus, Phillipsia, Griff ithides и
Bathyurus являются представителями этой группы.
Phacopida (фиг. XI. 13; см. также фиг. XI. 16). Факопиды очень разнооб-
разный отряд, хотя не такой большой, как отряд Ptychopariida. Факопиды
сущестсвовали с ордовика по девон. Они имели преимущественно пропарие-
вые лицевые швы, некоторые - гонатопариевые, а немногие даже опистопа-
риевые. Глабель разнообразной формы. Торакс состоял из восьми—девятнад-
цати сегментов. Пигидий обычно средних и крупных размеров, но у некото-
рых ранних предствителей - маленький. Род Calymene принадлежал одной
из групп факопид с гонатопариевыми швами и суживающейся впереди гла-
белью с четырьмя парами боковых долей и маленькими голохроическими
глазками. Роды Phacops и Dalmanites принадлежали другой группе Phacopi-
da, имевшей глабель, расширяющуюся впереди, шизохроические глаза и
пропариевые лицевые швы.
Lichida (фиг. XI. 14; см. также фиг. XI.6, фиг. XI. 16). Лихиды имели сред-
ние и крупные размеры; они просуществовали с начала ордовика по конец
девона. Пигидий обычно больше, чем цефалон; он обладал кинжаловидными
плеврами. Глабель широкая, а швы - опистопариевые. Панцирь нередко был
188
КАМЕННАЯ КНИГА
Фиг. XI. 12. Отряд Proetida (ордовик—пермь)
а-в — Proetus concinnus, х3.5: а - головной щит, б - хвостовой щит, в - головной
щит с фронтальной стороны, силур, Швеция, о-в Готланд; г — Phillipsia gemmu-
lifera, х3.5, реконструкция, ранний карбон, Англия; д — Gnffithid.es seminife-
rus, xl.7, реконструкция, ранний карбон, Англия (Treatise ... Part 0, 1959)
Griflithides
покрыт многочисленными бородавочками (туберкулы). Род Terataspis,
имевший шипастый и бородавчатый облик, являлся характерной формой
лихид (см. фиг. XI. 16).
Odontopleurida (см. фиг. XI. 16). Одонтоплевриды шипастые, относитель-
но мелкие трилобиты, известные из верхнекембрийских и нижнеордовикс-
ких пород. Пигидий короткий и очень шипастый. Торакс также с длинными
шипастыми сегментами в числе восьми-десяти. Глабель из двух или трех
пар долей; лицевые швы - опистопариевые. Роды Radiaspis и Leopaspis явля-
ются яркими примерами этих причудливых трилобитов.
Если проанализировать все группы трилобитов, то можно заметить следу-
ющее. Голохроические глаза дали начало шизохроическим. Пигидии увели-
чивались в размерах, изменяясь от микропигиевых к изопигиевым и даже
макропигиевым вариантам. Относительно простые неорнаментированные
трилобиты дали начало сильно шипастым и бородавчатым формам. Вместе с
тем в течение 350 млн. лет своей истории, несмотря на все эти преобразова-
ния, трилобиты недалеко отошли от исходного плана строения тела.
Древнейшие трилобиты принадлежат к Redlichiida. Задолго до окончания
кембрия к ним присоединились Agnostida, Corynexochida и Prychopariida. В
конце кембрия разразился крупный экологический кризис. Только несколь-
ко ветвей трилобитов перешли в ордовик, но уже в начале этого периода они
дали огромное разнообразие форм. Ордовикские трилобиты освоили много
новых экологических ниш, например, в рифовых сооружениях, в которых
они практически отсутствовали в кембрии. Следующий экологический кри-
зис наступил в конце ордовика. Силурийские и девонские трилобиты при-
ЖИВОТНЫЕ ИЗ ТРЕХ ЧАСТЕЙ: ТРИЛОБИТЫ
189
Ceraurus
Фиг. XI. 13. Отряд Phacopida (ордовик—девон)
а — Cerauruspleurexanthemus, xl .5, реконструкция, средний ордовик, США; б —
Flexicalymene senaria, х2, головной щит, средний ордовик, Северная Америка;
в, г - Calymene blumenbachi сбоку (е) и сверху (г), х1.3, силур, венлок, Англия;
д — Phacops altaicus, средний девон, Алтай; е - Dalmanites caudatus, х0.7, си-
лур, Англия; ж - Dipleura dekayi, xl, средний девон, США (Основы палеонтоло-
гии, том VIII, 1960; Treatise ... Part 0, 1959)
надлежали к близким ветвям; они не так разнообразны, как в раннем палео-
зое. После девона сохранились только представители отряда Proetida. В кон-
це перми трилобиты, как и многие другие животные, вымерли в результате
еще одного экологического кризиса.
190
КАМЕННАЯ КНИГА
Arctinurus
Akantharges
Фиг. XI.14. Отряд Lichida (ордовик—девон)
a —Arctinurus boltoni, длина до 15 см, силур, венлок, Нью-Йорк; б—Akantharges
gourdoni, xl, реконструкция, средний девон, Франция
ОСОБЕННОСТИ СТРОЕНИЯ И ОБРАЗ ЖИЗНИ ТРИЛОБИТОВ
Все трилобиты были морскими формами, и, насколько известно, они даже
не заходили в солоноватые устья рек. В океанах и морях трилобиты плавали
в толще воды и у ее повехности, ползали, скользили и прыгали по дну, лежа-
ли на нем и зарывались в песок или ил. По способу питания трилобиты варь-
ировали от детритофагов и фильтраторов до хищников и падалеедов.
Пути развития Olenellus. Род Olenellus из отряда Redlichiida имел доволь-
но примитивное строение. Вероятно, они и близкие ему рода дали начало
многим другим трилобитам. Только что вылупившийся Olenellus был кро-
хотным трилобитом округлой формы, который мог во взвешенном состоянии
дрейфовать в водах океана. Он продолжал так плавать и на мераспидной ста-
дии, но, достигнув голаспидной стадии, опускался на дно и ползал по нему в
мелководных участках обычно недалеко от побережья моря.
Нет лучшего района для изучения Olenellus, чем долина озера Louise в за-
падной Альберте, хотя род встречается по всей Северной Америке, Гренлан-
дии и Швеции. В западной Альберте осыпь склонов у подножия скал, увен-
чанных ледниками, представлена нижнекембрийскими кварцитами и
темно-серыми слюдистыми сланцами. Последние содержат панцири и от-
печатки трилобитов, в то время как кварциты богаты следами их жизне-
деятельности.
Olenellus был способен плавать, используя свои жаберные ножки, или
ползать с помощью эндоподитов. Плавая, он не оставлял следов, но при пол-
зании членистые эндоподиты внедрялись в песок, а шипы цефалона и туло-
ЖИВОТНЫЕ ИЗ ТРЕХ ЧАСТЕЙ: ТРИЛОБИТЫ
191
Фиг. XI.15. Донные (бентосные) трилобиты
1,2 — Trimerus и Ectenaspis, неподвижно ожидающие добычу; пигидии заглуб-
лены в осадок; 3 - Bumastus, продирающийся сквозь ил и вспахивающий его;
4 - Paraharpes, скользящий по субстрату; 5 - Flexicalymene, свернувшийся в
момент опасности (может также ползать по дну и плавать в толще воды)
вищных сегментов местами пропахивали острые бороздки. Часть следов ока-
менело и превратилось в фоссилии, как и углубления, несущие отпечатки эн-
доподитов и шипов. Некоторые углубления были в виде неглубоких ямок
неправильной формы. Возможно, в них обитали маленькие мягкотелые за-
рывающиеся черви, служившие пищей для трилобитов. Ямки могли пред-
ставлять также места отдыха и кладок трилобитов (см. фиг. XI.4).
Разнообразие ползающих и зарывающихся трилобитов. Ползание было и
оставалось в течение палеозоя основным способом передвижения трилоби-
тов, хотя облик и размер ползающих изменялись чрезвычайно (фиг. XI. 15).
Вид “Olenellus” vermontanus обладал длинным и узким телом, а ширина тела
Olenellus thompsoni почти равна его длине. В среднекембрийское время и поз-
же отмечено развитие многочисленных ползающих трилобитов, имеюших
широкое и уплощенное тело. У некоторых (например, Paradoxides) были ма-
ленькие пигидии и много туловищных сегментов; у других (например, Dike-
locephalus и Eobronteus) торакс был маленьким, а пигидий большим. Род
Arctinurus из силура имел короткий широкий цефалон, вытянутый и заост-
ренный впереди, и очень широкий торакс.
Isotelus и его сородичи могли ползать, но их широкий пигидий был спосо-
бен закапываться в ил, работая как борона при вспахивании.
Узкорылый Ectenaspis пошел еще дальше, на что указывает положение
хорошо сохранившихся ископаемых форм, где виден “хвост”, заглубленный
в мягкий субстрат, и голова, оставшаяся на поверхности. В таком положении
трилобит был способен хватать крошечных ползающих существ или мерт-
вые тельца, когда их проносило мимо.
Донные трилобиты: шипастые и с каймой-лимбом. Существует много те-
орий, объясняющих функции шипов у трилобитов. Раньше считали, что ши-
пы указывают на планктонный образ жизни, так как, увеличивая поверх-
ность тела, они почти не влияют на вес и, таким образом, препятствуют по-
гружению Недавно стали предлагать другие объяснения. Большинство'
шипастых трилобитов все-таки слишком крупные, чтобы шипы могли со-
здать им дополнительные условия для плавания (фиг. XI. 16). Шипы, несо-
мненно, временами предохраняли от хищников. Удлиненные боковые шипы
192
КАМЕННАЯ КНИГА
Фиг. XI. 16. Гладкий (а) и разнообразно-шипастые (б-ж) трилобиты
а - Cyclopyge, хб (отряд Ptychopariida), большеглазый свободно-плавающий
трилобит, ордовик, Европа и Северная Америка; б - Albertella helena, хО.6 (от-
ряд Corynexochida), ползающая форма, средний кембрий, Северная Америка;
в - Атрух длиной 15 см (отряд Ptychopariida), образ жизни неясен, ордовик,
Швеция; г — Deiphon forbesi, xl.3 (отряд Phacopida), вероятно, планктонная
дрейфующая форма, силур, Богемия; д - Radiaspis radiata длиной 2.5 см (отряд
Odontopleurida), девон, Германия; е - Terataspis grandis длиной 46 см (отряд Li
chida), преимущественно ползающий бентос, средний девон, Нью-Йорк; ж -
Ceratarges armatus длиной 3.8 см (отряд Lichida), вероятно, ползающая форма,
средний девон, Германия
ЖИВОТНЫЕ ИЗ ТРЕХ ЧАСТЕЙ: ТРИЛОБИТЫ
193
и брюшные шипы были весьма полезны для облегчения веса трилобита, ког-
да он отдыхал на мягком илистом субстрате. Брюшные шипы могли также
служить опорой, когда трилобит слегка приподнимался над субстратом.
В случае с одонтоплевридным Acidaspis шипы, очевидно, поддерживали це-
фалон в определенном специфическом положении, которое было эффектив-
ным, когда трилобит питался подножным кормом. Шипы также использова-
лись для зарывания.
Некоторые трилобиты, такие как представители подотряда Harpina (на-
пример, Harpes), имеют на головном щите широкую уплощенную кайму
(лимб), продолжающуюся назад в пару удлиненных щечных шипов. Такая
широкая внешняя кайма, как и удлиненные щечные шипы, могли рассредо-
точивать вес этого малоподвижного трилобита, используя широкую пло-
щадь, удерживая его, таким образом, от погружения (как это очень похоже
достигается на лыжах и снежной доске). Другое объяснение: кайма, прони-
занная кровеносными сосудами, служит для дыхания.
Свободно- и пассивно-плавающие трилобиты. Примером свободно-плаваю-
щих трилобитов может служить род Cyclopyge с громадными глазами, кото-
рые могли видеть во всех направлениях (см. фиг. XI. 16). Глаза кажутся ужас-
но большими относительно тела взрослого Cyclopyge, размеры которого не пре-
вышали 1 см. Некоторые исследователи думают, что трилобит Cyclopyge днем
прятался, а ночью бодрствовал, освещая тьму с помощью света, испускаемого
его округлыми органами, находящимися на верхней поверхности цефалона.
Кроме свободно плавающих были, вероятно, пассивно плавающие (дрей-
фующие), то есть планктонные трилобиты. Маленькие размеры, как у агно-
стид, могли способствовать такому образу жизни. Некоторые крошечные аг-
ностиды, вероятно, могли составлять часть планктона раннего палеозоя. Не-
которые плавающие трилобиты имели увеличенные глаза, а другие, как
Agnostus, были слепыми. Большие глаза полезны во многих отношениях:
они помогали в ночном видении, осуществляли широкий обзор и давали сте-
реоскопическое изображение. Слепота встречается у животных, ведущих
различный образ жизни. Слепыми могут быть формы, живущие в простран-
ствах, где отсутствует свет и видеть невозможно. Зарывающиеся формы
обычно тоже слепые, такими же бывают планктонные формы. Таким обра-
зом, надо аккуратно использовать информацию при реконструкции образа
жизни ископаемого. Необходимо осмыслить всю анатомию животного, ког-
да ведут “восстановление” повседневной жизни ископаемых форм.
Трилобиты-гигинты. Маленькие агностиды имеют тольк 0.6 см в длину,
подавляющее большинство трилобитов достигают 1.8-8 см. Некоторые фор-
мы, конечно, еще больше, но ряд трилобитов превосходят своих соплеменни-
ков настолько, что мы можем назвать их гигантами.
Первым гигантом был Paradoxides harlani, найденный в нижнекембрий-
ских слоях Массачузеттса; он достигал в длину 48 см, а его головной щит почти
равен 30 см в ширину. Другим гигантом был Isotelus, который достигал в дли-
ну 69-76 см. Более яркими, тем не менее, были Uralichas и Terataspis, имею-
щие инкрустированные панцири. Последний достигавший в длину 48-69 см,
был среди всех крупных трилобитов и наиболее шипастым (см. фиг. XI. 16).
ПРОБЛЕМАТИЧНЫЕ КЕМБРИЙСКИЕ АРТРОПОДЫ - “TRILOBITOIDEA”
Кембрийские породы Европы, Сибири, Северной Америки, Китая и Авст-
ралии содержат разнообразных членистоногих, когда-то называвшихся ра-
13 Каменная книга
194
КАМЕННАЯ КНИГА
Фиг. XI. 17. Anomalocaris nathorsti — среднекембрийское членистоногое из
сланцев Бёрджесс Канады (Gould, 1989)
рд — ротовой диск, описанный первоначально как медуза Peytoia (см. также
фиг. VII.3)
кообразными. Только некоторые принадлежат этой группе, но остальные
выглядят иначе, их родственные отношения среди членистоногих неясны.
Они могли развиваться в течение длительного времени “эволюционного экс-
перимента” в докембрии, а затем дать начало группам членистоногих, суще-
ствовавших в последующее время. В старых консервативных классификаци-
ях их считают родственными трилобитам и называют “трилобитоидами”.
Необходимо провести еще много исследований и полевых работ, прежде чем
появится понимание этих необычных и древних “членистых животных”. Ог-
ромное число экземпляров и видов найдено в среднекембрийских сланцах
Бёрджесс; тем не менее группа просуществовала по девон включительно.
Marrella, часто называемая кружевным крабом, имела очень узкое туло-
вище, четыре очень крупных шипа и много жабронесущих конечностей. Ее
считали только что полинявшим трилобитом, но эта интерпретация, по-ви-
димому, базировалась на двух парах перистых антенн. Несмотря на свой
нежный кружевной облик, Marrella часто встречается в сланцах Бёрджесс.
Sidneyia имела около 13 см в ширину, очень широкие головогрудь и туло-
вище, стебельчатые глаза и тельсон, подобный плавнику.
Эти и другие проблематичные кембрийские артроподы включают не толь-
ко плавающих хищников, но и зарывающиеся формы, то есть включают все
то разнообразие первых “эволюционных проб”, окончившихся преимущест-
венно вымиранием (фиг. XI. 17).
Глава XII
РАКООБРАЗНЫЕ
• Трилобиты напоминают нам раков, омаров и даже крабов, так как
все они имеют членистые конечности и панцири, что характерно
для артропод. И все-таки трилобиты и ракообразные принадлежат
разным ветвям типа Arthropoda. Раки, омары и крабы относятся к ракооб-
разным - группе, куда также входят креветки, усоногие рачки и менее изве-
стные существа, часть из которых представлена микроскопическими форма-
ми. Хотя большинство ракообразных живет в соленой или пресной воде, не-
которые обитают на суше, а часть даже стала паразитами.
Название “Crustacea” обязано присутствию панциря в виде твердой корки
(греч. crust - корка, кора), что характерно для этой группы, но не для всех ее
представителей. У некоторых ракообразных панцирь развит плохо или даже
отсутствует.
РАЗНООБРАЗНЫЕ РАКООБРАЗНЫЕ
Некоторые исследователи считают ракообразных классом; другие - над-
классом или подтипом. Несмотря на разный таксономический ранг, присуж-
даемый им, животные остаются по существу теми же самыми. Все они четко
отличаются от трилобитов в присутствии двух пар антенн (а не одной) и трех
пар конечностей, которые стали специализированными органами, рвущи-
ми, жующими и толкающими добычу в рот. Конечности редко сохраняются
у ископаемых форм. Тело ракообразных состоит из головы, грудного отдела
(торакс) и брюшка (абдомен); первые две части обычно в различной степени
сливаются, образуя головогрудь (цефалоторакс).
Ракообразные состоят из нескольких групп, но только Branchiopoda,
Ostracoda, Cirripedia и Malacostraca несомненно представлены в ископае-
мом состоянии.
Жаброногие, или Branchiopoda. Жаброногие представлены довольно
примитивными созданиями крошечных, маленьких и средних размеров от
2 мм до 10 см. Они встречаются, возможно, в кембрии, но достоверно с девона
по настоящее время.. Как и примитивные многочленистые трилобиты, они
имеют большое число сегментов и членистых ножек, несущих жабры, ис-
пользуемые для дыхания, передвижения и передачи пищи ко рту. Ножки
развиты только у грудных сегментов, а у брюшных они отсутствуют. Совре-
© 1997. О.Б. Бондаренко, перевод на русский язык и изменения.
13*
196
КАМЕННАЯ КНИГА
Conchostraca
(Раковинные листоногие)
Фиг. XII. 1. Представители отряда листоногих рачков (Phyllopoda) подкласса
жаброногих (Branchiopoda)
а - щитни, или нотостраки: реконструкция Tryops, х2, пермь, Оклахома; б- ра-
ковинные листоногие, или конхостраки: реконструкция Cyzicus, х10, поздний
карбон, Огайо. Род указывают повсеместно от девона—поныне
менные жаброногие редко живут в нормально соленых морях, засоленных
лагунах и соленых озерах, а все больше в пресноводных бассейнах, в том чис-
ле и временных, таких как лужи и прудики, остающиеся после дождя и таю-
щего снега.
Жаброногие отряда Phyllopoda имеют хитиновый спинной щит - кара-
пакс (Notostraca, или подотряд щитни) либо двустворчатую раковину (Con-
chostraca, или подотряд раковинные листоногие). Некоторые, тем не менее,
не имеют ни того, ни другого (отряд Anostraca), их остатки встречаются на-
чиная с юры.
Нотостраки, или щитни, довольно прогрессивная группа, так как
голова и туловище покрыты карапаксом, то есть хитиновым спинным щи-
том, отсюда название щитни. Они встречаются с карбона-поныне. Наиболее
известный род — Triops, в его состав включают палеозойские и мезо-
кайнозойские виды (фиг. XII. 1). На нескольких ископаемых экземплярах
сохранились следы железы, выделявшей карапакс.
Конхостраки, или раковинные листоногие, наиболее частые
ископаемые, встречающиеся с девона. Подавляющее большинство покрыва-
ют свое тело двустворчатой раковиной и поэтому выглядят как маленькие
двустворки, если не видеть жабронесущие ножки и антенны. Некоторые ис-
копаемые представители этой группы были морскими формами, но боль-
шинство обнаружены в пресноводных континентальных отложениях. Среди
современных листоногогих рачков наиболее известна Daphnia, она является
пресноводной формой, часто продаваемой в зоомагазинах как еда для аква-
риумных рыбок. Некоторые виды Cyzicus и близких родов встречаются в де-
вонских и более поздних отложениях и до сих пор обильны в современных
прудах. Громадное число раковин Cyzicus найдены в пресноводных сланцах
триасового возраста у Принстона в штате Нью-Джерси.
Ископаемые конхостраки очень важны как руководящие фоссилии в не-
морских разрезах, таких как девон Северного полушария, пермо-карбон обо-
их полушарий и мезозой Азии. В этих местах и для этого времени трудно
найти подходящие руководящие ископаемые, и тогда маленькие конхостра-
ки приобретают важное значение для стратиграфии.
РАКООБРАЗНЫЕ
197
Фиг. XII.2. Остракоды: отряды Bradoriida (ранний—поздний кембрий), Palaeo-
copida (ордовик—пермь, ?современность) и Podocopida (ордовик—современность)
а — отряд Bradoriida: “Bradoria" robusta, х8, ранний кембрий; б, в - Palaeocopi-
da: б - Paraechmina spinosa, х20, силур, венлок, от Онтарио до Мэриленд; в -
Glyptopleura, хЗО, карбон-пермь, Европа и Северная Америка; г—и - отряд
Podocopida: г — современная форма, увел. (Ан - антенна, Гл - глаз, Мо - отпечат-
ки мускула, Нж - ножки); д — Ranapeltis, хЗО, девон, Северная Америка; е -
Eocytheropteron, х35, мел-эоцен, повсеместно; ж,з~ Loxoconcha, снаружи и из-
нутри, х15, мел-современность, повсеместно; и - “Bradleya", х80, поздний
мел-современность, Европа, Африка, Северная Америка
Остракоды, или ракушковые рачки. Эти ракообразные так же, как и рако-
винные листоногие, имеют двустворчатые раковины, но известковые, тело
короче и без сегментации, число пар грудных ножек от одной до трех, но они
могут и отсутствовать. Одна пара конечностей - это ногочелюсти, другие па-
ры конечностей используются для ползания, плавания и рытья; самцы ис-
пользуют одну пару конечностей также для удержания самочек во время
спаривания.
Большинство видов имеют только один глаз, хотя некоторые имеют два
сложных глаза, но известны и слепые формы. Створки открываются и за-
крываются полностью и особенно хорошо у примитивных форм. Большинст-
во видов от 0.2 до 2 мм в длину, некоторые ископаемые превышают 80 мм; со-
временные планктонные формы достигают 50 мм.
Современные остракоды живут в морях, опресненных заливах и в прес-
ных водоемах, где они плавают, ползают, прикрепляются к водорослям или
зарываются в ил; несколько видов обитают в почвах тропических лесов. Ост-
ракоды, вероятно, возникли в морских условиях. Они внедрились в пресные
198
КАМЕННАЯ КНИГА
Фиг. XII.3. Остракоды: отряды Beyrichicopida (ордовик—пермь) и Leperditicop-
ida (ордовик—девон)
а-г — отряд Beyrichicopida: а — Drepanella crassinoda, х15, ордовик, от Виргинии
до Индианы; б - Eurychilina reticulata, х20, средний ордовик, США; в — Zygobey-
richia, х20, поздний силур-ранний девон, Европа и Северная Америка; г - Kirk
byella, Х16, силур-карбон, Северная Америка; д-ж — отряд Leperditicopida: ра-
ковинка остракоды в трех положениях, х2
воды в карбоне и там развивались в различных направлениях. Некоторые ос-
воили лесные почвы. Остракоды питаются водорослями, в том числе и диато-
меями, а также различными живыми и мертвыми животными. Остракоды,
по-видимому, были падалеедами уже в далеком геологическом прошлом, о
чем свидетельствует находка палеозойского “ракоскорпиона” из эврипте-
рид. Его конечности и тело, где когда-то находилась пищеварительная систе-
ма, были облеплены сотнями раковинок остракод. То, что картина не обрат-
ная (ракоскорпион поедал остракод), доказывается тем, что раковинки ост-
ракод целые, не разломанные и не раздавленные.
Остракоды линяют, как и другие членистоногие; некоторые виды по мно-
гу раз, пока достигнут своего окончательного облика и размеров.
Остракоды появились в раннекембрийских морях и обильны до сих пор.
Некоторые имеют гладкие раковины, но другие украшены ребрами, бугорка-
ми, ямками и бороздками, все это учитывается при определении системати-
ческого положения (фиг. XII.2, 3). Благодаря тому, что многие виды геогра-
фически были распространены широко, а существовали недолго, они явля-
ются хорошими руководящими ископаемыми (индекс-фоссилии).
Действительно, из всех ракообразных остракоды имеют наибольшее гео-
логическое значение, являясь одной из прекрасно документированных
групп беспозвоночных. Среди них самыми важными оказались морские бен-
тосные, а не планктонные формы, потому что последние имеют столь слабо
объизвествленные раковинки, что редко сохраняются в ископаемом состоя-
нии. В результате этого и потому, что у остракод нет планктонной личинки,
их можно использовать для внутрибассейновой, но не межконтинентальной
корреляции. Кроме того, остракод очень часто используют и при палеоэколо-
гических реконструкция, так как они существуют в определенных условиях
обитания.
Остракод обычно подразделяют на пять отрядов (Archaeocopida, Leperditi-
copida, Myodocopida, Beyrichicopida, Podocopida), основываясь на строении и
РАКООБРАЗНЫЕ
199
скульптуре раковины, конструкции замка, соединяющего две створки (если
он есть), степени объизвествления или фосфатизации раковины и на распо-
ложении рубцов на внутренней поверхности, оставленных мускулами, кон-
тролирующими открывание и закрывание створок.
Усоногие рачки, или Cirripedia. До 1829 года усоногие рачки относились
к моллюскам на основании того, что большинство из них имеет толстые изве-
стковые скелеты. Затем обнаружили, что они - ракообразные, которые на
молодых стадиях плавают или дрейфуют во взвешенном состоянии. После
линьки (от одной до трех) они строят двустворчатые раковинки, очень похо-
жие на раковинки остракод; затем они прикрепляются головой к субстрату,
обычно теряя двустворчатую раковинку, и секретируют другой наружный
скелет, состоящий из табличек, которые перекрывают друг друга или плотно
держатся вместе с помощью жесткой кожи. Пища подается ко рту ветвисты-
ми бахромчатыми ножками, которые напоминают колеблющиеся перышки
или усики, что и послужило причиной названия Cirripedia (Циррипедии),
что в переводе означает “бахромчатоногие” или “усоногие”. Эти ножки, рит-
мично двигаясь, хватают пищу, которую потом “слизывают” гребенчатые
придатки у рта. Утверждение, что ножки создают ток воды, который вносит
пищу в рот, оказалось несостоятельным. Некоторые усоногие стали парази-
тами, они питаются, вгрызаясь ртом внутрь, или прямо абсорбируют пищу
через покровы тела.
Наиболее древний усоногий рачок, Cyprilepas, найден прикрепленным к
лимбу панциря позднесилурийского “ракоскорпиона” из эвриптерид. Сур
rilepas относится к отряду Thoracica, единственной группе усоногих рачков,
хорошо представленной в ископаемом состоянии. Эта древняя форма имела
“стебель”, как современные усоногие, называемые “морскими гусями” или
“морскими уточками”. Вся конструкция рачка со стеблем внешне напомина-
ет гусиную голову с длинной шеей. Их называли так потому, что когда-то ду-
мали, что они связаны с древними дикими гусями. Современные виды при-
крепляются к субстрату длинными “стеблями”, чешуйчатыми или голыми,
но некоторые современные и древние роды имели короткие стебли, которые
полностью были покрыты табличками. Таблички на теле рачка подвижны,
они удерживаются вместе благодаря плотной коже. После смерти кожа сгни-
вает, поэтому ископаемые усоногие рачки обычно представлены разрознен-
ными табличками. Таблички довольно часто встречаются в мезозойских и
кайнозойских отложениях Донбасса, Крыма, Западной Европы и других ре-
гионов. Несколько “морских уточек” было обнаружено на внешней поверх-
ности раннемелового аммонита (фиг. XII.4).
Усоногие рачки, называемые “морские желуди”, получили свое название
за внешнюю форму, округлую или коническую (см. фиг. XII.46). В отличие
от морских уточек таблички у них плотно соединены. У морских желудей,
кроме того, нет стебля, они плотно прикрепляются всем основанием к субст-
рату, а таблички, которые покрывают тело, плотно соединены друг с другом,
хотя те, что образуют крышечку, свободно двигаются. Превосходные эк-
земпляры морских желудей были найдены в некоторых мезозойских и кай-
нозойских отложениях.
Усоногие рачки, хотя и появились в палеозое, по существу дивергировали
только в мезозое, а своего пика они достигли в кайнозое. Некоторые палеон-
тологи, обратив на это внимание, называют современное время “Веком усо-
ногих рачков”. Прослеженная закономерность, возможно, больше отражает
скудость ископаемых данных, чем факт усиления современного разнообра-
зия. Так, многие современные паразитические циррипедии не имеют твер-
200
КАМЕННАЯ КНИГА
Фиг. ХП.4. Усоногие рачки — Cirripedia (кембрий—современность)
а — Protobalanus, х10, систематическое положение дискуссионное, средний де-
вон, Нью-Йорк; б — морской желудь Balanus, прикрепленный к раковине гаст-
роподы, миоцен Северной Америки (род встречается с эоцена-поныне, повсеме-
стно); в — морская короткостебельчатая уточка Loriculina, мел, почти повсеме-
стно; г — пять морских уточек, прикрепленных к раковине раннемелового
аммонита Cheloniceras (Друщиц, Зевина, 1969)
дых образований и поэтому у них очень мало шансов сохраниться в ископае-
мом состоянии.
Усоногие рачки, вероятно, произошли от группы ракообразных, называе-
мых Maxillopoda, которые обладают пятью головными, шестью грудными и
пятью брюшными (абдоминальными) сегментами. От Maxillopoda произош-
ла и часть других членистоногих, включая миниатюрных водных копепод и
остракод. Следует знать и то, что Чарльз Дарвин, всемирно известный благо-
даря своей теории эволюции, в свое время написал многотомную моногра-
РАКООБРАЗНЫЕ
201
фию по ископаемым и ныне живущим усоногим рачкам. И хотя с тех пор
проведено много исследований, работа Дарвина оказалась столь полной и
тщательной, что до сих пор остается основополагающей по этой группе.
Твердые “мягкораковинпые”, или Malacostraca
Остракоды, может быть, являются наиболее многочисленными среди ис-
копаемых ракообразных, а усоногие рачки представляют собой наиболее вы-
соко специализированную группу, но малакостраки, или высшие ракообраз-
ные, — это те, о ком вспоминают прежде всего, когда говорят о ракообразных
вообще. Это крабы, креветки, омары и еще огромное разнообразие форм, ко-
торых нам никогда полностью не изучить.
Malacostraca (дословный перевод с греческого - “мягкораковинные”) -
неудачное название для высших ракообразных, так как оно отражает только
кратковременное состояние, наступающее сразу после линьки. В остальное
время большинство малакострак хорошо защищено раковиной или панци-
рем, который состоит из плотного хитина, часто отвердевшего и утолщенно-
го за счет известкового материала. Карапакс, или панцирь, представлен раз-
нообразно: от почти сплошного покрова головы и тела до почти полного его
отсутствия.
Для высших ракообразных характерно присутствие брюшных ножек и
фиксированное число сегментов: пять головных, восемь грудных и шесть
(реже семь) брюшных. Головные сегменты несут антеннулы, антенны и три
пары челюстей. Грудные передние сегменты с одной-двумя или тремя па-
рами челюстей, иногда преобразованных в клешни, а задние с ходильными
ножками. Многие высшие ракообразные имеют веерообразный хвост. На го-
лове обычно два сложных фасеточных глаза, реже глаза отсутствуют. Мала-
костраки впервые появляются в раннем кембрии и существуют по настоящее
время.
Филлокариды — Phyllocarida (фиг. XII. 5). Они являются важной группой
малакострак. Филлокариды имеют двустворчатую раковину, которая по-
крывает только фронтальную часть тела, в отличие от остракод, у которых
тело полностью заключено в раковину. Филлокариды (“листовидные кревет-
ки”) представлены относительно крупными ископаемыми размером от 8 мм
до 4 см, а не микроскопическими: таким образом, они отчетливо крупнее ос-
тракод и жаброногих.
Филлокариды, вероятно, являются наиболее примитивными малакостра-
ками, и формы, подобные им, несомненно, дали начало более совершенным
группам этого класса. Филлокариды впервые появились в раннем кембрии и
живут до сих пор. Два рода из сланцев Бёрджесс среднего кембрия, отнесены
к филлокаридам: Perspicaris и Canadaspis. Интересно, что Canadaspis явля-
ется наиболее примитивным ракообразным.
Десятиногие — Decapoda. Большинство ископаемых малакострак отно-
сятся к декаподам — ракообразным, имеющим десять ножек, осуществляю-
щих движение. Грудной отдел (торакс) состоит из шести сегментов, которые
объединены с головой в головогрудный панцирь. Глаза находятся на по-
движных стебельках. Хвостовая часть обычно оканчивается хвостовым
веером.
Декаподы впервые появляются в перми. Они редки в триасовых породах,
но в верхнеюрских литографских известковых сланцах южной Германии
они многочисленные и разнообразные. Ископаемые сохраняют панцирь, ан-
тенны, ножки, конечности брюшка и веерообразный хвост.
202
КАМЕННАЯ КНИГА
Фиг. ХП.5. Представители филлокарид подкласса высших ракообразных
(Malacostraca)
а — Caryocaris curvilata, х4, ордовик, граптолитовые сланцы, широко распрост-
ранен в Северной Америке; б - Dithyrocaris oceani, х0.5, поздний девон,
Нью-Йорк; в - Echinocarispunctata, xl, средний-поздний девон, США
Раки-кроты представлены тонкопанцирными зарывающимися формами.
Раки-отшельники имеют незащищенное брюшко, которое они прячут в ра-
ковинах других животных. Клешни раков и креветок часто находят в мело-
вых и палеоген-неогеновых отложениях (фиг. XII.6).
Шипастых омаров иногда называют лангустами. Современные виды име-
ют колючие панцири, массивные антенны, все десять ног используют для
хождения. Некоторые ископаемые имели длинные тонкие клешни; у других
передние конечности были очень длинными, а последний сегмент был по-
крыт волосовидными отростками, которые, по-видимому, использовались
для осязания. Большинство ископаемых шипастых омаров находят в мезо-
зойских породах Европы, лучшие экземпляры происходят из юрских лито-
графских известковых сланцев Германии. В них также обнаружены первые
настоящие омары (Егута), размером менее 5 см в длину с клешнями, похо-
жими на удлиненные плоскогубцы. Род Astacus, речной рак, был найден в
миоценовых, возможно, плиоценовых породах Айдахо и восточного Орегона.
Крабы являются наиболее распространенными современными ракообраз-
ными и более редкими, как ископаемые. Краба легко узнать по его широко-
му тяжелому карапаксу, короткому брюшку, который прячется под торак-
сом, коротким антеннам и по отсутствию веерного хвоста. Первые крабы по-
явились в средней юре Европы. Древние роды очень сходны с теми
современными формами, которые до сих пор сохранили черты, связываю-
РАКООБРАЗНЫЕ
203
Фиг. XII.6. Представители отряда десятиногих раков (Decapoda) подкласса
высших ракообразных (Malacostraca)
а - Antrimpos kiliani, х0.5, примитивная креветка из золенгофенских сланцев,
юра, Германия; б - Callianassa mortoni, х0.75, клешня роющего норы рака, по-
здний мел, от Нью-Джерси до Техаса; в - Palaeop alaemo п newberryi, х2, панцирь
рака сбоку, поздний девон Огайо-рацний карбон Айовы; г, д - панцирь — кара-
пакс краба с конечностями с верхней и нижней стороны из палеоген-неогено-
вых отложений Панамы, близкий современному тихоокеанскому съедобному
крабу Cancer magister
щие их с ракообразными, имеющими веерные хвосты. Много причудливых
крабов существовало в морях, которые покрывали Южную Европу, Север-
ную Азию и Северную Африку в течение эоценовой эпохи. Часть из них (как
и немногие современные крабы) сильно шипастые, они выглядели подобно
известковым водорослям. Некоторые роды и виды крабов являются руково-
дящими ископаемыми для палеоген-неогеновых формаций, хотя они встре-
чаются намного реже, чем моллюски. Восточный съедобный, или голубой
краб (Callinectes sapidus) обнаружен в плейстоценовых отложениях Масса-
чусетса и Нью-Джерси. Близкий западный съедобный краб (Cancer fissus) -
плиоценовое ископаемое Калифорнии.
Все основные группы декапод появились в юрское время. Все они продол-
жают существовать и сейчас, и многие достигли большого разнообразия и
широкого распространения в течение второй половины кайнозоя.
Глава XIII
ЧЛЕНИСТОНОГИЕ ОТ МЕЛКОВОДНЫХ
ДО СУХОПУТНЫХ И ЛЕТАЮЩИХ
• Хотя многие членистоногие в настоящее время являются наземны-
ми формами, самые древние представители типа были водными.
Первая вспышка разнообразия членистоногих (радиация), произо-
шедшая в докембрийские времена, разыгрывалась в мелководных эпиконти-
нентальных морях, где многие червеподобные группы дали начало различ-
ным артроподам. Членистоногие, очевидно, оставались водными, пока кон-
тиненты не покрылись растительностью, представленной настоящими
наземными растениями, а не только мхами и лишайниками. Эти растения
создали убежища и влажные участки, а также разнообразную пищу, открыв
для членистоногих новые богатые условия обитания, облегчив им решение
проблемы жизни в наземных условиях.
Процесс преодоления проблем был постепенным и медленным. Еще до
окончания кембрийского периода, потомки некоторых трилобитов проник-
ли в устья рек. В течение ордовика родственники этих животных внедрялись
выше по течению. Постепенно у них исчезли антенны, а вторая пара конеч-
ностей стала клешнеподобной. В настоящий момент нет уверенности, кто
точно явился предком хелицеровых, но очень соблазнительно думать, что
они произошли непосредственно от трилобитов.
Наземные растения, очевидно, появились в середине силура в венлокский
век, что примерно означает рубеж, соответствующий 420 млн. лет тому на-
зад. Прежде чем окончился силурийский период, то есть в интервале
420-405 млн. лет назад, водные хелицеровые стали осваивать сушу, о чем
свидетельствуют ископаемые остатки из Швеции, Шотландии, Англии и Се-
верной Америки. У других хелицеровых (не сухопутных) развивались осо-
бенности строения, позволяющие им жить в мелководных солоноватых во-
дах или в приливно-отливных зонах. Один из таких современных потомков
может жить вне воды целыми днями, совершая путешествия в глубь суши на
расстояние от нескольких метров до более чем 1.5 км.
Хелицеровые образуют огромную группу артропод, встречающуюся
уже в породах кембрия. До сих пор они являются весьма многочисленной
группой. Тело всех хелицеровых подразделяется на две части: просому
(слившиеся голова и грудь) и опистосому (абдомен). Название группы, тем
не менее, происходит от клешневидных конечностей (хелицеры), нахо-
дящихся перед ртом, что характерно для этих членистоногих. Паукооб-
разные (Arachnida) являются хелицеровыми, как и две другие группы:
© 1997. О.Б. Бондаренко, перевод на русский язык и изменения.
ЧЛЕНИСТОНОГИЕ ОТ МЕЛКОВОДНЫХ ДО СУХОПУТНЫХ
205
Хелицеры
Фиг. XIII.1. Строение современного мечехвоста Limulus polyphemus с верхней
(слева) и нижней (справа) стороны (подтип Chelicerata)
мечехвосты (Xiphosura) и эвриптериды (Eurypteridd), или водные “рако-
скорпионы”.
СТОЙКИЕ КСИФОСУРЫ, ИЛИ МЕЧЕХВОСТЫ
Современный скиталец мечехвост, или краб-подкова, знаменит тем, что
он так называемое живое ископаемое. Его называют Limulus, или мечехво-
стом; он существует, мало меняясь, с раннепермского времени. Уже в тече-
ние 280 млн. лет мечехвост имеет широкую головогрудь (просома), которая
напоминает по очертаниям подкову лошади, оканчивающуюся “щечными”
шипами разной длины (фиг. XIII. 1, 2). Как и головной щит трилобитов,
головогрудный панцирь мечехвоста прикрывает как голову, так и внут-
ренние органы (печень, желудок и кишку). За головогрудью идет шипас-
тый брюшной щит (называемый также абдоменом), за которым следует
“хвост” - тельсон.
У мечехвостов имеется два сложных и два простых глаза; рот расположен
центрально под головогрудью; имеется шесть пар конечностей, но без ан-
тенн. Первые две конечности клешневидные, они называются хелицеры. Ос-
тальные четыре пары используются для хождения. Они оканчиваются пин-
206
КАМЕННАЯ КНИГА
Фиг. XIII.2. Древние ископаемые мечехвосты (подтип Chelicerata)
a -Aglaspts spinifera длиной около 10 см, поздний кембрий, Висконсин; б-Bun-
odes, длиной около 10 см, специализированный орнаментированный мече-
хвост, силур, Европа; в - Pseudoniscus roosevelti, силур, Нью-Йорк; г - Prestwi-
chianella danae длиной около 6 см, пресноводный мечехвост, средний-поздний
карбон, Иллинойс; д - Palaeolimulus avitus длиной около 1.8 см, ранняя пермь,
Канзас
цетовидными щипчиками, которые режут пищу на кусочки, прежде чем она
попадет в рот. Шестая пара конечностей является специализированной, чья
основная функция, как у тягача, протащить мечехвоста вперед сквозь ил и
песок. Нижняя сторона брюшка (или абдомена) несет широкие придатки,
ЧЛЕНИСТОНОГИЕ ОТ МЕЛКОВОДНЫХ ДО СУХОПУТНЫХ
207
двигающиеся равномерно, когда животное плывет. Эти придатки также ис-
пользуются для дыхания, так как они несут жаберные “книжки”, состоя-
щие из 150—200 очень тонких листочков, от которых кислород воды посту-
пает в кровь.
Наиболее известный мечехвост - это краб-подкова, или Limuluspolyphe-
mus, живущий вдоль Атлантического побережья от Новой Шотландии и
далее на юг вдоль побережья Флориды, где начинается Гольфстрим. Мече-
хвост Limulus роет и вспахивает дно в поисках червей и других мелких
животных. Весной мечехвосты заходят в очень мелководные заливы, где
они откладывают яйца. Следы ползания мечехвостов легко увидеть во вре-
мя отлива, одни следы оставлены одиночками, а другие - парами, в кото-
рых маленький самец следует за хвостом-тельсоном своей более крупной
подруги. Эти следы могут быть как мелкими, так и глубокими; они часто
изгибаются и вьются в разных направлениях на значительных простран-
ствах, прежде чем закончатся неправильной ямкой, в которой откладыва-
ются яйца.
Древние родственники мечехвостов. Пока юный мечехвост Limulus
развивается, он линяет, растет и изменяет свой облик весьма значительно.
По мере развития он проходит стадии, сходные с теми стадиями, которые
прошли мечехвосты в течение палеозоя (см. рис. XIII.2). Первая стадия
напоминает род Aglaspis, позднекембрийского членистоногого с относи-
тельно коротким головным щитом (цефалоном), одиннадцатью туловищ-
ными сегментами и шестнадцатью парами ног, идущими после хелицер.
Род Aglaspis относится к одноименному отряду Aglaspida, существовавше-
му только в кембрии. Затем у мечехвоста Limulus наступают стадии, напо-
минающие палеозойские формы со все уменьшающимся числом сегмен-
тов. Силурийский Bunodes, например, имел девять сегментов; Belinurus
(девон-карбон) обладал восемью, a Euproops (средний-поздний карбон)
имел семь. Род Palaeolimulus из пермских пород Канзаса обладал уже
слившимся брюшком, хотя насечки в его срединной части являются
реликтами шести сегментов. Начиная от древнейшего мечехвоста Palaeo-
merus (ранний кембрий) к ныне живущему Limulus основные изменения
сводятся к укорачиванию туловища и к увеличению общих размеров чле-
нистоногого.
Большинство древних мечехвостов были морскими, но Belinurus и Eu-
proops внедрились в солоноватые лагуны и даже проникли в заболоченные
пространства. Особенно это относится к роду Euproops, он является редким,
но очень характерным ископаемым каменноугольных конкреций Mazon
Creek, Иллинойс, которые также содержат наземные растения, пресновод-
ных рыб и моллюсков.
Как было сказано, современный Limulus ползает по песчаному и песча-
но-илистому субстрату во время спаривания. Древние следы, оставлен-
ные мечехвостами, были обнаружены в формациях от ордовика по мел.
Одни из них, как, например, следы из девона и позднего карбона, были
когда-то приняты за следы конечностей амфибий; другие, из триаса Нью-
Джерси, были приписаны крошечным рептилиям. О наиболее драматиче-
ской истории повествуют следы на поверхности юрских сланцев Герма-
нии. Оказалось, что они оканчиваются у остатков мечехвостов, назван-
ных Mesolimulus. Вместо того, чтобы двигаться к побережью, там спари-
ваться и откладывать яйца, эти мечехвосты легкомысленно пытались
вернуться в соленые воды, но потеряли силы и погибли в своих неудач-
ных попытках.
208
КАМЕННАЯ КНИГА
Фиг. XIII.3. Эвриптериды и наземный скорпион (подтип Chelicerata)
а — Palaeophonus nuncius, достоверный сухопутный скорпион с жалящим “хвос-
том”, поздний силур, Швеция; б, в - водный ракоскорпион Eurypterus remipes
длиной около 20 см с верхней (б) и нижней (в) стороны, поздний силур, Нью-Йорк
ЭВРИПТЕРИДЫ, ИЛИ МОРСКИЕ РАКОСКОРПИОНЫ
Другая ископаемая группа, принадлежащая меростомным хелицеровым -
это эвриптериды, или водные ракоскорпионы.
Разнообразные виды эвриптерид, существовавших от ордовика по пермь в
Европе, Северной и Южной Америке, обнаружены в солоноватоводных и
пресноводных отложениях. Некоторые формы, известные из позднего силура
Западной Австралии, оставили “четкие следы хождения по сухой” земле - это
первое свидетельство о жизни, шагнувшей в наземные условия обитания.
В целом эвриптериды напоминают нам стройных агласпид с прямоугольны-
ми головами, расходящимися конечностями и панцирем, орнаментирован-
ным бугорками, чешуйками и ребрышками. Обычная длина эвриптерид от
10-20 до 43 см, но некоторые достигали в длину 1.8-2.1 м (см. фиг XIII.4).
Род Eurypterus имел шесть пар конечностей: первая пара - хелицеры, сле-
дующие четыре пары - короткие шипастые, используемые для хождения, и
ЧЛЕНИСТОНОГИЕ ОТ МЕЛКОВОДНЫХ ДОСУХОПУТНЫХ
209
Фиг. XIII.4. Молодые и взрослые звриптериды (подтип Chelicerata)
а, б — Stylonu.ru.s-. а - молодой экземпляр S. myops длиной 2.4 см, силур, венлок,
Нью-Йорк, б - взрослый экземпляр близкого вида длиной 1.37 м, силур, вен-
лок, Нью-Йорк (интервал существования рода - силур-девон Европы и Се-
верной Америки); в, г- Pterygotus: в - Р. sp., молодой экземпляр длиной 1.7 см,
г — взрослый экземпляр Р. buffaloensis длиной 1.8-2.1 м, поздний силур,
Нью-Йорк
шестая пара выступает наружу в виде лопат или весел (фиг. ХШ.З). Хелице-
ры были столь малы, что ходильные конечности должны были использовать-
ся для удержания пищи, но основания лопатовидных конечностей, несущие
зубцы, служили для перетирания. Последняя пара (шестая) конечностей
просомы были самые крупные и на концах сильно расширенными; они, по-
видимому, служили для плавания. Два крупных сложных глаза и два ма-
леньких простых находились на спинной стороне просомы.
Eurypterus отражает основной план строения эвриптерид. Но время шло, и
эти ракоскорпионы специализировались в различных и часто неожиданных на-
правлениях. У некоторых возникла почти треугольная просома, а первые шесть
сегментов брюшка стали очень широкими. Другие приобрели квадратную про-
сому, их первые четыре конечности удлинились, покрылись множеством тон-
ких шипиков, которые выглядели как ловчие сети для захватывания пищи.
14 Каменная книга
210
КАМЕННАЯ КНИГА
Ордовикский род, ошибочно принятый за граптолит и названный Megalograp-
tus, стал шипастым, и у него вместо острого возник клешневидный тельсон.
Stylonurus сочетал длинный тельсон и сужающийся абдомен с относитель-
но маленькой и очень четкой просомой, на которой сложные глаза придвину-
лись почти к центру (фиг. ХШ.4). Хелицеры (первая пара) были короткими.
Следующие конечности были ходильными, из них первая пара конечностей
была короткой и шипастой, вторая и третья пары были длинными с шипами,
расположенными, как зубцы гребенки, а две последние пары были еще длин-
нее и несли продольные длинные гребни. Эти последние конечности выгля-
дят совсем непохожими на те лопатовидные и весловидные конечности ос-
тальных эвриптерид, оканчивающихся острыми загнутыми коготками.
Stylonurus несомненно ходил на этих восьми конечностях, а шипастые из
них мог использовать для удержания пищи, так как хелицеры были корот-
кими. Силурийские виды были около 13 см длиной, но один вид из среднего
девона Нью-Йорка вырастал до 1.37 м. Но даже этот вид не достигал размеров
Pterygotus, который достигал в длину 1.8-2.1 м. Его сложные глаза были
крупными и находились на краю щита; тельсон представлял собой широкую
пластину; хелицеры были длинными, увенчанными зубовидными клешня-
ми и так сочленены, что могли доставать до рта. Pterygotus мог плавать так
же хорошо, как и ползать; он был активным охотником и наиболее опасным
членистоногим позднего силура и раннего девона.
Стадии роста и тип строения (габитус). Сброшенные панцири показыва-
ют, что эвриптериды проходили интенсивный метаморфоз. Очень молодой
Stylonurus myops, например, был коротким и широким с глазами на припод-
нятых овальных площадках и только с пятью сегментами между головогруд-
ным щитом и тельсоном. На последующих стадиях число сегментов увеличи-
валось, их окончания становились шиповидными, тельсон удлинялся и ста-
новился тонким.
Древние эвриптериды были морскими животными, но в силурийское вре-
мя они стали обитать в широких лагунах или заливах, где вода была опрес-
ненной или, наоборот, чрезмерно соленой. Огромное число молодых эврипте-
рид в сланцевых слоях Отисвилля (штат Нью-Йорк) свидетельствуют о том,
что это пространство служило удобным местом для размножения; оно вполне
сопоставимо с мелководными заливами, где в настоящее время Limulus
polyphemus откладывает яйца.
По мере того, как время шло, все большее число эвриптерид вселялись в оп-
ресненные заливы, а оттуда в реки и даже болота. Об этом свидетельствуют по-
гребенные эвриптериды, оказавшиеся в сланцах и известняках, переслаиваю-
щихся с углями таких районов, как Германия, Новая Шотландия и Централь-
ный Иллинойс США. Род Lepidoderma из среднего-позднего карбона штатов
Пенсильвания и Иллинойс длиной 20-23 см является чешуйчатой формой с
шиповидными сегментами. Он найден среди растений влажных лесов (будущие
угли) в сланцевых слоях, которые содержат также пресноводных двустворок и
гастропод. Последние эвриптериды (пермь Европы) также встречены в ассоци-
ации с ископаемыми лианами и деревьями болотистых лесных массивов.
СКОРПИОНЫ И ДРУГИЕ ПАУКООБРАЗНЫЕ
Другой многочисленной группой хелицеровых являются Arachnida, то
есть паукообразные, которые включают скорпионов, пауков, клещей и не-
сколько других групп.
ЧЛЕНИСТОНОГИЕ ОТ МЕЛКОВОДНЫХ ДО СУХОПУТНЫХ
211
Паук Eophrynus Многоножка Eophoberia
Фиг. XIII.5. Паук и многоножка
а — паук Eophrynusprestvici, средний-поздний карбон, Англия; б - многоножка
Euphoberia armigera, около 9 см в длину, средний-поздний карбон, Иллинойс,
округ Grundy
Одно время думали, что скорпионы произошли от эвриптерид, таких как
Pterygotus, чьи хелицеры были длинными мощными клешнями. На самом
деле хелицеры скорпионов короткие и невыразительные, а их клешни, буду-
чи первой парой ходильных ног, увеличены и изменены. Шестая пара конеч-
ностей никогда не имеет веслоподобного расширения, как это присуще боль-
шинству эвриптерид. Тельсон скорпиона стал чем-то вроде гиподермальной
иглы, способной вспрыснуть яд, выделяемый крупной железой у его основа-
ния. Органы дыхания представлены легкими, забирающими кислород из
воздуха, а не из воды.
Эти черты строения указывают на то, что скорпионы являются “племян-
никами и племянницами” эвриптерид, а не прямыми потомками. Скорпио-
ны весьма древняя группа, так как их полностью развитые экземпляры обна-
ружены в позднесилурийских породах Нью-Йорка и Швеции. Особенно ве-
ликолепные экземпляры происходят из каменноугольных отложений Mazon
Creek, округ Гранди, Иллинойс.
Скорпионы были более разнообразными в прошлом. В течение силура, де-
вона и карбона у скорпионов наблюдается наибольшее число типов строения.
Только одна из этих групп дала начало всем современным скорпионам.
Пауки-долгоножки. Пауки, как и скорпионы, дышат атмосферным возду-
хом, хотя большинство из них для этого используют не легкие, а трубочки -
трахеи, которые разветвляясь пронизывают все тело. Несколько водных ви-
дов существуют как в морских, так и пресных водах. Пауки не имеют клеш-
ней, их тело короткое, без тельсона. Несмотря на эти различия, пауки и скор-
пионы принадлежат одной группе, то есть Arachnida, которая также включа-
ет сенокосцев, сольпуг, фринов и несколько других групп. Тело у
паукообразных, кроме клещей , подразделено на уплощенную головогрудь
(цефалоторакс) и членистое брюшко. Головогрудь несет шесть пар конечнос-
тей, четыре из них - ходильные ноги. Антенны отсутствуют.
Пауки впервые обнаруживаются в средне-позднекаменноугольных поро-
дах (например, Palaeophorius), хотя их полнота развития и разнообразие
указывает, что они появились и эволюционировали задолго до этого времени
(фиг. XIII.5).
14*
212
КАМЕННАЯ КНИГА
Фиг. ХШ.6. Морской паук Palaeoisopus problematicus (Pycnogonida) с разма-
хом конечностей 18 см, ранний девон, Германия (рисунок Frank Knight)
Пауки были намного разнообразнее в прошлом, чем сейчас; только поло-
вина из шестнадцати (или около этого) установленных отрядов дожили до
сих пор; наибольшее разнообразие наблюдалось в позднем палеозое. Один во-
истину гигантский паук, Megarachne servinei, из позднего карбона Аргенти-
ны, обладал конечностями длиной более чем 50 см.
Другая характерная группа паукообразных — это Pycnogonida, или мор-
ские пауки, считавшиеся когда-то редкими. В последнее время думают, что
они произошли от древней морской группы паукообразных. Пикногониды
обладают маленьким брюшком, четырьмя парами ходильных ног, имеющих
разный облик. Третья пара приспособлена для ношения яиц во время инку-
бации. Размеры пикногонид изменяются от нескольких миллиметров до
40 см; они преимущественно бентосные хищники (фиг. ХШ.6). Группа впер-
вые появляется в раннем девоне Германии (Palaeoisopus и Palaeopantopus) и
в настоящее время исключительно многообразна в Антарктических водах,
откуда известно более одной сотни видов.
Наиболее древняя наземная форма из Arachnida - это паук Alkenia из ран-
него девона, он определенно древнее, чем первый наземный скорпион. В обе-
их группах (пауки и скорпионы) наблюдается тенденция слияния и редук-
ции сегментов, что с течением времени превратило их в довольно компакт-
ных животных.
Превосходно сохранившиеся арахниды известны из желваковидных мио-
ценовых пород Калифорнии, из олигоцена западной части Северной Амери-
ки (Oligocene Florissant insect beds) и палеогеновых (эоцен) янтарей Прибал-
тики. Янтарь, который является окаменевшей смолой деревьев, сохраняет
строение даже миниатюрных волосиков этих арахнид.
У НИР АМИИ
Другое крупное подразделение, иногда выделяемое в особый тип, — это те
членистоногие, которые включают всем известных многоножек, насекомых
ЧЛЕНИСТОНОГИЕ ОТ МЕЛКОВОДНЫХ ДО СУХОПУТНЫХ
213
и группу онихофор. Они принадлежат Uniramia, все представители которых
имеют конечности, состоящие из единственной ветви, а не из двух ветвей,
как у трилобитов. Конечности унирамий заканчиваются коготками.
Унирамии впервые появляются в кембрии и существуют поныне. Род
Aysheaia из среднего кембрия сланцев Бёрджесс, панцирь и кутикулярная
структура которой напоминает нам онихофор, является Uniramia (см.
фиг. XI.2). Вместе с ним часто встречаются фрагменты губок, возможно слу-
живших едой для этих членистоногих.
Кембрий и, вероятно, докембрий, как ясно показывает фауна из сланцев
Бёрджесс, были временами “экспериментов” во многих группах членистоно-
гих. У некоторых групп возник удачный план строения, но большинство дру-
гих вымерло в середине палеозоя из-за возрастания конкуренции с другими
беспозвоночными, включающими также и более успешных членистоногих.
Победители соревнования стали предвестниками будущего и дали начало
ныне живущим унирамиям, ракообразным и хелицеровым.
ОНИХОФОРЫ
Современные онихофоры являются небольшими (до 15 см) червеподобны-
ми беспозвоночными, которые живут в сырых влажных условиях, обычно в
пнях или под камнями, преимущественно в южном полушарии. Их иногда
называют бархатными червями. В отличие от червей, у онихофор настоящие
и многочленистые конечности с хитиновыми коготками, как у Aysheaia (см.
фиг. XII. 2). Из-за мозаичной природы строения онихофор помещают в раз-
личные систематические группы, но в большинстве случаев среди членисто-
ногих. У онихофор просто устроенная голова из трех сегментов и червеподоб-
ное сегментированное туловище (до 43 сегментов), не подразделенное на от-
делы. Многое из внутренней анатомии соответствует типу членистоногих, а
эмбриональные стадии онихофор очень сходны с многоножками из подтипа
трахейнодышащих. Нередко онихофор рассматривают как самостоятель-
ный подтип и даже как тип Onychophora, или Protracheata.
Онихофоры впервые появляются в кембрии и существуют поныне, явля-
ясь одной из групп животных, распространенных преимущественно на кон-
тинентах бывшей Гондваны.
МНОГОНОЖКИ
Многоножки представлены длинными ползающими существами, похо-
жими на гусениц, с множеством (до 375 пар) довольно коротких конечнос-
тей. Под названием многоножки, или Myriapoda, объединены четыре класса
подтипа трахейнодышащих. Группа, называемая тысяченожками, состоит
из форм, которые прячутся в почве или под омертвевшими стволами и опав-
шими листьями, где они питаются гниющим материалом или корнями. Они
обладают твердыми, блестящими экзоскелетами, одной парой антенн и дву-
мя парами коротких ножек у подавляющего числа туловищных сегментов,
за что их называют двупарноногими (Diplopoda).
Из известных тысяченожек наиболее древняя ископаемая форма происхо-
дит из пород верхнего палеозоя. Род Euphoberia из каменноугольной форма-
ции Мэзон Крик (Mazon Creek, США) является шипастой формой с относи-
тельно длинными ножками и сегментами, по-видимому, разделяющимися
214
КАМЕННАЯ КНИГА
на нижней стороне тела (см. фиг. XIII. 5). Другая форма из этой же формации
достигает в длину 20 см и является крупнейшей многоножкой мира.
У группы, называемой стоножками (Губоногие - Chilopoda), туловищные
сегменты обладают только одной парой ног, а те, что идут сразу после голо-
вы, превращены в ядовитые зубцы. В отличие от тысяченожек, они являют-
ся быстродвигающимися хищниками. В янтаре Прибалтики обнаружены не-
сомненные стоножки, и, вероятно, к ним также относятся ископаемые, обна-
руженные в среднекаменноугольной формации Mazon Creek.
Все многоножки являются наземными атмосфернодышащими членисто-
ногими с большим числом сегментов и конечностей. Они, как и онихофоры,
обладают отчетливой головой и однородно сегментированными туловищем,
не подразделенным на отделы. У них есть мандибулы, которые приспособле-
ны для кусания косо или поперек, что не характерно для остальных членис-
тоногих. Наиболее представительные формы многоножек - это гигантские
артроплевриды, которые достигали в длину 1.5-2 м (см. фиг* XI.2). Они су-
ществовали в болотистых лесах каменноугольного периода и, вероятно, бы-
ли растительноядными формами, так как внутри них обнаружены остатки
плауновидных растений. Артроплевриды исчезли после каменноугольного
периода, так как перестали существовать те влаголюбивые флоры, с которы-
ми они были тесно связаны по условиям обитания.
ПОРАЗИТЕЛЬНО УСПЕШНЫЕ НАСЕКОМЫЕ: ШЕСТИНОЖКИ
Насекомые являются самым большим и наиболее успешным классом чле-
нистоногих. Более того, они являются самой процветающей группой во всем
царстве животных. Мало того, что они живут в самых разнообразных усло-
виях, их около двух миллионов видов, что составляет около 70% всех видов
животных. Многие насекомые существуют в таком громадном и все увеличи-
вающемся количестве, что, кажется, не реагируют на яды и другие происки,
используемые человеком против них. Все-таки во многих других случаях
разрушение природы человеком привело к исчезновению некоторых видов
насекомых, часть насекомых находятся на грани исчезновения.
Название “insecta” происходит от латинского слова, означающего “разре-
занные на части”. Оно отражает характер строения тела, резко разделенного
на голову, грудь (торакс) и брюшко (абдомен). Голова состоит из шести сег-
ментов, грудь из трех, брюшко состоит самое малое из пяти сегментов и самое
большое из одиннадцати или двенадцати. Голова несет одну пару антенн и
три пары ротовых конечностей. К груди прикреплены три пары ног (всего
шесть), а брюшко вообще не имеет конечностей. Ноги состоят из восьми или
девяти члеников и обычно оканчиваются коготками. Юные формы, то есть
личинки, могут вести водный образ жизни и дышать легкими, но взрослые
особи дышат атмосферным воздухом с помощью разветвленных трубочек -
трахей, значительно более сложных, чем у пауков. Подавляющее большин-
ство взрослых насекомых имеет также плотный хитиновый экзоскелет.
Первичнобескрылые насекомые (Apterygota). Мы часто изображаем на-
секомых, как быстрых созданий с двумя или четырьмя крыльями, проводя-
щих свое основное время в полете, хотя они могут отдыхать или охотиться за
едой и на земле. Тем не менее огромное число насекомых является наземны-
ми членистоногими и почти нелетающими, да и сам класс начинался с бес-
крылых форм. Первичнобескрылые насекомые до сих пор продолжают су-
ществовать — это ногохвостки (Collembola) и щетинохвостки (Thysanura).
ЧЛЕНИСТОНОГИЕ ОТ МЕЛКОВОДНЫХ ДО СУХОПУТНЫХ
215
Фиг. XIIL7. Насекомые палеозойского и раннемезозойского возраста
а - Triassomachilis, триас; б — Rhyniella, вероятно современная форма, нечаянно
попавшая в “ловчую” трещину девонских пород и описанная когда-то как де-
вонское насекомое; в — Stenodictya lobata, х2, примитивное крылатое насекомое
с двумя крыловидными выростами впереди настоящих крыльев, карбон, Фран-
ция (с разрешения F.M. Carpenter); г - Adeloblatta Columbiana, таракан длиной
2.5 см, средний-поздний карбон, Иллинойс
Первые из них очень маленькие, длиной 0.2-10 мм, с короткими антеннами
и с простыми глазками, которые у почвенных форм отсутствуют. Ногохвост-
ки бегают на коротеньких ножках или прыгают с помощью вильчатых “хво-
стиков”, прикрепленных к брюшку. В современной классификации ного-
хвостки относятся к особому классу, а не к насекомым. Щетинохвостки, по-
видимому, более развитые, чем ногохвостки. Они хотя и маленькие (до 2 см),
но обладают длинными антеннами, фасеточными глазами и бегательными
ножками. В современной классификации они относятся к подклассу наруж-
ночелюстных насекомых. Эти существа не только древние, известные с ка-
менноугольного времени, но и являются выдающимся примером гибкости и
успеха насекомых. Казалось, что, будучи живыми ископаемыми, они обре-
чены на вымирание, но на самом деле они заполонили весь мир. Некоторые
216
КАМЕННАЯ КНИГА
Фиг. XIII.8. Ископаемые насекомые
а - крыло предка стрекоз Tipus gracilis длиной 14.5 см из перми Оклахомы (фо-
то F.M. Carpenter); б — муха, прекрасная сохранность в прибалтийском янтаре
(фотоГ.М. Carpenter); в-два термита в янтаре; оба насекомых с крыльями, сви-
детельствующими, что они попали в смолу во время весеннего сезона спарива-
ния (фото P.S. Tice из работы Buchsbaum “Животные без позвоночника”)
из них устроились в хорошо обогреваемых человеческих жилищах. И там,
оказавшись вне своей природной пищи, они буйно расплодились, используя
накрахмаленную одежду, книги, глянцевые журналы и всякие крошки, что
накапливаются в кухнях.
Ранние древнекрылые (Palaeoptera). Многие конечности членистоногих
трансформируются: одни ножки превращаются в клешни, другие ножки
служат челюстями и т.д. Крылья по сравнению с ними являются новыми
структурами, так как они возникли из четырех или шести выростов, кото-
рые появились у первых трех грудных сегментов. Это демонстрируют иско-
паемые, такие как Stenodictya из каменноугольных отложений Франции
(фиг. XIII. 7). Stenodictya имела два выроста на первом грудном сегменте и
настоящие крылья на втором и третьем. Выросты и крылья по существу по-
хожи, они очень тонкие, имеют разветвляющиеся жилки и обладают кро-
шечными волосиками, покрывающими их верхнюю поверхность.
Выросты превратились в крылья в течение каменноугольного времени,
так как насекомые, полностью готовые к полету, были обнаружены в отло-
жениях нижнего карбона. Этих крылатых насекомых называют древнекры-
лыми, или Palaeoptera, потому что характер жилкования является архаич-
ным, хоти и сложным, а крылья не складывались назад, когда их обладатели
отдыхали. Крылья просто висели, они имели набор мускулов, осуществляю-
щих их движения во многих направлениях. Современные поденки (Ephe
meroptera) очень похожи на этих первых летающих насекомых. Первые ис-
копаемые из этой группы известны из позднего карбона.
ЧЛЕНИСТОНОГИЕ ОТ МЕЛКОВОДНЫХ ДО СУХОПУТНЫХ
217
Фиг. XIII. 9. Protolindenia wittei, настоящая стрекоза с размахом крыльев око-
ло 10 см. Поздняя юра, Золенгофен, Германия (фото F.M. Carpenter)
Подавляющее большинство палеоптер имело размах крыльев 2.5-10 см,
но у некоторых он был намного больше. Некоторые виды Protodonata, часто
воспринимаемые прародителями стрекоз, имели размах крыльев 70-76 см и
тело длиной 30-38 см. Как и настоящие стрекозы, они были быстролетаю-
щими хищниками с шипастыми конечностями, используемыми для ловли
жертв. Протодонаты существовали от раннего карбона по пермь почти по-
всеместно. Исключительно хорошие экземпляры были найдены в тонко-
плитчатых пермских известняках у Elmo, около Abilene в Канзасе. Настоя-
щие стрекозы (СМоиа7а)появились в ранней перми и существуют поныне
(фиг. XIII.9).
В позднем палеозое было несколько отрядов древнекрылых, но только два
из них, Ephemeroptera и Odonata, существуют в настоящее время. Палеопте-
ры были задвинуты в тень более “продвинутыми” формами, такими как та-
раканы, кузнечики и близкими им формами.
Новокрылые насекомые с складывающимися крыльями (Neoptera).
Крылья оказались спасительным приобретением; они помогали своим обла-
дателям избежать пауков, скорпионов и мелких позвоночных, которые охо-
тились на земле. В то же время примитивные крылья, не способные склады-
ваться, создавали и неудобства, так как мешали палеоптерам прятаться под
сучками и листьями. От этого противоречия помогли избавиться крылья, ко-
торые приобрели способность складываться назад, давая тем самым возмож-
ность спрятаться в любом убежище или избежать опасности, улетев.
Несколько групп насекомых приобрели крылья, способные складывать-
ся, но в некоторых отрядах сохранились особенности строения палеоптер.
218
КАМЕННАЯ КНИГА
Фиг. XIII. 10. Ископаемые насекомые перми, триаса и олигоцена (фото
F.M. Carpenter)
а — крыло Clatrotitan andersoni, длиной 13.8 см (вымерший отряд титаноптера,
близкий к современным прямокрылым) с очень большим органом “стрекота-
ния”, триас, Новый Южный Уэлс, Австралия; б - древнекрылое Dunbaria fasci
ipennis шириной 3.7 см с остатками окраски на крыльях, ранняя пермь, Элмо,
Канзас; в — муха-ктырь (семейство Asilidae) длиной 2.5 см, сохранились волос-
ки-щетинки на ножках, олигоцен, Колорадо; г — верблюдка Raphidia mortua
длиной 1.7 см, олигоцен, Florissant, Колорадо; д - Lithosmylus columbianus,
длиной 3.0 см, сетчатокрылое из семейства осмилид, олигоцен, Florissant, Ко-
лорадо; е - бабочка Prodryas persephone шириной 5.3 см, с остатками окраски,
олигоцен, Florissant, Колорадо
ЧЛЕНИСТОНОГИЕ ОТ МЕЛКОВОДНЫХ ДО СУХОПУТНЫХ
219
Задолго до того, как закончился пермский период, возникли разные направ-
ления развития новокрылых насекомых, чьи крылья складывались назад,
одна пара над другой. Среди этих новых групп были таракановые (Blattodea,
или Blattoptera), чьи ископаемые остатки можно узнать благодаря ог-
ромным надкрыльям с характерным жилкованием, а также благодаря
широкой груди, которая у многих видов намного больше головы (см.
фиг. ХШ.7г). Некоторые средне-позднекаменноугольные тараканы достига-
ли в длину 7.6-10 см, но последующие виды были уже не такими большими.
Так как первые тараканы жили на влажной земле в болотистых лесах, их ос-
татки хорошо сохранились в илистых отложениях, и поэтому они непропор-
ционально многочисленны среди палеозойских насекомых.
Полный и неполный (простой) метаморфоз. Древние неоптеры, такие как
тараканы, проходят неполный метаморфоз. Это означает, что из яйца выхо-
дят крошечные личинки (нимфы), которые выглядят и живут почти так же,
как их полностью развитые родители. Тараканья личинка, например, меня-
ет свой покров несколько раз, пока станет половозрелой, но в течение роста
почти не меняется. Приобретенные две пары крыльев потом исчезают. Люди
знают, что маленькие тараканьи личинки (нимфы) бегают, как и их родите-
ли, едят то же самое и живут в тех же условиях.
В этом нет ничего плохого для самих таракановых. Доказательством слу-
жит тот факт, что тараканы остаются преуспевающими и многочисленными
формами более чем 280 млн. лет. Многие насекомые повышают свои шансы
выживания благодаря тому, что молодь живет в одних условиях, а взрослые -
в других. Это может происходить в течение неполного метаморфоза: личин-
ки стрекоз, например, ползают по дну водоемов, но взрослые стрекозы выхо-
дят на сушу и охотятся в воздушном пространстве. Диапазон условий обита-
ния намного больше, когда молодь имеет одно строение, взрослые - другое, а
промежуточная стадия, то есть куколка, имеет еще один тип строения. Такое
явление наблюдается у бабочек, у которых молодь (гусеница) в виде червеоб-
разной личинки ползает по растениям и питается их листьями. Через какое-
то время гусеница становится куколкой: безногой, без рта, заключенной в
панцирь, обеспечивающий защиту, пока идет реорганизация. Затем из ку-
колки выходит взрослая особь — бабочка, с четырьмя широкими крыльями,
конечностями, способными цепляться, и ротовыми приспособлениями, со-
сущими сок (нектар) цветов. Бабочки не могут вытеснить свою молодь, гусе-
ницу и куколку или отнять у них еду, так как у последних другие условия и
режим жизни. Метаморфоз бабочек является полным.
От древних тараканоподобных предков произошли три основные группы
(надотряды) насекомых: ортоптероиды, гемиптероиды и невроптероиды.
Надотряд ортоптероиды (Orthopteroidea) представлен отрядами: Blattoptera -
таракановые (поздний карбон-ныне); Orthoptera - прямокрылые, то есть
кузнечики, саранчовые и сверчки (поздний карбон-ныне); Dermaptera — ко-
жистокрылые, то есть уховертки (пермь-ныне); Isoptera - изоптеры, то есть
термиты (палеоген-ныне). Все они проходят простой метаморфоз и имеют
среди прочих характеристик следующую - на задних крыльях выделяется
большая лопасть.
Другой надотряд насекомых (Hemipteroidea) включает гемиптероидные
отряды: Mallophaga — пухоеды и власоеды (неоген-ныне); Anoplura - вши
(неоген-ныне); Thysanoptera — трипсы (юра-ныне); Hemiptera — полужест-
кокрылые, то есть клопы, цикады, тли и др. (пермь-ныне). Гемиптероидные
также проходят простой метаморфоз, но не имеют большой лопасти на зад-
них крыльях; кроме того, тонкие жилки на крыльях редуцированы.
220
КАМЕННАЯ КНИГА
Третий надотряд, невроптероиды (Neuropteroidea), или сетчатокрыловид-
ные, характеризуется полным метаморфозом с гусеницей и куколкой, сов-
сем непохожих на взрослых особей. В этот надотряд входят отряды: Ну-
menoptera — перепончатокрылые, то есть пчелы, осы, муравьи (триас-ныне);
Coleoptera - жесткокрылые, то есть жуки (ранняя пермь-ныне); Neuroptera -
сетчатокрылые (пермь—ныне); Mecoptera — скорпионницы (карбон-ныне);
Siphonaptera, или блохи (? мел Австралии; палеоген-ныне повсеместно);
Diptera — двукрылые, то есть комары, мухи и москиты (триас-ныне); Tri
choptera - ручейники (пермь-ныне) и Lepidoptera — чешуекрылые, то есть ба-
бочки (мел—ныне).
Преимущества, связанные с полным метаморфозом, были достигнуты по
крайней мере в пермское время, так как некоторые раннепермские насеко-
мые принадлежат двум отрядам, которые в настоящее время проходят ста-
дии гусеницы и куколки, прежде чем стать взрослыми. Они составляли тогда
только 5% известных видов, но их доля увеличивалась в течение последую-
щих эпох. В настоящее время более чем 80% насекомых мира проходят пол-
ный метаморфоз.
Современные фауны насекомых. Насекомые, которые существуют в насто-
ящее время, представляют собой смесь древних (тараканы и стрекозы) и более
современных ветвей, среди них преобладают невроптероиды и ортоптероиды.
Жуки появились в позднепермское время и стали многочисленными в на-
чале мезозойской эры. Описано около двух тысяч палеоген-неогеновых видов,
а плотные передние крылья встречаются в большом количестве в плейстоце-
новых асфальтах Калифорнии и в одновозрастных породах по всему свету.
Современные мухи сохранили только одну пару крыльев, что отражено в
названии Diptera или “Двукрылые”. Примитивные бабочки {Lepidoptera -
чешуекрылые) впервые появляются в раннем мелу. Чешуекрылые, найден-
ные совместно с разнообразными другими беспозвоночными и растениями в
олигоценовых озерных отложениях Florissant и повсюду в Колорадо (см.
главу XXXII), сохранили следы окраски, а также антенны, волосовидные от-
ростки (щетинки), которые покрывали брюшко и грудь. Примитивные ви-
ды, сходные с муравьями и пчелами (Hymenoptera — перепончатокрылые),
встречаются уже в триасе, но более специализированные формы, в том числе
и с социальным поведением, неизвестны в домеловое время.
В целом ископаемые насекомые являются весьма важной, имеющей боль-
шое значение группой, но они не обладают очарованием и красотой совре-
менных форм. Они содержат много сведений об эволюции своего класса, но
их находки известны только из некоторых формаций нескольких районов,
но и там они могут остаться незамеченными из-за ископаемых растений.
Иногда комбинация растений и насекомых хорошо различается, например,
по ископаемым листьям, имеющим наросты - галлы, или по листьям, сре-
занным или продырявленным определенными насекомыми.
Глава XIV
УЛИТКИ И ИХ КОРОЛЕВСТВО
• Название моллюски, или мягкотелые, произошло от латинского
слова “soft” — мягкий. Это второй по величине тип беспозвоночных,
представленный более чем 100 000 ныне существующих видов.
В повседневной речи моллюсков называют ракушками, а съедобных пред-
ставителей - “shellfish”. В действительности многие животные, имеющие
раковину, не являются моллюсками, в то же время некоторые моллюски ли-
бо утратили раковину, либо раковина стала внутренней. Для лучшего озна-
комления с данным типом необходимо сконцентрировать свое внимание в
первую очередь на строении мягкого тела, а затем уже на раковине.
Тело у моллюсков мягкое: его нижняя часть превратилась в мышечную
структуру, которая в основном служит для передвижения и поэтому называ-
ется ногой. Верхняя часть мягкого тела покрыта своеобразной мантией.
Мантия строит раковину. Раковина моллюсков является известковой,
преимущественно кальцитовой, могут содержаться слои родственного мине-
рала - арагонита, который легко поддается растворению.
Строение мягкого тела моллюсков существенно отличается от такового у
брахиопод. Моллюски являются подвижными животными и обычно не осе-
дают на одном месте. У них сквозная пищеварительная система: имеется и
рот, и анальное отверстие, а не “слепая кишка”, как у большинства брахио-
под, у которых отходы выводятся через единственное ротовое отверстие. Эти
преимущества сформировали животных, способных освоить множество раз-
личных, выгодных с точки зрения добычи пищи, экологических ниш. Мол-
люски, по сравнению с брахиоподами, более подвижны, в мезо-кайнозое они
стали господствующей группой. Моллюски, вероятно, являются потомками
свободноживущих червей.
МОНОПЛАКОФОРЫ
Моноплакофоры - группа блюдечкообразных морских моллюсков, кото-
рая, по-видимому, является довольно примитивной и не испытала торсион-
ный процесс (фиг. XIV. 1, 2). Они имеют псевдосегментированные мускулы,
жабры и внутренние органы. Нога у моноплакофор круглая и окружена ман-
тийной щелью. Это единственные современные раковинные моллюски с при-
знаками сегментации, видимо, именно они представляют собой предковую
© 1997. И. А. Михайлова, перевод на русский язык и изменения.
222
КАМЕННАЯ КНИГА
Фиг. XIV.l. Neopilina galatheae,
современная моноплакофора с
западного побережья Централь-
ной Америки
Внешний вид раковины со спин-
ной стороны, размер около 1 см в
длину (Frank Knight по Tasch,
1973)
Фиг. XIV.2. Scenella, монопла-
кофора из среднего кембрия Ка-
нады
Раковина изнутри, видно распо-
ложение сегментированных мус-
кулов U-образной формы; разме-
ры 1.2 см в длину (Frank Knight по
Tasch, 1973)
группу, возникшую от кольчатых червей или членистоногих. Первоначаль-
но моноплакофоры были встречены в отложениях кембрийской системы, а
теперь известны и в современных морских бассейнах, например Neopilina.
Так как эта группа первоначально была известна только в ископаемом состо-
янии, то, когда в 1957 году была обнаружена Neopilina, она была воспринята
как “живое ископаемое”.
ХИТОНЫ И БИВНЕВЫЕ РАКОВИНЫ
Самые ранние, наиболее просто устроенные моллюски фактически неиз-
вестны вследствие их принадлежности к группе мягкотелых животных, ко-
торые появились, развивались и вымерли в глубокой древности. Но мы мо-
жем составить о них верное представление, изучив хитонов — простейшие
формы среди живущих ныне моллюсков.
Хитоны, или боконервные (фиг. XIV.3). Научное название хитонов - Ат-
phineura вызвано спецификой их нервной системы, которая образует двой-
ную нервную цепочку вдоль всего тела. Современный хитон представляет со-
бой животное овальной формы с нечетко обособленной головой и подошвооб-
разной ногой, расположенной под мягким телом. Их размеры могут
достигать от 0.6 до 36 см в длину — такова длина гигантского Cryptochiton с
западного побережья Северной Америки. Во рту у хитона расположен шер-
шавый язык, снабженный радулой лентообразной формы с рядами острых
роговых зубов. Рот находится непосредственно в передней части животного.
Благодаря мускулам радула перемещается вперед и назад, позволяя живот-
ному соскребать водоросли со скал, по которым ползает хитон. Пищевари-
тельная система у них довольно проста, и раковина (отсутствующая у неко-
торых разновидностей) состоит из 7 или 8 отдельных пластин, налегающих
друг на друга. Жабры хитона в основном развиты достаточно хорошо.
Несмотря на принадлежность хитонов к примитивным моллюскам, их ос-
татки встречаются в отложениях позднего кембрия, например, Matthevia из
Северной Америки. Палеозойские особи отличаются тем, что их пластинки
УЛИТКИ И ИХ КОРОЛЕВСТВО
223
в
Фиг. XIV.3. Строение хитоиов
а, б — вид современного хитона: а — сверху, б — снизу, в — внутреннее строение
мягкого тела, г — три пластинки ископаемых хитонов
расположены друг за другом, тогда как у мезозойских и более молодых раз-
новидностей они черепицеобразно налегают друг на друга так же, как у со-
временных хитонов.
Скафоподы, или бивневые раковины (фиг. XIV. 4). Современные предста-
вители этой группы часто использовались американскими индейцами в ка-
Фиг. XIV.4. Две скафоподы в прижизненном положении, одна частично вскрыта
224
КАМЕННАЯ КНИГА
честве бусинок. Это достаточно странные, специфические моллюски, чьи ра-
ковины довольно тонкие, искривленные в форме конусов, открытых с обоих
концов. Большинство разновидностей не превышает 7.5 см в длину, кроме
одной из средне-верхнекаменноугольных (пенсильванских) отложений Те-
хаса, достигающей поразительной длины около 60 см. Современные скафо-
поды обитают в норках на морском дне от уровня малой воды до глубины
4500 м. Раковина в осадке располагается наклонно, задний конец возвыша-
ется над дном, а нога служит для передвижения и зарывания. Рот периоди-
чески углубляется в осадок; короткие щупальца вокруг него собирают пищу
в виде мелких животных. Анальное отверстие находится около верхнего
конца раковины; жабры почти редуцированы. Скафоподы известны начиная
с ордовика, но значительно чаще встречаются в кайнозойских отложениях,
например, род Dentalium.
БРЮХОНОГИЕ МОЛЛЮСКИ, ИЛИ ГАСТРОПОДЫ
Слово гастроподы означает брюхоногие. Улитки и их королевство обычно
имеют раковины простой структуры.
Мягкое тело гастропод (фиг. XIV. 5). Большинство ископаемых гастропод
представлено раковинами, но для их познания необходимо дать описание
современных форм. У типичных гастропод имеется нога, которая может до-
статочно далеко высовываться из раковины и втягиваться в нее обратно.
Мягкое тело либо вздутое, либо спирально свернутое, сверху покрытое ман-
тией. Голова, имеющаяся среди моллюсков только у улиток и головоногих,
снабжена двумя щупальцами, на каждом из которых с наружной части по-
средине между основанием и вершиной расположен глаз. Мантия сужается
к краю и образует с мягким телом карманообразную полость. У видов, обита-
ющих в воде, в этой полости находятся одна или две жабры, к которым вода
проникает сквозь сифон-желобок, имеющий форму трубки. После того как
жабры получают кислород, вода выходит через щель, разделяющую тело и
мантию. Гастроподы на личиночной стадии испытывают поворот верхней ча-
сти туловища вокруг оси (торсионный процесс). В результате этого анальное
отверстие нередко оказывается расположено над головным отделом.
Раковина гастропод. Существует большое разнообразие раковин гастро-
под, но все они выделены (секретированы) мантией. Снаружи раковина по-
крыта тонким слоем периострака, состоящего из рогового материала (проте-
ин), предохраняющего раковину от разъедания морской водой. Периострак
редко сохраняется у ископаемых форм и не развит у тех гастропод, чьи рако-
вины перекрыты широким отворотом мантии.
Раковины в большинстве случаев состоят из арагонита, расположенного в
виде серии слоев или призм. Внутренние слои, как правило, образованы
очень тонким слоем арагонита, чья структура параллельна внутренней по-
верхности раковины; она гладко “отполированная” или перламутровая. По-
скольку раковины в основном состоят из арагонита, они часто разрушаются
в процессе захоронения. Раковины с более грубыми фибрами и призмами не-
редко обламываются при извлечении ископаемых из известняка, мергеля
или сланца.
Большинство раковин улиток имеет форму свернутой спирали, но суще-
ствуют и исключения. Поэтому только эта характеристика является недо-
статочной. Улитки, произошедшие от несвернутых предков, встречаются в
кембрии, а возможно, появились и раньше. Некоторые примитивные пред-
УЛИТКИ И ИХ КОРОЛЕВСТВО
225
Фиг. XIV.5. Строение мягкого тела и раковины некоторых гастропод
а, б — схема строения современного Buccinum; в - Leptoptygma vlrgatum, архео-
гастропода, х0.5, средний-поздний карбон, долина Миссисипи, г — род Eotoma-
ria, археогастропода, Х1.3, средний ордовик, Восточная Канада и долина
р. Миссисипи. Род существовал с ордовика до силура, Евразия и Северная Аме-
рика, д - род Propilina, х4, моноплакофора из позднего кембрия-раннего ордо-
вика, Северная Америка и Азия
ставители, например беллерофонтиды, имеют спирально-плоскостную ра-
ковину, тогда как для большинства гастропод характерна раковина асим-
метричная. Не исключено, что симметричные формы даже не относятся к
улиткам. Среди палеонтологов существует мнение, что они близки моно-
плакофорам.
Крышечка. Раковина гастропод представляет прочную оболочку в виде
панциря, куда ее владелец всегда может спрятать свое мягкое и легко транс-
формирующееся тело. Некоторым группам не требуется более серьезной за-
щиты: к примеру, каури и конусообразные раковины (Conus) имеют такое
узкое щелевидное устье, что почти никто не может проникнуть внутрь. Но у
блюдечек (Patella) и других конических или слабо свернутых раковин устье
имеет такие же размеры, как сама раковина. Такие моллюски для обеспече-
ния собственной безопасности подползают на своей широкой мускулистой
ноге как можно ближе к скалам или другим объектам. Шероховатая область
мускульного отпечатка фиксирует размер и указывает на длину мускула, ко-
торый, сжимаясь, втягивает мягкое тело в раковину.
Многие гастроподы защищают себя с помощью крышечки, представляю-
щей роговую или известковую пластинку, расположенную над задней час-
тью ноги. Несмотря на то что крышечка выглядит абсурдно маленькой по
15 Каменная книга
226
КАМЕННАЯ КНИГА
сравнению с развернувшейся улиткой, она плотно закрывает устье ракови-
ны, когда животное втягивается внутрь.
Роговые крышечки редко встречаются в ископаемом состоянии, чаще со-
храняются более толстые и твердые известковые крышечки. Некоторые наи-
более утолщенные крышечки, известные у ископаемых, относятся к так на-
зываемым Maclurites из среднего и позднего ордовика.
Радула. Особым устройством пищеварительной системы гастропод явля-
ется расположенная во рту радула, представляющая собой серию рядов рого-
вых зубов. Это образование позволяет улитке полакомиться самыми разно-
образными блюдами (растительноядные и хищные гастроподы). Радула мо-
жет тереть, зацеплять, захватывать, проникать, сверлить и даже жалить
подобно змее. Радула является основой для классификации многих групп
улиток, она была найдена и у ископаемых.
Развитие и рост в течение жизни. Многие улитки вырастают до значитель-
ных размеров: одна современная особь достигает 60 см в длину. Раковины
многих форм, как ископаемых, так и современных, имеют линии нараста-
ния или складки, благодаря которым они выглядят достаточно старыми.
В действительности улитки растут быстро, а протяженность их жизни невели-
ка. Трубчатые улитки достигают 10-13 см в длину за 2-3 года; у рода Мигех,
имеющего толстую раковину, всего за 3 дня складки вырастают на 0.5—1 см.
Длительность жизни улиток составляет от 1 до 5 лет, хотя известны формы,
живущие до 20 лет.
АРХЕОГАСТРОПОДЫ
Несомненно, улитки эволюционировали с докембрия, но наиболее древ-
ние известные нам формы найдены в отложениях нижнего кембрия
(фиг. XIV. 6). Некоторые палеонтологи самой древней известной гастроподой
считают томмотский (ранний кембрий) род Aldanella из Сибири. Загадочное
кембрийское ископаемое - род Pelagiella, - видимо, является промежуточ-
ным между Aldanella и моноплакофорами: скорее всего они произошли от
Фиг. XIV.6. Некоторые археогастроподы
а, б—Bellerophon troosti, xl. 7, этот вид известен из ордовикских отложений вос-
тока США; род Bellerophon встречается с силура до триаса, повсеместно; в, г —
Salpingostoma boulli, ордовик, долина верховьев Миссисипи; д — Scenella reticu-
lata, х2, примитивный раннекембрийский моллюск
УЛИТКИ И ИХ КОРОЛЕВСТВО
227
единого предка. Род Aldanella может принадлежать к тем примитивным ар-
хеогастроподам, которые были закручены в плоскую, а не коническую спи-
раль. Отпечатки в тех местах, где прикреплялись мускулы, свидетельствуют
о том, что мягкое тело находилось в передней части раковины, а две пары
листообразных жабр — в задней. Жабры снабжены волосками, которые рас-
положены вдоль них в два ряда. У некоторых форм имеется только одна жа-
бра. Макушка отогнута назад. Археогастроподы, очевидно, медленно полза-
ли по илистому дну, питаясь морскими водорослями.
В греческой мифологии был герой, носивший имя Беллерофон, он отпра-
вился на битву на крылатом коне Пегасе. Такое название — Bellerophon —
было дано вымершему брюхоногому моллюску. Его раковина симметрично
закручена в одной плоскости, устье овальное. Наружная губа расширяется в
обе стороны и имеет в плоскости симметрии мантийную щель, при ее зарас-
тании образуется мантийная полоска. У нескольких родственных форм, ра-
ковина которых закручена неплотно, устьевая часть может быть еще более
расширена.
До сих пор существует неясность относительно места беллерофонтид в об-
щей системе гастропод. Некоторые палеонтологи относят их к археогастро-
подам, а другие специалисты считают, что эти формы более тесно связаны с
моноплакофорами. Очевидно, необходимо провести более тщательное иссле-
дование, чтобы взаимосвязи многих кембрийских моллюсков, включая и
беллерофонтид, стали более ясными. Беллерофонтиды представлены огром-
ным разнообразием форм и встречаются начиная с кембрия и кончая триа-
сом. Наиболее благоприятными для них были ордовик, силур и девон.
ПРЕУСПЕВАЮЩИЕ ПЕРЕДНЕЖАБЕРНЫЕ
Археогастроподы были представителями переднежаберных - одной из
трех наиболее крупных групп гастропод, которые встречаются с раннего
кембрия вплоть до нашего времени (фиг. XIV.7). Переднежаберные состав-
ляют преобладающее большинство палеозойских гастропод, а также значи-
тельную часть постпалеозойских сообществ. Их жабры и мантийная полость
располагаются около головы, устье направлено вперед, а вершина раковины
ориентирована назад. Пищеварительный тракт образует резкий изгиб, в ре-
зультате чего анальное отверстие оказывается почти над головой.
Несомненные переднежаберные, появившиеся в кембрийских морях, в
изобилии представлены и в наше время. Большинство форм данного класса
могут быть определены как морские улитки, раковины которых закручены
преимущественно по спирали. Они варьируют от Euomphalus, чья раковина
имеет форму спирали, закрученной почти в одной плоскости, до высококо-
нических (башенковидных), таких как Ectomarta и Turritella. Некоторые из
них гладкие, как фарфор высшего качества, другие имеют разнообразную
скульптуру в виде ребер и складок, шипов и игл; иногда в состав раковины
включены посторонние образования. Окраска раковин ныне живущих видов
варьирует от белой или серой до красной, зеленой и пурпурной, то есть
охватывает почти все цвета радуги.
Обычаи переднежаберных гастропод. Образ жизни и особенности обита-
ния этих форм столь же разнообразны, как и строение раковины. Современ-
ные литторины (род Littorina) и “собачьи прыщи” (род Thais) обитают в ли-
торальной зоне на скалах, которые осушаются во время отлива и омываются
волнами во время шторма. Оливы (масличные раковины) ползают в песча-
15*
228
КАМЕННАЯ КНИГА
Фиг. XIV. 7. Разнообразие переднежаберных гастропод
а — род Straparolus, археогастропода, широко распространенная с силура до
перми; б - Turritella, мезогастропода, этот вид известен из палеоцена и эоцена,
Мэриленд и Техас; в - Holopea symmetrica, археогастропода, х2.5, средний ор-
довик, восток Северной Америки; г - Helminthozyga vermicula, мезогастропода,
х20, средний-поздний карбон, Северная Америка; д - Vermicularia, мезога-
стропода, известная с миоцена по настоящее время, Северная и Южная Амери-
ка; е - Turbonopsis shumardi, археогастропода, средний девон, долина Мисси-
сипи; ж - Hormotoma whiteavesi, археогастропода, х0.8, ранний силур,
Нью-Йорк; з - Turritella, мезогастропода, внутреннее ядро, мел, Нью-Джерси;
и — Murex mississippiensis, неогастропода, xl.2, олигоцен, Миссисипи; к — позд-
немеловая гастропода, Нью-Джерси и Юта
УЛИТКИ И ИХ КОРОЛЕВСТВО
229
ном осадке, оставляя следы, почти идентичные найденным в пенсильван-
ских отложениях (средний-верхний карбон). Многие другие улитки полза-
ют по песчаному дну, присасываясь к подводным камням или передвигаясь
по коралловым рифам. Передвижение в основном осуществляется с помо-
щью единственной ноги, которая может иметь замечательные каналы, слу-
жащие присосками. Благодаря этому улитка быстро прикрепляется к ска-
лам или другим ракушкам. Род Strombus и некоторые другие переднежабер-
ные обладают когтеобразной крышечкой (оперкулюм), которая может
выбрасываться вперед и закрепляться в песке либо в иле. Мускулы ноги в
этом случае сокращаются, толкая животное вперед.
Современные переднежаберные гастроподы питаются преимущественно
водорослями, которые они соскребают с помощью радулы с каменистого
грунта. Однако “лунные” улитки и сверлильщики устриц проникают внутрь
других моллюсков и затем поедают их мягкое тело. Это очень древний способ
питания. Просверленные раковины моллюсков часто встречаются в третич-
ных и мезозойских отложениях, а просверленные раковины брахиопод изве-
стны начиная с позднего ордовика. Предполагается, что палеозойские хищ-
ники предпочитали брахиопод съедобным морским моллюскам, но мы не мо-
жем быть точно уверены, поскольку наши образцы не достаточно точно
определены. Большинство древних двустворок имеют слишком плохую со-
хранность, чтобы судить об этом.
Некоторые хищные гастроподы проникали внутрь двустворчатых моллю-
сков или морских уточек с помощью зуба или иглы, расположенных с на-
ружной стороны на кончике раковины. Род Strombus и некоторые другие яв-
ляются падалеядами (мусорщиками), питающимися мертвой рыбой и бес-
позвоночными .
Многие переднежаберные гастроподы с колпачковидной раковиной (“блю-
дечки”) большую часть суток проводят в спокойном безопасном месте. Ночью
они выползают за водорослями, служащими им пищей, а к утру возвращают-
ся назад. После возвращения на свое место блюдечки так крепко присасыва-
ются, что края их раковин по мере роста повторяют все неровности дна.
Ископаемые блюдечки могли как иметь, так и не иметь такую особен-
ность, но нет сомнения, что другие переднежаберные занимали в период по-
коя определенные места (“места отдыха”). Например, некоторые виды родов
Platyceras и Cyclonema присасывались к криноидеям так крепко, что остав-
ляли на них контуры своей раковины. Руки морских лилий образовывали
барьер, который эти улитки при движении к съедобным водорослям не мог-
ли преодолеть. Можно сделать вывод, что они питались отходами, вырабаты-
ваемыми лилиями.
Еще более необычен род Vermicularia. На ранней стадии его раковина
представляет собой вытянутую башенковидную спираль, похожую на рако-
вину рода Turritella. После спиральной стадии этот моллюск строит ракови-
ну в виде неправильно закрученной трубки, которая может ошибочно прини-
маться за известковые трубки кольчатых червей. Трубка рода Vermicularia
может быть свободна или же прикрепляется к скалам или другим ракови-
нам. Часто образуются скопления в виде общей запутанной массы. В пен-
сильванских отложениях (средний-верхний карбон) встречается род Hel-
minthozyga, который тоже утрачивает спиральную форму раковины.
Мезогастроподы и неогастроподы. Дополнительно к археогастроподам
среди переднежаберных гастропод выделяют две названные группы. Первая
из них - мезогастроподы - впервые появилась в ордовике и продолжает
существовать и в настоящее время (см. фиг. XIV. 7). Обособление мезогастро-
230
КАМЕННАЯ КНИГА
под основано на строении мягкого тела. Они имеют одну жабру и одно пред-
сердие. По строению радулы мезогастроподы подразделяются на более мел-
кие группировки. Форма раковины варьирует в значительной степени, хотя
многие мезогастроподы имеют раковины башенковидные. Эта группа незна-
чительно дивергировала в течение палеозоя по сравнению с мезозоем, осо-
бенно с меловым периодом. Интенсивное развитие мезогастропод началось в
мезозое и продолжилось в кайнозое.
Неогастроподы впервые появились в меловое время (фиг. XIV.8). Они об-
ладают жабрами, достаточно сходными с жабрами мезогастропод. Так же
как у некоторых семейств мезогастропод, у них имеется сифон и, как пра-
вило, вытянутый желобок в устьевой части раковины (тоже называемый
сифон). К данным двум группам принадлежат многие хорошо известные со-
временные морские гастроподы, такие как род Turritella с башенковидной
раковиной, вид Murex tribulus с многочисленными тонкими шипами (гор-
дость коллекционеров современных раковин), другие виды рода Мигех, а
также сильно ядовитый род Conus.
Дивергенция гастропод началась в раннем ордовике и продолжалась до кон-
ца девона. Гастроподы раннего палеозоя позднее были частично вытеснены дру-
гими группами животных и испытали массовое вымирание в конце перми, хотя
отдельные реликты просуществовали до конца триаса. К меловому периоду по-
степенно сформировался современный набор форм: увеличилось число неогаст-
ропод и уменьшилось число вновь появляющихся мезогастропод. Большинство
из них не подверглось вымиранию в конце мела. Господство гастропод,
достигнутое в кайнозое, сохраняется вплоть до настоящего времени.
ЗАДНЕЖАБЕРНЫЕ ГАСТРОПОДЫ: ПРОЦЕССЫ ДЕГЕНЕРАЦИИ
Род Acteon имеет яйцеобразную форму, разнообразные виды этого рода
встречаются начиная с формаций Pierre и Bearpaw (глава II) до современных
умеренных морей (см. фиг. XIV.8). Они являются типичными представите-
Фиг. XIV.8. Неогастроподы, заднежаберные гастроподы и хиолиты
a-Liopeplum, неогастропода, xl.5, этот вид происходит из верхнемеловых отло-
жений Техаса и Мэриленд; б — Busycon, неогастропода, этот вид происходит из
верхнемеловых отложений Теннесси, в -Acteon vaughani, заднежаберная гаст-
ропода, х8, поздний миоцен, Южная Каролина и Флорида; г, д - два экземпляра
Hyolithes, х0.9 (г), х2.7 (д), ранний кембрий, Северная Америка
УЛИТКИ И ИХ КОРОЛЕВСТВО
231
лями заднежаберных гастропод, которые, видимо, развились в основном в
процессе дегенерации. Некоторые из них, такие как Acteon, имеют хорошо
развитую раковину, мантию, одну пару жабр, расположенную позади серд-
ца. Другими словами, они сформировались как результат деторсионного
процесса и “выпрямления себя изнутри”. Раковина постепенно стала реду-
цироваться, у голожаберных она уже на эмбриональной стадии исчезает пол-
ностью. В ископаемом состоянии голожаберные неизвестны, но группа, к ко-
торой относится род Acteon, появилась в раннем карбоне и продолжает суще-
ствовать поныне. Некоторые виды являются руководящими ископаемыми
для меловых и третичных отложений; наибольшее разнообразие наблюдает-
ся в мезозое и кайнозое. Формы раковин варьируют в значительной степени,
и хотя у большинства они тонкие, хрупкие, у некоторых видов раковины
утолщены и поэтому сохраняются в ископаемом состоянии. Все названные
формы являются морскими.
Планктонные (“свободноплавающие”) птероподы. Птероподы преиму-
щественно безраковинные и мягкотелые, как и голожаберные, но у мень-
шинства имеются тонкие спирально закрученные низкоконические ракови-
ны. Голова у них нечетко выражена, а нога преобразована в пару крылооб-
разных плавников (птероподы означают крылоногие), с помощью которых
животные приспособились к обитанию в толще воды, войдя в состав планк-
тона. В основном они имеют маленькие размеры - менее 1 см. У ископаемых
птеропод раковина тонкая и хрупкая, у некоторых имеются крышечки для
закрывания устья.
Древнейшие птероподы встречены в отложениях эоцена и, так же как их
современные сородичи, являлись для своего времени составной частью мор-
ского планктона. Поэтому и раньше и сейчас они широко распространены
географически.
Хиолиты — это кембрийские ископаемые, одно время относившиеся к пте-
роподам (см. фиг. XIV. 8). Однако и сейчас у нас нет ясного представления от-
носительно занимаемого ими места в системе беспозвоночных. Они могут
принадлежать к моллюскам, но скорее всего их следует рассматривать в ка-
честве отдельного типа.
ДЫШАЩИЕ ВОЗДУХОМ ЛЕГОЧНЫЕ МОЛЛЮСКИ
Любой, кто бывает в лесу, знает, что некоторые улитки и слизни живут на
земле и дышат воздухом. Эту особенность разделяют пресноводные улитки,
которые вдыхают воздух, поднимаясь к поверхности воды. У обеих групп ут-
рачены жабры и дыхание осуществляется с помощью органа, похожего на
легкие, развивающегося из мантийной полости. Эти гастроподы являются
единственными моллюсками, сумевшими успешно освоить сушу.
Пресноводные легочные гастроподы обладают раковинами, достаточно
разнообразными по форме (фиг. XIV. 9). На голове у них имеются две пары
щупалец, у основания задней пары расположены простые глаза. Эти улитки
могли появиться уже в палеозое, хотя они впервые обнаружены только в ме-
зозойских отложениях. Некоторые легочные являются характерными иско-
паемыми для позднего мела, а одна группа характеризует формацию Morri-
son. В этой формации было найдено большое разнообразие огромных северо-
американских динозавров и ранних млекопитающих.
У многих пресноводных улиток сохранились раковины, которые могут
быть башенковидными или низкоконическими, тонкими или толстыми,
232
КАМЕННАЯ КНИГА
Фиг. XIV.9. Легочные гастроподы
Род Helisoma, х2. Эта пресноводная легочная форма обитает на Атлантическом
и Тихоокеанском побережье Пацифики и на северо-западе Аляски, известна
также в плиоцене и плейстоцене
хрупкими или прочными. Некоторые слизни полностью утратили раковину,
у других родов раковины редуцированы до тонкой пластинки, скрытой под
мантией.
Несмотря на то что легочные составляют незначительную часть ископае-
мых гастропод, у них есть своя собственная интересная история. Стивен
Джеу Гулд в качестве части своей дипломной работы в Колумбийском уни-
верситете в Нью-Йорке изучил большую коллекцию ископаемых легочных
гастропод, обитавших триста тысяч лет назад на Бермудских островах. Он
выявил некоторое количество незначительных изменений, которые произо-
шли на протяжении всего этого времени у различных популяций донных ма-
леньких улиток. Это обусловлено тем, что они неоднократно оказывались в
изоляции на небольших островах вокруг Бермуд из-за того, что в плейстоце-
не уровень моря то опускался, то поднимался во время теплых и холодных
(оледенения) периодов. Стивен'Джеу Гулд смог показать, что эволюционные
изменения у исследованных улиток происходили быстро (скачкообразно), а
не постепенно, что привело к новому взгляду на процесс эволюции. Благода-
ря этому ископаемые, принадлежащие к довольно незначительной группе,
привели к появлению важной научной идеи.
Глава XV
ДВУСТВОРКИ
• Улитки, о которых шла речь в предыдущей главе, самый распрост-
раненный класс среди моллюсков, к тому же единственный, освоив-
ший сушу. Двустворчатые моллюски, или двустворки, наиболее из-
вестны благодаря тому, что к ним относятся съедобные устрицы, гребешки,
мидии и другие.
И вновь названия становятся источником проблем, поскольку в Канаде и
США этих моллюсков называет пелециподы, а в Европе и остальных частях
света - пластинчатожаберные. Первое приведенное название означает топо-
роногие и основано на форме ноги. Второе название ламеллибранхиаты, или
пластинчатожаберные, отражает строение жабр, с помощью которых дышит
большинство моллюсков. Название Bivalvia, или двустворки, дано в связи с
тем, что их раковина состоит из двух створок. Наиболее древние двустворки
были обнаружены в ископаемом состоянии в отложениях нижнего кембрия.
В палеозое разнообразие двустворок не очень велико, но они достаточно ши-
роко представлены в мезозое. На сегодняшний день двустворки являются
господствующей группой беспозвоночных, имеющих прочную раковину и
обитающих в мелководной морской среде.
КТО ТАКИЕ ДВУСТВОРКИ?
Двустворки устроены по более простой и в то же время по более сложной
схеме, чем примитивные брюхоногие моллюски или хитоны. Упрощенность
проявляется в строении тела, не имеющего обособленной головы и радулы
(фиг. XV. 1). Раковина состоит из двух частей, или створок, которые могут
быть как равными, так и неравными и отличаться между собой по размерам,
форме и функциям.
Тело двустворок обычно симметричное, имеются правая и левая стороны
и соответственно правая и левая створки. В отличие от типичной улитки
мягкое тело не закручено и не высовывается в такой степени, как, например,
подошвообразная нога у блюдечка. Нога двустворок представляет продол-
жение нижней части туловища; у всех форм, за исключением сильно спе-
циализированных, она перестала быть широкой и плоской, а стала более уз-
кой, клинообразной или приобрела топоровидное очертание. Жабры нахо-
дятся в нижней, а не в верхней части тела и варьируют от собственно
© 1997. И.А. Михайлова, перевод на русский язык и изменения.
234
КАМЕННАЯ КНИГА
Жабры Мантия
а
Выводной
сифон
. Вводной
сифон
Периостракум
б
Фиг. XV. 1. Строение раковины и мягкого тела двустворчатых моллюсков
а — мягкое тело рода Unio, х0.8, удалена левая створка вместе с мантией; б — род
Tagelus, х0.8, олигоцен-современность, Северная и Южная Америка, Африка и
Европа; в — птериоморфная двустворка, х0.8, поздний мел, Алабама и Техас; г —
род Pteria, х0.8, триас—современность, повсеместно, современность — тепловод-
ные моря; д - гетеродонтная двустворка Eucrinatella, х0.8, мел-миоцен, Европа
и Северная Америка; е — таксодонтная двустворка Cucullaea vulgaris, х0.5, мел,
Северная Америка
пластинчатых - листообразных до складчатых или даже стержневидных.
Жабры закрыты двумя широкими лопастями мантии.
Различные способы питания. Двустворчатые моллюски являются фильт-
раторами и не могут употреблять в пищу крупные частицы. Они используют
ДВУСТВОРКИ
235
только микроскопические организмы, находящиеся в воде, из которой с по-
мощью жабр добывают и кислород и пищу. Это означает, что каждое живот-
ное должно всасывать воду в полость мантии, отфильтровывать пищу, уда-
лять песок или ил, передавать пищу ко рту, одновременно избавляясь как от
посторонних частиц, так и от воды, из которой уже извлечен кислород. Эта
задача выполняется по-разному, в соответствии с образом жизни различных
дву створок.
Мы наблюдаем этот процесс в простейшем виде у рода Solemya и его при-
митивных родственников, живших от девона и до настоящего времени. На
заднем конце мягкого тела мантия сворачивается в два неотчетливых канала
(сифона). Движение ресничек создает ток воды через нижний канал и доно-
сит ее до жабр. Там кровь отфильтровывает кислород и те частицы, которые
легко проходят сквозь тонкие жаберные мембраны. Одновременно с этим пи-
ща оседает на покрывающей жабры липкой слизи и доставляется ко рту.
Другое внутреннее “течение” направляет ил, песок и “прочие отходы”, а так-
же использованную воду в верхний канал (сифон); таким образом все пере-
численное выводится обратно в морскую среду.
У более высокоорганизованных двуствЬрок, особенно живущих в норках,
каналы образуют две самостоятельные более длинные трубки — истинные си-
фоны, которые иногда могут покрываться единой плотной кожистой оболоч-
кой. Когда двустворке угрожает опасность, продольные мускулы сифонов со-
кращают их и частично или полностью втягивают в раковину. Другие муску-
лы могут перекрыть трубки и таким образом защищают мантийную полость
от попадания в нее илистых или песчаных частиц. Следующая степень защи-
ты обеспечивается тем, что мускульные окончания краев мантии позволяют
так крепко сжать две лопасти мантии, как мы сжимаем губы. Створки могут
смыкаться и таким образом, что единственными отверстиями являются отвер-
стия для сифонов, а через щель между створками может высовываться нога.
На коралловых рифах обитает гигантский двустворчатый моллюск три-
дакна (Tridacna); во время отлива большинство раковин оказывается выше
уровня воды, а во время прилива вода их полностью покрывает. Когда вода
отходит, створки плотно смыкаются, когда возвращается, створки открыва-
ются и края мантии раскрываются в виде толстых разноцветных складок.
В складках мантии живут водоросли-симбионты, которые питаются, ис-
пользуя энергию солнечного света и углекислый газ, выделяемый моллюс-
ком. Между съедобными двустворками и водорослями установились тесные
симбиотические связи, выгодные для каждой стороны. Род Tridacna встре-
чается в четвертичных отложениях, возможно, у ископаемых моллюсков
также имелся симбиоз с водорослями.
Нет сомнений относительно способа питания некоторых древнейших мол-
люсков, сверливших дерево подобно современным корабельным червям.
Просверливая норку, они соскребают дерево, превращая его в порошок, ко-
торый заполнял бы их норки, если бы они его не употребляли в пищу Мы мо-
жем с уверенностью сказать, что ископаемые двустворки усваивали целлю-
лозу древесины так же хорошо, как их современные сородичи.
РАКОВИНА ДВУСТВОРОК
Большинство ископаемых двустворок имело раковину. Раковина совре-
менных двустворок, как и гастропод, состоит из трех слоев. Различной тол-
щины роговой периостракум (от тонкого до толстого) перекрывает сверху из-
236
КАМЕННАЯ КНИГА
вестковый - призматический - слой, в основном состоящий из призм каль-
цита. Этот слой хорошо сохраняется у таких ископаемых, как род
Inoceramus. Внутренний - пластинчатый - слой содержит тонкие листочки
или пластинки кальцита или арагонита, которые могут чередоваться с мате-
риалом, напоминающим периостракум. Если пластинки толстые и гладкие,
то они напоминают фарфор. Если же они тонкие, то становятся блестящими
и известны нам как перламутр или жемчуг. Несмотря на то, что жемчуж-
ный блеск нередко обнаруживают у ископаемых форм, однако сам
перламутр, как и сходная с ним фарфоровидная разновидность, не сохраня-
ется в процессе фоссилизации. Кроме этих трех типичных слоев у двуствор-
чатых моллюсков существуют по крайней мере еще три других. Комбина-
ции этих слоев и их число помогают в выявлении взаимоотношений этих
животных.
Онтогенетические изменения и структура раковины. Рост раковины дву-
створчатых моллюсков начинается с появления на личиночной стадии вы-
пуклой пластинки, разделяющейся далее на две ровные и почти круглые
створки. Эта форма может сохраняться на протяжении всей жизни. Ракови-
ны чаще всего становятся выпуклыми, формируя макушку, которая заметно
выделяется в начале расширения створок и сокращающейся кривизны. Ма-
кушки могут заканчиваться клювовидно, образуя оригинальные начальные
раковинки, иногда макушки стираются с возрастом.
Макушки могут находиться вблизи смычного края, вдоль которого соеди-
няются обе створки. Нередко каждая створка имеет под макушкой треуголь-
ную площадку - арею, обычно являющуюся наружной. Поверхность створок
может быть гладкой, видны только линии нарастания, свидетельствующие о
“периодах отдыха”, вызванных низкой температурой, недостатком пищи
или репродуктивной деятельностью. Чаще всего линии нарастания совпада-
ют со складками, концентрическими ребрами, бороздками, желобками или
иглами. Смычный край по бокам может переходить в ушки, наиболее разви-
тые у гребешков (роды Pecten, Chlamys и другие).
Раковины двустворок являются достаточно разнообразными по размерам:
некоторые не больше булавочной головки, но есть и очень крупные. Самая
большая, поистине огромная раковина рода Tridacna достигает в ширину 1.3 м
и весит около 225 кг. Род Haploscapha из позднего мела Канзаса, видимо, не
был столь тяжелым, но имел 1 м в ширину и 1.5 м в длину. Неизвестно,
сколько лет жили эти гиганты, но многие обычные современные двустворки
средних размеров живут от 8 до 18 лет.
Внутреннее строение. Раковина двустворок, как и многих других рако-
винных моллюсков, сформирована мантией. Мантия прикреплена к внут-
ренней поверхности обеих створок с помощью серии мелких мускулов, кото-
рые фиксируются на раковине в виде так называемой мантийной линии;
края мантии всегда остаются свободными. Мантийная линия хорошо пред-
ставлена у многих современных и ископаемых форм, хотя и не у всех видов
одинаково отчетливо. При наличии сифонов мантийная линия изгибается
внутрь (мантийный синус), глубина изгиба показывает относительные раз-
меры сифонов. Обладатели сифонов обитают в норках, наличие и величина
мантийного синуса дают нам представление о возможном образе жизни мно-
гих ископаемых. Однако это представление должно быть подтверждено фор-
мой раковины и другими признаками.
Двустворки имеют один или два мускула, которые выполняют функцию
закрывания раковины. Поскольку мускулы сводят створки вместе, их назы-
вают аддукторами. Широко известны употребляемые в пищу гребешки.
ДВУСТВОРКИ
237
Фиг. XV.2. Некоторые фолады
а — род Martesia, хЗ, юра-современность, широко распространен в неполярных
областях, б— рад Pholas, хО.6, мел-современность, Атлантика и Индо-Пацифика
В действительности мы едим только мускулы гребешков (роды Pecten,
Chlamys и другие).
Смычный край, зубы и лигамент (связка). У всех двустворчатых моллю-
сков створки соединяются вдоль смычного (или замочного) края. В большин-
стве случаев на этом крае на каждой створке располагаются выступы — зубы
и ямки между ними, зубы одной створки входят в соответствующие им ямки
другой створки. У основных групп двустворок зубы и разделяющие их ямки
расположены различно.
Створки раковины закрываются с помощью мускулов, а открываются ав-
томатически благодаря приспособлению, которое называется лигамент, или
связка. Связка представляет собой шероховатый, местами очень толстый
эластичный тяж, расположенный между створками в различных частях
смычного края. Когда раковина закрыта, упругий лигамент сжат, а когда
мускулы расслабляются, он расправляется, заставляя створки открываться,
а точнее - приоткрываться.
Странная особенность установлена у рода Pholas и близких ему форм, ко-
торые жили начиная с раннечетвертичного времени (фиг. XV.2). Род Pholas
утратил лигамент, передний мускул переместился на наружную поверх-
ность раковины, где он прикреплен к пластинам, изгибы которых противо-
положны изгибам створок. Когда передний мускул сокращается, то вместо
закрывания раковины происходит приоткрывание ее заднего конца.
Классификация двустворок. Палеонтологи всегда сталкивались с затруд-
нениями при классификации двустворчатых моллюсков и выяснении их
взаимосвязей друг с другом. Основной причиной этого затруднения является
то, что раковины двустворок имеют ограниченное число признаков, позволя-
ющих устанавливать родственные взаимоотношения. Это общая проблема
для большинства ископаемых. При классификации высших таксонов, таких
как подклассы, строение мягкого тела, и в частности строение жабр, являют-
ся наиболее важными признаками. Очевидно, что у ископаемых эти органы
не сохраняются. Кроме того, в процессе фоссилизации раковины часто ока-
зываются перекристаллизованы. В связи с этим иногда сложно определить
первоначальный состав раковины (кальцит или арагонит). К счастью, клас-
сификация на видовом, родовом и семейственном уровне не является боль-
шой проблемой, так как строение раковины обеспечивает нас достаточной
информацией.
Одна из классификаций, широко распространенных на сегодняшний
день, основывается на микроструктуре стенки раковины, строении смычно-
го края и ареи, а также особенностях мягких частей: жабр и желудка. По
238
КАМЕННАЯ КНИГА
а
Фиг. XV.3. Некоторые криптодонты (а) и птериоморфы (б-е)
а - род Solenomya, xl.7, девон-современность, широко распространен; б — Му а-
Una, х0.4, средний-поздний карбон, юго-запад США, род известен повсеместно
в карбоне и перми; в - Naiadites carbonarius с пучком биссусных нитей, xl .3, по-
здний карбон Швеции; г, д — род Ostrea, х0.9, мел-современность, широко рас-
пространен в неполярных морях; е — Chlamys (Lyropecten), х0.4, миоцен-плио-
цен, восток Пацифики и запад Атлантики
этим признакам выделяют шесть подклассов. Палеотаксодонты - маленькие
формы с листообразными жабрами (протобранхии), со смычным краем с
многочисленными зубами, расположенными параллельно друг другу (зубы
таксодонтного, или рядозубого, типа), и с арагонитовой раковиной. Они яв-
ляются инфауной, живущей и питающейся в осадке. Род Nucula - типичный
род данной группы - распространен начиная с ордовика и до нашего време-
ни. Это относительно примитивные двустворчатые моллюски.
С ордовика и до настоящего времени существуют и криптодонты
(фиг. XV.3). У них, как правило, зубы развиты недостаточно или отсутству-
ют полностью, либо около переднего края имеются простейшие зубы (дизо-
донтный тип). Криптодонты также живут в осадке и имеют арагонитовую ра-
ковину. Сегодня эту группу представляет только род Solemya, который до-
стигал своего пика в палеозойской эре.
Птериоморфы впервые появились в ордовике и живут в современном оке-
ане до сих пор. Это достаточно разнообразная группа, у взрослых форм
обычно развиты биссусные нити. Их раковины бывают и кальцитовыми и
арагонитовыми. Сюда входят такие известные представители, как мидии
(род Mytilus), гребешки (род Pecten), род Inoceramus, устрицы (роды Ostrea
и Exogyra).
Другая группа, просуществовавшая с ордовика до нашего времени, - па-
леогетеродонты (фиг. ХУЛ). Они обладают либо гетеродонтным (разнозу-
ДВУСТВОРКИ
239
Фиг. XV.4. Некоторые палеогетеродонты (а, б), шизодонты (в, г) и гетеродонты
(д-ж)
а - Archanodon catskillensis, хО.З, средний-поздний девон, Пенсильвания и
Нью-Йорк; б — род Unio, хО.6, этот экземпляр из позднего мела, Монтана, три-
ас-современность, Европа, Африка, Северная Америка; в, г-(?) род Pterotrigo-
nia, хО.6, поздний мел, Орегон; д — род Venus, хО.6, левая створка изнутри, оли-
гоцен—современность, Евразия, Северная Америка; е — род Venus, хО.6, этот эк-
земпляр из миоцена, прибрежная равнина Атлантики; ж — Venericardia
planicosta, хО.З, эоцен, Европа
бым), либо шизодонтным (расщепленнозубым) типом замка. Гетеродонты
имеют в основном два или три главных (кардинальных) зуба, расположен-
ных под макушкой, и длинные субгоризонтальные боковые (латеральные)
зубы, находящиеся впереди и сзади главных зубов. Шизодонтные зубы
известны только у одной группы, включающей среди современных морских
форм единственный род Neotrigonia - современное ископаемое, найденное на
Австралийском побережье. Близкие родственники неотригонии были изве-
стны в мезозойских отложениях Северной Америки и Европы задолго до то-
го, как в начале XIX века французские исследователи выловили современно-
го живого представителя этой группы на южном австралийском побережье.
В этот подкласс входят также пресноводные униониды.
Гетеродонты (разнозубые) впервые появились в ордовике и являются наи-
более изобилующими современными мелководными и глубоководными дву-
створчатыми моллюсками. Их смычный край с гетеродонтным или десмо-
донтным (связкозубым) типом замка, у последнего типа зубы очень малень-
кие или отсутствуют. Вместо отсутствующих зубов параллельно смычному
краю может располагаться связка. Почти все гетеродонты имеют арагонито-
вую раковину, они стали наиболее разнообразными представителями инфау-
ны. Животные защищены осадком; они собирают пищу с помощью удлинен-
240
КАМЕННАЯ КНИГА
ных сифонов, которые выступают над дном и фильтруют воду. Примерами
могут служить роды Му a, Pholas и Venus.
Другой группой зарывающихся и сверлящих двустворок являются Апо-
malodesmata. Они впервые встречены в триасовых отложениях и продолжа-
ют существовать до настоящего времени, например, род Pholadomya. У них
десмодонтное строение замка и арагонитовая раковина.
Геологическая история. Хотя первая известная двустворка (род Fordilia)
была обнаружена в отложениях нижнего кембрия Северной Америки и Евра-
зии, дивергенция двустворчатых моллюсков началась только в раннем ордо-
вике. До конца кембрия двустворки имели чрезвычайно маленькие размеры,
что справедливо и для всего класса моллюсков. Древнейшие двустворчатые
обитали в осадке, начиная с ордовика они существовали и в осадке (инфауна)
и на дне (эпифауна), освоив несколько типов обитания. Один из способов оби-
тания, давший двустворкам большое преимущество перед замковыми брахио-
подами, заключается в способности быстро зарываться в грунт при помощи
ноги. У некоторых зарывающихся форм к концу палеозоя развились удли-
ненные сифоны, позволявшие им погружаться в грунт достаточно глубоко,
сохраняя возможность добывать пищу из воды. Они успешно приспособи-
лись к обитанию в приливно-отливных условиях (частое взбалтывание осад-
ка волнами и присутствие под открытым небом во время отлива). Глубокое и
быстрое зарывание сделало их идеальными первооткрывателями гостепри-
имного дна такого типа, способствовало их расцвету, что подтверждается
широким распространением двустворок в течение последних 200 млн. лет.
Несмотря на этот прогресс, роды и виды двустворок в связи с длительнос-
тью своего существования не являются особо ценными для палеонтологов,
желающих определить возраст отложений. Но есть и исключения, напри-
мер, зональное подразделение карбона по неморским двустворкам; значение
рода Buchia для юры и раннего мела и, главное, подразделение меловых от-
ложений по иноцерамам. Двустворки чрезвычайно полезны для палеоэколо-
гических построений в связи с тем, что дают хорошее представление о их об-
разе жизни и условиях окружающей их среды.
Ростроконхи. Ростроконхи были маленькой палеозойской (с раннего кем-
брия до перми) группой моллюсков, которые имели небольшие размеры (они
составляли от нескольких миллиметров до нескольких сантиметров). Внеш-
не ростроконхи похожи на двустворок. Возможно, что их внутреннее строе-
ние напоминало внутреннее строение двустворок, несомненно они имели
функционирующую ногу. От типичных двустворок ростроконхи отличались
отсутствием на раковине характерного для двустворок смычного края, слои
раковины распространялись и на площадь связки. Однако у некоторых выс-
ших форм, например у рода Conocardium, наружный слой раковины не пере-
секал смычный край.
В связи с этим некоторые палеонтологи, в частности Дж. Пойета и Б. Рун-
негар, предполагают, что ростроконхи являются ступенькой в направлении
к двустворчатым моллюскам. Самостоятельный класс Rostroconchia обособ-
ляют, делая акцент не на их сходстве с двустворками, а, напротив, на отли-
чиях этой группы. Некоторые палеонтологи даже высказали предположе-
ние, что ростроконхи могли иметь предков среди моноплакофор. Возможно,
тогда же ростроконхи дали начало двустворкам и, видимо, скафоподам. По-
требуется провести более глубокое исследование, прежде чем мы сможем со-
ставить верное представление о разнообразии моллюсков, обитавших в кемб-
рии, а возможно, и ранее. Прекрасно сохранившиеся отпечатки, такие как
Mootwingee в среднем кембрии формации Куниган с запада Нового Южного
ДВУСТВОРКИ
241
Уэльса, могут пролить свет на эту очень интересную фазу, когда произошло
быстрое изменение в составе моллюсков, и смогут помочь удовлетворитель-
ному объяснению взаимоотношений ростроконхов.
Несмотря на то что двустворчатые моллюски существовали начиная с
кембрийского периода, они не были широко распространены до среднего ор-
довика. Для самой примитивной группы, называемой палеотаксодонты, ха-
рактерны удлиненные створки, на смычном крае расположены плохо разви-
тые зубы, либо зубы отсутствовали. Современные представители обладают
маленькими листообразными жабрами, как у гастропод и хитонов.
Форма раковины и действие ноги. Палеотаксодонты определяют основ-
ную модель, от которой могли неоднократно ответвляться потомки. Легко
производимые изменения укрепляли раковину, улучшали ее защитные
свойства или сокращали усилия для передвижения двустворки. Во время ор-
довикского периода у некоторых двустворчатых моллюсков раковины были
укорочены и утолщены, а на смычном крае добавлены зубы. Остальные дву-
створки в интервале с ордовика по девон были продолговатыми клинообраз-
ными с узкими низкими створками. Они могли перемещаться (скользить) по
илу с минимальным трением.
Мы часто рассматриваем ползание с точки зрения толкания, тогда как
двустворчатые моллюски приспособились к медленному тянущемуся пере-
движению (фиг. XV.5). Нога выбрасывается вперед и закрепляется в иле ли-
бо песке: сильные мускулы сокращаются и подтягивают раковину вперед.
Результатом этого является медленное прерывистое продвижение, но неко-
торые ползающие двустворчатые могут так быстро сокращать мускулы, что
просто подпрыгивают вперед. На такое движение способен род Cardium, так
же как и зарывающиеся двустворки, когда они находятся на поверхности
(фиг. XV.6). Острые ножевидные формы (роды Solen и Tagelus) иногда пры-
гают на 25-30 см.
Рытье норок и зарывание (фиг. XV. 7). Процесс зарывания двустворок
неоднократно усовершенствовался у различных групп. Раннюю стадию это-
го процесса мы наблюдаем у подрода Encrintella (Crassatellites'), который
устраивает норки в углублениях, поворачивая раковину из стороны в сторо-
ну. Род Venus продвинулся вперед: он зарывается вглубь до тех пор, пока
большая часть раковины не покроется осадком, при этом задняя часть рако-
вины и короткие сифоны остаются снаружи. Род Муа со своей тонкой рако-
б
Фиг. XV.5. Ползание двустворки по дну
а — обычная ползающая двустворка проталкивает свою ногу сквозь ил либо пе-
сок; б — кончик ноги начинает вытягиваться и принимать форму якоря; в — мус-
кулы ноги сокращаются, двустворка подтягивается вперед; г — через короткий
промежуток времени нога вытягивается вновь и процесс повторяется
16 Каменная книга
242
КАМЕННАЯ КНИГА
Фиг. XV.6. Некоторые свободно передвигающиеся и прикрепляющиеся двус-
творчатые моллюски
Для ископаемых форм образ жизни восстановлен в соответствии с ныне живу-
щими: а - плавающая Lima (L. reticulata, миоцен, Мэриленд); б — Cardium, для
передвижения, выкидывающий вперед ногу (С, coosense, миоцен-современ-
ность, Тихоокеанское побережье); в — лежащий на дне Cornellites (С. flabella,
девон, восточная часть США); г — цементирующаяся Anomia (A. micronema,
поздний мел, запад Северной Америки); д — прикрепляющийся биссусными ни-
тями Mytilus (М. conradinus, миоцен, от Нью-Джерси до Техаса); е — прикреп-
ляющаяся с помощью биссусаВyssonychia (В. radiata, поздний ордовик, восточ-
ная часть США); ж, з — две изогнутые формы, цементирующиеся к погибшим
раковинам: ж - Toucasia patagiata, ранний мел, Техас и Мексика, з — Monopleu-
ra texana, ранний мел, Техас и Мексика
виной зарывается полностью; при этом сифоны значительно превосходят ра-
ковину по длине. Раковина рода Муа, зарываясь, тоже поворачивается с од-
ной стороны на другую, прокладывая свой путь сквозь песчаный или
илистый осадок.
Род Масота также зарывается в илистый песок. Один из его видов - Ма-
сота nasuta - лежит на левом боку и направляет вверх своей длинный ввод-
ной сифон, который обшаривает песок, отыскивая мягкую пищу. Когда
больше ничего найти нельзя, сифон втягивается обратно и затем опять вы-
брасывается сквозь песок за новой порцией питательного ила. Короткий вы-
водной сифон выпускает отходы.
Многие двустворчатые моллюски способны зарываться очень быстро. Не-
которые из них обладают толстыми раковинами и живут в песке: хорошим
примером в данном случае является род Venus. Острые раковинки (роды
Solen, Ensis, Tagelus и Siliqua) зарываются в песок почти вертикально и име-
ют различной длины сифоны. Один из представителей рода Siliqua закапы-
вается на песчаных пляжах Тихоокеанского побережья. До тех пор пока мо-
ре спокойно, моллюск остается около поверхности, но, когда начинается
шторм или накатываются волны, Siliqua уходит вглубь, погружаясь в осадок
за пределы опасной зоны.
Сверлящие моллюски и корабельные черви. Ни один сверлящий моллюск
никогда не беспокоил свою отсутствующую голову различием между зарыва-
нием и сверлением. Под первым мы, главным образом, подразумеваем зака-
пывание с помощью ноги. Сверлильщики осуществляют свой способ, преодо-
левая породу, раковины, дерево, бетон и гранит.
ДВУСТВОРКИ
243
Фиг. XV.7. Типы зарывающихся двустворок
Образ жизни ископаемых форм восстановлен в соответствии с родственными им
современными видами: а — Crassatella, б - Tellina, в — Macoma, г — Mactra, д -
Ensis, е — Martesia, ж — Pholas. Сифоны обозначены стрелками, показывающи-
ми направление тока воды
Ранняя стадия видна у рода Pholadidea, который прикрепляется своей но-
гой к различным твердым поверхностям, а затем начинает продвигать рако-
вину вверх и вниз. Передняя часть раковины покрыта складками, с по-
мощью которых происходит соскабливание окружающей породы. Некото-
рые представители данного семейства продолжают рост и сверление в
течение всей жизни. Другие, достигая максимального размера, втягивают
ногу внутрь раковины и проводят остаток жизни только питаясь и ре-
продуцируя.
Род Lithophaga, современный сверлящий моллюск, прикрепляется пуч-
ком биссусных нитей и выделяет комплексный состав, растворяющий поро-
ду и ныне живущие кораллы, в которые он всверливается. Периостракум до-
вольно толстый, защищающий раковину.
“Корабельные черви” на самом деле не являются червями: это древоточ-
цы, чьи раковины чаще всего достаточно малы, скребущие органы также
стали маленькими; имеются две маленькие пластины, видимо, защищаю-
щие сифоны. Молодой корабельный червь появляется на свет весной, личин-
ка плавает неделю или две, а затем оседает на погруженные в воду бревна или
другие формы древесины. Там древоточец всверливается, образуя трубку, на
которую он изнутри наносит тонкий известковый слой. В местах, где древо-
точцы довольно многочисленны, они заполняют большую часть древесины
высверленными ими ходами, при этом настолько разрыхляя дерево, что оно
разрушается от столкновения с волнами.
Род Martesia близок роду Pholas, хотя и не имеет с ним явного сходства.
У них хорошо развитые раковины, которые они в раннем возрасте использу-
ют при сверлении. Позднее створки цементируются и прикрепляются к труб-
ке, окружающей сифоны. Раковины и трубки рода Martesia найдены и в ка-
честве ископаемых вдоль рек Two Medicine в Монтане, а также в других
формациях.
16*
244
КАМЕННАЯ КНИГА
Гребешки и другие плавающие моллюски. Как сказано выше, крылоно-
гие гастроподы (птероподы) плавают, взмахивая своими крыловидными “но-
гами”, а некоторые другие гастроподы плавают, используя свою расширен-
ную ногу в качестве плота. Некоторые двустворчатые также являются плов-
цами, но они для этого раскрывают и закрывают свои раковины, хлопая
створками.
Этот способ был освоен несколькими группами. Первыми представителя-
ми семейства пектенид были формы, имевшие прямой смычный край, заост-
ренные ушки и более выпуклую правую створку. Виды с равными створками
могли только ползать, а другие — лежать на морском дне. Со временем неко-
торые из их потомков стали прикрепляться к морским водорослям или рако-
винам биссусными нитями, выходившими через вырез под одним из ушек.
Другие открывали и захлопывали свои створки и тем самым быстро вытал-
кивали воду. Результатом этого было плавание резкими толчками; живот-
ные продвигались смычным краем вперед, но использовали они этот способ
только при необходимости быстрого удаления, а не в качестве постоянного
способа передвижения. По форме раковины выделяются позднепалеозой-
ские виды рода Aviculopecten, плававшие таким же образом, как род Pecten и
его сородичи в современных морях. Их мантия окаймлена щупальцами,
снабженными глазками, которые, возможно, предупреждают о надвигаю-
щейся опасности.
Еще один пловец - род Lima - известен и как ископаемое, и как современ-
ная форма, живущая ныне в Тихом и Атлантическом океанах. Лима имеет по
краю мантии множество длинных щупалец, хотя ее раковина часто достига-
ет всего 5 см в длину. Неплавающие представители оседают в ямках и выде-
ляют упругие волокна, сплетающие открытое с двух сторон гнездо. Создавае-
мое ногой течение входит в “гнездо” с одного конца, а отходы выбрасываются
через другой конец.
Закрепление биссусом и цементация. Пектениды были не первыми мол-
люсками, у которых развился биссус. Эта структура впервые появилась у ор-
довикского рода Byssonychia, у него на переднем крае имеется широкий же-
лоб, из которого выходили биссусные нити. Биссус имеется также у рода
Mytilus и у других морских мидий: у рода Anomia, у молодых экземпляров
рода Tridacna и у некоторых других групп. Представители семейства Pteri-
idae встречаются от девона до настоящего времени и включают в свой состав
как современных жемчужниц, так и древнейшие раковины, внешне очень
похожие на род Aviculopecten. К семейству Pemidae, также имеющему бис-
сус, относятся такие важные мезозойские роды, как Gervilliopsis и Inocera-
mus. Молодые двустворки многих типов для первоначального закрепления
также выделяют биссусные волокна.
Эти волокна формирует расположенная в ноге биссусная железа. Они вы-
тягиваются так, чтобы каждое из них закреплялось на своем месте. Некото-
рые “пряльщики”, такие как Mytilus, “передвигаются”, помещая новые бис-
сусные нити далеко вперед, а нити, выделенные ранее, при этом разлагают-
ся, и таким образом животное перемещается вперед. У рода Anomia, биссус
обызвествляется или превращается в роговую пробку, вокруг которой распо-
лагается правая створка. Такие раковины крепко прикрепляются на всю ос-
тавшуюся жизнь.
Устрицы являются типичным примером цементирующихся или сидячих
двустворчатых моллюсков (фиг. XV.8). Проведя короткое время на стадии
личинки в планктоне, род Ostrea прикрепляется к скале, раковине или
древесине, к которым она “цементируется” левой створкой; эта створка ста-
ДВУСТВОРКИ
245
Фиг. XV.8. Две позднемеловые устрицы: Exogyra costata (слева) и Gryphaea
mutabilis (справа). Показан способ их обитания на морском дне; Теннесси
новится крупнее, выпуклее, толще, чем правая, которая иногда не больше,
чем крышка, закрывающая полость, в которой находится мягкое тело. Род
Ostrea впервые появился в триасовых морях, существовал в изобилии в ме-
ловое время и широко распространен в настоящее время. И восточные и ти-
хоокеанские разновидности были найдены в плейстоценовых отложениях
Северной Америки. В недавнем прошлом вид Ostrea virginica дорастал до
40 см в длину, когда “белый человек” начал собирать и продавать моллюски
в качестве пищи.
АБЕРРАНТНЫЕ ОБИТАТЕЛИ МОРСКОГО ДНА
Устрицы не являются “нормальными” двустворками, поскольку они ле-
жат на одной из своих створок и она становится намного больше, чем другая.
Другие двустворки стали еще более необычными, обладая створками, закру-
чивающимися как у улитки, и даже иногда принимая форму “рожковых” ко-
раллов.
Роды Gryphaea и Exogyra (фиг. XV. 9). Впервые закрученные макушки
мы наблюдаем у родов Gryphaea и Exogyra - двух представителей устричного
семейства, появившихся в мелководных юрских морях. Как и все истинные
устрицы, они сначала строили гладкие симметричные створки, которые, как
казалось внешне, предназначались для ползания и зарывания. Однако вско-
ре эти молодые устрицы ложились на левую створку, которая затем начина-
ла расти намного быстрее правой. У ранних представителей рода Gryphaea
левая створка сначала принимала форму, подобную примитивной улитке, а
246
КАМЕННАЯ КНИГА
после дальнейшего роста становилась массивной с центральной закрученной
макушкой. В дальнейшем она освобождалась от своего прикрепления и сво-
бодно лежала на илистой поверхности, где ее вес и форма способствовали со-
хранению раковины в изначальном положении. Правая створка превраща-
лась в ровную пластину, которая могла подниматься и опускаться, приот-
крывая или закрывая раковину.
В течение позднего мела большое число видов Gryphaea утратили склон-
ность к закручиванию макушки и стали чаше- или шарообразными. Некото-
рые представители с возрастом отрываются от субстрата, к которому они
прикреплены, но остальные остаются прикрепленными в течение всей жиз-
ни. Раковины обоих типов представлены в изобилии в формациях Атланти-
ки и заливов береговой равнины Северной Америки, а вторая группа распро-
страняется и севернее до Теннесси.
Род Exogyra отличается от рода Gryphaea наличием на смычном крае не-
отчетливо выраженного зуба, очень узким углублением для связки и всегда
спирально закрученными макушками. Левая створка рода Exogyra в основ-
ном значительно больше и толще, чем у рода Gryphaea. У крупных предста-
вителей - некоторые из них достигают 15 см в высоту - створки имеют ради-
альные ребра и линии нарастания, распространяющиеся и на тонкие слои.
Левая створка у Exogyra варьирует до массивных структур, плотно закру-
ченных на макушке. Некоторые раковины, как и Gryphaea, полностью ото-
рвались от субстрата. Другие стали такими большими и тяжелыми, что по-
гружены в ил, наклоняясь таким образом, чтобы их правая, или верхняя,
створка находилась в горизонтальном положении. Удивительно, что не-
сколько створок были искривлены взаимодействием с соседними раковина-
ми. И хотя род Exogyra был распространен во многих местах, чаще всего он
не создавал плотные банки, что свойственно современным устрицам.
Двойные спирали и конусы. Хотя у рода Exogyra обе макушки спирально
закручены, макушка правой створки в сущности является плоской. У дру-
гих форм, створки которых были почти одинаковыми и закручивались в
форме спирали, раковина в целом выглядела так, как будто две улитки при-
а
Фиг. XV.9. Устрицы
а — Exogyra, х0.5, поздний мел, от Нью-Джерси до Мексики; б — Gryphaea
(размер до 16 см в длину), этот экземпляр из ранней юры Европы; в, г — Exogyra,
xl, поздний мел, почти повсеместно, за исключением Южной Америки и
Австралии
ДВУСТВОРКИ
247
Фиг. XV.10. Рудист Plagioptychus partschi, х0.5, мел Австрии
липли губами друг к другу. Достигая такой формы, некоторые роды развива-
лись дальше, и одна из створок — левая или правая — превращалась в массив-
ную рогообразную спираль или конусообразную структуру, которая лежала
или стояла вертикально на дне моря, в то время как другая створка станови-
лась чуть больше пластины-крышки, закрывающей мягкое тело сверху. Не-
которые из этих странных раковин являются важными меловыми руководя-
щими ископаемыми Техаса, Нью-Мексико и Мексики.
Наиболее отличаются от обычных двустворок рудисты, почти полностью
утратившие следы закручивания в спираль (фиг. XV. 10). Нижняя створка
стала конической, она представляет собой комплексную структуру, состоя-
щую из пластин и слоев. Внешне эти формы похожи на “складчатые” корал-
лы (ругозы). Они могли достигать 102 см в высоту. Верхняя створка умень-
шилась до размеров крышки, на внутренней поверхности которой располо-
жены очень длинные зубы, заходящие в углубления нижней створки.
Рудисты ограничены интервалом поздняя юра-мел и более распространены
в Европе, где они образуют биогенные тела более значительных размеров,
чем в Северной Америке.
ОТ СОЛЕНОЙ ДО ПРЕСНОЙ ВОДЫ
Двустворчатые моллюски впервые появились в море, и длительное время
большинство из них жило в этой среде. Однако некоторые формы перемести-
лись в солоноватые заливы и в течение девона адаптировались к обитанию в
пресной воде. Один широко известный род - Archanodon - включает формы с
хрупкими, тонкими створками, составляющими около 8 см в длину. Они оби-
тали в течениях, пересекающих великую дельту Gaitskill, и во время среднего
и позднего девона распространялись на Пенсильванию и Нью-Йорк.
248
КАМЕННАЯ КНИГА
Пока род Archanodon и его потомки ползали на дне солоноватоводных
бассейнов, родственные им формы, называемые Naiadites, развивали биссус
и некоторые прикреплялись к затопленным бревнам. Их раковины найдены
среди знаменитых пенсильванских папоротниковых слоев Mazon Creek Ил-
линойса, а также в Огайо и Новой Шотландии. В последнем регионе они
встречены совместно с другими родами, один из которых (“Anthracomya”)
похож на крошечный “лошадиный” хвост.
Большинство современных пресноводных двустворок принадлежат к ро-
дам Unio nAnodonta. Ископаемые из триаса Массачусетса, Пенсильвании и
Юго-Запада одно время относились ко второму из родов, но сейчас очевидно,
что они принадлежат к другим типам, выжившим только в Южной Америке.
Род Unio, видимо, развивался в триасе Европы или Азии, достигнув в позд-
ней юре Северной Америки, и с тех пор всегда процветал на этом континенте.
Эти ископаемые найдены в слоях, содержащих остатки гигантских юрских
динозавров, а также в меловых и более молодых отложениях. И хотя в неко-
торых случаях сохранились раковины, большинство образцов являются
лишь внутренними ядрами. Униониды представляют хороший поисковый
признак для тех палеонтологов, которые ищут наземных позвоночных.
Глава XVI
НОГИ ВПЕРЕДИ ГОЛОВЫ:
НАУТИЛОИДЕИ И ИХ СОРОДИЧИ
• Рыбаки с юго-западного побережья Тихого океана часто забрасыва-
ют сплетенные из бамбука сети для ловли моллюсков рода Nautilus.
У него красивая белая раковина с оттенком слоновой кости и крас-
новато-коричневыми полосами, глаза с булавочными “отверстиями”, фоку-
сирующими свет, и около 90 коротких рук-щупалец, использующихся для
передвижения и добычи пищи. Мягкое тело наутилуса варят или делают из
него суп, большую прибыль приносят и раковины.
Nautilus - единственный род из огромной группы себе подобных, дожив-
ший до настоящего времени. Он взял на себя роль активного охотника в от-
крытой воде. Наутилус и его родичи имели возможность вести такой образ
жизни в основном из-за плавучести их раковин и специализированной систе-
мы “реактивного движения”. Это происходило в то время, когда было огром-
ное множество “взвешенных” организмов, то есть были своеобразные “подве-
шенные кормушки”, которые уже в раннем ордовике служили добычей для
наутилоидей. Эти времена в сущности отличались от кембрия тем, что подоб-
ные “едоки” были главным образом приурочены к морскому дну.
НАУТИЛУС КАК ОБРАЗ ГОЛОВОНОГИХ МОЛЛЮСКОВ
Род Nautilus - это больше, чем необычная и красивая раковина: он также
является образцом целого класса морских моллюсков, известных как цефа-
лоподы, или головоногие. Их название, дословно означающее “головная но-
га”, становится понятным, когда мы распознаем, что их продолговатые щу-
пальца в действительности являются преобразованием высоко адаптирован-
ной ноги. Нога и голова располагаются таким образом, что щупальца
находятся перед глазами, а их основание окружает рот (фиг. XVI. 1).
Имеются и другие характерные черты, которые отличают цефалопод от
других классов моллюсков, в частности от улиток. Во рту у наутилуса сохра-
няется радула. Помимо этого имеются роговые челюсти, или клюв, исполь-
зующийся для раскусывания добычи. Голова, с ее воспринимающим мозгом
и усложненными глазами, развита достаточно хорошо. Мантия облекает
мягкое тело, в мантийной полости находятся четыре листообразные жабры
и открывается анус. Щупальца, или руки, выбрасываются вперед для захва-
та пищи и передачи ее ко рту, а также используются для ползания и закреп-
© 1997. И.А. Михайлова, перевод на русский язык и изменения.
250
КАМЕННАЯ КНИГА
Фиг. XVI.1. Современный головоногий моллюск — Nautilus, х0.15
а — прикрепившийся к дну руками или щупальцами; б — плывущий в обратном
направлении; в — раковина вскрыта, видны внутренние органы
ления на месте. Но если у наутилуса возникает необходимость быстрого пе-
редвижения, он заполняет мантийную полость водой, а затем с силой вытал-
кивает струю воды через специальную трубку-воронку. Воронка, как и
щупальца, также является частью некогда единой “ноги” предков и состоит
из двух скрепленных вместе лопастей, которые и образуют сужающуюся на
наружном конце трубку. Вырывающаяся из воронки струя воды позволяет
наутилусу плыть быстро, но не плавно, а толчками. В том случае, когда во-
ронка (гипоном) обращена вперед, ее обладатель плывет назад, а когда труб-
ка отгибается назад, то животное направляется вперед. Этот способ передви-
жения главным образом предназначен для ловли неосторожной рыбы и
другой пищи.
Многокамерная раковина. Раковина наутилуса напоминает большую
узкую улитку, закрученную в одной плоскости, но сходство это только
внешнее. Стенку раковины слагают три слоя. Верхний наружный конхио-
линовый слой, который не сохраняется в ископаемом состоянии, второй
слой раковины фарфоровидный, а третий слой - жемчужный, или перла-
мутровый. Эти слои сформированы мантией, два внутренних слоя образо-
ваны арагонитом.
Существует большое различие в строении раковины брюхоногих и голово-
ногих моллюсков. Раковина улитки неделима: если бы мы могли развернуть
ее, она бы представляла собой один неправильный, но сплошной конус. А ра-
ковина наутилуса обычно состоит из 30 или более камер, которые разделены
НОГИ ВПЕРЕДИ ГОЛОВЫ: НАУТИЛОИДЕИ И ИХ СОРОДИЧИ
251
между собой перламутровыми септами, или перегородками. Пока наутилус
растет, мантия надстраивает жилую камеру вперед. Далее мускулы фикси-
руют новую линию прикрепления мягкого тела к раковине, и животное
продвигается вперед. В результате этого процесса сзади остается свободное
пространство, которое отграничивается новой перегородкой. Таким образом
каждая септа была сформирована задней частью мантии, отделяя освободив-
шуюся часть раковины от жилой камеры.
Сифон и септальные трубки. Когда мягкое тело продвигается вперед, то
остается кожистая трубка — сифон. Перегородка вокруг сифона отгибается
в виде короткой, направленной назад трубки (некк). Не очень длинное
соединительное кольцо закрывает сифон снаружи. Сифон протягивается
через все камеры — от жилой камеры до самого начала раковины. Он хоро-
шо обеспечен кровеносными сосудами и, видимо, поставляет газ в камеры.
Кроме того, в камеры может быть привнесена или удалена из них так назы-
ваемая камерная жидкость. Раковина при этом становится либо тяжелее,
либо легче.
У наутилуса соединительные кольца тонкие, после смерти животного они
не сохраняются. У ископаемых соединительные кольца нередко утолщены
вторичными отложениями.
ПУТЬ ОТ ПРОСТОЙ УЛИТКИ до головоногих моллюсков
Исследователи часто с этим не согласны, и те из них, кто занимается клас-
сификацией цефалопод, не являются исключением. Многие имеющиеся
факты являются единичными для утверждения того, что предки этих су-
ществ незначительно отличались от колпачковидной улитки. Вероятно, они
имели крышечную раковину - домик с широкой присасывающейся ногой,
рот располагался в передней части тела, а жабры и анус — в задней. Счита-
лось, что эти моллюски закреплялись на плотном, но илистом дне, а не на
скалах, как современные блюдечки (например, Patella).
На протяжении веков потомки этих пионеров все более и более усовер-
шенствовали свое мягкое тело. Это означает, что их нога укоротилась, а пи-
щеварительный тракт изогнулся (стал петлевидным), в то время как рако-
вина стала конической или слегка рогообразной, чтобы вместить все более
“углубляющееся” мягкое тело. Позднее нога начала изгибаться кверху,
мантия приобрела способность секретировать несколько перегородок, одно-
временно формировался примитивный сифон. И на последней стадии наше-
го гипотетического ряда пищеварительный тракт изогнулся в виде
латинской буквы V, раковина и сифон стали продолговатыми. Нога продол-
жала расти около (вокруг) головы, подразделяясь на щупальца и две лопас-
ти, загибающиеся по направлению друг к другу. В месте их соединения об-
разовалась воронка, с помощью которой сформировавшийся головоногий
моллюск начинал плавать в тех случаях, когда ползание становилось для
него слишком медленным. Так и представлялась история происхождения
цефалопод, пока американский палеонтолог Э.Л. Йохельсон и его коллеги
не наткнулись на очень интересное маленькое ископаемое. Этим ископае-
мым был Knightoconus antarcticus из отложений верхнего кембрия с наибо-
лее невероятного, на сегодняшний день, места - Западной Антарктики. Это
была моноплакофора, с высокой, слегка изогнутой конической раковиной,
которая отличалась от подобных себе наличием около макушки нескольких
септ. Все, что было необходимо для того, чтобы отнести эту форму к наути-
лоидеям, — это развитие сифона и сифонального комплекса, столь характер-
252
КАМЕННАЯ КНИГА
ного для цефалопод. Поэтому головоногие моллюски, включающие в себя
наутилоидей, так же как брюхоногие моллюски, скорее всего произошли от
древних моноплакофор.
РАЗНООБРАЗНЫЕ И ДОЛГОЖИВУЩИЕ НАУТИЛОИДЕИ
Наутилус одновременно является и типичным головоногим моллюском, и
единственным дожившим до наших дней представителем подкласса Nauti-
loidea. И хотя все члены этого подкласса значительно отличаются друг от
друга, у них есть общая черта - наличие относительно простых перегородок,
которые в основном напоминают часовые стекла с вогнутой стороной или яв-
ляются относительно невысокими конусами (фиг. XVI.2). Каждая перего-
родка около воронки вогнута назад, формируя гипономический синус, а на
стыке со стенкой раковины образует перегородочную линию. Эта линия мо-
жет быть прямой (за исключением синуса), или может быть изогнута разно-
образным способом. У некоторых форм, таких как раннечетвертичная
Aturia, перегородки были сложно устроенными, подобными перегородкам
некоторых палеозойских гониатитов (аммониты).
Одно время самым древним известным представителем наутилоидей счи-
тался род Volborthella из нижнекембрийских отложений Северной Европы и
Канады. У этого рода имелась прямая известковая раковина длиной в не-
сколько миллиметров, однако отсутствовали перегородки и была радиаль-
ная, а не двусторонняя симметрия. Раковина имела пластинчатую структу-
ру, которую Йохельсон смог описать на основе новых хорошо сохранивших-
ся ископаемых, найденных в Норвегии. Специализируясь на ранних
моллюсках, Йохельсон установил, что семейство Volborthellidae и другие
группы, например Salterellidae, следует выделить в отдельный, отличаю-
щийся от моллюсков, новый тип, названный им Agmata.
Некоторые палеонтологи предположили, что древнейшим и простейшим
из всех наутилоидей является род Plectronoceras из отложений верхнего кем-
брия Китая. Раковина этого рода не прямая, а слегка изгибающаяся, с глубо-
кой жилой камерой и часто расположенными перегородками, вогнутыми
как стекла крошечных часов. Сифон располагается в краевой части ракови-
ны, а не в ее центре, как у Nautilus.
Строение мягкого тела этого и других ранних головоногих моллюсков не-
известно. Но мы можем с уверенностью утверждать, что они были очень по-
хожи на наутилуса с глазами, имеющими узкие щели, заканчивающиеся
крошечным отверстием, и с шероховатым капюшоном, закрывающим рако-
вину, когда мягкое тело, голова и щупальца втягивались внутрь. На щупаль-
цах почти всегда отсутствовали присоски, а сами щупальца длинные и менее
многочисленные, чем у наутилуса. По имеющимся наблюдениям, у некото-
рых видов число их всего 10.
Почему мы называем их “роговые раковины”? Название родов наутилои-
дей нуждается в комментариях, так как многие из них заканчиваются на
-ceras, что по гречески означает “рог”. Эти ископаемые нередко имеют сход-
ство с изогнутым или закрученным рогом, и первоначальное значение -ceras
было основано именно на этом сходстве. Со временем это окончание стало
привычным и распространилось на других головоногих моллюсков, имев-
ших наружные раковины. И хотя имеются исключения, можно говорить,
что почти любое ископаемое, чье название заканчивается на -ceras, является
головоногим моллюском.
НОГИ ВПЕРЕДИ ГОЛОВЫ: НАУТИЛОИДЕИ И ИХ СОРОДИЧИ
253
Вторичные
отложения
Соединительные
синус
кольца
Фиг. XVI.2. Древние наутилоидеи
а — схема строения прямой раковины головоногих моллюсков, слева и справа
показано продольное сечение, в центре — ядро; б — род Volborthella, хЗ, видимо,
наутилоидный моллюск, ранний-средний кембрий; в - род Plectronoceras, х4,
поздний кембрий; г - широкий сифон рода Cassinoceras с эндоконами; д — про-
дольный разрез рода Ascoceras, силур, Чехия; е- род Billingsites, Северная Аме-
рика; ж,з- сифон pojxaActinoceras; и — продольное сечение рода Endoceras, ши-
рокий сифон приближен к брюшной стороне и заполнен эндоконами
254
КАМЕННАЯ КНИГА
ХАРАКТЕР ОБИТАНИЯ И ИЗМЕНЧИВОСТЬ РАКОВИН
Примитивные головоногие моллюски могли быть прямыми, но, вероятнее
всего, были слегка изогнуты. Поскольку их камеры содержали только газ,
раковины были легче воды и намного легче, чем мягкое тело животного. Лег-
кий вес должен был обуславливать положение животного в воде: закрытый
конец направлен вверх, как у раковин на последней стадии предполагаемого
нами развития от брюхоногих моллюсков до цефалопод.
Это, вероятно, не вызывало никаких проблем, пока раковины были ма-
ленькими и короткими и достаточно уравновешивались весом мягкого тела.
Но со временем размеры раковин увеличивались и возникали сложности.
Крупные раковины, которые намного легче, чем вода, при расположении за-
крытым концом вверх должны были болтаться взад и вперед по прихоти лю-
бого волнения или течения. Это приводило к сокращению скорости движе-
ния, вызывая затруднения с сохранением направления движения. Это также
удерживало головоногих с такими раковинами от положения на правом боку.
Мы не уверены, что так происходило на самом деле, но так должно было
происходить. Наутилоидеи, неприспособленные для активного образа жиз-
ни, возможно, были вытеснены хищниками, голодом или другими неблаго-
приятными факторами и стали отклоняться от нормы. Их место заняли соро-
дичи, сумевшие преодолеть трудности (увеличение размеров), о которых мы
упомянули.
Имеется несколько теорий, объясняющих, как и почему развивались эти
новые, более удачно приспособившиеся наутилоидеи, но одни из них проти-
воречат другим. Поэтому мы опустим вопросы “почему?” и “как?” и остано-
вимся на фактических изменениях раковин и их сравнительных взаимоот-
ношениях, ориентации в толще воды и характере обитания.
Балласт и обитатели дна. Первые головоногие были присасывающимися
и ползающими, и некоторые из ранних изменений сохраняли и даже увели-
чивали их соответствие обитанию на дне, то есть бентосному образу жизни.
Они довольно заметны на так называемых ортоцераконах, чьи раковины бы-
ли прямыми, хрупкими и иногда очень большими. Один из ордовикских ро-
дов - Endoceras - достигал от 2.7 до 3.6 м в длину.
Такие раковины в том случае, когда они были пустыми, дрейфовали за-
крытым концом вверх, так как после смерти моллюска мягкое тело сгнива-
ло, а иногда выпадало на дно. Но редкие образцы с полосками, пятнышками
и зигзагообразной цветной каймой на верхней, или спинной, стороне ракови-
ны не оставляют сомнений, что ортоцераконовые прямые раковины при
жизни находились в горизонтальном положении, при этом воронка распола-
галась внизу, а голова — вверху. Это было осуществимо благодаря вторичным
отложениям на стенках раковины, перегородках и сифоне, которые в основ-
ном концентрировались около нижней (брюшной) стороны.
Сифональный балласт располагался тремя способами: кольцами, лучеоб-
разными пластинками и коническими слоями, называемыми эндоконы или
“внутренние конусы”. Последние иногда образовывались внутри широкого
сифона, находившегося около брюшной стороны животного или в непосред-
ственной близости от нее.
Род Actinoceras - одна из наиболее распространенных форм головоногих
моллюсков, у которых сифон имел систему радиальных лучеобразных от-
ветвлений. Род Endoceras — образец раковины прямой формы - обладал эн-
доконами, особенно развитыми в начальной части раковины. Короткий и
утолщенный род Cassinoceras отличается тесно расположенными перегород-
НОГИ ВПЕРЕДИ ГОЛОВЫ: НАУТИЛОИДЕИ И ИХ СОРОДИЧИ
255
Фиг. XVI.3. Характер обитания некоторых палеозойских наутилоидей
а — обитатель дна Gonioceras с уплощенной раковиной; б— Cyrtoceras, вероятно,
ползающий с помощью щупалец; в — Michelinoceras, видимо, ползал, но мог и
плавать; г - дрейфующий Mandaloceras с яйцеобразной раковиной; д — питаю-
щийся трилобитами Gigantoceras, который и ползал и плавал
ками, которые загибались вокруг сифона с массивными эндоконами. Устье
слегка наклонено вниз, будто тяжеловесный конус поднял его достаточно
высоко над дном — местом обитания животного.
Наиболее странным было приспособление, развитое в ордовике и силуре у
рода Ascoceras и его сородичей. Они начали с формирования хрупкой слабо
изогнутой раковины, не имеющей балласта (вторичных отложений), за ис-
ключением сифона, перемещавшегося все ближе и ближе к брюшной сторо-
не. Жилая камера начала удлиняться, а в течение позднего ордовика и раз-
растаться вширь, позволяя перегородкам сокращать содержащие газ каме-
ры, широко огибающие спинную область. Таким образом балластом для
животного становилось мягкое тело. Наконец, когда мягкое тело перемеща-
лось в увеличенную жилую камеру, оно закрывало сифон и позволяло отде-
литься оставшейся части раковины. В результате этого первоначально длин-
ная, остроконечная раковина преобразовывалась в короткую, приспособлен-
ную к более активному, чем в раннем возрасте, образу жизни.
Ползающий Gonioceras (фиг. XVI.3). Разнообразие приспособлений к
жизни на дне хорошо представлено на примере ордовикского рода Gonioceras.
Его раковина была очень низкой и широкой с умеренно конической
центральной частью и утонченными, заостренными по бокам “крыльями”.
256
КАМЕННАЯ КНИГА
Внутренняя поверхность раковины почти гладкая, а сифон не очень утяже-
лен; в течение взрослого периода жизни устье резко сужается до закрыва-
ния боковых краев. Это явление объяснялось как приспособление, предо-
храняющее животное от выталкивания мягкого тела из раковины моллюс-
ка, живущего среди морских водорослей и кораллов.
Эта идея сомнительна по двум причинам. Первая из них заключается в
том, что широкая раковина и сифон актиноцератидного типа способствуют
лучшему прикреплению (пропорционально своему весу), чем массивные
прямые конусы с широким устьем. Вторая: род Gonioceras, возможно, мог
немного ползать. Его широкая раковина напоминает по форме мелкую кам-
балу, лежащую на дне в ожидании пищи, но временами переплывающую с
одного места на другое. Возможно, так же поступал род Gonioceras. Когда он
чувствовал себя принужденным плыть, то немного проплывал, как бы сколь-
зя по воде и протягивая за едой щупальца.
Ползающие наутилоидеи с коническими и рогообразными раковинами.
И строение, и окраска говорят о том, что длинные прямые наутилоидеи пла-
вали по существу в горизонтальном положении и ползали. У некоторых ши-
роких и коротких раковин сохранились цветные полоски, распространен-
ные по всей поверхности. Эти полоски, форма раковины и отсутствие балла-
ста (вторичных отложений) не оставляют сомнений в том, что их обладатели
ползали, а раковины были ориентированы закрытым концом вверх.
Простейшие ползающие наутилоидеи - это роды Rizoceras и Cyrtoceras.
Первый имел коническую, а второй слегка изогнутую раковину, причем во-
гнутая сторона была брюшной. В конце концов один палеонтолог решил, что
обладатели этих раковин подобно улитке ползали по поверхности субстрата
при помощи рудимента первичной “гастроподной” ноги. Это делает данных
животных похожими на примитивных цефалопод, то есть это реликты, кото-
рым удалось сохраниться на протяжении большей части палеозойской эры.
Кажется более вероятным, что они были настоящими истинными наутилои-
деями с укороченными раковинами и мягким телом, которое специализиро-
валось на обеспечении использования любой прикрепляющей поверхности,
что необходимо для передвижения.
Изогнутые раковины не были ограничены только родом Cyrtoceras и по-
добными ему: они появлялись и в других семействах и даже в других отря-
дах. Такие раковины строились по двум противоположным схемам: одни — с
выпуклой (экзогастрические формы), а другие — с вогнутой (эндогастрические
формы) брюшной стороной. Первая схема не вызывает никаких проблем, осо-
бенно если сифон содержит вторичные отложения, служившие балластом. А
эндогастрические раковины с балластом или без него совершенно противопо-
ложны. Были ли их тела достаточно тяжелыми, чтобы они могли ползать с
направленным вверх концом раковины? Или, поскольку их раковины были
легче воды, их обладатели могли дрейфовать в толще воды и касаться дна
только щупальцами?
Причудливые ползающие и плавающие формы. Таковыми являются ко-
роткие изогнутые раковины, встреченные в отложениях силура и девона. У
некоторых из них, таких как род Protophragmoceras, жилая камера была
сжата, но открыта, а их обладатели, должно быть, вели образ жизни, похо-
жий на образ жизни рода Cyrtoceras.
Раковины типа Phragmoceras короче, а их камерная часть быстро перехо-
дит в большую, но существенно сжатую жилую камеру. Устье заужено - не
просто изначально узкое, а сжато в конечной устьевой части жилой камеры,
пока оно не приобретает почти I-образную форму, или еще более усовершен-
НОГИ ВПЕРЕДИ ГОЛОВЫ: НАУТИЛОИДЕИ И ИХ СОРОДИЧИ
257
ствовано. Даже если сифон достаточно массивный, животное не являлось
очень тяжелым. У рода Phragmoceras раковина слегка наклонена к задней
части, и задача уравновешивания в первую очередь ложится на мягкое тело.
Вероятно, Phragmoceras основную часть времени ползал, высовывая голову
со щупальцами и глазами из большей части жилой камеры, воронка у него
находилась в задней части. Свободные камеры наделяли животное способно-
стью держаться на воде, что было особенно необходимо в те моменты, когда
моллюск плыл. Так как воронка переместилась в заднюю часть, то при пла-
вании животное двигалось вперед, а не назад.
Такие формы, как род Hexameroceras, не были так резко закручены. Они
почти всегда дрейфовали, форма жилой камеры говорит о том, что глаза и ру-
ки располагались ниже, чем воронка.
Иной способ адаптации к дрейфующему образу жизни был встречен у
наутилоидей с короткой раковиной. Это роды Mandaloceras и Ovoceras из от-
ложений ордовика и девона. Раковины у них широкие и короткие с едва за-
метными следами изогнутости и устьем Y- и Т-образной формы. Глаза были
снабжены Y- и Т-образными перепонками. Некоторое расширение раковины
в устьевой части (в действительности - на брюшной стороне) обеспечивало
выход для воронки и рук (щупалец). Эти животные, вероятно, дрейфовали,
голова была при этом направлена вниз, время от времени они погружались
на дно и поднимались над ним под воздействием воды через воронку. Накло-
няя воронку в сторону задней части, они могли передвигаться вперед, слегка
отклоняясь при этом в сторону.
Это движение не могло быть очень быстрым, а положение, в котором
животные дрейфовали, показывает, что они искали пищу на дне. На основа-
нии того, что у этих форм было узкое устье, австрийский палеонтолог сделал
вывод, что эти и другие подобные им головоногие употребляли очень мелкую
пищу. Его оппоненты утверждают, что клюв мог высовываться даже из уз-
кой щели, а моллюск удерживал свою жертву руками. В непосредственном
процессе принятия пищи принимала участие радула, которая, возможно,
могла работать эффективнее, чем у рода Nautilus.
Закрученные, ползающие и плавающие наутилоидеи. Одна из теорий по-
лагает, что несомненные ранние цефалоподы были изогнуты таким же обра-
зом, как род Cyrtoceras, но их устье не было коротким и суженным. На сле-
дующей стадии раковины начали закручиваться: сначала неплотно, а затем
все более и более плотно, пока последние обороты не стали перекрывать друг
друга и даже полностью закрывать предыдущий оборот.
Эта теория, вероятно, является правильной, но пока еще нет фактической
серии ископаемых, которые бы ее подтвердили. Очевидно, что раннекемб-
рийский род Volborthella имел раковину почти прямую, а позднекембрий-
ский род Plectronoceras - довольно длинную и умеренно изогнутую. В морях
раннего кембрия обитали две группы плотно свернутых раковин, но отсутст-
вовали промежуточные формы. Три другие группы, или роды, появившиеся
в течение девона и карбона, видимо, имели предков с плотно свернутой рако-
виной. Как у них сформировался такой способ расположения оборотов, оста-
ется вопросом для рассуждений.
В зависимости от степени объемлемости оборотов и формы поперечного се-
чения свернутые спирально-плоскостные раковины получили различные на-
звания. Когда вопрос касается характера обитания, более важным является
размер мягкого тела, а также ширина и основная форма раковины. Такие го-
ловоногие, как роды Titanoceras и Solenocheilus, с широкими раковинами и
относительно крупным и тяжелым мягким телом, большую часть времени
17. Каменная книга
258
КАМЕННАЯ КНИГА
Фиг. XVI.4. Спирально свернутые наутилоидеи, некоторые с длинными шипами
а, б - Titanocerasponderosum, х0.35, поздний карбон, от Иллинойса до Небрас-
ки; в — Cooperoceras texanum, хО.6, пермь, Скалистые горы, Техас; г — Endolo-
bus, xl, карбон—пермь, Европа и Северная Америка; д — Solenocheilus (около
5 см), карбон—пермь, широко распространен; е—Acanthonautilus, х0.5; этот эк-
земпляр из России, род существовал в раннем карбоне Европы и Северной Аме-
рики; ж —Aturia, х0.5; этот экземпляр из эоцена Нью-Джерси; род существовал
с палеоцена по миоцен; широко распространен
должны были проводить, ползая по дну и плавая только на короткие дистан-
ции, и то лишь в случае угрожающей им опасности (фиг. XVI.4). Объемле-
мость оборотов раковины на оказывала заметного влияния на характер их
обитания.
НОГИ ВПЕРЕДИ ГОЛОВЫ: НАУТИЛОИДЕИ И ИХ СОРОДИЧИ 259
Формы подобные Eurystomites имели дисковидные раковины, легкие по ве-
су и длинные, но небольшие по объему жилые камеры, которые на конечной
стадии становились почти прямыми. Мягкое тело, чтобы обеспечивать бал-
ласт, очевидно, являлось достаточно тяжелым, а плавучесть раковины, скорее
всего, была такова, что устье располагалось под углом вниз. Эти животные
часть времени ползали, а часть — дрейфовали и могли плавать легче, чем Ti-
tanoceras. Хотя их высокими легкими раковинами управлять было труднее.
Род Nautilus имеет типичную для головоногих моллюсков спирально-пло-
скостную раковину. Мешковидное мягкое тело находится в короткой жилой
камере. Взрослый Nautilus незначительно тяжелее воды, всего настолько,
чтобы он без излишних усилий мог ползать, или дрейфовать, или свободно
“болтаться” над поверхностью дна, лишь в незначительной степени исполь-
зуя воронку. Сильная струя жидкости из воронки направляет Nautilus рако-
виной вперед, головной отдел при этом располагается сзади. Современные
формы в течение дня остаются на значительной глубине (около 550 м), а но-
чью поднимаются к поверхности воды. Вымершие животные могли иметь та-
кие же привычки. Поскольку на глубине 350 м вода могла раздавить ракови-
ну наутилуса, вероятно, он обладал способностью изменять давление газа в
камерах, что и позволяло животному выдерживать давление на больших
глубинах. Однако отсутствуют доказательства того, что газ достаточно уп-
лотнялся и увеличивал давление внутри воздушных камер, чтобы помочь
животному погрузиться, и, напротив, уменьшал это давление, когда ему не-
обходимо было всплыть.
Более приспособленными к плаванию были закрученные наутилоидеи с
очень узкой раковиной. Некоторые из них появились в пенсильванских мо-
рях, а другие присоединились к ним во время мезозойской и кайнозойской эр.
Многие из этих животных обладали гладкими раковинами, а края их перего-
родок были сильно изогнуты, в связи с чем перегородочная линия таких
форм была достаточно усложненной.
Некоторые аберрантные наутилоидеи. Большинство древних наутилои-
дей обладало гладкими раковинами со слабыми ребрами и линиями нараста-
ния, отражающими короткие перерывы роста. У некоторых развивались
кольцеобразные ребра, а иногда - целая система ребер. У Rotoceratida рако-
вины, кроме того, были украшены бугорками, шинами и пережимами.
Некоторые из этих скульптурных особенностей делали плавание невозмож-
ным и, вероятно, мешали и ползанию. Трудно поверить, что шипы у рода
Cooperoceras не запутывались в морских водорослях, или что морщины у ро-
да Ryticeras не затрудняли прикрепление или плавание. Второму роду
(Ryticeras) приходилось держаться вдали от маленьких кораллов, которые
могли зацепить и разрушить тонкие морщины.
Некоторые наутилоидеи, предки которых имели свернутые раковины, ос-
тавили свободное плавание в пользу дрейфования или ползания. Мы это
видим на примере ордовикского рода Schroederoceras. Сначала раковина это-
го рода плотно свернута, а затем она становится слегка изогнутой, и у ее хо-
зяина остается выбор между дрейфованием и ползанием. У силурийского
рода Lituites мы видим дальнейшее развитие этого процесса: меньшая часть
раковины спирально свернутая, а большая — прямая; устье суженное
(фиг. XVI. 5). Поскольку ни стенки раковины, ни сифон не были достаточно
весомыми, литуитесы должны были дрейфовать головой вниз, как и удли-
ненный Mandaloceras.
Другой силурийский типичный наутилоидный род Mitroceras был свер-
нут в спираль, как и улитки, хотя некоторые девонские роды в зрелой стадии
17*
260
КАМЕННАЯ КНИГА
/ /у I
Фиг. XVI.5. Прямые и согнутые наутилоидеи
а - Rutoceras, х0.5, девон, Европа и Северная, возможно, и Южная Америка;
б, в - Phragmoceras, х0.5, короткая, несоприкасаютцаяся спираль с узким усть-
ем, ранний силур, Европа и Северная Америка; г — Bickmorttes bickmoreanus, до-
стигающий в длину 23 см, ранний силур, Индиана; д, е — Hexameroceras , хО.6,
силур, Европа и Северная Америка; ж—Actinoceras, достигающий в длину 20 см,
на образце слева—ядро фрагмокона и сифона, в центре вскрыт фрагмокон, а спра-
ва — сечение сифона; средний ордовик-ранний силур, Евразия, Гренландия и Се-
верная Америка; з — Huronia bigsbyi, только сифон длиной до 13 см; ранний си-
лур, северный Мичиган; и — Lttuites lituus, х0.5, начальная часть спирально свер-
нута, затем, подобно Baculites, выпрямляется, ордовик, Северная Европа
НОГИ ВПЕРЕДИ ГОЛОВЫ: НАУТИЛОИДЕИИИХСОРОДИЧИ
261
не были свернуты. Эти наутилоидеи ползали, раковина наполовину возвы-
шалась над дном, хотя отсутствуют факты, подтверждающие, что их мягкое
тело имело широкое основание, подобное ноге улиток.
НАУТИЛОИДЕИ КАК ИСКОПАЕМЫЕ
Поскольку арагонитовая раковина наутилоидей быстро разрушается,
большинство принадлежащих к этой группе ископаемых представлено внут-
ренними ядрами. Они отражают форму раковины, разнообразие камер, по-
ложение, а иногда строение сифона. Перегородки наблюдаются в виде изо-
гнутых поверхностей или перегородочных линий, последние пересекают
разрушенные образцы и показывают, как перегородки соединялись со
стенками.
Естественно, что из этого правила существует множество исключений,
так как при разрушении самих раковин могло сохраниться внутреннее стро-
ение. Наиболее важными являются окремнелые ископаемые, у которых ара-
гонит был замещен кремнеземом (кварцем), окрашенным примесями типа
оксида железа. Подвергшиеся атмосферным явлениям карбонатные элемен-
ты могут растворяться в соляной или уксусной кислоте.
Для детального изучения ископаемых наутилоидей необходимы образцы,
показывающие, каким образом строилась раковина с ранних до зрелых ста-
дий. Многое можно узнать с помощью неполных образцов. Достаточно наблю-
дать форму раковины, чтобы распознать большинство родов и видов, некото-
рые могут быть определены по фрагментам сифона. Полезны и выветрелые об-
разцы, подвергшиеся атмосферным воздействиям, хотя у них почти никогда
не сохраняется скульптура наружной поверхности раковины.
Более сложным, чем идентификация видов, является определение родов.
Так, до 1925 года к роду Orthoceras (“прямой рог”) относились многие пря-
мые и изогнутые наутилоидеи, у которых сифон располагался около центра
и раковина, за исключением колец и линий нарастания, была гладкой. В род
Nautilus было включено большое разнообразие спирально свернутых рако-
вин, у которых сифон находился около центра, но несколько ближе к брюш-
ной стороне. К роду Gomphoceras относили короткие, изогнутые, не очень
крупные раковины, и т.д.
Такие роды были простыми, большинство из них определялось по внешне-
му сходству. Когда род Orthoceras был детально проанализирован, то в его со-
ставе оставили только несколько видов. Многие виды вошли в новые роды,
такие как Michelinoceras и Geisenocerina. Аналогичная ревизия постигла и
другие сборные группы, в результате этого наутилоидеи разделены сейчас на
бесчисленное число родов, распределенных по многим десяткам семейств и
одиннадцати отрядам.
В этом нет неправильности: классификация должна учитывать различия
и взаимосвязи, даже если они достаточно сложны. Правда, создается пробле-
ма для исследователей, которые могут оперировать современными названи-
ями, используя только доступные учебники, всегда полезную книгу Шайме-
ра и Шрока “Index Fossils of North America”, а также представленные здесь
роды. Другие образцы могут быть идентифицированы как виды, используя
публикации и коллекции, поэтому специалисты палеонтологи работают в
музеях, университетах и других институтах.
Самые ранние наутилоидеи, такие как род Plectronoceras, были маленьки-
ми (древнейшие - только около 10 мм в длину) со слегка изогнутой ракови-
262
КАМЕННАЯ КНИГА
ной (циртоконовый тип), что типично для кембрийских форм. В раннем ор-
довике появилось большое разнообразие форм, в основном они отличались
типами раковин и являлись более крупными; в палеозое господствовали
наутилоидеи с циртоконовыми (изогнутыми) и ортоконовыми (прямыми)
раковинами.
В среднем ордовике появилось девять из одиннадцати основных групп,
многие из которых существовали большую часть палеозоя. Последний из
возникших в позднем силуре или раннем девоне отрядов наутилоидей вклю-
чает в качестве одного из представителей и современный род Nautilus.
Большинство групп наутилоидей вымерло в конце палеозоя, и только от-
ряд Nautilida, продолжавший существовать в течение мезозоя, дожил до на-
стоящего времени. Возможно, вымирание наутилоидей было связано с широ-
ким распространением аммонитов, господствовавших в мезозойских морях.
КЛАССИФИКАЦИЯ НАУТИЛОИДЕЙ
Наутилоидеи и их ближайшие сородичи классифицируются некоторыми,
но не всеми палеонтологами на основе положения и строения сифона. При
этом выделяются три подкласса: Endoceratoidea, известный только в ордови-
ке и силуре; Actinoceratoidea, живший с ордовика до карбона, и Nautiloidea,
встречающийся с позднего кембрия до настоящего времени.
Эндоцератоидеи имели раковины прямые (ортоконы), реже изогнутые
(циртоконы). Для них характерен и наиболее широкий сифон, расположен-
ный около брюшной стороны раковины. Все актиноцератоидеи обладали
только прямыми раковинами. У них имелись короткие изогнутые перегоро-
дочные трубки, полукруглые соединительные кольца, а внутри сложно уст-
роенные радиальные каналы. Раковины наутилоидей бывают как прямые и
изогнутые, так и полностью свернутые в спираль, т.е. наблюдается наи-
большее разнообразие. Формы с прямыми раковинами существовали до три-
аса, но затем они уступили свое место спирально-плоскостным формам. Не-
давно появились сведения о том, что прямые ортоцератиды жили и в раннем
мелу (Doguzhaeva, Mutvei, 1994). Строение сифона у этой группы также до-
статочно разнообразно. У ранних наутилоидей перегородочные трубки соеди-
нены кольцами, утраченными у некоторых постпалеозойских форм.
Глава XVII
РОГА АММОНА
И ВНУТРЕННЕРАКОВИННЫЕ
ГОЛОВОНОГИЕ
• Название Ammonites было дано ископаемым раковинам, которые
имели спирально-плоскостные ребристые раковины. Оно означает
“рога Аммона”; происхождение названия связано с египетским богом
Аммоном, которого изображали с бараньими рогами на голове. На сегодняш-
ний день мы понимаем под названием аммоноидеи (Ammonites — это собствен-
но родовое название) подкласс головоногих моллюсков, имевший сложногоф-
рированные перегородки. Раковины аммоноидей варьируют от вздутых до уз-
ких и от гладких до невероятно скульптированных. Большинство из них
имело средние размеры, но некоторые огромны, например, меловой Pachydis-
cus seppenradensis, достигавший 1.7 м в диаметре. Недавно был встречен Diplo-
moceras maximus — гигант среди прямых аммоноидей (см. фиг. XVII.4). Когда
мягкое тело полностью втягивалось в жилую камеру, многие, а возможно, и
все представители этой группы могли закрывать свое устье шероховатой плас-
тинкой (анаптих) или двумя известковыми пластинками (аптихи). Мягкое те-
ло, глаза и руки, видимо, подобны таковым у наутилоидей, но, как установле-
но в последние годы, в действительности они имели наиболее близкое сходство
с колеоидеями, включая кальмаров и осьминогов. Радула аммонитов, с помо-
щью которой они питаются, имела, как у колеоидей, семь зубцов в ряду и че-
люсти, похожие на клювы осьминога, видимо, присутствовал чернильный ме-
шок. У наутилуса радула имеет в ряду тринадцать зубцов, и он отличается и от
аммонитов и от колеоидей отсутствием чернильного мешка; для аммонитов
также характерно краевое, преимущественно брюшное положение сифона,
выпуклые, а не вогнутые, как у наутилуса, перегородки. Таким образом, мяг-
кое тело аммонитов может реконструироваться, используя в качестве образца
модель кальмара и его сородичей, а не наутилуса, как считалось ранее.
СЕПТЫ (ПЕРЕГОРОДКИ) И ЛОПАСТНАЯ ЛИНИЯ
Аммониты появились в девоне, вероятно, они произошли от бактритоидей -
другого подкласса головоногих моллюсков, у которых преобладали прямые
раковины. Новая группа до среднего девона была достаточно консервативна.
Так называемые девонские гониатиты имели разнообразные спирально-плос-
костные гладкие раковины, варьирующие от широких до узких. Их название
объясняется наличием в лопастной линии заостренных лопастей (см. ниже).
© 1997. И.А. Михайлова, перевод на русский язык и изменения.
264
КАМЕННАЯ КНИГА
Складчатые перегородки (септы). Примитивные простые перегородки го-
ловоногих моллюсков с ровными краями были по площади лишь ненамного
больше поперечного сечения. Они имели прямые или слабоволнистые пере-
городочные линии. У аммонитов, так же как и у наутилоидей типа Aturia, пе-
регородки имели волнистый край, значит, по площади они значительно
превосходили площадь сечения оборота. Перегородочные линии станови-
лись при этом извилистыми или глубоко рассеченными. Этот процесс анало-
гичен заталкиваний кусочков бумаги в высокий стакан. Если площадь лист-
ка бумаги значительно больше, чем сечение стакана, то она при погружении
в стакан центральной части листка будет сжиматься и спрессовываться.
Это не более чем аналогия, поскольку не учитываются два фактора. Пер-
вый фактор: перегородки не заталкивались в раковину, а были сформирова-
ны мантией уже внутри нее и соответствовали поверхности заднего конца
мягкого тела (и мантии). Второй фактор: сжатие в нашем примере представ-
ляет собой случайный процесс, так как два одинаковых куска бумаги не мо-
гут быть втиснуты в стакан абсолютно идентичным способом. Складки (сед-
ла) и желобки (лопасти) мантии головоногих моллюсков являются характер-
ными особенностями, соответствующими определенным типам. Это ярко
выражено в строении лопастных (перегородочных) линий у аммонитов. По-
следние состоят из седел - складок, направленных вперед, и лопастей - же-
лобков, направленных назад. Для изучения аммонитов лопастную линию
зарисовывают, как бы распрямляя ее на плоскости, причем обычно только
одну половину.
Три типа лопастных линий (фиг. XVII. 1). Гониатиты жили и процветали
на протяжении около 150 млн. лет. Они появились в среднем девоне и дожи-
ли до конца палеозоя, в конце перми число их сильно сократилось и они
вымерли. В течение этого времени гониатиты значительно изменились, не-
которые из них стали очень вздутыми, почти шарообразными. Одна из от-
клоняющихся от нормы форм приобрела треугольные очертания, а другая
стала похожей на гипертрофированно увеличенную фораминиферу.
Фиг. XVII. 1. Аммониты с гоииатитовой (а), цератитовой (б) и аммонитовой (в)
лопастной линией
а — Goniatites, хЗ, б- Xenaspis, хЗ, пермь, Техас и Мексика; в - Hypophylloceras
ononense, ранний мел, Калифорния
РОГА АММОНА И ВНУТРЕННЕРАКОВИННЫЕ ГОЛОВОНОГИЕ
265
Возросла складчатость перегородок гониатитов, и их поверхность увеличи-
лась по площади, поэтому в лопастной линии, соответственно, возникли
новые седла и лопасти. Ни седла, ни лопасти не были рассечены, и их простое
строение характерно для так называемого гониатитового типа лопастной
линии.
Лопастная линия цератитов имела рассеченные лопасти и цельные нераз-
деленные седла. Аммониты с лопастной линией такого типа появились в
Миссисипи (ранний карбон) и просуществовали до конца триаса. Однако ам-
мониты с цератитовой лопастной линией вторично появились у позднемело-
вых родов, упростивших свои перегородки. Их называют псевдоцератиты
(ложные цератиты). Цератитовая лопастная линия была не у всех форм отря-
да Ceratitida, но имелась у большинства его представителей. После исчезно-
вения гониатитов цератиты разнообразно дивергировали в триасе и стали
вымирать только к концу этого периода.
Аммониты с аммонитовой лопастной линией появились в раннем триасе.
И седла, и лопасти были дополнительно разделены, образуя сложные фигу-
ры, подобные морозным узорам на стекле, или очертания сложно рассечен-
ных невероятно тонких элементов листьев папоротников. За этими измене-
ниями последовали и другие, затронувшие форму и скульптуру раковины и
сделавшие ее более сложной и нередко боле нелепой, чем самые необычные
раковины наутилоидей.
Высоко специализированные цератиты триаса вымерли к концу этого пе-
риода, оставив после себя продолжавшие существовать морфологически
консервативные типы. У их потомков вскоре стали развиваться более слож-
ные лопастные линии. Впоследствии, в позднем мелу, у некоторых форм про-
изошло вторичное упрощение и они вернулись к цератитовому типу. У неко-
торых из них лопастные линии в действительности были почти такими же
упрощенными, как у ранних гониатитов.
Почему лопастные линии стали рассеченными? Многие авторы пытались
объяснить, почему перегородки у аммонитов становились по краям складча-
тыми, образуя все более и более витиеватые лопастные линии. Одна теория
называет это ортогенетическим процессом — тенденцией к такому типу изме-
нений, которые приносили аммонитам заметную пользу. Другая теория -
теория селекции - предполагает, что головоногие моллюски со сложно уст-
роенными перегородками имели селекционные преимущества, так как
складчатые перегородки повышали прочность раковины. Эта идея под-
тверждается тем фактом, что многие гладкие, тонкие раковины аммонитов,
жившие на относительной глубине, имели сложные складчатые перегород-
ки, а раковины аммонитов, обитавших на мелководных участках, обладали
намного более простыми перегородками. Для большинства гладких рако-
вин, по сравнению с их современниками с разнообразной скульптурой
(ребра, шипы, бугорки), также характерны более усложненные перегород-
ки. С другой стороны, мы не знаем аммонитов с ровными краями перегоро-
док, подобными таковым у наутилоидей, живущих на достаточной глубине
для того, чтобы давление могло разрушить раковину. В то же время некото-
рые скульптированные раковины аммоноидей из мелководья обладали более
сложными лопастными линиями, чем гладкие глубоководные формы.
Другая особенность сопряжена с тем, что аммониты при появлении ново-
го варианта изменения перегородки и лопастной линии не меняли эти при-
знаки каждый раз. Напротив, новые детали появлялись после серии на-
чальных стадий развития, повторяющих лопастные линии более ранних
предковых родов. Если последняя из этих стадий оказывалась более выгод-
266
КАМЕННАЯ КНИГА
ной, чем у предков, то почему растущий аммонит повторял предваритель-
ные стадии? Это можно объяснить тем, что молодые и взрослые особи жили
в разных местах или тем, что более мелкие раковины часто были прочнее
более крупных.
Независимо от того, что было более выгодно - перегородки более слож-
ные или менее сложные, - очевидно, что существует большое разнообразие
форм с достаточно узким интервалом существования, что обусловило важ-
нейшую роль аммонитов в качестве биостратиграфического инструмента
исследования. Они особенно полезны для определения возраста мезозой-
ских отложений.
ФОРМА, СТРУКТУРА И УСЛОВИЯ ОБИТАНИЯ
Ранние аммоноидей были спирально-плоскостными, хотя у некоторых из
них обороты неплотно соприкасались друг с другом (фиг. XVII. 2). Палеозой-
ские и мезозойские представители этой группы развивались как типичные
плавающие и ползающие формы. Многие из них выглядели и, возможно,
жили, как современный Nautilus, у других были такие узкие обтекаемые ра-
ковины, что удивительно, каким образом в периоды отдыха они сохраняли
вертикальное положение. Видимо, для того, чтобы они не переворачивались,
в качестве опоры служили руки.
Ползающие аммониты - широкие гладкие гониатиты — появились в мис-
сисипских морях. В течение пенсильванского и юрского времени некоторые
роды приобретали широкие плоские раковины, как у медлительных водо-
емных улиток рода Helisoma, ранее известных как Planorbis. Некоторые
пермские, а также позднетриасовые и юрские ползающие формы были поч-
ти шаровидными. Другие, тоже, очевидно, ползающие типы, имели яйце-
видную либо округленно-треугольную форму или были чрезвычайно похо-
жи на “переросших” фораминифер. Эти типы появились в морях позднего
девона.
Ребра и другая скульптура. Мы помним, что свободно закрученные рако-
вины некоторых наутилоидей имели ребра-складки, образованные по типу
расширяющейся по периметру мантии. Когда каждая складка достигала
своей полной величины, мантия сокращалась до размера обыкновенной
стенки и затем опять расширялась вверх. Аммониты такого типа, как и Nau-
tilus, повторяли этот процесс, хотя у ископаемых часто сохраняется более
сглаженная скульптура.
Огромное число аммонитов имело ребра и пережимы, расположенные
вдоль или поперек оборотов, а иногда бугорки и даже шипы. Большинство
этих структур укрепляло раковину, так же как складки (гофрировка) дела-
ют твердой листовую сталь и барельефные украшения придают прочность се-
ребряному кофейнику или вазе. Многие таким образом украшенные и укреп-
ленные роды имели внешнее сходство с Nautilus’ом и, вероятно, были похо-
жими на него и по особенностям обитания. Другие роды имели киль и были
обтекаемыми, что свидетельствует об их способности к быстрому плаванию.
Однако большинство аммонитов обладало признаками, соответствующими
медленно ползающему образу жизни, например, род Zemistephanus, похо-
жий на невероятно широкую приземистую раковину улитки Helisoma, и род
Arcestes, имеющий почти шаровидный облик. Такая форма снижала ско-
рость движения; видимо, животные с подобным типом раковины почти все-
гда ползали.
РОГА АММОНА И ВНУТРЕННЕРАКОВИННЫЕ ГОЛОВОНОГИЕ
267
Фиг. XVII.2. Некоторые мономорфные аммониты
а - Prodromites gorbyi, х0.5, ранний карбон, центральная часть США; б -Arces-
tes, х2, видна цератитовая лопастная линия, широко распространен в среднем-
позднем триасе; в — Zemistephanus, юра, Британская Колумбия; г - Norman-
nites, средняя юра, Европа; д, е - Dufrenoya, xl, ранний мел, Европа, Северная
и Южная Америка, Африка; ж - Lytoceras, xl, юра-мел, Европа; з — Schloenba-
chia, xl, мел, Европа, Гренландия; и — Mortoniceras, xl, ранний мел, Африка,
Евразия, Северная и Южная Америка; к - Dipoloceras, xl, мел; л, м - Engono-
ceras, xl, мел, Европа, Северная Африка, Северная и Южная Америка
268
КАМЕННАЯ КНИГА
Ростры, ушки и воронка. И снова, подобно некоторым наутилоидеям, мно-
гие аммоноидеи в период зрелости сужают свое устье. У других форм развива-
лись брюшные (вентральные) выступы - ростры или боковые выросты - ушки.
Одна из теорий гласит, что сужение устья предотвращало “выпадение” мягко-
го тела из раковины. Это объяснение предполагает, что многие молодые жи-
вотные могли выталкиваться из своего широкого устья или что взрослые особи
становились все менее и менее прочно прикрепленными к стенкам раковины.
К сожалению, этому предположению нет подтверждения, как и тому, что все
молодые и взрослые аммониты очень прочно закреплялись в жилой камере.
Ростры и ушки означали, что их обладатели редко или вообще никогда не
плавали. Эта теория лишает киль этих животных функции рассекателя воды
и стабилизатора и присваивает ростру и ушкам незаслуженное значение. Ес-
ли бы эти образования причиняли животному неудобство, то как раз только
в случае ползания вперед, а не плавания назад с руками, которые подобно ру-
лю направляют животное. Узкая килеватая раковина аммонитов типа Di-
poloceras “имеет смысл” только для хороших пловцов. Возможно, некоторые
из названных структур были стабилизаторами.
Аптихи и анаптихи. В мезозойских отложениях очень часто вместе с ам-
монитами находят парные плоские известковые образования, похожие на
две створки. Редко они встречаются внутри жилых камер аммонитов и, веро-
ятно, выполняли роль крышечек и действовали как раскрывающиеся в цен-
тре старые салонные качающиеся двери. Это аптихи, очевидно, они могли за-
крывать устье аммонита, защищая тем самым мягкое тело раковины. Дру-
гая найденная в аммонитах структура - анаптих - образована из хитина или
какого-либо другого органического материала. Он представляет собой еди-
ную пластинку, часто имеющую форму бабочки. В исключительных случаях
они (преимущественно аптихи) были обнаружены сходящимися вместе и
могли выполнять функцию челюстей.
Гетероморфы — незакрученные аммоноидеи. У двух неплавающих аммо-
нитов, имевших треугольную раковину (роды Soliclymenia и Parawocklume-
ria), форма не менее нелепа, чем их названия. Другие роды утратили пра-
вильное спирально-плоскостное расположение оборотов и приобрели укло-
няющуюся форму раковины (фиг. XVII. 3). Разнообразие гетероморфных
раковин можно суммировать следующим образом.
Спирально-винтовые, высоко закрученные типы появились в позднем
триасе и были широко представлены в период со среднего (альб-сеноман) до
позднего мела. Они были похожи на улиток; возможно, ползали, а если и
плавали, то недолго и недалеко.
Раковины аммонитов с несоприкасающимися оборотами найдены в ниж-
неюрских и меловых отложениях. У некоторых обороты только не соприка-
сались друг с другом, а у других раковина в конце выпрямлялась. Первые две
группы могли либо ползать, либо плавать, так же как отдельные представи-
тели третьей группы. Остальные могли дрейфовать близко от дна, хотя это
лишь предположение.
Род Baculites после одного оборота строил прямую сжатую с боков ракови-
ну, заканчивающуюся необычно широким ростром. Некоторые специалисты
считают, что бакулиты дрейфовали и голова их была опущена вниз, другие —
что они ползали. Фрагменты с жемчужно-перламутровыми раковинами и
отличными лопастными линиями широко распространены в верхнемеловых
сланцах Pierre.
Раннемеловой род Hamulina и подобные ему формы первоначально росли
в виде прямого ствола, затем следовал коленообразный изгиб, и далее в обрат-
РОГА АММОНА И ВНУТРЕННЕРАКОВИННЫЕ ГОЛОВОНОГИЕ
269
Фиг. XVII.3. Гетероморфные аммониты девона (а, б), юры (в) и мела (г-ж)
а — Soliclymenia paradoxa, х2, поздний девон, Германия; б — Parawocklumeria,
xl.5, девон; в — Oecoptychius, xl.5, юра; г, д — Baculites, х10: г — начальная часть
раковины, д — поверхность перегородки, поздний мел, повсеместно; е - Pictetia
astieriana, х2, ранний мел, Западная Европа; ж - Hamites (Hamitella), имею-
щая 16.5 см в высоту; эта форма из меловых отложений Великой равнины
США; род встречается в Евразии, Африке и Северной Америке
ном направлении образовывался второй ствол. Мы можем представить мо-
лодых и совсем взрослых особей в виде ползающих и иногда плавающих жи-
вотных. В зрелом возрасте устьевая часть раковины выступает под и над ру-
ками. Но что делали эти животные, когда образовывали коленообразный
крутой изгиб?
Столь же загадочен образ жизни меловых Hamites и Macroscaphites. Они
считаются дрейфующими, а их форма определяла ориентацию в воде и на-
правление движения.
Род Scaphites и подобный по форме род Macroscaphites имеют аналогич-
ные проблемы, касающиеся характера их обитания и ориентации в воде. В
раннем возрасте они могли плавать и ползать, но в зрелости и старости были
плохо приспособлены к ползанию и, возможно, дрейфовали. Гетероморфы со
скафитидной формой раковины процветали и были самыми широко распро-
страненными ископаемыми в сланцах Pierre и других позднемеловых фор-
мациях.
Последняя из причудливых форм - Nipponites (и некоторые виды Helico-
ceras) — была найдена в меловых отложениях Японии. Это клубок неправиль-
ной формы, который, возможно, питался исключительно на дне. Род
Hamites включает виды с несоприкасающимися стволами (фиг. XVII.4) и та-
кие виды, которые начинают строить раковину из свернутых оборотов, далее
270
КАМЕННАЯ КНИГА
Фиг. XVII.4. Некоторые юрско-меловые гетероморфы
а — Hamites, 7.6 см в высоту, ранний мел, Африка, Евразия и Северная Амери-
ка; б — Macroscaphites yvani, 13 см в высоту, ранний мел (неоком), Европа; в -
Spiroceras bifurcati, средняя юра, байос, Европа; г - Hamulina astieriana, 30 см
в длину, ранний мел, Европа; д - Diplomoceras maximus, наиболее крупный ге-
тероморфный аммонит (3.5 м), поздний мел, Антарктида
образуют острый U-образный изгиб и затем формируют несколько оборотов
спирали, после чего прекращают закручивание и начинают изгибаться в об-
ратную сторону по направлению к макушке раковины.
МЕСТА ОБИТАНИЯ МЕЛОВЫХ АММОНИТОВ
Меловые отложения Техаса и северо-восточной Мексики изменяются от
рыхлых песчаников до сланцев и известняков, которые отлагались в раз-
личных местах: от дельт и приливно-отливных участков моря до рифов, об-
ширных мелководий и до глубин около 180 м. Эти отложения содержат ог-
ромное число аммонитов, раковины которых настолько слабо повреждены
и окатаны, что, очевидно, они были захоронены вблизи от места обитания
животных. Вместе с ними найдены гастроподы, обычные морские двус-
творки, в том числе устрицы, а также морские ежи, фораминиферы и мно-
гие другие формы. Ископаемые и отложения дают нам следующее
представление об условиях обитания аммонитов и других, живших вместе
с ними животных.
РОГА АММОНА И ВНУТРЕННЕРАКОВИННЫЕ ГОЛОВОНОГИЕ
271
1. Аммониты не обитали в пресных водоемах, течениях и в солоноватых
заливах дельт. Это неудивительно, поскольку ни один головоногий моллюск
не был найден в пресной воде или в пресноводных отложениях.
2. Аммониты редко жили около берега в мелкой, содержащей песок воде.
Такое место, возможно, было слишком мелким для этих моллюсков, вода по-
сле сильных дождей становилась здесь почти солоноватой, а в течение лета
было слишком тепло. Яйца современных головоногих в теплой воде погиба-
ют очень быстро.
3. Аммониты также избегали обширных мелководий, куда иногда забре-
дали огромные динозавры. Вода там имела различную глубину: от 2.5 до 6 м,
и волны часто достигали дна, поднимая легкую зыбь. Иногда морская вода
отходила, осушая илистое дно, покрывающееся ямками от капель дождя.
Крупные гастроподы и морские двустворки с толстой раковиной нашли та-
кое место очень гостеприимным для своего обитания, так же как и морские
ежи и бесконечное множество фораминифер Orbitolina. Те же самые факто-
ры, возможно, не допускали аммонитов к песчаному побережью.
4. Некоторые аммониты особенно процветали в местах, где дно было усея-
но пустыми раковинами двустворок, образующими залежи ракушняка. До
сих пор наиболее широко распространенными среди ракушников являются
узкие, но толстостенные раковины рода Dufrenoya.
5. Аммониты избегали коралловые и рудистовые рифы, приуроченные к
неглубоким и неспокойным водам. Было побережье, заселенное Gryphaea и
Exogyra, здесь процветали аммониты с узкой раковиной типа Engonoceras и
Dipoloceras, хотя другие формы встречались реже.
6. Эти аммониты с узкими раковинами наиболее изобиловали на глубинах
от 9 до 37 м. Температура и соленость в этих местах изменялись незначитель-
но, илистое дно никогда не обнажалось, самые огромные волны создавали
лишь легкие колебания, собиравшие устриц в гигантские валы. Такими же
многочисленными были другие моллюски и морские ежи.
7. Своего наибольшего разнообразия и многочисленности аммониты до-
стигли на глубинах от 37 до 180 м. Различным формам, вероятно, была необ-
ходима различная степень глубины и связанные с ней условия обитания.
Песчанистые известняки и сланцы содержат различные типы скульптиро-
ванных раковин, тогда как в мелководных отложениях обнаружены узкие
преимущественно гладкие формы. Гетероморфные представители, напри-
мер Scaphites, встречаются редко.
Чистые глинистые сланцы и известняки содержат огромное число разно-
образных аммонитов: гладких и ребристых, широких и узких, свернутых в
плотную спираль и свернутых частично. Самым идеальным местом обитания
для меловых аммонитов было бы карбонатное илистое дно на глубине более
37 м, где могли расти многочисленные водоросли, снабжавшие пищей огром-
ное число животных, включая и тех, которые питались самими аммонитами.
8. Море глубиной в 180 м занимало пространство, простирающееся к запа-
ду от того места, где сейчас располагается Мексиканский залив. Вода там бы-
ла слишком глубокой и темной для морских водорослей; поэтому брюхоно-
гие и двустворки встречались редко или вообще отсутствовали. Удивитель-
но, но около края этой впадины существовало огромное множество
аммонитов с гладкой раковиной. На большей глубине вода была холодной и
содержала мало кислорода, что являлось неблагоприятным фактором для
нормальной жизнедеятельности животных. Брюхоногие не могли жить на
такой глубине, а двустворки встречались достаточно редко, при этом они об-
ладали тонкой раковиной и значительно отличались от своих мелководных
272
КАМЕННАЯ КНИГА
сородичей. Наиболее преуспели аммониты: виды с широкими, гладкими ра-
ковинами оказались почти единственными животными, чьи останки сохра-
нились в ископаемом состоянии. Большинство из них встречено в ожелез-
ненных конкрециях и благодаря хорошему качеству образцов являются
весьма привлекательными. Наличие их там, возможно, означает, что они
обитали в водах выше данной впадины, а не в ней самой.
9. Сами аммониты служили некоторым животным в качестве добычи: и
позвоночным и беспозвоночным, включая рыб и огромных крабов. Класси-
ческий пример поедателей аммонитов - морские рептилии (мозозавры). На-
пример, один экземпляр аммонита рода Placenticeras из позднемеловых от-
ложений Южной Дакоты был сильно укушен мозозавром много раз, о чем
свидетельствуют следы зубов на раковине.
Имеется еще один факт: аммониты жили стаями и, исключая непригод-
ные места обитания, селились большими группами. Они обитали так же,
как и другие головоногие моллюски: от Volborthella до современного Nau-
tilus's. и кальмаров. Только осьминоги и каракатицы обитают иначе. Они в
одиночку рыскают в поисках пищи и в одиночку прячут свое легкое, неза-
щищенное тело в расщелинах скал. Некоторые аммониты, возможно, пи-
тались падалью и растительностью, и, вероятно, большинство из них жили
на самом дне или около него. Для некоторых аммонитов точно известен ха-
рактер пищи. Содержимое желудка рода Arnioceras свидетельствует о том,
что он предпочитал фораминифер и остракод. Род Hildoceras питался дру-
гими аммонитами, в то время как остальные употребляли в пищу морских
лилий.
Челюсти многих аммонитов не были предназначены для кусания, а зна-
чит, аммониты преимущественно не были хищниками. С помощью лопато-
образных нижних челюстей многие из них могли разворошить осадок не дне
и извлечь бентосные организмы, которые затем употреблялись в пищу. Не-
которые формы могли питаться также планктонными организмами и загла-
тывать микропланктон.
СТРЕЛООБРАЗНЫЕ ПЛОВЦЫ - БЕЛЕМНИТЫ
В то время как аммониты усовершенствовали свою раковину, некоторые
прямые головоногие моллюски сокращали (редуцировали) раковину до тех
пор, пока она не скрывалась под мантией. Таким образом возникли белемни-
ты - первые быстро плавающие головоногие.
Белемниты являются представителями еще одного подкласса цефалопод -
Coleoidea, который объединяет все формы, имеющие в мантийной полости
две жабры (и называющиеся двужаберные). К нему также относятся осьми-
ноги и кальмары. Все остальные головоногие (Endoceratoidea, Actinocera-
toidea, Nautiloidea, Bactritoidea и Ammo noidea) имеют или, вероятно, имели
четыре жабры и поэтому называются четырехжаберные.
Строение и характер обитания белемнитов. Некоторые ископаемые, у ко-
торых сохранились отпечатки мягкого тела, свидетельствуют о том, что бе-
лемниты выглядели и, предположительно, действовали как современные
кальмары. Мягкое тело у них торпедообразное, окруженное мантией, по бо-
кам имелись два горизонтальных плавника. Плавники поддерживали белем-
нитов в определенном положении в воде и при перемещении могли направ-
лять животное вверх или вниз (фиг. XVII.5). У них хорошо развита голова,
большие, круглые и темные глаза, вероятно фокусирующие свет с помощью
РОГА АММОНА И ВНУТРЕННЕРАКОВИННЫЕ ГОЛОВОНОГИЕ
273
хрусталика лучше, чем наутилус, через “булавочные” отверстия. У белемни-
тов, видимо, было десять рук, которые не были гладкими, как у наутилуса,
и обладали множеством крючков, повышавших эффективность при захвате
пищи. Ископаемые не проясняют ситуацию относительно характера пищи,
но их сходство с кальмарами позволяет предположить, что они питались
мелкой рыбой и лакомыми кусочками ракообразных типа крабов, креветок
и погибшими (ранеными) сородичами.
Как и большинство головоногих, белемниты плавали, всасывая воду в
мантийную полость и выталкивая ее через воронку. Когда воронка была на-
правлена вперед, они плыли назад, а когда воронка была повернута в сторону
задней части, белемниты двигались вперед. При опасности животные выпу-
скали через воронку из своего чернильного мешка, открывающегося в ман-
тийную полость, вязкую темную жидкость. Чернильная жидкость разлива-
лась в форме непрозрачного облака, которое, предположительно, ослепляло
противника, пока белемнит устремлялся прочь. А чернильная жидкость со-
временных кальмаров парализует органы обоняния, с помощью которых
охотятся главные враги кальмаров — рыбы. После того как рыбы выплывают
из ослепляющего облака, они еще долго мечутся бесцельно. Возможно, чер-
нильная жидкость белемнитов обладала аналогичным эффектом.
Раковина внутри мягкого тела. Белемниты, вероятно, эволюционировали в
тот момент, когда мягкое тело некоторых потомков ранних наутилоидей вы-
растало за пределы жилой камеры и начинало перекрывать мантией свою ра-
ковину. Раковина белемнитов состоит из ростра и фрагмокона. В верхней, или
спинной, области у них располагался продолговатый щит - проостракум, под-
держивающий и защищающий мягкое тело.
Длинные пустые раковины ранних белемнитов позволяли им держаться
на воде, затрачивая минимум усилий. А фрагмокон был очень тонким, поэто-
му животные достигали большой скорости, нередко сталкиваясь между со-
бой. Им был необходим балласт на конце мягкого тела, чтобы они могли рас-
секать воду, не переворачиваясь и не виляя.
Оба эти требования выполнялись ростром, крупной сплошной структу-
рой, расположенной сзади фрагмокона и частично вокруг него. Ростр состоит
Фиг. XVII.5. Форма и строение типичных белемнитов
а - ростр юрского белемнита, Вайоминг, Южная Дакота и Юта; б - схема строе-
ния белемнита; в — реактивный способ движения белемнита
18. Каменная книга
274
КАМЕННАЯ КНИГА
из радиальных волокон, чередующихся светлых и темных слоев. Темные
слои содержали большое количество органического материала, а светлые бы-
ли однородно известковыми. Оба слоя хорошо видны на расколотых или от-
полированных разрезах, свидетельствующих также об изменениях формы
ростра в процессе роста белемнита. На многих расколах наблюдаются нару-
шения и смещения слоев ростра - достаточное доказательство того, что бе-
лемниты во время стремительного движения назад часто сталкивались с дру-
гими объектами.
У некоторых белемнитов установлена четвертая внутренняя структура.
Это эпиростр, который не содержал призматических слоев и легко разрушал-
ся после смерти животного. Эпиростр обнаружен только у нескольких иско-
паемых родов.
Белемниты появились в миссисипских морях, но не стали изобилующими
до позднего триаса. Бесчисленное множество их плавало стаями в морях юр-
ского и мелового времени, а в конце последнего периода число белемнитов
резко сократилось. Последние ископаемые аберрантные фрагмоконы и рос-
тры исключительно редко встречаются в эоцене.
ОТ КОНИЧЕСКИХ РАКОВИН К ПЛАСТИНАМ-СЕПИОНАМ КАРАКАТИЦ
Преобразование внутреннего скелета до пластинок-сепионов было резуль-
татом того, что фрагмокон закручивался или менял свою структуру, ростр
уменьшался в размерах или редуцировался вообще.
Эти проявившиеся в юре изменения затем развивались по двум различ-
ным направлениям. Представители первого направления утратили проост-
ракум и ростр, сифон стал краевым, раковина - свернутой по типу крошеч-
ного Gyroceras, а мягкое тело и руки - короткими. Результатом таких изме-
нений было появление рода Spirilla. Это маленькое животное (около 10 см в
длину) единственный известный настоящий морской головоногий моллюск
с внутренней камерной раковиной. Спирулы (пелагическая форма) дрейфу-
ют и плавают в современных океанах. Некоторые из предков спирул, обитав-
шие из-за своих раковин и почти редуцированного ростра в мелких морях,
были найдены в третичных отложениях Северной Америки и Европы.
Вторая группа сепионов сохранила лишь небольшую часть ростра, но до-
бавился новый, очень широкий спинной проостракум и фрагмокон в виде по-
ристой арагонитовой пластинки, в которой перегородки - септы и другие
пластинки были соединены маленькими опорами. Таким образом появился
сепион, который дают канарейкам и другим домашним птичкам в качестве
источника карбоната кальция. Ископаемые сепионы и проостракумы найде-
ны в третичных отложениях Европы, а современная каракатица (Sepia) рас-
пространена по всему миру. Это короткое, овальное животное с двумя длин-
ными и восемью короткими руками. Каракатица может плавать и быстро (с
помощью воронки) и медленно, покачивая своими плавниками, располо-
женными по обе стороны мягкого тела.
КАЛЬМАРЫ И ОСЬМИНОГИ
После исчезновения белемнитов их место заняли кальмары, или теутиды.
Эти животные утратили фрагмокон и ростр, но сохранили роговое “перо”,
которое часто похоже на птичье и, возможно, является преобразованным
РОГА АММОНА И ВНУТРЕННЕРАКОВИННЫЕ ГОЛОВОНОГИЕ
275
спинным проостракумом белемнитов. Кальмары всегда были быстрыми
пловцами, и при захвате и удержании пищи по сравнению с крючками чрез-
вычайно важны присоски на руках. Кальмары появились в ранней юре и в
течение миллионов лет очень медленно развивались, а затем стали наиболее
широко распространенными современными головоногими. Они ведут образ
жизни, подобный таковому у белемнитов, и, вероятно, в начале третичного
времени вытеснили последних, так как употребляли ту самую пищу, от ко-
торой зависели белемниты, а также захватывая наиболее беспомощных и
наименее активных сородичей.
Осьминоги, как и предполагает их название, сократили число своих рук с
десяти до восьми, в то же время утратив все следы ростра и наружной рако-
вины. В связи с этим они почти никогда не встречались в ископаемом состо-
янии, но один вид с круглым телом и горизонтальными плавниками был най-
ден в позднемеловых отложениях Lebanon. Другая группа - аргонавты - ха-
рактеризуется наличием тонкой раковины, имеющейся только у самок.
В раковине происходит развитие яиц, иногда там находится и захваченная
женскими руками мужская особь значительно меньших размеров. Несколь-
ко таких раковин были обнаружены в третичных отложениях.
Согласно своему названию, современные бумажные кораблики (Argonau-
ta) плавают около поверхности теплых морей всего мира. В соответствии с
этим характер их обитания сильно отличается от такового у осьминогов, жи-
вущих вдоль скалистых берегов, и от гигантских кальмаров, местом обита-
ния которых являются темные, холодные глубины океана. К последним отно-
сится огромный Megateuthis, достигающий 17-20 м в длину и имеющий вес
от 450 до 1800 кг. Это единственный случай, когда современные представите-
ли превышают по размерам всех своих предков. Известно всего несколько ис-
копаемых кальмаров, первые из них встречены в раннетретичное время.
Интересный факт, что большинство головоногих, обладающих ракови-
ной, вымерли к концу мезозоя. Безусловно, в конце этой эры имел место
крупнейший биотический кризис, но некоторые палеонтологи предполага-
ют, что процесс вымирания был вызван возрастным соперничеством наруж-
нораковинных головоногих моллюсков с костистыми рыбами, которые пере-
мещались в более верхние слои моря. Цефалоподы, не обладавшие ракови-
ной, способны обитать на большей глубине, не опасаясь разрушения своей
раковины, и выжили потому, что на значительных глубинах конкуренция с
рыбами не была столь серьезной. Это интересное предположение заслужива-
ет дальнейшего рассмотрения.
Несмотря на вымирание аммонитов в конце мезозоя, следует помнить, что
на протяжении суммарного периода их жизни (девон-мел) они были самыми
разнообразными морскими животными. Сохранившиеся остатки раковин
позволяют детально подразделять осадочные отложения и проводить их кор-
реляцию на очень широких территориях.
18*
Глава XVIII
ПРЕИМУЩЕСТВЕННО СТЕБЕЛЬЧАТЫЕ
ИГЛОКОЖИЕ
• Расположенный около Le Grand (Айова) известняковый карьер на
протяжении многих лет был известен благодаря своим ископаемым.
И хотя некоторые слои не изобилуют ископаемыми, зато другие со-
держат хорошо сохранившиеся остатки, которые и рабочие карьера и иссле-
дователи называют морскими лилиями. Название Crinoidea (krinon - по-гре-
чески означает лилия) справедливо для данных ископаемых, напоминаю-
щих цветок на стебле.
Морские лилии знакомят нас с типом Echinodermata, или иглокожие жи-
вотные. Все представители этого типа являются морскими организмами. Из
большого количества классов иглокожих в настоящее время существуют
только шесть, среди них морские звезды, морские ежи, морские огурцы (го-
лотурии), а также криноидеи и их сородичи. У некоторых иглокожих имеет-
ся стебель, у других он отсутствует; многие имеют прочный скелет, другие —
мягкотелые. Иглокожие включают в свой состав и сидячих домоседов, и
дрейфующих, зарывающихся и ползающих представителей. Окраска совре-
менных особей представлена широким спектром от темного серо-зеленого до
голубого, пурпурного и ярко-красного цветов.
Несмотря на эти различия, для иглокожих характерен ряд общих основ-
ных признаков. Их мягкое тело и скелет в большинстве случаев обладают пя-
тилучевой симметрией. У некоторых, например у морских ежей, вторично
развивается двусторонняя симметрия. А у остальных представителей, таких
как вымершие гомалозои, отсутствуют даже следы радиальной симметрии.
Многие иглокожие имеют скелет, состоящий из табличек или игл кристал-
лического кальцита, которые под микроскопом могут быть пористыми или
сетчатыми и иногда образуют равносторонние ячейки. Эти элементы образо-
ваны мезодермой, которая также формирует и мускулатуру. Таблички могут
соединяться, но в основном они не соприкасаются, что означает возможность
увеличения их размеров на протяжении всей жизни животного.
Рост часто фиксируется соответствующими линиями или кольцами, рас-
положенными на табличках, а иглы с возрастом становятся длиннее и толще.
Другая важнейшая черта иглокожих - наличие водно-сосудистой или ам-
булакральной системы. Это взаимосвязанная система наполненных жидкос-
тью каналов и пузырей. Одним из основных элементов этой системы являют-
ся амбулакральные ножки, выходящие наружу и используемые для дыха-
ния, движения и питания. Амбулакральная система была обнаружена не у
© 1997. И.А. Михайлова, перевод на русский язык и изменения.
ПРЕИМУЩЕСТВЕННО СТЕБЕЛЬЧАТЫЕ ИГЛОКОЖИЕ
277
всех ископаемых групп иглокожих, но возможность ее существования не
исключена.
Иглокожие появились в начале палеозоя. Они настолько отличаются от
других групп беспозвоночных, что происхождение их определено нечетко.
К этому типу может быть отнесен род Tribrachidium (см. гл. IV) из докемб-
рийской эдиакарской фауны Южной Австралии, но не исключено, что он
принадлежит неизвестной вымершей группе, обладающей только внешним
сходством с иглокожими. Лишь данные эмбриологии могут немного прояс-
нить взаимоотношения этой группы. Полухордовые, включающие в себя
“желудевых червей” (Balanoglossus) и (Bhabdopleura) (см. гл. XX), и хор-
довые обладают похожими личинками и сходным способом образования
целома.
Иглокожие, как и некоторые другие типы, например членистоногие, зна-
чительно эволюционировали в течение кембрия и ордовика. После появления
более прогрессивных форм в связи с увеличивавшейся конкуренцией многие
группы начали вымирать, и лишь некоторые из них дали начало новым. Эф-
фективность дыхания, фильтрующего типа питания и лучшей защищеннос-
ти были критичными для выживания некоторых групп. Разнообразие игло-
кожих, измеряемое числом родов, появившихся за один миллион лет, возра-
стало в период с кембрия до карбона, далее упало до очень низкого уровня в
триасе и затем достигло своего пика в третичное время.
Несмотря на то что ископаемые остатки предоставляют нам мало инфор-
мации о происхождении иглокожих, они многое говорят об эволюционных
особенностях этой группы на протяжении 530-590 млн. лет. Ископаемые
свидетельствуют о том, что к концу ордовика строение тела большинства ос-
новных групп иглокожих усовершенствовалось. После чего их история со-
провождалась незначительными изменениями на фоне основного плана
строения этих организмов.
ВЫМЕРШИЕ СТЕБЕЛЬЧАТЫЕ ИГЛОКОЖИЕ
На протяжении большей части палеозоя в морской среде доминировали
разнообразные стебельчатые иглокожие. Разнообразие было значительно бо-
гаче, чем в настоящее время, и стебельчатые формы преобладали в сообщест-
ве иглокожих. Большинство некогда существовавших классов вымерло, и на
сегодняшний день наиболее дивергировавшими иглокожими являются мор-
ские звезды и морские ежи. Создается обманчивое впечатление, что данные
группы преобладали среди этого типа животных на протяжении предшест-
вующих 500 млн. лет.
Стебельчатые иглокожие подразделяются натри основные группы: цисто-
идеи (морские пузыри), бластоидеи (морские бутоны) и криноидеи (морские
лилии). Мы начнем рассмотрение с маленькой группы эокриноидей.
Эокриноидеи. Эти маленькие (6—7 см) ранние иглокожие чаще всего име-
ли конусообразный стебель, а у некоторых из них стебель вообще отсутство-
вал (фиг. XVIII. 1). Их вазообразная тека, или чашечка, образована тверды-
ми многоугольными табличками кристаллического кальцита с порами, рас-
положенными на границе соседних табличек. Чашечка обладала
несколькими простыми неветвящимися брахиолями, или ручками, отлич-
ными от нередко разветвленных брахиолей криноидей. В течение жизни бра-
хиоли с ресничками, возможно, удлинялись и доставляли пищу ко рту,
который располагался на вершине чашечки. Несмотря на незначительную
278
КАМЕННАЯ КНИГА
Фиг. XVIII.1. Карпоидеи (а, б), эокриноидеи (в) и цистоидеи (г-м)
а - Placocystites forbesianus, Xl, поздний силур, Англия; б - Mitrocystites mitra,
xl, средний ордовик, Чехия; в — Macrocystella marinae, xl.5, ранний ордовик,
Англия; г — Holocystites, хО.6, поздний ордовик-ранний силур, Европа и Север-
ная Америка; д, е — Pleurocystites filitextus, xl.3, средний ордовик, Онтарио и
Квебек; ж, з — Aristocystites bohemicus, Xl, ордовик, Чехия, род встречается в
Европе и Азии; и — ромбовые поры; к — двойные поры; л, м - Gomphocystites
bownockeri, х0.8, ранний силур, Огайо
дивергенцию, эокриноидеи просуществовали достаточно долго: с раннего
кембрия до силура.
Цистоидеи (Diploporita и Rhombifera). Эти животные, чье название означа-
ет пузыри, имели сжатое, яйцеобразное или шарообразное тело, защищенное
ПРЕИМУЩЕСТВЕННО СТЕБЕЛЬЧАТЫЕ ИГЛОКОЖИЕ
279
чашечкой, состоящей из табличек, пронизанных порами (см. фиг. XVIII. 1).
Некоторые роды непосредственно прикреплялись к другим объектам, у дру-
гих был короткий стебель, образованный круглыми, плоскими табличками.
Каждая чашечка имела в основании отверстие, через которое в стебель час-
тично заходили мягкие ткани. Отсутствие стебля, вероятно, признак прими-
тивности, и в случае, когда число табличек превышает двести и они распола-
гаются беспорядочно, можно в целом говорить о примитивности животного.
Небольшое число табличек, их определенная форма и закономерное располо-
жение являются специфической характеристикой.
Рот у цистоидей почти всегда находился в центре верхней стороны, иногда
от него в стороны расходились пищевые желобки (амбулакры); они были
прикрыты известковыми табличками. У других родов пищевые желобки от-
сутствовали; нередко брахиоли группировались вокруг рта в виде пучка или
протягивались вдоль обоих краев пищевых желобков. В этом случае брахио-
ли располагались на концах ответвлений пищевых желобков. Брахиоли все-
гда были короткими и тонкими. От них могли отходить дополнительные
придатки, называемые пиннулы. Видимо, пищевые желобки были снабже-
ны ресничками, двигавшимися взад и вперед и доставлявшими пищу ко рту.
На верхней стороне чашечки, на некотором расстоянии от рта, находился
анус, он был прикрыт мелкими табличками.
Цистоидеи представлены двумя основными группами (Rhombifera и
Diploporita). Различие между ними обусловлено расположением пор на
табличках чашечки. Видимо, через поры вода поступала в чашечку, и
обеспечивался газообмен. У одной группы поры создавали ромбовидные
структуры и открывались наружу либо сквозь щели, либо через маленькие
поры, находившиеся на соседних табличках. У второй группы поры разме-
щались парами, каждая пара находилась в пределах одной таблички. Эти
группы не обязательно были монофилетическими. Большинство из 22 се-
мейств цистоидей жили в период с кембрия до конца силура, лишь некото-
рые из них известны в девоне.
Цистоидеи представляли собой преимущественно прикрепленных, филь-
тровавших пищу животных. С помощью брахиолей они перемещали органи-
ческие частицы по направлению ко рту. Вероятнее всего, цистоидеи могли
несильно погружаться в осадок, обитали в морской воде на малых глубинах,
где и процветали. Некоторые формы значительно модифицированы. Так,
род Pleurocystites в действительности мог быть и подвижной формой. Он об-
ладал гибким стеблем и не имел якоревидного окончания. Стебель такой
формы мог работать как пропеллер и продвигать животное в воде недалеко от
дна волнообразными движениями.
Парабластоидеи. Парабластоидеи внешне похожи на бластоидей. Это
маленькие животные, например род Blastoidocrinus, были немного меньше
3 см в диаметре. Стебель был образован тонкими дисковидными членика-
ми, а чашечка состояла из большого числа табличек. Верхняя, или
оральная, сторона имеет вид пятиконечной звезды, в центре этой стороны
находится ротовое отверстие. По пяти лучам располагаются амбулакры.
Глубоко врезанные участки между ними называются интерамбулакры.
Противоположная аборальная сторона значительно отличается от ораль-
ной. На аборальной стороне расположены три или более поясов табличек и
серия крошечных интерамбулакральных табличек вдоль каждого из пяти
ответвлений пищевых желобков. Поры располагались вдоль края табли-
чек. Только два рода, каждый представлен одним видом, были известны из
отложений нижнего ордовика России и среднего ордовика востока Север-
280
КАМЕННАЯ КНИГА
Фиг. XVIII.2. Паракриноидеи (а, б) и бластоидеи (в—и)
а — Comarocystitespunctatus, xl, средний ордовик, Восточная Канада; б— Сапа-
docystis emmonsi, х4, средний ордовик, восток Северной Америки; в — реконст-
рукция Pentremites godoni, xl; г - строение теки Pentremites robustus; д -
Nucleocrinus vemeutli, х2, девон, долина Миссисипи; е — Tricoelocrinus wood-
mani, х12, ранний карбон, Индиана и Миссури; ж — Orophocrinus stelliformis,
xl.5, ранний карбон, долина Миссисипи; з - Timoroblastus coronatus, xl.5,
пермь. Восточная Индия, о-в Тимор; и — Stephanocrinus angulatus, х4, силур,
Нью-Йорк и Индиана
ной Америки. Их взаимоотношения с другими иглокожими изучены не до-
статочно хорошо.
Бластоидеи. Этих ископаемых часто называют “окаменелыми орехами”,
хотя Blastoidea означает “бутонообразные”. Они в большинстве случаев бы-
ПРЕИМУЩЕСТВЕННО СТЕБЕЛЬЧАТЫЕ ИГЛОКОЖИЕ
281
ли маленькими, иногда крошечными стебельчатыми иглокожими, чашечка
которых имела в диаметре менее чем 2.5 см. Бластоидеи жили с силура до
поздней перми; наиболее изобиловали в раннем карбоне. Выветрелые скаль-
ные выходы иногда содержат тысячи прекрасных образцов.
Чашечку бластоидей можно легко определить по форме: имеется пять ши-
роких пищевых желобков (амбулакров), которые глубоко прорезаны в цент-
ральной части, и радиальные крупные V-образные таблички с вырезом для
амбулакров (фиг. XVIII.2). Чашечка в основном состояла из этих крупных
табличек плюс некоторое число более мелких. Имеются значительные отли-
чия в расположении табличек от такового у морских лилий, в состав чашеч-
ки последних иногда входило огромное число дополнительных маленьких
табличек. Рот располагался на верхушке “бутона”, вокруг него находились
пять отверстий (спиракулей), одна из спиракулей одновременно служила
анальным отверстием. В течение жизни рот, так же как и амбулакры, при-
крывался маленькими подвижными табличками. Эти таблички у ископае-
мых форм чаще всего утрачены, так же как и многочисленные членики бра-
хиолей, располагавшиеся вдоль обеих сторон пищевых желобков и нередко
имевшие дополнительные ответвления.
Органы дыхания у бластоидей начинались рядами пор, расположенных
по обеим сторонам амбулакров. Многочисленные поры открывались в склад-
чатые структуры с тонкими стенками (гидроспиры), которые недалеко от рта
завершаются спиракулями. Гидроспиры — отличительная особенность блас-
тоидей. Вода проникала внутрь через множество пор, кислород усваивался
мягкими мембранами, окаймляющими гидроспиры, далее вода выходила
через спиракули.
Большинство бластоидей имели стебель, которым они закреплялись на
морском дне с помощью корневидных ответвлений. У некоторых форм сте-
бель утрачивался и чашечка располагалась непосредственно на дне или час-
тично погружалась в ил. У некоторых из этих бесстебельчатых родов, таких
как род Timoroblastus, развились крылообразные окончания или структуры,
напоминающие ножки старомодных жардиньерок.
Бластоидеи, как и многие подобные группы, не были особенно изобилую-
щими, за исключением определенных мест, в особенности в верхней части
криноидных рифогенных отложений. Силурийские бластоидеи известны
только в Северной Америке, тогда как девонские - по всему миру.
Паракриноидеи и эдриобластоидеи. Паракриноидеи появились, развива-
лись и вымерли в среднем ордовике. Вполне возможно, что в будущем они бу-
дут обнаружены как в более древних, так и в более молодых отложениях.
Они похожи на многие стебельчатые группы. Чашечка паракриноидей была
такой же, как и у цистоидей, руки — как у криноидей, а стебли похожи на
стебли бластоидей. Беспорядочное расположение табличек, а также система
пор отличны от таковых у бластоидей. Их однорядные амбулакры и пинну-
лы, или маленькие ответвления рук, позволяют обособлять эту группу от ци-
стоидей. Пиннулы являлись прекрасным улавливателем пищи.
Эдриобластоидеи были примитивной группой прикрепленных иглоко-
жих, которые имеют черты сходства с бластоидеями, цистоидеями или эдрио-
астероидеями (см. главу XIX). Они найдены только в среднем ордовике Кана-
ды и Австралии, но и там очень малочисленны. Это были животные ма-
леньких размеров, всего лишь около 2 см в диаметре. Чашечка имела
луковицеобразную или бутоновидную форму, она обладала пятилучевой
симметрией и состояла из двадцати табличек, расположенных в четыре
пояса. Каждый пояс образован пятью табличками: базальными, или основ-
282
КАМЕННАЯ КНИГА
ними, радиальными, дельтоидальными и оральными, или ротовыми.
В отличие от цистоидей таблички чашечки эдриобластоидей не имели пор, и
у них отсутствовали брахиоли. Между основными табличками беспорядочно
располагались дополнительные маленькие таблички, что отличает эдрио-
бластоидей от бластоидей. К этой группе относится только один вид Astrocys-
tites ottawaensis, его взаимоотношения с другими иглокожими на данный
момент не ясны.
Короноидеи. Некоторые роды, например Stephanocrinus, одно время отно-
сились к бластоидеям. Они были ограничены ордовиком и силуром. Короно-
идеи обладали спиралевидными, двурядными и разветвлявшимися брахио-
лями, не соединенными с полостью тела, напоминая брахиоли бластоподоб-
ных животных. Тека имела пирамидальный облик, напоминая корону, и
состояла из трех поясов табличек. Пять амбулакров сужались по направ-
лению к вершине чашечки. Внутри отростков, или зубцов, этой короны на-
ходились V-образные целомические каналы, характерные для бластоподоб-
ных животных, возможно использовавшиеся при дыхании. Короноидеи
имели стебель средней длины, он оканчивался прикрепительным дискооб-
разным расширением. Ближайшие родственники короноидей дали начало
бластоидеям.
РАЗНООБРАЗНЫЕ ПРОЦВЕТАЮЩИЕ КРИНОИДЕИ
Криноидеи, или морские лилии, представляют наиболее развитую группу
прекрасных стебельчатых иглокожих. В течение палеозойской эры они были
наиболее многочисленными, наиболее широко распространенными и наибо-
лее преуспевающими. Криноидеи появились в раннем ордовике. Род Echma-
tocrinus, обнаруженный в среднекембрийских сланцах Бёрджесс, некоторы-
ми палеонтологами причисляется к криноидеям; если это действительно
так, то появление данного класса следует отнести к более раннему периоду.
Хотя многие криноидеи вымерли к концу палеозоя, некоторые из них про-
должают существовать и в настоящее время. Область их распространения
моря и океаны, она простирается от освещенных солнцем мелководий и ри-
фов до темных, холодных вод глубиной около 4000 м и более. Некоторые
криноидеи достигают 15-18 м в длину; а руки у мелового рода имеют 1.2 м в
длину. Многочисленные сообщества обитали в мелководных морях девона и
карбона, где их остатки входили в состав карбонатных отложений. Одна из
таких серий в Скалистых горах (Альберта) суммарно составляет 60 м.
Большая часть карбонатных отложений с криноидеями состоит из разру-
шившихся стеблей и отдельных члеников, и только изредка встречаются ча-
шечки. Из этого можно сделать вывод, что взрослые криноидеи могли отры-
ваться от стеблей и самостоятельно дрейфовать на большие расстояния, осе-
дая после смерти животного на дно. Это также может означать, что чашечки
вскоре после смерти животного распадались на таблички, а членики стебля
какое-то время оставались соединенными друг с другом.
Основной план строения криноидей (фиг. XVIII. 3). Внешне они напоми-
нают эокриноидей и цистоидей; их также можно сравнивать с морскими
звездами, когда они лежат нижней стороной вверх, при этом анус и рот мор-
ских лилий были расположены на верхней - оральной - стороне чашечки.
Их руки всегда начинаются одной брахиальной табличкой. Вода проникает
внутрь тела через перфорированную табличку (как и у эдриоастероидей), да-
лее она проходит по кольцевому и радиальным каналам и наполняет трубча-
ПРЕИМУЩЕСТВЕННО СТЕБЕЛЬЧАТЫЕ ИГЛОКОЖИЕ
283
Фиг. XVIII.3. Строение морских лилий
а — реконструкция Eucalyptocrinites crassus, х0.5, видны чашечка, руки, сте-
бель и корневые образования, силур, центр США; б—е — чашечки: б — Botryocri-
nus, xl.5, силур-девон, Европа и Северная Америка; в - Phanocrinus, xl.6, кар-
бон, Африка, Европа и Северная Америка, г—е - три типа чашечек; ж — строение
стебля с узлами и междоузлиями; з-к — членики стеблей; л — современный Ап-
tedon c усиками - цирри. Условные обозначения табличек чашечки: В - ба-
зальные, IB— инфрабазальные, R — радиальные, X — анальные, AR - аниради-
альные, RA— радианальные
284
КАМЕННАЯ КНИГА
тые амбулакральные ножки, которые высовываются через поры на руках и
используются для дыхания. Собирают пищу и доставляют ее ко рту реснич-
ки, расположенные на руках вдоль пищевых желобков и их ответвлений.
Это основной план строения; от него имеются значительные отклонения,
с чем связано большое число различных терминов. Большинство из них мы
можем опустить, но необходимо отметить, что руки и чашечка вместе обра-
зуют крону, а стебель с корневидными образованиями или закрепляющимся
расширенным основанием называется нельма. Этот термин появился у сте-
бельчатых иглокожих и иногда используется при классификации всех игло-
кожих, имеющих или когда-либо имевших стебель, как, например, эдриоас-
тероидеи.
Корнеподобные выросты и иные образования. Видоизменение начинает-
ся в нижней части стебля. У многих криноидей имелись корнеобразные
структуры, враставшие в ил подобно тому, как корни наземных растений — в
почву. Стебель в основании может разветвляться, как бы приклеиваясь к ра-
ковинам, кораллам или другим твердым объектам. Такой фундамент иногда
разрушался волнами; так, верхнеордовикская формация в Цинциннати,
Огайо, содержит множество плоских тонких конкреций, в которых заключе-
но основание стебля криноидей. “Корешки” криноидей были также обнару-
жены в девонской окаменевшей древесине; можно предположить, что неко-
торые из этих иглокожих свисали вниз с дрейфующих стеблей растений.
Чашечки родов Lichenocrinus и, возможно, Cystoidosaccus были прикры-
ты сверху табличками. Эти таблички должны были закрывать мягкое тело.
Род Ancyrocrinus представляет собой другой тип, имевший в основании че-
тыре зубца, которые служили в качестве якоря. Корнеподобные образования
Scyphocrinttes формировали толстую, пластинчатую “луковицу”, разделен-
ную на секции (так называемые Lobolithus и Camarocrtnus). Лоболиты рас-
сматриваются в качестве плота, который позволял криноидеям дрейфовать
со свешенным вниз мягким телом; “луковица”, вероятно, могла обеспечи-
вать и закрепление на скалистом дне или на таких твердых объектах, как ра-
кушки, а также стебли других криноидей.
Стебель. На реконструкциях иногда демонстрируется, что криноидеи гра-
циозно покачивались на волнах или закручивались и изгибались в разные
стороны, чтобы достичь уровней, на которых изобилует пища и где кринои-
деи были недосягаемы для беспокоивших их течений. Такая картина не пол-
ностью учитывает природу стебля, который, как правило, состоит из соеди-
ненных друг с другом ровных члеников. Связки, расположенные между чле-
никами, допускают некоторую свободу движения. У некоторых родов
членики узкие со складчатыми или изогнутыми сочленовными поверхностя-
ми, позволяющими им наклоняться в одну или другую сторону. Членики
стебля рода Platycrinites имеют овальную форму, и, возможно, они могли
способствовать наклонам стебля в различных направлениях.
Форма члеников стебля в значительной степени варьирует: от круглой, эл-
липтической, четырехугольной, звездообразной и даже полукруглой. На по-
верхности имеются складки и разнообразные выемки для размещения
связок, основная функция которых должна была состоять в укреплении со-
единений члеников. Форма члеников у многих криноидей изменялась в про-
цессе роста. Одно из простейших изменений приводило к увеличению разме-
ров одних члеников по отношению к другим; в этом случае различаются
центральные узловые членики и расположенные между ними межузловые.
Боковые ответвления от стебля, отходящие только от центральных члени-
ков, называются усики.
ПРЕИМУЩЕСТВЕННО СТЕБЕЛЬЧАТЫЕ ИГЛОКОЖИЕ 285
В связи с тем, что стебли и их основание растут на протяжении всей жизни
животного, они должны быть обеспечены пищей и растворенными солями. В
центральной части членика или на фрагментах стеблей хорошо виден осевой
канал. Ширина и форма канала варьируют от больших до маленьких, от ци-
линдрических до звездчатых с пятью ярко выраженными секциями.
Основная функция стеблей - закрепление животного на дне. У рода Myelo-
dactylus имеется кончик стебля, который обвивается вокруг кораллов, кри-
ноидей или других подходящих объектов и прикрепляется к ним с помощью
одноветвистых усиков - цирри. Нижняя часть стебля преимущественно
утолщенная, но около середины, ближе к чашечке, стебель сужался и, как
правило, делал резкий изгиб, окружая собственно крону. За исключением
нескольких видов, стебель криноидей был достаточно длинным и распола-
гался почти вертикально, направляя свое мягкое тело вверх, у рода Myelo-
dactylus он, по всей вероятности, напоминал по форме знак вопроса.
Более значительными были изменения, при которых стебель утрачивал-
ся. Это происходило несколько раз у различных отрядов, при этом возника-
ли такие распространенные по всему миру формы, как современный род Ап-
tedon и позднемеловой род Uintacrinus. Первый из них первоначально фор-
мирует стебель, затем отрывается от него и прикрепляется к раковинам или
другим объектам с помощью пучка изогнутых усиков, временами отцепля-
ясь для плавания или медленного ползания, используя для этого руки. Род
Uintacrinus вел планктонный образ жизни, он не имел усиков, обитал стая-
ми вблизи поверхности, иногда заплывал на отмель, где и оседал, сохраняясь
в ископаемом состоянии в большом количестве.
Чашечка, или тека. Чашечка морских лилий состоит из двух частей: чаш-
ка и крышка (тегмен). Название первой части уже говорит само за себя: это
чашевидная, кубкообразная или вазообразная структура, образованная таб-
личками, имеющими определенную форму и расположенными у различных
родов в определенном порядке. Крышка варьирует от кожистого покрытия
чашки до крыши из толстых ротовых, или оральных, табличек, которые мо-
гут подниматься в виде высокого анального конуса или трубки или могут
формировать прочную крышу над ртом и пищевыми желобками.
Древнейшие и самые примитивные криноидеи (Inadunata) имели малень-
кие, жесткие, удлиненные чашечки, сложенные двумя поясами чередующих-
ся табличек (моноциклический тип), которые называются основные, или ба-
зальные (В), и радиальные (R). Первые прикреплялись к стеблю, а от вторых
отходили руки. Некоторые из этих ранних инадунат имели чашечку дицикли-
ческую, т.е. обладали третьим поясом табличек, расположенных ниже основ-
ных и называющихся нижнеосновные, или инфрабазальные. Особая таблич-
ка, анальная (X), находилась между двумя радиальными и выше таблички,
которая называется радиальной у одних родов и анирадиальной — у других.
У некоторых криноидей дополнительно к нижнеосновным были и иные
таблички, а кроме того, иногда таблички рук включались в состав чашечки.
При этом возрастало общее число табличек и дополнительно развивались но-
вые интербрахиальные таблички, которые не были соединены с анусом.
В связи с этим даже такие маленькие по размерам криноидеи, как роды Up-
erocrinus и другие, а также такие, как роды Ichthyocrinus и Scyphocrinites,
обладали огромным числом табличек чашечки, насчитывавшим, как прави-
ло, более ста. Палеозойские роды затем вымерли, а у большинства живот-
ных, обитавших в мезозое и кайнозое, размер чашечки и количество слагаю-
щих ее табличек сократились. Род Uintacrinus, однако, сохранил лукович-
ную чашечку, состоящую из огромного числа табличек.
286
КАМЕННАЯ КНИГА
Фиг. XVIII.4. Разнообразие рук морских лилий
Членики рук постепенно укорачиваются, и руки становятся двурядными,
одновременно возрастает число пиннул
Мы упомянули о криноидеях, стебли которых были настолько сильно изо-
гнуты или закручены, что могли обвивать чашечку. Эти уклоняющиеся ти-
пы были прямостоящими, а род Halystocrinus изгибался вниз ко дну, хотя
его чашечка при этом была направлена вертикально вверх, или лежал на
дне. При этом одна сторона была уплощенной, она являлась нижней сторо-
ной чашечки.
Меловой род Hemicrinus обладал коротким стеблем, прикрепленным к ос-
нованию широкой поверхностью, чашечка была повернута в сторону, в пра-
вый угол, а таблички и членики стебля сливались в единую твердую массу,
из которой высовывались толстые руки. Эта морская лилия могла обитать в
условиях сильных течений или в бурных водах, где волны достигали дна.
Некоторые изменения затрагивали и чашечку и крышку. Например, у ро-
да Eucalyptocrinites чашечка в нижней части была глубоко вогнута и стебель
как бы внедрялся в нее снизу. Крышка формировала трубку, затем расширя-
лась, вокруг располагалось десять перегородок, разделявших секции, в каж-
дой секции находились разветвленные руки.
Руки, или брахиоли. Руки криноидей значительно различаются по форме.
Они несут на себе окончания пищевых желобков и воднососудистой системы
и отличаются от рук большинства других стебельчатых иглокожих, у кото-
рых брахиоли представляют прямые отростки, отходящие от центральной
части мягкого тела. В основании рук криноидей находится отверстие, веду-
щее в полость тела, расположенную внутри чашечки. Руки криноидей обра-
зованы прямыми рядами соединенных брахиальных табличек. У относи-
тельно примитивных типов руки не ветвятся либо разветвляются один раз. У
рода Carabocrinus руки были короткими. У рода Petalocrinus таблички рук
резко увеличены в размерах и напоминают лепестки, загибающиеся назад по
направлению к основанию.
Остальные вариации могут быть суммированы следующим образом.
Многие криноидеи сохранили один ряд рук, при этом длина рук могла
значительно увеличиваться. У других криноидей с однорядными руками
брахиальные таблички приобрели клинообразную форму. Дальнейшее уко-
рачивание табличек и их сужение позволило им располагаться бок о бок друг
с другом таким образом, что остались видны только зигзагообразные швы.
В результате руки стали двурядными, сохранив однорядное начало
(фиг. XVIII.4). И однорядные, и двурядные руки имели боковые ответвле-
ния, или пиннулы.
У некоторых криноидей руки прекратили разветвляться, но возникли
длинные тонкие таблички, как, например, у рода Petalocrinus. У многих
ПРЕИМУЩЕСТВЕННО СТЕБЕЛЬЧАТЫЕ ИГЛОКОЖИЕ
287
криноидей руки ветвились многократно, становясь сложными и запутанны-
ми. Примерами такого типа служат сильно разветвленные руки мезозойских
и современных пентакринитид. Разветвления рук силурийских Crotalocrtnt-
tes соединялись в гибкие сети, которые либо образовывали широкие веерооб-
разные “вайи”, либо частично охватывали друг друга. Более значительные
изменения мы наблюдаем у рода Barrandeocrinus из силура северной Евро-
пы: пиннулы свободны только на кончиках, в нижней части они образованы
ровными смыкающимися табличками, последние формируют десять сек-
ций, изогнутых вниз и похожих на дольки апельсина. Рот погружен в углуб-
ление между сильно развитыми руками. Род Cornucrinus утратил свободные
руки, сохранив только амбулакры и пищевые желобки, которые в действи-
тельности были частью чашечки.
Иглы. Многие роды криноидей имели в чашечке толстые шероховатые
таблички; у других таблички перерастали в шипы, торчащие из чашечки,
крышки, рук, стебля и даже из длинных анальных трубок. У рода Ancyrocri-
nus были развиты шипы основания, служащие зубцами его “якоря”. А иглы
девонского рода Arthroacantha, известного в Европе и Северной Америке,
были с помощью связок прикреплены к маленьким бугоркам (туберкулам) и
благодаря этому могли шевелиться.
ОСНОВНЫЕ ГРУППЫ КРИНОИДЕЙ
Известно более шести тысяч видов ископаемых криноидей. В современ-
ных морях насчитывается 25 родов стебельчатых и 19 родов бесстебельчатых
морских лилий. Современные и ископаемые формы могут быть разделены на
четыре четко обособленные группы, часто называемые отрядами. Три отряда
в большинстве своем ограничивались палеозойской эрой, когда они форми-
ровали густые “криноидные сады”, а некоторые из них вели хищный образ
жизни, обитая преимущественно в подводных течениях. И только один из
четырех отрядов существует до сих пор.
Отряд Inadunata (фиг. XVIII. 5). Это относительно простые криноидеи;
они включают древнейших и примитивнейших членов класса, которые
просуществовали только до раннего триаса. Не исключена вероятность то-
го, что современный Hypocrinus может быть “современным ископаемым”,
дожившим до наших дней. Таблички чашечки плотно скреплены друг с
другом, крышка (тегмен) закрывает рот, пищевые желобки расположены
под ней. Руки однорядные или двурядные, без пиннул или с ними. У боль-
шинства родов они свободно или неплотно соединяются выше радиальных
табличек. Пятилучевая симметрия чашечки часто нарушается наличием в
радиальном поясе специальной радиальной таблички. Древнейшие инаду-
наты обладали вытянутыми, прямыми чашечками, а у более молодых па-
леозойских родов чашечка была расширена по бокам и крона стала похожа
на цветок.
Отряд Flexibilia (фиг. XVIII.6). Палезозойские криноидеи (ордовик-
пермь) с гибкой чашечкой. Соединения между табличками чашечки и крыш-
ки показывают, что они были достаточно свободно скреплены друг с другом
и оставались гибкими в течение всей жизни животного. В состав чашечки
входят первые (нижние) брахиальные таблички, а число интербрахиальных
и интеррадиальных табличек варьировало от нескольких штук до множест-
ва. В верхней части крышки располагались рот и открытые пищевые желоб-
ки. Руки были однорядными, не имели пиннул, ширина рук, в основном,
288
КАМЕННАЯ КНИГА
Фиг. XVIII.5. Инадунаты
а - Halysiocrinus, xl, стебель стелится по дну, средний девон-ранний карбон,
Северная Америка; б - Petalocrinus, xl.6, с необычным строением рук, ранний
силур, Северная Америка; в - Cyathocrinites, xl, с многократно ветвящимися
руками, силур-ранний карбон, возможно, пермь, повсеместно; г — Myelodacty-
lus, xl.5, с необычным строением стебля, ранний силур-девон, Европа и Север-
ная Америка; д - Carabocrinus с очень короткими руками, ордовик, Европа и Се-
верная Америка; е - Crotalocrinitespulcher с необычайным строением рук, х0.8,
поздний силур, о-в Готланд, Швеция
значительно превышала высоту. Как и у инадунат, имелись радиальная и
анальная таблички. Эти криноидеи, подобно инадунатам, имели специфиче-
ские черты, но были менее дивергировавшими.
ПРЕИМУЩЕСТВЕННО СТЕБЕЛЬЧАТЫЕ ИГЛОКОЖИЕ
289
Фиг. XVIH.6. Флексибилии
а - тека Sagenocrinites clarki с брахиальными и интербрахиальными табличка-
ми, силур, Теннесси; б — Eutaxocrinus whiteavesi, xl.5, средний девон—ранний
карбон, Европа и Северная Америка; в - Eutaxocrinus fletcheri, xl.l, ранний
карбон, Северная Америка; г - Taxocrinus, х0.5, девон-ранний карбон, Европа
и Северная Америка
Отряд Camerato, (фиг. XVIII. 7). Из всех палеозойских групп криноидей
наиболее разнообразными были камераты. Все таблички чашечки обычно
были утолщенными и жестко соединялись друг с другом. Тегмен (крышка)
закрывал и рот и пищевые желобки. В состав чашечки входили первое бра-
хиальные таблички и интеррадиальные таблички. Руки несли пиннулы и
были однорядными либо двурядными. Радиальная табличка отсутствовала.
Камераты существовали с ордовика до поздней перми, но наиболее изоби-
чующими были в раннем и среднем карбоне. Ранние камераты обладали боль-
шим сходством с инадунатами, от которых они, возможно, и произошли.
О т р я д Articulata (фиг. XVIII.8). У этой разнообразной группы стебельча-
тых и бесстебельчатых криноидей чашечки были преимущественно малень-
кими и состояли лишь из нескольких табличек. Они приобрели способность
передвигаться, а не оставаться навсегда прикрепленными. Это открывало ог-
ромные возможности, чем и объясняется успех рассматриваемой группы.
19. Каменная книга
290
КАМЕННАЯ КНИГА
Фиг. XVIII.7. Камераты
а — тека Scyphocrinites elegans, х0.5, с многочисленными рядами дополнитель-
ных табличек, поздний силур-ранний девон, Африка, Евразия и Северная Аме-
рика; б - лоболиты Scyphocrinites, х0.25, силур-девон, Евразия и Северная
Америка; в — Calliocrinus, xl, силур, Швеция; г - Uperocrinus с длинной тонкой
анальной трубкой, х0.7, ранний карбон, долина Миссисипи; д — Cactocrlnus,
xl.4, ранний карбон, Айова и Миссури; е - Eutrochocrinus christyi, ранний кар-
бон, долина Миссисипи и Нью-Мексико; ж -Lampterocrinus tennesseensis, xl.4,
ранний силур, Теннесси
ПРЕИМУЩЕСТВЕННО СТЕБЕЛЬЧАТЫЕ ИГЛОКОЖИЕ
291
Фиг. XVIII.8. Артикуляты
а - Marsupites, х0.8, поздний мел, Африка, Австралия, Евразия и Северная
Америка; б, в- современныйRhizocrinus, х10; г — фрагментPentacrinites,х0.7,
юра, Европа и Северная Америка; д — Hemicrinus astierianus, хЗ, поздний мел,
Европа
Таблички чашечки скреплялись гибкими соединениями. Очень гибкие и все-
гда однорядные руки с пиннулами; тегмен представляет кожистую перепон-
ку, пищевые желобки открытые.
Артикуляты, должно быть, появились в перми, когда остальные отряды
криноидей почти либо полностью вымерли, и на протяжении мезозоя и кай-
нозоя криноидеи представлены именно этим отрядом. Существует предполо-
жение, что артикуляты образуют не одну, а две группы, представители одной
произошли от Inadunata, а другой — были потомками Flexibilia. В мезозое и
в раннечетвертичное время артикуляты достигли некоторого разнообразия,
такие формы, как Uintacrinus socialis, были достаточно многочисленными
(фиг. XVIII.9). Экспансия артикулят началась в четвертичное время, в совре-
менных морях насчитывается более шестисот разнообразных видов. Распро-
странившись от мелководий до глубоководных пучин и от тропических до
близких к полярным морей, они сравнимы по значимости с камератами ран-
него карбона.
ПРОБЛЕМАТИЧНЫЕ НОМALOZOA
Воображение палеонтологов часто волнуют ископаемые, чьи взаимоотно-
шения не очевидны. Возможно, они привлекают к себе больше внимания,
чем того заслуживают. Двумя такими головоломными группами, безуслов-
но, являются конодонты и граптолиты; в равной степени или даже более про-
блематичны — Homalozoa, некоторые из них известны как кальцихордаты
или карпоидеи. Чаще всего их относят к иглокожим, но некоторые исследо-
ватели, использующие название Calcichordata, предположили, что они име-
ли сходство с хордовыми и тем самым с позвоночными животными.
Гомалозои в отличие от многих иглокожих не обладали радиальной сим-
метрией. У многих форм симметрия отсутствовала полностью. Некоторые
19*
292
КАМЕННАЯ КНИГА
Фиг. XVIII.9. Отпечатки вида Uintacrinus socialis, хО.З, обитавшего в поздне-
меловых морях Европы и Северной Америки. Этот образец найден в штате Ай-
ова (рисунок Е.П. Загозиной по фотографии из англоязычного оригинала)
карпоидей имели почти двусторонне-симметричный скелет, подобно таково-
му у многих хордовых.
Выделяют три основных группы карпоидей: стилофоры, гомостелеи и
гомойостелеи. Скелет гомостелей состоял из теки и стелы, которая представ-
ляла собой суженное окончание теки. Тека имела две непохожие стороны: од-
на плоская, на ней, по-видимому, лежало животное, противоположная сторо-
на - слегка выпуклая. Центральная часть теки состояла из множества ма-
леньких табличек (centralia), а периферийная область сложена большими
краевыми, или маргинальными, табличками (marginalia). Узкая хвостооб-
разная структура - стела - прикреплялась к теке и состояла из двойной се-
рии табличек, разделявшихся более мелкими табличками. Брахиоли отсут-
ствовали. Гомостелеи ограничены средним кембрием. Они, возможно, вели
сидячий образ жизни, заякориваясь стелой в морское дно и фильтруя пищу
из воды. А может быть, это были подвижные животные, питавшиеся органи-
ческим детритом. В настоящее время мы не знаем, какая модель наилучшим
образом объясняет их образ жизни.
Другой группой карпоидей являются стилофоры. Типичным представите-
лем стилофор является род Cothumocystis (фиг. XVHI.10). Он имел стебель и
по форме был похож на ботинок. Стилофоры обладали крупными краевыми
маргинальными табличками и мозаикой из маленьких центральных табли-
чек. Вдоль края теки располагалось несколько шипов. Около носка “ботин-
ка” находилась серия щелевидных пор. Некоторые палеонтологи считают,
что эти поры функционировали подобно жаберным щелям у позвоночных.
Не все исследователи согласны с такой интерпретацией, поэтому дискуссия
ПРЕИМУЩЕСТВЕННО СТЕБЕЛЬЧАТЫЕ ИГЛОКОЖИЕ
293
Фиг. XVIII. 10. Cothurnocystis (около 5 см в ширину), карпоидеи группы стило-
фор, ордовик, Европа
о систематическом положении стилофор продолжается. Эта группа встреча-
ется со среднего кембрия до позднего девона.
Третьей группой карпоидей являются гомойостелеи. Они очень похожи на
гомостелей, но в отличие от них имели одну брахиоль, а стела состояла из
двойного ряда табличек. Их тека была полностью сформирована серией ма-
леньких табличек. Данная группа существовала от кембрия (Невада) до ран-
него девона (Виктория, Австралия).
На основе эмбриологических исследований, а также работ о эволюции и
биохимии можно сделать вывод о родственных взаимосвязях иглокожих и
хордовых. Поэтому не исключено, что карпоидеи могут находиться в неизве-
стной области между этими двумя типами. Для успешного разрешения всех
противоречий необходимо дальнейшее изучение внутренней анатомии у тех
немногочисленных образцов, у которых она сохранилась, а также поиски но-
вых ископаемых.
Глава XIX
ПОДВИЖНЫЕ ИГЛОКОЖИЕ
Морские звезды ассоциируются у нас с криноидеями, которые от-
дыхают на соединенных с центральным диском (лучах), но рот
криноидей ориентирован вверх, а морских звезд вниз. Сейчас мы
переходим к тем группам иглокожих, у которых рот находится на нижней
стороне, либо у тех форм, которые напоминают собой “сосиску”, он распо-
ложен на одном из концов.
МОРСКИЕ ЗВЕЗДЫ И ИХ ПРОИСХОЖДЕНИЕ
Книги о современных животных в основном описывают две большие груп-
пы звездообразных иглокожих, одна из которых включает в себя настоящих
морских звезд (Asteroidea), а к другой относятся хрупкие змеевидные “звез-
ды” (Ophiuroidea). Палеонтологи добавляют к ним третью группу, Somaste-
roidea, и объединяют все три группы в отдельный класс, названный Stelleroi-
dea, или “звездообразные”.
Somasteroidea существовали преимущественно в Западной Европе с ран-
него ордовика до позднего девона. У них короткие толстые лучи и крупное
отверстие, к которому иногда приурочен рот, окруженный шероховатой ко-
жистой перепонкой. У края ротового отверстия начинались таблички, про-
низанные большими порами для выхода амбулакральных ножек; они про-
тягивались двумя рядами вдоль середины каждого луча. Множество рядов
стержнеобразных табличек, называемых virgalia, разветвлялось с каждой
стороны, завершая структуру, напоминающую руки и пиннулы кринои-
дей. Образцы прекрасной сохранности показывают, что верхняя поверх-
ность была покрыта шероховатой кожей, содержащей маленькие “спику-
лы”. Кожа на нижней стороне покрывала также и virgalia.
Если эти ископаемые означают то, что мы предполагаем, то морские звез-
ды являются потомками лежавших ртом вверх кембрийских криноидей.
Они утратили свой стебель и начали ползать с помощью абулакральных но-
жек, которые их предки использовали только для дыхания. “Спикулы” ор-
довикских Somasteroidea, вероятно, представляют собой характерную осо-
бенность этой группы, поскольку настоящие древнейшие морские звезды
имели таблички, напоминающие таблички криноидей. Чрезвычайно инте-
ресны уклоняющиеся формы, например офиоцистоидеи (фиг. XIX. 1).
© 1997. И. А. Михайлова, перевод на русский язык и изменения.
ПОДВИЖНЫЕ ИГЛОКОЖИЕ
295
Фиг. XIX.1. Офиоцистия “многоножка” Gillocystis, около 4 см в поперечнике,
ранний девой; Австралия, Виктория
Пути развития морских звезд
Это сходство в строении табличек морских звезд и криноидей проявилось
только у палеозойских морских звезд. Таблички большинства более поздних
Asteroidea соединены, и в основном они либо шероховатые, либо несут иглы.
Типичные Asteroidea имеют толстые лучи, в которые заходят доли желудка,
пищеварительные железы и репродуктивные (воспроизводительные) орга-
ны. Вода может всасываться и затем выталкиваться через кожу, а также про-
никать через пористую пластинку (мадрепорит), расположенную на верхней
стороне, около места соединения двух лучей. Через мадрепорит вода попада-
ет в каменистый канал, далее переходит в кольцевой и радиальные каналы,
подобные таковым у криноидей и эдриоастероидей.
Питание морских звезд. Морские звезды нащупывают еду или пробуют
ее с помощью специализированных амбулакральных ножек, расположен-
ных около окончания лучей. Более короткие ножки, тянущиеся вдоль хо-
рошо развитых амбулакров, используются главным образом для присасы-
вания или ползания со скоростью, изменяющейся от 2 до 30 см в минуту.
Однако часто животное часами лежит на одном месте, едва передвигая
лучами.
Морские звезды употребляют несколько видов пищи и добывают ее раз-
личными способами. Некоторые виды питаются органическими частицами,
дрейфующими вдоль морского дна, захватывая их пеленой слизи, которую
они затем заглатывают и переваривают. Когда такая пища становится скуд-
ной, выделение слизи прекращается, и желудок, представляющий собой пе-
репончатый мешочек, начинает через рот выворачиваться наружу, раство-
ряя относительно крупные морские водоросли и растущие на скалах малень-
кие морские водоросли или даже некоторые губки. Питающиеся улитками
морские звезды обычно заглатывают их целиком, проталкивая в рот с помо-
щью амбулакральных ножек. Обычные съедобные двустворки мидии и уст-
рицы втягиваются с раскрытыми створками посредством амбулакральных
ножек, после чего желудок окружает жертву и быстро переваривает содер-
жимое мягкого тела.
Ископаемые морские звезды (фиг. XIX. 2). Описанные способы питания
характерны для современных морских звезд. Похоже, что ископаемые име-
ли сходный тип питания, например, род Hudsonaster, живший со среднего
до позднего ордовика. Этот род имеет в ширину менее дюйма и короткие, как
бы обрубленные лучи. Его верхняя поверхность состоит из гладких, толстых
296
КАМЕННАЯ КНИГА
Фиг. XIX.2. Морские звезды (а, в, г), сомастероидеи (д) и офиуры (б, е-з)
а — Hudsonaster incomptus, х2, поздний ордовик, Огайо; б - Onychaster flexilis,
х0.9, хрупкая звезда, ранний-средний карбон, Индиана; в - Devonaster eucha-
ris, х0.9, средний девон; г — Metopaster, мел-миоцен, Европа, Северная Амери-
ка и Новая Зеландия; д — Villebrunaster thorali, ордовик, Франция; е — Loriolas-
ter mirabilis, ранний девон, Германия; ж — Cheiropteraster giganteus, х0.2, де-
вон, Германия; з -Aganastergregarius, х0.9, ранний-средний карбон, Индиана
табличек, расположенных, как и таблички чашечки криноидей, в опреде-
ленном порядке. Роды Palaeaster и Devonaster имеют более длинные и тон-
кие лучи и маленькие таблички между рядами более крупных. Маленькие
таблички, в отличие от более крупных, не располагаются в определенном по-
рядке.
Некоторые по существу примитивные морские звезды имели необычную
форму. Например, род Stuertzaster обладал широкими, лепестковидными
лучами и луковицеобразным диском, напоминающим грибы, которые назы-
ваются “земляные звезды”. Род Cheiropteraster еще более отклонялся от
ПОДВИЖНЫЕИГЛОКОЖИЕ
297
обычной нормы. У него было очень большое ротовое отверстие, а лучи отсут-
ствовали. Амбулакры были непосредственно соединены с мешковидным те-
лом, не имевшим табличек или спикул. Loriolaster имел маленький рот и
хорошо развитые в лучах амбулакры; у остальной части тела скелет от-
сутствовал. Этот род, как Cheiropteraster и некоторые другие необычные мор-
ские звезды, был обнаружен в плотных девонских сланцах Бунденбаха
в Германии.
Морские звезды редко встречаются в палеозое и еще реже в мезозойских и
кайнозойских отложениях. Большинство ископаемых принадлежит фор-
мам, развивавшим большие толстые таблички, которые могли быть и глад-
кими, и покрыты иглами. Они включают в себя как современные , так и вы-
мершие роды. Меловой род Metopaster, как и его современные сородичи,
имел массивные краевые таблички и резко укороченные лучи. Род Oreaster,
появившийся в юрское время, существует и в современных морях. У боль-
шинства ископаемых морских звезд лучи довольно тонкие, в то время как у
большинства известных современных видов хорошо защищенное мягкое те-
ло красного, желтого или голубого цвета с короткими толстыми лучами.
Офиуроидеи, или хрупкие звезды
Офиуроидеи (“змеевидные хвосты”) получили такое название в связи с
тем, что у большинства из них тонкие змеевидные лучи. Их популярное на-
звание вызвано чрезвычайной хрупкостью лучей. Формы, у которых лучи
разветвляются снова и снова, называются плетеными “звездами”.
Хрупкие “звезды” являются специализированными потомками Somastero-
idea, тело у которых стало дискообразным, а лучи более тонкими. Желудок не
простирался в лучи, рот был узкий, звездообразный. Лучи состояли из члени-
ков, подвижно сочлененных между собой и напоминающих позвонки позво-
ночных животных. У современных видов эти так называемые позвонки пред-
ставляют структуру из грубой ткани с широкими табличками и иглами, таб-
лички покрывает кожица. Животные ползают с помощью комплекса
изгибающихся движений, подталкивая себя одними лучами и подтягиваясь
другими. Лучи используются и для того, чтобы удерживать большие куски
пищи и переправлять их ко рту. Маленькие частицы проникают между амбу-
лакральными ножками. Род Опуchaster из раннего-среднего карбона “прили-
пал” к рукам или анальным трубкам криноидей и, вероятно, использовал их
отходы для питания. Эти отходы могли собираться с помощью слизи, подобно
той, которую выделяют, а затем заглатывают современные морские звезды.
Хрупкие “звезды” впервые появились в раннем ордовике, в отложениях
более поздних веков и эпох они встречаются достаточно редко. И хотя луч-
ший образец был найден в черных сланцах нижнего девона в Европе, хоро-
шие экземпляры обнаружены также в породах среднего ордовика в восточ-
ной части Северной Америки.
Эхиноидеи, или “морские разбойники”
Слово “urchin” означает “еж” (Erinaceus) и относится к хорошо всем зна-
комому округлому животному с множеством игл, распространенному от За-
падной Европы и Северной Африки до Китая и Новой Зеландии. Морские
ежи, или Echinoidea, являются не имеющими лучей и стебля иглокожими,
но обыкновенно обладающими большим числом игл, как и те млекопитаю-
щие, в честь которых они названы. Морские ежи, как и их наземные собра-
298
КАМЕННАЯ КНИГА
тья, имеют широкое распространение, встречаясь в морях всего мира и в от-
ложениях, варьирующих от нижнего палеозоя до четвертичных.
Панцирь и анатомия. Длинная геологическая летопись отражает разви-
тие панциря, который становился все более прочным - жестким. Панцири
(их не следует называть раковинами) примитивнейших морских ежей были
образованы маленькими пластинками, частично перекрывавшими друг дру-
га. Пластинки сочленялись в панцире подвижно и, как правило, распада-
лись после смерти животного. У прогрессивных типов пластинки срастаются
между собой; они обыкновенно утолщены либо укреплены вторичными отло-
жениями, иногда доходящими до половины панциря. Несколько родов, у ко-
торых пластинки остаются сравнительно тонкими, имеют массивные була-
вообразные иглы. И несмотря на то, что внутренняя структура пластинок яв-
ляется кружевной или сетчатой, многие из них достаточно прочны, чтобы
сохраниться при захоронении и фоссилизации.
Самые древние и примитивные морские ежи имели сигаровидную форму,
более поздние были уплощенными, субконическими, дискообразными или
приобретали такую неправильную форму, что адекватно описать ее могут
только рисунки. Нижняя (оральная) сторона, как правило, почти плоская, а
верхняя (аборальная) сторона в различной степени выпуклая. В центре
нижней стороны находится перистом (ротовое поле), которое при жизни жи-
вотного было покрыто кожистой оболочкой, упрочненной известковыми таб-
личками. У большинства правильных морских ежей (по старой классифика-
ции называемых Regularia) в центре верхней стороны имеется перипрокт
(анальное поле), внутри которого располагается анус (фиг. XIX.3). Вокруг
перипрокта находится пояс, включающий пять глазных пластинок, которые
завершают окончания абмулакров, и еще пять более крупных половых плас-
тинок с большими порами, сквозь которые выходят наружу половые продук-
ты. Таким образом, центральная часть верхней стороны сложена двумя по-
ясами пластинок, образующих вершинный щиток.
У большинства неправильных морских ежей (Irregularia в более ранних
классификациях) вершинный щиток преобразуется различным способом.
В центре верхней части панциря или около него остается собственно вершин-
ный щиток, а анус и перипрокт перемещаются по заднему интерамбулакру
назад. Анус может располагаться и на нижней (оральной) стороне, на срав-
нительно небольшом расстоянии позади рта.
Правильные и неправильные морские ежи имеют общее сходство, обладая
одной прободенной (одна из половых) мадрепоровой пластинкой, через кото-
рую вода проникает в систему каналов, ведущих к амбулакральным ножкам.
Ножки могут вытягиваться, как у криноидей, морских звезд и других игло-
кожих. Однако у морской звезды каждая ножка выходит из одной поры и
наполнена жидкостью из одного мускульного, сокращающегося мешочка.
А амбулакральные ножки морских ежей выходят наружу через пару пор, об-
разуя петлю, разъединяясь перед порами и соединяясь после них.
Амбулакры варьируют почти так же, как и панцирь в целом. Некоторые
амбулакры узкие и почти прямые, другие — изогнутые, а третьи — петалоид-
ные — лепестковидные. Некоторые из них могут протягиваться от перистома
до вершинного щитка, а другие ограничиваются частью верхней (абораль-
ной) поверхности. У новых морских ежей амбулакры состоят из двух рядов
пластинок, а некоторые палеозойские ископаемые имеют несколько рядов
более маленьких пластинок. Некоторые мезозойские и кайнозойские мор-
ские ежи за счет срастания маленьких простых амбулакральных пластинок
формируют более крупные, сложные амбулакральные пластинки.
ПОДВИЖНЫЕ ИГЛОКОЖИЕ
299
Фиг. XIX.3. “Морские разбойники”, или морские ежи
а—в — Hardouinja, поздний мел, Северная Америка; г, д — Holaster, мел—эоцен,
повсеместно; е, ж - Holectypus, юра-мел, Европа и Северная Америка; з —
Diplopodia, триас—мел, Евразия и Северная Америка; и — Salenia, мел-совре-
менность, Северное полушарие и Австралия, ныне повсеместно; к, л — Goniopy-
gus, юра-эоцен, Евразия, Северная Африка, Северная и Южная Америка;
м,н- цидароидная форма, палеоцен, Нью-Джерси
Панцири у всех морских ежей имеют бугорки, которые могут быть боль-
шими, хорошо заметными или слишком маленькими, чтобы различить их
без помощи лупы. В течение жизни панцирь был покрыт кожицей. Бугорки
300
КАМЕННАЯ КНИГА
служат для прикрепления игл, которые удерживаются на месте связками и
приводятся в движение шнуровидными мускулами. У примитивных родов
иглы размещены только на пластинках, расположенных между амбулакра-
ми, то есть на интерамбулакрах; на самих амбулакральных пластинках бу-
горки для игл появляются у мезозойских и кайнозойских морских ежей.
Морские ежи питаются с помощью комплексной системы, называемой
аристотелев фонарь. Он содержит пять заостренных зубов и целых тридцать
пять скелетных табличек, управляемых шестьюдесятью мускулами. Эти
таблички формируют пять идентичных кругов, служащих оболочкой и ры-
чагами и для зубов и для прикрепления мускулов. Стираясь на одном конце,
зубы растут на другом и, таким образом, медленно опускаются вниз сквозь
выемки (желобки) в аристотелевом фонаре.
Более пристального внимания требуют две другие группы структур, по-
скольку они очень важны для определения различных типов морских ежей,
нечасто встречающихся в ископаемом состоянии. Начнем с педицеллярий —
крошечных “клыкообразных” структур, двигающихся в различных направ-
лениях, захватывая или передвигая объекты, оседающие на поверхности жи-
вотного. Другими структурами являются таблички, палочки и крючочки из
кальцита, обнаруженные в кишечнике, репродуктивных гонадах, амбула-
кральных ножках и в некоторых других органах. Как и педицеллярии, они
настолько крошечные, что могут быть изучены только под микроскопом.
Мы уже упомянули о правильных и неправильных морских ежах
(фиг. XIX.4). До середины нашего века морских ежей подразделяли на эти
две группы. Вскоре после этого было обнаружено, что “неправильность”, воз-
можно, развивалась не один раз, и поэтому данная характеристика не может
использоваться для систематики животных. Классификация и выяснение
взаимоотношений различных группировок морских ежей должны опреде-
ляться на основании изучения комплекса анатомических деталей.
Новая классификация учитывает внешние характеристики и степень проч-
ности панциря, число рядов амбулакральных и интерамбулакральных пласти-
нок, тип амбулакральных пластинок, строение аристотелева фонаря, строение
гребней внутри панциря, структуры, окружающие перистом и поддерживаю-
щие челюстной аппарат морских ежей - аристотелев фонарь. В классификации
также используются строение вершинного щитка и форма панциря.
Выделяют две основные группы: перишэхиноидеи, живущие с ордовика до
настоящего времени, и эуэхиноидеи, появившиеся в позднем триасе и просу-
ществовавшие до настоящего времени. Рассмотрим обе группы более подробно.
Группа Perischechinoidea. Это примитивные представители Echinozoa,
включающие всех вымерших палеозойских морских ежей, а также ископае-
мых и современных цидароид. У палеозойских форм существует ряд харак-
терных особенностей по сравнению с более молодыми морскими ежами. Пан-
цирь обычно шаровидный и всегда правильный; известно всего несколько бо-
лее уплощенных форм. Перипрокт расположен в центре вершинного щитка.
Многие формы имеют гибкий панцирь и достигают значительных размеров.
В связи с гибкостью панциря ископаемые образцы достаточно часто разбиты
и деформированы. Амбулакральные пластинки всегда простые, даже если
они слагают много рядов в пределах одного поля.
Цидароиды были единственной палеозойской группой, пережившей пер-
мо-триасовый кризис и дожившей до наших дней. Возможно, они дали нача-
ло всем мезозойским и кайнозойским стволам морских ежей. Сами цидарои-
ды незначительно изменились после мезозоя и, так же как лингулидные бра-
хиоподы и дерево гинкго, похожи на “современных ископаемых”.
ПОДВИЖНЫЕ ИГЛОКОЖИЕ
301
е
Фиг. XIX.4. Позднемеловые и третичные морские ежи
а—в — Holaster, палеоцен, ныне повсеместно; г, д — Linthia, мел-плиоцен, повсе-
местно; е—з — Hemiaster, мел—современность, повсеместно; и, к — Rhopostoma,
палеоцен, Северная Америка; л—н — Heteraster, мел, Аравия, Средиземноморье,
Северная и Южная Америка
Цидароиды имеют крупные интерамбулакральные пластинки, в центре
каждой из них расположен один большой бугорок, его окружает кольцо мел-
ких, к бугоркам прикреплялись иглы. У них узкие, змеевидные амбулакры,
образованные маленькими пластинками, каждая из которых имеет одну па-
302
КАМЕННАЯ КНИГА
ру пор. Аристотелев фонарь устроен довольно просто. Типы игл представле-
ны достаточно разнообразно. Мелководные формы имеют булавовидные или
веретенообразные иглы, вероятно служившие стабилизаторами в неспокой-
ной воде. У глубоководных форм, обитающих на дне, иглы были тонкими и
продолговатыми. Цидароиды могли появиться в силуре, но несомненные на-
ходки известны начиная с девона.
Группа Euechinoidea. Все постпалеозойские морские ежи, за исключе-
нием цидароид, принадлежат к этой группе, которая впервые появилась в
позднем триасе. Все эуэхиноидеи имеют пять амбулакральных и пять интер-
амбулакральных полей, каждое из них состоит из двух рядов пластинок. На
амбулакральных пластинках может быть расположено более одной пары пор.
Как сказано выше, такие пластинки называют сложными. В отличие от пе-
ришэхиноидей, форма панциря может быть как правильной, так и неправиль-
ной. Более того, ежи с панцирем правильной формы могут иметь аристотелев
фонарь, но у неправильных форм он может быть утрачен. Эуэхиноидеи смогли
настолько разнообразить свой образ жизни по сравнению со своими предками,
что некоторые из них приспособились к зарыванию в мягкие грунты.
Среди эуэхиноидей выделяют четыре основные группы, объединяющие
около восемнадцати отрядов. Кратко рассмотрим эти группы.
Группа Diadematacea. Они впервые появились в поздней юре и сущест-
вуют в настоящее время. Амбулакральные пластинки сложные, реже про-
стые. Аристотелев фонарь имеет простое строение, как у примитивных цида-
роид, которые, возможно, и дали начало эуэхиноидеям. И хотя многие виды
обитают на большой глубине, наиболее запоминающимися являются рифо-
вые виды, имеющие удлиненные, ядовитые иглы, например род Diadema.
Группа Echinacea. Эхинацеи существуют от позднего триаса до настоя-
щего времени. Они являются правильными морскими ежами с более сложной
структурой, чем та, которая преобладает у диадематоидных форм. Они имеют
сложно устроенный околочелюстной пояс, амбулакральные пластинки более
чем с одной парой пор, аристотелев фонарь более сложного строения.
Группа Gnathostomata. Челюстные морские ежи являются неправиль-
ными морскими ежами, впервые появившимися в юре. Одна группа принад-
лежащая клипеастероидам, известна нам сегодня как песчаные доллары
(фиг. XIX.5). Они имеют простые амбулакральные пластинки с одной парой
пор, аристотелев фонарь и вершинный щиток размещены противоположно
друг другу. Клипеастероидеи в истории морских ежей появляются в палеоце-
новое время, то есть значительно позднее других групп. По многим признакам
их можно отнести к высоко специализированным формам. Половые и глазные
пластинки не образуют кольцо, а смыкаются на вершине панциря на месте
бывшего анального отверстия. Панцирь принимает необычные формы и чаще
всего укрепляется внутренними ребрами, табличками и столбиками из каль-
цита. Иглы у таких форм маленькие и достаточно многочисленные, в основ-
ном двух типов. Они имеют множество мелких, вспомогательных собираю-
щих пищу амбулакральных ножек, расположенных за пределами петалоидов.
Ранние клипеастероиды были крошечными и яйцеобразными с нечеткой
петалоидностью амбулакров. Более поздние роды, включая собственно род
Clypeaster, стали крупнее и толще с хорошо развитыми петалоидами и распо-
ложенным в центре нижней стороны ртом. Большинство клипеастероид яв-
ляются “песчаными долларами” - сплющенными животными с ровной
нижней поверхностью, на которой находятся собирающие пищу, обыкно-
венно разветвляющиеся, пищевые желобки. Иглы настолько короткие и
тонкие, что могут быть приняты за “шерстистый” покров.
ПОДВИЖНЫЕ ИГЛОКОЖИЕ
303
В связи с тем, что клипеастероиды обладали прочным панцирем, они изо-
билуют в ископаемом состоянии. В некоторых местах панцири почти запол-
няют рыхлые известняки, в других, когда дождь размывает отложения рых-
лых мергелей, сланцев или песчаников, они покрывают всю почву.
Группа Atelostomata. Все представители этой группы морских ежей
имеют неправильную форму панциря. У них отсутствует аристотелев фо-
нарь. Ротовое поле (перистом) и вершинный щиток часто противоположны
друг другу, а амбулакры нередко формируют петалоидную структуру. К дан-
ной группе принадлежат кассидулоиды, которые впервые появились в юре и
существуют до сих пор. Их лепестковидные амбулакры, образующие тупые
лепестки, ограничены верхней поверхностью панциря. Рот окружен двой-
ным кольцом лепестковидных пластинок; у всех других групп морских ежей
эта структура отсутствует.
Ателостоматами являются также спатангоиды и холастероиды, или серд-
цевидные морские ежи, так как их форма нередко напоминает сердце. Они
встречаются с ранней юры до настоящего времени. Спатангоиды появились
в раннем мелу и продолжают существовать до сих пор. Их амбулакры имеют
петалоидную, или лепестковидную, форму. Рот смещен вперед от центра, а
перипрокт располагается на почти вертикальной задней стенке, которая в
действительности является частью верхней - спинной - поверхности. Иглы
тонкие и “шелковистые” и могут быть как короткими, так и очень длинны-
ми. Некоторые роды имеют пояса бугорков (фасциолы); расположенные на
них микроскопические бугорки несли крошечные иглы.
Один из наиболее известных спатангоидных родов - это Micraster, широ-
ко распространенный в верхнемеловых отложениях Западной Европы. На-
чавшееся в 1899 году подробное изучение огромного числа относительно це-
лых образцов привело к выявлению эволюционных стадий видов рода
Micraster. Эти виды демонстрируют развитие от незарывающихся типов до
зарывающихся на мелководных участках, как, например, Micraster leskei, и
глубоко зарывающихся, как М. coranguinum. Эти представления обеспечи-
вают лучшее понимание функции панциря спатангоид, так же как и осно-
ванное на смене комплексов детальное расчленение мелководных морских
пород, слагающих меловые отложения Европы, включая Англию.
МЕСТА И ХАРАКТЕР ОБИТАНИЯ МОРСКИХ ЕЖЕЙ
Морские ежи имели достаточно разнообразные места обитания от рифов и
песчаных пляжей и мелководий до глубоких вод. Некоторые из них обитают да-
же в мелководных бухтах и на скалистых берегах, обнажаемых во время отлива.
Морские ежи ведут различный образ жизни. Почти все живут группами,
формируя “колонии”, насчитывающие дюжины, сотни или даже тысячи ин-
дивидуумов. Многие правильные морские ежи крепко “прилипают” к ска-
лам или закрепляются в щелях и трещинах с помощью игл и амбулакраль-
ных ножек, заканчивающихся присасывающимися дисками. Другие выка-
пывают норки, и не только в известняках и сланцах, но могут поселяться и в
гранитах, базальтах и твердом песчанике. Такие породы намного прочнее
известняков, и непонятно, как морские ежи могли в них просверливать себе
норки.
Другие правильные морские ежи обитают на илистой поверхности среди
скал и кораллов или на ровном твердом дне. Там они ползают с помощью ам-
булакральных ножек или используют иглы как ходули. Поскольку иглы с
304
КАМЕННАЯ КНИГА
помощью мускулов могут менять ориентировку, то и передвигаться ежи мо-
гут в любом направлении.
Пища этих правильных морских ежей является подходящей для харак-
тера их обитания и мест распространения. Некоторые из них питаются мор-
скими водорослями, покрывающими поверхность скал, другие улавливают
кусочки водорослей своими иглами и доставляют их ко рту с помощью амбу-
лакральных ножек. Морские ежи, обитающие на скалах, поедают губок,
гидроидов, мшанок и трубчатых червей, когда они могут их добыть, а виды,
живущие в расщелинах или зарывающиеся, ограничены пищей, заползаю-
щей в их ниши или дрейфующей вместе с водой. Педицеллярии схватывают
крошечные организмы и переправляют их ко рту, а некоторые обитающие
на дне морские ежи заглатывают ил и переваривают его, независимо от со-
держащейся в нем пищи.
Неправильные морские ежи обитают на илистом или песчаном дне, где
они ползают или зарываются. Многие сердцевидные ежи медленно ползают,
“зачерпывая” в рот ил и переваривая его органическое содержимое. Другие
обитают в норках на глубине 10—15 см, всасывая воду для дыхания через
трубчатое, выходящее не поверхность отверстие. Длинные амбулакральные
ножки также высовываются через это отверстие, собирая пищу и переправ-
ляя ее вниз, где она может быть доставлена ко рту.
Многие песчаные доллары ползают в тонком слое осадка, очень близко от
поверхности. А некоторые виды, обитающие в эстуариях, лежат на “ребре”,
располагаясь частично в воде, а частично вклиниваются в песок, при этом
всегда перпендикулярно направлению приливно-отливного потока. Когда
вода отступает, они закапываются, покрывая себя песком, и либо спокойно
отдыхают, либо ползут в поисках пищи. Они также зарываются при прибли-
жении морских звезд, питающихся морскими ежами, но другие животные
их не беспокоят.
Питание для песчаных долларов является комплексным процессом. Их
иглы, несомненно, покрыты кожей. Иглы, расположенные на верхней
части панциря, выделяют слизь и имеют двигающиеся назад и вперед рес-
нички. Последние создают течение, направляемое в сторону нижней по-
верхности и образующее маленькие водовороты, сталкивающие крошечные
организмы со слизью, которая их и поглощает. Слизь в свою очередь направ-
ляется к нижней брюшной поверхности, где она накапливается в пищевых
желобках и доставляется ко рту. И пока зубы не перетрут ее, песчаный дол-
лар фактически не ест.
Кажется вполне очевидным, что древние песчаные доллары питались так
же, как и их современные потомки, что большинство ископаемых сердце-
видных ежей зачерпывало пищу, ползая по илистому дну, а обитатели ри-
фов поглощали и растения, и маленьких животных. Но в дополнение к это-
му следует немного рассказать об особенностях питания специфических
эхинозоа.
МОРСКИЕ ОГУРЦЫ, ИЛИ ГОЛОТУРОИДЕИ
Морские огурцы представляют собой мягкотелых животных с кожным
покровом; их внешняя форма достаточно хорошо отражена в названии. Рот,
расположенный на переднем конце мягкого тела, окружен щупальцами.
Анус обычно находится на противоположном конце. У большинства голоту-
рий ряды трубчатых амбулакральных ножек протягиваются вдоль тела, а
ПОДВИЖНЫЕ ИГЛОКОЖИЕ
305
Фиг. XLX.5. Песчаные доллары
а - Abertella, х0.5, средний миоцен, Мэриленд; б - Clypeaster, хО.6, плиоцен,
Южная Калифорния, эоцен-современность, род распространен повсеместно;
в — Scutaster andersoni, х0.8, поздний олигоцен-ранний миоцен, Южная Кали-
форния; г-е- Mellita, х0.5, плейстоцен, Южная Каролина, Джорджия, Техас
20. Каменная книга
306
КАМЕННАЯ КНИГА
Х200
Х180
Х130
Фиг. XIX.6. “Спикулы” различных голотурий, или морских огурцов, Европа и
Северная Америка
крошечные пористые таблички (склериты) из кальцита или фосфата железа
могут выступать под кожей. Многие склериты имеют форму якоря, тогда как
другие — форму решетки или колеса. Рот окружен одним сплошным кольцом
табличек. Морские огурцы преимущественно лежат на дне или ползают по не-
му, заглатывая ил либо захватывая с помощью щупалец маленьких живот-
ные. Некоторые голотурии зарываются. Представители одной современной
группы (Elasipoda) плавают, вибрируя зонтиковидной перепонкой из кожи,
натянутой между продолговатыми щупальцами, и питаются планктоном.
Древнейшие голотурии, вероятно, появились в кембрии. Однако “спику-
лы”, несомненно принадлежащие этой группе, достоверно обнаружены в де-
вонских породах (фиг. XIX.6). Некоторые формы почти полностью покрыты
табличками, но у большинства ископаемых и современных форм скелет зна-
чительно редуцирован.
В настоящее время голотурии обитают в теплых мелководных и глубоко-
водных морях.
ВЫМЕРШИЕ ЭХИНОЗОИ: EDRIOASTEROIDEA И ДРУГИЕ
Другие группы эхинозоа встречаются в ископаемом состоянии значитель-
но реже, чем морские ежи. Все они на сегодняшний день являются вымерши-
ми и ограничены палеозоем. Экспериментальным периодом в развитии игло-
кожих был, главным образом, ранний палеозой.
Класс Edrioasteroidea (фиг. XIX. 7). Эдриоастероидеи являются малень-
кой, распространенной во всем мире группой, у которой имелись пять отдель-
ных, обычно извилистых, амбулакров и пять областей, соответствующих ин-
терамбулакрам. И амбулакры, и интерамбулакры были ограничены верхней
поверхностью животного и сходились в области рта, расположенного в центре.
Интерамбулакры покрыты серией гибких, беспорядочно расположенных таб-
личек. Изгибаясь, эти таблички позволяли некоторым животным передви-
гаться, а другие вели сидячий или прикрепленный образ жизни.
Амбулакральная система некоторым образом напоминает аналогичную
структуру у криноидей, но у эдриоастероидей отсутствовали брахиоли. Веро-
ятно, они имели родственные связи с морскими ежами, и некоторые ученые
считают их возможной предковой группой.
Древнейшим представителем эдриоастероидей был род Stromatocystites,
живший в раннем и среднем кембрии, то есть это одно из наиболее ранних иг-
локожих. Тека имела пятистороннюю форму, выделяются пять узких, пря-
мых амбулакров и между ними мозаика крошечных табличек в интерамбу-
ПОДВИЖНЫЕ ИГЛОКОЖИЕ
307
Фиг. XIX.7. Эдриоастероидеи
а — Edrioasterbigsbyi, xl.l, ордовик, Северная Америка; б - Foerstediscus splen-
dens, ордовик, Миннесота
лакрах. Это была свободно обитающая форма. Другие, более поздние эдриоа-
стероидеи тоже были свободными, но многие из них с помощью
маргинального кольца периодически прикреплялись к морскому дну, ска-
лам или другим беспозвоночным.
Группа Helicoplacoids. Огромное число ископаемых геликоплакоидей -
ранней и необычной группы иглокожих - обнаружено в нижнекембрийских
сланцах на вершине Белых гор восточной Калифорнии. Они также были най-
дены в породах этого возраста в других местах западной части Северной Аме-
рики. Это были радиально-симметричные, в основном чечевицеобразные
животные со спирально расположенными рядами табличек, образующих
гибкий панцирь. Вершинная и ротовая области располагались на противопо-
ложных концах. Подробности анатомии геликоплакоидей до сих пор нахо-
дятся в стадии изучения, например структура перистома и место расположе-
ния ануса.
Возможно, геликоплакоидей были свободными обитателями, не исключе-
но, что закрепленными или зарывающимися. Они могли и лежать на дне и
ползать на небольшие расстояния, как голотуроидеи.
Вместе с геликоплакоидеями известны другие группы ранних иглоко-
жих, такие как эдриоастероидеи, карпоидей и эокриноидеи. Разнообразие
иглокожих раннего кембрия свидетельствует о том, что дифференциация в
пределах этого типа, вероятнее всего, началась еще в докембрии. Род Tri-
brachidium из эдиакарской фауны Австралии некоторыми палеонтологами
признан как возможный предок иглокожих. Эти ранние ископаемые наво-
дят на мысль, что предки иглокожих могли быть свободно обитающими
формами.
Кроме геликоплакоидей и эдриоастероидей имеется ряд других малоиз-
вестных специфических палеозойских иглокожих, например обитавшие с
ордовика до девона офиоцистоиды и циклоцистоиды. Каждая из них более
подробно описана в “Treatise on Invertebrate Paleontology” под ред. Р.С. Му-
ра. Этот справочник, подробно освещающий группы ископаемых беспозво-
ночных, необходим каждому серьезному палеонтологу: и любителю, и про-
фессионалу. Совсем недавно классификация морских ежей была слегка
модифицирована в важной книге Эндрю Смита (1984). В ней также пре-
доставлено много новой информации об их анатомии, биологии и палео-
биологии.
20*
Глава XX
СЕТОЧКИ, ПРУЖИНКИ
И ПАЛОЧКИ: ГРАПТОЛИТЫ
Многие раннепалеозойские формации содержат маленькие “обуг-
ленные” или “серебристые” ископаемые, которые когда-то имели
склеропротеиновые внешние скелетики, близкие к роговым, а
внешне похожие на хитиновые. На поверхности породы они выглядят, как
будто кто-то нарисовал их карандашом или ручкой, за что они получили на-
звание “граптолиты”, то есть “разрисованные камни” (фиг. XX. 1).
Несмотря на то что граптолиты являются такой выдающейся группой, их
природа и систематическое положение долгое время были загадкой. Карл
Линней считал их “фальшивыми фоссилиями”, другие помещали их среди
растений, губок, книдарий (кишечнополостных) или мшанок. В 1948 году
польский ученый Р. Козловский объединил их с Rhabdopleura (крыложабер-
ные), современными червеобразными животными из полухордовых, чьи ис-
копаемые остатки были обнаружены начиная с ордовика.
Такая интерпретация систематического положения граптолитов была са-
мой неожиданной из всех. Несмотря на то, что крошечные (менее чем 0.5 мм
в длину) морские Rhabdopleura выглядят как неподвижные сидячие черви,
они имеют стержневидный выступ (дивертикулюм) выше рта, то есть зача-
ток хорды. Эти существа, как и большинство птеробранхий, к кому они отно-
сятся, строят колонии почкованием. Мускулистый стержень, или столон,
разветвляясь, соединяет каждую особь (или зооид) с другими на протяжении
всей колонии. Все особи и столон находятся в трубочках (фиг. XX.2). Когда
Rhabdopleura пребывает в спокойном состоянии, она высовывает свое тело с
щупальцами (лофофор), имеющими реснички, которые создают ток воды,
подающий ко рту пищевые частицы. Тело может быстро спрятаться обратно
в трубочку благодаря сокращению мускулистого стержневидного стебелька.
Трубочки или чашечки называются теками; они состоят из полуколец, со-
единенных по зигзагообразному шву. Теки образуют колонию, называемую
“рабдосома”. Между птеробранхиями и граптолитами столько поразитель-
ного сходства в строении скелетов, что их объединяют вместе в тип Hemichor-
data, или полухордовые.
Несмотря на то, что птеробранхии и граптолиты сходны, необходимо обра-
титься еще к одной группе, чтобы понять их соотношения с другими живот-
ными. Эта группа так называемых “желудевых червей”, или Enteropneusta
(кишечнодышащие), которые часто прячутся в приливных выбросах. Они
обладают длинным стройным телом, напоминающим червей, но в отличие от
© 1997. О.Б. Бондаренко, перевод на русский язык и изменения.
СЕТОЧКИ, ПРУЖИНКИ И ПАЛОЧКИ: ГРАПТОЛИТЫ
309
Фиг. ХХ.1. Граптолитовые сланцы
Черные сланцы раннего ордовика юго-восточной Австралии, содержащие “се-
ребристые” колонии Tetragraptus decipiens, Kiaerograptus antiquus, Clonograp
tus, Adelograptus victoriae (фото F.Coffa, с разрешения музея Виктории,
Мельбурн)
последних у Enteropneusta имеется дисковидное утолщение выше рта. Это
утолщение (пробосцис) является приспособлением для рытья. Сзади (ниже)
него находится область воротничка, и там находится спинной нервный тяж.
Такой тип нервного тяжа характерен для хордовых животных, включая и
позвоночных. В этом участке тела также находится массивное образование,
которое некоторые зоологи интерпретируют как зачаток опорного стержня
(нотохорда), также являющегося особенностью строения хордовых. Но не
все зоологи согласны с таким заключением. Пожалуй, наиболее важным и
примечательным является тип жабр, которыми обладают “желудевые чер-
ви”. Тип жабр очень сходен с жабрами ланцетника -Amphioxus, являющего-
ся рыбоподобным примитивным хордовым животным, близким к группе, ве-
дущей к позвоночным животным.
Ланцетники, “желудевые черви”, птеробранхии и группа, называемая
оболочниками (Tunicata), находятся по уровню развития между беспозво-
ночными и позвоночными. Граптолиты также находятся в этой “предрас-
светной” группе, так непохожей на родственников наших предков.
ДВЕ ГРУППЫ ГРАПТОЛИТОВ
Граптолитов обычно подразделяют на шесть отрядов. Четыре из них так
редки, что можно на них не останавливаться. Это донные ползающие или ин-
крустирующие формы с очень ограниченным распространением.
310
КАМЕННАЯ КНИГА
Фиг. ХХ.2. Rhabdopleura — современное полухордовое животное
а - зооид, сидящий в трубке длиной около 0.5 мм; б - кустистая колония, сте-
лющаяся по субстрату, ж — желудок, з - зигзагообразный шов, к - концевая
почка нового зооида, ки — кишка, лф — лофофор, п — полукольца, образующие
трубку, ид - приротовой диск, с - столон, ст - стержень-стебелек, сз - спрятав-
шийся зооид, т — тека
Дендроидеи — Dendroidea. Дендроидеи наиболее примитивные граптоли-
ты и наиболее сходные с Rhabdopleura. Тонкостенные теки растут группами
по три и соединены столонами. Каждая колония начиналась с узкоконичес-
кой теки (сикула), от острого кончика которой тянулось нитевидное образо-
вание - нема. Склеропротеиновая пластиночка на ее конце служила для при-
крепления к морскому дну или другим объектам, таким как водоросли или
раковины. Колонии дендроидей напоминают тонкие сеточки (фиг. XX.3).
Дендроидеи имеют три типа тек: столотеки, автотеки и битеки
(фиг. XX.4). Столотеки, или “почкующиеся особи”, представлены закры-
тыми трубочками, благодаря почкованию которых образуются все новые и
новые группы тек. Теки с открытыми устьями свидетельствуют о том, что
их обитатели едят, дышат и размножаются половым путем. Учитывая, что
одна из таких тек всегда крупная (автотека), а другая маленькая (битека),
считают, что в первой из них находилась женская особь, а во второй —
мужская.
Дендроидеи существовали с позднего кембрия по карбон, то есть длитель-
ность существования была более чем 250 миллионов лет. Некоторых дендро-
идей, таких как Inocaulis, иногда рассматривают как морские водоросли.
Граптолоидеи — Graptoloidea. Дендроидеи дали начало очень успешной
процветающей группе - граптолоидеям. Из них анизограптиды находятся
где-то между дендроидеями и настоящими граптолоидеями. Анизограптиды
потеряли прежнее разнообразие зооидов и сохранили только один тип тека.
Каждый зооид поэтому был способен почковаться и строить колонию, а так-
же размножаться половым путем.
Граптолоидеи являются наиболее многочисленными, разнообразными и
наиболее специализированными представителями своего класса и одними из
самых многочисленных в ископаемом состоянии (фиг. XX.5). Тем не менее,
их распространение ограничено интервалом от ордовика по ранний девон.
Большинство граптолоидей - бентосные формы, но некоторые стали планк-
тонными, широко распространенными в прошлых океанах мира. Благодаря
этому они являются хорошими руководящими ископаемыми для раннего-
среднего палеозоя, особенно для ордовика и силура.
СЕТОЧКИ, ПРУЖИНКИ И ПАЛОЧКИ: ГРАПТОЛИТЫ
311
Фиг. XX. 3. Сетчатые колонии дендроидей (поздний кембрий—карбон)
а - в ископаемом состоянии (К ore mag rapt us, Rhabdinopora), xl, б— реконструк-
ция прижизненного положения (Dictyonema)
Фиг. XX.4. Строение трех типов тек у дендроидий
а, б — триады тек, внешний вид (а) и разрез (б). As — стержень-стебелек, Av — ав-
тотека, Bt — битека, Dt — столотека
ВНЕШНИЙ ВИД, ОСОБЕННОСТИ СТРОЕНИЯ И ОБРАЗ ЖИЗНИ
Древние граптолиты строили маленькие колонии со стройными веточка-
ми, растущими вверх от основания, прикрепленного к раковинам, водорос-
лям или плотному илу. Еще до окончания кембрийского периода колонии
были представлены несколькими вариантами. Огромное разнообразие форм
возникло в течение ордовикского времени.
312
КАМЕННАЯ КНИГА
Фиг. ХХ.5. Граптолоидеи (ордовик—ранний девон)
а—в — ранние стадии развития граптолоидей: а - сикула, несущая круглое от-
верстие на стенке (перфорация), б — сикула и первая тека, возникшая на месте
перфорации, в — начало образования ветвей; г-е — двурядные и однорядные
граптолоидеи с различным’расположением и формой тек; ж — Cyrtograptus
murchisoni около 6 см в поперечнике, ранний силур, венлокский век, повсеме-
стно, кроме Южной Америки (рис. F. Knight по Moore, 1970); з — Spirograptus
turriculatus около 2 см длиной, ранний силур, Европа (рис. F.Knight по Moore,
1970). Е — центральный канал, F - нема
Dendropgaptus (из дендроидей) характеризуется ветвистой или кустистой
формой колонии (фиг. ХХ.6#)- Нема была представлена коротким стержнем
или почти отсутствовала; колонии прикреплялись к раковинам, водорослям
или к отвердевшему илу. Такое строение ограничивало широкое площадное
распространение личинок и взрослых форм. Несколько видов кустистых
дендроидей обладало длительным геохронологическим интервалом сущест-
вования, что снижает их стратиграфическую ценность, но их можно успеш-
но использовать в палеоэкологических реконструкциях.
Dictyonema (из дендроидей) имела вееровидную колонию, состоящую из
параллельных прутьев, разветвляющихся вновь и вновь и соединяющихся
СЕТОЧКИ. ПРУЖИНКИ И ПАЛОЧКИ: ГРАПТОЛИТЫ
313
Фиг. ХХ.6. Реконструкция образа жизни донных прикрепленных дендроидей
а - Inocaulis, xl, силур, Северная Америка; б - Dendrograptus, xl, кембрий-
карбон, почти повсеместно; в - Dictyonema, xl, поздний кембрий-ранний кар-
бон, повсеместно; г — Medusaegraptus, х0.2 (положение среди граптолитов дис-
куссионное), силур, Северная Америка и Европа
Фиг. XX .7. Реконструкция образа жизни планктонных граптолоидей
а — Glossograptus, xl, ордовик, почти повсеместно; б - Tetragraptus, xl, ордо-
вик, повсеместно; г — Didymograptus, xl.5, ордовик, повсеместно; в — Syndyo-
graptus, прикрепленный к водорослям, xl, поздний ордовик, Северная Америка
и Китай; д - Cyrtograptus, х0.5, силур, повсеместно, кроме Южной Америки
314
КАМЕННАЯ КНИГА
DENDROGRAPT1DAE
Dirty слегла
scrtulue
ANISOGRAPTIDAE
Adelograptus
virtoriae
Anisograptue
compactus
Clooograptus
rlgidus
DICHOGRAPTIDAE
Stgmagraptus
praecursw
Tetragraptus
approximate
decipiens
(Pendeograpius) fruticoeus (4 hr.)
(Pendeograpius) fruticose (4 hr.)
(Pendeograptus) pendens
sarra
Ptiylfograptus
typus
Zygograptus
junori
Didymograptus
(Didymograptellus) protobifidus
1SOGRAPTIDAE
isograptus
caducous australis
caducous imilatus
virtoriae difergens
virton'ae lunatus
victonae maximodivergens
virtoriae maximus
victoriae virtoriae
Oncograptus
upsiton
DIPLOGRAPTIDAE
Cliptograptus
auetrodenlatus
intersitus
toratiusculus
Diplograptus
decoratus
Orthograptus
quadrimucronatus
Climacograptus
bicornis
spiniferus
tubuliferus
DICRANOGRAPTIDAE
Dicellograptus
omatus
Dicranograptus
nicholsoni nicholsoni
RETIOLIDAE
OrthoretkMes
puJchernmus
ORDOVICIAN
LOWER MIDDLE UPPER
LANCE- FIELDIAN BENDIGOIAN BENDIGOIAN CASTLE- MAINIAN YAPEENIAN DARRIWIUAN GISBO- NIAN EASTONIAN BOLINDIAN
i $ h F Q ARENIGIAN ! Ш I CARADOCIAN ASHGILL
СЕТОЧКИ, ПРУЖИНКИ И ПАЛОЧКИ: ГРАПТОЛИТЫ
315
перемычками (см. фиг. XX.36, бе). Граптолиты рода Dictyonema или близ-
ких к нему форм вели бентосный или псевдопланктонный (плавучий) образ
жизни. Вееровидные колонии Dictyonema прикреплялись к субстрату или
водорослям дисковидной пластинкой, они также инкрустировали плотные
предметы или закреплялись в мягких грунтах с помощью отростков. Дру-
гие роды в этой группе, такие как Rhabdinopora, всю свою жизнь проводили
в открытом океане, свободно паря в воде.
Большинство ископаемых Diplograptus и Glossograptus (из граптолоидей)
представлены фрагментами, но несколько превосходных экземпляров най-
дены в виде пучков рабдосом, чьи немы оказались прикрепленными к цент-
ральному диску (фиг. XX.7). Диск, в свою очередь, был сверху соединен с
зонтиковидным вздутием, окруженным меньшими вздутиями шаровидной
или цилиндрической формы. Все сооружение представляло собой суперко-
лонию, или синрабдосому, которая плавала в палеозойских морях вместе с
волнами и течениями, как современные гидрозоа - сифонофоры, называе-
мые “португальскими корабликами”.
Неизвестно, какое количество граптолитов строило суперколонии, так
как большинство ископаемых форм представлено разрозненными фрагмен-
тами, чье прежнее строение невозможно определить. Несколько родов обра-
зуют ветвистые или спиральные колонии, которые прикреплялись к плава-
ющим предметам, свешиваясь вниз.
Развитие разнообразных колониальных форм и возникновение одной
или нескольких ветвей зависит от способа почкования. Если от первой теки-
сикулы возникают две ветви, называемые также рядами тек, колония (раб-
досома) называется двурядной (см. фиг. XX. 5). Если образуется только одна
ветвь - это однорядная колония. Направления, по которым ветви отходят в
стороны от сикулы также важны для формирования структуры колонии.
Если ряды тек растут в стороны вверх от сикулы, колония называется воз-
дымающейся. Если, наоборот, ветви свисают вниз, они являются висячими.
Палеонтологи, изучающие граптолиты, чтобы разграничить разные виды,
используют также такие характеристики, как форма тек и очертания усть-
ев, то есть апертур, наличие и характер приустьевых шипиков, число тек на
10 мм и т.д.
Изучая изменения граптолитовых комплексов в длинной последовательно-
сти пород в таких регионах, как юго-восточная Австралия, западная часть Се-
верной Америки, Британия, Шпицберген, Сибирь, Казахстан, Китай и др.,
можно выделить очень дробные подразделения (фиг. ХХ.8). Граптолиты пред-
ставлены разнообразными формами, которые быстро изменялись во времени,
одновременно широко распространяясь по акваториям, поэтому их широко
используют для корреляции отложений далеких регионов. Граптолиты чрез-
вычайно важны для определения возраста сильно складчатых и деформиро-
ванных черных сланцев, которые обычно не содержат других ископаемых.
Основное направление в эволюции граптолитов сводится к упрощению
строения, что облегчает вес колонии. Происходит редукция числа ветвей от
Фиг. ХХ.8. Временные интервалы некоторых дендроидей (Dendrograptidae,
Anisograptidae) и граптолоидей (Dichograptidae, Isograptidae, Diplograptidae,
Dicranograptidae, Retiolitidae), найденных в ордовикских породах Австралии.
Толстые черные линии показывают время распространения каждого вида. Ос-
новываясь на первом и последнем появлении, а также на одновременном суще-
ствовании этих разнообразных граптолитов, были установлены многие зоны и
ярусы в юго-восточной Австралии, такие как Lancefieldian и Chewtonian (схема
составлена I. Stewart и Р. Morrison)
316
КАМЕННАЯ КНИГА
многорядных до однорядных; стенки тек становятся тоньше; усиливается
плавучесть, что помогает держаться колониям в верхней толще воды, или
хотя бы над дном. Развитие плавучести и планктонный образ жизни привел
к тому, что колонии с висящими ветвями сменились колониями с воздымаю-
щимися ветвями.
В течение ордовика разнообразие граптолитов достигло пика. В ордовикс-
кое время отмечается похолодание, приведшее к образованию более четких
климатических зон и провинций подобно тому, как это наблюдается сейчас.
К концу ордовика мощное оледенение глобально понизило температуру,
включая и моря. Это довольно сильно подействовало на морские фауны, при-
ведя к исчезновению большинства групп граптолитов. Только несколько ви-
дов пережили этот крупный биологический кризис. Эти виды дали начало
гораздо более бедным силурийским фаунам, где доминировали монограпти-
ды, обладавшие однорядными колониями.
Местонахождения и сохранность. Граптолиты находят в кремнистых изве-
стковых и глинистых сланцах и в известняках. Чаще всего они встречаются в
черных глинистых сланцах (см. фиг. XX. 1). Есть много причин, почему так
происходит. Колонии граптолитов довольно нежные и хрупкие образования,
и поэтому песок быстро разбивает их на кусочки. Колонии также разрушают
трилобиты и более мелкие падалееды, которые обычно кишат в илистых осад-
ках. Однако большая часть черных сланцев образовалась в застойных услови-
ях, где вода содержала мало кислорода и очень много сероводорода за счет
медленно гниющей органики. Только немногие падалееды живут в таких ус-
ловиях, и поэтому отмершие граптолиты лежат нетронутыми. В процессе за-
хоронения они пиритизируются или замещаются серебристым слюдистым
минералом. Местом захоронения были также участки спокойной воды, где
механическое разрушение было небольшим. В условиях, описанных выше,
скорость седементации была низкой, поэтому высокая аккумуляция органи-
ческого материала сопровождалась небольшим содержанием осадка.
Глава XXI
“ЗУБЫ” БЕЗ ЧЕЛЮСТЕЙ:
КОНОДОНТЫ И КОНОДОНТОФОРИДЫ
• Название конодонты с греческого означает “конические зубы”. Ко-
нодонты загадочные морские ископаемые. Впервые они были описа-
ны Хр.Пандером в 1856 году из палеозойских отложений Россий-
ской Империи. Животные, ими обладавшие, называются конодонтофорида-
ми, или конодонтоносителями, но обычно их называют просто конодонтами.
Некоторые палеонтологи считают конодонтов позвоночными. Другие пале-
онтологи предполагают, что конодонты, возможно, стоят ближе к живот-
ным, имеющим лофофор, таким как брахиоподы, мшанки и форониды. Одно
из ископаемых из сланцев Бёрджесс среднего кембрия Британской Колум-
бии Odontogriphus, имеющий червеобразный облик с длиной тела около 6 см,
содержит конусовидные структуры, напоминающие конодонты. Эти конусы
лежат в основании каждого щупальца числом 20-25 и, очевидно, служили в
качестве внутренних опор для этого ловчего аппарата (фиг. XXI. 16). Odonto-
griphus был мягкотелым животным с правильными пережимами и отчетли-
вой головой.
Достоверный облик конодонтов был установлен благодаря сенсационной
находке отпечатка животного с конодонтовым аппаратом в отложениях ни-
жнего карбона Шотландии (см. фиг. XXI. 16, е). Отпечаток имел форму чер-
веподобного тела длиной около 7 см с головой, несущей две боковые лопасти.
В головном отделе на уровне глотки находился конодонтовый аппарат, состо-
ящий из маленьких зубовидных элементов различной формы, располагаю-
щихся в определенном порядке. До этой находки форма тела и расположение
“зубов” интерпретировали самым различным образом (см. фиг. ХХ1.1а-г).
Достоверные конодонты обнаружены в отложениях конца кембрия-начала
ордовика и далее на протяжении всего палеозоя и начала мезозоя по поздний
триас включительно.
Конодонты представляют собой микроскопические (0.1—5 мм) конические
зубовидные или уплощенные образования, чья функция не совсем понятна
(фиг. XXI.2). Иногда конодонты выглядят как крошечные зубы, они часто
встречаются симметричными парами или рядами, как и зубы позвоночных
животных. В отличие от позвоночных конодонтовые зубы не имеют жева-
тельных поверхностей и, вероятно, никогда не использовались для непосред-
ственного пережевывания пищи. Конодонты в основном состоят из сложного
соединения фосфата кальция (группа апатита) и очень небольшого количест-
ва органического белкового вещества.
© 1997.0.Б. Бондаренко, перевод на русский язык и изменения.
318
КАМЕННАЯ КНИГА
Фиг. XXI.1. Коиодоитоиосители: предполагаемые (а—г) и реальный (д, ё)
а—г — различные реконструкции предполагаемых конодонтоносителей (а, в—г —
Benton, 1987; б — Gould, 1989: род Odontogriphus, вид снизу, средний кембрий,
сланцы Бёрджесс, Канада); д, е — реальный конодонтоноситель Clydagnathus?
cf. С. cavusformis Rhodes длиной около 7 см, ранний карбон, Шотландия, Эдин-
бург (Briggs, Clarkson, Aldridge, 1983). к — конодонтовые элементы; М, Sc, Sb,
Pb — символы конодонтовых элементов
Известно три основных типа конодонтовых элементов: простые коничес-
кие и сложные - многозубчатые стержневидные и платформенные
(фиг. XXI.3). У сложных конодонтовых элементов обычно развиты различ-
ные выступы, некоторые из них сходны с выступами на наших коренных зу-
"ЗУБЫ" БЕЗ ЧЕЛЮСТЕЙ: КОНОДОНТЫ И КОНОДОНТОФОРИДЫ
319
Vjalovognathus
Фиг. XXI.2. Конодонтовые платформенные элементы в электронном микроскопе
а, б — N eogondolella clarki, вид сверху (а) и снизу (б), средний карбон, Morrowan
(башкирский век), Северная Америка (Р.Н. von Bitter, G.K. Merrill, 1977); в -
Vjalovognathus synguensis, вид сверху, ранняя пермь, болорский (кунгурский)
век, Юго-Восточный Памир (А.Н. Реймерс, 1991); г - Neostreptognathodus ре-
quopensis, ранняя пермь, артинский век, Южный Урал (коллекция А.Н. Рей-
мерса, Палеонтологический институт, № 4606)
бах. Платформенные конодонты имеют на верхней поверхности, как прави-
ло, разнообразный орнамент, что важно для их систематики.
Классификация конодонтов вызывает много проблем потому, что в коно-
донтовом аппарате животных содержатся зубы разных типов, как это наблю-
дается и в челюстях позвоночных. В прежнее время палеонтологи давали
каждому конодонтовому элементу свое латинское название. Открыв, что эти
элементы располагаются в определенном порядке и принадлежат конодонто-
вому аппарату одного животного, родовые и видовые латинские названия
стали давать аппаратам. Отдельные элементы аппарата получили буквенные
символы: М, S, Sa, Sb, Sc, Sd, P, Pa, Pb и т.д. (фиг. XXI.4a).
Конодонтоносители, вероятно, были свободно плавающими нектонными
животными. Многие виды распространены почти всесветно: в Северной Аме-
рике, Европе, Азии, Австралии. Конодонты являются превосходными руко-
водящими ископаемыми, особенно важными при обработке керна, благода-
ря своим миниатюрным размерам, массовости, узкому вертикальному ин-
320
КАМЕННАЯ КНИГА
Фиг. XXI.3. Отдельные конодонтовые элементы (сильно увел.)
а — конические элементы Protopanderodus, ордовик; б — стержневидные элемен-
ты; в, г - платформенные элементы: в - Kockelella, со стороны основания с ба-
зальной ямкой в центре, силур; г — Polylophodonta, вид сверху, поздний девон
(Treat. Invert. Paleont. Part W, 1981)
тервалу и широкому пространственному распространению. Тысячи этих
крошечных ископаемых сохраняются в маленьком кусочке керна. Особенно
большое биостратиграфическое значение конодонты имеют для среднего ор-
довика, девона и карбона.
Кроме биостратиграфического значения конодонты можно использовать
как индикаторы на нефть и газ. Дело в том, что окраска конодонтов в зависи-
мости от температуры диагенеза меняется от светло-янтарной прозрачной до
черной и белой непрозрачной (табл. XXL 1). Изменение окраски связано с по-
следовательным разрушением органической белковой составляющей по ме-
Таблица XXI. 1. Изменение окраски конодонтов от степени катагенеза
(Treatise..., Part W, 1981; Овнатанова, Петросянц, 1985)
Индекс окраски конодон- тов (ИОК) Цвет конодонтов Темпе- ратура, °C Тип скопления
1 светло-желтый, прозрачный 50-80 газ
р/2 подобный, светло-желтый 50-90 газ
2 желто-коричневый 60-140 нефть
3 темно-коричневый 110-200 конденсат
4 темно-коричневый, пятнистый 190-300 газ
5 темно-серый, почти черный 300-480 газ
6 черный с белесостью 360-550 Углеводороды раз-
61/2 белый с зеленоватым 480-720 рушаются, нельзя ожидать углеводо-
7 белый молочный родных скоплений
8 белый молочный, опоковидный 800
"ЗУБЫ" БЕЗ ЧЕЛЮСТЕЙ: КОНОДОНТЫ И КОНОДОНТОФОРИДЫ
321
а
Фиг. XXI. 4. Конодонтовые аппараты
а — реконструкция головного отдела животного, где показано положение коно-
донтовых элементов (М, Sb, Sc, Sd, Pb, Pa) в конодонтовом аппарате (R.Nicol,
1984); б — реконструкция конодонтового аппарата рода Prioniodus, представ-
ленного семью конодонтовыми элементами, ранний-средний ордовик (Treat. In-
vert. Paleont., Part W., 1981)
ре повышения температуры. В зависимости от температуры изменяется не
только цвет конодонтов, но изменяется и содержание нефти и газа в окружа-
ющих породах.
21 Каменная книга
Глава XXII
ОТ МОРСКИХ ЗВЕЗД ДО РЫБ,
ВЛАСТЕЛИНОВ ВОДЫ
• Ланцетник (Amphioxus) и целый зверинец других странных существ
являются связующим звеном между миром беспозвоночных, осо-
бенно иглокожих, и миром позвоночных животных. К этому проме-
жуточному звену между главными группами животных принадлежат “же-
лудевые черви” и крыложаберные, которые, как полагают, связаны с грапто-
литами и которые взятые все вместе относятся к полухордовым. К
беспозвоночным хордовым относятся также мешковидные морские живот-
ные - туникаты (урохордовые), взрослые особи которых не напоминают рыб,
а личинки внешне напоминают головастиков.
Хордовые — это животные, у которых на некотором этапе развития имеет-
ся нотохорд. У Amphioxus нотохорд представляет собой важную структуру
взрослого организма, являющуюся его главной опорой (фиг. XXII. 1). У рыб
и человека эта структура сильно редуцирована и присуща главным образом
эмбриональной стадии. Беспозвоночные хордовые особенно интересны, по-
тому что они свидетельствуют о том, что иглокожие и хордовые, вероятно,
имели общее происхождение. У иглокожих и хордовых сходно развитие ме-
зодермы. Мезодерма — это зародышевый листок, который залегает между
внешним слоем (или эктотодермой), развивающейся в кожу и в нервную си-
стему, и внутренним слоем (или эндодермой), который развивается в кишеч-
ник, и его придатки такие, как аппендикс. Мезодерма морской звезды, лан-
цетника и людей сходным образом формирует вырост в стенке кишечника.
Внутри этого выроста остается полость, внутри которой закладываются та-
кие органы, как легкие и печень. У червей, моллюсков и членистоногих ме-
зодерма закладывается совершенно другим путем и полости тела изначально
формируются как щели в толще ткани, а не как выросты.
Иглокожие и некоторые беспозвоночные хордовые также имеют очень по-
хожие личинки, которые довольно разительно отличаются от личинок дру-
гих беспозвоночных. Также у них похожи и белки плазмы крови, отличные
от белков всех других групп беспозвоночных. С другой стороны, некоторые
беспозвоночные хордовые имеют признаки, присущие позвоночным — жа-
берные щели, нотохорд, трубчатый спинной нервный тяж и так далее, прояв-
ляющиеся либо на юношеских либо на взрослых стадиях развития.
Хотя морские звезды и ежи с их лучевой симметрией совершенно не похо-
жи на примитивных позвоночных, таких как рыбы, существуют иглокожие
более близкие к позвоночным. Карпоидеи, упоминаемые в главе XIX, вклю-
© 1997. Д.Н. Есин, перевод на русский язык.
ОТ МОРСКИХ ЗВЕЗД ДО РЫБ. ВЛАСТЕЛИНОВ ВОДЫ
323
Фиг. XXII.1. Современный ланцентник (Amphioxus). Длина тела около 5 см
Фиг. XXII.2. Реконструкция внешнего вида Notocarpos garratti
а — спинная сторона, б — вид сбоку, в — брюшная сторона (по G.M. Philip)
чают виды, которые были двусторонне симметричными и по внешнему виду
походили на первых хордовых. Некоторые, такие как Notocarpos, даже
имели, по-видимому, отчетливые “голову” и “хвост”, были уплощены в
дорзовентральном направлении и являлись подвижными животными
(фиг. ХХП.2.). Некоторые палеонтологии, например Р.П.С. Джеффрис,
предполагают, что эта группа дала начало хордовым и, таким образом, всем
позвоночным животным, однако эта точка зрения оспаривается Г.М. Фили-
пом и другими.
21*
324
КАМЕННАЯ КНИГА
Первые позвоночные хордовые
Хотя ланцетник довольно похож на рыбу, у него отсутствуют многие чер-
ты, присущие настоящим позвоночным. Для начала, у него нет позвоночни-
ка. Кроме того, у него нет зубов и ему приходится процеживать воду для пи-
тания. Нервная система не имеет увеличенных участков, которые могли бы
выполнять роль мозга. У него нет и сердца, хотя система кровообращения по-
строена по позвоночному типу.
Первые остатки предполагаемых позвоночных обнаружены в верхнем
кембрии и раннем ордовике Северной Америки, Гренландии и Шпицберге-
на, т.е. в морских отложениях Северного полушария. Остатки этих прими-
тивных рыбообразных, часто называемые остракодермами или костнощит-
ковыми, фрагментарны и фосфатизированы. В минералогическом смысле
они сложены апатитом, совершенно как живая кость, но их микроструктура
отличается от микроструктуры кости современных хордовых.
Многие из этих чешуевидных ископаемых остатков объединены под на-
званием Anatolepis, но не все палеонтологи согласились бы с отнесением их к
позвоночным.
Среднеордовикские породы в центральной Австралии содержат некото-
рые из наиболее древних известных позвоночных и представлены отпечат-
ком целого бесчелюстного Arandaspis (фиг. XXII.3). Кость совсем не сохра-
нилась, но отпечаток дает представление как о внешнем, так и о внутреннем
строении этого животного, у которого имелись серии больших дорсальных и
вентральных пластин и один ряд более мелких пластин поверх жаберных от-
верстий. Небольшие чешуи покрывали если не все тело, то по крайней мере
его переднюю часть. Andreolepis был 12-14 см (около 5-5.6 дюймов) длиной,
и его тело в поперечнике было овальным, а не сплющенным в дорсовентраль-
ном направлении, как у многих древних щитковых бесчелюстных. Это древ-
нее животное, возможно, питалось органическими частицами и микроорга-
низмами и двигалось, извиваясь, как головастик, так как у него не было пар-
ных плавников для более совершенного плавания. Другие формы, такие как
Porophoraspis, также известны из отложений Австралии, но ни одна не со-
хранилась так полно, как Arandaspis. Совсем недавно в 1987-1988 гг. в Се-
верной и Южной Америке было найдено еще несколько полностью сохранив-
шихся экземпляров подобных животных.
В немного более молодых морских отложениях ордовика Скалистых гор
Северной Америки (песчаники Хардинг) обнаружены первые бесспорные ко-
стные остатки. Эта кость, называемая аспидином, состояла из четырех сло-
ев: базальный пластинчатый слой, за ним сетчатый слой, затем дентиновый
слой, а затем эмалевый слой. Многие из этих ископаемых представлены
фрагментарно, но обнаружена и почти полностью сохранившаяся форма
Asteraspis disiderata, костная броня которой была сложена серией неболь-
ших многоугольных пластин. У другой формы, Eriptychius, имелся внутрен-
ний скелет, сложенный кальцифицированным хрящем. Обе эти формы, как
и Arandaspis, были бесчелюстными фильтраторами.
ПРОЧИЕ ПТЕРАСПИДАМОРФНЫЕ ИЛИ КРЫЛОЩИТКОВЫЕ ФОРМЫ
Кроме ордовикских остракодерм, имеется ряд других, очевидно, родст-
венных силурийских и девонских бесчелюстных рыбообразных. У всех них
имеется не несколько, а единственное парное жаберное отверстие, парные
ОТ МОРСКИХ ЗВЕЗД ДО РЫБ, ВЛАСТЕЛИНОВ ВОДЫ
325
Фиг. ХХГГ.З. Arandaspis prionotolepis — ранний представитель бесчелюстных
из ранне-среднеордовикских отложений Центральной Австралии (реконструк-
ция F. Knight)
326
КАМЕННАЯ КНИГА
Фиг. XXII.4. Бесчелюстные, или агнаты (Agnatha). Длина животных от 6 до
30 см
носовые капсулы и относительно цельный головной щиток. У большинства не
было парных плавников. К этим бесчелюстным относились циатаспидные,
амфиаспидные, птераспидные, псаммостеидные и, возможно, телодонты.
Циатаспидные. Эта группа остракодерм впервые появилась в среднем си-
луре и существовала вплоть до раннего девона. Хотя древние формы встрече-
ны преимущественно в морских отложениях, многие представители в девоне
переселились в солоноватоводные и пресноводные водоемы. Циатаспиды по-
ходили на некоторых ордовикских остракодерм, так как их головной щиток
был сформирован из дорсальной и вентральной пластин, разделенных длин-
ными и относительно узкими брахиальными пластинами. Отдельные плас-
тины несли округленные дентиновые гребни, параллельные краям пластин.
Эти рыбообразные, также как и их непосредственные предки и ланцетник,
были, возможно, фильтраторами. Тело циатаспидных было слегка сплю’
щено, а глаза находились не сверху, а по сторонам головного щитка. Хвост
был почти симметричным (фиг. XXII.4). Многие поздние остракодермы, ко-
торые в девоне стали довольно разнообразными, возможно произошли от
этой группы.
Амфиастидные. Известны лишь из нижнего и среднего девона Сибири.
Эти высоко специализированные остракодермы имели почти цельный, срос-
шийся панцирь (из головного и грудного щитков). Они также почти полно-
стью утратили глаза,, что свидетельствует о том, что они жили в воде, перена-
сыщенной взвешенными частицами осадка, или вообще были грунтовыми
животными. У некоторых из них рот располагался на конце длинной трубки.
Птераспиды. Подобно амфиаспидным, эти остракодермы, возможно, про-
изошли от циатоаспидных. Как и у амфиаспидных, многие пластины были
сросшимися, но не сросшихся пластин имелось несколько больше. Птерас-
ОТ МОРСКИХ ЗВЕЗД ДО РЫБ, ВЛАСТЕЛИНОВ ВОДЫ
327
пидные также отличались присутствием костных выростов сверху и сбоку
панциря, которые, возможно, выполняли роль стабилизаторов при плава-
нии. Нижняя (вентральная) лопасть хвоста была значительно более длинной
по сравнению с верхней дорсальной (см. фиг. XXII.4). Довольно характерной
для этой группы была выдающаяся вперед ростральная пластина, которая
протягивалась далеко вперед рта, а у некоторых форм развивались даже уд-
линенные “рога” на боковом щитке. Это, несомненно, были самые странные
животные!
Псаммостеидные. Эти сплющенные в спинно-брюшном направлении ры-
бообразные, представляющие ветвь эволюции, противоположную той, что на-
блюдалась у циатаспидных, в раннем девоне обитали в прибрежных речных
водах, а затем некоторые переселились в океаны. Они существовали до конца
позднего девона и были широко распространены в Северном полушарии.
Некоторые псаммостеидные достигли значительных размеров, до двух мет-
ров. Как они в точности питались, не совсем ясно; хотя они были сплющены,
как многие придонные формы, но их рот был обращен кверху (см. фиг. XXII.4).
Телодонты. Телодонты, или целолепиды, представляют группу вымер-
ших бесчелюстных животных, которые очень полезны при определении воз-
раста отложений, но, как и в похожем случае с конодонтами, многое в их
строении остается неизвестным. Их тела были покрыты маленькими ромби-
ческими неперекрывающимися чешуями. По-видимому, эти чешуи не срас-
тались и, как у большинства других древних рыбообразных и рыб, внутрен-
ний скелет не обызвествлялся. Скорее всего он был построен исключительно
из хряща. Для определения возраста пород используются только расшлифо-
ванные чешуи. Тело телодонтов, обычно сохраняющееся в виде плоских от-
печатков, было, очевидно, немного сплющено в спино-брюшном направле-
нии, но сохраняло по-прежнему обтекаемую форму. Глаза располагались по
бокам головы, рот был впереди тела, а не на дорсальной или вентральной сто-
роне головы. Разделенные жаберные отверстия располагались вдоль боковой
стороны тела в отличии от единого дыхательного отверстия, характерного
для всех других птераспидоморф.
Телодоны также, по-видимому, имели разнообразные плавники, включая
пару пекторальных (боковых) плавников, маленький спинной плавник и ана-
льный плавник, лежащий впереди гипоцеркального хвостового с увеличен-
ной спинной лопастью. Чешуи телодонтов известны из отложений позднего
ордовика, и эта группа продолжала свое существование в девоне. Однако пока
не ясно, кто их ближайшие родственники. С одной стороны, глаза у них были
расположены по бокам головы и не было ростра, как у птераспидоморф (гете-
ростраки). С другой стороны, присутствие нескольких отдельных жаберных
щелей и парных плавников делают их похожими на цефаласпидоморф, что
будет рассмотрено далее. Для того чтобы понять их действительные родствен-
ные связи, необходимо найти более полно сохранившиеся экземпляры, одна-
ко это не умаляет их ценности для геологических исследований.
Цефаласпидоморфные
Еще одной большой группой бесчелюстных были цефаласпидоморфы.
Они отличались от гетеростраков по ряду признаков, например, тем, что они
имели носоглотку с одним отверстием (ноздрей) в середине сильно окостенев-
шего головного щитка. Эти рыбообразные всесторонне изучались шведской
школой палеоихтиологов, возглавляемой Эриком Стеншо. Стеншо в течение
нескольких десятилетий делал серийные пришлифовки многих экземпля-
328
КАМЕННАЯ КНИГА
ров этих древних животных и получил возможность проделать детальную
реконструкцию мозговых полостей и нервов мозга, а также изучить анато-
мию всего черепа.
Существуют три группы цефаласпидоморфов: анаспидные, остеостраки и
галеаспидные. Они довольно сильно различаются по форме тела, которая
приблизительно отражает их образ жизни.
Анаспидные. Эти существа имели торпедообразную форму тела и были
покрыты узкими, перекрывающимися чешуями. Их глаза располагались
по бокам головы. Задний конец нотохорда уходил в нижнюю лопасть хвос-
тового плавника, что сформировало обратно гетероцеркальный (или гипо-
церкальный) хвостовой плавник. Это, по мнению некоторых ученых, под-
талкивало животное вверх в момент движения. Во всяком случае оно суще-
ствовало хотя бы некоторое время в толще воды, а не было постоянно
привязано ко дну. Все это были относительно небольшие рыбообразные, у
которых рот располагался в передней части головы. Способы их питания
могли существенно различаться. Несколько (от 6 до 15) жаберных отвер-
стий диагонально пересекали бока этих животных, а позади этих жабр рас-
полагался шип. На каждом боку к этому шипу присоединялся чешуйчатый
плавник. Возможно, эти плавники развились из вентролатеральных плав-
никовых складок, которые имеются у примитивных хордовых, таких как
ланцетник.
Первые анаспиды появились в позднем силуре, например, такая “брони-
рованная” форма, как Birkenia (см. фиг. XXII.4). Более поздние формы, на-
пример позднедевонский Endeiolepis, были почти “голыми”, так как их че-
шуя почти совсем не окостеневала. Jamoytius тоже принадлежит к этой
группе, хотя когда-то его считали беспозвоночным хордовым. Также нео-
бычным является то, что остатки этого животного происходят из морских
отложений, хотя большинство анаспидных, вероятно, обитали в пресновод-
ных бассейнах.
Остеостраки. Наиболее известными и наиболее полно сохранившимися из
палеозойских бесчелюстных следует, скорее всего, считать остеостраков. То,
что они вели придонный образ жизни, ясно показывает строение их головно-
го щита, сплющенного в спинно-брюшном направлении.
У типичного, но довольно специализированного рода Cephalaspis голо-
вные пластинки формируют щиток, который был округлен спереди, сильно
выдавался назад в виде заостренных выростов и напоминал головные щиты
трилобитов (фиг. XXII.5). Два глаза были сближены и направлены вверх,
единая ноздря располагалась перед ними, а между глазами помещалось пи-
неальное отверстие, являвшееся, возможно, рудиментом второй пары глаз.
На нижней стороне головы располагался овальный рот, следом за ним - 10
пар жаберных отверстий. Ветвистые каналы протягивались от мозга к трем
спинным областям, покрытым маленькими пластинками. В настоящее вре-
мя полагают, что эти области, которые когда-то считались электрическими
органами, являются сенсорными полями, которые распознавали колебания,
давление и вибрацию воды.
Тело Cephalaspis заострялось в хвосту, где позвоночник загибался вверх и
поддерживал плоский плавник с хрящевыми лучами. Этот тип хвоста, назы-
ваемый гетероцеркальным, встречается у многих агнат и у примитивных че-
люстных рыб, а также у акул. Маленькие пластинки позволяли телу живот-
ного свободно изгибаться, а дорсальный плавник не давал слишком сильно
переваливаться из стороны в сторону. Два грудных плавника по бокам тела
ОТ МОРСКИХ ЗВЕЗД ДО РЫБ, ВЛАСТЕЛИНОВ ВОДЫ
329
Фиг. ХХП.5. Остеостраки, или костнопанцирные
а — Cephalaspis — остракодерма из Древнего Красного песчаника, длина тела око-
ло 18 см; б — головной щит Sclerodus из раннедевонских отложений Европы, на-
туральная величина
за головой служили для балансировки, однако, судя по их строению, они не
являлись настоящими плавниками (см. фиг. ХХП.5).
По-видимому, Cephalaspis достаточно хорошо плавал, хотя его хвост, воз-
можно, скорее тянул его ко дну, чем помогал двигаться вперед. Эти живот-
ные, очевидно, большую часть своей жизни существовали в озерах, реках и
эстуариях, где они питались, засасывая в рот, как пылесос, пищевые части-
цы, и задерживая их, когда они проходили через жабры. Так же питались и
другие остеостраки.
Остеостраки впервые встречаются в силурийских отложениях, но к этому
времени их разнообразие уже довольно высоко, что предполагает длитель-
ную предысторию. В этой группе впервые развилась ткань, имеющая ла-
куны для костных клеток, которую мы называем настоящей костью. На-
ружный скелет остростраков был сложно устроен, напоминая скелет гетеро-
страков. Элементы скелета были трехслойными: пластинчатый слой в
основании, слой, пронизанный кровеносными сосудами в середине, и тре-
тий, наружный слой, образованный нижней дентиноподобной зоной, и верх-
ней, эмалеподобной твердой зоной.
Хотя эта группа в раннем довоне заселила многие бентосные трофические
ниши, все остеостраки вымерли к концу этого периода.
Галеаспидные. Из нижнедевонских отложений Китая происходит третья,
отличная от других группа остракодерм, которая, вероятно, развивалась в
изоляции. Хотя эта группа имеет признаки, характерные как для гетерост-
раков, так и для остеостраков, скорее всего галеаспидные все же были ближе
к последним.
Галеаспидные внешне похожи на остеостраков. У них имелся сильно око-
стеневший головной щиток с глазами наверху и два полулунных канала вну-
треннего уха, ориентированные вертикально. Однако, по-видимому, у них
было две ноздри. Нижняя часть панциря была сложена мозаикой из малень-
ких пластин, а по бокам был ряд жаберных отверстий. Рот располагался сни-
зу, что свидетельствует о питании этих форм придонным бентосом. Однако в
отличие от остеостраков у них не было дорсальных сенсорных полей и хвост
был гипоцеркальный, а не гетероцеркальный.
330
КАМЕННАЯ КНИГА
Фиг. XXII.6. Минога — современный представитель бесчелюстных (а) и ее ро-
товое отверстие (б)
Круглоротые
Остракодермы были широко распространены в позднем силуре, но к кон-
цу девона их число уменьшилось и в конце концов они вымерли. Некоторые
ученые считают, что они исчезли бесследно, другие полагают, что они оста-
вили потомков в виде миног и миксин, объединенных под названием “круг-
лоротые” или “циклостомы” (фиг. XXII. 6). Это змеевидные, бесчелюстные
создания, лишенные конечностей, живущие и в пресной и в соленой воде. Их
кожа гладкая, у них только одна ноздря, и некоторые ведут паразитический
образ жизни, прикрепляясь к другим рыбам. Они проникают через кожу и
мясо и достигают полости тела, где они питаются соками тела их все еще жи-
вых жертв.
Возможно, такие анаспидные, как Jamoytus, дали начало современным
круглоротым, судя по их схожести в строении жабр и рта.
Достоверные находки циклостом также известны, одна из них - из средне-
пенсильванских (верхний карбон) пород серии Мэйзон Крик (штат Илли-
нойс). Moyomyzon из этого местонахождения имеет удивительно сходное с
современной миногой строение хрящевых структур в области рта. Однако у
Moyomyzon не было специализированных слившихся жаберных опорных
структур, как у современных миног. Hardistiella — древнейшее из известных
круглоротых, обнаружено в нижнем карбоне штата Монтана.
ПЕРВЫЕ ЧЕЛЮСТНЫЕ РЫБЫ - ПЛАКОДЕРМЫ
Существует несколько теорий, объясняющих происхождение челюстей у
позвоночных, но промежуточные звенья остаются неизвестными. По-види-
мому, настоящие челюсти представляли собой модифицированные жабер-
ные дуги и были важнейшим приобретением, позволившим гнатостомам (че-
люстноротым) опробовать огромный спектр типов питания, недоступных
для агнат (бесчелюстных).
Наиболее ранние из этих челюстноротых появились в позднем силуре и
обитали в морских и пресных водах. Их ранние находки довольно разнообраз-
ны, что предполагает их значительно более длительную историю развития.
ОТ МОРСКИХ ЗВЕЗД ДО РЫБ, ВЛАСТЕЛИНОВ ВОДЫ
331
Четыре группы челюстноротых рыб известны из нижнего девона: плако-
дермы с окостеневшим наружным скелетом, хрящевые рыбы (акулы, скаты
и химеры), костные рыбы и акантоды (“шипастые акулы”). Они так сильно
отличались друг от друга, что мы даже не уверены, что они произошли от
единой предковой группы. Возможно, челюсти и парные плавники, которые
так важны для улучшения маневренности, возникали в ходе эволюции не од-
нажды. Каменная летопись, однако, еще может предъявить более определен-
ные свидетельства.
Плакодермы
Плакодермы принадлежали к числу первых рыб, у которых имелись и
парные плавники, и челюсти, хотя ни одна из жаберных дуг не участвовала
в поддержке челюстей. Позвоночник у представителей этой группы был при-
митивным, поскольку нотохорд сохранялся в течение всей жизни, а позвон-
ки представляли собой Y-образные шипы, располагавшиеся сверху и снизу
него.
Плакодермы были довольно разнообразны в раннем девоне, может быть
потому, что это были первые позвоночные с челюстями, что давало им пре-
имущество над другими животными.
В ископаемом состоянии акантоды известны из более древних отложе-
ний, чем плакодермы, и может быть они являются первым челюстнороты -
ми, но плакодермы первыми получили широкое распространение. Плако-
дермы обычно имели сильно окостеневший экзоскелет. Костные покрытия
головы и груди были несросшимися и обычно подвижными по отношению
друг к другу. Их кость по структуре была очень похожа на трехслойную, со-
держащую лакуны кость, уже наблюдавшуюся нами у остеостраков. Много
групп плакодерм известны из девонских отложений, они очень важны для
биостратиграфии девона. Хотя в начале своей эволюции плакодермы были
некрупными животными, позже они стали достигать больших размеров.
Titanichthys - активная хищная артродира, остатки которой найдены в
верхнедевонских кливлендских сланцах штата Огайо, достигала шести ме-
тров (19.5 футов). В течение девона возникло большое разнообразие плако-
дерм, а затем они вымерли - единственная крупная группа рыб, не дожив-
шая до наших дней. Наиболее разнообразными среди плакодерм были арт-
родиры.
Артродиры (сочлененношеие). Некоторые из первых артродир (напри-
мер, Sigaspis) были полностью одеты в костную броню, включая хвост. Груд-
ной щиток протягивался далеко назад, а брюшная область была прикрыта
большими, подвижными чешуями. У этого рода, как и у других, ранних ар-
тродир, щитки ростра и головы имели скользящее сочленение, а не по прин-
ципу ключа и замка, как у более поздних форм. Головной и грудной щитки
артродир состояли из ряда больших пластин, которые оставались обособлен-
ными в течение всей жизни рыбы. Поскольку трудно судить о том, как эти
кости связаны с костями черепа более эволюционно продвинутых рыб (как,
например, форель или “золотая рыбка”), черепные и грудные пластины по-
лучили ряд различающихся названий. Например, у ранних артродир груд-
ной щиток состоит из большой медиальной дорсальной пластины, гранича-
щей с передней дорзолатеральной и латеральной пластинами. Названия пла-
стин в целом отражают их положение на теле.
Мозговая капсула была сильно окостеневшая, но сформирована эндохон-
дральной костью. Она первая преобразовалась в хрящ, который впоследст-
332
КАМЕННАЯ КНИГА
Фиг. ХХП.7. Некоторые средне- и позднедевонские артродиры
а — Dunkleosteus, длина тела около 9 м, б - Coccosteus, длина тела до 60 см. Оба
вида обитали в Северном полушарии до побережья Северной Африки
вии при внедрении костных клеток окончательно окостеневал. Kujdanoivias-
pis из раннего девона является очень важной формой, давшей много инфор-
мации о мозге этих рыб. Челюсти ранних артродир были сформированы
верхней небноквадратной костью, тоже эндохондральной, как и мозговая
капсула. Две пары дермальных верхнечелюстных костей присоединились к
этой кости и к мозговой капсуле. Верхнечелюстные кости были покрыты ма-
ленькими зубчиками, которые, вероятно, выполняли роль зубов. Нижнюю
челюсть поддерживала другая эндохондральная кость, а именно меккелев
хрящ. В верхней челюсти, как и в нижней, имелись кожная (инфрагнаталь-
ная) кость, которая осуществляла захват пищи или ее измельчение.
О более специализированных артродирах (как, например, Coccosteus)
(фиг. XXII. 76) известно больше, чем о примитивных формах. Однако ясно,
что главными направлениями в эволюции группы были уменьшение веса и
площади головного и грудного щитков и освобождение щечного соединения
из-под крыши черепа. Большая часть эндохондральных костей мозговой
капсулы и челюстей превратилась в хрящ. Это означало, что все меньшая
часть головы рыбы была защищена костью, что способствовало усовершенст-
вованию плавания и в целом облегчало скелет. Даже шипы уменьшились
или вообще исчезли, что тоже способствовало уменьшений) веса скелета.
Многие из продвинутых артродир стали высоко специализированными хищ-
никами, как, например, гигант Dunkleosteus (=Dinichthys), что означает
“ужасная рыба” (фиг. XXII. 7а).
Антиархи. Антиархи приспособились к придонному образу жизни и менее
активному передвижению. На протяжении большей части своей истории они
оставались покрытыми костями наружного скелета и внешне напоминали
современную кот-рыбу из Южной Америки, хотя они и не связаны с этой бо-
лее эволюционно продвинутой рыбой. Самые древние антиархи — это юнано-
лепиды из верхнего силура и нижнего девона Китая. Большинство антиар-
хов было сплюснуто в спинно-брюшном направлении, и их глаза были на-
правлены кверху, что отражает их придонный образ жизни и способ
питания, подобный действию пылесоса. У большинства антиархов имелись
ОТ МОРСКИХ ЗВЕЗД ДО РЫБ, ВЛАСТЕЛИНОВ ВОДЫ
333
Фиг. ХХП.8. Два скульптированных антиарха, напоминающих внешним ви-
дом остракодерм
сросшиеся шипы грудных плавников, которые действовали скорее как опо-
ры, нежели были каким-то образом связаны с плаванием. Один из хорошо
изученных антиархов - это Pterichthys (крылатая рыба) из Древнего Красно-
го песчаника (верхний девон) Шотландии (фиг. ХХП.8).
Pterichthys был настолько полно покрыт наружным скелетом, что некото-
рое время ошибочно принимался за краба. Костные пластины полностью
прикрывали его голову и морду, и толстые перекрывающиеся чешуи покры-
вали тело до хвоста. Возможно, сходство парных грудных плавников с кры-
льями послужило причиной для его родового названия. Причиной массовой
гибели этих животных являлась их неспособность переползать по суше из од-
ного водоема в другой. Это привело к тому, что многие хорошо сохранивши-
еся экземпляры из Древнего Красного песчаника погибли в результате сезон-
ного пересыхания водоемов вслед за половодьем.
Bothriolepis, чье название означает “ячеистая чешуя” вследствие харак-
терной скульптуры панциря, это еще один хорошо известный антиарх
(фиг. ХХП.8). Самые древние находки происходят из среднего девона Китая
и Австралии, но к позднему девону этот род получил широкое распростране-
ние по всему земному шару. Одно из наиболее знаменитых местонахождений
остатков Bothriolepis находится на полуострове Гаспе в Квебеке. Там рыбаки,
которые ловят современную сельдь, выкапывают и окаменевших антиарх из
слоев серого сланца и мягкого песчаника, обнажающегося вдоль морского
берега. Частой находкой здесь является Bothriolepis около 25 см (=10 дюй-
мов) длиной (некоторые особи достигали 1 м в длину). Это рыба с тупой голо-
вой и широким телом, заключенным в панцирь, покрытый замысловатыми
ячеями. Хвост и часть морды этой рыбы были свободны от панциря, покры-
тые панцирем грудные плавники были довольно длинными, а вдоль краев
несли ряды коротких шипов.
По всему миру обитало множество ботриолепид, панцирь которых отра-
жал разнообразие придонных способов существования. Одна австралийская
форма (В. gippslandiensis) имела костный гребень на макушке своего корот-
кого рыла, который мог служить стабилизатором в быстрых горных пото-
ках, где обитала эта рыба. Другая повсеместно распространенная панцирная
рыба - Groenlandaspis тоже имела такой гребень.
Филлолепиды. Филлолепиды представляют другую группу плакодерм,
которая, как полагают, была связана как с антиархами, так и с артродирами.
Phyllolepis был сильно сплющенной в спинно-брюшном направлении при-
334
КАМЕННАЯ КНИГА
донной рыбой с характерным рисунком головного и грудного щитков, напо-
минающим отпечатки пальцев, иначе говоря, скульптура представляла со-
бой ряд изогнутых гребней. Средние пластины щитков были очень больши-
ми, а боковые значительно меньше.
Филлолепиды обитали в пресноводных водоемах позднего девона, и их
анатомия и наружный панцирь довольно хорошо изучены, особенно у осо-
бей, найденных в Австралии.
Другие плакодермы. Несколько других групп плакодерм известны из де-
вонских отложений по всему миру. Петалихтиды были морскими рыбами,
которые существовали с раннего по поздний девон. Они тоже были бентосны-
ми или придонными, на что указывает положение глаз в верхней части голо-
вы и уплощенные щитки. Хотя они были во многих отношениях похожи на
большинство древних артродир, у них были очень короткие грудные плавни-
ки. У петалихтид появилось сочленение рыла с головой, которое было сфор-
мировано несколько иначе, чем у других артродир.
Acanthothoraci были раннедевонскими морскими плакодермами, которые
во многих отношениях были похожи на артродир. Однако, подобно антиар-
хам, носовые отверстия у них были направлены вверх. Птиктодонты отлича-
лись очень небольшим панцирем, удлиненным телом и очень длинным и уз-
ким хвостом. Они встречаются главным образом в морских средне- и верхне-
девонских отложениях и имеют пару необычных верхних и нижних зубов.
На самом деле они были довольно необычными среди плакодерм, в действи-
тельности больше напоминающими современных скатов или химер вплоть
до того, что самцы имеют класперы, соединенные с грудными плавниками,
которые главным образом использовались для удержания самки во время
копуляции (брачного обряда). Только акулы и их близкие родственники
(включая скатов) имеют подобные приспособления. Без сомнения, образ
жизни этих плакодерм, если они и в самом деле принадлежали к этой группе,
очень отличается от других. Ренаниды имели несколько больших костных
пластин, которые покрывали голову и рыло, а остальная часть тела была по-
крыта разнообразными мелкими пластинами и чешуями. Челюсти были схо-
жи с челюстями акул и имели изолированные зубчики. Многие из этих рыб,
например Gemuendina, были сравнительно небольшими, всего лишь около
30 см (12 дюймов) длиной. Они были главным образом сплющенными, бен-
тосными формами и напоминали скатов. Они встречаются в нижнедевон-
ских морских отложениях, но обычно их находки довольно редки. Стеншо-
еллиды (названные в честь знаменитого шведского палеоихтиолога Эрика
Стеншио), также как и псевдопеталихтиды, это очень плохо изученные ры-
бы из нижнедевонских морских отложений Германии. Все они были неболь-
шими и сплющенными в дорзовентральном направлении. Они внешне напо-
минали ренанид, и их челюсти были устроены сходным образом. Но они так
плохо изучены, что потребуется много времени, чтобы адекватно интерпре-
тировать их систематическое положение и родственные связи.
Плакодермы представляли собой первые результаты эволюционных экс-
периментов над “челюстным” и “плавниковым” образами жизни, которые
оказались довольно удачными. Только к концу девона они были вытеснены
хрящевыми и костными рыбами, господствующими сегодня.
Акантоды
Акантоды, или “шипастые акулы”, как их часто называют, появились в
раннем силуре и дожили до перми. Они являются наиболее древней группой
ОТ МОРСКИХ ЗВЕЗД ДО РЫБ. ВЛАСТЕЛИНОВ ВОДЫ
335
Фиг. ХХП.9. Раннедевонская акантода — Climatius. Длина тела 7—13 см. Эта
рыба имела настоящие челюсти и несколько пар плавников, поддерживаемых
плавниковыми шипами
челюстноротых позвоночных. Их название происходит от того, что они
имели много парных и непарных плавников, поддерживаемых плавнико-
выми шипами - ихтиодорулитами. У них было множество разнообразных
черт, напоминающих костистых рыб и акул, а одно время их даже относили
к плакодермам. Они напоминали некоторых из ранних представителей
костистых рыб, за исключением их шипов и гетероцеркального плавника.
Их голова была покрыта мозаикой из маленьких не перекрывающихся пла-
стин, а тело — очень маленькими правильно расположенными чешуями.
Как у многих из высших рыб, кость, называемая геомандибулярной, помо-
гала в присоединении небноквадратной кости (или верхней челюсти) к моз-
говой капсуле.
Акантоды были небольшими рыбами, в большинстве не более чем не-
сколько см длиной, хотя один род достиг длины 4-5 футов (около 1.5 м). У
них имелся гетероцеркальный хвостовой плавник с верхней лопастью более
длинной, чем нижняя. Также они имели увеличенные чешуи, находящиеся
в области жабр.
Climatius — типичная прогрессивная акантода. Она была небольшой (дли-
на тела взрослой особи колебалась в пределах 7.5-13 см) с тупой головой, с
хвостовым плавником, напоминающим акулий, и с твердыми чешуями, пол-
ностью покрывающими тело. Парные плавники были многочисленными, но
многие только слегка были больше шипов. Также имелись непарные спин-
ной и анальный плавники (фиг. ХХП.9).
Climatius жил в пресной воде, возможно, в позднем силуре и, совершенно
точно, в раннем довоне. Однако многие поздние формы увеличились в разме-
рах, и у них уменьшалась площадь, закрытая чешуями, и размер последних.
Acantodes из ранней перми имел тело удлиненной формы, частично
покрытое чешуей, не имел зубов и сильно отличался от девонских родствен-
ных форм.
АКУЛЫ И СКАТЫ: ХРЯЩЕВЫЕ РЫБЫ
В прошлом акул часто считали довольно примитивными рыбами, по-
скольку их черепа и скелеты были построены не из кости, а из хряща. У не-
336
КАМЕННАЯ КНИГА
Зуб Orodus (около 8 мм),
ранний карбон Сев. Америки
Зуб Petalodus (2.5 см),
карбон-пермь
Сев. Америки, Европы
и Гренландии
поверхность шести
зубов Orodus
Жевательная
Фиг. XXII. 10. Древние хрящевые рыбы и их зубные элементы
которых акул хрящ начал обызвествляться, но настоящая кость у них не раз-
вивалась, и у всех этих рыб (кроме скатов) имелся ряд открытых раздельных
жаберных щелей.
Хотя палеонтологи до сих пор обсуждают детали родственных взаимоот-
ношений акантод, акул и костистых рыб, эти рыбы, по-видимому, довольно
сильно отличаются от плакодерм. Одно отличие заключается в том, что у со-
временных рыб, включая акул, челюстная мускулатура помещается сбоку
небно-челюстной кости, в то время как у большинства плакодерм она нахо-
дится внутри. Имеются и другие отличия, но, очевидно, филогенетическое
древо всех этих групп и их родственные взаимоотношения будут воссозданы
очень нескоро.
Первые хрящевые рыбы в позднем силуре представлены находками ма-
леньких зубчиков, которые, как известно, встречаются в коже современных
акул. Эти ископаемые происходят из морских пород, и эти акулы (включая
скатов и химер, относящихся к эласмобранхиям) и родственные им брадио-
донты с незначительными исключениями так и остались жить в море.
На протяжении истории существования группы акулы претерпели две
главные вспышки диверсификации. Одна пришлась на девон, а вторая, дав-
шая начало современной морской фауне - на мезозой. Акулы всегда остава-
лись морскими хищниками, хотя некоторые представители этой группы
начали питаться планктоном и моллюсками. Зубы акул возникли путем пре-
образования чешуй, расположенных вдоль челюстей, и имеют разные очер-
тания коронки в зависимости от характера пищи.
В течение палеозоя существовали шесть или семь главных групп акул, и
их родственные взаимоотношения еще недостаточно ясны. Разнообразие
акул отражает эволюционные эксперименты с целью приобретения опти-
мальной формы тела, одна из которых послужила основой для всех совре-
ОТ МОРСКИХ ЗВЕЗД ДО РЫБ, ВЛАСТЕЛИНОВ ВОДЫ
337
Фиг. XXII.11. Гибодонт, кладоселяхия и их зубы
менных форм. Палеозойские группы включали кладоселахий, симмориид,
ксенакантид, евгенодонтид, скватинактид, петалодонтид, ородонтид и кте-
накантид. Мы кратко рассмотрим лишь три из этих групп.
Кладоселлахии. Это одна из наиболее известных примитивных акул. Ос-
татки этих форм встречаются в позднедевонских гальках из сланцев Клив-
ленд в Огайо. Этот род включал узкотелых, торпедообразных рыб до 4 футов
(немного более 1 м) длиной с широким, гетероцеркальным хвостовым плав-
ником (фиг. XXII. 11). У этого рода грудные плавники имели узкое основа-
ние, в то время как многие другие примитивные акулы часто имели широкое
основание плавника. Все эти черты указывают на то, что эти рыбы были спо-
собны быстро плавать и хорошо маневрировать. Верхняя челюсть срасталась
или была тесно связана с мозговой капсулой. Такая подвеска челюсти назы-
вается амфистильной.
У кладоселляхий не было анального плавника, но имелось два спинных.
Их плавниковые шипы были сложены дентином. Чешуи на коже отсутство-
вали, за исключением границ плавников и участков кожи вокруг глаз.
Ксенокантиды (плевроканты). Эти очень необычные представители акул
жили в пресной воде. Их зубы всегда легко узнать, поскольку у них было две
больших боковых коронки и одна маленькая посередине, которая перекры-
вала соседний зуб (фиг. XXII. 10). На основании, главным образом, изолиро-
ванных остатков зубов время существования этой группы оценивается при-
мерно с позднего девона до позднего триаса, а их географическое распростра-
нение от Северной Америки до Австралии и Антарктиды.
Некоторыми чертами строения ксенаканты напоминают кладоселляхий
и, возможно, происходят от них, но во многих отношениях они совершенно
уникальны и загадочны. Их спинной плавник был длинным, начинаясь сра-
зу позади головы и протягиваясь на всю длину тела. На брюшном плавнике
имелся класпер. У грудного плавника имелась длинная центральная ось, от
которой расходились небольшие лучи, создавая структуру, подобную перу.
Такие плавники называются актиноптеригоидными. Один большой плавни-
ковый шип часто прикреплялся к задней стенке мозговой капсулы.
22 Каменная книга
338
КАМЕННАЯ КНИГА
Фиг. ХХП.12. Зуб палеогеновой акулы-гиганта Carcharadon (высота зуба более
15 см, длина акулы около 25 м)
Ктенакантиды. Эта группа палеозойских акул, вероятно, явилась предко-
вой для всех современных акул. В целом они не слишком отличались от кла-
доселляхий. Главные отличия заключались в структуре плавников и плав-
никовых шипов. В шипах имелся дополнительный слой твердого вещества,
называемого витродентином, который, вероятно, формировался во внешнем
слое кожи. Ктенаканты также имели анальный плавник и зачаточное
трехчленное строение основания плавника, характерное для всех современ-
ных акул.
Ктенаканты появились в девоне и жили до триаса включительно. Однако
пик их расцвета приходится на карбон. Одной из первых групп, произошед-
ших от ктенакантов, были гибодонты (фиг. XXII. 11), эволюционировавшие
в карбоне в направлении современных акул.
У гибодонтов развились различные типы зубной системы, что свидетельст-
вует о разнообразии трофических адаптаций. Возможно, что многие из этих
рыб, равно как и другие палеозойские акулы, занимали трофические ниши,
которые впоследствии были заняты мезозойскими костистыми рыбами.
Современные акулы (неоселлахии). Современные акулы, скаты и химеры
являются доминирующей группой хрящевых рыб со среднего мезозоя до на-
ших дней. Они впервые появились в юре и быстро распространились по всему
миру. В ископаемом состоянии эти формы представлены отпечатками кожи
ОТ МОРСКИХ ЗВЕЗД ДО РЫБ, ВЛАСТЕЛИНОВ ВОДЫ
339
и тела, но от большинства сохранились только зубы, которые варьируют от
мелких зубчиков с многочисленными окончаниями до широких треуголь-
ных лезвий. Каждая акула имела в своих челюстях десятки и сотни зубов,
одновременно и старые зубы, поврежденные при питании или утраченные в
битвах, быстро замещались новыми.
Можно указать следующие различия между современными и примитив-
ными палеозойскими акулами, приобретенные в ходе эволюции:
— центры позвонков обызвествлены,
— более подвижные плавники,
— шипы редуцировались,
- верхняя челюсть стала подвижной по отношению к мозговой капсуле.
Все эти изменения привели к появлению хорошо маневрирующей рыбы с
более разнообразными возможностями использования челюстей. Например,
челюсти многих современных акул могут выдвигаться вперед относительно
черепной коробки, что позволяет отрывать куски от жертвы большого разме-
ра (например, от китов), не заглатывая всей жертвы целиком. У палеозой-
ских акул челюсти работали в основном как ножницы и движения подобные
тем, которые могут совершать челюсти современных акул, были для них не
доступны.
Некоторые акулы достигали огромных размеров, как, например, род Саг-
charodon (фиг. XXII. 12), который встречается до сих пор (это белая акула,
называемая еще акулой-людоедом). Большая белая акула может достигать
30-40 футов (9-12 м), а ее зубы 3 дюймов (около 8 см) в длину. Некоторые ви-
ды третичного периода имели зубы 6-8 дюймов (15-20 см) длиной, помещав-
шиеся в толстых челюстях, которые были шести футов (1.8 м) или более того
в ширину. Эти животные, таким образом, были, конечно, более массивными,
чем белые акулы, их голова была большего размера, и они, должно быть, до-
стигали 50 футов (15 м) в длину. Это были хищники, опасные даже для круп-
ных морских животных.
Скаты (известные также как батоиды) представляли собой акул, которые
превратились в уплощенных придонных рыб посредством значительного
увеличения грудных плавников, которые позволяли им практически “ле-
тать” в толще воды. Как у большинства придонных форм, их глаза перемес-
тились наверх. Большая часть плавников редуцировалась, а хвостовой плав-
ник превратился в хлыст.
Челюсти у этих рыб значительно отдалились от мозговой капсулы. Зубы у
скатов и химер обычно уплощенные, перетирающие. Поэтому эти рыбы мо-
гут питаться животными, которые заключены в твердую раковину (моллюс-
ки, ракообразные).
Самым древним батоидом является Squatina, чьи родственники, конечно,
существуют и поныне.
Цельноголовые, или химеры. Химеры являются заключительной и, воз-
можно, наиболее запоминающейся группой хрящевых рыб, которую мы рас-
смотрим.
Химер трудно связать с какой-либо другой группой акул. Они единственная
среди акул группа, у которой верхняя челюсть приросла к мозговой капсуле и
жабры прикрыты единой крышкой (как у костистых рыб). Зубы у них пред-
ставлены двумя рядами пластин в верхней челюсти и одним рядом в нижней,
возможно, функционирующих так же, как у скатов. Передние позвонки срас-
таются в единую структуру, называемую синаркуалярной, и к ней
прикрепляется дорсальный шип. Хвост превращен в тонкий хлыст. Как у акул,
у химер имеются класперы не только на грудном плавнике, но и на голове.
22*
340
КАМЕННАЯ КНИГА
Фиг. XXII. 13. Эоценовый обрыв, сложенный отложениями формации Грин Ри-
вер, штат Вайоминг. Отложения накапливались в озерной обстановке, за ис-
ключением верхней части разреза, сформированной речными отложениями.
Разрез содержит большое число ископаемых остатков рыб
Химеры, возможно, существовали еще в карбоне, если представители ро-
да Echinochimaera из нижнего миссисиппия в действительности принадле-
жат к этой группе. Кроме того, иниоптеригии из Пенсильвания Северной
Америки также могут относиться к этой группе рыб. Как бы то ни было, про-
исхождение этих форм и достоверная информация о происхождении химер
по-прежнему принадлежит к числу нерешенных проблем.
КОСТНЫЕ РЫБЫ
У перекрестка под названием Фоссил, недалеко от Кеммерера (штат Вай-
оминг), обнажаются обрывы мергелистых известняков и глинистых слан-
цев. Эти отложения эоценового возраста, которые накапливались в одном
из многочисленных озер, простиравшегося на территории юго-западного
Вайоминга и соседних областей Юты и Колорадо. Эти отложения, которые
являются частью формации Грин Ривер, знамениты встречающимися там в
большом количестве прекрасно сохранившимися ископаемыми рыбами
(фиг. XXII. 13).
Как мы имели возможность убедиться, кожа многих плакодерм и акул бы-
ла покрыта костными пластинами, однако их внутренний скелет был постро-
ОТ МОРСКИХ ЗВЕЗД ДО РЫБ, ВЛАСТЕЛИНОВ ВОДЫ
341
Фиг. XXII. 14. Лучеперые рыбы мезо-кайнозоя
а — костистая рыба Priscacara из эоценовых отложений формации Грин Ривер;
б— рентгеновский снимок ископаемой рыбы из красноцветных триасовых отло-
жений окрестностей Принстона, штат Нью-Джерси
ен из хряща. Группа рыб, к которым принадлежат находки в формации Грин
Ривер, имела такое же строение кожных покровов, а вот внутренний скелет
у этих рыб изменился. Их сложный внутренний скелет был построен из на-
стоящих костей, и все больше кожных костей добавлялось к черепу, челюс-
тям и плечевому поясу. Их хвостовые плавники изменились, и у более про-
двинутых форм хвост приобрел билатерально симметричную, или гомоцер-
кальную, форму (фиг. XXII. 14).
Известны две основные группы костных рыб: лучеперые, или актинопте-
ригии, и кистеперые, или саркоптеригии. Обе эти группы появились в ран-
нем девоне. Возможно, некоторые изолированные чешуи из позднего силура
также принадлежат к этим группам.
Актиноптеригии
Лучеперые рыбы названы таким образом потому, что они имеют парные и
непарные плавники, пронизанные тонкими костными лучами, которые от-
ходят прямо от тела, а не подобны оборкам по обеим сторонам костных или
мускульных лопастей.
Актиноптеригии из всех костных рыб наиболее близки к акантодам, но,
конечно, сильно отличаются по ряду черт, как, например, закономерности
смены зубов.
Примитивные лучеперые (хондростеи). Хондростеи претерпели наиболь-
ший расцвет в палеозое и раннем мезозое, но несколько видов дожили до на-
ших дней. Хондростеи, возможно, впервые появились в позднем силуре. Эти
и в действительности многие палеозойские формы относились к группе пале-
онисцид. Большинство были небольшими рыбами, хотя такие девонские
формы, как Cheirolepis, могли достигать двух футов (несколько больше 0.5 м)
в длину (фиг. XXII. 15).
Cheirolepis обладал тонким телом, тупорылой мордой и имел V-образный
гетероцеркальный хвостовой плавник, в котором нотохорд протягивался
прямо в верхнюю лопасть. У этой рыбы имелись непарные дорсальный и ана-
льный плавники и парные боковые плавники, все с широким основанием.
Череп состоял из нескольких гибких костных пластин, в которых поме-
342
КАМЕННАЯ КНИГА
Фиг. XXII. 15. Лучеперые рыбы палеозоя
Cheirolepis — примитивная девонская лучеперая рыба. Длина тела около 50 см
(слева). Высокотелый палеониск Amphicentrum из карбона Англии. Обитатель
спокойных вод, близкий по образу жизни к рифовым рыбам. Длина тела 15 см
(справа)
щались отверстия маленьких глаз. Более типичные палеониски имели боль-
шие глаза.
Тело палеонисков было покрыто рядами маленьких, ромбических чешуй,
которые были довольно толстыми. Они состояли из трех слоев, включая на-
ружный слой ганоина, похожего на эмаль. Эти чешуи, возможно, происхо-
дят от чешуй акантод, которые очень похожи на них (фиг. XXII. 15).
Челюсти этих рыб были длинными, с ножницеобразным прикусом, и со-
единялись позади мозговой капсулы. Начиная с момента появления, пер-
вые лучеперые рыбы развивались в нескольких разных направлениях. У не-
которых развивались большие глаза и более широкие туловища, а парные
плавники редуцировались до тех пор, пока не стали совершенно бесполез-
ными. У других тело расширялось в дорзо-вентральном направлении и ста-
новилось более тонким, чем у рыбы-солнце, которую они напоминали по
размерам и форме тела. У некоторых редуцировались кости черепа и позво-
ночного столба, так же как и число чешуй, вплоть до того, что оставалось
лишь несколько рядов больших костных пластин. Одним из результатов
развития в таком направлении являются практически лишенные чешуи
осетры (Acipenser), группа связанных с дном падалеядов, ныне живущих в
заливах, озерах и больших реках. Веслонос (Polydon), известный из мело-
вых отложений, а также живущий ныне в Миссисипи и в некоторых реках
Китая, утратил еще больше чешуй. У них осталось только несколько ма-
леньких чешуй у основания хвостового плавника. У обеих названных рыб
остались весьма примитивные плавники и гетероцеркальный хвост. Однако
некоторые рыбы, чьи потомки живут и сейчас, такие как африканский мно-
гопер (Polypterus), сохранили толстую броню блестящих чешуй. У этой ры-
бы имеются своеобразные парусообразные спиняые плавники и дифицер-
кальный хвост.
Это развитие примитивных “лучевых” плавников происходило на протя-
жении большей части интервала от девона до перми и сопровождалось при-
обретением разнообразных форм туловища.
ОТ МОРСКИХ ЗВЕЗД ДО РЫБ. ВЛАСТЕЛИНОВ ВОДЫ
343
Фиг. XXII. 16. Пять типов строения хвостового плавника
Lepisosteus — современная панцирная щука, возможно, также принадле-
жит к этим примитивным лучеперым, поскольку обладает сильно развиты-
ми чешуями и гетероцеркальным хвостовым плавником. Она встречается в
меловых отложениях практически по всему миру, и ее многочисленные че-
шуи ромбоидальной формы называются некоторыми коллекционерами “ис-
копаемыми финиками”. Их присутствие привело к открытию одного из са-
мых значительных местонахождений микрофоссилий ранних млекопитаю-
щих — Буг Крик Антиллз, около Форта Пек.
Неоптеригии, современные лучеперые
Помимо групп лучеперых, о которых шла речь выше (Palaeonisci, Chon-
drostei, Holostei и Polypteri), остальные объединяются в большую группу не-
оптеригий или Teleostei. Эта группа впервые появилась в позднем палеозое и
стала доминировать как в соленых, так и пресных водах в мезозое и кайно-
зое. Главные изменения при переходе от древних к современным лучеперым
происходили в костях и мускулатуре, связанных с питанием и движением. У
современных лучеперых максилла отделилась от остальных щечных костей,
добавились новые кости и увеличилась мускулатура, контролирующая дей-
ствие челюстей. Эти изменения позволили сильно увеличить диапазон от-
крывания рта и значительно усилить укус. Во многих случаях передняя
часть челюстей, в сущности, могла выдвигаться вперед. Многие формы вооб-
ще могли засасывать пищу в рот.
Продвинутые лучеперые рыбы также утратили многослойные чешуи. Хо-
тя последние и служили хорошей защитой, они были очень тяжелыми. С их
утратой рыбы приобрели способность к быстрому передвижению. Вновь воз-
никшие легкие чешуи обычно округлые, а не ромбические называются цик-
лоидные. Наряду с этими изменениями проявилась тенденция к приобрете-
нию цилиндрических катушковидных тел позвонков. Плавники стали более
подвижными, брюшные и грудные плавники часто занимают необычное по-
ложение, например, грудные плавники прямо над брюшными, что обеспечи-
вает хорошую маневренность. Хвостовой плавник демонстрирует тенденцию
к приобретению симметричных очертаний, возможно, как результат облег-
чения веса рыбы благодаря утрате чешуй и действию плавательного пузыря,
обеспечивающего хорошую плавучесть.
344
КАМЕННАЯ КНИГА
Фиг. XXII. 17. Примитивные неоптеригии
а — Semionotus — пресноводный костный ганоид триасового времени из Аризоны,
длина тела около 20 см; б — Dapedium — высокотелый костный ганоид из юрских
отложений, длина тела 36 см
Наиболее примитивных из современных лучеперых часто называют хря-
щевыми ганоидами (Chondrostei). Их взаимоотношения с другими таксона-
ми неясны, так что, возможно, наиболее правильным будет называть их про-
сто современными примитивными лучеперыми до тех пор, пока не появится
новая информация.
Semionotus был одной из таких лучеперых рыб, существовавших в триасе
(фиг. XXII.17а). Тело этой рыбы имело удлиненную форму и напоминало те-
ло форели. Однако другие семионитиды, такие как Dapedium были более вы-
сокотелыми, напоминая современных скалярий (фиг. XXII.176). У Dapedi-
um имелись тупые колышковидные зубы, с помощью которых эти рыбы дро-
били раковины моллюсков и других беспозвоночных.
Ильные рыбы (Amia) — это другие примитивные современные лучеперые.
Близкие родственники этих рыб впервые появились в пресноводных юрских
бассейнах. Amia появившиеся в позднему мелу были и поныне остаются
опасными хищниками. Они достигают более 30 дюймов (76 см) в длину и
имеют длинный спинной плавник и тупой хвостовой (не вполне симметрич-
ный), в котором есть несколько перевернутых позвонков. Эти рыбы в основ-
ном питаются мелкой рыбешкой, но также употребляют в пищу и разнооб-
разных беспозвоночных. Самцы устраивают на дне гнезда и охраняют икру,
отложенную самкой. Эти рыбы способны использовать воздушный пузырь
как легкое.
Разнообразные другие рыбы, такие как высокотелые и обладавшие мас-
сивными зубами пикнодонты (существовавшие с триаса до эоцена), мезозой-
ские макросемииды, а также некоторые другие рыбы занимали промежуточ-
ное положение между хрящевыми и наиболее продвинутыми костистыми
рыбами — телеостеями.
Teleostei - наиболее современные лучеперые. Позднетриасовые фолидо-
фориды являются древнейшими телеостеями, той группой актиноптеригий,
которая наиболее многочисленна и разнообразна сейчас, доминируя в вод-
ных биоценозах начиная с позднего мезозоя. Это золотые рыбки, морские
коньки, скалярии, форели и многие другие. С большой уверенностью можно
ОТ МОРСКИХ ЗВЕЗД ДО РЫБ. ВЛАСТЕЛИНОВ ВОДЫ
345
сказать, что телеостеи самые костистые из известных до настоящего времени
рыб. В отличие от хрящевых и мясистолопастных (Sarcopterygii) они имеют
полностью окостеневающий внутренний скелет, который дополнен много-
численными мелкими косточками, не связанными с позвоночным столбом.
Хвостовой плавник этих рыб гомоцеркальный (фиг. XXII. 16) и не продолжа-
ется в хвостовую лопасть, давая у окончания позвоночного столба начало
плавниковым лучам.
Чешуи не имеют даже следа ганоина или другого эмалеподобного слоя и
налегают друг на друга подобно черепице. У некоторых видов костистых рыб
чешуи могут быть сильно редуцированы или отсутствовать вообще.
Наиболее примитивными из всех современных телеостей, возможно, яв-
ляются остеоглоссоморфы, пресноводные рыбы, распространенные главным
образом в Южном полушарии. Только немногие из них встречаются в Юго-
Восточной Азии и Северной Америке. Впервые они появились на территории
Китая в юрское время.
Другая важная группа телеосетей - элопоморфы — также появились в по-
здней юре. Тарпаны и угри являются представителями этой, главным обра-
зом, морской группы, имеющей сложные начальные стадии развития, пред-
ставленные полупрозрачной личинкой - лептоцефалом.
“Бульдожий тарпан” Protheus, известный из меловых отложений Европы
и Северной Америки, принадлежит к этой группе. Американская форма об-
ладала тупой головой и челюстями, несущими ряд впечатляющих зубов, до-
стигая в длину 12 футов (3.7 м) и весил 500-700 фунтов (225-275 кг). Пита-
лись эти рыбы более мелкими собратьями и молодыми морскими рептилия-
ми, населявшими мелкие эпиконтинентальные моря Канзаса и Южной
Англии. Один огромный Protheus даже заглотил шестифунтового (1.8 м) со-
седа, но умер так и не переварив его. Это ясно из изучения скелетных остат-
ков, известных в ископаемом состоянии.
Другая группа телеостей включает клупеоморф, которые появились по
крайней мере в раннем мелу. Сельдь (Clupea) — типичный представитель этой
группы, так же как сардины и анчоусы.
Все другие телеостеи принадлежат к эутелеостеям, которые известны с по-
зднего мела. У многих имелась ячеистая кость и отмечается специализация
костей хвостового плавника. Обычно эутелеостей подразделяют на: лососео-
бразных (Salmoniformes), включающих лосося и форель; костнопузырных
(Ostriophisi), включающих карпа, карася, сома, пиранью, электрических уг-
рей и пр.; неотелеостей (Neoteleostei), которые представлены наиболее стран-
ными по форме тела костистыми, такими как удильщики, летучие рыбы,
скалярии и т.п.
Наравне с находками чешуй и даже целых скелетов большинства групп
телеостей в ископаемом состоянии можно встретить “ушные камешки” (или
отолиты) очень разнообразной формы. Полость внутреннего уха (саккулюс)
практически заполнена кристаллическим кальцитом. В результате форма
этих отолитов отражает форму саккулюса и специфична для каждого вида
рыб. Много отолитов можно встретить в тонком буровом керне, и поэтому
они чрезвычайно важны для определения возраста пород в дополнение к оп-
ределению рыбы, которой они принадлежали.
Другой вариант “слуха” реализован Ostriophisi. На передних позвонках
этих рыб имеются маленькие косточки, называемые косточками Вебера
Они соединены друг с другом и образуют цепочку, соединяя воздушный пу-
зырь с ушной областью; действуют они очень похоже на цепочку ушных ко-
стей млекопитающих, помогая усилить и передать звуковые колебания воз-
346
КАМЕННАЯ КНИГА
Фиг. XXII.18. Саркоптеригия
а — Holoptychus — саркоптеригия, обитавшая в среднем девоне-раннем карбоне
практически повсеместно, включая Антарктиду, длина тела рыбы 76 см; б— по-
перечный разрез зуба этой рыбы. Отчетливо видно его так называемое “лаби-
ринтодонтное” внутреннее строение
душного пузыря в жидкость внутреннего уха, которая действует на нервные
окончания, связанные с мозгом.
Конечно, костистых рыб следует считать наиболее удачливой группой
водных позвоночных, если учитывать их разнообразие и число особей.
МЯСИСТОЛОПАСТНЫЕ, ПРЕДТЕЧИ ОБИТАТЕЛЕЙ СУШИ
В противоположность актиноптеригиям, саркоптеригии, или мясистоло-
пастные, дожили до сегодняшнего дня в очень небольшом количестве. В эту
группу входят три вида современных двоякодышащих, встречающихся со-
ответственно в Южной Америке, Африке и Австралии, а также целакант ла-
тимерия (Latimeria).
При обращении к каменной летописи выделяются три основные группы
мясистолопастных рыб. Это двоякодышащие, целаканты, имеющие совре-
менных представителей и рипидистии, жившие только в палеозое. Все эти
группы появились в девоне. Двоякодышащие рыбы формируют отдельную
группу (Dipnoi), тогда как две другие группы иногда объединяют и называют
кистеперыми (Crossopterygii).
Кистеперые рыбы (фиг. XXII. 18) имеют плавники с мясистыми основани-
ями, включающие серию костей, отличную от костей в плавниках других
рыб. Именно от рыб этой группы произошли первые четвероногие - амфи-
бии. Так что если их самих и не так уж много сейчас, то их “отпрыски” — чет-
вероногие - преуспели в разнообразии жизненных стилей.
Двоякодышащие рыбы, или дипнои
Девонский род Dipterus (фиг. XXII. 19) был типичной двоякодышащей
рыбой, которая внешне несколько походила на кистеперую рыбу, но только
до тех пор, пока не открывала своего рта. Тогда и обнаруживалось, что у
многих представителей этой группы рыб существовала специализирован-
ная зубная система, с широкими веерообразными ребристыми зубами, ко-
торые свидетельствовали о том, что их хозяин мог дробить твердые объек-
ОТ МОРСКИХ ЗВЕЗД ДО РЫБ, ВЛАСТЕЛИНОВ ВОДЫ
347
Фиг. XXII. 19. Dipterus — девонская примитивная двоякодышащая рыба. Ее ос-
татки чрезвычайно широко распространены по миру
а — внешний вид; б — верхняя зубная пластинка; в — небная поверхность черепа
Фиг. XXII.20. Три вида специализированных двоякодышащих рыб
Два вида - вымершие, однако Ceratodus из раннего триаса очень близок к совре-
менной форме, живущей в Восточной Австралии
ты, такие как раковины беспозвоночных. У современных двоякодышащих
рыб эти зубы и мускулатура челюсти обеспечивает сильный прикус, кото-
рый и позволяет им разнообразно питаться, включая и питание твердыми
объектами. Череп этих рыб состоял из мозаики, сложенной маленькими
костями. Расположение этих костей сильно отличается от такового в любой
другой группе рыб.
Существенной чертой является то, что зубы и кости их черепа похожи на та-
ковые современных двоякодышащих рыб. Поскольку же мы много знаем об
образе жизни современных форм, мы можем предполагать, что ископаемые
рыбы были в этом похожи на современных. Хотя современные представители
имеют жабры, они могут дышать и атмосферным воздухом, получая кислород
из плавательного пузыря, преобразованного в мешковидное легкое.
348
КАМЕННАЯ КНИГА
Фиг. XXII.21. Eusthenopteron — позднедевонская кистеперая рыба (рипидис-
тия). Длина тела 1-1.5 м
Фиг. XXII.22. Osteolepis — кистеперая рыба (рипидистия) из средне-верхнеде-
вонского Древнего Красного песчаника Европы. Ее остатки часто встречаются
в отложениях этого возраста по всему миру. Чешуя этих рыб называется космо-
идная, так как содержит слой космина. Парные плавники содержали набор ко-
стей, близких к костям конечностей современных четвероногих
Но какую пользу легкие могут принести рыбе? В настоящее время легкое
отлично служит такой рыбе во время засухи, когда водоемы застаиваются
или полностью пересыхают. Африканские протоптерусы (Protopterus)
(фиг. XXII.20) и южно-американские чешуйчатники (Lepidosiren) в период
засухи выкапывают норы, где они впадают в спячку до тех пор, пока не изме-
нится погода. Представители пермского рода Gnathoriza, должно быть, тоже
вели себя так, поскольку их остатки найдены прямо в норах, которые они
использовали для пережидания периодов засухи.
Хотя самые первые двоякодышащие не имели веерообразных зубов и ха-
рактеризовались сильным окостенением скелета, в середине девона большая
часть их скелета заместилась хрящем и развилось особенное озубление. Че-
шуи тоже были сильно редуцированы. На протяжении своей истории радиа-
ция двоякодышащих происходила несколько раз, сначала в морской и прес-
ной воде, но в позднем палеозое и мезозое, несомненно, в пресной воде. Сего-
дня, однако, их редкие представители живут только в пресной воде на
южных континентах.
ОТ МОРСКИХ ЗВЕЗД ДО РЫБ, ВЛАСТЕЛИНОВ ВОДЫ
349
Фиг. XXII.23. Целаканты
а — Dipterus — пресноводный целакант из позднего триаса Нью-Джерси, имел в
длину всего лишь 10-13 см; б — Holophagus — морской целакант, обитавший в
позднетриасовое и юрское время на территории Европы и других регионов ми-
ра. Длина тела 60 см
Кроссоптеригии, или кистеперые
Рипидистии, возможно, самые важные для нас кистеперые, поскольку
когда-то в девоне они дали начало амфибиям и, следовательно, позже и опо-
средованно - нам. Рипидистии появились в начале девона и существовали до
ранней перми. В течение этого времени они были самыми главными пресно-
водными хищниками из всех хищных рыб.
Многие черты ранних рипидистий похожи на примитивных актиноптери-
гий. Кроме того, черепа некоторых рипидистий, как, например, Eus-
thenopteron, были очень похожи на черепа амфибий. Зубы рипидистий ха-
рактеризовались наличием складчатого дентина, который впоследствии дал
название группе древних амфибий - лабиринтодонты, многие из которых
имели такое же строение зубов (фиг. XXII. 18А). Так что мы имеем некоторое
представление о том, откуда произошла эта группа и кто был ее потомком.
Eusthenopteron и другие остиолепиформные рипидистии также имели ряд
черт, которые свидетельствуют о том, что они были предками амфибий
(фиг. XXII.21). К их числу относится слезный канал, возможно, связанный
с обитанием в приповерхностном слое воды, когда глазные и носовые отвер-
стия находились вне воды достаточно долго и нуждались в увлажнении. У
них развились внутренние ноздри (хоаны), которые предположительно поз-
воляли им дышать открытым ртом. Их мозговая капсула была разделена на
два отчетливых сегмента, а остатки подобного разделения обнаруживаются
на черепах первых амфибий, например ихтиостеги из Гренландии. Центры
позвонков как у Eusthenopteron, так и у Osteolepis довольно похожи на тако-
вые примитивных лабиринтодонтных амфибий тем, что у них имеются две
пары сегментов, дорсальный плевроцентр и вентральный интерцентр.
Как и в остальной своей анатомии, рипидистии сильно отличаются от дру-
гих рыб своими конечностями (фиг. XXII.22). Именно такая конечность да-
ла начало конечности четвероногих, способной поддерживать вес животного
вне воды. В грудном плавнике имелась большая проксимальная кость, впол-
не сравнимая с плечевой костью, и, кроме того, аналоги лучевой и локтевой
кости. Имелось запястье, но далее в дистальном направлении не было ника-
ких существенных опорных структур, так что пястье и кости фаланги паль-
350
КАМЕННАЯ КНИГА
цев, вероятно, впервые появились только у тетрапод. Очень похожим обра-
зом была устроена задняя конечность.
Выстоявшие целаканты
Рипидистии дали начало не только тетраподам, но также и другой группе -
целакантам, среди которых есть одна ныне живущая, хотя и редкая рыба -
латимерия (Latimeria).
Первые целаканты известны из отложений среднего девона
(фиг. XXII.23). Их связь с рипидистиями прежде всего основывается на стро-
ении мозговой капсулы, которая разделена на две части. Их тела были корот-
кими и у них обычно были высокие и несколько округлые головы. Лопасти
парных плавников были относительно короткими, а дифицеркальный хвост
имел трехраздельный плавник.
В позднем девоне существовали и морские и пресноводные формы, но
большинство мезозойских целакантов морские. До 1938 года считалось, что
эта группа вымерла в позднем мезозое. Но в 1938 году из глубин Индийского
океана у побережья Южной Африки был извлечен современный целакант,
названный латимерией. Этот экземпляр и еще несколько, добытых недавно
вблизи Мадагаскара, дали великолепный материал для представления о том,
как выглядели мягкие ткани представителей этой группы, известной до от-
крытия латимерии только в ископаемом состоянии. Это и в самом деле “жи-
вое ископаемое”, совсем как гинкго, впервые найденное живым в Китае.
Глава XXIII
ЗАЗЕЛЕНЕВШАЯ ЗЕМЛЯ
• История монер, низших животных и растений, которую мы рассмо-
трели в предыдущих главах, проходила в течение трех с лишним
миллиардов лет. В основном все эти организмы были морскими.
Пресноводные моллюски и наземные беспозвоночные появились значитель-
но позднее, через многие и многие десятки и даже сотни миллионов лет после
того, как их родственники заселили моря и океаны.
Нам мало что известно о возможных обитателях древней суши. Тем не ме-
нее, кое какие окаменевшие остатки донесли до нас скудные сведения о на-
земных обитателях того времени. Самые древние из таких окаменелостей от-
носятся к силурийским отложениям. Насекомые, сейчас значительно пре-
восходящие своим числом все остальные наземные организмы, появились не
ранее девонского времени.
К сожалению, сейчас мы ничего не можем сказать об обитателях докемб-
рийской и раннепалеозойской суши. Первые достоверные организмы появ-
ляются с силурийского периода.
ПРОБЛЕМЫ ПОЯВЛЕНИЯ НАЗЕМНЫХ ОРГАНИЗМОВ
Растения существовали миллиарды лет до того, как они вышли на сушу.
Много новых проблем вставало перед растениями и животными, живущими
в суровых условиях суши. Наземные растения решили некоторые из этих
проблем развитием специфического вещества кутин, который представляет
собой полимерную пленку, препятствующую потере воды. Они также обла-
дали устьицами - развитыми специализированными системами клеток эпи-
дермиса, располагавшимися на листьях. Устьица позволяли проникать
внутрь двуокиси углерода (СО2), необходимой для фотосинтеза, но не давали
воде выходить наружу. Система корней у наземных растений не только ук-
репляла их в грунте, но также поглощала воду и нитриды, необходимые для
роста и жизнеобеспечения растения. Высшие растения также образовали со-
судистые ткани для транспортировки воды и нитридов от корней к побегам и
листьям. Кроме этого, у них появились специальные механические ткани,
которые помогли им достичь крупных размеров. И наконец, наземные расте-
ния образовали органы размножения, споры и семена, защищенные от
высыхания.
© 1997. С.В. Наугольных, перевод на русский язык и изменения.
352
КАМЕННАЯ КНИГА
Фиг. XXIII. 1. Проптеридофиты.
а — Psilophyton, б - Gosslingia (по Schweitzer, 1984). Длина масштабной линей-
ки — 10 см
ПРЕДКИ НАЗЕМНЫХ РАСТЕНИЙ
Где и когда появились наземные растения, которые столь многочисленны
сегодня? Откуда они пришли? На этот вопрос легче ответить, глядя на ныне
живущие формы, а не ископаемые, поскольку остатки наземных растений из
кембрия, ордовика и раннего силура нам не известны. Наземные ландшафты
тогда были еще “безжизненными”, точнее они были освоены только бактери-
ями и, возможно, мхами и низшими грибами.
Многие данные указывают на то, что группой, которая дала начало назем-
ным растениям, были хлорофиты, т.е. зеленые водоросли. Как и наземные со-
судистые растения, хлорофиты имеют как хлорофилл-а, так и хлорофилл-б,
а также настоящий крахмал и целлюлозу в своих клеточных стенках. Эта
группа водорослей также охарактеризована тем, что именуется “чередовани-
ем поколений”. Это означает, что они проходят фазы полового и бесполого
ЗАЗЕЛЕНЕВШАЯ ЗЕМЛЯ
353
размножения, почти как современные папоротники. Все эти признаки, вмес-
те взятые, обособляют зеленые водоросли (хлорофиты) от всех остальных низ-
ших растений и делают их наиболее сходными с сосудистыми растениями.
ДРЕВНЕЙШИЕ НАЗЕМНЫЕ РАСТЕНИЯ
Толстостенные споры, по форме сходные со спорами современных наземных
растений, были обнаружены в кембрийских породах в Индии и России. Кроме
спор, известны и побеги, такие как алданофитон, предположительно принад-
лежавшие древнейшим наземным растениям. Алданофитон (Aldanophyton)
представлен множеством углефицированных отпечатков из Сибири с ветвями
до 13 см длиной. Он похож на небольшое плауновидное со спирально распола-
гающимися листьями. Побеги алданофитона достигали 8 мм ширины.
Однако до тех пор, пока не будут сделаны более многочисленные и лучше
сохранившиеся находки в кембрийских и ордовикских отложениях, палео-
ботаники будут сомневаться в том, рассматривать ли эти окаменелости как
остатки настоящих наземных растений или нет.
Когда наземные растения впервые появляются в геологической летописи,
они представляют собой смесь простых и более сложно организованных
форм, как бы намекающих на предыдущую историю, нам до сих пор не изве-
стную (фиг. XXIII. 1). Они не были сосудистыми растениями (другими слова-
ми, у них отсутствовали сосуды, переносившие нитриды и воду). Сосудистые
растения появляются только с конца силурийского периода.
Барагванатия (Baragwanathia) из Австралии - одно из этих ранних “сосуди-
стых” растений, предположительно позднесилурийского или раннедевонского
возраста. Барагванатии достигали довольно больших размеров, с ветвями до
26-28 см в длину и около 1-6 см в диаметре. Побеги были стройными и несли
большое количество тонких гибких листьев. В некоторых местах вдоль побега
или ветвей располагались спорангии, которые содержали споры. С их помо-
щью барагванатии размножались также, как папоротники делают это и по сей
день. Интересно, что по общему строению, как внутреннему, так и внешнему,
барагванатии напоминали современные ликоподиевые, или плауновидные.
В отличие от относительно сложно устроенной барагванатии, куксония
(Cooksonia) была очень просто организованным, небольшим растением, всего
несколько сантиметров в высоту. Ее побеги были тонкими и голыми, лишен-
ными листьев. Спорангии располагались на концах веточек. Куксония
встречается в силурийских отложениях Ирландии, но также попадается и в
более молодых породах, например в раннем девоне Уэльса и Австралии. Во
многих случаях это растение рассматривается палеоботаниками как наибо-
лее примитивное сосудистое растение. Об этом говорит, например, отсутст-
вие настоящих листьев и корней.
ДЕВОНСКИЕ БОЛОТА И ЛЕСА
Туристы, пользующиеся входом Серебряных Ворот в Йеллоустонский на-
циональный парк, проходят мимо интересного местонахождения древней-
шей наземной флоры. Для того чтобы его достигнуть, они поднимаются не-
сколько тысяч футов на автомобиле и двигаются, пересекая древнее, сейчас
обнаженное морское дно, наблюдающееся по берегам ледниковых озер. Око-
ло одного из них поднимается Медвежий Зуб, утес, сложенный морскими от-
23 Каменная книга
354
КАМЕННАЯ КНИГА
ложениями от кембрийского до девонского возраста. Недалеко от вершины
находится русло раннедевонской реки, прорезанной в более древних слоях в
те времена, когда северо-западный Вайоминг был низменностью, пересечен-
ной меандрирующими потоками.
Фотографы останавливаются “поснимать” озеро, а охотники за ископае-
мыми тем временем паркуют свои машины и карабкаются на южную сторону
скалы. Недалеко от ее основания они находят зеленовато-коричневые слан-
цы и песчаники, некоторые из которых пронизаны норками кембрийских
аннелид (кольчатых червей). Затем идут доломиты и известняки, содержа-
щие строматолиты, перекрывающиеся красными, темно-желтыми и серыми
речными отложениями, мощность которых достигает 45 м. Отложившиеся
недалеко от устья медленной реки, эти осадки содержат окаменелых эврип-
терид, остракодерм (агнат), рыб и множество наземных растений. Наиболее
обычные из них псилофиты (представители рода Psilophyton) — колючие без-
листные кустарники (см. фиг. XXIII. 1).
Растения, очень похожие на те, которые встречаются у скалы Медвежий
Зуб, произрастали в восточной Канаде в течение ранне- и среднедевонского
времени. Их остатки, обычно плохо сохранившиеся, могут быть встречены в
гаспинских песчаниках (песчаниках Гаспе), в юго-восточном Квебеке. Луч-
шие же образцы раннедевонских растений известны из Западной Европы. В
раннедевонскую эпоху там существовали укрытые между горными хребтами
долины, в которых росли грибы и множество более сложно устроенных рас-
тений. Наиболее хорошо известные виды девонских растений были найдены
в окремненных угольных болотах недалеко от поселка Рини, в Абердиншире
(Шотландия). Подпитывавшиеся кремнеземсодержащей водой из близлежа-
щих горячих источников, эти болота образовали окаменелости, сохранив-
шие тонкие структуры клеточной организации, хорошо заметные в тонких
срезах слоев породы. Риния (Rhynia), одно из этих растений, было простым.
Оно имело корнеподобные структуры (но не настоящие корни) и было лише-
но листьев. Известно несколько видов риний, отличающихся друг от друга
по размерам: от очень маленьких до достигавших около 50 см в высоту. Хор-
неофитон (Homeophyton), другое растение из Рини, является особенно важ-
ным, поскольку оно образовывало переходную форму между мхами (бриофи-
тами) и растениями типа риний, в которых спорофит (спороносное поколе-
ние) и гаметофит (поколение, продуцировавшее мужские и женские половые
клетки) были полностью не зависимы друг от друга. У хорнеофитона споро-
фит был гораздо больше по сравнению с гаметофитом.
Как риния, так и хорнеофитон относятся к риниопсидам, группе, существо-
вавшей только в силуре и девоне. Возможно, они родственны ныне живущему
незаметному псилотуму (Psilotum), встречающемуся в тропиках. Он также яв-
ляется растением, почти лишенным придатков: отсутствуют корни и листья с
проводящим пучком, а спорангии располагаются на концах коротких побегов.
ПОЗДНЕДЕВОНСКИЕ ЛЕСА
Возвращаясь в Северную Америку, мы обнаруживаем, что горы, подняв-
шиеся после того, как Протоатлантический океан закрылся, в течение позд-
недевонского времени были островами, простирающимися от Джорджии до
Ньюфаундленда. Эти горы были влажными, со многими потоками, которые
стекали в отступившее море. Там, где потоки обогащались соленой водой,
они откладывали слои осадков, как, например, Катсхилл Дельта в
ЗАЗЕЛЕНЕВШАЯ ЗЕМЛЯ
355
Нью-Йорке. Такие дельты, расширяясь, формировали между собой почти
бескрайние низменности, покрытые одними из самых первых лесов на Земле.
Существование таких лесов впервые было доказано в 1869 году, когда
“взбесившийся” горный ручей Шохари Крик, сломав несколько мостов, вы-
мыл глубокие рытвины и обнажил несколько ископаемых пней недалеко от
поселка Гильбоа в штате Нью-Йорк. Еще несколько фрагментов стволов бы-
ло обнаружено в 1897 году, а в 1920 коллекционеры открыли целый лесной
слой, располагавшийся несколько выше того, который был вымыт потоком.
Наконец в 1921 году администрацией Нью-Йорка было решено открыть два
карьера по добыче камня для строительства дамбы. Эти карьеры помогли бо-
лее полно узнать историю древних лесов Гильбоа.
Эосперматоптерис (Eospermatopteris). Наиболее многочисленные дере-
вья в лесах Гильбоа имели прямые высокие стволы до 12 м в высоту с расши-
ряющимися, как бы вспухшими, похожими на луковицы основаниями око-
ло 1 м в толщину. Корни были тонкими и гибкими, ремневидными, расходя-
щимися во всех направлениях, как корни современных пальм. Листья были
папоротниковидными, до 2.7 м в длину, а их кончики несли заостряющиеся
образования, напоминающие семена. Поэтому эти растения были отнесены к
группе, называемой семенными папоротниками, и получили название эос-
перматоптерис. Последующие исследования показали, что некоторые из
этих “семян” в действительности все еще содержали споры, в то время как
стволы и листья были идентичны остаткам, долгое время известным как ане-
врофитон (Aneurophyton). Аневрофитон и другие растения, такие как кал-
ликсилон, как правило, относятся к прогимноспермам. Они использовали
тот же способ размножения, что и папоротники, но древесина делала их бо-
лее сходными с голосеменными. Таким образом, они являлись очень важ-
ным переходным звеном между двумя крупными группами растений.
Калликсилон (Callixylon), встречающийся часто вместе с археоптерис (Аг-
chaeopteris) - другое растение, образовавшее как бы переходное звено между
папоротниками и настоящими голосеменными (растениями с “голыми”, но
настоящими семенами). Минерализованные экземпляры, относящиеся к
этой форме, довольно часто встречаются и представлены как крупными ство-
лами деревьев с диаметром до 1.5 м и более 8 м в высоту, так и мелкими побе-
гами и веточками. В поперечном сечении их древесина имела строение, ха-
рактерное для современных хвойных. Калликсилон Залесского (Callixylon
zalesskyi), обнаруженный во многих местонахождениях в северо-восточной
части США, имеет особенно большое значение для науки, поскольку точно
известно, что его спороносные папоротниковидные листья прикреплялись к
побегам с древесиной голосеменного типа. В действительности, папоротни-
ковидные вайи, прикреплявшиеся к калликсилону, были названы археопте-
рис мацилента (Archaeopteris macilenta) задолго до того, как они были найде-
ны в одной ассоциации с калликсилон. Поскольку археоптерис получил на-
звание ранее, в 1871 году, целое реконструированное растение, включавшее
в себя как побеги, так и листья, должно носить именно это название, по-
скольку сейчас все эти части были обнаружены в прикреплении одно к дру-
гому. Таким названием и стало археоптерис мацилента. Многие из позднеде-
вонских ископаемых древесин из северо-восточных частей Соединенных
Штатов принадлежат этой группе растений, которая доминировала в лесах
того времени.
Ранние лепидофиты. Гораздо менее обычными в болотистых лесах Гиль-
боа были два вида плауновидных растений, родственники которых сохрани-
лись и до нашего времени в виде широко известных селагинелл (Selaginella)
23*
356
КАМЕННАЯ КНИГА
и плаунов (Lycopodium). Плауновидные из Гильбоа были стройными деревь-
ями с поникшими, опущенными вниз ветвями. Один вид имел узкие листья
около 30 см в длину, а у другого вида они были более широкими и короткими
и находили друг на друга подобно чешуе. Они росли как на стволе дерева, так
и на его ветвях и, наверное, должны были делать дерево похожим на покры-
тое мхами. Шишки, располагавшиеся на кончиках некоторых ветвей, были
наполнены спорами, а не семенами.
Вопрос о спорах или семенах очень важен в классификации растений. Ес-
ли говорить об этом упрощенно, то спора - это единственная клетка, которая
может вырасти в новое растение. Такие споры есть у водорослей, грибов,
мхов и папоротников. У многих других растений, однако, две клетки долж-
ны вступить во взаимодействие и сформировать новую единственную клетку
до того, как начнется развитие растения. Эти “женящиеся” клетки могут
быть идентичны друг другу, как у плауновидных и некоторых водорослей,
или могут отличаться по размерам и форме, как у хвощей. Когда мы имеем
дело только с одной простой спорой, конечным продуктом всегда будет яв-
ляться новое растение, которое должно пройти начало своего жизненного пу-
ти без всякой помощи.
Семя - это уже новое растение, имеющее запасы пищи и находящееся в за-
щитной оболочке. Семя появляется после того, как две клетки - всегда раз-
ные - объединяются в частично развитое растение, называемое эмбрионом.
Этот эмбрион снабжен запасом пищи и заключен в покровные ткани или
скорлупу, обеспечивающие защиту от повреждений. Когда семя полностью
развивается, эмбрион внутри него начинает расти. Оно может лежать меся-
цы или иногда даже годы, прежде чем прорастет, разовьется в новое расте-
ние, которое в дальнейшем будет воспроизводиться половым путем.
Плауновидные или растения, очень близкие предкам плауновидных,
впервые появляются в позднем силуре или раннем девоне. Ряд форм, таких
как зостерофиллюм (Zosterophyllum), располагаются где-то между псилоп-
сидами или псилофитами и плауновидными. Многие из девонских плауно-
видных были небольшими травянистыми формами, не очень отличавшими-
ся по общей форме от маленького и незаметного нынеживущего плауна (Ly
copodium). Другие формы, такие как барагванатия (Baragwanathia), уже
упоминавшаяся в этой главе, достигали более крупных размеров. Эти плау-
новидные имели важную особенность, отличавшую их от других растений.
Они имели микрофиллы (“маленькие листики”) и адаксиальные (обращен-
ные к оси растения) спорангии, что означает, что структуры, содержавшие
споры, располагались на верхней поверхности листьев.
КАМЕННОУГОЛЬНЫЙ ПЕРИОД И КАМЕННОУГОЛЬНЫЕ ЛЕСА
Когда девонский период подошел к своему концу, начали сминаться в
складки горные породы в хребтах, протянувшихся от Ньюфаундленда до то-
го места, где сейчас располагается Северная Каролина, закрылся Прото-Ат-
лантический океан, приведя к активному горообразованию как в Северной
Америке, так и в Европе. Продукты разрушения североамериканских гор
Акадия образовали дельту Катсхилл, в которой были погребены ископаемые
леса Гильбоа. Затем началась раннекаменноугольная эпоха (в Северной Аме-
рике ее называют миссиссипской), продолжавшаяся 40 миллионов лет. В те-
чение большей части этого времени во внутренних районах Северной Амери-
ки были очень широко распространены мелкие моря, однако многие из
ЗАЗЕЛЕНЕВШАЯ ЗЕМЛЯ
357
восточных районов представляли собой заболоченные низменности. Ископа-
емые остатки показывают, что аневрофитоны с клубнеобразными стволами
вымерли вслед за исчезнувшими лесами Гильбоа. На их месте появились
плауновидные, вместе с папоротниками, птеридоспермами, а также растени-
ями, близкими современным хвощам. Для того чтобы дать картину жизни
каменноугольных растений, мы должны упомянуть основные подразделе-
ния этого периода в целом.
Эти подразделения несут различные названия: миссиссипий (раннекамен-
ноугольная эпоха), пенсильваний (средне- и позднекаменноугольная эпохи);
вместе они объединяются в каменноугольный период. Это было время, когда
моря то наступали, то отступали, особенно в Северном полушарии, а речные
потоки снова заполняли их новыми осадками. Другие реки располагались во
внутриконтинентальных бассейнах, которые опускались примерно с той же
скоростью, с какой реки заносили их продуктами разрушения возвышенных
участков суши. В результате появлялись многочисленные болота, которые
погребались под толщами отложений ила, гравия и песка.
Эти болота были истинным раем для растительной жизни, обусловленной
богатыми почвами, мягким и влажным климатом, а также высокой общей
влажностью. В результате растительность была столь обильной, что ее отми-
рающие остатки образовывали слои торфа, который затем превращался в
уголь по мере того, как погружался все глубже и глубже. Верхнекаменно-
угольные формации в восточной Пенсильвании, например, содержат три-
надцать слоев каменного угля, имеющих промышленное значение, один из
которых - Питтсбургский пласт - имеет около 2-4.3 м мощности. Разнообра-
зие различных листьев, побегов, стволов в этих отложениях поражает вооб-
ражение. Почти правильное повторение морских, болотных, угольных и реч-
ных отложений называется циклотемой. Эта “полосатая” последователь-
ность горных пород часто наблюдается в дорожных выемках, разрезающих
геологические напластования в восточной части США. Она напоминает нам
о том, насколько разные условия существовали в прошлом. Эти полосы со-
хранили свидетельства о менявшейся линии берега: морские трансгрессии, а
также те моменты времени, когда линия берега регрессировала (двигалась в
направлении от суши).
Кроме пластов угля, наземные растения пенсильванской эпохи оставили
нам многочисленные окаменелости. Некоторые из них сохранились в виде
пленки угля, оставшейся в тонкозернистом песчанике или сланце от разло-
жившихся листьев, побегов и корней. Другие представляют собой лишь от-
печатки в песчанике. Кроме этого, существует множество великолепных
окаменелостей анатомической сохранности, законсервировавшихся в стя-
жениях кальцита или “кол-боллах” (см. фиг. I, 2). Встречаются целые дере-
вья, сохранившиеся в угольных пластах, потому что в пустоты, образовав-
шиеся после выгнивания растительных тканей, просочился осадок. Такие
стволы встречены в Джоггинсе и Сиднее (Новая Шотландия). В некоторых
из этих стволов, которые были полыми или дуплистыми до того как они бы-
ли погребены осадком, встречаются прекрасно сохранившиеся скелеты
древних рептилий и амфибий, о которых в этой книге будет рассказано
далее.
Каждый, кто занимался определением пенсильванских растений, вскоре
обнаруживает, что они были очень широко распространенными. Многие ви-
ды, обильные в Новой Шотландии, Западной Вирджинии, Иллинойсе или
Канзасе, были впервые открыты и получили свои названия в Европе. Эти
широко встречающиеся растения оказывают большую помощь в сопоставле-
358
КАМЕННАЯ КНИГА
Фиг. ХХ1П.2. Хвойные (а) и руфлориевые (б, в) перми Ангариды
а - Timanostrobus, реконструкция стробила, вверху слева неуплощенная семен-
ная чешуя с укороченными семеножками, внизу слева - изолированное семя;
б— Bardocarpus, стробил с семенами, сохранившимися в прикреплении; в —
Pholidophyllum, уплощенный полисперм, на плоских семеножках видны рубцы
прикрепления семян (а - по Мейену, 1987, б, в - по Наугольных, 1996). Длина
масштабной линейки - 1 см
нии и корреляции геологических формаций. Кроме этого, поскольку расте-
ния довольно хорошо отражают окружающую их обстановку, их широкое
распространение говорит в пользу того, что климат и другие природные усло-
вия были довольно однообразными в то время. В отличие от Северной Амери-
ки и Европы, в Южном полушарии главные угольные месторождения имеют
не каменноугольный возраст. В Австралии, например, самые крупные пале-
озойские угольные бассейны образовались в пермском периоде.
ЗАЗЕЛЕНЕВШАЯ ЗЕМЛЯ
359
Фиг, ХХП1.3. Родственники хвощей из перми Ангариды
а - Annulina, б, г- Phyllotheca, в - Sachyogyrus, д - Equisetina. (а, г, д - по Рас-
сказовой, 1972; б, в - по Наугольных, 1996). Длина масштабной линейки - 1 см
К северо-востоку от экваториального пояса в каменноугольный и перм-
ский периоды располагался крупный массив суши, окаймленный Урало-
Монгольским геосинклинальным поясом. О существовании этого древнего
континента было известно уже давно, с начала нашего века. Назвали этот ма-
терик Ангаридой.
Одной из ярких особенностей Ангариды было то, что на этом материке
произрастала своеобразная, не похожая на другие растительность. Позд-
непалеозойская флора Ангариды во многом отличалась от других флор.
Наиболее характерны для ангарской флоры кордаиты, принадлежавшие
360
КАМЕННАЯ КНИГА
Фиг. ХХШ.4. Типичные каменноугольные растения
а - Lepidodendron, б - Catamites, в - Bothrodendron (Hirmer, 1927). Длина мас-
штабной линейки —1м
двум эндемичным семействам - руфлориевым (фиг. XXIII.26, в) и войнов-
скиевым.
Второй по частоте встречаемости группой высших растений Ангариды бы-
ли членистостебельные семейства черновиевых - дальние предки современ-
ных хвощей (фиг. XXIII.3).
НЕКОТОРЫЕ ТИПИЧНЫЕ ПЕНСИЛЬВАНСКИЕ РАСТЕНИЯ
(СРЕДНИЙ-ПОЗДНИЙ КАРБОН)
Ни одна книга, тем более — единственная глава, не может осветить все
многообразие пенсильванских растений. Тем не менее, мы кратко опишем те
роды, которые часто встречаются в каменноугольных отложениях Северного
полушария, для того чтобы дать общее представление о растительности ка-
менноугольных лесов и болот.
Плауновидные (ликоподиофиты). Современные плауновидные, как мы
уже знаем, включают в себя плауны, селагинеллы, шильники (изоэтес) и не-
которые другие растения. Большинство из них представляют собой неболь-
шие формы с многочисленными боковыми ответвлениями, размножающие-
ся спорами, содержащимися в шишкоподобных органах. В отличие от совре-
менных плауновидных, пенсильванские были большими деревьями, из
которых самым высоким был лепидодендрон (Lepidodendron, фиг. ХХШ.4).
ЗАЗЕЛЕНЕВШАЯ ЗЕМЛЯ
361
Фиг. ХХ1П.5. Stigmaria
а — аппендиксы, б — стигмы (рис. Наугольных по экземплярам из нижнего
карбона Кизеловского угольного бассейна, Урал). Длина масштабной, линейки —
10 см
Некоторые из видов, принадлежащих этому роду, достигали высоты около
30-40 м. Ствол был прямым и неветвящимся в своем основании и средней ча-
сти, но затем, выше, он делился надвое, образуя две больших ветви, которые
далее раздваивались снова и снова. Молодые побеги дерева были покрыты
кинжаловидными листьями, расположенными близко друг к другу на коре
дерева. Когда листья опадали со ствола и старых ветвей, они оставляли руб-
цы ромбической формы, образующие чешуйчатый рисунок на поверхности
коры, благодаря чему эти растения и получили название “чешуйчатых дере2
вьев” или “чешуедревов” (так переводится название “лепидодендрон”). Спо-
роносные шишки были длинными и росли либо на кончиках ветвей, либо
свешивались вдоль сторон фертильной ветви. Виды лепидодендрона — а их
насчитывается более сотни — хорошо определимы по форме и тонким дета-
лям строения рубцов от опавших листьев.
Основание стволов лепидодендронов разделялось на подземные части, ко-
торые образовывали органы, получившие специальное название корненос-
цев или корневых поддержек (фиг. XXIII.5). Такие корневые поддержки бо-
таники называют “боковыми абсорбционными придатками”, поскольку они
могут и не являться настоящими корнями. Сами корешки оставляли круг-
лые рубцы на корненосцах. Эти рубцы располагались гораздо более свобод-
но, чем рубцы от листьев на коре ветвей и стволов.
Стигмария (Stigmaria). До того, как была точно установлена природа
этих корненосцев, они были названы стигмариями (см. фиг. XXIII.5). Есть
сомнения относительно того, все ли стигмарии принадлежали лепидоденд-
ронам. Возможно, некоторые из них были подземными частями стволов дру-
гих плауновидных. Наиболее обычный вид стигмарий - S. ficoid.es - получил
свое название фикоидес (фигоподобный), поскольку он обладает некоторым
сходством с ветвями фигового дерева. Корненосцы Stigmaria постепенно су-
живались по мере удаления от ствола до 2.5 см в диаметре в самых дисталь-
ных частях. Сами корешки редко бывают в диаметре больше чем 0.6 см и
362
КАМЕННАЯ КНИГА
выглядят как узкие черные ленточки, сохранившиеся в сером сланце, кото-
рый был когда-то почвой.
Другим широко распространенным в пенсильванских лесах растением бы-
ла сигиллярия (Sigillaria). Сигиллярия была более стройным деревом, чем ле-
пидодендрон, но не таким высоким. Один из видов достигал около 2 м в диа-
метре у основания ствола, сужавшегося до 30 см на высоте 5.5 м и достигал об-
щей высоты около 12 м. Листья сигиллярий были тонкими, заостренными и
более длинными, чем листья лепидодендронов, обнаруживая некоторое сход-
ство с большими снопами травы. Когда листья опадали, они оставляли руб-
цы, располагавшиеся более редко на коре дерева. У некоторых видов кора
между листовыми рубцами была гладкой; у других она несла продольные ре-
бра, располагавшиеся вдоль ствола. Подавляющее большинство видов сигил-
лярий имело неветвящиеся стволы. Иногда они могли ветвиться у верхушек
один или два раза. Шишки (спороносные стробилы) около 10-20 см в длину,
прикреплялись к стволам и ветвям несколько ниже листьев.
Хвощи и их родственники. Современные хвощи произрастают как во
влажных, так и в сухих местах. Их прямые тонкие побеги обычно соединены
подземными корневищами и имеют очень жесткие покровы благодаря содер-
жащемуся в них большому количеству кремнезема. Каламитес (Catamites),
главный пенсильванский род, вырастал до 12 м в высоту (см. фиг. XXIII.46).
Стволы каламитов могли быть гладкими или, напротив, несли грубые про-
дольные ребра. Обычно встречаются экземпляры около 8 см в диаметре, но
иногда эти растения могли достигать 25-30 см в толщину. Древесные стволы
поднимались вверх от подземных корневищ, которые поддерживали расте-
ния, соединяясь друг с другом в узлах нижних частей побегов. Некоторые
виды не имели боковых ветвей, другие — несли боковые ветви, отходившие от
узлов. Пучки узких листьев также росли из узлов, особенно на молодых вет-
вях, часто сохраняющихся на достаточно крупных стволах.
Каламиты образовывали обширные джунгли по берегам рек и озер, а также
и в мелких болотах. Известны минерализованные побеги каламитов, встреча-
ющиеся в некоторых сланцах. Нередко можно найти углефицированные лис-
тья каламитов. Однако более обычны песчаниковые слепки, заместившие рас-
тительные ткани. На этих слепках хорошо видны продольные ребра, распола-
гающиеся вдоль междоузлий, и сами узлы, а также рубцы от опавших ветвей.
Тем не менее, скульптура этих слепков, как правило, не совпадает с той, кото-
рая существовала на поверхности стволов каламитов при жизни.
Аннулярия и астерофиллитес (Annularia, Asterophyllites). Это названия,
предложенные для листьев каламитов. Листья имели иглоподобную или
ланцетовидную форму, с единственной жилкой, и образовывали пучок или
мутовку вокруг ветви или ствола. Изолированные фрагменты корней кала-
митов описаны под другим родовым названием - астромиелон (Astromyelori).
Хвощевидные (или членистостебельные) имеют широкое стратиграфичес-
кое распространение, от девонского периода до наших дней. Но временем их
полного расцвета был, безусловно, поздний палеозой. Как показывают неко-
торые древние переходные формы, так же как хиения (Hyenia), предки чле-
нистостебельных близки древнейшим папоротникам (хиения имеет призна-
ки, характерные для обеих этих групп).
Многие важные эволюционные эксперименты мы можем видеть в средне- и
позднедевонской истории таких групп, как птеридофиты и ранние голосемен-
ные. Неудивительно, что есть несколько групп растений, которым довольно
сложно найти место в общей системе. Сфенофиллюм (Sphenophyllum) - одно
из таких “трудных” растений.
ЗАЗЕЛЕНЕВШАЯ ЗЕМЛЯ
363
Фиг. ХХШ.6. Sphenophyllum biarmicum Zalessky
Встречающиеся в отложениях кунгурского яруса (нижняя пермь) Среднего
Приуралья облиственные побеги этого представителя сфенофиллов, сохранив-
шиеся в прижизненном положении с корневыми придатками, указывают на то,
что растение произрастало в увлажненных экотопах (рис. Наугольных), Длина
масштабной линейки - 1 см
Сфенофиллюм был травянистым растением (фиг. ХХШ.6), имевшим мел-
кие листочки, как правило, треугольной формы, нередко рассеченные на не-
сколько лопастей. Произрастали они на заболоченных участках. Корни отхо-
дили от узлов, располагавшихся на тонких побегах. Первые сфенофиллы по-
являются в позднем девоне, достигают своего расцвета в позднем карбоне и
исчезают в поздней перми или раннем триасе. Считалось, что сфенофиллы -
родственники хвощей. Тем не менее, они имеют некоторые признаки, придаю-
щие им сходство с плауновидными. Сейчас они рассматриваются в качестве
уникальной, полностью вымершей группы (Sphenophyllales или Bowmanitales).
Настоящие папоротники. Первые папоротники также появились в сред-
нем девоне, в то время, когда прошло разделение сосудистых растений на все
основные группы. Предположительно они произошли от растений, не слиш-
ком-то отличавшихся от ринии - от тримерофитов. Интенсивное формообра-
зование среди папоротников происходило в течение каменноугольного пери-
ода, а затем, в раннепермскую эпоху, разнообразие папоротников уменьши-
364
КАМЕННАЯ КНИГА
лось. Позднее, в мезозое, они вновь испытали расцвет, вместе с цветковыми
растениями, и продолжают оставаться довольно многочисленными и разно-
образными и в настоящее время.
Пенсильванские папоротники были представлены различными формами
роста. Среди них были и растения, сходные с нынеживущим папоротником -
орлецом, широко распространенным в лесах средней полосы, но широко рас-
пространены и формы, являвшиеся настоящими деревьями. Впечатляющий
мараттиевый древовидный папоротник псарониус (Psaronius) встречался в
позднекаменноугольных болотах Северной Америки и Европы. Псарониус
имел высокий, слегка суживающийся к вершине стройный ствол, в верхней
части увенчанный кроной листьев (фиг. XXIII. 7), точно так же как и многие
древовидные папоротники, существующие и в настоящее время. Сложнопе-
ристые листья псарониусов (вайи), когда они встречаются отдельно от ство-
лов, часто называются по другому - пекоптерис (Pecopteris). Они достигают
3 метров в длину. Корни пекоптерисов очень напоминали корни современ-
ных папоротников.
Кордаиты и хвойные. Эти ископаемые растения напоминают нам, что “де-
ревья” - это не естественная группа, а всего лишь большие одревесневающие
растения, которые могут и не быть родственными друг другу. Большинство
деревьев каменноугольного периода принадлежали к плауновидным, но кор-
даитес (Cordaites) был голосеменным растением и мог быть родственником
девонского калликсилона (Callixylon). Сам по себе кордаитес был высоким
растением с относительно тонким стройным стволом. Ствол в верхней части
нес многочисленные облиственные ветви, образовывавшие крону растения.
Листья были широкими, мечевидными или ланцетовидными и менее заост-
ренными, чем, например, листья плауновидных. Судя по ископаемым остат-
кам, эти листья были широкими, плотными и кожистыми и достигали в дли-
ну от 20 до 100 см. Плоские сердцевидные семена прикреплялись к семенож-
кам фруктификаций, располагавшихся среди листьев.
Группа кордаитовых (Cordaitales или Cordaitanthales) — еще один хоро-
ший пример того, как отдельные части одного и того же растения по мере то-
го, как они встречались, получали свои собственные названия, поскольку
долго не удавалось установить, что это органы одного и того же растения.
В результате этого фруктификации еврамерийских (северо-американских и
европейских) кордаитов (т.е. их органы размножения, продуцировавшие се-
мена) получили название кордаитантус (Cordaitanthus), слепки внутренней
полости стволов были названы артизиями (Artisia), а корни стали называть-
ся амиелонами (Amyelon). Чуть позже, однако, было доказано, что все эти ор-
ганы были частями одного растения.
Кордаитовые были важной частью лесов в пенсильванское время. Расцвет
кордаитовых приходится на следующий, пермский период. Они имели поч-
ти глобальное распространение: их остатки известны на всех континентах,
за исключением Антарктиды. Кордаиты достигали довольно больших раз-
меров: до 30 м высоты. Однако средние их размеры были значительно мень-
ше — около 4.5 м или менее. Некоторые из кордаитов имели корневые под-
держки, как современные растения мангровых зарослей, указывающие на
произрастание представителей этих групп в болотистых условиях, в то время
как другие кордаиты имели неглубоко залегавшие корни, далеко распрост-
ранявшиеся по всем направлениям, почти так же как у современного эвка-
липта или резинового дерева.
Вальхия (Walchia) была другим древним голосеменным растением, суще-
ствовавшим в каменноугольных лесах, и относилась к хвойным. Возможно,
ЗАЗЕЛЕНЕВШАЯ ЗЕМЛЯ
365
Фиг. ХХШ.7. Древовидный каменноугольный папоротник Psaronius
а - отдельное стерильное перо (Pecopteris), б - фертильное (спороносное) перо
(Ptychocarpus), в - реконструкция целого растения (по Magdefrau, 1956, с
изм.), г — ствол (Caulopteris)
вальхия была переходной формой между кордаитами и настоящими хвой-
ными. Поскольку эта группа растений охарактеризована мозаичным набо-
ром признаков, встречающихся и у кордаитов, и у хвойных, решающее зна-
чение имеют признаки органов размножения (шишек, семян, пыльцы и др.),
с помощью которых можно установить действительные связи между предпо-
лагаемыми предками хвойных.
Вальхия и некоторые родственные ей формы сходны с нынеживущей ара-
укарией (Araucaria или “сосна острова Норфолк”), с ветвями, торчащими в
366
КАМЕННАЯ КНИГА
стороны от основного ствола. И то, и другое растения имели иглоподобные
листья, очень непохожие на листья кордаитовых.
Возникновение семян. Происхождение семян, случившееся где-то в сред-
нем палеозое, безусловно, одно из наиболее важных событий в истории на-
земных растений - такое же, как и появление амниотного яйца у рептилий.
Семена дали растениям независимость от воды или влажных местообитаний,
необходимых споровым растениям для размножения. Это позволило семен-
ным растениям начать колонизацию значительно более удаленных от воды
районов суши. Целый ряд групп растений, существовавших в карбоне, были
уже семенными (“предголосеменными”). Некоторые растения, прогимнос-
пермы, соединяли в себе признаки как папоротников, так и голосеменных.
Голосеменные сами по себе демонстрируют довольно широкое разнообразие
способов размножения, а также типов листьев, семян (фиг. XXIII.8) и древе-
сины. Это разнообразие нередко приводит к ошибкам, когда исследователь
сталкивается только с одной или несколькими изолированными частями
растения. Нередко тяжело определить даже то, к какой крупной группе от-
носится тот или иной фрагментарный остаток растения.
В общем разнообразии голосеменных угадывается две генеалогических
линии, одна - ведущая к цикадовым, и другая - к хвойным (иногда указыва-
ют еще и третью филетическую линию — к гинкговым). В то время как цика-
довые обладали большими сложноперистыми листьями и стволами с широ-
ким кольцом коры, для хвойных более характерны простые листья и ком-
пактные древесные стволы с хорошо развитой вторичной ксилемой.
Цикадофиты относятся к двум группам - цикадалес (Cycadales) и беннет-
титалес (Bennettitales). Предками цикадофитов и гинкгофитов были птери-
доспермы или “семенные папоротники” (Pteridospermales), также относив-
шиеся к настоящим голосеменным растениям.
Птеридоспермы. Каменноугольные и пермские отложения содержат
большое количество ископаемых листьев, которые когда-то считались лис-
тьями настоящих папоротников, а затем растениями, соединяющими при-
знаки папоротников и голосеменных. По формам роста некоторые птеридос-
пермы были сходны с древовидными папоротниками, в то время как другие
были более низкорослыми или образовывали лианы. Папоротниковидные
сложноперистые листья птеридоспермов, как правило, располагались на
стволе по спирали. Семена располагались прямо на листьях или листоподоб-
ных семеносных органах (фиг. XXIII.95) и никогда не были собраны в шиш-
ки, в отличие от других голосеменных, например кордаитов и хвойных.
Медуллоза (Medullosa) — один из наиболее хорошо известных медуллозо-
вых (или тригонокарповых) птеридоспермов, распространенных от раннека-
менноугольной эпохи до перми. Точно так же, как и у большинства других
растений, обсуждавшихся нами ранее, многие части медуллозы получили
свои собственные названия до того, как они были найдены в прикреплении к
одному растению. Черешки с оборванными листьями были названы миелок-
силон. Изолированные листья были отнесены к родам невроптерис (Neuropt-
eris) и алетоптерис (Alethopteris). Название кольпоксилон (Colpoxylori) — бы-
ло использовано для побегов. Для неспециалиста такая множественность на-
званий неудобна и неизбежно приведет к ошибкам. Однако это помогает
понять, насколько важны находки относительно полных растений, позволя-
ющих установить прижизненную взаимосвязь изолированных частей и дать
реконструкцию целого растения.
Цикадовые и беннеттиты (цикадоидеи). В настоящее время цикадовые со-
ставляют очень небольшую часть растительности Земного шара, однако в
ЗАЗЕЛЕНЕВШАЯ ЗЕМЛЯ
367
Фиг. ХХП1.8. Различные типы семян из пермских отложений Ангариды
а, в, ж, и — Samaropsis с оборванной крылаткой; д, з - Samaropsis с сохранив-
шейся крылаткой; е - Sylvella; б, г — Carpolithes (по Наугольных, 1996). Длина
масштабной линейки — 1 см {а—б, г—з) и 2 мм (в, и)
позднем палеозое и особенно в мезозое, они были гораздо более широко рас-
пространены и являлись очень важной группой растений. Расцвет цикадо-
вых, безусловно, приходится на юрский и начало мелового периода. Совре-
менные цикадовые напоминают своей формой низкорослые пальмы (пальмы
в действительности относятся к цветковым растениям). Это, как правило,
небольшие растения или низкорослые деревья. Семена и пыльценосные ор-
ганы размножения располагались в шишкоподобных образованиях (строби-
лах). Пыльца продуцировалась одним растением, в то время как шишки с се-
менами располагались на другом. Пыльцевые зерна цикадовых имеют ха-
рактерную лодочковидную форму.
Из каменноугольных отложений известно несколько проблематичных
растений, таких как лазиостробус (Lasiostrobus), и из пермских - спермопте-
рис (Spermopteris), которые могли быть ранними, может быть древнейшими
представителями цикадовых. Тем не менее, однако, только с триасового пе-
риода известны настоящие цикадовые, такие как Lyssoxylon, известные по
прекрасно сохранившимся стволам из формации Чипл (Аризона). Более
древние находки все-таки заставляют сомневаться, существовали ли настоя-
щие цикадовые в дотриасовое время.
368
КАМЕННАЯ КНИГА
Фиг. XXIII.9. Пельтаспермовые птеридоспермы перми Ангариды
б-д - Peltaspermum retensorium (Zal.) Naug. et Kerp, б — базальная часть вайи с
пельтоидами, собранными в кистевидные полиспермы, в - мужские фруктифи-
кации (синангии), г — общая архитектура вайи, д - семеносный диск с хорошо
заметными семенными рубцами, а, е - листья Psygmophyllum expansum
(Brongn.) Schimper (Наугольных, 1996). Длина масштабной линейки - 1 см
Беннеттиты, или цикадеоидеи, - другая группа растений, внешне сход-
ных с пальмами, родственная цикадовым, но существовавшая исключитель-
но в мезозойскую эру. Вильямсония (Williamsonia) — один из наиболее полно
изученных представителей этой группы. Великолепная реконструкция это-
го растения была предложена знаменитым индийским палеоботаником Са-
ни. Органы размножения беннеттитов очень напоминали цветы. Как беннет-
титы, так и цикадовые скорее всего произошли от птеридоспермов в поздне-
каменноугольную эпоху.
ЗАЗЕЛЕНЕВШАЯ ЗЕМЛЯ
369
Фиг. XXIII. 10. Гинкго и переходные формы
а — лист Ginkgo из нижнемеловых отложений Восточной Сибири; б—г - пермское
растение Kerpia macroloba Naug., родственное переходным формам между пель-
таспермовыми птеридоспермами и гинкговыми (по Наугольных, 1995). Длина
масштабной линейки — 1 см
Гинкгофиты. В течение мезозоя помимо цикадофитов существовало мно-
жество других семенных растений, некоторые из которых сохранились до
настоящего времени. Кейтония (Caytonia) и позднепалеозойский глоссопте-
рис (Glossopteris) не оставили потомков, но такая мезозойская группа, как
гинкговые (гинкгоалес, Ginkgoales, фиг. ХХШ.10), представлена и в совре-
менной флоре, правда, единственным видом - Ginkgo biloba. Это живое иско-
паемое сейчас произрастает в садах в некоторых местах Китая и Японии. По-
скольку это растение неплохо развивается после пересадки и искусственно
разводится, сейчас оно культивируется по всему миру и чувствует себя не-
плохо. Согласно одной из точек зрения, гинкговые произошли от пельтас-
пермовых птеридоспермов (см. фиг. XXIII.9), по другой - от кордаитов. Они
обладают многими признаками, придающими им сходство с другими голосе-
менными растениями.
Кроме гинкговых и цикадовых в мезозойских флорах доминировали
хвойные. В позднемеловую эпоху у всех этих групп голосеменных растений
инициативу перехватили цветковые или покрытосеменные растения, кото-
рые будут подробно описаны в главе XXXII.
РАЙ ИСЧЕЗ: ПЕРМСКАЯ ФЛОРА
Каменноугольные болота были настоящим раем для растительных орга-
низмов, но этот рай исчез, как только условия изменились. Это случилось в
конце пенсильванской эпохи и далее, в течение пермского периода. В это вре-
24 Каменная книга
370
КАМЕННАЯ КНИГА
мя началось оледенение, начали расти полярные ледниковые шапки. Это
привело к резкому падению уровня моря, поскольку много воды Мирового
океана перешло в замороженное состояние, в снег и лед полярных ледников.
Климат стал более холодным и сухим. Тем не менее в межледниковые эпохи
кое-где существовали и более теплые и влажные условия.
Эти изменения были более заметны в Южном полушарии. В Северном же
(за исключением Ангариды) заболоченные низменности существовали еще
миллионы лет. Поэтому мы видим, что раннепермские растения в этом реги-
оне остаются очень сходными со своими пенсильванскими предками. Однако
в Южном полушарии, где многие континенты были покрыты ледниками,
развился совершенно иной тип флоры.
Глоссоптерис и его родственники. Эта флора, произраставшая в суровых
условиях, содержит множество растений: настоящие папоротники, хвой-
ные, мелкие членистостебельные и некоторые другие. Но главным ее пред-
ставителем, безусловно, был глоссоптерис (Glossopteris), птеридосперм с
простыми ланцетовидными листьями, которые обильны в пермских и триа-
совых отложениях Южного полушария. Листья глоссоптерис имели одну
ложную “среднюю жилку”, состоящую из нескольких сближенных прово-
дящих пучков, и множество боковых жилок, образовавших характерное
сетчатое жилкование. У другого птеридосперма, родственного глоссоптерис -
гангамоптерис (Gangamopteris) — средняя жилка отсутствовала. Третий род -
невроптеридиум (Neuropteridium) — хотя и не относился к глоссоптеридам,
но тоже очень характерен для глоссоптериевой флоры Южного полушария.
Невроптеридиум имел развитый центральный стержень, к которому при-
креплялись лопастные перышки с широким основанием.
Глоссоптерис был деревом средних размеров и имел ствол с древесиной го-
лосеменного типа, относящийся к роду Araucarioxylon и достигавший 40 см
в диаметре.
Флора Хермит. Неплохую коллекцию пермских растений можно собрать
вдоль дороги, ведущей в Аризонский Большой Каньон. Флора Хермит сохра-
нилась в сланцах, образовавшихся из отложений дельты. В этих сланцах не-
редко встречаются слепки кристаллов соли и поверхности, покрытые трещи-
нами усыхания. Эти отложения образовались на равнине, орошаемой редки-
ми, но мощными дождями, после которых наступали более сухие сезоны, в
течение которых реки высыхали под палящими лучами солнца.
Болотные влаголюбивые растения не могли произрастать в таких услови-
ях. Вместо них красноцветные породы хермитских сланцев содержат водо-
рослевые маты, располагавшиеся на илистом дне, и остатки птеридоспер-
мов, обладавших кожистыми листьями, произраставшие по берегам редких
водоемов и в речных долинах. Кроме этого, в этих условиях существовали
толстостебельные виды сфенофиллов (Sphenophyllum), вальхии (Walchla) с
маленькими игольчатыми листьями и некоторые другие хвойные. Интерес-
но отметить тот факт, что общее видовое разнообразие этой флоры было до-
вольно низким, что указывает на более суровые условия, существовавшие в
этом районе, в отличие от других регионов Северной Америки, где встреча-
ются остатки пермских растений.
КУНГУРСКАЯ ФЛОРА ПРИУРАЛЬЯ
Другой очень интересной пермской флорой является флора кунгурского
яруса Среднего Приуралья. Нередко ее называют бардинской флорой - по
ЗАЗЕЛЕНЕВШАЯ ЗЕМЛЯ
371
р. Барде, располагающейся в предгорьях Урала. На берегах этой реки нахо-
дятся наиболее известные местонахождения кунгурских растений.
Интерес, который вызывает к себе бардинская флора, обусловлен в пер-
вую очередь тем, что в ее составе находятся многие растения, давшие начало
группам, широко распространенным уже в мезозое. Одной из таких групп
были пельтаспермовые птеридоспермы, обладавшие сложноперистыми лис-
тьями, похожими на листья папоротников, и грибообразными, пельтатными
семенными органами (см. фиг. XXIII.9).
Другим интересным бардинским растением была керпия (Kerpia) (см.
фиг. XXIII. 10б-г). Этот род, возможно, был близок переходным формам от
пельтаспермовых к гинкговым, группе, характерной уже для мезозойской эры.
ЗАВЕРШЕНИЕ ЭРЫ
Пермский период определенно был временем глобального изменения рас-
тительности, отмеченным также мощными климатическими изменениями.
Многие из групп, доминировавших в каменноугольном периоде, вымерли
или резко сократили свою численность. Такие группы, как хвойные, пельта-
спермовые птеридоспермы, глоссоптериды и другие голосеменные, получи-
ли широкое распространение.
В то время как многие группы птеридофитов исчезли в начале пермского
периода, одна из них, семейство осмундовых (Osmundaceae), получила даль-
нейшее развитие. События второй половины пермского периода связаны уже
с предвестием новой, мезозойской эры.
24-
Глава XXIV
ДРЕВНИЕ И СОВРЕМЕННЫЕ АМФИБИИ
• Слово “амфибия” означает “ведущий двойную жизнь”. Действи-
тельно, большинство представителей этой группы позвоночных
часть своей жизни проводит в воде, а часть на суше. Лягушки, на-
пример, обычно откладывают свою икру в воду. Вылупляющиеся из нее го-
ловастики дышат жабрами и вообще являются исключительно водными су-’
ществами. Со временем они подвергаются метаморфозу, приобретают легкие
и конечности, после чего вполне способны жить на суше.
Амфибии могут быть подразделены на три основные группы: лабиринто-
донты, лепоспондилы и лиссамфибии. Точные родственные связи этих
групп друг с другом, а также с рыбами, от которых они происходят, и с реп-
тилиями, которые являются их производной, не ясны. Лиссамфибии факти-
чески не могут рассматриваться в качестве естественной группы, так как ее
члены (лягушки, саламандры и др.), возможно, происходят от различных
групп.
Лабиринтодонты — древнейшая группа амфибий. Дословный перевод на-
звания одной из упомянутых выше групп амфибий, лабиринтодонтов, озна-
чает “лабиринтовые зубы”. Эти животные характеризуются сложно сверну-
той эмалью зубов. В результате при просмотре тонкого шлифа зуба кажется,
что зуб имеет лабиринтовую структуру (фиг. XXIV. 1).
Наиболее примитивные лабиринтодонты - ихтиостегиды, из которых
лучше всего известна позднедевонская Ichthyostega (Гренландия). Обладая
четырьмя короткими, толстыми, хорошо развитыми конечностями, крепки-
ми пальцами лап и сильными поясами конечностей, эти животные были спо-
собны выбираться на сушу. Но по сравнению с современными наземными
животными они, вероятно, имели несколько неуклюжий вид (фиг. XXIV. 2).
То, что они действительно могли выходить на сушу, подтверждают ископае-
мые следы, найденные в Австралии (фиг. XXIV.3). Однако эти ранние амфи-
бии все еще сохранили несколько признаков, характерных для их рыбных
предков: хвостовой плавник, систему боковой линии на поверхности черепа
и остаток предкрышечной кости, которая у рыб соединяет покрывающую
жабры крышечную кость с остальным черепом (фиг. XXIV.2, 4, 13). Вся
внешняя поверхность черепа и оперкулярного аппарата покрыта крупной
скульптурой в виде длинных костных гребней, типичной для более поздних
лабиринтодонтов. Обладая большими зубами и мощными челюстями, ихти-
остега, вероятно, была грозным хищником.
© 1997. В.К. Голубев, перевод на русский язык и изменения.
ДРЕВНИЕ И СОВРЕМЕННЫЕ АМФИБИИ
373
Фиг. XXIV.1. Поперечный шлиф половинки зуба лабиринтодонта. Хорошо
видны лабиринтоподобные складки эмали
Фиг. XXIV.2. Позднедевонская амфибия из Гренландии, Ichthyostega, имела
четыре лапы и хвостовой плавник. Ее длина несколько меньше 1 м
Тела позвонков ихтиостеги и других лабиринтодонтов состоят из двух ко-
стей — плевроцентра и гипоцентра (фиг. XXIV.4). У одной группы лабирин-
тодонтов — темноспондилов — в теле позвонка начинает доминировать гипо-
центр. В конце концов у некоторых форм плевроцентр полностью исчезает.
У антракозавров, другой группы лабиринтодонтов, наоборот, плевроцентр
начинает доминировать над гипоцентром. Последний постепенно уменьша-
ется, а в некоторых случаях совсем исчезает. Несколько забегая вперед, сле-
дует отметить, что рептилии и их потомки имеют позвонки, сформирован-
ные из плевроцентров древних амфибий. Именно по этой причине предпо-
лагается, что антракозавры являются группой, от которой произошли
рептилии.
Название “темноспондилы” переводится как “разрезанные позвонки” и
подразумевает сложное строение тел позвонков у этих амфибий. Данное на-
звание нельзя назвать очень удачным, т.к. позвонки других лабиринтодон-
тов также состоят из более чем одной кости. Как бы то ни было, темноспон-
дилы были наиболее преуспевшей группой амфибий, имевшей самое боль-
шое число семейств.
Карбон и ранняя пермь — время радиации темноспондилов: появляется
громадное количество форм, приобретается все большее число наземных
374
КАМЕННАЯ КНИГА
Фиг. XXIV.3. Позднедевонская следовая дорожка одного из древнейших четве-
роногих. Длина дорожки около 2 м. Слои Геноа Ривэ, Виктория, Австралия
адаптаций. Черепа темноспондилов становятся более высокими, а конечнос-
ти более развитыми. В качестве типичных представителей этой группы ам-
фибий можно привести 8-сантиметрового Amphibamus из первой половины
позднего карбона Европы и Северной Америки (фиг. XXIV. 5) и двух несколь-
ко более молодых формы из ранней перми Техаса - Eryops длиной около
1.5 м и Cacops длиной около 40 см. Эриопс и какопс, как и все лабиринтодон-
ты и лепоспондилы, были хищниками.
Eryops вел преимущественно наземный образ жизни. Будучи активным
хищником, он нападал как на более мелких амфибий, так и на рыб
(фиг. XXIV.6, 7,13). Насколько можно судить по хорошо развитым конечно-
стям и их поясам, Cacops, вероятно, был наиболее приспособленным к жизни
ДРЕВНИЕ И СОВРЕМЕННЫЕ АМФИБИИ
375
Кистеперая рыба (Eusthenopteron)
Фиг. XXIV.4. Изменение строения черепа, конечностей и позвонков прн пре-
вращении рыбы в амфибию
Идентичные кости черепа имеют одинаковую заливку. Идентичные кости пе-
реднего плавника, передней лапы и позвонков обозначены одинаковыми буква-
ми: Н - плечевая кость, I - гипоцентр, М - пястные кости, N - невральная дуга
позвонка, Р - плевроцентр, R - лучевая кость, U - локтевая кость
на суше среди всех известных в настоящее время темноспондилов. Треть всей
длины какопса приходилась на голову. И, несмотря на скромные размеры,
он должно быть был серьезным, кровожадным хищником (фиг. XXIV.8).
Эти ранние, преимущественно наземные темноспондилы часто имели пан-
цирь, который использовался в качестве защиты, а один из них, Platyhys-
trix, даже имел удлиненные остистые отростки позвонков - явление, неодно-
кратно встречаемое в разных группах рептилий, но более неизвестное у ам-
фибий.
Темноспондилы рассмотренного возраста объединяются вместе как рахи-
томы, что дословно означает животные с “рассеченным спинным хребтом”: в
их позвонках плевроцентр и гипоцентр развиты в равной степени
(фиг. XXIV. 13, вверху слева).
376
КАМЕННАЯ КНИГА
Фиг. XXIV.5. Amphibamus — темноспондил первой половины позднего карбо-
на Европы и Северной Америки. Длина 8 см
Фиг. XXIV.6. Череп Eryops. Длина 46 см. Окрестности Бейлора, Техас, США
Начиная с пермского времени прежние тенденции увеличения числа
адаптаций для жизни на суше сменились на противоположные во второй
группе темноспондилов, стереоспондилы (“прочные позвонки” - у этих ам-
фибий наблюдается постепенное укрупнение гипоцентра и уменьшение, а в
крайних случаях и полная редукция плевроцентра (фиг. XXIV. 13, вверху в
центре)). Тела и особенно головы этих животных со временем становились
все более и более уплощенными (фиг. XXIV.9). Их конечности упрощались,
а сами амфибии становились все более водными. Внешне они напоминали
крокодилов. В одной группе, метопозавры, глаза сместились далеко вперед,
почти к самому переднему краю морды (фиг. XXIV. 10). Metoposaurus из по-
зднего триаса Северной Америки, Европы и Азии никогда не выходил из во-
ДРЕВНИЕ И СОВРЕМЕННЫЕ АМФИБИИ
377
Фнг. XXIV.7. Eryops был крупной, мощной, активной амфибией раннеперм-
ского времени. Длина 1.5 м
Фиг. XXIV.8. Лабиринтодонт Cacops, чьи остатки были обнаружены в
красноцветных континентальных отложениях перми Техаса (США), достигал
40 см в длину
ды на сушу (фиг. XXIV.9, 13). Другой представитель рассматриваемой груп-
пы амфибий - Mastodonsaurus из триаса Европы. Это животное имело череп
метровой длины и тело длиной более 4 метров.
Еще одна группа стереоспондилов, брахиопоиды, характеризуется укоро-
ченным черепом. Это нашло отражение в ее названии, которое означает “ко-
роткомордые”. Один из самых последних лабиринтодонтов, Siderops, явля-
ется брахиопоидом. Его остатки происходят из раннеюрских отложений
Квинсленда.(Австралия) (фиг. XXIV.11, 12). Другие юрские формы извест-
ны из Китая, Монголии и Европейской России. Но самым молодым (геохро-
нологически) лабиринтодонтом, вероятно, является раннемеловая форма из
Австралии. Брахиопоиды, очевидно, большую часть своего времени прово-
дили лежа на дне пресноводных водоемов. Они пристально смотрели вверх в
ожидании случайной добычи, схватывая снизу любое проплывающее над ни-
ми животное.
Самыми водными из всех темноспондилов были плагиозавры. По крайней
мере один из них, Gerrothorax, имел жабры в течении всей своей жизни, а не
378
КАМЕННАЯ КНИГА
Фиг. XXIV.9. У позднепермского Metoposaurus тело и голова были широкими
и уплощенными в горизонтальной плоскости, а конечности маленькими и сла-
быми. Этот лабиринтодонт никогда не выходил из воды. Длина 2.4 м
Фиг. XXIV. 10. Шесть черепов и кости Metoposaurus, обнаруженные в отложе-
ниях небольшого водоема, который существовал в окрестностях Санта-Фэ
(Нью-Мексико, США) в триасовом периоде
только на стадии личинки1. Сохранение ювинильных структур во взрослом
состоянии называется неотенией. Это весьма распространенный способ, с по-
мощью которого животные приспосабливаются к изменению внешних усло-
вий. Все, что для этого требуется - это изменить механизм синхронизации
развития организма. Это можно наблюдать у современного аксолотля, ино-
гда встречающегося в зоомагазинах. Аксолотль может сохранять жабры всю
жизнь вместо того, чтобы резорбировать их.
1 — Позднепермские восточноевропейские брахиопоиды, двинозавры, также сохраняли жабры в
течении всей своей жизни (прим, перев.).
ДРЕВНИЕ И СОВРЕМЕННЫЕ АМФИБИИ
379
Фиг. XXIV.11. Череп Siderops из нижнеюрской формации Эвэгрин (Квин-
сленд, Австралия). Сидеропс — один из самых поздних лабиринтодонтов. Оче-
видно, изолированная от всего остального мира Австралия послужила убежи-
щем для этой амфибии, так же как и для многих других групп животных. Дли-
на черепа 49 см
Вытянутые черепа и длинные, иглоподобные зубы - характерные особен-
ности трематозавров, активных раннетриасовых рыбоядов. Некоторые из
них населяли океанские побережья, располагавшиеся в то время на террито-
рии Гренландии и Шпицбергена. Если это действительно так, то трематозав-
ры были очень необычными животными, т.к. подавляющее большинство (но
не все) современные амфибии не могут существовать в соленой воде. По всей
видимости, и ни одна из других вымерших амфибий не была к этому приспо-
соблена. Вполне возможно, что данные необычные места обитания тремато-
завров представляли собой дельты: районы впадения пресноводных рек в
океаны. В дельтах часто морские организмы встречаются совместно с прес-
новодными. Однако трематозавры, очевидно, вышли в моря и океаны вслед
за своей основной добычей, лучеперыми рыбами, разнообразие морских
форм которых непрерывно увеличивалось в течение триаса.
Возврат к жизни в воде у лабиринтодонтов был, вероятно, связан с увели-
чением в течение пермского времени количества хищных рептилий, кото-
рые, несомненно, более приспособлены к жизни на суше, чем амфибии. Один
из способов избежать конкуренции с рептилиями - покинуть сушу и стать
водными хищниками.
Другая группа лабиринтодонтов - антракозавры, или “угольные ящери-
цы”. Они получили такое название потому, что их остатки весьма обычны в
угольных отложениях карбона Европы. Они, конечно, не были ящерицами,
но греческое слово sauros (ящерица) часто используется в научных названи-
ях и рептилий, и амфибий. Другого более подходящего слова нет ни в грече-
380
КАМЕННАЯ КНИГА
Фиг. XXIV.12. Так, по мнению Фрэнка Найта, выглядел сидеропс
ДРЕВНИЕ И СОВРЕМЕННЫЕ АМФИБИИ
381
Ichthyostega, девон
Eryops, пврмь
Metoposaurus, триас
Фиг. XXIV.13. Так изменялись позвонки и небо лабиринтодонтов в процессе
эволюции
ском, ни в латинском языках, а именно на основе этих двух языков составля-
ются научные названия живых и вымерших организмов.
Антракозавры никогда не были столь разнообразными и многочисленны-
ми, как темноспондилы. Различные группы антракозавров, возможно, име-
ют общего позднедевонского или раннекаменноугольного предка, напоми-
навшего ихтиостегу. Почти все они исчезают к концу перми. Последние их
представители из группы хрониозухий, быстровианиды, известны из ранне-
го и среднего триаса только Европейской России.
Плевроцентр был основной костью в теле позвонка антракозавров. Но од-
на группа имела гипоцентр, который был почти так же хорошо развит и так
же образовывал полное кольцо вокруг хорды — гибкого опорного стержня,
характерного для примитивных позвоночных. Эта группа антракозавров по-
лучила название эмболомеры. Типичный ее представитель — длиннотелый
(более 5 м) водный Pteroplax из первой половины позднего карбона Велико-
британии (фиг. XXIV. 14). Как большинство эмболомеров, он имел малень-
кие конечности и даже повторно развитый дорсальный плавник. Будучи ис-
ключительно водным животным, он, однако, не приобрел обтекаемую форму
тела, как более поздние лабиринтодонты. Пятнадцатисантиметровый Diplo-
vertebron из позднего карбона Чехословакии имел очень необычный для эм-
382
КАМЕННАЯ КНИГА
Фиг. XXIV. 14. А это Pteroplax, 4.5-метровый антракозавр из позднего карбона
Великобритании
Фиг. XXIV.15. Позднекаменноугольный лабиринтодонт Diplovertebron имел
грациозное тело и не очень сильные лапы. Голова была средних размеров и не
так сильно уплощена, как у других лабиринтодонтов. Длина 15 см
боломера вид. Его тело было относительно более коротким, а лапы относи-
тельно более длинными. Вероятно, он не был чисто водным животным
(фиг. XXIV. 15).
Все другие антракозавры имели большой плевроцентр. Гипоцентр, если
присутствовал, то только в виде маленькой серповидной кости. Типичный
представитель - Seymouria, много лет рассматривавшаяся в качестве проме-
жуточной формы между амфибиями и рептилиями (фиг. XXIV. 16). Она по-
лучила свое название по городу Сеймур (Техас, США), около которого впер-
вые были найдены ее остатки. Строение посткраниального скелета сеймурии
имеет весьма рептилийный облик. Однако она сохраняет и амфибийные чер-
ты, например ушную вырезку в задней части черепа, отсутствующую у всех
рептилий. Этот близкий родственник пресмыкающихся даже сохраняет жа-
берные дуги в личиночном состоянии. Но ни одна рептилия, вылупившись
из яйца, их не имеет.
Существует и другая проблема рассмотрения Seymouria в качестве прямо-
го предка рептилий. Она состоит в том, что сеймурия появляется слишком
поздно, чтобы быть истинным предком: она существовала в течение ранней
перми, в то время как самые древние рептилии известны уже в первой поло-
вине позднего карбона. Как бы то ни было, сеймурия, вероятно, не очень от-
личается от настоящих предков рептилий.
В последние годы был детально описан другой антракозавр, который ка-
жется ближе к предкам рептилий. Это - Gephyrostegus из первой половины
позднего карбона Чехословакии. Он жил одновременно с самыми ранними
ДРЕВНИЕ И СОВРЕМЕННЫЕ АМФИБИИ
383
Фиг. XXIV.16. Seymouria (слева) могла бы рассматриваться в качестве пере-
ходной формы от амфибий к рептилиям, если бы не существовала так поздно.
Длина ее тела 51 см. Trimerorhachis (справа) - очень специализированный лаби-
ринтодонт. Обе амфибии из перми Техаса, США
Фиг. XXIV. 17. Личиночный “бранхиозавр” из позднего карбона и ранней пер-
ми Европы. Длина от 4 до 15 см
рептилиями и, таким образом, не был, конечно, их истинным предком. Од-
нако череп и остальной скелет очень сходны с таковыми примитивных реп-
тилий, отличаясь главным образом деталями строения шейных позвонков и
формой неба черепа.
Загадочные бранхиозавры. Многие позднекаменноугольные и ранне-
пермские амфибии были маленькими существами. Они получили название
бранхиозавры (фиг. XXIV. 17). Короткие черепа, длинные хвосты и пре-
имущественно хрящевой скелет - характерные особенности этих живот-
ных. Ископаемые остатки некоторых представителей свидетельствуют о
том, что бранхиозавры имели внешние жабры подобно современным амби-
стомам (или аксолотлям) и головастикам. Однако критическое изучение
этой группы в конце концов показало, что эти предполагаемые взрослые
особи в действительности были личинками диссорофоидных лабиринто-
донтов. Таким образом, бранхиозавры, как особая группа амфибий, никог-
да не существовали.
384
КАМЕННАЯ КНИГА
Фиг. XXIV. 18. Diplocaulus — лепоспондильиая амфибия с широким черепом.
Длина животного достигала 60—90 см
Аберрантные лепоспондилы. В отличие от лабиринтодонтов, лепоспонди-
лы, или амфибии с “красивыми позвонками”, характеризуются одной полой
костью, формирующей тело позвонка. С момента своего появления в раннем
карбоне и до полного исчезновения в конце перми эта группа была довольно
консервативной в своей эволюции.
Лепоспондилы - преимущественно маленькие животные. Самые крупные
из них достигали в длину до 1 метра, средний же размер не превышал 30 см.
Обычно они имели вытянутое тело с уменьшенными лапами и поясами ко-
нечностей, которые могли вообще отсутствовать. Некоторых лепоспондилов,
например Ophiderpeton из первой половины позднего карбона Европы и Се-
верной Америки, легко спутать с 75-сантиметровой змеей.
Одним из наиболее причудливых лепоспондилов был Diplocaulus из позд-
него Пенсильвания (поздний карбон) и ранней перми Северной Америки
(фиг. XXIV. 18). Он имел уплощенный череп и широкие выступающие за те-
ло “рога”. Крошечные лапки у этих амфибий висели рядом с длинным, но уп-
лощенным в горизонтальной плоскости телом. Голова Diplocaulus напомина-
ла массивный костный полумесяц или треугольник, который имел разнооб-
разную форму у разных видов. Глаза и ноздри были направлены вверх, но
маленький рот располагался на нижней стороне головы. Diplocaulus, вероят-
но, всю свою жизнь проводил в воде, часами и даже целыми днями непо-
движно покоясь на одном месте. Очнувшись, он поедал мелкую молодь рыб,
которую мог поймать около дна. Диплокаулюс дышал с помощью жабр, ко-
торые, вероятно, были внутренними, располагаясь в складках кожи позади
массивной головы.
Лиссамфибии: лягушки, саламандры и другие. Всех современных амфи-
бий объединяют в одну группу лиссамфибий (“гладкие амфибии” - подразу-
мевается гладкая, лишенную всяких чешуек кожа у этих животных). К ним
относятся лягушки, жабы, саламандры и червяги, или безногие амфибии.
Червяги — это тропические, редко встречающиеся, почти слепые формы, не
имеющие ни конечностей, ни их поясов. По своему образу жизни они на-
поминают больших земляных червей.
Многие годы считали, что лягушки происходят от лабиринтодонтов, а са-
ламандры и безногие амфибии от лепоспондилов. Однако разрывы в палеон-
тологической летописи этих групп настолько велики, что никто точно не зна-
ет, связаны ли лиссамфибии непосредственными родственными связями с
другими амфибиями или происходят от особого рыбного предка. Некоторые
ДРЕВНИЕ И СОВРЕМЕННЫЕ АМФИБИИ
385
Фиг. XXIV. 19. Кайнозойские амфибии
Слева, Andrias — позднемиоценовая саламандра из Европы, длина 60-120 см.
Справа, Palaeobatrachus — 8-сантиметровая лягушка из миоцена Северной
Америки
структуры вроде уникальной ослабленной зоны в основании зубов среди всех
позвоночных, известные только у темноспондила Doleserpeton, подкрепляют
теорию отделения лиссамфибий от других амфибий в допозднепермское время.
Лиссамфибии не известны в палеонтологической летописи до поздней
перми, хотя уже из средний перми имеется несколько позвонков, которые
определены как позвонки лягушек и саламандр. Самый древний бесспорный
представитель лиссамфибий - Triadobatrachus, лягушка, известная по пол-
ному скелета из нижнего триаса Мадагаскара.
Великолепно приспособленные к прыганью, современные лягушки отли-
чаются от всех других позвоночных самым сильно измененным скелетом
(фиг. XXIV.19, справа). Будучи несомненной лягушкой, Triadobatrachus
имеет несколько примитивных черт, что можно ожидать от самого раннего
представителя этой группы. В передней лапе имеются две (лучевая и локте-
вая) кости, а не одна, как у настоящих лягушек. Аналогично и для костей
задней конечности. Хвост состоял из серии отдельных позвонков - вполне
нормальное явление для позвоночного. У других лягушек хвост - это единая,
шипообразная структура, уростиль, которая сформировалась в результате
срастания примерно дюжины позвонков.
Ко времени появления Triadobatrachus лепоспондилы уже исчезли, а ла-
биринтодонты еще просуществуют некоторое время до раннего мела. Между
поздним триасом и поздней юрой амфибии практически исчезли с нашей
планеты. Известно только несколько видов, доживших до мелового периода.
Это только один из многих случаев, когда группа резко теряет разнообразие,
чтобы в будущем вновь достигнуть расцвета.
Сегодня лягушки известны на всех континентах, кроме Антарктиды, и до-
стигли многих отдаленных областей Земли, например Новой Зеландии. С
другой стороны, саламандры, не известны не только в Антарктиде (трудно
ожидать, что они могли бы выжить в суровых антарктических условиях), но
также в Австралии и в Новой Зеландии.
Большинство современных саламандр имеет длину несколько сантимет-
ров. Самая крупная из них - Andrias из Китая и Японии, который достигает
более 1 м в длину2. Миоценовый экземпляр Andrias из Европы сначала, в
2 - Палеоценовые саламандры Монголии и Канады достигали в длину более 1.5 м (прим, перев.).
25 Каменная книга
386
КАМЕННАЯ КНИГА
семнадцатом столетии, получил название Homo diluvii testis (“человек - сви-
детель потопа”), поскольку тогда считали, что это остатки бедного грешни-
ка, погибшего во время библейского потопа (фиг. XXIV. 19).
К сожалению, ископаемые остатки раннекаменноугольных тетрапод поч-
ти не известны нигде в мире. А это, вероятно, было время не только расхож-
дения лабиринтодонтов и лепоспондилов, но и отделения темноспондилов от
антракозавров внутри лабиринтодонтов (хотя данная дивергенция могла
произойти и раньше) и происхождения рептилий от антракозавров. Очевид-
но, ранний карбон - это самый важный период в истории амфибий, когда по-
является большинство групп, за исключением (?) лиссамфибий.
К счастью, с материалом по позднедевонским и позднекаменноугольным
наземным позвоночным дела обстоят значительно лучше, предоставляя нам
возможность увидеть мир тетрапод накануне и после этого важного этапа их
истории. Позднедевонские наземные позвоночные известны из восточной
Гренландии, юго-восточной Австралии, Канады и России, а позднекаменно-
угольные - из Европы и с востока Северной Америки. Однако, короткий спи-
сок регионов подчеркивает, что палеонтологическая летопись наземных по-
звоночных даже этих времен чрезвычайно неполна.
Глава XXV
МИРИАДЫ РЕПТИЛИЙ НА ЗЕМЛЕ
ВИСОЧНЫЕ ОТВЕРСТИЯ И КЛАССИФИКАЦИЯ РЕПТИЛИЙ
• Hylonomus, самое древнее позвоночное, которое относят к рептили-
ям. Его остатки происходят из раннего Пенсильвания (поздний кар-
бон) Джоггинса (Новая Шотландия, Канада). В отличие от большин-
ства амфибий эти животные не использовали воду в процессе воспроизводст-
ва, а откладывали яйца, сохранявшие жизнеспособность в наземных
условиях. Таким образом, у этих животных появилась настоящая возмож-
ность освоить сушу и начать использовать те ресурсы, которые прежде не бы-
ли доступны позвоночным1.
Покровный череп Hylonomus позади каждого глазного отверстия, или
глазницы, образует сплошную поверхность. У других рептилий в этой обла-
сти располагается одно или два отверстия (височные окна), имеющие различ-
ное положение и окруженные разными костями. Несмотря на то, что строе-
ние височных окон в деталях может сильно различаться, их положение (или
отсутствие) в черепе служит основой для выделения в составе рептилий че-
тырех основных групп, или подклассов.
Анапсиды. Эта наиболее примитивная группа рептилий объединяет су-
ществ, которые подобно Hylonomus не имеют никаких височных отверстий
(фиг. XXV.2). Позади каждой орбиты располагается пять покровных костей
черепа, которые плотно срастаются друг с другом. Черепа подобного строе-
ния известны у котилозавров и черепах* 1 2. Котилозавры вымирают к концу
юрского периода, а морские пресноводные и сухопутные черепахи процвета-
ют до сих пор (фиг. XXV. 1).
Синапсиды. Эта одна из древнейших, но не примитивная группа репти-
лий, исчезающая в ранней юре. Ее представители имели одно височное от-
верстие на каждой стороне черепа, располагающееся под заглазничной и че-
шуйчатой костями (фиг. XXV. 2). Синапсиды представлены пеликозаврами
и терапсидами, “зверообразными рептилиями” (см. главу XXXI).
© 1997. В.К. Голубев, перевод на русский язык и примечания. ,
1 — То. что Hylonomus был яйцекладущим, является не фактом, а предположением, которое осно
вано на том, что данное животное, вероятно, физиологически было рептилией. В действитель
ности, яйца не известны ни у одной палеозойской рептилии (прим, перев.).
2 - Анапсиды - сборная группа, включающая как примитивных рептилий, капториноморф, так и
черепах и их предков. Последних в настоящее время выделяют в отдельный класс Parareptilia.
Это наиболее молодая среди всех “рептилий” группа происходит от особых амфибийных пред-
ков, дискозаврисков, широко распространенных в ранней перми южной Евразии. Парарепти-
лии с рептилийной физиологией появляются где-то в середине перми (прим, перев.).
25*
388
КАМЕННАЯ КНИГА
Фиг. XXV. 1. Meiolania — гигантская рогатая черепаха. Ее остатки были обна-
ружены в четвертичных отложениях о. Лорд-Хау в Тихом океане (реконструк-
ция выполнена Фрэнком Найтом)
МИРИАДЫ РЕПТИЛИЙ НА ЗЕМЛЕ
389
Синапсид (Edaphosaurus)
Эвриапсид (Plesiosaurus)
Диалсид (Youngina)
Фиг. XXV. 2. Черепа представителей четырех основных групп (подклассов)
рептилий. Апапсиды (Limnoscelis) не имеют височных отверстий; диапсиды
(Youngina) имеют два височных отверстия; эвриапсиды (Plesiosaurus) и синаи-
сиды (Edaphosaurus) имеют по одному височному отверстию соответственно вы-
ше и ниже контакта заглазничной и чешуйчатой костей. Обозначения: PF — зад-
нелобная кость, РО — заглазничная кость, SQ - чешуйчатая кость, ST - надви-
сочная кость
Эвриапсиды. Животные этой группы имели только одно височное отвер-
стие на каждой стороне черепа, которое располагалось выше заглазничной и
чешуйчатой костей (фиг. XXV.2). К эвриапсидам относят исключительно
водных существ - нотозавров, плезиозавров, плакодонтов и ихтиозавров.
Диапсиды. Как следует из названия, череп имеет два височных отверстия
с каждой стороны: выше и ниже чешуйчатых и заглазничных костей
(фиг. XXV.2). С момента своего появления в позднем карбоне, диапсиды
представлены двумя основными группами: лепидозаврами (“чешуйчатые
ящерицы”) и архозаврами (“ящерицы-правители”). Первая группа включа-
ет ящериц, змей и мозазавров, вторая - динозавров, крокодилов, летающих
ящеров и их различных родственников. Уже более 200 млн. лет диапсиды
превосходят большинство других рептилий по изобилию, разнообразию и
размеру3.
Описать эволюцию всех рассмотренных четырех подклассов в,общем виде
не представляется возможным. Одни группы рептилий в процессе эволюции
увеличивались в размерах, в то время как другие, наоборот, превращались в
карликов. Некоторые группы резко меняли направления своего эволюцион-
ного развития. Многие семейства сокращали число костей как в черепе, так
и в остальном скелете. Другие, наоборот, добавляли много новых. А некото-
рые, особенно змеи, потеряли кости таких частей тела, как конечности, но
3 — Весьма вероятно, что синапсиды, с одной стороны, и диапсиды с эвриапсидами, с другой, про-
исходят от разных амфибийных предков. Однако это самые древние группы “рептилий”, и их
достоверные предки среди антракозавровых амфибий пока неизвестны (прим, перев.).
390
КАМЕННАЯ КНИГА
Фиг. XXV.3. Вертикально залегающие ископаемые стволы деревьев в форма-
ции Джоггинс (окрестности Камберленда, Новая Шотландия, Канада)
У многих стволов до их захоронения сгнила сердцевина и образовалась полость.
В настоящее время в этих полостях находят остатки скелетов амфибий и репти-
лий пенсильванского возраста
МИРИАДЫ РЕПТИЛИЙ НА ЗЕМЛЕ
391
Фиг. XXV.4. Пермские “щекастые ящеры” — парейазавры
Слева, череп Elginia, позднепермского парейазавра из Шотландии, длина 25 см.
Справа, парейазавр Bradysaurus, населявший Южную Африку в среднеперм-
ское время. Его длина достигала 2.4 м
увеличили количество позвонков и ребер. Ящерицы и их родственники осва-
ивали болота, пустыни, моря и даже деревья. Ихтиозавры, представители
эвриапсид, по своему облику были более рыбоподобны, чем некоторые спе-
циализированные рыбы. Несмотря на то, что сфенодонтиды за последние
150 миллионов лет не испытывали никаких серьезных изменений ни в стро-
ении, ни в форме, ни в размере, мы можем считать дивергенцию одной из ос-
новных особенностей истории рептилий. Далее мы проследим основные на-
правления эволюции этой группы позвоночных и оценим их результаты.
КОТИЛОЗАВРЫ4 И ДРЕВНИЕ ЛОВУШКИ
Залив Фанди между Нью-Брансуиком и Новой Шотландией (Канада) ши-
роко известен самыми большими приливами на земле. Менее известным, но
весьма важным для истории позвоночных является местонахождение иско-
паемых остатков Джоггинс (Новая Шотландия, Канада), расположенное на
берегу этого залива.
Три сотни миллионов лет назад, в раннем Пенсильвании (поздний карбон),
эта область представляла собой болотистую равнину, в которой накапли-
вались растительные остатки, впоследствии превратившиеся в громадные
толщи каменного угля. Болота были окружены обширными лесами плау-
новидных типа Sigillaria (см. главу ХХШ). Стволы этих растений достигали
до 0.9—1.2 м в диаметре и до 30 м в высоту. В отличие от современных де-
ревьев, у стволов плауновидных очень плотным и крепким был только внеш-
ний слой толщиной не более 10 см. Внутренняя же часть дерева была полой
или образована из материала, который очень быстро сгнивал. Время от вре-
мени основания деревьев заносились слоем осадка мощностью до 3 м. Дере-
вья умирали, их стволы обрушивались, а на поверхности оставались полые
пни, внутрь которых и попадали неосторожные рептилии и амфибии
(фиг. XXV.3).
4 - Термин котилозавры используется для группы, объединяющей примитивных рептилий (прим,
перев.).
392
КАМЕННАЯ КНИГА
Кости всех скелетов, обнаруженных внутри описанных пней, обычно от-
делены друг от друга. Данный факт, а также присутствие многочисленных
копролитов, указывает на то, что падение в эти естественные ловушки не
убивало животных. Они еще долго жили здесь, поедая друг друга5.
Из Джоггинса известно две рептилии. Одна из них - Hylonomus, упоми-
навшийся выше. Это животное было длиной около 20 см. Морфологически
оно является подходящим предком для более поздних рептилий: гаттерий,
ящериц, змей, крокодилов, черепах6, а также разнообразных исчезнувших
мезозойских групп. Внешне Hylonomus похож на гаттерию и примитивных
современных ящериц, но отличается от них прежде всего наличием монолит-
ного черепа анапсидного, а не двудырочного, диаспидного, типа. Позвонки,
конечности и пояса конечностей по существу такие же, как у современных
гаттерий и ящериц.
Парейазавры. Большинство котилозавров были маленькими животны-
ми, не существенно отличаясь по размеру от Hylonomus или несколько круп-
нее. Однако средне- и позднепермские парейазавры были исключением.
В средней перми7 эта группа была представлена большими, странными
животными (фиг. XXV.4, справа). Их остатки известны из южноафрикан-
ских пород данного возраста. Самые крупные парейазавры достигали 3 м в
длину, имели мощные лапы, бочкообразное тело и весили около 900 кг. Их
череп был массивным и шишковатым, кожу украшали костные пластины, а
зубы на челюстях были зазубрены подобно ножу для мяса8. Скелеты парейа-
завров обычно находят в позах, соответствующих естественному положению
животного. Вероятно, парейазавры утонули в болотной трясине, куда они за-
брели, поедая болотные растения9. Последние парейазавры, существовав-
шие в поздней перми, значительно лучше известны по находкам из России,
Шотландии (Великобритания) (фиг. XXV.4, слева) и Китая. К этому времени
в Южной Африке они уже стали достаточно редкими животными.
СИНАПСИДЫ - ПРЕДКИ МЛЕКОПИТАЮЩИХ
Значительно реже, чем Hylonomus, в стволах деревьев в Джоггинсе встре-
чаются остатки Protoclepsydrops, возможно, самого древнего представителя
синапсид. Он известен только по плечевой кости и нескольким позвонкам,
поэтому имеются некоторые сомнения относительно отнесения его именно к
синапсидам. Из другого местонахождения ископаемых стволов деревьев,
расположенного поблизости от предыдущего, во Флоренции (Новая Шотлан-
дия, Канада) известен Archaeothyris. Здесь вмещающие отложения имеют
5 - Найденные в Джоггинсе и близлежащих местонахождениях внутри окаменевших стволов ко-
стные остатки принадлежат мелким животным с цепкими лапками, которые, конечно же, лег-
ко могли вылезти из описанных “ловушек”. В данном случае мы имеем дело с уникальным,
очень редко (!) встречающимся типом захоронения древоживущих наземных позвоночных. Hy-
lonomus и другие найденные вместе с ним рептилии жили на сигилляриях и в огромных полос-
тях их стволов, охотясь на различных беспозвоночных. Где они жили, там и погибали, а их тру-
пы поедались их же собратьями (прим, перев.).
6 — 0 предках черепах см. одну из сносок выше (прим, перев.).
7 - По европейской схеме пермь подразделяется на две части. Средняя пермь, используемая амери-
канскими исследователями, соответствует нижней половине верхней перми. Соответственно,
среднепермское время соответствует первой половине поздней перми (прим, перев.).
8 - Предполагается, что парейазавры были фитофагами, т.е. растительноядными животными
(прим, перев.).
9 - К подобному типу захоронений относится местонахождение примитивных парейазавров-бра-
дизаврид на р. Вятка у г. Котельнич в Кировской области, Россия (прим, перев.).
МИРИАДЫ РЕПТИЛИЙ НА ЗЕМЛЕ
393
Фиг. XXV.5. Пеликозавр Dimetrodon из ранней перми Северной Америки имел
высокий “парус” на спине. Максимальная длина 3.0-3.3 м
несколько более молодой возраст, чем в Джоггинсе: поздний пенсильваний
(конец позднего карбона). Остатки Archaeothyris имеют хорошую сохран-
ность, пожалуй, самую лучшую для всех известных позднепенсильванских
синапсид.
Позднепенсильванские и раннепермские синапсиды представлены пели-
козаврами. Среди пеликозавров выделяется две группы хищников и две фи-
тофагов. Все они хорошо представлены в раннепермских отложениях цент-
ральной части севера Техаса (США).
Сегодня эта область представляет собой равнину, усеянную фермами и
пастбищами. Мескитовое дерево является основным деревом в этих местах;
самые крупные рептилии — гремучие змеи. Сильные дожди — редкое явле-
ние, что весьма благоприятно для охотников за ископаемыми. Потоки дож-
девой воды, мчащиеся по широким промоинам, обнажают слои разноцвет-
ных глин, красных мергелей и красноцветных песчаников, в которых часто
попадаются ископаемые остатки позвоночных.
В те времена, когда накапливались все эти отложения, северный Техас
представлял собой прибрежно-океанскую дельтовую равнину с постоянно
изменяющей свое положение в пространстве береговой линией. Внутрикон-
тинентальный климат мог быть засушливым, но районы дельт были сильно
увлажненными, что способствовало развитию здесь пышной растительнос-
ти, очень напоминавшей растительность более ранних “угольных” болот.
Древние хвощи Calamites (см. главу XXIII) по размерам напоминали малень-
кие деревья. Леса состояли из хвойных и плауновидных растений. Были ши-
роко распространены также настоящие папоротники. Среди семенных папо-
ротников преобладала Medullosa.
Наиболее ужасным из хищных пеликозавров, населявших те болота, был
сфенакодонт Dimetrodon (фиг. XXV.5). Кровожадный вид ему предавали не
394
КАМЕННАЯ КНИГА
Фиг. XXV.6. Пермские растительноядные пеликозавры
Слева вверху - Edaphosaurus с высоким “парусом” на спине. Справа вверху, че-
реп Edaphosaurus. Справа внизу, рыбояд Ophiacodon. Обе рептилии из перми
Техаса (США)
только двойные кинжаловидные клыки в передней части пасти, но и режу-
щие зубы, располагающиеся позади них (фиг. XXXI. 1, слева). В дополнение
ко всему этому на спине располагался крупный “парус”, сформированный
сильно удлиненными остистыми отростками спинных позвонков. Между
этими костными отростками была натянута богатая кровеносными сосудами
кожная перепонка. Вероятно, Dimetrodon использовал “парус” для ускоре-
ния разогрева тела, располагая его перпендикулярно лучам солнца. Хотя все
сфенакодонты были хищниками, подобными Dimetrodon, не все имели “па-
рус” на спине. Например, Ophiacodon, по размерам соизмеримый с Dimetro
don, такого спинного “паруса” не имел (фиг. XXV.6). Он был преимущест-
венно водным хищником и охотился главным образом за рыбой, а не за реп-
тилиями.
Несомненно, главным объектом охоты Dimetrodon был растительноядный
пеликозавр Edaphosaurus (фиг. XXV. 6). Он также имел “парус” на спине, по
строению несколько отличающийся от “паруса” диметродона: на длинных
остистых отростках позвонков располагаются многочисленные короткие го-
ризонтальные поперечные костные выросты. Кинжаловидные клыки отсут-
ствовали, и все зубы были похожи друг на друга.
Edaphosaurus по размерам был таким же, как и все рассмотренные выше
пеликозавры, — до 3.3 м в длину. Однако не все пеликозавры были такими
большими. Varanosaurus был более мелким офиакодонтным хищником, не
превышавшим в длину 1.5 м (фиг. XXV. 7). Еще более мелким был Myctero-
saurus, растительноядное животное длиной 60 см.
Следующий этап синапсидной истории - происхождение и радиация те-
рапсид. Большинство палеонтологов считает, что эти рептилии происходят
МИРИАДЫ РЕПТИЛИЙ НА ЗЕМЛЕ
395
от сфенакодонтных пеликозавров. Череп у терапсид стал более прочным, по-
скольку в нем появились дополнительные связи. Однако височное отверстие
на задне-боковой стороне черепа в большинстве случаев увеличилось в раз-
мерах. Это связано с развитием челюстной мускулатуры, которая во время
сжимания челюстей помещалась именно здесь, что позволяло сделать укус
более мощным.
По сравнению с пеликозаврами терапсиды характеризуются более про-
грессивным строением задних конечностей. У пеликозавров задние лапы
широко расставлены в стороны так, что бедренная кость, верхняя, самая
ближняя к телу, кость задней конечности, перемещается в горизонтальной
плоскости. Такая постановка конечностей называется латеральной. Она ха-
рактерна для многих древних и современных рептилий, например крокоди-
лов и ящериц. В данном положении тело оказывается расположенным близ-
ко к земле. Именно эта особенность рептилий лежит в основе другого назва-
ния данной группы наземных позвоночных - пресмыкающиеся. У терапсид
задние лапы были приведены в положение под тело и перемещались в верти-
кальной плоскости. Данный способ положения конечностей, называемый
парасагитальным, значительно более эффективен для быстрого передвиже-
ния. Однако передние конечности так и оставались латеральными у боль-
шинства терапсид и парасагитальными становятся только у некоторых наи-
более продвинутых форм в самом конце пермского времени.
Остатки самых ранних терапсид известны в среднепермских отложениях
Северной Америки, самой верхней части последовательности напластования
пермских континентальных пород на этом континенте10 11. К счастью, конти-
нентальные отложения нижней части верхней перми Европейской России,
синхроничные средней перми Северной Америки, также содержат остатки
примитивных терапсид. В Евразии, вероятно, и произошла начальная ради-
ация этой группы зверообразных рептилий11. Разрез пермских отложений
Восточной Европы сопоставляется с основанием более длинной последова-
тельностью напластования континентальных отложений Южной Африки,
где история терапсид прослеживается до позднего триаса. В Европе терапси-
ды доживают до средней юры, откуда известна всего одна форма. Таким об-
разом, эволюционная история синапсид весьма подробно отображена в пале-
онтологической летописи, чем не может похвастаться ни одна другая группа
позвоночных.
Большинство палеонтологов считают, что все терапсиды происходят толь-
ко от одной группы пеликозавров - хищных сфенакодонтов. Однако были
предположения, что некоторые из растительноядных терапсид могли быть
потомками казеидных пеликозавров. Среди позвоночных, как правило,
крупные, более продвинутые группы происходят именно от хищников, а не
от фитофагов. Обычно даже растительноядные формы в продвинутых груп-
пах происходят от хищников, а не от более примитивных фитофагов. Конеч-
но из этого правила имеются исключения. Поэтому нет ничего удивите льно-
10 — в действительности терапсиды не известны из Северной Америки. В данном случае речь, веро-
ятно, идет о наиболее терапсидообразном пеликозавре Тetraceratops. Широко известные севе-
роамериканские пеликозавры — это тупиковая ветвь развития синапсид (прим, перев.).
11 — Наибольшим разнообразием отличаются гондванские терапсиды. Скорее всего, именно этот
древний материк являлся местом происхождения и начальной радиации терапсид. Основные
группы терапсид из Гондваны попадают в Европу и Азию в самом начале поздней перми (или
самом начале средней перми по американской схеме). К сожалению, позднекаменноугольные
и раннепермские континентальные отложения с остатками тетрапод с территории бывшей
Гондваны в настоящее время не известны (прим, перев.).
396
КАМЕННАЯ КНИГА
Фиг. XXV.7. Некоторые некрупные животные, чьи остатки встречены в перм-
ских красноцветных отложениях Техаса (США)
Офиакодонт Varanosaurus, длиной 1.5 м; котилозавр Labidosaurus, длина 60 см;
продвинутая амфибия или очень примитивная рептилия Diadectes, длина 1.5-
1.8 м
го в том, что казеиды могли быть предками некоторых или даже всех расти-
тельноядных терапсид.
Наиболее многочисленной среди терапсид группой по количеству извест-
ных видов и родов являются аномодонты. В момент появления этой исклю-
чительно растительноядной группы в самом начале средней перми все ее
представители были маленькими существами длиной от 25 до 50 см. Воз-
можно, маленькие размеры ранних аномодонтов связаны с тем, что в это вре-
мя существовали такие крупные (более 2 м в длину) фитофаги, как диноце-
фалы (другая группа терапсид) и парейазавры. После исчезновения диноце-
фалов во второй половине поздней перми и перехода поздних парейазавров к
жизни в крупных водоемах аномодонты стали увеличиваться в размере, за-
нимая место крупных растительноядных животных в сообществе наземных,
неводных, тетрапод.
Одной из наиболее ранних групп аномодонтов были дицинодонты. Эти те-
рапсиды потеряли все резцы и нижнечелюстные клыки: на их месте образо-
вался роговой клюв. Верхнечелюстные клыки были крупными и длинными
(фиг. XXV.8, справа). Щечные зубы, располагающиеся позади клыка, при-
сутствовали лишь у некоторых дицинодонтов.
В поздней перми существовало три группы дицинодонтов. Все они исчез-
ли к концу этого периода. Из триаса известна только одна группа дицинодон-
тов, которая достигла всех уголков Земли и просуществовала до конца этого
периода (фиг. XXV.8, слева).
Одним из самых необычных и наиболее широко распространенных дици-
нодонтов был Lystrosaurus (фиг. XXV.9). Это животное имело сильно укоро-
ченную, опущенную вниз морду. В отличие от большинства дицинодонтов,
элементы конечностей состояли главным образом из хряща. Кроме того,
Lystrosaurus имел округлое, бочкообразное тело. Вероятно, эти животные по
МИРИАДЫ РЕПТИЛИЙ НА ЗЕМЛЕ
397
Фиг. XXV.8. Дицинодонты
Kannemeyeria - раннетриасовый дицинодонт из Южной Африки. Длина 1.8 м.
Ближайшие ее родственники известны также из России и Австралии. Справа,
череп самого Di.cyn.odon, верхняя пермь Южной Африки. Длина черепа 15 см
образу жизни напоминали современных гиппопотамов, которые большую
часть времени проводят в реках и озерах, но при этом способны кое-как пере-
двигаться и по суше. Lystrosaurus был одним из первых древних позвоноч-
ных, обнаруженных в Антарктиде, указывая тем самым, что в течение ран-
него триаса рептилии и амфибии этого континента были весьма сходны с ам-
фибиями и рептилиями Южной Африки. Сам Lystrosaurus, однако, имел
более широкое распространение, будучи обнаруженным также в России, Ин-
дии, Китае и Индокитае.
В начале триаса появилась еще одна группа рептилий, которые подобно
дицинодонтам утратили зубы и приобрели хорошо заметные клювы для пе-
реработки растительности. Это - ринхозавры, одна из многих диапсидных
групп. Может быть то, что дицинодонты больше никогда не восстановили
своего доминирующего положения в сообществах наземных тетрапод после
крупного кризиса в конце перми, было вызвано появлением в триасе ринхо-
завров, занявших нишу крупных наземных фитофагов. В противном случае
дицинодонты заняли бы эту экологическую нишу и восстановили бы свое бы-
лое разнообразие. Ринхозавры и дицинодонты исчезают из палеонтологиче-
ской летописи примерно к концу позднего триаса.
Диноцефалы не были столь долгоживущей группой терапсид, как аномо-
донты. Время их существования ограничивается только средней пермью,
когда они населяли Россию, Южную Африку и Китай. Типичные диноцефа-
лы — это крупные, длиной около 2-х метров животные с массивными черепа-
ми. Толстые покровные кости черепа, особенно поверх крошечного мозга,
наводят на мысль, что самцы могли устраивать брачные турниры, бодаясь
головами, подобно современному снежному барану-толсторогу.
Среди диноцефалов были и фитофаги, и хищники. Хотя некоторые дино-
цефалы действительно вели хищный образ жизни, очень трудно себе предста-
вить, как эти тяжелые, неуклюжие животные преследовали свою добычу.
Возможно, они были падалеядами или нападали на свои жертвы из засады.
Более мелкие и более грацильные горгонопсы могли быть активными
хищниками. Типичный для горгонопсов размер имел Lycaenops из поздней
перми Южной Африки, достигавший около 60 см в длину. Большие клыки
398
КАМЕННАЯ КНИГА
Фиг. XXV.9. Типичная для триаса Антарктиды и Южной Африки сцена
Хищная зверообразная рептилия Thrinaxodon стоит над только что убитым
проколофоном Procolophon. На заднем плане в воде затаился крокодилообраз-
ный текодонт Chasmatosaurus, подкарауливая трех растительноядных Lyst.ro-
saurus. Листрозавры, почуяв опасность, озираются по сторонам. Двое из них
смотрят на хасматозавра (реконструкция выполнена Питером Траслэ)
на обеих челюстях - характерная черта всех горгонопсов, причем у большин-
ства эти зубы были настолько длинными, что их можно назвать саблевид-
ными. Зубы позади клыков обычно очень маленькие, а у некоторых форм мо-
гут отсутствовать. Горгонопсы, очевидно, только отрезали куски мяса от сво-
ей добычи и проглатывали их не пережевывая.
Рогатый хищник - редкое явление, но восточноевропейский позднеперм-
ский горгонопс Proburnetia имел пару хорошо развитых костных рогообраз-
ных выростов в задней части черепа и костные возвышения над глазами.
Горгонопсы были доминирующими хищными терапсидами в течение по-
здней перми, но они вымирают в конце этого периода, не оставив никаких
потомков.
Более мелкая группа хищных терапсид - тероцефалы. Основная масса те-
роцефалов — хищники, некоторые тероцефалы были всеядными, и лишь от-
дельные формы, вероятно, были растительноядными, что в общем является
нормальным для образа жизни терапсид. Но развитие некоторых структур
у этих рептилий позволяет предположить наличие у них редких и уникаль-
ных адаптаций. Например, позади клыков у одной формы, которая, как
считают, была падалеядной, располагаются не зубы, а серия роговых зуб-
ных пластин.
Никакие другие терапсиды не были столь мелкими, как насекомоядные (?)
тероцефалы. Позади их клыков располагались крошечные, остроконечные
зубы, хорошо приспособленные для прокусывания кутикул жуков или тара-
канов (эти насекомые уже существовали в пермское время, см. главу XIV).
МИРИАДЫ РЕПТИЛИЙ НА ЗЕМЛЕ
399
Фиг. XXV.10. Зверообразная рептилия Cynognathus из среднего триаса Юж-
ной Африки. Длина около 1.5 м
Тероцефал Euchambersia, вероятно, имел ядовитый укус. Кость, окружа-
ющая основание верхнечелюстного клыка, покрыта маленькими характер-
ными лунками, которые могли бы быть связаны с ядовитой железой. На
внешней стороне клыка, или ядовитого зуба, располагается нисходящий же-
лобок, по которому яд мог стекать в рану жертвы после того, как Euchamber-
sia крепко сжимал челюстями добычу, пытаясь ее умертвить.
В самом конце поздней перми от тероцефалов происходят цинодонты (жи-
вотные с “зубами, как у собаки”). В конечном итоге, именно от цинодонтов в
позднем триасе возникают млекопитающие. В начале триаса существует по
крайней мере шесть семейств цинодонтов. Со временем цинодонты приобре-
тали все более млекопитающеподобный облик, причем каждая группа своим
особым путем.
Вместе с другими продвинутыми терапсидами цинодонты приобретают
вторичное небо. Оно было необходимо для отделения ротовой полости от но-
сового прохода. Более примитивные терапсиды не могли дышать и есть одно-
временно. Для многих животных, типа ящериц, и сегодня это не имеет осо-
бого значения, поскольку процессы внутри их тела протекают достаточно
медленно, и они могут просто прекращать дышать во время еды. Но для ак-
тивных животных, вроде нас с Вами, желательно, если ни необходимо,
иметь возможность есть и дышать одновременно. Вспомните, когда Вы про-
студились в последний раз и могли дышать только ртом. Насколько Вам бы-
ло трудно тогда есть пищу! Только наличие одного вторичного неба уже ука-
зывает на то, что цинодонты были весьма активными животными.
По прохождению кровеносных сосудов в черепе, насколько это может
быть восстановлено, цинодонты - скорее млекопитающие, чем рептилии.
Кроме того, относительные размеры ребер указывают, что цинодонты имели
диафрагму между легкими и нижележащей полостью тела. Наконец, кости
черепа гладкие и нигде не имеют ни тонких желобков, ни маленьких лунок,
за исключением самой передней части морды, где присутствуют глубокие
ямки. Эта особенность отмечается только у животных с хорошо развитыми
мясистыми губами и вибриссами, или сенсорными волосами.
Развитие у млекопитающих кровеносной системы с четырехкамерным
сердцем и появление диафрагмы связано с тем, что теплокровным животным
требуется более эффективные кровообращение и дыхание. Присутствие губ
позволяет перед тем, как проглотить пищу, обрабатывать ее, разжевывая на
400
КАМЕННАЯ КНИГА
маленькие части. Современные рептилии глотают пищу крупными кусками.
Поскольку цинодонты стали более млекопитающеподобными, они, вероят-
но, имели волосы и кормили детенышей молоком.
Позже цинодонты приобрели ряд других прогрессивных черт. Передние
конечности больше не были расставлены в стороны, а приобрели парасаги-
тальное положение, как это ранее произошло с задними конечностями. Они
были вытянуты под телом, при движении перемещаясь только в вертикаль-
ной плоскости, тогда как у более примитивных групп плечевая кость переме-
щается в горизонтальной плоскости. Продвинутые цинодонты также поте-
ряли костную перемычку, которая отделяла височное отверстие от глазницы
(фиг. XXXI.3).
Большинство семейств цинодонтов исчезает к концу триаса. Из ранней
юры известен Oligokyphus — маленькое животное длиной около 45 см. Он
имел высоко специализированную зубную систему. Клыки отсутствовали, а
на месте, где они могли бы располагаться, присутствует большой промежу-
ток в зубном ряду (диастема). Спереди от диастемы располагались резцы, а
позади сложные щечные зубы, каждый с многочисленными конусами или
бугорками. Строение коронки зуба подобно тому, что мы можем наблюдать
сегодня у животных, питающихся семенами или орехами.
Однако зубы Oligokyphus довольно необычны для цинодонта. Большинст-
во цинодонтов, вроде Cynognathus из среднего триаса Южной Африки, были
хорошо приспособленными хищниками (фиг. XXV. 10). От одного из таких
цинодонтов с хищной зубной системой в позднем триасе происходят первые
настоящие млекопитающие, взрыв разнообразия которых будет наблюдать-
ся в самом начале кайнозойской эры после длительного покоя во времена гос-
подства на земле “ужасных ящеров”, динозавров12.
12 — Конечно же “покой” был только внешним. Мезозойская эра - время основной радиации мле-
копитающих. Именно в это время появляются все основные группы млекопитающих (прим,
перев.).
Глава XXVI
ПРОЩАЙ, СУША
• Как только постоянно меняющая свой облик Земля предоставила
возможность четвероногим позвоночным преуспеть на суше, репти-
лии сразу стали активно развиваться. Тем не менее, вскоре после то-
го, как это случилось, некоторые из этих только что появившихся прогрес-
сивных животных вновь вернулись к жизни в воде. Возможно, что некото-
рые из них вообще никогда полностью не расставались с водой.
Мы уже знаем Ophiacodon, рыбоядного пермского синапсида, который,
вероятно, был хорошим пловцом, хотя также мог передвигаться и по суше
(фиг. XXV.6, справа). Наверняка, более водным был Mesosaurus, недолго су-
ществовавший род из ранней перми Южной Африки и Южной Америки
(фиг. XXVI. 1). Стройное существо длиной около 40 см, Mesosaurus имел че-
люсти, усаженные многочисленными иглообразными зубами. Во время по-
гони за ракообразными он, очевидно, использовал передние лапы как управ-
ляющие органы, перемещаясь в воде с помощью широких задних конечнос-
тей. Мезозавр имел интересное географическое распространение: его остатки
встречаются только в Бразилии (восток Южной Америки) и Южной Африке.
Во время существования Mesosaurus эти два материка еще были частями
единого континента Гондвана.
МОЩНЫЕ И НЕ ТАКИЕ УЖ КОНСЕРВАТИВНЫЕ ЧЕРЕПАХИ
Когда мы сегодня говорим о водных рептилиях, большинство из нас вспо-
минает черепах. Эти рептилии появляются к позднетриасовому времени и
весьма обычны во все последующие периоды. Они захватили болота, озера,
реки и моря и приобрели самую надежную броню, которую можно встретить
у четвероногих позвоночных. Большинство черепах придерживалось этой
своей ранней анатомической особенности, чем, вероятно, и объясняется их
успешное существование на протяжении вот уже 200 миллионов лет. Однако
за это время две отдельные группы черепах независимо друг от друга измени-
ли череп и шею уникальным для позвоночных образом. Это однозначно сви-
детельствует о том, что данная группа рептилий была далека от эволюцион-
ной статичности с триаса по настоящее время.
Черепахи произошли от неизвестных пермских анапсид, которые прово-
дили жизнь в тихих и, вероятно, болотистых водоемах, где их тела со време-
© 1997. В.К. Голубев, перевод на русский язык и примечания.
26 Каменная книга
402
КАМЕННАЯ КНИГА
Фиг. XXVI. 1. Mesosaurus — раннепермская водная рептилия из Южной Афри-
ки и Южной Америки. Длина 40 см
нем расширились, в то время как лапы превратились в ласты с кожистыми
перепонками между пальцами. На спине появился панцирь, или карапакс,
соединившийся с ребрами и позвонками. Карапакс состоял из костных плас-
тин, которые сформировались в коже. Сверху он был перекрыт чешуями, ко-
торые постепенно расширились в роговые щитки. Щитки также закрыли
вентральную часть панциря, или пластрон, который был соединен с кара-
паксом с обеих сторон. Части поясов конечностей были включены в плас-
трон. Зубы стали маленькими и в конце концов исчезли, одновременно на че-
люстях образовался роговой клюв.
Группы черепах. С момента своего появления черепахи эволюционируют
тремя основными группами. Типичным представителем одной группы, Pro
ganochelydia, является сам Proganochelys (фиг. XXVI.2, справа внизу). Ос-
татки представителей этого рода найдены в триасе Европы и Таиланда. Их
челюсти были уже беззубыми, но крошечные зубы остались на небе. Ни голо-
ва, ни хвост не могли втягиваться в панцирь, но они оба были защищены ши-
пами. Лапы, которые также оставались только снаружи от панциря, были
защищены длинными, заостренными по краям щитками.
Другие две группы черепах являются более продвинутыми, чем Proga-
nochelydia, во многих отношениях. На заре своей истории обе группы приоб-
ретают крепкие черепа, которые больше не имеют подвижных сочленений. В
то же самое время каждая группа независимо от другой развивала в черепе
своеобразные блоковые устройства, которые изменили направление дейст-
вия челюстных мускулов. Позже, вероятно в конце мезозойской эры, шеи у
представителей этих двух групп изменились так, что теперь голову можно
было втягивать назад под защиту панциря.
Современные члены одной из рассмотренных групп, плевродиры (“боко-
шейные”), втягивая свои головы в панцирь, изгибают длинные шеи вбок. Ис-
копаемые остатки из меловых отложений принадлежат формам, удивитель-
но сходным с черепахами, которые все еще живут в Южном полушарии. Все
дожившие до современности австралийские черепахи принадлежат к этой
группе.
Криптодиры (“скрытошейные”) включают всех остальных черепах. Со-
временные криптодиры глубоко втягивают голову, S-образно изгибая шею в
вертикальной плоскости между плечевыми пластинами. Однако не все позд-
некайнозойские криптодиры были способны это делать. Восточноавстралий-
ская Meiolania (фиг. XXV. 1) и аргентинская Niolamia, несомненно, испыты-
ПРОЩАЙ. СУША
403
Фиг. XXVI.2. Мезозойские черепахи
вали в этом большие затруднения, так как обе эти сухопутные черепахи име-
ли хорошо развитые рога!
Ранние криптодиры были обитателями болот. Они хорошо плавали, часто
грелись на солнце, забираясь на бревна или песчаные отмели. А прогулки по
суше были для них весьма обычным занятием. Некоторые из их потомков
стали значительно более водными, но так и остались обитателями рек и бо-
лот, другие же ушли жить в море. Одним из самым первых представителей
морских черепах был Archelon - гигант длиной более 3.7 м, с узкой головой,
громадным клювом, огромными ластами и весом приблизительно 2.7 т
(фиг. XXVI.2, слева). Archelon обитал в позднемеловых морях Южной Дако-
ты, Канзаса (США) и смежных регионов, которые кишели большими рыба-
ми и кровожадными рептилиями, близкими родственниками ящериц. Са-
мый прекрасный по сохранности скелет Archelon не имеет только одного зад-
него ласта. Возможно, он был откусан одним из этих хищников.
На примере черепах мы сталкиваемся с большим количеством тех контра-
стов и противоречий, которые характеризуют эволюцию рептилий. Черепа-
хи появляются как существа, развивающие панцирь и осваивающие воду.
Некоторые формы, типа современной каймановой черепахи, практически
такими и остаются. Другие же теряют панцирь или возвращаются к жизни
на суше, хотя ни одна черепаха никогда не делала сразу и то, и другое.
Archelon был одной из черепах, которые потеряли большую часть панци-
ря, закрывающей кости, и сохранили только жесткую кожу. Две другие
группы морских черепах повторили этот процесс, и тоже самое сделали
речные мягкокожие черепахи. Раннетритичные представители последней
группы имели панцири из тонких, усеянных ямками пластин; более поздние
26*
404
КАМЕННАЯ КНИГА
Фиг. XXVI.3. Мезо-кайнозойские черепахи
Слева — панцирь Taxochelys, североамериканской меловой морской черепахи,
сходной с современным логгерхедом; длина около 3.7 м. Справа, Testudo аттоп —
сухопутная раннетретичная черепаха из Египта; длина панциря составляет 1 м
формы потеряли всю свою броню, оставив только кожистый панцирь. Раз-
розненные кости третичных мягкокожих черепах настолько обычны на Го-
лых Холмах в районе Континентального Водораздела Северной Америки, в
центральных районах южного Вайоминга (США), что они образуют здесь
большую часть гальки. Очевидно и панцири, и скелеты быстро развалива-
лись на части после смерти животных, т.к. целые экземпляры редки.
Пресноводные черепахи сегодня часто выползают на сушу и, очевидно, дела-
ли это во все времена. Возможно, самая известная ископаемая пресноводная че-
репаха, Stylemys, обитавшая на Сырых Равнинах североамериканских штатов
Южная Дакота, Небраска и восточного Вайоминга в олигоцене. В миоценовое
время она становится менее обычной. Типичные экземпляры достигают в длину
от 13 до 20 см., однако имеются сообщения о панцирях длиной от 51 до 91 см.
Сухопутные черепахи - это наземные черепахи с колонообразными лапа-
ми, которые потеряли перепонки между пальцами. Большие панцири, полу-
чившие название Stylemys, могут в действительности принадлежать Testudo
(фиг. XXVI.3, справа), исключительно наземному роду, который включает
современных роющих и пустынных сухопутных черепах, а также гигант-
ские виды с островов Галапагос (Тихий океан) и Альдабра (Индийский оке-
ан). Сухопутные черепахи появляются в эоцене и уже в раннем олигоцене
стали весьма разнообразны. Первоначально получившие название Colos
sochelys, самые большие экземпляры Testudo были найдены в отрогах Гима-
лаев в горах Сивалик (Индия). Они имеют плейстоценовый возраст. Их пан-
цири достигали 2.2 м в длину, 1.5 м в ширину и 88 см в высоту и весили более
1 т. Это было неплохим достижением для наземных рептилий, чьи низкорос-
лые предки в свое время оставили сушу!
ИХТИОЗАВРЫ
Все 120 млн. лет своей истории ихтиозавры активно развивались.
Их наиболее примитивные представители впервые появляются в среднетри-
ПРОЩАЙ, СУША
405
Фиг. XXVI.4. Ихтиозавры
Cymbo&pondylus — примитивный ихтиозавр, чьи остатки обнаружены в среднем
триасе штата Невада (США); длина 12.2 м. Позвоночник в хвостовой части не
соглут вниз: возможно, не было никакого хвостового плавника. Ophthalmosau-
rus из юры Вайоминга (США) и Англии был высоко специализированным. Его
хвост напоминал хвост акулы, а зубы были очень маленькими; длина 3 м. Mix-
osaurus известен из Европы, Северной Америки, Шпицбергена и Индонезии
асовых морях Калифорнии, Шпицбергена, Европы и Индонезии. Это были
стройные существа со сжатыми, но почти прямыми хвостами, на верхней и
нижней стороне которых располагались невысокие плавниковые складки
(фиг. XXVI.4, вверху). Со временем позвоночник начал загибаться вниз. Од-
новременно с этим происходило увеличение верхнего плавникового лепестка
(фиг. XXVI.4, внизу справа). В конце концов хвост приобрел реверсирован-
ный гетероцеркальный вид, превратившись в мощный движущий в воде ор-
ган, сходный с таковым у самых быстрых акул (фиг. XXVI.4, внизу слева).
Вытянутые челюсти имели много зубов; передние ласты были больше зад-
них, хотя и у тех, и у других кости конечностей были укорочены, в то время
как пальцы были существенно удлинены за счет появления дополнительных
фаланг (гиперфалангия) (фиг. XXVI.5 и 6). Во многих случаях рядом с пер-
воначальными пальцами добавлялись ряды из большого количества допол-
нительных костей, в результате чего образовывались более широкие ласты.
Позвонки были короткие и многочисленные, а огромные выступающие глаза
406
КАМЕННАЯ КНИГА
Фиг. XXVI.5. Stenopterygius из нижней юры Германии; длина 1.6 м (фото
Фрэнка Коффы, любезно предоставленная музеем Виктории, Мельбурн)
Фиг. XXVI.6. Передний ласт Stenopterygius, на котором видны дополнитель-
ные фаланги на пальцах (гиперфалангия), но отсутствуют дополнительные
пальцы (гипердактилия). Длина ласта 24 см (фото Фрэнка Коффы, любезно пре-
доставленная музеем Виктории, Мельбурн)
укреплялись костными склеротическими кольцами. Ихтиозавры явно охоти-
лись при помощи зрения, поэтому их глаза были сконструированы так, что
могли фиксировать максимальное количество необходимой информации.
При реконструкции древних позвоночных нам приходится восстанавли-
ваться мягкие части их тела на основании тех свидетельств, которые предо-
ставляют кости скелета. Совсем другое дело — ихтиозавры. Темноцветные
юрские породы Германии содержат скелеты ихтиозавров, окруженные
ПРОЩАЙ, СУША
407
обугленными пленками, позволяющими увидеть контуры тел животных,
широкие ласты, хвост и дорсальный, напоминающий акулий, лишенный
костей плавник. Эти обугленные пленки также показывают, что кожа ихти-
озавров была гладкой, а на переднем крае ласт была усеяна редкими малень-
кими чешуями. Крошечные пигментные зерна свидетельствуют о том, что
“рыбо-ящерицы” (так переводится название этой группы) были темными
сверху и светлыми снизу. Такая окраска, увеличивающая их сходство с аку-
лами и дельфинами, имела маскировочное значение и помогала этим реп-
тилиям скрываться от проплывавшей ниже и выше них потенциальной
добычи.
Хотя ихтиозавры все еще дышали воздухом, они вели вполне водный об-
раз жизни. Они питались рыбой, кальмарами и белемнитами, остатки кото-
рых иногда находят между ребрами некоторых скелетов. Ихтиозавры, веро-
ятно, никогда не выходили на берег, поэтому не могли откладывать яйца и
вынашивали их внутри своего тела, став живородящими рептилиями. На не-
которых экземплярах видны скелеты маленьких ихтиозавров внутри боль-
ших, при этом нет никаких явных свидетельств, что первые были съедены
вторыми вместо белемнитов или рыбы. На других экземплярах зафиксиро-
ваны события, когда молодые животные, по-видимому, выходят из той обла-
сти, где хранились яйца. Возможно, самка ихтиозавра погибла во время “ро-
дов” или молодые особи вылупились вскоре после ее смерти. Такое иногда
случается у ящериц и змей, чьи яйца вскрываются внутри тела, после чего
молодые особи выходят наружу.
ОТ ПОЛЗАЮЩИХ ЖИВОТНЫХ ДО ПЛЕЗИОЗАВРОВ
В поздней перми остров Мадагаскар отделился от восточного побережья
Африки. На морском побережье, которое только что образовалось на запад-
ной части Мадагаскара, в те времена обитал Claudiosaurus, рептилия длиной
около 70 см. Это животное имело маленькую голову, умеренно длинную шею
и задние конечности, превышающие по размерам передние. Оно питалось
маленькими ракообразными, которых вылавливало в водах близлежащего
океана. После охоты, клаудиозавр выбирался на теплые камни и грелся на
солнце подобно современным тюленям.
Claudiosaurus — первый представитель весьма разнообразной группы но-
тозавров и плезиозавров, которые появятся позже и будут процветать в мезо-
зойскую эру. Он происходит от неизвестного пермского диапсидного предка
и, подобно ящерицам, теряет нижнюю перемычку нижнего височного окна,
приобретая таким образом эвриапсидную конструкцию черепа.
С начала триасового периода близкие родственники Claudiosaurus, ното-
завры, стали более водными, приобретя более приспособленный для жизни в
воде скелет. Подобно современным морским крокодилам, они, вероятно, вы-
ходили на берег всякий раз, когда им это было нужно (фиг. XXVI. 7, слева
внизу). Их кости были найдены в морских отложениях, связанных прежде
всего с древним океаном Тетис: в Южной Европе, Азии и Северной Африке.
Плезиозавры. Название этих потомков нотозавров означает “подобные
ящерицам” или “почти ящерицы”. Тем не менее, некоторые из этих существ
меньше всего были похожи на ящериц как внешним видом, так и своими раз-
мерами. Плезиозавры были морскими рептилиями, приспособленными к
жизни в открытом море, хотя они иногда наведывались в прибрежные боло-
та, эстуарии и реки. Одна группа этих рептилий имела короткие, уплощен-
408
КАМЕННАЯ КНИГА
Фиг. XXVI.7. Водные нотозавры, плезиозавры и плакодонты
ные головы, длинные шеи и толстые овальные туловища, укрепленные сни-
зу циновкообразным сплетением брюшных ребер (фиг. XXVI. 7, слева ввер-
ху; фиг. XXVI.8, справа). Их хвосты, хотя и довольно длинные, были
лишены плавниковой лопасти: они двигались в воде с помощью передних и
задних лап, которые превратились в веслоподобные ласты. Кости бедра и
плеча были широкими, обеспечивая достаточно места для прикрепления
мощных мускулов, смещавших ласты вниз и назад. Длина плезиозавров из-
менялась в пределах 2.4—15.2 м, а число позвонков шеи в основном варьиро-
вало от тридцати до восьмидесяти.
Вторая группа плезиозавров была сходна с первой по строению ласт и хво-
ста, но их туловища были более высокими, а челюсти более длинными и от-
носительно более тонкими (фиг. XXVI.8, слева). Характерная особенность -
короткая шея, насчитывавшая не более тридцати позвонков.
И длинношеие, и короткошеие плезиозавры появляются в раннеюрских
морях Европы. Они были весьма многочисленны здесь на протяжении
многих миллионов лет, но практически неизвестны в других районах
(фиг. XXVI. 7, слева вверху). Только в меловом периоде их ареал существен-
но расширяется, а сами они превращаются в очень крупных животных.
Австралийский Kronosaurus имел череп длиной почти 2.7 м. Эласмозавры,
населявшие моря на территории современной Великой Равнины в Северной
Америке, имели очень стройные хвосты и шеи длиной 7 м (фиг. XXVI.8,
справа). Такие вытянутые шеи и хвосты объясняют, почему плезиозавров
ПРОЩАЙ, СУША
409
Фиг. XXVI.8. Позднемеловые плезиозавры из западного Канзаса (США)
Elasmosaurus достигал в длину 12.5-15.2 м, а короткошеий Trimacromerum
всего 3 м
иногда сравнивают со змеями, продетыми сквозь тела черепах. Тем не менее,
ни один из известных плезиозавров не имел панциря, и хотя шея и была до-
статочно гибка, но даже самые длинношеие виды не могли вращать и изги-
бать ее так, как это делают змеи. Плезиозавры, очевидно, плавали на поверх-
ности воды или в приповерхностной ее части и редко ныряли. Находясь под
водой, они “летали” с помощью своих ласт так же, как это делают современ-
ные морские черепахи и пингвины. Основной их добычей были рыбы, белем-
ниты и кальмары, которых они ловили, резко бросаясь вперед всем телом,
или выбрасывая только голову. Жертвы, слишком большие, чтобы быть про-
глоченными целиком, разрывались на части резкими толчками и рывками
головы - способ, который часто используют современные крокодилы и алли-
гаторы.
Плакодонты. Меньшее родство с нотозаврами имеет другая группа мор-
ских эвриапсид: плакодонты. Подобно нотозаврам, они обнаружены только
в триасовых морских отложениях океана Тетис: южная Европа, северная
Африка и юго-западная Азия. Плакодонты были существами с короткими
тупыми головами, толстыми туловищами и пастообразными лапами (фиг.
XXVI.7, справа внизу). Они походили на современного ламантина или мор-
скую корову. Плакодонты, очевидно, плавали на мелководьях и поедали
моллюсков, раздрабливая их раковины широкими, низкими зубами, кото-
рые размешались на челюстях и костях неба (фиг. XXVI.7, справа вверху).
410
КАМЕННАЯ КНИГА
У Placodus ребра ближе к позвоночнику сильно утолщались, а сплетение
брюшных ребер (гастралии) защищало тело животного снизу. Гастралии,
возможно, также выполняли роль балласта, поддерживающего животного в
устойчивом положении во время всплывания к поверхности. У позднетриа-
сового Placochelys спина была скрыта панцирем.
АРХАИЧНЫЙ ХАМПСОЗАВР
Многие диапсиды осваивали жизнь в воде. К этой группе относятся все
другие рептилии, описываемые в данной главе. Наиболее консервативным
среди них был Champsosaurus. Обладая узкой мордой, он внешне напоминал
гавиала. Подобно последнему, Champsosaurus был очень эффективным “ры-
боловом”. Однако он отличался весьма примитивным строением черепа. Это
вполне можно было бы ожидать от триасовой рептилии, но хампсозавры на-
селяли Северную Америку и Европу с позднего мела до эоцена, а также изве-
стны из позднего мела восточной Азии. Они были одними из самых больших
позвоночных, которые пережили крупное вымирание в конце мелового
периода.
МОЗАЗАВРЫ - НАСТОЯЩИЕ ВОДНЫЕ ЯЩЕРИЦЫ
Раньше плезиозавров иногда описывали как “ужасные свирепые чудови-
ща”. Фактически, такие мрачные прилагательные более применимы к на-
стоящим водным ящерицам - мозазаврам.
Мозазавры - странные рептилии. А история их открытия не менее захва-
тывающая, чем они сами. В 1780 г. в подземных песчаных каменоломнях
под Питерсбергом, район города Маастрихт (Нидерланды), рабочие обнару-
жили окаменелый череп. Люди послали за армейским хирургом и натурали-
стом, который собирал ископаемые остатки из каменоломен. Он работал на-
столько искусно, что очень быстро извлекал из горных пород целые скелеты.
Доктора Хофманна вполне устроила стоимость, назначенная данному образ-
цу. Он также оплатил его транспортировку, а позже подготовил его к выстав-
ке. Но земля над каменоломнями принадлежала некому Кэнону Годдину, ко-
торый подал в суд, выиграл дело и стал обладателем данной находки.
В 1795 г. французская армия осадила Маастрихт. Поскольку данное иско-
паемое было уже широко известно в Европе, главнокомандующий приказал
своим канонирам поберечь ту часть города, где располагался дом Кэнона Год-
дина. Заподозрив причину этого “покровительства”, Кэнон Годдин спрятал
свое сокровище. После сдачи города он не выдал свою тайну, поэтому фран-
цузами было объявлено вознаграждение - шестьсот бутылок вина - всякому,
кто найдет этот скелет. Результат не заставил себя долго ждать: томимые
“жаждой” гренадеры вскоре нашли его. Находка была доставлена в Париж,
где сначала была описана как “недоразвитый кит” или “дышащая рыба”, а
после как “громадная ящерица с Мааса” (Маас - река, протекавшая рядом с
Питерсбергом). Британский автор перевел это многоречивое название в ла-
тинское М osasaurus.
Мозазавры были распространены по всему миру в позднем мелу, достиг-
нув максимального расцвета в морях, которые занимали районы, примыка-
ющие к современным Атлантическому океану и Мексиканскому заливу. Са-
мые прекрасные экземпляры происходят из меловых отложений западного
ПРОЩАЙ, СУША
411
Фиг. XXVI.9. Мозазавры — морские ящеры позднемелового возраста
Tylacosaurus (слева) из Канзаса (США) и Новой Зеландии был 6.7 м длиной
Канзаса (США) и достигают максимальной длины 9 м. Однако челюсти и зу-
бы, найденные в слоях глауконитового песка в Нью-Джерси (США), могли
принадлежать гигантскому мозазавру длиной около 14 м. Самый странный
представитель этой группы был Globidens, встречавшийся на юге Соеди-
ненных Штатов, в Южной Америке, Африке, Европе и Азии. Он, вероятно,
промышлял у морского дна, где собирал двустворчатых, брюхоногих и дру-
гих мало подвижных моллюсков. Их раковины он раздрабливал своими
бульбовидными зубами, которые почти полностью потеряли некогда острые
вершины.
Более типичные мозазавры, типа Tylosaurus из Канзаса (США) и Новой
Зеландии, были активными пловцами с длинным телом, с уплощенным хво-
стом и конечностями, превратившимися в широкие, перепончатые ласты с
хорошо развитыми пальцами (фиг. XXVI.9). Костные пластины закрыли
верхнюю часть головы. Шея, тело и хвост были покрыты чешуями такими
же, как у ящериц, - они хорошо сохранились на некоторых обугленных ис-
копаемых. Склеротические кольца укрепили глаза. Барабанная перепонка
состояла из толстого хряща. Каждая ветвь нижний челюсти была вооружена
шестнадцатью-восемнадцатью острыми зубами. В задней части каждой вет-
ви располагалась рычажная кость - квадратная кость. Квадратная кость
412
КАМЕННАЯ КНИГА
имела шарнирную связь с челюстью на одном конце, а на другом конце - та-
кое же сочленение с черепом (фиг. XXVI.9, справа вверху). Этот факт, а так-
же связка, посредством которой ветви челюстей спереди соединялись друг с
другом, свидетельствуют, что мозазавры, подобно змеям, были способны
опускать и широко раздвигать свои челюсти, чтобы проглотить слишком
крупные куски пищи.
Окаменелости из брюшной области некоторых скелетов показывают, что
рыба была главным объектом питания мозазавров. Достигая 6-9 м в длину,
они были единственными позвоночными, которые могли в те времена ловить
таких животных, как “бульдожий тарпон” (Portheus molossus), которые ве-
сили от 270 до 360 кг, и были так же опасны, как большинство морских реп-
тилий. В меню мозазавров входили также аммониты. Были обнаружены ра-
ковины аммонитов со следами прокусов, соответствующих зубам мозазав-
ров. Поскольку старые, изношенные зубы выпадали из челюстей и
заменялись новыми, мозазавры всегда были хорошо вооружены как для
борьбы с конкурентами, так и для ловли добычи.
ФИТОЗАВРЫ - ЖИВОТНЫЕ С ОШИБОЧНЫМ НАЗВАНИЕМ
Заходившие в реки мозазавры иногда встречали здесь крокодилов длиной
до 15.2 метров. Однако они появились слишком поздно, чтобы столкнуться с
существами, чьи привычки, внешний облик и строение скелета очень сильно
напоминают таковые крокодилов (фиг. XXVI. 10). Формально мы должны
этих псевдокрокодилов называть фитозаврами (“растительные ящерицы”)
не потому, что это название удачно описывает их образ жизни, а потому, что
один уже почти забытый профессор считал их растительноядными.
В действительности фитозавры были хищными рептилиями длиной от' 1.8 до
6.1 м. Их тело было защищено костным панцирем, перекрытым сверху рого-
выми чешуями. Череп походил на череп современного гавиала, крокодила с
выпуклым “лбом” и вытянутыми челюстями (фиг. XXVI. 11). Ноздри у гави-
ала, однако, находятся на его морде, тогда как у фитозавров они были сдви-
нуты близко к глазам, располагаясь на костном возвышении, так что они бы-
ли несколько приподняты над черепом (фиг. XXVI. 12).
Самые древние известные фитозавры обнаружены в среднем триасе Евро-
пы (фиг. XXVI. 10). Они распространяются по всему миру в течение позднего
триаса и еще сохраняются в ранней юре. Их кости были найдены в Европе и
Южной Америке, на противоположном от Нью-Йорка берегу реки Гудзон, в
Северной Каролине и во многих других местах североамериканского Юго-За-
пада. Эти рептилии жили в болотах и медленно текущих реках. Они плавали
с помощью своих сжатых хвостов, при этом над водой часто возвышались од-
ни только ноздри. Хотя они, несомненно, пожирали амфибий и других реп-
тилий, все же их основной добычей, вероятно, была рыба.
КРОКОДИЛЫ
Крокодилы образуют длительно существующий отряд диапсидных репти-
лий, который объединяет собственно крокодилов, гавиалов, аллигаторов,
кайманов и их ископаемых родственников. Первые крокодилы появляются
в позднем триасе Европы, Южной Америки и Восточной Африки. Хорошо
известный Ископаемый Лес в северо-восточной Аризоне является одним из
ПРОЩАЙ, СУША
413
Фиг. XXVI.10. Rutiodon — позднетриасовый фитозавр из Аризоны (США).
Длина черепа 107 см. Растения представлены цикадофитами, папоротниками и
араукариевыми. Крайнее справа дерево - Araucarioxylon, левее от него Wood-
worthia
тех мест, где были найдены ископаемые остатки наиболее примитивных кро-
кодилов. Яркие современные полупустыни сильно контрастируют с теми бо-
лотами, которые существовали здесь во времена Protosuchus. Протозух -
рептилия длиной около 76 см, с относительно короткой головой, большими
глазами и умеренной длины хвостом (фиг. XXVI. 13). Его спина, брюхо и
хвост были покрыты прямоугольными пластинами, которые, вероятно,
защищали его от зубов голодных фитозавров. Конечности имели типично
крокодиловый облик и могли использоваться как для ходьбы, так и для пла-
вания. Задние лапы были длиннее передних. Эта особенность, которую до
сих пор сохраняют крокодилы, была характерна также для большинства
динозавров.
Мезозухии. История крокодилов включает два этапа адаптивной радиа-
ции. Один из них охватывает юрский период, когда группа, называемая ме-
зозухиями, широко распространяется как по Новому, так и по Старому Све-
ту. Самые маленькие из них были меньше 30 см в длину и не имели почти ни-
какого панциря. Самыми большими были тонкомордные, похожие на
современных гавиалов существа длиной 4.6-6.1 м. Все они имели большие
верхние височные отверстия и плоские или слегка вогнутые сзади позвонки.
Эти позвонки и изолированные зубы - самые обычные ископаемые остатки
мезозухий.
Большинство мезозухий было активными хищниками, которые бродили
по побережьям и болотистым отмелям в поисках рыб или рептилий. Метрио-
414
КАМЕННАЯ КНИГА
Фиг. XXVI.11. Mestriosuchus — позднетриасовый европейский длинноморд-
ный фитозавр. Длина черепа 83 см
Фиг. XXVI.12. Череп Rutiodon, верхний триас Северной Америки. Длина 107 см
ринхиды были наиболее необычными из этих крокодилов, потому что их те-
ла были сильно изменены для жизни в море, где они конкурировали с ихти-
озаврами и мозазаврами (фиг. XXVI.14, слева). Это были гладкокожие, ли-
шенные панциря рептилии со стройными туловищами, вытянутыми
мордами и очень длинными хвостами, в которых позвоночник загнулся вниз
как у ихтиозавров, свидетельствуя таким образом о наличии терминального
плавника. Как и у ихтиозавров, верхняя лопасть хвостового плавника не
имела никаких костей. Ее присутствие достоверно известно по ископаемым
остаткам из тонкослоистых черных уплотненных глин, в которых сохрани-
лись также отпечатки тел. Большинство этих крокодилов плавали с помо-
щью хвоста. Их конечности, которые превратилась в ласты, служили в каче-
стве балансирующих органов. Большинство метриоринхид обитало в юрских
морях Европы и Южной Америки, но другие мезозухии, встречавшиеся в мо-
рях, реках и болотах Европы, Африки и Америки, доживают до эоцена.
Современные крокодилы. Следующая большая волна радиации крокоди-
лов началась в самом начале мелового периода (периода накопления писчего
мела). В пределах этого довольно длинного периода появляются три группы,
которые существует до сих пор: длиннорылые гавиалы, остромордные собст-
венно крокодилы и широкомордные аллигаторы (фиг. XXVI. 14, справа).
Кроме некоторых аллигаторов, все эти современные крокодилы, или эозу-
хии, не имеют спинного и брюшного панциря. Их височные отверстия ма-
ленькие, а позвонки вогнуты спереди, но выпуклы сзади. Они также значи-
тельно лучше решили проблему, связанную с расположением внешних нозд-
рей на длинном носе или морде. У фитозавров этой проблемы не было, так
как их ноздри располагались около глаз. У мезозухий кости верхних челюс-
тей и неба формируют трубу, которая располагается под истинной крышей
рта. Эта труба отводила воздух к отверстиям, располагавшимся ниже и поза-
ди глаз. У эозухий ноздри остались на морде, в то время как воздуховодная
труба протягивается до самой задней части черепа, где располагаются отвер-
ПРОЩАЙ, СУША
415
Фиг. XXVI. 13. Protosuchus — примитивный крокодил из ранней юры Аризоны
(США). Длина 76 см
Фиг. XXVI. 14. Мезо-кайнозойские крокодилы
Geosaurus - позднеюрский морской мезозухиевый крокодил (метриоринхид)
из Центральной Европы и Аргентины с ластами и акулоподобным хвостом.
Справа - череп Alligator thomsoni, пресноводной рептилии из позднего миоцена
Небраски (США). Длина 36 см
стия трахеи. Благодаря такому усовершенствованию, а также мембране, ко-
торая закрывала горло, эти рептилии, были способны дышать, не опасаясь,
что вода попадет в их легкие, даже если их пасть открыта.
Крокодил Deinosuchus, чьи остатки обнаружены в меловых отложениях
Европы и Северной Америки, имел череп длиной более 1.8 м и достигал в
длину от 13.7 до 15.2м. Значительно более крупный, чем современные виды
(современный индийский гавиал достигает 9 м в длину, в то время как мор-
ской крокодил, который распространен от Индии до Соломоновых островов
и северной Австралии, вырастает до 10 м в длину) Deinosuchus напоминал
метриоринхид, хотя он и не был так высоко специализирован для жизни в
воде. Его конечности все еще вполне пригодны в качестве опоры и для хож-
дения, а хвост, великолепный плавательный орган, не имеет никаких следов
плавника.
Хотя большинство крокодилов — это водные животные, среди них имеется
две группы, которые в кайнозойскую эру, по-видимому, процветали как на-
земные хищники. Обе группы имели уплощенные, кинжаловидные зубы,
напоминающие таковые больших хищных динозавров, типа Tyrannosaurus.
416
КАМЕННАЯ КНИГА
Такие зубы называются зифодонтными, что по древнегречески означает
“кинжаловидные зубы”. Пристихампсины (или зифодонты), одна из этих
групп, были эозухиями, которые просуществовали в Северном полушарии до
конца эоцена. В Африке они встречались еще в миоцене, а самый молодой
представитель известен из плейстоцена Австралии. Вторая группа — хищные
Sebecidae, которые процветали в Южной Америке до позднетретичного време-
ни, когда на этот континент по образовавшемуся сухопутному Панамскому
перешейку попадают плацентарные хищники. Себециды рассматриваются
как специфические мезозухий или помещаются в особую группу себекозухий.
Зифодонтные зубы известны также у другой, более ранней и не так широ-
ко распространенной группы крокодилов - хсисозухид. Их остатки происхо-
дят из юры Китая и мела Австрии.
В истории крокодилов выделяются два явления, которые поражают нас
наиболее сильно. Одно из них - это то, с какой очевидной легкостью эти реп-
тилии приобретают в процессе эволюции фитозавровую конструкцию тела, и
не однажды, а по крайней мере три раза, причем в других случаях это наблю-
дается у крокодилов и хампсозавров. Второе явление - выживание неспе-
циализированных животных и вымирание специалистов. Современные
крокодилы менее специализированы, чем ихтиозавры, плезиозавры, моза-
завры и многие исчезнувшие крокодилы. Однако эти более специализиро-
ванные формы исчезли, в то время как генерализованные крокодилы сохра-
нились. Подобно черепахам, они не являются реликтами, живущими на
грани вымирания, но представляют собой большую, процветающую группу.
Если люди преднамеренно не уничтожат их, они, вероятно, будут неплохо
себя чувствовать и через многие миллионы лет.
Глава XXVII
ЧЕШУЙЧАТЫЕ РЕПТИЛИИ
И ЯЩЕРОТАЗОВЫЕ ДИНОЗАВРЫ
Снова и снова мы убеждаемся, что важнейшие предковые организ-
мы РеДко производят сильное впечатление. Большинство из них бы-
ло небольшими, малозаметными и относительно неспециализиро-
ванными существами. Мало кто становился многочисленным; еще реже кто-
то переживал более чем одну или две эпохи. Как сказал один из экспертов,
организмам редко удается долгое время называться генерализованным пред-
ком. Животные либо вымирают, либо дают потомков, которые, насколько
это возможно с учетом анатомии, специализируются, стремясь выжить. Как
только новые приспособления закреплялись, потомки вытесняли своих при-
митивных предшественников, затем часто полностью вымиравших.
Из этого правила простоты и короткой жизни существовали несколько ис-
ключений. Диапсиды - одно из них. Они впервые появились в позднем карбо-
не Северной Америки, прошли всю пермь и лишь в конце триаса заместились
своими более прогрессивными потомками. Одним из самых ранних и наиболее
хорошо изученных диапсид был Petrolacosaurus, ящерицеподобное животное
с длинным хвостом, стройными конечностями и черепом длиной около 6 см.
Его челюсти были усажены заостренными зубами; другие зубы, короче и не та-
кие острые, были рассыпаны по небной поверхности пасти. Оба височных окна
были хорошо развиты, а пинеальное отверстие было относительно большим.
ЩИТКИ, ЧЕРЕПА И ЗМЕИ
Прогрессивные рептилии в триасе развивались по нескольким направле-
ниям. Одна из групп, лепидозавры, приобрела щитки и некоторые другие
приспособления. Среди них сфенодонтиды придерживались ползающего
стиля жизни и в процессе эволюции изменялись очень мало. Сфенодонтиды
дожили до наших дней в лице туатары или гаттерии из Новой Зеландии. Бо-
лее радикальные изменения по сравнению с исходными диапсидами произо-
шли у ринхозавров, приобретших мощные челюсти, притупленные зубы и
попугаеобразную морду, что позволило перейти к питанию грубой расти-
тельной пищей. Расцвет этой группы пришелся на триас и к концу этого пе-
риода ринхозавры вымерли. Лепидозавры-хампсозавры становятся вторич-
новодными рыбоядами и приобретают крокодилообразную форму. Они изве-
стны с триаса до эоцена (см. гл. XXVI).
© 1997. Ю.М. Губин, перевод на русский язык и изменения.
27 Каменная книга
418
КАМЕННАЯ КНИГА
Фиг. XXVII.1. Реконструкция черепа Megalania prisca, гигантской варанид-
ной ящерицы из плейстоцена Австралии. Длина черепа 74 см. Ниже череп со-
временного рода Varanus, его ближайшего родственника (фото Ф. Коффы, соб-
ственность музея Виктории, Мельбурн)
У другой, более важной группы лепидозавров исчезает одна из щечных
костей, а две другие уменьшаются в размерах, что приводит к появлению
настоящих ящериц (фиг. XXVII. 1). Ранние их представители были родст-
венны варанам и игуанам, встречающимся сейчас в тропиках. Они были
предшественниками хамелеонов и других специализированных ящериц
(фиг. XXVII.2).
Одна из групп раннемеловых варанов дала потомков, чьи лапы станови-
лись все меньше и меньше, а количество позвонков в крестце и ребер все
больше. Первым результатом этих изменений были длиннотелые коротконо-
гие ящерицы, ползавшие за счет изгибов тела. Позже эти абберантные яще-
рицы дали змей: ранние змеи близки современным боа и питонам, у которых
еще сохранились в скелете следы конечностей. Ранние змеи также теряют
остатки костной дуги позади глаз, так что на месте двух теменных окон оста-
ется единое широкое отверстие. Как у мозазавров, челюсти становятся сво-
бодно прикрепленными и соединенными двойным суставом. Это обеспечива-
ло широкое открытие пасти, при котором ее хозяин мог проглотить добычу
толще, чем он сам.
Наиболее древние змеи были неядовитыми, хотя фоссилии, которые мог-
ли принадлежать гадюкам, были встречены уже в меловых и палеоценовых
формациях. Первые достоверные остатки рептилий, принадлежавшие на-
стоящим гадюкам и кобрам, происходят из миоценовых отложений. Ямкого-
ловые, в том числе гремучие змеи, появились в плиоцене. Ядовитые змеи
также включают настоящих морских змей - гидрофид. Они встречаются от
Мадагаскара до Панамы, концентрируясь в основном в районе Австралии,
причем некоторые из них никогда не покидают водной среды, даже для раз-
множения, перейдя к живорождению в море.
ОТ ТЕКОДОНТОВ К ДИНОЗАВРАМ
Кроме лепидозавров, примитивные диапсиды дали начало другой группе
рептилий - архозаврам или “ящерицам-правителям”. Эта группа доминиро-
вала на суше со среднего триаса по конец мела. Наиболее известными их
ЧЕШУЙЧАТЫЕ РЕПТИЛИИ И ЯЩЕРОТАЗОВЫЕ ДИНОЗАВРЫ
419
Фиг. XXVII.2. Megalania (длина 5.5 м), пирующая на трупе Diprotodon (рису-
нок Ф. Найта, собственность музея Виктории, Мельбурн и Пионер Дизайн
Студио)
27*
420
КАМЕННАЯ КНИГА
Фиг. XXVII.3. Текодонты и строение таза двух отрядов динозавров
представителями были динозавры, но Archosauria также включали прими-
тивных текодонтов, от которых отошли все другие группы архозавров, лета-
ющих рептилий или птерозавров и единственную, дожившую до современно-
сти группу - крокодилов.
Вернемся в мир триаса. Эволюционирующие текодонты развили новые
черты в черепе, челюстях и тазовом поясе, которые и обеспечили успех архо-
заврам (фиг. XXVII.3). Перед каждым глазом в черепе появляется отверстие,
другое характерное отверстие развивается в нижней челюсти. Изменения в
тазе включают увеличение площади прикрепления мышц, углубление “кар-
мана” для головки бедра и присоединение дополнительных позвонков к тазу,
что усилило его связь с позвоночником. Все это было важным для перехода к
полувертикальному двуногому передвижению и увеличению размеров, в чем
многие текодонты и их потомки преуспели.
Одной из бипедальных текодонтов была Euparkeria, которая бегала на длин-
ных задних лапах, балансируя своим полуприподнятым телом с длинным уп-
ругим хвостом. Длина животного колебалась от 1 до 1.5 м, а рост — 25—37 см.
Этозавры относились к текодонтам, продолжавшим передвигаться на че-
тырех лапах. Впервые появившись в триасе (фиг. XXVII.4), они были среди
первых, покрытых тяжелым костным панцирем наземных позвоночных.
Другой четырехногой группой текодонтов были Phytosauria (см. гл. XXVI).
Два динозавровых отряда. В триасе текодонты дали начало наиболее фан-
тастической ветви рептилий — динозаврам.
Термин “динозавр”, что обозначает “ужасный ящер”, как полезен, так и
обманчив. Он полезен, так как приложим к колоссальному разнообразию ар-
хозавров, обитавших в мезозое и не имеющих другого обиходного имени в на-
ши дни. Он обманчив, так как предполагает, что все эти рептилии были ог-
ромными и свирепыми хищниками, относящимися к единой естественной
группе. В действительности, так называемые динозавры включали как мас-
сивных гигантов весом в 40 или 50 т, так и стройных грациозных животных
ЧЕШУЙЧАТЫЕ РЕПТИЛИИ И ЯЩЕРОТАЗОВЫЕ ДИНОЗАВРЫ
421
Фиг. XXVII.4. Typothorax, позднетриасовый этозавр из Северной Америки.
Длина тела 3.7 м
размерами меньше домашнего петуха. Хотя многие виды были хищниками,
еще больше их современников питалось только растительностью. К сказан-
ному надо добавить, что эти разнообразные рептилии образуют два самосто-
ятельных отряда, хотя и происходящих от общих ранних текодонтов.
Главные отличия между отрядами проявлялись в строении тазового ком-
плекса. У заурисхий, или “ящеротазовых”, эта структура остается трехлуче-
вой формы, присутствующей у ранних текодонтов и развивающейся у кроко-
дилов. У орнитисхий (“птицетазовых”) лобковая кость обычно имеет две вет-
ви, одна из которых протягивается вперед, а другая - назад, под седалищную
кость. Зубной ряд заурисхий идет вперед до переднего края челюстей, тогда
как у большинства орнитисхий останавливается не доходя до него. У всех ор-
нитисхий, за исключением нескольких родов, челюсти были покрыты рого-
вым чехлом. Эти признаки, а также характер зубов говорят, что все орнити-
схии были растительноядными, в том время как заурисхии включали при-
мерно по половине как плотоядных, так и растительноядных животных.
НАИБОЛЕЕ ДРЕВНИЕ ДИНОЗАВРЫ
Переход от текодонтов к динозаврам-заурисхиям был малозаметным и
произошел в досреднетриасовое время. Во второй половине этого периода за-
урисхии уже бродили по Европе, Азии и долинам востока Северной Амери-
ки. Они оставили редкие скелеты и бесчисленные следы на темно-красных
грязных песках, позже превратившихся в камень. Находимые с трудом те-
перь, эти следы были обычны в дни, когда триасовый “железистый песча-
ник” активно добывался для строительства церквей, тюрем и даже дренаж-
ных каналов под железными дорогами.
Триасовые динозавры. Строение костей и следы показывают, что многие
позднетриасовые динозавры были хищниками со стройными лапами,
длинной шеей, маленькой головой и длинным хвостом. Многие достигали
90-120 см длины, некоторых, длиной 2-2.5 м, уже можно назвать больши-
ми, а существа, достигавшие 5-6 м - были гигантами. Все они передвига-
лись на задних конечностях, которые имели три, редко четыре длинных
пальца и еще один настолько короткий, что он редко оставлял отпечаток на
земле. По этой причине их ископаемые следы вначале ошибочно принимали
за следы птиц.
Детальную картину позднетриасовой жизни позволяют восстановить на-
ходки из формации Чинли на юго-западе США. В те времена мы могли бы
увидеть здесь возвышенности, покрытые лесами шишконосных араукарие-
422
КАМЕННАЯ КНИГА
Фиг. XXVII.5. Coelophysis, примитивный динозавр из позднего триаса Север-
ной Аризоны и Нью-Мексико. Длина до кончика хвоста 2.4 м
вых “сосен”, чьи упавшие стволы смывались потоками вниз по склонам и от-
лагались на илистых отмелях озер, чтобы в конце концов образовать широко
известный Аризонский Окаменелый лес и другие, чуть менее красивые леса.
Большие папоротники росли под деревьями, маленькие цикадофиты запол-
няли открытые места, а хвощи образовывали заросли вдоль русел потоков
(см. главу XXIII). Климат был умеренным, хотя иногда облака пепла от вул-
канических извержений заполняли воздух.
Потоки, водоемы и мелкие озера были местом обитания мидий, двояко-
дышащих и ганоидных рыб и крупных амфибий. Панцирные текодонты до-
стигали в длину 3.5-4.5 м. На них охотились ящеротазовые динозавры дли-
ной до 6 м, передвигавшиеся в основном на двух задних лапах. В болотах ча-
сто встречались фитозавры до 14 м длиной, а в лесах на возвышенностях
обитал по крайней мере один род дицинодонтов (Placerias), имевший клыки
и клюв на челюстях. Он мог сталкиваться с более мелкими (до 2.5 м) ящеро-
тазовыми динозаврами - стройными двуногими существами, обладавшими
высокими узкими черепами и полыми костями конечностей. Подобно своим
предкам текодонтам, они хватали добычу передними конечностями и челю-
стями, а тело удерживали в равновесии с помощью вытянутых хвостов
(фиг. XXVII.5). Вес таких динозавров доходил до 20-23 кг, и на юго-западе
США они были представлены родом Coelophysis.
ЦАРСТВО ГИГАНТСКИХ ЗАВРОПОД
В начале юрского периода на западе США болота сменяются обширными
пустынями. Постоянно дующие по этой бесплодной земле ветры навевали пе-
счаные дюны, которые сегодня представляют собой толщу косослоистых пе-
счаников более 1100 м мощности. Туристы могут их наблюдать в каньонах,
арках, башнях, утесах в районе Моавской пустыни (штат Юта, США).
Хотя пустыни и были малопригодны для обитания динозавров, в литифи-
цированных юрских дюнах было обнаружено несколько их костей. Другие
ЧЕШУЙЧАТЫЕ РЕПТИЛИИ И ЯЩЕРОТАЗОВЫЕ ДИНОЗАВРЫ
423
Фиг. XXVII.6. Древнейшие динозавры
Слева - Plateosaurus, примитивный завропод из триаса Европы и Аргентины,
длина 4.9—6.1 м. Справа - Anchisaurus из восточных районов Северной Амери-
ки, был длиной 2.4 м
районы Северной Америки были более благоприятными, и в теплых и часто
болотистых низинах заурисхии процветали. Заурисхии подразделяются на
две группы: тероподы (“звероногие”) и завроподы (“ящероногие”). Названия
этих групп мало соответствуют действительности, но сами группы очень
важны. За несколькими исключениями, тероподы передвигались на двух
задних лапах с тремя опорными птицеподобными пальцами на каждой, име-
ли короткие передние лапы и были хищниками. Завроподы, напротив, пере-
двигались на четырех массивных конечностях и имели зубы, приспособлен-
ные для поедания мягких сочных растений.
Предки гигантов. Разделение ящеротазовых динозавров началось в триа-
се. Среди стройных рептилий, что бродили по долинам востока Северной
Америки, встречались более крупные с мощными задними лапами живот-
ные рода Anchisaurus. Это были прозавроподы или “первые ящероногие”.
Они имели очень маленькие головы, тупые зубы и пять пальцев на каждой
лапе, из которых четыре хорошо развиты (фиг. XXVII.6). Они могли пере-
двигаться либо в полуприподнятом положении, либо на четырех лапах, а пи-
тались как растениями, так и животными. Так, в рацион Thecodontosaurus,
видимо, кроме растений входили и его близкие родственники текодонты.
Прозавроподы были очень широко распространены в позднем триасе и
ранней юре, и их остатки известны со всех континентов, кроме Антарктиды.
Plateosaurus из Западной Европы и Аргентины достигал 6 м в длину и, хотя
и мог передвигаться на задних лапах, вероятно, проводил больше времени на
четырех лапах, чем Anchisaurus.
Самым большим прозавроподом был Euskelosaurus из позднего триаса
Южной Африки. Он достигал в длину 12 м и весил около двух тонн. Самое не-
обычное в его строении — отсутствие признаков, которые указывали бы на то,
что в недалеком прошлом он произошел от двуногих предков. Является об-
424
КАМЕННАЯ КНИГА
Фиг. XXVII.7. Череп, голова и левая задняя стопа Camarasaurus из позднеюр-
ской формации Моррисон Северной Америки. Взрослая рептилия достигала
12.2 м длины
щепризнанным, что все более поздние архозавры происходят от двуногих те-
кодонтов. Однако существование Euskelosaurus с настоящей четвероногой
постановкой конечностей так рано в геологической летописи архозавров поз-
воляет допустить, что завроподы происходят непосредственно от четвероно-
гих форм и никогда не имели двуногих предков.
Триумф завропод. Преимущественно более крупные, чем прозавроподы и
никогда не передвигавшиеся только на задних лапах, завроподы легко отли-
чаются по строению передних лап. У них на передней лапе есть только один
коготь, и он располагается на большом пальце. Напротив, прозавроподы
имели когти по крайней мере на первых трех пальцах.
Завроподы появились в ранней юре, но их расцвет начинается лишь в по-
здней юре. Они становятся многочисленными и разнообразными, одновре-
менно сильно увеличиваясь в размерах. Самые интересные и обильные на-
ходки позднемеловых завропод известны из североамериканской формации
Моррисон, Северной (Тендагуру, Танзания) и Восточной Африки и Китая
(Сычуань).
Формация Моррисон сложена глинами, мергелями и песчаниками, кото-
рые накапливались в болотах, мелких озерах и широких потоках, блуждав-
ших по низкой равнине. Цикадофиты, гинкго и хвойные растения формиро-
вали леса на фоне мелколесья, состоявшего преимущественно из папоротни-
ков. Хвощи и сочные растения в изобилии росли на болотах. Все это
указывает на то, что климат был умеренным и ровным, хотя и с чередовани-
ем влажных и сухих сезонов.
Один из обитавших там завропод, Camarasaurus, был относительно неспе-
циализированным животным, достигавшим 18 м длины во взрослом состоя-
нии, со стройными шеей и хвостом и короткими передними лапами — послед-
ний признак напоминает предков теропод (фиг. XXVII. 7). И задние и перед-
ние лапы были массивные, причем на последних имелся только один полный
палец.
ЧЕШУЙЧАТЫЕ РЕПТИЛИИ И ЯЩЕРОТАЗОВЫЕ ДИНОЗАВРЫ
425
Фиг. XXVII.8. Гигантские завроподы
Brachiosaurus (слева) п. Apatosaurus или бронтозавр (справа) были одними из са-
мых больших ящеротазовых динозавров. Оба жили в Колорадо и прилегающих
районах США в позднеюрское время. Brachiosaurus также обитал в Централь-
ной Европе. Уже когда этот рисунок был сделан, выяснилось, что череп Apato
saurus больше похож на Diplodocus, а не на Camarasaurus, как показано здесь
Apatosaurus, завропод, больше известный под ошибочным названием
Brontosaurus (“громовой ящер”) был гораздо крупнее и специализирован-
ной. Он достигал в длину 20—23 м, имел толстое короткое тело, гибкую шею
и хвост, который у основания был достаточно массивным, а кзади становил-
ся все тоньше, превращаясь в плетеобразный (фиг. XXVII.8). Лапы были
больше похожи на толстые колонны, как у слонов, и доходили в диаметре до
65-70 см. Когти были только на полностью развитых пальцах: на задних ла-
пах было по три массивных когтя, а на передних - по одному. Вес этой репти-
лии мог доходить до 27-32 тонн.
Apatosaurus часто сталкивался со своим родственником Diplodocus. Это
существо достигало в длину 24-27 м и 4 м в высоту от земли до самой верхней
точки кривой спины непосредственно впереди от тазового пояса. Почти одну
треть длины животного составляла шея, больше половины приходилось на
хвост, который к кончику приобретал стройную, плетеобразную форму.
Diplodocus был гигантом, весящим до 18 т, что приблизительно соответству-
ет весу пяти крупных слонов.
426
КАМЕННАЯ КНИГА
Фиг. XXVII.9. Череп Diplodocus длиной 60 см. Ноздри обозначены буковой N,
орбиты - Е
Фиг. XXVII.10. Череп Brachiosaurus длиной 76 см. Ноздри обозначены буквой
N, орбиты — Е
Спереди этого огромного тела располагалась невзрачная голова длиной
60 см. Ноздри открывались в единую лунку, находившуюся между и
несколько выше глаз, тупые зубы имели наклон наружу и, вероятно, ис-
пользовались для обдирания, а не откусывания растений (фиг. XXVII.9 и
XXVII. 10).
Мозг был маленьким гладким органом, весящим не более 100—150 г и по
размерам соизмеримым с мозгом кролика. Если взять отношение объема
мозга небольших рептилий к их телу и экстраполировать на размер завро-
под, то выяснится, что “ожидаемый” мозг для рептилии такого размера, как
Diplodocus, в пять раз больше, чем объем мозговой полости в ископаемых ос-
татках. Так что, вероятно, они не были среди самых интеллектуально раз-
витых динозавров. Однако завроподы, как группа, просуществовали более
100 млн. лет, в течение которых многие другие группы динозавров успели и
возникнуть и исчезнуть (фиг. XXVII.11 и XXVII.12). Поэтому остается счи-
тать, что умственных способностей завропод было вполне достаточно для их
типа жизни.
Diplodocus представлял собой малоподвижное существо, удовлетворявше-
еся умеренным количеством еды. Мозг был способен переработать зритель-
ЧЕШУЙЧАТЫЕ РЕПТИЛИИ И ЯЩЕРОТЛЗОВЫЕ ДИНОЗАВРЫ
427
Фиг. XXVII.il. Mamenchisaurus, самый длинношеий завропод из средней или
верхней юры Сычуаня, Китай (собственность музея Виктории, Мельбурн)
Фиг. XXVII.12. Mamenchisaurus (рисунок П. Траслз, собственность музея Вик-
тории, Мельбурн)
428
КАМЕННАЯ КНИГА
ную, обонятельную и звуковую информацию и передавать ее в нервные узлы,
расширения спинного мозга в районах плечевого и тазового поясов. Эти
нервные узлы координировали ходьбу и движения хвоста. Такие действия
плюс добывание продовольствия, спаривание и, возможно, миграции в ста-
дах заполняли жизнь Diplodocus и других завропод.
Еще больше был Brachiosaurus, остатки которого найдены на западе
Сев. Америки, в Африке и Португалии. В отличие от остальных завропод
этот гигант имел передние лапы длиннее задних, так что положение его
спины напоминало современного жирафа. Немного цифр: плечевая кость
Brachiosaurus достигала 210 см, высота тела на уровне плечевого пояса - 5
м 80 см; длина шеи - 8.5 м и голова находилась на высоте 12 м над землей.
Хотя хвост брахиозавра был намного короче, чем у Apatosaurus, общая
длина тела Brachiosaurus достигала 22 м 60 см. Массивное тело с ребрами
длиной до 2.6 м весило по крайней мере 37 т. При этом вес мозга составлял
только 200 г.
Все эти измерения сделаны на одном скелете брахиозавра, еще не достиг-
шем максимальных размеров. Есть находки костей более крупных, почти на
13%, особей. Подсчет показывает, что такие животные достигали 13м высо-
ты, 25 м длины и веса в 45 т.
Из формации Моррисон известны еще два, может быть, более крупных за-
вропода, однако их остатки весьма неполны. Меньший и лучше известный -
Supersaurus, длина которого могла доходить до 30 м. Ultrasaurus был еще
длиннее и его предполагаемый вес мог достигать 130 т (фиг. XXVII. 13 и
XXVII. 14). Если это было так, то эта рептилия была тяжелей современного
синего кита, до сих пор считавшегося самым тяжелым животным в истории
Земли.
Но даже Ultrasaurus выглядел бы маленьким на фоне Breviparopus, завро-
пода из Марокко, известного только по следам, и недавно обнаруженной фор-
мы с американского юго-запада. Его назвали Seismosaurus или “сотрясатель
Земли”; по размерам следов и расстоянию между ними впервые описавшие
их ученые предположили, что животное было 48 м длиной.
Скелет завропод. Завроподы были не только самыми большими рептили-
ями в мире; они также были самыми большими из наземных животных.
Проблемы, возникающие между размерами и окружающей средой были ре-
шены комплексом из костей, связок, мускулов, привычек и репродуктив-
ных органов.
Первой и наиболее очевидной проблемой было обеспечение каркаса для
тела. Ребра также были деталями каркаса, но наиболее важной, критичес-
кой частью здесь был позвоночник. Позвонки должны были быть большими
и крепкими, но не литыми, так как их монолитность приведет к такому уве-
личению веса, которое ноги не смогут выдержать. С превращением завропод
в гигантов их позвонки приобрели ажурное строение, и можно сказать, что
они стали по большей части состоять из полостей (фиг. XXVII. 15). В реально-
сти эволюция предпочла пренебречь костью, там где это возможно, а до-
бавить разнообразные перегородки и гребни, сохранявшие жесткость по-
звонка и противостоявшие нагрузкам. Как результат, была получена струк-
тура, сформированная связками и межполостными перегородками и больше
похожая ажурностью на современный стальной мост.
Однажды созданное тело необходимо было поддерживать и перемещать с
места на место. Для этой цели завроподы использовали длинные выпрямлен-
ные вертикально массивные ноги. Их размер можно оценить на примере пле-
чевой кости, Brachiosaurus, достигавшей в длину 2.1м. Бедренная кость, Ар
ЧЕШУЙЧАТЫЕ РЕПТИЛИИ И ЯЩЕРОТАЗОВЫЕДИНОЗАВРЫ
429
Фиг. XXVII.13. Модель передней лапы гигантского динозавра Ultrasaurus, за-
вропода из позднеюрской формации Моррисон, Колорадо, США (фото собст-
венность Дж.А. Дженсена)
atosaurus, была более массивной, длиной около 2 м и имевшей в обхвате на
уровне середины длины 90 см.
Тропы завропод. При обсуждении, могли ли даже такие массивные ноги
поддерживать огромное тело завропод, издавна используется интересный ар-
гумент. Те, кто утверждали, что это невозможно, указывали на длинные ко-
сти конечностей, оканчивавшиеся хрящом, и говорили, что последний был
бы разрушен весом более 20 т. Поэтому, мол, рептилии не могли передвигать-
ся по суше. Однако этой беды завроподы избежали бы в воде, где они могли
бы плавать, пуская в ход лапы и тяжелые хвосты.
Это предположение, но в его пользу говорят следы завропод, найденные в
раннемеловых породах на Пэлюкси Ривер (Глен Роуз, Техас, США). Оставив-
шие следы, вероятно, были близки к Apatosaurus и Diplodocus, хотя их зад-
ние лапы несли четыре когтя вместо трех, а на передних не было ни одного
(фиг. XXVII. 16). Эти существа, очевидно, пробирались в прибрежных боло-
тах раннемелового моря и оставляли следы на известковистом песке дна.
Здесь же были найдены раковины устриц и других морских животных. Сле-
ды были 96 см длиной и 66 см шириной, а расстояние между ними -3.7 м.
430
КАМЕННАЯ КНИГА
Фиг. XXVII.14. Лопатка Ultrasaurus на дне карьера Драй Мейса, Колорадо,
США (фото собственность Дж.А. Дженсена)
Большинство следов было оставлено животными, передвигавшимися на
четырех лапах, а их хвосты были приподняты над землей или дном. Один
из динозавров на некотором расстоянии оставил след своего хвоста, другой
шел на передних лапах, в то время как задняя часть тела плавала. Чтобы
повернуть, эта рептилия сделала толчок одной из задних лап наружу и
вверх. При этом когти вырыли на грунте глубокие желобки, сохранившие-
ся на окаменевшей породе. Вывод: завроподы жили в воде, в том числе на
глубинах.
Критики не подвергают сомнению эти факты, но дают им другое объясне-
ние. Кости конечностей, говорят эти критики, росли у завропод на протяже-
нии всей жизни. Поэтому, если их хрящевые окончания не заместились кос-
тью, то этот хрящ был достаточно крепок, чтобы вынести вес тела. Более ва-
жен еще один факт. Следовые дорожки Глен Роуз содержат следы разных
типов. У первого типа они глубокие, но не отчетливые, как будто бы влаж-
ные, мягкие осадки, составляющие края следа, обвалились вниз вскоре по-
сле прохода рептилий. Другие следы отчетливей, но относительно мельче,
как будто бы грунт был крепче и суше. Обычное отсутствие следов хвоста
ЧЕШУЙЧАТЫЕ РЕПТИЛИИ И ЯЩЕРОТАЗОВЫЕ ДИНОЗАВРЫ
431
Фиг. XXVII. 15. Детали строения завропод
Слева — головка бедра Camarasaurus. Поверхность торца кости при жизни жи-
вотного была покрыта более мягким хрящом. В центре - на позвонке из шеи Ap-
atosaurus видно, что перегородки и пластины сохраняли жесткость кости, а пу-
стоты уменьшают вес. Справа - человек 1.7 м ростом рядом с плечевой костью
Brachiosaurus (2.1 м), найденной в Колорадо, США
говорит просто о том, что завроподы большую часть времени пребывания на
суше держали свои хвосты приподнятыми над землей.
Кроме того, хотя скелеты завропод и массивны, они облегчены макси-
мально возможно. Если же завроподы были полностью водными, подобно
китам, исчезает необходимость в облегчении скелета. Основания лап завро-
под были относительно небольшими (по отношению к размерам) и из-за это-
го животные могли вязнуть в мягком илистом грунте болот. Эти лапы ка-
жутся лучше приспособленными к движению по твердому, плотному осно-
ванию. В дополнение ко всему, тела завропод высокие и неширокие,
подобно наземному слону, а не расширены, как у амфибиотического гиппо-
потама.
Таким образом, к решению этого спора мы сможем приблизиться, только
если удастся найти следы завропод в осадках, сформировавшихся либо на
суше, либо на мелководье, при глубине не более 50-60 см. Сейчас же мы мо-
жем только сказать, что завроподы, по крайней мере время от времени, бро-
дили по воде, их тела имели положительную плавучесть, а сами они умели
плавать.
Еще одна проблема активно обсуждается - тип размножения. В южной
Франции были найдены ископаемые яйца рептилий диаметром 30 см в тех
же отложениях, где были обнаружены остатки завропода Hypselosaurus. В
Аргентине полный скелет ювенильного прозавропода Mussaurus длиной
около 25 см был найден рядом с ископаемыми яйцами диаметром 2.5 см. Та-
кие ассоциации напрямую дают возможность предположить, что завроподы
откладывали яйца, а не были живородящими. Можно допустить, что, как со-
432
КАМЕННАЯ КНИГА
Фиг. XXVII. 16. Гигантские завроподы
Вверху -Apatosaurus, идущий на передних лапах и гребущий задними. Внизу -
Diplodocus, идущий по мелководью; его хвост подвсплыл
временные крокодилы, они защищали своих детенышей после вылупления
на свет.
Когда исчезли последние завроподы? Завроподы в северном полушарии
достигли своего расцвета в позднеюрское—раннемеловое время. Ранее счита-
лось, что после этого наступил упадок и лишь несколько разбросанных по
планете родов небольших размеров дожили до позднего мела. Последние от-
крытия показывают, что и большие завроподы добрались до конца периода.
Так выяснилось, что среди разнообразных позднемеловых завропод Арген-
тины присутствовали гиганты Chubutisaurus, достигавшие 23 м длины. В Се-
верной Америке были найдены остатки Alamosaurus, весившего до 27 тонн и
21м длиной. Однако в целом на планете разнообразие гигантских позднеме-
ловых завропод было меньше, чем в поздней юре.
ХИЩНЫЕ КАРНОЗАВРИДЫ
Завроподы обладали “большим” количеством мяса - от десятков кило-
граммов до десятков тонн. Благодаря этому богатству появилась возмож-
ность стать гигантами и хищниками.
Ранние, среднетриасовые, плотоядные динозавры были небольшими
трехпалыми бипедальными тероподами. К концу триаса некоторые из их по-
томков достигли 2.5-3.7 м длины и веса 70 кг и стали гораздо сильней, чем
самые прогрессивные ящерицы. Несомненно, в качестве добычи у них при-
сутствовали Plateosaurus, давшие начало завроподам.
ЧЕШУЙЧАТЫЕ РЕПТИЛИИ И ЯЩЕРОТАЗОВЫЕ ДИНОЗАВРЫ
433
Фиг. XXVII. 17. Три позднеюрских хищных динозавра. Они жили в районе со-
временных Скалистых гор на западе США. Самый крупный AUosaurus был
10.4 м длиной; Ceratosaurus был 5.3 м длиной, а самый маленький — Ornitho-
lestes длиной 1.8 м
Возможно, эволюция плотоядных динозавров шла впереди их жертв.
Древнейшие завроподы были найдены в ранней юре, тогда как полностью
сформированные карнозавры, видимо, обитали уже в позднем триасе Евро-
пы и Южной Африки. Однако фоссилии этого возраста настолько проблема-
тичны и фрагментарны, что могут принадлежать и большим хищным теко-
донтам. В раннеюрское время представители семейства, получившего назва-
ние мегалозавры, или “большие ящеры”, уже включали животных длиной
от 3.5 до 9 м с 30-сантиметровыми и более черепами и пильчатыми зубами,
которые загибались назад в процессе роста. Их передние лапы были короче,
чем у Plateosaurus, а задние, сильные и мощные, несли длинные изогнутые
когти. Мегалозавры, возможно, поджидали добычу в засаде, затем настига-
ли ее и убивали, переломив позвоночник. Копти удерживали добычу во вре-
мя поедания, как это делают в наши дни стервятники.
Среднеюрские карнозавры известны мало, а в позднеюрское время группа
становится многочисленной и разнообразной (фиг. XXVII. 17). Одним из са-
мых известных ее членов является Ceratosaurus, найденный недалеко от Ка-
ньон Сити в штате Колорадо, США. При ходьбе его высота достигала 2.1 м, а
длина от носа до кончика хвоста — 5.3 м. Голова была 51 см длиной и 33 см
высотой с саблеобразными зубами, поднимавшимися на 5 см над челюстями.
На носу был притупленный узкий рог и по небольшому выступу перед каж-
дым глазом. Подобно мегалозаврам, Ceratosaurus передвигался на задних
лапах, балансируя длинным хвостом.
28. Каменная книга
434
КАМЕННАЯ КНИГА
Мощный и свирепый охотник Ceratosaurus, однако, кажется маленьким
на фоне Allosaurus. Эта рептилия была 13 м длины, 2.4 м высоты с наклонен-
ной к земле головой и 4.3-4.6 м, когда поднимала тело и голову вверх. Ее го-
лова была 91 см длиной с 15-сантиметровыми пильчатыми зубами. Прикреп-
ленная к заднему концу черепа нижняя челюсть, опускаясь, открывала ог-
ромную пасть. Ею это чудовище могло наносить мощные ужасные удары и
использовать для отрывания больших кусков добычи.
В Национальном Динозавровом парке в штате Юта, США, остатки аллоза-
вра были найдены вместе с диплодоком и другими завроподами. На знамени-
том динозавровом местонахождении Боун Кэбин в штате Вайоминг, США,
кости Apatosaurus и других завропод были поцарапаны зубами Allosaurus,
найденными тут же. Очевидно, ужасный укус хищника был так силен, что
сломались несколько из его 15-сантиметровых зубов, а само животное ушло,
отказавшись от добычи. Можно, тем не менее, допустить, что многие древние
рептилии питались другим образом, чем Allosaurus.
Остается неясным еще один вопрос. Мог ли Allosaurus нападать на гигант-
ских беззащитных завропод или он был падалеядом? Одни эксперты говорят
одно, другие — второе, и все могут быть правы. Вполне возможно, что аллоза-
вры нападали на молодь гигантов или на таких животных, как Camarasau-
rus. Более того, по крайней мере один карнозавр размером с Allosaurus под-
крадывался к завроподам в прибрежном болоте, чьи отложения были
найдены около Глен Роуз в Техасе. Но даже ныряние на глубину не обяза-
тельно спасло бы жизнь завроподу, так как тероподы тоже были хорошими
пловцами.
Тираннозавриды, “ящеры-тираны”. В первой половине мелового периода
некоторые из теропод выделились в группу тираннозаврид, “ящериц-тира-
нов”, возможно самых крупных из всех сухопутных хищников.
Одним из ранних тираннозаврид был относительно стройный Gorgosau-
rus, который обитал во влажных сильно залесенных низинах на территории
нынешней провинции Альберта, Канада (фиг. XXVII. 18). Будучи в высоту
2.1м стоя, он имел низкий череп с удивительно длинными зубами, на перед-
них лапах у него было по два пальца, на задних по четыре с рудиментом пя-
того. Брюшные ребра защищали живот, что является примитивным при-
знаком.
Однако это было только начало. У более поздних видов Gorgosaurus идет
увеличение размеров: длина до 8.8 м, высота в приподнятом положении до
2.7 м, а низкий череп, вооруженный изогнутыми зубами, достигал 56 см дли-
ны. Брюшные ребра еще хорошо развиты, но рудиментарный пятый палец
на задних лапах исчезает. Рептилия, возможно, передвигалась наклонив-
шись вперед, как Ceratosaurus и Allosaurus, но могла и быстро бегать на ко-
роткие расстояния. Его передние лапы были маленькими и, видимо, беспо-
лезными. Они уменьшились настолько, что на них осталось лишь по два
пальца.
Позже всех появился Albertosaurus и его более известный родственник
Tyrannosaurus. Оба были массивными, приподнятыми динозаврами с самы-
ми крупными среди заурисхий головами. Tyrannosaurus был 14.3 м длиной,
5.5 м высотой и весил от 8 до 10 тонн. Его коленный сустав был на высоте
1.8 м над землей, кости на задних лапах 15—20 см длиной; череп достигал
130 см от носа до затылка, зубы представляли собой пильчатые лезвия
15-сантиметровой длины и 2.5 см ширины (фиг. XXVII.19 и XXVII.20). Пе-
редние лапы имели лишь по два пальца и были слишком малы, чтобы удер-
живать пищу.
ЧЕШУЙЧАТЫЕ РЕПТИЛИИ И ЯЩЕРОТАЗОВЫЕ ДИНОЗАВРЫ
435
Фиг. XXVII. 18. Хищные тероподы
Справа - страусоподобный Struthiominus из мела Альберты, Канада. Высота
животного 2.4—2.7 м. Слева — один из ранних видов Gorgosaurus высотой 2.1м
из мела Альберты
Соблазнительно представить, что добыча Tyrannosaurus по пропорциям
соответствовала его размерам. В действительности же этот гигантский хищ-
ник жил в конце позднего мела, через 50 миллионов лет после вымирания
Diplodocus и Brachiosaurus. Меньшие завроподы были редки. Добычу яще-
риц-тиранов составляли растительноядные рептилии 6-9 м длиной, которые
пробирались, плавали и бродили на низменностях запада Сев. Америки. Они
потеряли в размерах, но выиграли в количестве. Тираны редко оставались
голодными, они просто получили более разнообразный стол, чем их предше-
ственник Allosa urus.
Колючие хищники. Самыми странными среди всех теропод были спиноза-
вриды. Они жили в позднемеловое время в Европе и Северной Африке. Хотя
их находки неполные, однако можно сказать, что североафриканский Spino
saurus был крупным животным, у которого из спины торчали вверх длинные
невральные отростки-шипы, как у Dimetrodon, достигавшие 1.8 м длины.
Удивительно, как такие рептилии могли ходить на двух лапах и убивать ак-
тивную добычу. Может быть они затаивались в засаде и нападали, или они
питались падалью?
Тетраподы с сильно развитыми невральными отростками, такие как
пермские пеликозавры Dimetrodon, Edaphosaurus из Техаса, триасовый те-
кодонт Ctenosauriscus из Германии и североафриканские позднемеловые
динозавры Ouranosaurus и Spinosaurus все жили в приэкваториальных рай-
онах того времени. Это показывает, что “парус” на их спине мог быть полезен
для регулирования температуры тела, отдачи лишней энергии. Если на-
править кровь в кровеносные сосуды паруса, когда животное находится в
тени или даже, когда плоскость паруса параллельна лучам солнца, темпера-
тура крови будет понижаться. В случае, если температура животного слиш-
28*
436
КАМЕННАЯ КНИГА
Фиг. XXVII. 19. Tyrannosaurus, хищный динозавр 14.3 м длиной, 5.5 м высотой
и черепом длиной 127 см. Мел Монтаны, США
ком низкая, оно может нагреть себя на солнце быстрее, чем животное без па-
руса - просто поворачивая плоскость паруса перпендикулярно солнечным
лучам.
Этот “парус” мог также использоваться, чтобы зрительно увеличить раз-
меры животного в конкурентной борьбе между самцами в период размноже-
ния, либо для отпугивания более крупных хищников.
Длиннорукие тероподы. По крайней мере два теропода из позднего мела
Азии известны только по своим передним лапам. Но каким! (фиг. XXVII.21).
Каждая лапа 2.5 м длиной, и каждая вооружена тремя мощными костями.
Deinocheirus имел относительно стройные лапы, a Therizinosaurus - более
массивные. Так как полный скелет ни одного из этих животных еще не най-
ден, у нас нет полного представления, как они выглядели. Скорее всего, они
походили на Tyrannosaurus с сильно увеличенными передними лапами.
Ловцы птиц и похожие на птиц. Пока одни динозавры превращались в ги-
гантов, другие становились стройнее и проворнее. У некоторых даже лапы,
тело и шея становились похожими на современного страуса.
Эта тенденция началась с триасового Coelophysis или со стройного двуно-
гого Compsognathus, жившего в позднеюрских прибрежноморских лесах
Германии. Так как две трети их 76-сантиметровой длины составляли шея и
хвост, вес такого животного был не больше, чем крупного петуха. Пальцы
были длинными и имели когти, а маленькие челюсти были усажены остры-
ЧЕШУЙЧАТЫЕ РЕПТИЛИИ И ЯЩЕРОТАЗОВЫЕ ДИНОЗАВРЫ
437
Фиг. XXVII.20. Череп Tyrannosaurus длиной 127 см (фото Ф. Коффы, собствен-
ность музея Виктории, Мельбурн)
Фиг. XXVII.21. Передние лапы Deinocheirus, в остальном неизвестного теро-
пода из позднего мела Монголии. Каждая лапа 2.6 м длиной (фото Ф. Коффы,
собственность 3. КиеланЯворовской и музея Виктории, Мельбурн)
438
КАМЕННАЯ КНИГА
Фиг. XXVII.22. След небольшого динозавра из раннего мела Австралии
Длина отпечатка 20 см (фото Ф. Коффы, собственность музея Виктории, Мель-
бурн)
ми зубами. Compsognathus, видимо, обитал в подлеске и охотился на еще бо-
лее мелких рептилий. На самом деле, у одного из образцов в районе живота
нашли остатки такой пищи.
Ornitholestes, из позднеюрской формации Моррисон в штате Вайоминг,
США, был 1.8 м длины с крепким телом и относительно длинным не сгибаю-
щимся хвостом. Передние лапы превращаются в трехпалый хватающий ор-
ган, задние лапы и стопы становятся длинными, стройными, хорошо при-
способленными к бегу. Часто изображающийся охотящимся на птиц, Omit
holestes (что означает “грабитель птиц”), возможно, нападал в основном на
маленьких, проворных рептилий, таких как ранние ящерицы.
Deinonychus из раннего мела штата Монтана, США, был 2.4-4 м длиной и
имел большую голову. Это был необычный динозавр по многим причинам.
Во-первых, второй палец на каждой задней лапе был вооружен увеличенным
когтем. Используя его как крюк, Deinonychus был способен быстрым ударом
вспороть живот своей добыче или сопернику. В отличие от других динозав-
ров, он, возможно, обладал объемным зрением, так как поля зрения его глаз
перекрывались. Сами глаза были очень большими, поэтому он мог видеть в
темноте не хуже, чем днем. Возможно, он охотился ночью. По сравнению с
размерами тела, мозг у Deinonychus был значительно больше, чем у любого
другого динозавра, примерно того же размера, что у соответствующей ему со-
временной птицы, например страуса.
Позднемеловой монгольский родственник дейнонихуса, Adasaurus, имел
тазовый пояс более похожий на птиц, чем на типичных теропод
(фиг. XXVII.22). Лобковая кость взамен того, чтобы быть направленной вниз
и вперед, шла вниз и назад, параллельно седалищной кости.
ЧЕШУЙЧАТЫЕ РЕПТИЛИИ И ЯЩЕРОТАЗОВЫЕ ДИНОЗАВРЫ
439
Фиг. XXVII.23. Скелет Gallimimus длиной 4 м из позднего мела Монголии
Он является родственником своего известного североамериканского современ-
ника Struthiominus (фото Ф. Коффы, собственность 3. Киелан-Яворовской и му-
зея Виктории, Мельбурн)
Другим семейством небольших теропод с таким же устройством лобковой
кости были меловые сегнозавриды Монголии и Китая. В отличие от Deinony-
chus viAdasaurus, они, однако, имели четыре пальца на задних лапах, а не
три. Также, в отличие от Deinonychus, у них отсутствовал увеличенный “бо-
евой” коготь. Кроме того, сегнозавриды теряют передние зубы в пасти, а вза-
мен получают клюв.
Struthiomimus из позднего мела Северной Америки, приподнявшись, был
2.4—2.7 м высоты и по пропорциям напоминал страуса, сохраняя в то же вре-
мя типичные черты рептилий - тазовый пояс заурисхий, четыре лапы и
длинный хвост. Маленькая голова была похожа на страусиную, а беззубые
челюсти имели роговой клюв.
Жил Struthiomimus, очевидно, во влажных низинах и на песчаных равни-
нах, где бродил или быстро бегал на длинных задних лапах. Быстрый бег был
440 КАМЕННАЯ КНИГА
его средством защиты от врагов, так как тонкие трехпалые лапы были неспо-
собны к удару (в отличие от мощных лап страусов).
Орнитомимиды (это название семейства), видимо, были всеядными
(фиг. XXVII.23). Они могли легко достать фрукты с низких деревьев, их
клюв мог поймать маленьких рептилий; их рукоподобные передние лапы
могли выкопать яйца из кладок других животных на поверхность земли и
придерживать их, пока клюв разламывал скорлупу.
Овирапториды, как показывает название (“похитители яиц”), были также
любителями поедания яиц. По сравнению с орнитомимидами, их черепа бы-
ли короче и массивнее, но как и первые, они утеряли зубы. Обычно эти воры
убегали, но один из монгольских овирапторов был убит во время ограбления
кладки другого динозавра. Его разбитый череп сохранился среди окаменев-
шей скорлупы.
Глава XXVIII
ПТИЦЕТАЗОВЫЕ ДИНОЗАВРЫ
В 1822 г. жена Гидеона Мантелла, деревенского доктора из Англии,
нашла несколько окаменелых зубов в породах раннемелового возра-
ста. Ее мужу позже удалось собрать еще несколько зубов и костей,
которые эксперты из Англии и Франции относили то к носорогам, то к гип-
попотамам, то к рыбам. Сам доктор Мантелл думал, что зубы относятся к
рептилиям, похожим на современных ящериц - игуан, и поэтому назвал жи-
вотное Iguanodon. Прошли годы, прежде чем эти зубы и кости были опреде-
лены как остатки орнитисхий, птицетазовых динозавров.
ОРНИТОПОДЫ ИЛИ “ПТИЦЕНОГИЕ”
Iguanodon - это прогрессивный представитель подотряда, называемого
орнитоподами (“птиценогими”), хотя их ноги в действительности менее пти-
цеподобны, чем у некоторых теропод. Значительно более примитивное жи-
вотное, Camptosaurus, появившееся в Северной Америке и Европе в поздней
юре (фиг. XXVIII. 1), дожило до раннего мела и, возможно, добралось до Ав-
стралии. Оно было довольно неуклюжим, 2.1-5.2 м в длину, с низким и от-
носительно тяжелым черепом, передвигалось на двух ногах. Передние лапы
были пятипалыми, а задние четырехпалыми, заканчивавшимися копытооб-
разными когтями. Бедренная кость была изогнута, и, возможно, Camptosau-
rus иногда наклонялся и переходил на четырехногое хождение. Как и у всех
орнитисхий, у него была дополнительная кость, предзубная (praedentale), в
передней части нижней челюсти. И верхняя и нижняя челюсти впереди бы-
ли покрыты роговым чехлом, уплощенные, лезвиеобразные зубы занимали
задние две трети рта. Уровень челюстного сустава был ниже уровня зубов, в
отличие от карнозавров, у которых сустав находился на уровне зубного ряда
или даже выше. У карнозавров челюсти смыкались заходя друг за друга, как
ножницы, в то время как у Camptosaurus они не доходили друг до друга, как
у щипцов для орехов, что позволило разрезать только мякоть плода.
Впервые открытый в Европе, Iguanodon был также найден в Северной
Америке, Азии и Северной Африке. Разнообразные его виды были от 4.6 до
9.1 м в длину и достигали 4.6 м в высоту (фиг. XXVIII.2). Все они имели узкое
тело и сжатый с боков хвост, передвигались на задних лапах, имеющих три
больших и один очень маленький палец. На передних лапах было пять паль-
© 1997. В.М. Губин, перевод на русский язык и изменения.
442
КАМЕННАЯ КНИГА
Фиг. XXVIII.1. Юрская орнитисхия Camptosaurus длиной 5.2 м из Европы и
Северной Америки
цев, один из которых “большой”, стал костным шипом. Пока не были найде-
ны хорошо сохранившиеся черепа, некоторые исследователи считали, что
этот шип был на носу животного.
В угленосных районах, недалеко от Монса, Бельгия, раннемеловые пото-
ки прорезали глубокие (до 180 м) ущелья в карбоновых отложениях. Потоки
заполнили эти ущелья грязью, остатками растений и разнообразных живот-
ных, снесенных из окружающих болот. В 1878 г. шахтеры, прокладывая
туннель через такое древнее ущелье, обнаружили остатки двадцати трех
больших игуанодонов, а также крокодилов, черепах и рыб. Игуанодоны, по-
видимому, были представлены особями обоих полов одного вида, что позво-
лило изучить их анатомию исключительно детально.
Другое замечательное открытие было сделано в угольной шахте недалеко
от Седариджа, штат Колорадо, США. Там позднемеловой игуанодон прошел
по навозу, который сейчас превратился в уголь, а следы животного заполни-
лись песком, ставшим серым песчаником. Размеры следов (86 см в длину и
ширину) показывают, что их хозяин был значительно больше, чем любой
другой известный игуанодон.
Hypsilophodon, примитивный птицетазовый динозавр из раннего мела Ев-
ропы и Северной Америки был 1.5-1.8 м длиной и имел 15-сантимеровый че-
реп. Передние лапы были короткими, а спина и бока, возможно, несли тон-
кие щитки. Hypsilophodon — это один из наиболее хорошо изученных пред-
ПТИЦЕТАЗОВЫЕ ДИНОЗАВРЫ
443
Фиг. XXVIII.2. Iguanodon высотой 3.4 м и прорисовка костей его правой перед-
ней лапы. Iguanodon жил в Европе, Северной Америке, Азии и Северной Афри-
ке в раннемеловое время
ставителей группы примитивных, относительно маленьких орнитопод,
которые лишь слабо изменились с позднего триаса до позднего мела. Но вну-
три группы на ранних стадиях эволюции было несколько “экспериментов”.
Одним из них является позднетриасовый или раннеюрский Heterodontosau-
rus (“ящер с разными зубами”) из Южной Африки. В дополнение к щечным
зубам он имел хорошо выраженные клыки. В целом от этой в основном кон-
сервативной группы отошли не только игуанодонтиды, но и все другие груп-
пы орнитисхий.
Утконосые или гадрозавры. Эти рептилии отличались очень длинным че-
репом, который заканчивался беззубым расширением. При жизни оно было
покрыто роговым чехлом, использовавшимся для сбора мягких растений и
выискивания личинок в грязи между корнями. Пища перерабатывалась зу-
бами, сгруппированными в располагавшиеся перпендикулярно длинной оси
черепа “обоймы” (фиг. XXVIII.3). В каждой из таких обойм было по несколь-
ко налегавших друг на друга зубов, так что при стирании старого зуба на его
место выдвигался новый, а старый вываливался. Хотя “зубная” часть рта и
ограничена в размерах, в каждой ветви челюсти можно было насчитать от
двухсот до пятисот зубов, а всего во рту от восьмисот до двух тысяч зубов.
Примерно треть из них использовалась, остальные были в резерве.
Размеры гадрозавров варьировали от 3.7 до 12 м, в среднем около 9 м. Жи-
вотные передвигались в наклонной позиции и часто переходили на четыре
лапы. Тело было высоким, но нешироким, хвост — высоким и узким, а тон-
444
КАМЕННАЯ КНИГА
Фиг. XXVIII.3. Некоторые меловые утконосые динозавры
Edmontosaurus (слева) и Hadrosaurus (справа внизу), оба высотой 4.6-5.5 м,
жившие на востоке Северной Америки. Слева вверху — череп Edmontosaurus
длиной 1.1м; справа вверху - сечение через челюсть утконосого динозавра, по-
казывающее, как сменяются зубы; увел. х0.25
кая морщинистая кожа покрывалась не налегавшими друг на друга малень-
кими округлыми щитками. Они имели четыре пальца на передних и три
пальца на задних лапах; когти были копытообразными. Между пальцами пе-
редних и, возможно, задних лап были перепонки.
Бесспорно, утконосые могли как плавать, так и бродить по мелководью, и
гулять по суше. Это видно по их перепончатым лапам и высоким узким хво-
стам, которые могли использоваться как рулевое весло. Позвоночник был
усилен сильно кальцитизированными сухожилиями, позволявшими муску-
лам изгибать тело и хвост из стороны в строну. Эти сухожилия сохранились
на многих скелетах утконосых.
Остатки плодов, веток, семян и хвойных иголок в полостях тела некото-
рых гадрозавров говорят о том, что хотя они были хорошими пловцами, кое-
кто, по крайней мере, предпочитал диету из наземных растений. Возможно,
способность плавать первично была приспособлением, позволявшим сбе-
жать от хищников и лишь во вторую очередь перейти к питанию водными
растениями.
Тела у всех гадрозавров были однотипны, но череп - это особая история.
В целом он был длинным и высоким, слабо понижался сзади вперед и имел
ПТИЦЕТАЗОВЫЕ ДИНОЗАВРЫ
445
Фиг. XXVIII.4. Скелет Tsintaosaurus, гадрозавра из позднего мела Китая дли-
ной около 10 м (фото Ф. Коффы, собственность музея Виктории, Мельбурн, и
Института палеонтологии позвоночных и палеоантропологии, Пекин)
очень большие отверстия для ноздрей. Кости, окружавшие эти отверстия,
были тонкими.
Внутри общей схемы присутствовало широкое разнообразие форм. В ос-
новном имеются два типа черепа: не имеющие гребня (или имевшие литой
гребень) и те, у которых был пустотелый гребень. Разнообразие формы голо-
вы у гадрозавров, возможно, было полезно для различения представителей
своего вида, особенно в сезон размножения. Похожая ситуация наблюда-
ется в Африке сейчас, когда антилопы узнают своих сородичей по форме
рогов.
Гадрозавры, такие как Edmontosaurus из Северной Америки, не имевшие
гребня совсем и Tsintaosaurus из Китая, могли иметь свисающие складки ко-
жи по бокам головы (фиг. XXVIII.4, XXVIII.5, XXVIII.6). Они могли разду-
вать их как шары, примерно также, как раздуты горловые мешки у некото-
рых современных ящериц. Ярко раскрашенные, они делали этих динозавров
зрительно больше и свирепее. Животные также могли использовать эти кож-
ные складки для издавания звуков, подобно голосовым мешкам лягушек.
У обладавших пустотелым гребнем гадрозавров носовые каналы проходи-
ли через гребень (фиг. XXVIII. 7). Этот дополнительный объем носовых про-
ходов действовал как резонирующая камера, когда гадрозавры переклика-
лись друг с другом.
446
КАМЕННАЯ КНИГА
Фиг. XXVIII.5. Реконструкция скелета Tsintaosaurus, гадрозавра из позднего
мела Китая длиной около 10 м (рисунок П.Траслэ, собственность музея Викто-
рии, Мельбурн)
Гадрозавры произошли от игуанодонтид в позднемеловое время, когда
они проникли в Европу через Азию и через Северную Америку в Южную
Америку. Их остатки наиболее часто встречаются в отложениях русловых
потоков, существовавших в Восточной Азии и вдоль западного побережья
моря, которое покрывало Северную Америку от Техаса до Северной Канады.
В бедлендах Ред Дир Ривер, провинции Альберта, Канада, часть района, в
котором известны обильные остатки гадрозавров, была объявлена Заповед-
ником. Здесь было найдено много почти полных скелетов, некоторые с кон-
турами мягких тканей и отпечатками кожи. Эти животные, скорее всего,
умерли в воде, были снесены вниз по течению и захоронились вне досягаемо-
сти хищников. Здесь есть и отдельные кости скелетов, разломанные на кус-
ки, возможно, тираннозавридами.
В прекрасном местонахождении в Центральной Монтане, США, удалось
обнаружить настоящую “детскую” гадрозавров. Здесь остатки яиц, молоди и
взрослых особей Maiasaura были найдены захороненными группами вместе.
Поэтому можно предположить, что взрослые животные обеспечивали без-
опасность молоди после их вылупления из яиц.
Толстоголовые динозавры. Впервые эти рептилии появились в раннеме-
ловое время в Англии (фиг. XXVIII.8). Тем не менее, больше всего их извест-
но из позднего мела запада Северной Америки и Восточной Азии. Majungath-
olus, найденный на Мадагаскаре, показывает, что эти динозавры проникли
также и в Гондвану.
Stegoceras — один из самых изученных толстоголовых (фиг. XXVIII.9). Он
был безобидным вегетарианцем, 1.5-1.8 м длины с пятью короткими
ПТИЦЕТАЗОВЫЕ ДИНОЗАВРЫ
447
Фиг. XXVIII.6. Череп Tsintaosaurus длиной 1.3 м (фото Ф. Коффы, собствен
ность Института палеонтологии позвоночных и палеоантропологии, Пекин, и
музея Виктории, Мельбурн)
пальцами на передних и четырьмя длинными пальцами на задних лапах.
Стегоцерас сщипывал растения заостренными зубами, которые шли до пе-
редней части рта, так что клюва у него не было. Предчелюстная кость была
маленькой, а те, которые образуют поверхность черепа, слились вместе и
достигали 7.5 см толщины. В задней части головы имелось несколько ма-
леньких шипов.
Pachycephalosaurus (“толстоголовый ящер”) жил в самом конце мелового
периода, когда тираннозавриды были в своем зените. Его куполообразный
череп 66 см длиной и 25 см толщиной над мозгом был украшен шипами и
гребнями. Так как большая часть тела покрывалась только кожей, этот мас-
сивный череп для защиты был бесполезен. Скорее всего он использовался
как оружие турнирного боя между конкурирующими самцами, примерно
так же, как это делают современные горные бараны в сражениях за самок в
брачный сезон.
Самым маленьким из толстоголовых динозавров был Micropachycephalo
saurus (“крошечный толстоголовый ящер”), живший в Китае в позднем ме-
лу. Он был всего 51 см в длину.
Все толстогловые динозавры, упомянутые выше, имели куполообразные
черепа, однако Wannanosaurus, Hotnalocephale и Goyocephale из позднего ме-
ла Восточной Азии имели плоские головы. Было ли это следующим шагом в
развитии турнирного оружия или просто это другая группа, пока неясно.
448
КАМЕННАЯ КНИГА
Фиг. XXVIII.7. Три утконосых динозавра с полыми костными гребнями
Слева вверху -- голова Lambeosaurus длиной 81см. Справа вверху - голова Рага-
saurolophus. Показано, как располагался внутри гребня носовой проход. Внизу -
плавающий Cory thesaurus. На суше он передвигался как Edmontosaurus. Дли-
на животного 10 м
Покрытые броней и динозавры-дрендоуты. Две группы орнитисхий при-
обрели броню. Носившие пластины (Stegosauria) появились в средней юре
Европы и Китая. Закончилось их время в позднем мелу в Индии вместе с
“миниатюрным” стегозавром Dravidosaurus длиной 3 метра. Dravidosaurus
был “живым ископаемым”, изолированном на островном континенте, каким
была Индия в то время. В других местах стегозавры вымерли к концу ран-
него мела.
Наиболее изученным из группы и одним из самых известных динозавров
был Stegosaurus из поздней юры Северной Америки (фиг. XXVIII.10). Самые
крупные особи достигали 5.5-7.6 м длины, 3.7-4.0 м высоты и весили от 7 до
10 т. 40-сантиметровый череп был низким и уплощенным, с хорошо разви-
тым клювом и 90-100 маленькими зубами. Тело было высоким, но неширо-
ким; задние лапы длинные, передние — короткие, и три из пальцев на каж-
дой ноге имели копытообразное окончание. Широкие бока были покрыты
ПТИЦЕТАЗОВЫЕ ДИНОЗАВРЫ
449
Фиг. XXVIII.8. Голова Pachyce-
phalosaurus, череп длиной 66 см,
толщина лобных костей 23 см
Фиг. XXVIII.9. Stegoceras, при-
митивный “толстоголовый” с че-
репом длиной 19 см
грубой кожей, а шея, спина и часть хвоста несли два ряда костных пластин.
Кроме того, на конце хвоста было от двух до восьми длинных шипов.
На рисунке Stegosaurus изображен как мирное, малоподвижное живот-
ное, слишком низкое, чтобы бродить по болотам, и слишком слабо защищен-
ное, чтобы рисковать встречаться с хищниками типа Allosaurus. Эти пласти-
ны защищают только спину, хвост как оружие недостаточен, а бока открыты
для атаки. Скорее всего, пластины были нужны не для защиты. Они давали
возможность стегозавру быстро охлаждаться или -нагреваться, прогоняя
кровь через сеть кровеносных сосудов, испещеривших эти пластины. Сами
животные, вероятно, постоянно находились среди деревьев, чьи стволы и
ветви служили им укрытием.
Вслед за завроподами Stegosaurus с мозгом весом всего в 70 г был одним из
самых несмышленных в мезозое. Как и они, стегозавр перенес большую
часть мыслительной деятельности в большой ганглий спинного мозга в пле-
чевом поясе и еще один в тазовом районе. Последний весил 1.4 кг.
Хорошо защищенные анкилозавры. Систематически динозавры-дрендо-
уты образуют подотряд Ankylosauria. Появившись в поздней юре Европы,
они стали широкими, низкопосажнными фитофагами с короткими массив-
ными лапами и коренастыми, несущими копытца стопами. Один из наиболее
известных — Ankylosaurus был 5.2 м в длину, 1.8 м в ширину и немного более
1.2 м в высоту. 70-сантиметровый массивный череп не имел височных отвер-
стий; толстые костные пластины нависали над каждым глазом и ноздрей и
вытягивались наружу подобно ушам (фиг. XXVIII. 11). Ребра крепко прира-
стали к позвонкам, а спина и бока были перекрыты толстыми костными
щитками. Другие щитки покрывали хвост, который заканчивался костной
дубиной, хорошо прикрепленной к позвонкам. Челюсти были тяжелыми, а
зубы настолько малы, что были почти бесполезны.
Euoplocephalus достигал 5.3 м длины и 2.4 м ширины; лапы были корот-
кие и широко расставлены, поэтому ходить он мог только маленькими шаж-
ками вперевалку (фиг. XXVIII. 12). Кожа тела была усеяна костными шипа-
29. Каменная книга
450
КАМЕННАЯ КНИГА
Фиг. XXVIH.10. Stegosaurus, юрский динозавр из западных районов США
длиной 5.5—7.6 м. Эта рептилия известна по своему крестцовому нервному ган-
глию или “второму мозгу”. Справа вверху - крестцовый (или тазовый) ганглий.
Справа внизу — головной мозг, изображенный в том же масштабе
ми и наростами до 15 см длиной. Хвост заканчивался костной булавой и нес
два больших шипа, покрытых роговым чехлом.
В отличие от Ankylosaurus и Euoplocephalus, Panoplosaurus утратил кост-
ную булаву на конце хвоста. Этот недостаток восполнялся наличием ряда ос-
трых шипов вдоль обеих сторон тела. Самые большие, над плечами, были
отогнуты вперед, к шее.
Некоторые анкилозавры были найдены в песках эолового происхожде-
ния, что предполагает, что животные могли жить среди песчаных дюн, по-
едая растущие на них и рядом растения. Однако более вероятно, что анкило-
завры проводили большую часть своего времени во влажных лесах или боло-
тах, где ползали как гигантские причудливые черепахи, скользили и
выкарабкивались из грязи. Когда появлялась опасность, они ложились на
землю, прижимали к бокам лапы, а хвост был готов отразить атаку любого
существа. Даже Tyrannosaurus не мог пробиться через их броню или пере-
вернуть незащищенным животом вверх. То, что они на самом деле могли за-
щитить себя таким образом, подтверждают многочисленные находки тиран-
нозаврид со следами прижизненных поврежедний оснований лап, возмож-
но, от резких ударов хвостом анкилозавров.
ЯЙЦА, ВОРОТНИКИ И РОГА
В 1887 г. профессор О.С. Марш описал окаменевшие рога, найденные не-
далеко от г. Денвер, штат Колорадо, США, назвав их Bison alticornis. Двумя
годами позже сходные рога были обнаружены на черепах рептилий. Таким
образом, предполагаемый бизон превратился в цератопса или рогатого
динозавра.
ПТИЦЕТАЗОВЫЕ ДИНОЗАВРЫ
451
Фиг. XXVIII.il. Извлечение анкилозавра Saichania из позднемеловых отло-
жений Монголии. Обратите внимание на череп животного на противопложной
от людей стороне фотографии (фото В.Скаржински, собственность 3. Киелан-
Яворовской, Институт палеобиологии, Варшава)
Хотя эти существа впервые были найдены в Северной Америке, появились
они, возможно, сначала в Азии. По крайней мере один примитивный вид до-
жил там до позднего мела, в том месте, которое сейчас называется пустыней
Гоби. Это ископаемое было названо Protoceratops - дословно “первое рогатое
лицо” или “первый цератопс”.
Это имя не соответствует действительности, так как Protoceratops очень
редко имел рог и он не был первым цератопсом. Их корни уходят к Psittaco
saurus (“ящер-попугай”) из раннего мела, двуногому родственнику Hypsilo
phodon (фиг. XXVIII. 13). Проходят миллионы лет и потомки этого родона-
чальника становятся четвероногими, с коротким хвостом и лапами, массив-
ным телом и костным воротником позади головы (фиг. XXVIII. 14). Зубы
располагаются в ряд в желобке, а верхний клюв присоединяется к новой от-
дельной кости в передней части верхней челюсти.
Эти черты сохранились у Protoceratops, достигавшего 1.5-1.8 м в длину.
Его задние лапы пока еще длиннее, чем передние, все они имеют по пять
пальцев, в воротнике позади черепа есть два очень больших отверстия, на но-
су не было рога, а челюсти несли крючкообразные клювы. На каждой пред-
челюстной кости остаются по два цилиндрических зуба.
В период размножения самка Protoceratops устраивала гнездо в мягком
песке и откладывала туда дюжину или более 15-сантиметровых яиц. Песок
вокруг яиц иногда из-за сильных дождей уплотнялся и эмбрионы погибали,
не успев вылупиться. Их остатки окаменевали и в таком виде дошли до нас.
Кроме этого, известны находки скелетов Protoceratops от только что вылу-
пившихся детенышей до полноразмерных динозавров. Они показывают в
разной степени детальности развитие животных, которые жили и умерли
80 млн. лет назад.
29*
452
КАМЕННАЯ КНИГА
Фиг. XXVIII. 12. Три меловых панцирных динозавра из Северной Америки
Слева вверху - череп Ankylosaurus длиной 69 см. Справа вверху - Panoplosaurus
длиной 7 м с рядами шипов по бокам. Внизу — Euoplocephalus длиной 5.5 м
Цератопсы достигли Северной Америки задолго до того времени, когда сам-
ка Protoceratops откладывала яйца в монгольский песок. Они также стали боль-
шими животными с толстыми телами, свисающими хвостами и лапами, пре-
вратившимися в вертикальные колонны. На передних лапах было пять паль-
цев, на задних четыре и рудимент пятого, и все хорошо развитые пальцы несли
копытца. Рога росли на носу и над глазами, череп с воротником стал очень боль-
шим. Слившиеся между собой шейные позвонки и массивные мышцы, присое-
динявшиеся к основанию черепа, поддерживали тяжелую голову. Как и у ан-
килозавров, подвздошная кость увеличивается и отгибается наружу, формируя
пластину над тазобедренным суставом, однако кожа остается без щитков.
Это общий план строения, а сейчас мы коснемся их разнообразия.
В начале своей американской карьеры, цератопсы разделились на две
группы, длинноворотниковых и относительно коротковоротниковых. Ран-
ние длинноворотниковые имели короткие рога на носу, длинные над глазами
и отверстия в воротнике. Эти черты постоянно развиваются и достигают сво-
его зенита у Pentaceratops и Torosaurus с черепами до 2.6 м длиной и 60-сан-
тимеровыми рогами над глазами. Длина таких рептилий доходила до 6 м
(фиг. XXVIII. 15).
Менее последовательны изменения в коротковоротниковой группе. Они
начались с Monoclonius, имевшего высокий череп с длинным прямым рогом
на носу, более короткими над глазами и небольшими шишками вдоль их
края. У Styracosaurus шишки превратились в покрытые рогом шипы. У
Triceratops воротник стал цельным с морщинистой поверхностью, а рога над
глазами больше, чем рог на носу.
Triceratops отмечает как наивысшую точку в развитии цератопсов, так и
их закат. Разные виды этого рода были от 6 до 7.6 м в длину и 2.5-3 м в высо-
ПТИЦЕТАЗОВЫЕ ДИНОЗАВРЫ
453
Фиг. XXVHI.13. Первые цератопсы
Слева — Psittacosaurus длиной 1.2 м был очень примитивным рогатым динозав-
ром. Справа - Protoceratops длиной 1.5-1.8 м имел воротник, но был еще почти
безрогим
ту, а весили от 7.5 до 11 тонн (фиг. XXVIII. 16). Череп длиной 2.4 м нес 1-ме-
тровые рога над глазами и защищенный мозг весом почти 900 г, в двенадцать
раз больше, чем у Stegosaurus. Во время динозавров этих животных было
очень много: один сборщик во время пешеходных и верховых маршрутов по
отложениям формации Хелл Крик в Центральной Монтане в течение семи
летних сезонов видел остатки более чем пятисот черепов. Такую многочис-
ленность можно попытаться объяснить редкой комбинацией силы, воору-
женности, способности постоять за себя и подвижности. Рубцы на воротни-
ках показывают, что Triceratops часто дрались между собой, видимо, защи-
щая свою территорию и самок в брачный сезон. Крепкие лапы, сильная шея
и опасные рога помогали Triceratops отражать атаки хищников, надеяв-
шихся с помощью своих челюстей добраться до незащищенного брюха жи-
вотного. Тираннозавриды не были умственными гигантами и Triceratops,
бесспорно, знали достаточно, чтобы защищаться от этого врага, если, прав-
да, не бывали захвачены врасплох нападением с тыла.
ПАНИЧЕСКОЕ БЕГСТВО ДИНОЗАВРОВ
На наш взгляд, некоторые динозавры как будто созданы для скорости. То,
что они могли двигаться быстрее, чем требуют обстоятельства, показал ана-
лиз следовых дорожек.
В середине мела, там, где сейчас центральная часть штата Квинсленд (Ав-
стралия), смешанная группа орнитопод и маленьких теропод была на водо-
пое на берегу озера (фиг. XXVIII. 17). Всего здесь было не менее 130 особей,
размером от петуха до страуса эму. Внезапно появился большой (до 8 м в дли-
ну) теропод и медленно, со скоростью около 7 км в час, двинулся по направ-
лению к ним. Потом он повернул направо и на этом информация о его движе-
ниях исчезает, так как он вышел за район, где сохранились следы.
Через несколько мгновений орнитоподы и тероподы побежали в противо-
положном от места появления хищника направлении. Орнитоподы развили
454
КАМЕННАЯ КНИГА
Фиг. XXVIII. 14. Неполный скелет Рsittacosaurus, двуногого динозавра с попу-
гаеобразным клювом из раннего мела Центральной Азии. Длина полного ске-
лета была около 90 см (фото Ф. Коффы, собственность Института палеонтологии
позвоночных и палеоантропологии, Пекин, и музея Виктории, Мельбурн)
скорость 16 км в час, а маленькие тероподы — 12 км в час. Самыми медленны-
ми оказались маленькие животные, чьи следы были последними и часто на-
кладывались сверху на следы первых. Очевидно, в момент паники каждый
динозавр был сам за себя, и даже если обычно молодь и находилась под защи-
той, в данной ситуации такого не случилось. То же самое происходит в наши
дни, когда стадо зебр бывает внезапно испугано львицами.
ОБРАЗЦЫ АДАПТИВНОЙ РАДИАЦИИ
Динозавры представляют собой хорошие примеры адаптивной радиации,
при которой связанные между собой животные и растения изменяются, стре-
мясь выжить, в различных направлениях и под воздействием разнообразных
условий. Так, ранние заурисхии, первоначально двуногие хищники средних
размеров, превращаются в двуногих карликов и гигантских охотников, че-
тырехногих фитофагов, бывших самыми крупными животными, когда-либо
ходившими по поверхности Земли, и страусоподобных животных, утеряв-
ших зубы и приспособившихся к всеядности. Этими различными путями
они жили и процветали по крайней мере 130 млн. лет.
Орнитисхии также произошли от двуногих хищников, однако они оста-
лись растительноядными. В поздней юре и мелу у них сформировалось пять
адаптивных моделей.
1. Двуногая жизнь в лесах. Передние лапы обычно использовались для за-
хвата пищи, хотя могли использоваться и для ходьбы. Кроме бега у них не
было никакой защиты (Camptosaurus, Hypsilophodon, игуанодонтиды, пахи-
цефалозавриды и другие маленькие двуногие фитофаги).
2. Жизнь в лесах, обеспечивающая нормальные условия для больших
медленных четвероногих животных. Защита обеспечивалась броней и по-
движным, вооруженным шипами хвостом (Stegosaurus).
3. Жизнь в болотах, пресных или соленых водах, но с постоянным пере-
движением по суше. Пища попадала в рот с помощью клюва, как у утки. При
ходьбе использовались задние лапы, а хвост помогал при плавании. Так как
оружие и броня отсутствовали, безопасность обеспечивалась либо убегани-
ем, либо уплыванием (утконосые или гадрозавры).
ПТИЦЕТАЗОВЫЕ ДИНОЗАВРЫ
455
Фиг. XXVIII.15. Головы больших цератопсов или динозавров с клювами, шей-
ными воротниками и рогами
Слева вверху - Styracosaurus", слева внизу - Monoclonius, справа вверху - Pen-
taceratops, справа внизу - Chasniosaurus. Все из позднего мела Северной Амери-
ки; xl/30
4. Жизнь на возвышенностях и в болотах, где широкотелые рептилии бро-
дили вперевалку или ползали в грязи. Растительная пища срывалась рого-
выми челюстями; безопасность обеспечивалась костным панцирем и массив-
ным булавообразным хвостом, хотя часто таким широким, покрытым креп-
кой броней существам, чтобы чувствовать себя в безопасности от врагов,
было достаточно просто спокойно лежать (анкилозавры).
5. Активная жизнь в заросших растительностью низинах среди сухих
дельт и болот в стиле современного носорога. Для защиты используются пан-
цирь (воротник) и рога, подкрепленные мощью 8—11-тонного тела. Рога ис-
пользовались также для распознавания себе подобных (большие цератопсы).
БЫЛИ ЛИ ДИНОЗАВРЫ ТЕПЛОКРОВНЫМИ?
В течение многих лет к динозаврам относились как к большим, медли-
тельным, туповатым рептилиям, которые в конце концов заместились не-
большими, быстрыми, сообразительными млекопитающими. Затем группа
ученых поставила под сомнение такую картину. Со временем некоторые ста-
ли думать о динозаврах, как о больших, быстрых рептилиях, возможно, теп-
локровных. Может быть они могли бы и до сих пор доминировать на планете,
если бы не случилось чего-то внезапного, что привело к вымиранию диноза-
вров и позволило млекопитающим широко расселиться и занять их место.
456
КАМЕННАЯ КНИГА
Фиг. XXVIII.16. Triceratops, позднемеловой рогатый динозавр 6.1—7.6 м дли-
ной. Иногда на него нападал Tyrannosaurus. Слева вверху - череп с показанны-
ми мозгом (зачернено) и шейными мышцами
В процессе этих споров было сделано много серьезных попыток выяснить,
что же это были за животные. Стало ясно, что динозавры обладали таким ши-
роким спектром адаптаций, что невозможно говорить о их стиле жизни в це-
лом, как и о всех млекопитающих и о всех рептилиях.
Некоторые из динозавров были очень умны. Завроподы, например, имели
мозг в пять раз меньше, чем полагается рептилиям их размера. С другой сто-
роны, мозг небольшого теропода Stenonychosaurus был в шесть раз больше,
чем должно быть у такой рептилии, и такой же большой, как у млекопитаю-
щего или птицы близкого размера.
Теплокровные животные требуют в десять раз больше пищи в день, чем
холоднокровные. Увеличение активности дает преимущества теплокровным
животным, но за это надо платить. В случае завропод с их маленькими голо-
вами и относительно слабыми зубами, животным не хватило бы дневного
времени, чтобы обеспечить себя едой, если бы они были теплокровными. Па-
радоксально, но другой трудностью для гигантских завропод было бы избав-
ление от избыточного тепла. Киты, тех же размеров, способны делать это, но
они живут в воде, гораздо более эффективной для передачи тепла. Фактиче-
ски завроподы почти идеально приспособлены к тому, чтобы быть холодно-
кровными. При их размерах невозможно успеть заметно охладиться из-за
перепада дневных и ночных температур. Так что, если, греясь и питаясь на
солнце, в течение нескольких дней им удавалось поднять температуру до
уровня, при котором животные становились активными, в дальнейшем этот
уровень мог поддерживаться уже без вклада такого же большого количества
пищи и энергии.
Все свидетельствует о том, что завроподы были холоднокровными.
С другой стороны, есть не менее убедительные примеры, что по крайней ме-
ре некоторые маленькие хищные динозавры были теплокровными. Очевид-
но, что самые первые летающие птицы были теплокровными, так как полет
требует высокого метаболизма. Птицы произошли от небольших теропод,
так что если разделение на две группы не произошло на протяжении одного
ПТИЦЕТАЗОВЫЕ ДИНОЗАВРЫ
457
Фиг. XXVHL17. Карта следов динозавров из карьера Ларк около Винтона,
Квинсленд, Австралия. Они были найдены в позднемеловых отложениях фор-
мации Винтон и, возможно, оставлены большим тероподом (Tyrannosauropus),
орнитоподами поменьше (Wintonopus) и маленькими тероподами (Skartopus).
(собственность М. Вэйд, А. Талборна и музея Квинсленда, Брисбен)
поколения и теплокровность не была мгновенно приобретена птицами,
логично думать, что и некоторые небольшие тероподы тоже были тепло-
кровными.
458
КАМЕННАЯ КНИГА
ГИГАНТЫ ПРОТИВ КАРЛИКОВ
Мы проследили историю динозавров на протяжении позднего триаса, юр-
ского и мелового периодов, которые занимают около 165 млн. лет. В течение
этих лет несколько больших групп превратились в гигантов, в то время как
другие оставались средних размеров или даже становились карликами. Ка-
кие преимущества заставляли рептилий становиться все больше и больше и
какой штраф надо за это платить? Что получали их небольшие родственники
и что они теряли, если их размеры уменьшались?
Штрафы за гигантизм многократно обсуждались в статьях, книгах и даже
анимационных фильмах. Один из них - вес как таковой: большие животные
должны постоянно бороться против своего собственного тела и давящего
вниз его веса. Мы видели, как эти причины вынудили больших завропод
приобрести ажурные позвонки и массивные конечности, чтобы поддержать
их тело. На суше такие монстры, как Albertosaurus и Tyrannosaurus могли
приблизиться к пределам, при которых связки еще могут держать тело вме-
сте, а мускулы способны передвигать тонны крови, плоти и костей достаточ-
но быстро, чтобы обеспечить себя пищей.
Пища - достаточность пищи - предлагает другую проблему. Большие жи-
вотные еДят больше, чем маленькие, хищные или вегетарианцы, рыбы, мле-
копитающие или рептилии. Сегодня такие большие животные, как слоны и
тигры, не могут жить на маленьких островах: на них недостаточно пищи,
чтобы поддержать воспроизводящуюся популяцию. Примерно также ги-
гантские завроподы не могли жить в небольших лесах, а в больших лесах не
встречались стада завропод. Больших теропод всегда было меньше, чем их
жертв, и каждый из хищников, вероятно, специализировался на определе-
ной группе животных, защищая их от других агрессоров. Так побуждал
делать их инстинкт, но основой инстинкта была необходимость обеспечения
в питании.
Это была плата за увеличение размеров, но это также давало и преимуще-
ства, особенно растительноядным. Большие животные защищены от нападе-
ния своими размерами вне зависимости от их подвижности или наличия
средств защиты. Ни один большой теропод не будет настолько глуп, чтобы
напасть на взрослого Apatosaurus или Diplodocus, и даже аллозавр, возмож-
но, дважды подумает, прежде чем атаковать таких гигантов. Да и сам алло-
завр врагов не имел, конечно, за исключением других аллозавров, и поэтому
становясь взрослыми, они могли считать себя в безопасности.
Даже потребность в большом количестве пищи имела свои положитель-
ные стороны: конечно, 25-тонному динозавру нужно было больше еды, чем
25-килограммовому, но не в тысячу раз. Поэтому “затраты” на жизнь гиган-
та завропода были значительно меньше, чем на эквивалентный вес триасо-
вых динозавров. В то же самое время завропод получал преимущества в виде
размеров и долгой жизни, вероятно, более длинной, чем у любой современ-
ной рептилии.
Наконец, коснемся температуры. При своих гигантских размерах боль-
шие динозавры могли поддерживать почти постоянную температуру. Она
практически не успевала упасть ночью, на холодном ветру или в прохлад-
ные периоды, при их медлительности поддерживалась на одном и том же
уровне.
И наоборот. Мы привыкли обсуждать эффект холода и забываем, что со-
временные рептилии не менее чувствительны к перегреву. В летние месяцы
в горячий полдень они вынуждены скрываться в тени, а быстрый перегрев
ПТИЦЕТАЗОВЫЕ ДИНОЗАВРЫ
459
Фиг. XXVIII.18. Яйцо динозавра из меловых отложений Южного Китая
может привести к смерти. Но большие динозавры, с их относительно невысо-
ким отношением площади тела к его весу, поглощают минимальное количе-
ство солнечных лучей и, таким образом, избегают эффекта перегрева.
Приобретения и потери карликов обратны гигантам. Карлик не обременен
весом, со своими небольшими размерами он может обитать в местах, недо-
ступных гигантам. Ему надо значительно меньше пищи, что означает отсут-
ствие необходимости в ее разнообразии. С другой стороны, его может случай-
но убить любой гигант, даже не подозревающий о его существовании. Он по-
требляет относительно много пищи, из-за этого вынужден конкурировать с
множеством животных близкого размера. Можно уверенно сказать, что жиз-
ненный путь его значительно короче - несколько лет против десятилетий и
даже столетий. Наконец, никакой карлик не может сохранять постоянную
температуру или жить в одном температурном диапазоне. Он может быть бы-
стрым - для рептилии, при подходящих условиях, но он становится медли-
тельным при прохладной погоде и должен защищаться от слишком высокой
температуры и прямых солнечных лучей.
Мы не можем сказать однозначно, что хорошо и что плохо. Все, что мы мо-
жем сделать, это заявить, что один набор признаков давал возможность про-
цветать гигантам, а другой - их родственникам оставаться небольшими или
даже превращаться в карликов. Это совершенно не значит, что гигантов бы-
ло много, а карликов мало и для первых было больше шансов сохраниться в
ископаемом состоянии. И не так уж и важно, что гиганты вымерли; их сме-
нили меньшие рептилии, некоторые из них в свою очередь стали большими.
Этот процесс продолжался и шел вплоть до конца мелового периода, когда
динозавры вымерли, оставив после себя только своих потомков — птиц.
460
КАМЕННАЯ КНИГА
Фиг. XXVIII.19. Интригующая гипотеза, основывающаяся на представлениях
Дэйла Рассела, как мог бы выглядеть небольшой разумный двуногий динозавр,
если бы эти рептилии не вымерли в конце мезозойской эры
На переднем плане динозавроид, позади него Stenonychosaurus inegualis, близ-
кий по размеру к человеку тероподный динозавр (собственность Д. Рассела)
ЗАГАДКА ВЫМИРАНИЯ
Более чем сто лет назад палеонтологи узнали, что динозавры вымерли
относительно быстро в конце мезозоя. Существовало много попыток объясне-
ния этому. Некоторые считали, что климат стал слишком теплым, другие —
слишком холодным. Третьи объяснения допускали, что более сообразитель-
ные млекопитающие, жившие одновременно с динозаврами, так или иначе
выжили своих больших, но глупых современников.
Для начала надо вспомнить, что динозавры были не единственной груп-
пой, вымершей в конце мезозойской эры. В воздухе исчезли птерозавры;
плезиозавры и мозазавры исчезли в морях. Кроме того, среди беспозвоноч-
ных вымирают аммониты, иноцерамиды, много родов и видов брахиопод, иг-
локожих, кокколитов и пелагических фораминифер. Таким образом, выми-
рание динозавров было не изолированным событием, а только частью обшир-
ного феномена.
Однозначное объяснение, почему эти события произошли одновременно,
отсутствует. Ближе к концу мела климат становится более прохладным, хо-
тя некоторые места, такие как Альберта, Канада, были своеобразными пар-
никами даже зимой. Уровень моря понижался, что приводило к уменьше-
нию количества мелководных морей. Исходя из этих фактов, одна из теорий
допускает, что мог измениться химизм океана из-за поступления туда боль-
шого количества осадков. Изменение содержания элементов, в том числе
ПТИЦЕТАЗОВЫЕ ДИНОЗАВРЫ
461
Фиг. XXVIII.20. Голова динозавроида вблизи (собственность Д. Рассела)
ядовитых, могло привести к уменьшению численности многих морских ор-
ганизмов, особенно мельчайших, составляющих планктон. С их исчезнове-
нием уменьшается количество питающихся ими животных и, в свою оче-
редь, животных, охотившихся на этих животных, и так далее.
Другая теория, связанная с понижением температуры, особенно в районе
Северного полюса, напрямую не затрагивает динозавров. Но общее похоло-
дание позволило млекопитающим стать многочисленнее и больше. С измене-
нием климата это сообщество млекопитающих начинает распространяться
по направлению к экватору, вытесняя динозавров.
Используя очень чувствительные химические методы, удалось обнару-
жить, что в осадочных породах, переходных от позднего мела к раннему па-
леогену, наблюдается необычно высокая концентрация иридия. Редкий на
Земле, иридий, один из металлов группы платины, встречается в повышен-
ных концентрациях в метеоритах. Пытаясь объяснить это явление, предпо-
ложили, что большой метеорит, диаметром около 10 км, в самом конце мела
упал на Землю. Непосредственно от него могли пострадать только несколько
неудачников, оказавшихся в месте падения, однако поднявшееся пылевое
облако могло затенить Солнце. Это означало бы, что процесс фотосинтеза мог
прекратиться на срок от нескольких месяцев до нескольких лет, убивая
взрослые растения и не трогая их семян. Без обильной растительности в те-
чение нескольких лет сумели бы выжить только те фитофаги, кто питался се-
менами или сохранившимися листьями. Как следствие, крупные раститель-
ноядные животные исчезли бы, а сохранились только те небольшие формы,
которые тратили бы на поиски пищи меньше энергии, чем получали от нее.
462
КАМЕННАЯ КНИГА
(Если бы слон жил подобно мыши, собирая семена, он бы голодал.) Кроме то-
го, температура после падения метеорита стала бы ниже.
Другим эффектом от падения метеорита могло быть мгновенное повыше-
ние температуры в атмосфере, невыносимое для организмов, неспособных
найти временное укрытие.
Кометы обычно описываются как “грязные комья снега”, так как они со-
стоят из замороженной воды, двуокиси углерода, аммиака и метана. При-
ближаясь к Солнцу, эти вещества испаряются и формируют длинный хвост
кометы. При столкновении с Землей эти вещества, в том числе яды типа ам-
миака, оказались бы выброшенными в атмосферу. Мгновенный выпуск боль-
шого количества ядовитых газов также мог бы вызвать вымирания.
Вопросы “что если?” часто могут привести к интересным рассуждениям
относительно альтернативной истории или предыстории. Что было бы, ес-
ли бы динозавры пережили конец мела? Один ученый предположил, что
было бы, если бы из небольшого хищного динозавра сформировалось ра-
зумное двуногое существо (фиг. XXVIII.19 и XXVIII.20). Мы никогда не бу-
дем знать, могло бы это произойти или нет, но подумать об этом занятно.
Глава XXIX
РЕПТИЛИИ: ЛЕТАЮЩИЕ
И ПЛАНИРУЮЩИЕ
©Пока позднетриасовые и юрские динозавры превращались в гиган-
тов, в рептилийном мире появились два новых эволюционных на-
правления. Некоторые архозавры приобретают крылья и, осваивая
воздух, становятся теплокровными. У части из них появляются перья - они
становятся птицами. На других появляются волосы - они превращаются в
птерозавров.
КРЫЛАТЫЕ ПТЕРОЗАВРЫ
Первая ископаемая летающая рептилия была найдена в известняках, со-
держащих остатки насекомых, креветок и отпечатки медуз. В 1784 г. обра-
зец был описан кратко как “одно из тех позвоночных, которые в древние вре-
мена обитали в море”. В 1809 г. этот образец был переизучен великим фран-
цузским анатомом Ж. Кювье, по-видимому, среди живших в то время
больше всех знавшем о древних животных. Он определил, что ископаемое
является крылатой рептилией и назвал ее Pterodactylus или “крылатым
пальцем”. Группа рептилий, к которой она относится, сейчас называется
Pterosauria.
Давайте сравним птерозавров с птицами - они действительно выглядели и
функционировали не так, как другие позвоночные. У всех было короткое те-
ло и большие костные головы с мозгом, протягивающимся впереди до уровня
больших глаз со склеротическими кольцами. Хвосты были короткими или
длинными, крылья состояли из кожистой перепонки, натянутой на костный
каркас. Длинные кости крыльев были пустотелыми и наполнены воздухом,
к грудине крепились мощные летательные мышцы. Долго ожидавшееся под-
тверждение теоретическим предположениям о существовании у этих живот-
ных волосяного покрова появилось с находкой в позднеюрских отложениях
Казахстана небольшого, размером с голубя, рамфоринхоида Sordes.
Как и у летучих мышей и птиц, в крыльях птерозавров можно отчетливо
увидеть передние конечности, или “руки”. Мы можем определить плечевую
кость, кости предплечья, грудину. В старых книгах даже описывается пять
пальцев: гигантский удлиненный “мизинец” поддерживал большую часть
крыла плюс три коротких пальца, заканчивающихся когтями, и рудимен-
тарный большой палец. Сейчас представляется, что у большинства птероза-
© 1997. Ю.М. Губин, перевод на русский язык и изменения.
464
КАМЕННАЯ КНИГА
Фиг. XXIX.1. Птерозавры
Rhamphorhynchus (слева) был прогрессивным длиннохвостым птерозавром, до-
стигавшим 60 см длины. Dimorphodon (справа) обладал высоким и очень легким
черепом и был чуть больше метра длиной. Оба они жили на территории Европы
в юрское время
вров этот “большой палец” был только окостеневшим сухожилием, так что
“большим” был четвертый, а пятый был утерян еще на ранних стадиях эво-
люции летающих ящеров.
Длиннохвостые птерозавры. Хвосты, пальцы и зубы разделили птерозав-
ров на две группы. Более древние из них имели длинные хвосты, пятипалые
лапы и обычно хорошо развитые, хотя часто и немногочисленные зубы. Их
остатки найдены в позднетриасовых и юрских породах.
Dimorphodon жил в ранней юре. Как и полагается одному из древнейших
длиннохвостых птерозавров, он был еще достаточно примитивен. Хотя череп
и был легким, с тремя большими отверстиями с каждой его стороны, морда
еще была неудлиненной, в отличие от всех поздних представителей группы.
По зубам он больше напоминал неспециализированных текодонтов, чем ко-
го-нибудь из птерозавров.
Rhamphorhynchus был последним и наиболее развитым длиннохвостым
птерозавром и жил в позднеюрское время (фиг. XXIX. 1). У него был развит
большой вытянутый череп, вооруженный большими острыми и изогнутыми
зубами и хорошо приспособленный к охоте за рыбой. Длинный тонкий хвост
на конце расширялся, что, видимо, помогало рулить во время полета.
Короткохвостые птерозавры. Эти рептилии, скорее всего, являются по-
томками длиннохвостых, хотя их прямой предок еще не известен. В процессе
эволюции хвост постепенно укорачивается, исчезает пятый палец на перед-
них лапах, зубы сначала становятся стройней и многочисленней, а у поздних
птерозавров исчезают совсем. При этом морда приобретает форму клюва. Ко-
роткохвостые птерозавры появились в поздней юре и вымерли незадолго до
окончания мелового периода.
Ctenochasma из германской юры была примитивным безклювым животным
с коротким хвостом и головой длиной всего 15 см (фиг. XXIX.2). Зубы живот-
ного были тонкими и частыми (похожими на щетину), а в передней части чере-
па отсутствовало несколько костей. У Pterodactylus эта потеря была компенси-
рована: хотя кости и не появились вновь, те, что остались, удлинились и вос-
полнили потерю. Зубы на челюстях, видимо, исчезли, и появляется клюв.
РЕПТИЛИИ: ЛЕТАЮЩИЕ И ПЛАНИРУЮЩИЕ
465
Фиг. XXIX.2. Летающие рептилии с разными типами зубов
а — Pterodactylus — короткохвостый юрский птерозавр размером с голубя; б — че-
реп Ctenochasma с многочисленными длинными зубами, длина черепа около
15 см; е — череп Dtmorphodon длиной около 25 см
В течение поздней юры и раннего мела более двадцати видов Pterodactylus
обитали на пространстве от Африки до Северной Европы; некоторые из них до-
стигали размеров гуся, однако большинство было не крупнее обычного воробья.
С появлением короткохвостых птерозавров начинается увеличение разме-
ров крыльев. Например, у позднемелового Nyctosaurus с телом, как у голубя,
крылья были больше, чем у орла. Еще необычнее был Pteranodon, обитавший
в Северной Америке и Европе. Его гигантские крылья, достигавшие в разма-
хе 8 м, поддерживали тело всего 50 см длиной (исключая голову и хвост); а ве-
сило все животное не более 10 кг. Шея была длиннее чем тело, а череп дости-
гал 2 м, причем его длина зависела от того, развивался ли выступающий на-
зад костный гребень или нет. Нижняя челюсть была глубокой и представляла
собой верх “кармана”, как у пеликанов, в котором добыча сохранялась до то-
го, как быть проглоченной и переваренной (фиг. XXIX.3).
Quetzalcoatlus был самым крупным птерозавром, или, говоря другими
словами, самым крупным организмом среди тех, кто мог летать. Он был
найден в Техасе, США. К сожалению, от Кецалькоатля известно только не-
сколько фрагментов черепа, костей крыла и позвоночника. Тем не менее,
расчеты показывают, что животное весило около 86 кг и имело размах
крыльев до 12 м.
Одним из самых необычных птерозавров был Dsungaripterus из верхнего
мела Китая (фиг. XXIX. 5). На голове у него был гребень, но направленный не
назад, как у Pteranodon, а вперед, над глазами. Кроме того, его морда и ни-
жняя челюсть были загнуты вверх. Необычно?
Еще более своеобразным был Pterodaustro из верхней юры или раннего ме-
ла Аргентины. В нижней челюсти животного сидели длинные изогнутые па-
лочковидные зубы, причем их количество достигало 24 штук на один санти-
метр длины. Может быть, Pterodaustro использовал такие необычные зубы,
как современный черный ножеклюв или водорез (родственник чаек, встреча-
ющихся в наши дни в Северной Америке и Африке)? Пролетая, он мог погру-
жать нижнюю челюсть в воду и выцеживать из нее мелкую рыбу. Хотя чер-
ный водорез и не имеет настоящих зубов, гребнеобразная форма его нижней
челюсти дает возможность успешно охотиться именно таким образом.
Условия жизни крылатых рептилий. В старых книгах птерозавры изобра-
жались похожими на летучих мышей, неспособными к прогулкам и практи-
30. Каменная книга
466
КАМЕННАЯ КНИГА
Фиг. XXIX.3. Pteranodon и его череп. Этот короткохвостый беззубый меловой
птерозавр из Канзаса, США, имел размах крыльев до 8 м
Фиг. XXIX.4. Dimorphodon, рамфоринхоид из ранней горы Европы. Череп дли-
ной до 20 см (рисунок из работы К. Падиана, 1983)
чески беспомощными на земле. Считалось, что они могли взлетать только
спрыгивая с деревьев; часто они изображались прицепившимися лапами к
веткам или потолкам пещер, как это делают в наши дни летучие мыши.
Позже выяснилось, что по своим характеристикам птерозавры были более
птицеподобными, чем летучемышеподобными. В отличие от последних, их
задние конечности не присоединялись к мембране. Более того, на этих сво-
бодных от мембран лапах птерозавры могли гулять! У некоторых даже были
на лапах плавательные перепонки. Распределение летательных мышц было
более похожим на птиц, чем на летучих мышей. И птерозавры, возможно,
могли взлетать без затруднений как с земли, так и с поверхности воды.
Летающие ящеры охотились, активно используя зрение и значительно
меньше обоняние. На слепках мозга животных видно, что зрительные доли
сильно увеличены, в то время как отвечающие за обоняние уменьшены. Ма-
ленькие виды охотились на насекомых, ухватывая их на лету или сидящими
на деревьях. Большие птерозавры поедали рыб и водных беспозвоночных.
Остатки непереваренной пищи были найдены в районе глотки Pteranodon, в
то время как объемистый горловой мешок был пустым. Может быть это от-
рыжка? Некоторые птерозавры могли быть даже мусорщиками.
Птерозавры юры Германии жили на островах и заселенных побережьях,
вблизи от моря. Pteranodon также обитал в прибрежных районах и сильные
РЕПТИЛИИ ЛЕТАЮЩИЕ И ПЛАНИРУЮЩИЕ
467
Фиг. XXIX.5. Череп Dsungaripterus, примитивного птеродактилоида с сохра-
нившимися редкими зубами в челюстях. Длина черепа до 41 см, а крыла — до
3.5 м. Он был найден в позднеюрских (? раннемеловых) отложениях провинции
Синдзянь, Китай (фото Франка Коффы, собственность Института палеонтоло-
гии позвоночных и палеоантропологии. Пекин, и музея Виктории, Мельбурн)
ветры могли относить его на сотни километров от берега. Часть крылатых
рептилий, возможно, жила на озерах и охотилась на рыб.
Две опасности поджидали как маленьких, так и больших птерозавров. Во-
первых, это попасть в глотку более крупных летающих рептилий, или же, пла-
нируя, угодить в пасть мозазавров и плезиозавров во время их охоты за рыбой
и беспозвоночными. Вторая опасность - угроза повреждения крыльев. Поэто-
му усиление и упрочнение мембраны крыльев за счет спутанных внутренних
волокон внутри кожи было одним из эволюционных приобретений животных.
ПЛАНИРУЮЩИЕ ЯЩЕРИЦЫ
За 20-30 млн. лет до появления первых летающих птерозавров, другая
группа рептилий, ящерицы, стала осваивать воздух, однако они выбрали ме-
нее удачный путь. Вместо развития крыльев на измененных передних конеч-
ностях, ребра таких ящериц расширяются для поддержки мембраны, позво-
лявшей животным планировать с дерева на дерево. Эти ящерицы известны
только из триаса Нью-Джерси, США (Icarosaurus) и из пермско-юрских от-
ложений Англии (Kuehneosaurus, Kuehneosuchus). Известно лишь несколь-
ко групп летающих ящериц, исчезнувших, не дав потомков. Через сто девя-
носто миллионов лет другие ящерицы приспособились делать то же самое.
Это современный Draco, “летающий дракончик”, планирующая ящерица из
Юго-Восточной Азии.
Интересная рептилия, близкая к ящерицам, обитала в раннетриасовое
время в Средней Азии. У Sharovipterix тонкая кожистая перепонка соединя-
ла короткие передние и длинные задние лапы (у птерозавров наоборот). Ле-
тать эти животные, однако, не могли, но планировали не хуже ящериц, по-
крывая при этом расстояния в несколько десятков метров.
зо*
Глава XXX
ПТИЦЫ - ИЗОБРЕТАТЕЛИ ПЕРА
• Только птицы имеют перья. Это единственный признак, которым
обладают все создания, относимые к птицам. Перо - замечательная
структура, очевидно, возникшая из сильно измененных чешуй реп-
тилий. Они служат хорошим изолирующим покровом, помогающим сохра-
нять тепло, а структура пера такова, что отталкивает воду, чему помогает
смазка секретом копчиковой железы. Но самое важное назначение пера -
формирование из этих легких образований основы крыла и хвоста, необходи-
мых для полета. Полет - безусловный фирменный признак птиц, хотя неко-
торые из них стали нелетающими, пройдя через фазу полета.
ПРОИСХОЖДЕНИЕ ПТИЦ
Позднеюрские известняки в Германии, в которых было найдено много ле-
тающих рептилий — птерозавров также сохранили для нас другое позвоноч-
ное, названное археоптериксом {Archaeopteryx). Сначала в августе 1861 г.
было найдено единственное перо. Скелет самого археоптерикса в перьях был
найден месяцем позже, а еще один был раскопан в 1877 г. Сейчас известно
7 его экземпляров и то - первое перо. Из них один - частичный скелет, еще
два - без черепа, остальные 4 - практически полные с черепами, с ясными от-
печатками перьев крыла и хвоста. Интересно, что ни один из экземпляров
археоптерикса не показывает никакого другого оперения, что, возможно,
указывает на возникновение перьев в процессе эволюции прежде всего с по-
требностями полета.
Archaeopteryx значит - “древнекрыл”. До сих пор он - самое древнее из из-
вестных существ, имеющих перья. Археоптерикс найден только в лагунных
отложениях Южной Германии возрастом 150 млн. лет. Более сотни лет уш-
ло, чтобы найти эти несколько бесценных скелетов. Однако, если бы они бы-
ли без перьев, археоптерикс вполне мог бы быть классифицирован как реп-
тилия, а не птица.
Мозговая коробка черепа археоптерикса увеличенных размеров, поэтому
его мозг был заметно крупнее, чем мозг его рептильных предков, а швы меж-
ду отдельными костями черепа почти полностью сросшимися. Но в боль-
шинстве других особенностей это вымершее существо, размером с ворону,
было больше похоже на маленького бегающего динозавра, чем на птицу
© 1997. Е.Н. Курочкин, перевод на русский язык и изменения.
ПТИЦЫ - ИЗОБРЕТАТЕЛИ ПЕРА
469
Фиг ХХХ.1. Археоптерикс приготовился взмахнуть крыльями и планировать
с ветки цикадового. Длина около 46 см
(фиг. ХХХ.1). Например, костная основа хвоста состояла из длинного ряда
удлиненных цилиндрических позвонков, как у мелких тероподных диноза-
вров, а не была крайне редуцированной, как у современных птиц. И хвосто-
вые перья крепились у археоптерикса вдоль хвоста, а не поперек его укоро-
ченной костной основы (пигостиля), как у современных птиц.
Кости археоптерикса были плотными, а не полыми, как у большинства
птиц. Челюсти, очевидно, были покрыты плотным чешуйчатым покровом, а
не роговым клювом и вооружены рядом мелких конических зубов, сидевших
в альвеолах. Таз был совершенно динозавровый, хотя лобковые кости смот-
рели вниз и назад. Общая форма и пропорции костей конечностей более
сходны с таковыми крошечных тероподных динозавров, чем с кем-либо еще
(фиг. ХХХ.2).
Джон Остром, специалист по динозаврам из Йельского университета, изу-
чил археоптерикса довольно основательно и заключил, что его предками
должны быть тероподные динозавры. Не все палеонтологи согласны с этим,
некоторые заявляют, что предки были ближе к крокодилам или птицетазо-
вым динозаврам скорее, чем к ящеротазовым. Тем не менее, доказательства
Острома в пользу тероподных предков весьма основательны.
Летал, планировал или просто бегал археоптерикс до сих пор остается
предметом споров. Асимметрия его первостепенных маховых перьев предпо-
лагает, что он мог быть активным летуном. У современных летающих птиц
первостепенные маховые определенно асимметричны, то есть опахала на
разных сторонах от стержня пера имеют разную ширину. Это создает крылу
лучшие аэродинамические качества и помогает увеличить подъемную силу
при полете. Алан Федуччия и Сторрс Олсон - палеонтологи, изучающие птиц
в Соединенных Штатах Америки, - дополнительно обратили внимание, что
структура костей плечевого пояса — лопатки и вилочки, которые уникальны
у птиц, — обеспечивает все необходимое для полета у археоптерикса.
Долгое время археоптерикса считали непосредственным предком всех по-
зднейших птиц. Но в 1980-х годах была открыта новая группа птиц - энан-
470
КАМЕННАЯ КНИГА
Фиг ХХХ.2. Седьмой экземпляр археоптерикса (Archaeopteryx lithographica),
найденный в каменоломнях около г. Зольнгофена (Золенгофена), Германия, в
1992 г. и описанный в 1993 г. Поздняя юра (фото Е. Курочкина, благодаря лю-
безности д-ра П. Вельнхофера). Масштаб 1 см
циорнисы (Enantiornithes). Энанциорнисы являются непосредственными
родственниками археоптерикса, хотя не все специалисты согласны с таким
заключением. Они имели особенное строение черепа, позвонков, крыла и
задней конечности, из которых никак не могли произойти подобные же
структуры у настоящих птиц. Кроме того, сами настоящие птицы найдены
теперь и в раннемеловых отложениях - только немного более молодых, чем
отложения, в которых был обнаружен археоптерикс. Это также ставит под
вопрос возможность происхождения настоящих птиц от археоптерикса. Та-
ким образом, по новой гипотезе археоптерикс и его родственники энанциор-
нисы относятся к боковой и тупиковой линии в эволюции птиц.
ПРОТОАВИС
В 1986 г. в позднетриасовых отложениях штата Техас в США были найде-
ны остатки нового маленького существа размером с сороку. Нашедший и
изучающий его американских палеонтолог Шанкар Чаттерджи в 1991 г. дал
ему имя протоавис техасский (Protoavis texensis). По внешне тяжеловатым и
плотным костям протоависа складывается четырехлапое существо с удли-
ненными передними конечностями, с длинным хвостом из тонких позвонков
и с большой головой (фиг. XXX.3). Однако череп его оказался сложен необы-
чайно легко для ранних архозавров - из тонких пневматизированных кос-
ПТИЦЫ - ИЗОБРЕТАТЕЛИ ПЕРА
471
Фиг ХХХ.З. Реконструкция скелета протоависа (Protoavis texensis) — малень-
кого архозавра из позднего триаса США. Протоавис, вероятно, мог быть пред-
ком эволюционной линии настоящих веерохвостых птиц (по Ш.Чаттерджи,
1991. From Philosophical Transactions of the Royal Society of London, series B,
vol. 332, Number 1265, s. 277-346)
тей, с огромными глазницами и с огромной мозговой полостью. Также
протоавис обладал седловидными позвонками, длинным и тонким коракои-
дом, плечевой костью с двумя мыщелками, тазовыми костями с внутренни-
ми полостями для размещения крупных почек. Эти и другие прогрессивные
черты морфологии ставят протоависа среди весьма вероятных предков насто-
ящих птиц.
Многие палеонтологи не согласны с выводами Чаттерджи. Они считают,
что он ошибается в трактовке анатомии протоависа и приписывает ему кости
от разных животных. Но почти никто не видел воочию его скелета. Действи-
тельно, и скелет передней и задней конечности протоависа нельзя назвать пти-
чьими крылом и лапой. Но они обладают такими особенностями строения, из
которых эволюционно вполне могли возникнуть птичьи крыло и лапа.
ЭНАНЦИОРНИСЫ
Первые энанциорнисы были описаны в 1981 г. английским палеонтоло-
гом Сирилом Уокером из отложений позднего мела Аргентины. Строение их
скелета оказалось настолько необычным, что не все палеонтологи сразу пове-
рили в реальность этой новой группы птиц. Некоторые специалисты первое
время считали, что Уокер ошибся и описал под одним названием смесь кос-
472
КАМЕННАЯ КНИГА
Фиг ХХХ.4. Реконструкция скелета маленького древесного беззубого энанци-
орниса — нананциуса В. Алифанова (Nanantius valifanovi) из верхнего мела
Южной Гоби в Монголии (сохранившиеся в ископаемом состоянии кости зату-
шеваны). Масштаб 1 см. (По E.N. Kurochkin, 1996. A new enantiornithid of the
Mongolian Late Cretaceous, and a general appraisal of the infraclass Enantiorni-
thes (Aves). Moscow: Palaeontological Institute of RAS. Special Issue)
тей рептилий и птиц. Но вскоре выяснилось, что энанциорнисы обитали на
протяжении всего мелового периода и были найдены на всех континентах
мира, кроме Антарктиды. Они были очень разнообразны. Больше всего энан-
циорнисов обнаружено в раннемеловых отложениях Китая.
Среди энанциорнисов теперь известны маленькие зубастые древесные
формы, беззубые формы, крупные и маленькие бегающие наземные и, веро-
ятно, ныряющие птицы. Короче, энанциорнисы оказались наиболее разно-
ПТИЦЫ - ИЗОБРЕТАТЕЛИ ПЕРА
473
образными и, вероятно, доминировавшими в мелу среди птиц (фиг. XXX.4).
Но в конце мела энанциорнисы вымерли, как и динозавры.
Название энанциорнисы (Enantiornith.es) в переводе с греческого обозна-
чает “противоположные птицам”. Такое название полностью соответствует
их необычной анатомии. Внешне их кости очень похожи на кости настоящих
птиц. Но тонкие детали их строения показывают, что функции передней ко-
нечности как крыла, задней конечности как птичьего типа лапы и облегчен-
ного челюстного аппарата были достигнуты у энанциорнисов своим путем,
параллельным эволюционному пути настоящих птиц. Поэтому настоящие
птицы не могли произойти от энанциорнисов.
С другой стороны, в анатомии меловых энанциорнисов и юрского археоп-
терикса прослеживается много общеприобретенных унаследованных особен-
ностей строения, указывающих на их близкое родство друг с другом и с теро-
подными динозаврами непосредственно. Таким образом, эволюционная ли-
ния археоптериксы-энанциорнисы, скорее всего, произошла от общего
предка с тероподными динозаврами. Эволюционная же линия настоящих
птиц прямых родственных связей с тероподными динозаврами, очевидно, не
имела.
Не все ученые согласны с такой оценкой родственных отношений энанци-
орнисов. Так, американский специалист Люис Киаппи доказывает прямое
родство энанциорнисов и настоящих птиц. А всех птиц он считает прямыми
потомками тероподных динозавров.
ЗУБАТЫЕ МЕЛОВЫЕ ПТИЦЫ
Археоптерикс и энанциорнисы были успешным экспериментом эволюции
своего времени, но позднее они постепенно проиграли в конкуренции с на-
стоящими птицами. Но и среди настоящих птиц было не менее двух безус-
пешных эволюционных попыток завоевать воду и воздух. Этот опыт природа
совершала с птицами, у которых сохранялось древнее наследство настоящих
зубов в челюстях.
Хорошим примером таких птиц является гесперорнис (Hesperornis), что
значит “западная птица”, лучше всего известный из отложений, сформиро-
ванных большим внутренним морем, разделявшим Северную Америку попо-
лам в меловом периоде. Много частичных скелетов гесперорнисов дали Нио-
брарские меловые отложения штата Канзас и окружающих территорий в
США, а их отдельные кости находили севернее, вплоть до Канадской Аркти-
ки. Форма костей гесперорниса очень похожа на форму костей современных
птиц. Но он еще имел зубы, а его кости были толстостенными, так как это бы-
ла ныряющая птица. Гесперорнису не требовалось летать и его крылья были
сильно редуцированы (фиг. XXX. 5). В мелководных внутренних морях того
времени гесперорнисы были довольно крупными хищниками, так как неко-
торые виды достигали 1.5 метра в длину.
Более 100 лет считалось, что гесперорнисы обитали в мелу только в Север-
ной Америке. Но в 1990-х годах русский палеонтолог Лев Несов окончатель-
но подтвердил, что такие же крупные нелетающие гесперорнисы жили в по-
зднемеловое время в морских проливах на территории современных Север-
ного Казахстана, Центральной России и Южной Швеции.
Род Hesperornis — только один из многих гесперорнисообразных птиц, рас-
пространенных во всем мире. Их пресноводные формы были найдены в позд-
нем мелу Северной Америки, Канады и Монголии, хотя большинство связа-
474
КАМЕННАЯ КНИГА
Фиг ХХХ.5. Зубатые птицы позднего мела
Ихтиорнис (слева) и гесперорнис (справа). На других реконструкциях геспе-
рорниса изображают без хохла и со сплошной перепонкой между пальцами, а
не с лопастями на отдельных пальцах
но с морскими условиями. Гесперорнисы были похожи на современных га-
гар по форме тела и образу жизни, но не были связаны с ними родством.
В тех же отложениях Ниобрары, откуда происходит гесперорнис, найдена
небольшая летающая зубатая птица ихтиорнис (Ichthyornis), что означает
“рыбная птица”. Ихтиорнис имел большой киль на грудине и длинные кры-
лья, в отличие от гесперорниса, и, вероятно, вел успешную жизнь как лета-
ющий хищник над меловыми морями, по крайней мере в Северной Америке
(см. фиг. ХХХ.5).
МЕЗОЗОЙСКИЕ НАСТОЯЩИЕ ПТИЦЫ
Не позднее чем с самого начала мелового периода, уже появились птицы,
обладавшие признаками специализации современных птиц в строении ске-
лета. Во всяком случае некоторые из них еще сохраняли зубы, как, напри-
мер, хаоянгия (Chaoyangia) из раннего мела Китая. Уже в это время появи-
лись родственники современных неогнатических птиц, такие как китайский
ляонинорнис (Liaoningornis) и румынский евролимнорнис (Eurolimnornis).
Вместе с ними уже существовали и родственники современных палеогна-
тических птиц, каковыми были амбиортус (Ambiortus) из Монголии
(фиг. XXX.6) и отогорнис (Otogornis) из Китая. Немногие известные теперь
раннемеловые настоящие птицы жили одновременно с, пожалуй, более мно-
гочисленными и разнообразными энанциорнисами и даже - в одних и тех же
местах рядом с ними, что показывают раскопки в Монголии и Китае. Еще
совсем недавно некоторые палеонтологи не верили, что настоящие птицы
обитали на Земле так давно - 120-135 млн. лет назад (фиг. XXX. 7), в эпоху
расцвета динозавров. Считалось, что время появления и первого разнообра-
зия настоящих птиц приходится на кайнозойскую эру, на начало палеогена
(фиг. XXX.8). Теперь же установлено, что уже в позднемеловое время встре-
чались куликоподобные, гагароподобные, бакланоподобные и альбатросопо-
добные птицы, мало отличающиеся от их современных потомков.
ПТИЦЫ - ИЗОБРЕТАТЕЛИ ПЕРА
475
Фиг ХХХ.6. Частичный скелет (позвоночник, плечевой пояс, крыло) амбиорту-
са Дементьева (Ambiortus dement]evi), раннемеловой палеогнатической птицы
из озерных отложений неокома Центральной Монголии. В нижней части ри-
сунка на отпечатке и противоотпечатке концевой части крыла хорошо видны
когти пальцев крыла и отпечатки перьев. Масштаб 1 см. Оригинал
476
КАМЕННАЯ КНИГА
Фиг ХХХ.7. Раннемеловые перья птиц из местонахождения Байса, Забайка-
лье, Россия. На одном из образцов, показанных отпечатком и противоотпечат-
ком, сохранился характер распределения окраски. Одно маленькое пуховое пе-
ро указывает на терморегуляторные функции оперения уже у раннемеловых
птиц. Перья найдены в отложениях возрастом 120 млн. лет, что вместе с други-
ми находками раннемеловых перьев во всем мире указывает на широкое рас
пространение птиц, уже с начала мела. Масштаб 1 см (фото Е. Курочкина, Па
леонтологический институт РАН)
КАЙНОЗОЙСКИЕ “ЛЕТАЮЩИЕ МАШИНЫ’
Время расцвета настоящих птиц приходится на кайнозой. Настоящие
птицы имеют много общих особенностей, связанных с полетом, даже если не-
которые из них стали вторично нелетающими. Для того чтобы хорошо ле-
тать, важно иметь легкий вес и прочный скелет. Поэтому скелет птиц стал
легче, утратив зубы и приобретя полые кости, которые быстро в первые
2-3 месяца жизни достигают окончательных размеров и строения. Произош-
ло полное слияние ряда костей в крыльях и задних конечностях, так же как
и в черепе, что уменьшило вес, но увеличило их прочность.
ПТИЦЫ - ИЗОБРЕТАТЕЛИ ПЕРА
477
Фиг ХХХ.8. Пещерные отложения эоцена и олигоцена во Франции дали бога-
тые коллекции ископаемых птиц. Это местонахождение в Керси - одно из наи-
более известных (фото благодаря любезности Бернарда Зига и Сесиль Морер-
Шавире)
Скелет настоящей птицы имеет и другие прочные узлы, особенно в груд-
ной области, образованные слиянием костей и развитием маленьких крюч-
ковидных отростков на ребрах. Отросток одного ребра связывает его со сле-
дующим сзади ребром, что создает во время полета прочную “коробку” для
грудной полости. Такая конструкция защищает жизненно важные органы
птицы, такие как сердце и легкие, когда во время полета возникает мощное
давление на эту область.
Все эти характеристики птиц делают их уникальными. Поэтому палеон-
толог, изучая даже отдельные кости, во многих случаях может установить
их принадлежность к птицами и даже определить их вид, а часто и образ
жизни (фиг. XXX.9). Некоторые кости лучше определяются по сравнению с
другими. В ноге особенно хорош в этом отношении тарсометатарсус
(фиг. XXX. 10). Он формируется слиянием тарсальных и метатарсальных ко-
стей ступни. Тарсометатарсус имеет важное значение в познании ископае-
мых птиц. Он очень прочный и часто сохраняется в ископаемом состоянии.
478
КАМЕННАЯ КНИГА
Фиг ХХХ.9. Один из плотоптерусов (Plotopteridae) — пингвиноподобных птиц,
родственных пеликанам. Пингвины известны из Южного полушария. Плотоп-
териды встречались только в Северном полушарии, развившись из других
групп птиц, хотя они вели образ жизни, сходный с пингвинами. Этот экземпляр
из верхнемиоценовых морских отложений Японии. Ископаемые плотоптериды
известны из Северной Пацифики и были внешне похожи на пингвинов средних
и крупных размеров. (Фото благодаря любезности Й. Хасегавы)
Он также имеет сложное строение с большим количеством диагностических
признаков, позволяющих даже по фрагменту установить систематическую
принадлежность птицы.
Ископаемая летопись птиц раннего кайнозоя основана преимущественно
на изучении фрагментов их костей, хотя в последние десятилетия находят
все больше и больше отпечатков скелетов. Она неплохо отражает происхо-
дившие эксперименты, некоторые из которых заканчивались вымиранием
отдельных созданных эволюцией типов птиц, в то время как в других фило-
генетических стволах птиц происходила модернизация строения, что позво-
лило птицам стать к концу кайнозоя самым многочисленным по числу видов
классом позвоночных животных.
В общем, птицы того времени были промежуточны по своему строению
между известными сегодня семействами птиц. Если бы читатель смог отпра- I
виться в палеоцен или эоцен наблюдать птиц, он не смог бы легко поместить
увиденных птиц в современные семейства.
Пресбиорнис (Presbyornis) — одна из таких птиц, известная из эоценовых
отложений южной части штата Вайоминг (фиг. XXX. 11). У пресбиорниса
ПТИЦЫ - ИЗОБРЕТАТЕЛИ ПЕРА
479
Eudyptes-
пингвин
Diomedea-
альбатрос
Falco-
сокол
Cacatua-
попугай
Согсогах-
воробьиная
птица
Фиг XXX. 10. Скелет птицы с наиболее важными (затушеваны) для палеонтоло-
га костями. Разнообразие тарсометатарсусов (справа) иллюстрирует некоторые
различия между таксонами птиц. Тарсометатарсус часто сохраняется в ископае-
мом состоянии и может быть использован для определения семейств птиц и для
более точного определения, если он хорошо сохранился. (С изменениями по Rich
and Thompson, 1982, The Fossil Vertebrate Record of Australasia.) Масштаб 1 см
удивительным образом сочеталась мозаика признаков современных кули-
ков, уток и фламинго. Это была длинноногая околоводная птица с клювом,
очень похожим на утиный, с крыльями, как у куликов, с лапами, похожими
480
КАМЕННАЯ КНИГА
Фиг XXX.11. Отпечатки следов птиц из эоцена формации Грин Ривер штата
Юта, ширина 2.5—3.8 см. Много водных птиц, включая пресбиорниса (Presbyor-
nis), жили на и около больших озер, которые простирались через штаты Вайо-
минг, Юта и Колорадо в то время
Фиг XXX.12. Череп фороракоса (Phororhacos longissimus) из раннего миоцена
(сантакрусий) Аргентины, около 70 см длиной (фото благодаря любезности Ро
зендо Паскуаля, музей Ла-Платы, Аргентина)
одновременно на куличиные и фламинговые. Таким образом, куда бы чита-
тель поместил пресбиорниса в его определителе птиц эоцена? Конечно, и се-
годня мы знаем много птиц, занимающих промежуточное положение между
несколькими группами птиц. Такие промежуточные формы, как пресбиор-
нис, позволяют проникнуть внутрь семейственных связей птиц и рождают
идеи о родственных связях между крупными группами птиц.
Птицы раннего кайнозоя отличаются от современных также и в другом от-
ношении. Отдельные группы птиц, малочисленные сегодня, такие, как, на-
пример, журавлеобразные птицы (Gruiformes), были во много раз разнооб-
разнее в палеоцене и эоцене. Некоторые из них даже терроризировали мле-
копитающих того времени. Южноамериканская группа форорацид широко
ПТИЦЫ - ИЗОБРЕТАТЕЛИПЕРА
481
Фиг ХХХ.13. Диатрима (Diatryma) — нелетающая эоценовая птица из штата
Вайоминг, США, около 2.1 м высоты
радиировала в кайнозое, представляя основных наземных хищников в тече-
ние всего этого времени. Они имели огромный высокий клюв с порядочным
крючком на конце, такой же как у орлов, а голова была почти таких же
размеров, как у современной лошади (фиг. XXX.12). Когда наземные позво-
ночные пересекали Средне-Американский сухопутный мост, один из форо-
рацид — титанис (Titanis) прошел на север; огромные фаланги пальцев его ла-
пы были найдены в плейстоценовых отложениях штатов Флорида и Техас.
Тяжеловесные эоценовые диатримы (Diatryma) из Северной Америки и
Европы, несмотря на устрашающе огромный и крючковатый клюв, вели, по
всей видимости, мирную растительноядную жизнь, но их мощные ноги пред-
ставляли серьезное оборонительное оружие против хищных млекопитаю-
щих (фиг. ХХХ.13). Крылья у диатрим были редуцированы до состояния ма-
леньких обрубков.
Кроме журавлеобразных птиц и их родственников, были разнообразные
ржанкообразные (Charadriiiformes), особенно фламингоподобные формы,
такие как пресбиорнис. Более разнообразны, чем сегодня, были также куро-
образные птицы (Galliform.es) и совы (Strigi formes).
Некоторые группы того времени не оставили живущих ныне родст-
венников. Одной из таких групп являются костнозубые или ложнозубые
птицы - псевдодонторнисы, известные из Англии, Франции, Азербайджана,
Нигерии, США, Канады, о.Сеймур (Антарктика) и Новой Зеландии
(фиг. XXX. 14). У этих гигантских родственников пеликанов и бакланов не
было настоящих зубов, но у них на челюстях образовались ряды костных вы-
ступов, функционировавших как зубы, делая их вероятными великолепны-
31 Каменная книга
482
КАМЕННАЯ КНИГА
Фиг XXX.14. Реконструкция одной из костнозубых или ложнозубых птиц — псев-
додонторниса (Pseudodontomis), имевшего рассчитанный размах крыльев в 4.5 м,
Первые родственные псевдодонторнису формы известны из эоцена, а последние —
из плиоцена. Этот псевдодонторнис был найден в Новой Зеландии. (Рисунок вос-
произведен благодаря любезности компании Шелл, Новая Зеландия)
ми поверхностными рыболовами. Это были одни из самых больших, когда
либо известных, летающих птиц, существовавших от эоцена до плиоцена.
Однако самой большой летающей птицей была позднемиоценовая аргентин-
ская хищная птицы аргентавис (Argentavis) (фиг. XXX. 15). Аргентавис ве-
сил около 70 килограмм и имел размах крыльев около 7 метров!
МИР БЕЗ ПЕСНИ
Некоторые группы птиц, очень заметные и многочисленные сегодня, как,
например, певчие воробьиные, вероятно, даже не существовали в начале
кайнозоя. Хотя в эоцене Австралии в последнее время были найдены родст-
венники современной птицы-лиры, относящиеся к примитивным воробьи-
ным (Sub-Oscines). Однако настоящие певчие птицы (Oscines) не известны ра-
нее олигоцена. Таким образом, в эоцене было не слышно крика вороны или
утреннего хора жаворонков. Вместо них разнообразные маленькие древес-
ные птицы, которые могли только кричать или пищать, а среди них - примо-
буккониды - отдаленные родственники дятлов и пуховок, дальние родствен-
ники современных кукушек, птиц-мышей, козодоев (фиг. XXX. 16), стри-
жей, трогонов, зимородков заполняли пространство, ныне захваченное
певчими птицами. В последние несколько миллионов лет певчие птицы нео-
быкновенно успешно радиировали в сотни форм. Сегодня они составляют бо-
лее половины из 9000 видов всех птиц мира.
ПТИЦЫ - ИЗОБРЕТАТЕЛИ ПЕРА
483
Фиг XXX.15. Реконструкция аргентависа (Argentavis magnificens) из поздне-
го миоцена (уякерий) Аргентины и палеонтолог К.Кемпбелл (из статьи
Е. Курочкина, Наука и жизнь, 1990, № 5, с. 84-85; фото благодаря любезности
д-ра К.Кемпбелла, музей Естественной Истории округа Лос-Анджелес, США)
СОВРЕМЕННЫЕ ПТИЦЫ
Не позднее начала миоцена, около 22 миллионов лет назад, в ископаемой
летописи начали появляться птицы, очень похожие на сегодняшних
(фиг. XXX. 17). Эти ископаемые формы еще не относились к современным
видам, но были их близкими родственниками. Кроме них, все еще встреча-
лись некоторые архаичные птицы раннего кайнозоя, такие как ложнозубые
птицы и фламингоподоные палелёдусы. Но большую часть уже составляли
примитивные формы птиц из современных семейств и родов.
Современные виды начали появляться в позднем плиоцене. Около
1.8 миллионов лет назад, не позднее начала плейстоцена, большинство уже
составляли современные виды птиц. В ледниковый период авифауна была
смешанной из современных и вымерших видов (фиг. XXX. 18). Ископаемая
летопись птиц для этого интервала времени наиболее заполнена, по сравне-
нию с какой-либо предшествующей эпохой. В том числе, и общее число видов
птиц, известных для ледникового периода, много выше, чем для предшест-
вующих интервалов времени. Такой результат лишь отражает хорошую со-
хранность ископаемых остатков птиц из плейстоцена, но не показывает
31*
484
КАМЕННАЯ КНИГА
Фиг XXX.16. Скелет квиполлорниса (Quipollornis) — древнейшего совиного ля-
гушкорота, представителя современного семейства Aegothelidae, из миоцено-
вых озерных отложений штата Новый Южный Уэльс, Австралия
Длина черепа около 4 см. Это редкий случай полной сохранности птичьего ске-
лета в ископаемом состоянии, даже с сохранившимися отпечатками перьев, к
тому же, геологический возраст находки точно датирован, так как вскоре после
гибели птицы поток лавы покрыл озеро. Абсолютный возраст этих вулканичес-
ких отложений определен по радиоактивным изотопам как 13.7-17 млн. лет на-
зад (фото Ф. Коффы, благодаря любезности музея Виктории, Мельбурн)
действительного количества видов существовавших тогда, по сравнению с
прошлым или с настоящим.
Не позднее конца плейстоцена, около 10 тыс. лет назад, вымерли некото-
рые группы птиц, которые прошли из раннего кайнозоя в плейстоцен, такие
как гигантские михиранги или дроморнитиды (Dromornithidae) в Австра-
лии. В некоторых группах гигантские формы возникли только в плейстоце-
не. К ним относятся, например, гигантские большеноги (Progura) Австра-
лии, огромная катарта тераторнис (Teratornis) в Новом Свете (фиг. XXX. 19)
и кубинские плейстоценовые совы орнимегалониксы (Ornimegalonyx)
(фиг. XXX.20-21). Другие сформировались определенно раньше, как ново-
зеландские моа, мадагаскарские эпиорнисы или крупный маврикийский
дронт (фиг. ХХХ.22). Однако в конце концов эти огромные птицы также ис-
чезли. Во многих случаях еще остается открытым вопрос: что или кто послу-
жили причиной их вымирания? Климат или человек?
МОА, МИХИРАНГИ И ДРУГИЕ НЕЛЕТАЮЩИЕ ПТИЦЫ
Африканские страусы, казуары, нанду, и киви относятся к разнообразной
группе птиц, называемой - бескилевыми (Ratitae), хотя предпочтительнее
их называть палеогнатами (Palaeognathae). Некоторые представители пале-
ПТИЦЫ - ИЗОБРЕТАТЕЛИ ПЕРА
485
Фиг XXX.17. Shandongornis shanwanensis — перепелоподобная птица из мио-
цена провинции Шаньдун, Центральный Китай
Тысячи скелетов позвоночных и масса отпечатков растений сохранились там в
тонких озерных осадках, сформировавшихся в вулканической кальдере (благо-
даря любезности Института палеонтологии позвоночных и палеоантропологии
Китайской Академии наук, Пекин)
огнат были и остаются маленькими, такие как, например, киви, хотя другие
оказались самыми большими из когда-либо живших птиц. Интересно, что
современные палеогнаты распространены по фрагментам древней Гондваны:
киви и моа - в Новой Зеландии, нанду и тинаму - в Южной Америке, двупа-
лые страусы - в Африке, а казауры и эму - в Австралии. Эпиорнисы Мадага-
скара также относились к бескилевым птицам. Прерывистое распростране-
ние современных палеогнат можно объяснить широким распространением
их предков.
До недавних пор древние палеогнаты были совсем неизвестны. Длинный
ряд разнообразных палеогнат из палеоцена и эоцена Европы, Северной и
Южной Америки был открыт после 1981 г. Раннемеловые палеогнаты были
найдены в Китае, Монголии и в Европе. Стало известно, что некоторые пале-
огеновые палеогнаты имели нормальную грудину с килем и могли хорошо
летать. В то же время, уже с палеоцена появились довольно крупные бес-
килевые нелетающие формы, как, например, бразильский диогенорнис
486
КАМЕННАЯ КНИГА
Тераторнис
кряква
Кваква
кондор
Falcoruformes (2570)
Беркут
Зубчатоклювая
Журавль
Луговой балобан
Каракара
* Вымершие виды
Орел
Г риннелла
Воробьиная
пустельга
Американским
стервятник *
Канадская
казарка
индейка *
Белоголовый
орлан
Лабреискии
кондор
Болотный
лунь
Лабреи
аист
Полосатый
ястреб
Краснохвос-
канюк
Лентохвостый Плачущая
горлица
голубь
мохноногим
канюк
Крикливым
Шилоклювка зуёк Кроншнеп
Орел
Даггетта
Поганка
орокопут
илоклювыи
дятел
Калифорниискии
бегун
Виргинский Ла^Т^кая
филин совка
Ворон Сорока Рогатый
Cuculiformes 06
Crilumbilonnes
Калифорнийская
Фиг XXX. 18. Относительная представленность различных групп птиц, собран-
ных в битумных ямах Ранчо Ла-Бреа в Калифорнии плейстоценового возраста
Обилие хищных птиц и падалеядов^в ископаемом состоянии из этих древних би-
тумных отложений было много выше, чем в естественных популяциях, потому
что много хищников попадало в эти “ловушки” при охоте на других животных.
Этот пример хорошо показывает, насколько относительна ископаемая лето-
пись, не всегда ясно отражающая действительное обилие животных прошлого
(из работы Хильдегард Ховард, рисунок Драга Джельт)
(Diogenornis). В общем, древние палеогнаты были шире распространены и за-
метно разнообразнее современных их потомков.
Для всех современных бескилевых характерна сильная редукция крыль-
ев, в ответ на жизнь в изоляции без хищников на больших или малых остро-
вах. Большинство - всеядно, и их пища чрезвычайно разнообразна. Не все
ученые согласны, что бескилевые относятся к одной и той же родственной
линии в эволюции птиц.
ПТИЦЫ - ИЗОБРЕТАТЕЛИ ПЕРА
487
Фиг XXX. 19. Тераторнис Мериэма (Teratornis merriami) из битумных четвер-
тичных отложений Ранчо Ла-Бреа в Калифорнии
Эта птица весила около 14 кг и имела размах крыльев почти 4 м. Тераторнис -
близкий родственник аргентависа из Южной Америки (благодаря любезности
музея Естественной Истории округа Лос-Анджелес)
Эпиорнис и его родня. Эпиорнисы (Aepyornis) или “слоновые птицы” изве-
стны с Мадагаскара. Хотя несколько костных фрагментов из Северной Аф-
рики также были отнесены к ним, а скорлупа яиц с Канарских островов в Ат-
лантическом океане очень напоминает по своему строению эпиорносовую,
только ископаемые остатки с Мадагаскара без сомнений представляют этот
отряд птиц.
Эпиорнис (“птица-великан”) достигал высоты 3 м, это значит, что он был
заметно выше современного африканского страуса (фиг. XXX.23). Лучше
было бы его назвать “тяжеловесом”, так как из-за широкого туловища и мас-
сивных ног его вес достигал 600 кг - самый большой известный для птиц, что
заметно больше 100-160 кг для самцов современного африканского страуса.
У одного вида цевка была длиной 64 см, при диаметре широкого верхнего
конца 46 см и 15 см посредине кости. Бедро было короче, но толще, при такой
толщине оно, возможно, было частично полым. Вместе с костями ног сохра-
нились яйца эпиорнисов. Они достигали 30-33 см в длину при объеме около
7.3 л. Через много лет после гибели последнего эпиорниса люди нашли их в
болотах и использовали вместо мисок или кувшинов. Первые европейцы,
488
КАМЕННАЯ КНИГА
Фиг ХХХ.20. Кубинские плейстоценовые совы орнимегалониксы (Ornimegalo-
пух) — самые большие из известных сов
Орнимегалониксы относятся к типичным совам (Strigidae), но они имели ма-
ленькие крылья и уменьшенную грудину, что указывает на полную неспособ-
ность к полету. На рисунке метровый орнимегалоникс атакует хутию (Са-
promys), крупного кубинского грызуна, сохранившегося до наших дней (рису-
нок художника И. Разиной по реконструкции Е. Курочкина. Наука и жизнь,
1991, №6, с. 118-119)
столкнувшиеся с этими яйцами, применяли их на факториях в качестве со-
судов под ром. Позднее сами яйца стали товаром и были развезены по миру
до самых отдаленных уголков.
Моа — “ужасные” птицы. В то время как эпиорнисы процветали и вымер-
ли на Мадагаскаре, другая группа оперенных гигантов прошла сходный путь
эволюции в Новой Зеландии.
Эту вторую группу составляли моа, научное название которых - Dinorni
thiformes может быть грубо переведено с греческого как “ужасные птицы”. В
действительности они были травоядными, вероятно, питаясь различными
растениями. У моа не было других средств нападения или обороны, кроме их
мощных ног, чтобы ударить или чтобы убежать. Но защищаться моа было не
от кого. До появления в Новой Зеландии около 1000 лет назад маорийцев,
там не было ни одного наземного хищника.
ПТИЦЫ - ИЗОБРЕТАТЕЛИ ПЕРА
489
Фиг ХХХ.21. Ископаемые животные являются популярной темой у коллекци-
онеров марок. На этой кубинской марке изображен орнимегалоникс (Ornime-
galonyx) (благодаря любезности О. Арредондо)
Наиболее древние моа были обнаружены в плиоценовых глинах, возрас-
том около 3-4 млн. лет, на о.Южный. Эти моа не были гигантскими, в отли-
чие от многих более молодых плейстоценовых форм, таких как динорнис (Di-
no mis). Хотя весь список находок моа ограничен коротким интервалом вре-
мени, вероятная история их была во много раз более длинной, возможно,
растянувшись на 80 млн. лет. Приблизительно тогда от остальной Гондваны
отделилась Новая Зеландия и, похоже, на ней остались предки моа. 80 мил-
лионов лет изоляции вполне могли дать все разнообразие их известных форм
(фиг. XXX.24). К тому же, в кайнозое происходили неоднократные подъемы
и падения уровня моря, когда Новая Зеландия в течение времени разделя-
лась на группы островов изменявшихся размеров, формы и количества.
Остатки моа из сухих пещер показывают, что у них голова и большая
часть шеи были почти голые, как у эму. Клюв с острыми режущими краями
и содержание желудков нескольких моа предполагают, что многие из них
ели листья и побеги деревьев и кустарников, хотя другие могли питаться
травянистой растительностью. Высота различных форм была от 1 до 3.7 м,
вес доходил до 300 кг, а внешний вид менялся от грациозных длинноногих
созданий до бочкоподобных птиц с короткими ногами и толстыми голенями
и бедрами. Перья были тонкими и шелковистыми, очень похожими на пух
молодых современных птиц. Как у эму, казуаров и киви каждое перо имело
два стержня, а не один, как у африканского страуса и у американских нанду.
Окраска оперения изменялась от коричневой или каштановой с белыми или
черными пестринами до белой, как у кур породы плимут-рок.
Очень часто остатки моа находят в болотах, в таких, как в долине Пира-
мид Вэлли, в округе Северный Кентербери, на о.Южный в Новой Зеландии.
Там, на каждом гектаре находится, по крайней мере, 1700 скелетов. Подчас
кости ног у этих скелетов стоят вертикально в черной жирной грязи, очевид-
490
КАМЕННАЯ КНИГА
Фиг ХХХ.22. Дронт, или додо (Didus)
Реконструкция дронта с о. Маврикий в Индийском океане, истребленного евро-
пейскими моряками в XVI веке. Родственник голубей, огромный дронт имел
уменьшенные кости крыльев, увеличенные кости задних конечностей и сильно
редуцированный киль грудины. Все это было связано с утратой им способности
к полету (благодаря любезности Американского музея Естественной Истории,
Нью-Йорк)
Фиг ХХХ.23. Два вида эпиорнисов (Aepyornis) ростом 2.7—3 м
Слева - кости лапы и яйца одного из них. Черное пятно внизу - яйцо курицы в
том же масштабе
ПТИЦЫ - ИЗОБРЕТАТЕЛИ ПЕРА
491
Фиг ХХХ.24. Длинноногие и коротконогие моа послеледниковой Новой Зелан-
дии. Самые высокие достигали роста 3.6 м
но, в том же положении, в котором моа погибли, увязнув в древней трясине.
Вместе со скелетами попадаются кучки камней — гастролитов или желудоч-
ных камней — которые моа подбирали и накапливали в “верхнем желудке”.
С их помощью в желудке перетирались листья и ветки, составлявшие боль-
шую часть пищи моа. Однако за много лет в сухих пещерах были найдены
3 самых лучших экземпляра, у которых сохранились кожа, мускулатура и
перья, подходящие для научного исследования. Дальнейшее их изучение,
возможно, откроет, насколько вымершие моа родственны к другим группам
птиц.
Михиранги — возможно, не бескилевые. Моа и эпиорнисы были большими
птицами. Но, вероятно, самой большой из всех известных птиц, во всяком
случае, по весу, был дроморнис Стиртона (Dromonis stirtoni) из позднего ми-
оцена центральной Австралии. Он относился к одному из 8 известных видов
михирангов (Dromornithidae), размеры которых варьировали от близких к
современному эму до самых больших для птиц.
Михиранги отличались от других птиц некоторыми уникальными при-
знаками. Они имели маленькие копытообразные когтевые фаланги на паль-
цах лап и очень небольшой внутренний палец, приближающийся к состоя-
нию, известному для африканского страуса, где он полностью утрачен
(фиг. XXX.25, 26). Квадратная кость - маленькая косточка, связывающая
492
КАМЕННАЯ КНИГА
Фиг ХХХ.25. Жениорнис Ньютона (Genyornis newtoni)
Слепок скелета с реконструированным черепом из плейстоценовых озерных от-
ложений озера Коллабонна в Южной Австралии. Рост этого жениорниса около
1.8 м (благодаря любезности музея Виктории, Мельбурн)
верхнюю и нижнюю челюсти у птиц, рассказывает самые интересные вещи.
Она совсем не похожа на квадратную кость у палеогнат. Вместо этого она
сходна с квадратной костью фазанов, индеек и болыпеногов. Возможно, что
предковая для михирангов курообразная птица оказалась изолированной в
Австралии после отсоединения ее от Гондваны, как это случилось с предком
австралийских бескилевых — казуаров и эму.
Михиранги были разнообразны в миоцене, но затем к плейстоцену остал-
ся только один их вид. Самые последние датированные их находки происхо-
дят из болотных отложений возрастом 26 тыс. лет в Лэнсфильде, в юго-вос-
точной Австралии. Эти отложения по крайней мере на 10 тыс. лет перекры-
вают время появления в Австралии человека (фиг. XXX.27). Возможно,
человек и изменения климата вместе привели к концу эту группу птиц.
Многие виды михирангов, вероятно, были связаны с лесными местообита-
ниями, характерными для большей части Австралии до плиоцена. Когда с
распространением засушливых условий площади лесов стали сокращаться,
а лугов - расширяться, некоторые михиранги, очевидно, двинулись в лу-
говые местообитания, такие как, например, ильбандорнис (Ilbandornis), у
которого были длинные ноги и грацильные пропорции, что характеризует
бегающих животных открытых местообитаний. Но на этих лугах уже жили
ПТИЦЫ - ИЗОБРЕТАТЕЛИПЕРА
493
Фиг ХХХ.26. Реконструкция внешнего вида жениорниса Ньютона (Genyornis
newtoni)
Родственный ему дроморнис Стиртона (Dromornis stirtoni) из миоцена Австра-
лии, возможно, был самой большой птицей из всех живших на Земле, достигая
более 3.6 м высоты (рисунок Ф. Найта, благодаря любезности музея Виктории и
Пайониир Дизайн Студио)
494
КАМЕННАЯ КНИГА
Фиг ХХХ.27. Наскальная роспись австралийских аборигенов (полуостров
Кейп-Йорк, Квинсленд, Австралия) с мифическим человекоподобным Квинка-
ном, стоящим рядом с большой птицей. Эта птица заметно крупнее, чем совре-
менный эму, и может изображать вымершего михиранга жениорниса (Genyor-
nis) (фото К.Деннетт, благодаря любезности П. Трезиса)
Фиг ХХХ.28. В древних дюнных отложениях на о. Норфолк в юго-западной ча-
сти Тихого океана была собрана большая коллекция костей морских птиц,
живших до и после прибытия на остров полинезийского человека. Пожары по
вине человека, вероятно, привели к вымиранию некоторых видов на о. Нор-
фолк. Позднее по вине европейцев вымерли и другие виды птиц
ПТИЦЫ - ИЗОБРЕТАТЕЛИ ПЕРА
495
разнообразные кенгуру. Михиранги могли проиграть такую конкурентную
битву, после того как встретили этих “наглецов”, приведших их к вымиранию.
По другому сложилась судьба птиц на океанических островах. Там птицы
были единственной заметной группой наземных позвоночных. Тысячи и
миллионы лет изоляции привели к образованию многих необычных их
форм, часто утративших способность к полету. Человек принес на эти остро-
ва свиней, собак, крыс и других животных, и новые птичьи болезни. Люди
выжигали леса и распахивали луга, а также напрямую истребляли птиц. Все
это привело к вымиранию на островах за последнюю тысячу лет нескольких
сотен видов птиц (фиг. XXX.28).
ИТОГ
Птицы очень отличаются от других позвоночных своим перьевым покро-
вом. Возможно, перья впервые появились в связи с полетом. Птицы подня-
лись в воздух и освоили пространство, которое прежде успешно освоили
только насекомые и птерозавры. Отдельные птицы утратили способность к
полету или в связи с наземным образом жизни, как африканский страус и
эму, или в связи с совершенным освоением водной среды, как пингвины.
Действительный успех группы все-таки связан с воздухом, где теперь птицы
господствуют среди позвоночных животных.
Глава XXXI
ШЕРСТИСТЫЕ РЕПТИЛИИ
СО СЛОЖНЫМИ УШАМИ:
РАННИЕ МЛЕКОПИТАЮЩИЕ
В те времена, когда в небе парили крылатые ящеры, а в различных
водоемах плавали бескрылые птицы, представители одной группы
наземных позвоночных оставались маленькими и незаметными.
Начиная с позднего триаса они скрывались в лесах и чащах, избегая хищных
рептилий, и питались главным образом ночью. Таким образом, они заняли
нишу, которую не использовало большинство других животных, и медленно
развивались в качестве мелких членов наземного сообщества позвоночных.
ОТ РЕПТИЛИЙ К МЛЕКОПИТАЮЩИМ
Мы часто всех животных называем “зверями”. Однако в биологии и пале-
онтологии этим термином обозначена только одна группа животных, которую
мы лучше знаем под другим названием - млекопитающие. К млекопитаю-
щим относят всех теплокровных, покрытых волосами существ, имеющих же-
лезы, которые выделяют молоко для вскармливания детенышей. За редкими
исключениями они рождают живых детенышей, а не откладывают яйца.
Изменения в костях и зубах (фиг. XXXI.2). Те характерные признаки, ко-
торые наблюдаются в мягких тканях млекопитающих, обычно не могут быть
выделены на ископаемых остатках их предков. Но некоторые твердые эле-
менты тела млекопитающих, вроде костей и зубов, также отличаются от ко-
стей и зубов типичных рептилий. Мозговая коробка, например, намного
крупнее чем у рептилий и размещает мозг, чьи “мыслительные центры”, или
мозговые полушария, постоянно увеличивались в размере. Древняя костная
перегородка позади каждого глазного отверстия исчезла, хотя млекопитаю-
щие типа обезьян и лошадей заменили ее новой.
Внешние ноздри имеют одно общее отверстие в черепе. Внутренние носо-
вые отверстия перемещены в заднюю часть ротовой полости, посредством
развития вторичного костного неба. Последняя структура наводит нас на
мысль о небе крокодилов, но у млекопитающих она имеет независимое про-
исхождение и действительно позволяет дышать во время еды. Каждая ветвь
нижней челюсти состоит только из одной кости (зубная кость), а зубы имеют
несколько видов и выполняют разные функции: удерживание, разрывание,
резание и дробление. Коренные зубы в конце зубного ряда имеют два или
больше корней. Единый затылочный мыщелок, соединяющий у рептилий
© 1997. В.К. Голубев, перевод на русский язык и примечания.
ШЕРСТИСТЫЕ РЕПТИЛИИ СО СЛОЖНЫМИ УШАМИ
497
Фиг. XXXI. 1. Череп пеликозавра и плечевой пояс зверообразной рептилии и
млекопитающего
Слева — череп пеликозавра Dimetrodon с дифференцированными зубами, указы-
вающими на близкое родство с млекопитающими; длина черепа 33 см. Плече-
вой пояс зверообразной рептилии (в центре) и млекопитающего (справа)
череп с позвоночником, у млекопитающих разделен на две части, или мы-
щелки, по одному с каждой стороны спинного мозга.
Другие новообразования, типичные для скелета млекопитающих. Пле-
чевой пояс намного уже, чем у рептилий, и обычно теряет две кости
(фиг. XXXI. 1, справа). Подвздошная кость тазового пояса наклонена вперед.
В тазовом поясе ниже тазобедренного сустава располагается крупное отвер-
стие. Бедренная и плечевая кости более длинные и более стройные, чем у ти-
пичных рептилий. Головка бедра располагается сбоку, а не на самом конце
бедренной кости. Последний признак так же, как новые местоположения
крепления мускулов, связан с изменениями характера движения конечнос-
тей, которые у млекопитающих расположены близко к телу, а не расставле-
ны в стороны, как у большинства рептилий.
Родственные связи между рептилиями и млекопитающими. В главе XXV
мы проследили историю синапсидных рептилий. Самые древние синапсиды -
одни из самых ранних рептилий, которые появляются в позднем карбоне
вместе с котилозаврами, сборной группой, из которой выводят всех осталь-
ных рептилий. Наиболее примитивными среди синапсид были пеликозавры
(фиг. XXXI. 1, слева). По мере эволюции пеликозавров некоторые их группы
становились все более млекопитающеподобными. В поздней перми пелико-
завров сменили терапсиды. Среди терапсид были группы, которые продол-
жали приобретать все больше и больше признаков, характерных для млеко-
питающих. В конце концов, в позднем триасе появляются самые ранние на-
стоящие млекопитающие.
САМЫЕ РАННИЕ МЛЕКОПИТАЮЩИЕ
Млекопитающие могли возникнуть только однажды, от одного рептилий-
ного предка. Однако родственные связи некоторых примитивных групп
млекопитающих друг с другом до сих пор не ясны, поэтому вполне возмож-
но, что различные группы млекопитающих имеют разных рептилийных
предков.
32 Каменная книга
498
КАМЕННАЯ КНИГА
Структуры у типичных рептилий
Костная перемычка позади глаза
4 „ ' Один тип
Несколько костей в зубов
нижней челюсти
Структуры у типичных млекопитающих
Млекопитающие и
зверообразные
рептилии
Раздельные носовые
отверстия
Типичная рептилия
Одно общее отверстие
для ноздрей
Единый
затылочный мыщелок
Коренной зуб
млекопитающего
Внутренние ноздри
сдвинуты далеко
назад
Внутренние ноздри расположены
далеко впереди. Нет вторичного
костного неба
Бедренная
кость
Крупное вторичное
костное нёбо
Головка
бедра
Подвздошная
кость направлена Зуб
вверх рептилии
Головка
бедра
I
Бедренная
кость
Тазовый пояс и бедро
рептилии
Подвздошная костью
наклонена
вперед
Бедренная $
кость
Тазовый пояс и бедро
млекопитающего
Фиг. XXXI.2. Сравнение строения черепа, костей посткраниального скелета и
зубов рептилий, звероподобных рептилий и млекопитающих
Наиболее примитивные млекопитающие. В позднем триасе Лесото (Юж-
ная Африка), Западной Европы, Аризоны (США) и провинции Юннань (Ки-
тай) появляются морганукодонтиды (фиг. XXXI.4). У этих самых примитив-
ных известных млекопитающих главный контакт между нижней челюстью
ШЕРСТИСТЫЕ РЕПТИЛИИ СО СЛОЖНЫМИ УШАМИ
499
Cynognathus, типичный средне-
триасовый хищный цинодонт
из Южной Африки, который
имел широкую костную пере-
мычку позади глаза. Длина че-
репа 33 см
Bienotherium, позднетриас-
совый растительноядный
цинодонт из Китая не имеет
никаких следов костной пе-
ремычки позади глаза. Дли
на черепа 13 см
Ваипа, продвинутый раннетриасовый
тероцефал с неполной костной
перемычкой позади глаза.
Длина черепа 13 см
Фиг. XXXI.3. Степень развития костной перемычки позади глаза у различных
терапсид
и черепом уже располагался между зубными и чешуйчатыми костями. Дру-
гие маленькие кости все еще остались в нижней челюсти, но они не играли
большой роли в процессе жевания. Поскольку нижняя челюсть еще содер-
жит несколько костей, очевидно, специализированное среднее ухо млекопи-
тающих у этих животных еще не развилось. Сочленовные и квадратные кос-
ти, которые у морганукодонтид также были вовлечены в челюстное
сочленение, позже превратились в ушные кости - молоточек и наковальню,
что было очень важно для обострения слуха.
Эти ранние млекопитающие напоминали современных землероек. Подоб-
но землеройкам, они были маленькими активными хищниками, которые по
ночам среди опавших листьев разыскивали различных насекомых и их ли-
чинок, основную свою пищу. Активный поиск пищи был необходим, потому
что, будучи теплокровными, им, вероятно, в день необходимо было съесть
пищи в два-три раза больше веса собственного тела.
Несмотря на свой землеройкообразный внешний вид, морганукодонтиды
сохранили много примитивных признаков не только в строении челюстей.
Например, в тазе морганукодонтид вертлужная впадина, в которую входит
головка бедра, имеет широкое отверстие, как это наблюдается у терапсид.
У более продвинутых млекопитающих отверстие закрыто костной стенкой.
Мозг морганукодонтид также был более мелким, а его участки, отвечающие за
обоняние, развиты слабее, чем у сходных по размеру современных землероек.
Морганукодонтиды - самые древние и наиболее примитивные триконо-
донты. В течение юры и мела эта группа становится более продвинутой: мно-
гочисленные кости нижней челюсти исчезают, и остается только одна, зуб-
ная, кость. Название “триконодонт” означает “трехбугорчатый зуб” и отме-
чает тот факт, что обычно щечные зубы у этих животных имеют три хорошо
заметных конуса, располагающихся спереди назад вдоль одной линии. Боль-
шинство триконодонтов было маленькими, размером с землеройку, живот-
ными. Тем не менее, одним из самых крупных мезозойских млекопитающих
был меловой триконодонт, размером с домашнюю кошку.
В течение юры и мела от триконодонтов или близкородственных им форм
происходят ряд других групп млекопитающих. Некоторые из них исчезнут
к концу мезозойской эры, в то время как другие превратятся в современных
млекопитающих, включая и нас с Вами.
32*
500
КАМЕННАЯ КНИГА
Фиг. XXXI.4. Megazostrodon — позднетриасовый триконодонт из Лесото (Юж-
ная Африка). Длина около 15 см (рисунок Фрэнка Найта по реконструкции
Ф. Дженкинса и Ф.Р. Паррингтона)
Докодонты были группой всеядных, размером с крысу, млекопитающих,
чьи остатки известны только из средне- и позднеюрских отложений. Не смо-
тря на то, что докодонты, вероятно, не отличались большим разнообразием
(известно не очень много видов этих зверей), они были достаточно широко
распространены: от западных районов Северной Америки (США) до Запад-
ной Европы. Так как наиболее хорошо изученные местонахождения средне-
и позднеюрских млекопитающих располагаются именно в этих регионах, у
нас нет серьезных оснований считать, что докодонты не имели более широко-
го распространения и, соответственно, не были более разнообразными. Нам
необходим дополнительный палеонтологический материал, чтобы подтвер-
дить эти суждения или опровергнуть их.
Другая древняя группа млекопитающих, мультитуберкуляты, появилась
в поздней юре и просуществовала до раннего олигоцена (фиг. XXXI. 5).
В течение этого времени они широко распространятся по всему Северному
полушарию. Мультитуберкуляты очень разнообразная группа. Некоторые
из них были размером с мышь, в то время как другие становились более
крупными, достигая размеров современного бобра. Название данной группы
млекопитающих отражает наличие многочисленных конусов, расположен-
ных рядами на коренных зубах. Эта особенность сближает мультитуберку-
лят с тритилодонтными цинодонтами, с которыми, как считают, они состоят
в близком родстве. Череп мультитуберкулят был широким и несколько уп-
лощенным. Мощные резцы указывают, что их пища и образ жизни, вероятно,
очень напоминали таковые грызунов. Мультитуберкуляты вымирают веко-
ШЕРСТИСТЫЕ РЕПТИЛИИ СО СЛОЖНЫМИ УШАМИ
501
Фиг. XXXI.5. Ptilodus, палеоценовый мультитуберкулят из Северной Америки.
Длина 30 см (рисунок Фрэнка Найта по реконструкции Ф.А. Дженкинса и
Д.В. Крауса с изменениями)
рости после появления и ранней радиации грызунов. Возможно, это было
следствием конкуренции, возникшей между этими двумя группами.
Яйцекладущие однопроходные Австралии - наиболее примитивные со-
временные млекопитающие. Они представлены только двумя формами: во-
доплавающим утконосом и питающейся муравьями ехидной. Они также
сильно отличаются друг от друга, как выдра и еж. Этот факт наводит на пред-
положение о наличии большого количества промежуточных между ними
форм, которые исчезли к настоящему времени. Доплейстоценовая палеонто-
логическая летопись однопроходных чрезвычайно скудна: известно только
несколько костей и зубов из миоцена и единственная челюсть из раннего ме-
ла. Миоценовый однопроходной Obdurodon является бесспорным утконо-
сом, но отличается от ныне живущих совершенно беззубых во взрослом со-
стоянии форм наличием хорошо развитых зубов (фиг. XXXI.7). Steropodon,
раннемеловой однопроходной из Австралии, имеет зубы сходные с зубами
Obdurodon, но характеризующиеся намного более простыми корнями
(фиг. XXXI. 6). Его можно рассматривать как утконоса или в качестве про-
межуточной формы между утконосами и ехиднами. Когда мезозойская ис-
тория позвоночных Австралии станет известна несколько лучше, весьма
502
КАМЕННАЯ КНИГА
Фиг. XXXI.6. Замещенная опалом челюсть Steropodon из нижнемеловых опа-
ловых разработок у Лайтнин Ридж (Новый Южный Уэлс, Австралия)
Образец на нижнем снимке был снят в отраженном свете; на верхнем снимке ис-
точник света располагается за образцом и свет проходит через образец
Фиг. XXXI.7. Покрытый эмалью верхнечелюстной зуб Obdurodon, второго
древнейшего известного утконоса, из среднемиоценовых отложений севера
Южной Австралии
Современные взрослые утконосы не имеют зубов, а у молодых особей их корот-
коживущие зубы оразованы только дентином. Длина по горизонтали 6.5 мм
ШЕРСТИСТЫЕ РЕПТИЛИИ СО СЛОЖНЫМИ УШАМИ
503
Фиг. XXXI.8. Нижняя челюсть ShuQtherium, млекопитающего размером с зем-
леройку из средней или поздней юры Сычуаня (Китай). Длина челюсти 12 мм
вероятно, выяснится, что эти примитивные млекопитающие были более раз-
нообразными. Сходства в форме мозговой коробки и микроструктуры зубов
мультитуберкулят и однопроходных предполагают, что эти две группы мо-
гут быть близко родственными.
Примитивные, неспециализированные терии впервые появляются в позд-
нем триасе. Их остатки обнаружены в тех же самых отложениях в Мэндир-
Хиллсе (окрестностях Бристоля, Англия), из которых известны находки
морганукодонтид. Примитивные терии — группа, от которой в меловое время
происходят сумчатые и плацентарные млекопитающие. Хотя они и не увели-
чивали свой размер на протяжении мезозойской эры, это никак не свиде-
тельствует о том, что они вообще не эволюционировали. Как у триконодон-
тов, в нижней челюсти исчезают все кости кроме зубной. Две из этих упомя-
нутых выше нижнечелюстных костей вошли в последовательность из трех
маленьких костей, связывающих барабанную перепонку со слуховыми нер-
вами. Сочленовная и квадратная кости рептилий превратились, соответст-
венно, в молоточек и наковальню млекопитающих, став функционально свя-
занными с древней костью уха рептилий, стапесом, которая у млекопитаю-
щих получила название стремечко.
Это новообразование у млекопитающих, возможно, было связано с необ-
ходимостью увеличения остроты слуха. Поскольку самые ранние млекопи-
тающие были маленькими ночными животными, лучший слух давал пре-
имущество и в уходе от хищников, и при поиске добычи.
В отличие от триконодонтов зубы прогрессивных млекопитающих позво-
ляли намного более эффективно пережевывать пищу, что обеспечивалось
сложным характером смыкания бугров на верхних и нижних коренных зу-
бах (молярах). Однако эти изменения происходили не в одном единственном
направлении: в течение мезозойской эры млекопитающие много “экспери-
ментировали”.
Нижние щечные зубы у наиболее примитивных терий представляли собой
по существу простые треугольные столбы (тригониды), которые имели режу-
щие грани. По мере развития группы, на нижних молярах развивалось уг-
лубление, или бассейн (талонид), о которое новый конус (протокой) на верх-
них коренных зубах раздрабливал пищу. Среди териев, которые эволюцио-
нировали в сумчатых и плацентарных, в позднеюрское и раннемеловое
время на задней части моляров появляется талонид. Но челюсть средне- или
позднеюрского Shuotherium, найденная в Сычуане (Китай), имеет талонид
на передней части зуба (фиг. XXXI.8). Всех териев с талонидами позади три-
504
КАМЕННАЯ КНИГА
гонидов объединяют в янготериев, a Shuotherium был помещен в группу, на-
званную инотерии (зеркальные звери), поскольку их зубы являются как бы
зеркальным отражением друг друга. Происхождение названия этих групп
связано с китайской концепцией инь и янь: идеи неизбежного притяжения и
отталкивания противоположностей.
С развитием зубов, эффективных как в разрезании, так и в раздроблении
пищи, у териев появилась возможность радиировать в громадное количество
форм, объединяемых в сумчатых и планцентарных, с самым широким диа-
пазоном пищи.
Расцвет сумчатых. Примитивные древние терии где-то на одном из четы-
рех континентов - в Австралии, Антарктиде, Северной или Южной Амери-
ке, превращаются в сумчатых. Позже они распространяются по другим кон-
тинентам. Самые древние данные по сумчатым - позднемеловые ископаемые
остатки из обеих Америк. Но их отсутствие в палеонтологической летописи
Антарктиды и Австралии этого возраста, возможно, связано с отсутствием
находок, а не с тем, что этих животных на данных континентах тогда не бы-
ло. Некоторые продвинутые особенности, обычные для всех австралийских
сумчатых и неизвестные почти у всех американских, позволяют предполо-
жить, что сумчатые мигрировали в Австралию из Южной Америки через Ан-
тарктиду, а не в обратном направлении. Это произошло к концу мела или в
самом раннем кайнозое перед отделением Австралии от Антарктиды (и, сле-
довательно, от Южной Америки) и началом движения ее на север, к Азии1.
Хотя мы обычно считаем Австралию страной сумчатых, родиной кенгуру
и вомбатов, американский опоссум, вероятно, больше напоминает примитив-
ных сумчатых. Он также принадлежит к одной из древнейших американских
линий позвоночных: разрозненные кости и зубы мелких опоссумов обнару-
жены в меловых отложениях на западе Соединенных Штатов, которые также
содержат остатки динозавров. Подобно современным сумчатым, эти живот-
ные, очевидно, не имели эффективного механизма вынашивания эмбриона в
утробе матери. В результате детеныши рождались крошечными, слабо разви-
тыми существами. Затем они переползали в материнскую сумку, где, крепко
прикрепившись к соску, сосали молоко, продолжая свое развитие.
Хотя сумчатые Северной Америки имеют длинную историю, они никогда
не были особенно разнообразными на этом континенте. Многие из них исчез-
ли к концу мела, а в кайнозое известно только одно семейство, Didelphidae.
Дидельфиды вымирают в Северной Америке в миоцене, но в плейстоцене
вновь появляются. Правда они будут представлены здесь только одним ви-
дом, Didelphis marsupialis, - иммигрантом из Южной Америки.
Крупные дидельфиды - единственные представители сумчатых, которые
достигли Европы, но это произошло не раньше палеоцена. Они никогда не
были здесь особенно разнообразны и полностью исчезают в миоцене. На дру-
гих континентах история сумчатых еще более коротка: в Африке они были
известны в эоцене и олигоцене, а в Азии — только в олигоцене.
Именно в Австралии и Южной Америке сумчатые наиболее разнообразны
как в прошлые эпохи, так и в настоящее время.
1 - Глобальные палеозоогеографические реконструкции, выполненные Н.Н. Каландадзе и А.С. Ра-
утиан, свидетельствуют, что родиной сумчатых была южная часть Гондваны (Южная Америка,
Антарктида и Австралия), изолировавшаяся от всего остального мира в допозднеюрское время.
Вероятно, именно после этого события и происходит разделение примитивных териев на сумча-
тых и плацентарных. От рассмотренного блока материков Австралия отделяется в раннем ме-
лу. Несколько позже происходит кратковременное восстановление сухопутной связи между Се-
верной и Южной Америками. Именно тогда сумчатые попадают в Северную Америку, а прими-
тивные плацентарные - в Южную Америку (прим, перев.).
ШЕРСТИСТЫЕ РЕПТИЛИИ СО СЛОЖНЫМИ УШАМИ
505
Фиг. XXXI.9. Два южноамериканских сумчатых, напоминающих плацентарных
В Южной Америке сумчатые были основными хищниками среди назем-
ных позвоночных начиная с третичного периода и до плиоцена, когда форо-
ракоидные птицы сменили больших хищных сумчатых боргиенид. Thyla
cosmilus, близкого родственника боргиенид, при поверхностном и даже бо-
лее пристальном взгляде можно ошибочно принять за саблезубого тигра
Smilodon (фиг. XXXI.9, справа). Их сходство - превосходный пример кон-
вергентного развития: два вида происходят от совершенно разных предков,
но примерно одинаковые давления естественного отбора приводят к тому,
что эти виды приобретают большое внешнее сходство. Оставаясь главными
хищниками в Южной Америке на протяжении большей части кайнозойской
эры, некоторые сумчатые превратились в мелких растительноядов.
Возможно, в отдельные периоды сумчатые Южной Америки через Ан-
тарктиду проходили в Австралию, и наоборот. Единственные ископаемые,
свидетельствующие о том, что это действительно имело место, несколько че-
люстей и зубы с о. Сеймур около антарктического полуострова, выступающе-
го к Южной Америке2.
Но именно в Австралии сумчатые испытали самую широкую адаптивную
радиацию. За 60 млн. или около того лет, когда Австралия была изолирована
от остального мира океанскими барьерами, потомки опоссумоподобных сум-
чатых приобрели внешний вид и привычки, сходные с таковыми многих дру-
гих животных.
В среднем миоцене самые крупные известные австралийские наземные
млекопитающие были размером с овцу. Ими были дипротодонтиды, четы-
рехлапые листояды (фиг. XXXI. 11). По мере того, как кайнозойская эра под-
ходила к завершению, дипротодонтиды постепенно становились все крупнее
и крупнее. Наконец в плейстоцене появляется Diprotodon. Это животное бы-
ло размером с носорога и к тому же очень на него похожее, за исключением
2 — Глобальные палеозоогеографические реконструкции свидетельствуют о том, что Австралия от-
делилась от Южной Америки и Антарктиды до второй половины раннего мела и никогда после,
до настоящего времени включительно, эта связь не восстанавливалась. Именно поэтому здесь
отсутствуют плацентарные млекопитающие, известные в Южной Америке (прим, перев.).
506
КАМЕННАЯ КНИГА
Фиг. ХХХЫО.Череп Diprotodon, наиболее крупного из когда-либо живших
сумчатых (Юго-Восточная Австралия)
Этот череп длиной всего 66 см, тогда как самые крупные формы имели черепа в
полтора раза длиннее
одного признака - отсутствие рога (фиг. XXXI. 12). Несмотря на крупные
размеры, Diprotodon вовсе не был интеллектуальным гигантом: большая
внутренняя часть его головы была, очевидно, занята воздушными кармана-
ми, а не мозгом (фиг. XXXI. 10).
Другой группой австралийских сумчатых, чьи представители значитель-
но увеличились в размерах в позднем кайнозое, были кенгуру. Самые круп-
ные среднемиоценовые кенгуру были размером с маленьких кроликов. Поз-
же климат на континенте становится более засушливым, и эта группа испы-
тывает настоящий эволюционный взрыв: появляется много новых, крупных
форм. К концу плейстоцена самый большой кенгуру достигал приблизитель-
но 2.6 м в высоту (фиг. XXXI.13, 15).
Не все австралийские сумчатые, для которых было характерно отмечен-
ное увеличение размеров на протяжении позднего кайнозоя, были безопас-
ными фитофагами. Несмотря на то. что тиляколеониды происходят от расти-
тельноядных или всеядных форм, их последние премоляры были изменены
в крепкие режущие лезвия, занимающие четвертую часть длины всей челю-
сти. Самое большое из этих животных было размером с леопарда, а самое ма-
ленькое — с домашнего кота (фиг. XXXI. 14). Так же как и многие другие
большие сумчатые, они вымирают в конце плейстоцена.
Другие австралийские сумчатые, чьи древние представители известны по
ископаемым остаткам, — это вомбаты, коалы, поссумы и всеядные бандику-
ты. Из них только вомбаты имеют формы, сильно отличающиеся от ныне жи-
вущих видов. Некоторые вомбаты в плиоцене и плейстоцене достигали 80 см
в высоту и 2 м в длину, т.е. в двое выше и длиннее любого современного пред-
ставителя. Если они рыли подземные норы, как это делают их современные
родственники, то они были самыми большими среди всех когда-либо жив-
ших роющих млекопитающих.
Специфическое ответвление поссумов - эктоподонтиды. Имея многочис-
ленные бугорки на своих зубах, они, как считали, являются близкими родст-
венниками мультитуберкулят. Эти миоценовые и плиоценовые животные,
вероятно, питались твердыми орехами, подобно белкам и, возможно, были
на них очень похожи.
ШЕРСТИСТЫЕ РЕПТИЛИИ СО СЛОЖНЫМИ УШАМИ
507
Фиг. XXXI. 11. Palorchestes — близкий родственник Diprotodon, но сильно от-
личающийся от него размерами
Обладая большими когтями, эти сумчатые могли быть аналогами наземных ле-
нивцев. Они населяли Австралию в плиоцене и плейстоцене. Наиболее крупные
виды могли достигать 2-2.5 м в длину (художник Фрэнк Найт)
508
КАМЕННАЯ КНИГА
Фиг. XXXI.12. Взрослая самка Diprotodon (длина 2.5 м) стоит в растерянности,
в то время как ее детеныш барахтается в болоте, куда он угодил, желая утолить
жажду
26 тыс. лет назад, район озера Каллабонна (Южная Австралия) (художник
Фрэнк Найт)
ШЕРСТИСТЫЕ РЕПТИЛИИ СО СЛОЖНЫМИ УШАМИ
509
Фиг. XXXI.13. Sthenurus (длина 2 м), питающийся ветками и молодыми побе-
гами кенгуру из плиоцена и плейстоцена Австралии
Один палец на задней лапе был развит сильнее остальных, которые были сильно
редуцированы или вовсе отсутствовали. Аналогичное явление наблюдается у
лошадей, у которых, правда, пальцы редуцируются не только на задних, но и на
передних конечностях (художник Фрэнк Найт)
510
КАМЕННАЯ КНИГА
Фиг. XXXI. 14. Thylacoleo (длина 1.5 м) из плиоцена и плейстоцена Австралии
использовал свои сильно увеличенные последние премоляры в качестве
страшных режущих лезвий
Тиляколеониды — это одни из немногих хищных млекопитающих, которые про-
исходят от всеядных или растительноядных предков (художник Фрэнк Найт)
ШЕРСТИСТЫЕ РЕПТИЛИИ СО СЛОЖНЫМИ УШАМИ
511
Фиг. XXXI.15. Propleopus — вымерший гигантский крысиный кенгуру, чьи ос-
татки обнаружены в плейстоценовых отложениях Австралии
На рисунке кенгуру изображен пожирающим яйца страуса эму. Современные
кенгуру — это растительноядные животные, но Propleopus, возможно, был хищ-
ником (художник Фрэнк Найт)
МЛЕКОПИТАЮЩИЕ С ПЛАЦЕНТОЙ
Хотя сумчатые не откладывали яйца, немногие из них смогли приобрести
эффективный механизм вынашивания детенышей в утробе матери. Этот
барьер смогли преодолеть те меловые млекопитающие, которое соединили
512
КАМЕННАЯ КНИГА
Фиг. XXXI.16. Череп насекомоядного Asioryctes. Длина черепа 30 мм. Мел
Монголии
Фиг. XXXI.17. Asioryctes по внешнему виду и, возможно, по образу жизни на-
поминал современную землеройку. Длина черепа около 5 см
мембрану яйца со стенкой матки. В результате появился новый орган: пла-
цента, через которую питательные вещества и кислород передавались эмбри-
ону и одновременно удалялись вредные продукты его жизнедеятельности.
Эти два процесса теперь позволяли эмбриону проходить полное развитие,
приобретая более сложный мозг, чем у любого сумчатого. Занимательно, что
некоторые сумчатые (бандикуты) развили структуру, очень напоминающую
плаценту.
Ранние насекомоядные. Описанные события в эволюции млекопитающих
являются только предположением, основанным главным образом на уровне
организации ныне живущих животных. Дальнейшую историю плацентар-
ных продолжают позднемеловые ископаемые остатки из Монголии. Это уже
остатки бесспорных плацентарных млекопитающих. Некоторые из них бы-
ли специализированными зверями, другие, наоборот, были очень примитив-
ными насекомоядными. От них происходят разнообразные плацентарные
млекопитающие, которых мы видим сегодня. Эти примитивные насекомояд-
ные были неприметными существами, которые, вероятно, питались ночью
ШЕРСТИСТЫЕ РЕПТИЛИИ СО СЛОЖНЫМИ УШАМИ
513
или в сумерках насекомыми и их личинками, а также червями. По отно-
шению к телу их головы и хвосты были длинными, а лапы короткими
(фиг. XXXI. 16,17). Некоторые виды имели по крайней мере до сорока четы-
рех зубов, или одиннадцать на каждой стороне каждой челюсти. Эти зубы
были устроены по определенной схеме, или формуле: три резца спереди, да-
лее следуют один клык, четыре предкоренных и три коренных зуба. Многие
более поздние млекопитающие будут терять одни из них и изменять форму
других, лишь несколько специализированных групп, вроде китов и неполно-
зубых, будут значительно увеличивать их число. Судя по строению зубной
системы, а также другим признакам, меловые насекомоядные уже были го-
товы произвести большинство современные отрядов, которые они в конце
концов произвели и которые будут рассмотрены нами в главе XXXIII.
33 Каменная книга
Глава XXXII
ЗЕМЛЯ ЦВЕТЕТ
• Дорога от Беркалильо к Ацтеку в штате Нью-Мексико проходит че-
рез горный хребет Напаньенто и поворачивает к западу в долину
Сан-Хуан. Это засушливый район, представляющий собой холмы,
бедленды, каньоны, в обрывистых уступах которых видны выходы разно-
цветных глинистых сланцев. Индейцы, коренные жители этой области, сей-
час живут в нескольких резервациях, хотя руины доисторических индей-
ских городов и поныне сохранились в некоторых ущельях. Один из них,
Пуэбло-Бонито, служил когда-то убежищем более чем для тысячи людей.
ЛАРАМИЙСКАЯ РЕВОЛЮЦИЯ
Приезжающие сюда археологи устремляются к этим развалинам, в то вре-
мя как охотники за ископаемыми разыскивают расщелины и каньоны, в ко-
торых видны слои, рассказывающие нам о том, как закончился Век Репти-
лий и начался Век Млекопитающих. Эта стратиграфическая история начи-
нается с грубозернистых желтых песчаников позднемелового возраста,
относящихся к формации Ойо Аламо-Сандстоунз. Эти песчаники включают
кости и черепа рогатых, носивших шейные воротники динозавров, переме-
шанные с остатками древесины и чешуей ганоидных рыб. Как сами породы,
так и заключенные в них окаменелости рассказывают нам о влажной низ-
менности, где трупы гигантских пресмыкающихся переносились потоками
от одной речной отмели к другой и в конце концов быстро покрылись осад-
ком на одной из песчаных кос. То же происходило и с затонувшими стволами
деревьев.
Мы не знаем, сколько времени происходило накопление этих толщ, по-
скольку самые молодые отложения были разрушены эрозией. Это “несогла-
сие” является лишь одним из свидетельств ларамийской революции, горооб-
разовательной эпохи, ознаменовавшей конец мезозойской эры и продолжав-
шейся в течение начала кайнозоя. В действительности, ларамийская
революция была лишь кульминацией тектонических процессов, начавших-
ся в конце триасового периода и начале юрского и постепенно продолжав-
шихся в течение 65-75 миллионов лет. Во многих местах, таких, например,
как долина Сан-Хуан, эти процессы привели лишь к умеренному повыше-
нию уровня земли, одновременно съедаемому эрозией. В других же местах
© 1997. С.В. Наугольных, перевод на русский язык и изменения.
ЗЕМЛЯ ЦВЕТЕТ
515
Фиг. XXXII.1. Листья древнейших цветковых растений
1 — Magnoliaephyllum, 2 — дихотомически сложный лист, 3 - Liriophyllum, 4 —
лист платаноидного облика, 5 — Menispermites, 6 — Sapindopsis, 7 — Araliop-
soides, 8 - Araliaephyllum, 9— Vitiphyllum, 10 - Sapindopsis, 11 — Plantaginopsis,
12 - Rogersia, 13 - Acaciaephyllum, 14 - Quercophyllum, 15 — Ficophyllum, 16 —
Proteaephyllum, 17 - неописанный лист из нижнего мела Забайкалья (Мейен,
1987). Вне масштаба
ларамийский орогенез (горообразование) привело к возникновению мощных
горных цепей, простиравшихся от Кэйт-Хорка до Карибского моря и от Мек-
сики до Аляски. Жидкие, расплавленные граниты, поднимаясь к поверхно-
сти земли, создали тот горный ландшафт, который характерен для Северо-
Американского запада и горной цепи Анд в Южной Америке.
ПОЯВЛЕНИЕ ЦВЕТОВ
Эволюция наземных растений также отражает это событие, поскольку
они приспосабливались к новым природным условиям. Юрские динозавры
обитали среди гинкговых лесов и зарослей беннеттитов. Хорошим примером
последних может служить вильямсония (Williamsonia) - растение, принад-
лежавшее к группе, сходной с цикадовыми и вымершей в настоящее время,
одной из разновидностей шишкообразующих голосеменных. Когда-то доми-
нировавшие “семенные папоротники” (птеридоспермы) и мхи в это время
были уже очень незаметными и немногочисленными. Несмотря на общее
разнообразие растительности, мы, тем не менее, не смогли бы почувствовать
аромат цветов, к которому так привыкли сейчас.
Однако незадолго до массового вымирания динозавров в конце мелового
периода миллиарды цветковых растений заполнили земные леса. После по-
явления в раннем мелу, их все возраставшее разнообразие и численность бы-
ли подобны взрыву. В породах этого возраста известны листья, пыльца и цве-
ты, принадлежавшие как однодольным (цветковым растениям с одним эмб-
риональным “листком” и листьями с параллельным жилкованием), так и
зз*
516
КАМЕННАЯ КНИГА
Фиг. XXXII.2. Органы размножения примитивных цветковых
а — Calycites, б — Kenella, в, д — Archaeanthus, в - схема листовки, г — зрелое со-
брание листовок, д — побег с цветком. По Dilcher, Crane, 1984 из Мейен, 1987.
Длина масштабной линейки - 1 мм (а), 1 см (б, в), 3 см (г, д)
двудольным (растениям с двумя эмбриональными “листками” — долями и
сложноветвящимися жилками). Эти растения были как ветроопыляемыми,
так и опыляемыми насекомыми. Разнообразие типов организации цветко-
вых растений наталкивает нас на мысль, что история покрытосеменных
должна была начаться задолго до раннемеловой эпохи, однако пока нам не
удалось найти самых древних ископаемых покрытосеменных.
Древнейшими известными ископаемыми остатками цветковых являются
небольшие, простые листья с перистым жилкованием из нижнемеловых (не-
окомских) отложений Сибири (фиг. XXXI. 1), возраст которых составляет
около 120 миллионов лет. Немногим более молодые (барремские и аптские)
остатки цветковых, представленные целым рядом листьев различного обли-
ка, принадлежавших как однодольным, так и двудольным, происходят из
восточной части Северной Америки. Из этого района Северной Америки, а
также из некоторых других известны растения, напоминающие современ-
ные протейные, аралии, березы, фиговые деревья и тополя. Ближе к концу
мелового периода появляются родственники дубов, платанов, грецких оре-
хов, магнолий, лавров и многих других. Эти растения доминируют в место-
нахождениях на Аляске, Гренландии, в Европе и Азии. Они также известны
из альбских отложений Австралии.
Пыльца покрытосеменных, микроскопические мужские половые клетки
растений, которые многих из нас заставляют страдать от аллергии, “весен-
ней лихорадки”, также впервые встречаются в породах нижнемелового воз-
раста Англии в несколько более молодых отложениях в Западной Африке и
Северной Америки. Некоторые из этих пыльцевых зерен (такие как клавати-
полленитес - Clavattipollenites и лилиацидитес - Liliacidites) очень сходы с
пыльцой магнолиевых. Развитие разнообразия типов пыльцы аналогично
динамике разнообразия типов листьев цветковых.
ЗЕМЛЯ ЦВЕТЕТ
517
Древесины ранних цветковых растений, однако, более редки. Единствен-
ное исключение составляют находки стволов с плохо сохранившимися тка-
нями из аптских отложений Японии.
Плоды цветковых растений впервые появляются в нижнем мелу (баррем-
ский ярус), самые древние из них обнаруживают сходство с плодами ниссы
(Nyssa). Цветы также известны из пород этого возраста, однако древние и хо-
рошо сохранившиеся цветы происходят из нижней части верхнего мела (се-
номанский ярус) Северной Америки. Там же были найдены головчатые соп-
лодия с семенами и сложные соцветия (фиг. XXXII.2). Многие из этих цветов
довольно примитивны и выглядят сходными с родственниками роз (семейст-
во Rosaceae) и хамамелидовыми.
В близких по возрасту отложениях из Роуз Крик в штате Небраска на про-
тяжении последних нескольких лет было собрано много прекрасных окаме-
нелых цветов. Один из наиболее хорошо сохранившихся имеет средние раз-
меры около 3.5 см в диаметре. Этот цветок имел пять лепестков, пять тычи-
нок, несущих пыльцу (тоже сохранившуюся), пять пестиков и пять завязей.
В основании каждой завязи были небольшие возвышения, которые, возмож-
но, продуцировали нектар. С этим ископаемым цветком сходны цветы розы
и винограда. Тот факт, что пыльца этого цветка была исключительно мел-
кой, а также то, что растение имело органы, продуцировавшие нектар, за-
ставляет предположить, что растение опылялось насекомыми, а не ветром.
Комплексы растений позднего мела указывают на то, что климатические
условия были более жаркими и влажными (гумидными), чем те, которые
мы видим сейчас. Флора Медсан Боу из Центрального Вайоминга, сейчас
представляющего собой высокоширотную пустыню, включает в свой состав
широколистные покрытосеменные, такие как фикус (Ficus), магнолия
(Magnolia), лавры (Lauras), а также пальмы (Sabal) и красное дерево
(Metasequoia).
ТРЕТИЧНЫЕ ФЛОРЫ
С началом кайнозойской эры (включающей палеогеновый, неогеновый и
четвертичный периоды) в течение почти 65 миллионов лет в эволюции расте-
ний уже не происходило драматических событий. В отложениях палеоцено-
вой формации Форт-Юнион, обнажающейся в штатах Монтана, восточном
Вайоминге, а также на западе Северной и Южной Дакоты, встречаются скоп-
ления растительных остатков, иногда формирующих слои угля. Водоросли,
грибы, мхи, папоротники, хвойные, включая араукарию (Araucaria, сосна
острова Норфолк), а также гинкго произрастали здесь когда-то вместе с по-
крытосеменными, такими как пальмы, клены, ясени и даже хлебное дерево
(артокарпус, Artocarpus). Такой состав флоры указывает на теплый и влаж-
ный субтропический климат, сохранившийся и в эоцене. Хорошим приме-
ром эоценовых флор является флора лондонских глин в Англии, представля-
ющая собой смесь растений низин и возвышенностей, произрастающих сей-
час в Юго-Восточной Азии. Как и почему эти растения оказались все вместе
в одно время и в одном регионе, до сих пор удовлетворительно не объяснено.
Тем не менее в течение всего кайнозоя сохранилось три главных направле-
ния в развитии наземной растительности. Общее понижение температуры,
время от времени прерывалось более теплыми климатическими эпизодами.
Внутренние районы континентов постепенно высыхали, особенно начиная с
миоценовой эпохи. В некоторых случаях причиной этому служило поднятие
518
КАМЕННАЯ КНИГА
горных хребтов, как это было, например, на западе Северной Америки. В
других случаях, как в Австралии, иссушение климата стало результатом
движения материка к северу и глобальным сужением тропических поясов. В
третьих, это случилось вследствие чередования ледниковых и межледнико-
вых условий ближе к концу кайнозойской эры. Все эти изменения привели к
тому, что растительные комплексы изменялись и “мигрировали” или замы-
кались в некоторых районах в зависимости от меняющихся климатических
условий.
Степи получили широкое распространение в Северной Америке, Южной
Америке и Австралии, особенно в середине кайнозойской эры, поскольку
именно в это время на перечисленных материках началось иссушение кли-
мата. Тропическая растительность вторглась в Северную Австралию или
эволюционировала там в результате “водвижения” этой части материка в
тропический пояс. Леса хвойных деревьев заняли север и юг Северной Аме-
рики и Европы, в то время как огромные массы ледников то наступали, то
несколько уменьшались в размерах несколько раз в течение последних
1.8 миллионов лет.
Одна из наиболее известных третичных флор происходит из олигоценовых
отложений (слои Флориесант), обнажающихся недалеко от Колорадо
Спрингс, штат Колорадо. Сотни тысяч растительных остатков сохранились в
тонкослоистых вулканических пеплах, отложившихся на дне древнего озера.
Пальмы и многочисленные другие деревья составляли основу этой флоры,
включавшей большое количество видов, в настоящее время уже не встречаю-
щихся в Северной Америке (зелкова - Zelkova, или кики, айлантус - Allan
thus, или небесное дерево). Используя сведения о жизни современных родст-
венников этих растений, палеоботаники смогли установить, что климат,
существовавший в то время, был теплым субтропическим, с достаточно
обильными и частыми дождями, способствовавшими произрастанию об-
ширных лесов в речных долинах. Сосны и дубы доминировали на возвы-
шенностях.
Одним из самых интересных деревьев флоры Флориссат была Sequoia, или
красное дерево. Секвойи по настоящее время произрастают в некоторых ме-
стах Калифорнии. Гигантские секвойи достигают в высоту более 95 м. Сек-
войя - это хвойное, а не цветковое растение, нам оно интересно еще и потому,
что в прошлом секвойи имели очень широкое распространение, в отличие от
того, что мы видим сегодня. История секвойи начинается где-то в глубине
мелового периода в северном полушарии, лишь затем это растение рассели-
лось по всей Северной Америке.
Другое родственное растение, часто встречающееся вместе с остатками
секвойи, также может послужить темой для интересного и интригующего
рассказа. Это — метасеквойя (Metasequoia). Листва метасеквойи сходна с ли-
ствой секвойи, но отличается наличием пар “иголок”, которые противопос-
тавлены одна другой, в отличие от чередования, когда они последовательно
отходят от главной оси. Метасеквойи существуют на Земле уже более чем
70 млн. лет. Метасеквойи были известны нам только как ископаемые расте-
ния, и лишь позднее были обнаружены их нынеживущие потомки - “живые
ископаемые”. В 1948 году, среди рисовых полей в провинции Сычуань, в
Центральном Китае было обнаружено большое, ранее неизвестное хвойное.
Оно было несколько необычным, поскольку, подобно лиственнице сбрасыва-
ло свою листву за зиму, но это было несомненное хвойное. Эти растения по-
сле последующего изучения и сравнений оказались метасеквойями, неболь-
шие популяции которых сохранились в маленьком районе Китая, хотя ког-
ЗЕМЛЯ ЦВЕТЕТ
519
да-то ими были заселены Северная Америка, Европа, Азия (включая
арктическую) и Гренландия. Эти растения преуспевали в эпохи с мягким
климатом. Во время постепенного похолодания в течение третичного време-
ни северные границы распространения метасеквойи медленно, но неуклонно
сместились к югу. Вследствие иссушения климата метасеквойи вымерли во
многих других местах, сохранившись лишь в виде реликта только в запад-
ном Китае. Однако сейчас это растение произрастает в интродукции во мно-
гих районах мира.
ТРИУМФ ЦВЕТОВ
Сегодня цветковые растения, появившиеся в эпоху расцвета динозавров,
воистину царствуют в мире. За всю их историю существовало более четверти
миллиона видов, а это гораздо больше, чем число видов всех других групп на-
земных растений вместе взятых. Цветковые действительно превзошли все
остальные группы растений, как числом, так и разнообразием, и только па-
поротники сейчас распространены также широко.
Распространение покрытосеменных растений тесно связано с развитием
насекомых, птиц и млекопитающих. Они питаются нектаром цветковых
растений, одновременно способствуя их опылению, поедают листья и плоды
и помогают распространению семян. Таким образом они приспособились
друг к другу.
Глава XXXIII
ВЕЛИКОЕ РАЗНООБРАЗИЕ
ПЛАЦЕНТАРНЫХ
• Как Вы узнали в главе XXXI, плацентарные - это млекопитающие,
находящиеся на самой высокой и наиболее продвинутой стадии
развития. Они очень разнообразны, попытаемся хотя бы кратко
охарактеризовать основные группы класса Mammalia.
НЕ СОВСЕМ БЕЗЗУБЫЕ НЕПОЛНОЗУБЫЕ
Броненосцы. Современные броненосцы имеют острую мордочку, тело,
покрытое броней из костных пластин, укрепленных в коже и колышкооб-
разные коренные зубы, число которых от 14 до 50 штук. Пища броненос-
цев состоит, в основном, из пауков, многоножек, скорпионов, взрослых на-
секомых, личинок и червей. Броненосцы ловят их длинным, липким
языком.
Броненосцы относятся к отряду Edentata (буквально “беззубые”), кото-
рый включает наиболее примитивных, во многих отношениях, плацентар-
ных млекопитающих. Ископаемые остатки неполнозубых впервые обнару-
жены в палеоценовых отложениях Южной Америки. На этом континенте в
течение палеогенового и неогенового периодов они были очень разнообраз-
ны. Однако известны находки настоящего неполнозубого Ernanodon из позд-
него палеоцена Китая, и менее уверенно определяемого как неполнозубый
Eurotamandua из среднего эоцена Европы. Миграция отдельных представи-
телей отряда из Южной Америки - центра происхождения в Азию и Европу
возможна, однако вызывает множество вопросов.
Ранние неполнозубые были маленькими, длиннохвостыми животными,
не покрытыми броней; они достигали длины 38—46 см. Их потомки разви-
лись в пять хорошо отличающихся групп, разделяемых на бронированных и
“голых”. Бронированные неполнозубые включают броненосцев и глиптодон-
тов. К “голым” неполнозубым относятся муравьеды, наземные и древесные
ленивцы.
По систематике, предложенной Р. Кэрроллом (1993) все американские не-
полнозубые относятся к отряду Xenarthra (“с необычными сочленениями”),
делящемуся на три инфраотряда: панцирные (Loricata), ленивцы (Philosa) и
муравьеды (Myrniecophagidae). Панцирные, в свою очередь, делятся на бро-
неносцев и глиптодонтов, а ленивцы - на наземных и древесных.
© 1997. Т.В. Кузнецова, перевод на русский язык и изменения.
ВЕЛИКОЕ РАЗНООБРАЗИЕ ПЛАЦЕНТАРНЫХ
521
Фиг. XXXIII. 1. Реконструкция внешнего вида и отдельные скелетные элемен-
ты глиптодонтов
а-в - Glyptodon, приблизительно 2.7 м в длину. Это бронированное животное
переселилось в Северную Америку в течение плейстоцена; г — булавохвостый
Doedicurus жил только в Южной Америке. Его длина 3.7-4.6 м
Панцири броненосцев никогда не были полностью твердыми, они вклю-
чают несколько колец брони, которые подвижно соединены между собой, и
это позволяет телу сгибаться. Самый большой ныне живущий вид достига-
ет приблизительно 91 см в длину. Плейстоценовый представитель броне-
носцев - Chlamydotherium был по размерам близок к носорогу и весил около
1.8 тонн.
Глиптодонты. Глиптодонты или “вырезаннозубые” названы так по строе-
нию своих зубов. Отличительными особенностями этой группы являются:
низко посаженная голова; твердая “раковина”, сделанная из костных щит-
ков, сцементированных вместе, и хвост, заключенный в кольцевые ряды
пластин или в твердый костный футляр. В миоцене самые крупные предста-
вители глиптодонтов были всего 91 см в длину, а в плейстоцене они достига-
ли уже 3.7 м. Собственно род Glyptodon имел хвост, окруженный подвиж-
ными, накладывающимися друг на друга костными кольцами с шипами
(фиг. XXXIII. 1). У Sclerocalyptus кольца, защищающие хвост, были соеди-
нены между собой, а у Doedicurus хвост заканчивался массивной костной
“булавой” с шипами (фиг. XXXIII. 1). Мощные мускулы раскачивали хвост
из стороны в сторону, нанося удары, которыми можно сломать кости любого
хищника. Глиптодонты были особенно широко распространены в Аргентин-
ских пампасах в течение плейстоцена.
Ленивцы. Древесные ленивцы сегодня - вялые животные, которые висят
на ветках вниз головой, их ископаемые остатки встречаются довольно редко.
Наземные же ленивцы были разнообразными и очень специализированными
животными. Развитие наземных ленивцев началось в олигоцене, они были
522
КАМЕННАЯ КНИГА
Фиг. XXXIII.2. Наземные ленивцы
а, б — Mylodon, животное, имевшее 3.1 м в длину; в — Megatherium — крупней-
ший наземный ленивец, приблизительно 6.1 м в длину, реконструкция
листоядными и травоядными животными не крупнее 91 см в длину. Хотя ни
один из них не стал полностью беззубым, зубы сильно редуцировались.
Об о лигоценовых наземных ленивцах известно мало. Примитивный по
строению скелета миоценовый Hapalops был приблизительно 1.2 м длины,
включая хвост. На ногах имелось по пять пальцев с когтями, запястья и
лодыжки были искривлены так сильно, что животное ходило на внешней
стороне передней и задней ног. Череп низкий и относительно длинный с вы-
ступающей вперед нижней челюстью. Выступание вперед нижней челюсти
облегчало животныму сдирание листьев с веток.
Nothrotherium -плейстоценовый потомок Hapalops приблизительно 2.3 м
длиной. Ноги животного были массивны, один палец на задней ноге исчез, на
двух остались лишь рудименты когтей. На передней ноге было пять пальцев,
но первый и пятый были слишком маленькими, чтобы нести когти. Хотя
Nothrotherium появился в Аргентине, он затем преодолел Панамский пере-
шеек и распространился на север до Калифорнии. Представители этого рода,
очевидно, предпочитали сухие, гористые области, где питались листьями
юкки и другими морозоустойчивыми растениями, в основном из семейства
лилейных. Скелет, сохранивший сухую кожу и сухожилия, был найден око-
ло г. Аден в Новой Мексике. Отдельные кости, очищенные каменными ножа-
ми, а также экскременты, волосы и когти обнаружены в Гипсовой пещере на
востоке от Лас-Вегаса, штат Невада. Остатки имеют возраст 8.5-11 тыс. лет.
Эта находка показывает, что Nothrotherium жил в настоящую геологичес-
кую эпоху и был съеден древними американцами.
Megatherium также появился в Южной Америке, а затем распространил-
ся в течение плейстоцена на север до штатов Огайо и Каролина (США). Взрос-
лое животное достигало в длину 6.1 м и весило более 4.5 т (фиг. ХХХШ.2).
Череп был относительно маленький, с глубокими нижними челюстями, за-
ВЕЛИКОЕ РАЗНООБРАЗИЕ ПЛАЦЕНТАРНЫХ
523
канчивающимися желобообразным выступом, который использовался при
сдирании листьев с деревьев. Имелся широкий гребень, располагающийся
ниже каждого глаза, полностью окруженного костью. Шея была короткая, а
тело длинное и мощное с хвостом. Тазовый пояс состоял из двух костей, ко-
торые простирались вперед и лопатообразно расширялись. Такое своеобраз-
ное строение таза является приспособлением для поддержания внутренних
органов, когда Megatherium сидел вертикально. Кости передней и задней ног
были массивны, но передние ноги менее тяжелы. На каждой передней лапе
было по четыре когтя, а задняя лапа несла только один. Первые два пальца
задней ноги исчезли, а последние два сильно редуцировались. Очень боль-
шая пяточная кость и короткие фаланги пальцев несли большую часть веса
животного.
Mylodon и его близкие родственники были травоядными животными
3-4 м длиной и, вероятно, весили немного больше тонны. Они имели корот-
кие, широкие головы с широкой мордой и мелкие нижние челюсти. Все че-
тыре ноги были массивны; на передней имелось пять пальцев, три из кото-
рых с когтями, на задней - четыре пальца, из которых только второй и тре-
тий несли когти, пяточная кость очень крупная (фиг. ХХХШ.2).
Мы представляем себе ленивцев робкими и неуклюжими существами,
становившимися жертвами саблезубых тигров, львов, ягуаров и пещерных
медведей. Однако забываем про их когти и мощные мускулы на ногах. Эти
мускулы использовались в основном, чтобы валить деревья или отламывать
ветки, но они использовались и для защиты от врагов. Приподниматься и
размахивать лапами ленивцы могли свободно. Megatherium и Mylodon бы-
ли намного крупнее, чем самые огромные ныне живущие медведи.
ПРЫГУНЧИКОВЫЕ - МАЛЫШКИ С “ХОБОТОМ”
Макросцелиды - это животные с длинной, узкой мордой. Даже самый
большой прыгунчик длиной 31 сантиметр больше всего похож на землерой-
ку. Внешне они отличаются наличием длинного гибкого хоботка. Известны
с олигоцена до настоящего времени в Африке (фиг. XXXIII.3). Эти живот-
ные, по-видимому, из ныне живущих самые близкие родственники кроли-
ков. Если это так, то их возможный общий предок Anagale жил в Восточной
Азии в течение палеоцена.
ЗАЙЦЕОБРАЗНЫЕ, ИСТОРИЯ НЕКОТОРОГО УСПЕХА
Кролики, зайцы и пищухи впервые появляются в эоцене и олигоцене
Азии, Европы и Северной Америки. В плиоцене кролики и зайцы достигают
Африки, а в позднем плейстоцене - Южной Америки.
Даже самые ранние зайцеобразные имеют немного отличий от их ныне
живущих потомков. Даже пищухи, несмотря на их короткие уши, слабо от-
личаются от других зайцеобразных. Схожесть внешнего облика обусловлена
их адаптацией. С этой общей для всех формой тела они сумели успешно при-
способиться к жизни в различных окружающих условиях. На каждом участ-
ке суши, которого достигли зайцеобразные, они процветали. Переселившись
из Европы в Австралию и Новую Зеландию, они быстро приобрели статус
главного вредителя, успешно колонизируя эти области, вытесняя многих
местных животных.
524
КАМЕННАЯ КНИГА
Фиг. XXXIII.3. Современный представитель отряда прыгунчиковые. Хоботко-
вая собачка (Rhynchocyon). Длина тела 24-31 см, длина хвоста 19-26 см
(В.Е. Соколов, 1973)
АРХАИЧНЫЕ ХИЩНИКИ - КРЕОДОНТЫ
Известны с позднего палеоцена и раннего эоцена Северной Америки и Ев-
ропы. Охуаепа — один из самых ранних креодонтов. Это был коротконогий
хищник более 91 см в длину, его внешний вид напоминал современную росо-
маху (фиг. XXXIII.4). Некоторые ученые предполагают, что Охуаепа плава-
ли подобно выдре, ловили рыбу и мягкопанцирных черепах. Другие пред-
полагают, что они жили на суше и ели падаль или охотились из засады. Все
соглашаются, что их мозг был слабо развит и животное было не способно к
хитрости, которая является обычной для современных росомах, намного бо-
лее развитых хищников.
Patriofelis (в переводе “отец котов”) известен из среднего эоцена Северной
Америки и являлся родственником Охуаепа. Хотя он не был гигантом, но отно-
сился к крупным хищникам (фиг. XXXIII.4). Patriofelis весил не больше чер-
ного медведя, но имел череп почти такого же размера, как у льва. Нижние че-
люсти были тяжелыми и глубокими, с мощной мускулатурой, это придавало
им большую сокрушительную мощь. Зубы были приспособлены для поедания
мяса, но ноги слишком коротки для активной охоты. Такой контраст дает воз-
можность некоторым палеонтологам предположить, что Patriofelis также имел
привычки, подобные современным выдрам. Другие палеонтологи предполага-
ют, что эти животные охотились из засады или питались падалью, подобно со-
временным гиенам, которые, как известно, ловят иногда и живую добычу.
Одним из наиболее преуспевших креодонтов был Hyaenodon. Он появился
в позднем эоцене Европы, Северной Америки и Азии и сохранился на всех
трех континентах в течение олигоцена. В эту же эпоху род достиг Африки и
вымер там в раннем миоцене. Известно несколько видов гиенодонов. В ран-
нем олигоцене на западе Соединенных Штатов имеется, по крайней мере, че-
тыре вида, размер которых варьирует от размера мелкого койота до крупного
гризли (фиг. XXXIII.5).
Креодонты вымерли к концу миоцена и не дали начало никаким другим
группам млекопитающих. Ныне живущие млекопитающие, которых мы на-
зываем хищниками, совершенно другая группа - истинные хищные.
ВЕЛИКОЕ РАЗНООБРАЗИЕ ПЛАЦЕНТАРНЫХ
525
Охуаепа, жил в Северной Америке
в течение позднего палеоцена
и в Европе в течение раннего эоцена.
Длина тела 100 см
Patriofelis, ранний-средний эоцен,
жил только в Северной Америке
и был приблизительно 1.5 м длиной,
череп такого же размера, как у льва
Фиг. XXXIII.4. Два эоценовых креодонта
Фиг. XXXIII.5. Реконструкция внешнего вида и черепа гиенодонов
а—б - Hyaenodon horridus; а — реконструкция; б — череп; в - череп меньшего ви-
да Н. musteunus. Известны из олигоцена Северной Америки
ИСТИННЫЕ ХИЩНЫЕ - CARNIVORA
Истинные хищные, то есть члены отряда Carnivora, появляются в палео-
цене. До олигоцена все хищные были так сильно похожи, что были объеди-
нены в одно надсемейство Miacoidea. Все животные были маленькие, самое
большое достигало размера колли.
К концу раннего олигоцена с исчезновением мезонихидных кондилартр и
снижением численности креодонтов произошло увеличение количества
хищных. Хищные делятся на две основных группы: “группа собак” и
“группа котов”. “Группа собак” включает кроме собак медведей, енотов,
526
КАМЕННАЯ КНИГА
Фиг. XXXIII.6. Саблезубые кошки из олигоцена Северной Америки
a- Dinictis, длина тела от носа до основания хвоста 102 см; б - Hoplophoneus, не-
сколько большего размера и более тяжелого строения. Ископаемые остатки обо-
их найдены в олигоценовых отложениях местонахождения группы Белая река,
центральная часть США
скунсов и ласк. Кроме котов, “группа котов” включает гиен и вивер. Отличи-
тельной особенностью большинства членов “группы котов” является воз-
можность втягивать когти, в то время как большинство “группы собак” этого
не могут.
По данным Р. Кэрролла, отряд хищных делится на четыре надсемейства:
примитивные хищные (Miacoidea), кошачьи (Aeluroided), псовые (Arctoidea)
и моржовые (Otarioidea). К кошачьим относятся семейства: виверовые, ко-
шачьи и гиеновые. К псовым - семейства: псовые, енотовые, медвежьи, ку-
ньи. К моржовым - семейства: моржи, ушастые тюлени и вымершие инали-
арктиды. По ряду других систематик надсемейство моржовые и семейство
тюленей объединяются в отряд ластоногих (Соколов, 1979).
В истории развития кошачьих саблезубые формы появлялись дважды.
Ложные саблезубые коты появились в раннем олигоцене и вымерли в конце
миоцена. Представителями этой группы были Dinictis и Hoplophoneus
(фиг. ХХХШ.6). Истинные саблезубые коты, часть живущего сейчас се-
мейства кошачьих - Felidae, которое включает не только ласкового домаш-
него кота, а также львов и тигров. В это семейство входили такие живот-
ные, как Smilodon californicus, хорошо известный по тысяче экземпляров,
найденных в ямах смолы местонахождения Ранчо ла Бреа (около Лос-Анд-
желеса). Истинные саблезубые коты появились в начале миоцена и некото-
рое время (около эпохи) жили совместно с ложными саблезубыми.
Развитие саблезубости у млекопитающих, однако, было присуще не толь-
ко хищным. Thylacosmilus - плотоядный сумчатый из плейстоцена Южной
Америки, о котором мы рассказывали в главе XXXI, был также саблезубым.
Корни мощных клыков продолжались выше глаз. Некоторые креодонты то-
же были саблезубыми. Независимое возникновение саблезубости, по край-
ВЕЛИКОЕ РАЗНООБРАЗИЕ ПЛАЦЕНТАРНЫХ
527
ней мере четыре раза в истории развития млекопитающих, говорит о том, что
это весьма удачная адаптация для плотоядных животных. Почему ни один
из них не дожил до настоящего времени, остается загадкой.
К началу миоцена некоторые хищные начали жить в море. Они названы
ластоногими по форме их конечностей. Фактически, две различных группы
морских млекопитающих отдельно произошли от наземных хищников.
Группа моржовые и ушастые тюлени, к которой относятся моржи, морские
котики, морские львы и сивучи, одна из них. Они, очевидно, произошли от
животных подобных медведям, и их история развития сосредоточена вокруг
Тихоокеанского бассейна.
Другая группа — тюленевые - своим происхождением связана с куньими и
известна преимущественно из Атлантики и областей, примыкающих к Сре-
диземному морю.
ПАНТОДОНТЫ - РАСТИТЕЛЬНОЯДНЫЕ ПОТОМКИ ДРЕВНИХ ПЛОТОЯДНЫХ
Как мы узнали из главы XXXI, остатки некоторых самых ранних плацен-
тарных млекопитающих найдены в позднемеловых отложениях Монголии.
Среди них были плотоядные млекопитающие, от которых произошли крео-
донты и хищные, а также многие неплотоядные животные.
Coryphodon — один из хорошо известных в позднем палеоцене и раннем эо-
цене представителей отряда пантодонты (фиг. ХХХШ.7). Его толстые кости
хорошо противостоят разрушению, хорошо сохраняются и обычны в ископа-
емом состоянии. Некоторые виды были не больше тапиров, другие же веси-
ли, как волы, 725-820 кг и были более тяжело построены. Coryphodon имел
длинное тело и короткие пятипалые конечности, которые походили на ко-
нечности слона. Череп большой, широкий и тупой. Клыки длинные, и самцы
использовали их в схватках. По своему внешнему виду Coryphodon был по-
хож на карликового гиппопотама. Предполагают, что животное проводило
много времени в болотах.
Фиг. ХХХШ.7. Coryphodon, позднепалеоценовый и раннеэоценовый панто-
донт. Его остатки, найдены в Европе, Азии и Северной Америке. Он был прибли-
зительно 2.4 м в длину и, возможно, был обитателем болот
а — реконструкция; б — череп
528
КАМЕННАЯ КНИГА
Фиг. XXXIII.8. Barylambda, пантодонт. Остатки найдены в позднем палеоцене
Северной Америки, он был приблизительно 2.4 м длиной
а — реконструкция; б — череп; в — скелет левой стопы
Coryphodon был широко распространен, его ископаемые остатки найдены
в Северной Америке, Азии и в Европе. Другой же пантодонт - Barylambda
жил только в Северной Америке и только в течение позднего палеоцена
(фиг. XXXIII.8).
НАСЕКОМОЯДНЫЕ, ПРИМИТИВНЫЕ И СПЕЦИАЛИЗИРОВАННЫЕ
В течение длительного времени палеонтологи считали ныне живущих на-
секомоядных “живыми ископаемыми”, фактически неизмененными поздне-
меловыми предками всех других плацентарных млекопитающих. Однако со-
временные ежи, землеройки и кроты также имели кайнозойскую историю
развития и, конечно, более эволюционно продвинуты, чем их позднемеловые
или раннекайнозойские предки. Эти изменения столь значительны, что
предков современных насекомоядных часто помещают в отдельный отряд
Lipotyphla.
Мы думаем о ежах, как о маленьких безопасных животных, и в основном
это верно. Deinogalerix, по своим размерам тела не уступавший койоту, а по
размерам головы волку, был, конечно, исключением. Это животное жило на
территории современного полуострова Гаргано, который выступает в Адриа-
тическое море примерно в середине северо-восточного побережья Италии.
В позднем миоцене эта область была островом. В составе фауны, жившей на
острове, отсутствовали хищники размера Deinogalerix. И хотя это насекомо-
ядное не могло добывать себе пищу так же эффективно, как сходный по раз-
меру истинный хищник, оно выступало в этой роли, так как на острове не бы-
ло конкуренции.
ЛЕТУЧИЕ МЫШИ, ЕДИНСТВЕННЫЕ ЛЕТАЮЩИЕ МЛЕКОПИТАЮЩИЕ
Если бы не адаптация к полету, летучие мыши не так сильно отличались
бы от своих насекомоядных предков из позднего мела, как большинство дру-
гих плацентарных млекопитающих. Мы часто сравниваем их с птерозавра-
ВЕЛИКОЕ РАЗНООБРАЗИЕ ПЛАЦЕНТАРНЫХ
529
Фиг. ХХХШ.9. Icaronycteris, древняя летучая мышь эоцена из формации Зе-
леной Рекн (Вайоминг), Северная Америка. Длина 12 см (фото Don Baird)
ми, потому что у тех и у других были широкие крылья, покрытые очень тон-
кой кожей. Крыло птерозавра натягивалось от тела и ног к четвертому пальцу,
для поддержки другие пальцы не использовались. Летучие мыши избежали
этого дефекта, у них удлинились кости в “руке”, и четыре из пяти пальцев ис-
пользуются для усиления и растяжения крыла. Они также развили сильные
грудные мускулы и увеличили слуховые области мозга. Древние летучие мы-
ши, очевидно, ориентировались в полете, слушая эхо от издаваемых звуков,
поскольку многие из их потомков используют эхолокацию и сегодня.
Самые древние из известных летучих мышей были маленькими существа-
ми, они утратили несколько зубов, характерных для их предков и обладали
хорошо развитыми крыльями. Некоторые виды жили в лесу, другие пресле-
довали насекомых над реками и озерами. Охотясь над Зеленой рекой и озе-
ром эоценового возраста (юго-западный Вайоминг), летучие мыши время от
времени так низко пролетали над водой, что ударялись о нее. Они очень быс-
тро тонули и погружались на дно водоема, где кости, а иногда даже кожа со-
хранялись в мелкозернистом осадке.
В олигоцене группа раскололась на две части, одну составляли мелкие лету-
чие мыши, питающиеся насекомыми, а представители другой начали есть пло-
ды. Плодоядные постепенно увеличивали размер тела и черепа, самые круп-
ные представители, “летучие лисицы”, в настоящее время живут на островах
Индийского океана, в Австралии и в Японии. С олигоцена до настоящего вре-
мени большинство других ныне живущих отрядов млекопитающих очень
сильно изменились по своему составу и морфологии. Это не относится к лету-
чим мышам. Многие летучие мыши, жившие в олигоцене, отличаются только
в незначительных деталях от летучих мышей, летающих сегодня
(фиг. ХХХШ.9).
ШЕРСТОКРЫЛЫ ИЛИ КАГУАНЫ
Шерстокрылов называют еще летающими лемурами, однако это не насто-
ящие лемуры и они не летают, а планируют. Сегодня шерстокрылы живут в
34 Каменная книга
530
КАМЕННАЯ КНИГА
Юго-Восточной Азии и на Филиппинах. Их немногочисленные ископаемые
остатки известны из палеоцена и эоцена Северной Америки. Вместе с летучи-
ми мышами и приматами они, возможно, формируют одну ветвь развития в
древе млекопитающих - надотряд Archonta. Общей чертой всех представите-
лей надотряда является то, что по крайней мере их примитивные члены бы-
ли приспособлены к жизни на деревьях.
ЛЮДИ И ДРУГИЕ ПРИМАТЫ
Возможно потому, что мы сами являемся приматами, много усилий было
потрачено, чтобы распутать эволюционную историю этой группы. Однако
даже если бы мы принадлежали к другому отряду млекопитающих, прима-
ты очень показательны и удобны для изучения, потому что много раз различ-
ные их группы приспосабливались к сходным условиям существования. Та-
ким образом, они дают плодородную почву для изучения различных путей
приспособления животных к окружающей среде.
Примитивные приматы. Первыми плацентарными млекопитающими, ко-
торые влезли на деревья, были, вероятно, приматы палеоцена. Считается, что
в палеоцене они произошли от мультитаберкулат. Самые ранние из извест-
ных приматов - плезиадаписы (подотряд Plesiadapiformes) — были специали-
зированной ветвью развития. Они имели промежуток в зубном ряду и боль-
шие, вперед торчащие нижние резцы. По этим особенностям строения они
больше напоминают живущих сейчас белок или австралийских древесных
поссумов, чем других приматов. Никакие более поздние приматы не имеют
такую большую диастему, промежуток в зубном ряду между резцами и пре-
молярами.
Примитивные приматы карабкались по деревьям так же, как это делают
сегодня белки, то есть они вонзали в кору свои острые когти. В результате
эволюции карабканье по деревьям стало выполняться при помощи цепких
пальцев рук и ног, а когти приматов уменьшились до ногтей.
Глаза древних приматов не имели области перекрытия зрения, которая
имеется у большинства ныне живущих приматов и дает возможность видеть
в трех измерениях. У древних приматов область зрения каждого глаза была
направлена больше в стороны. С течением времени приматы стали более
визуально ориентирующимися животными, а значение обоняния умень-
шилось.
Известны остатки приматов из палеоценовых отложений Северной Аме-
рики и Европы. Однако весьма вероятно, что прямые предки современных
приматов жили в другом месте, возможно, в экваториальных областях, где
пышные тропические леса обеспечивали им любимые места обитания.
В эоцене появились животные, которые могут быть отнесены к наиболее
примитивным группам ныне живущих приматов, - это лемуровые и долго-
пяты. Вместе с вымершими представителями Adapiformes они объединяются
в единый подотряд полуобезьяны (Prosimi).
Лемуровые. Наиболее известный из этих примитивных приматов - Noth-
arctus. По внешнему виду он напоминал лемуров с относительно длинной
мордой (фиг. XXXIII. 10). В течение среднего эоцена был, вероятно, актив-
ным прыгуном, обитателем леса, современные аналоги которого известны на
юго-западе Вайоминга. Для успешного передвижения глаза у него были рас-
положены так, чтобы область перекрытия зрения давала животному воз-
можность видеть в трех измерениях. К лемуровым из ныне живущих прима-
ВЕЛИКОЕ РАЗНООБРАЗИЕ ПЛАЦЕНТАРНЫХ
531
Фиг. XXXIII.10. Эоценовые лемур и долгопят Северной Америки
а, б — Notharctus, примитивный эоценовый лемур: а - реконструкция, б - череп
8 см длиной; в — голова Tetonius, крошечного эоценового тарзиоида из Вайомин-
га, длина его черепа 3 см
тов относятся собственно лемуры, карликовые лемуры, руконожки, индри и
лори.
Долгопяты. Ныне живущий в Юго-Восточной Азии род долгопятов явля-
ется единственным представителем некогда разнообразной группы мелких
приматов - тарзиоидов, которые процветали в течение эоцена в Европе, Азии
и Северной Америке. Tetonius из раннего эоцена Северной Америки являет-
ся одним из немногих ранних тарзиоидов, которые известны по остаткам че-
репов. На крошечном черепе, только 3 см в длину, выделяется большая че-
репная коробка и огромные глазные орбиты (фиг. ХХХШ.10). Возможно, та-
кими большими глазами Tetonius мог видеть даже ночью.
Наиболее эволюционно развитые обезьяны и человек относятся к подотря-
ду человекоподобных, делящемуся на широконосых обезьян (обезьян Нового
Света) и узконосых обезьян (обезьян Старого Света).
Обезьяны Нового Света, или Южной Америки, появились в раннем олиго-
цене. Много споров велось относительно того, мигрировали ли они из Север-
ной Америки или из Африки, и до настоящего времени ученые не пришли к
единому мнению. Научное название для них — Platyrrhines, что переводится
как “широкие носы”. Название дано по факту, что их две ноздри располага-
ются далеко друг от друга и открываются вбок, по сравнению с носами обезь-
ян Старого Света, у которых ноздри располагаются рядом друг с другом и от-
крываются вперед. Большинство южноамериканских обезьян имеют по
тридцать шесть зубов и цепкий хвост, который активно используется при ла-
зании, за это их часто называют паукообразными обезьянами. Тридцать два
зуба только у маркозеток. Они потеряли последний четвертый коренной зуб
(“зуб мудрости”), но сохранили четыре предкоренных зуба, которые были ут-
рачены приматами Старого Света.
Ископаемые остатки нескольких обезьян были найдены в олигоценовых и
миоценовых отложениях Южной Америки, но намного больше остатков из-
вестно из отложений плейстоценовых пещер Бразилии. Хотя паукообразные
обезьяны теперь обитают и к северу от Мексики, их ископаемые остатки
вообще не известны в Центральной и в Северной Америке. Они были найде-
34*
532
КАМЕННАЯ КНИГА
ны только в очень молодых отложениях на Ямайке и в Доминиканской
Республике.
Обезьяны Старого Света. Из позднего эоцена Бирмы и раннего олигоцена
Африки известны самые ранние представители человекоподобных обезьян,
к которым относятся мартышки и человекообразные обезьяны, включая лю-
дей. Эти самые ранние из известных антропоидов были размером с малень-
ких ныне живущих обезьян, но настолько примитивны по своему строению,
что их нельзя относить к истинным обезьянам Старого Света. Возможно, они
могли быть общим предком всех более поздних антропоидов.
Человекообразные обезьяны возникли последними из антропоидов, они
появились в раннем миоцене. Около 20 млн. лет назад роль, которую в насто-
ящее время играют обезьяны в сообществе млекопитающих Африки, была
сыграна человекообразными обезьянами.
Предки ныне живущих человекообразных обезьян обитали в лесу. Напро-
тив, наши предки приспособились к жизни в саваннах и сухих степях, кото-
рые образовались на месте влажных лесов Африки в течение миоцена и пли-
оцена, поскольку климат стал более засушливым. Эти климатические изме-
нения повлияли на передвижение животных. Начало вырабатываться
прямохождение, положение тела стало более вертикальным и, в конечном
счете, они начали ходить только на нижних конечностях, освободив руки
для переноса орудий труда и для других действий.
ИЗОБИЛИЕ КОПЫТНЫХ
Большинство ископаемых остатков плацентарных млекопитающих мож-
но объединить в группу копытных. Не все копытные имеют копыта, но мно-
гие обладают ими, и главным образом это травоядные.
КОНДИЛАРТРЫ - УДИВИТЕЛЬНЫЕ КОПЫТНЫЕ
Это были удивительные животные, сочетавшие в строении скелета при-
знаки, характерные как для хищных, так и для растительноядных живот-
ных. Представители именно этого отряда считаются предками парнокопыт-
ных, непарнокопытных, хоботных и полностью вымерших южноамерикан-
ских копытных.
Наиболее примитивные копытные - арктоционидные кондилартры
появились в самом начале палеоцена. По внешнему виду это маленькие че-
тырехногие животные, являющиеся идеальными предками для более позд-
них копытных. В течение очень короткого по геологическим масштабам
времени, нескольких миллионов лет, арктоциониды дали начало многим
группам.
Phenacodus известен из позднего палеоцена и раннего эоцена Северной
Америки и раннего эоцена Европы (фиг. ХХХШ.11). По-видимому, он яв-
лялся предком лошадей и других непарнокопытных. На это указывает преж-
де всего строение лодыжки задней ноги. Некоторые виды были маленькими,
как лиса, другие достигали размера овцы. Имели длинную голову с глазами,
которые располагались в передней части черепа; длинные хвосты и пятипа-
лые конечности с маленькими копытами.
Кондилартры достигли Южной Америки. Раннеэоценовый Didolodus из
Аргентины был весьма похож на Phenacodus. В то время как кондилартры
ВЕЛИКОЕ РАЗНООБРАЗИЕ ПЛАЦЕНТАРНЫХ
533
Фиг. ХХХШ.11. Phenacodus — кондилартр, найденный в позднем палеоцене и
раннем эоцене Северной Америки и раннем эоцене Европы. Самый крупный
вид был около 1.7 м в длину
Северного полушария вымерли в конце эоцена, в Южной Америке они про-
должали существовать по крайней мере до среднего миоцена.
Тиллодонты. Многие потомки арктоционид относились к небольшим или
недолго просуществовавшим группам. Одной такой группой были тиллодон-
ты (отряд Tillodonta), обитавшие в эоцене в Северном полушарии. С больши-
ми долотоподобными резцами эти животные по внешнему виду напоминали
грызунов, с которыми, как раньше думали, они связаны по происхождению.
Однако тиллодонты — самостоятельный отряд растительноядных млекопи-
тающих. Trogosus, среднеэоценовый тилодонт, был гигантом размером с со-
временного черного медведя.
Трубкозубы. Капский трубкозуб, живущий в настоящее время в Африке,
является единственным продолжающим существование видом из отряда
трубкозубы (Tubulidentata). Его зубы имеют характерное трубчатое строение
жевательной поверхности, по которому и дано название отряда. Сначала
трубкозубы появляются в раннем миоцене Африки, а затем распространяют-
ся по Европе и Азии. Однако после плиоцена они остались лишь в Африке.
Возможно, представители этого отряда также произошли от кондилартр. Че-
тыре известных рода трубкозубов отличаются друг от друга главным образом
по степени приспособления их скелетов к поеданию термитов и муравьев, а
также к рытью и карабканью.
Уинтатерии. Другие потомки арктоционидных кондилартр - уинтатерии
(отряд Uintatheria) - имели очень своеобразное строение черепа. Uintatherium
среднеэоценового возраста достигал 1.5 м высоты в плече и весил как совре-
менный носорог, на которого он немного был похож. Длинная, низкая голова
несла три пары тупых костных рогов, которые были закрыты кожей и достига-
ли от 5 до 25 см высоты (фиг. ХХХШ.12). Самцы имели кинжалоподобные
клыки на верхних челюстях, которые могли быть использованы против круп-
ного хищника или против других самцов во время поединков за самку. Его по-
томок - Eobasileus — был еще крупнее, длина черепа достигала 1 м, тогда как
длина черепа Uintatherium была только 76 см. Eobasileus был почти
2.1м высоты, а клыки на 23 см торчали вниз из верхних челюстей
(фиг. ХХХШ.12).
534
КАМЕННАЯ КНИГА
Фиг. XXXIII. 12. Два эоценовых унитатерия Северной Америки
а - Uintatherium, достигал 1.5 м высоты в плече, длина черепа 76 см, б -
Eobasileus, высота около 1.5 м и череп 102 см длиной
Phenacodus
Caryphodon
Uintatherium
Свинья
Носорог
Фиг. XXXIII. 13. Головной мозг архаичных и современных млекопитающих.
Архаичные млекопитающие (верхний ряд) имели намного меньший головной
мозг в целом и его полушария, чем таковые у современных млекопитающих
(нижниц ряд), и поэтому были намного менее интеллектуальными животными.
Одна четвертая естественного размера
Наряду с такими достижениями в размере и вооружении Eobasileus имел
и очевидные слабости. Крупное животное нуждалось в большом количестве
пищи, a Eobasileus имел зубы, которые не были ни больше, ни лучше, чем зу-
бы Uintatherium. Мозг также был маленький, по крайней мере по современ-
ным стандартам. И у Eobasileus и у Uintatherium мозг был намного меньше,
чем у гиппопотама или носорога равного размера, с крошечными полушари-
ями (фиг. ХХХШ.13). Они были способны думать, но настолько смутно, что
ВЕЛИКОЕ РАЗНООБРАЗИЕ ПЛАЦЕНТАРНЫХ
535
инстинкты и рефлексы полностью управляли его жизнью, как это было и у
крупных динозавров.
Данное утверждение относится ко всем “архаичным” млекопитающим -
кондилартрам, креодонтам, а также к примитивным копытным. С непово-
ротливыми телами, управляемыми маленьким головным мозгом, они были
не подготовлены к конкуренции с новыми и более интеллектуально развиты-
ми животными.
ПАРНОКОПЫТНЫЕ ЖИВОТНЫЕ
К таким более интеллектуальным животным относятся парнокопытные,
млекопитающие с раздвоенными копытами. Они характеризуются тем, что
главная ось опоры ноги проходит между вторым и третьим пальцами. Та-
ким образом, тяжесть при опоре на ногу одинаково распределена между
двумя пальцами, которые поэтому и одинаково развиты. Такое строение
конечности контрастирует с конечностью непарнокопытных, у которых
главная ось опоры проходит только через один третий палец. В этом случае
третий палец самый большой на ноге и может быть даже единственным,
как у современных лошадей.
Хотя парнокопытные появились только в раннем эоцене, в конце этой же
эпохи они подверглись настоящему эволюционному взрыву, значительно
увеличившему их разнообразие. К концу эоцена появилось 20 семейств пар-
нокопытных и возникли все основные группы. Современных животных
можно отнести к трем основным группам: свинообразные (подотряд Suina),
мозоленогие (подотряд Tylopoda) и жвачные (подотряд Ruminant ia). По мне-
нию некоторых ученых, мозоленогие не относятся к парнокопытным, а обра-
зуют самостоятельный отряд. Современные свинообразные представлены
свиньями, пекари и бегемотами; мозоленогие - верблюдами и ламами; жвач-
ные — оленьками, жирафами, оленями, вилороговыми и полорогими, вклю-
чая домашний скот.
Ископаемых представителей парнокопытных значительно больше, чем
современных. Так, известно шесть вымерших семейств свинообразных, в два
раза больше, чем современных. Из ископаемых свинообразных хорошо
изучена “гигантская свинья” Dinohyus. Ископаемые остатки этого животно-
го найдены в миоценовых отложениях Северной Америки и, в частности, в
костеносных пластах местонахождения Агата Спрингс (фиг. XXXIII. 14). Со-
временные представители мозоленогих относятся всего лишь к одному се-
мейству - верблюдовые, тогда как еще пять семейств представлены исклю-
чительно вымершими формами. Вымершими являются и так называемые
примитивные парнокопытные, остатки которых известны с раннего эоцена
до позднего олигоцена.
Ореодонты. Полностью вымерший отряд. Oreodon, или по другому Мегу-
coidodon, был коротконогим, тупоголовым растительноядным животным,
имеющим по пять пальцев на передней ноге (первый палец был очень ма-
леньким) и по четыре пальца на задней (фиг. XXXIII. 15). Merycoidodon, ве-
роятно, жили стадами, на это указывает большое количество костей, зубов и
хорошо сохранившихся черепов по сравнению с ископаемыми остатками
других млекопитающих, собранных из формации Бэдленд (штаты Южная
Дакота и Небраска, США). Там же найден один скелет самки с костями заро-
дышей двух близнецов.
Leptauchenia имел короткие ноги, короткий хвост и крошечные копыта.
Эти копыта, да к тому же глаза и ноздри, перемещенные на самую верхнюю
536
КАМЕННАЯ КНИГА
Фиг. XXXIII. 14. Парнокопытные миоцена
Остатки обоих животных собраны из костеностных пластов местонахождения
Агата Спрингс
Фиг. ХХХШ.15. Три ореодонта, обычных в позднем олигоцене Южной Дакоты
а - Agriochoerus, тело длиной 1.2-1.5 м, б - Merycoidodon, 41 см высоты в плече,
найдено большое количество ископаемых остатков, в — Leptauchenia, плаваю-
щий с полностью погруженным телом
ВЕЛИКОЕ РАЗНООБРАЗИЕ ПЛАЦЕНТАРНЫХ
537
часть черепа, располагающийся на вершине морды рот указывают, что это
млекопитающее много времени проводило в воде (фиг. XXXIII. 15). Он мог
плавать, полностью погружая тело в воду. Другой ореодонт - Agriochoerus
имел очень длинный хвост, когтистые лапы и напоминал некоторых крео-
донтов (фиг. XXXIII. 15). Он, вероятно, мог залезать на деревья для защиты
от врагов. Когти использовал, в основном, для вырывания корней и луковиц
растений, так как его зубы были приспособлены для поедания растительной
пищи.
Ореодонты процветали в олигоцене и раннем миоцене (фиг. XXXIII. 14).
Их разнообразие постепенно уменьшилось в среднем и позднем миоцене и к
концу эпохи они полностью вымерли. Область распространения ореодонтов
была ограничена Северной и Центральной Америкой, их остатки не находят
южнее Панамского канала. Не ясно, почему такая распространенная группа
на континенте была неспособна мигрировать, когда другие семейства сумели
перейти по Берингийскому мосту даже в Азию.
История семейства верблюдовых (Camelidae). Сегодня мы считаем верб-
людов жителями пустынь Старого Света. Немного подумав, вспоминаем, что
ламы Южной Америки также члены семейства верблюдовых. Поэтому ка-
жется странным, что большая часть истории развития этого семейства про-
ходила в Северной Америке, и только к концу кайнозоя они мигрировали на
другие континенты.
Protylopus - примитивный верблюд. Первый известный член семейства
верблюдовых появился в эоцене. По систематике Р. Кэрролла он не относит-
ся к семейству верблюдовых, а принадлежит полностью вымершему семей-
ству оромерисиды. Этот патриарх - Protylopus - был коротконогим живот-
ным немного крупнее американского зайца, с удлиненным черепом и глаз-
ными впадинами, частично закрытыми костными выступами (фиг.
XXXIII.16). Челюсти несли сорок четыре зуба, клыки были похожи на рез-
цы, а щечные зубы имели очень низкие коронки. На передней ноге было че-
тыре пальца, а на задней — два и небольшие косточки - рудименты двух бо-
Фиг. XXXIII. 16. Два североамериканских представителя семейства верблю-
довых
Голова (а) и череп (б) Protylopus, поздний эоцен Северной Америки, самый древ-
ний из известных представителей семейства, длина черепа 12 см; в — Stenomy-
lus, раннемиоценовый верблюд Северной Америки, 69 см высоты в плече
538
КАМЕННАЯ КНИГА
Фиг. ХХХШ.17. Два вида Poebrotherium из олигоцена североамериканских
Больших равнин. Большой — 66 см высоты в плече
а, б— реконструкция, в — череп крупного вида
ковых пальцев. Копыта направлены подобно копытам оленя, это показыва-
ет, что пальцы не дополнялись мозолью, как у современных верблюдов.
Задние ноги длиннее передних и при беге животные, вероятно, скакали.
Poebrotherium. Потомки Protylopus бродили огромными стадами на запа-
де Северной Америки в течение олигоцена. Наиболее характерным был Poe-
brotherium, чьи кости найдены в отложениях фармации Бэдлэнд (Южная Да-
кота, США) и в некоторых других областях. Poebrotherium напоминал ламу,
был приблизительно 66 см высоты в спине и весил меньше овцы
(фиг. ХХХШ.17). Зубы походили на зубы Protylopus, на передних и задних
ногах было только по два пальца. Хвост, хотя и короткий, длиннее, чем у со-
временной ламы.
Мы обычно предполагаем, что все древние животные с одним родовым на-
званием выглядели похоже, поэтому описание внешнего вида можно отно-
сить ко всем родственным видам. Это не всегда правильно. Фактически, на-
ше описание Poebrotherium относится только к виду Р. labiatum, другой вид,
Р. wilsoni, имел более длинные и намного более стройные ноги, более корот-
кое тело и сильно изогнутую спину (фиг. XXXIII. 17). Искривление спины не
явилось результатом посмертных изменений скелета, позвоночник не вы-
прямлялся и у живых особей.
Верблюдов миоцена можно разделить на три группы, которые эволюцио-
нировали самостоятельными путями. Одна группа — мозоленогие, горбатые
животные, известные сегодня. Они могут быть расценены как основной
ствол в семействе верблюдов. Две другие группы были ветвями, которые
кратко развились и увяли.
Изящные газелиевые верблюды. Род Stenomylus символизирует одну из
увядших ветвей — маленькие тела и стройные ноги этих животных объясня-
ют название группы “газелиевые верблюды”. Stenomylus был немного выше
60 см в плече, с тонкой шеей. Его нижняя челюсть, на первый взгляд, несет
двенадцать резцов, однако на самом деле имеется всего лишь шесть резцов по
три с каждой стороны, но клыки и два премоляра также приняли форму рез-
цов и использовались для щипания травы (фиг. XXXIII. 16).
ВЕЛИКОЕ РАЗНООБРАЗИЕ ПЛАЦЕНТАРНЫХ
539
Фнг. XXXIII. 18. Три североамериканских верблюда
а — Oxydactylus, длинношеий верблюд миоцена, 137 см в плече; б - Аерусате-
lus, животное миоцена с такими длинными ногами и шеей, что получил про-
звище “жирафовый верблюд”; в, г — голова и череп Procamelus, длина черепа
30 см
Большинство ископаемых остатков Stenomylus было найдено в отложе-
ниях раннего миоцена местонахождения Агата Спрингс (запад штата Не-
браска). Карьер представляет часть древней “дневной поверхности”, где
огромное стадо “газелиевых верблюдов” провело ночь, как спят стада
современных лам на равнинах Патагонии. Под утро больные и старые жи-
вотные умерли и были засыпаны песком, принесенным ветром. Смерть но-
чью наступила для сотни или больше животных, чьи кости были потом
найдены.
Верблюды, такие же высокие как жирафы. Если Stenomylus походил на
газель, то его родственник Oxydactylus (это имя значит “длинный палец”)
принадлежит ветви, которая длинными ногами и шеей напоминает нам о
жирафах. Сам Oxydactylus был довольно маленьким, всего 137 см высоты в
плече, хотя, вытягивая шею, мог доставать листья на высоте 2.4 м над зем-
лей (фиг. XXXIII. 18). На каждой ноге имелось по два пальца с копытами, но
нет никаких следов мозолей.
540
КАМЕННАЯ КНИГА
Фнг. XXXIII. 19. Американские верблюдовые
а — Camelops, 2.1 м высоты в плече. Животное ледникового периода, оставался
в живых до недавнего времени, б - гуанако, самый негорбатый современный
верблюд, живущий в горах и на равнинах Южной Америки
Эта группа достигла расцвета в среднем и позднем миоцене. Род Аерусате-
lus (“высокий верблюд”), его крупные виды имели 3.7 м высоты и могли бы
соперничать с ныне живущими жирафами, но их тела не снижались к хвосту
(фиг. ХХХШ.18). Низкокоронковые, короткие зубы были приспособлены к
ощипыванию молодых побегов, на ногах имелись гвоздеобразные копыта и
мозоли, которые позволяли передвигаться по песчаной почве.
От Procamelus к современным верблюдам. В течение среднего и позднего
миоцена главная линия развития верблюдов характеризовалась представи-
телями рода Procamelus, который напоминал Poebrotherium, но имел более
длинную шею и был 1.2 м высоты в плече. Голова длинная и низкая; в перед-
нем отделе верхней челюсти зубы атрофировались (фиг. ХХХШ.18). Это по-
казывает, что животное захватывало траву между твердыми верхними дес-
нами и нижними резцами. Костная арка сзади орбиты полная, а ноги, подоб-
но ногам Aepycamelus, имели мозоли и копыта.
Потомки Procamelus расселились широко. Некоторые пошли в Мексику и
в юго-западном направлении пересекали Южную Америку в конце плиоце-
на, как только исчезли морские барьеры. Эти мигранты образовали несколь-
ко форм с хорошо развитыми мозолями на пальцах ног, включая современ-
ных гуанако и викунья (фиг. ХХХШ.19). Древние гуанако, приспособлен-
ные к жизни среди гор были приручены доисторическими индейцами. Их
домашних потомков - лам и альпака - разводят ради шерсти.
В плиоценовое время верблюды также достигли Азии и образовали два ро-
да: Camelus и Рагас amelus, которые в плейстоцене были распространены от
Китая до Европы и северной Африки. Последний род теперь является пол-
ностью вымершим, а от рода Camelus образовались два ныне живущих до-
машних вида. Одногорбый верблюд - дромадер, его родиной, возможно, бы-
ВЕЛИКОЕ РАЗНООБРАЗИЕ ПЛАЦЕНТАРНЫХ
541
Фиг. XXXIIL20. Некоторые современные парнокопытные
а — косуля (Capreolus capreolis), длина тела 100-150 см, высота в холке 65—100 см,
распространены в умеренном поясе Евразии; б — вилорог (Antilocarpidae ameri-
сапа), длина тела 113-140 см, высота в холке 88-105 см, обитает в центральных
областях Северной Америки; в - черная антилопа (Hippotragus nig er), длина те-
ла 188—267 см, высота в холке 100-160 см, распространены в Центральной и
Южной Африке; г - сайгак (Saiga tatarica), длина тела 110-146 см, высота в
холке 60-79 см, распространен в Восточной Европе, Средней и Центральной
Азии (рис. Е.П. Загозиной по книге В.Е. Соколова, 1979)
ла Африка. Это крупное животное 3-3.3 м длиной и 1.8-2.1 м высотой в пле-
чах. Двугорбый верблюд - бактриан, обитающий в Центральной Азии, не
такой большой, имеет более длинную шерсть и может выносить более холод-
ный климат, чем его одногорбые родственники.
Никто не знает, когда и где возникли горбы, так как эти заполненные жи-
ром структуры не оставляют никаких отпечатков на скелете. Многие авторы
предполагают, что горбы появились у верблюдов Старого Света; другие го-
ворят об их наличии у плейстоценовых верблюдов Северной Америки. Не-
которые из североамериканских верблюдов были такого же размера, как
542
КАМЕННАЯ КНИГА
дромадеры. Однако были и намного крупнее (4.6 м высотой) и намного бо-
лее тяжело построены, чем ранние жирафообразные верблюды. Остатки од-
ного рода — Camelops, вымершего в современную эпоху, череп с клочками
сухих мышц, были найдены в пещере около г. Филлморе (штат Юта)
(фиг. ХХХШ.19).
Жвачные. К отряду парнокопытных относится еще такая очень разнооб-
разная группа, как жвачные, различающиеся между собой строением рогов.
Наиболее разнообразным является семейство полорогих, к нему относятся
широко распространенные и хорошо знакомые нам животные: различные
антилопы, газели, сайгаки, козы, бараны и быки. Представителей двух
других семейств - оленьковые и оленевые (косули, лоси, олени, оленьки,
северные олени) можно еще встретить в лесах и в тундре. Жирафов
(семейство жирафовые) и вилорогов (семейство вилороговые) мы можем
увидеть только в зоопарках (фиг. ХХХШ.20).
Фиг. XXXIII.21. Эоценовые мезонихнды и современные хищные
а — Synoplotherium, собакоподобный мезонихид среднеэоценового возраста.
Длина тела около 1.2 м; б — голова гигантского мезонихида Andrewsarchus из
Монголии; в — голова аляскинского бурого медведя, самого большого ныне жи-
вущего хищного животного {б, в — нарисованы в одинаковом масштабе). Череп
медведя 46-48 см длиной; череп мезонихида 86 см длиной
ВЕЛИКОЕ РАЗНООБРАЗИЕ ПЛАЦЕНТАРНЫХ
543
Мезонихиды. Среди копытных выделяется особая группа - мезонихиды,
хищные копытные. По своему происхождению они связаны с арктоционид-
ными кондилартрами. Много лет их относили к креодонтам. Очевидно, что
это плотоядные животные, однако они не имеют признаков, присущих ис-
тинным плотоядным. Недавно было доказано, что мезонихиды — это часть
большого разнообразия парнокопытных.
Synoplotherium жил в течение среднего эоцена в Северной Америке. Это
был активный охотник, похожий на коротконогого волка с крупной головой
(фиг. ХХХШ.21). Andrewsarchus из позднего эоцена Азии имел череп 86 см
длиной, для сравнения: длина черепа аляскинского бурого медведя только
46-48 см (фиг. ХХХШ.21). Если говорить о размерах тела этого животного,
то его длина достигала 3.8 м, а высота около 1.8 м, когда оно стояло на четы-
рех лапах.
Современных мезонихид нет, однако, по палеонтологическим данным,
они дали начало очень разнообразной современной группе - китообразные.
МЛЕКОПИТАЮЩИЕ МОРЯ
Ранее мы выяснили, что рептилии, господствовавшие на Земле до млеко-
питающих, очень быстро образовали большое количество водных форм.
Освоение млекопитающими водных условий обитания также произошло
очень быстро, выделился даже особый отряд китообразных животных.
Самые древние остатки китов относятся к подотряду археоцета (Archaeo
ceti) и известны из отложений раннего эоцена Пакистана. Череп и зубы поз-
воляют предположить, что предками археоцетеса были мезонихиды, воз-
можно, питавшиеся рыбой и проводившие все больше и больше времени в во-
де. Кроме черепа этого самого раннего археоцета, другие части скелета не
известны. Вполне вероятно, что он мог выходить время от времени на сушу.
Среднеэоценовый археоцет был уже полностью водным животным. У него
заостряется рот, удлиняется тело, передние конечности приобретают форму
ласт, а задние настолько редуцируются, что полностью скрыты в теле. Щеч-
ные зубы имели двойные корни, а зубы в передней части рта были заострены
или гвоздеподобны. Ноздри передвинулись на верхнюю часть головы, при-
мерно на половину расстояния между ртом и глазами (фиг. ХХХШ.22).
Остатки Basilosaurus, также известного как Zeuglodon, найдены в поздне-
эоценовых отложениях южной части Атлантического побережья Северной
Америки. Basilosaurus достигал длины 16.8-21.3 м, имел относительно ко-
роткий череп и очень длинный хвост (фиг. ХХХШ.22). Хвостовые позвонки
были крупнее, чем позвонки короткого тонкого тела, их часто находят на
востоке Америки, в Африке, Европе и Новой Зеландии. Несколько малень-
ких, короткотелых китов-археоцет известно из олигоцена.
Позднее их место в мире заняли истинные киты, которые, вероятно, про-
изошли от неспециализированного археоцета. Ранние киты имели корот-
кие, коренастые тела, их ноздри были объединены в единое дыхало, которое
располагалось между или даже позади глаз. Некоторые рода обладали тре-
угольными зубами, подобными зубам акул, хотя их тела походили на
дельфинов. В течение олигоцена группа разделилась на зубатых (Odontoce-
ti) и усатых (Mysticeti) китов.
Зубатые киты, к которым относятся современные дельфиновые, нарвало-
вые, кашалотовые и клюворыловые, сохранили зубы и даже увеличили их
число в несколько раз относительно исходных - сорока четырех. Усатые ки-
544
КАМЕННАЯ КНИГА
Фиг. ХХХШ.22. Примитивные и специализированные киты — археоцеты
ты, наоборот, стали беззубыми и отцеживают мелких беспозвоночных плас-
тами “усов” - специализированной кожей, свисающей с крыши ротовой по-
лости. Усатые киты - самые крупные из всех ныне живущих животных. Со-
временный синий кит, например, достигает длины 30 м и весит 112.5 т.
Такие огромные динозавры достоверно не известны, хотя некоторые непол-
ные скелеты позволяют предположить, что в истории Земли могли сущест-
вовать такие гиганты (см. главу XXVII).
НЕПАРНОПАЛЫЕ КОПЫТНЫЕ
В конце палеоцена возникла группа копытных, у которых ось силы тяже-
сти проходила через третий центральный палец ноги. У этой группы, назван-
ной непарнокопытные, боковые пальцы ноги уменьшаются, а центральный
все больше развивается. В некоторых группах, подобно современной лоша-
ди, имеется только один, третий, функциональный палец ноги. Все, что ос-
тается от боковых пальцев, это рудименты костей.
ВЕЛИКОЕ РАЗНООБРАЗИЕ ПЛАЦЕНТАРНЫХ
545
Фиг. XXXIII.23. Непарнокопытные миоцена
а - Moropus, травоядное животное, чьи копыта стали когтями. Ископаемые ос-
татки халикотерия собраны из местонахождения Агата Спрингс, б - Dicerathe-
rium — миоценовый носорог, чьи остатки обильны в костеносных пластах около
г. Агата. Высота в плече приблизительно 102 см; два маленьких рога располага-
лись на носу
В дополнение к этому, скакательный сустав (лодыжка) задней ноги разви-
вался по пути уменьшения боковых движений. Такое развитие скакательно-
го сустава, - по сравнению с изменениями в суставе парнокопытных, сделало
непарнокопытных лучшими бегунами.
К отряду непарнокопытных относятся три подотряда. Первый подотряд -
анцилопода (Ancylopoda) - представлен исключительно вымершими живот-
ными; ко второму подотряду - гиппоморфа (Htppomorpha), - относятся брон-
тотерии и лошадиные и третий подотряд - цератоморфа (Ceratomorpha), объ-
единяет тапироидов и носорогообразных.
Халикотерии — когтистые непарнокопытные. Когда мы думаем о непар-
нокопытных, то представляем животных с копытами, ныне живущие лоша-
ди, тапиры и носороги все имеют их. Однако в отдельное семейство отряда не-
парнокопытных подотряда анцилопода были выделены животные, имею-
щие когти на ногах. Это - халикотерии. Когда впервые был найден череп, то
он был отнесен к непарнокопытным, а ноги - к гигантскому ящеру или не-
полнозубому. Более шестидесяти лет потребовалось, чтобы понять, что эти
части скелета принадлежали одному животному.
Одним из самых больших халикотериев был Moropus, скелеты которого
известны из костеностного пласта Агата Спрингс. Это существо имело голо-
ву, похожую на лошадиную, мощное тело и короткий хвост, как у тапира.
Передние ноги массивные, похожи на ноги носорога, а задние похожи на
ноги медведя, и на обоих мощные когти, которые являются отличительным
признаком группы (фиг. ХХХШ.23). Клыки и верхние резцы отсутствова-
ли. Щечные зубы были широкие и низкие, такими зубами можно было же-
вать только мягкую пищу. По одной из теорий, Moropus питался молодыми
побегами; сидел и тянул вниз ветви мощными передними ногами. Другая,
35 Каменная книга
546
КАМЕННАЯ КНИГА
Фиг. ХХХШ.24. Безрогие носороги
а — Hyracodon — быстро бегающий носорог, около 1.5м длиной. Лучше всего из-
вестен из отложений позднего эоцена—позднего олигоцена Северной Америки.
Возможно, достигал Европы в раннем олигоцене. Жил на сухих высокогорьях;
б - Metamynodon — болотный носорог, живший с позднего эоцена по поздний
олигоцен в Северной Америке и в раннем-позднем олигоцене в Азии. Длина те-
ла - 2.4—4.3 м
по мнению автора, более убедительная теория предполагает, что Moropus
питался в основном корнями и луковицами растений и использовал когти
для рытья. Они становились также оружием, когда на животное кто-то
нападал.
Носороги — рогатые и не очень. Сегодня носороги - большие, тяжелые жи-
вотные с крупными рогами. Но в раннем эоцене их предки были маленькими
и безрогими.
Некоторые носорогообразные типа гиракодона существовали в среднем
олигоцене Северной Америки. Это были маленькие животные без рогов со
стройными ногами, бегавшие по сухим нагорьям (фиг. ХХХШ.24). Metamy-
nodon, наоборот, был бочонкообразным животным 2.4-3 м длиной, близким
по внешнему виду к современному карликовому гиппопотаму, только более
крупный (фиг. ХХХШ.24). Он, несомненно, проводил большую часть време-
ни на реках и болотистых озерах, где взрослые особи крупного размера не бо-
ялись рыскающих крокодилов.
Сильно отличались от современных носорогов животные, которые жили в
олигоцене и миоцене. В 1911 году британский ученый, работавший на хол-
мах Балучистан (северо-запад Индии), нашел несколько костей существа,
которое, казалось, было носорогом, но по размеру крупнее любого известного
слона. Он назвал его Baluchitherium.
Намного больше костей этого животного было найдено Вальтером Гранге
из Американского музея естествознания, в 1922 году работавшим в Монго-
лии. Там он нашел три неполных скелета Baluchitherium, один из которых
включал череп. 365 фрагментов были собраны и отправлены в Нью-Йорк, где
ВЕЛИКОЕ РАЗНООБРАЗИЕ ПЛАЦЕНТАРНЫХ
547
Фиг. XXXIII.25. Buluchitherium, олигоценовый носорог из Азии. 5.5 м высоты
в плече
а — реконструкция; б — бедренная кость носорога в сравнении с человеком 1.8 м
роста
квалифицированный препаратор и его помощники потратили три месяца,
собирая фрагменты скелета. В итоге они восстановили самого большого носо-
рога и самое большое млекопитающее, которое нам известно из когда-либо
живших на суше.
В детском возрасте, когда гиппопотам кажется большим, а слон - гиган-
том, Baluchitherium кажется почти нереальным. Его голова была приблизи-
тельно 1.5м длины и выглядела нелепо маленькой на гигантском толстом те-
ле 5.5 м высоты в плечах (фиг. ХХХШ.25). Baluchitherium жил небольшими
стадами, питался листьями и молодыми побегами, поднимая голову и вытя-
гивая шею. Он мог легко объедать ветки деревьев на высоте 6.1м над землей.
Большинство ископаемых остатков носорогов найдено разрозненными и в
небольших количествах. Однако в 1877 году капитан Джеймс Кук, бойскаут
и работник фермы обнаружили около почты в г. Агата, штат Небраска, боль-
шое местонахождение ископаемых млекопитающих, содержащее остатки
тысяч индивидуумов. В раннем миоцене по сухим равнинам часто проно-
сились пыльные бури, в которых задыхались свиноподобные ореодонты
(фиг. XXXIII. 14), жавшиеся друг к другу для защиты. Потоки, превращав-
шиеся в наводнения после внезапных дождей, топили много животных и
уносили трупы умерших, лежащие на земле. Эти остатки отлагались на пес-
чаных отмелях, которые затем стали настоящими костными пластами, пере-
полненными сверху донизу остатками млекопитающих (фиг. XXXIII.26).
Скопления, найденные Джеймсом Куком, расположены на двух плоско-
вершинных холмах, по-видимому, песок, гравий и кости заполнили русло
древнего миоценового потока, который протекал приблизительно там же,
где и ныне существующая река Ниобрара. Никто не знает, сколько костей со-
держало местонахождение, потому что все, кроме двух маленьких холмов,
было разрушено эрозией. Но имеется четыре тысячи триста черепов и от-
35*
548
КАМЕННАЯ КНИГА
Фиг. XXXIII.26. Плита из костеностного пласта миоценового возраста место-
нахождения Агата Спрингс, штат Небраска. Она содержит приблизительно че-
тыре тысячи триста костей и черепов (фото предоставлено Американским музе-
ем Естественной Истории, Нью-Йорк)
дельных костей скелета в одной плите серо-желтого песчаника площадью
4.1 кв. м. На основании этих данных можно предположить, что один из двух
холмов должен содержать ископаемые остатки, принадлежащие приблизи-
тельно семнадцати тысячам скелетов. Этого достаточно, чтобы удовлетво-
рить потребности всех музеев на все оставшееся время.
Из тех семнадцати тысяч скелетов, по крайней мере шестнадцать тысяч
относятся к носорогам, названным Diceratherium (см. главу I, там изображен
отпечаток этого животного, сохранившийся в потоке лавы). Он весил не
больше среднего борова, хотя ноги у него были более длинные и более мощ-
ные (фиг. XXXIII.23). Самцы имели два маленьких рога, расположенных ря-
дом на носу, а самки были безрогими. Diceratherium обитал на раннемиоце-
новых равнинах запада Соединенных Штатов в стадах таких же больших,
какими были стада бизонов, насчитывавщие приблизительно 60 млн. особей
до их истребления европейцами.
Носороги известны в Северной Америке в течение всего миоцена и вымер-
ли лишь в среднем плиоцене. В Африке, Азии и Европе они продолжали про-
цветать весь плейстоцен.
Массивные бронтотерии. Полностью вымершим является семейство
бронтотерии, или титанотерии, подотряда гиппоморфа. Они были самыми
крупными жителями раннего олигоцена Азии и Северной Америки, до-
стигали длины 3.7-4.3 м, высоты 2.1-2.5 м и весили 4-5 т. По внешнему
виду походили на современных носорогов с мощными, крупными телами и
массивными ногами. Подобно носорогам, бронтотерии были непарнопалы-
ми. На задних ногах они имели по три пальца, несущие копыта, на перед-
них - по четыре. Голова была длинной, низкой и вогнутой. Два широких
тупых костных рога росли рядом на носу и были у самцов больше, чем у са-
ВЕЛИКОЕ РАЗНООБРАЗИЕ ПЛАЦЕНТАРНЫХ
549
Eotitanops,
51 см высотой,
ранний эоцен
Palaeosyops
97 см высотой
средний эоцен
Фиг. XXXIII.27. Этапы эволюции североамериканских бронтотериев
550
КАМЕННАЯ КНИГА
Фиг. XXXIII.28. Brontotherium platyceras, один из самых больших бронтотериев
Самки (б) были меньше самцов (д) и имели более короткие рога, в — череп самца
приблизительно 114 см длины
мок. Это было опасное оружие, подкрепленное весом их владельцев. Самцы
использовали их при защите от хищников и в поединках с конкурентами.
Самые древние раннеэоценовые бронтотерии известны из Азии. Они были
небольшими животными, всего 30-51 см высотой, с черепами без каких-ли-
бо следов от рогов. В среднем эоцене, когда достигли Северной Америки,
бронтотерии стали крупными животными, приблизительно 1 м высотой и у
них начали развиваться рога, в виде низких шишек выше глаз. Рога, также
как и размер тела, стали крупнее в позднем эоцене. Один из самых ранних
видов, известных в олигоцене, был 3 м длиной и 1.5 м высотой или такого же
размера, как крупный носорог. Настоящие гиганты, появившиеся несколь-
ко позже, были массивными животными с большими рогами и низкими, ши-
рокими черепами, под стать которым было и крупное тело (фиг. XXXIII.27).
Все бронтотерии, по-видимому, питались молодыми побегами и листья-
ми. Самый мелкий вид питался растениями немного выше травы. Гиганты
типа Brontops и Brontotherium (фиг. XXXIII.27, XXXIII.28) срывали листья
с высоких кустов и низких деревьев губами, их резцы были слишком малень-
кими для использования или вообще исчезли. Щечные зубы преобразова-
лись в крупные давящие и размалывающие зубы, вдвое шире в верхней че-
люсти, чем в нижней (фиг. XXXIII.29). Их размер показывает, что они могли
перетирать грубую пищу, хотя не могли долго противостоять песку и жест-
ким травам.
Мозг бронтотериев увеличился в размере, однако был значительно мень-
ше мозга современного носорога. Бронтотерии были менее интеллектуаль-
ны, они имели меньшее количество складок в мозжечке, и переднее части
ВЕЛИКОЕ РАЗНООБРАЗИЕ ПЛАЦЕНТАРНЫХ
551
Верхний (слева) и два нижних (справа)
коренных зуба Eotitanops, х0.8
Нижняя челюсть Broritotherium,
около 64 см длиной
Фиг. XXXIII.29. Отдельные коренные зубы и нижняя челюсть Brontotherium и
Eotitanops
больших полушарий были менее раздуты. По своим относительным разме-
рам мозг был маленьким (фиг. ХХХШ.27). 4-тонный бронтотерий имел
мозг, немного больший по размеру, чем два сжатых кулака человека. Совре-
менный носорог с мозгом подобного размера весит немного более 2 т. Некото-
рые ученые предполагают, что слабые умственные способности плюс зубы,
которые не могли справляться с устойчивой травяной диетой, объясняют,
почему бронтотерии вымерли в конце раннего олигоцена.
Лошади. Большинству из нас лошадь наиболее знакома из непарнокопыт-
ных. Самый ранний предок лошадей был обнаружен дважды, с сопутствую-
щей путаницей в именах. Первые ископаемые остатки найдены в Европе и
названы Hyracotherium, из-за сходства восстановленного внешнего облика
животного с кроликом или даманом Старого Света. Остатки из Америки бы-
ли богаче и лучше интерпретируемы; их описали как Eohippus или “лошадь
рассвета”. В научных работах это животное называют Hyracotherium, т.к.
это название было предложено первым и имеет приоритет. Название эогип-
пус знакомо и хорошо обосновано, однако является синонимом первого на-
звания и не используется в научной литературе.
Предок первых лошадей. Подобно наиболее перспективным предкам,
гиракотерии (эогиппусы) были маленькими и неспециализированными жи-
вотными. Некоторые виды имели всего 25 см высоты в плече и весили
3.6-4.1 кг, по размеру они были близки современным домашним кошкам.
Другие достигали 51 см высотой и весили приблизительно в четыре раза
больше. У всех была изогнута дугой спина и задняя часть тела поднята выше
передней, что придавало животным почти кроликоподобный облик
(фиг. XXXIII.30). Можно предположить, что из них получились бы прекрас-
ные домашние животные.
Такая идея опровергается, когда мы исследуем мозг гиракотерия. Он ма-
ленький, примитивный и, вероятно, не намного более развит, чем у фенако-
дуса. Череп походил на череп кондилартра расположением глаз, помещен-
ных близко к середине морды, и строением орбит, которые были открыты
552
КАМЕННАЯ КНИГА
Фиг. ХХХШ.ЗО. Hyracotherium или Eohippus (“лошадь рассвета”) из раннего
эоцена Европы и Северной Америки. Этот древнейший известный предок лоша-
дей был около 46 см длиной и 23 см высотой в плече
сзади (фиг. ХХХШ.34а). Имелось сорок четыре зуба примитивного строения
с маленькими промежутками между резцами и клыками, а также между
клыками и предкоренными зубами. Зубы перемалывающего типа низкоко-
ронковые, пригодные только для еды мягких листьев и мясистых плодов
(фиг. XXXIII.35а). На передних ногах было по четыре пальца, каждый с кро-
шечным копытом; на задних ногах только по три пальца, поэтому гиракоте-
рий относится к непарнокопытным. Большая часть веса животного приходи-
лась на подушечки подошв ног, как у собак, а не на копыта.
Разнообразие семейства лошадиных. Гиракотерии вымерли в Европе в те-
чение среднего эоцена. Однако они оставили потомков, названных палеоте-
рии, которые имели большие тела, короткие ноги, длинные шеи и большие
головы, подобно тапирам (фиг. ХХХШ.31). Копыта были хорошо развиты, и
некоторые виды выработали высококоронковые перемалывающие зубы с це-
ментом. Палеотерии распространились в Западную Азию и дожили до ранне-
го олигоцена. Самый поздний вид был такого же размера, как современные
носороги, и вдвое тяжелее любой современной лошади.
В Северной Америке гиракотерии дожили до конца эоцена. Их средне- и
позднеэоценовые потомки потеряли боковые пальцы на задних ногах, а
предкоренные зубы стали похожими на коренные. Эти варианты были удос-
тоены двух имен: Orohippus и Epihippus, но только ученые могут отличить их
от предкового рода.
Две примитивные трехпалые лошади. Большие изменения произошли в
эволюции лошадей в олигоцене. Они отражены в строении Mesohippus. Хотя
виды различались по размеру, типичный мезогиппус был около 51 см высо-
ты и 1 м длины, или приблизительно такого же размера, как собака колли
(фиг. ХХХШ.32). Череп узкий и довольно глубокий, с мордой лошадиного
типа, но с мелкой нижней челюстью. Глаза больше сдвинуты к задней части
морды, чем у гиракотерия; орбиты все еще открыты сзади, а мозг удивитель-
но большой для млекопитающего олигоцена (фиг. XXXIII.346). В строении
ВЕЛИКОЕ РАЗНООБРАЗИЕ ПЛАЦЕНТАРНЫХ
553
Фиг. XXXIII.31. Palaeotherium, родственник лошадей, живший в Европе со
среднего эоцена до раннего олигоцена. Высота около 1.2 м
Фиг. XXXIII.32. Mesohippus, трехпалая лошадь олигоценового возраста. Дли-
на тела около 102 см. Ископаемые остатки найдены в местонахождении Белая
река, штат Южная Дакота, США
зубной системы четко выделяется диастема - промежуток между клыками и
предкоренными зубами. Щечные зубы, все еще низкокоронковые, показы-
вают, что Mesohippus питался листьями и, возможно, мясистыми плодами
(фиг. ХХХШ.356).
Прогрессивные и архаичные черты также присутствуют в строении ног ме-
зогиппуса. Передние конечности потеряли один палец, его остаток - щепа -
скрыт под кожей. Средний палец каждой ноги стал самым большим, но еще
все три пальца имели хорошо развитые копыта. Большая часть веса живот-
ного все еще приходилась на подушку подошвы ноги, как у собаки и как у
гиракотерия. Задняя половина животного была дугообразно изогнута, зад-
554
КАМЕННАЯ КНИГА
Equus,
1.5 м высоты в плече. Лошади произошли
в Северной Америке в плиоцене
и вымерли там в конце плейстоцена.
В Азии, Африке и в Европе продолжают существовать
сейчас и расселены европейцами на всех континентах
itt.
да* Й!
Pliohippus,
первая однопалая лошадь.
Средний-поздний миоцен
Северной Америки
Mesohippus,
примитивная
трехпалая лошадь.
Ранний-средний олигоцен
Северной Америки
Merychippus,
продвинутая трехпалая лошадь,
102 см высотой.
Только один палец на ноге
соприкасается с землей.
Ранний-средний миоцен
Северной Америки
Hyracotherium или Eohippus,
самый древний из известных лошадей.
Ранний эоцен Европы, Азии
и Северной Америки
Фиг. XXXIII.33. Эволюционная линия лошадей
ВЕЛИКОЕ РАЗНООБРАЗИЕ ПЛАЦЕНТАРНЫХ
555
Hippidion, южноамериканская лошадь,
плейстоцен
Hyracotherium, ранний эоцен
Фиг. XXXIII.34. Изменение строения черепов в семействе лошадиных
Фиг. ХХХШ.35. Изменение строения коренных зубов лошадей. Коронка зуба
становится выше и эмаль (показанная черным) образует все более сложные
складки. Вид с жевательной поверхности и сбоку зуба
ние ноги были длиннее, чем передние. Mesohippus, очевидно, бегал, как
олень, подпрыгивая.
Mesohippus жил почти одновременно с Miohippus, который был большим
и несколько более лошадеподобным животным, его потомки разделяются на
четыре основные группы. Две из них сохраняют низкокоронковые зубы,
дающие возможность ощипывать молодые побеги, и используют для пере-
движения три пальца на ногах. Некоторые виды этих групп известны из
556
КАМЕННАЯ КНИГА
Азии и Европы. В Северной Америке к ним относится Megahippus (“боль-
шая лошадь”) по размеру близкая к африканскому носорогу, который 1.5 м
высотой, 3.4-3.7 м длиной и весит около 1.8 т. Такие гиганты контрастиру-
ют с третьей группой потомков Miohippus — Archaeohippus. Члены этой
группы эволюционировали в карликов не крупнее гиракотерия среднего
размера.
Прогрессивный Mery chippus. Четвертой среди потомков Miohippus была
группа, которую мы называем прогрессивной просто потому, что от нее про-
изошли современные лошади (фиг. XXXIII.33). Наиболее важный член этой
группы - мерикгиппус - появился в раннем миоцене и дожил до позднего ми-
оцена.
При сравнении Merychippus с более ранним родом выявляются следую-
щие отличительные черты строения скелета:
размер приблизительно такой же, как у шотландского пони. Высота в пле-
чах 1 м;
туловище лошадеподобно по своему строению, с почти прямой задней ча-
стью. Некоторые коренастые виды были медленными бегунами, другие -
стройны и быстры;
глаза располагались далеко сзади на голове, как у современных лошадей.
Орбиты замкнуты позади глаз костной пластиной;
кости предплечья плотно соединены, что исключает возможность боко-
вых движений. Часть малой берцовой кости редуцирована;
средний палец как передней, так и задней ног большой, с широким копы-
том, на которое приходился основной вес тела. Вторые и четвертые пальцы
ног очень коротки.
Изначально у мерикгиппуса были низкокоронковые зубы, но в процессе
эволюции они постепенно преобразовались в высококоронковые. Эмаль
предкоренных и коренных зубов сильно складчатая и чередуется с цементом
(фиг. XXXIII.35в). Зубы росли большую часть жизни животного, их рост
прекращался с формированием корня зуба только у животного среднего воз-
раста. Челюсти двигались из стороны в сторону, совершая размалывающее
движение, а также вверх и вниз. Высококоронковые зубы дали возможность
Merychippus отказаться от поедания молодых побегов и перейти на жесткую
траву. Широкое распространение травы в миоценовое время позволило трех-
палому пастбищному животному перейти жить в прерии и на равнины, где
длинные ноги и быстрый бег обеспечивали спасение от хищников. Потомки
Merychippus распространились в Северной и Южной Америке, Азии, Европе
и Африке в самом конце миоцена и в плиоцене. Одна из африканских форм
дожила до плейстоцена.
От трехпалых к однопалым. Дальнейшие изменения строения скелета
были отмечены у Pliohippus средне-позднемиоценового возраста. У некото-
рых видов маленькие боковые пальцы ног сильно укоротились, у других
они уменьшились до крошечных костей. Плиогиппус имел широкое тело и
размалывающие зубы (фиг. XXXIII.35г), которые имели высокую корон-
ку, клыки стали маленькими, а диастема расширилась. В конце плиоцена
эти изменения привели некоторых североамериканских Pliohippus к со-
временным лошадям (Equus) (фиг. ХХХШ.ЗЗ, ХХХШ.34г, ХХХШ.35д).
В Южной Америке другая ветвь Pliohippus дала начало коротконогим
лошадям рода Hippidion, чей череп имел удивительно длинные носовые
кости, которые поддерживались рядом хрящей, не сохранившихся в иско-
паемом состоянии (фиг. XXXIII.34в). Некоторые из этих животных были
ВЕЛИКОЕ РАЗНООБРАЗИЕ ПЛАЦЕНТАРНЫХ
557
распространены в пампасах, другие, очевидно, жили на восточных
склонах Анд.
Equus, современная лошадь. Хотя род эквус возник в Северной Америке в
плейстоцене, он быстро распространился по другим континентам и развился
в различные подрода и большое количество видов. Оставшиеся на родине жи-
вотные в среднем достигали 102 см высоты, были и близкие по размеру к со-
временной бельгийской лошади, которая достигает 165 см высоты в плече и
весит около тонны (0.9 т). Лошади в изобилии были распространены на Боль-
ших Равнинах центра Северной Америки. Все лошади Северной Америки
вымерли приблизительно в конце плейстоцена. Мигрировав в Азию в раннем
плиоцене (по нашему мнению, несколько позже - в самом конце плиоцена),
они быстро распространились в Африке и в Европе, где существуют до на-
стоящего времени. Намного позже уже одомашненные лошади были завезе-
ны европейцами в Северную Америку, и сейчас лошади живут на всех кон-
тинентах.
Снижение численности диких лошадей. Огромные стада лошадей жили в
Европе, Азии и Африке в течение плейстоцена. В Америках они были рас-
пространены от Аляски до Магеланова пролива. Лошади вымерли в Новом
Свете, затем стали редкими и совсем исчезли в Европе, оставаясь в большом
количестве только на равнинах Африки и некоторое время в Азии. Сейчас ос-
тавшиеся в живых дикие лошади представлены африканским ослом, тремя
видами зебр, тремя видами куланов и лошадью Пржевальского. Азиатские
виды могут сохраниться только в зоопарках. Домашние породы, по-видимо-
му, произошли от вымерших азиатских лошадей, а также от африканского
осла.
Такое практически повсеместное вымирание лошадей не совсем понятно.
Правда, люди уничтожили многие современные виды, другие страдали от
вытеснения с традиционных пастбищ домашними породами. Но эти факто-
ры не объясняют снижение количества видов, а затем и полное вымирание
лошадей в течение позднего плейстоцена в Северной Америке. Эту загадку
вымирания еще больше усугубляет тот факт, что домашние лошади Старого
Света одичали и стали распространенными в Северной и Южной Америке по-
сле того, как они были повторно завезены европейцами.
ЮЖНОАМЕРИКАНСКИЕ КОПЫТНЫЕ - ДЕТИ ИЗОЛЯЦИИ
Большую часть кайнозойской эры Южная Америка была полностью окру-
жена водой, как Австралия сегодня. Очевидно, некоторые копытные пересе-
лились на континент в палеоцене прежде, чем сухопутные связи с внешним
миром были полностью прерваны. От этих первых поселенцев произошло не-
сколько уникальных отрядов южноамериканских животных.
Нотоунгуляты. Подавляющее большинство южноамериканских копыт-
ных относится к отряду нотоунгуляты (Notoungulata). Их распространение
не ограничено этим континентом, но история развития связана именно с
Южной Америкой. Один род известен из Северной Америки, несколько ро-
дов — из палеоцена Азии. Все же в основном это южноамериканская группа.
Наиболее распространенным в кайнозое является один из последних нотоун-
гулят — Toxodon, плиоценовая и плейстоценовая форма. У него массивная го-
лова, мощное тело и трехпалые конечности, которые кажутся слишком ма-
ленькими, чтобы удерживать это огромное тело. Toxodon, возможно, был
полуводным животным или питался молодыми побегами подобно бронтоте-
558
КАМЕННАЯ КНИГА
Фиг. ХХХШ.36. Два плейстоценовых копытных Южной Америки
а — Toxodon ростом более 1.2 м; б - Macrauchenia ростом 1.5 м в плече
рию (фиг. ХХХШ.36). Другой род нотоунгулят жил, подобно креодонтам или
современным кроликам и даже походил на них внешне.
Литоптерны. Второй очень распространенной группой копытных Южной
Америки были литоптерны (Litoptern). Они, вероятно, были потомками при-
митивных копытных, очень похожих на Phenacodus. В более позднее время
некоторые из них стали странными длинноносыми существами (семейство
макроухениды), в то время как другие были более лошадеподобными (семей-
ство протеротерииды), чем лошади того же геологического возраста.
Это высказывание звучит парадоксально, но оно имеет анатомический
смысл. Термином “лошадь” мы обозначаем млекопитающее, которое имеет
один крупный палец на каждой ноге, закрытый копытом. Лошади миоцена
и плиоцена имели по три пальца на ноге, и хотя животные могли ходить
только на одном из них, два других присутствовали и имели копыта. Миоце-
новый же литоптерн Thoatherium имел однопалые конечности с остатками
двух очень маленьких боковых костей. С легким телом и длинными строй-
ными ногами это животное, вероятно, бегало также стремительно, как совре-
менная антилопа (фиг. XXXIII.37).
Второе семейство литоптерн — макроухениды — достигло расцвета в плио-
цене и плейстоцене и представлено родом Macrauchenia. Лучше всего извест-
ный вид этого рода имел широкое тело приблизительно 1.5 м высоты, с мощ-
ными ногами и тремя пальцами на конечностях. Шея и голова были бы похо-
жи на шею и голову верблюда, если бы не короткий, мускулистый хобот
(фиг. ХХХШ.36).
Другие южноамериканские копытные. Известно еще три отряда южно-
американских копытных. Все они были довольно крупными животными и
проявляли черты сходства внешнего облика с некоторыми неродственными
формами из других частей мира. Пиротериев можно сравнить со слонами, ас-
трапотериев с древними носорогами, а ксенонгулят с пантодонтами
(фиг. XXXIII. 7, XXXIII.8) и дицератами (фиг. XXXIII. 12). По сравнению с
нотоунгулятами и литоптернами к этим трем отрядам относится намного
меньше видов, а ксенунгуляты представлены всего двумя. Все они появи-
ВЕЛИКОЕ РАЗНООБРАЗИЕ ПЛАЦЕНТАРНЫХ
559
Фиг. ХХХШ.37. Thoatherium, “ложная лошадь”, 43 см высотой из миоцена
Южной Америки
а — реконструкция, б — скелет стопы задней однопалой конечности
лись в палеоцене или в эоцене и вымерли в конце миоцена. Напротив, ното-
унгуляты и литоптерны сохранились до плейстоцена.
ХОБОТНЫЕ - ДЛИННЫЕ НОСЫ И БИВНИ
История хоботных - это повторяющиеся миграции с эволюционными из-
менениями, которые превратили маленькое, непримечательное животное в
гиганта с бивнями и хоботом, весящего 5, а возможно, и 7 т.
Предки и нетипичные слоны. Одним из самых древних известных членов
отряда хоботных является Moeritherium, относящийся к самостоятельной
группе меритерии. По строению черепа и зубов он также близок к сиренам и
десмостилидам. Возможно, эта форма связывает все три отряда: хоботные,
сирены и десмостилиды. Название Moeritherium животному было дано пото-
му, что его ископаемые остатки были впервые найдены на берегу древнего
озера Мерис, чье дно теперь стало плодородной долиной в Египетской облас-
ти Фаюм. Moeritherium был приблизительно 1.5 м длиной и 64 см высотой в
плече, с длинным, мощным, округлым телом, крепкими и короткими пяти-
палыми конечностями. Низкий, широкий и тупой череп, сильно сдвинутые
вперед глаза и высоко расположенные ноздри напоминают морду тапира
(фиг. XXXIII.38). Зубы низкокоронковые перетирающего типа. Четыре рез-
ца большие и можно отметить, что рост бивней уже начался. Это животное,
возможно, было частично водным, так как форма его тела подобна форме те-
ла гиппопотама, а остатки очень часто находят в морских дельтовых отложе-
ниях.
Moeritherium не был путешественником, он жил на низменностях Север-
ной Африки в течение позднего эоцена и раннего олигоцена.
Группой, уклонившейся от общего пути развития хоботных, является по-
дотряд динотериоиды. Deinotherium был животным слоновьего телосложе-
560
КАМЕННАЯ КНИГА
Фиг. ХХХ1П.38. Moeritherium — древнейший из известных родственников сло-
новых. Приблизительно 60 см высотой в плече; жил в болотистых областях Се-
верной Африки в течение позднего эоцена и раннего олигоцена
Фиг. ХХХШ.39. Череп Deinotherium с нижней челюстью. Его длина 122 см.
Deinotherium был широко распространен в Старом Свете в миоцене, а в Африке -
в плейстоцене
ния и размера, но все же не истинным слоном. Один из миоценовых видов
имел низколобый череп приблизительно 122 см длиной, с квадратной челю-
стью и выступающими носовыми костями, которые поддерживали хобот.
Перетирающие зубы низкокоронковые, примитивного строения. Верхняя
челюсть была короткая, резцы отсутствовали, а нижняя челюсть изгибалась
наружу и вниз, имелось два коротких, изогнутых бивня (фиг. ХХХШ.39).
Deinotherium обитал в течение миоцена в Африке, Азии и в Восточной Евро-
пе. В плейстоцене он вымер в Европе и в Азии, но еще некоторое время про-
должал существовать в тропической Африке. Нет никаких данных, что
представители этой группы когда-либо достигали Америки.
Разнообразие мастодонтов. Богатый палеонтологический материал по хо-
ботным верхнего эоцена и нижнего олигоцена Северной Африки относится к
роду Palaeomastodon. Некоторые виды были тапироподобными животными
ВЕЛИКОЕ РАЗНООБРАЗИЕ ПЛАЦЕНТАРНЫХ
561
Фиг. ХХХШ.40. Эволюционные линии хоботных
36 Каменнай книга
562
КАМЕННАЯ КНИГА
0.9—1.2 м высотой, другие достигали высоты 1.8 м и весили пару тонн. Все
имели мощные, прямые ноги, высокие сзади черепа и короткие хоботы
(фиг. XXXIII.40). И верхние и нижние челюсти несли бивни, но верхние
бивни были длиннее и загнуты вниз. Зубы перетирающего типа с низкими
коронками, тупыми гребнями и несколькими корнями.
В течение последующих 5 млн. лет потомки Palaeomastodon достигли Ев-
ропы, пересекли Азию и распространились в Северной Америке, но осталось
немного окаменелостей по маршрутам их путешествий. Пока животные
путешествовали, они также эволюционировали, становясь слоноподобными
по размеру и форме, однако сохранили низкокоронковые гребенчатые зубы.
Некоторые сохранили четыре бивня, другие потеряли бивни на нижней
челюсти.
От палеомастодонтов произошло несколько групп хоботных. Одну из них
ученые долгое время называли мастодонтами. Название Mastodon ранее
применялось к целому ряду животных, которые жили в течение миоцена и в
более поздние эпохи. В процессе изучения животные получали другие родо-
вые названия, а название Mastodon стало относиться лишь к двум видам
плейстоценового и голоценового возраста. Позже было установлено, что для
этих видов родовое название Mammut более старое и по правилам зоологиче-
ской номенклатуры его следует использовать для научных целей. Таким об-
разом, название мастодон стало синонимом названия рода маммут, так же
как эогиппус синонимом родового названия гиракотерий.
Животное, описанное как Mammut americanum было крупным, его зубы
(которые часто находят в ископаемом состоянии) имели отдельные корни и
тупые гребни. Два крупных бивня были развиты только на верхней челюс-
ти, на нижней челюсти они исчезли или стали рудиментарны. Тело по-
крыто рыже-коричневыми волосами, и хотя на каждой ноге имелось по
пять пальцев, большая часть веса приходилась на эластичную подушку.
Самки были мельче, около 2.1 м, а крупные самцы достигали 2.9 м
(фиг. XXXIII.40).
Это животное по старинке называют американским мастодонтом. Оно
было распространено от Новой Англии и Флориды до Мексики, Калифорнии
и Аляски. Находки ископаемых зубов указывают, что мастодонты пересек-
ли северную Азию и достигли равнин Восточной Европы. В Северной Амери-
ке Mammut americanum жил и после Ледникового периода и являлся объек-
том охоты древнего человека. Животное, чьи остатки найдены в штате
Огайо, умерло всего восемь-девять тысяч лет назад.
Хотя мастодонты имели бивни и длинные хоботы, они не дали начало на-
стоящим слонам.
Происхождение гомфотериид. Вторая ветвь развития отряда хоботных,
берущая свое начало от палеомастодонтов, дала начало семейству гомфоте-
рии. Родоначальником этого семейства является род Phiomia. Он жил в Егип-
те в течение олигоцена, был 135 см высотой, с вытянутой мордой и хоботом,
длинной нижней челюстью и очень короткими бивнями (фиг. XXXIII.40).
Коренные зубы очень специфичны по своему строению, они разделены на
гребни и дополнительные бугры. Если бы не их размер, то они были бы очень
похожи на коренные зубы свиньи.
От Phiomia произошли Gomphotherium (фиг. XXXIII.40) и Trilophodon.
Имена здесь не очень важны. Эти рода в основном отличаются строением зу-
бов и нижних челюстей. Зубы стали более сложными, а нижние челюсти
более длинными - до 1.8 м. Верхние челюсти укоротились, а бивни, которые
они несли, стали длиннее, чем у Phiomia. Некоторые виды были только
ВЕЛИКОЕ РАЗНООБРАЗИЕ ПЛАЦЕНТАРНЫХ
563
Нижняя челюсть Gnathobelodon,
“слона-ложки’’ из позднего миоцена
Amebelodon, “слон-совок”,
нижняя челюсть длиной более 2 7м
Житель Северной Америки
в позднем миоцене
Platybelodon. “слон совок",
нижняя челюсть более 91 см длиной.
Ранний миоцен Африки.
Расселился в Азии и в Европе
в среднем миоцене, а в конце
X. среднего миоцена появился
в Северной Америке
Фиг. XXXIII.41. Копатели ила
1.8 м высотой, а другие достигали 2.4 м и были близки по размеру к слонам
из цирка.
Отклоняющиеся от нормы формы. Копатели ила. Род Gomphotherium,
очевидно, произошел в Африке, но вскоре расселился по Европе, Азии и
Северной Америке. С ним распространился родственный род, названный
Platybelodon. У представители этого рода нет верхних бивней, нижние че-
люсти сужены перед щечными зубами, но затем расширяются для разме-
щения двух широких, плоских бивней, образующих огромный “совок”
(фиг. XXXIII.41). Этим “совком” платибелодон вырывал корни из болот. Ко-
роткий хобот удерживал корни на “совке”, в то время как длинный язык от-
делял пищу от грязи и переправлял ее дальше в рот для жевания.
Остатки Platybelodon были обнаружены на Кавказе, находка его остатков
в начале 1920-х годов в Монголии стала неожиданностью. Затем близкие по
строению челюсти и черепа были найдены наряду с мастодонтами на северо-
западе Небраски и названы Amebelodon и Gnathobelodon. Амебелодон имел
чрезвычайно длинные нижние челюсти, несущие плоские бивни, более
длинные, но более узкие, чем у Platybelodon (фиг. XXXIII.41). У гнатобело-
дона не было нижних бивней, но нижняя челюсть сформировалась в виде во-
гнутой, тонко заостренной “ложки”, которая была очень удобна для корчева-
ния (фиг. XXXIII.41).
36*
564
КАМЕННАЯ КНИГА
Стегодон и настоящие слоны. Предком современных слонов, относящих-
ся к отдельному семейству слоновые, вероятно, был ранний Gomphotherium.
В Африке от него произошли потомки, чьи черепа стали короткими, но высо-
кими, а нижние челюсти укороченными. Их предкоренные зубы уменьши-
лись, а коренные увеличились в размере и на них развились эмалевые греб-
ни, частично закрытые цементом. Молодые зубы начали выдвигаться на-
клонно вместо роста прямо вверх или вниз, и только восемь могли
использоваться в одно время.
Первым результатом этих изменений был Primelephas, известный из позд-
него миоцена Восточной и Центральной Африки. В плиоцене и плейстоцене
Азии известны находки еще одного слона Stegodon, примитивного по строе-
нию зубов (фиг. XXXIII.42). Постепенные изменения в строении черепа,
нижних челюстей и зубов привели к образованию от Primelephas трех родов:
Mammuthus, Loxodonta и Elephas. Они распространились в Азии, Европе,
Африке и Северной Америке.
Мамонты {Mammuthus}. Эти вымершие слоны плиоцена и плейстоцена
были описаны под несколькими родовыми названиями, но теперь объеди-
нены в род Mammuthus, который не следует путать с Mammut. Черепа ма-
монтов скошены назад и более высокие, чем у локсодонтов. Бивни направ-
лены вниз и вперед в форме арки, очень большие. У старых самцов они ча-
сто изгибались так сильно, что загибались обратно к телу
(фиг. XXXIII.43). У некоторых примитивных европейских и азиатских ви-
дов, типа южного мамонта (Mammuthus meridionalis), коренные зубы со-
держат только десять—двенадцать пластин, а у прогрессивных видов коли-
чество пластин третьего коренного зуба от двадцати шести до тридцати.
Имелось по четыре копыта на задних ногах, но не ясно, по четыре или по
пять копыт было на передних ногах. Если их было пять, то одно копыто бы-
ло очень маленьким.
Самый большой североамериканский мамонт, названный Mammuthus im-
perator жил на юге и юго-западе Соединенных Штатов. Это один из двух ви-
дов, известных из смоляных ям, описанных в главе XXXIV. Другой, называ-
емый М. columbi, был распространен к северу, в умеренных областях. Ма-
монты этого вида или подвида достигали высоты 3.2 м в плече, в отличие от
крупных самцов М. imperator, достигавших высоты 4.1 м.
Хотя ископаемые остатки колумбийского мамонта и его родственников
превосходят численностью находки других американских видов, эти живот-
ные менее широко известны, чем Mammuthus primigenius. Этот мамонт рас-
пространился из Западной Европы в Азию и Северную Америку. Он был не
очень крупным животным; самцы достигали 2.7 м высоты в плече, а голова
была на 20-30 см выше. Бивни большие и сильно изогнутые, до 4.9 м в дли-
ну. Передние ноги длиннее задних, туловище сильно наклонено к хвосту
(фиг. XXXIII.43). Шерсть имела два цвета, подшерсток серого цвета сверху
прикрывался длинными, грубыми красновато-коричневыми волосами.
Шерсть защищала животное от дождя, снега, холода, и это объясняет, поче-
му мамонт был способен жить комфортно только в нескольких милях от лед-
ника.
Mammuthusprimigenius - один из тех редких ископаемых животных, чьи
мягкие части известны, т.к. замороженные тела целиком с волосами, кожей
и высушенной кровью были найдены на Аляске и в Сибири. Его внешний об-
лик запечатлен на картинах и в статуэтках, сделанных обитателями пещер
Европы, которые заманивали животных в ямы-ловушки и убивали камен-
ными кувалдами.
ВЕЛИКОЕ РАЗНООБРАЗИЕ ПЛАЦЕНТАРНЫХ
565
Elephas, современный азиатский слон.
Бивни относительно маленькие,
высота 2.4-3.0 м в плече; реконструкция
Коренной зуб
Коренной зуб
Loxodonta, современный африканский слон.
Этот род в плейстоцене также жил в Европе.
Высота от 0.9 до 4.3 м в плече; реконструкция
Stegodon производит впечатление
примитивного слона. Жил в Азии,
в плейстоцене;реконструкция
Фиг. ХХХШ.42. Три стадии эволюции слонов. Происходит увеличение разме-
ра тела и усложнение коренных зубов. У современных азиатских слонов
наблюдается редукция бивней
566
КАМЕННАЯ КНИГА
Фиг. ХХХПЕ43. Мамонт (Mammuthus primigenius). Этот вид был распростра-
нен в Европе, Азии и Северной Америке, достигал 2.7 м в плече
Локсодонты (Loxodonta). Эта группа появилась в плиоцене и включает со-
временных африканских слонов - Loxodonta africana и. их ископаемых род-
ственников, Череп локсодонта не очень высокий, щечные зубы относительно
низкие, с нерегулярными ромбовидной формы пластинами, количество ко-
торых редко больше десяти. Бивни длинные, но не сильно изогнуты, задняя
часть туловища ниже плечей и резко склонена к хвосту; уши очень большие.
Передние ноги длиннее задних, на передних ногах по четыре пальца, на зад-
них - только по три (фиг. XXXIII.42).
Некоторые локсодонты очень крупные, отдельные особи африканского
слона достигают 3.5 м высоты в плече, a Loxodonta antique из ледникового
периода достигал 4.3 м. С другой стороны, виды, которые населяли Мальту и
другие средиземноморские острова, были карликами, самый маленький был
немного больше 90 см высотой и весил не больше, чем крупная свинья
(фиг. ХХХШ.44). Они были намного меньше, чем современные африканские
лесные слоны-карлики, которые достигают высоты приблизительно 1.8 м.
Азиатские слоны (Elephas). Сегодня они обитают в юго-восточной Азии и
в восточной Индии. Единственный современный вид - Elephas maximus (ин-
дийский слон) знаком нам по зоопаркам и циркам. Ископаемые остатки ази-
атских слонов плиоценового и плейстоценового возраста известны также из
Африки и Европы. Череп очень короткий, глубокий и округлый на вершине,
часто с выступающим лбом. Бивни направлены вниз и меньше, чем у афри-
канских слонов. Многие самки не имеют бивней. Коренные зубы высокие и
широкие, последние имеют двадцать четыре сильно складчатые пластины.
Задняя часть туловища аркообразно изогнута позади плеч (фиг. XXXIII.42).
Ноги короткие и толстые, с пятью пальцами и эластичной подушкой, на ко-
торую приходится самая большая нагрузка при хождении, потому что она
ВЕЛИКОЕ РАЗНООБРАЗИЕ ПЛАЦЕНТАРНЫХ
567
Фиг. XXXIII.44. Реконструкция ископаемых карликовых слонов из плейсто-
ценовых отложений островов Средиземноморья (фото предоставлено музеем
Естественной Истории, Базель, Швейцария)
соприкасается с землей раньше и лучше, чем пальцы ноги. Имеется по пять
копыт на передних ногах и по четыре на задних, на пятом пальце тоже может
быть копыто, но очень маленькое. Головной мозг весит 4.5-5.0 кг и кажется
более развитым, чем у Loxodonta. Высота взрослого самца достигает
2.4-3.0 м в плече, а вес - 3.6-4.5 т.
ВОДНЫЕ РОДСТВЕННИКИ ХОБОТНЫХ
Так как Moeritherium являлся амфибиотическим животным, то возмож-
но, что от него или от его близких родственников произошли два отряда вод-
ных млекопитающих.
Сирены или водные растительноядные. По берегам Тетиса в раннем эоце-
не появились первые сирены. Они быстро распространились в теплых водах
всех океанов и крупных рек. В течение миоцена они достигли максимума
своего разнообразия, в это время жило около дюжины родов сирен. Сегодня
обитают только два рода: дюгони (Dugong) и ламантины (Trichechus). Третий
род - стеллерову морскую корову (Hydrodamalis) - человек полностью
истребил в восемнадцатом веке, всего лишь за двадцать семь лет после его
открытия.
Сирены - единственные млекопитающие, которые питаются исключи-
тельно водными растениями. Их передние конечности изменены в ласты и не
имеют ногтей. Задние конечности полностью редуцированы. Характерно,
что кости, особенно ребра, являются очень плотными и тяжелыми. Это поз-
воляет животному легко совершать глубокие погружения.
568
КАМЕННАЯ КНИГА
Фиг. ХХХШ.45. Palaeoparadoxia (десмостилид) из раннего миоцена Калифор-
нии. Длина скелета около 2.4 м (фото предоставлено D. Savage)
Десмостилиды, или “связанные столбы”. Много лет десмостилиды были
известны только по остаткам зубов, которые похожи на ряд коротких тру-
бок, как бы спаянных друг с другом, отсюда и название. Так необычны были
эти зубы, что ученые, первыми изучавшие их, предположили, что они при-
надлежат примитивным млекопитающим — мультитаберкулатам.
Впоследствии некоторый скелетный материал был найден в Японии и в
Калифорнии. Десмостилиды, как сейчас известно, были морскими млекопи-
тающими, напоминающими тюленей, с четырьмя хорошо развитыми конеч-
ностями (фиг. ХХХШ.45). Возможно, они плавали в мелкой воде и питались
моллюсками и ракообразными, раздавливая их между своими тяжелыми
столбообразными зубами.
Десмостилиды обитали только в циркум-тихоокеанской области. Они по-
явились там в раннем миоцене и вымерли к концу этой эпохи.
Фиг. XXXIII.46. Реконструкция панголина Eomanis из среднего эоцена Евро-
пы. Чешуи сохранились вместе со скелетом. Длина тела до 88 см, длина хвоста
до 80 см (Storch, 1978)
ВЕЛИКОЕ РАЗНООБРАЗИЕ ПЛАЦЕНТАРНЫХ
569
б Садовая соня
а Обыкновенный хомяк
д Мышь-малютка
е Четырехпалый тушканчик
Фиг. XXXIIL47. Современные мышеобразные
а — обыкновенный хомяк (Cricetus cricetus), длина тела 24—34 см, длина хвоста
3.5-6 см, распространен в Европе и в Западной Сибири; б - садовая соня
(Eliomys quercinus), длина тела до 14 см, длина хвоста до 12 см, распространена
в Европе, Северной Африке и на западе Малой Азии; в - полевка-экономка (Mi-
crotus oeconomus), длина тела 11-20 см, длина хвоста 2-9 см, распространена
на территории Европы, Северной Америки, Азии и Северной Африки; г - боль-
шая песчанка (Rhombomys opimus), длина тела 13-20 см, длина хвоста
13-16 см, встречается в южной Европе и Азии; д — мышь-малютка (Micromys
minutus), длина тела до 7 см, длина хвоста также до 7 см, распространена в Ев-
разии; е - четырехпалый тушканчик (Allactaga tetradactyla), длина тела
10-26 см, длина хвоста 16-31 см, встречается на юге Европы, в Азии и в Север-
ной Африке (рис. Е.П. Загозиной по книге В.Е. Соколова, 1977)
570
КАМЕННАЯ КНИГА
Фиг. XXXIII.48. Современные белкообразные и дикобразовые
а — белка-летяга (Glau.com.ys volans), длина тела 21-27 см, длина хвоста 9-18 см,
распространена в восточной части Северной Америки; б — азиатский бурундук
(Tamias sibiricus), длина тела 8-18 см, длина хвоста 6-14 см, распространен на
северо-востоке Европы и в Азии; в — полосатая белка (Funisciurus), длина тела
31 см, длина хвоста 10-20 см, встречается в Африке; г — шиншилла (Chinchilla
laniger), длина тела 22-38 см, длина хвоста 8-15 см, сохранилась только на се-
вере Чили; д - малайский дикобраз (Hystrix brachyura), длина тела 60-90 см,
длина хвоста 12—15 см. Обитает на отдельных островах Малазии (рис. Е.П. За-
гозиной по книге В.Е. Соколова, 1977)
ПАНГОЛИНЫ, ПАНЦИРНЫЕ МУРАВЬЕДЫ
Панголины сегодня — это покрытые панцирными пластинками специали-
зированные муравьеды, живущие в Африке и в южной Азии. В олигоцене
они также существовали в Северной Америке и в Европе. Где и от кого они
произошли, не известно. В течение 35 млн. лет существования они не сильно
изменились в своем внешнем облике (фиг. XXXIII.46).
ВЕЛИКОЕ РАЗНООБРАЗИЕ ПЛАЦЕНТАРНЫХ
571
Фиг. ХХХШ.49. Самый крупный современный грызун — водосвинка (Hydro-
choeridae hydrochaeris). Длина тела 100-130 см, высота в плечах 50 см. Распро-
странена в Южной Америке (рис. Е.П. Загозиной по книге В.Е. Соколова,
1977).
ГРЫЗУНЫ, БОЛЬШАЯ ИСТОРИЯ УСПЕХА
Наиболее успешным отрядом млекопитающих по числу родов и видов яв-
ляются грызуны. Их количество большее, чем всех других млекопитающих
вместе взятых. Перечислим лишь некоторые, наиболее знакомые нам семей-
ства: беличьи, летяговые, бобровые, хомяковые, полевки, песчанки, мыши-
ные, соневые, мышовки, тушканчиковые, дикобразовые, свинковые, нутри-
евые, шиншиловые и другие (фиг. XXXIII.47, XXXIII.48, ХХХШ.49).
Парадоксально, но происхождение такого многочисленного отряда и взаи-
моотношения семейств внутри неясны.
Несомненно, грызуны произошли от некого примитивного насекомоядно-
го в течение позднего мела или палеоцена. Однако самые древние известные
грызуны Paramys и Franimys из позднего палеоцена Северной Америки так
заметно отличаются от всех других плацентарных, что они дают не больше
ключей к разгадке, чем более поздние грызуны.
Сразу после своего появления грызуны быстро радиировали и рассели-
лись по всем континентам, кроме Антарктиды. Подобно приматам, они до-
стигли Южной Америки в раннем олигоцене и образовали формы, ограни-
ченные почти исключительно этим континентом, например, морские свин-
ки. Наиболее интересный представитель этой группы - гвинейская свинья.
Мы обычно представляем себе грызунов как млекопитающих маленького
или среднего размера, однако плейстоценовый кавиоморф из Аргентины -
Telicomys - был близок по размеру к носорогу. Из Южной Америки после об-
разования в плиоцене Панамского перешейка успешно эмигрировал только
один род - североамериканский дикобраз.
В Северной Америке, Европе, Азии и Африке развились свои главные
группы грызунов, и на каждом континенте имеются уникальные семейства.
На каждом из этих континентов грызуны очень разнообразны. Австралия,
572
КАМЕННАЯ КНИГА
напротив, имеет представителей только одного семейства - мышиные, кото-
рое включает крыс и мышей. Грызуны достигли Австралии только в плиоце-
не и не имели времени, чтобы также разносторонне развиться, как в других
местах.
Бобры сегодня все живут в воде, но в миоцене Северной Америки были и
наземные формы. Один из самых древних - Ceratogaulus. Название означает
“рогатый землекоп” и указывает на факт, что на черепе этих животных име-
лась пара рогов и что они копали норы, в которых жили. Это достоверно из-
вестно, потому что их ископаемые остатки были обнаружены в норах. Иско-
паемые норы Ceratogaulus имеют собственное название - Daemonelix или
“штопор дьявола”. Это спиральные норы, заполненные осадком, который в
конечном счете стал тверже, чем окружающая порода. Сегодня они часто мо-
гут быть найдены на западе Небраски, где эрозия частично их отпрепариро-
вала. Некоторые спирали нор уходят вниз на 2.4 м и имеют почти горизон-
тально расположенные комнаты на всю длину хода.
Глава XXXIV
ЖИВОТНЫЕ И ПТИЦЫ
ЛЕДНИКОВОГО ПЕРИОДА.
ГУБИТЕЛЬНОЕ ВЛИЯНИЕ ЧЕЛОВЕКА
• “Около Пуэбло (Лос-Анджелес) имеются более двадцати мест выхо-
да на поверхность источников жидкой нефти, смолы и т.д. Западнее
в середине большой равнины есть обширное озеро смолы с многими
мелкими лужами, в которых непрерывно образуются и лопаются пузыри. Во
время засухи животные, приходящие на водопой, увязали в смоле, не могли
вытащить ноги, и озеро поглощало их. После многих лет их кости окамене-
ли. Я собрал несколько экземпляров”.
Так испанский исследователь Жозе Лонгинес Мартинес описал залежи
смолы в Ранчо ла Бреа, которые он видел в 1792 г. Но современники не рас-
сматривали собранные Мартинесом кости как ископаемые, пока в 1906 г. не
были начаты палеонтологические раскопки. В течение десяти лет были со-
браны сотни тысяч костей разнообразных животных плейстоцена, или лед-
никового периода. Другие тысячи костей и сейчас остаются в смоле парка
Ханкок на западе Лос-Анджелеса.
ЧЕТВЕРТИЧНЫЙ ИЛИ ЛЕДНИКОВЫЙ ПЕРИОД
Перед тем как пойти дальше, мы должны разъяснить этот обманчиво про-
стой термин “ледниковый период”. В своем развитии Земля прошла через не-
сколько “возрастов”, в течение которых климат был холодным и ледники
были широко распространены в различных частях мира. Первый холодный
период отмечался в течение Гурона, около 2300 млн. лет назад. Имелись и
другие эпохи похолодания в конце протерозоя и в течение палеозоя. По-
следний период похолодания начался приблизительно 1.5 млн. лет назад и
длится до настоящего времени. Наличие ледника в течение четвертичного
периода - наиболее признанный и наиболее изученный факт в истории оле-
денений Земли. Однако, называя четвертичный период ледниковым, мы
забываем, что в течение периода было несколько эпох оледенения
(фиг. XXXIV. 1). Многим четвертичный ледниковый период кажется зимой,
которая сменяется катастрофическими наводнениями при таянии ледяных
пластов. Такая картина, однако, является слишком простой. В Северной
Америке, Азии и Европе выделяются четыре главных ледниковых эпохи и
более мелкая пятая. Между этими холодными эпохами были эпохи тепла и
таяния, в которые климат иногда был даже более теплым, чем сегодня. Так,
© 1997. Т.В. Кузнецова, перевод на русский язык и изменения.
574
КАМЕННАЯ КНИГА
Общая стратиграфичес кая шкала Хронологическая шкала (млн. л) Палео- магнитная шкала Восточно- европейская стратиграфичес- кая шкала Альпийская стратиграфичес- кая шкала Североамери- канская стратиграфичес- кая шкала
Система Отдел Раздел
ГОЛО'-’’ цен 0.01 Валдайское (оледенение) Вюрм (оледенение) Висконское (оледенение)
-0.1 Микулинское Рисе-Вюрм Сангамон
Московское \ (межледниковье) / у (межледниковье) /
-0.2 -0.3 -0.4 -0.5 (оледенение) Одинцовское (межледниковье) Днепровское (оледенение) Рисе (оледенение) Иллинойс (оледенение)
стоцен О ф X 2 Лихвинское (межледниковье) Миндель-Рисс (межледниковье) Ярмут (межледниковье)
еоплей LD Окское (оледенение) Миндель (оледенение) Канзас (оледенение)
К л m о т ф о с о X -0.6 -0.7 Л 7Q Гюнц- Миндель (межледниковье) Артон (межледниковье)
X X ф
о
(К СО X т X н о >х ф с; -0.9 -1.0 -1.1 -1.2 -1.3 -1.4 О 0.89 5 • Г юнц (оледенение) Небраска (оледенение)
ф” m ф □плейстоцен о со ла 01 Харамк Дунай Гюнц (межледниковье)
ф Матуяг
-1.5 Дунай (оледенение)
1.67 ’5 ш ч:
О 1.87
Фиг. XXXIV. 1. Хроностратиграфическая схема антропогеновых отложений
Восточная Европа, Западная Европа, Северная Америка и их корреляция
(по данным Стратиграфия СССР, четвертичная система, полутом 1, 1982;
К.К. Марков и др., 1965; решение бюро МСК, 1995)
ЖИВОТНЫЕ И ПТИЦЫ ЛЕДНИКОВОГО ПЕРИОДА
575
например, межледниковые отложения около города Торонто (Канада) содер-
жат остатки деревьев, которые теперь не растут севернее Центральной Мис-
сисипской долины (штаты Огайо и Пенсильвания).
Эпохи оледенения (гляциалы) длились приблизительно 200-300 тыс. лет,
а эпохи потепления (интергляциалы) — 10-16 тыс. лет. Оледенение начина-
лось постепенно, тогда как отступление ледников было быстрым. Последняя
ледниковая эпоха началась приблизительно триста тысяч лет назад и достиг-
ла своего максимального похолодания восемнадцать тысяч лет назад. Десять
тысяч лет назад последняя ледниковая эпоха закончилась. Если ритмичное
чередование эпох оледенений и потеплений сохранится и далее, то можно
ожидать, что следующая эпоха похолодания постепенно начнется в течение
ближайшей тысячи лет. В пределах главных эпох оледенения имелись ко-
роткие стадии потепления и, наоборот, стадии похолодания отмечались в пе-
риоды межледниковий.
Цикличный характер четвертичного периода, возможно, связан с регу-
лярными, незначительными изменениями в форме орбиты Земли относи-
тельно Солнца. Это вызывает изменения в количестве и распределении полу-
чаемого солнечного тепла. Колебания достаточно большие, чтобы сущест-
венно уменьшить количество растаявшего льда. Уменьшение таяния
вызывает накопление льда в течение года. Год за годом лед медленно растет,
пока его вес не становится таким большим, что из-за давления ледник начи-
нает течь.
Хотя изменения в форме орбиты Земли могут объяснять, почему ледники
наступали и отступали в течение четвертичного периода, они не объясняют,
что послужило толчком к первому оледенению. Ритмичные изменения орби-
ты Земли существовали и до четвертичного периода, но оледенений не проис-
ходило. Возможно, в течение кайнозоя температура Земли снизилась. И это
явилось причиной, что небольшие изменения в количестве получаемого сол-
нечного тепла смогли вызывать оледенения и потепления.
Для объяснения снижения температуры Земли в течение кайнозоя было
выдвинуто несколько гипотез. Одна из них основана на сокращении коли-
чества тепла, выделяемого Солнцем. К сожалению, никто не придумал,
как проверить это предположение. Другая гипотеза основывается на том,
что кайнозой и особенно поздний кайнозой - время интенсивного горооб-
разования. Обширная область Земли в этот период была поднята выше, в
более холодные климатические зоны, и это привело к всеобщему пониже-
нию температуры.
Сторонники третьей теории предполагают, что поскольку континенты пе-
реместились в результате тектоники плит, то изменилась циркуляция воз-
душных и океанических потоков. При уменьшении потока тепла от тропи-
ков к высоким широтам можно ожидать глобальное похолодание климата.
Кроме того, изменяющиеся относительные количества суши и моря в раз-
личных широтах повлияли на понижение температуры всей Земли. В част-
ности, уменьшилось соотношение отраженной и поглощенной энергии в сто-
рону увеличения отраженного количества. Возможно, все эти факторы вы-
звали последний ледниковый период.
В течение четвертичного периода имелось четыре главных центра конти-
нентального оледенения в Северном Полушарии: два в Северной Америке,
один в Европе и один в Азии (фиг. XXXIV.2). В период максимального разви-
тия ледников два ледника Северной Америки объединились и перекрыли
более 15.5 млн. кв. км. Ледниками был занят весь север континента, их
южная граница проходила по южной части Великих Озер. Однако большая
576
КАМЕННАЯ КНИГА
Фиг. XXXIV.2. Максимальное распространение льдов Северного полушария в
четвертичном периоде
Материковые льды, наземные, подземные и морские льды ( К.К. Марков и др.,
1965)
1 - современная граница морских льдов; 2 — древняя граница морских льдов;
3 — современные наземные материковые льды; 4 - древнее наземное максималь-
ное оледенение; 5 - современная граница многолетней мерзлоты и подземного
оледенения
часть Аляски и запад территории Юкон не подверглись оледенению. В этих
районах не было достаточного количества осадков, хотя температура была
очень низкая.
По той же причине, очевидно, Северо-Восточная Сибирь осталась свобод-
ной от ледников. Континентальные ледяные покровы были развиты в Севе-
ро-Западной Сибири и в Финно-Скандинавском районе. Эти две области оле-
денения при объединении перекрывали площадь в 9.6 млн. кв. км.
Сегодня континентальные ледяные покровы существуют только на остро-
ве Гренландия и в Антарктиде. Антарктическая ледяная шапка была един-
ственным обширным континентальным ледником в Южном полушарии и в
течение четвертичного периода. Это шапка, вероятно, была в три раза боль-
ше во время максимальной фазы развития ледников и, возможно, в три раза
ЖИВОТНЫЕ И ПТИЦЫ ЛЕДНИКОВОГО ПЕРИОДА 577
меньше десять тысяч лет назад, в течение самой теплой стадии современного
межледниковья.
ЖИЗНЬ НА ДРЕВНЕЙ ЛОС-АНДЖЕЛЕССКОЙ РАВНИНЕ
После небольшой геологической подготовки мы постараемся восстано-
вить условия жизни и население во время ледникового периода по отдель-
ным образцам, сохранившимся в залежах смолы местонахождения Ранчо ла
Бреа. При первом изучении окаменелости, найденные в смоле, были отнесе-
ны к раннему плейстоцену. Позже эта ошибка была исправлена, и сейчас их
возраст оценивается от двенадцати до сорока тысяч лет, то есть конец послед-
него (для Северной Америки - Висконсинского) ледниковья.
Рассматриваемая Лос-Анджелесская равнина во время ледникового пери-
ода не была покрыта ледником, самые близкие ледники располагались в
Сьерре Неваде (в 240 км). Ручьи текли к древней реке Лос-Анджелел, кото-
рая затем бежала к югу и впадала в залив, который теперь называется зали-
вом Сан Педро. Растительный покров равнины не был однороден. Можно бы-
ло выделить заросли вечнозеленого карликового дуба и дубовые рощи, рощи
можжевельника, кипариса и епископских сосен.
Мелкие млекопитающие, жившие на равнине, были очень похожи на со-
временных. Они включали мышей, земляных белок, кенгуровых крыс, аме-
риканских кроликов и американских зайцев. Однако остатки растений, най-
денные в отложениях этого возраста, говорят о более теплом климате и мень-
шем влиянии океана, чем сегодня. Сезоны умеренных дождей чередовались
с летней и осенней засухами.
Копытные и когтистые травоядные. Хотя мелкие млекопитающие нам
знакомы, большинство крупных кажутся необычными. Например, на рав-
нине обитало большое количество бизонов (Bison antiquus). Они были
крупнее современных представителей и достигали более 2.1м высоты в хол-
ке. У крупных самцов черепа от 69 до 81 см шириной между рогами, которые
менее изогнуты, чем у современного бизона (фиг. ХХХГУ.Зи).
Дикие лошади были меньшей численности, чем бизоны, и бродили по
равнине небольшими группами или стадами. Все они относились к одному
виду Equus occidentalis, до 1.47 см высоты в холке, с выпуклым лбом, мощ-
ным телом и относительно короткими ногами (фиг. ХХХГУ.Зк). Они были не
такие высокие как современные североамериканские лошади, и более
тяжелые.
Остатки верблюдов, найденные в ямах со смолой, говорят об их крупных
размерах - 2.1 м высоты в самой высокой задней части туловища
(фиг. XXXIV.Зз). Череп по строению близок к черепу ламы, а кости посткра-
ниального скелета похожи на таковые современного двугорбого азиатского
верблюда. На равнинах обитали также большие стада Вгеатегух, вилорогой
антилопы 48 см высотой в плече (фиг. XXXIV.36). У самцов рога были раз-
делены в два прямых шипа, один из которых намного больше другого. Самки
часто безрогие. По строению зубов вилорогая антилопа близка к совре-
менным, и можно сказать, что она ела такую же траву, как едят антилопы
сегодня.
Самые крупные млекопитающие, остатки которых найдены в залежах смо-
лы - это мастодонты и мамонты (фиг. XXXIV.4, XXXIV.5). Наиболее древний
из известных мастодонтов с Лос-Анджелесской равнины - Mammut ameri
сапит. Мамонты появились позже и представлены двумя видами: Mammuth-
37 Каменная книга
578
КАМЕННАЯ КНИГА
Фиг. XXXIV.3. Типичные млекопитающие и птицы, остатки которых найдены
в залежах смолы местонахождения Ранчо Ла Бреа. Поздний плейстоцен, около
20 тыс. лет назад
а - большой стервятник (Teratornis), с размахом крыльев 3-4 м; б - маленькая
вилорогая антилопа (Вгеатегух), ростом 51 см; в - наземный ленивец (No-
throtherium), 99 см высоты в спине; г — аист, ростом около 1.2 м; д — гигантский
короткомордый медведь (Arctodus), 127 см высоты в задней части туловища; е -
лев (Felts atrox), 127 см высоты в плече; ж - два императорских мамонта, 3-4 м
в высоту; з — верблюд, 2.1 м в плече; и - череп бизона; к — череп лошади
us imperator (фиг. XXXIV.Зж) и М. columbi (см. фиг. XXXIV.5). Однако ни
один из них не был многочисленен.
Самым обычным неполнозубым, ископаемые остатки которого наиболее
многочисленны в местонахождении Ранчо ла Бреа, является наземный лени-
вец Mylodon или Paramylodon. Он был распространен от Патагонии до штата
Орегон, а в восточном направлении до штата Небраска. Представители дру-
гого рода наземных ленивцев Nothrotherium бродили на равнине у подножья
близлежащих гор (фиг. XXXIV.Зе). Третий род, Megalonyx, также жил сре-
ди гор, но иногда посещал равнину, и его остатки были найдены в смоле.
Ископаемые остатки Megalonyx из Ранчо ла Бреа принадлежат виду, распро-
страненному в восточном направлении до Атлантического побережья. Этот
вид был описан в 1797 г. Томасом Джефферсоном (ставшим впоследствии
президентом США), который ошибся и отнес кости лапы с когтями к огром-
ному льву.
ЖИВОТНЫЕ И ПТИЦЫ ЛЕДНИКОВОГО ПЕРИОДА
579
Фиг. XXXIV.3. Окончание
Различные хищники. Там, где обитают травоядные животные, обяза-
тельно должны быть и хищные. На Лос-Анджелесской равнине они были
представлены ласками, скунсами, барсуками, лисами, койотами. Все отно-
сятся к современным видам или очень близки им. Там же обитал ныне вы-
мерший “страшный волк” — Cants dims, крупноголовое животное, но не та-
кой крупный, как современный лесной волк. Он являлся стайным живот-
ным и, вероятно, охотился группами, ел как свежее мясо так и падаль.
Кошачьи были представлены рыжей рысью, пумой или горным львом, Fe
Its (или Pantherd) atrox, а также саблезубыми формами. F. atrox мощное по-
хожее на льва животное было распространено от Аляски до Миссисипи и
Центральной Мексики. Самцы были крупнее самого большого тигра, а сам-
ки близки по размеру к современному ягуару (фиг. XXXIV.Зе). Этот лев был
активным охотником и охотился на лошадей, верблюдов, бизонов и других
травоядных.
Smilodon — последний из саблезубых тигров, был короткохвостым живот-
ным, намного более тяжелым, чем современный лев (фиг. XXXIV.6). Тело
массивное, ноги мощные, а мозг меньше, чем у Felis atrox или пумы. Клыки,
подобно клыкам других саблезубых тигров, ножеобразные, изогнутые, ис-
пользовались для разрезания или протыкания шкуры крупных животных
типа бизона или наземного ленивца, а не для кусания. Smilodon не бегал за
37*
580
КАМЕННАЯ КНИГА
Фиг. XXXIV.4. Зубы мастодонтов Северной Америки
а — верхний коренной зуб мастодонта Sertidentinus из Флориды; б - кончики
бивней двух американских мастодонтов, на них углубления, которые образова-
лись в результате истирания корнями растений при рытье мастодонтом земли;
в - второй левый нижний коренной зуб американского мастодонта
Фиг. XXXIV.5. Самец южного или колумбийского мамонта. Высота в плечах
около 3.2 м. Находки известны из многих местонахождений на юго-западе
США
ЖИВОТНЫЕ И ПТИЦЫ ЛЕДНИКОВОГО ПЕРИОДА
581
Фиг. XXXIV.6. Smilodon — последний саблезубый тигр. Приблизительно
102 см высотой в плече
а — череп с максимально открытой нижней челюстью; б — реконструкция
добычей, так как его ноги по своему строению не были приспособлены для
быстрого бега. Можно представить такую картину: животное затаилось и
ждет около рощи или водопоя, а затем набрасывается на наземного ленивца
или на какого-нибудь другого животного. Сильные передние лапы с когтями
хватают жертву сзади, а длинные клыки разрезают шкуру снова и снова, на-
нося рваные раны. Высоко помещенные ноздри позволяли смилодону ды-
шать, даже когда его рот был набит длинными, грубыми волосами наземного
ленивца.
Самый большой из хищников, остатки которых были найдены в смоле, -
гигантский короткомордый медведь Arctodus 127 см высотой в задней части
туловища и 229 см длиной (фиг. XXXIV.35). Для сравнения - большой
гризли 102 см высотой и 178 см длиной. Он был распространен от Юкона до
Пенсильвании и Техаса. Если плейстоценовые гиганты имели такие же при-
вычки, как современные медведи, и защищали свою территорию от других
медведей, то ни гризли, ни черный медведь не имели возможностей для ус-
пешного расселения на древней Лос-Анджелесской равнине.
Насекомые и птицы. В отличие от многих других ископаемых фаун фауна
Ранчо ла Бреа включает большое количество остатков насекомых и птиц.
Среди насекомых много жуков-трупоедов, которые питались и даже жили в
разложившихся трупах. Птицы известны очень разные: от гусей и аистов
более 1.2 м высотой (фиг. XXXIV.Зг) до дятлов, восковиц и крошечных воро-
бьев. Больше половины остатков птиц приходится на стервятников, орлов и
ястребов.
Многие ястребы принадлежали современным видам. Другие были близки
к видам, ископаемые остатки которых найдены в Центральной Америке. Во-
семь или больше видов американских грифов питались рядом с залежами
смолы, и их остатки найдены в ней. Один был сходен с современным кали-
форнийским кондором, но принадлежал к вымершему виду, который был
распространен от Каскадных гор до Флориды и Мексики. Другой близок к
кондору из Южной Америки, но имел более длинный клюв. Оба кажутся ма-
582
КАМЕННАЯ КНИГА
ленькими рядом с Teratornis, который был распространен от Калифорнии до
Техаса, Флориды и северной Мексики. Teratornis достигал роста 76 см, когда
садился, весил около 22.7 кг и имел размах крыльев 3.7 м (фиг. XXXIV.За).
Отверстия для ноздрей очень большие, поэтому ученые предполагают, что он
при охоте пользовался больше обонянием, чем зрением, то есть охотился по
запаху. Возможно, в своем поведении тераторнис отличался от других
стерв ятников.
ПОЧЕМУ И КАК ОНИ УМИРАЛИ?
Исследуя остатки различных животных, нам интересно, почему они най-
дены в ямах смолы. Что привело их к этим ямам? Как они попали в смолу, в
вещество, которое они должны избегать?
Ответы появляются, когда мы рассматриваем строение смоляных ям, ка-
ким оно было в позднем плейстоцене. Вопреки обычно поддерживаемому
представлению, в Ранчо ла Бреа нет окаменелостей в маленьких озерах, ко-
торые имели прослойку воды над глубоким бассейном смолы. Много окаме-
нелостей известно из ручьев и небольших речек, в которых липкая смола,
вытекающая на поверхность почвы, смешивалась с глиной, песком и грави-
ем. В некоторых местах в результате смешения образовывались пропитан-
ные асфальтом зыбучие пески.
Небольшие заводи речек были заселены улитками, водяными жуками и
клопами, это были места для корма гусей, уток и паганок. Цапли, журавли
и береговые птицы бродили на отмелях, и жабы приплывали, чтобы отло-
жить икру. Время от времени стада бизонов, антилоп или лошадей останав-
ливались у заводи на водопой.
Это картина сезона дождей, а в сухой период, который продолжался с
поздней весны до осени, потоки прекращали течь и сокращались до ряда изо-
лированных заводей. Они оставались единственными источниками воды.
Травоядные животные проходили десятки километров, чтобы достигнуть
их; большинство стояли и пили на отмелях, они были в безопасности. Другие
же останавливались на зыбучих песках, пропитанных липким асфальтом и
увязали. Здесь же добывали пищу хищники, нападая на увязших
беспомощных травоядных.
Имеется последовательная толща ископаемых остатков млекопитающих,
птиц и насекомых. Сначала к водопою приходили измученные жаждой тра-
воядные от кролика до императорского мамонта. Очень сильно желая пить,
они заходили в воду слишком глубоко и увязали в смоле. При отчаянных по-
пытках освободиться животное погружалось все глубже, в то же самое время
привлекая внимание бродящего рядом хищника. Для хищников это была
легкая добыча, но если они были неосторожны, то сами тоже увязали в смо-
ле. Трупы ели голодные волки, которые рычали и огрызались друг на друга
на разорванных тушах.
Ястребы, орлы и стервятники наблюдали борьбу или улавливали аромат
падали. Они снижались, ели мясо увязших животных и улетали, но когда не-
сколько птиц нападали на одну тушу, то разгорались поединки. Птицы тол-
кали и били друг друга крыльями и те, которые теряли равновесие, часто па-
дали в смолу. На заключительной стадии прибывали последние пожиратели,
это мясные мухи, откладывающие яйца в падаль, и жуки, питающиеся ею.
Многие из этих насекомых или падали в смолу, или были захоронены с остат-
ками туш при их погружении.
ЖИВОТНЫЕ И ПТИЦЫ ЛЕДНИКОВОГО ПЕРИОДА
583
Погружение происходило медленнее, чем можно было бы предположить.
Много костей в залежах смолы сильно выветрелые, другие погрызаны мы-
шами после того, как хищники обглодали их. Кости находились на поверх-
ности в течение недель или месяцев, возможно, большими грудами. Они при-
влекали все больше хищных млекопитающих и мелких птиц, остатки кото-
рых также найдены в смоле.
В результате такой необычной последовательности накопления ископае-
мого материала в захоронении имеется намного больший процент плотояд-
ных и падалеядных птиц и животных, чем обычно, давая нам редкую воз-
можность “бросить взгляд” на эти обычно плохо известные ископаемые
формы.
Местонахождение близкое по условиям захоронения к Ранчо ла Бреа,
имеется и в Евразии - это Бинагады (Азербайджан). По времени образования
оно немного древнее и отличается условиями накопления ископаемых остат-
ков. По мнению И.М. Громова, изучавшего это местонахождение, основная
масса костных остатков крупных млекопитающих и птиц была принесена
потоком, впадавшим в озеро, и скапливалась в области его конуса выноса.
Особенно большое количество трупов приходилось на период паводков. По-
сле спада воды на берегах и отмелях хищники и падалеядные птицы набра-
сывались на туши и часто гибли сами, увязая в песке и иле, пропитанном
смолой. При следующем половодье остатки трупов переносились водой, от-
лагались в ямах на дне озера и постепенно засыпались песком.
Образование асфальтовой корки на костях предотвращало их дальнейшее
разрушение. В песке, пропитанном смолой, хорошо сохранялись и остатки
насекомых, погибших в нефтяной пленке, покрывавшей отдельные части
озера. В Бинагадах были найдены не только кости ископаемых животных,
но и мягкие ткани, законсервированные в асфальте.
В позднем плейстоцене равнинная часть Апшеронского полуострова была
занята степями, а в предгорных областях - саваннами. Копытные и боль-
шинство хищников обитали в равнинной части полуострова и в низких
предгорьях. Возможно, лишь гиены и медведи жили выше в горах. Из круп-
ных растительноядных основную массу населения составляли сайгаки, ло-
шади, олени и бизоны, хоботные же полностью отсутствовали. Из хищников
волки, лисы и барсуки были многочисленны, а саблезубые кошки вообще не
встречены. Для Бинагадов характерно также большое количество остатков
грызунов.
Местонахождения Ранчо ла Бреа и Бинагады, близкие по времени и усло-
виям образования, показывают разницу в населении Лос-Анджелесской и
Апшеронской равнин в позднеплейстоценовое время.
ЖИВОТНЫЕ ЛЕДЯНОГО ФРОНТА (ПРИЛЕДНИКОВЬЯ)
Хотя животные, найденные в смоле, жили в ледниковом периоде, ни одно
из них никогда не видело ледника и не испытало метели. Другие же млеко-
питающие жили в приледниковой зоне всего в нескольких километрах от по-
стоянно двигающегося ледника и перемещались на север, когда ледник таял,
и на юг, когда он начинал расти снова.
Один из таких морозоустойчивых животных - овцебык, или мускусный
бык. В течение плейстоцена он был распространен в Азии и Европе. Корена-
стый 1.1-1.2 м высотой, его длинная шерсть не пропускала дождь и снег, а
волосяной покров (подшерсток) хранил тепло. Современный овцебык пред-
584
КАМЕННАЯ КНИГА
Фиг. XXXIV.7. Растительноядные “шерстистые” животные плейстоцена
а — Preptoceras, около 1.1—1.2 м высотой, жил в Калифорнии; б — шерстистый
носорог (Coelodonta), жил в Европе и на севере Азии
почитает холодную, ветреную погоду. Возможно, они могли обитать и в ме-
нее суровом климате, так как по находкам ископаемых остатков установле-
но, что эти животные жили и в Калифорнии (фиг. XXXIV.7). Северный ка-
надский олень (карибу) также животное Арктических бесплодных земель.
Он жил около фронта ледника в течение плейстоцена. Там они часто пас-
лись вместе с мамонтами и мастодонтами, хотя последние имели область
распространения, протягивающуюся далеко на юг до Флориды. Северные
канадские олени обитали в болотистых лесистых местностях, где иногда
увязали в грязи. Американские лоси также кормились на болотах
(фиг. XXXIV. 7), а горные козлы предпочитали сухие скудные земли, где
погода изменялась от прохладной до холодной. Все эти животные или близ-
кие к ним формы, предпочитающие скудные арктические земли, имели
циркумполярное распространение в течение плейстоцена, их остатки
встречаются в Азии, Европе и Северной Америке. Исключение составляют
мастодонты, но даже они произошли в позднем миоцене в Старом Свете, а в
плейстоцене вымерли.
Большинство млекопитающих плейстоцена жило в более благоприятных
регионах. В Северной Америке Большие Равнины и плато были заселены ог-
ромными табунами лошадей. Лошади были многочисленны и в прериях и
среди лесов на востоке. Представители большинства видов были близки по
размеру к современным североамериканским ковбойским лошадям. Однако
Equus giganteus из Техаса была больше самого крупного современного тяже-
ловоза, а Е. tau из Мексики ростом не превышала шотландского пони.
Тапиры, которые жили в Северной Америке начиная со среднего олигоце-
на до максимального оледенения были обычны на востоке от Миссисипи, но
после продвижения ледника далеко на юг полностью вымерли в Северной
Америке. Носороги исчезли из Северной Америки в конце плиоценовой эпо-
хи, хотя один из видов, с двумя длинными рогами, дожил до конца леднико-
ЖИВОТНЫЕ И ПТИЦЫ ЛЕДНИКОВОГО ПЕРИОДА
585
Фиг. XXXIV.8. Парнокопытные ледникового периода
а — Megaloceros — гигантский или большерогий олень, известен из Европы, Се-
верной Африки и Азии, высота в плече около 1.8 м; б - Cervalces — американ-
ский лось, был распространен от Нью-Джерси до Айовы
вого периода в Европе и Азии вместе с мамонтами. Подобно мамонту, этот но-
сорог имел волосяной покров из длинной, грубой шерсти желтовато-красно-
го цвета. Из-за этого животное получило название “шерстистый носорог”
(фиг. XXXIV. 7).
Североамериканские парнокопытные в плейстоцене были представлены
большим количеством видов, многие из которых живут и сейчас, а некото-
рые вымерли. Так, североамериканский лось Cervalces известен нам только
по ископаемым остаткам. Он был крупным животным с разветвленными ро-
гами и очень большими копытами, жил в лесах от Айовы до Нью-Джерси
(фиг. XXXIV.8). В то же время в Старом Свете обитал гигантский или боль-
шерогий олень - Megaloceros. Он вымер приблизительно десять тысяч лет
назад и действительно был гигантским, с размахом рогов от 2.1 до 3 м. Ске-
леты, найденные в болотных торфяниках, кажутся слишком тонкими, что-
бы удерживать эти огромные рога (фиг. XXXIV.8).
Самые древние бизоны известны из раннего плейстоцена Китая и Ин-
дии. В среднем плейстоцене они достигли Европы, а в позднем — Северной
Америки. В Северной Америке обитало несколько видов бизонов, одни
паслись в огромных стадах на Больших Равнинах и в прериях, другие оби-
тали в лесах на востоке континента. Так, Bison latifrons был распростра-
нен в лесных областях от Флориды до Орегона. Это огромное животное с
размахом слегка изогнутых рогов 1.5-1.8 м (фиг. XXXIV.9). Другой вид
бизона с юго-западных равнин, которого обычно называют Bison taylort, а
следует называть бизоном Фриггинса (фиг. XXXIV.9). Именно он был од-
ним из тех, кого выбили и съели доисторические американцы приблизи-
тельно одиннадцать тысяч лет назад. Кремневые наконечники копий были
найдены в 1926 г. среди костей этих вымерших бизонов около города Фол-
586
КАМЕННАЯ КНИГА
Фиг. XXXIV.9. Бизоны
а — бизон Фриггинса был истреблен древними людьми; б - Bison Latifrons, круп-
ный вид, с очень высоким горбом и размахом рогов более 1.8 м
сом (Нью-Мексико). У многих скелетов не было хвостов, предполагают,
что они были отрезаны. Подобные находки сделаны и в других областях,
где форсомскими людьми кроме бизонов были убиты верблюды, лошади,
мамонты. Эти ценные находки являются доказательством того, что древ-
ний человек активно охотился и убивал этих животных, а не просто жил
рядом с ними.
Некоторые бобры ледникового периода принадлежали к ныне живущему
виду и были распространены на большой части Европы, Азии, Соединенных
Штатов и северной Мексики. Гигантские бобры Castoroides немного крупнее
черных бобров и в длину достигали 2.4 м от носа до кончика хвоста. Они были
северными животными и населяли озера от Нью-Йорка до Небраски. Воз-
можно, эти бобры питались водными растениями вместо коры, хотя резцы у
них крупные. Мы ничего не знаем о том, строили ли они хатки и дамбы, как
это делают бобры сегодня.
Дикобразы, которые теперь являются животными гор и северных лесов,
мигрировали из Южной Америки и распространились по всем Соединенным
Штатам и Канаде. С этими колючими грызунами прибыли и наземные ле-
нивцы: Mylodon, Nothrotherium и Megatherium. Остатки первых двух были
найдены среди млекопитающих в ямах смолы. Представители рода Megath-
erium обитали на востоке Северной Америки между штатами Флорида и Юж-
ная Каролина. Они должны были часто сталкиваться с Megalonyx, который
расселился в этих же районах в течение плиоцена.
ЮЖНОЕ ПОЛУШАРИЕ
На Антарктиде ледяной щит сформировался в миоцене, а в течение плей-
стоцена только периодически увеличивался или уменьшался. На этом кон-
ЖИВОТНЫЕ И ПТИЦЫ ЛЕДНИКОВОГО ПЕРИОДА
587
Фиг. XXXIV. 10. Один из многих позднеплейстоценовых глиптодонтов Арген-
тины — Glyptodon clavipes
а — панцирь и броня хвоста 1 м длиной; б — скелет 2.7 м длиной. (Фото R. Pas-
cual, музей Ла Плата, Аргентина)
тиненте в плейстоцене никогда не было крупной области суши, не занятой
льдами, на которой наземные млекопитающие и птицы могли бы разви-
ваться.
Из трех других южных континентов только в Южной Америке да и то
только в южной части, было континентальное оледенение. Однако горные
ледники были развиты в Южной Америке, Австралии и Африке, а также в
Новой Зеландии и Новой Гвинее.
В то время как увеличение и уменьшение континентального ледяного щи-
та было главным результатом колебаний температуры в плейстоцене Ан-
тарктиды, на трех других южных континентах отмечалось чередование
588
КАМЕННАЯ КНИГА
Фиг. XXXIV.il. Скелет крупного наземного ленивца Glossotherium myloid.es
из позднего плейстоцена Аргентины
а - скелет, длина около 2.3 м; б - череп с фронтальной и боковой сторон (фото
R. Pascual, музей Ла Плата, Аргентина)
влажных и сухих климатических условий. Они коррелятивно связаны с на-
ступлением и отступлением ледников в Северном полушарии. Очевидно, су-
хие периоды соответствуют максимальному развитию ледниковых щитов, а
влажные - их отступлению и таянию.
В течение влажных стадий сухие озера в пустынях центральной Австра-
лии и в Сахаре заполнялись водой. Часто вода достигала такого высокого
уровня, что эти озера сообщались с морем. Сегодня вода заходит в них в ред-
ких случаях, а скапливающаяся там дождевая вода просто испаряется.
Фауна млекопитающих Африки в течение плейстоцена подверглась наи-
меньшим изменениям, чем в любой другой части мира. Хотя, конечно, жи-
вотные, которые там обитали, будут казаться нам странными сегодня. Од-
ним из наиболее необычных животных был оленерогий жираф Sivatherium,
с короткой шеей и двумя парами рогов. Передние рога - пара конических вы-
пуклостей, подобно рожкам у ныне живущих жирафов. Вторая пара - паль-
чатые рога - напоминали рога лося. Однако даже такое необычное животное
является членом семейства жирафовых, живущего и сейчас в Африке. Это
отражает общую картину для Африки, где многие рода и виды млекопитаю-
щих вымерли в течение плейстоцена, но они принадлежали семействам, к
ЖИВОТНЫЕ И ПТИЦЫ ЛЕДНИКОВОГО ПЕРИОДА
589
Фиг. XXXIV. 12. Вымерший сумчатый Diprotodon Австралии
а - скелет лапы; б - след
Найдены при раскопках на озере Каллабонна, юг Австралии. След первоначаль-
но отпечатался на грязи, а песок, нанесенный ветром заполнил и запечатал его.
Позже, после того как заполняющий песок стал крепким, как камень, произо-
шла эрозия вметающей породы и получился слепок следа. (Фото R. Tedford,
Американский музей Естественной Истории, Нью-Йорк)
которым относятся различные ныне живущие виды. Те семейства, которые,
подобно халикотериям, уже вымерли на других континентах или сильно
уменьшили свое разнообразие, оставались как реликты в Африке.
В Южной Америке в позднем плейстоцене произошло очень заметное
снижение разнообразия млекопитающих. Многие длительно существовав-
шие семейства, процветавшие в кайнозое, стали вымершими, например
глиптодонты. По меньшей мере десять родов крупных броненосцеподобных
млекопитающих жило в среднем плейстоцене северной Аргентины, и даже
в позднем плейстоцене их было еще восемь (фиг. XXXIV. 10). Теперь группа
стала полностью вымершей наряду с тремя семействами наземных ленивцев
(фиг. XXXIV.11), двумя семействами нотоунгулят и одним семейством ли-
топтерн. Все они длительное время существовали в кайнозое Южной Амери-
ки и в большом количестве даже жили в плейстоцене. Вероятно, их вымира-
ние можно объяснить отрицательным влиянием различных млекопитаю-
щих, в том числе и человека, пришедших из Северной Америки.
Меньшее количество семейств млекопитающих вымерло в течение плей-
стоцена в Австралии. Вымерли дипротодонты, палорхестиды и тилаколео-
ниды (см. главу XXXI). Эти три семейства австралийских сумчатых были
590
КАМЕННАЯ КНИГА
крупными животными (фиг. XXXIV.12). То, что много крупных млекопи-
тающих вымерло в течение плейстоцена, отмечается и на других конти-
нентах.
Это истинно не только для континентов, но и для островов, и относится не
только к млекопитающим, но и к птицам. На острове Мадагаскар слоновая
птица и гигантский лемур вымерли даже не в плейстоцене, а в течение по-
следней тысячи лет. То же самое произошло и в Новой Зеландии, где исчезла
бегающая птица — моа.
В обоих случаях снижение численности и полное уничтожение животных
произошло только после прибытия людей. Во всех областях Земли люди по-
явились раньше, чем произошло самое сильное плейстоценовое вымирание.
Губительное влияние людей на крупных животных не может быть объяснено
просто охотой. Скорее всего, эти вымирания вызваны изменением ландшаф-
та, например, применением широко распространенной практики выжига-
ния растительности.
Возможно, другим фактором, сыгравшим роль во всемирном исчезнове-
нии крупных млекопитающих и птиц к концу плестоцена, являются суро-
вые климатические условия. Однако ранние стадии оледенения были более
серьезны, чем последняя.
Вымирание происходило постепенно в течение всего плейстоцена, но оба
фактора, вероятно, сыграли роковую роль к концу эпохи. При воздействии
двух факторов одновременно результат давления на фауну был более серьез-
ным, чем при влиянии одного.
ЧЕЛОВЕК - ПРОДУКТ ЛЕДНИКОВОГО ПЕРИОДА?
Гоминиды - семейство, к которому относятся люди, оно отделилось от
высших узконосых обезьян где-то между 14 и 4 млн. лет назад. Более точно
сказать нельзя, так как никаких остатков человекообразных в течение этих
10 млн. лет (от 14 до 4 млн. лет) не найдено. Это, вероятно, произошло в
Африке, хотя некоторые ученые считают Азию местом происхождения
человека.
Самые древние из известных ископаемых остатков гоминид - остатки ав-
стралопитеков - датированы 3.7 млн. лет. С маленьким объемом мозга и тя-
желыми челюстями они, вероятно, более походили на необычных обезьян,
чем на людей (фиг. XXXIV. 13). Однако следы, сохранившиеся на вулканиче-
ском пепле в Лайтоли (Танзания) и скелетные остатки из нескольких место-
нахождений на юге и востоке Африки показывают, что они ходили верти-
кально.
Взрослые австралопитеки были высотой 1.3-1.5 м, ниже, чем большинст-
во людей, живущих сегодня. Они жили не в лесах, а в саваннах, где вместо
мягких плодов их пищу составляли жесткие клубни и твердые орехи. Это от-
ражено в строении зубов: резцы уменьшены, а эмаль на коренных зубах
сложноскладчатая. Своими короткими резцами они раскусывали жесткую
кожуру, а коренные зубы использовались для дробления и размалывания
грубой растительной пищи, найденной в саванне.
Род Ното, к которому мы принадлежим, вероятно, произошел от рода
Australopithecus около 2 млн. лет назад, и первым его представителем был
Homo habilis (человек умелый) (фиг. XXXIV. 14). Австралопитеки же не вы-
мерли с появлением рода Ното, а продолжали жить рядом, приблизительно
до одного миллиона лет назад.
ЖИВОТНЫЕ И ПТИЦЫ ЛЕДНИКОВОГО ПЕРИОДА
591
Фиг. XXXIV.13. Australopithecus robustus из позднего плиоцена Африки
а — череп, ширина 16 см; б - нижняя челюсть, длина 15 см (фото M.G. Leakey,
Национальный музей Кении)
Фиг. XXXIV. 14. Homo habilis (человек умелый) из позднего плиоцена Африки
а - череп, ширина 13 см; б— нижняя челюсть, длина 8 см (фото M.G . Leakey, На-
циональный музей Кении)
Около 1 млн. лет назад род Ното, а более точно его вид Н. erectus (человек
прямоходящий) расселился из Африки в Восточную и Южную Азию.
Хотя австралопитеки использовали простейшие орудия, только люди
смогли усовершенствовать и усложнить их. Более сложной была и социаль-
592
КАМЕННАЯ КНИГА
ная структура рода Ното. Вместо добывания еды индивидуально, люди
начали делать это коллективно и делить пищу друг с другом. Следующим
шагом в социальном развитии стало возникновение речи.
Следует отметить, что при эволюции гоминид выделяется общая тенден-
ция относительного увеличения и развития головного мозга. Очень любо-
пытный факт, что неандертальцы, самые ранние члены нашего вида — Ното
sapiens, имели головной мозг в среднем больше, чем наш. Кроме того, их ске-
леты показывают, что они были сильнее. Все же современный Homo sapiens
сменил их приблизительно сорок тысяч лет назад. Размер мозга не является
показателем эволюционной продвинутости. Она может быть связана с неуло-
вимыми путями работы нашего головного мозга. Хотя неандертальцы очень
напоминали нас, они могли вести себя совсем по-другому. Однако мы не мо-
жем этого выяснить, их просто нет в настоящее время.
Почему они вымерли, а возможно, ассимилировались с современным Но-
то sapiens, не выяснено. Это остается одной из миллионов загадок, которые
преподносит человеку природа.
ЛИТЕРАТУРА
(научно-популярные книги по палеонтологии
на русском языке)
Акимушкин И.И. Исчезнувший мир. М.: Молодая гвардия, 1982. 191 с.
Аугуста Й. Исчезнувший мир. М.: Недра, 1979.191 с.
Аугуста Й„ Буриан 3. Летающие ящеры и древние птицы. Прага: Артия, 1961. 104 с.
Аугуста Й., Буриан 3. Книга о мамонтах. Прага: Артия, 1962. 57 с.
Аугуста Й., Буриан 3. Ящеры древних морей. Прага: Артия, 1965. 68 с.
Аугуста Й., Буриан 3. По путям развития жизни. Прага: Артия, 1971. 51с.
Биологический энциклопедический словарь. М.: Советская энциклопедия, 1989.864 с.
Бодылевский В.И. Малый атлас руководящих ископаемых. 5-е изд. Л.: Недра, 1990.
263 с.
Бойко М.С., Габдуллин Р.Р., Рякин ДА. Методические рекомендации по курсу пале-
онтологии. Сборник тестов (Моск. гор. станция юных натуралистов). М.: Б.и.,
1996. 97 с.
Быстров А.П. Прошлое, настоящее и будущее человека. Л.: Медгиз, 1957. 314 с.
Балха Каменное Царство. Перевод Андрея Нартова. Санкт-Петербург, 1784. 146 с.
24 табл, (первое русскоязычное издание по палеонтологии).
Верещагин Н.К. Почему вымерли мамонты. Л.: Наука, 1979.195 с.
Верещагин Н.К. Записки палеонтолога; по следам предков. Л.: Наука, 1981. 166 с.
Вологдин А.Г. Земля и жизнь. 2-е изд. М.: Недра, 1976. 239 с.
Габуния Л.К. Следы динозавров (по материалам горы Сатаплия и данным литерату-
ры). М.: Изд-во АН СССР, 1958. 72 с.
Герман Т.Н. Органический мир миллиард лет назад. Лик докембрия СССР. Л.:
Наука, 1990. 50 с.
Давиташвили Л.Ш. Курс палеонтологии. М.-Л.: Госгеолиздат, 1949. 835 с.
Давиташвили Л.Ш. Причины вымирания организмов. М.: Наука, 1969. 440 с.
Джохансон Д., Иди М. Люси. Истоки рода человеческого. М.: Мир, 1984. 295 с.
Диксон Д. Динозавры: ил. энцикл. М.: Моск, клуб, 1994. 144 с.
Динозавры. Как живые! Б.м.: Фирма “Биз-энтерпрайз”, 1994. 30 с.
Ефремов ИА. Дорога ветров (гобийские заметки). Переизд. М.: Молодая гвардия,
1980. 416 с.
Ивахненко М.Ф., Корабельников В А. Живое прошлое Земли. М.: Просвещение,
1987. 255 с.
Ископаемые цветковые растения России и сопредельных государств (в 7-ми т.).
Л.-СПб.: Наука, 1974-1994. 457 с. (т. 1 - 1974. 124 с.; т. 2 - 1982. 216 с.; т. 3 -
1994. 117 с.).
История крупных млекопитающих и птиц Северной Евразии. СПб.: ЗИН, 1992.158 с.
Калмыков Н.П., Пискунов А.С., Хандуев В.Б. Ископаемые млекопитающие Музея
природы Бурятии. Улан-Удэ: БНЦ СО РАН, 1992. 72 с.
38. Каменная книга
594
КАМЕННАЯ КНИГА
Катастрофы и история Земли. Новый униформизм. М.: Мир, 1986. 471 с.
Кондратов А.М. Шанс для динозавра. 3-е изд. СПб.: Гидрометеоиздат, 1992. 288 с.
Коробков ИЛ. Палеонтологические описания. 3-е изд. Л.: Недра, 1978. 208 с.
Кремп Герхард О.У. Палинологическая энциклопедия. М.: Мир, 1967. 411 с.
Криштофович А.Н. Палеоботаника. 4-е изд. Л.: Гостоптехиздат, 1957. 650 с.
Крумбигелъ Г., Вальтер X. Ископаемые: сбор, препарирование, определение, ис-
пользование. М.: Мир, 1980. 333 с.
Крылов И.Н. На заре жизни. Органический мир докембрия. М.: Наука, 1972. 104 с.
Крымголъц Г.Я. Методика сбора и обработки палеонтолого-стратиграфического ма-
териала. Л.: Изд-во Ленингр. ун-та, 1954. 46 с.
Кузнецов С.С. Геологические экскурсии. М.: Недра, 1978. 170 с.
Кучинский М. Тропик динозавра. М.: Наука, 1982. 192 с.
Кэрролл Р. Палеонтология и эволюция позвоночных (в 3-х т.). М.: Мир, 1992, 1993.
863 с. (том 1, 1992. 280 с.; том 2, 1993, 278 с.; том 3, 1993. 310 с.).
ЛаппоЛВ. Следы былых биосфер, или рассказ о том, как устроена биосфера и что ос-
талось от биосфер геологического прошлого. 2-е изд. М.: Знание, 1987. 207 с.
Малахов АА. Занимательно о геологии. М.: Молодая гвардия, 1969. 240 с.
Мартинсон Г.Г. Загадки пустыни Гоби. 2-е изд. Л.: Наука, 1980. 149 с.
Мартинсон Г.Г. В поисках древних озер Азии. Л.: Наука, 1989. 156 с.
Мартинсон Г.Г. Что мы знаем о динозаврах? Л.: Недра, 1990. 93 с.
Мейен С.В. Из истории растительных династий. М.: Наука, 1971. 223 с.
Мейен С.В. Следы трав индейских. М.: Мысль, 1981. 159 с.
Микропалеонтология (Н.И. Маслакова, Т.Н. Горбачик, А.С. Алексеев и др.). М.:
Изд-во Моск, ун-та, 1995. 256 с.
Михайлова ИЛ., Бондаренко О.Б. Палеонтология. Часть 1, 2. М.: Изд-во Моск, ун-
та, 1997. 878 с.
Морозов П.Е. Методическое пособие по сбору и определению ископаемой фауны ме-
зозоя Подмосковья (Моск. гор. станция юных натуралистов). М.: Б.и., 1994. 34 с.
Морозов П.Е., Ильясов И.В., Кузьмина СЛ. Методические рекомендации по сбору и
определению ископаемой фауны Московской области (Моск. гор. станция юных
натуралистов). М_: Б.и., 1992. 33 с.
Моя первая энциклопедия: Динозавры. М.: “Персей”, "Вече” ACT, 1994. 200 с.
Музафаров В.Г. Определитель минералов, горных пород и окаменелостей (справ,
пособие). М.: Недра, 1979. 327 с.
Наставление по сбору и изучению ископаемых органических остатков. Вып. I-XI.
М.: Изд-во АН СССР, 1953-1982.464 с.
Невероятные существа. СПб.: Фонд “Ленингр. галерея”: АО “Норинг”, 1996. 48 с.
Немков Г.И. Историческая геология с элементами палеонтологии (пособие для учи-
телей). М.: Просвещение, 1980. 191 с.
Немков Г.И., Карский Б.Е., Лин Н.Г. и др. Краткий геологический словарь для
школьников. М.: Недра, 1989. 176 с.
Николае Т.Г. Долгий путь жизни: о возникновении и развитии жизни на Земле. М.:
Мир, 1986. 168 с.
Определитель ордовикских окаменелостей окрестностей Санкт-Петербурга для
учебной геологической практики. СПб.: СПб ГУ, 1993. 70 с.
Орлов ЮЛ. В мире древних животных. Очерки по палеонтологии позвоночных.
3-е изд. М.: Наука, 1989. 163 с.
Очев В.Г. Тайны пылающих холмов. Саратов: Изд-во Саратов, ун-та, 1976. 94 с.
Очев В.Г., Янин Б.Т., Барское И.С. Методическое руководство по тафономии позво-
ночных организмов. М.: Изд-во Моск, ун-та, 1994. 143 с.
Палеонтологический словарь. М.: Наука, 1965. 616 с.
Палеонтология и палеоэкология. Словарь-справочник. М.: Недра, 1995. 494 с.
ЛИТЕРАТУРА (научно-популярные книги по палеонтологии на русском языке)
595
Погоня за тенью. О реальных и фантастических водных животных. Сборник. М.:
Знание, 1992. 80 с.
Поплавская МД. Увлекательная палеонтология. Клав: Наукова думка, 1982. 128 с.
Проблемы доантропогеновой эволюции биосферы. М.: Наука, 1993. 314 с.
Ранняя история Земли. М.: Мир, 1980. 620 с.
РаупД., Стенли С. Основы палеонтологии. М.: Мир, 1974. 390 с.
Решетов В.Ю. Ковалевский В.О. (палеонтолог). Кн. для учащихся. М.: Просвеще-
ние, 1986. 126 с.
Розанов А.Ю. Что произошло 600 миллионов лет назад. М.: Наука, 1986. 95 с.
Рождественский АК. На поиски динозавров в Гоби. 3-е изд. М.: Наука, 1969. 293 с.
Ружечка Б., Диттлер К. О чем рассказывают окаменелости. М.: Наука, 1964. 71 с.
Симпсон Дж. Темпы и формы эволюции. М.: Мир, 1948. 369 с.
Симпсон Дж. Великолепная изоляция. М.: Мир, 1983. 255 с.
Следы жизнедеятельности древних организмов. М.: Наука, 1993. 125 с.
Современная палеонтология: методы, направления, проблемы, практическое прило-
жение. Том 1, 2. М.: 1988. 925 с.
Сребродольский Б.И. Биологическая минералогия. Киев: Наукова думка, 1983. 100 с.
Стратиграфический кодекс. Изд. 2-е. СПб.: Изд. Межвед. стратигр. ком., 1992.120 с.
Татаринов Л.П. Очерки по теории эволюции. М.: Наука, 1987. 222 с.
Трофимов Б.А. Кости дракона. М.: Знание, 1964. 47 с.
Фауна и экосистемы геологического прошлого. М.: Наука, 1993. 125 с.
Федонкин МА. Бесскелетная фауна венда и ее место в эволюции. Тр. Пин. Т. 226. М.:
Наука, 1987. 176 с.
Хаксли Джулиан. Удивительный мир эволюции. М.: Мир, 1971. 112 с.
Харленд У.Б. и др. Шкала геологического времени. М.: Мир, 1985. 140 с.
Циттель К. Основы палеонтологии (палеозоология). Ч. 1. Беспозвоночные. М.:
ОНТИ, 1934. 1056 с.
Черкесов В.Ю. Палеонтологический определитель. Л.-М.: Глав. ред. геол.-разв. и
геофиз. лит-ры, 1934. 204 с.
Чижевский Г. В дебрях времени. Палеонтологическая фантазия. М.: Детгиз, 1963.
158 с.
Энциклопедия для детей. Т. 4. Геология. М.: Аванта плюс, 1995. 624 с.
Яковлева И.Н., Яковлев В.Н. По следам минувшего. М.: Дет. лит., 1983. 319 с.
Янин Б.Т. Терминологический словарь по палеонтологии. М.: Изд-во Моск, ун-та,
1990. 134 с.
38-'
УКАЗАТЕЛЬ РУССКИХ НАЗВАНИИ
Австралопитек 589, 590
Агната 326, 328, 330
Агностиды 180,184, 186,193, 194
Азиатский бурундук 570
Азиатский слон 565, 566
Аисты 578, 581
Акантоды 331, 334, 335, 336, 341, 342
Акритархи 44, 79-81
Акротретиды143
Аксолотль 378, 383
Актинии 17
Актиноптеригии 341, 344, 346, 349
Актиноцератоидеи 262
Акула-людоед 339
Акуловые 18
Акулы 18, 329, 331, 334-337, 338, 339, 341,
405, 407
Алданофитон 353
Алетоптерис 366
Аллигатор 409, 412, 414
Аллогромиины 87
Аллозавр 434, 458
Альбатросоподобные 474
Альпака 540
Аляскинский бурый медведь 542, 543
Амбиортус 474
Амбиортус Дементьева 475
Амбистома 383
Амеба 78
Амебелодон 563
Американский заяц 537, 577
Американский кролик 577
Американский лось 584, 585, 585
Американский мастодон 561, 562, 580
Американский нанду 489
Американский стервятник 486
Амиелоны 364
Аммониты 31, 31, 38, 109, 252, 262-275,
264,412,460
Аммоноидеи 39, 44, 263, 265, 266, 268
Амфиаспидные 326
Амфибии 5, 6, 17, 18, 39, 56, 57, 207, 346,
348, 349, 372-386, 375, 384, 389-391,
396,397,412,422
Анапсиды 387, 389, 401
Анаспид 326
Анаспидные 328
Ангиоспермы 59
Аневрофитовые 60
Аневрофитон 355
Анемоны 120
Анизограптиды 310
Анкилозавр 449, 450, 451,452, 455
Аннелиды 163
Аннулярия 362
Аномииды 55
Аномодонт 396, 397
Антиархи 332, 333, 334
Антилопы 29, 445, 542, 558, 577, 578, 582
Антракозавры 373, 379, 381, 382, 386, 389
Антропоиды 532
Анцилопода 545
Анчоус 345
Аралия 516
Араукариевые 413, 421
Араукария 365
Арахниды 212
Аргентавис 482, 483
Аргонавт 275
Арктоционидные кондилартры 532, 533,
543
Арктоциониды 532, 533
Артизии 364
Артикуляты 291, 291
Артокарпус 517
Артродиры 331, 332, 333, 334
Артроплевриды 214
Артроподы 174,175,176,177,193,195, 204
Архаичные птицы 483
Археогастроподы 225, 226, 226, 227, 228,
229
Археоптериевые 60
Археоптерикс 9, 468, 469, 470, 473
Археоцетес 543
Археоцеты 543, 544
Археоциаты 44, 100-102,100,101,105,125
Архозавры 389, 424, 418-467, 470
Астерофиллитес 362
Астропотерии 558
Ателостоматы 303
Атиридины 153
Атлантическая устрица 54
Атрипины 151
Аулопориды122,122,125
Африканский многопер 342
Африканский осел 557
УКАЗАТЕЛЬ РУССКИХ НАЗВАНИЙ
597
Африканский протоптерус 347
Африканский слон 565, 566
Африканский страус 484, 487, 489, 491, 495
Афросальпингидеи 93, 104
Бабочки 29, 218, 219, 220
Баклан 481
Бакланоподобные 474
Бактерии 8, 37, 50, 56, 65, 66, 71,125
Бактриан 541
Бактритоидеи 263
Бакулиты 268
Бандикут 506, 512
Барагванатия 353
Баран-толсторог 397
Бараны 447, 542
Барсуки 579, 583
Бархатные черви 213
Батоиды 339
Бахромчатоногие 199
Бациллы 37
Бегемот 535
Беззамковые брахиоподы 141-143,144,145
Безногие амфибии 384
Белемниты 23, 31, 32, 44, 272, 273-275
Беличьи 571
Белка 506
Белк а-летяга 570
Белкообразные 570
Беллерофонтиды 225, 227
Белоголовый орел 486
Беннеттиталес 366
Беннеттиты 366, 368
Беркут 486
Бескилевые 484-486
Бескрылые насекомые 39
Беспозвоночные 5-7, 14-16, 21, 30, 39, 75,
145,155, 221, 229, 231, 233, 272, 277, 307
Бесчелюстные 22, 39, 60, 324-330, 325, 326,
326
Бесчерепные 61
Бизон Фриггинса 585, 586, 586
Бизоны 450, 548, 577, 579, 579, 580, 582,
583 585
Бластоидеи 44, 277, 279, 280, 280-282
Блохи 220
Боа 418
Бобровые 571
Бобры 500, 572, 586
Боконервные 222
Болотный лунь 483
Болотный носорог 546
Большая песчанка 569
Большеноги 492
Большерогий олень 585, 585
Большие ящеры 432
Большой стервятник 578
Боргиенида 505
Ботриолепиды 333
Брадизавриды 392
Брадиодонты 336
Бранхиозавр 383
Брахиозавр 427
Брахиопоиды 377, 378
Брахиоподы 5, 7, 11, 12, 22, 25, 26, 29, 31,
32, 33, 39, 44, 56, 60, 61, 122, 126, 138-
159,139-141,155-157, 221, 229, 300
Бриофиты 354
Бродячие черви 163
Броненосцы 520-521
Бронтозавр 425
Бронтопс 549
Бронтотерии 545, 548-551, 549, 550
Брюхоногие моллюски 31, 227, 233, 250,
252,254, 271
Бульдожий тарпон 23, 345, 412
Бурый медведь 542, 543
Быки 542
Быстровианиды 381
Вальхия 364, 365, 370
Варан 418
Варанидпая ящерица 418
Верблюдка 218
Верблюдовые 535, 537-542, 537, 540
Верблюды 13, 27, 535, 537, 538, 539, 540-
542, 577, 579, 579, 586
Верметиды 165
Виверовые 526
Виверы 526
Викунья 540
Вилорог541, 542
Вилорогая антилопа 578
Вилороговые 535, 542
Вильямсония 368, 515
Виргинский филин 486
Власоеды 219
Впутреннераковинные головоногие 263
Водорез 465
Водоросли 5, 11, 37, 48, 49, 52, 58, 59, 65,
69, 70, 73, 85, 101, 126, 158, 159, 161,
227, 229, 235, 244, 256, 259, 271, 295,
304,311,315
Водосвинка 571
Волки 543, 579, 582, 583
Вольвокс 93
Вомбат 504, 506
Воробьи 465, 581
Воробьиная пустельга 486
Воробьиные 482
Ворон 486
Ворона 468,482
Воротничковые жгутиконосцы 93
Восковица 581
Вши 219
Выдра 501
Высшие ракообразные 201, 202, 203
Высшие растения 39, 47, 59, 101
Гавиал 410, 412-416
Гагара 474
Гагароподобные 474
Гадрозавры 443, 444, 445, 446, 455
Гадюка 418
Газели 542
Газелиевые верблюды 538, 539
Галеаспидные 328, 329
Гангамоптерис 370
Ганоид 344
Ганоидные 422
Гастроподы 5, 11, 15, 17, 23, 25, 26, 30, 31,
33, 44, 81, 87, 162, 200, 210, 224,
225-227, 225, 228, 229, 235, 241, 244,
271
598
КАМЕННАЯ КНИГА
Гаттерия 392, 417
Гвинейская свинья 571
Гексакораллы 38, 119, 120,120, 126
Геликоплакоидеи 307
Гелиолитоидеи 123-125, 124
Гемиптероидные 219
Гемиптероиды 219
Герматипные кораллы 120
Гесперорнис 473, 474
Гесперорнисообразные птицы 473
Гетеродонты 234, 239, 239,
Гетероморфные аммониты 268-270, 269
Гетеростраки 327, 329
Гибодонтная акула 337
Гибодонты 337, 338
Гигантская свинья 535, 536
Гигантские большеноги 484
Гиганский бобр 586
Гигантский короткомордый медведь 579,
581
Гигантский лемур 590
Гигантский олень 585, 585
Гидра 57,105, 111
Гидрозоа 315
Гидроидные 109
Гидроидные медузы 109
Гидроидные полипы 106
Гидроиды 81,127, 304
Гидроконозоа 110,111
Гидрокораллы 102, 106, 106,107
Гидрофида 418
Гиеновые 526
Гиенодоны 524, 525
Гиены 526, 583
Гимнолематы 130
Гинкго 300, 369, 424
Гинкгоалес 369
Гинкговые 60, 366
Гинкгофиты 369
Гиппарионы 38
Гиппоморфа 548
Гиппопотамы 27, 397, 431, 441, 534
Гиракодон 546
Гиракотерии 551, 552, 555
Гистрихосферы 81
Гладыш 219
Глиптодонты 520, 521, 521, 587, 587, 588,
589
Глоботрунканиды 38
Глоссоптериевые 60
Глоссоптерис 369
Гнатобелодон 563
Головастики 328, 383
Головоногие моллюски 12, 47, 58, 58, 224,
249-252, 250, 253, 254, 257, 259, 263,
264,271-275
Голожаберные 231
Голосеменные 15, 39, 355, 356, 364—371
Голотурии 18, 276, 304, 306, 306
Голотуроидеи 304, 307
Голубой краб 203
Голубь 463, 465, 490
Гомалозои 276, 291
Гоминиды 38, 589-592
Гомойостелеи 292, 293
Гомостелеи 292, 293
Гомфотерии 562
Гониатиты 252, 263-266
Горгонопс 397, 398
Горный козел 584
Горный лев 579
Граптолиты 5, 9, 30, 33, 39, 44, 58, 60, 81,
137,210, 291, 308-311, 315, 316, 322
Граптолоидеи 310, 312, 313, 315
Гребенчаторотые 135
Гребешки 233, 236-238, 244
Гребневики 59
Гремучие змеи 393, 418,
Грецкий орех 516
Грибки 53
Грибы 15, 37, 48, 49, 52, 53, 59, 66, 160, 296
Гризли 581
Грифы 581
Громовый ящер 425
Грызуны 18, 18, 19, 38, 42, 488, 500, 501,
571,571,572, 583
Гуанако 540
Губки 5,17,25,26, 31,48, 53, 60, 75,82, 93-
96, 94—97, 100, 102, 105, 106, 115, 121,
162,163,169, 170, 213, 295, 304, 308
Губковые44, 93, 97, 103, 105
Губоногие 214
Гу боротые 136
Гуси 465,580, 581
Дазикладиевые 53
Двинозавр 378
Двоякодышащие 12, 60, 347, 422
Двугорбый верблюд 541, 577
Двужаберные 273
Двукрылые 220
Двуногие тероподы 22
Двупалые страусы 485
Двупарноногие 213
Двустворки 5, 11, 12, 22, 25, 25, 26, 26, 31-
33, 44, 54, 55, 157, 196, 229, 233-248,
241,243, 271, 275
Двустворчатые моллюски 5, 15, 31, 47, 58,
138, 139, 229, 233-241, 234, 242, 243,
244, 247,411
Дейнонихус 440
Декаподы 201, 203
Дельфин 407
Дельфиновые 543
Дендроидеи 310, 311, 312, 313, 315
Десмостилиды 559, 568, 568
Десятиногие 201, 203
Диапсидные рептилии 413
Диапсиды 389, 410, 417, 418
Диатомовые водоросли 9, 25, 38, 80, 83
Диатрима 481
Дидельфиды 504
Дикобраз 586
Дикобразовые 570, 571
Диметродон 394
Динозавроид 460, 461
Динозавры 1,9,11,12,17,19, 21, 22, 29, 36,
38,126, 231, 248,271, 389,400, 414, 416,
418, 426, 431, 432, 441-462, 468, 469,
473, 474, 504
Динорнис 489
Динотериоиды 559, 560
Динофлагелляты 44, 60, 79, 80, 81, 82, 120
Диноцефал 396, 397
УКАЗАТЕЛЬ РУССКИХ НАЗВАНИЙ
599
Диогенорнис 485
Диплодок 1,434
Диплокаулюс 384
Дипнои 346
Дипротодонтиды 505
Дипротодонты 589
Дискозавриск 387
Диссорофоидные лабиринтодонты 383
Дицинодонт 396,397, 422
Додо 22,62, 490
Докодонты 500
Долгопяты 530, 531, 531
Домашняя кошка 499, 506
Доядерные 50, 66
Древесные ленивцы 520, 521
Древесные птицы 482
Древнекрыл 468
Древнекрылые 216, 217, 218
Древоточец 244
Дромадер 540,542
Дроморнис Стиртона 491, 493
Дроморнитиды 484
Дронт 484, 490
Дубы 11, 29, 577
Дюгони 567
Дятлы 482, 581
Евгенодонтиды 337
Евролимнорнис 474
Ежи 501, 528
Ель 27
Енотовые 526
Еноты 525
Епископские сосны 577
Ехидна 501
Жаба 384, 582
Жаброногие 195, 196, 201
Жаворонок 482
Жвачные 535, 542
Жгутиковые 85
Жгутиконосные 79
Жгутиконосцы 82
Желе-рыба 105
Желудевые черви 277, 308, 309, 322
Жемчужница 244
Жениорнис Ньютона 492, 493
Жесткокрылые 220
Жирафовые 542, 588
Жирафы 427, 535, 542, 597
Жирафовый верблюд 539, 539, 540
Жуки 29, 220, 398, 582
Жук-плавунец 21
Жук-трупоед 581
Журавлеобразные птицы 480, 481
Журавли 486, 582
Завроподы 422, 423, 424, 425, 426, 427-432,
434,436,449,456,458
Заднежаберные гастроподы 230, 230, 231
Зайцеобразные 523
Зайцы 523
Замковые брахиоподы 141-143, 145, 153,
240
Заурисхии 421, 423, 435, 440, 454
Звери 5, 496, 500
Зверообразные рептилии 38, 387, 395, 399
Зебры 454, 557
Зеленые водоросли 25, 26, 50, 52, 53, 59, 60,
66
Зеленые одноклеточные протисты 52
Землеройки 499, 503, 512, 523, 528
Земляные белки 577
Земляные черви 384
Земноводные 56, 60
Зимородок 482
Зифодонт 416
Змеехвостки 26
Змеи 384, 389, 392, 407, 409, 412, 418,427
Золотая рыбка 331, 344
Золотистые водоросли 60
Зооксантеллы 85,120
Зоохлореллы 85
Зубатые киты 109, 543
Зубатые птицы 473, 474
Зубчатоклювая куропатка 486
Иглокожие 5, 7, 25, 44, 60, 61, 109, 162,
163,276-307, 322, 460
Игуана 418, 441
Игуанодонтиды 443, 446, 454
Игуанодонты 442
Известковые водоросли 29, 53, 203
Известковые губки 95, 96, 126
Измененноротые 131
Изоптера 219
Ильбандорнис 492
Ильная рыба 344
Императорский мамонт 579, 582
Инадунаты 285, 287, 288, 288
Иналиарктиды 526
Индейка 492
Индийский слон 566
Индри 530
Индрикотерий 57
Инотерия 504
Иноцерамы 240
Ихтиозавры 389, 405, 406, 407, 414, 416
Ихтиорнис 4 74
Ихтиостега 372, 373, 381
Ихтиостегиды 372
Кавиоморфы 571
Кагуаны 529, 530
Казеидный пеликозавр 395
Казеиды 396
Казуары 484, 485, 489, 492
Кайман 413
Какопс 374, 375
Кактусы 21, 27
Каламитес 362
Каламиты 362
Калифорнийская индейка 486
Калифорнийский бегун 486
Калифорнийский кондор 486
Калликсилон 355
Калликсилон Залесского 355
Кальмары 23, 263, 272-275
Кальпионеллиды 83
Кальцихордаты 291
Камераты 289, 290, 291
Канадская казарка 486
Канадский олень 584
Канарейка 274
600
КАМЕННАЯ КНИГА
Капский трубкозуб 533
Капториноморфы 387
Каракара 486
Каракатица 272, 274
Карась 345
Карибу 584
Карликовые слоны 567
Карликовый дуб 577
Карликовый лемур 531
Карнозавриды 432
Карнозавры 432-434, 441
Карп 345
Карпоидеи 44,278, 291-293, 307
Кассидулоиды 303
Катарта тераторнис 484
Каури 225
Кашалотовые 543
Кваква 486
Квиполлорнис 484
Кейтония 369
Кенгуровые крысы 577
Кенгуру 495,504, 506, 509
Керпия 371
Кецалькоатль 465
Киви 484, 485, 489
Кики 518
Кипарис 577
Кистеперые 60, 341, 375
Китообразные 543, 544
Киты 57, 339, 411, 427, 430, 456, 513
Киты-археоцеты 543, 544
Кишечнодышащие 308
Кишечнополостные 59, 105, 308
Клаватиполленитес 516
Кладоселахии 336, 337
Клаудиозавр 407
Клен 517
Клещ 210
Клипеастероидеи 302, 303
Клопы 219, 582
Клупеоморфа 345
Клюворыловые 543
Книдарии 51, 59, 60, 105, 308
Коала 506
Кобра 408
Кожистокрылые 219
Козодой 482
Козы 542
Койоты 579
Кокки 37
Кокколитофориды 38, 80, 80, 81
Кокколиты 80, 81, 460
Колеоидей 263
Колумбийский мамонт 564, 580
Кольпоксилон 366
Кольчатые черви 60, 77, 160, 161, 163-167,
169,177, 222
Кольчецы 163
Колючие хищники 436
Комары 220
Кондилартры 532-535, 533
Кони 11
Конодонтоморфы 44
Конодонтоносители 317, 318, 319
Конодонтофориды 317
Конодонты 9, 38,44, 291, 317,318, 320,321,
327
Конулярии 109, 110, ПО
Конхостраки 196
Копеподы 200
Копытные 532-559, 583
Корабельные черви 235, 243, 244
Коралловые водоросли 59
Коралловые губки 96
Кораллы 9, 11, 12, 14, 24, 25, 25, 26, 29-31,
44, 46-48, 58, 59 , 75 , 75, 76, 105, 106,
110-127, 129, 133, 135, 156, 158, 159,
163, 243, 245, 247, 256, 259, 284, 285,
303
Кордаитантус 364
Кордаитес 364
Кордаитовые 60, 364
Коринексохиды184
Корненожки 84
Короноидеи 282
Кистеперые рыбы 346, 348
Костистые рыбы 275, 336, 338, 339, 341,
344,345
Костнозубые птицы 481, 482
Костнопанцирные 329
Костнопузырные 345
Костнощитковые 324
Костные рыбы 331, 334, 340, 341
Косули 541, 542
Котилозавры 39, 387, 391, 392, 396, 497
Кот-рыба 332
Кошачьи 526, 579
Кошачья трава 56
Краб-подкова 205, 207
Крабы 18, 23, 174-176, 194, 195, 201-203,
203, 272, 273, 333
Кранииды 143
Красное дерево 518
Краснохвостый канюк 486
Красные водоросли 25, 26, 52, 60, 70, 125
Креветки 195
Кремневые губки 94, 95, 95, 97
Кремнероговые губки 95
Креодонты 524 -526, 525, 535, 537
Крикливый зуек 486
Криноидеи 23, 26, 44, 229, 276, 277, 281,
282, 284-291, 294-298,306
Криптодиры 402, 403
Криптодонты 238, 238
Криптостоматы 133
Крокодилы 22, 376, 389, 392, 395, 407, 409,
412-414, 415, 416, 420, 421, 431, 441,
469,496
Кролики 426, 506, 523, 582
Кроншнеп 486
Кроссоптеригии 348
Кроты 528
Крошечный толстоголовый ящер 447
Круглоротые 58, 330
Круглые черви 160, 161, 167
Кружевной краб 194
Крылатые насекомые 39
Крылатые ящеры 496
Крылатый палец 463
Крыложаберные 60, 308, 322
Крылоногие гастроподы 244
Крылощитковые 324
Крысиный кенгуру 511
Крысы 495, 500, 572
УКАЗАТЕЛЬ РУССКИХ НАЗВАНИЙ
601
Крышечный коралл 115, 116
Кряква 486
Ксенакантиды 337
Ксенаканты 337
Ксенонгуляты 558
Ксифосуры 205
Ктенакандиды 337
Ктенаканты 338
Ктеностоматы 135
Ктеностомы 135
Кузнечики 29, 217, 219
Куксония 353
Кукушка 482
Куланы 557
Кулик 479
Куликоподобные 474
Куличиные 480
Куньи 526, 527
Курица 489
Курообразные птицы 481, 492
Куторгиниды 143
Лабиринтодонты 39, 349, 372—386, 373, 377
Лабрейская совка 486
Лабрейский аист 486
Лабрейский кондор 486
Лама 535, 537-540
Ламантины 410, 567
Ламеллибранхиаты 233
Лангусты 202
Ланцетник 309, 322, 323
Ласки 526, 579
Ластоногие 526, 527
Латимерия 346, 349, 350
Легочные гастроподы 231, 232, 232
Лемуровые 530, 531
Лемуры 530, 531, 531
Ленивцы 13, 17, 23, 45, 520-523
Лентохвостый голубь 486
Леопард 506
Лепидодендрон 360, 361
Лепидозавры 389, 417, 418
Лепоспондилы 372, 374, 381, 384-386
Лептоцефал 345
Лесной волк 579
Летающие лемуры 529
Летающие птицы 456, 482
Летающие рептилии 420, 465, 467
Летающие ящеры 389, 464, 466
Летающий дракончик 467
Летучие лисицы 529
Летучие мыши 465, 466, 528, 529, 529, 530
Летучие рыбы 345
Летяговые 571
Лжемедузы 108, 109
Лилейные 522
Лима 244
Лингула 156
Лингулиды 26,143, 155, 157,158,
Лиссамфибии 372, 384-386
Листовидные креветки 201
Листоногие рачки 196
Листрозавр 398
Лисы 579, 583
Литистидные губки 95
Литоптерны 558, 559, 589
Литторина 227
Литуитес 259
Лихенарииды123, 125
Лихиды 187,188
Лишайники 59, 204
Логгерхед 404
Ложная лошадь 559
Ложнозубые птицы 481, 482, 483
Локсодонты 564, 566
Лопатоногие моллюски 31, 31
Лори 531
Лоси 542
Лососеобразные 345
Лосось 345
Лошади 27, 496, 509, 532, 535, 544, 545,
551-557, 554 , 555, 558, 577, 578, 579,
582-584, 586
Лошадиные 545, 555
Лошадь Пржевальского 557
Лошадь “рассвета” 551, 552
Луговой балоба 486
Лучеперые 341,341, 342, 343, 379
Львы 29,453, 523-526, 578, 579
Люди 17, 29, 36, 219, 322, 416, 487
Лягушки 372, 384, 385, 445
Ляонинорнис 474
Магнолиевые 517
Магнолия 517
Макросемииды 344
Макросцериды 523
Макроухениды 558
Малайский дикобраз 5 70
Малакостраки 201
Малиновка 24
Малощетинковые черви 163, 166
Маммалии 18, 56
Маммут 562
Мамонты 12, 13, 21, 57, 564, 566, 577, 584-
586
Маркозетки 531
Мартышки 532
Мастодонты 560-563, 577, 580, 584
Мегалозавры 432, 433
Медведи 525, 527, 542, 583
Медвежьи 526
Медузоподобные 107
Медузы 75, 75, 77, 105-109, 108
Медуллоза 366
Мезогастроподы 228, 229, 230
Мезогиппус 552-556
Мезозавры 401
Мезозухий 414, 415
Мезонихидные кондилартры 525
Мезонихиды 542, 543
Мерикгиппус 556
Меритерии 559
Мескитовое дерево 393
Метасеквойя 518, 519
Метопозавры 376
Метриоринхиды 414, 416
Мечехвосты 204, 205, 206, 207
Мидии 55, 233, 238, 244, 295
Миксины 58, 130, 330
Миллепоры 106
Миноги 58, 130, 330
Митилиды 26, 33
Михиранг жениорниса 494
602
КАМЕННАЯ КНИГА
Михиранги 484, 491, 492, 495
Млекопитающие 5, 6, 8, 9, 22, 29, 60, 231,
297, 392, 399, 400, 456, 458, 480, 498-
513, 520-592
Многоклеточные 39, 51, 73, 74, 75, 76, 78,
81, 93, 94
Многолучевые 104
Многоножки 169, 211
Многощетинковые черви 162,163,164,166,
167,168
Моа13,19,484,485, 489, 491, 590
Можжевельник 577
Мозазавры 23, 24
Мозоленогие 535, 538
Мокрецы 21
Моллюски 5,14, 22, 23, 33, 57, 60, 109,138,
221, 222, 224, 226, 227, 229, 231, 233,
235, 236, 240, 241, 243-245, 249, 251,
252,256, 271
Моллюскоподобные 138
Монера49, 49, 50, 51, 59
Мономорфные аммониты 267
Моноплакофоры 162, 221, 222, 222, 225,
225, 226, 227, 228, 240,251,252
Моргануконтиды 498, 499, 503
Моржи 526, 527
Моржовые 526, 527
Морская капуста 59
Морская корова 410
Морская черепаха 404, 409
Морские анемоны 17
Морские бутоны 277
Морские гребешки 55
Морские гуси 199
Морские ежи 11, 14, 26, 31, 31, 33, 39, 270,
271, 276, 277, 297-300, 299, 301, 302,
304-307
Морские желуди 199, 200
Морские звезды 23, 26, 61, 276, 277, 282,
294-298, 296, 304
Морские котики 527
Морские лилии 11, 23, 29, 61, 229, 272, 276,
277,281,282, 283, 285-287, 286
Морские львы 527
Морские огурцы 18, 276, 304, 306, 306
Морские пауки 212
Морские перья 75, 76, 129, 133
Морские пузыри 277
Морские разбойники 297, 299
Морские свинки 571
Морские уточки 199, 200
Морской конек 344
Морской салат 59
Морской целакант 349
Москиты 220
Мохноногий канюк 486
Моховидные 60, 127
Мультитаберкулаты 530
Муравьеды 520
Муравьи 220
Мускусный бык 583
Муха-ктырь 218
Мухи 29,216,218, 220
Мхи 59, 127, 204
Мшанки 5, 9, 11, 12, 25, 26, 31, 32, 39, 44,
58, 60, 126-137, 128, 136,158, 159, 304,
308,317
Мшистые 127
Мышеобразные 569
Мыши 462, 500, 572, 577
Мышиные 571, 572
Мышовки 571
Мышь-малютка 569
Мягкокожая черепаха 404
Мягкораковинные 201
Мясистолопастные 344, 346
Мясные мухи 582
Мяун 56
Наземные ленивцы 13, 15, 507, 520-523,
578, 578, 580, 586, 587, 588, 589
Наземные растения 53, 78, 79, 207, 351-
371, 514-519
Нананциус 472
Нанду 484, 485
Нарваловые 543
Наружнораковинные головоногие 275
Наружночелюстные насекомые 215
Насекомоядные 512, 513, 528, 571
Насекомые 5, 6, 15, 20, 21, 22, 29, 60, 61,
161, 169, 174-176, 212, 214, 215, 216,
217, 218, 219, 220, 398, 463, 466, 495,
499, 513, 529, 581, 582
Настоящая устрица 54, 55
Настоящие многоклеточные животные 94
Настоящие мхи 59
Настоящие птицы 470, 471, 473, 474, 476,
477
Наутилоидеи 44, 249, 251, 252, 253, 254,
255, 256, 257, 258, 259, 260, 261-266,
268,273
Наутилус 11,249-252, 263, 273
Неандерталцы 592
Небесное дерево 518
Невроптеридиум 370
Невроптерис 366
Невроптероиды 219, 220
Немертины 160,161
Неогастроподы 228, 229, 230, 230
Неогнатические птицы 474,
Неоптеригии 343, 344
Неоптеры 219
Неоселлахии 338
Неотелеостеи 345
Неотригония 239
Непарнокопытные 532, 535, 544-557, 545-
555
Непарнопалые 544-557
Неполнозубые 513, 520-523, 578
Неправильные морские ежи 298, 300, 302,
304
Низшие грибы 79
Низшие многоклеточные 104
Низшие черви 167
Низшие четвероногие 5
Нисса 517
Новокрылые насекомые 217, 219
Новые морские ежи 298
Ногохвостки 214, 215
Носороги 13, 15, 16, 16, 17, 28, 441, 505,
534, 545, 545, 546-548, 546-548, 550-
552, 584
Носорогообразные 545, 546
Нотозавр 389, 407, 408, 409
УКАЗАТЕЛЬ РУССКИХ НАЗВАНИЙ
603
Нотостраки 196
Нотоунгуляты 557-559, 587, 589
Нуммулитиды 90, 92
Нутриевые 571
Ныряющие птицы 472, 473
Обезьяны 496, 531, 532, 589, 590
Обезьяны Нового Света 531, 532
Обезьяны Старого Света 531, 532
Оболеллиды 143
Оболочники 59, 309
Обыкновенный хомяк 569
Овцебык 583
Овцы 538
Одногорбый верблюд 540
Одноклеточные водоросли 78, 81
Одноклеточные животные 81, 93
Однопроходные 501
Одонтоплевриды188, 193
Оленевые 542
Оленеллиды 177
Оленерогий жираф 588
Олени 535,542, 583
Оленьки 535, 542
Оленьковые 542
Оливы 227
Омары 176, 180, 195, 201, 202
Онихофоры 160, 161, 163, 168-170, 169,
176, 177, 213, 214
Орбитоиды 90
Орел Гриннема 486, 580-582
Орел Даггета 486
Ореодонты 535—537, 536, 547
Орлы 581, 582
Орнимегалоникс 484, 488, 489
Орнистомимиды 440
Орнитисхии 421, 441, 442, 443, 448, 454
Орнитоподы 441, 442, 453, 454
Ородонтиды 337
Оромерисиды 537
Ортиды 39, 145, 158
Ортоптероидеи 219, 220
Ортоцератиды 262
Осетр 342
Осмундовые 371
Остеоглоссоморфы 345
Остеостраки 328, 329, 329
Остиолепиформные рипидистии 349
Остракодермы 324, 326, 328, 329, 330, 333
Остракоды 25, 26, 44, 197, 198, 199—201,
272
Осы 220
Осьминог 263, 272-275
Отогорнис 474
Офиакодонт 396’
Офиоцистия 295
Офиоцистоидеи 294
Офиоцистоиды 307
Офиуроидеи 297
Офиуры 26, 296
Паганки 582
Палеледусы 483
Палеогетеродонты 238, 239
Палеогнаты 484-486, 492
Палеомастодон 562
Палеониски 342
Палеонисциды 341
Палеоптеры 217
Палеотаксодонты 238, 241
Палеотерии 552
Палорхестиды 589
Пальмы 27
Панголины 568, 570
Пантодоны 527, 527, 528, 528
Панцирная щука 343
Панцирные 520
Панцирные муравьеды 570
Папоротники 15, 21, 27, 60, 265, 355, 363,
364,393, 413, 422, 424, 519
Папоротниковидные 35
Парабластоидеи 279
Паракриноидеи 280, 281
Парарептилии 387
Парейазавры 391, 392, 396
Парнокопытные 532, 535-542, 545, 585,
585
Патериниды 143
Пауки 20, 210, 211, 212, 214, 217
Пауки-долгоножки 211
Паукообразные 204, 210, 212
Паукообразные обезьяны 531
Пахицефалозавриды 454
Певчие воробьиные 482
Певчие птицы 482
Пекари 535
Пектениды 244
Пелециподы 233
Пеликан 465, 481
Пеликозавры 3.9, 387, 393, 394, 395, 436,
497
Пентакринитиды 287
Пентамериды 145, 149, 149, 158
Первичнобескрылые насекомые 214
Первичнотрахейные 169
Первобытные люди 11
Переднежаберные гастроподы 227, 228, 229
Перепелоподобная птица 485
Перепончатокрылые 220
Перишэхиноидеи 300, 302
Песчанки 571
Песчаные доллары 302, 304, 305
Петалихтиды 334
Петалодонтиды 337
Печеночные мхи 59
Пикногониды 212
Пикнодонты 344
Пингвиноподобные птицы 478
Пингвины 22, 409, 478, 495
Пиранья 345
Пиротерии 558
Питон 418
Пищухи 523
Плагиозавры 377
Плакодермы 60, 330, 331, 333-336, 341
Плакодонты 389, 408, 410
Пластинчатожаберные 233
Платан 516
Платибелодон 563
Плаун 356
Плауновидные 15, 60, 214, 355, 356, 360-
362,391, 393
Плацентарные 38, 503-505, 512, 520-592
Плачущая горлица 486
604
КАМЕННАЯ КНИГА
Плевродиры 402
Плевроканты 336, 337
Плезиадаписы 530,
Плезиозавры 19, 389, 407, 408, 410, 416,
460, 467
Плеченогие 56
Плиогиппус 556
Плоские черви 60, 160
Плотоптериды 478
Плотоптерус 478
Поганка 486
Подвижные иглокожие 294
Поденки 216
Позвоночные 5—7, 14, 18, 21, 22, 30, 39, 44,
58, 59, 61, 160, 175, 217, 248, 272, 291,
292, 309, 322, 324-350, 372-513, 520-
592
Покрытосеменные 60, 515-519
Полевка-экономка 569
Полевки 571
Полорогие 535, 542
Полосатая белка 570
Полосатый ястреб 486
Полужесткокрылые 219
Полуобезьяны 530
Полухордовые 59, 277, 308, 310, 322
Пороносцы 93
Португальский кораблик 315
Поссумы 506, 530
Правильные морские ежи 298, 300, 303, 304
Пресбиорнис 478, 480, 481
Пресмыкающиеся 56, 60, 382, 395
Пресноводная черепаха 404
Пресноводные рыбы 207
Приапулиды160, 161, 162,167, 168
Прикрипленные иглокожие 281
Приматы 38, 530-532, 571
Примитивные многоклеточные 93
Примитивные парнокопытные 535
Примитивные хищные 526
Примитивный верблюд 537
Примобуккониды 482
Пристихампсины 416
Продуктиды148, 148
Проетиды 186
Прозавроподы 423, 424, 431
Прокариоты 37, 39, 50, 66, 72, 73, 74, 78
Проколофон 398
Простейшие 49, 56, 78, 166
Протеротерииды 558
Протисты 27, 51-53, 51, 56, 59, 78-81, 80,
83, 85, 90, 93, 94
Протоавис 470, 471
Протобранхии 238
Протодонаты 217
Протоптерусы 347
Прыгунчиковые 523, 524
Прямокрылые 218, 219
Псаммостеид 326
Псаммостеидные 326, 327
Псевдодонторнисы 481, 482
Псевдопеталихтиды 334
Псевдоцератиты 265
Псилотум 354
Псилофит 354
Псовые 526
Птераспидаморфные 324, 327
Птераспидные 326
Птераспиид 326
Птеридоспермы 15, 366, 368, 368, 370, 371,
515
Птериоморфы 54, 234, 238, 238
Птеробрапхии 308, 309
Птеродактилоид 467
Птерозавры 420, 460, 463, 464, 466-468,
495,528, 529
Птероподы 231, 244
Птиктодонты 334
Птихопарииды 185, 186
Птицетазовые динозавры 469
Птица-лира 482
Птица-мышь 482
Птицы 5, 6, 13, 17-19, 22, 23, 38, 55, 60, 61,
62, 438, 456, 457, 468-496, 469-472,
474-494, 581-583,590
Пузырчатопоровые 131
Пума 579
Пурпурные бактерии 51
Пуховка 482
Пухоеды 219
Пчелы 220
Радиолярии 9, 25, 44, 82, 84, 84, 85
Радиоциаты 104
Разнолучевые 104
Раки 195, 202, 203
Раки-кроты 202
Раки-отшельники 202
Раковинные листоногие 196, 197
Ракообразные 20, 25, 60,194-203,196-203,
213,401,407
Ракоскорпионы 198, 199, 205, 209
Ракушковые рачки 197
Рамфоринхоиды 463, 466
Растения 5-7, 11, 14, 15, 19, 21-23, 25, 26,
27, 28, 30, 48-50, 49, 51-54, 56, 59, 61,
62, 65, 66, 78, 79, 82, 351-371, 514-519
Расщепленные растения 50
Рахитомы 375
Редлихииды 184
Ренаниды 334
Рептилии 1, 5, 6, 14, 17, 18, 19, 21-23, 30,
33,57,207,272, 345, 372-376, 379, 382,
383, 386-468, 388-467, 496-513, 497,
498, 543
Ресничные 60, 83
Рецептакулиты 93, 97, 102-104, 102
Ржанкообразные 481
Ризоподы 84
Ризостомиды 109
Риниопсиды 60, 354
Риния 354
Ринхозавр 397, 417
Ринхонеллиды150, 150, 152,158
Рипидистии 346,348, 349, 350
Рогатый динозавр 450, 453
Рогатый землекоп 572
Роза 517
Ростроконхи 240, 241
Роталиины 89, 90
Ругозы 39,111-119, 112-118, 125, 132, 247
Рудисты 38, 247, 247
Руконожки 531
Ручейники 220
УКАЗАТЕЛЬ РУССКИХ НАЗВАНИЙ
605
Рыба-солнце 342
Рыбообразные 324, 326, 327
Рыбояд 379, 394, 417
Рыбы 5, 6, 9, 11, 14, 19, 21, 23, 26, 229, 250,
272, 273, 275, 324, 328, 330-350, 332-
349
Рыбы-хищники 273
Рысь 579
Саблезубые кошки 526, 526, 527, 583
Саблезубые тигры 505, 579, 581
Садовая соня 569
Сайгаки 541, 542, 583
Саламандра 384, 385, 372
Саранчовые 219
Саркодовые 60
Саркоптеригии 341, 346
Сверлильщики 229, 243
Сверлящие моллюски 243
Сверчки 29, 219
Свинковые 571
Свинообразные 535
Свиньи 23, 495, 534, 535, 549
Себекозухии 416
Себециды 416
Северный канадский олень 584
Северный олень 27, 542
Североамериканский дикобраз 571
Североамериканский лось 585
Сегнозавриды 440
Сеймурия 382
Секвойя 29, 518
Селагинелла 355
Сельдь 333
Семенные папоротники 59, 393, 515
Семионистиды 344
Семонтэостомиды 109
Сенокосцы 211
Сердцевидные морские ежи 303, 304
Серпула 165
Серпулиды 165
Сетчатокрыловидные 220
Сетчатокрылые 220
Сивучи 527
Сигиллярия 362, 392
Сидеропс 379, 380
Сидячие черви 163
Сикамора 29
Силикофлагелляты 80, 82
Симморииды 336
Синапсида 387, 389, 392, 393, 395, 401, 497
Синапсидные рептилии 497
Синезеленые водоросли 50, 66, 125
Синий кит 544
Сирены 559, 567
Сирингопориды 122, 125
Сифонофоры 106, 315
Скалярия 344, 345
Скат 331,334-336,338, 339
Скафоподы 25, 26, 223, 223, 224,240
Скватинактиды 337
Склерактинии 119, 120, 120
Склероспонгии 93, 96, 97, 98, 102
Скорпионницы 220
Скорпионы 208, 210-212,217
Скребни 160, 161
Скунсы 526, 579
Слизни 232
Слон-ложка 563
Слоновые 560, 564
Слоновая птица 590
Слон-совок 563
Слоны 11,17, 21,62, 559, 560, 563-567,565
Слоны-карлики 566
Соаниты 93, 104
Собаки 495
Собаковидные 22
Сова 481,484, 488
Совиный лягушкорот 484
Современная лошадь 481, 556, 557
Современные птицы 469, 473, 474, 486
Сольпуги 211
Сом 345
Сомастероидеи 296
Соневые 571
Сорока 470, 486
Сорокопут 486
Сосна 29
Сосудистые растения 44, 353
Сотовые табуляты 121, 121
Спатангоиды 303
Спермоптерис 367
Спинозавриды 436
Спирифер 158
Спирифериды 145, 152, 153, 155, 158
Спириферины 152
Спирорбис165
Спирула 274
Спонгиоморфиды 106, 126
Стебельчатые иглокожие 276, 277, 281, 282,
284,286
Стегодон 564
Стегозавры 448, 449
Стегоцерас 447
Стеклянные губки 32, 57,
Стеллерова морская корова 567
Стенолематы 130
Стеншоеллиды 334
Стервятники 433, 580-582
Стереоспондилы 376, 377, 381
Стигмария 361
Стиластерины 106
Стилофоры 292, 293
Стоножки 214
Страусы 438, 440, 453, 511
Страшный волк 579
Стрекающие 105
Стрекозы 21,216, 217, 219, 220
Стриж 482
Строматолиты 29, 30, 37, 39, 50, 65-71, 66-
70, 73, 125
Строматопораты 12, 93, 97, 98, 103, 126
Строфомениды145,146, 148,153, 155, 158
Сумчатые 38, 526, 587, 589, 589
Сумчатые волки 13
Сухопутная черепаха 404
Сфенакодонт 393-395
Сфенакодонтный пеликозавр 395
Сфенодонтиды 391, 417
Сфенофиллюм 362, 363
Сферактиноиды 104, 126
Сфинктозоа 93, 98-101, 98
Сцифоидные медузы 109
Сцифомедузы 109
606
КАМЕННАЯ КНИГА
Табулятоидеи 39, 121—125, 121, 122, 124,
125
Табулятоморфные кораллы 121, 124
Табулятоморфы 121
Табуляты121
Тапироиды 545
Тапиры 62, 545, 552, 584
Таракановые 219
Тараканы 217, 219, 220, 398
Тарзиоиды 531, 531
Тарпан 345
Тасманитиды 81
Тарсометатарсус 479
Текодонты 398, 418, 420, 421-424, 432, 436,
464
Текстуляриины 89
Телодонты 326, 327
Темноспондилы 373-375, 376, 381, 385, 386
Тентакулиты 170
Терапсидный пеликозавр 395
Терапсиды 387, 394-398
Тераторнис 486, 582
Тераторнис Мериэма 487
Теребратулиды 145, 153,154, 158
Терия 503, 504
Термиты 216, 219
Тероподный динозавр 460, 469, 473
Тероподы 22, 423, 424, 432, 434, 435, 436,
437, 440, 441, 453, 454, 457
Тероцефал 398, 399, 499
Тетрадииды123, 124, 125
Тетракораллы 112, 114, 119
Тетраподы 349, 386, 395, 396, 436
Теутиды 274
Тецидные брахиоподы 154, 158
Тигры 458, 526
Тилаколеониды 589
Тиллодонты 533
Тиляколесниды 506, 510
Тинаму 485
Тинтинниды 83, 83
Тираннозавриды 434, 435, 446, 450, 453
Титанис 481
Титанотерии 548
Тли 219
Толстоголовый динозавр 446, 447
Толстоголовый ящер 447
Тополь 516
Трахейнодышащие 169, 213
Трахейные 169
Трематозавр 379
Трепостоматы 131, 132
Трепостомные мшанки 132
Трепостомы 132, 133
Трехосные шестилучевые губки 95
Тригониды 503
Тридакна 235
Триконодонты 499, 500, 503
Трилобитоиды194
Трилобиты 5, 18, 22, 36, 39, 44, 60, 162,
174-195,192, 204, 205, 255, 316
Тримеропсиды 60
Трипсы 219
Тритилодонтные цинодонты 500
Трогон 482
Трубкозубы 533
Трубчатопоровые 130
Трубчатые табуляты 121, 122
Трубчатые черви 304
Туатары 417
Тубулипораты 131
Туникаты 328
Тушканчиковые 571
Тысяченожки 213, 214
Тюленевые 527
Тюлени 407, 526
Угорь 345
Удильщики 345
Узконосые обезьяны 531, 590
Уинтатерия 533, 534
Улитки 6, 11,12, 23, 26, 221, 224-227, 229,
231-233, 246, 249-251, 256, 259, 261,
266,268, 295, 582
Униониды 239, 248
Унирамии 60, 212, 213
Урохордовые 328
Усатые киты 543, 544
Усоногие рачки 25, 26, 195, 199, 200, 200,
201
Устрица виргинская 55
Устрицы 23, 24, 26, 54, 55, 141, 155, 229,
233, 238, 243, 244-246, 245, 246, 270,
271,295,429
Утка 479, 582
Утконос 501, 502
Утконосые 443, 444, 455
Утконосый динозавр 17, 447, 448
Уховертки 219
Ушастые тюлени 526, 527
Фавозитиды 121, 121, 123, 165
Фазан 492
Факопиды 186
Фенакодус 551
Фиговые деревья 516
Фигоподобный 361
Фикус 517
Филактолематы 129, 130
Филлокариды 201, 202
Филлолепиды 333, 334
Фитозавры 412, 413, 414, 415, 422
Фитофаги 449, 454, 461, 506
Фламинго 479
Фламинговые 480
Фламингоподобные 481
Флексибилии 289
Фолады 237
Фораминиферы 5, 9, 25, 26, 29, 38, 44, 64-
67, 70, 85, 86, 87, 88, 89-92, 137, 264,
266, 270-272, 460
Форамы 85
Форель 331,344, 345
Форониды 60, 127, 138, 317
Фороракоидные птицы 505
Фороракос 480
Форорациды 480, 481
Фрины 211
Фузулинины 89, 90, 92
Хализитиды 123-125, 125
Халикотерии 545, 545, 546, 587
Хамелеон 418
Хампсозавр 410, 416, 417
УКАЗАТЕЛЬ РУССКИХ НАЗВАНИЙ
607
Хаоянгия 474
Харовые 53
Харофиты 52, 53
Хасматозавр 398
Хвойные 59, 60, 358, 364, 365, 393, 424,
518
Хвощевидные 60
Хвощи 359, 360, 362, 393, 422, 424
Хейлостоматы 136
Хейлостомные мшанки 136
Хейлостомы 136
Хелицеровые 60, 204, 208, 210, 213
Хететиды 93, 96, 123, 125, 126
Хететоидеи 123, 124, 124
Химера 331, 334, 336, 338, 339
Хиолиты 162, 168, 170, 230, 231
Хитинозоа 44, 81, 82
Хитоны 25, 26, 222, 223, 233, 241
Хищные 525-527, 542
Хищные копытные 542, 543
Хищные млекопитающие 481
Хлебное дерево 517
Хлорофиты 53, 66
Хоботковая собачка 524
Хоботные 532, 559- 567, 561, 583
Холастероиды 303
Хомяковые 571
Хондростеи 341
Хонетиды 148
Хордовые 39, 61, 130, 162, 277, 291-293,
309,322-324
Хорнеофитон 354
Хризамеба 51
Хрониозухии 381
Хрящевые ганоиды 344
Хрящевые рыбы 60, 331, 334, 335, 336, 338,
339,344
Хсисозухиды 416
Цапля 582
Цветковые 38, 59, 515-519
Целаканты 346, 349, 349, 350
Цел о лепиды 327
Цельноголовые 339
Цепочечные табуляты 121-123, 125
Цератиты 265
Цератоморфа 545
Цератопс 450-452, 453, 455
Цефаласпидоморфные 327, 328
Цефалоподы 25, 249, 251, 252, 254, 256,
257,273, 275
Цианобионты 25, 26, 37, 44, 50, 51, 59, 60,
66, 67, 69-72, 98, 101
Цианофиты 50, 66
Циатаксонида 120
Циатаспидные 326, 327
Циатаспиид 326
Цидароиды 300-302
Цикадовые 21, 59, 60, 366, 367, 469
Цикадофиты 413, 422, 424
Цикады 219
Циклозоа 109
Циклостомы 330
Циклоцистоиды 307
Цинодонты 399, 400, 499
Циррипедии 199
Цистоидеи 277-279, 278, 281, 282
Чайка 465
Человек 5, 8, 12, 22, 54, 322, 246, 460, 484,
494, 531, 589
Ч. прямоходящий 591
Ч. разумный 55
Ч. умелый 590, 591
Человекообразные обезьяны 532, 590
Человекоподобные 494, 531, 532
Челюстноротые 39, 330, 331, 335
Челюстные морские ежи 302
Челюстные рыбы 329, 330
Черви 18, 23, 25, 26, 31, 31, 67, 75, 81, 109,
138, 160-173, 165, 169, 170, 177, 207,
213,243,322,513,520
Червяги 384
Черепаха 387, 388
Черная антилопа 541
Черные бобры 586
Черный медведь 581
Черный ножеклюв 465
Четырехжаберные 273
Четырехпалый тушканчик 569
Четвероногие 346, 348, 374, 401
Чешуекрылые 220
Чешуеносцы 100
Чешуйчатые рептилии 417
Членистоногие 9, 22, 26, 75, 108, 162, 163,
164, 166, 169, 174-220, 176, 194, 222,
272,322
Шерстистые мамонты 11, 27, 36
Шерстистый носорог 14, 584, 585
Шерстокрылы 529, 530
Шестилучевые кораллы 31, 119,
Шестиножки 214
Шизодонты 239
Шизокораллы 123
Шизофиты 50
Шилоклювка 486
Шиншилла 570
Шиншилловые 571
Шипастые акулы 331, 334
Шипоклювый дятел 486
Широконосые обезьяны 531
Шляпочные грибы 53
Штопор дьявола 18, 24, 572
Щегол 24
Щетинохвостки 214, 215
Щитковые бесчелюстные 324
Щитни 196
Эбридии 82
Эвриапсиды 387, 389, 391, 410
Эвриптериды 205
Эдриоастероидей 281, 282, 284, 295, 306,
307
Эдриобластоидеи 281, 282
Эквус 557
Эктоподонтиды 506
Эласмобранхии 336
Эласмозавр 409
Электрический угорь 345
Элопоморфы 345
Эмболомеры 381, 382
Эму 62, 453, 485, 489, 491, 492, 495, 511
Энанциорнисы 469-474
608
КАМЕННАЯ КНИГА
Эндоцератоидеи 262
Энтелодоны 23
Эогиппус 551
Эозоон 65
Эозухии 415, 416
Эокриноидеи 277, 278, 278, 282, 307
Эосперматоптерис 355
Эпиорнис 484,485,487, 488, 490, 491
Эриопс 374
Этозавр 420, 421
Эукариоты 37, 39, 52, 66, 72, 73, 78
Эуэхиноидеи 300, 302
Эхинацеи 302
Эхинозоа 304, 306
Эхиноидеи 44, 297
Южноамериканская лошадь 555
Южноамериканские копытные 532, 557-
559, 558
Южноамериканские чешуйчатники 347
Южный мамонт 564
Юкка 552
Юнанолепиды 332
Ядерные 66
Ямкоголовые 418
Янготерия 503
Ястребы 581, 582
Ящер 411
Ящерица-правитель 389, 418
Ящерица-тиран 434, 436
Ящерицы 17, 389, 392, 395, 399, 403, 407,
408, 410-412,418,432,439,441,445,467
Ящеротазовые динозавры 417, 425, 469
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИИ
Abertella 305
Acaciaephyllum 515
Acantharia 85
Acanthocephala 161
Acanthonautilus 258
Acanthothoraei 334
Acantodes 335
Acidaspis 183,193
Acipenser 342
Acrothele 155
Acrotreta 143
Acrotretida 143, 155
Acteon 230, 231
A.vaughani 230
Actinoceras 253, 254, 260
Actinoceratoidea 262, 272
Actinoptychus 80
Actinosalpinx uralensis 165
Actinozoa 58
Aculatostroma verrucosum 98
Adapiformes 530
Adasaurus 438, 439
Adeloblatta columbiana 215
Adelograptus victoriae 309
Aegothelidae 484
Aeluroidea 526
Aepycamelus 539, 540
Aepyornis 487, 490
Aganaster gregarius 296
Aglaspida 207
Aglaspis 207
A. spinifera 206
Agmata 252
Agnatha 60, 326
Agnostida 184,184, 188
Agnostus 184, 193
Agriochoerus 536, 537
Ailanthus 518
Ajacicyathus nevadensis 101
Akantharges 190
A. gourdoni190
Alamosaurus 432
Albertella helena 192
Albertosaurus 434, 435, 458
Aldanella 226, 227
Aldanophyton 353
Alethopteris 366
Alkenia 212
Allactaga tetradactyla 569
Alligator thomsoni 415
Allogromiina 89
Allosaurus 433, 434, 435, 436, 449
Alveolites 121,123
A.suborbicularis 121
Ambiortus 474
A. dementjevi 475
Amblysiphonella barroisi 99
Ambocoelia 153
A.umbonata 153
Amebelodon 563, 563
Amia 344
Ammonites 263
Ammonoidea 272
Amphibamus 374, 376
Amphibia 57, 60
Amphicentrum 342
Amphigenia elongate 154
Amphineura 222
Amphioxus 309, 322
Ampyx 192
Amyelon 364
Anabarites 166
Anagale 523
Anastrophia verneuili 149
Anatolepis 324
Anatosaurus 17
Anchisaurus 423
Ancylopoda 545
Ancyrochitina sp. 82
Ancyrocrinus 284
Andreolepis 324
Andrewsarchus 542, 543
Andrias 385
Aneurophytales 60
Aneurophyton 355
Angiospermopsida 60
Angochitina sp. 82
Animalia 54
Anisograptidae 314
Ankylosauria 449, 452
Ankylosaurus 449, 450, 452
Annelida 60, 76
Annelides 161,163
Annularia 362
Annulina 359
Anodonta 247
39. Каменная книга
610
КАМЕННАЯ КНИГА
Anomalocaris 108, 162
A. nathorsti 194
Anomalodesmata 240
Anomia 55, 242, 244
A. micronema 242
Anoplura 219
Anostr аса 196
Antedon 283, 285
Anthozoa 58,106, 110, 111
Anthracomya 248
Antilocarpidae americana 541
Antrimpos 203
A. kiliani 203
Apatosaurus 425, 428, 429, 431,432, 434, 458
Apterygota 214
Arabellites 167
Arachnida 204, 210-212
Araliaephyllum 515
Araliopsoides 515
Arandaspis 324
A. prionotolepis 325
Araucaria 365, 517
Araucarioxylon 371, 413
Area 31
Arcestes 266, 267
Archaean th us 516
Archaeata 102
Archaeoceti 543
Archaeoconularia 110
Archaeocopida 198
Archaeocyathi 60, 100, 102
Archaeocyathus regularis 101
Archaeodiscina 81
Archaeohippus 556
Archaeopteridales 60
Archaeopteris 355
A. macilenta355
Archaeopteryx 9, 38, 468
A. lithographica 470
Archaeospongium 104
Archaeothyris 392, 393
Archanodon 247, 248
A. catskillensis 239
Archelon 403
Archimedes 134, 135
A. wortheni 134
Archohelia vicksburgensis 120
Archonta 530
Archosauria 420
Arctinurus 180,183, 191
A. boltoni 190
Arctodus 578, 581
Arctoidea 526
Argentavis 482
A. magnificens 483
Argonauta 275
Aristocystites bohemicus 278
Arnioceras 272
Arthroacantha 287
Arthropleura 176
Arthropoda 76,108, 174, 195
Articulate 141, 142, 145,155, 289
Artisia 364
Artocarpus 517
Asaphus186, 187
A. expansus 187
Ascoceras 253, 255
Asilidae 218
Asioryctes 512
Astacus 202
Astarte 31
Asteroidea 294, 295
Asterophyllites 362
Asteropyge 180
Asterosalpinx asiaticus 165
Astraspis disiderata 324
Astrocyatites ottawaensis 282
Astromyelon 362
Astylospongia praemorsa 56
Atelostomata 302
Athyrididina 152
Athyris spiriferoides 153,157
Atrypa 55,141,156, 158
A. devoniana 57
A. nevada 57
A. nuntia 57
A. occidentalis 151
A. reticularis 55, 57, 151,157
A. sp. 151
Atrypidina 151, 152
Aturia 252, 258, 264
Aulacophyllum 117
A. sulcatum 117
Aulocaulis 122, 123
A. annectens 122
Aulocystis jacksoni 122
Aulopora 122,123, 129
Auloporida 122
Australopithecus 590
A. robustus 591
Aves 55, 60
Aviculopecten 244
Aysheaia 162, 169,169, 170,177, 213
A. pedunculate 176
Bacteria 60
Bactritoidea 273
Baculites 260, 268, 269
Balanoglossus 277
Balanus 200
Baluchitherium 546, 547, 547
Baragwanathia 353, 356
Bardocarpus 358
Barrandeocrinus 287
Barroisella 156
Barylambda 528, 528
Basilosaurus 57, 543, 544
Bathyurus 187
Bauria 499
Beachia 141, 158
Beinotherium 499
Belinurus 207
Bellerophon 226, 227
B. troosti 226
Bennettitales 366
Beyrichicopida 198
Bickmorites bickmoreanus 260
Bienotherium 499
Bija 111
Billingsella pepina 146
Billingsites 253
Birkenia 328
Bison alticornis 450
B. antiquus 577
B. latifrons 585, 586
B. taylori 585
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
611
Bivalvia 54, 233 Blastoidea 280 Blastoidocrinus 279 Blattodea 219 Blattoptera 219 Blothrophyllum 114 B. decorticatum 114 Bothriolepis 333 B. gippslandiensis 333 Bothrodendron 360 Botryocrinus 283 Bothryodictya ramosa 94 Bowmani tales 363 Brachiopoda 60, 138 Brachosaurus 425, 426, 428, 431, 433, 435, 436 Brachyspirifer 156 Bradleya 197 Bradoria robusta 197 Bradoriida 197 Bradysaurus 391 Branchiopoda 195, 196 Breameryx 577, 578 Breviparopus 428 Breviphyllum 112, 113 B. cliftonense 112 Brontops 549, 550 Brontosaurus 425 Brontotherium 549, 550, 551 B. platyceras 550 Brooksella 108 B. Cambria 108 Bryophyta 60 Bryozoa 60,127 Buccinum 225 Buchia 240 Bumastus 178, 180, 191 Bunodes 206, 207 Burgessia 162 Burgessochaeta 162 Busycon 230 Byssonychia 242, 244 B. radiata 242 Camptosaurus 441, 442, 454 Canadaspis 162, 201 Canadia 164,164 C. spinosa 164, 164 Canadocystis emmonsi 280 Cancer 203 C. fissus 203 C. magister 203 Caninia 116 Canis dirus 579 Cannopilus 80 Capreolus capreolis 541 Capromys 488 Capsulocyathus subcallosus 100 Carabocrinus 286, 288 Carcharodon 338, 339 Cardium 241, 242 C. coosense 242 Carinacyathys sp. 100 Carnivora 525-527 Carpolithes 36 7 Carybdeida 109 Caryocaris 202 C. curvilata 202 Cassinoceras 253, 254 Castoroides 586 Catenipora 123, 125 C. micropora 125 Caulopteris 365 Caytonia 370 Celpoxylon 367 Centrospira 84 Cephalaspis 328, 329, 329 Ceratarges 192 C. armatus 192 Ceratitida 265 Ceratodus 347 Ceratogauhis 18, 24, 572 Ceratomorpha 545 Ceratoporella nicholsoni 98 Ceratosaurus 433, 434, 435 Ceraurus 178, 180 C. pleurexanthemus 189 Cervalces 585, 585
Cacops 374, 377 Cactocrinus 290 Calamites 359, 360, 362, 393 Calceola 116 Cal cichordata 291 Calcispongiae 96, 96, 99 Callianassa 203 C. mortoni 203 Callinectes sapidus 203 Calliocrinus 290 Callixylon 355, 364 C. zalesskyi 355 Calycites 516 Calymene 187,189 C. blumenbachi 189 Camarasaurus 424, 425, 431, 434 Camarocrinus 284 Camarotoechia congregate 150 C. sappho 157 Camelidae 537-542 Camelops 540, 542 Camelus 540 Camerata 289 Camptosalpinx sibiriensis 165 Chaetetes 124 C. septosus 124 Champsosaurus 410 Chancelloria 104 Chaoyangia 474 Charadriiformes 481 Charniodiscus 7 5 C. longus 76 Chasmatosaurus 398 Chasmosaurus 455 Cheilostomata 130, 136 Cheirolepis 341, 342 Cheiropteraster 296, 297 C. giganteus 296 Chelicerata 60, 205, 206, 208, 209 Cheloniceras 200 Chilopoda 214 Chinchilla laniger 570 Chitinozoa 81 Chlamydotherium 521 Chlamys 31, 236, 237 Chlamys (Lyropecten) 238 Chlorellopsis 70, 71 Chlorophyta 60
39*
612
КАМЕННАЯ КНИГА
Choia 162
Chondrichthyes 60
Chondrophora 109
Chondrostei 343, 344
Choristites 57
Chrysamoeba 50, 51, 87
Chrysophyta 60
Chubutisaurus 432
Ciliophora 60, 83
Cirripedia 195, 199, 200
Clatrotitan andersoni 218
Claudiosaurus 407
Clavattipollenites 516
Clidastes 411
Climatis 335
Clonograptus 309
Clupea 345
Clydagnathus ? cf. cavusformis 318
Clypeaster 302, 305
Cnidaria 60,103, 105,107, 109,111
Coccolithus 80
Coccosteus 332
Coelodonta 584
Coelophysis 422, 436
Coleoidea 273
Coleoptera 220
Collembola 214
Collenia 68, 70
C. frequens 69
C. versiformis 69
Colossochelys 404
Colpoxylon 366
Comarocystites punctatus 280
Composite 156
Compsognathus 436, 438
Conchidium nysius 149
Conchostraca 196
Congregopora 106
C. nasiformis 106
Conifer ales 60
Conocardium 240
Constellaria 132, 133
C. constellate 133
Conularia 109, 110
C. undulata 110
Conulata 110
Conus 225, 230
Cooksonia 353
Cooperoceras 259
C. texanum 258
Cordaitales 60, 364
Cordaitanthales 364
Cordaitanthus 364
Cordaites 364
Cornellites 242
C. flabella 242
Cornucrinus 287
Coronata 109,110
Corynexochida 184,186,188,192
Corynotrypa 131
C. delicatula 131
Coryphodon 527, 527, 528, 534
Corythosaurus 448
Coscinocyathus dianthus 101
Cothonion 111
Cothurnocystis 292, 293
Cranaena 140
Crania 143
Crassatella 243
Craticularia cylindrica 97
Cricetus cricetus 569
Crinoidea 276
Crossopterygia 60
Crossopterygii 346
Crotalocrinites 287
C. pulcher 288
Crustacea 60, 195
Cruziana 171, 180
Cryptochiton 222
Cryptolithus 180, 186,187
C. tesselatus 187
Cryptostomata 130, 133, 134
Cryptozoon 70, 71
C. proliferum 70
Ctenidodinium 80
Ctenochasma 464, 465
Ctenosauriscus 435
Ctenostomata 130,135
Cubomedusae 109
Cucullaea vulgaris 234
Cyanobionta 50, 60
Cyathocrinites 288
Cycadales 60, 366
Cycadeoidales 60
Cyclococcolithina 80
Cyclogyra 86
Cyclomedusa davidi 107
C. plana 107,108
Cyclonema 229
Cyclopyge 192, 193
Cyclostomata 58, 130
Cyclotrypa 129
Cyclozoa 107, 109
Cylindroporella 52
Cymobospondylus 405
Cynognathus 399, 400, 499
Cyprilepas 199
Cyrtia 158
C. exporrecta 152
Cyrtoceras 255, 256, 257
Cyrtograptus 313
C. murchisoni 312
Cyrtospirifer disjunctus 152
Cystiphylloides 117
C. americanus 115, 117
Cystoidosaccus 284
Cystoporata 130,131,132
Cyzicus 196
Daemonelix 17,18, 24, 572
Dalmanites 187
D. caudatus 189
Dapedium 344
Daphnia 196
Dasycladaceae 103
Decapoda 201, 203
Deinocheirus 436, 437
Deinogalerix 528
Deinonychus 338, 438
Deinosuchus 415, 416
Deinotherium 559, 560, 560
Deiphon 192
D. forbesi 192
Dekayella 128, 132, 133
Delthyris 57
Dendrograptidae 314,315
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
613
Dendrograptus 312, 313
Dendroidea 310
Dentalium 224
Dermaptera 219
Devonaster 296
D. eucharis 296
Devonochonetes carinata 148
Diadectes 396
Diadema 302
Diadematacea 302
Diagoniella hindei 94
Diatryma 481
Diceratherium 15,16, 545, 548
Dichograptidae 314, 315
Dickinsonia 75, 75,108,163
D. costata 76
Dicranograptidae 314, 315
Dictyoclostus 148
Dictyonellidina 145, 155
Dictyonema 311, 312, 313, 315
Dictyospongia sceptrum 94
Dicynodon 397
Didelphidae 504
Didelphis marsupialis 504
Didolodus 532
Didus 490
Didymograptus 313
Dielasma 141
D. bovidens 154
Dikelocephalus 187, 191
D. oweni 187
Dimetrodon 393, 394, 435, 497
Dimorphedon 464-466
Dinichthys 332
Dinictis 526, 526
Dinobolus conradi 144
Dinohyus 535, 536
Dinomischus 162
Dinornis 489
Dinornithiformes 488
Dinorthis pectinella 146
Diogenornis 485
Dipleura 189
D. dekayi189
Diplocaulus 384
Diplodocus 1,425, 426, 428, 429, 433, 458
Diplograptidae 314
Diplograptus 315
Diplomoceras maximus 263, 270
Diplopoda 213
Diplopodia 299
Diploporita 278, 279
Diplovertebron 381, 382
Dipnoi 60, 346
Dipoloceras 267, 268, 271
Diprotodon 419, 505, 506-508, 589
Diptera 220
Dipterus 346, 34 7, 349
Discorbina 88
Dithyrocaris 202
D. oceani 202
Di tripula 167
Doedicurus 521, 521
Doleserpeton 385
Draco 467
Dravidosaurus 448
Drepanella crassinoda 198
Dromornis stirtoni 491, 493
Dromornithidae 484, 491
Dsungaripterus 465, 467
Dufrenoya 267, 271
Dugong 567
Dunbaria fasciipennis 218
Dunkleosteus 332
Dysodonta 55
Echinacea 302
Echinocaris 202
E. punctata 202
Echinochimaera 340
Echinodermata 60, 76, 276
Echinoidea 56, 297
Echinozoa 300
Echmatocrinus 162, 282
Ectenaspis 191
Ectomaria 227
Edaphosaurus 389, 394, 435
Edentata 520, 521
Edestus 336
Ediacaria 75
E. flindersi 108
Edmontosaurus 444, 445, 448
Edrioaster bigsbyi 307
Edrioasteroidea 306
Ehmaniella 180
Eichwaldia 145,155
Eif felia globosa 104
Elasipoda 306
Elasmojerea sequana 96
Elasmosaurus 409
Elephas 564, 565, 566
E. maximus 566
Elginia 391
Eliomys quercinus 569
Elliasopora 135
E. siluriensis 135
Elrathia 186
Emmonsia 123
Enantiornithes 470, 473
Encrintella (Crassatellites) 242
Endeiolepis 328
Endoceras 253, 254
Endoceratoidea 262, 273
Endolobus 258
Endothyranella 88
Engonoceras 267, 271
Ensis 242, 243
Enteletes hemiplicatus 146
Enteropneusta 308, 309
Entoprocta 128
Eobasileus 533, 534, 534
Eobronteus 191
Eocytheropteron 197
Eodiscus 184,184
E. punctatus 184
Eohippus 551, 552, 554, 555
Eomanis 568
Eophrynus prestvici 211
Eospermatopteris 355
Eospirifer 57
E. radiatus 152
Eotitanops 549, 551
Eo tomaria 225
Eozoon 64-66, 68, 70
E. canadense 64, 65
Ephemeroptera 216, 217
614
КАМЕННАЯ КНИГА
Epihippus 552
Epitheles hemisphaerica 68, 96
Equidae 38
Equisetina 359
Equisetopsida 60
Equus 554-557
E. giganteus 584
E. occidentalis 577
E. tau 584
Eridophyllum 118
E. seriale 118
Erinaceus 297
Eriptychius 324
Ernanodon 520
Erutaxocrinus whiteavesi 289
E. fletcheri 289
Eryma 202
Eryops 374,376, 377, 381
Eucalyptocrinites 286
E. crassus 283
Euchambersia 399
Eucrinatella 234
Eudea cribraria 96
Euechinoidea 302
Eumetazoa 94
Eunicites 167
E. atavus 164
Euomphalus 227
Euoplocephalus 449, 450, 452
Euparkeria 420
Euphoberia 211, 213
E. armigera 211
Euproops 207
Eurolimnornis 474
Eurotamandua 520
Eurychilina reticulata 198
Eurypterida 205
Eurypterus 208, 209
E. remipes 208
Eurystomites 259
Euskelosaurus 423, 424
Eusthenopteron 348, 349, 375
Eutaxocrinus fletcheri 289
E. whiteavesi 289
Eutrochocrinus christyi 290
Exoconularia 110
Exogyra 238,245,245, 246, 246, 271
E. costata 245
Fallotaspis 177, 184
F. typica 185
Favosites 121, 123,129
F. argus 123
Favositida 123
Felidae 526
Felis atrox 578, 579
Fenestella 135
F. retiformis 134
Fenestella s.l. 134
Fenestrata 130, 133, 134
Ficophyllum 515
Ficus 517
Filicales 60
Fistulipora 131
Fistuliporas.l. 132
Flabellotrypa rugulosa 131
Flexibilia 287, 291
Flexicalymene 191
F. senaria 189
Floridinella vicksburgica 136
Foerstediscus splendens 307
Foraminifera 85
Fordilla 240
Franimys 571
Funisciurus 5 70
Fursenkoina 88
Fusulinella 88
Fusulinina 87, 89
Galliformes 481
Gallimimus 439
Gangamopteris 370
Gastroconus 110, 111
Geisenocerina 261
Gemuendina 334
Genyornis 494
G. newtoni 492, 493
Geosaurus 415
Gephyrostegus 382
Gerrothorax 377
Gervilliopsis 244
Gigantoceras 255
Gigantoproductus 148
Gillocystis 295
Ginkgo 369
G. biloba 369
Ginkgoales 60, 369
Girvanella 70
Glaucomys volans 5 70
Globidens 411
Globigenna 88
Globigerinidae 90
Globivalvulina 88
Globorotalidae 90
Glossograptus 313, 315
Glossopteridales 60
Glossopteris 370
Glossotherium myloides 588
Glottidia 141,154
Glyptodon 521, 521
G. clavipes 587
Glyptopleura 197
Glyptosaurus 17
Gnathobelodon 563, 563
Gnathoriza 348
Gnathostomata 302
Gomphoceras 261
Gomphocystites bownockeri 278
Gomphotherium 561, 562, 564
Goniatites 264
Gonioceras 255, 255, 256
Goniophyllum 115, 116
Goniopygus 299
Gorgonia 129
Gorgosaurus 434, 435
Gosslingia 352
Goyocephale 447
Graptolithina 58, 60
Graptoloidea 58, 310
Graptozoa 58
Griffithides 187
G. seminiferus 188
Groenlandaspis 333
Gruiformes 480
Gryphaea 245, 245, 246, 246, 271
G. mutabilis 245
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
615
Guadalupia cylindrica 99
Guettardiscyphia stellata 97
Gymnolaemata 130
Gyroceras 274
Hadrophycus 68
H. immanis 68
Hadrophyllum 116
Hadrosaurus 444
Haimesiastraea conferta 120
Hallopora 132
Halysiocrinus 286, 288
Halysites 123,125
H. amplitubatus 125
Halysitida 123
Hamitella 268
Hamites 269, 269, 270
Hamptonia bowerbanki 94
Hamulina 268
H. astieriana270
Hamulus 166
Hapalops 522
Haploscapha 236
Hardistiella 330
Hardouinja 299
Harpes 193
Harpina 193
Hebertella sinuata 146
Helicoceras 269
Helicoplacoids 307
Helicoprion 336
Heliolitoidea 123
Heliophyllum 114,117
H. halli 114
H. obconicum 117
Heliosphaera 84
Helisoma 232, 266
Helminthozyga 229
H. vermicula 228
Hemiaster 301
Hemichordata 308
Hemicrinus 286
H. astierianus 291
Hemip ter a 219
Hemipteroidea 219
Hesperornis 473
Hesperorthis 155,156
Heteractinida 104
Heteraster 301
Heterodontosaurus 443
Heterorthis clytie 146
Hexabactron 102
Hexadoridium 84
Hexagonaria 118
H. percarinata 118
Hexameroceras 257, 260
Hildoceras 272
Hippidion 555, 556
Hippomorpha 545
Hippotragus niger 541
Holaster 299, 301
Holcorhynchia 155
Holectypus 299
Holocystites 278
Holopea symmetrica 228
Holophagus 349
Holoptychus 346
Holostei 343
Homalocephale 447
Homalozoa 291
Homo 590-592
H. diluvii testis 386
H. erectus 591
H. habilis 590, 591
H. sapiens 55, 592
H. sapiens sapiens 55
Homolophyllum 117
H.ungula 117
Hoplophoneus 526, 526
Hormotoma whiteavesi 228
Horneophyton 354
Hudsonaster 295
H. incomptus 296
Huronia bigsbyi 260
Hyaenodon 524
H. horridus 525
H. musteunus 525
Hyalospongia 57
Hydnoceras tuberosum 94
Hydra 51,112
Hydroconozoa 110
Hydrochoeridae hydrochaeris 571
Hydroconus 110, 111
Hydrodamalis 567
Hydrosamalis 567
Hydrozoa 102, 106,106,108,108
Hyenia 363
Hylonomus 387, 392
Hymenoptera 220
Hyolitha 170
Hyolithes 162, 230
Hypagnostus 184
H. parvifrons 184
Hypocrinus 287
Hypophylloceras ononense 264
Hypothyridina 158
H. venustula 150
Hyppothoa hyalina 136
Hypsilophodon 442, 451, 454
Hypsolosaurus 431
Hyracodon 546
Hyracotherium 551, 552, 554, 555
Hystrix brachyura 570
Icaronycteris 529
Icarosaurus 467
Ichthyocrinus 285
Ichthyornis 474
Ichthyostega 372, 373, 375, 381
Iguanodon 441, 443
Ilbandornis 492
Ildraites 167
Illaenus 186
Inadunata 285, 287, 291
Inarticulate 141, 142,155
Inocaulis 310, 313
Inoceramus 31, 236, 238, 244
Inordozoa 109
Insecta 60, 214
Irregularia 298
Ischadites iowensis 102
I. murchisoni 102
Isograptidae 314
Isoptera 219
Isotelus 191, 193
I. maximus 181
616
КАМЕННАЯ КНИГА
Jamoytus 328, 330
Johoia 162
Kannemeyeria 397
Kenella 516
Kerpia 371
K. macroloba 369
Khangailites 124
Kiaerograptus antiquus 309
Kirkbyella 198
Knightoconus antarcticus 251
Kockelella 320
Kootenia 180
Koremagraptus 311
Kronosaurus 408
Kuehneosaurus 467
Kuehneosuchus 467
Kujdanowiaspis 332
Kutorgina cingulata 144
Kutorginida 143,155
Labidosaurus 396
Lagenochitina estonica 82
Lambeophyllum 111, 112
L. profundum 112
Lambeosaurus 448
Lampterocrinus tennesseensis 290
Laqueus140,158
Lasiostrobus 367
Latimeria 346, 350
Laurus 517
Leanchoilia 162
Leopaspis 188
Leperditicopida 198
Lepidocarpon 15
Lepidocyclina 88, 90
Lepidodendron 360, 360
Lepidoderma 210
Lepidoptera 220
Lepidosiren 348
Lepisosteus 343
Leptaena 155
L. rhomboidalis 147
Leptauchenia 535, 536
Leptoptygma virgatum 225
Lermontovia dzevanowskii 185
Liaoningornis 474
Lichenaria 124
L. typa 124
Lichenariida 123
Lichenocrinus 284
Lichida 187,192
Liliacidites 516
Lima 242, 244
L. reticulata 242
Limnoscelis 389
Limulus 205, 207
L. polyphemus 205, 207, 210
Lindstroemella aspidium 144
Lingula 139, 141-143, 154,155,158
L. anatina 144
L. elderi 144
Lingulella 143
Lingulida 142,143, 155
Linthia 301
Liopeplum 230
Lipotyphla 528
Liriophyllum 515
Lithophaga 243
Lithorhizostomatida 109
Lithosmylus colupbianus 218
Lithostrotion s.l. 114
Litoptern 558
Littorina 227
Lituites 259
L. lituus 260
Lobolithus 284
Lorenzinites rarus 108
Loricata 520
Loriculina 200
Loriolaster 297
L. mirabilis 296
Louisella 162
Loxoconcha 197
Loxodonta 564, 565, 566, 567
L. africana 566
L. antique 566
Lumbriconereites 167
Lycaenops 397
Lycopodiopsida 60
Lycopodium 356
Lyginopteris 15
Lyropora 134
L. subquadrans 134
Lyssoxylon 367
Lystrosaurus 396, 397, 398
Lytoceras 267
Macgeea solitaria 117
Maclurites 226
Macoma 242, 243
M. nasuta 242
Macrauchenia 558, 558
Macrocystella marinae 278
Macroporella 52
Macroscaphites 269
M. yvani 270
Mactra 243
Magadina 159
Magnolia 517
Magnoliaephyllum 515
Maiasaura 446
Majungatholus 446
Malacostraca 195, 201, 202, 203
Mallophaga 219
Mamenchisaurus 426-428
Mammalia 60, 520-592
Mammut 562, 564
M. americanum 561, 562, 577
Mammuthus 564
M. columbi 564, 578
M. imperator 564, 577
M. meridionalis 564
M. primigenius 564, 566
Mandaloceras 255, 257, 259
Manteoceras 549
Marginalia burlingtonensis 148
Margochitina margaritana catenaria 82
Marrella 162, 194
Marsupites 291
Martesia 237, 243, 243
Martinopsis 142
Mastodon 562
Mastodonsaurus 377
Matthevia 222
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
617
Mawsonites spriggi 76, 108
Maxillopoda 200
Meandrina alveolus 120
M. baretti 120
Mecoptera 220
Mediospirifer 155
Medullosa 366, 393
Medusaegraptus 313
Megahippus 555
Megalania 419
M. prisca 418
Megaloceros 585, 585
Megalograptus 210
Megalonyx 578, 586
Megarachne servinei 212
Megateuthis 275
Megatherium 522, 522, 523, 555, 586
Megazostrodon 500
Meiolania 388, 402
Mellita 305
Melosiras.l. 80
Menispermites 515
Meristella barrisi 157
Meristina maria 153
Merychippus 554, 555, 556
Merycoidodon 535, 536
Mesohippus 552, 553, 553-555
Mesolimulus 207
Mesosaurus 401, 402
Mestriosuchus 414
Metamynodon 546, 546
Metasequoia 55, 517, 518
M. occidentalis 55
Metopaster 296, 297
Metoposaurus 376, 378, 381
Miacoidea 525, 526
Michelinoceras 58, 255, 261
Micraster 303
M. coranguinum 303
M. leskei 303
Microcyclus 116
Micromys minutus 569
Micropaehycephalosaurus 447
Microtus oeconomus 569
Miliolina 87, 89, 91, 91
Millepora 107
Miohippus 555, 556
Mitroceras 259
Mitrocystites mitra 278
Mixosaurus 405
Moeritherium 559, 560, 567
Mollaria 162
Mollusca 54, 57, 60
Monera 60
Monoclonius 452, 455
Monopleura texana 242
Monticulipora 132
Montlivaultia nattheimensis 120
Moropus 545, 545, 546
Mortoniceras 267
Mosasaurus 410
Moyomyzon 330
Mucrospirifer mucronatus 157
Multiplicisphaeridium 81
Murex 226, 230
M. mississippiensis 228
M. tribulus 230
Mussaurus 431
Mya240, 241,242
Myalina 238
Mycterosaurus 394
Myelodactylus 285, 288
Mylodon 522, 523, 578, 586
Myodocopida 198
Myriapoda 213
Myrmecophagidae 520
Mysticeti 543
Mytilus 55, 238, 242, 244
M. conradinus 242
M. edulis 55
Naiadites 248
N. carbonarius 238
Nanantius valifanovi 472
Nassellaria 84
Nautilida 262
Nautiloidea 252, 262, 272
Nautilus 249, 250, 252, 257, 259-262, 266,
272
Nemathelminthes 161
Nemertini 161
Neogondolella clarki 319
Neopilina 222
N. galatheae 222
Neoptera217
Neospirifer cameratus 152
Neostreptognathodus pequopensis 319
Neoteleostei 345
Neotrigonia 239
Nereites 171
Neuroptera 220
Neuropteridium 370
Neuropteris 366
Neuropteroidea 220
Niolamia 402
Nipponites 269
Normannites 267
Nostoc 51
Notharctus 530, 531
Nothrotherium 522, 578, 578, 586
Notocarpos 323
N. garratti 323
Notostraca 196
Notoungulata 557
Nucleocrinus verneuili 280
Nucula 31, 238
Nummulites 88, 90
Nususia 162
Nyctosaurus 465
Nyssa 517
Obdurodon 501, 502
Obolellida 143,155
Obovithyris 155
Odonata 217
Odontoceti 543
Odontogriphus 317, 318
Odontopleurida 188,192
Oecoptychius 268
Oenonites 167
Ogygopsis 180
O. klotzilSd
Oldhamia 52
Olenelloides armatus 182
Olenellus 177,179,180,181,184,190
O. thompsoni 185, 191
618
КАМЕННАЯ КНИГА
О. vermontanus 191
Oleneothyris 158
Olenoides 162, 180, 185
О. serratus 186
Olenus186
Oligochaeta 163
Oligokyphus 400
Omphyma 115
Ony ch aster 297
O. flexilis296
Onychophora 161,168, 177, 213
Opabinia 162
Ophiacodon 394, 401
Ophiderpeton 384
Ophiuroidea 294
Ophthalmosaurus 405
Orbitolina 271
Oreaster 297
Oreodon 535
Ornimegalonyx 484, 488, 489
Ornitholestes 433, 438, 439
Orodus 336
Orohippus 552
Orophocrinus stelliformis 280
Orthida 145,146, 155
Orthoceras 57, 58, 261
Orthop tera 219
Orthop ter oidea 219
Orthorhynchula linneyi 150
Oscinos 482
Osmundaceae 371
Osteolepis 348, 349
Ostracoda 195
Ostrea 55, 238, 238, 244, 245
Ostrea (Ostrea) lutraria 55
Ostrea virginica 55, 245
Ostreidae 55
Ostriophisi 345
Otarioidea 526
Otogornis 474
Ottoia 162
O. prolifica 168
Ottonosia 70
Ouranosaurus 435
Ovoceras 257
Oxyaena 524, 525
Oxydactylus 539, 539
Pachycephalosaurus 447, 449
Pachydiscus seppenradensis 263
Pachysphaera 81
P. transitans 182
Pagetia bootes 184
Paiutiida 111
Palaeaster 296
Palaeobatrachus 385
Palaeocopida 197
Palaeognathao 484
Palaeoisopus 212
P. problematicus 212
Palaeolimulus 207
P. avitus 206
Palaeomastodon 560, 561, 562
Palaeomerus 207
Palaeonisci 343
Palaeopalaemon 203
P. newberryi 203
Palaeopantopus 212
Palaeoparadoxia 568
Palaeophorius 211
P. nuncius 208
Palaeoptera 216
Palaeosyops 549
Palaeotherium 553
Palorchestes 507
Panapeltis 197
Panoplosaurus 540, 452
Panthera atrox 579
Paracamelus 540
Paraconularia 110
Paradoxides 184, 185, 191
P. paradoxissimus 185
P. harlani 193
Paraechmina spinosa 197
Paraharpes 179,191
Paramylodon 578
Paramys 571
Parareptilia 387
Parasaurolophus 448
Parasmilia 120
Paraspirifer 156, 158
Parawocklumeria 268, 269
Parazoa 94
Parvancornia 75
P. minchami 76
Patella 225, 251
Paterinida 143,155
Patriofelis 524, 525
Pecopteris 364, 365
Pecten 236—238, 244
Pelaginella 226
Pelecypoda 54
Pelomyxa 78
Peltaspermum retensorium 368
Pennatula 75
Pentaceratops 452, 455
Pentacrinites 291
Pentamerella arata 149
Pentamerida 148,155
Pentamerus laevis 149,149, 155
P. oblongus 149
Pentremites godoni 280
P. robustus 280
Peridinium 80
Perischechinoidea 300
Pernidae 244
Peronidella sp. 96
Perspicaris 201
Petalaxis 118
P. stylaxis 118
Petalocrinus 286, 288
Petalodus 336
Petalonamae 76
Petrolacosaurus 417
Peytoia 108, 193
Phacopida 178,179,187,189,192
Phacops 180, 187
P. altaicus 189
Phanocrinus 283
Phenacodus 532, 533,534, 558
Phillipsastraea 118
Phillipsia 187
P. gemmulifera 188
Philosa 520
Phiomia 561, 562
Pholadidea 243
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
619
Pholadomya 240
Pholas 237, 237, 240, 243, 243
Pholidophyllum 358
Phorarhacos lonissimus 480
Phoronida 60
Phragmoceras 256, 257, 260
Phylactolaemata 129
Phyllocarida 201
Phyllolepis 333
Phyllopoda 196
Phyllotheca 359
Phytosauria 420
Pictetia astieriana 269
Pikaia 162
Pirania 162
Placenticeras 272
Placerias 422
Placochelys 410
Placocystites forbesianus 278
Placodermi 60
Placodus 408, 410
Placosaurus 17
Plagioptychus partschi 247
Planorbis 266
Plantae 60
Plantaginopsis 515
Plasmoporas.l. 124
Platecarpus 411
Plateosarus 423, 432,438
Platybelodon 562, 563, 563
Platyceras 23, 229
Platycrinites 284
Platyhelminthes 60, 161
Platyhystrix 375
Platyrrhines 531
Platysolenites 163,164,166
Platystrophia 145,156,158
P. biforata 57
P. lynx 57
P. moritura 57
P. ponderosa 57
Plectronoceras 252, 253, 257, 261
Plesiadapiformes 530
Plesiochelys 403
Plesiosaurus 389, 408
Plethorhyncha speciosa 150
Pleurocystites 279
P. filitextus 278
Plicocyathus rosanovi 100
Pliohippus 554, 555, 556
Plotop teridae 478
Podocopida 197,198
Poebrotherium 538, 538, 540
P. labiatum 538
P. wilsoni 538
Polychaeta 163
Polydon 342
Polylophodonta 320
Polypora 129
Polypteri 343
Polypterus 342
Polyscyphia pseudocoeloptychium 97
Polystomellina 86
Polyzoa 127
Porifera 93,102
Porophoraspis 324
Portheus molossus 23, 412
Praecambridium 75
Prasopora 132
Preptoceras 584
Presbyornis 478, 480
Prestwichianella canae 206
Pretiosocyathus subtilus 101
Priapulida 161,167
Priapulopsis bicaudatus 168
Priapulus caudatus 168
Primelephas 564
Primocorallina 52
Prioniodus 321
Priscacara 341
Prismodictya prismatica 94
Proburnetia 398
Procamelus 539, 540
Procolophon 398
Prodromites gorbyi 267
Prodryas persephone 218
Productella 148
P. truncata 157
Productida 148
Proetida 186, 188,189
Proetus 187
P. concinnus 188
Profistulipora 130
Proganochelydia 402
Proganochelys 402
Progura 484
Promerycochoerus 536
Propilina 225
Propleopus 511
Prosimi 530
Proteaephyllum 515
Protheus 345
Protista 58, 79, 103
Protoavis texensis 470, 471
Protobalanus 200
Protoceratops 451, 452, 453
Protoclepsydrops 392
Protodonata 217
Protolindenia 217
Protomedusae 108, 109
Protopanderodus 320
Protophragmoceras 256
Protopterus 348
Protospongia erixo 94
Protosuchus 413, 415
Protozoa 50, 58, 65, 78, 84
Protracheata 161, 169, 213
Protylopus 537, 538
Prozeuglodon 544
Psaronius 364, 365
Pseudodontornis 482
Pseudoniscus roosevelti 206
Pseudosyringocnema eleganta 102
Psilophyton 352, 354
Psilotum 354
Psittacosaurus 451, 453,454
Psygmophyllum expansum 368
Pteranodon 465, 466
Pteria 234
Pterichthys 333
Pteridospermales 366
Pteriidae 244
Pteriomorpha 54
Pterobranchia 60
Pterocorys 84
Pterodactylus 463, 464, 465
620
КАМЕННАЯ КНИГА
Pterodaustro 465
Pteroplax 381, 382
Pterosauria 463
Pterotrigonia 239
Pterygotus 209, 210, 211
P. buffaloensis 209
Pterygotus sp. 209
Ptilodus 501
Ptychocarpus 365
Ptychopariida 178,179,182,185-188,192
Pugnoides calvini 150
Pustulina pustulosa 157
Pycnogonida 212
Quercophyllum 515
Quetzalcoatlus 465
Quipollornis 484
Radiaspis 188
R. radiata 192
Radiocyatha 104
Rafinesquina 158
R. alternata 147
Ranapeltis 197
Raphidia mortua 218
Ratitae 484
Receptaculita 102
Receptaculites 102
Redlichiida 182,184,185,186,188, 190
Regularia 298
Rensselaeria 158
R. marylandica 154
R. ovoides 154
Reptilia 57, 60
Retiolitidae 314
Retrorsirostra carleyi 146
Retziacea 152
Rhabdinopora 315
Rhabdopleura 308, 310, 277
Rhamphorhynchus 464
Rhipidomella penellope 157
Rhizocrinus 291
Phizophyllum 116
Rhizostomida 109
Rhizostomites 108
R. admirandus 108
Rhodophyta 60
Rhombifera 278, 279
Rhombomys opimus 569
Rhombopora 134
Rhopostema 301
Rhynchocyon 524
Rhynchonellida 150,155
Rhynia 354
Rhyniella 215
Rhyniopsida 60
Richthofenia 159
Rizoceras 256
Rogersia 515
Ropalonaria venosa 135
Rosaceae 517
Rostroconchia 240
Rotaliina87, 89,91,91
Rotoceratida 259
Ruminantia 535
Rusophycus 180, 181
Rutiodon 413,414
Rutoceras 260
Ryticeras 259
Sabal517
Sabellidae 163
Sabellidites 163,166
Sachyogyrus 359
Sagenocrinites clarki 289
Saichania 451
Saiga tatarica 541
Salenia 299
Salmoniformes 345
Salpingostoma boulli 226
Salterellidae 252
Saltoposuchus 420
Samaropsis 367
Sapindopsis 515
Sarcodina 60, 84
Sarcopterygii 345
Saturnalidae 84
Savroniella cincinnata 83
S. rugosa 83
Scaphites 269, 271
Scenella 162,222
S. reticulata 226
Schizambon canadensis 144
Schizocoralla 124
Schizophoria 156
Schloenbachia 266
Schroederoceras 259
Schuchertella woolworthana 147
Sclerocalyptus 521
Sclerodus 328
Sclerospongiae 93, 96—98, 99, 103, 121
Scutaster andersoni 305
Scyphocrinites 284, 285, 290
S. elegans 290
Scyphozoa 107, 108,108
Sebecidae 416
Seismosaurus 428
Selaginella 356
Semionotus 344
Sepia 274
Septastrea crassa 120
Sequoia 518
Serpula 164,166
Serpulidae 165
Sertidentinus 580
Seymouria 382, 383
Shandongornis shanwanensis 485
Sharovipteria 467
Shumardia pusilia 182
Shuotherium 503, 504
Siderops 377, 379
Siderospongia sirensis 95
Sidneyia 194
Sieberella 155
Sigaspis 331
Sigillaria 391, 362
Siliqua 242
Siphonaptera 220
Siphonophrentis 116, 117
S. gigantea 116,117
Sivatherium 588
Skartopus 457
Smilodon 505, 579, 581,582
S. californicus 526
Solemya 235, 238, 238
Solen 241, 242
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
621
Solenocheilus 257, 258
Solenopora 70
Soliclymenia 268
S. paradoxa 269
Somasteroidea 294, 299
Sordes 463
Spermopteris 367
Sphenophyllales 363
Sphenophyllum 362, 370
S. biarmicum 363
Sphenopsida 60
Sphinctozoa 98
Spinatrypa hystrix 151
Spinocyrtia granulosa 152,157
Spinosaurus 435
Spirifer 57,141
Spiriferida 150,155
Spiriferina 151
Spiroceras 58
S. bifurcati 270
Spirograptus turriculatus 312
Spirorbis 164,166
Spirula 274
Spongia 60, 93
Spongiata 93, 97, 100, 102,103
Spongiomorphida 107
Sporadoscinia sp. 98
Spriggia wadea 107
Spriggina 163, 177
S. flounders! 76
Spumellaria 84
Squamiferida 103
Squatina 339
Steginoporella magnilabris 136
Stegnammina 86
Stegodon 564, 565
Stegosauria 448
Stegosaurus 448, 449, 450, 453, 454
Stelleroidea 294
Stellispongia glomerata 96
Steloceras 446, 449
Stenodictya 216
S. lobata 215
Stenolaemata 130
Stenomylus 537, 538, 539
Stenonychosaurus 456
S. inegualis 460
Stenopora fibrosa 129
Stenopterygius 406
Stephanocrinus 282
S. angulatus 280
Stephanoscyphus 110
Steropodon 501, 502
Sthenurus 509
Stigmaria 361, 361
S. ficoides 361
Stomatopora 129, 131
S. parvipora 128
Straparolus 228
Streptelasma 112,115
S. corniculum 112
Streptindytes sp. 165
Stricklandia castellana 149
Strigidae 488
Striformes 481
Stromatocystites 307
Stroma toporata 98,103
Stromatospongia 98
Strombus 229
Stropheodonta demissa 147,157
Strophomena 142, 158
S. planumbona 147
Strophomenida 145,147, 155
Strophonella 142
Struthiomimus 435, 439, 440
Stuertzaster 296
Stylemys 404
Styliolinas.l. 170
Stylocoeniella armata 120
Stylonurus 209, 210
S. myope 209, 210
Styracosaurus 452, 455
Suina 535
Supersa'urus 428
Sydneyia 162
Sylvella 367
Synaptophyllum 118
S. simcoense 118
Syndyograptus 313
Synoplotherium 542, 543
Syringopora 122
S. retiformis 122
Syringoporida 122
Syringothyris 57
Tabulata 121
Tabulatom orpha 121
Tagelus 234, 241, 242
Tamias sibiricus 570
Tasmanites 81
Taxochelys 404
Taxocrinus 289
Taxodium occidentale 55
Teleostei 343, 344
Telicomys 571
Tellina 31, 243
Tentaculita 170
Tentaculites s.l. 170
Terataspis 178,188,192,193
T. grandis 192
Teratornis 484, 578, 582
T. merriami 487
Terebellidae 116
Terebratalia 154,156, 158, 159
Terebratulida 153,155
Testudo 404
T. ammon 404
Tetonius 531, 531
Tetraceratops 395
Tetradium 124
T. fibratum 124
Tetragraptus 313
T. decipiens 309
Tetrataxis 88
Tetraxonida 94, 95, 96
Textularia 87
Textulariina 86, 89, 91, 91
Thais 227
Thalamopora cribrosa 99
Thamnopora 123
Thecideidina 153, 155
Thecodontosaurus 423
Therizinosaurus 423, 436
Thoatherium 558, 559
Thoracesphaera 80
Thoracica 199
622
КАМЕННАЯ КНИГА
Thrinaxodon 398
Thylacoleo 510
Thylacosmilus 505, 526
Thysanoptera 219
Thysanura 214
Tillodonta 533
Timanostrobus 358
Timoroblastus 281
T. coronatus 280
Tipus gracilis 216
Titanichthys 331
Titanis 481
Titanoceras 257, 259
T. ponderosum 258
Tomopteria 75
Tomopteridae 164
Torosaurus 452
Torynifera setigera 153
Toucasia patagiata 242
Toxodon 557, 558
Trachylina 109
Trematis ottawaensis 144
Trepostomata 130,131,133
Triadobatrachus 385
Triarthrus 180
Triassomachilis 215
Triaxonida 94, 96, 97
Tribrachidium 75, 277, 307
T. heraldicum 76
Triceratops 452, 453, 456
Trichechus 567
Trichoptera 220
Tricoelocrinus woodmani 280
Tridacna 235, 236, 244
Trilobita 60, 177,184
Trilobitoidea 193
Trilophodon 562
Trimerella ohioensis 144
Trimeropsida 60
Trimerorhachis 383
Trimerus 180,191
Trinacromerum 409
Triops 196
Tritoechia 156
Trogosus 533
Tropidoleptus carinatus 146,157
Tryops 196
Tsintaosaurus 445-447
Tubulidentata 533
Tubuliporata 130,131
Tubuliporida 58
Tunicata 309
Turbinia 165,167
T. abbreviata 167
Turbonopsis shumardi 228
Turritella 227,228, 229, 230
Tylacosaurus 411
Tylopoda 535
Tylosaurus 411
Typothorax 421
Tyrannosauropus 457
Tyrannosaurus 415, 434, 435, 436, 437, 438,
450, 457, 457, 458
T. rex 11
Uintacrinus 285
U. socialis 291, 292
Uintatheria 533
Uintatherium 533,534, 534
Ultrasaurus 428, 429, 430
Unio 234, 239, 248
Uniramia 60, 213
Uperocrinus 290
Uralichas 193
Vaceletia 99
V. cripta 99
Valeriana officinalis 56
Varanosaurus 394, 396
Varanus 418
Vauxia 162
Venericardia planicosta 239
Ventriculites pedester 97
Venus 239, 240, 242
Vermes 108, 160
Vermetidae 165
Vermetus 167
Vermicularia228, 229
Villebrunaster thorali 296
Vinella 135
V. repens 135
Vitiphyllum 515
Vjalovognathus synguensis 319
Volborthella 252, 253, 257, 272
Volborthellidae 252
Volvox 93
Vulvulina advena 92
Waagenella cf. W. elongata 99
Walchia 270, 364
Wannanosaurus 447
Waptial62
Westergaardia 180
Whidbornella lachrymosa 148
Williamsonia 368, 515
Wintonopus 457
Wirgiana decussata 149
Wiwaxia 162
Woodworthia 413
Xenarthra 520
Xenaspis 264
Xenusion 177
Xiphosura 205
Yadjaicyathus astashkini 100
Youngina 389
Zelkova 518
Zemistephanus 266, 267
Zeuglodon 57, 543
Zittelispongia alcyonoidea 97
Zoophycos 171
Zoophyta93, 111
Zosterophyllum 356
Zygobeyrichia 198
Zygospira modesta 151
ОГЛАВЛЕНИЕ
Предисловие к англоязычному изданию книги 1989 года...........5
Предисловие к русскоязычному изданию..........................8
I О чем рассказывают ископаемые...............................11
II Фоссилии, породы и возраст..................................28
III Группы организмов, названия и отношения между ними..........48
IV Остатки древнейших организмов...............................63
V Разнообразие протистов......................................78
VI Губковые: настоящие и предполагаемые........................93
VII Простые кишечнополостные: тип Cnidaria, или стрекающие.....105
VIII Моховидные животные: тип Bryozoa...........................127
IX Брахиоподы-крепыши.........................................138
X Черви, следы жизнедеятельности и проблемы палеоихнологии .... 160
XI Животные из трех частей: трилобиты.........................174
XII Ракообразные...............................................195
XIII Членистоногие от мелководных до сухопутных и летающих......204
XIV Улитки и их королевство....................................221
XV Двустворки ................................................233
XVI Ноги впереди головы: наутилоидеи и их сородичи.............249
XVII Рога Аммона и внутреннераковипные головоногие..............263
XVIII Иглокожие: преимущественно стебельчатые....................276
XIX Подвижные иглокожие .......................................294
XX Сеточки, пружинки и палочки: граптолиты ...................308
XXI “Зубы” без челюстей: конодонты и конодонтофориды...........317
XXII От морских звезд до рыб, властелинов воды..................322
XXIII Зазеленевшая Земля.........................................351
XXIV Древние и современные амфибии..............................372
XXV Мириады рептилий на Земле..................................387
XXVI Прощай, суша...............................................401
XXVII Чешуйчатые рептилии и ящеротазовые динозавры................417
XXVIII Птицетазовые динозавры.....................................441
XXIX Рептилии летающие и планирующие ...........................463
XXX Птицы - изобретатели пера..................................468
XXXI Шерстистые рептилии со сложными ушами .....................496
XXXII Земля цветет................................................514
XXXIII Великое разнообразие плацентарных .........................520
XXXIV Животные и птицы ледникового периода. Губительное влияние
человека............................................................573
Литература..................................................593
Указатель русских названий..................................596
Указатель латинских названий................................609
Научно-популярное издание
Патриция Викерс Рич,
Томас Хьювитт Рич,
Милдред Адамс Фентон
КАМЕННАЯ КНИГА
Летопись доисторической жизни
Выпускающий редактор О.П. Кузина
Корректоры Т.И. Чернышова , Т.В. Благосова
Издательская лицензия 090151 от 06.06.96
Подписано в печать 24.11.97. Формат 70x108/16
Гарнитура "Школьная". Печать офсетная
Усл. печ. л. 54,6. Уч. изд. л. 55,9
Тираж 4000 экз. Тип. зак. 42
Международная академическая издательская компания “Наука
117864, Москва, Профсоюзная ул., 90
Типография издательства “Дом печати”
432601 г. Ульяновск, ул. Гончарова, 14
ОБЩАЯ
СТРАТИГРАФИЧЕСКАЯ ШКАЛА
ФАНЕРОЗОЯ
(Стратиграфический кодекс, 1992)
Эра тема Сис- тема Отдел и подотделы Ярус
КАЙНОЗОЙСКАЯ KZ Четвертичная Q (антропогеновая) О
Неогеновая N Плиоцен N2 I Верхний N2 Средиземноморье I Пьяченцский N2pia Занклский N2zan Восточный Паратетис Акчагыльский N2a Киммерийский N2k Понтский N,.2p Мэотический Щт Сарматский N,sr Конкский N,kn Караганский N, кг Чокракский N,ch
Нижний nJ
Миоцен | Верхний N, Мессинский Ntmes Тортонский N,‘tor
Средний N2 Серравальский N(srv Лангийский N, Ian
Нижний N] Бурдигальский N,bur Аквитанский N,aqt Тарханский N, t Коцахурский N(kc Сакараульский N,s Кавказский N, к
Палео- геновая -Р । Олигоцен ₽31 Верхний Хатский ₽3h
Нижний Рюлельский ₽3г
Ордовикская О Силурийская S Девонская D Каменноугольная С Пермская Р
250
Татарский P2t О Дорашамский P2dr Джульфинский P2d
Верхний Р2 Казанский P2kz Уфимский Р2и з1Й Паратети >ie районы) Мидийский P2md Мургабский Р2т Кубергандинский P2kb
Нижний Р, Кунгурский Р]к Артинский Р(аг Сакмарский P,s Ассельский Р,а ВОСТОЧН1 (южнь Бол орский Рф Яхташский PJa Сакмарский P^s Ассельский Р,а
«гои Верхний С3 Гжельский С3д Квсимовский С3к
Средний С2 Московский С2т Башкирский С2Ь
Нижний С, Серпуховский C,s Визейский C,v Турнейский С(t
3G0 Верхний D3 Фаменский D3fm Франский D3f
Средний D2 Живетский D2zv (D2g) Эйфельский D2ef
Нижний D, Л 1 п Эмсский D,e Пражский О, р Лохковский D, I
Верхний S2 Пржидольский S2p Лудловский S2ld
Нижний S| 440 Венлокский S,v(3,л Лландоверийский St )
Верхний Оз Ашгиллский O3as (О3а)
Средний О2
Нижний О,
— 505 ---
Карадокский О2к (О2с)
Лландейловский O2ld
Лланвирнский О21
Аренигский О,а
Тремадокский О
МЕЗОЗОЙСКАЯ MZ КАЙНОЗОЙСКАЯ KZ
- Палеогеновая Р г3 nehoe Верхний Приабонский ₽2р
Средний ₽| Бартонский Р2Ь Лютетский Р2|
Нижний Ипрский ₽2i
Палеоцен P, | Верхний Танетский ₽, t
Нижний ₽j ес . Монтский ₽, m Датский ₽,d
Меловая К Верхний К2 Маастрихтский К2т Кампанский К2кт (К2ср) Сантонский K2st Коньякский К2к(К2сп) Туронский K2t Сеноманский K2s (K2cm)
Нижний К1 1 ДЕ Альбский К1а< Аптский К, а Барремский К, Ьг Готеривский Kt (K,h) Валанжинский K,v Берриасский К,Ь
1 1 Триасовая Т | Юрская J г j Верхний Jg Титонский J3U Волжский J3V Кимериджский J3km Оксфордский J3o
Средний Jg Келловейский J2k (J2c) Батский J2bt Байосский J2b Ааленский J2a
— Нижний Jj Тоарский t ПлинсбахскийJ,p Синемюрский J,s Геттангский J(g (Jih)
Z1 □ Верхний Тз Рэтский T3r Норийский T3n Карнийский T3k
Средний Т2 Ладинский T2I Анизийский Т2а
Нижний Т1 Оленекский Т,о Индский T(i
ПАЛЕОЗОЙСКАЯ PZ Кембрийская G 505 Верхний €3 Батырбайский С3Ь Аксайский С3ак Сакский C3s Аюсокканский 633s
Средний 62 Майский С2т Амгинский С2ат
Нижний С, 570(530) Тойонский tn Ботомский С,Ь
Атдабанский at Томмотский С Л
ОБЩАЯ СТРАТИГРАФИЧЕСКАЯ ШКАЛА ДОКЕМБРИЯ
(Стратиграфический кодекс, 1991)
Акротема Эонотема Эратема Система
Фанерозой Палеозой Кембрий
:рхний протерозой РЯ2 Венд\/ | Верхний V2 —- 6501 15 Нижний V,
Г X S э *) э L Рифей R I Верхний рифей (каратавий) R3 1000150 Средний рифей (юрматиний) П2 1350120 Нижний рифей (бурзяний) R,
5 L OJ СО л сел 4. с.л
оелл + ел Нижний протерозой (карелий) PR, Верхний карелий PR? 1900 150—— Нижний карелий PR)
£Г Верхний архей AR ел + ел 2
1 i и К L Нижний архей AR