/
































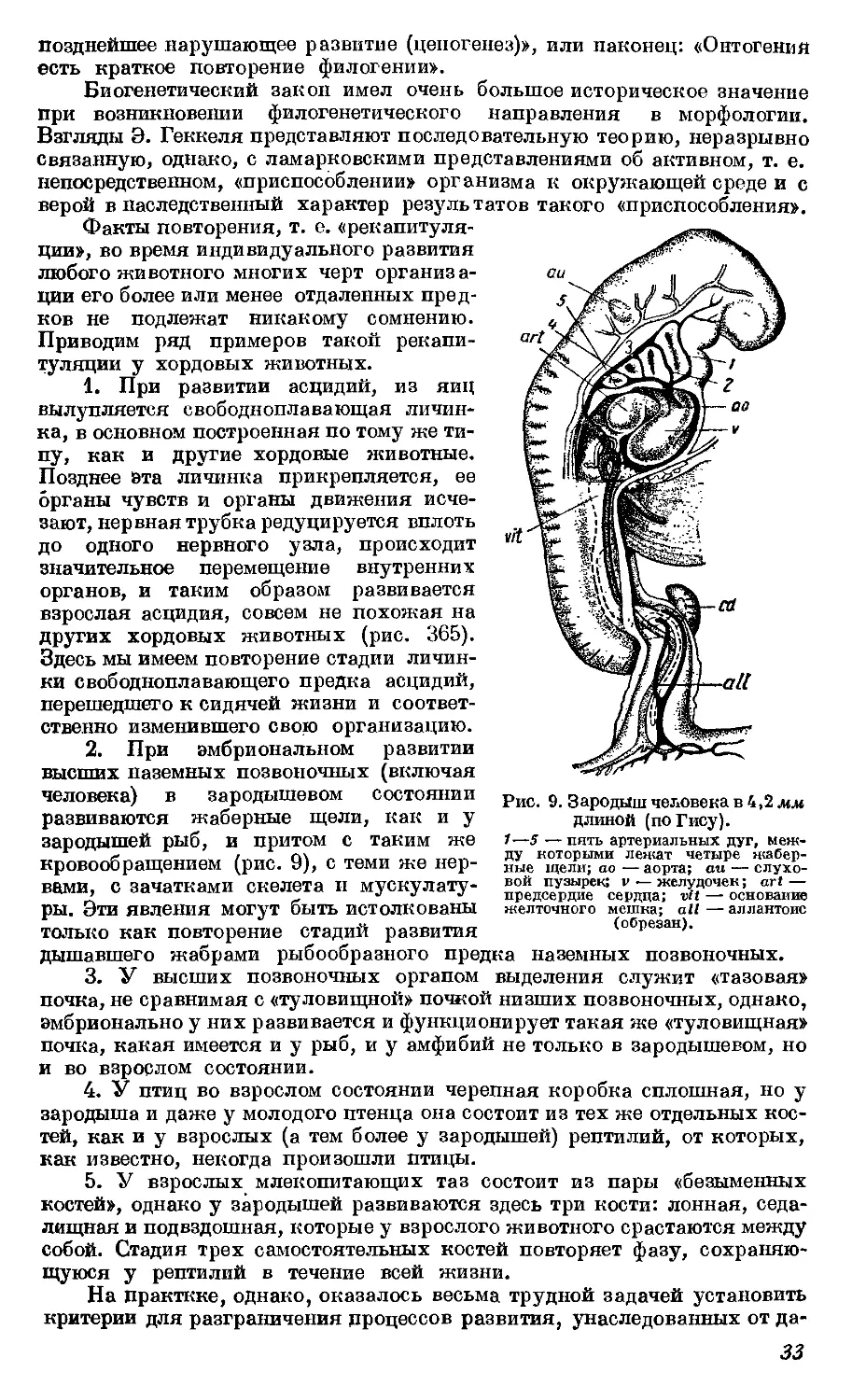








































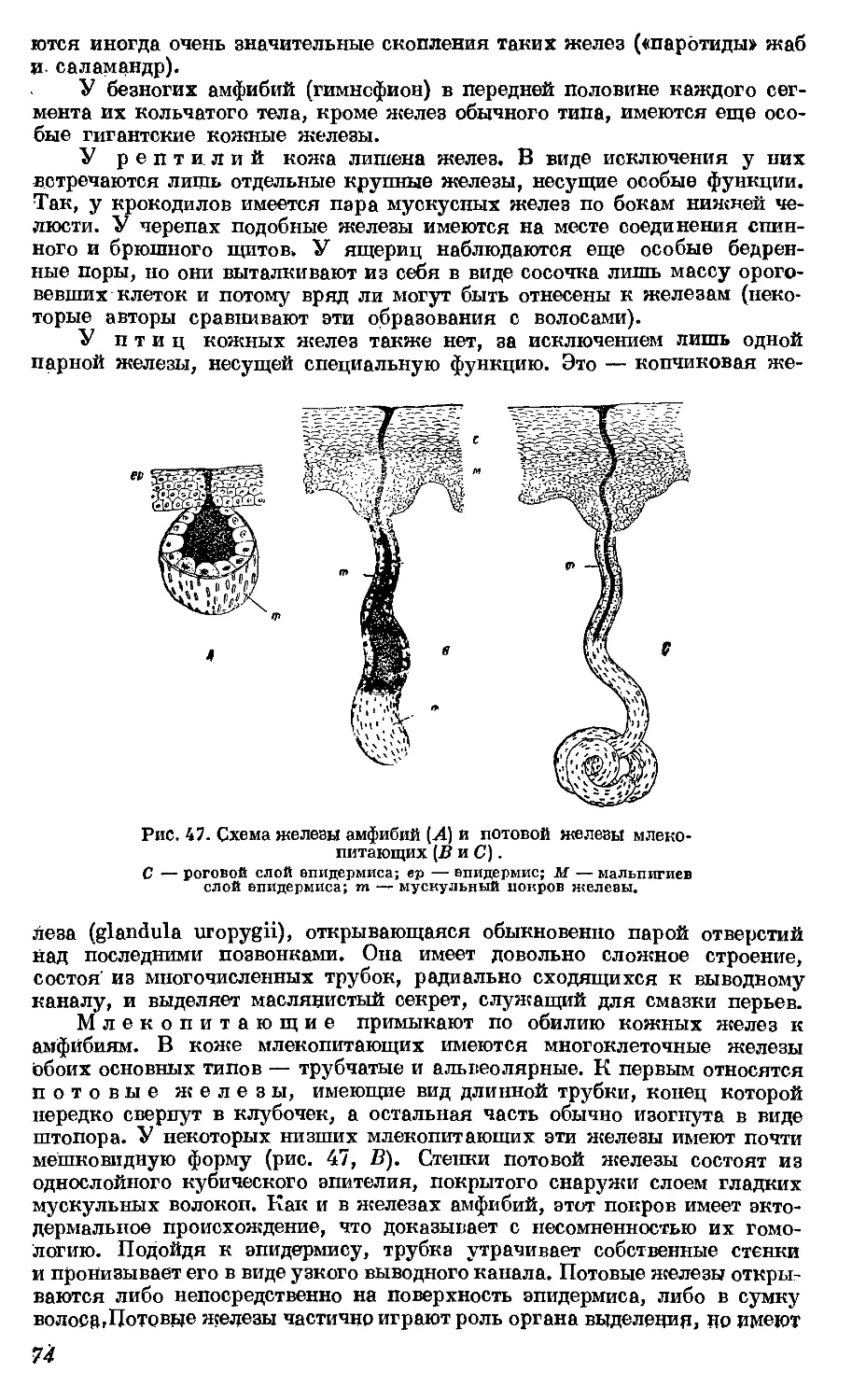






























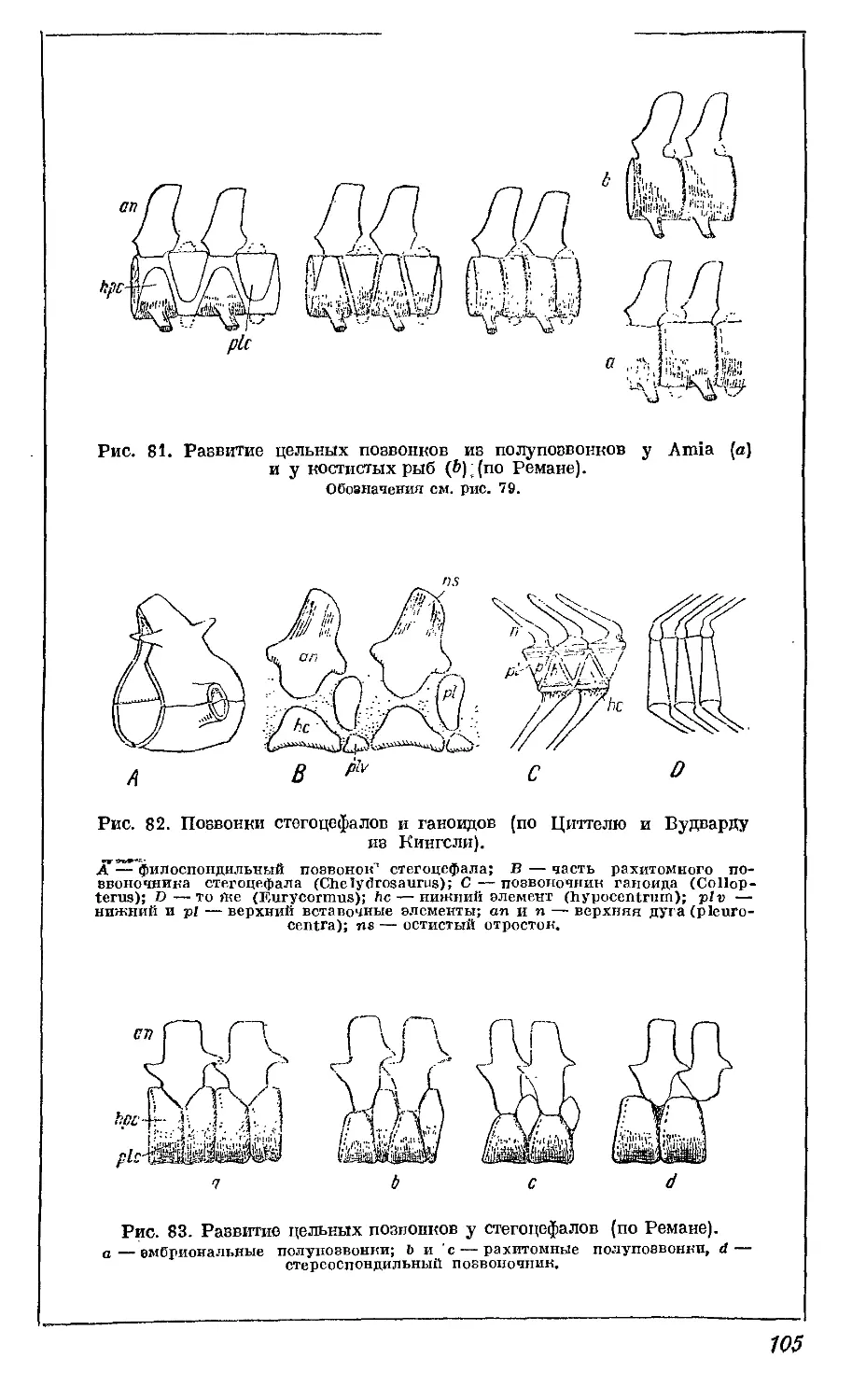













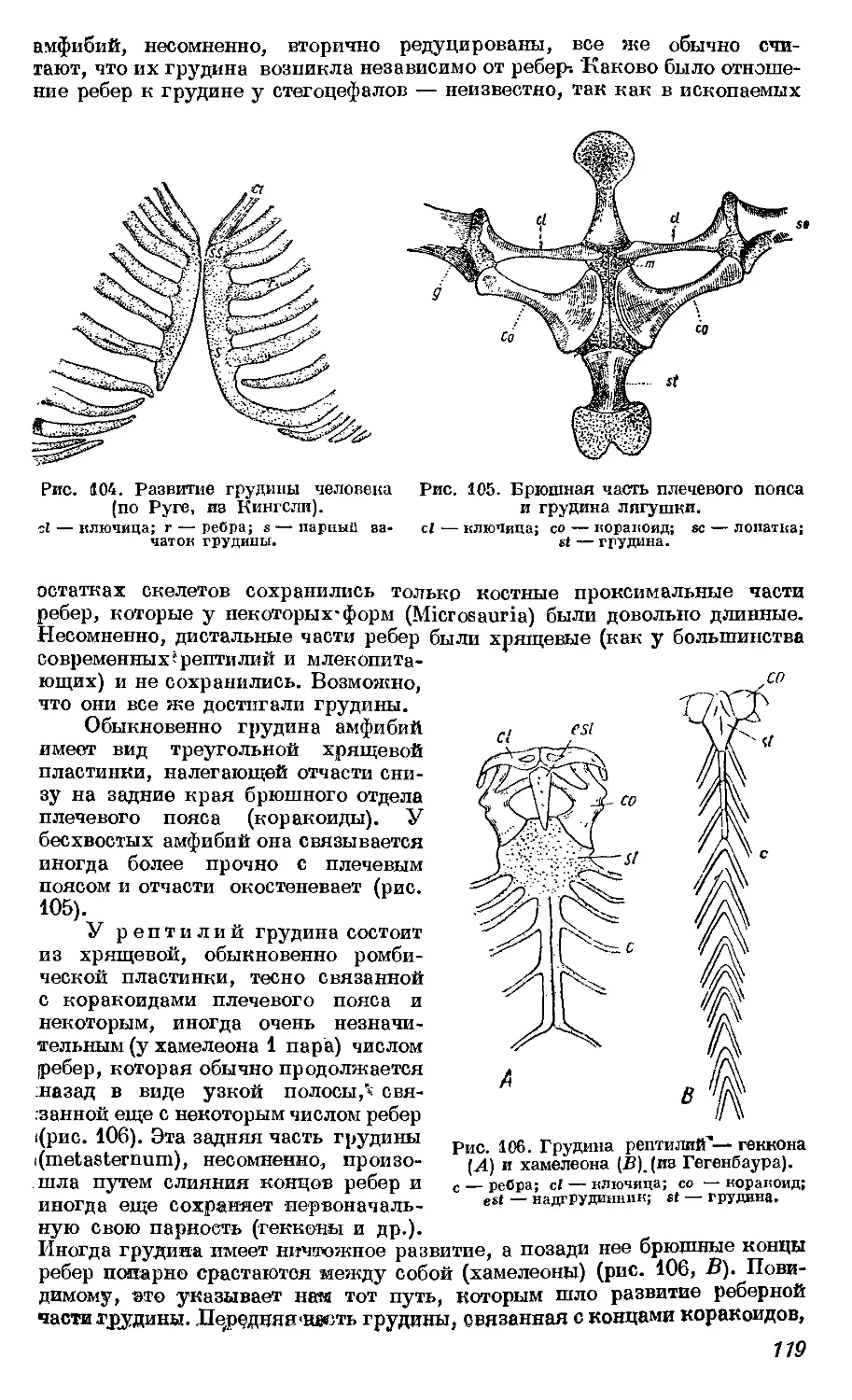








































































































































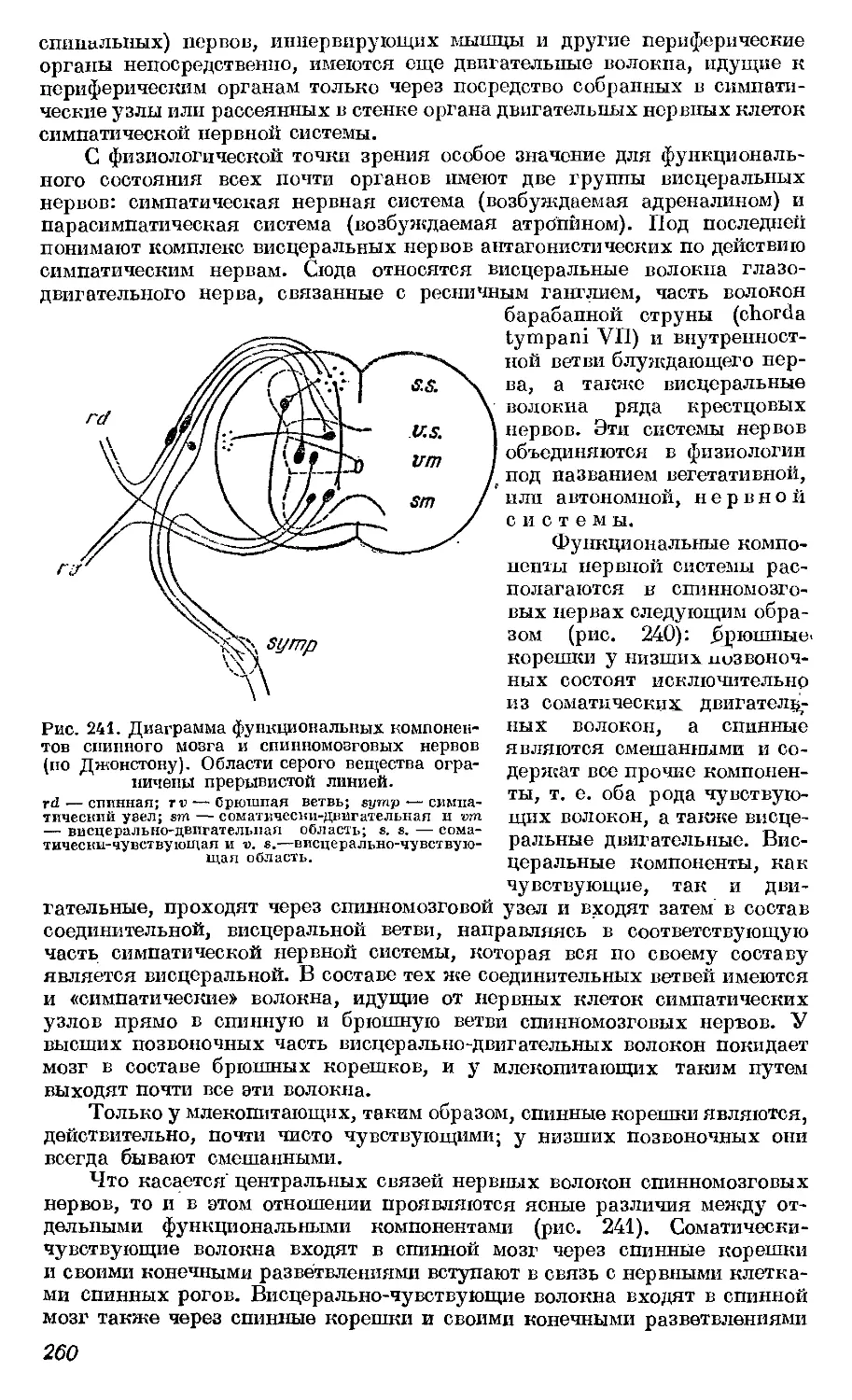













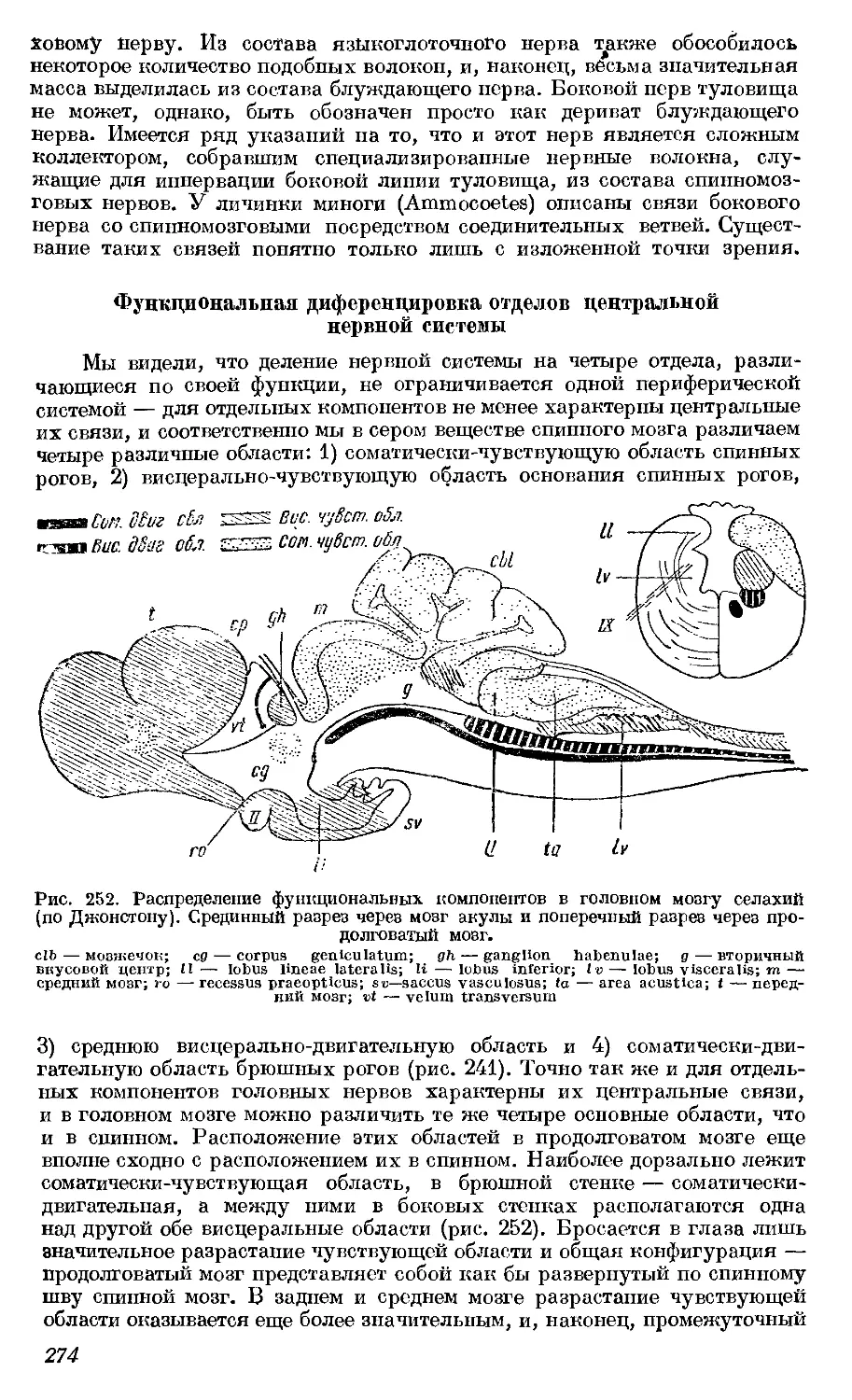













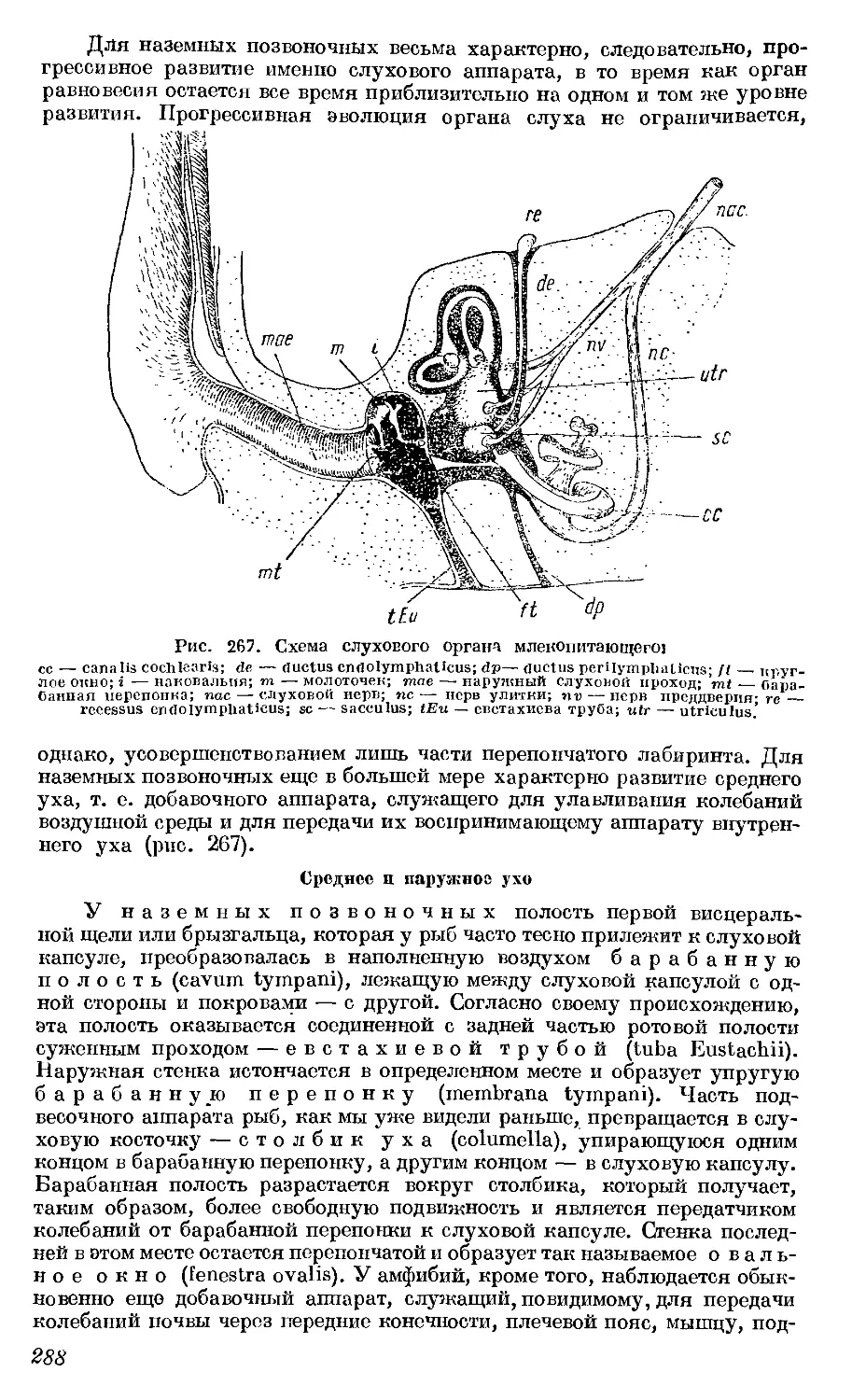















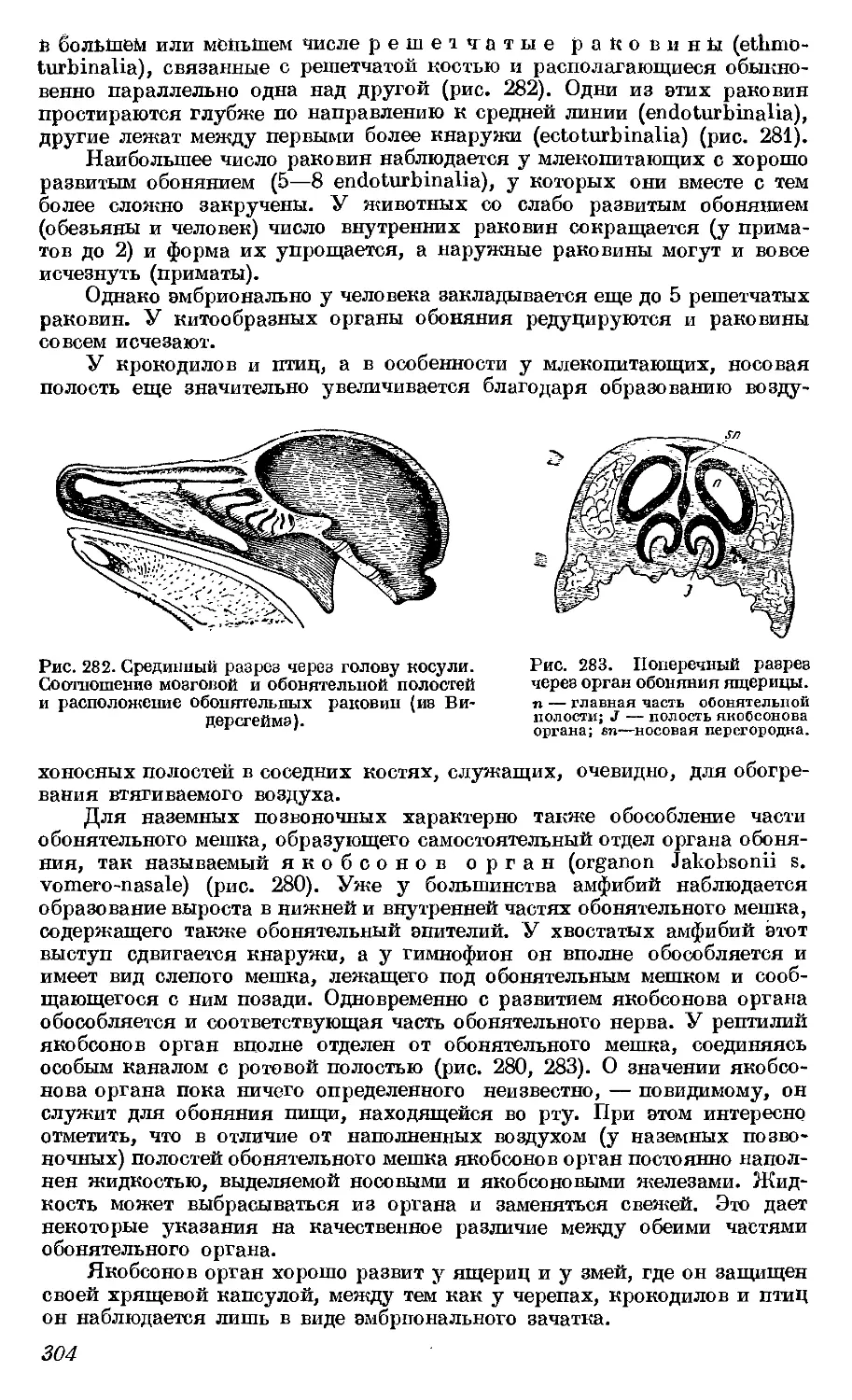












































































































































































































































Текст
И. И. ШМАЛЬГАУВЕН
ОСНОВЫ
СРАВНИТЕЛЬНОЙ АНАТОМИИ
позвоночных животных
ИЗДАНИЕ ЧЕТВЕРТОЕ
ИСПРАВЛЕННОЕ И ДОПОЛНЕННОЕ
Допущено
Министерством высшего образования СССР
в качестве учебника для биофаков
еосударственных у ниве рситетов
ГОСУДАРСТВЕННОЕ ИЗДАТЕЛЬСТВО «СОВЕТСКАЯ НАУКА»
Москва —1947
Книга представляет собой четвертое издание
известного руководства сравнительной анатомии
академика И. И. Шмальгаувена и рассчитана на
студента-биолога. В отличие от большинства дру-
гих подобных руководств в него включена до-
вольно значительная общая часть, содержащая
очерк истории сравнительной анатомии, опреде-
ление основных ее понятий и разъяснение основ-
ных принципов филогенетического преобразова-
ния органов. Кроме того, как единственный опыт
такого рода дан краткий очерк филогении с за-
ключительной главой о морфо-физиологических
закономерностях эволюции. Эта синтетическая
часть является логическим завершением курса.
ПРЕДИСЛОВИЕ
Настоящий курс рассчитан на студентов биологического факультета
универститетов. Предполагается, что студент, приступающий к изучению
сравнительной анатомии, получил некоторую общебиологическую подго-
товку и прослушал уже основной курс зоологии. Все же я считал не лиш-
ним дать вначале краткую характеристику хордовых и позвоночных жи-
вотных, а также ознакомить читателя с элементами их эмбриологии, по-
скольку это нужно для понимания сравнительной анатомии позвоночных.
Кроме того, я предпосылаю каждой главе краткий очерк по сравнительной
анатомии беспозвоночных для того, чтобы дать отправную точку для
сравнения и чтобы яснее охарактеризовать эволюцию соответствующих
органов позвоночных. Эти вводные очерки набраны мелким шрифтом.
Мелким шрифтом набрано также изложение строения органов по клас-
сам в тех случаях, когда такое изложение казалось мне необходимым; в
других случаях оно совершенно опущено, так же как и описание многих
анатомических особенностей отдельных групп, которое с успехом может
быть приурочено полностью к учебнику зоологии (например, совершенно
опущено описание органов, служащих для питания зародыша у живородя-
щих позвоночных).
Можно быть различного мнения о методах преподавания сравнитель-
ной анатомии и соответственно материал может распределяться по-раз-
ному. Есть сторонники рассмотрения строения позвоночных по классам,
как в известной «Зоологии» Паркера. Мне кажется, что одно не исключает
другого. Большой практикум, естественно, ведется и должен вестись по
зоологической системе. Однако в исследовательской практике разработка
основных проблем филогенетической морфологии всегда велась и ведется
по системам органов и отдельным органам. Вне этого порядка нельзя
получить цельного представления об эволюции этих систем и органов.
С другой стороны, анализ должен дополняться синтезом, и от эволюции
систем и органов мы должны вернуться к эволюции организмов в целом,
т. е. к общей филогении животных. В очень краткой форме это сделано
в настоящем курсе. Конечно, при таком рассмотрении все внимание фик-
сируется лишь на общих путях эволюции основных ветвей позвоночных.
Частные линии развития по пути «адаптивной радиации» форм в пределах
классов и отрядов, с их приспособлениями к тем или иным условиям
существования, с их иногда крайней специализацией, остаются в боль-
шинстве вне поля зрения. Я считаю, что этот материал, относящийся в
значительной мере к «экологической» морфологии, должен входить в
курс зоологии позвоночных.
При выборе материала я старался отвести первое место тем фактам,
которые имеют большое филогенетическое значение, характеризуя эво-
люцию больших групп. Особое внимание уделено скелету как системе
органов, наиболее ясно иллюстрирующей эволюцию позвоночных вообще
и отдельных классов и групп в частности.
3
Во втором издании по сравнению с первым, помимо внесения некото-
рых поправок и дополнений, значительно увеличено число рисунков.
Кроме этого, заново составлено введение, содержащее исторический
очерк развития сравнительной анатомии, разъяснение основных понятий
и принципов сравнительной анатомии, а также разбор методов филогене-
тического исследования. Далее, как опыт синтеза сравнительноанатоми-
ческих данных, к курсу прибавлен краткий очерк филогении позвоночных.
И 'здесь для цельности изложения этому очерку предпослан набросок
основных моментов филогении беспозвоночных. Наконец, в конце книги
разбираются некоторые основные закономерности эволюционного про-
цесса, поскольку о них можно говорить на основании филогенетических
исследований.
В третьем издании добавлено 15 рисунков, часть прежних заменена,
и внесены исправления, уточнения и немногие дополнения в тексте, в
особенности в описании осевого скелета и половой системы.
В четвертом издании введены изменения в изложении общих покро-
вов, изменена номенклатура некоторых костей черепа и добавлен ряд
рисунков.
И. Шмалъгаузен
ЧАСТЬ I
ВВЕДЕНИЕ
При изучении животных организмов мы их расчленяем на простран-
ственно органиченные части, несущие определенную функцию. Такие
более или менее сложные комплексы, служащие для тех или иных отправ-
лений, мы называем органами. Разлагая сложные органы на более простые,
подчиненные органы, или еще далее—на наиболее простые, элементарные
органы (а последние—на ткани и клетки, что представляет вместе с том
уже задачу гистологии), мы путем анализа получаем детальнейшую кар-
тину строения органов и их взаимоотношений в целом организме. Такое
исследование строения организма, т. е. его формы в самом широком смысле
этого слова, составляет задачу описательной анатомии. Другая задача
исследователя—изучить изменения этой формы как процесс — состав-
ляет предмет морфологии. При этом изучение тех преобразований в строе-
нии организма, которые наблюдаются в сравнительно короткое время
его индивидуального развития, начиная от яйца и до вполне закончен-
ного организма, составляет специальную задачу эмбриологии. Исследо-
вание же преобразований сходных органов у различных животных со-
ставляет предмет сравнительной анатомии. Путем синтеза сравнительная
анатомия создает ряды форм, позволяющие проследить шаг за шагом
постепенные изменения строения органов.
Главный метод сравнительной анатомии—метод сравнения, позво-
ляющий установить большее или меньшее сходство между органами раз-
личных животных и дающий возможность проследить постепенные их
преобразования. Для обозначения сходства общего строения и происхож-
дения, или морфологического сходства, служит термин «гомология» в
отличие от сходства приспособлений или физиологического сходства,—
аналогии.
Установление гомологии органов и составляет ближайшую задачу
сравнительной анатомии, как она была поставлена уже давно (в конце
XVIII столетия). Критерием гомологии является, таким образом, простое
сходство строения органов, причем чем более сходны между собой и соот-
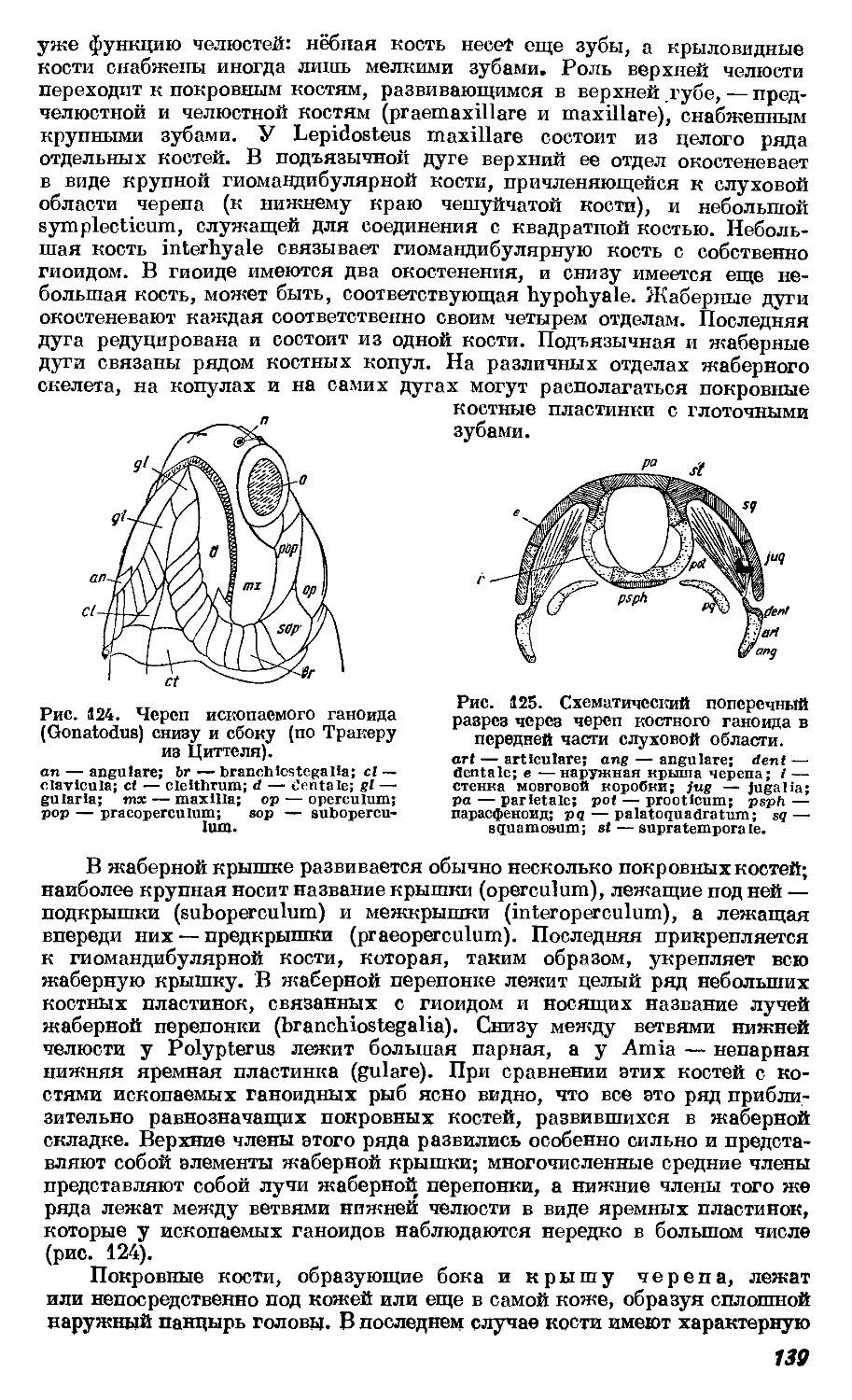
ветствующие организмы (что определяется сходством строения и взаимо-
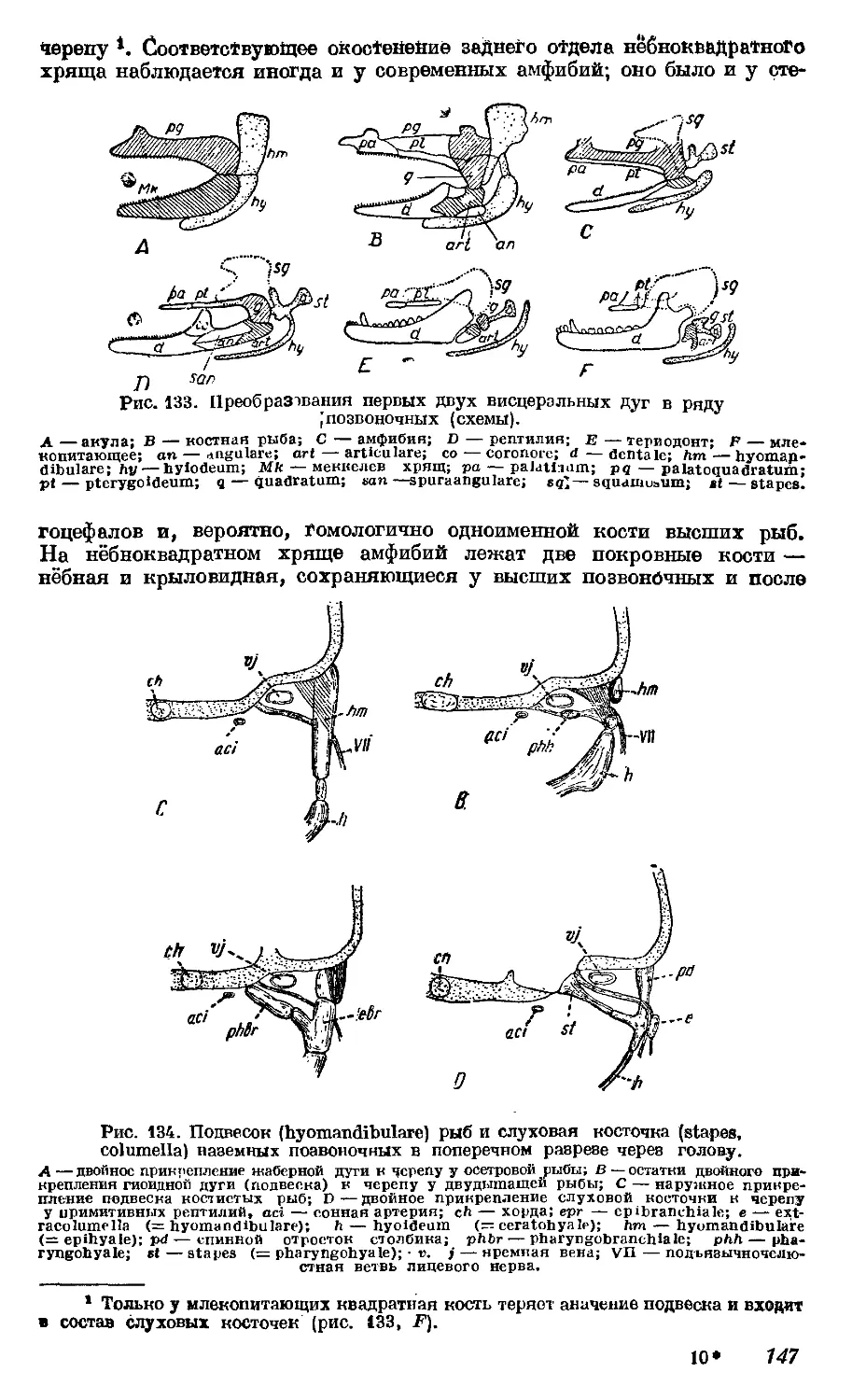
отношений других органов), или, как говорят, чем они «ближе», тем и
надежнее выводы, тем меньше риска принять аналогию за гомологию.
Отсутствие промежуточных форм, которые позволили бы провести срав-
нение и менаду более «далекими» организмами, отчасти восполняется
сравнением всех стадий индивидуального развития, так как зародыши
таких животных и вообще не вполне еще развитые формы обнаруживают
обычно большую степень сходства, чем взрослые. Поэтому в настоящее
время сравнительная анатомия не довольствуется изучением одних взрос-
5
лых организмов, но проводит детальное сравнение на всех стадиях их
индивидуального развития1. Таким образом, сравнительная анатомия
сливается со сравнительной эмбриологией в единую науку.
С установлением эволюционной теории задача сравнительной ана-
томии расширилась. Изменился и взгляд па значение сходства гомоло-
гичных органов. Если раньше гомология, т. е. морфологическое сходство
органов, объяснялось единством плана творения, то теперь оно объясня-
ется общностью происхождения от одного и того же органа более или менее
отдаленного предка, т. е. кровным родством организмов. Соответственно
и самое понятие гомологии органов несколько изменилось, и в современ-
ной науке гомологичными признаются сходно построенные органы, сход-
ство которых есть выражение их происхождения от одного и того же
органа общего предка.
Вместе с тем установление гомологий органов и изучение их преобра-
зований в ряду животных, т. е. построение сравнительноанатомических
рядов форм, не является уже конечной целью, а оказывается лишь сред-
ством для выяснения родственных отношений и, таким образом, средством
для изучения истории организмов, т. е. их филогении. Так как необхо-
димый при изучении всякой истории фактор—время—дается здесь лишь
путем сложных умозаключений, то некоторые выводы нуждаются в про-
верке данными палеонтологии, в которой историческая последователь-
ность дается более непосредственно и надежно. Отсюда вытекает необхо-
димость самого тесного контакта с наукой об ископаемых животных,
строение которых, само собой разумеется, составляет в равной мере и
предмет сравнительноапатомического изучения.
Изучение истории организмов, интересное и само по себе, становится
существенно необходимым, если мы желаем подойти ближе к тем процес-
сам преобразования форм, которые сопровождают историческое или фило-
генетическое развитие организмов, если мы желаем понять приспособления
органов и организмов, их прогрессивное и регрессивное развитие, смены
функций и т. д. И не менее необходимо это изучение, если мы желаем
найти удовлетворительное объяснение процессов индивидуального
(онтогенетического) развития. На стадии яйцеклетки очень далекие
животные нередко столь сходны между собой, что при современных сред-
ствах исследования они по внешности неотличимы; между тем в дальней-
шем, даже при идеально одинаковых условиях, из них постепенно разви-
ваются совершенно отличные организмы. Это различие в процессах формо-
образования обусловлено, конечно, индивидуальным строением яиц,
связанным с различным их происхождением и различной историей соот-
ветствующих организмов.
Можно прямо сказать, что ни в какой другой области знания важность
изучения истории не выступает с такой очевидностью, как в биологии,
и наиболее существенной особенностью живых организованных существ
является их историческая обоснованность.
Таким образом, изучение истории организмов животных—важнейшая
задача сравнительной анатомии и связанной с нею филогенетики. Можно
задачу понимать еще шире и требовать объяснения этой истории, т. е.
ставить своей целью каузальное изучение истории, но это составляет уже
в значительной мере предмет эволюционной теории, и здесь на первый
план выступает экспериментальный метод исследования. Нужно сказать,
что в этом направлении работа еще только развертывается.
1 Эмбриологический критерий гомологии, взятый сам по себе, ненадежен. Неко-
торые процессы индивидуального развития оказываются весьма изменчивыми, так
что нередко несомненно гомологичные органы развиваются у близких форм за счет
различных источников (меторизис), в различных местах (гетеротопия) или в иной
последовательности (гетерохрония).
б
ИСТОРИЯ СРАВНИТЕЛЬНОЙ АНАТОМИИ
Анатомия животных изучалась уже в глубокой древности. Натура-
листы-философы древней Греции обладали известными познаниями в этой
области. Особенно значительную работу проделал Аристотель (384 —
322 г. до н. э.), изучивший и описавший очень большое число разнообраз-
нейших животных (более 500). Аристотелю принадлежит заслуга создания
первой научной классификации. Он обращает особое внимание па един-
ство плана строения животных в пределах устанавливаемых им крупных
групп, в которых одни и те же органы различаются лишь по степени своего
развития. В строении организма он различает «гомогенные» части (т. е.
ткани) и «гетерогенные» (т. е. органы). Аристотель произвел также первые
наблюдения над эмбриональным развитием цыпленка, причем отмечает,
что общие черты организации выявляются раньше, чем специальные (много
позднее это было сформулировано в законе К. Бэра). Аристотель рас-
полагал все организмы (как и неорганические тела) по совершенству их
строения в один восходящий ряд (что в XVIII веке было развито далее
натуралистами, говорившими о «лестнице живых существ»). Как большую
заслугу Аристотеля следует отметить также и то обстоятельство, что он
изучал организмы в связи с окружающей средой, а органы — в связи с их
функцией, причем последняя рассматривалась как фактор, определяющий
строение органа. Вместе с тем, однако, по Аристотелю орган существует
для выполнения определенной функции и его развитие определяется во
всех деталях именно этой конечной целью. Такая установка приобрела
большое значение в построениях многих биологов-идеалистов и носит
название телеологической (по-гречески тзког.— цель, kfc-. —
учение).
В древнем Риме наибольшую известность получили компилятивные
сочинения Плиния (23—79). Анатомия древности завершилась работами
Галена (131—210), из которых черпали свои знания и средневековые
ученые. Гален был разносторонним врачом, детально изучавшим анатомию
и в частности анатомию нервной системы на животных, в особенности
на обезьянах как животных наиболее похожих на человека.
В средние века анатомия животных (как и все естествознание) была
в состоянии полного упадка и даже в эпоху Возрождения она не подня-
лась выше уровня знаний Аристотеля. Анатомия животных очень долго
играла чисто служебную роль. Путем вскрытия животных старались
познать строение человеческого организма, так как вскрытие челове-
ческих трупов не допускалось вследствие религиозных предрассуд-
ков.
Эпоха Возрождения не оставила без внимания и анатомию. Мы не мо-
жем не упомянуть здесь гениального Леонардо да Винчи (1452—1519)
как одного из пионеров в этой области. Работы Ронделета, Белона и,
в особенности, знаменитого анатома Везалия (1514—1564) значительно
пополнили знания по анатомии человека и животных. Итальянцы Фаб-
ричио и Северино возродили в своих исследованиях сравнительный метод,
введенный еще Аристотелем. Исключительно большое значение в истории
морфологии имело, однако, открытие действительного хода кровообраще-
ния и выяснение его механизма Гарвеем (1578—1657). Бартолини продол-
жил исследования Гарвея в области лимфатической системы. Мальпиги
(1628—1694) углубил исследования анатомов применением микроскопа и
произвел также ряд блестящих исследований по морфологии беспозво-
ночных. В этой области также выделяются необычайно тщательные иссле-
дования Сваммердама (1637—1680), которому принадлежит еще ряд
важных эмбриологических исследований. Собственно основателем срав-
нительной анатомии можно, пожалуй, считать сотрудника Бюффона —
Л. Добантона (1718—1800), давшего анатомическое описание скелета
7
и других органов целого ряда птиц и млекопитающих и проведшего
детальное их сравнение друг с другом и с органами человека.
На заре современного эмбриологического исследования имели немалое
значение также наблюдения и теоретические представления К. Бонне
(1720—1793), как ярого приверженца теории преформизма, и К. Ф.
Вольфа (1733—1734), как основателя теории эпигенеза. Интересные
сравнительноанатомические исследования принадлежат затем знаменитому
голландскому анатому П. Камперу (1722—1789), французу Ф. Вик д’Азиру
(1748—1794), а также английскому анатому Дж. Гейгеру и немецкому
И. Блюменбаху (1752—1840). Все указанные исследователи при сравне-
ниях всегда исходили из анатомии человека.
Однако сколько бы ни было сделано указанными учеными, все же
только Ж. Кювье (1769—1832) удалось поднять сравнительную анатомию
до уровня самостоятельной научной дисциплины. Из позвоночных им
впервые была подробно изучена анатомия рыб и в особенности детально
разработана анатомия млекопитающих, причем впервые основательно
изучены и скелеты многих ископаемых животных. Критический ум Ж.
Кювье удержал его от легковесных натурфилософских спекуляций того
времени, однако он же не дал ему подняться до высоты тех эволюционных
идей, которые уже в то время зарождались в свободных умах Ж. Ламарка
и др., но не имели еще достаточно солидной научной базы. Ж. Кювье
был твердо убежден в неизменяемости видов; он разделил все животное
царство на несколько типов, построенных каждый по особому плану, и
совершенно исключал возможность сравнения животных, принадлежащих
к разным типам. Однако было бы неправильно характеризовать Ж. Кювье
как консерватора и голого эмпирика. Учение о типах представляет собой
результат известных обобщений, не потерявших своего значения и дошед-
ших в измененной форме до настоящего времени. Ж. Кювье не только
является основателем двух новых научных дисциплин — сравнительной
анатомии и палеонтологии. Мы обязаны Ж. Кювье установлением заме-
чательного для своего времени и до сих пор недостаточно оцененного
«принципа корреляций», по которому каждый организм представляет
сббой целостную систему, причем пи одну часть этой системы нельзя
изменить, не вызвав изменения остальных. Все части организма зависят
в своих проявлениях p;.jr от друга, и эта взаимозависимость функций
определяет сосуществси ание частей внутри целого. Функциональное един-
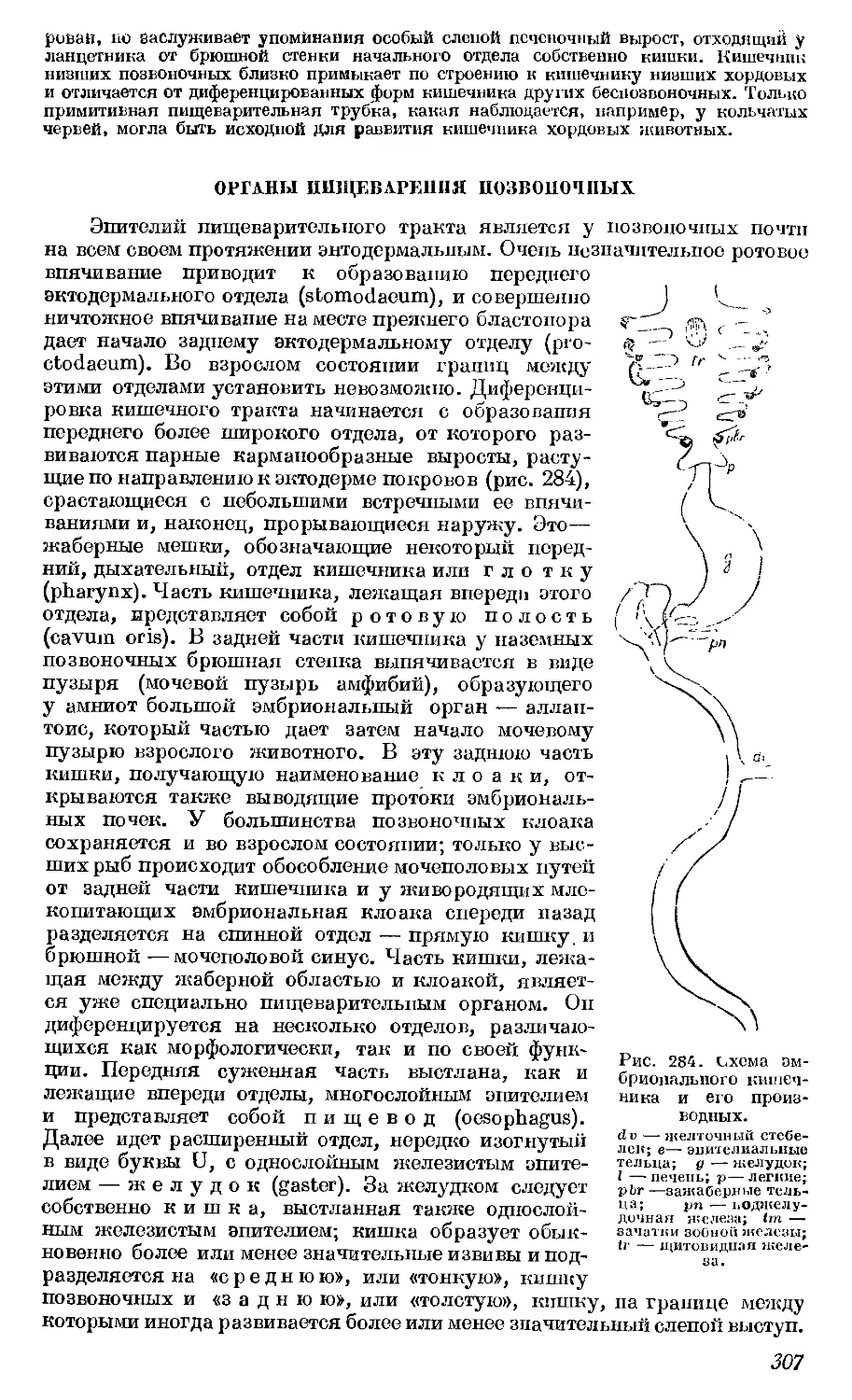
ство организма рассматривается как необходимое условие, которое только
и делает возможным существование живых существ. Форма и функция
связаны неразрывно, и именно функция определяет строение органа.
В этих представлениях НС. Кювье стоит значигельно выше своих совре-
менников, хотя^н и не мог освободиться от телеологических установок.
Изучение функций органов в глазах Кювье является средством для пони-
мания их строения.
Пониманием приспособления органа к 'несению известной функции
и исчерпывается, однако, «объяснение» его строения.
Из учеников Ж. Кювье следует назвать Г. Блэнвилля, И. Меккеля
и в особенности Г. Мильн-Эдвардса (1800—1885). Последний заслужил
большую известность своими исследованиями строения и развития различ-
ных беспозвоночных и особенно ракообразных, что привело его к уста-
новлению принципа диференциации, основанной на разделении труда
между различными частями организма. В этих работах мы видим еще
влияние идей Кювье. Дальнейшее развитие морфологии пошло, однако,
в целом по иному пути, именно по пути изучения «чистой» формы (неза-
висимо от функции данных частей).
Широким охватом материала и многими новыми идеями мы обязаны
современнику Кювье французскому зоологу Э. Ж. Сент-Илеру (1772 —
1844). Сравнение органов различных животных привело его к установле-
8
нию особой категории сходства, названного им «аналогией» (соответствует
тому, что теперь называют гомологией). Это сходство определяется постоян-
ством соотношений с другими органами, а не функцией и не формой, кото-
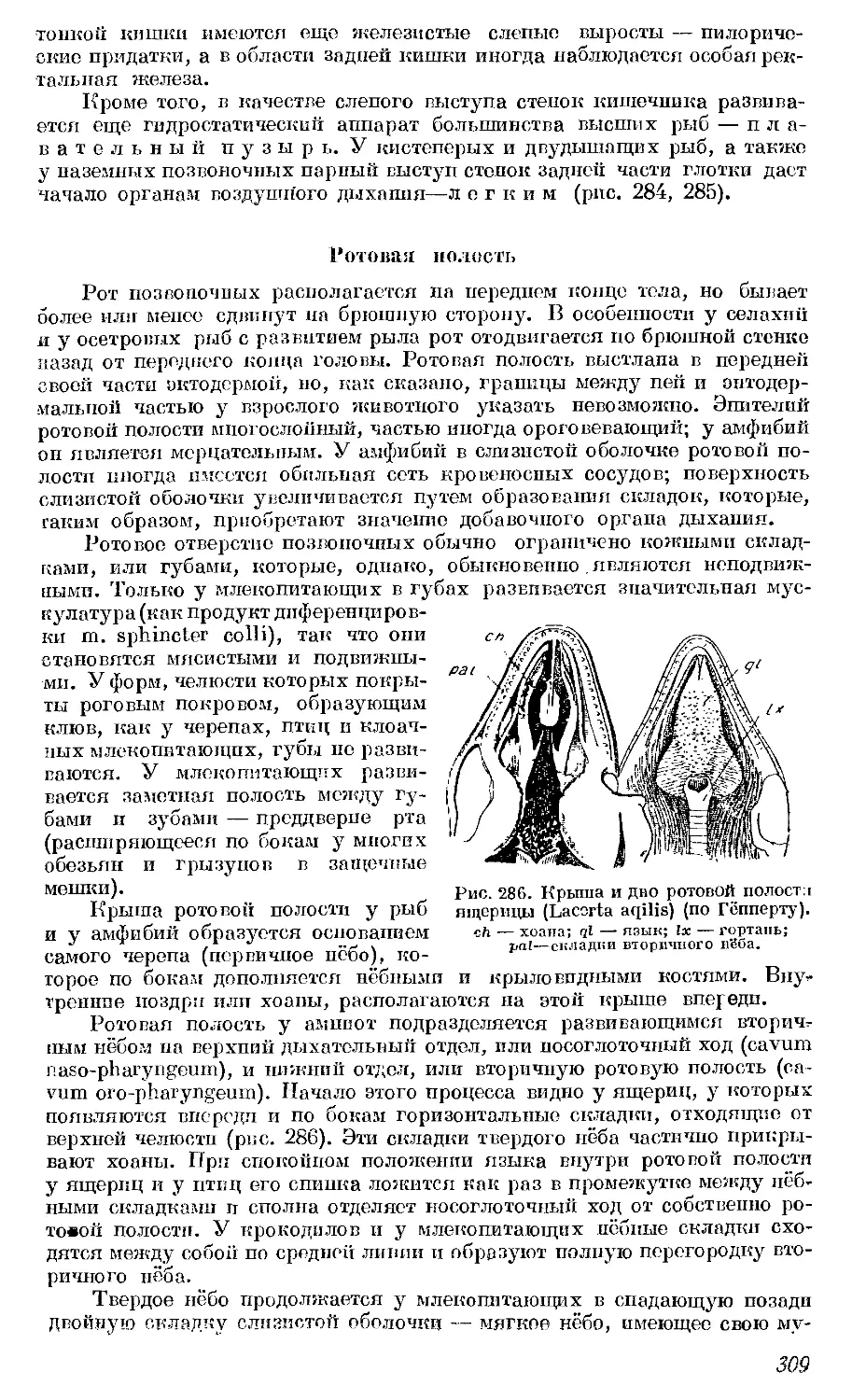
рые оказываются весьма изменчивыми. Интересен также его «принцип
уравновешивания органов» (principe de balancement des organes). Уравно-
вешивание понимается здесь как соотносительная компенсация в развитии
органов; при ограниченности «материала» прогрессивное развитие одного
органа связано с регрессом другого и, наоборот, атрофия какого-либо
органа приводит к разрастанию другого. Сент-Илер изучал также строение
зародышей различных позвоночных (между прочим, нашел провизорные
закладки зубов у зародышей беззубых китов) и проводит сравнение заро-
дышей высших позвоночных со взрослыми низшими (например, найдя
обособленные кости черепа у зародыша птицы, он провел их сравнение
с костями черепа взрослой рыбы).
Идеологическая установка Э. Ж. Сент-Илера в общем противоположна
установке Ж. Кювье. Если у Кювье функция определяет строение органа,
то у Сент-Илера строение органа определяет его функцию и изменения
последней зависят от изменений в строении органа. Таким образом,
изучение функций теряет для морфолога всякий смысл. Морфологисчекое
сходство объясняется «единством плана». Видоизменения этого плана
зависят от изменений в окружающей среде, причем последняя чисто физи-
чески меняет строение организма и его органов. Естественно, что согласно
такому взгляду природа не создает ничего нового, а только преобразует
уже существующее. Сравнение различных организмов, построенных по
одному плану, позволяет выделить общее, сходное, типичное, позволяет
отвлечься от обусловленных воздействием среды изменений, отклонений
от этого «типа». Таким образом создается представление об исходном,
обобщенном идеальном «типе», в котором реализованы все органы, все
возможности. В результате хотя Сент-Илер как-будто удачно избегает
телеологических установок Кювье и в вопросах преобразования организ-
мов стоит на противоположной, именно механистической позиции, хотя
Сент-Илер возвышается до признания принципа преобразования всех
организмов без ограничения, приближаясь в этом к основателям теории
эволюции, — все же он в своем учении скатывается к чисто формальному
Пониманию живых существ и, в конце концов, к идеалистической концеп-
ции единого плана строения. Освобожденная от изучения функциивчистая»
морфология оперирует с преобразованиями отвлеченных «идеальных»
форм. Этому особенно способствовало влияние романтической натурфило-
софии начала XIX столетия.
Таким образом, созданное Кювье учение о типах при унаследованных
от феодального строя общих идеалистических установках только и могло
привести к развитию идеалистической морфологии, кото-
рая достигла особого расцвета в Германии (Кильмейер, поэт Гёте, зоолог
Окэн). Для идеалистической морфологии особенно характерны следующие
представления: 1) «единство плана», лежащего в оснцре строения живых
существ, принадлежащих по меньшей мере к одному типу; 2) расположение
всех организмов в один ряд по восходящим ступеням совершенства, т. е.
по «лестнице живых существ»; 3) признание параллелизма между разви-
тием особи и восходящим рядом взрослых организмов; 4) представление
о повторении частей внутри организма.
Анатомы разрабатывали положения об единстве плана и о повторении
частей внутри целого. Наиболее яркое выражение получили эти тенденции
в позвоночной теории черепа, рассматривавшей голову как повторение
туловища с его позвонками, и в учении об «архетипе», развитое главным
образом английским анатомом Р. Оуэном (1804—1892) (рис. 1).
Оуэн был наиболее крупным сравнительным анатомом Англии, обра-
ботавшим огромный материал и введшим в науку ряд основных понятий,
9
являющихся и в настоящее время фундаментом сравнительной анатомии.
Так, Р. Оуэн различает две категории сходства менаду органами различных
животных—аналогию и гомологию. Гомологичным Оуэн называет «тот же»
орган у различных животных независимо от сходства или различия его
функций. Аналогичными он называет «разные» органы, несущие, однако,
у различных животных сходную функцию. Кроме «специальной гомологии»
он различает еще «общую гомологию», или отношение между органами
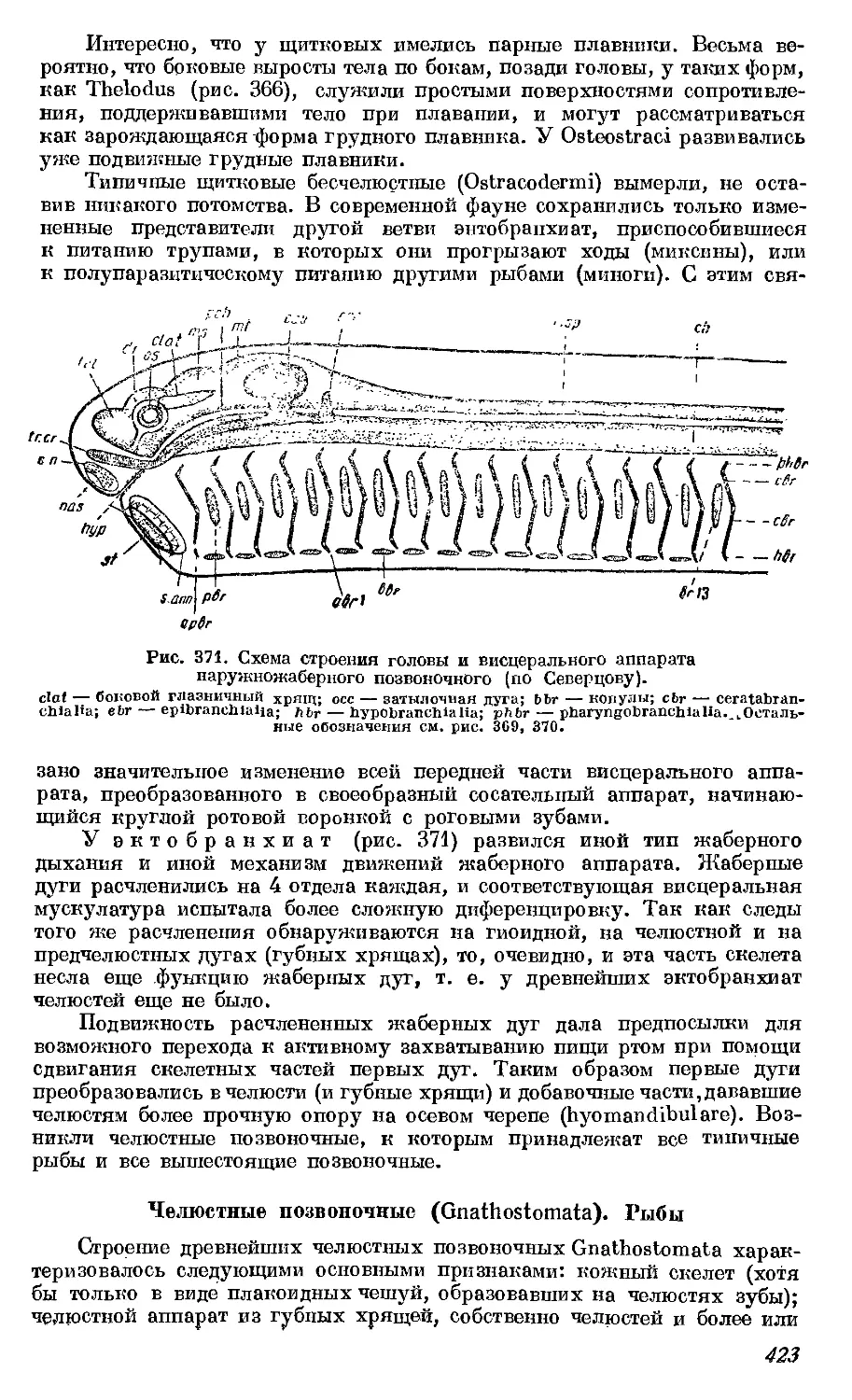
данного животного и соответствующего типа, и, наконец, «сериальную
гомологию», или сходство между последовательно повторяющимися орга-
нами одного и того же животного (например, между позвонками; то, что
теперь называется гомодинамией). Оуэн является последним типичным
представителем отживающего натурфилософского направления. Быстрый
Рис. 1. «Архетип» скелета позвоночного животного по Р. Оуэну. Гомодинамные
части обозначены одинаковой штриховкой. Черным цветом обозначены тела позвонков
и «придатки» ребер (гомодинами скелета парных конечностей).
ар — «придатки» ребер; ha — пюкние дуги («гемапофизы»); hs -— нижпие остистые отростки;
па — верхние дуги («нсурапофпзы»); pl — ребра («илеврапофизы»); n. sp. — верхние остистые
отростки.
расцвет буржуазного хозяйства требовал развития точного знания. Все
более выдвигалось значение не идеальных построений разума, а знания
фактов, изучения конкретных связей и их исторической обоснованности.
Одним из наиболее ранних представителей точного исследования является
И. Мюллер (1801—1858), детально изучивший анатомию низших позво
ночных и имевший большое влияние па дальнейшее развитие морфологии.
К. Зибольд и Ф. Станниус, кроме ряда исследований, оставили после
себя прекрасную сводку по сравнительной анатомии. Одновременно с
изучением взрослых форм подвигалось и развитие сравнительной эмбри-
ологии. Здесь следует назвать имена Г. Пандера (1794—1865), описавшего
зародышевые листки цыпленка, М. Ратке (1793—1860), открывшего жабер-
ные щели и жаберное кровообращение у зародышей птиц и млекопитающих,
а также смену органов выделения, и К. Бэра (1792—1876), давшего класси-
ческое описание эмбрионального развития цыпленка и по справедливости
считающегося основателем научной эмбриологии.
Новый и наиболее мощный толчок получило развитие сравнительно-
анатомической мысли с восторжествованием эволюционной идеи. Морфо-
логи начала XIX века не были эволюционистами. Однако накопленный
ими эмпирический материал послужил существенной основой для построе-
ния эволюционной теории. Впрочем сам Чарльз Дарвин не был морфологом
и далеко не использовал того фактического материала, который к тому
времени уже был накоплен. Наиболее энергичным проводником идей
Ч. Дарвина в Англии был Т. Гексли (1825—1895), заслуживший особую
известность сравнительноанатомическим исследованием черепа позвоноч-
ных, где он совершенно разбивает позвоночную теорию черепа, а следо-
вательно, и теорию «архетипа» Оуэна.
10
Широкий отклик нашло применение эволюционных идей в морфологии
у русских исследователей: А. О. Ковалевского (1840—1901), И. И.
Мечникова (1845—1916), А. В. Заленского (1845—1920), показавших на
эмбриологическом материале единство происхождения животного мира,
и В. О. Ковалевского (1842—1883), который своими классическими иссле-
дованиями скелета ископаемых копытных поло?кил начало эволюционной
палеонтологии. В Германии роль «апостола дарвинизма» выполнял зоолог
Э. Геккель (1834—1919), оказавший известное влияние и на дальнейшее
развитие сравнительной анатомии построением «родословных дерев», что
положило начало филогенетическому направлению, и
в особенности популяризацией «биогенетического закона». Этот закон был
уже в основном сформулирован Фр. Мюллером (1821—1897) как правило
краткого повторения каждым высшим организмом в течение его индиви-
дуального развития («онтогенеза» по терминологии Э. Геккеля) тех этапов,
которые прошли его предки в своем историческом развитии («филогенезе»).
Применение биогенетического закона дало опору эмбриологическому ме-
тоду в сравнительной анатомии.
Эволюционная идея вдохнула новую жизнь в сравнительную анатомию
и быстро довела ее до небывалого еще расцвета.
Учение о гомологии органов, являющееся основой сравнительной
анатомии, получило совершенно новое освещение. Если в идеалистической
морфологии сходство гомологичных органов являлось лишь выражением
единства плана, т. е., в конце концов, отражением единства идеи творца,
то теперь оно объясняется кровным родством организмов. Поэтому и
понятие гомологии получило новое определение. К. Гегенбаур дает такую
формулировку: «Гомологией (в тесном смысле слова) обозначают отноше-
ние между двумя органами одинакового происхождения, которые, сле-
довательно, развились из одного и того же зачатка и обнаруживают одина-
ковое морфологическое соотношение». Установление гомологий как основ-
ная задача сравнительной анатомии становится теперь средством для
определения родственных соотношений между организмами, т. е. для
установления их филогении.
Несмотря на коренное изменение взглядов на значение этого сход-
ства, несмотря на замену формальной классификации генеалогической,
все же многие представления идеалистической морфологии иногда пре-
красно уживались с новой, филогенетической. Представление об единстве
плана, об «архетипе» сохранилось в виде представления об исходной
форме как общем предке данной группы организмов. И эта исходная
форма строилась путем отвлечения от всех «приспособлений», обу-
словленных функцией. И в филогенетической морфологии нередко
оперировали с преобразованием идеальной «чистой» формы. В «общей
морфологии» Э. Геккеля (1866) еще чрезвычайно ясно видны, наряду с
материалистическими взглядами, и чисто идеалистические конструкции.
Такова, например, его «проморфология», как «кристаллография органи-
ческого», или «тектология», как учение о составе организма из индиви-
дуальностей разного порядка.
Наиболее крупным представителем филогенетического направления
в сравнительной анатомии был К. Гегенбаур (1826—1903), оставивший
ряд классических исследований в области морфологии главным образом
скелета низших позвоночных. Его «сравнительная анатомия позвоночных»
представляет собой и до сих пор наиболее солидную переработку всего
сравнительноанатомического материала на основе эволюционного учения.
В сравнительной анатомии беспозвоночных подобную работу пред-
принял А. Ланг (1855—1916). Из учеников Гегенбаура .следует назвать
М. Фюрбрингера и Г. Брауса. Хотя К. Гегенбаур признавал значение эмб-
риологического метода в сравнительной анатомии, но относился к нему
весьма критически. Другие исследователи, однако, широко воспользова-
11
лись им и достигли в этом направлении больших результатов. В Англии
Ф. Бальфур (1851—1882) оставил нам первое крупное сравнительно эмбрио-
логическое сопоставление в свете эволюционного учения. Целый ряд
сравнительноанатомических и эмбриологических описаний принадлежит
В. К. Паркеру. Эмбриологическому методу последовали в Англии также
А. М. Маршалл, Э. Рай Лэнкестер, Г. Керр, Мэк Брайд и другие современ-
ные ученые; в Германии—морфологи Р. Видерсгейм, А. Гетте, А. Дорн,
К. Рабль, А. Ланг, Е. Гаупп, Е. Коршельт и многие другие; в Голландии—
фан Вай; у нас —И. Мечников, А. Ковалевский, В. Заленский, А. Север-
цов и др. следовали также главным образом эмбриологическому методу.
По мере развития сравнительной анатомии все более выяснялась
недостаточность методов. Сравнение даже в соединении с эмбриологическим
методом не всегда давало достаточно надежные результаты, что вызвало
отрицательное отношение многих биологов ко всему филогенетическому
направлению в целом.
Новейшая сравнительная анатомия, однако, не без успеха пользуется
и данными палеонтологии для более прочного обоснования своих выводов
(Г. Осборн, В. Грегори, Е. Гудрич, П. Сушкин, А. Северцов). С другой
стороны, некоторые палеонтологи всецело посвящают себя морфологи-
ческим исследованиям и достигают иногда блестящих результатов (Д. Уот-
сон в Англии и в особенности Е. А. Стеншио и его школа в Швеции).
Другие исследователи пошли по пути искания совершенно новых
методов, в частности по пути эксперимента (В. Ру, Г. Браус и многие
другие). Задачи экспериментальной морфологии, однако, уже только
частично захватывают область сравнительной анатомии, почему мы на
этом направлении здесь не останавливаемся.
В настоящее время задачи сравнительной анатомии значительно рас-
ширились. После установления основных филогенетических связей есте-
ственно былоперейти к изучению морфологических закономерностей самого
эволюционного процесса, что и было сделано в основном трудами акад.
А. Н. Северцева и его школы. Созданное им направление эволюционной
морфологии выдвигает следующие основные проблемы: филогенетическое
изменение форм и функций органов и их соотношений с внешней средой,
соотносительное преобразование органов, типичные направления эволю-
ционного процесса.
Кризис сравнительной анатомии, наступивший к началу XX века,
имеет, несомненно, глубокие основания и указывает не на исчерпание
объекта исследования, как это обычно думают, а прежде всего на при-
менение неправильной методологии.
Путь исследователя-анатома, изучающего строение животных, —
это всегда путь анализа. Вырывая организм из его естественной среды
и расчленяя мертвое тело па части, органы и ткани, он забывает,
что организм есть органическое целое, которое по существу своему
является нерасчленимым. В попытках синтеза, без которого, конечно,
нет никакой пауки, анатом и филогепетик забывают, что целое не есть
простая сумма частей в живом организме, забывают о том, что «чистая»
форма вообще лишена всякой реальности и воссоздают, таким образом,
лишь безжизненные схемы. Ближайшей задачей является разработка
«биологической», или «синтетической» морфологии, намеченной еще
Ж. Кювье, с применением, однако, последовательной методологии мате-
риалистической диалектики, к чему в настоящее время уже приближаются
некоторые морфологи. Орган изучается как неотъемлемая часть целого
организма в неразрывной связи с его функцией. И в изучении истори-
ческого его становления преобразование формы изучается совместно с
преобразованием функции. Организм, неизбежно, конечно, расчленяемый
для анатомического описания, изучается в своих жизненных проявлениях
и восстанавливается как целое в связи с окружающей его средой. История
12
живых организмов реконструируется в неразрывной связи с историей
этой среды. На этом пути предстоит еще большая и многообещающая
работа.
ОСНОВНЫЕ ПОНЯТИЯ СРАВНИТЕЛЬНОЙ АНАТОМИИ
Организм и орган
Организм — живое существо, т. е. единственно реальный но-
ситель жизни, характеризующийся всеми свойствами живого. Всякий
организм является структурно сложной системой, непрерывно взаимо-
действующей с окружающей средой таким образом, что в «нормальных»
условиях поддерживает свою целостность и жизнь. Такое строение и
такие реакции мы называем целесообразными. Целесообразность не есть,
следовательно, внутреннее свойство живых организмов вообще, а выра-
жается только в соотношениях определенного организма с той средой,
которая исторически определила его развитие. Организмы характери-
зуются также своим химическим составом, в котором основное значение
имеют белковые тела. Физиологически они характеризуются обменом
веществ (ассимиляцией и диссимиляцией), ростом, размножением и раздра-
жимостью.
Интересующее здесь нас строение организма определяется, следова-
тельно, его историческим развитием в конкретных условиях меняющейся
среды и без учета этой истории и вне этой среды не может быть понято.
Органом (греч. opyavov — орудие) мы называем более или менее
обособленную часть организма, несущую определенную функцию. Органы
могут иметь довольно сложное строение, причем отдельные их части
могут нести ту или иную подчиненную функцию. Таким образом, можно
говорить об органах разного порядка, о расчленении и диференциации
органов. С другой стороны, различные органы могут объединяться и
совместно участвовать в выполпепии какой-либо сложной функции. Тогда
говорят об аппаратах и о системах органов. Связь между органами одной
системы является в основном физиологической связью (внутрисекреторная
система, мускулатура, органы чувств, иногда скелет), но может выражать-
ся и в морфологическом объединении (пищеварительная система, дыха-
тельный аппарат, нервная система, кровеносная, выделительная, иногда
половая, иногда скелет).
Различают органы постоянные и органы временные. Постоянные, или
дефинитивные, органы характеризуют зрелое животное. Времен-
ные, или провизорные, органы имеют переходящее значение в
течение известного периода развития у зародыша или у личинки, а после
этого исчезают, заменяясь другими образованиями. Примерами эмбрио-
нальных провизорных органов будут: хорда большинства позвоночных,
жаберные щели наземных позвоночных, первичная почка и зародышевые
оболочки высших позвоночных (амниот). Примерами личиночных органов
могут быть: железистый желобок пескоройки, наружные жабры личинок
амфибии, присоски, роговые челюсти и хвост головастиков бесхвостых
амфибий.
Рассматривая органы в их историческом развитии, мы можем говорить
о прогрессивных органах, которые увеличиваются в разме-
рах и становятся более сложными по своему строению (например, головной
мозг позвоночных, рис. 2), и о регрессивных органах, которые
уменьшаются в размерах и получают более простое строение. В первом
случае их функция усиливается, приобретая большее значение, во втором
случае значение органа падает. Если функция органа теряется, то это
морфологически выражается в редукции органа. Рудиментар-
ными (лат. rudimentum, зачаток) мы называем органы, утратившие
13
Рис. 2. Разрез через крышу черепа ящерицы с
изображением головного мозга (Ьп) и париеталь-
ного глаза (Т) (по Лехе).
CfecJe зйачейие и сохраняющиеся лишь в виде незначительных остатков
(рудиментов), положение, связи, развитие, а иногда и строение которых
указывают на их происхождение от более развитых органов непосредст-
венных предков данного организма, у которых они играли гораздо более
существенную роль. Примерами рудиментарных органов могут служить:
остатки глаз пещерного протея (рис. 7), теменной орган позвоночных,
хорошо развитый у некоторых ящериц (рис. 2), остатки задних конечно-
стей у некоторых змей, остат-
ки, таза у китообразных,
остатки плечевого пояса у
безногих ящериц, остатки
боковых пальцев — грифель-
ные косточки лошади, оста-
ток мигательной перепонки
глаза человека, ушные и
хвостовые мышцы человека
и т. п.
Строение органа объяс-
няется его функцией, его
связями с другими частями
(в пределах аппарата, систе-
мы органов и всего организ-
ма), и, следовательно, орган
не может пи в данном состо-
янии, ни в разрезе исто-
рического развития рассматриваться изолированно, вне этих связей, а
только как одна из специализированных, но вместе с тем соподчиненных
частей всего организма.
Метод сравнения и гомология органов
При сравнении различных организмов между собой обнаруживаются
нередко известные сходства в общем плане их строения и в строении
отдельных частей, отдельных органов. Это сходство может выражаться
главным образом в сходстве функций, не сопровождаясь глубоким морфо-
логическим сходством. Такое сходство называют аналогией (греч.
dvct dvabyoc — соответственный). Примером аналогичных органов могут
служить совершенно различные по происхождению органы дыхания:
жабры рака и жабры рыбы, трахеи насекомого и легкие позвоночного пли
различные органы движения; конечности членистоногого и позвоночного
животного или крыло бабочки и крыло птицы. Гомологией (греч.
брос, бцоХоуос — одинаковый) в широком смысле называют морфологи-
ческое сходство, т. е. сходство в строении и соотношениях частей,
основанное на происхождении от сходных зачатков, которые когда-то
несли и сходную функцию. Гомологичными могут быть органы, несущие
в настоящее время самые различные функции, -и само собой разумеется,
что эти различные функции связаны и с характерными различиями в
строении гомологичных органов. Гомологичными являются, например,
передние конечности наземных позвоночных: бегательная нога ящерицы,
крыло птицы, роющая лапа крота, хватательная конечность обезьяны,
плавательный ласт кита (рис. 3). Различия в форме этих органов связаны
с весьма различной их функцией, а глубокое сходство внутреннего строения
и соотношений с другими частями (расчленение конечности, плечевой пояс,
его связь с грудиной и позвоночником, ее мышцы, нервы, сосуды) объяс-
няется происхождением от одного и того же органа—ходильной конечности
далекого общего предка этих животных — примитивного наземного по-
звоночного (стегоцефала, котилозавра). Морфологическое сходство может
14
быть разного порядка, и соответственно различают несколько видов
ГОМОЛОГИИ.
Гомономия, или общая гомология Гегенбау-
р а — морфологическое сходство как соотношение между органами одного
и того же порядка, повторяющимися в теле одного и того же животного.
Это сходство, в свою очередь, подразделяется на разные тины в зависи-
мости от характера расположения органов.
1. Гомотипия (бцб; — одинаковый, тбтос — форма, строение) —
соотношение между
симметрично распо-
ложенными органа-
ми. При совершенной
двусторонней сим-
метрии гомотипич-
ные органы правой
и левой стороны мо-
гут быть весьма сход-
ными, образуя один
как бы зеркальное
изображение друго-
го: правая и левая
рука, правая и ле-
вая почка и т. п.
Однако правое и ле-
вое легкое человека
или, еще лучше, пра-
вые и левые яични-
ки и яйцеводы птицы
дают нам уже при-
меры возрастающих
различий. Симме-
трично расположен-
ные части и органы
н азываются анти-
мерам и.
2. Гомоди-
нами я (бро; —оди-
наковый, 85vaptc —
сила, значение), или
сериальная гомоло-
гия, — соотношение
между органами или
частями, располо-
женными последова-
тельно вдоль глав-
ной оси животного.
Рис. 3. Скелет передней конечности наземных позвоночных.
В верхнем ряду слева направо: саламандры, морской чере-
пахи, крокодила и птицы; в нижнем ряду: летучей мыши,
кита, крота, человека. Гомологичные кости обозначены
одинаковыми буквами (по Лехе).
Такие повторяющиеся вдоль главной оси части носят наименование м е-
т а м е р о в. Таким образом, гомодинамны сегменты тела членистоногих
со всеми их частями, в том числе конечностями, гомодинамны отдельные
позвонки позвоночных животных. В общем гомодинамны также передние
и задние конечности позвоночных.
3. Гомономия (Sjjloc — одинаковый, vopoc — разделенный) в
тесном смысле слова—соотношение между различными другими одноимен-
ными органами—лучами плавников, пальцами конечностей и т. п.
Гомология, или частная гомология Геген-
б а у р а,—соотношение между органами одинакового происхождения,
т. е. развивающимися из одного и того же органа общего предка данных
15
животных. Такие органы развиваются у разных животных из такого
же зачатка и обнаруживают сходные соотношения с другими органами.
Так как гомология покоится, таким образом, на родстве, то она устанав-
ливается с тем большей степенью достоверности, чем ближе родство иссле-
дуемых организмов, т. е. чем больше сумма морфологических сходств
между ними.
Гомология может быть полной, если сравниваемые органы хотя
и обнаруживают известные, быть может, даже глубокие преобразования,
но полностью сохраняют свои соотношения и не позволяют установить
выпадения или прибавления каких-либо частей извне. Полная гомология
имеется, например, между скелетом конечности ящерицы и примитивного
млекопитающего, между головным мозгом амфибии и рептилии.
Неполная гомология обнаруживается в существовании морфо-
логического сходства между частями сравниваемых органов. Такая
гомология может быть дефективной, если известная часть оргаяа
утеряна (например, сердце костистой рыбы по сравнению с сердцем хря-
щевой, заключающее все гомологичные части за исключением артери-
ального конуса, который у костистых рыб утерян; конечность лошади,
утратившая по сравнению с другими млекопитающими все пальцы за
исключением лишь одного среднего; нижняя челюсть млекопитающих,
утратившая по сравнению с рептилиями целый ряд костей сочленовной
области, и т. п.), или аугментативпой, если к органу приба-
вились известные части извне (например, ухо млекопитающих, в котором
по сравнению с низшими наземными позвоночными прибавились еще
две слуховые косточки, и наружное ухо, или ухо наземных позвоночных
вообще, у которых по сравнению с ухом рыб к полностью гомологичному
лабиринту прибавилось еще среднее ухо).
Гомойология (Плате) — сходство гомологичных органов, не-
сущих одинаковую функцию и поэтому развивающихся конвергентно
или параллельно. Гомойология — это, следовательно, аналогия гомоло-
гических органов. Примером гомойология может служить сходство между
ластами ихтиозавра, плезиозарва и кита.
Развитие (онтогенез и филогенез)
Развитие — процесс становления органических форм — не следует
понимать буквально, т. е. как развертывание чего-то уже существующего.
Хотя понятие развертывания и входит в наше понимание развития, но
так же, как и простой рост, не составляет его сущности. Характерным
для развития является преобразование форм с установлением новых соот-
ношений и выделением новых частей.
Различают индивидуальное развитие, или онтогенез (ov, бусос —
существо, yeysatc — возникновение, происхождение), т. е. процесс ста-
новления отдельной особи, и историческое развитие, или филогенез
(фбХоу — род), т. е. процесс становления вида.
Сравнительная анатомия имеет дело с филогенетическим развитием,
и здесь в особенности преобразование форм и новообразование частей
суть совершенно необходимые и в сущности исчерпывающие элементы
этого понятия.
При сравнении процесса исторического развития различных орга-
низмов мы наблюдаем их расхождение, т. е. уменьшение сходства, если
они развиваются в различной среде. Гомологичные органы таких орга-
низмов несут разные функции и становятся все более различными. Такое
соотношение в развитии различных организмов называется диверген-
цией, или дивергентным развитием.
В других случаях мы видим, что организмы, по существу, быть мо-
жет, весьма различные, развиваясь в сходной среде, приобретают все
16
большие сходства. Различные органы таких организмов могут приобре-
сти сходные функции и сходное строение, т. е. становятся «аналогичными»
(рис. 5). Такое соотношение в развитии разных организмов называется
конвергенцией. Наконец близко родственные организмы могут
в сходной среде развиваться конвергентно или параллельно, т. е. гомо-
логичные их органы, несущие сходные функции, приобретают и в даль-
нейшем развитии независимо друг от друга новые обусловленные фук-
цией аналогичные сходства. Такое сходство аналогичных и в то же время
гомологичных органов называется, как мы уже отмечали, гомойологией
(Плате).
ОСНОВНЫЕ ПРИНЦИПЫ ФИЛОГЕНЕТИЧЕСКОГО
ФОРМООБРАЗОВАНИЯ
Диференциация и интеграция
(лат. differentia — различие, integratio — возобновление)
. Основным принципом развития органических форм является уста-
новленный еще Мильн-Эдвардсом (1851) принцип диференциации, осно-
ванной на разделении труда. Диференциация представляет собой разде-
ление однородного на обособленные части, которые в силу различного
положения, различных связей и, следовательно, различных функций
приобретают и различное строение. Диференциация, т. е. морфологи-
ческое разделение, всегда связана, таким образом, с физиологическим
разделением труда между отдельными частями (рис. 4). Принцип дифе-
ренциации мы называем основным принципом всякого развития, так как
именно этим путем возникает все новое: гомогенные части с весьма общими
отправлениями распадаются на органы, несущие все более определенные,
более частные функции, и весь организм получает все более сложное
строение.
Так как, однако, диференциация, т. е. морфологическое обособление
частей, предполагает и разделение труда, т. е. специализацию частей,
а части, специализирующиеся для несения одной определенной функции,
теряют способность нести иные функции, то ясно, что в диференциро-
ванном организме отдельные части несут главным образом лишь некоторые
жизненно важные функции, а не все. С обособлением органов питания
другие части специализируются для несения иных функций, они теряют
способность самостоятельно захватывать твердую пищу и тем самым по-
падают в зависимость от органов питания, поставляющих им пищевой
материал в уже растворенном виде. С обособлением органов дыхания
общие покровы теряют эту функцию, становятся менее проницательными,
более прочными (функция защиты) и вместе с тем, как и другие органы,
становятся в зависимость от органов дыхания, поставляющих им кисло-
род (через кровеносную систему), и т. д. Обособляясь в отношении одной
специальной функции, эти части тем самым становятся в полную зависи-
мость от всех остальных частей, несупщх иные жизненно важные функции.
Таким образом, одновременно с диферециацией идет и соподчинение
частей целому организму; вместе с распадом организма на различные
части, с его морфофизиологическим расчленением, неизбежно идет и про-
тивоположный процесс увеличения взаимозависимости частей, восста-
новления гармоничного целого, или процесс интеграции. Процесс интег-
рации (Спенсер) необходимо, органически связан с процессом диферен-
циации и является вместе с ним основным принципом исторического раз-
вития организмов.
Морфофизиологические преобразования органов
Диференциация связана с разделением функций. Каждая часть по-
лучает главным образом лишь некоторые из прежних функций неразде-
ленного целого, однако, почти никогда не приходится говорить об одной
17
основная пасть
наружная нить
внутренняянт
первая антенна
вторая антенна
1-я малШа
?> 2-я. Maxilla
2-я
ходильные
3-я
4-я
5'Я
2-Я
3-Я
-Я
Заднепроходные
нови
Рис. 4. Диференциация конечностей речного рака, связанная с разделе-
нием _и сменой функций (по Кюкенталю).
вторая антенна с
СрюшнсО стороны
чешуя
отверстие
сяжновой
Железы
Жевательная
пластинва
palpus
wandieula
entopodit
exopodit
t-я челюстная нова
^entopodit
2-я челюстная нова
exopocif
' entopodit
3-я челюстная нова
exopodit
- 1-я '
4-Я
18
только функции известной части (принцип мультифункциональности).
Даже специализированные органы высших животных несут многие функ-
ции, хотя при атом очень заметно выделяется одна основная функция.
Таким образом, каждый орган несет, кроме характерной для него глав-
ной функции, еще ряд второстепенных и этим самым даются широ-
кие возможности преобразования органов (Ч. Дарвин). Мы различаем,
разные типы преобразований, и соответственно этому современные иссле-
дователи выделяют некоторые наиболее типичные формы таких измене-
ний.
Смена функций. При изменении условий существования
главная функция может терять свое значение и, наоборот, какая-либо
из второстепенных функций может приобрести значение главной. Про-
исходящая таким образом смена функций сопровождается и соответст-
венным изменением строения органа (принцип смены функций А. Дорна).
Так, например, в желудке позвоночного происходит, с одной стороны,
химическая переработка пищи выделением пищеварительных желез,
но, с другой стороны, также и механическая обработка. У птиц происхо-
дит диференцировка, т. е. разделение на два отдела: в одном из них со-
храняется прежняя главная функция (железистый желудок), в другом
происходит смена, главной функцией становится механическая обработка
пищи (мускульный желудок). Диференцировка конечностей рака также
дает примеры такой смены функций. Первоначально главной функцией
этих конечностей была плавательная, а второстепенными функциями—хо-
дильная, хватательная и даже дыхательная. При диференцировке голов-
ные и передние грудные конечности речного рака стали хватательными
и жевательными, задние грудные — ходильными, брюшные плаватель-
ные — частично служат и для вынашивания икры и для обмена воды на
органах дыхания. Почечные канальцы позвоночных служат не только
для выделения, но и для выведения половых продуктов. Эта последняя
функция становится главной в части органа, обособляющейся в вид©
придатка семенника. Конечности млекопитающих служат в основном
для хождения по земле, и это их главная функция, однако, кроме того,
эти конечности могут служить для лазания, для рытья земли, для пла-
вания, для хватания, для защиты и даже для нападения (хищники). Такая
мало специализированная конечность очень пластична, т. е. легко может
быть изменена в зависимости от условий среды. При измененных усло-
виях местообитания любая из второстепенных функций может стать на
место главной (у обезьяны конечность служит в основном для лазания,
у крота — для рытья, ласт тюленя — для плавания, передняя конечность
человека — орган хватания). Но даже крыло птицы — весьма специали-
зованный орган летания — может изменить свою функцию, как это видно
на примере пингвина, у которого оно преобразовалось в орган плавания
(рис. 5).
Расширение функций (Плате) сопровождает нередко
прогрессивное развитие, т. е. диференцирование органа, который по мере
своего расчленения приобретает некоторые новые функции. Так, напри-
мер, парные плавники рыб, возникшие как пассивные планирующие
органы, вызывающие лишь полезное при известных условиях сопротив-
ление воды, с приобретением своей мускулатуры и прогрессивным рас-
членением становятся в основном активными рулями глубины, но притом
еще и органами, поддерживающими равновесие, органами плавания и
у донных рыб иногда органами опоры и движения по дну. С переходом
позвоночных на сушу последняя функция опоры при ползании стала
главной, конечность значительно преобразовалась и при дальнейшем
прогрессивном развитии (диференцировании) приобрела целый ряд новых
функций (бегание, скакание, лазание, хватание, рытье и т. д.). Расшире-
ние функций увеличивает пластичность органа и всего организма, так
7Р
как открывает много новых возможных путей эволюции при дальнейшей
смене функций.
Сужение функций (Северцов), или уменьшение числа функ-
ций, составляет тип преобразований, противоположный предыдущему.
Таковы, например, преобразования конечностей копытных, которые по
мере своей специализации в направлении более совершенного выполнения
основной функции теряют свои второстепенные функции. Специализация
связана, следовательно, с решительным преобладанием главной функции,
а вместе с тем и с утерей
ти и ограничением возможности даль-
нейших преобразований (конечность
лошади, крыло летучей мыши, ласт
кита).
Усиление функций
(Плате) связано, например, с пере-
ходом гладкой мускульной ткани в
поперечнополосатую, с увеличением
секреторной части железы, с увеличе-
нием размеров органа или с его боль-
шей концентрацией. В последнем
случае эти преобразования частично
совпадают с тем, что Франц обозна-
чает как принцип централизации.
Активация функций
(Северцов) — преобразование пас-
сивных органов в активные. Напри-
мер, развитие подвижных плавников
рыб из малоподвижных кожных скла-
док; развитие подвижного ядовитого
зуба у змей, выдвигаемых когтей ко-
шек, подвижного таза лягушек и т. п.
Иммобилизация час-
тей (Северцов) — преобразование
активного органа в пассивный,
подвижного — в неподвижный. Сюда
Рис. 5. Крыло пингвина (.4), преобра-
зованное в ласт и приобретшее общее
сходство с плавником акулы (В) (по
Осборну)
относятся потеря подвижности первичной верхней челюсти в ряду позво-
ночных, образование крестца путем срастания позвонков у наземных
позвоночных и особенно у птиц.
Разделение функций (Северцов) сопровождается разде-
лением органа (например, мышцы, части скелета) на самостоятельные
отделы. Примером такого разделения может служить распадение непре-
рывного непарного плавника рыб на отделы. Такой распад связан и с
изменением функций отдельных частей — передние отделы (спинной и
анальный плавники) становятся рулями, направляющими движение
рыбы, задний отдел (хвостовой плавник) — двигательным органом.
Принцип диференцирования всего организма или отдельных его
органов — основной принцип всякого развития, сопровождающегося
Новообразованием. Связанный с ним принцип смены функций лежит
в основе всех процессов преобразования отдельных органов. Важно от-
метить здесь совершенно ясную взаимозависимость процессов расчленения
и преобразования функций и расчленения и преобразования частей орга-
низма (органы). В этих процессах развития форма и функция столь не-
разрывно связаны, что каждый наш привычный термин оказывается одно-
сторонним и неспособным выразить сущность явления (поскольку смена
функций является одновременно и сменой формы). Поэтому Северцов,
которому принадлежит наиболее полный современный анализ морфоло-
гических закономерностей эволюции, говорит всегда о морфофизиологи-
ческих преобразованиях.
20
Прогресс, регресс и специализация органов
В большинстве рассмотренных морфологических преобразований
органов мы выделяли как более важные и интересные такие преобразова-
ния, которые ведут к осваиванию новых функций или к более совершен-
ному выполнению прежних функций, т. е. дают для организма новые
возможности использовать жизненные средства окружающей среды или
же дают при той же затрате энергии больший эффект. Такие изменения
мы называем прогрессивными. Диференциация, расширение
и увеличение числа функций, усиление и активизация, а также обычно
и разделение функций и даже иммобилизация часто относятся к таким
прогрессивным преобразованиям.
Уменьшение числа функций при специализации предста-
вляет пример прогрессирования в отношении лишь некоторых и, в осо-
бенности, главной функции. Некоторые органы могут в результате такой
специализации достигнуть высокой степени совершенства (конечность
копытного, язык хамелеона, крыло птицы, органы высших чувств, многие
внутренние органы, как мозг, отдельные мышцы, легкие, сердце, почки
и т. п.). Однако большая специализация тех органов, которые обеспе-
чивают более непосредственную связь организма с внешней средой («экто-
соматические» органы Северцова: конечности, челюсти и зубы, покровы
и др.), ограничивает возможность смены функций при изменении среды,
т. е. делает организм менее пластичным. При специализации утеря второ-
степенных функций связана с регрессом соответствующих частей органа.
Таким образом, у копытных прогрессивная специализация конечностей
связана с редукцией ключицы, редукцией малой берцовой и локтевой
костей, редукцией боковых пальцев и многих мышц, нервов и сосудов,
связанных с этими частями.
Уменьшение числа функций может, однако, сопровождаться и общим
ослаблением органа, при котором и главная функция теряет свое значение.
В этом случае мы имеем регрессивное развитие органа, его
общую редукцию. В случае сохранения какой-либо незначительной вто-
ростепенной функции от такого органа надолго может сохраниться его
остаток — рудимент (остатки таза кита, к которым причленяются
мышцы penis). При полной потере всякого значения орган исчезает бес-
следно (органы боковой линии наземных позвоночных, конечности змей).
Регрессивное развитие отдельных органов (например, конечностей
змей) не предполагает обязательно общего регресса всей организации и,
наоборот, наблюдается постоянно и при прогрессивном развитии орга-
низмов (редукция чешуй у млекопитающих и волос у человека, редукция
пальцев в крыле птицы), поскольку в меняющихся соотношениях между
организмами и средой меняется и удельный вес отдельных органов и вообще
их функциональные соотношения внутри целого организма. Некоторые
органы и функции приобретают все большее значение и заступают место
других органов, которые в условиях новой среды оказываются излишними.
Таким образом, мы переходим к вопросам соотносительного развития
органов в эволюции организмов, при котором еще ярче выступает отно-
сительность понятий прогресса и регресса.
Соотносительное развитие органов
Анатом-морфолог имеет дело главным образом с преобразованиями
отдельных органов. Однако, в конкретной эволюции меняются не отдель-
ные органы, а целые организмы во всех своих частях. В организме все
части взаимосвязаны, и в исторических его преобразованиях мы уста-
навливаем не только изменения соотношений, но и соотносительные изме-
21
пения частей. При всех этих изменениях целостность и жизненность
организма сохраняются лишь при строгой координации изменений его
частей, органов. Как говорят о непрерывном приспособлении (адаптации)
организма к меняющейся среде, так же можно говорить и о взаимных
приспособлениях органов в изменяющемся организме или об их коадапта-
ции (коаптации). Эти коадаптации строятся на конкретном материале
индивидуальных наследственных изменений нормы реакции данного
организма, нередко со многими их генетическими связями, частью в
результате непосредственного морфогенетического или физиологического
взаимодействия между органами, частью через естественный подбор
наиболее жизненных соотношений при данных биологических условиях.
Из этогоследует.что понятие коадаптации охватывает различные процессы,
в основе которых лежат закономерности разного порядка. Отсюда вы-
текает необходимость расчленения этого понятия.
Уже Ж. Сент-Илер пытался выразить в своем принципе «уравно-
вешивания органов» наблюдаемые закономерные соотношения в развитии
различных органов одного организма. Еще более общий характер имеет
установленный Ж. Кювье принцип корреляций. Ж.. Кювье было^
однако, чуждо понимание истории.ррганизмов^его представление 6 кор-_
беляциях носило статический характер, и критерием корреляций являг
лось лишь постоянство сосуществования органов. Хотя мы и можем при
известных исследованиях подходить к коррёляцйям’со статической точки
зрения (хотя бы, например, при биометрической обработке материала),
но о статическом понимании корреляций в настоящее время не может
быть и речи. С установлением эволюционного учения в понятие корреляций
естественно были внесены элементы динамики, поскольку сосуществова-
ние обычно объясняется взаимозависимостью в развитии. Кроме того,
пришлось расчленить это понятие, различая корреляции физиоло-
гические, поскольку они выражаются в индивидуальном развитии
организма («индивидуальные» корреляции Плате), ифилетические,
поскольку они выражаются в его историческом (филогенетическом) раз-
витии. Были предложены и разные другие термины и были сделаны раз-
личные попытки дальнейшего расчленения понятия. Так как «сосущество-
вание» частей в организме определяется по Кювье взаимозависимостью
функций, то термин «корреляция» следует сохранить в смысле физиоло-
гической, индивидуальной корреляции (впрочем, примеры, приводимые
Кювье, несомненно, включают и филетические корреляции биологического
характера). Таким образом, сохраняя термин «корреляции» для индиви-
дуального развития организма (как это и принято в «механике развития»),
мы будем пользоваться для обозначения соотносительного развития
органов в филогенезе термином «координация». Термин «координация»
мы берем у А. Н. Северцова, однако, влагаем в него иное содержание,
разумея его не статически как сосуществование, а как закономерную
зависимость в филогенетических изменениях частей (органов).
Соотношение частей в индивидуальном и историческом развитии
организма может выразиться в форме замещения одного органа другим о
подобной функцией. Эта частная форма корреляций и координаций обозна-
чается как «субституция» органов и функций.
Корреляции индивидуальные или физиологические
Сюда мы относим взаимозависимости частей или органов при инди-
видуальном развитии организма. В этом случае мы имеем в виду экспе-
риментально устанавливаемые зависимости частей. Так как, однако, орга-
низм развивается как целое, то, в конце концов, все части взаимозависимы,
и, вероятно, можно было бы говорить о корреляции между всеми частями
организма. При таком расширении этого понятия оно, однако, теряет
свою определенность и становится совершенно ненужным. Мй будем го-
22
ворить о корреляциях только при наличии совершенно ясной связи между
развитием одной части и определением качества или количества другой
части. Эта связь может осуществляться непосредственно или через по-
средство третьей части.
Корреляции могут иметь различный характер, и соответственно
можно различать геномные корреляции, морфогенетические и эргонти-
ческие корреляции.
Геномные корреляции. Геномные, или идиоплазматиче-
кие (Плате), корреляции представляют собой соотношения, обусловленные
множественным действием одних и тех же наследственных факторов.
Геномные корреляции могут также покоиться на тесном сцеплении разно-
родных наследственных факторов и вообще на относительном постоянстве
факторов, содержащихся в одной и той же хромосоме (например, половой).
Таковы, например, геномные корреляции между хохлом кур, открытой
формой ноздрей и раздвоением гребня или корреляции между различными
половыми признаками (поскольку они не зависят от гормонов), например,
у насекомых.
Морфогенетические корреляции. Морфогенетиче-
скими корреляциями я назвал соотношения, обусловленные взаимо-
зависимостями эмбриональных процессов (т. е. теми взаимозависимо-
стями, которые исследуются «механикой развития»). Сюда относится
и принцип уравновешивания Ж. Сент-Илера, поскольку последний пред-
полагал наличие соотносительной компенсации в развитии органов (при
наличии ограниченного запаса образовательного материала прогрессивное
развитие одного органа связано с регрессом другого). Как показывают
данные механики развития, к морфогенетическим корреляциям относится
также большинство соотношений в величине и расположении отдельных
органов в организме и их частей внутри органа. Топографические соотно-
шения между хордой, миомерами туловища и центральной нервной систе-
мой являются примером такой типичной и весьма устойчивой корреля-
тивной зависимости. Связь между подвздошными костями таза и ребрами
крестцовых позвонков (развитие крестцовых ребер определяется контак-
том с подвздошными костями), соотношения между перепончатым лаби-
ринтом уха и слуховой капсулой, соотношения между глазным яблоком
и хрусталиком глаза—во всех этих случаях нарушение положения одного
компонента сказывается в виде изменения положения или даже в виде на-
рушения развития другого компонента. Эти зависимости чрезвычайно
сложны (развитие хорды и мезодермы определяет развитие мозга и глаза,
развитие мозга определяет развитие уха, развитие уха определяет развитие
капсулы, развитие глаза определяет развитие хрусталика, развитие хру-
сталика определяет развитие роговицы и т. д.), и мы в самом деле имеем
дело с целым рядом звеньев корреляционной цепи, в котором изменение
первого звена неизбежно влечет за собой изменение всех последующих. Из
более мелких признаков расового характера к таким морфогенетическим
корреляциям относится, например, зависимость между оперением ног у кур
и брахидактилией (укорочением 4-го пальца, зависящего от того, что ске-
летогенный материал его закладки частично используется на развитие
оперения).
Эргонтические1 (или функциональные) корре-
ляции. Первые два типа корреляций определяются зависимостями
эмбриональных функций, изучаемых физиологией наследственности и
физиологией развития. Эргонтические корреляции определяются зави-
симостями в дефинитивных функциях уже сформировавшегося, растущего
или взрослого организма и выражаются также в более или менее ясных
морфогенетических изменениях связанных таким образом частей.
1 От греческого epyov—работа.
23
Такие арготические (и притом положительные) корреляции имеются
между развитием нервных центров, нервов и периферических органов
(недоразвитие центров и нервов при удалении органов чувств, конечностей;
гиперплазия их при экспериментальном увеличении области иннервации,
например, при пересадке органа чувств или индукции лишней конечности).
Подобные же арготические корреляции имеются между развитием мышцы,
нерва и кровеносных сосудов, снабжающих мышцу, между развитием
мышцы и скелетных частей, к которым эта мышца прикрепляется.
При этом зависимость эта не только чисто количественного характера:
с функцией мышцы нередко связано развитие особых отростков и гребней
на костях, например, черепа. При ранней парализации мышцы путем
перерезки соответствующих нервов у молодого, еще растущего животного
эти отростки и гребни недоразвиваются. Эргонтические корреляции
имеются и между различными частями скелета (и определяют, между
прочим, во многом развитие суставов) и между различными внутренними
органами. Особенно пластичны кровеносные сосуды, развитие которых
всегда обнаруживает явную положительную корреляцию с развитием
органа. Во взрослом организме особенно ярко выделяются корреляции
между различными железами внутренней секреции и скелетом и еще
яснее между половыми железами и многими вторичными половыми при-
знаками. Кроме наиболее часто наблюдаемых положительных, возможны
и отрицательные корреляции, например, между гонадами и эпифизом,
секреция которой тормозит развитие половых желез.
Координации, или филетические корреляции
Сюда мы относим филогенетические взаимозависимости частей или
органов, устанавливаемых методом сравнения по соотносительным их
изменениям при эволюции организмов. Эти изменения могут быть одно-
именными или противоположными изменениями количественного харак-
тера (часто при динамических координациях), или координированными
оказываются различные изменения качественного характера. Во всяком
случае характерно, что при изменениях данных органов между ними
сохраняется более или менее постоянное соотношение. Зависимость может
быть непосредственной или косвенной; обе части могут в своем развитии
зависеть от третьей, или обе части могут зависеть от определенных условий
среды в широком смысле этого слова. Ясно, однако, что в понятие коорди-
нации не входит простое сосуществование частей (хотя бы и в развитии
организмов). Например, положение: «животные, покрытые шерстью, имеют
млечные железы и три слуховые косточки», хотя и отмечает определенное
постоянное сосуществование частей, но не выражает никакой зависимости
в развитии. Это сосуществование обосновано исторически — происхожде-
нием определенной группы организмов от одной общей формы.
Мы различаем три вида филетических корреляций; 1) топографические
2) динамические и 3) биологические.
1. Топографические координации (Северцов); к ним
близки тектонические (Домбровский). Они выражаются в согласованных
изменениях топографических соотношений между органами, между кото-
рыми непосредственной функциональной связи нет. Таковы очень многие
общие соотношения органов, характеризующие «план» строения целых
групп организмов. Стойкость этих соотношений определяется относитель-
ной стойкостью многих морфогенетических корреляций, на которых они
именно и базируются. Таковы топографические соотношения между
осевым скелетом (хордой и позвоночником) и спинным мозгом, между
формой и величиной черепной коробки и головным мозгом, перепончатым
и костным лабиринтом, между величиной глаз и формой черепа, между
прогрессивным развитием средних и редукцией крайних пальцев у ко-
пытных и т. п. Во всех этих случаях имеется взаимная обусловленность
24
формы и размеров частей, хотя и нет непосредственной функциональной
связи между, например, головным мозгом и окружающей его у высших
позвоночных вплотную черепной коробкой, повторяющей в общем форму
мозга. Здесь имеется зависимость менаду формообразовательными процес-
сами, о которых также можно говорить как о функциях, но во всяком
случае эти функции не имеют ничего общего со специфическими дефини-
тивными отправлениями центральной нервной системы или части скелета.
2. Динамические, или конструктивные, координации
выражаются в согласованных изменениях частей, непосредственно свя-
занных в своих функциях. Они определяются относительным постоянством
функциональных зависимостей (корреляций) между различными органами.
Особенно постоянны такие зависимости между специализированными
органами, имеющими строго определенную функцию, допускающую глав-
ным образом количественные изменения, как, например, координации
между нервными центрами и периферическими органами, координации
между скелетом, мышцами, их кровоснабжением и иннервацией вообще
или соотношения между вкусовыми органами рыб, их нервами и соответ-
ственными отделами мозга или между органом обоняния и обонятельными
долями, как это выясняется из многих сравнений (например, крупных
обонятельных долей мозга млекопитающих, имеющих высоко развитый
орган обоняния, и ничтожных по размеру обонятельных долей птиц, у
которых орган обоняния очень прост). Подобные простые соотношения
имеются и между уровнем развития крыла, грудной мышцы, ее крово-
снабжением и иннервацией и величиной киля грудины у птиц. Примером
несколько более сложной координации с явно качественными изменениями
в отдельных звеньях может служить зависимость между формой нижней
челюсти и формой ее суставной головки, развитием височной мышцы и
гребнями черепа у млекопитающих, координация между весом головы и
туловища и развитием остистых отростков передней части позвоночника
(холка является скелетной системой, противодействующей изгибу позво-
ночника на месте опоры в области передних конечностей).
Динамические координации наблюдаются также между самыми раз-
личными частями скелета, в частности между задней конечностью, тазом
и крестцом, как это выясняется из сравнения скакательной задней конеч-
ности, а также формы и положения таза бесхвостых амфибий и хвостатых.
Та же координация выявляется при сравнении задних конечностей,
формы таза и развития крестца у четвероногих рептилий и ходящих на
двух ногах птиц, и т. п.
Примером динамической координации между различными более отда-
ленными органами может служить соотношение между развитием органов
внутренней секреции и зависимых от них признаков, например, величины
тела и величины гипофиза (и то и другое достигало очень больших разме-
ров у некоторых динозавров).
3. Биологические координации выражаются в сопря-
женных изменениях органов, не связанных топографически и не обнару-
живающих также непосредственной функциональной зависимости. Эти ко-
ординации чрезвычайно многочисленны и разнообразны и обусловлены бо-
лее или менее непосредственно характерными условиями биологической об-
становки, с изменением которой все эти связи легко нарушаются (для
специализированного животного, однако, коренное изменение биологиче-
ской обстановки невозможно). Многие примеры корреляций Ж. Кювье
относятся к этой группе. Таковы соотношения между формой и структурой
коренных зубов жвачных копытных, частичным недоразвитием клыков и
резцов (образование диастемы), специализацией конечностей и диференци-
ровкой желудка. Они определяются растительным питанием и соответст-
вующим образом жизни. Такова связь между длиной шеи и длиной перед-
них конечностей у травоядных, а также связь между развитием мышц зад-
25
ней конечности и дыхательной мускулатурой у копытных как приспособле-
ние к быстрому бегу, связь между хвостовым плавником, ластами, формой
тела и подкожным жиром у китов, обусловленным жизнью в воде. Такова
же связь между диференцировной зубов и строением кишечника. При
крайней специализации обнаруживаются многие такие типичные соотно-
шения, как, например, связь между хватательными лапами и цепким
хвостом, а также подвижностью глаз и длинным выбрасываемым языком
хамелеона, приспособленного к ловле насекомых на тонких ветвях де-
ревьев.
Разные типы координации различаются как характером, так и проч-
ностью соотношений, устанавливаемых при филогенетических изменениях
компонентов такой координационной цепи.
Конечно, постоянство соотношений в изменениях органов, выражае-
мое устанавливаемой нами координацией, никогда не является абсолют-
ным. При более крупных изменениях биологической обстановки легко
происходит разрыв многих существующих (особенно, биологических)
координаций и установление новых.
Многие динамические (конструктивные) координации оказываются,
однако, гораздо более прочными, в особенности если они наблюдаются
между специализированными органами, не допускающими изменения
функции (нервный центр, нерв и мышца, орган чувств и нервный центр).
Точно так же весьма стойкими оказываются многие топографические
координации, которые покоятся на основных морфогенетических
соотношениях, при нарушении которых невозможно было бы развитие
данных органов.
План строения организмов, характерный для крупных групп живот-
ных (в особенности «типов»), и основывается как раз на таких стойких
координациях.
Замещение органов и функций
Особый вид взаимоотношений органов во время развития животных
(частная форма корреляций и координаций) был обозначен Клейненбергом
как «субституция» органов. Под субституцией органов понимают замеще-
ние в течение филогенетического \или онтогенетического) развития орга-
низма одного исчезающего органа другим, несущим подобную же функцию
и занимающим подобное же (гомотопная субституция) или — совсем иное
положение (гетеротопная субституция).
1. Гомотопная субституция. Примером гомотопной
субституции может служить замещение первичного осевого скелета, хорды,
хрящевым, а затем костным позвоночником в течение онтогенетического
и филогенетического развития позвоночных. Сюда же относится вообще
замещение хряща костью в скелете позвоночных. Дальнейшим примером
может служить замещение сочленовной кости зубной в нижней челюсти
млекопитающих.
2. Гетеротопная субституция. К атому типу соотно-
шений между органами близок тип филогенетических изменений, обозна-
ченный Северцовым как субституция функций. Гетеротопная субституция
обозначает замещение одного органа подобным или даже совершенно
другим органом, не становящимся на его место, но несущим физиологи-
чески или, по крайней мере, биологически равноценную функцию. Типич-
ными примерами являются: замена нефридий как органов выделения у
насекомых мальпигиевыми сосудами; замена волосяного покрова, защи-
щающего тело от потери тепла, слоем подкожного жира у китообразных;
замена хвостового плавника как локомоторного органа грудными плавни-
ками у скатов; замена задних ластов как органов движения хвостовым
плавником у китообразных. К этому же порядку соотношений относится
и обратная зависимость (координация) между развитием реберных дыха-
26
тельных мышц грудной клетки млекопитающих и развитием диафрагмы
(хотя полного замещения здесь не бывает). Такова же, например, замена
работы конечностей как оргапа движения у змеевидных ящериц и у змей
работой боковых мышц тела при боковых (служащих для движения)
изгибах тела, замена органа зрения своеобразным щупальцем и органом
обоняния у подземных роющих гимнофион (эти два последних примера
показывают гетеротопную субституцию лишь биологически равноценных
органов).
Возникновение органов и их исчезновение
Вопрос о возникновении новых органов и вопрос об исчезновении
рудиментарных органов, потерявших свое значение, относятся к более
трудным морфологическим проблемам эволюционной теории, которые
неоднократно служили поводом для критических выступлений против
дарвиновской теории естественного отбора. Поэтому вполне уместно
отдельно рассмотреть эти вопросы в связи с другими вопросами филогене-
тических преобразований организмов.
Мы говорили о непрерывной связи между формой и функцией; но
может ли ничтожный зачаток органа, начальная фаза его возникновения,
выполнять какую-либо функцию, может ли он иметь значение для жизни
особи и может ли он, следовательно, быть предметом естественного отбора?
Кроме того, как может существовать орган, утративший функцию, руди-
мент, не выполняющий никакой работы, если мы говорим, что связь формы
и функции неразрывна, и наоборот, почему такие рудименты все яге часто
бесследно исчезают, если они не несут никакой функции, безвредны и,
следовательно, такяге не могут быть предметом естественного отбора,
который признается основным фактором эволюции? Вот круг вопросов,
смущающих каягдого бролога, недостаточно знакомого с конкретным
материалом сравнительной анатомии и эмбриологии.
В самом деле вопрос о возникновении новых органов во всех конкрет-
ных хорошо изученных случаях разрешается без особого труда при учете
принципов диференциации и смены функций. Органы возникают истори-
чески не сами по себе как независимые зачатки, а путем постепенного
выделения и обособления от иного органа, несущего функцию более общего
характера, т. е. путем диференцировки уяге существующих органов или
частей организма.
Большинство органов при своем возникновении несет совсем иную и,
во всяком случае, неизмеримо более простую и общую функцию.
Конечность наземного позвоночного в зачаточном состоянии не могла
бы служить в качестве органа опоры и двиягения на суше (неверное возра-
жение, выставленное против теории Дарвина), но она и в самом деле
возникла не как орган опоры, а представляет собой результат преобразо-
вания (связанного со сменой функции) плавника, на котором ползал
рыбообразный предок наземного позвоночного. Однако, и эти плавники
в зачаточном состоянии не могут слуягить в качестве плавников. Они
тогда и не были плавниками в собственном смысле этого слова, они возник-
ли как небольшие кили по бокам брюшной поверхности тела, не произво-
дившие никаких собственных двиягений и игравшие роль поверхностей,
увеличивавших полезное сопротивление воды, подобно брюшной поверх-
ности самого тела, т. е. парные плавники возникали в качестве бокового
расширения брюшной стенки тела рыбы.
Такой слоягный орган чувств, как глаз многих высших ягивотных,
такяге не мог возникнуть сразу как таковой. Все органы чувств возникли
путем диференцировки покровов, эпителий которых обладает более или
менее значительной чувствительностью по отношению к разнообразным
раздражениям. Иногда в этом эпителии бывают рассеяны клетки, особо
чувствительные по отношению к световым раздраягениям (земляные черви).
27
Обычно светочувствительные клетки связаны с другими клетками, в кото-
рых отлагается пигмент. Последний поглощает световые лучи и, очевидно,
трансформирует световую энергию в тепловую. Концентрация чувствую-
щих клеток приводит к образованию глазных пятен в покровах (рис. 6).
Собственно о зрении в этих случаях говорить не приходится, это — свето-
чувствительные органы, более или менее позволяющие животному хорошо
оценивать потемнение или посветление при собственном перемещении из
света в тень и наоборот. Погружение покровов ниже общего уровня
(защита более чувствитель-
ных частей эпителия от меха-
нических или иных раздра-
жений наблюдается при раз-
витии всех органов чувств)
приводит к образованиюглаз-
ной ямки, замыкающейся за-
тем в пузырек, при условии
выработки максимальной
Рис. 6. Эволюция органа врения у моллюсков
(по Боасу).
1 — глазное пятно; 2 и г — глазные ямки; 4 и 5 —
глазные пузыри без хрусталика; в — глазной пузы-
рек с хрусталиком; с —кутикула; в — эпидермис; g —
стекловидное тело; I — хрусталик; а — чувствующие
клетки; st — зрительные палочки.
прозрачности передней его
стенки и покровов в данном
месте. Однако, вместе с тем,
благодаря изгибу чувствую-
щего эпителия, он теперь
различно освещается лучами
света, падающими в различ-
ных направлениях. Живот-
ное может теперь оценивать
до известной степени распре-
деление света и тени в окру-
жающей среде без собствен-
ного перемещения. В этом
имеется уже нечто принципи-
ально новое.
Чувствительность глаз-
ных пузырьков увеличи-
вается, если прозрачная
передняя стенка пузырька
или соответственный участок
покровов утолщается и по-
лучает известную выпук-
лость. В этом случае световые
лучи, преломляясь в утол-
щенной стенке, сходятся,
концентрируются, прежде
чем упасть на зрительный эпителий. На сетчатке глаза получается более
яркое световое пятно. Так как на сетчатку теперь падают сходящиеся
лучи, то вместе с их концентрацией на сетчатке охватывается более широ-
кое пространство окужающей среды. С увеличением выпуклости «хруста-
лика» чувствительность органа повышается. Происходит дальнейшая
интенсификация функции, основанная главным образом на количественных
изменениях. Эта интенсификация имеет свой возможный максимум —
наиболее яркое световое пятно получится на сетчатке тогда, когда кривизна
хрусталика станет такой, что ее главный фокус придется как раз на сет-
чатку (рис. 6, 6). Однако в это самое время сразу будет достигнут еще
один, так сказать, побочный результат — на сетчатку упадет изображение
окружающих предметов. Только теперь, при условии диференци рования
нервных волокон и клеток зрительного центра, становится возможным
настоящее зрение, т. е. оценка внешнего вида окружающих предметов,
28
и притом тем более совершенное, чем правильнее кривизна хрусталика.
Из постепенных количественных изменений возникло опять нечто каче-
ственно новое. Зрение это совершенствуется с приобретением подвижности
глаза, т. е. с развитием собственной мускулатуры путем выдиференцировкп
ее из окружающих тканей, с развитием аккомодационного аппарата и т. д.
В этих изменениях мы имеем достижение еще более широкого зри-
тельного охвата окружающего пространства не только в ширину, но и
в глубину (аккомодация).
У двустороннесимметричных животных, в том числе у позвоночных,
органы зрения обычно парные. Каждый глаз, расположенный у позво-
ночного сбоку головы, охватывает известную часть пространства по бокам
и впереди животного. С сокращением лицевых частей (челюсти), редук-
цией органа обоняния и разрастанием головного мозга у обезьян глаза
оказываются расположенными все более впереди. Угол между зритель-
ными осями глаз становится все более острым, и, наконец, в тот «момент»,
когда они становятся параллельно, т. е. когда оба глаза смотрят вперед
на одно и то же поле, зрительные представления совмещаются и получа-
ется опять нечто новое — стереоскопическое зрение с весьма совершенным
восприятием глубины.
Мы разобрали довольно подробно наиболее трудный пример воз-
никновения нового органа чрезвычайной сложности и установили ряд
этапов, знаменующих подъем органа на более высокую ступень и свя-
занных с приобретением совершенно новых качеств при полной, однако,
непрерывности самого процесса развития. Возникновение органа слуха
высших позвоночных, также весьма сложного органа, объясняется еще
проще, и мы можем утверждать, что этот орган образовался в результате
диференцировки и дальнейшего усложнения комплекса кожных органов
боковой линии низших родных позвоночных (см. во второй части), ко-
торый затем дополнился различными вспомогательными частями. Воз-
никновение различных других органов оказывается также вполне понят-
ным во всех случаях, когда вопрос достаточно хорошо изучен (в насто-
ящем курсе имеется много примеров такого рода). Всегда новые органы
возникают путем постепенной диференцировки в пределах имеющихся
уже органов или частей, обособляются от них и преобразовываются вместе
с обособлением и преобразованием их функций.
Вопрос о регрессивном развитии уже был отчасти освещен. Если,
однако, ставить вопрос в разрезе более широких биологических проблем,
то он не так прост, как это кажется. Когда орган теряет свое значение,
то он вовсе не всегда становится вредным и, следовательно, прямо не уни-
чтожается под действием естественного отбора. Чаще всего орган стано-
вится просто бесполезным. Существует предположение, что прекращение
действия естественного отбора ведет к сохранению независимых вариаций
отдельных частей, которые при скрещиваниях ведут ко все большему
нарушению соотношения частей, т. е. к ухудшению, ослаблению органа
(гипотеза панмиксии). Хотя эти соображения совершенно верны и реду-
цирующиеся органы, действительно, обнаруживают весьма большую и
разнообразную изменчивость всех своих частей, все же эта гипотеза не
может объяснить самого явления редукции и рудиментации. Однако,
если принять во внимание, что наличие эмбриональных вариаций отдель-
ных частей приводит к нарушению нормальных морфогенетических кор-
реляций и даже к их разрыву, то мы легко поймем, что нарушение тех
соотношений, которые являются условием нормального развития органа,
приводит к недоразвитию органа, к его остановке на некоторой эмбрио-
нальной фазе, вплоть до полного исчезновения самой закладки, если эти
нарушения коснулись основных зависимостей, определяющих реализацию
этой закладки. Относительно индиферентный («презумптивный») мате-
риал будущей закладки при этом, конечно, используется на построение
29
соседних органов. Таким образом, при редукции глаз у пещерных живот-
ных, например, у протея, наблюдается колоссальная изменчивость частей
не только взрослого глаза, но и глаза зародыша. Замедленное развитие
глазного бокала приводит обычно к недоразвитию хрусталика (рис. 7); весь
глаз останавливается на более или менее ранней эмбриональной стадии
развития (хотя в виде индивидуальной вариации иногда развивается
еще довольно полно).
Такая картина типичной редукции органа приводит нередко к обра-
зованию незначительного остатка органа — рудимента, не несущего ни-
какой функции.Стро-
Рис. 7. Разрез черев рудиментарный глав протея (Proteus
anguineus) (по Колю)
а> — стекловидное тело; п. ор. — зрительный нерв; pig — пиг-
ментный эпителий; rt^~ ретина; sc+ch —склера и сосудистая
оболочка.
го говоря, конечно,
нельзя сказать, что
рудименты не несут
никакой функции: по
меньшей мере руди-
мент остается дей-
ственным звеном в
какой-либо' цепи
морфогенетиче с к и х
зависимостей, опре-
деляющих нормаль-
ное формирование
других органов. G
этим связано иногда
довольно упорное со-
хранение «провизор-
ных» органов в эм-
бриональном разви-
тии животных (на-
пример, эмбриональ-
ная хорда и жабер-
ные щели позвоноч-
ных). Если же эти морфогенетические зависимости разрываются и
заменяются иными, то такой рудимент исчезает бесследно. Рудименты,
сохраняющиеся во взрослом организме, обычно несут хотя бы и мало
заметную, но определенную функцию, часто, правда, не имеющую ничего
общего с первоначальной функцией органа (рудимент непарного глаза,
эпифиза, а также многие рудименты жаберного эпителия являются желе-
зами внутренней секреции).
По способу исчезновения редуцирующихся органов Северцов разли-
чает собственно рудиментацию, сопровождающуюся редукцией на всех
стадиях развития и приводящую обычно к образованию типичного руди-
мента, и «афанизию», при которой происходит активное уничтожение
органа взрослого животного при нередко полной сохранности его у эмбри-
она (где он, очевидно, в этих случаях сохраняет свои морфогенетические
функции).
ЭМБРИОЛОГИЧЕСКИЙ МЕТОД В СРАВНИТЕЛЬНОЙ АНАТОМИИ
И ВОПРОС О СООТНОШЕНИЯХ МЕЖДУ ФИЛОГЕНЕЗОМ
И ОНТОГЕНЕЗОМ
Сравнительная анатомия при изучении строения животных пользу-
ется методом сравнения. Если мы обозначаем как задачу сравнительной
анатомии изучение истории организмов, то сравнению подлежат не только
взрослые формы, но и эмбрионы, так как вместе с первыми изменяются
и последние. При сравнении более далеких форм метод сравнения эмбри-
онов приобретает даже особенно большое значение, так как эмбрионы
30
обычно изменяются менее значительно, чем взрослые организмы, и обна-
руживают черты сходства и в таких частях, где у взрослых произошло
большое расхождение. Таким образом, использование эмбриологического
материала облегчает сравнение и установление гомологий у более далеких
форм и, во всяком случае, позволяет провести глубокое и детальное
сравнение и, следовательно, позволяет составить себе более ясную картину
филогенетических преобразований данных форм.
То обстоятельство, что эмбрионы более сходны между собой, чем соот-
ветствующие взрослые формы, и притом тем более, чем они моложе, было
отмечено еще знаменитым эмбриологом К. Бэром, который сформулировал
следующие положения: «Чем более ранние стадии развития мы исследуем,
тем более сходства находим мы между различными животными»... «Исто-
рия развития индивида есть история растущей индивидуальности во всех
отношениях». При этом развитии первоначально выявляются признаки,
наиболее общие, характерные для типа, позднее — менее общие, выявля-
ющие принадлежность животного к определенному классу, еще позднее —
признаки, характерные для отряда, семейства, рода, и, наконец, признаки,
наиболее специальные, характеризующие данный зародыш как предста-
вителя определенного вида и, в заключение, как отдельную особь. При
сравнении эмбрионального развития различных животных мы видим посте-
пенное возрастание различий — постепенное обособление, расхождение
этих форм друг от друга (рис. 8). Эти положения К. Бэра вполне сохра-
нили свое значение и до настоящего времени. Однако они были дополнены
и видоизменены в связи с установлением эволюционного учения.
Еще натурфилософы говорили о «параллелизме» между историей
индивидуального развития высших животных (особенно человека) и
«лестницей организмов». Уже Кильмейер (1793) доказывал, что высшие
животные проходят в течение своего индивидуального развития стадии,
сходные со взрослыми состояниями (т. е. со ступенями лестницы) низших
животных. В этом находила свое выражение натурфилософская идея
повторения. К. Бэр решительно выступил против таких сравнений заро-
дышей со взрослыми низшими животными.
Эволюционное учение осветило как значение того сходства за-
родышей, о котором говорил К. Бэр, так и значение тех фактов, которые
отмечались натурфилософами как параллелизмы. Это было вполне пра-
вильно понято уже Ч. Дарвином, впервые наметившим правильное раз-
решение проблемы, но особенно ясно выражено Ф. Мюллером. Эмбриональ-
ное сходство объясняется кровным родством. Родственные формы и раз-
виваются сходно, особенно вначале (рис. 8). Явления ж® параллелизма
зависят от того, что при эволюции животных их измененля затрагивают
преимущественно позднейшие фазы развития и что новые черты органи-
зации появляются обычно в конце, образуя как бы надстройку да прой-
денных уже этапах. Индивидуальное развитие по мере усложнения взрос-
лой формы все более удлиняется путем последовательного ряда надставок,
в своей хронологии повторяющих исторический путь эволюции взрослой
формы. Ф. Мюллер, правда, вносит известные ограничения: не всегда все
новое появляется в конце развития, иногда изменяются и более ранние
стадии, и, кроме того, «историческая летопись в индивидуальном разви-
тии постепенно стирается, по мере того как развитие пробивает себе все
более прямой путь от яйца к зрелому животному, и часто искажается
вследствие борьбы за существование, которой подвергаются свободно-
живущие личинки». Правило повторения признаков взрослых предков,
или правило «рекаиитуляции», было популяризировано Э. Геккелем в
его «биогенетическом законе»: «Ряд форм, которые пробегает индивиду-
альный организм во время своего развития от яйца до взрослого состо-
яния, есть краткое и сжатое повторение длинного ряда форм, который
прошли животные предки данного организма, или исходная форма его
3t
вида от древнейших времен так называемого органического творения до
настоящего времени», или «Развитие зародыша (онтогенез) есть сжатое
и сокращенное повторение развития рода (филогенез); при этом повто-
D
Рис. 8. Зародыш кролика (/), обезьяны (//) и человека (///) на равных стадиях
развития (по Кейбелю и Зеленке).
рение тем полнее, чем более постоянным наследованием сохраняется пер-
воначальное извлеченное развитие (палингенез), и, напротив, тем менее
полно, чем более через посредство различных приспособлений вносится
32
позднейшее нарушающее развитие (цепогенез)», или наконец: «Онтогения
есть краткое повторение филогении».
Биогенетический закон имел очень большое историческое значение
при возникновении филогенетического направления в морфологии.
Взгляды Э. Геккеля представляют последовательную теорию, неразрывно
связанную, однако, с ламарковскими представлениями об активном, т. е.
непосредственном, «приспособлении» организма к окружающей среде и с
верой в наследственный характер результатов такого «приспособления».
Факты повторения, т. е. «рекапитуля-
ции», во время индивидуального развития
любого животного многих черт организа-
ции его более или менее отдаленных пред-
ков не подлежат никакому сомнению.
Приводим ряд примеров такой рекапи-
туляции у хордовых животных.
1. При развитии асцидий, из яиц
вылупляется свободноплавающая личин-
ка, в основном построенная по тому же ти-
пу, как и другие хордовые животные.
Позднее эта личинка прикрепляется, ее
органы чувств и органы движения исче-
зают, нервная трубка редуцируется вплоть
до одного нервного узла, происходит
значительное перемещение внутренних
органов, и таким образом развивается
взрослая асцидия, совсем не похожая на
других хордовых животных (рис. 365).
Здесь мы имеем повторение стадии личин-
ки свободноплавающего предка асцидий,
перешедшего к сидячей жизни и соответ-
ственно изменившего свою организацию.
2. При эмбриональном развитии
высших наземных позвоночных (включая
Рис. 9. Зародыш человека в 4,2 мм
длиной (по Гису).
1 —5 — пять артериальных дуг, меж-
ду которыми лежат четыре жабер-
ные щели; ао — аорта; аи — слухо-
вой пузырек; v — желудочек; art —
предсердие сердца; vtt — основание
желточного мешка; all — аллантоис
(обрезан).
человека) в зародышевом состоянии
развиваются жаберные щели, как и у
зародышей рыб, и притом с таким же
кровообращением (рис. 9), с теми же нер-
вами, с зачатками скелета и мускулату-
ры. Эти явления могут быть истолкованы
только как повторение стадий развития
дышавшего жабрами рыбообразного предка наземных позвоночных.
3. У высших позвоночных оргапом выделения служит «тазовая»
почка, не сравнимая с «туловищной» почкой низших позвоночных, однако,
эмбрионально у них развивается и функционирует такая же «туловищная»
почка, какая имеется и у рыб, и у амфибий не только в зародышевом, но
и во взрослом состоянии.
4. У птиц во взрослом состоянии черепная коробка сплошная, но у
зародыша и даже у молодого птенца она состоит из тех же отдельных кос-
тей, как и у взрослых (а тем более у зародышей) рептилий, от которых,
как известно, некогда произошли птицы.
5. У взрослых млекопитающих таз состоит из пары «безыменных
костей», однако у зародышей развиваются здесь три кости: лонная, седа-
лищная и подвздошная, которые у взрослого животного срастаются между
собой. Стадия трех самостоятельных костей повторяет фазу, сохраняю-
щуюся у рептилий в течение всей жизни.
На практкке, однако, оказалось весьма трудной задачей установить
критерии для разграничения процессов развития, унаследованных от да-
33
леких предков, т. е. «палингенезов» (по терминологии Э. Геккеля), of
процессов, вторично искажающих этот исторический ход развития, т. е.
«ценогенезов». Между тем это разделение совершенно необходимо, так как
для филогенетических построений, очевидно, могут быть использованы
только первые. Поэтому теперь некоторые эмбриологи совершенно отри-
цают значение биогенетического закона в исследовательской практике. На-
рушения унаследованного от далеких предков палингенетического разви-
тия состоят не только из приспособлений самого зародыша или личинки
к условиям их существования, что выражается в присутствии у них особых
зародышевых оболочек, зародышевых органов питания, дыхания и выде-
ления или в присутствии специальных личиночных органов, которых пи
у каких взрослых предков никогда не было (собственно ценогенезы в
узком смысле этого слова), но также из ряда изменений, мало, невидимому,
связанных с внешними условиями развития зародыша. К такого рода
изменениям относятся: гетерохронии, т. е. изменение времени
закладки органа или части (например, ранняя закладка центральной
нервной системы или сердца у высших позвоночных), гетерото-
пии —изменение места закладки (сердце высших позвоночных заклады-
вается на желтке по бокам в виде двух далеко отстоящих отдельных час-
тей), затем упрощение или сокращение развития, связанное с выпадением
стадий («прямое» развитие пресноводного рака по сравнению со сложным
преобразованием личиночных форм у морских раков) и т. п.
Эти нарушения правильной повторяемости исторического хода раз-
вития данной формы в ее индивидуальном развитии были предметом це-
лого ряда специальных исследований (Кейбель, Менерт, Северцов), авторы
которых пытались внести известные поправки в биогенетический закон.
В настоящее время обычно допускают довольно далеко идущую рекапи-
туляцию в развитии отдельных органов, но вместе с тем отрицают палин-
генетическое значение последовательности возникновения отдельных
органов во всем организме. Наиболее обстоятельный разбор всего этого
вопроса сделан Северцовым, определившим границы применимости био-
генетического закона эволюционными процессами, которые являются
результатом последовательных изменений конечных фаз формообразо-
вания. Такие «надставки» развития Северцов называет «анаболиями».
Если эволюция организма протекает путем отклонения процессов формо-
образования на более ранних стадиях («девиации»), то рекапитуляция
наблюдается только в начале развития. Если же эволюция происходит
путем изменения эмбриональных зачатков («архаллаксисы»), то никакой
рекапитуляции в оптогенезе измененных таким образом форм не наблю-
дается. Таким образом, мы вместе с Северцовым различаем такие элементы
онтогенеза: 1) процессы, унаследованные от более или менее далеких
предков, или палингенезы, 2) эмбриональные уклонения от палингене-
тического хода развития, связанные с приспособлением самого зародыша
или личинки к их специфическим условиям существования, или ценоге-
незы, и 3) эмбриональные изменения, связанные с филогенезом взрослого
организма, или филэмбриогенезы. Последние могут осуществляться пу-
тем архаллаксиса, девиации или анаболии.
Вопрос о соотношениях между онтогенезом и филогенезом представ-
ляется в настоящее время в таком виде: поскольку взрослая форма в сущ-
ности неотделима от предшествующих стадий ее развития, мы под фило-
генезом всегда должны понимать исторический ряд последовательных
онтогенезов (а не только ряд взрослых форм). Следовательно, можно го-
ворить о филогенетическом развитии любой стадии эмбриона, личинки,
взрослого животного. Не только филогенез определяет онтогенез (Гек-
кель), но и онтогенез является базой для филогенеза (Северцов). Онто-
генез в основном составляется из процессов, в общем повторяющих собой
процессы индивидуального развития более или менее отдаленных пред-
54
Ков. Эмбрион всегда сходен только с эмбрионом, однако в его развитии
нередко явно выделяются фазы или частные процессы, которые у более или
менее далеких предков претворялись затем в характерный орган или
признак зрелого организма, а у наблюдаемого теперь потомка преобра-
зовываются в существенно иной орган, утративший прежнее значение,
или останавливаются в развитии и даже вовсе исчезают.
На ранних стадиях можно установить сходство и между очень дале-
кими формами, между тем как на более поздних стадиях эти формы все
более, расходятся. Это сходство более молодых зародышей между собой
указывает на то. что в своих видимых филогенетических изменениях
ранние стадии отстают от более поздних. Новые черты организации чаще
проявляются лишь на более поздних стадиях, в конце морфогенеза (ана-
болии). Однако это правильно лишь в общем. В частных случаях возможны
и отклонения развития на более ранних стадиях («девиации») и проявле-
ние нового признака сразу в самом начале морфогенеза («архаллаксис»).
Здесь рекапитуляция более далеких исторических фаз невозможна.
Исторически сложившийся «палингенетический» ход развития повто-
ряется в онтогенезах потомков с тем большим постоянством, чем сложнее
и теснее взаимная зависимость отдельных формообразовательных про-
цессов, составляющих эмбриогенез в целом (характерно в особенности
для ранних стадий развития всего зародыша, а также для начальных фаз
развития частных систем и органов), и чем более предшествующие морфо-
генетические процессы являются необходимой базой для последующих.
«Ценогенетические» и «филэмбриогенетические» изменения этого унасле-
дованного процесса наступают тем легче, чем рыхлей становится связь
между формообразованием отдельных частей внутри целого («самодифе-
ренцирование» органов на более поздних стадиях развития). Таким об-
разом, рекапитуляция есть выражение целост-
ности развивающегося организма, сохраняющейся
во всех его исторических изменениях в глубоких внутренних связях между
отдельными морфогенетическими процессами, между тем как нарушения
этой рекапитуляции указывают на относительную самостоятельность
развивающихся частей в индивидуальном развитии целого. Чем ярче
выражена целостность, т. е. неделимость развивающегося организма,
тем полнее проявляется и правило рекапитуляции (особенно у позвоноч-
ных и более всего у высших позвоночных).
ОРГАНИЗАЦИЯ ХОРДОВЫХ животных
Характеристика хордовых
Тип хордовых животных представляет собой группу животных,
характеризующуюся вполне определенным планом строения, выраженным
особенно ясно в зародышевой фазе развития. У всех хордовых имеется
внутренний осевой скелет в виде продольного эластического стержня —
спинной струны, или хорды. Передняя часть кишечника сообщается е
наружной средой посредством ряда парных отверстий, называемых жа-
берными щелями, так как у живущих в воде хордовых в них распола-
гаются жабры. Наконец, у всех хордовых животных центральная нервная
система является трубчатой. Кроме того, не менее характерно и располо-
жение органов. Вдоль спинной стороны тела располагается центральная
нервная система, под ней лежит хорда, а под хордой с брюшной стороны
тела помещается общая полость тела с кишечной трубкой (рис. 10). Кроме
того, хордовые животные принадлежат к двусторонне-симметричным
вторичнополостпым животным (Coelomata) и построены метамерно.
Приведенное определение требует, однако, пояснений. Далеко не
всегда все эти признаки бывают выражены у взрослого животного, и вся
характеристика целиком применима лишь к зародышам. Наиболее общим
35
Цризйаком взрослых животных является существовайие трубчатой цент-
ральной нервной системы, лежащей на спинной стороне. Этот признак
не удерживается лишь у взрослых оболочников, у которых нервная
система редуцируется и канал ее зарастает. Затем хорда у всех высших
хордовых замещается позвоночником, развивающимся на ее месте. Мета-
мерия выражается главным образом в мускулатуре и скелете и полностью
исчезает у взрослых оболочников. Наконец, жаберные щели зарастают
у наземных позвоночных с переходом их к легочному дыханию. Нои хорда,
и жаберггые щели всегда имеются у зародышей высших позвоночных.
Рис. 10. Схематический продольный разрез через тело ланцетника (по Боасу).
ап — анальное отверстие; Ы — жаберная часть нишечнина'; ch — хорда; int —
нитка; п — мозг; or — рот; р — оноложаберная полость; ро — атриальное
отверстие.
К хордовым животным относится большинство сложно построенных
высших животных, включая и человека. Сюда относятся некоторые так
называемые «беспозвоночные» (асцидии, сальпы, аппендикулярии) и ти-
пичные позвоночные животные—круглоротые, рыбы, земноводные, репти-
лии, птицы и млекопитающие. Очень просто организованным представи-
телем типа является ланцетник, анатомия и эмбриология которого дала
нам чрезвычайно много для решения вопроса о происхождении позвоноч-
ных и для выяснения их организации.
Организация ланцетника
Ланцетник (Amphioxus lanceolatus) — небольшое рыбообразное жи-
вотное с довольно узким и заостренным с обоих концов телом. Брюшная
поверхность более плоская, чем спинная, и продолжается по бокам в
особые «метаплевральные» складки, идущие от рта, вдоль большей
части туловища (рис. 11). Вдоль спины по средней линии идет складка,
имеющая значение непарного спинного плавника. На заднем конце тела
этот плавник значительно повышается и, огибая его, продолжается
на брюшную сторону тела. Эта более развитая часть плавника носит
название хвостового плавника. На брюшной стороне тела хвостовой
плавник впереди переходит в более низкий анальный плавник, кото-
рый идет вплоть до того места, где кончаются парные метаплевральные
складки. В этом месте находится атриальное отверстие называемое
иногда — жаберной порой (рис. 10). Несколько отступя от заост-
ренного переднего конца тела, лежит рот на дне чашевидного углубления,
края которого («ротовой вырост») обсажены 12—20 парами усиков, окру-
жающих рот в виде бахромы. Со спинной стороны ротового углубления
помещается мерцательный орган, вызывающий у живого животного непре-
рывный ток воды, направляющийся в ротовое отверстие и выходящий из
атриального отверстия. Заднепроходное отверстие помещается позади
атриального отверстия с левой стороны от анального плавника.
Покровы ланцетника состоят из однослойного цилиндрического
эпителия — эпидермиса и подстилающей его прозрачной основной пере-
понки. Эпидермис покрыт тонкой пористой кутикулой. Среди обыкно-
венных цилиндрических клеток находятся отдельные чувстующие клетки
36
с волосками и бокаловидные железистые клетки, выделяющие слизь на
его поверхность. Бесклеточная основная перепонка представляет собой
однородное вещество, выделенное клетками эпидермиса. Под основной
перепонкой помещается слой почтй бесклеточной студенистой ткани с
отдельными волокнами, ограниченный снизу кожным листком мезодермы,
сохраняющим характер эпителия.
Мускулатура ланцетника состоит из двух больших мышечных
масс, сплошь покрывающих бока
делены прослойками почти бес-
клеточной ткани (миосептами) на
целый ряд (около 60) следующих
друг за другом мускульных
сегментов, или миомеров. Так как
миосепты направлены не прямо
поперек тела, а косо под углом
к продольной оси, и сверх того изо-
гнуты в виде угла «)> направлен-
ного вершиной вперед, то миоме-
ры имеют вид полуконусов, вло-
женных друг в друга. Миомеры
правой и левой стороны не совпа-
дают, а расположены поочередно.
Мускульные волокна направлены
горизонтально вдоль длинной оси
тела. В спинной половине мус-
кульная масса значительно толще,
чем в брюшной.
Кроме этой боковой мышцы,
имеется еще сплошной слой попе-
речных мускульных волокон с
брюшной стороны тела в области
метаплевральных складок. Эта
поперечная мышца служит для
сжимания и опорожнения около-
жаберной (атриальной) полости
(рис. 11, mt).
Скелет. Вдоль всего тела
ланцетника, начиная от перед-
него конца его, тянется хорда,
представляющая собой плотный
и весьма эластичный стержень.
Наружная стенка хорды предста-
влена -очень прочной оболочкой;
внутренняя масса состоит из диско-
образных клеток с волокнистыми
стенками и вакуолями с жидким
содержимым. Оболочка хорды
представляет собой продукт выде-
животного. Боковые мышцы раз-
uf
Рис. 11. Схематический разрез через жа-
берную область Amphioxus lanceolatus.
Слева разрез прошел через вторичную жабер-
ную дужку, справа — через первичную; ао —
аорта; с — corium; ес — субэндостильный це-
лом; / — слой, образующий миокоммы (фас-
ции); fh — луч спинного плавника; g — гона-
ды; gl — петля сосуда около нефридиальной
трубки (glomerulus); к — жаберный сосуд, или
дужка; kd — кишечник; Id — перегородка ме-
жду целомической и околожаберной полостями;
т — миотом; те — спинной мозг; ml — попе-
речная брюшная мышца; п — нефридиальный
канал; of—целомические полости в метаплев-
рах; р — околожаберная полость; sc — субхор-
дальный целом; si—жаберная артерия, или
брюшная аорта; sk — скелетогенный слой;
uf — лимфатические полости в брюшной стен-
ке (по Бовери и Гатчеку из Шимкевича),
ления клеток хорды; остатки кле-
точных тел располагаются вдоль спинного и брюшного края хорды под ее
оболочкой. Остальной скелет ланцетника построен из опорных перепонок
и почти или совсем бесклеточной массы различной консистенции, которая
здесь заменяет типичную соединительную ткань других хордовых. Эта
ткань образует тонкий студенистый слой под эпидермисом, продолжается
в миосептах, разделяет в виде сагиттальной перегородки миомеры правой
и левой стороны и окружает хорду и центральную нервную систему. Мно-
гочисленные стержни более плотной студенистой ткани образуют скелет
37
непарных плавников; это так называемые лучи плавников. Более плотные
тонкие стержни волокнистого бесклеточного вещества образуют жаберный
скелет — продукт выделения прилежащих эпителиальных клеток. Ротовой
вырост поддерживается также своим скелетом в виде кольца, состоящего
из отдельных звеньев «хряшеподобной» ткани. Каждое звено дает от себя
отросток, служащий осью, поддерживающей соответствующий усик.
Нервная система. Центральная нервная система ланцетника
имеет вид стержня, содержащего узкий центральный канал, который
продолжается на спинной стороне в узкую срединную щель, доходящую
до поверхности органа (рис. 11). Таким образом, мозг имеет форму не
вполне замкнутой трубки, а скорее желобка с сомкнутыми краями. Обо-
собленного головного мозга у ланцетника нет, но центральный канал в
переднем конце мозга образует расширенную полость, которую сравни-
вают со средним (третьим) же-
Рис. 42. Мозговой пузырек ланцетника
в сагиттальном разрезе.
Ер — епендима; gg — мерцательная ямка
(слева); gz — нервная клетка; Jnf — инфун-
дибулярный орган; Р — пигментное пятно;
I — передняя пара нервов.
лудочком головного мозга позво-
ночных; на нижней поверхности
этого желудочка у молодого жи-
вотного имеется углубление, ко-
торое иногда сравнивают с ворон-
кой позвоночных. Здесь рано раз-
вивается значительная группа
чувствующих клеток (рис. 12) «ин-
фундибулярного» органа. Непо-
средственно позади передней
полости мозга срединная щель
расширяется на спинной стороне
органа, образуя вторую полость,
которую сравнивают с четвер-
тым желудочком позвоночных.
Передний конец мозга образует непарный выступ (обонятельная
лопасть), тесно связанный с обонятельной ямкой при помощи короткого
нерва. Под этим выступом в передней стенке мозга имеется скопление
пигмента, неправильно считавшееся органом зрения. Скопления пигмента
располагаются и вдоль центрального канала всего мозга, сопровождая
здесь небольшие бесспорно светочувствительные органы (глазки Гессе).
Таким образом, у ланцетника весь мозг оказывается светочувствительным.
При микроскопическом исследовании можно обнаружить, что главная
масса стенок мозга состоит из волокон, идущих вдоль органа; среди
этих волокон имеются отдельные очень толстые, так называемые гигант-
ские волокна. Клеточные элементы расположены исключительно по
внутренней поверхности органа, т. е. у стенки центрального канала и
вдоль срединной щели. Клетки, непосредственно выстилающие эту по-
верхность, образуют эпителий, (эпендима) и нервной функции не несут:
от них отходят отростки, проникающие сквозь всю толщу мозга и служа-
щие для его укрепления, т. е. играющие роль скелета. Между этими
клетками, обычно немного отступя от поверхности, расположены отдель-
ные более крупные нервные клетки, даюшие от себя нервные отростки.
Кроме обыкновенных ганглиозных клеток, в мозге ланцетника наблюдают-
ся еще правильно расположенные гигантские мультиполярные клетки, от
которых отходят упомянутые гигантские волокна.
Периферические нервы отходят от центральной нервной сисмемы в
определенном порядке. Соответственно каждому мускульному сегменту
с каждой стороны отходит по одной паре нервных корешков, по одному
спинному, отходящему одним стволом с боковой стороны мозга, и по
одному брюшному, отходящему отдельными пучками волокон по бокам
брюшной стороны мозга. Спинные и брюшные корешки каждой стороны
отходят не на одном уровне, а поочередно один за другим. Корешки
38
правой и левой стороны чередуются так же, как и соответствующие миоме-
ры, так что против спинного корешка одной стороны приходится брюшной
корешок противоположной, и наоборот. Спинные корешки продолжаются
в спинные нервы, которые дают чувствительные ветви к коже и к внутрен-
ним органам и двигательные ветви к мускулатуре внутренних органов
(кишечника, сосудов) и к поперечной мышце брюха. Таким образом,
спинные корешки и нервы по своему составу являются смешанными;
кроме чувствующих волокон, они содержат двигательные волокна для
определенной категории мышц, называемых висцеральными. Нервные
пучки брюшных корешков направляются непосредственно в прилежащий
миомер боковой мышцы, где распадаются на отдельные волокна, вступа-
ющие в связь с мускульными волокнами. Таким образом, брюшные ко-
решки являются чисто двигательными для миомеров туловища, или для
так называемой париетальной мускулатуры. Первые две пары нервов
Рис. 13. Передний конец тела ланцетника. Левая половина.
Ротовая чаша, окруженная усиками с мерцательным органом (с/1); парус (vet) с велярными
щупальцами; передняя часть глотки с жаберными щелями (bi); eft — хорда.
отходят от области головного мозга: передняя — от его переднего конца,
вторая — над желудочком. Эти «головные» нервы лежат симметрично
друг против друга и содержат чувствующие волокна, иннервирующие
передний (предротовой) конец тела.
Кроме мозговых нервов, имеются также симпатические сплетения
нервов с ганглиозными клетками в стенке кишечника. Они связаны посред-
ством соединительных ветвей со спинным мозгом.
Органы чувств ланцетника очень просты. Прежде всего во
всем эпидермисе рассеяны чувствующие клетки, располагающиеся обычно
парами. Местами такие клетки образуют скопления. Особенно многочислен-
ны чувствующие клетки на усиках, где они образуют чувствующие почки,
и вообще на переднем конце тела, а также п по краю паруса и на его
щупальцах. Эти клетки сравнивают с обонятельными. Имеются и более
сложные органы. На спинной стороне передней части тела, слева, имеется
ямка, выстланная мерцательным эпителием, к которой подходят 1 — 2
нервные веточки от первой пары нервов. Эта ямка (ямка Кёлликера)
считается органом обоняния (рис. 12). У личинки она сообщается с поло-
стью мозга посредством отверстия — невропора. Небольшие своеобразные,
светочувствительные органы располагаются вдоль всего мозга у централь-
ного его канала. Эти «глазки Гессе» состоят каждый из чувствующей
клетки с тонкими палочковидными образованиями на одной стороне, где ее
59
которые дыхательный ток воды выходит из глот:
околожаберную полость и затем выводится через
Рис. 14. Поперечный разрез через эндостиль ланцетника.
ао. i>— брюшная аорта; с—целомический канал; НЬ — первич-
ная жаберная перегородка; Zb—язычковая перегородка; gl —
группы железистых клеток.
покрывает в виде чашки или шапки другая, пигментная клетка. Свето-
чувствительность мозга ланцетника связана с прозрачностью его тела.
Кишечник начинается лежащим на дне ротовой чаши ротовым
отверстием, ограниченным круглой складкой слизистой оболочки, парусом
(velum), с собственной мускулатурой и особыми выростами — велярными
щупальцами, направленными внутрь глотки. Расположенный в ротовой
чаше мерцательный орган гонит воду в ротовое отверстие (рис. 13). Ротовая
чаша и весь кишечник ланцетника выстланы мерцательным эпителием.
Передняя, большая, часть кишечника представлена очень широкой труб-
кой — жаберной частью кишечника или глоткой (рис. 10). Глотка проре-
зана большим числом (свыше 100) косых жаберных щелей (рис. 13), через
;и в окружающую ее
атриальное отверстие
наружу. Жаберные
щели отделяются
друг от друга узки-
ми промежутками —
жаберными перего-
родками, по кото-
рым проходят кро-
веносные сосуды.
Кровь в жаберных
сосудах окисляется
кислородом, раство-
ренным в воде, омы-
вающей жаберные
перегородки. По-
следние поддержива-
ются скелетом, со-
стоящим из тонких,
но плотных бескле-
точных стержней,
соединенных между собой тонкими перекладинами в цельную решетку
(рис. 13). Таким образом, жаберные щели поддерживаются открытыми.
Скелетный стержень каждой второй перегородки несколько отличается
по форме от первой, и соответственно мы различаем расположенные
поочередно первичные и вторичные (или «язычковые») перегородки. Вдоль
первичной жаберной перегородки проходит целомический канал (рис. 11).
На дне глотки по срединной линии расположен желобок, выстланный
мерцательным эпителием, с четырьмя продольными рядами железистых
клеток и одним средним рядом клеток с длинными ресничками; это под-
жаберная борозда, или эндостиль (рис. 14). Впереди у паруса обе губы
поджаберной борозды расходятся направо и налево, охватывая глотку
изнутри в виде окологлоточных мерцательных лент, которые опять схо-
дятся па спинной сторо-пе глотки и продолжаются назад в наджаберную
борозду. Весь этот аппарат служит своеобразным органом для извлече-
ния из воды пищевых частиц. Слизь, выделяемая железистыми клетками
поджаберной борозды, гонится действием мерцательных ресничек по
желобку вперед, навстречу току воды; оседающие из воды пищевые час-
тицы прилипают к слизи и увлекаются ее током к окологлоточным лентам,
перегоняющим их к спинной борозде, в которой ток слизи направлен
назад, прямо в кишечник.
Таким образом, и дыхание, и питание ланцетника происходят почти
пассивно, действием одного только мерцательного эпителия. Собственно
кишка начинается довольно широким отделом, который, постепенно
суживаясь, кончается заднепроходным отверстием, снабженным коль-
цевой мышцей (сфинктером)/От начального отдела кишки с брюшной
40
vca ааа
ао.и и
Рис. 15. Схема кровообращения ланцетника.
ао. d — спинная аорта; ао. v — брюшная аорта; и. с. а — пе“
редняя и v. с, р — задние кардинальные вены; vp — ворот"
ная венц, печени; v. si — подкишечная вена.
стороны отходит направлонныйвпород слепой вырост в виде полого ме-
шочка. Это печеночный вырост, гомология которого с печенью позвоноч-
ных доказывается, между прочим, существованием на нем воротного
кровообращения (рис. 15).
Органы кровообращения представлены кровеносными
сосудами, несущими бесцветную кровь. С брюшной стороны, непосред-
ственно под глоткой, лежит сократимый поджаберный сосуд, несущий
венозную кровь вперед (рис. 15). От этого сосуда, называемого также
брюшной аортой (aorta ventralis), с обеих сторон отходят боковые ветви,
направляющиеся вверх в первичные жаберные перегородки. Эти жабер-
ные сосуды соединяются поперечными анастомозами с сосудами, лежа-
щими во вторичных жаберных перегородках, и затем изливают окислен-
ную уже кровь с каждой стороны в парный продольный сосуд, идущий
вдоль спинной стенки глотки. От этих наджаберных сосудов отходят
вперед в голову небольшие сонные артерии. Главная масса артериальной
крови направляется
назад, где оба над
жаберных сосуда
соединяются в один
непарный ствол —
спинную аорту (aorta
dorsalis), идущую по
средней линии тела
непосредственно под
хордой до заднего
конца тела, отда-
вая попутно ветви к мышцам, кишечнику и другим органам, где они рас-
падаются на сеть капилляров. Во всех этих органах кровь отдает свой
кислород, поглощает взамен углекислоту из тканей, извлекает всевоз-
можные продукы обмена и собирается в венозные сосуды. Кишечные капил-
ляры изливают кровь в так называемую подкишечную вену (vena subin-
testinalis), которая тянется непосредственно под кишкой вперед до пече-
ночного выроста, где она разбивается па оплетающую его сеть капилляров,
представляющую собой воротную систему печени. Отсюда кровь вновь
собирается в сосуд (vena hepatica), образующий небольшое расширение
на месте впадения в него с двух сторон парных вен, называемых кювьеро-
выми протоками. Дальше вперед брюшной сосуд продолжается в виде
уже рассмотренной брюшной аорты. Кювьеровы протоки составляются
из одной пары вен, несущих кровь от передней половины тела, и одной
пары вен, несущих кровь от задней половины тела. Эти сосуды, носящие
название передних и задних кардинальных вен, собирают кровь со стенок
тела по метамерно расположенным веточкам.^
Таким образом, сердца у ланцетника нет, и кровь перегоняется исклю-
чительно пульсацией самих сосудов, главным образом брюшной аорты
и оснований жаберных сосудов.
Полость тела (целом) выстлана плоским эпителием, клетки
которого снабжены каждая одним мерцательным волоском. У ланцетника
она представляет собой систему сравнительно небольших полостей, так
как она в значительной мере вытеснена развивающейся здесь другой
полостью — околожаберной, или атриальной, которая охватывает всю
жаберную часть кишечника снизу и сбоку. В жаберной области имеется
лишь пара продольных каналов, лежащих вдоль спинной части глотки
и сообщающихся с третьим каналом, лежащим под кишечником, узкими
каналами по первичным жаберным перегородкам (рис. 11). Позади глотки
целом вполне охватывает кишечник. Остатки целомических полостей
находятся в мотапловральных складках, а также окружают половые
железы.
41
Органы выделения ланцетника представлены парным ря-
дом отдельных коротких трубок (нефридиев), лежащих по бокам спинной
части глотки между продольными каналами полости тела и околожабер-
открывается несколькими отверстиями —
«нефростомами» в целом и одним вы-
водным отверстием в околожаберную
полость. Нефростомы усажены особы-
ми булавовидными клетками с тон-
кими внутренними канальцами и длин-
ным мерцательным волоском внутри
канальца—«соленоцитами» (рис. 16).
Нефридии расположены правильно
по одному против каждой вторичной
жаберной перегородки. Кроме того,
на верхней стенке ротового углубле-
ния слева имеется еще большая выде-
ной полостью. Каждая трубка
Рис. 16. Почечные канальцы
Amphioxus.
7 — целый каналец со многими нефро-
стомами и соленоцитами; Ьг — верхний
конец жаберной щели; р — отверстие
почечных канальцев в перибранхиаль-
ную полость (по Боверн); 2 — часть
стенки почечного канальца с сидящими
па ней 7 соленоцитами, сильно увеличен-
ная (по Гудричу из Гертвига).
лительная трубка — «нефридий Гат-
чека» со многими соленоцитами и
с большим сосудистым клубочком. По
своему строению нефридии ланцет-
ника поразительно сходны с нефри-
диями некоторых кольчатых червей.
Половые железы (гонады)
построены одинаково у самца, и
у самки. Они представляют собой парный ряд округлых тел, окруженных
целомическим эпителием и правильно метамерно (по одному на миомер)
расположенных в стенках тела по бокам околожаберной полости (рис. 11).
Половые железы вдаются в атриальную полость и открываются в нее
только при созревании половых продуктов, которые выводятся затем из
околожаберной полости через жаберную пору.
Развитие ланцетника
Отложенные яйца ланцетника одеты тонкой оболочкой и после выде-
ления редукционного тельца и оплодотворения начинают дробиться.
Дробление полное и почти равномерное. Бластомеры раздвигаются,
образуя в середине полость, и таким образом в результате дробления
яйца ланцетника развивается типичная шаровидная бластула (рис.
17, Л). Так как дробление все же не вполне равномерное, то и стенка
бластулы на одной стороне зародыша несколько толще, чем на другой.
Та половина зародыша, где помещаются более мелкие клетки, является
анимальной («животной»), так как здесь развиваются впоследствии органы,
характерные для животных, — нервная система и органы чувств. Про-
тивоположная половина, состоящая из более крупных клеток, является
вегетативной («растительной»), так как здесь уже с самого начала сосре-
доточены функции питания. Более крупные клетки вегетативного полюса
бластулы начинают вворачиваться внутрь, и таким образом, путем ти-
пичной инвагинации, получается двуслойный зародыш — гаструла
(рис. 17). Выстланная энтодермой гастральная полость зародыша сооб-
щается с наружной средой посредством бластопора. Эктодермаль-
ные клетки наружной стенки получают мерцательные жгутики, под
действием которых зародыш начинает двигаться, еще находясь внутри
яйцевой оболочки. Затем гаструла несколько вытягивается, отверстие
бластопора уменьшается, и одновременно начинается развитие централь-
ной нервной системы и образование среднего зародышевого листка.
Спинная сторона зародыша рано намечается: здесь стенка гаструлы
становится более плоской. Клетки эктодермы, образующие среднюю
42
часть спинной стороны, начинают опускаться; обособляясь от остальных
клеток, они дают начало нервной пластинке, клетки которой принимают
несколько более высокую цилиндрическую форму (рис. 18, Л). По краям
нервной пластинки остальная часть эктодермы приподнимается в виде
складок и зарастает над ней. Таким образом, нервная пластинка оказы-
вается лож а шей под эктодермальным покровом (рис. 18, В). Затем края
нервной пластинки приподнимаются, так что она принимает вид желобка
(рис. 18, С), и, наконец, смыкаются на спинной стороне (рис. 18, D).
Желобок превращается в нервную трубку с центральным каналом, про-
должающимся на спинной стороне в узкую щель. Нервные складки про-
должаются и за бластопор, до которого доходит нервная пластинка.
Рис. 17. Эмбриональное развитие ланцетника (по Серфонтену и Гатчеку).
А — бластула; В, С, D — гаструлнция; Е, F — образование мезодермы, хорды
и нервной системы; ап — анимальный полюс; Ър — бластопор; ch — хорда;
сп — нервнокишечный канал; сое/ — целомические мешки; g — гастральная
полость; mes— складка меводермы; п —нервный канал; пр — невропор; red-—
вегетативный полюс.
Когда складки срастаются между собой, они покрывают сверху, кроме
нервного желобка, и бластопор, который тогда открывается в полость
нервной системы (рис. 17, Е). Таким образом, на заднем гонце зародыша
устанавливается прямая связь между кишечной полостью и полостью
мозга посредством нервнокишечного канала (canalis neu-
roentericus). На переднем конце тела складки смыкаются позже всего, и
здесь нервный канал открывается наружу большим сначала отверстием —
п европором (neuroporus) (рис. 17, Е, F). На этом месте развивается
обонятельная ямка, на дне которой долгое время еще остается открытый
невропор.
Одновременно с развитием центральной нервной системы начинается
диференцировка и в энтодерме. На спинной ее стороне справа и слева
начинают развиваться парные продольные выступы в виде глубоких
складок — закладки мезодермы (рис. 18), являющейся, следова-
тельно, согласно общему происхождению с первичной энтодермой, «энто-
мезодермой». Между этими закладками средняя часть спинной стенки
первичной кишечной полости начинает утолщаться и изгибаться в про-
дольный желоб, который отшнуровывается от кишки и превращается в
плотный продольный стержень, лежащий над кишкой и непосредственно
под нервной трубкой. Это — зачаток хорды (рис. 18). Лежащие по
бокам от закладки хорды продольные складки первичной энтодермы
постепенно отделяются от места своего образования. Стенки этих складок
представляют собой мезодерму (мезобласт), а полости их—вторичную
полость тела. Оставшаяся часть энтодермы образует собственно кишечник,
который сначала является замкнутым (сообщаясь позади только с нервной
трубкой). Боковые складки мезодермы разрастаются по бокам тела заро-
43
дыша и начинают разделяться со спинной стороны на следующие друг
за другом мешковидные участки (рис. 17, Е). Спинные мешочки развива-
ются не все одновременно, а последовательно спереди назад, так что число
их все увеличивается (рис. 17, Е, F). По мере своего образования они
обособляются от остальной мезодермы и образуют ряд толстостенных
пузырьков, называемых сомитами. Таким образом, зачаток мезо-
дермы оказывается разделенным на спинной отдел, состоящий из мета-
мерного ряда сомитов, располагающихся по бокам от хорды и нервной
Рис. 18. Поперечные разрезы через зародыши ланцетника, показываю-
щие развитие мезодермы, хорды и нервной системы.
ch — хорда; coel — целомический мешок (в D — полость тела); rn.es — закладка
мезодермы; ту — полость сомита; п— нервная трубка; пр— нервная пла-
стинка; npl — нервный желобок.
трубки, и брюшной отдел, называемый боковой пластинкой
и лежащий в виде сплошного двуслойного образования по бокам от кишеч-
йика (рис. 18, D, справа). Каждый отдел получает свой участок полости
первоначальной складки, т. е. зачатка вторичной полости тела.
От нижнего внутреннего угла сомита вырастает плоская складка,
которая располагается между хордой и нервной системой, с одной сто-
роны, и остальной частью сомита — с другой (рис. 19). Эта складка
носит название с к л е р о т о м а; составляющие ее клетки образуют
затем ткань, окружающую хорду и нервную трубку, а также ткань
плавниковых лучей. Вся остальная часть сомита носит название
м и о т о м а. Его полость — миоцель — отделяет наружную его стенку,
кожный листок, от внутренней, или мускульного листка. Наружный
листок сохраняется у взрослого лапцетника в виде тонкого кожного
клеточного слоя, продуцирующего соединительную ткань, лежащую под
эпидермисом. Клетки внутреннего листка миотома превращаются в про-
44
Рис. 19. Поперечный
разрез через личинку
ланцетника.
ch — хорда; сое/ — полость
тела; go —гонотом; int —
кишка; ту — миотом; п —
мозг; sc — склеротом.
ДольнЫо мускульное волокна и дают начало миомёрам, иЛи мускульным
сегментам боковой мышцы взрослого животного (рис. 19).
В боковой пластинке, или спланхнотоме, внутренняя полость (спланх-
ноцель) отделяет прилежаший к кишечнику кишечно-волокнистый, или
висцеральный, листок от наружного, кожно-волокнистого, или периеталь-
ного листка. В отличие от спинного отдела брюшная часть мезодермы
лишена сегментации. Полости боковых пластинок образуют по бокам
кишечника непрерывную парную полость тела, или целом взрослого
животного. Сошедшиеся между собой пад и под кишкой внутренние
листки боковых пластинок представляют собой спинной и брюшной мезен-
терии, в которых в виде продольных каналов развиваются спинной (спин-
ная аорта) и брюшной (подкишечная вена и брюш-
ная аорта) кровеносные сосуды. Мезентерии ис-
чезают, и парная полость тела сливается в одну
непарную (рис. 19). У взрослого ланцетника, как
было сказано, целомическая полость сильно со-
кращена. Кишечно-волокнистый, или висцераль-
ный, листок боковой пластинки дает между про-
чим начало мускулатуре кишечника. За счет кле-
ток боковой пластинки развивается и поперечная
мышца ланцетника. Таким образом, вся вис-
церальная мускулатура ланцетника развивается
за счет брюшного отдела мезодермы — боковых
пластинок, менаду тем как вся париетальная
мускулатура происходит за счет спинного отдела
мезодермы — миотомов. Следовательно, наблю-
даемое у взрослого животного различие в иннер-
вации этих мышц вполне соответствует разли-
чию в эмбриональном их развитии.
Наконец, от стенок полости тела в виде сле-
пых выростов развиваются нефридиальные ка-
нальцы, которые позже открываются в околожа-
берную полость. От миотомов развиваются еще половые железы. От ниж-
него их конца отделяется по мешочку, содержащему целомическую по
происхождению полость (рис. 19). В наружной стенке этого «гонотома>
образуется утолщение эпителия, разрастающееся в половую железу (го-
наду), вполне заполняющую полость мешочка.
Одновременно с описанными процессами диференцировки мезодермы
происходит и дальнейшее развитие энтодерма льны х органов. На переднем
конце тела, слева, развивается эктодермальное углубление, на дне кото-
рого энтодерма срастается с эктодермой и затем здесь появляется неболь-
шое сначала, но быстро растущее отверстие. Это — рот, который у лан-
цетника закладывается асимметрично (рис. 20) и представляет невидимо-
му по своему происхождению переднюю жаберную щель левой стороны.
На брюшной стороне таким же образом появляется другое маленькое
отверстие, которое затем перемещается на правую сторону. Это — первай
жаберная щель левой стороны, которая существ}ет непродол-
жительное время и затем зарастает. В той же области развивается и
вырост переднего конца кишечника, лежащий с правой стороны, который
превращается затем в большой замкнутый мешок, называемый колбо-
видной железой. Он представляет собой чисто личиночный орган, исче-
зающий у взрослого животного (рис. 20). По мере того как личинка вы-
тягивается в длину, позади первой щели на брюшной стороне развивается
один за другим целый ряд (14) жаберных щелей левой стороны, которые
по мере развития перемещаются на правую сторону (рис. 20). Вслед за
этим на той же правой стороне тела закладывается в той же последова-
тельности ряд жаберных щелей (8) правой стороны, лежащий со спинной
45
сторони от первого. Затем нижний ряд передвигается обратно на-брюхо
и после этого на левую сторону (причем число щелей сокращается до 8
вследствие зарастания первой и пяти задних). Теперь расположение
жаберных щелей становится вполне симметричным. Рот передвигается
на брюшную сторону.
Жаберные щели расположены строго метамерно. Число щелей опять
увеличивается вследствие появления новых щелей позади старых. Затем
со спинной стороны каждой щели вырастает вниз в виде «язычка» пере-
Рис. 201 Личинки ланцетника сбоку. Развитие рта и жаберных щелей (по Уилли).
а. с —головная полость; ап — задний проход; ch —хорда; g — нолбовидная железа; int —
кишка; п —нервная трубка; пр— невропор; о — глазок; рр — предротовая ямка; st — рот;
1, 2, 3, 4 — вакладки жаберных щелей.
городка, которая делит щель вдоль на две. Следовательно, число щелей
увеличивается еще вдвое. Развившиеся так перегородки и представляют
собой вторичные, или язычковые, перегородки взрослого ланцетника.
Наконец, на брюшной стороне непосредственно над жаберными
отверстиями личинки развивается пара продольных метаплевральных
складок, ограничивающих с боков эктодермальный желобок, в который
открываются жаберные щели. Края желобка срастаются между собой,
ограничивая таким образом некоторую полость, идущую вдоль брюшной
стенки и представляющую собой зачаток околожаберной по-
лости. Позади остается отверстие, представляющее собой атриальное
отверстие. Околожаберная полость затем сильно разрастается в стороны,
охватывая с боков глотку. Таким образом, околожаберная (атриальная)
полость ланцетника выстлана эктодермой, т. е. наружными покровами,
и ограничивает собой часть наружной среды.
Развитие ланцетника дает нам крайне упрощенную, можно сказать,
схематическую картину того, что наблюдается и у других хордовых.
Особенностью, свойственной исключительно ланцетнику, является лишь
асимметричное развитие рта и жаберных щелей. Такая асимметрия на-
блюдается у донных рыб (камбаловых) в связи с тем, что они лежат на
одном определенном боку. Временная асимметрия ланцетника объяснима
принятием предположения, что и предки ланцетника обладали такой же
биологической особенностью — они лежали на левом боку ртом вниз,
а жаберные щели помещались на верхнем правом боку. Когда эти формы
стали зарываться в песок, подобно современному ланцетнику, те жаберные
щели покрылись заросшими над ними кожными складками, образовав-
шими атриальную полость, и симметрия восстановилась.
Общая организация позвоночных
Позвоночные — свободно перемещающиеся двусторонне-симметричные
хордовые животные с более или менее значительным внутренним скеле-
том и хорошо развитыми и весьма характерными органами чувств.
46
На heреднем конце тела, обыкновенно несколько с брюшной стороны,
Помещается рот (рис. 21). Впереди рта или чаще па спинной стороне
помещаются парные обычно ноздри. Над ртом помещаются парные органы
зрения, а позади них — парный же орган слуха. Позади рта распола-
гается парный ряд жаберных щелей, которых нет у наземных форм.
В задней половине тела с брюшной стороны помещается отверстие клоаки
или реже отдельные заднепроходное, мочевое и половое отверстия.
Рис. 21. Схема позвоночного (рыбы).
А и В — границы головы, туловища и хвоста. С части туловища кожа снята, чтобы показать
мускулатуру; аи — положение органа слуха; Ьг — жаберные щели; И — каналы боковой ли-
нии; m — миотомы; п — ноздря; па, nc, nd — анальный, хвостовой и спинные плавники; пр,
nv — парные грудной и брюшной плавники; spj — брызгальце.
Рис. 22. Схематические поперечные разрезы черев голову, туловище и хвост
позвоночного (рыбы) (по Паркеру).
ае и at — наружное и внутреннее жаберные отверстия; а. Ьг — жаберная дуга; aod, aov —
спинная и брюшная аорта,- Ьг —жаберный мешок; с и с' — верхнее и нижнее ребро; ей
гемальный канал; сп — нервный канал позвоночника; ст — осевой череп; d — мочеполовой
канал; g — половая железа; /Ьг — жаберные лепестки; med—головной мозг; п — спинной
мозг; г — почка; rad — луч плавника; вс — кардинальная вепа; vert — позвоночник; Ы —
боковая вена; vsf — подкишечнан вена.
Тело поделено по крайней мере на три отдела: голову, туловище
и хвост. В голове расположены головной мозг и важнейшие органы чувств,
защищенные черепом, рот и глотка с поддерживающим их висцеральным
скелетом (рис. 22). В туловище располагается общая полость тела с орга-
нами пищеварения, выделения и размножения. Хвост снабжен у рыб
сильной мускулатурой и является главным органом движения; у наземных
позвоночных он играет менее существенную роль. У наземных позвоночных
47
обособляется еше один отдел — шея, и голова вместе с тем приобретает
известную подвижность. На теле позвоночных имеются придатки, служа-
щие органами движения, — конечности. У рыб они представлены непар-
ными и парными плавниками. Непарные плавники расположены в сагит-
тальной плоскости (вертикально) и поделены обычно па 1—2 спинных,
Рис. 23. Схема скелета низшего позвоночного (рыбы).
а. Ъг. 3 — третья жаберная дуга; a. hy — подъязычная дуга; а. тп — челюстная дуга; а. п
и а. Л — верхние и нижние дуги позвоночника; а. pect — плечевой пояс; ch — хорда; с.
аи — слуховая капсула; с. ol — обонятельная капсула; сг. п — осевой череп; cr. v — вис-
церальный скелет; па, пс, nd — непарные (анальный, хвостовой, спинные) плавники; п. р,
п, v — парные плавники (грудной и брюшной); pelv — тазовый пояс; rad — лучи плавников.
хвостовой и анальный (рис. 21). Парные плавники, в числе двух пар,
располагаются по бокам тела; передняя пара, грудные плавники, лежит
тотчас позади головы, вторая пара, брюшные плавники, — обычно не-
посредственно впереди клоаки. У наземных позвоночных имеются две
пары пятипалых конечностей в виде сложных рычагов, служащих для
хождения.
Наружные покровы позвоночных состоят из эпителиальной
надкожицы — многослойного эпидермиса и подстилающей его волокнистой
кожи собственно, или дермиса.
с. аи pact
Рис. 24. Схема первичного скелета наземного позвоночного.
— челюстная дуга; с. аи, с. ol — слуховая и обонятельная капсулы; cr, п, cr. v — осе-
вой череп и висцеральный; I. а, е. р — передняя и задняя конечности; hy — подъязычный
аппарат; Ц — подвздошная кость; pect, pelv — плечевой и тазовый пояса; st — грудина.
Мускулатура тела представлена боковыми мышцами, поде-
ленными соединительнотканными перегородками на миомеры. У наземных
позвоночных метамерия теряется, и строение мускулатуры много сложнее.
Кроме этой париетальной мускулатуры, имеется еще и висцеральная
мускулатура челюстей и жаберного аппарата (или его производных).
Скелет позвоночных нередко представлен обширными кожными
окостенениями. Сверх того, всегда имеется внутренний скелет (рис. 23),
состоящий из хорды, идущей от середины головы (от подмозговой железки)
назад вдоль всего тела, и из более или менее развитого хрящевого или
костного скелета.
Вокруг хорды располагается осевой скелет, представленный в туло-
вище и хвосте позвоночником, который в простейшем случае состоит из
парного ряда метамерно расположенных дуг, ограничивающих спинномоз-
говой канал (рис. 23); в более развитом состоянии он состоит из метамер-
ного ряда позвонков с ребрами, тела которых более или менее вытесняют
48
хорду. В черепе различают два отдела — осевой и висцеральный. Осевая
часть черепа, представляющая собой как бы продолжение позвоночника,
имеет вид коробки, защищающей головной мозг и некоторые органы
чувств (органы обоняния и слуха). Висцеральная часть черепа состоит из
ряда висцеральных дут, из которых первая носит название челюстной
дуги, вторая — подъязычной, а следующие — жаберных. У наземных
позвоночных эти дуги подвергаются значительным преобразованиям.
Непарные плавники имеют свой скелет в виде ряда стержней — лучей
плавников. Парные плавники имеют скелет, в общем сходный со скелетом
непарных плавников, но он укрепляется в теле животного посредством
поясов конечностей — плечевого и тазового. У наземных позвоночных
скелет парных плавников преобразуется в скелет пятипалой' конечности
(рис. 24). У низших позвоночных внутренний скелет состоит из хряща;
у высших рыб и наземных позвоночных хрящ замещается костью.
Рис. 25. Схема нервной системы позвоночного (рыбы).
<11 — промежуточный мозг; gn — спинномозговые ганглии; hy — подъязычный нерв; I, о —•
обонятельная доля; т — средний мозг; ml — продолговатый мозг; mt — мозжечок; т. sp —•
спинной мозг; nl —боковой нерв; п. sp — спинномозговые нервы; pl. Ъг—плечевое сплете-
ние; t—передний мозг; 1 —X—головные нервы.
Центральная нервная система позвоночных (рис. 25) состоит
из двух, ясно различимых частей — головного мозга и спинного. Головной
мозг состоит из пяти отделов — переднего, промежуточного, среднего,
заднего и продолговатого мозга, содержащих соответствующие расширения
внутренней полости — желудочки. У высших позвоночных преоблада-
ющее развитие получает передний мозг. Периферическая нервная система
состоит из 10—12 пар головных нервов и из метамерного ряда спинно-
мозговых нервов. Из головных нервов часть представляет собой нервы
высших органов чувств (1-я, 2-я и 8-я пара нервов), часть представляет
собой чисто двигательные нервы (3-я, 4-я , 6-я и 12-я пары нервов), а все
остальные представляют собой смешанные нервы, снабженные ганглиями
и содержащие, кроме чувствующих волокон, и двигательные для мышц
висцерального аппарата (5-я, 7-я, 9-я, 10-я и 11-я пары нервов). Спинно-
мозговые нервы отходят в каждом сегменте тела с каждой стороны двумя
корешками: одним — брюшным, чисто двигательным, для париетальной
мускулатуры, и другим — спинным корешком, снабженным ганглием,
содержащим смешанные волокна (кроме чувствующих, еше и двигатель-
ные) для висцеральной мускулатуры кишечника, сосудов и пр. У всех
позвоночных, за исключением миног, оба корешка соединяются вместе
для образования смешанного нерва. Значительная часть висцераль-
ных нервов соединяется в особое сплетение, образующее симпа-
тическую нервную систему, обладающую и своими особыми нервными
узлами.
Органы чувств у позвоночных очень высоко развиты. Кроме
49
свооодных окончании нервов, в покровах у высших позвоночных имеются
особые осязательные пятна и тельца. Органами Вкуса являются концевые
почки, которые располагаются главным образом в ротовой полости.
У живущих в воде низших позвоночных имеются еще особые органы
боковой липни (сейсмосенсорные органы), служащие для восприятия
грубых колебаний и движения воды. У всех позвоночных имеются парные
органы обоняния (сросшиеся у круглоротых в непарный орган) в виде
мешковидных полостей со складками чувствующего эпителия. Парный
орган зрения построен очень сложно. Глаза- имеют вид полого шара,
стенка которого изнутри выстлана нервным и воспринимающим аппара-
том — ретиной, а снаружи состоит из скелетной оболочки — склеры.
Внутренняя полость шара выполнена прозрачным стекловидным телом,
а в передней ее части помещается особое светопреломляющее сферическое
тело — хрусталик. Парное ухо представляет собой комбинацию органа
Рис. 26. Схема внутренних органов позвоночного (рыбы).
с— сердце; ch—хорда; cl—клоака; с. о—ротовая полость,- ср—околосердечная полость; сгЬ—го-
ловной мозг; с»—брюшная полость; de—желчный проток; д— половая железа; ga—желудок;
hep—печень; int—кишка; т. ер— спинной мозг; od—яйцевод; oes — пищевод; р—плава-
тельный пузырь; ph — глотка; рп—поджелудочная железа; г—почка; spl—селезенка; (Л—
щитовидная железа.
слуха с органом равновесия и состоит из перепончатого мешочка с тремя
полукружными каналами, наполненными жидкостью с плавающими в ней
отолитами. Это внутреннее ухо дополняется у наземных позвоночных
добавочным аппаратом для передачи звуковых колебаний, для чего служит
система слуховых косточек, расположенных в полости среднего уха.
Полость тела (целом) выстлана брюшиной и поделена на
передний отдел — околосердечную полость, в которой помещается сердце,
и задний — брюшную полость, в которой располагаются все важнейшие
внутренние органы (рис. 26). Брюшная полость не вполне поделена бры-
жейками (мезентериями) на правую и левую половины. У млекопитающих
развивающаяся диафрагма отделяет еще особый грудной отдел от собствен-
но брюшного.
Органы пищеварения и дыхания начинаются рото-
вой полостью, выстланной многослойным эктодермальным эпителием.
Для захватывания пищи рот вооружен очень твердыми образованиями —
зубами. Ротовая полость ведет в глотку (рис. 26), боковые стенки которой
прорезаны рядом жаберных щелей, ведущих наружу, образуя иногда
заметное расширение — жаберный мешок. Жаберные перегородкп между
последовательными жаберными щелями поддерживаются скелетными
жабернымп дугами и несут на поверхностях, обращенных внутрь жабер-
ных люшков (передней и задней), особые сложные складки слизистой
оболочки — жабры. У наземных позвоночных жабры исчезают, а в* ка-
честве органов дыхания имеются наполняемые воздухом мешковидные
выросты глотки — легкие. Глотка ведет в несколько суженный отдел —
50
Пищевод, за которым следует обыкповсппо расширенный желудок и затем
собственно кишка. Кпптсчпик постоянно снабжен двумя крупными пи-
щеварительными железами: печенью п поджелудочной железой (рис. 26).
Кровеносная система состоит пз замкнутой системы
сосудов, сводящихся в общем к продольному брюшному сосуду, на пути
которого имеется сердце и в котором кровь движется вперед, и к спинному
сосуду, в котором кровь течет назад. Сердце низших позвоночных состоит
из четырех последовательных отделов, разделенных клапанами: венозной
пазухи, предсердия, желудочка и артериального конуса. Оно несет исклю-
чительно венозную кровь, которая направляется вперед в брюш-
ную аорту (рис. 27); от последней направо и палево отходят в жаберные
перегородки приносящие жаберные артерии, несущие кровь в жабры,
откуда она по окислении собирается в выносящие жаберные артерии,
соединяющиеся затем в непарную спинную аорту (или сразу, пли сначала
Рис. 27. Схема кровообращения низшего позвоночного (рыбы).
а. с — сонная артерия; a. caud — хвостовая артерия; а. сое—нишечпая артерия; aff—прино-
сящие жаберные артерии; a.g — половая артерия; a. И — подвздошная артерия; ао. d —спин-
ная и ао. о— брюшная аорты; a. mes — брыжеечная артерия; а. г — почечная артерия; а.
fd — подключичная артерия; d. с—шовьеров проток; eff— выносящие жаберные арте-
рии; о. с. а —передняя и v. с. р— задняя кардинальные вены; v. caud — хвостовая всна;р.
h — печеночная вена; v. I — боковая вена; v. р. h — воротная вена печени; v, р. г — ворот-
ная вена почехх; v. scl — подключичная вена.
в парные наджаберные сосуды) (рис.. 27). Последняя отдает парную ветвь
в голову (сонные артерии) и затем идет назад непосредственно под хордой,
снабжая по пути артериальной кровью все органы брюшной полости.
Венозная кровь от кишечника собирается в крупную вену (рис. 27),
которая направляется в печень, где опа распадается на капилляры, обра-
зуя воротную систему печени. Из печопи кровь по печепочпой вене направ-
ляется в венозную пазуху сердца. Из хвоста кровь течет по хвостовой
вене к почкам, где эта вена распадается на капилляры, образуя воротную
систему почек. От почек кровь собирается в парные стволы, лежащие по
бокам спинной аорты, — задние кардинальные вены, собирающие крогь
от боковых стенок тела. Из головы кровь возвращается по парной перед-
ней кардинальной воне (яремной). Обе пары кардинальных вей, соеди-
няясь вместе над сердцем, образуют парные кювьеровы протоки, впада-
ющие в венозную пазуху.
У наземных позвоночных (рис. 28), в связи с исчезновением жабер-
ного дыхания, одна пара жаберных артерий преобразуется в парную
дугу аорты, несущую кровь из брюшной аорты непосредственно в спип-
ную, а другая пара преобразуется в легочные артерии, несущие ве-
нозную кровь для окисления в легкие. Артериальная кровь возвраща-
ется по легочным венам назад в сердце, и таким образом устанавливается
второй, «малый», круг кровообращения. Сердце получает теперь, кроме
венозной, и артериальную кровь, и для разделения этих двух токов в
его основных двух камерах, сначала в предсердии, а затем н в желудочке,
Я
развиваются перегородки. Этими перегородками сердце разделяется на
левую, артериальную, и правую, венозную, половины. Из «двукамерного»
сердце становится «четырехкамерным».
В венозной системе также происходят изменения: задние кардиналь-
ные вены замещаются у наземных позвоночных непарной нижней полой
веной (рис. 28, Р. с. I.). У высших позвоночных (птиц и млекопитающих)
воротная система почек исчезает и исчезает одна из дуг аорты.
Органы выделения позвоночных представлены парными,
довольно компактными органами — почками, состоящими из большого
числа более или менее извитых канальцев (нефридиев), открывающихся у
низших позвоночных в’оронками в полость тела. Канальцы связаны каж-
дый с фильтрационным аппаратом в виде камеры, заключающей клубо-
Рис. 28. Схема кровообращения наземного позвоночного.
а. ао — дуга аорты; а. с. в — наружная и а. с. i — внутренняя сонные артерии; a. caud —
хвостовая артерия; а. сое — кишечная артерия; a. g. — половая артерия; а. I —подвздошие
артерия, а. тез — брыжеечная артерия; ао. d — спинная и ао. v — брюшная аорты; а. р —
легочная артерия; а. г — почечная артерия; a. scl — подключичная артерия; к. а — брюш-
ная вена; v. caud — хвостовая вена; о. с. а — передняя и о. с. р — задняя кардинальные ве-
ны; к. с. г —задняя полая вена; в. р — легочная вена; v. р. h — воротная вена печени; к. р.
г — воротная вена почек.
чек кровеносных сосудов, и изливают свое выделение в парные протоки,
впадающие обыкновенно в клоаку.
Половые органы представлены парными половыми желе-
зами (яичниками или семенниками), выводные протоки которых разви-
ваются в связи с органами выделения и обычно впадают в клоаку.
Кроме того, у позвоночных всегда имеются еще особые беспроточныо
железы — органы внутреппей секреции К таким орга-
нам относятся непарные подмозговая и надмозговая железки, щитовид-
ная и зобная железы, а также околопочечные тела.
Классификация позвоночных
Подтип позвоночных делится на следующие шесть классов: бесчелюстные (Ag-
natha), рыбы (Pisces), земноводные (Amphibia), пресмыкающиеся (Reptilia), птицы
(Aves) и млекопитающие (Mammalia). Первые два класса объединяют водных позво-
ночных, остальные — наземных. Принимают и некоторые более крупные подразделе-
ния. Так, первому классу бесчелюстных позвоночных (Agnatba) противополагается
все прочие — челюстные позвоночные (Gnathostomata, челюстноротые). Первые три
класса — бесчелюстных, рыб и земноводных — объединяют в группу Anamnia по
отсутствию у них амниона и других зародышевых оболочек, кроме яйцевых, проти-
вополагая их всем высшим — пресмыкающимся, птицам и млекопитающим, составляю-
щим группу Amniota, у которых такие оболочки имеются. Иногда рыб и амфибий
объединяют под названием ichthyopsida (рыбообразных), а рептилий и птиц — под
названием Sauropsida (ящерообразных).
I класс Agnatha. (бесчелюстные) характеризуется отсутствием челюстей, непарной
ноздрей, отсутствием парных конечностей и типичными жаберными мешками с мери-
диональным расположением внутренних жаберных лепестков (Entobranchiata). Скелет
хрящевой. Хорда постоянная. Сюда относятся в современной фауне рыбообразные
животные с удлиненными цилиндрическим телом и ртом в виде присасывательной
52
воронки, круглоротые (Cyclostomata), т. е. миноги и миксины (Petromyzontes, Myxi-
noidei). Кроме того, к бесчелюстным относятся ископаемые панцырные, или щитковые
Ostracodermi).
II класс Pisces (рыбы) характеризуется присутствием парных и непарных плавни-
ков. Жаберные лепестки сидят снаружи на жаберных дугах (Ectobranchiata). Сердце
венозное, «дву камерное*. В коже обычно имеются дентиновые или костные чешуи.
Сюда относятся в качестве подкласса Elasmobranch!! (сростножаберные) с хря-
щевым скелетом (Chondrichthyes— хрящевые рыбы) и дентиновыми чешуями в коже.
Жаберные щели обычно открываются непосредственно наружу, и впереди нормальных
щелей имеется рудиментарная щель — брызгальце. В сердце имеется артериальный
конус с несколькими рядами клапанов и в кишке складка — спиральный клапан.
Подкласс включает селахий (Selacbii), или поперечноротых, т. е. акул (Squaloidei)
и скатов (Batoidei) и цельноголовых (Holocephala), т. е. химер и близкие к ним формы.
У всех остальных рыб имеется настоящая кость (Osteichthyes—костные рыбы). В ка-
честве второго подкласса сюда относятся Teleostomi (завершенноротые), обладающие
уже отчасти костным скелетом и вторичными костными челюстями. Жаберные щели
прикрыты жаберной крышкой. В подкласс входят хрящевые ганоиды (Chondrostei)
с современными осетровыми рыбами, костные ганоиды (Holostei), кистеперые ганоиды
(Crossopterygii), многоперые (Polypteroidei) и костистые рыбы (Teleostei). К последним
относится громадное большинство современных рыб, не имеющих ни брызгальца, ни
артериального конуса, ни спирального клапана. Третий подкласс Dipnoi (двудышащие)
содержит в современной фауне лишь несколько видов, характеризующихся, кроме жа-
берного дыхания, еще и легочным и соответствующими изменениями в кровообра-
щении.
III класс Amphibia (амфибии, или земноводные) характеризуется личиночным
дыханием посредством жабер, в то время как взрослое животное дышит легкими (и
кожей). Сердце трехкамерное, несущее слева артериальную и справа венозную кровь.
Конечности пятипалого типа. У современных форм кожа голая. Сюда относятся хвоста-
тые амфибии (Urodela), включающие саламандр, тритонов и т. п., бесхвостые амфибии
(Anura), содержащие лягушек и жаб, и наконец гимнофионы, или червяги (Gymnophio-
na s. Apoda), безногие змееобразные животные, ведущие подземный образ жизни.
Кроме того, к классу амфибий относятся ископаемые стегоцефалы (Stegocephali),
имевшие иногда панцырь из костных чешуй.
IV класс Reptilla (рептилии, или пресмыкающиеся) характеризуется роговым
покровом ив чешуй или щитков. Дышат исключительно легкими. Сердце трех-, четы-
рехкамерное с обеими дугами аорты. Сюда относятся черепахи (Chelonia) с коротким
телом, защищенным панцырем, состоящим из спинного и брюшного щита, ископаемые
котилозавры (Cotylosauria), имевшие еще некоторые черты, общие со стегоцефалами,
и ископаемые вверезубые (Theriodontia), приближавшиеся к млекопитающим. Эти
формы объединяются вместе с некоторыми другими ископаемыми в группу Synapsida,
характеризующуюся присутствием одной только скуловой дуги (как у млекопитающих).
Ко второй группе Diapsida, характеризующейся развитием двух височных дуг (которые
впоследствии могут и исчезнуть), относятся хоботоголовые (Rhynchocephalia), очень
примитивные ящерицеобразные рептилии с единственным современным представителем
гаттерией (Hatteria), крокодилы (Crocodilia) и многие ископаемые формы, как дино-
завры, птерозавры и др. Несколько обособленное положение занимают чешуйчатые
(Squamata), объединяющие ящериц и змей.
V класс Aves (птицы) характеризуется видоизменением передних конечностей
в крылья, общим покровом из перьев и четырехкамерным сердцем с одной правой
дугой аорты. Температура тела постоянная. К птицам относятся: ископаемые Archae-
ornithes с единственными представителями археоптериксом и археорнисом, которые
обнаруживают известные черты сходства с рептилиями; бескилевые птицы (Ratitae),
куда относятся современные страусы, казуар и киви, утратившие способность летать,
и килевые птицы (Carinatae), к которым относятся все прочие птицы.
VI класс Mammalia (млекопитающие) характеризуется покровом из волос,
присутствием млечных желез и четырехкамерным сердцем с одной левой дугой аорты.
Температура тела постоянная. Сюда относятся яйцекладущие клоачные (Monotremata),
современными представителями которых являются только ехидна и утконос, сумчатые
(Marsupialia), живородящие млекопитающие, которые донашивают своих детенышей
в особой сумке, и все типичные звери (Eutheria), обладающие плацентой (Placentalia).
Последние содержат следующие отряды: насекомоядные (Insectivora), наиболее при-
митивная группа, включающая ежей, кротов, землероек; рукокрылые (Chiroptera),
куда входят летучие мыши; шерстокрылы (Galeopitheci); неполнозубые (Edentata),
включающие ленивцев, муравьедов, броненосцев и панголина; грызуны (Rodentia)
с зайцами, белками, бобром, сонями, тушканчиками и мышами; креодонты (Creodontia)
ископаемая группа примитиввых хищников; хищные (Carnivora): медведи, волки,
куницы, выдры, кошки и гиены, а также и водные: котики, моржи и тюлени; китообраз-
ные (Cetacea) с зубатыми китами, как кашалоты и дельфины, и беззубыми китами;
кондилартры (Condylarthra), ископаемая группа примитивных копытных, непарно-
копытные (Perissodactyla), включающая тапиров, носорогов, лошадей; парнокопытные
(Artiodactyla) с бегемотом, свиньями и жвачными, к которым относятся: верблюды,
53
позвоночные) Ichthyopterygia
Таблица 1
Клазсы Подклассы Отряды 11 подотряды
Л n a m n i a Agnatha (бесчелюст- ные) Pisces (рыбы) Amphibia (земно- водные) Reptilia (пресмы- кающиеся) ( I Ostracodermi -j- < Cyelosiomata Arthrodira 4- Elasmobranchii ( Actino- Teleostomi )Р”ГУ& " Crosso- {pterygii Dipnoi Stegocophali 4- , • Liss'jmphibia < Anapsida -J- Synapsida < f Anapspida 4- Osteostraci 4- Heterostraci 4- Potromyzontes Myxinoidei Arthrodira 4- Acanthodei4- Pleuropterygii 4* Ichthvotomi 4- { IIS’S" Holocephali Chondrostei Polypteroidei Holostei Teleostei Crossopterygii Monopneumones Dipneumones Temnospondyli 4- Lepospondyli — Stereospondyli 4- Phyllospondyli 4- Gymnophiona Urodela Anura Cotylosauria — Pelycosauria 4- rr, . „ , f. Theriodontia -f- iheromorpha , jjiCyno^ontia 4- Chelonia Ichthyosiuria — Sa uro pterygia -j- Piacodontia -4 I Protorosauria 4- Prosauna Rhynchoceptlalia Бесщитковые Костнощитковые Разнощптковые Миноги Млксины II ан цы рн ые акул с о б р аз и ы е Акулы Скаты Целы;оголовые, химеры Хрящэкостные Миогоиерые ганопды Костные ганоиды Костистые рыбы Кистеиерые Однолег очные двудышащпе Двулегочные двудышащие Темпоопондильные стегоц 'фалы Микровавры Лабиринтодонты Бранхиозавры Гимпсфионы, червяги Хвостатые амфибии Бесхвостые амфибии Котилозавры Пеликозавры Териодонты (зверог.убы.с; Дицинодоиты Черепахи Ихтиозавры Плезиозавры Плоскозубые Первичные ящеры Х.сботогологые
Tetrapoda s. Cheiropterygia (наземные
Diapsidn
a n i о t a Aves (птицы) Parapsida Archaeorn Seornithes P rototheri Meta- llic ria " 'thes a Marsu- pial ia
< Mammalia (млеко- питающие) < Eutheria s. Placen- talia Eden- tata
Ungu- lata
Pterosauria -}
Pseudosuchia -f-
Parasuchia -j-
Crocodilia
_i_ J Saurischia 4-
Dinosauna + | Qrnithischia +
Rhamphoryn-
choidea —
Pterodacty-
loidea -p
Чап am at a / Lacertilia
bquamata | Ophidicl
Saururae -|-
Ratitae
Carinatae
Allotheria — , Monotremata
Triconodonta
Polyprotodontia
Diprotodontia
Pantothsria -j-
Insectivora
Chiroptera
Galeopitheci
Creodontia+ { psC™ rtodi +
Carnivora {
( Arch-ieoceti
Cctacea < Odontoceti
( Mystacoceti
Rodentia
Xenarthra
Pholidota
Condylarthra —
Artiodactyla
Perissodactyla
Amblypoda
Proboscides
Sirenia
Hyracoidea
Tubulidcnt ata
r, . . I Pros i mi is
Primates |
I Псевдозухпп (ложные крокодилы)
Паразухип
Крокодилы
Ящэротазые динозавры
1 Пгицетазые динозавры
Рамфоринхи
Птеродактили
Ящерицы
Змеи
Ящерохвостые птицы
I Бескилевые птицы
I Килевые птицы
Клоачные млекопитающие
Первичные сумчатые
Плотоядные сумчатые
Растительноядные сумчатые
Первичные плацентарные
Н юекомоядные
I Рукокрылые
I Шерстокрылы
Креодонты
Хищные
Ластоногие
Первичные киты
Зубатые киты
Беззубые киты
Грызуны
Неполнозубые
Панголины
! Первичные копытные
Парнокопытные
Непарнокопытные
Амблпподы
Слоны
Сирены
! Даманы
’ Трубксзубы
( Полуобезьяны (лемуры)
i Обезьяны
олени, антилопы, быки, козы, бараны и жирафы; даманы (Hyracoidea); слоны (Probos-
cidea); сирены (Sirena ), включающие морских дюгоней и ламантинов; полуобезьяны,
или лемуры (Prosimiae), — группа очень примитивных обезьян; обезьяны (Simiae),
к которым относятся «широконосые» обезьяны Нового Света — игрунки (уистити)
и ревуны, «узконосые» обезьяны Старого Света — мартышки, павианы и высшие
«человекообразные» бесхвостые обезьяны — гиббон, орангутан, горилла и шимпанзе.
Классификация позвоночных, принятая в этой книге, представлена
на табл. 1.
Эмбриональное развитие позвоночных
Зрелое яйцо позвоночных одето обыкновенно тонкой прозрачной
оболочкой, пронизанной многочисленными порами (zona radiata в. pel-
lucid а). Если эта яйцевая оболочка отличается значительной плотностью
и проникновение сперматозоида через пору было бы невозможно, то для
этого имеется особое отверстие (micropyle), что наблюдается у круглоро-
тых, ганоидных и костистых рыб. Кроме такой первичной оболочки, раз-
вивающейся еще в яичнике, обыкновенно имеются также вторичные оболоч-
ки — продукты выделения железистых стенок яйцеводов, которыми яйцо
Рис. 29. Полное и неравномерное дробление лягушечьего яйца (сбоку);
цифры обозначают число шаров дробления в каждой стадии [(по Эккеру
из Гертвига).
окружается во время прохождения вдоль последних. К таким оболочкам
относятся: студенистая оболочка рыб и амфибий, «белок> яйца селахий,
рептилий и птиц, «роговая> скорлупа селахий, кожистая скорлупа реп-
тилий и пропитанная известью скорлупа некоторых рептилий и всех птиц.
Количество желтка в яйце позвоночных весьма различно; отчасти в
зависимости от этого меняется и величина яйца. Яйцо млекопитающих
очень мало и не содержит заметного количества желтка; наоборот, огром-
ные яйца селахий, многих рептилий и птиц заключают очень большой
запас желтка. Желток всегда концентрируется на одном, вегетативном
полюсе, который, вследствие большего его удельного веса, поворачива-
ется вниз.
Характер дробления зависит от количества и распределения желтка
в яйце. У миног, у двудышащих рыб и у большинства земноводных желтка
в яйце сравнительно мало («голобластические» яйца), и дробится оно
по типу полного и неравномерного дробления. Первые две плоскости
дробления лежат вертикально (по главной оси) и делят яйцо последова-
тельно на два и четыре бластомера одинаковой величины. Третья плоскость
дробления лежит горизонтально и отделяет четыре меньшие клетки ани-
мального полюса от четырех больших клеток вегетативного, и эта разница
в величине клеток верхнего и нижнего полюсов во время дальнейшего
хода дробления не уменьшается (рис. 29). В результате дробления разви-
56
вается довольно типичная бластула, стенка которой неравномерна,
соответственно чему сегментационная полость сдвинута ближе к анималь-
ному полюсу (рис. 31); У ганоидных рыб дробление вегетативного полюса
так сильно запаздывает, что пока первые четыре борозды доходят до конца,
на анимальном полюсе уже успевает развиться множество мелких клеток.
Здесь наблюдается как раз переход от полного дробления к частичному,
Рис. 30. Три стадии дробления куриного яйца (по Келликеру). Изображен тольк
зародышевый диск.
дискоидальному, которое характерно для большинства позвоночных.
Яйца миксин, селахий, костистых рыб, гимнофион, рептилий и птиц при-
надлежат к типу «меробластических> яиц, в которых дробится лишь срав-
нительно бедная желтком протоплазма анимального полюса (рис. 30),
а’перегруженная желтком большая часть яйца совсем не дробится. В ре-
зультате дробления на анимальном полюсе образуется так называемый
зародышевый диск, под которым развивается и щелевидная сегментаци-
Рис. 31. Бластула и гаструляция яйца тритона (по Гертвигу).
ап — апикальный полюс; Ьр — бластопор; d — дорзальная губа бластопора; cd — эктодерма;
ent— эндодерма; g — гастральная полость; mes—зачаток мезодермы; р — желточная пробка;
в—сегментационная полость; v—вентральная губа бластопора; veg—-вегетативный полюо.
онная полость (рис. 33, Л). Таким образом, и здесь различима стадия бла-
стулы с сильно сдвинутой и сжатой полостью дробления внутри, но в то
время как верхняя ее стенка состоит из клеток зародышевого диска, ниж-
няя представлена огромной массой нераздробившейся части яйца, так
называемого «желтка>. Последняя содержит, однако, кроме переполнен-
ной желтком протоплазмы, также и ядра, которые в частях, прилежащих
к сегментационной полости, довольно энергично делятся и в дальнейшем
принимают участие в построении зародыша, а также в передаче ему пи-
тательного материала. Такие клеточные элементы разного, впрочем, зна-
чения носят название мероцитов.
57
У низших, клоачных млекопитающих дробление также частичное
и дискоидальное, но у высших, в связи с их живородностью, развивается
питание зародыша непосредственно за счет матери, и запас желтка в япце
исчезает. Заметное количество желтка еще наблюдается в яйце у сумчатых,
по и у них уже дробление вернулось к типу полного и равномерного,
которое характерно для всех типичных млекопитающих. В результате
этого дробления развивается первоначально плотная кучка клеток, ко-
торая затем путем их расхождения и выделения жидкости в образовавшу-
юся таким образом внутреннюю полость превращается в пузырь — бла-
стулу.
Рис. 32. Схематические медиальные разрезы через зародыши
тритона на последовательных стадиях гаструляции, иллюстрирую-
щие ход перемещения клеточного материала.
Черным обозначен материал будущей нервной пластинки, белым —эпидер-
миса, мелкими редкими точками — энтодермы, мелкими густыми точка-
ми — мезодермы и крупными точками — хорды. Крестиком отмечен ма-
териал вегетативного полюса. Стрелки показывают направления дви-
жений (по Фогту из Балинского).
Образование первых двух зародышевых листков идет путем г астру-
ля ц и и, которая наиболее типично выражена у низших позвоночных
с голобластическими яйцами, как, например, у земноводных. У послед-
них наблюдается при этом ясно выраженная инвагинация, т. е. впячи-
вание части клеток внутрь полости бластулы (рис. 31).
Это впячивапие происходит, однако, не на вегетативном полюсе, а В
некоторой пограничной зоне по экватору, в задней части яйца. Впячнва-
пие идет наиболее энергично в области спинной губы бластопора (рис. 31,
d). Ввертывающиеся здесь внутрь клетки экваториальной зоны бластулы
ложатся затем под клетками анимального полюса и образуют крышу
первичной кишки, или гастральную энтодерму (называемую также эпто-
мезодермой или хордо-мезодермой, так как она содержит материал будущих
зачатков хорды и мезодермы).
58
Образуется и гастральная полость. Крупные бластомеры вегетатив-
ной половины не вворачиваются внутрь, по поворачиваются па месте, так
что их первоначальная парулшая поверхность обращается внутрь га-
стральной полости, т. о. в конце концов они занимают такое же положение,
какое должны были бы занять, если бы произошла типичная инвагинация
(рис. 32). Клетки пограничной зоны все более надвигаются на клетки ве-
гетативного полюса и начинают их обрастать. Своими краями они обра-
зуют как бы кольцевую складку, которая постепенно под ней стягивается.
Таким образом, гаструляция идет здесь одновременно и путем инвагинации
и путем эпиболии. Край обрастания, который обозначает границу между
обрастающими анимальными и обращаемыми вегетативными клетками,
представляет собой край бластопора, т. е. его губы. Вспомним, что губы
бластопора представляют собой место перехода клеток эктодермы в клетки
энтодермы. Таким образом, бластопор имеет форму кольца, но кольцо
это не ограничивает отверстия, а окружает массу вегетативных клеток.
По мере того как кольцо постепенно стягивается под клетками вегета-
5
Рис. 33. Две последовательные стадии гаструляции у селахий. Сагиттальный разрев
через зародышей акулы (по Циглеру).
Ьр — бластопор; ecl — энтодерма; ent — энтодерма; g — гастральная полость; s — полость
дробления.
тивного полюса, оно принимает в то же время все более резкие очертания.
Выступающие между губами бластопора набитые желтком клетки веге-
тативного полюса известны под названием желточной пробки. Наконец,
губы бластопора вполне сходятся, покрывая собой желточную пробку.
Бластопор получает при этом вид продольной щели (сравнимой с «пер-
вичной бороздой»). Верхняя часть этой щели впоследствии закрывается
нервными складками ц превращается в нервно-кишечный канал (сапаИн
neuroenterici.’s), а нижняя ее часть дает начало заднему проходу (рис. 34)..
Нередко гаструляция у амфибий осложняется тем, что сегментационная
полость пе исчезает, а гастральное впячиванне прорывается в нее, и таким
образом обе полости сливаются в одну. Вегетативные клетки затем пере-
мещаются по внутренней стенке и выстилают дно полости, которая после
этого становится идентичной первичнокпшечпой полости. Как бы то пи
•было, но мы имеем здесь, несомненно, процесс измененной инвагинации,
соединенной с эпиболией, в результате которого однослойный зародыш
превращается в двухслойный: он покрыт снаружи эктодермой и содержит
внутри одну гастральную полость, выстланную энтодермой и сообщаю-
щуюся с наружной средой посредством отверстия — бластопора. Таким
образом, стадия гаструлы здесь ясно выражена.
По существу таким же образом протекает гаструляция и у селахий.
На заднем крае зародышевого диска происходит типичная инвагинация
(рис. 33), между тем как весь остальной диск своими краями постепенно
обрастает нераздробленяую часть яйца («желток»). И здесь, кроме инва-
гинации, наблюдается и эпиболия. Образование гастральной полости про.
59
текает вполне типично, и с ее разрастанием сегментационная полость
вытесняется. Нижней стенкой гастральной полости первоначально яв-
ляется нераздробленный «желток>.
У высших позвоночных (амниот) дело происходит немного сложнее.
Посредине зародышевого диска наблюдается утолщенная область, нося-
щая название зародышевого щита. В задней части этого зародышевого
шита образуется небольшая продольная «первичная бороздка> на утол-
щенной полосе бластодермы или «первичной полоске>. При образовании
энтодермы эта полоска оказывается местом перехода эктодермальных кле-
ток в энто-мезодермальные, образующиеся путем непрерывного переме-
щения клеточных масс снаружи внутрь (рис. 39). На переднем конце пер-
вичной бороздки происходит настоящая инвагинация, приводящая к
образованию гастральной полости и покрывающей ее сверху энто-мезо-
дермы. Инвагинация не происходит лишь у млекопитающих, у которых
Рис. 34. Сагиттальный разрез через зародыш амфибии (по Кейбелю).
ап — анальное отверстие; сп — нервнокишечный канал; ch — хорда; сое/—
полость тела; ер — эктодерма; int — кишка; т — мезодерма; п — нерв-
ная трубка.
процесс гаструляции наиболее изменен. В этом случае наблюдается, од-
нако, врастание сплошно й массы клеток, представляющей собой такую
же гастральную энтодерму (энто-мезодерму).
На этой стадии развития первичная бороздка высших позвоночных
обнаруживает известное сходство со стадией сомкнутого щелевидного
бластопора зародыша амфибий. Таким образом, можно сказать, что пер-
вичная бороздка обладает признаками бластопора, а первичная полоска
представляет собой его сомкнутые губы.
Однако у высших позвоночных наблюдается и процесс обрастания
«желтка> краями зародышевого диска, но этот процесс происходит позже
и независимо от гаструляции. Следовательно, у высших позвоночных
наступило разделение одного связного процесса (инвагинации, соединен-
ной с эпиболией) на два независимых.
Бластопор, развивающийся на месте инвагинации, превращается
и здесь при замыкании нервных складок в нервно-кишечный канал. Однако
не вся энтодерма образуется путем инвагинации. Часть энтодермы обра-
зуется под зародышевым диском на поверхности желтка и носит название
60
желточной энтодермы. Здесь имеется и Полость, но это —
не гастральная полость, а сегментационная. Когда гастральное впячи-
вание достигает этой полости, оно прорывается в нее, и гастральная по-
лость соединяется с сегментационной. Дно этой соединенной полости об-
разовано нераздробленной массой желтка, которая содержит на своей
поверхности многочисленные клеточные элементы, идущие в дальнейшем
на образование энтодермы. Энтодерма кишечника образуется за счет жел-
точной энтодермы, а хорда и мезодерма — за счет гастральной. В этом
процессе образования энтодермы нельзя не видеть дальнейшего измене-
ния в том же направлении, какое намечается уже у амфибий, когда га-
стральная полость прорывается в сегментационную, а часть энтодермы
образуется простым
поворачиванием и
перемещением кле-
ток вегетативного по-
люса.
По сравнению с
ланцетником процесс
образования первых
зародышевых лист-
ков позвоночных
Рис. 35. Стереограммы переднего конца зародышей амфи-
бий на двух последовательных стадиях развития: Разви-
тие нервной системы, хорды и мезодермы.
ch — хорда; сое/ — полость тела; ect — эктодерма; ent — энто-
дерма; int — кишка; тл — мезодерма; п — нервная трубка; пр—
нервная пластинка.
которая дает начало эпителиальной стенке кишеч-
оказывается, таким
образом, довольно
сильно измененным.
Большая часть эн-
тодермы диференци-
руется на месте, и это
именно та энтодерма,
ника и ее производным. Другая, более активная, часть первичной энто-
дермы, образующаяся путем инвагинации (гастральная энтодерма), хотя
и входит иногда на короткое время в состав стенок первичной кишки,
образуя ее крышу, в дальнейшем диференЦируется, однако, только на
хорду и мезодерму. У высших позвоночных образование последних частей
идет быстрее, процесс инвагинации обособляется от остальных гаструля-
ционных процессов (образование желточной энтодермы, эпиболия) и пред-
ставляет собственно процесс непосредственного образования совместного
зачатка хорды и мезодермы. Так как этот материал крыши первичной
кишки (т. е. часть первичной энтодермы) представляет совместный зачаток
хорды и мезодермы, то его нередко и обозначают как «хордомезодерма»
(или, по происхождению, как «энтомезодерма»).
После образования энтодермы собственно, а также хордомезодермы,
начинается дальнейшая диференцировка в пределах последней. Полоса
клеток, лежащая посредине вдоль спинной стороны крыши первичной
кишки (т. е. хордомезодермы), утолщается и затем отшнуровывается.
Это — зачаток хорды (рис. 34, 35).
Образование мезодермы, которое у ланцетника происходит путем
обособления части крыши первичной кишки по бокам зачатка хорды, у
позвоночных происходит иначе. Хотя зачаток мезодермы после обособле-
ния занимает совершенно такое же положение по бокам зачатка хорды
(рис. 35), но он образуется относительно раньше, чем у ланцетника,
именно во время гаструляции, а не после ее окончания. С этим связаны
и различия первой ее диференцировки. В то время как у ланцетника
мезодерма появляется в виде выступов на уже сформировавшейся первич-
ной кишке, у позвоночных она выявляется одновременно с гаструляцией
вокруг бластопора и особенно по бокам от спинной губы бластопора.
Гаструляционными движениями этот клеточный материал выносится впе-
ред и ложится впереди бластопора в крыше первичной кишки, по бокам
61
зачатка хорды, в виде* парной массы, напоминающей складки (рис. 35).
Наиболее ясно все это видно у амфибий.
Позади бластопора обе складки переходят одна в другую, так что он
вполне окружен развивающейся мезодермой, и здесь последняя переходит
с одной стороны в энтодерму, а с другой стороны — в эктодерму, так что
Рис. 36. Поперечгые разрезы через зародышей селахий (по Кейбелю).
Л — образование осевой и краевой [ мезодермы; В — их соединение; ch — хорда;
ect — эктодерма; ent — энтодерма; mes — мезодерма; mg — осевая мезодерма; тр —•
краевая мезодерма; п — нервный желобок.
блазтопор оказывается местом соединения всех трех зародышевых листков,
без видимых границ переходящих здесь друг в друга (рис. 38, Л).
У селахий наблюдается образование мезодермы в' виде кольцевой
сю ауки по краю зародышевого диска. В области инвагинации эта краевая
ect
Рис. 37. Поперечные разрезы через зародышевы" диск ящерицы (Gecco)
(по Биллю из Шимкевича)!
А — более ранняя стадия; В — более поздняя; ect — эктодерма (не затушевана); ent —-жел-
точная энтодерма; ch — зачатрк хорды; rn.es—зачаток мезодермы; nisp — но’кно-волокиист» й
и rn.se— лишенно-волокнистые листки ес.
мезодерма переходит внутрь и ложится в виде как бы складок, по бокам
от зачатка хорды, образуя так называемую осевую мезодерму (рис. 36).
Так как края зародышевого диска селахий сравнимы с губами бластопора,
то, следовательно, здесь наблюдается в сущности то же самое, что и у
амфибий. Губы бластопора являются и здесь местом образования и соеди-
нения всех трех зародышевых листков.
Точно так же и у высших позвоночных происходит образование
мезодермы в области первичной полоски в виде парной массы врастающих
внутрь клеток (рис. 38), которая перемещается вперед и ложится по
бокам хорды, образуя здесь зачаток осевой мезодермы (рис. 37). Это дает
62
нам лишнее доказательство в пользу того, что первичная полоска в дей-
ствительности представляет собой сомкнутые губы бластопора, в которых
Рис. 38. Сравнение образования мезодермы в области бласто.пора
у амфибий (Л) и в области первичной полоски у млекопитающих
(В) (по Кейбелю).
Ьр — бластопор; ms — мезодерма; рг — первичная бороздка.
соединяются все три зародышевых листка. Из сказанного видно, что
осевая (гастральная) и краевая (перистомальная) мезодермы, различаясь
по положению, представляют собой только различные фазы одного и того
Рис. 39. Перемещения материала (гаст-
руляционные движения) при образова-
нии мезодермы в окружности первичной
полоски (бластопора) у млекопитаю-
щего.
а.т — рсевая мезодерма; / —передняя губа;
р,— первичная бороздка (бластонор); ps —
перистомальная мезодерма.
Рис. 40. Поперек! ые разрезы через
зародыш тритона (ио Гертвпгу из
Шпмкевича).
А — башке кзади; В — ближе кпереди,
ect — эктодерма; ent — энтодерма; Coel —
полость тела; тр .— наружный; mt—
внутренний листок мезодермы; ch — хорда;
ту — сомиты; п — зачаток нервной систе-
мы, углубляющийся (на В) в виде желобка
II, ограниченного с боков валиками nt.
же образования, так как краевая мезодерма постепенно превращается
в осевую (рис. 39).
По краю самого диска у амниот образования мезодермы не происходит.
В дальнейшем мезодерма разрастается, образуя сплошной листок
между эктодермой и энтодермой. В ее пределах происходит затем даль-
нейшая диференцировка (рис. 40). Спинная часть мезодермы, лежащая
по бокам от хорды, правильно сегментируется, и в каждом сегменте по-
является небольшая полость. Остальная часть мезодермы, лежащая по
бокам кишечника, не сегментируется. Сегментированная часть мезодермы
представляет собой ряд с о м и т о в, а песегментированная часть — бо-
ковые пластинки. Полость, заключающаяся в боковых пластин-
ках, представляет собой целом, или вторичную полость тела. И в этих
процессах обнаруживается большое сходство с ланцетником.
Одновременно с образованием мезодермы идет и развитие нервной
системы путем образования утолщенной нервной пластинки вдоль
спины зародыша, которая затем образует жолоб и замыкается в трубку
(рис. 35, 36, 40).
Количество желтка в яйце оказывает значительное влияние и на
внешнюю форму зародыша.
Рис. 41. Медианный разрез через зародыш птицы. Обособление зароды-
ша и образование амниона и аллантоиса (по Кейбелю).
all—закладка аллантоиса; ат. а—передняя и ат. р—задняя складки амниона;
ап •— закладка заднего прохода; с — сердце; ch—хорда; сп—нервно-кишечный
канал; coel — полость тела; ect — эктодерма; ent — энтодерма; тр— париеталь-
ный и ш» — висцеральный листки мезодермы; п — нервная трубка; st — рото-
вое впячивание.
Если этого желтка не особенно много, то брюшная стенка зародыша
оказывается просто несколько вздутой вследствие того, что нижняя
стенка кишечника представлена массой набитых желтком более крупных
клеток (рис. 34). Если желтка много, то он лишь постепенно обработается
краями зародышевого диска, а самое тело зародыша образуется в неболь-
шой средней части диска. По мере того как развиваются зародышевые
листки и закладываются первые органы на тело зародыша, впереди обо-
собляется голова в виде вздутия, а позади в виде выступа начинает вы-
растать хвост (рис. 41). Передняя и задняя части тела постепенно обособля-
ются от лежащего снизу «желточного мешка», состоящего из массы желтка,
окруженной всеми тремя зародышевыми листками. Зародыш перетяжкой
все яснее обособляется от желточного мешка, так что у многих позво-
ночных (селахий, рептилий и птиц) последний оказывается подвешенным
к зародышу посредством тонкого стебелька. Интересно, что такое обра-
зование желточного мешка происходит и у млекопитающих, но так как
желток последними утрачен, то желточный мешок выполнен лишь бел-
ковой жидкостью. Во время дальнейшего развития зародыша желток по-
степенно потребляется, и желточный мешок, сокращаясь, втягивается
внутрь и, наконец, совершенно исчезает.
У амфибий в задней части кишечника развивается выступ брюшной
ее стенки, который впоследствии дает начало мочевому пузырю. Такая
закладка образуется и у высших позвоночных (рис. 41, all), но у них она
разрастается в большой зародышевый пузырь, служащий не только ре-
зервуаром для жидких выделений, но и органом зародышевого дыхания.
Это — так называемый алл а п i о и с,, который потом подвергается
64
Рис. 42. Четыре стадии развития вародышевых
оболочек птицы (схема по Боасу из Шимкевича).
ek — энтодерма; еп — энтодерма; ш — мезодерма; ат —
амнион; ат'— енладни, за счет которых образуются
амнион и сероза — s; al — аллантоис; Ы — желточный
пузырь; t — •по'ЛОТЗТБ'-тшшечника
обратному развитию и отбрасывается. У плацентных млекопитающих ал-
лантоис принимает участие в развитии детского места, и только небольшая
часть его идет затем па образование дефинитивного мочевого пузыря.
Для высших позвоночных характерно еще образование особых приспо-
соблений, защищающих зародыш от вредного влияния толчков. У них
развивается система выполненных жидкостью зародышевых обо-
лочек, посреди которых располагается зародыш. К этой системе
относится и упомянутый уже аллантоис, состоящий из энтодермы с ме-
зодермальным покровом снаружи. Кроме того, вокруг зародыша рано
развивается полая внутри кольцевая складка эктодермы с подстилающим
ее слоем мезодермы (рис.
41, ат. а и ат. р). Эта
складка поднимается во-
круг^ зародыша и срас-
тается над ним (рис. 42,
ат).
Так как складка со-
держит полость и является
двойной, то она дает на-
чало двойному покрову.
Внутренний покров,
развивающийся вокруг те-
ла зародыша и состоящий
из слоя эктодермы, обра-
щенного к зародышу, и
слоя мезодермы, высти-
лающего ее снаружи, но-
сит название ал. н и о н а.
Наружная степка той*
же амниотической склад-
ки, срастаясь над амнио-
мом, образует наружную
оболочку зародыша, со-
стоящую из эктодермы
снаружи и мезодермаль-
ной подстилки, — это
серозная о б о л о ч-
к а. Сначала [сероза обособлена лишь в области зародыша, но по
мере расщепления «внезародышевой» мезодермы, покрывающей желточный
мешок, она превращается в полную оболочку, окружающую все яйцо.
Между телом зародыша и амнионом помещается замкнутая, выполненная
жидкостью амниотическая полость. Между обеими стенками амниоти-
ческой складки, а затем между, амнионом и серозой, имеется выстланная
мезодермой полость, которая является продолжением полости тела и
носит название внезародышевого целома. Аллантоис, разрастаясь, ложит-
ся во внезародышевом целоме между амнионом и желточным мешком,
с одной стороны, и серозой — с другой (рис. 42). Наружная стенка аллан-
тоиса покрывается богатой сетью кровеносных сосудов и служит для
газового обмена между зародышем и наружной средой. У млекопитающих
эта стенка аллантоиса срастается с серозой (хорион), образующей на своей
поверхности многочисленные сосочки. Последние врастают в углубления
слизистой оболочки матки, и таким образом получается орган, связующий
зародыш с телом матери,—плацента, или детское место, служащее не
только для дыхания, но и для питания зародыша посредством осмоза
через стенки близко соприкасающихся кровеносных сосудов.
Зародышевые оболочки являются примером чисто эмбриональных
приспособлений.
65
Йе менее часто наблюдаются йриспособления развивающегося орга-
низма к особым условиям его существования в тех случаях, когда этот
организм развивается не под защитой различного рода оболочек, а вы-
нужден вести вполне самостоятельное существование. В таких случаях
личиночного развития обычно наблюдаются специально личиночные ор-
ганы и приспособления. Различного рода личиночные органы нередко
наблюдаются у костистых рыб и в особенности у амфибий. Примером
такого рода органов являются наружные жабры, наблюдающиеся в виде
ветвистых выростов у личинок многоперых ганоидов (Polypterus), неко-
торых двудышащих (Protopterus и Lepidosiren) и у всех амфибий. У амфи-
бий преобразования, сопровождающие переход от личиночной формы
к взрослой, так резко выражены, что их приходится отмечать как мета-
морфоз. Ясно выраженный метаморфоз наблюдается и у миног (Petromy-
zon), личинка которых, известная под названием пескоройки, столь
значительно отличается от взрослой формы, что раньше ее относили к
особому роду — Ammocoetes.
Мы проследили образование зародышевых листков позвоночных и
развитие некоторых специально зародышевых органов. С развитием орга-
нов взрослого животного мы познакомимся при описании последних.
Теперь остается лишь указать на обшую судьбу зародышевых листков.
Наружный листок — эктодерм а — представляет собой первич-
ные покровы животного, служащие для связи его с внешней средой.
И в дальнейшем эта роль остается за эктодермой и ее производными.
Путем диференцировки отдельные ее части получают ту или иную спе-
циальную функцию, но в общем за счет эктодермы развиваются органы
защиты и даже нападения и органы восприятия раздражений со стороны
внешнего мира. Таким образом, весь эпидермис и его производные—же-
лезы, роговые образования кожи — развиваются за счет эктодермы.
Эктодерма принимает такяге участие и в развитии некоторых чешуй рыб
(плакоидной чешуи), а такяге и зубов. Ротовая полость образуется путем
впячивания эктодермы по направлению к переднему концу кишки
(рис. 41), после чего следует срастание и прорыв ее в полость кишки.
Подобным же образом развивается и задний проход. И здесь обычно на-
блюдается впячивание эктодермы, которое соединяется затем с энтодер-
мальной частью кишки. Таким же образом эктодерма принимает участие
и в образовании жаберных щелей. Наконец, за счет эктодермы у позво-
ночных развиваются вся нервная система и воспринимающий аппарат
органов чувств (за возмоягным исключением лишь органов вкуса).
Вдоль спинной стороны зародыша образуется полоса утолщенной
эктодермы — нервная пластинка, края которой приподнимаются в виде
складок или валиков, образуя нервный яголоб, который затем замыкается
в нервную трубку. Нервная трубка представляет собой зачаток всей
центральной нервной системы. На переднем конце нервные валики смы-
каются позже, и внутренняя полость трубки сообщается некоторое время
с наруягной средой посредством невропора. Развитие нервной системы
идет, следовательно, в общем таким же путем, как и у ланцетника. Вдоль
спинной стороны нервной трубки из боковых частей нервной пластинки
(нервных складок) развивается парный продольный выступ — гангли-
онарная пластинка, за счет которой образуются затем нервные узлы.
Такая ганглионарная пластинка развивается и в области головы, но
здесь к ганглиям присоединяются еще клеточные элементы, происходящие
непосредственно из определенных утолщений эктодермы, называемых
плакодами. Периферические нервы образуются за счет отростков нервных
клеток.
Орган обоняния образуется из утолщения эктодермы в передней
части головы, которое дает затем парное мешковидное впячивание, пре-
вращающееся в обонятельный мешок. Орган зрения развивается в качестве
66
парного выступа боковых стенок передней части нервной трубки. Выступы
имеют сначала вид пузырей, а затем наружная их стенка вворачивается
внутрь, так что зачаток глаза приобретает форму бокала, связанного
с мозгом тонким стебельком. Степки бокала дают начало наиболее сущест-
венным частям глаза — ретине и пигментной оболочке; в стебельке раз-
вивается зрительный нерв. Хрусталик глаза образуется за счет эктодермы
непосредственно. Орган слуха образуется в виде парного мешковидного
впячивания эктодермы, замыкающегося затем в так называемый слуховой
пузырек; последний представляет собой*зачаток внутреннего уха. Органы
боковой линии развиваются в виде утолщений эктодермы и с эпидермисом
кожи связи не теряют, хотя иногда и погружаются в более или менее
глубокие каналы.
За счет эктодермы образуется также отчасти и мезенхизма. Именно
ганглионарная пластинка головы (развивающаяся из нервных складок)
дает начало массе мезенхимы (мезэктодерме), которая мигрирует к боковым
стенкам глотки и идет затем здесь на построение висцерального скелета.
В области туловища ганглионарная пластинка образует мезенхиму, иду-
щую на построение скелета непарных плавников рыб.
Энтодерма дает начало эпителию органов пищеварения, т. е.
прежде всего эпителиальной выстилке всего кишечника, за исключением
лишь части ротовой полости и незначительной части задней кишки. В ка-
честве слепого выроста брюшной стенки кишечника развивается печень.
На такой стадии развития закладка печени позвоночного весьма напоми-
нает печеночный вырост ланцетника. Впоследствии этот вырост сильно
ветвится и превращается в сложную трубчатую, а затем, путем образо-
вания анастомозов, в сетчатую железу взрослого животного..Подобным
же образом из нескольких выростов кишечника развивается и поджелу-
дочная железа. В качестве полого выроста спинной стенки кишечника
развивается плавательный пузырь рыб.
Органы дыхания, т. е. точнее эпителий этих органов, развиваются
также за счет энтодермы. У рыб в области глотки развивается ряд парных
мешковидных выпячиваний, растущих по направлению к эктодерме,
откуда к ним направляются встречные впячивания. Оба листка срастаются,
и затем жаберный мешок прорывается наружу. Эпителий жаберных
лепестков рыб и амфибий развивается в эктодермальной части мешков.
Органы воздушного дыхания — легкие — развиваются как парные меш-
ковидные выросты брюшной стенки задней части глоточной области.
Наконец, за счет энтодермы в той же глоточной области развиваются
и две железы внутренней секреции — зобная железа и щитовидная же-
леза. Последняя развивается в виде непарного выступа на дне глотки,
который у круглоротых имеет вид желобка с железистыми стенками,
строение которых поразительно напоминает подглоточную борозду (эндо-
стиль) низших хордовых.
Мезодерма дает начало огромному числу органов и, кроме того,
участвует и в образовании органов, развивающихся из других листков,
образуя их скелет, соединительную ткань, мускулатуру и сосуды. От
внутреннего нижнего угла сомита выступает масса клеток, выполняющих
затем промежуток между сомитом, с одной стороны, и хордой и нервной
системой — с другой. Эта масса носит название склеротома, и за ее счет
развиваются позвоночник, череп, скелет плавников и соединительноткан-
ные перегородки между миомерами и их отделами. Остальная часть сомита
представляет собой миотом. Наружная стенка миотома (кожный листок)
расплывается на отдельные клетки, образующие соединительнотканную
часть кожи. Внутренняя его стенка утолщается и дает миомеры и прочую
париетальную мускулатуру взрослого животного.
Часть промежуточная между миотомом и боковой пластинкой (сте-
белек сомита) дает начало почечным канальцам. Утолщения спинной
67
стенки боковой пластинки представляют собой зачатки половых желез.
Сами боковые пластинки с их полостями дают начало полости тела и ее
перитонеальной выстилке. Разрастаясь вокруг кишки, их стенки сраста-
ются между собой над ней и под ней, образуя спинной и брюшной мезен-
терии.
Прилежащая к кишке часть висцерального листка дает, кроме того,
начало соединительной ткани слизистой ее оболочки, кровеносным и
лимфатическим сосудам и кишечной мускулатуре. Париетальный листок
также дает, кроме собственно бркЯшшы, еще и многочисленные соедини-
тельнотканные элементы и пр. Кровеносные сосуды развиваются также за
счет мезодермы, причем главные стволы (подкишечный и надкишечный)
и сердце образуются в виде продольных каналов в брюшном и спинном
мезентериях.
Эмбриология хордовых дает нам много примеров постепенной и
последовательной диференцировки органов. Эта онтогенетическая дифе-
ренцировка во многих случаях, бесспорно, является ясным повторением
филогенетической диференцировки, происходившей у более или менее
отдаленных предков изучаемой формы. Таким образом, мы имеем много
прекраснейших иллюстраций биогенетического закона. Не говоря уже
об эмбриональном развитии асцидий, ясно обнаруживающих во время
своего онтогенеза более совершенную организацию своих свободно пла-
вавших предков, напомню закладку щитовидной железы у круглоротых,
которая повторяет процесс ее развития путем преобразования эндостиля
низших хордовых; напомню факт закладки хорды и жаберных щелей у
высших позвоночных, у которых они никакой роли не играют, — факт,
объяснимый исключительно как повторение признаков живших в воде
рыбообразных их предков.
Изучение онтогенеза хордовых и, в частности, позвоночных живот-
ных имеет поэтому очень большое значение для установления генезиса
органов и для изучения вопроса о родственных соотношениях между
различными группами позвоночных.
При изучении сравнительноанатомического материала его принято
излагать по системам органов, распределяемых по функциональному
принципу.
В этой книге принято деление на следующие системы:
1) система общих покровов,
2) скелет,
3) мускулатура и электрические органы,
4) нервная система,
5) органы чувств,
6) органы пищеварения,
7) органы дыхания,
8) органы кровообращения,
9) общая полость тела,
10) органы выделения,
11) органы воспроизведения,
12) органы внутренней секреции.
ЧАСТЬ II
СРАВНИТЕЛЬНАЯ АНАТОМИЯ ОТДЕЛЬНЫХ СИСТЕМ
ОРГАНОВ
ОБЩИЕ ПОКРОВЫ
Покровы беспозвоночных
Сношения животного с внешней средой осуществляются через посредство экто-
дермы. Поэтому восприятие раздражения, чувствительность и защита тела от вредных
воздействий представляют собой основные функции эктодермы, которые у низших
животных и остаются целиком за ней. Специализация отдельных клеток и целых
участков эктодермального покрова для восприятия тех.или иных раздражений приво-
дит к высокой диференцировке, которая будет рассмотрена в главе о нервной системе.
Здесь мы ограничимся главным образом изучением покровов как органов защиты.
Главная роль в этом смысле у беспозвоночных почти всегда остается за эктодермой
и ее производными. Подстилающие эктодерму мезодермальные клетки обыкновенно
не образуют заметного слоя, который был бы ясно отграничен отсоединительной тка-
ни остального тела, но иногда они уже могут быть обозначены как соединительно-
тканная часть кожи. Таким образом, в коже (cutis) различают поверхностный
эктодермальный слой—- а п и д е р м и е..(epidermis) — и подстилающий его мезодер-
мальный слой — соединительнотканную часть кожи, или кориум (corium s. dermis).
У беспозвоночных эпидермис однослойный, чаще Btero цилиндрический. У низ- *
ших форм он часто снабжен мерцательными волосками. У турбеллярий он реснитча-
тый. Реснички характерны и для личинок беспозвоночных (за исключением членисто-
ногих), где нередко, кроме общего ресничного покрова, наблюдаются определенные
полосы эпителия с очень длинными ресничками — мерцательные шнуры. Такой рес-
ничный покров является наиболее примитивной формой органов движения.
У ктенофор склеенные ряды ресничек превратились в своеобразные весельные пла-
стинки. Иногда, особенно у сидячих форм, реснички сидят на выростах около рта и
служат для привлечения пищевых частиц помощью вызываемого ими тока воды. Это
наблюдается у коловраток, а также у мшанок и плеченогих, у которых «щупальца» и
«руки» снабжены таким мерцательным покровом.
У других животных поверхность эпителия обыкновенно покрывается более или
менее толстой кутикулой. Кутикулярная оболочка образует нередко уже у
кишечнополостных (гидроидные полипьЛдовольно прочный покров. У многих червей
имеется значительная кутикула на всей поверхности. У членистоногих эпителий обычно
теряет цилиндрическую форму и образует на своей поверхности толстый слой очень
прочной кутикулы, которая получает название хитина. Значение эпителия сво-
дится теперь к продукции все новых слоев хитина, и такой эпителий, лежащий под
толстым хитиновым покровом, получает название гиподермиса. Хитин слоист и обычно
пронизан канальцами, в которые проникают отростки клеток гиподермиса. Подвижность
животного обеспечивается тем, что хитиновый покров на промежутках между сегмен-
тами и на суставах конечностей остается тонким, между тем как на сегментах и чле-
никах достигает значительной толщины. Нередко хитин пропитывается известью и
достигает еще большей твердости (ракообразные). Благодаря тому, что тело животного
заключено теперь в неподатливую твердую оболочку, рост его ограничен довольно
узкими пределами, по достижении которых старый хитиновый покров целиком отсла-
ивается, обнажая тонкий и мягкий слой молодого хитина. Такая периодическая линька
Всегда сопровождает рост животного. Взрослое животное часто вовсе не растет.
Покровы моллюсков образуют очень твердую, пропитанную известью рако-
вину не на всей поверхности, так что остальная часть тела, остающаяся непокры-
той, может выдвигаться и втягиваться под защиту раковины. Такая раковина состоит
69
ив нескольких слоев различной структуры (поверхностный, призматический, перла-
мутровый) и росту животного не препятствует, так как и сама она увеличивается вслед-
ствие непрерывного отложения новых слоев по переферии краевыми частями мантии
животного.
Очень своеобразна оболочка туникат. Весьма разнородная по своей консистен-
ции, то студенистая, то хрящеподобная, она представляет собой продукт выделения
зктодермального зпителия, но в нее после проникают мезодермальные клетки и даже
сосуды, так что она приобретает вид опорной ткани.
В эпидермисе обычно наблюдаются рассеянные железистые клетки. В некоторых
случаях такие одноклеточные железы достигают значительной величины, и тогда
их тело погружается в подлежащую массу ткани, так что в эпидермисе остается только
суженный их выводной канал (пиявки). Наблюдаются и многоклеточные слизистые
кожные железы у аннелид и у моллюсков, а также железы специального значения,
как, например, слюнные, паутинные, ядовитые и пахучие железы ракообразных,
многоножек и особенно пауков и насекомых. Возможно, что и органы дыхания много-
ножек и насекомых — трахеи — представляют собой результат видоизменения кож-
ных желез.
Соединительнотканный слой, кожи лишь редко достигает заметного развитии,
как, например, у кольчатых червей. У головоногих моллюсков в этом слое располо-
жены многочисленные х ром ато форы — очень подвижные ветвистые клетки,
содержащие пигмент. При сокращении многочисленных отростков пигмент концен-
трируется в теле клетки. Этой сократимости хроматофоров головоногие обязаны спо-
собностью менять свою окраску.
Очень мощный кожный слой соединительной ткани с перепутанными волокнами,
идущими в различных направлениях, имеется у иглокожих. У последних в этом слое
развиваются более или менее обширные обызвествления, образующие обычно мощный
кожнЫй скелет, состоящий из тесно связанных друг с другом пластинок (у го-
лотурий имеются лишь отдельные известковые тельца различной формы).
Очень часто у беспозвоночных наблюдается тесная связь кожи с подлежащей
мускулатурой. Особенно характерна такая связь для червей, у которых таким образом
развивается так называемый кожно-мускульный мешок.
В связи с кожей у беспозвоночных стоит обравование еще некоторых органов.
Ветвистые выросты покровов, обильно снабженные кровеносными сосудами, явля-
ются нередко в виде жабер, как, например, у кольчатых червей. Затем кутикулярные
щетинки тех же кольчатых червей, приводимые в движение специальными мускулами,
служат у них органами движения. У многощетинковых червей пучки таких щетинок
располагаются по бокам на специальных метамерных выростах тела — параподиях,
которые и являются наиболее примитивной формой парных конечностей.
У ланцетника кожа очень слабо диференцирована и состоит ив однослойного
зпидермиса, подостланного незначительным количеством соединительной ткани.
Покровы позвоночных
У позвоночных животных всегда ясно различимы оба слоя кожи:
верхний, вктодермальный — эпидермис и нижний, мезодермальный —
кориум (рис. 43). Эпидермис позвоночных всегда имеет вид много-
слойного эпителия, нижний слой которого состоит из цилиндрических
клеток, сохраняющих до известной степени эмбриональный характер^
У эмбрионов из однослойной эктодермы первоначально развивается
двуслойный эпителий, нижний цилиндрический слой которого позже,
Путем клеточных делений, дает начало новым слоям клеток. И у взрослого
животного этот нижний слой находится в деятельном состоянии, продув
цируя все новые слои клеток эпидермиса. Верхний слой плоских клетоК
у рыб и личинок амфибий образует на своей поверхности тонкую пористую
каемку подобно таковой ланцетника. Такая тонкая каемка характерна
для мерцательного эпителия, а так как у личинок ланцетника, а также
у личинок амфибий и молодых эмбрионов костистых рыб действительна
наблюдается покров из мерцательных ресничек, то каемка эпидермиса и
считается остатком общего мерцательного покрова. Настоящей кутикулы,
служащей наружным скелетом, у позвоночных никогда не развивается.
У низших. позвоночных.нпидермис отграничен от подстилающей его
соединительной части кожи бесструктурной основной перепонкой, которай
Является совместным продуктом выделения обоих компонентов кожи.
Между клетками эпидермиса обыкновенно прекрасно видны протоплаз-
матические связи в виде межклеточных мостиков. По межклеточным
10
пространствам происходит циркуляция питательных жидкостей, полу-
чающих только таким путем доступ к клеткам эпидермиса. В эпидермисе
низших позвоночных наблюдаются отдельные железистые клетки различ-
ных типов, иногда в огромном количестве.
У наземных позвоночных эпидермис-диференцируется на два слоя:
нижний, содержащий неизмененные клетки—мальпигиев слой* и верхний—
роговой слой (рис. 44). В роговом слое клетки испытывают особого рода
перерождение: они переполняются капельками жироподобного вещества—
элеидина, которое постепенно уплотняется, в то время как сами клетки
становятся все более плоскими; ядро перестает быть различимым, и клетка
превращается в твердую роговую чешуйку. По мере того как поверхност-
Рис. 43. Схематический разрез через кожу
рыбы.
Ь — бокаловидные клетки; bas — основной
слой епидермса; со — кожа собственно; ер —
впидермис; g — зернистые клетки; к — колбо-
видные клетки; п — нервы; v — сосуды.
ные слои ороговевших клеток
изнашиваются и сбрасываются,
Рис. 44. Разрез через кожу человека
(из Видерсгейма).
Со — кожа собственно; ' ср — сосудистые
сосочки; В— сальные железы; F—под-
кожный жир; G — сосуды; Н — волос;
N — нервы; пр — нервные сосочки; SC —
роговой слой; SB — потовые железы;
SIX — их выводные протоки; SM — маль-
пигиев слой.
роговой слой непрерывно возобновляется снизу за счет клеток
мальпигиева слоя. Сбрасывание поверхностных клеток совершается
путем постепенного шелушения или путем линьки, т. е. отделения це-
лого слоя. Последнее наблюдается обычно у змей и ящериц, у кото-
рых верхний слой иногда сходит в виде целого чехла.
Клетки эпидермиса, в особенности более глубоких его слоев, нередко
переполнены красящими веществами — пигментами, обусловливающими
окраску кожи животного, так же как и пигмент, заключающийся в спе-
циальных мезодермальных клетках.
Соединительнотканная часть кожи, кориум позвоночных, разви-
вается за счётТаджнйГо листка миотома зародыша. У лайцётника этот
листок сохраняет еще характер эпителия, который выделяет бесклеточную
опорную ткань студенистого характера, подстилающую эпидермис. У
позвоночных в этой ткани появляются и клеточные элементы вследствие
распада кожного листка на отдельные клетки — мезенхиму. Однако у
личинок низших позвоночных (крутлоротых, амфибий) мезодермальные
клетки ложатся еще непрерывным слоем, подобно эпителию, под эктодер-
мой, и затем выделяют на своей поверхности основное вещество с соедини-
тельнотканными волокнами (рис. 45). По мере ррста этого промежуточного
71
слоя в него включаются затем и клеточные элементы. Таким образом
развивается соединительнотканная часть кожи. Волокна кожи у низших
позвоночных располагаются весьма правильно—чередующимися слоями с
взаимно перпендикулярным направлением волокон, идущих под углом
к оси тела (рис. 43). Слои горизонтальных волокон пронизываются пучками
вертикальных волокон, вместе с которыми в кожу входят питающие ее
сосуды и нервы. У высших позвоночных (птиц и млекопитающих) волокна
кожи неправильно спутаны (рис. 44). У птиц кожа вообще очень тонка,
но у млекопитающих оба слоя кожи отличаются значительной толщиной,
в особенности у некоторых форм. Если эпидермис очень толст, то питание
его осуществляется при помощи сосочков кориума с сосудами и нервами,
глубоко в него вдающихся снизу.
И в мезодермальной части кожи встречается пигмент, но здесь он кон-
центрируется всегда в специальных, обыкновенно очень сложно разветв-
ленных пигментных клетках (рис. 45). Кроме обыкновенных пигментных
клеток разных цветов, различают еще иридоциты, наполненные кристал-
ликами гуанина, которые придают серебристый блеск многим рыбам.
Пигментные клетки обычно лежат в кориуме, но они могут переползать
и в межклеточные
Рис. 45. Разрез через кожу личинки тритона.
со — закладка кориума; ер — двуслойный эпидермис с желе-
зистыми (лейдиговыми) клетками; т — клетки кориума pig—
пигментные клетки; ас — подкожная соединительная ткань.
пространства эпидер-
миса.
Под вижность
представляет вообще
характерную особен-
ность пигментных
клеток. Иногда осо-
бо развитая подвиж-
ность пигментных
клеток связана со
способностью живот-
ного менять свою
окраску. Такие клет-
ки носят название хроматофоров. Изменение окраски достигается в этих
случаях либо перемещением зерен пигмента из отростков в тело клетки
и наоборот, либо сокращением всего хроматофора. Способность менять
окраску наблюдается у некоторых рыб (камбалы, скорпены и др.), амфи-
бий (древесницы и др.) и рептилий. Наиболее известным примером этой
способности являются хамелеоны, большие хроматофоры которых лежат
довольно глубоко и снабжены многочисленными отростками, доходящими
непосредственно до эпидермиса. Пигмент может перемещаться по отрост-
кам под самый эпидермис и может передвигаться обратно вглубь кожи
в тело клетки. Такие перемещения стимулируются подходящими к хрома-
тофорам нервами.
Кожные железы
Клетки эпидермиса обнаруживают способность к секреторной деятель-
ности, выражающейся в выделении их протоплазмой разнообразных
жидких веществ, в особенности слизи. Такие железистые клетки
рассеяны в эпидермисе низших позвоночных иногда в колоссальном коли-
честве, как, например, у круглоротых. У миксин эпидермис
состоит не менее как на половину всей своей толщины исключительно
из железистых клеток. И у круглоротых и у р ы б имеются железистые
клетки различного типа. Наиболее распространены обыкновенные бока-
ловидные слизистые клетки, открывающиеся непосредственно на поверх-
ности эпидермиса (рис. 43). Кроме того, встречаются еще в середине
эпидермиса шарообразные клетки с зернистым содержимым и большие
72
колбовидные клетки, сидящие на ножке, доходящей до основания эпидер-
миса, где она покоится в основном слое цилиндрических клеток. Эти
клетки выделяют, невидимому, серозную жидкость не непосредственно
на поверхность эпидермиса, а в межклеточные пространства. Типичных
многоклеточных кожных желез у рыб не встречается. Железистые углубле-
ния эпидермиса при основании различного рода игл и шипов служат
у некоторых рыб в качестве ядоотделительных желез. Сложные образо-
вания железистого характера представляют собой и органы свечения
некоторых глубоководных рыб (некоторые акулы из сем. Spinacidae и
костистые из сем. Scopelidae и др.). В таких органах различают централь-
ную массу клеток железистого характера, окруженных отражательной
соединительнотканной оболочкой с кристаллами гуанина и покрытых со
стороны, обращенной внутрь тела, пигментной оболочкой. Спереди к
железистой части органа примыкают часто прозрачные массы клеток,
играющие роль линзы, а к последней — сбоку еще особый рефлектор,
состоящий из вогнуто-
го отражательного зер-
кальца, покрытого сза-
ди пигментом.
У личинок зем-
новодных эпидер-
мис также содержит
большое количество же-
лезистых клеток (рис.
45), но у взрослых
животных последние ис-
чезают и заменяются
многоклеточными желе-
зами. Опи развиваются
Рис. 46. Разрез через вожу амфибий.
по — кориум; d — выводной проток железы; ер— эпидер-
мис; gl — железистые клетки; тп — мускульный покров
железы; pig — пигментные клетки.
за счет клеток основно-
го слоя эпидермиса,
которые погружаются в
соединительнотканную
часть кожи и путем раз-
множения дают начало
шаровидной группе клеток, лежащей под эпидермисом. Около вре-
мени метаморфоза внутри такого зачатка железы появляется полость,
которая продолжается в виде узкого выводного канала, проходящего
через эпидермис на наружную поверхность последнего. Клетки распо-
лагаются в один слой по стенке железы. Многоклеточная железа взрослого
животного имеет, следовательно, вид простого мешка или пузыря, стенка
которого состоит из одного слоя железистых клеток (рис. 46). Сама железа
лежит в кориуме и узким горлышком связана с эпидермисом, через ко-
торый проходит ее выводной капал. Снаружи эпителий железы покрыт
слоем меридионально расположенных гладких мускульных клеток экто-
дермального происхождения, и такие мускульные клетки вообще рассеяны
в кориуме земноводных (явление среди позвоночных вообще совершенно
исключительное). Основная функция кожных желез амфибий заключа-
ется в поддержании кожи во влажном состоянии, что является весьма
существенным ввиду слабого развития рогового ее слоя. Увлажнение
кожи является также условием возможности кожного дыхания, которое
имеет у амфибий довольно большое значение. Наконец, секрет кожных
желез имеет значение и в смысле защиты организма от проникновения
паразитов и иногда даже от нападения более крупных врагов. Эти железы
бывают двух родов — обыкновенные слизистые и другие с каким-то зер-
нистым содержимым. Секрет последних часто бывает ядовитым. На неко-
торых местах тела, в особенности по бокам задней части головы, образу-
73
ются иногда очень значительные скопления таких желез («паротиды» жаб
и саламандр).
У безногих амфибий (гимнсфион) в передней половине каждого сег-
мента их кольчатого тела, кроме желез обычного типа, имеются еще осо-
бые гигантские кожные железы.
У рептилий кожа лишена желез. В виде исключения у них
встречаются лишь отдельные крупные железы, несущие особые функции.
Так, у крокодилов имеется пара мускусных желез по бокам нижней че-
люсти. У черепах подобные железы имеются на месте соединения спин-
ного и брюшного щитов. У ящериц наблюдаются еще особые бедрен-
ные поры, но они выталкивают из себя в виде сосочка лишь массу орого-
вевших клеток и потому вряд ли могут быть отнесены к железам (неко-
торые авторы сравнивают эти образования с волосами).
У птиц кожных желез также нет, за исключением лишь одной
парной железы, несущей специальную функцию. Это — копчиковая же-
Рис. 47. Схема железы амфибий (Л) и потовой железы млеко-
питающих (В и С).
С — роговой слой епидермиса; ер — епидермис; М — мальпигиев
слой епидермиса; т — мускульный покров железы.
лева (glandula uropygii), открывающаяся обыкновенно парой отверстий
Над последними позвонками. Она имеет довольно сложное строение,
состоя' из многочисленных трубок, радиально сходящихся к выводному
каналу, и выделяет маслянистый секрет, служащий для смазки перьев.
Млекопитающие примыкают по обилию кожных ягелез к
амфибиям. В конге млекопитающих имеются многоклеточные железы
обоих основных типов — трубчатые и альвеолярные. К первым относятся
потовые железы, имеющие вид длинной трубки, конец которой
нередко свернут в клубочек, а остальная часть обычно изогнута в виде
штопора. У некоторых низших млекопитающих эти железы имеют почти
мешковидную форму (рис. 47, В). Стенки потовой ягелезы состоят из
однослойного кубического эпителия, покрытого снаружи слоем гладких
мускульных волокон. Как и в ягелезах амфибий, этот покров имеет экто-
дермальное происхождение, что доказывает с несомненностью их гомо-
логию. Подойдя к эпидермису, трубка утрачивает собственные стенки
и пронизывает его в виде узкого выводного канала. Потовые ягелезы откры-
ваются либо непосредственно на поверхность эпидермиса, либо в сумку
волоса,Потовые железы частично играют роль органа выделрцир, до имеют
74
также значение для терморегуляции организма. Потовая железа разви-
вается в виде плотного выроста основного слоя эпидермиса, растущего
вглубь кориума (рис. 48, Л). Позже в этом выросте появляется канал.
Обычно закладка потовой железы связана с закладкой волоса. Потовые
железы имеются у всех млекопитающих за редкими лишь исключениями
(китообразные, сирены; у мышей имеются только подошвенные железы).
Иногда они приобретают форму разветвленных трубок (у медведей, гип-
попотама). У многих млекопитающих имеются местные скопле-
ния более слояшых трубчатых желез, представляющих Собой модифи-
кацию потовых. Секрет потовых желез — это продукт выделения, т. е.
результат жизнедеятельности железистых клеток, Составляющих их
стенки. Обыкновенно он очень жидкий, но иногда слизистый (гиппопотам)
или белковый, иногда густой и вязкий (ушная сера человека) и иногда
даже жирный; в некоторых случаях цветной (красный у гиппопотама и
у самца большого кенгуру, у которого он окрашивает шерсть в крас-
ный цвет).
Рис. 48. Развитие потовой железы (Л) и развитие млечной железы (/?),
со — иориум; ер — эпидермис.
К типу альвеолярных желез относятся сальные железы.
Это гроздевидные железы, состоящие обыкновенно из нескольких ло-
пастей, связанных с сумками волос (рис. 64), в которые они выделяют свой
жирный секрет. Развиваются сальные железы в виде выростов от влага-
лища волос. Эти железы не имеют мускульного покрова. Эпителий их
многослойный, и клетки его при образовании секрета испытывают жировое
перерождение, разрушаются и выталкиваются. Таким образом, эти же-
лезы совсем иного типа, чем потовые. У млекопитающих встречается,
кроме обычных сальных желез, много специализированных желез того
же типа. Такие железы встречаются иногда и независимо от волос, в осо-
бенности на местах перехода в слизистую оболочку (на губах, на конъюнк-
тиве, перианальные и препуциальные железы многих животных, мускус-
ная железа кабарги, цибетиновая железа виверры и многие другие).
Млечные железы типичных млекопитающих по своему стро-
ению являются также сложными гроздевидными железами и долгое время
считались продуктом преобразования сальных желез. Сравнению с пос-
ледними благоприятствует и богатый жиром секрет млечных желез. Однако
в настоящее время доказано с достаточной достоверностью, что млечные
железы — результат видоизменения трубчатых, потовых желез. Секрет
Последних иногда также содержит и белки и жиры.
У клоачных млекопитающих (Monotremata) млечные железы пред-
ставляют собой очевидное видоизменение потовых; это — длинные труб-
чатые мешки с гладким мускульным покровом, которые открываются в
сумки волос на особом железистом поле. В те же сумки одновременно
открываются и обыкновенные сальные железы. Следует еще отметить,
75
что стенки млечных желез состоят из однослойного эпителия, клетки
которого при секреции не разрушаются. Железистое поле помещается
у ехидны по обеим сторонам брюха в особой сумке, которая развивается
только во время беременности и служит для вынашивания яйца, а потом
и детеныша. Секрет железы стекает по волосам железистого поля и сли-
зывается детенышем.
У живородящих млекопитающих железистое поле ограничивается
очень небольшой областью покровов, выпяченной в виде сосца. Сами
млечные железы построены здесь сложнее и принадлежат к альвеоляр-
ному типу. Тем не менее у сумчатых (Marsupialia), если и не на самих же-
лезах, то на их протоках сохраняется гладкий мускульный покров, дока-
зывающий существование генетической связи этих желез с млечными
железами клоачных млекопитающих, а также и с потовыми железами.
В отношении выводных протоков различают два типа сосцов: 1) так назы-
ваемые «истинные» сосцы, на вершине которых открываются более или ме-
нее многочисленные выводные протоки желез отдельными отверстиями,
и 2) «ложные» сосцы, внутри которых имеется общий выводной канал
значительного диаметра, открывающийся на вершине сосца одним отвер-
Рис. 49. Расположение выводных протоков млечной железы
на железистом поле у клоачных млекопитающих (Л), на
«истинном» сосце (В) и на «ложном» сосце (С) живородящих
млекопитающих.
стием (рис. 49). Первый тип сосцов встречается у большинства сумчатых,
полуобезьян, обезьян и человека. Второй тип характерен для хищных
и копытных.
У ехидны млечные органы закладываются очень рано в виде парного
утолщения эпидермиса на брюхе. Это — зачатки железистого поля, на
которых затем закладываются многочисленные волосы. От зачатков во-
лос в виде боковых выростов развиваются трубчатые железы, которые
вскоре достигают гораздо большего развития, чем обыкновенные потовые.
У зародышей живородящих млекопитающих по бокам брюха разви-
вается парная полоска утолщенного эпителия — млечная линия. Па пос-
ледней обособляются млечные бугорки, которые затем уплощаются и,
погружаясь одновременно вглубь собственно кожи (кориума), превра-
щаются в так называемые млечные точки. За счет такого плотного эпите-
лиального зачатка развиваются стенки сосцов и сами железы. Последние
образуются в виде выростов, погружающихся вглубь кориума, весьма
сходно с развитием потовых желез (рис. 48, В). У сумчатых по краям
развивающегося сосца наблюдаются все переходы между обыкновенными
потовыми и типичными млечными железами. У сумчатых, а иногда у выс-
ших млекопитающих (и у человека) наблюдается также связь зачатков
млечных желез с сумками рудиментарных волос — та же связь, которая
характерна для взрослых клоачных.
Все эти факты, вместе взятые, позволяют в настоящее время утвер-
ждать, что млечные железы всех млекопитающих представляют собой
преобразованные потовые железы.
как и секрет потовых_жйдаа*.^схь результат,жизнедеятельности клеток,
й не следствие их разрушения, как в сальных железах.
76
Расположение и число желез и сосцов весьма различны. У плодовитых
форм они располагаются в два ряда вдоль всей брюшной поверхности тела
от подмышечной области до паховой (у одного сумчатого, опоссума Didel-
phys henseli, до 25 сосцов — рис. 50; у насекомоядного, тенрека Cente-
tes~, — 22 сосца). Редукция’числа сосцов сопровож-
дает уменьшение числа одновременно рождаемых
детенышей. Обычно наблюдается 1—8 пар. У
копытных имеются 1—2 пары сосцов в паховой обла-
сти. У полуобезьян имеется 1 пара на груди и сверх
того часто 1 пара на брюхе. У летучих мышей,
обезьян и человека только 1 пара на груди. Но в
этих случаях имеются указания на происхождение
от форм более плодовитых, и у человека, например,
эмбрионально закладывается парный ряд в 5 сос-
цов. В виде аномалии лишние сосцы наблюдаются
и у взрослого человека (гипертелия).
Защитные образования кожи
Чешуи рыб
В коже низших позвоночных почти всегда
имеются твердые скелетные пластинки, образую-
Рис. 50. Расположе-
ние сосцов у опоссу-
ма (Dldelphys) (ио
Томасу).
щие нечто вроде панцыря, защищающего тело
животного. Такие пластинки лежат в соединитель-
нотканной части кожи и носят название чешуи.
У рыб встречаются три основных рода чешуй,
различающихся как по форме, так и по материалу, из которого
они построены. Это — плакоидные, ганоидные и костные чешуи. Все
эти виды чешуй связаны, однако, переходными формами и, не-
сомненно, представляют результат видоизменения одного, наиболее
Рис. 51. Расположение плакоидных
чешуй в коже акулы (Scyllium). В
одной чешуе показано расположе-
ние канальцев и отмечены зуб (d)
и основная пластинка (bas).
альных направлениях отходит
примитивного типа—плакоиднойчешуи.
Плакоидная чешуя со-
стоит из лежащего в коже собственно
(кориуме) довольно широкого основа-
ния и сидящего на нем шипа, острие
которого выдвигается через эпидермис
наружу (рис. 51, 52). Шип имеет раз-
личную форму: изогнутого назад про-
стого конуса, плоского трезубца и т. д.
Основание чаще всего является в
виде ромбической пластинки (рис. 51).
Внутри чешуи имеется полость, вы-
полненная богатой кровеносными со-
судами мякотью (pulpa). Плакоидная
чешуя состоит из очень твердого веще-
ства — дентина, представляющего со-
бой соединение органического веще-
ства с солями извести. Дентин еще
тверже, чем кость, и в отличие от пос-
ледней клеточных элементов не содер-
жит. От внутренней полости в ради-
множество тонких разветвленных ка-
нальцев, которые пронизывают всю толщу дентина плакоидной чешуи.
Иногда вершина шипа покрыта тонким колпачком еще более твердого
вещества, в котором нет ясных канальцев. Это вещество еще богаче из-
вестью и носит название эмали. В канальцы дентина проникают отростки
клеток, выстилающих внутреннюю полость. Благодаря образователь-
77
Hoti деятельности этих клеток, Носящих наименование склеробластов,
происходит нарастание массы дентина.
Такие чешуи залегают в коже селахий, покрывая все их тело мелкими
шипиками (рис. 51). На челюстях подобные же плакоидные чешуи крупнее
и имеют вид зубов. Сходство в строении этих органов так велико, что
нередко й обычную плакоидпую чешую называют кожным зубом. По мере
изнашивания старые плакоидные чешуи сбрасываются (так же, как и
зубы на челюстях) и заменяются вновь развивающимися.
Плакоидная чешуя развивается на границе эктодермы и мезодермы
(рис. 52). Нижний слой эктодермы приобретает форму колпачка, в который
снизу в виде сосочка входит масса мезодермальных клеток. Клетки основ-
ного слоя эпидермиса, образующие стенку колпачка, приобретают высокую
цилиндрическую форму, а находящиеся внутри колпачка мезодермальные
клетки располагаются частью непосредственно под эпидермисом, образуя
сплошной слой, напоминающий эпителий и выстилающий полость колпачка
Рис. 52. Развитие плакоидной чешуи.
Ь — основная пластинка; d — дентин шипа; р — сосочек; sc —
склеробласты.
изнутри. Клетки этой мезодермальной выстилки носят название склеро-
бластов. Первая закладка самой плакоидной чешуи обнаруживается в виде
утолщения основной перепонки, подстилающей эпидермис. Таким образом,
закладка происходит как раз на границе между эктодермой и мезодермой.
Образующийся слой твердого вещества представляет собой эмаль в том
случае, если оно появляется как результат образовательной деятельности
основного слоя эпидермиса. У селахий, однако, чаще зачаток плакоидной
чешуи появляется сразу в результате совместной деятельности основного,
слоя эпидермиса и слоя мезодермальных склеробластов. Этот первона-
чально развивающийся слой покрывает затем верхушку зуба и очень
тверд; вообще он очень сходен с эмалью, отличаясь от нее главным образом
лишь присутствием тончайших канальцев. В настоящее время это вещество
считают особой разновидностью дентина. Во всяком случае этот витроден-
тин, как его называют, и по своему составу и по происхождению очень
близок к эмали. Дальнейшее нарастание вещества развивающейся.пла-
коидной чешуи происходит изнутри и, следовательно, за счет образова-
тельной деятельности слоя склеробластов. Последние образуют на своей
наружной поверхности все новые массы дентина, и растущий таким обра-
зом шип пробивается через эпидермис наружу. Деятельность эктодермы
отступает теперь совсем на задний план, и значение ее, повидимому,
главным образом формирующее. В дальнейшем развитие плакоидной
чешуи заканчивается уже полностью в пределах мезодермы, где склеро*
бласты, располагаясь в два слоя, выделяют между ними дентиновую пла-
стинку, служащую основанием чешуи. Мезодермальный сосочек оказы-
вается теперь внутри чешуи, образуя его мякоть. Таким образом, плакоид-
ная чешуя является вполне сформированной.
78
эм, эта ткань действительно близка
же встречаются в небольшом коли-
Heftiyfc селахий состоит в основном из дентина, по последний встре-
чается в нескольких модификациях. Эмаль, или более плотный витроден-
тин, весьма сходный с эмалью, покрывает шип в виде колпачка. Главная
масса шипа состоит из типичного дентина, пронизанного тонкими ветвя-
щимися канальцами, в которые проникают протоплазматические отростки
склеробластов. Основание плакоидной чешуи состоит обычно из дентина,
содержащего сложную систему каналов, которые являются продолжением
центральной полости и содержат также мякоть с сосудами и склеробласта-
ми, отростки которых проникают в канальцы дентина. Такой дентин
похож на кость с ее гаверсовыми каналами и носит название трабекуляр-
ного дентина, или вазодентина. В наружном слое основной пластинки
заключаются также характерные для кости колагенные волокна, которые
здесь не обызвествлены. Таким образ
к настоящей кости. Однако в ней все
честве характерные для дентина ка-
нальцы , в которые проникают отрост-
ки склеробластов. Такую ткань, со-
ставляющую главную массу круп-
ных чешуй некоторых скатов (Raja),
называют иногда псевдодентином
(Третьяков). Наконец, иногда ден-
тин принимает слоистое строение и,
включая клеточные элементы, ста-
новится еще более сходным с ко-
стью.
Второй род чешуй — ганоид-
ные чешуи — встречается лишь
у немногих современных «ганоидных»
рыб (у единственных представителей
многоперых — африканских Poly-
pterus и Calamoichthys и одного из
двух родов Holostei — американской
панцырной щуки — Lepidosteus), но
были широко распространены среди
ископаемых. Ганоидные чешуи имеют
вид ромбических очень толстых и
твердых пластинок, тесно соединяю-
щихся друг с другом (особого рода сочленение) в один сплошной панцырь,
покрывающий все тело животного. У различных рыб ганоидная чешуя бы-
вает разная. Ее основание составляется во всяком случае из настоящей ко-
стной ткани характерного слоистого строения, называемой изопедином.
Снаружи она состоит из очень твердого эмалеподобного вещества, которое
может иметь различное строение (рис. 53 и 54). Наиболее примитивным
видом ганоидной чешуи является «космоидная» чешуя, в которой наружный
слой содержит систему полостей, выполненных мякотью. От нее пуч-
ками отходят тонкие канальцы, содержащие только отростки клеток
(склеробластов, находящихся в мякоти). Таким образом, наружный слой
имеет структуру, характерную для дентина. Такое видоизменение дентина
называется космином. На шлифах можно видеть, что космин предста-
вляет собой как бы комплекс большого числа сросшихся между собой
плакоидных чешуй (рис. 53, В). Космоидные чешуи имеются у многих
ископаемых кистеперых рыб и у двудышащих. У некоторых рыб ганоидная
чешуя имеет первоначально сходное же строение, но затем по мере роста
чешуи на ней сверху развиваются один за другим слои плотного дентина,
получающего название ганоина. На разрезе через такую чешую видна
основная масса слоистой кости — изопедина, средняя масса — космина и
наружная масса слоистого дентина — ганоина (рис. 54). Таков тип чешуй
Рис. 53. Происхождение ганоидйой че
шуи (С) путем срастания ряда плане ид
ных чешуй с кожным окостене ние
(Л, В).
d — ганопн; о—изопедин.
79
ископаемых хрящекостных рыб (Palaeoniscidae) и современных мног.опе-
рых (Polypterus, Calamoichthys). Наконец, у других рыб средняя масса
космина редуцируется, и вся чешуя с самого начала имеет слоистое
строение. По мере ее роста концентрически нарастают снизу все новые
слои изопедина, а сверху все новые слои ганоина (рис. 53, С). Такая
чешуя имеется у современной панцырной щуки (Lepidosteus).
Рис. 54. Ганоидная чешуя Eurynotus (по Гудричу).
А — шлиф через часть чешуи при большом увеличении; В — общий вид
чешуи с поверхности; С — поперечный разрез через чешую, а — перед-
няя часть чешуи, покрываемая предыдущей; ар — сочленовный от-
росток; с— канальцы в слое космина; g— слои ганоина; Л—система
горизонтальных каналов зубной мякоти; i —слои изопедина; s— на-
ружная поверхность.
Ганоидная чешуя развивается вся за счет мезодермы, и, следовательно,
настоящей амали на ней нет. На поверхности ганоидной чешуи, однако,
развиваются и многочисленные шипики, вполне идентичные шипам пла-
коидпых чешуй, которые потом иногда спадают. Изучение шлифов гано-
идных чешуй различных ископаемых рыб показало также с достаточной
ясностью, что ганоидная чешуя представляет собой продукт срастания
Рис. 55. Разрез через кожу костистой рыбы.
ер — эпидермис; s — костная чешуя
довольно большого числа плакоидных чешуй. Новоприобретением гано-
идной чешуи является лишь настоящее костное вещество (изопедин),
образующее нижнюю половину чешуи. Это костное вещество рассматри-
вают как кожное окостенение, с которым срослось известное число пла-
коидных чешуй (рис. 53), но можно также предполагать, что сам дентин,
составлявший основания атих плакоидных чешуй, превратился путем
включения клеточных элементов из остеодентина в настоящую кость.
Во всяком случае участие плакоидных чешуй в образовании ганоидной
80
чешуи не возбуждает никаких сомнений. Ганоидные чешуи являются
постоянными органами и смене не подлежат.
Третий род чешуй — костные чешуи — имеется у всех совре-
менных костистых рыб и встречается также и у ганоидных рыб. Костные
чешуи состоят из простой, обычно круглой и тонкой пластинки костного
вещества, лежащей в кориуме. Обычно такие чешуи помещаются в кожных
складках и черепицеобразно налегают друг на друга, так что снаружи
виден только задний край каждой чешуи (рис. 55). Этот задний край бы-
вает гладким, и тогда чешуя называется циклоидной, или он зазубрен,
и тогда "чешую называют ктеноидной. Как и ганоидные чешуи, костные
чешуи развиваются под эпидермисом за счет мезодермальных склеробла-
стов. Костные чешуи связаны целым рядом переходных форм с ганоид-
ными чешуями ископаемых рыб. Кроме типичных ромбических ганоидных
чешуй, известны и круглые циклоидные чешуи того же строения, и на
ряде переходных форм можно проследить постепенное утончение этой
чешуи и редукцию верхнего ганоинового слоя. На костной циклоидной
чешуе современной ильной рыбы (Amia calva) имеется лишь тонкий
поверхностный орнамент из ганоина. Тонкий покров из ганоина имелся
еще и на циклоидной чешуе древнейших ископаемых костистых рыб.
Покровный слой чешуй современных костистых рыб обнаруживает также
известное сходство с ганоином. Дальнейшее утончение циклоидной чешуи
приводит у некоторых современных форм к утрате всяких каналов, а
иногда даже к утрате костных телец, так что от нее у большинства костис-
тых рыб остается лишь одна костная пластиночка без всяких полостей.
Костные чешуи, как и ганоидные, смене не подлежат. Своеобразные
чешуи осетровых рыб состоят также из настоящей кости.
Однако и у современных костистых рыб (сомовые) иногда еще встре-
чаются настоящие плакоидные чешуи с шипом, состоящим из дентина
с мякотной полостью внутри; вершина его выдвигается через эпидермис
наружу и покрыта колпачком из эмали. Основание шипа подвижно со-
членяется с лежащими под ним кожными костями.
Таким образом, все виды чешуй рыб оказываются генетически друг
с другом связанными.
Роговые чешуи
Основное значение рогового покрова наземных позвоночных состоит
в предохранении кожи от высыхания и ее защите от повреждений. Значи-
тельное утолщение и видоизменение рогового слоя приводят у высших
позвоночных к образованию особых органов, служащих для защиты
тела от механических повреждений и несущих, таким образом, функцию
наружного скелета, или к развитию более сложного покрова, сокраща-
ющего также теплоотдачу тела животного. Образования первого рода
распространены в особенности у рептилий, у которых они образуют
нечто вроде панциря из ч е ш у й и щитков; второе наблюдается
у птиц и млекопитающих (перья и волосы).
У рептилий утолщение рогового слоя происходит небольшими обыкно-
венно участками, между которыми он остается более тонким, благодаря
чему сохраняется подвижность животного (рис. 56). Таким образом
развиваются чешуи в виде бугорков — бугорчатые чешуи, представля-
ющие собой наиболее примитивную форму чешуй (гекконы, хамелеоны,
гаттерия) или в виде пластинок, налегающих своим задним краем друг
на друга, — черепицеобразные чешуи, характерные в особенности для
ящериц и змей (Squamata). Иногда чешуи преобразуются в шипы
и различного рода сложные выросты. На неподвижных частях тела, на-
придТ'* в» голове у всех рептилий, а также на спинном и брюшном щите
че£о °К СТСр’ ,ва10ТСЯ более крупные плоские роговые пластинки — щитки.
IJr1 , сходясь л'ИНОГда наблюдаются соответствующие им окостенения
Р°Д°1е^ 81
в мезодермальной части кожи; такие костные пластинки наблюдаются
иногда у ящериц (гекконы и особенно сцинки) и очень хорошо развиты,
Рис. 56. Схемы роговых чешуй (по Боасу, ивменено).
А — бугорчатые чешуи. В — шитки, С и D — черепицеобравные чешуи; с — роговой слой
и m — мальпигиев слой эпидермиса; os — окостенения.
кроме черепах (щиты), у крокодилов, где костным пластинкам соответ-
ствуют также плоские роговые щитки. У других позвоночных роговые
чешуи обыкновенно не образуют общего покрова тела: у птиц они покры-
вают только нижние части ног,
а у млекопитающих встреча-
ются только в виде исключе-
ния, особенно на хвосте (боб-
ры, крысы и др.; рис. 56).
Перья
У птиц тело покрыто ро-
говыми образованиями более
сложного строения — перья-
м и. Наиболее крупные и
сложные контурные перья со-
стоят из прочной оси,’поддер-
живающей легкую пластинку
сложного строения — опахало.
Основание оси образовано
полым внутри цилиндром—ОТИ-
gpM». конец которого погружен
в мешочек кожи (сумку). Осталь-
ная часть ствола представляет
собой стержень, несущий по
бокам по ряду тонких пластин-
чатых веточек — бородок, несу-
щих в свою очередь на обращен-
ных друг к другу сторонах
Рис. 57. Схема строения пера птицы
(по Бючли).
8 — ствол; Ва — бородки; rd — бородочки; г —
крючочки.
веточки второго порядка, или
бородочки; последние снабже-
ны крючочками, посредством
которых все бородочки проч-
но между собой сцеплены,
образуя одну цельную пла-
стинку (опахало) (рис. 57). Перо такого устройства оказывается вйзду-
хоупорным образованием, соединяющим значительную прочность и упру-
гость с необычайной легкостью — свойства, весьма важные для лекция.
Такие контурные перья определяют форму птицы; особенно они развиты
на крыльях (маховые перья) и на хвосте (рулевые). Кроме того, наблюда-
82
ются еще перья с менее прочным стержнем, бородки которых крючками
не сцеплены, — это пуховые перья или, если стержень очень короток,
так что бородки отходят все вместе как бы кисточкой, — пух. К сумке
пера подходят пучки гладких мышц, сокращением которых перо может
приподыматься. Перья периодически сбрасываются и заменяются новыми.
Развитие пера. Молодой зачаток пера появляется в виде
скопления мезодермальных клеток, приподнимающих эпидермис в форме
бугорка, который затем начинает
расти назад (рис. 58). На этих ста-
диях развития зачаток пера вполне
сходен с зачатком чешуи. Затем
кожа, окружающая основание зачат-
ка, углубляется, образуя влагалище
пера, а сам зачаток состоит из цен-
тральной массы мезодермы (мякоть
пера), окруженной довольно тол-
стым эпидермисом. Поверхностный
слой эпидермиса состоит из плос-
ких клеток, глубокий — из цилин-
дрических. В эпидермисе образу-
ются продольные утолщепия, кото-
рые более или менее глубоко вда-
ются в мякоть пера и затем вполне
С
Рис. 58. Разрез черев три последова-
тельные стадии развития пера.
с — роговой слой; т — мальпигиев слой;,
у—зачатой влагалища Пера; р— сосо-
чек пера.
отделяются от поверхностного слоя,
образующего чехлик на молодом пере _
вают и дают бородки эмбрионального пуха, которые свободно распа-
даются на коротком стержне, когда покрывающие чехлик сбрасывается.
Мезодермальная мякоть пера отстает от эпи-
в виде сосочка у
обильно снабжается
питающими
(рис. 59). Сами утолщения орогове-
расту-
Рис. 59. Развитие бородок вмбрионального пуха.
А и В—поперечные разрезы через развивающееся перо; С—продольный
разрез; с — роговой слой; М — мальпигиев слой; р — сосочек пера; s —
бородки.
дермиса сохраняется
его основания. Сосочек
кровеносными сосудами,
щее перо.
Эмбриональный пух заменяется затем дефинитивным пером, которое
вырастает под ним на том же сосочке и постепенно его сталкивает. Разви-
тие такого пера идет по существу таким же образом с той разнипей, что
утолщения, дающие начало бородкам, идут не прямо вдоль зачатка.
Одно7 продольное утолщение развивается сильнее и представляет собой
зачаток стержня (рис. 60). По бокам последнего зачатки бородок идут
косо, сходясь легкой спиралью к стержню, и таким же образом зачатки
бородочек развиваются под некоторым углом к первичным бородкам;
83
Когда чехлик спадает и молодое перо постепенно выдвигается, то бородки
оказываются перисто расположенными по бокам стержня.
По своему эмбриональному развитию перо обнаруживает на ранних
стадиях такое большое сходство с развитием роговой чешуи, что перо
рассматривают как весьма усложненную чешую. Доказательством в
щользу происхождения пера от чешуи могут служить факты существования
переходных форм между чешуей и перьями на ногах у некоторых птиц
(страусы), а также факт обратного упрощения пера при его редукции
у пингвинов, у которых перья имеют форму перистозазубренных чешуй.
Кроме того, перья пингвинов линяют особым образом — старое перо
сбрасывается в виде чехлика с раз-
вившегося под ним нового, подобно
чешуям рептилий.
Волосы
Если для рептилий характерен
покров из роговых чешуй, для птиц—
покров из перьев, то для млекопита-
ющих не менее характерен волосяной
покров, служащий прежде всего, как
отчасти и перья птиц, для защиты
тела животного от потери тепла.
Волосы — тоже роговые образо-
вания довольно сложного строения
6’
Рис. 60. Стереограмма развиваю-
щегося контурного пера (ив Гессе),
г — зачаток стержня; в — бородки.
(схема по Видерсгейму).
Ар — мышцы, подымающие волос; Со — кожа
собственно; F — волокнистая сумка волоса; Fb —
жировав ткань; GH — пограничная перепонка;
HBD — сальные железы; HP — сосочек волоса
с сосудами; М — сердцевина; О — кутикула
волоса; R — корковое вещество; Sc — роговой
слой эпидермиса; Sch — стержень волоса; SM —
мальпигиев слой эпиде рмиса; WS. WS'— наруж-
ное и внутреннее влагалища волоса.
(рис. 61). В волосе различают нижнюю, сидящую глубоко в коже вздутую
часть — луковицу и выступающий наружу длинный цилиндрический
стержень. Нижняя часть волоса и луковица, образующие вместе «корень»
волоса, сидят в «сумке», состоящей из наружной соединительнотканной
части и внутреннего эпидермического влагалища (рис. 61, VE51). Различают
наружное влагалище волоса, представляющее собой продолжение эпи-
дермиса с его слоями (слой цилиндрических клеток обращен кнаружи,
а внутри лежат слои плоских клеток), и внутреннее влагалище, образу-
ющее как бы заворот эпидермиса на поверхность волоса. Во внутреннем
влагалище различают слой Генле, слой Гексли и прилежащий к самому
волосу слой плоских клеток — «кутикулу» влагалища. В промежуток
84
между волосом и его влагалищем открываются протоки сальных желез,
служащих для смазывания поверхности»волоса. Луковица волоса состоит
из индиферентных клеток, путем размножения которых волос непрерывна
растет. В луковицу снизу вдается богатый сосудами сосочек кориума,
служащий для питания растущего волоса. Стержень волоса состоит и»
следующих частей: 1) наружной кожицы, или «кутикулы» волоса, из
плоских черепицеобразно налегающих друг на друга клеток, 2) корко-
вого вещества, в котором клетки вытянуты вдоль оси волоса, и 3) сердце-
винного вещества, клетки которого вытянуты поперек. В сердцевине
иногда заключаются пузырьки воздуха, в корковом веществе — пигмент.
Клетки волоса ороговевают. Когда рост волоса прекращается, луковица
его зарастает, ороговевает и
спадает, а на его месте в той
же сумке развивается новый
сосочек и на нем — новый
волос.
Волосы сидят наклонно,
и к их сумкам подходят
пучки гладких мышц (mu-
sculi arrectores pilorum), слу-
жащие для приподнимания
волос (иннервируются сим-
патическими нервами). У
млекопитающих различают
несколько родов волос.
Обыкновенно более длинные
и жесткие волосы образуют
шерсть (ость), а расположен-
ные между ними более ко-
роткие и мягкие — подшер-
сток, который особенно раз-
вивается в зимнем наряде.
Более толстые волосы обра-
зуют щетину (свиньи) и иглы
(ежи, дикобразы и др.). По-
следние обычно снабжены
Рис. 62. Развитие волоса. Четыре последова-
тельные стадии (ив Гессе).
со — мезодермальная часть кожи; ер — эпидермис;
f — зачаток влагалища волоса; g — зачаток саль-
ной железы; р—сосочек волоса;;.. pl— зачатов
волоса.
поперечнополосатыми мыш-
цами (подкожной мускулатуры). Осязательные волосы («vibrissae» ^наблю-
даются на щеках и других местах; чаще всего они образуют на верхней
губе млекопитающих, в особенности у хищных, так называемые усы.
Эти волосы обладают также поперечнополосатыми мускулами, их влага-
лища обильно снабжены нервами (ветвями тройничного нерва) и окру-
жены венозными лакунами. Впрочем, волосы вообще обладают довольно
богатой иннервацией и могут служить частью общего осязательного
аппарата.
Развитие волоса в отличие от развития чешуи и пера начи-
нается в эпидермисе. Некоторая группа клеток приобретает более высокую
цилиндрическую форму и начинает деятельно размножаться (рис. 62).
Эпидермический зачаток погружается глубоко в мезодермальную часть
кожи, и затем в массе его клеток начинается диференцировка, — наруж-
ные слои дают начало наружному и внутреннему влагалищу (а также
и сальным железам, образующимся в виде боковых выростов зачатка),
а глубокая масса клеток превращается в зачаток собственно волоса.
Снизу в зачаток вдается питающий его мезодермальный сосочек. Клетки
зачатка волоса начинают ороговевать, и по мере того как волос нарастает
снизу за счет деятельности луковицы, конец его пробивается через влага-
лище наружу.
55
Таким образом, по своему развитию во—с не напоминает нам каких-
либо других роговых образований кожи, — если происхождение волоса
от роговой чешуи и должно считаться весьма вероятным, то пока мы не
имеем еще доказательств в пользу такого взгляда. Все другие воззрения
на происхождение волоса являются также лишь гипотезами, иногда
{весьма мало обоснованными. Наиболее популярна гипотеза Маурера,
согласно которой волосы развились путем ороговения из органов
боковой линии. Молодые зачатки волос, действительно, обнаружи-
вают известное сходство с органами боковой линии, и в некоторых случаях
клетки последних с прекращением их деятельности ороговевают (при
метаморфозе у амфибий). Однако эта гипотеза встречает слишком большие
затруднения вследствие того, что у рептилий, от которых несомненно
произошли млекопитающие, органов боковой линии уже не было.
Если, однако, вопрос о происхождении волоса млекопитающих пока
еще является неразрешенным,
Рис. 63. Кожа хвоста мыши с че-
шу ями (sg) и группами волос (tr)
между ними (по Плате).
то одно можно установить с несомнен-
ностью — предшественником волосяного
покрова был покров из чешуй. Это до-
казывается не только тем, что у многих
млекопитающих еще встречаются роговые
чешуи, в особенности на хвосте (многие
сумчатые, насекомоядные и грызуны), а
иногда закладываются провизорные че-
шуи, которые после исчезают (на хвосте у
мадагаскарского насекомоядного, тенрека
Centetes), но также и расположением во-
лос у зародышей многих млекопитающих.
В тех случаях, когда одновременно с
волосяным покровом имеются и чешуи, волосы располагаются правиль-
ными группами, обыкновенно по три волоса, из которых средний, са-
мый крупный, — позади каждой чешуи (рис. 63). У млекопитающих,
не имеющих чешуй, сохраняется все же расположение волос правильно
чередующимися группами, как-будто бы они находились позади чешуй. У
зародышей млекопитающих эти группы обычно состоят из трех во-
лос, из которых средний — более крупный. Позже число волос в каж-
дой группе увеличивается. И у человеческого зародыша в четыре
месяца волосы располагаются ясными поочередными группами по 3 и 5
волос, из которых средний — более крупный. Эти группы ясно указы-
вают на расположение чешуй у отдаленных предков млекопитающих.
У всех млекопитающих во время эмбриональной жизни развивается
сплошной волосяной покров — lanugo foetalis, который сменяется затем
постоянным. У человека lanugo на пятом и шестом месяце покрывает
все тело и все лицо (в виде аномалии может сохраниться и у взрослого).
Вообще тело человека и во взрослом состоянии сплошь покрыто
волосами (исключая ладони и подошвы ног), как и кожа большинства
млекопитающих, но волосяной покров здесь значительно редуцирован.
Физиологическую роль волосяного покрова — защищать тело животного
от потери тепла — в некоторых случаях берет на себя вообще утолщение
эпидермиса (слоны, носороги) или особо развитый подкожный слой жира
(киты). В этих случаях волосяной покров редуцируется и у китов, напри-
мер, почти совсем исчезает. Редуцируется волосяной покров и у чело-
века. Произошла эта редукция, вероятно, тогда, когда предки человека
жили в теплых странах с ровным климатом.
Местные роговые образования, когти, рога
Кроме такого общего покрова, каким являются чешуи рептилий,
перья птиц и волосы млекопитающих, у наземных позвоночных наблю-
даются еще местные утолщения рогового слоя различного значения.
86
I ак, i . челюстях черепах, птиц и клоачных млекопитающих имеется
толстый роговой покров, образующий клюв и заменяющий отсутствую-
щие у этих животных зубы.
Роговые утолщения на нёбе млекопитающих дают так называемые
нёбные пластинки. Сильное развитие последних приводит у
беззубых китов к образованию больших расщепленных роговых пластинок,
свешивающихся с нёба вглубь ротовой полости, служа особого рода
цедилкой; вто так называемый китовый ус.
Рис. 64. Продольный разрез черев ноготь (Л), коготь (В) и копыто (С) (по Боасу).
Ь — подушечка пальца; п — ногтевая пластинка; ph — конечная фаланга пальца; в —•
подошвенная пластинка; v — когтевой валик.
На концах пальцев уже у амфибий замечается некоторое утолще-
ние рогового слоя, которое иногда имеет вид настоящих когтой (Dactyle-
thra из бесхвостых и Onychodactylus из хвостатых амфибий). У рептилий
и птиц все пальцы, как правило, вооружены когтями, охватывающими
их концы в виде чехлика. Когти по происхождению являются орга-
нами, защищающими концы пальцев ог повреждений; в дальнейшем они
становятся вспомогательными органами при передвижении животного
и несут различные связанные с этим функции, и, наконец, у млеко-
С
Рис. 65. Развитие рогов у овцы (по А. Брандту).
1 — лобная кость; 2 — окостенение (os cornu); 3 — рог.
питающих они иногда превращаются в органы активной защиты жи-
вотного и даже в орудия нападения. Когти более или менее заострены
на концах и слегка изогнуты. Верхняя стенка когтя состоит из более
твердого рогового вещества (когтевая пластинка); нижняя стенка не-
сколько мягче (подошвенная пластинка). Наибольшего развития дости-
гают когти у млекопитающих, у которых диференцировка на когтевую и
подошвенную пластинки очень резко выражена (рис. 64, В). Так как
верхние слои когтя оказываются наиболее твердыми, а нижние, мягкие,
слои изнашиваются быстрее, то верхний край всегда сильнее выступает,
и таким образом коготь всегда остается острым. Особенно развиты когти
87
у хищных, у которых они нередко (у кошек) движением конечной фа-
ланги могут втягиваться внутрь особой сумки на тыльной стороне
пальца. Основание когтевой пластинки покрыто кожной складкой— ногте-
вым валиком, а снизу конец пальца вздут в виде подушечки. Ноготь
представляет собой плоский коготь, подошвенная пластинка которого
Рис. 66. Развитие рогов оленей (схемы
по Нитше).
1 — развитие первого рога; 2 —кожа сбро-
шена, начало резорбции кости; За. ЗЬ —
первый рог сбрасывается; 4, 6, 6 — аажи-
вление раны и развитие второго рога; 7—
развитие второго рога закончено, кожа
сброшена. Сот—cerium; Kpld—epidermis.
сильно редуцирована (рис. 61, Л).
Копыто — это также видоизменен-
ный коготь, утолщенная когтевая
пластинка которого образовала стен-
ку копыта, подошвенная пластин-
ка — подошву копыта, а подушечка
вдается в подошву в виде «стрелки»
(рис. 64, С). Копыто служит наиболее
прочным защитным покровом концов
пальцев.
На костных выростах лобных
костей у млекопитающих нередко
сидят рога. У полорогих копыт-
ных — это толстые роговые чехлы,
имеющие постоянный характер (за
исключением Antilocapra, у которой
такие рога периодически сбрасыва-
ются). Костные выросты, на которых
сидят рога, образуются при участии
особого кожного „окостенения (os
cornu), которое прирастает к черепу
(рис. 65). У других копытных роль
кожного покрова рогов отступает на
задний план по сравнению со значе-
нием самой кости. У жираф костяные
рога одеты мягкой кожей. У оленей
только молодые рога одеты кожей с
волосами, но затем кожа отмирает и
спадает (рис. 66). Рога оленей, имею-
щиеся обычно только у самцов, пе-
риодически сбрасываются вслед-
ствие разрушения кости у основания
рога. Затем к периоду половой деятельности у самца вырастают новые
рога, обыкновенно несколько более сложной формы.
Кожный скелет
На различных частях тела ганоидных рыб отдельные кожные чешуи
срастаются между собой или разрастаются, образуя более крупные пла-
стинки. Такие пластинки, располагаясь в ряд по бокам и вдоль спины
осетровых рыб, образуют так называемые жучки. Подобные же разрос-
шиеся чешуи покрывают область плечевого пояса и голову. Здесь на
децо^вижных частях тела кожные чешуи разрастаются, еще .больше-и
обрязуют~к. о ж н ы е к о с т и? У древнёй1пйх~рыб~кожные кости головы
часто еще сравнительна невелики, имеют не вполне установившееся рас-
положение и весьма многочисленны (как отчасти и у современных осе-
тровых рыб, а также и у упомянутой уже панцырной щуки Lepidosteus).
У более высоко стоящих форм кожные кости получают весьма по-
стоянное расположение. Располагаясь сначала непосредственно в самой
коже, они обнаруживают ту же структуру, что и кожные чешуи данной
формы, т. е. у рыбы с типическими ганоидными чешуями (как совре-
менные Polypterus и Lepidosteus) и кожные кости плечевого пояса
88
Рис. 67. Врюшной панцирь стего-
цефалов (по Креднеру и Циттелю).
А — Branchlosaurus; В — подроб-
ности того же; С — Archegosаигиз;
D — Petrobates
и череда оонаруживают то же строение из нижнего костного слоя и верх-
него слоя типичного ганоина. На таких костях обычно развиваются и
шипы. Пока кости располагаются 'в самой коже на поверхности обычно
имеется та же скульптура, которая характерна для кожных чешуй
данной формы. Одновременно с исчезновением ганоинового покрова на
чешуях он редуцируется и на кожных костях, так что у рыб с костными
чешуями и кожные кости состоят из одного только костного вещества.
У некоторых костистых рыб как в чешуях, так и в этих костях вторично
исчезают и каналы, и клеточные элементы.
Появившись в коже, кожные кости, однако, у более высоко стоящих
форм погружаются глубже и вступают в более тесную связь с частями
внутреннего скелета — черепом и плече-
вым поясом, образуя на нем так называе-
мые покровные кости.
Аналогично образованию кожных
костей на наружной поверхности тела
происходит образование подооных же ко-
стей и внутри ротовой полости7~3дёсь бо-
лее или менее обширные костные пластин-
ки развиваются в связи с основаниями
зубов, которые, как мы видели, представ-
ляют собой лишь специализированные
плакоидные чешуи. Такие кости, назы-
ваемые по своему происхождению з у б-
ными костями, также могут утра-
тить свою первоначальную связь со сли-
зистой оболочкой ротовой полости, по-
гружаются глубже и, вступая в связь с
частями внутреннего скелета, образуют
покровные кости дна черепа и челюстей.
В некоторых случаях даже у наземных
позвоночных такие кости еще ясно обна-
руживают свое происхождение в связи с
зубами, как это наблюдается во время
эмбрионального развития их у амфибий.
У наземных позвоночны
следы покрова из кожных чешуй. Панцырь из чешуй циклоидного
типа имелся у многих ископаемых амфибий — стегоцефалов, иногда на
всем теле, иногда только на брюхе (рис. 67). Из современных земновод-
ных остатки кожных чешуй сохранились лишь у гимнофион в виде очень
маленьких костных пластинок, лежащих глубоко в коже (в задней поло-
вине каждого кожного кольца).
Но если у современных наземных позвоночных, как правило, общий
чешуйчатый покров исчез, то у всех у них сохранились его производные:
покровные кости черепа и плечевого пояса являются постоянными остат-
ками кожного панцыря рыб, точно так же как находящиеся в ротовой
полости зубы являются остатками древнейшего покрова из плакоидных
чешуй. У некоторых рептилий сохраняются, сверх того, еще несомнен-
ные остатки брюшного панцыря стегоцефалов. Это так называемые брюш-
ные ребра (gastralia) гаттерии, крокодилов и многих ископаемых рептилий
(рис. 397, 403), представляющие собой, несомненно, кожные кости. По
своему расположению «брюшные ребра» очень сходны С косыми рядами
удлиненных чешуй брюшного панцыря некоторых стегоцефалов (рис.
67, D). Те же элементы приняли, невидимому, участие и в образовании
брюшного щита черепах.
Кроме таких унаследованных от рыб кожных окостенений, у назем-
ных позвоночных наблюдаются и новые окостенения. Так, встречаются
89
х лишь редко сохраняются
кожные окостенения у некоторых амфибий (например, на голове) и у
рептилий (костные пластинки под роговыми чешуями некоторых ящериц
и в коже крокодилов). Очень сильно развиты кожные окостенения у че-
репах, где они, срастаясь с частями внутреннего скелета, дают начало
прочному панцырю из двух больших щитов — спинного (сагарах) и брюш-
ного (plastron). Спинной щит образован рядом непарных спинных плас-
тинок (neuralia), сросшихся с остистыми отростками позвонков, и пар-
ным рядом больших реберных пластинок (costalia), сросшихся с соответ-
ствующими ребрами; край щита образован особыми краевыми пластин-
ками (marginalia). Брюшной щит состоит из одной непарной кости (endo-
plastron) и четырех больших парных пластинок (epi-, hyo-, hypo- и xyphi-
plastron). Из этих костей непарная кость (endoplastron) и передняя пар-
ная (epiplastron) представляют собой кожные кости, тесно связанные у
других позвоночных с грудиной и плечевым поясом — надгрудинник
(episternum) и ключицы (claviculae). И у млекопитающих встречаются
в виде исключения новые кожные окостенения. Примером очень значи-
тельного их развития является панцырь броненосцев.
Из двух слоев, составляющих покровы животных,—эктодермального
эпидермиса и подстилающего его соединительнотканного слоя — у бес-
позвоночных явно преобладает значение однослойного эпидермиса и его
производных, причем выделяемая им кутикула является наружным ске-
летом животного.
У позвоночных эпидермис всегда многослойный и кутикулы никогда
не выделяет; большое значение приобретает мезодермальный кориум.
Эпидермис рыб содержит многочисленные отдельные железистые
клетки. Многоклеточные железы развиваются лишь в коже наземных
позвоночных; у амфибий — это простые мешковидные органы, а у млеко-
питающих наблюдаются железы двух типов—трубчатые потовые железы,
гомологичные железам амфибий, и гроздевидные сальные, представляю-
щие собой новое приобретение млекопитающих. Млечные железы — это
преобразованные потовые железы.
Кожный скелет позвоночных в простейшем случае представлен плако-
идными чешуями, построенными из дентина и развивающимися на гра-
нице между эктодермой и мезодермой в результате образовательной дея-
тельности мезодермальных склеробластов. Верхушка плакоидной чешуи
бывает покрыта слоем эмали, образуемой эктодермой. В результате сра-
стания известного числа плакоидных чешуй с кожным окостенением раз-
вилась ганоидная чешуя, которая через редукцию дентинового слоя пре-
образовалась у высших рыб в костную чешую. Прогрессивное развитие
плакоидных чешуй на челюстях связано со сменой функции и привело к
образованию зубов позвоночных.
Диференцировка эпидермиса приводит у наземных позвоночных
к образованию более или менее значительного рогового слоя. Утолщения
последнего па концах пальцев — когти и их видоизменения — копыта
и ногти. Па челюстях у беззубых форм развивается клюв, а на лбу у
некоторых млекопитающих — рога. В образовании рогов могут принять
участие и кожные окостенения, которые у оленей образуют весь рог.
За счет того же рогового слоя у рептилий развивается общий покров из
чешуй, продуктом усложнения которых являются перья птиц и волося-
ной покров млекопитающих. У птиц и млекопитающих образуется лег-
кий густой покров, прекрасно защищающий животное от потери тепла.
Путем срастания чешуй между собой, а также разрастания отдельных
чешуй возникли на поверхности тела (в области головы и плечевого пояса)
кожные кости, которые, погружаясь под покровы, вступают в тесную
связь с внутренним скелетом. Подобным же образом в ротовой полости
развились покровные кости как окостенения внутри слизистой оболочки,
90
связанные с основаниями зубов. Следовательно, зубы и покровные кости
являются у наземных позвоночных единственным остатком кожного пан-
циря рыб.
ВНУТРЕННИЙ СКЕЛЕТ
Скелет беспозвоночных
Наиболее примитивной формой внутреннего скелета должна считаться основ-
ная перепонка, находящаяся у кишечнополостных между эктодермой и энтодермой
и выделяемая клетками обоих листков совместно. У медув эта перепонка достигает
вначительной мощности, образуя бесклеточную массу, — так называемую галлерту
их зонтика. Галлерта имеет иногда студенистый характер, а иногда почти достигает
плотности хряща. Гомогенная у низших медуз, она переполняется у высших тонкими
волокнами. Наковец, у последних в галлерте появляются и разветвленные клетки
(мезенхима), и таким образом бесклеточная опорная масса превращается в студенистую
соединительную ткань. У кишечнополостных встречаются и другие виды скелета.
В щупальцах иногда имеется плотная скелетная ось из клеток энтодермальвого про-
исхождения. Еще большее значение имеют у кишечнополостных известковые отложе-
ния клеток мезенхимы и «роговой» или известковый скелет, развивающийся в экто-
дермальных складках подошвы коралловых полипов (являющийся, следовательно,
наружным).
У губок скелетные образования представляют собой продукт выделения клеток
мезенхимы и состоят из известковых, или кремневых игл определенной формы
и расположения, или ив переплетающихся «роговых» (спонгиновых) волокон. Очень
сильного развития достигают известковые отложения в теле иглокожих. Они возни-
кают также в виде правильных структур как продукт выделения мезодермальных кле-
ток. Этот скелет является в главной своей массе кожным.
У других беспозвоночных преобладающее значение имеет, как мы уже видели,
наружный кутикулярный скелет. Хитиновый скелет членистоногих является не только
органом защиты, но служит также местом прикрепления их сложно диференцирован-
ной поперечно-полосатой мускулатуры. Встречаются и другие структуры, имеющие,
несомненно, значение очень примитивного внутреннего скелета и генетически свя-
занные с основной перепонкой и галлертой кишечнополостных. У червей обычно на-
блюдается система пластинок, тяжей и пограничных перепонок, развивающихся не-
редко всюду, где соприкасаются два органа (и, в частности, в диссепиментах). Эти
перепонки имеют характер бесклеточной опорной массы и играют ту же роль, что со-
единительная ткань у позвоночных животных. Но встречается иногда специфическая
скелетная ткань клеточного строения, например, в жабрах некоторых аннелид, и на-
конец, как высшая форма такой ткани — настоящий хрящ. Последний наблюдается
иногда у червей и в особенности у моллюсков, но только у головоногих моллюсков
из него построены более значительные скелетные образования — нечто вроде хряще-
вого черепа, защищающего нервные узлы, глаза и органы слуха, особый спинной
хрящ, хрящи у основания плавников и др.
У ланцетника, однако, за исключением хорды нет клеточных скелетных образо-
ваний. Все органы отделяются друг от друга бесклеточным или почти бесклеточным
опорным веществом, выделяемым соседними епителиями. Более прочная «хитинопо-
добная» или «хрящеподобная» бесклеточная масса образует жаберный скелет, раз-
вивающийся путем уплотнения основной перепонки, и лучи непарного плавника.
Скелет позвоночных
Центральная скелетная ось хордовых животных — хорда — состоит
из • совершенно особой ткани. Материалом, из которого строится весь
остальной внутренний скелет позвоночных, является волокнистая соеди-
нительная ткань, хрящ и кость. Все эти ткани появляются в тех же мес-
тах, где у ланцетника имеется содержащая волокна бесклеточная или почти
бесклеточная опорная масса: осевой скелет развивается вокруг хорды
и центральной нервной системы, висцеральный скелет — на-месте жабер-
ной решетки и скелет конечностей — в виде хрящевых лучей в плавни-
ковых складках. Таким образом, бесклеточное опорное вещество фило-
генетически является предшественником клеточных тканей позвоночных,
и нужно думать, что путем включения клеточных элементов более рыхлая
ткань ланцетника превратилась в типичную плотную соединительную
ткань, а более прочная «хрящеподобная» ткань — в настоящий хрящ.
Еще более твердый скелетный материал появляется у низших рыб
сначала только в коже. Как плакоидная чешуя представляет собой наи-
91
более примитивный вид кожного скелета, так и составляющий ее дентин
представляет собой древнейший твердый скелетный материал позвоноч-
ных. Развитие плакоидной чешуй показывает нам, что дентин появляется
путем преобразования опорной перепонки, подстилающей эпидермис,
и в дальнейшем нарастает в результате образовательной деятельности
мезодермальных склеробластов. Позже развитие может перейти целиком
в пределы мезодермы. Дентин, как и другие виды древнейших скелетных
тканей, является бесклеточным образованием. Существование переход-
ных форм между дентином и костью, а также история происхождения
ганоидной чешуи (нижняя половина которой является костной) путем
слияния плакоидных, позволяют нам думать, что во время дальнейших
филогенетических преобразований дентин посредством включения колла-
генных волокон, а затем и клеточных элементов превратился в кость. По-
следняя, будучи окружена со всех сторон мезодермой, находилась в луч-
ших условиях питания и роста, вследствие чего постепенно вытеснила
и заместила дентиновый скелет (типичный дентин нарастает и питается
только со стороны мякоти). Кость вначале возникла также непосредственно
под эктодермой в связи с основной перепонкой (как костные чешуи неко-
торых рыб), но затем постепенно эмансипировалась от нее и окончательно
погрузилась в мезодерму, клетки которой приобрели способность выделять
костное вещество не только в самой коже, но и в других местах. Во всяком
случае и типичная кость появляется раньше всего в коже, а лишь затем
и во внутреннем скелете.
Способность выделять известь возникает, впрочем^ и в мезодерме
внутреннего скелета довольно рано, но у низших рыб приводит не к обра!-
зованию кости, а лишь к появлению обызвествлений. Отложения извести
довольно широко распространены в хрящевом скелете селахий, где они
имеют обыкновенно очень правильную форму. В позвонках они имеют
вид цилиндров, снабженных иногда правильными радиальными гребнями,
а в черепе—вид многогранных пластинок лучистого строения. Эти отло-
жения извести не изменяют, однако, характера ткани.
С появлением настоящей кости во внутреннем скелете значение хряща
падает, и он постепенно вытесняется и замещается костью. Филогенети-
чески это происходило двояким путем: 1) отдельные кожные кости
погружались вглубь и иногда вступали в тесную связь с хрящевым ске-
летом в качестве покровных костей; главная роль переходила к кости,
состоящей из более прочного материала, а лежащий под ней хрящ терял
свое значение и редуцировался; 2) отдельные склеробласты погружались
вглубь и входили в состав перихондрия хрящевого скелета, где на поверх-
ности хряща стали выделять слой кости. Кость как более прочный мате-
риал развивалась прогрессивно и иногда даже врастала внутрь хряща,
а хрящ отставал в развитии и редуцировался. Первым путем развились
так называемые «вторичные», или, лучше, покровные кости (ossa inyes-
tientia); вторым- путем появились «первичные», или замещающие, кости
сразу во внутреннем (первичном) скелете (ossa substituentia). Во время
эмбрионального развития различие в происхождении костей обоего рода
всегда ясно обнаруживается.
Покровные кости развиваются в соединительной ткани
в результате образовательной деятельности остеобластов (представля-
ющих собой те же склеробласты), располагающихся сплошным слоем
сверху и снизу вокруг развивающейся костной пластинки (так же как и
костной чешуи). По мере роста кости отдельные остеобласты включаются
в кость, становясь костными тельцами. Покровная кость развивается не-
зависимо от первичного (хрящевого) скелета и с хрящом непосредственно
никогда не связана. Если покровная кость вторично вступает в очень тес-
ную связь с первичным скелетом, то она все же всегда отделена от хряща
по крайней мере, надхрящницей.
92
Замещающие кости развиваются под надхрящницей непо-
средственно на хряще за счет образовательной деятельности остеобластов
надхрящницы, которая тогда получает название надкостницы. Отдель-
ные ее клетки по мере роста кости включаются в нее, превращаясь в кост-
ные тельца. Это — тип перихондрального окостенения. Иногда масса
остеобластов с сосудами врастает внутрь самого хряща, разрушая его
и выделяя на его месте пластинки костного вещества. Это — позднейший
тип эндохондрального окостенения, встречающийся чаще у высших по-
звоночных.
Таким образом, мы имеем два критерия для распознавания покровной
и замещающей кости: 1) эмбриологический критерий — покровная (или
вторичная) кость развивается в соединительной ткани независимо от
хряща, между тем как замещающая (или первичная) кость развивается
под перихондрием на месте хряща (первичного скелета), который она вы-
тесняет; 2) сравнительно-анатомический критерий — покровная кость
есть результат перемещения кожной кости и может быть прослежена
назад до чешуй или зубов, из которых она произошла, между тем как
замещающая кость возникает сразу во внутреннем скелете.
С появлением кости во внутреннем скелете значение хряща, как ска-
зано, падает, и он постепенно редуцируется, замещаясь костью. У высших
позвоночных внутренний скелет является вполне костным и хрящ сохра-
няется только в тех местах, где требуется более гибкий материал. Таким
образом, филогенетически соединительнотканный внутренний скелет за-
мещается хрящевым, а последний — костным. Во время онтогенетиче-
ского развития эта последовательность повторяется, и мы у высших позво-
ночных различаем последовательно стадии перепончатого (соединительно-
тканного), хрящевого и, наконец, костного скелета.
Кости замещающие появляются на месте первоначального хряща.
Таким образом, происходит замена одного материала другим без нару-
шения общей формы данного скелетного образования. Поэтому хрящевой
скелет вместе с возникающими в нем замещающими окостенениями объеди-
няют под названием первичного скелета. Между тем присоединяющиеся
к внутреннему скелету извне покровные кости образуют нередко совер-
шенно новые структуры и поэтому объединяются под названием вторич-
ного скелета. Так, например, в хрящевом черепе позвоночных развива-
ется целый ряд замещающих костей, но развивающийся таким образом
костный череп повторяет форму хрящевого. С другой стороны, развиваю-
щиеся на том же черепе покровные кости дают начало совершенно новому
образованию — вторичной, наружной крыше черепа.
В качестве скелетного материала хрящ и кость имеют каждый свои
особенности. Хрящ растет всей массой, хотя питание внутренних его час-
тей затруднено, он гибок, но не столь прочен, как кость. Пластичность
и приспособляемость хряща очень велики, и расчленение и слияние хря-
щевых элементов происходят очень легко. Кость растет только с поверх-
ности; питание кости обеспечено системой кровеносных сосудов и более
тесной связью клеточных элементов. Кость представляет собой материал
очень прочный, но неподатливый, и поэтому костные скелетные образо-
вания характеризуются множественностью элементов, более или менцр
подвижно между собой связанных. Костные элементы реже срастаются
и не так легко расчленяются, представляя собой более консервативный
материал.
Общий обзор скелета позвоночных1
Скелет позвоночных выполняет в жизни животных двойную функцию.
С одной стороны он защищает животное и его органы от различных
механических воздействий, с другой — служит опорной частью двига-
1 Общий обзор скелета составлен Б. С. Матвеевым.
93
тельной системы организма. Двигательные функции, выполняемые ске-
летными элементами и мускулатурой, разделяются на соматические
(передвижение тела в пространстве) и висцеральные (двигательные
функции, связанные с питанием и дыханием).
Соответственно этому скелет позвоночных животных состоит из трех
разделов: 1) осевого скелета туловища, представленного хордою и позво-
ночником, 2) скелета органов движения — у рыб плавников, у наземных
позвоночных скелета передних и задних конечностей и их поясов, 3) ске-
лета головы — черепа, состоящего из осевого отдела — нейрокраниума
(мозговой череп) и висцерального — спланхнокраниума (челюстной и
жаберный аппарат).
У бесчерепных скелет представлен лишь хордою, окруженною бес-
клеточной, студенистою тканью, и плотными волокнистыми тяжами, под-
держивающими плавники и жаберные щели. У круглоротых, низшего
класса черепных позвоночных, скелет сохраняет наиболее примитивное
состояние. Скелет туловища представлен хордою, одетой плотным соеди-
нительнотканным футляром с зачаточными хрящевыми верхними дуж-
ками. Лишь слабо намечается разделение осевого скелета на туловищный
и хвостовой отделы. Парных плавников нет, а непарные плавники под-
держиваются нерасчлененными хрящевыми нитями. В голове осевой череп
и висцеральный скелет еще не составляют единого черепа. Осевой череп
сохраняет перепончатую крышу. Наравне с примитивными чертами череп
усложнен приспособлением к паразитическому питанию (губные хрящи).
Висцеральный скелет сохраняет почти полную самостоятельность как
скелет переднего отдела пищеварительной трубки, связанный с дыхатель-
ной функцией жаберного аппарата.
Круглоротые принадлежат к бесчелюстным позвоночным (Agnatha).
Все остальные позвоночные составляют особую группу челюстноротых
позвоночных (Gnathostomata), имеющих челюстной аппарат для активного
захватывания добычи.
У всех рыб, независимо от структуры ткани скелета (хрящ или кость),
имеется ряд общих черт строения, отражающих их первично водный образ
жизни. Скелет туловища представлен хорошо расчлененным у большин-
ства рыб хрящевым или костным позвоночником. Имеется ясное деление
позвоночника на два отдела: туловищный с ребрами, охватывающими по-
лость тела с ее органами, и хвостовой отдел — локомоторный с гемальным
каналом позади полости тела. В плавниках появляется самостоятельная
система скелетных лучей двойного происхождения. Основание плавни-
ков поддерживается хрящевыми или костными лучами, принадлежащими
к внутреннему скелету. Кнаружи от них развиваются скелетные лучи
кожного происхождения (эластиновые или костные). Парные плавники
имеют кроме того пояса конечностей, связывающие правый и левый плав-
ники в единую скелетную систему.
Чед>е п рыб неподвижно соединен с позвоночником. У большинства
рыб (у всех, кроме химеровых и двудышащих) висцеральный скелет по-
движно соединяется с мозговым черепом при помощи подвеска. Однако
оба отдела черепа уже составляют единое целое. Передние висцеральные
дуги (челюстная и подъязычная) преобразованы в челюстной аппарат для
активного захватывания пищи. Остальные висцеральные дуги выполняют
дыхательную функцию. У всех костных рыб (Osteichthyes) в связи с ды-
хательной функцией висцерального скелета, развивается жаберная крышка
(Operculum).
У всех_наземных позвоночных (амфибий, рептилий, птиц и млеко-
питающих) в связи с переходом от водного к наземному образу жизни,
наблюдается ряд обцщх черт строения. Скелет непарных плавников пол-
ностью исчезает. Передние и задние конечности, предназначенные для
хождения по суше, подвижно расчленены на ряд отделов. Плечевой пояс
94
теряет связь с черепом и укрепляется грудиной. Тазовый пояс прочно
причленяется к позвоночнику в его крестцовом отделе. В черепе наземных
позвоночных произошли следующие преобразования: ^развитие по-
движного сочленения черепа с позвоночником; 2) редукция~жабёрного
скелета й Преобразование его в подъязычный аппарат; 3) срастание че-
люстного аппарата с основанием черепа (аутостилия); 4) обособление
среднего уха и слуховых косточек, играющих роль звукопроводящего
аппарата; 5) соединение обонятельных полостей с ротовой или глоточной
полостью через задние ноздри (хоаны). Указанные выше общие признаки
наземных позвоночных типично представлены в скелете хвостатых амфибий.
Скелет высших классов наземных позвоночных (рептилий, птиц и
млекопйтающй5Г),~ббъедйняемых в общую группу амниот (Amniota), также
имеет ряд общих черт строения, отражающих, независимо от образа жизни
животного, их более высокую активность, по сравнению с первыми назем-
ными позвоночными — амфибиями.
Осевой скелет их туловища расчленяется на пять отделов: 1) ш е fi-
ll ы и—подвижный,"'передний отдел с редуцированными ребрами, 2) г руд-
ной отдел с ребрами, соединяющимися с грудиной в грудную клетку, сокра-
щением которой осуществляются дыхательные движения, 3) пояснич-
ный отдел (не всегда ясно выраженный) с приросшими к позвонкам руди-
ментарными ребрами, 4) крестец, соединенный с тазовым поясом,
5) подвижный хвостовой отдел.
Череп амниот окостеневает полностью, грань между покровными
и основными окостенениями полностью стирается. Межглазничная пере-
городка в черепе сужена и мозговая полость оттеснена назад. Вторичные
челюсти (maxilla и praemaxilla) разрастаясь образуют в дне черепа костный
свод — твердое нёбо, отодвигающее хоаны назад по направлению к гло-
точной области.
Конечности становятся мощными органами передвижения на суше.
В особенности птицы и млекопитающие при беге высоко приподнимают
свое тело над уровнем почвы (рис. 409 и 411).
У птиц весь скелет отражает приспособление к полету. Преобразо-
вание передних конечностей в крылья и двуногое хождение на задних ко-
нечностях связано с облегчением и перестройкой всего скелета: облег-
ченный череп с громадными глазницами и клювом соединяется с очень
подвижной шеей. Мощно развитая грудная клетка с килем на грудине
причленяется к неподвижному грудному отделу позвоночника. Хвостовой
отдел укорочен. Мощные ноги причленяются к большому тазу и получают
прочную опору на усложненном крестце.
У млекопитающих независимо от образа жизни фиксируется посто-
янное число шейных позвонков (семь) и относительно постоянное соотно-
шение между скелетом конечностей и позвоночником (12—18 грудпых по-
звонков, 5—7 поясничных, 2—4 крестцовых и неопределенное число хво-
стовых). Череп характеризуется мощным развитием обонятельной об-
ласти и мозговой коробки, появлением в ушной области барабанной кости
(tympanicum) и сложным звукопроводящим аппаратом из трех косточек.
Резко выражена разнозубость и самостоятельное причленение нижней
челюсти через венечный отросток к слуховой области черепа у основания
своеобразной скуловой дуги.
Хорда
Первоначальным осевым скелетом позвоночных является хорда, ко-
торая у низших форм лишь дополняется отдельными элементами дуг,
а у других позвоночных более или менее вытесняется развивающимися
телами позвонков.
Хорда позвоночных на ранних стадиях развития представляется
клеточным шнуром энто-мезодермального происхождения, лежащим под
95
центральной нервной системой. Клетки хорды получают вид плоских
дисков и располагаются весьма правильно подобно монетному столбику.
На наружной поверхности хорды появляется тонкая бесструктурная
оболочка, представляющая собой продукт выделения ее клеток. Эта обо-
лочка называется наружной эластической оболочкой (elastica externa).
Затем внутри клеток хорды появляются крупные вакуоли, заполненные
жидкостью, а ядра с окружающей их протоплазмой постепенно оттесня-
ются к наружной поверхности. Границы между отдельными клетками
исчезают, так что можно различить лишь отдельные ядра, располагающи-
еся главным образом в периферическом слое протоплазмы, и внутреннюю
ег
Рис. 68. Часть поперечного
разреза через хорду личин-
ки речной миноги (по Гуд-
ричу).
ct — соединительная ткань;
el — наружная эластичная
оболочка; fs — волокнистая
оболочка; nt — вакуолизиро-
ванная ткань хорды.
крупноячеистую массу.
Под наружной эластической оболочкой, которая имеется и у ланцет-
ника, у позвоночных развивается затем еще другая оболочка, волокнистого
строения, которая достигает обыкновенно значительной толщины (рис. 68).
Эта волокнистая оболочка нарастает изнутри
и представляет собой, несомненно, продукт
выделения периферического синцития, который
по общему виду обнаруживает известное сход-
ство с эпителием и носит название эпителия
хорды.
Наконец, иногда внутренний слой оболочки
хорды оказывается также бесструктурным, и
тогда эта часть ее получает название внутренней
эластической оболочки.
В таком виде хорда развивается у всех
позвоночных. Но в некоторых случаях насту-
пают дальнейшие изменения. У хрящевых рыб,
а также и у двудышащих, во время их разви-
тия в наружной эластической оболочке хорды
появляются отверстия, и через эти отверстия
проникают внутрь волокнистой оболочки
мезодермальные клетки из окружающей хор-
ду скелетогенной ткани. Эти клетки прини-
мают веретеновидную форму и переполняют
всю волокнистую оболочку. Таким образом,
у некоторых рыб бесклеточная оболочка хор-
ды путем вторичного включения мезодер-
мальных клеточных элементов становится особого рода клеточной волок-
нистой тканью. В данном случае мы имеем еще один пример возникновения
скелетной ткани из бесклеточных вначале продуктов выделения, т. е. сна-
чала появляется основное вещество, а затем уже в него проникают клетки.
Вселившиеся в оболочку хорды мезодермальные клетки не остаются без-
деятельными, —. они гомогенизируют волокнистое вещество и превра-
щают его в основное вещество хряща. Таким образом, у селахий внутри
оболочки хорды развиваются хрящевые тела позвонков, в которых затем
происходят еще отложения извести. У цельноголовых и у двудышащих
особых тел позвонков не развивается, но у первых развиваются многочис-
ленные известковые кольца внутри оболочки хорды.
В качестве постоянного органа хорда функционирует лишь у немногих
низших позвоночных — у круглоротых, у цельноголовых, осетровых и
двудышащих рыб. Во всех этих случаях оболочка хорды обладает зна-
чительной толщиной и прочностью. Скелетная роль хорды основывается
на прочности и упругости ее оболочек, а также и на прочности стенок
внутренних вакуолей одновременно со значительным внутренним давле-
нием, т. е. на принципе, имеющим гораздо большее применение в тканях
растений (тургор).
У других позвоночных хорда вытесняется развивающимися телами
96
Иозвонков, Которые обычно перетягивают хорду посредине, так что остатки
последней сохраняются главным образом между телами позвонков. В этом
случае у взрослого животного хорда сохраняется в виде четкообразного
шнура внутри позвоночника, как это наблюдается у большинства рыб.
У земноводных хорда, наоборот, наиболее вытесняется между телами по-
звонков развивающимися здесь межпозвоночными хрящами, и остатки
хорды нередко сохраняются только внутри тел. У млекопитающих остатки
хорды сохраняются между телами соседних позвонков внутри межпозво-
ночных хрящей в виде их студенистого ядра (nucleus gelatinosus s. nucleus
pulposus).
Позвоночник
Элементы собственно позвоночника развиваются в «скелетогенной»
соединительной ткани, обволакивающей хорду и центральную нервную
систему. Источником этой ткани эмбрионально является мезодерма скле-
Рис. 69. Две последовательные стадии диференцировки мезодермы и
образования мезенхимы у позвоночных. Схематические поперечные
разрезы.
ао — аорта; ch — хорда; coet — полость тела; cut — кожный листок мезодермы;
f—вакладка парного и/' — непарного плавника; gen—половая складка;
gn —ганглионарная пластинка; int — кишка; mb —мускульная почка парно-
го и mb' — непарного плавника; msc — мускульная пластинка миотома; ту —-
миотом; п — нервная трубка; nph — закладка почечного канальца; set — скле-
ротом; st — подкишечная вена; som —париетальный и spl — висцеральный
листки; sm — скелетогенная мезенхима конечности.
ротомов (рис. 69). У зародыша осевая мезенхима непрерывно продолжа-
ется в сагиттальную соединительнотканную перегородку между миото-
мами правой и левой стороны, в горизонтальную перегородку между
спинным и брюшным отделами миомеров, в более или менее вертикальные
перегородки между отдельными миомерами и, наконец, в слой соединитель-
ной ткани между брюшиной и боковыми мышцами. Расположение этой
системы осевых оболочек и перегородок совпадает с расположением их
у ланцетника, у которого они состоят еще из почти бесклеточной во-
локнистой массы (рис. 70). У позвоночных это типичная соединительная
ткань. В этих перегородках у позвоночных развиваются основные
элементы осевого скелета (рис. 71).
У круглоротых, и именно у миног, появляется лишь парный ряд
небольших хрящиков, сидящих на хорде в соединительнотканном слое
97
Рис. 70. Расположение опорных перегородок у ланцетника (Л) и у позво-
ночных (В).
md — спинной отдел боковой мышцы и mv — брюшной отдел; sh — горизон-
тальная и ss — сагиттальная перегородки.
Рис. 71. Схема элементов осевого скелета и их отношения к главным
соединительнотканным перегородкам (по Гудричу).
ас — стенка брюшной полости; bd — верхняя дуга; bv — нижняя дуга; tw —
стенка тела; dr — верхнее (добавочное) ребро; i — кишка; id, iv — передние,
вставочные дуги; т— поперечная перегородка (миосента); ms— брынакейка;
msd, msv — сагиттальные спинная и брюшная перегородки; nes — мозговая
трубка; s — оболочка хорды; рг — шпкнес (плевральное) ребро; sp — верхний
остистый отросток; ts — горизонтальная перегородка.
Рис. 72. Схема позвоночника миноги.
аа — передняя и ар — задняя дуги; ch — хорда; dn — спинной и т —брюш-
ной нервные корешки; ms —• миосента.
98
Рис. 73. Схема эмбрионального позвоноч-
ника селахий.
аа — передняя и ар — задняя верхние дуги; av —
нижние дуги; с — ребро; ch — хорда; dn — спин-
ной и ж — брюшной нервные корешки; mt —
миосепта.
йо бокам спинного мозга. Это — очень тонкие стержни, довольно пра*
вильно чередующиеся с выходами корешков спинномозговых нервов
(рис. 72). Так как последние отходят поочередно (брюшной и затем спин-
йой), то, следовательно, на каждый сегмент тела (определяемый грани-
цами миотома, сегментальными
Йится по две пары элементов,
В передней части каждого сег-
мента ле/кит передняя (крани-
йльная) дуга, располагающаяся
непосредственно позади выхо-
да брюшного нервного ко-
решка; в задней части сегмен-
та лежит задняя (каудальная)
дуга, расположенная непосред-
ственно позади спинного ко-
решка.
У низших рыб наблюдается
по существу то же самое распо-
ложение верхних дуг. У села-
хий верхние дуги закладыва-
ются также в виде двух пар
хрящей в каждом сегменте. Они
также правильно чередуются с
выходами спинномозговых нер-
вов. Передняя и задняя дуги каждого сегмента в общем еще равнозна-
чащи, но в то время как задняя дуга сидит непосредственно на
хорде, передняя дуга до хорды не доходит (рис. 73). Брюшной
нервный корешок выходит позади задней дуги (иногда сквозь нее), спин-
ной корешок выходит в середине сегмента позади передней дуги. Кроме
верхних дуг, у селахий закладываются и нижние дуги также в
кровеносными сосудами и т. д.) прихо-
называемых верхними дугами.
Рис. 74. Последовательные стадии образования позвонка
селахий (поперечные разрезы).
ai, as — нижняя и верхняя дуги; с — ребро; ph — гемальный от-
росток; pl — боковой отросток позвонка; рп-—нервная дуга; «р —
остистый отросток (dorsospinale).
числе двух пар хрящей в каждом сегменте. Обе пары нижних дуг сидят
непосредственно на хорде.
Следовательно, у низших позвоночных элементы позвоночника воз-
никают в виде двух нар верхних дуг и двух пар нижних дуг в каждом
метамере тела (первичная_диплпеппттдылия). В дальнейшем у селахий
в задней половине каждого сегмента наступает охрящевение внутри
оболочки хорды, и таким образом развивается тело позвонка в виде
хрящевого кольца. Основания задних пар дуг разрастаются вокруг обо-
лочки хорды и дополняют тело позвонка еще вторым, наружным слоем
хряща. В результате развивается цельный хрящевой позвонок, на ко-
99
тором верхние и нижние дуги задней пары являются в виде отростков
(рис. 74). Верхние и нижние дуги передней части сегмента остаются в виде
самостоятельных хрящей, вклинивающихся между соседними позвонками.
Это— так называемые вставочные пластинки (intercalaria) взрослых села-
хий (рис. 75). Подобным же образом развиваются элементы позвоночника
и у цельноголовых рыб, с той лишь разницей, что у них сильнее разви-
вается передняя дуга каждого сегмента, так что у взрослого животного
задняя дуга является в виде вставочного хряща (следовательно, послед-
няя гомологична собственно верхней дуге позвонка селахий, а не вста-
вочной пластинке).
Рис. 75. Схема позвоночника селахий.
ап — верхние дуги; с — ребра; ch — хорда; 1с — вставочные
пластинки; pl — боковые отростки позвонка.
У других низших позвоночных точно так же первыми элементами
позвоночника являются верхние и нижние его дуги, закладывающиеся
обычно тоже в числе двух пар в каждом сегменте. Однако передние дуги
каждого сегмента сразу закладываются в виде гораздо менее значительных
хрящей, чем задние дуги. У взрослых ганоидных рыб первые сохраняются
в виде вставочных хрящей между позвонками, но уже не играют столь
заметной роли, как у селахий (рис. 87). У костистых рыб и у земноводных
во время их развития наблюдаются иногда только рудиментарные закладки
вставочных хрящей (передних дуг), которые у взрослых животных исче-
зают бесследно. У высших позвоночных в каждом сегменте закладывается
уже только одна пара верхних и одна пара нижних дуг. Однако и у высших
позвоночных соотношения некоторых частей в области нижних дуг дают
ясные указания на первичную двойственность (диплоспондилмю) эле-
ментов позвоночника.
За счет верхних и нижних дуг, являющихся первоначальными эле-
ментами позвоночника, развиваются и другие его части. Попарным слия-
нием концов верхних дуг (neurapophyses) над спинномозговым каналом
образуются непарные верхние остистые отростки (processus
spinosus neuralis). Нижние дуги разрастаются в стороны в виде боковых
отростков (parapophyses). Примыкая к боковым отросткам, в миосептах
развиваются ребра (рис. 71). Ребра позвоночных возникают, как и другие
элементы осевого скелета, в виде охрящевений, расположенных в системе
соединительнотканных перегородок, связанных с одной стороны с хордой
и с другой стороны с боковыми мышцами тела. Первично ребра занимают
положение по бокам общей полости тела,* между брюшиной и боковыми
мышцами, вдоль внутреннего края миосепт (рис. 71, рг). Позднее они,
однако, нередко перемещаются по миосепте вглубь брюшной мускулатуры
и иногда почти достигают горизонтальной перегородки (у взрослых села-
хий). Эти ребра получили название нижних ребер в отличие от подобных
100
добавочных образований, развивающихся у высших рыб и у амфибий
сразу в горизонтальной перегородке, отделяющей брюшную мускулатуру
от спинной, и получивших название верхних ребер (рис. 76, 77). Крите-
рием для распознавания ребер обоих типов служило до сих пор только
положение ребер. Так как нижние ребра нередко значительно переме-
щаются к спине и погружаются вглубь мускулатуры и, кроме того, у на-
земных позвоночных в связи с диференцировкой боковых мышц исчезает
горизонтальная перегородка, этот критерий теряет свое значение.
Рис. 76. Соотношения элементов осевого скелета в области туловища (Л, В) и хвоста
(С) у хрящевой (Л) и у костной рыбы (В, С).
ап — нервные дуги; ah — гемальные дуги; cd — верхнее и е — нижнее реОро; coel —
полость тела; md — спинной и иго — Орюшной отделы мускулатуры; sp — верхний и sp'—
нижний остистые отростки.
Ребра наземных позвоночных обыкновенно обозначались как верхние
ребра. В последнее время введен, однако, новый, эмбриологический
критерий (Емельянов), позволяющий провести более надежную гомологию
между ребрами рыб и наземных позвоночных. По этим данным, ребра
РиС. 77. Соотношения элементов осевого скелета в области туловища и хвоста у прими-
тивного гипотетического наземного позвоночного (Л) и у амниот (В, С).
Обоаначения см. на рис. 76.
амниот, как и ребра рыб, закладываются вблизи хорды и растут отсюда
в дистальном направлении, охватывая полость тела и проникая затем
более или менее глубоко в миосепты. Добавочные, верхние, ребра много-
перых ганоидов (Polypteridae) и костистых рыб развиваются в горизон-
тальной перегородке на ее пересечении с миосептами вдали от осевого
скелета и растут затем по направлению к последнему. При этом у кости-
стых рыб добавочные ребра развиваются обычно без хрящевой стадии
путем окостенения соединительной ткани.
Ребра большинства современных амфибий вообще заметно редуци-
рованы. По эмбриологическим данным, ребра Urodela и Anura относятся
101
к верхним ребрам. У Urodela рудименты нижних ребер прирастают к
позвонкам. У безногих амфибий, гимнофион, ребра нижние, как и у
амниот. Ребра возникают в миосептах боковой мышцы и дают последним,
очевидно, более прочную опору при передвижении рыбы посредством
боковых изгибов тела. Эта функция ребер исчезает у наземных позво-
ночных. в связи с чем и стоит их редукция у амфибий, особенно у форм
с укороченным телом (бесхвостые). Вместе с тем у амниот, начиная с
рептилий, ребра получают в грудной области новую функцию опоры при
дыхательных движениях и соответственно развиваются и преобразу-
ются как скелет грудной клетки. В поясничной области они редуциру-
ются, и только у змей в связи с установлением новой формы движения —
ползания посредством боковых изгибов тела — восстанавливается локо-
моторное значение ребер на всем протяжении туловища (как это было у
рыб).
Добавочные (верхние) ребра имеют, очевидно, иное значение. Ди-
стальный конец ребра образует иногда как у рыб, так и у амфибий изгиб
или даже развилку, защищающую нерв и канал боковой линии. Кроме
того, прочное соединение дистальных концов этих ребер с кожным панцы-
рем у многоперых рыб (Polypterus) и направление эмбрионального развития
от покровов к позвоночнику позволяют думать, что они служили также
связью между кожным и осевым скелетом. Редукция добавочных ребер
у наземных позвоночных обусловлена в таком случае утратой кожного
скелета, а также органов боковой линии.
Итак, у позвоночных наблюдается образование двоякого рода ребер —
верхних и нижних. Верхние ребра располагаются по миосептам и в гори-
зонтальной соединительнотканной перегородке, между спинным и брюш-
ным отделами боковой мышцы. Нижние, или плевральные, ребра перво-
начально развиваются также по миосептам, но глубже, в соединительной
ткани между брюшиной и брюшным отделом боковой мышцы, непосред-
ственно охватывая общую полость тела (рис. 76). У многоперых рыб
(Polypteridae) имеются одновременно ребра обоего рода (рис. 89); для
большинства рыб характерны нижние ребра; которые иногда, особенно
у хрящевых рыб (Elasmobranchii), перемещаются внутрь самих миосепт
в толще брюшной мускулатуры.
Нижние края нижних дуг иногда (селахии и осетровые рыбы) не-
сколько выступают, охватывая с боков спинную аорту (рис. 74). В хвосто-
вой области позвоночника нижние дуги разрастаются не в бока, а вниз,
охватывая с боков продолжение спинной аорты — хвостовую артерию и
вену (рис. 76 и 77). Здесь нижние дуги являются в виде гемальных
отростков позвонков (haemapophyses).
Под хвостовой артерией и веной гемальные отростки каждой пары
смыкаются и срастаются с образованием нижнего остистого
отростка (processus spinosus haemalis). При переходе от туловищной
области позвоночника к хвостовой можно видеть, что боковые отростки,
широко расходящиеся в туловище, начинают сходиться под все более
острым углом, пока не сомкнутся в хвосте, образуя гемальный канал,
являющийся в сущности лишь сокращенной в своих размерах общей
полостью тела. У некоторых рыб (Lepidosteus, Polypterus) нижние, ребра,
постепенно сходясь, срастаются между собой, принимая, таким образом,
участие в образовании гемального канала и нижних остистых отростков.
Верхние ребра в ограничении гемального канала никогда участия не
принимают— они имеются только в туловищной области, или, иногда
в начале хвостовой области видны еще соответствующие им небольшие
боковые отростки.
Тело позвонка (centrum) развилось в различных группах позво-
ночных совершенно независимо и далеко не всегда одинаково. Наиболее
примитивные представители различных групп позвоночных имеют нередко
102
постоянную хорду с сидящими па ней элементами позвонков в виде
верхних и нижних дуг. У круглоротых, цельноголовых, осетровых и
двудышащих рыб тел позвонков еще нет, хотя последние группы имеют
в остальных отношениях прекрасно развитый и даже отчасти костный по-
звоночник. У селахий имеются, как мы видели, хрящевые тела позвонков,
развивающиеся внутри оболочки хорды и дополняемые хрящевыми же
продуктами разрастания оснований дуг. У современных костных ганоидов
(Holostei), у многоперых ганоидов (Polypteridae) и у костистых рыб (Те-
leostei), так же как и у всех современных наземных позвоночных, имеются
уже костные тела позвонков, образующие одно неразрывное целое с первич-
ными их элементами — дугами,
являющимися в виде отростков
позвонков (рис. 89, 90). Так же
как и тело хрящевого позвонка
селахий, тело костного позвонка
высших рыб имеет форму вогну-
того с двух сторон цилиндра (за
одним лишь исключением —
Lepidosteus, тело позвонка ко-
торого опистоцельно, т. е. во-
гнуто сзади и выпукло спереди),
т. е. является амфицельным. В
вогнутостях двух соседних позвон-
ков (интервертебрально) помеща-
ются остатки хорды, которая в
области тела позвонка сильно
перетянута, т. е. имеет четковид-
ную форму (рис. 90).
В развитии тел позвонков у
высших рыб оболочка хорды
никакой роли не играет: костное
Рис. 78. Развитие позвонка аксолотля
(из Кингсли).
сс — хрящ внутри хорды; i — суставная щель
в межпозвоночном хряще <с; ch — хорда; c/is—
оболочка хорды; v — тело позвонка (кость
черного цвета).
тело развивается вокруг хорды с
ее оболочками (перихордально).
Костное тело позвонка разви-
вается в виде тонкого сначала
цилиндра (рис. 78), независимо от
хряща в соединительной ткани.
Таким образом, костное тело позвонка появляется сразу в виде
одного цельного образования.
Данные эмбрионального развития показывают, что у низших
животных (селахий) тело позвонка развивается частью в результате
обрастания хорды основаниями хрящевых верхних и нижних дуг, т. е.
из нескольких парных частей.
У высших же рыб и у наземных позвоночных тела позвонков разви-
ваются сразу как цельное кольцевидное тело вокруг оболочки хорды,
независимо от сидящих на ней дуг. История ископаемых форм, как рыб,
так и амфибий, показывает, что все же костное тело позвонка первона-
чально составлялось из нескольких частей, располагавшихся на хорде
соответственно основаниям верхних и нижних дуг, а также вставочным
хрящам. Участие элементов дуг в образовании этих частей, т. е. элементов
тел, выражалось в различной степени. В редких случаях тела образо-
вались исключительно путем простого разрастания оснований дуг вокруг
хорды (Pycnodonti). Гораздо чаще вокруг хорды развивались самостоя-
тельные окостенения, дававшие дугам позвонков более прочную опору
на хорде. Таким образом, в каждом сегменте развивалось иногда до
четырех пар окостенений, получивших названия эпицентров (со-
ответствующих основаниям верхних дуг, т. е. basidorsalia), гипоцен т-
ров (соответствующих основаниям нижних дуг, т. е. basiventralia) и
плевроцентров (верхних и иногда нижних, соответственно вста-
вочным хрящам, interdorsalia и interventralia). Наиболее постоянными
оказываются затем в каждом сегменте два окостенения как результат
попарного соединения гипоцентров и плевроцентров — непарный гипо-
А
Рис. 79. Схема полупозвонков, состоящих из гипоцентров
и плевроцентров, с соответствующими дугами (по Ремане).
Л — поперечный раврев на уровне гипоцентра; В— то же на
уровне плевроцептра; С —вид позвоночника сбоку; ап — верхняя
дуга; av — нижняя; hpc — гипоцентр; pic — плевроцентр.
центр, дававший опору нижним дугам и ребрам, и непарный верхний
плевроцентр, лежавший между верхними дугами (рис. 79 и 80).
В дальнейшем окостенение захватывало и дуги позвонков, которые
тогда прочно соединялись с названными элементами тела в одно целое.
У позднейших позвоночных шло затем разрастание отдельных элементов
Рис. 80. Позвонок костного гаиоида Osteorhachis (по'Гудричу).
Л — вид сбоку и сзади; В — вид спереди; ап — верхняя дуга;
ch — место прохождения хорды; he — нижняя дуга (гипоцентр);
pic — вставочная дуга (плевроцентр); г — боковой отросток (при-
членсние ребра).
и их последующее соединение, приведшее к образованию цельных костных
тел позвонков, связанных с их дугами и отростками. Этот процесс в различ-
ных группах позвоночных протекал по-разному.
У большинства рыб произошло срастание эпицентров и гипоцентров
в одно цельное кольцо, а плевроцентры редуцировались. Однако у неко-
торых ископаемых костных ганоидов преобладающее развитие получили
гипоцентры (hypocentrum) и верхний промежуточный элемент (pleuro-
centrum dorsale), которые образовали два полукольца, поочередно
104
Рис. 81. Развитие цельных позвонков из полу позвонков у Amia (а)
и у костистых рыб (Ъ) (по Ремане).
Обозначения см. рис. 79.
Рис. 82, Позвонки стегоцефалов и ганоидов (по Циттелю и Вудварду
из Кингсли).
А — филоспондильный позвонок"' стегоцефала; В — часть рахитомного по-
ввоночника стегоцефала (Chelydrosaurus); С—позвоночник ганоида (Collop-
terus); D —то Л:е (Eurycormus); he — нижний элемент (hypocentrum); plv —
нижний и pl — верхний вставочные элементы; ап и п — верхняя дуга (pleuro-
centra); ns — остистый отросток.
А
Рис. 83. Развитие цельных позвонков у стегоцефалов (по Ремане).
а — эмбриональные полупозвонки; b и с — рахитомные полуповвонки, d —
стереоспондильный позвоночник.
105
А
охватывающие хорду снизу и сверху и вклинивающиеся между соседними
полукольцами (рис. 79, 80, 81). Иногда каждое из полуколец разрастается
вокруг всей хорды, так что в каждом сегменте развивается по два полных
кольца, т. е. по два тела позвонка: одно, связанное с верхними и нижними
дугами, и другое — без дуг (диплоспондилия в телах) (рис. 81, 82, D).
То же самое наблюдается и у одного современного представителя костных
ганоидных рыб (Amia calva), но только в хвостовой области позвоночника.
Совершенно независимо от рыб про-
изошло развитие тел позвонков у назем-
ных позвоночных, и также из несколь-
ких элементов в каждом сегменте, что
приводило и кобразованию полупозвон-
ков (рис. 83, 84). У темноспондилышх
стегоцефалов можно проследить про-
грессивное развитие гипоцентров и эпи-
центров и редукцию плевроцентров (рис.
82, В и 83).
« У бранхиозавров, стоявших ближе к
современным амфибиям, имелись лишь
топкие костные пластинки, соответству-
ющие эпицентрам и гипоцентрам, т. е.
каждый позвонок состоял из двух пар
окостенений (рис. 82, А). У современных
амфибий эти окостенения образуют одно
неразрывное целое. С другой стороны, у
высших наземных позвоночных (Amniota)
тела позвонков образованы, повидимому,
главным образом за счет верхних вста-
вочных элементов (pleurocentra dorsalia), с которыми срастались
основания верхних дуг, вследствие чего нижние дуги у них причленяются
между телами позвонков (интервертебрально) (рис. 85).
Во всяком случае совершенно ясно, что тела позвонков, действи-
тельно, представляют собой сравнительно позднее приобретение, развив-
Рис. 84. Два туловищных позвон-
ка емболомерного стегоцефала
Eogyrinus (по Ватсону).
с — головка ребра; dp — попереч-
ный отросток; hp — гипоцентр (тело
поввонка); па — верхняя дуга; pl—
плевроцептр (промежуточное тело
позвонка); prz, ptz — суставные от-
ростки; « — бугорок ребра.
Рис. 85. Развитие позвонков амниот (по Ремане, с изменениями).
а — исходный эмбриональный тип; ь — позвонки котилозавра (сейму-
рии); с—типичные позвонки рептилий; ап—верхние дуги; с — прй-
члепспис головой ребра; 1с —• интерцентр (гипоцентр); pit — поперечный
отросток (причлепснис бугорка ребра).
шееся частью в связи с разрастанием оснований дуг позвоночника, частью
путем окостенения в самой скелет огенпоп ткани вокруг хорды, независимо
в различных группах позвоночных животных. Соответственно перво-
начальной множественности элементов дуг (2 пары верхних и 2 пары
нижних дуг) и тело позвонка состояло первоначально из нескольких
(до 4 пар) частей, причем у одних позвоночных преобладающее развитие
получили одни элементы, у других — другие, но в результате у всех
современных позвоночных развилось одно пельное цилиндрическое тело
позвонка в каждом сегменте.
Прогрессивное развитие позвоночника находится в явной связи со
значением мускулатуры и осевого скелета при локомоторных движениях
тела и в особенности хвостовой области. У плохо плавающих химер,
двудышащих и осетровых сохраняется еще постоянная хорда, а у лучших
пловцов — акул и костистых рыб с коротким и сильным хвостовым плавни-
ком — имеется всегда вполне развитый позвоночник.
Рис. 86. А — часть поввоночника акулы (Lanina) из области туловища; В — попереч-
ный разрез через середину позвонка (из Гудрича).
с — тело позвонка; са — иввестиовые кольца; dr — отверстие для спинного нервного ко-
решка; ha — нижняя дуга; in, iv — верхняя и нижняя вставочиые дуги; И — продольная
спинная свявка; па — верхняя дуга; пс — спинномозговой канал; nt — хорда; гсы — ради-
альные обыввествления; vr — отверстие для брюшного нервного корешка; w — межпозвоноч-
ная связка.
Первоначальная множественность элементов тел связана со множе-
ственностью дуг позвонков и обеспечивала прочность и гибкость позво-
ночника (основой которого оставалась еще хорда) без наличия сочленений
между его частями. Разрастание элементов тел внутри сегмента и развитие
сочленений на границе сегментов позволило соединить значительную
подвижность позвоночника с прочностью целостных позвонков.
Позвоночник низших позвоночных
У круглоротых, и именно
имеется, как мы видели, в зачаточном
парный ряд небольших хрящевых
верхних дуг (по две пары в каждом
сегменте). Эти хрящи лежат в соеди-
нительнотканной стенке спинномоз-
гового канала и играют, несомнен-
но, крайне незначительную роль.
Главное значение имеют еще соеди-
нительнотканные оболочки и перего-
родки., У миксин хрящ в позвоночнике
только у миног, позвоночник
состоянии, представляя собою
Рис. 87. Позвоночник осетровой
рыбы.
ап — верхние и av —нижние дуги;
ch — хорда; Гс — вставочные хрящи;
sp — остистые отростки.
отсутствует.
У хрящевых рыб имеется
уже вполне развитый хрящевой по-
звоночник (рис. 86). Позвоночник
селахий состоит из ряда хрящевых
позвонков с двояковогнутыми, амфицельными, телами, верхними и ниж-
ними дугами и со вставочными хрящами, правильно чередующимися
с верхними и нижними дугами. Кроме того, у некоторых форм имеется
ряд непарных хрящевых пластинок, замыкающих спинномозговой канал
сверху. Спинномозговой канал оказывается, таким образом, прекрасно
107
защищенным системой тесно связанных менаду собой хрящевых пластинок,
менаду которыми (или внутри которых) имеются лишь небольшие отверстия
для выхода корешков спинномозговых нервов. Стенки его образованы
верхними дугами (точнее — дугами задней пары), верхними вставочными
пластинками (дугами передней пэры) и замыкающими пластинками.
Нижние дуги имеют различный вид в туловищной и в хвостовой области.
В туловищной области они образуют боковые отростки с причленяющи-
мися к ним нижними ребрами и небольшие гемальные выступы по бокам
от спинной аорты. В хвостовой области ребер нет, а боковые отростки и
гемальные выступы вместе образуют гемальные отростки, ограничи-
вающие гемальный канал и продолжающиеся в непарный нижний
остистый отросток. Внутри тел. позвонков наблюдаются отложения
извести в виде цилиндров, иногда с радиальными продольными гребнями
(рис. 86, В).
У цельноголовых позвоночник лишен тел позвонков, но
в остальном построен сходно.
У осетровых рыб позвоночник менее развит, чем у селахий:
тел позвонков у них еще не имеется, так что элементы его сидят непо-
средственно на постоянной хорде (рис. 87). Верхние дуги вполне охва-
тывают спинномозговой канал и тесно соприкасаются своими верхними
концами, к которым причленяется небольшой верхний остистый отросток.
К небольшим боковым отросткам нижних дуг причленяются нижние
ребра, лежащие непосредственно под брюшиной. Имеются и небольшие
гемальные выступы, охватывающие спинпую аорту. В хвостовой области
гемальные выступы и боковые отростки смыкаются для образования
гемального канала. Гемальные дуги продолжаются под гемальным кана-
лом в непарный нижний остистый отросток. У осетровых имеются и не-
большие вставочные хрящи, вклинивающиеся между основаниями верхних
и нижних дуг. В позвоночнике осетровых появляются первые окосте-
нения — обычно окостеневают ребра и остистые отростки и нередко
также концы верхних дуг.
У современных костных ганоидов и у костистых
рыб имеются вполне развитые костпые тела позвонков двояковогнутой
(амфицельной) формы (рис. 90). Исключение составляет лишь костный
ганоид Lepidosteus с опистоцельными позвонками. У другого ганоида,
Arnia calva, позвонки амфицельные. У этой формы основания дуг являются
хрящевыми, и между ними вклиниваются еще вставочные хрящи (рис. 88).
В хвостовой области этой рыбы имеется по два тела позвонка на каждый
сегмент: переднее тело несет вставочные хрящи (передние дуги), а заднее
тело — верхние и нижние дуги (задние дуги).
У всех других рыб с костным скелетом лишь крайне редко наблю-
даются во взрослом состоянии остатки вставочных хрящей, и весь позво-
ночник состоит только из одних цельных костных .позвонков. Как
и у других рыб, наблюдается диференцировка менаду двумя отделами
позвоночника — туловищным и хвостовым.
Типичный туловищный позвонок состоит из цилиндрического тела
и его отростков (рис. 90). Тело глубоко вогнуто спереди и сзади и имеет
небольшое отверстие, через которое проходит редуцированная четковид-
ная хорда. Верхние отростки охватывают с боков спинномозговой канал,
над которым они срастаются, образуя верхний остистый отросток. Между
верхними остистыми отростками соседних позвонков остаются значитель-
ные пространства, затянутые лишь соединительной тканью, через которые
выходят корешки спинномозговых нервов. Основания верхних дуг снабже-
ны суставными отростками, посредством которых соседние позвонки
сочленяются друг с другом (zygapophyses). Боковые отростки выдаются
более или менее значительно в бока и несут нижние ребра. Но, кроме
нижних ребер, у многоперых ганоидов (Polypteridae) имеются одновремен-
705
Ио й верхние ребра, лежащие в соединительнотканной перегородке между
спинным и брюшным отделами боковой мышцы тела (рис. 89) и связывающие
нозвоночник с кожным панцырем. В передней части туловища верхние
Рис. 88. Три позвонка молодой
Amia calva из области перехо-
да туловища в хвостовой от-
дел. Позвонок хвостовой обла-
сти имеет два тела (по Шауин-
сланду).
па — верхние дуги; in — верхние
вставочные хрящи; па — нижние
дуги; iv — нижние вставочные
хрящи.
Рис. 89. Позвонки многоперо-
го [ганоида (Polypterus) (по
Гегенбауру).
А — ив передней и В — ив задней
части туловищной области; С —
хвостовой поввонок; ci — нижнее
ребро; cs — верхнее ребро; pl —
боковые отростки; sp — верхний
и sp' — нижний остистые от-
ростки.
своей части развиваются не на хрящевой
Рис. 90. Позвоночник костистой рыбы в сагит-
тальном разрезе (С). Отдельные позвонки из туло-
вищной (Л) и хвостовой (В) области.
ап — верхние дуги; ah — гемальные дуги; с — реб-
ра; ср — тела поввонков; ра — сочленовные отрост-
ки; pl — боковые отростки; sp — верхний и sp' —
нижние остистые отростки.
ребра крупнее, чем нижние, в задней части — соотношение обратное.
У некоторых костистых рыб наблюдаются, кроме лижних ребер, верхние,
которые, однако, в большей
основе, а прямо в соедини-
тельной ткани. Кроме того,
у костистых рыб в связи с
позвоночником имеется еще
более или менее сложная
система тонких «межмышеч-
ных> косточек, развиваю-
щихся так же как окостене-
ния соединительной ткани.
В типичном хвостовом
позвонке боковые отростки
сходятся, образуя гемальный
канал. Вместе с тем иногда
сходятся и нижние ребра,
срастающиеся внизу для
образования нижнего остис-
того отростка (Polypterus,
Lepidosteus и некоторые костистые рыбы). Таким образом, хвостовой
позвонок получает симметричную форму с верхними и нижними дугами и
остистыми отростками (рис. 89, 90).
У двудышащих рыб тел позвонков нет, и хрящевые осно-
вания верхних и нижних дуг сидят непосредственно на постоянной хорде.
Верхние части верхних дуг и длинные расчлененные остистые отростки,
а также и ребра, принадлежащие к типу нижних ребер, окостене-
вают.
109
Йозвоиочники наземных позвоночных
У амфибии между телами позвонков развиваются особые межпозво
ночные хрящи, которые более или менее перетягивают хорду между
телами, так что последняя нередко сохраняется в виде остатков только
внутри тел (интравертебрально) (рис. 91). В хрящах между телами позвон-
ков развивается сферическая суставная щель, обращенная у одних форм
своей вогнутой стороной вперед, у других — назад. Так как межпозво-
ночные хрящи
плотно сращены с передними и задними поверхностями
тел позвонков, то последние получают
соответственно вогнутую спереди — п р о-
СА
цельную, или вогнутую сзади — о п и-
стоцельную форму. Первая форма
наблюдается у большинства бесхвостых
амфибий, вторая — у высших хвостатых
амфибий. У низших хвостатых амфибий
межпозвоночные хрящи развиты слабее,
и позвонки сохраняют амфицельную
форму. Предполагается иногда, что меж-
позвоночные хрящи представляют собой
сросшиеся между собой остатки вставоч-
ных элементов, или плевроцентров.
Костные верхние дуги, образую-
щие одно целое с костными телами
Рис. 92. Поперечный разрез через IV [груд-
ной позвонок саламандры (по Гёпперту).
ап — невральная дуга; av — позвоночная арте-
рия; с — ребро; ch — хорда; к — костная пла-
стинка на границе между верхней дугой и боко-
вым отростком; ph — гемальный отросток; pl —
боковой отросток; pt — поперечный отросток.
Рис. 91. Позвоночник хвостатых
амфибий Ranidens (слева) и Spelerpes
(справа) в горизонтальном разрезе
(из Видерсгейма).
ch — хорда; R — тело позвонка; СК —
хрящ внутри хорды IvK — межпо-
звоночный хрящ.
позвонков, ограничивают спинномозговой капал и, срастаясь над ним,
образуют крайне незначительный верхний остистый отросток. У основа-
ния дут имеются крупные суставные отростки.
Боковые отростки разделяются у хвостатых амфибий на два вы-
ступа, верхний и нижний; верхний располагается па верхней дуге,
с которой он и срастается (рис. 92). Таким образом, вместо простого бо-
кового отростка рыб у наземных позвоночных имеется Два выступа —
один на теле позвонка и другой на верхней дуге (рис. 96). Первый по-
степенно редуцируется, а второй, получивший название поперечного отро-
стка (diapophysis), имеет преобладающее значение. Положение поперечных
отростков на верхних дугах позвонков характерно для наземных
йозвоночпых.
Ребра хвостатых амфибий в отличие от других позвоночных принад-
лежат к типу верхних ребер и образуют у своего основания развилку,
обе ветви которой соответствуют обоим выступам бокового отростка
110
в качестве нсзиачи-
(рис. 77, 92). Ребра современных амфибий отличаются весьма незначи-
тельной длиной и до грудины никогда не доходят. У некоторых стегоце-
фалов ребра обладали значительно большей длиной, охватывая с боков
брюхо, что дает основание предполагать, что у них имелись и нижние
ребра, которые у современных амфибий (за исключением гимнофион)
редуцировались.
У бесхвостых амфибий эта редукция пошла гораздо дальше, чем
/у хвостатых, и самостоятельные ребра наблюдают
тельных рудиментов лишь у немногих форм и то
лишь на двух-трех позвонках (рис. 93). У других
бесхвостых рудиментарные ребра сращены в
одно целое с поперечными отростками.
В хвостовой области имеются типичные ге-
мальные дуги с небольшими нижними остистыми
отростками и одновременно в передних, по край-
ней мере, позвонках наблюдаются и боковые
отростки. У бесхвостых амфибий хвостовые по-
звонки срастаются в одну длинную кость — так
называемый уростиль (рис. 93).
У всех наземных позвоночных, начиная с
амфибий, диференцируются еще две новые обла-
сти позвоночника. Передние позвонки приобре-
тают оолее значительную подвижность, и ребра
их становятся рудиментарными. Это шейный
отдел позвоночника, состоящий _у амфибий
только из одного первого позвонка, снабженно-
го спереди парой суставных поверхностей для
сочленения с затылочными буграми черепа. Затем
на границе между туловищной и хвостовой обла-
стью развивается еще один отдел позвоночника,
образующий опору для тазового пояса. Это
крестцовый отдел, к ребрам позвонков которого
причленяются подвздошные кости (ossa ilei) таза.
У амфибий этот отдел состоит также только из
одного позвонка (рис. 93). У хвостатых амфибий
можно ясно видеть, что подвздошные кости при-
членяются именно к концам самостоятельных
крестцовых ребер. У бесхвостых эти ребра, как и
ребра туловищной области, срослись в одно це-
лое с поперечными отростками, которые здесь
обладают очень значительной величиной.
Некоторые низшие рептилии имеют
еще двояковогнутые тела позвонков (гаттерия,
гекконы и многие ископаемые формы), и иногда
между телами позвонков сохраняются и остатки хорды, но у громадного
большинства современных форм имеются процельные позвонки, и хорда
совершенно исчезает. В некоторых случаях тела соседних позвонков
связаны между собой посредством межпозвоночных дисков или менисков,
как это наблюдается у гаттерии и у крокодилов. У гаттерии и у гекконов
между телами шейных позвонков, а также между телами задних туловищ-
ных позвонков и в начале хвостовой области снизу вклиниваются еще
особые элементы, носящие неудачное наименование межцентров (inter-
centra). Ребра соответствующей области нередко причленяются головкой
к этим элементам (рис. 94). В хвостовой области к ним причленя-
ются гемальные дуги (рис. 95). У крокодилов и у ящериц такие же
«межцентры» прирастают к позвонкам, образуя особые нижние их
отростки. Эти элементы, соответствующие описанным ранее гипоцен-
Рис. 93. Позвоночник
бесхвостой амфибии (Dis-
coglossus) (из Видерс-
гейма).
ап — верхняя дуга перво-
го позвонка; рг — сочле-
новный отросток; sp —
остистый отросток; ir —
поперечные отростки туло-
вищных позвонков; tre —
поперечные отростки хво-
стовой области (уростиль—
со); г — ребра; s — крест*
новый позвонок.
777
трам, указывают па первоначальную множественность вяемонтов по-
звоночника.
Рис. 94.
А — схематический продольный разрез через три первых шейных позвонка примитивной
рептилии, показывающий образование атласа (Л') и эпистрофея с его зубовидным отростком
(Z>Z'); Л — гипоцентр (нижняя дуга); pZ — тело (плевроцентр); nt — хорда; В — два грудных
позвонка примитивной рептилии; с — головка ребра; па — верхняя дуга; pz — передний
суставной отросток; t — бугорок ребра; tp — поперечный отросток (по Гудричу).
Верхние дуги смыкаются над спинномозговым каналом и образуют
хорошо развитый, сжатый с боков, верхний остистый отросток (рис. 95).
От дуг отходят вперед и назад суставные отростки, служащие для сочле-
нения позвонков друг с другом. Боковые отростки в некоторых случаях
двойные — у крокодилов от верхней дуги отходят довольно значитель-
ные поперечные отростки и на теле позвонка имеются также небольшие
Рис. 96. Два хвостатых позвонка
аллигатора (из Кингсли).
с — тело; ha — нижняя дуга; hs—
нижний остистый отросток; ап — верх-
няя дуга; ns — верхний остистый от-
росток; poz, prz — задние и передние
сочленовные отростки; t — поперечный
отросток.
Рис. 96. Передний и задний груд-
ные позвонки аллигатора (из Геген-
баура).
с — ребро; ср — тело; ра — суставной
отросток; pt — поперечный отросток;
sp — остистый отросток.
выступы (рис. 96). Ребра амниот относятся, по новейшим данным (Емель-
янов), к типу низших ребер. Они образуют у своего проксимального конца
развилку, нижняя ветвь которой сочленяется с упомянутым выступом на
теле позвонка, а верхняя ветвь — с концом поперечного отростка. Такое
двойное сочленение ребра с позвонком наблюдается у крокодилов в шей-
ном отделе и в начале грудной области (рис. 97). То же самое наблюдается
и у многих ископаемых рептилий. Нужно думать, что у рептилий верхний
поперечный отросток образовался разрастанием бокового отростка. И у
других современных рептилий, в особенности у гаттерии и у гекконов,
772
наблюдаются нередко ясные следы прежнего существования такого двой-
ного сочленения ребра с позвонком, и, таким образом, юно должно счи-
таться примитивным признаком наземных позвоночных. По сравнению с
млекопитающими, у которых также имеется двойное сочленение ребра
с позвонком, нижнюю ветвь
развилки ребра можно назвать
его голоькой (capitulum), а
верхнюю ветвь — бугорком
(tuberculum). Двойное соеди-
нение ребра с позвоночником
является особенностью назем-
ных позвоночных: оно дает
необходимую поддержку вну-
тренностям, допуская в то же
время свободные дыхательные
движения при активном уча-
стии ребер.
Ребра имеются на большин-
стве и иногда па всех позвонках
предкрестцовой области. У
крокодилов, например, и на
первом шейпом позвонке (ат-
ласе) имеется уже самостоятель-
ное ребро (рис. 98). У других рептилий первые позвонки обыкновенно
лишены. ребер, по следующие шейные позвонки обладают свободными
ребрами (только у черепах их рудименты прирастают к позвонкам),
области лишь несколько
меньшей длиной. В задней
части грудной области
имеется всегда (и у кроко-
дилов) простое причлепе-
ние ребра только к по-
перечному отростку (рис.
Иногда в последних
туловищных позвонках
ребра 1 редуцируются и
срастаются с поперечны-
ми отростками. В этом
случае различают еще одну
область позвоночника, не
имеющую самостоятель-
ных ребер, — пояснич-
ную.
В наиболее развитых
ребрах грудной области
различают по два отдела:
проксимальный — позво-
ночный отдел, который
Рис. 97. Схема ребра амниот; вид спереди
и сбоку (по Ремане).
cap — головка; с», ct, cs — позвоночный, проме-
жуточный и грудинный отделы ребра; ри —
крючковидный отросток; st — грудина; tub —
бугорок ребра.
отличающимися от ребер следующей грудной
Рис. 98. Первые четыре шейные позвонка кроко-
дила (по Гудричу).
с — головка ребра; hp — гипопентр (нижняя дуга);
пц — верхняя дуга; ра — проатлас (его верхняя дуга);
pl — тело позвонка; prz, ptz — передний и задний
суставные отростки; г — ребро; t—бугорок ребра; 1р—
поперечный отросток. Нервная дуга первого позвонка
(атласа) и проатласа на левой стороне удалены для того,
чтобы показать тело первого позвонка (рГ), образующее
зубовидный отросток.
всегда является костным, и дистальный — грудинный отдел, который
обыкновенно остается хрящевым (у крокодилов и он окостеневает).
Между этими основными отделами у примитивных форм (особенно у
млекопитающих) имеется еще небольшой промежуточный отдел (рис. 97).
Иногда позвоночный отдел ребер снабжен сзади небольшим крючко-
видным отростком (processus uncinatus), налегающим сверху па следующее
позади ребро (гаттерия, крокодилы) (рис. 97). Все указанные отделы
ребра имеют свои самостоятельные хрящевые закладки и независимые
113
•центры окостенения ’(если грудинный отдел вообще етистенежает; чащй'
здесь наблюдается лишь обызвествление).
В хвостовой области позвоночника у рептилий имеются настоящие
костные гемальные дуги с нижними остистыми отростками, обладающие ха-
рактерной формой буквы Y, но эти дуги причленяются не к телам позвон-
ков, а между телами (промежуточное их положение дает указания на
происхождение тел позвонков из вставочных элементов, т. е. плевроцентров
позвоночника) (рис. 95).
Как правило, в хвостовой области имеются и остатки ребер, прира-
стающие к телам и образующие здесь поперечные отростки позвонков.
Для высших наземных позвоночных (Amniota) весьма характерны
изменения, происходящие в области первых шейных позвонков (рис. 94г
98). Тело первого позвонка не срастается^с^йбТвбтстеующйми-дугами и
связками соединяется с телом~второго позвонка (у змей, птиц и млекопи-
тающих вполне прирастает), образуя его зубовидный отросток (processus
odontoideus). Второй позвонок оказывается, таким образом, состоящим
из тела второго позвонка с его дугами, из связанного с ним тела первого
позвонка и из вклинивающегося между этими телами гипоцентра. Этот
позвонок получает название эпистрофея. Первый позвонок лишен
своего тела и состоит из одних дуг, которые замыкаются снизу сраста-
ющимся с ними передним'Типоцентром (т. е. нижней дугой). Таким образом,
этот позвонок получает форму кольца, в котором горизонтальная связка
отделяет спинномозговой канал от того места, куда входит связанное со
вторым позвонком тело, т. е. зубовидный отросток. Первый позвонок,
атлас, довольно свободно может вращаться на зубовидном отростке
эпистрофея. У некоторых ископаемых крокодилов очень хорошо видно
происхождение атласа и эпистрофея из частей темноспондильного позво-
ночника. У этих форм вращение атласа еще не было возможно и первое
ребро причленялось как раз на границе между нижней дугой (гипоцентром)
атласа и его «телом» (плевроцентром), т. е. зубовидным отростком. У позд-
нейших крокодилов причленепие ребра перемещается вниз на дугу атласа.
Впереди первого позвонка иногда наблюдаются остатки еще одного
позвонка, лежавшего между ним и черепом. Это так называемый проатлас
(proatlas), тело которого прирастает к телу атласа и также входит в состав;
зубовидного отростка эпистрофея, а верхние дуги вклиниваются между
атласом и затылочной частью черепа и наблюдаются в виде самостоятель-
ной кости у взрослых крокодилов, гаттерии и хамелеонов (рис. 98).
На передней поверхности атласа снизу имеется суставная ямка, дно1
которой образовано зубовидным отростком, для сочленения с единствен-
ным у рептилий затылочным бугром черепа.
Шейная область позвоночника рептилий содержит различное число
позвонков, обладающих, как правило, самостоятельными, хотя и неболь-
шими ребрами. Наиболее диференцирован этот отдел у черепах, где руди-
ментарные шейные ребра прирастают к позвонкам, и шея обладает зна-
чительной подвижностью. Первым грудным позвонком считается тот по-
звонок, ребро которого доходит до грудины. В общем грудная область
позвоночника состоит из различного числа позвонков с самостоятельными
ребрами. Поясничная область характеризуется значительными попереч-
ными отростками, представляющими результат прирастания к ним руди-
. ментарных ребер. Эта область у рептилий редко бывает выражена. Крест-
цовая область служит для причленения подвздошных костей таза (рис. 99)
и состоит у современных рептилий всегда из двух позвошсов (у ископаемых
рептилий крестцовых позвонков бывало и больше). Подвздошные кости
причленяются и здесь, как у амфибий, к ребрам крестцовых позвонков
(рис. 99), но только эти ребра срастаются в одно целое с поперечными
отростками, которые соответственно выделяются своей мощностью. За-
кладку самостоятельных крестцовых ребер можно еще обнаружить во
114
время эмбрионального развития. Хвостовая область состоит из различного
и иногда значительного числа позвонков. Для этой области характерно
существование гемальных дуг, причленяющихся интервертебрально
(рис. 95).
Так как границы меягду отдельными областями определяются отно-
шением к грудине и тазовому поясу, то у форм, лишенных конечностей,
у которых исчезают и грудина и тазовый пояс, как, например, у змей,
границы между отделами становятся неясными. Общее число позвонков
у форм с длинным телом, каковы безногие ящерицы и змен, может быть
очень велико (до 400).
У птиц позвоночник морфологически очень сходен с позвоноч-
ником рептилий, но он своеобразно специализирован. В связи с потреб-
ностью в прочной опоре для летательных мышц и для задних конечностей,
Рис. 99. Крестцовые позвонки крокодила (Л) со спинной сто-
роны и ящерицы, игуаны (В) — с брюшной (по Ремане).
Ц — подвздошная кость; г — крестцовые ребра; s — крестец.
которые одни поддерживают тело, вся туловищная часть позвоночника
становится неподвижной, и многие позвонки срастаются между собой,
образуя более или менее крупные комплексы позвонков. В противопо-
ложность этому шейный отдел приобретает очень большую подвижность,
и обладает нередко значительной длиной в зависимости от увеличения
числа позвонков, входящих в состав шеп. Изгибание шеи (в вертикальной
плоскости) в виде буквы S является средством для перемещения центра
тяжести птицы во время полета. Тела позвонков соединяются здесь между
собой седловидными поверхностями (гетероцельная форма позвон-
ков) с менисками между ними, которые позволяют производить свобод-
ное пзгибапие шеи во всех направлениях, но не допускают вращения.
Атлас и эпистрофей специализировались для совершения вращательных
движений. Зубовидный отросток вполне срастается с телом эпистрофея.
Шейные ребра рудиментарны и прирастают к позвонкам за исключением
нескольких последних, которые могут остаться свободными. Так как
ребра соединяются с позвонком двойным сочленением и таким образом
прирастают головкой к телу, а бугорком — к поперечному отростку, то
между приросшим ребром, телом и поперечным отростком остается про-
странство, через которое проходит позвоночная артерия. Таким образом,
получается картина продырявленного поперечного отростка.
В грудной области сильно развитые ребра имеют также двойное со-
членение с позвонком. У птиц оба отдела ребра — позвоночный и грудин-
ный — окостеневают. Первый обладает костными же крючковидными
отростками (processus uncinati), как и у крокодилов. Эти отростки нале-
гают каждый на следующее позади ребро, что способствует достижению
большей устойчивости всей грудной клетки. Несколько грудных позвон-
ков срастаются обыкновенно между собой в один неподвижный комплекс.
115
Самое замечательное в позвоночнике птиц — это огромный сложный
S
Рис. 100. Крестец
молодого голубя (по
Бючли).
S — первичные крест-
цовые позвонки с реб-
рами.
крестец (synsacrum), состоящий из значительного числа сросшихся между
собой позвонков. В этом крестце, однако, лишь два позвонка выделяются
своими крестцовыми ребрами, и этим обнаруживается генетическая связь
с рептилиями (рис. 100). Эти два позвонка носят название истинных крест-
цовых позвонков. Впереди них лежит некоторое
число позвонков без ясных ребер (точнее, руди-
ментарные ребра образуют здесь одно целое с
поперечными отростками) — это позвонки пояснич-
ной области, впереди которых в составе крестца
имеются иногда еще и грудные позвонки с типичны-
ми грудными ребрами. Позади двух истинных крест-
цовых позвонков в состав крестца входит еще более
или менее значительное число позвонков хвостовой
области. В этой области число свободных позвон-
ков у современных птиц очень незначительно, и
последние позвонки’ почти7 всегда срастаются между
собой в плоскую вертикальную пластинку — коп-
чик (os coccygis), служащий для поддерживания
рулевых перьев. У ископаемой птицы Archaeopte-
ryx было значительное число свободных хвостовых
позвонков, как у рептилий.
У млекопитающих поверхности тел
позвонков плоские, и между ними залегают меж-
позвоночные диски, или мениски. Верхние дуги
продолжаются в довольно длинный (особенно
в грудной области) верхний остистый отро-
сток. У основания верхних дуг имеются суставные
отростки. Ребра млекопитающих сохраняют двойное причленение к позвон-
кам. В шейной области они рудиментарны и срастаются с позвонками,
как и у птиц, образуя продырявлен-
ные поперечные отростки (рис. 101, /Л
102). В грудной области ребра причле- Z7 ап
няются головкой между двумя соседни- /
ми телами позвонков и бугорком к й \ Р^
поперечному отростку. Они не обра-
зуют более ясной развилки, как у .
нижестоящих форм, так как верхняя С ср С J
ветвь развилки укорачивается и те-
перь, действительно, может быть на-
звана бугорком (рис. 102). Гебро и у
млекопитающих состоит из двух или
даже трех отделов, из которых грудин-
ный обыкновенно остается хрящевым (у
клоачных и неполнозубых он также
Рис. 101. Третий шейный’ позвонок
молодой ехидны (по Флоуеру).
ап — верхняя дуга; с — ребро; ср —
тело позвонка; pl — боковой и pt —
поперечный отростки.
окостеневает). Поперечные отростки занимают типичное положение на
верхних дугах и только в поясничной области сидят иногда на телах.
Кроме того, вЧтоясничной области (и на задних позвонках грудной)
нередко (в особенности у грызунов и у неполнозубых) развиваются до-
вольно значительные добавочные отростки, известные под названием
метапофиз и анапофиз.
Число шейных позвонков у млекопитающих за очень редкими исклю-
чениями ограничено семью (у ламантина и у одного ленивца — 6, у пан-
голина — 8 и у другого ленивца — 9). Длина шеи зависит не от числа
позвонков, а от их длины. Почти отсутствующая шея китов состоит из
7 совершенно плоских позвонков, которые нередко между собой сраста-
ются. Длинная шея жирафы содержит также лишь 7 позвонков, но только
116
очень длинных. Первые 2 позвонка представлены атласом и эпистрофеем
типичного строения. В кольцевидном атласе брюшной отдел (гипоцентр)
нередко имеет свой центр окостенения (рис. 103). Зубовидный отросток
у клоачных представлен еще особой костью, но у живородящих млеко-
питающих вполне срастается с телом эпистрофея (однако сохраняет свое
самостоятельное окостенение у зародыша).
Число грудных позвонков вариирует (9—24); чаще всего их около 13.
Ребра передних грудных позвонко
чаются свободно. У китообразных
pt ап
в доходят до грудины, последние кон-
до грудины доходит только одно или
во всяком случае немногие ребра.
Число поясничных позвонков также
различно (2—9). В состав их попереч-
ных отростков входят и рудиментар-
ные ребра, что во многих случаях
доказано эмбриологически.
Крестец состоит из небольшого
числа сросшихся позвонков (1—10;
максимальное число у неполнозу-
бых), чаще всего 2—4, из которых
Рис. 102. Схема шейного (Л) и грудно-
го (В) позвонков млекопитающих (из
Вебера).
ап — нервная дуга; ! с — ребро (заштри-
ховано); cap — головка ребра; ср — тело
позвонка; ра— суставные отростки; pt —
поперечные отростки; tub — бугорок реб-
ра; sp — остистый отросток.
Рис. 103. Атлас и эпистрофей молодо-
го носорога (из Вебера)
at — атлас; ер — эпистрофей; о — вубо-
видный отросток (тело атласа); v — ниж-
нее окостенение кольца атласа (гипо-
центр).
лишь 1—2 позвонка являются истинными, так как в их боковых частях,
служащих для сочленения с подвздошными костями тазового пояса, наблю-
даются самостоятельные окостенения, соответствующие крестцовым реб-
рам. Следующие позади позвонки, если и обнаруживают иногда закладки
ребер, то во всяком случае они не принимают участия в подвздошно-крест-
цовом соединении. Эти позвонки являются, следовательно, лишь прирос-
шими к крестцу хвостовыми позвонками.
Хвостовой отдел позвоночника состоит из различного числа позвон-
ков (3—46) в зависимости от длины хвоста. В этой области нередко наблю-
даются типичные нижние дуги с нижними остистыми отростками в виде
буквы ¥, которые, как и у рептилий, причленяются между телами позвон-
ков (интервертебрально).
Человек и по строению своего позвоночника занимает высшее
место среди млекопитающих, в частности среди высших обезьян. Шейны#
отдел у человека состоит из тех же 7 позвонков; из них на двух последних
эмориологически доказана закладка шейных ребер. Особенностью шейных
позвонков человека является раздвоенный обыкновенно остистый отросток.
Грудных позвонков 12, как и у оранга (у гориллы и у шимпанзе их 13),
но известны случаи нахождения и тринадцатого ребра. Форма грудной
клетки — сплющенная со спинной и брюшной стороны, бочонкообразная
117
и у человека, и у человекообразных обезьян. Поясничных позвонков 5,
причем доказана закладка ребер, срастающихся с поперечными отростками
(в виде аномалии свободное ребро наблюдалось на первом поясничном
позвонке и у взрослого человека). Крестцовых позвонков 5, из которых 2
истинных. В боковых массах крестца (massae laterales) эмбрионально
наблюдаются особые окостенения, соответствующие закладке крестцовых
ребер. Хвостовая область позвоночника рудиментарна и в связи с этим
сильно вариирует. У взрослого человека наблюдается 3—6 позвонков
(обычно 4), а эмбрионально до 9. У человекообразных обезьян хвоста
нет, и число хвостовых позвонков близко к тому, какое имеется у
человека. У оранга 3 позвонка, а у других обычно 4. Для позвоночника
человека в целом характерен его изгиб (намечающийся, впрочем, и
у обезьян), особенно вогнутая поясничная область и резкий изгиб назад
на границе крестца, где последний образует своим передним краем вы-
втуп внутрь — мыс (promontorium).
Грудпна
Грудина представляет собой систему элементов скелета, связывающих
брюшные концы ребер грудной области между собой и с плечевым поясом.
Первоначальное ее значение, очевидно, состояло в укреплении пояса,
необходимом ввиду того, что конечности являются органами опоры всего
тела наземного позвоночного. Таким образом, в тазовом поясе развилось
причленение к позвоночнику, вызвавшее образование крестца, а в плечевом
поясе произошло общее увеличение его поверхности и установилась
связь с ребрами посредством грудины. Естественно, что настоящая грудина
имеется только у наземных позвоночных и вторично исчезает у тех форм,
у которых исчезли конечности (змеи). У птиц она значительно развивается
соответственно большой роли передней конечности (крыла) и в особенности
вследствие увеличения поверхности прикрепления сильных грудных
мышц.
Грудина низших наземных позвоночных — амфибий развивается из
парных зачатков, возникающих в миосептах брюшной стенки тела, т. е.
в местах, соответствующих нормальному положению концов нижних
ребер (которые у амфибий редуцировались). Так же развивается и пред-
грудинник. Это дает нам возможность считать и грудину амфибий и даже
предгрудинник продуктом соединения брюшных концов ребер, дававших
опору поясу конечностей. С другой стороны, возможно, что грудина
амфибий диференцировалась на месте за счет возникших в миосептах
охрящевений, подобных тем, которые имеются в брюшной стенке Necturus.
Сходные охрящевения могли дать начало и грудинным отделам ребер
наземных позвоночных, которые, как известно, имеют самостоятель-
ную закладку и, быть может, вторично соединились с позвоночными
отделами ребер. Однако и в этом случае мы должны признать существо-
вание генетической связи грудины с ребрами.
У высших наземных позвоночных — млекопитающих грудина разви-
вается в виде парных зачатков, образующихся путем слияния брюшных
концов ребер (рис. 104) (иногда парные полоски хряща развиваются
раньше, а затем с ними сливаются брюшные концы ребер). Парные
полоски хряща сливаются затем в одну непарную пластинку, вырастающую
и вперед от брюшных концов первой пары ребер по направлению к эле-
ментам плечевого пояса, а иногда и назад от концов последней пары.
Позже, во время окостенения грудины, иногда замечаются еще следы
парного ее происхождения в появлении парных центров окостенения
(некоторые млекопитающие).
У амфибий грудина непарная и развивается совершенно неза-
висимо от ребер, которые до нее никогда не доходят. Хота нижние ребра
118
амфибий, несомненно, вторично редуцированы, все же обычно счи-
тают, что их грудина возникла независимо от ребер-. Каково было отноше-
ние ребер к грудине у стегоцефалов — неизвестно, так как в ископаемых
Рис. 104. Развитие грудины человека
(по Руге, из Кингсли).
с! — ключица; г — ребра; s — парный ва-
чаток грудины.
Рис. 105. Брюшная часть плечевого пояса
и грудина лягушки.
cl — ключица; со — коракоид; sc — лопатка;
st — грудина.
остатках скелетов сохранились только костные проксимальные части
ребер, которые у некоторых-форм (Microsauria) были довольно длинные.
Несомненно, дистальные части ребер были хрящевые (как у большинства
современных?рептилий и млекопита-
ющих) и не сохранились. Возможно,
что они все же достигали грудины.
Обыкновенно грудина амфибий
имеет вид треугольной хрящевой
пластинки, налегающей отчасти сни-
зу на задние края брюшного отдела
плечевого пояса (коракоиды). У
бесхвостых амфибий она связывается
иногда более прочно с плечевым
поясом и отчасти окостеневает (рис.
105).
У рептилий грудина состоит
из хрящевой, обыкновенно ромби-
ческой пластинки, тесно связанной
с коракоидами плечевого пояса и
некоторым, иногда очень незначи-
тельным (у хамелеона 1 пара) числом
ребер, которая обычно продолжается
:назад в виде узкой полосы,* свя-
занной еще с некоторым числом ребер
|(рис. 106). Эта задняя часть грудины
i(metasternum), несомненно, произо-
шла путем слияния концов ребер и
иногда еще сохраняет первоначаль-
Рис. 106. Грудина рептилий'— геккона
(Я) и хамелеона (В), (из Гегенбаура).
с — ребра; cl — ключица; со — порапоид;
est — надгрудиннип; st — грудина.
ную свою парность (гекконы и др.).
Иногда грудина имеет ничтожное развитие, а позади нее брюшные концы
ребер попарно срастаются между собой (хамелеоны) (рис. 106, В). Неви-
димому, это указывает нам тот путь, которым шло развитие реберной
части грудины. Передняя >ц£еть грудины, связанная с концами коракоидов,
119
вероятно, возникла раньше в виде независимого хряща между ними и
концами первой пары ребер и может быть гомологична всей грудине
амфибий.
У птиц грудина развита очень сильно и состоит из большой костной
пластинки, связанной впереди с коракоидами, а сбоку — с костными же
брюшными концами ребер (рис. 107). По средней линии па ней имеется
Рис. 107. Грудина птицы (цесарки)
(из Гегенбаура).
с — ребра; со — коракоид; st — гру-
дина.
Рис. 108. ’ Грудина лемура
(по Боасу).
с — ребра; cl — ключицы; т — руко-
ятка грудины; х — мечевидный отрос-
ток.
большой плоский непарный выступ — киль (carina), служащий для
прикрепления грудных мышц (рис. 180, В). У нелетающих птиц (Ratitae и
некоторых других) гребень редуцируется.
Грудина млекопитаю
Рис. 109. Плечевой пояс и грудина
клоачного млекопитающего (утконо-
са) (из Видерсгейма).
с! — ключица; со — коракоид; est —
надгрудивник; gl — сочленовная ямка;
рсо — прокоракоид; sc — лопатка; st —
грудина.
Ввиду того, что передняя и.
рептилий обнаруживает тесную
щ и х состоит из нескольких отделов.
ПереднийТрасишренный ; отделу связан с
первой парой ребер и с ключицами, если
таковые имеются (рис. 108). Он но-
сит название рукоятки (manubrium).
Следующий отдел связывает между
собой концы нескольких пар ребер и
носит наименование тела (corpus).
Этот отдел очень часто состоит из
целого ряда костных сегментов,
лежащих по одному между соседними
парами ребер. И у человека этот
отдел, одновременно с наступающим
окостенением сегментируется па 4—5
сегментов, которые затем уже сра-
стаются. Позади тела выдается более
или менее значительный мечевидный
отросток (xiphisternum), несомненно,
происшедший, как и тело, путем слия-
ния брюшных концов известного числа
ребер, которые затем утратили связь
с грудиной.
повидимому, первичная часть грудины
связь с концами коракоидов, весьма
интересно провести сравнение с грудиной клоачных млекопитающих,
которые одни только обладают хорошо развитыми коракоидами. Ока-
зывается, что у клоачных млекопитающих, впереди несомненной рукоятки,
связанной с первым ребром, имеется у молойого животного еще один
120
хрящевой отдел грудины — предгрудинник (prosternum), лежащий между
концами коракоидов. Очевидно, это первичная, передняя часть грудины
ч (гомолог грудины амфибий). Позже этот хрящ редуцируется и отчасти
вытесняется развивающейся здесь костью (рис. 109, est). У живородящих
млекопитающих наблюдались закладки небольших хрящей впереди
рукоятки, которые впоследствии обычно с ней срастаются. Это остатки
предгрудинника, который редуцировался в связи с редукцией коракоидов.
Надгрудиииик
С грудиной наземных позвоночных довольно тесно связана покровная
кость, характерная для современных рептилий (рис. 106, Л). Этот над-
грудинник (episternum) имелся и у стегоцефалов в виде типичной
кожной кости, обычно ромбической формы, нередко с характерной для
последних скульптурой. У современных амфибий этой кости нет вовсе.
У большинства современных рептилий она имеет форму буквы Т,
непарная ветвь которой налегает на грудину, а боковые ветви связаны
с ключицами (откуда другое название этой кости — межключица, inter-
clavicula). У черепах она вместе с ключицами вошла в состав их брюшного
щита (endoplastron). У безногих рептилий она, понятно, редуцируется;
у живородящих млекопитающих ее также нет, но у клоачных име-
ется кость типичной для надгрудинника формы, к обеим боковым ветвям
которой прилежат ключицы (рис. 109). Эта кость по своей форме и поло-
жению столь типична, что должна считаться гомологом надгрудинника
рептилий. Однако в то время как последний представляет собой типичную
покровную кость, надгрудинник клоачных врастает в лежащий под ним
предгрудинный хрящ (prosternum) и отчасти его вытесняет.
Повидимому, здесь перед нами один из случаев превращения покров-
ной кости в кость, замещающую хрящ. Другой аналогичный и совершенно
несомненный пример такого же превращения представляет ключипа —
кость, явно покровная по своему происхождению, по отчасти вытесняющая
у млекопитающих лежащий под нею прокоракопдпый хрящ.
Брюшные ребра,
Позади грудины у некоторых рептилий наблюдается еще парный
ряд удлиненных костей, известных под названием брюшных ребер (gastra-
lia). Это чисто кожные окостенения, с ребрами, понятно, ничего общего
не имеющие. По своему происхождению это остатки брюшного панцыря
стегоцефалов (рис. 67). Эти образования пользовались большим распро-
странением среди ископаемых рептилий (рис. 397, 403), из современных
же форм они сохранились у гаттерии и у крокодилов.
Череп
Диференцировка мезодермы в области головы
Мы уже отмечали, что у позвоночных спинной отдел мезодермы сег-
ментируется и образует сомиты, а брюшной отдел, так называемая боковая
пластинка (спланхнотом), остается несегментированным. Сомиты отде-
ляют внизу на своей внутренней стороне склеротомы, дающие начало
осевому скелету (рис. 110). Остальные части сомитов — миотомы —
сохраняют у низших позвоночных свою сегментацию пожизненно в миоме-
рах боковой мышцы. Боковая пластипка ограничивает несегментирован-
ную полость тела.
В области головы мезодерма также сегментируется. В спинном ее
отделе развиваются сомиты головы, которые также диференцируются
на склеротомы и миотомы (рис. 110). В брюшпом отделе боковая пластинка
головы (в отличие от туловища) тоже сегментируется, хотя этот процесс
происходит здесь вторично и обусловлен развитием жаберных щелей.
121
Развивающиеся жаберные мешки растут по направлению к эктодерме,
оттесняя на своем пути мезодерму. Таким образом, жаберные мешки
делят боковую пластинку на участки, лежащие между ними, т. е. в жабер-
ных перегородках (рис. 111). Эти участки мезодермы дают начало висце-
ральной мускулатуре.
Рис. 110. Источники образования осевого скелета. Схематические поперечные разрезы
через туловище (Л) и голову (В) зародыша позвоночного.
on — закладка дуг позвоночника; ch — хорда; сое!—полость тела; сгп—закладка occBoiti
части черепа; его —закладка висцерального скелета; gn— ганглионарная пластинка; ту —
многом; п — мозг; ini — кишка; ph — глотка; set — склеротом.
Головные сомиты развиваются весьма типично у наших позвоноч-
ных. Различают сомиты передней части головы, лежащие впереди от
закладки уха, — предушпые сомиты, которых развивается 3 пары (по
Рис. 111. Сегментация мезодермы в области головы. Стереограмма (согласно Матвееву).
ahc — передняя головная полость; аи — слуховой пузырек; сег — головной {мозг; ch —
хорда; сое! —полость тела; hy — подъязычная перегородка (зачаток дуги); ini—кишка;
it — боковая пластинка; md — челюстная перегородка (зачаток дуги); ту—миотом; п —
мозг; ос — глазной бокал; scl — склеротом; st — рот. 7, 2, 3 — жаберные перегородки; I—IX —
головные миотомы.
более новому толкованию их 5 пар), и лежащие позади — заушные сомиты,
которых развивается от 4 до 6 пар. Средний из трех предушных сомитов
неразрывно связан с той частью боковой пластинки, которая лежит в
области закладки челюстной дуги, и потому получил название челюстного
сомита (это, невидимому, двойной сомит) (рис. 111). Передний сомит
722
соответственно называется предчелюстным, а третий — зачелюстным
(также двойной сомит). И в области головы каждый сомит диференци-
руется на внутренний отдел, который затем расплывается в массу скеле-
тогенной мезенхимы — склеротом и наружную часть — миотом; склеро-
томы развиваются в голове даже сильнее, чем в туловище (рис. 110);
наоборот, миотомы здесь разрастаются значительно меньше и частью
имеют явно рудиментарный характер.
Из предушных миотомов развиваются глазные мышцы. Из первого —
три прямые мышцы глаза (верхняя, внутренняя и нижняя) и нижняя
косая, из второго — верхняя косая и из третьего — наружная прямая.
Заушные миотомы обычно принимают участие в развитии подъязычной
мускулатуры и у круглоротых сохраняются полностью в виде ряда миоме-
ров взрослого животного. У остальных позвоночных заушные миотомы
большей частью исчезают бесследно, и лишь редко 1—2 последних миотома
сохраняются в затылочной области взрослого животного.
Во всяком случае можно принять как правило, что у типичного
взрослого позвоночного в области головы миомеров нет; факт же существо-
вания миотомов во время эмбрионального развития показывает, что
у предков позвоночных голова была построена метамерно, подобно туло-
вищу, как это наблюдается в настоящее время у ланцетника (у которого
миомеры доходят до переднего конца головы). У типичных позвоночных
метамерия головы утеряна, и причину этого указать нетрудно. Концентра-
ция важнейших органов чувств на переднем конце тела вызвала прогрес-
сивное развитие центральной нервной системы — головного мозга. Про-
грессивное развитие органов чувств и головного мозга представляет
наиболее характерный процесс, действительно определяющий в общих
чертах всю эволюцию позвоночных животных. Важное значение этих орга-
нов и одновременно большая их нежность вызвала необходимость в спе-
циальной их защите, и такой защитой явилась черепная коробка.
С развитием черепа передний конец тела потерял, однако, свою
гибкость, и мускулатура его в виде миомеров боковой мышцы стала
излишней.
Значение черепа не только в том, чтобы защищать известный ряд
органов. Позвоночные — в основе хищники. В передней части головы,
в области рта, у них развились сильные органы захватывания пищи —
челюсти (а у круглоротых — сложный губной аппарат). Функциониро-
вание челюстей требует известной опоры для них и значительной мускула-
туры. Черепная коробка служит для прикрепления этой муску-
латуры и дает опору для самих челюстей. Наконец, значительной трате
энергии подвижного хищника должен соответствовать и усиленный обмен
веществ, связанный с интенсивностью окислительных процессов. Это
определило и прогрессивное развитие органов дыхания — жаберного
аппарата, с его мускулатурой и скелетом, который разви вился
также в области головы и для которого опорой является та же череп-
ная коробка.
Череп. Скелет головы, или череп (cranium), естественно распа-
дается на два отдела — осевой и[висцеральный,
неурокраниум, представляетсобоУ KaKJjLHRQ&ogKgHHgjpceBoro скелета
туйовища и так же, как и последний, служит для защиты центральной
нервной системы. Это — коробка, заключающая в себе головной мозг,
органы обоняния и слуха, и служащая отчасти защитой и органу зренид.
Вис ц_е р а л ь н ы й череп, или спланхнокраниум, представляет
скелет передней части кишечника. Он состоит из ряда висцеральных
дуг, из которых передние преобразованы в органы захватывания
пищи, а все остальные служат для поддерживания органов дыхания. Оба
отдела черепа развиваются независимо друг от друга и различным об-
разом.
123
Развитие осевого черепа
На внутренней стороне головных сомитов, как сказано, развиваются
склеротомы, вполне соответствующие склеротомам туловища. Склеротомы
головы развиты даже сильнее, так как они поставляют значительный
клеточный материал, потребный для развития всей черепной коробки.
Как и в туловище, склеротомы расплываются в массу мезодермальных
клеток (мезенхиму), которые распределяются около хорды и вокруг
центральной нервной системы, образуя соединительнотканную оболочку,
Рис. 112. Хрящевые закладки осевого черепа (схема).
<fn — верхние дуги позвоночника; ch — хорда; pch— парахордалии; st — глазничные хря-
щи; tr — трабекулы.
окружающую головной мозг и развивающиеся органы чувств (рис. 110).
Это стадия перепончатого черепа, за которой в дальнейшем следует стадия
хрящевого черепа, а у большинства позвоночных — и стадия костного
черепа. Развитие хряща идет не сразу во всей скелетогенной ткани. По-
следняя и вообще неравномерно распределена — она образует местные
сгущения, в особенности под головным мозгом (дно перепончатого черепа)
Рис. ИЗ. Стереограмма головы зародыша акулы. Развитие хрящевого черепа (по
Северцеву).
а. ьг — жаберные дуги; a. hy — подъязычная дуга; ат — челюстная дуга; с. аи — слухо-
вая капсула; ch — хорда; h — гипофпв; int—кишка; т — средний мозг; п — спинной
мозг; па—-ноздря; ос —глаз; peh— парахордалии; рг—первичный передний мозг; si —
глазничные хрящи; tr —трабекулы.
и в этих сгущениях начинается охрящевение от некоторых довольно
постоянных центров. Таким образом, у позвоночных развиваются перво-
начально лишь определенные элементы хрящевого черепа. Таких постоян-
ных элементарных хрящей имеется две пары, лежащие обе в основании
черепа.
По бокам хорды, образуя непосредственное продолжение позвоноч-
ника, развивается пара удлиненных околохордовых хряще й—
парахордалии (parachordalia cranii) (рис. 112). Впереди хорды, под осно-
ванием переднего мозга, развивается другая пара удлиненных хрящей —
черепные балки, или трабекулы (trabeculae cranii, рис. 112)»
724
Кроме этих хрящей, образующих основание черепа, наблюдается иногда
еще пара хрящей в боковых его стенках, в задней части глазниц. Это так
называемые глазничные, или сфенолатеральиые, хрящи черепа (cartilagines
orbitales s. sphenolaterales). Наконец, более или менее независимо от этих
элементов черепа развивается хрящ в соединительнотканных оболочках
(капсулах), окружающих органы чувств. Слуховые капсулы развиваются
в непосредственной близости (а иногда и в связи) с передним отделом
парахордалий, с которыми они во всяком
случае вскоре срастаются (рис. 113). Обо-
нятельная капсула развивается обычно сразу
в связи с передним концом трабекул (рис. 114).
Наконец, капсула органа зрения остается
всегда независимой (и подвижной) и обра-
зует склеру глаза, которая у громадного
большинства позвоночных является хрящевой,
а иногда частью окостеневает.
Парахордалии занимают такое же
положение по отношению к хорде, как и по-
звоночник, и в задней их части можно иногда
заметить следы подобной же сегментации с
указанием даже на существование двух пар
элементов в каждом сегменте (рис. 112, 113).
Именно здесь наблюдались у хрящевых рыб
выступы, соответствующие по своему положе-
нию верхним концам дуг позвоночника, и
между ними правильно располагаются выходы
затылочных нервов. Одним словом, имеются
ясные указания на образование задней части
парахордалий путем срастания известного чис-
ла дуг позвоночника. Такие дуги носят на-
звание затылочных, и вся задняя часть парахор-
далий обозначается как затылочный их отдел.
Передняя часть парахордалий обозначается по
отношению ее к слуховой капсуле как слухо-
вой отдел. Хотя в этом отделе ясных следов сег-
ментации не наблюдалось, он образует непо-
средственное продолжение затылочного отдела,
занимает такое же положение по отношению к
хорде и, очевидно, имеет такое же происхожде-
ние. Вся эта область черепа вследствие при-
сутствия в ней хорды носит название хордаль-
ной части черепа в отличие от передней, или
прехордальной, части черепа. Надо прибавить,
что сегментация мезодермы и образование головных сомитов наблюдаются
именно в хордальной части черепа, которая, следовательно, обнаружи-
вает следы метамерного строения по целому ряду признаков.
Довольно точной границей между хордальной и прехордальной
частями черепа служит подмозговая железка, или гипофиз (hypophysis
cerebri, рис. 113), которая лежит как раз впереди переднего конца хорды.
Она обозначает границу между обоими отделами черепа и в таких
случаях, когда передний конец хорды редуцируется. В прехордальной
части черепа располагаются трабекулы, в которых следов сегмента-
ции не обнаружено, и глазничные хрящи. В строении этой части черепа
мы не можем установить сходства со строением осевого скелета туловища.
Очевидно, это скелет ипого происхождения.
Если хордальная часть черепа представляет продукт преобразования
осевого скелета передней части тела соответственно изменившимся по-
725
Рис. 114. Расположение
хрящевых вакладок осево-
го черепа акулы. Вид со
спинной стороны. Заклад-
ки обведены ковтурами
черепа и головы взрослого
животного.
с. аи — слуховая капсула,
ch — хорда; со—обонятельная
капсула; h — гипофиз; ос —
глав; р. аи — слуховая, р.
о — обонятельная, р. осс —
ватылочная и р. orb —
глазничная области черепа;
pch — парахордалии; st —
глазничные хрящи; tr — тра-
бекулы.
треёностям (развитие головного мозга и органа слу&а, прикрепление
висцерального аппарата и мышц), то прехорлальпая часть черепа пред-
ставляет новообразование, развившееся в связи с развитием переднего
мозга и соответствующих органов чувств.
Во время дальнейшего развития черепа задние концы трабекул
срастаются с передними концами парахордалий. Последние срастаются
между собой под и над хордой, совершенно ее обволакивая. Трабекулы
также срастаются между собой, и таким образом развивается общая хря-
щевая пластинка, представляющая основание или дно черепа. В этой
пластинке довольно долго (а иногда и пожизненно) сохраняется непарное
отверстие впереди переднего конца хорды — основная фонтанель (fenestra
Рис. 115. Эмбриональный хрящевой череп лосося со спинной (Л) и с
брюпшой (В) стороны (по Гудричу).
ас — слуховая капсула; bh, bbr — непарные связующие элементы
висцерального скелета (basihyale, basibranchialia); ebr — ceratobran-
chialia; ch — ccrotohyale; fh — основная фонтанель; hbr — hypobranchi-
alia; hm — hyomandibulare; hp — hypohyale; mk — меккелев хрящ; nt —
хорда; par— парахордалии; pq — palataquadratum; tr — трабекулы;
vf — отверстие для блуждающего нерва.
basicranialis anterior). У взрослого животного на этом месте во всяком
случае остается ямка, в которой лежит гипофиз (турецкое седло челове-
ческой анатомии).
С хрящевым основанием черепа с самого начала уже тесно связаны
обонятельная капсула впереди и слуховая — в области прежних пара-
хордалий. Глазничные хрящи (cart, orbitales) также рано срастаются
с основанием черепа, которое постепенно разрастается вверх, по бокам
головного мозга, образуя боковые стенки черепа. Передний конец осно-
вания черепа разрастается в виде вертикальной пластинки между обо-
нятельными капсулами, образуя носовую перегородку. Наконец, хрящ
боковых стенок черепа разрастается над головным мозгом, образуя крышу
черепа. Последняя почти никогда не бывает полной (рис. 115,116). Обыкно-
венно более или менее значительная часть ее не охрящевевает вовсе, а
остается перепончатой. Такие пространства в хрящевом черепе носят
название фонтанелей; они или затянуты очень прочной соединительно-
тканной перепонкой (хрящевые рыбы), или прикрываются сверху очень
рано развивающимися покровными костями. Иногда в верхней стенке
перепончатого черепа развивается только хрящевой мостик, лежащий
сверху между слуховыми капсулами (tectum synoticuin) (рис. 116, t. sot.).
У высших позвоночных в связи с ранним развитием костей хрящ не
только рано редуцируется, но нередко и закладывается уже в сокращен-
126
I
"С.осе
ном объеме (рис. 117). Фонтанели, имеющиеся и в черепе низ1пих позво-
ночных, получают здесь большие размеры, а расположенные в боковых
стенках черепа отверстия для выхода нервов также значительно разраста-
ются, так что весь хрящевой череп вместо сплошной коробки приобретает
вид решетки, составленной
•из сложной системы пере-
кладин. Более значительные
!массы хряща. сохраняются
обычно лишь в слуховых и
обонятельных капсулах.
Первичный череп взрос-
лого животного имеет вид
Коробки со вздутой передней
'частью — обонятельной об-
ластью черепа, сжатой сред-
ней частью — глазничной
областью, за которой сле-
дует опять вздутая — слухо-
вая область, и, наконец, по-
зади череп замыкается заты-
лочной областью (рис. 114).
По строению осевого че-
репа различают два основ-
ных типа черепов: 1) пла-
тибазальный тип — черепа
с широким основанием, и
2) тропибазальный тип —
черепа с узким основанием
(рис. 118). В платибазальных
черепах стенки глазниц, ши-
роко раздвинуты, и между
ними продолжается мозго-
вая полость черепа. К этому
типу относятся черепа круглоротых, селахий и амфибий. В тропибазаль-
ных черепах стенки глазниц сильно сближены, мозговая полость вытесне-
на, и весь мозг располагается позади. Обычно стенки глазниц даже
вполне срастаются между собой, образуя тонкую межглазничную пере-
городку (septum interorbitale).
осе.
Рис. 116. Хрящевой череп личинки тритона. Вид
со спинной стороны (по Гаупу).
с. аи — слуховая капсула; с. осе — затылочные бу-
гры; ch — хорда; т — меккелев хрящ; осе — заты-
лочная область р. ant — пределазничный отросток
черепа; р. asc — processus ascendens; р. ot — pro-
cessus oticus пёбноквадратного хряща; t. sot — заты-
лочное перекрытие; I — VII -— отверстия головных
нервов.
Развитие и строение висцеральной части черепа
Висцеральная часть черепа, или просто висцеральный скелет, разви-
вается независимо от осевого черепа за счет мезенхимы эктодермального
происхождения (из ганглионарной пластинки; рис. 110) в виде ряда не-
зависимых друг от друга дужек, состоящих сначала из сгущенной мезен-
химы, которая затем превращается в хрящ (рис. 113). Висцеральные дуги
имеют правильное метамерное расположение и отделены друг от друга
висцеральными щелями. Эта метамерия висцерального скелета, или бран-
хиомерия, не находится, однако, в полном соответствии с метамерией
спинного отдела мезодермы, или с мезомерией. Висцеральные дуги и вис-
церальные щели обыкновенно не" вполне совпадают с сомитами головы
даже и на ранних стадиях развития.
Первые две висцеральные дуги получают особенно значительное
развитие и дают начало челюстной и подъязычной дуге взрослого жи-
вотного. Челюстная дуга расчленяется на два отдела, подъязычная — на
два-четыре отдела. Следующие за ними дуги расчленяются на четыре
отдела каждая и дают начало жаберным дугам взрослого животного
(рис. 119).
727
P bpt
Рис. i 17. Хрящевой череп ящерицы, вид сбоку (по Гауну).
с. ии — слуховая капсула; с. nas — обонятельная капсула; col — столбик уха; р.
asc — processus ascendens, р. pt — processus pterygoideus нёбнонвадратного хряща;
p. bpt — processus basiplerygoideus основания черепа; q — quadratum; s. io — меж-
глазничная перегородка; t. sol — затылочное перекрытие.
Рис. 118. Положение головного мозга и платибазаль-
ном (Л) п тропнбазальпом (В) черепе.
Рог рай
Рис. 119. Череп акулы (Mustelus) (по Гудричу).
Ъ. Ьг — basibranchialc; ь. hy — basihyae; с. Ъг — ceratobranchialia; ер. Ьг —
epibrachialia; hm—hyomandibidarc; hy — hyoideum; lb—губные хрящи; МЛ—
меккелев хрящ; рай — слуховая; ро — обонятельная; р. осе — затылочная и р.
от — глазничная области черепа; ph. Ьг — pharyngobranchialia; pq — пёбпонвадрат-
ный хрящ; г — rostrum; i—3 — жаберные дуги; II—X — отверстия для выхода голов-
ных нервов.
128
В висцеральном скелете, впереди челюстной дуги, нередко, в особен-
ности у хрящевых рыб, наблюдаются еще так называемые губные хрящи,
до двух пар в верхней губе и иногда одна пара в нижней (рис. 119). По
своему развитию и положению эти хрящи представляют собой, невиди-
мому, рудименты передних висцеральных дуг. Задняя пара верхних губ-
ных хрящей иногда соединяется в углах рта с парой нижних хрящей,
и тогда эти элементы и по форме напоминают висцеральную дугу, расчле-
ненную на два главных отдела — верхний и нижний. Передняя пара верх-
них губных хрящей не имеет соответствующей пары нижних хрящей и
представляет собой остаток одного только верхнего отдела висцеральной
дуги, нижний отдел которой исчез бесследно. Эти передние висцеральные
дуги носят наименование предчелюстных дуг.
Челюстная (arcus mandibularis) и подъязычная дуга
(arcus hyoideus) состоят у хрящевых рыб (селахий) из двух отделов каж-
дая. Верхний отдел челюстной дуги называется нёбноквадрат-
ным хрящом (palatoquadratum), нижний отдел — меккеле-
вым хрящом (cartilago Meckeli). Оба эти отдела служат собственно
челюстями только у низших рыб. Верхний отдел подъязычной дуги носит
название подъязычночелюстного, или гиомандибулярного,
хряща (hyomandibulare), а нижний отдел — подъязычного, или гио-
идного хряща собственно (hyale). Жаберные дуги (arcus branchiales)
состоят из четырех отделов каждая: в верхней половине—pharyngobran-
chiale сверху и epibranchiale снизу и в пижней половине — ceratobran-
chiale сверху и hypobranchiale снизу. Висцеральные дуги правой и левой
стороны соединяются на брюшной стороне тела между собой посредством
ряда непарных элементов — copulae. Передний непарный элемент свя-
зывает между собой нижние концы подъязычной дуги и носит название
basihyale, а следующие связывают концы жаберных дуг и носят название
basibranchialia (рис. 119 и 121).
Число жаберных дуг различно у различных позвоночных. У наиболее
примитивных форм оно доходит до семи (круглоротые и низшие предста-
вители селахий), обычно же их у рыб пять. У наземных позвоночных
жаберные дуги теряют свое значение, и остатки их значительно преобра-
зуются, но все же обычно можно установить закладки четырех жа-
берных дуг. Если у рыб число жаберных дуг сокращается, то редуциру-
ются всегда задние дуги. И у рыб с нормальными пятью дугами нередко
находили рудименты шестой дуги. Это Доказывает, что позвоночные жи-
вотные произошли от форм, обладавших большим числом жаберных дуг,
из которых последние редуцировались. Мы видели, что и в передней части
висцерального аппарата произошла редукция предчелюстных дуг. Таким
образом, сокращение числа дуг происходило с обоих концов висцераль-
ного аппарата. Если мы вспомним, какое число жаберных щелей наблю-
дается у низших хордовых (и у круглоротых оно иногда еще доходит
до 14), то мы поймем, какое значительное сокращение испытал висцераль-
ный аппарат у позвоночных. Все висцеральные дуги развиваются вполне
сходно за счет одного и того же скелетогенного материала и имеют сходное
расположение, т. е. гомодинамны. Первые дуги, однако, отличны от сле-
дующих по функции и по расчленению. Тем не менее имеются указания
на то, что и эти дуги некогда были расчленены точно так же, как и типич-
ные жаберные дуги. Гиоидная дуга нормально обладает своим непарным
элементом — basihyale — подобно жаберным дугам. Оба отдела подъязыч-
ной дуги у хрящевых рыб по положению вполне соответствуют двум сред-
ним отделам жаберных дуг (epibranchiale и ceratobranchiale), которые
и вообще являются наиболее крупными. Над гиомандибулярным хрящом
у’цельноголовых рыб имеется еще один отдел гиоидной дуги, вполне соот-
ветствующий По положению верхнему элементу жаберных дуг. У селахий
находили рудиментарный хрящик между гиомандибулярным хрящом
129
и слуховой областью черепа. Наконец, и под нижним концом гиоида соб-
ственно находили небольшие хрящи, представляющие остатки нижнего
отдела гиоидной дуги. Таким образом, над гиомандибулярным хрящом,
представляющим собой epihyale, найдены остатки pharyngohyale, а под
гиоидом собственно, представляющим собой ceratohyale, найдены остатки
hypohyale. Следовательно, имеются ясные указания на первоначальное
расчленение подъязычной дуги на те же типичные отделы, как и жабер-
ных дуг. Челюстная дуга отличается от жаберных дут более заметно, но
найденные остатки копулы и некоторые указания на прежнее существо-
вание нижнего парного элемента — hypomandibulare, а также нахожде-
ние рудимента pharyngomandibulare у селахий, заставляют предпо-
лагать, что и челюстная дуга была расчленена так же, как и жаберные
дуги, и два ее отдела представляют собой epimandibulare и ceratomandi-
bulare. Одинаковое расчленение дуг делает вероятным предположение
об одинаковой функции, которой должны были обладать эти дуги. Естест-
венно, что все дуги по своей функции были первоначально жаберными.
Доказательством справедливости этого предположения является факт
развития между челюстной дугой и подъязычной такой же висцеральной
щели, как и следующие жаберные. Эта висцеральная щель сохраняется
и во взрослом состоянии у низших рыб (у большинства селахий, осетровых
и кистеперых рыб) в виде так называемого брызгальца (spiraculum), и
в ней находится даже рудиментарная жабра. Кроме того, и впереди
этой щели находили иногда эмбриональные закладки рудиментарных
висцеральных щелей. Все это вместе взятое доказывает нам, что все вис-
церальные дуги были у предков современных позвоночных некогда жа-
берными дугами, несли ту же функцию и были так же расчленены. Мы
теперь знаем, что у ископаемых щитковых бесчелюстных (Ostracodermi),
действительно, все висцеральные дуги были еще снабжены жабрами и
несли одну и ту же функцию. Затем у челюстных позвоночных одна из
передних (но не самая передняя) дуг изменила свою функцию, стала слу-
жить сначала для удерживания добычи во рту, а затем и для ее захваты-
вания. Соответственно этой смене функции преобразовался и сам орган,—
сохранились лишь два средних отдела дуги, но они развились очень сильно
и, превратившись в челюсти, получили сильную мускулатуру и прочное
прикрепление к черепу. Подъязычная дуга видоизменилась в связи с
преобразованием челюстной. Верхняя половина подъязычной дуги стала
служить специально для более прочного укрепления челюстной дуги,
подвешивая задние углы челюстей к черепу. Верхний ее отдел — pha-
ryngohyale — редуцировался, а следующий отдел — epihyale, — наобо-
рот, развился очень сильно, дав начало гиомандибулярному хрящу, слу-
жащему у огромного большинств рыб подвеском для челюстей. Нижняя
половина подъязычной дуги сохранила значение связующего звена между
челюстным и жаберным аппаратом.
Мы видели, что первоначально висцеральные дуги совершенно не-
зависимы от осевого черепа. Позднее между ними устанавливается
связь посредством связок или непосредственным причленением. Эта связь
является, следовательно, всегда вторичной. Подъязычная дуга, и именно
гпомандибулярный хрящ, получает особенно прочное прикрепление к
слуховой области черепа.
Верхний отдел челюстной дуги прикрепляется наиболее сложно:
своим передним концом у низших позвоночных он подвешивается к обо-
нятельной или глазничной области черепа либо посредством связки, либо
посредством особого отростка, а своим задним концом, нередко, посред-
ством связки или особых отростков, к основанию черепа или к слуховой
капсуле. Сверх такой непосредственной связи, задний край челюстной
дуги прикрепляется у большинства рыб еще при помощи гиомандибу-
лярного хряща.
130
По отношению первых висцеральных дуг к осевому черепу различают
йесколько типов строения черепа (рис. 120).
В гипотетическом исходном типе и челюс'гная, и гиоидная дуги под-
вешивались к осевому черепу своими верхними концами независимо друг
от друга — это тип протостиль н о г о черепа. У наиболее древних
ископаемых селахий и у наиболее примитивных современных форм (Нер-
tanchus, Hexanchus и Chlamydoselachus), так же как п у костных гано-
идов, наблюдается тройная связь челюстной дуги с черепом: верхний
отдел челюстной дуги прикрепляется впереди посредством одного отростка
к дну черепа и позади посредством другого отростка к слуховой его об-
ласти, и сверх этих непосредственных связей задний угол челюстей прочно
Рис. 120. Отношение передних висцеральных дуг к осевому черепу.
А — амфистильный череп акулы (Heptanchus); В — гиостильный череп
акулы; С — амфистильный череп костного ганоида (-Amia); D — аутостиль-
ный череп навемного позвоночного (лягушка), hm — hyomandibulare; hy —
hyoideum- ih —- interhyale; Mk — меккелев хрящ; md — нижняя челюсть;
ot — processus oticus; ph — processus palatobasalis; pq — palatoquadratum.
связан связками с гиомандибулярным хрящом. Такой тип строения че-
репа носит название амф и стильного (рис. 120, А, С). У неко-
торых рыб (цельноголовых и двудышащих) и у наземных позвоночных
челюстная дуга связана (и даже срастается) только непосредственно с
осевым черепом, впереди с обонятельной областью и позади со слуховой,
и иногда верхний отдел челюстной дуги (palatoquadratum) даже вполне
сливается с осевым черепом (рис. 120, D). В этом случае гиомандибуляр-
ный хрящ пе играет никакой роли в укреплении челюстей. Это тип а утр-
стильного черепа, развивающийся вторично из амфистилии или
гиостилии. У огромного большинства рыб верхний отдел челюстной дуги
связан непосредственно с осевым черепом только впереди, задний же ко-
нец нёбноквадратного хряща подвешивается к черепу только посредством
сильно развитого гиомандибулярного отдела подъязычной дуги, служа-
щего подвеском для челюстей (рис. 120, В). Этот тип строения черепа
называется гиостильным. При крайнем развитии гиостилии
челюсти висят исключительно на гиомандибулярном хряще (осетровые).
Различные типы строения черепа выработались в связи с особен-
ностями строения аппаратов захватывания пищи и дыхания. В амфистилии
131
сбг
Рис. 421. Жаберный скелет акулы (Не-
ptanchus) со спинной стороны (по Геген-
бауру).
ЬЬт — basibranchiale; bh — basihyale; cb —
ceratobranchiale; cbr — cardiobranchiale
(задняя копула); ер — epibranchiale; h_____
hyoideum; hbr — hypobranchiale; ph—
pharyngobranchiale; 7—7 — жаберные
дуги.
ботан английским
находит свое выражение связь обеих функций первых двух висцеральных
дуг — первичной, дыхательной (участие в боковых дыхательных движе-
ниях), и новой, хватательной. В гиостилии вта связь еще не нарушена,
однако одновременно с прогрессивным развитием новых, более совершен-
ных форм дыхательных движений, при которых главная роль переходила
к активным движениям жаберной крышки, связанной с подъязычной
дугой, в атом типе устанавливается большая свобода в движениях задней
части аппарата. Наконец, в аутости-
лии имеется полное разделение обеих
функций и максимальное укрепле-
ние челюстной дуги, совершенно
обособившейся от дыхательных дви-
жений гиобранхиального аппарата.
Теории происхождения черепа
Сходство черепа с позвоночни-
ком, выражающееся прежде всего в
отношениях к центральной нервной
системе, уже давно направило науч-
ную мысль на более детальное срав-
нение этих частей скелета и привело
еще в конце XVIII столетия к попыт-
ке рассматривать череп как преобра-
зованную переднюю часть позвоноч-
ника. Этот взгляд, известный под
названием позвоночной теории че-
репа, был высказан сначала поэтом-
натуралистом Гёте (опубликован мпо-.
го позднее) и натурфилософом Оке-
ном и затем уже к середине прошло-
го века детальнейшим образом разра-
1 его учении об «архетипе» (рис. 1). По-
звоночная теория рассматривала костный череп позвоночных как комплекс
небольшого числа (3—5) полных позвонков. В костях черепа находили
гомодинамы тел, верхних дуг, остистых отростков и т. д. Так как изучался
главным образом скелет млекопитающих как наиболее совершенных
животных (т. е. более полно выражающих основную «идею») по взглядам
того времени, то и части позвонков пытались находить прежде всего в
черепе млекопитающих. Нужно сказать, что в расположении костей че-
репа у последних, действительно, обнаруживается некоторая правиль-
ность, которая заставляет для чисто описательных целей и теперь нередко
прибегать к посегментпому распределению костей.
Сравнение же костей черепа низших позвоночных с костями черепа
млекопитающих позволило распределить таким же образом и первые.
Так, различали затылочный, теменной, лобный и носовой сегменты,
в каждом из которых имеется непарная кость у основания (тело), парные
кости по бокам (дуги) и непарная или парные кости сверху. Однако уже
Гексли показал (в шестидесятых годах прошлого столетия), что эта сег-
ментация — чисто вторичная, не имеющая никакой почвы в черепе низших
позвоночных. В черепе низших позвоночных, Tait же как и в эмбриональ-
ном хрящевом черепе позвоночных вообще, нет никаких указаний на
существование каких-либо сегментов, соответствующих костным позвон-
кам позвоночной теории. Таким образом, последняя оказалась неверной.
В основе ее лежала, однако, по существу верная мысль о происхож-
дении черепа из метамерно расположенных частей.
В строении висцерального аппарата, именно низших позвоночных,
метамерия выражена совершенно ясно. Исходя первоначально (в согласии
132
о позвоночной теорией) из сравнения метамерно расположенных висце-
ральных дуг с ребрами позвоночника, Гегенбаур вывел заключение,
что осевая часть черепа представляет собой результат полного слияния
известного числа позвонков, по крайней мере в своей задней части, в
которой было найдено продолжение хорды. Метамерному расположению
висцеральных дуг в этой области соответствует и подобное ясе расположе-
ние нервов. Последовавшее затем детальное изучение эмбрионального
развития головы показало, что и эта теория неправильна. Прежде всего,
исходное сравнение висцеральных дуг с ребрами оказалось неверным.
Висцеральные дуги хотя и развиваются как образования, метамерно
расположенные, но независимо от осевого скелета и иным путем.
Однако было найдено, что сама мезодерма сегментируется в голове
так же, как и в туловище, и за счет развивающихся таким образом сомитов
(точнее, их склеротомов) строится осевой череп. Таким образом, сам
материал оказывается сегментированным. Хрящи, развивающиеся в
хордальной части черепа (парахордалии), занимают по отношению к
хорде положение, вполне сходное с элементами позвоночника в туловище,
и в своей задней части обнаруживают даже следы сходной сегментации
(Северцов). Кроме того, в задней (затылочной) части черепа наблюдается
иногда и прямое прирастание дуг позвонков (затылочные дуги) и даже
целых позвонков. Целый ряд затылочных дуг с сидящими на них остистыми
отростками имеется в черепе осетровых рыб. Одна или две затылочные
дуги наблюдаются в черепе многих позвоночных. У костистых рыб нередко
и во взрослом черепе ясно различим приросший передний туловищный
позвонок, а у двудышащих рыб с затылочной областью черепа связана
даже пара крупных «головных» ребер.
Таким образом, благодаря трудам целого ряда ученых, как Бальфур,
фан Бай, Дорн и др., постепенно развилось то эмбриологическое учение
о метамерии головы, которое в существенных частях было уже изло-
жено. По этому учению, голова представляет собой преобразованный
передний конец тела, который раньше был построен так же, как и все
остальное тело. Как и у ланцетника, он поддерживался хордой и был
правильно сегментирован. С прогрессивным развитием органов чувств
и головного мозга, а также с преобразованием передних жаберных дуг в
челюстной аппарат, элементы позвонков, находившиеся в хордальной
части головы, срослись между собой и дали начало осевой части черепа.
Таким образом, хордальный отдел черепа является результатом пре-
образования передней части осевого скелета. Лежащая впереди хорды
прехордальная часть черепа представляет, невидимому, новообразование,
развившееся в связи с разрастанием переднего мозга. Появившись в
результате преобразования передней части осевого скелета хордальная
часть черепа продолжает у низших позвоночных нарастать путем дальней-
шего присоединения элементов позвоночника к затылочной его области.
У рыб это возможно вследствие отсутствия у них подвижного затылочного
сочленения.
Хрящевой череп низших позвоночных
Осевая часть черепа круглоротых находится на очень низкой
ступени развития. Дно черепа обладает значительной основной фонтанелью,
продырявленной отверстием для своеобразного назогипофизарного канала.
Крыша черепа остается почти на всем своем протяжении перепончатой.
Имеется’лишь небольшой мостик в области слуховых капсул. У миксин
не только крыша, но и бока черепа остаются перепончатыми. Весь осевой
череп круглоротых развивается из парахордалий, и настоящих трабекул
у них еще не имеется (Северцов). Череп круглоротых особенно примитивен
и в том отношении, что в нем нет еще затылочной области. Череп кончается
133
слуховой капсулой, позади которой свободно выходят 9-я и 10-я пары
головных нервов (языкоглоточный и блуждающий), которые у всех прочих
позвоночных выходят через особые отверстия в затылочной области черепа.
Капсулы органов чувств обладают еще значительной самостоятельностью.
Слуховые капсулы заметно выступают в виде шаровидных вздутий. Обо-
нятельные капсулы связаны с остальным черепом лишь соединительной
тканью.
Дно черепа продолжается вперед в виде большой непарной хрящевой
пластинки, которая налегает своим передним концом на подобную же
переднюю пластинку. Обе эти пластинки прикрывают со спинной стороны
сложно развитый губной аппарат. У круглоротых нет челюстей, и передняя
часть висцерального скелета частично срослась с осевым черепом, а в
остальном преобразована в очень сложно построенный сосательный
аппарат, состоящий из известного числа так называемых губных хрящей
(очевидно, гомологов предчелюстных дуг). Собственно жаберный скелет
у миксин редуцирован, а у миног представлен решеткой из семи неправиль-
но изогнутых и нерасчлепенных жаберных дуг, связанных менаду собой
горизонтальными перекладинами под хордой, наджаберными отверстиями,
под жабершдми отверстиями и с брюшной стороны.
У селахий хрящевой череп достигает высокой степени развития.
Осевой череп представляет хрящевую коробку с широким основанием
(платибазальпый тип), несколько сжатой глазничной областью, вздутую
впереди и позади соответственно положению обонятельных и слуховых
капсул (рис. 118, Л). Позади слуховой области имеется небольшая заты-
лочная область, а впереди обонятельной области череп продолжается
в вырост — rostrum (рис. 119). Впереди часть крыши черепа остается
перепончатой (верхняя фонтанель). В боковых стенках черепной коробки
имеются отверстия для выхода головных нервов, большинство которых
лежит в глазничной области, где помещается крупное отверстие для
зрительного нерва и позади от него — отверстия для нервов глазпых
мышц (3-й, 4-й и 6-й пар), а также для ветвей тройничного и лицевого
нервов. Позади слуховой капсулы, в затылочной области черепа, нахо-
дятся отверстия для языкоглоточного и блуждающего нервов. Под этими
отверстиями помещается обыкновенно еще несколько очень маленьких
отверстий для выхода затылочных нервов.
Висцеральный скелет селахий состоит из хрящевых типично расчле-
ненных дуг, верхние концы которых обычно соединяются между собой
связками, а нижние концы — хрящевыми копулами. Челюстная дуга
состоит из нёбноквадратного хряща, служащего верхней челюстью, и
меккелева хряща, служащего нижней челюстью. Передние концы нёбно-
квадратных хрящей правой и левой стороны у селахий соединяются.
У громадного большинства селахий пёбноквадратный хрящ соединяется
с черепом только в передней своей части посредством отростка (processus
palatobasalis) или связки, задняя же его часть поддерживается прочным
соединением с гиомандибулярным хрящом (гиостилия, рис. 120, В),
хотя иногда здесь наблюдаются и связки, подвешивающие задний угол
челюстей непосредственно к черепу. Только у немногих примитивнейших
акул (Notidanidae) пёбноквадратный хрящ соединяется также со слуховой
областью черепа посредством особого заднего отростка (processus oticus),
так что череп имеет амфистильное строение (рис. 120, А). Снаружи к
элементам челюстной дуги часто прилежат остатки предчелюстных дуг —
губные хрящи, располагающиеся в губных складках. Подъязычная дута
состоит из двух отделов — верхнего, гиомапдибулярного хряща,*служа-
щего подвеском для челюстей и при пленяющегося к слуховой области
черепа, и нижнего, гиоидного. Оба эти отдела соответствуют двум средним
элементам типичной висцеральной дуги, представляя^собой epihyale и
ceratohyale. У большинства селахий имеется 5 жаберных дуг. Только у
134
особо примитивных форм их бывает больше: у Heptanchus — 7, у Нехап-
chus и у Chlamydoselachus — 6 (рис. 121). Жаберные дуги расчленены
на четыре отдела каждая (pharyngobranchiale, epi-, cerato- и hypobran-
chiale). Подъязычная и жаберные дуги правой и левой стороны соединяются
между собой и со следующими парами рядом копул; из них передняя
представляет собой basihyale, а следующие — basibranchialia. У боль-
шинства селахий копулы задних дуг сливаются в одну большую пластинку,
прикрывающую сердце со спинной стороны (cardiobranchialie). Кроме
етих существенных элементов висцерального скелета, имеются еще и иные.
Передняя стенка гиомандибулярной висцеральной щели — брызгальца —
поддерживается небольшим спиракулярным хрящом. На элементах подъ-
язычной дуги, так же как и на средних членах жаберных дуг, вдоль
ваднего их края сидит ряд жаберных лучей, поддерживающих жаберные
складки.
Рис. 122. Череп осетра сбоку по удалении покровных костей (из Видерсгейма).
Хрящ обозначен точками.
ЬЬг — basibranchialia; с — ребра; ch — хорда; сЬг — ceratobranchiale; ebr — epibran-
chiale; hm — hyomandibulare; hy — гиоид; th— Interhyale; md—нижняя челюсть; n —нозд
ря; па — верхние дуги; о — глазница; ph.br — pharyngobranchialia; ps — парасфеноид
р<г — нёбноквадратный хрящ; г — rostrum; sy — symplecticum.
У цельноголовых осевой череп, вследствие большой вели
чины глаз, имеет сильно сжатую глазничную область, стенки которой
соединились для образования тонкой межглазничной перегородки (тро-
пибазальный тип). Нёбноквадратный хрящ вполне прирос к осевому
черепу, и подъязычная дуга челюстей не подвешивает (аутостилия). В связи
с этим подъязычная дуга обнаруживает такое же строение, как и типич-
ные жаберные дуги. Лучи подъязычной дуги очень сильно развиты и под-
держивают большую кожную складку, прикрывающую не только бли-
жайшую жаберную щель (как это наблюдается на жаберных складках
селахий, которые прикрывают напосредственно позади лежащую щель),
но и все остальные, образуя наиболее примитивную форму жаберной
крошки. У цельноголовых весь жаберный скелет располагается под че-
репной коробкой, а не позади нее, как у селахий.
У хрящевых ганоидов череп с узким основанием и имеет
вид сплошной хрящевой коробки без всяких фонтанелей. В затылочной
области черепа наблюдается прирастание большого числа элементов по-
звоночника (рис. 122). В висцеральном скелете наблюдаются уже при-
знаки, характерные для высших рыб. Верхние концы жаберных дуг под-
вешены к основанию осевого черепа двойными pharyngobrachialia (верх-
ними и нижними). Имеется резко выраженная гиостилия, и челюсти висят
исключительно на гиоидном подвеске. Нижний конец гиомандибулярного
хряща путем отчленения дал начало новому элементу — symplecticum,
который служит специально для прикрепления заднего угла челюстей.
С гиоидом собственно гиомандибулярный хрящ связан небольшим про-
135
межуточным элементом — interhyale. Существование этих элементов
весьма характерно для высших рыб, равно как и появление кости в ка-
честве скелетного материала.
Окостенений первичного скелета еще немного. Некоторые элементы
висцерального скелета одеты перихондральным костным чехлом. Кроме
того, у старых осетровых рыб наблюдаются
Рис. 123. Крыша черепа
осетра.
с. lat — каналы органов бо-
ковой линии; fr — лобные
кости; п — передняя и задняя
ноздри; о — глазница; ор —
жаберная крышка; orb — ог-
bltalia; ра— теменные кости;
рра — заднетеменные; st —
supratemporale; sp — брыз-
гальце.
неправильно ограниченные окостенения в хря-
ще осевого черепа. Гораздо большее значение
имеют покровные кости. Основание черепа
осетровых рыб подостлано длинной непарной
костью — парасфеноидом — и лежащим впере-
ди нее под рылом «сошником» (рис. 122). Свер-
ху череп покрыт большим числом кожных
костей, образующих сплошной панцырь, в ко-
тором имеются лишь отверстия для глаз, для
ноздрей и для брызгальца (рис. 123). По свое-
му развитию и строению и по характерной
скульптуре эти кожные кости представляют та-
кие же разросшиеся чешуи, как и расположен-
ные рядами вдоль спины и по бокам тела осе-
тровых крупные «жучки». Некоторые из этих
костей по своему расположению вполне срав-
нимы с наиболее постоянными покровными
костями черепа высших рыб. Имеется у осе-
тровых рыб и кожное окостенение в жабер-
ной крышке (operculum) и покровные кости
на челюстной дуге (palatinum, pterygoideum,
quadrato-jugale и dentale), а также и кость,
неправильно сравниваемая со вторичной верх-
ней челюстью.
По всем этим признакам осетровые зани-
мают положение, переходное к высшим рыбам
с хорошо развитым костным скелетом. У иско-
паемых хрящевых ганоидов система покровных
костей черепа была развита выше, чем у совре-
менных осетровых, и нередко вполне сходна по
своему составу с покровными костями черепа
высших рыб.
У высших рыб число покровных костей черепа сокращается, отдель-
ные кости разрастаются, погружаются глубже под кожу и вступают в
более тесную связь с первичным черепом.
Костный череп
У высших рыб и у наземных позвоночных первичный череп частью
окостеневает, т. е. его хрящ вытесняется развивающейся на его месте за-
мещающей («первичной») костью, частью дополняется покровными костя-
ми, которые более или менее тесно к нему прилегают. В результате зна-
чение хряща падает, и он у высших позвоночных вполне вытесняется
костью. Расположение костей черепа у позвоночных в общем довольно
постоянно. Наиболее типичны следующие кости.
Осевой череп. Покровные кости. Крыша черепа состав-
ляется из парных теменных костей (parietalia), лежащих обыкно-
венно над слуховой областью черепа, парных лобных костей (fron-
talia), покрывающих глазничную область сверху, и парных носовых
костей (nasalia), покрывающих обонятельную область. На боковой стенке
слуховой капсулы лежат парньщ надвисочная (supratemporale)
136
и чешуйчатая кость (squamosiim). Сверх того нередко впереди
лобных костей имеются 1—2 пары переднелобных (praefron-
talia) и позади них заднелобные (postfrontalia) и задне-
глазничные (postorbitalia). Бока черепа позади глазниц (щеки или
височная область) покрываются щечными, или скуловыми, костями
(jugalia). Под глазницами и впереди них наружный костный череп огра-
ничивается расположенными в верхней губе челюстными и пред-
челюстными костями (maxillare и praemaxillare). Дно черепа
подстилается у низших позвоночных большой непарной костью — пара-
сфеноидом (parasphenoideum), а под обонятельной областью лежит
парный по происхождению сошник (vomer). Последние кости имеют
зубное происхождение.
Замещающие кости. В осевом черепе окостеневают глав-
ным образом бока и основание, тогда как крыша черепа, прикры-
тая покровными костями остается на большом протяжении перепон-
чатой.
В затылочной области черепа развиваются непарная
основная затылочная кость (os basioccipitale), парные б о-
ковые затылочные (exoccipitalia или occipitalia lateralia) и
непарная верхняя затылочная (supraoccipitale). Первые
кости окружают затылочную дыру (foramen occipitale); последняя обычно
лежит выше, входя отчасти в состав крыши черепа. В качестве окосте-
нений стенок слуховых капсул развиваются парные около-
ушные кости — ossa periotica. Из них наиболее постоянна перед-
н е у ш н а я (prooticum), обычно продырявленная отверстием для выхода
лицевого нерва. Позади нее развивается снизу заднеушная (opis-
thoticum) и сверху верхнеушная (epioticum). Впереди слуховых
капсул в глазничп о-в исочной области черепа развива-
ются у основания непарные кости.— основная клиновидная
кость (os basisphenoideum) и передняя клиновидная
(praesphenoideum) и по бокам парные кости — боковые клино-
видные (later os phenoidea) и глазничн о-к линовидные (ог-
bitos phenoidea).
В обонятельной области находится иногда непарная
средняя обонятельная кость (mesethmoideum) и парные
боковые обонятельные (exethmoidea), а иногда единая р е-
шетчатая кость (ethmoideum).
Висцеральный скелет. Покровные кости. Вторичные
челюсти (ossa praemaxillaria и maxillaria) развиваются как покров-
ные кости в верхней губе и, быть может, первоначально лежали на верх-
них губных хрящах, т. е. на предчелюстных висцеральных дугах. С че-
люстной дугой вторичные челюсти не связаны. На верхнем отделе ч е-
люстной дуги — нёбноквадратном хряще — лежат снизу по край-
ней мере две покровные кости — нёбная кость (palatinum) впереди
и крыловидная кость (pterygoideum) на средней его части.
На нижнем отделе челюстной дуги — меккелевом хряще — ле-
жат две большие кости зубного происхождения: пластинчатая
(spleniale) на внутренней его стороне и зубная (dentale) на наруж-
ной стороне. Есть основние думать, что последняя генетически не отно-
сится к челюстной дуге, а подобно вторичным верхним челюстям разви-
лась в нижней губе как покровная кость на нижнем губном хряще. В зад-
ней половине меккелева хряща снаружи и снизу лежит угловая
кость (angulare) и иногда над ней еще надугловая (supraangulare)
и с внутренней стороны коясно-сочленовная (goniale s.
dermarticulare). На других висцеральных дугах покровных костей не раз-
вивается; близкое отношение к подъязычной дуге имеют лишь кости жа-
берной крышки (ossa opercularia) и жаберной перепонки (branchios-
137
tegalia) высших рыб, в качестве покровных костей, развившихся в кож-
ной складке на лучах гиоидной дуги.
Замещающие кости. В челюстной дуге обычно окостене-
вают только задние отделы нёбноквадратного и меккелева хряща. Окосте-
нением первого образуется квадратная кость (os quadratum) и
окостенением второго — сочленовная кость (articulare). В подъ-
язычной и в жаберных дугах обыкновенно каждый отдел имеет свое
собственное окостенение, сохраняющее название соответствующего отдела,
но иногда наблюдается и развитие нескольких костей в одном отделе
(например, в гиоиде высших рыб).
Костный череп высших рыб
У костных ганоидов и у костистых рыб черепная коробка окостене-
вает в значительной мере. У низших их представителей между отдельными
окостенениями сохраняются еще заметные массы хряща, но у высших
форм хрящ вытесняется разрастающимися костями, сохраняясь главным
образом лишь в обонятельной области. Осевой череп высших рыб имеет
в общем узкое основание (тропибазальный тип), стенки глазниц у ганоидов
сближены, а у большинства костистых рыб имеется лишь тонкая межглаз-
ничная перегородка.
В затылочной области у костных ганоидов имеется основ-
ная затылочная. (basioccipitale) и боковые затылочные (exoccipitalia)
кости, последние с отверстиями для 9-й и 10-й пар нервов. В слуховой
области костных ганоидов развиваются: переднеушная (prooticum) с отвер-
стиями для 5-й и 7-й пар нервов, заднеушная (opisthoticum) и верхнеуш-
ная (epioticum). В глазнично-височной области в основании черепа по-
зади непарная основная клиновидная (basisphenoideum) и парная боковая
клиновидная (laterosphenoideum), а впереди парная же глазнично-клино-
видная (orbitosphenoideum). Других типичных замещающих костей в осе-
вом черепе костных ганоидов не имеется. Однако две парные покровные
кости — передне- и заднелобные — иногда врастают более или менее
глубоко в хрящ.
Череп костных ганоидов принадлежит к типу амфистильных черепов.
У обоих современных представителей Holostei — Amia и Lepidosteus —'
верхний отдел челюстной дуги прикрепляется к черепу не только впереди,
в обонятельной области, но и позади — к основанию черепа на границе
между глазничной и слуховой областью посредством особого отростка
па крыловидных костях. У некоторых ископаемых рыб у основания
черепа на основной клиновидной кости имелись особые выступы (proc,
basipterygoidei), к которым причленялись указанные отростки крыло-
видных костей челюстной дуги. У многоперых ганоидов — Polypterus —
имеется просто очень широкая связь крыловидных костей с основанием
черепа. Сверх того, и гиомандибулярный отдел подъязычной дуги служит
подвеском для челюстей. Передние концы palatoquadrata между собой
не соединяются.
В задней части нёбноквадратного хряща развиваются обыкновенно
две замещающие кости: квадратная (quadratum) — снизу и задняя кры-
ловидная (metapterygoideum) — сверху. В средней части нёбноквадрат-
ного хряща имеется не более двух покровных костей — наружная и
внутренняя крыловидные (ectopterygoideum и entopterygoideum) и впе-
реди нёбная кость смешанного происхождения (dermo- autopalatinum).
Меккелев хрящ в задней своей части замещается сочленовной костью
(articulare) и покрывается целым рядом покровных костей: снаружи
зубной костью (dentale), изнутри пластинчатой’(spleniale), которые обе
несут зубы, и, кроме того, позади и снаружи угловой и надугловой
костью (angulare и supraangulare). Верхний отдел челюстной дуги теряет
уже функцию челюстей: нёбная кость несет еще зубы, а крыловидные
кости снабжены иногда лишь мелкими зубами. Роль верхней челюсти
переходит к покровным костям, развивающимся в верхней .губе, — пред-
челюстной и челюстной костям (praemaxillare и maxillare), снабженным
крупными зубами. У Lepidosteus maxillare состоит из целого ряда
отдельных костей. В подъязычной дуге верхний ее отдел окостеневает
в виде крупной гиомандибулярной кости, причленяющейся к слуховой
области черепа (к нижнему краю чешуйчатой кости), и небольшой
symplecticum, служащей для соединения с квадратной костью. Неболь-
шая кость interhyale связывает гиомандибулярную кость с собственно
гиоидом. В гиоиде имеются два окостенения, и снизу имеется еще не-
большая кость, может быть, соответствующая hypohyale. Жаберные дуги
окостеневают каждая соответственно своим четырем отделам. Последняя
дуга редуцирована и состоит из одной кости. Подъязычная и жаберные
дуги связаны рядом костных копул. На различных отделах жаберного
скелета, на копулах и на самих дугах могут располагаться покровные
Рис. 125. Схематический поперечный
разрез через череп костного ганоида в
передней части слуховой области.
art — articulate; ang — angulare; dent —
flentale; e —наружная крыша черепа; i —
стенка мозговой коробки; jug — Jugal ia;
pa — parietale; pot — prooticum; psph —
парасфеноид; pq— palatoquadratum; sq—
squamosum; st — supratemporale.
Рис. 124. Череп ископаемого ганоида
(Gonatodus) снизу и сбоку (по Тракеру
из Циттеля).
ап — angulare; Ьг — branchicstegalia; cl —
clavicula; ct — cleithrum; d — Centale; gl —
gularia; na — maxilla; op — operculum;
pop — praeoperculum; sop — subopercu-
lum.
В жаберной крышке развивается обычно несколько покровных костей;
наиболее крупная носит название крышки (operculum), лежащие под ней —
подкрышки (suboperculum) и межкрышки (in ter operculum), а лежащая
впереди них — предкрышки (praeoperculum). Последняя прикрепляется
к гиомандибулярной кости, которая, таким образом, укрепляет всю
жаберную крышку. В жаберной перепонке лежит целый ряд небольших
костных пластинок, связанных с гиоидом и носящих название лучей
жаберной перепонки (branchiostegalia). Снизу между ветвями нижней
челюсти у Polypterus лежит большая парная, а у Amia — непарная
нижняя яремная пластинка (gulare). При сравнении этих костей с ко-
стями ископаемых ганоидных рыб ясно видно, что все это ряд прибли-
зительно равнозначащих покровных костей, развившихся в жаберной
складке. Верхние члены этого ряда развились особенно сильно и предста-
вляют собой элементы жаберной крышки; многочисленные средние члены
представляют собой лучи жаберной перепонки, а нижние члены того же
ряда лежат между ветвями нижней челюсти в виде яремных пластинок,
которые у ископаемых ганоидов наблюдаются нередко в большом числе
(рис. 124).
Покровные кости, образующие бока и крышу черепа, лежат
или непосредственно под кожей или еще в самой коже, образуя сплошной
наружный панцырь головы. В последнем случае кости имеют характерную
139
скульптуру и такой же покров ганоина, как и кожные чешуи. Весь этот
наружный панцырь значительно шире, чем осевой череп, и прилегает не-
Рис. 126. Крыша черепа кост-
ного ганоида (схема).
eth— ethmoldeum; fr— лобные
кости; jug — щечные кости; тх —
maxilla; п — носовая кость;
orb — глазничные кости; ор —
крышка; ра— теменная кости;
ртх — praemaxilla; рра — зад-
нетеменные; pop — предкрышка;
prf — переднелобная и ptf —
заднелобная кости; sq — над-
височная.
посредственно к последнему только свер-
ху; на боках же между ними вдвигаются
части висцерального скелета (челюстная
дуга и подвесок) и висцеральная мускула-
тура (рис. 125).
Характерны следующие кости крыши
черепа: теменные (parietalia), лобные (fron-
talia), носовые (nasalia) и непарная верх-
няя обонятельная (supraethmoideum); кроме
того, в обонятельной области лежат отчасти
врастающие в хрящ переднелобные (praefron-
talia) и позади глазниц в начале слуховой
области, также отчасти врастающие в хрящ,
заднелобные (postfrontalia); позади послед-
ней на верхней части слуховой капсулы
лежат надвисочная (supratemporale) и чешуй-
чатая кости (squamosum) и, наконец, по зад-
нему краю крыши черепа лежит попереч-
ный ряд обычно парных костей — заднете-
менные, или табличные кости (postparietalia
s. tabularia). По бокам черепа между глаз-
ницей и жаберной крышкой располагается
большее или меньшее число щечных костей.
Передние кости этой группы окружают глаз-
ницу и носят название глазничных костей
(orbitalia), а лежащие позади них кости на-
зываются щечными, или скуловыми костями
(jugalia). У Lepidosteus имеется очень большое число таких костей, у
Polypterus — одна большая. Предкрышечная кость по своему происхожде-
нию относится, невидимому, также к
этой группе костей, а не к костям жа-
берной крышки. Все эти кости обра-
зуют у наиболее примитивных форм
(Lepidosteus, Polypterus и многие иско-
Рис. 127. Череп ископаемой кистеперой рыбы (Osteolepis); вид сверху (Л) и сбоку (В)
(по Гудричу). Каналы боковой линии показаны пунктиром.
а — angulare; d— dentale; е — ethmoldeum, сросшееся с ростральным щитом; ft — срос-
шиеся лобные кости с отверстием для пинеального органа; ioc — подглазничный канал;
it — Intertemporale; j — iugale; I — lacrymale; Ig. vg— боковые и нижние яремные пла-
стинки (Jugularia); mdc — нижнечелюстной канал; mg —средняя яремная пластинка; тх —
maxillare; п — ноздря; па — nasalia, входящие в состав рострального щита; о — глазница;
ор — operculum; р —отверстие пинеального органа; pop —praeoperculum; ptf — postfron-
tale; pto — postorbitale; rs — ростральный щит; so — supraorbitale; soc — надглазничный
канал; sop — suboperculum; ppa — postparietale; sq — squamosum (quadratojugale?); st—
supratemporale (pteroticum); stc — теменной канал; sqc— яремный канал; tb—tabulate;
toe — затылочный канал.
140
паемые ганоиды), как сказано, совершенно сплошной наружный панцкрЬ
головы (рис. 126, 127).
Дно черепа подостлано большим парасфеноидом (parasphenideum)
и парным сошником (vomer), лежащим под обонятельной областью черепа
и несущим зубы. У Polypterus и парасфеноид покрыт мелкими зубами.
При сравнении древнейших рыб с современными можно заметить,
что число покровных костей первоначально было велико и распределение
их не было точно фиксировано (как отчасти и у современных осетровых).
Впоследствии определенные кости разрастались, а другие исчезали,
но некоторые кости сохранялись с большим постоянством и унаследо-
ваны большинством современных позвоночных. Далеко не всегда можно
указать, почему та или иная кость отличается особенным постоянством,
но в отношении некоторых костей это можно установить.
Щечные кости были значительно распространены среди ископаемых
рыб (рис. 127), из современных же рыб они сохранились лишь у костных
ганоидов, у которых к ним прикрепляется часть жевательной мышцы,
почему, очевидно, они и сохранились (рис. 125). На голове у рыб наблю-
дается очень правильное распределение каналов боковой линии, которые
в области туловища проходят через ряд чешуй, а в области головы — через
покровные кости черепа, служащие им защитой (рис. 126, 128). Главный
Рис. 128. Череп трески сбоку. Положение органов боковой линии и их иннервация
(по Гудричу).
ап — ваднесочленовная кость; art — сочленовная; Ьс — n. buccalis VII; d — вубная кость;
/г — лобная кость; hm — hyomandibulare; hmf — п. hyomandibularis VII; top — меж-
крышка; 1 —слезная кость; tat—n. lateralia X; Io — косточки канала боковой линии;
tnx — maxilla; mpt — metapterygoideum; n — носовая кость; op — крышка; pop — пред-
крышка; pmx — praemaxilla; pi — переднелобные кости; pst — posttemporale; q — quadra*
turn; toe — верхнезатылочная кость; sop— подкрышка; so — n. ophtalmicus super! icia lis
УП; sob — косточки подглааничного канара боковой линии.
канал проходит по надвисочным, лобным и носовым костям (надглазничный
канал); отходящий от него подглазничный канал проходит по глазничным
костям; гиомандибулярный канал — по предкрышке и по угловой и зубной
костям нижней челюсти и поперечный затылочный канал — по ряду
заднетеменных костей (postparietalia, tabularia). Все эти кости отличаются
очень большим постоянством, зависящим, невидимому, отчасти именно
от их связи с органами боковой линии. Только заднетеменные кости исче-
зают у большинства костистых рыб, но сохраняются, иногда, их остатки
в виде небольших трубчатых косточек, окружающих надвисочный канал.
Мы несколько подробнее остановились на черепе костных ганоидов
ввиду того, что он построен наиболее типично и имеет большое значение,
141
Являясь ИСХОДНЫМ и для костистых рыб, и для наземных позвоночных,
между тем как череп костистых рыб обнаруживает признаки известной
специализации.
В черепе костистых рыб стенки глазниц обычно между собой
срастаются, образуя межглазничную перегородку, и орбитосфеноиды пре-
вращаются в непарную кость. У большинства же эта кость редуцируется,
и вся межглазничная перегородка низводится до тонкой перепонки.
Осевой череп окостеневает еще полнее, чем у костных ганоидов, и число
замещающих костей еще больше (рис. 129).
Рис. 129. Схема черепа костистой рыбы. Висцеральный скелет отделен от черепной
коробки.
alsph— laterosphenoideum; ang — retroarticulare; art—articulare; bo—basioccipitale"
bsph — bosisphenoideunT; terbr —ceratobrancljialia; dent — dentale; eeth — ectoetlimoidcuinl
ecpt— ectopterygoideum; enpt— entopterygoideum; eo— exocdpitalc; epbr — epibranchalia;
epot — epioticum; fr — frontale; hm — hyomandibulare; Льг — hypobranehiale; hy—подъязычная
дуга; thy — tinterliyale; meth — mesethmoideum; md —челюстная дуга; mpt — metapterygoide-
um; mx — maxilla; n — nasale; opot — opisthoticum; osph — orbitosphenoideum; pa —
parietale; pat.— palatinum; phbr — pliaryrigbbranchialc; pmx — rraernaxilta; prr>f —.proofi-
cum; plot—pterptlciim.; psph —paraspKenoIdeum; q— qjiadratum; so — supraoccipitale;
ephot — sphenoticum; sp — symplecticum; v — voineir; I—V — жаберные дуги.
От глазниц отходят назад под основание черепа обыкновенно парные
глубокие каналы, куда проникают четыре прямые мышцы глаза — каналы
глазных мышц, или миодомы.
В затылочной области, кроме основной и боковых затылочных костей,
имеется еще верхняя затылочная кость (supraoccipitale), развившаяся,
невидимому, в результате прирастания остистого отростка одного из
позвонков. В слуховой области, кроме передне- и верхнеушной, имеются
еще две смешанные кости, лежащие над первой — клиновидноушпая
(sphenoticum) и позади нее крыловидноушная (pteroticurn). Обе эти
кости занимают то же положение, что и покровные заднелобная и падви-
сочная кости ганоидных рыб, и представляют, невидимому, результат
врастания этих костей в хрящ верхней стенки слуховой капсулы. В
глазнично-височной области имеется основная клиновидная и боковые
клиновидные кости. Глазнично-клиновидные кости имеются только у
низших костистых рыб. В обонятельной области имеется непарная средняя
обонятельная кость (mesethmoideum) и парная боковая (ectoelhmoideum).
Эти кости также смешанного характера и являются результатом врастания
в хрящ переднелобных костей и верхней обонятельной. Первая начинает
врастать в хрящ уже у костных ганоидов, вторая .наблюдается в виде
покровной кости еще у низших костистых рыб. Дно черепа подостлано
742
Seth
pk
pa-'
scS.
pep-"
-pt/
'РРЧ
sf)u
парасфеноидом й непарным у костистых рыб сошником, который начинав®
врастать в хрящ.
Висцеральный аппарат в общем весьма сходен с таковым костных
ганоидов. Череп костистых рыб относится к гиостильному типу, так рак
пёбноквадратный отдел челюстной дуги непосредственно соединяется с
черепом только впереди, в обонятельной области. Верхними челюстями
служат вторичные кости верхней губы — praernaxillare и maxillare.
Нёбноквадратный хрящ окостеневает позади, образуя квадратную и
заднюю крыловидную кости, и покрывается в средней своей части покров-
ными наружной и внутреннейкрыловидными костями; впереди развивает-
ся смешанная нёбная кость,
несущая зубы. В нижней поло'
вине челюстной дуги позади
развиваются сочленовная и зад-
несочленовная (retroarticulare)
кости, а впереди зубная. Ни
пластинчатой, ни надугловой
костей у костистых рыб нет.
В гиоидной дуге имеются те же
окостенений, что и у костных
ганоидов (hyomandibulare, sym-
plecticum, interhyale и hyale
с несколькими окостенениями).
В жаберных дугах окостеневают
все отделы. Последняя, пятая
жаберная дуга состоит из одного
только ceratobranchiale, жабер
не имеет, а несет так называе-
мые нижнеглоточные зубы.
Pharyngobranchialia второй,
третий и четвертой дуг соеди-
няются между собой и несут
«верхнеглоточные» зубы. На
копулах также нередко нахо-
дятся зубные пластинки. Жа-
берная крышка состоит из ти-
пичных костей: operculum, sub-
operculurn, inter operculum и
praeoperculum. Жаберная пе-
репонка поддерживается кост-
ными лучами (branchiostegalia).
Нижних яремных пластинок
у современных костистых рыб
нет (за исключением одного лишь сем. Elopidae).
Из покровных костей крыши и боков черепа редуцируются задне-
теменные кости, сохраняющиеся в виде самостоятельных элементов лишь
у низших костистых рыб (рис. 130), исчезают чешуйчатые кости и щечные
пластинки. На боках черепа сохраняется, кроме челюстных костей, лишь
кольцо орбитальных косточек (рис. 128). Вторичная крыша черепа со-
ставляется из парных теменных, лобных и носовых костей. Верхняя
обонятельная кость, имеющаяся еще у низших костистых рыб (рис. 130),
врастает в хрящ и получает название средней обонятельной кости (те-
sethmoideum). Переднелобные кости врастают в хрящ и получают назва-
ние наружных обонятельных костей (ectoethrnoidea). Заднелобные и над-
височные кости врастают в хрящ слуховой капсулы и получают названия
клиновидно-ушных и крыловидноушных костей (sphenoticum и pteroticum).
Все эти кости по положению соответствуют названным покровным костям
зос
Рис. 130. Череп карпа сверху; положение
органов боковой линии (по Гудричу).
еро — eploticum; />р — operculum; sbo, sob — ог-
bltalla; pop — praeoperculum; ppo — postparle-
talla; ptf — postfrontalia; pf — praefrontaBa;
seth — supraethmoideum. остальные обозначения
см. рис. 129.
ййЖе стоящих рыб и развиваются у костистых рыб как смешанные кости,
т. е. закладываются в качестве типичных покровных костей вне хряща и
его перихондрия, но затем окостенение захватывает и последний, и кость
начинает вытеснять хрящ. То же самое явление замечается отчасти и при
развитии сошника и некоторых других костей у высших костистых. Мы
толкуем это как превращение покровной кости в кость, замещающую
хрящ, — явление, вообще говоря, довольно редкое. По другому толко-
ванию здесь произошло срастание покровной кости с костью замещающей.
Однако у низших рыб мы на их месте находим одну покровную кость.
Череп двудышащих рыб отчасти примитивен, отчасти весьма
своеобразно специализирован. Он имеет широкое основание и состоит в
значительной мере из хряша, дополненного почти исключительно покров-
ными костями. У древнейших форм имелось очень большое число костей
крыши- черепа, которое у современных форм значительно сокращено.
Своеобразным является присутствие больших непарных костей крыши
черепа, сравнение которых с костями других рыб несколько затруднено.
С осевым черепом срастается несколько позвонков с остистыми отрост-
ками и даже ребрами. В хрящевой черепной коробке развивается лишь
одна пара замещающих костей — боковые затылочные кости. Снизу
череп подостлан парасфеноидом и парным сошником с зубами.
Нёбноквадратный хрящ вполне срастается с основанием черепа
(аутостилия), и лежащие на нем крыловидные кости (pterygoidea) несут
большие сложные нёбные зубы. На меккелевом хряще имеются
такие же сложные зубы на пластинчатых костях (splenialia). Снаружи
на нижней челюсти лежат угловая и незначительная зубная кость, не
несущая зубов. Гиомандибулярный хрящ рудиментарен; он связан с
слуховой капсулой и соединяется связками не только с большим гиоидом,
нои с задней частью нёбноквадратного хряща, указывая этим на прежнюю
роль его в качестве подвеска. Вторичных челюстей у двудышащих нет.
Жаберный аппарат отчасти редуцирован в связи с появлением легочного
дыхания. Кости жаберной крышки также отчасти редуцированы. Покров-
ная кость, лежащая на слуховой капсуле и продолжающаяся вниз на
задний отдел нёбноквадратного хряща, представляет, повидимому, резуль-
тат срастания чешуйчатой кости с предкрышкой и в высшей степени
напоминает «чешуйчатую» кость амфибий, с черепом которых череп дву-
дышащих вообще в некоторых отношениях сходен.
Череп наземных позвоночных н его окостенения
Череп наземных позвоночных отличается от черепа рыб главным
образом изменениями в области висцерального скелета, чзсг^.находится
в связи с утратой жаберного дыхания. Предъязычная и жаберные дуги
редуцируются и частью преобразуются в органы другого значения.
Челюстная дуга срастается с основанием черепа, давая типичную ауто-
стилию. Подвесок преобразуется в слуховую косточку. Жаберная крышка
совершенно исчезает.
В осевом черепе изменения гораздо менее значительны. Характерным
является развитие подвижного сочленения с позвоночником посредством
затылочных бугров и известное облегчение черепа вследствие частичной
редукции его. наружной крыши.
По своему составу череп наземных позвоночных очень близок к черепу
костных ганоидов. У низших их представителей — земноводных — в
осевом черепе сохраняется еще много хряща и число замещающих костей
невелико, напоминая в этом отношении череп кистеперых ганоидов.
У более высоко стоящих форм хрящ почти вытесняется костью.
В черепе наземных позвоночных развиваются следующие з вме-
щающие кости.
744
В затылочной области — основная, боковые и верхняя затылочные
кости (basi-, ex-, supraoccipitalia). В слуховой области — передняя,
задняя и верхняя ушные кости (pro-, epi-, opisthotica). У млекопитающих
здесь развивается одна околоушная кость с несколькими окостенениями.
В глазнично-височной области у основания — основная и у высших форм
передняя клиновидные кости (basi-, praesphenoideum). По бокам от них —
парные — боковая клиновидная и глазнично-клиновидная (latero-, orbi-
tosphenoidea). В обонятельной области у низших наземных позвоночных
сохраняется хрящ, а у высших развивается решетчатая кость с несколь-
кими окостенениями (ethmoideum).
Как видно, большинство костей — те же самые, что имеются и у
костей приобретены
4J
костных ганоидов. Впрочем, некоторые из этих
наземными позвоночными независимо и, следователь-
но, не гомологичны одноименным костям рыб. Так,
например, латеросфеноиды отсутствуют у низших
наземных позвоночных и, по всей вероятности,
представляют собой новоприобретение высших форм.
Таким же новоприобретением являются тем более
и те кости, на месте которых и у земноводных, и
у ганоидных рыб имеется только хрящ. Сюда отно-
сятся верхняя затылочная и обонятельные кости,
которые, несомненно, не гомологичны одноименным
костям костистых рыб.
Что касается покровных костей че-
репа, то наиболее характерны для наземных позво-
ночных следующие кости крыши черепа: теменные
(parietalia), лобные (frontalia), носовые (nasalia) и
иногда передне- и заднелобные (prae-и postfrontalia);
по бокам черепа в височной области чешуйчатые
(squamosa) и задпеглазничные (postorbitalia), а у
примитивных форм также надвисочные (supratempo-
ralia) и под ними квадратпоскуловые (quadratoju-
galia) и скуловые (jugalia). Под глазницами и впере-
ди от них челюстные (maxillaria) и предчелюстные
(praemaxillaria) кости образуют нижний край кры-
ши черепа. Дно черепа подостлано самостоятель-
ным парасфеноидом только у амфибий. Под обо-
нятельной областью черепа у всех наземных по-
звоночных лежит парный сошник (vomer), срастаю-
щийся у высших форм в одну кость. Все это —
кости, характерные и для черепа высших рыб.
И для покровных костей черепа наземных позвоночных приходится
повторить сказанное при описании черепа рыб: число костей у более
примитивных форм в общем выше, а у высших форм сокращается. Череп
амфибий, казалось бы, противоречит этому, так как состоит из сравни-
тельно небольшого числа покровных костей, в особенности у бесхвостых;
изучение же ископаемых амфибий — стегоцефалов — убеждает нас в том,
что череп современных амфибий испытал значительную редукцию.
Строение черепа стегоцефалов является, несомненно, исходным для
всех современных наземных позвоночных и вместе с тем позволяет без
особого труда провести сравнение между черепом наземных позвоночных
с одной стороны, и рыб — с другой (рис. 131, 132).
Прежде всего следует отметить, что покровные кости черепа стего-
цефалов составляли такой же сплошной наружный панцырь головы,
как и у ганоидных рыб, причем кости его были также тесно связаны с
кожей. Мы уже видели, что поверхность типичных кожных костей обычно
отличается характерной скульптурой. Такая скульптура наблюдается и
145
Рис. 131. Череп сте-
гоцефала (Tremato-
saurus) с отпечатками
каналов боковой ли-
нии (контуры по Ис-
ке лю).
fr — frontale; jug — ju-
gale; mx — maxilla;
n — nasale; pa — pa-
rietale; pmx — praema-
xilla; po — postorbita-
le; ppa — postparietale;
prf—praefrontale; ptf —
postfroitale; qj— qua-
dratojugale; sq— squa-
mosum; st— supratem-
porale.
на крыше черепа стегоцефалов (рис. 142). Состав этой крыше также вполне
сходен с составом крыши черепа костных ганоидов и в особенности некото-
рых ископаемых кистеперых (рис. 387). Кроме обычных костей, имеются и
характерные специально для ганоидов — поперечный ряд костей позади
теменных и кости, покрывающие височную область черепа, или щечные
кости, которые у наземных позвоночных имеются в числе двух пар и
получают название скуловых костей, так как входят в состав скуловых
дуг. У стегоцефалов, как и у рыб, череп подостлан большим парасфенои-
дом и парным сошником.
Интересно, что на черепе некоторых стегоцефалов сохранились и
отпечатки каналов боковой линии головы, причем положение их в общем
соответствует положению у высших рыб (рис. 131). Таким образом, на-
ружная крыша черепа построена у стегоцефалов так же, как и у костных
ганоидов. Разумеется, есть и различия, и прежде всего это выражается в
отсутствии у первых жаберной крышки.
Рис. 132. Череп стегоцефала Orthosaurus (по Ватсону).
ап — angulare; d — dentalc; ju — jugale; la — lacrymale; ptsp — postspleniale; дз — qua-
dratojugale; san—supraangulare;, smx — septomaxillare; sp! —spleniale; sq — squamosum.
В висцеральном скелете наземных позвоночных произошли по сравне-
нию с рыбами крупные преобразования, значение которых установить
нетрудно. Возникли они в связи с переходом к жизни на суше, а так как
низшие из наземных позвоночных — амфибии — в личиночном состоянии
живут в воде и дышат жабрами, то значительная часть этих преобразова-
ний происходит во время их индивидуального развития и без труда
может быть прослежена.
В области челюстной дуги для наземных позвоночных характерна
непосредственная связь нёбноквадратного хряща с осевым черепом. На
черепе амфибий можно видеть, что эта связь устанавливается в тех же
местах, как и у костных ганоидов: впереди — с обонятельной областью
черепа и позади — с основанием черепа на границе между глазничной и
слуховой областью черепа (точнее — со стенкой камеры тройнично-лице-
вых ганглиев). Различие по сравнению с костными ганоидами лишь в том,
что сочленение заменилось у амфибий срастанием и, главное, что гиоман-
дибулярный отдел подъязычной дуги потерял у последних значение под-
веска. Таким образом, аутостилия наземных позвоночных развилась
вторично из амфистилии костных ганоидов (рис. 120, С, D). То же самое
наблюдается и у двудышащих рыб, у которых гиомандибулярный хрящ
вторично утратил значение подвеска.
Нёбноквадратный отдел челюстной дуги, утрачивающий значение
челюсти уже у высших рыб, у наземных позвоночных окончательно
перестает нести эту функцию (рис. 133). В связи с этим у многих
амфибий и у всех высших наземных позвоночных передняя часть нёбно-
квадратного хряща редуцируется, а задняя окостеневает в виде квадратной
кости, служащей исключительно для подвешивания нижней челюсти к
146
черепу *. Соответствующее окосГейейие заднего отдела небнокваДраТноГо
хряща наблюдается иногда и у современных амфибий; оно было и у сте-
Рис. 133. Преобразования первых двух висцеральных дуг в ряду
(позвоночных (схемы).
Л —акула; В — костная рыба; С — амфибия; D — рептилия; Е — териодонт; F — мле»
копитающее; ап — angulare; art — articulare; со—сотового; d—dentale; Лш—hyoman-
dibulare; hy — bylodeum; Mk — меккелев хрящ; pa — paJatinum; pq — palatoquadratum;
pl — pterygoideum; q — quadratum; san —spuraangulare; eq*— aquamusum; st — stapes.
гоцефалов и, вероятно, Гомологично одноименной кости высших рыб.
На нёбноквадратном хряще амфибий лежат две покровные кости —
нёбная и крыловидная, сохраняющиеся у высших позвоночных и после
Рис. 134. Подвесок (hyomandibulare) рыб и слуховая косточка (stapes,
columella) наземных позвоночных в поперечном разрезе через голову.
А—двойное прикрепление жаберной дуги в черепу у осетровой рыбы; В —остатки двойного при-
крепления гиоидной дуги (подвеска) к черепу у двудытащей рыбы; С — наружное прикре-
пление подвеска костистых рыб; D—двойное прикрепление слуховой косточки к черепу
у примитивных рептилий, асг — сонная артерия; ей — хорда; ерг — epibranchiale; е — ext-
racolumella (= hyomandibulare); h — hyoideum (— ceratohyale); hm — hyomandibulare
(=. epihyale); pd — спинной отросток столбика; phbr — pharyngobranchiale; phh — pha-
ryngohyale; at—stapes (= pharyngohyale); v. j — яремная вена; VH — подьязычночелю-
стная ветвь лицевого нерва.
1 Только у млекопитающих квадратная кость теряет значение подвеска и входит
в состав слуховых косточек (рис. 133, F).
10*
147
редукции переднего отдела того хряща, на котором они первоначально
возникли (рис. 133). Эти кости также соответствуют одноименным костям
костных ганоидов.
В нижнем отделе челюстной дуги задний конец меккелева хряща око-
стеневает, образуя сочленовную кость, которая редуцируется у совре-
менных амфибий, а остальная его часть окружена покровными костями —
зубной, угловыми и пластинчатой костью. Все это те же кости, что име-
ются и у костных ганоидов.
В подъязычной дуге наземных позвоночных интересны преобразо-
вания ее верхнего отдела (рис. 133, 134, 135). Вполне естественно, что
с потерей значения подвеска гиомандибулярный отдел редуцируется,
Рис. 135. Соотношения между элементами первых двух висцеральных дуг, их нервами
и сосудами у рыб (.4), примитивной рептилии (/?), ящерицы (С), млекопитающего (D)
(по Гудричу, изменено). Нервы обозначены черным, вены — густыми точками, арте-
рии — поперечными штрихами.
а — articulare; ап — angulare (= tympauicum); br — processus basalis; ch — ceratohyale;
cht — chorda tympani; dpr— processus dorsalis; / — лицевой нерв; g — goniale; hm — hyo-
mandibuiare; hn — подъязычночелюстная ветвь лицевого нерва; i — incus ( — quadratum);
ic — внутренняя Сонная арерия; tmd—внутренний нижнечелюстный нерв VH; jv — яремная
вена; ер — epipterygoideum ( =alisphenoideum); т — malleus ( = articulare); тк — меккелев
хрящ; md и тх — ни:кне- и верхнечелюстная ветви тройничного нерва; тп — рукоятка
молоточка; орг — processus oticus; or — глазничная артерия; р — нёбный отдел квадратного
хряща (кости); phh— pharyngohya ie; pn — нёбный нерв VH; q— квадратный отдел;
sp —брызгальце; st — stapes; sta — arteria stapedia-; tr — тройничный нерв; ty — барабан-
ная перепонка.
как это наблюдается и у двудышащих рыб. У наземных позвоночных он,
однако, приобрел новую функцию и существенно преобразовался. Поло-
жение его между жаберной крышкой и стенкой слуховой капсулы сде-
лало возможным приобретение значения посредника при передаче зву-
ковых колебаний внутреннему уху. Эта функция облегчалась еще тем,
что полость первой висцеральной щели — брызгальца — при переселе-
нии животного на сушу наполнилась воздухом и, представляя собой упру-
гое пространство, способствовала колебательным движениям наружной
стенки под влиянием доходящих до нее звуковых волн. Таким образом,
полость брызгальца преобразовалась в барабанную полость, а гиоман-
дибулярная кость — в слуховую косточку, так называемый столбик (colu-
mella). При этом достигается более совершенная передача самых слабых
колебаний внутреннему уху, благодаря тому, что стенка слуховой кап-
745
Сулы истончилась в том месте, где в нее упирается слуховая косточка,
И превратилась в затянутое перепонкой овальное окно. При эмбриональ-
ном развитии многих наземных позвоночных наблюдалось, что столбик
уха развивается из верхнего отдела подъязычной дуги (рис. 136), а в не-
которых случаях у примитивных рептилий (гаттерия и гекконы) он даже
и пожизненно сохраняет связь с гиоидом
(рис. 135, В, С).
Однако столбик уха представляет собой до-
вольно сложное образование и состоит из двух
частей, имеющих значение самостоятельных отде-
лов. Упирающийся в овальное окно внутренний
элемент (stapes) соответствует верхнему элементу
гиоидной дуги — pharyngohyale, рудимент кото-
рого эмбрионально наблюдается у стенки слухо-
вой капсулы и у двудышащих рыб (рис. 134).
Более крупный наружный^отдел столбика — ext-
racolumella, упирающийся в барабанную пере-
понку и связанный иногда с гиоидом, соответ-
ствует следующему элементу гиоидной дуги—
epihyale, т. е. собственно .гиомандибулярному
хрящу. Интересно, что наружный отдел столби-
ка иногда причленяется к квадратной кости (не-
которые амфибии и рептилии, в особенности ис-
копаемые пеликозавры и териодонты). В этом
нетрудно усмотреть следы прежней связи подвеска
с челюстной дугой, т. е. остаток амфистилии.
С другой стороны, иногда сохраняются и следы
связи extracolumella с боковой стенкой слуховой
Рис. 136. Слуховой стол-
бик эмбриона геккона
(Platidactylus) (по Верс-
люису).
af — arteria facaHs (stape-
dis); is —• interealare; exc —
extracolfimella; hy — гио-
ид; at — stapes; par —
вакладка processus parotl-
cus черепа.
капсулы и с чешуйчатой костью, т. е. как раз с той областью, где причле-
няется подвесок у высших рыб, посредством особого спинного отростка
(processus dorsalis, in tercal аге) (рис. 135, 136).
Нижний отдел подъязычной дуги преобразуется у наземных по-
звоночных в так называемые передние или гиоидные рожки подъязыч-
ной кости. Остальные отделы висцерального аппарата сохраняются частью
в виде подъязычной кости с ее рожками, представляющими собой остатки
жаберных дуг, частью в виде гортанных хрящей. За этими преобразова-
ниями проследить нетрудно, в особенности у амфибий. Это лишнее дока-
зательство происхождения наземных позвоночных от живших в воде и
дышавших жабрами предков, которые были близки к костным ганоидам.
Череп амфибий
Осевой череп амфибий имеет широкое основание (платибазальный
тип) и состоит в значительной части из хряща. Имеется крупная верхняя
фонтанель (рис. 137), закрытая сверху лобными и теменными костями,
а у хвостатых амфибий имеется, сверх того, еще и большая нижняя фон-
танель, прикрытая снизу парасфеноидом. У основания сильно вздутой
слуховой капсулы имеется небольшое «овальное» окно (fenestra
ovalis, рис. 137), в которое упирается слуховая косточка, столбик. У края
этого окна иногда обособляется часть стенки самой капсулы (к которой
прикрепляется часть мышцы, поднимающей лопатку), образуя добавоч-
ную слуховую косточку, носящую нерациональное название operculum
(не смешивать с крышкой рыб). Было высказано предположение, что эта
добавочная слуховая косточка амфибий служит специальным аппаратом
для передачи колебаний почвы на внутреннее ухо через передние конеч-
ности, плечевой пояс и мышцу, поднимающую лопатку (m. levator sca-
pulae), которая одним пучком волокон прикрепляется к operculum. В глаз-
749
ничной области значительная часть стенки черепа остается хрящевой
и хрящевой же остается и вся обонятельная капсула, выделяющаяся на
черепе в виде значительного вздутия.
В осевом черепе современных амфибий костей немного (рис. 138).
В затылочной области имеется пара больших боковых затылочных костей,
снабженных суставными буграми для сочленения с позвоночником. Для
амфибий характерно присутствие именно двух затылочных бугров. В
боковых затылочных костях имеются отверстия для выхода блуждающих
нервов. У стегоцефалов, кроме этих
костей, наблюдалась еще и основная
затылочная кость, а также иногда и
верхняя затылочная кость. В слухо-
вой области имеется только большая
переднеушная кость с отверстием
или вырезкой для выхода тройнично-
го и лицевого нервов. У стегоцефалов
имелась также заднеушная кость.
В глазничной области лежат большие
Рис. 138. Череп лягушки снизу (по Эк-
керу и Видерсгейму). На одной стороне
черепа покровные кости удалены. Хрящ
обозначен пунктиром.
саи — слуховая капсула; вое — exoeeipi-
talia; eth — хрящ обонятельной области;
Ьр —frontoparietale (край); тх— maxilla;
о — глазница; pal—- palatinum; ртх — ргае-
maxilla; pro — prooticum; ps — parasphe-
noideum; pt — Pterygoideum; pq — нвбно-
квадратный хрящ; qj — quadrate jugale;
eph — sphenethmeideum; в — vomer; II—
отверстие для зрительного нерва; V — от-
верстие для тройничного нерва.
Рис. 137. Хрящевой череп молодой
лягушки (по Гаупу из Кингсли).
fov—овальное окно; тп—меккелев хрящ;
pq — нёбноквадратннй хрящ; сп — ноеовая
капсула.
парные у хвостатых амфибий, глазничпоклиновидные кости (рис. 139),
которые у бесхвостых срастаются в одну кольцевидную кость, охватываю-
щую всю черепную коробку в передней части глазничной области (рис.
138). Эта кость носит название клиновиднообонятельной кости (sphene-
thmoideum), так как она частью зарастает и в область обонятельных
капсул. Последние остаются в остальном хрящевыми. У стегоцефалов,
однако, иногда имелась и основная клиновидная кость.
Крыша черепа составляется из следующих покровных костей: темен-
ные, лобные (срастающиеся у бесхвостых в одну парную лобнотеменную
кость), носовые, а у хвостатых амфибий, сверх того, еще переднелобные
(иногда в числе двух пар) (рис. 139). В слуховой области черепа лежит
чешуйчатая кость (squamosum), которая одной своей ветвью разрастается
вниз, покрывая и заднюю часть (pars quadrata) нёбноквадратного хряща.
Нижний край крыши черепа впереди глазниц образован вторичными
челюстями, а позади глазниц у бесхвостых амфибий — тонкой квадратно-
скуловой костью (quadratojugale), образующей по краю рта костный
мостик, который связывает задний конец челюстной кости с нижним
концом задней части нёбноквадратного хряща. У хвостатых амфибий
задний конец челюстной кости оканчивается свободно (рис. 139). Дно
черепа подостлано большим парасфеноидом, который у неко-
торых хвостатых амфибий еще покрыт многочисленными зубами. Под
обонятельной областью лежит парный сошник, несущий зубы.
Наружная стенка черепа у современных амфибий не образует сплош-
ного панцыря, но у стегоцефалов такая сплошная наружная крыша
150
имелась (стегальный череп), и отсутствие ее у современных форм есть
результат редукции. Впрочем, и в одной группе современных амфибий —
у гимнофион — имеется такая же наружная крыша черепа. Однако
число костей у них не больше, чем у других амфибий, и, вероятно, мы
имеем здесь случай вторичного разрастания тех же костей, что имеются
и у современных хвостатых амфибий. У последних область осевого черепа
позади глазниц (височная область) обыкновенно совершенно обнажена,
но в некоторых случаях наблюдаются незначительные остатки наружного
покрова в виде костных мостиков или височных дуг, покрывающих ату
область черепа. У бесхвостых амфибий имеется, как уже сказано, мостик,
Рис. 439. Череп хвостатой амфибии (Amblystoma) сверху и снизу
(по Видерсгейму из Кингсли)
f — frontale; тх — maxilla; n — nasale; os — orbitosphenoideum; of — prooticum; p— parie-
tale; pm — praemaxilla; prf — praefrontale; ps — parasphenoideum; pf — pterygoideum;
sg — squamosum; up — vomeropalatinum.
идущий вдоль края рта (нижняя височная дуга, образованная квадратно-
скуловой костью). Подобный мостик имеется в виде исключения и у
одной из хвостатых амфибий (Tylototriton), у которой, как и у некоторых
других тритонов, наблюдается еще мостик в верхней части височной об-
ласти (верхняя височная дуга). На месте верхней дуги у амфибий обычно
имеется направленный вперед отросток чешуйчатой кости и связка.
Висцеральный скелет амфибий составляется из шести висцеральных
дуг. Верхний, нёбноквадратный отдел челюстной дуги, окончательно
утратил функцию верхней челюсти. Последним указанием на ату функцию
служат лишь зубы, имеющиеся на нёбной кости у хвостатых амфибий.
Небноквадратный хрящ прирастает к черепу в обонятельной области
своим передним концом и своим задним концом к основанию черепа у
переднего края слуховой капсулы (рис. 137). Обычно весь нёбноквадрат-
ный отдел остается хрящевым, и только задняя его часть иногда окосте-
невает в области причленения нижней челюсти (квадратная кость). Более
обширную область занимает это окостенение только у гимнофион, у кото-
рых, как и у ископаемых стегоцефалов, имеется крупная квадратная
кость. На нёбноквадратном хряще лежат снизу две покровные кости —
крыловидная и нёбная (рис. 138). Последняя кость, как сказано, у хвоста-
тых амфибий снабжена зубами и срастается с сошником (рис. 139).
Нёбноквадратный хрящ сохраняется на всем своем протяжении лишь
у бесхвостых амфибий (и в виде исключения у Ranides); у хвостатых
амфибий передняя часть его редуцируется и имеет тогда вид более или
менее значительного крыловидного отростка (processus pterygoideus) на
заднем или квадратном его отделе (рис. 140). В некоторых случаях и
крыловидный отросток исчезает, и тогда от нёбноквадратного хряща
остается лишь его pars quadrata, служащая подвеском для нижней челюсти..
Нижний отдел челюстной дуги сохраняется обыкновенно на всем своем
151
протяжении в виде меккелева хряща. Иногда наблюдается небольшое
окостенение в заднем его конце, на месте сочленения с квадратной костью
(сочленовная кость). У бесхвостых амфибий регулярно окостеневает
Рис. 140. Нёбноквадратный хрящ у бес-
хвостых амфибий (Л) и редукция его
у хвостатых (В, С).
передний конец меккелева хряща,
образуя маленькую подбородочную
кость, наблюдающуюся и у неко-
торых костных ганоидов. Вся осталь-
ная часть хряща покрыта покров-
ными костями — зубной снаружи и
пластинчатой и покровной сочленов-
ной (goniale) изнутри.
У стегоцефалов имелись, кроме
того, угловая, надугловая, венечная
и другие покровные кости нижней
челюсти.
Верхний отдел подъязычной ду-
ги представлен слуховой косточкой,
столбиком, который одним концом
упирается в овальное окно слуховой
капсулы, а другим концом у бесхвос-
тых амфибий в барабанную перепон-
ку. У хвостатых и у безногих амфибий
барабанной перепонки нет, барабан-
ная полость также исчезла; сама слу-
ховая! косточка (columella) нередко
редуцируется и иногда вполне заме-
няется добавочной косточкой (opercu-
lum), о которой речь была выше. За-
служивает внимания, что у хвостатых
амфибий нередко сохраняется прямая связь столбика с задней частью
нёбноквадратного хряща. У гимнофион короткая и массивная слуховая
косточка также причленяется к квадратной кости. Интересно, что здесь
она продырявлена отверстием для прохождения лицевой артерии (art.
Рис. 141. Подъязычный аппарат аксолотля (Л) и молодой лягушки (В)
(ив Видерсгейма).
ЬЬг — basibranchialia; chy — гиоидные рожки; со. h —тело; pr. thy — задние рожки-
сЫ — 2—4 — ceratobrahchialia; hbr — hypobranchia На; hph — hypohyale; ch — ceratohyale.’
facialis s. stapedia), так ate как это иногда наблюдается в столбике репти-
лий и в стремени млекопитающих. Нижняя часть гиоидной дуги вместе
с жаберными дугами (функционирующими у личинки) образует у амфибий
так называемый подъязычный аппарат (рис. 141). Последний у хвостатых
152
и у безногих амфибий наименее изменен и состоит из нескольких (3—5)
дуг, соединенных копулами. Первые дуги нередко расчленены на два
отдела (hypo- и ceratobranchialia). Верхний конец гиоида у хвостатых
амфибий связан связкой с концом нёбноквадратного хряща, с которым
связан и столбик уха, но непосредственной связи его с последним у амфи-
бий во взрослом состоянии не имеется.
У бесхвостых амфибий подъязычный аппарат представлен хрящевой
пластинкой с передними и задними «рожками» (рис. 141, В). Передние
рожки представляют собой остаток гиоида и прирастают верхним концом
к стенке слуховой капсулы. Кроме этих бесспорных остатков висцераль-
ных дуг, предполагается, что и гортанные хрящи амфибий (боковые
хрящи и их производные) представляют собой преобразованную пятую
пару жаберных дуг.
Череп стегоцефалов (рис. 131, 132, 142, 389) отличается
от черепа современных амфибий прежде всего присутствием сплошной
наружной крыши из большого сравнительно числа покровных костей,
ртх
ртх
psph
Рис. 142. Череп стегоцефала (Branchiosaurus) сверху и снизу
(по Креднеру и Фритчу).
pal — palattaum; р/ — praefrontale; psph — parasphenoideum; pl — pterygoideum;
v — vomer. Остальные обозначения, как на рис. 131.
среди которых имелись и щечные кости (jugalia и quadratojugalia) и
заднетеменные или табличные (postparietalia s. tabularia). Эти особенности,
как уже сказано, сближают череп стегоцефалов с черепом костных ганои-
дов. Присутствие особого отверстия в сагиттальном шве черепа, где,
очевидно, помещался теменной глаз, сближает этот череп отчасти с черепом
некоторых ископаемых ганоидов, имевших подобное отверстие, а также
и с черепом многих рептилий, для которых череп стегоцефалов также
оказывается исходным. У стегоцефалов нередко имелось и кольцо окосте-
нений в склере глаза. То же самое наблюдается и у некоторых ископаемых
ганоидов, и у многих рептилий и птиц. Число окостенений хрящевого
черепа было у стегоцефалов значительно выше. На нёбе имелись также
эктоптеригоиды (ossa transversa рептилий). На нижней челюсти имелись
многочисленные покровные кости, как и у рептилий. Во всем этом стего-
цефалы оказываются связующим звеном между рыбами и высшими назем-
ными позвоночными. Череп современных амфибий оказывается значительно
редуцированным. Палеонтология не дает материала для того, чтобы, мы
могли шаг за шагом проследить эту редукцию, но все же среди много-
численных стегоцефалов с массивными черепами встречаются и такие
(Branchiosauria), черепа которых по своей легкости и общей структуре
приближаются к черепу современных амфибий (рис. 142).
Череп рептилий
Череп рептилий относительно не столь широк, как у амфибий, он
имеет узкое основание и тонкую межглазничную перегородку, которая
да окостеневает, а остается хрящевой или перепончатой. Хрящ даже
153
ембрионально никогда не образует цельной коробки, а имеет вид системы
перекладин, разделенных более или менее обширными перепончатыми
пространствами (рис. 117). Более значительные массы хряща окружают
лишь ухо и орган обоняния. В слуховой капсуле, кроме овального окна,
появилось еще небольшое «круглое» окно (fenestra rotunda),
также затянутое перепонкой. Череп взрослого животного окостеневает
довольно полно и во всяком случае дополняется большим числом покров-
ных костей, так что хрящ играет заметную роль лишь в обонятельной
области.
В затылочной области развиваются все четыре затылочные кости
(основная, боковые и верхняя) (рис. 144). В боковых затылочных, кроме
Рис. 143. Череп гаттерии (Hatteria)
сбоку и сверху (по Циттелю).
ап—angulare; art—articulare; bo — basioccipitale;
со — coronare; col — columella; den — dentale;
eo— exoccipitale; ept — epipterygoideum; fr —
frontale; j-—• jugale; mx — maxilla; n — nasale;
pa — parietale; pfr— praefrontale; pmx — prae-
maxilla; pro — prooticum; pt — pterygoi-
deum; po — postorbitale; ptf — postfrontale;
«-—quadratum; qj— quadra to jugale; san—
supraangulare; so — supraoccipitale; sq—squa-
mosum; spl — spleniale.
ртт
Рис. 144. Череп гаттерии (Hatteria),
снизу и сзади. Нижняя челюсть изнутри
(по Циттелю).
Ьо — basioccipitale; bs — basisphenoideum;
ch — хоаны; opo — opisthoticum; pal — pa-
latiium; tr — transversum; v—vomer;
Остальные обовначения, как на рис. 143.
отверстий для блуждающего нерва, имеются еще отверстия для подъязыч-
ного нерва. Под затылочной дырой располагается единственный у рептилий
(и птиц) затылочный бугор, в образовании которого принимают участие
основная и боковые затылочные кости. В стенке слуховой капсулы разви-
ваются три кости — передняя, верхняя и задняя ушные кости, из которых
только переднеушная (рис. 143) остается у рептилий самостоятельной;
верхнеушная срастается с верхней затылочной костью, а заднеушная —
с боковой затылочной (за исключением черепах, у которых она остается
154
самостоятельной, рис. 155). Пр бокам слуховая капсула дает сильно
выдающиеся боковые отростки (proc, parotici), в которых располагаются
заднеушные кости (рис. 144). Эти отростки упираются в наружную крышу
черепа, и в этом месте происходит причленение квадратной кости, которая
получает, таким образом, прочную опору на черепе. Это одна из наиболее
характерных особенностей черепа рептилий. Впереди основной затылочной
кости в основании черепа лежит основная клиновидная кость с особыми
боковыми отростками (processus basipterygoidei), к которым у примитивных
рептилий причленяются крыло видные кости (рис. 144). У ящериц основная
Рис. 145. Схематическое изображение черепа
крокодила (по Гудричу)
а — articulare; ап — angulare; d — dentale; fr — fron-
tale; if — нижняя височная яма; j — jugale;
I — lacrymale; mx—maxillare; na — nasale; о — глав-
нипа; pa — parietale; pm — praemaxillare; po — post,
orb it ale; prf — praefrontale; q — quadratum; qj —
quadrate jugale; sa — supraangulare; sf — верхняя
височная яма; sq —squamosum.
an
Рис. 146. Череп ящерицы (Amei-
va) сбоку (из Гертвига).
d — dentale; ept — epipterygoideum;
sf — supratemporale. Остальные odo-
вначения, как на рис. 148.
клиновидная кость часто срастается с основной затылочной и с передне-
ушными костями. Парных костей глазничной области обычно не имеется.
Только у крокодилов и динозавров имеются латеросфеноиды и орбито-
сфеноиды да у ящериц имеются небольшие окостенения межглазничной
перегородки, сравниваемые с орбитосфеноидами. В обонятельной области
у рептилий окостенений нет.
У рептилий имеется большое число покровных костей черепа. Крышу
черепа образуют теменные, лобные, носовые, переднелобные, слезные,
заднелобные, заднеглазничные, чешуйчатые, иногда надвисдчные, квадрат-
носкуловые, скуловые, челюстные и предчелюстные кости (рис. 143, 145).
Теменные кости обычно (исключая черепах) срастаются в одну непарную
кость, в середине которой у ящериц и у гаттерии имеется отверстие для
теменного глаза. Надвисочные кости (supratemporalia) встречаются только
у ящериц. Под основанием черепа у ящериц закладывается еще вполне
типичный парасфеноид, который после срастается с основной клиновид-
ной костью. У взрослого животного остатки передней части парасфеноида
имеют вид направленного вперед отростка («клюва», rostrum) базисфе-
ноида (рис. 144). Под обонятельной областью лежит парный сошник,
несущий у гаттерии еще по одному зубу. У черепах он срастается в одну
непарную кость, а у крокодилов покрывается снизу развивающимся
вторичным твердым нёбом.
Нёбноквадратный хрящ окостеневает в задней своей части, образуя
os quadratum. Далее впереди имеется еще одно окостенение, развиваю-
щееся в восходящем отростке нёбноквадратного хряща у ящериц и у
гаттерии. Это столбчатая кость (epipterygoideum), соединяющая у взрос-
лого животного крыловидную кость с теменной (рис. 146). У гаттерии
эпиптеригоиды близко прилегают к стенке черепа (рис. 143), а у ископае-
мых териодонтов уже входят в ее состав. Квадратная кость или подвижно
соединяется с боковыми отростками черепа (proc, paroticus) и с чешуйча-
155
той костью (стрептостилия) или вклинивается между другими костями
и оказывается неподвижной (монимостйлия). Первое характерно для
ящериц и змей, между тем как неподвижное соединение наблюдается у
гаттерии, черепах и крокодилов. Весьма подвижное соединение у репти-
лий, несомненно, выработалось вторично. У змей эта подвижность очень
велика, и сама чешуйчатая кость также подвижно соединена с черепом
(рис. 147). Это специальное приспособление к возможно более широкому
раскрыванию рта. Из покровных костей челюстной дуги на дне черепа
лежат крыловидные и нёбные кости (рис. 144). Эти кости сравнительно
слабо еще связаны с дном черепа у ящериц, а у змей образуют подвижную
систему костей, снабженных зубами. Задним концом крыловидные кости
достигают квадратных костей, а впереди соединяются с нёбными. Внутрен-
ним краем они соединяются с базиптеригоидными отростками основной
клиновидной кости (исключая змей). Кнаружи от крыловидных костей
ап
Рис. 147. Схематическое изображение черепа змеи (по Гудричу),
п — ноздря; ро/ — posfrontale. Остальные обозначения, как на рис. 145.
у большинства рептилий (исключая черепах) лежат еще покровные попе-
речные кости (transverse), которые соединяют крыловидные кости с
задним концом челюстных (рис. 151, В). Эти кости имелись и у некоторых
стегоцефалов и сравниваются с наружными крыловидными костями (ecto-
pterygoidea) рыб. У змей челюстные кости становятся очень подвижными,
и поперечные кости служат рычагом, передающим движение квадратной
и крыловидной кости на челюстную. У крокодилов и черепах, наоборот,
вся эта система костей очень прочно связана с другими костями, и впервые
вполне входит в состав дна черепа (рис. 151).
Хотя стрептостилия и представляет собой результат вторичного
изменения и именно приспособления для схватывания живой, притом
весьма подвижной, добычи (особенно насекомых), все же и монимостилия,
несомненно, также развилась вторично. Первоначально верхний конец
квадратной кости соединялся довольно рыхлой связью с боковыми отрост-
ками (proc, parotici) черепа посредством вставочного элемента (inter-
calare), представляющего собой остаток части прежнего подвеска (hyornan-
dibulare). Кроме того, птеригоиды подвижно соединялись е базиптери-
гоидными отростками основания черепа. Сам череп мог несколько изги-
баться в области теменных костей, и таким образом при открывании рта
происходило некоторое выдвигание вперед крыловидных костей и при-
поднимание верхней челюсти вместе с передней частью черепа (первично-
кинетический тип черепа).
В нижнем отделе челюстной дуги имеется всегда хорошо развитая
сочленовная кость. Хорошо развиты и все покровные кости нижней
челюсти — зубная, пластинчатая, угловая, надутловая и еще особая
венечная кость (согопаге), образующая венечный отросток нижней челюсти
(processus coronoideus); имеется и кожная сочленовная кость (goniale),
которая у ящериц срастается с сочленовной (рис. 146, 148).
156
Верхний отдел подъязычной дуги Представлен xopotao развитой слухо-
вой косточкой, имеющей форму длинного столбика (columella) (рис. 144).
В этом столбике различим отдельно окостеневающий внутренний отдел,
замыкающий овальное окно слуховой
капсулы — стремя (stapes), — и ж. наруж-
ный отдел — extracolumella, который свя-
зан j е барабанной перепонкой, t соеди-
няется иногда с квадратной костью и
Рис. 149. Подъязычный аппарат
ящерицы (по Гаупу-Циглеру).
Ьг — 1—2 — жаберные дужкн; egl —
entoglossum; hy — гиоидные рожки.
Рис. 148. Развитие костей нижней челюсти
рептилии (Sceleporus) (из Кингсли).
аг — articulare; d — dentale; g — goniale; mk — мек-
келев хрящ, Остальные обовначения. как на
рнс. 143.
с гиоидом (гаттерия, гекконы) и сохраняет иногда также дорзальную
связь со слуховой капсулой (proc, dorsalis, intercalare) (рис. 135).
Остальная часть висцерального скелета представлена подъязычным
аппаратом, который состоит из
непарного тела (соответствует
сросшимся копулам) и трех пар
рожков, соответствующих ос-
таткам гиоида и двух жабер-
ных дуг (рис. 149). У крокоди-
лов имеется только одна пара
рожков.
На черепе рептилий можно
проследить развитие образова-
ний, характерных и для черепа
вышестоящих позвоночных —
вторичного твердого нёба и ску-
ловых дуг.
Крыша ротовой по-
лости образована у низших
наземных позвоночных дном
самого черепа (рис. 150, Л)
е его парасфеноидом и сошни-
ками и нёбноквадратным хря-
щом с его покровными ко-
стями — крыловидными и нёб-
ными. Наружный край этого
первичного твердого
нёба образован предчелюст-
ными и челюстными костями.
В
Рис. 150. Образование вторичного твердого
нёба у рептилий. Схематический парамедиаль-
ный разрез через череп.
ch. рг —первичные и ch. sec — вторичные хоаны;
d. nas. ph — носоглоточный ход; pal. pr — пер-
вичное и pal. sec — вторичное твердое нвбо;
polf — обонятельный отдел; р. resp — дыхатель-
ный отдел.
Внутренние ноздри, первичные хоаны, открываются непосредственно у
этого наружного края между челюстной костью и сошником (рис. 144).
У ящериц нёбо устроено еще по этому типу, но на челюстных костях у
них можно уже заметить небольшие горизонтальные выросты, идущие
вдоль внутреннего края этих костей. Такие выросты носят название
нёбных отростков. У черепах они разрастаются и образуют костный
помост, отделяющий впереди верхнюю часть ротовой полости, где поме-
щаются первичные хоаны, от остальной части. Этот помост получил
157
Название вторичного твердого нёба (рис. 150, В). Сошникй
срастаются в одну непарную кость, которая делит отгороженное прост-
ранство ротовой полости на правое и левое. Отверстия, лежащие по бокам
сошника и ведущие в пространство, отгороженное вторичным твердым
нёбом, являются вторичными хоанами, а само отгороженное пространство
представляет правый и левый носоглоточные ходы (ductus nasopharyngei)
Рис. 151. А — череп черепахи (Trionyx), В — крокодила (Alligator)
снизу (из Кингсли и Шиплей и Мэк Брайда).
eh — хоаны, Обозначение костей прежнее.
У черепах и сам сошник своей нижпей частью участвует в образовании
твердого нёба (рис. 151), и это участие сошника наблюдается иногда еще
у крокодилов. Однако в большинстве случаев у последних сошник вы-
тесняется с поверхности и совершенно покрыт нёбными отростками других
костей, являясь лишь перегородкой между правым и левым носоглоточ-
ными ходами.
Рис. 152. А — череп стегоцефала (по Иекелю) и В — ископаемой рептилии
(Seymouria) (по Броили). Обозначения см. рис. 131.
Дальнейшим разрастанием твердого нёба вторичные хоаны отодви-
гаются все далее назад. Этот процесс постепенного отодвигания хоан
может быть прекрасно прослежен палеонтологически в ряду ископаемых
паразухий и крокодилов. Значение такого отодвигания, очевидно, кро-
ется в потребности обеспеченного дыхания при более энергичном обмене
158
йеЩеств у высших наземных позвоночных. Легко понять, что у форм, обла-
дающих хоанами в передней части крыши ротовой полости, дыхание пре-
рывается на все время, пока принимаемая пища находится во рту; между
тем как у форм с позади лежащими вторичными хоанами дыхание пре-
рывается лишь на короткое время самого акта глотания. У современных
крокодилов вторичное твердое нёбо очень сильно развито и простирается
вдоль почти всего основания черепа. Оно образовано сошедшимися по
средней линии горизонтальными нёбными отростками межчелюстных,
челюстных, нёбных и даже крыловидных костей, в области которых
под основной клиновидной костью открываются вторичные хоаны
(рис. 151).
Наружная крыша черепа имела вид совершенно непре-
рывного панцыря не только у стегоцефалов, но и у древнейших рептилий.
Эта крыша была иногда еще покрыта
характерной для кожных костей скульп-
турой и имела в некоторых случаях даже
буквально тот же состав, что и у стегоце-
фалов, так что череп их сверху не отличим
(рис. 152). У позднейших рептилий, как и
у амфибий, этот скелет облегчается путем
частичной его редукции. Богатство из-
ученных ископаемых форм рептилий дает
возможность шаг за шагом проследить ход
этой редукции. Оказывается, что редукция
наружной крыши черепа шла двумя не-
зависимыми путями, ведущими к черепам
современных высших наземных позвоноч-
ных (у амфибий эта редукция шла также
независимо) (рис. 153). У одних рептилий
частичная редукция наружной крыши
привела сразу к образованию двух отвер-
стий в височной ее области. У древнейших
форм эти отверстия невелики (рис. 153,
С), но у позднейших разрастаются, так
что остатки наружной крыши сохраняются
лишь в виде узких мостиков, покрываю-
щих височную область черепа (рис. 143).
Таким образом, стегальный че-
реп со сплошной крышей превращается
в витальный череп с височными
дугами. В височную область черепа теперь
можно проникнуть не только сзади (задняя
височная яма) и снизу (нижняя височная яма), как и в стегальном чере-
пе, но и сбоку и сверху через образовавшиеся отверстия, носящие назва-
ния боковой и верхней височных ям. Горизонтальный костный мостик,
лежащий между верхней и боковой височными ямами, называется верх-
ней височной дугой; мостик, лежащий под боковой височной ямой и
представляющий собой остаток нижнего края крыши черепа, носит на-
звание нижней височной дуги.
Передний вертикальный мостик, ограничивающий глазницу сзади,
называется глазнично-височной дутой, а задний мостик, представляющий
остаток заднего края наружной крыши черепа позади верхней височной
ямы, носит название задней височной дуги. Что касается состава височных
дут (рис. 143), то верхняя дута образована заднеглазничной костью и
чешуйчатой, соответствующие части которых при образовании височной
дуги получают вид горизонтальных отростков этих костей. Нижняя ви-
сочная дуга составляется за счет скуловой и квадратноскуловой костей.
Рис. J53. Стегальный (Л), моно-
зигальный [В) и дизигальный (С)
черепа ископаемых рептилий.
А — Labidosaurus; В — Cynogna-
thus; С — Palaeohatterla.
159
йта дуга связывает квадратную кость з челюстной. Состав височных дуг
может быть пояснен следующей диаграммой:
(frontale) (parle! ale)
верхняя височная яма
postorbitale------------------squamosum
глазница боковая височная яма
(maxillare)---------jugale---------------quadratojugale-------(quadratum)
aent
Рис. 154. Черен морской черепахи (Che-
lone) сбоку (по |Видерсгейму). Обозначе-
ния см. рис. 143
По этому типу шло развитие височных дуг у большинства рептилий
(группа Diapsida). Из современных рептилий обе дуги, и верхняя и ниж-
няя, имеются у гаттерии и у крокодилов (дизигальные черепа). У других
рептилий та или другая дуга редуцируется, и таким образом развивается
череп с одной височной дугой—
верхней или нижней (монозигаль-
ные черепа). У птиц, происходя-
щих от рептилий с двумя височ-
ными дугами, исчезла верхняя
височная дуга и имеется только
одна нижняя (рис. 157). Иначе шла
редукция крыши черепа у чешуй-
чатых (группа Parapsida), у ко-
торых образование одной только
верхней височной ямы сопрово-
ждалось образованием вырезки,
развивавшейся от нижнего края
крыши черепа. У современных
ящериц имеется только верхняя
височная дуга (рис. 146), но и
она нередко редуцируется и является неполной. У змей и некоторых
ящериц исчезли обе дуги (рис. 147), и височная область черепа является
совершенно обнаженной.
Рис. 155. Череп черепахи (Trionyx) сбоку (по Раблю). Обозначения
см. рис. 143, 144.
Наконец, еще в одной группе рептилий частичная редукция наружной
крыши черепа привела к образованию вырезки в задней верхней части
височной области (рис; 153, В). Разрастание этого отверстия, называемого
также верхней височной ямой, приводит к образованию единственной
височной (скуловой) дуги, покрывающей височную область черепа (рис. 154,
155). Такие черепа с первично единственной височной дугой характерны
для некоторых ископаемых рептилий и для современных черепах (группа
160
Synapsida), а также и для млекопитающих. По своему составу эта ску-
ловая дуга ясно отличается и от верхней, и от нижней височных дуг,
так как она составляется из элементов, характерных для обеих. Задняя
ее часть состоит из отростка чешуйчатой кости и из квадратноскуловой
кости, а передняя часть — из скуловой кости (рис. 155). Височная яма,
разрастаясь вперед, обычно соединяется с глазницей. В некоторых слу-
чаях у черепах и у млекопитающих и эта дуга редуцируется и является
неполной, а иногда и вовсе исчезает, и тогда височная область черепа
совершенно обнажается.
У некоторых ископаемых рептилий в крыше
черепа развивалось еще одно окно впереди глаз-
ниц — предглазничная яма (динозавры) (рис. 401).
Ф* so
Рис. 156. Череп мо-
лодого аиста снизу
(ив Зеленки). Обовна-
чения см рис, 143,144.
ppm — нёбный отрос-
ток челюстной кости,
г—rostrum;
Рис. 157. Череп цыпленка сбоку (по Боасу).
as — laterosphenoideum; е — ethmoideum; I — lacrymale;
остальные обозначения прежние.
Черен птиц
Череп птиц отличается от черепа рептилий главным образом своей
конфигурацией — большими орбитами с тонкой межглазничной перего-
родкой, сильно вздутой черепной коробкой позади орбит и вытянутой в
виде клюва передней частью черепа (рис. 157). С таким черепом до извест-
ной степени сходны черепа некоторых ископаемых рептилий, именно псев-
дозухий (рис. 408) и динозавров, которые и по другим признакам близки
к птицам, а также и черепа летавших птерозавров. Если сходство первых
по своей глубине является выражением действительного их родства с
птицами, то во вторых мы имеем интересный пример сходства в резуль-
тате приспособления к одинаковым условиям существования, т. е. пример
схождения признаков (конвергенции).
Состав черепа птиц почти в точности тот же самый, что и у рептилий,
хотя это и трудно установить на взрослом черепе вследствие того, что у
птиц большинство костей срастается между собой, так что границ отдель-
ных костей обнаружить нельзя. Это срастание костей обеспечивает проч-
ность черепа при относительной тонкости и легкости отдельных компо-
нентов.
В затылочной области черепа развиваются все четыре затылочные
кости (рис. 156). Как и у рептилий, имеется один непарный затылочный
бугор. Последний, как и затылочная дыра, лежит на нижней стороне
черепа, так как ось последнего у птиц стоит перпендикулярно к оси
позвоночника. В связи со значительным разрастанием крыши черепа (боль-
767
Шой мозг) слуховые капсулы сдвинуты вниз. В слуховой капсуле разви-
ваются все три ушные кости, которые рано срастаются между собой и
с соседними костями (верхнеушная с верхней и заднеушная с боковой
затылочной костью) в одну сплошную массу. В глазничновисочной об-
ласти черепа у основания развиваются основная и передняя клиновид-
ные кости и в глазницах парные боковые клиновидные (рис. 157) и глаз-
нично-клиновидные кости (как у крокодилов). В передней части межглаз-
ничной перегородки лежит обонятельная кость (ethmoideum), которая
развивается из пескольких цен-
тров окостенения. Под дном че-
репа и у птиц развивается руди-
ментарный парасфеноид, который
затем срастается с базисфеноидом,
причем передняя часть его обра-
зует выступающий вперед клюв
Рис. 159. Подъязычный аппарат
голубя.
Ь. ы 1, Ъ, Ъг 2 — basibranchialia 1 и 2;
bhy — basihyale; chy — гиоидные рожки;
cbr., ер. br — части первой жаСерной дуги.
Рис. 158. Дно черепа и твердое нёбо страуса
нанду (Rhea americana) как тип палеогната
(по Пикрафту).
bpt — базиптеригоидное сочленение; pbpt—ба-
зиптеригоидный отросток; prsph — парасфе-
ноид; v — сошник.
(rostrum) (рис. 156). Сошник развивается еще как парная кость, но
затем срастается в одну кость, обнаруживающую обычно следы двой-
ственности и у взрослого животного. Крыша черепа состоит из сле-
дующих покровных костей: теменные, лобные, носовые, чешуйчатые,
переднелобные (рис. 157); вдоль нижнего края впереди большая пред-
челюстная кость, сросшаяся в одну непарную кость, образующую все
почти надклювие; позади нее небольшие челюстные и затем очень тонкие
скуловые и квадратноскуловые кости, образующие нижнюю височную
дугу. У птиц нередко также хорошо развито вторичное твердое нёбо,
образованное сошедшимися между собой нёбными отростками предче-
люстных и челюстных костей.
Что касается висцерального скелета, то квадратные кости соединяются
с осевым черепом подвижно, как и у ящериц (стрептостилия); они сочле-
няются с задними концами крыловидных костей, а последние — с концами
нёбных (рис. 156). На месте соединения крыловидных костей с нёбными
они обычно прилежат к клюву базисфеноида, вдоль которого они могут
передвигаться («пеогпаты»). Так как передние концы нёбных костей
связаны с челюстными, то вся эта подвижная система костей может толкать
челюсти снизу вперед, и надклювие при этом приподнимается, так как
162
оно связано с черепом лишь со спинной стороны тонкими и гибкилЫ
частями носовых костей. Однако у низших птиц («палеогнаты») вместо
этой системы имеется соединение внутренних краев крыловидных костей
с базиптеригоидными отростками основной клиновидной кости, как мы
это видели и у рептилий (рис. 158). В нижней челюсти меккелев хрящ
весьма полно окостеневает с образованием двух костей — подбородочной
впереди и сочленовной позади. Сверх того, развивается и весь ряд покров-
ных костей, характерных для рептилий, — зубная, пластинчатая, угловая,
надугловая и венечная (рис. 157). Все кости нижней челюсти срастаются
затем между собой в одно целое (также и в симфизе, правая челюсть с
левой). Слуховая косточка — столбик уха — вполне сходна с таковой
у рептилий. Подъязычный аппарат сильно упрощен и состоит из длинного
тела, в котором можно различить несколько частей, из ничтожных остатков
гиоида в виде небольших выступов на нем и из длинных рожков (рис. 159),
соответствующих первой паре жаберных дуг (как у крокодилов). Все
эти части окостеневают.
Череп млекопитающих
Череп млекопитающих имеет широкое основание (платибазальный
тип); однако это результат вторичного изменения тропибазалыгого черепа
нижестоящих форм, обусловленный разрастанием мозговой коробки.
У эмбрионов млекопитающих наблюдаются иногда еще следы межглаз*
ничной перегородки.
Хрящевой череп млекопитающих эмбрионально представлен
иногда довольно полно. Почти сплошной хрящ наблюдается нередко не
только в стенке слуховой и обонятельной капсул и в основании черепа,
но и на его боках. В крыше черепа, кроме обонятельных капсул, хрящ
наблюдается только позади, в затылочной области, в виде хрящевого
мостика, связывающего слуховые капсулы над затылочной дырой. В
слуховой капсуле имеются оба окна — овальное окно, в которое упирается
слуховая косточка, и круглое окно, затянутое только перепонкой. В
обонятельной капсуле уже на хрящевой стадии развиваются более или
менее закрученные выросты боковых стенок, так называемые обонятель-
ные раковины. В костном черепе млекопитающих наблюдается
весьма характерный процесс сокращения числа костей путем их срастания
в некоторые типичные комплексы. В некоторых, случаях отдельные кости
настолько утратили свою индивидуальность, что сохранились лишь в
виде отдельных центров окостенения сложной кости, но так как несколько
центров окостенения встречается нередко и в простых костях, то устано-
вить морфологический состав таких сложных костей иногда оказывается
делом нелегким (например околоушная кость).
В затылочной области черепа боковые затылочные кости образуют
обычно совместно с основной затылочной костью парные затылочные
бугры. Последние образовались из простого бугра рептилий путем редук-
ции его средней части, как это видно у некоторых ископаемых форм и у
примитивнейших млекопитающих. Уже у ископаемых териодонтов разви-
вался двураздельный затылочный бугор, а у современной ехидны оба
затылочных бугра связаны между собой хрящевой полосой, и суставная
щель в них одна общая. Затылочная дыра лежит у примитивнейших форм
позади черепа в плоскости, почти перпендикулярной к основанию, а у
форм с сильно развитым мозгом перемещается вниз, ложась у человека
почти в одной плоскости с основанием черепа. Затылочная дыра ограни-
чена основной и боковыми затылочными костями; верхняя затылочная
кость обычно лежит выше. Почти всегда все четыре затылочные кости
срастаются в одну затылочную кость (рис. 160). В слуховой
капсуле развивается несколько окостенений, из которых главное сравни-
вают, с предушной (prooticum), а заднее — с заднеушной костью (opistho-
163
Рис. 160. Череп собаки сверху, снизу, сбоку и сагиттальный распил (по Видерсгейму).
В. осс — основная затылочная кость; Cho — хоаны; С. осс — затылочные бугры; Eth и
Eth' — решетчатая кость: For. т— затылочная дыра; Jg —скуловая кость; Jm —- межче-
люстная кость; L — слезная кость; М — челюстная кость; Mated — наружный слуховой
проход; N—носовая кость; Р— теменная кость; Ра! — нёбная кость; Pet—каменистая
кость; Pjt — скуловой отросток; Pt —крыловидная кость; Sph — алисфеноид; Sph' — бази-
сфеноид; Sph" — пресфеноид; Sq-—чешуйчатая кость; Sq., осс — верхняя затылочная кость;
Т — барабанная кость; Vo — сошник.
164
ticum s. mastoideum). Эти окостенения очень рано срастаются в одну
каменистую кость (os petrosum) и не образуют характерного
для рептилий бокового отростка. Заднее, наружное окостенение стенки
капсулы образует сосцевидную часть каменистой кости, которая у человека
выступает в виде сосцевидного отростка (processus mastoideus). С этой
сложной околоушной костью вступают в более или менее тесную связь
еще две покровные кости. Одна новая, барабанная кость (tympa-
nicum) (рис. 160), ограничивающая снаружи полость среднего уха, оче-
видно, гомологичная угловой кости нижней челюсти рептилий, и другая —
большая чешуйчатая кость (squamosum). У человека все эти кости сраста-
ются в одну височную кость (os temporale). В глазнично-височной
области развиваются две непарные кости у основания черепа — основная
клиновидная и передняя клиновидная. Закладывается и рудиментарный
парасфеноид, который у эмбриона дает иногда еще выступающий вперед
клюв (rostrum). К основной клиновидной кости по бокам прирастают
эпиптеригоиды, получающие здесь название височных крыльев
(alae temporales s. alisphenoidea) (уже у некоторых рептилий эпиптери-
гойды начинают входить в состав стенки черепа). К передней клиновидной
кости по бокам прирастают глазнично-клиновидные кости, образующие
глазничные крылья клиновидной кости (alae orbitales s. orbi-
tosphenoidea) (рис. 160).
У высших обезьян и у человека височные крылья превышают своими
размерами глазничные и получают название больших крыльев (alae
magnae). Височные крылья, образующие, следовательно, одно целое с
базисфеноидом, снабжены отростками, с которыми связаны крыловидные
кости. У высших обезьян и у человека последние прирастают к клино-
видной кости и образуют внутреннюю пластинку нисходящих крыловидных
ее отростков. У некоторых млекопитающих и у человека основная клино-
видная кость с ее височными крыльями прирастает к передней клино-
видной кости с ее глазничными крыльями, образуя одну клиновидную
кость. В основании глазничных крыльев имеется отверстие для выхода
зрительного нерва. Между обоими крыльями через особое отверстие,
имеющее у человека форму щели, проходят нервы глазных мышц и глаз-
ничная ветвь тройничного нерва. Остальная часть последнего проходит
через отверстия в височных крыльях или позади них.
Впереди клиновидных костей в обонятельной области черепа из
нескольких окостенений развивается решетчатая кость (os
ethmoideum). Отдельно окостеневает средняя часть, образующая носовую
перегородку (lamina- perpendicularis), и боковые части, которые скоро
с нею срастаются. Боковые части обращены к черепной полости решетча-
тыми пластинками (lamina cribrosa), которые продырявлены у утконоса
одним отверстием каждая, а у других млекопитающих — многочислен-
ными отверстиями для прохождения обонятельного нерва. Боковые
части решетчатой кости имеют ячеистое строение («лабиринты» с воздуш-
ными полостями), и от них внутрь носовой полости свешиваются пластин-
чатые выросты, или обонятельные раковины (ethmoturbi-
nalia), имеющие весьма различное и иногда очень сложное строение.
Верхняя из этих раковин окостеневает самостоятельно, прилежит к
носовой кОКИ и иногда с нею срастается (nasoturbinale). Впереди и ниже
решетчатой ftocTii располагается еще одна большая и самостоятельно
окостеневающая нижняя раковина (maxillotmjbinale), которая иногда
прирастает к челюстной кости. Под носовой перегородкой лежит непарный
сошник, простирающийся позади до пресфеноида.
Крыша черепа млекопитающих образована теменными и
лобными костями. Теменные кости нередко срастаются между собой
(у клоачных, многих сумчатых и копытных). Между ними и верхней
затылочной костью наблюдается еще особая покровная м е ж т е м е н-
165
пая кость (interparietale), которая развивается как парная кость
или даже как две пары костей. Иногда она остается самостоятельной,
но обыкновенно срастается с соседними костями — с теменными (грызуны
и жвачные) или, гораздо чаще, с верхней затылочной костью (хищные,
обезьяны и человек). Межтеменные кости гомологичны заднетеменным
(табличным) костям котилозавров и стегоцефалов. Лобные кости иногда
также срастаются между собой (насекомоядные, рукокрылые, обезьяны
и человек). На обонятельной области черепа лежат носовые кости. У пе-
реднего края глазницы помещается небольшая слезная кость
(lacrymale), продырявленная отверстием для прохождения слезного ка-
нала. На слуховой области черепа лежит чешуйчатая кость, которая
иногда срастается с окостенениями слуховой капсулы в одну височную
кость, образуя ее чешую (squama). С прогрессивным развитием мозговой
полости и оттеснением слуховой капсулы вниз чешуйчатая кость прини-
мает участие в образовании стенок мозговой полости. Чешуйчатая кость
обладает суставной ямкой для причленепия нижней челюсти. Непосред-
ственное сочленение последней с чешуйчатой костью черепа (а не с квадрат-
ной костью) представляет собой наиболее характерную особенность черепа
млекопитающих. Чешуйчатая кость несет направленный вперед скуловой
отросток, который вместе со скуловой костью (jugale s. zygomaticum) и
одноименным отростком челюстной кости образует единственную у мле-
копитающих скуловую дугу, прикрывающую височную область черепа.
С разрастанием мозговой полости связано исчезновение боковых ушных
отростков (proc, parotici). При этом оттесняется назад и исчезает также
задняя височная яма (следы которой сохранились у однопроходных в
виде височного канала в заднем корне скуловой дуги). Другая покровная
кость слуховой области черепа, барабанная кость (tympanicum),
представляет особенность черепа млекопитающих. У низших форм она
имеет вид неполного кольца, на которое натянута барабанная перепонка
(клоачные, сумчатые и многие насекомоядные). У прочих млекопитающих
опа заметно крупнее и образует наружную стенку барабашюй полости,
нередко вздутую в виде пузыря (грызуны, хищные и др., рис. 160); раз-
растаясь, она охватывает в виде трубки и наружный слуховой проход.
Этакость иногда остается вполне самостоятельной(низшие млекопитающие)
чаще более или менее полно прирастает к каменистой кости. По своему
происхождению барабанная кость представляет результат преобразования
угловой кости (angulare) нижестоящих позвоночных (рис. 162).
Височная область чарепа, ^как сказано, прикрыта единственной у
млекопитающих скуловой дугой, составленной из скуловой кости
и скуловых отростков чешуйчатой и челюстной кости. Обычно у млеко-
питающих глазница не отграничена от височной ямы, но иногда (копытные,
обезьяны) отросток скуловой кости разрастается позади глазниц и сходится
с подобным заднеглазничным отростком лобных костей, дополняя, таким
образом, костное кольцо вокруг орбиты. У обезьян этот процесс идет даль-
ше, и глазница вполне отгораживается от височной ямы костной стенкой.
Вторичные верхние челюсти образуют переднюю
нижптою часть черепа. Предчелюстные или межчелюстные кости млеко-
питающих отличаются отсутствием переднего отростка, ограничивающего
ноздри спереди и с внутренней стороны. Редукция этого отростка, имеюще-
гося у всех нижестоящих позвоночных, сделала у млекопитающих возмож-
ным развитие наружного носа. Межчелюстные кости срастаются у обезьян
и у человека в одно целое с челюстными костями. Эти кости закладываются,
однако, всегда в виде самостоятельных костей, положение которых обозна-
чается у взрослого резцами, сидящими всегда на межчелюстных костях.
Челюстные кости образуют остальную, ббльшую часть верхней челюсти и,
между прочим, участвуют также и в образовании наружной стенки носовой
полости.
166
Твердое нёбо млекопитающих образовано за счет нёбных
отростков межчелюстных, челюстных и нёбных костей, которые сходятся
на средней линии и образуют здесь шов. Нёбные кости, как и крыловидные,
стоят вертикально и ограничивают хоаны с боков. Крыловидные кости
у большинства млекопитающих являются вполне самостоятельными
костями, но у обезьян и у человека срастаются с клиновидной костью,
образуя с каждой стороны внутреннюю пластинку нисходящего, крыло-
видного ее отростка.
В нижней челюсти млекопитающих (mandibula), развива-
ющейся как покровная кость на меккелевом хряще, не удалось
до сих пор обнаружить сложного состава, и большинство исследователей
принимает, что она составляется из одной только зубной кости. Задняя
ее часть изогнута вверх и образует нередко почти прямой угол с передней
частью, несущей зубы. На восходящем заднем колене нижней челюсти
помещается сочленовный отросток, снабженный головкой для сочленения
с чешуйчатой костью. Впереди этого отростка находится венечный отросток
для прикрепления височной мышцы, а снизу обращенный назад угловой
отросток (proc, angularis).
Интересно отметить зависимость между формой челюстей, положением
и формой челюстного сустава и способом питания млекопитающих. У
насекомоядных и хищных усиление челюстей сопровождалось лишь
ограниченным развитием жевательных движений. Челюсти остаются в
основном хватательным органом, снабженным у млекопитающих режу-
щими зубами. Соответственно суставная поверхность имеет глубоко
цилиндрическую форму и располагается на одном уровне с рядом зубов.
У всеядных и растительноядных форм жевательная способность усили-
вается вследствие перемещения челюстного сустава вверх. Развивается
задний, «восходящий» отдел нижней челюсти, благодаря которому движе-
ния последней приобретают скользящий (вперед при закрывании рта)
характер, с сильным дробящим действием в области коренных зубов.
Суставная поверхность приобретает более плоскую форму, допускающую
более свободные движения во всех направлениях, и зубы становятся
тупобугорчатыми или складчатыми с широкой плоской коронкой.
У млекопитающих, как сказано, имеется совершенно иное сочлене-
ние нижней челюсти с черепом, чем у других позвоночных. У млекопи-
тающих квадратная кость не служит подвеском для нижней челюсти,
и в составе последней нет костей, характерных для ее задней половины.
Изучение эмбрионального развития дало, однако, возможность установить
существование соответствующих частей челюстной дуги и у млекопи-
тающих. У последних, так же как и у нижестоящих позвоночных,
развивается и пёбноквадратный хрящ и меккелев хрящ
(рис. 161). Первый значительно редуцирован, но он все же окосте-
невает. Передняя его часть, соответствующая эпиптеригоиду, входит
в состав стенки черепа и вследствие своей связи с клиновидной
костью получила название алисфеноида; задняя часть,, собственно квад-
ратная кость, занимает в сущности такое же положеште, как у репти-
лий. Но развивающаяся таким образом кость не служит подвеском,
как у последних, а дает слуховую косточку, так называемую нако-
вальню (incus), которая, следовательно, гомологична квадратной
кости (рис. 161). Меккелев хрящ, так же как у рептилий, окосте-
невает в своей утолщенной задней части, но развивающаяся таким обра-
зом кость не остается в составе нижней челюсти, как у других позво-
ночных, а дает другую слуховую косточку — молоточек (malleus).
На этой косточке найдена и небольшая покровная кость, срастающаяся
с ней и соответствующая покровной сочленовной кости (goniale) нижней
челюсти низших наземных позвоночных. Она образует «длинный» отросток
МОДОточка (proc, folii в. longus mallei) (рис. 135). Нужно прибавить, что
767
и отношение нервов (chorda tympani VII) к слуховым косточкам млеко-
питающих вполне сходно с их отношением к слуховой косточке и нижпей
челюсти рептилий, так что гомология этих частей теперь не подлежит
сомнению. Верхний отдел подъязычной дуги, образу-
ющий у рыб подвесок, а у амфибий, рептилий и птиц слуховую косточку —
столбик, и здесь входит в состав слуховых косточек. Как и столбик, он
запирает овальное окно слуховой капсулы. Это — стремя (stapes),
продырявленное, как и столбик некоторых других позвоночных (гимно-
фионы и некоторые рептилии), отверстием, через которое у низших мле-
Рис. 161. Череп зародыша броненосца Tatusia (из Вндсрсгсйма).
a. ty—annulus tyrapanicus; аи— слуховап капсула; Ъ. hy — basihyale; с. hy — eeratohy-
ale; cr— перстневидный хрящ; d — dentale e. hy — epihyale; en — ноздря; eo — exoecipi-
tale; f—лобная кость; h. hy — hypohyale; i — jugale; in — наковальня; Ic — слезная
косточка; mh— меккелес хрящ; ml—молоточек; тх— maxilla; n — носовая кость; ос.
С-—затылочный бугор; р — теменная кость; -ра— нёбная кость; рх— praemaxilla; so —
supraoccipitale; sq—squamosum; st — стремя; st. m — musculus stapedialis; th — щито-
видный хрящ; tr — трахея. Римские цифры обозначают отверстия для выходных нервов.
копитающих (насекомоядные) проходит лицевая артерия (arteria facialis
s. stapedia) (рис. 135). У других позвоночных, в том числе и у человека,
эта артерия наблюдается по крайней мере эмбрионально. Таким образом,
у млекопитающих имеется цепь из трех слуховых косточек, сочленяющихся
друг с другом: стремя запирает овальное окно, молоточек связан с бара-
банной перепонкой, а наковальня служит связующим звеном между пими.
Связь этих элементов между собой не есть нечто повое, приобретенное
млекопитающими. Мы видели, что у амфибий, а иногда и у рептилий,
наблюдается связь столбика с квадратной костью, и мы толковали ее
как остаток прежней связи подвеска (hyomandibulare) с челюстной дугой
(palatoquadratum). С другой стороны, квадратная кость регулярно сочле-
няется с сочленовной костью. Следовательно, новым является не взаимная
связь этих костей, а другое — обособление заднего конца нижней челюсти
и развитие нового челюстного сустава.
Нижний отдел подъязычной дуги представлен и у
млекопитающих передними рожками (cornua anteriora) подъязычного
аппарата. Эти рожки имеют часто несколько отдельных окостенений,
соединенных связками, и прикрёплены к слуховой капсуле (рис. 161).
Верхнее окостенение у человека и у оранга прирастает к каменистой
кости п образует ее шиловидный отросток (processus styloideus). Тело
подъязычной кости представляет собой остаток копулы, а задние рожки —
преобразованную первую жаберную дугу. Вторая и третья жаберные
дуги соединяются для образования щитовидного хряща гортани, впервые
168
появляющегося у млекопитающих. Происхождение этого хряща из двух
жаберных дуг можно проследить эмбриологически и даже сравнительно-
анатомически на низших млекопитающих, у которых срастание обеих
дуг еще неполное (рис. 313). Четвертая жаберная дуга вместе с пятой
образуют, невидимому, остальные гортанные хрящи, а быть может являют-
ся также источником для образования трахеальных хрящей.
Таким образом, череп млекопитающих довольно легко выводится из
черепа ниже стоящих форм. Состав черепа в общем тот же самый, но
довольно заметно изменилась общая его конфигурация. Это
изменение находится в тесной связи с развитием головного мозга, а сле-
довательно и с увеличением объема мозговой полости черепа. Увеличение
мозговой полости приводит к сближению передней ее стенки с обонятель-
ными капсулами. При этом межглазничная перегородка редуцируется,
а глазницы отодвигаются на бока черепа. Эмбрионально иногда еще
наблюдается совершенно типичная межглазничная перегородка (например
у обезьян). Остатки межглазничной перегородки сохраняются у млеко-
питающих в их передней клиновидной кости и в задней части носовой
перегородки. Таким образом, череп млекопитающих является преобразо-
ванным тропибазальным черепом, который лишь вторично приобрел
сходство с платибазальными черепами. С тем же прогрессивным разви-
тием мозга связано и изменение в относительном расположении носовой и
мозговой полости. Первоначально они располагаются одна позади другой,
у млекопитающих же разрастающаяся мозговая полость все более надви-
гается на носовую (рис. 282), так что при высшем ее развитии у человека
она совершенно покрывает последнюю сверху. За всеми этими изменениями
проследить нетрудно, так как они развиваются постепенно и не связаны
с полным морфологическим перераспределением существующих частей.
Другое дело — преобразования в области челюстной
дуги; здесь мы имеем, действительно, нечто совершенно новое и притом
трудно выводимое из строения ниже стоящих форм. Если у млекопитающих
имеется иное сочленение нижней челюсти с черепом, чем у других позво-
ночных, то развитие этого нового сочленения нельзя себе представить
иначе, как допустив существование переходной стадии, когда одновременно
функционировали два сочленения челюсти: старое соединение — сочле-
новной кости с квадратной и новое сочленение — зубной кости с чешуй-
чатой. Такое предположение всегда казалось мало вероятным и служило
препятствием к признанию изложенной уже теории Рейхерта о происхож-
дении слуховых косточек, заставляя искать иных путей для объяснения
различий, наблюдающихся между млекопитающими и другими позвоноч-
ными. Теория Рейхерта связана со следующими допущениями. Для того,
чтобы развилось новое сочленение, нужно, чтобы зубная кость и чешуй-
чатая близко сходились в области челюстного сустава; между тем в боль-
шинстве случаев у рептилий их разделяет целый ряд других костей.
Нужно также допустить, что в течение некоторого времени функциони-
ровало двойное сочленение или по крайней мере сочленение двойного
состава (старое — между сочленовной и квадратной костью и новое —
между зубной и чешуйчатой). Затем части челюстей, входившие в состав
первоначального сочленения, должны были совершенно обособиться от
главной их массы, соединенной в новом сочленении. Все зти предпо-
сылки долгое время не находили фактов для своего подтверждения,
и только в последнее время палеонтология дала нам целый ряд если не
прямых, то косвенных доказательств правильности теории. Были найдены
хорошо сохранившиеся черепа рептилий, которые по своему строению,
а также и по начинающейся диференцировке зубной системы очень близки
к черепам млекопитающих (кроме черепа, и в остальном скелете имеется
много признаков, характерных для последних, так, например, весьма
сходны пояса конечностей). У высших представителей этой группы Theri-
169
odontia зубы были диференцированы на резцы, клыки и коренные и
имелся двураздельный затылочный бугор. В черепе Theriodontia имеются
верхняя височная яма и единственная скуловая дуга такого же состава,
как и у млекопитающих. В отличие от других рептилий у них венечный
отросток нижней челюсти имеет огромные размеры и представляет собой
восходящий отросток зубной кости. Дальнейшая эволюция подобных
форм могла итти следующим образом: от основания венечного отростка
зубная кость разрасталась постепенно еще далее назад до сочленовной
области. С другой стороны, и чешуйчатая кость разрасталась вдоль квад-
ратной кости вниз вплоть до области причленения нижпей челюсти.
Таким образом, в состав сочленения нижпей челюсти с черепом вошли,
кроме первичных костей (articulare и quadratum), еще и соответствующие
покровные кости. У некоторых рептилий (черепахи, крокодилы, гаттерия)
снаружи; В — задняя часть ниж-
Рис. 162.
А—нижняя челюсть териодонта Cynognathus вид ___________ . ____ _____ _____
ней челюсти териодонта, вид изнутри; С — задняя часть нижпей челюсти и слуховые кос-
точки позднего эмбриона сумчатого Perameles, вид снаружи; D — задняя часть нижпей
челюсти и слуховые косточки новорожденного опоссума Didelphys, вид изнутри (из Гудри-
ча), ас — хрящ па вторичном суставе; ап —angulare; аг — articulare; d — dentale, д — goni-
ale; i — incus; m—malleus; mb— меккелев хрящ; sa—supraangulare; st —stapes; ty—
tympanicum (angulare).
в состав сочленения уже входят и покровные кости, именно надугло-
вая кость (supraangulare) внизу и квадратноскуловая (quadratojugale)
вверху, и, кроме того, весьма близко к сочленению подходит зубпая
кость внизу и чешуйчатая вверху. Этим даны все условия для возможной
диференцировки единого сочленения на два: внутреннее — между квад-
ратной и сочленовной костью и наружное — между зубной костью и
чешуйчатой Ч
На обособление заднего отдела нижней челюсти имеются также
указания, и во всяком случае в ряду Theriodontia ясно наблюдается ре-
дукция этого отдела (сочленовной и угловой костей), с одной стороны
(рис. 162), и квадратной кости — с другой. Эта редукция показывает
нам совершенно ясно, что у высших териодонтов старое (внутреннее)
1 По Домбровскому, это наружное сочленение развивалось между надугловой
и квадратноскуловой костями, которые у рептилий иногда принимают участие в че-
люстном сочленении, а у млекопитающих, может быть, срослись с зубной и чешуй-
чатой костями.
170
сочленение сочленовной кости с квадратной во всяком случае уже теряло
свое прежнее значение. Кроме того, у териодонтов в челюстном сочленении
безусловно участвовали и покровные кости (рис. 414, 415). У Регшосу-
nodon, по Сушкину, в состав сочленения входили надугловая и квадратно-
скуловая кости. У других териодонтов в сочленении, очевидно, участво-
вали, кроме того, также зубная и чешуйчатая кости (рис. 162). У терио-
донта Ictidosaurus описано уже прямо второе сочленение нижней челюсти
непосредственно с чешуйчатой костью (Брум).
Таким образом, теория происхождения слуховых косточек млеко-
питающих, бывшая долгое время чисто эмбриологической теорией, под-
тверждена и палеонтологическими находками, установившими действи-
тельное существование по крайней мере некоторых переходных стадий.
Что касается черепа так называемых высших млекопитаю-
щих,!. е. обезьян и человека, то эти черепа представляют собой совер-
шенно последовательный ряд изменений, связанных главным образом с
прогрессивным развитием их головного мозга. Увеличивающаяся мозго-
вая полость, с одной стороны, сдвигает затылочную дыру к основанию
черепа, так что она у человека ложится в плоскости, почти параллельной
последнему и, с другой стороны, впереди надвигается на носовую полость,
покрывая ее сверху, так что решетчатая пластинка ложится горизонтально.
Параллельно этому идет изменение положения лицевых частей черепа по
отношению к мозговой коробке. Ось, проходящая через лицевые части у
низших млекопитающих, лежит приблизительно на продолжении оси,
проходящей через основание черепа; у высших форм лицевая ось откло-
няется вниз, образуя очень тупой угол с осью основания черепа; это
отклонение особенно значительно у обезьян, а у человека угол прибли-
жается к прямому. Череп человека отличается от черепа обезьян
значительно большим объемом мозговой полости и более слабым разви-
тием челюстей. Объем мозговой полости у человекообразных обезьян
достигает 400—560 см3, у ископаемого австралопитека (рис. 429) достигал,
быть может, 650 см3, у ископаемого питекантропа был около 900 см3,
а у человека — около 1500 см3, хотя объем нормального черепа иногда
падает до 1000 см3. Таким образом, мы здесь имеем довольно последова-
тельный ряд форм, и для морфолога безразлично, будем ли мы смотреть
на промежуточную форму, на питекантропа, как на человекообразную
обезьяну или как на обезьяноподобного человека. По новейшим данным,
именно находкам остатков очень близкого существа — синантропа, поль-
зовавшегося уже огнем и изготовлявшего простейшие орудия, и на
питекантропа следует смотреть как на очень примитивного человека.
У человека черепная коробка округлая и гладкая и в этом отношении
гораздо более напоминает черепа детенышей и самок человекообразных
обезьян, чем черепа взрослых самцов. Эти последние снабжены довольно
крупными гребнями, в особенности большим сагиттальным гребнем для при-
крепления височной мышцы, что находится в связи со значительной силой
их челюстей. У других обезьян с более слабыми челюстями, особенно у
небольших обезьян, черепная коробка обыкновенно округлая и гладкая.
У человека, несомненно, произошла значительная редукция челюстей, и
последние этапы этой редукции еще можно проследить. И у некоторых
рас человека, особенно на древнейших ископаемых черепах, можно видеть
более крупные и заметно выступающие вперед челюсти (прогнатизм). Редук-
ция и отступание альвеолярной части нижней челюсти, связанное с более
слабым развитием зубов, приводят у большинства современных рас к
образованию выступающего вперед подбородка. Одновременно происхо-
дит и редукция характерного обезьяньего признака — сильно высту-
пающих надбровных дуг, которые в некоторых случаях более заметны
и у современного человека, а на ископаемых человеческих черепах иногда
еще соединяются между собой, образуя один сплошной валик над орбитами
ц над переносицей.
Скелет конечностей
Непарные плавники
Непарные плавники рыб развиваются в виде одной непрерывной
кожной складки, идущей по средней линии тела вдоль спины, огибающей
его задний конец и продолжающейся на брюшной стороне вплоть до
заднепроходного отверстия. Нередко наблюдается участок такой складки
и впереди анального отверстия (преанальный плавник). В таком виде
непарный плавник зародыша рыбы до известной степени сходен с плавни-
ком ланцетника, который на брюшной стороне продолжается и вперед от
анального отверстия. В дальнейшем у большинства рыб определенные
участки этой складки развиваются дальше и дают начало дефинитивным
плавникам взрослой рыбы, между тем как промежуточные части сначала
отстают в своем развитии, а затем редуцируются и исчезают бесследно.
Таким образом обычно развиваются 1—2 спинных плавника, хвостовой
Рис. 163. Развитие мускулатуры в плавниках (схемак
А — область вакладок спинного и брюшного плавника сбоку; В — поперечный разрез;
coel — полость тела; mb — мускульная почка брюшного плавника, делящаяся на спинную
и брюшную; ту—многом; nd — спинной и nv — брюшной плавник; pv — брюшной
отросток миотома.
плавник, состоящий из спинной и брюшной лопасти, и анальный плавник,
лежащий позади анального отверстия. В этих плавниках развивается
своя мускулатура и свой скелет.
Зачатками мускулатуры служат так называемые мускульные почки,
т. е. эпителиальные каплеобразные выросты концов миотомов, обыкно-
венно заключающие в себе небольшую полость (рис. 163). Мускульные
почки закладываются метамерно — по одной на каждом миотоме. Они
вытягиваются в длинные выросты, врастающие в плавниковую складку,
и затем теряют связь с миотомом; полость в них исчезает, границы клеток
теряются, и вслед затем каждый такой вырост преобразовывается в пучок
продольных мускульных волокон. Мускульные почки закладываются не
только в области дефинитивного плавника, но нередко и во всей промежу-
точной области. Такие промежуточные почки частью перемещаются потом
в тот или другой плавник, частью же имеют совсем рудиментарный харак-
тер и рассасываются (абортивные почки). Так как мускульные почки
развиваются одинаково на миотомах правой и левой сторон, то в резуль-
тате в каждом плавнике у его основания имеется парный'ряд мускульных
зачатков (рис. 163, В). Клетки мезенхимы, наполняющие плавниковую
складку и происходящие из ганглионарной пластинки, начинают стя-
гиваться и образуют клеточные сгущения в виде стержней, расположен-
ных как раз между мускульными зачатками каждой пары (рис. 163* В).
Затем в этих стержнях сгущенной мезенхимы происходит охрящевение,
и таким образом развиваются хрящевые лучи непарных плавников (га-
dialia). Только скелет хвостового плавника высших рыб развивается
172
Рис. .164. Спинные плавники акул (Л—Heptan-
chus, С — Acanthias) и ската (В — Raja)
(из Гудрича);
иначе. Его источником является осевая мезенхима и он должен рассматри-
ваться как производное от позвоночника.
Лучи непарных плавников развиваются, таким образом, метамерно
(по одному на каждую пару мускульных почек, а следовательно и на
каждый миотом) и совершенно независимо от осевого скелета. Если к
этому прибавить, что каждый мускульный пучок иннервируется веточкой
нерва того метамера, к которому он относится по происхождению, то
будет ясно, что такой плавник имеет метамерное строение. У взрослого
животного метамерное строение плавника, однако, маскируется или даже
теряется вследствие того, что его элементы теснее сдвигаются и при этом
хрящевые лучи нередко сливаются, особенно своими основаниями, в более
или менее крупные плас-
тинки. Такое явление на-
зывается концентрацией
плавника. Особенно зна-
чительная концентрация
плавников наблюдается у
рыб с хрящевым скелетом
(селахий и цельноголо-
вых), и это явление, не-
сомненно, находится в свя-
зи с сравнительно малой
прочностью скелетного ма-
териала (рис. 164). С при-
обретением более прочного
скелетного материала —
кости — такая концентра-
ция плавников оказывает-
ся излишней. Таким обраеом, наиболее измененные формы скелета
плавников наблюдаются у низших рыб, в особенности в тех случаях,
когда он играет особо значительную роль в качестве органа опоры, как,
например, при наличии крупного шипа в передней части плавника (рис.
164, С). Чаще всего у основания плавника наблюдаются крупные хряще-
вые пластинки, а дистально на них сидят менее измененные хрящевые
лучи. У высших рыб плавниковые лучи окостеневают и сохраняют свою
самостоятельность.
Развитие непарных плавников из одной непрерывной складки, так
же как и закладка мускульных почек в области промежуточной между
плавниками, . доказывает совершенно ясно, что отдельные плавники
большинства современных рыб — результат разделения одного общего
непарного плавника, который окаймлял почти все тело. Метамерная
закладка мускулатуры и скелета служит указанием на то, что этот непре-
рывный плавник был построен метамерно, в точном соответствии с сег-
ментацией туловища. Разделению общего плавника на отделы соответ-
ствовало, несомненно, и физиологическое разделение функций. В то
время как спинные и анальный плавники получили значение килей и
отчасти рулей, служащих для направления движения и придания ему
устойчивости, хвостовой плавник стал служить главным органом, вызы-
вающим это движение (локомоторным органом).
У круглоротых имеется еще один непрерывный плавник.
Деление на отделы, наблюдающееся только у миног (два спинных и хво-
стовой плавники), является исключительно наружным, так как в самом
деле здесь имеется совершенно непрерывный ряд мускульных пучков и
хрящевых лучей; последние в промежутке между выступающими лопа-
стями плавника только значительно короче. Скелет состоит из большого
числй длинных и тонких хрящевых лучей (supradorsalia), поддерживающих
всю лопасть плавника вплоть до наружного ее края (рис. 165, Л).
173
У с е л а х и й плавниковые лучи имеют вид гораздо более мас-
сивных хрящевых стержней (radialia), обыкновенно расчлененных на три
отдела каждый, но они значительно короче и входят обыкновенно только
в основание лопасти плавника. Главная часть последней поддержи-
вается вторичным кожным скелетом, состоящим из большого числа тонких
э л а с т о и ди ио вы х нитей (elastotrichia) (рис. 165, В). Эти нити—
Рис. 165. Сколет непарных плавников круглоротых (Л), селахий (В),
осетровых рыб (С) и костных (Z>) (схемы).
ch—хорда; ап — верхние дуги; sp — остистые отростки; s — кожные лучи (lepidotrichiaj);
г — лучи внутреннего скелета (птсрпгпофоры).
продукт деятельности мезодермальных клеток, и развиваются непосред-
ственно под эктодермой.
У ганоидных и костистых рыб также развиваются
подобные лучи (actinotrichia), но они редуцируются и замещаются
кожными же, но костными лучами (lepidotrichia). Последние
имеют такое же происхождение, как кожные кости вообще, т. е. представ-
ляют собой измененные кожные чешуи, и подобно последним могут иметь
ганоиновый покров и иногда снабжены на своей поверхности шипиками
(рис. 166, В). На лопасти плавника кожные чешуи располагаются пра-
вильными рядами на правой и левой сторонах (рис. 165, С). Члены каждого
ряда принимают удлиненную форму и, сочленяясь между собой, образуют
правую и левую половины членистого плавникового луча. Такие членистые
лучи бывают гибкими (мягкими) и нередко на конце расщепляются, но
иногда отдельные членики каждого луча срастаются между собой, и тогда
лучи являются цельными и твердыми (жесткими, колючими). У низших
ганоидов не наблюдается соответствия между числом кожных! лучей и
распределением лучей внутреннего скелета: первые имеются в значительно
большем числе и непосредственно со вторыми не связаны (рис. 165, С).
У высших ганоидов и у костистых рыб число кожных лучей сокращается,
так что на каждый луч внутреннего скелета приходится лишь один луч
наружного, который и вступает с ним в прочную связь (рис. 165, D).
Основание кожного луча охватывает своей правой и левой половиной
дистальный членик внутреннего луча с боков и прочно с ним соединяется
(рис. 166, В}. Дистальный членик луча внутреннего скелета принимает
174
шаровидную форму и очень подвижно сочленяется с остальной частью
луча. Луч внутреннего скелета становится носителем кожного луча
Рис. 166. Поперечные разрезы черев непарный плавник селахии (Л)
v и костного ганоида (В) (схемы).
atr — actinotrichla; etr — elastotrichia; Itr—постные лучи (lepidotrichla); mr—musculus
radlalis; mrp — глубокая и mrs — поверхностная лучевая [мышца; г — птеригиофор. т. е.
первичный луч.
плавника всегда прикрепляются к основаниям
(птеригиофором). Мышцы
кожных лучей.
Особого внимания заслуживает
хвостовой плавник рыб (рис. 167).
Последний состоит из ’ спинной и
брюшной лопастей, которые первона-
чально располагались симметрично
на заднем конце тела, так что хорда
проходила между ними посредине.
Это — первично симметричная, ийи
протоцеркальная, форма
хвостового плавника, как она наблю-
дается у круглоротых и во время
онтогенетического развития у всех
рыб (рис. 167, Л). У последних эта
стадия является, однако, только пре-
ходящей, — задний конец хвоста
изгибается вверх, и брюшная лопасть
начинает увеличиваться в размерах.
Таким образом, получается несим-
метричная, или гетероцеркаль-
ная, форма хвостового плавника
(рис. 167, В), которая чрезвычай-
но характерна для всех низших
рыб (селахии, цельноголовые, хря-
щевые ганоиды). Для высших рыб,
однако, и эта стадия оказывается
преходящей. Брюшная лопасть у ос-
нования хвостового плавника все
увеличивается и достигает размеров
Рис. 167. Скелет хвостового плавника.
А — протоцеркальная форма; В — ге-
тероцеркальная форма (осетр); С — го-
моцеркальная форма (костистая рыба);
D — дифицеркальная форма.
ah, ап — гемальные и невральные дуги;
ch — хорда; sp, sp — верхние и нижние
остистые отростки; г — лучи плавника;
vert — позвонки.
175
изогнутого вверх конца хвоста с его спинной и брюшной оторочкой
Таким образом, развивается наружно симметричная, или гомоцер-
кальная, форма хвостового плавника, которая представляет собой
лишь скрытую гетероцеркию (рис. 167, С).- При редукции конца осевого
скелета, который продолжается в верхнюю половину хвостового плав-
ника, такая наружная симметрия становится и внутренней. Вторичная
симметрия может развиться и обратным отклонением осевого скелета
вниз к средней линии тела, и развитием спинной лопасти хвостового
плавника до размеров брюшной. Таким образом, развиваются вторично
симметричные, или дифицеркальные, формы хвостового плав-
ника, как они наблюдаются у некоторых костных ганоидов, двудышащих
рыб и иногда у костистых (рыс. 167, D). Стадия гетероцеркального хвосто-
вого плавника пройдена всеми современными рыбами. Гетероцеркия
связана с неравномерным распределением масс в теле рыбы и неодина-
ковым распределением сил сопротивления воды около продольной оси при
движении ее вперед. Гетероцеркия заменяется симметрией, хотя бы
только наружной, одновременно с приобретением плавательного пузыря,
который у высших рыб компенсирует вес тела в воде.
Скелет хвостового плавника первоначально состоит из двух рядов
(спинного и брюшного) таких же независимых от осевого скелета хрящевых
лучей, каки в других непарных плавниках. Однако впоследствии этот пер-
вичный скелет редуцируется и вытесняется остистыми отростками позвон-
ков, которые у высших рыб служат вторичным внутренним скелетом хво-
стового плавника. У костистых рыб нижние остистые отростки разрастаются
здесь в довольно широкие пластинки (гипуралии) (рис. 167, С). Сами
плавниковые лопасти поддерживаются в хвосте кожным скелетом того
же типа, что и в других плавниках.
Непарные плавники наблюдаются и у водных амфибий, рептилий
и млекопитающих, однако в них не имеется ни собственной мускулатуры,
ни твердого скелета.
Парные плавники
У рыб имеются парные плавники в числе двух пар. Передняя пара
находится непосредственно позади головы и носит название грудных
плавников, а задняя пара — брюшных. Только у некоторых рыб с очень
длинным телом парные плавники могут вторично исчезнуть (у угрей нет
задних плавников, а у мурен нет и передних).
Как было указано выше, у рыб главным органом поступательного
движения в воде является хвост, а парные плавники служат, как и не-
парные, для регуляции движения. У хрящевых рыб парные плавники
располагаются в горизонтальной плоскости и служат несущими поверх-
ностями и рулями глубины, а также отчасти органами опоры на дне.
У костных рыб выделяются два типа строения плавников. У большинства
костных лучеперых рыб (Actinopterygii) парные плавники располагаются
в вертикальной или наклонной плоскости и способны совершать повороты,
являясь исключительно органами регулировки плавания.
Такой тип плавника имеют хрящевые и костные ганоиды и костистые
рыбы. У многоперых, кистеперых и двудышащих рыб, ведущих придон-
ный образ жизни и живущих в зарослях, парные плавники имеют широ-
кое мясистое основание и служат органами опоры.
Парные плавники рыб развиваются в виде двух пар горизонтальных
складок по бокам тела; эти кожные складки между собой не соединяются
(некоторая связь между передней и задней парой наблюдается только у
скатов), но они вначале занимают относительно более широкое простран-
ство, которое после заметно сокращается. Мускулатура парных плавни-
ков развивается также в виде каплеобразных набуханий эпителиальных
нижних концов миотомов, т. е. в виде ряда мускульных-почек (рис. 163).
176
В отличие от непарных плавников на каждом миотоме развиваются у села-
хий по две мускульные почки. У ганоидов и у костистых рыб развивается,
как и в непарных плавниках, по одной почке на каждом миотоме. Мус-
кульные почки развиваются не только на тех миотомах, которые лежат
вполне в пределах плавниковой складки, но иногда и на целом ряде бли-
жайших промежуточных миотомов. Эти мускульные почки развиваются
медленнее и имеют нередко рудиментарный характер. Некоторые из них
позже еще перемещаются в закладку плавника и принимают там участие
в образовании его мускулатуры, другие рассасываются (абортивные
почки). Это обстоятельство, так же как и факт закладки плавника в виде
более широкой складки, которая затем сокращается, указывает на то,
что парные плавники раньше занимали большее пространство, а затем
стали более узкими (сконцентрировались). В особенности это заметно
при развитии брюшного плавника, который первоначально, очевидно,
простирался далеко вперед. В некоторых случаях у селахий наблюдается
развитие мускульных почек и на всем протяжении между грудным и брюш-
ным плавником (Pristiurus). Это делает вероятным предположение, что
парные плавники развились, как и непарные, путем разделения одной
первоначально непрерывной боковой складки, расположенной в области
туловища (от головы до анального отверстия).
Каждая мускульная почка отделяется от соответствующего миотома
и делится на две почки — спинную и брюшную, которые затем врастают
в плавниковую складку, где они ложатся у ее основания — одна вдоль
спинной стенки, а другая вдоль брюшной (рис. 163, В). Затем каждый та-
кой мускульный зачаток превращается в пучок продольных мускульных
волокон. Таким образом, в зачатке плавника развивается двойной ряд
мускульных пучков — спинной и брюшной.
Мезенхима, наполняющая плавниковую складку, происходит из бо-
ковой пластинки зародыша. Она концентрируется соответственно каждой
паре мускульных зачатков, образуя клеточные сгущения в виде стержней,
расположенных как раз между спинным и брюшным мускульными зачат-
ками (рис. 163, В). Таким образом, не только мускулатура, но и скелет,
развивающийся в соответствии с ней, имеют по существу метамерное
распределение, причем на каждый сегмент туловища приходится у села-
хий по два луча скелета и по два мускульных пучка, а у ганоидов и кости-
стых рыб по одному сегменту плавника. Во время дальнейшего развития
зачатки лучей скелета охрящевевают, причем развивающиеся таким образом
хрящевые лучи обычно оказываются с самого начала связанными между
собой хрящевой пластинкой, тянущейся вдоль основания плавника.
От переднего конца этой пластинки внутрь тела врастают хрящевые вы-
росты, дающие начало поясу конечности. Скелет вполне развитого плав-
ника низших рыб состоит обычно из 1—3 крупных хрящевых пластинок,
лежащих у основания, и из целого ряда членистых хрящевых лучей,
соединяющихся с наружным их краем. В этом виде скелет парного плав-
ника весьма сходен с некоторыми концентрированными формами непар-
ных плавников (рис. 169,170). Существенное различие заключается лишь в
том, что скелет парных плавников обладает более прочной опорой в теле
животного в виде пояса конечностей. Последний имеет в передней конеч-
ности вид дужки, охватывающей тело с боков, а в задней конечности —
вид пластинки, лежащей в брюшной стенке тела.
Происхождение конечностей
Общее сходство в строении скелета (также и мускулатуры и нервов)
парных и непарных плавников и огромное сходство их эмбрионального
развития заставляют нас думать, что оба рода конечностей имеют сходное
происхождение. Непарные плавники представляют собой, несомненно,
777
Рис. 468. Три стадии образования парных
и непарных плавников (схемы).
результат разделения одной непрерывной вначале плавниковой складки
(рис. 168). Последняя представляла собой вначале простую кожную
складку, лишенную и мускулатуры, и особого скелета. Такая складка
служила лишь для увеличения известных поверхностей, важных для раз-
личных функций, связанных с передвижением животного в воде. Затем
в ней развился свой внутренний скелет в виде метамерного ряда незави-
симых хрящевых стержней, чем было достигнуто более прочное укреп-
ление этой складки. И, наконец, когда тело животного становилось менее
гибким, недостающая подвижность самого тела была заменена активностью
плавников. Соответственно этому от каждого миотома обособился мускул,
вступивший в связь с кожей плавниковой складки. В подвижной и гибкой
хвостовой области мускулатура плавника развивается гораздо позже
путем преобразования частей
самих миотомов. Разделение
общей плавниковой складки на
отделы сопровождалось и раз-
делением функций. Задняя ее
часть сохраняет роль главного
органа движения и становится
хвостовым плавником. Муску-
латурой этого органа является
осевая мускулатура. Передние
части приобретают известную
самостоятельную подвижность
и получают значение рулей для
направления движения, стано-
вясь спинными и анальными
плавниками.
По видимому, и парные плавники возникли в виде непрерывных бо-
ковых кожных складок, лишенных вначале и особого скелета, и собст-
венной мускулатуры (рис. 168). Эти складки служили для увеличения
брюшной поверхности тела и представляют результат ее разрастания по
бокам. Небольшие складки такого рода развиваются иногда и вторично
у современных селахий. Функциональное их значение сходно со значе-
нием непарной складки. С увеличением размеров складки понадобился
свой скелет для ее укрепления. Скелет возник в виде метамерного ряда
независимых хрящевых стержней (в одной группе низших рыб по два на
каждый сегмент, как у современных селахий, у других рыб — по одному).
От соответствующих миотомов обособились затем мышечные пучки, ко-
торые и прикрепились к коже плавниковой складки, располагаясь сверху
и снизу от каждого луча внутреннего скелета. Таким образом развилась
парная боковая плавниковая складка с метамерным распределением ске-
лета и мускулатуры (и нервов). Наблюдения над низшими рыбами пока-
зывают, что парные плавники служат у них, главным образом, в качестве
рулей глубины. Это значение могла иметь и простая кожная складка при
достаточно гибком туловище животного; с утратой же общей подвижности
эта последняя могла быть заменена лишь подвижностью самой складки.
Легко понять, что в качестве рулей глубины наибольшее значение имели
конечные части складки — передняя и задняя, как наиболее удаленные
от точки вращения. Поэтому значительное развитие получили лишь пе-
редняя часть складки, которая дала начало грудному плавнику, распо-
ложенному непосредственно позади головы, и задняя часть складки, ко-
торая дала начало брюшному плавнику, расположенному первоначально
непосредственно впереди анального отверстия. Средняя часть складки
не получала заметного развития и рано редуцировалась (рис. 168). Таким
образом, указанное первичное значение боковых складок вполне объяс-
няет тот факт, что у всех современных рыб развились только две пары
178
плавников. Кроме факсов эмбрионального развития парных Плавников,
теория боковой складки (или теория метамерного происхождения конеч-
ностей) опирается, помимо общего сходства с непарными плавниками,
еще и на некоторые другие сравнительно-анатомические данные. Прежде
всего, зачаточные боковые складки имеются у ланцетника, хотя генети-
ческая связь парных плавников рыб с метаплевральными складками лан-
цетника и не доказана. Боковые складки сравнимые с зачатком плавников
имелись и у некоторых ископаемых бесчелюстных. Более существенно
то обстоятельство, что в скелете парных конечностей, особенно у прими-
Рис. 469. Три стадии образования скеле-
та парного плавника. А — гипотетическая
исходная форма; В — примитивный брюш-
ной и С — грудной плавник (схемы).
bpt — basipterygium; mpt — metapterygium;
n — отверстия для нервов; pct — плечевой
иояс; г — лучи
Рис. 170. Тазовый пояс и брюшные плав-
ники хрящевого гаиоида (Л — Scaphir-
hynchus) и акулы (В — Chlamydosela-
chus) (по Раутенфельду и Гудричу).
bpt — basipterygium; п — отверстия для
нервов; pv — тазовый иояс; г — Лучи.
тивных рыб, можно найти еще указания на происхождение основных их
хрящей, а также и пояса конечностей путем слияния оснований лучей
плавника (рис. 169). У некоторых древнейших ископаемых селахий зна-
чительное число лучей сочленялось непосредственно с поясом, и ясно
видно, что основные хрящи представляют собой результат слияния основ-
ных члеников лучей (Cladoselache, Cladodus, Symmorium). В заднем плав-
нике, который вообще обнаруживает более примитивное строение, это
иногда наблюдается и у современных рыб. Так, у очень примитивной
акулы Chlamydoselachus большое число лучей брюшного плавника при-
членяется непосредственно к заднему поясу. Последний имеет вид широ-
кой пластинки, лежащей в брюшной стенке тела, и снабжен двойным ря-
дом отверстий для прохождения нервов (рис. 170). Это обстоятельство
объяснимо, если принять предположение, что здесь имелись раньше про-
179
должения хрящевых лучей, между которыми проходили нервы; оснований
лучей затем срослись между собой и дали начало пластинке — поясу,
продырявленному отверстиями для нервов на месте прежних промежутков
между лучами. Однако еше более примитивными оказываются брюшной
плавник и задний пояс хрящевых ганоидов (рис. 170, А). У последних
не только нет никаких основных хряшей в самом плавнике, где имеется
лишь ряд самостоятельных лучей, но и пояс находится еще только на
стадии своего образования путем слияния основных члеников лучей плав-
ника — в задней его части наблюдаются еще самостоятельные основные
члены лучей, а в передней они более или менее полно срастаются между
собой, оставляя небольшие отверстия для прохождения нервов. У высших
рыб пояса конечностей хорошо развиты, а в самом плавнике лучи продол-
жают сохранять свою самостоятельность, хотя они здесь значительно
редуцированы в связи с сильным развитием кожного скелета (рис. 172).
Таким образом, у высших рыб в парных плавниках, как и в непарных,
лучше, чем у низших, сохраняется примитивный признак — независи-
мость лучей, и это, казалось бы, парадоксальное явление находит себе
объяснение в свойствах самой скелетной ткани. Кость представляет собой
более прочный материал, который гораздо менее пластичен, чем хрящ,
и потому костные элементы консервативнее хрящевых. Кроме того, ко-
нечность высших рыб представляет собой более слабый орган, чем плавник
селахий, и соответственно она менее концентрирована.
Изложенная теория метамерного происхождения конечностей разра-
ботана целым рядом авторов.
Кроме того, был высказан и другой взгляд на происхождение парных
конечностей. По теории архиптеригия, разработанной Гегенбауром, пар-
ные конечности позвоночных представляют собой результат преобразо-
вания двух жаберных дуг с их жаберными лучами. Собственно дуга дала
пояс конечности, а жаберные лучи дали скелет свободной конечности.
Задняя конечность по этой теории испытала значительное перемеще-
ние назад. Наиболее примитивной формой скелета является, по Геген-
бауру, архиптеригий, т. е. скелетная ось, несущая два ряда лучей, как
это наблюдается, например, у двудышащих рыб. Против этой теории го-
ворят, однако, следующие факты: метамерная закладка мускулатуры и
скелета (за счет нескольких сегментов, а не одного); закладка плавника
в виде горизонтальной, а не вертикальной складки и горизонтальное
положение плавников у низших рыб, заменяющееся вертикальным у выс-
ших, а не наоборот; развитие пояса конечности от скелета плавника, а
не наоборот; положение пояса снаружи от туловищной мускулатуры,
а не внутрь от нее, как лежит жаберный скелет, и, наконец, тот факт,
что мускулатура конечностей является париетальной (развивается от
миотомов), а не висцеральной (развивающейся от боковых пластинок),
и иннервируется рядом спинномозговых нервов, а не блуждающим. Таким
образом, современные данные говорят решительно в пользу теории бо-
ковой складки. Однако некоторые новейшие экспериментальные данные
дают как-будто указания на независимое от скелета свободной конечности
образование плечевого пояса. В таком случае последний, быть может,
имеет висцеральное происхождение, а его связь со скелетом боковой склад-
ки установилась позднее. В пользу этого может быть истолковано наличие
висцеральной мышцы плечевого пояса m. trapezius уже у низших рыб.
В скелете парных конечностей различаются, как уже указывалось,
внутренняя их опора — пояс и собственно скелет свободной конечности.
Передний пояс рыб
Передний пояс селахий представляет хрящевую дугу, охватываю-
щую тело с боков и с брюшной стороны, где правая и левая его полови-
180
ны соединяются между собой (у ископаемого Pleuracanthus этого соедине-
ния не было). Пояс лежит поверхностно и снабжен снаружи и сбоку
выступом, служащим для причленения свободной конечности (плав-
ника). В этом месте хрящ наиболее массивен и пронизан обыкновенно двумя
каналами для прохождения нервов конечности. Место причленения плавни-
ка является границей между спинным и брюшным отделами пояса, кото-
рые по сравнению с вышестоящими формами называют лопаточным
(pars scapularis) и коракоидным (pars
coracoidea) (рис. 169, С). С этим поя-
сом в общем сходен и первичный
пояс хрящевых ганоидов, имеющий
вид хрящевых дужек, лежащих по
бокам тела, но не соединяющихся
между собой на брюшной стороне.
Этот первичный пояс покрыт, од-
нако, уже несколькими кожными
костями, примыкающими к нему сна-
ружи. На спинном отделе пояса лежит
большая кость — cleithrum, а на
брюшном отделе также крупная
кость, гомологичная ключице выс-
ших позвоночных — clavicula. Пер-
вая кость соединяет вторичный пояс
с наружной крышей черепа при
посредстве еще одной кожной кости,
Рис. 171. Передний пояс двудышащей
рыбы (Ceratodus) и миогоперого ганоида
(Polypterus) с брюшной стороны (из
Гегенбаура).
cl — clavicula; clt — cleithrum; 0 — су-
ставный бугорок.
лежащей над ней (supracleithrale).
а вторая кость соединяется на брюшной стороне тела с одноименной
костью другой стороны. Обе кости обладают снаружи скульптурой, ха-
рактерной для типичных кожных костей.
Рис. 172. А — правая половина плечевого пояса с грудным плавником костного га-
лоида (Amia), вид изнутри; В — правая половина первичного скелета плечевого
пояса и грудного плавника костистой рыбы (Salmo), вид изнутри (по Гудричу).
сП — cleithrum; со — cpracoideum; de — дистальные хрящики; тсо — mesocoracoideum;
mt—basale metaptcrygii; pelt — postcleithra 1c; pt — posttemporale; sc — scapula; sett —
supracleithrale; rp, r — лучи; sf — отверстие для нервов.
У двудышащих рыб первичный плечевой пояс также является хря-
щевым и по своему полному развитию напоминает пояс селахий, но кортц
вторичного пояса (clavicula и cleithrum) уже утратили характер кожных
костей и тесно прилежат к хрящу (рис. 171).
У костных ганоидов и у костистых рыб наблюдается значительная
редукция первичного пояса, которая, очевидно, связана с прогрессивным
181
развитием вторичного. Редко первичный пояс остается еще хрящевым
(Amia) (рис. 172, А), обычно он окостеневает в виде двух сравнительно
небольших костей, лежащих в области причленения скелета свободной
конечности. Кость, лежащая со спинной стороны, носит наименование
лопатки (scapula), а кость, лежащая с брюшной стороны, — коракоида
(coracoideum) (рис. 172, В). Первичный пояс сам по себе играет, однако,
у высших рыб лишь ничтожную роль, — он служит связью между плавни-
ком и вторичным поясом, который и является собственно действительной
опорой конечности. Вторичный пояс высших рыб (рис. 172, 4) представлен
крупной парной костью, соответствующей разросшейся cleithrum ганоидов,
с которой, быть может, срослась и ключица. Последней в виде самостоя-
тельной кости во всяком случае нет. Обе cleithra соединяются на брюшной
стороне тела между собой, а на спинной стороне прикрепляются к крыше
черепа посредством двух других покровных костей (supracleithralia и
posttemporalia). Таким образом, плечевой пояс укрепляется у высших
рыб почти неподвижно.
Задний иояс рыб
У селахий задний пояс имеет вид пластинки, лежащей в брюшной
стенке тела и связывающей оба брюшных плавника между собой. В этой
пластинке имеются обыкновенно 1—2 пары отверстий для прохождения
нервов. В некоторых случаях наблюдается целый ряд таких отверстий
(рис. 170). История развития показывает, что
непарная пластинка селахий — результат сра-
стания парных зачатков, Г развивающихся от
оснований брюшных плавников. Иногда на по-
ясе, над местом причленения плавника, возвы-
Рис. 173. А — тазовый пояс и брюшной плавник ископаемого кистеперого ганоида
(Eusthenopteron) (по Гудричу); В — тазовый пояс и брюшной плавник костистой
рыбы (Salmo).
I — лепидотрихии; р и — таз; г — лучи; s — чешуя; d. г — ножные лучи.
шается небольшой спинной отросток (processus iliacus). У цельноголо-
вых задний пояс состоит из двух независимых половин и замечателен
по своей форме: он разрастается не только на брюшную сторону, но
и на спинную, так что в нем, как и в переднем поясе, можно различать
брюшной отдел, лежащий под местом причленения свободной конеч-
ности (pars ischiopubica), и спинной отдел, лежащий над ним (pars
752
iliaca). Последний соответствует сильно развившемуся спинному отростку
(proc, iliacus) пояса селахий.
У высших рыб задний пояс развит очень слабо и представлен обычно
лишь небольшой парной пластинкой. У хрящевых ганоидов он находится
на стадиях образования, и в задней своей части представлен иногда
еще независимыми основными члениками лучей скелета брюшного плав-
ника (рис. 170, Л). Постепенное прирастание этих основных члеников
приводит к образованию цельной хрящевой пластинки треугольной
формы, продырявленной одним или несколькими отверстиями для про-
хождения нервов. У костных ганоидов и у костистых рыб эта парная
пластинка окостеневает (она обычно неправильно обозначается как основ-
ной элемент свободной конечности) (рис. 173).
Скелет грудного плавника
Скелет грудного плавника селахий состоит из нескольких более
крупных хрящевых пластинок, лежащих в суженном основании плавника
и вдоль заднего его края (рис. 169, С). Иногда имеется лишь один такой
основной хрящ, но типичным считается обыкновенно существование трех
хрящей (basalia propterygii, mesopterygii и metapterygii), из которых
наибольшее значение имеет задний, который тянется вдоль заднего края
плавника (basale metapterygii). К наружному краю этих хрящей при-
крепляется ряд расчлененных хрящевых лучей, которые у акул не доходят
до края плавника. Дистальная часть лопасти последнего поддерживается
такими же кожными образованиями (эластоидиновыми нитями), как и в
непарных плавниках. Иногда внутренний скелет грудного плавника
является более измененным и состоит из большого числа многоугольных
хрящевых пластинок. Скелет грудного плавника хрящевых ганоидов
состоит из небольшого сравнительно числа хрящевых лучей, которые
частью причленяются непосредственно к поясу, частью сидят на небольшом
основном хряще, лежащем у заднего края плавника. Скелет этот сохра-
няет, следовательно, известные примитивные признаки, но вместе с тем
он развит сравнительно слабо и поддерживает только основание плавника.
Это находится в очевидной связи с прогрессивным развитием вторичного
скелета плавника, состоящего из кожных костных лучей (lepidotrichia).
Последние вполне сходны с кожными лучами непарных плавников, имеют,
несомненно, такое же происхождение из чешуй и являются главной
опорой лопасти плавника. У костных ганоидов и у костистых рыб первич-
ный скелет плавника частью окостеневает. У ганоидов наблюдаются
иногда еще основные хрящи, к которым причленяются окостеневшие
лучи (рис. 172, А). У костистых рыб первичный скелет состоит только
из небольших окостеневших лучей, сидящих в небольшом числе (чаще
всего около пяти) непосредственно на костях первичного пояса и примы-
кающего к ним снаружи ряда небольших хрящиков (рис. 172, Б). Вторич-
ный скелет состоит из довольно значительного числа сильно развитых
кожных лучей. Кожные лучи парных плавников являются членистыми,
и только передние лучи иногда превращаются в твердые колючки.
Таким образом, у высших рыб наблюдается редукция первичного скелета
плавника и функциональное замещение его вторичным, кожным скелетом.
У двудышащих рыб, наоборот, имеется прекрасно развитый внутрен-
ний скелет, поддерживающий почти всю лопасть плавника за исключением
лишь наружного ее края, где эта роль переходит к тонким кожным лучам.
Внутренний скелет плавника остается хрящевым и имеет своеобразное
строение: он состоит из длинной членистой оси, несущей у Ceratodus
два ряда боковых членистых лучей. Этот скелет соединяется одним лишь
основным членом своей оси с поясом и .в этом отношении до известной
степени сходен с конечностью, наземных позвоночных.
183
Скелет брюшного плавника
Рис. 174. Брюш-
ной плавник и
часть таза двуды-
шащейрыбы Neo-
ceratodus) (по
Давыдову и
Кингсли).
ах — ось плавни-
на; pH—подвздош-
ный отросток;
ргт — пояс; г —
лучи.
для хождения,
В скелете
Скелет брюшного плавника рыб имеет вообще более простое строение,
чем скелет грудного плавника. У селахий он состоит из одного основного
хряща (b as ale metapterygii), тянущегося вдоль внутреннего (заднего)
края плавника, с рядом сидящих на нем лучей. Иногда некоторое число
передних лучей причленяется непосредственно к поясу
(рис. 170, Б). Особенно примитивен скелет брюшного
плавника у хрящевых ганоидов, у которых он состоит из
некоторого числа лучей, непосредственно причленяющих-
ся к элементам не вполне сформировавшегося заднего
пояса (рис. 170, Л). У костных ганоидов и у костистых
рыб первичныйскелетплавника совершенно рудиментарен
и состоит иногда из нескольких незначительных косточек
(рис. 173, В), а у высших форм и вовсе исчезает. Хорошо
развит лишь вторичный кожный скелет, лучи которого
у костистых рыб в случае полной редукции первичного
скелета причленяются непосредственно к заднему поясу.
Скелет брюшного плавника двудышащих рыб вполне
сходен со скелетом грудного плавника и состоит также
из длинной членистой оси с расположенными в два ряда
(у Neoceratodus) боковыми членистыми лучами (рис. 174).
Весь этот скелет у двудышащих остается хрящевым.
Конечности наземных позвоночных и их происхождение
Непарные плавники наблюдаются только у живу-
щих в воде личинок амфибий и у тех представителей
высших позвоночных, которые вторично переселились
в воду (многие хвостатые амфибии, некоторые репти-
лии, главным образом ископаемые, и некоторые мле-
копитающие, как, например, китообразные). Однако во
всех этих случаях лопасть плавника представляет собой
лишь более или менее прочную кожную складку, которая
лишена внутреннего скелета и мускулатуры. Парные
конечности у этих же форм нередко имеют форму плав-
ников (ласты), но такие плавники непосредственно не
сравнимы с рыбьими плавниками, так как они всегда
являются результатом вторичного преобразования ти-
пичной конечности наземного позвоночного. Последняя
представляет собой сложный рычаг, предназначенный
т. е. для перемещения тела животного на суше.
парных конечностей наземных позвоночных мы также
различаем собственно свободную конечность и пояс, лежащий в самом
туловище и служащий для нее опорой.
Пояса конечностей имеют у наземных позвоночных вид дуги, охва-
тывающей тело снизу и с боков. В общем такой пояс сходен с поясом
парного плавника рыбы (рис. 175). Имеется некоторое различие в причле-
нении свободной конечности: у рыб для этого имеется выступ на поясе,
а у наземных позвоночных —- сочленовная ямка. Наиболее сходны пояса
передних конечностей, которые у наземных позвоночных отличаются
лишь большей шириной как спинного, так и брюшного отделов. Это увели-
чение поверхности поясов связано с прогрессивным развитием мускула-
туры конечностей (и особенно задней конечности, выполняющей при дви-
жении на суше большую работу). Пояса задних конечностей у наземных
позвоночных развиты значительно выше, чем у большинства рыб, имеют
хорошо развитый спинной отдел (pars iliaca), который причленяется к
184
ребрам (первоначально к одной паре ребер) позвоночника. Такое причле-
нение служит для более прочного укрепления пояса и представляет
характерную особенность наземных позвоночных (некоторая аналогия
имеется лишь в прикреплении переднего пояса к позвоночнику у скатов).
Скелет свободной конечности наземных позвоночных состоит из причле-
няющегося головкой к поясу довольно длинного проксимального отдела—
stylopodium, который несет два также довольно крупных элемента, со-
ставляющих следующий отдел конечности — zeugopodium, несущий уже
собственно лапку, autopodium (рис. 176). В последней различают основа-
ние, состоящее из значительного числа мелких элементов — basipodium,
Рис. 175. Сравнение скелета типичного плавника
рыбы и пятипалой конечности при одной и той же
ориентировке. А — скелет брюшного плавнйка
рыбы и В -— скелет задней конечности наземного
позвоночного; С — скелет грудного плавника и
D — скелет передней конечности примитивного
наземного позвоночного.
civ — clavicula; clt — cleithrum; co — coracoideum;
f — fibula; fe — femur; h—humerus; V.— ileum;
is — ischium: mpt — metapterygium; p —pubis; p. co—
pars coracoidea; p. sc — pars scapularis; pr. Ц — pro-
cessus iliacus; sc —scapula; и — ulna; I—V — пальцы,
Рис. 176. Схема строения
пятипалой конечности.
С — Centralia; F — fibula; Fe —
femur; f — fibulare; H — hume-
rus; i— intermedium; m — medi-
an»; P — praepolex; pis — pisi-
forme; R—radius; r—radiale;
T — tiba; t — tibialc; U — ulna;
и — ulnare; 1—5 — basalia dis-
talia 1—5 пальцев; 6 — postmi-
nimus; I—V — metabasalia 1 —
5 пальцев.
и дистальную часть — acropodium, состоящую из более длинных эле-
ментов, образующих правильные ряды или лучи, значительная часть
которых обособлена друг от друга и образует собственно пальцы.
В передней конечности stylopodium получает название плечевой
кости (humerus), оба элемента zeugopodium, или предплечья, назы-
ваются лучевой (radius) и локтевой костью (ulna); первая лежит
вдоль переднего (внутреннего) края конечности, вторая — вдоль заднего
(наружного) края. Autopodium передней конечности представляет собой
к и с т ь, ее основание — basipodium — носит название запястья (carpus).
Дистальная часть кисти •— acropodium — состоит из соответствующих
755
пальцам основных члеников, образующих пястье (metacarpus), и собственно
пальцев, состоящих каждый из нескольких члеников, или фаланг (pha-
langes digitorum).
В задней конечности stylopodium является в виде бедренной
кости (femur). Передний (внутренний) элемент zeugopodium, или голени,
представляет собой большую берцовую кость (tibia), а
задний (наружный) элемент — малую берцовую кость (fibula).
Autopodium задней конечности представляет собой ступню, и ее
основание — basipodium — носит название предплюсны (tarsus). Ди-
стальная часть ступни — acropodium задней конечности — состоит
из соответствующих пальцам основных члеников, образующих плюсну
(metatarsus), и собственно пальцев, состоящих из нескольких фаланг
(phalanges) каждый.
Наиболее сложно строение основания лапки (basipodium), которое
состоит из трех рядов косточек — проксимального (basalia proximalia),
среднего (basalia medialia) и дистального (basalia distalia). Проксимальный
ряд нормально состоит из трех косточек. Две крайние косточки примы-
кают к концам обоих элементов zeugopodium и получают соответствующие
названия: в передней конечности передняя косточка — radiale и задняя —
ulnare; в задней конечности передняя — tibiale и задняя — fibulare.
Между ними лежит промежуточная косточка — intermedium. Средний
ряд состоит из одной или нескольких центральных косточек — Centralia.
Дистальный ряд состоит из косточек, примыкающих к костям следующего
отдела. Эти косточки обозначаются цифрами соответственно пальцам —
basalia (carpalia, tarsalia) distalia 1—5.
Элементы передней и задней конечностей можно сопоставить сле-
дующим образом:
Передняя конечность Задняя конечность
Humerus Stylopodium Femur
radius, ulna zeugopodium tibia, fibula
radiale, intermedium, ulnare ) ( tibiale, intermedium, fibulare
centralia, medialia > basipodium < centralia, medialia
carpalia distalia 1, 2, 3, 4, 5 ) ( tarsalia distalia 1, 2, 3, 4, 5
metacarpalia I, II, III, IV, V I .. f metatarsalia I, II, III, IV, V
1 phalanges, / acropodium | phalanges
Строение скелета конечности наземных позвоночных, или пятипалой
конечности, как ее называют, обнаруживает известное сходство со строе-
нием скелета некоторых форм плавников рыб. Это сходство выражается
в общем сходстве пояса конечностей и в лучевом расположении дистальных
элементов свободной конечности (рис. 175). Так как наземные позвоночные,
несомненно, произошли от рыбообразных форм, то сама собой напраши-
вается мысль, что и парные их конечности представляют собой преобра-
зованные парные плавники.
Первые попытки провести сравнение между пятипалой конечностью
и плавником рыбы произведены давно, и теория происхождения конеч-
ностей имеет не менее долгую историю, чем сама сравнительная анатомия.
Старые анатомы не проводили еще различия между первичным, внутрен-
ним, скелетом и вторичным — кожным, и поэтому пальцы наземных
позвоночных сравнивались непосредственно с кожными лучами плавни-
ков высших рыб. Такое сравнение с современных точек зрения, разумеется
недопустимо. Натурфилософы (Окен и др.) полагали, что весь скелет
позвоночных представляет собой результат диференцировки его осевого
скелета, и соответственно видели в скелете парных конечностей преобра-
зованные ребра. Так как и висцеральные дуги считались преобразован-
ными ребрами, то, следовательно, согласно этому взгляду, пояс конечности
является гомодинамом висцеральной дуги и вся конечность — образо-
756
ванием мономерным. Таким образом, натурфилософы до известной степени
предвосхитили те мысли, которые легли в основу теории архиптеригия
Гегенбаура. Согласно последнему взгляду конечность наземных позво-
ночных представляет собой преобразованный архиптеригий, т. е. древней-
шую форму плавника с рядовым расположением лучей на определенной
оси.
Изучение истории развития конечностей наземных позвоночных
привело в общем к тем же результатам, как и ее исследование у рыб.
Пятипалая конечность закладывается в виде небольшой горизонтальной
складки и на ранних стадиях имеет в общем форму плавника. Мускулатура
конечности развивается в виде эпителиальных выростов — мускульных
почек — на целом ряде миотомов (обычно около 5, иногда до 8). Мускуль-
ные почки врастают в конечность и здесь расплываются в массу клеток,
располагающихся в виде сплошного слоя на спинной и на брюшной сто-
роне конечности, образуя зачатки спинных и брюшных мышц (разги-
бателей и сгибателей) конечности. Мезенхима концентрируется посредине
между этими мускульными зачатками и образует закладку скелета, в
котором затем наступает охрящевение соответственно отдельным элемен-
там взрослой конечности. В большинстве случаев все элементы конечности
затем окостеневают. Таким образом, в развитии конечностей наземных
позвоночных наблюдается сходство с развитием плавников рыб. То обсто-
ятельство, что мускульные почки быстро сливаются в общую массу кле-
ток, не может служить препятствием для сравнения, так как это наблю-
дается и у высших рыб. В некоторых случаях и у рыб (у двудышащих),
и у наземных позвоночных (у амфибий) развитие является еще более
измененным, и вместо врастания мускульных почек в зачаток конечности
происходит простое вселение клеток, отделяющихся от тех же нижних
концов миотомов.
Таким образом развитие пятипалой конечности идет в общем тем же
путем, как и развитие парных плавников, и если мы для скелета последних
принимаем независимое его происхождение из метамерного ряда лучей,
следовательно, то же самое мы должны принять и для конечности назем-
ных позвоночных. Во всяком случае, сходство в развитии парных конеч-
ностей рыб и наземных позвоночных является бесспорным доказательством
того, что пятипалые конечности последних представляют' собой преобра-
зованные парные плавники рыб. Если, однако, происхождение этих
конечностей от плавников является совершенно несомненным, то остается
все же еще вопрос — каков же тип того плавника, из которого развилась
пятипалая конечность. На этот вопрос разные авторы давали различные
ответы.
Все современные авторы сравнивают конечность наземных позво-
ночных с типичным рыбьим плавником, обладавшим лишь не особенно
большим числом лучей, как это наблюдается у большинства высших рыб.
Первичный скелет плавников костистых рыб, как мы видели, редуцирован
и функционально замещен кожным скелетом, так что, естественно, отпа-
дает как объект для сравнения. Таким образом, с пятипалой конечностью
сравнимы лишь плавники ганоидов и двудышащих (Ceratodus). Мы не
будем входить в рассмотрение взглядов отдельных авторов, укажем только,
что современные исследователи обычно выводят пятипалую конечность
из плавника с неясно выраженной осью, которая лежала вдоль заднего
края конечности (metapterygium). В общем эти взгляды довольно близки
к взглядам Гегенбаура, если только не смотреть на архиптеригий как на
древнейшую форму плавника и совершенно отбросить теорию его проис-
хождения от висцерального скелета.
В настоящее время можно считать несомненным, что пятипалая
конечность развилась не путем усложнения, а путем постепенного упро-
щения из более богато расчлененной конечности (А. Н. Северцов). Все
187
пятого пальца по заднему
Рис. 177. Полная схема строе-
ния основания кисти.
г — centrale; Н — humerus; i —
intermedium; т — medialia; рр—
praepollex; pls— pisiforme; R—
radius; r — radiale; U — ulna;
u—ulnare; 1—5—carpalia dista-
lia; 6 — postniinimus; I — V —
metacarpalia.
элементы конечности морфологически равнозначащи и представляют
собой отчасти результаты слияния лучей (члены оси конечности), отчасти
продукт их расчленения. Лучевое расположение элементов конечности
ясно выражено в особенности у хвостатых амфибий, по крайней мере,
во время развития конечности. В проксимальной части конечности разли-
чима ось, проходящая вдоль заднего края и состоящая из трех более
крупных элементов (humerus, ulna и ulnare в передней конечности и femur,
fibula и fib lare — в задней). Эта ось несет сбоку и на конце лучи. Первый
боковой луч представлен рядом — radius, radiale, carpale 1, metacarpale 1
и первый палец; второй боковой луч рядом — intermedium, centrale,
carpale 2, metacarpale 2 и второй палец; третий луч — carpale 3, meta-
carpale 3 и третий палец, а четвертый и пятый лучи, имеющие подобный
же состав, занимают конечное положение. Но, кроме этого, наблюдаются
еще и остатки лишних лучей — элементы, лежащие впереди первого
пальца и напоминающие иногда рудиментарный палец (praepollex в пе-
редней конечности и praehallux—в задней), и элементы, лежащие позади
краю конечности (postminimus) (рис. 176).
Нужно допустить, что в конечности назем-
ных позвоночных произошло уменьшение
числа лучей, которых раньше было не менее
семи. В области основания кисти (basipodium)
нередко находят рудиментарные элементы,
обычно прирастающие к соседним элемен-
там, и вообще у наземных позвоночных
весь этот отдел подвергается редукции—он
сокращается в размерах, в особенности
у высших форм, и многие его элементы сра-
стаются между собой. Таким образом, строе-
ние основания кисти (стопы) становится
проще, число элементов уменьшается, а ино-
гда и весь этот отдел прекращает свое су-
ществование, прирастая к соседним отделам:
(задняя конечность птиц). Нахождение мно-
гочисленных рудиментарных элементов в этой
области у низших наземных позвоночных
показывает, что и у них произошло уже за-
метное упрощение строения. В дальнейшем
у наземных позвоночных исчезает собствен-
но весь средний ряд элементов основания
кисти, состоящий, кроме обычной централь-
ной косточки (centrale proximale), перво-
начально еще из целого ряда дистальных
центральных косточек (centralia distalia
s. medialia), входивших в состав всех лучей
конечности (рис. 177). Следовательно, скелет плавника при преобразова-
нии в пятипалую конечность испытал сокращение числа лучей и упроще-
ние в строении известного среднего отдела, который вообще затем сокра-
щается в размерах. С другой стороны, история развития показывает, что
длинные проксимальные и дистальные кости конечности закладываются
в виде небольших сравнительно хрящевых элементов, приблизительно
равнозначащих всем прочим. И у низших наземных позвоночных, в осо-
бенности у некоторых ископаемых форм, эти длинные элементы все же
относительно много короче, чем у высших (рис. 190, 191). Таким образом,
можно думать, что эти проксимальные и дистальные элементы в пятипалой
конечности развились прогрессивно по сравнению с исходным плавником.
Что длина некоторых костей конечности, действительно, связана с функ-
цией конечности наземного позвоночного, видно из того, что при обратном
188
переселений Последних й воду и превращении пятипалой конечности в
ласт наступает обратное укорочение этих костей. Все элементы конечности
рновь становятся приблизительно равнозначащими, как это произошло,
например, у китообразных и, в особенности, у ископаемых водных репти-
лий (ихтиозавры и плезиозавры). У этих форм наблюдается и увеличение
Числа элементов конечности (числа фаланг) и даже вторичное увеличение
числа лучей. Все это является косвенным указанием на то, что процессы,
Сопровождавшие преобразование рыбьего плавника в пятипалую конеч-
ность, действительно, обусловлены сменой его функции.
Таким образом, исходным для развития пятипалой конечности был
плавник с не очень большим числом лучей (не менее семи), но богато
р асчлененный.
Сильно расчлененным является плавник одной из двудышащих рыб
(Ceratodus), и у основания его мы находим лишь один элемент, причле-
няющийся непосредственно к поясу и соответствующий Stylopodium пяти-
палой конечности.- Однако скелет этого плавника является гораздо более
специализированным, чем это можно допустить для исходной формы.
Известное сходство с конечностью наземных позвоночных обнаруживает
скелет плавника многоперых ганоидов (Polypterus), но этот скелет явля-
ется очень мало расчлененным. Гораздо большее и, можно сказать, пора-
зительное сходство с пятипалой конечностью обнаруживает скелет плав-
ника некоторых ископаемых представителей кистеперых (Crossopterygii),
как, например, скелет плавника Eusthenopteron (рис. 173, 4). Прокси-
мальная часть этого скелета почти идентична скелету пятипалой конеч-
ности, дистальная его часть, к сожалению, неизвестна, — очевидно, она
была хрящевой и потому не сохранилась. Еще большее сходство обнару-
живает скелет плавника Sauripterus, в котором к тому же на переднем
(нижнем) крае кожные лучи редуцировались, что, очевидно, указывает
на иную его функцию (рис. 388). Нужно прибавить, что эти представи-
тели кистеперых и по другим признакам очень близки к наземным позво-
ночным, так что сходство в строении их конечностей объясняется, дей-
ствительно, близким родством этих форм.
Плечевой пояс наземных позвоночных
Плечевой пояс наземных позвоночных отличается от первичного
пояса рыб, главным образом, большей шириной как брюшного, так и спин-
ного отдела (рис. 175). У рыб, как мы видели, передний пояс прикрепля-
ется к черепу посредством ряда покровных костей (supracleithrale и post-
teniporale). У наземных позвоночных такое прикрепление по необходи-
мости исчезло одновременно с развитием подвижности головы. Вместе
с тем понадобилось иное средство для укрепления пояса, и так как послед-
ний лежит кнаружи от туловищной мускулатуры и с внутренним скелетом
не связан, то единственным способом для его укрепления оказалось общее
увеличение поверхности его опоры. Таким образом, и спинной (pars scapu-
laris), и брюшной отдел (pars coracoidea) значительно расширяются, в
особенности в своей дистальной части, приобретая вид треугольных пла-
стинок, соединяющихся своими вершинами.
У амфибий особенной шириной отличается брюшной отдел пояса,
на который опирается значительная часть тела (рис. 178). У высших по-
звоночных здесь устанавливается через посредство грудины соединение
с целым рядом ребер, и таким образом эта часть пояса приобретает новую
и еще более прочную опору. Вместе с тем большая ширина брюшного от-
дела оказывается излишней, и наступает его редукция, так что нередко
остается лишь одна кость (коракоид или ключица), причленяющаяся к
грудине. Спинной отдел пояса, наоборот, и у высших позвоночных отли-
чается значительной шириной (рис. 181).
189
В спинном отделе первичного пояса развивается одно окостенение —
лопатка (scapula), которая первоначально занимает лишь прокси-
мальную часть отдела, между тем как более широкая дистальная его часть
остается хрящевой и получает название надлопаточного хряща (cartilago
suprascapulare). У высших позвоночных собственно лопаточное окосте-
нение разрастается обыкновенно на весь спинной отдел пояса. В брюшном
отделе первичного пояса развивается обыкновенно одно окостенение,
которое получает название вороновидной кости, или коракоида
(coracoideum). В этом отделе пояса нередко наблюдается образование
окон, затянутых лишь соединительной тканью (рис. 180, 4). Такие окна
отделяют более массивную заднюю, собственно коракоидную, часть, в
которой и располагается указанное окостенение, от передней, прокора-
Рис. 178. Плечевой пояс хвостатой амфи-
бии (Л) и бесхвостой (жабы — В).
cl — clavicula; сП — cleithrum; со — coracoi-
deum; ерс — epicoracoidcum; rjl — суставная
ямка; рсо — procoracoideum; sc — scapula;
st — грудина.
коидной, части, которая может
оставаться хрящевой (cartilago
procoracoideum), или на нее рас-
пространяется окостенение от ко-
ракоида, или, наконец, здесь на-
блюдается самостоятельное око-
стенение, получающее название
прокоракоида (os proco-
racoideum) (рис. 181).
Кроме костей первичного по-
яса, у наземных позвоночных на-
блюдаются в поясе и покровные
кости, которые никогда, однако,
не играют такой « значительной
роли, как у рыб. Покровная кость
спинного отдела пояса — cleith-
rum, которая является наиболее
крупной костью пояса рыб, у на-
земных позвоночных редуцирует-
ся. У стегоцефалов и у некоторых
древнейших ископаемых репти-
лий эта кость еще имеется и зани-
мает типичное положение на наружной поверхности лопаточного отдела
пояса (рис. 179). В некоторых случаях на ней наблюдается и типичная
для кожных костей скульптура. Из современных форм cleithrum сохра-
няется только у бесхвостых амфибий в виде костной пластинки, лежащей
на наружной поверхности надлопаточного хряща (рис. 178, В). У других
наземных позвоночных эта кость исчезла бесследно. Покровная кость
брюшного отдела пояса — ключица (clavicula), — наоборот, сохра-
няется в виде значительной кости у большинства наземных позвоночных.
У стегоцефалов она нередко имела вид широкой пластинки с характерной
скульптурой (рис. 179, А). Она лежит обычно в передней части пояса на
прокоракоидном хряще, но иногда вполне обособляется от внутреннего
скелета и располагается впереди первичного пояса (рептилии) (рис. 180, А).
У стегоцефалов, кроме небольшой коракоидной кости в
плечевом поясе, имелись и обе покровные кости — clavicula и cleithrum,
обладавшие нередко характером типичных кожных костей. Пояс хвоста-
тых амфибий отличается значительной шириной как лопаточного,
так, в особенности, коракоидного отдела (рис. 178). Оба отдела имеют вид
широких хрящевых пластинок. Брюшной отдел вырезкой подразделяется
на переднюю — прокоракоидную и большую заднюю — собственно ко-
ракоидную часть. В проксимальной части лопаточного отдела развива-
ется небольшое окостенение (scapula), которое нередко разрастается на
область сочленовной впадины, захватывая отчасти и коракоидный отдел
пояса, в котором самостоятельных окостенений нет (за исключением одной
190
Только формй — Siren, у которой наблюдается небольшая коракоиднай
кость). Брюшной отдел пояса правой и левой стороны налегает один на
другой, а позади на них налегает еще хрящевая пластинка, представля-
ющая собой грудину.
Полное отсутствие покровных костей на плечевом поясе у хвостатых
амфибий объясняется, очевидно, их вторичной редукцией.
У бесхвостых ам фи б и й в брюшном отделе пояса
имезтся затянутое перепонкой окошко, отделяющее переднюю, или про-
коракоидную, его часть, остающуюся хрящевой, от задней, окостенева-
Рис. 179. А — плечевой пояс стегоцефала (ре-
ставрация) и В — ископаемой рептилии (Pelyco-
sauria) (по Каве).
civ— clavicula; clt — cleithrum; co — coracoideum;
rest — episternum; sc — scapula; st — sternum.
Рис. 180. A — плечевой пояс
ящерицы с брюшной стороны (ив
Гегенбаура), В — плечевой пояс
и грудина чайки сбоку (ив Зелен-
ки). Обозначения см. рис. 179.
ющей — собственно коракоидной части (рис. 178, В). В проксимальной
части спинного отдела имеется хорошо развитая лопаточная кость, а на
широком надлопаточном хряще лежит пластинчатая покровная кость —
cleithrum. В брюшном отделе пояса имеется хорошо развитая коракоид-
ная кость, а на прокоракоидном хряще лежит тонкая покровная кость —
ключица. У более примитивных бесхвостых амфибий пояса правой и- ле-
вой стороны налегают друг на друга своими хрящевыми брюшными частями
(так называемый эпикоракоидный хрящ), как у хвостатых амфибий
(рис. 178), но у специализированных форм наступает прочное соединение
их между собой по средней линии. Вместе с тем наступает и более прочное
соединение с алиментами грудины (рис. 105).
У рептилий также нередко, в особенности у низших форм, в
поясе сохраняется еще значительное количество хряща: в спинном от-
деле — надлопаточный хрящ, а в брюшном отделе, впереди — прокора-
коидный й по внутреннему (брюшному) краю — эпикоракоидный хрящ
(рис. 180, Л). Имеются оба типичных окостенения первичного пояса —
лопатка и коракоид, но нередко (у ящериц) они в области сочленовной
ямки срастаются между собой в одну кость. У ящериц имеется типичная
покровная ключица, которая на большей части своего протяжения отде-
лена от первичного пояса и лежит впереди прокоракоида. Внутренние
концы ключиц соединяются с покровной костью грудины — надгрудин-
ником (episternum). Внутренние края коракоидного отдела пояса (эпико-
ракоидный хрящ) соединяются впереди между собой, а позади с грудиной.
Так как последняя у рептилий уже связана с концами ребер, то плечевой
пояс приобретает таким образом более прочную опору на остальном внут-
191
рением скелете. У крокодилов нет ни ключиц, ни прокоракоидов, и пояс
состоит только из лопатки и коракоида собственно. У черепах имеется
хорошо развитый коракоид и, кроме того, от лопатки на брюшную сторону
отходит большой акромиальный отросток, самостоятельно окостенева-
ющий и неправильно называемый прокоракопдом. Ключицы, так же как
и связанный с ними надгрудинпик, совер-
шенно обособились от пояса и вошли
у черепах в состав брюшпого щита (epi-
plastralia и endoplastron).
У птиц плечевой пояс полностью
окостеневает. Лопатка изменяет свою фор-
му и положение, — она тянется в виде
узкой и длинной, слегка изогнутой («са-
блевидной») кости разад вдоль спины.
Коракоиды хорошо развиты и соединяют-
ся с грудиной. У некоторых низших птиц в
широком коракоиде имеется окно (страус),
отделяющее передний прокоракоидпый
отдел, который у других форм является
в виде рудиментарного отростка, а у
огромного большинства птиц и вовсе исче-
Рис. 181. А—правая половина пле-
чевого пояса молодого клоачного
(утконоса) (А) — молодой обезья-
ны (В) (по Боасу), рсо — ргосо-
racoideum; остальные обозначе-
ния см. рис. 179.
зает. Ключицы птиц срастаются между
собой своими внутренними концами, об-
разуя характерную для птиц вилочку
(furcula) (рис. 180, В).
В плечевом поясе клоачных
м л е к о п и т а ю щ и х можно разли-
чить еще все существенные части типичного пояса наземных позво-
ночных (рис. 181, Л). Кроме лопаток, имеются хорошо развитые корако-
иды, упирающиеся своими нижними концами в грудину, и впереди от
коракоидов еще самостоятельные кости — прокоракоиды. Лопатки снаб-
жены особым выступом (acromion), к которому примыкает ключица, со-
Рис. 182. Левая половина плечевого пояса териодонтов и млекопитающих; А — Dicy-
nodon; В — Cynognathus; С — утконос; D — кролик. Хрящ обозначен точками,
ас — акромион; с — коракоид; ерг — коракоидпый отросток; ес — прокоракоид; д — су-
ставная ямка; sc — лопатка; sp — гребепь лопатки.
единяющаяся с передним краем надгрудипника. Акромиальный выступ
появляется и у некоторых рептилий (черепахи) и был хорошо развит у
близких к млекопитающим Theriodontia (рис. 182). У живородя-
щих млекопитающих лопатка снабжена гребнем (spina
scapulae), продолжающимся в хорошо развитый ,о р о м и о н. Кора-
коиды редуцируются и прирастают к лопатке, образуя на ней особый
коракоидный отросток (processus coracoideus). Этот отросток
всегда развивается в виде самостоятельной кости, но до грудины никогда
не доходит (лишь у зародышей сумчатых коракоиды иногда достигают
192
грудины). Прокоракоидов у живородящих млекопитающих Не наблюда-
ется (хотя в коракоиде иногда имеется два центра окостенения). Ключицы
у млекопитающих имеют характер смешанной кости, так как их внутрен-
ний конец отчасти замещает хрящ. У млекопитающих, конечности кото-
рых приспособлены к разнообразным движениям, наблюдаются обычно
хорошо развитые ключицы (многие" сумчатые, насекомоядные, грызуны,
рукокрылые и обезьяны). У млекопитающих, передние конечности кото-
рых специализировались для производства однообразных движений, как
у быстро бегающих и плавающих форм, они совсем редуцируются (копыт-
ные, некоторые хищные, китообразные, сирены). У человека ключица
очень хорошо развита, а лопатка отличается значительной шириной ди-
стальной (спинной) ее части и большим развитием гребня. Коракоидный
отросток довольно долго (до 12—15-летнего возраста) остается в виде
самостоятельной кости.
Тазовый пояс наземных позвоночных
Тазовый пояс наземных позвоночных отличается от пояса рыб силь-
ным развитием спинного его отдела, который только намечается у низших
рыб в виде небольшого отростка (processus iliacus) (рис. 175). Этот спин-
ной, или подвздошный, отдел пояса прикрепляется к ребрам крестцовых
позвонков, и таким образом тазовый
пояс, а следовательно, и задние конеч-
ности получают прочную опору на
осевом скелете. Брюшной отдел пояса
представлен широкой и первоначально
сплошной, как и у рыб, пластинкой,
прободенной лишь отверстием для про-
хождения
нерва (canalis obturatorius)
А
Рис. 183. Тазовый пояс стегоцефалов (А, В),
саламандры (С) и ископаемых рептилий
(D — Procolophon и Е — Diademodon) сбоку
(ив Боаса).
il — ilium; is — ischium; p—pubis.
Рис. 184. Тазовый пояс саламандры
с брюшной стороны (Л) и лягушки
Сбоку (В) и с брюшной стороны (С).
ас — acetabulum; ер — cartilage ypsiloi-
des; рр — pars pubis. Остальные об-
означения, как на рис. 183. (А—из
Видерсгейма).
(рис. 184). Правая и левая половины пояса соединяются между со-
бой по средней линии широким сращением (symphysis). Спинной
отдел пояса окостеневает у всех наземных позвоночных, образуя удлинен-
ную подвздошную кость (os ilium). В брюшном отделе тазового пояса
первоначально окостеневает лишь задняя часть, образующая седалищную
кость (os ischium) (рис. 184), а затем и передняя, где развивается лобковая
193
кость (os pubis) (рис. 183). Иногда все три кости срастаются затем между
собой. Начиная с рептилий, в сплошной первоначально брюшной пластинке
появляется окно, затянутое перепонкой (fenestra ischio-pubica). У иско-
паемых котилозавров его, однако, еще не было, а у некоторых примитив-
ных форм, как гаттерия, это окно относительно невелико и окна правой
и левой стороны разделены хрящом или даже костью (рис. 183,185). У дру-
гих рептилий окно увеличивается в размерах^ и окна правой и левой сто-
роны тогда разделены лишь тонкой связкой (ящерицы и крокодилы).
Canalis obturatorius наблюдается первоначально независимо от окна,
185). Но у черепах и млекопитающих окно
canalis obturatorius — foramen obturatum.
Сочленовная ямка (acetabulum) поме-
щается на месте схождения всех трех
костей.
У стегоцефалов наблюдают-
ся нередко уже все три окостенения
(рис. 183). У современных амфи-
бий в брюшном отделе пояса (pars
ischio-pubica) обычно имеется только одно
седалищное окостенение (os ischium), а
впереди него сохраняется хрящ (рис. 183,
184). Однако нередко окостенение захва-
тывает и этот лобковый отдел пояса,
причем иногда здесь наблюдается само-
кнаружи и впереди него (рис.
образуется путем разрастания
Рис. 185. Тазовый пояс гаттерии
(Hatteria) (по Видерсгейму).
ас — acetabulum; fip — fenestra
ischiopubica; fo—canalis obturato-
rius; * — proc, hypoiscliiadicus, Ос-
тальные обозначения см. рис. 183.
стоятельное окостенение, вскоре срастаю-
щееся с седалищной костью. Вполне самостоятельная лобковая кость
(os pubis) имеется только у одной бесхвостой амфибии (Dactylethra).
Нужно думать, что это окостенение, как и многие другие, утеряно
амфибиями вторично. Форма пояса различна у хвостатых и у бесхвостых
амфибий (рис. 184). У хвостатых подвздошный отдел представлен костью
умеренной длины, а седалищно-лобковый отдел имеет вид широкой пла-
стинки, продырявленной лишь отверстием для нерва. У бесхвостых амфи-
бий подвздошные кости значительно вытянуты в длину, а седалищно-лоб-
ковый отдел сильно сжат с боков, так что вся нижняя часть таза приобре-
тает вид вертикального диска, в середине которого находится сочленов-
ная ямка (вертлужная впадина). По средней линии впереди от брюшного
отдела пояса наблюдается у многих хвостатых амфибий (и Dactylethra из
бесхвостых) совершенно самостоятельный хрящ в виде расширенного
или расщепленного впереди стержня (cartilago ypsiloides) (рис. 184, Я).
У рептилий в тазовом поясе всегда хорошо развиты все три
кости, сходящиеся вместе в области вертлужной впадины (рис. 185).
В брюшной части пояса, между лобковой и седалищной костью, у совре-
менных рептилий имеется окно (fenestra ischio-pubica), сравнительно не-
большое у гаттерии, крупное у ящериц и, в особенности, у крокодилов.
Независимо от этого имеется и отверстие для нерва (canalis obturatorius),
которое у крокодилов соединяется с окном, а у черепах разрастается и
образует подобное же окно. Обе половины пояса связаны между собой
по средней линии хрящом, который при сильном развитии окон редуци-
руется и остается в виде связки между ними. Впереди этот хрящ продол-
жается иногда в непарный отросток (cartilago epipubis). Подобный отрос-
ток наблюдается иногда и у амфибий (Menobranchus), у двудышащих и га-
ноидных рыб (Polypterus). Таз крокодилов отличается большой величи-
ной окон, направленными вперед лобковыми костями и необычайной для
современных рептилий шириной подвздошных костей. У ископаемых
динозавров подвздошные кости были еще шире, а лобковые и седалищные
кости были еще более раздвинуты. Кроме того, лобковые кости иногда
были разделены па- две ветви — переднюю (praepubis) и заднюю (post-
754
pubis). Последняя направлена назад параллельно седалищным костям
(рис. 186, Л).
Тазовый пояс птиц отличается очень большой шириной подвздош-
ных костей (рис. 186), которые соединяются и срастаются с большим
числом позвонков, образующих сложный крестец. Все это связано с хожде-
нием на двух ногах. Другой особенностью птичьего таза является поло-
жение лобковых костей, которые направлены назад параллельно седа-
лищным. Наконец, у птиц ни лобковые, ни седалищные кости не образуют
между собой сращений. Нижние концы этих костей широко раздвинуты,
и это представляет необходимое приспособление для кладки крупных
яиц, снабженных твердой скорлупой. Лобковые кости по своему положению
напоминают postpubis некоторых динозавров, с тазом которых птичий
Рис. 18G. Тазовый попе динозавра
(А — Camptos4urus) и молодых птиц
Tinamu (В), Rhea (С) сбоку (по Боасу);
рр — postpubis. Остальные обозначения
см. рис. 183.
таз обнаруживает вообще очень боль-
шое сходство (рис. 186). Эмбриональ-
но лобковый отдел таза птиц напра-
влен первоначально вниз, а затем
повертывается все более назад. На-
блюдающийся у птиц передний от-
Рис. 187. Левая половина тазового пояса
клоачного млекопитающего (Ornithorhyn-
chus) (из Боаса).
ас — acetabulum; т — os marsupialc, Осталь-
ные обозначения см. рис 183.
росток (processus pectineus) представляет собой, быть может, рудимент
(рис. 186, р) передней ветви лонного отдела таза динозавров.
У млекопитающих, кроме типичных трех костей таза, разви-
вается еще одна небольшая вертлужная кость (os acetabulare), лежащая
в передней части вертлужной впадины. В образовании этой впадины
лобковая кость обыкновенно участия не принимает. У взрослого живот-
ного все кости таза срастаются между собой на каждой стороне в одну
безыменную кость (os innominatum). Foramen obturatum
окружено co всех сторон костью. У большинства млекопитающих имеется
широкое сращение, соединяющее и лобковый, и седалищный отделы между
собой (но у насекомоядных, у хищных и, в особенности, у обезьян и у
человека сращение имеется лишь в передней, лонной части пояса). У
ехидны и у сумчатых с лобковыми костями связана впереди пара костей,
служащих для укрепления сумки (ossa marsupialia) (рис. 187). Для мле-
копитающих по сравнению с другими позвоночными характерно иное
положение таза — довольно длинные подвздошные кости обращены вперед,
лонные невелики, а седалищные разрастаются назад. Таз приобретает,
благодаря этому, наклонное положение, с осью, направленной вниз и
назад (вентрокаудально). Вертлуг лежит позади крестца (рис. 187). Это
795
изменение положения связано с переходом от ползания к хождению и
беганию.
Особенностью человеческого таза являются его положение (сращение
обращено не столько к брюшной стороне, сколько вниз, т. е. морфологи-
чески назад по оси, так что длинная ось симфиза образует с осью тела
значительный угол) и широко разрастающиеся в бока подвздошные
кости, которые поддерживают внутренности при вертикальном положении
тела.
Скелет свободной конечности
Пятипалая конечность уже с самого начала отличается от типичного
рыбьего плавника по своему положению относительно остального тела
(рис. 175). В то время как у рыб передний край плавника у высших форм
становится верхним, так как горизонтальное прикрепление плавника
заменяется вертикальным, у наземных позвоночных (как и у двудышащих)
произошел обратный поворот, и передний край конечности становится
нижним. Когда рыба откидывает свой плавник назад, она прижимает
его к телу спинной поверхностью, между тем как наземное позвоночное
при аналогичном движении прижимает к телу брюшную (ладонную)
поверхность конечности. Впрочем, соединение конечности наземных
позвоночных с поясом столь подвижно, что допускает самое различное ее
положение. У низших форм проксимальная часть конечности (stylopodium)
нормально направлена почти прямо в сторону перпендикулярно к телу,
следующий отдел (zeugopodium) направлен в передней конечности слегка
вперед, а в задней назад и, наконец, сама лапка (autopodium) направлена
в сторону и вперед. У высших форм все эти изгибы выражены резче.
Проксимальный отдел конечности прижимается к телу и направлен в
передней конечности (плечо) вниз и назад, а в задней (бедро) — вниз и
вперед. Следующий отдел направлен в передней конечности (предплечье)
вниз и вперед, а в задней конечности (голень) — вниз и назад. Таким
образом, эти отделы образуют между собой угол, направленный в передней
конечности вершиной (локоть) назад, а в задней — вершиной (колено)
вперед (рис. 175). Сама лапка (кисть, ступня) лежит горизонтально и
направлена прямо вперед, так что морфологически передний ее край
(первый палец) обращен внутрь. Таким образом, вся конечность оказы-
вается расположенной в сагиттальной плоскости. Различие в расположенья
отделов передней и задней конечности определяется различной их функ-
цией: при передвижении животного передняя конечность тянет тело, а
задняя толкает.
Проксимальный конец пятипалой конечности в отличие от рыбьего
плавника соединяется с поясом шаровидной головкой, образующей с
сочленовной ямкой пояса очень подвижный сустав. Из двух элементов
следующего отдела первоначально, невидимому, более развитым был
задний (ulna, fibula), но нередко сильнее развивается передний (radius,
tibia), между тем как у специализированных форм задний (fibula) иногда
и вовсе редуцируется. Локтевая кость снабжена на своем проксимальном
конце локтевым выступом (olecranon), служащим для прикрепления мышц.
Кости этих отделов конечностей у высших позвоночных относительно
гораздо длиннее, чем у низших форм. В основании самой кисти у низших
наземныхпозвоночных наблюдается нередко очень большое число правиль-
но расположенных элементов, но у высших форм происходит сокращение
числа костей вследствие срастания некоторых элементов. Относительные
размеры этого отдела у высших форм меньше, чем у низших (ср. рис. 190
и 197).
В дистальных частях конечности у наземных позвоночных крупных
преобразований общего значения не наблюдается. По сравнению с репти-
лиями у всех других позвоночных наблюдается сокращенное число фаланг.
196
Если прогрессивное развитие пятипалой конечности, несомненно, сопро
вождается уменьшением числа фаланг,то также несомненно, что и редукция
конечностей связана с уменьшением числа фаланг. Этим объясняется,
повидимому, незначительное число фаланг у хвостатых амфибий.
Конечность хвостатых амфибий построена в высшей сте-
пени типично и содержит обыкновенно все существенные элементы схемы
(рис. 188). Проксимальные кости сравнительно еще коротки, а основание
лапки (basipodium) относительно значительно крупнее, чем у высших
позвоночных. Число фаланг сравнительно невелико (обычно 2, 2, 3, 3, 2),
Передняя конечность имеет всегда только 4 пальца, причем недостает
пятого пальца, который следует считать редуцированным. Задняя конеч-
ность имеет обычно 5 пальцев. Однако
Рис. 188. Задняя конечность хво-
статой амфибии (Spelerpes) (из
Видерсгейма).'
с — centrale; F — fibula; Fe — fe-
mur; f — fibulare; i — intermedium;
m,— mediate; T —tibia; t—tibiale;
2 — tarsalia distalia 1 — 2; 3 — 5 —
tarsalia distalia; I—V—metatarsalia.
иногда конечность заметно редуцируется,
в особенности, у постоянножаберпых ам-
фибий, и число пальцев вторично сокра-
щается до трех или даже двух (Amphiuma,
Рис. .189. Основание ступни хвостатой амфибии
(Ranldens). Обозначения см. рис. 188.
Proteus). Одновременно с этим наблюдается и значительное упрощение
строения основания лапки, так что число его элементов может сократиться
до трех (Proteus). В запястье нормальной конечности имеются следующие
элементы: radiale, intermedium, ulnare, centrale proximate, 1—2 centralia
distalia (medialia), carpale distale 1 + 2 и carpalia distalia 3 и 4. Как видно,
кроме обычной центральной косточки, имеется еще один или два элемента
среднего ряда (medialia). Один из дистальных элементов является комплекс-
ным — результат сращения первого и второго. Задняя конечность очень
сходна с передней, отличаясь от нее лишь несколько большей полнотой.
Нормально имеется пять вполне развитых пальцев, а иногда еще незна-
чительные х'ящики, рассматриваемые как рудименты лишних лучей
(рис. 189). Кроме обычной центральной косточки, наблюдается еще от
одного дс трех элементов среднего ряда (medialia 1, 2, 3). Первые два
дистальных элемента предплюсны срастаются между собой, образуя такой
же комплекс, как и в передней конечности. Таким образом, в предплюсне
нормально имеются следующие элементы: tibiale, itermedium, fibulare,
197
central© proximale, 1—3 Centralia distalia (medialia), tarsale distale 1 + 2
и tarsalia 3, 4 и 5.
В конечности хвостатых амфибий прекрасно сохраняется первона-
чальное правильное лучевое расположение элементов, и такое же правиль-
ное расположение наблюдалось и у стегоцефалов, конечность которых
по своему строению очень близка к конечности хвостатых амфибий.
У стегоцефалов также наблюдается несколько элементов среднего ряда
в основании лапки, а вместо комплекса первых двух дистальных элементов
Рис. 190. Задняя конечность стего-
цефала Trematops 'по Уиллистону).
Обозначения см. рис. 188.
Рис. 191. Передняя конечность сте-
гоцефала Eryops (по Майнеру).
Обозначения см. рис. 177.
иногда имелись самостоятельные косточки (tarsalia distalia 1 и 2) (рис.
190, 191). Состав конечности хвостатых амфибий можно выразить следу-
ющими диаграммами.
с с с
b b b Ь Ь b b
а а а а а а а а а
I 11 ! III IV 1 II III IV V
1 + 2 3 4 1 + 2 3 4 5
(т2) in1 (m2) (m3)
с с
г u t I
i
R U T F
H Fo
Конечность б есхвостых амфибий весьма значительно изме-
йена вследствие приспособления к особому способу передвижения скачка-
ми. Чрезвычайно развиты и специализированы, в особенности, задние,
конечности. В передней конечности лучевая и локтевая кости срастаются
между собой. Вполне развиты только последние четыре пальца. Первый
палец имеется, но совершенно рудиментарен. В основании кисти interme-
dium срастается с lilnere, a centrale с radiale, Элементы среднего ряда
198
эмбрионально закладываются (в числе трех), ио затем прирастают к
дистальным элементам. Таким образом, первоначально довольно большое
число элементов сокращается у примитивных бесхвостых амфибий до 7;
но у более специализированных форм срастаются между собой еще и
дистальные элементы, начиная с последних, так что число костей запястья
падает до 5, а иногда даже до 3. В задней конечности бесхвостых прокси-
мальные элементы отличаются большой длиной; берцовые кости сраста-
ются менаду собой. Все пять пальцев прекрасно развиты и, кроме того,
имеется значительный рудимент переднего пальца (praehallux), состоящий
из нескольких члеников. Две проксимальные кости предплюсны — tibiale
и fibulare — чрезвычайно сильно вытянуты в длину и сращены своими
концами; они образуют добавочный рычаг конечности. Другие проксималь-
1Рис. 192. Передняя конечность гат-
терии (Hatteria) (по Гаусу из Север-
цова).
— centrAIe; г — intermedium; т — me-
diate; В — radius; г — radiale; pis —pi-
siforme; V—ulna; u—ulnare; 1—a —
.carpalia distalia; I—V — metacarpajia,
Рис. 193. Передняя конечность примитивной
рептилии Ophiacadon (по Уиллистону),
о! — локтевой отросток; остальные обозначения
см. рис. 102.
ные элементы исчезли, а дистальные элементы редуцированы и представ-
лены лишь двумя-тремя небольшими косточками.
В конечностях рептилий проксимальные кости заметно
удлинены, а область основания лапки обычно значительно сокращена.
Число фаланг больше, чем у других позвоночных, и нормально выра-
жается следующей формулой: 2, 3, 4, 5, 3 (рис. 192), падая только у чере-
пах до 2, 3, 3, 3, 3. Обе конечности имеют по пяти пальцев. У змееобразных
ящериц и у змей конечности редуцируются и у большинства змей совер-
шенно исчезают, что находится в связи с изменением способа передвижения.
Передняя конечность сохраняет в общем примитивное строение и
содержит следующие элемиты запястья: radiale, intermedium, centrale,
ulnare и carpalia distalia 1—-;<> (рис. 192,193). Однако в эмбриональной конеч-
ности нередко наблюдается еще два-три элемента среднего ряда. Кроме
того, и у взрослого живо,тцрго обычно имеется еще один элемент, лежащий
снаружи (или с ладонной поропы) PT ulnare и носящий название г о р о -
199
ховидной косточки (pisiforme). Гомолог этой косточки наблю-
дается иногда эмбрионально и у амфибий и представляет собой, невиди-
мому, остаток заднего луча. Эмбрионально у рептилий наблюдается и
остаток переднего луча в виде элемента, лежащего кнаружи от radiate
(radiale externum). У гаттерии (рис. 192), у большинства ящериц (рис. 194)
и у черепах передняя конечность построена вполне типично, и только
у крокодилов наблюдается дальнейшее упрощение. У последних две
Рис. 194. Основание передней (Л) и задней (В) конеч-
ности молодого геккова (Ascalabotes) (по Северцеву).
tpr — tarsale proxhnale, Остальные обозначения
см. рис. 177 и 188.
проксимальные кости
radiale и ulnare значи-
тельно увеличены, а
другие элементы сокра-
щаются и срастаются
между собой.
В задней конечно-
сти рептилий наблю-
дается значительное уп-
рощение строения пред-
плюсны, связанное от-
части с развитием ме-
зотарзального сустава
между проксимальным
и дистальным рядами
костей (рис. 194, В).
Проксимальные кости
прочно связаны связка-
ми с концами берцовых
костей и срастаются между собой, а дистальные кости прочно связаны с
основаниями костей плюсны и иногда прирастают к ним. У черепах наблю-
даются иногда еще самостоятельные fibulare и centrale, наряду с компле-
ксом tibiale + intermedium, у других черепах и у крокодилов здесь име-
ются две проксимальные кости (tibiale + intermedium + centrale и fibu-
lare), а у большинства черепах и у всех ящериц только один большой
комплекс — tarsale proximate (рис. 194, В). Из дистальных элементов
у рептилий tarsalia distalia 4 и 5 постоянно срастаются между собой,
образуя так называемый кубоид. У ящериц первые два дистальных эле-
мента (tarsalia distalia 1 и 2) срастаются с соответствующими костями
плюсны (metatarsalia 1 и 2) (рис. 194). Строение конечности рептилий
можно выразить следующими диаграммами, где знаком 4-обозначены
постоянные срастания, а знаком — характерные для многих форм (в
особенности, для ящериц).
ь С b d с b е d с b С b ь С Ь d с b е d с b с b
а а а а а а а а а а
I II III IV V I II III IV V
1 2 3 4 5 1 2 3 4 + 5
с ш1 с
г i и pis t 4-i — f
R и Т F
Н Fe
Конечности птиц изменены чрезвычайно сильно. Передняя конеч-
ность преобразована в крыло, и вся дистальная часть ее редуцирована.
У птиц сохраняются лишь три первых пальца с рудиментарными фаланга-
ми. Запястье состоит только из двух проксимальных костей (radiale,
сросшиеся с intermedium, и ulnare, сросшееся с pisiforme) (рис. 195, Л).
Дистальные элементы у зародыша также закладываются (рис. 196, Л),
200
но впоследствии срастаются с основаниями пястных костей, которые
также срастаются между собой в одну кость — carpometacarpus.
Задняя конечность птиц представляет собой в высшей степени спе-
циализированный орган, но построенный, однако, по тому же типу,
как и конечность рептилий. Малая берцовая кость (fibula) редуци-
руется и прирастает к большой берцовой. Проксимальные элементы
предплюсны срастаются между собой (рис. 196, В), как и у рептилий,
но затем прирастают к берцовой, образуя tibiotarsus; дистальные элементы
редуцируются и срастаются между собой и с элементами плюсны, образуя
одну длинную цельную кость — цевку (tarsometatarsus). Число пальцев
обычно равняется четырем, и
Рис. 195. Скелет крыла ворона (Л) и ноги
молодого цыпленка (В) (по Боасу).
ста — carpometacarpus; tmt — tarsometatarsus; Tt—
tibiotarsus; 1—3 — пальцы. Остальные обозначения
прежние.
первый палец противопоста-
влен остальным (рис. 195,В).
Конечности ископаемых ди-
Рис. 196. Скелет конечностей за-
родыша пингвина (А — передней,
В — задней) (по Шеудеру из
Зеленки).
1—4 — пальцы. Остальные обозна-
чения прежние.
нозавров по многим признакам занимают промежуточное положе-
ние между типичными конечностями рептилий и конечностями птиц
(рис. 406).
Конечности млекопитающих отличаются обычно очень зна-
чительной длиной проксимальных отделов, но в общем сохраняют довольно
примитивное строение (рис. 197). Обычно имеются все пять пальцев и не-
редко также рудименты переднего луча (praepollex в передней конечности и
praehallux в задней). Постоянным элементом передней конечности является
и pisiforme. Число фаланг у млекопитающих сокращенное и выражается
следующей формулой: 2, 3, 3, 3, 3. Общая форма конечности весьма разно-
образна и находится в зависимости от приспособления ее ко всевозможным
специальным функциям (рис. 3). У роющих форм конечности коротки
и очень широки (кроты, броненосцы и др.). У плавающих форм конечности
преобразуются в ласты, длинные кости также укорачиваются, а число
фаланг увеличивается (киты, сирены). У летающих млекопитающих
чрезвычайно удлиняются пальцы (2—5), на которых натянута летательная
перепонка (рукокрылые).
Большинство млекопитающих при хождении опирается па всю
кисть и ступню (стопоходящие), но быстро бегающие формы, как, например,
высшие хищники, некоторые грызуны и пр., опираются только на концы
пальцев (пальцеходящие). У лучших бегунов концы пальцев при атом
201
защищены копытами, и животное опирается только на концы средних
пальцев (третий и четвертый у парнокопытных или третий у непарно-
копытных); крайние пальцы редуцируются (рис. 198), и у высших форм
остаются только два одинаково развитых средних пальца (жвачные),
основания которых (metacarpalia, metatarsalia) срастаются между собой
в одну кость, или, наконец, остается лишь один третий палец (лошади)
(рис. 420). У лазающих млекопитающих развивается хватательная ко-
нечность, которая характеризуется противоположением первого пальца
всем остальным. В передней конеч-
ности вместе с тем развивается спо-
собность лучевой кости вместе с
кистью вращаться вокруг локтевой
кости так, что рука из первичного
пронированного положения (ргопа-
tio) ладонной стороной вниз может
переходить в супинированное (supi-
tr.m
Рис. 197. Скелет передней (Л) и задней (В) конечностей лазающего примитивного,
млекопитающего (опоссум, Didelphys marsupialis) (по Верслуису).
ol — локтевой отросток; ph и рр — praehallux и pracpollex; tub. с — пяточный бугор;
tr. т. и tr —большой и малый трохантср. Остальные обозначения прежние.
natio), ладонью вверх (при параллельном положении костей предплечья);
Передняя конечность млекопитающих довольно примитивна. Нижний
конец плечевой кости часто пронизан отверстием (foramen entepicondylare),
которое наблюдается иногда и у рептилий. Лучевая и локтевая кости
обычно одинаково развиты, но иногда последняя редуцируется. Локтевая
кость снабжена значительным проксимальным выступом (olecranon). В
запястье имеются все типичные элементы: radiale, intermedium, ulnare,
pisiforme, centrale distale (mediate) и carpalia distalia 1, 2, 3 и 4 + 5
(рис. 197 и 199, Л). Характерным является срастание двух последних
дистальных элементов между собой. Эмбрионально находили и закладку
centrple proximate, которое затем срастается обычно с intermedium.
Centrale distale часто срастается с radiale. Задняя конечность несколько
203
более изменена. Проксимальный конец бедренной кости несет 2—3 выступа
(troch an teres) для прикрепления мышц. В голени малая берцовая обычно
развита слабее большой и иногда редуцируется и прирастает к последней.
Рис. 198. Скелет задней конечности парнокопытных; гиппопотама, свиньи, оленька,
оленя и быка (по Лейтгардту). Обозначения прежние.
позади большой пяточный выступ и
Рис. 199. Кисть и ступня человека (из
Гегепбаура). Обозначения прежние
(рис. 188, 177)
В коленном сочленении развивается сухожильная кость — коленная
чашечка (patella). У млекопитающих имеются только две довольно зна-
чительные проксимальные кости предплюсны (рис. 197 и 199, В). Наружная,
пяточная кость (calcaneus) имеет
представляет собой fibulare. Вну-
тренняя, таранная кость (astra-
galus) представляет, повидимому,
комплекс intermedium ]- centrale.
В голеностопном сочленении уча-
ствуют почти исключительно tibia
и astragalus. Tibiale, повидимому,
обычно редуцируется (согласно
другому взгляду, она входит в со-
став таранной кости). У млекопи-
тающих имеется также крупная
кость среднего ряда — ладьевид-
ная кость (naviculare), соответ-
ствующая centrale distale (media-
le), с которой иногда, повиди-
мому, срастается и tibialo. Из
дистальных элементов предплюсны
последние два, как и у рептилии,
срастаются между собой и обра-
зуют кубоид (tarsalia distalia 4 1- 5).
Конечнозтп человека построены вполне типично (рис. 199).
Передняя конечность является хватательной, как у обезьян. В ней
найдены и закладки центральных косточек, из которых одна срастается
с intermedium, а другая — с radicle (из высших обезьян центральная
203
косточка остается самостоятельной только у орапга). Задняя конечность
человека, несомненно, вторично утратила особенности хватательной
конечности обезьян (рис. 430). Первый палец потерял способность проти-
вопоставляться остальным и получил очень значительное развитие, что
связано с прямохождением человека.
В анатомии человека принята своя номенклатура, которая в прила-
гаемой таблице сопоставлена со сравнительноанатомической.
Carpus
radiale
intermedium (+ centrale
ulnare
centrale distale (mediate)
pisiforme
carpale distale 4
carpale distale 2
carpale distale 3
carpalia distalia 4 + 5
Naviculare (scaphoideum)................
Lunatum (semilunare)....................
Triquetrum (cuneiforme).................
Centrale................................
Pisiforme (accessorium).................
Multangulum majus (trapezium)...........
Multangulum minus (trapezoideum) . . .
Capitatum (magnum)......................
Hamatum (uncinatum).....................
pr.)
Tarsus
Talus (astragalus)...........
Calcaneus....................
Naviculare (scaphoideum) . .
Cuneiforme 4 (entocuneiforme)
Cuneiforme 2 (mesocuneiforme)
Cuneiforme 3 (ectocuneiforme)
Cuboideum....................
intermedium + centrale pr.
( + tibiale?)
fibulare
centrale distale (-|- tibiale?)
tarsate distale 4
tarsale distale 2
tarsale distale 3
tarsalia distalia 4 + 5
Свод важнейших фактов и положений морфологии скелета
Если хорда как постоянный орган и сохраняется еще у некоторых
низших позвоночных (круглоротые, цельноголовые, осетровые и двуды-
шащие), то у огромного большинства их она более или менее полно вы-
тесняется развивающимися телами позвонков. Основой для развития
осевого скелета является система соединительнотканных оболочек и пе-
регородок, которая затем дополняется хрящом как более совершенной
формой скелетной ткани, и, наконец, последний вытесняется еще более
прочной костью. У высших позвоночных во время их эмбрионального раз-
вития наблюдается последовательная смена тех же трех стадий разви-
тия скелета.
Позвоночник в наиболее примитивном виде состоит из одних лишь
дуг, сидящих на постоянной хорде. У низших рыб в каждом сегменте
развиваются две пары верхних и две пары нижних дуг. Обычно задние
пары получают преобладающее развитие, а передние редуцируются,
сохраняясь у низших рыб в виде вставочных хрящей. Все части позвонков
развиваются за счет этих первичных элементов, а также особых окосте-
нений вокруг хорды. Боковые отростки представляют собой продукт
разрастания нижних дуг. У позвоночных развивается два рода ребер и
боковых отростков, различающихся по своему положению и иногда
наблюдаемых совместно. В туловище низших наземных позвоночных
имеются верхние ребра, у Polypterus и костистых рыб одновременно обе
пары ребер, а у остальных позвоночных — нижние, лежащие первона-
чально непосредственно под брюшиной. В связи с первичными дугами
позвонков развиваются и тела позвонков, и соответственно множествен-
ности первых и тела первоначально составлялись из нескольких частей,
что прослежено палеонтологически. У селахий тела развиваются внутри
оболочки хорды, а также путем разрастания и слияния оснований дуг
вокруг хорды. Таким образом развились цельные позвонки, состоящие из
тела, на котором первичные дуги с их производными имеют вид отростков.
204
У высших рыб и у наземных позвоночных тела развиваются в онтогенезе
независимо от дут. У последних боковые отростки располагаются на верх-
них дугах и развивается двойная связь ребра с позвонком.
У рыб имеются лишь две области позвоночника — туловищная о
ребрами и хвостовая с гемальным каналом. У всех наземных позвоночных
появляются еще две новые области: впереди подвижная шейная, а на
границе между туловищной и хвостовой областью—крестцовая. Послед-
няя служит для укрепления тазового пояса, подвздошные кости которого
сочленяются с ребрами истинных крестцовых позвонков. Для всех амниот
характерно преобразование первых двух позвонков в атлас и эпистрофей,
образующие особый вращательный сустав. Число шейных позвонков
ограничивается у млекопитающих семью.
Грудина развивается только у наземных позвоночных. Первично она
состоит из пластинки, связывающей концы коракоидов с передними
ребрами. Эта грудина нарастает затем вследствие срастания между собой
концов более или менее значительного числа ребер.
Надгрудинник представляет собой кость, кожную по происхождению.
Он наблюдается у стегоцефалов и рептилий, сохранившись и у клоачных
млекопитающих, где вступил в тесную связь с хрящом первичной
грудины.
Мезодерма головы сегментируется в спинной своей части так же,
как и в туловище, образуя ряд головных сомитов. Метамерное распределе-
ние париетальной мускулатуры головы у взрослых позвоночных теряется,
так как первые три миотома превращаются в глазные мышцы, а следующие
редуцируются. Эта потеря метамерии обусловлена развитием черепа,
благодаря которому голова утратила свою гибкость. Мезодерма скле-
ротомов служит материалом, из которого строится осевая часть черепа,
окружающая и защищающая головной мозг и высшие органы чувств. Пере-
пончатый вначале череп превращается затем в хрящевую коробку, развитие
которой начинается от пары хрящей, лежащих по бокам переднего конца
хорды (парахордалии), пары хрящей, лежащих у основания переднего
мозга (трабекулы), и иногда еще пары глазничных хрящей. В заднем,
хордальном, отделе черепа наблюдаются иногда еще следы сегментации
и нередко происходит дальнейшее прирастание элементов позвоночника
к затылочной его области. Хордальная часть черепа представляет собой,
несомненно, результат преобразования передней части осевого скелета
(позвоночника). Прехордальная часть черепа представляет собой ново-
образование. Кроме указанных хрящей, развиваются еще хрящевые
оболочки — капсулы вокруг органов чувств. Обонятельная капсула
прирастает к переднему концу трабекул, а слуховая — к переднему
отделу парахордалий. Слиянием хрящей между собой образуется основа-
ние черепа, от которого затем хрящ разрастается по бокам от мозга и,
наконец, появляется и в крыше.
Только у низших позвоночных осевой череп сохраняет вид хрящевой
коробки. Уже у хрящевых ганоидов в качестве скелетного материала
появляется и кость: разросшиеся кожные чешуи образуют на голове
сплошной наружный покров из многочисленных и не вполне еще постоян-
ных костей; появляются и покровные кости, подстилающие череп снизу
(парасфеноид и сошник). У высших рыб число покровных костей черепа
сокращается; они приобретают довольно постоянное расположение, и,
кроме того, появляются и замещающие кости, которые почти полностью
вытесняют хрящ, в особенности в задней половине черепа. Боль-
шинство характерных костей черепа высших рыб имеется и у наземных
позвоночных.
Различают два вида кости — покровную, или «вторичную», кость,
развивающуюся прямо в соединительной ткани и представляющую собой
погрузившееся вглубь кожное окостенение, и замещающую, или «первич-
205
ную», кость, развивающуюся на хряще или внутри его (во всяком случае
под перихондрием) и вытесняющую последний.
Из высших рыб наиболее типично построен череп костных ганоидов
и кистеперых, у которых имеется еще значительное количество хряща,
но вместе с тем развиваются и все типичные окостенения. Покровные
кости крыши черепа образуют сплошной панцырь, совершенно покрыва-
ющий его снаружи и с боков, оставляя лишь отверстия для глаз.
Для черепа наземных позвоночных характерно развитие затылоч-
ного сочленения с позвоночником посредством затылочных бугров —
двух у амфибий, одного у рептилий и птиц и вторично парного у млеко-
питающих. По своему составу череп наземных позвоночных очень близок
к черепу костных ганоидов и кистеперых, и у древнейших наземных по-
звоночных (стегоцефалов) он обладал такой же сплошной наружной кры-
шей из кожных костей, в которой имелись лишь отверстия для глаз. Для
наземных позвоночных характерна также более или менее значительная
редукция наружной крыши черепа. Эта редукция произошла независимо
у амфибий и у рептилий. У древнейших ископаемых рептилий наружная
крыша черепа была сплошная, как у стегоцефалов (стегальный череп).
Редукция этой крыши в височной области приводит к образованию височ-
ных ям и системы височных дуг между ними (зигальный череп). В одной
группе рептилий развились две височные ямы и соответственно две гори-
зонтальные височные дуги. В другой группе развилась одна верхняя ви-
сочная яма и только одна скуловая дуга, сохранившаяся и у современных
черепах. От этой группы унаследовали свою единственную скуловую дугу
и млекопитающие. В черепе последних наблюдается образование харак-
терных комплексов костей и заметное изменение общей конфигурации
в связи с прогрессивным развитием головного мозга и соответственным
увеличением черепной коробки.
Висцеральная часть черепа развивается независимо о! осевой части
за счет экто-мезенхимы ганглионарных пластинок в виде ряда следующих
друг за другом висцеральных дуг. Все висцеральные дуги представляют
собой первоначально равнозначащие образования; они развились в жа-
берных перегородках, несли одинаковую функцию, были единообразно
расчленены на четыре отдела и соединялись на брюшной стороне между
собой посредством ряда непарных копул. Одна из передних висцеральных
дуг изменила свою функцию и сохранила лишь два средних отдела в виде
первичных челюстей — нёбноквадратного хряща и меккелева хряща.
Впереди лежащие дуги редуцировались, и их остатки, невидимому, со-
хранились у хрящевых рыб в виде губных хрящей. Позади лежащая гио-
идная дуга служила для укрепления челюстной дуги и сохранила в пол-
ной мере также лишь два средних отдела, из которых верхний превратился
в подвесок для челюстей. С усовершенствованием отдельных жабер число
жаберных щелей и дуг сокращалось также и сзади. У большинства рыб
осталось пять типичных жаберных дуг.
С появлением кости висцеральный аппарат частью окостеневает,
частью покрывается покровными костями.
Значение верхнего отдела челюстной дуги падает с появлением вто-
ричных челюстей, развивающихся в верхней губе, независимо от челюст-
ной дуги. Уже у высших рыб зубы обычно несет только нёбная кость, а
у наземных позвоночных нёбноквадратный хрящ окончательно теряет
значение верхней челюсти. Соответственно передняя часть нёбноквадрат-
ного хряща редуцируется, а задняя, окостеневающая в виде квадратной
кости, служит лишь подвеском для нижней челюсти. Покровные кости,
развившиеся на нёбноквадратном хряще, сохраняются в составе дна че-
репа. У некоторых рептилий и у птиц они иногда до известной степени
подвижны, но обычно у высших форм соединяются неподвижно с другими
костями. Скелет крыши ротовой полости образован у высших форм вто-
206
ричным твердым небом, развивающимся за счет небных отростков Пред-
челюстных и челюстных костей, к которым могут присоединиться нёбные
отростки нёбных и иногда крыловидных костей. Квадратная кость служит
у наземных позвоночных подвеском для нижней челюсти и может при-
обрести известную подвижность. Верхний отдел подъязычной дуги с уте-
рей значения подвеска редуцируется, но не исчезает, а приобретает новую
функцию передачи звуковых колебаний и преобразуется в слуховую
косточку — столбик. Для млекопитающих характерно развитие нового
причленения нижней челюсти к черепной коробке. Квадратная кость
теряет значение подвеска и преобразуется в другую слуховую косточ-
ку млекопитающих — наковальню. Сочленовная кость нижней челюсти
обособилась от зубной и преобразовалась в третью слуховую косточку —
молоточек. Взаимная связь слуховых косточек пе является новой, это
остаток связи подвеска рыб с квадратной костью п последней — с сочле-
новной. Нижний отдел подъязычной дуги и жаберный скелет преобразу-
ются у наземных позвоночных в подъязычный аппарат, передние
рожки которого образованы гиоидом, и в гортанные хрящи.
Скелет непарных плавников возник в виде метамерного ряда хряще-
вых стержней в первоначально непрерывной непарной складке. Путем
разделения этой складки образовались затем отдельные плавники. Пер-
вичный хрящевой скелет дополняется кожными образованиями, разви-
вающимися в лопасти плавника.
Скелет парных конечностей возник также в виде метамерного ряда
хрящегых стержней в непрерывных первоначально парных боковых склад-
ках. Путем развития конечных частей складок и редукции промежуточ-
ной области образовались передние и задние конечности. Хрящевые лучи
слиллсь своими основаниями, и путем разрастания этой основной части
скелета плавника в стенках тела развились пояса конечностей.
Передний, или плечевой, пояс имеет вид дужки, охватывающей тело
с боков, и снабжен выступом для причленения скелета плавника у рыб
или ямкой для причленения скелета свободной конечности у наземных
позвоночных. С развитием значительного вторичного пояса из кожных
костей, прикрепляющихся верхним концом к черепу, первичный пояс
высших рыб значительно редуцируется. У наземных позвоночных хорошо
развит первичный пояс, спинной и брюшной отделы которого дистально
значительно расширяются. Из покровных костей пояса у наземных по-
звоночных сохраняется только ключица. У млекопитающих весь брюшной
отдел первичного пояса редуцируется, и остаток коракоида прирастает
к лопатке.
Задний, или тазовый, пояс является у рыб парной по происхождению
пластинкой, лежащей в брюшной стенке тела. Редко он снабжен восхо-
дящим отростком. Последний у наземных позвоночных сильно развит и
соединяется с ребрами крестцовых позвонков, образуя подвздошный отдел
пояса.
Скелет парных конечностей низших рыб представляет собой гибкую
пластинку, поддерживаемую 1—3 основными хрящами, которые тесно
связаны между собой и несут более или менее значительное число членис-
тых лучей первичного скелета. В парных плавниках рыб наблюдается
такое же развитие вторичного кожного скелета, как и в непарных плав-
никах, и со значительным его развитием у высших рыб первичный скелет
заметно редуцируется.
Конечности наземных позвоночных произошли от плавников с сильно
расчлененным скелетом, обладавшим одним только основный! элементом.
Прочное соединение элементов скелета нарушается и заменяется подвиж-
ным соединением посредством суставов. Число лучей уменьшается до
пяти. Проксимальные элементы скелета конечности сильно увеличива-
ются в размерах, а средние элементы уменьшаются, нередко срастаются
207
й теряют первоначальное лучевое расположение. Последнее сохраняется
у высших форм лишь в дистальных частях конечностей.
Различные отделы конечности соединяются между собой подвижными
сочленениями и образуют вместе сложный рычаг, различно изогнутый в
передней и задней конечностях.
МЫШЕЧНАЯ СИСТЕМА
МУСКУЛАТУРА БЕСПОЗВОНОЧНЫХ
Наиболее примитивный вид мускулатуры наблюдается у кишечнополостных
животных, у которых мускульные злементы еще не обособлены, а представлены лишь
особыми сократимыми мускульными отростками эпителиальных клеток. В эктодерме
эти отростки образуют сплошной слой продольных волокон, а в энтодерме — слой
кольцевых. У высших кишечнополостных наблюдается уже известная диференци-
ровка, а у медуз мускульные отростки приобретают даже поперечную полосатость,
но все же они еще не являются самостоятельными элементами.
У плоских червей мускулатура вполне обособлена от эпителия и образует непо-
средственно под эктодермой сплошную массу, состоящую ив наружных кольцевых,
внутренних продольных и иногда еще косых гладких мускульных волокон. Вся эта
масса тесно связана с кожей и образует так называемый кожно-мускульный мешок.
Кроме того, и паренхима тела пронизана многочисленными мускульными волокнами.
У кольчатых червей кожно-мускульный мешок состоит ив непрерывного слоя коль-
цевых волокон, лежащих непосредственно под эктодермой, и слоя продольных воло-
кон, который обычно разделен на четыре продольные мышцы и развивается ва счет сте-
ной полости тела (соматоплевры). Кроме того, наблюдаются особые мускульные пучки,
прикрепляющиеся н щетинкам, мышцы в перегородках полости тела и, наконец, слой
кольцевых волокон окружает кишечник, обусловливая его перистальтику. Точно так
же имеется слой мускульных клеток и в стенках кровеносных сосудов. Все эти мышцы
развиваются также ва счет мезодермы. У более специализированных червей, а также
и у моллюсков кожно-мускульный мешок нередко дает начало более или менее круп-
ным мышцам специального значения, но почти все эти мышцы, как и у аннелид, отно-
сятся к гладким. Поперечно-полосатая мускулатура имеется лишь иногда в продоль-
ной мышце аннелид и в сердечной мышце моллюсков. У головоногих моллюсков почти
вся мускулатура является поперечно полосатой.
У членистоногих развивается типичная поперечнополосатая мускулатура,
которая прикрепляется к наружному их скелету и образует сложную систему обособ-
ленных мышц. У некоторых раков и насекомых даже и мускулатура внутренностей
состоит ив поперечнополосатых мускульных волокон. Сравнение мускулатуры у раз-
личных животных показывает ясно, что поперечнополосатые мышцы не отличаются
от гладких ни по своему происхождению, ни по общей функции. У одних животных
вся мускулатура, как «произвольная», так и «непроизвольная», является гладкой, у
других и та, и другая может быть поперечнополосатой. Можно только сказать, что
поперечнополосатые мышцы способны к более быстрым и внергичным сокращениям и
могут в связи с повышением интенсивности работы возникнуть нв месте гладких мышц
в любом органе.
Гораздо более существенным оказывается деление всей мускулатуры на сомати-
ческую, которая обусловливает движение тела, и висцеральную, которая производит
движения внутренних органов (кишечника, сосудов и пр.). Это деление может быть
проведено у всех животных, хотя обычно между этими группами мышц нет заметных
различий в их происхождении и иннервации. Но у хордовых животных различие между
соматической и висцеральной мускулатурой оказывается уже гораздо более глубоким.
У ланцетника вся соматическая мускулатура развивается за счет внутренней
стенки сомитов, которые по сравнению с нижестоящими вторичнополостными живот-
ными являются выростами спинной стенки полости тела (соматоплевры). У взрослого
животного эта поперечнополосатая мускулатура является в виде продольной боковой
мышцы, поделенной студенистыми перегородками на ряд сегментов (миомеров), соот-
ветствующих эмбриональным миотомам. и иннервируется она исключительно брюш-
ными спинномозговыми нервами. Гладкая мускулатура сосудов и кишечника разви-
вается ва счет остальной мезодермы, т. е. боковых пластинок. Но, кроме того, ва счет
последних развивается еще одна мышца, состоящая ив слоя поперечнополосатых во-
локон, охватывающих снизу околожаберную полость, — так называемая поперечная
мышца (musculus transversus).. Эта висцеральная мускулатура иннервируется нерв-
ными волоннами, выходящими ив мозга в-составе спинных спинномозговых нервов.
МУСКУЛАТУРА ПОЗВОНОЧНЫХ
У позвоночных, как и у ланцетника, ясно выражено деление на сома-
тическую (париетальную) и висцеральную мускулатуру, которая раа-
208
йятся и по своему развитию, и по иннервации. Последняя является очень
важным критерием для суждения о происхождении той или иной мышцы.
Дело в том, что каждое мускульное волокно связано с определенным нерв-
ным волокном и образует вместе с ним одну физиологическую единицу.
Таким образом, и целая мышца неразрывно связана с определенным нер-
вом. Эта связь устанавливается с пер вых моментов появления мускульных
элементов, что всегда сопровождается и диференцировкой особых нервных
элементов (эпителиальномускульные клетки кишечнополостных также
уже связаны посредством отростков с нервными отростками нервных
клеток) и не может нарушаться ни при каких филогенетических преобра-
зованиях. Всякое нарушение этой связи привело бы к прекращению функ-
ции всего аппарата. Поэтому когда одна мышца в результате известной
диференцировки разделяется на несколько новых мышц, то и ее нерв де-
лится на соответствующее число ветвей. Если несколько мышц соединя-
ется для образования одной, то последняя иннервируется несколькими
нервами. Если мышца развивается прогрессивно, то увеличивается и
нерв; если мышца редуцируется, то одновременно исчезает и соответству-
ющий нерв. Мышца, развивающаяся из определенного мускульного сег-
мента, иннервируется всегда нервом данного сегмента, и т. д.
Соматическая мускулатура позвоночных вся по-
перечнополосата, развивается за счет мускульного листка миотома и ин-
нервируется нервами, волокна которых выходят из мозга в составе брюш-
ных корешков спинномозговых нервов (и соответствующих им двигатель-
ных головных нервов). Висцеральная мускулатура по-
звоночных является частью гладкой, частью поперечнополосатой. Она
развивается за счет остальной мезодермы, т. е. боковой пластинки (в виде
исключения иногда и за счет эктодермы), и иннервируется висцеральными
нервами, волокна которых у низших позвоночных, как и у ланцетника,
выходят в составе спинных корешков (и соответствующих им смешанных
головных нервов). Поперечнополосатая висцеральная мускулатура не
может быть выделена в качестве особой системы, так как она представ-
ляет собой лишь результат более высокой диференцировки известных
участков этой мускулатуры.
ВИСЦЕРАЛЬНАЯ МУСКУЛАТУРА
Часть гладкой мускулатуры позвоночных развивается иногда еще
непосредственно за счет основного слоя эктодермы. Таким образом
развиваются гладкие мускульные волокна кожи амфибий, которые про-
низывают ее в различных направлениях и образуют тонкий сплошной
покров на их кожных железах. Такой же покров из продольных гладких
мускульных клеток эктодермального происхождения характеризует и
потовые железы млекопитающих (и млечные железы клоачных). Наконец,
за счет ретинального листка радужины развиваются мускульные волокна
радужины, образующие кольцевой замыкатель (m. sphincter pupillae) и
радиальный расширитель зрачка (m. dilatator pupillae). У птиц эта му-
скулатура оказывается поперечнополосатой.
Все эти мышцы иннервируются висцеральными нервами симпати-
ческой системы.
Вся остальная мускулатура развивается за счет мезодермы.
Гладкая мускулатура кожи развивается за счет элементов кожного листка
сомита. Мускулатура кишечника развивается за счет висцерального листка
боковых пластинок, а мускулатура мочеполовых органов и сосудов — за
счет мезенхимы, происходящей из боковых пластинок, или частью (со-
суды кожи) из мезенхимы самой кожи. Мускулатура кожи состоит из во-
локон различного направления, которые лишь редко дают начало более
или менее обособленным мускульным пучкам (к влагалищам перьев у
209
птиц — m. arrectores plumarum и к сумкам волос у млекопитающих —
ш. arrectores pilorum). Мускулатура сосудов образована непрерывным
слоем кольцевых мускульных волокон, которые на капиллярах ложатся
продольно. На различных внутренних органах наблюдаются обычно слои
кольцевых и продольных волокон. Стенки кишечника окружены также
внутренним слоем кольцевых волокон и наружным слоем продольных, со-
вместное действие которых вызывает перистальтические движения кишки.
В области желудка имеется еще слой косых волокон. Вся эта муску-
латура иннервируется висцеральными нервами главным образом симпа-
тической нервной системы.
Мы видели, что гладкая мускулатура замещается иногда поперечно-
полосатой. Это наблюдается, например, в радужине глаза птиц. То же
самое произошло на известном участке брюшного кровеносного сосуда,
в котором мускульная стенка развилась очень значительно и стала попе-
речнополосатой. Таким образом развилось сердце позвоночных. Такая
замена гладкой мускулатуры поперечнополосатой представляет у позво-
ночных довольно редкое явление, но имеется, однако, еще целая область,
где подобный процесс наблюдается не как исключение, а как правило.
Это — передняя часть кишечника, висцеральная мускулатура которой
является поперечнополасатой у всех позвоночных и простирается назад
на более или менее значительную часть пищевода. Иногда эта мускула-
тура разрастается и на область желудка или даже на самую кишку, где
она тогда образует наружный мускульный покров поверх обычного глад?
кого (линь, вьюн). Иногда и стенка плавательного пузыря приобретает
поперечнополосатую мускулатуру (костные ганоиды).
Особенно значительно развивается поперечнополосатая висцераль-
ная мускулатура в области глотки, где она вступает в связь с распола-
гающимся здесь висцеральным скелетом и становится произвольной. Эта
мускулатура подвергается у позвоночных значительной диференцировке,
образует большинство мышц головы и представляет собственно висцераль-
ную мускулатуру в тесном смысле этого слова (точнее — висцеральную
мускулатуру висцерального аппарата). Эта мускулатура развивается,
как и остальные мышцы кишечника, за счет мезодермы боковых пласти-
нок и иннервируется рядом висцеральных нервов головы, которые иннер-
вируют вообще соответствующую область кишечника. Задний из этих
нервов — блуждающий нерв — иннервирует и значительную позадиле-
жащую часть кишечника, по меньшей мере до желудка включительно
(у низших позвоночных иногда и весь кишечник).
Висцеральная мускулатура висцерального аппарата и ее производные
Висцеральная мускулатура висцерального аппарата представляет
Собой результат диференцировки первоначально сплошной массы коль-
цевых волокон, охватывающих переднюю часть кишечника. У низших
позвоночных большая ее часть состоит еще из совершенно непрерывной
массы поперечных волокон, охватывающих весь висцеральный аппарат
снизу и с боков и образующих общий его сжиматель — mus-
culus constrictor superficialis (рис. 200, 201, 155). От этой мышцы уже у
круглоротых и у селахий обособляются отдельные более глубокие мус-
кульные пучки, вступающие в более тесную связь с отдельными висце-
ральными дугами. В переднем отделе висцерального аппарата часть этих
мышц подвергается более сложной диференцировке, в особенности у кру-
глоротых, в связи с развитием их сложного губного аппарата. Висцераль-
ная мускулатура круглоротых покрыта снаружи целым рядом миотомов
боковой мышцы, разделенных здесь на спинные и брюшные участки,
между которыми располагаются наружные жаберные отверстия. У рыб
соматическая мускулатура ташке отчасти покрывает висцеральный аппа-
210
рат, образуя продольные мускульные ленты с брюШной, а иногда и со
спинной стороны, но никогда не имеет здесь столь значительного развития,
как у круглоротых (рис. 200). У низших рыб (селахий) еще очень
хорошо развит общий сжиматель висцерального аппарата (ш. constrictor
super! icialis) в виде слоя мускульных волокон, охватывающих весь аппа-
рат сбоку и снизу. В этой непрерывной мышце различают лишь известные
Рис. 200. Схема соотношений между париетальной и висцеральной мускулатурой.
А — передний нонен, тела сбоку; В — поперечный разрез через голову; аЪг — жаберная дуга»
аи __ ухо; Ъг — жаберный мешок; d— спинной нерв (смешанный); hyp — подъязычный
нерв; md— спинной и mv—-брюшной отдел боновой мышцы; ту — миотом; па—ноздря;
п — мозг; ос — глаз; ог. os —; нижняя и верхняя косая мышца; par — париетальная (подъ-
язычная) мускулатура; re, ri, rif,rs — наружная, внутренняя, нижняя и верхняя прямые
мышцы глаза; sp — брызгальце; v — брюшной нерв (двигательный); ^isc — висцеральная
мускулатура.
области, как, например, спинную часть во сти висцеральных щелей,
и брюшную, лежащую снизу (рис. 201). здо существеннее деление
ее на области иннервации, которые совпадают с общим сегментальным
делением всего аппарата. Впереди, в области челюстной дуги вплоть до
брызгальца, мышца иннервируется тройничным нервом (5-я пара), далее
позади, в области подъязычной дуги от брызгальца и до первой жаберной
щели, мышца иннервируется лицевым нервом (7-й пары), затем в области
первой жаберной Дуги, вплоть до второй экаберной щели, она иннерви-
руется языкоглоточным нервом
(9-й пары), и, наконец, вся зад-
няя часть мышцы представляет
собой область блуждающего
нерва (10-й пары). В передней
части сжимателя наблюдается
обособление крупных мускуль-
ных масс, прикрепляющихся
к обоим отделам челюстной
дуги. Часть, прикрепляющаяся
к верхнему отделу челюстной
дуги, лежит глубже и предста-
вляет собой мышцу, подни-
мающую нёбноквад-
ратный хрящ (m. levator
palatoquadrati, V) (рис. 201)1.
Часть, прикрепляющаяся к
нижнему отделу, представляет
нижнюю челюсть (ш.
I pg sda spir bri
ил CSS
Рис. 201. Висцеральная мускулатура акулы
(Heptanchus) (по Феттеру).
adm—m. adductor mandibular; Ы I — первая
жаберная щель; css—- m. constrictor superficialis;
im — m. intermandibularis; Ipq — m. lavator pa-
latoquadrati; spir — брызгальце; tr—m. trapezius.
собой мышцу, приводящую
adductor mandibulae) (рис. 201). Эти
мышцы иннервируются тройничным нервом. Брюшная часть сжима-
теля, лежащая снизу между ветвями нижней челюсти, представляет собой
межчелюстную мышцу (m. intermandibularis, V, VII). Перед-
няя часть последней иннервируется тройничным, а задняя — лицевым
1 Римские цифры при названии мышц обозначают головной нерв, иннерви
рующий данную мышцу.
211
нервом. Кроме того, и в области жаберного аппарата обособляются неко-
торые более глубокие мышцы, прикрепляющиеся к элементам жаберных
дуг: mm. adductores arcuum, связывающие оба средние отдела каждой дуги
(epi- и ceratobranchialia), и mm. arcuales dorsales, соединяющие верхние
концы соседних дуг (epi- и pharyngobranchialia). Наконец, позади жабер-
ного аппарата со спинной стороны наблюдается обособленная часть об-
щего сжимателя, которая небольшими пучками прикрепляется к послед-
ним жаберным дугам, а главной своей массой прикрепляется к переднему
краю спинного отдела плечевого пояса; это — трапециевидная
мышца (m. trapezius, XI) (рис. 201). У высших рыб мускулатура вис-
церального аппарата несколько сложнее диференцпрована, и, между
прочим, обособляются особые мышцы подвеска (m. adductor hyomandi-
bulae, VII) и жаберной крышки (т. opercularis, VII, который в свою
очередь разбивается на систему отдельных мышц).
У земноводных иу других наземных позвоночных в связи
с развитием аутостилии и неподвижным прикреплением верхнего отдела
челюстной дуги к черепу, естественно, должна была редуцироваться и
мышца, поднимающая нёбноквадратный хрящ. Эта мышца сохраняется
только на стрептостильных черепах в виде mm. levatores pterygoidei
(у ящериц и у змей) и mm. orbito-quadrati (у птиц). Остатки этой мышцы,
но уже с иной функцией, наблюдаются и у амфибий в виде мышцы, подни-
мающей глаз (hi. levator bulbi, V). Мышца, приводящая нижнюю челюсть
(ш. adductor mandibulae), распадается на более поверхностную мышцу,
отходящую от элементов скуловой дуги, — жевательную мышцу
(ш. masseter, V) и более глубокую, отходящую от крыши черепа и за-
полняющую височную яму черепа, — височную мышцу (m. tem-
poralis, V). У высших наземных позвоночных также
различимы обе эти мышцы, причем первая (m. masseter) отходит от ску-
ловой дуги и прикрепляется к наружной стороне нижней челюсти, а
вторая (ш. temporalis) помещается в височной яме и прикрепляется к
венечному отростку челюсти. Кроме того, обособляется еще более глубокая
мускульная масса, отходящая от крыловидной кости и прикрепляющаяся
к внутренней поверхности нижней челюсти, — это крыловидная
мышца (m. pterygoideus, V), которая у млекопитающих делится еще
на наружную и внутреннюю. Между ветвями нижней челюсти у наземных
позвоночных сохраняется передняя часть общего сжимателя в виде меж-
челюстной мышцы (m. intermandibularis, V, VII). У млекопитающих
развивается в особенности передняя часть этой мышцы, принадлежащая
к области тройничного нерва, и получает название подбородочно-подъ-
язычной мышцы (m. mylohyoideus, V, VII).
В области мускулатуры лицевого нерва у наземных позвоночных
вместо мышц подвеска и жаберной крышки рыб наблюдается довольно
крупная мышца, прикрепляющаяся к заднему углу нижней челюсти.
Так как она действует на челюсть позади точки опоры (сустава), то при ее
сокращении челюсть опускается, так что она является антагонистом
приводящей мышцы. Это мышца, опускающая нижнюю че-
люсть (m. depressor mandibulae, VII). У млекопитающих она, очевидно,
перенесла свое прикрепление далее вперед (на зубную кость), и у высших
форм к ней впереди присоединяется еще часть, обособившаяся от меж-
челюстной мышцы и иннервируемая тройничным нервом. Эта часть мышцы
имеет вид особого переднего брюшка так называемой двубрюшной мышцы
(m. digastricus mandibulae, V, VII).
Задняя часть поверхностного сжимателя той же области иннервации
(VII) разрастается у рептилий в довольно крупную мышцу, охватывающую
шею снизу и с боков, — сжиматель шеи (ш. sphincter colli, VII).
У млекопитающих эта мышца делится на более глубокий слой, сохра-
няющий прежнее название (m. sphincter colli), и поверхностный слой,
272
располагающийся непосредственно под кожей (platysma myoides). Волокна
этих слоев имеют косое направление и перекрещиваются между собой.
Эти мышцы разрастаются на всю область головы и дают начало сложной
системе подкожной лицевой мускулатуры. От более
глубокого сжимателя шеи обособляются здесь мышцы рта (m. orbicularis
oris, m. buccinator и др.), а от более поверхностной платизмы — мышцы
ушные и окружающие глазницу (m. auriculares, т. orbicularis oculi и др.).
У обезьян и у человека эти мышцы особенно сложно диференцированы
и образуют так называемую мимическую мускулатуру (рис. 202). При
общем высоком развитии этой мускулатуры у человека отдельные ее
мышцы подвергаются редукции, как, например, ушные мышцы, имеющиеся
обычно лишь в виде нефункционирующих рудиментов.
Мышцы собственно жаберного аппарата, принадлежащие к области
языкоглоточного и блуждающего нервов, с утратой жаберного дыхания
Рис. 202. Лицевая мускулатура и нервы обезьяны (Propithecus)
(но Руге из Видерсгейма).
у наземных позвоночных, естественно, редуцируются, но частью сохра-
няются в виде мышц подъязычного аппарата, глотки и, в особенности, в
виде гортанных мышц. Трапециевидная мышца окончательно теряет
связь с жаберными дугами и становится уже исключительно мышцей
плечевого пояса. Она разрастается у наземных позвоночных и особенно у
млекопитающих, где она отходит не только от шейной и грудной области
позвоночника, но и от самого черепа вплоть до сосцевидного его отростка
и прикрепляется не только вдоль гребня лопатки и ее акромиального
отростка, но и к ключице и даже к грудине. Эта нижняя часть мышцы
иногда обособляется и получает название грудино-ключично-
сосцевидной мышцы (m. sterno-cleido-mastoideus, XI), между
тем как верхняя часть сохраняет прежнее название (m. trapezius s. cucul-
laris, XI). Иннервирующая эту мышцу задняя часть блуждающего нерва
обособляется у высших наземных позвоночных и становится самостоятель-
ным головным нервом 11-й пары (n. accessorius).
213
Соматическая мускулатура
Вся соматическая (париетальная) мускулатура позвоночных разви-
вается исключительно за счет внутреннего, мускульного листка миотома
(следовательно, за счет лишь небольшого участка первоначальной сома-
топлевры, выростом которой является сомит). Мускульный листок миотома
значительно утолщается, клетки его становятся цилиндрическими и теряют
свои границы, так что в каждом сегменте развивается один сплошной
синцитий со многими ядрами. На внутренней поверхности мускульного
синцития начинается образование мускульных волоконец (фибрилл),
и вслед за тем эта поверхность увеличивается вследствие образования
складок, врастающих внутрь синцития и делящих его, в конце концов,
на целый ряд горизонтальных мускульных лент. По всей поверхности
мускульных лент развиваются фибриллы, между тем как внутри остается
первоначально неизмененная саркоплазма с ядрами. Однако вслед за
этим происходит дальнейшее увеличение поверхности путем разделения
каждой мускульной ленты на целый ряд мускульных волокон, содержащих
по периферии мускульные фибриллы. В промежутки между мускульными
лентами, а затем и между волокнами врастает соединительная ткань,
служащая для обособления мелких мускульных единиц и несущая кро-
веносные сосуды для их питания. Таким образом, из эпителиальных
стенок миотомов развивается мускульная ткань взрослого животного.
У высших позвоночных эти процессы протекают несколько проще.
Наряду с процессами гистогенеза наблюдаются и морфологические
изменения. Нижний конец миотома образует эпителиальный вырост,
который растет вниз, охватывая стенку полости тела снаружи (рис. 69, В).
Эти брюшные отростки миотомов разрастаются до взаимного соприкосно-
вения; во внутренней их стенке также идет развитие продольных мускуль-
ных волокон, и таким образом развивается брюшной отдел миомеров
взрослого животного. В результате всех этих пропессов эпителиальные
миотомы, лежащие в спинной части тела зародыша, превращаются в
миомеры боковой мышцы, охватывающие тело взрослого животного со
всех сторон, хотя спинные их отделы всегда остаются более массивными.
Глазные мышцы
Глазные мышцы развиваются из трех (пяти) предушных головных
миотомов, которые распадаются на мезенхиму, образующую затем зачатки
отдельных мышц. У всех позвоночных развиваются четыре прямые
мышцы, отходящие все вместе от дна глазницы с задней ее стороны и
прикрепляющиеся к верхнему, переднему (внутреннему), нижнему и
заднему (наружному) краю глазного яблока, и две косые мышцы,
отходящие от передней стенки глазницы и прикрепляющиеся к верхней
и нижней стенке глазного яблока (рис. 203). Из первого (предчелюстного)
миотома головы развиваются три прямые мышцы — верхняя (ш.
rectus superior, III), внутренняя (m. rectus internus, III) и н и ж-
н я я (m. rectus inferior, III) инижняя косая мышца (m. obli-
quus inferior, III). Все эти мышцы иннервируются глазодвигательным
нервом (3-й пары), являющимся нервом этого сегмента (рис 203). Из вто-
рого челюстного миотома, являющегося, невидимому, двойным, разви-
вается одна только верхняя косая мышца (m. obliquus superior,
IV), иннервируемая блоковым нервом (4-й пары), а из третьего (также
двойного) миотома — наружная прямая мышца (m. rectus exter-
nus, VI), иннервируемая отводящим нервом (6-й пары).
У наземных позвоночных от наружной' прямой мышцы обособляется
более глубокая порция, идущая прямо к основанию глазного яблока;
это — мышца, втягивающая глазное яблоко (m. retractor
214
bulbi, VI), иннервируемая, согласно своему происхождению, отводящим
нервом. При втягивании глазного яблока внутрь глазницы отчасти пас-
сивно происходит смыкание век над глазом и в особенности третьего
века или мигательной перепонки, которая обычно лишена собственной
мускулатуры. У рептилий и птиц от
вающей глазное яблоко, развивается
перепонки (m. nicticans). У мле-
копитающих прямые мышцы
глаза располагаются тесное вокруг
зрительного нерва, но в общем здесь
имеются те же мышцы, что и у ниже-
стоящих форм. Верхняя косая мышца
получает у млекопитающих свое-
образное положение: опа удлиня-
ется, причем место ее отхода пере-
носится от переднего края глазницы
в глубь последней; однако при этом
положение ее конечной части фикси-
руется у переднего края глазницы
посредством сухожильного ушка, об-
разующего блок, через который пе-
рекидывается мышца на своем пути,
и, таким образом, несмотря на зна-
чительное перемещение места при-
крепления, действие мышцы остается
дистальной части мышцы, втяги-
специальпая мышца мигательной
0
Рис. 203. Глазные мышцы (схема).
oi — нижняп и os — верхняя косые мыш*
цы; гс — наружная, ri — нижняя, rit —
внутренняя и rs — верхняя прямые мыш-
цы; III, IV, VI — нервы.
в сущности тем же. Мышца, втягивающая глазное яблоко, имеется и
у большинства млекопитающих, но у обезьян она редуцируется. У выс-
ших обезьян и у человека лишь редко наблюдаются остатки этой мышцы.
Кроме того, у млекопитающих иногда от верхней прямой мышцы обо-
собляется еще порция волокон, образующая мышцу, поднимающую
верхнее веко (m. levator palpebrae superiors, имеющую иногда и иное
происхождение) и иннервируемую глазодвигательным первом.
Подъязычная мускулатура
Полный ряд заушных миотомов головы сохраняется во взрослом
состоянии только у круглоротых, где они разделяются па спинной и
брюшной отделы, образуя наджаберную и поджаберную мускулатуру.
У рыб и у наземных позвоночных передние заушные миотомы совершенно
исчезают, а задние, затылочные, миотомы редко сохраняются в виде мало
измененных остатков в области затылка, большей яге частые образуют
вместе с передними туловищными миотомами особую наджаберную и
поджаберную, или подъязычную, мускулатуру. Наджаберная мускула-
тура наблюдается в виде системы небольших мышц между верхними
концами жаберных дуг только у низших рыб. Она иннервируется заты-
лочными нервами (n. spino-occipitales). Поднгаберная, или подъязычная,
мускулатура имеется, наоборот, у всех позвоночных.
Развивается подъязычная мускулатура за счет брюшных отростков
затылочных, а иногда и передних туловищных миотомов, которые растут
вниз, огибая сзади жаберный аппарат, и затем вперед, ложась в виде
сплошной массы под висцеральным аппаратом. У взрослого животного
здесь имеется диферепцированная па отдельные пучки масса продольных
мускульных волокон, нередко поделенная сухожильными перегородками
(inscriptiones tendineae) па отдельные сегменты. Эта мускулатура пред-
ставляет собой непосредственное продолжение вперед брюшной мускула-
туры туловища, и сегментация ее у низших позвоночных — остаток
прежнего деления на миомеры (рис. 200); однако соответствие этой сегмен-
275
со а
тации с первоначальной обычно утрачивается. У низших рыб (рис. 204)
подъязычная мускулатура идет от плечевого пояса вперед и расщепляется
на пучки, идущие к отдельным висцеральным дугам, — mm. согасо-
arcuales (m. coraco-branchiales, m. coraco-hyoideus и m. coraco-mandibu-
laris). У земноводных подъязычная муску-
латура представляет непосредственное
продолжение прямой мышцы брюха. Она
распадается, как и у всех других назем-
ных позвоночных, на задний участок,
идущий от плечевого пояса и грудины до
подъязычного аппарата — m. sterno-hyoi-
deue (от грудины) и m. omo-hyoideus (от
пояса), и передний участок, идущий от
подъязычного аппарата до подбородка,—
подбородочно-подъязычная мышца (m. ge-
nio-hyoideus). От последней у наземных
позвоночных обособляется и собственная
мускулатура языка — подборо-
дочно-язычная мышца (m. genio-glossus),
служащая для выдвигания языка, и
подъязычно-язычная мышца (m. hyoglos-
sus), служащая для его втягивания.
Вся подъязычная мускулатура иннер-
вируется соответственно своему происхо-
ждению комплексом затылочных и иногда
передних спинномозговых нервов, обра-
зующих так называемый подъязычный
нерв (n. hypoglossus), который только
у высших позвоночных становится типич-
ным головным нервом (12-й пары).
Мускулатура туловища
У низших позвоночных вся мускула-
тура туловища состоит, как и у ланцет-
ника, из ряда миомеров правой и левой
стороны, которые вместе образуют так
называемые боковые мышцы.
Боковая мышца каждой стороны раз-
делена горизонтальной соединительноткан-
ной перегородкой на спинную и брюшную мышцы (перегородки нет
только у миног), и соответственно каждый миомер поделен на спинной
и брюшной отделы (рис. 200). Каждый миомер первоначально иннерви-
руется двигательной ветвью одного спинномозгового нерва — спинной
отдел—спинной веткой; а брюшной отдел—брюшной веткой. Вертикаль-
ные миосепты, служащие перегородками между отдельными миомерами,
изгибаются, образуя на границе между спинным и брюшным отделами
угол, направленный вершиной вперед. Затем верхние и нижние концы
миотомов изгибаются вперед, и таким образом каждая миосепта образует
ломаную линию в виде буквы W, средний угол которой лежит на гори-
зонтальной перегородке и направлен вершиной вперед. Однако миосепты
не остаются вертикальными и по отношению к сагиттальной плоскости —
они изгибаются в каждом отделе, образуя конусы, направленные посредине
вершиной вперед. Таким образом миомеры взрослого животного образуют
довольно сложную систему конусов, вкладывающихся друг в друга и
имеющих в поперечном разрезе вид концентрических колец. Первона-
чальное продольное направление волокон уже у рыб начинает изменяться
Рис. 204. Подъязычная мускула-
тура акул (Heptanchus) (по^Фет-
теру).
тс — myocomma; md — нижняя че-
люсть; со. а — m. coracoarcuales;
со. Ы — т. coracobranchiales; со,
hy — т. coracohyoideus; со. md —m.
coracomandibularis; со — плечевой
пояс; Ьг 1—7—жаберные дуги;
hy — подъязычная дуга.
Q16
в области брюшного отдела мускулатуры. В боковых стенках брюха
волокна принимают косое направление, причем нередко оно различно
на различной глубине. По средней линии брюха сохраняется продольное
направление волокон. Таким образом, здесь намечается уже некоторая
диференцировка внутри самих миомеров. Эта диференцировка, образуя
посредине прямые и по бокам косые мышцы брюха, приводит к обособле-
нию различных слоев брюшной мускулатуры.
Спипная мускулатура наземных позвоночных
У наземных позвоночных прикрепление миосепт точно приспосабли-
вается к отдельным сегментам скелета, что связано с развитием подвиж-
ного сочленения между позвонками.
У амфибий туловищная мускулатура состоит еще из ряда независимых
миомеров, подразделенных, как и у рыб, горизонтальной септой на спин-
ной и брюшной отделы. У амниот горизон-
тальная септа исчезает и граница между
спинной и брюшной мускулатурой стирается.
Начиная с рептилий, миомеры частью раз-
биваются на отдельные мускульные пучки,
сохраняющие метамерное распределение и
связывающие между собой отдельные части
соседних позвонков, частью соединяются ме-
жду собой для образования более крупных
мышц (рис. 205). Таким образом развиваются
остистые мышцы (mm. inters pinales), соеди-
няющие верхние остистые отростки, попереч-
ные (mm. intertransversales), соединяющие
между собой поперечные отростки, попереч-
но-остистые (mm. transversospinales). От по-
перечных отростков к ребрам идут попереч-
нореберные мышцы (mm. transversocostales).
От передних позвонков к голове идут пря-
мые и косые мышцы головы (mm. recti et
obliqui capitis). Вдоль всей спины в углу
между остистыми и поперечными отростками
лежит длинная мышца спины (m. longissimus
dorsi), передняя часть которой может обо-
собиться в виде длинной мышцы головы
(m. longissimus capitis). По бокам над реб-
рами лежит система mm. ileo-ccstales. На-
конец, позади тазового пояса диференци-
руется особая хвостовая мускулатура (ча-
стью за счет брюшной мускулатуры), остат-
ки которой наблюдаются и у бесхвостых
форм (включая человека).
В результате этой сложной диференци-
ровки от первоначальной метамерии спинной
мускулатуры у высших позвоночных сохра-
няются только следы в виде метамерного
Рис. 205. Спинная мускула-
тура млекопитающего (из Верс-
луиса). Плечевой пояс с его
мускулатурой удален.
с — ребра; сг — череп; er. sp. —
m. erector spinae; ilc — m. ileo-
costalis; lc, Id — m. longissimus
dorsi, spl — m. splenitis; tr.sp —
m. transversospina lis.
распределения отдельных мышц, связываю-
щих между собой последовательные позвонки. Значение спинной мускула-
ТУРЫ, которая у рыб является важнейшей частью локомоторного аппарата,
у наземных позвоночных в общем гораздо меньше. Вместо этого разви-
вается весьма сильно мускулатура конечностей.
217
Брюшная мускулатура наземных позвоночных
У хвостатых амфибий и в брюшном отделе боковой мышцы туловища
сохраняется деление на миомеры. Однако и у низших форм наблюдается
ясная диференцировки этой мускулатуры на отделы и слои, различаю-
щиеся направлением волокон (рис. 206, 208). Как и у высших рыб, посре-
дине брюха наблюдается обособление продольной мускульной полосы,
сохраняющей продольное направление своих волокон. Это прямая
мышца брюха (m. rectus abdominis), по средней линии которой
имеется соединительнотканная перегородка (linea alba), отделяющая,
как и у рыб, мускулатуру правой и левой стороны. В боковых стенках
брюха направление волокон меняется, причем различно на различной
глубине: в поверхностных слоях волокна направляются косо назад к
брюху (рис. 205, 207), в более глубоких они идут косо назад и к спине, а
в самых глубоких слоях принимают приблизительно поперечное направ.
Рис. 206. Поверхностная брюшная мускулатура тритона (по Мауреру).
h. Ъг—поджаберная мышца; тс — ммосепта; о. е. s. — m, obliquus externus superficialis;
r. i. — m. rectus internus r. s — m. rectus superficialis.
ление. У низших амфибий все три слоя еще связаны друг с другом, и ди-
ференцировка выражена лишь в различном направлении волокон, но у
более высоко стоящих форм наступает полное обособление этих слоев
друг от друга и соответственное образование наружной косой
мышцы брюха (m. obliquus externus), внутренней косой
мышцы брюха (m. obliquus internus) и поперечной мышцы
(т. transversus abdominis). Лежащие по средней линии продольные волокна
прямой мышцы также вполне обособляются от соседних мышц. У высших
амфибий, как и у прочих наземных позвоночных, миосепты исчезают,
и мускульные волокна в косых и в поперечной мышце идут без перерыва
на всем их протяжении. Следы прежней сегментации сохраняются, однако,
еще в наружной косой мышце некоторых примитивных млекопитающих
и в особенности в прямой мышце брюха даже у высших форм (inscripti-
ones tendineae). У обезьян, в особенности у человекообразных, и у чело-
века прямая мышца брюха значительно уменьшается и состоит лишь из
незначительного числа сегментов.
С значительным развитием ребер у высших наземных позвоночных
(амниот) брюшные мышцы разделяются в грудной области на отдельные
участки, лежащие между ребрами. Это м е ж р е б е р н ы е мышцы,
наружная и внутренняя (mm. intercostales externus et internus), являю-
щиеся непосредственным продолжением косых мышц брюха и играющие
большую роль при совершении дыхательных движений (рис. 207). В по-
ясничной области им соответствует квадратная поясничная мышца (т.
quadratus lumborum), продолжающаяся и под межреберными мышцами
и примыкающая сбоку к поперечной. Впереди ребер та же мускулатура
является в виде лестничных мышц (mm. scaleni). У млекопитающих на-
ружная и внутренняя межреберные мышцы дают начало еще новым про-
дуктам диференцировки — задним зубчатым мышцам, верхней и нижней
(mm. serrati postici superior et inferior), которые специализируются в
качестве дыхательных мышц, служащих для расширения грудной клетки.
У сумчатых с брюшной стороны от прямой мышцы брюха обособляется
еще особая мышца, отходящая от сумчатых костей и доходящая впереди
218
до грудины. Это пирамидальная мышца (га. pyramidalis), которая имеется
и у низших плацептных млекопитающих (насекомоядные), а у прочих
наблюдается лишь в виде рудимента, связанного с лонными костями..
Вся эта сложная диференцировка, очевидно, обусловлена сменой
функции брюшной мускулатуры. Если у рыб она, как и спинная муску-
латура, несла функцию локомоторную, то
теперь с изменением способа передвижения
и приобретением легочного дыхания она
служит для сжимания брюшной полости и,
в частности, для совершения дыхательных
движений.
Мускулатура конечностей
Мускулатура непарных плавников раз-
вивается в виде метамерного ряда каплео-
бразных эпителиальных выростов спинных
или брюшных концов миотомов (рис. 163).
Эти мускульные почки врастают в основание
закладки плавника, теряют связь с миото-
мами и превращаются в целый ряд синци-
тиев, в которых затем идет образование
мускульных волокон. Таким образом, соот-
ветственно каждому лучу внутреннего ске-
лета развивается с каждой стороны по одной
первичной лучевой мышце (m. radialis).
В более концентрированных плавниках лу-
чевые мышцы тесно сближаются между собой,
образуя более или менее сплошной слой
с каждой стороны скелета. У высших рыб
отдельные лучевые мышцы диференцируются
на глубокие и поверхностные.
Мышцы плавников прикрепляются все-
гда к лучам кожного скелета. Иннервирует-
ся эта мускулатура числом спинномозговых
нервов, равным количеству миотомов, при-
нявших участие и их рзавитии.
Мускулатура парных плавников закла-
дывается также в виде ряда мускульных
почек, вырастающих от нижних концов
миотомов (по две от каждого миотома у се-
лахий и по одной у высших рыб) (рис. 163).
Мускульные почки делятся каждая на верх-
нюю и нижнюю вторичные почки, которые
врастают в основание закладки плавника со
спинной и брюшной его стороны и превра-
щаются в такие же первичные лучевые мыш-
цы (mm. radiales), как и в непарных плав-
никах. Лучевые мышцы тесно сближа-
Рис. 207. Брюшная мускула-
тура гаттерии (по Мауреру).
ici — внутренние межреберные
мышцы; o. l. s. —поверхностная
наружная косая мышца; pct —
грудная мышца; pst— брюшное
«ребро»; st — грудина; с—ребра;
г — прямая мышца.
ются между собой и образуют довольно сплошной мускульный слой на
спинной (m. levator s. abductor) и на брюшной (т. depressor s. adductor)
стороне плавника. Отдельные пучки этого слоя довольно точно соответ-
ствуют лучам внутреннего скелета, но прикрепляются, как и в непарных
плавниках, к лучам кожного скелета. У высших рыб наблюдается упро-
щенное развитие мускулатуры — мускульные почки быстро теряют свои
контуры и расплываются в массу мезенхиматозных клеток, образующих
сплошной слой миогенпой бластемы на спинной и на брюшной стороне
27Р
плавника. В пределах спинной (отводящей) и брюшной (приводящей)
мышц плавника наблюдается и некоторая диференцировка, по крайней
мере на поверхностные и глубокие мышцы.
Мускулатура конечностей на* земных позвоночных раз-
вивается также из ряда мускульных почек, которые быстро теряют свои
контуры и расплываются в массу мезенхиматозных клеток, образующих
сплошной слой на спинной (тыльной) и на брюшной (ладонной) стороне
зачатка конечности. В пределах этих мускульных зачатков происходит
затем диференцировка, причем мускульные зачатки передней конечности
образуют проксимальные выросты, разрастающиеся в области грудного
пояса и даже дальше на спину и на грудь, образуя мышцы плечевого
пояса. Мышцы, развивающиеся из этих первичных мускульных зачат-
ков, гомологичны мышцам парных плавников и составляют первич-
ную мускулатуру конечностей, которая (так же как и мышцы
плавников) иннервируется сплетением известного числа спинномозговых
Рис. 608. Поверхностные мышцы передней части тела саламандры
(по Фюрбрингеру ив Кингсли).
а — anconeus; bi — humero-bra g chialis; cue — trapezius; dtr— dorsotrachealisj dg— de-
pressor mandibulae; as— dorsalis scapulae; eo — obliquus externus; id — latissimus dorsit
is— levator scapulae; m — masseter; mh — intermandibularis; mhp —mylohyoideus poste-
rior; pc — pectoralis; ph — procoracohumeralis; та—rectus abdominis; «pc—supracoracoideus
нервов, плечевым в передней конечности (plexus brachialis) и пояснично-
крестцовым — в задней (plexus lumbosacralis). Из спинного зачатка мус-
кулатуры, гомологичного отводящей мышце плавника, развивается группа
разгибателей пятипалой конечности, а из брюшного зачатка, гомологич-
ного приводящей мышце плавника, развивается система сгибателей. Кроме
этой первичной мускулатуры, в качестве продуктов дальнейшей диферен-
цировки миотомов грудной области развивается еще особая вторич-
ная мускулатура плечевого пояса, служащая для передвижения
самого пояса и иннервируемая непосредственно спинномозговыми нер-
вами (пп. thoracici superior et inferior).
Из спинного мускульного зачатка развиваются’ в передней конеч-
ности следующие первичные мышцы (рис. 208): спинные мышцы плече-
вого пояса, идущие к плечевой кости, — дельтовидная (m. del-
toideus scapulae) впереди, спинная мышца (т. dorsalis scapulae)
над лопаткой, подлопаточная (m. subsc apularis) и широкая
мышца спины (m. latissimus dorsi) позади; в пределах свободной
конечности — трех-, четырехглавая мышца (m. anconeus),
идущая от лопатки, коракоида и плеча к локтевому отростку, и все раз-
гибатели конечности (mm. extensores), включая разгибатели паль-
цев. У лазающих млекопитающих, особенно у приматов, в передней ко-
нечности за счет разгибателей развиваются еще мышцы, поворачивающие
конечность ладонью вверх (mm. supinatores).
Из брюшного мускульного зачатка развиваются в передней конеч-
ности следующие первичные мышцы (рис. 208): брюшные мышцы плече-
вого пояса — коракоидно-плечевая (m. coraco-brachialis)
220
впереди и грудная мышца (m. pectoralis) позади; Последняя
отходит не только от пояса, грудины и ребер, но и дальше позади от брюш-
ной фасции. В пределах свободной конечности коракоид н о - лу-
че в а я мышца (m. coraco-radialis) и все сгибатели (mm.
flexores) конечности. У лазающих млекопитающих в хватательной перед-
ней конечности за счет сгибателей развиваются еще мышцы, поворачи-
вающие конечность ладонью вниз (mm. pronatores).
Вторичная мускулатура плечевого пояса состоит из значительной
мышцы, развивающейся впереди пояса — мышцы, поднимающей
лопатку (m. levator scapulae), и передних зубчатых мышц
(mm. serrati antici superficialis et profundus), развивающихся позади
Рис. 209. Поверхностная мускулатура туловища и конечностей собаки
(по Мартину).
ап — m. anconeus; Ы/ — m. biceps femoris; dlt — т. deltoideus; ex. d—m, extensor digitorum;
fl. d — m. flexor di itorum; sen — m. gastrocnemicus; gl— m. glutaeus; Id — m. latissi-
mus dorsi; o. e.—m. obliquus externus abdominis; pct — m. pectoralis; rm.— rectus
abdominis; slclm — m. sterno-cleidomastoideus; tr — m. trapezius; i> — m. sartorius.
пояса. Мышца, поднимающая лопатку, появляется уже у селахий, но ее
нет у высших рыб, у которых пояс неподвижно прикреплен к черепу. У на-
земных позвоночных она идет от ребер и от поперечных отростков шей-
ных позвонков к внутренней поверхности лопатки. Кроме этих париеталь-
ных мышц, в связь с передним поясом вступает (уже у селахий) и еще
одна, висцеральная мышца — трапециевидная (m. trapezius),
которая, повидиму, также должна быть отнесена ко вторичной мускула-
туре пояса.
У птиц сильно развиваются мышцы передней конечности — ив осо-
бенности колоссальная грудная мышца, которая является главной лета-
тательпой мышцей.
У млекопитающих (рис. 209) очень значительно разраста-
ется вдоль всей спины широкая мышца спины (m. latissimus dorsi) и
сильно развивается и диференцируется грудная мышца (m. pectoralis
major et minor); кроме того, сильно разрастается и диференцируется
трапециевидная мышца (m. trapezius s. cucullaris, т. sterno-cleido-mastoi-
deus). Коракоидно-плечевая мышца диференцируется далее, и с редук-
цией коракоида частично переносит место своего отхода на лопатку.
Вместе с коракоидно-лучевой мышцей она принимает участие в образо-
вании характерной для млекопитающих двуглавой мышцы (m. biceps
brachii). Кроме того, у млекопитающих получают большое значение
зубчатые мышцы. У животного, поднимающего свое тело над землей,
оно как бы висит между лопатками именно на зубчатых мышцах.
221
В задней конечности нет вторичной мускулатуры пояса, так как
последний прикреплен к позвоночнику и более или менее фиксирован в
своем положении. Первичная мускулатура составляется из следующих
главнейших мышц: приводящие мышцы (mm. adductores)
и гребенчатая (m. pectineus) лежат впереди, лобковобед-
ренная (m. pub of ет oralis) — с внутренней стороны конечности,
грушевидная (m. pyriformis s. caudofemoralis, т. е. хвостовая
мышца низших форм) — сзади, и ягодичная (m. glutaeus s. ileofe-
moralis амфибий), диференцпрованная у млекопитающих на несколько
мышц, — сзади и снаружи. Мышцы, ограничивающиеся пределами сво-
бодной конечности, разделяются на разгибатели (m. extensores),
которые представляют собой результат диференцировки первичной спин-
ной мускулатуры (m. quadriceps femoris и др.), и сгибатели (т.
flexores), представляющие собой результат диференцировки первичной
брюшной мускулатуры (m. biceps femoris и др.). Четырехглавая мышца
(m. quadriceps femoris) идет от подвздошной, лонной и бедренной костей
через колено к большой берцовой. В ее сухожилии развивается колен-
ная чашечка (patella).
Сложная диференцировка мускулатуры конечностей находится в
очевидной связи с изменением их функций при переходе от водной жизни
к наземной. Для выполнения функции рулей или весел достаточна была
сравнительно простая мускулатура, служащая для изменения положения
лопасти плавника, являющейся простой упругой пластинкой. С превра-
щением конечности в сложный рычаг, поддерживающий и передвигающий
тело животного на суше, естественно связано обособление большого числа
мышц, точно координированное действие которых обусловливает разно-
образные движения наземных позвоночных. Мускулатура туловища соб-
ственно отступает на задний план, а мышцы конечностей занимают пре-
обладающее положение, в значительной мере покрывают туловищную
мускулатуру и даже частично ее вытесняют (m. latissimus dorsi и т. рес-
toralis).
Диафрагма
Диафграмма представляет собой мускульную перегородку, отделя-
ющую грудную полость с легкими и сердцем от брюшной полости. Эта
перегородка имеется только у млекопитающих и является важной дыха-
тельной мышцей.'Первый зачаток ее обнаруживается в виде поперечной
соединительнотканной перегородки (septum transversum), развивающейся
в передней части тела (в шейной области) непосредственно позади сердца
в связи с задней стенкой перикардия. Одновременно с перемещением
сердца и перегородка перемещается назад. В эту перегородку врастает
затем брюшная туловищная мускулатура в виде пары мышечных складок,
растущих от брюшной стенки тела. У взрослого животного или вся диа-
фрагма является мускульной (с радиальным расположением волокон) или
середина ее состоит из соединительной ткани. Сквозь диафрагму проходят
пищевод и крупные кровеносные сосуды. Диафрагма выпукла спереди
и становится при сокращении более плоской вследствие чего грудная по-
лость увеличивается и воздух всасывается в легкие. Таким образом,
диафрагма является важнейшей дыхательной мышцей млекопитающих.
Иннервируется диафрагма специальным нервом (n. phrenicus),
отходящим от шейного сплетения спинномозговых нервов, и такая иннер-
вация также ясно указывает на перемещение диафрагмы назад, что нахо-
дится, невидимому, в связи с процессом обособления шеи у наземных по-
звоночных.
Диафрагма представляет собой чрезвычайно важное приобретение
млекопитающих, связанное с развитием более совершенного дыхания,
а следовательно и более интенсивного обмена веществ в организме. Интен-
222
сивное окисление является источником высокой температуры тела, обу-
словливает возможность освобождения гораздо большего количества
энергии и, таким образом, связано с общим повышением жизнедеятель-
ности.
Подкожная мускулатура
Кроме описанной уже гладкой мускулатуры кожи, у наземных по-
звоночных нередко развивается вторичная связь между поверхностными
слоями поперечнополосатой мускулатуры тела и кожей. Такая мускула-
тура может совершенно обособиться и располагается тогда или в подкож-
ной соединительной ткани, или даже в самой коже, образуя систему кож-
ной поперечнополасатой мускулату-
ры. Зачатки такой мускулатуры на-
блюдаются и у бесхвостых амфибий;
сильнее она развита у рептилий,
где прикрепляется к кожным чешу-
ям, и у змей образует довольно слож-
ную систему отдельных мышц, частью
идущих от ребер к чешуям; эта му-
скулатура служит для перемещения
животного помощью координирован-
ных движений кожных чешуй.
Гораздо большего развития до-
стигает кожная мускулатура у мле-
копитающих, где она нередко обра-
зует почти сплошной подкожный
слой, покрывающий тело животного
(рис. 210). В области туловища это
сильно разросшиеся и обособившиеся
поверхностные слои двух мышц пле-
чевого пояса — широкой мышцы
спины (m. latissimus dorsi) и груд-
ной мышцы (т. pectoralis). В обла-
сти шеи и головы это сильно разрос-
шиеся поверхностные слои висце-
ральной мышцы — сжимателя шеи
(m. sphincter colli, VII). Очень силь-
но развита подкожная мускулатура у
клоачных млекопитающих (рис. 210),
где она покрывает все тело (panni-
culus carnosus). Сильно развита она
также у броненосцев и ежей, у ко-
торых она служит, можду^ прочим,
для свертывания их тела в шар. При
помощи этой мускулатуры животные
встряхивают свою кожу, чтобы из-
бавиться от насекомых и т. п. У ежей
Рис. 210. Кожная мускулатура
ехидны (по Руге).
с! — отверстие клоаки; тт — млечная
железа; ms — сумка; pct — дериват груд-
ной мышцы; sph. с VII—m. sphincter colli
VII; sph. cl —m. sphincter cloacae; sph.
m — m. sphincter niarsupii.
и дикобразов передвижение игл производится пучками той же попереч-
нополосатой мускулатуры. Осязательные волосы также снабжены попе-
речнополосатой мускулатурой, развивающейся от лицевой мускулатуры.
Последняя образует в области шеи и головы так называемую платизму
и мимическую мускулатуру, о которой речь была выше. Эта мускулатура
иннервируется лицевым нервом и особенно сильно развита у обезьян
(рис. 202) и у человека, у которых туловищная подкожная мускулату-
ра исчезает.
223
Для позвоночных животных весьма характерно, Следовательно, рез-
кое деление всей мышечной системы на соматическую и висцеральную
мускулатуру, различные по происхождению и по иннервации.
Особо высоко диференцируется висцеральная мускулатура передней
части кишечника в связи с развивающимся здесь висцеральным аппара-
том. Эта поперечнополосатая мускулатура, являющаяся сначала в виде
простого общего сжимателя всей жаберной области, распадается затем
на целый ряд отдельных мышц, приводящих в движение те или иные части
висцерального аппарата. У наземных позвоночных с утратой жаберного
дыхания особое значение приобретают лишь мышцы челюстей и некото-
рые поверхностные мышцы, образующие шейную и лицевую мускула-
туру млекопитающих.
Соматическая мускулатура служит первоначально только для пере-
движения животного в воде при помощи волнообразных изгибаний тела
и поделена в связи с атим на целый ряд отдельных сегментов. С утратой
передней частью тела своей гибкости и образованием головы миомеры
здесь, естественно, редуцировались за исключением тех, которые при-
обрели иную функцию в качестве глазных мышц и продольной мускула-
туры висцерального аппарата. С изменением способа передвижения у
наземных позвоночных и туловищная мускулатура теряет свою сегмен-
тацию и диференцируется на сложную систему мышц различного значе-
ния, связанных частью с осевым скелетом и его производными и с поясом
конечностей, частью служащих в качестве брюшного пресса и дыхатель-
ных мышц. Мускулатура конечностей, развившаяся первоначально в
качестве сегментального образования, теряет постепенно свою сегмен-
тацию уже у рыб, но испытывает особо сложную диференцировку лишь
у наземных позвоночных в связи с изменением функций конечностей.
Наконец, у высших наземных позвоночных за счет поверхностных слоев
мышц плечевого пояса иногда развивается еще более или менее значитель-
ная кожная поперечнополосатая мускулатура.
Электрические органы
Электрические органы, встречающиеся только у скатов и у костистых
рыб, представляют собой почти всегда видоизмененную поперечнополо-
сатую мускулатуру, которая специализировалась для производства элек-
трических разрядов. Такая специализация оказалась возможной, так
как мышечное сокращение и вообще всегда сопровождается освобожде-
нием небольшого количества электрической энергии. Эти органы имеют
иногда весьма значительный объем (у электрического угря более трети
веса всего тела) и у разных рыб различное происхождение.
У некоторых скатов (Raja) это слабый еще парный орган, развива-
ющийся из участка боковой мышцы и связанный переходами с неизмен-
ной мускулатурой; иннервируется он спинномозговыми нервами соответ-
ствующей области. У других, «электрических», скатов (Torpedo) имеется
очень сильный электрический орган по бокам головы, представляющий
собой видоизменение висцеральной мускулатуры и иннервируемый вис-
церальными нервами (7-й, 9-й и 10-й пары). Также очень сильные органы
имеются в брюшной половине тела у электрического угря (Gymnotus
electricus), где oini иннервируются, согласно своему происхождению,
большим числом спинномозговых нервов. У других костистых рыб наблю-
даются более слабые мускульные электрические органы (Mormyrus, Gym-
narchus), сходные по общему положению (хвостовая область) и происхож-
дению с органами угря. У Astroscopus электрические органы расположены
в орбитах глаз и развиваются из глазных мышц.
Электрические органы состоят из целого ряда электрических пласти-
нок, представляющих собой видоизмененные мускульные волокна с их
224
нервными окончаниями (концевыми пластинками) и отделенных друг от
друга прослойками студенистой соединительной ткани (рис. 211). В элек-
трических пластинках более слабых органов сохраняются еще следы по-
перечной полосатости в части, противоположной месту вхождения нерв-
ного волокна. У электрического ската электрические пластинки собраны
столбиками, образующими вертикальные призмы, и каждый орган состоит
из большого числа (около 400) таких призм. К каждой пластинке подхо-
дит снизу нервное волокно и распадается на фибриллы, образующие тон-
кую сеть в нижней ее половине (соответствующей концевой пластинке).
Верхняя половина электрической пластинки представляет собой преобра-
Рис. 211. Схема тонкого строения электрического органа; две электрические пластинки
в окружающей соединительной ткави; поперечный раврев (по Верслуису). Нервы
показаны черным цветом, сосуды — белые.
е — центральный слой пластинки; п — нерв; pl. el. — электрический слой; sf —соедини-
тельнотканные волокнистые оболочки; v — сосуды.
зованное мускульное волокно. Ее верхняя стенка образует множество
сосочков, между которыми проникают многочисленные разветвления
кровеносных сосудов. Нервные волокна подходят к электрическим пла-
стинкам с одной определенной стороны во всем органе (нижней у Torpedo),
и эта сторона пластинки, а также и всего органа оказывается электро-
отрицательной (у Torpedo — нижняя). Разряд органа происходит «про-
извольно» через посредство электрических нервов и всегда в определен-
ном направлении.
Совершенно особый электрический орган имеется у электрического
сома (Malapterurus electricus),—он охватывает все тело животного и пред-
ставляет собой орган кожного происхождения. Иннервируется этот орган
парой колоссальных простых нервных волокон, независимо отходящих
от мозга между вторым и третьим спинномозговыми нервами. Эти волокна
связаны с парой огромных многополюсных нервных клеток.
НЕРВНАЯ СИСТЕМА
НЕРВНАЯ СИСТЕМА БЕСПОЗВОНОЧНЫХ
В простейшем виде нервная система представляет собой сеть клеточных элемен-
тов, связанных между собой длинными отростками, функциональное вначение которых
состоит в восприятии и передаче раздражений внешней среды. Естественно, что для
225
несения этой функции специализируются именно эктодермальные клетки как Непо-
средственно соприкасающиеся с окружающей средой. Первоначально, очевидно, лю-
бая клетка могла и воспринимать раздражение, и отвечать на него известным образом.
Но затем некоторые эктодермальные клетки становятся особо раздражимыми, «чув-
ствующими» клетками/Раздражение, воспринятое эктодермальной чувствующей клет-
кой, может быть в виде особого возбуждения передано соседним клеткам через посред-
ство обычных протоплазматических связей и вызвать известную реакцию, как, напри-
мер, сокращепие,?У губок, не имеющих еще особой нервной системы, все координиро-
ванные двигательные реакции осуществляются, очевидно, путем передачи возбуждения
исключительно по протоплазматическим связям (плазмодесмам).
Но уже у низших кишечнополостных такие особо чувствительные эктодермаль-
ные клетки теряют непосредственную связь с поверхностью и погружаются глубже
обычных клеток покровов, а связи их с другими клетками развиваются в длинные
отростки. Таким образом, в эктодерме появилась система более глубоких чувствующих
«нервных» клеток, которые воспринимают раздражения внешней среды уже не прямо,
а через посредство поверхностных эктодермальных клеток. Нервные клетки соединя-
ются посредством длинных и тонких отростков — нервных волокон — как между
собой, так и с другими эктодермальными же клетками, образуя одну цельную систему.
Так как эпителиальные клетки кишечнополостных снабжены мускульными отрост-
ками (эпителиально-мускульные клетки), то каждая такая клетка может реагировать
на раздражение сокращением своего мускульного отростка. Нервные волокна связаны
не непосредственно с мускульными отростками, а именно только с эпителиальной
частью клетки. Возможно, что последнюю поэтому следует рассматривать как двига-
тельный нервный элемент простейшего вида.сЦепь, состоящая из чувствующей эпите-
лиальной клетки (рецептора), нервной клетки и мышечного волоййа или железы
(эффектора), представляет собой простейший путь возбуждения, ведущий от раздраже-
нМГк" реакции на это раздражение. Это и есть то, что физиологи называют рефлек-
торной дугой.
Такая нервная система простейшего строения, состоящая из клеточных элемен-
тов и волокон, равномерно распределенных по всему телу в виде сети, носит наимено-
вание разлитой, или диффузной, нервной системы. У кишечнополостных подобная
же нервная система развивается и в энтодерме, где она служит для восприятия раз-
дражений со стороны содержимого кишечной полости. У других животных развива-
ется единая эктодермальная нервная система, воспринимающая возбуждения как со
стороны покровов с их органами чувств, так и со стороны кишечника и других внутрен-
них органов. Таким образом, нервная система становится аппаратом, связующим
организм со средой и различные части организма между собой в одно целое.
У большинства кишечнополостных наблюдаются и в наружном слое
эпителия еще многочисленные чувствующие клетки, непосредственно связанные с длин-
ными нервными волокнами и образующие местами значительные скопления чувству-
ющего эпителия. У медуз чувствующий эпителий входит также в состав довольно вы-
соко диференцированных органов чувств. Такие чувствующие клетки являются уже
как бы второй генерацией чувствующих элементов, если считать связанные с ними
и лежащие глубже нервные клетки также за чувствующие клетки, лишь утратившие
непосредственную связь с внешней средой.
Локализация чувствующих клеток на определенных местах сопровождается и
концентрацией нервных элементов, образующих здесь более густую сеть. Таким об-
разом развивается нервное сгущение на ротовом диске коралловых полипов. У медуз
наблюдается распределение чувствующего эпителия по краю зонтика, где развиваются
их органы чувств. Соответственно этому и нервные элементы концентрируются в тех
же местах и образуют кольцевой ствол, состоящий из массы волокон с большим числом
нервных клеток. Такой ствол уже может рассматриваться как центральная нервная
система, а пучки волокон, отходящие от него к органам чувств, образуют перифери-
ческие нервы.
Таким образом, концентрация нервных клеток приводит к образованию нерв-
ных центров, а соединение нервных волокон дает периферические нервы. Эта концен-
трация обусловлена прежде всего локализацией чувствующих клеток, т. е. развитием
органов чувств. С появлением двигательных нервных элементов последние также
образуют нервные центры в связи с теми или иными группами мышц.
Нервные центры могут иметь различную форму. У примитивных животных цен-
тральная нервная система имеет обычно вид сплошных стволов, состоящих из массы
волокон и клеток. Распределение нервных стволов находится в согласии с общим пла-
ном строения животного. У животных с лучевой симметрией и нервная система по-
строена соответствующим образом. У иглокожих имеется кольцевой ствол с отходя-
щими от него радиальными нервами.
У животных с двухсторонней симметрией наблюдается диференцировка между
передним концом тела, где развиваются важнейшие органы чувств, и задним. Соот-
ветственно положению органов чувств и центральная нервная система в голове разви-
вается сильнее и образует так называемый головной мозг в виде более или менее зна-
чительной нервной массы, от которой обыкновенно отходит несколько продольных
стволов назад вдоль всего туловища. У некоторых червеобразных вся нервная система
226
еще связана с эктодермой (ще'гинкочелЮстныс, кишечнодьппащие), а у других (немер-
тины) можно проследить постепенное ее обособление и погружение в глубь тела.
Довольно примитивная нервная система имеется у ресничных червей.
На переднем конце тела сосредоточена парная нервная масса (головные узлы), от ко-
торой отходит множество нервных волокон вперед к коже и к парным органам зрения.
От головных узлов назад отходит обыкновенно пара значительных продольных нерв-
ных стволов, от которых отходят многочисленные периферические нервы. Иногда на-
блюдается и большее число продольных нервных стволов, но пара средних, лежащих
с брюшной стороны, отличается более значительной мощностью. Все эти стволы за-
ключают в себе, кроме нервных волокон, и множество нервных клеток и являются,
таким образом, частями центральной нервной системы.
Обычно головные узлы соединяются между собой не только со спинной стороны,
но и с брюшной стороны от глотки, образуя таким образом сплошное нервное кольцо
вокруг глотки. Такое окологлоточное кольцо имеется иу круглых червей.
У последних от него отходит несколько продольных нервных стволов, из которых
спинной и брюшной являются наиболее крупными. У немертин нервная система
построена по тому же типу сплошных стволов и состоит из пары надглоточных (голов-
ных) узлов, двух боковых нервных стволов и одного спинного, соединенных между
собой поперечными анастомозами.
У более высоко организованных животных наблюдается обыкновенно некото-
рая дальнейшая диференцировка в пределах самой центральной нервной системы.
В нервных стволах появляются местные более значительные поверхностные скопле-
ния нервных клеток, образующие более или менее значительные вздутия — нервные
узлы, или ганглии. Таким образом развивается система нервных узлов, соединенных
между собой продольными комиссурами, которые состоят главным образом из волокон,
но содержат обычно и отдельные нервные клетки.
У членистых животных концентрация нервных клеток продольных стволов про-
исходит соответственно каждому сегменту тела и обусловлена, повидимому, главным
образом развитием сегментально расположенных конечностей (параподии кольчатых
червей). В этих случаях нервная система приобретает обыкновенно вид парной цепочки,
состоящей из правильного ряда связанных между собой нервных узлов.
У кольчатых червей центральная нервная система состоит из пар-
ного надглоточного узла (головной мозг), от которого отходят нервы к чувствующему
аппарату головы и, в частности, к органам зрения, и парного брюшного ствола, обра-
зующего «брюшной мозг». Надглоточные узлы связаны между собой спинной комис-
сурой, а с брюшными стволами — окологлоточным кольцом. Оба брюшных нервных
ствола снабжены н каждом сегменте утолщениями, от которых отходят поперечные
комиссуры, связывающие каждую пару узлов между собой. В стороны о т каждого
узла отходят также правильно расположенные сегментальные периферические нервы.
Таким образом, брюшной мозг нередко приобретает вид парной цепочки с попереч-
ными комиссурами в каждом сегменте (вид веревочной лестницы). Но очень часто обе
цепочки сильно сближены и нередко соединяются между собой по средней линии,
образуя одну непарную брюшную цепочку, во внутреннем строении которой, однако,
обычно сохраняются ясные следы парного происхождения. В брюшном мозге нервные
волокна занимают центральную часть, а нервные клетки лежат на поверхности. Попе-
речные комиссуры составляются из волокон, отходящих от нервных клеток одной сто-
роны и переходящих затем через узел другой стороны в периферический нерв (пере-
крест нервных волокон).
Центральная нервная система членистоногих построена по тому же
типу. У примитивных ракообразных, у первичнотрахейных и у многоножек наблю-
дается по существу такая же парная брюшная цепочка, как и у кольчатых червей.
С развитием гетерономной сегментации у высших форм происходит диференцировка
и в области нервной системы. При слиянии сегментов в области головы, груди и т. п.
соответствующие нервные узлы также сливаются между собой. В тех сегментах, в ко-
торых исчезают конечности, редуцируются и нервные узлы. Наконец, наблюдается
и перемещение узлов в другие сегменты и их слияние в большие нервные массы. В связи
со значительным развитием органов зрения происходит обособление специальных
зрительных узлов. Непосредственно под глоткой, в нижней части окологлоточного
кольца, образуется подглоточный увел путем слияния нескольких пар узлов, обслу-
живающих ротовые конечности. Наконец, в области груди у высших раков, у пауков
и насекомых наблюдается сближение ганглиев, обслуживающих ходильные конеч-
ности, между тем как лежащие позади узлы совсем исчезают. Это сближение нередко
приводит и к полному слиянию всех брюшных узлов в одну большую ганглиозную
массу (пауки, мухи).
У моллюсков центральная нервная система развивалась иначе. У при-
митивнейших форм имеется окологлоточное нервное кольцо, от которого назад отхо-
дит пара брюшных нервных стволов и пара боковых. У большинства моллюсков про-
изошла диференцировка в пределах этой системы, приведшая к образованию парного
надглоточного (головного) узла и нескольких пар туловищных узлов (ножные, плев-
ральные, париетальные и висцеральные), соединенных продольными комиссурами
между собой и с надглоточными узлами. У высших форм все узлы сближаются между
собой вокруг глотки.
227
Совершенно особый тип центральной нервной системы свойствен хордовым
животным и отчасти кишечнодышащим, которых иногда и причисляют к ним. У к и-
шечнодышащих имеется в общем очень примитивная нервная система в виде
сплошного нервного сплетения, связанного еще с эктодермой. Более значительного
развития достигает нервный слой в хоботке. В туловище выделяются в особенности
два непарных продольных ствола — один спинной и один брюшной, которые соеди-
няются между собой нервным кольцом у основания ошейника. Продолжение спин-
ного нервного ствола в ошейнике развито значительно выше и вполне отделено от по-
кровов. Замечательно, что эта часть центральной нервной системы имеет форму трубки,
полость которой у некоторых видов открыта и впереди, и позади. В глубоких частях
трубки помещаются нервные клетки, а по периферии располагаются только волокна.
Трубчатая форма центральной нервной системы представляет один из наиболее
характерных признаков хордовых животных. У некоторых низших хордовых нерв-
ная трубка наблюдается, правда, только эмбрионально и ватем дегенерирует, заме-
няясь одним простым спинным нервным узлом (оболочники), но это явление находится
здесь в связи с общими явлениями регресса в органиэации этих форм.
НЕРВНАЯ СИСТЕМА ХОРДОВЫХ ЖИВОТНЫХ И ЕЕ ПРОИСХОЖДЕНИЕ
У хордовых вся центральная нервная система представлена нервной
трубкой, лежащей со спинной стороны животного. Передний конец трубки
обыкновенно расширен и образует головной мозг, между тем как задняя
цилиндрическая часть трубки получает название спинного мозга. У заро-
дыша центральная нервная система закладывается в виде продольной
полосы утолщенной эктодермы, нервной пластинки, которая углубля-
ется, образуя жолоб, замыкающийся затем в трубку. Передний конец
трубки остается довольно долго открытым и это отверстие носит название
невропора. Задний конец трубки у зародыша сообщается с полостью
кишки (нервно-кишечный канал).
Закладка центральной нервной системы в виде полосы утолщенной
эктодермы наблюдается нередко и у беспозвоночных, но у них она не за-
мыкается в трубку. Расположение нервных элементов у хордовых также
иное: нервные клетки помещаются в центральной части органа, а волок-
на — в периферической.
Мы видели, что нервная система возникла, повидимому, путем-обо,-
собления чувствующих эктодермальных клеток, которые погружались
глубже под защиту поверхностною впиавлии. У предков хордовых жи-
вотных, вероятно, имелась продольная спинная полоса чувствующего
эпителия, которая вся целиком углубилась сначала в виде открытого жо-
лоба, а затем образовала замкнутую трубку. Аналогичное явление наблю-
дается на органах боковой линии позвоночных: эти органы также погру-
жаются сначала в открытую борозду, которая затем замыкается в канал.
Что нервная трубка хордовых животных первоначально имела значение
органа чувств, видно из того, что у низших хордовых последние находятся
еще в самом мозге. Отоцист и глазок личинок асцидий развиваются в
стенках вздутой передней части нервной трубки. У ланцетника весь мозг
является светочувствительным, так как внутри его стенок имеются осо-
бые чувствующие клетки, частью окруженные пигментными клетками и
образующие так называемые глазки Гессе. Обонятельная ямка ланцет-
ника развивается в связи С невропором. V"позвоночных животных органы
зрения развиваются всегда за счет стенок самого мозга, а орган обоняния
по своему происхождению, быть может, до известной степени связан с
невропором. Во всяком случае он развивается за счет эктодермального
утолщения, непосредственно прилежащего к переднему концу нервной
пластинки. Наконец, органы боковой линии и органы слуха, так же как
и нервные узлы, развиваются за счет эктодермы, непосредственно приле-
жащей к краям нервной пластинки1. Эпителий, выстилающий централь-
1 Имеются и еще более непосредственные указания на первоначальную чувст-
вующую функцию нервной трубки: в нижней стенке промежуточного мозга, именно
в сосудистом мешке рыб, а также в эпендиме вокруг центрального канала спинного
мозга позвоночных также найдены чувствующие клетки.
228
ный канал нервной трубки, первноначально является мерцательным и
вызывает движениями своих ресничек ток воды, входящий через невропор
и направленный назад через нервно-кишечный канал наружу. Это обсто-
ятельство освещает до известной степени значение втих отверстий. Как
и раньше, на сталии нервной пластинки и открытого жолоба, мерцатель-
ные реснички производят постоянный обмен воды, что было необходимо,
<<ли этот орган служил для химического ее испытания (обоняние, вкус).
Это объясняет нам и возникновение органа обоняния в связи с певропором.
Наконец, постоянный обмен воды мог иметь огромное значение в смысле
осуществления более интенсивного газового обмена (дыхания) нервной
ткани, что было чрезвычайно важно при слабой диференцировке органов
дыхания и кровообращения.
Таким образом, центральная нервная система хордовых животных
занимает, невидимому, особое положение и непосредственно с централь-
ными органами беспозвоночных не сравнима. Тогда как последние обра-
зовались путем концентрации злементов субэпителиальной нервной сети,
мозг хордовых развился путем обособления от эктодермы особой чувствую-
щей пластинки. Весьма вероятно, что пол такой чувствующей пластинкой
имелось уже значительное нервное сплетение. Во время дальнейшей вво-
люции хордовых животных периферические части чувствующей пластинки
диференцировались в качестве более совершенных органов чувств: перед-
няя часть углубилась в виде парной ямки и дала начало органу обоняния,
а краевые части — клеткам периферических нервных узлов и в связи
с ними органам боковой линии (и органу слуха). Средняя часть пластинки
погрузилась глубже, свернулась в трубку и сохраняла еще некоторое
время значение светочувствительного аппарата, но затем вта роль перешла
к особым выростам передней части трубки, которые высоко специализи-
ровались в качестве органов зрения. Остальная часть трубки утратила
значение чувствующего органа и превратилась в центральный нервный
аппарат, связывающий все органы чувств, возникшие на периферии пла-
стинки, в единую систему, и служащий для выработки координированного
ответа на те или иные воспринятые воздействия внешней среды. Возможно,
что этот новый центральный аппарат находился в лучших условиях, чем
старый, вследствие более обильного снабжения кислородом, по крайней
мере на стадиях пластинки желобка и открытой с двух сторон трубки,
когда в нем действием мерцательных ресничек происходила непрерывная
смена воды. В особенности хорошо снабжалась свежей водой передняя
часть мозга через широкий невропор, и это создало условия, необходимые
для прогрессивного развития как самого головного мозга, так и связан-
ных с ним органов чувств. Находившееся же под чувствующей пластин-
кой нервное сплетение утратило свое значение. Когда пластинка сверну^
лась в трубку, оно оказалось на ее поверхности1.
Строение центральной нервной системы ланцетника еще в высшей
степени примитивно. Мозг ланцетника по существу представляет собой
жолоб с тесно сомкнутыми краями. Спинная щель, так же как и централь-
ный канал, ограничена морфологически наружной поверхностью нервной
пластинки, которая, как и вся поверхность тела, выстлана эпителием.
Клетки нервного эпителия диференцированы — ббльшая их часть явля-
ется просто эпителиальными клетками, снабженными лишь особыми
радиальными отростками, имеющими значение скелета для всего органа,—
это клетки эпендимы; другие, более крупные, клетки являются чувству-
ющими (глазки Гессе), и, наконец, третьи, также довольно крупные,
клетки являются собственно нервными клетками. Таким образом, нерв-
1 У позвоночных, действительно, имеется нервное сплетение неизвестного зна-
чения на поверхности мозга (А. Немилов). Весьма возможно, что и вся симпатическая
нервная система позвоночных унаследована ими от бесповвоночпых животных.
229
ные клетки лежат здесь, подобно чувствующим, еще на самой поверх-
ности, между тем как нервные клетки даже низших беспозвоночных
лежат уже глубже. Это является лучшим доказательством того, что нерв-
ная трубка хордовых животных не гомологична центральной нервной
системе низших животных, а представляет собой более новое приобретение,
развившееся из особого органа чувств (чувствующей пластинки).
Сравнение чувствующего аппарата позвоночных с таковым беспозво-
ночных показывает, что у первых произошло дальнейшее обособле-
ние чувствующих клеток от эктодермы и превращение их в нервные
клетки особых периферических узлов, лежащих по пути чувствующих
нервов. У большинства беспозвоночных чувствующие клетки лежат в ряду
других клеток эпителия покровов, от которых они отличаются лишь своей
формой, присутствием чувствующего волоска и — главное — тем, что они
А
Рис. 212. Соотношения между чувствующими клетками и центральной нервной
системой (заштрихована) (по Ретциусу ив Эдингера).
Л — чувствующий эпителий дождевого червя; В — улитки; С — обонятельный эпителии
позвоночных; D — эпидермис позвоночных; —слуховой эпителий позвоночных; ер — эпи-
дермис; дас —ганглиозная клетка из спирального ганглия улитки уха; gsp — ганглиозная
клетка спинномозгового ганглия; 1о — обонятельная доля мозга; пс — центральная нерв-
ная система.
продолжаются в длинные отростки — нервные волокна, которые своими
конечными разветвлениями вступают в связь с дендритами нервных
клеток (рис. 212, Л). У некоторых высших беспозвоночных чувствующие
клетки иногда погружаются глубже под эпителий, в котором тогда оста-
ется лишь периферический отросток клетки (рис. 212, В). У Позвоночных
животных в эпителии покровов совсем нет чувствующих клеток описанного
типа, а имеются лишь так называемые свободные окончания нервов (т. е.
конечные разветвления нервных волокон, являющихся периферическими
отростками чувствующих нервных клеток, расположенных в узлах) (рис.
212, D). Таким образом, клетки чувствующих нервных узлов позвоночных
представляют собой первичночувствующие клетки типа беспозвоночных,
т. е. продукт недавнего еще обособления чувствующих элементов от
эктодермы. У ланцетника обособленных нервов узлов нет, а имеются
лишь рассеянные ганглиозные клетки в спинном мозге и по ходу спинного
корешка.
У позвоночных имеются также особые чувствующие клетки в эпите-
лии, но эти клетки не имеют нервного отростка, а охватываются конечными
разветвлениями чувствующего нервного волокна (рис. 212, Е). Такие
клетки типа позвоночных можно назвать вторичночувствующими клетками.
Однако в двух важнейших органах чувств позвоночных — в органе зрения
230
и в органе обоняния — чувствующие клетки обладают нервными отрост-
ками, которые своими конечными разветвлениями вступают в связь с
дендритами нервных клеток (рис. 212, С). Эти органы чувств связаны не
с периферическими узлами, как другие, а непосредственно с центральной
нервной системой. Орган зрения и развивается как часть центральной
нервной системы, а орган обоняния по своему происхождению также,
вероятно, связан с нервной пластинкой. Эти органы представляют собой
остаток унаследованного от беспозвоночных первичночувствующего аппа-
рата, другая часть которого дала начало собственно нервной системе.
Точно так же первичпочувствующие клетки встречаются у позвоночных и
внутри самого мозга — в сосудистом мешке промежуточного мозга рыб
(saccus vasculosus) и в выстилке центрального канала спинного мозга.
В общем мы различаем, следовательно, такого рода чувствующие
клетки: 1) первичпочувствующие клетки, расположенные в эпителии
покровов или погруженные под покровы, но сохраняющие чувствующий
отросток в этом эпителии, 2) гомологичные им чувствующие нервные
клетки (невроны) узлов со свободными окончаниями, разветвленными
между клетками эпителия или охватывающими вторичночувствующие
клетки и 3) вторичночувствующие клетки, не имеющие нервного отростка.
Этот последний тип чувствующих клеток встречается только у позво-
ночных.
Нервные клетки (невроны) центральной нервной системы разли-
чаются по положению и по характеру связей. Мы различаем невроны
трех основных типов: 1) чувствующие невроны узлов, которые лежат
полностью вне центральной нервной системы, 2) ассоциативные невроны
мозга, которые лежат полностью внутри центральной нервной системы
и 3) двигательные невроны, тело которых лежит в центральной нервной
системе (или в симпатических узлах), а неврит входит в состав нервов
и проходит вне центральной нервной системы к эффектору — мышце
или железе.
В
Д
Е
D
Рис. 213. Последовательные стадии развития цен-
тральной нервной системы. Схематические попереч-
ные разрезы.
ер — эпидермис; дп — ганглионарная пластинка.
Развитие нервной системы позвоночных
На ранних стадиях развития зачаток большей части нервной системы
имеет вид неоднократно упоминавшейся нервной пластинки, т. е. полосы
утолщенной эктодермы, лежащей на спинной стороне зародыша и огра-
ниченной с обеих сторон
небольшими валиками —
нервными складками (рис.
213). Последние в перед-
ней части зародыша рас-
ходятся, ограничивая бо-
лее широкое пространство,
соответствующее будуще-
му головному мозгу, впе-
реди которого нервные ва-
лики сходятся между со-
бой, образуя поперечный
головной валик. Нервная
пластинка вместе с нерв-
ными складками дает на-
чало спинному и головно-
му мозгу, органам зрения
и большинству перифери-
ческих узлов и нервов. Эктодерма, непосредственно прилежащая к
нервной пластинке, дает начало обонятельному эпителию и чувствую-
щему эпителию органов боковой линии и слухового органа, их нервным
узлам и нервам. Одни только вкусовые органы развиваются в эпи-
телиальной выстилке передней части кишечника.
231
Нервная пластинка сворачивается затем в жолоб, нервные складки
смыкаются над ним, и таким образом жолоб превращается в трубку
(рис. 213). Нервные складки срастаются раньше всего в области задней
части головы, и отсюда этот процесс продолжается назад и вперед. Позади
некоторое время сохраняется отверстие, ведущее в кишечник (canalis
neuroentericus), а впереди сохраняется отверстие, ведущее наружу (пеиго-
porus). Когда нервная пластинка превратилась в нервную трубку, то
клетки, составлявшие первоначально нервные складки, оказываются в
промежутке между трубкой и сомкнувшейся над ней эктодермой (рис. 213).
Эти клетки образуют затем парные боковые выросты, лежащие вдоль
спинной стенки нервной трубки (рис. 214). Это ганглионарные пластинки,
за счет которых идет затем образование спинномозговых и большей части
головных нервных узлов. В области закладки органа слуха ганглио-
нарная пластинка прерывается. В области переднего мозга она также
исчезает.
Рис. 214. Нервная трубка и
ганглионарная пластинка (gn)
вародыша позвоночного (из
Кингсли)
Рис. 215. Невромеры в головном мозге вародыша
форели (по Хиллу). Вид сбоку.
аи — ухо; ос—глав; ту — миотомы; рт и т — грани-
цы переднего, среднего и продолговатого мовга; 1—
11 — невромеры.
У переднего края нервной пластинки по бокам будущего невропора
появляется парное утолщение эктодермы, которое затем углубляется и
образует обонятельные ямки. По бокам будущего продолговатого мозга
также развивается пара эктодермальных утолщений, которые затем
углубляются, отделяются от эктодермы и образуют замкнутые слуховые
пузырьки.
Обыкновенно уже на стадии нервной пластинки и затем во время
образования нервной трубки можно заметить, что последняя легкими
перехватами поделена на целый ряд последовательных сегментов, или
невромеров (рис. 215). Ганглионарная пластинка образует также выступы
соответственно каждому сегменту нервной трубки (рис. 214). Эта сегмен-
тация, или невромерия, есть выражение общей метамерии тела позвоноч-
ного животного и вполне ясно выражена в области спинного мозга и задней
части головного мозга, где невромеры правильно чередуются с сомитами
(рис. 215). В передней части головного мозга невромеры также наблю-
даются, но они здесь менее ясны, весьма кратковременны и маскируются
ранним появлением других перехватов и вздутий, соответствующих уже
дефинитивным отделам головного мозга.
Соответственно большей ширине передней части нервной пластинки
вся передняя часть нервной трубки оказывается с самого начала заметно
вздутой. В дальнейшем, вследствие неравномерного роста различных
частей, появляются новые вздутия, разделенные перехватами (рис. 216).
Таким образом, сначала намечается вздутие в передней части головного
мозга, которое перехватом отделяется от задней части (рис. 217). По-
следняя лишь очень постепенно переходит в заднюю цилиндрическую
часть нервной трубки, соответствующую спинному мозгу. Переднее
вздутие сравнивают с мозговым пузырьком низших хордовых и называют
232
иногда первичным головным мозгом (archencephalon). Вскоре, однако,
это вздутие перетягивается еще па два отдела. Таким образом, в переднем
конце нервной трубки развиваются три вздутия, разделенные перехва-
тами,— мозговые пузыри, образующие первичный передний
мозг (prosencephalon), средний мозг (mesencephalon) и пер вич-
Рис. 216. Развитие отде-
лов головного мозга.
Схемы.
А — стадия трех пузырей
(с глазными пузырями ос);
В — стадия пяти отделов
(с глазными бокалами ос);
d — diencephalon; т —me-
sencephalon;ml —myel-
encephalon; mt — mctence-
phalon; p — telencephalon.
ный задний мозг (rhombencephalon)
(рис. 216, A, 218). Вслед за этой стадией трех
мозговых пузырей дальнейшая диференцировка
приводит к образованию пяти дефинитивных
отделов головного мозга (рис. 216, В). От перед-
него мозгового пузыря впереди развивается
простой или парный выступ, представляющий
собой собственно передний мозг (telencepha-
lon), который у большинства позвоночных обра-
зует так называемые полушария большого мозга,
полости которых получают название боковых
желудочков. Задняя часть переднего пузыря обра-
зует промежуточный мозг (dieiiceplia-
lon), полость которого называется третьим желу-
дочком. Боковые стенки этого отдела выпячивают-
ся в виде глазных пузырей. Средний
пузырь образует средний мозг (mesence-
phalon), полость которого обыкновенно имеет вид
узкого капала (aquaeductus Sylvii s. iter), связы-
вающего полости третьего и следующего, четвер-
того, желудочков. Задний пузырь образует
в передней своей части задний мозг (rneten-
cephalon), крыша которого выступает в виде
поперечной складки и получает название м о з-
ж е ч к a (cerebellum), и позади продолго-
ватый мозг (myelencephalon), с полостью
еп
Рис. 217. Схематический медиальный разрез через эм-
бриональную нервную трубку (по Купферу).
с. пей — canalis neurentericus; ch — хорда; еп —первичшяй
мозговой пузырь; г — воронка; med — нервная трубка; пр —•
невропор; tp — tuberculum postering.
четвертого желудочка, переходящего непосредственно в центральный
канал спинного мозга.
Головной мозг никогда не сохраняет формы прямой трубки, отделы
которой следуют друг за другом; быстрый рост отдельных частей, отчасти
брюшной и в особенности спинной стенки приводит к образованию ха-
рактерных изгибов (рис. 219). Наиболее значителен передний, или
теменной, изгиб, который развивается в области среднего мозга
вследствие быстрого роста его крыши и крыши переднего мозга. В этом
233
месте эмбриональный мозг как бы складывается пополам своей брюшной
стенкой, причем передний мозг оказывается подогнутым. Менее значи-
телен задний, или затылочный, изгиб, который развивается в задней
части продолговатого мозга и имеет то же направление (вогнутостью
на брюшную сторону). Третий изгиб располагается между обоими пре-
дыдущими в области заднего мозга и имеет обратное направление, т. е.
образует на брюшной стороне выпуклость, — это средний изгиб, или
Рис. 218. Схематический медиальный разрез через эмбриональный
мозг на стадии трех первичных пузырей (по Купферу).
ch— хорда; chi — перекрест зрительных трактов; i —воронка; It— концевая пластинка
med — спинной мозг; ms— средний пузырь; mt — задний пузырь; рг — передний пузырь.
рпр — processus neuroporicus; го — recessus opticus.
изгиб моста. Все эти изгибы особенно резко выражены в эмбриональном
мозге, а затем более или менее сглаживаются, за исключением высших
позвоночных, у которых они и во взрослом состоянии хорошо выражены.
Соотношение между первичной сегментацией мозга, или невромерией,
и дефинитивными отделами установлено не вполне точно. Повидимому,
первичный передний мозг развивается за счет трех первых невромеров,
Рис. 220. Схема расчленения эмбрионально-
го мозга. Срединный разрез (по Купферу).
ch — перекрест зрительных трактов; сЫ—моз-
жечок; ср — задняя комиссура; са — верхняя
комиссура; d—промежуточный мозг; ер—эпи-
физ; inf — воронка; 11 — концевая пластинка;
т — средний мозг; ml — продолговатый мозг;
mt — задний мозг; t — конечный мозг;
го —recessus praeopticus; tp — tuberculum
posterius.
Рис. 219. Изгибы головного мозга.
Срединный разрез головного мозга за-
родыша свиньи (из Кингсли).
с — изгиб среднего мозга; ch — хорда;
pl — сосудистое сплетение; hy — гипофиз;
i — воронка; т — средний мозг; п — за-
тылочный изгиб; р — изгиб моста; di —
промежуточный мозг; t — передний мозг.
средний мозг — за счет четвертого и пятого, задний мозг — за счет шестого,
а продолговатый мозг — за счет различного числа следующих невромеров,
начиная с седьмого.
Границы между отделами являются частью совершенно условными
(рис. 220). Границей между передним мозгом и промежуточным является
небольшое углубление брюшной стенки впереди перекреста зрительных
234
трактов (recessus praeopticus) и свешивающаяся со спинной стенки эпите-
лиальная поперечная складка (velum transversum), наблюдающаяся у
всех позвоночных, по крайней мере, эмбрионально. Границей менаду
промежуточным мозгом и средним служит небольшой выступ нижней
стенки впереди переднего изгиба (tuberculum posterius) и пучок поперечных
нервных волокон— так называемая задняя комиссура (comissura posterior),
лежащая в спинной стенке мозга. Средний мозг отделен от заднего ясным
перехватом — перешейком (isthmus), в области которого со спинной
стенки мозга внутрь вдается складка, образующая передний мозговой
парус (velum medullare anterius).
Стенки мозга утолщаются также неравномерно (рис. 223, 224). В
различных отделах головного мозга наблюдается утолщение различных
частей стенок, и эти местные утолщения собственно и характеризуют
отделы мозга взрослого животного. Передняя стенка мозга остается
посредине сравнительно тонкой и образует так называемую концевую
пластинку (lamina terminalis) (рис. 220), ограничивающую спереди
полость третьего желудочка. По бокам от нее у большинства позвоночных
развиваются парные выросты — полушария переднего мозга.
Но даже и в тех случаях, когда передний мозг остается непарным, наблю-
даются парные утолщения нижней части его передней стенки, образующие
обонятельные доли (lobi olfactorii), и парные же утолщения
его дна (рис. 224, 225), образующие так называемые полосатые
тела (corpora striata). У высших позвоночных значительно утолщается
крыша переднего мозга, носящая название мантии (pallium). В про-
межуточном мозге развиваются сначала парные выросты, дающие начало
глазным пузырям, а затем происходит утолщение боковых стенок, обра-
зующих зрительные бугры (thalami optici). Крыша проме-
жуточного мозга на большей части своего протяжения остается тонкой,
состоя из одного лишь слоя плоских.эпителиальных клеток, на котором
развивается переднее сосудистое сплетение. В позади
лежащих отделах дно мозга имеет довольно значительную толщину, и,
кроме того, наблюдается утолщение крыши среднего мозга, образующего
парные вздутия — зрительные доли (lobi optici, corpora quadrigemina),
и значительное утолщение крыши заднего мозга, образующее мозже-
чок (cerebellum). В области продолговатого мозга утолщаются главным
образом боковые стенки, между тем как крыша остается совершенно
тонкой, состоя из одного лишь плоского железистого эпителия, на кото-
ром развивается заднее сплетение кровеносных сосу-
дов. Наконец, и в области спинного мозга разрастаются главным образом
боковые стенки, вследствие чего центральный канал вытягивается перво-
начально в вертикальную щель. Спинная и брюшная стенки — «пластинки
дна и покрышки» — остаются тонкими. Боковые стенки, разрастаясь
еще больше, спускаются затем ниже брюшной стенки нервной трубки,
которая оказывается, таким образом, на дне глубокой брюшной
щели спинного мозга (fissura inferior). В брюшную щель врастает соеди-
нительная ткань с кровеносными сосудами. На спинной стороне спинного
мозга по средней линии сходным путем происходит образование более
узкой спинной щели, и наблюдается также врастание соедини-
тельной ткани с сосудами (fissura superior).
Кроме процессов общего формообразования (морфогенез), для пони-
мания нервной системы имеют существенное значение и процессы развития
микроскопических структур (гистогенез).
Стенка нервной пластинки и нервной трубки состоит первоначально
из цилиндрического эпителия, ядра которого лежат на разном уровне,
клетки частью занимают всю толщу эпителия, частью неправильно распре-
делены между другими. Затем более высокие цилиндрические клетки
располагаются радиально вокруг полости трубки; на своей внутренней,
235
а также и на наружной поверхности трубки они выделяют пограничную
перепонку. Ядра этих клеток располагаются частью у основания, не-
посредственно на внутренней поверхности эпителия, и сохраняют это
положение и во взрослом мозге. Дистальные части тела таких клеток
по мере утолщения стенок мозга вытягиваются в длинные радиальные
волокна (рис. 221). Этот слой клеток образует внутреннюю эпителиаль-
ную выстилку всех полостей взрослого мозга и носит название эпен-
димы (ependyma). Отростки этих клеток имеют значение опорных обра-
зований. Расположенные между цилиндрическими клетками более корот-
кие клетки деятельно размножаются и частью превращаются в ветвистые
клетки, образующие невроглию (neuroglia) (рис. 221, В), частью
преобразуются в нервные клетки (рис. 221, Л). Первые образуют
Рис. 221. Поперечные разрезы через спинной мозг млекопитающих на двух
стадиях развития (из Боннета по Ретциусу и Ленгоссеку).
А — молодой эмбрион летучей мыши; показана часть клеток эпендимы и некоторые нервные
клетки; В — эмбрион человека; справа показаны только некоторые волокна эпендимы, сле-
ва —только клетки эпендимы и глии; линией отмечена граница серого вещества; d— спинной
корешок нерва; ер — волокна эпендимы; gl — клетки глии; дп — ганглий; пс — комиссу-
ральная нервная клетка; пт — двигательная нервная клетка; v — брюшной корешок нерва^
своими многочисленными отростками сложную сеть, служащую опорой
для всего мозга. Вторые имеют вполне сходное происхождение и разли-
чаются лишь по своей дальнейшей диференцировке и функции. И те и
другие удаляются более или менее значительно от центральной полости
мозга. Эмбриональные нервные клетки (невробласты) деятельно размно-
жаются только на ранних стадиях развития; во взрослом мозге размно-
жение нервных клеток наблюдается редко. Преобразования невробласта
начинаются развитием отростка, неврита, который вытягивается в длин-
ную тонкую нить, образующую осевой цилиндр нервного волокна.
Позже развиваются другие отростки, дендриты, которые более или менее
обильно ветвятся в веществе стенок мозга. Нервные волокна выходят на
периферию мозга и здесь входят в состав периферических нервов или
растут более или менее далеко вдоль его поверхности, входя затем
в другие отделы мозга (рис. 221, Л). Масса волокон, развивающихся
на поверхности мозга, образует (вместе со связывающими их клетками
глии) его «белое» вещество, между тем как нервные клетки распола-
гаются внутри органа, в его «сером» веществе, которое заключает в себе,
кроме нёрвных клеток и их отростков, также и клетки невроглии. Пери-
ферическое вещество мозга является белым вследствие присутствия
миэлиновой оболочки на нервных волокнах в этой области. Нервные
отростки одних клеток растут по направлению" к периферии (центро-
бежно) и образуют двигательные пути мозга и двигательные перифери-
ческие нервы; отростки других клеток растут по направлению к выс-
шим центрам (центростремительно) и образуют чувствующие пути мозга,
236
Однако не во всех частях мозга нервные клетки сохраняют центрайь-
ное положение недалеко от внутренней повезти. В различных отделах
головного мозга часть нервных клеток перемещается по направлению к
периферии и иногда образует значительный слой серого нервного вещества
даже на наружной поверхности мозга. У низших позвоночных значительная
часть нервных клеток сохраняет еще связь с внутренней выстилкой мозго-
вых полостей посредством особого центрального отростка, между тем как
тело клетки иногда уже заметно перемещается к периферии. Интересно
отметить, что у рыб клетки, наиболее удаленные от внутренней полости,
оказываются вместе с тем и наиболее высоко диференцированными, — их
дендриты являются более многочисленными и образуют более сложную
и широкую систему ветвлений.
Нервные клетки располагаются в сером веществе мозга не равно-
мерно, а образуют местные скопления, которые называют ядрами. Среди
ядер мы различаем первичные ядра, в которых начинаются (в двигатель-
ных ядрах) или кончаются (в чувствующих ядрах) волокна перифери-
ческих нервов (соответственно двигательных или чувствующих), и вто-
ричные, или ассоциационные, ядра, которые вставляются между чувству-
ющими и двигательными ядрами, связываясь с ними волокнами, входя-
щими в состав центральных нервных путей.
Развитие периферической нервной системы
Ганглионарная пластинка образует, как сказано, в каждом сегменте
утолщения, которые при энергичном размножении составляющих их
клеток растут, подвигаясь вниз по бокам от спинного мозга. Таким образом
развивается метамерный ряд лопастей, лежащих против промежутков
между сомитами. Лопасти эти содержат материал, за счет которого обра-
зуются зачатки спинномозговых узлов. Посегментные скопления клеток
обособляются от спинной поверхности мозга, подвигаясь далее к брюшной
стороне. Часть клеток отделяется, перемещается по направлению к спин-
ной аорте и образует здесь парный ряд ганглиев симпатической нервной
системы х. Клетки этих ганглиев дают затем отростки, развивающиеся
частью по направлению к различным внутренним органам (двигательные и
чувствующие симпатические нервные волокна), частью по направлению
к спинномозговому узлу (чувствующие волокна соединительной ветви).
Пройдя через узел, волокна соединительной ветви врастают в мозг, входя
в состав спинных нервных корешков.
Клетки спинномозговых узлов приобретают веретеновидную форму и
образуют затем каждая два отростка (рис. 221, Л). Один, центральный,
отросток развивается немного раньше на спинном конце клетки и растет
по направлению к спинной части боковой поверхности мозга, входя в
состав спинных нервных корешков. Эти центральные отростки рассматри-
вают как невриты; они врастают в мозг, где образуют на его поверхности
пучки центростремительных или чувствующих нервных волокон, конечные
разветвления которых вступают в связь с дендритами нервных клеток
мозга. Другой, периферический, отросток развивается немного позднее
и растет к периферии, где образует конечные чувствующие разветвления.
Эти периферические отростки сравнивают с дендритами нервных клеток
мозга. Периферические отростки клеток спинномозговых узлов соеди-
няются вместе для образования чувствующих компонентов спинномозго-
вых нервов. Клетки спинномозговых узлов меняют свою веретеновидпую
форму, в особенности у высших позвоночных, на колбовидную, униполяр-
ную, развивающуюся из биполярной вследствие сближения и срастания
основных частей обоих нервных отростков.
1 По более новым данным ганглии симпатической системы образуются из невро-
бластов, мигрирующих из брюшной стенки мозга.
237
Двигательные нервы развиваются несколько Позже и растут непосред-
ственно от центральной нервной системы. От крупных невробластов
брюшной части мозга развиваются нервные отростки, которые выходят из
мозга, слагая брюшные нервдые корешки (рис. 221, А), растут по направле-
нию к той или иной мышце и образуют двигательные компоненты спинно-
мозговых нервов. Эти волокна иннервируют исключительно сомати-
ческую мускулатуру (т. е. миотомы и их производные). Кроме того, невро-
бласты боковых частей мозга образуют также двигательные волокна,
выходящие у низших позвоночных через спинные корешки и узлы, а
у высших форм — через оба корешка спинномозговых нервов. Эти волокна
иннервируют висцеральную мускулатуру (гладкую и поперечнополосатую)
и железы или непосредственно, или через посредство симпатической
нервной системы.
Как в центральной нервной системе, так и в ганглионарной пластинке
не все клетки дают начало нервным элементам. Часть клеток ганглионар-
ной пластинки идет на образование скелетогенной мезенхимы (участву-
ющей в построении зачатков висцерального скелета и скелета непарных
плавников). Другие клетки выходят из ганглионарной пластинки, а
позднее также из брюшной части мозга и располагаются вдоль нервов,
образуя затем шванновскую оболочку их волокон. Таким образом клетки
шванновской оболочки по происхождению сходны с клетками невроглии,
что доказывается еще и тем обстоятельством, что в центральной нервной
системе шванновская оболочка исчезает, а нервные волокна окружаются
клетками глии. Позднее вокруг нервов развиваются еще и соединительно-
тканные оболочки, которые у места выхода нервов из мозга переходят
в твердую мозговую оболочку.
В области головы развитие периферической нервной системы идет в
сущности таким же образом — с той лишь разницей, что, кроме гангли-
онарной пластинки, в образовании нервных узлов принимают участие и
непосредственно части эктодермы .(.рис. 222).. Ганглионарная пластинка
сравнительно слабо ра’зйита в задней части продолговатого мозга, опа
совсем прерывается в области'уха и отсутствует в переднёй“частй головного
мозга. Слабому развитию ганглионарной пластинки в отдельных областях
соответствует либо недоразвитие периферических узлов, либо присутствие
и иных источников их образования.
По бокам головы, над каждой висцеральной щелью, развивается
утолщение эктодермы, из которого выползают затем клетки, присоеди-
няющиеся к прилежащим зачаткам узлов головных нервов. Эти утолщения
эктодермы носят название на д ж а бе р н ы х (эпибранхиалышх) пла-
код (рис. 222). Клетки, происходящее-из этих плакод, соединяющей
с клетками, происходящими от ганглионарной пластинки, и образуют
вместе с ними узлы головных нервов 7-й, 9-й и 10-й пар (лицевого, языко-
глоточного и блуждающего), от которых отходят типичные висцеральные
нервы. Было высказано предположение, что наджаберные плакоды пред-
ставляют собой остатки особых висцеральных органов чувств, чувствую-
щие клетки которых затем соединились с узлами обычного (спинномозго-
вого) типа.
Кроме того, по бокам головы, несколько выше, развиваются еще
особые утолщения эктодермы, которые носят название боковых
(дорзо-латеральных) плакод (рис. 222). Иногда это одно непрерывное
продольное утолщение, иногда оно состоит с самого начала из 3—4 неза-
висимых частей. Во всяком случае, некоторая средняя часть утолщенной
эктодермы, лежащая в области ганглия лицевого нерва, обособляется и
погружается вглубь, образуя ямку. Эта ямка представляет собой зачаток
слухового пузырька. Клетки, обособляющиеся от его стенки, ложатся
у ганглия лицевого нерва и образуют ганглий слухового нерва (8-й пары),
который целиком развивается исключительно за счет плакоды. Клетки
238
клеток, сама плакода раз-
Рис. 222. Поперечный разрез че-
рез голову зародыша акулы в об-
ласти 3-й жаберной щели (tr) (по
Фрорипу).
ch — хорда; dl — боковая плакода;
ер — наджаберная плакода; дп —
ганглий блуждающего нерва; X —
зачаток десятого нерва.
этого ганглия развивают затем периферические отростки по направлению
к чувствующему эпителию слухового пузырька и центральные отростки,
врастающие в мозг. Слуховой пузырек, его ганглий и нерв развиваются,
таким образом, за счет боковой плакоды. Остальные части боковой пла-
коды развиваются только у рыб и амфибий. Передняя часть утолщенной
эктодермы, лежащая впереди слуховой плакоды, отделяет от себя клетки,
ложащиеся также у ганглия лицевого нерва и образующие более или
менее тесно связанную с ним массу нерв!
растается дальше вперед в виде полосы
над глазом и под глазом и дает ветвь,
растущую вниз, по направлению к нижней
челюсти. В этих полосках развиваются
через известные промежутки небольшие
чувствующие ямки — сейсмосенсорные ор-
ганы или органы боковой линии. Затем по-
лоски обыкновенно погружаются вглубь,
образуют жолоб, который обычно замы-
кается в канал. Таким образом разви-
ваются надглазничный, подглазничный и
подъязычно-челюстной каналы боковой
линии головы. Нервные клетки, развив-
шиеся за счет этой части плакоды, дают,
кроме центральных отростков, образую-
щих часто обособленные корешки лице-
ввого нерва, периферические отростки,
растущие вдоль эктодермальных полосок
и образующие нервы этих каналов боковой
линии (n. ophtalmicus superficialis VII, n.
buccalis VIIh n. mandibularis externusVII).
Клетки, выползающие из задней, заслу-
ховой части плакоды, образуют часть
ганглия языкоглоточного нерва и зна-
чительную, обычно ясно обособленную,
нервную массу, лежащую со спинной стороны ганглия блуждающего
нерва (рис. 222). Сама плакода постепенно разрастается назад, образуя
узкую полосу утолщенной эктодермы вдоль средней боковой линии тела.
В этой полосе развиваются также сейсмосенсорные органы и она также
обычно замыкается в канал боковой линии, идущий назад вдоль всего
туловища. Нервная масса, лежащая над ганглием блуждающего нерва,
представляет собой ганглий боковой линии. Центральные отростки его
клеток врастают в мозг, образуя корешок бокового нерва, а- перифери-
ческие отростки растут вдоль боковой линии, вступают в связь с разви-
вающимися органами чувств и составляют боковой нерв, который у
взрослого животного тесно связан с блуждающим нервом (n. lateralis X).
В области уха ганглионарная пластинка, как сказано, вовсе не
развивается, а впереди него она хорошо выражена и образует здесь два
ганглия тройничного нерва (5-й пары), которые развиваются только
за счет этого источника.
Остается еще упомянуть, что все чисто двигательные нервы области
головы, т. е. нервы 3-й, 4-й, 6-й и 12-й пар, развиваются совершенно
таким же образом, как и двигательные нервы брюшных корешков спин-
ного мозга, от крупных нервных клеток брюшной стенки мозга.
Наконец, первые две пары головных нервов являются нервами совер-
шенно особого типа. Зрительный нерв развивается в глазном стебельке
за счет отростков нервных клеток ретины. Последняя представляет собой
по происхождению часть стенок самого мозга, имеет сложное строение и
является не просто чувствующим эпителием, а вместе с тем и сложным
239
йервйЫм Центром. Таким образом, зрительный нерв Является не Перифе-
рическим нервом, а центральным трактом. Как и сам мозг, он содержит
опорные элементы (невроглия) и окружен мозговыми оболочками. Обо-
нятельный нерв является также нервом особого типа, не сравнимым
с другими нервами позвоночных, так как составляется из волокон, вы-
растающих непосредственно от чувствующих клеток обонятельного эпи-
телия по направлению к мозгу, где они входят в обонятельную луковицу.
Спинной мозг
Спинной мозг простирается у низших позвоночных вдоль всего туло-
вища до конца хвоста. Он лежит непосредственно над хордой, впереди
без резкой границы переходит в головной мозг, а позади постепенно
утончается. У низших позвоночных иногда наблюдается плоская ленто-
видная форма мозга, как у круглоротых (и у химер), но чаще поперечное
сечение его имеет у рыб округлотреугольную
Рис. 223. Продолговатый
мозг и мозжечок остера (по
Джонстону).
(с расширенным основанием) или, у наземных
позвоночных, овальную или круглую форму.
Внутренняя полость спинного мозга предста-
влена узким центральным каналом, выстлан-
ным эпителием — эпендимой, клетки которого
продолжаются в длинные радиальные волокна,
доходящие до периферии органа. Между мас-
сивными боковыми стенками глубоко врезы-
ваются со спинной и брюшной стороны раз-
деляющие их очень узкие спинная и брюшная
борозды, заполненные соединительной тканью.
Брюшная борозда более заметна и в особенно-
сти сильно развивается у наземных позво-
ночных.
У рыб спинной мозг обычно вполне рав-
номерно развит и позади очень постепенно
утончается. В некоторых случаях сильное раз-
витие периферических нервов вызывает увели-
чение соответствующих нервных центров, на-
ружно выражающееся в образовании местных
утолщений стенок мозга; так, у морского пе-
туха (Trigla) развиваются пять парных вздутий
сЫ —мозжечок; lat—кореш-
ки бокового нерва VII; I.
aclat — lobus acustleo-latera-
Us ; I. visl — lobus vlsceraBs;
V — X — головные нервы.
спинной стенки передней части спинного мозга
соответственно отходу пяти нервов, идущих
в передние увеличенные и способные к самосто-
ятельным движениям лучи грудного плавника.
У наземных позвоночных значительное усиленно парных конечностей,
поддерживающих все тело животного, ведет обыкновенно к образованию
утолщений спинного мозга в области отхода нервов, входящих в состав
плечевого и поясничного сплетений. Такие утолщения особенно зна-
чительны у птиц, у которых соответственно сильному развитию задних
конечностей сильно расширена крестцовая область спинного мозга, в кото-
рой и центральный канал расширяется, образуя нечто вроде желудочка
ромбической формы (sinus rhomboidalis). Еще более значительное расши-
рение имелось в крестцовой области у динозавров, массивное тело которых
опиралось главным образом на задние конечности. У стегозавра площадь
поперечного сечения крестцового мозга превышала таковую головного
мозга в 10 раз. Имеются расширения и у млекопитающих в шейной и
поясничной областях позвоночника.
У рЫб мозг простирается обычно вдоль всего тела. У наземных позво-
ночных задний конец спинного мозга нередко редуцируется, как, напри-
240
мер, у бесхвостых амфибий. У млекопитающих хвостовой отдел тела
низводится до степени сравнительно незначительного придатка, и в связи
с этим находится редукция заднего конца спинного мозга, который кон-
чается обычно уже в поясничной области и никогда не заходит далее
крестцовой. Однако этот конец мозга не исчезает бесследно — от него
остается рудимент в виде конечной нити (filum terminale). Так как ре-
дукция спинного мозга сопровождается его укорочением, то нервы, идущие
к задним частям тела, получают все более наклонное положение и про-
ходят до своего выхода значительный путь по спинномозговому каналу
вдоль заднего конца мозга, образуя сложный пучок — так называемый
конский хвост (cauda equina).
Серое вещество спинного мозга занимает центральное положение и
состоит из звездчатых опорных элементов (клеток глии) и нервных клеток
с их отростками. Последние более или менее удалены от центрального
канала. Более крупные нервные клетки лежат со спинной стороны цен-
трального канала и вступают в связь с чувствующими волокнами спинных
корешков, а еще более крупные клетки лежат с брюшной стороны серого
вещества по бокам от центрального канала, это — двигательные клетки,
нервные отростки которых составляют брюшные корешки спинномозговых
нервов.
Уже у рыб скопление двигательных нервных клеток образует заметные
парные выступы серого вещества, так называемые брюшные рога.
У наземных позвоночных развиваются подобные же спинные рога
серого вещества, содержащие чувствующие нервные клетки. Таким обра-
зом, у высших позвоночных серое вещество спинного мозга имеет в попе-
речном разрезе приблизительно форму буквы Н. У млекопитающих
развиваются еще более или менее заметные боковые выступы — боко-
вые рога серого вещества (рис. 241).
Белое вещество спинного мозга, состоящее только из нервных волокон,
окруженных здесь мизлиновой оболочкой (за исключением круглоротых),
занимает всю периферию и делится вдающимися в него рогами серого
вещества на так называемые столбы: спинные столбы между спинными
рогами, брюшные столбы между брюшными рогами и боковые столбы
по бокам мозга. Спинные столбы составляются из чувствующих нервных
волокон, восходящих к головному мозгу, брюшные столбы содержат
двигательные волокна, нисходящие от головного мозга, а боковые столбы
содержат волокна различного значения как центростремительные, так и
центробежные, а также и короткие пути, связывающие различные части
спинного мозга.
Головной мозг
Продолговатый мозг (myelencephalon s. medulla joblongata) по своему
строению больше других отделов приближается к спинному, от которого
он ясно не отграничен. Полость центрального канала здесь чрезвычайно
сильно расширяется, образуя ромбический четвертый желу-
дочек (рис. 224, 225), который простирается вперёд и в область заднего
мозга, где затем резким сужением — перешейком (isthmus) —
отделяется от полости среднего мозга. Боковые стенки нервной трубки
в области продолговатого мозга сильно разрастаются в стороны, растя-
гивая совершенно тонкую эпителиальную ее крышу, покрывающую
полость четвертого желудочка. Кровеносные сосуды мягкой мозговой
оболочки образуют на этом эпителии заднее сосудистое спле-
тение, которое вместе с ним вдается многочисленными складками в
полость желудочка (plexus chorioideus posterior) (рис. 233). Эпителий
эпендимы имеет здесь железистый характер. Сильное развитие боковых
стенок продолговатого мозга находится в связи с значительным развитием
головных нервов, обслуживающих висцеральный аппарат и органы
247
боковой линии и слуха, которые все отходят от этой части мозга. И в
продолговатом мозге, как и в спинном, чувствующая область серого
вещества располагается со спинной стороны органа, который здесь явля-
ется как бы развернутым, так что наиболее дорзальные его части ока-
зываются наиболее раздвинутыми в стороны. Эта чувствующая область
Рис. 224. Схема головного мозга рыбы.
А — срединный разрез; В, С, D, Е — поперечные разрезы через передний, промежуточный
средний и продолговатый мозг; F — мозг сврху.
ch — хиазма; сЫ — мозжечок; cst — полосатые тела; di — промежуточный мозг; ер — эпи-
физ; hy — гипофиз; if —воронка; 1у — нижние доли; 1о — обонятельные доли; та — средний
мозг; ml —продолговатый мозг; pt — задний мозг р — теменной орган; t — передний мозг;
ih — зрительные бугры; III, IV — третий и четвертый желудочки.
продолговатого мозга особенно сильно развита у рыб и нередко вдается
внутрь полости четвертого желудочка двумя продольными валиками
(рис. 223), из которых нижний представляет собой висцеральную лопасть
(lobus visceralis), а верхний — лопасть боковой линии (lobus acustico-
lateralis), являющуюся центром не только для органов боковой линии,
Рис. 225. Схема головного мозга наземного позвоночного (обозначения
см. на рис. 224).
А
но и для органа слуха. Висцеральная лопасть в некоторых случаях не-
обычайно разрастается в связи с значительным развитием органов вкуса
и образует два больших полушария, являющихся иногда самыми круп-
ными отделами мозга (карповые, сомовые рыбы). У электрического ската
разрастается лежащая более медиально (вентрально) двигательная область
продолговатого мозга и образует пару крупных электрических долей.
242
У наземных позвоночных редукция жаберного аппарата и органов боковой
линии приводит к некоторому сокращению названных областей, причем
от лопасти боковой линии, естественно, остается одна только слуховая
область (area acustica). В основании продолговатого мозга помещаются
двигательные ядра большинства головных нервов (от 5-й до 12-й пары).
Белое вещество спинного мозга продолжается и на поверхности
продолговатого мозга, образуя более или менее значительные массы
продольных волокон, которые наружно в особенности ясно различимы
у млекопитающих. Брюшные столбы продолжаются вперед в виде пары
постепенно расширяющихся валиков, так называемых пирамид, в
самом начале которых наблюдается перекрест волокон, переходящих
сюда частью и из боковых столбов спинного мозга. Последние также
ясно видны в области продолговатого мозга и у высших обезьян впереди
переходят в заметное вздутие — оливы (olivae), которым внутри
соответствуют ядра серого вещества. Спинные столбы образуют в самом
начале продолговатого мозга небольшие вздутия, соответствующие также
ядрам серого вещества, и впереди постепенно сглаживаются. В передней
части продолговатого мозга, вдоль края крыши четвертого желудочка,
лежат тяжи продольных волокон, переходящие затем на мозжечок; это
так называемые веревочные тела, особенно выраженные у мле-
копитающих (corpora restiformia). От продолговатого мозга отходит
большая часть головных нервов, именно отводящий нерв (6-й пары),
с брюшной стороны, и все висцеральные нервы (5-й, 7-й, 9-й и 10-й пар;
первые из них у млекопитающих лежат в области моста), а также и слу-
ховой нерв (8-й пары) — с боков. Подъязычный нерв (12-й пары) нередко
связан уже со спинным мозгом.
Задний мозг (metencephalon) представляет собой у низших
позвоночных лишь часть крыши нервной трубки в области четвертого
желудочка, впереди от заднего сосудистого сплетения. Это так называемый
мозжечок (cerebellum), который у круглоротых и у амфибий имеет
вид небольшой поперечной складки, ограничивающей спереди крышу
четвертого желудочка (рис. 224, 225). У некоторых рыб это также еще не-
значительное образование, между тем как у других он разрастается и
прикрывает частью соседние отделы мозга — продолговатый мозг позади
(рис. 230) и иногда средний мозг впереди. Впереди мозжечок переходит
в поперечную складку, более или менее глубоко вдающуюся во внутрен-
нюю полость и служащую границей между задним мозгом и средним; это
передний мозговой парус (velum medullare anterius s.
valvula cerebelli) (рис. 231); иногда подобная же складка развивается и
позади мозжечка (velum medullare posterius). Нередко уже у рыб наблю-
дается увеличение поверхности мозжечка путем образования многочис-
ленных складок, благодаря чему развивается сложная система извилин.
У амфибий впервые на наружной поверхности мозжечка появляется серое
вещество определенной структуры, которое образует так называемую
кору мозжечка. У рептилий эта кора развивается сильнее, и мозжечок
становится более выпуклым вследствие разрастания верхней поверхности.
У птиц и у млекопитающих увеличение поверхности коры мозжечка со-
провождается образованием глубоких и сложных складок, заметных сна-
ружи в виде системы борозд и извилин между ними (рис. 235, 236). Так
как изнутри кора подостлана белым веществом, проникающим во все про-
межутки между складками серого вещества, то на разрезе через мозжечок
видна очень сложная картина распределения белого вещества (на сагит-
тальном разрезе вид разветвленного дерева—arbor vitae). У птиц и у мле-
копитающих наблюдается еще и наружное обособление более крупных
отделов мозжечка. В средней части наблюдается образование ряда пра-
вильных поперечных борозд, придающих этой части вид «червячка» (vermis);
по бокам развиваются у птиц небольшие парные выступы (flocculi), а у
243
млекопитающих, в особенности у высших форм, часто значительные полу-
шария (hemisphaerae), на которых сбоку иногда имеются и flocculi. Кроме
серой коры, в стенках мозжечка имеются еще парные массы глубокого
серого вещества, образующие так называемое зубчатое ядро (nucleus den-
tatus). Мозжечок имеет значение важного центра, ведающего функцией
сохранения равновесия путем регуляции мускульного тонуса. Вместе
с тем, он оказывается основным органом регуляции функционального
состояния всех органов при посредстве вегетативной нервной системы.
Мозжечок связан с другими отделами мозга значительными пучками
волокон, образующими позади упомянутые уже веревочные тела, или
задние ножки мозжечка (corpora restiformia s. crura cerebelli
ad medullam), идущие по бокам крыши четвертого желудочка. Впереди
мозжечок связан посредством передних ножек мозжечка
(brachia conjunctiva s. crura cerebelli ad cerebrum) co средним мозгом;
между передними ножками помещается передний мозговой парус. Наконец,
у млекопитающих с развитием моста в передней части продолговатого
мозга развиваются и значительные пучки волокон, соединяющие полу-
шария мозжечка с мостом, или средние ножки мозжечка
(crura cerebelli ad pontem).
Кроме мозжечка, к заднему мозгу относят еще и варолиев мост
(pons Varolii), который впервые выделяется только у млекопитающих в
виде поперечной массы волокон, соединяющих средние ножки мозжечка.
Развивается этот отдел за счет передней части продолговатого мозга и
достигает особой мощности только у высших млекопитающих.
Средний мозг (mesencephalon) представляет собой у низших
позвоночных довольно крупный отдел мозга (рис. 224). Его крыша (tectum
opticum) вздута в виде парных зрительных долей (lobi optici),
в сером веществе которых кончаются волокна зрительных трактов. По-
лость среднего мозга у низших позвоночных довольно объемиста и продол-
жается в зрительные доли в виде парных выступов, образующих зритель-
ные желудочки (рис. 224). У наземных позвоночных (и у двудышащих
рыб) средний мозг теряет значение зрительного центра и заметно умень-
шается в размерах. Боковые стенки его утолщаются и внутренняя полость
сильно сжимается, превращаясь в узкий канал — сильвиев водопровод
(aquaeductus Sylvii) (рис. 225). У млекопитающих крыша среднего мозга
подразделяется поперечным перехватом, так что «двухолмие» низших
форм превращается в «четверохолмие» (рис. 236).
В основании среднего мозга имеются некоторые ядра, из которых
особое значение ассоциационного центра имеют «красные ядра» млеко-
питающих. Здесь же помещаются и двигательные ядра глазодвигательных
нервов. Кроме того, здесь проходят большие парные массы нервных
волокон, соединяющих передний мозг с продолговатым и спинным мозгом.
Эти массы продольных волокон образуют так называемые ножки боль-
шого мозга (pedunculi cerebri s. crura), которые наружно особенно
ясно выделяются у млекопитающих.
В области среднего мозга отходят две пары головных нервов — гла-
зодвигательный нерв (3-й пары) с брюшной стороны и блоковый (4-й пары)
со спинной стороны между крышей среднего мозга и мозжечком.
Промежуточный мозг (diencephalon) представляет со-
бой небольшой отдел мозга, который снаружи виден только у животных
с примитивным мозгом (круглоротые, двудышащие, земноводные) (рис. 229,
232); у других позвоночных он сверху закрывается соседними отделами,
и только снизу видны известные выступы стенок промежуточного мозга
(рис. 230, 235). В этом отделе в высшей степени утолщаются боковые стен-
ки, в которых имеются значительные массы серого вещества, образующие
ври тельные бугры (thalami optici) (рис. 224, 225). Дно проме-
жуточного мозга остается сравнительно тонким, а крыша на значитель-
244
ной части своего протяжения и вовсе не содержит нервного вещества;
она представлена простым эпителием, с которым срастается мягкая моз-
говая оболочка, сосуды которой образуют здесь переднее сосу-
дистое сплетение (plexus chorioideus anterior) (рис. 233), мно-
гочисленными складками свешивающиеся во внутреннюю полость. По-
следняя имеет, благодаря утолщению боковых стенок, форму вертикаль-
ной щели (рис. 224, 225). Это — третий желудочек, передняя
часть которого может проникать более или менее далеко в передний мозг,
где он спереди ограничивается концевой пластинкой (lamina
terminalis). В задней части крыши промежуточного мозга имеется парное
скопление нервного вещества, образующее нес им метр и чные
г а н г л и и (ganglia habenulae), связанные между собой верхней комис-
сурой (comissura superior s. habenularis). Непосредственно позади послед-
ней наблюдаются еще особые непарные придатки, имеющие обыкновенно
железистый характер (рис. 224, 225). В некоторых случаях, однако, эти
образования имеют строение органов зрения. Круглоротые обладают двумя
такими органами, имеющими вид пузырьков, соединенных нервами с кры-
шей мозга (рис. 268). Передний, меньший, пузырек носит название т е-
м е н н о г о (париетального) органа; его нерв связан с верхней ко-
миссурой. Задний, больший, пузырек представляет собой эпифиз, или
пинеальный орган (epiphysis cerebri s. glandula pinealis); его
нерв связан с задней комиссурой, лежащей в крыше мозга па границе
среднего мозга. У некоторых рыб и у большинства ящериц имеется хорошо
развитый теменной орган глазоподобного строения. У других позвоноч ,ых
сохраняется обыкновенно только эпифиз, причем он функционирует уже
не в качестве органа чувств, а приобретает значение железы внутрен-
ней секреции (рис. 232, J33).
Нижняя стенка промежуточного мозга образует направленный на-
зад непарный полый выступ — воронку (infundibulum), к передней
(нижней) стенке которой примыкает другая железа внутренней секреции —
под мозговая железка (hypophysis cerebri s. glandula pitui-
taria) (рис. 233). Впереди воронки нижняя стенка промежуточного мозга
образует серый бугор (tuber cinereum), который спереди ограничен пере-
крестом зрительных трактов. В задней стенке воронки парная масса се-
рого вещества образует сосковидные тела (corpora mamilla-
ria). У низших позвоночных на воронке имеются довольно значительные
тонкостенные полые выросты — задний непарный, оплетенный крове-
носными сосудами, сосудистый мешок железистого характера
(lobus posterior s. saccus vasculosus) (рис. 231) и парные боковые — ниж-
ние доли (lobi inferiores) (рис. 230). У наземных позвоночных эти
образования редуцируются и присоединяются к подмозговой железке,
образуя ее нервную заднюю лопасть. Нервные образования дна проме-
жуточного мозга (нижние доли, сосковидные тела и серый бугор) нередко
объединяют под названием hypothalamus.
В нижней стенке промежуточного мозга по бокам продолжаются нож-
ки большого мозга (pedunculi cerebri), идущие к переднему мозгу. Впереди
масса поперечных волокон образует перекрест зрительных
трактов (chiasma), поднимающихся отсюда в зрительные бугры,
охватывая по пути ножки большого мозга. У млекопитающих зрительные
тракты можно проследить до заднего края бугров, где имеются две пары
небольших возвышений — коленчатые тела (corpora geniculafa la'eral’a
et medialia), связанные пучками волокон также с четверохолмием. Между
обоими зрительными буграми у млекопитающих происходит посредт не
сращение, получившее нерациональное название серой или мягкой ко-
миссуры.
Передний мозг (telencephalon) представляет собой всегда
значительный передний отдел мозга, который, постепенно прогрессируя,
245
достигает у высших позвоночных огромных размеров, покрывая сверху
большую часть остального мозга. По средней (сагиттальной) плоскости
он спереди ограничен концевой пластинкой (lamina terminalis), до кото-
рой внутри доходит и полость среднего желудочка, отграниченная от по-
лости третьего желудочка промежуточного мозга у низших позвоночных
поперечной эпителиальной складкой, свешивающейся с крыши мозга —
поперечным парусом (рис. 231). Так как в области переднего мозга утол-
щаются не боковые стенки, а основание в виде больших выпуклых парных
масс, образующих основные ганглии, или полосатые тела (cor-
pora striata), то и полость желудочка имеет здесь более сложную форму:
продолжаясь у низших позвоночных в виде вертикальной щели, лежащей
между обоими полосатыми телами, вплоть до концевой пластинки, она
вверху, по бокам переходит в парные полости, покрывающие сверху вы-
пуклые полосатые тела (боковые желудочки) (рис. 234). У низших позво-
ночных бока и крыша переднего мозга, образующие мантию, или
плащ (pallium), остаются сравнительно тонкими и повторяют в общем
форму лежащих под ними полосатых тел, от чего весь передний мозг ка-
жется разделенным на два полушария. В передней стенке переднего мозга
развивается парное выпячивание более или менее крупных размеров, в
которое может заходить и продолжение полости боковых желудочков
(рис. 224); это обонятельные доли (lobi olfactorii), которые
могут быть различно диференцированы. Иногда обонятельные доли явля-
ются сидячими, располагаясь всей своей массой на передней стенке мозга,
иногда значительно вытягиваются в длину и нередко диференцируются
на основную часть (собственно обонятельную долю), более или менее длин-
ный стебелек (pedunculus olfactorius) и обонятельную
луковицу (bulbus oefactorius) (рис. 229, 234). В задней части крыши
переднего мозга, непосредственно впереди от поперечного паруса, наблю-
дается продолжение того же сращения тонкой эпителиальной крыши с
мягкой мозговой оболочкой (tela chorioidea anterior), которое покрывает
полость промежуточного мозга; здесь эта крыша дает у многих позвоноч-
ных вверх непарный вырост, невидимому, железистого характера, назы-
ваемый парафизом (paraphysis). У двудышащих рыб и у наземных позво-
ночных передняя стенка мозга вдвигается посредине между обоими поло-
сатыми телами назад в виде вертикальной складки, разделяющей перед-
ний мозг на два обособленных теперь полушария (haemisphaerae
cerebri) (рис. 225, F), в которую проникают и мозговые оболочки, образую-
щие здесь прочную перегородку. С развитием обособленных полушарий,
Полости боковых желудочков впереди отделяются друг от друга, соеди-
няясь позади мо и роевыми отверстиями (foramina Monroi)
с передним концом среднего желудочка (рис. 225, F, III).
Серое вещество переднего мозга, образующее большие парные скопле-
ния в обонятельных долях и в полосатых телах, имеется, однако, и в кры-
ше мозга, где оно сплошным слоем выстилает полости желудочков (epis-
triatum). Поверхность переднего мозга у низших позвоночных содержит
белое вещество. Только у высших рыб нервное вещество совершенно ре-
дуцируется в крыше переднего мозга.
В концевой пластинке располагается довольно значительная п е-
редняя комиссура (commissura anterior), связывающая между
собой оба полосатых тела (рис. 231).
У двудышащих рыб и у амфибий довольно значительно развиваются
полушария; их верхняя стенка — мантия (pallium) — остается еще срав-
нительно тонкой; но в концевой пластинке, рядом с передней комиссурой,
над ней появляется еще одна комиссура (commissura pallii anterior s.
hippocampi), связывающая между собой мантию обоих полушарий (рис.
227, 233). Часть нервных клеток этой мантии перемещается по направле-
нию к поверхности и располагается внутри белого вещества. У рептилий
246
передний мозг разрастается еще больше, но и здесь величина его зависит
от развития больших полосатых тел в его основании. Мантия остается
еще тонкой, но в ней впервые появляется поверхностное серое вещество
определенной структуры, представляющее собой кору большого мозга.
Эта кора в большей своей части играет, однако, лишь роль высшего
обонятельного центра и сравнима только с незначительной ее частью у
млекопитающих, носящей название гиппокампа. Поэтому мантию низших
наземных позвоночных обозначают как первичную мантию,
или зачаток гиппокампа (archipallium s. primordium hippocampi), хотя
в боковых ее частях и имеется уже область, соответствующая вторичной
мантии млекопитающих (рис. 226).
У млекопитающих передний мозг получает колоссальное развитие
(рис. 236), постепенный ход которого можно ясно проследить, переходя
от низших форм к высшим. Обонятельные доли обычно хорошо развиты.
Рис. 226. Развитие вторичной мантии и гиппокампа у млекопитающих
(по Эдингеру).
д___змея В — сумчатое млекопитающее Ilyysiprimnus; апр — первичная мантия;
’ * hip — гипнонами; пр — вторичная машия.
Полосатые тела помещаются в виде довольно значительных вздутий па
дне передней части боковых желудочков («переднего рога») и продолжа-
ются назад по бокам от зрительных бугров в виде постепенно суживаю-
щихся «хвостатых тел» (corpora caudata). Но главной частью мозга явля-
ется здесь мантия, отличающаяся чрезвычайным развитием коры. Эта
кора имеет огромное значение в качестве, комплекса ассоциационных
центров, в которых сосредоточиваются все высшие, психические функции
мозга. Мы видели, что соответствующая область имеется и у низших по-
звоночных, но даже у рептилий она сравнительно еще ничтожна. Так как
эта область помещается в боковых частях мантии, то при значительном
ее разрастании верхние части мантии отодвигаются к средней плоскости,
где становятся медиальной стенкой полушарий. Здесь эта прежняя, пер-
вичная мантия (archipallium), глубоко вдавливается внутрь полости бо-
ковых желудочков в виде складок, получающих теперь название гиппо-
кампа (hippocampus в. cornu Ammonis) (рис. 226), который сохраняет
свою функцию высшего обонятельного центра. На медиальной поверх-
ности полушарий складка гиппокампа отмечена проникающей в нее глу-
бокой бороздой (fissura hippocampi). Нижняя часть первичной мантии,
отделенная от гиппокампа развитием новой коры, остается на нижней
поверхности полушарий в виде так называемых грушевидных долей, также
сохраняющих свою обонятельную функцию. Сильно разрастающаяся
вторичная мантия (neopallium) диференцируется,, разделяясь
247
на области, носящие название долей. Раньше всего у большинства млеко-
питающих развивается глубокая сильвиева борозда (fis-
sura Sylvii) (рис. 238), идущая на наружной поверхности снизу, косо,
вверх и назад и отделяющая переднюю лобную долю от нижней
височной. Сильвиевым бороздам по положению в глубине мозга
соответствуют полосатые тела. Выше и впереди от сильвиевой борозды
развивается также глубокая поперечная роландова борозда
(sulcus centralis s. Rolandi), отделяющая переднюю, лобную долю от верх-
ней, теменной (рис. 238). Наконец, у обезьян появляется еще попе-
речная борозда (sulcus transversus), отделяющая позади небольшую з а-
тылочную долю. Кроме этих крупных подразделений, у высших
сра
млекопитающих разра-
стающаяся кора обра-
зует еще многочислен-
ные более или менее
глубокие складки, отме-
ченные снаружи прони-
кающими в них бороз-
дами. Этим достигается
значительное увеличе-
ние поверхности коры
большого мозга. Непра-
вильно располагающие-
ся пространства между
этими бороздами носят
Рис. 227. Передние комиссуры позвоночных. Схемати-
ческие поперечные разрезы через передний мозг рыбы
(Л), амфибии или рептилии (В) клоачного (С) и жи-
вородящего млекопитающего (Z)).
apt — archipallium; npl — neopallium; ca — commissura
anterior; cpa — commissura pallii anterior; oc— corpus cal-
losum; cst — corpus striatum; f — fornix.
у крупных животных, чем у небольших.
cst
D
название извилин. Сис-
тема борозд и извилин
особенно богато разви-
та у высших млекопи-
тающих с высоким
интеллектом (рис. 238,
239) и гораздо сложнее
В области боковых желудочков происходит диференцировка. Одно-
временно с оформлением височной доли обособляется и соответствующая
часть желудочка в виде нижнего рога (cornu inferius), в полость
которого с медиальной стороны вдается гиппокамп. Остальная, верхняя
часть желудочка проникает вперед, в лобную долю, и носит название
переднего рога (cornu anterius), в полость которого снизу и
снаружи вдается полосатое тело. Наконец, с развитием затылочных долей
развивается и проникающий в них задний вырост желудочков, носящий
название заднего рога (cornu posterius), в который у приматов
с медиальной стороны вдается утолщение, называемое птичьей шпорой
(calcar a?is).
Параллельно изменениям в расположении серого вещества переднего
мозга у млекопитающих происходят и преобразования в области передних
комиссур (рис. 227, 228). Собственно передняя комиссура, связывающая
между собой полосатые тела, остается без изменения в концевой пластинке;
лежащая же над ней комиссура мантии в связи с прогрессивным разви-
тием последней увеличивается и одновременно изменяет свою форму.
Еще очень небольшая у клоачных млекопитающих, она у сумчатых почти
достигает величины передней комиссуры и при этом изгибается в виде
колена, обращенного выпуклостью назад (рис. 228). Верхняя ветвь этого
колена служит комиссурой между корой вторичной мантии и получает
название мозолистого тела (corpus callosum), между тем как
нижняя ветвь представляет собой так называемый свод (fornix), заклю-
чающий волокна, связывающие между собой первичную мантию, т. е.
248
оба гиппокампа (рис. 227, 228) У более высоко стоящих млекопитающих
эти образования развиваются еще более и, в особенности, мозолистое
тело достигает весьма зна-
чительной мощности.Часть
концевой пластинки, за-
ключенная менаду мозоли-
стым телом и сводом, рас-
тягивается в чрезвычайно
тонкую треугольную плас-
тинку (septum pellucidum),
в которой путем расщепле-
ния получается вертикаль-
ная щелевидная полость,
носящая неудачное назва-
ние пятого желудочка
(рис. 237).
Одновременно с раз-
витием этих крупных ко-
миссур передняя комис-
сура отступает на задний
план и представляет собой
у высших млекопитающих
сравнительно ничтожное
образование.
У млекопитающих от
переднего сосудистого
Рис. 228. Диференцировка передних комиссур у мле-
копитающих. Срединные разрезы через концевую
пластинку (заштрихована) (по Джонстону). А —
рептилия; В — клоачное; С — сумчатое; D — ле-
тучая мышь; Е — высшее млекопитающее.
са — передняя комиссура; ср — комиссура мантии;
сс — моаолистое тело; ch — комиссура гиппокампа;
Мрр — гиппокамп.
сплетения проникают пар-
ные ветви вперед через монроевы отверстия внутрь боковых желудоч-
ков, где они в виде боковых сосудистых сплетений ложатся в нижних
рогах вдоль гиппокампа.
Обзор строения головного мозга в различных классах
Головной мозг круглоротых сравнительно весьма примитивен. Все его
Отделы лежат в одной плоскости и не налегают друг на друга. Сравнительно крупные
Обонятельные лопасти едва обособлены от переднего мозга и внутри содержат полости.
В небольшом переднем мозге преобладают полосатые тела, между тем как крыша явля-
ется тонкой, чисто эпителиальной. Промежуточный мозг у миног хорошо виден сверху,
где на нем ясно выделяются крупные несимметрические ганглии (g. habenulae) и оба
глазоподобных органа (теменной и пинеальный). Снизу воронка связана с сосудистым
мешком. Средний мозг слабо развит: крыша его посредине является эпителиальной,
как и в передних отделах мозга, и на ней также развивается сосудистое сплетение
(plexus chorioideus medius). Мозжечок совершенно зачаточен и имеет вид валика, огра-
ничивающего ромбическую ямку спереди. Продолговатый мозг довольно длинный
и постепенно переходит в спинной. Головной мозг миксин по сравнению с мозгом ми-
ног сильно укорочен и сжат.
Для селахий (рис. 229) весьма характерно значительное развитие перед-
него мозга, в крыше которого имеется и нервная ткань, выстилающая полость желу-
дочка (epistriatum). Последняя является непарной и, таким образом, передний мозг
еще не поделен на два полушария, хотя он и подразделяется поверхностной бороздой
на симметрические половины. Обонятельные доли очень велики, иногда являются
вполне сидячими, иногда диференцированы на примыкающие к переднему мозгу соб-
ственно доли (lobi olfactorii), на промежуточные стебельки (pedunculi olfactorii) и на
прилежащие к обонятельным мешкам луковицы (bulbi olfactorii). Промежуточный мозг
снабжен со спинной стороны эпифизом, а с брюшной стороны имеются хорошо разви-
тые нижние доли (lobi inferiores) и сосудистый мешок (saccus vasculosus). Средний
1 К этой комиссуре гиппокампа (psalterium) присоединяются еще продольные
волокна, идущие от обонятельной лопасти к гиппокампу и от последнего к сосцевид-
ным телам (образуя «столбы» свода) и частью к несимметрическим ганглиям. Таким
образом, свод представляет собой сложное образование. С редукцией гиппокампа у
высших млекопитающих и его комиссура становится лишь незначительным образо-
ванием в своде.
249
мозг хорошо развит и поделен на два полушария (lobi optici). Очень хорошо развит
также мозжечок, прикрывающий нередко не только переднюю часть продолговатого
мозга, но и часть среднего. Поверхность мозжечка образует нередко сложную систему
извилин. Продолговатый мозг то удлиненный, то более сжатый.
Головной мозг ганоидов и костистых рыб (рис. 230) имеет у различ-
ных представителей весьма разнообразное строение, но все же можно подчеркнуть
некоторые общие им характерные черты. Прежде всего, это незначительные относи-
тельные размены всего головного мозга и, в.частности, небольшая величина переднего
мозга, крыша которого или содержит самые незначительные количества нервной ткани
V.tr
Рис. 229. Мозг акулы (Scymnus lichia). А — сбоку, В — сверху, С — снизу.
В первом рисунке (Л) вписан контур желудочков.
с bl —мовжечон; са, ср. cs— передняя, задняя, верхняя комиссуры; di — промежуточный
Мозг; ер — эпифиз; hyp—гипофиз; I. olf— обонятельная доля; olf— обонятельная лу-
ковица; т— средний мозг; ml —продолговатый мозг; nt—конечный нерв; tch — сосудй
стое сплетение; sv — сосудистый мешок; г. tr—поперечный парус; I—XI •—головные нервы
(ганоиды) или не содержит ее вовсе, являясь тогда чисто эпителиальной (это так на-
вываемая развернутая форма переднего мозга костистых рыб). Главная масса перед-
него мозга представлена полосатыми телами. Полость переднего мозга является
неразделенной и дает парный вырост в небольшие сидячие обонятельные доли.
Промежуточный мозг снабжен большими нижними долями и сосудистым мешком
(рис. 230, 231). Весьма характерно для ганоидов и костистых рыб сильное раз-
витие среднего мозга, который образует у них самый большой отдел головного мозга.
Хорошо развитый мозжечок (за исключением многоперых ганоидов) налегает на
250
продолговатый мозг. Он увеличивается еще колоссально развитым передним моз-
говым парусом, образующим так называемый клапан мозжечка (valvula cerebelli),
вдающийся в виде сложной складчатой массы внутрь полости среднего мозга (рис. 231).
В продолговатом мозге сильно развиты висцеральные доли, которые у некоторых ко-
стистых рыб (например, у карповых и сомовых) выдаются в виде больших лопастей
или полушарий, значительно превышающих по своим размерам другие отделы голов-
ного мозга. Это сильное развитие висцеральных долей находится в связи с чрезвы-
чайно сильным распространением органов вкуса, которые у костистых рыб наблюда-
ются в покровах на всем теле.
Рис. 230. Головной мозг форели сверху (Л), снизу (В) и сбоку (С).
С переднего мозга снята крыша (из Видерсгейма).
сЫ — cerebellum; ch — chiasma; с; st — corpora striata; ep — эпифиз; hyp — гипофиз;
li — lobi inferiores; lo — lobi olfactorii; m — mesencephalon; ml — myelencephalon;
ep —корешки спинномозговых нервов; to-—saccus vasculosus; t—telencephalon; I—
X— корешки головных нервов.
Головной мозг двудышащих рыб отличается значительными разме-
рами переднего мозга, вполне поделенного па два полушария. Крыша полушарий
содержит нервное вещество, клеточный слой которого выстилает полости желудочков.
Обонятельные доли — сидячие. В промежуточном мозге имеются хорошо развитые
нижние доли. Средний мозг развит сравнительно слабо, а мозжечок — совершенно
ничтожный.
У амфибий передний мозг имеет сравнительно еще более крупные размеры
(рис. 232). Его мантия, как и у двудышащих, состоит из глубокого слоя нервных кле-
ток и поверхностного слоя'нервных волокон. Полушария вполне обособлены друг от
друга и содержат полости боковых желудочков. Обонятельные доли лишь очень слабо
отграничены от переднего мозга. Изгибы головного мозга выражены очень слабо. Про-
межуточный мозг ясно виден сверху вследствие слабого налегания соседних отделов
(рис. 232). Воронка хорошо выражена, но сосудистый мешок и нижние доли редуци-
251
ровались (рис. 233). Средний мозг начинает терять значение зрительного центра, но
образует еще довольно крупный отдел головного мозга. Мозжечок у амфибий совер-
шенно зачаточный и имеет вид небольшого валика у переднего края ромбоидальной
ямки (рис. 232, 233).
В головном мозге рептилий имеются прекрасно развитые полушария боль-
шого мозга (рис. 234), характерной особенностью которых является поверхностный
Рис. 231. Срединный разрез через головной мозг форели (из Галлера).
са— commissura anterior; ср—commissura posterior; cs — commissura superior (hahenu-
laris);ier — epiphysis; lo — lobus opticus; I. olf— lobus oifactorius; ro — recessus praeopti-
cus; vt — velum transversum; Mu. cbl — valvula cerebelli Остальные обозначения, каи
на рис. 230.
слой серого вещества определенной структуры, образующий уже настоящую кору
большого мозга (cortex cerebri). Толщина мантии все же еще сравнительно незначи-
тельна, и главную массу переднего мозга, как и у низших форм, образуют полосатые
тела. Обонятельные доли— иногда сидячие,(чаще, однако, диференцированы на стебли
и луковицы (рис. 234). Мозговые изгибы у рептилий прекрасно выражены.
Рио. 232. Головной мозг лягушки сверху (Л) и снизу (В) (из Видерсгейма).
2 — n. hypoglossus; di — diencephalon, Остальные обозначения см. рис. 230.
Сильное развитие переднего мозга, налегающего отчасти на задние отделы, при
заметных еще размерах среднего мозга приводит к тому, что промежуточный мовг
сверху не виден. Снизу выдается воронка, к которой сзади прилежит гипофиз. Сверху
видны лишь придатки промежуточного мозга — эпифиз — и нередко прекрасно раз-
витый теменной орган (рис. 268). У гаттерии и у ящериц теменной орган имеет вид
глазоподобного пузырька, верхняя прозрачная стенка которого образует утолщение
в виде линзы, а задняя стенка имеет вид ретины с внутренним слоем чувствующих
252
клеток, изолированных особыми пигментными клетками, и с наружным слоем нерв-
ных волокон и клеток. Теменной орган помещается в теменном отверстии черепа под
прозрачными покровами и связан с мозгом посредством нерва. Средний мозг репти-
Рис. 233. Срединный разрез через мозг саламандры (ив Галлера).
clb— мозжечок; i—infundibulum; d — diencephalon; ер—epiphysis; tch — сосудистое
сплетение. Остальные обозначения см. рис. 231.
лий еще хорошо развит. Мозжечок развивается довольно значительно и иногда при-
крывает значительную часть продолговатого мозга. Иногда намечается уже и деление
среднюю часть и боковые лопасти.
на несколько утолщенную
Рис. 234. Головной мозг алли-
гатора сверху (из Видерсгей-
ма).
Ьо — bulbus olfactorius; ро — ре-
dunculus olfactorius. Остальные
обозначения см. рис. 230.
еда » У птиц передний мозг достигает уже
относительно огромных|размеров (рис. 235). Однако
это и здесь обусловлено^' еше необычайно сильным
развитием полосатых тел. Обонятельные доли
очень малы, что связано со слабым развитием
органов обоняния, и тесно примыкают к переднему
мозгу. Мозговые изгибы выражены очень резко.
Средний мозг хорошо развит (соответственно высо-
кому развитию органа зрения), и обе его доли
сильно сдвинуты на бока мозга вследствие разраста-
ния полушарий большого мозга назад вплоть до со-
прикосновения с мозжечком (рис. 235). Последний
очень велик, прикрывает значительную 'часть про-
долговатого мозга и имеет сложное строение. Боль-
шая средняя часть мозжечка, снабженная попереч-
ными бороздами, образует «червячок», а по бокам
имеются небольшие боковые выступы (flocculi).
Головной мозг млекопитающих ха-
рактеризуется чрезвычайным развитием полушарий
(рис. 236) переднего мозга, причем это развитие
Рис. 235. Головной мозг голубя сбоку
(из Видерсгейма).
if — воронка; 1,2 — спинномозговые нервы.
Остальные обозначения прежние.
обусловлено уже разрастанием крыши, а не полосатых тел, которые оказываются
сравнительно незначительными. Эта крыша представляет собой главным образом пео-
pallium, развившийся из боковых частей мантии нижестоящих форм, и содержит
253
Bticinne чувствующие (зрительные, слуховые и осязательные) и двигательные центры
и, что особенно важно, ассоциационные
деятельностью. Первичная, обонятельная м
Рис. 236. Головной мовг кролика сверху
(из Видерсгейма).
hm — полушарие мозжечка; v — червячок.
Остальные обозначения прежние.
центры, связанные с высшей нервной
ития, archipallium, оттесненная 1Лэвным
образом вверх и внутрь, дала начало
гиппокампу (рис. 226), частью же сдви-
нута к основанию мозга, где осталась
в виде грушевидной доли (lobus pyrifor-
mis).
Поверхность полушарий большого
мовга у низших форм гладкая, песте-
пенно усложняется с развитием сложной
системы борозд (рис. 238). Развиваю-
щаяся раньше всего глубокая сильвие-
ва борозда отделяет переднюю, лобную
долю полушарий от нижней — височной.
Развивающаяся выше и впереди от силь-
виевой поперечная роландова борозда
отделяет лобную долю от верхней —
теменной. Наконец, у обезьян еще
одна, поперечная борозда, отделяет
позади небольшую затылочную долю.
Кроме зтих наиболее важных борозд,
развивается у многих млекопитающих
с высокоразвитым интеллектом (как
копытные, хищные и приматы; однако
особенно у китообразных) еще боль-
шое число’других борозд, отделяющих
друг от друга’ множество извилин коры
большого мовга (рис. 238). Соответ-
ственно обособлению лобвой, височной
и затылочной долей обособляются и
части боковых желудочков, получаю-
щие названия переднего, нижнего и
заднего рогов. Вместе с развитием коры
большого мозга прогрессивно разви-
вается и соответствующая комиссура,
которая, начиная с сумчатых млекопи-
тающих, диференциру’ется па два отде-
ла: большое мозолистое тело (corpus cal-
losum), связывающее собственно кору
обоих полушарий между собой, и ле-
жащий под ним свод (fornix), в котором располагаются волокна, связывающие
остаток первичной мантии — оба гиппокампа (рис. 228, 237).
Обонятельные лопасти обык-
Рис. 237. Срединный разрез черев головной
мозг человека (ребенка) (ив Видерсгейма).
call — мозолистое тело; / — свод; Са и Ср — пе-
редняя и задняя комиссуры; FM — монроево
отверстие; hyp — гипофиз- сЫ —мозжечок; т—
средний мовг; ml—продолговатый мозг; Р—мост;
med — спинной мозг; То — зрительный бугор;
t — передний мовг; ер — эпифиз;
новенно хорошо развиты. Изгибы
мовга ясно выражены (рис. 237).
Рис. 238. Извилины мозга человека
(из Видерсгейма).
FS — сильвиева борозда; сЫ — мозже-
чок; Lf — лобная доля; Lo — затылоч-
ная доля; Lp —теменная доля; ml— про-
долговатый мовг; Ро — задняя попереч-
ная борозда; med — спинной мовг; Т —
височная доля; В — центральная (ро-
ландова) борозда.
254
Промежуточный мозг сверху не виден. Эпифиз представляет собой небольшой
железистый орган; гипофиз также невелик. Доли крайне незначительного среднего
мозга подразделяются поперечной бороздой на четыре бугра, образующие четверо-
холмие (corpora quadrigemina). Мозжечок очень велик и диференцирован на
среднюю часть — червячок — и боковые полушария, на которых сбоку и снизу часто
tuasma
opt
Dypamida
Oliva
Рис. 239. Мозг человека снизу (по Эдингеру).
—5-'fundi bulum
- Corp
maniill
Pedunculus
Cere-
bellum
сидят небольшие придатки (flocculi) (рис. 236). Оба полушария мозжечка связаны
между собой массой поперечных волокон, лежащих в нижней стенке заднего мозга и
образующих характерный для млекопитающих варолиев мост (pons Varolii) (рис. 239).
В продолговатом мозге по бокам крыши четвертого желудочка обособляются продоль-
ные пучки волокон, продолжающихся затем в задние ножки мозжечка — веревочные
тела (corpora restiformia), а у основания продольные же волокна образуют выступающие
валики — пирамиды.
Мозговые оболочки
Центральная нервная система ланцетника одета сплошным слоем
соединительной ткани. У позвоночных же развиваются специальные обо-
лочки (meninges) за счет надхрящницы /тли надкостницы окружающего
скелета (черепа и позвоночника). Первоначально развивается, таким обра-
зом, простая первичная оболочка (meninx primitive), оде-
вающая мозг и несущая кровеносные сосуды.
В спинном мозге между ней и выстилающей окружающий спинно-
мозговой канал надхрящницей, или надкостницей, имеется полость, вы-
полненная у рыб главным образом жировой тканью, а у наземных позво-
ночных — жидкостью, с сетью соединительнотканных перемычек, подве-
шивающих таким образом спинной мозг внутри канала. Первичная обо-
лочка спинного мозга расщепляется у наземных позвоночных на покры-
вающую мозг богатую сосудами мягкую мозговую оболочку (pia mater
primitive) и твердую оболочку (dura mater), разделенные лим-
фатической щелью. У млекопитающих, наконец, мягкая мозговая обо-
лочка диференцируется еще на два слоя, разделенные лимфатической
полостью со сложной сетью нежных перекладин, на прилежащую к мозгу
255
сосудистую вторичную мягкую оболочку (pia mater secun-
daria) и нежную, ячеистую, богатую лимфой паутинную о б о -
лочку (arachnoidea).
В области головного мозга диференцировка протекает сходно, но
твердая мозговая оболочка тесно прилегает к надкостнице и у млеко-
питающих с нею срастается. У млекопитающих твердая оболочка обра-
зует две большир складки: продольную «серповидную» складку головного
мозга (falx cerebri), врезывающуюся между полушариями большого мозга,
и поперечный «шатер» (tentorium), вдающийся между большим мозгом и
мозжечком. Богатая сосудами мягкая оболочка у всех позвоночных
срастаются с утонченной эпителиальной крышей промежуточного и про-
долговатого мозга и образует вдающиеся в мозговую полость сложные
системы складок с сосудистыми сплетениями (plexus chorioidei). Кроме
того, она проникает и во все борозды между извилинами коры мозга.
Наконец, кровеносные сосуды вместе с соединительной оболочкой вра-
стают также внутрь самого мозп.
. Диференцировка оболочек идет, как упомянуто, путем образования
лимфатических щелей, ведущих к их расщеплению. Этот процесс понятен,
так как прилежащие к мозгу части оболочек несут функцию питания
мозга и содержат обильную сеть кровеносных сосудов, с прогрессивным
развитием которых, сопровождающим рост мозговой деятельности, уве-
личивается и общее количество лимфы, оттекающей по поверхностным
частям оболочек.
Периферическая нервная система
Периферическая нервная система составляется из нервов, т. е. из
путей, связывающих центральную нервную систему с периферическим
органами, и немногих расположенных на этих путях периферических
узлов. Тесная функциональная связь между этими органами, естественно,
приводит к тому, что и эволюция самих путей определяется эволюцией,
соответствующих периферических органов. Рост какой-либо мышцы неиз-
бежно сопровождается прогрессивным развитием ее двигательного нерва
(а также и соответствующих двигательных ядер мозга), и, наоборот, ре-
дукция мышцы влечет за собой редукцию данного нерва. Всякое переме-
щение, диференцировка или, наоборот, слияние различных мышц всегда
сопровождается и соответственными процессами в иннервирующих их
нервах. Прогрессивное развитие органа чувств точно так же определяет
развитие соответствующего чувствующего нерва (а также и центра в мозге)
и т. д. Таким образом, изменения в области иннервации ясно определяют
ход эволюции периферической нервной системы.
Что касается гистологического состава периферической нервной си-
стемы, то следует только отметить, что у круглоротых вся периферическая
нервная система состоит еще из безмякотных нервных волокон, между
тем как у рыб и у вышестоящих позвоночных нервы, отходящие от цен-
тральной нервной системы, состоят исключительно из мякотных волокон,
а безмякотные волокна органичиваются симпатической нервной системой.
По месту отхода различают нервы спинномозговые и головные; однако,
как мы увидим в дальнейшем, морфологически между ними резких разли-
чий не имеется, тем более что существуют и переходные между ними
затылочные нервы. В связи с периферическими нервами стоит еще особая
система ганглиев и нервов, снабжающих внутренние органы, так назы-
ваемая симпатическая нервная система.
Спинномозговые нервы отходят от мозга двумя кореш-
ками каждый — спинным (или «чувствующим») и брюшным (двигатель-
ным). У ланцетника корешки отходят от мозга поочередно, причем спинной
нерв проникает в перегородку между мышечными сегментами, а брюшной
состоит из пучка волокон, направляющихся непосредственно в соответству-
256
Ющин миомер. Точно так же и у миног оба корешка продолжаются в неза-
висимые друг от друга нервы. В отличие от ланцетника спинной корешок
снабжен здесь, как и у всех позвоночных, ганглиозным утолщением,
образующим спинномозговой узел. У всех позвоночных, за исключением
миног, оба спинномозговых корешка соединяются вместе для образования
одного смешанного нерва.
Чередование спинных и брюшных корешков у большинства рыб еще
ясно видно, но у высших позвоночных оно теряется. После соединения
обоих корешков спинномозговой нерв делится на три главные ветви —
спинную (г. dorsalis), снабжающую кожу п мышцы спины, брюш-
ную (г. ventralis), идущую в боковые и брюшные стенки тела, и висце-
ральную, или соединительную (г. visceralis s. communicans),
которая спускается к верхней стенке общей полости тела в области аорты,
где соединяется с симпатической
нервной системой (рис. 240).
Спинномозговые нервы имеют
правильное метамерное располо-
жение, в точности соответствуя
мускульным сегментам и правиль-
но чередуясь с элементами позво-
ночника. Однако в распределении
мелких разветвлений этих нервов
такой правильности уже не заме-
чается; особенно это касается тех
частей тела, в которых теряется
первоначальная метамерия. Таким
образом, и здесь видна та зависи-
мость между органом и его нервом,
о которой мы- уже говорили в
начале-этой главы. Прежде всего
это касается конечностей, органов
метамерного происхождения, рано
теряющих свою первичную сег-
ментацию. Уже в области непар-
ных плавников рыб между иннер-
вирующими их ветвями спинномозговых нервов развиваются анастомо-
зы, соединяющиеся иногда в непрерывный продольный ствол у осно-
вания плавника (и. collector). •
В еще большей мере это замечается в области парных плавников,
где, наряду с простыми продольными анастомозами, образующими кол-
лектор, наблюдаются еще и более сложные нервные сплетения
(plexus). Таким образом, у позвоночных развиваются два крупных спле-
тения спинномозговых нервов, служащих для иннервации парных конеч-
ностей: плечевое сплетение (plexus brachialis) в области передней
конечности и п о я с н и ч н о-к рестцовое (plexus lumbo-sacralis) в
области задней.
Число нервов, входящих в состав этих сплетений, зависит от числа
метамеров, вошедших в состав конечности — у рыб, в частности, у хря-
щевых рыб, оно может быть очень велико (особенно у скатов), у наземных
позвоночных оно изменяется от 3 до 7 в каждом сплетении. У наземных
позвоночных от плечевого сплетения обособляется впереди шейное
сплетение, с которым связан и подъязычный нерв.
С перемещением конечности может перемещаться и соответственное
сплетение, причем с одной стороны элементы выпадают из сплетения, с
другой стороны вновь включаются, как это наблюдается, например, при
перемещении передних конечностей назад в связи с обособлением шеи
у наземных позвоночных.
Рис. 240. Отхождение и состав спинномоз-
говых нервов. Схема.
d —- спинной корешок; dn — ганглий; т—мовг;
/с — соединительная ветвь; rd — спинная
ветвь; rv— брюшная ветвь; v— брюшной
корешок. Условные обозначения компонентов
см. рис. 242.
257
С и м п а т и ч е с к а я нервная с и с т е м а состоит из системы
узлов и серых безмякотиых (большей частью) волокон, иннервирующих
внутренние органы, снабжая их как чувствующими волокнами, так и дви-
гательными (к гладкой мускулатуре кишечника и различных других
органов, к мускульной стенке сосудов, к поперечнополосатой мышце
сердца, а у птиц к поперечнополосатой мышце радужины глаза, а также
секреторные волокна к разным железам). Кроме того, у млекопитающих
симпатические нервы сопровождают и все двигательные нервы мозга
вплоть до их окончаний на поперечнополосатых мускульных волокнах.
Симпатические двигательные волокна, выходящие из мозга, отли-
чаются от других висцерально-двигательных волокон тем, что они не
доходят непосредственно до области своей иннервации, а кончаются
всегда в симпатических ганглиях, где вступают в связь с двигательными
нервными клетками, от которых уже отходят безмякотные волокна к
мышце или железе.
У круглоротых симпатическая система состоит из отдельных ганглиев,
лежащих по бокам аорты и связанных соединительными веточками (rami
communicantes) со спинномозговыми нервами. Продольных соединений
между этими ганглиями нет, но периферические нервы соединяются на
различных внутренних органах в сплетения, в которых также распо-
лагаются и ганглиозные образования. Такие сплетения развиваются в
стенках кишечника, сердца, в мочевых органах и половых железах.
У рыб также имеются хорошо развитые периферические сплетения сим-
патической системы, но продольных соединений между ганглиями и у них
обыкновенно нет. Только у костистых рыб и у двудышащих, как и у на-
земных позвоночных, ганглии симпатической системы соединяются между
собой по бокам от позвоночника продольным пограничным ство-
лом (truncus sympaticus). Этот пограничный ствол продолжается у
хвостатых амфибий, как и у названных рыб, назад в хвостовой канал.
Симпатическая нервная система продолжается впереди и в области
головы вплоть до тройничного нерва, образуя ряд симпатических ганглиев,
частью более или менее самостоятельных (g. ciliare и g. sphenopalatinum),
частью тесно связанных с ганглиями головных нервов. Симпатические
стволы и нервы в области головы обыкновенно идут совместно с ветвями
головных нервов, и только местами симпатический ствол идет самостоя-
тельно (якобсонова комиссура, идущая вдоль стенки слуховой капсулы
и связывающая 7-ю пару нервов с 9-й)‘. Имеются в области головы и
периферические симпатические ганглии (g. submaxillare).
Из периферических нервов симпатической системы следует отметить
более крупные сердечные (rr. cardiaci) и кишечные нервы
(пн. splanchnici), отходящие в передней части пограничного ствола.
Головные нервы отходят большей частью от боковых частей
головного мозга, иногда несколькими корешками, но никогда в их распо-
ложении не наблюдается той правильности, которая характеризует спин-
номозговые нервы. Ближайший анализ показывает, что большая часть
их в действительности построена по тому же типу, что и спинномозговые;
однако ни один из головных нервов не соответствует всему спинномоз-
говому нерву, а лишь отдельным спинным или брюшным корешкам,
между которыми в области головы нет соединения. В этом отношении
головные нервы, быть может, сохраняют более примитивное строение,
как оно наблюдается в спинномозговых нервах ланцетника и миног.
Таким образом, одни головные нервы, лишенные ганглиев, чисто
двигательные, отходящие большей частью от брюшной поверхности
мозга, соответствуют брюшным корешкам спинномозговых нервов (нервы
3-й, 4-й, 6-й и 12-й пар); другие головные нервы, снабженные ганглиями
и значительной частью чувствующие, отходят от боковых частей мозга и
соответствуют спинным корешкам спинномозговых нервов (нервы 5-й,
258
7-й, 8-й, 9-й и 10-й пар). Наконец, как мы уже видели раньше, две пары
нервов — обонятельные и зрительные — занимают совершенно особое
положение и с другими нервами не сравнимы. Зрительные нервы пред-
ставляют собой центральный тракт, т. е. часть самого мозга, а обонятель-
ный нерв — комплекс отростков первичночувствующих клеток.
Таким образом, в периферической нервной системе области головы
замечаются некоторые особенности, отличающие ее от нервной системы
области туловища. Одни из этих особенностей сравнительно легко объ-
яснимы: так, например, перемещение места отхода спинных корешков на
бока мозга, очевидно, обусловлено значительным развитием спинной
части самого мозга.
Другие особенности связаны с эволюцией периферических органов
области головы. Для того чтобы эта связь была понятна, нам нужно пред-
варительно ознакомиться с функциональным значением различных частей
нервной системы.
Функциональные компоненты нервной системы
Периферические нервы, как известно, могут быть разделены на две
основные группы: чувствующие нервы, передающие возбуждение от из-
вестного чувствующего поля по направлению к центральной нервной
системе (в центростремительном направлении), и двигательные нервы,
передающие возбуждение от центральной нервной системы к периферии,
т. е. к мышце, сосуду, железе и т. п. (в центробежном направлении). Каж-
дая из этих групп может быть в свою очередь разделена на два отдела,
отличающиеся не только по иннервируемым ими периферическим органам,
но и по своим центральным связям, это — соматические и висцеральные
нервы. Первые иннервируют собственно стенки тела с его кожей и муску-
латурой; вторые — внутренние органы.
Таким образом, мы различаем следующие группы нервов:
1. Соматические чувствующие нервы, идущие от
органов, воспринимающих физическое раздражение, как давление, тепло
или холод, звуковые колебания среды и свет, т. е. от кожных и сейсмо-
сенсорных органов (боковой линии), органов слуха и зрения. Эти органы
связывают организм с внешней средой, и раздражения их вызывают те
или иные ощущения, на которые животное может реагировать произ-
вольными движениями.
2. Висцеральные чувствующие нервы идут от
органов восприятия химического раздражения (от растворенных или
взвешенных в окружающей среде или во внутренних полостях частиц
различных веществ), т. е. от нервных окончаний в области кишечника и
других внутренних органов, от специальных органов глотки и ротовой
полости, т. е. органов вкуса, которые, однако, иногда наблюдаются и на
наружной поверхности тела, и, наконец, от органов обоняния. Раздра-
жение этих нервов вызывает часто рефлекс без всяких ощущений или с
мало диференцированными ощущениями (от внутренних органов), но
нередко связано с ясно выраженными специфическими ощущениями
(вкус и обоняние).
3. Соматические двигательные нервы иннерви-
руют париетальную мускулатуру тела, т. е. миотомы и их производные
(мышцы спины и брюха, мышцы конечностей, подъязычную мускулатуру
и глазные мышцы).
4. Висцеральные двигательные нервы иннерви-
руют висцеральную мускулатуру, т. е. всю гладкую мускулатуру (сосу-
дов, кишечника и различных внутренних органов), поперечнополосатую
сердечную мышцу, поперечнополосатую мускулатуру передней части
кишечника и висцерального аппарата (моторные волокна), а также раз-
личного рода железы (секреторные волокна). Кроме мозговых (церебро-
259
спинальных) нервов, иннервирующих мышцы и другие периферические
органы непосредственно, имеются еще двигательные волокна, идущие к
периферическим органам только через посредство собранных в симпати-
ческие узлы или рассеянных в стенке органа двигательных нервных клеток
симпатической нервной системы.
С физиологической точки зрения особое значение для функциональ-
ного состояния всех почти органов имеют две группы висцеральных
нервов: симпатическая нервная система (возбуждаемая адреналином) и
парасимпатическая система (возбуждаемая атропином). Под последней
понимают комплекс висцеральных нервов антагонистических по действию
симпатическим нервам. Сюда относятся висцеральные волокна глазо-
двигательного нерва, связанные
i£S.
sm
symp
Рис. 241. Диаграмма функциональных компонен-
тов спинного мовга и спинномовговых нервов
(по Джонстону). Области серого вещества огра-
ничены прерывистой линией.
rd — спинная; rv— брюшная ветвь; symp— симпа-
тический увел; sm — соматичесни-двмгательная и vm
— висцерально-двигательная область; а. а. — сома-
тическп-чувствующая и v. а.—висцерально-чувствую-
щая область.
ит
с ресничным ганглием, часть волокон
барабанной струны (chorda
tympani УП) и внутренност-
ной ветви блуждающего нер-
ва, а также висцеральные
волокна ряда крестцовых
нервов. Эти системы нервов
объединяются в физиологии
под названием вегетативной,
или автономной, нервной
системы.
Функциональные компо-
ненты нервной системы рас-
полагаются в спинномозго-
вых нервах следующим обра-
зом (рис. 240): .брюшные'
корешки у низших позвоноч-
ных состоят исключительно
из соматических, двигатель-
ных волокон, а спинные
являются смешанными и со-
держат все прочие компонен-
ты, т. е. оба рода чувствую-
щих волокон, а также висце-
ральные двигательные. Вис-
церальные компоненты, как
чувствующие, так и дви-
узел и входят затем в состав
гательные, проходят через спинномозговой
соединительной, висцеральной ветви, направляясь в соответствующую
часть симпатической нервной системы, которая вся по своему составу
является висцеральной. В составе тех же соединительных ветвей имеются
и «симпатические» волокна, идущие от нервных клеток симпатических
узлов прямо в спинную и брюшную ветви спинномозговых нервов. У
высших позвоночных часть висцерально-двигательных волокон покидает
мозг в составе брюшных корешков, и у млекопитающих таким путем
выходят почти все эти волокна.
Только у млекопитающих, таким образом, спинные корешки являются,
действительно, почти чисто чувствующими; у низших позвоночных они
всегда бывают смешанными.
Что касается центральных связей нервных волокон спинномозговых
нервов, то и в этом отношении проявляются ясные различия между от-
дельными функциональными компонентами (рис. 241). Соматически-
чувствующие волокна входят в спинной мозг через спинные корешки
и своими конечными разветвлениями вступают в связь с нервными клетка-
ми спинных рогов. Висцерально-чувствующие волокна входят в спинной
мозг также через спинные корешки и своими конечными разветвлениями
260
вступают в связь с клетками, лежащими в основании спинных рогов.
Соматически-двигательные волокна отходят от моторных клеток брюшных
рогов и покидают спинной мозг в составе брюшных корешков. Висцераль-
но-двигательные волокна отходят от моторных клеток средней зоны
спинного мозга, которая у млекопитающих выступает в виде боковых ро-
гов, и покидают спинной мозг у низших позвоночных в составе спинных
корешков, а у высших более или менее значительной частью — в составе
брюшных корешков (рис. 241).
Таким образом, и в сором вещество спинного мозга можно различить
четыре различные функциональные области: соматически-чувствующую
область спинных рогов, висцоралыю-чувствующую область основания
спинных рогов, висцерально-двигательную среднюю область и сомати-
чески-двигательную область брюшных рогов (рис. 241).
В составе головных нервов имеются все те же компоненты, что и в
спинномозговых нервах. Как сказано уже раньше, головные нервы срав-
нимы лишь с отдельными спинными и брюшными корешками спинномоз-
говых нервов, между которыми здесь нет соединения. Анализ состава
головных нервов убеждает в этом. Чисто двигательные головные нервы
(движущие глазом и подъязычный) являются именно соматически-дви-
гательными, так как иннервируют мускулатуру, развивающуюся из
миотомов, так же, как и брюшные корешки спинномозговых нервов.
Смешанные головные нервы (тройничный, лицевой, языкоглоточный и
блуждающий) содержат все остальные функциональные компоненты,
подобно спинным корешкам спинномозговых нервов. Чисто чувствующий
слуховой нерв, оказывается, не имеет самостоятельного значения, так
как лишь вторично обособляется от лицевого, а висцерально-двигательный
добавочный нерв развивается лишь у высших позвоночных путем обособ-
ления задней порции блуждающего нерва. Об особом положении обоня-
тельного и зрительного нервов уже говорилось.
Таким образом, все головные нервы, за исключением лишь первых
двух пар, по своему составу сходны с отдельными корешками спинномоз-
говых нервов. Однако в относительном развитии отдельных компонентов
замечаются крупные отличия от последних, и это определяется эволю-
цией периферических органов в области головы. Прежде всего бросается
в глаза ничтожное развитие брюшных, соматически-двигательпых нервов.
Это находится в связи с редукцией миотомов в области головы (что в свою
очередь связано с развитием черепа), дающих начало лишь незначитель-
ным глазным мышцам. Затем не менее характерно чрезвычайно значитель-
ное развитие смешанных нервов и, в особенности, висцеральных их ком-
понентов, что связано с эволюцией передней части кишечника (хвататель-
ной и дыхательной), в области которой развивается висцеральный аппарат
со своим чувствующим аппаратом и значительной мускулатурой. Нако-
нец, в связи с развитием некоторых специальных органов чувств разра-
стаются и соответствующие компоненты головных нервов. Так, развитие
органов вкуса в слизистой оболочке ротовой полости и глотки сопровож-
дается разрастанием соответствующих специальных висцерально-чув-
ствующих компонентов в составе .лицевого, языкоглоточного и блужда-
ющего нервов. Развитие сейсмосенсорных кожных органов сопровожда-
ется специализацией и разрастанием части соматически-чувствующих
нервных волокон, которые затем обособляются в виде особых крупных
отделов лицевого, языкоглоточного и блуждающего нервов, известных
под названием нервов боковой линии. Наконец, развитие органа слуха
в результате специализации группы сейсмосенсорных органов сопро-
вождается полным выделением иннервирующих его волокон в особый
слуховой нерв, обособляющийся от лицевого нерва.
Таким образом, состав головных первов изменился главным образом
количественно, но вместе с тем произошла и новая диференцировка и спе-
261
циализация известных частей в связи с развитием новых периферических
органов в области головы. Что касается центральных связей головных
нервов, то и здесь мы не найдем принципиальных отличий от спинномоз-
говых, но вместе с тем увидим дальнейшее подтверждение изложенным
положениям о взаимной зависимости между эволюцией периферических
органов, их нервов и соответствующих отделов центральной нервной
системы. Отложив рассмотрение этого вопроса до заключительной части
этой главы, перейдем к систематическому описанию головных нервов.
Головные нервы
Обычно различают двенадцать пар головных нервов, которые выхо-
дят из черепа через особые отверстия в боковых его стенках. У низших
позвоночных постоянно имеется лишь десять пар вполне типичных го-
ловных нервов, если не считать небольшого концевого нерва.
О. Концевой нерв (n. terminalis) отходит от переднего конца
переднего мозга у концевой его пластинки. Это небольшой соматически-
чувствующий нерв, снабженный небольшим ганглием и идущий к сли-
зистой оболочке органа обоняния (рис. 229). Обонятельной функции он
не несет. У наземных позвоночных (амфибий, рептилий, млекопитающих)
концевой нерв разветвляется в слизистой оболочке якобсонова органа.
I. Обонятельный нерв (n. olfactorius) занимает особое
положение, отличаясь от всех прочих нервов позвоночных тем, что состав-
ляется из волокон, представляющих собой отростки чувствующих клеток
обонятельного эпителия. Входя в обонятельные доли головного мозга,
они оканчиваются там конечными разветвлениями, вступающими в связь
с дендритами особых нервных клеток. Как было уже сказано, обонятель-
ные доли иногда диференцируются на два отдела, связанные между собой
посредством более или менее длинного стебелька, — собственно обоня-
тельную долю и прилежащую к обонятельному мешку луковицу (рис. 244).
Обонятельный стебелек (pedunculus olfactorius) нередко совершенно не-
правильно обозначается как обонятельный нерв. Последний представляет
собой комплекс чувствующих волокон, идущих от обонятельного эпителия
к обонятельной лопасти (луковице) (рис. 242). Эти нервные волокна отли-
чаются обыкновенно самой незначительной длиной в тех случаях, когда
луковица тесно прилежит к обонятельному мешку (селахии, ганоиды,
некоторые костистые рыбы и некоторые ящерицы). В других случаях
обонятельный нерв длиннее, а сидячая обонятельная доля не обнаружи-
вает указанной диференцировки (некоторые костистые рыбы, амфибии,
некоторые ящерицы, черепахи) (рис. 232). Обонятельный нерв относят
к висцерально-чувствующему отделу.
II. Зрительный нерв (n. opticus), или тракт, отходит
от дна промежуточного мозга, перекрещивается под его основанием с
одноименным нервом противоположной стороны (перекрест зрительных
нервов, chiasma nervorum opticorum) и направляется в глаз, где распро-
страняется по внутренней поверхности сетчатки. У млекопитающих пе-
рекрест неполный, так как часть волокон остается на той же стороне. В осо-
бенности много прямых волокон в составе зрительных трактов животных,
обладающих стереоскопическим зрением (высшие приматы, человек).
Как было уже сказано, волокна зрительного нерва развиваются как от-
ростки клеток глазного бокала внутри глазного стебелька. И тот, и дру-
гой развиваются из стенок переднего мозга (см. развитие органа зрения).
Сетчатка представляет собой не просто чувствующий эпителий, а сложный
центр с нервными и опорными клетками, а глазной нерв — центральный
тракт, поддерживаемый элементами глии и окруженный мозговыми обо-
лочками. Зрительный тракт относят к соматически-чувствующему отделу.
III. Г лазодвигательпый нерв (n. oculomotorius.
(рис. 242, 244, 247) отходит от брюшной поверхности среднего мозга и
262
иннервирует три прямые мышцы глаза — верхнюю, пижиюю и внутреннюю
(переднюю) — и нижнюю косую мышцу, т. о. то мышцы, которые развива-
ются из предчелюстного миотома, сегментальным нервом которого он,
следовательно, и является. По своему составу это •— соматичоски-двига-
тольпый нерв (соответствующий брюшному корешку спинномозгового
нерва). Эмбрионально наблюдается связь этого перва с первым тройнич-
ным (V, 1, и. ophthalmicus profundus), который, очевидно, является спин-
ным корешком того же сегмента. Часто группа ганглиозных клеток всту-
пает в связь с глазодвигательным первом, образуя ресничный узел
(g. ciliaro), большей частью относящийся к висцеральной нервной системе.
sow т «км vis т «к» soiTi S lat
ш n 11111 m VIS
Рис. 242. Головные нервы рыбы. Схема с обозначением функциональных
компонентов I—X — первая по десятую пары головных нервов.;.
с х — n. cardiacus; сЫ — мозжечок; hy — hypoglossus; ini. X — n. intestinalis; m — mesen-
cephalon; ph —r. pharyngeus X; m. X — r. mascularis; pl. br. — plexus brachialis; prt — r.
praetrematicus; pit — r. posttrcmaticus; r. hy VH — r. hyoideus; г. IX — r, lateralis; r. md.
I-—r mandibularis trigemini; rmdi, md. e VH — ramus mandibularis internus и externus
facialis; r. buc. VH —r, buccalis; г. тх. V —r. maxillaris; r. o. pr — r, ophthalmicus pro-
fundus; ros — r. ophthalmicus supcrficiahs V; r. os. VH—r, ophthalmicus superficialis
VII; r, pa VII и IX—r, palatines VII и IX; s—spiraculum; 1—5 —жаберные щели; som.
m. vise. m. — соматически- и висцерально-двигательные нервы; som, s. rise, s—соматически!
и висцерально-чувствующпс первы; lat — нервы боновой линии,
Таким образом, к глазодвигательному нерву вторично примешиваются
висцеральные волокна. Наконец, у млекопитающих в составе корешка
глазодвигательного нерва наблюдаются еще висцеральные двигательные
волокна, идущие к ресничной мышце (m. cillaris) и к мышце радужины
(m. iridis). Невидимому, эти волокна по своему происхождению отно-
сятся к тройничному нерву, передние ядра которого близко примыкают
к ядрам глазодвигательного нерва и представляют собой результат того
же процесса вторичного включения висцеральных элементов, который
наблюдается и в брюшных корешках спинномозговых нервов.
IV. Блоковый нерв (n. trochlearis) (рис. 242, 244, 247) отходит
от спинной поверхности мозга на границе между средним и задним мозгом
и направляется к верхней косой мышце глаза, которую он иннервирует.
Верхняя косая мышца развивается из второго (челюстного) миотома го-
ловы, и, следовательно, блоковый норв является сегментальным нервом
этого миотома. Отхождение этого нерва со спинной стороны мозга есть
результат вторичного перемощения. Ядра блокового нерва помещаются
в брюшной стенке мозга, сейчас же позади ядер глазодвигательного нерва;
отсюда волокна его поднимаются в боковых стенках мозга вверх, пере-
крещиваются в крыше мозга с нервом противоположной стороны и выхо-
дят затем на границе среднего и заднего мозга. По своему составу это —
соматичоскп-двигатольный нерв. Во время эмбрионального развития
263
обнаруживаются иногда следы связи его со вторым тройничным нервом
(V, 2, n. trigeminus II), который, очевидно, является спинным корешком
того же сегмента. Иногда в составе блокового нерва наблюдаются и чув-
ствующие волокна, относящиеся по своему происхождению к тройнич-
ному нерву.
VI. О т в о д я щ и й и е р в (n. abducens) (рис. 242, 244, 247) отхо-
дит от брюшной поверхности продолговатого мозга обыкновенно двумя,
а иногда и несколькими корешками и направляется к наружной прямой
(отводящей) мышце глаза. С обособлением мышцы, втягивающей глазное
яблоко (m. retractor bulbi) у наземных позвоночных и мышцы мигательной
перепонки (m. nicticans) у рептилий и птиц отводящий нерв иннервирует
и эти мышцы. Названные мышцы развиваются из третьего головного мио-
тома. По своему составу отводящий нерв является соматически-двига-
тельным первом, который вполне сохранил характерные черты брюшных
корешков. Отводящий нерв обнаруживает следы связи с лицевым нервом.
Все три пары по priori, движущих глаз, отличаются весьма незначительной
величиной соответственно величине глазных мышц. В тех случаях, однако, когда
глаз и его мышцы имеют крупные размеры (например у луны-рыбы, Orthagoriscus
mola), и соответствующие нервы выделяются своей величиной.
V. Тройничный нерв (n. trigeminus) представляет собой,
как вцдно по его развитию, результат соединения двух совершенно само-
стоятельных нервов, обладающих каждый своим особым ганглием. И у
взрослого животного ганглий тройничного нерва нередко ясно обнару-
живает двойственность своего происхождения. Нервы, составляющие
Рис. 243. Головные нервы акулы. Вид сбоку (из Гудрича).
Ьс — n. buccalis VII; dg — еппннап ветвь языкоглоточного нерва; dr — спинные корешки
dv—сиипиан ветвь блуждающего нерва; е—губной хрящ; д—n. glossopharyngeus; hm—hyo-
mandibularc;. Im — n. hyoniandibularis VII; , hy — u. liypoglossus; ll —n. lateralis vagi
(X); me — меккелев хршц; md — n.»inanbibularis — n. mandibularis VII; nc — обо-
нятельная капсула; opl— зрительный нерв; otf—n. otieus VII; pg — palatoquadratum;
prl>—предо:абернал ветвь; pro — н. ophthalmicus profundus V; ptb — зажаберпая ветвь;
r, v — ростральный хрящ; sop — и. ophthalmicus; superficialis V'+VII; ет — брюшной коре-
шок; во — н. visccralis (intcstinalis) X.
тройничный нерв, обозначаются как первый, или глубокий глазничный
(n. ophthalmicus profundus s. n. trigeminus I), и второй, или собственно
тройничный нерв (n. trigeminus II).
Vr Глубокий глазничный нерв (n. ophthalmicus
profundus) (рис. 242) отходит от передней части боковой поверхности про-
долговатого мозга и снабжен своим ганглием (g. profundi). В качестве
вполне самостоятельного нерва он сохраняется только у многоперых га-
ноидов (Polypterus); у других позвоночных он более или менее тесно со-
единяется со вторым тройничным нервом и тогда является лишь одной из
трех главных ветвей тройничного нерва (рис. 246). Этот нерв покидает
264
череп через осооое отверстие, проходит через глазницу между глазными
мышцами, отдав предварительно поверхностную ветвь (n. ophthalmicus
supcrficialis V), идущую над глазом совместно с одноименной ветвью ли-
цевого нерва (рис. 242, 244). Обе эти ветви чисто соматически-чувству-
ющие и идут к коже
передней части го-
ловы.
V2. Собствен-
но тройничный
нерв (n. trigemi-
nus II) (рис. 242, 243,
244) отходит от боко-
вой стенки продолго-
ватого мозга совмест-
но с глазничным нер-
вом и имеет свой осо-
бый ганглий (g. tri-
gemini s. g. Gasseri).
По выходе из черепа
через особое отвер-
стие он делится на
две крупные ветви—
верхнечелюст-
ную (ii. maxillaris)
и нижнечелюст-
ную (n. mandibu-
lar is). Верхнечелюст-
ная ветвь проходит
под глазом вдоль
верхней челюсти и
иннервирует кожу
передней части голо-
вы, нёбо и верхнече-
люстные зубы. По
своему составу — это
чувствующая ветвь,
содержащая главным
образом соматические
(кожные), но обыкно-
веппо также и висце-
ральные волокна.
Нижнечелюстная
ветвь отдает крупную
двигательную ветвь
к жевательной мус-
кулатуре (m. levator
palatoquadrati, т.
adductor mandibulae
Рис. 244. Головной мозг и нервы акулы (Carcharias)
(по Принстону из Кепгсли).
Ъо — обонятельная луковица; Ьг 1—4 — жаберные нервы;
hm — n. hyomanrtibularis VII; I —n. lateralis X; w.d— n. man-
dibularis V; rax — n. maxillaris V; os — n. ophthalmicus
supcrficialis VII; ol — pedunculus olfactorius; pal —n. palati-
nus VII; po — y. posttrematicus; pr — r. praetrematicus;
v — t. intestinalis X; tel —передний мозг; di—промежуточ-
ный мозг; ras — средний мозг; rat — мозжечок; ral — продол-
говатый мозг.
и его производные и m. intermandibularis)
и направляется затем вдоль нижней челюсти, где снабжает кожу,
слизистую оболочку ротовой полости и нижнечелюстные зубы (рис: 246).
У млекопитающих этот нерв отдает еще чувствующую ветвь к языку
(п. lingualis V) (рис. 246). Таким образом, собственно тройничный нерв
является смешанным нервом: он содержит, кроме двигательных, еще и
чувствующие волокна и притом главным образом кожные, но обычно
также висцеральные.
VII. Лицевой нерв (n. facialis) отходит от боковых стенок
продолговатого мозга непосредственно позади тройничного нерва и не-
265
редко в связи с ним (у рыб и амфибий) несколькими корешками, со-
единяющимися затем в сложном ганглии. Верхня ячасть ганглия имеет са-
мостоятельное значение и представляет собой ганглий группы нервов боко-
вой линии; нижняя часть представляет собственно ганглий лицевого
нерва (g .facialis s. g. geniculi). Таким образом, и здесь следует у низших
позвоночных различать два нерва — нерв боковой линии . (n. lateralis
VII) и собственно лицевой.
Боковой нерв делится на три ветви: 1) поверхностный глаз-
ничный нерв (n. ophthalmicus superficialis VII), идущий совместно
с одноименной ветвью тройничного нерва и иннервирующий надглазнич-
ный канал сейсмосенсорных органов (и ампулы верхней стороны рыла
у селахий) (рис. 242, 243), 2) щ е ч н ы й нерв (n. buccalis VII), идущий
обыкновенно совместно с верхнечелюстной ветвью тройничного нерва и
Рис. 245. Головные нервы наземного позвоночного. Схема;
1—ХП — двенадцать пар головных нервов; chi — chorda tympanx VII; hy — n. hypoglos-
sus; г. с. X — r. cardiacus X; rhy VII — r. hyoideus; Tint. X — r. pneumogastricus; r. I. V и
X — г. lingualis V и IX; rmd V — r. mandibularis; г. mx. V — r. maxillaris; r. o. pr — r.
ophthalmicus V; rph — r. pharyngeus IX и X. Остальные обозначения прехкние.
иннервирующий подглазничный канал сейсмосенсорных органов (и ампулы
нижней стороны рыла у селахий) (рис. 242) и 3) наружный нижне-
челюстной нерв (n. mandibularis externus), который идет сна-
чала совместно с подъязычно-челюстной ветвью лицевого нерва вдоль
подвесочного аппарата и переходит затем на наружную поверхность ниж-
ней челюсти (рис. 258). Этот нерв иннервирует гиомандибулярный и ниж-
нечелюстной каналы сейсмосенсорных органов. Все эти ветви являются
по своему функциональному составу соматическими и связаны с водными
органами чувств, так что у наземных позвоночных естественно исчезают
вместе с исчезновением этих органз в.
Собственно лицевой нерв дает: 1) чисто чувствующую (висцеральную)
нёбную ветвь (n. palatinus) к слизистой оболочке нёба (рис. 242,
244), 2) небольшие чувствующие ветви к передней стенке брызгальца у
селахий (г. praetrematicus VII) и 3) большой смешанный подъязыч-
н о-ч елюстной нерв (n. hyomandibularis), идущий позади брыз-
гальца вдоль гиоидной дуги (рис. 242, 244) и отдающий значительные
двигательные ветви к висцеральной мускулатуре этой области (n. hyoi-
deus VII) и чувствующую ветвь, переходящую на внутреннюю стенку
нижней челюсти (n. mandibularis internus), где опа иннервирует слизи
266
тола, получают значительное развитие и
Рис. 246. Схема головных нервов кошки. (Ниж-
няя челюсть отвернута) (по Миварту из Кингсли).
cl — chorda tympani; d — n. alveo laris; g—ганглий;
io— n. infraorbitalis; I —n. lingualis V; li, Is—
n. laryngaus X; md — n. mandibularis V; mx—
n. inaxillaris V; о — n. opthhalmicus V; t — язык.
стую оболочку ротовой полости с ео вкусовыми органами. Двигательные
ветви иннервируют, между прочим, мышцы подвеска и жаберной крышки
у рыб и часть межчелюстной мышцы, мышцу, опускающую пижпюю че-
люсть, и поверхностную шейную мускулатуру у наземных позвоночных.
У млекопитающих получает особенно значительное развитие лицевая
мускулатура, иннервируемая в этом случае крупной специальной ветвью
того же нерва (рис. 246). Чувствующая ветвь (n. mandibularis internus)
проходит у наземных позвоночных через барабанную полость и получает
название барабанной струны (chorda tympani), сохраняющей
в общем ту же область иннервации (рис. 245) в слизистой оболочке рото-
вой полости.
Лицевой нерв является, между прочим, одним из нервов специальных
висцеральных органов чувств, именно органов вкуса. С сильным разви-
тием последних у костистых рыб, у которых органы вкуса располагаются
и на наружной поверхности
особые ветви лицевого нерва
(рис. 276).
Таким образом, собствен-
но лицевой нерв является
типичным смешанным нер-
вом, содержащим все три
компонента (висцеральные,—
чувствующие и двигатель-
ные, — и соматические чув-
ствующие) со значительным,
однако, преобладанием вис-
церальных.
У наземных позвоноч-
ных из состава лицевого
нерва, естественно, выпадают
нервы боковой линии, в зна-
чительной степени редуци-
руются чувствующие компо-
ненты вообще, и с другой
стороны, как упомянуто,
получают значительное пре-
обладание двигательные ком-
поненты, в особенности, у
млекопитающи х.
VIII. Слуховой нерв (n. acusticus) по своему развитию пред-
ставляет собой обособившуюся часть предыдущего нерва и именно его
специального соматически-чувствующего отдела, т. е. нерва боковой ли-
нии (n. lateralis VII). Этот нерв снабжен отдельным ганглием и делится
на ветви, соответствующие отделам воспринимающего аппарата внутрен-
него уха (рис. 242, 245).
IX. Языкоглоточный п е р в (n. glossopharyngeus) отхо-
дит от боковой стенки продолговатого мозга несколькими корешками,
снабжен своим ганглием (g. petrosum) и выходит из черепа сейчас же
позади слуховой капсулы, у высших позвоночных обыкновенно совместно
с блуждающим нервом. У рыб наиболее дорзальные его корешки отно-
сятся к группе боковых нервов, снабжены иногда обособленным ганглиом
и дают небольшую ветвь к сейсмосенсорным органам головы.
Главная часть нерва является типичным висцеральным нервом.
Этот нерв делится у рыб и водных амфибий на следующие три главные
ветви (рис. 242): 1) небную (n. palatinus IX), чисто висцерально-чув-
ствующую ветвь, идущую к слизистой оболочке нёба, 2) п р е д ж а б е р-
н у ю (г. praetrematicus), висцеральпо-чувствующую ветвь, идущую
267
вдоль подъязычной дуги по передней стенке первой жаберной щели,
иЗ)зажаберную (г. posttrematicus) смешанную ветвь, идущую
вдоль первой жаберной дуги. Последняя ветвь является наиболее значи-
тельной и снабжает, между прочим, и висцеральную мускулатуру соот-
ветствующей области (первой жаберной дуги). У наземных позвоночных
он иннервирует глотку с ее мускулатурой и является главным вкусовым
нервом [язычный нерв (n. lingualis IX)] (рис. 245, 246).
Как и в других типичных висцеральных нервах, в языкоглоточном
нерве преобладающее развитие получают висцеральные его компоненты,
между тем как соматически-чувствующие (кожные) редуцируются, a v
высших рыб даже вполне исчезают, за исключением специализированных
волокон, иннервирующих сейсмосенсорные органы. Последние исчезают
у наземных позвоночных.
X. Блуждающий нерв (n. vagus) отходит многочисленными
корешками от боковой стенки продолговатого мозга и обладает своим
ганглием (g. vagi) (рис. 242, 243, 244). Этот нерв делится у рыб и у водных
амфибий на два крупных отдела: 1) б ок о вой н е р в (n. lateralis X),
идущий назад вдоль боковой линии туловища и снабжающий соответствую-
щие органы чувств, и 2) ж а б е р н о-к и ш е ч н ы й нерв (n. branchio-
intestinalis), иннервирующий весь жаберный аппарат, начиная со второй
щели, а также и внутренние органы. Первый нерв имеет самостоятельное
значение и иногда сохраняет свой особый ганглий и вообще свою самостоя-
тельность и у взрослого животного. Второй нерв проходит сначала вдоль
спинной части жаберного аппарата, где против каждой щели, начиная
со второй, отдает по одной жаберной ветви, которая сейчас же
делится на три типичные ветки (рис. 242):
1) г л о т о ч и у ю (г. pharyngeus), чисто висцерально-чувствующую,
идущую к слизистой оболочке глотки,
2) преджаберную (г. praetrematicus), также висцерально-
чувствующую ветвь, идущую вдоль передней стенки соответствующей
щели, и
3) з а ж а б е р н у ю (г. posttrematicus), смешанную ветвь, которая
содержит, кроме чувствующих волокон, еще и двигательные для висце-
ральной мускулатуры соответствующей дуги. Позади места отхождения
последней жаберной ветви главный ствол отдает еще двигательные ветви
к висцеральной мышце плечевого пояса (m. trapezius) и продолжается
в виде кишечной в е т в и (г. intestinalis), направляющейся внутрь
и назад для иннервации внутренностей и именно сердца (n. cardiacus),
пищевода, желудка и легких у наземных позвоночных (n. pneumogas-
tricus), плавательного пузыря рыб, кипит и других внутренностей.
Жаберно-кишечный отдел блуждающего нерва представляет собой
большой комплекс типичных висцеральных нервов. Его соматически-
чувствующие компоненты редуцируются (особенно у высших рыб), а
висцеральные, наоборот, чрезвычайно разрослись соответственно разви-
тию жаберного аппарата. В составе жаберных ветвей блуждающего нерва
имеются и специализированные висцерально-чувствующие компоненты,
иннервирующие вкусовые органы.
У наземных позвоночных с редукцией жаберного аппарата реду-
цируются и собственно жаберные нервы, от которых остаются главным
образом лишь ветви, иннервирующие глотку и гортань сих мускулатурой,
Боковой нерв, естественно, совершенно исчезает у наземных позвоночных
(рис. 245, 245).
XI. Добавочный нерв (n. accessorius) является в виде
вполне самостоятельного нерва лишь у млекопитающих, хотя различим
и вообще у амниот (рис. 245, 246). По своему происхождению это обособ-
ленные задние корешки блуждающего нерва, волокна которых и у рыб
можно проследить до ветви, иннервирующей висцеральную трапециевид-
268
ную мышцу плечевого пояса. У рептилий область задних корешков блуж-
дающего нерва разрастается назад вплоть до первых спинномозговых
нервов. Эти корешки соединяются между собой в продольный стволик,
направляющийся вперед на соединение с ганглием блуждающего нерва
(рис. 245). Таким образом, у рептилий добавочный нерв теснейшим обра-
зом еще связан с блуждающим, п то же самое относится к птицам. Только
у млекопитающих он является вполне самостоятельным нервом (хотя
между ним и блуждающим нервом имеется соединительная ветвь). Зад-
ние корешки добавочного нерва простираются у млекопитающих еще
дальше назад, вдвигаясь посредине между спинными и брюшными кореш-
ками вплоть до уровня пятого — седьмого шейного нерва. Этот нерв
содержит только висцерально-двигательные волокна и иннервирует у
млекопитающих трапециевидную мышцу и грудино-ключично-сосце-
видную.
XII. Подъязычный нерв (n. hypoglossus) является типич-
ным головным нервом лишь у амниот и особенно у млекопитающих, где
он отходит 2—3 корешками от брюшной степкн задней части продолго-
ватого мозга. По выходе через затылочную область черепа эти корешки
Рис. 247. Схема сомачически-двигательных головных нервов.
plcbr — plexus cervicobrachialis; hy — hypoglossus. Остальные обозначения прежние.
соединяются в один ствол, связанный обыкновенно соединительными
веточками с передними спинномозговыми нервами (шейное сплетение —
plexus cervicalis) и направляющийся затем к подъязычной мускулатуре
и в язык, мускулатуру которого он иннервирует (рис. 246). У нижестоящих
позвоночных связь со спинномозговыми нервами оказывается гораздо
более тесной (рис. 243), а в некоторых случаях (у амфибий) подъязычный
нерв в действительности представлен лишь 1—2 спинномозговыми нервами.
По своему развитию подъязычный нерв в действительности всегда
представляет собой комплекс нервов, принадлежащих по своему типу
к спинномозговым и иннервирующим затылочные и передние туловищные
миотомы. В этих нервах прежде всего редуцируются спинные корешки
вместе с их ганглиями (которые могут быть обнаружены эмбрионально,
а в виде рудиментов наблюдаются иногда и у взрослых животных), но
затем вместе с редукцией передних заушных миотомов редуцируются и
иннервирующие их брюшные корешки и, таким образом, большее или
меньшее число передних нервов бесследно исчезает. Следующие нервы
сохраняются в виде более или менее значительных брюшных корешков,
частью покидающих череп через особые отверстия в затылочной его об-
ласти (затылочные нервы), частью выходящих позади черепа через заты-
лочную дыру или между элементами первых позвонков. Эти нервы соеди-
няются в общий ствол, огибающий жаберную область сзади и иннерви-
рующий затем мускулатуру, развившуюся из соответствующих (заты-
лочных и иногда первых туловищных) миотомов — подъязычную муску-
269
латуру всех позвоночных и мускулатуру языка наземных форм. У селахий
имеется обыкновенно 4—5 затылочных нервов, выходящих через отдельные
отверстия в черепе и ио выходе соединяющихся в коллектор, образующий
вместе со следующими спинномозговыми нервами общее шейпо-плечевое
нервное сплетение (plexus cervicobrachialis). Волокна коллектора вскоре,
однако, вновь отделяются от общего сплетения и направляются вперед
к подъязычной мускулатуре (рис. 247). Таковы же в общем соотношения
первых нервов спинномозгового типа и у всех рыб (число затылочных
нервов обычно ниже, чем у селахий). У амфибий затылочные нервы совсем
редуцируются, и в состав шейного сплетения входят лишь настоящие
спинномозговые нервы. Наконец, у амниот, с обособлением шейной об-
ласти и перемещением передних конечностей далее назад, шейное спле-
тение (pl. cervicalis) обособляется от плечевого (pl. brachialis), и в первом
получает преобладание значительный ствол, состоящий исключительно
из соматически-двигательных волокон передних корешков и предста-
вляющий собой подъязычный нерв.
Таким образом, мы видим, что в головных нервах можно более или
мепее ясно обнаружить тот же состав, что и в спинномозговых нервах:
одни из них сходятся в одном отношении со спинными, другие — с брюш-
ными корешками последних. Тем не мепее ясно бро-
Рис. 248. Схема жа-
берного нерва (со-
гласно Северцеву).
дп. v — висцеральный
ганглий; г. ph — г.
pharyngeus; г. prt. — г.
praetrematicus г. prt i.—r
praetrematicus internus;
r ptt. —г. posttremati-
cus.
саются в глаза и различия, которые не органичи-
ваются относительным развитием тех или иных
компонентов в связи с развитием соответствующих
периферических органов области головы, на что уже
было указано, по в особенности касаются еще рас-
положения отдельных головных нервов. В то время
как в области туловища спинномозговые нервы рас-
полагаются метамерно в соответствии с сегментацией
мускулатуры и скелета, в голове такой правильности
непосредственно обнаружить невозможно. Но все
же имеются известные данные, позволяющие вывести
заключение, что и головные нервы располагались
первоначально метамерпо и утратили это располо-
жение лишь впоследствии в связи с исчезновением
сегментации мускулатуры и скелета головы.
Прежде всего наблюдается известная правиль-
ность если не в отходе, то в ветвлениях висцераль-
ных нервов. Каждая жаберная ветвь висцерального
нерва делится на три ветви: 1) глрточную (г. pharyn-
geus), 2) преджаберную (г. praetrematicus) и 3) за-
жаберную (г. posttrematicus) (рис. 248), причем
первые две ветви являются висцерально-чувствую-
щими, а последняя — смешанной. Это касается
прежде всего типичных жаберных нервов: блуждаю-
щего и языкоглоточного. Нёбная ветвь последнего
является гомодинамом глоточной ветви. Однако и
лицевой нерв ветвится таким же образом: его чув-
ствующая нёбная ветвь соответствует глоточной, а
смешанная подъязычно-челюстная является зажа-
берпой ветвью; кроме того, у селахий имеется и
небольшая преджаберпая ветвь (рис. 249). Следо-
вательно, лицевой нерв относится таким же образом к первой висцераль-
ной щели — брызгальцу, как языкоглоточный к первой жаберной щели,
а жаберные ветви блуждающего нерва.— к остальным щелям. Лицевой
нерв — это висцеральный нерв брызгальца. Это сравнение можно про-
270
вести и дальше. Тройничный нерв имеет две висцеральные ветви, из ниХ
передняя, верхнечелюстная, является чувствующей, а задняя, нижне-
челюстная, смешанной (рис. 249). Если принять, что в состав ротового
отверстия вошла висцеральная щель, находившаяся впереди от первой
(брызгальца), то тройничный нерв является висцеральным нервом этой,
назовем ее ротовой, щели. Верхнечелюстная ветвь сравнима с нреджабер-
ной, а нижнечелюстная ветвь — с зажаберной ветвью типичного висце-
рального нерва.
Следовательно, в ветвлениях висцеральных нервов сохранилось еще
метамерное расположение, находящееся в связи и согласии с метамерным
строением иннервируемого им висцерального аппарата (с бранхиомерией)
(рис. 249). Еще яснее была выражена первоначальная метамер ая висце-
Рис. 249. Распределение висцеральных нервов головы рыбы.
Обозначения см. рис. 242.
ральных нервов у ископаемых бесчелюстных позвоночных (Ostracodermi),
у которых и глубокий глазничный нерв был типичным висцеральным
нервом предротовой дуги (рис. 250).
Кроме этих указаний, имеются еще и другие. Прежде всего двига-
тельные, брюшные головные нервы эмбрионально являются сегменталь-
ными нервами определенных миотомов головы: нервы глазных мышц для
предушных миотомов, а затылочные нервы — для заушных. Таким обра-
зом, в этой области метамерия нарушается лишь некоторым смещением
и соединением отдельных корешков, а также редукцией некоторых ко-
решков между корешками отводящего нерва и затылочными корешками
подъязычного (рис. 247). При правильном сегментальном строении
Сег- мент Миотом Брюшной нерв Спинной нерв Висцеральная щель
1 2 3 4 5 I П-а П-Ь Ш-а Ш-Ь n. oculomotorius n. trochlearis » » n. abducens » » п. ophthalmicus prof. n. trigeminus » » n. facialis n. glossopharyn- Ротовая щель Брызгальце 1-я жаб. щель
6 IV Редукция » » gens n. vagus 2-я » »
7 V МИОТОМОВ » » » » 3-я » »
8 VI и соответ- ствующих VII нервов (n. hypoglossus) n. vag.is, шп цер. 4-я » »
9 » » комп. To же 5-я » »
10 11 VIII IX » » » » » » 6-я » » 7-я » «
271
головы каждому брюшному корешку должен соответствовать спинной,
но в области головы нет соединения, характерного для спинномозговых
нервов, и поэтому нет точного критерия, который позволил бы нам отнести
Рис. 250. Головной мозг и нервы щитковой бесчелюстной рыбы Cephalaspis
(по Стеншио).
VII е — электрический нерв; di—промежуточный мозга; д. gass— ganglion gasserl V;o.
prof — ganglion n. ophthalm. profundi V; hab. I. и halt. r — левый и правый ganglion
habenulae; mes — средний мозг; mt — мозжечок; nil —n. lateralis X; nopht. lat. — n. oph-
thalmicu superf. VII; r. int— intestinaiis X; r. rise—передние висцеральночувствующие ветви
жаберных нервов; spi. v., spi. d.— брюшные и спинные корешки; tel — передний мозг;
I—X — головные нервы.
известный спинной нерв к тому или иному метамеру. Иногда, впрочем,
некоторая связь эмбрионально наблюдается, и на основании таких наблю-
дений можно вывести нижеследующее предварительное распределение
нервов по сегментам головы.
272
В таблице (см. стр. 271) второй и третий миотомы показаны двойными,
как это является вероятным по новейшим данным, и тройничный нерв
показан на двух метамерах, так как имеются. данные, заставлляющие
предполагать, что в состав верхнечелюстной ветви вошла еще рудиментар-
ная зажаберная ветвь лежащего впереди предротового пли предчелюстного
сегмента.
Блуждающий нерв, имеющий огромную область иннервации, несом-
ненно, является сложным нервом (рис. 249). По своему развитию он
соответствует, невидимому, двум полным сегментальным спинным кореш-
кам, к которым затем присоединяются висцеральные компоненты из целого
ряда лежащих позади сегментов. Мы знаем, что висцеральные элементы
ганглиев типичных висцеральных нервов развиваются в значительной
мере за счет особых наджаберных плакод. Ганглий блуждающего нерва
Рис. 251. Нервы боковой линии. Обозначения см. рис. 242.
составляется из элементов целого ряда таких плакод (начиная с плакоды
второй щели), которые иногда дают начало и целому ряду самостоятель-
ных наджаберных ганглиев, лежащих в стволе блуждающего нерва (крут-
лоротые) или у основания соответствующих жаберных нервов (скаты).
Таким образом, полимерная природа блуждающего нерва выступает
совершенно ясно. Блуждающий нерв представляет собой коллектор из
одного или двух полных спинных корешков и висцеральных компонентов
целого ряда следующих. В последних метамерах наравне с брюшными
корешками, входящими в состав подъязычного нерва, наблюдаются также
и рудиментарные спинные корешки с ганглиями, указывающие, что
последние не вошли полностью в состав блуждающего нерва.
Развитие блуждающего нерва показывает, что этот типичный головной
нерв постепенно разрастался, захватывая все новые области позади и
не только весь висцеральный аппарат собственно, по и большинство
внутренних органов.
При развитии подъязычного нерва виден обратный процесс: этот
нерв составляется из типичных спинномозговых нервов и перемещается
затем в голову, что, очевидно, находится в связи с нарастанием головы
за счет туловища, с прогрессивной цефализацией передней части послед-
него.
Наконец, следует еще отметить, что в связи с развитием органов бо-
ковой линии часть кожных нервов специализировалась и стала обособ-
ляться в особую систему боковых нервов (рис. 251). Таким образом, от
лицевого нерва обособилась значительная масса специализированных
волокон и дала начало главным боковым нервам головы, а также и слу-
273
Хойому Нерву. Из состава языкоглоточного нерва также обособилось
некоторое количество подобных волокон, и, наконец, весьма значительная
масса выделилась из состава блуждающего нерва. Боковой нерв туловища
не может, однако, быть обозначен просто как дериват блуждающего
нерва. Имеется ряд указаний на то, что и этот нерв является сложным
коллектором, собравшим специализированные нервные волокна, слу-
жащие для иннервации боковой линии туловища, из состава спинномоз-
говых нервов. У личинки миноги (Ammocoetes) описаны связи бокового
нерва со спинномозговыми посредством соединительных ветвей. Сущест-
вание таких связей понятно только лишь с изложенной точки зрения.
Функциональная диференцировка отделов центральной
нервной системы
Мы видели, что деление нервной системы на четыре отдела, разли-
чающиеся по своей функции, не ограничивается одной периферической
системой — для отдельных компонентов не менее характерны центральные
их связи, и соответственно мы в сером веществе спинного мозга различаем
четыре различные области: 1) соматически-чувствующую область спинных
рогов, 2) висцерально-чувствующую область основания спинных рогов,
Рис. 252. Распределение функциональных компонентов в головном мозгу селахий
(по Джонстону). Срединный разрез через мозг акулы и поперечный разрез через про-
долговатый мозг.
clb— мозжечок; со — corpus geniculatum; gh— ganglion habenulae; g—вторичный
вкусовой центр; ll— lobus lineae lateralis; li — lobus inferior; Iv— lobus visceralis; m —
средний мозг; го — recessus praeopticus; sv—saccus vasculosus; ta — area acustica; t — перед-
ний мозг; vi — velum transversum
3) среднюю висцерально-двигательную область и 4) соматически-дви-
гательную область брюшных рогов (рис. 241). Точно так же и для отдель-
ных компонентов головных нервов характерны их центральные связи,
и в головном мозге можно различить те же четыре основные области, что
и в спинном. Расположение этих областей в продолговатом мозге еще
вполне сходно с расположением их в спинном. Наиболее дорзально лежит
соматически-чувствующая область, в брюшной стенке — соматически-
двигательная, а между ними в боковых стенках располагаются одна
над другой обе висцеральные области (рис. 252). Бросается в глаза лишь
значительное разрастание чувствующей области и общая конфигурация —
продолговатый мозг представляет собой как бы развернутый по спинному
шву спинной мозг. В заднем и среднем мозге разрастание чувствующей
области оказывается еще более значительным, и, наконец, промежуточный
274
и Передний мозг, представляют собой огромную чувствующую массу и
совсем не содержат двигательных ядер. Рассмотрим это распределение
н сколько ближе (рис. 252).
е 1. Сомати чески -д вигате л ьная область головного
мозга ограничивается ядрами 12-й, 6-й, 4-й и 3-й пар нервов, которые
располагаются на дне главным образом в продолговатом и частью в сред-
Рис. 252-а Схема головного мозга насекомоядных и низших приматов. Вид с левой
стороны. Рисунки показывают нарастание преобладания центров зрения над цент-
рами обоняния.
I _ Прыгунчик [Macroscclididae]; II—Тупайя [Tupajidac]; III—Долгопят [Tarsioidea ];
ly—Игрунок [Callirichidae ], Области мозга: 1 —обонятельная; й—двигательная;
3 — осязательная; 4 — зрительная; 5 — слуховая; 6 — переднелобпал.
нем мозге. Эта область головного мозга по своему объему очень незначи-
тельна.
2. В и с ц е р а л ь п о-д питательная область головного
мозга исчерпывается ядрами (11-й), 10-й, 9-й, 7-й и 5-й пар нервов, распо-
лагающимися на дне продолговатого мозга, по бокам соматических ядер.
Эта область продолговатого мозга развита довольно значительно соответ-
ственно развитию висцеральной мускулатуры головы.
Огромных размеров достигает эта область у электрического ската, висцераль-
ная мускулатура которого частью преобразована в электрический орган. Соответ-
ствующая двигательная область продолговатого мозга образует здесь крупные «элек-
трические доли».
3. Висцерально-чувствующая область распола-
гается в боковых стейках продолговатого мозга. Висцерально-чувствую-
щие волокна 10-й, 9-й, 7-й и 5-й пар нервов вступают в связь с нервными
клетками этой области. Соответственно развитию висцерального аппарата
у рыб эта область более или менее выступает в виде валиков, вдающихся
в полость четвертого желудочка, и обозначается тогда как висцеральная
лопасть (lobus visceralis). Особенно разрастается эта область при значитель-
ном развитии вкусовых органов у костистых рыб, у которых висцеральные
лопасти иногда выдаются наружу в виде пары значительных полушарий
(у карповых). Наконец, для обонятельных нервов имеется специальная
чувствующая область в виде обонятельной лопасти переднего мозга.
4. Сомати чески- чувствующая область распола-
гается в спинной части боковых стенок продолговатого мозга, в мозжечке
и крыше среднего мозга. Соматически-чувствующие волокна 10-й, 9-й,
8-й, 7-й и 5-й пар головных нервов вступают своими конечными разветвле-
275
ниями в связь с нервными клетками этих частей мозга. У рыб и водных
амфибий это — также весьма значительная область, размеры которой
стоят в связи с развитием органов боковой линии. В продолговатом мозге
рыб соответствующая область выступает внутрь полости четвертого же-
лудочка в виде продольного валика, лежащего над висцеральной ло-
пастью и получающего название лопасти боковой линии (lobus lineae
lateralis).
Известная часть этой лопасти является специально слуховой областью
(area acustica). У наземных позвоночных с редукцией органов боковой
линии редуцируется и соответствующий центр, так что вся эта область
оказывается представленной главным образом лишь слуховой областью
да незначительными центрами общего кожного чувства.
Что касается органа зрения, то, как уже упоминалось, сама ретина
содержит не только воспринимающий аппарат, но и элементы, соответ-
ствующие нервному центру.
Роль головного мозга не ограничивается, однако, простой передачей
возбуждений от чувствующих центров к двигательным ядрам. При простей-
ших рефлексах путь, действительно, оказывается здесь таким же простым,
как и в спинном мозге в аналогичных случаях. Для головного мозга,
однако, в особенности характерно существование высших центров,
до которых восходит возбуждение от перечисленных уже первичных
чувствующих областей.
От клеток соматически-чувствующей области спинного мозга волокна
восходят по проводящим путям спинных столбов спинного мозга до ядер
спинных столбов в продолговатом мозге; от последних, как и от нервных
клеток всей соматически-чувствующей области продолговатого мозга
(лопасть боковой линии рыб), отходят новые волокна, восходящие далее
до мозжечка, до крыши среднего мозга, а у высших позвоночных — и до
коры большого мозга. Таким образом, мозжечок и средний мозг являются
не только первичными, но одновременно и высшими чувствующими цент-
рами.
Для органа зрения высшим центром является у низших позвоночных
также крыша среднего мозга, хотя коллатерали зрительных путей идут
и в зрительные бугры промежуточного мозга. У высших позвоночных
роль высшей зрительной области переходит к коре большого мозга, куда
волокна восходят через зрительные бугры (именно corpora geniculata).
От клеток висцерально-чувствующей области спинного мозга волокна
восходят по проводящим путям боковых столбов спинного мозга так же,
как и волокна, идущие от клеток висцерально-чувствующей области
продолговатого мозга (висцеральная лопасть) до заднего мозга, где име-
ются высшие висцеральные и, в частности, вкусовые центры (в боковых
стенках заднего мозга у низших позвоночных и в самом мозжечке у высших
позвоночных; в связи с ними стоит клапан мозжечка, который получает
такое развитие у костистых рыб). От заднего мозга третичные висцераль-
ные пути восходят еще до промежуточного мозга, где кончаются главным
образом в нижних долях и в сосцевидных телах. Отсюда волокна восходят
и до большого мозга. От обонятельной доли переднего мозга волокна
идут далее в полосатые тела, в первичную крышу переднего мозга, т. е.
грушевидную долю и гиппокамп млекопитающих, и вступают в связь
также и с центрами промежуточного мозга.
Из этой довольно сложной картины, детальное рассмотрение которой
составляет уже предмет физиологии, можно вывести известные заклю-
чения и о происхождении различных отделов головного мозга. Продолго-
ватый мозг представляет собой по своему строению тот же спинной мозг,
лишь модифицированный в связи с развитием висцерального аппарата
и специальных органов чувств (вкуса, боковой линии и слуха). Задний
и средний мозг построены в сущности по тому же плану — они содержат
276
те же компоненты и распределение их в общем то же. Крыша этих отделов
представляет собой еще продолжение первичной чувствующей области, но
она сильно разрослась и приобрела также и новую функцию: она связы-
вается с различными другими центрами, как низшими, так и высшими,
и приобретает значение координационных центров.
В первичном переднем мозге совсем пет двигательных ядер п нет
первичной чувствующей области; это — комплекс высших центров, пост-
роенный иначе, чем лежащие позади отделы головного мозга, и обнару-
живающий в своем развитии явную связь с двумя органами высших
чувств—органом обоняния и органом зрения.
Таким образом, изучение головного мозга приводит нас к тому же
выводу, как и изучение периферической нервной системы, мускулатуры,
скелета и прочих органов головы. Задняя, хордальная, область головы —
это результат преобразования передней части туловища; передняя, про-
хордальная, область представляет собой новообразование. Первичный
передний мозг представляет собой образование особого рода, развив-
шееся, вероятно, путем разрастания известного участка чувствующей
области переднего конца мозга в связи с развитием органа обоняния.
Собственно передний мозг и является у всех низших позвоночных лишь
комплексом обонятельных центров. Между этим отделом и остальным
мозгом естественно возник первый, древнейший координационный аппарат,
каковым является промежуточный мозг. Позже и в крыше следующей
части мозга стали развиваться координационные центры (среднего и
заднего мозга). У наземных позвоночных с изменением среды в значитель-
ной мере теряют свое значение органы восприятия химического раздра-
жения и, наоборот, приобретают огромное значение органы зрения,
слуха и осязания; соответственно происходит значительное развитие
высших соматических и, в частности, зрительных центров. Значение
высшего координационного аппарата переходит к большому мозгу, к
соматической области его коры, которая и получает колоссальное развитие
у млекопитающих. Висцеральная (обонятельная) область коры (первичная
кора) отступает на задний план и сохраняется главным образом в гип-
покампе.
ОРГАНЫ ЧУВСТВ
ОРГАНЫ ЧУВСТВ ОБЩЕГО ЗНАЧЕНИЯ
Раздражимость как основное свойство живых организмов присуща вообще про-
топлазме, но с диференцировкой клеток сложного организма наступает специализация
и в этом отношении. Естественно, чю такая специализация в особенности наблюдается
среди клеток покровного эпителия, кот орый связывает организм с внешней средой, но
отчасти она происходит и среди клеток эпителия кишечной полости, который подверга-
ется различного рода раздражениям со стороны воспринимаемой пищи. Уже у кишеч-
нополостных среди индиферентных клеток эпителия, которые, конечно, и сами являются
раздражимыми, наблюдаются отдельные клетки, отличающиеся своей формой и тесной
свяэью с нервной системой. Такие чувствующие клетки нередко снабжены плотными
отростками, несколько выступающими над общей поверхностью тела. В некоторых
местах тела — в области рта, а также на выступающих частях, каковы, например,
щупальца, эти чувствующие клетки образуют более значительные скопления.
Из таких скоплений чувствующих клеток и возникли простейшие органы чувств.
Чувствующие клетки эпителия первоначально являются просто особо раздражимыми
клетками и воспринимают раздражения различного рода (апэлективные органы).
Точно так же и простейшие органы чувств первоначально имеют лишь самое общее
значение. Однако различное положение этих органов уже само по себе определяет их
фактическое значение Для организма. Чувствующие клетки в области ротового от-
верстия, естественно, возбуждаются пищевыми веществами и специализируются в
качестве органов восприятия химического раздражения (вкуса). Скопления чувствую-
щих клеток на различных периферических частях тела, в особенности на подвижных
выростах, определяют их значение в качестве органов восприятия механического
раздражения (осязания). Таким образом, уже само положение определяет точнее
функциональное значение органа. Строение органа усложняется — чувствующие
клетки располагаются тем или иным характерным образом, приспосабливаясь для
277
восприятия определенных раздражений, и окружаются индиферентными опорными
клетками, защищающими их от всяких иных раздражений (элективные органы).
Чувствующая часть opiana погружается нередко ниже общего уровня поверхности
тела, а из индиферентных клеток эпителия, нередко и из окружающих иных тканей,
строится добавочный аппарат различного рода, приводящий к чувствующему эпителию
лишь раздражения определенного рода (адекватные). Таким образом произошла
специализация органов чувств, так что каждый из них проявляет свою особенную
«специфическую энергию», определяемую строением органа и его связями с нервной
системой.
Рис. 253. Чувствующая поч-
ка дождевого червя (из
Гессе)
ер — эпителий; п — нерв.
Различного рода раздражения воспринимаются
у беспозвоночных животных рассеянными в покровах
отдельными чувствующими клетками, снабженными
выступающими наружу плотными отростками (щетин-
ками, волосками) и продолжающимися в нервный от-
росток, вступающий в связь с нервной клеткой опре-
деленного нервного центра (рис. 212, Л). У кольчатых
червей наблюдаются и более совершенные кожные
органы — группа чувствующих клеток окружается
более длинными обкладочными клетками эпидермиса
и образует так называемые бокаловидные органы,
служащие, очевидно, органами восприятия химического
раздражения, и боковые органы, воспринимающие,
повидимому, механическое раздражение (рис. 253).
У моллюсков также имеются, кроме отдельных чув-
ствующих клеток, и специализированные органы более
сложного строения (эстеты, осфрадии). У членистоногих отдельные чувствующие
клетки или группы таких клеток снабжены отростками, входящими в подвижную у
основания или неподвижную хитиновую щетинку разнообразной формы. Такие органы
специализируются в качестве «осязательных» или «обонятельных» щетинок. У лан-
цетника имеются в эпидермисе отдельные чувствующие клетки со щетинками, образую-
щие на ротовых щупальцах значительные скопления. Кроме того, обонятельная ямка
ланцетника является уже более сложным органом.
Наравне с особыми чувствующими клетками, однако, уже у низших червей
имеются многочисленные свободные окончания нервов в покровном эпителии — нерв-
ные волокна кончаются многочисленными разветвлениями, проходящими по межкле-
точным пространствам эпителия.
У позвоночных животных отдельных первичпо-чувствующих клеток в
покровах нет, а свободные нервные окончания (рис. 212, D} имеют вид многочисленных
разветвлений волокон, доходящих до самых наружных клеток эпидермиса (за исклю-
чением роговых). У позвоночных имеются, кроме того, и особые кожные органы,
составляющиеся из комплексов чувствующих и опорных клеток.
Эти чувствующие клетки являются, однако, клетками новой генерации: они не
продолжаются непосредственно в нервное волокно, а оплетаются конечными развет-
влениями чувствующих нервных волокон, исходящих от клеток нервных узлов
(рис. 212, Е).
СПЕЦИАЛИЗИРОВАННЫЕ ОРГАНЫ ЧУВСТВ
Органы восприятия механического раздражения
Кожные органы. Простейшим воспринимающим аппаратом
у позвоночных являются свободные окончания нервов
(рис. 254), широко распространенные как по всей коже, так и в кишечнике
и в различных других органах. Раздражение свободных окончаний нервов
в эпителии кожи, в зубной мякоти человека связано с ощущением боли х.
У позвоночных имеются, кроме того, и специальные концевые органы,
состоящие из чувствующих, «осязательных» клеток, отдельных или чаще
собранных вместе в характерные группы.
Осязательные клетки происходят из эпидермиса и имеют вид плоских
или эллипсоидальных тел, оплетенных сетью нервных волокон, к которым
с одной стороны примыкает конечная пластинка в виде блюдца («осяза-
тельный мениск»), в свою очередь состоящая из густой сети нервных во-
локонец. Такие осязательные клетки нередко располагаются еще в самом
1 Раздражение свободных окончаний нервов во внутренних органах также мо-
жет быть связано с чувством боли. Чаще, однако, оно связано с менее определенными
ощущениями (голод, жажда) или вызывает рефлекс без ощущения.
278
эпидермисе (меркелевские клетки) (рис. 255). Группы подобных осяза-
тельных клеток, лежащих под эпидермисом, носят название осязательных
пятен, а если они окружены соединительнотканной оболочкой, то их
называют осязательными тельцами.
Осязательные тельца имеются уже у хрящевых рыб, хотя гораздо
большее значение они приобретают у наземных позвоночных. У бесхвостых
амфибий осязательные пятна распространены на всей коже. У рептилий
они нередко покрыты более тонким эпидермисом и очень правильно рас-
полагаются на заднем крае каждой чешуи. У птиц имеются более сложно
построенные осязательные
тельца, состоящие из груп-
пы осязательных клеток,
окруженных соединитель-
нотканной оболочкой. У
птиц распространение ося-
Рис. 254. Свободные окончания
нервов в коже саламандры (из
Кингсли).
Рис. 255. Осязательные клетки Меркеля (Л), осяза-
тельные тельца Грандри (В) и Фатер-Пачини (С).
п — мякотное и п*— бсзмякотное нервное волокно.
зательиых телец ограничивается языком и восковицей клюва-
У млекопитающих наблюдаются изолированные осязательные клетки на
местах, лишенных волос, или группы подобных клеток (тельца Меркеля),
лежащие непосредственно под эпидермисом или во .влагалищах волос,
имеются также более сложно построенные органы, состоящие из централь-
ной массы клеток, окруженной соединительнотканной оболочкой, в которых
нерв распадается на множество веточек, образующих спутанный клубочек
(тельца Краузе и Мейсснера). Такие тельца лежат обыкновенно под
эпидермисом в сосочках кожи (рис. 255, В).
Наиболее сложными кожными органами, невидимому, общего харак-
тера, являются колбовидные тельца высших позвоночных (амниот). Эти
тельца имеют то удлиненную (фатер-пачиниевы тельца) (рис. 255, С),
то более сжатую форму (тельца Гербста) и состоят из центрального
нервного стволика, окруженного слоем эпителиальных клеток, между
которыми проникает масса боковых веточек центрального нерва. Другое
нервное волокно, подходящее к тому же тельцу, распадается на сеть
волоконец, оплетающих эпителиальные клетки снаружи. Все это образо-
вание окружено оболочкой, состоящей из многочисленных пластинчатых
соединительнотканных клеток. Эти тельца располагаются глубоко в коже,
а у птиц и у млекопитающих они находятся, кроме того, и в различных
внутренних органах — в брыжейках, мышцах, сухожилиях, надкостнице,
в плевре и околосердечной сумке, в головке копулятивного органа и т. п.
Не подлежит сомнению, что эти органы чисто эктодермального про-
исхождения лишь вторично погружаются в глубь тела у высших по-
звоночных.
Роль органов осязания у млекопитающих играют также волосы,
в сумке которых имеются осязательные клетки и сложная сеть свобод-
275
вых нервных окончаний. Некоторие волосы являются весьма совершен-
ными органами осязания, как, например, «усы» хищных (vibrissae).
Сейсмосенсорные органы (боковой линии). Сейсмосен-
сорные органы представляют собой специализированные кожные органы,
в высшей степени характерные для рыб и живущих в воде амфибий.
Органы эти возникают на определенным образом расположенных экто-
Рис. 256. Орган боковой линии
амфибий в разрезе.
h — опорные клетки; п — нерв;
s — чувствующие клетки.
дермальных утолщениях путем кле-
точной диференцировки небольших
участков, дающих начало отдельным
чувствующим холмикам, как их назы-
вают. В каждом органе (рис. 256)
имеется два рода клеток—грушевид-
ные вторичночувствующие клетки,
занимающие более центральное по-
ложение и не доходящие до основа-
ния органа, и длинные цилиндри-
ческие опорные клетки, идущие
через весь орган от его поверхности
до самого основания. Чувствующие
клетки снабжены на своем свободном
конце плотной щетинкой и оплетают-
ся у основания конечными разветвле-
ниями нервных волокон.
У круглоротых и у амфибий эти органы лежат на поверхности (рис.
256) или погружаются лишь в неглубокие ямки. У рыб они погружаются
в ямки, желобки или каналы (ри. 257).
Сейсмосенсорные органы располагаются обыкновенно очень пра-
вильно. Своеобразна их локализация у круглоротых, где они распола-
Рис. 257. Продольный разрез черев канал боковой линии костистой рыбы.
п i — боковой нерв; s. о — сейсмосенсорные органы; s —• чешуя; as — трубчатая часть
чешуи, через которую проходит навал.
гаются на туловище в три продольных ряда с каждой стороны. У амфибий
нервные холмики располагаются по определенным линиям на голове
и в два-три продольных ряда вдоль туловища. Для рыб, как упомянуто,
весьма характерно образование сплошных каналов, в которых через извест-
ные промежутки располагаются отдельные органы. У низших акул
(Chlamydoselachus) и у химер вместо каналов имеются еще открытые
желобки, между тем как у всех прочих селахий, у ганоидов и костистых
рыб имеются замкнутые каналы, погружающиеся более или менее глубоко
в кожу. Такие каналы открываются через известные промежутки отвер-
стиями наружу. У рыб с костным скелетом каналы боковой линии про-
ходят в чешуях и в покровных костях, где они нередко обра-
зуют сложные разветвления. У костистых рыб боковая линия туловища
280
ясно видна благодаря наружным отверстиям, лежащим в каждой чешуе
по этой линии (рис. 257).
Кроме системы поверхностных каналов, у рыб наблюдаются и от-
дельные сейсмосенсорные органы или группы органов, погруженные
в болеэ или менее глубокие ямки. Таковы нервные мешочки ганоидов
и слизевые каналы селахий (ампулы Лоренцини). Последние предста-
вляют собой глубокие трубки, оканчивающиеся вздутием (ампулой), в
котором и располагаются органы такого же строения, как и в боковых
каналах. Слизевые каналы селахий иногда ветвятся, образуя сложные
гроздья, и располагаются в области головы, главным образом на рыле.
У рыб в типических случаях различимы следующие сейсмосенсорные
каналы (рис. 258): 1) н а д г л а з н и ч н ы й канал, идущий над
глазом вперед на верхнюю сторону рыла, иннервируется поверхност-
Рис. 258. Каналы боковой линии головы и их иннервация у трески
(но Колю из Кингсли).
ь — П. buccalis VII; dl и vl — n. lateralis X; h — n. hyomandibularis VII; os — n. ophthal-
micus superficialis VII; I — Соковой главный, lim — подъязычно-челюстной, io — подглазнич-
ный, so — надглазничный и st — задиетеменной каналы.
ной глазничной ветвью лицевого нерва; 2) п о д г л а з н и ч н ы й к а-
н а л, идущий над глазом вперед на нижнюю сторону рыла, иннерви-
руется щечной ветвью того же нерва; 3)подъязычн о-ч елюстной
канал, идущий вдоль подвеска (у рыб с костным скелетом по пред-
крышке) на нижнюю челюсть, иннервируется подъязычно-челюстной
ветвью лицевого нерва и 4) главный боковой канал, идущий
назад до конца тела, вдоль так называемой боковой линии, иннервируется
боковой ветвью блуждающего нерва; кроме этого, обыкновенно имеется
небольшой поперечный заднетеменной канал, соединяющий
главные боковые каналы обеих сторон между собой в затылочной области,
который иннервируется боковой ветвью блуждающего нерва. Небольшая
часть главного канала, образующего продолжение бокового канала туло-
вища в области головы, иннервируется еще веточкой языкоглоточного
нерва.
Функциональное значение органов боковой линии состоит в восприя-
тии легких движений и колебаний воды небольшой частоты. Воспринимая
разницу между давлением токов воды в различных частях тела, рыба
может ориентироваться не только относительно скорости и направления
водяных течений, но и относительно движений собственного тела, а также
может воспринимать отраженные водяные токи от посторонних предметов
и таким образом ориентироваться в пространстве.
281
Особое положение занимает инфундибулярный орган,
располагающийся в стенке сосудистого мешка промежуточного мозга
рыб. Он состоит из группы первичночувствующих клеток, снабженных
пучками волосовидных окончаний. Высказывалось предположение, что
этот орган воспринимает давление в желудочках нервной системы, и дает
рыбе возможность рефлекторно регулировать глубину своего положения
в воде.
Органы равновесия и слуха
У наземных позвоночных, а повидимому и у многих других животных функции
восприятия звуковых раздражений и восприятия своего положения в пространстве
соединяются в одном органе. Поэтому во многих случаях трудно бывает решить во-
прос, имеем ли мы дело со слуховым органом или со статическим. Раньше органы,
заключающие в себе «слуховые» камешки и чувствующие клетки, всегда обозначались
как слуховые. Теперь в результате целого ряда экспериментов чаще приходят к вы-
воду, что у водных животных мы имеем дело обычно с органами чувства равновесия.
Органами чувства равновесия являются, например, «слуховые» щупальца ме-
дуз, представляющие собой укороченные щупальца, эктодермальные клетки которых
отделяют слуховые камешки, или статолиты. Чувствующие клетки с волосками лежат
в эктодерме самого щупальца или у его основания. Такие щупальца могут погружаться
в ямки или замыкаться в пузырьки. Изменение положения тела вызывает изгибание
щупальца в ту или иную сторону под влиянием тяжести статолита и давление на чув-
ствующие волоски соответственной стороны.
Чаще наблюдаются эктодермальные «слуховые» ямки, или замкнутые пузырьки
(статоцисты), наполненные жидкостью (эндолимфой). Внутренняя стенка этих ямок,
или пузырьков, выстлана чувствующим эпителием с волосками, на которые давит ле-
жащий во внутренней полости статолит. При изменении положения тела статолит,
естественно, давит на волоски других клеток.
У десятиногих раков эти органы имеют вид открытых ямок, помещающихся на
основном членике первой пары усиков. Роль статолитов у них играют песчинки, ко-
торые при каждой линьке выбрасываются вместе с внутренним хитиновым покровом
органа и заменяются новыми.
Доказано с достаточной точностью, что все эти различные по происхождению
органы являются органами равновесия, хотя и нельзя отрицать возможности воспри-
ятия звуковых колебаний теми же органами, так как ясно, что колеблющийся стато-
лит (отолит) должен раздражать чувствующие волоски, на которые он опирается.
Однако у наземных беспозвоночных имеются органы, несомненно, восприни-
мающие звуковые колебания. Эти органы построены на иных принципах. У насекомых
наблюдаются иногда так называемые хордотональные органы, существенной частью
которых является натянутая под покровами струнообразная связка. Чувствующие
клетки соединяются одним концом с покровами и другим концом — со связкой, при-
ходящей в состояние колебаний, когда до нее доходят звуковые волны из окружающей
среды. Иначе построены тимпанальные органы, имеющиеся на голенях передних
ног у кузнечиков и сверчков (а также у муравьев и термитов) и по бокам первого брюш-
ного членика у саранчи и у цикад. Подобные органы имеются и в различных местах
тела у разных бабочек. Здесь тонкая кутикулярная перепонка натянута на прочной
хитиновой рамке: под этой перепонкой, играющей роль барабанной перепонки, лежит
трахейный пузырь; между перепонкой и трахейным пузырем располагаются группы
чувствующих клеток.
Слуховой орган позвоночных является прежде всего органом равно-
весия; он содержит статолиты, опирающиеся на волоски чувствующих клеток и раз-
дражающие их при всяком изменении положения тела, а также при изменении скоро-
сти движения. Но этот орган построен гораздо сложнее, чем соответствующие органы
беспозвоночных, и у наземных позвоночных дополняется аппаратом, несомненно, слу-
жащим для восприятия звуковых колебаний.
Слуховой орган позвоночных
Слуховой орган позвоночных развивается, как и органы боковой
линии, из утолщения эктодермы (боковой плакоды) в задней части головы.
Этот зачаток уха углубляется в виде ямки, которая затем принимает вид
пузырька, первоначально сообщающегося с наружной поверхностью
посредством канала, а потом обычно вполне отшнуровывающегося от
эктодермы (рис. 259). В дальнейшем слуховой пузырек диференцируется
на два отдела: верхний и нижний, отделенные друг от друга некоторым
перехватом. В верхнем отделе (pars superior) образуются три кармано-
образные складки, располагающиеся в трех взаимно перпендикулярных
плоскостях; в середине каждой складки обе стенки слипаются и затем
282
рассасываются, так что остается лишь периферическая часть складки
в виде полукружного канала (рис. 259). Остальная часть верхнего отдела,
Рис. 259. Последовательные стадии развития внутреннего уха.
А — слуховая плакода; В — ямка; С и Л — пузырек в разрезе; Е, F — образование no iv
кружных каналов. Схемы: с — зачаток улитки; cs — полукружный капал; ге______ rpce«s»e ... ”
dolympbaticus; s — sacculus; и — urliculus. leccsbus ел-
связывающая полукружные каналы между собой, получает название
овального мешочка (utriculus). В нижнем отделе (pars inferior) образу-
ется более или менее значительный полый выступ (lagena), который у
млекопитающих вытягивается в спи-
рально завитой канал улитки (canalis
cochlearis). Остальная часть нижнего
отдела получает название круглого
мешочка (sacculus). Оба мешочка
вместе носят название преддве-
рия (vestibulum).
Канал, соединяющий первона-
чально полость слухового пузырька,
а затем преддверие (обыкновенно на
границе между обоими мешочками)
с наружной средой, или эндолим-
фатический проток (duc-
tus endolymphaticus s. aquaeductus
vestibuli), обыкновенно замыкается
на своем конце и сполна отшнуровы-
вается от эктодермы.
Развившееся таким образом из
слухового пузырька довольно слож-
ное образование выполнено жидко-
стью — эндолимфой — и предста-
вляет собой наиболее существенную
часть органа равновесия и слуха
позвоночных — перепончатый
(с а б и р и н т, или внутреннее ухо
лрис. 260). Перепончатый лабиринт
вободно лежит в полости слуховой
Рис. 260. Схема лабиринта позвоноч-
ного (из Зеленки).
са, се, ср — передний, наружный и задний
полукружные каналы; ст — cristae acusti-
сае; de — ductus endolymphaticus:? —la-
gena; nil — macula lagenac; mn — macula
neglecta; ms — macula sacculi; mu — ma-
cula utriculi; ol — отолиты; s — sacculus.
283
капсулы, внутренние стенки которой повторяют у высших позвоночных
сложную форму эпителиальных стенок внутреннего уха, образуя так
называемый наружный, или скелетный, лабиринт (хрящевой
или костный). Между перепончатым и наружным лабиринтом имеется
щелевидное пространство, выполненное жидкостью — перилимфой.
У наземных позвоночных полость наружного лабиринта сообщается с
лимфатическими полостями головы
Рис. 261. Перепончатый лабиринт цель-
ноголовой рыбы (Chimaera monstrosa).
Вид с внутренней стороны (по Ретциусу).
аа, ае, ар — передняя наружная и задняя
ампулы; с — кожа; са, се, ср — передний, на-
ружный и задний полукружные каналы; сг —•
слуховой гребень; de — ductus endolympha-
ticus; mn — macula neglecta; ms — macula
sacculi; VII — слуховой нерв; s — sacculus;
и — utriculus.
при помощи особого перилимфати-
ческого протока (ductus perilym-
phaticus).
Перепончатый лабиринт низ-
ших позвоночных состоит из обоих
мешочков — овального (utri-
culus) с полукружными кана-
лами и круглого (sacculus),
разделенных перехватом (ductus
utriculo-saccularis), по близости
которого от круглого мешочка
отходит эндолимфатический про-
ток (ductus endolymphaticus).
Эндолимфатический проток у хрН-
щевых рыб открывается наружу
на спинной поверхности головы
(рис. 261). Это остаток первичной
связи слухового органа с наруж-
ной средой. У всех других позво-
ночных связь эта прерывается, и
эндолимфатический проток окан-
чивается слепо, обыкновенно не-
большим вздутием (sacculus endo-
lymphaticus) х. Полукружные
каналы (canales semicirculares)
лежат в трех взаимно перпенди-
кулярных плоскостях. Различают
приблизительно горизонтальный
наружный и два вертикаль-
ных канала — передний и
задний) (рис. 260, 261). На
одном конце каждого из каналов
имеется вздутие — ампула, в
котором помещается гребень чув-
ствующего эпителия. Ампулы на-
ружного и переднего каналов ле-
жат рядом друг с другом на пе-
реднем их конце, ампула заднего
канала—на заднем его конце. Внутренние колена обоих вертикальных
каналов сливаются между собой (образуя общий sinus utriculi).
1 Иногда эндолимфатический мешочек продолжается в значительные выросты
и каналы, как у Protopterus над продолговатым мозгом, у амфибий в области голов-
ного мозга. У бесхвостых амфибий слившиеся между собой мешки продолжаются на-
зад в длинный отросток, лежащий над спинным мозгом и дающий еще боковые ветви,
ложащиеся над спинномозговыми ганглиями. Все эти выросты содержат эндолимфу
с многочисленными мелкими статолитами. У гекконов подобные выросты распростра-
няются в области затылка и шеи под кожей. У костистых рыб, обладающих веберовым
аппаратом (Ostariophysi, к которым относятся карповые, сомовые и др.), оба эндолим-
фатических мешочка соединяются между собой под мозгом и продолжаются позади
в непарный вырост, связанный через окружающую его перилимфатическую полость
с косточками веберова аппарата.
284
Эпителий, образующий стенку перепончатого лабиринта, йе весь
носит характер чувствующего эпителия — таковой имеется лишь в неко-
торых местах, образующих так называемые слуховые пятна
(maculae acusticae),чувствующие клетки которых снабжены короткими
волосками, и вдающиеся во внутреннюю полость в виде пластиночек
слуховые гребни (cristae acusticae), чувствующие клетки кото-
рых снабжены длинными волосками. Слуховые пятна имеются в овальном
мешочке (macula acustica utriculi п, за исключением млекопитающих
еще небольшая macula neglecta) и в круглом мешочке (macula acustica
sacculi). Слуховые гребни располагаются в ампулах полукружных ка-
налов (рис. 260).
Чувствующий эпителий состоит из цилиндрических опорных клеток
и грушевидных чувствующих клеток, снабженных волосками. Основания
Рис. 2G2. Левый лабиринт костистой рыбы
и лягушки (В) (ив Гессе).
I — macula lagenae; р. b. — papilla basilaris;
чувствующих клеток густо оплетаются конечными разветвлениями воло-
кон слухового нерва. Таким образом, чувствующий аппарат органа слуха
вполне сходен с таковым органов боковой линии. Напомню, что им также
соответствуют и общие цен-
тры в головном мозге. К слу-
ховым пятнам и гребешкам
подходят ветви слухового
нерва, который снабжен ган-
глием, лежащим в непосред-
ственной близости с пере-
пончатым лабиринтом.
В эндолимфе взвешено
обыкновенно множество ото-
кониев (мелких ото- или
статолитов), придающих ей
молочный вид, а кроме того
имеются нередко и отдельные
более крупные камешки (со-
стоящие из кристалликов уг-
лекислой извести, склеенных
органическим веществом).
Всякое изменение положения
(А)
I — macula lagenae; р. b. — papilla basilaris; s —
macula sacculi; и — macula utriculi (обведены пунк-
тиром)
тела вызывает перемещение эндолимфы и
отокониев, раздражающих при этом чувствующие волоски слуховых пятен
и гребешков. Таким образом, слуховой орган низших позвоночных (в осо-
бенности его полукружные каналы с ампулами) является прежде всего
органом чувства равновесия. Казалось бы естественным, что и звуковые
колебания могут привести в движение отоконии, и, таким образом, трудно
отрицать возможность восприятия, по крайней мере, различного рода
шумов. Экспериментально, однако, доказано, что рыбы в большинстве
случаев глухи.
У костных ганоидов и у костистых рыб отоконии сливаются в крупные
«отолиты» (статолиты), лежащие по одному в овальном мешочке (recessus
utriculi), в круглом мешочке (sacculus) и в особом выступе последнего
(lagena).
У большинства рыб (за исключением химер) (рис. 261) па круглом
мешочке наблюдается более или менее значительный выступ (lagena)
(рис. 260, 262), в котором тогда имеется н отдельный слуховой сосочек
выступа (papilla acustica lagenae), обособляющийся от слухового пятна
мешочка. II от слухового нерва одновременно обособляется ветвь выступа
(n. lagenae). Слуховой сосочек этого выступа и является зачатком соб-
ственно слухового аппарата. У наземных позвоночных этот выступ разви-
вается прогрессивно. У а м ф и б и й от слухового сосочка выступа обо-
собляется отдельный основной слуховой сосочек (papilla
acustica basilaris), и соответствующая часть стенки выступа укрепляется
285
на скелетной рамке, образуя особую основную перепонку (рис. 262, В).
У рептилий выступ мешочка развивается сильнее, а у кроко-
дилов превращается в довольно длинный и несколько изогнутый канал
улитки (canalis cochlearis) (рис. 263). Часть стенки, укрепленная на ске-
летной раме, — основная перепонка — вытягивается в длину, так же
как и расположенный на ней основной сосочек. На слепом конце канала
различим и первоначальный
сосочек выступа (рис. 263, В).
Вместе с обособлением основ-
ного сосочка обособляется и
специальная ветвь слухового
нерва (г. basilaris) со своей
ганглиозной массой. Так как
основная перепонка прикре-
пляется своими краями к
стенкам скелетного лабирин-
Рис. 263. Левый лабиринт черепахи (Л) и кро- та> то ЯСНО, что перилим-
кодила (В) (по Гессе). Объяснения см. рис. 262. фатическое пространство ме-
жду каналом перепончатой
улитки и скелетной делится на два отдела — верхний (или лестницу
преддверия) и нижний (или лестницу барабанной полости). Воспри-
нимающий аппарат усложняется, и па основном сосочке развивается еще
особая упругая кроющая пластинка, и эта часть органа приобретает боль-
шое сходство с кортиевым органом млекопитающих (рис. 265). Очень
сходно построен, но еще несколько выше развит слуховой орган птиц
(рис. 264, Л), а также однопроходных
млекопитающих, у которых также
имеется изогнутый канал улитки,
отделяющийся от мешочка значитель-
ным перехватом в виде узкого кана-
ла (canalis reunions). У живородящих
Рис. 265. Разрез через оборот улитки
лабиринта морской свинки (по Шнейде-
ру из Кингсли). Кость заштрихована.
Is-—спиральная связка; г — перепонка
Рейсснера; sg — спиральный ганглий; sm,
st, sv— scalae media, tympani, vestibuli.
Рис. 264. Левый лабиринт птицы (Л) и
млекопитающего (В] (из Гессе). Объясне-
ния см. рис. 262.
млекопитающих слуховой орган достигает высшей степени своего разви-
тия. Канал улитки значительно вытягивается в длину и при этом изги-
бается по спирали, образуя от 1% До 5 оборотов (рис. 264, В). Конечный
слуховой сосочек выступа (papilla lagenae) исчезает1, а основной сосочек
преобразован в сложно построенный кортиев орган. Основная
перепонка (membrana basilaris), т. е. нижняя стенка канала улитки,
прикреплена своим внутренним краем к спиральному выросту костной
1 Есть еще у утконоса.
286
улитки (lamina spiralis ossea), по которому проходит к кортиеву органу
нерв улитки (и. cocblearis), имеющий и свою обособленную ганглиозную
массу (g. spirale). Наружный край основной перепонки прикреплен по-
средством спиральной связки (ligamentum spirale) к наружной стенке
костной улитки (рис. 265). Основная перепонка содержит на всем своем
протяжении множество очень упругих волокон, натянутых, подобно стру-
нам, между спиральным костным выростом и спиральной связкой (число
их у человека определяют в 16—20 тысяч). Длина этих волокон посте-
пенно увиличивается с увеличением диаметра канала улитки от ее осно-
вания к вершине (у человека от 0,064 до 0,480 мм). Предполагают, что
эти волокна, подобно струнам разной длины, резонируют в унисон со
Рис. 266. Кортиев орган (по Лавдовскому из Видерсгейма).
В—основная перепонка; Ва— клетки столбов; С—кроющаяТперелонка; По1. Lo—lamina
spiralis ossea; Ls— спиральная связка; N, N‘, N1—слуховой нерв с ганглием; G — чувст-
вующие.; клетки; К — псрсионка Рейсснера; Sm — исшил улитки (средняя лестница).
звуками различной высоты (теория Гельмгольца). Кортиев орган состоит
из опорных и чувствующих клеток (рис. 266): Из опорных клеток обра-
щают на себя внимание два ряда особых клеток столбов; последние накло-
нены друг к другу и ограничивают нечто вроде туннеля в середине органа
(кортиев канал). Чувствующие клетки короче опорных и располагаются
в один ряд кнутри от клеток столбов и в несколько рядов кнаружи от mix,
вместе с опорными (дейтерсовыми) клетками (рис. 266). Чувствующие
волоски погружаются в особую кутикулярную кроющую пере-
понку (membrana tectoria), выделяемую эпителиальными клетками
аксиальной части канала. Канал улитки (seal a media) граничит
своей основной перепонкой с перилимфатической нижней или бара-
банной лестницей (scala tympani) (рис. 265) и своей верхней
стенкой — перепонкой Рейспера — с лестницей преддверия
(scala vestibuli). Оба перилимфатические канала переходят друг в друга
на верхушке улитки. Перепончатая улитка сообщается с остальной частью
лабиринта очень узким соединительным каналом (canalis reunions), и
перехват между обоими мешочками имеет также характер узкого канала
(ductus utriculo-saccularis), от которого отходит эндолимфатический про-
ток, оканчивающийся слепым вздутием в области мозговых оболочек.
Слуховые пятна и слуховые гребни иннервируются ветвями нерва
преддверия (и. vestibularis) с его ганглием. Таким образом, у млекопи-
тающих произошло глубокое разделение слухового нерва на две части —
нерв улитки и нерв преддверия (рис. 267).
287
Для наземных позвоночных весьма характерно, следовательно, про-
грессивное развитие именно слухового аппарата, в то время как орган
равновесия остается все время приблизительно на одном и том же уровне
развития. Прогрессивная эволюция органа слуха не ограничивается,
Рис. 267. Схема слухового органа млекопитающего!
сс — canalis cochlearis; de — ductus endolymphaticus; dp— ductus perilymphalicns; ft — круг-
лое окно; i — наковальня; m —- молоточек; mae — наружный слуховой проход; mt — бара-
банная перепонка; пас — слуховой нерв; пс — нерв улитки; ш> — нерв преддверии; тс —
recessus endolymphaticus; sc — sacculus; tEu — евстахиева труба; utr — utriculus.
однако, усовершенствованием лишь части перепончатого лабиринта. Для
наземных позвоночных еще в большей мере характерно развитие среднего
уха, т. е. добавочного аппарата, служащего для улавливания колебаний
воздушной среды и для передачи их воспринимающему аппарату внутрен-
него уха (рис. 267).
Сродное ц наружное ухо
У наземных позвоночных полость первой висцераль-
ной щели или брызгальца, которая у рыб часто тесно прилежит к слуховой
капсуле, преобразовалась в наполненную воздухом барабанную
полость (cavum tympani), лежащую между слуховой капсулой с од-
ной стороны и покровами — с другой. Согласно своему происхождению,
эта полость оказывается соединенной с задней частью ротовой полости
суженным проходом — евстахиевой трубой (tuba Eustachii).
Наружная стенка истончается в определенном месте и образует упругую
барабанную перепонку (membrana tympani). Часть под-
весочного аппарата рыб, как мы уже видели раньше, превращается в слу-
ховую косточку — столбик уха (columella), упирающуюся одним
концом в барабанную перепонку, а другим концом — в слуховую капсулу.
Барабанная полость разрастается вокруг столбика, который получает,
таким образом, более свободную подвижность и является передатчиком
колебаний от барабанной перепонки к слуховой капсуле. Огенка послед-
ней в этом месте остается перепончатой и образует так называемое о Баль-
ное окно (fenestra ovalis). У амфибий, кроме того, наблюдается обык-
новенно еще добавочный аппарат, служащий, повидимому, для передачи
колебаний почвы через передние конечности, плечевой пояс, мышцу, под-
288
нимающуй) лопатку, и обособленную часть слуховой капсулы, образу-
ющую добавочную слуховую косточку (operculum). Со вторичным пере-
ходом к водной жизни хвостатые амфибии теряют барабанную полость
и перепонку, и в слуховом аппарате наблюдаются особые приспособления
для передачи колебаний водной среды. У гимнофион, ведущих роющий
образ жизни, барабанная полость также исчезла, и то же самое наблю-
дается у роющих змей.
У рептилий и птиц в стенке слуховой капсулы появляется
второе перепончатое окно, так называемое круглое окно (fenestra
rotunda), отделяющее перилимфатическое пространство от барабашюй
полости. Это, несомненно, важное приобретение, сделавшее возможной
гораздо более совершенную передачу колебаний барабанной перепонки
через столбик и овальное окно на перилимфатическое пространство, а
следовательно, и на внутреннее ухо, для чего раньше значительным пре-
пятствием служила несжимаемость жидкостей. У крокодилов и птиц ба-
рабанная полость разрастается в окружающих костях, образуя в них
воздухоносные полости. Барабанная перепонка погружается несколько
ниже общего уровня кожи, так что образуется зачаточный наружный слу-
ховой проход, у края которого иногда наблюдается небольшая кожная
складка, представляющая собой зачаточную ушную раковину (ящерицы,
крокодилы, птицы).
Среднее ухо млекопитающих отличается главным образом
большей сложностью передаточного аппарата (рис. 267). Из трех слухо-
вых косточек млекопитающих (о происхождении их см. в главе о черепе)
наружная — молоточек (malleus) — прикреплена своей рукоят-
кой к барабанной перепонке, а другим концом сочленяется со второй
косточкой — наковальней (incus), — которая своим коротким
отростком прикреплена помощью связки к стенке барабанной полости,
а длинным отростком, играющим роль рычага, сочленяется с третьей,
внутренней косточкой — стременем (stapes), — которая замыкает
собой овальное окно. Таким образом, передача колебаний барабанной пе-
репонки происходит по цепи из трех косточек. Особая мышца, прикрепля-
ющаяся к рукоятке молоточка, натягивает барабанную перепонку
(m. tensor tympani), а другая, мышца стремени (m. stapedius), наклоняет
стремя и натягивает перепонку овального окна. Рефлекторно сокраще-
нием этих мышц уменьшается амплитуда колебаний всего аппарата при
более сильных звуках.
Барабанная полость и у млекопитающих образует выступы в сосед-
них костях (tympanicum, squamosum, mastoideum, occipitale). У млеко-
питающих, наконец, достигает значительного развития и наружный
слуховой проход и, кроме того, позади прохода развивается
вырост — наружная ушная раковина (concha), которая соби-
рает звуковые волны и направляет их в проход (рис. 267). Ушная раковина
имеет хрящевой скелет и приводится в движение своей особой мускула-
турой (лицевой).
Слуховой орган позвоночных по своему развитию, по иннервации
и по строению чувствующего аппарата не сравним с подобными органами
беспозвоночных, но вполне сходен с органами боковой линии и, следо-
вательно, представляет собой такое же новое приобретение позвоночных,
как и последние. Можно считать вполне доказанным, что внутреннее ухо
представляет собой специализированный определенным образом комплекс
сейсмосенсорных органов. С образованием кристалликов извести в по-
лости этих органов обозначилась новая их функция — органов равно-
весия. Образование более крупных статолитов, повидимому, послужило
уже толчком к приобретению еще новой функции — органов слуха соб-
ственно. Однако эта функция могла приобрести большое значение только
289
у йаземных позвоночных, у которых, благодаря упругой воздушной среде,
оказалось возможным выработать более совершенный слуховой орган.
Постепенное усовершенствование улавливающего и передающего аппа-
рата (наружное и среднее ухо) точно так же характеризует наземных
позвоночных, как и усовершенствование собственно воспринимающего
звук аппарата внутреннего уха. Это выражается в прогрессивном разви-
тии выступа мешочка, с его основным сосочком, и их преобразовании в
канал улитки с кортиевым органом.
Органы зрения
В некоторых случаях установлено, что восприятие светового раздражения воз-
можно и без специальных органов зрения. В этих случаях, очевидно, чувствующие
клетки общего значения возбуждаются также и световыми лучами. Как правило, од-
нако, у животных, реагирующих на световое раздражение, наблюдается особый све-
точувствительный аппарат. В простейшем случае это — рассеянные светочувстви-
тельные клетки, встречающиеся, например, во множестве в покровах дождевых чер-
вей (а также в соединительной ткани и в мозге). Эти клетки заключают в себе особое
внутреннее тело, но не связаны с какими-либо скоплениями пигмента.
Гораздо чаще, однако, светочувствительные органы обладают пигментом, заклю-
чающимся или в самих чувствующих клетках или в особых пигментных клетках. У не-
которых ресничных червей наблюдаются интересные глазки, состоящие из чувству-
ющей клетки, продолжающейся на конце, обращенном к свету, в нервный отросток
и снабженной на противоположном конце краевым слоем из чувствующих палочек,
погруженных в особую пигментную клетку, имеющую вид чаши.
Подобные же главки имеются и в спинном мозге ланцетника. Чувствующие клетки
других животных снабжены обыкновенно палочковидными выростами, содержащими
окончания нервных волоконец, и соединяются между собой в более или менее значи-
тельные группы, образующие сетчатку, или ретину, причем в одних случаях свето-
чувствительные клетки обращены своими палочками к свету (прямые глаза), в других
случаях — в противоположную сторону (обращенные глаза). В редких случаях группы
светочувствительных клеток лежат на одном уровне с окружающим эпителиальным
покровом, образуя главные пятна (медузы). Обыкновенно соответственный участок
покровов углубляется и образует глазную ямку (рис. 6). Таким образом, орган зрения
предохраняется от грубых механических раздражений, оставаясь допустимым для све-
товых. Нередко утолщенная прозрачная кутикула доставляет органу еще более на-
дежную защиту. Если края глазной ямки смыкаются между собой, то образуется пу-
зырчатый глав. Такое постепенное развитие органа врения можно проследить в различ-
ных группах животного царства, особенно у кольчатых червей и у моллюсков, у кото-
рых и дальнейшая эволюция глав в высшей степени наглядна (рис. 6).
Прозрачные клетки передней стенки глазного пузырька превращаются в особое
светопреломляющее тело — хрусталик, который, подобно двояковыпуклой линзе,
отбрасывает изображение предметов внешней среды на сетчатку (в некоторых случаях
линза развивается как утолщение кутикулярного покрова). В органе зрения, как ука-
зано, большую роль играет пигмент, который изолирует каждую светочувствитель-
ную клетку от всяких побочных лучей за исключением лишь тех, которые падают на
нее в строго определенном направлении; без такой оптической изоляции образное зре-
ние было бы совершенно невозможно. Дальнейшее усовершенствование пузырчатого
глава сводится к увеличению числа зрительных клеток, к приобретению диафрагмы —
зрачка, устраняющего боковые лучи, к введению особого аккомодационного аппарата,
при помощи которого глаз устанавливается на дальние или близкие предметы («наво-
дится на фокус»), и к приобретению подвижности самого глава. Аккомодация дости-
гается обыкновенно или перемещением хрусталика или изменением его кривизны.
У беспозвоночных небольшие глазки иногда имеются в очень большом числе.
Соединением большого числа таких глазков в группу с одним общим полем зрения,
составляющимся ив тесно соприкасающихся полей отдельных глазков, изолированных
друг от друга пигментом, образуются сложные глаза. Образование таких сложных
глав наблюдается уже у кольчатых червей, у моллюсков и у многоножек, но особенно
характерны они для ракообразных и насекомых, обладающих весьма совершенными
сложными главами, приспособленными для образного, мозаичного зрения.
Парный орган зрения позвоночных обнаруживает известное общее сход-
ство с самыми совершенными органами беспозвоночных — с глазами головоногих мол-
люсков; однако при ближайшем рассмотрении оказывается, что расположение элемен-
тов ретины у них является обратным, хрусталик у позвоночных развивается в покро-
вах, а не в передней стенке главного пувырька, и, таким образом, сходство между этими
органами ьокоится лишь на аналогии. Обратное расположение элементов ретины на-
блюдается иногда и у беспозвоночных (некоторые моллюски и различные паукообраз-
ные) и развивается в результате впячивания передней стенки главного пузыря внутрь,
так что образуется бокал с двойными стенками. В втом случае ретина развивается из
290
передней стенки, палочки врительных клеток которой обращены внутрь, т. е. от света
к пигментному слою, развивающемуся в задней стенке. Процесс впячивания наружной
стенки глазного пузыря обусловлен образованием хрусталика, который в этих случаях
всегда развивается за счет покровов. Однако глаза позвоночных отличаются и от об-
ращенных глаз беспозвоночных тем, что развиваются как выросты из центральной
нервной системы, а не путем обособления от покровного эпителия, как глаза беспозво-
ночных. Таким образом, парные глаза позвоночных занимают вполне изолированное
положение и представляют собой новое их приобретение.
На вопрос о происхождении органов зрения позвоночных проливает некото-
рый свет факт светочувствительности самой центральной нервной системы у ланцет-
ника. У последнего в спинном мозге, вблизи центрального канала, имеются отдельные
светочувствительные клетки (глазки Гессе), очень сходные с таковыми ресничных чер-
вей. Светочувствительная клетка имеет на своем внутреннем конце краевой слой, со-
стоящий из множества палочек, которым клетка погружается в пигментную клетку,
имеющую форму чаши. Наружный, обращенный к свету конец клетки, продолжается
в нервный отросток. Сходные клетки имеются и в спинной стенке передней части нерв-
ной трубки. Эти глазки Гессе являются, таким образом, по своему типу обращенными
органами зрения, как и парные глаза позвоночных. Наконец, в переднем конце мозга
имеется значительное пигментное пятно, которое, однако, по экспериментальным дан-
ным не имеет вначения органа зрения. У личинок асцидий имеется небольшой, но не-
сколько более совершенный глазок, развивающийся также в стенках мозговой трубки
(пузырька).
У позвоночных мы различаем более или менее рудиментарные непарные органы
зрения и в высшей степени совершенные парные глаза.
Непарные глазные пузырьки позвоночных
Непарные глаза позвоночных располагаются по средней линии на
спинной поверхности головы в числе не более двух (рис. 268). Эти органы
развиваются как полые выросты крыши промежуточного мозга, которые
постепенно отшнуровываются и превращаются в лежащие один позади
другого пузырьки, соединенные с мозгом посредством ножки. У большин-
ства позвоночных эти органы дают затем образования железистого харак-
тера, но у круглоротых и у многих рептилий здесь развиваются насто-
ящие органы зрения (рис. 268). Передняя стенка глазного пузырька ста-
новится прозрачной, а задняя стенка превращается в сетчатку, от клеток
которой по направлению к мозгу развиваются нервные отростки, образу-
ющие более или менее длинный нерв. У круглоротых развиваются два
таких глазных пузырька: передний является вполне рудиментарным,
это теменной орган (парапинеальный или париетальный), зад-
ний лежит над первым непосредственно под прозрачными покровами и
представляет собой пинеальный орган (эпифиз). Последний
имеет прозрачную переднюю стенку и светочувствительную заднюю. В пиг-
ментированной сетчатке имеются светочувствительные клетки с палочко-
видными окончаниями, обращенными внутрь пузыря, цилиндрические
опорные клетки и ганглиозные клетки, залегающие в основном слое ре-
тины и продолжающиеся в волокна нерва. Последний снабжен у стенки
пузыря ганглием и идет к задней комиссуре в крыше промежуточного
мозга йот туда в сильно развитый правый несимметрический ганглий (g. ha-
benulae). Нерв переднего пузырька также вначале снабжен ганглиозной
массой, и волокна его продолжаются через верхнюю комиссуру до левого
несимметрического ганглия. Таким образом, имеются некоторые указания
на то, что эти органы первоначально были парными.
Пинеальный орган имеет вид лежащего под кожей пузырька, свя-
занного с особым нервом и у бесхвостых амфибий. У других позвоночных
пинеальный орган совершенно теряет характер органа зрения и превра-
щается в железу внутренней секреции (gl. pinealis s. epiphysis cerebri).
Теменной орган получает иногда глазоподобное строение
или, по крайней мере, вид пузырька у высших рыб (особенно у Amia и у
двудышащих), но обычно он редуцируется. Только у некоторых р е пти-
л и й, именно у гаттерии (Hatteria) и у большинства ящериц, париеталь-
ный орган имеет вполне характер пузырчатого глаза. Передняя его стенка
291
утолщена и ооразует хрусталик, задняя пигментирована и представляет
многослойную ретину. Светочувствительные клетки обращены палочко-
Рис. 268. Схемы париетального и пинеального
органов миноги, лягушки и ящерицы
(по Студничке).
с — ножа; сг — нрыша черепа; ср и се — задняя
и верхняя комиссуры; ер — эпифиз; п. ра и п.
pi — nervus parietalis и n, pinealis; pa и рг —
париетальный и пинеальный орган; par — пара-
физ; v tr — поперечный парус.
видными окончаниями во вну-
треннюю полость, а обратный
конец их продолжается в нерв-
ное волокно. Между светочув-
ствительными клетками распо-
лагаются длинные пигментные
клетки. В лежащем под свето-
чувствительными клетками слое
нервных волокон располагают-
ся и ганглиозные клетки сет-
чатки. Нерв направляется в
верхнюю комиссуру (com. ha-
benularis) крыши промежуточ-
ного мозга. Экспериментально
доказано, что теменной орган
ящериц, действительно, спосо-
бен воспринимать световые
раздражения.
Спинные глазные пузырьки
позвоночных принадлежат, сле-
довательно, к типу прямых
пузырчатых глаз; однако отглаз
беспозвоночных они отличаются
своим развитием от централь-
ной нервной системы.
Возможно, что это — орга-
ны, парные по происхождению,
которые вторично сместились
к средней линии. Судя по суще-
ствованию теменного отверстия
в черепе многих ископаемых
рыб (Crossopterygii) и стегоце-
фалов, можно думать, что рань-
ше эти органы имели более
значительное распространение.
Парный орган зрения
позвоночных
Глаза позвоночных закла-
дываются на очень ранней ста-
•дии, когда зачаток центральной
нервной системы имеет еще вид
утолщенной пластинки. По бо-
кам передней, расширенной
части этой пластинки можно
заметить зачатки глаз в видеы пар пигментных пятен, которые затем углу-
бляются, образуя пару ямок. Когда нервная пластинка замкнется в трубку,
зачатки глаз оказываются в области боковых стенок первичного перед-
него мозга (рис. 269). Здесь они все более выпячиваются в виде пары пу-
зырей, соединительная часть которых постепенно суживается. Таким
образом из стенок мозга развиваются глазные пузыри, соеди-
ненные с боковыми стенками промежуточного мозга посредством полого
стебелька. По мере развития спинной области мозга глазные сте-
бельки оттесняются все более на брюшную сторону (рис. 269). Затем на*
292
ружная часть стенки глазного пузыря впячивается внутрь полости по-
следнего, и таким образом глазной пузырь превращается в бокал
с двойной стенкой. Бокал этот имеет не совсем правильную форму,
так как, вследствие более быстрого роста его спинной части, глазной сте-
белек лежит ближе к нижнему его краю и нижняя стенка частью также
вдавливается внутрь, образуя здесь жолобообразную сосудистую вырезку,
или «хориоидальную» щель (рис. 270). По мере приближения глазного
бокала к покровам по бокам головы эктодерма образует утолщение, ко-
торое погружается под покровы навстречу глазному бокалу сначала в
виде ямки, а затем замкнутого пузырька, представляющего собой зача-
ток хрусталика глаза (рис. 269). Хрусталик вполне отделяется
Рис. 270. Пластическое изображение
глазного бокала с хрусталиком (из
Гертвигэ). fsh — сосудистая щель,
I — хрусталик.
Рис. 269. Схемы развития глаза.
А, В, С — поперечные равревы (согласно Джон-
стону); D — горизонтальный разрез; с — рого-
вица; г — радужина; I — хрусталик; р — пиг-
ментная оболочка; г — сетчатка; set — склера.
от эктодермы и ложится в отверстии глазного бокала. Внутренняя стенка
хрусталика сильно утолщается вследствие удлинения ее клеток, прини-
мающих цилиндрическую, а затем даже волокновидную форму, так что
полость хрусталика совершенно вытесняется. Это — волокнистое ядро
хрусталика. Наружная стенка остается тонкой и образует так называ-
емый эпителий хрусталика. Рост хрусталика происходит в результате
размножения клеточных элементов в области экватора на границе обеих
стенок. Отверстие глазного бокала суживается впереди хрусталика, вслед-
ствие роста его краев, и образует зрачок глаза (рис. 269, 2)). Внутренняя,
более толстая стенка бокала представляет собой будущую сетчатку,
а тонкая, наружная стенка дает начало пигментной оболочке глаза. Обво-
лакивающая глаз мезодерма дает начало наружным оболочкам глаза:
сосудистой оболочке, служащей для питания сетчатки, и склере, пред-
ставляющей собой скелет глаза. Эта скелетная оболочка продолжается
и впереди хрусталика, где она срастается с покровами, образуя прозрач-
ную роговицу (в которой, следовательно, различают наружную эпидер-
мическую часть — эпителий роговицы, и внутреннюю — соединительно-
тканную) (рис. 269, D). Мезодермальные клетки проникают, однако, и
внутрь глазного бокала через упомянутую сосудистую вырезку и обра-
зуют здесь эмбриональные кровеносные сосуды. Позднее эти клетки при-
нимают также участие в отделении студенистого вещества, выполняющего
полость бокала и возникающего вначале в результате отделительной дея-
тельности клеток самой ретины. На более поздних стадиях развития со-
судистая вырезка зарастает. В сетчатке происходит сложная диференци-
ровка клеточных элементов, и от нервных ее клеток развиваются отростки,
врастающие по глазному стебельку в мозг и образующие зрительный
тракт («нерв»). Клетки самого стебелька дают элементы глии. Края глаз-
293
ного бокала впереди от хрусталика вместе с прилежащей соединительной
тканью образуют радужину. Мезодермальные клетки, лежащие между
радужиной (и хрусталиком) и роговицей глаза, расходятся к стенкам и
оставляют свободную полость с жидкостью — переднюю камеру.
Таким образом развиваются наиболее существенные части глаза. По-
смотрим теперь, каково строение вполне сформированного глаза (рис. 272).
Сетчатка (retina) (рис. 271) имеет очень сложное строение и
содержит, кроме чувствующих клеток, еще нервную ткань и опорные
элементы. Чувствующие клетки лежат на наружной (обращенной от света)
стороне и несут каждая на своем наружном конце плотное воспринима-
Рис. 271. Схематический разрез через сетчатку глаза (по Гессе).
Т — внутренняя пограничная перепонка; 2 — слой нервных волокон; 3 — слой ганглиозных
клеток; 4 — внутренний сетчатый слой; 5 — слой биполярных клеток («внутренний ядер-
ный»); в — наружный сетчатый слой; 7 — слой зрительных клеток («наружный ядерный»);
8 — наружная пограничная перепонка; 9 — палочки и колбочки; 10 — пигментный эпите-
лий; 11 — опорная клетка; 12, 13 — центробежные и центростремительные волокна зри-
тельного тракта; I, II, III — пояса трех последовательных яевронов сетчатки.
ющее окончание — палочку или более короткую и вздутую у осно-
вания колбочку. Эти палочки и колбочки проходят через погранич-
ную перепонку сетчатки и погружаются в лежащий снаружи пигментный
слой. Тела чувствующих клеток с их ядрами образуют так называемый
наружный клеточный слой. В следующем далее наруж-
ном сетчатом слое располагаются дендриты — здесь происходит
соединение конечных разветвлений нервного отростка чувствующих кле-
ток с дендритами нервных клеток следующего, внутреннего кле-
точного слоя. Нервные клетки этого слоя соединяются опять при
помощи отростков во внутреннем сетчатом слое со сло-
ем ганглиозных клеток, покрытым изнутри слоем исхо-
дящих от них нервных волокон, составляющих далее зритель-
ный тракт. Последний проходит сквозь сетчатку и другие оболочки на
наружную поверхность глаза; естественно, что на месте прохождения
зрительного тракта нет воспринимающих элементов и это место является
слепым пятном глаза. Сетчатка, а также и зрительный тракт
содержат, кроме нервных элементов, и опорные клетки, соответствующие
элементам невроглии1.
1 Воспринимающие элементы сетчатки (палочки и колбочки) распределены у
различных животных различно. У круглоротых и у селахий различия между палоч-
ками и колбочками еще мало выражены, а у рептилий обычно имеются одни колбочки.
У млекопитающих преобладают палочки, а у ночных и водных млекопитающих кол-
294
Снаружи ретину покрывает пигментная оболочка, раз-
вивающаяся из наружной стенки глазного бокала и служащая для опти-
ческой изоляции отдельных воспринимающих элементов (рис. 271, 272).
Далее кнаружи лежит мезодермальная сосудистая оболочка
(chorioidea) глаза с богатой сетью кровеносных сосудов (рис. 272). В зад-
ней стенке сосудистой оболочки имеется нередко особый белый слой кле-
ток, наполненных мелкими кристалликами извести, — серебристая обо-
лочка (argentea) большинства рыб. Внутренняя поверхность сосудистой обо-
лочки иногда образует блестящий слой — зеркальце (tapetum),
состоящее из плоских клеток, содержащих мелкие кристаллики, или из
I
Рис. 272. Схематический горизонтальный разрез через глаз человека
(из Паркера).
с — роговица; ch—сосудистая оболочка; d—твердая оболочка зрительного тракта; ес—
эпителий роговицы; ecj — эпителий конъюнктивы; i — радужина; I — хрусталик; os — зуб-
чатый край; р — пигментная оболочка; рс — ресничное тело; г — сетчатка; sc — склера;
о — j — зрительная ось; II — зрительный нерв.
тонких эластических волокон. Этот слой отражает световые лучи обратно
на сетчатку и обусловливает свечение глаз в темноте у селахий и у млеко-
питающих. В этом случае пигментная оболочка, вследствие редукции в
пей пигмента, оказывается прозрачной. На переднем крае глазного бо-
кала ретина редуцирована и представлена топким эпителиальным слоем,
покрытым продолжением пигментного слоя. Вместе с продолжением со-
судистой оболочки этот передний край бокала образует кольцевую пере-
городку, ограничивающую сзади переднюю камеру глаза — р а д у-
ж и п у (iris) (рис. 272). Соединительнотканный передний слой радужины
может содержать пигмент, обусловливающий темную окраску глаз.
бочек обыкновенно нет вовсе. У человека в средней части сетчатки имеются одни кол-
бочки. Колбочки служат, повидимому, для восприятия цветов при дневном освещении,
а палочки воспринимают лишь свет различной интенсивности.
295
Радужина ограничивает отверстие — зрачок (pupilla), который может
расширяться и сужаться благодаря присутствию в ней радиальных (m. dila-
tator pupillae) и кольцевых (т. sphincter pupillae) мускульных волокон
эктодермального происхождения (развивающихся за счет клеток самого
бокала). Обе мышцы радужины состоят обычно из гладких мускульных
волокон. Непосредственно позади радужины стенки глазного блока
образуют обычно выступающий внутрь складчатый кольцевой валик,
выполненный продолжением сосудистой оболочки, который содержит
у наземных позвоночных массу меридиональных, радиальных и кольцевых
мускульных волокон, составляющих ресничный мускул (m. ciliaris). Это
так называемое ресничное тело (corpus ciliare), играющее боль-
шую роль при аккомодации глаза у наземных позвоночных. По краю
ресничного тела прикрепляется соединительнотканная капсула, одеваю-
щая хрусталик (сумка хрусталика). Между роговицей и зрачком поме-
щается, как уже упомянуто, выполненная «водянистой влагой» перед-
няя камера глаза; между радужиной и хрусталиком находится
небольшая кольцевидная задняя камера. Внутренняя полость
глазного бокала выполнена студенистым стекловидным телом
с небольшим количеством волокон и клеток.
Кнаружи от всех описанных частей глаза лежит прочная капсула —
склера, состоящая обычно из хрящевой ткани, но у млекопитающих
волокнистая. У рептилий и птиц в передней части склеры развивается
кольцо окостенений (склеротикальное кольцо, имевшееся также у сте-
гоцефалов и у некоторых ископаемых рыб). Впереди хрусталика склера
образует вместе с эпителием покровов прозрачную роговицу. На
глазном тракте склера продолжается в виде грубо волокнистой оболочки,
переходящей дальше в твердую мозговую оболочку. Лежащая под ней
мягкая оболочка переходит в сосудистую оболочку глаза и в мягкую
оболочку головного мозга. Эти факты являются лишней иллюстрацией
того, что орган зрения позвоночных представляет часть центральной
нервной системы.
Описанные выше части образуют глазное яблоко (bulbus
oculi), которое более или менее подвижно лежит в углублении — орбите —
боковой стенки черепа. С глазным яблоком связаны придаточные части
глаза.
Мускулатура, движущая глаз, состоит из описанных ранее четырех
прямых и двух косых мышц глаза, являющихся очень постоянными;
кроме того, у наземных позвоночных (за исключением змей и приматов)
имеется мышца, втягивающая глазное яблоко внутрь орбиты (m. retractor
bulb i), ay амфибий челюстная по происхождению мышца служит для под-
нятия глазного яблока (m. levator bulbi). У высших наземных позвоноч-
ных поднятие глазного яблока нередко достигается помощью гладкой глаз-
ничной мышцы (m. orbitalis), иннервируемой симпатическими волокнами.
Передняя стенка глаза прикрыта участком прозрачной кожи, прини-
мающей участие в образовании роговицы. Вокруг этого участка разви-
ваются обыкновенно еще складки кожи, или веки (palpebrae), выстланные
изнутри эпидермисом, образующим, как и на роговице, конъюнктиву
глаза. У рыб имеется обыкновенно только одно кольцевидное веко, у
наземных позвоночных различают верхнее и нижнее веки, а, кроме того,
у большинства позвоночных имеется еще тонкое прозрачное третье веко —
мигательная перепонка (membrana nicticans), лежащая
глубже других век и задергивающая глаз от переднего (внутреннего) угла.
Верхнее и нижнее веки особенно хорошо развиты у млекопитающих.
Мигательная перепонка наблюдается у акул, у амфибий и особенно у
рептилий и птиц. Она имеется и у млекопитающих, но нередко редуци-
руется. У обезьян, как и у человека, рудимент ее образует во внутреннем
углу глаза полулунную складку (plica semilunaris).
296
Веки подвижны, особенно у высших позвоночных, и снабжены тогда
собственной мускулатурой (m. levator palpebrae superioris, m. depressor
palpebrae inferioris) различного происхождения. У амниот, особенно у
млекопитающих, имеется мышца, поднимающая верхнее веко и проис-
шедшая, невидимому, путем обособления от верхней прямой мышцы
глаза (иннервируется глазодвигательным нервом), или ее заменяет гладкий
мускул (m. palpebralis), иннервируемый симпатическим нервом. Имеется
также кольцевая мышца, замыкающая верхнее и нижнее веки (m. orbi-
cularis oculi) и представляющая собой часть подкожной лицевой муску-
латуры. Мигательная перепонка задергивается обыкновенно пассивно при
втягивании глазного яблока внутрь орбиты действием ретрактора. Но
у рептилий и птиц от последней мышцы обособляется специальная мышца
мигательной перепонки (m. nicticans). У некоторых ящериц и у змей
оба века прозрачны, сращены’ между собой и образуют постоянное перед-
нее прикрытие роговицы глаза.
У наземных позвоночных в области век развиваются различные же-
лезы, смачивающие своим выделением переднюю стенку глаза. Одна
группа крупных желез развивается в области нижнего века у переднего
(внутреннего) угла глаза; это гардерова железа, выделяющая
жирный секрет; другая группа — слезная железа (glandula lacry-
malis) — развивается у заднего угла. Слезная железа у млекопитающих
передвигается под верхнее веко. Гардерова железа редуцируется у обезьян
и у человека. Жидкое водянистое выделение слезных желез стекает по
конъюнктиве по направлению к переднему (внутреннему) углу глаза,
откуда по слезному каналу отводится в носовую полость.
Глаза рыб обладают плоской роговицей и шаровидным хрусталиком (рис.’273)-
Не имеется пи мышц зрачка, пи ресничной мышцы. В полость глаза вдается’от сосу-
дистой оболочки особый серповидный отросток
(processus falciformis), прикрепляющийся своим
конечным вздутием к сумке хрусталика. Этот от-
росток содержит гладкие мускульные волокна,
при сокращении которых хрусталик перемещается
внутрь (ш. retractor lentis). С противоположной
стороны связка служит антагонистом, оттягива-
ющим хрусталик к прежнему месту. Таким обра-
зом, аккомодация достигается здесь перемещением
самого хрусталика.
У амфибий роговица выпуклая, а хру-
сталик имеет вид двояковыпуклой линзы. В ра-
дужине и в ресничном теле имеется гладкая
мускулатура. Аккомодация достигается переме-
щением хрусталика под действием особой мыш-
цы, притягивающей хрусталик вперед, к роговице
(m. protractor lentis), а также иногда давлением
ресничной мышцы1.
У рептилий и птиц в стекловид-
ное тело вдается характерный вырост с сосудами,
который у птиц имеет вид складчатой пластинки.
К аккомодации этот своеобразный вырост, или
гребень (pecten), не имеет никакого отношения.
Аккомодация достигается не только перемещением
хрусталика, но, в особенности, изменением его
формы под давлением ресничного тела, содержа-
щего (как и радужина зауропсид) поперечнополо-
сатую мускулатуру. При сокращении ресничного,
мускула выпуклость хрусталика увеличивается.1
Глаза птиц относительно очень велики.
Рис. 273. Глаз костистой рыбы
(из Видерсгейма).
Ад — серебристая оболочка; Со —
роговица; Ср — campanula! lialleri;
Cv — стекловидное тело; Ir — раду-
жина, L—хрусталик; Тл — сосудистая
оболочка; Ор — зрительный тракт;
Os—твердая оболочка; Рг — серпо-
видный отросток; Rt — сетчатка;
.Sc — склера; ТЕ —пигментный эпи-
телий; Тр — зеркальце; VK — пере-
дняя камера.
У млекопитающих склера глаза волокнистая. Нередко имеется отра-
жательное зеркальце (tapetum). Кроме подвижных верхнего и нижнего век, края ко-
торых обрамлены волосами— ресницами, часто имеется и мигательная перепонка.
1 Так как ресничное тело представляет собой кольцеобразный валик позади
хрусталика, то при его сокращении хрусталик отодвигается от ретины ближе к ро-
говице.
297
Имеется хорошо развитая слезная железа, которая у млекопитающих помещается в
области верхнего века, и, кроме того, часто имеется еще гардерова железа (ее нет у
приматов). По краям век у основания ресниц открываются древовидно разветвленные
мейбомиевы железы, которые представляют собой измененные сальные железы. Акко-
модация глаз млекопитающих достигается сокращением ресничной мышцы. Последняя,
однако, не действует непосредственно на хрусталик. Хрусталик в спокойном состо-
янии растянут сумкой хрусталика и имеет более плоскую форму (установка зрения
вдаль). При сокращении ресничной мышцы ослабляется натяжение сумки, и хрусталик
становится более выпуклым.
Парные органы зрения позвоночных развиваются как выросты из
центральной нервной системы. Поэтому нужно думать, что эти глаза
возникли не непосредственно в покровах, как глаза беспозвоночных, а
путем диференцировки центральной нервной системы. Что такая диферен-
цировка, действительно, возможна, доказывает факт нахождения свето-
чувствительных клеток в мозге дождевых червей и, в особенности, кон-
центрация подобных элементов в мозге ланцетника. Когда и каким обра-
зом возникли глаза позвоночных, нам в точности неизвестно, но общая
картина все же более или менее ясна. Весьма вероятно, что вся центральная
нервная система позвоночных первоначально представляла собой чувст-
Рис. 274. Схемы, иллюстрирующие происхождеение парных глаз позвоночных
(по Бовери).
I — хрусталик; т — миотомы; п — мовг; ос — глазки Гессе; р — пигментные клетки.
вующую пластинку, которая, между прочим, была и светочувствительной.
Можно предположить, что в пределах такой пластинки наступила затем
диференцировка, приведшая к образованию глазных пятен, а затем и
глазных ямок. Когда нервная пластинка погрузилась глубже и затем
замкнулась в трубку, глазные ямки оказались в виде выступов на этой
нервной трубке. В пользу такого взгляда говорят факты из истории эмбри-
онального развития. Несколько иначе рисуется эта картина на основании
сравнительноанатомических данных. Согласно этому взгляду, органы
зрения позвоночных образовались филогенетически немного позже, на
стадии, когда нервная пластинка уже замкнулась в трубку (Бовери,
Северцов) (рис. 274). У ланцетника центральная нервная система имеет
уже вид трубки, но вся она еще является светочувствительной. У предков
позвоночных произошла иная диференцировка. С увеличением размеров
тела и уменьшением прозрачности тканей мозг попадает в менее выгодные
условия для восприятия световых раздражений. В эволюции позвоночных
эти неблагоприятные моменты были устранены, так как некоторые участки
мозга с большим количеством светочувствительных Элементов стали вы-
пячиваться в виде пузырей, приближаясь, таким образом, к покровам,
которые в этих местах сохранили или даже усилили свою прозрачность.
Утолщение эктодермы образовало светопреломляющее тело — хрусталик,
приобретение которого представляло - собой огромный шаг вперед, так
как сделало возможным образное зрение. По мере приближения глазных
пузырей к покровам кривизна хрусталика должна была увеличиваться.
Так как глазные пузыри развивались от мозга, то наибольшее значение
для восприятия световых раздражений имела близкая к покровам наруж-
ная стенки глазных пузырей. Выпуклая форма этой стенки была небла-
гоприятна для образного зрения, так как хрусталик отбрасывал внутрь
сферическое вогнутое изображение, вогнутость которого увеличивалась
вместе с ростом кривизны хрусталика, т. е. по мере приближения глазных
пузырей к покровам. Поэтому передняя (наружная) стенка глазных пузырей
вдавливалась внутрь, и таким образом развились глазные бокалы, внутрен-
няя (передняя) стенка которых представляет собой сетчатку Ч
Светочувствительные клетки мозга развились из поверхностного
эпителия нервной пластинки, и чувствующие их выросты, естественно,
были направлены наружу. С развитием нервной трубки они оказались
внутри последней, в эпителии, выстилающем внутреннюю полость, куда
были направлены и чувствующие выросты. При образовании глазных
пузырей чувствующие клетки сохранили свое положение, и в передней
пх стенке, давшей начало сетчатке, чувствующие выросты были направлены
внутрь полости пузырей, т. е. от света. Нервные волокна, расположенные
в мозге поверхностно, оказались на наружной поверхности глазных
пузырей и, следовательно, на передней (внутренней) поверхности сетчатки.
В задней стенке глазного бокала чувствующие и нервные элементы реду-
цировались, но зато сохранились пигментные клетки, которые в передней
стенке исчезли.
Дальнейшая эволюция парных органов зрения сводится к усовер-
шенствованию светопреломляющих сред, развитию скелетной оболочки
и приобретению мускулатуры, служащей для движения глазного яблока,
ряда позвоночных происходит постепенное усовершенствование акко-
модации, и наблюдается приобретение подвижных век, слезпых желез
и других придаточных частей глаза.
Органы восприятия химического раздражения
У большинства беспозвоночных химическая раздражимость не локализуется
на определенных местах тела. У кишечнополостных, у червей и у моллюсков хими-
чески возбудимые клетки рассеяны по всему телу, хотя в области головы они нередко
образуют более значительные скопления. У ресничных червей и у немертин на перед-
нем конце тела наблюдаются иногда мерцательные ямки («обонятельные») с большим
количеством чувствующих клеток. У кольчатых червей в покровах весьма распрост-
ранены группы первичночувствующих клеток, образующие так называемые чувству-
ющие почки (рис. 253).
Наиболее высоко диферепцированы органы химического чувства у членистоно-
гих. У ракообразных это — небольшие выросты, располагающиеся обычно на первой
паре усиков, в которых лежат отростки первичночувствующих клеток. Хитиновый
покров этих выростов очень топок. У насекомых различают отдельные органы восприя-
тия раздражений от жидких и газообразных веществ, т. е. органы вкуса и обоняния.
Органы вкуса расположены внутри пищевода и иногда на ротовых частях. Они пред-
ставляют собой конические выступы, содержащие продолжение чувствующей клетки
и сидящие в углублениях. Органы обоняния располагаются главным образом на антен-
нах (усиках). Это также конические выступы, сидящие иногда в ямках, иногда открыто
и содержащие отростки первичночувствующих клеток.
1 Непарные спинные глаза находились ближе к покровам и сохранили пузыр-
чатое строение.
299
Органы вкуса позвоночных
Органами вкуса у позвоночных являются вкусовые почки. Последние
состоят из длинных цилиндрических вторичночувствующих клеток, снаб-
женных тонким и коротким чувствующим волоском (рис. 275). Между
чувствующими клетками располагаются не менее длинные опорные клетки.
Вкусовая почка пронизывает всю толщу эпидермиса; под ней соединитель-
нотканная часть кожи образует сосочек. Нервные волокна проникают в
почку и оплетают своими конечными разветвлениями чувствующие клетки.
Вкусовые почки представляют собой единственные диференцирован-
ные органы чувств, которым приписывают эктодермальное происхождение.
У рыб они первоначально возникают в пищеводе, глотке и энтодермальной
части ротовой полости. У костистых рыб эктодермальные зачатки пере-
мещаются и в область эктодермы, где затем распространяются по на-
ружной поверхности всего1 тела. Таким образом, у костистых рыб вку-
совые почки встречаются в различнейших местах тела: в ротовой полости,
в глотке, на жаберных дугах,г'в покровах всего тела, на плавниках, но,
в особенности, на губах и на'’’усиках. У наземных позвоночных распро-
странение вкусовых почек ограничи-
вается ротовой полостью и отчасти
глоткой.
Рис. 275. Разрез через вкусовую
почку:
k — опорные клетки; п — нерв;
s — чувствующие клетки.
Рис. 276. Вкусовые ветви лицевого псрва
сома Amieurus (по Еррику).
Прекрасно развиты вкусовые почки и у млекопитающих, у которых
они располагаются главным образом на сосочках языка, но встречаются
и на небе, в зеве и на надгортаннике. На языке вкусовые почки поме-
щаются на рассеянных по всей его поверхности грибовидных сосочках
(papillae fungiformes), на лежащих в задней части языка нескольких
крупных вальковатых (papillae circumvallatae) и на парном листовидном
сосочке (papilla foliata), тянущемся позади вдоль края языка у большин-
ства млекопитающих и состоящем из ряда параллельных складочек со
вкусовыми почками по бокам.
Вкусовые почки иннервируются у низших позвоночных всеми типич-
ными висцеральными нервами, т. е. лицевым, языкоглоточным и блуждаю-
щим. Вкусовые почки, располагающиеся на наружной поверхности тела,
иннервируются передним из вкусовых нервов, именно лицевым, что ука-
зывает на то, что эти органы, действительно, переместились на наружные
покровы из ротовой полости. Соответственно значительному развитию
этих органов у костистых рыб колоссально развиваются и специальные
ветви лицевого нерва, распространяющиеся тогда по всему туловищу,
образуя главный боковой ствол, параллельный боковой ветви блуждаю-
щего"нерва (рис. 276). Отметим, что в этом случае чрезвычайно разраста-
ются^ соответствующие висцеральные центры головного мозга.
У млекопитающих вкусовыми нервами являются: барабанная струна
(chorda~tympani VII) и язычная ветвь языкоглоточного нерва (г. lingua-
lis IX).
300
Органы oooiniHiui позвоночных
Орган обоняния позвоночных животных развивается в виде парного
утолщения эктодермы на переднем конце головы (обонятельные плакоды),
которые затем погружаются глубже, образуя сначала простые обонятель-
ные ямки. В простейшем случае он н у взрослого животного представлен
парными ямками, выстланными чувствующим эпителием и располагающи-
мися па переднем конце головы Ч
Обонятельный эпителий состоит из длинных тонких первичночув-
ствующих клеток и не менее длинных опорных клеток. Обонятельные
клетки почти волокновидны с веретено видным вздутием па месте ядра
(рис. 212, С). На своем периферическом конце они снабжены пучком
чувствующих волосков, между том как проксимальные нх концы про-
должаются в волокна обонятельного нерва, которые своими конечными
разветвлениями вступают в связь с дендритами нервных (митральных)
клеток обонятельной доли, образуя характерные клубочки. Опорные
клетки цилиндрической формы нередко несут мерцательные реснички.
Обонятельные ямки, пли мошки, защищаются снаружи скелетной
капсулой, срастающейся с передней частью мозговой коробки.
Чувствующая поверхность эпителия обонятельных мешков увели-
чивается путем образования более или менее сложной системы складок, в
которые от стенок обонятельной капсулы могут вдаваться поддерживаю-
щие их скелетные выросты, или раковины.
Наружное отверстие мешка представляет собой ноздрю.
Уже у низших позвоночных наблюдаются приспособления к обновле-
нию воды в обонятельном мешке, которые приводят к образованию раз-
личного рода обонятельных ходов.
У к р у г л о р о т ы х орган обоняния вторпчо соединяется с гипо-
физарной трубкой и превращается в непарный орган, открывающийся
наружу непарной «ноздрей», пли, вернее, назогппофизарпым отверстием
па спинной стороне головы. При сжатии и растяжении гипофизарного
мошка по гипофизарному каналу через «ноздрю» и орган обоняния прохо-
дит ток воды. У миксин гипофизарная трубка открывается в ротовую
полость. Таким образом, достигается постоянное обновление воды в
органе обоняния. Увеличение поверхности обонятельного эпителия до-
стигается образованием много численных радиальных (меридиональных)
складок.
У всех других позвоночных обонятельный орган но имеет никакого
отношения к гипофизу, остается парным и открывается наружу парной
ноздрей. У селахий обонятельные ямки лежат на нижней стороне
рыла, впереди рта. От каждой ноздри идет ко рту желобок, прикрытый
нередко складкой кожи (рис. 277). Если представить себе, что такая
складка кожи срастается своим краем с покровами, то желобок превра-
тится в трубку, ведущую от обонятельного мошка ко рту. Такой процесс,
действительно, наблюдается эмбрионально у большинства позвоночных.
У зародышей наземных позвоночных обонятельные ямки первона-
чально бывают ограничены так называемыми носовыми отростками и
продолжаются в желобок между внутренним носовым и верхнечелюстным
отростками (рис. 278). Этот желобок, соединяющим обонятельную ямку
с ротовой полостью, потом через сращение его краев превращается в
канал, открывающийся наружу одним отверстием в области первичной
1 Хотя орган обоняния рыб служит для восприятия раздражений от веществ,
растворенных в воде, мы его обозначаем как орган обоняния ввиду его полной гомо-
логии с обонятельным органом наземных позвоночных, у которых ои служит для
восприятия раздражения от газообразных веществ или от веществ, взвешенных в воз-
духе. Кроме того, следует отметить, что какая-то качественная разница между орга-
нами вкуса и обоняния у рыб. несомненно, имеется.
301
обонятельной ямки на спинной поверхности головы — наружная нЭзДря —
и другим отверстием в ротовую полость — внутренняя ноздря, или хоана.
У рыб тоже развиваются две пары ноздрей, но у них обе пары
располагаются снаружи и притом обыкновенно на спинной стороне го-
ловы. У костистых рыб вода входит в переднюю ноздрю и выходит через
заднюю. Только у двудышащих рыб обе пары ноздрей перемещаются
Рис. 277. Голова акулы снизу
(Scyllium) (по Видерсгейму)
N — ноздри; М — рот; HSO — боковая
линия.
на брюшную поверхность: передняя
пара открывается под верхней губой, а
задняя — в ротовую полость (хоаны).
Рис. 278. Голова вародыша высшего позво-
ночного снизу (из Шимкевича).
Ад — глав; А — наружный и J — внутренний
носовые отростки; Ь — складка века; М — пер-
вичная ротовая полость; NP — обонятельная
ямка; О — верхнечелюстной отросток; S — лоб-
ный отросток; Т — слезоносовой желобок.
У рыб многочисленные складки обонятельного эпителия распола-
гаются обыкновенно радиально по отношению к продольной оси (рис. 279).
У наземных позвоночных ближайшая ко рту пара
ноздрей переместилась в ротовую полость, образовав хоаны (choanae),
Рис. 279. Поперечный разрез через обо-
нятельную область головы акулы (из Ши-
плей и Мэк Брайда).
ol — обонятельный мешок; п — ноздря;
sn — носовоя перегородка.
Рис. 280. Диференцировка обонятель-
ного мешка у ящериц.
о. Jac — якобсонов орган.
между тем как другая пара остается на спинной поверхности головы в виде
наружных ноздрей (иares). Образовавшийся таким образом
носо-ротовой канал приобретает большое значение в качестве дыхатель-
ного пути. Таким образом, приспособление к обмену воздуха в органе
обоняния привело к усовершенствованию органов дыхания. Орган обо-
няния одной своей частью приобрел новую функцию, и это в значительной
мере определило ход его дальнейшей эволюции. Уже у амфибий обо-
нятельный мешок начинает подразделяться на собственно обонятель-
1 Черев орган обоняния у Protopterus направляется дыхательный ток воздуха
во время спячки в период засухи.
302
и у ю (внутреннюю) область, выстланную обонятельным эпителием
(pars olfactoria), и дыхательную (наружную) область, выст-
ланную простым эпителием (pars respirator!а). У рептилий обоня-
тельная область лежит выше, а дыхательная помещается ниже и диферен-
Рис. 281. Поперечный разрез че-
рез орган обоняния птицы (Л) и
млекопитающего (В).
а — воздушные полости; ect— ecto-
turbinalia; ent —endoturbinalia;
t — turbinale; sn — septum nasi.
цируется еще на три отдела — передний отдел, или преддверие
(vestibulum), открывающиеся ноздрей наружу и выстланное плоским
эпителием, средний отдел, соединенный с собственно обонятельной об-
ластью и выстланный цилиндрическим мерцательным эпителием, и задний
отдел, открывающийся хоанами в полость рта, или носоглоточный
ход (ductus nasopharyngeus) (рис. 280). Последний отдел получает
у всех высших позвоночных значительное
(первичные хоаны) вместе с верхней частью
ротовой полости отделяются от остальной
части ротовой полости развивающимися
горизонтальными нёбными отростками,
позади которых открываются тогда носо-
глоточные ходы парным обычно отвер-
стием, или вторичными хоанами (рис. 150).
Этот известный нам уже процесс разви-
тия вторичного нёба отодвигает внутрен-
ние носовые отверстия все далее назад и
иногда до задней части черепа (у кроко-
дилов). Значение такого отодвигания хоан
как можно ближе к глотке (и гортани),
очевидно, заключается в том, что оно обес-
печивает возможность свободного дыхания
в то время, когда пища находится во рту.
Это в особенности важно для высших по-
звоночных с более интенсивным обменом
веществ и в свою очередь дало возмож-
ность млекопитающим приобрести способ-
ность разжевывать пищу.
Прогрессивное развитие обонятельной
поверхности у наземных позвоночных
привело так же, как и у рыб, к образова-
нию сложной системы складок. Однако
только у личинок амфибий эти складки
правильно радиально расположены, во-
обще же имеют вид более или менее закру-
ченных горизонтальных выростов. Эти выросты поддерживаются отрост-
ками скелетных стенок носовой полости, в которых могут появиться и
самостоятельные окостенения, получающие название обонятельных
раковин. Уже у рептилий в связи с наружной стенкой носовой по-
лости развивается слегка завитой выступ, челюстная раковина, которая
отчасти подразделяет носовую полость на верхний, обонятельный, и
нижний, дыхательный, отделы. У птиц, кроме того, развивается еще
одна раковина впереди, в преддверии, не имеющая обонятельного значения,
а служащая, очевидно, для обогревания и увлажнения вдыхаемого воз-
духа, и другая раковина — позади, над челюстной раковиной. Эта верх-
няя, или носовая, раковина и является собственно единственной настоя-
щей обонятельной раковиной птиц (рис. 281, Л). У млекопитающих
имеется впереди нижняя, или челюстная, раковина (maxil-
loturbinale) (рис. 282), связанная с челюстной костью и выстланная про-
стым эпителием (иннервируется тройничным нервом). Обыкновенно име-
ется также носовая раковина (nasoturbinale), лежащая впереди
в верхней части носовой полости и связанная с носовой костью (рис. 282).
Собственно обонятельными являются у млекопитающих лежащие позади
303
й большем или мОньШем числе решетчатые райовинЬ (ethmo-
turbinalia), связанные с решетчатой костью и располагающиеся обыкно-
венно параллельно одна над другой (рис. 282). Одни из этих раковин
простираются глубже по направлению к средней линии (endoturbinalia),
другие лежат между первыми более кнаружи (ectoturbinalia) (рис. 281).
Наибольшее число раковин наблюдается у млекопитающих с хорошо
развитым обонянием (5—8 endoturbinalia), у которых они вместе с тем
более сложно закручены. У животных со слабо развитым обонянием
(обезьяны и человек) число внутренних раковин сокращается (у прима-
тов до 2) и форма их упрощается, а наружные раковины могут и вовсе
исчезнуть (приматы).
Однако эмбрионально у человека закладывается еще до 5 решетчатых
раковин. У китообразных органы обоняния редуцируются и раковины
совсем исчезают.
У крокодилов и птиц, а в особенности у млекопитающих, носовая
полость еще значительно увеличивается благодаря образованию возду-
Рис. 282. Срединный разрез через голову косули.
Соотношение мозговой и обонятельной полостей
и расположение обонятельных раковин (из Ви-
дерсгеймэ).
Рис. 283. Поперечный разрез
через орган обоняния ящерицы.
п — главная часть обонятельной
полости; J — полость якобсонова
органа; «п—носовая перегородка.
хоносных полостей в соседних костях, служащих, очевидно, для обогре-
вания втягиваемого воздуха.
Для наземных позвоночных характерно также обособление части
обонятельного мешка, образующего самостоятельный отдел органа обоня-
ния, так называемый якобсонов орган (organon Jakobsonii s.
vomero-nasale) (рис. 280). Уже у большинства амфибий наблюдается
образование выроста в нижней и внутренней частях обонятельного мешка,
содержащего также обонятельный эпителий. У хвостатых амфибий этот
выступ сдвигается кнаружи, а у гимнофион он вполне обособляется и
имеет вид слепого мешка, лежащего под обонятельным мешком и сооб-
щающегося с ним позади. Одновременно с развитием якобсонова органа
обособляется и соответствующая часть обонятельного нерва. У рептилий
якобсонов орган вполне отделен от обонятельного мешка, соединяясь
особым каналом с ротовой полостью (рис. 280, 283). О значении якобсо-
нова органа пока ничего определенного неизвестно, — невидимому, он
служит для обоняния пищи, находящейся во рту. При этом интересно
отметить, что в отличие от наполненных воздухом (у наземных позво-
ночных) полостей обонятельного мешка якобсонов орган постоянно напол-
нен жидкостью, выделяемой носовыми и якобсоновыми железами. Жид-
кость может выбрасываться из органа и заменяться свежей. Это дает
некоторые указания на качественное различие между обеими частями
обонятельного органа.
Якобсонов орган хорошо развит у ящериц и у змей, где он защищен
своей хрящевой капсулой, между тем как у черепах, крокодилов и птиц
он наблюдается лишь в виде эмбрионального зачатка.
304
У млекопитающих также обычно имеется хорошо развитый якобсонов
орган в виде длшшин парном труоки, окруженной хрящевом капсулой
(якоосомов храму, лежащем медналвно мод органом ооонянмя н снаожае-
мои осооом ветвью оооннтельного нерва. Позади якоосонов орган кон-
чается слепо, а впереди переходит в неоно-носовой, ими стемсонов, канал
(d. назорашьншз), которым открывается на неое (проходя через Зогашша
шсыпа твердого неоа). У приматов атот орган редуцируется.
Остается еще отметить, что у наземных позвоночных анмтелнн обо-
нятельном полости мрмооретает характер слизистой ооолочкм, поддержи-
ваемом во вмамшом состоянии секретом оокаловмдных клеток и сложных
желез. У амфиоин м рептилии имеются железы, лежащие дорзалыю у
наружном (рис. 263) и вентрально у внутреннем стенок ооонмтельного
мешка. У млекопитающих, кроме маленьких, рассеянных повсюду в но-
совом целости ооуменовых желез, имеется еще крупная стенсонова железа,
лежащая снаружи у основания носовой полости и открывающаяся в
преддверие.
13 носовую полость открывается у наземных позвоночных также
слезноносовом канал.
Орган обоняния рыб представлен парными мешками, выстланными обонятельным
зшпедпем, иььерхиоыь кихирою увеммчаваеюя uyicM оиразивоимя мниючислеиилмх
и ииыкииввиио радиальных складыч (рис. х/У). Парные ииаДрн у селахии иоиощиклсн
ИОСредсхьим нОиерхнисгиШ'О Ячелиииа си pi им (рис. У ьысшах рыО имею юн две
иары_11иадреи» лежащих из иншшии итириие гилоиы (1 ильки у дьудышащах — иа
брюшном).
У амфибий имеется пара наружных низдрен и лара хиаи. Намечается днфереицм-
ровка посипим иилисш на дыхалельныи и оиоиячелюныи отделы. Имеются илсани-
ьисовий каИаЛ и ьиАих’ОчнсЛенные женены. ЛкиииОНиь оргии начинав! иоосиОлЛхься,
в оиоиеьнисти у гимнофион.
У рентнлш! и шиц ооосибляегся еще особое преддверие, и удлиняется носогло-
точный ход в Связи с развитием вторичного неоа (рис. хъир носовые ракошшы илаои
развиты. JiHOOCOHOB vpiан вполне Оиииоилен у ящериц и у вмеи, а у других реиыыам
и у мтмц редуцируется.
У млекопитающих орган обоняния достигает высшей степени своего развитии,
410 выражается, в осиивнности, в сложном развитии оооияхельпых раковин. 1 кроме
нередиих, нижнечелюстной и порхнем, носовой раковин ммвеюя oo.iee млн минее
зьачительное число решетчатых раковин (ряс. zt>xj. лкоосонов оргаи имеется у ооль-
шинства млекопитающих в качесхье ьнилпе ооосиоленво! о органа, открывающегося
в риювую полость. У приматов ают орган редуцируется. млекоишающпх разня-
вается еще мяснехый наружный нос, поддерживаемым носовыми костями и хрящами.
У некоторых форм наружным нос удлиняется в хоиот, сиаижевпын сложной муску-
латурой (земЛериШШ, свиньи и оииоенно слоны).
Орган обоняния позвоночных появляется в виде пары простых обо-
нятельных ямок, выстланных изнутри цилиндрическим анителием с пер-
вично чувствующими клетками. Весьма вероятно, что атот орган чувств
является самым древним органом чувств позвоночных ииерьоначальни имел
вид чувствующен пластинки, тесни примыкающей, а может оыть, и свя-
занной с передней частью нервной пластинки. С ооразоьаннем углуолешш
понадобились присносооления для смены виды в атих органах, и у рыо
они сводятся к иоразованню отдельных вводных и выводных отверстии.
1 аким ооразоА! возникли две нары ноздрей. & наземных позвоночных
одна нара ноздрей переместилась внутрь ротовой полости и получила
название хоан. Носовая полость стала частью дыхательных путей, кото-
рые в дальнейшем более или менее ооосоОляются от сооственно оооня-
телыюи ооласти.
Увеличение обонятельной поверхности приводит у рыб к образованию
многочисленных мелких радиальных складок, а у наземных позвоночных
развиваются оолее крупные складки, иногда очень сложной (рормы, под-
держиваемые скелетными выростами стенок ооонятельнои капсулы —•
оионятельными раковинами. Наибольшей сложности достигает система
М5
раковин у млекопитающих. Увеличение поверхности происходит, впрочем,
и в дыхательной области и также отчасти сопровождается образованием
складок, поддерживаемых раковинами, отчасти же осуществляется путем
разрастания носовой полости в соседних костях. Значение этого процесса
исчерпывается, повидимому, необходимостью предварительного увлажне-
ния и обогревания втягиваемого воздуха. В нижней части обонятельного
органа у наземных позвоночных обособляется добавочный, также обоня-
тельный, отдел, остающийся в связи с ротовой полостью, — якобсонов
орган. Наконец, у всех наземных позвоночных в стенках обонятельного
органа развиваются многочисленные железы, увлажняющие слизистую
оболочку.
ОРГАНЫ ПИЩЕВАРЕНИЯ
ОРГАНЫ ПИЩЕВАРЕНИЯ БЕСПОЗВОНОЧНЫХ
У всех животных главная часть пищеварительного тракта развивается из эмбрио-
нальной первичной кишки (archenteron), т. е. из энтодермы, к которой затем присо-
единяются более или менее значительные эктодермальные впячивания, образующие
начальный и конечный отделы кишечного канала. К энтодермальному и частью к экто-
дермальному эпителию пищеварительного тракта присоединяется, начиная с червей,
еще мезодермальный покров с мускулатурой и кровеносными сосудами. Ротовое от-
верстие помещается обычно впереди, на брюшной поверхности тела, и развивается
у большинства беспозвоночных (первичноротовые) из эмбрионального бластопора.
Заднепроходное отверстие лежит обычно на заднем конце тела и развивается заново
путем образования впячивания навстречу заднему слепому концу кишки и прорыва его
внутрь последней. У иглокожих и у хордовых за счет бластопора развивается, однако,
ваднепроходное отверстие, а ротовое отверстие образуется заново в виде эктодермаль-
ного впячивания, прорывающегося в передний конец кишки (вторичноротовые). В
некоторых случаях у членистоногих (Peripatus) наблюдалось развитие за счет вытя-
гивающегося в длину бластопора обоих наружных отверстий кишечника.
У кишечнополостных вся пищеварительная полость является собственно полостью
первичной кишки, хотя и у них к этому уже иногда присоединяется (у коралловых
полипов и ктенофор) эктодермальное впячивание, образующее глоточную трубку.
Пищеварительная полость является слепым мешком, ротовое отверстие которого
(соответствующее бластопору) служит в одно и то же время и для выбрасывания непе-
реваренных частиц. Пищеварительная полость у кишечнополостных нередко дает
радиальные выросты или продолжается в более сложную систему периферических
каналов, служащих, очевидно, для распределения питательных материалов по всему
телу. У низших червей (ресничных червей и сосальщиков) кишечный канал также не
имеет ваднепроходного Отверстия и продолжается во множество разветвлений, кончаю-
щихся слепо. Эктодермальная глотка сильно развита. Начиная с высших червей,
в пищеварительном тракте имеется, кроме переднего эктодермального отдела (stomo-
daeum), еще такой же задний отдел (proctodaeum). Таким образом развивается кишеч-
ник, состоящий из трех отделов — передней кишки, средней (mesenteron) и задней.
В пределах этой пищеварительной трубки происходит затем диференцировка,
которая особенно значительна у членистоногих и у головоногих молюсков. У насе-
комых, например, пищеварительный тракт состоит из пищевода, зоба, жевательного
желудка, железистого желудка или средней кишки, извитой задней кишки и прямой
кишки с ректальными железами. Ив всех этих отделов только средняя кишка является
энтодермальной, и, следовательно, эктодермальная часть здесь значительно превы-
шает первичную часть кишки. К этому нужно еще прибавить, что для измельчения
пищи у ротового отверстия развиваются сложно устроенные ротовые части (челюсти,
жвала и т. д.)., представляющие собой главным образом преобразованные конечности.
Очень высоко диференцирован также кишечный канал у головоногих моллюсков, у
которых он состоит ив глотки, пищевода, зоба, желудка и извитой кишки со значитель-
ным слепым отростком. У головоногих есть и приспособления для измельчения пищи —
клювообразные челюсти и зубчатка (radula) в глотке. Кроме того, у головоногих
имеются две настоящие пищеварительные железы, называемые печенью и поджелу-
дочной железой и изливающие свой секрет в основание слепой кишки.
В высшей степени своеобразна диференцировка кишечника у хордовых
животных. Прежде всего, в отличие от членистоногих почти весь кишечный канал
оказывается эктодермальным, между тем как эктодермой выстланы лишь небольшой
начальный и ничтожный конечный его отделы. Наиболее характерной особенностью
является, однако, Преобразование более или менее значительной передней части
кишечника н орган дыхания: стенки обширного у низших хордовых глоточного мешка
прорезаны вначительным числом жаберных отверстий, н стенках которых происходит
окисление крови ва счет кислорода, протекающего через эти отверстия наружу дыха-
тельного тока воды. В остальном кишечник у низших хордовых часто мало диференци-
306
роваи, но заслуживает упоминания особый слепой печеночный вырост, отходящий у
ланцетника от брюшной стенки начального отдела собственно кишки. Кишечник
низших позвоночных близко примыкает по строению к кишечнику низших хордовых
и отличается от диференцированных форм кишечника других беспозвоночных. Только
примитивная пищеварительная трубка, какая наблюдается, например, у кольчатых
червей, могла быть исходной Для раввития кишечника хордовых животных.
ОРГАНЫ ПИЩЕВАРЕНИЯ ПОЗВОНОЧНЫХ
Эпителий пищеварительного тракта является у позвоночных почти
на всем своем протяжении эктодермальным. Очень незначительное ротовое
впячивание приводит к образованию переднего
эктодермального отдела (stomodaeum), и совершенно
ничтожное впячивание на месте прежнего бластопора
дает начало заднему эктодермальному отделу (рго-
ctodaeum). Во взрослом состоянии границ между
этими отделами установить невозможно. Диференци-
ровка кишечного тракта начинается с образования
переднего более широкого отдела, от которого раз-
виваются парные карманообразные выросты, расту-
щие по направлению к эктодерме покровов (рис. 284),
срастающиеся с небольшими встречными ее впячи-
ваниями и, наконец, прорывающиеся наружу. Это—
жаберные мешки, обозначающие некоторый перед-
ний, дыхательный, отдел кишечника или г л о тку
(pharynx). Часть кишечника, лежащая впереди этого
отдела, представляет собой ротовую полость
(cavum oris). В задней части кишечника у наземных
позвоночных брюшная стенка выпячивается в виде
пузыря (мочевой пузырь амфибий), образующего
у амниот большой эмбриональный орган — аллан-
тоис, который частью дает затем начало мочевому
пузырю взрослого животного. В эту заднюю часть
кишки, получающую наименование клоаки, от-
крываются также выводящие протоки эмбриональ-
ных почек. У большинства позвоночных клоака
сохраняется и во взрослом состоянии; только у выс-
ших рыб происходит обособление мочеполовых путей
от задней части кишечника и у живородящих мле-
копитающих эмбриональная клоака спереди назад
разделяется на спинной отдел — прямую кишку, и
брюшной —мочеполовой синус. Часть кишки, лежа-
щая между жаберной областью и клоакой, являет-
ся уже специально пищеварительным органом. Он
диференцируется на несколько отделов, различаю-
щихся как морфологически, так и по своей функ-
ции. Передняя суженная часть выстлана, как и
лежащие впереди отделы, многослойным эпителием
и представляет собой пищевод (oesophagus).
Далее идет расширенный отдел, нередко изогнутый
в виде буквы U, с однослойным железистым эпите-
лием — желудок (gaster). За желудком следует
собственно кишка, выстланная также однослой-
ным железистым эпителием; кишка образует обык-
новенно более или менее значительные извивы и под-
Рис. 284. схема эм-
брионального кишеч-
ника и его произ-
водных.
dv —• желточный стебе-
лек; е— эпителиальные
тельца; д — желудок;
I —печень; р—легкие;
pbr —зажаберные тель-
ца; рп — поджелу-
дочная железа; 1т —
зачатки зобной железы;
tr — щитовидная желе-
за.
разделяется на «среднюю», или «тонкую», кишку
позвоночных и «з а д н ю ю», или «толстую», кишку, па границе между
которыми иногда развивается более или менее значительный слепой выступ.
307
пой выстилки, покрытой слоем
/1
Эпителий желудка и собственно кишки является однослойным цилий-'
дрическим. У личинки миноги, а также у Protopterus из двудышащих
вся кишка выстлана, как и у ланцетника, мерцательным эпителием. У ми-
ноги, у рыб и у амфибий в различных частях кишечника наблюдаются
остатки мерцательного покрова.
Стенки кишки составляются прежде всего из внутренней эпителиаль-
соединительпой ткани с кровеносными
сосудами. Это — слизистая обо-
лочка кишечника (mucosa). Слизи-
стая оболочка покрыта более рыхлой
подслизистой соединительной
тканью (submucosa) и затем мускуль-
ным слоем (muscularis), состоящим
обыкновенно из внутреннего слоя коль-
цевых волокон и наружного слоя про-'
дольных. Поперечнополосатая мускула-
тура наблюдается обычно только в на-
чальной части кишечника (ротовая по-
лость, глотка и частью пищевод) и
в конечной. Все остальные части пи-
щеварительного тракта имеют гладкую’
мускулатуру. Результатом сокраще-
ний этой мускулатуры являются ритми-
ческие движения стенок кишечника,
или перистальтика, помощью которой
происходит перемешивание содержимого
кишечника и его передвижение. Соеди-
нительная ткань и мускулатура ки-
шечника развиваются из висцерального
листка (splanchnopleura) боковой плас-
тинки. Из того же источника разви-
вается и наружный покров лежащей
в брюшной полости части пищеваритель-
ного тракта (часть пищевода, желудок
и кишка собственно), так называемая
серозная оболочка, предста-
вляющая собой не Что иное, как часть
перитонеальной выстилки брюшной по-
лости. Так как эта выстилка образуется
благодаря схождению по средней линии
висцеральных листков парных боковых
пластинок, то естественно, что кишеч-
ник оказывается укрепленным на двой-
ных листках перитонеума — брыжейках
(mesenterium). Брюшной мезентерий
большей частью исчезает, а спинной
сохраняется в виде брыжейки взрослого1
животного, подвешивающей кишечник и подводящей к нему сосуды и
нервы (разветвления блуждающего нерва и нервы симпатической системы).
В связи с пищеварительным трактом у позвоночных стоит довольно’
значительная система желез. В ротовой полости у наземных позвоночных
развиваются более или менее сложные слюнные железы (рис. 285),
В начальном отделе кишки собственно развивается очень крупная железа
в виде полого непарного выступа, лежащего в брюшном мезентерии, —
печень (рис. 284, 285).^Де менее постоянной является и другая круп-
ная железа, развивающаяся в этой же области кишки из нескольких за-
чатков, — поджелудочная железа. Иногда у рыб в начале
Рис. 285. Схема кишечника чело-
века (из Видерсгейма).
А — задний проход; Са, Cd, Ct — ча-
сти ободочной (толстой) кишки; Dd—
тонкая кишка; Gls — слюнные железы;
Glth — щитовидная железа; Gl. thy —
зобная железа; Lb — печень; Lg — лег-
кие; Мд — желудок; Ое — пищевод;
Ра — поджелудочная железа; Ph —
глотка; Р»— червеобразный отросток,
R — прямая кишка; Z — диафрагма.
308
топкой кишки имеются еще железистые слепые выросты — пилориче-
ские придатки, а в области задней кишки иногда наблюдается особая рек-
тальная железа.
Кроме того, в качестве слепого выступа стенок кишечника развива-
ется еще гидростатический аппарат большинства высших рыб — пла-
вательный пузырь. У кистеперых и двудышащих рыб, а также
у наземных позвоночных парный выступ стопок задней части глотки дает
начало органам воздушного дыхания—л о г к и м (рис. 284, 285).
Ротовая полость
Рис. 286.
ящерицы
Крыша и дно ротовой полост:t
(Lacerta aqilis) (по Гепперту).
ch — хоана; ql — язык; lx — гортань;
pal—складки вторичного и’ёба.
и крыловидными костями. Ену-
Рот позвоночных располагается па переднем конце тела, но бывает
более или менее сдвинут на брюшную сторону. В особенности у селахий
и у осетровых рыб с развитием рыла рот отодвигается по брюшной стенке
назад от переднего конца головы. Ротовая полость выстлана в передней
своей части эктодермой, но, как сказано, границы между пей и энтодер-
малыюй частью у взрослого животного указать невозможно. Эпителий
ротовой полости многослойный, частью иногда ороговевающий; у амфибий
он является мерцательным. У амфибий в слизистой оболочке ротовой по-
лости иногда имеется обильная сеть кровеносных сосудов; поверхность
слизистой оболочки увеличивается путем образования складок, которые,
таким образом, приобретают значение добавочного органа дыхания.
Ротовое отверстие позвоночных обычно ограничено кожными склад-
ками, или губами, которые, однако, обыкновенно являются неподвиж-
ными. Только у млекопитающих в губах развивается значительная мус-
кулатура (как продукт диференциров-
ки rri. sphincter colli), так что они
становятся мясистыми и подвижны-
ми. У форм, челюсти которых покры-
ты роговым покровом, образующим
клюв, как у черепах, птиц и клоач-
ных млекопитающих, губы но разви-
ваются. У млекопитающих разви-
вается заметная полость между гу-
бами и зубами — преддверие рта
(расширяющееся по бокам у многих
обезьян и грызунов в защечные
мешки).
Крыша ротовой полости у рыб
и у амфибий образуется основанием
самого черепа (первичное нёбо), ко-
торое по бокам дополняется нёбными
треннпе ноздри или хоаны, располагаются на этой крыше впереди.
Ротовая полость у амниот подразделяется развивающимся вторич-
ным нёбом па верхний дыхательный отдел, или носоглоточный ход (cavum
naso-pharyngeum), и нижний отдел, или вторичную ротовую полость (ca-
vum oro-pharyngeum). Начало этого процесса видно у ящериц, у которых
появляются впереди и по бокам горизонтальные складки, отходящие от
верхней челюсти (рис. 286). Эти складки твердого нёба частично прикры-
вают хоаны. При спокойном положении языка внутри ротовой полости
у ящериц и у птиц его спинка ложится как раз в промежутке между нёб-
ными складками и сполна отделяет носоглоточный ход от собственно ро-
товой полости. У крокодилов и у млекопитающих нёбные складки схо-
дятся между собой по сродней линии и образуют полную перегородку вто-
ричного нёба.
Твердое нёбо продолжается у млекопитающих в спадающую позади
двойную складку слизистой оболочки — мягкое нёбо, имеющее свою му-
309
скулатуру и отграничивающее ротовую полость от глотки. Мягкое нёбо
продолжается справа и слева в нёбноглоточные дуги (arcus palato-pharyn-
gei), а посредине от него у некоторых высших обезьян и у человека све-
шивается выступ — язычок.
На твердом нёбе млекопитающих имеются обычно плотные попереч-
ные валики, называемые нёбными пластинками. Эпителий
нёбных пластинок нередко обладает толстым роговым слоем, который
иногда значительно разрастается (роговые пластинки сирен и китовый
ус беззубых китов, образующий своеобразную цедилку). Повидимому,
нёбные пластинки помогают перетиранию пищи языком. У человека они
также закладываются и хорошо видны у новорожденных, но затем реду-
цируются.
В ротовой полости позвоночных развивается целый ряд органов спе-
циального значения: зубы, язык, ротовые железы.
Зубы
У позвоночных животных края рта, а нередко и часть внутренней
стенки ротовой полости снабжены зубами, обыкновенно конической формы,
служащими первоначально для удерживания во рту захваченной добычи,
а у высших позвоночных — нередко и для размельчения пищи.
Развитие зубов начинается с появления эктодермального утолщения,
вдающегося более или менее глубоко в соединительнотканную часть кожи.
Это утолщение у селахий, у рептилий и особенно у млекопитающих имеет
вид сплошного выроста, который носит название зубной пластин-
Рис. 287. Развитие зуба (с его
заместителем) у млекопитающего
(по Гертвигу).
ds — зубная пластинка; ео — эмале-
вый орган; р — мякоть зуба; pq —
зачаток постоянного зуба.
Рис. 288. Схема зуба низшего позвоночного (Л —
без корня) и млекопитающего (В — с корнем)
(по Шимкевичу).
1 — эмаль; 2 — дентни; 3 — полость зубной мякоти;
4 — кость (цемент).
В
к и (рис. 287, 289). В основной части последней происходит образование
зачатков отдельных зубов. Мезодермальные клетки образуют местные
сгущения — сосочки, вдающиеся более или менее глубоко в эктодермаль-
ное утолщение. Клетки основного слоя эпителия располагаются в виде
колпачка, покрывающего мезодермальный сосочек. Этот колпачок со-
стоит из цилиндрических клеток, образующих на своей внутренней по-
верхности твердое вещество — эмаль и поэтому носит название эма-
левого органа (рис. 287), Наружные клетки мезодермального
310
сосочка располагаются правильным слоем, напоминающим эпителий, и
образуют на своей наружной поверхности также очень твердое вещество —
дентин. Эти наружные клетки сосочка получают название одон-
тобластов. По мере увеличения массы дентина развивающийся
таким образом зуб постепенно принимает свою окончательную форму
и проталкивается наружу, прорезывая эпителий. Во вполне сформиро-
ванном зубе мы различаем очень твердый богатый известью поверхност-
ный слой эмали (substantia adamantine) (рис. 288), состоящий из
призматических волокон, расположенных вертикально к поверхности зуба,
и покрытый топким слоем органического вещества — зубной кутикулой.
Главная масса зуба состоит из дентина (substantia eburnea), обла-
дающего призматическим строением с многочисленными ветвистыми ка-
нальцами (рис. 288), в которые проникают отростки прилежащих к нему
в сосочке клеток (одонтобластов). Наконец, у млекопитающих основание
зуба покрывается еще снаружи слоем костного вещества, носящего здесь
название цемента. Зубной сосочек сохраняется внутри зуба в виде
зубной мякоти (пульпы), содержа-
щей нервы и сосуды, питающие
зуб-
Внутренняя часть дентина
содержит иногда каналы с крове-
носным сосудами (вазодентин) и
весьма сходна с костью, в кото-
рую он иногда и переходит у ос-
нования (рис. 288). Эмаль иногда
развита очень слабо (рыбы и реп-
тилии) или даже вовсе отсутствует
(некоторые рыбы и немногие мле-
копитающие).
По мере изнашивания зуба он
Рис. 289. Схематический разрез через
край челюсти акулы.
ds — зубная пластинка; d — зуб; р — зубная
мякоть; cs — плакоидиая чешуя.
расшатывается и выпадает, а ему
на смену приходят новые, кото-
рые у низших позвоночных раз-
виваются в течение всей жизни
(рис. 289). Такую неограниченную
смену зубов обозначают как поли
щих происходит ограничение числа
лишь две последовательные зубные
это — дифиодонтизм (рис. 287).
ф и о д о и т и з м. У млекопитаю-
смен зубов, так что наблюдаются
системы (молочная и постоянная),
Наконец, в некоторых случаях
у млекопитающих прорезывается лишь одно, никогда не сменяющееся
поколение зубов, что обозначается как м о п о ф и о д о п т и з м.
Форма зубов в простейшем случае является конической, и число зу-
бов может быть очень значительным. У селахий наблюдаются очень раз-
нообразные формы зубов — в виде гребней, трехзубцев и т. д.; в некото-
рых случаях они превращаются в плоские дробящие пластинки (особенно
у скатов). У других рыб и у низших наземных позвоночных имеются обык-
новенно простые конические зубы. Иногда в таких зубах наблюдается ра-
диальная складчатость дентина, придающая зубу более или менее сложное
внутреннее строение (Lepidosteus, многие ископаемые кистеперые ганоиды,
стегоцефалы и некоторые рептилии). У млекопитающих происходят ди-
ференцировка зубов, увеличение размеров и одновременно ограничение
их числа. Особенно сложную многобугорчатую или складчатую форму
приобретают задние зубы млекопитающих.
У селахий зубная система на наружных краях челюстей свя-
зана непрерывными переходами с общим покровом из плакоидных чешуй
(рис. 289). Это обстоятельство, так же как и большое сходство строения
и развития зубов ср строением и развитием плакоидной чешуи, доказы-
311
пяет нам, что зубы представляют собой не что иное, как расположенные
в ротовой полости плакопдные чептчи, которые соответственно требова-
ниям новой функции лишь выше диференцированы.
Зубы первоначально располагаются па различных частях стенок
потовой полости, хотя наибольшего развития достигают всегда на челюстях.
У некоторых акул почти вся слизистая оболочка рта и глотки покрыта
небольшими зубами (Chlamydoselachus, Galeus, Mustelus). У высших рыб
все кости ротовой полости могут быть снабжены зубами — они могут быть
на пёбе, на языке, а также и в глотке. Остатки такого общего зубного
покрова наблюдаются иногда и у амфибий, по у высших наземных позво-
ночных зубы имеются только на челюстях.
В некоторых случаях зубная система совсем редуцируется, и тогда
обыкновенно развивается функционально заменяющий ее роговой клюв
или другие образования (чепепахи, птицы, клоачные млекопитающие,
некоторые неполнозубые и беззубые кпты).
У круглоротых настоящих зубов пет гонге. Так пэзьтвэемые зубы, распо-
лагающиеся в ротовой воронке и ня языке, представляют собой бугорки ороговевшего
эпителия.
У р ы б имеются обыкновенно многочисленные мелкие зубы. У селахий они
сравнительно крупны и бывают ланцетовидными, вазубпепными или в виде плоских
пластинок (главным образом у скатов). У костистых рыб [в частности, у харяпинид
(Characinidae)] иногда тэвже'наблюдаются довольно крупные пифереппировапные вубы,
а в некоторых случаях происходит слияние нескольких зубов в крупные пластинки.
Зубы сипят в слизистой оболочке или прирастают к костям. У ганоидных и костистых
рыб зубы располагаются-)™ челюстях, а иногда и ня всех вообще костях ротовой по-
лости (па предчелюстных, челюстных, нёбпых. крыло-
Рис. 290. Поперечный раз-
рез через нижнюю челюсть
геккона. Зуб и его заме-
б стители (по Гегенбауру).
d — dentale; ds —вубная пла-
стинка; mk — меккелев хрящ;
р —• вубная мякоть.
видных, зубных и пластинчатых, а также на сошниках,
на парасфеноиде) и па языке. Сверх того, развиваются
зубы и в глотке, где они образуют так называемые
верхнеглоточные и нижнеглоточные вубы, сидяшие пт
верхних и иижпгх элементах жаберных дуг(яя Ph агу г-
роЪгапсЫаИя 2—4 и на cerafobranchiale 5-й дуги).
Глоточные вубы постигают иногда высокой степени
диференцировки, особенно v капповых рыб.
У двудышащих пыб вубы, закладывающиеся в виде
отдельных конических зубов, срастаются затем между
собой, образуя крупные зубные пластинки. Таким
Рис. 291. Схемы отношений зубов к челюстным костям
у плевродонтных (я), акродонтпых (Ь) и текодонт-
ных (с) рептилий (по Видерсгейму).
образом, получается пара нёбных зубных пластинок (соответствеппоГпёбпокрыловид-
ным костям) и пара нижнечелюстных на внутренней стенке челюсти (соответственно
пластинчатым костям). Кроме того, имеется пара1 зубов на сошниках. Зубные кости
у двудышащих рудиментарны и зубов не несут,’а челюстных костей у них нет вовсе.
У амфибий имеется много мелких конических зубов, сидящих на челюстях,
предчелюстных, челюстных, зубных и иногда на пластинчатых костях, на сошниках
и нередко на нёбных костях: в некоторых случаях вубы имеются и на покровной кли-
новидной кости (на парасфеноиде). У бесхвостых амфибий вубы редуцируются и
сохраняются обычно только в верхней челюсти и на сошниках. У некоторых жаб
зубов нет вовсе.
Зубы р е.птилий также обычно простой конической формы, сидят на челюстях,
но иногда имеются я на нёбных и крыловидных костях (у многих змей и ящериц),
а у гаттерий (Hatteria) в молодости имеется по одному зубу на сошниках. В огромном
312
большинстве случаев все аубы одинаковой конической формы (гомодонтная зубная
система), ио иногда наблюдается дифереппировна на аубы различной формы (гетсро-
доптная зубная система) — передние, сравнительно небольшие, напоминающие реппы,
следующие большие конические, похожие на клыки, и вадиие бугорчатые или иногда
трехзу^чатые. Такая диференпнровка намечается у некоторых современных яптерпп,
но в особенности она характеризует зубную систему ископаемых териодонтов (Theri-
ot! ontia).
Зубы рептилий прирастают или сбоку к внутренней поверхности челюстных
костей (рис. 290) — п левродоптиые зубы, или своим основанием к краю
челюстей—а кродон тине зубы, или, наконец,"опи не прирастают, а сипят в ячей-
ках, или альвеолах —-текодонтиые зубы, как это наблюдается у крокодилов
и некоторых ископаемых рептилий (Theriodontia и др.) (рпс. 291). V ядовитых вмей
произошла своеобразная специализация отдельных «ядовитых" вубов: иногда они
имеют желобок, проходящий вдоль передней стопки, — бороздчатые зубы, или желобок
изменяется замкнутым каналом, открывающимся у основания и па вершине зуба, —
трубчатые зубы. ГГо борозде или по каналу протекает яд, выделяемый ядовитой желе-
зой. Смена зубов у рептилий обычно постоянная и неограниченная (рпс. 290) (то же
самое относится и к ядовитым зубам змей). У черепах вубов пет вовсе, и челюсти одеты
роговым чехлом (эмбрионально зубы у черепах иногда еще закладываются) .
j У современных птиц зубная система также редуцирована и заменена роговым
клювом, но у археоптерикса, а также у меловых птиц имелись многочисленные кони-
ческие зубы, сидевшие у некоторых форм в отдельных ячейках (текодонтиые вубы).
У млекопитающих зубы сидят только в челюстях, в отдель-
ных’ячейках, и высоко диферешш рованы (гетеродонтная система)(рис.292).
Передние зубы р е з ц ы (incisivi) получили долотовпдвую форму и при-
способлены спецпалыго для гахгатыгапг.я и разрезания пгщп сле-
Рис. 292. Зубы собаки сбоку и снизу (по Видерсгейму).
с -— клыки; г — резцы; т — коренные; рт — ложноноренные зубы.
дующие за ними зубы сохранили свою коническую форму, но нередко имеют
значительные размеры — это клыки (canini), служащие для разры-
вания пищи; задние зубы, лежащие по бокам челюстей, приобретают более
сложную бугорчатую или складчатую форму и служат для перетирания
пищи — это коренные зубы, из которых передние называются ложно-
коренным и (praemolares), а задние — истинными коренными
(molares); последние имеют обыкновенно более сложную форму,чем первые.
С диференцировкой и прогрессивным развитием отдельных зубов общее
число их у млекопитающих уменьшается и уменьшается также число
смен. Число зубов у примитивных млекопитающих выражается следу-
ющей зубной формулой в которой зубы' одной стороны верхней
челюсти обозначены над чертой, а зубы нижней челюсти — под ней; пер-
вая цифра обозначает число резцов, вторая — клыки, третья — ложно-
коренные и последняя —- истипнокоренные зубы. Это исходное для боль-
575
шинства млекопитающих1 число зубов (44) в вышестоящих группах по-
степенно сокращается еще более и, например, у многих обезьян и у че-
ловека падает до 32, выражаясь формулой 2.1.2.3 для каждой челюсти.
Если в зубной системе частичная редукция приводит к образованию пе-
рерыва (diastema), лишенного зубов, то говорят об открытой зубной си-
стеме в отличие от замкнутой.
У китообразных наблюдается обратный процесс — возвращение всех
зубов к первоначальной простой конической форме, уменьшение их вели-
чины и одновременно значительное увеличение их числа (дельфины и
другие зубатые киты). Этот процесс дедиференцировки зубной системы,
развития вторично гомодонтной системы из гетеродонтной, прекрасно
иллюстрируется рядом ископаемых китообразных.
Число смен зубов у млекопитающих крайне ограничено, и обычно
имеются лишь два поколения зубов: молочное и постоянное,
причем сменные зубы развиваются на дне ячеек с внутренней стороны от
первых. Однако даже и этой однократной смене подлежат не все зубы:
истинные коренные не имеют молочных предшественников. Имеются
ясные Доказательства того, что дифиодонтизм млекопитающих развился
из полифиодонтизма нижестоящих позвоночных. У некоторых млеко-
питающих были обнаружены зачатки лишних поколений зубов, которые
не прорезываются. Зачатки, найденные кнаружи от молочных зубов,
естественно, представляют собой рудименты предмолочного поколения,
а зачатки, имеющиеся с внутренней стороны от постоянного поколения,—
недоразвитые остатки последующего, постдефинитивного поколения.
Зубы млекопитающих обладают обыкновенно, как и зубы других
позвоночных, ограниченным ростом; своеобразным, однако, является
формирование настоящего корня, т. е. сидящего в ячейке челюсти
суженного основания, иногда в виде нескольких ветвей, с тонким каналом
внутри, через который вглубь зубной мякоти проникают нервы и сосуды.
Корень отделен иногда ясным перехватом — шейкой — от верхней части,
коронки зуба, и покрыт костью — цементом, между тем как коронка
обычно покрыта эмалью (рис. 288). Образование корня связано всегда
с прекращением роста зуба. Иногда, однако, высокодиференцированные
и благодаря особенностям своей функции сильно стирающиеся зубы
обладают длительным ростом. В этом случае корня не образуется, и
зубной сосочек проникает в широко раскрытое основание зуба; эмалевый
орган получает форму кольца, одевающего растущее основание зуба.
Тачовы резцы грызунов, высокие призматические зубы копытных и т. п.
Процесс постепенного усложнения формы коренных зубов млекопи-
тающих в настоящее время с исчерпывающей полнотой прослежен на
ископаемых. Прежнее воззрение, что коренные зубы млекопитающих обра-
зовались путем слияния известного числа простых конических зубов,
должно было уступить место так называемой тритуберкулярной теории
палеонтологов Копа и Осборна. По этой теории сложная форма коренных
зубов млекопитающих развилась путем постепенного преобразования
простых конических зубов, как это иногда наблюдается и у рептилий
(особенно у ископаемых териодонтов) (рис. 293).
Первоначально имелся простой ряд конических зубов (гаплодонт-
н а я стадия), которые в верхней и нижней челюстях чередуются (как это
наблюдается и теперь у хищных и многих других млекопитающих). У осно-
вания конического зуба имелся кольцевой валик (cingulum), от которого
исходило образование добавочных вершин зуба. Сначала впереди и позади
главной вершины (на месте соприкосновения с зубом противоположной
1 У сумчатых исходная зубная формула несколько полнее и, повидимому, может
быть выражена следующим образом: 5 . 1 . 5 . 5 для каждой челюсти. У ископаемых
Pantotheria (юра) число истинных коренных зубов доходило ДО 7 (Amblotherium),'
314
челюсти) развились небольшие добавочные вершины (протодонт-
н а я форма). Эти добавочные вершины вырастали до величины главной,
и таким образом получился зуб с тремя одинаковыми зубцами в одной
плоскости (трехзубчатая, или триконодонтная форма). Средняя
вершина носит название протоконуса в верхней челюсти и протоконида —
в нижней; передняя добавочная вершина — параконус и параконид; зад-
няя — метаконус и метаконид. Добавочные вершины одной челюсти
входили в промежуток между главной и добавочной вершиной другой
челюсти. Затем, по Осборну, происходило смещение главной вершины
в одну сторону, а добавочных — в другую: в верхней челюсти средняя
Рис. 293. Образование основных форм коренных зубов млекопитающих. Схемы.
А — гаплодоптпая форма; В, С — протодонтпая; D—трехвубчатая форма; Е___трехбу-
горчатая форма; F — остробугорчатая форма; G — четырехбугорчатая форма; Н --- склад-
чатый и I — лунчатый зубы.
вершина (protoconus) перемещается внутрь, а передняя и задняя — кна-
ружи, между тем как в нижней челюсти средняя вершина (protoconid)
сдвигается кнаружи, а передняя и задняя — внутрь (рис. 293) J. Теперь
три вершины зуба располагаются по углам треугольника (трехбугор-
чатая, или тритуберкулярная форма). Такая форма зубов наблюдается
и у некоторых современных сумчатых, насекомоядных и полуобезьян.
При дальнейшем усложнении сначала на зубах нижней челюсти (которая
в этом процессе идет все время впереди верхней), а затем и на зубах верхней
развивается позади у основания коронки выступ — пятка (talonid в
нижней челюсти и talon в верхней) с бугорком на ней (hypoconid, hypo-
conus). Такие зубы встречаются у многих современных млекопитающих,
и эта форма лежит во всяком случае в основе различных типов зубов
насекомоядных и хищных. Если вершины такого зуба соединены между
собой острым режущим краем, то такой зуб получает название режу-
щего (векториальный или секодонтный зуб). Кроме того, различают
остробугорчатые и тупобугорчатые (бунодонтные)
формы зубов. Дальнейшее усложнение строения зуба выражается в том,
что между главными буграми развиваются различные добавочные бугорки.
Пижний бугор на пятке может достигнуть величины других трех
1 По некоторым новейшим данным, собственно перемещения вершин не было,
а трехбугорчатый тип зуба развился самостоятельно ив того же простого конического
(Грегори). Особым, развившимся независимо типом зубов является также многобу-
горчатый зуб некоторых мезозойских млекопитающих (Multituberculata).
315
бугров, спускающихся до уровня бугра пятки, и тогда мы получим ч е-
тырехбугорчатый зуб (квадритуберкулярный) с правильным
расположением бугров по углам четырехугольника (рис. 293). Другие
формы бунодонтных зубов развиваются в результате дальнейшего увели-
чения числа бугров.
Из того же четырехбугорчатого зуба выводятся, однако, и складча-
тые (лофодонтные) зубы. Передние и задние бугры соединяются попарно
между собой и образуют поперечные гребни (передний — proto-
loph и задний — metaloph); это — билофодонтные зубы, какие наблю-
даются, например, у тапира. Затем оба гребня соединяются между собой
еще гребнем вдоль наружной стенки зуба (ectoloph); такая форма зуба
со складками в виде буквы П наблюдается, например, у носорогов. Зуб
лошади представляет собой результат усложнения того же типа, как
это может быть прослежено палеонтологически (рис. 422). Из прими-
тивного билофодонтного типа через увеличение числа поперечных греб-
ней выводятся складчатые зубы слонов, грызунов и некоторых других
млекопитающих.
Из четырехбугорчатого типа выводятся и лунчатые зубы
жвачных через изменение формы бугров, которые вытягиваются и изги-
баются под углом (селенодонтные зубы) (рис. 293).
Наконец, следует еще указать, что при наиболее высокой диферен-
цировке зубы травоядных млекопитающих (подверженные значительному
стиранию) приобретают все более продолжительный рост, корни у них
не развиваются, и соответственно меняется форма зубов — короткие, бра-
хиодонтные зубы с ограниченным ростом преобразуются в высокие, ц и-
линдрические, гипселодонтные зубы с постоянным (точнее —
продолжительным) ростом, причем бугорки вытягиваются в длинные
призмы, а в промежутках между призмами откладывается цемент. Палеон-
тология дает нам прекрасные ряды форм, иллюстрирующие этот процесс.
Специальная морфология зубной системы различных отрядов млекопитающих
изучается более подробно в курсах зоологии позвоночных; здесь мы можем ограни-
читься лишь некоторыми замечаниями.
У клоачных млекопитающих зубы редуцировались и функционально
замещены роговым клювом.
У утконоса н молодости развиваются, однако, еще зубы многобугорчатого типа,
которые потом выпадают.
У сумчатых исчезла смена зубов (меняется лишь опин ложнокоренной).
Характерна диференцировка зубов: у травоядных (Diprotodontia) преобладающее
значение получают два средних резца, особенно сильно развивающиеся в нижней
челюсти, где другие резцы исчезают: у хищных (Polyprotodontia) имеется до пяти
мелких резцов с каждой стороны в каждой челюсти.
У насекомоядных имеется очень примитивная зубная система с остро-
бугорчатыми коренными, которые иногда являются еще трехбугорчатыми (Centetes,
Chrysochlorip) или даже трехзубчатыми (Hemicentetes). У рукокрылых зубная
система также очень примитивна, и коренные зубы близки к трехбугорчатому типу.
Зубы не сменяются (монофиодонтизм).
У неполнозубых зубная система редуцируется — эмаль на зубах не
развивается, и обыкновенно совсем исчезают резцы и клыки, а иногда (у панголина и
у муравьеда) зубов нет вовсе.
У грызунов развивается лишь пара средних резцов (небольшой второй
резец имеется в верхней челюсти заячьих), которые получают крупные размеры и
обладают постоянным ростом, причем эмалевый покров развивается только спереди.
Между резцами и коренными имеется широкая пиастема. Коренные имеют складчатое
строение, и число их прогрессивно сокращается. У некоторых грызунов зубная формула
10 0 2
падает до ~~ (Hydromys).
Для хищных характерно значительное развитие клыков и одного режущего
коренного зуба (последний ложнокоренпой в верхней челюсти и передний истинно-
коренной — в нижней), который получает название плотоядного, или хищного (рис.
292). У высших форм вубы, лежащие позади плотоядного, редуцируются.
У ластоногих остробугорчатые коренные вубы испытывают регрессивное
развитие, часто вторично возвращаются к примитивному трехзубчатому типу и теряют
смеяу.
316
Зубы к и т о о 6 р а а й ы х увеличиваются в числе и возвращаются и простой
конической форме (причем эмаль иечеваст). Смены зубов нет. У высших форм иеззубых
китов вубы совершенно редуцируются (хотя у зародышей еще закладываются).
У копыт н ы х сильно развиваются коренные зуоы> ирисиосоолеииые к пере-
тиранию растительной пищи. Многобугорчатые коренные наолюдаются у низших
всеядных парнокопытных. Лунчатые вуоы характерны для высших .парнокопытных,
у которых ооыкповенно редуцируются верхние резцы. У непарнокопытных коренные
вубы лофодонтного типа. У лошадей зубы лунчатоскладчатого типа и достигают на-
ибольшей сложности.
У хоботных пара верхних резцов образует огромные бивни с постоянным
ростом (у ископаемых слонов иногда и в нижней челюсти). Коренные зубы складчатые
(прекрасный палеонтологический ряд иллюстрирует их происхождение из бугорчатых
вубов и увеличение числа поперечных складок от 2 до 1о у африканского слона или
даже до 27 у мамонта и у Индийского слона) (рис. 426), причем одновременно в каждой
челюсти функционирует лишь по одному оольшому вуоу с каждой стороны. Всего
развивается 6 коренных вубов, которые прорезываются последовательно спереди назад,
постепенно выдвигаются вперед и сменяют друг друга (горивоатальван смена).
У сирен ревцы или редуцируются, или пара их (у дюгоня) превращена в
большие бивни с постоянным ростом. Клыки и ложнокорепьые редуцируются. Корен-
ные вубы в увеличенном числе, или с постоянным ростом (дюгонь), или с горизонталь-
ной сменой (ламантин).
У полуобезьян и у о бег ь ян вубная система мало дпфереяцировапа
и число вубов сокращается, доходя у узконосых обезьян до человеческой формулы:
2.1.2.3b каждой челюсти. Коренные зубы верхние •— четырсхоугорчаты, нижние —
пятибугорчаты.
Зубы позвоночных, возникшие как чисто кожные защитные образо-
вания (плакоидная чешуя), которые, между прочим, имелись п по краям
рта, приобрели более значительные размеры на челюстях, где они играют
роль вспомогательных органов схватывания и удерживания дооычи. Уже
у примитивных позвоночных нередко их форма меняется в связи с измене-
нием характера пищи; примерами могут служить режущие ланцетовидные
зубы хищных акул и плоские дробящие зубы скатов и некоторых акул,
питающихся моллюсками. Обычно, однако, зубы имеют простую кони-
ческую форму и сидят первоначально на всех костях ротовом полости,
а затем ограничиваются одними челюстями. Такие зубы имеют неограни-
ченное число смен. У млекопитающих зубы испытывают значительную
диференцировку, так что развиваются, по крайней мере, три основных
типа зубов, различающихся и по форме и по функции: резцы, клыки и
коренные зубы. Зубы сидят в отдельных ячейках, основание их сужается,
образуя корень. Отдельные зубы увеличиваются в размерах, коренные
зубы получают очень сложную форму, но число зубов значительно сокра-
щается, и одновременно падает число смен, так что развивается не более
двух поколений зубов, из которых одно только второе — постоянное —
характеризует взрослое животное. С увеличением продол?кительности
функционирования таких зубов, особенно при значительном их стирании,
и продолжительность роста увеличивается, так что иногда зубы растут
в течение всей жизни. Эта эволюция зубной системы находится, очевидно,
в связи с приобретением способности предварительно разжевывать пищу,
что в свою очередь привело к быстрому и совершенному ее усва-
иванию, характеризующему энергичный обмен веществ и более интен-
сивную жизнедеятельность, как она наблюдается у млекопитающих.
Можно сказать, что в эволюции последних вместе с приобретением воло-
сяного покрова и живородности, с развитием коры большого мозга далеко
не последнюю роль играла и диференцировка зубной системы.
Язык
На дне ротовой полости у позвоночных имеется непарный выступ —*
язык (lingua), который в наиболее простом виде, у рыб, является складкой
слизистой оболочки, поддерживаемой передним непарным элементом вис-
церального скелета (basihyale). У рыб в языке пет своей мускулатуры,
и он движется лишь вместе с вентральной частью висцерального аппа-
317
Рис. 294. Язык ящерицы—
варана (Monitor) (по Видерс-
гейму).
Ь — вход в гортань; М —
нижняя челюсть; Z — язык;
Zs — влагалище языка.
рата.У наземных позвоночных язык приобрел згачение подвижного органа,
служащего не только для удерживания и перемещения пищи в ротовой
полости, но и для ее захватывания, испытания, измельчения, глотания,
а также для питья и для обособления и замыкания дыхательных путей.
Строение языка амфибий в высшей степени разнообразно. В неко-
торых случаях он представляет собой орган, построенный лишь немногим
выше, чем у рыб (повидимому, здесь, однако, имеется вторичное упро-
щение); в большинстве же случаев это гораздо более совершенный орган,
имеющий, повидимому, и более сложный состав — во всяком случав
в дополнение к складке, покрывающей переднюю часть подъязычного
аппарата (язык рыб), впереди от последнего на дне ротовой полости
развивается еще особое железистое поле, которое затем входит в состав
языка. В языке наземных позвоночных, начиная с амфибий, появляется
собственная мускулатура, являющаяся продук-
том обособления и диференцировки подъязыч-
ной мускулатуры. Мышца, выдвигающая язык
вперед [подбородочноязычная (m. genioglossus)],
и мышца, втягивающая язык обратно [подъ-
язычноязычная (m. hyoglossus)], обособились
от подбородочноподъязычной (geniohyoideus)
и от грудиноподъязычной мышцы (m. sterno-
hyoideus). От этих же мышц, повидимому, обо-
собилась и мускульная масса самого языка.
Соответственно своему происхождению муску-
латура языка иннервируется подъязычным
нервом.
Вкусовые органы языка иннервируются
у наземных позвоночных языкоглоточным
первом.
У амфибий передняя часть языка изобилует
железами, выделяющими клейкую слизь, по-
могающую при ловле насекомых и другой
добычи. В некоторых случаях язык может при
этом выбрасываться далеко вперед. У бесхвос-
тых амфибий язык обыкновенно прикреплен
к^краю нижней челюсти, а свободный конец
откидывается назад к глотке; при ловле до-
бычи этот задний конец выбрасывается вперед.
У амфибий, живущих постоянно в воде, язык
редуцируется (постоянножаберные хвостатые—
Perennibranchiata и безъязычные бесхвостые—
Aglossa).
Язык высших позвоночных представляет собой, повидимому, морфо-
логически более сложное образование, — если опираться на данные
истории развития. Он развивается у амниот из непарного зачатка, лежа-
щего в области второй и третьей жаберных дуг, к которому присоеди-
няется затем еще пара валиков, лежащих по бокам, впереди от подъязыч-
ной дуги. Между этими зачатками располагается закладка щитовидной
железы. Задний зачаток дает начало корню языка, передние, парные
зачатки впоследствии сливаются между собой и дают начало телу языка.
В иннервации языка у рептилий принимает участие, кроме двух
ранее названных нервов, еще тройничный нерв. Форма и степень подвиж-
ности языка у рептилий в высшей степени различны. У черепах
и крокодилов он мало подвижен, а у большинства ящериц и у змей он
имеет удлиненную форму, на конце обыкновенно глубоко раздвоен и
отличается чрезвычайной подвижностью и чувствительностью (рис. 294).
В этом случае в основании языка имеется кольцевая складка, образующая
318
влагалище языка. Подъязычный аппарат образует скелет языка,
причем передний непарный его элемент (basihyale) вытягивается в длин-
ный отросток, поддерживающий переднюю часть языка (os entoglossum).
Обе пары рожков подъязычного аппарата служат местом прикрепления
мышц, движущих языком. Особенно длинный червеобразный язык имеется
у хамелеонов, у которых он на конце снабжен железистым расширением.
У птиц обычно тонкий и малоподвижный
язык лишен внутренней мускулатуры, заострен
впереди и отличается сильным развитием рогового
слоя, который в задней части языка снабжен рого-
выми сосочками. У дятлов и у колибри очень длинный
язык далеко выдвигается при помощи подъязычного
аппарата, задние рожки которого огибают сзади
череп и ложатся на нем сверху, доходя до ноздрей.
У попугаев широкий мясистый язык весьма подвижен.
Язык млекопитающих развивается в об-
щем сходно с языком рептилий. Собственная мус-
кулатура достигает наибольшего развития, так что
язык оказывается необычайно подвижным нанесет
разнообразные функции, в особенности выполняет
важную роль при акте глотания, а также в ка-
честве вкусового органа (рис. 295). Язык млекопи-
тающих чрезвычайно богат железами и снабжен
сосочками различной формы и значения (см.
описание органов вкуса), частью чувствующими (со
вкусовыми почками), частью ороговевшими. Вообще
ороговение языка иногда бывает довольно значитель-
ным, как, например, у хищников. Очень длинный
и клейкий язык наблюдается у муравьедов.
У некоторых сумчатых, полуобезьян и обезьян
Рис. 295. Язык сум-
чатого млекопитаю-
щего (Phascolomys
wombat) стремя валь-
коватыми сосочками
(по Бючли).
— гортань; р. v. —
papilla vallata.
под языком имеется выступ (sublingua), который Гегенбаур сравнивал
с языком нижестоящих форм, полагая, что собственно язык млекопитаю-
щих представляет собой новообразование. Этот взгляд не получил до-
статочного подтверждения.
Ротовые железы
У рыб и у водных амфибий нет сложных ротовых желез. Последние
представляют собой приобретение наземных позвоночных, у которых они
служат первоначально для смачивания слизистой оболочки ротовой
полости и в особенности для смачивания пищи, но впоследствии, у мле-
копитающих, выделяемая слюна содержит не только слизь, но и серозную
жидкость, в состав которой входят, между прочим, и пищеварительные
ферменты, расщепляющие углеводы: птиалин, превращающий крахмал
в мальтозу, и фермент мальтаза, превращающий мальтозу в глюкозу.
У наземных амфибий имеется обыкновенно непарная межно-
совая железа (gl. internasalis s. intermaxillaris) трубчатого строения,
выделяющая слизистый секрет, и нёбные железы. У рептилий, кроме
этих желез, имеются еще особые подъязычные железы (gl. sublinguales),
губные железы (верхние — gl. labiales superiores и нижние — gl. labiales
inferiores) и зубные. У ядовитых змей именно пара задних верхнезубных
желез преобразована в сложную трубчатую ядоотделительную железу.
Выводные протоки этих желез находятся в связи с ядовитыми зубами.
Собственной мускулатурой ядоотделительные железы не обладают, но
они прикрыты частью жевательной мышцы (m. masseter), при сокращении
которой содержимое железы выдавливается и изливается по протоку,
к борозде или каналу ядовитого зуба.
319
Птицы обладают железами на нёбе и подъязычными железами,
которые особенно развиты у зерноядных птиц.
У млекопитающих ротовые железы достигают наивысшего
развития. Кроме многочисленных мелких слизистых желез, как губные
железы (gl. labiates), щечные (gl. buccales), нёбные (gl. palatinae) и языч-
ные (gl. Imguales), вполне, повидимому, сравнимых с одноименными
железами амфибий и рептилий, имеется еще несколько крупных так
называемых слюнных желез. Это: 1) подъязычная железа
(gl. sublingualis) с несколькими протоками (иногда один бартолиниев
проток), которая иногда является еще чисто слизистой, 2) з а д н е я з ы ч-
н а я (gl. retrolmgualis), 3) подчелюстная (gl. submaxillaris)
железа, частью слизистая и частью серо'зная, с протоком (вартоновым),
открывающимся у нижних резцов, и 4) околоушная (gl. parotis) —
серозная железа с протоком (стеноновым), открывающимся у коренных
зубов верхней челюсти. Из этих желез первые три представляют собой,
повидимому, результат диференцировки подъязычной железы рептилий,
а околоушная железа — новое приобретение млекопитающих, развив-
щееся из щечных желез.
Кроме того, у различных млекопитающих встречаются и некоторые
другие ротовые железы, как молярные железы копытных и глазничные
железы собак.
В связи с ротовой полостью у наземных позвоночных наблюдаются
и различные лимфоидные органы. У амфибий, рептилий и птиц
имеются отдельные лимфатические узелки в различных местах стенки
ротовой полости и в особенности позади хоан. У млекопитающих имеются
крупные лимфоидные образования в различных местах в области зева —
миндалины (tonsillae). НёбнЫе миндалины (tonsillae palatinae)
лежат у человека и у антропоидов между нёбно-глоточными и нёбно-
язычными дугами, по бокам зева. У других млекопитающих они лежат
в мягком нёбе.
Глотка
Участок пищеварительного тракта, расположенный между ротовой
полостью и пищеводом, носит наименование глотки (pharynx). Стенки
ее выстланы многослойным эпителием и снабжены обильной поперечно-
полосатой мускулатурой. В ее области развивается весь жаберный аппа-
рат, а также важные органы внутренней секреции — щитовидная железа,
эпителиальные тельца и зобная железа (рис. 284).
У рыб мы, следовательно, относим к глотке область жаберных щелей
(см. описание органов дыхания). У наземных позвоночных в области
глотки, так же как иногда и в ротовой полости, происходит соединение
двух путей — пищеварительного и дыхательного, которые позади разде-
ляются. Кроме гортани, открывающейся в задний конец глотки, впереди
в нее открываются еще отверстия евстахиевых труб, а у высших амниот
обыкновенно и носоглоточные ходы.
Пищевод
Пищевод — отдел кишечной трубки, следующий непосредственно за
глоткой и служащий уже исключительно для проведения пищи. От сле-
дующего за ним отдела он резко не отграничен (только у млекопитающих
устанавливается внешняя граница), но в общем отличается от него внут-
ренней выстилкой многослойным эпителием, подобно глотке и ротовой
полости, а отчасти и положением вне брюшной полости и, следовательно,
отсутствием серозной оболочки, что особенно характерно для высших
позвоночных. В его стенках пищеварительных желез не имеется, и внут-
ренний слой эпителия может даже ороговевать. Длина пищевода бывает
320
Оё—
Рис. 296. Перед-
няя кишка птицы
(по Видерсгейму).
DM — железистый
желудок; Ig — зоб;
М D — средняя ки-
шка; ММ — мус-
кульный желудок;
Ое — пищевод.
различной, и у наземных Позвоночных зависит от длины шеи, достигай
очень большой величины у птиц. Внутренняя стенка образует обыкно-
венно многочисленные продольные складки и часто
обильно снабжена слизистыми железами. За слизистой
оболочкой следует мускульный слой, который в перед-
ней части состоит из поперечнополосатых волокон,
в некоторых случаях образующих всю мускулатуру
пищевода.
У большинства птиц пищевод образует заметное
расширение — зоб, обильно снабженный железами,
секрет которых служит для предварительного размачи-
вания пищи, а иногда имеет и пищеварительное зна-
чение (рис. 296). У голубей во время насиживания
эпителий зоба утолщается, клетки его испытывают
затем частью жировое перерождение и образуют таким
образом жидкое выделение, служащее для вскармлива-
ния птенцов.
Желудок
Желудок представляет собой обычно расширенный
отдел кишечника, который наружно вполне ясно от-
граничен от пищевода только у млекопитающих, но и
в этом последнем случае граница оказывается до извест-
ной степени условной, так как физиологическая гра-
ница между этими отделами может не совпадать с на-
ружно видимой.
Физиологически желудок характеризуется как орган, в котором
проходит первая фаза пищеварения в кислой среде. Стенка его выстлана
слизистой оболочкой с сетеобразным расположением складок. Она обла-
дает однослойным цилиндрическим эпителием и снабжена специальными
трубчатыми пищеварительными железами. С этой точки зрения у некото-
рых низших позвоночных, как и у ланцетника, желудка нет вовсе (кругло-
ротые, цельноголовые, двудышащие и некоторые костистые рыбы, в част-
ности, карповые). У других рыб, так же как и у наземных позвоночных,
имеется и наружно более или менее ясно различимый желудок. Он иногда
вытянут в длину или изогнут в виде буквы U, так что оба его колена
ложатся параллельно. В общем форма его находится в тесной связи с
общей формой тела животного: он вытянут в длину у животных с удли-
ненным телом, ложится поперек и приобретает мешковидную форму
у животных с более коротким телом. Передняя стенка желудка в послед-
нем случае является вогнутой и образует так называемую малую кри-
визну (curvatura minor), а задняя, выпуклая стенка образует большую
кривизну (curvatura major). Часть желудка, ближайшая к пищеводу,
называется кардиальной (лежащая у сердца), часть, прилежащая к собст-
венно кишке — пилорической. Последняя отграничена от кишки муску-
листой кольцевой складкой слизистой оболочки (пилорический клапан).
У рыб желудок обыкновенно неясно отграничен, хотя иногда
принимает форму довольно объемистого слепого позади мешка.
У амфибий и рептилий желудок обособляется гораздо
яснее (рис. 299), особенно у крокодилов, у которых и в пределах самого
желудка наблюдается дальнейшая диференцировка: за большим мешко-
видным железистым отделом со значительной мускулатурой следует
ясно обособленная пилорическая часть.
У птиц подобная диференцировка приводит к полному обособле-
нию двух отделов — переднего (кардиального) едва вздутого железистого
321
желудка и отделенного от него ясной перетяжкой мускульного желудка
(ри. 296). Чрезвычайно толстые мышечные стенки последнего имеют вид
двух дисков с сухожильными центрами, от которых отходят радиальные
волокна. Внутренняя стенка мускульного желудка снабжена железами,
выделяющими твердую рогоподобную внутреннюю выстилку, имеющую
иногда бугорчатую поверхность. Эта твердая оболочка вместе с заглаты-
ваемыми камешками служит для перитирания пищи (что чезвычайно
важно при отсутствии зубов у .птиц). Наибольшего развития достигает
мускульный желудок у зерноядных птиц. Иногда позади мускульного
желудка имеется небольшой пилорический желудок.
Желудок млекопитающих характеризуется значительным
развитием и диференцировкой желез. Отличают разнящиеся несколько
по своему гистологическому строению й йо характеру ^Лекрета кардиаль-
Рис. 297. Схемы желудка человека (Л), собаки (В), крысы (С), хомяка (Р), лошади
(Е), жвачного (F) с обозначением железистых областей (по Оппелю из Гессе).
ое — пищевод. Многослойный эпителий пищеводного характера обозначен горизонтальными
штрихами, область кардиальных желез — косыми, область дойных — пунктиром, область
пилорических — крестиками.
ные железы, донные и пилорические. Из них последние наиболее харак-
терны. Кардиальные железы выделяют слизистый, щелочной секрет;
донные железы — кислый. В донных железах различают железистые
клетки двух типов: кубические, главные клетки, выделяющие профер-
менты — пепсиноген и прохимозин, и большие округлые, покровные
клетки, рассеянные среди главных и выделяющие соляную кислоту.
Кроме того, донные железы выделяют еще один фермент — липазу. Пило-
рические железы выделяют также пепсиноген и прохимозин.
Несмотря на довольно резкое наружное обособление желудка от
пищевода, во внутреннем строении мы нередко наблюдаем совсем иные
границы между отделами. Так, например, у клоачных млекопитающих
он выстлан таким же плоским многослойным эпителием, как и пищевод.
У других млекопитающих нередко ближайшая к пищеводу часть желудка
выстлана многослойным эпителием, и, таким образом, в желудке млеко-
питающих можно различать пищеводную область и область собственно
желудочную, которая в свою очередь распадается на участки, характе-
ризующиеся специфическими для них железами, — кардиальную часть,
дно и пилорическую область (рис. 297).
Области желудка могут быть отделены друг от друга перетяжками,
и тогда желудок принимает сложную форму. Иногда обособляется пище-
водный отдел (у хомяка) (рис. 297, D), в других случаях обособляются
различные другие области желудка (область дна и пилорическая область
у китообразных). Наибольшей сложности достигает желудок у жвачных
(рис. 297, F), у которых он, как известно, поделен на четыре отдела:
322
объемиСткй р у б е ц с многбчисленными прочными сосочками, Сетка
с ячеистой внутренней поверхностью, к н и ж к а с листовидными продоль-
ными складками и, наконец, удлиненный сычуг с более низкими про-
дольными складками. Первые три отдела выстланы многослойным эпите-
лием, и только сычуг представляет собой настоящий желудок железистого
характера (с донными и пилорическими железами) х. У верблюдов желудок
построен несколько проще, но и своеобразно: рубец имеет особые выступы
в стенке — водяные клетки, снабженные цилиндрическим эпителием и
кардиальными железами. В сетке также имеются водяные клетки и кар-
диальные железы. Книжка от сычуга не обособлена. В водяных клетках
у верблюдов надолго задерживаются запасы воды.
Кишка собственно
Кишка собственно вся выстлана слизистой оболочкой с цилиндри-
ческим однослойным эпителием, содержащим, кроме слизистых бокало-
видных клеток, также нередко и трубчатые пищеварительные железы.
В этом отделе пищеварительного тракта происходит, так же как и в же-
лудке, химическая обработка пищи (но в щелочной среде) при посредстве
пищеварительных ферментов, выделяемых железами, т.е. переваривание
пищи, и, кроме того, всасывание веществ, переведенных в растворенное
состояние. Впрочем, в некоторых случаях в кишечном эпителии позво-
ночных наблюдается и внутриклеточное пищеварение (у селахий и у
амфибий), причем клетки вы-
пускают амебоидные отрост-
ки и захватывают пищевые
частицы внутрь.
Значение поверхности
кишечного эпителия при про-
цессах всасывания делает по-
нятным существование раз-
личных приспособлений к рис. 298. Спиральный клапан ската (Raja)
удлинению пути, проходимо- (по Майеру из Кингсли).
го пищей, и к увеличению
всасывающей поверхности путем образования складок и ворсинок.
У низших позвоночных это достигается образованием одной большой
складки, идущей по спирали вдоль значительной части кишки и назы-
ваемой спиральным клапаном. У миног такая складка невысока и тянется
вдоль всей кишки, образуя едва заметный спиральный изгиб. У селахий
складка глубоко врезывается в просвет кишки и образует более или
менее крутые обороты, напоминая витую лестницу (рис. 298). Внутренние
края клапана изгибаются обыкновенно так, что обороты его более или
менее охватывают друг друга, приобретая вид вложенных друг в друга
конусов. В некоторых случаях обороты клапана очень отлоги, а свободный
край его сильно закручивается на себе самом. У ганоидов и у двудышащих
рыб клапан развит в общем слабее и у костных ганоидов (Lepidosteus)
может быть назван рудиментарным. У костистых рыб спирального кла-
пана нет (исключение — Chirocentrus). При существовании спирального
клапана кишка имеет сравнительно незначительную длину, но большой
диаметр; пища проходит по такой кишке спиральный путь вдоль клапана.
У высших рыб, а также у наземных позвоночных вместо этого наблю-
дается удлинение самой кишки, которая при этом образует более или
1 По своему эмбриональному развитию, однако, все отделы сложного желудка
вместе гомологичны простому желудку и представляют, следовательно, результат
диференцировки последнего. Железы исчезают в передних «пищеводных» отделах
вторично (по исследованиям Герке).
323
м
—А
—а
РиС. 299. Кишечник лягуш-
ки (по Видерсгейму).
А — впадение прямой кишки
в клоаку (Cl); В — средняя
кишка; Du — начало средней
кишки; НВ—мочевой пузырь;
М — гкелудок; М z — селезен-
ка; Ое — пищевод; Ру —
привратник; R — толстая
кишка.
менее значительные извивы (рис. 299). У различных позвоночных длина
кишки весьма различна, и это находится нередко в очевидной связи с
характером пищи; как общее правило, можно принять, что у травоядных
форм эта длина более значительна.
Увеличение всасывающей поверхности кишки достигается, кроме
того, еще и образованием складок и ворсинок. У низших рыб наблю-
даются обыкновенно продольные складки слизистой оболочки, обеспе-
чивающие главным образом возможность растяжения кишки. У высших
рыб, а также у амфибий и у рептилий развивается неправильная сеть
многочисленных перекладин, придающих внутренней выстилке кишки
иногда ячеистое строение. Края перекладин
нередко оказываются неровными, зазубренны-
ми, и если зазубрины глубоко врезываются,
то таким образом развиваются неправильные
ряды многочисленных, более или менее высо-
1 ких сосочков. В этом можно усмотреть первые
признаки появления кишечных ворсинок,
которые массами покрывают всю внутреннюю
поверхность слизистой оболочки кишки птиц 1
и млекопитающих и придают ей бархатистый
вид.
Кишка позвоночных обыкновенно более
или менее ясно подразделяется еще на отделы
(рис 299). Передняя ее часть, в которую впа-
дают протоки печени и поджелудочной железы,
обособляется у высших позвоночных под на-
званием двенадцатиперстной к и ш-
к и (duodenum), в которой наблюдаются же-
лезы (бруннеров ы), близкие к пилори-
ческим железам желудка. Обыкновенно и у
низших позвоночных различают, кроме главной
части, «средней» кишки, содержащей у на-
земных позвоночных многочисленные либер-
кюновы железы, еще заднюю, или
толстую, кишку, которая у некоторых ганои-
дов и костистых рыб, а в особенности у назем-
ных позвоночных отграничена кольцевой склад-
кой (valvula ileo-colica). У высших позвоноч-
ных на границе между тонкой и толстой
кишкой имеется обыкновенно более или менее
значительный слепой вырост (coecum).
Толстая кишка, вообще довольно короткая, до-
стигает у млекопитающих более значительной
очередь подразделяться на собственно толстую
Внутренняя поверхность толстой кишки
длины и может в свою
(colon) и прямую (rectum).
обыкновенно гладкая, и задняя ее часть получает название клоаки
(cloaca), если в нее открываются протоки почек и половых желез. Брюш-
ная стенка клоаки у амфибий образует мешковидный вырост — мочевой
пузырь, который у зародышей высших наземных позвоночных приобретает
большое значение в качестве аллантоиса, служащего не только резерву-
аром для мочи, но и эмбриональным органом дыхания. У рептилий, птиц
и млекопитающих он сильно разрастается, но к концу
жизни большая его часть дегенерирует и лишь очень
часть дает мочевой пузырь взрослого животного.
эмбриональной
небольшая его
1 У птиц кишечные ворсинки развиты особенно сильно, что, очевидно, связано
с относительно малой длиной кишки.
324
На протяжении кишки у различных позвоночных наблюдаются раз-
личного рода железистые выросты. В переднюю часть средней кишки у
всех позвоночных впадают протоки двух крупных пищеварительных
желез — печени и поджелудочной железы. В этой же области у высших
рыб имеется нередко большее или меньшее число (от одного до двухсот
и более) слепых выростов, называемых пилорическими придатками (ap-
pendices pylorici) и служащих, повидимому, только для увеличения вса-
сывающей поверхности. У селахий в передней части задней кишки име-
ется пальцеобразный железистый придаток — ректальная железа, функ-
ция которой неизвестна.
В результате деятельности всех этих желез кишечный сок содержит
слизь щелочной реакции со многими пищеварительными ферментами (у
млекопитающих различают эрепсин, затем липазу, амилазу, инвертазу,
мальтазу, лактазу и, кроме того, энтерокиназу, которая активирует трип-
синоген поджелудочной железы, превращая его в трипсин).
Кишечник круглоротых имеет вид недиференцированной, почти прямой
трубки со слабо развитым спиральным клапаном (у миног).
Кишечник низших рыб (селахий, ганоидов и двудышащих) имеет нередко
еще вид почти примой трубки, во в этом случае он снабжен сильно развитым спираль-
ным клапаном (рис. 298), который у высших форм редуцируется. С редукцией клапана
сама кишка удлиняется и образует многочисленные извивы. Внутренняя стенка кишки
снабжена системой складок, на которых наблюдается иногда образование сосочко в.
У хрящевых рыб и у двудышащих задняя часть кишки образует клоаку, которая у
ганоидов и костистых рыб исчезла.
В кишечнике амфибий наблюдается ясное деление на среднюю кишку и
короткую и широкую заднюю. Всегда имеется клоака, с передней стенкой которой
связан мочевой пузырь (рис. 299).
У рептилий задняя кишка отграничена от средней складкой слизистой
оболочки (клапан) и, кроме того, отмечена небольшим слепым выступом (за исклю-
чением крокодилов). Всегда имеется клоака, подразделенная у крокодилов кольцевой
складкой на два отдела: urodaeum и proctodaeum.
У п т и ц довольно длинная кишка образует многочисленные сложные и очень
правильные извивы (петли, спирали). Задняя кишка обладает более значительным
диаметром и ясно обособлена от средней кишки. На границе между ними имеется
обыкновенно пара иногда довольно длинных слепых отростков. Конечный отдел кишки
служит клоакой, на спинной стенке которой у многих птиц имеется слепой вырост
(фабрициева сумка) неизвестного значения. Этот вырост развивается из
эктодермы (проктодеума) и у взрослой птицы дегенерирует.
Кишечник млекопитающих достигает значительной длины и ясно ди-
фереяцирован на отделы (рис. 285). Начальный отдел средней кишки, образующий
петлю позади желудка и содержащий особые железы (бруннеровы), носит назва-
ние двенадцатиперстной кишки (duodenum). Следующий отдел, обра-
зующий многочисленные извивы и обладающий либеркюновыми железами,
представляет собой собственно тонкую кишку (jejunum). Очень значительного
развития достигают кишечные ворсинки. Задняя кишка отделена от средней клапаном;
она очень длинна, особенно у растительноядных форм, образует также характерные
извивы и отличается довольно значительным диаметром. Она подразделяется на
толстую кишку (colon) и п р я м у ю (rectum). Толстая кишка часто образует
последовательные выступы (hanstra), отделенные друг от друга перехватами, и, кроме
того, продольная мускулатура ограничена немногими мускульными лентами (taenia),
разделяющими ряды выступов.
Только у низших, яйцеродных млекопитающих задний участок кишки предста-
вляет собой клоаку. У сумчатых имеется рудиментарная клоака в виде нмкп, окру-
женной общим сфинктером. Но у всех высших млекопитающих, начиная с сумчатых,
имеются отдельные заднепроходное и мочеполовое отверстия, разделенные более или
менее значительной промежностью.
В высшей степени характерен для млекопитающих единичный слепой вырост
в начале толстой кишки—так называемая слепая кишка (coecum). В редких слу-
чаях она отсутствует (у некоторых насекомоядных, медведей, гиппопотамов, неполно-
зубых и китообразных), иногда имеет незначительные размеры, как у клоачных,
многих сумчатых и у хишных. Наоборот, большие и иногда очень большие размеры
имеет1 слепая кишка у травоядных сумчатых и у некоторых грызунов. Вообше можно
сказать, что у травоядных форм слепая кишка длиннее, в особенности в тех случаях,
когда имеется простой желудок (лошадь). У многих грызунов (заяц), у некоторых
хищных, у полуобезьян и у обезьян на конце слепой кишки имеется еще более тонкий
слепой вырост—червеобразный придаток (appendix), имеюший, как полагают неко-
325
торые, значение рудиментарного органа (т. е. рудимента дистальной части слепой
кишки, которая испытала сокращение). Богатство аденоидной ткани указывает,
однако, на его значение в качестве лимфатической железы.
Пищеварительные железы
Печень
Ри
Рис. 300. Поджелудочная железа и желчные
протоки лягушки (по Видерсгейму).
Dey — пузырные протоки; Dft, Db1 — печеночные про-
токи; De — желчный проток; G—желчный пузырь; L,
Ll, L‘, L* — печень (приподнята); М — желудок;
Р — поджелудочная железа; Ру — привратник.
Печень представляет собой огромную железу, совмещающую в себе
несколько совершенно различных функций. Она является пищевари-
тельной железой, выделяя внутрь кишечника желчь, которая, благодаря
своей щелочной реакции, нейтрализует желудочный сок, а кроме того,
эмульгирует жиры, активирует липазу поджелудочного сока и, следо-
вательно, способствует расщеплению жиров, растворяет жирные кислоты
и возбуждает перистальтику кишки. Вместе с тем печень — чрезвычайно
важный кровеочистительный орган, нейтрализующий вредные продукты
распада и образующий,
между прочим, мочевину
и мочевую кислоту, и,
наконец, она служит за-
пасным резервуаром для
углеводов, которые отла-
гаются в ней в виде гли-
когена.
Развивается печень из
направленного вперед сле-
пого выроста брюшной
стенки кишечника, лежа-
щего в брюшном мезенте-
рии (рис. 284). Зачаток
печени древовидно развет-
вляется, приобретая все
более сложное строение.
У круглоротых и у села-
хий эти ветви сразу полые,
у прочих позвоночных они
развиваются сначала в ви-
де компактных клеточных
тяжей, которые лишь по-
зднее к анали зи ру ются.
Между разветвлениями
проникает мезенхима с
кровеносными сосудами,
и общая масса железы бы-
стро увеличивается. По
способу своего развития
печень представляет со-
бой, таким образом, сложную трубчатую железу/ В дальнейшем, однако,
между разветвлениями развиваются многочисленные анастомозы, и пе-
чень приобретает характер сетчатой железы. -
Печень взрослого животного лежит позади сердца в области желудка.
Она представляет собой большой двулопастный или многолопастный орган,
содержащий сложную сеть желчных капилляров, располагающихся между
печеночными клетками и соединяющихся в более крупные желчные ходы,
впадающие, наконец, в выводные печеночные протоки (duc-
tus hepatici). Печеночные протоки соединяются обыкновенно в один ка-
нал, снабженный слепым выростом — желчным пузырем (vesica
fellea), служащим резервуаром, в котором скопляется желчь (рис. 300).
Узкая шейка пузыря носит название пузырного протока (duc-
326
tus cysticus), а общий проток от пузыря до впадения в начальный отдел
средней кишки обозначается как желчный проток (ductus chole-
dochus). Иногда, кроме этого, имеются еще печеночные протоки (d. hepato-
enterici), впадающие в кишку независимо от пузыря и общего протока.
В некоторых случаях желчный пузырь отсутствует.
Печень получает для своего питания артериальную кровь через пе-
ченочную артерию, но, кроме того, получает большое количество венозной
крови через воротную вену печени (v. portarum hepatis).
Форма печени находится в соответствии с общей формой тела живот-
ного и, кроме того, зависит от положения и формы соседних органов.
У млекопитающих она впереди вплотную примыкает к диафрагме. Обычно
она подразделена на две лопасти — правую и левую, которые в свою оче-
редь могут распадаться на различные по величине доли.
Общие размеры печени у низших позвоночных (рыб и амфибий) отно-
сительно больше, чем у высших. У плотоядных Животных печень крупнее,
чем у травоядных.
Так как печень развивается в брюшном мезентерии, то она и у взрос-
лого животного оказывается подвешенной на остатках этого мезентерия,
известных под названием связок, к брюшной стенке тела (lig. suspen-
sorium hepatis) и к кишечнику (lig. hepato-entericum), а у млекопитающих
и к диафрагме.
Как показывает история развития, печень возникла из простого же-
лезистого выступа брюшной стенки кишечника, который, постепенно
усложняясь, превратился сначала в ветвистую трубчатую железу, а затем
в сетчатую.>Сравнение подтверждает этот вывод; У ланцетника нет печени
в собственном смысле этого слова, но имеется направленный вперед же-
лезистый слепой вырост брюшной стенки кишечника, в стенке которого
обильно разветвляется венозный сосуд, несущий кровь от кишечника и,
очевидно, соответствующий воротной вене печени. Несомненно, этот вы-
рост представляет зачаточную печень позвоночных. У миксин печень со-
храняет еще первичный характер ветвистой трубчатой железы (у миног
желчные ходы и протоки зарастают и печень сохраняется лишь как же-
леза внутренней секреции). У всех других позвоночных она имеет характер
сетчатой железы
Поджелудочная железа
Поджелудочная железа представляет собой в высшей степени важную
пищеварительную железу. Выделяемый ею сок содержит различные фер-
менты, расщепляющие белки и углеводы. Кроме того, она является, од-
нако, и железой внутренней секреции.
Развивается поджелудочная железа из ряда небольших почечек,
образующих затем несколько зачатков в начале средней кишки в той же
области, где закладывается и печень. Чаще всего наблюдается один спин-
ной вырост и два брюшных, которые затем соединяются все вместе в спин-
ном мезентерии (рис. 301).
У взрослого животного поджелудочная железа представляет собой
то более компактный, то состоящий из рассеянных долек орган, несколько
выводных протоков которого (ductus pancreatici) впадают в начальный
отдел средней кишки иногда независимо друг от друга, иногда соединяясь
предварительно в один проток или впадая в общий желчный проток
(рис. 300).
Поджелудочная железа имеет строение сложной альвеолярной же-
лезы и располагается обыкновенно в петле начального отдела средней
кишки (двенадцатиперстной кишки птиц и млекопитающих). В некото-
рых случаях она скрыта в стенках кишечника (у миног и у двудышащих),
рассеяна в виде мелких долек в сцинном мезентерии (у некоторых кости-
327
стых рыб) или внедрена в ткань печени (у миксин и некоторых костистых
рыб). У млекопитающих это довольно компактный орган, лежащий в
петле двенадцатиперстной кишки, в которую он впадает обыкновенно
двумя протоками — брюшным, главным (ductus Wirsungi), и спинным,
добавочным (d.Santorini). У парнокопытных сохраняется иногда только
последний.
Сок поджелудочной железы содержит целый ряд пищеварительных
ферментов: трипсиноген (дающий с энтерокиназой трипсин), эрептазу,
диастазу, липазу. Между ветвями соб-
Рис. 301. Развитие поджелудоч-
ной железы (по Дзингу из Кинг-
сли)
с — желчный проток; d — двенадца-
типерстная кишка; dp — спинная и
гр — брюшная закладка поджелу-
дочной железы; / — тонкая кишка;
s — желудок.
ственно поджелудочной железы лежат
группы клеток, снабженные широкими
кровеносными капиллярами — островки
Лангерганса. Эти островки играют роль
железы внутренней секреции, имеющей
большое значение в углеводном обмене.
Плавательный пузырь
У рыб с костным скелетом (ганоиды,
костистые) в брюшной полости имеется
пузырь, наполненный газом и служащий
важным гидростатическим аппаратом.
Плавательный пузырь хрящевых и
костных ганоидов и костистых рыб закла-
дывается сначала в виде складки, а затем—
полого выроста спинной стенки кишечника, располагающегося в спинноч
мезентерии. У ганоидов и низших костистых рыб («открытопузырных»)
он в течение всей жизни сохраняет свою связь с кишечником посредст-
вом узкого воздушного канала (ductus pneumaticus), впадающего в пище-
вод, а иногда в желудок со спинной стороны. У других рыб («закрыто-
пузырных») воздушный канал зарастает, и пузырь оказывается совершенно
замкнутым.
Плавательный пузырь располагается в спинной части брюшной по-
лости непосредственно под позвоночником и почками и прикрыт брюшиной
только снизу. Общая форма пузыря бывает различной — обыкновенно
он тянется вдоль всей спинной стенки брюшной полости, нередко узким
перехватом поделен на два отдела, следующие друг за другом. Иногда
он снабжен многочисленными выростами, из которых большое значение
имеют наблюдающиеся у многих костистых рыб парные отростки, про-
ходящие вперед в голову, где они доходят до перепончатых окон слуховой
капсулы (тресковые) или даже проходят внутрь последней, касаясь пе-
репончатого лабиринта, как у сельдевых. В других случаях наблюдается
иная связь плавательного пузыря с ухом — цепь из трех косточек (аппа-
рат Вебера), представляющих собой видоизмененные части передних по-
звонков (ребра и верхние дуги), соединяется с одной стороны со спинной
степкой плавательного пузыря, с другой стороны с особым непарным вы-
ростом внутреннего уха1. Значение этой связи прежде всего в передаче
всяких изменений объема плавательного пузыря (в частности наступаю-
щих вследствие изменения внешнего давления при вертикальных пере-
мещениях рыбы) внутреннему уху.
Так как эти рыоы принадлежат большей частью к открытопузырным
(карповые и сомовые), то, кроме регуляции деятельности газовых желез,
можно предположить и возможность рефлекторного выпускания газа
при увеличении объема пузыря.
1 Правый и левый эндолимфатические протоки соединяются поперечным каналом
под мозгом и дают назад пару мещковидных выростов и непарный эндолимфатический
ринус,
328
Таким образом, рыбой, быть может, воспринимаются изменения давления, а
следовательно, и изменения глубины, что, несомненно, должно иметь большое значение
в особенности для морских костистых рыб, которые неизбежно связаны с определенной
глубиной обитания. Однако как раз у морских рыб связь между пузырем и ухом наблю-
дается сравнительно редко, а наиболее совершенная форма этой евнзи — веберов
аппарат — встречается только у пресноводных рыб.
С другой стороны, как раз у рыб с веберовым аппаратом (Ostariophysi) непосред-
ственно позади плечевого пояса воздушный пузырь нередко подходит вплотную к коже,
образуя род барабанной перепонки. Это делает возможным предположение о передаче
звуковых колебаний через посредство плавательного пузыря, что, повидимому, и дока-
зано новейшими опытами}
Стенка плавательного пузыря выстлана изнутри эпителием (обыкно-
венно плоским) и покрыта слоем соединительной ткани, обыкновенно
с большим количеством эластических волокон (у сельдевых иногда наблю-
даются охрящевения, а у карповых и сомовых — иногда даже окостенения).
В соединительнотканном слое имеются иногда гладкие мышечные волокна1.
Соответственно своему происхождению плавательный пузырь иннер-
вируется блуждающим нервом, снабжается кровью от кишечной артерии
(a. coeliaca mesenterica) и отдает венозную кровь в варотную вену.
У хрящекостных и костных ганоидов воздушный проток обыкновенно
довольно широкий и короткий, так что, очевидно, эти рыбы в состоянии
выпускать избыточный газ из пузыря через проток; нужно допустить,
что они в состоянии также заглатывать и пропускать в пузырь атмосферный
воздух. Но у костистых рыб воздушный канал длинный и узкий, и если
при сокращении стенок пузыря в некоторых случаях часть газа и может
быть через него удалена, то вряд ли возможно наполнить пузырь этим
путем. Экспериментально установлено, что и закрытопузырные рыбы
обладают способностью регулировать количество газа в пузыре, и при
искусственном его удалении пузырь довольно быстро вновь наполняется
газом. Ясно, что это наполнение может итти только путем диффузии из
кровеносной системы.
Так как из кровеносной системы могут диффундировать главным Образом лишь
кислород и углекислота, то во вновь наполненном газом пуэыре содержится значи-
тельно больше кислорода, чем в атмосферном воздухе, и совершенно ничтожное
количество азота. Так как и при удалении избытка газа всегда удаляется главным
образом кислород, то со временем в пузыре накапливается, наоборот, заметное
количество азота. У глубоководных рыб вообще преобладает кислород, достигающий
99% объема.
Стенки плавательного пузыря, действительно, обильно снабжаются
кровью, причем нередко наблюдаются местные сплетения сети тонких
артерий («чудесная сеть» — rete mirabile), являющиеся на внутренней
стенке пузыря в виде «красных пятен» или вдающиеся во внутреннюю
полость в виде «красных те л», которые оказываются особенно зна-
чительными у закрытопузырных, у которых поверхность этих тел покрыта
особым цилиндрическим железистым эпителием. Это уже настоящие газо-
отделительные железы.
Удаление избытка газа происходит чаще всего (во всяком случае у
всех закрытопузырных рыб) путем его всасывания обратно в кровь при
посредстве сосудистых сплетений. Повидимому, эту функцию несет сосу-
дистое сплетение в спинной стенке пузыря на так называемом «овале».
Таким образом, содержание газа в пузыре может регулироваться.
Это чрезвычайно важно, так как плавательный пузырь представляет
собой гидростатический аппарат переменного объема. При погружении
рыбы на большую глубину плавательный пузырь сжимается, благодаря
увеличенному давлению; удельный вес рыбы увеличивается, и она будет
1 Иногда имеется еще наружный покров поперечнополосатой соматической муску-
латуры (Trigja, Zeus).
329
уже пассивно погружаться еще глубже. При поднятии рыбы ближе к
поверхности наступит растяжение пузыря, и рыба, вследствие уменьшения
ее удельного веса, начнет всплывать. В атом случае в действительности
наблюдается регулирование количества газа в пузыре, его всасывание
при увеличении объема и, наоборот, секреция газа при увеличении внеш-
него давления и сжатии пузыря. Эта регуляция представляет собой ре-
флекторный акт на изменение давления, воспринимаемое иногда через
посредство лабиринта (особенно при наличии веберова аппарата).
Плавательный пузырь представляет собой очень важное приобрете-
ние высших рыб, которое создало возможность развития костного скелета.
Последний без такого аппарата чрезмерно повысил бы удельный вес тела,
который и у хрящевых рыб заметно выше единицы и требует постоянной
затраты мускульной анергии для того, чтобы рыба оставалась на известном
уровне. Высшие рыбы могут находиться в воде во взвешенном состоянии
без всякой затраты сил. Однако это приобретение ведет и к неудобствам—
рыба связана с определенной глубиной и может перемещаться в вертикаль-
ном направлении лишь в ограниченных пределах. Быть может, поэтому
иногда у превосходных пловцов, какова обыкновенная макрель, плава-
тельный пузырь может отсутствовать. Очень часто плавательный пузырь
редуцируется у донных форм (камбалы и др.).
Значение гидростатического аппарата имеют нередко и легкие у
водных наземных позвоночных и особенно у водных хвостатых амфибий,
у которых они иногда вовсе утрачивают значение органа дыхания.
С другой стороны, и плавательному пузырю рыб иногда приписы-
вается значение вспомогательного органа дыхания. У костных ганоидов
(Lepidosteus, Amia) внутренние стенки пузыря являются ячеистыми, и
довольно широкое отверстие пузыря снабжено мышцами, служащими
для его сжатия и растяжения. Эти рыбы часто поднимаются на поверх-
ность и заглатывают воздух, пропуская его, очевидно, в плавательный
пузырь. И у некоторых костистых рыб наблюдается то же самое. У одной
из мормирид (Mormyridae) — Gymnarchus кровоснабжение пузыря осу-
ществляется ветвями наджаберных артерий, а вена впадает прямо в кювь-
еров проток, как у двудышащих рыб.
У кистеперых рыб (Polypterus) и у двудышащих плавательный пу-
зырь является обыкновенно парным органом и открывается на брюшной
стороне пищевода щелевидным отверстием, снабженным своей мускула-
турой. Очевидно, это органы иного происхождения, чем спинной плава-
тельный пузырь других рыб. У двудышащих рыб стенки его являются
ячеистыми, и он, несомненно, имеет значение добавочного органа дыхания.
Ввиду близости этих органов по их развитию, положению и кровоснаб-
жению к легким наземных позвоночных*мы вернемся к их рассмотрению
в главе об органах дыхания.
ОРГАНЫ ДЫХАНИЯ
ОРГАНЫ ДЫХАНИЯ БЕСПОЗВОНОЧНЫХ
Газовый обмен между организмом животного и окружающей средой может про-
исходить на всей поверхности тела (диффузное дыхание), если его покровы хоть сколь-
ко-нибудь проницаемы для газов. У очень мелких животных (даже у некоторых мелких
членистоногих), у которых наружная поверхность относительно больше, чем у более
крупных *, обыкновенно нет особых органов дыхания, как нет их и у животных с очень
вялой жизнедеятельностью, например, у дождевых червей, у некоторых амфибий.
В большинстве случаев, однако, диффузное дыхание оказывается недостаточным,
и усиление газового обмена достигается увеличением поверхности в определенных
1 Как известно, объем тела возрастает пропорционально кубу радиуса, а поверх-
ность — лишь пропорционально квадрату радиуса.
330
местах тела и приспособлениями к постоянному обновлению среды на этих местах.
Таким образом развивается местное дыхание, ограниченное особыми органами.
Газовый обмен состоит в поглощении атмосферного кислорода ив окружающей
среды и в выделении в нее углекислоты, образующейся в организме в результате
окислительных процессов. Кислород, поглощенный путем диффузии из воздуха не-
посредственно или иэ воздуха, растворенного в воде, разносится затем по всему телу
или опять-таки путем диффузии или при посредстве циркулирующих в теле соков
(кровеносной системы). Во всяком случае кислород, в конце концов, через тканевые
жидкости, омывающие отдельные клетки, передается последним. Процесс окисления
происходит во всех клетках организма и сопровождается выделением углекислоты
в те же тканевые жидкости, передающие их далее наружной среде. Это — процесс
внутреннего, или тканевого, дыхания. В результате окислительных процессов происхо-
дит освобождение известного количества энергии, которое характеризует жизнедея-
тельность организма х.
Основные принципы, по которым строятся органы дыхания животных, сводятся
к следующему: 1) проницаемость стенок этих органов для газов (чему, в особенности,
благоприятствуют их тонкость и влажность), 2) возможно большая поверхность и
3) постоянное обновление внешней среды.
Соответственно среде, в которой обитает животное, и органы дыхания бывают
построены неодинаково, так что различают органы водного дыхания, или жабры,
и органы воздушного дыхания — легкие и трахеи.
В качестве жабер у беспозвоночных функционируют специальные придатки,
увеличение поверхности которых достигается образованием сложных разветвлений
или складок. По своему положению жабры чрезвычайно разнообразны. У кольчатых чер-
вей очень часто наблюдается диффузное дыхание, особенно у пресноводных. У много-
щетинковых червей имеются нередко ветвистые жабры, обильно снабжаемые кровью и
покрытые мерцательным эпителием, вызывающим постоянный, обмен окружающей
воды. Эти жабры располагаются иногда метамерно по бокам тела или, у сидячих ане-
лид, на переднем конце.
У ракообразных роль жабер играют обыкновенно пластинчатые или разветвлен-
ные придатки конечностей. У молюсков жабры могут иметь различное строение. Перво-
начальный тип жабер — это перисто разветвленный вырост стенок тела, распола-
гающийся по бокам заднего прохода в полости мантии и покрытый мерцательными
ресничками, действием которых вызывается обновление воды. У пластинчатожа-
берных моллюсков жабры приобрели вид пластинок, продырявленных множеством
щелевидных отверстий.
У хордовых животных для дыхания приспособлен передний отдел кишеч-
ника, продырявленный более или менее многочисленными отверстиями или щелями,
ведущими наружу или непосредственно, или через особую околожаберную полость.
Дыхательный ток воды поддерживается у низших форм действием мерцательных рес-
ничек, а у высших форм (у позвоночных)—мускульными дыхательными Движениями.
Органы воздушного дыхания беспозвоночных представляют собой нередко пре-
образованные органы водного дыхания, т. е. жабры, как, например, легкие моллюсков,
а, повидимому, и легкие паукообразных, представляющие собой полости, открыва-
ющиеся дыхальцем, внутри которых, подобно листкам книги, располагаются много-
численные параллельные тонкие пластинки. Но каково бы ни было строение этих ор-
ганов, они при воздушном дыхании погружаются вглубь, располагаясь в особых по-
лостях, где их поверхность может поддерживаться во влажном состоянии непрерывным
отделением жидкости, или может быть достигнуто чрезвычайное утончение их стенок,
что одинаково способствует быстрой диффузии газов.
: Совсем особое положение занимают трахеи навемных членистоногих. Эта — слож-
ная система воздухоносных трубок, начинающихся парными дыхальцами на боках
сегментов. Трахеи сильно ветвятся, и тончайшие конечные их разветвления проникают
не только во все органы, но и в ткани и даже нередко в клетки. Трахеи развиваются
on- покровов ивыстланы тонким покровным эпителием(«гиподермисом») со слоем хитина,
образующим спиральные утолщения, отсутствующие лишь в "самых мелких конечных
разветвлениях. Эти утолщения поддерживают каналы открытыми. Между утолщени-
ями слой хитина может быть чрезвычайно тонким, так что газы свободно через него
диффундируют. Атмосферный воздух проникает в трахеи не только путем диффузии,
но и при помощи дыхательных движений. У насекомых нередко трахеи местами расши-
ряются в крупные пузыри, которые растягиваются и сжимаются при дыхательных
движениях (периодическое сжатие и растяжение брюшка) и просасывают атмосферный
воздух через трахеи. Трахейная система отличается от всех других типов органов ды-
хания в том отношении, что доставляет всем частям тела непосредственно атмосферный
воздух, так что дыхание внешнее здесь почти совпадает с дыханием тканевым.
1 Некоторые животные, именно паразиты кишечника, могут обходиться, пови-
димому, без непосредственного доступа кислорода, черпая необходимый им кислород
из всасываемых органических соединений и пользуясь энергией, освобождаемой при
расщепценци последние.
331
ОРГАНЫ ДЫХАНИЯ позвоночных
Жаберные щели и жабры
У позвоночных, как и у низших хордовых, органы водного дыхания
развиваются в переднем отделе кишечника. Эмбрионально в области
глотки развиваются парные карманообразные выросты эктодермальной
стенки, растущие по направлению к покровам, откуда им навстречу разви-
ваются соответствующие эктодермальные впячивания. Внутренние вы-
росты сходятся с наружными впячиваниями, их стенки на месте сопри-
косновения срастаются, и затем происходит прорыв. Таким образом,
развивается парный ряд висцеральных мешков, открывающихся наружу
посредством щелей. Висцеральные щели (или, по крайней мере, их карма-
нообразные закладки) развиваются первоначально у всех позвоночных,
но у наземных позвоночных они затем зарастают и бесследно исчезают.
Рис. 302. Схематические горизонтальные разрезы через голову костистой рыбы (Л)
и акулы (В); Слева положение клапана при вдохе, справа — при выдохе (по Воско-
бойникову).
kbl —жаберные лепестки; hkl —гиоидный клапан; ml — клапан крышки; mhh — ротовая
жаберная полость; nhh — наружные жаберные полости; ор — жаберная крышка.
У низших позвоночных в стенках этих висцеральных мешков разви-
ваются жабры. У круглоротых жаберные мешки вздуваются и приобре-
тают вид круглых (или точнее — чечевицеобразных) полостей с суженными
в виде каналов входной и выводной частями. На эктодермальной стенке
этих мешков слизистая оболочка образует меридиональные жаберные
складки или лепестки.
У рыб жаберные мешки имеют вид щелевидных пространств, лежащих
между жаберными перегородками (рис. 302). В последних эмбрионально
имеется участок полости тела в виде канала, и развивается свой скелет
в виде висцеральных дуг, лежащих во внутренней части перегородки
(внутрь от целомической полости), развивается мускулатура в наружной
части перегородки и артериальные дуги, дающие начало жаберным со-
судам. На стенках жаберных мешков и именно в той их части, которая
332
имеет эктодермальное происхождение, развиваются складКи слизистой
оболочки — жабры1.
Первая висцеральная щель значительно сокращается, постепенно
зарастая снизу, так что от нее остается лишь спинная ее часть, которая у
большинства селахий и осетровых рыб и у многоперых ганоидов (Polyp-
terus) сохраняется на спинной стороне головы в виде «брызгальца», а у
других рыб совсем зарастает. У наземных позвоночных за счет полости
этого висцерального мешка развивается барабанная полость уха. Следу-
ющие висцеральные мешки образуют у рыб собственно жаберные щели.
Число жаберных щелей может быть различно. Наибольшее число
щелей наблюдается у круглоротых, где оно у одного вида (Bdellostoma
polytrema) достигает 14 (а эмбрионально у Bdellostoma Stouti до 17 ще-
лей), но обычно равняется 7. У наиболее примитивных акул число жабер-
ных щелей (не считая брызгальца) достигает 7 (Heptanchus) или 6 (Нехап-
chus, Chlamydoselachus). Обычно у рыб имеется 5 жаберных щелей, а
иногда это число сокращается еще больше.
Таким образом у современных рыб, очевидно, произошло сокращение
числа жаберных щелей. Это сокращение происходит в направлении сзади
наперед, т. е. всегда сначала исчезают последние щели, как это доказы-
вают факты нахождения различных рудиментов в задней части жабер-
ного аппарата, (особенно остатки лишних жаберных дуг) и эмбриональные
закладки рудиментарных жаберных мешков. Такие рудименты найдены
и у акул с семью жаберными щелями (Heptanchus). Однако несомненно,
что жаберный аппарат редуцировался и спереди. Брызгальце
(spiraculum) представляет собой такую же висцеральную щель, как
и следующие за ней жаберные щели, и нельзя сомневаться, что и оно
несло раньше ту же самую функцию. Строение скелета, мускулатуры,
иннервация брызгальца, закладка артериальных дуг и впереди (в че-
люстной дуге), и позади (в подъязычной) от нее дает достаточно ясные
указания. Наконец, в брызгальце селахий и других рыб наблюдается
и рудиментарная жабра — она лежит на передней его стенке и носит
название «ложножабр ы», так как кровоснабжение ее отличается
от кровоснабжения других жабер (она снабжается не венозной кровью
от брюшной аорты, как истинные жабры, а артериальной кровью
от первой выносящей жаберной артерии). Следовательно, мы имеем в
брызгальце настоящую рудиментарную жаберную щель. Кроме того,
имеются указания на то, что впереди брызгальца имелись еще преспи-
ракулярные щели, которые исчезли, очевидно, в связи с развитием че-
люстного аппарата. Таким образом, жаберный аппарат позвоночных
испытал сокращение не только сзади, но и спереди.
У низших хордовых число щелей гораздо больше, чем у позвоночных.
У ланцетника их свыше 100, и если даже принять во внимание факт вторич-
ного увеличения их числа вследствие разделения каждой щели язычковой
перегородкой на две и вообще развитие ряда новых щелей после неко-
торой паузы (на стадии 14 первичных щелей), то все же ясно, что эволюция
жаберного аппарата шла у позвоночных по пути уменьшения числа жабер-
ных щелей при одновременном их усовершенствовании как органов ды-
хания. Это усовершенствование выразилось в увеличении дыхательной
поверхности путем образования складок слизистой оболочки, т. е. в обра-
зовании жаберных лепестков. Как показывает история развития, перво-
начальной формой жабер являются нитевидные выросты наружной стенки
жаберных дужек, которые затем путем образования боковых веточек
превращаются в жаберные лепестки (Северцов).
1 Эктодермальное их происхождение доказывается в особенности ясно в тех слу-
чаях, когда закладка жаберных лепестков происходит раньше, чем образовался про-
рыв перегородки между эктодермальной и эктодермальной частями жаберного мешка
(Acipenser).
333
Жабры рыб представляют собой Пластинчатые выросты слизистой
оболочки, поделенные на отдельные лепестки с многочисленными мелкими
поперечными складочками, обильно снабженные кровеносными сосудами.
Такие выросты располагаются на передней и задней стенках каждого
жаберного мешка и носят название полужабер (рис. 302, 303). Обе полу-
жабры одной перегородки составляют целую жабру. Если не считать
ложножабру, о которой уже была речь, то первая полужабра лежит на
передней стенке первого жаберного мешка, т. е. на задней стенке подъ-
Рис. 303. Вскрытый первый^жаберный, мешок акулы (Acanthias)
с полужаброй,£первой идуги (по Нирштрасу).
Ьг. 1 —nejBaa жаберная дуга; hbr. 1 —передняя полужабра; hy —гмоид (перереван);
pct — Грудной плавник; sp — брывгальце.
язычной дуги (гиоидная полужабра). На задней стенке последней щели
полужабры у современных рыб не бывает.
Жаберные перегородки содержат, кроме скелета и мускулатуры, при-
носящие и выносящие жаберные артерии, распадающиеся в лепестках
на сеть капилляров х.
У селахий каждая перегородка продолжается снаружи в кожную
складку, прикрывающую следующую позади жаберную щель (рис. 302, В).
Первоначальная функция этих складок — защита жабер от попадания
посторонних тел извне. Однако они принимают также участие в дыхатель-
ных движениях в качестве разрежаю-
щего насоса, и при прогрессивном
их развитии это участие становится
все более активным. Особенно хорошо
развиты жаберные складки у аку-
лы — Chlamydoselachus (рис. 304),
у которой в особенности выдается сво-
ей величиной первая складка, отхо-
дящая от подъязычной дуги, причем
Рис. 304. Голова акулы (Chlamydose-
lachus). Жаберные складки; / — склад-
ка подъязычной дуги.
правая и левая соединяются снизу
между собой. Жаберные перегородки и складки поддерживаются хряще-
выми лучами, отходящими от висцеральных дуг. У цельноголовых рыб
передняя складка получает особенно значительное развитие и прикры-
вает все жаберные щели. Это так называемая жаберная крышка, которая
у этих рыб поддерживается сильно развитыми хрящевыми лучами подъ-
язычной дуги. Так как теперь функция защиты всех жабер перешла к
одной передней складке, получившей преобладающее развитие, то естест-
1 Повидимому, в жаберных лепестках кровь циркулирует даже просто в межкле»
точных пространствах.
334
Ёенйо, что .следующее складки редуцировались, причем
лись и несущие их жаберные перегородки.
Жаберный аппарат
всех высших рыб защищен
снаружи жаберной
крышкой (operculum),
поддерживаемой собствен-
г
ным \ : костным скелетом
(покровные кости, развив-
шиеся на лучах подъязыч-
ной дуги). Твердая соб-
ственно крышка i продол-
жается на брюшной сторо-
не в кожную складку
с лучеобразными, обыкно-
отчасти
D
сократи-
Рис. 305. Поперечные разрезы через жаберные дуги
и жабры акулы (Л), химеры (В), осетра (С), костис-
тых рыб (Д, Е) (по Боасу).*
Ъ — жаберная дуга; с — перегородка, s — жаберная
тычинка.
венно, кожными окостене-
ниями — в жаберную
перепонку. Жабер-
ная крышка снабжена
мускулатурой и активно
участвует в дыхательных
движениях, получая у многих рыб [весьма важное значение разрежаю-
щего насоса, просасывающего воду через жаберный аппарат (рис. 302,Л).
Рис. 306. А — жаберные лепестки кос-
тистой рыбы в их нормальном положе-
нии; стрелки показывают направление
дыхательного тока воды; В — вид жа-
берной решетки между лепестками двух
соседних дуг (по Воскобойникову).
kb — жаберные лепестки; hf — вторичные
лепесточки.
На жаберных перегородках не толь-
ко исчезли кожные складки, Hoin
сами перегородки испытывают зна-
чительное сокращение, как это можно
проследить в ряду форм (рис. 305).
У костистых рыб сохраняется лишь
основная часть перегородки, где ле-
жат костная жаберная дуга, крове-
носные сосуды и т. д. Та часть перего-
родки, которая отграничивала друг
от друга полужабры одной дуги,
исчезла, и полужабры из сидячих
превратились в свободно свисающие
«гребенчатые жабры». Каждая полу-
жабра состоит из свободных, при-
крепленных только своим основа-
нием, ланцетовидных жаберных ле-
пестков (рис. 305, Е). Последние
образуют на своей поверхности вто-
ричные лепесточки, располагающие-
ся в соседних лепестках поочередно.
Концы полужабер последовательных
жаберных дуг при жизни держатся
сомкнутыми (рис. 306, Л), и вода про-
цеживается при дыхательных движе-
ниях через сложную систему щелей,
лежащих между вторичными лепе-
сточками (рис. 306, В). Дыхательная
поверхность жабер достигает, таким
образом,весьма значительной величи-
ны. Жаберные лепестки защищаются
снаружи жаберной крышкой, а изну-
три — так называемыми жаберными
тычинками, т. е. рядом отростков, отходящих от жаберных дуг по направ-
335
лению к соседней дуЬё, Перегораживающих внутренние жаберные отвср*
стия (рис. 302, 303). Эти тычинки образуют цедилку, препятствующую
прохождению пищевых частиц из полости глотки в жаберный аппарат.
С редукцией жаберных перегородок и с заменой хряща костью в жаберных
дугах объем каждой дуги уменьшился, и весь жаберный аппарат скон-
центрировался, заняв сравнительно небольшую область в задней части
головы.
Жаберный аппарат круглоротых занимает сравнительно очень большой
объем (около */в тела) и состоит из парного ряда типичных жаберных мешков с мери-
диональным расположением складок. Каждый мешок сообщается довольно узким
внутренним каналом с полостью глотки (или обособленной ее частью) при помощи
отдельных небольших внутренних жаберных отверстий и подобным же наружным ка-
налом с наружной средой — наружные жаберные отверстия. У личинки миноги жа-
берные мешки широко сообщаются с глоткой. У взрослой миноги соответствующая
часть глотки обособляется в замкнутую позади дыхательную трубку, лежащую под
остальной частью, служащей для проведения пищи (пищевод); жаберных мешков име-
ется 7 пар, открывающихся самостоятельными наружными жаберными отверстиями.
У рода Bdellostoma (из миксин) сохраняются и во взрослом состоянии более прими-
тивные соотношения — внутренние отверстия жаберных мешков открываю ся непо-
средственно в глотку; наружные отверстия также самостоятельны. Число жаберных
мешков и отверстий различно у различных видов и достигает у одного вида 14 (В. poly-
trema). У настоящих миксин (Myxine) наружные каналы жаберных мешков удлинены,
и все соединяются позади между собой, открываясь на каждой стороне одним общим
жаберным отверстием. Позади последнего мешка у всех миксин слева имеется еще ка-
нал, соединяющий полость кишечника непосредственно с жаберным отверстием (duc-
tus oesophago-cutaneus).
Механизм дыхания состоит у круглоротых в периодическом сжатии и расшире-
нии всей жаберной области (а следовательно, и отдельных жаберных мешков), причем
вода может всасываться и выбрасываться через жаберные отверстия. Когда кругло-
ротые присасываются своим ртом, то у миног вода всасывается и удаляется через жа-
берные отверстия, а у миксин она всасывается через ноздрю и гипофизарный канал.
Когда миксина погружается всей передней частью тела внутрь тела животного (обык-
новенно трупа), которым она питается, то, очевидно, для вдыхания служит пищеводно
кожный канал.
У рыб максимальное число настоящих жаберных щелей равняется 7 (Heptan-
chus), но обычно имеется 5. У селахий наружные жаберные отверстия имеют вид уз-
ких поперечных щелей, лежащих по бокам головы и открывающихся непосредственно
наружу. Внутренние отверстия имеют форму еще более длинных щелей, а жаберные
мешки сводятся к щелевидным пространствам между соседними жаберными перего-
родками. Жабры, как сказано, сидячие, а жаберные щели прикрываются кожными
складками, отходящими от лежащих впереди жаберных перегородок. В брызгальце
имеется рудиментарная челюстная полужабра, называемая ложножаброй, так как
она снабжается не венозной, а уже окисленной кровью. При дыхании селахии наби-
рают воду через рот (при расширении ротовой полости и глотки), причем жаберные
складки автоматически замыкают щели, и выталкивают ее затем через жаберные щели
наружу (при сжатии соответствующих полостей). У некоторых акул и в особенности
у скатов брызгальце имеет значительные размеры и служит для всасывания воды;
обратно вода из брызгальца не выталкивается, так как этому препятствует имеющийся
в нем клапан.
У цельноголовых жаберный аппарат построен и функционирует в общем, как
и у селахий, но все жаберные щели прикрыть! большой кожной складкой, отходящей
от подъязычной дуги; в связи с этим жаберные перегородки несколько сокращены.
У всех рыб с костным скелетом имеется такая же большая складка с костным
скелетом из нескольких крупных костей в своей верхней части [собственно жаберная
крышка (operculum)] и обыкновенно многими более мелкими костями в своей нижней
части, загибающейся на нижнюю поверхность головы между ветвями нижней челюсти
[жаберная перепонка (membrana branchiostegi)]. Жаберные перегородки редуциру-
ются заметно уже у ганоидов, но особенно у костистых рыб, и жабры свободно свисают
теперь в общую жаберную полость, прикрытую снаружи жаберной крышкой. Таким
образом, отдельные у низших рыб жаберные мешки сводятся у высших рыб к одним
только внутренним щелям между соседними дугами. Последние снабжены выростами,
жаберными тычинками, образующими цедилку, задерживающую пищевые частицы
и случайные тела в полости глотки (жаберные щетки).
С развитием жаберной крышки передняя (гиоидная) полужабра редуцируется
и перемещается под самую крышку, где она получает название оперкулярной
жабры (осетровые рыбы). У осетровых в брызгальце сохраняется и ложножабра
(челюстная полужабра). У костных ганоидов и у костистых рыб наружное отверстие
брызгальца зарастает, а ложножабра сохраняется иногда еще под подвеском (hyoman-
dibulare) или перемещается под жаберную крышку и ложится рядом с гиоидной полу-
556
жаброй (Lepidosteus), образуя, как и последняя, оперкулярную жабру. У костистых
рыб также имеется оперкулярная полужабра, представляющая собой, однако, челюст-
ную ложножабру; гиоидная полужабра здесь вполне редуцировалась. У пелагических
личинок костистых рыб ложножабра является иногда в роли «глазной» жабры, снаб-
жающей артериальной кровью глаза, которые у них относительно очень велики.
Дыхание рыб производится посредством периодического сжатия и растяжения
ротовой полости1 (нагнетающий насос), а также при помощи активных движений жа-
берной крышки (разрежающий насос).
Вода всасывается через ротовую полость, причем автоматически прижимаются
оторочка жаберной крышки и жаберная перепонка. Выталкивается вода через жабер-
ный аппарат при поднятии жаберной крышки, причем изнутри к зубам прижимается
складка слизистой оболочки, замыкающая рот. Кроме того, гибкая оторочка жаберной
Рис. 307. Наружные жабры моло-
дой хвостатой амфибии (Amphiu-
ma), частью покрытые жаберной
складкой (из Кингсли).
крышки при поднятии последней скользит по поверхности плечевого пояса и произво-
дит всасывающее действие. Вода при этом просасывается черев жаберный аппарат
(рис. 302). Это просасывание производится при сомкнутых жаберных лепестках, при-
чем вода протекает через тончайшую систему жаберных складочек, дающих большую
поверхность для поглощения кислорода (Воскобойников).
У некоторых костистых рыб наблюдаются значительные изменения в строении
жаберного аппарата. У пучкожаберных (морских игл и коньков) жаберные лепестки
расположены пучками. Нередко четвертая жабер-
ная Дуга теряет свои жабры, и пятая жаберная
щель зарастает. Иногда редуцируются жабры и
на третьей дуге. Оперкулярная щель иногда
сокращается до размеров небольшого отверстия.
Но гораздо замечательнее различные доба-
вочные приспособления для сохранения воды,
для воздушного дыхания, развивающиеся в связи
с жаберным аппаратом рыб (см. описание доба-
вочных органов дыхания).
У наземных позвоночных,
как уже упомянуто, закладывается, как и у рыб,
до шести висцеральных мешков. Из первого мешка, соответствующего брызгальцу,
развивается барабанная полость; следующие жаберные мешки прорываются
еще обыкновенно наружу, но затем исчезают, давая лишь начало некоторым произ-
водным их зпителия (зачаткам зобной железы и другим телам, о которых речь будет
в главе об органах внутренней секреции). Только у амфибий развиваются
настоящие жабры в качестве личиночных органов дыхания. У хвостатых амфибий —
зто наружные жабры (рис. 307), а у бесхвостых развиваются затем и внутренние (но
все же зктодермальные жабры своеобразного, правда, строения (древовидно развет-
вленные) (рис. 308). Закладывается у амфибий и жаберная крышка в виде простой
кожной складки, прикрывающей основания наружных жабер. У личинок бесхвостых
амфибий жаберная складка разрастается, прикрывая весь жаберный аппарат, а
оперкулярная щель сокращается до степени небольшого отверстия, обыкновенно
срастающегося с таковым другой стороны и располагающегося обычно несиммет-
рично на одном боку. У взрослых амфибий жаберные щели совершенно зарастают,
за исключением лишь явно неотеничных хвостатых амфибий, у которых могут со-
храниться во взрослом состоянии 2—3 жаберные щели одновременно с наружными
жабрами (Perennibranchiata) или только одна (четвертая) жаберная щель (Derotre-
mata).
Наружные жабры
Кроме внутренних жабер, сидящих в жаберных мешках, у позвоноч-
ных наблюдаются иногда также и наружные жабры. У эмбрионов селахий
под этим названием известны длинные нитевидные, иногда разветвленные
продолжения жаберных лепестков, торчащие из брызгалец и жаберных
щелей наружу. Подобные нитевидные личиночные жабры наблюдаются
иногда у костистых рыб (Gymnarchus, Heterotis).
Иного рода наружные жабры имеются у личинок кистеперых ганоидов
(Polypterus), у которых пара больших перисто разветвленных жабер си-
дит на верхней части подъязычной дуги. Подобные перисто разветвленные
наружные жабры имеются и у личинок двудышащих рыб (Protopterus
и Lepidosiren), где они лежат в количестве 4 пар по бокам головы, непо-
1 Растяжение достигается опусканием дна ротовой полости с брюшной частью
жаберного аппарата и расширением боков вследствие поднятия нёбной дуги, опира-
ющейся на подвесок; сжатие происходит при поднятии дна ротовой полости действием
жаберной мускулатуры и опускании нёбной дуги, причем подвесок притягиваете^
к черепу.
337
средствейно позади жаберной крышки. Эти жабры затем редуцируются,
но у Protopterus вполне исчезает только передняя, а три задние ложатся
одна над другой и сохраняются в качестве рудиментарных образований
в течение всей жизни. Эти жабры снабжаются кровью от 2-й, 3-й и 4-й
артериальных дуг.
У личинок амфибий развиваются подобные же наружные жабры
на задних (верхних) концах )трех первых жаберных дуг и снабжаются
Рис. 308. Схема соотношения наружных и
внутренних жабер у головастика лягушки
(по Мауреру ив Кингсли).
at> и еЬ — приносящие и выносящие жаберные
артерии; h — сердце; о — ухо; ph — глотка;
та — корень аорты.
кровью от соответствующих арте-
риальных дуг (рис. 307). Наруж-
ные жабры амфибий могут иметь
форму простых выростов с одно-
рядным расположением боковых
ветвей (бесхвостые амфибии)
(рис. 308) или состоят из оси
с двурядным расположением вет-
вей (перистые жабры щележабер-
ных амфибий и гимнофион)
(рис. 307), или с несколькими
рядами ветвей (многие хвостатые
амфибии), или, наконец, они дре-
вовидно разветвлены (постоянно-
жаберные амфибии). Обновление
воды на наружных жабрах дости-
гается периодическими их движе-
ниями при помощи особой муску-
латуры, а также деятельностью
мерцательного покрова.
Наружные жабры личинок амфибий, несомненно, гомологичны со-
ответствующим личиночным органам двудышащих рыб. И те, и другие
органы эктодермальны по своему развитию и нередко противополагались
внутренним жабрам, которые раньше считались энтодермальными обра-
зованиями. В настоящее время существуют гораздо более многочисленные
данные в пользу теории эктодермального происхождения жабер как
амфибий, так и рыб.
Не совсем ясно еще отношение наружных жабер ко внутренним, но
существуют указания в пользу того, что наружные жабры развились из
жаберных лепестков верхнего конца дуг в результате их перемещения па
наружную поверхность.
Добавочные органы дыхания
У многих высших рыб имеются различные приспособления для ды-
хания атмосферным воздухом. Эти приспособления обеспечивают рыбе
возможность пребывания в испорченной воде с недостаточным содержа-
нием кислорода, как это наблюдается часто в водоемах со стоячей водой,
особенно в тропическом климате. Нередко, кроме того, такие рыбы обла-
дают способностью продолжительного пребывания вне воды.
Так называемые «наджаберные органы» (у Ophiocephalus
и Periophthalmus) представляют собой углубления, лежащие со спинной
стороны в области первой жаберной щели, с обильно снабжаемым кровью
утолщением слизистой оболочки. У некоторых тропических костистых
рыб (Anabas, Trichogaster, Osphromenus, Polyacanthus) имеется лаби-
ринтовый аппарат, состоящий из сложной системы складок
слизистой оболочки, помещающейся в особом углублении в спинной части
жаберной полости. Лабиринт поддерживается специальным постным
выростом соответствующей жаберной дуги. Этот орган помещается на
верхнем конце первых двух жаберных дуг и снабжается венозной кровью
338
асе
Рис. 309. Добавочные дыхательные орга-
ны (асе) сома (Clarias) (по Бриджу).
Ьг — жабра.
под эпителием.
от соответствующих жаберных артерий. У некоторых сомовых (Clarias,
Heterobranchus) имеются ветвистые придатки на верхних концах 1—2 жа-
берных дуг, сидящие также в углублениях жаберной полости (рис. 309).
Во всех таких органах задерживается вода, увлажняющая их стенки, а
также жаберный аппарат.
У одного индийского сома — Saccobranchus (сходно и у Amphipnous) —
имеются длинные мешковидные выросты спинной части первой жаберной
щели, идущие назад вдоль позвоночника до самого хвоста. Эти мешки
наполняются водой, которая ис-
пользуется для смачивания жабер-
ных лепестков, когда сом нахо-
дится в воде.
У многих сомовых наблю-
дается кишечное дыха-
ние — рыба заглатывает атмос-
ферный воздух и пропускает его
вдоль всего кишечного канала и
затем через заднепроходное отвер-
стие наружу. Такое же кишечное
дыхание имеется и у обыкновен-
ного вьюна (Cobitis fossilis), сред-
няя кишка которого необычайно
богата кровеносными капилляра-
ми, залегающими непосредственно
Наконец, в качестве добавочного органа специально воздушного
дыхания служит иногда и плавательный пузырь. Такова
его роль, например, у костных ганоидов (Lepidosteus и Amia), у которых
стенки пузыря при увеличении внутренней поверхности приняли ячеи-
стый характер. Это — непарный пузырь, открывающийся каналом на
спинной стороне пищевода.
У многоперых ганоидов (Polypterus) и двудышащих рыб плавательный
пузырь парный и сообщается с брюшной стороной пищевода, закладываясь
непосредственно позади жаберной области. У Polypterus стенки его глад-
кие, и он, очевидно, не играет заметной роли в качестве органа воздуш-
ного дыхания. Но у двудышащих он обладает ячеистыми стенками и,
несомненно, имеет в этом смысле большое значение. Он снабжается кровью
от четвертой наджаберной артерии и возвращает окисленную кровь не-
посредственно в сердце (в венозную пазуху). У Protopterus и Lepidosiren
плавательный пузырь необычайно сходен с легкими низших наземных
позвоночных и по общему своему виду, и по строению, и по кровоснаб-
жению. У Ceratodus плавательный пузырь непарный, но в остальном схо-
ден с пузырем других двудышащих, так что можно думать, что и он имеет
парное происхождение. Короткий и широкий воздушный канал откры-
вается у всех этих форм щелевидным отверстием на брюшной стенке пи-
щевода. Это отверстие напоминает гортанную щель наземных позвоночных
и снабжено мускулатурой, служащей для ее замыкания. Легкие двуды-
шащих функционируют обычно в качестве добавочного органа дыхания.
Во время засухи Protopterus и Lepidosiren зарываются во влажный ил
и дышат исключительно легкими.
Так как плавательный пузырь других рыб представляет собой с са-
мого начала непарный орган, развивающийся в спинном мезентерии и
притом далее позади, в области пищевода или даже желудка, то нужно
полагать, что он не гомологичен парному пузырю двудышащих рыб и
приобретен в обеих группах рыб (Polypteroidei и Dipnoi, с одной стороны,
и Chondrostei, Holostei и Teleostei—с другой) независимо.
В обоих случаях орган возник, очевидно, сходным путем — рыба
заглатывала воздух, и он задерживался сначала просто в складках, а
339
затем в углублениях, давших Начало слепым выростам. Но в одном слу*
чае этот процесс имел место в самом кишечнике, в спинной его части, и
в результате его возник непарный спинной плавательный пузырь боль-
шинства рыб. В другом случае воздух, очевидно, задерживался в послед-
них жаберных мешках, в которых, таким образом; развились сразу пар-
ные выступы, давшие начало парному плавательному пузырю многоперых
ганоидов и двудышащих рыб. Весьма возможно, что этот орган с самого
начала служил добавочным органом воздушного дыхания. Так как пар-
ный плавательный пузырь и в других отношениях сходен с легкими
наземных позвоночных, то он вполне заслуживает названия легких.
И у наземных позвоночных, именно у амфибий, бывают различные
добавочные органы дыхания. Хотя основным органом дыхания у них слу-
жат легкие, в качестве добавочных органов сохраняются иногда жабры,
и кроме того, в особенности сама кожа поддерживается во влажном со-
стоянии и обильно снабжается кровью. Нередко и слизистая оболочка
ротовой полости образует многочисленные складки и является также в
роли добавочного органа дыхания. Наконец, у личинок амфибий иногда
развиваются и различные иные органы дыхания (например, хвост у Hylo-
des martinicensis).
Легкие позвоночных
Легкие позвоночных представляют собой парные мешки, открыва-
ющиеся посредством более или менее длинного канала на брюшной стенке
глотки гортанной щелью, снабженной своей мускулатурой. В простейшем
виде у некоторых амфибий эти органы в высшей степени сходны с парным
плавательным пузырем кистеперых ганоидов и двудышащих рыб.
Развитие легких
Легкие закладываются у низших наземных позвоночных в виде пар-
ного выроста на брюшной стенке глотки позади последнего жаберного
Рис..31О. Закладка легких у амфибий
(по Гётте), Горизонтальный разрез
через молодую личинку.
Ьг — вакладка наружной жабры; с—по-
лость тела; 7 — вакладка легких; ph—
полость глотки; st — ротовая ямка;
1—в — висцеральные мешки.
мешка (рис. 310). Во время дальней-
шего развития эти зачатки растут на-
зад, вытягиваясь в мешковидные обра-
зования, и в то же время обособляется
соединяющий оба зачатка участок глот-
ки. Последний дает начало непарному
дыхательному пути (гортани и трахее),
а парные мешки врастают в брюшную
полость, покрываясь при этом брюшин-
ной выстилкой (плеврой), и преобра-
зуются в более или менее сложно
построенные органы. Внутренняя по-
верхность легочных мешков постепенно
увеличивается путем развития все более
сложной сети перекладин, подразделя-
ющих ее на множество ячеек.
У высших наземных позвоночных
дыхательный путь обыкновенно закла-
дывается раньше легочных мешков, и
развитие начинается, таким образом,
с обособления непарного жолоба на
брюшной стенке глотки позади закла-
док жаберных мешков. На этом жолобе
закладываются затем сами легкие в ви-
де парного уже выступа. Кроме того,
и в самО1М легком раньше всего закла-
340
даваются дыхательные пути — бронхи и
процесс развития легкого представляет
-образования новых ветвей,
снабженных коночными взду-
тиями (рис. 311). .
У низших наземных по-
La
:звоночных дыхательные пути г /
•еще слабо развиты и состоят
нередко (у низших хвостатых 7^212
.амфибий и у бесхвостых) из (У\Л о
небольшой каморы, связан-
пой с обоими легочными меш-
ками и открывающейся в
глотку посредством гортан-
ях разветвления, и поэтому
собой картину постепенного
ной щели. В других случаях
{у многих хвостатых амфибий
Рис. 311. — Две стадии развития легкого свиньи
(с брюшной стороны) (по Флинту из Кингсли).
и у рептилий) от переднего
участка, или гортани, обосо-
бляется более или менее
Ь — жаберный мешок; dt I, г — спинные, боновые и
брюшные бронхи; ое — пищевод; I — трахея; р — ва-
ч ток легких.
длинная дыхательная трубка, или трахея. Стенка легочных мешков
в некоторых случаях, очевидно, в результате вторичного упрощения
(у постоянножаборпых амфибий) оказывается гладкой, но обычно уже
у бесхвостых амфибий и особенно у рептилий приобретает совершенно
губчатый характер.
Происхоагдеиие легких
Сходство легких низших наземных позвоночных с плавательным пу-
зырем некоторых рыб ужо давно навело исследователей на мысль о гомо-
логии этих образований. В такой общей форме это распространенное мне-
ние встречает, однако, немалые затруднения. Мы видели, что плавательный
пузырь большинства рыб является непарным органом, развивающимся
в спинном мезентерии. Он снабжается артериальной кровью кишечных
сосудов (a. cooliaca) и отдает венозную кровь частью в кардинальные,
частью в воротную вону почони. Эти факты, несомненно, говорят против
указанной теории. Однако у некоторых рыб наблюдается парный плава-
тельный пузырь, сообщающийся с брюшной стенкой пищевода и притом
далее впереди. Этот орган снабжается, как и легкие наземных позвоноч-
ных, кровью от четвертой пары жаберных артерий и отдаст ее прямо
в сердце (в венозную пазуху у двудышащих и в прилежащую к ней часть
печеночной воны у Polyptcrus). Совершенно ясно, что мы имеем здесь дело
с образованиями того же рода, как и легкие.
Таким образом, вышеуказанную гипотезу о происхождении легких
можно принять с известными ограничениями — легкие наземных позво-
ночных представляют собой результат дальнейшей специализации (как
органа дыхания) легочного пузыря многопсрых ганоидов и двудышащих
рыб.
Опираясь на факт закладки легких у амфибий в виде парных мешко-
видных выростов позади последней пары жаберных мешков (рис. 310),
Гётте высказал предположение, что легкие представляют собой результат
преобразования пары жаберных мешков. Эта теория может быть сближена
с первой, если допустить, что и плавательный пузырь имеет такое же про-
исхождение. Таким’образом, некоторые авторы полагают, что плава-
тельный пузырь рыб и легкие наземных позвоночных развивались не-
зависимо (диворгонтно) из пары последних жаберных мешков.
В настоящее время можно считать что теория Гётте о происхождении
легких наиболее согласуется с фактами. Что же касается вопроса о про-
исхождении плавательного пузыря рыб, то мы можем принять только
341
для парного пузыря многоперых ганоидов и двудышащих рыб такое же
его возникновение, как и для легких. В таком случае отпадает также
необходимость принятия вполне независимого развития этих органов.
Легкие наземных позвоночных представляют собой специализированный
парный ^плавательный пузырь. Последний возник путем преобразования
из пары жаберных мешков.
Процесс возникновения этих органов представить себе нетрудно:
в задних жаберных мешках задерживался заглатываемый атмосферный
воздух; так как это было связано с известными выгодами — облегчался
удельный вес рыбы и вместе с тем доставлялся известный запас кислорода,
то в соответствующих местах образовались мешковидные выросты, слу-
жившие уже специально для заглатывания воздуха. Такие мешковидные
выросты могли сразу иметь значение добавочных органов дыхания.
Дыхательные пути
Дыхательные пути представляют собой по существу более или менее
длинную трубку, помощью которой легкие сообщаются с полостью глотки.
В отличие от дыхательных частей самих легких дыхательные пути выст-
ланы цилиндрическим мерцательным эпителием.
У некоторых амфибий, как упомянуто, легочные мешки сообщаются
с гортанной щелью посредством небольшой камеры. По бокам гортанной
щели в ее стенках залегают парные гортанные хрящи, связанные с мыш-
цами, служащими для сжимания и расширения гортанной щели. Уже
у многих хвостатых амфибий и в особенности у форм с длинным телом
(Siren, Amphiuma) наблюдается диференцировка этой соединительной
камеры на передний отдел с гортанными хрящами и мускулатурой, гор-
тань (larynx), и задний отдел, трахею (trachea), в виде более или
менее длинной трубки, в стенках которой залегают небольшие хрящики.
Начиная с рептилий, трахея позади разделяется соответственно обоим
легочным мешкам на две ветви—бронхи (bronchi), поддерживаемые также
хрящами. В самих легочных мешках по мере усложнения строения их
стенок также происходит диференцировка на центральные части, служа-
щие продолжением воздушных путей и выстланные также цилиндрическим
мерцательным эпителием, и собственно дыхательные части — ячеи, или
легочные пузырьки, выстланные плоским однослойным эпителием
(рис. 315).
Гортань
Органами опоры для начального отдела дыхательной трубки являются
гортанные хрящи, представляющие продукт преобразования последних
жаберных дуг. У млекопитающих к ним присоединяются еще и лежащие
впереди дуги.
В гортани некоторых хвостатых амфибий (Necturus, Proteus)
наблюдается лишь пара хрящей (cart, laterales), лежащих по бокам гор-
танной щели и продолжающихся отчасти и вдоль дыхательной трубки
(рис. 312). К этим хрящам прикрепляются мышцы-расширители, отхо-
дящие от задних висцеральных дуг. Однако эта простота строения пред-
ставляет собой, повидимому, результат редукции. У других амфибий
.имеются, по крайней мере, две пары хрящей: передняя пара, ограничи-
вающая гортанную щель, носит название черпаловидных хря-
щей (cart, arytaenoideae); задняя пара тянется отчасти вдоль трахеи
и распадается у многих амфибий на пару хрящей, лежащих позади черпа-
ловидных, и на целый ряд небольших трахеальных хрящиков. У бес-
хвостых задняя пара гортанных хрящей. образует один кольцевидный
(«перстневидный») хрящ (cart, cricotrachealis), имеющий довольно сложную
форму (рис. 312).
342
У бесхвостых амфибий черпаловидные хрящи поддерживают две
боковые складки слизистой оболочки, лежащие на внутренней поверх-
ности гортани, — голосовые связки. Кроме боковых мышц,
расширителей гортани (mm. dilatatores laryngis), имеются еще мускулы-
сжиматели (mm. constrictores laryngis) 'в спинной и брюшной стенках
гортани.
Рис. 312. Гортанные хрящи хвостатых амфибий (Л — Necturus,
В — Salamandra) и лягушки (С) (из Видерсгейма).
cl — боновые хрящи; аг — черпаловидные; ст — перстневидный.
Гортань рептилий содержит те же самые хрящи. Перстне-
видный хрящ (cart, cricoidea) представляет собой неполное позади
кольцо, которое обособляется от трахеальных хрящей; спереди на нем
сидит пара черпаловидных хрящей. Как и у амфибий, гортань тесно
связана с подъязычным аппаратом и имеет свою висцеральную муску-
латуру. Пара складок служит у гекконов й хамелеонов голосовыми
связками.
Сходное строение имеет и гортань
птиц, у которых голосовых связок
не имеется.
Гораздо выше развита гортань у
млекопитающих, у которых
она является голосовым аппаратом.
Кроме черпаловидных и перстневид-
ного хрящей, приравниваемых обычно
к одноименным хрящам низших форм,
имеется еще щитовидный хрящ
(cart, thyreoidea), лежащий с брюшной
стороны впереди от кольцевидного хря-
ща, и сидящий на нем еще далее впе-
реди в непарной складке слизистой
оболочки надгорт ан ник (epiglot-
tis). Гортань млекопитающих связана
теснейшим образом с подъязычным
Рис. 313. Жаберный аппарат безно-
гой амфибии (Ichthyophis) и подъ-
язычный аппарат и гортань клоач-
ного млекопитающего (Ornithoryn-
chus) (из Галлера).
hy — подъязычная дуга; с — нопула;
tr — трахея; I—III — жаберные дуги.
аппаратом, в особенности у клоачных, обнаруживающих, очевидно,
весьма примитивные соотношения. Кольцевидный хрящ тесно связан
еще с трахеальными хрящами, а щитовидный хрящ у клоачных явст-
венно обнаруживает свой состав из трех пар хрящей, несомненно,
представляющих собой первые три пары жаберных дуг (рис. 313), и обра-
зует одно целое с подъязычным аппаратом. У живородящих млекопитаю-
щих также нетрудно установить происхождение щитовидного хряща на
343
основании истории его развития — здесь в его состав входят лишь две
пары жаберных дуг, именно вторая и третья, между тем как передняя
их пара прилежит к щитовидному хрящу спереди, нс теряя своей само-
стоятельности; это задние рожки подъязычного аппарата. Впереди от
щитовидного хряща, также с брюшной стороны, лежит надгортанник,
происхождение которого, однако, иное и с висцеральным аппаратом не
связано (рис. 314). Он развивается в складке сли-
Рис. 314. Гортань косу-
ли; вид с левой стороны
(из Видерсгейма).
аг —черпаловидный и сг—
перстневидный хрящи;
ctr — хрящи трахеи; ер —
надгортанник; th — щито-
видный хрящ; tr—трахея.
зистой оболочки впереди от гортанной щели.
Такая складка наблюдается в зачаточном виде
и у некоторых рептилий, в особенности у яще-
риц. Надгортанник в виде клапана прикрывает
гортанную щель спереди во время акта глотания.
Между щитовидным хрящом, с одной сторо-
ны, и черпаловидными — с другой, тянутся бо-
ковые складки слизистой оболочки с эластиче-
ской тканью внутри (голосовые связки), которые
при передвижении последних хрящей могут натя-
гиваться и ослабляться, в зависимости от чего
меняется высота тона звуков, издаваемых при
их колебании. Впереди голосовых связок нередко
имеется еще вторая пара складок слизистой
оболочки (верхние, или ложные, голосовые связ-
ки), а между обеими парами складок по бокам
имеются более или менее глубокие ямки — мор-
ганьевы желудочки, сильно развивающиеся у не-
которых обезьян (особенно у человекообразных),
где они выпячиваются между щитовидным и
кольцевидным хрящами и играют роль резонато-
ров (гортанные мешки). Между гортанными хрящами наблюдаются иногда
и непарные гортанные мешки (у некоторых обезьян, китообразных, парно-
копытных и др.).
У китообразных надгортанник и черпаловидные хрящи вытягиваются
вперед в виде трубки, вдающейся в хоаны позади мягкого нёба. Это при-
способление обеспечивает дыхание животного и во время глотания пищи.
Подобное приспособление имеется у новорожденных сумчатых.
Трахея
Трахея имеется уже в виде довольно длинной трубки у некоторых
хвостатых амфибий, у которых ее стенка укрепляется обыкно-
венно неравномерно разбросанными хрящиками, иногда соединяющимися
по бокам между собой. Эти хрящи обнаруживают известную связь с гор-
танными хрящами, и потому им приписывают такое же висцеральное
происхождение (предположительно за счет пятой жаберной дуги), как и
Последним. У амфибий и рептилий можно отметить различные степени
диференцировки перстневидного и трахеальных хрящей, образующих
у них еще одну цельную систему.
У рептилий трахея хорошо развита, причем длина ее нахо-
дится в зависимости от общей длины тела и, в особенности, от длины шеи;
у ящериц она наименее значительна. Просвет трахеи поддерживается
открытым благодаря укреплению ее стенок помощью большого числа
прекрасно развитых хрящевых трахейных колец, которые, впрочем, у
многих рептилий являются неполными (со спинной стороны). Нижний
конец трахеи разветвляется на два коротких бронха, поддерживаемых
также хрящевыми кольцами.
В связи с удлинением шеи у птиц и трахея достигает значительной
длины. Трахейные кольца обыкновенно полные и иногда окостеневают.
344
Трахея нередко удлиняется еще более, нем шея, и образует тогда более
или менее сложные изгибы, помещающиеся иногда под кожей туловища или
внутри гребня грудины (у лебедей, журавлей, цапель). Такая длинная
трахея играет роль резонатора, и то же значение имеют и наблюдающиеся
на ней иногда вздутия. Голосовым аппаратом птиц является не гортань,
а так называемая нижняя гортань (syrinx), лежащая на границе
между трахеей и бронхами. Нижняя часть трахеи вздувается, образуя
резонатор — так называемый барабан. Источником звуковых колебаний
являются различно устроенные перепонки, располагающиеся частью в
области бронхов, частью уже в трахее; кроме того, нередко наблюдается
складка с костным или хрящевым скелетом — козелок (pessulus), —
вдающаяся в полость барабана от места схождания внутренних стенок
бронхов. Нижняя гортань имеет свою более или менее сложно дифереп-
цировапную мускулатуру, сокращением различных мышц которой ме-
няется натяжение отдельных перепонок и, следовательно, регулируется
высота звука. Эта мускулатура развивается (в отличие от висцеральной
мускулатуры гортани) путем обособления от подъязычной муску-
латуры.
Трахея млекопитающих обладает значительной длиной, под-
держивается хрящевыми кольцами (обыкновенно, неполными) и делится
на два явственных бронха, поддерживаемых такими же кольцами.
Легкие собственно
Легкие двудышащих рыб служат лишь добавочным органом дыхания,
так как нормально эти рыбы дышат жабрами. Только в период засухи,
когда животное зарывается в засыхающий ил (Protopterus) или когда
вода портится, легочное дыхание приобретает важное значение. Легкие
многоперых ганоидов (Polypterus) имеют гладкие стенки и, таким образом,
вряд ли играют заметную роль лаже в качестве добавочных органов
дыхания. То же самое касается, однако, и некоторых амфибий.
У постояннож'абериых амфибий (Necturus, Proteus) легкие имеют
вид таких же удлиненных гладкостенных мешков, как и у Polypterus, и при-
том также несимметрично развиты (левое длиннее правого) (рис. 315).
И здесь они, очевидно, имеют главным образом значение гидростати-
ческого аппарата. И не только у амфибий, сохраняющих жабры, но и
у многих других легочное дыхание имеет еще ничтожное значение. Во
многих случаях преобладающую роль играет диффузное кожное дыхание,
а также дыхание посредством слизистой оболочки ротовой полости и
глотки.
. В коже имеется весьма развитая сеть капилляров, получающая
венозную кровь по кожным артериям, отходящим от сердца вместе с
легочными. У некоторых хвостатых амфибий, и притом у наземных форм,
наблюдается даже исключительно такое кожное дыхание, и легкие совер-
шенно атрофируются (Desmognathus, Spelerpes, Plethodon). Кожное дыха-
ние имеет большое значение даже у бесхвостых амфибий, у которых
легкие развиты уже довольно высоко.
У большинства хвостатых амфибий внутренняя поверхность легких
увеличена путем образования сети перекладин, ограничивающих систему
ячеек, более значительно развитую в проксимальной части легких (рис.
315, В). В Перекладинах имеются гладкие мускульные волокна. У бес-
хвостых амфибий легкие имеют вид мешков с довольно сложной системой
ячеек, так как внутри отдельных более крупных ячей развивается система
вторичных перекладин, ограничивающих систему более мелких ячеек
(легочных '[пузырьков, или альвеол). Таким образом, стенка легочного
мешка может принять почти губчатый характер. Края перекладин, вдаю-
щихся во внутреннюю полость, выстланы цилиндрическим эпителием,
345
и таким образом центральная часть легочного мешка имеет характер
дыхательного пути.
Механизм дыхания у амфибий своеобразен. Животное набирает
воздух через ноздри в ротовую полость при опускании дна последней,
а затем при поднятии его ротовая полость сокращается, и воздух нагне-
тается в легкие (причем ноздри замыкаются особыми клапанами) через
расширенную гортанную щель. При сокращении брюшных мышц воздух
выходит из легких обратно в ротовую полость. Ясно, что земноводное
может дышать только с
закрытым ртом.
У высших наземных
позвоночных (амниот) с
развитием довольно мощ-
ного рогового покрова ко-
жи легкие приобретают
значение единственного
органа дыхания и соот-
ветственно диференциро-
ваны заметно выше. Лег-
Рис. 315. Схемы легких хвостатых амфибий (Л —
Necturus; В — саламандра) и рептилий (С — яще-
рица; D — черепаха или крупная ящерица).
Ьг — бронх; ipbr — внутрилегочный бронх; tr— трахея.
кие разрастаются впе-
ред в виде выступов по бо-
кам от бронхов, так что
последние открываются не
в передний конец лег-
кого, а на внутренней его стенке (это связано отчасти и с передви-
жением сердца, охватывающего бронхи своими дугами, назад в брюш-
ную полость). Стенки легких приобретают губчатый характер, а цен-
тральная полость диференцируется на систему внутрилегочных дыха-
тельных путей.
У рептилий легкие иногда еще довольно примитивны. У гат-
терии (Hatteria) они имеют простые ячеистые стенки и весьма сходны
с легкими амфибий, отличаясь от них главным образом $ем, что верхушка
легкого уже несколько выступает вперед (рис. 315, С). У ящериц в легком
появляются отдельные более крупные перекладины, отходящие от спин-
ной и брюшной поверхностей и делящие легкое на отделы. Эти перекла-
дины глубоко врезываются в полость легкого, оставляя у высших форм
свободным лишь сравнительно узкий центральный ход (рис. 315, D).
Так как края перегородок покрыты цилиндрическим мерцательным
эпителием, то ограничиваемый ими центральный ход получает значение
дыхательного пути, лежащего на продолжении бронха. В стенках этого
внутрилегочного продолжения бронха у черепах и крокодилов появляются
и охрящевения. Множество отверстий ведет из внутрилегочного бронха
в лежащие вокруг него отдельные камеры легкого. Стенки перегородок
являются ячеистыми, так же как и наружные стенки легочного мешка.
В отдельных камерах легкого перекладины также разрастаются, врезы-
ваясь во внутреннюю полость (рис. 315, D). Внутренние края этих пере-
кладин также покрываются мерцательным эпителием, и таким образом
и в'боковых камерах обособляются побочные дыхательные пути — внутри-
легочные бронхи второго порядка, ведущие в более мелкие подразделения
легкого, обладающие также ячеистыми стенками. Таким образом, легкое
приобретает сложный губчатый характер, и от центрального внутриле-
гочного бронха отходят боковые ветви — бронхи второго порядка, деля-
щиеся иногда еще дальше на бронхи третьего порядка (рис. 316). Стенки
мелких подразделений легкого оказываются ячеистыми. В некоторых
случаях и в стенках бронхов второго порядка появляются хрящи.
У ящериц и особенно у змей в задней части легких перекладины
бывают слабо развиты, а иногда стенки являются и совсем гладкими.
346
У гекконов, некоторых ящериц и особенно у хамелеонов легкие продол-
жаются в тонкостенные придатки — легочные мешки.
Дыхание рептилий происходит при периодическом расширении и
сжатии брюшной полости, а следовательно, и легких при помощи брюшной ,
а также и реберной мускулатуры.
У высших наземных позвоночных процесс обособления дыхательных
путей внутри самого легкого идет гораздо дальше, и в результате этого
развивается сложная система бронхов различного порядка.
Рис. 316. Схема строе-
ния легкого черепахи
(Thalassochelys) (по
Милави).
Ы I — IV—бронхи с 1-го
по 4-й порядок.
Рис. 317. Схема строения
легкого птицы (из Кингсли).
а — место соединения экто-
бронхов с воздушными мешка-
ми; ь—бронх; в—энтобронхи;
ес — эктобронхи; т —средний
бронх; р—парабронхи.
Рис. 318. Схема [воздуш-
ных мешков птицы
(из Галлера).
abd—брюшные мешки; cerv—
шейные, clav — межключич-
ный; thop — грудные мешки.
Легкие птиц представляют собой губчатые и притом мало растя-
жимые органы, помещающиеся в спинной части'грудной полости. Бронхи
входят в каждое легкое с брюшной стороны и продолжаются в виде глав-
ного, или среднего, внутрилегочного бронха до заднего конца легкого, г ;е,
однако, не заканчиваются слепо, а продолжаются вне легкого в полость
тонкостенного бронха воздушного мешка. От главного бронха отходят
вторичные бронхи, из которых обыкновенно четыре пронизывают все
легкое и продолжаются за его пределами, как и главный бронх, в виде
воздушных мешков (рис. 317). Кроме того, известное число вторичных
бронхов (10—14) кончается слепо. Между вторичными бронхами распо-
лагаются соединяющие их многочисленные, приблизительно параллель-
ные, анастомозы—парабронхи (рис. 317). От последних отходит множество
слепых радиально расположенных каналов (бронхиоли) с ячеистыми
стенками. Воздушные мешки располагаются не только в брюшной полости,
по разрастаются во все части тела, проникая своими разветвлениями
и внутрь костей (пневматизация костей). Различают следующие воздуш-
ные мешки (рис. 318): пара шейных мешков, от которых отходит
пара подмешёчных, непарный межключичный мешок, лежащий
впереди вилочки, пара переднегрудных, пара заднегруд-
н ы х- в пара брюшных. Пневматичными у птиц являются обыкно-
венно следующие кости: плечевая, коракоид, грудина и ребра, кроме того,
иногда бедро, вилочка, лопатка и другие кости. Воздушные мешки имеют
большое значение, уменьшая удельный вес птицы, а с другой стороны,
играют значительную роль при акте дыхания. Они снабжаются кровью
через посредство небольших сосудов, отходящих непосредственно от
спинной аорты. Соответствующие вены впадают в нижнюю полую вену.
347
При спокойном положении птицы дыхателыше движения состоят
в периодическом сжатии и растяжении грудной полости, совершаемых
главным образом при участии реберной мускулатуры, причем грудина
ритмически поднимается и опускается. Эти движения вызывают растя-
жение и сжатие воздушных мешков, благодаря чему воздух просасывается
через легкие. При полете поднятие и опускание крыла вызывают растя-
жение и сжатие подмышечных мешков, периодически сокращающаяся
грудная мышца давит на выросты грудных мешков, и таким образом
автоматически происходит непрерывное вентилирование легких. Следует
отметить, что, благодаря тому, что воздушные мешки при дыхании птицы
играют роль мехов, главная масса остаточного воздуха ограничена воз-
душными мешками, а в легких птиц происходит гораздо более совершен-
ная вентиляция, чем у других позвоночных. В связи с таким совершен-
ным дыханием и окисление происходит более энергично, и освобождается
значительное количество тепла.
Рис. 319. А — схема строения легкого
млекопитающего (ив Гертвига}. В—схема-
тический разрез черев легкое человека.
ad — альвеолярный ход; Ь—бронх; Ы—брон-
хиола; е — конечные пузырьки с альвеолами.
Птицы обладают постоянной тем-
пературой тела, притом очень вы-
сокой (превышающей температуру
тола млекопитающих).
В легких млекопитаю-
щих бронхи делятся на довольно
большое число вторичных бронхов
(рис. 319), а последние—на б ро н-
х и третьего и четвертого поряд-
ков, кончающиеся тонкостенными
трубочками — бронхиолями,
которые на конце вздуты, образуя
«воронку», или конечный пу-
з ы р е к с ячеистыми стенками.
Иногда не только конечное взду-
тие, но и сам канал покрыт
альвеолами и получает тогда
название альвеолярной трубки.
Бронхи выстланы изнутри цилин-
дрическим мерцательным эпите-
лием и поддерживаются собствен-
отделах из более или менее
ным скелетом, состоящим в начальных
правильных хрящевых колец, а в дальнейших ветвях — из небольших
хрящиков. Бронхиоли собственного скелета не имеют и выстланы куби-
ческим эпителием. Газовый обмен происходит в легочных альвеолах,
которые выстланы плоским эпителием и снабжены богатой сетью капил-
лярных сосудов.
Легкие млекопитающих нередко подразделяются на отдельные доли.
Они помещаются в грудной полости, которая отделена мускулистой диа-
фрагмой от брюшной. Акт дыхания совершается при помощи периоди-
ческого расширения и сжатия грудной полости вследствие: 1) расширения
и сжатия грудной клетки в результате изменения положения ребер при
помощи реберной мускулатуры и 2) сокращения и расслабления диафрагмы,
имеющей форму купола. Воздух при этом периодически всасывается и
удаляется из легкого (непосредственно он сменяется лишь в бронхах;
в мелких разветвлениях и в легочных пузырьках обмен происходит только
путем диффузии). При выдыхании известную роль играет и эластичность
легочных пузырьков и присутствие гладких мускульных волокон.
Легкие позвоночных, возникшие, повидимому, как добавочные органы
дыхания у рыб в виде мешковидных выростов последней пары жаберных
348
меткой, сохранили это значение у двудышащих рыб. У паземпых Позво-
ночных они постепенно приобрели значение единственного органа дыхания.
У амфибий они еще не играют столь большой роли и немногим отличаются
от легких двудышащих. Эволюция легких выражается в обособлении
более или менее сложных дыхательных путей и в увеличении дыхательной
Поверхности путем развития все более сложной сети перекладин, прида-
ющих стенкам легких губчатое строение. Более крупные перекладины
разбивают легкое на отдельные участки, внутри которых идет обособление
центрального дыхательного пути и периферической ячеистой части, в
которой в свою очередь идет такая же диференцировка. Таким образом,
в конце концов, развивается сложная система разветвлений внутриле-
гочных бронхов, завершающихся конечными пузырьками с ячеистыми
стенками (с легочными альвеолами).
ОРГАНЫ КРОВООБРАЩЕНИЯ
ОРГАНЫ КРОВООБРАЩЕНИЯ БЕСПОЗВОНОЧНЫХ
Обмен веществ в тканях и клетках животного организма возможен лишь в том
случае, если происходит постоянная смена окружающей их среды, т. е. непрерывная
доставка необходимых для них питательных веществ и кислорода и столь же непре-
рывное удаление продуктов разрушения и углекислоты. Таким образом, питание,
дыхание и выделение, т. е. основные жизненные функции клеток, немыслимы без по-
стоянного перемещения веществ внутри самого организма. В простейшем случае, у
низших животных, проведение питательных веществ и кислорода от места их воспри-
ятия к месту потребления, а также проведение экскретов от клеток к наружной среде
происходит путем диффузионных токов в тканевых жидкостях. У низших червей (ту-
беллярий) и у кишечнополостных передача питательных веществ отдельным частям
тела облегчается тем, что пищеварительная полость дает от себя многочисленные вы-
росты или каналы, по которым пищевые вещества доставляются по возможности ближе
к месту потребления. Движение жидкостей в паренхиме тела первоначально совершенно
неправильно; но затем обозначаются определенные пути, по которым происходит более
свободная циркуляция.
Таким образом, повидимому, в паренхиме обособились кровеносные сосуды, пред-
ставляющие собой систему каналов, которые могут местами переходить в более или
менее обширные полости. Содержимое сосудов — кровь — представляет собой, однако,
не просто межтканевую жидкость, замкнутую в определенную систему каналов. Изу-
чение строения и истории развития кровеносной системы показывает, что КрОвЬ пред-
ставляет собой жидкую соединительную ткань, ограниченную стенками из плотной
соединительной ткани, киотни которой- частью приобрели значительную сократимость.
С разрастанием полостей половых желез и образованием вторичной полости тела более
крупные кровеносные сосуды попали частью в промежутки между стенками полости
тела. Здесь за счет последних мог развиться более значительный мускульный слой
сосудов.
Содержимое сосудов — кровь — представляет собой белковую жидкость с пла-
вающими в ней обыкновенно амёбообразными клетками. В жидкости переносятся как
питательные вещества к местам потребления, так и экскреты к органам выделения.
Кровяные клетки несут различные функции, но главным образом удаляют различные
плотные или нерастворимые органические вещества, разрушенные остатки клеток,
а также посторонние тела, микроорганизмы и т. д., причем пользуются своей подвиж-
ностью и способностью к внутриклеточному пищеварению^Почти всегда кровь служит
и для перенесения кислорода от органов дыхания ко всем клеткам организма. Это до-
стигается благодаря присутствию в крови особых белковых веществ, способных хими-
чески связывать кислород, но вместе с тем легко его и отдающих при пониженном его
парциальном давлении в окружающей среде. Обыкновенно это пигменты: красный —
гемоглобин, синий — гемоцианин и др., но иногда это вещество бесцветное. У беспо-
звоночных они почти всегда растворены в жидкости, т. е. в кровяной плазме, но иногда
(немертины, некоторые аннелиды), как и у позвоночных, заключаются в кровяных
клетках.
Движение крови по сосудам в простейшем случае крайне нерегулярно и дости-
гается главным образом давлением со сторовы сокращающейся мускулатуры тела и
кишечника. Обычно же наблюдается правильное движение крови по определенным
путям, что достигается различным образом, но чаще всего благодаря сократимости
стевок самих сосудов. Иногда части сосудов, обладающие более значительной муску-
латурой, принимают на себя роль органов, движущих кровь по сосудам, и тогда по-
лучают название сердец.
Кровеносная система называется замкнутой, если кровь движется только по со-
судам, ограниченным собственными стевками (кольчатые черви, хордовые), и незамк-
349
йУ’Гой, если сосуды местами прерываются щелевидными пространствами, лишенными
собственных стенок (лакуны, синусы), что наблюдается гораздо чаще.
Кровь, насыщенная кислородом, называется артериальной, а кровь, отдавшая
кислород и поглотившая углекислоту из тканей, называется венозной. Если кровь
окрашена гемоглобином, то артериальная кровь имеет яркокрасный цвет, а венозная —
более темный. У ланцетника кровь бесцветная, а у позвоночных окрашена в красный
цвет. Кровь позвоночных состоит из бесцветной кровяной плазмы со взве-
шенными в ней клетками различных типов. Различают: 1) эритроциты, или красные
кровяные шарики, содержащие гемоглобин — небольшие плотные диски (у млекопи-
тающих безъядреные), 2) лейкоциты, бесцветные подвижные амёбообразные клетки,
и 3) тромбоциты, очень маленькие плоские клетки, играющие известную роль при
свертывании крови.
а
Рис. 320. Схемы кровообращения кольчатого червя, ланцетника или
низшей рыбы и наземного позвоночного.
abr — жаберные артерии; яс — сонная артерия; яо. d —спинная аорта; яо. в — брюшная
аорта; ар — легочная артерия; d—спинной сосуд, d.с — кювьеров протон; v — брюшной
сосуд; а. с. а. и -в. с. р. — передняя и задняя кардинальные вены; гс — вадняя полая вена;
vh — печеночная вена; vp — легочная вена; vph — воротная вена печени; pci — подкишеч-
ная вена.
Стенки сосудов состоят из внутренней тонкой эндотелиальной выстилки (intima),
среднего эластического слоя с гладкими мускульными клетками (media) и наружного
соединительнотканного слоя (adventitia).
Если имеется сердце или, по крайней мере, главный пульсирующий сосуд, то
различают артерии — сосуды, несущие кровь от сердца, рассчитанные на более зна-
чительное кровяное давление и потому обладающие более значительными мускуль-
ными и эластическими слоями, и вены — сосуды, несущие кровь к сердцу, более тон-
костенные и снабженные часто клапанами, регулирующими направленна тока крови.
Артерии и вены соединяются между собой в случае аамкнутой системы сетью сосудов
очень мелкого калибра — капиллярами, тонкие стенки которых состоят только иа
андотелия, покрытого слоем продольных мускульных клеток, охватывающих сосуды
своими ветвями. Весь обмен веществ между кровью и тканями происходит путем осмоса
череа стенки капилляров. Иногда сосуд сразу распадается на сеть капилляров, кото-
рые аатем сразу же вновь собираются в выносящей сосуд: такие сети носят название
«чудесных сплетений» (retepnirabile).
Наиболее примитивная кровеносная система имеется у немертин, у которых она
состоит главным образом ив трех продольных сосудов: спинного и двух боковых. Из
350
спинного сосуда кровь течет вперед в голову, где он посредством сосудов, охваТУваЮ-
щих мовг, соединяется с двумя боковыми сосудами, по которым кровь течет навад.
У аннелид имеется замкнутая кровеносная система, состоящая из спинного со-
суда, по которому кровь течет вперед в голову, и брюшного сосуда, по которому кровь
течет навад (рис. 320).~Оба продольных главных сосуда соединяются между собой по-
перечными анастомозами, идущими по стенкам кишечника, где они образуют обильную
сеть разветвлений (висцеральные сосуды), и по стенкам тела, где они отдают ветви к
боковым придаткам, а также к жабрам, если последние имеются (париетальные со-
суды). Кровь движется благодаря пульсации сократимого спинного сосуда,играющего,
следовательно, роль сердца, а также иногда одной или нескольких пар поперечных
сосудов, которые тогда образуют заметные расширения.
У членистоногих кровеносная система незамкнутая и состоит из длинного сокра-
тимого спинного сосуда —сердца, продолжающегося в короткие артериальные стволы,
изливающие кровь в щелевидные пространства между органами. Обратно кровь сте-
кает в участок полости тела, в котором лежит сердце, и всасывается последним черев
парные отверстия, снабженные клапанами.
У моллюсков кровеносная система также незамкнутая, но, кроме артериальных
сосудов, имеются и венозные, а сердце обнаруживает деление на тонкостенные отделы—
предсердия (по числу соответствующие числу жабер), куда впадают вены, и толсто-
стенный желудочек, от которого отходят артерии.
У хордовых животных расположение сосудов очень сходно с расположением
их у аннелид (рис. 320). Имеются продольные брюшной и спинной сосуды, связанные
между собой анастомозами как в стенках кишечника, так и в стенках тела. Так как
передняя часть кишечника преобразована в дыхательный аппарат, то здесь происхо-
дит окисление крови, и кишечные сосуды этой области получают название жаберных.
Сердце представляет собой расширение брюшного сосуда.
Кровообращение у оболочников своеобразно в том отношении, что у них сердце
периодически сокращается то в одном, то в другом направлении, а соответственно ме-
няется и направление кровяного тока.
У ланцетника кровь бесцветная и течет по замкнутой системе сосудов, но обо-
собленного сердца здесь нет (см. рис. 45). Имеются два крупных кровеносных сосуда|
брюшной сосуд, по которому кровь течет вперед, и спинной сосуд, по которому она
течет навад. Оба сосуда связаны между собой поперечными анастомозами, образующими
сеть капилляров в стенках кишечника. В этом схематизированном виде кровеносная
система ланцетника вполне сходна с таковой кольчатого червя, с той лишь разницей,
что ток крови в сосудах имеет обратное направление и пульсирует не спинной, а брюш-
ной сосуд. Однако дальнейшая диференцировка вполне специфична<Брюшной сосуд
прерывается посредине системой капилляров (на печеночном выросте), делящей его
на пульсирующий передний отдел, лежащий под жаберной областью кишечника, —
поджаберный сосуд, и задний отдел, идущий вдоль брюшной стенки собственно
кишки, — подкишечную вену. Поджаберный сосуд, называемый также брюшной аор-
той (aorta ventralis), гонит венозную кровь вперед, и отдает по пути боковые ветви,
идущие вверх в жаберные перегородки. Ив последних окисленная кровь собирается
в пару сосудов, идущих вдоль спинной стенки глотки. От этих-наджаберных сосудов
отходят небольшие сосуды вперед в голову, а позади они соединяются в непарный ствол,
идущий под хордой навад вдоль всего тела, отдавая по пути ветви к мышцам, кишеч-
нику и другим органам; это — спинная аорта (aorta dorsalis), главный артериальный
ствол, по которому кровь течет навад. От стенок кишечника кровь собирается в не-
парный брюшной сосуд — подкишечную вену, которая несет кровь вперед к печеноч-
ному выросту, где она распадается на оплетающую его сеть капилляров, представля-
ющих собой так называемую воротную систему печени. Отсюда кровь вновь собирается
в сосуд, образующий небольшое расширение на месте впадения в него пары так назы-
ваемых кювьеровых протоков, и продолжается вперед в виде уже рассмотрен-
ной брюшной аорты. Кювьеровы протоки составляются из пары вен, несущих кровь
от передней половины тела, и пары вен, идущих от задней половины. Эти сосуды, но-
сящие название передних и задних кардинальных вен, собирают кровь со стенок тела
по метамерно расположенным веточкам.
Таким образом, у ланцетника имеется высокодиференцированная система кро-
веносных сосудов, но нет обособленного сердца, и кровь перегоняется пульсацией
самих сосудов, в особенности брюшной аорты и оснований жаберных сосудов.
ОРГАНЫ КРОВООБРАЩЕНИЯ ПОЗВОНОЧНЫХ
У низших позвоночных (рис. 320, 27) кровеносная система по су-
ществу вполне сходна с таковой ланцетника и отличается от нее главным
образом присутствием сердца. Последнее состоит из двух камер —
тонкостенного предсердия и мускулистого желудочка, к
которым позади присоединяется еще мешковидная венозная пазуха, а
впереди мускулистая трубка — артериальный конус. Все эти отделы несут
351
венозную кровь и располагаются последовательно (венозная пазуха,
предсердие, желудочек и артериальный конус), отделяясь друг от друга
клапанами, препятствующими изменению направления тока крови.
От сердца вперед ведет крупный артериальный ствол — брюш-
ная аорта, от которой по бокам отходят приносящие жаберные
артерии, идущие по жаберным перегородкам и распадающиеся на
систему капилляров в жаберных лепестках, где происходит окисление
крови. Из жабер артериальная кровь течет по выносящим жаберным ар-
териям к спинной стенке глотки, где опа собирается или непосредственно
в непарный ствол — спинную аорту, или предварительно в пар-
ные наджаберные сосуды, сливающиеся позади в спинную аорту. От над-
жаберных сосудов вперед в голову отходит пара сонных артерий
(a. carotides). Но спинной аорте, лежащей непосредственно под хордой,
кровь течет назад вдоль всего тела, отдавая по пути ветви как к стенкам
тела, так и к внутренним органам и, в частности, к передним конечностям
(a. subclaviae), к кишечнику (a. coeliaco-mesenterica), к почкам (a. renales),
к половым органам (a. gemtales), к задним конечностям (а. Шасае), и про-
должается позади в виде хвостовой артерии (a. caudahs).
Венозная кровь из стенок кишечника собирается в сосуды, направ-
ляющиеся к печени (v. portaruni hepatis), где они распадаются на капил-
ляры, образую воротную систему печени. Из печени кровь
по печеночным венам течет в венозную пазуху,сердца. В последнюю впа-
дают и парные к ю в ь е р o_bjj—(ductus Cuvieri), несущие
венозную кровь от стенок тела и некоторых органов. Из хвоста венозная
кровь направляется по хвостовой вене (v. caudalis) к почкам, где эта вена
распадается па капилляры, образуя так называемую воротную си-
стему почек (v. portarum renarum). Почечные вены (v. renales),
направляются вперед, образуя парные задние кардинальные
вены (v. cardinales posteriores), лежащие по бокам от спинной аорты и
принимающие в себя также сосуды, идущие от половых органов (v. geni-
tales), и метамерпые вены, несущие кровь от стенок тела. На уровне сердца
задние кардинальные вены соединяются с парными же передними
кардинальными (v. cardinales anteriores s. jugulares), или ярем-
ными венами, несущими кровь из головы, и образуют кювьеровы протоки,
впадающие в венозную пазуху. От задних конечностей кровь возвраща-
ется по боковым венам (v. laterales), лежащим в брюшной стенке тела и
направляющимся к кювьеровым протокам, принимая в себя по пути вены,
идущие от передних конечностей (v. subclaviae).
У наземных позвоночных (рис. 28, 320) выключается жаберное крово-
обращение и взамен его устанавливается непосредственное сообщение по
одной паре дуг от брюшной аорты к спинной. С приобретением легочного
дыхания устанавливается новый, добавочный крут кровообращения, разви-
вающийся отчасти за счет задней пары жаберных сосудов, которые дают
начало легочным артериям (a. pulmonales), несущим веноз-
ную кровь к легким. Окисленная кровь возвращается по легочным
венам (v. pulmonales) назад в сердце. Последнее получает теперь,
кроме венозной, и артериальную кровь, и для разделения этих двух токов
в его основных камерах, сначала в предсердии, а затем в желудочке, раз-
виваются перегородки. Этими перегородками сердце разделяется на ле-
вую, артериальную, и правую, венозную, половины. Из «двукамерного»
сердце становится «четырехкамерным». В венозной системе также проис-
ходят изменения — задние кардинальные вены замещаются уна земных
позвоночных непарной н и жнедй"1гЬ^6йве н о й (v. cava inferior).
Парные боковые вены соединяются"в~непарную брюшную (v. abdo-
minalis). У высших позвоночных воротная система почек исчезает, и исче-
зает одна из дуг аорты.
352
Характерным для позвоночных является еще обособление системы
тонкостенных лимфатических сосудов, образующих особо
значительную сеть в кишечнике, в подкожной соединительной ткани, в
мозговых оболочках и служащих посредниками между тканями и веноз-
ными сосудами. Лимфатические сосуды несут бесцветную жидкость с
амёбообразными клетками (лимфоцитами) — лимфу; они наполняются
путем диффузии жидкости из тканей и из кишечника и соединяются в
более крупные стволы (образующие иногда большие мешковидные расши-
рения), снабженные клапанами и впадающие, в конце концов, в вены.
Мелкие разветвления лимфатических сосудов образуют сеть, частью окан-
чиваются слепо, но также посредством небольших отверстий сообщаются
с полостью тела. Через посредство лимфатической системы в кровеносную
систему возвращается жидкость, просачивающаяся из капиллярных со-
судов в ткани, а также передаются питательные жидкости, всасываемые
в стенках кишечника.
Развитие сердца и сосудов
У рыб и амфибий сердце развивается в области задней части головы
непосредственно под глоткой. В брюшном мезентерии оба листка расхо-
дятся, и между ними и энтодермой появляются отдельные мезодермаль-
ные сосудообразовательные клетки, образующие здесь сплошную клеточ-
ную полосу. Клетки закладки располагаются затем в один слой, ограни-
чивающий трубчатую полость. Этот слой представляет собой зачаток вну-
тренней стенки сердца (эндокардия) и прилежащих частей крупных со-
судов. Прилежащие стенки полости тела утолщаются, и мезенхима, раз-
вивающаяся за их счет, дает мускулатуру сердца (миокардий), а остав-
шаяся часть стенок образует перикардий, выстилающий околосердечную
полость, которая обособляется от остальной полости тела. Правый и левый
участки околосердечной полости здесь рано сливаются между собой, окру-
жая сердце со всех сторон.
У амниот сердце развивается из двух закладок, широко раздвинутых
на стадии, когда энтодерма еще распластана на поверхности желтка. Обо-
собляются отдельно левая и правая перикардиальные полости, и во вда-
ющихся в них изнутри выступах появляются зачатки правой и левой
половин сердца. По мере того как энтодерма замыкается в кишечную
трубку, обе закладки сходятся между собой под кишкой, где образуется
вентральный мезентерий (мезокардий). На продолжении сердечной трубки
впереди и позади из подобных же мезодермальных зачатков развиваются
крупные кровеносные сосуды.
Сердце закладывается сначала в виде прямой трубки, которая затем
быстро растет и изгибается в виде буквы S, причем задняя половина за-
чатка, остающаяся более тонкостенной, сдвигается на спинную сторону
и перемещается далее вперед, образуя предсердие, между тем как передняя
часть остается на брюшной стороне, образуя желудочек, мускульная
стенка которого сильно утолщается. Прилежащий к предсердию сзади
тонкостенный отдел сердечной трубки образует венозную пазуху. Впереди
желудочка у низших позвоночных развивается узкая, но мускулистая
трубка — артериальный конус. Все отделы сердца отделяются друг от
друга перехватами. Затем эндокардий образует складки или клапаны,
разграничивающие отделы сердца друг от друга; кроме того, большее или
меньшее число клапанов развивается на протяжении артериального ко-
нуса. У высших позвоночных развиваются еще перегородки, разделяющие
обе главные камеры сердца вдоль — на правую и левую половины.
От сердца вперед развивается непарный артериальный ствол — брюш-
ная аорта, от которой по жаберным перегородкам отходят крупные пар-
ные сосуды — артериальные дуги или дуги аорты (рис. 321),
353
охватывающие глотку и соединяющиеся на спинной ее стороне в спинную
аорту, отдавая вперед в голову сонные артерии. Число артериальных дуг
соответствует числу висцеральных дут и обычно у рыб равняется 6—7,
из них две первые называются по своему положению челюстной и подъ-
язычной. У рыб остатки первых двух дуг могут сохраняться в виде незна-
чительных сосудов, а следующие артериальные дуги постепенно разделя-
ются на приносящие и выносящие жаберные артерии с сетью соединяющих
их капилляров. У наземных позвоночных первые две дуги также рано
Рис. 321. Преобразование дуг аорты у различных позвоночных
(вид с брюшной сторовы) (по Боасу из Кингсли).
А — исходное расположение у зародыша; В — рыба (двудышащая); С — хвостатая'’амфибия}
D — лягушка; Е—змея; F — ящерица; G — птица; Н— млекопитающее; с — желудочно-ки-
шечная артерия; da — спинная аорта; d. с. — ductus carotid; db — боталлов проток; ес и
ic — наружная и внутренняя сонные артерии; р — легочная артерия, s — подключичная
артерия; v. а — брюшная аорта. Черным обозначены артерии, несущие венозную кровь,
Исчезнувшие сосуды обозначены пунктиром.
атрофируются, а следующие испытывают характерные преобразования
(рис. 321). Третья пара дут (идущая по первой жаберной дуге) теряет свою
связь со спинной аортой и продолжается вперед в сонные артерии. Эмбри-
ональная связь этой дуги со спинной аортой носит название сонного (duc-
tus caroticus) протока и иногда (у змей и ящериц) сохраняется и у взрос-
лого животного. Четвертая пара дуг получает значительное развитие и
дает начало собственно дугам аорты (или корням аорты) взрослого
животпого. Эти дуги симметрично развиты у амфибий и рептилий. У птиц
левая дуга атрофируется, и сохраняется только правая. Умлекопитающих
во в^рбслом состоянии сохраняется только леван._дуга аорты (рис. 321).
Пятая пара дут исчезает и только у хвостатых амфибий сохраняется во
взрослом состоянии в виде незначительного протока. Шестая дуга дает
от себя легочные артерии jt теряет связь со спинной аортой. Эмбриональ-
ная < гязь легочных артерий со спинной аортой носит название боталлова
протока и сохраняется во взрослом состоянии у хвостатых амфибий, у
гаттерии и у черепах. Седьмая дуга у наземных позвоночных не развива-
ется вовсе.
354
У зародыша кровь течет из брюшной аорты по артериальным дугам
в спинную аорту, откуда разносится по всему телу и, между прочим, к
кишечнику и по желточным артериям (a. omphalo-mesen-
tericae) — к стенкам желточного мешка (рис. 322). От последнего кровь
возвращается по желточным венам (v. omphalo-mesentericae)
сначала прямо в венозную пазуху, а затем, с развитием воротной системы
печени, в последнюю. В стенках желточного мешка происходит не только
извлечение питательного материала, но и насыщение крови кислородом.
У высших позвоночных (амниот) с развитием аллантоиса к последнему
переходит роль эм-
брионального органа
дыхания (а у живо-
родящих млекопи-
тающих он служит
и для извлечения пи-
тательного материа-
ла из плаценты).
Аллантоис снабжает-
ся, как и задняя киш-
ка, выростом которой
он является, спе-
циальной ветвью об-
щей подвздошной ар-
терии (a. iliaca com-
munis)— пупочны-
ми артериями
(a. allantoidis s. um-
bilicales). Вены, не-
Рис. 322. Схема зародышевого кровообращения высшего
позвоночного (ив Кингсли).
аа — дуги аорты; al — аллантоис; ап — задний проход;
са, cv — хвостовая артерия и вена; da — Спинная аорта;
de — проток Кювье; h — сердце; ha — подвздошная ар-
терия (пупочная); / — яремная вена; ота, omv — желточ-
ная артерия и вена; ре — задняя кардинальная вена;
si — подкишечная вена; st — краевой синус; va — брюш-
ная аорта; у — желток; ys — желточный стебелек.
сущие артериальную кровь из стенок аллантоиса, получают наимено-
вание пупочных вен. Они несут кровь в сердце сначала прямо
через кювьеровы протоки, а потом вместе с желточной веной через
воротную систему печени. Вдоль нижней стенки кишки у зародыша
тянется подкишечная вена, которая позади продолжается в
хвостовую вену, а впереди принимает в себя желточную вену. На ее пути
к сердцу лежит печень, по мере развития которой вена распадается на
сеть капилляров, образуя воротную систему печени. С развитием карди-
нальных вен хвостовая вена теряет связь с подкишечной веной и соеди-
няется с кардинальными посредством сети капилляров в ткани почек,
образуя воротную систему почек. У наземных позвоночных за счет задних
частей кардинальных вен развивается задняя полая вена, которая впереди
развивается независимо,
Сердце;
Главную толщу стенок сердца составляет его мускульный сдой,
или миокарлип. который изнутри выстлан однослойным плоским эндо-
телием — эндокардием, а снаружи — соединительнотканным слоем с
таким же плоским целотелием, составляющим перикардий. Внутренняя
стенка сердца не является гладкой, а образует сеть перекладин, или тра-
бекул, которая особенно сильно развита в желудочке, где нередко вну-
тренняя стенка имеет совершенно губчатый характер.
Вены впадают в сердце обыкновенно в косом направлении, так что
при сокращении стенок сердца их просвет спадается, и кровь но. может
поступить из него обратно в вены. Впрочем, устья вен могут, кроме того,
быть защищены и клапанами. Клапаны имеются и на границе между отде-
лами сердца. Между венозной пазухой и предсердием имеется пара с и п у-
а тд) .и а л_ь н ы х к л а п а ц о в. Между предсердием и желудочком
атриовентрикулярное отверстие снабжено обыкновенно также двумя п е-
355
репончатыми клапанами, и, наконец, в артериальном ко-
нусе или вообще на границе между желудочком и артериальным стволом
имеется большее или меньшее число карманообразных кла-
панов.
Сердце рыб (рис. 323) несет только венозную кровь и состоит из двух
главных камер — предсердия (atrium) и желудочка (ven-
triculus). В этом «двукамерном» сердце имеются, однако, еще два отдела:
тонкостенный мешок — венозная пазуха (sinus venosus), позади
предсердия (рис. 324) и мускулистая трубка — артериальный
конус (conus arteriosus) — впереди желудочка, у начала артериаль-
Рис. 323. Схематический продольный раврев черев сердце различных рыб: А — села-
хий; В — ганоида (Amia); С — костистой рыбы (по Боасу). В В и С предсердие не
изображено.
а — предсердие; b — артериальная луковица; с — артериальный конус; k — клапаны;
а — венозная пазуха; t — брюшная аорта; v — желудочек.
кого ствола (рис. 323). Эти отделы самостоятельно пульсируют и обладают,
как и главные камеры сердца, поперечнополосатой мускулатурой.
Венозная пазуха имеется у всех низших позвоночных, а артериальный
конус отсутствует у круглоротых и у~КЙсших рыб. У селахий и у
ганоидов в артериальном конусе имеется несколько поперечных
рядов карманообразных клапанов, края которых подвешены сухожиль-
ными тяжами к стенке конуса (рис. 325). Из костных ганоидов у Lepidos-
teus имеется еще очень хорошо развитый конус с большим числом клапа-
нов, но у Amia имеются лишь три поперечных ряда клапанов (рис. 323).
У костистых рыб иногда наблюдаются еще два поперечных
ряда клапанов (Albula и др.), но конус их, во всяком случае, совсем ру-
диментарен, и обычно от него остается лишь один ряд клапанов на гра-
нице между желудочком и артериальным стволом. Последний уже у Amia
образует у основания эластическое вздутие — артериальную
луковицу (bulbus arteriosus), которая у костистых рыб получает
значительное развитие (рис. 323, В, ).
У двудышащих рыб в связи с появлением легочного кро-
вообращения наблюдается ряд интересных изменений. Прежде всего в
предсердии появляется перегородка, отходящая от брюшной его стенки
и не сполна подразделяющая его на правую й левую части. Эта перего-
родка вдается отчасти и в желудочек через атриовентрикулярное отвер-
856
стие. В правую половину поступает вся венозная кровь, а в левую —
артериальная кровь, приносимая по легочной вене из легких. В артери-
альном конусе имеются также приспособления к разделению артериаль-
ного и венозного токов крови. В артериальном конусе у цератода (Сега-
todus) один идущий по спирали продольный ряд клапанов получает пре-
обладающее развитие и дает, таким образом, спиральную перегородку.
У Protopterus эти клапаны вполне сливаются между собой в непрерывную
складку. Благодаря спиральному изги-
бу этой перегородки артериальная
кровь из левой половины сердца пере-
ходит в конусе на брюшную сторону,
от которой отходят две передние
пары дуг, несущие кровь в голову по
сонным артериям (у Ceratodus эти
дуги являются жаберными сосудами,
Рис. 324. Сердце и брюшная аорта
акулы (Mustelus) со спинной стороны
(по Нирштрасу).
ао — брюшная аорта; at — предсердие;
d. с — кювьеров проток; а. о — веноз-
ная пазуха; т — желудочек; о. Л —
печеночные вены; 2—в жаберные^ ар<
тернн.
Рис. 325. Вскрытое сердпе акулы (Alopiae)
спереди (по Гудричу).
at — предсердие; a v — клапаны артериального
конуса (с); Ь — bulbus aortae; de — кювьеров
проток; > «а — атриовентрикулярное отверстие;
vn — желудочек.
но у Protopterus жабры здесь редуцируются, и дуги непрерывны). Веноз-
ная кровь из правой половины сердца переходит в конусе на спинную
сторону, от которой отходят две последние пары жаберных артерий, не-
сущие кровь к жабрам и легким, где она окисляется.
У а м ф и б и й (рис. 326) сердце также является «трехкамерным»,
так как имеется перегородка (обыкновенно с отверстиями), подразделя-
ющая предсердие на правую — венозную и левую — артериальную по-
ловины. Имеется венозная пазуха, соединяющаяся с правым предсердием
отверстием, защищенным двумя клапанами. Атриовентикулярное отвер-
стие снабжено двумя клапанами, сидящими с брюшной и со спинной сто-
рон, свободные края которых укреплены сухожильными и мышечными
тяжами на стенке желудочка. Последний не поделен, но обладает чрезвы-
чайно развитой сетью мускульных перекладин, глубоко врезывающихся
во внутреннюю полость и, несомненно, препятствующих свободному сме-
шению артериальной и венозной крови. Артериальный конус снабжен
клапанами у основания и на конце, а кроме того обладает длинным про-
дольным спиральным клапаном (напоминающим клапан у Protopterus).
Правый и левый антериальные стволы поделены горизонтальной пере-
557
городкой на (брюшной отдел, от которого отходят передние пары дуг (сон-
ные и системные дуги аорты), и спинной отдел, от которого отходит задняя
пара дут (6-я), несущая кровь в легочные артерии. Таким образом, здесь
наблюдается такое же распределение крови, как у двудышащих рыб.
Артериальная кровь из легочных артерий поступает в левую половину
сердца, поворачивает вдоль спирального клапана в конусе на брюшную
сторону и идет по передним дугам в сонные артерии и спинную аорту.
Венозная кровь поступает из венозной пазухи в правую половину сердца,
поворачивает в конусе на спинную сторону и направляется по последним
Рис. 326. Вскрытое сердце лягушки
(ио Керру).
а — вонд, пропущенный в легочный канал (ср.);
Ь — вонд, пропущенный в канал системной дуги
аорты (са); с — вонд, пропущенный в сонный
канал (сс); at. s. и at. d. — левое и правое предсер-
дии; con — артериальный конус; s. ар. — гори-
вонтальная перегородка (отделяющая легочную
артерию, отмеченную аондом a); s. at. — перего-
родка предсердий; s. « — отверстие венозной
пазухи; ». at. ®п — атриовентикулярные клапаны;
в. р — отверстие легочной вены; в. sp — спи-
ральный клапан артериального конуса.
Рис. 327. Рис. 327. Сердце крокоди-
ла со спинной стороны (по Резе).
а. са — об11хая сонная артерия; A. d. и
A. s. — правая и левая дуги аорты; А.
т.— arteria mesenterica; ар — легочные
артерии; at. s. и at. d. — левое и пра-
вое предсердия; as — правая и левая
подключичные артерии; v. сс — венеч-
ная вена; d. с. d. и d. с. s. — передние
полые вены; vp — легочные вены; г.
с. г. — задняя полая вена.
дугам в легочные и кожные артерии для окисления. Так как, однако, это
разделение токов крови не является совершенным, то у амфибий имеются
и иные приспособления для направления токов крови по определенному
руслу, с чем мы познакомимся при изучении артериальной системы.
В сердце рептилий (рис. 327) наблюдается более совершенное
разделение обоих токов крови. Кроме полной перегородки предсердий,
имеется еще перегородка, делящая желудочек на две половины.
Перегородка эта, правда, обычно неполная — она отходит от брюш-
ной стенки и не доходит до спинной. Однако при сокращении (систоле)
желудочка она может дойти до спинной стенки, и таким образом дости-
гается полное разобщение обеих полостей. Только у крокодилов имеется
полная перегородка, отделяющая левый желудочек от правого, так что
сердце становится «четырехкамерным».
358
Венозный синус редуцируется и входит в состав правого предсердия.
У гаттерии он еще ясно различим извне, а у других рептилий он может
быть найден в правом предсердии в виде незначительного отдела, огра-
ниченного двумя характерными клапанами. Артериальный конус реду-
цирован и, невидимому, вошел в состав стенки желудочка, но от него
остался один поперечный ряд клапанов, которые отделяют полость же-
лудочка от артериального ствола.
Артериальный ствол, поделенный у амфибий продольными перего-
родками, разделяется здесь сполна на три сосуда, самостоятельно отхо-
дящие от желудочка. Один сосуд отходит от правой половины желудочка
и делится на обе легочные артерии (задняя пара дуг), остальная часть
ствола разделяется до основания на правый и левый сосуды и образует
обе системные дуги (корни) аорты. Из дуг аорты правая дуга отходит от
левой половины желудочка и получает артериальную кровь, а левая —
от правой половины желудочка, вблизи свободного края перегородки,
и получает смешанную кровь. Оба корня аорты перекрещиваются, таким
образом, у основания друг с другом. На месте отхождения дуг аорты и
легочных артерий имеются карманообразные клапаны.
Перегородка предсердий делит и атриовентрикулярное отверстие
на два, причем каждое отверстие получает по половине первоначальных
клапанов (брюшного и спинного), которые срастаются вдоль перегородки
между собой. Таким образом, каждое атриовентрикулярное отверстие по-
лучает по одному перепончатому клапану на внутренней стороне. У кро-
кодилов в правом атриовентрикулярном отверстии на наружной стенке
развивается еще мускулистая складка, играющая также роль клапана
(мускульный клапан).
У пти ц в правом атриовентрикулярном отверстии перепончатый кла-
пан редуцируется, а мускульный клапан развивается сильнее и является
здесь единственным замыкательным клапаном. В левом атриовентрику-
лярном отверстии перепончатый клапан подразделяется на три отдельные
створки.
Венозный синус у птиц совершенно редуцирован, и вены открыва-
ются непосредственно в правое предсердие. Не только предсердия, но и
желудочки сполна разделены (сердце «четырехкамерное»). Правый более
тонкостенный желудочек в значительной мере охватывает собой толсто-
стенный левый желудочек. От правого желудочка общим стволом отходят
обе легочные артерии, а от левого отходит единственная здесь правая дуга
аорты, получающая чисто артериальную кровь, между тем как левая дуга
совершенно атрофировалась.
Таким образом, у птиц, наконец, наступило полное разделение арте-
риального и венозного токов крови не только в самом сердце (как у кро-
кодилов), но и в сосудах.
У млекопитающих сердце так же, как и у птиц, четырех-
камерное. И у млекопитающих венозная кровь вполне отделена от арте-
риальной не только в самом сердце, но и в сосудах. Исчезнувшей дугой
здесь является, однако, не левая, а правая. Брюшная аорта берет начало
от левого желудочка, а общий ствол легочных артерий—от правого.
Венозный синус вполне слит с правым предсердием, а его клапаны сохра-
нились у впадения нижней полой и венечной вен.
Правый атриовентрикулярный клапан разделился на три створки
(valvula tricuspidalis), а левый — на две (valvula bicuspidalis). Свободные
края этих клапанов прикреплены посредством сухожильных нитей (chor-
dae tendineae) к мускульным выступам (mm. papillares) внутренней стенки
соответствующего желудочка (у клоачных их не имеется).
359
Артерии
У рыб из артериальных дуг зародыша развиваются жаберные сосуды,
приносящие и выносящие, с соединяющей их сетью капилляров в'жабер-
ных лепестках (рис. 328). Так как пятая жаберная дуга жабры не несет,
то жаберные сосуды развиваются только в подъязычной дуге и в четырех
первых жаберных дугах. Таким образом, у низших позвоночных рыб име-
ется пять приносящих жаберных артерий, из которых первая носит на-
звание гиоидной. У селахий в каждой жаберной дуге имеется, по две выно-
сящие жаберные артерии, соответствующие каждой полужабре. Обе вы-
носящие жаберные артерии каждого мешка соединяются между собой
в один сосуд, несущий кровь прямо в спинную аорту.
Рис. 328. Жаберное кровообращение у рыб и у амфибий.
А — схема, показывающая соотношение между приносящими и выносящими сосудами у се-
лахий. aod и ооо — спинная и брюшная аорты, перереаанные поперек; о/ — приносящая и
е/ — выносящая жаберные артерии, соединенные капиллярами в жаберных лепестках,
В — схема, показывающая соотношение между жаберными сосудами у ганоидных рыб (обе
схемы по Северцову). Значительная часть приносящей жаберной артерии (выше уровня
соединения обеих выносящих в одну) являются вдесь новообразованием, С — общая схема
жаберного кровообращения костистой рыбы; D—схема жаберного кровообращения личинки
хвостатой амфибии и Е—взрослой саламандры; а и е — приносящие и выносящие артерии
наружных жабер (Ьг); се и ci — наружная и внутренняя сонные артерии; pl — сосудистое
сплетение, ив которого! развивается сонная железа; р — легочная артерия.
От первой (гиоидной) выносящей артерии отходит сосуд к ложножабре
брызгальца (приносящая артерия ложножабры). Выносящай артерия
ложножабры образует переднюю сонную артерию (а. са-
rotis anterior), которая снабжает артериальной кровью глаз и мозг. От
верхнего конца выносящей гиоидной артерии отходит задняя сон-
ная артерия (a. carotis posterior), снабжающая другие части го-
ловы. Таким образом, мозг и глаза получают артериальную кровь, окислен-
ную дважды — в гиоидной полужабре и в ложножабре брызгальца. И у
других рыб, сохраняющих гиоидную полужабру и ложножабру брыз-
360
гальца, хотя бы в виде оперкулярных жабср, наблюдается сходное крово-
обращение в области головы.
Важнейшим отличием высших рыб от селахий является присутствие
в каждой дуге одной лишь выносящей жаберной артерии. Эти артерии
обычно соединяются сначала в продольные наджаберные стволы, слива-
ющиеся позади для образования спинной аорты. Однако у двудышащих
рыб, как и у селахий, имеется по две выносящие жаберные артерии в каж-
дой дуге, соединяющиеся сначала попарно между собой соответственно
каждой дуге, а затем впадающие в спинную аорту. У ганоидных рыб на-
блюдаются переходные формы, причем в начале каждой дуги имеется по
две выносящие артерии, которые затем соединяются в одну (рис. 328, А, В).
От брюшной части системы выносящих артерий у рыб отходит еще
вперед челюстная, или язычная, артерия и брюшная артерия, несущая,
Рис. 329. Жаберное кровообращение двудышащей рыбы Protopterus
(из Гудрича).
аГ— приносящие жаберные артерии; abh — приносящая гиоидная артерия; afg — прино-
сящая артерия наружной жабры; ар — легочная артерия; <ю. о и со. d — брюшная и спин-
ная аорты; са — сонная артерия; coel — a. coeliaca; ct — нории аорты; de — кювьеров про-
ток; efg — выносящая артерия наружной жабры; ед — наружная жабра; eph — выносящая
гиоидная артерия; М — сердце; та — язычная артерия; р — легкое; ар — легочная вена;
vc — задняя полая вена. Две первые жаберные дуги лишены жабер.
между прочим, артериальную кровь для питания стенок самого сердца.
От спинной части системы выносящих артерий у рыб отходит еще в голову,
кроме упомянутых уже внутренних сонных артерий, пара наружных сон-
ных артерий (негомологичных одноименным артериям наземных позво-
ночных).
Вдоль всей спинной аорты к стенкам тела отходят метамерно распо-
ложенные веточки. От задних частей корней аорты или от передней части
спинной аорты отходят последовательно следующие более крупные арте-
рии: кишечная (a. coeliaca), снабжающая кровью желудок (a. gas-
trica), печень (a. hepatica) и селезенку, подключичные (a. sub-
claviae), идущие в грудные плавники, и брыжеечные (a. mesen-
tericae), снабжающие кровью собственно кишку и отходящие в большем
или меньшем числе. Далее позади 'спинная аорта дает парные и часто
многочисленные ветви к половым органам (a. genitales), к почкам (a. re-
nales) и две крупные подвздошные артерии (a. iliacae), снаб-
жающие кровью заднюю часть кишечника и брюшные плавники. После
отхода этих артерий спинная аорта продолжается назад в виде хво-
стовой артерии (a. caudalis). У двудышащих рыб (а также у ки-
степерых ганоидов) от последней пары выносящих жаберных артерий
361
отходят легочные артерии к легким (рис. 329). Когда жабры функциони-
руют, то эти артерии получают артериальную кровь, и легочное дыхание
оказывается ненужным. Если же в жабрах происходит недостаточное
окисление, то легочные артерии получают венозную кровь, которая оки-
слявтся в легких и возвращается по легочным венам прямо в левую по-
ловину сердца. Благодаря существованию системы перегородок эта арте-
риальная кровь лишь отчасти смешивается с венозной, но главным образом
направляется в передние две (т. е. в третью и четвертую) артериальные
Дуги, которые у Protopterus даже непосредственно соединяются со спинной
аортой, так как первые две дуги здесь лишены жабер (рис. 329). От второй,
третьей и четвертой жаберных артерий отходят у личинок Protopterus
и Lepidosiren ветви к наружным жа-
Рис. 330. Артериальная система лягуш-
ки (по Паркеру, ивменено).
— спинная аорта; ci — внутренняя
сонная артерия; cut — кожная; с. шее —
кишечно-брыжеечная; деп— половая; да—
желудочная; hp — печеночная; И — под-
вздошная; int — кишечная; 1д — язычная
(нар. сонная); осс — затылочная; pulm —
легочная; геп — почечные; — под-
ключичная; vert — позвоночная.
брам (остатки их, так же как и остат-
ки наружных жабер, сохраняются
и у взрослого Protopterus).
У амфибий в личиночном
состоянии наблюдается кровообра-
щение, очень близкое к таковому
рыб и, в частности, двудышащих рыб.
От короткого артериального ствола
отходят три приносящие жаберные
артерии, идущие по трем первым
жаберным дугам в наружные жабры,
откуда окисленная кровь по вынося-
щим жаберным артериям течет в спин-
ную аорту (рис. 328). Четвертая
(шестая) артериальная дуга является
непрерывной, так как здесь жабра не
развивается и дает от себя легочную
артерию (a. pulmonalis) в развиваю-
щееся легкое. Ко времени метаморфо-
за начинает разрастаться анастомоз,
соединяющий у основания каждой
жабры приносящие сосуды с вынося-
щими, и таким образом восстанав-
ливаются эмбриональные артериаль-
ные дуги (рис. 328). Первая из этих
дуг (собственно это третья дуга) те-
ряет связь со спинной аортой, но со-
храняет свою связь с сонной артерией,
На ее пути у личинки имеется «чу-
десное» сплетение сосудов (rete mira-
bile), которое превращается в муску-
листое вздутие с перегородками вну-
три — так называемую сонную же-
лезу. Вторая дуга (четвертая) сильно разрастается и становится главным
стволом, соединяющим брюшную аорту со спинной, — системной дугой
аорты. Третья дута сохраняется еще в виде незначительного сосуда у
хвостатых амфибий и гимнофион (рис. 328), но исчезает бесследно у бес-
хвостых (рис. 330). Четвертая дуга дает начало легочным артериям,
сохраняя еще у хвостатых амфибий свою первоначальную связь со спин-
ной аортой посредством боталлова протока, который у бесхвостых исче-
зает.
От первой дуги отходят, как сказано, сонные артерии, разделяющиеся
на внутреннюю и наружную сонные артерии (а. са-
rotides interna et externa). От последней дуги, кроме легочных артерий,
отходит также большая кожная артерия, разветвляющаяся
362
в коже всего тела, где у амфибий также происходит окисление крови
(рис. 330).
Все другие артерии отходят от системных дуг и от спиниой аорты,
образующейся в результате их соединения. От самих дуг отходят впереди
из
Рис. 331. Сердце ящерицы (Lacerta)
(из Видерсгейма).
А — предсердия; Ао — аорта; ар, vp — легочная
артерия и вена; a. s. — подключичная артерия;
са — сонная артерия; ci — нижняя полая вена: tr —
ствол аорты; t>. j — яремная вена; t>. s —подклю-
чичная вена; V — желудочек; 1—2—Дуги аорты.
челюстные артерии и далее подключичные артерии и позвоночные
(a. vertebrales), тянущиеся вдоль всего позвоночника у основания попе-
речных отростков и дающие от себя метамерно расположенные веточки
к стенкам тела. Позвоночные артерии соединяются анастомозами со спин-
ной аортой и развиваются в ре-
зультате образования продоль-
ного ствола, соединяющего ме-
тамерные боковые ветви спин-
ной аорты. От спинной аорты
отходят кишечные артерии (или
отдельно ее ветви — желудоч-
ная, печеночная, кишечные) и
иногда в большом числе отдель-
пые брыжеечные, половые, по-
чечные, подвздошные и седалищ-
ные (нередко соединенные вме-
сте). У хвостатых амфибий
аорта позади продолжается в
хвостовую артерию.
Распределение по сосудам
артериальной и венозной крови
достигается не только существо-
ванием системы перегородок,
Ci Я\
о чем уже говорилось, но и
некоторыми добавочными при-
способлениями. Артериальный
ствол отходит от правой стороны
желудочка и потому получает в
первую очередь венозную кровь,
которая идет по пути наименьшего сопротивления в сравнительно широкие
и короткие лёгочные и кожные артерии. При наполнении этой системы
кровью давление в ней возрастает, и следующая порция смешанной крови
направляется через дуги аорты в спинную аорту. По мере наполнения и
этой системы уже чисто артериальная кровь из левой половины желудочка
направляется под большим давлением в сонные артерии, обладающие срав-
нительно ничтожным диаметром. Сопротивление движению крови в этой
системе возрастает благодаря существованию упомянутой сонной железки.
У рептилий артериальный ствол разделен до основания (рис. 331,
332), так что легочные артерии отходят одним общим стволом от правой
стороны желудочка независимо от остальной части, поделенной в свою
очередь на правый и левый стволы (отходящие от левой и правой сторон
желудочка). Обе сонные артерии оказываются в связи с правым стволом,
который получает артериальную кровь и продолжается в правую дугу
аорты. От последней отходят обыкновенно и обе подключичные артерии.
Таким образом, левая дуга аорты, несущая смешанную кровь, развита
несколько слабее. На месте ее соединения с правой для образования спин-
ной аорты от нее отходят кишечные артерии (a. coeliaco-mesenterica)
(рис. 332, А). У некоторых рептилий сохраняются еще и во взрослом со-
стоянии остатки и других артериальных дуг. У змей и ящериц сохраня-
ется связь между сонными артериями и корнями аорты посредством сон-
ного протока (остаток третьей артериальной дуги) (рис. 331). У некоторых
ящериц сохраняются остатки артериальных дуг, следующих за систем-
ными (остатки пятой артериальной Дуги). У черепах наблюдается во взрос-
363
A
ао
Рис. 332. Сердце и дуги аорты крокодила (Л)
и птицы (£). Схемы (по Боасу из Шимкевича).
а — правое, а* — левое предсердия; и и if — пра-
вый и левый желудочки; 1 и 1' — сонные артерии;
2 и 2' — правая и левая дуги аорты, причем
левая делится на сосуды, отходящие к кишечни-
ку — т, и на тонкий ствол — с, вместе с правой
дугой образующий нисходящую аорту — ао; 4
и 4' — легочные артерии.
лом состоянии боталлов проток1, соединяющий легочные артерии с кор-
нями аорты (остаток шестой артериальной дуги). У примитивной совре-
менной рептилии — гаттерии — сохраняются во взрослом состоянии и
сонный и боталлов протоки. .В остальном у рептилий имеются обычные со-
суды—от корней аорты или от подключичных артерий отходят позвоночные
артерии, которые дают боковые
ветви вдоль мозга; позади от
кишечных артерий отходит по-
следовательно несколько задних
брыжеечных (a. mesentericae ро-
steriores), почечные и половые
(a. renales, a. genitales) и артерии
к задним конечностям и хвос-
товой области (a. iliacae, а.
ischiadicae, a. caudalis).
У птиц совершенно ис-
чезает левая дуга аорты, кото-
рая, как мы видели, и у репти-
лий уже развита несколько
слабее (рис. 332, 333). У птиц,
таким образом, наступило пол-
ное разделение артериального
и венозного токов крови, и все
органы тела снабжаются исклю-
чительно артериальной кровью.
У млекопитающих
также имеется лишь одна Дуга
аорты, но здесь это — левая
дуга (рис. 333), отходящая от
^евого желудочка. Очевидно,
редукция левой дуги
исходный тип симметричной кровеносной системы был иной, чем у со-
врёменных рептилий, у которых уже намечается
Рис. 333. Схемы артериальных дуг. Вид со спинной стороны. А — у молодой
ящерицы, В — у птиц; С — у млекопитающих (по Гессе).
aod — спинная аорта (отрезана); се — и ci — а. сагоШ externa и interna; р — a. pulmo-
nalis; scl — a. subclavia.
отходящей от правого желудочка. Остатком правой дуги у млеко-
питающих является только правая подключичная артерия, нередко само-
стоятельно отходящая от артериального ствола.
1 Боталлов проток имеет у водных черепах определенное значение; при долгом
пребывании под водой, когда второй круг кровообращения в связи с прекращением
дыхания пропускает лишь немного крони, избыток ее проходит черев боталлов проток
и поддерживает, таким образом, нормальное кровяное давление в артериальной системе
тела. Таково же и вначение связи между обеими дугами аорты посредством foramen
Panizzae у крокодилов.
364
Таким образом, у млекопитающих не произошло разделения артери-
ального ствола на правый и левый, что мы видели у рептилий. Отход глав-
ных артерий весьма различен у различных млекопитающих. Иногда под-
ключичные и сонные артерии отходят самостоятельно, или сонные артерии
отходят одним общим стволом, в других случаях одна из них или обе вместе
соединяются с одной или обеими подключичными артериями. Остальные ар-
терии обычные: позвоночные артерии, отходящие обыкновенно от подклю-
чичных, кишечные, брыжеечные, почечные половые и подвздошные. Послед-
няя разделяется на внутреннюю подвздошную артерию (a. iliaca interna)
и бедренную (a. femoralis). Седалищная артерия (a. ischiadica) может
отходить самостоятельно или является ветвью подвздошной.
Вены
Имеющаяся у ланцетника подкишечная вена всегда закладывается
у зародышей позвоночных, но затем ее путь прерывается на местах разви-
тия обеих воротных систем. С развитием воротной системы почек хвосто-
вая вена теряет связь с подкишечной веной и через почки вступает в
связь с парными кардинальными венами, идущими по спинной стенке
брюшной полости (рис. 334)<С развитием воротной системы печени все
кишечные сосуды соединяются в воротную вену, а сама подкишечная
вена у низших позвоночных (круглоротых и селахий) может сохраниться
в виде небольшого сосуда, идущего вдоль спирального клапана.
У рыб хвостовая вена (v. caudalis), подходя к почкам,
разделяется на две приносящие воротные вены почек, которые распадаются
в последних на капилляры (рис. 334). Почечные вены соединяются с каж-
дой стороны в продольный ствол, направляющийся вперед — задние
кардинальные вены (v. cardinales posteriores), собирающие
также венозную кровь из стенок тела по метамерным сосудам. На уровне
сердца задние кардинальные вены соединяются с передними кар-
динальными, или яремными, венами (v. cardinales ante-
riores s.( v. jugulares), несущими кровь из головы, и образуют вместе
короткий поперечный ствол — кювьеров проток (d. Cuvieri),
направляющийся прямо в венозную пазуху сердца (рис. 334). Из задних
конечностей подвздошные вены (v. iliacae) направляются
впервд в брюшной стенке тела, образуя боковые вены (v. later ales),
которые впадают в кювьеровы протоки, приняв в себя по
пути подключичные вены (v. subclaviae) от грудных плавни-
ков (см. рис. 27).
Это симметричное расположение венозных сосудов нарушается
уже у высших рыб. У костистых рыб преобладающей становится правая
кардинальная вена, собирающая кровь и из задней половины левой почки,
между тем как левая значительно короче и тоньше.
У многоперых ганоидов (Polypterus) и у двудышащих рыб появляется
новый непарный венозный ствол — задняя (или нижняя) полая
вена (v. cava posterior, s. inferior). Эта вена характерна для наземных
позвоночных и развивается в своей задней части за счет кардинальных
вен (у двудышащих за счет правой кардинальной вены, а у наземных
позвоночных — за счет обеих) и впадает непосредственно в венозную
пазуху. Почечные сосуды впадают теперь в заднюю полую вену, а карди-
нальные вены собирают кровь по метамерным сосудам из боковых стенок
тела. С образованием задней полой вены в нее же у ее основания впадает
и печеночная вена.
У двудышащих рыб обе подвздошные вены дают от себя ветви на со-
единение с воротной веной почек, а затем направляются в брюшной стенке
тела вперед и соединяются в непарный ствол — брюшную вену (v. abdo-
minalis), несущую кровь в правый кюрьеров проток. Эта брюшная вена
365
должна рассматриваться как результат соединения ооеих боковых вен
низших рыб. Подключичные вены принимают в себя большие боковые
кожные вены и впадают независимо в кювьеров проток. Легочная вена
(v. pulmonalis) идет от легких у Polypterus к основанию печеночной вены, а
р у двудышащих входит в венозную
пазуху, по стенке которой она про-
должается до левой половины пред-
сердия.
У амфибий венозная систе-
ма очень похожа на таковую двуды-
шащих рыб. Общая схема выражается
в следующем (рис. 335). Хвостовая
вена разделяется на две воротные
вены почек, с которыми соединяется
внутренняя ветвь подвздошной вены.
От почек кровь собирается в заднюю
(нижнюю) полую вену, которая при-
нимает в себя печеночную вену и
Рис. 335. Венозная система хвостатой
амфибии. Кардинальные сены раздвинуты.
аЪЛ — брюшная вена; с — задняя полая псп»;
ср — задние кардинальные йены; caud — хво-
стовая вена; cut —- кожная вена; de — кювьс-
ров проток; И —- подвздошные вены; з'е и ji —
наружная и внутренняя яремные вены; ph —
воротная вена печени; рг — воротная иена
почек; set — подключичная вена.
Рис. 334. Схема кровеносной
системы костистой рыбы.
А — предсердие; Ао — спинная аорт?>
_4s — подключичная артерия; С — сонные
артерии; Са — передняя кардинальная
вена; Ср — задняя кардинальная вена;
D — кишка; НС — кювьеров проток; К —
жаберные артерии; КА — брюшная аорта;
L — печень; N — почка; Sv — венозная
павуха; Ус — хвостовая вена; V — желу-
дочек; vh — печеночные вены; Ур — ворот-
ная вена печени; Ург— воротная вена по-
чек; Vs — подключичная вена.
впадает в венозную пазуху. Кардинальные вены сохраняют симметричное
расположение и принимают в себя вены из стопок тела; позади они связаны
с заднем полон веной. У бесхвостых кардинальные вены исчезают. Наруж-
ные ветви подвздошных вен принимают в себя вены, идущие от задней
кишки (клоаки и мочевого пузыря) и соединяются в непарную б р ю in н у то
вену (v. abdominalis s. epigastrica), которая у амфибии впадает по в
кювьеров проток, а в воротную вону печени. Последняя
собирает кровь от всего кишечника.
366
Из головы кровь возвращается по передним кардинальным, или
яремным, венам, которые соединяются с задними кардинальными для
образования кювьеровых протоков. Подключичные вены
вместе с большими кожными венами (v. cutaneae magnae) впадают
также в кювьеровы протоки. Кожные вены имеют большое
значение в связи с существованием кожного дыхания.
Легочные вены (v. pulmonales) впадают у наземных позвоноч-
ных непосредственно в левое предсердие.
У высших наземных позвоночных (амниот) воротная система почек
понемногу теряет свое значение. У рептилий хвостовая вена вместе
с ветвью подвздошной вены дает еще воротную вену почек, и то же самое
относится к птицам (рис. 336), но у
непосредственную связь с задним
концом нижней полой вены, и таким
образом главная масса крови про-
ходит мимо почек. У млекопитаю-
щих воротная система почек совсем
исчезла.
них эта воротная вена вступает в
Рис. 337. Развитие передних вен млеко-
питающих (из Кингсли). А — стадия раз-
вития; В — вены взрослого животвого.
а — непарная вена (v. azygos); с — венеч-
ная вена сердца; е, i —наружная и вну-
тренняя яремные вены; ha — v. hemiazy-
gos; it — подввдошные вены; in—*бевымен-
ная вена; р — задняя полая вена; рс —
задняя кардинальная; рис — передняя
(верхняя) полая вена; s — подключичная
вена; si — верхние межреберные вены;
г — почки.
Рис. 336. Воротное кровообращение почек
у крокодила (А) и у птицы (В)
(по Гегенбауру).
М — брюшные вены; cd — хвостовая; сг —
бедренная; ст — копчико-брыжеечная; И. г —
внутренняя подведошная; is — седалищная;
vct с — задняя полая вена.
Соединением ветвей хвостовой вены и подвздошных образуется брюш-
ная вена, которая иногда является еще парной (крокодил) (рис. 336, Л).
У млекопитающих эта вена развивается как парный сосуд, но
является чисто эмбриональным образованием, давая начало пупочным
венам зародыша. Задняя полая вена получает преобладающее значение,
и одновременно падает значение задних кардинальных. Главная масса
крови, идущей по к ювьеровым протокам, получается теперь
от яремных и подключичных вен, и поэтому границы кювьерова протока,
обозначенные раньше местом соединения обоих пар кардинальных вен,
у высших позвоночных теряются. Крупный ствол, образующийся слиянием
яремной п подключичной вен, получает название ле р е д п с й (верхней)
iT ст л о"й Вчгтгта7~^адппё~ тГардипальные вены являются незначитель-
ными сосудами, впадающими в передние полые пены, и получают название
позвоночных вен (v. vertebrales); эти вены принимают в себя
сегментальные межробсрныс вены.
Для млекопитающих характерно асимметричное расположение главных
венозных стволов (рпс. 337). Прежде всего асимметрично развиты остатки
кардинальных вен (позвоночные вены), которые здесь получают назва-
367
ние непарных вен (правая — v. azygos, левая — v. hemiazygos). У неко-
торых млекопитающих редуцируется одна или другая из этих вен, но
у большинства, так же как и у человека, обе соединяются между собой
поперечным анастомозом, после чего левая непарная вена (v. hemiazygos)
теряет свою связь с левым кювьеровым протоком. Непарная вена несет
теперь всю кровь из стенок тела, полученную по межреберным венам
(v. intercostales), в правую переднюю (верхнюю) полую вену.
Передние полые вены иногда еще развиты симметрично, но обыкно-
венно и здесь получают преобладание сосуды правой стороны (рис. 337).
Обе передние полые вены соединяются между собой поперечным анасто-
мозом (безыменной веной), после чего левая полая вена редуцируется.
Кровь из левых подключичной и яремной вен идет по безыменной вене
(v. innominate) в правую переднюю полую вену, в которую впадает и
и непарная вена (v. azygos). От левого кювьерова протока сохраняется
лишь небольшой сосуд, венечная вена (v. coronaria), несущий кровь
из стенок самого сердца (рис. 337).
Лимфатическая система
Первоначально единая кровеносная система диференцируется уже
у низших позвоночных животных на замкнутую систему кровеносных
сосудов и на лимфатическую систему (рис. 338). Последняя не представляет
собой замкнутой системы сосудов — ее разветвления находятся в связи
с межклеточными пространствами кожи, кишечника и других органов
и сообщаются также при помощи небольших отверстий с общей полостью
тела. Стенки мелких капиллярных лимфатических сосудов состоят из
одного эндотелия. Более крупные сосуды и синусы имеют, кроме того,
соединительнотканную оболочку с эластическими и гладкими мускульными
волокнами. Крупные сосуды снабжены часто клапанами, препятствую-
щими обратному току лимфы. Проксимальные концы сосудов открываются
в вены.
Очень сильно развита сеть лимфатических сосудов в подкожной
клетчатке, где они у низших позвоночных приобретают характер крупных
мешков (лимфатические синусы, достигающие особо значительного разви-
тия у амфибий). Значительная сеть лимфатических сосудов имеется также
в стенках кишечника, в оболочках мозга, в мышцах и, наконец, в стенках
крупных сосудов, особенно артерий, а также в стенках самого сердца.
Из сети лимфатических сосудов лимфа собирается в более крупные
протоки, из которых наиболее крупным является парный первоначально
туловищный проток, идущий вдоль аорты и получающий
лимфу, между прочим, из лимфатического синуса в брыжейке. В последнем
сходятся лимфатические сосуды кишечника («млечные» сосуды) и других
внутренностей, а также сосуды задних конечностей. Туловищный проток
изливает свое содержимое в яремный лимфатический мешок, соединяю-
щийся с венами в области основания подключичной и яремной вен. В тот
же яремный мешок изливается и лимфа из головного протока,
идущего параллельно яремной вене, и из локтевого протока, несущего
лимфу из передней конечности.
У млекопитающих туловищные протоки обыкновенно развиты асим-
метрично.
Движение лимфы в сосудах поддерживается благодаря периодическому,
их сжатию давлением соседних органов, мышц и т. п. при содействии
клапанов. Однако, кроме того, в особенности у низших позвоночных
имеются еще расширенные пульсирующие участки сосудов, называемые
лимфатическими сердцами. Они наблюдаются в различных местах как у
рыб, так и у наземных позвоночных. Особенно многочисленны лимфа-
тические сердца у амфибий. У хвостатых амфибий имеется целый ряд
368
лимфатических сердец, лежащих метамерно под кожей на протяжении
бокового лимфатического сосуда. У гимнофион лимфатическая система
построена метамерно и снабжена очень большим числом боковых сердец
(до 100 пар). У бесхвостых амфибий одна пара сердец лежит в тазовой
области по бокам уростиля и другая пара — между поперечными отрост-
ками в начале спины. У рептилий и птиц имеется еще лимфатическое
сердце в области таза (рис. 338), а у млеко-
питающих таких сердец совсем нет.
В связи с лимфатическими сосудами
находятся многочисленные скопления адено-
идной ткани (сетчатая соединительная ткань
с большим количеством лейкоцитов), так
называемые лимфатические желе-
зы, в которых лимфатические сосуды раз-
биваются на сеть, теряя собственные стенки.
В точности функция этих образований не-
известна, но здесь главным образом проис-
ходит размножение лимфоцитов (лейкоцитов).
Лимфатические железы наблюдаются в
особенно большом количестве в стенках по-
лости тела (в брыжейках) и в стенках кишеч-
ника, но имеются и в других местах. Из
крупных лимфатических желез низших по-
звоночных следует назвать так называемые
«жировые тела» амфибий, связанные с по-
ловыми железами. У наземных позвоночных
наблюдаются еще довольно крупные лимфа-
тические железы в ротовой полости. У высших
позвоночных они развиваются преимущест-
венно в глотке, где получают название гло-
точных миндалин, в особенности пара круп-
ных миндалин по бокам зева, а у млекопи-
тающих, сверх того, на языке
Самым крупным лимфоидным органом
позвоночных является селезенка, от-
личающаяся, однако, от лимфатических же-
лез тем, что не находится в открытом
сообщении с лимфатическими сосудами, а
обильно снабжается только кровью. У рыб
она иногда состоит из отдельных скоплений
Рис. 338. Схема лимфатиче-
ских сосудов .птицы (по Шим-
кевичу).
a. cl —'подключичная артерия;
а. г — седалищная артерия; а.
т—a. mesenteries; ао — аорта;
d. th — грудной лимфатический
проток; L —лимфатические серд-
ца; V. са — передняя полая
вена; «. Ь — бедренная вена.
лимфоидной ткани, но нередко сконцентри-
рована в одном большом компактном теле. У наземных позвоночных это
всегда плотный орган, лежащий в брыжейке вблизи желудка. В се-
лезенке, кроме образования лейкоцитов, наблюдается и распадение
красных кровяных ^париков.
Кровеносная система позвоночных построена в основе своей по тому
же типу, как и кровеносная система низших хордовых и даже высших
червей, состоя по существу из брюшного и спинного сосудов, соединенных
анастомозами в стенках кишки и в стенках тела. У позвоночных она
обнаруживает более высокое развитие, во-первых, в обособлении сердца,
представляющего собой участок брюшного сосуда с особо развитой мус-
кульной стенкой и разделяющегося на четыре отдела, расположенные
последовательно, во-вторых, в дальнейшем развитии системы сосудов,
которая диференцируется на собственно кровеносные сосуды и лимфати-
ческие. Кишечные анастомозы в передней, жаберной, части кишки дали
начало жаберным сосудам, которые у наземных позвоночных превра-
369
щаются в дуги аорты, непосредственно передающие кровь из брюшного
сосуда в спинной. Брюшной .сосуд в двух местах прерывается образова-
нием сосудистых сплетений — в почках и в печени — и таким образом
преобразуется в воротные системы. В дополнение к брюшному сосуду
развиваются парные венозные стволы в спинной стенке тела — карди-
нальные вены, которые уносят кровь из почек и из стенок тела.
У наземных позвоночных в связи с приобретением легочного дыхания
развивается второй круг кровообращения, и сердце получает теперь,
кроме венозной, еще артериальную кровь.
Дальнейшая эволюция выражается в развитии приспособлений для
разграничения венозного и артериального токов крови как в самом сердце,
так и в сосудах. Сердце становится «четырехкамерным» с левой артериаль-
ной и правой венозной половинами, из которых каждая состоит из двух
камер (предсердия и желудочка). Передний и задний отделы сердца рыб —
венозная пазуха и артериальный конус — исчезают. Артериальный ствол
разделяется до основания на легочную артерию, отходящую от правого
желудочка и дуги аорты. Последние у высших наземных позвоночных
получают асимметричное развитие, причем у птиц имеется только правая
дуга, у млекопитающих — левая.
В венозной системе у наземных позвоночных появляется непарная
задняя полая вена, а задние кардинальные более пли менее редуцируются.
Остатки последних — позвоночные вены — получают у млекопитающих
асимметричное расположение. Асимметрия развивается у млекопитающих
и в расположении передних венозных сосудов, причем и здесь разви-
ваются преимущественно сосуды правой стороны (правая верхняя полая
вена).
ПОЛОСТЬ ТЕЛА
ПОЛОСТЬ ТЕЛА БЕСПОЗВОНОЧНЫХ
У кишечнополостных животных, так же как и у низших червей, имеется лишь
одна полость тела — кишечная полость, которая в этих случаях обыкновенно снабжена
выростами или даже переходит в периферическую систему каналов. У турбеллярий
все пространство между кишечником и покровами выполнено мезенхимой (паренхи-
мой), в которой лежат сложно построенные половые органы в виде системы полостей
и выводных каналов, а также и органы выделения. У других червей, кроме кишечной
полости и половых органов, появляется еще общая полость тела, в которой помещаются
эти органы. Так, например, у круглых червей имеется обширная полость между стен-
кой кишки и наружной стенкой тела. Эта полость не имеет эпителиальной выстилки,
представляет собой результат расхождения клеток мезенхимы и носит название пер-
вичной полости тела. У высших червей (например у кольчатых) между кишечником
и стенкой тела также имеется полость, но эта полость, обычно поделенная на правую
и левую половины, выстлана плоским эпителием, и в ней нет особых половых
полостей: половые продукты развиваются в ее собственных стенках. Это — вторичная
полость тела, или целом. Сравнение с бесполостными червями показывает, что вторич-
ная полость тела представляет собой результат разрастания полостей половых желез
(теория гоноцеля). Вторичная полость тела имеется, кроме высших червей, у кишечно-
дышащих и у иглокожих. Она имеется и у членистоногих, но редуцируется здесь и
вступает в связь с кровеносной системой. Редуцируется целом и у моллюсков, у ко-
торых он сохраняется лишь в виде околосердечной полости и полостей половых желез.
В отличие от высших представителей «беспозвоночных» вторичная полость тела играет,
однако, большую роль у всех хордовых животных.
Вторичная полость тела развивается иногда путем расхождения листков мезо-
бласта, образующегося из мезодермальных телобластов, в своем возникновении
связанных с зачатком энтодермы (кольчатые черви, моллюски), но в других случаях —
как полые выступы первичной кишки. Это особенно ясно у щетинкочелюстных
червей, у кишечнодышащих и у ланцетника, но одинаково справедливо и для игло-
кожих, а также для брахиопад.
Эти наблюдения позволили вывести заключение, что общая полость тела пред-
ставляет результат обособления периферических частей (выростов) кишечной полости
(теория энтероцеля). Так как, однако, имеются определенные указания на первичную
половую функцию вторичной полости тела, то нужно думать, что в обособившихся
частях кишечной полости именно и происходило развитие половых продуктов, как
это в действительности наблюдается у многих кишечнополостных.
370
Общая полость тела может иметь метамерное строение, подразделяясь на ряд
следующих друг ва другом камер, соприкасающиеся стенки которых образуют по-
перечные перегородки, или диссепименты. Внутренние стенки правого и левого участ-
ков полости тела покрывают снаружи кишку и срастаются как над ней, так и под
ней, образуя спинной и брюшной мезентерии, на которых кишка подвешивается внутри
целома. Внутренняя стенка вторичной полости тела носит название кишечного, или
висцерального, листка; наружная стенка’ называется кожным, или париетальным,
листком. За счет последней развивается значительная часть мускулатуры.
В стенках целома, как уже упомянуто, развиваются половые железы, и его
полость сообщается у высших червей с наружной средой сегментальными каналами,
служащими органами выделения, и подобными же каналами, служащими для выве-
дения половых продуктов.
Число отделов полости тела может быть очень велико у полимерных червей
(аннелиды), в других случаях число сегментов меньше, и нередко имеются два или
чаще три последовательных участка, разделенных диссепиментами — олигомерные
формы(брахиоподы, мшанки, щетинкочелюстные черви, кишечнодышащие и иглокожие).
У ланцетника целом развивается путем обособления парных мешковидных
выростов кишечника, которые сразу имеют метамерное расположение. Полость тела
подразделяется затем на спинной участок — полость мускульных сегментов (миоцель),
который по происхождению представляет собой продукт разрастания некоторого
участка париетального листка, и брюшной участок — полость боковой пластинки,
или полость дефинитивного целома. Эта полость теряет свою метамерию, так как
диссепименты исчезают, и таким образом развиваются сплошные правая и левая по-
лости тела, которые затем вследствие частичного исчезновения мезентериев (особенно
брюшного) сливаются в одну непарную полость.
У позвоночных общая полость тела развивается независимо от кишечной
полости, но во всяком случае ограничивающие ее мезодермальные стенки развиваются
в связи с энтодермой. В отличие от ланцетника общая полость тела с самого начала
закладывается сплошной, несегментированной. Мезодерма позвоночных рано дифе-
ренцируется на спинной отдел, образующий метамерный ряд первичных сегментов,
или сомитов, и остающийся несегментированным брюшной отдел — боковую пластинку.
Обе части остаются в течение некоторого времени связанными при помощи «стебельков»
сомитов, дающих затем начало почечным канальцам. Обе боковые пластинки разра-
стаются по бокам кишечника вплоть до взаимного соприкосновения над кишкой и
под кишкой, где соединяющиеся между собой висцеральные листки правой и левой
пластинок образуют упомянутые уже спинной и брюшной мезентерии, укрепляющие
кишку внутри полости тела. И у позвоночных в сгенках полости тела развиваются
половые железы, и в связи с ними возникают органы выделения и выводящие пути
половых желез.
ПОЛОСТЬ ТЕЛА ПОЗВОНОЧНЫХ
Общая полость тела позвоночных охватывает кишечный канал и выст-
лана плоским эпителием (целотелием), который у круглоротых является
мерцательным. От внутренних (висцеральных) стенок правой и левой
полостей у взрослого животного сохраняется серозная выстилка кишеч-
ника, и других внутренних органов и спинная брыжейка (мезентерий),
подвешивающая в виде двойного листка кишку к спинной стенке полости.
От брюшного мезентерия сохраняются лишь незначительные остатки в
области печени. Париетальный листок образует выстилку наружной
стенки полости тела.
У всех позвоночных передняя часть полости тела, заключающая
в себе сердце, — околосердечная, или перикардиальная, по-
лость обособляется от задней части, или брюшной полости. У низших
позвоночных между этими отделами иногда сохраняется еще связь посред-
ством особых каналов. Выстилка околосердечной полости носит название
околосердечной сумки, или перикардия, а выстилка брюшной по-
лости называется брюшиной, или перитонеумом.
Околосердечная полость, расположенная сначала впереди брюшной,
подвигается у наземных позвоночных назад и вдавливается в брюшную
полость, от которой вновь отделяется у млекопитающих, с образованием
поперечной перегородки — диафрагмы. Последняя отделяет от собственно
брюшной полости всю переднюю часть — грудную полость,
содержащую посредине сердце, окруженное околосердечной полостью, и
по бокам пару плевральных полостей с легкими. Выстилка
плевральных полостей носит название плевры.
371
С образованием извилин кишки брыжейка получает очень слож-
ное расположение. У млекопитающих от желудка свешивается большая
двойная складка мезентерия желудка и мезентерия толстой кишки, обра-
зующая так называемый большой сальник.
У большинства рыб полость тела сообщается с внешней средой посред-
ством брюшных пор (pori abdominales) — коротких парных каналов,
лежащих в области клоаки, или заднего прохода. Кроме того, у низших
позвоночных полость тела сообщается еще с внешней средой через посред-
ство воронок выделительных канальцев, а у самок всех позвоночных —
посредством яйцеводов. Последние иногда редуцируются и принимают
характер коротких выводных каналов — половых пор (лососевые и неко-
торые другие рыбы).
ОРГАНЫ ВЫДЕЛЕНИЯ
ОРГАНЫ ВЫДЕЛЕНИЯ БЕСПОЗВОНОЧНЫХ
Органы выделения служат для выведения из организма растворенных в воде
конечных продуктов обмена — солей и азотистых соединений. У некоторых низших
животных это удаление вредных или просто негодных веществ производится всей
поверхностью тела, как у кишечнополостных, у которых, таким образом, можно
говорить о диффузном выделении>У иглокожих выделение производится, повидимому,
через посредство воднососудистой системы.
Однако в громадном большинстве случаев для выделения имеются специальные
органы, которые бывают построены различно в зависимости от того, имеется ли обо-
собленная общая полость тела, принимающая участие в экскреции, или нет.хВ послед-
нем случае наблюдаются так называемые протонефридии; в первом случае обычно
имеются сообщающиеся с полостью тела метанефридии.
Протонефридии представляют собой систему каналов, обычно разветвляющихся
в паренхиме всего тела. Очень тонкие разветвления этих каналов замыкаются особыми
клетками, снабженными мерцательным жгутом, входящим внутрь канальца^ Такая
концевая клетка является собственно выделительным аппаратом, и соответственно
всасывающая ее поверхность увеличена обыкновенно путем образования различных
выростов/Внутри этих клеток нередко наблюдаются многочисленные вакуоли, ука-
зывающие на их деятель ност1>.~£Иерцательный жгут находится в непрерывном коле-
бательном движении и вызывает в канальце ток окружающей его жидкости по напра-
влению к более крупным каналам и, следовательно, наружу/Канальцы очень тонки,
и их полости представляют собой внутриклеточные полости немногих очень вытянутых
вдоль канала клеток.
Главные выводные стволы служат отчасти резервуарами для выделяемой жид-
кости и открываются наружу одним, парой, а иногда и несколькими отверстиями.
Протонефридии имеются у бесполостнц^цдоских червей, у первичнополостных немер-
тцц, а также у личинок высппгЗс'чёрвей и моллюсков.
С появлением вторичной полости тела эти органы изменяются. Уже у личинок
кольчатых червей протонефридии более или менее вдаются внутрь первичной полости
тела. Стенки нефридиальных каналов омываются со всех сторон жидкостью и также
могут принять на себя активную роль в выделении веществ; в связи с этим стенки
канальцев утолщаются. Наоборот, концевая клетка в значительной мере теряет функ-
цию специально экскреторного органа, поверхность ее уменьшается, и значение ее
сводится главным образом к поддержанию тока жидкости колебаниями длинного
волосовидного жгутика/Такие небольшие концевые клетки снабжены своим очень
тонким внутриклеточным канальцем, в котором колеблется жгутик, и носят наимено-
вание соленоцитов/Соленоциты покрывают внутренние концы сравнительно коротких
и толстых нефридиальных каналов иногда в огромном количестве/Соленоциты с их
канальцами имеют, повидимому, значение главным образом фильтраторного аппарата,
между тем как сам нефридиальный канал несет экскреторную функциюдТакая изме-
ненная форма протонефридиев наблюдается и у многих взрослых кольчатых червей;
однако в большинстве случаев у молодых щетинконогих червей протонефридии про-
рываются» во вторичную полость предыдущего сегмента. Таким образом возникает
другая форма нефридиальных каналов — метанефридии.
Метанефридии, или просто нефридии, начинаются расширенным концом с от-
верстием в полости тела — так называемой мерцательной воронкой, или нефра-
стомом, эпителий которого снабжен мерцательными ресничками, вызывающиими
ток жидкости, направленный внутрь канальца (иногда при этом сохраняются и остатки
соленоцитов с их мерцательными жгутиками)л У щетинконогих червей воронка укре-
пляется в задней стенке каждого сегмента и ведет в канал, проходящий через диссе-
пимент в следующий отдел полости тела, где образует многочисленные извивы и на-
372
правляется затем к выводному отверстию в боковой стенке тела, образуя предвари-
тельно тонкостенный резервуар — конечный пузырек. ^'Извитая трубка нефридиаль-
ного канала — по крайней мере средняя, явно железистая ее часть — несет экскре-
торную функцию. Фильтрация жидкостей происходит через воронку.
Нефридиальные каналы располагаются метамерно в строгой зависимости от
метамерии вторичной полости тела. Так как последняя сама является частью экскре-
торных органов и фильтрация жидкостей происходит, очевидно, через ее стенки, то
каждая камерадедрма. должна иметь свой нефридиальный канал. У членистоногих
и моллюсков вторичная полость редуцируется, а соответственно и нефридии испы-
тывают значительные изменения.
Для нас здесь интересно отметить, что выделительные органы хордовых
животных построены именно по типу нефридиев.
У ланцетника нефридиальные канальцы располагаются в жаберной
области соответственно каждой паре первичных жаберных щелей (в области язычковой
перегородки) и имеют вид коротких трубок, изогнутых коленом и открывающихся
одним концом в околожаберную полость (представляющую собой часть наружной
среды), а другим коленом при помощи нескольких отверстий в общую полость тела,
которая у ланцетника тянется над жаберной областью кишечника в виде пары про-
дольных каналов.«.Число нефридиальных трубок может достигать 100 пар. Внутри
стенок полости тела по соседству с нефридиями кровеносные сосуды образуют густую
сеть капилляров в виде клубочка (glomerulus), через которую, очевидно, происходит
фильтрация жидкостей внутрь полости тела, откуда они извлекаются нефридиальными
трубками. Таким образом, нефридиальные трубки ланцетника принадлежат к типу
метанефридиев. Интересным является, однако, тот факт, что края целомических отвер-
стий нефридиев покрыты множеством типичнейших соленоцитов, придающих им пора-
зительное сходство с нефридиями некоторых щетинконогих червей (сем. Phyllodocidae)
(рис. 16). Таким образом, нефридии ланцетника занимают еще до известной степени
промежуточное положение между протонефридиями и метанефриднями.
Почки низших позвоночных состоят ив комплекса канальцев, открываю-
щихся в общую полость типичными мерцательными воронками, и располагаются
вначале (у эмбриона) строго метамерно.
Это делает весьма вероятным предположение, что и у позвоночных мы имеем
сегментальные органы типа метанефридиев, подобные тем, с которыми мы познакоми-
лись у кольчатых червей.
ОРГАН ВЫДЕЛЕНИЯ ПОЗВОНОЧНЫХ
Почки
Почки позвоночных представляют довольно компактный парный орган,
состоящий из большого числа извитых выделительных канальцев, соеди-
няющихся с общим выводным протоком, который открывает обыкновенно
н заднюю кишку (клоаку)/Почки высших позвоночных, однако, несколько
отличаются от почек низших позвоночных; они более компактны, лежат
далее позади, в тазовой области, канальцы их построены сложнее и лишены
нефростомов.
В эмбриональном развитии высших позвоночных наблюдается после-
довательная смена трех различных выделительных органов. Эти три поко-
ления почек носят названия предпочки, или головной почки (pro-
nephros), первичной, или туловищной, почки (mesonephros) и
вторичной, или тазовой, почки (metanephros). У низших позво-
ночных постоянной почкой является орган, вполне сходный с провизорной
первичной почкой высших позвоночных, а у круглоротых имеются остатки
органа выделения, сходного с предпочкой. Таким образом, эмбриональной
смене трех различных органов выделения у высших форм соответствует
и филогенетическая смена этих органов в ряду позвоночных, но эта смена
носит несколько иной характер, представляясь скорее в виде постепенной
эволюции, связанной с прогрессивным развитием определенных участков
органа.
Так, переходя от низших позвоночных к высшим, можно проследить
весьма постепенное усложнение строения почечных канальцев (рис. 339).
В наиболее простом виде каждый почечный каналец состоит из извитой
трубки, открывающейся мерцательной воронкой (нефростомом) в полость
т'ела, а другим концом впадающей в продольный выводной канал. Такая
373
трубка вполне сходна с метанефридиальным канальцем кольчатых червей
и отличается от него главным образом лишь тем, что он открывается не
непосредственно наружу, а в особый выводной канал (что, впрочем,
иногда наблюдается и у аннелид). В полости тела вблизи от воронки
нередко наблюдается вздутие стенки со сплетением кровеносных сосудов
внутри — так называемый наружный сосудистый клубок (glo-
mus), являющийся фильтраторным аппаратом. Такие почечные канальцы
наблюдаются только в предпочке (рис. 339, Л).
Прогрессивное усложнение почечного канальца выражается отчасти
в его удлинении, а главное в том, что фильтраторный аппарат развивается
в связи с самим канальцем — появляется слепой выступ канальца, вну-
Рис. 339. Схемы почечных канальцев.
А — предпочки при наружном клуСке (gl); В — предпочки или первичной почки о внутрен-
ним клуСочком; С —первичной иди вторичной почки; D—почки млекопитающиго; М—маль-
пигиево тельце; Н — петля Генле; nsJ — нефростом; ts — извитой каналец; tr — прямые
канальцы.
тренняя камерка, в которую вдается сосудистое сплетение, или клубочек
(glomerulus). Это образование носит название мальпигиева тельца.
Такие канальцы наблюдаются иногда уже в предпочке, но в особенности
они характерны для первичной почки (рис. 339, В .
Дальнейшая эволюция нефридиального канальца состоит в исчезно-
вении мерцательной воронки. Довольно длинный и извитой каналец
начинается теперь мальпигиевым тельцем. Канальцы такого строения
наблюдаются весьма часто в первичной почке (взрослого животного),
а также и во вторичной почке (рис. 339, С). Наконец происходит довольно
сложная диференцировка самого нефридиального канальца. От мальпи-
гиева тельца идет так называемая извитая трубочка, затем тонкая и
длинная петля Генле, состоящая из двух параллельных колен, потом
более короткая извитая соединительная часть и, наконец, прямая
собирательная трубочка, впадающая уже в мочеточник (рис. 339, D).
Эта наиболее сложная форма нефридиального канальца встречается
только во вторичной почке млекопитающих.
Таким образом, резкой границы между различными поколениями
почек на основании строения их канальцев установить нельзя.
В почечных канальцах отдельные части различаются по своей функции.
Фильтраторная функция (фильтрация растворов солей и органических
веществ, за исключением белков) принадлежит сначала самой полости
тела и осуществляется через посредство наружного сосудистого клубка.
Выделяемая жидкость удаляется через мерцательную воронку нефридия.
Затем, с развитием внутренней капсулы с ее клубочком, фильтраторная
функция переходит к этой части самого канальца (к мальпигиеву телу),
274
а нефростом после этого редуцируется. Функция более или менее извитых
канальцев почки, стенки которых образованы кубическим эпителием, со-
стоит в обратном всасывании воды, сахара, хлоридов и двууглекислого нат-
рия, благодаря чему выделяемая моча становится более концентриро-
ванной. В основном она содержит мочевину и другие вредные про-
дукты обмена.
Мы видели, что по строению нет резкой разницы между предпочкой,
первичной почкой и вторичной; можно лишь сказать, что типичный ка-
налец метанефроса построен сложнее, чем типичный каналец мезонефроса,
а этот последний сложнее, чем типичный каналец пронефроса. Посмотрим,
нет ли резкой разницы между ними по их эмбриональному развитию.
Предпочка (pronephros) закладывается на очень ранней стадии
развития в виде парного ряда отдельных зачатков. Источником для их
образования являются стебельки сомитов или нефротомы, т. е. суженные
части мезодермы, связывающие в течение некоторого времени сомит с
боковой пластинкой (рис. 340). От наружной (париетальной) стенки
стебелька сомита развивается вырост по направлению к эктодерме. Сомит
Рис. 340. Развитие предпочки (по Феликсу ив Кингсли). Две стадии.
с — целом; d — предпочечный каналец и проток; ту — сомит; h — боковая пластинка;
т — стебелек сомита; п — воронка; so — соматический и sp—висцеральный листки боковой
пластинки.
вполне отделяется от стебелька, и последний оказывается связанным
только с боковой пластинкой. Вырост вытягивается в трубочку, сохраняю-
щую свою связь с боковой пластинкой. Такая трубочка представляет
собой уже каналец предпочки, сообщающийся с полостью тела посред-
ством воронки, или нефростома. Эти канальцы развиваются в передней
части туловища обыкновенно на небольшом числе сегментов. У миног за-
кладывается до 6 канальцев, у селахий—3—4, а иногда, у скатов — до 8,
у осетровых рыб — 7 канальцев, у безногих амфибий — до 12, у других
амфибий, у костных ганоидов и у высших позвоночных —2—3 канальца.
Согласно источнику своего развития канальцы, разумеется, расположены
метамерно.
Предпочечные канальцы вступают затем в связь с выводным протоком,
который у амфибий, осетровых, двудышащих и лососевых рыб образуется
прямо как складка соматоплевры. Этот способ образования следует счи-
тать первичным (Машковцев). У селахий и у амниот канальцы разраста-
ются своими наружными концами назад, сливаются между собой, образуя
общий продольный канал — предпочечный (пронефрический)
проток, который затем самостоятельно растет назад вдоль стенки
полости тела, между ней и эктодермой, вплоть до клоаки или до наружной
стенки тела; после срастания со стенкой клоаки выводной канал проры-
вается наружу. Во всяком случае, предпочечный проток растет назад
в столь тесной связи с соматоплеврой, что можно считать, что он разви-
вается за счет последней. В некоторых только случаях наблюдается,
повидимому, довольно тесная связь растущего конца протока, по крайней
мере позади, с эктодермой (иногда у млекопитающих).
375
В спинной части полости тела, между спинной брыжейкой и воронками
развиваются сосудистые клубки (отдельные у миног), которые могут
срастись в один общий наружный клубок (у двудышащих и у амфибий).
Участок полости тела с сосудистым клубком и нефростомами может обо-
собиться от остальной части общей полости и получает тогда название
наружной камеры предпочки (амфибии).
В некоторых случаях ближайшая к нефростому часть стебелька
вздувается, дает боковой выступ, в который затем врастает сосудистый
клубочек. Таким образом, в самом канальце развивается внутренняя
камера с клубочком (ганоиды и гимнофионы, у которых, впрочем, отдель-
ные капсулы затем сливаются в одну общую внутреннюю камеру пред-
почки с одним общим сосудистым клубком).
Кровь приносится в клубочки короткими сегментальными артериями,
отходящими от спинной аорты, и уносится кардинальными венами.
Предпочка представляет собой обыкновенно чисто провизорный орган,
который иногда играет значительную роль в качестве личиночного выде-
лительного аппарата (ганоиды, костистые рыбы, двудышащие, амфибии).
Во взрослом состоянии остатки предпочки в измененном, однако, виде
сохраняются только у круглоротых, где они стоят в связи с перикарди-
альной полостью. Существует предположение, что вся почка некоторых
миксин (BdeUostoma) представляет собой предпочку, тянущуюся здесь
вдоль всего тела.
Предпочка обнаруживает известную связь с половыми органами —
за счет ее воронок (или одной воронки) развивается у большинства позво-
ночных воронка яйцевода, служащая для выведения женских половых
продуктов из полости тела наружу.
Канальцы первичной почки (mesonephros) развиваются
немного позже и в общем очень сходно. В сегментах, следующих позади
предпочки, иногда развиваются такие же наружные выросты стебелька
сомита (нефротома), которые дают начало канальцам, растущим по на-
правлению к предпочечному протоку, срастаются с ним своими концами
и прорываются в него. Начиная с этого момента, предпочечный проток
получает название первичнопочечного протока, так
как он служит теперь выводным каналом и для первичной почки. Вну-
тренний конец канальца представляет собой стебелек сомита, который,
сохраняя свою первоначальную связь с полостью тела, превращается в
нефростом (дорзальная связь с сомитом рано прерывается). Такое развитие,
почти идентичное с развитием предпочечных канальцев, наблюдается у
селахий и у осетровых. Здесь можно указать лишь на следующие различия:
первичнопочечные канальцы развиваются позади предпочечных, несколько
позже (как и вообще позадилежащпе части сегментальных органов) и
прорываются в готовый уже проток. Все эти различия не выходят за пре-
делы различий, наблюдающихся между отдельными частями метамерно
построенных органов, и у гимнофион, например, задние канальцы самой
предпочки иногда прорываются в готовый уже предпочечный проток.
Однако иногда наблюдается развитие передних первично почечных каналь-
цев в тех же сегментах, в которых уже имеются предпочечные канальцы,
причем эти канальцы располагаются тогда над последними (дорзально
от них) или позади них.
У других позвоночных развитие первичнопочечных канальцев пред-
ставляется более измененным. Иногда стебелек сомита предварительно от-
деляется от боковой пластинки и превращается в полый пузырек, из кото-
рого затем вырастает каналец (гимнофионы). В других случаях нефротом
.обособляется сначала в виде плотного комочка, в котором лишь потом
появляется полость (многие амниоты). Иногда, наконец, нефротомы сра-
стаются в сплошную • «нефрогенную» полосу (некоторые амниоты) или
расплываются в мезенхиму (ганоиды, амфибии). В этом случае форми-
376
руются сначала метамерно расположенные плотные комочки, превраща-
ющиеся в пузырьки, из которых уже кнаружи вырастают канальцы. Во
всех этих случаях развивающиеся канальцы первоначально не связаны
с полостью тела, но такая связь впоследствии устанавливается — прок-
симальные их концы прирастают к соматоплевре и прорываются в целом,
образуя таким вторичным путем нефростомы.
Каким бы образом ни развивались первичнопочечные канальцы, они
первоначально располагаются строго метамерно по одному на каждый
сегмент. Эта метамерия сохраняется пожизненно только у миксин из рода
Bdellostoma.<y всех других позвоночных число канальцев увеличивается
благодаря тому, что на каждом канальце, со спинной стороны внутреннего
его конца, развиваются добавочные канальцы путем своего рода почко-
вания. Так этот процесс идет у селахий, у осетровых рыб и у гимнофнон,
у которых собственно метамерное строение этим не нарушается, так как
канальцы располагаются правильными сегментальными группами, откры-
вающимися у селахий совместно в первичнопочечный проток. У других
позвоночных развитие вторичных канальцев идет за счет «нефрогенной»
ткани, оставшейся неиспользованной при развитии первичных канальцев.
Эти канальцы позднейшей генерации прорываются либо в первичные ка-
нальцы, либо в первичнопочеч-
ный проток (в некоторых слу-
чаях они прорываются в особые
выросты первичнопочечного про-
тока, о чем речь будет в даль-
нейшем). Таким образом, мета-
мерное строение органа, разу-
меется, исчезает.
При каждом первичнопочеч-
ном канальце развивается в виде
выроста своя внутренняя кап-
сула (боуменова капсула), в
которую врастает сосудистый
клубочек (рис. 341).> Кровенос-
ный сосуд, выходящий из такого
мальпигиева тельца, направляется, однако, не прямо к кардинальной
вене, а распадается предварительно на вторую сеть капилляров, охваты-
вающую стенку канальца, а затем уже собирается в сосуды, несущие кровь
в заднюю кардинальную вену (а у наземных позвоночных позже — в зад-
нюю полую вену). Сами канальцы удлиняются и принимают извитую форму.
Нефростомы первичнопочечных канальцев сохраняются пожизненно
у некоторых селахий и у амфибий, у других позвоночных они исчезают,
и связь нефридиев с полостью тела прерывается.
Первичнопочечный проток у селахий постепенно расщепляется вдоль
на два канала, из которых один, получающий название вольфова канала,,
остается в связи с канальцами первичной почки и служит ее выводным
протоком, а другой — мюллеров канал — сохраняет свою связь с остат-
ками предпочки и становится у самки яйцеводом. Процесс расщепления
начинается впереди и оттуда постепенно распространяется назад до кло-
аки. У других позвоночных такого расщепления обыкновенно не наблю-
дается, но все же развивается новый мюллеров канал, идущий
параллельно первичнопочечному, или вольфову каналу, слу-
жащему выводным протоком первичной почки (рис. 342).
У амниот и первичная почка представляет собой орган преходящего
значения, замещаемый затем вторичной, или тазовой, почкой.
Канальцы вторичной почки (metanephros) развиваются,
сходно с добавочными, вторичными канальцами первичной почки за счет
особого заднего участка той же «нефрогенной» ткани.
Рис. 341. Схема мальпигиева тельца
(из Кингсли).
а — артерия; Ъ — боуменова капсула; gl — клу-
бочек; п— воронка; t — каналец; v — вынося-
щий сосуд.
377
Вместе с тем, однако, из задней части вольфова протока развивается
боковая ветвь, врастающая в нефрогенную ткань. Эта трубка представ-
ляет собой мочеточник (ureter); она разделяется на конце на неко-
торое число конечных разветвлений. Канальцы вторичной почки растут
навстречу этим разветвлениям и прорываются в них. У млекопитатитиу
на месте отхождения ветвей мочеточника образуется расширение — по-
чечная лоханка. Ветви мочеточника превращаются в прямые собиратель-
ные трубочки почки, а все остальные отделы почечных каналов развива-
ются независимо за счет каналь-
Рис. 342. Схема предпочки и первичной
почки и их отношений к половым железам
у анамний.
А — индиферентная эмбриональная стадия;
В — самка; С — самец; cl — клоака; д — по-
ловая желева; md—первичнопочечный проток;
тп — первичная почка; М—мюллеров канал;
о» — яичник; od — яйцевод; рп — предпочка;
t — семенник; vu — мочевой пувырь; W —
вольфов проток.
цев. Почечные канальцы растут
в длину и диференцируются у
млекопитающих на мальпигиево
тельце, извитой каналец, петлю
Генле и соединительный каналец
(рис. 339, D). Вторичнопочечные
канальцы начинаются прямо маль-
пигиевым тельцем, и воронки на
них никогда не бывает; в располо-
жении их, как и в расположении
вторичных канальцев первичной
почки у амниот, метамерии нико-
гда не замечается. Новообразова-
ние вторичнопочечных канальцев
наблюдается у рептилий и птиц
в течение всей жизни и только у
млекопитающих (за исключением
клоачных) ограничивается эмбрио-
нальным периодом.
Таким образом, вторичная поч-
ка отличается от первичной почки,
в которой воронки также могут не
развиваться, только тем, что ее
канальцы прорываются не непо-
средственно в первичнопочечный
проток, а в его вырост — моче-
точник.
Однако и в этом отношении нет столь резкой границы между первич-
ной и вторичной почками, как это могло бы казаться (рис. 343). У селахий
и у костистых рыб в задней части почки обособляются отдельные выводные
каналы, развивающиеся как выросты первичнопочечного протока.
У гимнофион даже на всем протяжении первичной почки добавочные ка-
нальцы прорываются не прямо в первичнопочечный проток, а группами
в особые выросты из него. Эти выросты имеют небольшие размеры и рас-
полагаются правильно мётамерно, по одному на сегмент (у Hypogeophis
на 50 сегментах), и в каждый вырост может впадать до 20 канальцев.
У птиц также развиваются подобные выросты, в которые прорываются
вторичные канальцы, но только в немногих задних сегментах, и распо-
лагаются они неметамерно. Из этих выростов задний получает скоро пре-
обладающее значение, сильно растет и превращается в дефинитивный
мочеточник с канальцами вторичной почки. Иу млекопитающих
иногдащв_виде индивидуальной вариации закладывается несколько моче-
точников, из которых только задний" развивается затем сильнее'"ЕГ дает
начало дефинитивному.
Таким ебра'збм, не только по строению, но и по эмбриональному раз-
витию нет резкой разницы между тремя поколениями почек. Ясно разли-
чаясь своими типичными чертами, они вместе с тем связаны непрерывными
переходами.
378
Несмотря на это, все же совершенно несомненным является самый
факт замещения предпочки, вполне обособленного органа, функциони-
рующего у личинок рыб и земноводных, первичной почкой взрослого жи-
вотного, независимым органом, лежащим позади предпочки. Совершенно
также несомненен факт замещения первичной, туловищной почки амниот,
функционирующей не только у зародышей, но иногда и у молодого живот-
ного (ящерицы, клоачные и лекото'рые сумчатые млекопитающие), совер-
шенно иным органом — вторичной почкой, лежащей в области таза.
Каковы же генетические
взаимоотношения между эти-
ми органами — возможно ли
независимое возникновение и
функциональное замещение
друг друга такими близкими
органами вместо столь, каза-
лось бы, естественной посте-
пенной прогрессивной эволю-
ции одного и того же органа?
На этот вопрос отвечают
различно. Некоторые авторы
полагают, что предпочка,
первичная и вторичная почка
представляют собой органы,
действительно независимые
по своему происхождению,
которые в течение эволюции
позвоночных последователь-
но замещали друг друга. Этот
взгляд наименее согласуется
с фактами, так как он не
объясняет нам поразительно-
го сходства в строении и раз-
витии почечных канальцев и
связи их выводных протоков.
Согласно другому взгляду у
позвоночных первоначально
была одна почка, тянувшаяся
вдоль всей полости тела (ho-
lonephros). Постепенно эта
почка диференцировалась на
переднюю часть, которая раз-
вивалась раньше, функцио-
нировала уже у зародыша
и сохранила более прими-
Рис. 343. Схема почечных канальцев амниот
и их отношение к половым желееам.
А — пндиферентная исходная стадия; В — самка;
С — самец; ер — epididymis, epoophoron; д — гона-
да; h — стебельчатая гидатида; М—мюллеров
канал; md — первичнопочечный проток; mn —meso-
nephros; mtn — metanephros; ot — яичник; os —
воронка яйцевода; pn —pronephros; pa — paradidy-
mis и paroophoron'; r — прямая кишка; sug —моче-
половой синус; t— семенник; и — матка; ит —uterus
masculinus; иг — мочеточник; W — вольфов канал;
си — мочевой пувырь.
тивное строение, и заднюю часть, которая прогрессировала и дала
начало более совершенному органу взрослого животного (теория единой
почки). Эта теория довольно хорошо согласуется с фактами и в особен-
ности хорошо объясняет соотношения между почками различных кругло-
ротыхДУ Bdellostoma совершенно одинаковые почечные канальцы разви-
ваются вдоль всей полости тела; передний отдел этой почки остается в
связи с околосердечной полостью и у взрослого животного, но обособля-
ется от большей задней части, лежащей в брюшной полости. Передняя
часть представляет собой предпочку, задняя считается первичной почкой,
которая у Bdellostoma сохраняет еще свою первоначальную метамерию
и во взрослом состоянип.>У других круглоротых метамерное строение
первичной почки затем нарушается. Против теории единой почки (защи-
щаемой Седжвпком, Фильдом, Брауером и др.) выставлено, однако, одно
379
довольно существенное возражение — она не объясняет тех случаев,,
когда передние канальцы первичной почки закладыаются в тех же сегмен-
тах, в которых располагаются задние канальцы предпочки.
Третья теория может быть названа теорией смены поколений каналь-
цев (Рюккерт, Феликс). По этой теории прогрессивное развитие органов
выделения выражается в появлении новых, более совершенных канальцев,
развивающихся путем обособления (отщепления, почкования) на спинной
стороне старых. Первая генерация вторичных канальцев, развившихся
в предпочке (за исключением лишь передней ее части), имела такое же
метамерное расположение, как и предпочечные канальцы, и представляет
собой первичную почку. Дальнейшее развитие вторичных канальцев при-
вело к утере первоначальной метамерии. Последователи этой теории
считают, что у Bdellostoma вся почка представляет собой еще предпочку.
Предпочка, следовательно, первоначально тянулась вдоль всей полости
тела.>Позднее, с развитием более совершенных первичнопочечных ка-
нальцев в области брюшной полости, предпочечные канальцы здесь
редуцировались и большей частью даже перестали закладываться. Теперь
передние канальцы, развивающиеся раньше других и удовлетворяющие
еще потребностям организма на таких ранних стадиях, стали специаль-
ным эмбриональным (или личиночным) органом.
Эта теория объясняет нам факты нахождения канальцев обоих пер-
вых поколений почек — предпочки и первичной почки — в одних и тех
же сегментах тела, а также и разницу в положении развивающихся ка-
нальцев (первичнопочечные канальцы занимают несколько более дор-
зальное положение по сравнению с предпочечными, а вторичные канальцы
первичной почки лежат также дорзально от первичных).
По Машковцеву предпочечные канальцы развиваются в передней,
а первичнопочечные — в задней части сегмента (основой для такого раз-
вития является первичная диплоспондилия, выражающаяся не только
в двойственности закладки склеротомов, но и в двойственном характере
стебелька сомита). Задние канальцы первоначально, вероятно, несли
половую функцию. Позднее, с перемещением гонад в дорзальную стенку
целома, половые канальцы преобразовались в выделительные канальцы
первичной почки, а предпочечные утратили свое значение и редуцирова-
лись, за исключением только их фильтраторного аппарата (внутренних
камерок с сосудистыми клубочками), сохранившегося в виде боуменовых
капсул в связи с первичнопочечными канальцами.
Что касается происхождения вторичной почки, то этот вопрос в на-
стоящее время- можно считать вполне разрешенным. Мы видели, что у
гимнофион вторичные канальцы первичной почки могут прорываться не
только в другие канальцы или в первичнопочечный проток, но и в специ-
альные выросты последнего, которые у гимнофион располагаются мета-
мерно. Из этих выростов у предков амниот прогрессивно развивались
задние и в особенности самый задний вырост, который стал на конце вет-
виться и вступил в связь с очень большим числом вторичных канальцев
(рис. 343). Передние выросты одновременно теряли свое значение и реду-
цировались, а задний вырост с его комплексом канальцев развился в боль-
шой орган, обособившийся в качестве вторичной почки с ее мочеточником.
С этой точки зрения почка гимнофион представляет по своему строению
смешанный орган, так как она содержит в себе, кроме первичнопочечных
канальцев, большое число метамерно расположенных выростов с комплек-
сами вторичных канальцев, которые все сравнимы со вторичной почкой.
Для почек позвоночных в высшей степени характерна их связь с
половыми органами. Прежде всего эта связь обнаруживается иногда
(осетровые по Машковцеву) уже в закладке половых желез, развивающихся
за счет набуханий эпителия первичных воронок первичной почки. Затем
у самок остатки предпочечных канальцев вместе с обособляющимся от
380
первичнопочечного протока мюллеровым каналом дают начало яйцеводу
с его воронкой, открывающейся в полость тела (см. рис. 342). У самца
развивается иная связь •— мужская половая железа связывается через
соединительные канальцы непосредственно с канальцами первичной
почки, которые тогда служат для выведения семени через вольфов
проток наружу (см. рис. 342)1. Таким образом, первичная почка несет
кроме выделительной функции еще другую: она служит для выведения
половых продуктов — по меньшей мере у самца. В связи с этим первичная
почка обыкновенно диференцируется на два отдела — передний, половой,
•служащий специально для выведения половых продуктов и теряющий
свою выделительную функцию, и задний, специально выделительный,
который явно прогрессирует.
У амниот, с обособлением в этой задней области более совер-
шенной вторичной почки, выделительный отдел первичной почки реду-
цируется, но половой отдел сохраняется в виде придатка семенника. Воль-
фов проток становится теперь уже специально семепроводом (рис. 343).
Таким образом, мы видим, что почки позвоночных совмещают две различ-
ные функции — выделительную и выведения половых продуктов.
Нефридии кольчатых червей также иногда совмещают обе функции,
но все же первоначально, повидимому, они разделяются между различ-
ными образованиями. У щетинконогих червей кроме нефридиальных
каналов имеются нередко в тех сегментах, в которых расположены поло-
вые железы, еще сегментальные органы другого рода — генитальные
канальцы, открывающиеся в полость тела довольно широкой воронкой
и служащие.для выведения половых продуктов. Иногда нефридии сраста-
ются с генитальными канальцами, образуя смешанные «нефромиксии».
Что почечные канальцы ланцетника представляют собой настоящие
нефридии, об этом уже говорилось. Эти органы не имеют никакого отно-
шения к половым органам и, следовательно, резко отличаются от по-
чечных канальцев позвоночных. Последние по своему развитию как-будто
ближе к генитальным канальцам, чем к нефридиям червей. В настоящее
время вопрос о происхождении почек позвоночных нельзя считать раз-
решенным. Несомненно, это — комплекс сегментальных каналов. Но
существует предположение, что не нефридии, а генитальные канальцы
или, быть может, нефромиксии, дали начало почкам позвоночных и что,
следовательно, первичная их функция — половая, к которой позже при-
соединилась выделительная. После приобретения этой новой функции
старые органы выделения (нефридии, как у ланцетника) могли редуци-
роваться. С этой точки зрения, следовательно, предпочка представляет
собой ряд генитальных канальцев. Они распределены метамерно соответ-
ственно первоначально метамерному расположению половых желез (как
у ланцетника) и соединены между собой общим продольным выводным
кацалом (предпочечный протоком). Этот канал представляет собой гоно-
дукт, образование которого связано (как и во многих других случаях)
с приобретением внутреннего оплодотворения и копулятивного аппарата.
Почки круглоротых сохраняют в некоторых отношениях весьма при-
митивное строение, особенно у Bdellostoma, где имеется метамерное расположение
канальцев. Почка круглоротых поделена на два отдела, между которыми известное
число канальцев дегенерирует. Несколько передних канальцев сохраняет свои воронки,
которые открываются в околосердечную полость, — это предпочечные канальцы.
Большое число задних канальцев образует почку, тянущуюся в виде длинной парной
ленты, лежащей по бокам от спинной аорты вдоль всей брюшной полости.
У Bdellostoma эта почка сохраняет свою метамерию и относится некоторыми
авторами также к предпочке. У других миксин и у миног здесь метамерия теряется,
благодаря образованию добавочных канальцев следующей генерации, и почка тогда
1 Иногда, впрочем, и яйцевод самок развивается за счет воронок и продольного
канала первичной почки (ганоиды и костистые рыбы).
381
уже, несомненно, должна быть обозначена как первичная почка. Почечные канальцы
круглоротых теряют во взрослом состоянии свои нефростомы и начинаются прямо
мальпигиевыми телами.
У хрящевых рыб предпочка всегда рано дегенерирует, и функционирую-
щим органом выделения служит первичная почка, сохраняющая у некоторых форм
свои нефростомы. Почка диференцирована, и суженный передний отдел служит у
самца для проведения половых продуктов в первичнопочечный проток, функционирую-
щий в качестве семепровода. Расширенный задний отдел почки служит выделительным
органом, в котором обособляется несколько отдельных протоков, впадающих в осно-
вание первичнопочечного протока. Этим самым строение задней части первичной
почки становится близким к строению тазовой почки. Первичнопочечный проток на
конце расширяется и образует мочевой синус, впадающий вместе с синусом другой
стороны на общем сосочке в клоаку.
У рыб с костным скелетом предпочка является чисто личин очным
органом и иногда заключает довольно большое число канальцев (до 11 у Amia), которое
Рис. 344. Почкп ящерицы (Л) и млекопитающих (В, С). В — С)хема продольного
разреза; С — почка косули (из Видерсгейма].
Са — сосочек; М — сердцевинное вещество; R — корковое; В — бертпниевы столбы;
N —почка; Рг—пирамиды; Рр— лоханка; SG — собирательные канальцы;,Ur—мочеточник.
сильно редуцируется у костистых рыб. У взрослой рыбы предпочка превращается в
лимфиодный орган. Выделительным органом взрослой рыбы является первичная
почка, канальцы которой обыкновенно не сохраняют нефростомов (за исключением
лишь осетровых рыб и Amia). Задняя часть первичной почки состоит из канальцев,
впадающих в особый вырост первичнопочечпого протока, и потому сравнима с тазовой
почкой. Путем образования выроста из мочевых протоков, соединяющихся на конце
между собой, развивается особый резервуар — мочевой пузырь.
Предпочка амфибий представляет собой важный и хорошо развитый личи-
ночный орган. Число каналов обычно, правда, невелико (2—3), но у гимнофион дости-
хает 12 и больше. Обыкновенно имеется наружная камера с общим клубком. Пред-
лочка функцинирует вплоть до метаморфоза. Органом взрослого животного является
первичная почка, которая сохраняет свои нефростомы в течение всей жизни. У бес-
хвостых амфибий эти нефростомы, однако, теряют связь с канальцами и сообщаются
с венозной системой.
У хвостатых амфибий и у гимнофион почка имеет удлиненную форму и Дифе-
ренцирована на узкий передний половой отдел, который у самца служит для прове-
382
дения семени, и широкий падкий — выделительный (рис. 347). У бесхвостых амфибий
почка имеет более компактный вид и не обнаруживает такой ясной диференцировки.
Однако и здесь замечаются различные приспособления, ведущие к обособлению
половых выводных каналов от мочевых, вплоть до полного иногда отщепления от
почек передних или иногда задних канальцев, служащих тогда исключительно для
проведения семени. Вольфов канал служит у самца многих амфибий одновременно и
мочевым протоком и семепроводом.- Обыкновенно он снабжен у самца на заднем своем
конце расширением, служащим резервуаром для семени — семенным пузырьком.
Однако канальцы главного, выделительного, отдела почки обычно впадают в особые
ветви первичнопочеччного протока и в этом отношении приближаются к постоянной
почке амниот. У некоторых хвостатых амфибий наступает, таким образом, полное
обособление мочевого протока и семепровода (например у тритонов). В брюшную
стенку клоаки у амфибий впадает большой мешок — мочевой пузырь, слу-
жащий резервуаром для мочи.
Урептилий и птиц предпочка является чисто' эмбриональным органом
и вообще развита весьма слабо. Первичная почка представляет собой важный, но
также эмбриональный орган, который у многих ящериц функционирует еще неко-
торое время после вылупления (у молодого животного). Во всяком случае она после
дегенерирует (остатки ее сохраняются у рептилий в виде так называемых желтых тел),
за исключением лишь переднего, полоного отдела, который у самца сохраняется в виде
придатка семенника (epididymis) вместе с его вольфовым протоком,
служащим теперь уже специально семепроводом (vas deferens). Вторичные почки
вполне обособляются от первичных' и представляют собой сравнительно небольшие
компактные органы, лежащие в тазовой области.
Иногда почка приобретает дольчатое строение (у птиц обыкновенно три дольки,
у змей большое число лопастей). Мочеточник идет вдоль почки и дает внутрь почки
боковые разветвленные мочевые протоки, с которыми соединяются уже мочевые ка-
нальцы. Эти боковые протоки имеют, таким образом, гребенчатое расположение
(рпс. 344, А).
С брюшной стенкой клоаки связан мочевой пузырь, который, однако, у змей,
у крокодилов и птиц редуцируется, что находится в связи с тем, что их выделения,
состоящие главным образом из мочевой кислоты, теряют жидкую консистенцию и
приобретают вид белой кашицеобразной массы.
У млекопитающих предпочка является совсем рудимен-
тарным образованием. Первичная почка представляет собой важный
эмбриональный орган, который, однако, у клоачных и у сумчатых функ-
ционирует и в молодом возрасте (до половой зрелости), но затем дегене-
рирует. Нефростомы обыкновенно вовсе не развиваются. Постоянная
почка представляет собой компактный орган характерной бобовидной
формы с выпуклой наружной стенкой и вогнутой внутренней, в углубле-
ние которой входят мочеточник и кровеносные сосуды. На разрезе через
почку уже простым глазом можно легко различить зернистый наружный
слой — корковое вещество — и радиально исчерченное сердцевинное
вещество. В первом располагаются извитые канальцы и мальпигиевы
тельца, во втором — прямые собирательные трубочки (рис. 344, В). Для
млекопитающих характерно веерообразное расположение последних (в
отличие от гребенчатого их расположения у рептилий и птиц). Моче-
точник (ureter), войдя в почку на внутренней ее стороне, образует
сейчас же расширение — почечную лоханку (pelvis). Соби-
рательные трубочки соединяются группами, открывающимися каждая
одним отверстием на сосочке, вдающемся внутрь лоханки. Такая группа
собирательных трубочек образует вместе так называемую пирамиду серд-
цевинного вещества (рис. 344). Корковое вещество проникает в проме-
жутки между отдельными пирамидами, так что последние ясно видны на
разрезе через почку уже простым глазом. Число сосочков соответствует,
таким образом, числу пирамид. Иногда имеется лишь один большой со-
сочек, на котором открываются главные ветви мочеточника. В этом слу-
чае все сердцевинное вещество почки собрано в одну пирамиду. Такую
почку называют неразделенной. Отдельным пирамидам почки соответ-
ствует нередко и наружное ее деление на отдельные дольки (рис. 344, С).
Такое дольчатое строение наблюдается у некоторых копытных, хищных
и обезьян, но в особенности оно характерно для слонов и для водных мле-
копитающих — ластоногих и китообразных, у которых дольки резко
383
обособлены. У других млекопитающих дольки тесно связаны и почка
приобретает вид вполне компактного цельного органа.
У клоачных млекопитающих мочеточники впадают в обособленный
отдел клоаки — мочеполовой синус, как раз против отверстия мочевого
пузыря. У живородящих млекопитающих мочеточники впадают уже не-
посредственно в мочевой пузырь, откуда моча выводится через мочепо-
ловой синус наружу (см. рис. 353). У самца мочеполовой синус продол-
жается в мочеполовой канал, проходящий вдоль копулятивного органа.
Моча млекопитающих жидкая и содержит главным образом мочевину
подобно моче амфибий и рыб.
Таким образом, почки позвоночных представляют собой комплекс
выделительных канальцев, которые первоначально имели метамерное
расположение и открывались мерцательной воронкой в полость тела,
а другим концом — в продольный выводной канал. Эти канальцы вполне
сравнимы с сегментальными органами кольчатых червей (и притом, быть
может, скорее с генитальными канальцами, чем с нефридиями). Новым при-
обретением позвоночных является выводной канал, развитие которого,
быть может, связано с приобретением внутреннего оплодотворения, так
как почечные канальцы одновременно служили и для выведения половых
продуктов.
Эволюция органов выделения позвоночных происходила постепенно
путем эмбрионального отщепления от уже существующих канальцев но-
вых, более совершенных. Комплекс первоначальных, сравнительно просто
построенных канальцев представляет собой предпочку и сохранился
только в передней части тела у зародышей. Он развивается раньше дру-
гих почечных канальцев и потому обособился и специализировался в
качестве зародышевого выделительного органа. В лежащих позади сег-
ментах предпочечные канальцы редуцировались, а комплекс добавочных
канальцев следующих генераций представляет собой первичную почку.
Первоначально канальцы новой генерации располагались по одному
на каждый сегмент, за исключением лишь самых передних, и это не нару-
шало метамерного строения органа. Дальнейшие генерации канальцеве
возникавшие путем отщепления или почкования, давали начало сегмен-
тально расположенным группам (селахии). Впоследствии правильно,
расположение вторичных канальцев совершенно исчезает. Эти канальцы
первичной почки построены сложнее — они представляют собой длинные
извитые трубки, всегда несущие на конце мальпигиево тельце (фильтра-
торный аппарат). Нефростомы сохраняются у некоторых примитивных
форм и во взрослом состоянии, но чаще всего впоследствии исчезают.
Наконец, наступило дальнейшее усложнение — выводной проток
стал давать от себя боковые выросты, и новые генерации канальцев стали
вступать в связь с этими выростами. Самый задний вырост получает у
высших наземных позвоночных преобладающее значение (мочеточник);
он на конце разветвляется, и в конечные его разветвления впадает очень
большое число вторичных почечных канальцев, которые все начинаются
прямо мальпигиевыми тельцами (воронок нет). Эта часть органа обособ-
ляется и становится вторичной, или тазовой, почкой. Более сложная,
вторичная почка развивается медленнее и начинает функционировать
позднее, и потому ранее развивающаяся первичная почка специализи-
руется теперь в качестве зародышевого органа выделения.
Обособление отдельного органа — первичной почки — от зароды-
шевой предпочки есть видимое выражение разделения их функций у
низших позвоночных по различным периодам жизни, и обособление вто-
ричной почки от первичной у высших позвоночных обусловлено тем же
фактором.
384
Таким образом у позвоночных возникла картина последователь-
ной смены трех поколений почек.
Почечные канальцы позвоночных с самого начала совмещают в себе
две функции: выделительную и выведения половых продуктов. У различ-
ных позвоночных наблюдаются различные способы разграничения этих
функций: обычно у самки остатки предпочки со своим выводным каналом
(мюллеровым), обособляющимся от первичнопочечного протока, стано-
вятся яйцеводом, а у самца передняя часть первичной почки приспо-
собляется для выведения семени и при редукции первичной почки у
амниот сохраняется в виде придатка семенника.
ОРГАНЫ ВОСПРОИЗВЕДЕНИЯ
ОРГАНЫ ВОСПРОИЗВЕДЕНИЯ БЕСПОЗВОНОЧНЫХ
Половые продукты животных развиваются за счет особых первичных половых
клеток, возникновение которых не приурочено к определенному зародышевому листку.
Хотя половые органы обычно развиваются в мезодерме, первичные половые клетки
обособляются гораздо раньше — часто раньше, чем развивается третий зародышевый
листок, и оказываются тогда нередко в энтодерме, или обособляются даже еще раньше,
на первых стадиях дробления. Такое раннее обособление первичных половых клеток
в индивидуальной жизни, несомненно, является отражением такой же ранней фило-
генетической диференцировки клеток животного организма на соматические и половые.
Эта диференцировка предшествовала всякой иной.
У низших животных образование половых элементов иногда не связано с опре-
деленной частью тела, с определенным органом. У губок первичные половые клетки
рассеяны в паренхиме тела и превращаются или в яйца, или в группы сперматозоидов.
Зрелые половые продукты выпадают в какой-либо из каналов через прорыв его стенки
и затем выводятся наружу. Таким образом, здесь не имеется ни половых органов, ни
особых выводных путей для половых продуктов. Такое дифузное образование половых
продуктов наблюдается иногда и у кишечнополостных. У гидры в различных местах
тела в эктодерме развиваются или отдельные яйца или группы сперматозоидов. У
других кишечнополостных роль половых желез играют группы эктодермальных (у
гидромедуз) или эктодермальных (у сцифомедуз, коралловых полипов, гребневиков)
клеток, которые располагаются уже в определенных местах (на желудочном стебельке,
радиальных каналах и т. п.). Такие гонады не имеют никакого добавочного аппарата
и не обладают выводящими каналами — они при созревании половых продуктов
прорываются наружу или в гастральную полость.
У других животных половые железы развиваются в мезодердщи обыкновенно
связаны с особыми выводными каналами. У круглых червей Половая железа имеет
вид парной или непарной трубки, диференцированной на несколько отделов: у самки —
на яичник, яйцевод, матку и влагалище, а у самца — на семенник, семепровод и семен-
ной пузырек.
У плоских червей часть женской половой железы обособляется и вырабатывает
не яйца, а клетки, набитые питательным материалом, которые затем присоединяются
к зрелой яйцеклетке, это — желточник. Кроме того, наблюдаются еще особые белковые
железы. Яичник обыкновенно продолжается в яйцевод, который диференцируется на
различные отделы, как матка, влагалище, образует резервуары для хранения семени
и т. п. Одним словом, в связи с яичником и его яйцеводом развивается более или менее
сложный добавочный аппарат. У членистоногих половые железы, имеющие нередко
вид разветвленных трубок, связаны также с наружной средой посредством собственных
выводных путей. У моллюсков половая железа нередко связана с очень сложно дифе-
ренцированными выводными путями, снабженными различного рода добавочными
железами. При этом железа нередко является гермафродитной, т. е. служит для выра-
ботки как женских, так и мужских половых продуктов (многие брюхоногие моллюски).
Для нас, однако, гораздо интереснее не эта своеобразная диференцировка поло-
вого аппарата в различных группах беспозвоночных, а то индиферентное состояние,
которое наблюдается у червей с хорошо развитой вторичной полостью тела. У коль-
чатых червей половые железы развиваются как набухания стенок полости тела. Поло-
вые продукты выпадают из железы в полость тела (где они обыкновенно еще дозревают)
и выводятся из последней наружу через генитальные канальцы (или целомодукты,
нередко связанные с нефридиями).
У ланцетника половые железы имеют вид одинаковых у обоих полов
мешочков, которые заключены в участках полости тела и располагаются метамерно
в боковых стенках тела. Зрелые половые продукты выпадают в околожаберную полость
через прорыв ее стенки.
385
Органы восцропзввдышя позвоночных
Половые железы
Рис. 345. Схема строения
семенника молодой ля-
гушки (по Битши).
ат — семенной пузырек;
сс — центральный тяж;
eg — половой тяж.
Половые железы (гонады) позвоночных развиваются в виде парного
набухания стенки полости тела по бокам спинной брыжейки, между ней
и нефростомами первичной почки. У селахий и у осетровых рыб утолщения
образуются прямо в стенках отдельных первичных воронок и соединяются
затем в общий зачаток железы (Машковцев; рис. 346).
Этот парный зачаток состоит из продольной складки утолщенного
эпителия с внутренней массой соединительной ткани и носит название
половых (генитальных) складок. Потом половые складки вдаются глубже
в полость тела, обособляются от стенки тела и оказываются, таким обра-
зом, подвешенными на брыжейке внутри полости
тела (брыжейка яичника носит название mesova-
rium, брыжейка семенника — mesorchium). Пер-
вичные половые клетки, или гоноциты, обосо-
бляются у позвоночных очень рано. С диферен-
цировкой зародышевых листков они обычно выяв-
ляются в составе энтодермы, но затем активно
перемещаются и попадают в генитальные складки.
Эпителий половой железы, содержащий в себе
первичные половые клетки, врастает затем в
соединительнотканную строму в виде многокле-
точных шнуров, которые потом разбиваются на
округлые комплексы клеток — фолликулы.
Дальнейшее развитие половой железы идет
различно в особях различного пола.
В яичниках фолликулы распадаются
таким образом, что каждый в конце концов содер-
жит одну крупную яйцевую клетку, окруженную
многочисленными мелкими фолликулярными клет-
ками, служащими для питания яйца и участвую-
щими вместе с последним в образовании яйцевой
оболочки (zona pellucida s. radiata). У млекопи-
тающих внутри фолликула позднее развивается
полость, наполненная серозной жидкостью, и
такой фолликул получает название граафова пу-
зырька. При созревании яйца стенка фолликула
лопается, и яйцо выпадает в полость тела. На
месте лопнувшего фолликула на поверхности яич-
ника развивается рубец из особого рода клеток—
так называемое желтое тело (corpus luteum),
являющееся периодическим органом внутренней секреции. Новообра-
зование яиц происходит у низших позвоночных в течение всей жизни,
а у высших — только эмбрионально или в первое время жизни.
У самцов низших позвоночных (круглоротые, селахии) сохраняется
еще до известной степени фолликулярное строение и в семенниках, но
отдельные фолликулы приобретают вид настоящих пузырьков и всту-
пают в связь с канальцами, соединяющими семенник с первичной почкой
(рис. 345). У всех других позвоночных пузырьки вытягиваются в семе н-
н ы е трубочки (tubuli seminiferi), которые уже соединяются с семе-
выносящими канальцами. Степки семенных пузырьков и семенных тру-
бочек содержат кроме первйчных половых клеток также индиферентные,
или сертолиевы, клетки, служащие, невидимому, и для питания разви-
вающихся половых элементов. Наконец, из первичной почки (и именно
от эпителия мальпигеевых телец) врастают в семенник клеточные «поло-
вые» тяжи, вступающие в связь с семенными пузырьками или с семенными
386
трубочками. Подобное же врастание половых тяжей от мальпигиевых
телец первичной почки наблюдали и в яичнике. Здесь эти тяжи участвуют
в образовании сердцевинного вещества яичника.
Полозые щ.о гоки
Мы видели, что во время эмбрионального развития у некоторых
позвоночных первично почечный проток расщепляется вдоль на два канала,
из которых один сохраняет связь с первичной почкой, — это вольфов
канал, а другой обособляется от нее, — м ю л л е р о в канал. Послед-
ний развивается иногда и иным путем. У высших позвоночных (амниот)
Рис. 346. Развитие гонады и выводных протоков
у самца Стерляди. Черным обозначены первичные
и точечками — вторичные канальцы мезонефроса
(по Машковцеву).
д — гонада; ma и тс, — первичный и вторичный ка-
нальцы; md — первичнопочечный проток; тп — первич-
ная почка; od — мезонефрический яйцевод; рп — пред-
почка.
он развивается, например, в продольной! утолщении стенки иервичнопо-
чечного протока. По существу этот способ развития очень близок к
первому и представляет результат его упрощения.
Передний конец мголлерова канала срастается с воронкой предпочки
(одной сильно разросшейся или образовавшейся путем соединения не
скольких) и образует яй-
цевод, открывающийся в
полость тела широкой мер-
цательной воронкой, а
другим концом впадающий
в клоаку. Нужно думать,
что передние канальцы
предпочки служили пер-
воначально не только для
выделения мочи, но и для
выведения женских поло-
вых продуктов, а затем спе-
циализировались на этой
функции и вместе с тем
обособилась и соответст-
вующая часть выводного
протока (рис. 342). У хря-
щевых и костных ганоидов,
а такяге у костистых рыб
яйцевод имеет, однако,
другое происхождение. У
этих рыб 2—3 воронки
первичной почки срастают-
ся между собой и соединяются с продольным каналом почки. Послед-
ний впадает в первичнопочечный проток. У высших рыб место впаде-
ния отодвигается назад, и таким образом происходит обособление и этого
«мезонефрического» яйцевода от первичнопочечного протока (Машковцев).
В этом случае яйцевод развивается в общем весьма сходно с семепрово-
дом самцов (рис. 346).
Яйцеводы могут затем диференцироваться на различные отделы, и
задние их участки срастаются у млекопитающих в довольно значительные
непарные образования. У самца мюллеров канал более или менее полно
редуцируется, а выводные пути половых органов развиваются иным
путем.
Как уже упомянуто, от мальпигиевых телец первичнопочечных
канальцев развиваются полоски утолщенного эпителия стенок полости
тэла по направлению к семеннику. Эти половые тяжи врастают в семенник
и вступают в связь с семенными пузырьками или трубочками. Затем в
половых тяжах появляется просвет, и они превращаются в канальцы,
соединяющие трубочки семенника с почечными канальцами, — это с е-
мевыносящие канальцы (vasa efferentia). Семя проводится
25*
387
Рис. 347. Схемы мочеполового аппарата хвостатых
амфибий: А — самца; В — самки (пс Шпенгелю).
а — семевыносящие канальцы; дг—половой отдел почки;
tn — мюллеров канал; ot—воронка яйцевода; о о — яич-
ник; г — выделительная часть почки; ts — семенник;
®е — vasa efferentia; w — вольфов канал; + — краевой
канал почки.
по этим канальцам и по первичнопочечным канальцам в вольфов проток
(рис. 343). Последний служит, следовательно, не только мочевым протоком,
но и с е м е п р о в о д о м (vas deferens). По выходе из семенника семе-
выносящие канальцы соединяются между собой продольным краевым
каналом семенника, а перед входом в почку — подобным же продольным
краевым каналом почки,
который развивается путем
соединения эпителия мезо-
нефрических воронок.
Таким образом, пер-
вичная почка и вольфов
канал несут первоначаль-
но одновременно две функ-
ции. Впоследствии у раз-
личных позвоночных про-
исходит более или менее
совершенное разделение
этих функций между раз-
личными частями почек и
более или менее полное
обособление выводящих
путей. Обычно для выве-
дения семени специализи-
руются лишь некоторые
канальцы, чаще всего в
передней части почки. Эти
канальцы теряют свои
мальпигиевы тельца. Та-
ким образом, почка дифе-
ренцируется чаще всего
на передний, половой, от-
дел и задний, — выдели-
тельный (рис. 347). Перед-
няя часть вольфова канала
служит тогда исключи-
тельно для выведения семе-
ни и приобретает харак-
тер сильно извитой трубки.
В заднем, выделительном,
отделе почки могут обосо-
биться свои выводные каналы, впадающие лишь в самый задний конец
вольфова канала (рис. 347).
У высших позвоночных (амниот) первичная почка совершенно реду-
цируется. Задний, выделительный, ее отдел сохраняется иногда в виде
незначительного образования, прилежащего к семеннику (paradidymis).
Передний, половой, отдел вместе с сильно извитой частью вольфова канала
образует придаток семенника (epididymis) (рис. 343, С). У
самки оба отдела первичной почки редуцируются, и их рудименты могут
быть иногда обнаружены в складке брюшины между яичником и яйцеводом
(paroophoron — рудимент выделительного отдела и epoophoron — руди-
мент полового) (рис. 343, В).
У форм с внутренним оплодотворением в связи с клоакой и половыми
протоками обыкновенно развиваются различно построенные наружные
части, предназначенные для проведения семени в половые пути самки.
Эти совокупительные органы могут иметь различное происхождение.
Половые железы круглоротых представляют собой обыкновенно непарны
орган, тянущийся над кишечником вдоль всей брюшной полости. Семенник сохраняет
388
фолликулярное строение и не имеет выводных протоков (так же, как и яичник). У
миксин происходит очень поздняя и ^совершенная диференцировка полов, так что
половая железа обнаруживает следы гермафродитизма, выраженные весьма различно
у отдельных особей. Задняя часть железы молодого животного представляет собой
семенник, передняя — яичник. При наступлении половой зрелости развивается либо
мужская, либо женская часть железы, так что у самки в задней части яичника имеется
рудиментарный семенник, а у самца в передней части семенника — рудиментарный
яичник.
Зрелые половые продукты выпадают у круглоротых в общую полость тела, а
оттуда выводятся через половые поры, помещающиеся у миноги на стенках особого
мочеполового синуса. У миног яйца мелкие, а у миксин — крупные с плотной («ро-
говой») оболочкой.
У селахий половые железы обыкновенно парные (у некоторых скатов разви-
вается только левая). Половые продукты у самки выводятся посредством яйцеводов,
представляющих собой мюллеровы каналы. Передние концы их обыкновенно соеди-
няются между собой и образуют общее внутреннее отверстие. Яйцеводы диференци-
рованы на несколько отде-
лов. В передней части яй-
цевода, где происходит
оплодотворение, железистые
стенки выделяют белок,
обволакивающий яйцо вна-
чительным слоем. Далее
позади имеется железистое
утолщение стенок яйцевода,
представляющее собой скор-
луповую железу, выделяю-
щую вокруг яйца «роговую»
скорлупу. Наконец, задний
отдел яйцевода образует
расширение, называемое
маткой, в котором у живо-
родящих форм развиваются
зародыши. Яйца селахий
очень велики и превосходят
в этом отношении всех дру-
гих позвоночных (собствен-
но яйцо без оболочек может
достигать 14—22 см; у Не-
xanchus яйца достигают
9—И еле).
Семенники сохраняют
фолликулярное строение и
связаны посредством попе-
речных семевыносящих ка-
нальцев (vasa efferentia) с
канальцами передней части
первичной почки. Вдоль се-
менника канальцы соединя-
ются продольным краевым
каналом. Таким образом,
здесь передняя часть почки
является половым отделом.
Сильно извитой вольфов ка-
нал является семепроводом.
Рис. 348. Схемы мочеполовых органов самок рыб
(по Гудричу из Кингсли).
А — двудышащая рыба (Protopterus); В — многоперый
ганоид (Polypterus); С, D —костные ганоиды (С —Amia)
и (D — Lepidosteus); Е — большинство костистых рыб;
F — лососевые рыбы; cb — клоачный мочевой пувырь;
cl — клоака; / -— воронка яйцевода; др— половое отверстие
(сосочек); т — первичная почка; о — япчпик; od — яйцевод;
г — прямая кишка; s — мочеполовой синус; up — мочевое
отверстие (сосочек); идр — мочеполовое отверстие (сосо-
чек); w — вольфовы протоки.
Он обособляется от выделительной части почки, обладающей вторичными выводными
протоками. На заднем своем конце он расширяется в мешочек — семенной пузырек
(vesiculum seminalis), впадающий в мочеполовой синус, который также может быть
снабжен парным выростом, служащим резервуаром для сохранения семени.
У рыб с костным скелетом иногда наблюдаются весьма типичные
соотношения между половыми железами и органами выделения. В других случаях
развиваются своеобразные отношения между половой и выделительной системами,
причем у высших представителей (у костистых рыб) половые пути совершенно обо-
собляются от мочевых. У двудышащих рыб имеются парные яичники, и яйца выво-
дятся наружу при помощи длинных яйцеводов (мюллеров канал) (рис. 348, Л). Яичники
ганоидов представляют собой парные органы в виде довольно длинных складчатых
лент. Яйцеводы представляют собой мезонефрические продольные каналы, которые
более или менее полно срастаются с яичниками. У осетровых рыб, многоперых (Poly-
pterus) и у одного представителя костных ганоидов — ильной рыбы (Amia) — короткие
яйцеводы открываются широкой воронкой в полость тела и тесно прилегают к соот-
ветствующим яичникам (рис. 348). У другого представителя костных ганоидов —
389
яйцевод сросся с
(юа
1(3*^
IL-J Лч>
яичником, так что
9Р
up
Рис. 349. Схемы мочеполового
папцырпой щуки (Lepidostens) и у костистых рыб отдельного яйцевода нет, а мешковид-
!.ыг яичники непосредственно продолжаются в короткий канал, открывающийся
"1 >-.ы ч. мочеполовом пли отдельном половом сосочке (рис. 348, D, Е^. Во время
. .1 ' поп mi.того развития у самки ятих рыб половые складки образуют желобок и
< ем свесышютя в трубку, канал которой, тяким образом, представляет собор
о'- ылрнвую ч сп, обще й полости тела. Стенки яичника образуют зятем многочислен-
ны нт 1к’п. свешивающиеся во внутреннюю полость. Зрелые яйца выпадают внутрь
пос.|е.1!1< й
Только у лососевых (Salmonidae) и некоторых других костистых рыб (Muraenidae,
Monnyridae. Cobilis) яичник не имеет мешковидной формы (хотя иногда снабжен
продольным желобком), и яйца выпадают в общую полость тела. В последних случаях
яйца выводятся наружу через половые поры, открывающиеся вместе позади заднепро-
ходного отверстия. Половые поры нередко связаны с воронковидными углублениями
брюшины, а иногда (у лососевых — Osmerus, Mallotus) с парой коротких яйцеводов
(рис. 348, F).
Во всех других случаях особых яйпеводов, как сказано, не имеется, и сравнение
с ганоидами позволяет вывести заключение, что здесь короткий мезонефрический
его воронка соединилась с краевым желобом,
а затем и с внутренней
полостью яичника. Отноше-
ния, существующие у лосо-
севых и некоторых других
костистых рыб, представля-
ют собой скорее вторичный
возврат к первоначальному
состоянию, чем примитив-
ный признак.
У самца наблюдается
более или менее полное обо-
собление выводящих путей
от органов выделения.
У хрящевых и костных га-
ноидов имеются еще весьма
примитивные соотношения
(рис. 349. _-1). Кроме много-
численных семевыносящих
канальцев, имеются и оба
соединяющие их продоль-
ные канала (краевой канал
семенника и краевой канал
ночки). Подобные же при-
митивные
блюдаюгся
из двудышащих,
м н о го числе и н ые
сящие канальцы ведут через
первичную почку в вольфов
проток. Но у других дву-
дышащих наблюдается ди-
ференцировкя, причем в ка-
честве половых путей спе-
ция л из и ру ются почечные
канальцы задней части поч-
Lepidosiren семевыносящие
-О
.-о
^0
:О
аппарата самцов рыб
(по Гудричу из Кингсли).
А — ганоид (Amia и Lcpidosteus); В — костистые рыбы;
С — многиперый гапонд (Polypterus); D — двудаппащая
(Protopterus); Е — мочеполовые отверстия самки лососе-
вой рыбы; а — заднепроходное отверстие; о.р — брюшная
пора; cb —клоачный пузырь; е — vasa efferentia; др—по-
ловое отверстие (сосочек); I — краевой капал почни; It —
краевой капал семенника; т— первичная почка; md—яйце-
вод; г — прямая кишка; ге — мальпигиево тельце; s — мо-
чеиоловой синус; t— семенник; и, up — мочевое отверстие;
идр —мочеполовое отверстие; v— семепровод; w — вольфов
проток.
соотношения на-
и у Cera tod из
И здесь
Семены но-
ки (а не передней, как у большинства позвоночных).
канальцы имеются лишь в небольшом числе (5—6) и проходят через задние почечные
канальцы: у Protopterus связь поддерживается при помощи одного лишь заднего ка-
нальца, который вполне обособился от остальной почки и приобрел значение самостоя-
тельного выводного пути (рис. 349, D}. Таким образом, семепровод здесь развился из
кразвого канала первичной почки, соединяющегося лишь в самом конце с первично-
почечным протоком.
Совершенно независимо подобный же процэсс имел место у многоперых ганоидов
и у костистых рыб. У Polypterus имеются еще оба продольные краевые канала, со-
единенные между собой при помощи семезыпосящих канальцев, но краевой канал
почки вполне обособился от последней и самостоятельно впадает в мочеполовой синус
(рис. 349. С). Таким же образом у костистых рыб произошло обособление полового
протока, так что семенник как бы .продолжается в свой выводной канал, открывающийся
в мочеполовой синус (рис. 349. В) или даже непосредственно наружу.
Большинство рыб с костным скелетом является яйцекладущими и имеет сравни-
тельно мелкие яйца, но известны и живородящие формы среди различных групп кости-
стых рыб.
У амфибий наблюдаются такие же взаимоотношения между половыми
железами и почками, как у селахий (рис. 347, 350).
390
Яичники представляют собой удлиненные парные органы, подвешенные на бры-
жейках к спинной стенке брюшной полости. Зрелые яйца выпадают в полость тела,
откуда выводятся наружу по очень длинным извилистым яйцеводам (мюллеровым
каналам), открывающимися отдельными воронками в переднюю часть полости тела
и впадающим позади в клоаку. Железистые стенки яйцеводов выделяют студенистую
массу, обволакивающую яйца во время их прохождения по яйцеводу. В задней, рас-
ширенной части яйцеводов у бесхвостых происходит формирование комков и шнуров,
в виде которых они откладывают яйца.
Оплодотворение у бесхвостых амфибий наружное, а у хвостатых амфибий и гим-
пофион — внутреннее. Яйца обыкновенно мелки.
Семенники имеют вид овальных тел, подвешенных на спинных брыжейках. Иногда
они разделены на несколько самостоятельных отделов, а у гимнофион состоят из целого
ряда четковидно расположенных телец, связанных между собой продольным каналом.
Семевыносящие канальцы связывают семенник с почечными канальцами (с мальпи-
гиевыми тельцами), соединяясь предварительно между собой с помощью продольного
краевого канала.
Рис. 350. Освобождение полового протока от почечных протоков
у хвостатых амфибий (по Машковцеву)
А — аксолотль; Б — Triton; С — Salamandrlna; D — Spelerpes; Е —Desmognathus; md — по-
чечные протоки; I — краевой капал почки; I — семенник; vd — семепровод.
У хвостатых амфибий почка диференцируется на передний половой и задний
выделительный отделы, причем канальцы последнего более или менее обособляются
от вольфова канала, служащего семепроводом (рис. 347). У бесхвостых такой дпфе-
ренцировки не наблюдается, и вольфов канал несет обе фукнции на всем своем протя-
жении. Задняя его часть обыкновенно вздувается, образуя семенной пузырек. Однако
у некоторых форм семя выводится только через передние канальцы или даже один
передний каналэп почки, который вполне обособляется от остальной части почки. Одно-
временно и вольфов канал расщепляется спереди назад, образуя отдельный от почки
семепровод (Discoglossidae). Таким образом, здесь в передней части почки происходит
процесс, аналогичный тому, который наблюдается в задней части почки у двудышащих,
кистеперых и костистых рыб, т. е. и здесь краевой канал первичной почки является
семепроводом, но он соединяется с вольфовым каналом не позади почек, а впереди
(рис. 350). Сперматозоиды многих хвостатых амфибий склеиваются в пачки — спер-
матофоры, которые при спаривании схватываются краями клоаки самки.
У амфибий с половыми железами связаны особые лимфоидные органы, иногда
очень сложной формы — так называемые жировые тела, которые развиваются за счет
зачатка половой железы. Повидимому, эти органы содержат известные запасы пита-
тельных веществ, так как весной они подвергаются сильной редукции, а после икро-
метания постепенно восстанавливаются1.
У высших позвоночных (амниот) сохраняются типичные взаимоот-
ношения между половыми железами и органами выделения. С развитием вторичной
почки первичная почка редуцируется, и ее остатки сохраняются в связи с половыми
органами — передний, половой их отдел представлен спутанными семевыносящими
трубочками, которые вместе с сильно извитым вольфовым каналом образуют у самца
1 У жаб (Bufo) передняя часть половых желез (семенники и яичники) имеет стро-
ение, сходное с ранней стадией развития яичника (биддеров орган). Функция этого
стэрилыюго участка половой железы г. точности неизвестна, но, так же, как и жировые
тела, он периодически редуцируется (г.есной) и затем опять регенерирует. Повидимому,
это орган внутренней секреции. При кастрации биддеров орган дает начало настоящему
яичнику.
391
придаток семенника (epididymis), а остаток вадней, выделительной, части почек со-
храняется в виде незначительного рудимента (paradidymis). У самки обе части почки
редуцируются и сохраняются иногда в складке брюшины между яичником и яйцево-
дом в виде незначительных рудиментов (epoophoron и paroophoron) (рис. 343).
У рептилий и птиц парные половые железы иногда взаимно смеща-
ются, особенно у форм с длинным змеевидным телом, у которых иногда замечается и
асимметричное их развитие, причем у змей и некоторых ящериц левый яичник развит
слабее правого. У птиц, в связи, очевидно, с огромной величиной их яиц, развивается
только один левый яичник, а правый совсем редуцируется. Точно так же развивается
и один левый яйцевод.
Яйцеводы у рептилий и у птиц диференцированы на отделы. Передняя их часть
выделяет у черепах, крокодилов и птиц белок, обволакивающий яйцо значительным
слоем; стенка следующей позади части яйцевода выделяет кожистую скорлупу и про-
питанную известью скорлупу у птиц и некоторых рептилий.
Оплодотворение у рептилий и птиц внутреннее, причем у рептилий и у некоторых
птиц имеются совокупительные органы. Яйца отличаются крупными размерами.
Рис. 351. Мочеполовой аппарат
самца утконоса (ив Видерс-
гейма).
В — мочевой пувырь; С1 — клоа-
ка; Ер — epididymis; Gp — по-
ложение головки члена во вла-
галище; Но—семенник; m—мыш-
цы; г — почка; Рр — praeputium;
rt — прямая кишка; вид — моче-
половой синус; Ur —мочеточник.
Vd — vas deferens; Ve — vas effe-
rens.
Половые железы млекопитающих развиваются, как и у дру-
гих позвоночных, впереди от вторичных почек, но затем постепенно
перемещаются назад в тазовую область (вследствие того, что поддержи-
вающие их складки брюшины растут медлен-
нее, чем само тело животного). Яичники
остаются у всех млекопитающих в задней
части брюшной полости, между тем как се-
менники сохраняют это положение в течение
всей жизни только у клоачных и некоторых
плацентных млекопитающих (некоторые на-
секомоядные и неполнозубые, слоны, даманы,
китообразные и сирены). У других млекопи-
тающих семенники выпадают из брюшной
полости наружу в особый мешок — мошон-
ку, увлекая за собой обволакивающий их
перитонеальный покров (descensus testiculo-
rum) (рис. 353). Мошонка (scrotum) предста-
вляет собой выпятившуюся часть стенки
тела с ее мышцами и связками и заключает
в себе участок полости, выстланный брю-
шиной.
Канал, соединяющий полость мошонки
с остальной полостью тела, носит название
пахового канала и может оставаться откры-
тым. В этом случае семенники погружаются
в мошонку лишь периодически во время
течки (грызуны, насекомоядные, рукокры-
лые, некоторые обезьяны) и после этого
втягиваются обратно в брюшную полость
помощью особого мускула (m. cremaster),
обособившегося от поперечной брюшной
мышцы. В других случаях паховой канал
зарастает, и семенники постоянно лежат
в мошонке. У сумчатых мошонка лежит впереди копулятивного органа,
у плацентных млекопитающих — позади.
У млекопитающих от клоаки обособляется передняя ее часть, свя-
занная с мочевым пузырем, в качестве мочен ол о в о г о с ину с а
(sinus urogenitalis, рис. 351, имеющийся также уже у черепах), в который
впадают и половые протоки (яйцеводы и семепроводы, а у клоачных и
мочеточники). У живородящих млеко питающих клоака исчезает, и моче-
половой синус открывается независимо наружу, образуя у самки неболь-
шой отдел, так называемое преддверие влагалища (рис. 352,
353), и вытягиваясь у самца в длинный мочеполовой канал.
392
Для млекопитающих в высшей степени характерен процесс посте-
пенного срастания яйцеводов v(m ю л л е р о в ы х каналов), ведущий к
образованию более или менее значительны к нииггрИкх" отделов (рис. 352).
У клоачпых оба яйцевода
совершетноразделены и самостоятельно
еще
Рис. 352. Диференцировка яйцеводов у млекопитающих (схема).
А — клоачное; В — молодая двуутробка; С — взрослое сумчатое; О — плацентное млеко-
питающее с двойной маткой; Е — с двурогой и F — с простой маткой; cl — клоака; od —
яйцеводы; г — прямая кишка; sug — мочеполовой синус; иг — мочеточники; ut — матка:
vag — влагалище; v. и — мочевой пузырь.
впадают в мочеполовой синус. Задняя часть каждого яйцевода расширена
и образует, так называемую, матку (стенка которой выделяет рогоподоб-
ную скорлупу яйца). У сумчатых оба яйцевода могут оставаться незави-
симыми или срастаются на некотором протяжении. Задние отделы яйце-
Рис. 353. Схематическое изображение мужского мочеполового аппарата черепахи
(Л), клоачпого млекопитающего (В) и живородящих млекопитающих (С, Л), вид сбоку.
ап — задний проход; cl — клоака; с. ид — мочеполовой канал; р — penis; г — почка;
вид — мочевой синус; t—семенник; иг — мочеточник; _ vd—семевыносящий канал;
v. и — мочевой пузырь.
водов образуют пару длинных влагалищ, которые нередко срастаются
своими передними концами. От сросшегося переднего конца влагалищ
назад направлен слепой мешок (рис. 352, С), который иногда позади
срастается с мочеполовым синусом и прорываете, в него; таким образом
развивается так называемое третье влагалище. Остальная часть яйцеводов
диференцируется на собственно яйцеводы, или фаллопиевы трубы, от-
393
крывающиеся в полость тела широкой воронкой с бахромчатым краем, и
матки (uterus), в которых происходит развитие зародыша.
У плацентных млекопитающих задние концы мюллеровых протоков
всегда сливаются в непарное влагалище (vagina). Остальная часть
яйцеводов может остаться парной на всем своем протяжении, и тогда имеется
двойная матка (uterus duplex) с парным отверстием во влагалище (боль-
шинство грызунов, слоны и др.) (рис. 352, D), или срастание в большей
или меньшей степени захватывает матку. Если матки сливаются только
Рис. 354. Мужской мочеполовой аппарат
ежа (по Видерсгейму).
cd — пещеристые тела; ер — epididymis;
д. с — куперовы железы; gl — головка
члена; дрр — препуциальные железы; рр—
препуциум; рг — предстательная железа;
г — почка; te — семенник; иг — мочеточ-
ник; rd — семевьтносящий канал; v. s. —
семенной пузырек; vu — мочевой пузырь.
Рис. 355. Женский мочеполовой аппарат
ежа (по Видерсгейму).
adr — околопочечные тела; д — железы; о —
воронка яйцевода; od — яйцевод; г — почка;
t —прямая кишка; иг — мочеточник; ut —
матка; vg—влагалище; vu—мочевой пузырь.
позади, между тем как передняя часть остается подразделенной продоль-
ной перегородкой, то говорят о двуразделыюй матке (uterus bipartitus),
как это наблюдается у некоторых грызунов, хищных, свиней и др. У дру-
гих хищных и копытных, у китообразных и насекомоядных срастание
идет еще дальше, и следы парного происхождения выражаются лишь в
существовании парных выростов, связанных с фаллопиевыми трубами;
это — двурогая матка (uterus bicornis) (рис. 352, Е). Наконец, у некоторых
рукокрылых, у обезьян и у человека произошло полное срастание обеих
маток в одну простую матку (uterus simplex) (рпс. 352, F). Наблюдаются
у млекопитающих, разумеется, и все переходы между этими типичными
формами.
Семенники представляют собой компактные овальные тела, связанные
с придатком, состоящим из извилистых семенных протоков с чрезвычайно
извилистым вначале вольфовым каналом (рис. 356), образующим семепро-
вод. Выходя из придатка, семепровод идет в виде ровного канала до впа-
дения в мочеполовой синус. Последний вытянут в виде канала, снабжен-
ного значительной мускулатурой, и носит название мочеполового
канала (canalis urogenitalis s. urethra) (рис. 353). В переднюю его часть
нередко открываются рудименты мюллеровых каналов, которые своим
394
основанием сливаются между собой, как и у самки (так называемая муж-
ская матка). Мочеполовой канал продолжается в канал копулятивного
органа (рис. 353).
В связи с половыми протоками развиваются у млекопитающих раз-
личного рода добавочные железы, особенно сильно развитые у самцов
(рис. 354). С семепроводами перед впадением их в мочеполовой канал
связана у многих млекопитающих пара мешковидных желез, носящих
наименование семенных пузырьков, которые достигают осо-
бенно значительного развития у некоторых грызунов и насекомоядных.
Основание мочеполового канала охватывается железистой массой, обра-
зующей предстательную железу (gl. prostata), в ткань кото-
рой погружен упомянутый рудимент яйцеводов. Кроме того, почти у всех
млекопитающих имеются парные куперовы железы, лежащие
по бокам мочеполового канала и обладающие поперечнополосатой мускуль-
ной оболочкой.
Гомологи этих желез имеются и в преддверии у самки, где они из-
вестны под названием бартолиниевых желез (рис. 355).
Копулятивные органы
Органы совокупления позвоночных могут иметь различное проист
хождение. У самцов селахий копулятивные органы представляю
собой преобразованные задние (внутренние) части брюшных плавников.
Такие парные органы поддерживаются хрящевым скелетом и снабжены
на своей внутренней поверхно-
сти желобком, по которому семя
стекает из клоаки самца. Во
время совокупления эти органы
вдвигаются в клоаку самки.
У костистых рыб
совокупительный аппарат на-
блюдается редко. У живородя-
щих Cyprinodontidae таковым
служит преобразованный задне-
проходный плавник. Этот орган
служит для перенесения склеен-
ных в пачки сперматозоидов
(сперматофоров) и для прикре„
пления их к половому отвер-
стию самки.
Из а м ф и б и й органы
совокупления имеются только
у гимнофион, у самцов которых
Tubuti
contort/
-Cactus deferens
Ductus
aberrans
Титса
albuginea
Ductus
epididymidis
nets
'Tubulus
rectus
Epithet.
Pnc. 356. Схема строения семенника и его
придатка у млекопитающего (человека)^(по
Эберту).
Caput epididyrmdis
длинная клоака может выворачиваться
и во время полового акта вводится в клоаку самки.
У рептилий наблюдаются копулятивные органы двух различных
типов. У змей и у ящериц — это пара мешков, лежащих под кожей у
заднего края клоаки. Эти мешки отходят от стенки клоаки и могут выдви-
гаться наружу. По внутренней стенке выпяченного мешка проходит
бороздка для стекания семени.
У черепах, крокодилов и у некоторых птиц на передней стенке
клоаки (рис. 357) имеется непарный копулятивный орган (penis). Он
Представляет собой утолщение стенки клоаки, поддерживаемое волок-
нистой тканью, содержащей наполняемые кровью полости (corpus fibro-
sum). Вдоль этого утолщения проходит продольный желобок, служащий
для стекания семени и охваченный эректильной губчатой тканью (corpus
spongiosum).
395
Дистальный конец органа, на котором желобок кончается ямкой,
может выдвигаться из клоаки действием особой мускулатуры. Из птиц
совокупительный орган имеется только у бескилевых (страусов и киви),
у пластинчатоклювых, некоторых куриных и у тинаму.
Выше всего развиты копулятивные органы млекопитающих.
Это непарный орган, развивающийся у зародыша в передней стенке клоаки
в виде небольшого вначале выступа — клоачного бугорка. В передней
Рис. 357. Продольный средний раврез черев клоаку самца крокодила (ив Иле).
cd — coprodaeum; cf —волокнистое тело члена; сое! — полость тела; dd—впадение семевыно-
сящего канала; int — кишка; Pd — proctodaeum; ud— urodaeum; иг— впадение мочеточ-
ника,
части клоачного бугорка развивается «половой бугорок», на нижней
поверхности которого образуется продольный половой желобок, ограни-
ченный по бокам половыми складками (рис. 358). Вокруг всех этих частей
развивается кольцеобразная складка — половой валик. Дальнейшее раз-
витие этих индиферентных частей полового аппарата идет различно у
особей разного пола — у самца половые валики срастаются между собой
Рис. 358. Развитие наружных половых органов человека (по Феликсу)»
А — индиферентное состояние у эмбриона 28 мм длиной; В — эмбрион женского пола в 32,5
мм длиной; С — эмбрион мужского пола в З1/, месяца; ап—анальное отверстие; cl — кли-
тор; gl. р—головка члена; 1т — большие губы; ид — первичное мочеполоваое отверстие;
р — член; вег — Мошонка.
по средней линии и образуют стенки мошонки, а у самок многих обезьян
и у женщин — так называемые большие губы. У самца половой бугорок
сильно растет и дает половой член (penis); у самки он слабо развивается
и превращается в так называемый клитор (рис. 358). Небольшие кожные
складки (половые складки) по краям половой бороздки дают у самки
многих млекопитающих малые губы, а обособившийся от клоаки моче-
половой синус дает широкое преддверие влагалища. У самца края половых
складок срастаются над мочеполовым синусом и над половой бороздкой,
идущей от него вперед по половому бугорку, и таким образом развивается
длинный мочеполовой канал. Подобный капал развивается и в клиторе
396
у самок некоторых млекопитающих (некоторые насекомоядные и хищные
грызуны и полуобезьяны).
У клоачных млекопитающих копулятивный орган
сидит внутри клоаки, окруженный складкой слизистой оболочки (вла-
галищем), и может выдвигаться наружу и втягиваться обратно в клоаку
(рис. 359). Этот орган усовершенствован по сравнению с рептилиями
и птицами в том отношении, что бороздка замкнулась в канал, служащий
для проведения семени. Канал начинается отверстием у основания
члена в мочеполовом синусе и кончается на вершине. При замыкании
отверстия, ведущего из синуса в клоаку, семенная жидкость протекает
не в клоаку, а по каналу копулятивного органа.
У живородящих млекопитающих клоака исчезает,
и между мочеполовым синусом и каналом копулятивного органа насту-
пает постоянное и непрерывное сообщение, так что он служит не только
для выведения семени, но и для мочеиспускания (рис. 353, 359). Половой
Рис. 359. Схемы мужских мочеполовых органов: / — клоачных; II — сумчатых и
III — плацентных млекопитающих (по Веберу из Кингсли).
а — задний проход; Ъ — мочевой пузырь; с — клоака; си — пещеристое тело канала;
ср — пещеристые тела члена; cd — куперова железа; р — промежность; ро — предстатель-
ная железа; г — прямая кишка; з — лобковая кость; t — семенник; и — мочеточник;
v — семпвод; ад — семенной пузырек; ит, т — брюшные мышцы.
член помещается теперь снаружи, но окружается складкой кожи, обра-
зующей его влагалище. У сумчатых, многих грызунов и насекомоядных
половой член расположен еще очень близко от заднепроходного отверстия,
и влагалище члена окружено общим сфинктером вместе с заднепроходным
отверстием (остаток клоаки). С прогрессивным развитием промежности
расстояние между этими частями увеличивается. Первоначально, у сум-
чатых член обращен верхушкой назад, но у других млекопитающих он
поворачивается верхушкой вперед, ложится горизонтально под покровами
брюшной поверхности (или висит свободно, как у человека) (рис. 359).
Копулятивный орган поддерживается парным или иногда непарным эрек-
тильным волокнистым телом (corpus fibrosum), и непарным также эректиль-
ным губчатым телом (corpus spongiosum), окружающим канал и образую-
щим на конце члена вздутую головку разнообразной формы. У сумчатых
конец члена часто раздвоен соответственно двойному влагалищу самки.
У сумчатых, хищных, ластоногих, китообразных, грызунов, руко-
крылых, полуобезьян и некоторых обезьян в передней части волокнистого
тела копулятивного органа развивается еще кость (os penis). Подобная
кость наблюдается и в сильно развитом клиторе у самок некоторых млеко-
557
питающих. Влагалище копулятивного органа образует часто вокруг его
головки кожную складку (praeputium), под которой открываются иногда
с со рые препуциальные железы (рис. 342), представляющие собой специа-
лизированные кожные железы, обладающие обычно ароматическим вы-
делением (у бобра — «бобровая струя»),
ОРГАНЫ ВНУТРЕННЕЕ С1.КРЕЦИН
Всякая ткань, любой орган тела выделяет известные продукты обмена,
которые не сразу выводятся наружу, а некоторое время циркулируют
внутри тела. Эти вещества обычно не являются индгферентными, а ока-
зывают либо известное общее действие на организм или воздействуют тем
или иным образом специфически на определенные ткани и органы. Любой
орган, выделяя известные вещества, оказывает некоторое влияние на дру-
гие органы, и, таким образом, кроме нервной связи, устанавливается
некоторая химическая связь между органами всего тела. Такие деятель-
ные (возбуждающие или тормозящие) химические вещества носят назва-
ние гормонов, и процесс их отделения называется внутренней секрецией,
или инкрецией. Внутренняя секреция в широком понимании является
побочной функцией любого органа, хотя бы, например, мышцы, но в не-
которых случаях ее значение в этом смысле и специфическое действие
резко бросаются в глаза наравне с главной функцией (например, печень,
поджелудочная железа, половые железы) или, наконец, мы видим органы
или обособленные части органов, специально приспособленные для такой
функции, каковы, например, у позвоночных щитовидная и зобная железы,
гипофиз п эпифиз, околопочечные органы. Эти органы особых протоков
не имеют; железа обильно промывается кровью, увлекающей выделяемые
ею гормоны. Надо сказать, что эти органы еще не изучены с достаточной
полнотой и пока известны главным образом у позвоночных.
Производные эпителия глотки
От эпителия жаберной области кишечника развиваются следующие
органы внутренней секреции: щитовидная железа, зобная железа и эпи-
телиальные тельца.
Щито видная железа представляет собой орган фолликуляр-
ного строения, располагающийся с брюшной стороны глотки. Развивается
она в виде непарного выступа брюшной стенки глотки. У личинки миноги
(Ammocoetes) эта закладка занимает довольно-большую область (от второй
до четвертой щели) и имеет вид продольного желобка, стенка которого
выстлана мерцательным эпителием и содержит четыре ряда железистых
клеток. Этот желобок продолжается в мерцательную бороздку, разбива-
ющуюся впереди от первой жаберной щели на две ветви, охватывающие
глотку с боков и соединяющиеся затем на спинной стороне глотки в над-
жаберную бороздку, идущую назад в пищевод. Сходство этого железистого
желобка с эндостилем низших хордовых животных бросается в глаза,
и в их гомологии не может быть сомнений. Однако затем ко времени мета-
морфоза желобок постепенно отшнуровывается от эпителия глотки и пре-
вращается в замкнутый пузырек, эпителий которого образует в группу
отдельных фолликулов. У других позвоночных закладка имеет вгд про-
стого углубления, лежащего между первой и второй парами жаберных
щелей. Это углубление постепенно отшнуровывается и дает начало полому
пузырьку, стенка которого разрастается в виде трубочек или плотных
клеточных шнуров, распадающихся затем на отдельные фолликулы. По-
следние являются сначала однослойными, а затем становятся многослой-
ными. Внутри пузырьков происходит накопление особого содержащего
398
иод белкового вещества, называемого «коллоидом». Совокупность фолли-
кулов обрастает массой соединительной ткани.
У селахий щитовидная железа располагается впереди брюшной аорты,
между ветвями нижней челюсти. У костистых рыб она охватывает брюш-
ную аорту в области передних дуг. У двудышащих намечается.разделение
ее на две половины. У амфибий щитовидная железа является парной и
лежит в области подъязычного аппарата (у хвостатых — позади второй
дуги, а у бесхвостых — под задними рожками). Щитовидная железа реп-
тилий непарная, но часто поделена на две лопасти и располагается под
трахеей, а у птиц пара желез лежит у основания бронхов (рис. 361).
У млекопитающих щитовидная железа двулопастная, но иногда также
распадается на пару отдельных желез. Обыкновенно она располагается
в области гортанных хрящей.
Выделяемый в кровь продукт железистой деятельности щитовидной
железы млекопитающих представляет собой азотистое соединение, содер-
жащее иод и сильно действующее на обмен веществ. Этот гормон в настоя-
Рис. 360. Производные эпителия глотки у ската (Raja, Л), лягушки (7?)
и курицы (С) (по Вердену из Кингсли).
е — эпителиальные тельца; дт— остатки жабер; р — зажаберное тельце; tm — зобная железа;
tr — щитовидная железа; I—VI—жаберные мешки.
щее время получен синтетически (тироксин). При недостатке гормона
щитовидной железы в организме наблюдаются весьма серьезные заболе-
вания (кахексия).
Щитовидная железа есть, следовательно, результат преобразования
поджаберного железистого желобка (эндостиля) низших хордовых,
который у позвоночных изменил свою функцию и превратился в железу
внутренней секреции.
Зобная железа представляет собой парный, по происхожде-
нию, лимфоидный орган, лежащий у рыб в жаберной области, а у назем-
ных позвоночных — обыкновенно по бокам шеи. Развивается зобная
железа в виде плотных выступов эктодермального эпителия спинной
части жаберных мешков (рис. 360). Эти выступы отшнуровываются и сли-
ваются на каждой стороне между собой, образуя полосу с узкой полостью
и толстыми стенками, от которой вырастают многочисленные округлые
ветви. Впоследствии она прорастает соединительной тканью и превращается
в парный, иногда лопастный лимфоидный орган, физиологическое зна-
чение которого еще не достаточно изучено, но, очевидно, его секреция
оказывает влияние па рост организма, особенно его скелета, и на развитие
половых желез.
У рыб зачатки зобной железы наблюдаются иногда на всех жаберных
мешках (рис. 360), и развившаяся таким образом железа сохраняет свое
первичное положение на спинной стороне жаберного аппарата. У амфибий
399
зобная железа развивается из меньшего числа зачатков, и иногда (у бес-
хвостых) лишь одна пара жаберных мешков (вторая) участвует в образо-
вании железы, а на других мешках имеются только рудиментарные за-
кладки. Зобная железа располагается у амфибий в области заднего угла
нижней челюсти. У гимнофион она развивается на всех мешках и состоит
из отдельных бранхиомерно расположенных долей. У рептилий и птиц
зобная железа развивается за счет двух-трех пар жаберных мешков и
лежит по бокам шеи, вытягиваясь у крокодилов и птиц в длинный ленто-
видный орган (рис. 361). У млекопитающих эта железа развивается с
брюшной стороны за счет трех пар жаберных мешков (главным образом
за счет второй) и располагается в грудной полости впереди от сердца,
Рис. 361. Положение щитовидной и вобной желев у ящериц (Л), птиц (В}
и млекопитающих (С) (по Шимкевичу).
с — сердце; са — сонная артерия; spe — супраперикардиальные тела; t — трахея; tm — зоб-
ная железа; tr — щитовидная железа; vj — яремная вена; 1х —гортань.
непосредственно над грудиной. Этот орган достигает своего наибольшего
развития у молодого животного (у человека в возрасте около 14 лет) и
обыкновенно после наступления половой зрелости постепенно умень-
шается в размерах, хотя нормально всегда имеется и у взрослого живот-
ного. У млекопитающих, кроме «грудной» зобной железы, иногда разви-
вается еще и другая — «шейная», впереди первой, за счет эктодермы
покровов шеи (некоторые сумчатые, обыкновенный крот, грызуны, ко-
пытные, полуобезьяны).
Кроме того, за счет эпителия жаберных мешков у наземных позво-
ночных образуются еще так называемые эпителиальные тельца
(околощитовидные железы), о функциональном значении которых еще
мало известно. Эпителиальные тельца развиваются у наземных позво-
ночных из брюшной стенки двух-трех жаберных мешков (у амфибий
только после метаморфоза) (рис. 360). У млекопитающих эти зачатки
сдвинуты закладкой зобной железы на спинную сторону и развиваются
обычно на третьем и. иногда четвертом жаберных мешках. У млекопитаю-
щих они нередко тесно прилежат к щитовидной железе или даже с нею
срастаются. Околощитовидные железы регулируют кальциевый обмен
организма и особенно влияют на развитие костей.
Наконец, позади последней пары жаберных мешков по бокам глотки
наблюдается эпителиальное утолщение, соответствующее, повидимому,
паре рудиментарных жаберных мешков и обособляющееся потом в виде
скопления замкнутых фолликулов с «коллоидным» содержимым. Эти
образования носят название зажаберных (пост- или ультимо-
бранхиальных) телец (рис. 360); у амфибий они лежат в области гор-
танной щели. У взрослых млекопитающих зажаберные тельца обычно
также прилежат к щитовидной железе. Функция их неизвестна.
400
Железы, связанные с мозгом
Гипофиз (hypophysis cerebri s. glandula pituitaria) представляет
собой железистую массу, прилежащую к воронке промежуточного мозга,
и состоит из множества клеточных шнуров или трубок, залегающих
в соединительнотканной строме.
У круглоротых гипофиз развивается в виде непарного впячивания
эктодермы над верхней губой, которое по мере роста последней отодви-
гается на спинную сторону и соединяется с органом обоняния. Дистальная
часть трубки ложится под мозгом и образует у миног довольно крупный
гипофизарный мешок, а у миксин он прорывается в глотку. Так как
имеется достаточно оснований думать, что рот у предков позвоночных
помещался далее впереди (прежде всего на это указывает существование
предротовых частей висцерального аппарата), то некоторые авторы (Бирд,
Купфер) предполагают, что гипофизарная трубка и представляет собой
остаток первичного рта позвоночных (palaeostoma).
У других позвоночных гипофиз возникает в виде выпячивания экто-
дермального же эпителия крыши ротовой полости и растет также по
направлению к основанию промежуточного мозга. Этот зачаток потом
вполне обособляется от эпителия ротовой полости и ложится у конца
воронки. Только у хрящевых рыб во взрослом состоянии сохраняется
остаток первоначальной связи с эпителием рта. Здесь от стенки эпите-
лиального мешочка вырастает множество ветвящихся трубочек, которые
затем распадаются на клеточные тяжи и группы. Таким образом, разви-
вается железистый орган, который уже у рыб диференцируется на два
отдела: брюшную, главную, лопасть и спинную, прилежащую к воронке,
промежуточную лопасть.
У высших позвоночных (амниот) задняя часть воронки, соответствую-
щая сосудистому мешку и нижним долям низших позвоночных, реду-
цируется, теряет свои нервные элементы и превращается в клеточную
массу (исключительно из невроглии), прилежащую к гипофизу. Это —
задняя, нервная, лопасть гипофиза. Передняя, железистая, часть гипо-
физа диференцируется в свою очередь уже у рыб на два отдела (передний
и промежуточный). Эти части являются важными железами внутренней
секреции, действие которых изучено, однако, главным образом, на мле-
копитающих. Передняя, брюшная или главная часть, выделяет в кровь
гормоны, регулирующие рост костей, и при повышении ее деятельности
сверх нормы наблюдается его нарушение: гигантский рост или (у взросло-
го) акромегалия. Промежуточная, или спинная, часть передней лопасти
гипофиза выделяет инкрет в желудочек промежуточного мозга. Этот гор-
мон оказывает, через щитовидную железу, возбуждающее влияние на
развитие и функцию половых органов, на гладкую мускулатуру и млеч-
ные железы. При удалении этой части гипофиза наблюдается отложение
жира и недоразвитие половых органов.
Эпифиз развивается в виде выроста крыши промежуточного
мозга. По своему происхождению это орган зрения, который у кругло-
ротых еще сохранил до известной степени глазоподобное строение. В ре-
дуцированном состоянии он имеется под кожей головы и у бесхвостых
амфибий. У млекопитающих он превращается в железистый орган —
шишковидную железу (grandula pinealis). От конца выроста развиваются
многочисленные эпителиальные тяжи, распадающиеся на отдельные фол-
ликулы, связанные соединительной тканью. У человека гормоны шишко-
видной железы оказывают тормозящее влияние на деятельность половых
желез и развитие вторичных половых признаков (при удалении или
прирожденных дефектах ее наблюдается преждевременная половая зре-
лость). У взрослого человека железистая ткань редуцируется и прорастает
соединительной тканью.
401
Околопочечные органы
Под названием околопочечных органов известны два рода различных
по происхождению органов— надпочечные и межпочечпые тела, которые
у наземных позвоночных соединяются в один парный орган.
Надпочечные тела (corpora suprarenaies) образуются за счет
зачатков симпатических нервных узлов, от которых обособляются комп-
лексы клеток, получивших название хромафиновых (так как они интен-
сивно окрашиваются солями хромовой кислоты в желтый или коричневый
цвет). У рыб они образуют обыкновенно ряд парных, иногда довольно
ясно метамерно (у селахий) расположенных тел, лежащих вдоль спинной
поверхности первичных почек в тесной связи с их сосудами. У амфибий
они без особого порядка прилежат к межпочечным телам, а у амниот
тесно с ним сливаются, образуя пару околопочечных органов сложного
состава. Отдельные хромафиновые клетки могут также оставаться внутри
симпатических ганглиев, и, с другой стороны, в надпочечных телах встре-
чаются обычно также и нервные клетки.
У млекопитающих часть, гомологичная надпочечным орга-
нам рыб, образует внутреннюю массу компактного околопочечного ор-
гана — его мозговое вещество. Последнее состоит из округлых
комков упомянутых уже хромафиновых клеток, густо оплетенных кро-
веносными сосудами, и представляет собой очень важную железу внутрен-
ней секреции. Хромафиновая ткань встречается, однако, не только в
околопочечных органах, но, кроме того, и в отдельных тельцах —«пара-
ганглиях». Продукт ее секреции не только выделен в чистом виде, но даже
получен синтетически; это — адреналин (C9H1SNO3), который регулирует
возбуждение симпатической нервной системы и действует на органы,
иннервируемые последней, т. е. на гладкую мускулатуру (в частности,
действует на кровеносные сосуды как сосудосуживатель), и, очевидно,
играет большую роль в регуляции кровообращения. Кроме того, адре-
налин возбуждает процесс расщепления гликогена в печени.
Межпочечпые тела (corpora interrenales) развиваются в виде
метамерно расположенных утолщений перитонеального эпителия, которые
потом отшнуровываются и ложатся в виде парного ряда между внутрен-
ними краями обеих почек по бокам спинной брыжейки. Затем эти зачатки
обыкновенно сливаются в парный или непарный тян?, лежащий между
почками в области спинной брыжейки. Клетки межпочечной системы
характеризуются своими липоидными включениями.
У наземных позвоночных они, как сказано, соединяются с надпо-
чечными телами в один орган, тесно связанный с почками. У амфибий
они лежат на внутренней (у хвостатых) пли брюшной (у бесхвостых)
поверхности последних, у рептилий — вблизи половых органов. У м л е-
к опита ющих околопочечные органы довольно компактны и при-
лежат к переднему концу почки. Гомолог меж по чечных тел образует
у млекопитающих периферическое корковое вещество около-
почечных органов, которое также обильно снабжается кровью. Клетки
межпочечных органов содержат значительное количество липоидных
веществ. Функция межпочечных тел менее изучена, но это также гажная
железа внутренней секреции, удаление которой смертельно. Возможно,
что эти органы служат, между прочим, также и для связывания различных
токсических веществ.
Инкреторные части половых желез
Огромное значение в половой жизни и, в частности, для развития
вторичных половых признаков имеют гормоны, выделяемые в кровь
половыми железами, как семенниками (мужские половые гормоны), так
402
и Яичниками (женские половые гормоны). Значение этих гормонов ясне
выявляется при опытах кастрации позвоночных, а также при трансплан-
тации половых желез на особей другого пола. По Штейнаху, эта секреция
относится к деятельности межуточной ткани половых желез, развиваю-
щейся за счет мезодермальной стромы. Другие исследователи приписы-
вают эту деятельность половому эпителию или самим половым клеткам.
Женский половой гормон, вызывающий течку млекопитающих, накапли-
вается в фолликулах созревающих яиц и отсюда поступает в кровь. Этот
гормон — фолликулин, или фолликулостерон, накапливается в пла-
центе и в моче беременного животного, из которой он был извлечен в кри-
сталлическом виде и, наконец, недавно получен синтетически (С18Н22О2).
В яичниках высших позвоночных наблюдается, кроме того, еще другой,
периодически развивающийся секреторный аппарат. После выпадения
зрелого яйца на поверхности яичника образуется рубец в виде бугра из
железистых клеток; у млекопитающих клетки этого рубца усиленно
размножаются и дают начало довольно крупному образованию, — это
так называемое желтое тело (corpus luteum s. follicuJare), обычно
ярко окрашенное особым пигментом — липохромом — в оранжевый цвет.
Этот пигмент заключается в жире, который накапливается в клетках
желтого тела, называемых лютеиновыми клетками. У млекопитаю-
щих гормоны, выделяемые желтым телом, играют большую роль. Они
вызывают изменение слизистой оболочки матки, способствующее имплан-
тации оплодотворенного яйца, и, кроме того, препятствуют созреванию и
выделению дальнейших яйцевых клеток и, наконец, быть может, пара-
лззуют мускулатуру стенок матки в течение начального периода бере-
менности. Если оплодотворения не произошло, то желтое тело быстро
редуцируется; в противном случае оно редуцируется лишь во второй по-
ловине беременности. Во время беременности, как известно, созревание
дальнейших яиц прекращается. Многие незрелые фолликулы редуци-
руются и превращаются также в группы лютеиновых клеток, подоб-
ные желтым телам поверхности яичника. Гормон желтого тела «лю-
теостерон» недавно получен синтетически (С21Н оО2). Мужские поло-
вые гормоны имеют близкое химическое строение и также недавно синте-
зированы. Это — «андростерон» (С19НзоО2) и более сильный гормон «те-
стостерон» (С18Н28О2).
ЧАСТЬ III
ФИЛОГЕНИЯ ПОЗВОНОЧНЫХ
И ЕЕ ЗАКОНОМЕРНОСТИ
ФИЛОГЕНИЯ ПОЗВОНОЧНЫХ
Палеонтологический метод в филогении
Мы познакомились с филогенетическими преобразованиями отдель-
ных органов и целых систем органов; однако в конкретной эволюции
изменяются не отдельные органы, а целые организмы, и, таким образом,
для восстановления подлинной истории организмов надо произвести
синтез полученных данных с использованием всех доступных нам методов.
Дело в том, что при установлении филогении всегда есть опасность увле-
чения абстрактными формами, обобщенными «типами», лишенными всякой
жизненности, так как необходимое для этого восстановление истории
функций и истории связей организма с окружавшей их ранее средой
является задачей весьма нелегкой. Кроме сравнения целых организмов и
изучения их в связи с их естественной обстановкой, необходимым
является и возможно более полное использование палеонтологических
данных. Палеонтология дает нам возможность не только восполнить
анатомические данные материалом, касающимся строения ископаемых
форм (в этом мы не имеем принципиально нового метода), она дает нам,
кроме того, знакомство с конкретными организмами, жившими в прежние
эпохи в определенной биологической обстановке. Об этой обстановке
можно судить, с одной стороны, по типу строения ископаемых животных
и, с другой стороны, по остаткам сопутствующих им организмов и по
характеру самих осадочных пород. Все эти данные приводятся для неко-
торого прошедшего времени, в котором, действительно, некогда разви-
вались предки современных нам животных. К сожалению, однако, палеон-
тология лишь исключительно редко, да и то для ограниченных отрезков
времени, дает нам действительно полные ряды форм в их естественной
последовательности. Так как только особо счастливый случай может
сохранить даже твердые части скелета позвоночных от полного разруше-
ния, то палеонтологические данные являются всегда неполными. При
огромном количестве совершенно вымерших животных лишь крайне
редко попадаются единичные представители таких форм, которые, действи-
тельно, являются прямыми предками каких-либо современных животных.
Обычно находимые остатки лишь приближаются по своему строению к
этим предкам, оказываясь в самом деле более или менее уклонившимися
представителями вымерших боковых ветвей. Таким образом, несмотря
на данную в палеонтологии хронологическую последовательность форм,
восстановление их истории является все же делом нелегким и требует
в свою очередь также полного учета всех данных морфологии.
404
Естественно, что чем далее в глубь веков уходит история, тем менее
полны и палеонтологические данные и, следовательно, тем большие труд-
ности стоят перед изучением филогении живших тогда организмов. Так
как все основные типы животного царства во всем своем многообразии
имелись уже в начале палеозойской эры, то полная история типов вообще
на основании палеонтологических данных не восстановима; она относится
к более раннему времени, от которого не сохранилось почти никаких
следов организмов. Только история самого молодого типа, именно хордо-
вых животных, или, точнее, позвоночных (так как ископаемые остатки
низших хордовых неизвестны), может быть до некоторой степени просле-
жена по дошедшим до нас остаткам. В этом случае мы, действительно,
можем прежде всего установить известную последовательность появления
представителей различных классов позвоночных и можем в некоторых
случаях проследить все наиболее существенные этапы филогенетических
преобразований отдельных ветвей. В особенности это относится к более
молодой группе позвоночных — млекопитающим, преобладающая масса
которых относится к третичному периоду. Сохраняются в основном,
конечно, твердые части, т. е. скелет, и притом наиболее полно скелет
более крупных животных. Поэтому нас не должно удивлять, что в палеон-
тологии наиболее хорошо изучены скелеты самых крупных и вместе с тем
специализированных стегоцефалов, рептилий и особенно млекопитаю-
щих. Геологическая последовательность появления отдельных классов
позвоночных рисуется в следующем виде. Наиболее примитивные позво-
ночные, относящиеся к бесчелюстным формам (Agnatha), известны, на-
чиная с силура. Начиная с силура, известны также остатки зубов и ши-
пов низших рыб (Elasmobranchii). В девоне сразу появляется очень боль-
шое количество разнообразных рыб, относящихся к селахиям, цельно-
головым (Holocephali), хрящекостным (Chondrostei), кистеперым (Cros-
sopterygii) и двудышащим (Dipnoi) рыбам. Костные ганоиды (Holostei)
появляются с начала мезозоя (в триасе) и костистые рыбы (Telostei) с
середины мезозоя, достигая расцвета только в третичный период. Первые
наземные позвоночные — стегоцефалы — появляются уже начиная с
верхнего девона. Стегоцефалы достигают расцвета в каменноугольный
и пермский периоды, а до триаса доходят лишь наиболее специализиро-
ванные формы. В конце каменноугольного периода появляются рептилии,
которые уже в перми представлены весьма разнообразными формами.
Максимального развития рептилии достигают в мезозое, к концу
которого их начинают вытеснять млекопитающие. Птицы появляются
.в виде наиболее примитивных представителей в юре (Archaeornis), в виде
зубастых птиц — в мелу и основных современных групп — в третичном
периоде. Млекопитающие появляются еще раньше. В триасе они во
всяком случае уже имелись, и надо думать, что время их возникновения
относится еще к пермскому периоду. В течение мезозоя они, однако,
представлены лишь мелкими примитивными формами. Образование
современных отрядов относится к третичному периоду. Таким образом,
последовательность появления отдельных классов позвоночных вполне
совпадает с нашими общими представлениями о филогении позвоноч-
ных, полученными на основании данных сравнительной анатомии и
эмбриологии. Теперь нам остается перейти к более детальному рас-
смотрению материала.
Эволюция многоклеточных животных
Попытки восстановить историю организмов начались сейчас же после
установления эволюционного учения. Уже Ламарк пытался выяснить
родственные соотношения между различными группами организмов и
изобразить их графически в виде таблицы с указанием родственных свя-
зей, т. е. в виде «родословного древа». Гораздо более широкой была попытка
405
Э. Геккеля охватить весь фактический материал того времени и дать пол-
ную филогению живых существ (как животных, так и растений). Хотя
и эта попытка кажется нам теперь мало совершенной и геккелевское ро-
дословное древо встретило очень много возражений, но все же оно легло
в основу позднейших филогенетических построений.
Мы здесь не будем разбирать вопросов филогении беспозвоночных животных,
однако, вкратце коснемся основных моментов эволюции многоклеточных, чтобы яснее
определить положение хордовых во всей системе животного царства. О происхождении
многоклеточных (Metazoa) ничего определенного нельзя сказать. Ни в настоящем,
ни в прошедшем нет никаких организмов, которые бы связывали их с простейшими.
Разнородная группа Mesozoa объединяет ряд вторично упрощенных, деградировавших
организмов неизвестного происхождения, которые ни в коем случае не могут пролить
свет на интересующий нас вопрос. Гипотезы о происхождении Metazoa построены на
основании эмбриологических данных. Однако, так как у низших Metazoa, именно у
разных кишечнополостных, уже первые стадии развития и, в частности, процессы
гаструляции оказываются различными, то ученые по-разному представляют себе их
происхождение. Нзиболее популярна геккелевская теория гастреи. по которой много-
клеточные животные развились из шарообразной колонии простейших, подобной Vol-
vox. Разделение трудя привело к диференцировке на два полушария, одно из них взяло
на себя локомоторную функцию, а другое — питательную. Питающее полушарие
ввернулось затем внутрь, и таким образом получился организм, построенный по типу
двуслойного бокала — гастрея. Эмбриональные стадии бластулы и гаструлы пред-
сгшляют собой, по Геккелю, поз горение филогенетических стадий плайей и гастреи.
Т и: как современные кишечнополостные и особенно гидрополипы построена в осноз-
н 1М ио типу гастреи, то гипотету Геккеля можно в основном считать вероятной, однако,
с не (вторыми О'о юрками. До юль ю вероятно, что Metazoa развились из ш трообразной
(или из бесформенной) коло.ши простейших, подобной Volvox. О днако. утверждать,
что это была колония жгутиковых инфузорий в современном смысле этого слова, а
тем более говорить о представителях Volvocales, нет никаких оснований (хотя бы по-
тому, что колонии Volvocales составляются из гаплоидных особей). Кром° того, переход
однослойного полого шара в двуслойный организм типа гастреи мог итти и не пу:ем
инвагинации, как предполагал Геккель, а путем диференцировки и разделения кле ок
на два слоя (теория «планулы» Лэнкестера) или путем миграции отдельных клеток
внутрь полости шэра (теория «фагоцителлы» Мечникова) Отверстие—первичный
рот такого организма — могло образоваться вследствие неполного замыкания клеток
при развитии зародыша и рот мог первоначально служить для выделения наружу кле-
ток, служащих для размножения (теория «генитогаструлы» Заленского). Наконец,
многие особенности развития и строения губок заставляют думать, что по своему про-
исхождению они вряд ли тесно связаны с кишечнополостными, а скорее всего пред-
ставляют ветвь организмов (Parazoa), самостоятельно развившихся из каких-то про-
стейших.
Из всего сказанного ясно, что о происхождении многоклеточных животных мы
можем высказывать только различные предположения. К этому можно прибавить,
что некоторые исследователи считают, что мы вообще не вправе говорить о происхож-
дении многоклэточных из колоний простейших и что скорее можно предполагать
их возникновение из простых недифереицированных организмов путем вторичной
разбивки тела на обособленные участки, т. е. путем приобретения клеточного
строения.
Каково бы, однако, ни было конкретное происхождение многоклеточных, мы
должны отметить самый факт возникновения многоклеточного строения (безразлично —•
из колонии или путем расчленения простого тела) как чрезвычайно важный этап, узло-
вой пункт, знаменующий собой подъем на более высокую ступень организации, бла-
годаря тем возможностям разделения функций между (до известной степени обособ-
ленными) элементами тела, которые привели в дальнейшем к сложнейшим диферен-
цировка м.
Перной такой диференпировкой (имеющейся уже в колониях Volvox) является
разделение на соматические и половые элементы.
Кто) ой диференцировкой, связанной с разделением функций питания и движе-
ния, является образование двуслойного организма с внутренним и наружным листами
тела. Эго разделение ведет к рационализации принятия пищи — принятие нищи непо-
средственно внутрь отдельных клеток (внутриклеточное пищеварение) заменяется
принятием ее внутрь некоторой полости тела, где она может подвергаться более или
мепее значительной предварительной обработке при помощи выделяемых сюда фер-
мешов, может быть измельчена и частично растворена. В измельченном и растворенном
виде пищевой материал захватывается затем клетками энтодермы, в которых может
происходить и дальнейшее внутриклеточное пищеварение. Таким образом становятся
доступными для использования в качестве питательного материала гораздо более круп-
ные частицы органических веществ и целые организмы.
Дальнейшим важным моментом в прогрессивной диференцировке многоклеточ-
ных является образование внутреннего остова из продуктов выделения эктодермаль*
406
вых и, может быть, эктодермальных клеток, образующих пограничную между ними
перепонку или более значительную студенистую массу (галлерта медуз), в которую
мигрируют и разрозненные клетки эктодермы. Таким образом возникает здесь средний
слой мезенхимы. Это образование внутреннего остова позволило увеличить пищева-
рительную поверхность первичной кишки путем образования системы выростов и
каналов в промежутке между внутренней полостью и наружными покровами.
Довольно неясен вопрос о происхождении обширной и разнородной группы чер-
веобразных организмов. Кишечнополостные — зто в основном прикрепленные орга-
низмы с радиальной симметрией, которая у высших свободноплавающих форм пере-
ходит в двулучевую (ктепофоры). Переход к более активным формам передвижения —
к ползанию по субстрату или к активному плаванию — естественно, всегда связыва-
ется с развитием двусторонней симметрии. Животное движется в определенном направ-
лении, которое становится главной осью. Вокруг последней симметрично распола-
гаются различные органы. Тело вытягивается вдоль зтой главной оси и при активных
формах движения ориентируется всегда определенным образом. Нижняя поверхность
тела несет иные функции (ползание на субстрате или поддержка тела в воде) и стано-
вится более плоской — брюшной в отличие от более выпуклой — спинной. Конкретно
это-т переход можно себе мыслить по-разному. Большой популярностью пользуется
гипотеза Ланга о происхождении червей от ползающих ктенофор, подобных известным
Ctenoplana kowalevskii и Coeloplana metschnikovi. У зтих форм уже развился мерца-
тельный покров на брюшной стороне тела, а весельный аппарат начинает редуциро-
ваться. У Coeloplana развивается система сложно разветвленных гастральных каналов,
а гребни исчезают. Сидячая кгенофора Tj ilfiella tristoma еще интереснее. Гребни име-
ются лишь в молодости и затем погружаются и превращаются в особые мерцательные
каналы, что как будто согласуется с предположением Крумбаха о происхождении
протонефридий червей из весельного аппарата ктенофор. Наиболее интересно строение
рта. Как и у других ктенофор, он имеет вытянутую в виде щели форму. Tj ilfiella при-
крепляется своим ртом к субстрату и приподнимает углы рта (т. е. концы щели), ко-
торые продолжаются в небольшие трубки. Хотя, несомненно, имеются извесгные черты
сходства между ресничными червями и ктенофорами в вентральном положении рото-
вого отверстия, в строении кишечника, в дорзальном положении аборального органа
ктенофор и в соответственном положении головного ганглия и статоциста у морских
турбеллярий, а также в следах радиальной симметрии у последних (четырехлучевое
расположение бластомеров и в особенности зачатка мезенхимы, следы радиального
расположения нервных стволов), все же на подобные организмы, конечно, нельзя
смотреть как на переходные формы. Вполне мыслим подобный же переход от какого-
либо иного представителя кишечнополостных (по Седжвику — от сцифополипа). Од-
нако, указанные примеры ценны в том отношении, что показывают реальную осущест-
вимость подобных мыслимых переходов. Вопрос осложняется, правда, еще тем, что
группа червеобразных организмов весьма разнородна, и вполне возможно, что она
имеет полифилетическое происхождение. Известное сходство ранних стадий развития,
а также личиночных форм позволяет, однако, предполагать существование некоторой
генетической связи между ними. Говорить о происхождении различных червей в более
конкретной форме в настоящее время еще не представляется возможным.
Не подлежит сомнению, что основные черты строения червей связаны с переходом
к активному передвижению (двусторонняя симметрия), которое позволило перейти
и к более активным формам питания и к дальнейшей его рационализации. Первым
существенным моментом в этой эволюции является приобретение анального отверстия.
Этот процесс, очевидно, связан с изменением формы, т. е. с удлинением первичного
рта и с его диференцировкой на передний (ротовой) и задний (анальный) участки, по-
добно тому как это наблюдается у Tjalfiella tristoma. Средняя часть ротовой щели
срослась, и, таким образом, пищеварительная полость оказалась связанной.с внешней
средой посредством переднего отверстия, через которое при движении вперед пищевой
материал попадал внутрь кишечника, и посредством заднего отверстия, из которого
выбрасывались неиспользованные остатки. В таком устройстве мы видим дальнейшую
рационализацию пищеварительного аппарата, так как существование единого отвер-
стия первичного рта ведет к тому, что при опорожнении гастральной полости одновре-
менно с выбрасыванием непереваримых остатков пищи выбрасывается и вполне при-
годный,, но еще не использованный более свежий пищевой материал. Теперь, с образо-
ванием второго отверстия, устанавливается непрерывный ток пищевого материала
в одном определенном направлении, что обеспечивает его более равномерное исполь-
зование. Это использование было, очевидно, тем более полным, чем более (с удлине-
нием самого тела) удлинялась и образовавшаяся таким образом кишечная трубка.
Второе усовершенствование состояло в изменении системы распределения всо-
савшегося уже пищевого материала по различным частям тела. Первоначально про-
исходила непосредственная его подача в разные части через каналы и выросты самой
пищеварительной полости (турбеллярий). Теперь всосавшийся жидкий пищевой ма-
тер.ццл поступает в околокишечные межтканевые пространства, образующие «первич-
нэ䧻 полость тела,.и при сокращениях кожно-мускульного мешка, сопровождающих
движения животного, эта жидкость перемещается и распределяется весьма равномерно
по всему телу. С обособлением особых продольных, а затем и поперечных каналов,
переходивших далее обычно в межтканевые пространства, ток жидкости принимал.
407
более правильный координированный характер. Путем такой диференциации посте-
пенно образовалась кровеносная система как посредник между органами пищеварения
и остальным телом. Естественно, что движение жидкостей в этой системе способство-
вало и передаче растворенных в ней газов: кислорода от поверхностных частей тела
к более глубоким и, наоборот, углекислоты от тканей организма к его поверхности,
и этим намечалась вторая роль кровеносной системы как посредника в газовом обмене.
Наконец, и жидкие продукты распада, естественно, поступали в ту же межтканевую
жидкость и затем в систему сосудов, из которой они тем или иным путем выводились
через органы выделения наружу.
Весьма важным моментом в эволюции червей является также развитие ограни-
ченной собственными стенками вторичной полости тела. Образование этой полости
дало максимальную независимость движениям кишечника, освободив их, вместе с
тем, от движений кожно-мускульного мешка. С развитием собственной мускулатуры
кишечника ва счет стенок вторичной полости тела передвижение пищевого материала
производится путем перистальтических движений кишки и не зависит от движений
самого тела червя.
Наконец, немалое значение имела в дальнейшей эволюции и локализации дыхания
в ограниченных частях общих покровов. Местное увеличение дыхательной поверхности
привело к образованию специальных дыхательных органов — жабер. Это обусловило
не только возможность повышения интенсивности газового обмена, а следовательно
и обмена энергий и веществ вообще, т. е. общего повышения жизнедеятельности, но,
вместе с тем, освободило большую часть покровов от функции дыхания, позволило раз-
вить более плотную (и вместе с тем мало проницаемую) кутикулу и дало тем самым
более прочную защиту тела животного от разного рода повреждений, от проникновения
паразитов и т. п. Развитие плотной кутикулы (хитина), предохранявшей организм и
от потери воды, позволило животному перейти от жизни в воде к жизни на суше.
Не подлежит никакому сомнению, что членистоногие произошли от высших
червей со вторичной полостью тела и с метамерно расчлененным телом. По целому ряду
признаков они обнаруживают явное сходство с кольчатыми червями. Нервная система
построена совершенно по тому же типу парной брюшной нервной цепочки. Метанефри-
дии, имеющиеся еще в виде сегментальных органов у интереснейшего «переходного»
организма, представителя первичнотрахейных — Peripatus, — сохраняются у рако-
образных в их антенной и раковинной железах, по у трахейных замещаются совсем
иными выделительными органами — мальпигиевыми сосудами. Параподии червей
преобразовались в членистые ножки. Целом вторично редуцируется. Мы не можем
быть уверены, что ракообразные, с одной стороны, и трахейные — с другой, развились
из одного ствола, однако, все же несомненно, что это близко родственные ветви. Пауко-
образные, бесспорно, родственны ракообразным и, очевидно, произошли от древнейших
Merostomata (Giganthostraca). Низшими из настоящих трахейных являются много-
ножки, сохранившие еще в общем гомономную сегментацию тела, как у Peripatus,
но обладающие уже мальпигиевыми сосудами, подобно насекомым. Если у Peripatus
трахеи представляются в виде сегментально расположенных пучков неветвящихся
трубочек (сравнимых с кожными железами), то у многоножек имеются уже более сложно
разветвленные пучки трахей, открывающиеся, однако, наружу сегментально распо-
ложенными стигмами. У некоторых многоножек (Scutigera) появляются и сложные
глаза, как у насекомых, а в группе Symphylla (Scolopendrella) личинки обладают
вначале тремя парами ходильных ножек на грудном сегменте, как насекомые. Не под-
лежит сомнению, что насекомые, в частности Apterygota, связаны с многоножками
весьма близким родством. На происхождение органов летания насекомых бросают
свет ископаемые Palaeodictyoptera (из каменного угля), у которых, кроме ряда при-
митивных признаков, как довольно однородная сегментация тела, нескладывающиеся
крылья (как у стрекоз) на 2-м и 3-м грудных сегментах, были еще горизонтальные
крыловидные выросты на 1-м грудном сегменте и небольшие подобные выросты и на
всех сегментах брюшка.
Происхождение моллюсков также в общем довольно ясно. По своему эмбриональ-
ному развитию (типичное спиральное дробление) и по строению личинки — veliger,
на ранних стадиях вполне сходной с трохофорой, они, бесспорно, тесно примыкают
к высшим червям, очевидно, утратившим в этой ветви свою сегментацию (впрочем,
быть может, ее и не было). Имеются и формы до известной степени «переходные». У чер-
веобразных Solenogastres нервная система состоит еще из окологлоточного кольца с
отходящими от него парой брюшных (ножных) стволов, сравнимых с брюшной цепочкой
аннелид, и парой спинных стволов. У хитонов имеется еще участок целома около ки-
шечника, а у соленогастров полости гонад сообщаются с перикардиальной, и половые
продукты выводятся через последнюю и черев метанефридиальные каналы.
Мы здесь ни в какой мере не можем входить в более детальное рассмотрение
вопросов происхождения- отдельных групп беспозвоночных, но лишь отметим, что,
как это и можно было ожидать, при переходе к высшим формам родственные отношения
становятся все более ясными. Рассмотренные ветви высших беспозвоночных все ко-
ренятся в группе высших червей и вместе с ними образуют один крупный отдел
генетически тесно между собой связанных форм. Это первичиоротовые (Protostomia),
отличающиеся не только тем, что у них рот развивается на месте эмбрионального
бластопора, но также и рядом других признаков. Мезодерма у них развивается почти
408
всегда из телобластов (исключение — брахиоподы): кровеносная система незамкнутая;
эпидермис однослойный; нервная система лестничного типа и обычно вполне обособлена
от эпидермиса; кутикула хорошо развита (часто—хитин). Морские личинки обычно
построевы по типу трохофоры, снабжены протонефридиями; мезенхима эктодермаль-
ного происхождения. Второй большой отдел полостных животных носит название
вторичноротовых (Deuterostomia). У них на месте эмбрионального бл'астопора разви-
вается не ротовое, ча анальное отверстие; полость тела (целом) развивается в виде
мешкообразных выростов первичной кишки (у позвоночных это изменяется, очевидно,
вторично); кровеносная система в основном замкнутая; эпидермис часто многослойный
(щетинкочелюстные
Af
Рис. 362. Кишеч-
нодышащее Bala-
noglossus.
Af—заднепроходное
отверстие; Вд— жа-
берная область;
Вг — жаберные ще-
ли; К — ошейник;
В — печеночные вы-
росты; Е — хоботок.
и позвоночные); центральная нервная система занимает обычно
спинное положение, никогда не бывает построена по типу лес-
тницы и обнаруживает тесную связь с эпидермисом (у позво-
ночных эмбрионально); хитина нет; тело трехчленистое или
(у хордовых) с мускульной сегментацией. Морские личинки
хотя по внешности иногда и напоминают трохофору (у кишеч-
нодышащих и у иглокожих), но не имеют протонефридий, и
мезенхима у них эктодермальная. Эти оба больших отдела
достаточно резко разграничены, хотя в группе щупальцевых
(Tentaculata), относимых к первичноротовым, обнаруживаются
некоторые признаки как-будто промежуточного характера
(трехчленистое тело; у брахиопод — энтероцель).
В основе отдела вторичноротых лежат также червеобраз-
ные организмы, однако их связь с прочими червями не может
быть так просто установлена, и некоторые авторы принимают их
независимое происхождение от кишечнополостных. Наиболее
примитивными современными представителями этого отдела
являются щетинкочелюстные — Chaetognatha (Sagitta, Spadella)
с телом, разделенным на три отдела, которым соответствуют и
три парных отдела целома, где кишка укрепляется на сагитталь-
ной переогродке — брыжейке. У щитинкочелюстпых имеется,
между прочим, многослойный
эпидермис и поперечнополо-
сатая мускулатура. Две круп-
ные ветви этого отдела вто-
ричноротых связываются у ос-
нования интереснейшими орга-
низмами «переходного» харак-
тера — кишечнодышащими
(Enteropneusta), с наиболее из-
вестным их представителем в
лице Balanoglossus (рис. 362).
О родстве этих организмов
с хордовыми животными мы
еще будем говорить в дальней-
шем, а здесь отмечаем черты
организации, связывающие их
с другой ветвью вторичноро-
тых — иглокожими (Echinoder-
mata), с которыми их нередко
и объединяют под названием
Hydrocoelia, или Ambulacralia.
Самое название показывает
наиболее характерную особен-
ность организации, которая
Рис. 363. Личинка Balanoglos-
sus — торнария.
ап — заднепроходное отверстие;
st — рот; hy — закладка полости
хоботка (гидроцели).
естественно объединяет эти по
внешности весьма различные формы. Из двух-трех пар развивающихся целомических
полостей, симметрия которых бывает сильно нарушена, развивается впереди один
непарвый отдел — гидроцель, дающий начало в высшей степени своеобразной и
характерной для иглокожих воднососудистой, или амбулакральной, системе, свя-
занной отверстием с наружной средой. Наполняющая эту систему вода может
перекачиваться в набухающие от этого амбулакральные ножьи, являющиеся органа-
ми движения. У Balanoglossus из переднего, непарного отдела целома развивается
также гидроцель, связанный порой с окружающей средой; он дает начало гораздо
более просто построенной полости хоботка, который при наполнении водой набухает
и является органом, помогающим червеобразному животному зарываться в песок.
Кроме того, личинка Balanoglossus — торнария (рис. 363) — в высшей степени сходна
с двустороннесимметричной личинкой иглокожих (dipleurula). В родстве этих форм не
может быть никаких сомнений, несмотря на крайне значительное их расхождение.
Иглокожие первоначально перешли к сидячей жизни; в связи с этим они вторично
приобрели радиальную симметрию и прочный кожный панцырь. После этого они
частично вновь перешли к активному перемещению и активному питанию, но в совер-
шенно своеобразной форме.
409
Вторичная ветвь вторичноротовых животных, наоборот, переходила ко все более
активным формам движения — к быстрому плаванию, переходила от пассивных
форм питания к активному и при прогрессивном развитии органов движения, органов
дыхания, органов чувств (и нервной системы) и органов захватывания пищи достигла
высших форм диференцировки — сложнейшей организации высших позвоночных
животных. В основе этой эволюции лежит весьма важное приобретение, несомненно,
внаменующее собой подъем организма хордовых животных на более высокую ступень.
МЫ уже говорили о развитии более активных форм движения. У хордовых животных
это было достигнуто развитием мускульной сегментации, метамерии, существенно
отличающейся от метамерии высших червей. Развившаяся в стенках целома про-
дольная мускулатура оказывается разделенной на ряд мускульных сегментов — мио-
меров. Следующие вдоль тела спереди назад последовательные сокращения мускульных
сегментов поочередно правой и левой сторон приводят к волнообразным боковым
изгибам тела и, вместе с тем, к новой, весьма эффективной форме движения. Хордовые
животные в основном представляют собой быстро плавающих активных морских
хищников, что и явилось биологическим основанием их прогрессивной эволюции.
Происхождение хордовых
Вопрос о происхождении позвоночных животных стал па строго науч-
ную почву только со времени эмбриологических исследований А. О. Ко-
валевского над асцидиями и ланцетником. Эти исследования показали
такое огромное сходство асцидий на стадиях эмбрионального и личиноч-
ного развития с позвоночными, что в их близком родстве не оставалось
никаких сомнений. После работ Ковалевского положение асцидий в си-
стеме вполне определилось, и их пришлось соединить вместе с ланцетни-
ком и позвоночными в новый тип хордовых животных. Это, однако, не
разрешило вопроса о происхождении позвоночных, а только лишь ото-
двинуло его вглубь и заставило тем острее поставить вопрос о проис-
хождении хордовых животных. Естественно, что мысль исследователей
неизменно возвращалась к разнородной группе червей, центральное поло-
жение которой заставляло искать здесь и корней высших представителей
животного царства — позвоночных.
Исследования Семпера по истории развития выделительной системы
позвоночных показали, что она возникает в виде метамерного ряда каналь-
цев, в общем весьма сходных с метанефридиями высших червей. Это на-
толкнуло Семпера, а вслед за ним и Дорна, на мысль о происхождении
позвоночных от кольчатых червей. В пользу этой теории приводился целый
ряд данных: и те и другие принадлежат к двустороннесимметрмчпым живот-
ным со вторичной полостью тела в метамерным расчленением целого ряда
органов. Общее расположение органов также сходно, если представить
себе червя перевернутым на спину. Однако имеется и ряд препятствий
для такого сравнения. В частности, таким припятствием является совсем
иной тип центральной нервной системы. Если даже сравнивать спинной
мозг позвоночных с брюшной нервной цепочкой аннелид, то головной
мозг, представленный у последних надглоточными ганглиями, связанными
с остальной нервной системой окологлоточным кольцом, оказался бы
совершенно несравнимым с головным мозгом позвоночных. Гипотеза Дорна
о том, что первично рот у позвоночных проходил сквозь головной мозг,
не имеет под собой никакой почвы, а другие вспомогательные гипотезы
также не могут спасти положения. Впрочем, Неф еще недавно (1926) пы-
тался воскресить теорию Дорна и доказать, что первично рот открывался
на месте невропора и что первичный желудочек головного мозга представ-
ляет собой гомолога ротовой полости, которая через воронку сообщалась
с кишкой. На первый взгляд большую опору находит в эмбриологических
фактах теория Дельсмана, который предполагает, что находившийся
на месте невропора рот вел в длинный эктодермальный пищевод, сохра-
нившийся у позвоночних в виде нервной трубки, переходящей позади
через нервнокишечный канал в кишку. Эта теория, однако, совершенно
не считается с полной невозможностью такой смены функций у живого жи-
410
вотного и, кроме того, наталкивается на ряд других трудностей. В насто-
ящее время мы должны считать такие теории отжившими свой век. Мы
знаем, что основные особенности эмбрионального развития и общий план
строения позвоночных совпадают с особенностями эмбрионального раз-
вития и строения низших хордовых, а в некоторых весьма важных момен-
тах также и с общими признаками вторично ротовых животных вообще.
Таким образом, и вопрос о происхождении позвоночных понимается
ПозвонсчнЬ/е
БесчерепнЫе
Оболочники
1(ишечнодЬ/ша-
щие
Иглоко
Жие"
КордобЫе
[первичнЫе бесчерепное)
асекоА
Мноеошетиникслх
МалошетиниковЬе | } Пе
Ишаки
ОлигомернЫе
червеобразное
НомеркинЫ
HeMamodbi-^
РесничнЬ/е черви.
Пиявки
Пауки
Мечехво
пус /cmbi
„ , Ракоскорпион!»
Первична / / р
трахейнЫе /
/Пербичнбе
S раки
ПолимернЫе
черви
СосалЬщики
Лентеи,П
Ресничное
инфузории
СаркоШе
Споровики-
HuHSHOCUbl
[крвичнЫе черви
[Pilateralia] Гребневики
коралловые полип!»
Сцифомедузы
Гjdpoudh
Кишечнополостные
/ /Г№и
Первичнее биченосцЬ
шире — как вопрос о происхождении хордовых, а этот последний естест-
венно связывается с вопросом о происхождении вторичноротовых живот-
ных. В этой связи приобретает совершенно исключительный интерес уже
упомянутая нами группа кишечнодышащих и в особенности типичный их
представитель — Balanoglossus, который обнаруживает столь значительное
сходство с хордовыми животными, что некоторые авторы причисляют его
прямо к ним (Бетсон, 1886; Уилли, Мастерман и др.).
Balanoglossus (рис. 362) — червеобразное животное с телом, расчле-
ненным на три отдела — хоботок, короткий ошейник и длинное туловище.
В каждом из этих отделов имеется своя целомическая полость, развиваю-
щаяся, как и у низших хордовых, как энтероцель. Передняя полость —
непарная полость хоботка — сообщается с наружной средой посредством
одной или двух пор и играет, как уже упомянуто, роль гидроцеля.
Вторая полость помещается в ошейнике, а третья образует собственно
всю брюшную полость в области туловища. В туловище Balanoglossus
411
намечается, однако, еще и дальнейшая сегментация, выражающаяся в
повторности гонад, жаберных щелей и «печеночных» выступов кишечника.
Интересно отметить, что и у ланцетника два первых сомита развиваются
несколько иначе, чем следующие — именно как типичные «энтероцель-
ные» мешки, обособляющиеся от переднего конца первичного кишечника.
Передний мешок первоначально закладывается, как непарное образова-
ние (как у Balanoglossus), затем делится на два и левая полость (как у
иглокожих) открывается отверстием наружу. На месте этого отверстия,
сравнимого с порой хоботка Balanoglossus, развивается затем ямка Гат-
чека. Второй мешок ланцетника сравним, очевидно, с полостью ошейника.
Третий сомит развивается в виде сплошной складки по бокам крыши пер-
вичной кишки, которая затем уже вторично сегментируется на целый
ряд дефинитивных сегментов тела. Интересно отметить,’ что и у позвоноч-
ных животных первые три головных сомита развиваются иначе, чем сле-
дующие за ними «заушные» сомиты. Быть может они и являются остатком
первичной сегментации исходных форм, близких к кишечнодышащим
червям. Наиболее существенным признаком, связывающим Balanoglossus
с хордовыми, является его дыхательный аппарат. Передняя часть кишеч-
ника открывается наружу парным рядом небольших жаберных щелей,
напоминающих по своей U-образной форме личиночные жаберные щели
ланцетника. Жаберные щели поддерживаются открытыми, благодаря
присутствию эластического скелета, который имеет вид решетки, сходной
с жаберной решеткой ланцетника. Рот лежит на брюшной стороне тела
впереди ошейника. Кишечник имеет вид почти прямой трубки, снабжен-
ной в своей средней части парными «печеночными» выростами, и укреплен
в перегородке между правой и левой полостью тела. От переднего конца
кишечника отходит вперед в хоботок слепой отросток, сравниваемый
иногда с зачаточной хордой. Кровеносная система состоит в основном
из спинного и брюшного продольных сосудов. Сходство с хордовыми жи-
вотными существенно увеличивается еще строением нервной системы.
Она в общем устроена весьма просто, тесно связана с эпидермисом и мало
концентрирована. В непрерывном нервном сплетении, лежащем в покро-
вах, выделяются. более заметно лишь спинной и брюшной тяжи, соеди-
ненные кольцевидной комиссурой на границе ошейника с туловищем.
Однако в области ошейника спинной ствол вполне обособлен от эпидер-
миса и получает трубчатый характер, напоминая нервную трубку хор-
довых животных. Таким образом, мы находим у кишечнодышащих целый
ряд признаков, общих с хордовыми животными, и, так как это касается
таких признаков, которые характерны только для этого типа, мы должны
признать их родство совершенно несомненным.
Этим, однако, не исчерпывается вопрос о происхождении хордовых
животных. Если даже и связывать кишечнодышащих с такими вторично-
ротовыми червями, как щетинкочелюстные, то все же встает вопрос о
происхождении этих червеобразных форм. Как уже было сказано, между
группами вторичноротовых и первично ротовых непроходимой пропасти
нет. Прежде всего, щупальцевые (Tentaculata) занимают до известной
степени промежуточное положение. Различия в развитии мезодермы и
образовании вторичной полости тела не имеют решающего значения.
В обоих случаях мезодерма образуется из зачатка, общего с энтодермой,
и при телобластическом ее развитии у первичноротовых дело сводится
лишь к более раннему обособлению образовательного материала и соот-
ветственно уменьшению числа клеток зачатка, как это характерно и для
других закладок при детерминированном развитии. Вопрос о рте также
не имеет принципиального характера, так как вряд ли можно сомневаться,
что как ротовое, так и анальное отверстия представляют собой остатки
частей первичного рта, т. е. эмбрионального бластопора, который разде-
лился на передний — вводной и задний — выводной участки.
412
Однако все же группы первичноротовых и вторичноротовых обосо-
бились друг от друга весьма рано, и их общими предками являются какие-
то весьма еще примитивные черви, которых характеризовать точнее не
представляется возможным. Иногда этих общих предков ищут среди еще
более примитивных форм — представителей кишечнополостных (Седжвик,
Шимкевич), а среди них, естественно, обращают особое внимание на кте-
нофор (Зедерштрем, Третьяков).
полного отсутствия проме-
V
а
Рис. 364. Торнария, превра-
щающаяся во вврослое жи-
вотное.
а — задний проход; Е — хобо-
ток; ес — полость хоботка; ed •—
выступ кишечника («хорда»);
ер —пора хоботка; К—ошейник;
he — полость ошейника; kp—пора
ошейника; hs — закладка первой
жаберной щели; т — рот; В —
туловище; тс — полость тулови-
ща; sp — аборальный орган.
Соотношения между низшими хордозымл и позвоночными
Как и в других ветвях царства животных, при переходе к геологи-
чески более молодым формам соотношения между различными группами
становятся все более ясными. Вопрос о соотношениях между низшими
хордовыми и позвоночными в общем совершенно ясен, хотя и не поддается
вполне конкретному обсуждению, вследствие
жуточных форм. Оболочники ясно обнаружи-
вают принадлежность к хордовым животным
в своем эмбриональном и личиночном разви-
тии, ланцетники во взрослом состоянии пред-
ставляют как бы живую схему типичного
хордового животного. Однако может возник-
нуть вопрос (и он, действительно, был по-
ставлен А. Дорном): не произошли ли оболоч-
ники и лацнетник путем обратного упроще-
ния, путем деградации от вышестоящих по-
звоночных животных? Тот же самый вопрос
был поставлен и по отношению к низшим
современным позвоночным — круглоротым,
которых А. Дорн также склонён рассматри-
вать как формы, возникшие путем деградации
от настоящих рыб. В случае правильности
этих воззрений возникла бы действительно
целая пропасть между позвоночными (и
именно рыбами, как низшими их представи-
телями) и беспозвоночными.
Мы можем начать рассмотрение этого
вопроса с кишечнодышащих, так как и к
ним применима указанная точка зрения.
Нервная система Balanoglossus хотя и обна-
руживает сходство с нервной системой позво-
ночных присутствием нервной трубки в ошей-
нике, но в остальном весьма отлична; она
тесно связана с покровами и мало концен-
трирована — это бесспорные признаки при-
митивной организации, и такая нервная
система никак не могла возникнуть из глу-
боко централизованной нервной системы
позвоночных. Кроме того, расчленение поло-
сти тела лишь на три отдела связывает ки-
шечнодышащих с мало членистыми червями
и с иглокожими (рис. 364). Наконец, с последними их связывают и ори-
гинальные функциональные соотношения части полости тела с внешней
средой («гидроцель»), ведущие к образованию своеобразного локомоторного
аппарата (хоботок кишечнодышащих, амбулакральная система иглокожих).
Эта связь необъяснима, если допустить, что кишечнодышащие произошли
от позвоночных путем деградации. То же самое следует сказать о личиноч-
ной форме «торнарии», которая обнаруживает очень большое сходство с
413
личиночными формами иглокожих (рис. 363). Кишечнодышащие могут
рассматриваться лишь как примитивные животные, ведущие свое начало
от того же самого ствола, от которого ответвились иглокожие, с одной
стороны, и хордовые —с другой (мы, конечно, не рассматриваем совре-
менных кишечнодышащих как прямых предков позвоночных).
Несколько сложнее на первый взгляд обстоит дело с оболочниками
и, в частности, с асцидиями. Эти последние обнаруживают, действительно,
Рис. 365. Схематическое изображение хвостатой
личинки (Л) и ее превращения в сидячую асцидию
(В, С) (из Паркера и Гасвелла). adh — присоски;
atr — околожаберная полость; ch — хорда; сП —
мерцательная ямка; end — эндостиль; с — сердце;
пеиг — нервная трубка; п. gn — нервный узел;
or — рот; г — прямая кишка; sens — мозговой пу-
зырек; sig — жаберьые отверстия; stol — столон;
t — хвост.
прикрепляется при помощи трех сосочков,
нем конце тела, к какому-либо подводному
ряд признаков регрессив-
ной эволюции, которая еще
яспо отражается па ходе
их индивидуального раз-
вития.
Ранние стадии эмбрио-
нального развития асци-
дий обнаруживают очень
большое сходство с подоб-
ными же стадиями разви-
тия ланцетника, как это
впервые было установлено
А. Ковалевским (1866—
1867). Нервная система
также закладывается в ви-
де пластинки, края кото-
рой заворачиваются, обра-
зуя жолоб, замыкающийся
затем в нервную трубку.
Под нервной трубкой за
счет энтод< рмальных кле-
ток развивается хорда, а
по бокам от нее — мезо-
дермальные полоски, даю-
щие, между прочим, нача-
ло мускулатуре. Рот раз-
вивается как эктодермаль-
ное впячивание. растущее
навстречу эктодермальной
кишке. От стенки послед-
ней развиваются жаберные
выступы, прорывающиеся
отверстиями в эктодер-
мальную околожаберную
(атриальную) полость.
В результате полу-
чается свободноплавающая
личинка со всеми основны-
ми признаками хордового
животного (рис. 365). Од-
нако эта личинка вскоре
развивающихся на перед-
предмету. Вслед за при-
креплением начинается превращение личинки в сидячую асцидию, причем
перестройка ее организации имеет во многом характер регрессивного
метаморфоза (рис. 365). Хвост, с его мускулатурой, хордой и нервной
системой, исчезает. Исчезают также имеющиеся в мозгу личинки органы
чувств, и вообще вся нервная трубка редуцируется, так что от нее оста-
ется лишь один небольшой нервный узел. Все тело поворачивается внутри
выделенной им оболочки, и ротовое отверстие принимает конечное поло-
414
Жепие. Кроме многочисленных жаберных отверстий, открывающихся
в обширную атриальную полость, не сохраняется почти никаких призна-
ков, которые вскрывали бы принадлежность этих животных к хордовым,
и неудивительно, что до исследования А. Ковалевского их положение
в системе было совершенно неопределенным (чаще всего их относили к
моллюскам). Таким образом, регрессивные изменения выражены у асци-
дий весьма ярко, и, очевидно, этот-регресс связан с переходом к сидячему
образу жизни, при котором и мускулатура, и органы чувств с высоко-
развитой нервной системой утратили свое значение.
Можно ли, однако, предполагать, что асцидии произошли путем деге-
нерации от позвоночных? В пользу такого предположения у нас нет ни-
каких данных. Наоборот, имеется ряд указаний, определенно противо-
речащих этой гипотезе А. Дорна. Поджаберный слизистый желобок —
эндостиль, имеющийся и у оболочников и у ланцетника (рис. 14), пред-
ставляет собой орган захватывания взвешенных в воде пищевых частиц
(при пассивном питании этих животных, непрерывно процеживающих
воду через передний отдел кишечника) и, конечно, как специальное при-
способление мог бы также развиться вторично. У позвоночных имеется,
однако, гомолог этого образования. У личинок круглоротых он имеет
еще характер железистого желобообразного углубления с таким же рас-
положением слизистых и мерцательных клеток эпителия, как у низших
хордовых, и играет, несомненно, ту же роль аппарата для улавливания
взвешенных в воде пищевых частиц. И здесь ток слизи движется вдоль
поджаберного желобка вперед до окологлоточного мерцательного кольца
и затем вдоль спинного мерцательного тяжа назад в пищевод. Этот под-
жаберный желобок, однако, позднее совершенно обособляется от эпи-
телия глотки и превращается в беспроточную щитовидную железу. У
других позвоночных щитовидная железа также всегда закладывается
как углубление эпителия дна глоточной области кишечника и, следова-
тельно, несомненно представляет результат преобразования эндостиля
низших позвоночных. Возможность обратного перехода совершенно исклю-
чается. Никакая дегенерация не могла привести к тому, чтобы из обособ-
ленной беспроточной щитовидной железы получилось желобообразное
углубление, тянущееся вдоль дна глотки. Наоборот, использование же-
лезистого эпителия эндостиля после утраты им его функции у низших
хордовых, когда они перешли к активному захватыванию пищи, для
внутрисекреторных функций представляется вполне естественным и по-
нятным.
Таким образом, несмотря на несомненное наличие дегенеративных
изменений, происшедших в организме оболочников, мы ни в коем случае
не можем итти так далеко, чтобы допустить их происхождение путем де-
генерации от позвоночных. Оболочники, несомненно, представляют собой
деградировавшую, в связи с приобретением сидячего образа жизни, бо-
ковую ветвь низших хордовых животных, которые в некоторых отноше-
ниях были еще очень мало диференцированы. По всей вероятности, исход-
ные формы были довольно близки к современному ланцетнику.
Что касается последнего, то в нем мы тем более не можем видеть де-
градировавшее позвоночное. Ланцетник представляет собой как бы живую
упрощенную схему хордового животного. Конечно, схема могла бы по-
лучиться и путем редукции, при исчезновении всех признаков высшей
организации и специальных приспособлений. Однако в действительности
это не так. У ланцетника имеется ряд признаков, несомненно, примитив-
ной организации, многие из которых не могут быть выведены из строения
позвоночных.
Здесь прежде всего следует упомянуть об эндостиле. Для последнего
сохраняет значение все сказанное об эндостиле оболочников и о свое-
образных органах чувств ланцетника, которые также не могли произойти
415
путем деградации из органов чувств позвоночных. Особенно примитивны
много численные расположенные в стенках мозга, так называемые, глазки
Гессе. Каждый такой глазок состоит из чувствующей клетки с нервным
волокном, снабженной каемкой палочковидных образований на одной
стороне, где ее покрывает в виде шапки другая, пигментная, клетка. Эти
глазки чрезвычайно сходны с примитивнейшими глазками некоторых
плоских червей. Затем выделительная система ланцетника представлена
парным рядом самостоятельных канальцев — нефридиев. Трудно допу-
стить, чтобы общий выводной проток почек позвоночных исчез и сама
почка распалась на метамерный ряд независимых канальцев. Менаду
тем, обратный процесс при прогрессивном развитии органа является
вполне понятным. Так же трудно допустить, чтобы при дегенерации орга-
низма его выделительная система редуцировалась и заменилась другой,
более простой выделительной системой. Дело в том, что нефридии ланцет-
ника во многом своеобразны и, притом, именно примитивны. Они снаб-
жены большим числом особых выделительных клеток, называемых со-
леноцитами (это — булавовидные клетки с.тонкими внутренними каналь-
цами и длинным мерцательным волоском внутри канальца). Выделитель-
ные канальцы ланцетника, с их соленоцитами (рис. 16) поразительно
сходны с нефридиями некоторых кольчатых червей.
Наконец, в высшей степени примитивным является и гистологическое
строение ланцетника, тело которого состоит почти исключительно из
эпителиев и лишено еще настоящей соединительной ткани.
Таким образом, принцип дегенерации не может быть принят как ру-
ководящий принцип в эволюции низших хордовых. Это, конечно, не зна-
чит, что эти животные не подверглись регрессивным изменениям. Наобо-
рот, мы знаем достоверно, что оболочники представляют собой формы,
весьма значительно деградировавшие. Мы только подчеркиваем, что
низшие хордовые не могли произойти путем дегенерации от позвоночных
животных, а только от гораздо более просто построенных хордовых жи-
вотных, которые были еще лишены признаков организации, характерных
для позвоночных.
Более полно мы осведомлены в настоящее время об эволюции низших
позвоночных. Положение круглоротых в системе позвоночных в насто-
ящее время далеко не так неопределенно, как это было еще недавно и осо-
бенно во времена А. Дорна. Выяснению этого положения особенно спо-
собствовали работы академика А. Н. Северцова и шведского палеонтолога
Э. А. Стеншио (Е. A. Stensio).
По Северцову, череп круглоротых имеет крайне примитивное стро-
ение — он состоит из одной только хордальной части, и притом без заты-
лочного отдела. Капсулы обонятельного и слухового органов также при-
митивны и мало связаны с черепной коробкой. Жаберный скелет миног
весьма своеобразен, образуя цельную решетку. Впереди от него висце-
ральный скелет представлен рядом более или менее измененных преджа-
берных дут, обнаруживающих те же характерные для круглоротых осо-
бенности. Этот преджаберный скелет ни в коем случае не мог произойти
путем редукции из челюстного скелета остальных позвоночных, так как
оп сохранил характерные черты жаберной решетки круглоротых, что
указывает на его преобразование непосредственно из жаберных дуг типа
круглоротых. Собственно жабры круглоротых занимают иное положение,
чем у челюстных, —> они представлены эктодермальными жаберными
мешками, развивающимися с внутренней стороны от жаберных дут. Так
как у челюстных позвоночных жабры иного типа и развиваются перво-
начально как эктодермальные жаберные нити, сидящие на жаберных
дугах снаружи, то ясно, что собственно жабры приобретены обеими груп-
пами позвоночных независимо друг от друга.
416
Дальнейшее прогрессивное развитие дыхательного аппарата выра-
зилось в увеличении дыхательной поверхности. Это увеличение означает
усовершенствование каждой отдельной жаберной щели, что сделало воз-
можным заметное сокращение общего числа этих щелей. И у круглоротых
и у челюстных позвоночных эволюция пошла по этому пути. Таким образом
ясно, что все эти процессы шли в обеих группах независимо и что жабер-
ный аппарат круглоротых так же нельзя вывести из такового челюстных,
как и, наоборот, жаберный аппарат челюстных — из диференцированного
жаберного аппарата круглоротых (Северцов).
Во всяком случае, нельзя допустить, чтобы могла произойти редук-
ция дыхательного аппарата рыб, с его эктобранхиями, и чтобы вслед за
этим произошел новый прогресс, связанный с развитием энтЬбранхий.
Исходными могли быть лишь формы еще индиферентные, т. е. со многими
щелями, не имевшими жаберных лепестков.
Круглоротые сохранили также много признаков примитивной орга-
низации: в составе всех висцеральных нервов у них имеются и сомати-
ческие чувствующие (кожные) компоненты, которые у высших позвоноч-
ных исчезают, за исключением лишь нервов специального значения (орга-
нов боковой линии) и кожных компонентов тройничного нерва. Отсутствие
соединений между спинными и брюшными корешками спинномозговых
нервов у миног (как у ланцетника) также иногда считается примитивным
признаком.
У личинки миноги, так называемой пескоройки, имеется поджаберная
слизистая бороздка, подобно низшим хордовым, печень имеет еще труб-
чатый характер. К этому можно прибавить еще очень большое число при-
знаков как взрослых круглоротых, так и особенно личиночных, которые
с большой долей вероятности следует считать примитивными, но которые
все же могли бы произойти и путем редукции.
На этих фактах мы останавливаться не будем, так как приведенного
выше бесспорного материала вполне достаточно.
Таким образом, ясно, что и круглоротые не произошли путем деге-
нерации от челюстных позвоночных: челюстей у них никогда не было, и
общая организация исходных форм была гораздо более примитивной.
Картина соотношений между низшими хордовыми животными и позво-
ночными становится довольно ясной — современные их представители
оказываются остатками примитивных хордовых животных, имеющих
общее происхождение с непосредственными предками позвоночных. Од-
нако большинство этих форм настолько изменено, что не дает возможности
представить себе происхождение позвоночных более конкретно. Несом-
ненно, что из низших хордовых наименее измененным является ланцетник,
и нужно думать, что позвоночные в своей эволюции проходили некогда
стадию развития, относительно весьма близкую к современному ланцет-
нику.
Бесчелюстные позвоночные (Agnatha)
Из низших позвоночных круглоротые, несомненно, подверглись
значительным изменениям, главным образом в связи с их своеобразным
способом питания. На их историю бросают, однако, некоторый свет и пале-
онтологические данные. Здесь нельзя не упомянуть о замечательных
исследованиях шведского палеонтолога Стеншио. В результате этих иссле-
дований оказалось, что давно известные верхнесилурские и девонские
щитковые «рыбы — Ostracodermi (рис. 366, 367), которых по признаку
отсутствия челюстей и раньше пытались объединить с современными
круглоротыми, представляют собой формы, действительно чрезвычайно
близко к ним стоящие. Стеншио изучил ’в деталях строение различных
представителей семейства Cephalaspidae (путем тщательного препариро-
вания иглами под бинокулярным микроскопом, а также методами изго-
417
топления полных серий разрезов и реконструкции по этим разрезам).
'Гак как у многих бывших в его распоряжении экземпляров скелет был
в превосходной сохранности и ясно выделялся своим цветом из окружа-
ющей породы, явилась возможность восстановить не только внешнюю
форму костей с отпечатками различных каналов, но и все внутренние
полости и форму многих мягких частей.
Оказывается, у этих животных был не только наружный костный
панцырь, но и внутренний скелет был покрыт слоем костной ткани. Основ-
ные нервы и сосуды головы были окру-
жены тонкими костными чехликами.
Таким образом, по форме полости че-
репной коробки, с ее каналами, уда-
лось полностью воспроизвести и форму
мозга с отходящими от него нервами
и дальнейшие разветвления нервов,
а также и расположение артерий и вен.
Удалось также установить в общих
чертах и строение висцерального ске-
лета, расположение и важнейшие ветви висцеральных нервов и поло-
жение сосудов соответствующей области.
Сходство организации с современными миногами оказалось порази-
тельным; однако у Cephalaspidae не было специализированного губного
аппарата миног и передняя часть головы была гораздо более похожа на
соответствующие части личинки миног — пескоройки. Как и у миног,
висцеральный скелет был слит с черепной коробкой, хотя жаберный ап-
Рис. 366. Бесчелюстное щитковое
Thelodus из верхнего силура.
Рис. 367. Бесчелюстное панцырное щитковое Cephalaspis
ив нижнего девона.
парат не простирался так далеко назад, как у миног. Как и у миног, име-
лись непарное назо-гипофизарное отверстие, ведущее в орган обоняния,
и гипофизарный канал, кончающийся слепым мешком под основанием
мозга. Положение гипофизарного мешка указывает на то, что предобо-
нятельная часть черепа развивалась, как и у миног, в верхней губе и
имеет, очевидно, висцеральное происхождение. В ухе имелись лишь оба
вертикальных полукружных канала; горизонтальный канал отсутство-
вал, как он отсутствует у современных миног.
Весьма сходно и строение головного мозга, причем ganglia habenulae
асимметричны — правый значительно крупнее и лежит частью позади
левого, как и у современных круглоротых. Имелся, однако, большой
двулопастный мозжечок, которого нет у миног. Весьма замечательна
и периферическая нервная система (рис. 250): 1) спинномозговые нервы
правой и левой стороны отходят поочередно; 2) в спинных и затылочных
нервах спинные и брюшные корешки не соединяются между собой для
образования смешанного нерва; 3) имеются специальные висцеральные
ветви жаберных нервов, идущие к особым органам чувств (вкуса) в глотке;
4) жаберные нервы не имеют преджаберной ветви; 5) во всех черепных
висцеральных нервах имеется чувствующая кожная ветвь; 6) глубокий
глазничный нерв (n. ophthalmicus profundus V) имеет значение самостоя-
тельного нерва; 7) жаберные нервы располагаются с внутренней стороны
жаберного скелета; 8) выход и расположение висцеральных нервов по-
добны таковым у современных миног (рис. 250).
418
Ёо всех этих признаках, а также и в кровеносной системе видйо
совершенно поразительное сходство с современными миногами. Весьма
важно, что при всем этом сходстве у Cephalaspidae имелись, повидимому,
грудные плавники, опиравшиеся на хорошо развитый плечевой пояс.
Еще важнее, однако, то, что имеется и большое число, признаков,
по которым Cephalaspidae примитивнее круглоротых. Мы уже упомянули,
что у них не было специализированного ротового сосательного аппарата
современных круглоротых и
нает личинку, миноги.
Совершенно однотип-
ный жаберный аппарат
доходил до самого пе-
реднего конца головы.
Имелось всего 10 жабер-
ных мешков, по форме
напоминающих скорее
мешки пескоройки (рис.
368), чем мешки взрос-
лой миноги, и открыва-
лись они отдельными
наружными жаберными
отверстиями на брюш-
ной поверхности голов-
ного щита. Жабры, с
их нервами и сосудами,
лежали с внутренней
стороны от скелета, и
притом на всех жабер-
ных перегородках, не
устройство передней части головы напоми-
Рис. 368. Жаберный аппарат бесчелюстного панцирного
щиткового (по Стеншио).
А — горизонтальный разрез через голову Kiaeraspis (видна
нижняя часть сверху). По бокам ротовой полости видны
жаберные мешки. В — головной щит Cephalaspis снизу.
Ы — жаберные отверстия; sp — спиракулярный жаберный
мешок; st — рот.
исключая и самых передних. По иннервации
можно установить, что имелись и спиракулярный и один преспира-
кулярный жаберные мешки. Передняя висцеральная перегородка иннерви-
ровалась глубоким глазничным нервом (n. ophthalmicus profundus V)
и может быть рассматриваема как гомолог предчелюстной дуги. Следую-
щая перегородка, гомолог челюстной дуги, иннервировалась тройничным
нервом, а третья перегородка — лицевым нервом.
При этом следует отметить, что ветвления всех висцеральных нервов
вполне единообразны (рис. 250). Все нервы лишены преджаберной ветви,
но имеют специальную переднюю висцеральную ветвь, идущую к вку-
совым органам глотки. Тройничный нерв не делится на две челюстные
ветви (верхне- и нижнечелюстную ветви других позвоночных), но по
своему ветвлению вполне подобен другим висцеральным нервам. Глубокий
глазничный нерв также сохранил первичное строение последних, включая
и висцерально-двигательный компонент.
Если к этому добавить, что у Cephalaspidae в течение всей жизни
функционировала головная почка, то получается картина чрезвычайно
примитивного позвоночного животного. Сама картина по существу не
содержит ничего неожиданного для морфолога — это именно схема
первичного позвоночного; однако выполнение этой картины, осуществле-
ние такой простой схемы в столь специализированном организме предста-
вляет собой большую неожиданность для большинства филогенетиков,
привыкших мыслить схемами, лишенными следов жизни в реальной
обстановке.
В щитковых обычно видят крайне специализированную ветвь низших
позвоночных. Эта специализация выражается прежде всего в крайнем
развитии наружного костного панцыря. Затем у Cephalaspidae в боковых
лопастях головного щита имелись значительные электрические органы.
419
Конечно, нельзя утверждать, что миноги произошли именно от этих
форм, а не от неизвестных пока, менее специализированных форм той же
группы. Однако, если принять во внимание, что кожный скелет имелся
во всяком случае во всей группе щитковых (Ostracodermi), то мы должны
допустить, что, по крайней мере, этот признак более высокой организации
современными круглоротыми утерян вторично.
Наиболее примитивные щитковые образуют группу Thelodonti (рис.
336), у которых кожный скелет состоял из многочисленных равномерно
разбросанных плакоидных чешуй. К сожалению, эти рыбообразные су-
щества еще мало изучены. От этих примитивных форм произошли, оче-
видно, и более специализированные щитковые с мощным панцырем, подоб-
но описанным Cephalaspidae, входящим в группу костнощитковых (Osteo-
straci). Все это были донные формы. К высшим костнощитковым близко
примыкает группа бесчелюстных, перешедшая к нектонному образу
жизни и обладавшая соответственно веретеновидной формой тела и зна-
чительно редуцированным кожным панцырем (Anaspida). Грудных плав-
ников у анаспид не было. Весьма вероятно, что круглоротые произошли
именно от анаспид.
Во всяком случае, несомненные факты говорят в пользу происхожде-
ния современных миног от форм, близких к древнейшим щитковым Osteo-
straci, отличавшихся целым рядом признаков весьма примитивной орга-
низации, через повышение организации (прогрессивная диференцировка
висцерального аппарата, прогрессивное развитие выделительной системы),
сопровождавшееся специализацией (сосательный губной аппарат) и
частичной редукцией (мозжечок, костный скелет).
Что касается второй группы современных круглоротых — миксин,
то они по вентральному положению назогипофизарного канала и суще-
ствованию лишь одной пары общих наружных жаберных отверстий ока-
зываются более специализированными формами. Не подлежит никакому
сомнению, что у миксин, наряду с дальнейшей специализацией, были
и довольно значительные регрессивные изменения, которые охватили
в значительной мере скелет, особенно осевой и висцеральный (за исклю-
чением язычного аппарата), а также органы зрения и слуха.
Если учесть сказанное, то становится ясно, что филогения низших
позвоночных животных может быть восстановлена на основании всей
суммы сравнительноанатомического, эмбриологического и палеонтоло-
гического материала лишь путем сложных умозаключений, т. е. неизбежно
в абстрактной — до известной степени — форме. Ни одна из ныне суще-
ствующих форм не может считаться прямым предком современных живот-
ных вообще, а ископаемой формы, которую можно было бы считать пред-
ком позвоночных, пока не найдено. Поэтому приходится итти по пути
искусственной реконструкции строения организма, от которого берут
начало позвоночные животные, путем отбрасывания у современных при-
митивных форм всех признаков специализации. Такая реконструкция
приводит к обобщенной форме, лишенной всяких признаков специали-
зации, т. е. живущей в неточно определенной среде и, следовательно,
нереальной. И в отдаленные времена предки современных позвоночных
имелись всегда во множестве форм, приспособленных к тем или иным
частным условиям среды (например бентонные, нектонные и планктонные
формы, береговые и пелагические и многие другие). Однако путем отвле-
чения от этих частных приспособлений можно было бы и для этих форм
вывести обобщенный тип такого предка, как это и делается в систематике
современных форм при описании признаков рода, семейства, отряда и т. д.
Наши реконструкции предков, сделанные на основании суммы знаний
эмбриологии и анатомии современных форм, в лучшем случае прибли-
жаются к такому обобщенному типу реальных предков. Несмотря на то,
что в этом случае мы оперируем с абстракциями, такого рода попытки
420
имеют все же большое значение, так как подводят итоги нашим знаниям
в этой области и позволяют выделить ряд бесспорных процессов, игравших
руководящую роль в эволюции данной группы. Вопрос об эволюции
низших позвоночных в последнее время весьма полно разработан Север-
цовым, и мы здесь даем в основном изложение его выводов.
Примитивные бесчерепные, давшие начало позво-
ночным животным, несомненно, были весьма близки к современному
ланцетнику. Это были свободноплавающие животные с мускульной мета-
мерией, простиравшейся до переднего конца тела, имевшие околоротовое
кольцо с усиками и питавшиеся планктоном. В отличие от ланцетника
хорда не выступала вперед от мозга, число жаберных отверстий было
10—20 и атриальной полости не было. От этих форм произошли, с одной
стороны, современные бесчерепные; пройдя сначала через стадию донной
жизни, с которой было связано временное приобретение асимметрии, они
перешли затем к жизни в донном песке. Последнее связано с восстановле-
нием двусторонней симметрии, с образованием атриальной полости, уве-
Рпс. 369. Схема строения головы и висцерального аппарата первичного
черепного животного (по Северцову).
оЬг 1—13 — жаберные дуги; арЬг — преджаберные дуги (арЬг 1 — гомолог гиоидной дуги);
рЬг — преджаберные щели; Ъг 1—13 —• жаберные щели; ch — хорда; di — промежуточный
мозг; ер — эпифиз; hyp — гипофиз; st — рот; mbr 1—12 — жаберные мышцы; mhy — гио-
идная мышца; msp — спинной мозг; ms — средний Мозг; mt — задний мозг; пав — обоня-
тельные ямки; п. vise — висцеральные нервы; sp — гомолог брызгальца; int X — n. Inte-
sttaalis vagi; tel — передний мозг; I—X — головные нервы.
личением числа жаберных отверстий и заострением переднего конца
тела, в котором разрослась вперед хорда. С другой стороны, развилась
прогрессивная ветвь первичных черепных животных (Рго-
tocraniata), давших начало всем собственно позвоночным. У этих первич-
ных черепных животных (рис. 369) были уже парные органы высших
чувств (зрения и слуха), обособленное сердце и зачаточный череп с вис-
церальным скелетом. Кожа была голая, но с многослойным эпидермисом.
Зубов и челюстей не было, и питание было, как и у бесчерепных, пассив-
ное. Череп составлялся только из парахордалий и слуховых капсул.
Обонятельные капсулы были независимы. У них имелись околоротовое
кольцо и до 17 нерасчлененных хрящевых жаберных дужек. Из перед-
них миотомов головы уже развились глазные мышцы. Висцеральная
мускулатура имела сегментальное строение (m. constrictor и m. adductor).
В головном мозге прежняя невромерия (около 10 сегментов) уже уступила
место дефинитивному делению на непарный передний мозг (из одного
сегмента) с зачатками парных полушарий, промежуточный мозг (из двух
сегментов), средний мозг (из двух сегментов) и задний мозг (из пяти
сегментов). Нервы отходили от мозга обособленными спинными и брюш-
ными корешками и располагались правильно метамерно, причем тройнич-
ный нерв составлялся из трех независимых висцеральных нервов челюст-
ной и предчелюстных дуг.
421
От этих первичных черепных животных произошли две неравноцен-
ные ветви. Одна из них пошла по пути специализации, сохранив многие
примитивные черты, — это группа Entobranchiata, получившая свое
название благодаря тому, что здесь эктодермальные жаберные лепестки
развились в виде продольных вкладок внутрь от жаберного скелета на
самых жаберных мешках. Другая ветвь пошла по пути прогрессивного
развития и дала начало всем высшим формам — это группа Ectobranchiata,
названная так вследствие развития у них эктодермальных жаберных
нитей, а затем лепестков па наружной стороне жаберных дут.
У энтобранхиат (рис. 370), которые первоначально сохра-
няли пассивный способ питания, жаберные дуги соединились в непрсу
Рис. 370. — Схема строения головы и висцерального аппарата
внутреннежаберного позвоночного (по Северцеву).
с. апп — ротовой кольцевидный хрящ; саи — слуховая капсула; cda — передний спинной
хрящ; cept — наджаберные, связи скелетной решетки; chpt — поджаберные связи; сп — обо-
нятельная капсула; csch— субхордальная и со—брюшная связь; os — глаз; pch — пара-
хордалии; tr. сг — черепные балки. Остальные обозначения см. рис. 369.
рывную решетку, доходившую до переднего конца головы. Благодаря
вторичному сближению и соединению обонятельных ямок с отверстием
гипофиза образовалась непарная ноздря, характерная для всей этой
группы. По морфологическим признакам уже очень давно наметилось
разделение этой группы на две ветви — в одной из них разрослась верх-
няя губа и ноздри отодвинулись на спинную сторону, а гипофиз оканчи-
вался слепым мешком (Osteostraci, Petromyzontes), в другой разрослась
обонятельная часть черепа и ноздри отодвинулись на брюшную сторону,
гипофиз сообщался каналом с пищеводом (Heterostraci и Myxinoidei).
В каждой из этих ветвей произошло разделение биологических типов:
донных форм, которые сохранили пассивное питание (Heterostraci и
Osteostraci), и нектонных форм, которые перешли к активному питанию
трупами, а затем также к полупаразитизму (Myxinoidei и Petromyzontes).
Первые быстро специализировались, и у них очень рано развился зна-
чительный костный панцырь. Это давало им хорошую защиту, хотя и за
счет потери подвижности, что для донных форм с пассивным питанием
большой роли не играло. Вторые сохранили значительную подвижность,
и у них если и были уже твердые скелетные образования, то во всяком
случае они бесследно редуцировались. Здесь может явиться вопрос об
основательности последнего предположения. Однако тот факт, что кожный
скелет уже имелся у древнейших известных нам позвоночных из верхнего
силура (Thelodus) в виде плакопдной чешуи, заставляет думать о чрезвы-
чайно раннем ее приобретении общими предками, т. е. первичными череп-
ными животными. В таком случае мы должны допустить, что современные
круглоротые произошли от форм, имевших кожный скелет хотя бы только
в виде плакоидной чешуи. Это нас не должно удивлять, так как редукция
кожного панцыря, несомненно, произошла и в других группах по мере
развития все большей подвижности.
422
Интересно, что у щитковых имелись парные плавники. Весьма ве-
роятно, что боковые выросты тела по бокам, позади головы, у таких форм,
как Thelodus (рис. 366), служили простыми поверхностями сопротивле-
ния, поддерживавшими тело при плавании, и могут рассматриваться
как зарождающаяся форма грудного плавника. У Osteostraci развивались
уже подвижные грудные плавники.
Типичные щитковые бесчелюстные (Ostracodermi) вымерли, не оста-
вив никакого потомства. В современной фауне сохранились только изме-
ненные представители другой ветви энтобрапхиат, приспособившиеся
к питанию трупами, в которых они прогрызают ходы (миксины), или
к полупаразитическому питанию другими рыбами (миноги). С этим свя-
Рис. 371. Схема строения головы и висцерального аппарата
наружножаберного позвоночного (по Северцову).
clat — боковой глазничный хрящ; осс — затылочная дуга; ЬЬг — копулы; сЬг — ceratabran-
chlalia; ebr— epibranehlalja; hbr — hypobranchia На; phbr — pharyngobranchlalia._uOeTanb-
ные обозначения см. рис. 369, 370.
зано значительное изменение всей передней части висцерального аппа-
рата, преобразованного в своеобразный сосательный аппарат, начинаю-
щийся круглой ротовой воронкой с роговыми зубами.
У экто бр ан хи ат (рис. 371) развился иной тип жаберного
дыхания и иной механизм движений жаберного аппарата. Жаберные
дуги расчленились на 4 отдела каждая, и соответствующая висцеральная
мускулатура испытала более сложную диференцировку. Так как следы
того же расчленения обнаруживаются на гиоидной, на челюстной и на
предчелюстных дугах (губных хрящах), то, очевидно, и эта часть скелета
несла еще функцию жаберных дуг, т. е. у древнейших эктобранхиат
челюстей еще не было.
Подвижность расчлененных жаберных дуг дала предпосылки для
возможного перехода к активному захватыванию пищи ртом при помощи
сдвигания скелетных частей первых дуг. Таким образом первые дуги
преобразовались в челюсти (и губные хрящи) и добавочные части, дававшие
челюстям более прочную опору на осевом черепе (hyomandibulare). Воз-
никли челюстные позвоночные, к которым принадлежат все типичные
рыбы и все вышестоящие позвоночные.
Челюстные позвоночные (Gnathostomata). Рыбы
Отроение древнейших челюстных позвоночных Gnathostomata харак-
теризовалось следующими основными признаками: кожный скелет (хотя
бы только в виде плакоидных чешуй, образовавших на челюстях зубы);
челюстной аппарат из губных хрящей, собственно челюстей и более или
423
менее связанной с ними гиоидной дуги с соответствующей мускулатурой;
вполне сформированная черепная коробка с обонятельной и слуховой
капсулами; особый скелет, состоявший из хорды с хрящевыми дипло-
спондильными верхними и нижними дугами.Челюстные позвоночные имели,
кроме непарных, также и парные плавники с соответствующими поясами.
Рис. 372. Папгыриая акулообразная рыба Coccostcus из нижнего девона
(по Иекелю).
Парный орган обоняния открывался парными ноздрями наружу на пе-
реднем конце головы. Жаберных щелей было не более 8. В брызгальце
была еще функционирующая жабра.
Из конкретных представителей древнейших челюстных позвоночных
особый интерес представляют рыбы, имевшие также значительный пан-
цырь из кожных костей — Arthrodira (рис. 372). У этих рыб, живших
еще в девонском периоде, по исследованиям Стеншио, хрящевой череп
был покрыт перихондральным окостенением и плотно прилегал к цент-
Рис. 373. Череп панцирной акуло-
образной рыбы Macropetalichthys
сверху (по Стеншио).
d. end — канал для эндолимфатического
протока; elf — обонятельная капсула:
orb — главница; par — отверстие для
теменного органа. Позади глазниц
выступают слуховые капсулы. Сверху
видны отпечатки каналов боковой линии.
ральной нервной системе и связанным
с ней органам, что дало возможность
восстановить в весьма значительной
степени строение всей головы.
У наиболее полно исследованного
представителя этих рыб — Macropeta-
lichthys — строение головы до деталей
сходно со строением головы современ-
ных хрящевых акулообразных рыб
(Elasmobranchii). Как и у последних,
черепная коробка (рис. 373) имела
широкое основание, и вообще она по
форме вполне сходна с черепной ко-
робкой современных хрящевых рыб и
отличается от нее лишь присутствием
сплошного перихондрального окостене-
ния как на внешней, так и на внутрен-
ней поверхности хряща. Положение
обонятельных капсул также сходно, и
ноздри открывались на брюшной по-
верхности головы. Внутреннее ухо с его
полукружными каналами совершенно
сходно с таковым хрящевых рыб, и, как
у последних, эндолимфатический про-
ток открывался наружу на спинной
поверхности головы. Нёбноквадратный
хрящ не соединялся непосредственно
с обонятельной областью черепа, а,
очевидно, висел на связке, как и у хрящевых рыб. Зубы похожи на
акульи. Расположение органов боковой линии также сходно. Наконец,
и головной мозг с отходящими от него нервами (рис. 374) поразительно
сходен с таковым у хрящевых рыб, так же как и расположение крове-
носных сосудов головы.
424
п
Рис. 374. Головной мозг Масго-
petalichthys сверху (по Стеншио).
die — промежуточный мозг; lolf —
обонятельная лопасть; тез — сред-
ний мозг; met — мозжечок; myl —
продолговатый мозг; п — обоня-
тельная ямка; par—теменной орган;
tel — передний мозг; I—X — голов-
ные нервы.
скелета. Активность в нахо-
Одним словом, не подлежит никакому сомнению, что эти древнейшие
рыбы находятся в ближайшем родстве с современными акулообразными
рыбами. Однако, в отличие от последних, у них имелся хорошо развитый
костный скелет и притом не только кожный панцирь (откуда их наимено-
вание — Stegoselachii), но и окостенения
внутреннего хрящевого скелета. Все это
делает весьма вероятным, что костная
ткань у современных хрящевых рыб ис-
чезла вторично. Вспомним, что и у древ-
нейших настоящих акулообразных рыб—
Acanthodii—имелись поверхностные окос-
тенения на частях висперального скелета
(рис. 376). Имелись кости также и у
древнейших химер. Так как мы привыкли
считать присутствие костной ткани при-
знаком более высокой организации, то мы
должны признать, что в этом отношении
хрящевые рыбы деградировали. Это,
конечно, не значит, что их развитие из
древнейших Arthrodira и в остальном имеет
регрессивный характер. Головной мозг,
прогрессивное развитие которого вообще
играет руководящую роль в эволюции
позвоночных, при всем его сходстве, все
же развит у современных акул и скатов
гораздо выше, чем у Arthrodira (рис. 374).
Это касается именно переднего мозга.
Таким образом, приходится признать,
что древнейшие позвоночные очень рано
приобрели костную ткань и костный ске-
лет, который у некоторых форм, живших,
вероятно, в прибрежной зоне прибоя,
очень быстро достиг значительного разви-
тия. Приобретенная таким образом защита
имела, однако, и свои отрицательные
стороны — костный панцырь, несомненно,
значительно уменьшал подвижность жи-
вотного. Многие такие формы, очевидно,
переходили к менее активной жизни на дне
морей, как об этом можно судить по плос-
кой форме их тела, характерной для
бентонных рыб.
Прогрессивное развитие подвижности
у наиболее хищных представителей этих
первичных челюстных животных было
возможно лишь при редукции костного
ждении добычи связана, однако, и с высоким развитием органов чувств,
а прогрессивное развитие последних связано с прогрессивным развитием
головного мозга. У первой ветви высокоразвитых хищников особое значе-
ние приобрел орган обоняния и связанный с ним передний мозг.
Хрящевые рыбы (Ghondrichthyes). Развитие современных хрящевых
рыб, таким образом, наметилось через прогрессивное развитие органа
обоняния и переднего мозга и одновременную редукцию первых окосте-
нений (за исключением лишь покрова из плакоидных чешуй, который,
однако, тоже иногда редуцируется, у некоторых скатов и у Holocephala).
История хрящевых рыб довольно ясна. Примитивнейшими их предста-
вителями мы должны признать уже упомянутых Acanthodei, остатки
425
которых известны, начиная с девона (рис. 375). У Acanthodei имелись
еще остатки окостенений в виде тонких костных пластинок (без костных
телец) на поверхности хряща (рис. 376). Передний край плавников был
защищен крупными шипами, как у многих современных акул. Шипы
по бокам брюха указывают на существование у древнейших представи-
телей группы (Climating)
до 7 пар плавников (рис.
375). Здесь, следовательно,
парная боковая складка
предков челюстных живот-
ных разделилась па боль-
шее число плавников. У
более молодых представи-
телей той же группы
Рис. 375. Акулообразная рыба Climating с 7 боко- Acanthodei (Acanthodes)
выми шипами, соответствующими, очевидно, парным имелись лишь две пары
плавникам (по Вудварду). плавников, как у всех
других рыб. Своеобразен
еще кожный скелет из мелких квадратных табличек, состоящих из слои-
стого дентина, весьма сходного с ганоином (рис. 377, А). Такая чешуя
занимает как бы промежуточное положение между плакоидной чешуей
хрящевых рыб и ганоидной чешуей рыб с костным скелетом (рис. 377, В).
Ископаемые Acanthodei, несомненно, близки, с одной стороны, к
акулообразным рыбам (Elasmobranchii) и, с другой стороны, к прими-
Рис. 376. Челюсть акулообразной рыбы Acanthodes с окостенениями
(по Иекелю).
а, он Ь — окостенения нёбноквадратного хряща; с, d — окостенения меккелева хряща.
тивнейшим рыбам с костным скелетом — ископаемым хрящекостным
(Palaeon iscidae).
щ ележаборные (Elasmobranchii) представлены также, уже
начиная с девона, некоторыми древнейшими вполне акулообразными
формами отряда Pleuropterygii с известным представителем Cladodus
(Gladoselache) (рис. 378), у которых, как у современных акул, задние
части брюшных плавников у самцов были преобразованы в копулятивный
орган.
Несколько более специализированы пресноводные Ichthyotomi с пред-
ставителем Pleuracanthus каменноугольного и пермского периодов. Эти
рыбы обладали плавниками с членистой осью (типа «архиптеригия»),
которые, очевидно, служили органами опоры при ползании по дну.
Наконец, уже очень рано, еще в палеозое, появляются и настоящие
поперечноротые (Plagiostomi), разбившиеся на две биологические группы:
426
нектонные формы представлены (в основном) акулами, а бонтонные —
скатами.
Одна ветвь морских хрящевых рыб, перешедшая также рано к донной
Рис. 377. Шлиф через чешую А — Acanthodes (девон), В — Cheirolepis
(по Гудричу).
д — слоистый лептин (гапоип); г — изопсдин; <М — дептиповыс канальцы;
жизни (еще в палеозое), специализировалась на питании твердой пищей —
у них развились крупные зубные пластинки, верхняя челюсть срослась
с черепной коробкой (аутостилия), и таким обра-
зом получился необычайно сильный челюстной
аппарат, легко раздавливающий раковины молюс-
ков. Это — цельноголовые (Holocephali), наибо-
лее известными представителями которых явля-
ются химеры. Интересно отметить, что у древней-
ших представителей цельноголовых также имелись
большие костные пластинки. У современных форм
не только окостенения, но и плакоидные чешуи
кожи редуцировались.
Хотя частичная редукция костной ткани за-
мечается у всех древнейших позвоночных (у хря-
щевых, у двудышащих, у кистеперых, у хряще-
костных рыб и даже у амфибий), однако она
в большинстве случаев не привела к полной
утрате кости. Костная ткань щитковых давала
этим животным известную защиту, хотя и огра-
ничивала их подвижность (что при питании
планктоном не имело значения). Редукция кост-
ной ткани была связана с переходом к активному
питанию и, следовательно, с требованием большей
подвижности. Однако редуцировался не только
кожный панцырь, но и окостенения внутреннего
скелета. Это объясняется, с одной стороны, важно-
стью уменьшения удельного веса рыбы, а также
мало совершенной формой первых окостенений.
Окостенения эти были сплошными и охватывали
хрящевой скелет и, в частности, череп непрепыв- „ „„„
г ’ г, гн Рис. зу8. Ископаемая
ным костным покровом. Этим самым ограничивал- акула Cladodus. Вид сни-
ся рост этих частей. Очевидно, окостенения раз- зу (по Иекелю).
вивались лишь поздно — молодое животное имело
только хрящевой скелет, а с образованием кости дальнейший рост, по край-
ней мере, черепа прекращался. Частичная редукция костного скелета при-
427
вела прежде всего к тому, что сплошные костные пластинки разоились
на отдельные участки. Этим была достигнута возможность значительного
роста окостеневших частей, а следовательно, и заключенного в черепной
коробке головного мозга. В результате стало возможным и более раннее
развитие окостенений у молодых рыб, что, несомненно, имело большое
значение в их борьбе за существование.
Относительно высокий удельный вес рыбы с костным скелетом мог
быть уменьшен путем заглатывания воздуха в кишечник. С развитием
плавательного пузыря, вполне уравновешивавшего тело рыбы в воде,
явилась опять возможность прогрессивного развития окостенений нового
типа (состоявших из многих отдельных элементов, в особенности на че-
репе). Это и происходило у геологически более молодых рыб с костным
скелетом (Holostei, Teleostei).
Костные рыбы (Osteichthyes) характеризуются чешуями, содержа-
щими костную ткань, костными челюстями, развившимися частично
(вверху) на губных хрящах, присутствием жаберной крышки с костным
скелетом и двумя парами ноздрей.
Рис. 379. Ископаемая хрящекостная рыба Palaconiscus (дсвОи) (по Вудварду).
Они появляются в девоне сразу в виде нескольких ветвей. Из низших
форм к ним наиболее приближаются Acanthodei, в особенности по строению
чешуй, однако вряд ли все-таки в последних можно видеть прямых предков
высших рыб. Древнейшие из известных нам примитивных рыб с частично
окостеневшим скелетом относятся к семейству Palaeoniscidae (рис. 379),
которое более или менее непосредственно связывается с высшими формами.
Современные осетровые, несомненно, очень примитивны по всей своей ор-
ганизации и во многом близки к хрящевым рыбам (строение черепа, сим-
физ нёбноквадратного хряща, примитивные позвоночник и плавники, при-
митивные ромбические чешуи и очень примитивный покров из чешуеобраз-
ных кожных костей на черепе). Между тем в ископаемом состоянии близкие
формы известны только с начала мезозоя, и их возникновение недоста-
точно известно. Поэтому Северцов не без основания предполагает, что
осетровые произошли от особой ветви весьма примитивных рыб, остатков
которых до сих пор не найдено. Всю эту группу Северцов называет Chond-
rosteoidei и противопоставляет другой группе Holosteoidei, от которой
произошли все высшие формы. Из ископаемых форм к первой группе
Chondrosteoidei относятся, по мнению Стеншио, и Saurichthyidae, отчасти
приближающиеся к осетровым, но, с другой стороны, все же родственные
и древнейшим Palaeoniscidae. У Saurichthyidae костный скелет, несо-
мненно, подвергся некоторой редукцип. То же самое допускается иногда
и для осетровых, которые приспособились к донной жизни и подверглись
некоторой специализации.
Во второй группе — Holosteoidei — имеется, по крайней мере, одна
весьма богатая формами прогрессивная ветвь, начинающаяся от пале-
озойских Palaeoniscidae (Heterocerci) и ведущая к костистым рыбам. У этих
рыб была постоянная хорда, однако дуги позвонков и ребра окосте-
невали. Хвост гетероцеркальный. В непарных плавниках костяные лучи
428
Рис. 381. Ископаемая костная рыба Caturus (верх-
няя юра); чешуя не изображена (по Вудварду).
имелись в количестве, превышающем число лучен внутреннего скелета
(птеригиофоров); впереди они были защищены рядами чешуй — фулькрами
(иногда и на парных плавниках). У древнейших девонских форм имелась
иногда очень мелкая ромбическая чешуя (Cheirolepis), напоминающая
чешую Acanthodei (рис. 377, В). У других нередко была чешуя в виде
крупных ромбических (Palaeoniscus), иногда очень высоких (Cheirodus)
пластинок. Эти рыбы достигли полного расцвета в каменноугольном и
пермском периодах. Только в мезозое они уступают место высшим формам.
Некоторые имели очень высокое тело (Platysomidae), подобно некоторым
современным костным
рыбам (главным обра-
зом рифовым). Следую-
щую ступень той же
ветви образуют «кост-
ные ганоиды» — Holos-
tei, появляющиеся в
пермском периоде и до- Рис. 380. Ископаемая костная рыба Eugnathus (юра)
стигающие в мезозое (по Вудварду).
полного расцвета,с боль-
шим богатством форм. Holostei чрезвычайно сходны с Palaeoniscidae.
Различия сводятся главным образом к тому, что у Holostei появляются
зачатки тел позвонков в виде небольших вначале «плевроцентров» (interdor-
salia) и «гипоцентров» (basiventralia), которые у высших форм образуют
полупозвонки (рис. 79, 80) или, наконец, полные амфицельные тела по-
звонков. Кроме того, хвостовой плавник обнаруживает переходы от гете-
роцеркии ко вторичной симметрии (гомоцеркии) через редукцию его дор-
зальной лопасти. Чешуя ганоидная, ромбическая, переходящая у позд-
нейших форм в циклоидную. И здесь это были частью быстро плавающие
рыбы с веретено видным
телом (например, Lepido-
tus), частью увертливые
высокие формы (например,
Dapedius). В одной неболь-
шой ветви сохранились
массивные ромбические че-
шуи, внутренний скелет
сильно окостенел, и цель-
ные тела позвонков очень
рано совершенно вытесни-
ли хорду. Это — семейство Aspidorhynchidae, ведущее к современным
панцирным щукам — Lepidosteidae.
Кроме того, среди ископаемых Holostei особо выделяется прогрес-
сивное семейство Eugnathidae. Ромбическая чешуя древнейших его пред-
ставителей (Eugnathus, рис. 380) постепенно округляется (Caturus, рис. 381)
и истончается (Eurycormus). Небольшие элементы тел позвонков (Eugna-
thus) разрастаются (Caturus) и образуют, по крайней мере, в хвостовом
отделе полупозвонки (Eurycormus). В чрезвычайно близком семействе
Amiidae, один из представителей которого Amia calva дожил до наших
дней, чешуи уже вполне округлые (циклоидные), развиваются цельные
тела позвонков (в хвосте еще полупозвонки) и, кроме того, редуцируются
фулькры на плавниках.
Те же процессы, которые характерны для Holostei и особенно для
указанной их ветви, привели и к образованию современных кости-
стых рыб, связанных теснейшим родством с ископаемыми Eugnathidae
и Amiidae. Они имеют то же самое полное развитие тел позвонков, кото-
рые приобрели амфицельную форму (как у Amiidae), тонкую костную
циклоидную чешую, теряющую верхний слой ганоида; хвост становится
429
вполне гомоцеркальным. Кроме того, исчезают фулькры на плавниках
и редуцируются яремные пластинки между ветвями нижней челюсти.
Представители древнейшего ископаемого семейства костистых рыб —
Leptolepidae (рис. 382) — известны начиная с конца триаса. Их цикло-
идная чешуя имела еще топкий слой ганоина. Представители другого
семейства — Elopidae — известны с мела и дожили до наших дней (Elops,
Megalops) в тропиках; у них сохранилась еще одна непарная яремная
пластинка. Представители современного семейства сельдевых (Clupeidae)
известны тоже начиная с мелового периода.
Другие процессы, приводящие к современным костистым рыбам, —
редукция артериального конуса сердца и спирального клапана кишки —
на ископаемом материале не могут быть прослежены, но следы этих органов
сохранились еще у некоторых современных форм. Именно артериальный
конус имеется еще у Albula
(представители Albulidae из-
вестны начиная с мела) и
Osteoglossum, а также у Me-
galops (Elopidae), а спираль-
ный клапан имеется у Chiro-
centrus (представители из-
Рис. 382. Ископаемая костистая рыба Leptolepis вестны с мела) и в рудимен-
(верхняя юра); чешуя не изображена (по Будвар- тарнОм виде у лососевых
ду'‘ (Salmonidae).
Мы не можем утверждать, что все костистые рыбы развились из од-
ного ствола. Ужо у Amiidae мы видели ряд параллельных им изменений.
Точно так же и костистые могли развиться из разных представителей
Eugnathidae и Amiidae в виде нескольких параллельных ветвей.
Другая прогрессивная группа рыб с костным скелетом появляется в
виде очень древних девонских форм, связанных несомненным родством как
с древнейшими Palaeoniscidae (особенно Cheirolepis), так и с Holostei,
образующих, однако, вполне самостоятельную ветвь своеобразных рыб.
Это — группа кистеперых рыб в широком смысле этого слова
(Holosteoidei crossopterygii — по терминологии Северцова), заключающая
в себе собственно кистеперых рыб (Crossopterygii), многоперых (Polypteri-
dae) и двудышащих (Dipnoi). Общим признаком, выделяющим эту группу,
является устройство парных плавников, которые имеют узкое основание,
обеспечивающее их поворотливость, и мускулистую центральную часть,
прикрытую чешуей, в которой помещался богато расчлененный внутрен-
ний скелет. Расчлененный внутренний скелет и диференцированная му-
скулатура, помещавшаяся в самом плавнике (а не в туловище, у его осно-
вания, как у других рыб с костным скелетом— Actinopterygii), давали
возможность более сложных движений плавниковой лопасти. Все эти
рыбы пользовались плавниками не только при плавании, но и в качестве
органа опоры при ползании по дну водоемов, а также при продвижении
при их помощи в зарослях водорослей и водяных растений. Другим общим
признаком для этих рыб является высокоразвитый плавательный пузырь,
по происхождению, невидимому, парный, развивающийся в брюшной
стенке задней части глоточной области, очевидно, за счет разрастания
назад пары задних жаберных мешков. Брюшной плавательный пузырь,
несомненно, гомологичен легким наземных позвоночных и нередко (во
всяком случае у двудышащих) в самом деле функционирует как легкие.
Эта группа рыб в некоторых отношениях очень примитивна. У них
имелась постоянная хорда (исключая Polypteridae), а внутренний скелет
во многих частях, особенно в черепе, сохранял много хряща. У прими-
тивнейших девонских Osteolepidae (рис. 383) чешуя была ромбическая,
как и у современных Polypteridae, однако у всех прочих, не исключая
и древнейших девонских представителей этой группы, чешуя была уже
430
Круглой, «циклоидной» формы. По своей структуре йто был очень прими»
тивный род чешуи. Составляется она из основной костной части (изопедин)
и внешней дентиновой, обнаруживающей следы происхождения путем
слияния плакоидных чешуй (космин). Характерного для ганоидных чешуй
верхнего покрова из слоистого дентина (ганоина) здесь не было (за исклю-
чением Polypteridae). Такая чешуя носит наименование космоидной чешуи.
Как видно из сказанного, одна из- ветвей этой группы, именно ветвь,
ведущая к современным многоперам (Polypteridae), занимает в ней со-
вершенно обособленное положение. Особое строение имеет и скелет их
плавников. Остальные представители этой группы сразу, еще в девоне,
распадаются на две неравно-
ценные ветви — собственно кис-
теперых и двудышащих.
Из кистеперых (Cros-
sopterygii) наиболее прими-
тивным семейством являются
Osteolepidae (девон и каменный
уголь, рис. 383), у которых
расположение костей черепа
челюсти) было еще весьма сходно с Palaeoniscidae (особенно Cheirolepis).
Чешуя у них была ромбическая, основание плавников еще мало суженное
и центральная лопасть короткая. Другое семейство весьма примитивных
форм — Rhizodontidae (девон и каменный уголь) — также имело парные
плавники с довольно короткой лопастью, но чешуя была уже циклоидная
(рис. 386). Зубы обычной конической формы имели у основания радиаль-
ную складчатость. Это семейство кистеперых рыб представляет особый
интерес, так как более других приближается к низшим наземным позво-
Рис. 383. Ископаемая кистеперая рыба
Osteolepis (девон) (по Тракеру).
(включая rostralia и кости нижней
Рис. 384. Ископаемая двудышащая рыба Dipterus (девон) (по Тракеру).
ночным (стегоцефалам). Из других представителей кистеперых Holopty-
chiidae (девон) очень рано специализировались. Их парные плавники
обладали длинной лопастью. Особый интерес представляют зубы, дентин
которых имел весьма сложную складчатую структуру. Более позд-
ние и наиболее специализированные кистеперые относятся к семейству
Coelacanthidae (пермь и весь мезозой). К этому семейству относится един-
ственный известный представитель кистеперых в современной фауне —
Latimeria. У этих рыб все плавники имели своеобразную округлую форму;
дифицеркальный хвост отличался особой концевой лопастью; стенка пла-
вательного пузыря окостеневала.
Двудышащие (Dipnoi) также известны, начиная с девона, и
сразу легко отличимы, хотя древнейшие их представители (Dipteridae)
довольно близки к кистеперым (Osteolepida) по общей форме- тела, по
расположению плавников и форме эпицеркальпого хвоста, по располо-
жению покровных костей на переднем конце головы, по присутствию
splenialia и postsplenialia на нижней челюсти, по сложному строению
оперкулярного аппарата (operculum, suboperculum, одна средняя ярем-
ная, одна пара больших яремных и парный ряд мелких боковых яремных
пластинок) и, наконец, по строению парных плавников, имевших, впрочем,
уже длинную среднюю лопасть (рис. 384). Все же и у древнейших двуды-
431
Р°
Рис. 385. Дно черепа (нёбо)
Dipterus (по Гудричу).
ра — парасфеноид; р — кры-
ловидные кости; pt — нёбные
вубные пластинки.
Шащих имеется ряд признаков своеобразной специализации. Кости крыши
черепа получают несколько отличное расположение, постепенно реду-
цируются щечные кости, наружные ноздри перемещаются вниз под верх-
нюю губу (это связано с редукцией челюстных костей). Главная особен-
ность, сразу выделяющая двудышащих из других в общем сходных с ними
рыб, — это развитие сложных зубных пластинок (рис. 385) с большими
гребнями на них, образующихся в результате срастания между собой
зубов внутреннего ряда. Вместе с тем, зубы наружного ряда и несущие их
кости, т. е. основные кости челюстей других позвоночных (maxilla, ргае-
maxilla и dentale), редуцируются. Редуцируются также части первичной
челюсти: нёбные и наружные крыловидные
кости. Вверху развиваются нёбные зубные
пластинки на внутренних крыловидных костях
(рис. 385), и эти последние соединяются между
собой впереди посредством симфиза. Внизу
зубные пластинки развиваются на пластинча-
тых костях (splenialia).
Все это преобразование зубной системы
связано с переходом к иной форме питания:
полупресноводная и пресноводная рыба стала
питаться водорослями и водяными растениями,
раздавливая их между зубами, причем вместе
с растениями съедались и сидящие на них
животные (в особенности моллюски). Нижняя
челюсть становится короче и получает более
солидную опору на осевом черепе, благодаря
прирастанию к нему квадратной кости. Одно-
временно подъязычночелюстной хрящ (hyoman-
dibulare) теряет функцию подвеска и редуцируется. От наиболее прими-
тивного семейства Dipteridae развитие двудышащих шло по меньшей
мере в трех направлениях, причем для всех, направлений характерна
частичная редукция костного скелета и, в особенности, редукция окосте-
нений первичного черепа. Одна ветвь ведет через Scaumenacia к Phanero-
pleuron (разрастание спинного плавника и слияние его с хвостовым).
У некоторых форм происходили редукция зубных пластинок и вторичный
их распад на отдельные зубы (Uronemus и Concho рота). Другая ветвь
ведет через Ctenodus (каменный уголь) и Sagenodus (каменный уголь,
пермь) к современному Ceratodus, зубные пластинки которого известны,
начиная с триаса. Современные Protopterus и Lepidosiren представляют
собой конечное звено особой более измененной ветви, происхождение
которой не так ясно.
Почти все рыбы из группы кистеперых вымерли. В современной
фауне известен лишь один представитель семейства Coelacanthidae —
Latimeria, единственный экземпляр которой добыт в 1938 г. у южных
берегов Африки. Из двудышащих в настоящее время также имеются
лишь остатки (Ceratodus в Австралии, Protopterus в Африке и Lepidosi-
ren в Южной Америке). Однако общее этим рыбам присутствие добавоч-
ного органа дыхания непосредственно атмосферным воздухом, именно
легких, обеспечило представителям этой группы возможность выхода
на сушу. В самом деле, одна из ветвей, начинающаяся среди примитивных
представителей кистеперых рыб, дала уже очень рано (в девоне) начало
мощному стволу, из которого произошли все наземные позвоночные.
Происхождение наземных позвоночных
Одним из наиболее интересных вопросов филогенетики является
вопрос о происхождении наземных позвоночных. В общей форме вопрос
432
этот в настоящее время вполне ясен. Уже из истории индивидуального
развития любого наземного позвоночного и, в частности, из онтогенеза
амфибий следует, что они произошли от рыбообразных позвоночных,
дышавших жабрами. Данные сравнительной анатомии и, особенно, палеон-
тологии позволяют в значительной мере конкретизировать наши представ-
ления о переходе позвоночных от водной жизни к жизни на суше.
Прежде всего отметим, что хотя переход из одной среды в другую
сопровождался глубочайшей перестройкой всей организации животного,
мы все яге можем выделить некоторые процессы, имевшие руководящий
характер и обусловившие возможность такого перехода. За этим с необхо-
димостью следовали и все другие преобразования. Такими руководящими
процессами были: 1) переход к легочному дыханию и 2) переход к передви-
жению (ползанию) при помощи парных конечностей, служивших при этом
органом опоры; кроме того, можно отметить еще один важный позднейший
процесс: 3) преобразование органа слуха в связи с восприятием колебаний
воздушной среды (иногда вторично — колебаний почвы).
Легочное дыхание, несомненно, развилось сначала как
дополнительное дыхание у рыбы, дышавшей жабрами. Мы знаем, что в
жарких странах при сильном нагревании пресноводных бассейнов содер-
жание кислорода в воде падает, чему способствует еще усиление процессов
гниения различных органических остатков. Поэтому у тропических
пресноводных рыб очень часто имеются добавочные органы дыхания
(лабиринтовые органы, мешковидные выросты жаберной области неко-
торых сомовых, особенно Saccobranchus). Эмбриональное развитие легких
у амфибий указывает на то, что легкие развились как пара мешковидных
выпячиваний в задней части жаберной области (от пятого жаберного мешка).
Подобные же органы являются плавательным пузырем у кистеперых и
двудышащих рыб. Они имеют ячеистые стенки и, несомненно, являются
также добавочным органом воздушного дыхания, т. е. легкими. Как и у
наземных позвоночных, они снабжаются кровью от четвертой пары вы-
носящих жаберных артерий, дающих от себя ветвь — легочную артерию.
Таким образом, легочное дыхание установилось еще у жившего в воде
животного и обусловило возможность его выхода на сушу. При переходе
на сушу жабры, конечно, потеряли свою функцию, и легочное дыхание
стало основным. Замена жаберного дыхания легочным привела прежде
всего к крупным изменениям в кровообращении. Жаберное кровообраще-
ние вообще теряет свое значение, капилляры исчезают, и устанавливается
более непосредственная связь между брюшной и спинной аортой. Таким
образом, уже у Protopterus первые две жаберные артерии являются в роли
дуг аорты (рис. 329). У наземных позвоночных эту функцию несет только
вторая пара жаберных артерий; остальные жаберные артерии редуци-
руются, за исключением первой, сохраняющей связь с сонной артерией, и
четвертой, являющейся в роли легочной артерии. Окисленная кровь
из легких возвращается в венозную систему, и таким образом сердце
получает смешанную кровь. В связи с этим развиваются приспособления
для разделения токов крови — артериального от венозного. Уже у дву-
дышащих легочная вена проходит вдоль стенки венозной пазухи и впа-
дает непосредственно в предсердие. В предсердии у двудышапщх появля-
ется также и продольная перегородка (правда, неполная). В желудочке
смешению крови препятствуют многочисленные мускульные перекладины,
вдающиеся внутрь полости. В артериальном конусе такяге развивается
перегородка из продольного ряда клапанов (намечается у Ceratodus),
которые у Protopterus и у амфибий сливаются в один продольный спираль-
но изогнутый клапан. Венозная кровь переходит внутри конуса справа
на спинную сторону, где отходят третья и четвертая жаберные артерии,
несущие кровь к жабрам и к легким. Артериальная кровь переходит
слева на брюшную сторону и затем в первые две дуги, несущие кровь
433
Дальше в сонные артерии и в спинную аорту. Все эти преобразования
связаны с приобретением легочного дыхания. Последнее, как сказано,
возникло еще у тех рыб, от которых произошли наземные позвоночные.
Преобразования в кровеносной системе происходили постепенно — в
основном начались еще у рыб (причем у новейших двудышащих произошли
большей частью независимо от предков амфибий), но совершенно закончи-
лись полным разделением сердца на артериальную и венозную поло-
вины только у высших наземных позвоночных (крокодилов, птиц и
млекопитающих).
Сходство кровеносной системы амфибий с таковой двудышащих рыб
увеличивается еще тем, что у последних также развиваются задняя полая
вена, несущая кровь от почек непосредственно в венозную пазуху (карди-
нальные вены начинают редуцироваться), и непарная брюшная вена,
несущая кровь из конечностей в правый кювьеров проток. Утрата жабер-
ного дыхания отозвалась, конечно, не только на кровообращении. Крупные
изменения произошли в висцеральном скелете. Челюстной аппарат,
участвующий обычно в дыхательных движениях при жаберном дыхании,
с утратой последнего мог фиксироваться более прочно на черепе. Нёбно-
квадратный хрящ (palatoquadratum), который и у костных ганоидов
(включая кистеперых) имеет обычно самостоятельное причленение к слу-
ховой капсуле, прирос к черепу в этой области. В связи с этим подъязычно-
челюстной хрящ (hyomandibulare) утратил значение подвеска и в значи-
тельной мере редуцировался, сменив при этом функцию: поддерживая
еще связь с квадратной костью, он стал передавать звуковые колебания
(повидимому, первоначально от сидящей на нем жаберной крышки) к
слуховой капсуле. Весь жаберный аппарат подвергается значительной
редукции и преобразуется в довольно еще значительный у хвостатых
амфибий подъязычный аппарат и гортань, с ее хрящами и мышцами.
Конечности также должны были подвергнуться радикальной
перестройке. Непарные плавники, включая хвостовой, утеряв на суше
свою функцию, полностью редуцировались, и только у живущих в воде
личинок развиваются простые кожные складки без скелета и муску-
латуры. Парные плавники сменили свою фукцию в основном еще до пе-
рехода на сушу. Жившие в прибрежных мелких водах, а также в пресной
воде рыбы пользовались своими плавниками как органами опоры при
ползании по дну или при проползании в зарослях водяных растений, как
это делают и современные двудышащие и кистеперые. Эта смена функции
была связана с прогрессивным расчленением скелета свободной конеч-
ности и с усложнением строения ее мускулатуры и обусловила возмож-
ность выползания животного на сушу. Дальнейшие изменения связаны
именно с этим переходом. Перемещение тела на суше требовало гораздо
большей мускульной силы и более прочной опоры для конечностей. По-
этому прогрессивно развивались не только скелет и мускулатура конечно-
стей, но и пояса конечностей. Однако наружный плечевой пояс рыб был
прочно связан с костями черепа. Это ограничивало возможность движений
как самого пояса, так и головы. Между тем, для первых наземных позво-
ночных, которые были на суше весьма мало поворотливыми, подвижность
пояса, в особенности головы, имела очень большое значение. Поэтому эта
связь нарушилась, и постепенно развился более подвижной шейный отдел
тела. Кости наружного пояса, не связанные с мышцами (cleithra, supra-
cleithralia, posttemporalia и отчасти claviculae), редуцировались или
погружались вглубь под кожу (ключицы). Наоборот, первичный пояс,
дающий поверхности для прикрепления мускулатуры, разрастался, осо-
бенно на груди и на спине, в широкие пластинки, получившие опору на
ребрах. Устанавливались также сложные мускульные связи с соседними
частями (вторичная плечевая мускулатура). Тазовый пояс тоже разрастал-
ся, особенно в спинной части (ileum), которая также получила опору на
434
ребрах позвоночника. Необходимость облегчения веса тела на суше йрй-
вела к редукции кожного скелета вообще, и в частности быстро исчеа
покров из кожных чешуй, которые немного дольше сохранились лишь
на брюшной поверхности тела.
Органы чувств. Полость брызгальца, замкнутая снаружи
и у высших рыб, естественно, наполнилась воздухом. Существование
легко сжимаемого воздушного пространства под подвижной подъязычно-
челюстной костью (hyomandibulare), утратившей свою функцию, но свя-
занной еще с остатками жаберной крышки, дало благоприятные условия
для передачи различных колебаний, в том числе звуковых колебаний
воздушной среды, с жаберной крышки на подъязычночелюстную кость
и отсюда на слуховую капсулу, к которой она прикреплялась (через
посредство небольшой pharyngohyale). Полость брызгальца разрасталась
вокруг подъязычночелюстной косточки (образуя барабанную полость), и
движения последней становились более свободными. Жаберная крышка
редуцировалась, и на ее месте осталась упругая кожная пластинка (ба-
рабанная перепонка). Таким образом, развился добавочный аппарат для
передачи звуковых колебаний воздушной среды — среднее ухо, состоящее
из барабанной полости с заключенной в ней слуховой косточкой — стол-
биком (columella).
На суше были благоприятные условия и для дальнейшего развития
органа зрения, а в связи с большим значением этих органов чувств про-
грессивно развился и головной мозг.
Таковы основные процессы, характеризующие переход водных по-
звоночных к наземной жизни. Это логически вытекает из всей суммы
наших знаний по сравнительной анатомии и эмбриологии низших наземных
позвоночных. Однако только сравнение с рыбами и использование палеон-
тологического материала может нам помочь представить весь этот переход
в более конкретной форме и установить более точно исходные формы и
последовательность отдельных этапов этих сложных преобразований.
Предки наземных позвоночных
Сходство в строении костного скелета с высшими рыбами и, в осо-
бенности, сходство покровных костей черепа (например, такая в высшей
степени характерная рыбья кость, как парасфеноид) и плечевого пояса
заставляет искать предков наземных позвоночных именно среди высших
рыб с костным скелетом. Костистые рыбы при этом, однако, сразу отпадают
как слишком специализированные — отсутствие артериального конуса
в сердце, отсутствие клоаки, развернутая форма переднего мозга (с эпи-
телиальной крышкой), частичная редукция костей крыши черепа (щечные
кости), непарный сошник, полное развитие костных позвонков, отсутствие
ключиц, положение челюстей в губах и многое другое противоречит
всякой попытке вывести наземных позвоночных из костистых рыб. Многое
из сказанного сохраняет свою силу и в применении к костным ганоидам —
Holostei. Таким образом, для сравнения остаются только кистеперые и
двудышащие, у которых еще имелись артериальный конус, туловищная
почка с нефростомами, типичная связь половых желез с канальцами
почек и др. У тех и других имеются также и легкие, снабжаемые кровью
от четвертой жаберной артерии, и намечаются некоторые дальнейшие изме-
нения в кровеносной системе.
У двудышащих начинается деление сердца на левую — артериальную
и правую — венозную половины, появляются задняя полая вена и не-
парная брюшная. У двудышащих головной мозг похож на мозг амфибий.
У них нёбноквадратный хрящ прирос к осевому черепу, как у амфибий.
Подъязычночелюстной хрящ утратил функцию подвеска и редуцируется.
Конечности начинают служить органами опоры при ползании. Имеются,
435
однако, и крупные отличия: у двудышащих, и не только у современных,
но и у ископаемых, имеется в высшей степени специализированная зубная
система (сложные зубные пластинки), располагающиеся на нёбе (на птери-
го:’дах) и на внутренней стороне нижней челюсти. Это и является отли-
чительным признаком ископаемых двудышащих. Одновременно кости,
несущие у других рыб, а также у наземных позвоночных зубы наружного
ряда, редуцируются (верхнечелюстные, межчелюстные и нёбные вверху и
зубные внизу). Однако как раз эти кости получают у наземных позвоноч-
ных преобладающее значение, и, следовательно, в отношении развития
челюстей и зубной системы двудышащие, с одной стороны, и наземные
позвоночные — с другой, испытали резко расходящуюся (дивергентную)
Рис. 386. Ископаемая кистеперая рыба Eusthenopteron
(верхний девон) (по Вудварду).
эволюцию. Точно так яге у двудышащих развивается своеобразный покров
кожных костей черепа, весьма отличный от крыши черепа наземных позво-
ночных. У двудышащих рыб имеются ребра только нижнего типа, а у
низших наземных позвоночных были и верхние ребра. Все эти обстоятель-
ства (а также и ряд других) не позволяют выводить наземных позвоночных
от двудышащих рыб.
Остаются только еще кистеперые рыбы. У современных кистеперых
(Latimeria) не обнаруживается большого сходства с наземными позво-
ночными, а многоперые (Polypteridae) занимают, как мы видели, поло-
жение совершенно особой ветви, весьма отличной от известных нам
ископаемых форм. Если же привлечь к сравнению древнейших Osteolepida
и, в особенности, Rhizodontidae (рис. 386), то мы можем установить весьма
многие черты сходства с наземными позвоночными и, в особенности,
с древнейшими их представителями — стегоцефалами. Пожалуй, наиболее
интересным является то обстоятельство, что у кистеперых (Osteolepis,
Dictyonosteus и др.) были, кроме наружных, еще и внутренние ноздри,
или хоаны. Это указывает, очевидно, на большую роль, которую у них
играло легочное дыхание. У кистеперых рыб имелась такая же вторич-
ная крыша черепа в виде сплошного панцыря из покровных костей,
покрывавших не только дорзальную поверхность, но и обе стороны
головы (рис. 387). Единственные отверстия в этом панцыре — орбиты
глаз по бокам, ноздри впереди и небольшое теменное отверстие (для
теменного глаза) сверху, между лобными костями. Состав этого панцыря
и расположение костей весьма сходны с составом и расположением
костей черепа у стегоцефалов. Различие заключается лишь в отсутствии
у наземных позвоночных жаберной крышки и в ином положении темен-
ного отверстия, которое у стегоцефалов помещалось далее позади, между
теменными костями.
Так же как и у наземных позвоночных, у кистеперых рыб на брюшном
отделе плечевого пояса имелись покровные кости — ключицы (которых
нет у Holostei и у Teleostei). Особенно интересно поразительное сходство
строения скелета парных конечностей. У Eusthenopteron (рис. 173) и
особенно у Sauripterus имелся богато расчлененный внутренний скелет
плавника (рис. 388). Узкое основание этого плавника поддерживается
436
одним элементом, сравнимым с плечом (бедром), сочленяющимся посред-
ством округлой головки с поясом. Далее лежат два элемента, сравнимые
с лучевой и локтевой (или берцовыми) костями и несущие до десятка лучей.
Рис. 387. Крыша черепа. А — кистеперой рыбы Eusthenopteron
(по Стеншио) кВ — стегоцефала Dolichopareias (по Быстрову).
sp — каналы боковой линии; fr — лобная кость; it — intertemporale; mx — челюстная кость;
п — ноздря; па — носовая кость; о — глазница; ор — жаберная крышка; р — пинеальное
отверстие; ра — теменная кость; ртх — предчелюстная кость; ро — заднеглазничная кость;
рра — заднетеменные кости; г—ростральные кости; sq— чешуйчатая кость; st — надвисоч-
ная кость.
Все эти части гораздо короче и относительно шире, чем в пятипалой
конечности, но в плавнике это и не могло быть иначе (при вторичном
переходе от наземной жизни к водной из пятипалой конечности с длинными
Рис. 388. Скелет плавника кистеперой рыбы Sauripterus (по Грегори).
— ключица; clt—cleithrum; со — коракоид; sc — Лопатка; sclt—supracleithrale.
ht и, г —вероятные гомологи плечевой, локтевой и лучевой костей наземных позвоночных.
костями вновь образуются ласты с укороченными, но расширенными
частями скелета). Общий план строения этих плавников совпадает с пла-
ном строения пятипалой конечности. Кроме того, в плавнике Sauripterus
457
кожные лучи уже редуцировались, по крайней мере, в передней (нижней)
его части. Далее следует напомнить, что у современных кистеперых,
кроме нижних ребер, имеются и верхние ребра типа низших наземных
позвоночных. Наконец, не лишено интереса и то, что у.ископаемых Rhi-
zodontidae имелась радиальная складчатость дентина у основания зуба.
Такая радиальная складчатость наблюдалась нередко и у стегоцефалов и
достигала большой сложности, с одной стороны, у более специализи-
рованных кистеперых сем. Holoptychiidae и, с другой стороны, также
у более специализированных стегоцефалов, образующих группу Laby-
rinthodontia. Здесь в двух ветвях, — с одной стороны, у рыб, с другой —
у амфибий — на основе особенности, унаследованной от общих предков,
развились независимо и параллельно весьма сходные сложнейшие струк-
туры. В представителях Rhizodontidae мы имеем, таким образом, макси-
мальное приближение к строению стегоцефалов.
Можно ли, однако, смотреть на эти известные нам формы кистеперых
как на непосредственных предков наземных позвоночных? На это прихо-
дится ответить отрицательно хотя бы потому, что они оказываются совре-
менниками древнейших стегоцефалов. Последние теперь известны уже из
верхнедевонских отложений. Однако слишком еще мало изучены нижнеде-
вонские кистеперые. Среди них, наверное, были и несколько’ более при-
митивные формы, которые стояли еще ближе к прямым предкам стегоце-
фале в,чем изученные Eusthenopteron и Sauripterus. Не подлежит сомнению,
что именно такие формы, относящиеся к древнейшим кистеперым, дали
начало всему стволу высших позвоночных. Что же касается двудышащих,
то мы не закрываем глаз и на их значительное сходство с амфибиями
в некоторых отношениях, но мы безусловно должны признать в них бо-
ковую, своеобразную специализированную ветвь, происшедшую, очевидно,
от тех же примитивных кистеперых. Таким образом, путем изучения
остатков кистеперых рыб, палеонтология твердо приближается к разре-
шению одной из интереснейших проблем филогенетики. Углубляя же
изучение древнейших стегоцефалов, которые обнаруживают признаки,
унаследованные от рыбообразных предков, палеонтология приближается,
с другой стороны, к разрешению той же проблемы происхождения назем-
ных позвоночных.
Происхождение амфибий
Ископаемые стегоцефалы во многом отличаются от современных
амфибий. Мы их, однако, относим к одному классу, так как они, по сути,
были амфибиями: они развивались в воде и в личиночном состоянии дыша-
ли жабрами. У многих стегоцефалов был кожный панцырь из чешуй, покры-
вавших иногда все тело, как у рыб, или чаще только брюшную поверх-
ность. Чешуи округлой формы редуцировались у других стегоцефалов
(рис. 67), принимали удлиненную форму, располагались более редкими
косыми рядами на брюхе и, наконец, совсем исчезали. У стегоцефалов
(т. е. «покрытоголовых») голова была защищена сверху и с боков сплошным
панцырем из многочисленных покровных костей (стегальный тип черепа),
подобно тому как мы это видели у кистеперых рыб. Эти кости лежали
еще в коже, как видно по характерной их скульптуре (рис. 142). Между
теменными костями было отверстие для теменного глаза. У многих стего-
цефалов в крыше черепа имелись отпечатки каналов боковой линии (рис.
131). Расположение каналов вполне сходно с их расположением у кисте-
перых рыб, и это позволяет вполне точно произвести сравнение отдельных
костей черепа. У стегоцефалов имелись здесь многие кости, которые в
дальнейшем у наземных позвоночных редуцировались или утратили
самостоятельное значение. Так, например, характерные для кистеперых
(и Holostei) покровные кости затылочной области, образующие один
поперечный ряд, имеются и у стегоцефалов, где они палеонтологами обо,-
438
значились как postparietalia и tabularia (в них, так же как и у рыб, прохо-
дит поперечный заднетеменной канал боковой линии).
обычно имелись только борозды
в костях, в которых лежали
каналы боковой линии, между
тем как у рыб последние прохо-
дят внутри костей и только от-
крываются наружу отверстия-
ми.
Интересно, что у описанных
недавно верхнедевонских стего-
цефалов — Ichtyostegidae — ор-
ганы боковой линии лежали
в каналах костей, как у рыб,
и сообщались с наружной сре-
дой отверстиями в костях кры-
ши черепа (рис. 389). Кроме
того, у Ichthyostegidae еще не
было овального окна, и слухо-
вая косточка упиралась просто
в углубление в стенке слуховой
капсулы. У них была и другая
оригинальная особенность—не
только передние ноздри, но и
задние лежали на нижней сто-
роне черепа под верхней губой
между челюстными и предчелю-
стными костями (напоминая их
положение у двудышащих).
У стегоцефалов костное первич-
ное нёбо было образовано сош-
ником, нёбными и крыловид-
У стегоцефалов
Рис. 389. Крыша черепа примитивного стего-
цефала Ichthyostega (верхний .девон) (по Севе—
Седербергу).
enpt — ветвь энтоптеригоида на квадратной^нос-
ти; /г — лобная; гг — межростральная; /и — ску-
ловая; 1а — слезная; тх — верхнечелюстная;
па — носовая; ра — теменная; ртх — предчелю-
стная; pop — предкрышна; рра — заднетеменная;
pro, pto — передне- и заднеглазничная; qj —
нвадратноскуловая; sq — чешуйчатая; so — над-
глазничные; st— надвисочная; ta — tabulare; гос,
jc, soc — подглазничный, яремный и надглазнич-
ный каналы органов боковой линии.
ними костями, из которых вну-
тренние крыловидные у некоторых примитивных форм разрастались
в виде широких пластинок, соединявшихся впереди посредством шва
Рис. 390. Нёбо кистеперого гапоида Eusthenopteron (Л) и стегоцефала
Loxomma (В), (по Ватсону).
ос — basioccipitale; ecpt и tr — ectopterygoideum (transversum); mx — maxilla; pal — palali-
num; pmx — praemaxilla; pt — pterygoideum; » — сошник. •
(рис. 390). Такие широкие энтоптеригоиды были иногда и у кистеперых
рыб (рис. 390), а для двудышащих как раз характерно образование
439
соединения — симфиза этих костей в передней их части. У других, более
поздних стегоцефалов твердое нёбо значительно редуцировано, птериго-
иды получают трехветвистую форму; передние ветви сокращаются и
широко раздвигаются к краям нёба, оставляя между собой большое нёб-
ное окно. У некоторых примитивных стегоцефалов (Eogyrinus, Palaeo-
gyrinus, Loxomma) птеригоиды позади причленялись к базиптеригоидным
отросткам основной клиновидной кости (рис. 390), как у кистеперых,
с одной стороны, и у рептилий — с другой. У вышестоящих форм это
сочленение заменялось соединением посредством шва с задней расширен-
ной частью парасфеноида. Жаберной крышки у стегоцефалов, конечно
не было; интересно, однако, отмстить, что, по крайней мере, у одной из
Рис; 391. Скелет примитивного стегоцефала Eogyrinus (Л) (каменный уголь)
и более специализированного Eryops (й) (пермь) (по Грегори).
Ichtyostegidae найдена несомненная небольшая предкрышечка (ргаео-
perculum), в которой проходит и соответствующий канал боковой линии
(гиомандибулярный канал) (рис. 389). Жаберные дуги имеются и у личи-
нок современных амфибий. Они имелись и у личинок стегоцефалов. У
высших их представителей они были приблизительно такой же формы,
как у современных амфибий — дорзальные их концы широко расходились
и, очевидно, поддерживали наружные жабры. Однако у некоторых более
примитивных палеозойских стегоцефалов (Archegosaurus, Branchiosaurus)
жаберные дуги сближались между собой со спинной стороны и следова-
тельно несколько более напоминали жаберные дуги рыб.
Весьма интересны конечности и, особенно, пояса конечностей стего-
цефалов. У некоторых древнейших стегоцефалов в отличие от современных
амфибий не только в задних, но и в передних конечностях было 5 пальцев
[по Уатсону, у Diplovertebron формула фаланг 2, 3, 3, 3, 4; у Trematops
(рис. 190) число фаланг, повидимому, было больше]. Длинные кости
конечностей были еще относительно очень короткими (рис. 391, 191),
На плечевом поясе, кроме ключиц, имеющихся и у других наземных
позвоночных, были также cleithra — характерные покровные кости спин-
ного отдела пояса рыб. У современных бесхвостых амфибий также еще
имеются остатки cleithra в виде небольших пластинчатых костей, лежащих
на надлопатках (рис. 178). Однако иногда у древнейших стегоцефалов
(Eogyrinus) (рис, 391), кроме того, имелись еще и другие покровные
кости supracleithralia и posttemporalia (рис. 392), которые, как и у
440
рыб, прикреплялись к крыше черепа (к концам tabularia). Покровные
кости плечевого пояса были вообще очень широки, а первичный пояс
относительно невелик, что напоминало обычные у рыб соотношения
(рис. 392). У таких животных шеи совсем не было и голова не имела само-
стоятельной подвижности (как и у рыб). Единственный здесь затылочный
бугор был глубоко вогнут, т. е. сустава не было, как и у рыб. Ключицы
лежали у стегоцефалов обычно в коже и имели вид широких пластинок
с характерной скульптурой. Между ними на груди лежал подобный же
по скульптуре надгрудинник (episternum s. interclavicula). Весьма инте-
ресен также тазовый пояс некоторых примитивнейших стегоцефалов
(Embolomeri) (рис. 391). Подвздошные кости не имели причленения к
крестцовым ребрам, но у них был длинный задний отросток и они налегали
всей своей поверхностью на
крестцовые ребра сверху (по-
добно тому как лопатка ле-
жит на грудных ребрах) и
укреплялись только при по-
мощи мышц. У примитивных
стегоцефалов окостеневали не
только седалищные и под-
вздошные кости, но и лонные.
Позвоночник был у различ-
ных стегоцефалов весьма раз-
Рис. 392. Плечевой пояс кистеперой рыбы Eust-
henopteron (Л), примитивного стегоцефала Eogy-
rinus (В) и более специализированного стегоце-
фала Cacops (С) (по Ватсону).
Clav—ключица; Clei—cleithrum; Р. Тет. — posttem.
porale; Sc — первичный пояс; ё. cl —supraclei-
thrale.
лично развит (рис. 82,83, 84).
У наиболее примитивных
форм были полупозвонки
(Embolomeri), у других—ра-
хитомные позвонки (Temno-
spondyli), состоящие из от-
дельных элементов тела (hy-
pocentra, pleurocentra и основания верхних дуг). Эти же оба типа по-
звонков встречаются и у различных костных ганоидов. У других стего-
цефалов вставочные элементы (pleurocentra) редуцировались и образо-
вались цельные позвонки путем срастания вокруг хорды верхних дуг
с гипоцентрами (hypocentra). Этот же тип позвонков перешел и к совре-
менным амфибиям.
У древнейших стегоцефалов ребра были довольно длинные, а у позд-
нейших они значительно укорачивались (как у современных амфибий).
В некоторых отношениях особый интерес представляют и зубы.
Как у кистеперых рыб Rhizodontidae, и у примитивных стегоцефалов
в основании зуба имелись радиальные складки дентина, которые ребрами
вдавались внутрь полости пульпы. У более специализированных «лаби-
ринтодонтов» эта складчатость развивалась очень сильно и придавала
зубу сложное строение (как и у кистеперых Holoptychiidae). У дру-
гих, более мелких стегоцефалов, например, у Branchiosauria, зубы имели
гладкую коническую форму с большой пульпой. Эта последняя форма
зуба перешла и к современным амфибиям.
Стегоцефалы жили частью в воде, частью на суше. Многие прими-
тивные формы были, действительно, земноводными в полном смысле
этого слова, а многие позднейшие формы нередко полностью возвра-
щались назад к жизни в воде. Примитивные Embolomeri (Eogyrinus и др.),
о которых мы говорили, были настоящими водными животными (длинное
тело с плавательным хвостовым отделом, очень слабые конечности), однако
это были все же амфибии, а не рыбы, и нужно считать, что они уже вторично
вернулись в воду. Вполне сухопутными животными они, однако, несом-
ненно, никогда не были, так как иначе у них не могли бы сохраниться
каналы и, следовательно, органы боковой линии. То же самое, но еще
441
в большей мере относится к Ichthyostegidae, у которых каналы боковой
линии сохранили вполне рыбий характер (рис. 389). Однако некоторые
и притом многие примитивные стегоцефалы вели, несомненно, вполне
наземную жизнь и обладали сравнительно хорошо развитыми конечно-
стями (например, Diplovertebron). Наоборот, высшие лабиринтодонты
(из триаса), частью очень крупные, все вторично вернулись к жизни
в воде.
Стегоцефалы развивались в различных направлениях и дали начало
довольно разнообразным формам. Наиболее примитивные из них имели
в общем ящерообразную форму тела. Более крупные рахитомные стего-
цефалы были иногда по форме тела и головы похожи на крокодилов (Arche-
gosaurus) или обладали укороченным широким телом, напоминая огромную
лягушку (Cacops). Иногда череп стегоцефалов расширялся в виде больших
боковых выростов (Diplocaulus). У многих даже и крупных водных ла-
биринтодонтов конечности были чрезвычайно малы. Некоторые стего-
цефалы обладали длинным телом с рудиментарными ножками (Microbra-
chis) или были совсем змееобразны (Dolichosoma). Высшими из них были
пермские и триасовые лабиринтодонты, имевшие в общем довольно мас-
сивный скелет.
Уже среди древнейших стегоцефалов появляются также и более
легкие небольшие саламандрообразные формы, наиболее известным пред-
ставителем которых является Branchiosaurus. Укороченные ребра, отсут-
ствие лонных окостенений, хрящевое запястье и предплюсна, тонкие
тела позвонков, окружавшие постоянную хорду, крупная широкая го-
лова, уменьшенное число костей черепа, большие глазницы и большие
окна в твердом нёбе — все это указывает на значительную редукцию
костного скелета в этой группе и вместе с тем на несомненное приближение
к хвостатым амфибиям. Последние представляют собой довольно древнюю
группу. Уже для среднего карбона известна неотеничная форма Cocytinus,
которую теперь относят к древнейшим Urodela. Крыша черепа близкой
к ней пермской амфибии Lysorophus содержит то же самое небольшое
число костей, какое имеется и у современных амфибий. Эта форма также
считается представителем хвостатых амфибий (Urodela).
Древнейшие остатки бесхвостых амфибий известны (Protobatrachus
из нижнего триаса Мадагаскара) только начиная с мезозоя. Эта группа
представляет результат приспособления к особому способу передвиже-
ния прыжками, что было связано с весьма значительным удлинением и
усилением конечностей.
Гимнофионы имеют, быть может, происхождение, независимое от
стегоцефалов (Aistopodidae?).
Происхождение рептилий
Стегоцефалы вымерли. В воде они не могли выдержать конкуренции
с рыбами, а на суше их вытеснили рептилии. Впрочем, стегоцефалы не
просто все вымерли, так как некоторые из них сохранили связь главным
образом с пересыхающими водоемами такого типа, которые были мало
пригодны для рыб, и преобразовались в современных амфибий, а другие
вполне порвали с водной средой и дали начало высшим наземным позвоноч-
ным, в первую очередь рептилиям. Стегоцефалы, даже и вполне сухопут-
ные, были связаны с водой своим размножением — они откладывали яйца
в воду и здесь они проходили личиночные стадии своего развития. Полное
освобождение от воды стало возможным с приобретением более прочного
рогового покрова, предохранявшего кожу от высыхания, и в особенности
с развитием более плотных оболочек яиц, под загцитой которых зародыш
за счет увеличенных запасов желтка мог развиваться достаточно длитель-
ное время вне воды. Запас желтка должен был быть столь значительным,
442
чтобы развивающееся за его счет молодое животное, только что вылупив-
шись, могло самостоятельно добывать себе пищу. Таким образом, от
стегоцефалов произошли путем дивергентного развития две новейшие
ветви: с одной стороны, — современные амфибии, в результате приспо-
собления к довольно узким условиям существования, т. е. в результате
специализации, при сохранении многих примитивных черт организации,
но также и при явных регрессивных изменениях, по крайней мере, в ске-
лете; с другой стороны — рептилии, завоевавшие затем различные области
суши, вернувшиеся также в воду и везде побеждавшие исходные формы.
Кроме более совершенного приспособления к жизни на суше (роговой
покров кожи и крупные яйца с прочными оболочками), рептилии отли-
чаются и рядом признаков более высокой организации по сравнению с
амфибиями. Так, у них имеет место более совершенное разделение арте-
риального и венозного токов крови в сердце. Значительно выше развиты
легкие. Более совершенна выделительная система (тазовая почка), которая
обособилась от половой. Значительно выше развиты головной мозг и
органы чувств. Наконец (что нетрудно проследить и на ископаемом мате-
риале), прогрессивно
развился костный ске-
лет вообще и, в част-
ности, наступило значи-
тельное усиление конеч-
ностей и укрепление их
поясов. Ясно обособи-
лась шея, развилось
подвижное соединение
головы с позвоночником
(атлас и эпистрофей), и
все животное стало го-
раздо более поворотли-
вым и подвижным. Раз-
вилась система зароды-
шевых оболочек. Вот
наиболее характерные
изменения, поднявшие
Рис. 393. Костное нёбо котилозавра (Seymouria (Л)
и эмболомерного стегоцефала (Baphetes (В) (по
Ватсону).
рептилий на более вы-
сокую ступень органи-
зации по сравнению с
Ьос — basioccipitale; ch — первичные хоаны; ecpt и tr—ecto-
pterygoideum (tranasversum); rnx—maxilla; pal — palatinum;
pmx — praemaxilla; pt — pterygoideum; q — quadratum;
sph — basisphenoideum; tr — transversum; v — vomer.
амфибиями.
Мы теперь не можем сказать точно, когда наступили именно эти
изменения. Мы знаем только скелеты и можем отметить, что морфоло-
гические изменения скелета, сопровождавшие переход от стегоцефалов
к рептилиям, были постепенными, и мы не можем провести резкой грани
между теми и другими. Не подлежит лишь сомнению, что предки рептилий,
действительно, находятся среди типичных стегоцефалов и именно среди их
более примитивных представителей. С одной стороны, у некоторых стего-
цефалов были уже ясно выражены признаки, характерные собственно
для рептилий. Так, например, у древнейших стегоцефалов череп цмел
относительно узкое основание, и передний мозг располагался над меж-
глазничной перегородкой. Парасфеноид в этом случае был иногда так же
слабо развит, как и у рептилий (Loxomma). Хрящевой череп окостеневал
полнее, в частности, имелись все затылочные кости (supra-, basioccipitale,
exoccipitalia), включая основную, которые вместе с боковыми образовали
один (по составу тройной) затылочный бугор (Loxomma, Trimerorachis
и др.), как и у низших рептилий (котилозавры). У некоторых примитив-
нейших стегоцефалов (Ichtyostegidae, Loxomma) птеригоиды впереди
сходились между собой по средней линии, так что у них не было характер-
44?
ного для всех амфибий большого нёбного окна, и дно черепа было впе-
реди почти совершенно прикрыто первичным твердым нёбом, как у репти-
лий (рис. 390, 393). Имелась и основная клиновидная кость с базиптериго-
идными отростками, к которым причлонялпсь птеригоиды (Eogyrinus,
Loxomma и др.). Окостеневали эпиптеригоиды и квадратные кости
(epipterygoidea, quadrata). Имелось большое число костей нижней челюсти.
Все эти признаки у позднейших стегоцефалов (и у современных амфибий)
исчезли, но они весьма характерны для рептилий. У некоторых примитив-
ных стегоцефалов были и типичные амфицельные позвонки (M'Crosauria).
С другой стороны, древнейшие рептилии — котилозавры (Со-
tylosauria) — обладали многими чертами организации, характерными
для стегоцефалов (рис. 393, 394). Позвонки были амфицельные, как и у
некоторых современных форм (гекконы, хамелеоны), был только один
крестцовый позвонок, как у амфибий,
Рис. 394. Скелет примитивного
котилозавра Seymouria (пермь)
(по Уиллистону).
череп сочленялся с позвоночником посред-
ство и одного затылочного бугра, сидящего
на основной и боковых затылочных кос-
тях, как и у современных черепах. Наи-
более, однако, интересно, что у котило-
завров сплошная наружная крыша черепа
была того же самого состава и с такой же
скульптурой, указывающей на положение
в коже, как и у стегоцефалов. Особенно
интересны черепа пермских Seymouria
(рис. 152 и 393) и Kotlassia, которые во-
обще не отличаются от черепа стегоцефа-
ла. Имелись и теменное отверстие, и ха-
рактерные для стегоцефалов ушные вы-
резки между tabularia и squamosa (где
помещалась барабанная перепонка). Пояса
конечностей также сохранили еще то же
строение, как и у стегоцефалов. На груд-
ном поясе, кроме ключиц, были cleithra
(на лопатке), а подвздошные кости таза
причленялись к боковым отросткам одного
только крестцового позвонка (как у ам-
фибий). Конечности были примитивны,
однако развиты выше, чем у стегоцефалов,
и число фаланг такое же, как у боль-
шинства рептилий (2, 3. 4, 5, 3). Позво-
ночник такого же строения как у типичных рептилий. Котилозавры были
вполне сухопутными животными, и каналов боковой линии у них не было.
Парасфеноид уменшился в размерах, и впервые начинала обособляться
ясная шея. На зубах иногда видны еще следы радиальной складчатости
дентина. Таким образом, мы имеем в некоторых древнейших стегоцефалах
и в примитивнейших котилозаврах формы, теснейшим образом связанные
между собой. Здесь почти стираются грани между амфибиями и репти-
лиями, и, в конце концов, можно сказать, что древнейшие рептилии отли-
чались от стегоцефалов только тем, что они стали уже вполне наземными
животными. Первые их представители — котилозавры — быстро рас-
селились в новой биологической обстановке (где условия для их размно-
жения первоначально были весьма благоприятны) и затем при возрастав-
шей между ними конкуренции развивались дивергонтно, образовав огром-
ное количество разнообразнейших форм. Некоторые из них рано специ-
ализировались и быстро были вытеснены затем более прогрессивными
формами, другие развивались медленнее и более гармонично и дали
начало новым более совершенным организациям. Уже некоторые котич
444
лозавры приобрели характер своеобразных, крайне уклоняющихся форм.
Таковы, например, парейазавры — большие, неуклюжие животные с
крайне массивным скелетом, поднимавшие свое тело на коротких и тол-
стых ногах.
Дальнейшая эволюция рептилий шла по пути выработки более по-
движных форм. Конечности увеличивались в размерах (рис. 193), крестец
состоял не меньше, чем из двух позвонков; вообще же скелет становился
более легким, не теряя при этом своей прочности. В первую очередь это
касается черепа, крыша которого испытывает частичную редукцию вслед-
ствие образования височных ям (замена тяжелого сплошного перекрытия
системой балок). Образование височных ям шло в различных филогене-
тических ветвях по-разному.
Синапсиды. У некоторых рептилий образовалось одно окно в крыше
черепа — одна боковая височная яма, ограниченная снизу одной только
нижней височной дугой. Это — группа Synap-
sida. У других сразу развились два окна
в крыше черепа — верхняя височная яма и
боковая височная яма, отделенные друг от друга
верхней височной дугой; снизу крыша черепа
замыкалась нижней височной дугой. Это —
группа Diapsida, имевшая две скуловые дуги.
Наиболее примитивные представители первой
группы образуют ряд Procolophonia (триас).
Они похожи еще на котилозавров, по имели
височную яму, которая впереди сливалась
с глазницей. Наиболее известный представитель
Sclerosaurus имел еще остатки кожного пан-
цыря. Также очень древние, частью своеоб-
разно специализированные формы, имевшие
одну отдельную височную яму, образуют отряд
Pelycosauria. Известным представителем этих
рептилий является Dimetrodon (пермь) — хищ-
ник с большими зубами п чрезвычайно длин-
ными остистыми отростками позвонков, кото-
рые, очевидно, поддерживали своеобразный
гребень на спине животного.
Черепахи. Дожившими до наших
дней представителями синапсид являются че-
репахи, которые имеют также весьма древнее
происхождение. Первая совершенно типичная
черепаха, еще обладавшая рудиментами зубов,
известна уже в триасе (Triassochelys). Однако,
кроме того, известна еще более древняя перм-
ская рептилия, имеющая явно переходный ха-
рактер, — Eunnotosaurus (рис. 395). Это была
с зубами, с короткими и широкими ребрами, которые лежали прямо под
кожей спины и образовывали нечто вроде спинного щита. Брюшного щита
еще не было. Черепахи представляют собой по происхождению наземных
и притом, вероятно, роющих рептилий, которые вторично вступали иногда
в более тесную связь с водой (наши болотные черепахи) и иногда перехо-
дили к вполне водной жизни (пресноводные Trionychoidea, морские Che-
lonidae и Spliargidae). В последнем случае панцырь редуцировался.
Ихтиозавры. Во многих ветвях рептилий наблюдается вто-
ричное возвращение в водную среду. Так, в группе Synapsida, кроме мор-
ских черепах, развились некоторые типично морские животные. Удлинен-
ные ящерообразные Mesosauria (известный представитель Stereosternum)
были уже морскими животными с длинным плавательным хвостом. От
Рис. 395. Ископаемая при-
митивная черепаха Eunno-
tosaurus (пермь) (пОВатсону).
ящерицеобразная форма
445
них произошли дельфинообразные ихтиозавры (Ichthyosauria) с верете-
новидным телом, спинным плавником, ластами и высоким гипоцеркаль-
ным хвостовым плавником. Это были прекрасно плавающие живородящие
морские животные.
Плезиозавры. Несколько иначе по форме пошло приспособ-
ление к морской жизни в другой ветви — у плезиозавров (Sauropterygia),
Рис. 396. Скелет примитивного плезио-
вавра Lariosaurus (триас) (из Циттеля).
происшедших, вероятно, также от
мезозавров. У ихтиозавров локомо-
торным органом служил хвост (как
и у рыб), а ласты только регулирова-
ли направление движения. Между
тем у плезиозавров эволюция пошла
по пути прогрессивного развития лас-
тов, служивших гребными органами.
И здесь исходные формы были ящеро-
образными (рис. 396), но более позд-
Рис. 397. Скелет специализированного
Plesiosaurus (нижняя юра) (ив Циттеля).
ние типичные плезиозавры имели короткое бочонкообразное тело с колос-
сальными ластами и длинной шеей (рис. 397). У плезиозавров были (как
остаток кожного панцыря) очень хорошо развитые брюшные ребра. Как
ихтиозавры, так и плезиозавры жили в течение мезозоя, в конце которого
бесследно вымерли.
Наконец, и еще одна вымершая ветвь, связанная теснейшим родством
с плезиозаврами, также приспособилась к жизни в воде, вероятно, в при-
брежной зоне (в области прибоя). Это черепахообразные Placodontia,
имевшие на неое и на нижней челюсти позади широкие дробящие зубы
446
(очевидно, они питались моллюсками), большие ласты, а также спинной
и брюшной панцирь из многих кожных окостенений. Эти своеобразные
животные были чрезвычайно похожи на современных морских черепах
(Sphargys), и в этом сходстве мы имеем яркий пример схождения
признаков в эволюции, т. е. так
(рис. 434).
Териодонты. Наконец,
группы Synapsida развилась одна
называемого конвергентного развития
остается еще отметить, что из той же
крупная прогрессивная ветвь вполне
сухопутных животных,
которые приподнимали
свое тело над землей и
переходили от обычного
для рептилий ползания
к хождению и бегу на
высоких ногах(рис.412).
Начало этого процесса
можно усмотреть и у
котилозавров, из кото-
рых парейазавры ходи-
ли, приподнимая свое
тело над землей. Но это
были крайне неуклю-
жие, тяжелые формы.
Другая ветвь более по-
движных животных, по-
видимому, ближе свя-
занная с пеликозаврами
(рис. 398), пошла явно
по пути общего подъема
Рис. 398. Череп пеликовавра (схема) (по Гудричу),
в — articulare; ап — angulare; d — dentale; /г — frontale;
J — jugale; I — lacrymale; mx — maxillare; n — ноздря;
па — nasale; о — глазница; pa — parictale; pm — praema-
xillare; po—postorbitalc; pof — postfrontale; ppa — postpa-
rletale; prf — praefrontale; q — quadratum; qj — quadrato-
jugale; sa— supraangulare; sp — spleniale;sg—squamosum;
st — supratemporale; t — tabulare.
своей организации на
более высокую ступень (териодонты и млекопитающие). Общим призна-
ком для «зверезубых» (Theriodontia) было также усиление челюстей
и диференцировка зубной системы. Более примитивные их представители
известны уже в пермском периоде. Средняя часть тройного затылочного
бугра у них редуцировалась и, таким образом, развивался парный заты-
лочный бугор, сидящий главным образом на боковых затылочных костях,
как у млекопитающих.
В связи с усилением конечностей происходили изменения в поясах,
В тазовом поясе подвздошные, седалищные и лонные кости неподвижно
соединялись, а иногда и срастались между собой в одну кость. Крестец
увеличивался путем присоединения дальнейших позвонков (всего до 4
крестцовых позвонков). В плечевом поясе прогрессивно развивалась ло-
патка, которая приняла характерную треугольную форму. На ней раз-
вился акромиальный отросток для прикрепления мышц и с ней прочно
были связаны коракоиды (рис. 182). Вторичное твердое нёбо развилось
более значительно (особенно у позднейших форм), чем у других рептилий
этой группы, заметно отодвигая хоаны назад. Развился также длинный
наружный слуховой проход. Верхнечелюстные кости становились более
высокими и прочно вклинивались своими восходящими частями между
другими костями крыши черепа (рис. 399, 400). На нижней челюсти, и
именно на зубной кости, развился очень большой венечный отросток,
служащий для прикрепления мощных жевательных мышц. Во всех этих
признаках имеется очень большое сходство с млекопитающими; еще более
бросается в глаза это сходство, если обратить внимание на интересную
диференцировку зубной системы (рис. 400, 414). В углах рта развивались
большие клыкообразные зубы, а далее по бокам челюстей коронка зубов
получала у более специализированных териодонтов сложную трехзуб-
447
чатую (у хищных) или даже плоскую (у травоядных) форму, напоминающую
коренные зубы млекопитающих.
От териодонтов очень рано обособилась сильно специализированная
ветвь аномодонтов, которая кончается оригинальными формами (Dicyno-
dontia) с редуцирующейся зубной системой, от которой оставалась только
пара бивней вверху. Некоторые из них возвращались к жизни в воде и
по виду напоминали современных сирен (Lystrosaurus).
Диапсиды. В другой группе рептилий (Diapsida), как было сказано,
образовались сразу две височные ямы и две скуловые дуги. Наиболее
примитивными их представителями являются мало еще изученные Рго-
torosauria (пермь) и Rhynchocephalia (с триаса), к которым относится и
современная Hatteria. В этой группе имеется, кроме нескольких незна-
Рис. 399. Череп териодонта (схема) (по Гудричу).
ip — interparietale (postparietale); sm—septomaxillare; pf— отверстие для теменного
органа. Остальные обозначения см. рис. 398.
чительных, также одна большая ветвь, которая от мало еще специализи-
рованных псевдозухий привела при явно прогрессивной эволюции к боль-
шому разнообразию различных высших форм (крокодилы, птерозавры,
динозавры, птицы).
Псевдо зу хи и и крокодилы. Указанная ветвь начи-
нается с ископаемых триасовых псевдозухий (Pseudosuchia, ложные кро-
кодилы), вполне сухопутных рептилий с хорошо развитыми конечностями.
Среди них были и длинноногие формы, лазающие и скачущие на деревьях
(Scleromochlus) (рис. 402). Характерно было строение черепа, в котором,
кроме обеих височных ям, имелась также еще большая предглазничная яма
(рис. 401). Чрезвычайно близкие к псевдозухиям не менее древние формы
(также триас) приспособились к жизни в воде и в высшей степени напо-
минали крокодилов. Это параллельный настоящим крокодилам отряд
паразухий (известный представитель Phytosaurus s. Belodon). На-
стоящие крокодилы (Crocodilia) связаны с ними близким родством
и развивались немного позднее — начиная с юры. Характерной особен-
ностью их черепа является соединение обоих носовых отверстий в одно.
У крокодилов сильно развивалось твердое нёбо, в образовании которого
принимали участие не только предчелюстные и челюстные кости, но так-
же и нёбные, а у высших форм и птеригоиды. В результате этого хоаны
постепенно отодвигаются все далее назад. Унаследованное от псевдозухий
предглазничное окно в течение эволюции крокодилов постепенно исче-
зает.
Птерозавры. От сухопутных псевдозухий произошли не только
водные рептилии, но и летающие. Летающие ящеры (Ptcrosauria) разви-
лись сразу в виде двух параллельных ветвей, которые летали по-разному.
448
Рамфоринхи [Rhamphorhynchus (юра)] имели очейь длинный хвост, иг-
равший роль руля, и узкие длинные крылья, на которых животное пла-
нировало. У птеродактилей [Pterodactylus (юра), Pteranodon (мел)] хвост
редуцировался, и животное двигалось, ударяя широкими крыльями о
воздух. Крыло было образовано кожной складкой, отходившей от боков
тела; передний край ее поддерживался чрезвычайно длинным четвертым
пальцем передней конечности. Летательный орган был, следовательно,
Рис. 400. Череп [териодонта Lycosuchus7 (нижний
триас) (по Иекелю). Обовначения прежние.
Рис. 401. Череп псевдозухий (схема) (по Гудричу).
if — нижняя височная яма; а/ — верхняя височная яма;
pl — нредглавничная яма. Остальные обовначения см.
рис. 398.
Рис. 402. Реставрация псевдо-
зухии Scleromochlus (пр Абе-
лю).
построен совершенно иначе, чем у птиц. Наряду с этим небезынтересно
отметить целый ряд параллельных изменений в разультате независимого
друг от друга приспособления этих форм к полету. В обеих ветвях разви-
валась прочная грудная клетка, причем грудные позвонки срастались
между собой. Развивалась большая грудина с килем для прикрепления
грудных мышц. Имелся сложный крестец. Кости были с воздушными по-
лостями, как у птиц. Развивалась сидевшая на длинной шее почти под
прямым углом округлая черепная коробка с предглазничной ямой и длин-
ный клюв, у высших форм беззубый. В отличие от птиц, лопатка причле-
нялась на широком пространстве к сросшимся между собой остистым
отросткам грудных позвонков; ключиц не было.
Динозавры. Повидимому, птерозавры произошли от псевдо-
зухий, лазавших по деревьям. По всей вероятности, от таких же лаза-
ющих форм через вторичный возврат на землю возникла и большая группа
динозавров. Дело в том, что все древнейшие динозавры ходили на двух
ногах, а у некоторых из них (древнейшие Saurischia) была хватательная
передняя конечность с сильно развитым вторым пальцем, а в задней конеч-
ности первый палец противопоставлялся всем остальным (как у птиц).
449
В связи с хождением на Двух ногах сильно развивался таз, широкие под-
вздошные кости которого на большом протяжении соединялись с крест-
Рис. 403. Скелет ящеротазого диновавра Compsognathus (по Гейльману).
цом, состоявшим из большого числа позвонков (до 10). Динозавры разви-
вались в виде двух параллельных ветвей, которые с самого начала ясно
Рис. 404. Передняя конеч-
ность динованра Ornitholestes
(Л) и первоптицы Archaeornis
(В) (по Гейльману).
друг от друга отличались. Одна ветвь пред-
ставляет собой в основе хищников, имевших
нормальное для рептилий строение таза
с длинными направленными вперед лонными
костями и направленными назад седалищны-
ми костями, которые соединялись в брюш-
ном симфизе [ящеротазые (Saurischia)]
Рис. 405. Задняя конечность диновавра Compsogna-
thus (1), первоптиц Archaeopteryx (2) и Archaeornis
(3) и голубя (4).
(рис. 403). Другая ветвь представляет травоядных животных с сильно
измененными челюстями, причем в нижней челюсти впереди в симфизе
развивалась непарная кость — praedentale. У этих животных таз
450
йМел совершенно своеобразное строение (рис. 186). От направленных
вперед лонных костей, которые своими брюшными концами не соединя-
лись, а, наоборот, широко открывали вход в таз (приспособление к не-
сению крупных яиц), отходил назад параллельно удлиненным седалищ-
ным костям длинный заднелонный отросток (proc, postpubis),, что при-
давало этому тазу внешнее сходство с птичьим тазом [птицетазые (Ornith-
ischia)].
Ящеротазые. В основании этой группы стоят небольшие бе-
Рис. 406. Зад-
няя конечность
динозавра Ог-
nithomimus (по
Ломбе).
гающие и скачущие на задних ногах животные с очень легким скелетом —
Coelurosauria (рис. 403). Кости у них были с воздушными полостями.
Задние ноги с длинной плюсной (рис. 405, 406), с противопоставленным
первым пальцем и рудиментарным пятым напоминали птичьи (однако
первый палец был значительно редуцирован). Небольшая
хватательная передняя конечность с тремя пальцами
также напоминала конечность первоптицы (рис. 404).
Легкий череп — с округлой мозговой коробкой. Наибо-
лее известные представители — Compsognathus и Ornitho-
lestes (юра). Отсюда развитие этой ветви динозавров
ведет к более крупным формам (Pachypodosauria) и далее
к гигантским двуногим хищникам [Theropoda, предста-
витель — Tyrannosaurus (мел)]. Из тех же более крупных
форм, через вторичный возврат к хождению на четырех
ногах, развились самые большие травоядные динозав-
ры — Sauropoda, отдельные представители которых до-
стигали гигантских размеров: бронтозавр — около 18 м
длиной, диплодок — более 22 м. Но были и еще более
крупные формы, поскольку описан отдельный шейный
позвонок такого динозавра в 111 см длиной и 72 см высо-
той. Это самые крупные позвоночные, жившие когда-либо
на суше (из водных позвоночных киты достигают иногда
еще больших размеров). Это прогрессивное увеличение
размеров тела мы можем рассматривать так же как осо-
бого рода специализацию, которая сопровождалась умень-
шением пластичности и привела к вымиранию при изме-
нившейся биологической обстановке (прогрессивное раз-
витие млекопитающих).
Птицетазые. Исходные формы и здесь ходили
на двух ногах. Некоторые их представители достигали
также очень больших размеров, например, игуанодоны [Iguanodon (мел)},
достигавшие 10 м в длину и 5 м в высоту. Высшие представители этой
ветви также вторично вернулись к хождению на четыгех ногах. Таковы
стегозавры [Stegosaurus (юра)], у которых спина была защищена двумя
рядами огромных треугольных костяных пластинок, и оригинальнейшие
цератопсиды [Triceratops (мел)], напоминавшие огромных носорогов.
Все эти растительноядные формы обнаруживают крайнюю степень спе-
циализации.
Динозавры, как и птерозавры, все бесследно вымерли в конце мезо-
зоя, не оставив после себя никаких иных форм.
Ветвь диапсид, начинающуюся псевдозухиями, мы назвали прогрес-
сивной ветвью, так как она дала начало многим весьма разнообразным
высшим формам, которые решительно господствовали в течение всего
мезозоя. Это широкое распространение и широкая радиация форм, не-
сомненно, указывают на какие-то преимущества, имевшиеся у них в борь-
бе за существование. В настоящее время нам трудно точно охарактери-
зовать условия жизни этих животных, однако мы можем отметить, что
такими преимуществами должны были быть некоторые весьма важные
изменения внутренней организации. Судя по современным крокодилам
451
(и их сравнению с птицами), мы можем предполагать, что уже у йсевдо-
зухий развилось четырехкамерное сердце, а в дальнейшем произошло
полное разделение венозной и артериальной крови и в самой аорте (через
редукцию левой дуги). Несомненно, это имело очень большое значение.
Но, быть может, еще большее значение имело прогрессивное развитие
легких. Уже у крокодилов имеются весьма совершенные легкие, а у не-
которых ящериц легкие продолжаются в систему воздушных мешков
(хамелеон). Пневматизация костей у птерозавров и у динозавров (притом
уже у низших) указывает на сильное развитие системы воздушных меш-
ков, проникавших, как и у птиц, внутрь костей. Это указывает и на су-
ществование более совершенного дыхания птичьего типа. Кроме того
пневматизация костей приводила к большей легкости, а следовательно,
и подвижности животного. Интенсивное дыхание позволило повысить
интенсивность обмена, активность животного, и позволило развивать
гораздо больше мускульной энергии. Все это обеспечило этим формам
длительное процветание. Однако головной мозг у них был относительно
очень слабо развит, и это не могло не иметь значения при возникновении
конкуренции с не менее подвижными и активными млекопитающими,
имевшими перед ними также еще и преимущества живорождения. Все
же основная ветвь диапсид не вся вымерла — сохранились и доныне в
специальной (амфибиотической) биологической обстановке крокодилы
и, кроме того, как мы увидим дальше, возникла еще одна более высокая
ветвь с хорошо развитым мозгом, завоевавшая новую воздушную среду,
из которой она вытеснила птерозавров. Это — птицы, находящиеся в
настоящее время в периоде полного процветания.
Парапсиды. Осталось сказать несколько слов еще об одной ветви
рептилий, которая занимает особое положение. Обычно их относят к той
же группе диапсид. Однако у древнейших их представителей была только
верхняя височная яма. Единственная верхняя скуловая дуга этих реп-
тилий образовалась, повидимому, не через редукцию нижней дуги, а
путем образования вырезки в нижнем крае стенок черепа. Такие формы
называют парапсидными. Эта ветвь вела все время скромное существо-
вание, питаясь главным образом насекомыми и другими мелкими живот-
ными, и приспосабливалась к тем или иным частным условиям среды,
не повышая заметно общего уровня своей организации. Это — чешуй-
чатые гады (Squamata), которые мало известны в мезозое, но имеют, не-
сомненно, весьма древнее происхождение. Древнейшим их представи-
телем является Araeoscelis, пермская ящерицеобразная форма с длинными
ногами. Главной морфологической особенностью чешуйчатых гадов явля-
ется устройство их черепа, в котором квадратная кость и прилегающие
к ней впереди птеригоиды становились все более подвижными (развитие
стрептостилии). От мезозойских сухопутных ящериц неизвестно почти
никаких остатков. Однако они, несомненно, существовали и обособляв-
шиеся от них отдельные ветви неоднократно возвращались к жизни в воде.
Известно довольно большое число частью очень крупных морских ящериц
с длинным телом и плавательными конечностями (ластами). Триасовые
талаттозавры (Thalattosauria), длиной до 2 м, меловые долихозавры (Doli-
chosauria) и меловые же мозазавры (Mosasauria=Pythononiorpha), до-
стигавшие иногда огромных размеров (до 12 м длиной), представляют
собой хотя и родственные, но в разное время независимо и параллельно
развивавшиеся формы. Исходными для них были ящерицы, близкие к
современным варанам. Более удлиненные формы ящериц развивались
и на суше. Удлинение тела связывалось с измененным способом передви-
жения — путем волнообразных боковых изгибов тела, при которых ко-
нечности теряли свое значение. Таким образом развились, опять-таки
в результате независимой и параллельной эволюции, различные безногие
змееобразные ящерицы. Подобным же образом раньше, еще в начале ме-
452
лового периода, развились и настоящие змеи. Подвижность и гибкость
тела обеспечивались у них развитием сложных сочленений между позвон-
ками (зигосфены и зигантры). Подвижность квадратной кости (стрепто-
стилия) и челюстей достигла максимального развития и позволяет змеям
чрезвычайно широко растягивать рот при заглатывании крупных жи-
вотных.
Происхождение птиц
Птицы обнаруживают в своей организации много общих черт с реп-
тилиями. В коже как и у рептилий, нет желез (кроме некоторых особо спе-
циализированных, как, например, копчиковая железа), но зато чрезвы-
чайно развиты роговые образования — перья, которые, как мы знаем,
представляют собой преобразованные чешуи. Не только история разви-
тия, но и сравнительноанатомические данные указывают на это достаточно
определенно. На ногах у страусов видны переходы от чешуи к перу и точно
так же перья у пингвинов при редукции вновь приобретают чешуеобраз-
ную форму. Череп птиц имеет один затылочный бугор, как у рептилий.
Парасфеноид срастается с основной клиновидной костью и образует вы-
дающийся вперед клюв (rostrum). Имеются и латеросфеноиды и орбито-
сфеноиды, как у крокодилов. Имеется типичная нижняя скуловая дуга
и, кроме височных ям, имеются также предглазничные. Квадратная кость
подвижная (стрептостилия), причем имеется механизм передачи ее дви-
жений через птеригоиды на надклювье. В позвоночнике характерны длин-
ный подвижной шейный отдел с атласом и эпистрофеем, сложный крестец,
в котором, однако, выделяются- (своими ребрами) два истинных крест-
цовых позвонка. На ребрах имеются крючковидные отростки (proc, unci-
nati), как у крокодилов. В задних конечностях функционирует мезотар-
зальное сочленение, как у рептилий. Сердце четырехкамерное, как у
крокодилов. Единственная, правая дуга аорты (которая и у рептилий
развита сильнее левой) выходит из левого желудочка, как у рептилий.
Сходно также строение тазовой почки и многое другое. Таким образом,
птицы по своему строению, несомненно, родственны рептилиям. Однако
вместе с тем они стоят значительно выше рептилий. Гораздо выше развит
головной мозг, особенно полушария большого мозга. Выше развиты ор-
ганы зрения и слуха (зачаток улитки, как и у крокодилов). Кроме того,
птицы гораздо более активны и в этой своей активности очень мало зависят
от температуры внешней среды. Рептилии способны к быстрым движениям
только при достаточно высокой температуре. У птиц покров из перьев
защищает тело от потери тепла, продуцируемого организмом. Таким об-
разом у них стало возможным установление постоянной и притом довольно
высокой температуры тела (выше, чем у млекопитающих). Эта высокая
температура связана с весьма интенсивным обменом, который обусловлен,
между прочим, и весьма совершенным дыханием. Легкие птиц, благодаря
своему устройству и связи с воздушными мешками, вентилируются лучше,
чем у каких бы то ни было иных позвоночных. К этому нужно прибавить,
что у птиц впервые наступает и полное разделение артериальной и веноз-
ной крови не только в сердце, но и в артериях. Немалое значение в эво-
люции птиц имело, также увеличение размеров яиц, связанное с измене-
нием формы таза. Благодаря большим яйцам вылупившийся детеныш
имеет уже сравнительно крупные размеры, а заботы птиц о своем потом-
стве обеспечивают и относительно большое их выживание.
Таким образом, происхождение птиц от рептилий было связано,
очевидно, с общим подъемом их организации на более высокую ступень.
Кроме того, нельзя не отметить, что на всей их организации отразилось
приспособление к жизни в специальной среде и к особому способу пере-
движения — летанию в воздухе. Эти изменения мы можем обозначить
кдк приспособительные (адаптивные). Общее облегчение веса птиц и, в
453
особенности, их скелета в значительной мере связано с развитием системы
воздушных мешков, проникающих также внутрь костей и «пневматизи-
рующих» их. Кости черепа очень тонки, но зато рано срастаются между
собой, образуя тонкую и вместе с тем прочную черепную коробку. Тяже-
лые зубы редуцировались и заменены легким роговым клювом. Передние
конечности преобразовались в крылья, причем пальцы в значительной мере
редуцировались, а пястные кости срослись между собой. Прогрессивное
развитие летательной грудной мускулатуры связано с увеличением разме-
ров грудины и развитием на ней гребня (к которому прикрепляется груд-
ная мышца), а также с более прочным укреплением плечевого пояса и
грудной клетки. Ключицы срослись между собой, лопатки ложатся вдоль
спины. Ребра окостеневают не только в спинном, но и в грудном отделе
и укрепляются помощью крючковидных отростков. Кроме того, грудные
позвонки неподвижно между собой срастаются. С приспособлением к
полету связана и редукция хвостового отдела позвоночника, замененного
коротким копчиком, укрепляющим рулевые перья хвоста. Так как у
птиц для хождения служат только задние конечности, несущие всю тя-
жесть тела, то и они развиваются прогрессивно. Ноги удлиняются и
укрепляются благодаря срастанию берцовых костей между собой и сра-
станию плюсневых костей, образующих длинную цевку. Проксимальные
предплюсневые кости срастаются с берцовой, а дистальные — с плюсной.
Вместе с тем развивается и тазовый пояс. Подвздошные кости таза раз-
растаются в ширину и укрепляются путем срастания на большом протя-
жении с позвоночником — развивается большой сложный крестец, в
состав которого входит значительное число как поясничных, так и хвосто-
вых позвонков.
Таким образом, в эволюции птиц одинаково сказался и общий подъем
организации на более высокую ступень, и весьма совершенное приспо-
собление к специальным условиям их существования. В этих условиях
они оказались наиболее совершенными животными, которые быстро
вытеснили летающих ящеров и до сих пор не имеют еще серьезных конку-
рентов.
Подходя к конкретизации вопроса о происхождении птиц, мы прежде
всего можем отметить, что из ныне живущих рептилий к ним наиболее
близки крокодилы (строение сердца, строение уха, присутствие нижней
скуловой дуги, боковые клиновидные кости черепа, крючковидные отрост-
ки ребер и многое другое). Если же принять во внимание ископаемые
формы, то по весьма многим признакам к птицам гораздо ближе дино-
завры (рис. 403). Все исходные формы динозавров бегали на двух ногах
как и птицы; кроме того обычно у них были легкие, пневматизированные
кости. В черепе, кроме верхней, была и нижняя скуловая дуга, была
также предглазничная яма, и даже общая форма черепа нередко напоми-
нала форму птичьего черепа (особенно череп Coelurosauria). У динозавров
была довольно длинная шея, как у птиц, и особенно сходно построенные
конечности. В передних конечностях первые три пальца развиты сильнее
других, а иногда четвертый и пятый пальцы совсем редуцируются (Coelu-
rosauria), как и у птиц (рис. 404). В задних конечностях плюсневые кости
удлиняются и прочно связываются (а иногда и срастаются, как, например,
у Ceratosaurus) между собой, а также с редуцированными дистальными
элементами предплюсны (рис. 405). Проксимальные элементы пред-
плюсны прочно связываются с берцовой костью; при этом на астрагале
(astragalus) развивается отросток, восходящий вверх вдоль поверхности
берцовой кости (рис. 406, t). Интересно отметить, что у молодого стра-
уса, а также у гоацина развивается подобный же восходящий отросток
как предварительная фаза укрепления проксимального отдела пред-
плюсны перед его полным прирастанием к берцовой кости. В .тазовом
поясе также широко разрослись подвздошные кости и увеличивается
454
состав кресца. Кроме того, у «птицетазых» динозавров и таз по форме
был похож на птичий, поскольку лонные кости одной своей частью
(postpubis) были обращены назад и вытянуты вдоль седалищных костей.
Передний отдел лонных костей (praepubis) не имел лонного сращения.
Однако, наряду с большим сходством, нельзя не отметить и некоторых
различий. Так, например, у динозавров не было ключиц. Если по строению
таза к птицам как-будто приближались Ornithischia, то по строению
конечностей и черепа к ним, несомненно, ближе стояли Saurischia (и
именно Coelurosauria, особенно, например, Ornitholestes, Compsognathus).
Однако и у последних редукция первого пальца задней конечности обыч-
но шла даже дальше, чем у птиц, что связано с их приспособлением к
быстрому бегу. У некоторых хорошо бегающих форм первый палец и
вовсе исчезал (так же как и у некоторых бегающих птиц). G другой сто-
роны, передние конечности древнейших целурозавров были довольно
Рис. 407. Скелет псевдозухий Ornithosuchus (по Гейльману).
S—четыре кожные чешуи.
слабы. Все это вместе взятое заставляет думать, что предками птиц были
хотя и очень близкие к Coelurosauria, но все же несколько более прими-
тивные формы, чем известные нам динозавры вообще. Кроме того, имеет
значение еще и соображение иного рода. Наиболее примитивные дино-
завры бегали на двух ногах, поэтому переход к летанию можно было бы
себе представить как приспособление ко все более далеким планирующим
прыжкам с разбега. Это предполагает, однако, очень быстрое бегание.
Но при этом первый палец задней конечности никак не мог сохраниться
в противопоставленном положении, а должен был бы совсем редуциро-
ваться. G другой стороны, гораздо легче могло пойти приспособление
к летанию через выработку планирующих прыжков с ветвей деревьев,
как это происходит у многих других позвоночных [из амфибий совре-
менная «летающая» лягушка (Rhacophorus), из ящериц дракон (Draco
volans), из млекопитающих сумчатый летун (Petaurus), шерстокрыл
(Galeopithecus), летающая белка (Pteromys), начало этого процесса у
некоторых обезьян]. В пользу именно такого происхождения полета
птиц говорит факт существования противопоставленного первого пальца,
особенно же строение ископаемой первоптицы (Archaeornis), которая,
несомненно, еще и не летала как следует, а только планировала. С этой
точки зрения нельзя не обратить внимания на формы, исходные и для
самих динозавров, именно на псевдозухий (рис. 407), среди которых были,
повидимому, формы, лазавшие по деревьям [и скакавшие на ветках
(рис. 402) подобно современному долгопяту Tarsius],
У псевдозухий череп был не менее похож на птичий, чем у Coeluro-
sauria (рис. 408). Кроме височных ям, имелась и предглазничная. Имелась
длинная гибкая шея. В плечевом поясе были и ключицы. В тазовом поясе
были широкие подвздошные кости; седалищные кости были направлены
прямо назад, а лонные оригинальным образом изгибались вниз (рис. 407),
т. е. занимали такое положение, как ранние хрящевые закладки лонного
отдела пояса у птиц. Задняя конечность также напоминала птичью и
455
первый палец был иногда хорошо развит. Так же как у динозавров и как
у археорниса, у них имелись брюшные ребра. Интересно еще отметить
своеобразную форму чешуй. У Euparkeria чешуи были вытянуты в длину
спереди назад и вдоль оси чешуи проходило продольное ребро, от кото-
рого отходили в стороны и назад перисто
Рис. 408. Сравнение черепов псевдозухий Aeto-
saurus (-4)„ и Euparkeria {В} (триас), первоптицы
Archaeornis (С) (юра) и голубя (D) (по Гейль-
ману).
расположенные боковые ребра
(рис. 407), напоминая до из-
вестной степени структуру
пера.
Вряд ли теперь можно
сомневаться, что птицы про-
изошли от лазающих репти-
лий, связанных близким род-
ством с псевдо зухиями. С
этими же формами находи-
лись в близком родстве и
динозавры, особенно Coeluro-
sauria, однако эти последние
формы представляют боко-
вую ветвь, вернувшуюся к
беганию по земле (подобно
тому как и от летающих птиц
вторично через приспособле-
ние к беганию на земле про-
изошли бескилевые птицы).
Особенно ясно выявляется
родство этих форм, если при
сравнении исходить не от со-
временных птиц, а от иско-
паемого археорниса. В этом
случае создается впечатле-
ние, что последний стоит
ближе к псевдо зухиям, чем
к современным птицам. Древ-
нейшие птицы известны
только по остаткам двух
экземпляров, найденных в
юрских сланцах. В настоя-
щее время их относят к раз-
ным родам. Первоначальное
название археоптерикса (Ar-
chaeopteryx) сохранено за
экземпляром, хранящимся в
Лондоне. Другой экземпляр,
хранящийся в Берлине, те-
перь относят к другому роду
и называют археорнис (Ar-
chaeornis). Этот последний
находится в гораздо лучшем состоянии и изучен полнее. Сходство его
скелета со скелетом некоторых Coelurosauria так велико, что их, наверное,
отнесли бы к одной группе, если бы не самые настоящие птичьи перья,
которые покрывали его тело и отпечатки которых прекрасно сохранились.
Именно присутствие покрова из перьев, имевших на крыльях характерное
расположение маховых перьев, и заставляет смотреть на это животное как
на птицу. В строении скелета имеется, конечно, большое сходство с совре-
менными птицами (рис. 409), но все же в некоторых отношениях он ближе
к скелету псевдозухий и динозавров. Общая конфигурация черепа, по-
жалуй, ближе к последним (рис. 408). Задние конечности вполне птичьи.
456
Таз птичий; седалищные кости позади расходились, однако направленные
назад лонные кости соединялись между собой в лонном сращении. В отли-
чие от птиц грудные позвонки между собой не срастались, имелся длинный
хвост из большого числа позвонков (18—20), грудина была слаба (неви-
димому, не окостеневала) и не имела киля. Все три пальца передней
конечности имели полное число фаланг (2, 3, 4) и были снабжены когтями;
кости пястья (metacarpus) между собой не срастались. Имелись брюшные
ребра (как у псевдозухий и у динозавров). Из этого сопоставления уже
ясно, что эта птица была по сути ближе к рептилиям — у нее отсутство-
вало многое из того, что характерно для птицы, как летающего организма.
Рис. 409. Сравнение скелет первоптицы (Archaeornis) и голубя (по Гейльману).
с — carpajia; cl — ключица (вилочка); со — каракоид; d — пальцы передней конечности;
/ — Седро; fi — fibula; h ~ плечо; i — ileum; is — ischium; me — nietacarpalia; mi — nieta-
tarsalia; py — копчик; p — pubis; r — radius; s — лопатка; st — грудина; tl—tibia; tm — tarsonie-
tatarsus; It — tiblotarsus; u — ulna; v — брюшные ребра; I—IV—пальцы вадней конечности.
Кости у археорниса не были пневматизированы. Грудные мышцы не
могли быть сильно развиты, так как на грудине не было киля. Свободные
кости пястья (metacarpalia) не могли давать достаточной опоры маховым
перьям. На костях и не видно следов их прикрепления, как это имеется
на пястье современных птиц. Длинный хвост также мог служить только
для планирования (хорошо летающие птицы имеют короткий хвост). Судя
по всем этим данным, археорнис не мог летать по-настоящему. Присут-
ствие когтей на пальцах крыла указывает на их роль во время лазания.
Это была лазающая и, может быть, скачущая на деревьях птица, которая
совершала только длительные планирующие прыжки с ветки на ветку
457
или с дерева на дерево. 1аким образом, это — настоящая переходная
форма, которая соединяет в себе как чисто птичьи признаки, так и при-
Рис. 410. Контуры черепа и головной
мозг меловой птицы (Ichthyornis) и
современной крачки (Sterna)*, с — по-
лушария большого мозга; ol — обо-
нятельные доли.
знаки, характерные для исходных реп-
тилий. Насколько велики изменения,
приведшие к настоящим птицам, видно
по сопоставлению скелетов на рис. 409.
Настоящие птицы почти современ-
ного типа известны, начиная с меловых
отложений. Они отличаются от совре-
менных собственно только еще присут-
ствием зубов. Однако неверно было бы
предполагать, что дальнейшая эволюция
птиц состояла исключительно в адап-
тивной радиации, т. е. в приспособле-
нии к различным местным (климати-
ческим и экологическим) условиям.
Имеются некоторые указания и на про-
должавшееся еще общее повышение
уровня организации. Так, например,
несомненно, происходило дальнейшее
прогрессивное развитие головного моз-
га, как это видно из сопоставления
их объема у меловой и у современной
птицы (рис. 410).
В своей эволюции птицы сразу раз-
бились на большое число отдельных
ветвей, но в общем морфологические
различия этих ветвей не столь значи-
тельны, чтобы здесь нужно было на
этом останавливаться.
Происхождение млекопитающих
Млекопитающие представляют собой значительно менее специали-
зированных животных, чем птицы, однако их связь с рептилиями не так
легко бросается в глаза. По богатству кожных желез они напоминают
скорее амфибий. Происхождение волосяного покрова млекопитающих
неясно, и, в частности, было высказано предположение, что волосы пред-
ставляют собой результат преобразования органов боковой линии амфибий.
Череп имеет широкое основание (как и у амфибий), между тем как у
рептилий черепа тропибазальные (т. е. с узким основанием). Для млеко-
питающих характерно присутствие парного затылочного бугра (как у
амфибий), в то время как у рептилий имеется один непарный бугор. У ти-
пичных млекопитающих, как и у амфибий, нет надгрудинника и, наконец,
число фаланг пальцев у млекопитающих невелико (2, 3, 3, 3, 3), напоминая
их число у амфибий. У млекопитающих имеется только одна левая дуга
аорты, но она выходит из левого желудочка (у рептилий дуги аорты пере-
крещиваются и левая дуга выходит из правого желудочка). Среднее ухо
и слуховые косточки млекопитающих построены совсем иначе, чем у
рептилий, и трудно себе представить, как они могли развиться из соот-
ветствующих частей уже высоко диференцированного уха рептилий. У мле-
копитающих функционирует голеностопный сустав (между берцовыми
костями и предплюсной), между тем как у рептилий сустав занимает
межтарсальное положение, и- т. д. Между современными рептилиями
и млекопитающими, действительно, целая пропасть. Немного ближе
стоят, быть может, черепахи — у них имеется единственная скуловая
дуга, по своему составу напоминающая дугу черепа млекопитающих,
455
формула фаланг пальцев та же, что у млекопитающих, а затылочный
бугор если и не парный, то, по крайней мере, тройной, в образовании
которого одинаково принимают участие и основная, и боковые затылоч-
ные кости. Однако серьезно сближать млекопитающих с черепахами
нельзя, хотя бы, например, потому, что черепахи беззубы, между тем
как прогрессивное развитие зубной системы представляет одну из наиболее
характерных черт в эволюции млекопитающих. Неудивительно поэтому,
что в поисках источников происхождения млекопитающих морфологам
в свое время пришлось обратиться даже к амфибиям (Гексли). В настоящее
Рис. 411. Сравнение положения ног у примитивной рептилии (Seymouria, Л)
и примитивного млекопитающего (Didelphys, В) (по Грегори).
время мы, однако, обладаем достаточными палеонтологическими данными,
чтобы не заниматься предположениями, а подойти к вопросу о происхож-
дении млекопитающих во всеоружии большого материала по изучению
скелета ископаемых форм.
Отметим, однако, прежде всего те основные вехи, по которым шло
прогрессивное развитие этой группы.
У млекопитающих, как и у птиц, достигнуто значительное увеличение
активности, связанное с прогрессивным развитием органов дыхания и
кровообращения (при полном разделении артериального и венозного
токов крови в сердце и аорте) .и с прогрессивным развитием конечностей
(рис. 411). Эта активность и здесь мало зависит от температуры среды,
так как у млекопитающих развился изолирующий покров из роговых
образований кожи — волос, защищающий тело от потери тепла. Таким
образом у млекопитающих установилась постоянная температура тела.
459
В данном случае мы имеем ту же интенсификацию всех жизненных про-
цессов, как и у птиц. Притом и здесь прогрессивно развивались органы
чувств — орган зрения (быть может, и не так сильно), орган слуха, с его
улиткой и кортиевым органом, а также орган обоняния, со сложнейшей
системой обонятельных раковин, которые достигли здесь высшей степени
совершенства. Кроме того, мы имеем еще и значительную рационализацию
питания, благодаря которой сделались доступными очень многие новые
пищевые материалы, особенно растительные, которые при этом весьма
хорошо используются. Это связано с прогрессивной диференцировкой
зубной системы, при которой стало возможным весьма совершенное
измельчение даже очень твердой пищи. Диференцировка зубной системы
представляет один из наиболее характерных процессов в эволюции мле-
копитающих. Наконец, немалое значение в борьбе млекопитающих за
существование сыграло развитие живорождения и различных форм заботы
о потомстве и прежде всего кормление детенышей молоком. Благодаря
этому животное вступало в жизненную борьбу уже значительно более
окрепшим, и гибель молоди прогрессивно сокращалась. Естественно, что
забота о потомстве предполагает значительное развитие инстинктов, а
может быть даже и интеллекта. Высокое развитие органов чувств, органов
движения и органов дыхания предполагает и весьма совершенную коорди-
нацию. Таким образом, у всех млекопитающих прогрессивно развивался
головной мозг и особенно кора большого мозга, достигая у них максималь-
ной относительной величины и максимальной сложности строения.
Все это вместе взятое означает весьма значительный подъем этих
животных на более высокую ступень организации и обеспечивает возмож-
ность успешной конкуренции с нижестоящими формами во всякой обста-
новке. Победив рептилий на суше, некоторые млекопитающие ушли в воду,
вытеснив рептилий и оттуда. Наконец, млекопитающие делают многие
попытки завоевать воздух и в одной ветви небезуспешно начинают конку-
рировать с птицами.
Указанные процессы можно себе представить в большинстве случаев
как возникающие на базе организации некоторого примитивного, назем
ного позвоночного, стоящего, быть может, на грани между амфибиями
и рептилиями. У современных форм во всяком случае не видно даже и
начала каких-либо изменений, которые бы вели дальше по намеченным
нами путям. Однако иной состав нижней челюсти млекопитающих и
непосредственное ее причленение к черепу являются совершенно непо-
нятными и не могут быть выведены из тех отношений, которые имеются
у рептилий. Точно так же своеобразное строение среднего уха с его слу-
ховыми косточками не может быть выведено из строения среднего уха
рептилий.
Эмбриологические исследования уже давно показали, что слуховые
косточки млекопитающих развиваются не только из верхнего конца
гиоидной дуги, как у рептилий и птиц, а еще и из части челюстной дуги.
Со столбиком рептилий сравнима только одна косточка — стремя, другие
две косточки представляют собой новоприобретение млекопитающих.
Наковальня развивается из задней части верхнего отдела челюстной дуги,
который у других позвоночных дает начало квадратной кости, служащей
у наземных позвоночных подвеском. Молоточек развивается из заднего от-
дела меккелева хряща, который у других позвоночных дает сочленовную
кость нижней челюсти (articulare). Таким образом, и состав нижней
челюсти, и все сочленение нижней челюсти с черепом иное, чем у рептилий.
Должна была произойти замена одного сочленения другим. Представить
себе такую замену чрезвычайно трудно, так как одно время должно было
существовать двойное прикрепление к черепу. Ясная картина эмбриональ-
ного развития с трудом поддавалась филогенетической интерпретации.
Описанною в течение последних десятилетий Брумом (Broom) перм-
ские териодонты из отложений Карру в Южной Африке и примыкающие
460
К ним пермские же териодонты, найденные АмалиЦким на Северной Двине,
проливают, однако, достаточный свет на вопрос о происхождении млеко-
питающих и, в частности, на вопрос о преобразовании их челюстей и о
происхождении слуховых косточек. Эти замечательные животные (рис. 412)
представлены довольно разнообразными формами, начиная от небольших
недавно найденных животных величиной с крысу и кончая довольно круп-
ными зверями. Для всех териодонтов характерны относительно довольно
крупные размеры черепа. Они бегали, держась высоко на ногах, как мле-
Рис. 412. Реставрация скелета териодонта Cynognathus (по Грегори),
с — шейные ребра; сг — 7 шейных позвонков:. th — 17 грудных позвонков;
I — 4 поясничных позвонка; а — 4 крестцовых поввонка.
Рис. 413. Скелет кисти (Л) и стопы (В) прими-
тивного териодонта Ьусаепорв по Бруму. Обозна-
чения обычные.
копитающие, и по целому ряду признаков были к ним весьма близки
(мы имеем в виду териодонтов в тесном смысле этого слова, т. е. тероце-
фалий и цинодонтов, но не уклоняющихся аномодонтов или дицинодонтов,
которых мы здесь касаться не будем).
Длинные ноги териодонтов по составу скелета (особенно запястье
и предплюсна) были вполне сходны с ногами млекопитающих (рис. 413).
Число фаланг сокращено до 2, 3, 3, 3, 3, причем у примитивных терио-
донтов видны пути этого сокращения (рис. 413). Имелся большой локтевой
отросток (olecranon). Пояса
конечностей были также сход-
ны. Кости тазового пояса
прочно связывались между
собой. Коракоиды сокраща-
лись в размерах, а на ло-
патке у высших териодонтов
развивался небольшой гре-
бень и акромиальный отро-
сток. Позвоночник похож на
позвоночник млекопитающих
и в состав крестца входило
более двух позвонков (до
четырех). Череп довольно
большой, гладкий (без скуль-
птуры) (рис. 399, 400, 414).
Заднетеменные кости (postpa-
rietalia и tabularia) стегоцефа-
лов и котилозавров срослись с образованием характерной для млекопитаю-
щих межтеменной кости. Как сказано выше, у низших териодонтов, как и
у котилозавров, был тройной затылочный бугор, который у высших форм
через редукцию средней его части превратился в двойной. Единственная
скуловая дуга (рис. 399) имела тот же состав, как и у млекопитающих.
У них имелся длинный наружный слуховой проход. Челюстные кости
велики и высоко поднимались. Эпиптеригоиды начинали входить в состав
стенки черепа (алисфеноиды млекопитающих) (рис. 414). Хоаны разде-
лялись парными сошниками, которые вклинивались между нёбными
461
костями и Доходили позади до крЫловиДнЫх. Имелись еще небольШиб
поперечные кости (transversa=ectopterygoidea). У позднейших териодон-
тов развивалось вторичное твердое нёбо такого же состава, как и у млеко-
питающих (нёбные отростки межчелюстных, челюстных и нёбных костей).
Все это весьма сближает териодонтов с млекопитающими. Еще интереснее,
однако, челюсти. Зубы сидели в отдельных ячейках, число их было срав-
нительно невелико, но они имели относительно крупные размеры и под-
вергались лишь однократной смене. Зубная система была диференциро-
вана и, по крайней мере, в углах рта развивались более крупные, клы-
кообразные зубы, которых у низших форм бывало до трех пар в каждой
челюсти (рис. 400). У высших форм число зубов сокращалось, и они
Рис. 414. Череп примитивного териодонта Scymnognathus (Л) и более
специализированного Ictidopsis (В) (по Бруму).
alsp — алисфеноид; ап — angulare; art — articulare; dn — dentale; ju — jugale; la — la-
crymale; mx — maxillare; na — nasale; pa — parictale; pmx — praemaxillare; po — post-
orbitale; pof — postfrontale; prf — praefrontale; pro — prooticum; qj — quadratojugale;
su* — septomaxillare; sq—squamosum; san — supraangulare.
диференцировались далее на резцы (4—6 пар), клыки (одна пара) и корен-
ные зубы (около 9—12 пар). Последние сохраняли всегда простой корень
и имели у низших форм простую коническую коронку; однако у высших
териодонтов развиваются трехзубчатые зубы (Cynognathus) (рис. 412, 414)
или даже более сложные формы коронок с одной главной вершиной и
несколькими добавочными (Permocynodon) или, наконец, у вторично
травоядных форм плоские жевательные коронки (Sesamodon). Во всем
этом имеется, как видно, очень большое сходство с млекопитающими.
Очевидно, со специализацией зубной системы связаны и изменения
в строении нижней челюсти. Во всяком случае, в ряду форм, ведущих от
пеликозавров (рис. 398) к териодонтам и аномодонтам, происходило
прогрессивное разрастание зубной кости (dentale) и постепенная редукция
462
Рис. 415. Область причленения нижней
челюсти к черепу у аномодонта Oude-
nodon; вид сзади (по Бруму). В обра-
вовании сустава для нижней челюсти
участвует, кроме квадратной кости (со-
членяющейся с articulare), также ква-
дратноскуловая (сочленяющаяся с supra-
angulare). Стремя своим дистальным
концом причленяется в квадратной
кости.
Ьос — basioccipitale; ехос — exoccipitale;
q — quadratum; qi — quadrato jugale; sq—
squamosum; st — stapes; ty—предполага-
емое, положение барабанной перепонни.
костей задней половины челюсти. Уже у пеликозавров зубная кос-ТЬ,
разрастаясь назад, вытеснила венечную. У териодонтов зубная кость
становится еще крупнее и выше, образуя большой венечный отросток,
высоко поднимающийся под скуловой дугой (рис. 399, 400, 414). Сочленов-
ная кость вместе с лежащей на ней угловой сокращаются в размерах,
становятся тоньше и так же, как и квадратная кость, явно теряют значение
основной опоры челюстей (рис. 162). У некоторых териодонтов они были
такими тонкими и незначительными, что в этом смысле не могли играть
никакой роли (например, у Cynognathus). Кроме того, связь угловых
костей с зубной была не особенно тесной (рис. 414, 162) и, быть может,
здесь имелось подвижное соединение. В таком случае уже у этих животных
челюсть должна была иметь и иное более прочное соединение с черепом.
Сушкин описывает у Permocynodon, кроме внутреннего соединения между
articulare и quadratum, еще и наруж-
ное между supraangulare и quadra-
tojugale. Брум также описывает вто-
рое соединение между нижней че-
люстью и височной костью у Ictido-
sauria. Таким образом, у териодонтов
рядом со старым и кнаружи от него
зарождалось уже характерное для
млекопитающих новое сочленение ме-
жду зубной костью (с которой, быть
может, срослась и надугловая кость)
и височной (с которой, вероятно,
срослась и квадратноскуловая). Кро-
ме того, у некоторых териодонтов
имелись и все характерные связи и
прочие предпосылки для образования
слуховых косточек млекопитающих.
У териодонтов столбик имел отвер-
стие для артерии (как и стремя млеко-
питающих) и упирался своим дисталь-
ным концом только в квадратную
кость (рис. 415); с последней была по-
движно связана и сочленовная. Лежащая на последней угловая кость была
тонкая и имела снизу вырезку, в которой, вероятно, располагалась ба-
рабанная перепонка (рис. 162). Столбик уха у териодонтов во всяком
случае не имел непосредственной связи с барабанной перепонкой (то же
самое было у пеликозавров и даже у некоторых котилозавров, у которых
столбик упирался только в квадратную кость). Мы знаем, что барабанная
полость и особенно барабанная перепонка млекопитающих не вполне го-
мологичны таковой современных рептилий и птиц — они развиваются
значительно ниже. У териодонтов барабанная перепонка располагалась
под задним углом нижней челюсти, т. е. как раз там, где она развивается
и у млекопитающих. Во всем этом мы не можем не видеть указаний на
ближайшее родство этих форм. Конечно, это не значит, что мы такую
форму, как Cynognathus, которая в общем очень похожа на млекопитаю-
щих, рассматриваем как прямого их предка.
У териодонтов имеются некоторые признаки специализации, указы-
вающие на известное уклонение от прямого пути, ведущего к млекопитаю-
щим. Так, например, нижние клыки входили в отверстия в верхней че-
люсти. Коренные зубы всегда имели простой корень и т. п. Повидимому,
и в строении мозга были большие отличия. Кроме того, большинство
териодонтов было животными крупного размера, между тем как первые
млекопитающие были очень небольшими. Однако нельзя и недооцени-
вать значения того сходства, о котором мы здесь говорили. Необратимость
463
эволюции нельзя понимать в том смысле, что признаки специализаций
не могут исчезнуть, крупные формы не могут вторично измельчать и т. п.
Несомненно, прямые предки млекопитающих находились в близком
родстве с териодонтами, и весьма вероятно, что их следует искать среди
более Примитивных представителей териодонтов. Мелкие формы менее
известны лишь потому, что условия для их сохранения в ископаемом
состоянии неизмеримо менее благоприятны, а более примитивные формы
обычно бывают более мелкими. Все же и в этом отношении палеонтология
идет вперед. В 1927 г. Сушкин описал небольшого териодонта Permocy-
nodon, о котором мы здесь уже говорили, длина черепа которого была
всего только около 9 см и который, несомненно, более других предста-
вителей приближается к млекопитающим. Наконец, в 1929 г. Брум дал
предварительное описание совсем маленьких териодонтов Ictidosauria,
величиной с крысу (Karromys), из триасовых отложений Южной Африки,
которых он прямо характеризует как недостававшее звено между репти-
лиями и млекопитающими (рис. 414, В).
Эволюция млекопитающих
Настоящие млекопитающие известны начиная с триаса, однако в
течение мезозоя они большой роли в природе не играли. От большинства
мелких, несомненно, примитивных форм сохранились только разрознен-
ные зубы и обломки челюстей, причем относительно древнейших форм
обычно неизвестно даже, были ли это уже млекопитающие или еще близкие
к териодонтам рептилии. Все же известны и состоящие из одной зубной
кости целые нижние челюсти, принадлежащие, несомненно, зарождаю-
щимся млекопитающим (рис. 416), у которых коренные зубы имели одну
Рис. 416. Нижние челюсти юрских млекопитающих (по Осборну).
I, Prototherla, Allotheria: Plagiaulax (73), Bolodon (76), II. MarsupIaBa. Trlconodontla: Am-
philestesU). Triconodon (4), Phascolotherium (3), Spalacotherium (7), Peralestes (3). III.
Placentalia, Pantotheria: Amphitherium (2), Peramus (6). Leptocladus (10). Amblotherium
(11). Achyrodon (13), Perarspalax (9). Phascolostes (12). Stylodon (14), Kurtodon (16).
главную вершину и небольшие добавочные впереди и позади, а корень
только начинает подразделяться. Таковы Dromatherium и Microconodon из
верхнего триаса Америки и Tribolodon из триаса Южной Африки, несо-
мненно, примитивнейшие представители настоящих млекопитающих,
образующие группу первичнозубых — Protodonta.
Однако одновременно с такими примитивными формами в том же
триасе имелись уже и более крупные и вместе с тем высокоспециализи-
рованные млекопитающие, образующие своеобразную группу Allotheria
(Multituberculate), характеризующуюся большими коренными зубами с
многобугорчатой коронкой. Впереди имеется только одна пара бивне-
464
образных резцов, отделенная большой дийстемой от указанных коренных
зубок. Наиболее известные представители: Tritylodon из триаса Южной
Африки, Plagiaulax из горы п мела и далее Polymastodon из третичного
периода. Далее у третичных представителей той ?ке группы Multituber-
culate резцы редуцируются, а четыре пары многобугорчатых коренных
зубов также находятся на пути к исчезновению, челюсти принимают
плоскую клювообразную форму (Desmostylus, череп которого достигал
90 см в длину). Наконец, у современного утконоса (Ornithorhynchus)
закладывается и пара передних зубов и 4 пары многобугорчатых коренных,
однако они рано выпадают и заменяются роговым покровом челюстей,
образующим своеобразный клюв. Таким образом, ясно восстанавливается
ряд форм, ведущий от древнейших Multituberculate к современным кло-
ачным животным (Prototheria, Monotremata) с единственными
их представителями — утконосом и ехидной.
fes В юре намечается обособление и еще одной группы млекопитающих
(Metatheria, Marsupialia), которая, впрочем, не выделяется столь высокой
специализацией. Остатки юрских млекопитающих вообще очень скудны,
так как примитивные их формы были мелкие, и от них сохранились только
зубы и в лучшем случае нижние челюсти. Остатки зубов мезозойских
млекопитающих позволили восстановить ход их постепенного усложнения,
как это изложено в разобранной в соответствующем месте теории Копа —•
Осборна. Строение и эволюция зубов мезозойских млекопитающих по-
зволяют вывести заключение, что по роду пищи они были насекомоядными,
но это еще очень мало говорит об организации этих животных. Находки
обломков, а иногда и целых нижних челюстей показывают, однако,
начало разделения на две группы, в одной из которых задний угловой
отросток нижней челюсти загибался внутрь. Так как этот загиб угла
нижней челюсти характерен для современных плотоядных сумчатых
(Polyprotodontia), то и эти остатки относят к сумчатым животным.
Наиболее примитивными их представителями являются юрские Tricono-
dontidae с типично трехзубчатыми коренными зубами. К ним примыкают
сумчатые «крысы» (Didelphyidae) как низшие представители современных
полипротодонтов, имеющие трехбугорчатые коренные зубы. В ископаемом
состоянии они известны начиная с мела и попадаются довольно часто
в третичных отложениях Европы. Растительноядные дипротодонты пред-
ставляют собой более молодую боковую ветвь, которая развивалась
только в Австралии и частично перешла в Южную Америку.
Сумчатые представляют собой обособленную ветвь примитивных мле-
копитающих; она не дала начала высшим формам, но выработала своеоб-
разную форму заботы о потомстве (вынашивание детенышей в сумке),
которая давала им в свое время известное преимущество. Мы знаем, что
и плацента у сумчатых утеряна вторично (рудименты ее имеются у Рега-
meles и др.). Соответственно различным местным условиям эта группа
распалась на довольно большое число разнообразно специализированных
форм: известны лазающие (кускус — Phalanger; опоссум — Didelphys),
скачущие (кенгуру — Macropus, сумчатый тушканчик — Antechinomys),
роющие (сумчатый крот — Notoryctes), плавающие (Chironectes), плани-
рующие (сумчатый летун — Petaurus), формы насекомоядные и даже
специально питающиеся муравьями (мурашеед — Myrmecobius), расти-
тельноядные, напоминающие грызунов дипротодонты, частью довольно
крупные и сильные хищники (сумчатый волк — Thylacinus). Здесь мы
имеем пример типичной «адаптивной радиации» форм по термипо тогии
Осборпа.
Наконец третья ветвь — плацентных млекопитающих
(Eutheria s. Placentalia) — развивалась медленнее других и в течение всего
мезозоя представлена мелкими диференцированными формами. Вначале,
в юре, они едва могут быть отграничены от возникающих в это время
465
сумчатых. Формы нижних челюстей, у которых угол не был загнут внутрь,
считаются принадлежащими плацентным млекопитающим, — таковы
мезозойские Pantotheria (Amblotherium и др.), у которых коренные зубы
были трехбугорчатыми (рис. 416). Эти индиферентные формы были по
роду пищи насекомоядными, и не без основания считается, что современ-
ные насекомоядные являются наименее измененными их потомками.
В последнее время в меловых отложениях Монголии (американская
экспедиция Эндрюса 1925 г.) найдены первые более полные черепа неболь-
ших (величиной с крысу) примитивных млекопитающих. Эти черепа
отнесены Грегори к двум весьма близким друг к другу семействам Zalamb-
dalestidae и Deltatheriidae (рис. 417). Из них первые имели по паре уве-
личенных передних резцов, рудиментарные клыки и режущие трехбу-
горчатые коренные зубы и приближаются к наиболее примитивным совре-
Рис. 417. Черепа меловых млекопитаю-
щий из Монголии (по Грегори). При-
митивное насекомоядное Zalambdalestes
(Л) и приближающийся к хищникам
Deltatheridium (В).
менным насекомоядным, каковы ма-
дагаскарский тенрек и южно африкан-
ский золотой крот (группа Zalambdo-
donta), вторые обладали выдающими-
ся клыками и острорежущими трех-
бугорчатыми коренными зубами,
чем явно приближаются к древней-
шим представителям хищных млеко-
питающих (креодонтам). Очевидно,
в меловом периоде только начинается
расхождение более примитивных
мезозойских плацентных млекопи-
тающих на более диференцированные
ветви, ведущие к современным насе-
комоядным и древнейшим хищникам.
Таким образом, история мезозой-
ских млекопитающих ясно показы-
вает несостоятельность широко рас-
пространенного мнения, что клоач-
ные млекопитающие представляют
наиболее примитивную группу, от
которой некогда произошли сумча-
тые, что от сумчатых развились пла-
центные млекопитающие и притом сначала насекомоядные, а от последних
уже и прочие. Это неверно не только по отношению к современным формам,
по и по отношению к ископаемым. Млекопитающие, произошедшие от
примитивных териодонтов, очевидно, еще в конце перми сразу разбились
на две ветви. Одна из них пошла по пути ранней специализации (Multi-
tuberculate), привела к современным клоачным и находится на пути
к вымиранию, а другая в общем, очевидно, прогрессивная ветвь (Proto-
donta и Triconodonta), представленная мелкими формами в триасе,
начинает в юре разбиваться в свою очередь на две ветви. Одна из этих
ветвей пошла -довольно быстро по пути приспособительной эволюции
и дала начало большому числу разнообразных сумчатых (Marsupialia).
Другая ветвь сохраняла «насекомоядный» характер (Pantotheria); однако
это не значит, что мы их должны без оговорки отнести к одной группе
с современными насекомоядными. Это были еще индиферентные формы,
которые в меловом периоде начинали в свою очередь разбиваться, по край-
ней мере, на две ветви (происхождение дальнейших ветвей еще не просле-
жено) собственно насекомоядных (Insectivora), которые сохранили очень
многие черты примитивной организации, и древнейших хищных (Carnivora
creodontia), которые в дальнейшем пошли по пути прогрессивной эво-
люции.
466
Насекомоядные (Insectivora)
Современные насекомоядные представляют хотя и специализирован-
ных в известных отношениях, но все же наиболее примитивных млеко-
питающих. Некоторые из их резцов получают иногда преобладающее раз-
витие (особенно передние резцы верхней челюсти), другие часто редуци-
руются, но в остальном имеется обычно очень примитивная зубная систе-
ма с нормальной формулой • < сзцы часто конической формы,
а коренные зубы имеют иногда примитивное строение зубов юрских
млекопитающих. Так, у тенрека (Centeles), золотого крота (Ghrysochloris),
Potamogale, щелезуба (Selenodon) имеются острые трехбугорчатые корен-
ные зубы, а у близкого к тенреку мадагаскарского же Hemicentetes лож-
нокоренные зубы даже еще трехзубчаты. В большинстве случаев, однако,
коренные зубы насекомоядных имеют все же трехбугорчато-секторпаль-
ное строение (т. е. имеется еще добавочный бугорок на «пятке»). И среди
насекомоядных видна богатая «адаптивная радиация» форм. Известны
формы, лазающие по деревьям (Tupaja), скачущие тушканчике образные
формы (прыгунчик — Macroscelides), роющие—крот (Talpa) и золотой
крот (Ghrysochloris), землеройки (Sorex), плавающие европейские выху-
холи (Myogale) и африканская (Potamogale), оригинальные, благодаря
своему покрову из игл, ежи (Erinaceus) и тенрек (Centetes). Насекомояд-
ные приспосабливались и к передвижению в воздухе. Близкий к пим
шерстокрыл (Galeopithecus) представляет лазающее животное, ко-
торое вторично приспособилось к растительной пище и совершает далекие
планирующие прыжки, благодаря сильному развитию кожной складки,
начинающейся впереди от шеи и простирающейся назад до конца хвоста.
Наконец, от насекомоядных произошли и настоящие летающие живот-
ные — рукокрылые (Chiroptera). Крыло развилось из кожной
складки, отходящей от боков тела и растягиваемой при помощи сильно
развитой передней конечности с длинными пальцами. Рукокрылые рас-
падаются на две группы, одна из которых сохранила насекомоядный
характер (летучие мыши), а другая (летучие собаки) перешла на расти-
тельную пищу—фрукты.
Хищные (Carnivora)
Как мы видели выше, обособление хищных млекопитающих началось
еще в меловом периоде, и у древнейших их представителей зубная система
в общем такая же, как у насекомоядных; несколько яснее только высту-
пают клыки и трехбугорчато-секториальные зубы приобрели режущий
характер. Хищные представляют собой группу млекопитающих с ярко
выраженным прогрессивным развитием, связанным с дальнейшей диферен-
цировкой конечностей (переход от стопохождения к пальцехождению) и,
особенно, с весьма значительным развитием головного мозга. Их специали-
зация выражается главным образом в зубной системе —в преобладающем
развитии одного из коренных зубов, редукции передних ложнокорепных
(образование диастемы) и задних коренных при одновременном укорочении
и вместе с тем значительном увеличении мощности челюстей и их мышц.
С последним связаны величина скуловой дуги и развитие гребней черепа,
служащих для прикрепления височной мышцы. У древнейших хищных
еще не было особого «хищного» зуба (Oxyclaenidae, Mesonychidae).
Затем начинается преобладание одного из задних зубов, образующего
с зубом противоположной стороны режущие ножницы (рис. 418). В сем.
Hyaenodontidae такое особое развитие получает второй коренной зуб
в верхней челюсти и третий коренной в нижней. В сем. Oxyaenidae таким
зубом является первый истиннокоренной в верхней челюсти и второй —
в ишкпей. У современных хищных режущий «хищный» зуб развивается
467
еще далее впереди, и именно таковым становится последним лойШокореи*
ной (Р4) в верхней челюсти и первый истиннокоренной (Мх) в нижней.
Формы, имевшие хищный зуб далее позади (Pseudocreodi) или не имевшие
его вовсе (Acreodi), все вымерли и объединяются под названием креодон-
тов (Creodontia). Хищные современного типа (Eucreodi) распадаются
на две ветви — наземных (Fissi-
Рис. 418. Развитие плотоядного зуба у крео-
донтов (I и II) и у современных хищни-
ков (III)
pedia) и водяных (Pinnipedia)
хищных. Наземные хищные раз-
вились от третичного (эоцен) семей-
ства Miacidae (представитель
Vulpavus) путем расхождения в
двух направлениях. Одна из вет-
вей — Arctoidea — ведет к совре-
менным собакам (Canidae), енотам
(Procyonidae), медведям (Ursidae)
и куницам (Mustelidae). Другая
ветвь — Aeluroidea — ведет к со-
временным виверрам (Viverridae),
гиенам (Hyaenidae) и кошкам
(Felidae). Последние развивались
по двум линиям. С одной стороны,
в семействе Machairodontidae очень
быстрая специализация при редук-
ции нижних клыков и колоссаль-
ном увеличении верхних (особенно
Smilodon) привела так же быстро
и к вымиранию. С другой сторо-
ны, в семействе настоящих кошек
Felinae специализация шла мед-
леннее, достигла не столь крайних
степеней, и они в современной
фауне представляют высших хищ-
ников.
Водяные хищные, или л а с-
т о н о г и е, обособились от неко-
торых Arctoidea и развились сразу
в виде трех параллельных ветвей,
I — Hyaenodon; II — Oxyaena; III — Canls
lupus (волк) (по Абелю). Хищный вуб ва-
штрихован.
ведущих к современным тюленям
(Phocidae), нерпухам (Otariidae)
и моржам (Trichechidae).
И среди хищных ясно видна их адаптивная радиация. Это выражается
в том, что указанные подразделения могут быть охарактеризованы не
только по морфологическим признакам, но и по биологическим. Приспо-
собление к жизни в воде достигло у ластоногих довольно высокой степени,
но намечается в у некоторых других хищных (например, у выдры Lutra,
Latax из куниц). Куницы представляют формы, лазающие по деревьям.
Собаки наиболее приспособлены к быстрому бегу, и у них редуцируются
ключицы. Медведи вторично возвращаются к растительной пище, и их
коренные зубы приобретают тупобугорчатый характер.
Китообразные (Cetacea)
Китообразные, несомненно, произошли от древнейших хищных, т. е.
от креодонтов, в результате их приспособления к жизни в воде. Это послед-
нее шло, однако, иным путем, чем у ластоногих. В то время как у послед-
них хвост редуцировался, а органом движения в воде служат задние
конечности, у китообразных, наоборот, хвост приобрел локомоторную
468
функцию, на нем развился широкий горизонтальный плавник, а задние
конечности редуцировались. У древнейших (эоценовых) китов была еще
нормальная зубная формула и режущие коренные зуоы.
Носовые отверстия помещались па переднем конце черепа (Protocetus)
или перемещались немного назад на середину рыла (Zeuglodon). Передняя
часть черепа вытягивается вперед, и зубы раздвигаются шире, особенно
резцы. У Zeuglodon коренные зубы сплющены с боков и зазубрены. От
этих древнейших китов произошли две ветви: одна из них начинается
такой формой, как Patriocctus (олигоцен), имевшей еще сплющенные
коренные зубы с семью зубцами каждый, и ведет через распад коренных
зубов па небольшие отдельные зубы и через дальнейшую их редукцию
к современным беззубым китам. Другая ветвь начинается также примы-
кающей к древнейшим китам переходной формой Agorophius (миоцен)
через. Squalodon (миоцен) к современным зубастым китам. И в этом слу-
чае шло увеличение числа зубов (у Squalodon формула - г~ ),
которые приобрели простую коническую форму. Ноздри отодвигались
дальше назад и помещаются у современных китов высоко па голове.
Киты являются примером наиболее совершенного приспособления
высшего позвоночного животного к жизни в воде. С этим приспособлением
связаны очень значительные изменения не только в скелете, об одном
котором и можно было судить по ископаемым остаткам, по и во всей их
•организации. Приспособление к дыханию наружным воздухом , не огра-
ничилось только перемещением ноздрей на верхнюю часть головы. Бес-
препятственное дыхание обеспечено также даже во время принятия пищи
своеобразным устройством гортани, которая вытягивается в трубку
(поддерживаемую черпаловидными хрящами и надгортанником), вдви-
гающуюся в хоаны. Волосяной покров терял в воде свое значение тепло-
изолирующего слоя и заменился слоем подкожного жира. Форма тола
приобрела максимальную обтекаемость, как и у хорошо плавающих рыб.
Передние конечности преобразовались в большие ласты, и, кроме того,
наново развивались горизонтальный хвост и вертикальный спинной
плавник.
Грызуны (Rcdentia)
Грызуны представляют собой млекопитающих со своеобразно спе-
диализированной зубной системой. В ней получила преобладающее
развитие лишь одна пара передних резцов, обладающих постоянным
ростом, за которой следует большая диастема. Коренные зубы — тупо-
бугорчатые, складчатые и иногда призматические, представляют ряд
прогрессивного приспособления к растиранию твердой пищи. Грызуны
представляют собой древнюю группу млекопитающих, происхождение
которых не достаточно выяснено, так как ископаемые третичные формы
уже вполне диференцированы. По всей вероятности, они родственны
древнейшим насекомоядным. Это группа и теперь вполне процветающих
форм; она также довольно широко радиирует. Мы знаем формы лазающие
(белки, сопи), планирующие (летяга), плавающие (бобры), напоминающие
ежей (дикобразы), быстро бегающие (зайцы), роющие (слепыш, гоффор),
скачущие (тушканчики) и множество различных мышей.
Неполнозубые (Edentata)
Неполнозубые состоят, по меньшей мере, из двух независимых групп
узко специализированных млекопитающих, у которых зубная система
более или менее редуцируется. Xenarthra охватывают броненосцев, ле-
нивцев и беззубых муравьедов. Pholidota содержат своеобразных беззубых
469
панголинов, И те, и другие представляют остатки древних ветвей млеко-
питающих, которые очень рано достигли крайней специализации и нахо-
дятся на пути к полному вымиранию.
Коиытные (Ungulata)
Копытные представляют собой очень богатую формами группу мле-
копитающих, приспособленную к растительной (редко смешанной) пище,
у которых конечности служат исключительно для передвижения по земле.
Древнейшие копытные были и по черепу с его зубами и по строению
конечностей очень близки к древнейшим хищным (креодонтам и, в част-
ности, к семейству Oxyclaenidae) и представляют, очевидно, ветвь послед-
них, которая переходила на растительное питание. Направление эволю-
ции копытных определяется родом пищи и способом передвижения, т. е.
полностью идет под знаком адаптации к некоторым определенным условиям.
Примитивнейшие копытные были пятипалыми и стопоходящими; конеч-
ности были довольно коротки. Однако они быстро переходили к полустопо-
хождению, при котором метаподии (плюсна и пястье) приподнимаются
над землей и становятся все более круто, так что животное опирается
на нижнюю поверхность фаланг и дистальные концы метаподий, и затем
к пальцехождению, при котором метаподии вовсе не касаются земли,
а животное опирается только на концевые фаланги. Тяжесть тела при
этом падает главным образом на более длинные средние пальцы и средние
метаподии, которые соответственно получают все более мощное развитие,
в то время как боковые теряют свое значение и редуцируются. Рядовое
расположение костей запястья и предплюсны постепенно заменяется
поочередным, при котором вклинивающиеся между собой косточки полу-
чают более прочную взаимную связь. Происходит срастание локтевых и
берцовых костей, а также и костей метаподиумов (пястья и плюсны)
между собой. Благодаря этому конечность высших копытных получает бо-
лее прочную опору.
Древнейшие копытные имели нормальный состав зубной системы
3.1.4. 3
|3. 1.4.3
, однако с удлинением челюстей у них развиваются промежутки
между резцами и клыками и между клыками и коренными зубами. Верхние
резцы иногда вовсе редуцируются; часто редуцируются также верхние
клыки, в то время как нижние принимают характер и положение резцов.
В редких случаях некоторые резцы развиваются прогрессивно и прини-
мают характер бивней. Резцы первоначально были конические, как у
хищных, а затем стали долото видными. Коренные зубы были первона-
чально трехбугорчатыми, как у древнейших креодонтов, но у них рано
развивались добавочные бугорки. В основном исходная форма может
быть обозначена как четырехбугорчатая. Число бугров может возрастать
путем образования добавочных, но во всяком случае зубы имеют у всеядных
форм тупобугорчатую коронку (бунодонтный зуб). У более ясно выра-
женных растительноядных копытных бугры принимают форму луночек
(селенодонтные зубы) или, сливаясь между собой, дают складки (лофо-
донтные зубы). Низкая вначале коронка становится все более высокой,
и у более специализированных копытных имеются высокие призмати-
ческие коронки коренных зубов с весьма длительным ростом. Одновре-
менно со специализацией копытных обычно шло и увеличение общих
размеров тела, в результате которого копытные представляют в общем
группу самых крупных наземных млекопитающих. Таким образом, эво-
люция копытных в основном является приспособительной, однако, не-
сомненно, прогрессивное развитие головного мозга поднимало и эту
группу млекопитающих постепенно на более высокую ступень, как это
видно из сравнения мозга третичных п современных копытных.
470
Древнейшие копытные образуют группу Protungulala, или Condy-
lar thra, заключающую небольших пятипалых полу стопоходящих живот-
ных с черепом, напоминающим череп креодонтов, и с подобными же трех-
бугорчатыми зубами. Наиболее известным их представителем является
Phenacodus из нижнего эоцена Америки (рис. 419). От этих исходных
форм эволюция пошла по очень многим независимым ветвям. Большинство
этих ветвей имело кратковременное существование, — как и всегда,
быстрая специализация приводила к быстрому вымиранию. Таковы,
например, ископаемые Amblypoda, которые дают прекрасные палеонто-
Рис. 419. Скелет примитивного копытного Phenacodus (по Осборну).
логические ряды форм, начинающиеся небольшими примитивными фор-
мами, как Pantplambda, со стопоходящими конечностями и трехбугорча-
тыми зубами (нижние коренные, однако, уже лофодонтны) и ведущие
через тяжеловесных Coryphodon к неуклюжим своеобразно специализи-
рованными формам, как Dinoceras. При крупных размерах тела этих
животных поражает, однако, ничтожная величина их головного мозга.
Мы не можем здесь описать, хотя бы и вкратце, эволюцию всех отдель-
ных ветвей копытных. Остановимся поэтому только на некоторых более
важных линиях. Упомянем, что по новейшим данным современный труб-
козуб, или капская свинья (Orycteropus), относившийся раньше к непол-
нозубым, представляет конечное звено одной из ветвей, начинающейся от
древнейших копытных.
Парнокопытные (Artiodactyla)
Парнокопытные настолько обособлены от прочих копытных, что
не исключается возможность их независимого происхождения от древней-
ших креодонтов. Весьма еще примитивные ископаемые, парнокопытные
образуют семейство Dichobunidae. Общей особенностью парнокопытных
является преобладающее развитие двух средних пальцев (рис. 198),
именно третьего и четвертого. Некоторые из них сохраняют сравнительно
мало измененные четырех- илп трехпалые конечности и тупобугорчатыо
зубы; в современной фауне они представлены свиньями п гиппопотамами.
У других специализация идет дальше и коренные зубы имеют бугорчато-
лунчатое строение. Сюда относятся вымершие третичные Anthracotheriidae
и Anoplotheriidae, имевшие четырех- или трехпалые конечности. Третья,
группа содержит формы с лунчатыми коренными зубами и, кроме неко-
торых ископаемых четырехпалых форм (например, Oreodontidae), заклю-
чает в себе всех наиболее специализированных современных парнокопыт-
ных, каковы верблюды, оленьки, жирафы, олени и полорогие. Все это
двупалые формы с длинными ногами, у которых метаподии обоих пальцев
471
срастаются, резцы верхней челюсти редуцируются, а клыки нижней
челюсти присоединяются к резцам. У жираф развиваются небольшие
костные ронжи на лобных костях, у оленей — более или менее длин-
ные, периодически сбрасываемые костные рога (обычно только у самцов),
а у полорогих постоянные роговые рога, сидящие на костных отростках
лобных костей (обычно у обоих полов). Наиболее богата формами группа
полорогих, которая в настоящее время находится в состоянии полного
расцвета.
Непарнокопытные (Pcrissodactyla)
Непарнокопытные характеризуются тем, что у них преобладающее
развитие получил третий палец (рис. 420). У дрегнейших форм имелась
полная зубная формула и коренные зубы с низкой коронкой. У поздней-
ших развивалась диастема между передними зубами и коренными, которые
приобретали складчатое или лунчато-складчатое строение и высокую
призматическую форму. Из современных форм наименее изменены тапиры,
у которых передняя конечность четырехпала, а задняя трехпала; нижние
коренные с двумя поперечными складками, а верхние имеют, кроме того,
еще наружную продольную связь. У носорогов передние зубы более
или менее редуцированы, а коренные имеют две изогнутые поперечные
Рис. 420. Филогенетический ряд изменений строения передней
конечности лошади (по Осборну).
а—Eohippus (нижний эоцен); Ь—Orohippus (средний эоцен); с—Mesohippus (нижний олигопен)’
d—Miohippus (верхний олигоцен); е — Hypohippus (средний миоцен); /—Hipparion (верхний
миоцен и нижний плиоцен); д — Pliohippus (верхний миоцен и нижний плиоцен).
складки и наружную продольную. Известно большое число ископаемых
носорогов, образующих прекрасные ряды последовательных форм. Не-
которые носороги достигали высокой степени специализации, как, напри-
мер, вымерший Elasmotherium со сложно складчатыми призматическими
коренными зубами. Одна ветвь привела к колоссальным животным,
ходившим на очень высоких ногах. Описанный акад. А. Борисяком индри-
котерий (Indricotherium) был самым крупным наземным млекопитающим,
по своей вышине (более 4 м) значительно превышающим слонов.
К непарнокопытным относятся также большие неуклюжие титано-
терии, у которых на голове развивались странные парные выросты. Пре-
красные ряды этих своеобразных животных описаны Осборном.
Наиболее специализированная ветвь непарнокопытных представлена
современными лошадьми. В основном эта ветвь развилась в Северной
Америке, и ее история прослежена на ископаемом материале с исклю-
чительной полнотой (рис. 420, 421, 422). В основе этой ветви лежат очень
примитивные небольшие формы (величиной с кролика), которые могут
472
быть отнесены к примитивнейшим копытным (Condylarthra). Таков
Hyopsodus, который, очевидно, стоял очень близко у основания лоша-
диного ряда. Последний начинается бесспорно древнейшей небольшой
четырехпалой лошадкой Eohippus в Америке и Hyracotherium в Европе
(из нижнего эоцена) и ведет в Америке через Orohippus к трехпалым
Mesoliippus, Merychip-
pus, к однопалому РИо-
hippus и к современ-
ному роду Equus. Здесь
названы только основ-
ные формы; на самом
деле известно большое
число их, лежащих
частью - в прямом ряду
предков современной
лошади, частью обра-
зующих множество вы-
мерших боковых ответ-
влений. От этого основ-
ного ряда американских
форм, повидимому, не-
однократно происходи-
ло вселение лошадиных
в Европу, где они обыч-
но затем вымирали.
Так, лошадь, близкая
к Mesohippus, предста-
влена в Европе родом
Anchitherium, а форма,
близкая к Merychippus,
сильно размножилась в
Европе, дав начало ши-
роко известной форме
трехпалой лошади —
Hypparion. Во всем
лошадином ряду можно
отметить большое число
•последовательных изме-
Рис. 421. Изменения величины и строения черепа
в ряду лошадей.
А — Eohippus (нижний эоцен); 'В — Mesohippus (средний
олигоцен); С — Protohippus (нижний плиоцен); D — Equus
(соврем.).
нений, в которых ясно
выражается все боль-
шая специализация,
все более совершенное
приспособление к жиз-
ни в степи, к быстрому бегу и к питанию травой. Коренные зубы
из низких бугорчатых становятся складчатыми и призматическими
(рис. 422), конечность становится трехпалой (Mesohippus, Miohippus,
Hypohippus), боковые пальцы укорачиваются (Merychippus) и совсем
редуцируются (рис. 420), так что развивается вполне однопалая форма
(Pliohippus, Equus). Одновременно происходит редукция малой берцовой
кости и ее прирастание к большой, а также редукция нижней части
локтевой кости и прирастание ее верхней части с локтевым отростком
к лучевой кости. Общие размеры тела все более увеличиваются (рис.
421). Со временная , лошадь из Америки вселилась в Европу, а вслед затем
в Америке вымерла.
Интересно отметить, что в то время, когда в Северной Америке раз-
вивалась лошадь, в Южной Америке развивалась параллельная ветвь
копытных, независимо начинающихся также от форм, едва отличимых
473
от Phenacodus. Ветвь эта через редукцию боковых пальцев привела также
к однопалой форме (Thoatherium). Это рано вымершая группа Litopterna
Рис. 422. Преобразования верхних коренных зубов у предков лошадей
(по Осборну).
1 — Hyracotlicrium; 2 — Pachynolophus; 3 — Anehilophus; 4. — Mesohippus;
S — Anchitherium,
Слоны (Proboscidea)
Прекрасно прослеженная палеонтологами история слонов представ-
ляет еще один типичный ряд форм, идущих все далее по пути приспособ-
ления к определенным условиям существования через довольно односто-
роннее изменение своей организации (т. е. идущих по пути специализа-
ции). Наиболее интересны здесь своеобразные изменения зубной системы.
Из резцов получает преобладающее развитие вторая пара, вырастающая
в огромные бивни; остальные резцы и клыки исчезают. Из коренных раз-
виваются молочные ложнокоренные и истиннокоренные. У древнейших
Рис. 423. Череп примитивного хобот- Рис. 424. Череп примитивного мастодонта
ного Moeritherium (верхний эоцен Palaeomastodon (нижний олигоцен Египта)
Египта) (по Эндрью). (по Эндрью).
форм они все имеются одновременно, но позднее размеры коренных зубов
все более увиличиваются, так что все вместе они не поместились бы на
относительно короткой челюсти. Развитие задних зубов все более запаз-
дывает, и у слонов, таким образом, развивается своеобразная горизон-
тальная смена зубов, состоящая в том, что позадилежащие зубы проре-
зываются лишь постепенно по мере изнашивания передних и выдвига-
ются на их место. У современных слонов с каждой стороны обеих челю-
стей нормально имеется лишь один большой коренной зуб и, только если
он уже сильно изношен, за ним появляется следующий, который начи-
нает одновременно функционировать. Последний истиннокоренной зуб
(Л/3) развивается только в глубокой старости.
Исходной формой для слонов является меритерии (Moeritherium) из
эоцена Египта (рис. 423), у которого была почтй полная зубная система.
Однако в нижней челюсти уже редуцировались третий резец, клык и пер-
вый ложнокоренной зуб. Вторая пара резцов выступает в виде небольших
бивней. Коренные зубы четырехбугорчатые. Меритерий был болотным
474
животным. похожим на тапира, высотой около одного метра. У палеома-
стодонта (Palaeomastodon •— олиго цен) впереди остаются только бивни
(рис. 424). Верхние бивни крупнее, однако внизу сама челюсть сильно
вытянута вперед. Коренные зубы увеличиваются и становятся шестибу-
горчатыми. Строение черепа показывает, что у палеомастодонтов был
уже довольно значительный хобот. Отсюда развитие слонов начинает
расходиться в нескольких направлениях. Прежде всего обособилась не-
большая ветвь слонов, у которых развивались только нижние бивни,
изгибавшиеся вниз, а верхние редуцировались —- Dinotherium. У всех
остальных слонов, наоборот, все сильнее развивались верхние бивни,
а нижние редуцировались и нижняя челюсть сокращалась. Эти формы,
в свою очередь, начинали разбиваться на две ветви. В бунодонтных зу-
бах первичных слонов бугры с самого начала располагались парами,
между которыми затем появились промежуточные бугры, и таким образом
коронка оказалась покрытой поперечными рядами бугров (бугорчатыми
гребнями), которые могут более или менее полно сливаться в поперечные
складки (лофы). В одной ветви слонов зубы сохраняют явно бугорчатый
характер. Эта ветвь начинается от Bunolophodon с тремя-четырьмя по-
перечными рядами бугров и ведет к «мастодонтам», которые были в тре-
тичном периоде широко распространены и представлены большим коли-
чеством довольно уже специализированных форм с большим хоботом,
с четырьмя-пятью поперечными рядами бугров на коренных зубах, ко-
торых одновременно бывает не более трех в каждой челюсти с каждой
стороны. Верхние бивни достигали большой длины и были направлены
вперед. Нижние бивни постепенно сокращались и, наконец, совсем ре-
дуцировались (рис. 425). У других мастодонтов более ясно была выражена
складчатость коренных зубов, а бугорчатость постепенно сглаживалась.
Число складок, вначале небольшое (3—4 складки у Mastodon на каждом
зубе), постепенно возрастало (рис. 426); нижние бивни начали исчезать,
верхние расти в длину и изгибаться. У Stegodon было от 6 до 12 попереч-
ных складок, а у настоящих слонов рода Elephas, которые, появившись
в конце третичного периода, имели большое распространение в четвер-
тичном периоде, число складок было еще больше. У современного афри-
канского слона число это сравнительно невелико, достигая на последнем
коренном зубе 10, у ископаемых Elephas meridionalis было до 14 складок,
у Elephas antiquus до 20 складок, а у мамонта (Elephas primigenius),
так же как и у современного индийского слона, до 27 на последнем корен-
ном зубе (передние зубы всегда имеют меньшее число складок). Слоны
представляют прекрасный ряд форм, иллюстрирующий нам постепенный
ход их специализации (рис. 426). Следует еще упомянуть, что одновременно
с последней шло и неуклонное увеличение общих размеров тела, и в со-
временных слонах мы имеем самых крупных наземных млекопитающих.
К слонам, невидимому, довольно близка по своему происхождению
и небольшая ветвь, ведущая к современным даманам (Hyracoidea),
наиболее примитивные формы которых были найдены в том же эоцене
Египта, где найдены и остатки меритерия. Там же были найдены и древ-
нейшие сирены (Eotherium), имевшие еще задние конечности и зуб-
ную систему нормального состава. И череп, и зубы были весьма сходны
с черепом и зубной системой меритерия. Во время дальнейшей эволюции
передние зубы редуцировались, а в коренных зубах, получивших склад-
чатое строение (пара поперечных складок, как у тапира), выработалась
горизонтальная смена зубов, как у слонов, причем одновременно функци-
онировало обычно 5—6 зубов. При этом, однако, общее число зубов было
сильно увеличено (до 30 коренных зубов). Волосяной покров у сирен ре.
дуцировался. Передние конечности преобразовались в ласты, а задние
конечности исчезли (палеонтологические остатки дают прекрасный ряд
форм, иллюстрирующих постепенный ход редукции таза), па хвосте
475
развился горизонтальный плавник, как у китов, к которым раньше сирен
и причисляли. Однако по строению очень многих органов сирены, несом-
ненно, родственны копытным. По строению мозга они близки к слонам.
Сирены представляют собой результат самостоятельного приспособления
Рис. 425. Филогенетический ряд масто-
донтов (по Абелю).
А — Bunolophodon angustidens (средний
и верхний миоцен); В — Bunolophodon
longirostris (нижний плиоцен); С—Buno-
lophodon arvernensis (средний и верхний
плиоцен),
Рис. 426. Эволюция головы и коренных
зубов в ряду хоботных.
1 — Moeritherium (верхний эоцен); 2 — Palaeo-
mastodon (нижний олигоцен); з — Trilopho-
don (миоцен); 4 — Mastodon (дилювий);
5 — Stegodon (плиоцен); 6 — Elephas
(соврем.).
еще одной группы млекопитающих к жизни в воде. Исходные формы для
этой группы были, очевидно, родственны тому же меритерию, который
стоит в основании ряда слонов. Здесь мы имеем также пример довольно
односторонней специализации (питание водорослями), ведущей эту группу
к явному вымиранию.
Приматы (Primates)
Эволюция многих рассмотренных нами групп и в особенности послед-
них (копытные) шла в значительной мере под знаком специализации, т. е.
приспособления к сравнительно узким условиям существования. Что же
касается последнего отряда — приматов, то их эволюция представляет
ряд в общем неглубоких изменений, которые в большинстве не могут быть
охарактеризованы как далеко идущая специализация. Вместе с тем здесь
особенно ярко выражено то, что вообще подняло млекопитающих на весьма
высокую ступень, именно прогрессивное развитие головного мозга; со-
кращение числа рождаемых одновременно детенышей (обычно до одного),
высокие формы заботы о потомстве и, наконец, стадная жизнь с соответ-
ственным развитием инстинктов. Приматы сохранили ряд примитивных
признаков: они пятипалы, стопоходящи, большой палец противопостав-
лен другим, всегда имеется ключица, имеется отдельная центральная
476
йосточка в запястье. Ё связи с всеядным питанием зубная система по спе-
циализирована и коронные зубы обычно четырех-пятибугорчаты. Млечные
железы и соски сокращаются в числе и, как правило, располагаются на
груди. Из признаков, особенно характерных для приматов, можно отме-
тить замену когтей па пальцах ногтями и сокращение числа резцов до
2 13 3
двух пар в каждой челюсти (зубная формула 2 ^’3*3 )• Вследствие соеди-
нения лобных и скуловых костей позади орбит развивается все более пол-
ная костная перегородка, отделяющая глазницу от височной ямы. В ос-
Рис. 427. Скелет лазающего насекомоядного Tupaja (по Грегори).
Рис. 428. Череп примитивной ископаемой полу-
обезьяны Notharctus (средний эоцен) (по Гре-
гори).
тальном довольно ясно выражается приспособленность к жизни на деревь-
ях, хотя некоторые представители приматов вторично переходят к жизни
на земле в открытых местностях. С жизнью па деревьях связано преобла-
дающее развитие органа зрения и редукция органа обоняния. Последняя
приводит к сокращению предглазничной части черепа, к сближению орбит
и их повороту вперед, благодаря чему у высших приматов устанавлива-
ется стереоскопическое зрение. По своему происхождению приматы, не-
сомненно, связаны с древнейшими лазающими насекомоядными [из со-
временных, очевидно, наибо-
лее близка лазающая Tupaja
(рис. 427), у которой орбита
еще отделена цосторбиталь-
пой дугой от височной ямы
(зубная формула *-g--g-)]
Во всяком случае ствол
приматов очень рано расще-
пился на три крупные ветви:
полуобезьян (Lemuroidea),
обезьян широконосых, Ново-
го Света (Platyrhina) и обе-
зьян узконосых, Старого Све-
та (Katarhina). Все три ветви
развивались независимо друг
от друга, и ни в коем случае не приходится говорить о происхождении
высших обезьян от полуобезьян, хотя последние, бесспорно, сохранили
многие признаки низшей организации и, следовательно, стоят ближе
к исходным формам (рис. 428).
У полуобезьян, по крайней мере частично, сохраняются
на пальцах кэгти (по меньшей мере на втором пальце); на первом, а обык-
новенно и на последних пальцах имеются ногти. Нижние резцы сидят
вместе с клыками очень косо в челюсти. Полуобезьяны — сумеречные
или даже ночные лазающие животные, у которых задние конечности, как
правило, длиннее передних.
477
У^ широконосых обезьян иногда имеются еще Когти,
именно у игрунок (однако первый палец всегда с ногтем). Барабанная
кость сохраняет примитивную кольцевидную форму. Хвост длинный,
иногда хватательный. Как современные, так и ископаемые представи-
тели этой ветви ограничены в своем распространении только Южной Аме-
рикой.
У узконосых обезьян на всех пальцах всегда имеются
ногти. Барабанная кость ограничивает длинный наружный слуховой
проход. Хвост короткий, иногда редуцированный. Рождают единовре-
Челобек
ЧелобекообрознЬю I Arctoidea)
(Anlropoidea).. ' Л (Aeluroidea)
Узконосое
обезЬянЬ/
Jybdfnbie
kumbi
I ЬеззубЬю
I / kumbi
i ХтцнЬ/е /
Л (Carnivora) | /
в Ц ПерЬичнЬе kurnbi
\ kpeodoumbi // (Archeoceti)
''X ‘PseudncreodiЙ
ПриматЬ/ \ Il
(Primates) \ I
НасекомояднЬ1е
(Jnsecliuora) '
Рукокрылые
(Chiroptera) —
Ppbi3uHbi
(Roaeatia]'
Сумчатые
,, . (Marsupialia),
ллоа.чн1не
(Moyotremata) —-----------
HenaptfokonbiiTinbie
(Perissodoctyla)
(drfilypoda
Пер/Ыхе шценМе
// (Panlol/teria]
HapHokonbimubie
(Artiodactyla)
СлонЬ/
(Pro6oscidea)
Awwfibi
(Hyracoidea)
СиренЬ/
(5/renia)
RepSwHbtE korbmnbte
/'(ConduLarthra)
Г —ТрубкозуЬл
НелолнозуСЬчз
(Xenarthra) ПангалннЬч
----- (Phot i data)
flepHuwhe млекопитающие
Рис. 429. Родословное древо млекопитающих. Двойной линией обозначены пути раз-
вития прогрессивных ветвей, ведущих к приматам, хищным и копытным.
менно только одного детеныша. Зубная формула еще несколько более
2 12 3
сокращенная, именно ‘ „-'у , причем клыки выдаются более, чем со-
седние зубы, особенно у самцов. Как современные, так и ископаемые пред-
ставители узконосых обезьян ограничены в своем распространении только
Европой, Азией с островами Малайского архипелага и Африкой.
Известно довольно много ископаемых остатков обезьян, однако в
большинстве случаев это лишь обломки челюстей или даже отдельные
зубы. По этим остаткам история приматов или, точнее, их зубов изучена
довольно хорошо, однако не дает достаточно полной картины преобразо-
Л78
вания их организации. Впрочем, как видно уже из краткой характеристики
основных трех ветвей приматов, эти преобразования касались довольно
незначительных деталей.
Особый интерес представляет история человекообразных
обезьян, которые отличаются от прочих обезьян Старого Света ре-
дукцией хвоста, отсутствием седалищных мозолей и щечных мешков,
бочонкообразной грудью. Передние конечностп у них значительно длин-
нее задних. Спускаясь на землю, они ходят в наклонном положении, опи-
раясь главным образом на задние конечности (отчасти и на тыльную по-
верхность передних). Орбиты глаз направлены прямо вперед, так что у
них вполне развито стереоскопическое зрение. Головной мозг достигает
максимального разви-
тия, и особенно разра-
стается область ассоци-
ационных центров коры
большого мозга.
В ископаемом со-
стоянии человекообраз-
ные обезьяны, еще не-
большой величины, из-
вестны начиная с оли-
гоцена Египта. Родона-
чальной формой являет-
ся, очевидно, форма,
близкая к Parapithecus,
которая соединяет в се-
бе черты насекомоядно-
го и примитивной обе-
зьяны. Найденный там
же Propliopithecus пред-
ставляет собой настоя-
Рис. 430. Череп ископаемой человекообразной обе-
зьяны Australopithecus africanus (по Абелю).
щую, но самую малень-
кую и наиболее примитивную человекообразную обезьяну. Эта форма свя-
зана, повидимому, с Pliopithecus из миоцена Европы и Индии, которая в
свою очередь приближается к современным гиббонам. В миоцене же Европы
и Азии были широко распространены более крупные человекообразные
обезьяны, по происхождению близкие к Propliopithecus и получившие
родовое название Dryopithecus. Различают много видов этого рода, ко-
торые изучены главным образом по обломкам челюстей с зубами и по этим
признакам приближаются к различным современным человекообразным
(рис. 433). Некоторые из них приближались к современной горилле (через
более молодую ископаемую форму — Palaeopithecus), другие прибли-
жались к современному шимпанзе (Dryopithecus giganteus, Dryopithecus
rhenanus), третьи, наконец, наиболее приближались к ряду предков че-
ловека (Dryopithecus darwini). Из находок, еще более приближающихся
к предкам человека, следует упомянуть о найденном недавно .австрало-
питеке (Australopithecus africanus), описанном по черепу молодого живот-
ного (рис. 430). Это, несомненно, наиболее высокостоящая из известных
нам человекообразных. Положение затылочного отверстия указывает
на более вертикальное положение тела, чем у антропоидов. Челюсти с
вертикальным положением резцов и едва выдающимися клыками похожи
на человеческие. Прогнатизм слабо выражен, как нередко и у человека.
Глазницы круглые, и расстояние между ними небольшое. Надбровных
выступов нет, и объем черепной коробки выше, чем у какого-либо иного
антропоида (объем 390 см3 у молодого животного и до 500 см3 у взрослого).
Нахождение остатков черепа взрослого австралопитека подтвердило его
положение в ряду предков человека.
479-
ОСНОВНЫЕ ЭТАПЫ ЭВОЛЮЦИИ ПРЕДКОВ ЧЕЛОВЕКА
7 примитивная костная рыба, плавающая при помощи боковых
волнообразных изгибов тела; 2—кистеперая рыба, лопастные парные
плавники которой служат органом опоры при ползании по дну и
выползании на берег; 3 примитивное земноводное (стегоцефал) с
пятипалой конечностью; 4 примитивная рептилия (котилозавр),
живущая и размножающаяся на суше; 5—звероподобная рептилия
(териодонт); 6 лазающее примитивное млекопитающее (опоссум);
7 ~ примитивная обезьяна (лемур); 8—человекообразная обезьяна
(шимпанзе); 9 -примитивный человек (неандерталец).
Человек связан ближайшим родством с человекообразными обезь-
янами, и совершенно правильно будет, если мы скажем, что человек про-
изошел от человекообразных обезьян. Однако само собой разумеется,
что мы не будем искать предков человека среди современных нам челове-
кообразных, которые проделали свою историю, отличную от ^человече-
ской. Предками человека могли быть только третичные человекообразные.
В стремлении отдалить предков человека назад в глубь веков мы, однако,
не можем итти так далеко, как, например, Вуд Джонс, пытавшийся вы-
вести человека от полуобезьяны (долгопят — Tarsius), и даже не можем
последовать за Осборном, который
думал, что у человека могли быть
Рис. 431. Скелет вадней конечности обевьян
и человека (по Грегори).
лишь более далекие общие с ан-
тропоидами предки. Нет основа-
ния предполагать и «деградацию»
человека от человекообразной обе-
зьяны (Ранке), так как третичные
антропоиды еще не обладали спе-
циализацией современных. Но все
же мы должны принять существо-
вание и некоторых регрессивных
изменений. Не подлежит, напри-
мер, сомнению, что человек про-
изошел от формы, лазавшей по де-
ревьям (с этим связана и редукция
Рис. 432. Слепок ступни горной горил-
лы (Gorilla beringei) (по Экели).
органа обоняния), с противопоставленным первым пальцем (рис. 431).
Однако способность лазать человеком утрачена, и соответственно утеряна
и способность противопоставлять большой палец ноги остальным. Что
этот процесс мог произойти довольно быстро, видно по строению ноги
недавно найденной горной гориллы (рис. 432), которая также утра-
тила способность лазать по деревьям. С хождением на двух ногах связано
прогрессивное развитие большого пальца ноги и прогрессивное удлинение
задних конечностей вообще (рис. 431). Наоборот, передние конечности
укорачивались. Большой палец руки у хорошо лазающих обезьян реду-
цировался. У человека этот палец имеется, и, конечно, он был и у предков
человека, а не развился заново. Однако нет оснований отрицать воз-
можности некоторого его вторичного усиления, как это, несомненно,
имело место в задней конечности.
С вертикальным хождением человека связано, кроме изменений в
480
строении конечностей, и приобретение характерного изгиба позвоночника,
а также плоская форма грудной клетки и положение затылочного отвер-
стия. Форма черепа изменялась, с. одной стороны, благодаря прогрессив-
ному разрастанию мозговой коробки, и с другой — в связи с ослаблением
и укорочением челюстей. Укорочение альвеолярной части челюстей при-
вело к образованию выступающего вперед подбородка. С тем же ослабле-
нием челюстей связано и исчезновение гребней па черепе (если они были
у предков человека; у небольших антропоидов гребней могло и не быть).
Хождение на двух ногах освободило передние конечности, превратило
их в хватательный орган и сделало возможным их чрезвычайно разнооб-
разное использование. Становление человека связано с использованием
Рис. 433. Родословная человекообразных обезьян и человека по палеонтологическим
данным (по Грегори с изменениями).
передних конечностей для изготовления орудий труда из различных ес-
тественных материалов. Эти искусственные орудия заменяли ему более
сложные естественные органы и дали возможность пользоваться такими
средствами защиты, нападения, постройки защищенного жилища, заго-
товления запасов и приготовления пищи, которых нет ни у одного из
животных. Так как именно это и имело решающее значение в подъеме
человека на высшую ступень, то естественно, что мы считаем критерием
при оценке остатков человекообразных существ факты нахождения вместе
с ними изготовленных ими орудий.
Найденные уже давно (1882) на Яве остатки человекообразного су-
щества, названного питекантропом (Pithecanthropus erectus) (черепная
крышка, бедро и кусочек челюсти), долго были предметами споров. Форма
бедра указывала на прямохождение, гладкая форма черепа при сравни-
481
тельно большой величине животного и объем мозга более 900 ел8 ставили
ато существо почти рядом с человеком. Новая находка черепной крышки
и нижпей челюсти питекантропа показала несколько меныппй объем
головного мозга (около 800 см3), но вполне подтвердила его положение
в ряду ближайших предков человека. Наконец, находки (1928 и 1936)
в Китае многих остатков существа, названного синантропом (Sinanthropus
pekinensis), бросают дальнейший свет и на значение питекантропа. Си-
нантроп был чрезвычайно похож на питекантропа: черепная крышка
очень сходной формы (узкая височная область и сильно выступающие
надбровные дуги) и очень близкой емкости (около 1000 см3). Челюсть не
имеет выступающего подбородка и вообще напоминает челюсть гейдель-
бергского человека. Вместе с этими остатками найдены простейшие ка-
менные орудия, обработанные рога оленя и следы употребления огня.
Таким образом мы в синантропе, а следовательно, вероятно, и в питекан-
тропе имеем перед собой уже настоящего человека, история происхожде-
ния которого в настоящем очерке более детально не рассматривается.
МОРФО-ФИЗИОЛОГИЧЕСКИЕ ЗАКОНОМЕРНОСТИ ЭВОЛЮЦИИ
Приспособление
Из приведенного краткого обзора филогении позвоночных видно,
насколько паши обычные представления в самом деле упрощают необы-
чайно сложный процесс эволюции и насколько относительное значение
имеют все наши основные понятия. Распространенное представление об
эволюции немногим отличается от лестницы живых существ натурфило-
софов и от градаций Ламарка. Мы не можем говорить об эволюции вообще
как о непрерывном процессе усовершенствования или даже усложнения
организации. Эволюция представляет непрерывную цепь изменений,
происходящих на различных этапах с различной скоростью и в различных
направлениях. Самое представление об усовершенствовании имеет весьма
условное значение, так же как и понятие о приспособлении. Мы говорим
о высших, более сложно построенных организмах и о низших, или более
простых, говорим об адаптации и о специализации как о приспособлении
к определенной более или менее ограниченной среде. Но все эти опреде-
ления весьма условны. Конкретные организмы, даже и «примитивные»,
бывают всегда приспособлены к определенной среде и могут быть в неко-
торых отношениях весьма совершенными и даже специализированными
(например, конкретные осетровые, амфибии, насекомоядные). С другой
стороны, высшие формы могут быть во многих отношениях весьма прими-
тивными (из рыб — кистеперые и двудышащие, из рептилий — кроко-
дилы, из млекопитающих — приматы). Хотя мы в дальнейшем и будем
говорить об эволюции путем общего подъема организации (ароморфоз)
и путем приспособления к известной среде (адаптация), но и в этом мы
имеем чисто условные разграничения, так как по существу всякая эволю-
ция является приспособительной и протекает всегда в конкретных усло-
виях среды. Однако некоторые приспособительные изменения имеют более
общий характер и дают организму гораздо более широкие возможности, не
ограничивая его именно данной средой, а оставляя его весьма пластичным
(например, развитие органов чувств, прогрессивное развитие головного
мозга, усовершенствование органов питания или дыхания); в этих случаях
мы и будем говорить о повышении организации. Другие приспособитель-
ные изменения вносят, наоборот, ограничение в смысле возможности
существования только в данной строго определенной среде (приспособле-
ние конечностей к бегу и всей организации к жизни в степи у многих
копытных, приспособление конечностей и других органов к жизни на
ветвях деревьев у ленивца или к жизни в воде у кита); в этом случае мы
482
можем говорить о приспособительной эволюции в узком смысле этого
слова.
Следовательно, вся эволюция любого животного протекает в связи о
изменением окружающей среды и она является, в сущности, зависимой,
т. е. приспособительной. Различия появляются лишь в зависимости от
того, насколько широка и разнообразна среда, в которой живет и разви-
вается организм. Эта зависимость изменений от изменений внешней среды
проявляется на некоторых частях организма особенно ярко и является
понятной и ясной. Северное называет такие органы, которые по своей
функции имеют непосредственное отношение к внешней среде, экзосома-
тическими (конечности, зубы, органы чувств и т. п.). При всех изменениях
среды в первую очередь изменяются экзосоматические органы. Однако
в целостном организме невозможно никакое изолированное изменение
отдельной части. С экзосоматическими органами более или менее непосред-
ственно связаны и другие, эндосоматические органы, которые с изме-
нением первых также неизбежно должны в большей или меньшей степени
измениться. В этих изменениях приспособительный характер выражается
не столь ясно, однако в сущности и все изменения внутренних органов
есть результат их взаимного приспособления, а следовательно, и резуль-
тат приспособления к меняющейся среде.
Прогресс и регресс
Относительность наших понятий особенно ясно выявляется на таких,
казалось бы, диаметрально противоположных понятиях, как прогресс
и регресс. Прежде всего во избежание бесконечных противоречий нам
приходится вслед за Северцевым строго разграничить понятие прогресса
биологического и прогресса морфо-физиологического.
Биологический прогресс означает победу в борьбе за
существование и выражается: 1) в увеличении численности особей данного
вида или данной группы видов, 2) в более широком ее расселении, т. е.
в захвате новых мест обитания, и 3) в распадении на подчиненные систе-
матические группы. Противоположное понятие — биологический регресс
означает: 1) уменьшение абсолютного числа особей данной группы, 2) су-
жение ареала обитания, 3) сокращение числа видов и разновидностей,
входящих в состав данной группы, благодаря их постепенному вымиранию.
Биологический прогресс может быть результатом как усложнения, так
и упрощения организации, т. е. он может совпадать с морфо-физиологи-
ческим прогрессом, но может с-ним и не совпадать.
Прогрессивная эволюция, т. е. морфо-физиологический
прогресс — ароморфоз (Северцов), биологически определяется усложне-
нием условий существования и означает усложнение организации, т. е.
прогрессивную диференцировку организма, сопровождающуюся расчле-
нением функций; последнее связано и с возникновением новых частей,
новых органов, а следовательно, и новых функций. Это усложнение может
в основном касаться немногих органов, другие могут сравнительно мало
измениться, а третьи могут, наоборот, потерять свое значение и редуци-
роваться. Таким образом, понятие морфо-физиологического прогресса
организма вовсе не исключает возможности регресса отдельных частей
или органов. Прогрессивное развитие низших позвоночных сопровожда-
лось прогрессивным развитием костного скелета, состоящего из высшей
формы скелетной ткани. Однако этот прогресс неизбежно сопровождался
регрессом хряща, и без этого регресса не был бы возможен. Приобретение
млекопитающими волосяного покрова мы обозначили как важное приобре-
тение, имевшее большое значение в их эволюции, так как это сделало воз-
можным установление постоянной температуры тела. Однако развитие
волосяного покрова неизбежно было связано с редукцией прежнего
483
покрова из чешуи. Точно так же прогрессивное развитие кровеносной
системы птицы или млекопитающего было связано с редукцией одной
дуги аорты и т. п.
Приспособительная эволюция, или адаптация (Се-
верцев), в более узком значении этого слова (в широком смысле этого
слова всякая эволюция приспособительна), определяется изменением
условий существования и означает изменение организации соответственно
измененной среде, без заметного ее усложнения. Приспособление может
быть связано и с возникновением новых органов и с исчезновением старых.
Такого рода преобразования проявляются наиболее ярко при переходе
в значительно отличающуюся среду (например, из воды на сушу или
наоборот). Переход от кистеперой рыбы к стегоцефалу сам по себе не
означал еще заметного прогресса, заметного усложнения. Одни органы
развивались прогрессивно (легкие), другие редуцировались (жабры),
третьи преобразовывались соответственно новым требованиям (крове-
носная система, конечности). Точно так же и переход млекопитающего
к летанию в воздухе (летучие мыши) или плаванию в воде (киты) не озна-
чает общего усложнения организации, общего прогресса. Развитие черепах
или змей не связано с общим прогрессом. Одни органы развивались про-
грессивно, а другие, утерявшие в новой среде свое значение, редуциро-
вались.
Хотя всякая эволюция является приспособительной, но чем более
условия существования ограничиваются некоторыми узкими пределами,
тем ярче бросается в глаза приспособление к этим строго ограниченным
условиям. В этом случае обычно сохраняются многие примитивные черты,
нередко имеется налицо даже вторичное упрощение организации (в связи
с упрощением условий существования), но зато другие органы могут
достигать весьма высокой степени диференцировки. В таких случаях
говорят о специализации.
Такими специализированными организмами являются, например,
круглоротые, из рыб —• двудышащие, из амфибий — гимнофионы, из
рептилий — хамелеоны, из млекопитающих --- ленивцы, муравьеды, си-
рены и многие другие.
Регрессивная эволюция биологически определяется
крайним упрощением условий существования и выражается морфоло-
гически понижением, упрощением организации при регрессе и исчезно-
вении ряда органов, что связано с ослаблением, упрощением и выпадением
соответствующих функций. К регрессу ведут однообразные условия
существования на больших глубинах, в пещерах (постоянная температура,
отсутствие света), в особенности переход к сидячей жизни и еще более —
паразитизм. Так, например, у асцидий переход к сидячей жизни связан
с утратой органов движения, органов чувств, крайним упрощением нерв-
ной системы и т. п. У паразитических червей, раков, моллюсков вся
организация может быть упрощена до полной невозможности определить
систематическое положение организма на основании его строения. У пара-
зитирующего на крабах усоногого рака Sacculina не сохранилось вообще
ничего от прежней организации, кроме покровов и половых желез. И од-
нако даже при крайней степени регресса он не охватывает всего организма.
И у паразитов некоторые части развиваются прогрессивно —и это прежде
всего органы прикрепления паразита к телу хозяина и затем органы
размножения. Таким образом, прогресс и регресс органов не исключают,
а, наоборот, дополняют друг друга при всех изменениях организмов в
тзчение их эволюции.
484
Ароморфоз и адаптация как основные направления эволюционного
процесса
Если подойти с нашими определениями понятий прогрессивной и
регрессивной эволюции к конкретному филогенетическому материалу, то
можно отметить некоторые довольно определенные закономерности. Каж-
дая новая большая группа животных возникает путем прогрессивной
эволюции, т. е. ароморфоза, поднимающего ее организацию на более
высокую ступень, а затем переходит на путь адаптации или иногда Даже
регресса (Северцов). Эта закономерность имеет глубокие основания.
Всякая группа животных есть результат эволюции форм, вытеснивших
в борьбе за существование какие-то иные формы, т. е. имеющих перед
ними какие-то преимущества. Если это касается большой группы живот-
ных, живущей в разных условиях, то это были преимущества общего
характера, имевшие значение во всех этих условиях, т. е. эти преиму-
щества, действительно, поднимали организм на более высокую ступень,
и эволюция имела прогрессивный характер. Всякое значительное усо-
вершенствование организации, дававшее организму большие преиму-
щества в борьбе за существование (ароморфоз), приводило к увеличению
его численности, более широкому расселению и распаду на местные формы.
Увеличение численности приводит к обострению внутривидовой борьбы,
к расхождению признаков, к приспособлению отдельных форм к тем или
иным более частным условиям, т. е. к адаптации, которая может выра-
жаться и в специализании, и даже в общем регрессе при оттеснении данной
формы в крайне ограниченную среду.
Так, в ряду позвоночных путем ароморфоза возникли челюстные
позвоночные, распавшиеся затем при явлениях адаптации на ряд различ-
ных форм (акулы, скаты, химеры). Путем ароморфоза возникли костные
рыбы с плавательным пузырем, жаберной крышкой и костными лучами
плавников и затем быстро распались на ряд форм, изменявшихся далее
путем адаптации (хрящекостные рыбы, костные ганоиды, кистеперые,
двудышащие, амфибии). Путем ароморфоза возникли рептилии с более
развитыми конечностями, органами чувств и мозгом, с защищенным!
покровами, более совершенными лешими и т. д., и затем в результате
адаптации к различным условиям среды они разбились па множество
различных форм. Путем ароморфоза возникли, очевидно, и псевдозухий,
давшие затем при явлениях адаптации начало целому ряду форм (кроко-
дилы, динозавры, птерозавры, птицы). Путем несомненного ароморфоза
возникли млекопитающие с их высокоразвитым мозгом, диференциро-
ванной зубной системой, длинными и сильными конечностями, постоянной
Температурой тела, живорождением и т. п. И млеко питающие в резуль-
тате адаптации к различным условиям среды разбились на многочисленные
формы, живущие в разной среде и пользующиеся разной пищей (насеко-
моядные, хищные, грызуны, рукокрылые, китообразные, копытные, при-
маты). В пределах менее значительных групп можно говорить о тех же
самых закономерностях. Так, бесхвостые амфибии обособились в резуль-
тате известного ароморфоза (значительное развитие конечностей, легких),
а затем путем адаптации дали начало весьма разнообразным формам.
Воробьиные птицы обособились в результате некоторого ароморфоза, за
которыми последовала адаптация. Хищные млекопитающие образовались
в результате явного ароморфоза и затем дали начало многим весьма
разнообразно адаптированным формам. То же самое можно сказать о
приматах и т. д.
Адаптивная радиация (Осборн). Обычно адаптация не-
которой группы животных идет не в одном направлении, а во многих,
определяемых различными условиями среды, в которые попадают те или
иные группы особей. В таком случае мы имеем расхождение признаков
485
в разные стороны, или адаптивную радиацию форм (Осборн), как это
и видно на приведенных выше примерах (и на очень многих других при-
мерах, отмеченных нами в очерке филогении позвоночных). Чем более
значителен ароморфоз, приведший к образованию данной группы, тем
более значительна и разнообразна обычно и следующая за этим ароморфо-
зои адаптивная радиация форм.
Дивергенция, конвергенция и параллелизм в эволюции
Направление эволюции явно определяется той средой, в которой
развивается организм. Так как эта среда для разных организмов не может
быть вполне одинаковой и так как разные организмы реагируют неоди-
наково даже на одинаковое изменение среды, то направление эволюции
определяется не только средой, но и строением развивающегося организма
(т. е. его предыдущей историей, поскольку это строение есть ее результат).
Дивергенция, или расхождение признаков, есть процесс при-
обретения различного строения в результате развития родственных орга-
низмов (сходных в силу сходной истории) в различной среде.
Дарвин предполагал, что эволюция идет путем расхождения призна-
ков в результате вымирания промежуточных форм, оказывающихся
в менее благоприятных условиях существования при прогрессивной
адаптации отдельных групп. Чем дальше подвигается адаптация (или
прогрессивная эволюция), тем сильнее размножаются попадающие в более
благоприятные условия новые формы, тем быстрее они получают преобла-
дание над промежуточными, тем быстрее идет изменение и расхождение
признаков. Чем более различна среда, в которой живут и развиваются
отдельные группы особей родственных между собой организмов, и чем
дольше длится это расхождение, тем более ярко бывает выражена их
дивергенция. Так, например, невелико расхождение между речной фо-
релью и озерной, между болотной лягушкой и водяной; значительно
больше расхождение между выдрой и куницей, между белкой и сусликом
(несмотря на весьма близкое родство); еще больше расхождение между
ежом и летучей мышью, между собакой и тюленем, между слоном и сире-
ной или между крокодилом и птицей. Все это — организмы, связанные
несомненным родством.
При дивергенции морфологическое сходство объясняется родством
организмов, а различия в строении организмов и гомологичных их органов
выработались в процесс независимой их эволюции в разной среде после
их обособления друг от друга (рис. 435).
Конвергенция, или схождение признаков, есть процесс при-
обретения сходного строения в результате развития неродственных (раз-
ных вследствие длительно различной истории) организмов в сходной среде.
Под неродственными организмами здесь следует понимать организмы,
связанные лишь более далеким родством и сильно разошедшиеся по своей
организации. При конвергенции различные по происхождению и, следо-
вательно, негомологичные или если и гомологичные, то весьма разные
органы могут вторично приобрести сходное строение, т. е. стать анало-
гичными. Чем более специфические требования предъявляет к организму
среда и чем длительнее и полнее приспособление к этой среде, тем больше
может быть и сходство, тем ярче выражается конвергенция (рис. 435).
Сходством, основанным на конвергенции, будет, например, редукция
глаз у пещерных животных (это сходство ясно определяется здесь специ-
фическими условиями среды — отсутствием света) или сходство ластов
у водных рептилий и млекопитающих. Нас здесь, однако, интересует
не столько сходство органов, сколько конвергентное сходство целых
организмов. Так, например, у хорошо плавающих позвоночных разви-
вается и сходная общая форма тела, сходное расположение плавников,
486
как это видно из сравнения хорошо плавающей акулы (Lamna), костистой
рыбы (макрели), рептилии (ихтиозавра) и млекопитающего (дельфина).
Сходством, основанным на конвергенции, будет и сходств? между
неродственными «голенастыми» болотными птицами, как журавли и
Л/,
Сумчатке ПлацекшнЬз
чАе \ //
Тер'Юионтк
/7(
ПтииА
diobi I Ячирстазке Anoialptx
I I Rrnuu^rmsbit
|| / /динозабрЬ!
I'/ /,Рамфоринхи.
J уТ^Втсрйдактипи.
J heromurpha
Черепахи
ttxmuo;
ПсеМозу.
езмхАА
РЬосАА,
SyridpAh.
i ^hyKhecepbaba.
а | / ро!сго'/р^'
i//
Dia-.'ii'da Че'-ЩйчатЬл
bpa:;x.uC3a5pbt
бим > , с f ион bi
Kcmu.ioaafybi
PlukpOSaSpbt
/ jia6upuHmoSonml>i
Kocmucmbie pkfrbi
(TbleosieiJ
Атю/dei
LepidosizoiJf
Kocmhbie хучеперк
IHclostei) \
Anospida
\
Круглоротые \
ICyclostomain) \
Чер<
У-.рящекостнЬ1е
(Chondrosleoidei)
Acanlhodi
ПриучиглиЬнЬз стегоцефалк
Высшие
кистоперые
/'(Crossopteryg/t)
Jd6ydbiw.aw,ue
/Cipnoi)
Костике kuemerepbie
/ (Crcssopleryqu)
jp Многоперые
/' iPntypteroitleL)
-.Hribie котомке
IHolcs'tb.dei)
Ak(jroo5p03hb
Jhelcdontt
Шишковые
lOstracedemi
le.nioi.imbie
ВнутреннеA<-i6epnbie l:aP‘-l''':<n>Haf.epnbie
(BndwranchiatapZ Aclolranchiaba)
n 4
< upouuHbie чесепнЬ'е
Cec'iepenubie
(Acrunia Г4-
Xcpldihe (Chorc^cu >
(Cronulla)
Рис. 434. Родословное древо хордовых. Двойной линией обозначен эволюционный
путь от низших хордовых к птицам и млекопитающим.
аисты, сходство между хамелеоном (Chameleo) и древесной агамой (Lyrio-
cephalus), сходство между ископаемыми Placodontia и морскими чере-
пахами (рис. 436), между кареттой (Chelone) и кояшетой черепахой (Sphar-
487
gis), сходство между сумчатым волком (Thylacinus) и настоящим (Canis),
водяным опоссумом (Chironectes) и выдрой (Lutra), сходство между подзем-
ными роющими млекопитающими: сумчатым кротом Австралии (Noto-
ryctes), обыкновенным кротом (Talpa), золотым кротом Африки (Chry-
sochloris) и даже грызуном (Siphaeus). Приспособление различных млеко-
2
Д,/а.Ъсде/ tfjdbfge/
A'jabcd/ A'jabfgf^
Д,/аЪс/ A2/abf/ А, /аЬс/
Д/аЪ/
A^abf/
А/аЪ/
I Н III
иис. 435. Схема, поясняющая различные соотношения в эволюции отдельных ветвей;
I — дивергентное развитие, при котором в связи с расхождением в разную среду потомки
одной и той же формы А, характеризующейся признаками а, Ь, приобретают различные но-
вые признаки (cd п /д), 11 — параллельное развитие, когда родственные формы, характери-
зующиеся признаками а, Ь, но несколько разошедшиеся (признаки с и /), в связи с пребыва-
нием в сходной среде приобретают независимо друг от друга дальнейшие сходные признаки
(д. е), 111 — конвергентное развитие, при котором различные организмы (А и В), характери-
зующиеся разными признаками (ab и mn), в связи с пребыванием в сходной среде приобре-
тают сходные признаки (де).
питающих к планированию также приводит к развитию конвергентного
сходства между сумчатым летуном (Petaurus), шерстокрылом (Galeopi-
thecus), белкой летягой (Pteromys) и т. д. Во всех этих случаях можно
говорить о неродственных формах, поскольку то сходство, о котором
здесь идет речь, даже в своих начальных фазах при-
Рис. 436. Конвергентное сходство морской черепахи (отряд Chelonia) и ископаемых
плоскозубых рептилий (отряд Placodontia). Слева — Archelon (верхний мел), справа —
PJacochelys (триас).
обретено независимо. Так, например, у каретты и у кожистой
черепахи были, конечно, когда-то общие предки, но это были наземные
черепахи, которые еще на суше развивались дивергентно и дали начало
разным сухопутным формам. Эти разные сухопутные черепахи незави-
симо друг от друга стали затем переходить к жизни в море, и, следова-
488
тельно, все эти сходства приобретены совершенно независимо друг от
друга.
Морфологический параллелизм, или параллельная
эволюция, есть процесс приобретения сходного строения в результате
развития родственных (сходных в силу сходной истории) организмов
в сходной среде. Параллелизм можно, пожалуй, также определить как
конвергенцию близко родственных организмов. Несомненно, что при
суждении о родстве двух форм возможен известный произвол в оценке
близости этого родства, а следовательно, естественно возникают известные
трудности при попытках провести строгую грань между конвергенцией
и параллелизмом. В действительности на родственных организмах эти
понятия сливаются, так же как на них сливаются и понятия гомологии и
аналогии. Все же понятие параллельного развития не является излишним,
так же как не является лишним и понятие гомойологии, включающей и
гомологию и аналогию одновременно. Мы будем говорить о параллелиз-
мах в тех случаях, когда сходные структуры хотя и развиваются в двух
разных организмах независимо друг от друга (как при конвергенции),
однако на базе весьма сходных в силу родства, т. е. вполне гомологичных,
структур (рис. 435). Несколько примеров позволят нам яснее уловить
смысл такого разграничения.
Двудышащие и амфибии связаны не слишком близким родством —
они давно обособились от примитивных кистеперых рыб как независимые
друг от друга ветви. И те и другие живут в биологически сходной среде
(высыхающие водоемы) и во многом весьма сходны. Далеко не все эти
сходства основаны на родстве: и аутостилия и сходство в строении сердца
и сосудов приобретены независимо друг от друга, т. е. представляют
результат конвергенции. Однако нужно принять во внимание, что сходство
в строении сердца и сосудов есть неизбежный результат развития легоч-
ного дыхания, а легкие как добавочный орган дыхания есть общее при-
обретение, унаследованное от кистеперых рыб, у которых они уже были,
так же как была и легочная артерия. Tait как сходные структуры
здесь развились на базе особенностей, уна-
следованных от общих предков, то мы здесь вправе
говорить о параллелизмах в развитии кровеносной системы у двудышащих
и у амфибий. Еще проще объяснить независимое развитие сложной склад-
чатости дентина в зубах кистеперых рыб и стегоцефалов. Так как это
строение зубов развивается в непосредственно между собой не связанных
семействах высших кистеперых (Holoptychiidae) и высших стегоцефалов
(Stereospondyli), то мы должны здесь, казалось бы, говорить о конверген-
ции. Однако и у примитивных кистеперых (Osteolepida, Rhizodontidae)
и у примитивных стегоцефалов (Rhachitomi, Embolomeri), безусловно
связанных между собой ближайшим родством, у основания зубов была
радиальная складчатость дентина. Так как слоясная складчатость зубов
Holoptychiidae и Stereospondyli развилась на основе простой радиальной
складчатости, унаследованной от общих предков, то мы и здесь можем
говорить о параллелизме данных структур.
Если два родственных организма развиваются параллельно по мно-
гим признакам, мы с тем большим правом говорим о параллельной эво-
люции этих организмов. Примерами такой, во многих отношениях парал-
лельной, эволюции могут служить обе основные ветви динозавров •— яще-
ротазые и птицетазые, у которых параллельно шло увеличение общих
размеров тела, увеличение крестца, возврат к хождению на четырех
ногах и многое другое. Параллельно развивались обе ветви птерозавров —
птеродактили и рамфоринхи, которые, начав планировать, разделились
и после этого на унаследованной общей основе все более приспосаблива-
лись к летанию (пневматизация скелета, развитие крыльев, грудины,
клюв черепа и многое другое). Если же сравнивать птерозавров и птиц,
489
то правильнее будет говорить только о конвергентном сходстве, так как
хотя это и родственные организмы, но даже первые моменты перехода
к планированию у них безусловно независимы. Все сходство связано со
способом передвижения — летанием. Но как раз этот способ передвиже-
ния приобретен птерозаврами и птицами независимо друг от друга: органы
летания развились на различной базе (у птерозавров — кожная складка,
поддерживаемая одним пальцем, у птиц — маховые перья, поддержива-
емые кистью) и являются как таковые лишь аналогичными (хотя сами
конечности в общем гомологичны). Примерами параллельного развития
будут и многие сходства между териодонтами и млекопитающими (ди-
ференцировка зубов, акромиальный отросток лопатки и пр.). Наконец,
примером такого же развития может служить и образование однопалой
формы с призматическими зубами в ряду лошадей в Северной Америке
и в ряду Litopterna в Южной Америке. Параллельно развивались тюлени,
котики и моржи. Сходно шло развитие зубастых китов и беззубых и др.
Параллельная эволюция имеет очень большое значение и притом
тем большее, чем ближе родство сравниваемых форм. Как правило, боль-
шинство близких ветвей во всех группах животных всегда развивается
параллельно, если только эти ветви не получились в результате перехода
в резко различную среду. Дело в том, что первоначальное расхождение,
являющееся необходимым условием всякого разделения на обособленные
формы (подвиды, виды), обычно ограничивается сравнительно небольшим
экологическим или географическим разграничением, а затем эти обособ-
ленные формы (после ослабления конкуренции между ними) продолжают
развиваться дальше в сходной в общем биологической обстановке, и-
естественно, это развитие сходных организмов в сходной среде идет по
существу параллельно по большинству признаков. Таким образом, пара,
лельная эволюция представителей небольших систематических групп
составляет общее правило, имеющее лишь редкие исключения. Однако
при разборе эволюции таких близких организмов на практике, действи-
тельно, оказывается весьма трудной задачей разграничить сходство,
основанное на родстве, и параллелизмы. Такое разграничение возможно
лишь на основании тщательного изучения истории этих форм.
Монофилия и полифилия
С вопросами конвергентной и параллельной эволюции тесно связан
и вопрос о морфологическом или полифилетическом происхождении тех
или иных групп организмов или даже всех животных, всех растений
или всех организмов вообще. Под монофилией понимают, развитие из
одного общего корня, т. е. от однородных предков; под полифилией —
развитие из нескольких корней, т. е. от различных предков. Большинство
эволюционистов, начиная с Дарвина, считает, что эволюция идет главным
образом путем дивергенции и естественные группы организмов (таксо-
номические единицы систематики) развиваются путем расхождения из
одного ствола, т. е. монофилетически. Конечно, при этом не предпола-
гается, как это иногда утверждают, развитие от одной пары исходных
форм, а только от группы скрещивающихся между собой особей (т. е.
разновидности, вида). Некоторые новейшие аволюционисты предпола-
гают, что в результате конвергентного развития сходство между организ-
мами разного происхождения может сделаться столь значительным, что
мы вынуждены будем относить такие сходные организмы к одной таксоно-
мической единице. Сходство, основанное на конвергенции, может стать
неотличимым от сходства, основанного на родстве. Если это так, то есте-
ственно, что в одни и те же систематические группы будут попадать орга-
низмы самого различного происхождения, и тогда эти группы окажутся
по дифилетическими. Так это и бывает на самом деле. Группа бескилевых
49Q
птиц (Ratitae) является, повидимому, полифилетической, так как, веро-
ятно, объединяет бегающих птиц разного происхождения, сходство кото-
рых связано именно с утратой способности летать. Однако если это будет
доказано, то тем самым будет доказана неестественность данной системати-
ческой группы и необходимость переработки систематики птиц. Когда
будет доказана генетическая связь отдельных бескилевых птиц с кури-
ными, журавлями и т. д., то их и нужно будет объединить с соответствен-
ными группами, так как мы стремимся к естественной классификации,
которая должна совпасть с классификацией генетической, т. е. основан-
ной на общности происхождения. Так это и производится по мере накоп-
ления более точных знаний.
Не так давно говорили о птицах «плавающих», «лазающих», «хищных»,
«голенастых» и т. п. Уже самые названия показывают, что здесь выдвига-
лось сходство биологическое, которое, конечно, во многом могло покоиться
на конвергенции. Все эти группы при ближайшем исследовании оказа-
лись полифилетпческими и были разбиты (аисты были отделены от журав-
лей, совы от «дневных» хищных и т. д.) и наоборот — многие внешне не-
сходные формы были объединены в новые «естественные», т. е. монофи-
летические, группы. Таким образом, полифшлия в указанном выше смысле
втого слова не может считаться принципом эволюционного процесса, а
только лишь выражением несовершенства нашей классификации, которая
постепенно приближается к естественной и вместе с тем к генетической.
С другой стороны, следует отметить, что трудность для установления ге-
нетической классификации вовсе не так велика. В прежней систематике
нередко пользовались лишь отдельными признаками чисто внешнего ха-
рактера. При использовании знания строения организма (особенно «эндо-
соматических» органов) все эти трудности отпадают, так как конвер-
генция никогда не бывает глубокой. Конвергентное
сходство развивается в зависимости от сходной среды и касается главным
образом эктосоматических органов, т. е. внешности (в широком смысле).
Более глубокие реакции разных организмов, хотя бы и в одинаковой среде,
не могут быть одинаковыми, и как бы по внешности ихтиозавр ни был
похож на дельфина, первый по всей своей организации представляет на-
стоящую вполне типичную рептилию, а второй — не менее типичное мле-
копитающее. Таким образом, мы определенно должны признать монофи-
летическое происхождение достаточно хорошо изученных, т. е. верно
оцененных систематиками, групп животных. Палеонтологические данные
достаточно ясно указывают на происхождение от одного корня всех хищ-
ников, всех копытных, в частности парнокопытных, носорогов, слонов
и т. д., т. е. как раз наиболее хорошо изученных групп.
Иногда, однако, под полифилией понимают нечто иное — не резуль-
тат конвергентного развития разных организмов, а следствие параллель-
ной эволюции многих форм, объединяемых в одну естественную группу.
Б результате параллельной эволюции сходства могут быть и очень зна-
чительными, и они бывают тем более глубокими, чем более близким род-
ством эти организмы связаны. Однако близкое родство как раз и означает
происхождение из одного корня, и, следовательно, в этих случаях име-
ется именно монофилия, а не полифилия. Не следует погашать монофи-
лию слишком узко. Конкретные организмы эволюируют во множестве
особей, среди которых идет постепенная диференцировка, и вся эта
масса, как правило, продолжает развиваться в сходной (в общем) био-
логической обстановке и, следовательно, параллельно. Поэтому все те
отдельные стволы, ветви, родственные линии и т. д., о которых мы гово-
рили в очерке филогении, представляют собой не ветви и не отдельные
линии, а пучки ветвей, бесконечное количество параллельных (в общем)
линий, которые мы только для простоты и наглядности изображаем в
виде простых ветвей. В основе таких ветвей, пусть даже в глубине
491
ствола, лежит всегда масса, а не единичная особь, но только это — масса
свободно между собой скрещивающихся особей. Видообразование всегда
начинается с некоторой дивергенции, а какое бы большое значение мы
ни придавали параллелизмам, эволюция в основе своей представляет
процесс постепенной дифференциации. Следовательно, естественные группы
организмов по своему происхождению всегда монофилетичны.
Направленность эволюции
То обстоятельство, что в группах близко родственных форм эволюция
обыкновенно идет хотя и независимо, но параллельно, явно указывает
на существование определенных путей, т. е. известной направленности
эволюционного процесса. Это давало повод говорить об ортогенезе, номо-
генезе, законе инерции и т. п. Существование самого факта направлен-
ности не может, однако, служить основанием для принятия каких-то осо-
бых внутренних упорядочивающих сил, направляющих эволюцию по за-
ранее предначертанным путям, как это себе иногда представляют. Направ-
ление эволюционного процесса определяется для каждого данного мо-
мента всегда комплексом внутренних и внешних факторов, сложнейшим
образом взаимодействующих друг с другом. Факт широкого распростра-
нения параллелизмов служит явным указанием на значение внутренних
факторов в эволюции, так как настоящие, глубокие параллелизмы наблю-
даются только у близко родственных форм. В этом случае направление
эволюции сходно в значительной мере, благодаря сходству унаследован-
ной, исторически сложившейся структуры этих организмов. Конечно,
эти параллелизмы развиваются только в сходной среде, однако вместе
с тем они возможны только для близких организмов. Более далекие орга-
низмы даже в тождественной среде развиваются по-разному, и их конвер-
генция выражается самое большее в тех чертах организации, которые
имеют непосредственное отношение к этой среде (экзосоматические ор-
ганы) и никогда не приводит к глубокому сходству. Далекие организ-
мы реагируют по-разному даже в одинаковой среде. Наоборот, существова-
ние одинаковой реакции у разных организмов лучше всего доказывает их
глубокое родство, общее сходство их исторически сложившейся структуры.
Вместе с тем, параллельное развитие даже самых близких организмов
возможно только в сходной среде, и в этом также явно выражается зна-
чение среды как фактора, определяющего направление эволюционного
процесса.
В разной среде даже одни и те же организмы развиваются по-разному,
т. е. дивергентно (и яснее всего это сказывается, конечно, на тех органах,
которые наиболее непосредственно связаны с этой внешней средой, т. е.
на органах экзосоматических). Направление эволюционного процесса
определяется в одинаковой мере как унаследованной структурой орга-
низма, т. е. внутренними факторами, так и средой, в которой развивается
организм. Если эта среда относительно постоянна или в течение длитель-
ного периода времени меняется в одном определенном направлении (на-
пример, постепенный переход к более сухому, континентальному климату
с расширением области степей), то и эволюция некоторых организмов
может довольно долго продолжаться в одном определенном направлении
(развитие лошадиного ряда).
Неограниченность эволюции и вымирание
Палеонтологи вскрывают перед нами все большее —: можно сказать,
бесконечное — число ветвей животного царства. Все эти ветви, прогрес-
сивно развиваясь, приспособлялись к условиям своего времени, посте-
пенно достигали широкого распространения и большой численности, но
492
все же, как неизменное правило, это процветание Длилось недолго и
всегда заменялось упадком, приводившим к полному вымиранию. Это
дало повод для проведения аналогии между жизнью вида (в смысле фило-
генетической ветви) и жизнью особи. Палеонтологи часто говорят
о молодости, зрелости и старости отдельных форм и даже целых больших
групп животного царства. Проведением таких аналогий пытаются «объяс-
нить» систематическое отмирание ветвей филогенетического древа. Путь
аналогий, обладая известным эвристическим значением, не является,
однако, надежным. В данном случае закономерности филогенетического
развития, конечно, совершенно несравнимы с закономерностями инди-
видуального развития, и указанная аналогия является крайне поверх-
ностной. Если смерть является необходимым результатом индивидуальной
жизни, ее неизбежным логическим завершением, то ни в коем случае этого
нельзя сказать про жизнь видов (как филогенетических ветвей). Одни
виды — и таковых, действительно, большинство — в своих преобразо-
ваниях заходят в тупик и вымирают. Однако другие — пусть немногие —
развиваются дальше и дают начало новому многообразию форм, вытесняя
с жизненной арены угасающие формы. В целом мы не видим уменьшения
жизни на земле, не видим уменьшения ее разнообразия и не видим в вы-
мирании необходимого следствия филогенетического развития. Посто-
янно наблюдающееся вымирание целых групп организмов имеет, конечно,
свои основания, и эти основания следует искать не только во внешней
среде, но и внутри самих организмов. Непосредственной причиной выми-
рания является, очевидно, естественный отбор в результате борьбы за
существование, которая начинает принимать неблагоприятный для дан-
ного вида характер. Если эта борьба в прошлом приводила к процветанию,
а теперь приводит к вымиранию, то это указывает на то, что характер борь-
бы в процессе эволюции меняется, так как, с одной стороны, меняется
биологическая обстановка, но с другой стороны — меняется и сам орга-
низм. Ошибочно было бы искать причины вымирания в одной только среде
или в самих только организмах. Вымирание есть, несомненно, результат
своеобразгого соотносительного развития организма и среды. Это свое-
образное соотносительное развитие повторяется весьма закономерно в
отдельных ветвях филогенетического древа. Суть его, очевидно, заклю-
чается в прогрессивном приспособлении к более узким условиям сущест-
вования, т. е. в специализации. Специализированные формы и образуют
всегда концы ветвей. Это не значит, что не существует иных путей разви-
тия, кроме специализации; однако неспециализированные организмы,
поднимаясь все выше, располагаются в стволах и в основаниях ветвей
филогенетического древа.
Первым указавшим на значение неспециализированных форм в про-
грессивной эволюции был известный американский палеонтолог Коп.
Дальнейший анализ путей эволюционного процесса был проведен Осборном
и Северцовым. Мы уже ознакомились с ароморфозами, т. е. явлением
общего подъема организации на более высокую ступень. Такие ароморфозы
ведут не к вымиранию, а к прогрессивному видообразованию и процве-
танию. Мы не видим никаких ограничений для возможности дальнейших
ароморфозов и, следовательно, не видим никаких оснований для предпо-
ложений об ограниченности эволюции вообще. Однако в отдельных вполне
оформившихся филогенетических ветвях дело, очевидно, обстоит иначе —
здесь мы видим все далее идущее обособление отдельных ветвей при их
прогрессивной адаптации. Диференциация форм (адаптивная радиация)
ведет к прогрессивному обособлению и к приспособлению отдельных
групп к все более узким условиям существования, к все большей специ-
ализации. В этих ветвях мы неизменно видим вслед за периодом подъема,
ведущим к максимальному процветанию, следующий период упадка, за
которым быстро следует полное вымирание. Причины вымирания обычно
493
Искали во вйешней среде. Кювье говорил о грандиозных катастрофах.
Позднейшие палеонтологи говорили об изменениях климата, о вторжении
новых форм и т. п. Все эти изменения внешней среды, несомненно, имели
огромное значение, но они не имели характера катастроф, и в то время
как одни формы погибали, другие, наоборот, закалялись в борьбе, разви-
вались прогрессивно и переходили в фазу процветания. Следовательно,
дело не только во внешних факторах, но и в самих организмах. Италь-
янский зоолог Роза полагал, что таким внутренним фактором, веду-
щим к вымиранию, является прогрессивная редукция изменчивости —
процесс, будто неизбежно сопровождающий всякую эволюцию. Австрий-
ский палеонтолог Абель говорит о противоположном процессе уве-
личения изменчивости, ведущего к дегенерации. Мы не видим в этих
явлениях общего приципа, достаточного для объяснения всех фактов,
и предполагаем, что в основе процесса вымирания лежат те же самые про-
тиворечия, которые являются движущей силой эволюции. Как во внутри-
видовой борьбе создаются более совершенные формы через уничтожение
менее жизненных, так и в межвидовой борьбе возникают все новые более
совершенные ветви, вытесняющие и заступающие место старых, менее
совершенных. Все новое создается ценой уничтожения старого. Вопрос
о более или менее жизненных, о более или менее прогрессивных ветвях
решается теми сложными соотношениями между организмом и средой,
которые определяют пути их развития.
В отдельных ветвях мы можем отметить одну закономерность, на ко-
торую мы уже неоднократно обращали внимание: чем быстрее идет про-
грессивное развитие какой-либо ветви по пути адаптации, чем быстрее
она достигает расцвета, тем быстрее она идет и к вымиранию. Примерами
могут служить быстрая эволюция вымерших стегоцефалов и медленное
развитие современных амфибий, быстрая специализация птерозавров
и медленная эволюция птиц, ранняя специализация териодонтов и мед-
ленная — млекопитающих. Среди последних можно отметить раннюю
специализацию и вымирание прототерий (Multituberculate — Monotre-
mata) и медленную эволюцию плацентных млекопитающих; быструю
специализацию вымерших махайродонтид и более медленное развитие
кошек, или быструю специализацию вымерших литоптерн и более мед-
ленную специализацию современной лошади и т. д.
Совершенно ясна связь между процессом специализации, ее темпом
и процессом вымирания. Быстрая специализация нередко ведет, к образо-
ванию «инадаптивных», негармонических форм. Такая специализация
заводит в тупики, закрывающие пути для дальнейших преобразований,
как это показано на эволюции различных копытных В. Ковалевским.
Формы, развивающиеся более медленно, чаще идут по пути более совер-
шенной адаптации и вытесняют первые. Однако и эти формы, хотя и мед-
леннее, но также идут к вымиранию, и причину этого мы усматриваем в
самой специализации. Форма, приспосабливающаяся к определенным
более узким условиям существования — к определенной почве, климату,
к определенной пище, — тем самым получает преимущества в борьбе за
существование. Специализация означает для данной формы ослабление
борьбы, иногда это почти уход от борьбы с другими формами. Не имеющая
конкурентов форма значительно возрастает в численности. Вместе с тем,
однако, возрастает внутривидовая борьба, ведущая к все более совершен-
ной специализации, к все более точной приспособленности организма к
строго определенным условиям внешней среды. Все уклонения уничто-
жаются. Изменчивость «идеально» приспособленной формы падает до
минимума, и вместе с тем падает до минимума и пластичность данной фор-
мы. Нередко наблюдаемое прогрессивное увеличение общих размеров
тела мы рассматриваем тоже как специализацию, дающую организму в дан-
ный момент известные преимущества в борьбе за существование (облегче-
494
ние йепосредственной борьбы с врагами, более экономный обмен веществ).
Однако и увеличение размеров ведет к уменьшению пластичности вида,
так как связано с удлинением жизненного цикла, с ослаблением темпа
размножения, а следовательно, и с уменьшением темпа самой эволюции.
Благодаря специализации организмы достигают процветания, однако
это процветание, связанное с размножением и обострением внутривидовой
борьбы, неизбежно ведет к уменьшению пластичности. При сравнительно
константных условиях среды такие специализированные организмы могут,
как бы застыв на известном уровне, сохраняться в течение многих геоло-
гических периодов почти неизменными (брахиоподы Lingula и Crania
с силура доныне, Discina даже с кембрия, двудышащая рыба Ceratodus
с триаса доныне, близкие к современным Hatteria представители Spheno-
dontidae с транса).
При всяких изменениях (и, конечно, не только при изменениях кли-
мата, почвы и т. д.,но в особенности при изменениях биологической обста-
новки) среды сейчас же сказывается недостаток пластичности специали-
зированных организмов. Чем более специализированы организмы, тем
более они отстают от темпа изменений среды и тем быстрее они идут в
этом случае навстречу полному вымиранию. Преимущества специализа-
ции — это преимущества настоящего времени. Однако те же самые пре-
имущества являются источником гибели в дальнейшем. Мелкие неспе-
циализированные млекопитающие мезозоя не были многочисленны, не
играли большой роли и развивались весьма медленно. Господами поло-
жения были огромные рептилии, жившие на суше, в воде и в воздухе,
специализированные в различных направлениях. Все приемущества были,
казалось бы, на стороне рептилий. Но обстановка менялась, рептилии
быстро вымерли, и началась адаптация, размножение и специализация
многочисленных форм млекопитающих. Таким образом, более универсль-
ные неспециализированные формы, не обладая нигде преимуществами
специализированных, находятся всегда в условиях бесконечно тяжелой
межвидовой борьбы с врагами, но в этой борьбе непрерывно совершен-
ствуются, меняются. Такие неспециализированные формы после периода
ароморфоза дают от себя множество боковых ветвей (адаптивная радиа-
ция), которые с большей или меньшей скоростью специализируются, а
в результате рано или поздно идут к вымиранию. Все же основные стволы
неспециализированных форм развиваются все дальше и вновь дают бо-
ковые ветви еще более совершенных специализированных форм, побеж-
дающих прежние, но обладающих в свою очередь ограниченным сущест-
вованием. В целом эволюция безгранична, но эта безграничность дости-
гается ценой постоянного ограничения, т. е. непрерывного уничтожения
множества ветвей, уступающих свое место новым, более совершенным.
Необратимость эволюции
Раньше нередко высказывалась мысль об обратимости, о цикличности
эволюционного процесса. И в настоящее время есть сторонники таких
взглядов, говорящие о специализации и деспециализации, о старении и
омоложении видов. При этом предполагается, что одно направление эво-
люционно'го процесса может заменяться обратным процессом.
Иногда, быть может, и можно говорить о некоторой деспециализации,
об обратном развитии и т. д.; однако нужно сразу отметить, что это никак
не следует понимать буквально в смысле возвращения в исходное состо-
яние.
Известный бельгийский палеонтолог Долло первый ясно высказал
это положение, получившее название закона необратимости эволюции,
или просто закона Долло. Абель так выражает сущность этого закона:
1) орган, редуцированный в течение филогенетического развития, ни-
495
когда вновь не достигает прежнего состояния; совершенно исчезнувший
орган никогда не восстанавливается; 2) бели при приспособлении к новому
образу жизни (например, при переходе от хождения по земле к лазанию
на деревьях) исчезают органы, имевшие при прежнем образе жизни боль-
шее функциональное зна-
Рис. 437. Череп третичной птицы Odontopteryx
с роговыми зубами (по Оуэну).
чение, то при вторичном
возвращении к прежнему
образу жизни эти органы
никогда не восстанавли-
ваются; на их месте по-
являются другие органы.
Поясним этот закон
некоторыми примерами.
В эволюции млекопитаю-
щих коренные зубы посте-
пенно усложнялись, и простая, первоначально коническая, коронка
становилась трехзубчатой, трехбугорчатой и многобугорчатой. В част-
ности, у примитивных китов (Patriocetus) были коренные зубы со
сложной семибугорчатой коронкой (все бугры расположены в один
ряд). История эмбрионального раз-
вития беззубых китов показывает,
что эти сложные зубы в процессе их
редукции первоначально распались
на отдельные конические зубы (со-
ответственно отдельным буграм), а
Рис. 439. Рудименты первичного (ер, пу,
hyp, х) и вторичного (а, Ь, с) панцыря
у современной кожистой морской чере-
пахи Sphargis.
Рис. 438. Часть вторичного панцыря из
кожных костей у ископаемой морской че-
репахи Cosmochelys (по Эндрью). Подоб-
ный панцырь закладывается и у молодых
современных кожистых черепах Sphargis.
затем уже совсем редуцировались. Путь редукции не есть, таким
образом, путь обратного развития, и получившиеся при редукции простые
конические зубы ни в какой мере не равноценны исходным формам кони-
ческого зуба предков млекопитающего. У птиц когда-то были зубы, но
потом они исчезли, и вместо них на челюстях развился роговой клюв.
У птиц могут вторично опять развиться зубы, как у третичной птицы
Odontopteryx (рис. 437), однако это уже роговые зубы совсем иною проис-
хождения. У морских черепах, плавающих в открытом море, костный
панцырь редуцируется. На палеонтологическом материале можно про
следить шаг за шагом такую редукцию (ряд Thalassemys — Toxochelys —
496
Archelon).Такая редукция панциря произошла и у далеких предков
морской кожистой черепахи. Остались лишь небольшие расширения
ребер, остатки краевых окостенений спинного щита и небольшие кости
брюшного щита (меловая Protosphargis). Затем эти черепахи вновь стали
жить в прибрежной зоне морского прибоя, и, очевидно, в связи с этим
у них вновь развился панцирь из кожных костей. Однако у них не проис-
ходило восстановления редуцированных костей старого панциря, не
расширялись вновь ребра, не расширялись кости брюшного панциря
и краевые пластинки спинного. В коже развилась целая мозаика новых
кожных окостенений, образовавших совершенно новый костный панцырь
(эоценовая Cosmochelys) (рис. 438). Теперь эти черепахи вновь вернулись
к жизни в открытом море, и этот панцырь второй генерации так же реду-
цируется, как редуцировался первый. У молодой современной кожистой
черепахи (Spbargis) имеется закладка такой же мозаики кожных костей
панцыря, как и у ископаемой Cosmochelys, однако позднее эти кости
редуцируются, и у взрослой Sphargis имеются рудименты кожных панцы-
рей двух генераций, лежащие одни над другими (рис. 439).
Из этих примеров ясно видво содержание закона Долло. Нельзя
сказать, что возможность обратных преобразований совершенно исклю-
чается. Никогда, однако, развитие не идет вспять по прежним путям,
и никогда оно не приводит it восстановлению уже пройденных этапов
и прея,-них структур. Таким образом, мы видим, что из парного плавника
рыбы при переходе к жпзнп на суше развилась пятипалая конечность,
а при обратном переходе наземного позвоночного к жизни в воде хотя
и восстанавливается опять плавникообразная конечность — ласт, но все
же это не прежний плавник, а нечто новое, своеобразное. Зако» необра-
тимости эволюции имеет огромное принципиальное значение. Организм,
переживший некоторую историю, уже не есть прежний организм, и при
возвращении в прежнюю обстановку он не мояют реагировать так, как
он реагировал на эту обстановку раньше. В процесс эволюции организм
становится, действительно, иным, и новые его качества определяют вместе
с тем иные его реакции на изменение среды. Это одно из лучших доказа-
тельств того, что направление эволюционного процесса определяется
не только средой, но и унаследованной внутренней структурой самого
организма (в широком смысле этого слова). Всякий организм имеет всегда
свое историческое обоснование; эта история ни при каких обстоятельствах
не моя,-ет быть вычеркнута и имеет огролщое значение в определении
направления дальнейшей эволюции.
УКАЗАТЕЛЬ ЛИТЕРАТУРЫ
Список этот не претендует на полноту. В основном он содержит лишь некоторые
классические работы, сводки и более новые специальные исследования, еще не вошед-
шие в сводки. Немного полнее приведены не слишком специальные работы, напеча-
танные на русском языке.
I. История морфологии и основные понятия сравнительной
анатомии
1. Cuvier G., Lemons d‘anatomie согпрагбе recueillies et publiees par Durniril
et Duvernoy, 5 t., Paris, 1798—1805.
2. О ken L., Lehrbuch der Naturphilosophie, 1809—1811.
3. Geoffrey Saint-Hilaire E., Philosophic anatomique, Paris, 1818.
4. Meckel J. F., System der vergleichenden Anatomie. 6 Bde, Halle, 1821—1833.
5. В a e г К. E. von, Ueber die Entwicklungsgeschichte der Tiere. Konigsberg,
1828—1837.
6. Rathke H., Anatomisch-philosophische Untersuchungen uber den Kiemen-
apparat und das Zungenbein, Riga und Dorpat, 1832.
7. Cuvier G., Recherches sur les ossements fossiles, Paris, 1834—1836.
8. Stannius H.u. Siebold v., Lehrbuch der Zootomie, 2 Bde, Berlin, 1848.
9. О wen R., On the Archetype and homologies of the vertebrate skeleton, London,
1848.
10. Milne-Edwards H., Lemons sur la physiologic et 1‘anatomie сотрагёе
de 1‘homme et des animaux, 14 vol., Paris, 1857—1880.
11. Huxley T. H., On the theory of the vertebrate skull {the croonian lecture),
Proc. Roy. Soc. London, vol. 9, 1858.
12. Owen R., on the anatomy of vertebrates, 3 vol., London, 1866—1868.
13. Haeckel E., Generelle Morphologie der Organismen, 2 Bde, Berlin, 1866.
14. Gegenbaur C., Grundriss der vergleichenden Anatomie, Leipzig, 1874.
15. Dohrn A., Der Ursprung der Wirbeltiere und das Prinzip des Functions-
wechsels, Leipzig, 1875. Дорн А., Принцип смены функций, M., 1937.-
16. Б а л ь ф у р и Форстер, Элементы эмбриологии, СПБ, 1880.
17. Борзенков Я. А., Чтения по сравнительной анатомии. Учен. вап.
Моск, ун-та, отд. ест. ист., вып. 4, 1884.
18. Н а е с k е 1 Е., Systematische Phylogenie, 3 Bde, Berlin, 1894—1896.
19. Gegenbaur C., Vergleichende Anatomie der Wirbeltiere, 2 Bde, Leipzig,
1898—1901.
20. M e h n e r t E., Biomechanik erschlossen aus dem Principe der Organogenese,
Jena, 1898.
21. С e в e p ц о в A. H., Этюды по теории эволюции, Киев, 1912; переиэдано,
Берлин, 1921.
22. С е в е р ц о в А. Н., Современные задачи эволюционной теории, Москва,
1914.
23. R u s s е 1 Е. S., Form and Function, London, 1916.
24. Broman L, Das sogenannte biogenetische Grundgesetz und die moderne
Erblichkeitslehre, Munchen, 1920.
25. Бэр К. Э., Избранные работы, Гиз, Ленинград, 1924.
26 Nordenskiold Е., Geschichte der Biologic, Jena, 1926.
27. Dombrowski. Br., Ein Versuch der Analyse einiger Korrelationen,
Zschr. f. Anat. u. Entw., Bd. 79, 1926.
28. Franz V., Ontogenie und Phylogenie. Abhandl. z. Theorie d. organischen
Entwicklung, H. 3, Berlin, 1927.
29. Sewertzoff A. N., Ueber die Beziehungen zvdschen der Ontogenese und
Phylogenese der Tiere, Jen. Zschr. Naturwiss., Bd. 63, Jena, 1927.
30. В e e r G. R. de, Embryology and evolution, Oxford, 1930.
4PS
91. Северцов a. H., Дегенерация органов и субституция, Ивв. Акад,
наук СССР, 1930.
32. Hermann Friedrich, Kritische Studien zum Wesen deg Begrifs der
Homologie. Erg. d. Anat. u. Entw. Bd. 29, 1932.
33. S e w e r t z о f f A.N., Studien fiber die Reduktion der Organe der Wirbelticre,
Zool, Jahrb. Anat., Bd. 53, 1931.
34. S e w e г t z о f f A. N., Morphologische Gesetzmassigkeiten der Evolution,
Jena, 1931.
35. E ж и к о в И., К теории рекапитуляции, Зоол. жури., 12, 1933.
II. Руководства по сравнительной анатомии (со сравнительной
эмбриологией) и зоологии (с преобладанием сравнительноанатоми-
ческого материала)
По сравнительной анатомии (со срапнителыюй эмбриологией)
l. Vogt С. u. Jung Е. Lehrbuch der praktischen vergleichenden Anatomie,
2 Bde, Braunsschwcig, 1883—1894.
2. О p p e 1 A., Lehrbuch der vergleichenden mikroskopischen Anatomie def
Wirbeltiere, 1—VIII, Leipzig, 1896—1914.
3. G e g e n b a u r C., Vergleichende Anatomie der Wirbeltiere, 2 Bde, Leipzig,
1898—1901.
4. Ellenberger W. u. Baum H., Handbuch der vergleichenden Ana-
tomie der Haustiere, Berlin, 1900.
5. Her twig O., Handbuch der vergleichenden und experimentellen Entwick-
lungslehre der Wirbeltiere, 3 Bde (6 книг), Jena, 1901—1906.
6. Wiedersheim R., Vergleichende Anatomie der Wirbeltiere, Jena, 1909.
7. Go o dri ch E. S., Vertebrata craniata I (Cyclostomes and Fishes), Lankes-
ter R., A Treatise on Zoology, IX, London, 1909.
8. Bfitschli O., Vorlesungen fiber vergleichende Anatomie, 2 Bde, Berlin,
d910 (1921).
9. L a n g A., Handbuch der Morphologie der wirbellosen Tiere, Jena, 1912.
10. M ar t i n P., Lehrbuch der Anatomie der Haustiere, 3 Bde, Stuttgart, 1912—
1919.
11. Гессе и Д оф л ей и, Строение и жизнь животных. 1. Тело животного
как самостоятельный организм, СПБ, 1913.
12. Бгочли О., Лекции по сравнительной анатомии (вып. 1, Введение, кожа,
скелет), П., 1917.
13. Ш и м к е в и ч В., Курс сравнительной анатомии позвоночных животных,
Л., 1922.
14. Ihle J., van Kampen Р., Nierstrasz Н., Versluys J.,
Vergleichende Anatomie der Wirbeltiere, Berlin, 1927.
15. G о o d г i c h E. S., Studies on the structure and development of vertebrates,
London, 1930.
16. Bolk L., Go p pert E., Kallius E., Lubosch W., Handbuch
der vergleichenden Anatomie der Wirbeltiere, I—VI, Berlin, 1932—1937.
17. Bocker H. Einffihrung in die vergleichende biologische Aantomie der
Wirbeltiere. I, II, Jena, 1935—1937.
По зоологии (co сравнительной анатомией)
18. В г о n h G. Н., Klassen und Ordnungen des Tierreichs. VI. Wirbeltiere. Осо-
бенно: Hoffmann С. K., Amphibien, Leipzig, 1873—1878; Hoffmann С. K.,
Reptilien, 1887—1890; Rauther M. etc., Pisces, I, 1901—1924; II, 1927.
19. Delage J. et H erouard E„ Trait ё de zoologie concrfete (серия непол-
ная); особенно VIII, Les Ргосогйёв, 1898.
20. П a p к e p T. Дж. иГасвель В. А., Руководство к воологии, ч. 2,
М., 1899.
21. Ray Lankaster Е., A treatise on Zoology. (IX Goodrich E. S.,
Fishes, 1909).
22. The Cambridge natural history, 10 томов, ив них первые 6 относятся к беспозво-
ночным (VII — Harmer, Herdman, Bridge, Hemichordata, Tunicata, Amphi-
oxus, Fishes, 1910).
23. Kfikenthal W. u. Krumbach Th., Handbuch der Zoologie, Berlin-
Leipzig, 1923 (еще не вакончепо).
П1. Монографии и сводки
1. В а е г К. Е„ Ueber die Entwicklungsgeschichte der Tiere, KSnigsberg, 1828—
1837.
2. D u g 6 s A., Recherches sur l’ost6ologie et la myologie des Batraciens, Paris,
1835.
499
9. М й 11 e r J., Vergleichende Anatomle der Myxinoiden, Abhandl. d. Preuss.
Akad. d. Wissensch., Berlin, 1835—1845.
4. M tiller J., Ban und Grenzen der Ganoiden, Abhandl. d. Preuss. Akad. d.
Wissensch., Berlin, 1846.
5. Goette A., Entwicklungsgeschichte der Unke, Leipzig, 1875.
6.H uxley T. H., On Ceratodus Forsteri with observations on the classification
of fishes, Proc. Zool. Soc. London, 1876.
7. Balfour F. M., A monograph on the development of Elasmobranch fishes,
London, 1879.
8. D о h r n A., Studien zur Urgeschichte des Wirbeltierkorpers I—XXV, Mit-
teilungen a. d. Zoolog. Station Neapel, 3—18, 1880—1907.
9. Ftirbringer M., Untersuchungen zur Morphologic und Systematik der
Vogel, I, II, Amsterdam, 1888.
10. P о 1 1 a r d H. B., On the anatomy and phylogenetic position of Polypterus,
Zoolog. Jahrb. Anat., 5, 1892.
11. Willey A., Amphioxus and the ancestry of vertebrate, New York, 1894.
12. Dean B., Fishes living and fossil, New York, 1895.
13. Ecker A. u. Wiedersheim R., Anatomic des Frosches (E. Gaupp).
8-Bde, Leipzig, 1896, 1899, 1904.
14. S c h a и i n s 1 a n d H. Beitrage zur Etwicklungsgeschichte und Anatomic
der Wirbeltiere, I—III (Sphenodon, Callorhynchus, Chameleo), Zoologica (C. Chun.),
Stuttgart, 1903.
15. О s b о r n H. F., The Reptilian subclasses Diapsida and Synapsida, Mem.
Amer. Museum Nat. History, 1, 1903.
16. Gregory W. K., The orders of Teleostomous fishes, Ann. New York Acad.
Sci. 17, 1907.
17. Gregory W. K., The orders of Mammals, Bull. Amer. Mus. Nat. Hist.,
27, New York, 1910.
18. С e в e p ц о в Л. H., Исследование об эволюции низших позвоночных, I, If,
III, Русск. арх. анат. гистол., эмбриологии. 1, 2, Л., 1917, 1925.
19. Н е i 1 m a n n G., The origin of birds, London, 1926.
20. Weber M., Die Saugetiere, 2 Bde, Jena, 1927.
21. Franz V., Morphologic der Akranier, Erg. d. Anat. u. Entw. gesch., 27, 1927.
IV. Работы, касающиеся частей организма, систем органов и отдельных
органов
1. Общие покровы
1. HertwigO., Ueber den Bau und die Entwicklung der Placoidschuppen
Und der Zahne der Selachier, Jen. Zschr. f. Naturwiss., Bd. 8. 1874.
2. В о a s J. E. V., Ein Beitrag zur Morphologic der Nagel, Krallen, Hufe und
Klauen der Saugetiere, Morph. Jahrb., Bd. 9, 1884.
3. Львов В., Сравнительное исследование и описание волоса, щетины, иглы
у млекопитающих и пера у птиц. Учен. зап. Моск, ун-та, отд. ест., вып. 4, 1884.
4. Davies H.R., Die Entwicklung der Feder und ihre Beziehungen zu anderen
Integumentgebilden, Morph. Jahrb., Bd. 15, 1889.
5. В о a s J. E. V., Zur Morphologie der Wirbeltierkralle, Morph. Jahrb., Bd.
21, 1894.
6. M a u r e r F., Die Epidermis und ihre Abkommlinge, Leipzig, 1895.
7. К e i b e 1 F., Ontogenie und Phylogenie von Haar und Feder, Erg. d. Anat. u.
Entwickl. Gesch., Bd. 5, Wiesbaden, 1895.
8. К r a u s e W., Die Entwicklung der Haut und ihrer Nebenorgane, O. Her-
twig, Handbuch d. vergl. u. exp. Entw. Lehre der Wirbeltiere, Bd. 2., T. 1, Jena, 1906.
9. Goodrich E. S., On the scales of fish, living and extinct., Proc. Zool. Soc.
London, 1907.
10. S e w e r t z о f f A. N., The development of the scales of Acipenser ruthenus,
Journ. Morph, a. Phys., vol. 42, 1926.
11. Sewer tzof f A. N., Die Entwicklung der Knochenschuppen von Polypterus
delhesi, Jen. Zschr. f. Naturw. Bd. 67, Jena, 1932.
12. M а т в e e в Б. С., Об эволюции кожных покровов позвоночных путем
эмбриональных изменений, Зоол. журн., И, 1932.
2. Скелет (включая морфологию головы и конечностей в целом)
а. Позвоночник и грудина
1. Goppert Е., Untersuchungen zur Morphologie der Fischrippen, Morphol.
Jahrb. 23, 1895.
2. G 6 p p e r t E., Die Morphologie der Amphibienrippen, Fschr. f. C. Gegenbaur,
1896.
500
3. Schauinsland H., Die Entwicklung der Wirbelsaule nebst Rippen und
Brustbein. O. Hertwig, Handbuch d. Vergl. u. exp. Entw.-lehre., Bd. 3, T. 2, 1905.
4. Кравец Л. П., История развития грудины и epistem’ального аппарата
млекопитающих. Труды сравн.-анатом, инет. Моск, унив., вып. 5, 1905.
5. Емельянов С. В., Развитие ребер и соотношение их с позвоночником,
I, II, III, IV. Русек, зоолог, журн., 5, 6, 8, 1925, 1926, 1928.
6. Matveiev В. Die Entwicklung der vorderen Wirbel u. des Weber’schen
Apparates bei Cyprinidae. Zool. Jahrb., Anat., 51, 1929.
7, Еме лья нов С. В., Развитие ребер наземных позвоночных, изд. Акад,
наук СССР, Труды лаборатории эволюционной морфологии, I, 1933.
8. Gadow Н. Е., The evolution of the vertebral column. Cambridge, 1933.
9. Emelianov S. W., Die Morphologie der Fischrippen Zool. Jahrb. Anat.,
61, 1936.
10. E m e 1 i a n о v S. W., Die Morphologie der Tetrapodenrippen, Zoolog. Jahrb,
Anat., 62, 1936.
б. Морфология головы в целом
1. W i j h е J. W. van, Ueber die Mesodermsegmente und die Entwicklung der
Nerven des Selachierkopfes, Verhandl. Akad. Vetensc. Deel. 22, Amsterdam, 1882.
2. R a b 1 C,, Theorie des Mesoderms, Morph. Jahrb., 15, 1889.
3. Froriep A., Entwicklungsgeschichte des Kopfes, Erg. Anat. u. Entw.
gesch., Bd. 1 u. 3, 1891, 1893.
4. Северцов A. H., Развитие затылочной области низших позвоночных
в связи с вопросом о метамерии головы, Учен. зап. Моск, ун-та, отд. ест. ист., вып.
11, 1895.
5. Северцов А. Н., Очерки по истории развития головы позвоночных,
I. Метамерия головы электрического ската. Учен. зап. Моск, ун-та, отд. ест. ист.,
вып. 15, 1900.
6. Кольцов Н. К., Развитие головы миноги, Учен. зап. Моск, ун-та, отд.
ест, ист., вып. 16, 1901.
7. Schauinsland Н., Beitrage zur Entwicklungsgeschichte u. Anatomie
der Wirbeltiere. I, II, III (Sphenodon, Callorhynchus, Chameleo). Zoologica (C. Chun),
Stuttgart, 1903.
8. Filatoff D., Die Metamerie des Kopfes von Emys lutaria., Morph. Jahrb.,
37, 1907.
9. G r e i 1 A., Entwicklungsgeschichte des Kopfes und des Blutgefaszsystems von
Ceratodus Forsteri (S e m о n, Zoologische Forschungsreisen in Australien, Bd. I). Jena,
1908, 1913.
10. Luther A., Beitrage zur Kenntnis von Muskulatur und Skelett des Kopfes
des Haies Stegostoma tigrinum und der Holocephalen. Acta Societatis Scient. Fenni-
cae., T. 37, 1909.
И. Б e л о г о л о в ы й Ю. А., Сегментальное положение границы черепа у
Sauropsida, Учен. зап. Моск, ун-та, отд. ест. ист., вып. 28, 1911.
12. М а т в е е в Б. С., К вопросу о мезомерии головы селахий, Дневник эоол.
отд. О. Л. Е. А. и Э., нов. сер., т. II, № 5, 1915.
13. Северцов А. Н., Исследование об эволюции низших позвоночных,
I, II, III, Русск. арх. анат., гист. и эмбр., I, III, 1917, 1925.
14. Матвеев Б. С., Строение эмбрионального осевого черепа низших рыб
(к вопросу о сегментальном строении черепа), Бюлл. Моск, о-ва исп. прир., 1925.
15. Sewertzoff A. N., The head skeleton and muscles of Acipenser ruthenus,
Acta Zoologica, IX, 1928.
См. также в списке монографий работы Бальфура, Дорна.
в. Череп
1. Huxley Т. Н., On the theory of the Vertebrate skull, Proc. Roy. Soc.
London, 9, 1858.
2. Parker W. K., On the structure and development of the skull of the
common fowl, Philos. Trans. Roy. Soc. London, 1869.
3. P a r k e r W. K., On the structure and development of the skull in the Frog;...
in the Batrachia, I, II, III, Philos. Trans. Roy. Soc. London, 1871, 1876, 1881.
4. Gegenbaur C., Untersuchungen zur vergleichenden Anatomie der Wir-
beltiere, III. Das Kopfskelett der Selachier, Leipzig, 1872.
5. P a r k e r W. K., On the structure and development of the skull in the pig;
in Mammalia, II, Edentata, III, Insectivora,’ Philos. Trans. Roy. Soc. London, 1874,
1884, 1885.
6. W i e d e r s h,e i m R., Das Kopfskelett der Urodelen, Leipzig, 1877
7. P a r k e r W. K., On the structure and development of the skull in the common
Snake;... in the Lacertilia, Philos. Trans. Roy. Soc. London, 1878, 1879.
8. Parker W. K., On the structure and development of the skull in sharks
and skates, Trans. Zool. Soc. London, 10, 1878.
501
9. Parker W. К. On the structure and development of the skull in Sturgeonsi...
in Lepidosteus osseus, Philos. Trans. Roy. Soc. London, 1882.
10. P a г к e r W. K., On the skeleton of the Marsipobranch fishes, I, II, Philos.
Trans. Roy. Soc. London, 1883.
11. Parker W. K., On the structure and development of the skull in the Cro-
codilia. Trans. Zool. Soc. London, 1883.
12. Sagemehl M., Beitrage zur vergl. Anatomie der Fische I—IV, Morphol.
Jahrb., 9, 10, 17, 1884, 1885, 1891.
13. Wi n si о w G. M., The chondrocranium in the Ichthyopsida, Turfts College
Studies, 5, 1898.
14. С ушиин П. IL, К морфологии скелета птиц. I. Череп Tinnunculiis
alaudarius. Учен. вап. Моск, ун-та, отд. ест. ист., вып. 14, 1898.
15. S е w е г t z о f f A. N., Die Entwicklung des SelachierschSdels. Festschr. f.
Kupffer, Jena, 1899.
16. Sewer tzoff A. N., Zur Entwicklungsgeschichte des Ceratodus Forsteri,
Anat. Anz., 21, 1902.
17. Fflrbringer K., Beitrage zur Morphologie des Skelettes der Dipnoer,
Jenaische Denkschrift, IV, 1904.
18. G a u p p E., Die Entwicklung des Kopfskeletts, O. Hertwig, Handbuch d.
vergl. u. exp. Entw. Lehre, Bd. 3, T. 2, 1905.
19. Кравец Л. П., Развитие хрящевого черепа Ceratodus, Труды сравн.
анат. ин-та Моск, ун-та, 8, 1910.
20. V е i t О., Entwicklung des Primordialcranium von Lepidosteus, Anat. H.
44. 1911.
21. Шмальгаувен И. И., К вопросу о происхождении аутостилии у
двудышащих и у наземных позвоночных, Русск. зоол. жури., 3, 1922.
22. Stadtmflller Fr., Studien am Urodelensch&del. I. Zur Entwicklungsge-
schichte des Kopfskeletts der Salamandra maculosa, Zschr. Anat. u. Entw. gesch., 75, 1924.
23. С e в e p ц о в A. H-, Развитие покровных костей черепа Amia calva, Бюлл.
Моск, о-ва исп. прир. 1925.
24. Sewer tzoff A. N., Beitrage zu einer Theorie des KnochenscMdels
der Wirbeltiere, Anat. Anz., 60, 1925—1926.
25. S e w e r t z о f f A. N., Studies on the bony skull of fishes, Quart. J. Microsc.
Sci., 70, 1926.
26. De Beer G. R. The developement of the vertebrate skull. Oxford, 1937;
Костный череп амниот
27. F u c h s H., Untersuchungen fiber Ontogcnie und Phylogenie der Gaumenbil-
dungen bei den Wirbeltieren, 1, II, Zschr. f. Morph, u. Anthrop., 10, 11, 1907, 1908.
28. Versluys J.. Das Streptostylie Problem und die Bewegungen im Schadel.
bei Sauropsiden, Zool. Jahrb. Anat. Suppl., 15. 1912 (Festschr. f. Spengel, 2).
29. Boas J., Dio Schlafenuberdachung und das Palatoquadratum in ihrem Ver-
hSltniss zum ubrigen Schadel bei den Dipnoern und den terrestren Wirbeltieren, Morph.
Jahrb., 49, 1914.
30. L a k j e r T., Studien fiber die Gaumenregion bei Sauriern im Vergleich nlit
Anamniern und primit. Sauropsiden, Zool. Jahrb., Anat., 49, 1927.
Висцеральный аппарат
31. G a u p p E., Das Hyobranchialskelett der Wirbeltiere, Erg. d. Anat. u. Entw.
gesch., 14, 1905.
32. В о с к о б о й и и к о в М. М., Развитие висцерального скелета костистых
рыб. Зап. Киевск. о-ва естеств., 1909.
33. В о с к о б о й н и к о в М. М., Очерки по бранхиомерии позвоночных, I,
П. III. Зан. Кисвск. о-ва естеств., 24, 1914.
34. К р и в е ц к и й А., К вопросу о морфологии влементоз гиоидной дуги
селахий- Русск зоол. жури., 2, 1917.
35. Боголюбский С. Н.. К сравнительной морфологии подъязычного
аппарата амфибий. 1—IV. Русск. зоол. жури., 4. 5. 6, 1924, 1925, l^ve.
36 W о s к о b о i n i к о f f М. МД Der Apparat der Kiemen "timing bei den
Fischen Zoolog Jahrb., Amt.. 55 1932.
37. tony F., Evolution de I’appareil hyo-branchial Marseil e, 1933.
Слуховые косточки и среднее ухо
38- Reichert. не Visceralbogen der Wirbeltiem im a 'zemeinen und
deren Metamorphose bei den Vogein and Saugetieren. Mullers Arch. f. An .t u. Phvsiol.,
1837
39. Ga up p E., Ontogenese und Phylogenese des schalleitenden Apparates bei
den Wirbeltieren, Erg. d. Anat. u Entw.. 8 1898
40. Versluys J., Die mittlere und aussbre Ohrsph8re det Lacertilia und Rhyn-
chdcephalia, Zool, Jahrb., Anat., 12, 1899.
502
41. Сушкин П. П., Заметит о черепе, 1, Биол. жури. 4., Москва, 1910.
42. Gaupp Е., Die Reichert’sche Theorie, Arch. f. Anat. n. Physiol., Anat. Abt.,
Snppl., 1912.
43. Goodrich E. S., The chorda tympani and middle ear in reptiles, birds
and mammals. Quart. J. Mier. Sci., 61, 1915.
44. Домбровский Б. А., Срапнительноанатомический очерк проводя-
щего ввук аппарата земноводных, Зап. Киевск. о-па естеств., 26, 1917.
45. S chmal hausen I. I., Der Suspensorialapparat der Fische nnd das
Problem der Gehorknochelchen, Anat. Anz., 55, 1923.
46. К 1 a a u w. van der C. J., Bau und Entwicklung der Gehorknochelchen,
Erg. d. Anat. u. Entw.-gesch., 1924.
47. Домбровский Б. А., Строение слухового аппарата у ящериц; у
змей, ящериц и крокодилов; у птиц, Русек, воол. жури., 4, 5, 1924, 1925.
48. D о m b г о w s к у Br., Der Suspensorialapparat der Ichthyopsiden, Zschr.
f. Anat. u. Entw.-gesch., 82, 1927.
г. Конечности
1. G e g e n b a u r C., Untersuchungen zur vergleichenden Anatomie der Wir-
beltiere. 1. Carpus und Tarsus, 2. Schultergiirtel d. Wirbeltiere, Brustflosse der Fische,
Leipzig, 1864, 1865.
2. D о h r n A., Die paarigen und unpaaren Flossen der Selachier, Mitteil. Zool.
St. Neapel, 5, 1884.
3. Wiedersheim R., Das Gliedmassenskelett der Wirbeltiere, Jena, 1892.
4. M о 11 i e r S., Die paarigen Extremitaten der Wirbeltiere, I, II, III, Anat.
H., Bd. 3, 5, 8, 1893, 1895, 1897.
.5 . Emery C., Beitrage zur Entwicklungsgeschichte u. Morphologie des Hand-
und Fuszskeletts der Marsupialier; Hand- und Fuszskelett von Echidna hystrix (Semon,
Zoolog. Forsch. Reisen in Australien, I, II, III), Jen. Dkschr., 5, 6, 1897, 1902.
6. Braus H., Die Entwicklung der Form der Extremitaten und des Extremi-
tatenskelette. O. Hertwig. Handbuch d. vergl. u. exp. Entwicklungslehre d. Wirb-, Bd.
3, T. 2, Jena, 1904.
7. Goodrich E. S., Development, structure and origin of fins of fishes.
Quart. J. Mier. Sci., 1906.
8. Se wertz of f A. N., Studien uber die Entwicklung der Muskeln, Nerven
h. des Skelettes der Extremitaten der niederen Tetrapoda, Бюлл. Моск, о-ва испыт.
прир., 1908.
9. Дерюгин К. М., Строение и развитие плечевого пояса и грудных плав-
ников у костистых рыб. Труды С.-Петерб. о-ва естеств., 39, 1909.
10. Rabi С., Bausteine zu einer Theorie der Extremitaten der Wirbeltiere, I.
Leipzig, 1910.
11. Ш мал ьгау вен И. И., Непарные плавники рыб и их фплогенениче-
ское развитие, Зап. Киевск. о-ва естеств., 1913.
12. Ш м а л ь г а у в е н И. И., Развитие конечностей амфибий и их значение
в вопросе о происхождении конечностей наземных позвоночных, Учен. зап. Моск, ун-та,
отд. ест. ист., вып. 37, 1915.
13. Шмальгаувен И. И., К вопросу о морфологическом значении не-
парного плавника Dipnoi, Русск. воол. журн., 1, 1916.
14. Шмальгаувен И. И., О функциональном значении плавников рыб,
Русск. воол. Htypii., 1 1916.
15. К р Ы ж а н о в с к и й С., Развитие скелета парных плавников у Pristi-
urus melanostomus, Русск. воол. журн. 1, 1916.
16. Ш м а л ь г а у в е н И. И., О конечностях Ranidens sibiricus Kessl, Русск.
воол. журн., 2, 1917
17. Шмальгаузен И. И.. О покронных костях плечевого пояса амфибий,
Русск. воол. HtypH., 2, Г 17.
18. Steiner Н . Die Entwicklung des VogelfiJgelskeletts, Acta Zoologica,
3, 1922.
19. Васнецов В В., О функции mesocoracoide'.im у костистых рыб, Русск,
воол. HtypH., 3, 1922.
20. К р ы ж а н о в с • и й С. Г., Развитие парных плавников Acipenser, Amia,
Lepidosteus и вопросы тео ли конечностей. Труды научно-иссл. ин га зоол. Моск,
ун-та, 1, 1925. *
21. Sewertzoff A. N-, Die МогрЬс1ог'е лег Brustflosse der Fische, Jen.
Zschr. F. Naturw., 62, 1926
22. Шестакова Г. С., К развитию к; ыла Rhea sp., Бюлл. Моск, o-ва псп.
прир., 1926.
23. Васнецов В. В, Равпитие нерпичьи» с плечевого пояса и скелета груд-
ного плавника Lepidosteus osseus, Русск. зоол. журн., 7, 1927.
24. Holmgren N., On the origin of the tetrapod limb., Acta Zoologica, 14,
1933.
25. Sewertzoff a. N ., Evolution der BauchflossBn der Fische, Zoolog.
Jahrb, Anat., Б8, 1934. й
503
3. Мускулатура (и электрические органы)
1. Edgeworth F. The cranial muscles of vertebrates, Cambridge Univ, press,
1935.
2 Fiirbringer M., Zur vergleichenden Anatomie der Schultermuskeln, 1,
2, Jen. Zschr. f. Naturw., 7, 8, 1873, 1874; 3. Morphol. Jahrb., 1, 1876; 4, 5, Jen. Zschr.
f. Naturw., 34, 36, 1900, 1902.
3. Vetter B., Untersuchungen zur vergleichenden Anatomie der Kiemen-und
Kiefermuskulatur der Fische, Jen. Zschr. f. Naturw., 8, 12, 1874, 1876.
4. Ruge G., Ueber die Gesichtsmuskulatur der Halbaffen, Morphol. Jahrb.,
11, 1886.
5. Maurer F-, Aufbau und Entwicklung der vcntralen Rumpfmuskulatur
der Urodelen Amphibien, Morphol. Jahrb., 18, 1891.
6. Ruge G., Die Hautmuskulatur der Monotremen und Hire Beziehungen zu
dem Masupial- und Mammarapparat., Jen. Denkschr., 6, 1895.
7. Иванцов H. А., Микроскопическое строение электрического органа у
Torpedo. Учен. зап. Моск, ун-та, отд. ест. ист., вып. И, 1895.
8. Maurer F., Die ventrale Rumpfmuskulatur ciniger Reptilien, Festschr,
f. Gegenbaur, 1. Leipzig, 1896.
9. Ruge G., Ueber das peripherischc Gcbiet des Nervus facialis bei Wirbeltieren,
Festschr. f. Gegenbaur., 3, Leipzig, 1897.
10. Allis E., The cranial muscles and cranial and first spinal nerves of Amia
calva, J. Morphol., 12, 1897.
11. Driiner L., Studien zur Anatomie der Zungenbein, Kiemenbogen und
Kehlkopfmuskeln der Urodelen, Zoolog. Jahrb. Anat., 15, 19, 1901, 1904.
12. Maurer F., Die Entwicklung des Muskelsystems und der elektrischen
Organe, O. Hertwig, Handbuch d. vergl. u. exp. Entw. lehre., Bd. 2, T. 1, 1904.
13. Luther A. Untersuchungen fiber die vom N. trigeminus innervierte Mus-
kulatur der Selachier; der Ganoiden und Dipneusten; der Amphibien, Acta Soc. Sci.
Fennicae, 36, 41, 44, 1908, 1913, 1914.
14. Garten S., Die Produktion von Elektrizitat. Winterstein, Handbuch d.
vergl. Physiologic, Bd. 3, 1910—1914.
15. Lubosch W., Vergleichcnde Anatomic der Kaumuskcln der Wirbeltiere,
Jen. Zschr. f. Naturw., 53, 54, 1914, 1917.
16. Рылкова E. В., Развитие плечевой мускулатуры амфибий, Дневгик
воол. отд. О. Л. Е. А. и Э., Нов. сер. 3, № 3, 1915.
17. L a k j е г Т., Studien uber die Trigeniinus-versorgte Kaumuskulatur der
Sauropsiden, Kopenhagen, 1926.
18. Dombrowski Br., Ein Versuch d. Klassifikation der Brust- und Bauch-
muskeln der Amnioten, Anat. Anz., 70, 1930.
См. также работы, касающиеся морфологии головы, особенно работы Лютера
и Северцева, а также работы по развитию конечностей, особенно Северцов, 190g.
4. Нервная система
1. S е г г е s, Anatomie comparee du cerveau dans les quatre classes des animaux
vertebres, I, II, Paris, 1821—1826.
2. GorOnOwitsch N., Das Gehirn und die Cranialnervc n von Acipenser
ruthenus, Morphol. Jahrb., 13, 1888.
3. Burckhardt R., Der Bauplan des Wirbeltiergehims, Morphol. Arbeiten
(Schwalbe), 4, 1894.
4. P i n k u s F., Die Himnerven des Protopterus annectens, Morphol. Arbeiten
(Schwalbe), 4, 1894.
5. Strong O. S., The cranial nerves of Amphibia, J. Morphol., 10, 1895.
6. Fiirbringer M., Ueber die spino-occipitalen Nerven der Selachier und
Holocephalen, Festschr. f. C. Gegenbaur, 3. Leipzig, 1897.
7. Neal H. V., The segmentation of the nervous system in Squalus acan-
thias, Bull. Mus. Comp. Zool. at Harvard College, 31, 1898.
8. Herrick C. J., The cranial nerves of the bony fishes, J. Comp. Neurology,
8, 11, 1898, 1901.
9. Hill Ch., Primary segments of the Vertebrate head, Anat. Anz., 16, 1899.
10. Johnston J. B., The nervous system of Vertebrates, Philadelphia,
1906.
11. Kupffer K. von, Die Morphogenie des Centralnervensystems, O.
Hertwig, Handbuch d. vergl. u. exp; Entw.-lehre, Bd. 2, T. 3. Jena, 1906.
12. Белоголовый IO. А., К развитию головных нервов у птиц. Учен,
зап. Моск, ун-та, отд. ест. ист., вып. 23, 1908.
13. Е "d i n g e r L., Vorlesungen liber den Bauder nervosen Zentralorgane
des Menschen und der Tiere, 2 Bde. Leipzig, 1911.
14. Эдингер, Введение в учение о строении нервной системы, СПБ, 1911.
15. Herrick С.- J., IntroductionЧо Neurology, Philadelphia, 1916.
504
16. Kappers A riens, C. Die vergleichende Anatomie des Nervensystems
der Wirbeltiere und des Menschen I, II, Haarlem, 1 920, 1921.
17. Kuhlenbeck H., Vorlesungen iiber das Zentralnervensystem der Wir-
beltiere, Berlin, 1927.
18. Kappers A., Huber C., Crosby E. The comparative anatomy
of the nervous system of vertebrates including man. New York, the Macmillan Co. 1936.
См. также список работ по морфологии головы в целом, особенно работы van
Wijhe, Froriep, Кольцова, Белоголового, в списке монографий работы Дорна и Север-
цева, в списке по органам чувств работу Митрофанова и в списке по мускулатуре ра-
боту Allis.
5. Органы чувств
1. Schulze F. Е., Ueber die Sinnesorgane der Seitenlinie bei Fischen und
Amphibien, Arch. f. mikr. Anat., 6, 1870.
2. Retzius, Das Gehororgan der Wirbeltiere, I, II, Stockholm, 1881, 1884.
3. Beard J., The system of branchial sense organs and their associated ganglia
in Ichthyopsida, Quart. J. Mier. Sci., 1885.
4. Митрофанов П. И., Исследования над развитием позвоночных живот-
ных. Происхождение нервов и закладка боковых органов, Варшава, 1892.
5. Hesse R., Die Sehorgane des Amphioxus, Zschr. f. wiss. Zool., 63, 1898.
6. К a 1 1 i u s E., Sehorgan, Erg. d. Anat. u. Entw. gesch., 12, 14, 1902, 1904.
7. Studnicka F. K., Die Parietalorgane. Oppel, Lehrbuch d. vergl. mik-
roskop, Anat. d. Wirbeltiere. T. 5. Jena, 1905.
8. Froriep A., Die Entwicklung des Auges der Wirbeltiere, O. Hertwig, Hand-
buch d. vergl. u. exp. Entwickl-lehre, Bd. 2, T. 2. Jena, 1906.
9. Krause R., Entwicklungsgeschichte des Gehororgans, O. Hertwig, Hand-
buch d. vergl. u. exp. Entw-lehre., Bd. 2, T. 2, 1906.
10. Peter K., Die Entwicklung des Geruchsorgans u. Jacobsonschen Organs,
O. Hertwig, Handbuch d. vergl. u. exp. Entw.-lehre, Bd. 2, T. 2, 1906.
11. Новиков M., Исследование о теменном глазе ящериц, Учен. зап. Моск,
ун-та, отд. ест. ист., вып. 27, 1910.
12. Третьяков, Д. К., Органы чувств речной миноги, Одесса, 1916.
13. Broman J., Das Organ vomero-nasale Jacobsoni — ein Wassergeruchs-
organ, Anat, Hefte, Bd. 58, 1919.
14. Plate L., Allgemeine Zoologie und Abstammungslehre, II. Die Sinnes-
organe der Tiere. Jena, 1924.
15. Chranilov V. S., Beitrage zur Kenntniss des Weber’schen Apparates
der Ostariophysi, 1, 2. Zoolog. Jahrb. Anat., 49, 51, 1927, 1929.
6. Органы пищеварения
Зубы
1. Osborn H. F., Evolution of Mammalian molar teeth, New York, 1907.
2. Kiikenthal W., Zahne, Handworterbuch der Naturwissenschaften, Bd.
10, 1915, новое издание, 1934.
3. В о 1 k L., Odontological essays I—V, J- of Anat., 55, 56, 57, 1921, 1922.
Ротовая полость
4. Gopport E., Die Entwicklung des Mundes und der MundhChle etc., O.
Hertwig, Handbuch d. vergl. u. exp. EntwickI.-lehre, Bd. 2, T. 1, 1902.
5. Kallius E., Beitrage zur Entwicklung der Zunge, I, II, Anatom. Hefte 1,
Bd. 16, 28, 31, 1906—1908.
6. G 6 r s E., Beitrage zur Entwicklung der Zunge, Anatom. Hefte, I. Bd. 52,
1915.
7. Sewertzoff S. A., Die Entwicklungsgeschichte der Zunge bei den Rep-
tilien, Acta Zoologica, 10, 1929.
Кишечник
8. Oppel A. Lehrbuch der vergleichenden mikroscopischen Anatomie der
Wirbeltiere. 1. Der Magen, Jena, 1896; 2. Schlund und Darm, 1897; 3. Mundhohle,
Pancreas, Leber. 1900.
9. Maurer F. Die Entwicklung des Darmsystems, O. Hertwig, Handbuch d.
vergl. u. exp. Entwickl.-lehre Bd., 2, T. 1, Jena, 1902.
10. J а с о b s h a g e n E., Untersuchungen iiber das Darmsystem der Fische und
Dipnoer, Jen. Zschr. f. Naturw., 47, 49, 53, 1911, 1913, 1915.
11. Jacobshagen E., Zur Morphologie des Oberflachenreliefs der Rumpf-
.darmschleimhaut der Amphibien (1); der Reptilien (2), Jen. Zschr., 53, 56, 1920.
505
7. Органы дыхания
Жабры
1. Sewertzoff A. N., Die Entwicklung der Kiemen und Kiemenbogen
geffisse der Fische, Zschr. f. wiss. Zool. 121, 1922.
2. Woskoboinikoff M. M., Der Apparat der Kiemenatmung bei den Fi-
schen, Zoolog. Jahrb. Anat., 55, 1932.
3. Крыжановскпй С. Г., Органы дыхания личинок рыб и псевдобранхия,
изд. Акад, наук СССР, Труды лабор. эволюц. морфол., I, 1933.
4. К г у z а п о v s к у S. G., Die Pseudobranchie (Morphologie und biologische
Bedeutung), Zool. Jahrb., Anat., 58, 1934.
См. также D о h r n, 1886, в списке монографий.
Добавочные органы дыхания рыбиплавательный пузырь
5. Bridge Т. W., Fishes., The Cambridge Natural History, vol. 7. London,
1904.
6. Rauther M., Die akzessorischen Atmungsorgane der Knochenfische, Erg.
u. Fortschr. d. Zool., 2, 1910.
7. Rauther M., Zur vergleichenden Anatomie der Sehwimmblase der Fische,
Erg. u. Fortschr. d. Zool. 5, 1922.
8. Wasnetzow W. V., Zur Frage fiber die Morphologie der Sehwimmblase,
Anat. Anz., 66, 1928.
Происхождение легких
9. GOtte A., Ueber den Ursprung der Lungon, Zool. Jahrb., Anat., 21, 1905.
10. MakuschokM., Zur Frage fiber die phylogenetische Entwicklung der
Lungen bei den Wirbeltieren, Anat. Anz., 39, 42, 46, 1911, 1912, 19! i.
11. M a k u s c h о k M., Ueber genetische Beziehung zwischen Sehwimmblase
und Lunge, Anat. Anz., 44, 1913.
Гортань
12. Geppert E., Der Kehlkopf der Amphibian und Reptilien, I, II, Morph.
Jahrb., 26, 28, 1898, 1900.
13. Go p pert E., Beitrage zur vergleichenden Anatomie des Kehlkopfes
und seiner Umgebung mit besonderer Berucksichtigung der Monotremen, S e mon,
Zoolog. Forsch. Reisen in Australian, III, 1901.
14. GOppert E., Die Entwicklung der luftfuhrenden Anhfinge des Vorder-
dams, O. Hertwig, Handbuch d. vergl. u. exp, Entwicklungslehre, Bd. 2., T. 1, Jena,
1902.
Легкие амниот
15. M i 1 a n i A., Beitrage zur Kenntnis der Reptilienlunge, Zool. Jahrb.,
Anat., 7, 10, 1894, 1897.
16. Moser F., Beitr8ge zur vergleichenden Entwicklungsgeschichte der
Wirbeltierlunge, Arch. f. mikroskop. Anat., 60, 1902.
17. Wolf S., Zur Kenntnis von Bau u. Funktion der Reptieienlunge, Zool.,
Jahrb., Anat., 57, 1933.
8. Органы крозооЗрэщенпя
1. Boas J. E. V. Ueber die Arterienbogen der Wirbeltiere Morph. Jahrb.
13, 1888.
2. T a n d 1 e r J., Zur vergleichendon Anatomie der Kopfarterien bei den
Mammalia. Anat., H. 1, Bd. 18, 1901—1902.
3. Tandler J. Zur Entwicklungsgeschichte der Kopfarterien bei den Mam-
malia, Morph. Jahrb., 30, 1902.
4. G r e i 1 A., Beitrage zur vergleichenden Anatomie u. Entwicklungsgeschichte
des Herzens und des Truncus arteriosus der Wirbeltiere, Morph. Jahrb., 31, 1903.
5. HOchstetter F., Entwicklung des Blutgefaszsystems, O. Hertwig,
Handbuch d. vergl. u. exp. Entwicklungslehre, Bd. 3, T. 2, 1906.
6. G r e i 1A., Entwicklungsgeschichte des Kopfes und des Blutgefaszsystems von
Ceratodus Forsteri, Semon. Zool. Forsch. Reisen in Australien, I, Jena, 1908, 1913.
7. Allis E. Ph., The pseudobranchial and carotid arteries in the Gnatho-
atome fishes, Zool. Jahrb., Anat., 27, 1908.
См. также монографии и руководства, в осЬбёшюсти Goodrich Е. 8., 1909
и 1930.
506
9. Полость тела, органы выделения и размножения
1. Semper G., Das Urogenitalsystem der Plagiostomen und seine Bedeu-
tung fur die iibrigen Wirbeltiere, Arbeiten a. d. Zool.-Zoot. Institut d. Univ. Wurz-
burg, 2, 1875.
2. Spengel J. W., Das Urogenitalsystem der Amphibien, Arbeiten a.
d. Zool.-Zoot. Institut d. Univ. Wurzburg, 3, 1876.
3. Balfour F. M., On the origin and history of the urogenital organs of
Vertebrates, J. of Anat. and Physiol., 10, 1876.
4. Hoffmann С. K., Zur Entwicklungsgeschichte der Urogenitalorgane
bei den Reptilien, Zschr. f. wiss. Zool., 48, 1889.
5. Rabi C., Ueber die Entwicklung deS Urogenitalsystems der Selachier,
Morph. Jahrb., 24, 1896.
6. Филатов Д. П., История развития выделительной системы у амфибий,
Бюлл. Моск. О-ва исп. прир., 1904.
7. Felix W. u. Buhler, Die Enfwicklung der Harn- und Geschlechtsorgane,
O. Hertwig, Handbuch d. vergl. u. exp. Entw.-lehre, Bd. 3., T. 1, Jena, 1904—1906.
8. Broman I., Ueber die Entwicklung und Bedeutung der Mesenterien und
der Korperhohlen bei den Wirbeltieren, Erg. d. Anat. u. Entw-gesch., 15, 1905.
9. Maschkowzeff A., Zur Phylogenie des Urogenitalsystems der Wir-
beltire etc. I, II, Zool. Jahrb. Anat., 48, 1926.
10. Maschkowzeff A., Der Genitalapparat der Acipenseridae, Zool.
Jahrb., Anat., 58, 1934.
11. Maschkowzeff A., Zur Phylogenie der Geschlechtsdriisen und der
Geschlechtsausfiihrgange bei d. Vertebrata, Zool. Jahrb. Anat., 59, 1935.
Копулятивные органы
12. Boas J. E. V., Zur Morphologie der Begattungsorgane der amnioten
Wirbeltiere, Moph. Jahrb., 17, 1891.
13. Fleischmann A., Morphologische Studien liber Kloake und Phallus
der Amnioten. 1—16, Morphol. Jahrb., 30—36, 1902—1907.
14. Gerhardt U., Der gegenwartige Stand der Kenntnisse von den Copu-
lationsorgane der Wirbeltiere, insbesondere der Amnioten, Erg. u. Fortschr. d. Zool,
1, 1908.
15. Henneberg B., Beitrag zur Entwicklung der Susseren Genitalor-
gane beim Sauger, Anat. Hefte, I, Bd. 50, Bd. 55, 1914, 1917.
10. Органы внутренней секреции
1. Maurer F., Schilddriise, Thymus und Kiemenreste der Amphibien, Moph.
Jahrb., 13, 1888.
2. Reese A. M., Structure and development of the thyroid gland in Pet-
romyzon. Proc. Acad. Nat. Sc., Philadelphia, 54, 1902.
3. Maurer F., Die Entwicklung des Darmsystems (die Kiemenspalten und
Hire Derivate), O. Hertwig, Handbuch d. vergl. u. exp. Entw.-lehre, Bd. 2, T. 1, 1902.
4. Poll H., Die vergleichende Entwicklungsgeschichte der Nebennierensysteme
der Wirbeltiere, O. Hertwig, Handbuch d. vergl. u. exp. Entw.-lehre, Bd. 3, T. 1, 1905.
5. Fritsche E., Die Entwicklung des Thymus bei Selachiern, Jen. Zschr.,
1910.
V. Филогения позвоночных
Обзоры и сводки, включая руководства по палеонтологии (2 и 7.— популярные книги)
1. Борисяк А., Курс палеонтологии, I, II, Москва, 1905—1906.
2. Гюнтер К., Происхождение и развитие человека, I, II, атлас, СПБ, 1909.
3. Abel О.. Die Stamme der Wirbeltiere, 1919.
4. Franz V., Geschichte der Organismen. Jena, 1294.
5. Давиташвили А. Ш., Курс палеонтологии, 1941.
6. H eider К., Entwicklungsgeschichte und Morphologie der Wirbellosen
(перепечатка из Kultur der Gegenwart, 1913). Berlin, 1928.
7. Дерюгин и Догель, Эволюция беспозвоночных и позвоночных
животных. Сборник «Человек и природа», т. V. Л., 1928.
8 N а е f A.. Phylogenie der Tiere. Baur u. Hartmann, Handbuch der Verer-
bungsuissenschaft, Bd. 3 (Lief. 13), Berlin, 1931.
9. Грегорн В. К., Эволюция лица от рыбы до человека. М., 1934.
10. Zittel К. A., Grundziige der Palaontologie. II. Vertebrata. Berlin,
1911. Русский перевод, 1935.
11. Ромер. Палентология позвоночных, 1939.
12. Kuhn, О, Die Pylogenie der Wirbeltiere. Jena, 1938.
См. также руководства, монографии и сводки по зоологии и сравнительной ана-
томии.
507
Основные источники
13. L a n g A., Der Ban von Gunda segmentate und die Verwandschaft der Plathel-
minthen mit Coelenteraten und Hirudineen. Mitt. Zool. St. Neapel, 3, 1881.
14. В a t e s о n W., The ancestry of the Chordata, Quart. J. Mier. Sci, 26, 1886.
15. H u b r e ch t, Die Abstammung der Anneliden und Chordaten und die Stel-
lung der Ctenophoren und Plathelminthen im System. Jen. Zschr. f. Naturw., 39, 1905.
16. W i 1 1 e у A., Amphioxus and the ancestry of Vertebrate. New York. 1894.
17. С e в e p ц о в A. H., Исследование об эволюции низших позвоночных I, II,
III, Русск. арх. анат., гист. и эмбр., 1 и 3, 1917—1925.
18. Stensio Е., А-son, The dowtonian and devonian vertebrates of Spitzbergen.
I Cephalaspidae. Norske Vidensskaps Ac., Oslo, 1927.
19. Stensio E. А-son, On the head of Macropetelichthyidae. Field Museum
of Nat. Hist. Publ. 232, Chicago, 1925.
20. Pollard H. B., On the anatomy and phylogenetic position of Polypterus.
Zool. Jahrb. Anat., 5, 1892.
21. Gregory W. K., The orders of Teleostomous fishes. Ann. of the New York
Ac. of Sci., 17, 2, 1907.
22. G r e g о г у W. K., Present status of the problem of the origin of the Tetrapoda.
Ann. New York Ac. Sci., 26, 1915.
23. S e w e r t z о f f A. N., Der Ursprung der Quadrupeda. Palaentol. Zschr., 8,
1926.
24. W a t s о n D. M. S., The evolution and origin of the Amphibia (Croonian lectu-
re), Philos. Trans. Roy. Soc., London, Ser. B., vol. 214, 1926.
25. Save-Soderberg G., Preliminary note on Devonian Stegocephalians
from east Greenland, Komiss. of. viedensk. Undersagelser i Gronland, 94, 1932.
26. О s b о r n H. F., The reptilian subclasses Diapsida and Synapsida, Mem. Amer.
Mus. Nat. Hist., 1. 1903.
27. Williston S. W., The phylogeny and classification of Reptiles, J. Geol.,
25, 1917.
28. N о p c s a F r. B., Die Familien der Reptilien, Fortschr. d. Geol. u. Paleontol.,
Berlin, 1923.
29. S w i n t о n W. C., The Dinosaurs, London, 1934.
30. Heilmann G., The origin of birds. London, 1926.
31. G r e g о г у W. K., The orders of Mammals, Bull. Amer. Mus. Nat. Hist., 27,
1910.
32. О s b о r n H. F., Age of Mammals, New York, 1910.
33. В г о о m R., On the origin of Mammals (Croonian lecture), Philos. Trans. Roy.
Soc., London, Ser. B., vol. 206, 1914.
34. Broom B., The Mammal-like reptiles of South Africa and the origin of mam-
mals, London, 1932.
Происхождение человека
35. Гексли Т., Положение человека в ряду органических существ, СПБ.,
1864.
36. Видерсгейм Р., Строение человека со сравнительноанатомической
точки зрения, М., 1900.
37. Ранке, Человек, СПБ., 1904.
38. Л е х е В., Человек, его происхождение и эволюционное развитие, М., 1913.
39. К л а а ч, Происхождение и развитие человеческого рода, «Вселенная и
человечество», изд. «Просвещение», СПБ.
40.Энгельс Фр., Диалектика природы. Роль труда в процессе очеловечения
обезьяны.
41. Gregory W. К. а. Н el 1 m an М., The dentition of Dryopithecus and the
Origin of Man, Anthropol. Papers of the Amer. Mus. Nat. Hist., 28, 1926.
42. A b e 1 O., Die Stellung des Menschen im Rahmen der Wirbeltiere, Jena,
1931.
43. Жуков Б. Происхождение человека, M., 1931.
44. Вишневский Б. Н., Происхождение человека.
45. Clark W. Е, G., Early forerunners of man, London, 1934.
46. Gr e g о г у W. K., Man’s place among the Anthropoids, Oxford, 1934.
47. Вейнерт Г. Происхождение человечества. M.—Л., 1935.
48. Бунак, Нестурдс,Рогинский. Антропология. 1941.
См. также книги Э. Геккеля.
VI. Морфологические закономерности эволюции
1. С о р е Е. D., The origin of the fittest (Essays on evolution), London, New
York, 1887.
2. Давиташвили А. Ш., Развитие идей и методов, в палеонтологии после
Дарвина, 1940.
508
3. D о 11 о L., Les lois de 1’evolution. Bull. Soc. Belg. Geol., 7, Bruxelles, 1893.
4. Willey A., Convergence in evolution, London, 1911.
5. Северцов A. H., Современные задачи эволюционной теории, Москва,
1914.
6. Д е п е р е Ш., Превращения животного мира, П., 1915.
7. Osborn Н. F., The Origin and evolution of life, London, 1918.
8. H e s s e R., Die Stufenleiter der Organisationshohe der Tiere, Sitzungsher.
Preuss, Akad. Wiss., Mat. Naturw. KI., Berlin, 1929.
9. Abel O., Palaobiologie und Stammesgeschichte. Jena, 1929.
10. Sewerzoff A. N., Morphologische Gesetzmassigkeiten der Evolution,
Jena, 1931.
11. С e в e p ц о в A. H., Главные направления эволюционного процесса, М.
1934.
12. Северцов А. Н., Морфологические закономерности эволюции, изд.
Академии наук СССР, 1938.
13. Шмальгаузён И. И., Организм как целое в индивидуальном и истори-
ческом развитии, изд. Академии наук СССР, 1938.
14. Ш м а л ь г а у з е н И. И. Пути и закономерности эволюционного про-
цесса. 1939.
ПМЕНИОИ УКАЗАТЕЛЬ
А
Абель 492, 494
Амалицкий 461
Аристотель 7
Б
Бальфур Ф. (Fr. Balfur) 12, 133
Белон 7
Бирд 401
Блэквилль Г. 8
Блюменбах И. 8
Бовери 298
Борисяк 472
Брауер 379
Браус Г. И, 12
Брум (Broom) 171, 460
Бюффон 7
Бэр К. (К. Ваег) 10, 31
Бэтсон 411
В
Бай 133
Вейсман 29
Вик д’Авир Ф. 8
Видерсгейм Р. 12
Воскобойников М. 337
Вуд Дж. 480
Г
Гаупп Е. 12
Гегенбаур К. 11, 15, 133, 180, 319
Геккель 9. (Е. Haeckel) 11, 31, 33, 406
Гексли Т. (Т. Huxley) 10, 132
Гельмгольц 287
Гентер Дж. 8
Гёте В. 9, 132
Гётте А. 12, 341
Грегори В. 12, 315, 466
Гудрич Е. 12
д
Дарвин Чарльз 10, 19, 31, 486
Дельсман 410
Добантон Л. 7
Долло 485
Дорн А. 12, 19, 133, 410, 413
3
Заленский В. 11, 12, 406
Зедерштрем 413
Зибольд К. 10
510
К
Кампер П. 8
Кейбель 34
Керр Г. 12
Кильмейер 9, 31
Клейненберг 26
Ковалевский А. О. 11, 414
Ковалевский В. О. 11, 494
Коп 314, 493
Крумбах 407
Купфер 401
Кювье Ж. (J. Cuvier) 8, 9, 12, 22, 494
Л
Ламарк Ж. 8, 405, 482
Ланг А. 11, 406
Лэнкестер. См. Рай Лэнкестер
м
Маршалл А. М. 12
М астерман 411
Машковцев 3S0
Меккель И. 8
Менерт 34
Мечников 11, 406
Мильн-Эдвардс Г. 8, 17
Мюллер И. (I. Muller) 10
Мюллер Фр. (Fr. Muller) И, 33
И
Немилов А. 229
Неф 410
О
Окэн 9, 132
Осборн Г. 12, 314, 465, 485,
Оуэн Р. (R. Owen) 9, 10, 132
п
Паркер В. К. 12
Плате 16, 19, 22, 23
Р
Рабль К. 12
Рай Лэнкестер 9. 12, 406
Ранке 480
Ратке М. 10
Рейхерт 469
Роза 694
Ру В. 12
Рюккерт 380
С
Сваммердам 7
Северино 7
Северцов А. Н. 12, 20, 22, 24,
416, 421, 483
Седжвик 379, 413
Семпер 410
Сент-Илер Э. Ж. 8, 9, 22, 23
Спенсер 17
Станниус Ф. 10
Стеншио Э. А. (Е, A- Stensio) 12,
Сушкин П. П. 12, 463
т
Третьяков 413
У
Уатсон 440
Уотсон Д. 12
Уилли (Willey) 411
ф
Фан-Вай (Van Wijhe) 12, 133
34, 133 Феликс 380
’ Фильд 379
Франц 20
Фюрбрингер М. 11
Ш
416, 424 Шимкввич В. М. 413
Штейнах 402
э
Эндрюс Э. 466
511
предметный указатель
А
Агама древесная 487
Адаптация 22, 482, 485, 494
— значение в ее эволюционном про-
цессе 485
Адаптивная радиация 485
Адекватные раздражения 277
Адреналин 402
Аисты 487
Акромион 192
Акулы 53, 54, 426
Аллантоис 64, 307
Алисфеноиды 462
Альвеолярная трубка 348
Амилаза 325
Амнион 65
Ампулы Лоренцини 281
Амфибии См. также Земноводные 53
— артерии их 362
— венозная система их 366
— взаимоотношения между их поло-
выми железами и почками 390
— головной мозг их 251
— грудина их 118
— жаберный аппарат их 337
— желудок их 321
— значение наружных жабер у их ли-
чинок 338
— зубы их 312
— кишечник их 325
— копулятивные органы их 395
— легкие их 344
— наружные жабры их 337
— органы обоняния у них 305
— позвоночник их 110
— почка 382
— происхождение их 438
— = ребра их 110
— ротовые железы их 319
— сердце их 357
— скелет свободной конечности их 197
— способность менять окраску у них 72
— тазовый пояс их 194
— трахея у них 344
— череп их 148
— язык их 318
Амфистилия 131
Амфицельные позвонки 103
Анаболии 34
Аналогичные органы 5, 10, 14
Анапофизы 116
Анатомия описательная 5
— сравнительная 5, 7
Аннелиды, кровеносная система их 351
Анамодонты 448
Антилп ы 56
512
Антимеры 15
Анэлективные органы 277
Аорта брюшная 351
— дуги ее 353
— корни ее 354
— спинная 352
Апендикулярии 36
Аромбрфоз 482, 493
— значение его в эволюционном про
цессе 485
Артериальная кровь 350
— луковица 356
Артериальные дуги 353
Артериальный конус 351, 356
Артерия (и) 360
— бедренная 365
— брыжеечные 361
— жаберная 352
— желточные 355
- г- кишечные 361
— кожная 362
— легочная 352, 362
— подвздошная 361
— подключичные 361
— позвоночные 363
— пупочная 355
— седалищная 365
— сонная 352, 360
— хвостовая 361
Архаллаксис 34
Археорнис 53, 456
Археоптерикс 53, 456
Архетип 10
Архиптеригий 180
Ассоциативные невроны 231
Асцидии 36
— регрессивные изменения их 415
— эмбриональное развитие их 414
Атлас 114
Аутостилия 131
Афанизия 30
Б
Барабанная кость 165
— лестница 287
— перепонка 289
— струна 267
Бараны 56
Бегемот 53
Бедренная артерия 365
— кость 186
Безыменная Вена 365
— кость 195
Белка летающая 455
Белки 53, 469
«Белое» вещество мозга 236
Берцовые кости 186
Бескилевые птицы 53, 490
Беспозвоночные, кровообращение их 349
и сл.
— мускулатура их 208
— нервная система их 225
— органы дыхания их 330
--- пищеварения их 306
— — чувств их 277
— покровы их 69
— полость тела их 370
— скелет их 91
— эпидермис их 69
Бесчелюстные позвоночные 53, 54
----геологическая эпоха их появле-
ния 405
---эволюция их 417 и сл.
Бесчерепные, примитивные 421
Билофодонтные зубы 315
Биогенетический закон Геккеля 33
Биологические координации 25
Биологический прогресс 483 и сл.
Блоковый нерв 263
Блуждающий нерв 239, 268
«Бобровая струя» 398
Бобры 53, 469
Бокал глазной 293
Боковая линия, лопасти ее 276
---Органы ее 50, 280
— пластинка 44
— складка. См. Теория боковой складки
Боковой нерв 268
Боковые вены 365
— желудочки 246
— затылочные кости 137
— клиновидные кости 137
— обонятельные кости 137
— плакоды 238
— рога 241
Большие губы 396
— крылья 165
Большой мозг, извилины его 254
---ножки его 244
Бородочки пера 82
Борозды большого мозга 254
Броненосцы 56, 469
— панцыри их 90
Бронхи 342, 347
Бронхиоли 347
Брыжеечные артерии 361
Брыжейка 308, 372
Брызгальце 333
— преобразование его у наземных позво-
ночных в барабанвую полость 148
Брюхо, косые мышцы его 218
— прямые мышцы его 218
Брюшина 371
Брюшная аорта 352
— вена 352, 366
— мускулатура 218
— полость 50
Брюшной плавник, скелет его 184
Брюшные воздушные мешки 347
— поры 372
— ребра 89
— рога 241
Бунодонтные формы зубов 315
Быки 56
В
Вазодентин 79, 311
Варолиев мост 244
Веберов аппарат 328
Веки 297
Вена (ы) 365
— боковые 365
— брюшная 366
— безыменная 368
— венечная 368
— кардинальные 352, 365
— кожные 367
— легочные 367
— подвздошные 365
— подкишечная 355
— подключичные 365
— позвоночные 367
— полая 352, 365, 367
— почечные 352
— хвостовая 352, 365
— яремные 365
Венечная вена 368
Венозная кровь 350
— пазуха 356
Венозные стволы, асимметрическое распо-
ложение их у млекопитающих 367
Верблюды 53
Веревочные тела 243
Вертлуг 195
Вертлужная впадина 194
Верхиеушная кость 137
Верхние дуги 99
— ребра 101
Верхняя затылочная кость 137
Виверры 468
Вилочка 192
Височная дуга задняя 160
— кость у человека 165
— мышца 212
Височные дуги 159
— крылья 165
— ямы 159
Висцеральная лопасть мозга 242
— мускулатура 209
----- висцерального аппарата 210
— часть черепа, развитие и строение
ее 127
Висцеральные двигательные нервы 258
— нервы, метамерия их 271
— чувствующие нервы 259
Висцерально-двигательная область голов-
ного мозга 275
— чувствующая область головного моз-
га 275
Висцеральный аппарат, висцеральная му-
скулатура его 210
— скелет 137
— развитие и строение его 127
Витродентин 79
Вкус, органы его у позвоночных 300
Вкусовые нервы 300
— почки 300
Влагалище 394
— языка 318
В незародышевый целом 65
Внутреннее ухо 282
Внутренние органы, роль энтодермы в их
развитии 66
Внутренняя косая мышца живота 218
— секреция 398
-----органы ее 52
Водяной опоссум 488
Воздушные мешки у птиц 347
Волки 53, 488
Волосы, происхождение их 84
— развитие их 85
513
Волосы, строение их 84
Волосяной покров млекопитающих 86
----редукция его 86
---- у человека 86
Вольфов канал 377, 387
Воронка мозга 245
Вороновидная кость 190
Воротная система печени 352
Воспроизводительные органы беспозво-
ночных 365
------позвоночных 386
Вставочные пластинки позвоночника села-
хий 100
Вторичная мантия мозга 247
— почка 373
----происхождение ее 380
Вторичное твердое небо 158
Вторичнопочечные канальцы 377
Вторичноротовые 409
Вторичные верхние челюсти 166
— кости 92. См. также Покровные кости
— челюсти 137
Вторичный скелет 96
Выделительные органы беспозвоночных
372
------позвоночных 52, 373
------эволюция их 384
Выдры 53, 468, 488
Высшие рыбы, костный череп их 138
Вымирание 492
Выхухоли 467
Г
Гады чешуйчатые 452
Газовый обмен 330
Ганглионарная пластинка, роль ее в обра-
зовании периферической нервной си-
стемы 237
Ганоидная чешуя 79
----генетическая связь ее с плакоид-
ной 80
Ганоидные рыбы, непарные плавники их
174
Ганоиды 53, 429
— головной мозг их 250
— кистеперые 53
— костные 53, 54, 429
— хрящевые 53, 428
Ганоин 79
Гаплодонтная форма зубов 314
Гардерова железа 297
Гастральная мезодерма 63
Гастрея 406
Гаструляция у амниот 60
— у земноводных 58
— у млекопитающих 60
— у селахий 59
Гаттерия 53
Гемальные отростки позвонков 102
Ген, плейотропное действие его 23
Генетические корреляции (Плате) 23
Генитальные складки 386
Генитогаструла 406
Гетеродонтная зубная система 313
Гетеротопия 6, 34
Гетерохрония 6, 34
Гетероцельные позвонки 111
Гетероцеркальная форма хвостового плав-
ника 175
Гетероцеркия 175
Гиббоны 56
574
Гягантише волокна
Гиены 53
Гимнофионы 53, 54, 442
Гиомандибулярный хрящ 129
Гиостилия 131
Гиоидный хрящ 129
Гипертелия 77
Гиподермис 68
Гиппокамп 247
Гипотеза панмиксии 29
Гипофиз 125, 401
Гипоцентры 429
Гипуралии 176
Гладкая мускулатура, замена ее попе-
речнополосатой 210
Глаз амфибий 297
— возникновение его 28
— млекопитающих 297
— мускулатура его 296
— развитие главных частей его 293
— рептилий 297
— рыб 297
— строение его 297
Глазки Гессе 39, 228 , 416
Глазнично-височная дуга 160
— клиновидные кости 137
Глазничные кости 140
— крылья 159
Глазничный нерв, глубокий 264
поверхностный 266
Глазное яблоко 296
Глазной бокал 293
Глазные мышцы 214
— пузыри 291
Глазодвигательный нерв 262
Глотка 307, 321
Голень 186
Голобластические яйца 56
Головная почка 373
Головной мозг 49, 233, 241
----висцерально-двигательная и вис-
церально-чувствующая области его 275
----высшие центры его 276
----границы между отделами его 233
---- значение прогрессивного развития
его для эволюции позвоночных 241
---- изгибы его 233
----первичный 233
-----соматически-двигательная область
его 275
-------чувствующая область его 275
----строение его 241, 249
----------у амфибий 251
-----------у ганоидов 250
-----------у двудышащих 251
-----------у костистых рыб 250
---------- у круглоротых 249
-----------у млекопитающих 253
---------- у птиц 253
-----------у рептилий 252
-----------у селахий 249
Головные нервы 258 , 262
Головоногие молюски, скелетная ткань
их 91
------хроматофоры их 70
Голосовые связки 343
Гомодинамия 15
Гомодонтная зубная система 313
Гомойология 16
Гомология 5, 10, 14, 15
— аугментативная 16
Гомология дефективная 16
— неполная 16
Гомология полная 16
— частная 15
Гоманомия 15
Гомотипия 15
Гомотопная субституция 26
Гомоцеркальная субституция 26
— форма хвостового плавника 176
Гонады 42, 386
Гоноциты 386
Горилла 56, 480
Гормон (ы) 398
— гипофиза 401
— желтого тела млекопитающих 403
— половых желез 402
— шишковидной железы 401
Гороховидная косточка 199, 200
Гортань 342
— нижняя 345
Гоффер 469
Гребенчатая мышца 222
Грудина 118
— млекопитающих 120
— у амфибий 118
— .у птиц 120
— у рептилий 119
Грудино-ключично-сосцевидная мышца 213
Грудная мышца 221
— полость 371
Грудной плавник, скелет его 183
Грушевидная мышца 222
Грызуны 53, 55, 469
— зубы их 316
— эволюция их 469
Губные железы 320
д
Даманы 56, 475
Двенадцатиперстная кишка 324
Двигательные невроны 231
Двойная матка 394
Двубрюшная мышца 212
Двудышащие 431, 53, 489
— геологическая эпохаихпоявления405
— головной мозг их 251
— филогения их 431
— череп их 144
Двураздельная матка 394
Двурогая матка 394
Девиации 35
Дегенерация 414
Дельтовидная мышца 220
Дельфины 53, 487
Дентин 77, 311
— трабекулярный 78
Детское место 65
Дефинитивные органы 13
Диапсиды, происхождение их 448
Диафрагма 222, 371
Дивергенция 17, 486
Дикобразы 469
Динамические координации 24
Динозавры 449
— происхождение их 450
— скелет свободной конечности их
201
Дипротодонты 465
Диференциация См. также Принцип
диферепциации 17
Дифиодонтизм 311
Дифицеркальные формы хвостового плав-
ника 176
Диффузная нервная система 226
Диффузное дыхание 330
Добавочный нерв 268
Долгопят 455
Долихозавры 452
Дорсолатеральные плакоды 238
Дыхание диффузное 330
— кишечное 339
— механизм его у амфибий 345
-------у fрептилий 346
Дыхание тканевое 331
— у беспозвоночных 331
Дыхательные органы 50, 332
— пути 342
Дыхательный аппарат 331
Дюгони 56
Е
Евстахиева труба 228
Ежи 53, 467
Еноты 468
Ехидны 53
•дл
iilw
Жаберная крышка 336
Жаберно-кишечный нерв 268
Жаберные артерии 352
— дуги 129
— перегородки ланцетника 40
— тельца 400
— щели 36
---число их 333
Жаберный скелет ланцетника 38
Жабры наружные 337
— позвоночных 332
— рыб 336
Жабы 53
— половые железы их 391
Жвачные 53
— лунчатые зубы их 316
Жевательная мышца 212
Железы бартолиниевы 395
— в эпидермисе беспозвоночных 70
— гардерова 297
— губные 319
— заднеязычная 320
— зубная 319
— кожные 72
— колбовидная 45
— копчиковая 74
— куперовы 395
— млечные 75
— нёбные 320
— околоушная 320
— пищеварительные 326
— поджелудочная 327
— подчелюстная 320
— подязычная 320
— половые 387
— потовые 74
— предстательная 395
— препуциальные 395
— ректальная 325
— ротовые 319
— сальные 75
— слезная 297
— слюнные 320
— шишковидная 401
— щечные 320
515
Железы (ы) щитовидная 398
— ядоотделительные зубные 319
— язычные 320
Железистые клетки эпидермиса 72
значение их у земноводных 73
у рыб 72*
Желтое тело 386, 403
Желточные артерии 355
Желудок 307, 321
Желудочки зрительные 245
—"полушарий мозга 241, 246
— сердца 351. 356
Желчный проток 327
— пузырь 326
Живородящие млекопитающие, копулятив-
ный орган их 397
Жирафы 56, 472
— рога их 88
Желток 57
— влияние количества его па внешнюю
форму зародыша 64
— - количество его в яйце позвоночных 56
Желточная пробка 57
— энтодерма 61
Журавли 487
Жучки 88
3
Заднеглазничные кости 137
Заднегрудные воздушные мешки 347
Заднеколобные кости 137
Заднеушная кость 137
Заднеязычная железа 320
Задняя дуга позвоночника 99
Задний мозг первичный 233, 243
Зайцы 469
Закрытопузырные рыбы 328
Замещающие кости 137
-развитие их 93
Запястье 185
Зародыш, влияние на его внешнюю форму
количества желтка в яйце 64
Зародышевые оболочки высших позво-
ночных 65
Зародышевый диск 56
— щит 60
Затылочная дыра 137
— кость 137. 138, 163
Затылочные кости 137
Зачелюстной сомит 123
Зверезубые 53. См. также Териодонты
Землеройки 53, 467
Земноводные 53, 54
Зеркальце (глаза) 295
Зигальпый череп 159
Змеи 53, 55, 453
Зобная железа 399
Золотой крот 467, 488
Зрачок 296
Зрение, орган его у позвоночных 50
Зрительные бугры 235, 244
— доли 235, 244
— желудочки 244
Зрительный нерв 262
----- развитие его 239
Зубная пластинка 310
*— система, эволюция ее 317
Зубные кости 89, 137
Зубчатые мышцы 218
Зуб (ы) 310
— акродонтные 313
516
Зуб (ы) амфибий 312
— билофодонтные 316
— бунодонтные 315
— гаплодонтные 314
— генетическая связь их с плакоидной
чешуей 89
— гетер одонтные 313
— гомодонтные 313
— квадритуберкулярные 316
— кожные* 78
— коренные 313
— ложнокоренные 313
— лофодонтные 316
— лошади 317
— лунчатые 316
— млекопитающих 313
— остробугорчатые 315
— плевродонтные 313
— протодонтные 315
— птиц 313
— режущий 315
— рептилий 312
— секодонтный 315
— секториальный 315
— складчатые 316
— смена их 311, 314
— текоцонтные 313
— тровоядных млекопитающих 317
— трехбугорчатые 315
— трехзубчатые 315
— триконодонтные 315
— тритуберкулярные 315
— тупобугорчатые 315
— формирование корней их у млеко-
питающих 314
— четырехбугорчатые 316
И
Иглокожие 409
— кожные, скелет их 70
Иглы 85
Игрунки 56
Игуанодоны 451
Идиоплазматические корреляции (Плате)
22
Известь, значение возникновения у мезо-
дермы способности выделять ее 92
Извилины большого мозга 248
Изопедин 80, 431
Иммобилизация частей 20
<'Инадаптивные>> формы 493
Инвагинация у земноводных 60
Иннертаза 325
Индивидуальные корреляции (Плате) 22
Индрикотерий 472
Инкреция 398
Интеграция 17
Инфундибулярный орган 281
Иридоциты 72
Истинные кресцовые позвонки 117
— сосцы 76
Ихтиозавры 446
— дельфинообравные 446
— происхождение их 446
К
Казуар 53
каменистая кость 165
Каналы боковой линии 280
— полукружные 284
Капская свинья 471
Кардинальные вены 365
---- задние 352
— — передние 352
Каретта 487
Карманообразные клапаны 356
Каудальная дуга позвоночника 99
Кахексия 399
Кашалоты 53
Квадратная кость 138
— поясничная мышца 218
Квадритуберкулярный зуб 316
Кенгуру 465
Киви 53
Кистеперые ганоиды 53.
— — значение наружных жабер у их
личинок 66
— рыбы 431
— — геологическая эпоха их появле-
ния 430
----значение их в происхождении на-
земных позвоночных 436
Кисть 185
Китовый ус 87
Китообразные 53, 468
— зубы их 314, 317
— эволюция их 468
Киты 53, 55, 468
Кишечник 323
— слизистая оболочка его 308
Кишечная артерия 361
Кишечнодышащие черви 409
Кишечное дыхание 339
Кишечнополостные, мускулатура их 208
— нервные клетки их 226
Кишечные артерии 361
— ворсинки 324
— нервы 258
Кишечный зек 325
Кишка 323
Клапаны сердца 355
Классификация позвоночных 53
Клиновидные кости 138
Клитор 396
Клоака 307, 324
Клоачные млекопитающие, зубы их 316
----копулятивные органы их 397
----млечные железы их 75
— — эволюция пх 465
Ключица 190
Коадаптация 22
Коаптация 22
Когтевая пластинка 87
Когтевой валик 87
Когти 87
Кожистая черепаха 487
Кожная артерия 362
Кожно-сочленовная кость 137
Кожные вены, большие 367
— железы 72
----ядоотделительные 73
— зубы 78
— кости 80
•— окостенения у черепах 90
— орга: ы чувств 278
Кожный скелет 88
----у иглокожих 70
Козелок 344
Козы 56
Колбовпдпа железа 46
Колбовидпые осязательные тельца высших
животных 279
Коллоид 399
Кольчатые черви, роль их кутикулярных
щетинок 70
Комиссура задняя 235
— передняя 246
Конвергентное развитие 486
Конвергенция 17, 486
Кондцлартры 53
Кондициональные факторы 25
Конечная нить 241
Конечноротые 53
Конечности позвоночных 47. См. также
свободная конечность
— диференцировка их мускулатуры 222
— мускулатура их 219
— наземного позвоночного, возникнове-
ние их 27, 184
-------- развитие их 434
— скелет их 172
Конский хвост 241
Конструктивные координации 24
Концевая пластинка 235, 245
Концевой нерв 262
Концентрация плавника 173
Координации 24, 25
— биологические 25
— динамические 25
— конструктивные 26
— тектонические 24
— топографические 24
Копулятивные органы 395
Копчик 116
Копыта 88
Копытные зубы их 317
— эволюция их 470
Коракоиды 190
Коракоидный отросток 192
— отдел пояса 181
Коракоидно-плечевая мышца 220
Корреляции 22. См. также Принцип кор-
реляций
— геномные 23
— идиоплазматические 23
— индивидуальные 22
— морфогенетические 23
— физиологические 22 и сл.
— филетические 22, 24
— функциональные 23
Коренные зубы, истинные 313
----процесс усложнения формы их у
млекопитающих 313
Кориум 69
Кортнев орган 287.
Космин 79, 431
Космоидная чешуя 79
Костистые рыбы*53, 54
---- головной мозг их 250
— — позвоночник их 108
Костная ткань, частичная редукция ее у
древнейших позвоночных 427
— чешуя 81
Костные ганоиды 53
----геологическая эпоха их появления
405
------позвоночник их 108
— лучи 174
— рога жирафы 88
— рыбы 53
Костные рыбы, геологическая эпоха их по-
явления 405
—- — непарные плавники их 174
517
Костные рыбы, филогения их 428
— чешуи 81
— череп 136
---высших рыб 138
Кость (и) 92
— бедренная 186
— безыменная 195
— берцовые 186
— верхнеушная 137
— верхняя затылочная 137
— височная 165
— вороновидная 190
— вторичная 92
— глазнично-клиновидные 137
— глазничные 140
— заднеглазничные 137
— заднелобные 137
— заднеушная 137
— замещающая 93, 137, 138
— затылочная 137, 163
— зубные 89, 137
— каменистая 165
— квадратная 137
— клиновидные 137
— кожно-сочленовная 137
— кожные 88
— крыловидная 137
— ладьевидная 203
— лобковая 194
— лобная 136
— локтевая 185, 202
— лучевая 185, 202
— межтеменная 165
— надвисочная 136
— надугловая 137
— небная 137
— носовая 136
— обонятельная 137
— околоушные 137
— первичная. См. вамещающие кости
92
— переднелобные 137
— переднеушная 137
— пластинчатая 137
— плечевая 185
— пневматичные 347
— покровная 92, 136
— предчелюстные 137
— пяточная 203
— решетчатая 137, 165
— седалищная 194
— слезная 166
— скуловые 137
— сочленовная 138
— средняя обонятельная 137
— таранная 203
— теменные 136
— угловые 137
— филогенетический процесс вытесне-
ния и замещения ею хряща 92
— челюстные 137
— чешуйчатая 137
— щечные 140
Котики 53, 488
Котилозавры, происхождение их 444
Кошки 53, 468
Краевая мезодерма 68
Краниальная дуга позвоночника 99
Креодонты 53, 468
Крестец 115
Кровеносная система 51, 349
---замкнутая 351
---эволюция ее 370
----------у наземных позвоночных
433
Кровообращение, органы его у ланцет-
ника 41
Кровь 349
Крокодилы 53, 448
— происхождение их 448
Крот (ы) 53, 467, 488
— золотой 488
— сумчатый 465, 488
Кроющая перепонка кортиева органа 287
Круглое окно 154, 290
Круглоротые 53, 54, 417
— головной мозг их 249
— жаберные мешки их 333
жаберный аппарат их 336
— зачатки позвоночника у них 97
— вубы их 312
— кишечник их 325
— непарные плавники их 173
— орган обоняния у них 301
— половые железы их 388
— почки их 381
— череп их 133
Круглый мешочек уха 284
Крыловидная кость 137
— мышца 212
Ктенофоры 69
Куницы 53, 468
Куперовы железы 395
Кускус 465
Кутикула 69
Кювьеровы протоки 352, 367
------ланцетника 41
Л
Лабиринт, перепончатый 283
— скелетный (наружный) 284
Лабиринтовый аппарат у костистых рыб
338
Ладьевидная кость 204
Лактаза 325
Ламантины 56
Ланцетник 36, 415
— атриальная полость его 41
— брюшная аорта его 41
— временная асимметрия его 47
— жаберные перегородки его 41
— жаберный скелет его 38
— кардинальные вены его 41
— кишечник его 40
— кожа его 70
— кровеносная система его 41, 351
— кювьеровы протоки его 41
— лучи плавников его 37
— мускулатура его 37
— нервная система его 38, 229
— нефридиальные канальцы его 373
— нефридии его 42, 415
— обонятельная лопасть его 39
— околожаберная полость его 41
— организация его 36
— органы выделения у него 42
кровообращения у него 41
-чувств у него 39
— и периферические нервы его 3&
— поджаберная борозда его 40.
— покровы его 36
— половые железы его 42
— развитие его 42
518
Ланцетник, скелет его 37
— соленоциты его 42
— спинная аорта его 41
— хорда его 37
— целом его 41
— эндостиль его 40'
Ластоногие 468
— зубы их 316
Легочные артерии 352, 362
— вены 352, 367
— мешки у рептилий 347
Легкие позвоночных 340, 345
— происхождение 341
--- развитие их 340
.Легочное дыхание, развитие его у назем-
ных позвоночных 433
Лейкоциты 350
•Лемуры 56
•Ленивцы 53, 469
Лестница преддверия 287
Летун сумчатый 488
Летучие мЫш и 53, 467
— собаки 467
Летяга 469, 488
Лимфа, движение ее в сосудах 368
Лимфоидные органы у наземных 320
Лимфатическая система 368
Лимфатические железы 369
— сердца 368
— сосуды 353
Лимфоциты 353
Липаза 325
Лицевой нерв 265
---- образование узлов его 239
Личиночные органы и приспособления
65
Лобковая кость 193
Лобково-бедренная мышца 222
Лобные кости 136
Ложножабры 333
Ложнокоренные зубы 313
Ложные сосцы 76
Локтевая кость 185
Лопасть боковой линии 242, 276
— висцеральная 242
Лопатка 190
Лопаточный отдел пояса 181
Лофодонтные зубы 316
Лошади 53, 473
— вубы их 317
Луковица волоса, строение ее 85
Лунчатые зубы жвачных 316
Лучевая кость 185
— мышца первичная 219
Лучи жаберной перепонки 139
Лягушки (а) 53
— «летающая» 455
м
Макрель 487
Малые губы 396
Мальпигиев слой 70
Мальпигиево тельце 374
Мальтаза 325
Мамонт 475
Мантия мозга 235, 246
Мартышки 56
Матка 394
Матка двойная 394
двураздельная 394
— двурогая 394
— простая 394
Медведи 53, 468
Межключнца 117
Межключичный воздушный мешок 347
Межпозвоночные хрящи 104
Межпочечные тела 402
Межреберные мышцы 218
Межтеменная кость 165
Межцентры 107, 111
Межчелюстная мышца 212
Мезодерма, диференцировка ее в области
головы 122
— краевая (перистомальная) 68
— образование ее у высших позвоноч-
ных 68
у позвоночных 61
у селахий 62
— осевая (гастральная) 63
— роль ее в эмбриональном развитии
позвоночных 67
Мезокардий 353
Меритерий 475
Меккелев хрящ 129
--развитие его у млекопитающих
Меробластические яйца 56
Мероциты 57
Метамерное происхождение конечностей
178
Метамеры 15
Метанефридии 372
Метапофизы 112
Меторизис 6
Мигательная перепонка 296
Миксины 53
Мимическая мускулатура 213
Миндалины 320
Миноги 53
— зачатки позвоночника у них 97
— личиночные формы их 66
Миокардий 353
МиомерЫ 37
Миосепты 37
Миотом 44
Млекопитающие 53, 55, 56, 454
— артерии их 364
— асимметрическое расположение у них
главных венозных стволов 367
— бронхи их 348
— венозная система их 367
— волосяной покров их 84
— геологическая эпоха их появления
405
— головной мозг их 253
— гортань их 343
— грудина их 120
— дифиодонтизм пх 314
— зубная формула их 314
— зубы их 313
— кишечник их 325
— кишечный сок их 325
— копулятивные органы их 397
— легкие их 348
— органы обоняния у них 304
— плацентные 465
— позвоночник их 116
— половые железы 392
— почки их 383
— происхождение их 458
— процесс усложнения формы их корен-
ных зубов 315
— ротцвые железы их 320
— сердце 359
— скелет свободной конечностй их 201
519
Млекопитающие, скуловая кость их 166
— сочленение нижней челюсти их с че-
репом 167
— тазовый пояс их 195
— твердое нёбо их 167
— трахея их 344
— формирование корней на их зу-
бах 313
— череп их 163
— число смен зубов у них 313
— эволюция их 464
— язык их 319
Млечная линия 76
Млечные железы 75
-генетическая связь их с потовы-
ми 76
--расположение и число их 77
— сосуды 368
— точки 76
Многоклеточные животные, эволюция их
405
Мозазавры 452
Мозговой парус 243
Мозговые оболочки 255
Мозжечок 233, 243
— ножки его 244
Мозолистое тело 248
Молоточек уха 167, 289
Моллюски, кровеносная система их
351
— нервная система их 227
— покровы их 69
• — происхождение их 408
— строение их жабер 331
Монимостилия 156
Монофилия 490
Монофиодонтизм 311
Монкроевы отверстия 246
Моржи 53, 468
Морские черепахи 488
Морфогенетические корреляции 23
Морфологический параллелизм 489
Морфология 5
Морфо-физиологические преобразования
органов 17
Морфо-физиологический прогресс 483
Мочевой пузырь 383
Мочеполовой канал 384, 395
— синус 384, 392
Мочеточник 378
Мошонка 392
«Мужская матка» 394
Муравьеды 53, 316, 459
Мускулатура беспозвоночных 208
-— брюшная 218
— гладкая 209 и сл.
— висцеральная 209
— конечностей 219 и сл.
----- диференцировка ее 221
— ланцетника 37
— мимическая 223
— париетальная 209
— подбородочно-язычная 216
— подкожная 223
— подъязычная 215
— позвоночных 48 , 208
— поперечнополосатая 210
— соматическая 209, 214
— спинная 217
— туловища 216
— хвостовая 21.7
— языка 216
Мышечная система 208
520
Мыши 53
Мышца (ы) брюха 218
— височная 212
— глазная 214
— гребенчатая 222
— грудная 221
— грудино-ключично-сосцевидная 213
— грушевидная 222
— двубрюшная 212
— дельтовидная 220
— жевательная 212
— зубчатые 218
— квадратная поясничная 218
— коракоидно-плечевая 220
— крыловидная 212
— лестничные 218
— лобково-бедренняа 222
— лучевые 219
— межреберные 218
— межчелюстная 211
— остистые 217
— пирамидальная 219
— подбородочно-подъязычная 212
— подбородочно-язычная 216
— подлопаточная 220
— поперечные 217
— поперечно-остистые 217
— поясничный 220
— приводящие 222
— разгибательные 220, 222
— сгибательные 222
— спинная 217
— - трапециевидная 221
— четырехглавая 220, 222
— ягодичная 222
Мюллеров канал 377
И
Наднисочнан кость 136
Надгортанник 340
Надгрудиннпк 121
Наджаберные органы 338
— плакоды 238
Надлопаточный хрящ 190
Надпочечные тела 402
Надугловая кость 137
Наземные позвоночные, брюшная муску-
латура их 218
------мускулатура конечностей их 219
— — органы обоняния у них 302
------плечевой пояс их 189
------позвоночник их НО
------предки нх 435
------происхождение пх 432
--------— конечностей их 184
------роговой слой пх 81
-----спинная мускулатура пх 217
-----тазовый пояс их 193
------череп их 144
Наковальня уха 167
Насекомоядные 53, 55, 467
— зубы их 316
— эволюция их 467
Насекомые, происхождение их органов
летания 408
Нёбная кость 137
Нёбноквадратный хрящ 129
Нёбные железы 320
— миндалины 320
— пластинки 87, 310
НёврОбласты 236
Невроглия 236
Невромерия 234
Невроны 231
Немертииы, кровеносная система их
350
Необратимость эволюции 495
Непарнокопытные 53, 55, 472
— зубы их 317
— эволюция их 472
Неполнозубые 53, 55, 469
— зубная система тх 316
Нерв (ы) блоковый 263
— блуждающий 268
— верхнечелюстной 265
— висцеральные 259
— вкусовые 300
— глазничный 264
— глазодвигательный 262
— головные 262,
— добавочный 268
— жаберно-кшиечнЫй 268
— зрительный 262
— кишечные 258
— концевой 262
— лицевой 265
— нёбный 266
— нижнечелюстной 265
— обонятельный 262
— отводящий 264
— подъязычно-челюстной 266
— подъязычный 269
— сердечные 258
— слуховой 267
— соматические 259
— спинномозговые 256
— тройничный 264
— щечный 266
— язычный 268
— языкоглоточный 267
Нерпухи 468
Неспециализированные формы, значение
их в прогрессивной эволюции 493
Неурокраниум 123
Нефридиальный каналец 372
---эволюция его 374
Нефридии 373
-— ланцетника 42, 415
Нефридий Гатчека 42
Нефростом 373
Нефростомы ланцетника 42
Нижнечелюстной нерв, наружный 266
Нижние доли 245
— дуги позвоночника, закладывание их
у селахий 99
— ребра 91
Нижний остистый отросток 100
Нижняя гортань 345
Низшие позвоночные, позвоночник их 97
---- развитие у них элементов позво-
ночника 94
— рыбы, геологическая эпоха их появле-
ния 405
Ноготь 87
Ножки большого мовга 244
Ноздри 301
Носовая раковина 303
Носовые кости 136
Носоглоточный ход 303
Носороги 53, 472
О
Обезьяны 56, 478
— Зубы их 317
Обезьяны человекообразные 479
— череп их 171
Обмен веществ в животном организме
349
— газовый 330
Оболочки мозга 255
Оболочники 35
— кровообращение у них 351
Обоняние, органы его у позвоночных
50
Обонятельная кость 137
— лопасть ланцетника 38
— луковица 246
Обонятельные доли 246
— мешки (ямки) 301
— плакоды 301
— раковины 165, 303
Обонятельный нерв 262
— — развитие его 240
— стебелек 246
— эпителий 301
«Овальное» окно 149, 289
Овальный мешочек уха 283
Одотонтобласты 311
Околожаберная полость ланцетника
41
Околопочечные 402
Околосердечная полость 50, 371
— сумка 371
Околоушная железа 320
Околоушные кости 137
Околохордовые хряши 99
Окостенение 92
Олени 56, 471
Оливы мозга 243
Онтогенез (онтогенетическое развитие) 6,
16
— соотношение между ним и филогене-
зом 30, 34
— элементы его 34
Опахало 82
Оперкулярная жабра 336
Опистоцельные позвонки 110
Опоссум 465, 488
Оранг-утан 56
Орган (ы) 5, 13
— боковой линии 280
— — — функциональное значение их
281
— вкуса 300
— внутренней секреции 398
— возникновение и исчезновение их
27
— восприятия механического раздраже-
ния 279
---химического раздражения 299
•— воспроизведения 385
— выделения 372
---ланцетника 42
--- эволюция их 384
— дефинитивные 13
— дифференциация их 17
— дыхания 330, 332
---добавочные 338
— эктосоматические 21
— замещение их 27
— зрения 290
---позвоночных, происхождение
его 291
— -----эволюция его 298
—1 — развиие его 298, 435
•--развитие его у беспозвоночных
290
527
Органы (ы) интеграция их 17
— инфундибулярный 281
— кожные 278
— копулятивные 395
— морфологическое сходство (гомология
их) 5
— морфофивиологическое преобравова-
— морфофивиологическое преобразова-
ние их 17
— наджаберные 338
— кровообращения 349, 351
— обоняния 301
----амфибий 302, 305
---- круглоротых 301
----млекопитающих 303, 305
----навемных поввоночных 302
---- птиц 304
----развитие дыхательных функций
его 301
----рептилий 303, 305
— — рыб 302, 305
----селахий 301
----эволюция его 305
— околопочечные 402
— париетальный 245, 291
— пинеальный 291
— пищеварения 50, 306
— принцип уравновешивания их 22
— равновесия 282
— слуха 282
— рудиментарные 13
— соотносительное развитие их 21
— специализация их 21, 278
— субституция их 26
— чувств 49, 277
----возникновение их 27
----значение прогрессивного развития
их для эволюции поввоночных 435
----ланцетника 40, 229
----раввитие их у наземных позвоноч-
ных 435
— экзосоматические 483
— эндосоматические 483
Огранивм 13
Осевая мезодерма 63
Осевой череп 123, 136
----развитие его 124
Осетровые рыбы 53, 429
----окостенение в их позвоночнике
103
----происхождение их 428
---- фонтанель черепа 122
Основная перепонка кортиева органа
286
Остистые мышцы 217
— отростки 100
Остробугорчатые вубы 315
Островки Лангерганса 328
Осязательный мениск 278
Отводящий нерв 264
Открытопузырные рыбы 328
Отоконии 285
Отолиты 285
П
Павианы 56
Полингенев 34
Палеомастодонт 475
Палеонтологический метод в филогении
404
Панголин 53, 55, 316, 470
522
Панмиксия 29
Панцыриые «рыбы» 423
Панцырь броненосцев 90
— стегоцефалов 89
Паравухии 448
Параллелизм морфологический 489
Параллельная эволюция 489
Парапинеальный орган 291
Параподии 70
Паропсицы, происхождение их 452
Парасфеноид 137
Парафиза 246
Парахордалии 125
Париентальный орган 292
Парнокопытные 53, 55, 471
— эволюция их 471
Паротиды 74
Парус мозговой 243
Паукообразные, происхождение их 408
Паутинная оболочка мовга 256
Первичная почка 373
Первичнопочечный проток (каналец) 376
Первичное твердое нёбо 157
Первичноротовые 408
Первичные кости 81
Первичный скелет 92
Переднегрудные воздушные мешки 347
Переднелобные кости 137
Переднеушная кость 137
Передний мовг, первичный 233
---полушария его 235
--- строение его 245
Передняя дуга позвоночника 99
Перепончатые клапаны сердца 355
Перепончатый лабиринт 283
Перешеек мовга 233, 241
Перикардиальная (околосердечная) полость
371
Перикардий 371
Перилимфа 284
Перистомальная мезодерма 63
Перитонеум 371
Периферическая нервная система 49,
256
------- развитие ее 237
-------роль ганглионарной пластинки
в ее образовании 236
Периферические нервы ланцетника 38
Перихондральное окостенение 93
Перстневидный хрящ 343
Пескоройка 67
Печеночные протоки 326
Печень 308, 326
— воротная система ее 352
— раввитие ее 327
Перья 82
— пингвинов 84
— происхождение их 84
— раввитие их 83
Пигментная оболочка глава 295
Пигменты 72
Пилорические придатки 325
Пингвины, перья их 84
Пинеальный орган 292
Пирамида сердцевидного вещества почек
383
Пирамидальная мышца 219
Пирамиды мовга 243
Питекантроп 481
Пищеварительные железы 326
Пищеварение, органы его у позвоночных
50, 306
Пищевод 307, 320
Плавательный пувырь 328
---как добавочный орган дыхания 330
Плавник (и) брюшной 184
— вовникновение их 177 и сл.
— грудной 183
— концентрация его 173
— непарные 172
— парные 176
— хвостовой 175
Плакоды боковые (дорсолатеральные) 238
— наджаберные (эпибронхиальные) 238
Планула эпибронхиальные 238
Плакоидная чешуя 77
---развитие ее 78
Пластинчатая кость 137
Платибазальный череп 127
Плацента 65
Плацентные млекопитающие, эволюция их
465
Плевральная полость 371
Плевральные ребра 101
Плевродонтные зубы 313
Плевроцентры 429
Плезиозавры, происхождение их 446
Плейотропное действие гена 23
Плечевая кость 185
Плечевое сплетение 257
Плечевой пояс амфибий 189
---генетическая связь его покровных
костей с кожным панцырем рыб 89
---млекопитающих 192
---наземных позвоночных 189
--- птиц 192
--- рептилий 191
---стегоцефалов 190
Плюсна 186
Подбородочно-подъязычная мышца 212
— язычная мышца 216
Подвздошная артерия 361
Подвздошные вены 365
Поджаберная борозда ланцетника 40
Поджелудочная железа 308, 327
Подкишечная вена 355
Подключичные артерии 361
— вены 365
Подкожная мускулатура 213, 223
Подлопаточная мышца 220
Подмозговая железка 245
Подошвенная пластинка (когтя) 87
Подшерсток 85
Подчелюстная железа 320
Подъязычная дуга 129
— железа 320
— мускулатура 216
Подъязычно-челюстной нерв 266
Позвонки, гемальные отростки их 102
— гетероцельные 113
— истинные кресцовые 116
— опистоцельные 103, 110
— процельные 104, 110
— рептилий 111
Позвонок, развитие его 103
— туловищный 104
— хвостовой 105
Позвоночник 97
— амфибий ПО
— млекопитающих 116
— наземных позвоночных 110
— низших позвоночных 107
— птиц 115
— - развитие его элементов у низших по-
ввбночных 107
— хрЯщбвоЙ 99
Позвоночник человека 117
— элементы его 97
Позвоночная теория черепа 132
Позвоночные артерии 363
— вены 367
— бесчелюстные 417
— геологическая последовательность по-
явления отдельных классов их 405
— жабры их 322
— классификация их 53 и сл.
— легкие их 340
— мускулатура их 208
— наземные 189, 217, 432, 435
— непарные глазные пузырьки их
291
— низшие 133
— общая организация их 47
— органы вкуса у них 300
----воспроизведения у них 386
----выделения у них 372
----дыхания у них 332
----кровообращения у них 351
---- обоняния у них 301
----пищеварения у них 307
----чувств у них 278
— парные органы зрения у них 292
— покровы их 70
— половые протоки их 387
— почки их 372
— происхождение их 410
— развитие дыхательного аппарата их
422
----нервной системы их 231
----сердца и сосудов их 353
— скелет их 91
— слуховой орган их 282
— соотношение между ними и низшими
хордовыми 413
— филогения их 404
— челюстные 423
— эмбриональное развитие их 56
и сл.
Покровные кости 92, 136
----развитие их 95
Покровы ланцетника 36
— общие 69
— позвоночных 48
Полая вена, вадняя (или нижняя) 365
----нижняя 352
----передняя (верхняя) 368
Полипротодонты 465
Полифилия 490
Полифиодонтизм 311
Половой член 396
Половые железы, инкреторные части их
402
----ланцетника 42
— органы позвоночных 52, 386
-------связь их с почками 380
— поры 372
— протоки позвоночных 387
— складки 386
Полорогие 471
Полосатые тела мозга 235, 246
Полость тела 50, 370
----беспозвоночных 370
---- грудная 371
---- перикардиальная 371
---- плевральная 371
----позвоночных 371
Полукружные каналы 284
Полуобезьяны 56, 477
— вубы их 315
523
Полушария большого мозга 252
— переднего мозга 235
Поперечная борозда мозга 248
Поперечные мышцы 217
Поперечнополосатая мускулатура, замена
ею гладкой 210
Поперечноротые 53, 426
Поперечный отросток позвоночника 103
Постбранхиалные тельца 400
Потовые железы 74
----значение их 74
Почечная лоханка 383
Почечные вены 352
Почки вкусовые 300
•— позвоночных 373
----связь пх с половыми органами 380
Пояс рыб 180
Поясничная мышца, квадратная 218
ПОяснично-крестцовое сплетение 257
Правило рекапитуляции 35
Преддверие влагалища 392
Предгрудинник 121
Предплечие 185
Предплюсна 186
Предпочка 375, 381
ПредпОчечный проток 375
Предсердие 351, 356
Предстательная железа 395
Предчелюстной сомит 123
Предчелюстные дуги 129
— кости 137
Препуциальные железы 398
Пресмыкающиеся 53. См. Рептилии
Приводящие мышцы конечности 222
Придаток семенника 388
Признаки, расхождение их 486
— схождение их 486
Приматы, эволюция их 476
Принцип диференциации (Мильн-Эвардс)
8, 17
— интеграции (Спепсер) 17
— корреляции (Кювье) 8, 22
— смены функций (Дорн) 19
— уравновешивания органов (Сент-
Илер) 9, 12
— централизации (Франц) 20
Приспособительная эволюция 483
Приспособление 482
Проатлас 114
Провизорные органы 13
Прогресс 21, 483
Прогрессивная эволюция 483
----значение для нее неспециализиро-
ванных форм 493
Прогрессивные органы 13
Продолговатый мозг 233, 241
Прокоракопд 190
Промежуточный мозг 233
----строение его 244
«Проморфология» Геккеля 11
Пронефрический проток 375
Протей, редукция глаза у него 30
Протодонтная форма зубов 315
Протонефрид 372
Протостилия 131
Протоцеркальная форма хвостового плав-
ника 175
Процельные позвонки 106
Прыгунчик 467
Прямая кишка 325
— мышца брюха 216
Псевдовухии, происхождение их 448
Псевдо дентин 79
524
Птеригиофор 129
Птеродактили 449, 490
Птерозавры 449, 489
Птицетавые 451, 489
— происхождение их 451
Птицы 53
— артерии их 364
— бескилевые 490
— бронхи их 347
— геологическая эпоха появления их 405
— головной мозг их 253
— гортань их 343
— грудина их 120
— желудок их 321
— зубы их 313
— кишечник их 325
— копулятивные органы их 395
— легкие их 347
— органы обоняния у них 305
— позвоночник их 115
— половые железы их 392
— почка их 383
— происхождение их 453
— сердце их 359
— скелет свободной конечности их 200
— тазовый пояс их 195
— трахея их 344
— череп их 161
— язык их 319
«Птичья шпора» мозга 248
Пузырпый проток 428
Пульпа зуба 311
Пупочные артерии 355
Пух 83
Пуховые перья 83
Пястье 186
Пяточная кость 203
Р
Равновесие, орган его у позвоночных 50
Радиация адаптивная 485
Радужина 295
Развитие 16
— дивергентное 16, 486
конвергентное 486
— ланцетника 42
— - прогрессивное 483
— регрессивное 21, 29, 486
Разлитая нервная система 226
Раковина моллюсков 69
Ракообразные, дыхание пх 331
Роландова борозда мозга 248
Рамфоринхи 449, 490
Расхождение признаков 486
Ребра 101, 113
— амфибий 11Q
— брюшные 89, 121
— плевральные 101
— рептилий 113
Ревуны 56
Регресс 21, 483
Регрессивная эволюция 484
Регрессивные органы 13
Редукция органов 13, 30
Режущий зуб 315
Резцы 313
Рекапитуляция 35
Ректальная железа 325
Рептилии, артерии их 363
венозная система их 367
— геологическая эпоха пх появления 405
— головпой мозг их 241
Рептилии, гортань их 343
— грудина их 119
— желудок их 321
— зубы их 312
— кишечник их 325 .
— копулятивные органы у них 395
— легкие их 346
— органы обоняния у них 305
— позвоночник их 114
— половые железы их 392
— почки их 383
— происхождение их 442
— ребра их 113
— ротовые железы их 319
— сердце их 358
— скелет свободной конечности их 199
— способность менять окраску у них 72
— трахея их 344
— череп их 159
— язык их 318
Ресничное тело 296
Ресничный покров, значение его 69
Рефлекторная дуга 226
Решетчатая кость 137
Решетчатые раковины 304
Рога 86
— серого вещества мозга 241
Роговица 296
Роговой покров, значение его у наземных
позвоночных 81
— слой 70
Роговые образования 81
Ротовая полость 309
Ротовые железы 319
Рудименты 21, 30
Рудиментарные органы 13
Рудиментация 30
Рукокрылые 53, 55, 467
— зубная система их 316
Рукоятка грудины 120
Рыбообразные 53
Рыбы 53, 54, 423
— артерии их 362
— вены их 365
— взаимоотношения между их половыми
железами и почками 389
— жаберный аппарат их 332
— желудок их 321
— задний попе их 182
— зубы их 312
— кишечник их 323
— копулятивные органы у пих 395
— костные 405, 428
— низшие 405
— органы обоняния у них 305
— первые формы окостенения у них 427
— передний пояс их 180
- — почка их 382
— сердце их 356
— развитие их 423
— способность менять окрску у пих
72
— хрящевые 425
С
Саламандры 53
Сальные железы 75
Сальпы 36
Свиньи 53
Свободная конечность, скелет ее 196
Свод мозга 248
Седалищная артерия 365
— кость 193
Секодоптпый зуб 315
Секториальный зуб 315
Селахии 53
— головной мозг их 249
— зубная система их 312
— копулятивные органы их 395
— непарные плавники их 174
— органы обоняния их 301
— половые железы их 250
— череп их 134
— чешуя их 79
Селезенка 369
Семенные пузырьки 395
— трубочки 386
Ссмявыносяшие канальцы 387
Семяпровод 388, 394
Сердечные первы 258
Сердце 355
— двукамерпое 356
• — развитие его 353
— четырехкамерпое 359
«Серое» вещество мозга, образование его237
Серозная оболочка 65, 308
«Серповидная» складка головного мозга 256
Сетчатка 294
Сильвиев водопровод 244
Сильвиева борозда 248
Симпатическая нервная система 49, 258
Синантроп 482
Сипапсиды, происхождение их 445
Сипуатриальпые клапаны 335
Сирены 56, 475
— зубы их 317
Скаты 53, 224
Скелет беспозвоночных 9J
— брюшного плавника 184
— висцеральный 127, 137
— вторичный 92
— грудного плавника 183
— конечностей 172
— морфология его 204
— позвоночных 91
— свободный конечности 196
Скелетный (наружный) лабиринт 284
Складчатые зубы 316
Склера глаза 296
Склеробласты 79
Склеротом 44
Скулэвая дуга, образование ее v рептилий
160
— кость млекопитающих 166
Скуловые кости 166
Слезная кость 166
Слепая кишка 325
Слепое пятно 294
Слепыш 468
Слизистая оболочка кишечника 308
Слоны 56
—• эволюция их 474
Слух, возникновение его 29
Слуховой нерв 267
---образование его ганглия 238
— орган амфибий 285
---млекопитающих 286
---наземных позвоночных 285
— — позвоночных 50, 282
--- птиц 286
---рыб 285
Слуховые гребни 285
— капсулы 137
— пятна 285
525
Слюнные железы 308, 320
Собаки 468
Соленоциты 372, 416
— ланцетника 42
Сом электрический 225
Соматическая мускулатура 209, 214
Соматически-двигательная область голов-
ного мозга 275
— чувствующая область головного мозга
275
Соматические двигательные нервы 259
Сомит 44, 122
Сонная артерия 360
Сони 53, 469
Сосковидные тела мозга 245
Сосудистые сплетения мозга 241, 245
Сосудистый мешок 245
Сосуды лимфатические 353
— развитие их 353
Сосцевидный отросток черепа у человека1б5
Сосцы 76
— расположение и число их 77
Сочленовная кость 137
Сошник 137
Специализация органов 21
Специальная гомология 10
Спина, широкая мышца ее 220
Спинная аорта 352
---ланцетника 41
— мускулатура наземных позвоночных
217
— мышца 220
— струна 35
Спинной мозг 240
Спинномозговые нервы 256
Спинные рога мозга 241
Спланхнокраниум 123
Спланхнотом 45
Спланхноцель 45
Сплетения нервные 257
Сравнительная анатомия, эадачи 5
— — история 7
Средняя обонятельная кость 137
Среднее ухо млекопитающих 289
---наземных позвоночных 288
---развитие его 435
Средний мозг 233
---строение его 244
Сростножаберные 53
Статолиты 285
Стегальный череп 159
Стегозавры 451
Стегоцефалы 53, 438
— геологическая эпоха их появления 405
— происхождение их 441
— тазовый пояс их 194
— череп их 145, 153
Столбик уха 148
Страусы 53
Стремя уха 157, 168
Стрептостилия 156
Ступня 186
Субституция гетеротопная 26
— гомотопная 26
— органов 24, 26
— функций 24, 26
Сумчатые 53, 55, 455
— «крысы» 465
— вубы их 316
— эволюция их 465
Сумчатый волк 465, 488
— крот 465, 488
— летун 455, 465, 488
526
1’
Тазовая почка 373
Тазовый пояс наземных позвоночных 193
Талаттозавры 452
Тапиры 53
Таранная кость 203
Твердое нёбо млекопитающих 167
Текодонтиые зубы 313
«Тектология» Геккеля 11
Тектонические координации (Домбровский)
24
Телеологическое направление в биологии
Тело (грудины) 120
Тельца Гербста 279
— колбовидные 279
— фатер-пачинеивы 279
Теменной изгиб мозга 233
— орган 245, 291
Теменные кости 136
Тенрек 467
Теория архиптеригия 180
— боковой складки 178
— гастреи 406
— ганитогаструлы 406
— дегенерации 414
— единой почки 379
— метамерного происхождения конеч-
ностей 178
— планулы 406
— смены поколений канальцев 380
— фагоцителы 406
— энтероцеля 370
Териодонты 460
— происхождение их 447
Тероцефалии 461
Тироксин 399
Титанотерии 472
Тканевое дыхание 330
Толстая кишка 324
Тонкая кишка 324
Топографические координации (Северцов)
24
Трабекулы 125
Трабекулярный дентин 79
Травоядные млекопитающие, зубы их
317
Трапециевидная мышца 212, 221
Трахеи наземных членистоногих 331
Трахейная система 331
Трахея 342, 344
Трехбугорчатая форма вубов 315
Трехзубчатая форма вубов 315
Триконодонтная форма вубов 315
Трипсин 325, 328
Трипсиноген 325, 328
Тритоны 53
Тритуберкулярная теория образования
коренных зубов у млекопитающих 314
— форма зубов 315
Тройничный нерв 264
-образование ганглия его 239
Тромбоциты 350
Тромпибазальный тип черепа 127
Трубкозуб 471
Туловище, мускулатура его 216
Туловищная почка 373
развитие ее 376
Туловищный поввонок, строение его 103
— проток 368
Турбелярии, эпидермис их 69
Тушканчики 53, 469
Тюлени 53, 468, 490
У
Угловая КОСТЬ 137
Угорь электрический 224
Узконосые обезьяны 56, 478
Уистити 56
Улитка уха, канал ее 286
Ультимобронхиальные тельца 400
Уравновешивание органов 22. См. также
Принцип уравновешивания органов
Уростиль 106
Усы 86
Утконос 53, 465
— развитие его вубов 316
Ф
Фабрициева сумка 325
Фагоцителла 406
Фаланги пальцев 186
Фаллопиевы трубы 393
Фатер-пачиниевы тельца 279
Физиологические корреляции 23
Филитические корреляции 24
Филогенез 17, 30
— соотношение между ним и онтогене-
зом 30
Филогенетика 10
Филогенетическое направление 11
— формообразование 17
Филогения 11
— позвоночных 404
Филэмбриогенез 34
Фолликулы 386
Формообразование, филогенетическое 17
Функции, активизация их 20
— вамещение их 26
— разделение их 20
— расширение их 19
— расчленение их 483
— смена их 19
— субституция их 22
— сужение их 20
— усиление их 20
Функциональные корреляции 23
X
Хамелеон (ы) 452, 487
— способность их менять окраску 72
Хвост, мускулатура его у рыб 47
Хвостатое тело мозга 247
Хвостовая вена 352, 365
— мускулатура 217
Хвостовой плавник 175
— позвонок, строение его 100
Химеры 53, 427
Хитин 69
Хитиновый скелет членистоногих 91
Хищные 53, 55, 467
— зубы их 316
— эволюция их 467
Хоаны 302
Хоботные, зубы их 317
Хоботоголовые 53
Хорда 37
— позвоночных 95
Хордовые, выделительные органы их 373
— дыхание их 331
— кровеносная система их 351
—- низшие 410
— организация их 35
Хордовые происхождение их 410
— нервная система их 38
— эволюция их 411
Хордомезодерма 61
Хориоидальная щель 293
Хроматофоры 70
— у головоногих моллюсков 70
Хрусталик глаза 293
Хрящ (и) 93
— гиоидный 129
— гиомандибулярный 129
— головоногих моллюсков 70
— межпозвоночные 104
— меккелев 129
— надлопаточный 190
— нёбнокавдратный 129
— околохордовые 124
— перстневидный 343
— сфенолатеральные 125
— филогенетический процесс вытеснения
и замещения его костью 93
— черпаловидные 342
— щитовидный 343
Хрящевой позвоночник 103
— череп низших позвоночных 133
---у эмбрионов млекопитающих 163
Хрящевые ганоиды 53
---череп их 135
— рыбы, позвоночник их 99
---филогения их 425
Хрящекостные рыбы, геологическая эпоха
их появления 405
ц
Целом 45, 370
— внезародышевый 64
— ланцетника 41
— позвоночных 50
— развитие его 371
Целурозавры 455
Цельноголовые 53, 4CS
— геологическая эпоха их появления 427
— череп их 135
Цемент 311
Ценогенез 33, 36
Централизация 17
Центральная нервная система позвоночных
49
-------функциональная дифереици-
ровка ее отделов 274
Цератопсиды 451
Циклоидная чешуя 81
Цинодонты 462
Ч
Человек, волосяной покров его 86
— позвоночник его 117
— происхождение его 481
— скелет свободной конечности его 203
— череп его 171
Человеческий таз, особенность его 196
Человекообразные обезьяны 56, 479
Челюсти вторичные 166
Челюстная дуга 129
— раковина 303
Челюстной сомит 123
Челюстные кости 137
— позвоночные 53
---развитие их 423
Червеобразный придаток 325
527
Червеобразные организмы, происхожде-
ние их 407
Черви, дыхание пх 331
— кишечнодышащие 409
— кожно-мускульный мешок их 70
— мускулатура их 208
— нервная система их 227
— щетинкочелюстные 409
Червяги 53
Череп 121
— амфибий 149
Череп амфистильный 131
— аутостильный 131
— взрослого животного, первичный 127
— висцеральный 123, 127
— высших рыб 138
— генетическая связь его покровных
костей с кожным пангырем рыб 89
— гиостильный 131
• — двудышащих рыб 144
— зигальный 159
— • конфигурация его у млекопитающих
1G9
— костный 136
— млекопитающих 163
• образование крыши его 165
— наземных позвоночных 144
— обезьяны 171
— осевой 124
— платибазальный тип его 127
— протостильный 129
— птиц 161
• —• развитие и строение висцеральной
части его 127
— редукция наружной крыши его 158
п сл.
— рептилии 153
— связь его образования с развитием
органов чувств и головного мозга 123
— стегальный 159
— стегоцефалов 153
— теория происхождения его 132
— тропибазальный тип его 127
— хрящевой 133, 163
— человека 171
Черепные балки 124
— первичные 421
Черепаха кожистая 487
Черепахи 53
— кожные окостенения их 89
— морские 487
— происхождение их 445
Черпаловидные хрящи 342
Четырехбугорчатый зуб 316
Четырехглавая мышца 220
Чешуйчатая кость 137
Чешуйчатые 53
Чешуйчатый покров, производные его 89
Чешуи рыб, генетическая связь всех видов
их 77
— у рептилий 81
Чешуя ганоидная 79
— космоидная 79
— костная 81
— ктеноидная 81
— плакоидная 77
— селахий 77
— циклоидная 81
Членистоногие, кровеносная система их
351
— мускулатура их 208
— нервная система их 227
— происхождение их 408
528
Членистоногие хитиновый скелет пх 69
Чувствующие клетки 230
Чудесная сеть 350
HI
«Шатер» мозга 256
Шванновская оболочка 238
Швейные воздушные мешки 347
Шерстокрылые 53, 55, 455, 467
Шерсть 84
Шея, значение ее у наземных позвоноч-
ных 47
Шимпанзе 56, 479
Широкая мышца спины 220
Широконосые обезьяны 56, 478
Шишковидная железа 401
щ
Щележаберпые, развитие их 426
Щетина 85
Щотинкочелюстпые черви 409
Щечные железы 320
—- кости 139
Щечный нерв 266
Щитки у рептилий 81
Щитковые 417, 423
Щитовидная железа 398
э
Эволюционный процесс, основные направ-
ления его 483
Эволюция, морфо-физиологические зако-
номерности ее 482
— направленность ее 492
— необратимость ее 495
— неограниченность ее 482
— определяющие ее направление фак-
торы 486
— параллельная 486, 489
— приспособительная 484
— прогрессивная 483
— - регрессивная 484
Экзосоматические органы 483, 496
Эктодерма, основные функции ее 67
— роль ее в развитии эпидермиса и его
производных 67
Эластическая оболочка хорды 96
Эластоидиновые нити 174
Элеидин 72
Элективные органы 278
Электрические органы 224
— скаты 225
Электрический сом 225
— угорь 225
Эмалевый орган 310
Эмаль 78, 311
Эмбриологическая теория черепа 133
Эмбриологический метод в сравнительной
анатомии 30
Эмбриология 6
Эндокардий 353
Эндолимфа 283
Эндолимфатический проток 283
Эндостиль ланцетника 40
Эндохондральное окостенение 92
Энтерокиназа 325
Энтероцель 370
Энтодерма, роль ее в развитии внутренних
органов 68
Энтомезодерма 60
Эпендима 236
Эпибронхиальные плакоды 238
Эпидермис 69
— железистые клетки его 72
— поввоночных 70
— роль эктодермы в его развитии 67
Эпистрофей 114
Эпителиальные тельца 400
Эпифив 401, 245
Эрепсин 325
Эритроциты 350
Я
Ягодичная мышца 222
Ядоотделительные кожные железы 73
— вубные железы 319
Ядра (серого вещества мозга) 237
Язык 317
— мускулатура его 216
Языкоглоточный вер 267
— образование узла его 238
Язычные железы 320
Яйца голобластические 56
— меробластические 56
Яйцеводы 392
Яйцекладущие клоачные 53
Яйцо, влияние количества желтка в нем
на внешнюю форму зародыша 64
Якобсонов орган 304
Ямка Келликера 39
Яремные вены 365
Ящерицы, змееобразные 452
Ящерообразные 452
Ящеротазые 451, 489
— происхождение их 451
INDEX
А
Acanthodei 425, 426
Acetabulum 194
Acromion 192
Acropodium 185
Actinopterygii 430
Actinotricha 174
Aeluroidea 468
Agnatha 52, 54, 405 , 417
Agorophius 469
Aistopodidae 442
Alae magnae 165
— orbitales s. orbitosphenoidea 165
— temporales s. alisphenoidea 165
Albula 430
Albulidae 430
Allotheria 464
Amblypoda 471
Ambulacralia 409
Amia calva 81, 106 , 429
Amiidae 429, 430
Amniota 52, 55, 106
Ammocoetes 67
Amphibia 53, 54
Amphioxus lanceolatus 36
Amphipnous 339
Amphitherium 466
Anarnnia 52, 54
Anaspida 426
Anchiterium 473
Anoplotheriidae 471
Antechinomys 465
Antbracotheriidae 471
Antilocarpa 88
Anura 53, 54
Aorta ven trails 41, 351
— dorsalis 41, 351
Apoda 53, 54
Appendices pylorici 325
Aquaeductus Sylvii 233, 244
— vestibuli 283
Arachnoidea 256
Araeoscelis 452
Arbor vitae 243
Archegosaurus 440, 442
Archelon 497
Archencephalon 233
Archaeopteryx 456
Archaeonris 405, 456,
Archaeornithes 53
Archipallium s. primordium hippocampi 247
Arctoidee 468
Arcus branchialis 129
— hyoideus 129
— mandibularis 129
— palato-pharingei 310
Area acustica 274
Argentea 295
Artoria (ae) allantoidis s. umbilicales 355
— carotides interna et externa 362
— carotis 352
---anterior 360
---posterior 360
— caudalis 352, 361, 364
— coeliaco-mesenterica(ae) 352, 361, 363
— femoralis 365
— gastrica 361
— genitales 352, 361
— hepatica 361
— iliaca communis 355
— — interna 365
— iliacae 352, 361, 364
— ischiadica 364
— mesenteri.cae 361
— omphalo-mesentericae 355
— pulmonales 352, 362
— renales 352, 361
— subclaviae 352, 361
— vertebrales 363
Arthrodira 424, 425
Artiodactyla 53, 55, 471
Aspidorhynchidae 429
Astragalus 204
Atrium 356
Australopithecus africanus 179
Autopodium 185, 196
Aves 53, 55
В
Balanoglossus 409, 411, 412, 413
Basale metapterygii 183,
Basalia 186
Basibranchialia 129
Basipodium 185
Basiventralia 429
Batoidei 53, 54
Bdellostoma polytrema 333
Belodon 448
Brachia conjunctiva s. crura cerebelli ad
cerebrum 244
529
Branchi osauria 153
Branchiosaurus 440, 442
Branchiostegalia 139
Bronchi 342
Bufo 391
Bulbus arteriosus 350
— oculi 296
— olfactorius 246
Bunolophodon 475
C
Cacops 442
Calamoichthys 79
Calcaneus 204
Calcar avis 248
Canales semicirculares 284
Canalis cochlearis 283, 286
— neuroentericus 232
— obturatorius 194
— urogenitalis s. urethra 394
Canidae 468
Canini (dentes) 313
Canis 488
Capitulum 113
Сагарах 90
Carina 120
Carnivora 52, 55, 467
— creodontia 466
Carpalia 186, 197
Carpus 185
Cartilagines arytaenoideae 342
— sphenolaterales 125
Cartilage cricoidea 443
— cricotrachealis 342
— epipubis 194
— Meckeli 123
— palatoquadratum 129
— procoracoideum 190
— suprascapulare 190
— thyreoidea 343
— ypsiloides 194
Caturus 429
Cauda equina 241
Cavum tympani 288
— naso-pharyngeum 309
— oro-pharyngeum 309
— oris 307
Centetes 316, 467
Centralia 186, 197
Cephalaspidae 418 и сл.
Ceratodus 67, 189, 432, 495
Cerebellum 233, 243
Cetacea 53, 55, 468
Chaetognatha 409
Chameleo 487
Cheirodus 429
Cheirolepsis 429, 431
Chelone 487
Cheolnia 53, 54
Chelonidae 445
Chiasma 245
Chirocentrus 430
Chironectes 465, 488
Chiroptera 53, 55, 467
Chlamydoselachus 135, 179
Choanae 302
Chondrichthyes 53, 425
Chondrostei 53, 495
Chondrosteoidei 428
Chorda tympani 267
Chorioidea 295
530
Chrysochloris 467, 488,
Cladodus 426
Cladoselache 426
Clavicula 90, 190
Cleithrum 181
Cloaca 325
Clupeidae 430
Cobitis fossilis 339
Coecum 325
Coelacanthidae 431
Coelurosauria 455
Colon 325
Columella 157, 288, 435
Comissura anterior 246
— palii anterior s. hippocampi 246
— posterior 235
— superior s. habenularis 245
Compsognathus 451
Concha 289
Conchopoma 432
Condylarthra 53, 55, 471
Conus arteriosus 356
Copulae 129
Coryphodon 471
Corium s. dermis 69
Cornu anterius, inferius et posterius 248
— Ammonis 247
Corpora caudata 247
— geniculata lateralia et medialia 245
— interrenales 402
— mamillaria 245 '
— quadrigemina 235
— restiformia 243
— striata 235, 246
— suprarenales 402
Corpus callosum 248
— ciliare 296
— fibrosum 395, 397
— luteum 386, 403
— spongiosum 395, 397
— sterni 120
Costalia 90
Cotylosauria 53, 444 и сл.
Crania 495
Cranium 123
Creodontia 53
Cristae acusticae 285
Crocodilia 53, 448
Crossopterygii 53, 405, 431
Crura cerebelli ad pontem 244
Crura cerebri 244
Cutis 69
Cyclostomata 53, 54
Cynognathus 463
I)
Dactylethra 194
Deltatheridae 466
Dermis (corium) 69
Descensus testiculorum 392
Desmostylus 465
Deuterostomia 409
Diapsida 53, 160, 448
Dicynodontia 448
Didelphyidae 465
Didelphys 459, 465
Diencephalon 233, 244
Dimetrodon 445
Dinoceras 471
Dinotherium 475
Diplocaulus 442
Diplovertebron 440
Dipnoi 58, 405, 481, 482
Dipteridae 432
Discina 495
Discoglossidae 391
Diprotodontia 316
Dolichosauria 452
Dolichosoma 442
Draco volans 455
Dromatherium 464
Dryopithecus 479
Ductus caroticus 354
— choledochus 327
— cuvieri 352, 365
— cysticus 327
— endolymphaticus s. aquaeductus
vestibuli 283
— hepatici 326
— hepato-enterici 327
— nasopalatini 305
— nasopharyngens 303
— pancreatici 327
— perilymphaticus 284
— pneumaticus 328
— santorini 328
— utriculo-saccularis 284
— Wirsungi 328
Duodenum 324
Dura mater 255
E
Echinodermata 409
Ectobranchiata 53, 423
Ectoturbinalia 304
Edentata 53, 55
Elasmobrachii 53, 405, 424, 426
Elasmotherium 472
Elastotrichia 174
Eiephas 475
Elopidae 430
Elops 430
Embolomeri 441, 489
Endoplastron 90
Endoturbinalia 422
Enteropneusta 409
Entobranchiata 52, 422
Eogyrinus 440
Eotherium 475
Eohippus 473
Ependyma 236
Epidermis 69
Epididymis 388, 392
Epiglottis 343
Epiplastron 90
Epistemum 90, 121
Epistriatum 246
Epoophoron 388, 392
Epiphysis cerebri s. glandula pinealis
245
Equus 473
Erinaceus 467
Ethmoturbinalia 165, 304
Eucreodi 468
Eugnathidae 429
Eugnathus 429
Eunnotosaurus 445
Eurycormus 429
EusthenopterOn 198, 436
Eutheria 53, 55
Extracolumella 148, 157
F
Falx cerebri 256
Felidae 468
Femur 186
Fenestra basicranialis anterior 126
— ischio-pubica 194
— ovalis 149, 289
— rotunda 154, 288
Fibula 186, 196
Fibulare 186, 196
Filum terminale 241
Fissipedia 468
Fissura hippocampi 247
— Sylvii 248
Flocculi 244
Foramen entepicondylare 202
— ocdpitale 137
Foramina Monroi 246
Furcula 192
G
Galeopitheci 53, 55, 467
Galeopithecus 455, 467, 488
Ganglia habenulae 245
Ganglion cilire s. sphenopalatinum 258
— spirale 287
— Gasseri s. trigemini 265
— geniculi s. facialis 266
— petrosum 267
Gastralia 89
Glandula (ae), buccalis (es) 820
— labiales 319
— lacrymalis 297
— linguales 320
— palatinae 320
— parotis 320
— pinealis 401
— pituitaria 245, 401
— prostata 393
— rectolingualis 320
— sublingualis(es) 320
— submaxillaris 32
— uropygi 74 о
Gnathostomata 53,
Gymnarchus 224J
Gymnophiona s. Apoda 53, 54
Gymnotus electricus 224
H
Hatteria 53, 291, 448
Haustra 325 >•
Hemicentetes 316, 467
Heptanchus 131, 135
Heterocerci 428
Hexanchus 131, 135
Hippocampus s. Cornu Ammonia 247
Holocephali 53, 405, 427
Holonephros 379
Holoptychiidae 431, 438
Holostei 53 , 405, 429, 435
Holosteoidei 428
— crossopterygii 430
Humerus 186
Hyaenodontidae 467
Hydrocoelia 409
Hyoplastron 90
Hyopsodus 473
531
Hypophysis cerebri s. glandula pituitaria
425, 245, 401
Hypoplastron 90
Hypothalamus 245
Hypparion 437
Hyracoidea 56, 475
Hyracotherium 473
I
Ichtidosauria 464
Ichtyopsida 53
IchtyOsauria 54 , 446
Ichthyostegidae 439, 442
Ichtyotomi 426
Iguanodon 451
Incisivi 313
Incus 167, 289
Indricotherium 472
Infundibulum 245
InsectivOra 53, 55, 467
Intercalate 449, 456
Intercalaria 100
Intercentra 411
Interclavicula 421
Interdorsalia 429
Interhyale 439
Intermedium 186, 197
Interoperculum 439
Iris 295
Isthmus 233, 244
J
Jejunum 325
Jugalia 440, 453
К
Karromus 454
Katarhina 477
1
Labyrinthodontia 438
Lagena 283
Lamna 487
Lamina cribrosa 465
— spiralis ossea 287
— perpendiXularis 465
— terminalis 235, 845,
Lanugo foetalis 86
Larynx 342
Latax 468
Latimeria 436
Lepidosiren 67, 472
Lemuorides 477
Lepidosteidae 429
Lepidosteus 79, 103
Lepidotrichia 474
Leptolepidae 430
Ligamentum hepato-entericum 327
— spirale 287
— suspensorium hepatis 827
Lingua 317
Lingula 495
LitOpterna 474, 490
Lobi inferiores 245
— olfactorii 235, 246
— optici 235, 244
Lobus acustico-lateralis 242
— posterior s. saccus vasulosue 245
— visceralis 242
Loxomma 440, 448
532
Lutra 468, 488
Lyriocephalus 487
LysOrophus 442
M
Machairodontidae 468
Macropetalichthys_424
Macropus 465/
Macroscelides 467
Macula(ae) acustica sacculi 285
----utriculi 285
— acusticae 285
— neglecta 285
Malapterurus electricus 225
Malleus 167
Mammalia 53, 55
Mandibula 167 ч
Manubrium sterni 420
Marginalia 90
Marsupialia 53, 55, 466
Mastodon 475
Maxilloturbinale 165, 272
Medialia 186, 498
Megalops 430
Membrana basilaris 286
— nicticans 296
— tectoria 287
— tympani 288
Meninges 255
Meninx primitive 255
Merychippus 473
Mesencephalon 233, 244
Mesenterium 308
Mesohippus 473
Mesonephros 372
Mesonychidae 467
Mesorchium 386
Mesosauria 445
Mesovarium 386
Metacarpalia 186
Metacarpus 186
Metanephros 372
Metasternum 119
Meta tarsalia 186
Metatarsus 186
Metazoa 405, 406
Metencephalon 233, 243
Micidae 468
Microbrachis 442
Microconodon 464
Micropyle 56
Microsauria 444
Moeritherium 474
Molares 313
Monotremata 53, 75, 465
Mosasauria 452
Multituberculata 315, 465
Musculus(i) adductor hyomandibulae 212
----mandibulae 211, 212
----adductores 222
— anconeus s. quadriceps 220
— arcuales dorsales 212
— auriculares 213
— biceps brachii 221
— buccinator 213
— ciliaris 296
— constrictor superficialis 210
— coraco-arcuales 216
— coraco-brachiales(is) 246, 220
----mandibularis 216
----radialis s. biceps 221
— cremaster 892
Musculus (i) deltoideus scapulae 220
— depressor mandibulae 212
----s. adductor 219
— digastricus mandibulae 212
— dorsalis scapulae 220
— extensores 220
— flexores 221
— genio-glossus 216, SIS'
----hyoideus 216, 318
— gluteus s. iliofemoralis 222
— hyoglossus 216, 318
— ileo-cqstales 217
— ° intercostales externus et intemus 218
— intermandibularis 211
— interspinales 217
— intertransversales 217
— latissimus dorsi 220, 221
— levator bulbi 212, 297
----palatoquadrati 211
----palpebrae superioris 215» 297
----scapulae 221
----s. abductor 219
— levatores pterygoidei 212
— longissimus capitis 217
----dorsi 217
— masseter 212
— mylohyoideus 212
— nicticans 215, 297
— obliqui capitis 217
— obliquus etxernus (abdominis) 218
----inferior 214
----internus (abdominis) 218
----superior 214
----omo-hyoideus 216
— Orbicularis oculi 213, 297
----oris 213 ’
— orbito-quadrati 212
— pectineus 222
— pectoralis 221
— «— major 221
----minor 221
— pronatores 221
— pterygoideus 212
— pubofemoralis 222
— pyramidalis 219
— pyriformis 222
— quadratus lumborum 218
— radiales(is) 219
— recti capitis 217
— rectus abdominis 218
----externus 214
----inferior 214
----internus 214
----superior 214
— retractor bulbi 214, 297
— scaleni 218
— serrati antici superficialis et profun-
dus 221
----pistici superior et inferior 218
— sphincter colli 212, 223
— stenocleido-mastoideus 213, 221
— sterno-hyoideus 318
— subscapularis 220
— supinatores 220
— temporalis 212
— transverso-costales 217
----spinales 217
— trapezius s. cucularis 213, 221
Mustelidae 468
Myelencephalon s. medulla oblongata 233,
.241
Myogale 467
Myrmecobius 465
Myxinoidei 58, 54
К
Nares 802
Nasoturbinale 165
Neopallium 247
Nervus(i) abducens 264
— accessorius 268
— (i) acusticus 267
— branchiointestinalis 268
— buccalis 265
— cardiacus(i) X 258, 268
— facialis 265
— glossopharyngeus 267
— hyoideus VII 266
— hyomandibularis 266
— hypoglossus 269
— intestinalis 268
— lateralis 239, 265,
— lingualis 265, 268
— mandibularis 265
----externus 266
— internus 266
— maxillaris 265
— oculo-motorius 262
— olfactorius 262
— ophtalmicus profundus s. n. trigemi-
nus I 264
----superficialis 239, 265
— opticus 262
— palatinus 266
— praetrematicus 267,
— posttrematicus 268, 270
— pharyngeus 270
— splanchnic! 258
— terminalis 262
— trigeminus 264
— trochlearis 263
— vagus 268
— vestibularis 287
Neuralia 89
Neurapophyses 100
Neuroglia 236
Neuroporus 4'3, 232
Notidanidae 134
Notoryctes 465, 488
Nucleus dentatus 244
— gelatinosus s. nucleus pulposus 97
0
Oesophagus 307
Olivae 243
Olecranon 196, 202
Operculum 139, 335
Oreodontidae 471
Organon Jakobsonii s. vomeronasale 304
Ornithischia 451
Ornitholestes 451
Ornithorhynchus 465
Orohippus 473
Orycteropus 471
Os(sa) acetabulare 195
— angulare 137
— articulate 138
— basioccipitale 137
— basisphenoideum 137
— branchiostegalia 137
— coracoideum 190
— cornu 87
533
Os (ва) dentale 137
— ectopterygoideum 138
— entopterygoideum 133
— epioticum 137
— epipterygoideum 155
— ethmoideum 137, 165
— ethmoturbinalia 165
— exethomoideum 137
— exoccipitalia 137
— frontalia 136
— goniale 138, 137, 156
— ileum 193
— innominatum 195
— interparietale 166
— investientia 92
— ischium 193
— jugalia 137, 153
— lacrymale 166
— laterosphenoidea 137
— marsupialia 195
— mastoideum 165
— maxillare(ia) 137
— maxilloturbinale 165
— mesethmoideum 137
— metapterygoideum 138
— nasalia 136
— nasoturbinalia 165
— naviculare 203
— opercularia 137
— opisthoticum 137
— orbitalia 140
— orbitosphenoidea 137
— palatinum 137
— parasphenoideum 137
— parietalia 136
— penis 397
— periotisa 137
— petrosum 165
— pisiforme 200
— postfrontalia 137
— postorbitalia 137
— postrarietalia 153
— praefontalia 137
— praemaxillare 137
— praesphenoideum 137
— procoracoideum 190
— prooticum 137
* — pteroticum 142
— pterygoideum 137
— pubis 194
— quadratojugalia 145, 153
— quadratum 138
— sphenethmoide.um 150
— sphenoticum 142
— spleniale 137
— squamosum 137
-substituentia 92
— supraangulare 137
— supraoccipitale 137
— supratemporale 136
— temporale 165
— transversa 156
— tympanicum 165, 166
Ostariophysi 329
Osteichthyes 53, 428
Osteoglossum 430
Osteolepida 489
Osteolepidae 430
Osteostraci 422, 423
Ostracodermi 53, 54, 417, 423
Otariidae 468
Oxyaenidae 467
Oxyelaenidac 467
554
P
Pachypodosauria 451
Palaeodictyoptera 408
Palaeogyrinus 440
Palaeomastodon 475
Palaeoniscidae 80, 426, 428
Palaeoniscus 428, 429
Palaeopithecus 479
Palaeostoma 401
Pallium 233, 246
Palpebrae 296
Pannicuius camosus 223
Pantolambda 471
Pantotheria 314, 466
Papilla acustica basilaris 285
------- lagenae 286
Papillae circumvallatae 300
— foliata 300
— fungiformes 300
Parachoraalia cranii 124
Paradidymis 388, 392
Paraphysis 246
Parapithecus 479
Parapophyses 100
Paroophoron 388, 392
Patella 203
Pathriocetus 469, 496
Pedunculi cerebri s. crura 244
Pedunculus olfactorius 246
Pelvis renarum 383
Pelycosauria 445
Penis 355, 395
Peripatus 408
Perissodactyla 53, 55, 472
Permocynodon 463
Pessulus 345
Petaurus 455, 465, 488
Petromyzon 67
Petromyzontes 54
Phalanger 465
Phalanges digitorum 186
Phaneropleuron 432
Pharyngohyale 147
Pharynx 307, 320
Phenacodus 471
Phocidae 468
Pholidota 469
Phytosaurus s. Belodon 448
Pia mater primitiva 255
-------secundaria 256
Pinnipedia 468
Pisces 53, 54
Pisiforme 200
Pithecanthropus erectus 481
Pithonomorpha 452
Placentalia 53, 55, 465
Placodontia 446, 487
Plastron 90
PlatyrhinB 477
Platysma myoides 213
Platysomidae 429
Plesiosaurus 446
Pleuracanthus 426
Pleuropteryqii 54
Plexus brachialis 257, 270
— cervicalis 269
— chorioideus anterior 245
-------prosterior 241
— lumbo-sacralis 257
Plica semilunaris 296
Pliohippus 473
Polymastodon 465
Polyprotodontia 316, 465
Polypteridae 430
Polypteroidei 53, 54
Polypterus 80
Pons Varolii 244
Pori abdominales 372
Posttemporale 182
Potamogale 467
Praechallux 199
Praemolares 313
Praeoperculum 139
Praeputium 389
Primates 476
Primordium hippocampi (archipallium) 247
Proatlas 114
Proboscides 56, 474
Processus basipterygoidei 155
— coracoideus 192
— iliacus 193
— mastoideus 165
— odontoideus 114
— oticus 134
— palatobasalis 134
— paroticus 155
— pectineus 195
— spinosus haemalis 102
----neuralis 101
— styloideus 168
— uncinati 115
Procolophonia 445
Proctodaeum 307
Procyonidae 468
Pronephros 373,
Propliopithecus 479
Prosencephalon 233
Prosimiae 56
Prosternum 121
Protocetus 469
Protocraniata 421
Protodonta 454
Protopterus 67, 432, 433
Protorosauria 448
Protosphargis 497
Protostomia 408
Prototheria 465
Protungulata 471
Pseudosuchia 448
Pteranodon 449
Pterodactylus 449
Pteromys 455
Pterosauria 448
Pulpa 77
Pupilia 296
S
В
Radlale 186, 197
Radialia 172
Radius 185, 196
Raja 224
Rami cardiaci 258
Ranidens 151
Ratitae 53, 55
Recessus praeopticus 235
Rectum 235
Reptilia 53, 54
Rete mirabile 329, 350
Retina 294
Rhachitomi 489
Rhacophorus 455
Rhamphorhynchus 449
Rhizodontidae 430, 436, 438, 441
Rhombencephalon 233
Rhynchocephalia 53, 448
Rodentia 53, 55
Rostrum 155, 162
Saccobranchus 339
Sacculina 484
'Sacculus 283
— endolymphaticus 284
Saccus vasculosus 245
Sagitta 409
Salmonidae 430
Saurichthyidae 428
Sauripterus 189, 436
Saurischia 449
Sauropoda 451
Sauropsida 52
Sauropterygia 54, 446
Scala tympani 287
— vestibuli 287
Scapula 190
Scaumenacia 432
Scleromochlus 448
Sclerosaarus 445
Scrotum 392
Selachii 53, 54
Septum interorbitale 127
— pellucidum 248
Sesamodon 462
Seymouria 444
Simiae 55, 56
Sinanthropus pekinensis 482
Sinus rhomboidalis 240
— urogenitalis 392
— venosus 356
Siphaeus 488
Sirenia 55, 56
Solenodon 467
Sorex 467
Spadella 409
Sphargidae 445
Spina scapulae 192
Spiraculum 130
Squalodon 469
Squaloidei 53, 55
Squamata 53, 55, 452
Stapes 148, 168
Stegocephali 53, 54
Stegodon 475
Stegosaurus 451
Stereospondyli 489
Stereosternum 445
Stomodaeum 307
Stylopodium 186, 196
Sublingua 319
Suboperculum 139
Substantia adamatina 311
— eburnea 311
Sulcus centralis s. Rolandi 248
— transversus 248
Supracleithrale 182
Symphysis 193
Symplecticum 135
Synapsida 53, 161, 445 и сл.
Syrinx 345
535
т Uterus simplex 394
Taenia 325 Talpa 467, 488 Tapetum 295 Tarsalia 186, 197 Tarsius 455, 480 Tarsus 186 Tectum opticum 244 — synoticum 216 Telencephalon 233, 245 Teleostei 53, 405 Teleostomi 53, 54 Temnospondyli 441 Tentaculata 409, 412 Tentorium 256 Thalami optici 235, 244 Thalassemys 496 Thalattosauria 452 Theriodontia 53, 54 Thoatherium 474 Thylacinus 465, 488 Tibia 186 Tibiale 186, 197 Tonsillae 320 — palatinae 320 Torpedo 225 Toxochelys 496 Trabeculae cranii 124 Trachea 342 Trematops 440 Triassochelys 445 Tribolodon 464 Triceratops 451 Trichechidae 468 Triconodonta 466 Triconodontidae 465 Trimerorachis 443 Trionychoidea 445 Tritylodon 465 Trochanteres 203 Truncus sympaticus 258 Tuba Eustachii 288 Tuberculum 235 — posterius 235 Tubuli seminiferi 386 Tupaja 467, 477 Tylototriton 151 Tympanicum 166 Tyrannosaurus 451 Utriculus 283 V Vagina 394 Valvula bicuspidalis, tricuspidalis 359 — cerebelli 243 Vas deferens 388 Vasa efferentia 387 Velum 40 — medullare anterius s. Valvula cere* belli 243 posterius 243 — transversum 235 Vena(ae) abdominalis s. epigastrica 352,366 — azygos 368 — cardinales anteriroress. jugulares352, 365 posteriores 352, 365 —> caudalis 352, 365 — cava inferior 352, 365 posterior 365 — coronaria 368 — cutaneae magnae 367 — epigastrica 366 — genitales 352 — hemiazygos 368 — hepatica 41 — iliacae 365 — innominata 368 — intercostales 368 — jugulares 352, 365 — laterales 352, 365 — portarum hepatis 352 renarum 352 — pulmonales 352, 367 — renales 352 — subclaviae 352, 365 — subintestiualis 41 — vertebralis 367 Ventriculus 356, Vermis 243 Vesica fellea 326 Vesiculum seminalis 389 Vestibulum 283, 303 • Vibrissae 85 Viverridae 468 Volvocales 406 Vomer 137 Vulpavus 468
u X
Ulna 186, 196 Ungulata 470' Ureter 378, 383 Urethra 394 Urodela 53, 54, 442 Uronemus 432 Ursidae 468 Uterus 394 — bicornis 394 — bipartitus 394 — duplex 394 Xenarthra 469 Xiphipl astron 90 Xiphistemum 120 z Zalambdalestidae 466 Zalambdodonta 466 Zeuglodon 469 Zeugopodium 186, 196 Zona radiata s. pellucida 56
СОДЕРЖАНИЕ
Предисловие ................................. 3
Часть I. ВВЕДЕНИЕ
История стравцитрльной анатомии.......................................... 7
Основные понятия сравнительной анатомии..................................13
Организм и орган..................................................13
Метод сравнения и гомология органов. .............................14
Развитие (филогенез и онтогенез) . ...............................16
Основные принципы филогенетического формообразования.....................17
Диференциация и интеграция........................................17
Морфофизиологические преобразования органов.......................17
Прогресс, регресс и специализация органов.........................21
Соотносительное развитие органов .................................21
Корреляции индивидуальные или физиологические.................22
Координации, или филетические корреляции......................24
Замещение органов и функций...................................26
Возникновение органов и их исчезновение...........................27
Эмбриологический метод в сравнительной анатомии и вопрос о соотношениях между
филогенезом и онтогенезом ..........................................30
Организация хордовых животных............................................35
Характеристика хордовых ..........................................35
Организация ланцетника'...........................................36
Развитие ланцетника...............................................42
Общая организация позвоночных.....................................47
Классификация позвоночных.........................................52
Эмбриональное развитие позвоночных................................56
Часть II. СРАВНИТЕЛЬНАЯ АНАТОМИЯ ОТДЕЛЬНЫХ СИСТЕМ ОРГАНОВ
Общие покровы........................................................ 69
Покровы беспозвоночных............................................ 69
Покровы позвоночных................................................70
Кожные железы..................................................72
Защитные образования кожи......................................77
Кожный скелет..................................................88
Внутренний скелет.....................................................91
Скелет беспозвоночных..............................................91
Скелет позвоночных.................................................91
Общий обзор скелета позвоночных................................93
Позвоночник.............................................. .... 97
Позвоночник низших позвоночных ............................107
Позвоночник наземных позвоночных............................НО
Грудина..................• . . . . ......................118
Надгрудинник .............................................121
Брюшные ребра..............................................121
Череп......................................................•
Диференцировка мезодермы в области головы...............
Раввитие осевого черепа...................................
Развитие и строение висцеральной части черепа...........
Теории происхождения черепа.............................
121
121
124
127
132
537
Хрящевой череп низших позвойочйых ........ 488
Костный череп............................................. 436
Костный череп высших рыб....................................438
Череп наземных позвоночных и его окостенения................444
Череп амфибий...............................................149
Череп рептилий..............................................153
Череп птиц................................................ 461
Череп млекопитающих.........................................163
Скелет конечностей.............................................172
Непарные плавники...........................................172
Парные плавники............................................176
Происхождение конечностей...................................177
Передний пояс рыб...........................................180
Задний пояс рыб.............................................482
Скелет грудного плавника....................................183
Скелет брюшного плавника ...................................184
Конечности наземных позвоночных и их происхождение . . . 184
Плечевой пояс наземных позвоночных..........................189
Тазовой пояс наземных позвоночных...........................193
Скелет свободной конечности.................................196
Свод важнейших фактов и положений морфологии скелета .... 204
Мышечная система......................................................208
Мускулатура беспозвоночных.........................................208
Мускулатура позвоночных............................................208
Висцеральная мускулатура...................................... 209
Висцеральная мускулатура висцерального аппарата и ее производные 210
Соматическая мускулатура.......................................214
Глазные мышцы............................................ 214
Подъязычная мускулатура....................................215
Мускулатура туловища.......................................216
Спинная мускулатура наземных позвоночных...................217
Брюшная мускулатура наземных позвоночных...................218
Мускулатура конечностей....................................219
Диафрагма..................................................222
Подкожная мускулатура......................................223
Электрические органы...........................................224
Нервная система.......................................................225
Нервная система беспозвоночных.....................................225
Нервная система хордовых животных и ее происхождение...............228
Развитие нервной системы позвоночных...........................231
Развитие периферической нервной системы........................237
Спинной мозг...................................................240
Головной мозг..................................................Z41
Обзор строения головного мозга в различных классах .... 249
Мозговые оболочки..............................................255
Периферическая нервная система................................256
Функциональные компоненты нервной системы......................259
Головные нервы............................................262
Функциональная диференцировка отделов центральной нервной системы 274
Органы чувств.........................................................277
Органы чувств общего значения.....................................277
Специализированные органы чувств................................ 278
Органы восприятия механического раздражения ....... 278
Органы равновесия и слуха.....................................282
Слуховой орган позвоночных................................282
Среднее наружное ухо......................................288
Органы зрения.................................................290
Непарные глазные пузырьки позвоночных.....................291
Парный орган зрения позвоночных...........................292
Органы восприятия химического раздражения.....................299
Органы вкуса позвоночных......................................300
Органы обоняния позвоночных...................................301
538
Органы пищеварения.................................................. 306
Органы пищеварения беспозвоночных...............................306
Органы пищеварения позвоночных..................................307
Ротовая полость................................................309
Зубы.......................................................310
Язык.................................................... 317
Ротовые железы............................................' 319
Глотка........................................ •...............320
Пищевод . • , • ...............................................320
Желудок........................................................321
Кишка собственно...............................................323
Пищеварительные железы.........................................326
Печень.....................................................326
Поджелудочная железа.......................................327
Плавательный пузырь............................................328
Органы дыхания........................................................330
Органы дцхания беспозвоночных.....................................330
Органы дыхания позвоночных...................................... 332
Жаберные щели и жабры.............................................332
Наружные жабры.............................................337
Добавочные органы дыхания.................................... 338
Легкие позвоночных.............................................340
Развитие легких................................... 340
Происхождение легких.......................................341
Дыхательные пути...............................................342
Гортань....................................................342
Трахея.....................................................344
Легкие собственно .............................................345
Органы кровообращения............................................... 349
Органы кровообращения беспозвоночных.............................349
Органы кровообращения позвоночных................................351
Развитие сердца и сосудов..................................... . 353
Сердце ....................................................355
Артерии....................................................360
Вепы.......................................................365
Лимфатическая система..........................................368
Полость тела..........................................................370
Полость тела беспозвоночных.......................................370
Полость тела позвоночных..........................................371
Органы выделения.....................................................-Г72
Органы выделения беспозвоночных...................................372
Органы выделения позвоночных...................................... 373
Почки .........................................................373
Органы воспроизведения....................................................385
Органы воспроизведения беспозвоночных.................................”385
Органы воспроизведения позвоночных.................................386
Половые железы..................................................386
Половые протоки.................................................387
Копулятивные органы............................................3115
Органы внутренней секреции............................................ 398
Производные эпителия глотки . .................................398
Железы, связанные с мозгом......................................401
Околопочечные органы ..........................................402
Инкреторные части половых желез................................402
Часть III. ФИЛОГЕНИЯ ПОЗВОНОЧНЫХ И ЕЕ ЗАКОНОМЕРНОСТИ
Филогения позвоночных...................................................404
Палеонтологический метод в филогении............................404
Эволюция многоклеточных животных................................405
Происхождение хордовых..........................................410
Соотношение между низшими хордовыми и позвоночными..............413
Бесчелюстные позвоночные (Agnatha)..............................417
Челюстные позвоночные (Quathostomata). Рыбы.....................423
539
Происхождение наземных позвоночных..........................432
Предки наземных позвоночных.............................435
Происхождение амфибий.......................................438
Происхождение рептилий......................................442
Происхождение птиц..........................................453
Происхождение млекопитающих.................................458
Эволюция млекопитающих......................................464
Морфо-физиологические закономерности эволюции......................482
Приспособление 482
Прогресс и регресс..........................................483
Ароморфоз и адаптация как основные направления эволюционного про-
цесса ......................................................485
Дивергенция, конвергенция и параллелизм в эволюции..........486
Монофилия и полифилия............................. . . . . 490
Направленность эволюции.....................................492
Неограниченность эволюции и вымирание.......................492
Необратимость эволюции......................................495
Указатель литературы............................................. 498
Именной указатель..................................................510
II р о д м е т н ы й у к а з а т е л ь.............................512
Index..............................................................529
Отв. редактор М. М. Местергази
А05663 Подписано к певати 20/VII 1947 г. Тираж 10 000. Объем 34 п. л., Формат 70x1087,,.
Цена 30 р. переплет 2 р. Заказ № 1780.
3-я типография « Краетый пролетарий» треста «Полиграфкнига» ОГИЗа при Совете Министров
СССР. Москва, Краснопролетарская, 10.