















































































































































































































































































Автор: Савельев С.В.
Теги: анатомия анатомия человека сравнительная анатомия антропология зоология анатомия животных позвоночные учебник для вузов издательство геотар-мед
ISBN: 5-9231-0045-2
Год: 2001




СРАВНИТЕЛЬНАЯ
АНАТОМИЯ
НЕРВНОЙ СИСТЕМЫ
ПОЗВОНОЧНЫХ
Учебник
для вузов
С.В. Савельев
СРАВНИТЕЛЬНАЯ
АНАТОМИЯ
НЕРВНОЙ СИСТЕМЫ
ПОЗВОНОЧНЫХ
Допущено Министерством образования Российской
Федерации в качестве учебного пособия для студентов
высших учебных заведений, обучающихся по направлению
подготовки бакалавров и магистров «Биология», а также
биологическим и медицинским специальностям подготовки
дипломированных специалистов
Учебник
для вузов
МОСКВА
ГЭОТАР-МВД
2001
УДК 611.835 (075.8)
ББК 28.706я73
С12
Рецензенты:
Академик РАМН, зав. кафедрой анатомии человека ММА им. И.М. Сече-
нова профессор М.Р. Сапин
Зав. кафедрой высшей нервной деятельности МГУ им. М.В. Ломоносова про-
фессор В. В. Шульговский
Научный редактор:
Профессор кафедры зоологии позвоночных МГУ им. М.В. Ломоносова
Ф.Я. Дзержинский
Савельев С.В.
С12 Сравнительная анатомия нервной системы позвоночных. — М.: ГЭО-
ТАР-МЕД, 2001. - 272 с.: ил. - (XXI век).
ISBN 5-9231-0045-2
Изложены основы сравнительной анатомии центральной и периферической
нервной системы позвоночных. Рассматриваются основные принципы структур-
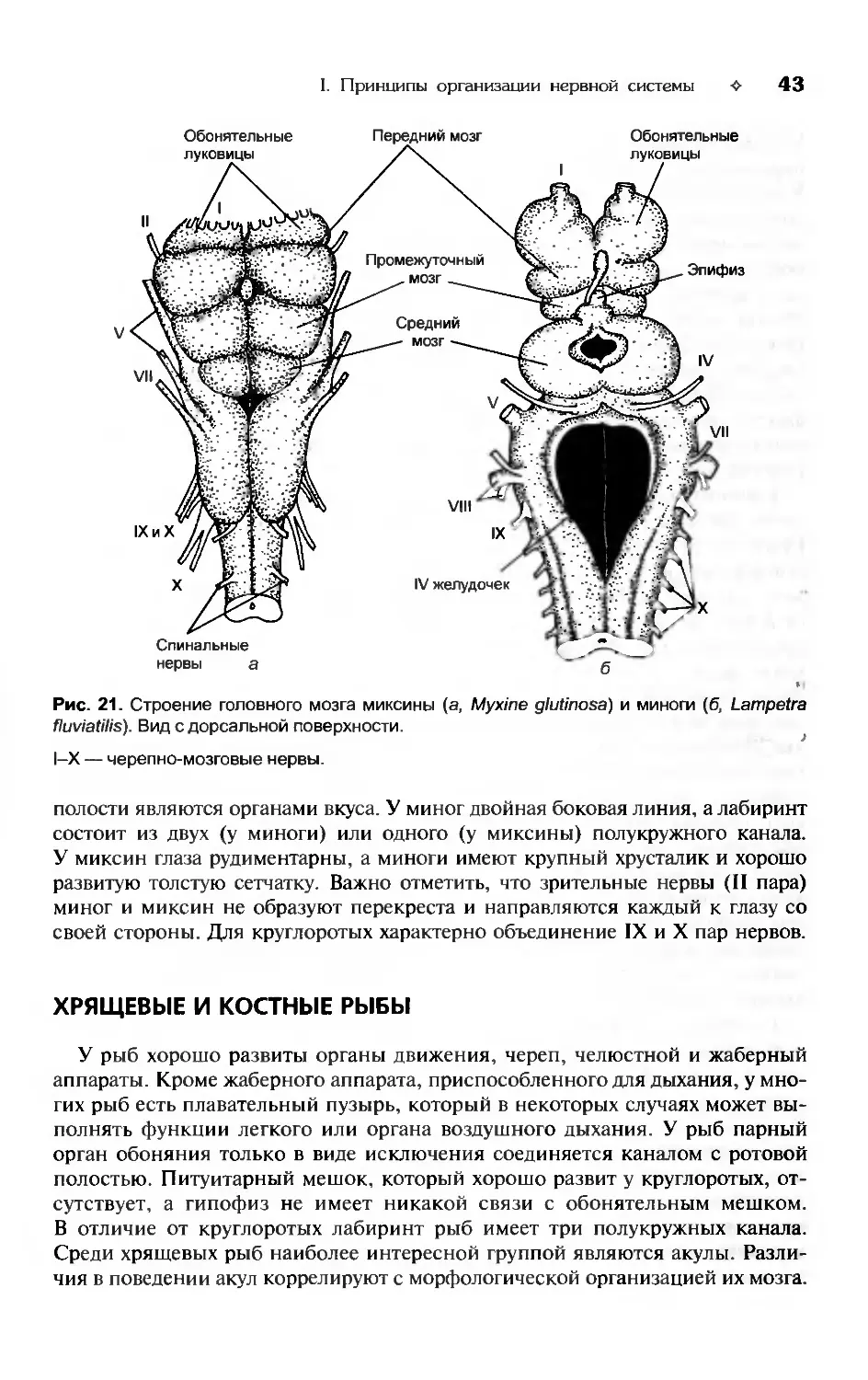
ной организации и направления эволюции нервной системы позвоночных и бес-
позвоночных животных. Описано эмбриональное развитие нервной системы и
даны основы сравнительной эмбриологии мозга позвоночных. В сравнительно-
анатомическом плане рассмотрены спиннои мозг, периферическая нервная си-
стема, черепно-мозговые нервы, продолговатый, задний, средний, промежуточ-
ный и передний мозг. Отдельная глава посвящена особенностям строения орга-
нов чувств позвоночных.
Книга предназначена для студентов, изучающих зоологию, ветеринарию, пси-
хологию, физиологию и анатомию, а также для преподавателей биологических
дисциплин.
УДК 611.835 (075.8)
ББК 28.706я73
Напечатано в Российской Федерации.
Права на данное издание принадлежат издательскому дому «ГЭОТАР-МЕД». Воспроизве-
дение и распространение в каком бы то ни было виде части или целого издания не могут
быть осуществлены без письменного разрешения издательства.
ISBN 5-9231-0045-2
© Издательский дом «ГЭОТАР-МЕД», 2001
©Савельев С.В., 2001
ПРЕДИСЛОВИЕ
Этот учебник подготовлен по материалам лекций, прочитанных на кафедре зоо-
логии позвоночных МГУ им. М.В. Ломоносова в 1997—1999 гг. К сожалению, объем
книги не позволяет в полной мере осветить проблему сравнительной анатомии
позвоночных. Поэтому доминируют телеграфный стиль и предельно краткое из-
ложение самого необходимого материала. Для учебника по столь обширному пред-
мету это единственный разумный выход. Экономические соображения заставили
меня отказаться от микрофотографий и полутоновых рисунков. Их функции вы-
полняют схемы и линейные рисунки. Примеры описания реальной работы отде-
лов головного мозга различных позвоночных предельно сокращены и даются в
скупом изложении. Все эти жертвы принесены на алтарь одной цели — созданию
предельно краткого и по возможности максимально полного учебника по сравни-
тельной анатомии нервной системы позвоночных. Небольшой учебник заполнен
многочисленными поясняющими схемами и рисунками реальных объектов. При-
мерно половина объема книги — рисунки. Такой подход основывается на моем
многолетнем опыте преподавания анатомии нервной системы. Простой рисунок с
объяснением запоминается намного лучше, чем несколько страниц очень хорошо
написанного текста. Практически все схемы, приведенные в учебнике, были де-
сятки раз нарисованы на аудиторной доске во время лекций. Они лишены некото-
рых подробностей, которые, как показывает опыт, впечатляют слушателей боль-
ше, чем сама суть рассматриваемого вопроса. Наиболее сложные вопросы строе-
ния нервной системы изложены подробнее, чем тривиальные проблемы, но не
выходят за рамки учебного курса сравнительной анатомии, читаемого на кафедре
зоологии позвоночных. При всех перечисленных упрощениях иллюстративного
материала и текста я старался сохранить существо рассматриваемых проблем и в
доступной форме изложить фундаментальные принципы строения нервной систе-
мы животных различных видов.
Считая, что логичное теоретическое построение поможет осмыслить анатоми-
ческий материал, я не избегал кратких экскурсов в эволюционную морфологию
нервной системы.
Учебник состоит из четырех взаимозависимых частей, которые могут изучаться
и по отдельности. В первой части изложены принципы структурной организации
нервной системы и основные теории происхождения нервных клеток. Для полно-
ценного понимания эволюции проведено сопоставление стратегии эволюции нерв-
ной системы позвоночных и беспозвоночных. В двух кратких обзорах изложены
общие планы строения нервной системы и их принципиальные структурные раз-
личия. Это своеобразное введение в нейробиологию.
Второй раздел посвящен сравнительной эмбриологии нервной системы. Эта
глава, с моей точки зрения, имеет большое дидактическое значение, поскольку
связное изложение процессов формообразования нервной системы в учебниках не
распространено. Практически отсутствует и информация по сравнительно-анато-
мическим особенностям развития мозга позвоночных. Этот пробел сравнительной
анатомии нуждается как в научной, так и в педагогической разработке.
Сравнительная анатомия нервной системы половозрелых животных начинается
с третьей главы. И зложение построено по классическому принципу, который пред-
полагает рассмотрение нервной системы по отделам. В одной главе объединены
спинной мозг и периферическая нервная система. Такой подход мне видится ло-
гичным, поскольку периферическая нервная система лучше запоминается студен-
тами при ее целостном описании вместе со спинным мозгом. В отдельную главу
вынесена сравнительная анатомия черепно-мозговых нервов. Этот раздел являет-
ся наиболее трудным для изучения, но в традиционных руководствах просто от-
сутствует. Поэтому я позволил себе подготовить оригинальную разработку проб-
лемы, которая позволяет студентам изучить данный вопрос без дополнительных
усилий и времени.
Сравнительная анатомия головного мозга сосредоточена в четырех главах. От-
дельная глава посвящена заднему, продолговатому мозгу и мозжечку. Эти двига-
тельные отделы мозга представляют собой целостный морфофункциональный комп-
лекс, который невозможно разделять по правилам формальной логики. Этот под-
ход не исключает выделение столь сложного анатомического объекта, как мозжечок,
в самостоятельную подглавку. Отдельные главы посвящены среднему, промежу-
точному и переднему мозгу.
Что касается приема изложения, то я предпочел начинать главу с внешнего
анатомического вида отдела мозга у различных позвоночных, затем давать его
сравнительно-анатомическое описание и рисунки тушью гистологических срезов.
Завершается описание информацией о системе нервных связей отдела с другими
частями мозга. Такая схема подачи нейроанатомического материала, как показал
опыт преподавания, пока приносит наилучшие результаты.
Сравнительная анатомия головного мозга лишается своей привлекательности и
внутренней логики без рассмотрения особенностей строения органов чувств. Этот
раздел формирует ясные представления об источниках и формах сенсорной ин-
формации, которую перерабатывает мозг позвоночных. Органы чувств классифи-
цированы по типу чувствительности на механорецепторы, хеморецепторы и ре-
цепторы электромагнитных полей и излучений. С этих позиций кратко рассмот-
рена сравнительная анатомия органов обоняния, вкуса, фоторецепторов,
элекгрорецепторов, рецепторов инфракрасного излучения, органов боковой ли-
нии и слуха. Изложены принципы организации соматической чувствительности.
В разделе даются некоторые основы знаний о физиологии сенсорных органов,
которые необходимы для понимания направления морфологической эволюции
периферических анализаторов.
Побудительной причиной создания данного учебника явилось полное отсутствие
в нашей оригинальной литературе курса сравнительной анатомии нервной системы.
До сих пор единственной книгой такого плана является упрощенное изложение
руководства А. Каперса 30-х годов, сделанное Е.К. Сеппом в 1949 г. Однако послед-
нее переиздание этой книги было сделано почти 40 лет назад. Небольшие главы по
анатомии нервной системы в общих руководствах носят элементарный характер и
часто содержат устаревшие сведения. Их обычно недостаточно даже для вводных
лекций курса зоопсихологии.
Поэтому у данной книги двойная задача. С одной стороны, удовлетворить по-
требность в сведениях по сравнительной анатомии нервной системы у студентов,
изучающих зоологию, ветеринарию, поведение животных и нейрофизиологию.
С другой стороны, она может стать первичным руководством для слушателей спе-
циальных курсов по нейробиологии.
Считаю долгом выразить благодарность профессору кафедры зоологии позво-
ночных Ф.Я. Дзержинскому, который предоставил мне возможность начать чте-
ние курса сравнительной анатомии нервной системы в МГУ им. М.В. Ломоносова
и провел огромную работу по научному редактированию учебника.
С. В. Савельев
I. ПРИНЦИПЫ ОРГАНИЗАЦИИ
НЕРВНОЙ СИСТЕМЫ
Нервная система — это объединение специализиро-
ванных клеток, которые воспринимают, обрабатывают,
хранят и используют информацию о внешней среде и
внутреннем состоянии организма. Именно этим функ-
циям подчинено строение нервных клеток. Нервные
клетки имеют ряд особенностей строения, которые от-
личают их от других клеток организма. Нейроны не-
одинаковы. Они различаются по размеру, форме ветвле-
ния дендритов и аксонов, выделению различных хими-
ческих веществ и физиологической активности. Однако
они являются и характерными структурными элемента-
ми нервных систем всех видов многоклеточных живот-
ных. Нейроны объединяются в сети и специализиро-
ванные структуры: ганглии или мозг.
У нервных клеток — нейронов — обычно можно вы-
делить три характерные области: клеточное тело, денд-
риты и аксон (рис. 1). Тело содержит ядро и биохими-
ческий аппарат синтеза молекул, необходимые для жиз-
недеятельности клетки. Обычно тело нейрона имеет
округлую, веретеновидную или пирамидальную форму.
Дендриты представляют собой тонкие отростки, кото-
рые многократно ветвятся в непосредственной близо-
сти от тела клетки. Вокруг него образуется ветвистое
дерево. Дендриты формируют ту основную физическую
поверхность, на которую поступают идущие к данному
нейрону сигналы. Аксоны распространяются далеко от
тела клетки. Их длина варьирует от 1 мм до 1 м, что
позволяет аксонам выполнять функции линий связи
между телом клетки и далеко расположенным opi аном-
мишенью или отделом мозга. По аксону проходят сиг-
налы, генерируемые в теле данной клетки. Аксон отли-
чается от дендритов как по строению, так и по свой-
ствам своей наружной мембраны. Большинство аксонов
длиннее и тоньше дендритов и имеет отличный от них
характер ветвления. Отростки дендритов в основном
6
❖
Сравнительная анатомия нервной системы позвоночных
Рис. 1. Строение нейронов, клеток глии и оболочек отростков нервных клеток. В централь-
ной нервной системе функции шванновских клеток выполняет олигодендроглия.
группируются вокруг клеточного тела, тогда как отростки аксонов располага-
ются на конце волокна, в том месте, где аксон взаимодействует с другими
нейронами или органами-мишенями.
Для взаимодействия между нейронами и клетками других тканей организ-
ма у нервных клеток существуют специализированные участки мембраны, рас-
положенные как на теле клетки, так и на ее отростках. Эти участки мембраны
имеют характерное строение и называются синапсами. Через них происходят
основной обмен информацией внутри нервной системы и взаимодействие
нейронов с другими органами и системами организма. Нейрон может иметь от
1000 до 10 000 синапсов и получать информацию примерно от 1000 других
нейронов. В типичном случае синапсы образуются между аксонами одной
клетки и дендритами другой. Однако существуют и другие типы синаптиче-
ских контактов: между аксоном и аксоном, аксоном и телом клетки, дендри-
том и дендритом, дендритом и телом клетки.
Происхождение нервных клеток в процессе эволюции животных довольно
спорно. Существует две основные точки зрения на эту проблему. Наибольшее
распространение получила точка зрения братьев Гертвигов (О. Hertwig,
R. Hertwig, 1878—1882). Их гипотеза состоит в том, что нервные и мышечные
клетки происходят из эктодермального эпителия. Они возникают независимо
друг от друга и вступают во взаимодействия уже вторично (рис. 2, а—в).
В соответствии с гипотезой Гертвигов источником всех нервных клеток явля-
ется первичная чувствительная клетка, возникшая из эктодермы и получив-
шая возможность воспринимать раздражения и генерировать и проводить воз-
буждение. Специализированные отростки этих клеток вступают в связь с не-
зависимо возникшими мышечными клетками. По мнению авторов этой
гипотезы, мышечные волокна возникли из клеток, сходных с эпителиально-
мышечными клетками кишечнополостных. Такой простейший нервно-мышеч-
ный комплекс развивается по пути дифференциации нервного аппарата, что
I. Принципы организации нервной системы
❖
7
п
Рис. 2. Две гипотезы происхождения нервных и мышечных клеток.
а-в — гипотеза братьев Гертвигов; г-е — гипотеза Клейненберга-Заварзина. По гипотезе
Гертвигов нервные и мышечные клетки происходят независимо из эктодермы, а затем объе-
диняются в нервно-мышечный комплекс. По представлениям Клейненберга-Заварзина из
эктодермы возникает клетка, обладающая сенсорными и моторными функциями одновре-
менно. Затем она дифференцируется на две системы: нервную и мышечную. Штриховка —
мышечные клетки или волокна, точки — чувствительные или нервные клетки.
приводит к возникновению вставочных нейронов или ганглиев (см. рис. 2, в).
Нервная система подразделяется на чувствительные клетки, лежащие в экто-
дерме, и вставочные нейроны, обладающие интегративной функцией. Послед-
ние являются предвестниками централизации обработки сенсорной информа-
ции и возникновения мозга.
Другая точка зрения на происхождение нервных клеток сформировалась в
работах Клейненберга и Заварзина (Н. Kleinenberg, 1872; А.А. Заварзин, 1941).
Ее суть сводится к тому, что нервная и мышечная системы рассматриваются
как единое и нераздельное целое. Считается, что обе системы возникли одно-
временно в виде такого образования, которое не имело дифференцированных
мышечных и нервных компонентов. С точки зрения этой гипотезы первичны-
8
Сравнительная анатомия нервной системы позвоночных
❖
ми были клетки, выделившиеся из эктодермы и обладавшие как нервными,
так и мышечными функциями (рис. 2, г, д). Часть клетки обладала способнос-
тью к чувствительности, а другая — к сокращению. В гипотезе подчеркивается
первичность морфофункциональной связи между нервными и мышечными
клетками. В дальнейшем, по мнению авторов, произошла дифференциация
чувствительно-сократимых клеток на чисто сенсорные и моторные, а общ-
ность их происхождения стала основой нервно-мышечных взаимодействий
(рис. 2, е).
Нейроны — это морфологические единицы, кирпичики, из которых пост-
роена нервная система как позвоночных, так и беспозвоночных животных.
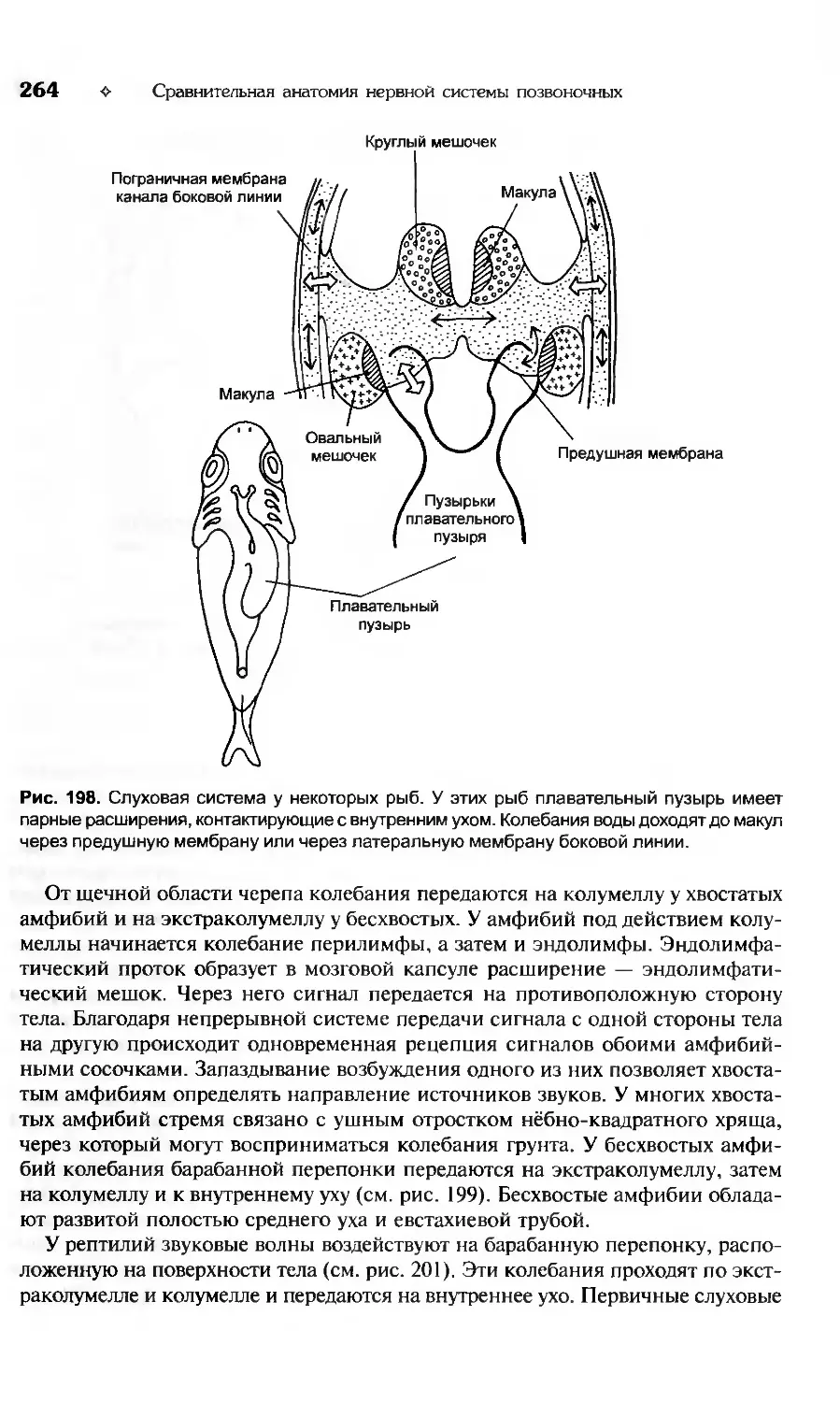
Сходство строения и функций нейронов объединяет все группы организмов,
обладающих нервной системой. Нейрон является минимальной единицей в
анализе нейробиологических процессов. Ионы и молекулы, находящиеся в
клетке, равно как и органеллы, обеспечивающие функционирование нейро-
нов, сходны с аналогичными структурами в других органах и тканях организ-
ма животных и человека. Их не стоит принимать за низший уровень организа-
ции нервной системы, как нельзя учитывать открывание и закрывание дверей
купе пассажирского поезда при составлении расписания поездов на конкрет-
ном участке железной дороги. Как поезд является единицей в расписании
движения поездов, так и нейрон представляет собой минимальную единицу в
организации нервной системы. Нервные клетки объединены в нервные систе-
мы различным образом. В простейшем случае эти элементы распределены
вполне равномерно по всему телу животного или по большей его части. Это
диффузное распределение нервных клеток. Являясь наиболее простым, оно в
то же время оказывается филогенетически наиболее старым. В этом случае
клетки расположены однородно и снабжены многочисленными одинаковыми
отростками, которые объединены в общую сеть (рис. 3, а). Протяженных и
организованных путей в такой сети нет, отростки клеток связывают только
соседние нейроны. В сеть входят волокна, идущие от воспринимающих кле-
ток эпителия или рецепторов, а из сети отходят двигательные волокна, окан-
чивающиеся на поверхности мышечных клеток.
Другой вариант представляет собой компактную организацию нервной си-
стемы. В этом случае клеточные тела передаточных и двигательных нейронов
собраны в небольших зонах тела, которые называются нервными ганглиями
или узлами. Для беспозвоночных характерны скопления нервных клеток, не
содержащие внутреннюю полость (см. рис. 3, а, б). Внутренняя зона ганглия
состоит из нервных клеток, окруженных нейропилем, представляющим собой
сплетение отростков дендритов этих клеток. В ганглиях расположены чувст-
вительные нейроны или оканчиваются их аксоны, что позволяет ганглиозным
клеткам получать информацию с периферии тела беспозвночного. Аксоны дви-
гательных нейронов, выходящие из ганглиев, образуют более или менее длин-
ные проводящие пути, связывающие с нервными центрами органы дыхания,
пищеварения, железы и мышечные клетки.
Для хордовых характерно появление более сложной структуры организации
нервной системы. Наряду с сетями и ганглиями у позвоночных формируется
центральная трубчатая нервная система. Возникает головной и спинной мозг с
полостью внутри, которая называется спинномозговым каналом или мозговы-
I. Принципы организации нервной системы
❖
9
Рис. 3. Схема основных морфологических типов организации нервной системы.
а — диффузный тип строения, характерный для кишечнополостных; б— ганглиозный тип
организации нервной системы, который характерен для червеобразных, моллюсков и чле-
нистоногих; в — центральная трубчатая нервная система хордовых, в которой присутству-
ют все типы интеграции нейронов.
ми желудочками (рис. 3, в). Размеры центральной трубчатой нервной системы
позвоночных позволяют разместить между чувствительным и двигательным ней-
роном большое количество вставочных нейронов, которые обрабатывают полу-
чаемую информацию. От качества переработки этой информации зависит слож-
ность поведения животного или интеллектуальная деятельность человека.
Эти общие принципы организации нервной системы характерны как для
беспозвоночных, так и для позвоночных животных. Однако существует ряд
принципиальных различий, которые следует подчеркнуть.
Во-первых, строение нейронов нервной системы беспозвоночных и позво-
ночных существенно различается как по размерам, так и по форме (рис. 4).
Во-вторых, для большинства беспозвоночных характерно отсутствие поло-
сти внутри мозга — мозговых желудочков. Исключением являются головоно-
гие моллюски, головные ганглии которых формируют внутренние полости.
В-третьих, у беспозвоночных стратификация нейронов распространена зна-
чительно менее, чем у позвоночных.
Эти морфологические различия приводят к тому, что эволюция нервных
систем позвоночных и беспозвоночных животных происходила в рамках очень
несхожих ограничений. Для беспозвоночных с их маленькой нервной систе-
мой было бы невозможным доминирование регуляционного эмбрионального
развития нервной системы над генетически детерминированным. Это связано
с тем, что при регуляционном развитии судьба эмбриональной клетки вероят-
ностна и }ависит от межклеточных взаимодействий. В таком случае требуются
некоторый переизбыток нейробластов, их конкурентное поведение при диф-
ференцировке и программированная гибель клеток. Такой переизбыток эмб-
рионального материала практически невозможен у беспозвоночных. В резуль-
тате возникла ситуация, когда нервная система развивается преимущественно
по детерминационному типу. Это означает, что число нервных клеток, их свя-
зи и даже ветвление дендритов предопределены генетически. Если попытат ься
выразить соотношение детерминированных и регуляционных событий в раз-
10
❖
Сравнительная анатомия нервной системы позвоночных
Рис. 4. Принципы объединения нейронов позвоночных и беспозвоночных.
а — типы нейронов; б — диффузная сеть, характерная для обеих групп; в — «параллель-
ные ганглии»; а — корково-нейропильный тип организации; О — островковая или ядерная
форма концентрации нервных клеток в нейропиле; е — периферический ганглий позво-
ночного; ж— кора, подкорковые ядра и стенка мозговых желудочков.
витии нервной системы беспозвоночных количественно, то оно составит при-
мерно 7:3. Понятно, что при таком соотношении практически не остается
места для морфологической индивидуализации нервной системы. Более того,
I. Принципы организации нервной системы
11
❖
достаточно жесткая детерминированность организации нервной системы со-
здает возможность эффективной передачи по наследству разнообразных форм
поведения. Поэтому мы встречаем среди беспозвоночных преимущественно
инстинктивное поведение, его незначительную индивидуализацию и отсут-
ствие «высоких» психологических свойств. Не менее важным морфологиче-
ским и функциональным отличием беспозвоночных является система нейро-
гормональной регуляции поведения. Нервная и эндокринная системы образу-
ют интегрированные нейроэндокринные комплексы. У наиболее эволюционно
«продвинутых» видов беспозвоночных в рефлекторные реакции и обработку
сенсорной информации входит нейрогормональный этап. Удельный вес ней-
рогормонального этапа в работе головных ганглиев может различаться, но его
значение у беспозвоночных несопоставимо выше, чем у позвоночных живот-
ных. Для большинства высших беспозвоночных характерно формирование
специального нейрогормонального органа, расположенного позади головных
ганглиев. Эти образования носят название нейрогемального органа, а при
интеграции с головными ганглиями или нервами — нейрогемальной зоны.
Наличие этого центра приводит к тому, что поведение животного при любой
активизации головных ганглиев начинает контролироваться выделяемыми гор-
монами. Следовательно, оно становится более генерализованным, програм-
мируемым и предсказуемым. Соотношение между влиянием нейрогормональ-
ных и нервных комплексов на поведение специально не изучали. Однако при-
близительная оценка показывает, что у высших беспозвоночных поведение
примерно на 85% контролируется нейрогормонами, тогда как у позвоночных
доля этого влияния не достигает 50%.
Таким образом, сопоставляя историческое развитие нервной системы, не-
обходимо отметить, что наибольшего результата эволюция достигла у бес-
позвоночных животных. Их нервная система очень небольшого размера, об-
ладает развитыми сенсорными входами и имеет практически полный набор
программ поведения, находящийся под контролем генетических или нейро-
гормональных процессов. Эта компактность, экономичность и детермини-
рованность делают нервную систему беспозвоночных совершенным инстру-
ментом для решения стандартных задач и коллективных действий. Учитывая
однотипность и предопределенность реакций нервной системы, легко по-
нять согласованность миграций, половую и суточную активности высших
беспозвоночных. Совершенство нервной системы беспозвоночных позволи-
ло им освоить огромные пространства и быть самой распространенной груп-
пой животных на планете.
Однако существует и обратная сторона медали. За компактность, эконо-
мичность и эффективность нервной системы беспозвоночным пришлось за-
платить индивидуализацией поведения. Беспозвоночные практически не об-
ладают внутривидовой изменчивостью строения нервной системы и как след-
ствие индивидуальными особенностями поведения. Они совершенны в
предусмотренных природой случаях и беспомощны в нестандартной ситуа-
ции. Из высших беспозвоночных получились бы идеальные солдаты, но не
появился бы ни один генерал. Там, где начинаются индивидуальность и твор-
чество, царят морфологическая изменчивость и случайность.
12 ❖ Сравнительная анатомия нервной системы позвоночных
Нервная система позвоночных построена на принципах вероятностного
развития, дублирования, избыточности и индивидуальной изменчивости. Это
не означает, что в мозге позвоночных нет места генетической детерминации
развития или нейрогормональной регуляции. Все эти слагаемые поведения
присутствуют и в организации нервной системы позвоночных, однако они
играют несоизмеримо меньшую роль, чем у беспозвоночных. При развитии
нервной системы позвоночных действуют законы формообразования, кото-
рые детерминируют первичную региональную экспрессию генов и морфоге-
нез (С.В. Савельев, 1993). Одновременно присутствует и огромная переизбы-
точность эмбриональных нейробластов. Строгой детерминации развития каж-
дого отдельного нейрона, как у беспозвоночных, нет. Судьба каждой клетки
носит вероятностный характер и зависит от тех коммуникативных взаимодей-
ствий, в которые она вступит во время своего индивидуального развития и
дифференцировки. Достаточно высокая случайность судьбы каждого нейро-
бласта отражается в огромных масштабах гибели клеток нервной системы.
При развитии головного мозга млекопитающих нормальная гибель клеток обыч-
но составляет 0,2—0,4%. К концу метаморфоза бесхвостых амфибий гибель
клеток достигает 85% от общего числа нейробластов спинного мозга. Такие
потери клеток для беспозвоночных были бы невосполнимы, тогда как у по-
звоночных эти события составляют нормальный морфогенез. Вероятностное
развитие нервных клеток позвоночных подтверждено в многочисленных экс-
периментах (С.В. Савельев, 1993). Достаточно упомянуть способность нервной
системы позвоночных к компенсации искусственно вызванной гибели клеток.
Если у эмбриона амфибии на стадии нервной пластинки случайным образом
(не в одном месте) разрушить 40% клеток, то зародыш разовьется в нормаль-
ный организм немного меньшего размера, чем обычная особь. Следовательно,
нервная система позвоночных уже в эмбриональный период закладывается с
«переизбытком» клеток, поскольку их судьба не столь строго детерминирова-
на, как у беспозвоночных. Таким образом, известные различия в развитии
нервной системы позвоночных и беспозвоночных животных позволяют назвать
онтогенетическое развитие позвоночных регуляционным, а беспозвоночных —
детерминационным. Это не означает полного отсутствия детерминационных
явлений у позвоночных и регуляционных процессов у беспозвоночных. Однако
явное превалирование одного процесса над другим совершенно очевидно.
Различные стратегии развития нервной системы позвоночных и беспозво-
ночных отражаются и на основных принципах ее морфологической организа-
ции. Если нервная система компактизована из-за размеров и массы беспозво-
ночных, то у позвоночных нет физических препятствий для ее увеличения.
Отсутствие ограничения на размер нервной системы позволяет достигать го-
ловному и спинному мозгу позвоночных крупных размеров и массы до 10 кг.
Однако различия в размерах мозга не исключают сходство гистологической
организации между позвоночными и беспозвоночными.
Нервные клетки в обеих группах не имеют принципиальных различий в
цитологической организации (см. рис. 4), хотя некоторые особенности в строе-
нии отростков и тел клеток могут быть предметом исследования сравнительной
цитологии. В самом простом случае нервные клетки образуют диффузную сеть,
характерную для обеих групп (см. рис. 4, б). В ганглиях беспозвоночных и
I. Принципы организации нервной системы
13
❖
нервной трубке позвоночных клетки и их отростки расположены закономерным
образом. К первому типу организации нервных клеток беспозвоночных следует
отнести их способность формировать «параллельные ганглии» (см. рис. 4, в). Эти
структуры характеризуются однонаправленным вытягиванием тел нейронов и
формированием контактов между параллельно лежащими отростками.
Второй тип организации нейронов наиболее характерен для беспозвоноч-
ных и носит название дифференцированного нейропиля (см. рис. 4, г). Он
состоит из тел клеток, расположенных в корковом слое, и нейропиля, состоя-
щего из переплетенных отростков нейронов. Это наиболее распространенный
принцип строения туловищных и головных ганглиев.
Третьим типом концентрации нервных элементов у беспозвоночных явля-
ется нейропиль с островками тел клеток (см. рис. 4, д). Локальные концентра-
ции тел клеток беспозвоночных сходны с ядрами головного и спинного мозга
позвоночных. Они характеризуются компактным расположением тел нейро-
нов внутри нейропиля, что позволяет клеткам эффективнее обмениваться
информацией между собой.
Для позвоночных характерно наличие периферических ганглиев, окружен-
ных соединительнотканной оболочкой (см. рис. 4, е). Тела клеток в таких ганг-
лиях расположены по всему объему структуры и не имеют поверхностной лока-
лизации, как у беспозвоночных. Нейропиль, состоящий из переплетенных от-
ростков клеток, не выражен. Корковые структуры позвоночных характеризуются
многослойным расположением тел клеток (см. рис. 4, ж). Они обычно разделе-
ны пучками волокон, которые позволяют выделять как горизонтальные слои
коры, так и вертикальные колонки. Для позвоночных специфично участие раз-
нообразных типов клеток в формировании коры, тогда как для беспозвоночных
это довольно редкое явление. Принципиальным отличием этих групп является
наличие мозговых желудочков у позвоночных, которые окружены специализи-
рованными эпендимными клетками. Эти клетки сохраняют свои эмбриональ-
ные свойства и могут участвовать в регенерации мозга. У постоянно растущих
животных, таких, как неотенические виды хвостатых амфибий, акулы и некото-
рые рептилии, эпендимные клетки спинного мозга могут пролиферировать и
формировать отростки у половозрелых животных. Для некоторых певчих птиц
характерны сезонная пролиферация эпендимных клеток и их миграция в «пев-,
ческие» центры мозга самцов. Таким образом происходит формирование мор-
фологического субстрата для усложнения песенного репертуара. Сохранение
эпендимными клетками способности к пролиферации является подтверждени-
ем регуляционного принципа организации мозга позвоночных. Подобная «ре-
зервная избыточность» для беспозвоночных неизвестна.
Эти данные свидетельствуют не только о глубоких различиях, но и о сход-
стве развития и организации нервной системы позвоночных и беспозвоноч-
ных. Реконструируя пути усложнения морфологической организации мозга,
можно предположить следующую последовательность событий. На первом этапе
исторического развития нервной системы произошло появление чувствитель-
ных элементов из клеток эктодермального зачатка (рис. 5, а, б). Специализи-
рованные клетки эктодермы одновременно обладали сенсорными и эффек-
торными функциями. Они рецептировали сигнал, проводили его к эффектор-
ным органам и запускали их реакцию. Эти клетки были связаны между собой
14
Сравнительная анатомия нервной системы позвоночных
Рис. 5. Реконструкция основных этапов усложнения морфологической организации нервной
системы.
а — эктодерма без специализированных чувствительных элементов: б — эктодерма, со-
держащая нервные клетки, связанные между собой отростками; в — диффузная нервная
сеть, не содержащая выраженных центров; г — «параллельные ганглии» беспозвоноч-
ных; д — ганглии без выраженного нейропиля; е — ганглии с нейропилем; ж — головной
ганглий беспозвоночного с грибовидными телами, которые являются стратифицированны-
ми структурами и островковыми (ядерными) скоплениями тел нейронов; з — нервная трубка,
образовавшаяся за счет выхода отростков нейронов на наружную поверхность нервого
тяжа; и — нервная трубка с ядрами и прижелудочковым расположением нейронов; к —
стратифицированные структуры позвоночных коркового типа.
I. Принципы организации нервной системы
15
❖
и формировали непрерывную сеть, которая не имела выраженных центров
(рис. 5, в). Такой тип организации нервной системы мы встречаем у кишечно-
полостных. При появлении более сложных поведенческих задач элементы нерв-
ной системы стали объединяться в небольшие скопления. По-видимому, это
происходило двумя путями. С одной стороны, формировались «параллельные
ганглии» (рис. 5, г), для которых характерны синаптические контакты между
телами клеток. Этот примитивный тип концентрации нервных элементов ха-
рактерен для свободноплавающих кишечнополостных.
С другой стороны, появились скопления нейронов с наружным расположе-
нием тел клеток и нейропилем из переплетенных отростков внутри ганглия
(рис. 5, е). Этот тип организации ганглиев оказался достаточно эффективным
и сохранился до настоящего времени у большинства беспозвоночных. Такой
ганглий обладает рядом преимуществ, которые имеют особое значение для
животных с незамкнутой кровеносной системой. Тела его нейронов располо-
жены преимущественно на наружной поверхности, что позволяет поддержи-
вать довольно высокий уровень метаболизма: снабжать питательными веще-
ствами, кислородом и отводить токсичные продукты экскреции клеток. Ней-
ропиль, находящийся внутри ганглия, дает возможность обмениваться
сигналами и формировать генерализованный ответ на разнообразные сенсор-
ные сигналы. По-видимому, из этой формы концентрации нервных элемен-
тов возникли головные и туловищные ганглии высших беспозвоночных
(рис. 5, ж), ганглии и нервная трубка позвоночных (рис. 5, д, з). В головных
ганглиях беспозвоночных сложились два основных типа гистологических струк-
тур: островковые скопления тел клеток и стратифицированные грибовидные
тела. Островковые скопления тел клеток беспозвоночных практически иден-
тичны подкорковым и стволовым ядрам позвоночных. Организация грибо-
видных тел напоминает слоистое расположение клеток в коре млекопитаю-
щих. Однако грибовидные тела беспозвоночных не имеют упорядоченных вер-
тикальных связей между нейронами и колончатой организации. Тем не менее
стратификация нейронов в грибовидных телах предполагает сходство меха-
низмов обработки информации в ассоциативных центрах позвоночных и бес-
позвоночных животных.
По-видимому, нервная система по шоночных возникла из ганглиев беспо-
звоночных с нейропилем из переплетенных отростков (см. рис. 5, е). Трубча-
тая нервная система сформировалась в результате выхода отростков нейронов
из внутренней полости ганглия (см. рис. 5, е, з). Это событие привело к фор-
мированию нейропиля из отростков нервных клеток с наружной стороны нерв-
ной трубки. На следующем этапе формирование новых нервных центров про-
исходило преимущественно вокруг желудочков. Это привело к возникно-
вению центрального серого вещества. Часть клеток выселялась из
прижелудочковой зоны и формировала структуры ядерного или стратифици-
рованного типа во внешнем нейропиле (рис. 5, и, к). У высших позвоночных
центральное серое вещество практически отсутствует, а основные нервные
центры мозга представлены сложными ядрами и корковыми структурами раз-
личных типов. В дальнейшем цефализация позвоночных развивалась по прин-
ципу количественного наращивания анатомического представительства ана-
лизаторных систем и ассоциативных центров. В отличие от беспозвоночных
16
Сравнительная анатомия нервной системы позвоночных
❖
нервная система трубчатого типа при наличии замкнутой кровеносной систе-
мы может бесконечно увеличиваться в размерах. Это позволило позвоночным
достигнуть крайне высокого уровня развития умственных способностей.
Подводя итог краткому обзору основных принципов анатомической и ги-
стологической интеграции нервных клеток у беспозвоночных и позвоночных
животных, необходимо сделать несколько общих выводов. Во-первых, нейро-
ны в обеих группах имеют сходное строение, но различаются по размерам. Во-
вторых, интегративные взаимодействия между нейронами осуществляются в
сходных гистологических образованиях: ганглиях, ядрах и стратифицирован-
ных структурах. Эти образования встречаются как у беспозвоночных, так и у
позвоночных животных. Основными морфологическими отличиями позвоноч-
ных являются система мозговых желудочков и организация кровеносной сис-
темы. Эффективная система кровоснабжения позволяет поддерживать высо-
кий уровень метаболизма нервной системы. Обширная система мозговых же-
лудочков и сосудистое сплетение обеспечивают осмотический и водный баланс
головного и спинного мозга. Эти морфологические особенности позвоночных
обеспечивают лабильность, высокие адаптационные возможности и большие
размеры нервной системы. Нервная система беспозвоночных более консерва-
тивна, имеет небольшие размеры и не обладает столь широкими адаптацион-
ными возможностями, как у позвоночных. Зато она имеет огромный набор
генетически детерминированных программ поведения, четко запрограммиро-
ванное ра шитие и эффективную систему гормональной регуляции физиоло-
гической активности. При небольших размерах беспозвоночных это позволяет
добиваться впечатляющих успехов в конкуренции с другими видами.
Таким образом, сравнение организации позвоночных и беспозвоночных
отражает различные принципы развития нервной системы, что дает представ-
ление о возможных вариантах ее строения и путях эволюции.
ОБЩИЙ ПЛАН СТРОЕНИЯ НЕРВНОЙ СИСТЕМЫ
БЕСПОЗВОНОЧНЫХ
Основной особенностью строения кишечнополостных является радиальная
симметрия. Форма тела кишечнополостных в основном напоминает чашу,
иногда уплощенную или удлиненную с простой полостью внутри, сообщаю-
щейся с внешней средой единственным отверстием, одновременно выполня-
ющим функции рта и ануса. Органы у кишечнополостных отсутствуют, что
говорит о тканевом уровне организации этих животных. У них существует
только два клеточных слоя со студенистой мезоглеей между ними (рис. 6).
Почти у всех кишечнополостных нервная система представлена диффуз-
ной сетью нейронов с отростками, лишенными оболочки. В наиболее чистом
виде мы встречаем ее у прикрепленных форм (см. рис. 6). У всех кишечнопо-
лостных нервная сеть залегает в слое неклеточного вещества — мезоглее, рас-
положенном между двумя слоями клеток, составляющих стенку тела. Из на-
ружного слоя — эктодермы — нервная сеть получает информацию от хеморе-
цепторных сенсорных ямок, светочувствительных глазков, сенсорных
тактильных клеток и гравитационных рецепторов — статоцитов. Сигналы от
I. Принципы организации нервной системы
❖
17
Рис. 6. Строение нервной системы кишечнополостных на примере пресноводной гидры.
Для гидры характерна диффузная нервная система с плотным расположением нейронов
на подошве и у ротового отверстия. Увеличен фрагмент стенки тела гидры с закрашенны-
ми нервными клетками, которые расположены в мезоглее и на границе гастродермы и
эпидермиса.
этих рецепторов распространяются по нервной сети и идут к сократительным
эпителиальным, железистым и пищеварительным клеткам гастродермы — внут-
реннего слоя. У этого общего для кишечноплостных типа организации суще-
ствуют многочисленные отклонения.
У актиний нервная сеть не везде одинакова. Нервные клетки встречаются
чаще у оснований щупалец и по краю ротовой площадки. Еще более неодно-
родна нервная система у свободноплавающих кишечнополостных. Нервная
сеть медуз ограничена нижней поверхностью зонтика и желудочного стебель-
ка. У гидромедуз по краю зонтика формируется двойное нервное кольцо, со-
единенное нервами с органами равновесия, хеморецепторами и сенсорными
тактильными клетками. Централизация нервной системы особенно заметна у
сцифоидных медуз. Их нервное кольцо состоит из многочисленных нервных
клеток, сконцентрированных в 8 краевых тельцах. Эти данные свидетельству-
ют о том, что уже на уровне кишечнополостных происходит концентрация
нервных клеток в ганглиоподобные структуры.
Различия в организации нервной системы реализуются в поведении кишеч-
нополостных. При сильном раздражении гидры или актинии происходит бы-
строе сокращение всего тела животного. Это общая генерализованная реакция
диффузной нервной системы. Интересно отметить, что если у актинии выре-
зать участок тела или отделить щупальце, то на протяжении суток оно будет
продолжать реагировать на раздражения, как и в теле животного. Следова-
тельно, диффузная нервная система представляет собой однородное образова-
ние, не имеющее более или менее важных зон или центров. Даже такая просто
устроенная нервная система может быть причиной весьма целесообразного
18 ❖ Сравнительная анатомия нервной системы позвоночных
поведения животного. Говоря о гидромедузах, необходимо отметить, что их
нервная система уже отличается от исходного диффузного типа. Кольцевые
нервные тяжи, проходящие по краю зонтика, одновременно включают не-
сколько нейронов и их отростков. Рассеянные тела нейронов имеют отростки,
вытянутые параллельно друг другу, что придает диффузной нервной системе
более сложную форму, носящую название медуллярной.
Для сцифоидных медуз характерны скопления нервных клеток, близкие по
строению с ганглиями. Если удалить все 8 краевых скоплений нервных кле-
ток, то сокращения исчезнут, а раздражение будет вызывать только одноразо-
вое сжатие колокола медузы. Нервная система кишечнополостных, организо-
ванная довольно просто с анатомической точки зрения, является основой до-
вольно сложного и недостаточно исследованного поведения. У актиний
вырабатывается устойчивое привыкание к однотипному механическому раз-
дражению. Попытки обмануть актиний, подбрасывая пропитанную соком рыбы
бумажку, приводят к выработке довольно устойчивого рефлекса. Через неко-
торое время актинии начинают отбрасывать ложную приманку, что говорит о
существовании кратковременной памяти у кишечнополостных. Для кишечно-
полостных характерна и ассоциативная память. Если собрать актиний в при-
роде и поместить в аквариум, то они расположатся в соответствии со своим
исходным положением. Следовательно, каждая особь запоминает, в каком на-
правлении было ориентировано ее ротовое отверстие.
ЧЕРВЕОБРАЗНЫЕ
К червеобразным относят животных, которые очень сильно различаются по
размерам, форме тела, внутренней организации и образу жизни. Червя можно
определить как билатерально-симметричное животное с мягким телом, длина
которого в 2—3 раза и более превышает ширину. В отличие от двухслойной
организации кишечнополостных у червей возникает третий зародышевый слой
клеток — мезодерма. Она расположена между эктодермой (наружным слоем
клеток) и энтодермой (внутренним слоем клеток). Этот признак характерен
для всех других вышестоящих многоклеточных животных. Появление третьего
слоя клеток определяет появление дифференцировки тела червей на внутрен-
ние органы. У большинства червей легко можно обнаружить выделительные
органы и органы размножения.
Для червей характерна билатеральная симметрия. Это означает, что у чер-
веобразных есть хорошо выраженный передний (головной) и хвостовой кон-
цы тела и сагиттальная плоскость, которая разделяет их на две более или ме-
нее симметричные половины тела. Наличие переднего конца тела, определя-
ющего направление движения червя, привело к концентрации рецепторных
нервных элементов в относительно небольшом участке тела — головной зоне.
У большинства червей на головном конце тела имеется группа вкусовых кле-
ток и светочувствительных элементов. Отростки этих клеток оканчиваются в
головных скоплениях нейронов, носящих название ганглиев (рис. 7).
Конгломерат клеток, расположенных в головной части червя, может быть
назван мозгом, поскольку выполняет характерные для него функции: получает,
I. Принципы организации нервной системы
❖
19
Рис. 7. Строение нервной системы
плоского червя. У плоского червя
лестничная нервная система. В уз-
лах лестницы находятся ганглии,
которые соединены между собой
поперечными комиссурами.
обрабатывает и передает к эффекторным органам информацию, поступаю-
щую от органов чувств. В качестве эффекторных органов у червей выступают
мышечные клетки, которые позволяют осуществлять несложные движения
ползания, плавания или демонстрируют про-
стейшие ориентировочные рефлексы. Нужно
отметить, что описанные выше особенности
строения червей относятся к свободноживу-
шим формам, тогда как у паразитических
форм наблюдается редукция или полное от-
сутствие органов чувств, нервных клеток и
органов движения.
У плоских червей нервная система центра-
лизована менее всего. Два боковых нервных
ствола тянутся у них вдоль всего тела от перед-
него конца к заднему (см. рис. 7). Боковые
стволы соединены поперечными нервными
пучками, а в зонах пересечений расположены
тела нейронов. В общем виде нервная систе-
ма плоских червей напоминает лестницу. По-
этому удачное название «лестничная нервная
система» сохранилось до настоящего време-
ни. Нервная система плоских червей распо-
ложена ближе к брюшной стороне тела, так
как на ней сильнее развита мускулатура и она
постоянно раздражается, соприкасаясь с суб-
стратом, по которому ползет червь. К перед-
нему концу тела размеры нервных стволов уве-
личиваются. В головной части червя располо-
жен мозг, который состоит как из брюшных,
так и из спинных скоплений нервных клеток.
Мозг плоских червей еще не играет принципиальной роли в поведении жи-
вотного. При разрезании планарий на несколько кусков каждый из них дви-
гается как целое животное, продолжает жить и регенерирует в целую особь.
У некоторых морских червей головное скопление нервных клеток играет зна-
чительно более важную роль. При разрезании такого червя пополам его пе-
редняя половина тела двигается более правильно, чем задняя.
Значительно сложнее организована нервная система у кольчатых червей.
Это обусловлено более сложным строением их тела и поведением свободно-
живущих форм. Тело кольчатых червей разделено на сегменты, называемые
метамерами. Каждый из них содержит ряд органов, характерных для всего
червя. Сегментация тела червя обусловлена развитием целома — внутреннего
отсека, заполненного жидкостью и выстланного мезотелием — производным
мезодермы. Кроме этого, кольчатые черви обладают сквозной пищеваритель-
ной системой, идущей от рта к анусу, и цепочкой нервных ганглиев, объеди-
няющих все сегменты червя.
У кольчатых червей формируется ярко выраженная центральная нервная
система. Клеточные тела нейронов сконцентрированы в отдельных сегментах
20 ❖ Сравнительная анатомия нервной системы позвоночных
так, что формируют два ряда нервных ганглиев. Ганглии каждого сегмента
связаны поперечными волокнами между собой и продольными волокнами с
ганглиями соседних сегментов. Ганглии сближены и образуют брюшную нерв-
ную цепочку. Головное скопление нервных клеток у кольчатых червей транс-
формировалось в настоящий мозг, расположенный над пищеводом (рис. 8).
Ганглии имеют организацию, характерную для большинства беспозвоноч-
ных. Тела нервных клеток расположены на наружной поверхности, а отростки
клеток и синаптические соединения — внутри ганглия. У кольчатых червей
имеются многочисленные рецепторы, специализирующиеся на восприятии
чувства равновесия, света и химических веществ. В примитивной форме при-
сутствует соматосенсорная система, которая позволяет воспринимать сигна-
лы, приходящие с поверхности тела червя Кроме этого, кольчатые черви
имеют специализированные рецепторы для осязания, ощущения давления и
болевые рецепторы.
Многообразие рецепторов, большой мозг и весьма совершенная мускулату-
ра позволяют червям вести разнообразную жизнь. Среди червей мы встречаем
роющие, плавающие и ползающие формы. Они используют все виды расти-
тельной и животной пищи. Черви могут вести паразитический образ жизни,
могут быть активными хищниками или вступать в симбиоз со свободноживу-
щими животными. Некоторые полихеты (многощетинковые кольчецы) явля-
ются комменсалами морских звезд (используют хозяина как источник пищи).
Полихеты находят своих хозяев в результате развитой хеморецепции. Они пред-
почитают здоровых и активных морских звезд. Если в Т-образном лабиринте
им предложить чистую морскую воду и воду, в которой жила морская звезда,
то они предпочтут последнюю. Если морская звезда была повреждена, то по-
лихета предпочтет чистую морскую воду.
Для червей характерны все типы рефлексов, описанные для кишечнополо-
стных. Однако организация нервной системы червей позволяет им проявлять
более сложное поведение. Червеобразные способны к ассоциативному и ус-
ловнорефлекторному обучению в Т-образном лабиринте. Обыкновенный дож-
девой червь использует 10—12 разнообразных приемов затаскивания опавшего
Рис. 8. Головные ганглии и нервы морского кольчатого червя — немертины.
I. Принципы организации нервной системы
21
❖
листа к себе в норку. Он отказывается от попытки использовать в пищу лист,
только перепробовав все возможные способы.
Это разнообразие поведения червей обусловлено усложнением организации
нервной системы и появлением нервных клеток, специализирующихся на вос-
приятии сигналов различной природы, которые позволяют червям вести сво-
бодноподвижный и хищнический образ жизни. Однако максимального разви-
тия нервная система беспозвоночных достигла у моллюсков и членистоногих.
ЧЛЕНИСТОНОГИЕ И СХОДНЫЕ С НИМИ ГРУППЫ
Для этой обширной группы животных характерно наличие по всей длине
тела конечностей, которые иннервируются парными ганглиями, организован-
ными в брюшную нервную цепочку. В головной части нервные клетки объе-
динены в крупные ганглии, называемые мозгом. У этих животных головной
мозг выполняет основные и незаменимые функции по интеграции жизнедея-
тельности организма. Развитие головного мозга членистоногих явилось ре-
зультатом длительной цефализации нервной системы. Цефализация — увели-
чение размеров нервных центров в головной части животного — обусловлена
б
Рис. 9. Строение нервной системы членистоногих различных систематических групп
а — муравей; б — майский жук; в — краб; г — телифон хвостатый (Thelyphonus caudatus;
паукообразные). Нервная система телифона вписана в контуры тела.
22
Сравнительная анатомия нервной системы позвоночных
концентрацией сенсорных органов на переднем конце тела червей, членисто-
ногих и сходных с ними животных (рис. 9).
У членистоногих обнаружена сегментация мозга, вызванная появлением спе-
циализированных отделов, в которых происходит переработка информации,
получаемой от сенсорных органов. У насекомых с хорошо развитым мозгом
протоцеребрум получает информацию от глаз, дейтоцеребрум — от антенн с
развитыми хеморецепторами, а тритоцеребрум — от передней части пищева-
рительного тракта. Наиболее важным результатом цефализации нервной сис-
темы является возникновение грибовидных тел, которые расположены в про-
тоцеребруме членистоногих и некоторых высших червей. Грибовидные тела,
по-видимому, участвуют в интеграции информации, поступающей от различ-
ных сенсорных систем. Они крайне сложно организованы. В шляпке грибо-
видного тела насекомых может располагаться от 3 до 12 слоев нейронов, раз-
меры которых не превышают 2—3 мкм. Ножка грибовидного тела состоит из
входящих и выходящих из шляпки волокон (рис. 10).
Грибовидные тела не выполняют функции непосредственной переработки
информации, поступающей от слуховых, зрительных или химических рецеп-
торов. В них происходит анализ уже первично обработанных сигналов от ди-
стантных анализаторов и брюшной нервной цепочки. По сути дела грибовид-
ные тела выполняют ассоциативные функции, позволяющие высшим беспо-
звоночным принимать решения на основе анализа информации, поступающей
от всех систем организма.
Такая сложная организация мозга членистоногих используется для контро-
ля самых разнообразных форм поведения. Примером может служить распо-
знавание пищи падальной мухой. Она исследует пищу при помощи хеморе-
цепторов, расположенных на дистальных сегментах ног. Контакт с потенци-
альной пищей вызывает прохождение нервных импульсов к подпищеводному
ганглию и активацию его нейронов. В свою очередь эти нейроны возбуждают
двигательные нейроны, которые стимулируют выворачивание хоботка и нача-
ло всасывающих движений. При разрушении мозговых ганглиев поиск пищи
Рис. 10. Надглоточный нервный узел перепончатокрылого с грибовидными телами Показа-
ны расположение тел нервных клеток и основные связи между центрами головного ганглия.
I. Принципы организации нервной системы
23
подавляется у аннелид и большинства насекомых. Необходимо учитывать, что
рецепторы у насекомых намного чувствительнее, чем у других животных. На-
пример, комары находят теплокровных животных, двигаясь по градиенту дву-
окиси углерода или аминокислот. Положительная реакция на аминокислоты
выявляется у комаров при разведении 1:10 000 000.
На уровне головного мозга осуществляется главный контроль рефлексов и
спонтанной активности ганглиев брюшной нервной цепочки. Он регулирует
включение и выключение рефлекторных реакций
низших центров, обладающих собственным аппа-
ратом для организации конкретной деятельности.
Разрушение церебральных ганглиев у большинства
беспозвоночных сопровождается гиперактивностью.
Не менее важным свойством сложного мозга явля-
ется способность сохранять информацию. Боль-
шинство насекомых обладает хорошей памятью.
Наряду с мозгом и брюшной нервной цепоч-
кой у членистоногих для управления организмом
используются нейроэндокринные органы. Основ-
ные из них расположены позади головного мозга
и соединены с ним посредством нервного ствола.
Нейрогормоны секретируются нервными оконча-
ниями, локализованными в кардиальных телах
нейронов мозга, или клетками самого кардиаль-
ного тела. Вырабатываемые в этих органах гормо-
ны необходимы для перестройки организма чле-
нистоногого во время метаморфоза — перехода от
стадии личинки к взрослому животному (имаго).
В этот момент значительным изменениям подвер-
гается центральная нервная система насекомых
(рис. 11). Так, например, у почти безногих личи-
нок дровосека грудные узлы практически одина-
ковы по размерам с брюшными и незначительно
отличаются по функциям. Среди этих ганглиев по
размерам и функциям выделяется только голов-
ной мозг. У взрослого жука с развитыми ножны-
ми и крыловыми мышцами разница между узла-
ми очень значительна. Во время метаморфоза про-
исходит объединение нескольких грудных ганглиев
личинки в один крупный ганглий взрослого жука.
Усложнение функций, связанных с полетом и дви-
жением конечностей, определяет увеличение раз-
меров управляющих ганглиев. Необходимо отме-
тить, что условия жизни личинки могут изменить
поведение взрослого насекомого. Если личинки
плодовой мушки дрозофилы были вскормлены на
среде, содержавшей перечную мяту, то взрослые
мухи обнаруживают положительный хемотаксис по
Рис. 11. Строение нервной
системы личинки и взрослого
комара.
а—личинка; б — комар после
метаморфоза. Во время мета-
морфоза происходит слияние
грудных и брюшных ганглиев,
что приводит к усилению це-
фализации нервной системы.
24 ❖ Сравнительная анатомия нервной системы позвоночных
отношению к этому растению. Эти данные свидетельствуют о сохранении
нервной системой информации, полученной на личиночной стадии разви-
тия. Учитывая, что нервная система в период метаморфоза подвергается весьма
значительным изменениям, запоминание «личиночного» опыта можно свя-
зать только с грибовидными телами или ассоциативными зонами церебраль-
ных ганглиев.
Значение мозга для животных с таким сложным поведением, как у члени-
стоногих, трудно переоценить. Даже в простых опытах по удалению головных
ганглиев легко заметить глубокие изменения поведения. Жесткокрылые (жуки),
раки и крабы после разрушения мозга начинают безостановочно ползать, од-
нообразно очищать поверхность тела или совершать непрерывные жеватель-
ные движения. Стрекозы замирают на месте и не совершают никаких движе-
ний. Утрата головного мозга приводит к потере целесообразности поведения
высших членистоногих, хотя автономные движения сохраняются. Сохранение
способности осуществлять непрерывные движения при удаленном мозге оп-
ределяется автономной активностью ганглиев брюшной нервной цепочки.
Грибовидные тела мозга насекомых оказывают тормозящее влияние на актив-
ность движений. Локальное разрушение участков мозга вне грибовидных тел
не влияет на активность движений, тогда как повреждение грибовидных тел
вызывает усиление ходьбы и прыжков. Под контролем головных ганглиев на-
ходятся наиболее сложные поведенческие реакции, например пение насеко-
мых. У сверчка контроль пения осуществляется грибовидными телами, цент-
ральным телом протоцеребрума и вторым грудным ганглием. Раздражая раз-
личные участки грибовидных тел и волокон, связывающих перечисленные
ганглии, можно вызывать или прекращать песню сверчка. Кроме этого, стре-
котание можно вызвать или затормозить, раздражая глаз, антенны, церки, тим-
панальные органы или половой аппарат. Это показывает, что интеграция сиг-
налов, приходящих от всех рецепторов, происходит в грибовидных телах, что
и определяет тип песни сверчка.
Необходимо подчеркнуть, что органы чувств членистоногих хорошо развиты
и могут быть специализированы так же, как и у позвоночных. Примером может
служить способность совок (Heliothis armgera, Mythimna separata, Amphipyra perflua)
к акустическому анализу ультразвуковых сигналов (Д.Н. Лапшин, 1995; Y. Xue
et al., 1996). Эти бабочки способны к эхолокационной ориентации среди систе-
мы сферических преград. Они могут определять препятствие на расстоянии от 7
до 12,5 см и совершать разнообразные маневры их облета. Эти наблюдения
показывают, что нервная система членистоногих достаточно развита для обра-
ботки и продолжительного хранения довольно сложной информации, которая
необходима для формирования ассоциативного поведения.
Таким образом, у высших членистоногих мы сталкиваемся с новым каче-
ством нервной системы. Мозг членистоногих выполняет настолько важные
функции по интеграции поведения, что его утрата становится невосполни-
мой. Если удаление любого участка нервной системы плоских червей не при-
водит к гибели животного, го для членистоногих губительна утрата любого
ганглия. Такая уязвимость нервной системы является результатом ее центра-
лизации и дифференциации строения головного мозга и ганглиев брюшной
1. Принципы организации нервной системы
25
❖
нервной цепочки. У насекомых мы встречаем крайне сложное поведение. До-
статочно упомянуть широко распространенную заботу о потомстве и суще-
ствование общественных насекомых. К ним относятся термиты, муравьи, осы,
пчелы и ряд других насекомых, которые живут большими группами. Внутри
таких сообществ различные особи выполняют специфические функции, что
повлекло за собой внутривидовую специализацию нервной системы. Это вер-
шина эволюции нервной системы беспозвоночных, поскольку выполнение
общественных функций в других сообществах животных не приводит к мор-
фологической трансформации нервной системы.
МОЛЛЮСКИ
Среди беспозвоночных особое место занимают моллюски. Это довольно
большая группа животных, включающая в себя несколько классов и около
80 000 видов. Рассмотрим основных представителей этой группы, коими явля-
ются брюхоногие (улитки), двустворчатые (мидии) и головоногие (кальмары и
осьминоги) моллюски. У брюхоногих моллюсков тело обычно заключено в
раковину, в большинстве своем они представляют собой малоподвижных жи-
вотных; двустворчатые, как правило, вовсе неподвижные, прикрепленные орга-
низмы, а головоногие моллюски являются свободноживушими формами и
весьма активными хищниками с практически полностью утраченной ракови-
ной. Моллюски обладают крайне разнообразной нервной системой. У мало-
Висцеральные
ганглии
Буккальные ганглии
Плевральные
ганглии
Педальные
ганглии
Церебральные
ганглии
Париетальные
ганглии
Рис. 12. Строение нервной системы брюхоногого моллюска, вписанной в контур тела. Пе-
рекрещенность нервных тяжей возникает при развороте тела в период формирования ра-
ковины. Вид сверху.
26
Сравнительная анатомия нервной системы позвоночных
Рис. 13. Строение нервной системы каракатицы, вписанной в контур тела.
подвижных и примитивных форм она организована, как у плоских червей, а
у головоногих моллюсков она наиболее сложная среди беспозвоночных
(рис. 12—14). Наиболее типично организована нервная система у брюхоногих
моллюсков (см. рис. 12). Головная часть нервной системы брюхоногих мол-
люсков состоит из четырех пар головных ганглиев, которые сгруппированы
вокруг пищевода. Парные ганглии соединяются между собой комиссурами, а
с ганглиями, расположенными вдоль оси тела, — коннективами, как у червей
и членистоногих. Буккальный ганглий иннервирует рот и передний отдел пи-
щеварительного канала, церебральный — глаза и щупальца, плевральный и
педальный — «ногу». Отдельно расположены абдоминальный ганглий, иннер-
вирующий органы висцерального мешка, и генитальный ганглий, лежащий
вблизи органов размножения. Несмотря на простоту организации нервной
системы брюхоногих моллюсков, они способны обучаться — вырабатывать
условные рефлексы на внешние воздействия. Например, морской заяц (Aplysia)
при длительной стимуляции жаберной пластинки и хвоста способен вырабо-
тать кратковременный условный рефлекс, сохраняющийся несколько часов.
Специальная программа обучения, приводящая к структурным изменениям
нервной системы, позволяет возникнуть долговременной памяти, которая под-
держивает выработанный навык в течение нескольких недель.
Нервная система головоногих моллюсков организована намного сложнее
(см. рис. 13, 14). Это связано в первую очередь с потерей раковины и хищни-
I. Принципы организации нервной системы
27
Центральный ганглий
Ротовой ганглий
Церебральные
ганглии
Глоточный
ганглий
Ротовой ганглий
Брахиальный ганглий
Церебральные
ганглии
Глаз и
зрительный
ганглий
Педальный ганглий Висцеральный ганглий
б
Рис. 14. Головные ганглии осьминога.
а — вид сверху; б — вид слева. Участок с точками — ротовая полость и пищевод.
Висцеральный
ганглий
ческим образом жизни. Большинство головоногих моллюсков — подвижные
животные. Их движение осуществляется за счет выбрасывания воды через си-
фон по принципу реактивного движения. Это позволяет головоногим мол-
люскам активно охотиться и быстро избегать опасности. Головной мозг ось-
минога приближается к мозгу рыбы сходных размеров и состоит примерно из
150—180 млн клеток.
Головные ганглии нервной системы осьминога сосредоточены, как и у дру-
гих моллюсков, вокруг пищевода (см. рис. 14). Ганглии увеличены, а попереч-
ные комиссуры сильно укорочены. Слияние головных ганглиев приводит к
формированию крупного мозга. Из органов чувств наибольшее развитие по-
лучили глаза. Нервные клетки специализированных зрительных долей мозга
имеют различную морфологическую структуру, нехарактерную для остальных
беспозвоночных, что указывает на использование зрительных центров в каче-
стве ассоциативных областей. Крупными долями мозга осьминога являются
брахиальные и педальные узлы, управляющие головными щупальцами и рото-
вой воронкой. Висцеральные ганглии иннервируют внутренности и содержат
в себе автономный дыхательный центр с обособленной локализацией субцен-
тров вдоха и выдоха. Буккальные ганглии определяют работу ротового аппара-
та. Сложность манипуляций, выполняемых щупальцами осьминога, не усту-
пает способностям к манипулированию у хоботных (слонов) и человекообраз-
ных обезьян. Это достигается в результате интегративной работы центральной
28 ❖ Сравнительная анатомия нервной системы позвоночных
части мозга, контролирующей все поведение осьминога. Однако центральные
мозговые ганглии не являются однородным образованием.
Существует морфологическая дифференцировка, связанная с контролем конк-
ретных функций. Первая пара центральных ганглиев контролирует движение «рук»
осьминога во время поедания добычи. Вторая и третья пары центральных ганглиев
через контроль педальных и брахиальных ганглиев управляют передвижением жи-
вотного при помощи щупалец, осязанием и изменением направления при плава-
нии. Во второй и третьей парах ганглиев находятся центры контроля окраски ось-
минога, которая постоянно изменяется, маскируя животное. При перерезке связей
между этими ганглиями изменение окраски осьминога нарушается. Левая полови-
на тела окрашивается иначе, чем правая. Для головоногих моллюсков контроль за
окрашиванием тела имеет принципиально важное значение для реализации поло-
вого поведения. Перед спариванием у каракатиц самец становится полосатым, а
самка, увидя его, принимает аналогичную пигментацию.
В жизни большинства головоногих моллюсков принципиальную роль игра-
ет зрение. Их глаза очень похожи на глаза позвоночных животных. У них есть
роговица, радужная оболочка, сетчатка и хрусталик. У моллюсков наведение
на резкость осуществляется за счет приближения или удаления хрусталика от
сетчатки. Аналогичным образом происходит аккомодация хрусталика у анам-
ний и змей (см. рис. 174). В области оптимальной зрительной чувствительно-
сти у каракатиц на один квадратный миллиметр приходится 150 000 зритель-
ных рецепторов, тогда как у человека — только 65 000. Среди моллюсков наи-
лучшее зрение имеет осьминог. Осьминог обладает некоторыми особенностями
восприятия зрительных стимулов, поскольку он имеет горизонтально-щеле-
вой зрачок и страдает астигматизмом. Эти анатомические особенности глаза при-
водят к тому, что осьминоги лучше различают вертикальные решетки, немного
хуже — косые и хуже всего — горизонтальные (W.R.A. Muntz и J. Gwyther, 1989).
Разрешающая способность глаз головоногих моллюсков сопоставима с тако-
вой млекопитающих. Столь совершенное зрение позволяет осьминогам разли-
чать предметы различной формы при одинаковых размерах. У них после 3—4
предъявлений вырабатываются устойчивые рефлексы на сложные зрительные
стимулы, подкрепляемые пищей. Головоногие способны к переделкам навы-
ков, которые заключаются в использовании предварительно выработанных
условных рефлексов для решения новых экспериментальных задач.
Наряду со зрением важным дистантным анализатором головоногих мол-
люсков является обоняние. Они могут различать очень низкие концентрации
веществ, растворенных в воде. Осьминог способен обнаружить при помощи
рецепторов на присосках хинин в концентрации, которая в 100 раз меньше
ощущаемой человеком с помощью вкуса. Головоногие моллюски могут на-
учиться различать большое количество зрительных образов, тактильных и хи-
мических раздражителей. Они умеют строить дома из камней, обладают хоро-
шей зрительной памятью и легко обучаются.
Мозг головоногих моллюсков представляет собой наиболее сложную си-
стему интеграции поведения среди беспозвоночных. Только самые высокоор-
ганизованные насекомые обладают аналогичными способностями решать экс-
периментальные задачи, использовать память и предварительный индивиду-
альный опыт. Однако эти свойства нервной системы в наибольшей степени
выражены у позвоночных животных.
I. Принципы организации нервной системы
29
❖
ПРОИСХОЖДЕНИЕ НЕРВНОЙ СИСТЕМЫ ХОРДОВЫХ
Начиная изучать позвоночных животных, необходимо понять основные
принципы их эволюции и историческую связь с беспозвоночными. Основным
признаком позвоночных животных является наличие хорды — гибкой стерж-
невидной структуры, которая проходит продольно через дорсальную часть тела.
Она является эволюционным предшественником позвоночного столба и не-
посредственно связана с другими важными признаками — дорсальным полым
нервным тяжом и глоточными жаберными щелями (рис. 15). В нервной систе-
ме позвоночных выделяют трубчатую центральную нервную систему и пери-
ферическую нервную систему, состоящую из ганглиев и нервов. Это разделе-
Рис. 15. Основные последовательные этапы возникновения примитивных хордовых.
а — прикрепленное кишечнополостное с радиальной симметрией; б — свободноплаваю-
щее животное с радиальной симметрией; в — придонное билатерально-симметричное
животное с дорсальным жаберным аппаратом; а — «ильное» животное, которое возникло
из придонной формы в результате поворота тела на 90е; д — примитивное хордовое с
вторичной билатеральной симметрией тела; е — свободноплавающее хордовое живот-
ное. Стрелками показана последовательность эволюционных событий.
30
Сравнительная анатомия нервной системы позвоночных
❖
ние условно, так как периферическая нервная система в значительной степе-
ни состоит из отростков нейронов, расположенных в центральной нервной
системе. Центральная нервная система подразделяется на спинной и голов-
ной мозг. Головной мозг расположен в мозговом черепе, а спинной мозг за-
щищен невральными дугами позвонков, образующими вокруг него сегменти-
рованный чехол. В центральную нервную систему входят нервные волокна,
идущие от различных чувствительных рецепторов. Волокна, выходящие из
центральной нервной системы, передают сигналы от мозга к внутренним орга-
нам и мускулатуре животного.
У наиболее низкоорганизованных хордовых можно заметить признаки орга-
низации нервной системы, сближающие их как с позвоночными, так и с
беспозвоночными животными. Бытует несколько точек зрения на возникно-
вение хордовых, которые имеют равные права на существование. Наиболее
известны гипотезы возникновения хордовых от членистоногих, паукообраз-
ных, немертин, кольчатых червей, иглокожих, гребневиков и кишечнодыша-
щих (В. Шимкевич, 1907; Р. Гессе, 1913; А.Н. Северцов, 1914; Н.А. Холодков-
ский, 1914; В.Н. Беклемишев, 1944; Р.А. Ливанов, 1955; Н. Kuhlenbeck, 1967;
R. Jefferies и D. Levis, 1978; R. Norchcutt и C. Gans, 1983; А.В. Иванов, 1976, 1991;
В.В. Малахов, 1977, 1980, 1991; Р. Кэрролл, 1992). Все эти теории с разной
степенью доказательности отвечают на один вопрос о механизме возникно-
вения трубчатой нервной системы позвоночных из ганглиозной нервной си-
стемы беспозвоночных. Рассмотрим наиболее логичную, с моей точки зре-
ния, последовательность преобразования нервной системы в процессе эво-
люционного формирования хордовых. Допуская, что кишечнополостные
являются безусловными предками основных групп животных, мы получим
гипотетическое существо, изображенное на рис. 15, б, 16, о. Внутри живот-
ного проходит пищеварительный канал, который представляет собой гастро-
васкулярную полость кишечнополостных, открытую спереди ротовым отвер-
стием. У этого организма еще нет билатеральной симметрии, и он мог пред-
ставлять собой какого-либо гидроида, перешедшего к свободноподвижному
образу жизни. Он мог свободно перемещаться в трехмерной водной среде за
счет сокращений щупалец, всего тела или пассивно, вместе с током воды.
Основным результатом перехода к такому образу жизни стало появление око-
логлоточного нервного кольца.
Животное с радиальной симметрией не нуждалось в усложнении организа-
ции, пока не перешло к придонному образу жизни. При передвижении по дну
неизбежно должны были возникнуть спинная и брюшная поверхности тела и
появиться признаки билатеральной симметрии. При переходе к активному
движению по субстрату у предка хордовых возникла необходимость в усиле-
нии метаболизма. Это произошло за счет формирования на спинной стороне
кишечной трубки щелей, которые выполняли роль жаберного аппарата
(см. рис. 15, в). Замена радиальной симметрии на билатеральную с неизбеж-
ностью привела к возникновению парных нервных тяжей, идущих вдоль тела
(см. рис. 15, г, 16, б). Вполне естественно, что ползание привело к формирова-
нию переднего (головного) скопления нервных клеток — головных ганглиев.
Эти ганглии стали выполнять функции нервного центра, в котором концент-
рировались сенсорные и ассоциативные нейроны. У такого животного два
I. Принципы организации нервной системы
❖
31
ж
Рис. 16. Строение нервной системы предковых форм хордовых. В верхнем ряду показаны
четыре этапа централизации нервной системы. Нервная система вписана в контур тела
гипотетических предков хордовых.
а — диффузная нервная система свободноплавающего животного с радиальной симмет-
рией и окологлоточным нервным кольцом; б— придонное червеподобное животное с плос-
ким телом и двойной нервной цепочкой, расположенной по краям тела; в — асимметрич-
ное «ильное» животное, возникшее из придонной формы после поворота тела на 90°; его
нервная система состоит из двух нервных цепочек: дорсальной и вентральной. Дорсаль-
ная нервная цепочка увеличивается в размерах, а ганглии сливаются; г — вторично-сим-
метричное свободноплавающее животное с дорсальной нервной трубкой и сегментиро-
ванной вентральной нервной цепочкой, из которой возникли вегетативные нервные ганглии.
В нижнем ряду приведено схематическое изображение сечений тела гипотетических пред-
ковых форм хордовых: д — радиально-симметричное животное; е — придонное животное
с билатеральной симметрией, мускулатурой, сегментированным телом, асимметричной
кровеносной системой, кишечными отверстиями для дыхания и двумя тяжами нервных
ганглиев; ж— вторично-билатерально-симметричное животное с нервной трубкой и вент-
ральными вегетативными ганглиями, возникшими в результате исчезновения продольных
волокон «нижнего» тяжа ганглиев.
нервных головных ганглия переходят в широко расставленные нервные тяжи,
расположенные по бокам тела животного. В центре проходит пищеваритель-
ный канал, дорсальнее и вентральнее которого идут поперечные комиссуры,
соединяющие левый и правый нервные тяжи (см. рис. 15, г, 16, б). Такое стро-
ение нервной системы характерно для многих плоских червей, хотя надо под-
черкнуть, что описываемое животное является исключительно гипотетической
32
о
Сравнительная анатомия нервной системы позвоночных
моделью. При ползании по горизонтальной поверхности у животных самых
различных систематических групп возникают сходные приспособления: пучки
мышечных волокон, расположенные вдоль тела и имеющие сегментарное строе-
ние. Такая мускулатура необходима для создания волнообразных продольных
движений тела, позволяющих двигаться головой вперед. Последовательно распо-
ложенные сегменты мускулатуры требуют согласования сокращений, что осуще-
ствляется посегментно расположенными латеральными ганглиями (см. рис. 16, б).
Каждый мышечный сегмент приобрел собственный ганглий, соединенный с со-
седними ганглиями при помоши отростков нервных клеток. На этом этапе эво-
люции нервной системы произошло принципиальное событие, приведшее к дор-
сальному положению центральной нервной системы хордовых. Гипотетическое
животное с двумя боковыми нервными тяжами развернулось на 90° — встало
на «ребро». Одна боковая поверхность животного стала брюшной, а другая —
спинной (см. рис. 15, г, 16, в).Вполне справедлив вопрос о причинах такого
поворота. Реальной причиной может быть изменение структуры морского грун-
та, на котором обитало это гипотетическое животное.
Если поверхность дна постепенно покрывалась илом или животное обитало
на границе интерстициали, то наиболее выгодно стало располагаться на ребре.
В таком положении животное могло скрывать в илистых осадках большую часть
тела, оставляя ротовое отверстие над поверхностью дна, и не затрачивало зна-
чительной энергии для маскировки и фиксации своего положения в грунте.
Возможно и другое объяснение. Если эти животные обитали в прибрежных
зонах моря, то становится актуальной проблема закрепления в грунте или быст-
рого погружения в него. Это могло быть связано с приливами и отливами,
морскими течениями и подвижностью придонной воды мелководных морей.
В результате этого поворота один из боковых нервных тяжей стал брюш-
ным, а другой — спинным. Животное оказалось асимметричным, так как но-
вая ось симметрии совпала с горизонтальной плоскостью тела ползающего
червеобразного предка. Поворот плоского червеобразного животного на ребро
приводит к нескольким важным результатам. На верхнем (спинном) крае тела
началось слияние ганглиев, составляющих один из тяжей нервной системы
(см. рис. 15, д, 16, в). Это связано с тем, что при проталкивании тела в или-
стом грунте основное сопротивление приходится на переднюю часть тела жи-
вотного, а задняя часть тела и хвост довольно пассивны. Поэтому координа-
ция движений должна быть максимальной в головной части животного, что и
приводит к слиянию ганглиев в передней части дорсального нервного тяжа.
Увеличение именно дорсального нервного тяжа обусловлено тем, что он
находился на границе ила и воды, в максимальном рецепторном контакте с
внешней средой, тогда как вентральный тяж был по сути дела депривирован и
изменил свои функции. На дорсальном тяже сохранились асимметричный гла-
зок и тактильные рецепторы. Затем, после появления нервной трубки, диффе-
ренцировались боковые парные глаза и парные дистантные хеморецепторы.
Вентральный тяж в головной зоне сохранил химическую и тактильную чувст-
вительность, которая стала основой для развития органов чувств ротовой и
жаберной области позвоночных.
Таким образом, центральная нервная система хордовых произошла из бо-
кового (левого или правого) нервного ствола червеподобного животного, ко-
I. Принципы организации нервной системы
❖
33
торое совершило поворот на ребро. Нервная система хордовых никогда не
была в вентральном положении и является гомологом одного из боковых нерв-
ных стволов билатерально-симметричных беспозвоночных.
В результате объединения спинных ганглиев сформировалось протяженное
скопление нервных клеток, не разделенное участками нервных волокон. Ти-
пичный ганглий беспозвоночного состоит из тел клеток, расположенных по
его периферии, и центральной части, образованной из переплетения отрост-
ков нервных клеток — нейропиля (см. рис. 16, б, е). При слиянии ганглиев
произошло объединение нейропилей, тогда как сами нейроны остались на
периферии нервного тяжа. Движение спинномозговой жидкости между отрост-
ками нейронов было слишком медленным для обеспечения метаболизма цент-
ральной нервной системы активного животного. Поэтому отростки вышли на
наружную поверхность, освободив пространство — внутреннюю полость — для
движения спинномозговой жидкости (см. рис. 16, ж). При этом симметрич-
ные комиссуры, идущие к брюшному нервному стволу, стали брюшными ко-
решками спинного мозга, а дорсальные корешки возникли из чувствитель-
ных окончаний верхнего нервного тяжа. Следовательно, нервная трубка явля-
ется гомологом слившихся ганглиев одного из нервных стволов беспозвоночных,
внутри которых шизоцельно сформировалась полость — будущие мозговые
желудочки и спинномозговой канал.
Совершенно другие изменения должны были произойти с брюшным нерв-
ным тяжом. Он не исчез и не редуцировался. У ползающего на ребре «ильно-
го» животного брюшной нервный тяж сегментировался. Сегментация заклю-
чалась в том, что ганглии сохранили свое строение, но межганглионарные
продольные связи стали иннервировать внутренние органы. Отростки нейро-
нов, расположенных в вентральных ганглиях, стали преимущественно лока-
лизоваться в кишечном канале, стенках сосудов и выделительной системе, а
не в соседних ганглиях. При этом ганглии брюшного нервного ствола не утра-
чивали комиссуральных связей со спинным нервным тяжом (см. рис. 15,
д, е, 16, в, г). Вентральные головные скопления нервных клеток стали нейраль-
ной основой для совершенствования функций передней части пищеваритель-
ного канала и развития ротовой хеморецепции.
Таким образом, ответ на поставленный вопрос о происхождении вегетатив-
ных ганглиев позвоночных довольно ясен. Вегетативные ганглии хордовых
возникли в результате разделения одного из первично-латеральных стволов
лестничной нервной системы беспозвоночных на отдельные ганглии, которые
сохранили свои комиссуральные связи с противоположным нервным тяжом.
Следовательно, поворот на 90° уплощенного червеподобного животного
привел к возникновению нервной системы, характерной для большинства хор-
довых: дорсальная нервная трубка и вентральные вегетативные ганглии. Ось
симметрии изменилась, но это не повлекло за собой катастрофических по-
следствий. Наоборот, исходя из этой модели, можно объяснить происхожде-
ние нервной трубки и вегетативных ганглиев. Нервная трубка хордовых гомо-
логична одному из тяжей нервной системы уплощенных червеобразных бес-
позвоночных предков, тогда как вегетативные ганглии гомологичны второму
боковому нервному тяжу этих животных.
Таким образом, рассмотрев становление нервной системы гипотетическо-
го предка хордовых, можно предположить основные события в филогенезе
2—5818
34
о
Сравнительная анатомия нервной системы позвоночных
нервной системы, которые привели к формированию дорсальной нервной труб-
ки и вегетативной нервной системы хордовых.
1. Самой древней предшественницей нервной системы хордовых была диф-
фузная нервная сеть свободноплавающих кишечнополостных.
2. Появление парных нервных стволов произошло при переходе свободно-
плавающего кишечнополостного к придонному образу жизни. В результате
возникли первая ось симметрии, брюшная и спинная стороны животного.
Скорее всего таким животным было плоское червеобразное существо, ползав-
шее по дну.
3. Одиночный дорсальный нервный ствол возник после поворота червеоб-
разного предка хордовых на 90°. Это могло произойти при переходе к ильному
существованию на «ребре». В этом случае один из нервных тяжей оказался
расположенным дорсально, а второй — вентрально. Из верхнего тяжа сфор-
мировалась центральная нервная система, а из нижнего — периферическая
нервная система, иннервирующая внутренние органы.
4. Полость нервной трубки возникла после слияния ганглиев дорсального
нервного ствола из нейропиля, который освободился от отростков нервных
клеток. Они вышли на внешнюю поверхность слившихся ганглиев, сформи-
ровав слой белого вещества вокруг тел клеток. Причинами этих преобразова-
ний были сенсорно-привилегированное положение дорсального тяжа и необ-
ходимость усиления обменных процессов в мозге.
5. Сегментация центральной нервной системы хордовых возникла в соответ-
ствии с количеством мышечных сегментов, иннервируемых из двух нервных
стволов червеобразного предка. Вентральные корешки спинного мозга сформи-
ровались из комиссур, а дорсальные (смешанные) нервы возникли из чувстви-
тельных и моторных нервов латеральной части тела червеобразного предка.
Описанное выше животное могло стать предком современных хордовых
после повторного перехода к активному плаванию в трехмерной водной среде
(см. рис. 15, е, 16, г). Для сохранения механической стабильности трубчатой
нервной системы возникла хорда и изменился внешний вид животного. Сле-
ды описанных событий можно выявить при внимательном изучении личиноч-
ных и взрослых форм головохордовых и круглоротых.
о
НЕРВНАЯ СИСТЕМА ЛИЧИНОЧНО-ХОРДОВЫХ
Это большая группа морских животных, имеющих на стадии личинки вы-
раженную спинную струну — хорду, над которой расположена нервная трубка
(рис. 17, а). Личинки этих животных всегда свободноплавающие и всегда снаб-
жены хвостом. Наиболее характерным представителем этой группы является
асцидия. Свободноплавающие личинки асцидий по общему виду немного на-
поминают головастиков лягушек, которые обладают овальным телом и длин-
ным, сжатым с боков хвостом. По оси хвоста идет хорда, представляющая
собой столбик студенистого вещества, одетый одним слоем клеток. Над хор-
дой расположена нервная трубка, а по бокам — мышечные волокна. В теле
дичинки нервная трубка утолщается, а в головной зоне приобретает вид пу-
зырька. Это чувствительное образование является предшественником мозга и
I. Принципы организации нервной системы
35
Рис. 17. Головной участок нервной системы свободноплавающей личинки и прикрепленг
нал взрослая асцидия.
а — личинка асцидии, вид сверху и сечение; б — строение взрослой асцидии. У личинки
нервная система представлена трубкой с асимметричным головным расширением, содер-
жащим фоторецептор, и двумя нервными стволами. Один ствол лежит выше нервной трубки,
а другой — ниже хорды. Для взрослой формы характерен один крупный нервный ганглий с
отходящими от него сенсорными и моторными нервами.
п
содержит в себе рецепторы. Личинка асцидии может воспринимать свет с по-
мощью светочувствительного глазка и узнавать о направлении гравитацион-
ного поля благодаря наличию статоциста. Активно перемещающееся живот-
ное имеет глотку, сердце, околосердечную полость и выделительную систему,
открывающуюся наружу одиночным отверстием. В таком состоянии большин-
ство личинок находится только несколько часов. Затем они прикрепляются к
какому-либо предмету при помощи специальных сосочков и превращаются во
взрослое оседлое животное.
Такое превращение носит название регрессивного метаморфоза. Он заклю-
чается в увеличении количества глоточных дыхательных отверстий, полном
исчезновении хвоста и хорды. Наибольшим изменениям подвергается нервная
система. Исчезают хвостовой отдел нервной трубки, глазок и статоцист. Вме-
сто головного пузырька и туловищной нервной трубки формируется одиноч-
ный ганглий, характерный для нервной системы беспозвоночных (рис. 17, б),
который лежит между ротовым и анальным отверстием. Он вытянут в спинно-
брюшном направлении и формирует несколько нервных стволов. На брюш-
ной стороне нервного узла расположена субнейральная железа, которую гомо-
логизируют с гипофизом позвоночных. Таким образом, из активной свобод-
ноплавающей личинки с хорошо развитыми органами чувств, хордой и сложцо
организованной нервной системой возникает сидячее взрослое животное, у
которого органы, указывающие на родство с позвоночными, изменяются до
36
Сравнительная анатомия нервной системы позвоночных
❖
неузнаваемости. Большую часть тела оседлой формы занимает пищеваритель-
ный тракт U-образной формы с обширной глоточной полостью Описанный
жизненный цикл не является универсальным для всех оболочников. В некото-
рых примитивных родах этой группы хвостатой стадии развития нет, а у дру-
гих развитие личинки происходит в родительском организме. Этому способ-
ствует гермафродитизм (двуполость) взрослых оболочников.
Личиночно-хордовые объединяют в себе признаки нервной системы позво-
ночных и беспозвоночных животных. Эта важная в эволюционном отношении
группа показывает единство происхождения позвоночных и беспозвоночных
животных, что имеет существенное значение для понимания исторического раз-
вития нервной системы. Следующей по уровню организации группой хордовых
являются бесчерепные или головохордовые, которые обладают всеми основными
признаками хордовых. Они имеют во взрослом состоянии хорошо развитые хор-
ду, спинной мозг и жаберные щели (рис. 18). Единственным представителем группы
является ланцетник — небольшое животное с прозрачным телом, которое заост-
рено с обоих концов и сжато с боков. Нервная система ланцетника расположена
над хордой, между мышцами правой и левой сторон тела.
Центральная нервная система имеет вид трубки, которая в поперечном раз-
резе представляет собой треугольник с полым центральным каналом. Сама
трубка состоит преимущественно из продольных нервных волокон и тел кле-
ток, сгруппированных вокруг центрального канала. Периферическая нервная
система состоит из нервов, отходящих от центральной нервной системы. Они
делятся на две группы: первая образована несимметричными парами нервов,
отходящих от передней части нервной трубки, называемой мозгом, вторая
состоит из большого числа спинных нервов, отходящих от спинного мозга
(см. рис. 18, б). Первая пара головных нервов отходит от переднего края нерв-
ной трубки, а вторая — от его спинной стороны. Обе пары нервов имеют
многочисленные нервные узлы, которые размещены в головной части ланцет-
ника. Спинные нервы распределены посегментно, и соответственно располо-
жению миомеров нервы правой и левой сторон отходят поочередно, а не друг
против друга. В каждом сегменте с каждой стороны тела есть два нерва: спин-
ной, отходящий одним корешком от верхней стороны спинного мозга, и брюш-
ной, отходящий многочисленными отдельными волокнами. Спинные нервы
идут к коже и к мышцам, являясь как чувствительными, так и двигательными.
Брюшные нервы исключительно двигательные и идут к миомерам.
В передней части нервной трубки центральный канал незначительно рас-
ширяется, образуя большую полость — желудочек передней части нервной
трубки, а несколько позади него формируется спинное расширение. Это обра-
зование, как и вся нервная трубка, покрыто тонкой соединительнотканной
оболочкой. Передний верхний отдел нервной трубки выдается небольшим
полым мешком (асимметричная ямка Келликера), который соединяется с ор-
ганом обоняния и носит название средней обонятельной доли. На его заднем
конце с брюшной стороны есть небольшое углубление, называемое воронкой.
Органы чувств ланцетника представлены асимметричной обонятельной
ямкой, открывающейся наружу на левой стороне головы. Этот хеморецептор-
ный орган имеет полость, которая открывается через нейропор в центральный
канал нервной трубки. Органы зрения ланцетника весьма своеобразны. В го-
I. Принципы организации нервной системы
37
а
Рис. 18. Клетка Рохда (Rohde). Внешний вид и центральная нервная система ланцетника.
а — центральная нервная система, вид сверху: I—VI — головные нервы; б — сечение тела
взрослого ланцетника: 1 — клетки Жозефа (Joseph); 2 — ламеллярные фоторецепторные
клетки; 3 — клетки, секретирующие катехоламины; 4 — нейроэндокринные клетки; 5 —
пролактин-иммунореактивные клетки; 6 — кальцитонин-иммунореактивные клетки; 7 —
катехоламинергические нейроэндокринные клетки; 8 — серотонинергические клетки; 9 —
клетки воронки; 10 — клетки радиальной глии (по Н. Obermuller-Wilen, 1984).
ловной части они представлены непарным глазом, который выглядит как пиг-
ментное пятно. Небольшие светочувствительные клетки расположены в тол-
ще нервной трубки на всем ее протяжении. Каждая из них частично окружена
пигментной клеткой. Такая пара клеток представляет собой отдельный про-
стой глазок, способный улавливать различия в освещенности. Органы осяза-
ния представлены нейронами, иннервирующими ротовые щупальца, а орган
вкуса — небольшой ямкой Гатчека, которая расположена на верхней стороне
предротовой воронки.
ПОЗВОНОЧНЫЕ ЖИВОТНЫЕ
К позвоночным относят обширную группу животных, включающую в себя
круглоротых, рыб, амфибий, рептилий, птиц и млекопитающих. Обособление
этой группы связано с тем, что все перечисленные животные обладают еди-
ным планом анатомического строения. У них имеются развитый позвоноч-
ник, череп, головной и спинной мозг. Именно внутри этой группы животных
нервная система достигла своего максимального развития. Сложнее всего орга-
38
Сравнительная анатомия нервной системы позвоночных
❖
низован мозг млекопитающих и человека. Он обладает способностью не толь-
ко получать, обрабатывать, хранить и передавать информацию, но и генери-
ровать мысли о том, чего еще не было в природе.
Центральная нервная система позвоночных организована по общим прин-
ципам. В ней выделяют головной и спинной мозг, который расположен над
позвоночником или хордой. Однако если головная часть ланцетника была
намного меньше, чем спинная, то у большинства позвоночных это соотноше-
ние носит обратный характер. Головной мозг позвоночных в большинстве
случаев намного больше спинного. Если принять величину спинного мозга за
единицу, то относительная величина головного мозга будет следующая:
пятнистая саламандра (хвостатые амфибии) — 0,9
травяная лягушка (бесхвостые амфибии) — 1
бык (млекопитающие) — 1,5
карп (костные рыбы) — 2
петух (птицы) — 2
кролик (млекопитающие) — 2
броненосец (млекопитающие) — 2,8
еж (млекопитающие) — 4
кошка (млекопитающие) — 4
летучая мышь (млекопитающие) — 6
макака (млекопитающие) — 8
дельфин (млекопитающие) — 18—23
человек (млекопитающие) — 26
Интересно отметить, что у кошки и макаки приблизительно равная масса
тела и спинного мозга (7,5 г). Масса же головного мозга сильно различается.
У макаки он в среднем равен 60 г, а у кошки — 28—30 г. Таким же образом
крупная собака, горилла и человек с близкими массами тела и размерами спин-
ного мозга обладают различной средней массой головного мозга: 135, 430,
'' 1350 г соответственно (рис. 19).
Нетрудно понять причину этих различий. Масса спинного мозга и его раз-
мер непосредственно зависят от количества иннервируемых мышц и числа
рецепторов, воспринимающих сигналы, идущие от тела животного. Чем боль-
ше размер тела животного, тем больше удельный вес спинного мозга в цент-
ральной нервной системе. По-видимому, зауроподы — самые крупные среди
четвероногих травоядных динозавров — обладали спинным мозгом, во много
раз большим по массе, чем головной мозг. С другой стороны, очевидно, что
размер головного мозга не может определяться только периферическими ана-
лизаторами и иннервацией мускулатуры головы. Главная часть головного моз-
га позвоночных состоит из ассоциативных нейронов, которые не имеют ника-
кой непосредственной связи с органами, воспринимающими раздражение, или
с двигательными системами организма. Основная задача ассоциативных обла-
стей головного мозга заключается в интеграции всего разнообразия сигналов,
приходящих из внешней среды и от внутренних систем организма. Анализ
этой информации и использование предыдущего опыта определяют поведе-
*ние животного и его реакции на внешнее раздражение.
Таким образом, более совершенное развитие ассоциативных зон нервной
системы является критерием приспособляемости и выживания конкретного
I. Принципы организации нервной системы
❖
39
Рис. 19. Внешний вид головного мозга различных позвоночных животных. Вид сверху,
а — рыбы; б — рептилии; в — птицы; г — сумчатые млекопитающие; д — хищные; в —
приматы; ж — человек.
организма. У высших позвоночных наиболее сложное поведение объясняется
развитием ассоциативных центров головного мозга. Спинной мозг сам по себе
вызывает лишь рефлекторные реакции, протекающие более или менее одно-
образно. Модификация этих реакций, их оттенки или полное подавление не-
которых рефлексов — та нервная деятельность, которая сопровождается явле-
ниями, называемыми памятью, рассудком и волей, — происходит в ассоциа-
тивных центрах головного мозга.
Для нервной системы позвоночных животных характерен ряд специфиче-
ских характеристик. Центральная нервная система состоит из спинного и го-
ловного мозга. Головной мозг включает в себя пять основных отделов: перед-
ний, промежуточный, средний, задний и продолговатый мозг (рис. 20). Каж-
дый из отделов головного мозга исторически связан с конкретными
дистантными анализаторами — хеморецепторами, фоторецепторами, тактиль-
ными или слуховыми системами анализа внешней и внутренней среды орга-
низма. Как правило, конкретные рецепторы расположены на некотором рас-
стоянии от мозга и соединены с ним посредством нервов. Нервы, относящие-
ся непосредственно к мозгу, носят название черепно-мозговых нервов. Всего
их насчитывается 12 пар. Черепно-мозговые нервы разделяются на сенсор-
ные, моторные и смешанные. Сенсорные нервы передают информацию от
внешних и внутренних рецепторов в головной мозг, а моторные проводят
эффекторные сигналы к мышцам.
40 ❖ Сравнительная анатомия нервной системы позвоночных
Внутреннее строение центральной нервной системы характеризуется нали-
чием полостей — мозговых желудочков, которые образуют систему сообщаю-
щихся цистерн, заполненных спинномозговой жидкостью (см. рис. 20). У низ-
ших позвоночных в переднем мозге расположена одна полость, которая носит
название переднего мозгового желудочка (I). Для более высокоорганизован-
ных позвоночных с парными полушариями переднего мозга характерно суще-
ствование двух полостей (I, II), которые носят название латеральных желудоч-
ков (см. рис. 20, б). Строение переднего мозга многих млекопитающих и чело-
века характеризуется наличием разветвленных латеральных желудочков
переднего мозга. Желудочки формируют два или три рога, которые распрост-
раняются в лобную, височную и затылочную доли полушарий переднего моз-
га. Эти полости носят название рогов латеральных желудочков (см. рис. 20, в).
Место схождения трех рогов носит название треугольника или звезды лате-
ральных желудочков. Латеральные желудочки соединяются между собой и с
III мозговым желудочком при помощи монроева отверстия.
III желудочек расположен на уровне промежуточного мозга и переходит в
сильвиев водопровод. У низших позвоночных III желудочек и сильвиев водопро-
вод образуют довольно широкую полость часто одинакового сечения на всем
своем протяжении. Однако у рептилий и млекопитающих сильвиев водопровод
значительно сужается. У человека его диаметр около 3—4 мм. Сильвиев водопро-
вод переходит в IV желудочек, который лежит внутри заднего и продолговатого
мозга. Стенки заднего и значительной части продолговатого мозга развернуты в
латеральном направлении, что приводит к формированию широкой полости IV
желудочка и истончению дорсальной стенки нервной трубки. IV желудочек сужа-
ется в каудальном направлении и переходит в центральный канал спинного моз-
га. Центральный канал проходит вдоль всего спинного мозга и иногда формирует
расширение в поясничном отделе. Это расширение у млекопитающих носит на-
звание люмбального желудочка, а у птиц — люмбосакральной полости. Люмбо-
сакральная полость часто заполняется у птиц желатинозной субстанцией, плохо
проницаемой для спинномозговой жидкости. Надо отметить, что центральный
канал спинного мозга у многих млекопитающих зарастает с возрастом на уровне
плечевого пояса и грудного отдела позвоночника.
Внутри желудочков головного мозга и центрального канала спинного мозга
проз гкает спинномозговая жидкость. Она образуется в сосудистых сплетениях
за счет фильтрации плазмы крови. В полость желудочков из кровеносного
русла проникают только вода и небольшое количество ионов. Сосудистых спле-
тений два (см. рис. 20). Одно расположено в латеральных желудочках и рост-
ральной части III желудочка, а второе — в IV желудочке. Надо отметить, что у
круглоротых существует сосудистое сплетение среднего мозга (plexus chorioideus
mesencephali), которое выходит на наружную поверхность мозга. Переднее со-
судистое сплетение обычно занимает 1/3 латеральных желудочков и 1/2 длины
III желудочка. У млекопитающих оно проникает только в боковые рога лате-
ральных желудочков. Заднее сосудистое сплетение обычно занимает большую
часть крыши IV желудочка. У млекопитающих оно похоже на букву М. Инте-
ресно отметить, что у приматов ножки буквы М высовываются в специальные
отверстия (отверстия Люшка) на наружную поверхность мозга около выхода
слухового (VIII) нерва, а центральный изгиб сосудистого сплетения просовы-
1. Принципы организации нервной системы
41
Передний рог
Монроево
отверстие
Сильвиев
водопровод
Переднее
сосудистое
сплетение
Боковой рог
Задний рог
Сильвиев
водопровод
Заднее
сосудистое
сплетение
=1 Задний
ЕЕЗ мозг
Продолговатый
Liililil МОЗГ
Передний
мозг
Промежуточный
мозг
Средний
мозг
Рис. 20. Строение головного мозга позвоночных.
а — организация, характерная для хрящевых и костных рыб, б—для амфибий, рептилий
и птиц, в — для млекопитающих. I-IV — желудочки мозга. Передний мозг (telencepnalon),
промежуточный мозг (diencephalon), средний мозг (mesencephalon), задний мозг
(metencephalon), продолговатый мозг (myelencephalon).
вается над мозговыми оболочками через асимметричное отверстие (апертура
Мажанди). Эти отверстия являются своего рода страховочными клапанами,
позволяющими быстро компенсировать резкие перепады давления спинно-
мозговой жидкости. Сосудистое сплетение может иметь значительные морфо-
логические различия у разных видов. Так, у круглоротых и двоякодышащих
переднее сосудистое сплетение может покрывать своими складками практи-
чески всю дорсальную поверхность переднего, промежуточного и среднего
мозга. Заднее сосудистое сплетение разрастается и закрывает собой как зад-
ний, так и продолговатый мозг. Мозг сверху выглядит как разросшаяся вино-
градная гроздь. У осетровых в развитом заднем сосудистом сплетении часто
осуществляется кроветворение. Для многих костистых рыб характерен перед-
ний мозг, вывернутый внутренними поверхностями желудочков наружу.
В этом случае сосудистое сплетение в переднем мозге практически отсутству-
ет. Однако основная схема анатомического строения остается неизменной.
42 ❖ Сравнительная анатомия нервной системы позвоночных
БЕСЧЕЛЮСТНЫЕ (КРУГЛОРОТЫЕ)
Как следует из названия группы, этих позвоночных объединяет отсутствие
челюстного аппарата. Типичными представителями являются речная и ручьевая
миноги. Кроме них в группу входят миксины — морские полупаразитические
животные или падальщики. Круглоротые являются длиннотелыми первично-
водными позвоночными, которые отличаются от рыб ротовой присоской без
челюстей, непарным органом обоняния, отсутствием парных конечностей.
Нервная система миноги имеет ряд своеобразных черт, выделяющих ее из
всех известных позвоночных животных. Спинной мозг миноги уплощен и на-
поминает по форме ленту. Его спинные корешки чередуются с брюшными и
не соединяются с ними для образования смешанных нервных стволов. Симпа-
тическая нервная система развита слабо и представлена несколькими неболь-
шими узелками по бокам спинной аорты, в стенках сердца, в почках и поло-
вых железах. Первичный задний мозг миноги хорошо развит, а обширная по-
лость IV желудочка занимает большую часть дорсальной поверхности этого
отдела (рис. 21). По размерам он практически равен среднему, промежуточно-
му и переднему вместе взятым. Интересно отметить небольшие размеры моз-
жечка, который выглядит, как небольшая поперечная лента у переднего края
IV желудочка. Для животного, передвигающегося в трехмерной водной среде,
такое строение мозжечка нехарактерно. Средний мозг, представленный пар-
ными зрительными долями, дифференцирован слабо. Его крыша открыта, как
и полость IV желудочка заднего мозга. Поверхность крыши состоит из тонко-
го слоя недифференцированных нервных клеток и сосудистого сплетения,
свешивающегося в полость сильвиева водопровода.
На верхней стороне промежуточного мозга находится два асимметричных
утолщения — хабенулярные ганглии, из которых правое намного больше ле-
вого. В правый хабенулярный ганглий входит тонкий нерв от пинеального
органа (эпифиза). Парапинеальный орган (теменной) посылает нерв в левый
ганглий и по своему строению аналогичен пинеальному органу. Он содержит
светочувствительные рецепторы, пигмент и несовершенный хрусталик. Это
третий глаз миноги, позволяющий воспринимать изменения освещенности.
Под эпифизом расположен вырост промежуточного мозга, связанный с левым
хабенулярным ганглием. Он носит название парапинеального органа. Еще ниже
расположен дорсальный мешок промежуточного мозга, который связан с обо-
ими хабенулярными ганглиями. На нижней стороне промежуточного мозга
расположен гипофиз. Передний мозг состоит из двух грушевидных выпячива-
ний. Парные задние утолщения являются собственно передним мозгом, а пе-
редние — обонятельными луковицами. У круглоротых орган обоняния непар-
ный, хотя снабжается парным обонятельным нервом (I пара). У миног ноздря
ведет через короткий носовой проход к обонятельному мешку, расположенному
впереди головного мозга. От обонятельного мешка отходит кзади слепой вы-
рост — назогипофизарный мешок, который прободает основание черепа и
оканчивается под передним краем хорды. Стенка этого мешка прилежит к
основанию промежуточного мозга и дает в развитии начало гипофизу. У мик-
син этот мешок открывается в глотку. Осязательными органами служат нервные
окончания в коже, а луковицеобразные группы нервных окончаний в ротовой
1. Принципы организации нервной системы
43
Рис. 21. Строение головного мозга миксины (a, Myxine glutinosa) и миноги (б, Lampetra
fluviatilis). Вид с дорсальной поверхности.
I—X — черепно-мозговые нервы.
полости являются органами вкуса. У миног двойная боковая линия, а лабиринт
состоит из двух (у миноги) или одного (у миксины) полукружного канала.
У миксин глаза рудиментарны, а миноги имеют крупный хрусталик и хорошо
развитую толстую сетчатку. Важно отметить, что зрительные нервы (II пара)
миног и миксин не образуют перекреста и направляются каждый к глазу со
своей стороны. Для круглоротых характерно объединение IX и X пар нервов.
ХРЯЩЕВЫЕ И КОСТНЫЕ РЫБЫ
У рыб хорошо развиты органы движения, череп, челюстной и жаберный
аппараты. Кроме жаберного аппарата, приспособленного для дыхания, у мно-
гих рыб есть плавательный пузырь, который в некоторых случаях может вы-
полнять функции легкого или органа воздушного дыхания. У рыб парный
орган обоняния только в виде исключения соединяется каналом с ротовой
полостью. Питуитарный мешок, который хорошо развит у круглоротых, от-
сутствует, а гипофиз не имеет никакой связи с обонятельным мешком.
В отличие от круглоротых лабиринт рыб имеет три полукружных канала.
Среди хрящевых рыб наиболее интересной группой являются акулы. Разли-
чия в поведении акул коррелируют с морфологической организацией их мозга.
44
Сравнительная анатомия нервной системы позвоночных
❖
У активных хищников хорошо развито обоняние и высокоспециализирован
передний мозг, который служит центром обработки обонятельных сигналов.
У относительно неспециализированных форм размеры мозга сходны, но наи-
более развиты ромбэнцефалон и мозжечок, которые являются координаци-
онным центром двигательной активности (рис. 22). Наибольшую вариабель-
ность развития демонстрирует передний мозг акул и скатов. У пелагических
акул, ведущих активный поиск пищи в верхних слоях воды, передний мозг
обычно выглядит, как монополушарное образование (см. рис. 22, а). Для
придонных форм акул характерно наличие парных полушарий (см. рис. 22, б).
Особое строение имеет передний мозг у скатов. Парные обонятельные цент-
ры, которые формируются вокруг монополушарного переднего мозга, соз-
дают иллюзию его трехполушарной организации (см. рис. 22, в). Особняком
стоит строение мозга химер, которые имеют чрезвычайно развитый продол-
говатый мозг (см. рис. 22, г).
У многих костистых рыб наиболее развиты зрительные отделы мозга. В ка-
честве примера следует рассмотреть мозг костистой рыбы — карпа (рис. 23).
При поверхностном вгляде на мозг карпа становится ясно, что основной струк-
турой является средний мозг. В средний мозг приходит вторая пара черепно-
мозговых нервов, идущих от глаз. Зрительные нервы у карпа перекрещивают-
ся и идут на противоположную сторону: от правого глаза к левой половине
среднего мозга, а от левого глаза — к правой. Мозжечок крупный по сравне-
нию с другими отделами мозга и почти полностью закрывает полость IV желу-
дочка. Передний и промежуточный мозг небольших размеров. Промежуточ-
ный мозг можно идентифицировать только по выходу эпифиза впереди пар-
ных выпячиваний среднего мозга (см. рис. 23). Нижняя часть промежуточного
мозга формирует воронку и гипофиз. Из строения мозга совершенно ясно, что
поведение карпа основано на хорошо развитом зрении и реализуется через
координацию движений, которая осуществляется крупным мозжечком. С дру-
гой стороны, связанный с обонянием передний мозг развит плохо, что гово-
рит о незначительном использовании хеморецепции. Для координации дви-
жений карпа большую роль играет анализ положения тела в пространстве, о
чем свидетельствует развитый лабиринт с тремя полукружными каналами. Они
позволяют карпу точно оценивать угловое ускорение движения. Органы ли-
нейного ускорения карпа представлены мешочками, содержащими крупные
отолиты, которые являются кальцифицированными выделениями клеток.
Таким образом, строение мозга карпа отражает основные чувствительные
механизмы, которые он использует для анализа окружающей среды. У рыб с
другой экологией и способом добывания пищи строение мозга может сущест-
венно отличаться. Примером может служить мозг клюворылых (см. рис. 185).
У клюворылых электрическим органам, иннервируемым продолговатым моз-
гом, соответствуют особые «электрические полушария» мозжечка, которые по
размерам превышают передний мозг. Они закрывают собой не только продол-
говатый мозг, но и все рострально лежащие отделы. У латимерии передний,
промежуточный и средний мозг развит слабо. У нее относительно небольшой
мозжечок и непропорционально крупные двигательные отделы продолговато-
го мозга (см. рис. 23, а).
I. Принципы организации нервной системы
45
в
Рис. 22. Строение головного мозга акул, скатов и химер (без обонятельных луковиц).
а — длиннокрылая акула (Calcharhinus longimanus)-, б — молодой (20 см) черноморский
катран (Squalus acanthias); в — гладкий скат (Raja batis)-, г — европейская химера (Chimaera
monstrosa). I-X — черепно-мозговые нервы, зачернен IV желудочек.
46
❖
Сравнительная анатомия нервной системы позвоночных
Рис. 23. Строение головного мозга рыб.
а—латимерия (Latimeria chalumnae)-, б — рыба-луна (Mola mola); в—карп (Cyprinus carpio)',
г — бурый протоптер (Protopterus annectens). I-X — черепно-мозговые нервы, зачернен
IV желудочек, нл — нерв латеральной линии.
I. Принципы организации нервной системы
47
<•
Особенности строения тела некоторых рыб могут приподнести неожидан-
ные нейроанатомические сюрпризы. Примером может служить строение цент-
ральной нервной системы рыбы-луны (см. рис. 23, б). Этот вид из отряда
скалозубообразных не имеет хвоста, питается зоопланктоном, откладывает до
300 млн икринок и ведет пассивный образ жизни. Отсутствие хвоста привело
к укорочению спинного мозга, а неразвитость обоняния — к редукции перед-
него мозга. У рыбы-луны гипофиз лежит ростральнее переднего мозга, а са-
мыми крупным отделом является средний мозг с развитыми зрительными не-
рвами (см. рис. 23, б). Основной объем головного мозга занят нейрогормо-
нальными и зрительными центрами, что полностью соответствует незатейлевой
биологии рыбы-луны.
У двулегочников более сложная биология, которая включает в себя циклы
спячки и активности, постройку гнезда, переход на различные типы дыхания и
всеядность. У бурого протоптера мозг отражает многогранность его поведения
(см. рис. 23, г). В его мозге хорошо развиты полушария переднего мозга, сред-
ний мозг и двигательные отделы продолговатого мозга. Поверхность IV желу-
дочка протоптера практически закрыта сосудистым сплетением, которое фор-
мирует своеобразные «виноградные грозди». Аналогичное сплетение прикрыва-
ет промежуточный и передний мозг, но на рис. 23, г не показано.
Многие рыбы проявляют сложные формы поведения. Они узнают и защи-
щают своего полового партнера, проявляют продолжительную заботу о потомст-
ве, строят и охраняют свое гнездо, обладают способностью к обучению. Евро-
пейский лосось обучается выпрыгивать из воды и проскакивать через подве-
шенное в воздухе кольцо. Известны случаи обучения рыб нажиманию на рычаг
для получения червяка. Реакция сохранялась месяцами, но без подкрепления
быстро угасала.
Рыбы являются первичноводными животными, приспособленными к пла-
ванию. Одна из наиболее важных черт их строения, накладывающая отпеча-
ток на все их поведение, — отсутствие рычажных конечностей. Поэтому инст-
рументальное поведение играет ограниченную роль в их жизни.
АМФИБИИ (ЗЕМНОВОДНЫЕ)
Амфибии внешне отличаются от рыб тем, что имеют рычажные конечности
вместо парных плавников. В личиночном состоянии почти все амфибии ды-
шат жабрами, которые у ряда видов сохраняются в течение всей жизни
У взрослых животных обычно имеются легкие. Личинки амфибий развивают-
ся в водной среде и обладают хрящевым скелетом, тогда как большинство
взрослых животных обитают на земле и имеют скелет из костной ткани. Пере-
ход от личиночной стадии к взрослому животному часто связан с выходом на
сушу, что приводит к значительным изменениям скелета и внутренних органов.
У лягушек исчезает хвост, развиваются легкие вместо жабр и формируются ко-
нечности. Этот процесс носит название метаморфоза. Некоторые земноводные
становятся половозрелыми на личиночной стадии. Это явление лучше всего
изучено у аксолотля и носит название неотении. Современные земноводные
обладают гладкой кожей, которая легко пересыхает. Амфибии размножаются
48
Сравнительная анатомия нервной системы позвоночных
❖
преимущественно в воде и не имеют тазовых почек. Поэтому они нуждаются
во влажной среде и чаще всего обитают поблизости от водоемов. В эту группу
входят бесхвостые (лягушки и жабы), хвостатые (саламандры и тритоны) и
безногие амфибии — червяги (червеобразные роющие земноводные).
Нервная система амфибий имеет много сходных черт с рыбами, но обладает
и рядом особенностей. Хвостатые и бесхвостые амфибии приобрели конечно-
сти, что повлекло за собой изменение организации спинного мозга. Спинной
мозг лягушки короткий и оканчивается тонкой конечной ниточкой. В соот-
ветствии с числом позвонков у лягушки существует только 10 пар спинных
нервов (рис. 24). II и III пары сливаются и образуют плечевое сплетение, про-
водящее нервы в передние конечности. VII, VIII, IX и X пары соединяются
для образования крестцово-поясничного сплетения, которое иннервирует зад-
ние конечности. Такая специализация нервов спинного мозга впервые встре-
чается у амфибий и отсутствует у рыб. Есть еще одно важное отличие в стро-
ении спинного мозга. У лягушки I спинной нерв выполняет функцию XII
нерва и иннервирует мышцы языка.
Головной мозг лягушки отличается от мозга рыб очень небольшим моз-
жечком, более крупным передним мозгом и хорошо заметным промежуточ-
ным мозгом (см. рис. 24). Парные полушария переднего мозга имеют спере-
ди крупные обонятельные луковицы, которые срастаются между собой по
средней линии у бесхвостых амфибий. Промежуточный мозг расположен сразу
за полушариями и имеет два выроста, соответствующих эпифизу и парафизу.
Нижний мозговой придаток промежуточного мозга — гипофиз — хорошо
развит. Средний мозг амфибий представляет собой наиболее крупную струк-
туру нервной системы. У большинства амфибий он выглядит как парные
полусферы, в которые входят зрительные тракты — продолжение зрительных
нервов. Позади среднего мозга расположен крайне маленький мозжечок,
который представляет собой небольшое утолщение передней стенки IV же-
лудочка. Полость IV желудочка открыта. Все черепно-мозговые нервы хоро-
шо заметны, но XII пара отходит от центральной нервной системы вне моз-
гового черепа и поэтому рассматривается как спинномозговой нерв. Доба-
вочному (XI) нерву соответствует ветвь блуждающего нерва (X). Блуждающий
нерв иннервирует внутренние органы и гортань (производное жаберного
аппарата рыб). У амфибий парасимпатические ветви блуждающего нерва
начинают иннервировать легкие. Языкоглоточный нерв (IX) иннервирует
полость рта, глотку и язык. Он редко выходит из черепа отдельно, чаще всего
сливается с блуждающим нервом.
У бесхвостых амфибий тройничный нерв (V) может сливаться после выхода
из мозга с лицевым и слуховым нервами (VI—VIII). Зрительный нерв (II) обра-
зует на нижней стороне мозга срастание, напоминающее букву X, со сложным
перекрестом волокон, тогда как у рыб это простое перекрещивание нервов.
Обонятельный нерв (I) состоит из двух пучков волокон, которые у безногих
амфибий разделяются и входят в мозг раздельно. Один из них идет от воздуш-
ного обонятельного мешка, а второй — от хеморецепторов якобсонова органа,
который у амфибий выполняет функции органа водного обоняния.
Основные органы чувств амфибий изменены в соответствии с приспособ-
лением к жизни в новой среде. У личинок и постоянно живущих в воде видов
I. Принципы организации нервной системы
49
Рис. 24. Строение головного мозга амфибий.
а — суринамская пипа (Pipa pipa)-, б — аксолотль (Ambystoma tigrinum); в — травяная ля-
гушка (Rana temporaria); г — молодая (90 мм) червяга (Chtonerpeton viviparum). I-XII —
черепно-мозговые нервы, зачернен IV желудочек, во — вомероназальный нерв, нл — нерв
латеральной линии.
50 ❖ Сравнительная анатомия нервной системы позвоночных
имеется сложная боковая линия, состоящая из нескольких рядов чувствитель-
ных луковиц, которая позволяет воспринимать колебания воды. У наземных
видов эти органы заменены специализированными осязательными пятнами.
Органами вкуса являются луковицеобразные тельца, разбросанные в слизи-
стой оболочке нёба и языка. Носовые мешки, имеющие две связанные каме-
ры, обладают способностью к хеморецепции как воздуха, так и воды благода-
ря расположенным в них рецепторным клеткам. Они открываются ноздрей
наружу и специальным отверстием — хоаной — в ротовую полость. Так как
вдыхание и выдыхание воздуха происходят обычно через ноздри, то носовая
полость амфибий не только является обонятельным органом, но и входит в
состав дыхательных путей.
Глаза амфибий позволяют различать цвета предметов и движущиеся объек-
ты. Земноводные обладают хорошим зрением. Хрусталик их глаза сзади более
выпуклый, чем спереди, и в обычном состоянии установлен для зрения вдаль.
Аккомодация глаза достигается сокращением лежащей вокруг хрусталика мыш-
цы, перемещающей его по направлению к роговице. У некоторых земновод-
ных глаза невелики или рудиментарны и спрятаны под кожей.
Орган слуха включает среднее ухо, которое впервые встречаются у амфи-
бий. Снаружи он ограничен барабанной перепонкой, за которой лежит боль-
шое пространство — барабанная полость, соединенная с ротовой полостью
короткой евстахиевой трубой. В барабанной полости расположено стремя,
передающее колебания барабанной перепонки на специальное окно внутрен-
него уха. Колебания воздуха вызывают движение барабанной перепонки, ко-
торое передается посредством стремени к эластичному окну внутреннего уха,
обладающего механочувствительными клетками. Соединение евстахиевой трубы
с ротовой полостью позволяет выравнивать давление воздуха в барабанной
полости с атмосферным. Понятно, что такой орган слуха не отличается высо-
кой чувствительностью и не позволяет воспринимать высокие звуки. Тем не
менее возможность воспринимать колебания воздуха, а не воды впервые воз-
никла у амфибий.
Симпатическая нервная система хвостатых амфибий состоит из большого
симпатического нерва, простирающегося в виде цепи узелков, соединенных
волокнами, от головы через туловище в хвостовой отдел тела. Там, где основ-
ные концы ребер раздвоены и охватывают позвоночную артерию, существует
добавочный ствол симпатического нерва, связанный с большим симпатиче-
ским нервом.
Адаптация амфибий к наземному существованию повлекла за собой изме-
нение поведения этих животных. Амфибии в большей степени, чем рыбы,
отличаются индивидуальным поведением, которое координируется централь-
ной нервной системой. Они проявляют различные формы заботы о потомстве
и способность к обучению. Земноводные могут проходить простые лабирин-
ты. В этом отношении жабы несколько превосходят лягушек, а тритоны и
саламандры с такой задачей не справляются.
Переход к наземному образу жизни повлек за собой изменение организа-
ции центральной нервной системы и поведения земноводных. У наиболее
высокоорганизованных амфибий становление поведения происходит в про-
цессе индивидуального развития. Оно основано на врожденных стереотипах,
I. Принципы организации нервной системы
51
❖
которые тем не менее могут быть незначительно изменены в результате лич-
ного опыта конкретного животного. Это принципиальный шаг в эволюции
нервной системы, поскольку способность к обучению и индивидуальному из-
менению поведения является мерой интеллектуальных способностей живот-
ных и человека.
РЕПТИЛИИ (ПРЕСМЫКАЮЩИЕСЯ)
При первом взгляде на пресмыкающихся становится заметно, что они от-
личаются от амфибий структурой кожи. Она всегда образует твердые роговые
чешуи, иногда с кожными окостенениями внутри. Обычно конечностей две
пары, хотя встречаются виды с одной передней парой или только с одной
зачаточной парой конечностей. У змей и безногих ящериц ног нет вовсе.
К ныне живущим рептилиям принадлежат ящерицы, хамелеоны, змеи, чере-
пахи и крокодилы. Эта обширная группа животных освоила огромные терри-
тории благодаря своей независимости от воды в период размножения.
Нервная система у рептилий развита значительно лучше, чем у амфибий.
Спинной мозг имеет цилиндрическую форму с плечевым и поясничным утол-
щениями, соответствующими конечностями. Сзади спинной мозг продолжа-
ется, постепенно сужаясь, в начало хвостового отдела позвоночника.
Головной мозг у рептилий, особенно у крокодилов и черепах, более широк
и массивен, чем у амфибий. Для мозга рептилий характерно изгибание вниз в
области продолговатого мозга и стремление крупных полушарий переднего
мозга надвинуться на лежащие позади промежуточный и средний мозг (рис. 25).
Полушария переднего мозга без резких границ переходят в обонятельные лу-
ковицы, от которых начинаются обонятельные нервы (I). Увеличение разме-
ров полушарий переднего мозга связано с появлением зачатков коры — одно-
го или нескольких слоев клеток, расположенных под поверхностью мозга.
В этой зоне переднего мозга осуществляется вторичный анализ информации,
поступающей от органов обоняния. Промежуточный мозг почти совсем зак-
рыт сверху передним мозгом, так что на поверхность высовывается только
париетальный (парапинеальный) орган. У многих ящериц этот орган сохраня-
ет строение глаза. Как и другие позвоночные, рептилии имеют воронку, пере-
ходящую в гипофиз. Средний мозг представлен в виде хорошо развитого дву-
холмия, которое выполняет функции зрительных центров. Однако у змей зри-
тельное двухолмие несколько редуцируется и впервые появляются небольшие
парные задние бугорки — среднемозговые центры слуха (у рыб они обслужи-
вали органы боковой линии). Двигательные отделы мозга рептилий представ-
ляют собой крупное образование с почти закрытым IV желудочком. Он при-
крыт сверху мозжечком, который может иметь вид полоски, треугольника,
овала или круга, как у крокодилов. У последних можно выделить центральный
и два боковых отдела мозжечка.
Черепно-мозговые нервы представлены 11 парами, которые преимуществен-
но обособлены друг от друга. Нервы, часто сливающиеся с тройничным и блуж-
дающим нервами, выходят из мозга рептилий по отдельности. Только у змей
лицевой нерв иногда тесно связан с узлом тройничного нерва. Обонятельный
52
Сравнительная анатомия нервной системы позвоночных
Рис. 25. Строение головного мозга рептилий
а — геккон (Gekko gekko); б—сетчатый питон (Python reticularis)', в — хамелеон (Chamaeleo
lateralis)', г — красноухая черепаха (Pseudemys skripta). I-XII — черепно-мозговые нервы,
зачернен IV желудочек, во — вомероназальный нерв.
I. Принципы организации нервной системы
53
нерв (I) снабжает своими ветвями обонятельный мешок и якобсонов орган.
Зрительные нервы (II) образуют в основании черепа перекрест, называемый
хиазмой. При этом у яшериц волокна одного зрительного нерва проходят сквозь
щель другого. Лицевой нерв (VII) представляет небольшую слаборазвитую ветвь.
Блуждающий нерв (X) является одним из наиболее длинных черепно-моз-
говых нервов. Это обусловлено тем, что сердце, желудок и легкие сильно уда-
лены от головы. К блуждающему нерву прилежит добавочный нерв (XI), а
подъязычный нерв (XII), начинаясь несколькими корешками от передней ча-
сти спинного мозга, выходит из черепа через одно или несколько специаль-
ных отверстий.
Симпатическая нервная система представлена у рептилий симпатическим
нервом, который в голове образует соединение с ветвями многих черепных
нервов. У крокодилов в шейном отделе он разделяется на два тяжа. Один из
них лежит в позвоночном канале, образованном раздвоенными основаниями
ребер, а другой — на нижней поверхности шейных позвонков. У змей шейная
и туловищная части симпатического нерва слабо развиты.
Органы чувств рептилий обладают рядом уникальных особенностей, что свя-
зано с ороговением покровов. Несмотря на ороговение, в чешуйках кожи мно-
гих рептилий найдены осязательные пятна, а под чешуйками существует разви-
тая сеть осязательных телец с нервными окончаниями. Огромное значение в
качестве осязательного органа имеет язык, который у многих змей и ящериц
постоянно находится в движении. Вкусовыми органами служат луковицеобраз-
ные тельца в слизистой стенке языка и нёба. Орган обоняния у рептилий пред-
ставлен двумя системами: основной (обонятельный эпителий носовых мешков)
и добавочной (якобсонов орган). Носовая полость парная, сообщается с рото-
вой и глоточной полостями через хоаны. У ящериц и змей часто встречается
парная полость, лежащая книзу от носовой и открывающаяся особым отверсти-
ем в ротовую полость. На дне полости расположены хемочувствительные кар-
маны — якобсонов орган, в который рептилии вкладывают раздвоенный на
конце язык. Слуховой аппарат рептилий состоит из внутреннего и среднего, а
иногда и из зачатков наружного уха. Лабиринт более развит, чем у амфибий, а у
крокодилов уже появляются признаки улитки. Глаза рептилий обычно несколь-
ко крупнее по отношению к голове, чем у амфибий. Глазное яблоко шаровид-
ной формы, а вокруг роговицы у многих ящериц и черепах формируется скле-
ротикальное кольцо из костных пластинок. Аккомодация у большинства репти-
лий достигается при помощи мышцы, расположенной вокруг хрусталика.
Сокращение этого мускула сжимает хрусталик, делая его толще, и тем самым
дает возможность рассматривать близко расположенные предметы. Только у
змей аккомодация происходит за счет перемещения хрусталика, как у амфибий.
Кроме того, у некоторых змей имеются терморецепторы очень высокой чув-
ствительности. Они могут улавливать инфракрасные волны от мыши, темпе-
ратура тела которой выше, чем температура окружающей среды, с расстояния.
15 см. Так как терморецепторы расположены в лицевых ямках, гремучие змеи
могут определять направление источника излучения.
В колбочках рептилий и птиц находятся специальные цветные масляные
капли, которые сужают кривые спектральной чувствительности рецепторов и
уменьшают их взаимное перекрытие. Масляные капли в 2—5 раз увеличивают
54
Сравнительная анатомия нервной системы позвоночных
❖
число различимых цветовых оттенков у этих животных. Многие черепахи об-
ладают хорошим цветовым зрением. В сетчатке одного из исследованных ви-
дов (Pseudemys scripta) было найдено несколько типов фоторецепторов, отве-
чающих за восприятие цвета и освещенности: палочки, красные колбочки,
красно-зеленые двойные колбочки, одиночные красные и зеленые колбочки.
Все эти фоторецепторы работают в различных условиях освещенности фона и
объекта, что позволяет черепахам использовать 4 механизма цветовосприятия.
Поведение рептилий построено на врожденных реакциях, которые значитель-
но индивидуализируются в процессе жизни. Надо отметить, что продолжительность
приобретения индивидуального опыта у рептилий довольно велика. Известно,
что крупные черепахи живут 150—250 лет, змеи и ящерицы — 7—25 лет, крокоди-
лы — 70—80 лет. У рептилий очень большую роль играет обучение. Многие из
них легко приручаются и быстро осваивают лабиринт. Черепахи могут узнать
фигуру, нарисованную сплошной линией, даже если она очерчена пунктиром
или повернута на небольшой угол. Для ящериц характерно некоторое прогно-
зирование развития событий. Это позволяет им преследовать добычу уже пос-
ле того, как она скрывается из виду. Интересно отметить, что водяные ужи,
обученные правильно проходить Т-образный лабиринт, после линьки все за-
бывают. Аналогично ведет себя шипохвост, который, привыкнув к новой тер-
ритории, забывает ее после кратковременного охлаждения.
Для рептилий характерна иерархическая организация компактных групп.
Доминирующей игуане уступают место не только на своей территории, но
также и на соседних, когда она направляется на охоту в общие нейтральные
зоны. Иерархия у гигантских черепах Галапагосских островов выражается в
строго установленном порядке следования друг за другом во время миграции.
Групповая организация рептилий является одной из сложных форм обществен-
ного поведения, которая была достигнута у холоднокровных животных.
ПТИЦЫ
Птицы отличаются от остальных позвоночных тем, что они имеют крылья,
покрыты перьями и способны поддерживать постоянную температуру тела. Это
яйцекладущие животные, которые по строению центральной нервной системы
весьма сходны с рептилиями. Птицы являются высокоспециализированными
животными, приспособленными к обитанию в воздушной среде, и почти каж-
дая часть их организма адаптирована к столь необычным условиям. Приспособ-
лениями к палету являются хорошая теплоизоляция, оперение, превращение
передних конечностей в крылья, видоизменение грудины и плечевого пояса для
крепления больших мышц крыла, а тазового пояса с задними конечностями —
для поддержания массы тела на земле, а также усовершенствование дыхатель-
ной системы, которая обеспечивает наивысшую температуру тела среди всех
животных. Кроме этого, у птиц отсутствуют зубы, утрачены левая дуга аорты и
правый яичник с яйцеводом. Этих особенностей строения достаточно для полу-
чения ясного представления о том, что птицы являются высокоспециализиро-
ванной группой позвоночных. С другой стороны, организация современных
птиц в наиболее существенных чертах довольно однообразна.
I. Принципы организации нервной системы ❖ 55
Строение мозга у птиц значительно менее разнообразно, чем у рыб, амфи-
бий и рептилий. Головной мозг заполняет полость черепа и имеет округлую
форму при крайне малой длине (рис. 26). Самый крупный из отделов головно-
го мозга передний мозг. Он состоит из двух полушарий, которые имеют глад-
кую поверхность или слабо обозначенные продольные углубления (см. рис. 26).
Эти углубления не являются бороздами, а отражают границы слоев неостриа-
тума. Только у попугаев можно заметить небольшое морфологическое обособ-
ление височной доли полушарий. Основной объем полушарий занимают под-
корковые ядра, тогда как кора имеет рудиментарное строение и занимает не-
большую часть верхней стенки мозга. Роль коры млекопитающих выполняет
неостриатум, который развивается из гомологичного участка стенки полуша-
рия. Полушария переднего мозга простираются назад до контакта с мозжеч-
ком. Поэтому парные выпячивания среднего мозга до такой степени сдвинуты
наружу, что лежат по бокам.
Следствием разрастания переднего мозга назад, а мозжечка вперед является
то, что промежуточного мозга снаружи совсем не видно, хотя его можно опре-
делить по выросту эпифиза. Эпифиз у птиц развит незначительно, тогда как
гипофиз имеет достаточно крупные размеры. Средний мозг сильно развит, но
из-за своего бокового расположения имеет нехарактерную внешнюю морфо-
логию. Мозжечок состоит из массивной средней части (червя), пересекаемой
обычно 9 извилинами, и двух небольших боковых долей, которые гомологич-
ны клочку мозжечка млекопитающих.
Рис. 26. Внешнее строение головного мозга птиц.
а — обыкновенная индейка (Meleagris gallopavo); б — серый гусь (Anser anser); в — бра-
зильский ябиру (Jabiru myeterica); г — обыкновенная зеленушка (Chloris chloris).
56
Сравнительная анатомия нервной системы позвоночных
❖
Продолговатый мозг относительно мал, его нижняя сторона в еще большей
степени, чем у рептилий, образует изгиб вниз, а в области промежуточного
мозга наблюдается изгибание вверх. Спинной мозг птиц очень хорошо развит.
Он образует большие утолщения в плечевой и поясничной области, откуда
отходят нервы передних и задних конечностей. В поясничном утолщении верх-
няя стенка спинного мозга расходится и центральный канал расширяется в
ромбовидный синус, покрытый сверху только мозговыми оболочками. Спин-
номозговые нервы соединяются у птиц корешками еще в канале позвоночно-
го столба и выходят между дугами или через отверстия слившихся позвонков.
В плечевом сплетении обычно участвуют 4 нерва, реже — 3, а у страусов —
только 1 нерв. Для управления задними конечностями у птиц существует два
сплетения: поясничное и седалищное, или крестцовое. Поясничное сплетение
обычно состоит из 3, а у страусов может включать и 5 нервов. Седалищное
сплетение у всех птиц состоит из 4 крестцовых нервов.
Черепно-мозговые нервы птиц представлены 12 парами, которые начина-
ются отдельно. Обонятельный нерв (I) идет от нижней поверхности обоня-
тельной доли, которая спереди и снизу примыкает к полушариям переднего
мозга. Зрительные нервы (II) после хиазмы плавно переходят в средний мозг,
а блуждающий нерв, как и у рептилий, идет из черепа далеко назад, иннерви-
руя сердце, легкие, пищевод и желудок. В отличие от рептилий добавочный
нерв (XI) представлен тонкой веточкой блуждающего нерва, а подъязычный
(XII) нерв имеет корешки, отходящие как от продолговатого, так и от спинно-
го мозга.
Симпатическая нервная система птиц построена по общему для всех по-
звоночных принципу. Однако отличительной чертой ее строения у птиц яв-
ляется расположение шейной части большого симпатического нерва, часто
называемого пограничным стволом. Он лежит в канале, образованном дву-
корневыми началами поперечных отростков. Участок этого нерва прилежит
к сонным артериям, а далее кзади симпатический нерв находится в канале,
образуемом головками и бугорками ребер, и только в поясничной области он
лежит свободно.
Строение органов чувств птицы несколько отличается от такового осталь-
ных животных. Осязание многих птиц (кулики, утки) сосредоточено в челю-
стном аппарате, в котором расположены специализированные механорецеп-
торы (тельца Гранди и Хербста). Осязательные тельца расположены в коже у
основания крупных перьев и сконцентрированы в восковице, которая по-
крывает основание клюва. Совы обладают особыми осязательными перьями,
расположенными вокруг клюва. У дятлов, дроздов, попугаев и глухарей ося-
зательные функции выполняет язык, весьма богатый нервными окончания-
ми. У птиц с хорошо развитым языком вкусовые рецепторы играют роль в
выборе пищи. Зрение птиц очень хорошо развито. Глаз способен к аккомо-
дации и обладает рядом морфологических особенностей, позволяющих пти-
цам эффективно ориентироваться, успешно охотиться и издалека определять
качество пищи. У птиц отлично развит слух. Орган слуха состоит из внутрен-
него, среднего и зачатков наружного уха. Птицы могут воспринимать сигна-
лы различной частоты. В этом отношении они намного превосходят многих
млекопитающих.
I. Принципы организации нервной системы
57
❖
Поведение птиц крайне сложно и разнообразно. Они заботятся о потомст-
ве, совершают перелеты на расстояния в тысячи километров, имеют сложное
брачное поведение, строят гнезда или специальные устройства для выведения
птенцов, обучают молодых птиц и сохраняют способности к обучению на про-
тяжении всей жизни. Птицы обладают хорошей памятью и могут осуществ-
лять несложную инструментальную деятельность. Они легко обучаются ре-
шать простые задачи и подражать человеческому голосу. «Слова», произноси-
мые птицами, являются эмоциональным символом настроения, в котором они
их запомнили, а не результатом рассудочной деятельности.
В отличие от всех рассмотренных выше групп животных птицы любят иг-
рать друг с другом или с различными предметами. Мозг птиц обеспечивает
возможность предвидения вероятных событий. Они рассчитывают свое пове-
дение на несколько ходов вперед, что уже характерно для высших млекопита-
ющих и человека.
МЛЕКОПИТАЮЩИЕ
Млекопитающие являются высшими из позвоночных животных. Они тепло-
кровны и выкармливают своих детенышей молоком. Все млекопитающие всю
жизнь дышат воздухом при помощи легких, хотя многие из них — водные жи-
вотные. Тело млекопитающих в большинстве случаев четко разделяется на го-
лову, туловище, хвост и четыре конечности. У китообразных и сиреновых раз-
виты только передние конечности, а у некоторых приматов и человека отсут-
ствует хвост. В группу млекопитающих входят однопроходные (утконос и ехидна),
сумчатые (американские опоссумы, бандикуты, вомбаты и кенгуру) и плацен-
тарные звери (грызуны, китообразные, копытные, хищные, насекомоядные,
приматы и т.д.). Масса головного мозга у млекопитающих очень различается
(табл. 1).
Таблица 1. Масса головного мозга млекопитающих
Вид Масса головного мозга, г
Мышь (грызуны) 0,4
Крыса (грызуны) 1,6
Морская свинка (грызуны) 4,8
Кролик (зайцеобразные) 9,3
Кошка (хищные) 31,0
Собака (хищные) 65,0
Макака-резус (приматы) 88,0
Корова (парнокопытные) 340,0
Шимпанзе (приматы) 355,0
Горилла(приматы) 940,0
Человек (Homo sapiens) 1350,0
Дельфин (китообразные) 1550,0
Слон (хоботные) 6075,0
Финвал (китообразные) 7200,0
Кашалот (китообразные) 9200,0
58 ❖ Сравнительная анатомия нервной системы позвоночных
Как видно из табл. 1, мозг млекопитающих может различаться по массе в
10 000 раз. Это разнообразие массы мозга соответствует разнообразию разме-
ров животных. Однако каждый вид весьма неоднороден. Он состоит из подви-
дов и пород, которые имеют существенные различия как по размерам тела,
так и по объему мозга.
Примером таких различий могут служить собаки. Их мозг может различаться
по массе в несколько раз. Самая маленькая масса мозга собаки, которая была
описана в литературе, составляла 41,5 г, а самая большая — 212 г (Н.И. Петро-
павловский, 1904). Ниже представлена внутривидовая изменчивость массы мозга
собак (в граммах):
Минимальная масса мозга — 41,5
Максимальная масса мозга — 212,0
Средняя масса мозга беспородных собак — 73,2
Средняя масса мозга породистых собак — 92,8
Средняя масса мозга сук — 76,9
Средняя масса мозга кобелей — 85,0
Весьма показательно отношение массы мозга и массы тела. Минимальное
отношение 0,2 выявлено у крупных собак, масса тела которых составляет
28—44 кг. Для собак массой менее 2,5 кг оно увеличивается до 2,9. Следова-
тельно, масса мозга по отношению к массе тела уменьшается с увеличением
размеров животного. Эта закономерность не наблюдается в период развития
щенков. У щенков и взрослых собак одной и той же породы отношение
массы мозга к массе тела практически одинаково. Это отличает индивиду-
альное развитие мозга собак от такового обезьян и человека. У обезьян и
человека плод рождается с большим мозгом и маленькой массой тела, а за-
тем масса тела увеличивается намного быстрее, чем масса мозга. Головной мозг
подвержен половому диморфизму. У самок масса мозга обычно на 10—12 г мень-
ше, чем у самцов. Тем не менее суки обычно мельче, чем кобели, что замет-
но влияет на относительную массу мозга и тела. У сук относительная масса
мозга больше, чем у кобелей.
Весьма любопытны различия породистых и беспородных собак. Средняя
масса мозга в выборках беспородных собак колеблется от 42 до 118 г, а в
группе породистых — от 41,5 до 212 г. Искусственно выведенные породы име-
ют большую амплитуду изменчивости головного мозга, чем беспородные со-
баки. По-видимому, у одичавшей в Австралии дикой собаки Динго мозг имеет
еще большую однородность по массе, чем у беспородных городских собак.
Возврат к природным формам уменьшает искусственно выведенное разнооб-
разие. Надо отметить, что у породистых собак относительная масса мозга по-
чти в два раза меньше, чем у беспородных.
Центральная нервная система у млекопитающих развита больше, чем у ка-
кой-либо другой группы животных. Основными отличиями строения мозга мле-
копитающих от мозга рептилий и птиц являются крупные парные полушария
переднего мозга, которые у высших зверей формируют борозды и извилины
Крупные полушария млекопитающих исторически возникли из боковой части
стенки переднего мозга рептилий в результате разрастания поверхностного
скопления клеток, которое носит название плаща или коры. У высших млеко-
I. Принципы организации нервной системы
59
❖
питающих новая кора состоит из 6 четко выделяемых слоев, толщина которых
у высших приматов доходит до 100 клеток. Общее число нейронов в коре
переднего мозга человека составляет 11 млрд клеток. В коре млекопитающих
продублированы почти все основные функции нервной системы на уровне
заднего, среднего, промежуточного и переднего мозга.
Кора подразделяется на поля — зоны, в которых представлен конкретный
периферический анализатор или моторная система. Примером может служить
затылочная кора человека, которая ответственна за зрение и дублирует функ-
ции среднего мозга низших позвоночных. Наличие коры позволяет млекопи-
тающим точнее и детальнее анализировать сигналы, приходящие из внешней
среды и от внутренних органов, что лежит в основе сложного поведения и
мышления человека. Однако только улучшение анализа рецепторных сигна-
лов не может быть достаточным условием для становления сложного поведе-
ния. Необходима глубокая интеграция всей информации, поступающей в мозг.
Эти функции выполняет кора за счет связей между расположенными в ней
нейронами. Подсчитано, что на каждый мотонейрон приходится до 2000 вста-
вочных нейронов, которые позволяют согласовывать активность отдельных
клеток. Таким образом, увеличение размеров коры и формирование ее слои-
стой структуры позволили повысить качество обработки поступающей в мозг
информации и интегрировать активность отдельных его структур. На этом
морфологическом субстрате возникло новое качество мозга — способность к
синтетической деятельности.
Полушария переднего мозга млекопитающих разрастаются вбок и кзади,
покрывая собой промежуточный мозг, часто также средний мозг, а у человека
и продолговатый мозг (рис. 27). Полушария переднего мозга имеют непра-
вильную овальную форму и более или менее расширены сзади. У многих мле-
копитающих, принадлежащих к самым разнообразным группам (насекомояд-
ные, грызуны, зайцеобразные), полушария гладкие, без признаков борозд и
извилин. У большинства высших млекопитающих в коре формируются бороз-
ды и извилины (см. рис. 27). Однако извилины бывают в различной степени
развиты даже в пределах одного отряда. Низшие приматы практически лише-
ны борозд и извилин, тогда как у высших приматов их строение значительно
сложнее (рис. 28). С увеличением размеров полушарий непосредственно свя-
зано появление мозолистого тела — обширной зоны волокон, соединяющих
левое и правое полушария. Это образование характерно для мозга млекопита-
ющих. Промежуточный мозг полностью скрыт полушариями переднего мозга.
В нем хорошо развит эпифиз, а воронка, как и у всех позвоночных, заверша-
ется крупным гипофизом. Он выполняет преимущественно нейроэндокрин-
ные функции, в частности для регуляции сезонной, суточной и половой ак-
тивности млекопитающих.
Средний мозг представляет собой плотное небольшое тело в виде двух пар
возвышений, которые носят название четверохолмия. У рептилий и амфибий
крыша среднего мозга состоит из двух возвышений, связанных с сетчаткой.
У млекопитающих они отчасти сохраняют свои функции и расположены в
передней зоне четверохолмия. Появление задних бугорков среднего мозга связа-
но с развитием слухового аппарата млекопитающих. Задние бугорки среднего
мозга являются слуховыми центрами и отражают уровень развития слуховой
60
❖
Сравнительная анатомия нервной системы позвоночных
Рис. 27. Разнообразие внешнего строения головного мозга млекопитающих. У низших мле-
копитающих головной мозг совершенно лишен борозд и извилин, тогда как у китообразных
их может быть значительно больше, чем у человека.
а — кролик; б — кенгуру; в — волк; а — дельфин (афалина).
системы у конкретного вида. По соотношению бугорков четверохолмия мож-
но судить о значении для животного слуха и зрения. У копытных и хищных
наиболее развиты передние бугорки четверохолмия, которые связаны со зре-
нием, а у китообразных, рукокрылых и утконоса преобладают задние бугорки.
Однако крыша среднего мозга у зверей занимает второстепенное положение в
обработке как зрительной, так и слуховой информации. Роль ведущих цент-
ров начинают выполнять специализированные поля новой коры.
Задний мозг млекопитающих хорошо развит и имеет крупный мозжечок.
В отличие от рептилий мозжечок млекопитающих состоит из трех частей: цент-
ральной части, или червя, и двух боковых полушарий. У сумчатых, грызунов и
насекомоядных размер червя не уступает каждому из полушарий, тогда как у
приматов и хищных по размерам доминируют полушария мозжечка. Нужно
подчеркнуть, что мозжечок не является ни сенсорным, ни моторным образо-
ванием. Он стоит в стороне от главных входных и выходных путей и служит
структурой, координирующей двигательную активность с нервной деятельно-
стью других отделов головного мозга.
Внутри головного мозга находится сложная система мозговых желудочков.
Полость спинного мозга переходит в ромбовидную ямку продолговатого моз-
га, которая носит название IV мозгового желудочка. Его передний край при-
I. Принципы организации нервной системы
❖
61
Рис. 28. Разнообразие латеральной и медиальной поверхностей полушарий переднего
мозга обезьян. Слева — наружная латеральная поверхность левого полушария; справа —
медиальная поверхность правого полушария, левое полушарие удалено.
а — лемур (Lemur variegatus); б — капуцин (Cebus apella)-, в — павиан (Cynopithecus niger)',
г — орангутанг (Pongo pygmaeus).
крыт у большинства млекопитающих мозжечком. Кпереди от мозжечка IV
желудочек переходит в сильвиев водопровод — канал, проходящий под сред-
ним мозгом и ведущий в III желудочек. III желудочек представляет собой
довольно узкое пространство между внутренними стенками промежуточного
мозга. Спереди через монроевы отверстия он сообщается с двумя боковыми
желудочками. I и II желудочки лежат внутри полушарий переднего мозга. Впе-
реди III мозгового желудочка вследствие местного срастания полушарий вок-
руг треугольного пространства образуется так называемый пятый мозговой
желудочек. Этот мозговой желудочек свойствен только млекопитающим.
Черепно-мозговые нервы представлены 12 парами. Обонятельный (I) нерв
обычно выходит из более или менее крупной обонятельной луковицы в виде
отдельных небольших волокон.
Только у утконоса он представляет целый ствол, а у дельфинов и китов
отсутствует. Зрительные нервы (II) образуют сложный перекрест, а тройнич-
ный нерв (V) обычно выходит из черепа отдельными ветвями, которые разли-
чаются по функциям. Лицевой нерв (VII) млекопитающих сильно развит, так
как является двигательным нервом мимической мускулатуры. Блуждающий,
языкоглоточный и добавочный нервы (IX—XI) выходят из черепа совместно.
Блуждающий нерв, как и у птиц, простирается далеко в туловище, иннерви-
руя внутренние органы.
62 ❖ Сравнительная анатомия нервной системы позвоночных
Симпатическая нервная система хорошо развита и образует много сплете-
ний. Самое крупное из них находится в передней части брюшной полости
позади желудка. Большой симпатический нерв образует парную цепь ганглиев
по обеим сторонам позвоночника, которая продолжается до области тройнич-
ного нерва и формирует ганглии для его первой, второй и третьей ветвей.
В шейной области ганглии немногочисленны, нервы формируют редкую сеть.
Органы чувств у млекопитающих более развиты, чем у всех остальных позво-
ночных. Для осязания служит кожа, богатая свободными нервными окончани-
ями и специализированными рецепторами тепла, холода, вибрации и давления.
У млекопитающих хорошо развит орган обоняния. В большинстве случаев он
состоит их двух частей: обонятельного эпителия, приспособленного для вос-
приятия большинства запахов, и якобсонова органа, который используется для
рецепции половых запахов. Слуховая система развита благодаря усложнению
строения улитки и развитию коры. У млекопитающих улитка впервые закручи-
вается в спираль, которая у китообразных делает 1,5 оборота, у человека — 2,5,
у кошки — 3, у свиньи — 4, а у некоторых грызунов — даже 5 оборотов. Способ-
ностью воспринимать электрические сигналы обладают водные позвоночные
животные. Ответы электрорецепторов клюва утконоса были обнаружены при
изменении напряженности электрического поля. Утконос может воспринимать
не только напряженность, но и частотные характеристики электромагнитного
поля. Эти особенности позволяют ему находить пищу в непрозрачной воде.
Зрение млекопитающих позволяет им различать цвет и форму объектов намно-
го лучше, чем животным других классов. Это достигается как сложностью стро-
ения глаза, так и наличием корковой части зрительного анализатора. Цветовым
зрением обладает большая часть млекопитающих, хотя у видов, ведущих суме-
речный образ жизни, цветовое восприятие понижено.
Сложность поведения млекопитающих не нуждается в подробном иллюст-
рировании. Большинство млекопитающих могут прогнозировать результат своих
действий, выражать эмоции, играть, устанавливать иерархические отношения
в группе, обучаться и решать сложные экспериментальные задачи. Коллектив-
ные действия волков, крыс и копытных, забота о потомстве, межвидовой аль-
труизм и заметные индивидуальные особенности поведения конкретных осо-
бей приближают млекопитающих к человеку. Морфологические различия между
человеком и высшими млекопитающими не носят принципиального характе-
ра. Мозг писателя А. Франса весил 1020 г, а мозг средней гориллы — 940 г
Извилин у дельфинов примерно вдвое больше, чем у человека, при равной
массе тела. Толщина коры ряда грызунов намного больше, чем у человека и
обезьян, хотя мозг полностью лишен извилин. Тем не менее только челове-
ческий мозг способен создавать то, чего до него не было в природе.
II. СРАВНИТЕЛЬНАЯ ЭМБРИОЛОГИЯ
НЕРВНОЙ СИСТЕМЫ
Проблема происхождения нервных клеток и их пред-
шественников имеет большое значение для понимания
причин возникновения разнообразия форм и типов орга-
низации мозга. По существу вопрос о происхождении
клеток нервной системы является одной из сторон про-
блемы детерминации формообразования головного моз-
га. На процессы детерминации развития нервной си-
стемы существует по меньшей мере два взгляда: с од-
ной стороны, это представления о мозаичности развития
нервной системы, а с другой — регуляционная модель
морфогенеза мозга.
Мозаичный тип развития предусматривает детерми-
нацию судеб потомков клеток-предшественниц на про-
тяжении всего онтогенеза мозга вплоть до его оконча-
тельной дифференцировки. Согласно мозаичной моде-
ли детерминация начинается на стадиях развития,
представляющих собой 32- и 64-клеточные бластулы
(морулы). На этих стадиях выделяются один или два
первичных бластомера, которые станут в дальнейшем
предшественниками всех клеток нервной системы.
Пройдя одно или два деления, бластомеры формируют
первичную группу клеток-предшественниц. Каждая из
них дает начало одному клону клеток (т.е. группе кле-
ток общего происхождения). На этой стадии развития
вся нервная система состоит из нескольких закономерно
расположенных клонов клеток. Клоны не смешиваются
друг с другом и имеют выраженные границы. В этих
границах заложена структурализация мозга на поздних
эмбриональных и дефинитивных стадиях. Однако в про-
цессе реального развития мозга существуют миграции
клеток, которые учитываются в мозаичной модели мор-
фогенеза.
Для объяснения этих явлений вводится представле-
ние о компартментах. Компартмент клеток отличается
от клона только тем, что клетки, расположенные на его
64 ❖ Сравнительная анатомия нервной системы позвоночных
границах, могут перемешиваться с клетками соседних компартментов. Несмотря
на перемешивание, клетки в компартменте на 80% состоят из потомков одной
клетки-предшественницы. Компартмент, по мозаичной модели развития, яв-
ляется не просто совокупностью топологически объединенных потомков од-
ной клетки, но и функциональной единицей. При формировании нервной
трубки клетки компартмента создают определенную область мозга или функ-
циональную структуру. Это может быть любая из областей мозга, в которой
клетки объединены анатомически, функционально и по происхождению. Счи-
тается детерминированным не только сам компартмент, но и входящие в него
клетки. Следовательно, мозаичная модель развития мозга предусматривает то-
пографическую детерминацию и его функциональную дифференцировку.
Мозаичная модель успешно объясняет и образование первичного паттерна
нервных связей. Известно, что первые нервные связи устанавливаются между
самыми удаленными друг от друга структурами. На основе первичных нервных
связей формируются вторичные, которые объединяют между собой близко рас-
положенные структуры. На самых поздних стадиях развития и дифференциров-
ки мозга возникают связи внутри отдельных структур, которые являются тре-
тичными. Эта схема сильно упрощена, но отражает дифференцировку мозга у
низших позвоночных. Последовательность перечисленных событий дифферен-
цировки хорошо объяснима, если допустить, что первые связи устанавливаются
между самыми дальними «родственниками» клеток из разных компартментов.
Тогда сначала возникнут связи между компартментами, наиболее удаленными
друг от друга. На втором этапе установятся связи между соседними компарт-
ментами, а затем между клетками внутри одного компартмента. Таким образом,
допустив, что связи в мозге устанавливаются в той же последовательности, в
какой формируются дочерние нейробластомеры, клоны и компартменты, мы
получим целостную и формально непротиворечивую модель раннего эмбрио-
нального формообразования и дифференцировки головного мозга.
Другая точка зрения предусматривает значительно меньшую детерминиро-
ванность судеб отдельных клеток. Основой регуляционной модели развития
мозга являются мультифункциональность клеток-предшественниц и наличие
самой общей детерминации нейральных зачатков. В этом случае предусматри-
вается детерминация только направления нейральной дифференцировки тка-
ни, а не судеб отдельных клеток. Судьба клеток нейрального зачатка зависит
от того, в какие коммуникационные отношения они вступят друг с другом в
процессе эмбрионального формообразования. Регуляционная модель подра-
зумевает, что судьба клетки в известной степени случайна. Она изменится,
если клетка переместится в нейральном зачатке. Следовательно, судьба клет-
ки в каждый конкретный момент времени зависит от ее топографического
положения и коммуникационных взаимодействий с соседями. Таким обра-
зом, параметром, ограничивающим степень изменчивости клетки, является ее
топологическая принадлежность к той или иной части нейрального зачатка.
Например, нейроэпителиальная клетка нервной пластинки, расположенная в
центре зачатка презумптивного среднего мозга, не может стать клеткой пере-
днего или заднего мозга, но внутри среднего мозга она может дифференциро-
ваться в любой нейрон. Безусловно, на стадии гаструлы эти возможности у
клеток значительно шире. С точки зрения регуляционной модели судьба клеток
II. Сравнительная эмбриология нервной системы ❖ 65
зависит от тех формообразовательных процессов, в которых они участвуют.
Каждая клетка нейрального зачатка детерминирована настолько, насколько
она топологически связана с развитием конкретных структур. В регуляцион-
ной модели судьба клетки зависит от нескольких постоянно изменяющихся
параметров. С одной стороны, это топография клетки в развивающемся зачат-
ке, а с другой — весь комплекс межклеточных взаимодействий. Согласно ре-
гуляционной модели, любая клетка может начать пролиферировать или выйти
в дифференцировку, если ее позиционная информация будет соответствовать
началу этого процесса. В этом случае первичный паттерн нервных связей ус-
танавливается не по генеалогическому принципу, а между клетками, реализу-
ющими одинаковую позиционную информацию.
В основе различий мозаичной и регуляционной моделей лежит отношение
к продолжительности детерминированного развития. Мозаичная модель под-
разумевает, что судьба клетки предопределена уже на стадиях дробления и
гаструлы. Все остальное время развития клетка реализует полученную инфор-
мацию. Другая ситуация в регуляционной модели. Здесь предполагается, что
на непродолжительное время детерминирована только исходная группа кле-
ток, которая будет дифференцироваться в головной мозг, а конкретная ин-
формация о дифференцировке структур мозга появляется во время взаимо-
действия клеток в развитии. Причем информация о дальнейшем развитии воз-
никает дискретными порциями. Клетка на ранних стадиях развития еще «не
знает» своей окончательной судьбы, а обладает программой действий только
на несколько часов. Следовательно, регуляционная модель отличается от моза-
ичной прерывистостью детерминации.
Таким образом, к настоящему времени сложились две концепции развития
нервой системы: мозаичная и регуляционная. Обе модели имеют свои поло-
жительные и отрицательные стороны, однако окончательный ответ на вопрос
о том, какая из них в большей степени отражает реальный механизм развития
нервной системы, дали только прямые наблюдения за судьбами клеток нерв-
ной системы.
ПЕРВИЧНАЯ ДИФФЕРЕНЦИРОВКА
КЛЕТОК НЕРВНОЙ СИСТЕМЫ
Попытки определить эмбриональную локализацию клеток-предшественниц
предпринимались давно. Первые удачные эксперименты по маркировке жи-
вых клеток нервной системы были проведены Фогтом (W. Vogt, 1925, 1929) и
Карпентером (Е. Carpenter, 1932, 1937). Однако проследить судьбу отдельных
бластомеров от ранних стадий дробления до органогенеза не удавалось. Ос-
новной трудностью исследований было отсутствие хорошего витального мар-
кера. Маркер должен быть нетоксичен, хорошо сохраняться в клетке и выдер-
живать большое разведение. Первой удачной попыткой промаркировать от-
дельные бластомеры на стадии 32 клеток является исследование японских
авторов (О. Nakamura и К. Kishiama, 1971; О. Nakamura, 1978). Они вводили
внутриклеточно витальный краситель в поверхностные бластомеры зароды-
шей шпорцевой лягушки (Xenopus laevis). Меченые бластомеры и их потомки
3—5818
66 ❖ Сравнительная анатомия нервной системы позвоночных
сохраняли метку длительное время. Это позволяло исследовать меченые по-
томки бластомеров на 23-й стадии развития, которая у этого объекта соответ-
ствует периоду формирования хвостовой почки. Исследователи определяли
топографическое распределение меченых клеток на тотальных анатомических
разрезах, поскольку применяемый ими способ мечения исключал использова-
ние гистологических процедур. Тем не менее они показали, что бластомеры,
из которых формируется нервная система, локализованы преимущественно в
дорсальных частях бластулы (рис. 29).
Наиболее важным шагом в исследовании клеток-предшественниц было при-
менение Хирозе и Джекобсоном метода внутриклеточной маркировки нейро-
бластов (Y. Hirose и М. Jacobson, 1979—1984). Эти авторы начали свои иссле-
дования с анализа судеб отдельных бластомеров, меченных на ранних стадиях
дробления. Первые работы показали, что уже на 16-клеточной стадии строго
определенные клетки дают столь же строго детерминированные зачатки. Была
обнаружена высокая степень детерминации отдельных бластомеров и их по-
томков в формировании определенных органов зародыша и структур головно-
го мозга. В каждом эксперименте потомки меченых одинаковых бластомеров
занимали одно и то же положение в головном мозге. На основании хорошо
воспроизводимых экспериментов Джекобсон создал карту локализации кле-
ток-предшественниц на различных стадиях дробления зародышей шпорцевой
лягушки (рис. 30).
Сетчатка
Головные ганглии
Спинной мозг
Отоцисты
Дорсальная часть мозга
Обонятельная плакода
Нервный гребень
Хрусталик
Сетчатка
Головные ганглии
Дорсальный спинной мозг
Обонятельная плакода
Нервный гребень
Дорсальный мозг
Хрусталик
Отоцисты
Головные ганглии
Нервный гребень
Вентральный мозг
Вентральный
спинной мозг
Сетчатка
Головные ганглии
Нервный гребень
Мозг, спинной мозг
Обонятельная плакода
Хрусталик
Нервный гребень
Дорсальная часть
спинного мозга
Сетчатка
Нервный гребень
Головные ганглии
Дорсальный спинной мозг
Отоцисты
Хрусталик
Головные ганглии
Нервный гребень
Головной мозг
Спинной мозг
Отолиты
Нервный гребень
Дорсальная часть мозга
Дорсальный
спинной мозг
Рис. 29. Схема представительства в отделах нервной системы потомков бластомеров,
меченных на 16-клеточной стадии развития шпорцевой лягушки (по S. Moody, 1987). Полу-
жирным шрифтом отмечены отделы нервной системы, которые преимущественно состоят
из потомков меченого бластомера.
II. Сравнительная эмбриология нервной системы
❖ 67
Детерминативный характер мозаичного развития головного мозга был столь
очевиден, что в 1982 г. Джекобсон отверг существование индукционного вли-
яния хордомезодермы на нейроэктодермальный зачаток нервной системы.
С его точки зрения, в индукционных вза-
имодействиях нейроэктодермы и хордо-
мезодермы нет необходимости, посколь-
ку первая уже детерминирована на 64-
клеточной стадии. Отказавшись от идеи
существования индукционных процес-
сов, Джекобсон создал клональную мо-
дель развития головного мозга, сущность
которой состоит в следующем. Отдель-
ный бластомер является клеткой-пред-
шественницей, которая, пройдя несколь-
ко пролиферативных циклов, формирует
группу потомков — клон клеток. В про-
цессе развития потомки клеток одного
клона не смешиваются и составляют ог-
раниченные части морфологических
паттернов (рис. 31).
Паттерн, по Джекобсону, — это по-
томки клеток одного клона, которые со-
0 -mesencephalon A -rhombencephalon
О - prosencephalon □ - medulla spinalis
Рис. 30. Схема зародыша шпорцевой ля-
гушки на 32-клеточной стадии развития и
представительство потомков меченых
бластомеров в отделах нервной системы
(по М. Jacobson, 1985; S. Moody, 1987;
Н. Takasaki, 1987). Темные значки — по-
томки бластомера постоянно встречают-
ся в отделе мозга; светлые значки — по-
томки бластомера встречаются в отделе
не всегда.
ставляют отдел или структуру головно-
го мозга. С его точки зрения, на позд-
них стадиях развития различные клоны
дают различные типы клеток. Форми-
рование морфологических и гистологи-
ческих паттернов зависит от того, как
клетки в различных областях эмбриона
становятся запрограммированными и
вступают на различные пути развития.
Он считает, что программирование
включает в себя иерархическую серию переключений, которые приводят к
ограничению клеточной судьбы. При этом иерархическое переключение кле-
ток является в первую очередь результатом клеточных линий (последователь-
ных поколений потомков одной клетки-предшественницы), клеточного поло-
жения и в последнюю очередь результатом межклеточных взаимодействий
В 1984 г. Джекобсон признал роль индукционных процессов в формирова-
нии нервной системы, но включил их в свою модель как одно из второстепен-
ных звеньев в системе межклеточных взаимодействий. В его модели интерес-
на интерпретация глубинных источников ограничения клеточных судеб. Дже-
кобсон полагает, что клональное ограничение судеб клеток происходит
первично, из наследования бластомерами цитоплазматических компонентов,
которые регионально локализованы в яйце зародыша. Следовательно, модель
предполагает неравноценное распределение между бластомерами цитоплаз-
матических компонентов. По мнению Джекобсона, именно в этот момент клетки
приобретают индивидуальные качества, которые являются основой для меж-
68
❖
Сравнительная анатомия нервной системы позвоночных
Рис. 31. Представительство меченных на 512-клеточной стадии бластомеров шпорцевой
лягушки в нервной системе. Бластомеры и серийные срезы мозга с маркированными до-
черними клетками имеют одинаковые номера (по М. Jacobson, 1985).
клеточных взаимодействий. Межклеточные взаимодействия в свою очередь
приводят к прогрессивному ограничению судеб клеток и окончательной де-
терминации клеточных типов.
Джекобсон и Хирозе создали карты судеб отдельных бластомеров, которые
позволили установить локализацию клеток-предшественниц нервной систе-
мы на ранних стадиях дробления. Продолжая исследования, Джекобсон про-
анализировал судьбу отдельных бластомеров на 128-, 256-, 512-клеточной ста-
дии. Он показал, что границы между клонами значительно менее четкие и
воспроизводимые, чем представлялось ранее. К 1987 г. модель развития нерв-
ной системы Джекобсона претерпела ряд существенных изменений. Он пред-
положил, что границы клонов, а следовательно, и детерминации начинают
определяться у зародышей эмбрионов шпорцевой лягушки на 512-клеточной
стадии. Образование границ приводит к формированию морфологических ком-
партментов, которые являются популяцией потомков групп исходных клеток.
Компартменты в свою очередь формируют определенные структуры или отде-
лы головного мозга. Джекобсон допускал, что компартменты могут переме-
шиваться на своих границах, а их клетки могут участвовать в образовании
одной и той же структуры.
II. Сравнительная эмбриология нервной системы
69
❖
Понятно, что ответить на вопрос о степени перемешивания клеток из двух
соседних клонов с помощью только одного красителя нельзя. Для решения
этой задачи был применен метод маркировки бластомеров с помощью виталь-
ных люминесцентных красителей. В бластомеры шпорцевой лягушки, при-
надлежащие двум соседним клеточным клонам, вводились красители, кото-
рые дают красную и желто-зеленую люминесценцию. Положение меченых
клеток показало, что разделение клонов имело место в 46% случаев, тогда как
интенсивное перемешивание клеток наблюдалось в 54% случаев. Таким обра-
зом, введение контрастных красителей в бластомеры соприкасавшихся кло-
нов не выявило унимодального распределения меченых потомков. Следова-
тельно, положение модели Джекобсона о клональной ограниченности судеб
потомков меченых бластомеров не подтвердилось. Однако данные этих авто-
ров согласуются с гипотезой ограниченного развития компартментов в не-
рвной системе. Эта гипотеза подразумевает, что потомки клеток-предшествен-
ниц дают первично ограниченные клоны нейроэпителиальных клеток, кото-
рые в дальнейшем формируют морфологические компартменты. Компартменты
в свою очередь отвечают за формирование определенных структур мозга. От-
дельные компартменты могут частично перемешиваться друг с другом, но их
морфологическая органоспецифичность остается неизменной. Иначе говоря,
потомки клеток-предшественниц нервной системы могут формировать только
нервную систему и никакой другой орган. Таким образом, тканеспецифич-
ность отдельных бластомеров сочетается с полипотентностью их потомков уже
внутри нервной системы.
Для экспериментатора вопрос звучит следующим образом: могут ли потом-
ки одного бластомера с равной вероятностью появляться или исчезать в опре-
деленных структурах головного мозга разных зародышей? Если это так, то
детерминация клеток-предшественниц носит общий характер и определяется
межклеточными взаимодействиями на относительно поздних стадиях разви-
тия. Ответы на эти вопросы дали работы Муди (S. Moody, 1987), в которых
использовалось пероксидазное маркирование отдельных бластомеров шпор-
цевой лягушки на 32-клеточной стадии. Муди установила, что большинство
органов состоит из потомков клеток, происходящих от разных бластомеров
Потомки одного бластомера хотя и смешиваются с соседними клетками, но
все-таки имеют более или менее характерное положение в дефинитивном за-
родыше. Однако вариации в локализации меченых клеток были достаточно
часты и повторялись в каждом исследованном эмбрионе. Эти различия явля-
ются нелинейными и могут быть разбиты на несколько групп. В первую груп-
пу Муди включила случаи, когда две дочерние клетки могут давать равное
количество клеток в один орган. Для второй группы характерно такое распре-
деление клеток, когда потомки одной дочерней клетки могут составлять боль-
шую часть органа, который формируется из материнского бластомера. Для
третьей группы характерна локализация всех потомков дочерней клетки в орга-
не, возникшем из материнскогоо бластомера на 16-клеточной стадии.
Принадлежность нейроэпителиальной клетки к одной из групп еще не яв-
ляется детерминацией развития, поскольку потомки одних и тех же бластоме-
ров демонстрируют поливариантность своего распределения в органе. На рис. 30
показано распределение потомков бластомеров в головном мозге на стадии
70
о
Сравнительная анатомия нервной системы позвоночных
хвостовой почки, меченных на 32-клеточной стадии. Из этих данных видно,
что ряд бластомеров неизбежно преобразуется в клетки нервной системы.
Потомки других бластомеров могут с равной вероятностью локализоваться в
различных органах. Появление этих клеток в нервной системе непредсказуемо
и зависит от комбинации межклеточных взаимодействий, в которые они всту-
пят на поздних стадиях развития зародыша. Например, по данным Джекобсо-
на и соавт., потомки вентральных бластомеров на 32-клеточной стадии не
входят в нервную систему. Муди же показала, что они могут включаться в
состав спинного мозга зародыша, однако их включение является вероятност-
ным процессом: у одного зародыша они могут дифференцироваться в мото-
нейроны, а у другого вообще отсутствуют в спинном мозге.
Сходные исследования были проведены на рыбах и мышах. Важно, что в
них, как и в работах Муди, показана только общая направленность развития
отдельных бластомеров. Детальную детерминацию развития нервной системы
установить не удалось. По-видимому, путь жесткой детерминации развития
характерен только для беспозвоночных. Действительно, у пиявок и нематод
развитие бластомеров и их потомков полностью детерминировано. Можно пред-
положить, что в процессе эволюции нервной системы позвоночных первым
возник детерминативный тип развития, на что указывает детерминация судеб
бластомеров асцидии, которая сходна с морфогенезом беспозвоночных.
Подводя итог обзору происхождения предшественников нейроэпителиаль-
ных клеток, можно сказать, что нервная система развивается из 8 ростродор-
сальных бластомеров (см. рис. 29, 30) на 32-клеточной стадии развития заро-
дышей. Остальные бластомеры анимальной половины зародыша также при-
нимают участие в формировании нервной системы, однако их участие носит
вероятностный характер. Потомки почти любого бластомера из дорсальной
половины зародыша могут дифференцироваться в нервные клетки, если ока-
жутся в соответствующей системе межклеточных взаимодействий. Из вент-
ральных бластомеров могут формироваться участки нервной системы, но этот
процесс носит индивидуальный характер.
ФОРМИРОВАНИЕ НЕЙРОЭПИТЕЛИЯ
Нейроэпителий образуется из потомков клеток-предшественниц. Это уча-
сток эктодермы, который является предшественником нервной ткани. Начи-
ная со стадии дробления эти клетки пролиферируют, а их потомки формиру-
ют небольшую зону на анимальном полюсе зародыша (рис. 32). Она выглядит
как сектор сферы, расположенный между анимальным полюсом и дорсальной
губой бластопора.
Почти у всех видов хвостатых амфибий нейральный зачаток организован
одинаково (см. рис. 32, а). Весьма вероятно, что на стадии ранней гаструлы в
нейроэпителии хвостатых амфибий существует клональная ограниченность
между потомками отдельных клеток-предшественниц. С началом гаструляци-
онных движений пластов зародыша начинается активная перегруппировка
клеток презумптивного нейроэпителия. Он растягивается в краниокаудальном
направлении и сжимается по направлению к оси зародыша. Краниальная гра-
II. Сравнительная эмбриология нервной системы
О
71
Рис. 32. Локализация нейральной эктодермы на последовательных стадиях гаструлы (а, б)
и ранней нейрулы (в) у хвостатых амфибий. Участок с точками — локализация нейро-
эктодермы.
* — анимальный полюс.
ница презумптивного нейроэпителиального пласта достигает анимального по-
люса зародыша, а каудальная опускается к бластопору. Необходимо отметить
более активную перегруппировку клеток по направлению к бластопору, что
приводит к неравномерности перераспределения материала нейроэктодермы.
Гаструляционная инвагинация мезодермы вызывает вытягивание каудальной
области нейрального зачатка и как следствие сжатие к средней линии (см.
рис. 32, в). Основную морфогенетическую нагрузку выполняет дорсальная губа
бластопора, через которую мигрирует хордомезодерма. Миграция хордомезо-
дермы — ведущее событие гаструляции, которое определяет морфогенез дру-
гих систем, в том числе и нейроэпителия.
В период гаструляции происходит первичная эмбриональная индукция,
которая морфологически состоит в том, что инвагинирующая вовнутрь заро-
дыша хордомезодерма вступает в контакт с нейроэпителием. Во время контак-
та между двумя эмбриональными пластами клеток происходит активация ней-
роэпителия, что дает начало нейральной дифференцировке. Первичная эмб-
риональная индукция хорошо изучена как морфогенетический процесс, но ее
химическая природа до настоящего времени не известна. Первичная эмбрио-
нальная индукция является не однозначным процессом, а совокупностью ин-
тегрированных морфогенетических реакций как индуктора, так и индуцируе-
мой ткани.
К моменту начала нейруляции формируется нейроэпителий, который со-
храняет свою организацию до замыкания нервной трубки. В нейроэпителии
позвоночных выделяют апикальную и базальную стороны пласта (рис. 33).
Апикальной стороной называется наружная поверхность эктодермы, базаль-
ной — ее внутренняя поверхность, которая обращена к хордомезодерме. На
ранних стадиях развития клетки нейроэктодермы одновременно связаны с
обеими поверхностями пласта. К окончанию гаструляции организация ней-
рального зачатка видоспецифично изменяется. У большинства хвостатых ам-
фибий и большей части млекопитающих мембраны клеток нейроэпителия
выходят одновременно на базальную и апикальную поверхности. Однако у
бесхвостых амфибий и ряда рептилий нейроэпителиальные клетки выходят на
поверхность только одной из сторон пласта.
72
о
Сравнительная анатомия нервной системы позвоночных
а б
в
Рис. 33. Типы организации нейроэпителия у позвоночных.
а — монослойный нейроэпителий; б— нейроэпителий смешанного типа; в — двухслойный
нейроэпителий. Нейроэпителий различается по закрепленности клеток на базальной и апи-
кальной поверхностях пласта.
Расположение клеток в пласте определяет существование двух основных
типов организации нейроэпителия: многослойного и однослойного. Оба типа
нейроэпителия встречаются у хорошо изученных видов амфибий. Многослой-
ным нейроэпителием обладают представители родов Xenopus, Hyta, а одно-
слойным — большинство видов Urodela (см. рис. 33). До начала нейруляции
монослойный пласт состоит из клеток одного типа, которые, как правило,
имеют одинаковую форму. В монослойном нейроэпителии клетки закреплены
одновременно на базальной и апикальной поверхностях пласта. Связь с одной
из мембран могут утрачивать клетки, находящиеся в пролиферативном цикле.
Базальный конец клетки в монослойном эпителии может менять свою лока-
лизацию в процессе пролиферации. Во время деления связь клетки с базаль-
ной стороной пласта утрачивается, а отросток втягивается в округляющуюся
клетку. Клетка проходит телофазу, и отростки дочерних клеток вновь устрем-
ляются к базальной поверхности пласта. Возвращается ли отросток дочерней
клетки в зону локализации материнского отростка, неизвестно.
Для многослойного эпителия характерна закрепленность клеток только на
одной из поверхностей пласта (см. рис. 33, в). Когда клетка расположена в
середине пласта, она не закреплена ни на одной из его сторон. При такой орга-
низации нейроэпителия клетки отличаются большим разнообразием форм и
высокой степенью гетерогенности взаимодействий. Клетки многослойного ней-
роэпителия могут смещаться в горизонтальной плоскости относительно друг
друга, что исключено для однослойного нейроэпителия. Тангенциальная под-
вижность клеток в многослойном пласте может сочетаться с радиальной мигра-
цией клеток от внешней до внутренней поверхности пласта. Такая встречная
интеркаляция клеток эпидермального и гиподермального слоев нейроэпителия
была выявлена у шпорцевой лягушки сразу после нейруляции (рис. 34, е).
Между двумя крайне различными формами организации нейроэпителия су-
ществует широкий спектр переходных форм, которые сочетают в себе призна-
ки, характерные как для однослойного, так и для многослойного нейроэпите-
лия. Смешанные типы организации нейроэпителия встречаются среди пред-
ставителей рода Triturus и у ряда бесхвостых амфибий. Возможно, что этот тип
организации нейроэпителия распространен гораздо шире, но специальных
исследований до сих пор не проводилось. Для смешанной организации ней-
роэпителия характерно наличие трех типов клеток, которые различаются по
положению в пласте (см. рис. 33, б). К первому типу следует отнести клетки,
одновременно выходящие на банальную и апикальную поверхности пласта.
II. Сравнительная эмбриология нервной системы
О
73
Рис. 34. Три основных типа нейруляции.
а-в — нейруляция у акул и хвостатых амфибий в монослойном нейроэпителии; г-е — ней-
руляция у бесхвостых амфибий в двухслойном нейроэпителии; ж-и— нейруляция у кости-
стых рыб. Основная причина нейруляции в монослойном нейроэпителии связана с изме-
нением формы клеток. В двухслойном нейроэпителии изменение формы клеток сопряжено
с проникновением клеток гиподермы к апикальной мембране эпидермального пласта, ко-
торый помечен точками. У костистых рыб клетки нейроэктодермы мигрируют между клет-
ками хордомезодермы и формируют нервный тяж, который трансформируется в нервную
трубку схизоцельным путем.
Второй тип составляют клетки, выходящие только на апикальную поверхность
пласта; третий тип включает клетки, связанные с базальной поверхностью.
Каждый из перечисленных трех типов организации эмбрионального нейро-
эпителия имеет ряд особенностей, которые накладывают отпечаток на весь
морфогенез нервной системы. В первую очередь структура нейроэпителия влия-
ет на становление формы мозга, которая начинается с нейруляции.
НЕЙРУЛЯЦИЯ
Нейруляция — это период развития, когда нервную систему можно впервые
выделить как анатомический зачаток, который отличается по струг туре от дру-
гих тканей зародыша. До начала нейруляции нейроэпителий можно локализо-
вать только на основании экспериментов по витальной маркировке клеток-
предшественниц, так как он неотличим от окружающих его клеток эктодермы.
У большинства видов позвоночных, за исключением костистых рыб, нейрут,
ляция включает в себя несколько более или менее выраженных периодов. Она
74
о
Сравнительная анатомия нервной системы позвоночных
начинается с изменения формы клеток, расположенных в зоне нейрального
зачатка. Изменение формы клеток приводит к увеличению толщины нейро-
эпителия, что выделяет его из окружающего презумптивного эпидермиса. При
утолщении нейроэпителия появляются медуллярные валики, которые ограни-
чивают нервную пластинку. Валики поднимаются над поверхностью эпидер-
миса зародыша и замыкаются, образуя нервную трубку (см. рис. 34).
Этот несложный эмбриологический процесс формирования нервной труб-
ки издавна привлекал пристальное внимание исследователей. Начиная с Гиса
(W. His, 1892—1894) предпринимались попытки объяснить природу наблюдае-
мого процесса в сворачивании нервной трубки, но до настоящего времени
вопрос остается открытым.
Лучше всего исследовано формирование нервной трубки у цыпленка, бес-
хвостых и хвостатых амфибий, крыс и мышей (В. Burnside, 1971, 1973; Т. Schro-
eder, 1970; Y. Schoenwolf et al., 1988; J. Smith и Y. Schoenwolf, 1988). Несмот-
ря на видимое анатомическое единство органогенеза, нельзя игнорировать
особенности нейруляции, характерные для животных с различными типа-
ми организации нейроэпителия. Поэтому имеет смысл рассмотреть нейру-
ляцию хотя бы в трех крайних формах организации нейроэпителия: в одно-
слойном, двухслойном и смешанном нейроэпителии. Различия в нейруля-
ции у животных носят принципиальный характер, что отражается на форме
мозга взрослых животных. Эмбрионы многих амфибий, рептилий, птиц и
млекопитающих обладают в процессе нейруляции монослойным нейроэпи-
телием, что делает этот вариант морфогенеза наиболее распространенным
среди позвоночных.
Нейруляция в монослойном эпителии начинается с утолщения нейроэпи-
телия в зоне будущей нервной трубки (см. рис. 34, а—в). По данным Джекоб-
сона (A. Jacobson, 1981), у тритона со стадии гаструлы (13) до стадии ранней
нейрулы (15) нейроэпителиальные клетки увеличиваются в длину почти на
50%, что сопровождается уменьшением апикальной и базальной поверхностей
клетки. На этой стадии развития клетки нейроэпителия имеют столбчатую
организацию, их апикальные и базальные участки по форме не отличаются
друг от друга. После начала вытягивания клеток (поляризации) нейроэпите-
лий можно выделить как анатомический предшественник нервной системы.
Если давать строгое определение, то только с началом поляризации клеток-
предшественниц можно говорить о появлении нейроэпителия — специфиче-
ской нейральной структуры. До этой стадии развития целесообразнее имено-
вать область локализации клеток-предшественниц нейральным или нейроэм-
бриональным полем. Как только клетки нейроэмбрионального поля эктодермы
начинают поляризоваться, клетки презумптивного эпидермиса уплощаются,
уменьшая толщину эмбриональной эктодермы. Необходимо отметить, что по-
ляризация клеток происходит неравномерно. В одном месте нервной пластинки
клетки вытягиваются в большей степени, а в другом — в меньшей. Поданным
Бернсайда, у Taricha torosa (хвостатые амфибии) длина клеток в передней час-
ти нервной пластинки на 20% больше, чем в задней. После начала нейруляции
среди веретеновидных клеток появляются колбовидные клетки. Колбовидные
клетки отличаются от веретеновидных клеток тем, что у них сжат апикальный и
расширен базальный участок мембран. Поданным Шенвольфа и Фрэнкса (1984),
II. Сравнительная эмбриология нервной системы ❖ 75
полученным на цыпленке, только около 10% веретеновидных клеток нервной
пластинки становятся колбовидными. Колбовидные клетки распределены в
сворачивающейся нервной пластинке неравномерно. Они находятся в непар-
ной средней зоне нервной пластинки, расположенной над зачатком хорды, и
в парных дорсолатеральных зонах. Между этими зонами нейроэпителиальные
клетки остаются веретеновидными. Механизмы, которые трансформируют
нейроэпителиальные клетки из веретеновидных в колбовидные, точно не из-
вестны. Нейроэпителиальные клетки могут становиться колбовидными в ре-
зультате апикального сжатия, базального расширения или в результате одно-
временного проявления обоих этих процессов. Базальное расширение клеток
нейроэпителия происходит одновременно с ретракцией апикального участка
цитоплазмы. Когда веретеновидные клетки трансформируются в колбовидные,
их ядра двигаются в сторону базального конца клетки. Эта миграция ядер вызы-
вает формирование базального расширения клеток. Аналогичные процессы на-
блюдали и в нервной трубке. Было показано, в частности, что миграция ядер
при изгибании нейроэпителиального пласта всегда направлена в сторону рас-
ширяющегося участка мембраны клетки.
Возникновение колбовидной формы клеток зависит от пролиферативного
цикла. Для большинства позвоночных характерна апикальная (прижелудочко-
вая) пролиферация нейробластов. Независимо от видовой специфичности ядра
клеток в монослойном нейроэпителии делятся преимущественно по апикаль-
ной поверхности пласта, причем телофаза латеральных клеток продолжается
дольше, чем медиальных. Поэтому их дочерние клетки находятся в базальном
районе меньшее время, чем клетки медиальной области. В результате этого в
латеральной зоне стенки нервной трубки формируется меньше колбовидных
клеток, чем в медиальной. Ядра клеток синтезируют ДНК, пока они непод-
вижны. Следовательно, основной причиной регионарной специфичности в
расположении колбовидных клеток являются продолжительность различных
фаз пролиферации и длина клеток.
Подводя итог обзору вероятных механизмов сворачивания нервной трубки
в однослойном эпителии, надо отметить, что основной и доказанной причи-
ной нейруляции является поляризация клеток — вытягивание вдоль длинной
оси за счет формирования пула парааксиальных микротрубочек. Вторым по
значимости процессом, приводящим к замыканию нервной трубки, является
возникновение колбовидных клеток. Хотя колбовидность клеток и имеет ме-
сто, ее роль в формировании нервной трубки недостаточно ясна. Можно толь-
ко констатировать наличие колбовидных клеток во время нейруляции, но яв-
ляются ли эти клетки причиной или следствием сворачивания нервной пла-
стинки, сказать нельзя. Открыт и вопрос о причинах зональных различий в
поляризации клеток нервной пластинки, а также о механизмах, интегрирую-
щих согласованные действия отдельных нейроэпителиальных клеток.
У ряда видов позвоночных животных нейроэпителий многослоен. Детально
изучена нервная пластинка шпорцевой лягушки (Xenopus laevis), которая состо-
ит из двух слоев эктодермы: эпидермального и гиподермального (см рис. 33, в,
34, г—е). Эпидермальный слой лежит снаружи и на поздних стадиях развития
включается в дно и внутренние стенки нервной пластинки. Гиподермальный
(подлежащий) слой пигментирован значительно сильнее, за что получил на-
76
о
Сравнительная анатомия нервной системы позвоночных
звание «темной» эктодермы. Гиподермальный слой дает производные нервной
системы: дно нервной пластинки и латеральные стенки нервной трубки. Кро-
ме того, из этого слоя дифференцируются клетки нервного гребня и пигмен-
тированный слой эпидермиса. В отличие от однослойного эпителия клетки
эпидермального и гиподермального слоев могут двигаться относительно друг
друга, что накладывает на нейруляцию шпорцевой лягушки ряд специфиче-
ских особенностей. У нее происходит не сворачивание нервной пластинки, а
постепенное наползание клеток, расположенных в зоне валиков, на клетки в
медиальной зоне нервной пластинки (см. рис. 34, г—е). Поверхностные слои
клеток скользят к средней части нервной пластинки по внутренним и слива-
ются по средней линии. Фактического подъема медуллярных валиков нет. Их
заменяет миграция поверхностно расположенных клеток эпидермального слоя.
Когда нервная пластинка у шпорцевой лягушки замыкается, формируется очень
маленькая полость внутри трубки — желудочек мозга. Просвет желудочка спин-
ного мозга меньше, чем диаметр одной клетки. Только спустя некоторое вре-
мя после окончания нейруляции формируется достаточно обширная полость
головного и спинного мозга. По сути дела можно сказать, что нервная трубка
возникает за счет наползания и последующей интеркаляции (встречной миг-
рации) клеток эпидермального и гиподермального слоев нейроэктодермы.
Из приведенного описания нейруляции ясно, что этот процесс осуществля-
ется гораздо более сложными механизмами, чем в однослойном нейропителии.
В клетках эпидермального слоя нейроэктодермы (Xenopus и Hyla) были найдены
высокоупорядоченные группы микрофиламентов, сходные по организации с
апикальными микрофиламентами однослойного нейроэпителия. Однако апи-
кальное сжатие не объясняет механизмы миграции клеток и независимых дви-
жений эпидермального и гиподермального слоев клеток. Как и в однослойном
эпителии, инициация замыкания нервной пластинки сопровождается тем, что
кубоидальные клетки начинают деформироваться в колбовидные. Поляризация
клеток обусловлена сборкой микротрубочек, а ретракцию апикального участка
клеток связывают с активностью микрофиламентов. Остается неясной роль ги-
подермальной части нейроэпителия, так как перечисленные особенности орга-
низации характерны для эпидермального слоя клеток. Открыт вопрос и о при-
чинах интеркаляции клеток эпидермального и гиподермального слоев нейро-
эпителия при формировании мозговых желудочков. Перечисленные особенности
нейруляции двухслойного нейроэпителия характерны и для нервных пластинок
смешанного типа, когда пласт организован из клеток, связанных одновременно
с одной или двумя мембранами. Но в таких нейроэпителиальных пластах про-
цесс нейруляции зависит от процентного соотношения между клетками этих
двух типов. Если больше половины клеток связаны с обоими поверхностями
пласта, то нейруляция пойдет по типу, характерному для однослойного нейро-
эпителия. В том случае, если преобладают клетки, связанные только с одной из
поверхностей пласта, будет преобладать миграционный тип нейруляции, харак-
терный для многослойного нейроэпителия.
Особняком стоит нейруляция у костистых рыб. Сворачивания нейроэпите-
лия в нервную трубку не происходит. Вместо этого начинается миграция ней-
роэпителиальных клеток от дорсальной эктодермы в вентральном направле-
нии (см. рис. 34, ж—и). Мигрировавшие клетки формируют нейроэпителиаль-
II. Сравнительная эмбриология нервной системы
О
77
ный тяж без внутренней полости, который представляет собой головной и
спинной мозг. Из центральной зоны тяжа клетки начинают мигрировать к
внешней стенке, что приводит к формированию нервной трубки. Следова-
тельно, возникновение полой нервной трубки у костистых рыб происходит по
шизоцельному типу. Шизоцельное образование мозговых желудочков и спин-
номозгового канала является крайней формой специализации морфогенеза
костистых рыб. Для многих видов характерны переходные варианты нейруля-
ции, которые сочетают в себе признаки нейруляционного сворачивания мо-
нослойного нейроэпителия с симптомами миграционного механизма переме-
щения нейроэктодермы. Так, у миног (Petromyzon marinus) и американского
чешуйчатника (Lepidosiren paradoxa) нейруляция происходит за счет миграции
двух плотно прижатых слоев нейроэпителиальных клеток, которые вертикаль-
но опускаются между клетками хордомезодермы (рис. 35, а). Затем эта двух-
слойная пластинка начинает расходиться и формировать полость. Возникает
нервная трубка с монослойными стенками.
Выпадает из общей схемы нейруляция у ланцетника. Эмбрион ланцетника
обладает обыкновенным монослойным нейроэпителием, описанным выше
(рис. 35, г). В начале нейруляции все события развиваются по традиционной
ж
Рис. 35. Нейруляция у миноги и американского чешуйчатника (а—в) у ланцетника (г-ж).
78
о
Сравнительная анатомия нервной системы позвоночных
схеме: утолщается нейроэпителий и формируются нервные валики. Клетки в
краях эктодермы, которая окружает нервную трубку, начинают пролифери-
ровать и образуют тонкий пласт, покрывающий весь нейральный зачаток
(см. рис. 35). Нервная пластинка остается открытой при полностью сформи-
рованной эктодерме. Замыкание нервной трубки происходит под эктодермаль-
ным слоем клеток.
В некоторых случаях нейруляция проходит параллельно с некоторыми нео-
бычно ранними процессами органогенеза, что делает ее черезвычайно видо-
специфичной. Примером может служить развитие одной из лягушек ( Eleuthero-
dactylus nubicola). У этого вида к концу нейруляции уже хорошо заметны почки
конечностей, которые сформировались вокруг нервной трубки. Такая гетерох-
рония уникальна для морфогенеза, но ее влияние может существенно сказать-
ся на анатомичесом строении нервной трубки.
Смыкание нервных валиков происходит не одновременно по всей длине
эмбриона. Существуют значительные различия по времени замыкания нервной
трубки в разных зонах головного и спинного мозга. Если у некоторых кости-
стых рыб и амфибий нейруляция может завершаться за 25—50 мин, то у выс-
ших млекопитающих и человека этот процесс растягивается на 7—10 дней.
Более того, сравнительная нейроэмбриология показывает, что анатомическая
сущность этих процессов может сильно различаться.
В самом тривиальном случае, который характерен для большинства позво-
ночных, нейруляция начинается со смыкания медуллярных валиков нервной
пластинки на расстоянии 1/3 от каудального края эмбриона (рис. 36). Возник-
шая зона смыкания валиков начинает распространяться в ростральном на-
правлении. Таким образом, нервная трубка первоначально формируется на
уровне спинного мозга и только затем — на уровне головного. Этот вариант
нейруляции наиболее известен, поскольку его просто наблюдать у амфибий.
Однако формирование нервной трубки происходит по-разному даже у млеко-
питающих (рис. 37). В начале нейруляции нервная трубка обыкновенно замы-
кается только в одном месте. У млекопитающих это обычно граница среднего
и заднего мозга. От этого места начинается движение нейруляционных волн,
которые распространяются в ростральном и каудальном направлениях. По мере
их продвижения нервные валики смыкаются, а нервная трубка закрывается
эктодермой. У человека и высших обезьян существует третья (вентральная)
нейруляционная волна, которая движется от рострального края нервной пла-
стинки навстречу ростральной волне нейруляции (рис. 37, б). Ростральная и
вентральная волны обычно короче каудальной и замыкают свои участки нерв-
Рис. 36. Формирование нервной трубки у Urodela. Две нейруляционные волны распрост-
раняются в ростральном и каудальном направлениях.
11. Сравнительная эмбриология нервной системы
О
79
Рис. 37. Сворачивание нервной трубки у эмбриона мыши (а) и человека (б). Передний
конец тела отмечен звездочкой. При нейруляции у млекопитающих возникает несколько
волн нейруляции (стрелки) и видоспецифических нейропоров (у).
п — передний нейропор; з — задний нейропор (нейруляция мыши по К. Sakai, 1989).
ной пластинки за несколько десятков часов. В зоне их движения формируется
средний, промежуточный и передний мозг, а там, где они сходятся, возникает
передний нейропор. Каудальная волна вызывает нейруляцию в заднем, про-
долговатом и спинном мозге (рис. 37, б). При ее смыкании формируется кау-
дальный нейропор, который часто продолжается в нервно-кишечный канал.
Нейропор и нервно-кишечный канал сохраняются относительно долго, а за-
тем зарастают. Время существования отверстий и их количество имеют высо-
кую видоспецифичность. У одних видов они существуют долго, тогда как у
других закрываются вскоре после нейруляции. У человека полноценного не-
рвно-кишечного канала не возникает. В целом нервно-кишечный канал су-
ществует дольше, чем нейропор, что делает нервную трубку открытой систе-
мой. У многих видов позвоночных нейруляция проходит намного сложнее,
чем описано выше. Детальное изучение сворачивания нервной трубки у мыши
80 О- Сравнительная анатомия нервной системы позвоночных
выявило по меньшей мере 8 морфологических фаз нейруляции (К. Sakai, 1987,
1989). На рис. 37. а приведена модифицированная схема этого автора, иллюс-
трирующая фазы формирования нервной трубки. Анализ нейруляции у мыши
позволил автору описать несколько нейропоров — зон открытой нервной труб-
ки, ограниченных смыкающимися нервными валиками. Кроме рострального
нейропора и нервно-кишечного канала, он обнаружил переднемозговой и зад-
немозговой нейропоры.
Заканчивая краткий обзор типов нейруляции у позвоночных, можно ска-
зать, что мы не располагаем достаточным количеством сравнительных данных
и можем только приблизительно оценить вклад этого морфогенетического
процесса в особенности анатомического строения нервной системы полово-
зрелых животных. Тем не менее ясно, что события, связанные с замыканием
нервной трубки, несут в себе запас топологической информации, которая ис-
пользуется в дальнейшем морфогенезе нервной системы.
ФОРМИРОВАНИЕ ГОЛОВНОГО МОЗГА
Начиная с работ Бэра (Н. Baer, 1837), Станиуса (S. Stanius. 1854) и Н.Н. Мик-
лухо-Маклая (1870) сложилась широко распространенная схема развития эмб-
рионального мозга, которая, не претерпев почти никаких изменений, сохра-
нилась до настоящего времени. Согласно этой схеме, развитие формы мозга
складывается из нескольких этапов. На первом этапе после нейруляции форми-
руется гомогенная нервная трубка со значительным изгибом в головной облас-
ти (рис. 38, а). Вторая стадия заключается в разделении головной части нервной
трубки на два отдела: prosencephalon и rhombcephalon. На третьем этапе prosen-
cephalon расчленяется на mesencephalon и telendiencephalon (рис. 38, б). Форми-
руются три мозговых пузыря: передний, средний и задний. Затем из этих трех
мозговых пузырей развиваются пять. Передний мозговой пузырь дифферен-
цируется на парные полушария переднего мозга и промежуточный пузырек
diencephalon; каудальный мозговой пузырь разделяется на задний (metencepha-
lon) и продолговатый (myelencephalon) пузыри. Таким образом, мозг приходит
к пятипузырной стадии, которая морфогенетически закреплена в существова-
нии пяти дефинитивных отделов мозга (рис. 38, в). Эта схема развития мозга
широко используется для сравнительно-эмбриологических работ и построе-
ния эволюционных концепций. Однако среди исследователей до настоящего
времени не существует единого мнения о последовательности дифференци-
ровки отделов мозга, а работы Таккет (F. Tuckett и Y.M. Morris-Kay, 1985)
поставили под сомнение вопрос об образовании мозговых пузырей за счет
поперечных перетяжек нервной трубки.
Кроме традиционных представлений, существует идея и о нейромерном про-
исхождении мозговых пузырей (рис. 38, г~ё) (В. Bergqwist, 1952, 1956; Н. Kallen,
1954). Согласно этой модели, в развитии мозга выделяются четыре периода.
На раннем этапе формообразования мозг состоит из двух частей: archencephalon
и deutoroencephalon Границей между этими отделами является SIP, которая
одновременно выполняет функции нейромерной границы. SIP выделяется на
протяжении всего онтогенеза и сохраняется у взрослых животных. В основе
II. Сравнительная эмбриология нервной системы
81
Рис. 38. Три гипотезы сегментации мозга после нейруляции.
а-в — наиболее известная схема расчленения нервной трубки за счет мозговых бороздок;
г-е — нейромерная модель сегментации мозга (по В. Bergqwist, 1956, с изменениями);
ж-и — модель развития мозга, сочетающая перемещение мозговых пластов и подразделе-
ние на основные отделы (С.В. Савельев, 1993). Межмозговые бороздки: SIP — sulcus inter-
cephalicus posterior, SIA — s. i. anterior, Sil — s. i. intermedius. Стрелками отмечены основ-
ные направления движения пластов клеток.
образования SIP лежит влияние хорды, которая подстилает rhombencephalon,
тогда как prosencepalon развивается над прехордальной пластинкой. Различ-
ные влияния структур, подстилающих эти отделы, приводят к формированию
на их границе теменного изгиба — каудального выпячивания воронки и опре-
деляют форму мозговой части гипофиза. Существование этого морфогенети-
ческого этапа признают сторонники как классической, так и нейромерной
схемы развития мозга.
На первом этапе эмбрионального развития мозга, с точки зрения нейро-
мерной модели, мозг разделяется на два крупных нейромерных блока:
prosencephalon и rhombencephalon. Любые возникающие после нейруляции по-
перечные перетяжки мозга рассматриваются как нейромеры 1-го порядка, хотя
они могут разделять далеко стоящие друг от друга отделы. Быстрая интерней-
ромерная фаза II, которая характеризуется образованием перетяжек в нейро-
мерах 1-го порядка, переходит в постнейромерный период. На постнейромер-
ной стадии принято выделять 11 нейромеров, которые включают в себя весь
головной мозг. Из первых двух нейромеров образуется передний мозг, третий,
четвертый и пятый нейромеры дают промежуточный мозг, шестой и седьмой —
средний мозг, а нейромеры с восьмого по одиннадцатый формируют задний
мозг (рис. 38, г~е). С точки зрения авторов нейромерной гипотезы развития
82 ❖ Сравнительная анатомия нервной системы позвоночных
мозга, каждый нейромер должен закладывать основу для зон головного мозга,
с которыми будут связаны черепно-мозговые нервы. При этом констатируют-
ся независимая от соседних нейромеров дифференцировка и миграция клеток
в области нейромеров. Нейромеры рассматриваются как морфогенетически и
анатомически обособленные субъединицы.
Существует морфогенетическая модель сегментации мозга, сформулирован-
ная в результате реконструкции развивающегося мозга сибирского углозуба
(Salamandrella keyserlingii) (Amphibia, Urodela). У этой амфибии головной конец
нервной трубки вначале образует два пузыря (prosencephalon и rhombencephalon),
затем четыре (prosencephalon разделяется на передний, промежуточный и сред-
ний мозг) и, наконец, пять пузырей (rhombencephalon разделяется на задний и
продолговатый мозг) (рис. 38, ж—и). Подобная последовательность формообра-
зования отличается от общепринятых представлений о морфогенезе мозга по-
звоночных, согласно которым головной отдел нервной трубки проходит двух-,
трех- и пятипузырные стадии. Однако эти данные не следует распространять
на все виды позвоночных, поскольку у двоякодышащих и костистых рыб изве-
стны двух-, четырех- и пятипузырные стадии морфогенеза мозга. У близко-
родственного сибирскому углозубу вида (Hynobius nebulosus) описаны двух-,
трех-, четырех- и пятипузырные стадии развития мозга (R. Sumi, 1926). Одна-
ко для сравнительной анатомии важно, что различные варианты морфогенеза
во всех случаях завершаются одинаковым результатом — образованием пяти
мозговых пузырей. Одновременно с этим в rhombencephalon сибирского угло-
зуба появляются пять поперечных вздутий — нейромеров. Они исчезают пос-
ле разделения rhombencephalon на задний и промежуточный мозг. Таким об-
разом, развитие мозга млекопитающих и некоторых амфибий указывает на
существование смешанного пути сегментации. В развитии мозга присутствуют
ростральные миграции, разделение мозга на постоянные отделы и нейромерная
организация двигательных отделов мозга. По всей видимости, наиболее близка
к реально существующим процессам схема, изображенная на рис. 38, ж—и.
На первом этапе мозг разделяется на два пузыря, в которые может входить
любая комбинация закладок будущих мозговых отделов. Затем начинаются
интенсивные миграции нейроэпителиальных клеток в ростральном направле-
нии, что приводит к закладке переднего мозга. Движение клеток завершается
после сегментации мозга на основные отделы. Независимо от расчленения
мозга постоянными бороздками задний мозг сегментируется временно суще-
ствующими нейромерами. Исчезновение нейромеров и начало дифференци-
ровки завершают период активного формообразования.
Рассматривая выделение основных отделов головного мозга, надо отметить,
что, несмотря на общую схему сегментации, существует масса видоспецифи-
ческих особенностей. Даже в пределах одной систематической группы можно
выделить видоспецифические характеристики образования мозговых границ,
которые отражаются на преобразованиях формы мозга после нейруляции. На
рис. 39 представлена внешняя форма мозга после замыкания нервной трубки
у трех видов хвостатых амфибий. Представители одной группы имеют не толь-
ко различную внешнюю форму мозга, но и различную последовательность его
сегментации (рис. 40). Сравнительно-анатомический анализ этого процесса
иллюстрирует определенную несостоятельность представлений о сходстве ран-
II. Сравнительная эмбриология нервной системы ❖ 83
в
Рис. 39. Рисунки, сделанные по восковым пластическим реконструкциям развития мозга у
трех видов хвостатых амфибий после нейруляции.
а — сибирский углозуб; б — иглистый тритон; в — обыкновенный тритон. Реконструкции
охватывают период от окончания нейруляции до начала активного питания личинок. За-
метно, что даже внешняя форма развивающегося мозга у представителей одной система-
тической группы сильно различается.
них стадий развития головного мозга у различных видов позвоночных. По
сути дела нет ни одного мозгового пузыря, который бы всегда соответствовал
конкретному отделу мозга, как это представлялось ранее. Даже двухпузырная
стадия не является строго детерминированной сегментацией. У сибирского
углозуба средний мозг объединен в общий пузырь с передним, а у обыкновен-
ного тритона — с задним мозговым пузырем (см. рис. 40). Трехпузырная ста-
дия может присутствовать, как у иглистого тритона, или заменяться сразу че-
тырехпузырной, как у сибирского углозуба. Такое изменчивое развитие голов-
ного мозга тем не менее завершается одинаково — формированием пяти
основных пузырей, которые дифференцируются в основные отделы мозга. Даль-
нейшее развитие формы мозга зависит от локализации зон пролиферации и
дифференцировки нейробластов.
84
Сравнительная анатомия нервной системы позвоночных
Рис. 40. Схема сегментации головного мозга на отделы у четырех видов хвостатых амфи-
бий: Hynobius nebulosus(no R. Sumi, 1926), Pleurodeles waltlii, Triturus vulgaris, Salamandrella
keyseriingii.
ОБРАЗОВАНИЕ НЕРВНЫХ СВЯЗЕЙ
Дифференцировка нейроэпителиальных клеток мозга включает в себя мно-
жество аспектов, среди которых кинетика пролиферации, стимуляция к выхо-
ду в дифференцировку и установление паттерна нейрональных связей являют-
II. Сравнительная эмбриология нервной системы
о
85
Рис. 41. Первичная дифференцировка нейробластов у амфибий.
а — схема поперечного среза головного мозга в зоне дифференцировки нейробластов;
латеральная поверхность головного мозга сибирского углозуба (б), иглистого тритона (в),
обыкновенного тритона (г). Слева — появление очагов дифференцировки, посередине —
вылупление личинок, справа — переход к активному питанию. Заштрихованы очаги диф-
ференцировки.
ся ключевыми. Эмбриологические исследования показывают, что нейробла-
сты дифференцируются в головном мозге в различное время, причем в неко-
торых структурах и после окончания эмбрионального периода развития. Гете-
рохронность этого процесса создает условия для появления центров первич-
ной дифференцировки нейробластов. Сравнительная анатомия очагов
дифференцирующихся нейробластов хорошо разработана на амфибиях. У хво-
статых амфибий первые дифференцированные группы клеток появляются на
латеральных поверхностях нервной трубки (рис. 41). Обычно зоны дифферен-
цирующихся нейробластов появляются одновременно в различных областях
головного мозга в виде быстро расширяющихся очагов. В таком очаге из про-
лиферации выходит только 15—20% клеток. Выход клеток в дифференцировку
быстро нарастает, что приводит к формированию непрерывной зоны диффе-
ренцирующихся клеток и их отростков (см. рис. 41).
В 30-х годах Когхиллом и Хэрриком (Y. Coghill, 1930; С. Herrick, 1937) были
установлены некоторые общие принципы образования нервных связей. Ими
было обнаружено, что существуют закономерности движения отростков меж-
ду центрами первичной дифференцировки. Если клетка дифференцируется в
непосредственной близости от центра первичной дифференцировки, то ее отро-
86 ❖ Сравнительная анатомия нервной системы позвоночных
сток растет по направлению к этому центру. Если клетка лежит на некото-
ром удалении от таких дифференцировочных зон, ее отросток движется по
границе между двумя этими центрами. Не менее важным результатом было
выявление этими авторами изменения направления движения отростков в
зависимости от активности ближайшего центра дифференцировки. Отрос-
ток клетки растет в сторону зоны с максимальной дифференцировочной ак-
тивностью. Следовательно, центр с максимальной и продолжительной ак-
тивностью должен собрать максимум первичных нервных связей. У амфибий
такой областью является наружная поверхность мозга в зоне IV желудочка,
которая дифференцируется первой и надолго сохраняет свое превалирова-
ние над остальными структурами головного мозга. В раннем эмбриогенезе
головного мозга существует явное колебание дифференцировочной активнос-
ти в различных областях головного мозга. По мнению Когхилла (1930), разни-
ца в скоростях дифференцировки нейробластов может вызывать образование
реципрокных связей.
Дальнейшие исследования показали, что сложившаяся зона дифференци-
рованных нейробластов расширяется по поверхности мозга медленно, образуя
связи преимущественно внутри себя (С.В. Савельев, 1987). К моменту вылуп-
ления личинок хвостатых амфибий около 70—80% площади поверхности го-
ловного мозга покрыто волокнами.
Таким образом, топология дифференцированных клеток и их отростков обус-
ловлена процессами формообразования, в основе которых лежат различные
проявления клеточной активности. В период дифференцировки клетки выхо-
дят на поверхность пласта и образуют отростки, причем отростки нейробла-
стов двигаются по поверхности пласта, состоящего только из торцов поляри-
зованных клеток. Поэтому зоны дифференцировки и распространение нервных
отростков топографически совпадают. Это наблюдение позволяет сделать один
важный вывод. По-видимому, растущий аксон предпочитает двигаться только
по неоднородному субстрату. Таким образом, модель Когхилла—Хэррика мо-
жет быть применена при условии, что отростки нейробластов выбирают для
движения наиболее гетерогенную поверхность пласта. Тогда последователь-
ность событий будет выглядеть следующим образом: 1) в различных отделах
мозга появляются зоны дифференцировки нейроэпителиальных клеток; 2) эти
зоны обладают различной дифференцировочной активностью; число диффе-
ренцированных клеток нарастает быстро или медленно и меняется в процессе
эмбрионального развития; 3) отростки нейроэпителиальных клеток двигаются
от зон невысокой дифференцировочной активности к областям высокой диф-
ференцировочной активности по поверхности пласта с максимальной гетеро-
генностью, что ограничивает свободу движения нервных отростков.
Эта модель пригодна для моделирования топологии дифференцировки только
на хвостатых амфибиях, поскольку у остальных позвоночных этот процесс
происходит значительно сложнее. Так, например, у эмбрионов приматов и
человека дифференцировка нейроэпителиальных клеток нервной трубки на-
чинается еще до окончания нейруляции. Нейробласты выходят в дифферен-
цировку в зоне первого слияния нервных валиков и образуют связи с соседни-
ми областями по мере замыкания медуллярных валиков.
II. Сравнительная эмбриология нервной системы ❖ 87
ФОРМИРОВАНИЕ ПЕРИФЕРИЧЕСКОЙ
НЕРВНОЙ СИСТЕМЫ И АНАЛИЗАТОРОВ
Нейруляция, сегментация головного мозга и первичная дифференцировка
происходят параллельно с развитием периферической нервной системы и ана-
лизаторов. Практически одновременно с возникновением нервной трубки
формируются группы нервных клеток, носящие название клеток нервного греб-
ня, или нейромезенхимы. Они возникают из клеток, расположенных на гра-
нице нервной пластинки и окружающей ее эктодермы (рис. 42, а). После от-
деления от зачатка нервной трубки и эктодермы клетки нервного гребня миг-
рируют от нервной трубки в вентральном направлении и участвуют в
формировании многих дериватов различных органов (рис. 42, б).
Из нервного гребня формируется большая часть периферических нервов и
ганглиев. В периферические нервы входят как отростки нейронов, расположен-
ных в периферических ганглиях, так и отростки центральных нейронов (рис. 42,6).
Основные производные клеток нервного гребня:
нейроны ганглиев дорсальных корешков спинного мозга,
ганглии симпатической нервной системы,
ганглии парасимпатической нервной системы,
хромаффинные клетки надпочечников, секретирующие гормоны,
шванновские клетки,
часть мозговых оболочек,
клетки хрящей висцеральных дуг,
пигментные клетки (исключая пигментные клетки сетчатки и центральной
нервной системы),
одонтобласты,
рецепторы сосудов,
часть нейрокраниума,
специализированные капсулы чувствительных нервных окончаний.
Определенная часть периферической нервной системы формируется из эк-
тодермальных плакод (рис. 43). Эктодермальные плакоды не связаны с клет-
ками нервного гребня и являются самостоятельными закладками. Из них фор-
мируются сенсорные волокна, ганглии и специфические образования дистант-
ных анализаторов. У эмбрионов позвоночных животных принято выделять две
группы эктодермальных плакод: дорсолатеральные и эпибрахиальные. Плако-
ды представляют собой зародышевые утолщения эктодермы. После инвагина-
ции происходит отделение зачатка и расползание будущих невромастов к зо-
нам дифференцировки.
Большая часть производных эктодермальных плакод формируется под индук-
ционным влиянием закладки центральной нервной системы или ее производ-
ных. Так, слуховой пузырек возникает после индукционного контакта между зад-
ним мозговым пузырем и эктодермой. Хрусталик инвагинирует и отделяется от
эктодермы под влиянием глазного пузыря или бокала, а обонятельный эпителий
формирует полость после индукционного влияния полушарий переднего мозга.
88 ❖ Сравнительная анатомия нервной системы позвоночных
ганглиев
Рис. 42. Эмбриональная закладка (а) и основные направления миграции клеток нервного
гребня (б).
Рис. 43. Производные эктодермальных плакод и схематическое изображение их закладок.
Дорсолатеральные плакоды расположены в верхней части зародыша а эпибрахиальные —
в нижней.
II. Сравнительная эмбриология нервной системы
89
Эктодермальные плакоды:
дорсолатеральные плакоды:
органы боковой линии (механорецепторы и электрорецепторы),
слуховой пузырек (вестибулярный аппарат),
Vll, VIII, IX, X головные нервы и ганглии;
эпибрахиальные плакоды:
V2, V3, VII, IX, X головные нервы и ганглии,
обонятельная плакода,
предглазничная плакода,
хрусталиковая плакода.
Периферические анализаторы формируются у позвоночных из нервной си-
стемы, эктодермальных плакод, клеток нервного гребня или имеют смешан-
ное происхождение. Зрительный анализатор позвоночных закладывается в
основном из двух источников: нервной трубки и хрусталиковой плакоды. Фор-
мирование глаза начинается сразу после замыкания нервной трубки. На гра-
нице между зачатком переднего и промежуточного мозга возникает парное
выпячивание глазных пузырей (рис. 44, а, б). Глазные пузыри достигают
эктодермы и индуцируют закладку хрусталика. Участок эктодермы прогиба-
ется навстречу глазному пузырьку, который трансформируется в глазной бо-
кал (рис. 44, в). Затем закладка хрусталика отделяется от эктодермы в виде
пузырька (рис. 44, г). Ножка глазного бокала трансформируется в зрительный
нерв, наружная стенка бокала — в пигментный эпителий, а внутренняя, более
толстая — в сетчатку (рис. 44, д).
Сходным образом происходит закладка органа обоняния. У всех позвоноч-
ных (кроме круглоротых и рыб) переднемозговой пузырь формирует парные
выпячивания (рис. 45, а). Они приходят в соприкосновение с головной экто-
дермой и индуцируют возникновение обонятельной плакоды (рис. 45,6). Цент-
ральная часть плакоды инвагинирует по направлению к полушарию и форми-
рует обонятельную полость (рис. 45, в, г).
Индукция эктодермы со стороны закладки заднего мозга приводит к фор-
мированию слухового пузырька. Он возникает из центральной части дорсола-
теральной плакоды. Индукция происходит в конце нейруляции, когда лате-
ральные стенки формирующейся нервной трубки вступают в контакт с приле-
жащим эпидермисом (рис. 46, а). Затем нервная трубка отходит от эктодермы,
Рис. 44. Стадии формирования сетчатки и хрусталика (а-д)
90
❖
Сравнительная анатомия нервной системы позвоночных
Рис. 45. Последовательные стадии индукции и формирования обонятельного мешка (а-а).
а индуцированный участок прогибается вовнутрь (рис. 46, б). Края прогнувшего-
ся участка смыкаются, и слуховой пузырек отделяется от эпидермиса (рис. 46, в).
Затем начинаются морфогенетические трансформации пузырька, которые за-
вершаются образованием полукружных каналов — рецептора углового ускоре-
ния, мешочков, содержащих гравитационный рецептор, и улитки — слухового
рецептора.
Периферические анализаторы соединены с головным и спинным мозгом с
помощью нервов. Эти связи носят различный характер. Иногда можно наблю-
дать прямое соединение рецепторной клетки с головным мозгом, как в случае
с обонятельным рецептором. Рецепторные клетки обонятельного и вомерона-
зального эпителия имеют чувствительные выросты, представляющие собой ви-
доизмененные дендриты. Аксоны этих клеток проникают непосредственно в
головной мозг и оканчиваются в обонятельных луковицах. Один и тот же ней-
рон является и сенсорным, и передающим сигналы. Аналогично устроены и
другие соматические рецепторные клетки. Тела чувствительных нейронов ле-
жат в ганглиях дорсальных корешков спинного мозга, а отростки тянутся к
дерме и мускулатуре и входят в спинной мозг. По-видимому, это самый древ-
ний тип соединения сенсорных нейронов и центральной нервной системы.
Другие анализаторы соединены с мозгом через систему ганглиев или специа-
лизированных групп клеток, которые выполняют функции предварительной
фильтрации и кодирования сенсорного сигнала. В качестве примера можно
Рис. 46. Последовательные стадии формирования слухового пузырька из дорсолатераль-
ной плакоды (а-е).
II. Сравнительная эмбриология нервной системы ❖ 91
рассмотреть волосковые клетки внутреннего уха, которые не имеют отростков
и не могут генерировать сигналы. Их чувствительность основана на модулиро-
вании сигнала, который поступает от эфферентного волокна и, проходя через
клетки, считывается афферентным волокном. При этом и афферентные, и
эфферентные клетки лежат в ганглии VIII нерва. В зрительной системе вспо-
могательная группа клеток приближена к первичным фоторецепторам и орга-
низована в сетчатку. Сенсорный сигнал, полученный палочками или колбоч-
ками, обрабатывается «на месте», в непосредстванной близости от короткоак-
сонного рецептора. В обычном случае сенсорная или вторично-чувствующая
клетка лежит в поле рецепции, а передающий нейрон — в сенсорном ганглии.
Примером является гломусная клетка каротидного тела, чувствительная к на-
пряжению кислорода в крови. Она иннервируется чувствительными нервны-
ми окончаниями сенсорных клеток, лежащих в каменистом ганглии IX (язы-
коглоточного) нерва. Следовательно, организация соединений анализаторов и
мозга довольно разнообразна, что требует ее специального рассмотрения.
Нужно отметить, что большая часть чувствующих и часть двигательных нервов
образованы отростками нейронов, расположенных в периферических гангли-
ях, лежащих вне головного и спинного мозга. С другой стороны, двигательные
нервы, состоящие из отростков мотонейронов, начинаются от клеток, распо-
ложенных в базальной пластинке спинного и головного мозга. Следовательно,
противопоставление периферической и центральной нервной системы носит
достаточно условный характер.
Все события, связаннные с эмбриональным развитием и дифференцировкой
нервной системы, происходят на фоне непрерывной пролиферации нейробла-
стов. На ранних этапах развития нервной трубки пролиферация сосредоточена
в специальных зонах нервной трубки (рис. 47). После окончания нейруляции
нейробласты активно пролиферируют на всем протяжении латеральных стенок
нервной трубки, тогда как в дорсальных (planum dorsale) и вентральных (planum
ventrale) зонах нервной трубки размножения клеток практически не происхо-
дит. Такая ситуация сохраняется непродолжительное время. Латеральная стен-
ка нервной трубки разделяется продольными бороздами (sulcus limitans). Выше
Planum ventrale
Рис. 47. Пролиферация клеток стенки нервной трубки сосредоточена в латеральных зо-
нах, которые разделяются граничной бороздкой (sulcus limitans).
92
❖
Сравнительная анатомия нервной системы позвоночных
Рис. 48. Пролиферативный цикл нейробластов в стенке нервной трубки.
нее располагается область чувствующих нейронов (planum alare), а ниже — зона
двигательных нейронов (planum basale). Такое строение центральной нервной
системы в большей степени характерно для спинного мозга, тогда как в голов-
ном мозге эти закономерности выражены не столь отчетливо.
Пролиферация нейробластов имеет ряд особенностей, которые придают ей
морфогенетический смысл, уже упоминавшийся при описании сворачивания
нервной трубки. Пролиферация происходит преимущественно в прижелудоч-
ковой зоне нервной трубки. В ее основе лежит морфогенетическая подвиж-
ность как самих нейробластов, так и их ядер (рис. 48). В наиболее обычном
случае деление начинается с вытягивания отростков рапировидных нейробла-
стов. После того как клетка устанавливает связь между наружной и внутрен-
ней поверхностью стенки мозга, начинается миграция ядра. Ядро нейробласта
начинает движение от прижелудочковой зоны стенки мозга к наружной зоне.
Немного не доходя до наружного сегмента клетки, ядро останавливается и
начинает обратное движение (см. рис. 48). В верхней фазе миграции ядра
происходит синтез ДНК. Возвращение ядра к прижелудочковой зоне происхо-
дит уже с двойным число хромосом. После достижения ядром прижелудочко-
вой зоны стенки мозга начинается втягивание наружного отростка клетки.
Затем клетка округляется и вступает в телофазу. Дочерние клетки начинают
формировать отростки, которые выходят к наружной поверхности пласта, и
пролиферативный цикл повторяется. При выходе в дифференцировку нейро-
бласты утрачивают связь с прижелудочковой поверхностью стенки нервной
трубки и мигрируют по окружающим их клеткам к наружной стенке мозга
(см. рис. 48). У наружного края пласта они образуют отростки, которые рас-
пространяются по наружной поверхности нервной трубки. Самый наружный
слой, состоящий исключительно из отростков нейробластов, носит название
краевого слоя (маргинального). Зона расположения тел клеток, вышедших из
пролиферации, носит название покровного слоя (мантийного).
Таким образом, эмбриональная стенка мозга проходит несколько фаз раз-
вития, которые отражают пролиферационные и миграционные периоды. На
II. Сравнительная эмбриология нервной системы
93
Зоны первичной
пролиферации
Рис. 49. Формирование вторичных зон пролиферации из мигрировавших из прижелудоч-
кового слоя нейробластов.
первом этапе стенка мозга состоит из монослойного эпителия. Затем он пре-
вращается в псевдомногослойный за счет подвижности ядер в делящихся клет-
ках. В это время происходит формирование краевого слоя, состоящего из на-
ружных отростков рапировидных нейробластов, в которые не «доходят» миг-
рирующие ядра. После начала дифференцировки и миграции нейробластов
формируется мантийный слой, состоящий из тел мигрировавших клеток.
Нужно отметить, что часть нейробластов удлиняется и связывает прижелу-
дочковый и краевой слои мозга. Эти клетки носят название клеток радиаль-
ной глии, или таницитов. По ним осуществляется миграция нейробластов как
в стратифицированные образования (кора переднего мозга, крыша среднего
мозга), так и в специализированные ядра.
При развитии мозга пролиферация может происходить и вне прижелудоч-
ковой зоны мозга. Вторичные очаги деления клеток образуются около приже-
лудочковых зон с повышенной пролиферацией (рис. 49). Они возникают бла-
годаря тому, что мигрировавшие клетки сохраняют способность к пролифера-
ции. Если пики пролиферации в прижелудочковой зоне возникают регулярно,
то может сформироваться несколько зон вторичной пролиферации. При этом
первоначально возникшие центры пролиферации окажутся лежащими лате-
ральнее, чем очаги пролиферации, возникшие позднее. В этом виден опреде-
ленный мозаицизм морфогенеза нервной системы.
III. СПИННОИ МОЗГ И ПЕРИФЕРИЧЕСКАЯ
НЕРВНАЯ СИСТЕМА
Сравнение морфологической организации спинного
мозга позвоночных следует начинать с обзора внешне-
го строения. Спинной и головной мозг составляют вме-
сте центральную нервную систему. Граница между эти-
ми крупными отделами нервной системы достаточно
условна, поскольку отростки клеток из головного мозга
проникают в спинной и наоборот. Традиционно при-
нято считать, что граница между спинным и головным
мозгом проходит на уровне перекреста пирамидных во-
локон. У человека и большинства млекопитающих эта
зона перекреста волокон хорошо заметна на вентраль-
ной поверхности продолговатого мозга. Все, что распо-
ложено каудальнее перекреста (задний край ромбовид-
ной ямки), принято считать спинным мозгом. Спин-
ной мозг лежит в полости позвоночного канала, который
образован невральными дугами позвонков. Спинной
мозг многих рыб, хвостатых амфибий, рептилий и птиц
распространяется до конца позвоночного канала, тогда
как у бесхвостых амфибий и млекопитающих занимает
только часть его полости.
Спинной мозг в сечении напоминает овал или круг с
неровной поверхностью. У многих животных на его дор-
сальной поверхности имеется продольная бороздка, кото-
рая называется верхней срединной бороздкой ffissura media-
na superior/ а на вентральной — аналогичная нижняя сре-
динная борозда (fissura mediana inferior). Спинной мозг
имеет внутри полость, которая называется центральным
каналом. Строение спинного мозга сильно варьирует в
различных систематических группах позвоночных (рис. 50).
Он может быть длинным и однородным, как у змей, или
быть не длиннее головного мозга, как у рыбы-луны.
Разнообразие внешней формы спинного мозга опре-
деляется функциональной нагрузкой на эту часть нерв-
ной системы. У четвероногих животных в спинном мозге
обычно хорошо заметны шейные и поясничные утол-
щения, в которые входят нервы от конечностей. Пояс-
III. Спинной мозг и периферическая нервная система
95
Рис. 50. Организация спинного мозга различных позвоночных. Вид с дорсальной поверх-
ности. Масштаб = 1 см (по R. Nieuwenhues et al., 1997).
ничные утолщения у птиц и некоторых млекопитающих настолько увеличе-
ны, что стенки спинного мозга расходятся и формируют полость. Эта полость
у птиц заполнена липидами и называется ромбовидным синусом (sinus lumbo-
sacralis, s. rhomboideus/ На латеральных поверхностях поясничного утолще-
ния птиц находится маргинальное ядро Гофмана—Келликера (nucleus marginalise
(рис. 52, 6). Аналогичное расширение поясничного утолщения у млекопитаю-
щих носит название конечного желудочка (ventriculus terminalis/
Достаточно часто встречаются разнообразные утонченные нити, которыми за-
канчивается спинной мозг (см. рис. 50). В различных группах позвоночных они
имеют различную форму, длину и происхождение. У костистых рыб спинной
мозг заканчивается утоньшением, переходящим в специфический эндокринный
орган, называемый урофизом (рис. 51, в). Для хвостатых амфибий, рептилий и
птиц характерна не очень длинная конечная нить, которая возникает в результате
билатерального сжатия развивающегося спинного мозга. У бесхвостых амфибий,
насекомоядных, рукокрылых и приматов длина конечной нити определяется ве-
личиной укорочения спинного мозга в спинномозговом канале (рис. 51,6).
Самый первый взгляд на сечения спинного мозга различных животных по-
казывает, что распределение серого и белого вещества неодинаково (рис. 52).
96
❖ Сравнительная анатомия нервной системы позвоночных
D
□
□
□
□
□
□
□
□
□
О
□
0
О' .
О О
« О
□
□
□
□
□
□
□
□
□
о
О
□
□
о
в
У миног и миксин серое вещество спинного моз-
га слабо дифференцировано (см. рис. 52, л).
Для ланцетника характерно дорсальное распо-
ложение тел нейронов (см. рис. 52, к). В неко-
торых случаях они могут локализоваться в цен-
тре нервной трубки, пересекая своими отрост-
ками полость центрального канала.
В остальных группах животных можно вы-
явить закономерное распределение серого и бе-
лого вещества на разрезе спинного мозга.
В большинстве случаев серое вещество распо-
ложено вокруг центрального канала в виде
классической бабочки, характерной для боль-
шинства чмниот. Верхние «крылья» серого ве-
щества принято называть дорсальными (спин-
ными) чувствующими рогами, а нижние —
вентральными (брюшными) двигательными ро-
гами спинного мозга. Спинные рога обычно
содержат мелкие вставочные нейроны, тогда
как брюшные — крупные моторные. В туло-
вищном отделе хвостатых амфибий слабо раз-
виты спинные рога, а у черепах — брюшные.
У многих амниот между спинным и брюшным
Рис. 51. Организация каудальной
зоны спинного мозга.
а — хвостатые амфибии, репти-
лии, птицы; б — бесхвостые ам-
фибии, насекомоядные, рукокры-
лые, приматы; в — костистые
рыбы обладают урофизом.
а
Рис. 52. Поперечные срезы спинного мозга различных позвоночных.
а — кот; б — американский опоссум; в — зеленая мартышка; г, д — голубь; е — крокодил;
ж— аксолотль; з — озерная лягушка; и — крокодиловый сарган; к—ланцетник; л — мик-
сина. f— маргинальное ядро.
III. Спинной мозг и периферическая нервная система
97
рогом формируется небольшой выступ серого вещества, называемый боковым
рогом и содержащий висцерально-двигательные нейроны (рис. 53).
Распределение серого вещества в спинном мозге неодинаково, что особен-
но заметно у тетрапод (см. рис. 53). Оно отражает количество входящих и
выходящих спинномозговых нервов. Через спинномозговые нервы осущест-
вляется соматическая чувствительность, висцеральная чувствительность, со-
матически-двигательная и висцерально-
двигательная иннервация. Соматическая
чувствительность включает в себя кожную,
сухожильную, связочную и мышечную чувст-
вительность. Висцеральная чувствитель-
ность включает вкусовую рецепцию и сиг-
налы от внутренних органов. Соматически-
двигательная иннервация обслуживает
скелетные мышцы, а висцерально-двига-
тельная — железы, глоточную, лицевую ми-
мическую и челюстную мускулатуру, глад-
кую мускулатуру кожи, сосудов и внутрен-
них органов. A icoJojCcxuOU ?
В состав спинного мозга традиционно
включают ганглий дорсального корешка
спйнногб мозга или спинальный ганглий, в
котором расположены тела афферентных
(чувствительных) соматических и висцераль-
ных нейронов. Эти клетки связаны со спин-
ным мозгом коротким отростком, который
оканчивается в дорсальных рогах серого ве-
щества. Вентральная часть спинного мозга
Рис. 53. Изменение диаметра спин-
ного мозга и формы очертаний се-
рого вещества в шейном, грудном,
спинном, поясничном и хвостовом
отделах у гориллы (слева) и черепа-
хи (справа).
состоит из эфферентных (двигательных) во-
локон. Они начинаются от клеток, распо-
ложенных в вентральных рогах спинного
мозга. Не у всех позвоночных встречается
четкое разделение корешков спинного моз-
гагаГмоторные и сенсорные. У многих пер-
вичноводных позвоночных и амфибий вис-
церальные эфферентные волокна выходят из спинного мозга как через дорсаль-
ные, так и через вентральные корешки. Для ланцетника, миног и миксин
характерен преимущественный выход висцеральных эфферентных волокон че-
рез дорсальный корешок спинного мозга. Нужно отметить, что слияния дор-
сальных и вентральных корешков спинного мозга (рис. 54) у ланцетника и
круглоротых не происходит. Они в виде самостоятельных нервов достигают
иннервируемых органов. Для большинства высших позвоночных характерен
выход дорсальных и вентральных корешков спинного мозга на одном уровне.
У ланцетника, миног, миксин и акул дорсальные корешки спинного мозга
чередуются с вентральными. Они лежат на разных уровнях, обозначая грани-
цы миотомов. Эфферентный соматический (моторный) корешок выходит из
спинного мозга в центре миотома, а афферентные (чувствительные) и висце-
4—5818
98
Сравнительная анатомия нервной системы позвоночных
Дорсальный
корешок
Ганглий дорсального Вентральный
корешка корешок
Соединительная
ветвь (ramus
communicans)
Симпатическая
цепочка
Висцеральный
ганглий Печень
Кожные
рецепторы
и железы
Рис. 54. Организация спинномозговых нервов и их переключений у млекопитающих.
Коллатеральный
ганглий
Кишечник
Спинальные
нервы
Симпатический
ганглий
ральный эфферентный — в промежутках между миомерами. Таким образом,
наиболее примитивными характеристиками морфологической организации спи-
нальных нервов можно считать чередование дорсальных и вентральных кореш-
ков, их самостоятельность и наличие висцеральных эфферентных волокон в
дорсальных корешках спинного мозга.
* Спинной мозг выполняет ряд автономных функций и интегрирует свою
активность с головным мозгом. Эффективность этой интеграции определяет-
ся нервными связями, которые организованы в виде восходящих и нисходя-
щих путей спинного мозга.
v Восходящие и нисходящие волокна спинного мозга организованы таким
образом, что восходящие локализуются преимущественно в дорсальной части
спинного мозга, а нисходящие — в вентральной (рис. 55).
Восходящие пучки волокон направляются в головной мозг и оканчиваются
в пяти основных центрах. Дорсальный и вентральный спинномозжечковые
тракты (tractus spinocerebellaris dorsalis, ventralis) в составе задних ножек входят
и оканчиваются в мозжечке. По этому тракту в мозжечок поступает проприо-
цептивная информация от мускулатуры. Из стройного и клиновидного пучков
(fasciculus gracilis, fasciculus cuneatus) волокна приходят в моторные центры
заднего и продолговатого мозга. По ним поступает информация о положении
тела в пространстве и позиционировании конечностей. По латеральному спи-
ноталамическому тракту (tractus spinothalamicus lateralis) волокна из спинного
мозга проникают в таламус, куда передают информацию о болевых ощущени-
ях и изменениях температуры. Вентральный спиноталамический тракт (tractus
spinothalamicus ventralis) также оканчивается в таламусе, но через него переда-
ется в мозг информация о тактильной чувствительности животного. Через спи-
нотектальный тракт (tractus spinotectalis) в крышу среднего мозга поступает
проприоцептивная информация от шеи и плечевого пояса.
III. Спинной мозг и периферическая нервная система
99
Восходящие волокна
Fasciculus
septomarginalis
Fasciculus
interfascicularis
Fasciculus
cuneatus
Нисходящие волокна
Fasciculus
gracilis
Tractus
reticulospinalis
ventralis
Fasciculus
proprius
Tractus
corticospinalis
lateralis
Tractus
rubrospinalis
Tractus
reticulospinalis
lateralis
Tractus
corticospinalis
Tractus
sulcomarginalis
Tractus
tectospinalis
Tractus
vestibulospinalis
ventralis
Tractus
spinocerebellaris
dorsalis
Tractus
spinothalamicus
lateralis
Tractus
spinocerebellaris
____ventralis
Tractus
spinotectalis
Tractus
spinothalamicus
ventralis
Рис. 55. Организация восходящих и нисходящих волокон спинного мозга на примере мле-
копитающих.
По спиноретикулярному тракту (tractus spinoreticularis) в ретикулярную фор-
мацию заднего мозга приходит информация о болевых ощущениях и от внут-
ренних органов. Надо отметить, что информация в восходящие пучки посту-
пает через дорсальные корешки спинного мозга и несколько раз переключает-
ся на вставочных нейронах по пути от спинного мозга к головному (рис. 56).
Нисходящие волокна расположены преимущественно в вентральной поло-
вине спинного мозга. Основная часть нисходящих волокон спинного мозга
начинается в заднем или среднем мозге, хотя для млекопитающих характерны
пучки волокон, передающие сигналы к эффекторным органам от коры перед-
него мозга. По латеральному и вентральному кортико-спинальному тракту
(tractus corticospinalis lateralis, ventralis) в спинной мозг проникают волокна
моторных нейронов, которые расположены в коре. Они осуществляют прямой
контроль первичных моторных нейронов, управляющих передними и задними
конечностями.
Руброспинальный тракт (tractus rubrospinalis) начинается в красном ядре и
тегментуме среднего мозга. По нему проходят нисходящие моторные сигналы
в спинной мозг. По тектоспинальному тракту (tractus tectospinalis), берущему
начало от крыши среднего мозга, проходят волокна, несущие информацию от
зрительной и слуховой систем к конечностям и туловищу животного. Боковой
и нижний пучки ретикулоспинального тракта (tractus reticulospinalis lateralis,
ventralis) начинаются в ретикулярной формации заднего мозга. Через них осу-
ществляется рефлекторный контроль за кинестезией. Вестибулоспинальный
100
❖
Сравнительная анатомия нервной системы позвоночных
Рис. 56. Организация восходящих волокон
спинного мозга млекопитающих. Дорсаль-
ные пучки восходящих волокон несут непе-
рекрещенные волокна, тогда как вентраль-
ные — преимущественно перекрещенные.
Рис. 57. Организация нисходящих волокон
спинного мозга млекопитающих.
тракт (tractus vestibulospinalis) берет свое начало в вестибулярных ядрах задне-
го мозга. По его волокнам проходят сигналы, управляющие рефлексами позы
с помощью осевой мускулатуры и мускулатуры конечностей. Нисходящие во-
локна переходят на противоположную сторону, как и восходящие волокна.
Перекресты образуются как на уровне продолговатого мозга, так и в непо-
средственной близости от нейронов-мишеней (рис. 57).
Нужно отметить, что передача как восходящих, так и нисходящих сигналов
неодинакова у животных, принадлежащих даже к одной систематической груп-
пе. Примером могут служить нисходящие пирамидные пути млекопитающих,
которые начинаются в коре головного мозга и заканчиваются на первичных
моторных нейронах спинного мозга (рис. 58).
Для кролика и опоссума характерно наличие нескольких проприоспиналь-
ных вставочных нейронов, через которые проходит сигнал, для кошачьих —
одного вставочного нейрона, а для приматов и человека специфичен моноси-
наптический моторный путь. Благодаря этому при повышении уровня орга-
низации нервной системы происходит сокращение скорости проведения сиг-
нала по спинному мозгу, что повышает эффективность двигательно-моторных
координаций.
Восходящие и нисходящие волокна у высших позвоночных объединяются при
выходе из спинного мозга в корешки, а затем в спинальные нервы (см. рис. 54).
III. Спинной мозг и периферическая нервная система ❖ 101
Следовательно, спинальные нервы содержат
как чувствительные, так и моторные волок-
на. Спинальные нервы довольно быстро раз-
деляются на несколько ветвей, которые зако-
номерно распределяются в органах и мускула-
туре. Примером такого распределения может
служить топологическая организация двигатель-
ных и чувствительных соматических волокон
(рис. 59). Эти волокна иннервируют мускула-
туру конечностей и тела животного.
У некоторых позвоночных существует объе-
динение спинальных нервов вдоль тела в виде
спинальных нервных сплетений. По сути дела
эта взаимосвязь отдельных спинальных нер-
вов позволяет максимально эффективно со-
гласовывать автономные периферические ре-
акции, в которых принимают участие мини-
мум два сегмента тела животного. Признаки
такой интеграции можно увидеть уже у акул
и костных ганоидов (рис. 60). Структурой, вы-
полняющей эти функции, является собираю-
щий нерв (п. collector), который у акул может
образовывать небольшое сплетение на уровне
анального плавника. У костных ганоидов та-
кого сплетения, как правило, не возникает.
Рис. 58. Организация пирамид-
ных нисходящих путей млекопи-
тающих.
а — кролик, опоссум; б — кошка;
в — приматы.
Рис. 59. Нервы спинного мозга, обслуживающие мускулатуру тела и конечностей (двига-
тельная и чувствительная соматическая иннервация). Схема условно отражает распреде-
ление нервов, существующее у эмбрионов млекопитающих.
102
❖
Сравнительная анатомия нервной системы позвоночных
Рис. 60. Спинальные сплетения в различных группах позвоночных.
Собирающий нерв выявлен и у некоторых костистых рыб. Однако он продол-
жается только в передней половине тела и не формирует сплетение. По-види-
мому, этот вариант объединения спинальных нервов следует принимать за
исходный тип. Для большинства позвоночных, имеющих конечности, харак-
терны два крупных сплетения: плечевое (plexus cervicitobrachialis) и пояснич-
но-крестцовое (plexus lumbosacralis). Функциональная роль такого объедине-
ния определяется необходимостью интегрировать работу мускулатуры конеч-
ностей на всех возможных уровнях анатомического строения. Такой тип
объединения спинальных нервов характерен преимущественно для анамний,
тогда как у амниот встречаются еще два типа обособленных сплетений. Для
млекопитающих обычно разделение plexus cervicitobrachialis на шейное (plexus
cervicalis) и плечевое (plexus brachialis).
Между плечевым и поясничным сплетением иногда появляется туловищ-
ное сплетение. Нужно отметить, что среди позвоночных встречаются исклю-
чения, у которых спинальные сплетения практически отсутствуют. Неболь-
шое плечевое сплетение найдено у Sirenidae, и никаких признаков сплетения
нет у большинства представителей Apoda. По-видимому, у этих животных спи-
нальное сплетение утрачено вторично.
На этом основные внешние анатомические различия спинного мозга и его
нервов исчерпываются, тогда как принципиальные особенности внутреннего
строения помогут пролить свет на организацию периферической нервной си-
стемы. Периферическая нервная система состоит из соматического и висце-
рального компонентов. Соматический компонент произошел из афферентов и
эфферентов, которые связаны с покровами, связками, сухожилиями и муску-
латурой. Эта система позволяла древним позвоночным отвечать на внешние
раздражения адекватными двигательными реакциями. У высших позвоночных
управление сложными соматическими функциями имеет обширное предста-
III. Спинной мозг и периферическая нервная система
103
вительство в коре переднего мозга. Висцеральный компонент отсутствует в
коре, но широко представлен в спинном мозге, а его высшие центры сосре-
доточены в промежуточном мозге на уровне гипоталамуса. Висцеральный
компонент периферической нервной системы имеет афферентные входы от
внутренних органов и кровеносных сосудов и эфферентные выходы к попе-
речнополосатой мускулатуре жаберных дуг, гладких мышц сосудов, мускула-
туре глотки, челюстного аппарата, мимической мускулатуре и железам. Су-
ществует предположение, что эти два компонента периферической нервной
системы возникли независимо друг от друга. Затем на основе соматического
компонента сформировалась центральная нервная система, подчинившая вис-
церальный компонент. В качестве доказательства приводится способность
многих внутренних органов автономно отвечать на различные раздражения.
Наиболее наглядным примером является автономная рефлекторная актив-
ность кишечника при прохождении через него пищи. Действительно, висце-
ральный компонент значительно более независим от головных центров, чем
соматический. Однако маловероятен даже непродолжительный процесс не-
зависимого эволюционирования пищеварительной и двигательной систем жи-
вотных. По-видимому, это представление возникло на основе ложного ощу-
щения автономности работы внутренних органов и «невозможности» их цент-
рального контроля.
Структурная организация периферической нервной системы построена по
иерархическому типу и отражает морфофункциональные взаимоотношения с
головным и спинным мозгом.
Эта классификация отражает и морфологические различия в строении каж-
дого из компонентов этой системы. Соматические двигательные волокна обычно
проводят импульсы по аксону единственного нейрона, который начинается в
спинном мозге, а оканчивается в иннервируемой мышце (рис. 61). Эфферен-
Парасимпатическая
104
❖
Сравнительная анатомия нервной системы позвоночных
Соматическая Висцеральная Парасимпатический
Рис. 61. Организация серого вещества спинного мозга и периферической иннервации.
тный висцеральный путь обычно состоит из двух нейронов. Нейрон, располо-
женный в спинном мозге, называется преганглионарным нейроном, а его от-
росток — преганглионарным волокном. Преганглионарные волокна имеют хо-
рошо развитую миелиновую оболочку (см. рис. 61). Преганглионарные волок-
на оканчиваются на постганглионарных нейронах, у отростков которых имеется
плохо развитая миелиновая оболочка. Отростки постганглионарных нейронов
оканчиваются в иннервируемых эффекторных органах.
Организация висцерального пути различается в симпатической и парасим-
патической частях. К симпатической системе (пояснично-грудной) относят
волокна, выходящие преимущественно из поясничного и грудного отделов
центральной нервной системы. У симпатических путей обычно бывают корот-
кое преганглионарное и длинное постганглионарное волокна. Постганглио-
нарные нейроны расположены в ганглиях симпатического ствола, шейных,
чревных и брыжеечных ганглиях. Из этих ганглиев постганглионарные волок-
на направляются к органам-мишеням (рис. 62). Нужно отметить, что преган-
глионарное волокно симпатического нейрона выделяет ацетилхолин, а пост-
ганглионарный нейрон — адреналин и его метаболический предшественник
норадреналин. Выделение этих медиаторов мобилизует организм животного и
стимулирует его физиологическую активность.
Для парасимпатической нервной системы характерно довольно длинное
преганглионарное волокно, которое оканчивается в ганглии, расположенном в
самом внутреннем органе или в непосредственной близости от него (см. рис. 62).
Постганглионарное волокно обычно короткое и почти полностью расположе-
но в самом иннервируемом органе. При передаче сигнала парасимпатическим
нейроном выделяется ацетилхолин, который действует, почти как антагонист
адреналина и норадреналина.
Ill. Спинной мозг и периферическая нервная система
105
Ресничный
тракт
Рис. 62. Организация автономной нервной системы млекопитающих. Пунктирная линия —
пути парасимпатической, непрерывная линия — симпатической нервной системы.
Следует уточнить, что эти условно антагонистические системы иннервиру-
ют одни и те же органы только у млекопитающих. Эта двойная иннервация
является значительным эволюционным достижением, которое возникло дале
ко не сразу. Большинство позвоночных имеет двойную иннервацию только у
некоторых органов, тогда как основная их часть иннервируется или симпати-
ческой, или парасимпатической системой. В таком случае противопоставле-
ние двух систем не совсем корректно.
У млекопитающих совмещена симпатическая и парасимпатическая системы,
которые действуют на внутренние органы по-разному. При воздействии на же-
лезы обе системы часто выполняют необычные для них функции. Симпатичес-
кая нервная сеть при воздействии на поджелудочную, слюнную и слезную же-
лезы ингибирует их секрецию, а парасимпатическая нервная система ее стиму-
лирует. При этом парасимпатическая система не влияет на потовые железы,
тогда как симпатическая вызывает обильное потоотделение. В пищеваритель-
ном тракте симпатическая сеть усиливает тонус сфинктеров и уменьшает под-
вижность стенки кишечника, а парасимпатическая действует наоборот. Симпа-
тическая система вызывает высвобождение глюкозы из печени, расслабление
желчного и мочевого пузыря, преганглионарными нейронами стимулирует ак-
тивность надпочечников. Парасимпатическая система вызывает сжатие желч-
106 ❖ Сравнительная анатомия нервной системы позвоночных
ного пузыря и гладкой мускулатуры мочевого пузыря, что сопровождается рас-
слаблением сфинктеров последнего. На сердце эти системы влияют прямо про-
тивоположным образом. Симпатическая система увеличивает частоту сердеч-
ных сокращений и их интенсивность. При этом расширяются коронарные арте-
рии. Парасимпатическая система замедляет сердцебиение и вызывает сужение
коронарных артерий. Симпатическая система немного расслабляет ресничную
глазную мышцу, а парасимпатическая сокращает сфинктер зрачка и цилиарную
мышцу (аккомодация). Симпатическая система вызывает расширение бронхов
легких и сокращение селезенки, брюшной полости и кожи. Парасимпатическая
система, оставаясь нейтральной к коже и брюшной полости, вызывает сужение
просветов бронхов и расслабление селезенки. При влиянии на половые органы
парасимпатическая система вызывает эрекцию как пениса, так и клитора. Сим-
патическая система запускает механизм эякуляции у самцов, но у самок ее роль
достоверно не известна. В целом можно констатировать, что симпатическая
система стимулирует метаболизм животного, тогда как парасимпатическая его
заметно угнетает. Столь эффективная система автономной регуляции активно-
сти внутренних органов характерна для млекопитающих, тогда как у других
позвоночных ситуация выглядит иначе.
Первые признаки автономной нервной системы можно обнаружить у лан-
цетника. Из дорсальных корешков спинного мозга ланцетника выходят нервы,
направляющиеся к диффузным сплетениям нервных клеток, лежащих во внут-
ренних органах. Специализированных ганглиев в органах нет. Переключение
происходит непосредственно на отдельных клетках. По-видимому, в такой орга-
низации можно увидеть признаки парасимпатической системы, но однознач-
ные выводы о проведении такой гомологии кажутся преждевременными. Фраг-
ментарная организация автономной нервной системы встречается уже у кругло-
ротых. У них симпатическая цепочка отсутствует, но коллатеральные ганглии
собирают часть висцеральных волокон, которые расходятся от них во внутрен-
ние органы. У миксин головные автономные волокна имеются только в блужда-
ющем (X) нерве. Предполагается, что спинальные автономные волокна могут
выходить у миксин в составе вентральных корешков, но их распределение в
теле плохо изучено. Для речной миноги характерно наличие автономных воло-
кон в блуждающем (X), лицевом (VII) и языкоглоточном (IX) нервах, которые
иннервируют жаберный аппарат. Есть непроверенные данные, что у речной
миноги автономные нервы выходят как через дорсальные, так и через вентраль-
ные корешки спинного мозга. Они направляются к почкам, гонадам, кровено-
сным сосудам, пищеварительному тракту и другим внутренним органам.
У хрящевых рыб автономная система весьма хорошо развита. Для акул
характерны отсутствие коллатеральных ганглиев, расчлененность симпатиче-
ской цепочки и несвязанность с ней черепных нервов (рис. 63). Блуждаю-
щий нерв гомодинамен V, VII и IX гипобрахиальным нервам. Блуждающий
нерв типичных акул рассматривается как коллектор таких нервов. Парасим-
патической частью является только внутренностная ветвь (ramus splanchnicus),
которая идет к внутренним органам. Нужно отметить, что симпатические
ганглии пластиножаберных сливаются с хромаффинными телами, происхо-
дящими из клеток нервного гребня. В ряде черепных и туловищных нервов
симпатической системы переключение происходит в самом иннервируемом
органе, как в парасимпатической системе. Однако большая часть симпати-
III. Спинной мозг и периферическая нервная система
107
Рис. 63. Организация автономной нервной системы акулы.
ческих нервов имеет короткие преганглионарные волокна, которые оканчи-
ваются в симпатических ганглиях, лежащих вдоль спинного мозга, и длин-
ные постганглионарные отростки, достигающие иннервируемых органов. Дви-
гательной иннервации кожи у пластиножаберных еще нет, но кровеносная
система уже иннервирована.
У костистых рыб симпатический нервный ствол непрерывен и связан с че-
репными нервами (рис. 64). Производные клеток нервного гребня (хромаф-
финные тела) выделяются из симпатических ганглиев в отдельные структуры
Для большинства костистых рыб характерно появление коллатерального ганг-
лия. Свидетельством значительного развития автономной нервной системы
костистых рыб можно считать появление волокон, снабжающих кожу. У кос-
тистых рыб получила дальнейшее развитие двойная иннервация головы туло-
вищными постганглионарными волокнами. В голову костистых рыб начина-
ют проникать туловищные волокна, которые иннервируют саму ротоглоточ-
ную полость и желудок. Автономные волокна костистых рыб, выходящие из
спинного мозга в туловищной части и иннервирующие внутренние органы,
имеют короткое преганглионарное волокно и длинный постганглионарный
отросток. Они явно принадлежат к симпатической системе. В каудальной ча-
сти туловища часть волокон имеет длинное преганглионарное и короткое пост-
ганглионарное волокна, которые расположены в стенке мочевого пузыря. Пе-
реключение происходит в самом сплетении мочевого пузыря, что сходно с
парасимпатической нервной системой млекопитающих.
108
❖
Сравнительная анатомия нервной системы позвоночных
Рис. 64. Организация автономной нервной системы костистых рыб.
Нужно отметить, что у многих костистых рыб обнаружены коллатеральные
ган1лии, а часть волокон спинальных нервов выходит из центральной нервной
системы в составе головных нервов. Эти признаки сближают строение авто-
номной нервной системы костистых рыб и амфибий.
У амфибий и других тетрапод организация автономной нервной системы сходна
с приведенным выше базовым описанием. Для них характерны парная симпати-
ческая нервная цепочка, идущая вдоль спинного мозга, и многочисленные колла-
теральные ганглии, которые распределены между внутренними органами (рис. 65).
Для большинства хвостатых амфибий характерно наличие выходов моторных
волокон автономной системы как через дорсальные, так и через вентральные
корешки спинного мозга. У бесхвостых амфибий, рептилий, птиц и млекопитаю-
щих все моторные волокна выходят через вентральные корешки спинного мозга.
Рассматривая автономную нервную систему рептилий, птиц и млекопитаю-
щих, следует уточнить классификацию ганглиев симпатической части автоном-
ной нервной системы. Обычно принято выделять два симпатических ствола ган-
глиев, лежащих в непосредственной близости от спинного мозга (рис. 66, 67).
Их традиционно называют тораколюмбальными, или паравертебральными,
симпатическими ганглиями, подчеркивая парность симпатических стволов.
Кроме них, выделяют непарные ганглии: верхний и нижний брыжеечные и
чревный, или брюшной, ганглии. Эти ганглии непарны и часто называются
превертебральными. Большая часть симпатических ганглиев удалена от ин-
нервируемых органов, но есть и исключения. Этим исключением являются
относительно небольшие симпатические ганглии, расположенные в непосредст-
III. Спинной мозг и периферическая нервная система
109
Рис. 65. Организация автономной нервной системы амфибий.
венной бди «ости от половых органов. К ганглиям сигнал приходит по очень
длинным симпатическим преганглионарным волокнам, а передается к поло-
вым органам по коротким волокнам. В этом случае анатомическое строение
симпатической системы сходно с таковым парасимпатической. Симпатичес-
кими эффекторными органами млекопитающих являются гладкие мышцы
сосудов, органов брюшной полости, волос, зрачка и выделительных органов.
Они иннервируют сердце, потовые, слюнные и пищеварительные железы,
клетки подкожной жировой клетчатки, печени и канальцы почек.
У млекопитающих ганглии парасимпатической системы расположены только
в области головы и вблизи органов таза. Во всех остальных случаях парасим-
патические клетки распределены в толше и на поверхности сердца, легких и
желудочно-кишечного тракта. Такие скопления клеток парасимпатической
системы носят название интрамуральных ганглиев.
Преганглионарные парасимпатические волокна, обслуживающие глазные
мышцы и железы, расположенные в головной части животного, выходят из
центральной нервной системы в составе глазодвигательного (III), лицевого
(VII) и языкоглоточного (IX) нервов. К органам брюшной и грудной области
парасимпатические волокна выходят в составе блуждающего нерва (X). Тазо-
вые органы обслуживаются нервами, выходящими из крестцового отдела. Па-
расимпатическая нервная система млекопитающих иннервирует гладкую муску-
латуру и железы желудочно-кишечного тракта, легкие, предсердия, слюнные
железы, выделительные и половые органы. Парасимпатические эффекторные
волокна в виде исключения снабжают артерии полового члена, клитора, ма-
лых половых губ и мозга.
110
❖
Сравнительная анатомия нервной системы позвоночных
Глотка
Брюшной ганглий
Желудок
Сердце
Легкие
Печень
Почка
Брыжеечный
ганглий
IX
X
Кишечник
Селезенка
Гонады
Толстая
кишка
Нервное
сплетение
конечностей
Мочевой
пузырь
Нервное
сплетение
конечностей
Клоака
Рис. 66. Организация автономной нервной системы рептилий.
Необходимо уделить внимание афферентным волокнам автономной не-
рвной системы. Висцеральные афференты обычно идут в составе крупных
периферических нервов. В спинной мозг они часто входят совместно с сома-
тическими афферентными волокнами. В таком случае тела висцеральных чув-
ствительных волокон лежат в спинальных ганглиях. Однако большая часть
афферентных волокон от брюшной и грудной полостей попадает в централь-
ную нервную систему в составе блуждающего нерва (X). Поэтому иногда
используется нестандартное анатомическое название висцеральных афферен-
тов — вагальные афференты. Большинство афферентов являются классичес-
кими механорецепторами, которые воспринимают или давление в просвете
артерии, или степень наполнения мочевого пузыря. Кроме этого, они могут
воспринимать pH среды, электролитный состав жидкостей в тканях внутрен-
них органов или болевые раздражения.
Нужно отметить, что особенности морфологической организации симпати-
ческой и парасимпатической систем позволяют оценить масштабы их влия-
ния на организм животного. Симпатические постганглионарные нейроны об-
ладают обширной зоной иннервации, что приводит к генерализованным реак-
циям животного. Суммарный эффект от действия симпатической нервной
Ш. Спинной мозг и периферическая нервная система
111
Рис. 67. Организация автономной нервной системы млекопитающих.
системы выражается в торможении активности многих внутренних органов и
стимуляции сердечной и скелетных мышц. Этот эффект известен в физиоло-
гии как подготовка организма к поведению типа «борьба и бегство». Парасим-
патическая система действует намного более локально, поскольку ее постган-
глионарные нейроны обладают значительно меньшими зонами иннервации.
Они сосредоточены в отдельных органах и оказывают в основном регулирую-
щее действие на функцию органа-мишени.
Суммируя взаимодействие органов животного с автономной нервной си-
стемой, необходимо подчеркнуть четыре основные закономерности.
1. Если орган иннервируется парасимпатической или симпатической систе-
мой, то их воздействия носят разнонаправленный характер, что повышает
эффективность его регуляции.
2. Симпатическая и парасимпатическая системы могут оказывать на тот
или иной орган или тормозящее, или возбуждающее влияние.
3. У многих животных в одном органе превалирует симпатическая или па-
расимпатическая система.
4. Для низших позвоночных характерно управление многими органами с
помощью только одной из систем.
IV. ЧЕРЕПНО-МОЗГОВЫЕ НЕРВЫ
Черепно-мозговые нервы позвоночных исторически
связаны со спинномозговыми нервами и органами
чувств, расположенными в голове. Специализирован-
ные чувствительные и моторные черепно-мозговые нер-
вы имеют в соответствии с разными теориями различ-
ное происхождение. Весьма рационально представле-
ние о том, что большая часть нервов, выходящих из
продолговатого и среднего мозга, может быть гомоло-
гизирована с дорсальными или вентральными кореш-
ками спинного мозга. Для этого предположения есть
все основания, поскольку расположение чувствитель-
ных и моторных столбов спинного мозга сохраняется в
продолговатом мозге (рис. 68).
Моторные столбы расположены в вентральной части
спинного мозга, а чувствительные — в дорсальной. На
уровне IV желудочка ситуация изменяется, поскольку дор-
сальные края нервной трубки оказываются развернуты-
ми в латеральных направлениях (см. рис. 68). Моторные
центры ствола мозга оказываются приближенными к ме-
диальной оси мозга, а сенсорные — к латеральному краю.
Картина топологической преемственности продолжения
столбов спинного мозга в головной нарушается появле-
нием вентральных мозговых ядер, которые возникают в
результате вентральной миграции клеток при формиро-
вании вторичных зон пролиферации среди базальных пуч-
ков волокон. Дополнительную путаницу вносят и спе-
циализированные центры, связанные с особой висцераль-
ной чувствительностью. По сути дела это выделившиеся
висцеральные двигательные и висцеральные чувствитель-
ные центры, которые дополняются специализированными
ядрами особой соматической чувствительности.
При достижении головного мозга спинномозговые
столбы утрачивают свою непрерывность. Они подразде-
ляются на ядра, которые отделены друг от друга группами
IV. Черепно-мозговые нервы
113
Особый соматический
*>У« I чувствительный столб
Висцеральный
чувствительный столб
£♦'*•1 Соматический
jLJ чувствительный столб
Особый висцеральный
чувствительный столб
Соматический
моторный столб
Висцеральный
моторный столб
Особый висцеральный
моторный столб
И
Рис. 68. Распределение висцеральных и соматических центров в спинном, продолговатом
и среднем мозге. В нижней части рисунка гипотетическая схема, отражающая предковое
состояние топологического распределения гомологичных центров в спинном, заднем и про-
долговатом мозге; боковая проекция. При переходе из спинного мозга в головной распре-
деление столбов сохраняется, что предопределяет топологию выхода моторных и сенсор-
ных нервов.
нервных волокон. Расчленение столбов на ядра, обслуживающие конкретные
нервы, не приводит к их кардинальному смещению относительно обшей схе-
мы, приведенной на рис. 68. Действительно, даже у млекопитающих ядра 111,
IV, VI и ХП соматических моторных нервов лежат вдоль медиальной оси сред-
него, заднего и продолговатого мозга. В пределах висцерального двигательно-
го столба расположены слюноотделительные (парасимпатические) ядра VII и
IX нервов, а также парасимпатический компонент III и X нервов. Аналогично
укладываются в общую схему центры локализации висцерально-чувствитель-
ного одиночного ядра IX и X нервов, которые выглядят как продолжение вис-
церального чувствительного столба спинного мозга. В несколько увеличенном
соматическом чувствительном столбе на уровне среднего мозга расположены
сенсорные ядра V нерва.
Однако возникают и дополнительные центры, обслуживающие специаль-
ные чувствительные нервы. По-видимому, эти дополнительные центры могли
сформироваться на основе специализации и морфологического увеличения
размеров основных четырех столбов спинного мозга. К ним относят специ-
альные моторные и сенсорные ядра как соматического, так и висцерального
114
❖
Сравнительная анатомия нервной системы позвоночных
Чувстви-
тельное
ядро
тройничного
нерва (V)
Коленчатый
ганглий
лицевого
нерва (VII)
Верхний и
нижний
каменистые
(g. petrosum)
ганглии
языкоглоточного
нерва (IX)
Чувствительные
ядра лицевого (VII),
языкоглоточного (XI)
и блуждающего (X)
нервов
Обонятельная
плакода (I)
Ганглий
(g. semilunare)
тройничного
нерва (V)
Сетчатка
(II)
а
Нижний
(g. nodosum)
ганглий
блуждающего
нерва (X)
Верхний ганглий
(g.julare) блуждающего
нерва (X)
Ядро
блокового
нерва (IV)
Моторное ядро
лицевого нерва (VII)
Моторное ядро
языкоглоточного
нерва (IX)
Моторное ядро
подъязычного
нерва (XII)
Моторное ядро
отводящего
нерва (VI)
Моторное ядро
блуждающего
нерва (X)
Моторное ядро
мандибулярной
ветви тройничного
нерва (V)
Моторное
ядро
добавочного
нерва (XI)
Рис. 69. Эмбриональная закладка черепно-мозговых нервов и их ганглиев.
а — формирование сенсорных нервов и ганглиев; б — формирование моторных нервов и
ганглиев.
Ядро
глазодвигательного
нерва (III)
происхождения (см. рис. 68). У млекопитающих в особых столбах соматиче-
ской чувствительности расположены зрительные центры крыши среднего мозга
(II нерв), улитковые и вестибулярные ядра (VIII нерв). С особой висцераль-
ной чувствительностью связаны вкусовые ядра VII и IX нервов. Особые вис-
церальные двигательные центры включают в себя ядра V, VII, IX, X и XI
нервов. Таким образом, общая топология распределения ядер в среднем, зад-
нем и продолговатом мозге соответствует гипотетической модели, представ-
ленной на рис. 68. Сходство расположения ядер черепно-мозговых нервов и
столбов спинного мозга особенно заметно у эмбрионов приматов (рис. 69).
Как видно на рис. 69, закладка сенсорных нервов происходит из дорсолате-
ральных, а моторных — из вентральных зон мозга. Определенную дисгармо-
нию в распределение закладок черепно-мозговых нервов вносят обонятель-
ный и зрительный нервы, которые являются особыми чувствительными не-
рвами и начинаются от обонятельной плакоды и ганглиозных клеток сетчатки
IV. Черепно-мозговые нервы
❖
115
соответственно. Поэтому при классификации черепно-мозговых нервов при-
нято выделять несколько групп нервов, различающихся как по историческому
происхождению, так и по принадлежности к соматической или висцеральной
части периферической нервной системы.
ОСОБЫЕ СОМАТИЧЕСКИЕ ЧУВСТВИТЕЛЬНЫЕ НЕРВЫ
К особым соматическим чувствительным нервам относят обонятельный нерв
(n. olfactorius; I), зрительный нерв (n. opticus; II) и слуховой нерв (n. acusticus;
VIII) (рис. 70). Лицевой нерв (n. facialis; VII), языкоглоточный нерв (n. glos-
sopharyngeus; IX) и блуждающий нерв (n. vagus; X) отнесе-
ны в эту категорию условно, поскольку в их составе име-
ются волокна латерального нерва боковой линии (у водных
позвоночных). Добавочный нерв (n. accessorius; XI) отно-
сить в эту группу не стоит. У водных обладателей иннерви-
руемой им трапециевидной мышцы он очень мал и входит
в состав X нерва как моторная веточка.
У низших позвоночных принято выделять передний и
задний нервы боковой линии (n. lateralis anterior, n. lateralis
posterior). Передний нерв входит в головной мозг обычно с
лицевым нервом. Это связано с тем, что он получает сен-
сорные волокна от органов боковой линии головы. Глаз-
ничная, щечная и гиомандибулярная ветви переднего нер-
ва боковой линии идут независимо от лицевого нерва, но
объединяются под общей оболочкой перед вхождением в
мозг. Задний нерв боковой линии может проникать в мозг
в составе как языкоглоточного, так и блуждающего нерва.
Задний нерв боковой линии включает в себя преимущест-
венно волокна, идущие от боковой линии, которая тянется
вдоль всего тела, и небольшое количество нервов от невро-
мастов задней части головы.
Рис. 70. Особые со-
матические чувст-
вительные нервы.
ЖАБЕРНЫЕ НЕРВЫ
Большую часть черепно-мозговых нервов составляют жаберные, или дорсаль-
ные, нервы. Название «жаберные, или дорсальные, нервы» сформировалось по
двум причинам. Во-первых, считается, что они произошли из гомологов дорсаль-
ных корешков спинного мозга. Жаберные нервы содержат чувствительный вис-
церальный и соматический компоненты, а также висцерально-двигательные во-
локна, что подтверждает их гомологию с дорсальными корешками спинного
мозга. Висцерально-двигательные, как и висцерально-чувствительные, волок-
на в некоторых жаберных нервах могут отсутствовать. Во-вторых, эти нервы
сформировались для обслуживания жаберной системы, что предопределяет их
сегментарность и единство анатомического плана строения. У современных
позвоночных следы этого сегментарного строения выражены достаточно ясно
116
Сравнительная анатомия нервной системы позвоночных
(рис. 71). Все черепно-мозговые жаберные нервы имеют характерное анатоми-
ческое отличие — ганглий, который служит подтверждением их происхождения.
У низших позвоночных обычно сохраняется примитивное строение жаберных
нервов. Из головного мозга они выходят в виде двух сливающихся корешков:
сенсорного и моторного. Гипотетический сенсорный корешок включает в себя
дорсальную ветвь — соматические сенсорные волокна (до сих пор не найденные)
и вентральную ветвь — висцеральные сенсорные волокна (рис. 72). Эти две ветви
переходят в сенсорный ганглий, после которого разделяются. Соматические во-
локна поднимаются дорсально, а висцеральные разделяются на три ветви: гло-
точную, прецщелевую и защелевую. Моторный корешок проходит мимо ганглия
и сливается с задней жаберной ветвью висцерального сенсорного нерва.
К жаберным нервам принято относить терминальный, или концевой, нерв
(n. terminalis; 0), глубокую глазничную ветвь тройничного нерва [n. trigeminus
(ramus ophthalmicus profundus); VJ, собственно тройничный нерв (n. trigeminus;
V2,V3), лицевой нерв (n. facialis; VII), языкоглоточный нерв (n. glossopharyn-
geus; IX), блуждающий нерв (n. vagus; X), добавочный нерв (n. accessorius; XI).
У некоторых видов висцерально-двигательные компоненты жаберных нервов могут
содержать глазодвигательный нерв (n. oculomotorius; III). Жаберные нервы неод-
нородны по составу, что позволяет разделить их на пять основных групп.
Общий соматический чувствительный компонент присутствует в терминаль-
ном нерве (n. terminalis; 0), глубокой глазничной ветви тройничного нерва
[n. trigeminus (ramus ophthalmicus profundus); VJ, собственно тройничном нер-
ве (n. trigeminus; V2. V3), лицевом нерве (n. facialis; VII), языкоглоточном нерве
a
Рис. 71. Распределение жаберных черепно-мозговых нервов.
а — предковая гипотетическая форма; б — распределение жаберных нервов у современ-
ных челюстноротых, s — брызгальце.
IV. Черепно-мозговые нервы ❖ 117
Сенсорный
корешок
Сенсорный
ганглий
Дорсальная ветвь
(соматическая
чувствительная
не выявлена)
Задняя жаберная ветвь
висцерального
чувствительного и
моторного нервов
Передняя
жаберная
ветвь
Моторный
корешок
висцерального
нерва
Глоточная ветвь
— висцерального
чувствительного
нерва
Жаберная щель
Рис. 72. Возможные компоненты жаберных нервов у рыб. Висцеральный чувствительный
нерв обозначен точками, соматический сенсорный — штриховкой.
(n. glossopharyngeus; IX), блуждающем нерве (n. vagus; X) и добавочном нерве
(n. accessorius; XI) (рис. 73). В VII и IX нервах общий соматический компо-
нент может отсутствовать или присутствовать в очень небольшом количестве.
Общий висцеральный чувствительный компонент и особый висцеральный чувст-
вительный компонент присутствуют в одних и тех же четырех жаберных нервах:
лицевом нерве (n. facialis; VII), языкоглоточном нерве (n. glossopharyngeus; IX),
блуждающем нерве (n. vagus; X) (рис. 74). К особой висцеральной чувстви-
тельности принято относить нервы, обслуживающие органы вкуса. Они вхо-
дят в состав VII, IX и X нервов. К специальным висцеральным чувствитель-
Рис. 73. Нервы, содержащие общий сома- Рис. 74. Нервы с общим и особым висце-
тический чувствительный компонент. ральным чувствительным компонентами
118
Сравнительная анатомия нервной системы позвоночных
Рис. 75. Нервы, содержащие особый вис- Рис. 76. Нервы, содержащие висцеральный
церальный двигательный компонент. двигательный компонент.
ным нервам относят обонятельный нерв (n. olfactorius; I), который формиру-
ется из отростков первично чувствующих клеток обонятельного эпителия.
Особый висцеральный двигательный компонент включает пять жаберных нер-
вов: собственно тройничный нерв (n. trigeminus; V2 3), лицевой нерв (n. facialis;
VII), языкоглоточный нерв (n. glossopharyngeus; IX), блуждающий нерв (n. vagus;
X) и добавочный нерв (n. accessorius; XI) (рис. 75).
Выделение этого компонента обусловлено тем, что наряду с обычной глад-
кой висцеральной мускулатурой в голове позвоночных присутствует особая
поперечнополосатая висцеральная мускулатура. Эта мускулатура обслуживает
висцеральные компоненты скелета и иннервируется особыми висцеральными
компонентами жаберных нервов. Нужно отметить, что этот компонент не сто-
ит рассматривать в качестве части автономной системы, поскольку он имеет
центральные механизмы контроля.
К «настоящей» автономной нервной системе относится общий висцераль-
ный двигательный компонент жаберных нервов. В него входят лицевой нерв
(n. facialis; VII), языкоглоточный нерв (n. glossopharyngeus; IX), блуждающий
нерв (n. vagus; X) и добавочный нерв (n. accessorius; XI) (рис. 76).
Для многих видов характерно наличие висцеральных двигательных волокон
в глазодвигательном нерве (n. oculomotorius; III), которые связаны с реснич-
ным ганглием.
ВЕНТРАЛЬНЫЕ НЕРВЫ
Соматические двигательные и соматические чувствительные нервы относят к
вентральным нервам, поскольку считается, что они произошли из гомологов
вентральных корешков спинного мозга. Они выходят и входят в головной мозг
IV. Черепно-мозговые нервы
119
в составе четырех черепно-мозговых нервов: глазодвигатель-
ном нерве (п. oculomotorius; III), блоковом нерве (n. troch-
learis; IV), отводящем нерве (n. abducens; VI) и подъязыч-
ном нерве (n. hypoglossus; XII) (рис. 77). Соматические чув-
ствительные волокна идут в составе двигательных нервов и
осуществляют как проприоцептивные функции, так и пере-
дачу сигналов от сухожилий и связок.
Таким образом, О, I, II и VIII пары черепно-мозговых
нервов являются чувствительными нервами, V, VII, IX, X —
смешанными нервами, содержащими как моторные, так и
чувствительные волокна, HI, IV, VI, XI и XII — двигатель-
ными нервами. В табл. 2 представлены черепно-мозговые
нервы млекопитающих и высокоорганизованных млекопи-
тающих. В некоторых случаях расположенные рядом череп-
но-мозговые нервы объединяются, что будет рассмотрено
ниже (рис. 78).
Черепно-мозговые нервы очень изменчивы в различных
группах позвоночных. Терминальный нерв отсутствует у
круглоротых и птиц, подъязычный не входит в состав чере-
па у рыб, добавочный нерв не отделен у низших позвоноч-
ных от блуждающего, а подъязычный нерв входит в состав
черепа (за счет трех зачатков позвонков, приросших к чере-
пу) У пресмыкающихся.
Рис. 77. Нервы, со-
держащие особый
висцеральный дви-
гательный компо-
нент.
Зрительный (II)
Лицевой (VII)
Слуховой (VIII)
Обонятельная
луковица
Отводящий (VI)
Языкоглоточный (IX)
Блуждающий (X)
Добавочный (XI)
Подъязычный (XII)
Рис. 78. Черепно-мозговые нервы млекопитающих на примере волка.
Глазо-
двигательный (III)
Блоковый (IV)
Тройничный (V)
120
❖
Сравнительная анатомия нервной системы позвоночных
Таблица 2. Черепно-мозговые нервы млекопитающих
Нерв Место выходы из мозга Функция
Обонятельный (1) Обонятельная луковица Сенсорный вход от обоня- тельных рецепторов
Зрительный (II) Зрительная хиазма Сенсорный вход от гангли- озных клеток глаза
Глазодвигательный (III) Перед мостом, на медиаль- Моторный выход к четырем ном крае ножки мозга из шести наружных мышц глазного яблока
Блоковый (IV) Дорсально. позади четверо- Моторный выход к перед- холмия ней косой мышце глазного яблока
Тройничный (V) Передний край варолиева Моторный выход к жева- моста, латерально тельным мышцам, основ- ной сенсорный вход от лица
Отводящий (VI) В борозде между мостом и Моторный выход к наруж- пирамидой ной прямой мышце глазно- го яблока
Лицевой (VII) Впереди и латерально от Основной выход к мышцам оливы лица, вход от вкусовых ре- цепторов
Слуховой (VIII) Латерально от оливы Сенсорный вход от слухо- вой и вестибулярной систем
Языкоглоточный (IX) В борозде позади оливы Вход от каротидного тела, вкусовых рецепторов, мотор- ный выход к мышцам зева, гортани и слюнным железам
Блуждающий (X) Позади языкоглоточного нер- Моторный выход к мышцам ва в борозде позади оливы сердца, легких и кишечника
Добавочный, состоящий Верхние корешки: позади Моторный выход к трапе-
из двух частей: верхних блуждающего нерва, ниж- циевидной мышце
корешков и нижних ние корешки: между перед-
корешков (XI) ними и задними корешка- ми шейных нервов
•Подъязычный (XII) Между пирамидой и оливой Моторный выход к мышцам языка
Концевой, или терминальный, нерв (nervus terminalis; 0). Этот нерв выявлен у
всех позвоночных, за исключением круглоротых и птиц. Считается, что он
возник из чувствительного дорсального корешка, гомологичного корешкам
спинного мозга (см. рис. 71). Для этих выводов существуют веские доказа-
тельства в виде сенсорных ганглиев терминального нерва. На сегодняшний
день известно четыре варианта анатомической организации терминального
нерва (рис. 79). У акул этот нерв начинается от различных учатвков переднего
мозга, включая терминальную пластинку. Однако для Hexanchus griseus изве-
стен выход терминального нерва в зоне вхождения обонятельного тракта в
IV. Черепно-мозговые нервы
121
а б в
Рис. 79. Терминальный нерв у различных позвоночных.
а — организация нерва у большинства акул, вид с вентральной поверхности мозга: б —
локализация терминального нерва yAmia, Dipnoi и Lepisosteus, вид с вентральной поверх-
ности: в — терминальный нерв, входящий в слившийся с передним мозгом чувствитель-
ный ганглий у Hexanchus griseus, вид с дорсальной поверхности.
мозг (см. рис. 79). Поэтому он может встречаться в литературе под названием
предобонятельного нерва (n. praeolfactorius). У этого вида акул терминальный
нерв не имеет видимого ганглия, который, судя по всему, сливается с пере-
дним мозгом. У других видов акул терминальный нерв имеет выраженный
ганглий. В некоторых случаях этот нерв распадается на две ветви, каждая из
которых имеет собственный чувствительный ганглий (см. рис. 79, о). Поскольку
нервы начинаются от терминальной пластинки или от медиальной поверхно-
сти переднего мозга, то они оказываются лежащими медиальнее обонятель-
ных нервов и трактов. У Am io, Dipnoi и Lepisosteusтерминальный нерв начина-
ется от вентральной зоны переднего мозга в непосредственной близости от
зрительной хиазмы. Такое положение нерва привело к возникновению еще
одного названия — «предзрительный нерв» (n. praeopticus). В этих группах
позвоночных нерв идет латеральнее обонятельного нерва и формирует оди-
ночный чувствительный ганглий У приматов и человека терминальный нерв
существует преимущественно в эмбриональном состоянии. Этот нерв являет-
ся чувствительным и иннервирует эпителий носовых полостей.
Обонятельный нерв (nervus olfactorius; I). Обонятельный нерв является чув-
ствительным. Он состоит из отростков рецепторных клеток обонятельного эпи-
телия, которые оканчиваются в обонятельных луковицах. Обонятельные луко-
вицы могут быть приближены к органам обоняния или сращены с головным
мозгом. У амфибий, рептилий и млекопитающих к обонятельному нерву при-
соединяется вомероназальный нерв (nervus vomeronasalis). Он начинается от ре-
цепторных клеток обонятельного эпителия вомероназального органа и окан-
чивается в собственной вомероназальной луковице (рис. 80). Она сливается с
обонятельной луковицей и анатомически не выделяется только у приматов и
122
❖
Сравнительная анатомия нервной системы позвоночных
Обонятельный
эпителий
Обонятельный
нерв
Обонятельная
луковица
Вомероназальная
луковица
Вомероназальный
эпителий
Вомероназальный
нерв
Рис. 80. Организация обонятельного и вомероназального нервов. Вид мозга с дорсальной
поверхности.
человека. У амфибий и некоторых рептилий вомероназальный нерв лежит лате-
ральнее обонятельного нерва, тогда как у большинства млекопитающих он
расположен медиальнее.
У низших позвоночных обонятельный нерв образует множественные пуч-
ки. У круглоротых, которые имеют только один обонятельный мешок, парные
обонятельные нервы собраны в компактные тяжи при входе в мозг. Обычно от
обонятельного органа отходит множество обонятельных волокон. Они доволь-
но самостоятельно достигают обонятельной луковицы и входят в нее. Иногда
волокна сливаются в 2, 3 пучка и более, которые входят в обонятельную луко-
вицу самостоятельно. От обонятельной луковицы начинается тракт, который
обычно один (рис. 81).
Для млекопитающих характерен выход обонятельного нерва через проды-
рявленную пластинку (lamina cribrosa) решетчатой кости. В этом случае обо-
нятельного нерва как такового нет, а есть многочисленные пучки раздель-
ных волокон.
Зрительный нерв (nervus opticus; II). Зрительный нерв является чувствитель-
ным нервом, берущим свое начало от ганглиозных клеток сетчатки глаза. Этот
черепно-мозговой нерв устроен достаточно однотипно. Участок от глаза до
хиазмы носит название зрительного нерва, а после хиазмы — зрительного тракта
(рис. 82). Для костистых рыб такое многообразие названий носит достаточно
условный характер, поскольку зрительный нерв от правого глаза переходит на
левую сторону мозга и наоборот без каких-либо морфологических изменений.
Хиазма у этих позвоночных является просто местом прохождения одного зри-
тельного нерва над другим. В этом случае происходит полный перекрест зри-
тельных волокон. Зрительный нерв правого глаза может проходить под не-
рвом от левого глаза и наоборот, что является, как правило, видоспецифичес-
кой анатомической особенностью. Для большинства хрящевых и костистых
рыб, амфибий и рептилий характерен полный перекрест зрительных волокон.
Однако для рыб и рептилий известны крайне своеобразные формы организа-
IV. Черепно-мозговые нервы ❖ 123
Рис. 81. Организация обонятельного нерва у различных позвоночных
а — акула, рыбы; б — амфибии, большинство рептилий, птицы, утконос; в, г — млекопита-
ющие. Участок с точками — обонятельная луковица, зачернен обонятельный нерв.
ции зрительной хиазмы, которая выглядит как сложное переплетение отдель-
ных пучков зрительных волокон (см. рис. 82). Для млекопитающих характерно
наличие хиазмы, в которой происходит только частичное перекрещивание зри-
тельных волокон. Часть волокон переходит на противоположную сторону мозга,
а часть остается на своей. Соотношение перекрещенных и неперекрещенных
волокон меняется в зависимости от относительной величины бинокулярного
поля зрения. Так, у зайцев и кроликов перекрещенных волокон больше, чем
неперекрещенных, а у приматов их количество примерно равно. Поскольку
зрительный нерв состоит из отростков ганглиозных клеток, его диаметр кор-
релирует с размером сетчатки, а строение хиазмы дает достоверную информа-
цию о степени бинокулярности зрения животного.
Зрительный тракт проникает в головной мозг на границе промежуточного
и среднего мозга. У низших позвоночных, рептилий и птиц он оканчивается
Рис. 82. Организация зрительного нерва и хиазмы у различных позвоночных.
а, е — млекопитающие; б — костистые рыбы; в — сельдевые; г, д — амфибии, большин-
ство рептилий и птиц.
124
Сравнительная анатомия нервной системы позвоночных
преимущественно в крыше среднего мозга, а у млекопитающих — как в ядрах
таламуса, откуда переключается в кору переднего мозга, так и в крыше сред-
него мозга.
Глазодвигательный нерв (nervus oculomotorius; III). Этот соматический мо-
торный нерв иннервирует четыре из шести наружных мышц глаза, которые в
развитии формируются из первого черепного сомита. В эту группу входят ниж-
няя, верхняя и передняя прямые и нижняя косая мышцы (m. recti: superior,
inferior, internus; m. obliquus inferior). В составе глазодвигательного нерва про-
ходят автономные волокна, иннервирующие мускулатуру ресничного тела и
радужки глаза. Они обеспечивают зрачковые рефлексы и аккомодацию глаза
(рис. 83). У большинства позвоночных автономная ветвь глазодвигательного
нерва проходит через ресничный ганглий. Обычно глазодвигательный нерв
одиночный и симметрично начинается от вентролатеральной поверхности сред-
него мозга. Однако имеется ряд интересных исключений как у низших, так и
у высших позвоночных. Для круглоротых характерна иннервация нижней пря-
мой мышцы волокнами, идущими в составе отводящего нерва (VI). У некото-
рых видов акул и чешуйчатников (Lepidosireri) глазодвигательный нерв, по-
видимому, может содержать чувствительные волокна, которые иннервируют
оболочки глаз и надхрящницу стенки глазницы. Наличие чувствительных во-
локон в этом нерве предполагается у бесхвостых амфибий. Считается, что они
могут иннервировать слизистую оболочку глаза.
Ьлоковый нерв (nervus trochlearis; IV). Этот соматический моторный нерв
иннервирует только верхнюю косую мышцу глаза (m. obliquus superior), кото-
рая формируется из второго предушного сомита. Он покидает дорсальную по-
верхность среднего мозга низших позвоночных позади тектума, а у млекопи-
тающих — ниже задних (слуховых) бугорков четверохолмия (рис. 84). Этот
нерв начинается от контралатеральных ядер, поднимается дорсально и обра-
зует перекрест. Поэтому он резко отличается от других двигательных нервов,
выходящих из мозга вентрально. У амфибий выявлена дополнительная ветвь,
начинающаяся ростролатеральнее основного ствола нерва. Есть сообщения
Рыбы, амфибии,
рептилии
Рептилии,
млекопитающие,
птицы
К нижней, верхней и
передней прямой и
нижней косой мышцам
Ресничный ганглий
К гладкой мускулатуре
ресничного тела и
радужки глаза
Рис. 83. Организация глазодвигательного нерва. Вид с вентральной поверхности
К нижней, верхней и
передней прямой и
нижней косой мышцам
IV. Черепно-мозговые нервы
125
Рис. 84. Организация блокового нерва позвоночных. Вид мозга с дорсальной стороны.
о наличии у рыб и амфибий чувствительных компонентов нерва, которые ин-
нервируют оболочку глаза (conjunctiva oculi). По-видимому, этим объясняется
наличие дополнительной передней ветви.
Тройничный нерв (nervus trigeminus; V). Это смешанный нерв, который обес-
печивает соматическую чувствительность головы и дает двигательные волокна
к челюстным мышцам. Тройничный нерв начинается от переднего края про
долговатого мозга несколькими крупными корешками, которые соединяются
в полулунный (тройничный) ганглий (ganglion semilunare). Из ганглия выхо-
дят три ветви: глубокая глазничная ветвь V] (ramus ophthalmicus profundus),
верхнечелюстная ветвь V2 (ramus maxillaris) и нижнечелюстная ветвь V3 (ramus
mandibularis).
Глубокая глазничная ветвь (V,) является соматическим чувствительным нер-
вом, который иннервирует покровы этмоидной части головы позвоночных.
Чувствительные волокна расположены в веках, слезных железах, слизистой
оболочке носовой полости и коже. Верхнечелюстная ветвь (V2) считается го-
мологом преджаберной ветви, а нижнечелюстная ветвь (V3) — гомологом за-
жаберной ветви типичного жаберного нерва. Обе ветви включают в себя сома-
тические чувствительные волокна, иннервирующие ротовую полость и боль-
шую часть поверхности головы. Чувствительные волокна нижнече. постной ветви
(V3) иннервируют покровы ниже разреза рта и нижнечелюстные зубы, а во-
локна верхнечелюстной ветви (V2) преимущественно связаны с верхней лице-
вой частью головы. Нижнечелюстная ветвь (V3) содержит волокна, иннерви-
рующие челюстную мускулатуру. Таким образом, тройничный нерв является
составным нервом, который не имеет выраженного висцерального чувстви-
тельного компонента.
Тройничный нерв возник в результате слияния двух жаберных нервов с
собственными ганглиями: глубокого глазничного нерва (n. ophthalmicus pro-
fundus, g. profundi) и верхненижнечелюстного нерва (n. maxillomandibularis s. tri-
geminus proprius, g. maxillomandibulare) (рис. 85).
126
Сравнительная анатомия нервной системы позвоночных
Некоторые amniota
Круглоротые, акулы
Многоперы
Млекопитающие
Рис. 85. Организация тройничного нерва позвоночных. Вид на мозг с вентральной поверхности.
У многоперов сохранилась эта древняя конструкция, хотя у остальных по-
звоночных оба ганглия сливаются и формируют полулунный ганглий
(g. semilunare). У рыб есть подчиненная веточка верхнечелюстной ветви трой-
ничного нерва (п. ophthalmicus superficialis), гомологичная лобной ветви чело-
века. Для круглоротых и акул характерно наличие четырех ветвей тройнично-
го нерва. Четвертая ветвь выходит из головного мозга отдельно от основного
ствола на границе заднего и среднего мозга и не входит в полулунный ганг-
лий. Поэтому ее часто называют таламическим, или среднемозговым, нервом
(п. thalamicus, s. n. mesencephalicus). Логичнее называть ее поверхностной глаз-
ничной ветвью тройничного нерва — ramus ophthalmicus superficialis (Vo).
У некоторых костистых рыб поверхностная глазничная ветвь проходит через
тройничный ганглий и выходит из него вместе с глубокой глазничной ветвью.
В таких случаях глубокая глазничная ветвь снабжена специальным ганглием,
который носит название ганглия глазничной ветви — g. ophthalmicus. У много-
перов поверхностная глазничная ветвь отсутствует, но тройничный нерв вы-
ходит из мозга двумя корешками: глубокой глазничной ветвью и слившимися
нижнечелюстной и верхнечелюстной ветвями (см. рис. 85). Для некоторых
амниот известен выход из мозга нижнечелюстной ветви (V3) в виде самостоя-
тельного нерва, который не проходит через ганглий. Эта веточка содержит
преимущественно двигательные волокна, иннервирующие мускулатуру челю-
стной (висцеральной) дуги. У млекопитающих нижнечелюстная ветвь (V3) фор-
мирует хорошо заметную чувствительную ветвь языка (ramus lingualis), кото-
рая может начинаться непосредственно от полулунного ганглия.
Отводящий нерв (nervus abducens; VI). Этот соматический моторный нерв
иннервирует только заднюю прямую мышцу глаза (т. rectus extemus), которая
образуется из третьего предушного сомита и контролирует отведение глаз на-
зад или вбок. У наземных позвоночных возникает дополнительная мышца,
IV. Черепно-мозговые нервы
❖
127
отвечающая за втягивание глаза (m. retractor bulbi). Она также развивается из
третьего сомита и иннервируется отводящим нервом. Эта же мышца контро-
лирует движение мигательной перепонки (третьего века) у рептилий и птиц.
У латимерии этот нерв иннервирует крупную парную
подчерепную мышцу (m. subcephalicus). Эта мышца
стелется по двублочной мозговой капсуле, при сокра-
щении опуская передний блок (etmosphenoideum) вместе
с верхней челюстью (рылом). При такой вариабельнос-
ти отводящий нерв выходит из мозга довольно однотип-
но — из вентральной поверхности заднего мозга, бли-
же к медиальной оси (рис. 86).
Лицевой нерв (nervus facialis; VII). Этот смешанный
нерв включает как чувствительные, так и двигательные
волокна. Исторически он сформировался как типичный
жаберный нерв, связанный с брызгальцем (рис. 87). По-
этому его чувствительные компоненты разделены на
висцеральные и соматические. Висцеральные чувстви-
тельные ветви иннервируют слизистую оболочку (от-
носящуюся к подъязычной дуге части глотки), ротовую
полость и вкусовые луковицы [нёбная ветвь (г. palatinus;
VII), (г. mandibularis intemus; VII) (s. chorda tympani)].
Ветвление лицевого нерва происходит медиальнее брыз-
Рис. 86. Организация от-
водящего нерва позво-
ночных. Вид мозга с вент-
ральной поверхности.
Брюшной
корешок
Рыбы, водные
амфибии
Спинной
корешок
Глазничная
ветвь
Щечная
ветвь
Коленчатый
ганглий
Барабанная
струна
Костистые
рыбы
Наземные
амфибии
Глоточная
ветвь
Защелевая
ветвь
VII
Змеи
Рис. 87. Организация лицевого нерва позвоночных. Вид мозга с вентральной поверхности.
V, VII, VIII — черепно-мозговые нервы.
128 ❖ Сравнительная анатомия нервной системы позвоночных
гальца, от него назад идет толстая гиомандибулярная ветвь (г. posttrematus),
впереди от брызгальца — тонкая рудиментарная (г. prespiracularis; VII) и доволь-
но толстая нёбная ветвь (г. palatinus; VII), которая сформировалась в результа-
те гипертрофии глоточной ветви (г. pharyngeus). Соматические чувствитель-
ные компоненты VII нерва (части предушного латерального нерва) включают
в себя три ветви: поверхностную глазничную (г. ophthalmicus superficialis; VII),
щечную (г. buccalis; VII) и гиомандибулярную, которая продолжается в виде
наружной нижнечелюстной ветви (г. mandibularis externus; VII). Висцеральные
двигательные волокна иннервируют мускулатуру, принадлежащую гиоидной
дуге. Лицевой нерв у амфибий и рептилий часто объединяется с тройничным
нервом, что приводит к значительным проблемам его анатомического выделе-
ния и анализа отдельных ветвей (см. рис. 87). У костистых рыб часто встреча-
ется разделение лицевого нерва на четыре практически равных корешка пря-
мо при выходе из мозга.
Кроме этих ветвей, у многих костистых рыб существуют особые вкусовые
(спинные) ветви VII нерва (г. lateralis; VII), которые распространяются от че-
репа назад, проходят вдоль остистых отростков позвонков и оканчиваются в
покровах и плавниках. Для большинства бесхвостых амфибий характерно сбли-
жение лицевого нерва с тройничным и слуховым нервами. У многих змей
встречается слияние тройничного и лицевого нервов, что маскирует распреде-
ление двигательных и чувствительных корешков. У рыб гиомандибулярная
ветвь составляет большую часть этого нерва. Эта ветвь проходит позади брыз-
гальца и иннервирует мускулатуру гиоидной дуги. От нее отходит крупный
пучок волокон, называемый оперкулярной или жаберной ветвью (ramus
opercularis), который идет к жаберной крышке. В этой ветви содержатся как
висцеральные двигательные, так и чувствительные волокна. У млекопитаю-
щих из мускулатуры гиоидной дуги формируются лицевые мышцы. Этот факт
и послужил причиной названия нерва. Для защелевой ветви тетрапод харак-
терно наличие крупной барабанной струны, которая у зауропсид проходит
через полость среднего уха и поворачивает вперед, давая чувствительные во-
локна во вкусовые луковицы и нижнюю челюсть.
Слуховой нерв (nervus acusticus; VIII) (n. vestibulocochlearis). Этот чувстви-
тельный нерв идет от внутреннего уха. Между внутренним ухом и мозгом рас-
положен ганглий, который состоит из двух частей: преддверной (g. vestibulare)
и улитковой (g. cochleare). Эти ганглии могут быть разделены и располагаться
на одноименных нервах: преддверном нерве (n. vestibularis) и улитковом нерве
(п. cochlearis). Эти ганглии могут лежать и в самом внутреннем ухе в непосредст-
венной близости от рецепторов. Улитковый нерв связан со слуховой систе-
мой, а преддверный — с сенсорной системой рецептора углового ускорения
(полукружными каналами) и рецептором линейного ускорения (гравитацион-
ным рецептором). Такая организация нерва характерна для большей части
амфибий, рептилий, птиц и млекопитающих (рис. 88). Однако у рыб и водных
амфибий улитковый нерв практически не выражен, хотя во внутреннем ухе
есть невзрачный сосочек (papilla neglecta), которому приписывают функции
слуховой системы.
Языкоглоточный нерв (nervus glossopharyngeus; IX) (рис. 89). Этот жаберный
нерв является как чувствительным, так и двигательным висцеральным нервом.
IV. Черепно-мозговые нервы
129
Как жаберный нерв он выходит из мозга от-
дельным корешком и имеет характерный ка-
менистый ганглий (g. petrosum). У млекопи-
тающих, птиц, рептилий и большинства ам-
фибий из ганглия обычно выходит одна ветвь,
а у рыб — две ветви; передняя ветвь (ramus
praetrematicus) и задняя ветвь (ramus posttrema-
ticus). Передняя вервь дает волокна к пере-
дней стенке жаберного мешка, глотке и сли-
зистой стенке нёба, а задняя — к мышцам
первой жаберной дуги. Часть волокон задней
ветви языкоглоточного нерва рыб и водных
амфибий связана с электрорецепторными
органами и боковой линией. У рыб и амфи-
бий он может включать в себя соматические
чувствительные волокна. Языкоглоточный
нерв сформировался в результате специали-
зации жаберного нерва, обслуживавшего первую типичную жаберную щель.
Он иннервирует часть висцеральной поперечнополосатой мускулатуры глот-
ки, гортани, надгортанник и слюнные железы. Висцеральные чувствительные
волокна этого нерва расположены в ротовой полости и языке и обслуживают
вкусовые почки, лежащие в корне языка и отвечающие (у приматов) за вос-
приятие горького.
У птиц вкусовая чувствительность не очень хорошо развита, поэтому чувст-
вительные волокна этого нерва выполняют преимущественно осязательные
функции.
Рис. 88. Организация слухового
нерва позвоночных. Вид на мозг с
вентральной поверхности.
а — слуховой ганглий; б — вести-
булярный ганглий.
Первичноводные
позвоночные
Рис. 89. Организация языкоглоточного нерва позвоночных. Вид на мозг с вентральной по-
верхности.
5—5818
130
Сравнительная анатомия нервной системы позвоночных
❖
Блуждающий нерв (nervus vagus; X) (рис. 90). Это крупный жаберный нерв,
содержит все виды висцеральных волокон и особые чувствительные волокна
боковой линии, которые принадлежат соматической системе. У низших вод-
ных позвоночных он иннервирует покровы всех жаберных щелей, кроме пер-
вой, покровы глотки, включая вкусовые почки, поперечнополосатую мускула-
туру жаберных дуг начиная со второй. Блуждающий нерв имеет крупный ярем-
ный ганглий (g. jugulare). От этого ганглия у низших позвоночных начинаются
две ветви: жаберно-кишечная (ramus branchio-intestinalis) и латеральная [ramus
lateralis (ramus splanchnicus)[. Каждая жаберная веточка формирует сочетание
волокон, описанное выше (см. рис. 72). У акул сначала (почти самостоятель-
но) отходит первая жаберная ветвь, затем — латеральный нерв, после — третья
и четвертая жаберные ветви. Позади этого продолжение жаберно-кишечной
ветви представляет собой внутренностную ветвь.
У рыб блуждающий нерв обслуживает значительное количество вкусовых
почек, а у млекопитающих — только почки корня языка, ответственные за
рецепцию горького. Благодаря этому нерву возникает рвотный рефлекс при
попадании горького (сигнал о ядовитости пищи) на корень языка. Наличие
специальной внутренностной ветви характерно для всех позвоночных. Эта ветвь
иннервирует желудок, кишечник, легкие и сердце. Через эту ветвь проходят
висцеральные чувствительные волокна и парасимпатические волокна авто-
номной системы. У тетрапод она начинается непосредственно от ганглия, а у
низших позвоночных выделяется только в качестве небольшой ветви основ-
ного ствола (см. рис. 90). У личинок и водных амфибий латеральная ветвь
выходит из головного мозга в виде отдельного корешка, который обладает
собственным ганглием.
Некоторые
амфибии
Первичноводные
позвоночные
Ганглий
X нерва
Латеральная
Ганглий
латеральной
ветви
Ганглий
X нерва
Латеральная
ветвь
Млекопитающие
Жаберная
ветвь
Ганглий
X нерва
ветвь
Рис. 90. Организация блуждающего нерва позвоночных. Вид на мозг с вентральной поверх-
ности.
IV. Черепно-мозговые нервы
О
131
Добавочный нерв (nervus accessorius;
XI) (рис. 91). Этот нерв содержит вис-
церальные двигательные волокна и
считается выделившимся моторным
компонентом X нерва. Действитель-
но, у всех позвоночных, за исключе-
нием млекопитающих, добавочный
нерв выходит из мозга в составе ко-
решков блуждающего нерва и может
начинаться от шейной части спинно-
Млекопитающие
Амфибии,
рептилии
Рис. 91. Организация добавочного нерва по-
звоночных. Вид на мозг с вентральной по-
верхности.
го мозга, как у птиц. Поэтому уверен-
ное анатомическое выделение XI нер-
ва возможно только у млекопитающих
и некоторых видов рептилий.
Подъязычный нерв (nervus hypoglossus; XII). Это соматический двигательный
нерв. У низших позвоночных, включая амфибий, подъязычный нерв находится
вне черепа и рассматривается как первый спинномозговой нерв. Однако у ис-
копаемых форм амфибий доказано его внутричерепное положение. У большин-
ства животных он выходит из продолговатого или спинного мозга нескс " экими
брюшными корешками, волокна которых сливаются в общий нерв (рис. 92).
Рис. 92. Организация подъязычного нерва позвоночных. Вид на мозг с вентральной поверх-
ности.
132 ❖ Сравнительная анатомия нервной системы позвоночных
У птиц этот нерв начинается частично от продолговатого, а частично от
спинного мозга. Этот нерв иннервирует мышцы, формирующиеся из соми-
тов затылочной области. Производные затылочных миотомов расположены у
рыб вокруг жаберной области и формируют поджаберную или подъязычную
мускулатуру, а у млекопитающих — и мышцы языка. Поэтому у Anamnia он
иннервирует дно ротовой полости, а у Amniota — язык и гипобрахиальную
мускулатуру.
V. продолговатый и задним мозг
Продолговатый и задний мозг позвоночных является
сосредоточением чувствительных и двигательных цент-
ров. Они содержат сенсорные и моторные ядра череп-
но-мозговых нервов, моторные ядра восходящих и ни-
сходящих путей, мозжечок, а также нервные волокна,
соединяющие спинной и головной мозг. Между задним
и продолговатым мозгом взрослого животного нет рез-
кой границы. Принцип такого подразделения двигатель-
ных отделов пришел из эмбриологии. У млекопитаю-
щих в эмбриональный период границы между этими
отделами довольно четкие, что и привело к появлению
двойного названия. Наиболее часто употребляют назва-
ние «ромбовидный мозг» (rhombencephalon), или ствол
головного мозга. Это название сложилось под влияни-
ем как внешнего вида отдела, так и его IV желудочка,
который у млекопитающих имеет ромбовидную форму.
Граница между ромбовидным и спинным мозгом мле-
копитающих проходит по нижнему краю перекреста пи-
рамидных пучков волокон. У низших позвоночных та-
кой границей считают место, где центральный канал
спинного мозга принимает вид круга. Ростральной гра-
ницей ромбовидного мозга является воображаемая, а
иногда и реальная линия, которая начинается от перед-
него края мозжечка и заканчивается позади сосцевид-
ных тел. Такое положение границ отдела характерно для
млекопитающих, хотя у других позвоночных ситуация
иная (рис. 100).
Задний и продолговатый мозг в целом повторяет стро-
ение спинного мозга (см. рис. 68). Расположение мо-
торных и сенсорных столбов спинного мозга сохраня-
ется, но полость IV желудочка и мозжечок придают сис-
теме дополнительные особенности. Моторные столбы
спинного мозга трансформируются в ретикулярную фор-
мацию, которая является основным моторным центром
отдела.
134 ❖ Сравнительная анатомия нервной системы позвоночных
У некоторых рептилий, птиц и млекопитающих IV желудочек с дорсальной
поверхности не виден. Его закрывают червь и парные полушария мозжечка.
У многих костистых рыб (рис. 93, в) на дне желудочка возникают парные или
одиночные бугорки, которые могут выпирать из полости выше краевых сте-
нок заднего мозга. В некоторых случаях желудочек закрывается не мозжеч-
ком, а разрастающимися латеральными стенками заднего и продолговатого
мозга (рис. 93, д). Рассмотреть форму IV желудочка можно только после уда-
ления складчатой эпендимной пленки, которая является дорсальной стенкой
большей части ромбовидного мозга. Эта стенка является гематоэнцефаличе-
ским барьером между мозговыми желудочками и задним сосудистым сплете-
нием. Сосудистое сплетение ромбовидного мозга очень хорошо развито как у
низших, так и у высших позвоночных. Однако максимального развития оно
достигает у круглоротых, двоякодышащих и некоторых хрящевых рыб. У осет-
ровых кровообращение в сосудистом сплетении ромбовидного мозга столь хо-
рошо развито, что в нем происходит образование клеток крови. Сосудистое
сплетение выступает в качестве кроветворного органа.
Рис. 93. Внешний вид головного мозга различных животных
а — минога; б — колючая акула; в — карп; г — травяная лягушка; д — геккон; е — сетчатый
питон; ж— голубь; з — кролик; и — собака. Участок с точками — задний и продолговатый
мозг, зачернен IV желудочек.
\J. Продолговатый и задний мозг О- 135
Ромбовидный мозг может составлять от 14 до 95% от общей массы головно-
го мозга, что зависит от уровня развития нервной системы животного. Общая
тенденция такова, что чем выше развитие мозга и сложнее поведение живот-
ного, тем меньшую долю составляет ромбовидный мозг. У птиц и приматов
эта тенденция нарушается, что обусловлено чрезмерным развитием мозжечка,
который необходим для сложной координации движений. Нужно отметить,
что в процессе эволюции мозжечок неоднократно достигал высокого развития
даже у первичноводных животных, но затем значительно редуцировался у ам-
фибий и рептилий (рис. 94, 95).
Рис. 94. Сагиттальные сечения головного мозга различных животных.
а — минога; б—колючая акула; в — латимерия;г — травяная лягушка. Участок с точками —
задний и продолговатый мозг. Ill, IV — желудочки мозга.
136
❖
Сравнительная анатомия нервной системы позвоночных
Рис. 95. Сагиттальные сечения головного мозга различных животных.
а — черепаха; б — голубь; в — собака. Участок с точками — задний и продолговатый мозг.
Ill, IV — желудочки мозга.
Сагиттальные сечения головного мозга различных животных покашвают,
что анатомические соотношения между продолговатым, задним мозгом, моз-
жечком и сосудистым сплетением IV желудочка изменяются. Причины таких
анатомических различий связаны как с моторными, так и с сенсорными цент-
рами. С одной стороны, на размер двигательного отдела мозга влияет форма
активности животного. У первичноводных позвоночных продолговатый и зад-
ний мозг развиты слабее, чем у наземных позвоночных. Более сложно разви-
тый задний мозг тетрапод обусловлен появлением конечностей и переходом к
наземному образу жизни. С другой стороны, развитие мозжечка связано с
\/. Продолговатый и задний мозг ❖ 137
адаптацией животных к ориентации и активным движением в трехмерной среде.
У акул и костистых рыб мозжечок развит во много раз больше, чем у амфибий.
Водные позвоночные двигаются в трехмерной среде, а большинство амфибий —
в двухмерной. Поэтому у земноводных мозжечок оказался вторично упрощен.
Не меньшее влияние на размер и форму ромбовидного мозга оказывает орга-
низация черепно-мозговых нервов позвоночных. Их представительство в виде
моторных и сенсорных центров на уровне ромбовидного мозга иногда оказы-
вает решающее влияние на форму отдела и IV желудочка. Примером могут
служить гигантские вкусовые или электрорецепторные ядра костистых рыб.
Поскольку у животных черепно-мозговые нервы развиты в различной сте-
пени, то и их мозговые центры могут быть различного размера и формы,
что и отражается на анатомическом строении ромбовидного мозга (рис. 96).
Существуют достаточно надежные морфофункциональные корреляции между
анатомическим строением ромбовидного мозга и развитостью сенсорных и
Рис. 96. Латеральные проекции ядер черепно-мозговых нервов в среднем, заднем и про-
долговатом мозге различных животных.
а — акулы; б— костистые рыбы; в — амфибии; а — рептилии; д — птицы; е — млекопита-
ющие. Ill—XII — ядра черепно-мозговых нервов. Ядра, выделенные внутри общего конту-
ра, морфологически не разделены (по A. Kappers, 1921).
138 ❖ Сравнительная анатомия нервной системы позвоночных
моторных ветвей черепно-мозговых нервов. Кроме этого, часть ядер черепно-
мозювых нервов появляется только у рептилий, птиц и млекопитающих.
Как видно, у низших позвоночных ядра черепно-мозговых нервов зани-
мают положение, соответствующее сенсорным и моторным столбам спинно-
го мозга (см. рис. 96, а, б). По мере усложнения мозга, а особенно с выходом
на сушу и появлением теплокровности топологические соответствия между
спинным и ромбовидным мозгом маскируются новыми ядрами черепно-моз-
говых нервов. При этом происходит дифференциация ядер, которые перво-
начально были представлены обобщенными структурами. Примером может
служить разделение общего ядра лицевого (VII), языкоглоточного (IX) и блуж-
дающего (X) нервов на три независимых компонента. У хрящевых рыб все
три ядра этих нервов слиты в общую структуру и могут быть разделены толь-
ко при использовании специальных маркеров (см. рис. 96, а). В связи с раз-
витием сложной чувствительности у костистых рыб ядро VII нерва выделяет-
ся в отдельную структуру, а два оставшихся мозговых центра IX и X нервов
остаются объединенными. Только у птиц происходит анатомическое разде-
ление ядер IX и X нервов (рис. 96, д). Однако нужно учитывать, что мышцы
певчей (нижней) гортани птиц иннервируются подъязычным нервом. У мле-
копитающих уже невозможно заподозрить существование связи между ядра-
ми IX и X нервов, поскольку IX нерв отделен от блуждающего X нерва уве-
личившимся центром подъязычного (XII) нерва (см. рис. 96, е).
В самом сложном случае, у млекопитающих, черепно-мозговые нервы в
ромбовидном мозге представлены 30 ядрами, которые разделяются на мотор-
ные и сенсорные центры. Они, как правило, имеют достаточно обособленные
входы, которые часто называют порциями нервов, сенсорными или моторны-
ми ветвями или даже корешками.
В составе ромбовидного мозга млекопитающих находятся ядра начиная с
глазодвигательного (III) (основная часть ядра лежит в среднем мозге) и кон-
чая XII нервом; всего 14 сенсорных и 16 моторных ядер.
Глазодвигательный нерв (nervus oculomotorius; III) представлен тремя мотор-
ными ядрами:
1) крупноклеточным латеральным ядром (nucleus lateralis);
2) мелкоклеточным латеральным ядром (Эдингера—Вестфаля) (nucleus lateralis);
3) крупноклеточным медиальным ядром (Пеля) (nucleus medialis).
Блоковый нерв (nervus trochlearis; IV) представлен только одним моторным
ядром:
ядром блокового нерва (nucleus nervus trochlearis).
Тройничный нерв (nervus trigeminus; V) на входе в мозг разделен на две пор-
ции: большую и малую. Большая порция является сенсорной и начинается от
полулунного (гассерова) ганглия. Она формирует в мозге три сенсорных ядра
тройничного нерва:
1) главное ядро моста (nucleus principalis pontinus);
2) ядро нисходящего корешка (nucleus radicis descendentis);
3) ядро одиночного тракта (частично) (nucleus tractus solitarii).
Малая порция тройничного нерва начинается из двух моторных ядер:
1) ядра нисходящего среднемозгового корешка (nucleus radicis descendentis,
mesencephalicae);
\J. Продолговатый и задний мозг ❖ 139
2) жевательного ядра моста (nucleus masticatorius pontinus).
Отводящий нерв (nervus abducens; VI) представлен одним моторным ядром:
ядром отводящего нерва (nucleus nervus abducentis).
Лицевой нерв (nervus facialis; VII) начинается из моторного ядра:
ядра лицевого нерва (nucleus nervi facialis).
Слуховой нерв (nervus acusticus; VIII) входит в мозг в виде двух слившихся
нервов. Улитковый нерв (nervus cochlearis) начинается от спирального улитко-
вого ганглия и оканчивается в двух сенсорных ядрах:
1) дорсальном ядре (слуховом бугорке) (nucleus dorsalis s. tuberculi acustici);
2) вентральном ядре (nucleus ventralis).
Преддверный нерв (nervus vestibularis) начинается от вестибулярного ганг-
лия и оканчивается в четырех сенсорных ядрах ромбовидного мозга:
1) угловом ядре (Бехтерева) (nucleus angularis);
2) гигантоклеточном ядре (Дейтерса) (nucleus magnocellularis);
3) вестибулярном ядре нисходящего корешка (Роллера) (nucleus radicis
desendentis vestibularis);
4) треугольном ядре (Швабе) (nucleus triangularis).
Языкоглоточный нерв (nervus glossopharyngeus; IX) имеет двигательный и сен-
сорный компоненты, которые связаны с четырьмя ядрами в мозге. Сенсорный
компонент берет свое начало в каменистом ганглии и, войдя в мозг, формиру-
ет два ядра:
1) дорсальное ядро латеральной порции блуждающего нерва (nucleus dorsalis
vagi lateralis);
2) ядро одиночного тракта (nucleus tractus solitarii).
Моторная порция языкоглоточного нерва начинается также в двух ядрах:
1) вентральном (двойном) ядре блуждающего нерва (nucleus ventralis vagi
s.ambiguus);
2) нижнем слюноотделительном ядре (nucleus salvatorius inferior).
Блуждающий нерв (nervus vagus; X) является смешанным нервом и фор-
мирует четыре ядра, которые составляют значительную часть продолгова-
того мозга. Сенсорная порция этого нерва начинается от яремного (узлова-
того) ганглия и, войдя в мозг, оканчивается в тех же ядрах, что и языкогло-
точный нерв:
1) дорсальном ядре латеральной порции блуждающего нерва (nucleus dorsalis
vagi lateralis);
2) ядре одиночного тракта (nucleus tractus solitarii).
Моторная порция блуждающего нерва начинается также в двух ядрах:
1) вентральном (двойном) ядре блуждающего нерва (nucleus ventralis vagi
s. ambiguus);
2) дорсальном ядре медиальной порции блуждающего нерва (nucleus dorsalis
vagi medialis).
Добавочный нерв (nervus accessorius; XI) является двигательным нервом и
берет свое начало в двух ядрах:
1) мозговом ядре добавочного нерва (nucleus nervi accessorii cerebralis), ко-
торое является каудальной частью вентрального (двойного) ядра блуждаю-
щего нерва;
140
Сравнительная анатомия нервной системы позвоночных
❖
2) спинальном ядре добавочного нерва, которое состоит из дорсальной и
вентральной частей (nucleus dorsalis nervi accessorii spinalis, nucleus ventralis nervi
accessorii spinalis).
Подъязычный нерв (nervus hypoglossus; XII) является смешанным нервом, в
котором на основании мозгового представительства часто выделяют самосто-
ятельную промежуточную порцию — промежуточный нерв (nervus intermedius).
Двигательная порция начинается в двух ядрах:
1) ядре подъязычного нерва (nucleus nervi hypoglossi);
2) верхнем слюноотделительном ядре (nucleus salivatorius superior).
Часть волокон проприоцептивные. Они начинаются в коленчатом ганглии,
а приходят в ядро X нерва:
ядро одиночного тракта (nucleus tractus solitarii).
Таким образом, черепно-мозговые нервы вносят значительный вклад в фор-
мирование ромбовидного мозга. В зависимости от развитости тех или иных
ядер продолговатый и задний мозг приобретают складки, выпячивания и про-
гибы, которые могут служить анатомическим эквивалентом адаптивных изме-
нений черепно-мозговых нервов.
Кроме ядер черепно-мозговых нервов, в ромбовидном мозге млекопитаю-
щих расположено несколько комплексов ядер, которые являются вторичными
сенсорными и моторными центрами, принадлежащими как к восходящим и
нисходящим путям черепно-мозговых нервов, так и к спинному мозгу. Эти
центры можно объединить в несколько групп по топографическим и функци-
ональным признакам.
Восходящие волокна из спинного мозга в составе нежного и клиновидного
пучков формируют каудальный комплекс продолговатого мозга, включающий
два ядра с одноименными названиями:
1) ядро нежного пучка (nucleus fasciculi gracilis);
2) ядро клиновидного пучка (nucleus fasciculi cuneati).
К ядрам дугообразного комплекса моста относят два скопления:
1) дугообразные ядра (nuclei arcuati);
2) ядра моста (nuclei pontis).
В состав оливарного комплекса входят шесть ядер:
1) нижнее оливное ядро (nucleus olivaris inferior);
2) дорсальное добавочное оливное ядро (nucleus olivaris accessorius dorsalis);
3) медиальное добавочное оливное ядро (nucleus olivaris accessorius medialis);
4) верхнее оливное ядро (nucleus olivaris superior);
5) ядро трапециевидного тела (nucleus corporis trapezoidei);
6) ядро латеральной петли (nucleus lemnisci lateralis).
Кроме перечисленных ядер, в состав ромбовидного мозга входят ядра моз-
жечка, которые будут рассмотрены ниже. Локализация и форма центров ром-
бовидного мозга отражают эволюционный уровень развития двигательных сен-
сорных и моторных функций (рис. 97—99).
Приведенные сечения ромбовидного мозга различных позвоночных дают
представление о связи между уровнем развития двигательной активности жи-
вотных и изменениями в морфологической организации нервной системы.
Ромбовидный мозг играет огромную роль как центр управления двигатель-
ной активностью. Через него проходят или оканчиваются все двигательные
V. Продолговатый и задний мозг
141
❖
сенсорные и моторные волокна. Эти структуры можно разделить на две груп-
пы: восходящие и нисходящие пути соматической чувствительности и анало-
гичные пути от мышечных, сухожильных и связочных рецепторов (рис. 100).
Восходящие пути соматической чувствительности входят в ромбовидный
мозг как со стороны спинного мозга, так и через корешки головных нервов.
В процессе эволюции у позвоночных сформировалось два восходящих пути
соматосенсорной системы: латеральный и дорсальный. Под латеральным при-
Рис. 97. Сечения заднего мозга на уровне мозжечка (по R. Nieuwenhuys et al., 1997, с
изменениями).
142
Сравнительная анатомия нервной системы позвоночных
Ambystoma tigrinum
Columba livia
Testudo hermanni
Stratum Purkinje
N. vestibularis
ventrolateralis
N. vestibularis
tangentialis
N. vestibularis
descendens
N. nervi V
N. magnocellularis
Oliva
superior
N. reticularis
medius
N. cochlearis
lamina ris
IV
N. motorius
nervi VII
N. raphes
N. reticularis
gigantocellularis
N. reticularis
parvocellularis
paragigantocellularis
cochlearis
Рис. 98. Сечения заднего мозга на уровне мозжечка.
нято понимать спиноталамический путь млекопитающих и рептилий, спино-
тектальный путь у амфибий и птиц и спиноретикулярный у первичноводных
животных. Название зависит от центра, в котором оканчиваются восходящие
волокна. Латеральный соматосенсорный путь существует у всех позвоночных
и является наиболее древним образованием. Через этот путь передаются боле-
вые и тактильные сигналы, поступает информация о температуре тела, а у
тетрапод — и от некоторых суставов. Он начинается от чувствительных нейро-
нов, лежащих в чувствительных ганглиях дорсальных корешков спинного мозга
(см. рис. 100). Отростки этих нейронов входят в спинной мозг, переходят на
противоположную сторону и образуют восходящий латеральный пучок воло-
V. Продолговатый и задний мозг
143
Didelphis virginiana
N. prepositus
hypoglossi
N interpositus
anterior
N. lateralis
cerebelli
N. fastigii
Flocculus
Ss
N vestibularis
N. motorius
nervi VII
N. cochlearis
ven trails
N. cochlearis
dorsalis
N. reticularis
gigantocellularis
N. reticularis
gigantocellularis
pars ventralis
N. spinalis nervi V
N. reticularis
parvocellularis
Stratum Purkinje
Рис. 99. Сечения заднего мозга на уровне мозжечка (по R. Nieuwenhuys et al., 1997, с
изменениями).
кон, называемый спиноталамическим трактом. Этот тракт разрыхляется в ром-
бовидном мозге и формирует многочисленные коллатерали к ядрам ретику-
лярной формации. Однако основной мишенью этого восходящего пути у мле-
копитающих является таламус, а у рыб и амфибий — крыша среднего мозга.
В таламусе млекопитающих волокна спиноталамического тракта переключа-
ются в соматосенсорную кору.
У рептилий впервые появляется филогенетически новый путь соматосен-
сорных волокон. Они проводят сигналы от проприорецепторов и тактиль-
ных рецепторов глубоких слоев покровов. Именно из этих волокон форми-
руются задние или дорсальные столбы белого вещества спинного мозга: кли-
новидный и нежный пучки. Эти волокна оканчиваются в ядрах задних столбов,
которые носят одноименные названия: «клиновидное ядро» и «нежное ядро»
Эти ядра расположены у самого каудального края продолговатого мозга. Выхо-
дящие из ядер задних столбов волокна переходят через среднюю линию и фор-
мируют путь медиальной петли (лемнисковый), который оканчивается в тала-
мусе. Этот путь у рептилий и млекопитающих обычно называют лемниско-
144
Сравнительная анатомия нервной системы позвоночных
Клиновидное ядро
Новая кора
Ретикулярная формация
Ядра V нерва
Лицо -»
Латеральный
тракт
Касание,
температура,
боль
Новая кора
Таламус
Медиальный лемниск
Ядро2
Нежное ядро
Задние столбы
Нейроны сенсорных
ганглиев Спинной мозг
Проприорецепция,
тактильная чувствительность,
давление, вибрация
Рис. 100. Восходящие пути ромбовидного мозга.
вой системой. От волокон лемнискового пути отходят коллатерали в ядра
ретикулярной формации.
Коллатеральные волокна из старого и филогенетически новых соматосен-
сорных путей оканчиваются почти во всех центрах ретикулярной формации
ромбовидного мозга. Нейроны этой структуры формируют собственные вос-
ходящие волокна, которые входят в восходящую ретикулярную систему. Эта
система оканчивается в таламусе у млекопитающих и в среднем мозге у рыб и
амфибий. Волокна ретикулярной восходящей системы участвуют в запуске
механизмов пробуждения и стимулируют неспецифическую активность мозга
животных.
Второй группой восходящих систем ромбовидного мозга тетрапод являются
волокна, идущие от мышечных и суставных рецепторов конечностей. Восхо-
дящие пути от передних и задних конечностей существенно различаются. Чувст-
вительные волокна, идущие от передней конечности, начинаются от сенсор-
ного нейрона, лежащего в ганглии задних корешков спинного мозга. Отростки
этого нейрона входят в спинной мозг и затем раздваиваются. Одни коллатерали
участвуют в местных нейронных взаимодействиях, а другие формируют восхо-
дящий путь. Местные спинномозговые связи организуют работу сегментар-
ных автономных рефлексов, тогда как восходящие волокна принимают учас-
тие в координации движений. Волокна от передней конечности поднимаются к
ядрам задних столбов на границе спинного и продолговатого мозга. У млекопи-
\/. Продолговатый и задний мозг
145
о-
тающих от клиновидного и нежного ядер этот восходящий путь продолжается
через медиальный лемниск и оканчивается в таламусе, где волокна переклю-
чаются на кору У ни зших тетрапод волокна переключаются в передней части
спинного мозга и формируют основные связи со средним мозгом.
Восходящие волокна от задней конечности также начинаются от сенсорных
нейронов задних корешков. Волокна этих нейронов раздваиваются в спинном
мозге, формируя сегментарные рефлекторные сети и восходящие пути. От зад-
ней конечности формируется два основных пути. Один путь начинается от
спинальных нейронов, расположенных в ядрах специализированного столба
(Кларка). Они формируют спинно-мозжечковый восходящий пучок волокон,
который переключается в ядрах мозжечка и оканчивается в корковом мозжеч-
ковом слое. У млекопитающих на пути к ядрам мозжечка спинно-мозжечко-
вый пучок дает коллатерали к ретикулярным ядрам ствола, которые в составе
медиального лемнискового пути достигают таламуса. От таламуса начинаются
восходящие волокна в сенсомоторную кору.
У низших тетрапод, лишенных неокортекса, восходящий путь несколько иной.
Волокна начинаются от предшественников специализированного столба (Кларка)
и оканчиваются практически во всех ядрах таламуса. По пути в таламус они
дают многочисленные коллатерали в ретикулярную формацию ромбовидного
мозга, мозжечок, крышу среднего мозга и ядра тройничного нерва. От рост-
ральных ядер таламуса начинаются две группы волокон, которые оканчиваются
в стриатуме и плаще переднего мозга. Для первичноводных животных характер-
но окончание волокон от мускулатуры туловища в ретикулярных ядрах, в ядре
VIII нерва, тегментуме и крыше среднего мозга. Только единичные волокна
проникают в таламические центры и некоторые ядра стриатума.
Нисходящие волокна ромбовидного мозга в основном обслуживают двига-
тельные системы организма. Управление движением позвоночных построено
по иерархическому принципу: чем выше уровень двигательного центра, тем
сложнее и генерализованнее сокращение мускулатуры организма. На самом
нижнем уровне этой системы лежат сегментарные двигательные пути, которые
обслуживают самые простые рефлекторные реакции и не выходят за границы
спинного мозга. Однако моторные нейроны спинного мозга, выполняя про-
стые функции, направляют «копии» выполненых команд в вышележащие цен-
тры. Такая обратная связь моторных центров носит название реафферентации и
позволяет контролировать параллельную активность моторных нейронов.
Высшие моторные центры, формирующие нисходящие волокна, у млеко-
питающих начинаются в моторной коре, проходят через базальные ганглии,
мозжечок, ретикулярную формацию ромбовидного мозга и оканчиваются на
моторных нейронах спинного мозга. Высшим моторным центром у птиц явля-
ется стриатум, у рептилий, амфибий и рыб — крыша среднего мозга. У всех
позвоночных животных нисходящие двигательные системы продублированы
параллельными путями. Одновременно существуют «длинный» и «короткий»
нисходящие моторные пути. При прохождении команды по «длинному» пути
происходят последовательные переключения волокон во всех перечисленных
выше структурах. В «коротком» пути осуществляются прямые связи между
высшими центрами и мотонейронами спинного мозга. Понятно, что «корот-
кий» путь быстрее, но менее интегрирован с сопредельными двигательными
146
0-
Сравнительная анатомия нервной системы позвоночных
центрами. «Короткие» пути сформировались для обслуживания быстрых реф-
лекторных реакций животных, тогда как сложная двигательная активность об-
служивается «длинными» путями (рис. 101).
Моторные ядра нисходящих двигательных путей расположены в ромбовид-
ном мозге в виде нескольких центров. Наиболее обширен комплекс ретикуляр-
ных ядер. Они имеют относительно небольшие размеры и распределены в сред-
ней части ствола. Из ретикулярных ядер начинаются прямые волокна ретику-
лоспинального тракта, оканчивающиеся на моторных нейронах спинного мозга.
Это наиболее древний центр управления движением. Именно в ретикулярных
ядрах моста расположены те крупные нейроны, которые управляют движением
хвоста у рыб и амфибий. Эти клетки называют мюллеровскими и маутнеров-
скими нейронами соответственно. По-видимому, они являлись предтечами мо-
торных центров, на что указывает большое количество (4—10) мюллеровских
клеток у миног и миксин. По паре маутнеровских клеток имеется у костистых
рыб, хвостатых амфибий и личинок лягушек. Эти клетки, возбуждаясь, затор-
Двигательная кора
Красное ядро
Ядра моста
Верхние бугорки
четверохолмия
Пирамидные
клетки коры
Таламус
Двигательная кора
Базальные ядра
расное ядро
Мозжечок
Вестибулярное ядро
Пирамидные
клетки коры
Ретикулярные
ядра моста
1 ll(llIVVVi
Мотонейроны
спинного мозга
Рис. 101. Нисходящие пути ромбовидного мозга млекопитающих. Непрерывная линия
«длинный», пунктирная — «короткий» путь нисходящих волокон. Оба варианта прохожде-
ния волокон могут присутствовать как в пирамидной, так и в экстрапирамидной системе
управления движением. У низших позвоночных нисходящие пути начинаются от крыши
среднего мозга и ретикулярной формации.
Бульбарные
ретикулярные ядра
V. Продолговатый и задний мозг
147
❖
маживают друг друга и одновременно быстро передают импульс, запускающий
сокращение мускулатуры на одной стороне хвоста. Этими клетками обеспечи-
ваются быстрые реакции избегания у имеющих хвост водных видов позвоноч-
ных. Есть сведения, что при помощи этих клеток исходный сигнал передается
на конец спинного мозга, что позволяет рыбам плыть хвостом вперед. У тетра-
под из ретикулярных ядер также начинаются волокна, оканчивающиеся на мо-
торных нейронах спинного мозга. В ретикулярных ядрах даже млекопитающих
еще можно встретить гигантские нейроны, сходные по цитологическому строе-
нию с маутнеровскими клетками амфибий. Они осуществляют контроль за сег-
ментарной активностью спинного мозга при движении конечностей.
На основании объединения ядер вестибулярных нервов и центров ретику-
лярной формации сформировался комплекс вестибулярных ядер. Эти ядра
являются источником информации для стимуляции активности мюллеровских
и маутнеровских нейронов. У млекопитающих вестибулярный комплекс со-
стоит из четырех ядер, два из которых формируют нисходящие пучки воло-
кон. Латеральное вестибулярное ядро имеет смешанное строение и включает в
себя крупноклеточное ядро Дейтерса. Из ядра Дейтерса и от других клеток
латерального ядра начинаются нисходящие волокна вестибулоспинального
тракта, которые оканчиваются на мотонейронах, иннервирующих работу мышц
разгибателей конечностей. В составе продольного нисходящего пути участву-
ют волокна, берущие свое начало от медиального вестибулярного ядра. Ниж-
нее и медиальное ядра млекопитающих в нисходящих путях прямого участия
не принимают. Они дают волокна к мозжечку.
У амфибий, рептилий, птиц и млекопитающих появляется еще один мор-
фологически обособленный источник нисходящих волокон — красное ядро
(мезенцефалическое). Во всех трех группах этот центр на разрезах свежего
мозга выглядит как красноватое пятнышко, что и предопределило название
ядра. В составе ядра выделяются крупные клетки, которые сходны по строе-
нию с гигантскими клетками ядра Дейтерса. Нисходящие от красного ядра
волокна формируют руброспинальный тракт, а коллатеральные связи оканчи-
ваются в ядрах ретикулярной формации. Наибольшее развитие этого тракта
произошло у рептилий и птиц, тогда как у млекопитающих он менее выражен
по сравнению с другими нисходящими путями. У млекопитающих, рептилий
и птиц через красное ядро осуществляется управление работой мышц сгибате-
лей конечности. У хвостатых амфибий красное ядро выполняет аналогичные
функции, но оно структурно объединено с ядром медиального (промежуточ-
ного) продольного пучка [nucleus (intersticialis) fasciculus longitudinalis medialis].
Красное ядро бесхвостых амфибий уже отделено от ядра медиального про-
дольного пучка, но анатомически выделяется крайне плохо. У двоякодыша-
щих рыб и латимерии красного ядра выявить не удалось. У млекопитающих к
этой группе центров подключаются кора и подкорковые ядра. Поскольку нис-
ходящие двигательные пути исторически описывали преимущественно у мле-
копитающих, то возникло не совсем логичное разделение нисходящих волокон
на две большие группы: пирамидный и экстрапирамидный нисходящие пути.
Причина такой классификации состоит в том, что на вентральной поверхности
продолговатого мозга млекопитающих находятся вытянутые бугорки, которые
носят название пирамид. Они являются наружными выпячиваниями нисходя-
148 ❖ Сравнительная анатомия нервной системы позвоночных
щих двигательных волокон. Все центры, волокна которых проходят через них,
относят к пирамидной системе, а те. которые идут в составе других нисходя-
щих пучков, приписывают к экстрапирамидной системе. К пирамидной сис-
теме принято относить ретикулярную формацию, вестибулярные ядра, крас-
ное ядро, подкорковые двигательные ядра и моторную кору. К экстрапира-
мидной системе относят двигательные ядра продолговатого мозга, верхние
бугорки четверохолмия и некоторые бульбарные ядра (см. рис. 101). Нужно
от метить, что первоначально у примитивных хордовых сформировалась пер-
вичная пирамидная система, но, естественно, без коры и базальных ганглиев.
Интегрирующей двигательный центр сформировался в среднем мозге. Таким
образом, экстрапирамидная система возникла как высший ассоциативный центр
головного мозга рыб, амфибий и рептилий. Функции неокортекса в этих группах
выполнял средний мозг, что привело к формированию новых путей, управля-
ющих моторными нейронами спинного мозга. При переходе к высшим репти-
лиям, птицам и млекопитающим осуществлялось параллельное «достраива-
ние» древнего, пирамидного, и нового, экстрапирамидного, путей. У репти-
лий «достройка» сохранила следы двойственности Часть волокон от обеих
систем оканчивается в стриатуме, а часть — в плаще переднего мозга. Для
птиц «достройка» завершилась на уровне подкорковых центров, а у млекопи-
тающих — в коре. Представительство обеих систем сохранилось и на корко-
вом уровне. Следовательно, экстрапирамидный и пирамидный нисходящие
пути представляют собой две параллельные системы управления двигательны-
ми нейронами спинного мозга. Они обладают многочисленными взаимными
связями на уровне спинного мозга, ретикулярной формации, таламуса и коры,
что интегрирует их влияние на моторную активность животных.
МОЗЖЕЧОК
Мозжечок занимает особое положение как в сравнительной анатомии, так и
в сенсомоторном управлении двигательной активностью. С анатомической точ-
ки зрения это весьма разнообразная структура мозга. Мозжечок формируется в
виде небольшой складки у переднего края заднего мозга. Он является произ-
водной структурой заднего мозга и образует с ним основные связи. Существует
устойчивая точка зрения, что организация мозжечка отражает эволюционный
уровень рассматриваемой группы животных (см. рис. 94. 103). В самом общем
виде эта закономерность действительно имеет место, но многочисленные ис-
ключения не позволяют ее использовать 1 ак инструмент анатомического ана-
лиза. Примером может быть организация мозжечка у акул. Мозжечок кошачьей
акулы (Pristiurus melanostomus) состоит из двух небольших складок, а у лососевой
акулы (Lamna ditropis) он примерно в 20 раз больше, с мощно развитыми склад-
ками поверхности. У костистых рыб мозжечок может достигать колоссального
развития и составлять до 85% от объема головного мозга.
Пример с акулами показывает, что даже в одной систематической группе
мозжечок может очень различаться. Мозжечок у миног неразличим макроско-
пически. Он идентифицируется только гистологически — по характерному кле-
точному составу (рис. 102, а). Для акул и костистых рыб характерно огромное
\J. Продолговатый и задний мозг
❖
149
д
Рис. 102. Сагиттальные сечения мозжечка различных животных.
а — минога; б— кунья акула; в — амия; г — форель; б — латимерия; е — травяная лягуш-
ка; ж— аллигатор; з — зеленая мартышка. На сагиттальных сечениях заметно, что размер
мозжечка не всегда соответствует эволюционному положению группы.
разнообразие форм и размеров мозжечка, в котором отражаются особенности
биологии каждого вида. У латимерии мозжечок состоит из двух небольших скла-
док и организован намного проще, чем у многих видов костных и хрящевых
рыб. У большинства хвостатых и бесхвостых амфибий мозжечок крайне незна-
чительного размера, что отражает незатейливую моторную координацию зем-
новодных. Небольшое увеличение размеров мозжечка наблюдается у рептилий.
Однако только у птиц в связи с развитием сложных форм двигательной актив-
150 ❖ Сравнительная анатомия нервной системы позвоночных
ности мозжечок заметно усложняется. Максимального развития он достигает у
млекопитающих, что обусловлено развитием корково-мозжечковых связей.
Формирование перекрещивающихся пучков волокон, идущих от мозжечкового
комплекса к коре, приводит к возникновению моста, который радикально ме-
няет внешний вид мозга млекопитающих (см. рис. 96).
Мозжечок является частью двигательной системы, но не имеет прямых свя-
зей с чувствительными клетками и мускулатурой. Он является своеобразной
надстройкой над восходящими и нисходящими путями и центрами ромбовид-
ного мозга. В функции мозжечка входят сенсомоторная координация, управ-
ление системами поддержания равновесия и регуляция мышечного тонуса.
Это не означает, что у животных с небольшим мозжечком перечисленные
функции редуцированы. Для двигательных систем характерна морфофункцио-
нальная параллельность обеспечения функций. Но при небольшом мозжечке
уровень интеграции двигательной активности становится ниже.
Мозжечок у акул, костистых рыб, птиц и млекопитающих имеет складча-
тую поверхность. Неглубокие складки обычно называют бороздками, а глубо-
кие — долевыми бороздами. Последние отделяют друг от друга морфофункцио-
нальные отделы мозжечка. Площадь поверхности мозжечка может достигать
огромных размеров. У человекообразных обезьян и многих копытных размеры
поверхности мозжечка могут быть больше, чем у коры переднего мозга (рис. 103).
На «развертках» поверхности мозжечок выглядит как довольно однородная
структура. На самом деле он состоит из трех основных частей, которые отра-
жают его эволюционное становление (рис. 104).
Рис. 103. Геометрические «развертки» поверхности мозжечка млекопитающих. По срезам
были сделаны графические реконструкции поверхности мозжечка, которые отражают его
реальные размеры (по R. Nieuwenhuys et al., 1997).
V. Продолговатый и задний мозг
151
Lobus anterior
Архицеребеллум
в
Рис. 104. Соотношение основных топографических отделов мозжечка у рептилий (а), птиц
(б) и млекопитающих (в). Сходная маркировка применена для одинаковых отделов моз-
жечка в различных группах позвоночных (по A. Kappers, 1921, с изменениями).
Анатомическую эволюцию мозжечка относительно точно отражает его раз-
деление на архицеребеллум, или старый мозжечок (archicerebellum), древний
мозжечок, или палеоцеребеллум (paleocerebellum), и новый мозжечок, или нео-
церебеллум (neocerebellum). Однако существует старая морфологическая тра-
диция выделять центральную (осевую) часть, называемую червем, и парные
полушария мозжечка. На самом деле это только анатомические складки еди-
ной поверхности мозжечка. Тем не менее их анатомическая номенклатура со-
вершенно необходима, поскольку каждый из структурных отделов мозжечка
точно соответствует масштабам представительства двигательных центров.
Наиболее древней частью мозжечка является старый мозжечок, или палео-
церебеллум (paleocerebellum). У млекопитающих его называют нижней, или
задней, долей мозжечка (lobus inferior). Он состоит из небольшой нодулярной
доли (uvula, nodulus) и латеральных ответвлений — миндалины и клочков моз-
жечка (tonsila, flocculus, paraflocculus). Палеоцеребеллум отделяется от неоце-
ребеллума глубокой препирамидальной бороздой (fissura secunda). К архице-
ребеллуму приходят волокна от вестибулярных центров. Размеры этой струк-
152
Сравнительная анатомия нервной системы позвоночных
❖
туры и анатомическое развитие довольно точно отражают уровень организа-
ции вестибулярного аппарата животного.
Архицеребеллум занимает переднюю долю (lobus superior) и отделен от ос-
тального мозжечка первичной бороздой (fissura prima). В архицеребеллуме вы-
деляют язычок (lingula), которому соответствует в полушарии уздечка мозжеч-
ка, центральную дольку (lobulus centralis), которой соответствует крыло цент-
ральной дольки (ala lobuli centralis), и холмик (montriculus), состоящий из
верхушки (culmen) и ската (declive), которые продолжаются в полушария в
виде четырехугольных долей (lobuli quadrangulares). В четырехугольной доле
выделяют переднюю и заднюю части (pars anterior, pars posterior) (рис. 105).
В архицеребеллуме оканчиваются преимущественно волокна восходящих
путей спинного мозга, которые идут в составе спинно-мозжечковых трактов
По ним доставляется информация от мышечных рецепторов. Нужно отме-
тить, что вентральная часть спинно-мозжечкового тракта присутствует у всех
позвоночных, а дорсальная впервые появляется у рептилий. У птиц и млеко-
питающих она получает дальнейшее развитие, что приводит к значительному
увеличению ее размеров по сравнению с вентральным трактом.
Старая номенклатура
Новая номенклатура
Vinculum lingulae
Ala lobuli centralis
Fissura
prima
Lingula
Cu men
Pars posterior
Dechve
Lobulus gracilis
Pyramis
Uvu a
Tonsi a
Nodulus
Paraflocculus
Flocculus
Foccu us
Lobulus
simplex
Lobulus
anterior
Lobulus
ansiform is
crus II
Lobulus
paramed ianus
Paraflocculus
dorsalis
Paraflocculus
ventralis
Lobulus ansiformis
crus I
Lobulus
centralis
Fohum
vermis
Fuber
vermis
Lobu us biventer
lateralis, medians
Lobuli semilunares
supenores
Lobuli semi unares
mferiores
Рис. 105. Основные отделы мозжечка млекопитающих по разным номенклатурам
Fissura
secunda
V. Продолговатый и задний мозг
153
❖
Неоцеребеллум впервые появляется только у рептилий. У млекопитающих
он занимает медиальную или заднюю долю мозжечка (lobus posterior). В пере-
дней части неоцеребеллума расположены лист червя (folium vermis) и бугор
червя (tuber vermis). Листу червя в полушариях соответствуют нижние и верх-
ние полулунные доли (lobuli semilunares supriores, lobuli semilunares inferiores),
а бугру червя — тонкая долька (lobulus gracilis). К неоцеребеллуму относят и
пирамиду червя (pyramis), которой в полушариях соответствует двубрюшная доль-
ка (lobulus biventer). У приматов и человека она расположена на нижней повер-
хности и обычно относится к архицеребеллуму (нижней доле), что вносит пута-
ницу в номенклатуру. К неоцеребеллуму приходят волокна от ядер моста и
нижних олив продолговатого мозга.
Приведенное выше описание мозжечка относится к его организации у мле-
копитающих, у которых он принимает наиболее сложную форму. Однако у ам-
фибий он выглядит крайне упрощенно даже по сравнению с гаковым акул и
костистых рыб. которые имеют развитый вестибулярный аппарат и тонко орга-
низованную координацию последовательных сокращений туловищной муску-
латуры. У первичноводных позвоночных, амфибий, рептилий и птиц полуша-
рия мозжечка практически отсутствуют. Они представлены только небольшими
латеральными выпячиваниями флоккулонодулярной доли. По сути дела в этих
группах мозжечок состоит из одного червя, в котором выделяются перечислен-
ные выше отделы. Большую часть мозжечка низших позвоночных составляет
развитая срединная часть, которую называют телом мозжечка (corpus cerebelli).
В этой зоне происходит интеграция сигналов от мышечных рецепторов, сен-
сорных входов и вестибулярной системы. Только у млекопитающих появляются
развитые полушария, что опосредовано развитием коры переднего мозга.
Мозжечок низших позвоночных соединен с задним мозгом непрерывной
пластинкой, в которой пучки волокон анатомически не выделяются. У млеко-
питающих эти пучки формируют три пары столбообразных структур, которые
носят название ножек мозжечка (pedunculi cerebelli). Через ножки осуществ-
ляются связи мозжечка с остальной центральной нервной системой. Задние
ножки состоят преимущественно из проприоцептивных волокон, которые
приходят из спинного мозга. Средние ножки состоят из волокон, соединяю-
щих мозжечок и кору переднего мозга, а передние ножки сформированы из
эфферентных волокон, соединяющих мозжечок и средний мозг.
У рыб, амфибий и большей части рептилий мозжечковые волокна непо-
средственно соединены со средним, ромбовидным и спинным мозгом. Для
крокодилов, птиц и млекопитающих характерно соединение мозжечка с дру-
гими центрами мозга через специализированные ядра нисходящих и восходя-
щих путей (рис. 106). Однако у высших позвоночных древние связи мозжечка
сохраняются наравне с вновь образованными путями. Примером может слу-
жить связь коры мозжечка с латеральным вестибулярным ядром. Между чер-
вем мозжечка и латеральным вестибулярным ядром существует прямая связь у
всех позвоночных животных начиная с круглоротых. Вестибулярно-мозжеч-
ковые связи определяют способность животных координировать движения тела,
что является основной функцией архицеребеллума. У птиц и млекопитающих
происходит расширение представительства структур, вовлеченных в вестибу-
лярные регуляции. Это приводит к появлению связей между латеральным
154
Сравнительная анатомия нервной системы позвоночных
Передний мозг
Красное ядро Вестибулярное ядро
Рис. 106. Основные нисходящие и восходящие связи мозжечка.
а — восходящие волокна; б — нисходящие волокна.
вестибулярным ядром и медиальной зоной коры червя, которые осуществляют-
ся через ядро шатра (nucleus fastigii). Аналогичные изменения происходили в
процессе эволюции и со связями между красным ядром и мозжечком. У низших
тетрапод связи между красным ядром и червем мозжечка осуществляются без
посредников. Однако при развитии коры млекопитающих в эти связи вовлека-
ются прилежащие к червю зоны, что приводит к появлению специализирован-
ного центра переключения — пробковидного ядра (nucleus emboliformis). Даль-
нейшее развитие неоцеребеллума привело к возникновению зубчатого ядра
мозжечка (nucleus dentatus). Через это ядро осуществляется третий путь комму-
никаций между красным ядром и мозжечком. Появление новых и более мощ-
ных путей не редуцирует древние связи между структурами. Они сохраняются в
качестве быстрых упреждающих и настраивающих волокон, которые подготав-
ливают мозговые центры к приему основного потока информации.
Наряду с системой дублирования древних связей мозжечка у млекопитаю-
щих возникают и совершенно новые пучки волокон. Наиболее значительным
приобретением мозжечка в связи с развитием полушарий неоцеребеллума яв-
ляется зубчатое ядро. Оно получает волокна от различных участков полуша-
рий мозжечка и передает сигналы в вентролатеральное ядро таламуса (nucleus
ventrolateralis). В этом центре происходит интеграция сенсомоторных сигна-
лов с активностью корковых центров переднего мозга. Эволюция мозжечка не
V. Продолговатый и задний мозг
155
ограничивается параллельным дублированием нисходящих и восходящих пу-
тей. Практически во всех случаях происходит формирование коллатеральных
связей и установление новых последовательных путей. Примером может быть
зубчатое ядро, которое у птиц и рептилий передает основное количество воло-
кон к красному ядру, а у млекопитающих — к вентролатеральному ядру тала-
муса. Красное ядро и коллатеральные связи мозжечка с ретикулярными ядра-
ми ствола позволяют поддерживать мышечный тонус и осуществлять рефлек-
торные реакции. Связи с вестибулярным центром позволяют осуществлять
контроль за положением тела в пространстве, а таламические связи предопре-
деляют тонкие сенсомоторные координации.
Все эти процессы осуществляются за счет сложной системы межклеточных
взаимодействий на уровне коры мозжечка (cortex cerebelli). Основные принципы
организации коры мозжечка сходны в различных трупах позвоночных. Сложнее
всего организована и лучше всего исследована кора у млекопитающих (рис. 107).
Кора мозжечка разделена на три слоя: гранулярный слой (самый глубокий),
слой клеток Пуркинье и молекулярный слой (поверхностный).
Информация в мозжечок приходит по трем основным типам волокон. Наи-
более обширный поток информации приходит по мшистым волокнам, кото-
рые начинаются в ядрах моста, а оканчиваются на отростках клеток-зерен.
Волокна, идущие от ядер нижних олив, носят название лиановидных, или
лазящих, волокон. Эти волокна получили свое название из-за того, что, по-
добно лианам, оплетают отростки клеток Пуркинье. Каждое лиановидное во-
локно образует синапсы только на одной клетке Пуркинье. Большое количе-
ство волокон приходит в кору мозжечка из голубого ядра, но они распределя-
ются по всей поверхности коры, не отдавая предпочтение определенному типу
клеток. Нисходящие волокна коры мозжечка начинаются от клеток Пуркинье
и направляются к ядрам нисходящего пути мозжечка или к центрам в других
Рис. 107. Основные принципы строения коры мозжечка млекопитающих.
а — слой клеток-зерен; б — слой клеток Пуркинье; в — молекулярный слой; г — астроцит;
д — клетки-зерна; е — моховидные волокна; ж — клетки Гольджи; з — клетки Пуркинье;
и — корзинчатые клетки; к — звездчатые клетки; л — лиановидные волокна; м — белое
вещество мозжечка.
156 ❖ Сравнительная анатомия нервной системы позвоночных
отделах головного мозга. Внутри коры существует строгая упорядоченность
связей между клетками. Аксоны клеток-зерен приходят в молекулярный слой,
разветвляются на два Т-образно расположенных волокна, которые тянутся в
противоположных направлениях. Они оканчиваются на клетках Пуркинье,
отростки которых лежат в плоскости, перпендикулярной аксонам клеток-зе-
рен. Интегративные функции выполняют корзинчатые клетки, отростки ко-
торых оплетают большие группы клеток Пуркинье. Аналогичные функции
выполняют звездчатые клетки и клетки Гольджи.
Общая схема организации мозжечка первичноводных позвоночных имеет
ряд специфических отклонений, что позволяет реконструировать происхож-
дение этой структуры. Наиболее интересной особенностью мозжечка кости-
стых рыб является наличие заслонки (valvula). Это переднее разрастание моз-
жечка, которое может проникать в полость III желудочка или выходить на
дорсальную поверхность мозга. У некоторых видов (Gnathonemus petersii} за-
слонка мозжечка принимает столь большие размеры, что может скрывать боль-
шую часть переднего и заднего мозга. Заслонка получает те же афферентные и
эфферентные волокна, что и мозжечок. Однако в заслонку большее количе-
ство волокон поступает из среднего мозга, а в тело мозжечка — из заднего и
продолговатого мозга.
В мозге некоторых костистых рыб непосредственно под мозжечком распо-
ложены специфические центры, которые обслуживают электрорецепторы.
У рыб с развитыми электрорецепторными системами мозжечок может быть
выражен значительно меньше, чем электрорецепторные ядра {Clarias anguillaris,
Xenomystus notopterus). Электросенсорные центры обладают связями, сходны-
ми с афферентными и эфферентными системами мозжечка. Их строение час-
то напоминает крайне упрошенный вариант организации коры мозжечка. По-
видимому, вполне вероятно предположение о происхождении мозжечка из
электрорецепторных центров, которые утратили свое функциональное значе-
ние. На первом этапе придонные животные, обитавшие в непрозрачном иле,
приобрели развитую систему электрорецепторов. Последние позволяли им хо-
рошо ориентироваться и разыскивать пищу в мутном придонном слое. При
придонном существовании потребность в интеграции двигательной активно-
сти и работе вестибулярного аппарата относительно невелика. Однако при
переходе к активному плаванию в толще воды возникла необходимость в сен-
сомоторной интеграции, но отпала необходимость в электрорецепции. Основ-
ными дистантными анализаторами стало зрение, обоняние и органы боковой
линии. В это время на структурной основе органов электрорецепции сформи-
ровалась интегрирующая надстройка над сенсомоторной системой — мозже-
чок. После утраты электрорецепторных входов мозжечок стал эволюциониро-
вать исключительно как аналитическая надстройка над сенсомоторной и вес-
тибулярной системами.
VI. СРЕДНИМ МОЗГ
Средним мозгом принято называть участок централь-
ной нервной системы, расположенный между ростраль-
ным краем мозжечка и эпифизом. На латеральной и
вентральной поверхностях мозга его границами явля-
ются мост и складка, образованная мамиллярными те-
лами (у низших позвоночных — задний край воронки).
У млекопитающих средний мозг окружает самый узкий
желудочек — сильвиев водопровод (aqueductus cerebri,
Sylvii), который у низших позвоночных представлен
широким «зрительным желудочком» (ventriculus opticus).
На дорсальной поверхности средний мозг виден не у
всех позвоночных. У первичноводных животных он
может быть прикрыт разросшимся мозжечком, а у мле-
копитающих и птиц — каудальным краем полушарий
переднего мозга (рис. 108—110).
Средний мозг исторически сложился как первичный
фительный и сейсмосенсорный центр. У всех позво-
ночных, за исключением млекопитающих, зрительные
волокна, идущие от ганглиозных клеток сетчатки, окан-
чиваются преимущественно в ростральной части кры-
ши среднего мозга. У первичноводных и амфибий дор-
сальная стенка среднего мозга полностью занята зри-
тельным центром, который называется крышей среднего
мозга, или зрительной крышей (tectum mesencephali,
opticum). У многих низших позвоночных зрительные
центры формируют парные полушария, которые могут
доминировать над другими мозговыми отделами, если
зрение для данного вида является ведущей системой аф-
ферентации (см. рис. 108—110). У млекопитающих и
человека дорсальная стенка состоит из четырех бугор-
ков, что отразилось и на ее названии — «пластинка чет-
верохолмия» [lamina quadrigemina (tecti)J. Передние бу-
горки — зрительные, а задние — слуховые.
Соотношение между передними и задними бугорка-
ми млекопитающих может быть эффективным крите-
158
Сравнительная анатомия нервной системы позвоночных
Рис. 108. Внешний вид головного мозга различных животных.
а — минога; б—колючая акула; в—карп; г—травяная лягушка; д — геккон; е — сетчатый
питон; ж — голубь; з — кролик; и — собака. Участок с точками — средний мозг.
рием оценки превалирования слуховой или зрительной системы. Если на
дорсальной поверхности мозга видно, что передние (верхние) бугорки раз-
виты более, чем задние, то можно уверенно предполагать преимущественное
развитие зрения. Такая картина наблюдается у копытных, многих хищных и
приматов. Если доминируют задние (нижние) бугорки, то более развитой
системой афферентации является слух. Это особенно ярко выражено у дель-
финов и летучих мышей, которые обладают развитым слухом в высокочас-
тотном диапазоне. У рептилий четверохолмие выражено значительно мень-
ше, что затрудняет морфофункциональную оценку развития слухового и зри-
тельного анализаторов. Задние бугорки рептилий расположены не в одной
плоскости с передними, а подвернуты в складку, образованную средним моз-
гом и мозжечком. Поэтому визуальная оценка относительных размеров этих
структур затруднена.
Дорсальная поверхность среднего мозга у низших позвоночных преимущест-
венно отражает организацию и степень развития зрительных центров. Это свя-
зано с тем, что зрительные волокна после зрительного перекреста поднимают-
VI. Средний мозг
159
Пинеальный орган Сосудистое сплетение в крыше среднего мозга
Средний мозг
Передний мозг
Гипофиз
б
Рис. 109. Сагиттальные сечения головного мозга различных животных.
а — минога; б—колючая акула; в—латимерия; г—травяная лягушка. Участок с точками —
средний мозг. Ill, IV — желудочки мозга.
ся от вентральной поверхности промежуточного мозга и оканчиваются в пере-
дней части крыши среднего мозга. У большей части рыб глаза хорошо разви-
ты, а основным анализаторным центром является средний мозг. Зрительные
центры доминируют над остальным мозгом и являются наиболее сложно орга-
низованной системой (рис. 109, 113). Поскольку зрение рыб и амфибий явля-
ется ведущей системой афферентации, то и основные ассоциативные центры
расположены в среднем мозге. Это связано с тем, что вентральная часть сред-
него мозга является продолжением двигательных центров ствола мозга.
160
Сравнительная анатомия нервной системы позвоночных
Сосудистое сплетение
Пинеальный орган
Сосудистое сплетение
Передний
мозг
б
Мозжечок
Гипофиз
в
Рис. 110. Сагиттальные сечения головного мозга различных животных.
а — черепаха; б — голубь; в — собака. Участок с точками — средний мозг. Ill, IV — желу-
дочки мозга.
Ствол фактически является непрерывной структурой, которая плавно пере-
ходит из продолговатого мозга в задний и оканчивается на границе промежу-
точного и среднего мозга. На основе интеграции зрительного анализатора с
другими сенсорными входами формируется центр, который выполняет слож-
ные ассоциативные функции, аналогичные таковым в коре переднего мозга
млекопитающих. Эта область расположена в ростровентральной части средне-
го мозга. Она является центром принятия решений у первичноводных позво-
ночных, амфибий и многих рептилий.
В среднем мозге позвоночных расположены основные слуховые центры.
Они возникли на основе мезенцефалического центра сейсмосенсорной чувст-
вительности рыб и личинок амфибий. При выходе позвоночных на сушу эта
VI. Средний мозг
161
система преобразовалась в орган слуха. Мозговое представительство этой сис-
темы у низших позвоночных носит название полукруглого валика (вздутия)
(torus semicircularis), а у млекопитающих — нижних, или задних, бугорков чет-
верохолмия (colliculi inferiores). Основные морфологические этапы развития
полукруглого валика легко оценить по сагиттальным сечениям среднего моз-
га, которые немного смещены от оси симметрии (рис. 111).
У латимерии полукруглый валик морфологически еще не выражен. Это прос-
то задний участок крыши среднего мозга, который с дорсальной поверхности
выглядит как единое целое. Только на внутренней поверхности крыши имеется
небольшая складка, обозначающая начало полукруглого валика (см. рис. 111, а).
Рис. 111. Парасагиттальные сечения головного мозга представителей основных групп по-
звоночных.
а — латимерия; б — амфибии; в — рептилии; г — птицы; д — млекопитающие. Участок с
точками — зрительный отдел, с крестиками — слуховой отдел среднего мозга, стрелки —
направление роста полукруглого валика
6—5818
162 ❖ Сравнительная анатомия нервной системы позвоночных
Для амфибий характерно увеличение размеров полукруглого валика, что опос-
редовано преобразованием центров боковой линии в мозговое представитель-
ство слуховой системы. Однако внешних анатомических изменений практи-
чески не видно. Только после рассечения мозга вдоль сагиттальной оси стано-
вится понятна причина отсутствия внешних изменений.
Дело в том, что III желудочек амфибий не узок, как сильвиев водопровод
млекопитающих, а представляет собой довольно обширную полость. Поэтому
полукруглый валик разрастается не в дорсальном, а в ростровентральном на-
правлении, выбухая в полость III желудочка (см. рис. 111,6). Таким образом,
значительное количественное приращение полукруглого валика не отражается
на форме дорсальной поверхности среднего мозга амфибий.
Для рептилий характерно появление полукруглого валика в виде выпячива-
ния на дорсальной поверхности мозга в мозжечково-среднемозговой складке
(см. рис. 111, в). В этом случае название полукруглого валика меняется на
нижние, или задние, бугорки (colliculi superiores). Вся крыша среднего мозга
начинает называться пластинкой четверохолмия, или четверохолмием (lamina
quadrigemina tecti, corpora quadrigemina), а зрительная крыша (tectum opticum)
становится верхними, или передними, бугорками (colliculi inferiores). Однако
у рептилий полость III желудочка достаточно велика и значительная часть
задних бугорков формирует внутрижелудочковое выпячивание. У рептилий
мы видим обе анатомические стратегии развития полукруглого валика мозга:
наружу мозга и внутрь. Обе эти стратегии оказались реализованными.
У птиц задние бугорки четверохолмия на дорсальной поверхности среднего
мозга отсутствуют. Полукружный валик развивался в полость III желудочка,
что привело к редукции наружной поверхности этого отдела (см. рис. 111, г).
В результате такого развития у птиц оказывается резко сужены дорсальные по-
лости III желудочка, а задние бугорки на поверхности мозга представлены толь-
ко узкой полоской в мозжечково-среднемо <говом перегибе. Зрительная специ-
ализация птиц привела и к особому развитию передних бугорков четверохол-
мия. Из небольших выпячиваний на поверхности мозга у рептилий они
превратились в огромные структуры — зрительные доли (lobi optici). Они на-
столько велики, что видны при рассмативании мозга и сверху, и сбоку, и снизу.
У млекопитающих была реализована иная стратегия развития задних бугор-
ков. Они увеличивались в дорсальном направлении, что было обусловлено
сужением просвета III желудочка за счет количественного увеличения восхо-
дящих в передний мозг волокон от двигательных центров заднего, продолго-
ватого и спинного мозга (рис. 111,6). В результате сформировалось четверо-
холмие, развитие которого может быть первичным критерием для оценки пре-
валирования слуховой или зрительной системы.
Участок среднего мозга, лежащий под крышей среднего мозга, имеет общие
принципы строения у всех позвоночных (рис. 112).
Вентральнее крыши среднего мозга расположен тегментум (tegmentum
mesencephali), который является прямым продолжением ретикулярной фор-
мации продолговатого мозга. Тегментум подразделяется, как и в спинном,
заднем и продолговатом мозге, на чувствительную и моторную зоны (tegmentum
sensorium, motoricum) (см. рис. 112). В сенсорной зоне расположены полу-
круглый валик и ядра латеральной петли, которые являются промежуточными
VI. Средний мозг
163
Tractus opticus medialis
Рис. 112. Фронтальное сечение среднего мозга позвоночных. Участок с точками — ядра и
среднемозговые центры, со штриховкой — пучки волокон.
центрами между слуховыми ядрами заднего и среднего мозга. В моторной
зоне лежат двигательные ядра черепно-мозговых нервов и нисходящие и вос-
ходящие спиноцеребральные волокна. Наиболее стабильным пучком волокон
является продольный медиальный пучок (fasciculus longitudinalis medialis), встре-
чающийся у всех позвоночных. Для амниот характерно появление специали-
зированного красного ядра (nucleus ruber), которое формируется в связи с
развитием конечностей и расположено в моторной зоне тегментума. В мотор-
ной зоне среднего мозга расположено двигательное ядро блокового нерва (IV).
У большинства позвоночных этот нерв переходит на противоположную сторо-
ну мозга и выходит наружу из под тектума.
Если с рис. 112 провести умозрительную операцию по расчленению тектума
и тегментума, то станет понятно анатомическое происхождение среднего моз-
га. Соединение дорсальных краев сенсорной зоны тегментума позволит полу-
чить копию сечения спинного мозга. При этом все зоны окажутся на своем
месте. Таким образом, в среднем мозге относительно новообразованным отде-
лом является тектум, который возник за счет разрастания края сенсорного
тегментума. На это указывают и специализированные пучки волокон, встре-
чающиеся у всех позвоночных: медиальный и латеральный зрительные тракты
(tractus opticus medialis, lateralis).
Перечисленные общие принципы организации среднего мозга значительно
маскируются многообразным морфологическим строением тектума (рис. 113).
На рис. 113 видно, что у амфибий, рептилий и птиц он организован значи-
тельно сложнее, чем у млекопитающих. Эта сложность проявляется в наличии
большого количества специализированных нейронов и стратифицированнос-
ти (многослойности) тектума, что обеспечивает более эффективную перера-
ботку информации. У многих видов рыб, амфибий и рептилий тектум имеет
большее количество слоев, чем кора переднего мозга человека (рис. 114—116).
164
❖
Сравнительная анатомия нервной системы позвоночных
Амфибии
Рептилии
Рис. 113. Поперечные срезы крыши (верхних бугорков) среднего мозга позвоночных. Имп-
регнация азотнокислым серебром (по R. Cajal).
Привычное умозаключение о более сложной организации мозга млекопитаю-
щих здесь не подходит. Существуют две причины наблюдаемых различий.
С одной стороны, бугорки крыши среднего мозга млекопитающих утрачи-
вают функции высшего центра обработки слуховой и фительной информа-
ции. Этим центром становится кора переднего мозга, куда чере s таламические
ядра поступает основной сенсорный поток информации. Крыша среднего мозга
млекопитающих играет второстепенную роль, которая связана с видоспеци-
фическими рефлексами.
В «домлекопитающий» период основные зрительные и слуховые функции
были сосредоточены в среднем мозге. В нем осуществлялись ассоциативный
анализ информации и основная синтетическая активность мозга. Средний
мозг являлся «интеллектуальной» надстройкой над остальными структурами
центральной нервной системы. По сути дела он выполнял функции коры
переднего мозга млекопитающих. В нем складывались видоспецифические и
наследуемые формы универсальных и скоординированных реакций, которые
являются основой жизнедеятельности. В связи с развитием переднего мозга
млекопитающих врожденные реакции среднего мозга позволили коре «не
VI. Средний мозг
165
N. nervi oculomotorii
Рис. 114. Поперечные срезы среднего мозга (по R. Nieuwenhuys et al., 1997, с изменениями),
заниматься» примитивными формами видоспецифических ответов на внеш-
ние сигналы.
Основная задача развивавшейся коры состояла в объединении афферент-
ной информации различных систем. Поэтому за средним мозгом млекопита-
ющих сохранились как анализаторные, так и рефлекторные функции, осво-
бождающие кору от контроля за такими простыми реакциями, как старт-реф-
лекс (непроизвольная реакция на резкое слуховое или зрительное раздражение).
В процессе эволюции среднего мозга можно обнаружить две основные стра-
тегии развития зрительного анализатора и среднего мозга. Зрительная система
приобретала дополнительные возможности за счет развития или среднего мозга,
166
❖
Сравнительная анатомия нервной системы позвоночных
principalis
magnocellularis
nuclei laminares
n.
n.
Rana
esculenta
Griseum
superfi ciale
Tupinambus
teguixin
Tectum mesencephali
Griseum
periventriculare
N. tegmenti:
anterodorsalis
anteroventralis
N. profundus
N. nervi oculomotorii
Griseum
centrale
Torus
semici rcularis:
n. laminares
n. centralis
N. profundus
pars rostralis
Griseum
centrale
mesencephali
N. mesencephalicus
nervi trigemini
Stratum
griseum
N. nervi
oculomotorii:
dorsalis,
ventralis
N. interstitialis
N. ruber Area ventralis
tegmenti
Рис. 115. Поперечные срезы крыши среднего мозга позвоночных (по R. Nieuwenhuys et al.,
1997, с изменениями).
или периферической части анализатора — сетчатки. Сравнительный морфоло-
гический анализ показывает, что даже у представителей одного отряда хвоста-
тых амфибий можно выявить оба варианта решения одной проблемы (рис. 117).
На пропорционально выполненных схемах животных из одной размерной груп-
пы видно, что в одном случае, у R.sibiricus, проблема эволюции зрительного
анализатора решалась за счет увеличения площади поверхности сетчатки и
количества нервных волокон, приходящих в крышу среднего мозга. У Р. waltlii
VI. Средний мозг
❖
167
Corvus
monedula
Didelphis
virginiana
Colliculus
superior:
stratum zonale
stratum griseum
superficiale
stratum griseum
intermedium
stratum griseum
profundum
N. nervi
oculomotorii
dorsolateralis
N. mesencephalicus
nervi trigemini
N nervi oculomotorii:
dorsomedialis —
ventromedialis —
N. semilunaris
N. tegmentalis
pedunculopontinus
N. mesencephalicus
lateralis, pars dorsalis
N. isthmi:
parvocellularis,
magnocellularis
Macaca
mulata
Griseum
centrale mesencephali
N. oculomotorius
N parabigeminalis
N. interpeduncularis
N. mesencephalicus
nervi trigemini
N. Edinger-Westfal
Substantia nigra:
pars reticulata
pars compacta
N. ruber (magnocellularis)
Colliculus superior:
stratum zonale
stratum griseum
superficiale
stratum griseum
intermedium
stratum griseum
profundum
Griseum centrale
mesencephali
N. parabigeminalis
N. Edinger-Westfal
N. oculomotorius
N. interpeduncularis
N. branchii
colliculi inferioris
N. tegmentalis
pedunculopontinus
Nuclei pontis
Рис. 116. Поперечные срезы среднего мозга позвоночных (по R. Nieuwenhuys et al., 1997, с
изменениями).
и T. cristatus превалируют объемы сетчатки и волокна ганглиозных клеток, до-
стигающие объема тектума. Несколько иначе решалась проблема зрительной
адаптации у O.fischeri, S. salamandra и A. tigrinum. У этих видов увеличен объем
как сетчатки, так и серого вещества крыши среднего мозга. Таким образом,
анатомические различия, наблюдаемые в строении крыши среднего мозга хво-
статых амфибий, преимущественно связаны с вариациями адаптации зритель-
ной системы. В одном случае недостаток поступления зрительной информа-
ции компенсировался при помощи простого увеличения размеров глаза, в дру-
168
❖
Сравнительная анатомия нервной системы позвоночных
в
г
Рис. 117. Схематическое изображение головного мозга и сетчатки хвостатых амфибий и
количественные отношения сетчатки и крыши среднего мозга.
а — Ambystoma tigrinum', б — Onychodactylus fischeri; в — Triturus cristatus; г — Pleurodeles
waltlii', d — Ranodon sibiricus; e — Salamandra salamandra. Рядом с сетчаткой — округлен-
ные средние величины объема и площади поверхности, рядом с крышей среднего мозга —
объем серого вещества
гом — количеством несенсорных элементов сетчатки, в третьем — объемом
сетчатки и серого вещества среднего мозга.
С другой стороны, крыша среднего мозга у рыб, амфибий и рептилий вы-
полняет не только зрительные и слуховые функции. Она является интегратив-
ным центром поведения животных. В крыше среднего мозга рыб, амфибий и
рептилий сосредоточены четыре из пяти основных сенсорных центра (рис. 118).
Их присутствие в среднем мозге организовано по стратификационному типу.
Это означает, что каждое из представительств одной из сенсорных систем за-
нимает более или менее определенный уровень в слоистой организации текту-
ма. Эти «поля» могут частично или полностью перекрываться, а их вертикаль-
ная интеграция является основой для выработки генерализованного ответа на
весь комплекс внешних раздражений. По сути дела строение тектума напоми-
нает сложенную колоду карт, каждая из которых без условностей взаимодей-
ствий с другими картами (правилами игры) не имеет смысла.
Закономерности организации крыши среднего мозга, выявленные у амфи-
бий, характерны и для рептилий. Более того, у рептилий выявлено точное
топологическое представительство каждой из сенсорных систем (рис. 119).
Таким образом, крыша среднего мозга состоит из «полей», расположенных
друг над другом. Организация среднего мозга позволяет интегрировать работу
VI. Средний мозг
169
Рис. 118. Строение головного мозга и крыши среднего мозга аксолотля (A tigrinum) с нане-
сенными на ее поверхность областями представительства различных анализаторов.
Дорсальная поверхность
Рис. 119. Представительство соматотопических проекций в крыше среднего мозга репти-
лии (Iguana iguana) с нанесенными на ее поверхность областями представительства ре-
цепторов от различных частей тела (по N. Gaither и В. Stein, 1979, с изменениями).
170
Сравнительная анатомия нервной системы позвоночных
❖
всей нервной системы у рыб, амфибий и рептилий, что отразилось на системе
связей этого центра.
У большинства первичноводных позвоночных средний мозг выполняет функ-
ции принятия генерализованных решений. Основой для такой активности слу-
жит разнообразная сенсорная информация, которая поступает в средний мозг
от внешних и внутренних анализаторов. Для первичноводных позвоночных ос-
новой для принятия решения являются сенсомоторная и зрительная информа-
ция, сигналы от вестибулярной системы, органов боковой линии и обонятель-
ной системы. Только последняя не представлена прямыми связями в крыше
среднего мозга (рис. 120). Практически все основные моторные ядра заднего и
продолговатого мозга взаимодействуют с крышей среднего мозга у круглоро-
тых, хрящевых и костистых рыб, амфибий и рептилий. У хрящевых и костных
рыб и амфибий они образуют прямые связи, идущие как от первичных мотор-
Nucleus reticularis
superior
Nucleus octavus
ascendens
Сетчатка
Nucleus intermedius
area octavolateralis
Nucleus dorsalis -
area octavolateralis
Nucleus reticularis
medius
Nucleus reticularis
isthmi
Nucleus ruber
Nucleus cerebelli
Lemniscus lateralis
Задний мозг
Nucleus descendens
nervi trigemini
Nucleus reticularis
inferior
Крыша среднего
мозга
Рис. 120. Основные волокна, восходящие в крышу среднего мозга акулы.
Передний мозг
Nucleus tegmentalis
lateralis
— Контур мозжечка
VI. Средний мозг < 171
ных центров, так и от вторичных ядер, обслуживающих мозжечок. Нужно отме-
тить, что у круглоротых таких прямых связей намного меньше, чем у акул (рис.
121). Это связано с тем, что исторически более древним вариантом поступления
сигналов в крышу среднего мозга является ретикулярный путь. По-видимому,
первоначально сигналы проходили через ядра рострального края ретикулярной
формации — покрышку среднего мозга и только затем поступали в тектум. Этот
путь хорошо выражен у круглоротых и представлен небольшим количеством
волокон практически у всех позвоночных. Однако уже у акул система сенсомо-
торно-тектальных связей значительно изменилась. Основные моторные центры
установили прямые связи с тектумом, а тегментно-тектальные моторные пути
стали иметь вспомогательный характер. У амфибий прямые сенсомоторно-тек-
тальные связи достигли максимального развития, а крыша среднего мозга стала
Tractus tectobulbaris
Продолговатый
и задний мозг
Thalamus
dorsalis
Thalamus
ventralis
Fasciculus
tectotegmentalis
Nucleus reticularis
superior
Nucleus reticularis
medius
Nucleus tegmentalis
lateralis
Commissura tecti
Commissura ansulata
Nucleus intercollicularis
Контур
мозжечка
— Nucleus fasticuli
longitudinals medialis
Nucleus raphes
superior
Крыша среднего
мозга
Nucleus reticularis
inferior
Nucleus reticularis
isthmi
Рис. 121. Основные волокна, нисходящие из крыши среднего мозга колючей акулы.
Fasciculus predorsalis
172
Сравнительная анатомия нервной системы позвоночных
высшим ассоциативным центром. Она стала выполнять функции, аналогичные
роли коры переднего мозга млекопитающих.
У рептилий интеграция сенсорных входов в крыше среднего мозга стала
уменьшаться. За тектумом продолжала сохраняться львиная доля ассоциатив-
ных функций, но восходящие сенсорные волокна вновь стали менять свой
путь. Возникли промежуточные центры переключений, поскольку часть воло-
кон стала направляться в передний мозг — новый интегративный центр. По
сути дела восходящие связи вновь стали проходить через дополнительные ядра,
как у круглоротых (рис. 122). Разница состоит в том, что дополнительные
сенсомоторные ядра рептилий перенаправляют сигналы в промежуточный и
передний мозг, а ядра круглоротых — в средний. Развитие переднего мозга
рептилий привело к радикальному переключению восходящих волокон не толь-
ко от сенсомоторных центров, но и от сетчатки. Часть как перекрещенных,
так и неперекрещенных в хиазме зрительных волокон направляется не в кры-
шу среднего мозга, а в латеральное коленчатое тело (corpus geniculatum laterale)
таламуса. Часть выходящих волокон латерального коленчатого тела направля-
ется в тектум, а часть — в передний мозг. Схожие преобразования, но не-
Сетчатка
Nucleus intercollicularis
Lemniscus medialis
Lemniscus spinalis
Продолговатый
и задний мозг
Nucleus isthmi pars
magnocellularis
Corpus
geniculatum
laterale
Крыша среднего
мозга
мозг
Striatum
Substantia nigra
Locus coeruleus
Nucleus
commissuralis
(anterior)
Nucleus descendens
nervi trigemini
Nucleus funiculi
dorsalis
Nucleus cochlearis
angularis
Рис. 122. Основные волокна, восходящие в крышу среднего мозга рептилий Светлые тре-
угольники — ядра, не имеющие прямых связей с тектумом.
Передний
VI. Средний мозг
173
❖
сколько в меньших масштабах претерпевает и слуховая система. Часть воло-
кон, минуя нижние бугорки четверохолмия, направляется в медиальное ко-
ленчатое тело (corpus geniculatum mediale), а оттуда в передний мозг. Форми-
рование вокруг среднего мозга обходных путей восходящих волокон свиде-
тельствует о постепенной утрате этим центром интегративных функций.
В нем сохраняются наборы автоматических врожденных реакций организма,
которые были получены на ранних этапах эволюции позвоночных.
Нужно отметить, что у птиц крыша среднего мозга еще выполняет довольно
много интегративных функций, хотя несравненно большее значение начина-
ют иметь структуры стриатума переднего мозга.
У млекопитающих, как у птиц и рептилий, связи среднего мозга и сенсор-
ных систем обслуживают врожденные автоматические реакции. Сложные ас-
социативные функции млекопитающих принимают на себя специализирован-
ные поля коры полушарий переднего мозга.
Описанные преобразования восходящих связей среднего мозга аналогичны
таковым нисходящих путей (см. рис. 121, 123). У низших позвоночных крыша
среднего мозга является местом принятия решения и как следствие источни-
Stnatum
Nucleus reticularis inferior
Nucleus isthmi pars
magnocellularis
Tractus
reticulospinalis
Corpus geniculatum
laterale
Tractus tectobulbaris
ventralis
Крыша среднего мозга
Nucleus rotundus
Fasciculus predorsalis
Nucleus reticularis medius
Продолговатый и задний мозг
Thalamus
ventralis
Thalamus
dorsalis
Рис. 123. Основные волокна, нисходящие из крыши среднего мозга рептилий.
Ростродорсальный зачаток
коры переднего мозга
(зрительное поле)
174 ❖ Сравнительная анатомия нервной системы позвоночных
ком генерализованного двигательного ответа. Поэтому у первичноводных по-
звоночных и амфибий превалируют прямые нисходящие двигательные пути,
которые оканчиваются в двигательных центрах заднего и продолговатого моз-
га. При помощи этих связей осуществляется контроль за моторной активно-
стью. В другие отделы мозга нисходящие волокна направляются в крайне не-
значительном количестве (см. рис. 121). Эта ситуация изменяется у рептилий,
птиц и млекопитающих. В этих группах животных значительная часть воло-
кон направляется из среднего мозга в таламус, стриатум и кору переднего
мозга. Присутствие этих связей хорошо заметно еще у рептилий (см. рис. 123).
Снижение количества нисходящих связей между крышей среднего мозга и
двигательными центрами объясняется появлением относительно автономных
стриарно-ретикулярных (у птиц) и кортико-ретикулярных (у млекопитающих)
связей. Новые центры перетнего мозга принимают на себя функции тектума и
формируют новые двигательные пути. Таким образом, крыша среднего мозга
была первой специализированной ассоциативной структурой, которую заме-
нил бурно эволюционирующий передний мозг.
VII. промежуточный мозг
Промежуточный мозг в соответствии с названием
занимает небольшой участок между средним и пере-
дним мозгом (рис. 124). У низших позвоночных про-
межуточный мозг заметен с дорсальной поверхности,
но, начиная с рептилий, он практически полностью
закрывается полушариями переднего мозга (рис. 125,
126). На дорсальной поверхности остаются только ча-
сти эпифизарного комплекса. Ростральный край про-
межуточного мозга ограничен терминальной пластин-
кой (lamina terminalis), которая находится между полу-
шариями переднего мозга. Ее вместе с передней
комиссурой (commisura anterior) относят к переднему
мозгу. Дорсокаудальной границей промежуточного
мозга обычно считается задняя комиссура (commissura
posterior), а вентрокаудальной — сосцевидные тела (cor-
pora mamillaria) у млекопитающих. Эта граница выра-
жена даже у рыб, поскольку на задней стенке воронки
находятся гомологи сосцевидных тел.
Особый интерес представляет строение дорсальной и
вентральной стенок промежуточного мозга (см. рис. 125,
126). Дорсальная стенка обычно не содержит нейронов,
но имеет ряд специфических образований, которые при-
нимают участие в нейроэндокринных регуляциях и вод-
но-солевом обмене нервной системы. Самой заметной
дорсальной структурой промежуточного мозга является
эпифизарный комплекс (рис. 127). В наиболее сложном
случае он включает в себя эпифиз, или пинеальный орган
(epiphysis cerebri, corpus pineale, glandula pinealis), парапи-
неальный орган, или теменной глаз (organum parapinealis),
и парафиз (paraphysis), или дорсальный сосудистый ме-
шочек (sacculus vasculosis). У низших позвоночных все эти
структуры встречаются одновременно.
У большей части взрослых птиц и млекопитающих
обычно сохраняется только эпифиз (corpus pineale), а па-
рафиз (corpus parapineale) исчезает в поздний эмбрио-
176
❖
Сравнительная анатомия нервной системы позвоночных
Рис. 124. Мозг различных позвоночных. Вид с дорсальной поверхности.
а—минога; б—колючая акула; в — карп; г—травяная лягушка; д — геккон; е — сетчатый
питон; ж— голубь; з — кролик; и — собака. Участок с точками — промежуточный мозг.
нальный период. Однако в эмбриональном развитии парафиз присутствует у
некоторых анамний и у всех змниот. У гаттерии и большинства ящериц имеется
теменной глаз, который сохраняется наряду с эпифизом (рис. 128). У миноги
парафиз посылает нерв к левому хабенулярному ганглию (у гаттерии к право-
му), а эпифиз — к правому. Эти особенности строения эпиталамуса круглоро-
тых послужили основой для создания фоторецепторной гипотезы возникнове-
ния пинеального комплекса. Согласно гипотезе парафиз является остатком пра-
вого глаза, а эпифиз — левого. Причины тупиковости этого пути видят в
неинвертированности фоторецепторов простых глазков современных рептилий.
По-видимому, наиболее примитивным типом организации пинеального
комплекса следует считать концентрацию светочувствительных элементов в
дорсальной стенке головного расширения центрального спинномозгового ка-
нала у ланцетника (см. рис. 128, а). Анатомическое обособление комплекса
началось с появления непарного выпячивания крыши среднего мозга, распо-
ложенной между хабенулярной (пинеальной, поводковой) комиссурой (commi-
ssure habenularum) и задней комиссурой (commissura posterior) (см. рис. 128, б).
VII. Промежуточный мозг
177
Пинеальный
Сосудистое сплетение в крыше
Пинеальный
комплекс
Мозжечок
Аденогипофиз
Sacculus vasculosis
Нейрогипофиз
Пинеальный
г
Рис. 125. Сагиттальные сечения головного мозга.
а — минога; б — колючая акула; в — латимерия; г — травяная лягушка. Участок с точками
— промежуточный мозг. Ill, IV — желудочки мозга.
Примером такого обособления является дорсальный холмик пиявкоротых мик-
син (Bdellostoma). Одиночное выпячивание комплекса широко распростране-
но среди пластиножаберных, цельноголовых и двоякодышащих. Пинеальный
комплекс в этих группах представляет собой трубчатый асимметричный вы-
рост крыши среднего мозга. Следующим этапом исторического развития ком-
плекса стала дифференцировка первичного пузыря на два образования: пара-
пинеальный орган и эпифиз (см. рис. 128, в).
Бинарная система у всех позвоночных остается асимметричной. Эволюция
парапинеального органа реализуется в пределах трех вариантов. Парапинеаль-
ный орган может полностью исчезать у крокодилов, некоторых птиц и боль-
шинства млекопитающих. Если он сохраняется, то может трансформировать-
ся в светочувствительный элемент или принимать на себя часть нейроэндо-
кринных функций, характерных для эпифиза.
Эпифиз, как правило, значительно крупнее парапинеального органа и со-
храняется в виде самостоятельной анатомической структуры практически у
178
❖
Сравнительная анатомия нервной системы позвоночных
Сосудистое сплетение
переднего мозга
Пинеальный Мозжечок
комплекс
Мозжечок
Нейрогипофиз
Аденогипофиз
Сосудистое
сплетение
Пинеальный
орган
Сосудистое сплетение
III желудка
Сосудистое
сплетение
Передний
мозг
Воронка
Нейрогипофиз
Сосудистое
сплетение
Аденогипофиз
Эпифиз
Рис. 126. Сагиттальные сечения головного мозга.
а — черепаха; б—голубь; в — собака. Участок с точками — промежуточный мозг. Ill, IV —
желудочки мозга.
Сосцевидные
Нейрогипофиз Аденогипофиз тела
в
Передник
озг
всех амниот. Исключение составляют аллигаторы и некоторые виды червяг.
У аллигаторов весь пинеальный комплекс заметно редуцирован, а у червяг
парапинеальный орган превалирует над эпифизом. Если парапинеальный орган
сформировался преимущественно как париетальная светочувствительная си-
стема, то эпифиз является типичным нейроэндокринным органом. По сути
дела это тандем, состоящий из железы — эпифиза и регулятора ее нейроэн-
докринной активности — пинеального глаза.
Для активных днем животных характерен следующий циркодианный цикл
активности пинеального комплекса. Днем, когда свет воздействует на пине-
альный глаз, эпифиз получает сигналы, стимулирующие выработку серотони-
на. В светлое время суток синтез и секреция серотонина возрастают в эпифизе
примерно в 5 раз. При снижении освещенности выработка серотонина резко
уменьшается, в эпифизе начинает синтезироваться мелатонин. Синтез этих
производных индола связан между собой. Накопленные за светлое время су-
VII. Промежуточный мозг
179
Нерв теменного глаза
Эпифиз
Парафиз
Передняя
комиссура
Хабенулярная
комиссура
Зрительная
хиазма
Терминальная
пластинка
Теменной глаз,
или парапинеальный орган
Epithalamus
Thalamus dorsalis
Tha lamus ventralis
Hypothalamus
Воронка
Sacculus vasculosis
Задняя
комиссура
Аденогипофиз
Нейрогипофиз
Рис. 127. Сагиттальное сечение через промежуточный мозг и терминальную пластинку с
обозначением основных анатомических структур, которые могут встречаться у позвоноч-
ных в разных сочетаниях.
ток запасы серотонина подвергаются последовательному ацетилированию и
метилированию и превращаются в мелатонин. Уровень секреции мелатонина
возрастает в темное время суток примерно в 10 раз (рис. 129). Мелатонин и
серотонин действуют разнонаправленно. Мелатонин влияет на пигментацию
кожи, тормозит развитие половых функций у молодых животных и дает цир-
кодианный антигонадотропный эффект. Иначе говоря, он вызывает генерали-
зованное снижение активности животного. Серотонин не оказывает такого
угнетающего влияния на организм. Как нейромедиатор катехоламинового ряда
он оказывает как тормозящее, так и возбуждающее действие. Под его влияни-
ем находятся уровень бодрствования, восприятие сенсорных сигналов и эмо-
циональное состояние. Предельно упрощая ситуацию, можно сказать, что ме-
латонин снижает, а серотонин стимулирует активность животного. Нужно от-
метить, что у позвоночных с сумеречной активностью весь описанный выше
процесс происходит в обратном порядке. Днем, когда животное неактивно,
вырабатывается мелатонин, а в ночное время — серотонин.
Механизм регуляции суточной активности вполне понятен у животных,
имеющих пинеальный глаз (см. рис. 129), который может непосредственно
передавать сигналы к пинеалоцитам — основным нейросекреторным клеткам
эпифиза. В дневное время под влиянием отростков фоторецепторов и проек-
ционных нейронов происходит стимуляция синтеза серотонина, а ночью —
180
❖
Сравнительная анатомия нервной системы позвоночных
Рис. 128. Сагиттальное сечение через пинеальный комплекс.
а-в — объяснение в тексте, х — Glyptolepis groenlandica; z — Rattus rattus. Участок co
штриховкой — хабенулярная комиссура, с точками — задняя комиссура.
мелатонина. Однако у большей части рептилий, птиц и млекопитающих пине-
ального глаза нет и механизм регуляции суточной активности эпифиза совер-
шенно другой. Он также построен на световой регуляции суточной активно-
сти, как и у животных, имеющих пинеальный глаз.
Информация об изменении светового дня поступает не от парапинеального
органа, а непосредственно от зрительной системы (рис. 130). Основным ис-
точником информации является небольшой пучок волокон ретиногипотала-
мического тракта, который начинается от ганглиозных клеток сетчатки и окан-
чивается в супрахиазматическом (надзрительном) ядре (nucleus suprachiasmaticus,
supraopticus). От этого ядра начинаются два независимых пути к эпифизу. Один
обеспечивается волокнами, начинающимися в супрахиазматическом ядре и
оканчивающимися в гипоталамической, септальной и преоптической облас-
тях (см. рис. 130). Из этих центров выходят волокна, направляющиеся в эпи-
физ непосредственно или через уздечку (habenula) промежуточного мозга. Вто-
рой путь также начинается от супрахиазматического ядра и в составе медиаль-
ного пучка нисходящих волокон достигает шейного отдела спинного мозга.
VII. Промежуточный мозг ❖ 181
Нерецепторная клетка в
Рис. 129. Разрез через пинеальный комплекс промежуточного мозга. Парапинеальный орган
представляет собой неинвертированный глазок.
Рис. 130. Основные структуры и связи головного мозга млекопитающих, регулирующих
суточный ритм активности эпифиза при утрате пинеального глаза.
182 ❖ Сравнительная анатомия нервной системы позвоночных
Затем из зоны локализации симпатических мотонейронов спинного мозга
выходит несколько волокон, оканчивающихся в верхнем шейном ганглии. От
верхнего шейного ганглия начинаются симпатические волокна, которые окан-
чиваются непосредственно в эпифизе. Такая иннервация мозга симпатичес-
кой системой является уникальной в эволюции позвоночных и вызвана необ-
ходимостью замены функций исчезнувшего пинеального глаза. Механизм ре-
гуляции суточной активности эпифиза, по-видимому, построен на том, что в
дневное время пинеалоциты стимулируются через гипоталамическую область,
а в ночное — через симпатические волокна верхнего шейного ганглия. Основ-
ным водителем ритма является супрахиазматическое ядро, которое метаболи-
чески активно в период бодрствования животного.
Все образования пинеального комплекса мозга относятся к эпиталамусу
(epithalamus) (см. рис. 127). В ростральной части эпиталамус у низших позво-
ночных образует парафиз. Сосудистое сплетение, расположенное на границе
переднего и промежуточного мозга, иногда имеет огромные размеры. Оно не
только свешивается в просвет желудочков, но и выходит на поверхность мозга.
Для двоякодышащих характерна ситуация, когда сплетение закрывает собой
большую часть дорсальной поверхности головного мозга. Нужно отметить, что
парафиз может являться частью сплетения или трансформироваться в структу-
ру, аналогичную сосудистому мешочку (sacculus vasculosis) нейрогипофиза. Под
пинеальным комплексом в эпиталамусе находятся хабенулярные ганглии (ядра)
(ganglion habenulae). У круглоротых и костистых рыб они асимметричны и полу-
чают афферентную иннервацию от переднего мозга. Нисходящие волокна из
хабенулярных ганглиев направляются в составе пучка Мейнерта (отогнутый пу-
чок) (fasciculus retroflexus) в межножковое ядро (nucleus interpeduncularis), кото-
рое расположено в основании среднего мозга. Эти ганглии осуществляют пере-
дачу обонятельных сигналов в ствол мозга. Вентральнее эпиталамуса (epithalamus)
расположен зрительный бугор, или таламус («брачное ложе» для полушарий
переднего мозга) (thalamus). Он состоит из двух частей: дорсального таламуса
(thalamus dorsalis) и вентрального таламуса (thalamus ventralis) (см. рис. 127).
Непосредственно под ним лежит гипоталамус (hypothalamus), который образует
воронку, переходящую в нейрогипофиз. Ростральнее воронки лежит зрительная
хиазма, а каудальнее (у млекопитающих) — сосцевидные тела. Гипоталамус яв-
ляется основным нейроэндокринным отделом мозга, а его размеры коррелиру-
ют с выраженностью гормонально-зависимого поведения.
Гипоталамус превалирует над другими отделами мозга у некоторых рыб, ам-
фибий и рептилий. У рыб он может образовывать своеобразные выпячивания
воронки — нижние доли (lobi inferiores). Для мозга рептилий обычно используется
другое название разросшихся стенок промежуточного мозга — «латеральные доли»
(lobi laterales). Воронка сужается в вентральном направлении и переходит в нейроги-
пофиз, а у рыб — и в сосудистый мешочек (sacculus vasculosis). С нейрогипофи-
зом непосредственно соединен аденогипофиз, который происходит из дорсаль-
ной части ротового эпителия в раннем эмбриогенезе.
Эпиталамус, таламус и гипоталамус являются стабильными образованиями
и присутствуют во всех группах позвоночных, хотя каждый из отделов может
быть выражен в большей или меньшей степени (рис. 131). Особую роль в
эволюции промежуточного мозга играет таламус. Существует выраженная
VII. Промежуточный мозг
183
Рис. 131. Сагиттальные сечения промежуточного мозга различных животных.
а — минога; б—лопатонос; в — латимерия; г — аксолотль; б—черепаха; е —опоссум (по
R. Nieuwenhuys et al., 1997).
эволюционная закономерность изменения количественных соотношений меж-
ду дорсальным и вентральным таламусом. У анамний вентральный отдел та-
ламуса развит более, чем дорсальный, и представляет собой преимуществен-
но прижелудочковую массу клеток. Небольшой латеральный компонент со-
ответствует вентральной части латерального коленчатого тела млекопитающих
(рис. 132—134), что было доказано при помощи ретроградного транспорта
различных маркеров. В процессе эволюции размеры вентральной части тала-
муса уменьшаются, а дорсальной — увеличиваются. У млекопитающих пре-
валируют ядра дорсального таламуса, а не вентрального. Для низших позво-
ночных характерно развитие вентрального таламуса. Это обусловлено тем,
что дорсальная часть таламуса преимущественно связана с развитием восхо-
дящих путей от октаволатеральной, зрительной и сенсомоторных систем к
коре полушарий переднего мозга.
184
Сравнительная анатомия нервной системы позвоночных
Ganglion habenulae
Нейрогипофиз
N. dorsalis
hypothalami
Thalamus dorsalis
Thalamus ventralis
- Аденогипофиз
N. ventralis
hypothalami
Corpus geniculatum
laterale
Thalamus ventralis,
pars medials
Ganglion habenulae
N. medius hypothalami
Thalamus ventralis,
pars ventromedialis
Эпифиз
Thalamus dorsalis,
pars dorsomedials
N. suprachiasmaticus
N. preopticus
hypothalami
в
Рис. 132. Срезы промежуточного мозга различных животных.
а — минога; б— колючая акула; в—многопер (по R. Nieuwenhuys et al., 1997, с изменениями).
У низших позвоночных высшие сенсорные и ассоциативные центры сосре-
доточены в среднем мозге, а дорсальный таламус является скромным интегра-
тором между средним мозгом и обонятельной системой переднего мозга. Дор-
сальный таламус состоит из нескольких небольших ядер, которые осуществ-
ляют передачу сенсорных сигналов в передний мозг. Переднее ядро таламуса
(nucleus anterior thalami) получает волокна из сетчатки глаза, а дорсолатераль-
ное и латеральное ядра дорсального таламуса (nucleus dorsolateralis, nucleus
lateralis) являются полифункциональными. Они получают непрямые зритель-
ные волокна, которые начинаются от крыши среднего мозга. Центральное
таламическое ядро (nucleus centromedianus) получает сигналы из соматосен-
сорной и слуховой систем и от механорецепторов боковой линии, которые посту-
пают от среднего мозга (torus semicircularis). Слуховые, зрительные и сенсомо-
торные сигналы от ретикулярной формации заднего мозга передаются через
дорсальный таламус в передний мозг, где интегрируются с обонятельной ин-
VII. Промежуточный мозг
185
Эпифиз
Ganglion
habenulae
Thalamus dorsalis
pars dorsalis
Thalamus dorsalis,
pars ventralis
N. ventromedials
thalami
Pars dorsalis
hypothalami
Pars ventralis
hypothalami
Рис. 133. Срезы промежуточного мозга различных животных.
а — латимерия; б — аксолотль; в — озерная лягушка (по R. Nieuwenhuys et al., 1997, с
изменениями).
формацией. Переднее ядро дорсального таламуса слабо развито у анамний. Оно
представляет собой небольшое прижелудочковое клеточное скопление, тогда
как задние ядра развиты намного лучше и имеют выраженные границы из пуч-
ков волокон. У амниот развитие дорсального таламуса связано в основном с
увеличением и дифференцировкой переднего ядра, которое прогрессирует бла-
годаря развитию восходящих афферентных связей в кору переднего мозга.
Описанные выше различия в организации ядер дорсального таламуса при-
вели к гипотезе их двойственного происхождения (Y. Butler, 1995). В соответ-
ствии с этой гипотезой ядра дорсального таламуса разделяются на две группы:
коллоталамус (collothalamus) и лемноталамус (lemnothalamus).
Это разделение предлагается автором на основании происхождения связей
ядер дорсального таламуса. С его точки зрения, коллоталамус преимуществен-
186
Сравнительная анатомия нервной системы позвоночных
N. entopeduncularis
anterior
N. rotundus
Tectum
N. intersticalis
striae terminalis
N. habenularis medialis
N. habenularis lateralis
N. dorsomedialis
thalami
N. geniculatus lateralis,,
pars dorsalis
N. dorsolateralis
thalami
N. lateralis hypothalami
N. ventralis hypothalami
N. dorsolateralis
hypothalami
N. ventolateralis
thalami
N. ventrolateralis
thalami
N. geniculatus lateralis,
pars ventralis
N. habenularis medialis
N. habenularis lateralis
N. ovoidalis
subrotundus
N. dorsolateralis
posterior thalami
Stratum cellulare:
externum,
internum
N. lentiformis
N. dorsomedialis
posterior thalami
N. superficialis
parvocellularis
thalami
N. geniculatus
lateralis,
pars dorsalis
Pars ventralis
N. medialis
dorsalis
thalami
Pulvinar
thalami
Corpus
geniculatum
laterale
N. reticularis
thalami
Hypothalamus
6
N. habenularis
lateralis
Cauda nuclei
caudati
N. posterior
thalami
Corpus
geniculatum
mediate
St
Substantia
nigra N. ruber N. intersticialis N. centromedianus
N. lateralis
posterior
thalami
N
в
Рис. 134. Схемы срезов промежуточного мозга.
а — тегу (Tupinambis teguixin)-, б—голубь; в — макака резус (по R. Nieuwenhuys et al., 1997,
с изменениями).
VII. Промежуточный мозг
187
но связан с крышей среднего мозга, а лемноталамус — с лемнисковым путем,
включая зрительный тракт. У анамний из лемноталамического зачатка фор-
мируются ростральные (передние) ядра дорсального таламуса, а из коллотала-
мического — каудальные (задние) и центральные ядра, которые занимают
прижелудочковое положение. Этот подход позволяет непротиворечиво гомо-
логизировать ядра дорсального таламуса в различных группах позвоночных
(рис. 135). В пользу такого разделения дорсального таламуса свидетельствуют
гомологичность дорсокаудального ядра таламуса акул и костистых рыб, круг-
лого ядра (nucleus rotundus) рептилий и птиц и латеральных ядер комплекса
подушки (pulvinar) таламуса млекопитающих. Однако ситуация не так ясна,
как может показаться. Дело в том, что отдельные прижелудочковые скопления
клеток анамний гомологизируются с целыми комплексами ядер амниот. Сле-
довательно, четкое пошаговое прослеживание эволюционных пребразований
отдельных ядер невозможно. Это хорошо видно на примере с лемноталами-
ческим передним ядром, которое у анамний состоит из непрерывного превен-
трикулярного матрикса. Это ядро дает у рептилий, птиц и млекопитающих
большую группу ядер. По сути дела этот подход предлагает проводить рацио-
нальную гомологизацию не отдельных ядер, а целых таламических комплек-
сов, которые имели общих предшественников у низших позвоночных. Дей-
Рептилии (диапсиды)
Млекопитающие
Дорсолатеральное
зрительное
ядро
Дорсомедиальное
и переднее
дорсолатеральное
ядра
Медиальное
ЯДРО
Передняя
группа ядер
лухова
зона
лкт
мкт
Вентральная
группа ядер
Комплекс
задних ядер
Z Ростромедиальная
зона
Медиальная и
интраламинарная
группы ядер
Амфибии
Медиальное Подушка
заднее
ядро
Латеральная
зрительная зона
Исходное
состояние
Соматосенсорная
(полисенсорная)зона
Зрительная
зона
Предшественники
амниот
Рис. 135. Схема предполагаемой эволюции ядер дорсального таламуса.
Незакрашенные ядра — лемноталамус, с точками — коллоталамус.
лкт — латеральное коленчатое тело, мкт — медиальное коленчатое тело (переработано
по У. Butler, 1995, с изменениями).
188 ❖ Сравнительная анатомия нервной системы позвоночных
ствительно, развитие неокортекса у млекопитающих и стриатума у птиц при-
вело к резкому увеличению размеров таламических структур. Внутри таламуса
стали выделяться специализированные ядра, которые обеспечивали передачу
сигналов к конкретным центрам переднего мозга.
Это привело к разделению первично однородного прижелудочкового скоп-
ления клеток на группы ядер. Одновременно с этим происходила экспансия
полушарий переднего мозга в каудальном направлении. Дорсальный таламус
млекопитающих постепенно становился важнейшим центром переключения вос-
ходящих путей в неокортекс полушарий переднего мозга. Из схемы на рис. 135
становился понятно исторически различное происхождение латерального ко-
ленчатого тела, отвечающего за переключение зрительных волокон в кору боль-
шого мозга, и медиального коленчатого тела, отвечающего за переключение
слуховых волокон.
Нужно отметить, что у костистых рыб в каудальной части промежуточного
мозга находится несколько ядер, носящих название клубочкового комплекса
(nuclei glomiformis). Они происходят из мигрирующих от задней части средне-
го мозга групп прижелудочковых клеток. Через эти ядра осуществляется пере-
дача механосенсорной информации от боковой линии и электросенсорных
органов в передний мозг.
Таким образом, дорсальный таламус промежуточного мозга, по-видимому,
является смешанным образованием. Комплексы его ядер формируются как
непосредственно из зачатка промежуточного мозга, так и за счет миграции из
среднего мозга. Вполне допустимо, что лемноталамический комплекс возник
из древних ядер латеральной петли, которые дифференцировались по мере
становления коры переднего мозга. Коллоталамус явно сформировался под
влиянием рострального края среднего мозга. Будучи «вынесенным вперед»
суммирующим работу тектума центром, коллоталамус оказался идеальным цен-
тром для передачи информации как на базальный, так и на корковый уровень
переднего мозга. Его расцвет пришелся на становление мозга рептилий.
У млекопитающих он сохранился как важнейший центр переключения слухо-
вых и соматосенсорных сигналов. Соматосенсорные сигналы у тетрапод, а
особенно у млекопитающих, играют огромную роль в координации движений.
Поэтому соматосенсорная область превратилась в наиболее заметное образо-
вание промежуточного мозга.
Вентральный таламус представляет собой обособленную внутрижелудочко-
выми бороздками часть таламуса (см. рис. 127). Вентральный таламус осуще-
ствляет контроль за моторными функциями. Это происходит двумя основны-
ми способами. У низших позвоночных вентральный таламус является одним
из главных моторных центров. Он сам интегрирует моторную активность и
генерирует управляющие сигналы. У рептилий, птиц и млекопитающих он
становится промежуточным комплексом ядер на пути нисходящих волокон из
двигательных центров переднего мозга.
У костистых рыб вентральный таламус представляет довольно развитый
центр, сопоставимый по разнообразию сенсорного представительства с кры-
шей среднего мозга. В него поступают сигналы из ретикулярной формации
ствола, спинного мозга, сенсорных ядер тройничного нерва, слуховой части
среднего мозга, сетчатки, тектума и мозжечка. Нисходящие связи распределе-
VII. Промежуточный мозг
189
❖
ны аналогичным образом, хотя к ним и добавляются отдельные волокна из
базальной области переднего мозга. По сути дела вентральный таламус кости-
стых рыб является одним из вариантов ассоциативного центра ядерного типа,
который смог бы объединять все сенсорные входы и формировать генерализо-
ванную программу действий. Однако он стал специализированным моторным
центром, а ассоциативные функции сосредоточились в развитой крыше сред-
него мозга. У амфибий и рептилий основную роль начинают играть связи
вентрального таламуса с ядрами дорсальных столбов спинного мозга. Вент-
ральный таламус становится все более «моторным», а нисходящие из передне-
го мозга волокна уменьшают его самостоятельность. Это приводит к тому, что
у теплокровных он утрачивает роль самостоятельного моторного центра и транс-
формируется в промежуточную станцию на пути двигательных волокон от
переднего мозга в двигательные центры продолговатого и спинного мозга.
Под вентральным таламусом расположен гипоталамус (см. рис. 127). Он об-
разует развитые латеральные выпячивания стенки мозга и полый стебелек, ко-
торый носит название воронки. В каудальной части воронка заканчивается ней-
рогипофизом, плотно соединенным с аденогипофизом. Часть стенки воронки
может образовывать тонкостенный сосудистый мешочек (sacculus vasculosis) ней-
рогипофиза (см рис. 125, 126). Гипоталамус у акул и костистых рыб может
образовывать выпячивания, которые дифференцируются в крупные парные
нижние доли промежуточного мозга. У латимерии и рыбы-луны (Mola mold)
гипофиз достигает такого размера, что при взгляде на мозг с дорсальной поверх-
ности он выступает ростральнее полушарий переднего мозга. Основные типы
строения гипофиза приведены на рис. 136. Гипофиз, являясь составной струк-
турой, имеет сложную историю исследования, что отразилось на путанице в
названиях его отдельных частей. Аденогипофиз происходит из кармана Ратке,
который представляет собой складку дорсальной стенки ротовой полости. Эта
складка находится в контакте с мозгом и в процессе эмбриогенеза отделяется от
ротовой полости. Из кармана Ратке формируются три части аденогипофиза:
бугорковая часть (pars tuberalis), промежуточная часть (pars intermedia) и дис-
тальная (периферическая) часть (pars distalis). В нейрогипофиз входят средин-
ное возвышение (eminentia mediana), воронка (infundibulum) и нейромозговая
часть (pars nervosa). Эта номенклатура сложилась под влиянием эмбриологичес-
ких исследований. В литературе чаше можно встретить чисго анатомическое
разделение гипофиза на переднюю и заднюю доли (anterior, posterior lobe). При
этом к задней доле относят промежуточную и нейромозговую части, а к перед-
ней — бугорковую и дистальную. Понятно, что при таком разделений гипофиза
на доли воронка и срединное возвышение не учитываются.
Перечисленные структуры гипофиза не всегда встречаются одновременно.
У круглоротых, пластиножаберных, двоякодышащих, кистеперых и костных рыб
отсутствует бугорковая часть аденогипофиза. При этом аденогипофиз акул может
образовывать многочисленные ростральные выросты (lobus ventralis на рис. 136),
которые не встречаются у других позвоночных и могут быть больше самого
гипофиза. Бугорковая часть аденогипофиза присутствует только у амфибий, птиц
и млекопитающих. Проксимальная и ростральная части дистального аденогипо-
физа четко анатомически дифференцируются у миноги, хрящевых и костных
рыб. У миксин, амфибий, рептилий, птиц и млекопитающих их признаки есть,
190
♦
Сравнительная анатомия нервной системы позвоночных
_ Eminentia
Птицы mediana
Млекопитающие
форма
Рис. 136. Сагиттальные сечения гипофиза различных позвоночных. Передний конец тела
находится слева
но четкой гомологизации провести невозможно. Промежуточная доля аденоги-
пофиза идентифицируется у всех перечисленных групп, за исключением мик-
син. Нейрогипофиз четко выявляется у всех позвоночных. Однако срединное
возвышение отсутствует у миксин, а сосудистый мешочек характерен только
для хрящевых и костистых рыб, многоперов и латимерии. У двоякодышащих,
круглоротых, амфибий, рептилий, птиц и млекопитающих он отсутствует.
Гипоталамус играет основную роль в гормональной регуляции организма,
поддержании гомеостаза, полового и пищевого поведения. Все эти функции
осуществляются благодаря специализации нервных клеток этого отдела и своеоб-
разной системе кровобращения базальной части промежуточного мозга (рис. 137).
Клетки нейрогипофиза не синтезируют и не секретируют гормоны. Гормоны
доставляются в нейральную долю гипофиза по отросткам нейросекреторных
клеток, которые расположены в вышележащих областях гипоталамуса. Синтез
гормона осуществляется в гипоталамусе, а его секреция в кровеносное русло
происходит в нейрогипофизе.
VII. Промежуточный мозг
191
Рис. 137. Сечение гипофиза млекопитающих с системой сосудов. Стрелками обозначено
направление течения крови. Передний конец тела находится слева.
Отростки нейронов трех ядер гипоталамуса: супрахиазматического, преоп-
тического и паравентрикулярного ядра (n. suprachiasmaticus, n. preopticus,
n. paraventricularis) — играют основную роль в секреции гормонов нейральной
доли гипофиза. По отросткам нейронов этих ядер в нейральную долю направ-
ляются два основных гормона: окситоцин и вазопрессин (антидиуретический
гормон). Окситоцин стимулирует сокращения гладкой мускулатуры молочных
желез и матки, а вазопрессин влияет на почечные канальцы, вызывая обрат-
ное всасывание воды. Его дополнительное действие проявляется в стимуля-
ции сокращения гладкой мускулатуры артериол, что вызывает повышение ар-
териального давления. Отростки нейросекреторных клеток ядер гипоталамуса
оканчиваются на капиллярах сосудистой системы нейрогипофиза. Гормоны
попадают в кровь и разносятся по организму животного.
Аденогипофизарная система регуляции выделения гормонов построена по
иному принципу. Отростки секреторных нейронов, расположенных в гипотала-
мусе, оканчиваются не в аденогипофизе, а в сосудистом сплетении срединного
возвышения (см. рис. 137). Капиллярная сеть срединного возвышения снабже-
на аксонами нейронов, в которых накапливаются гипоталамические пептиды.
При гормональной или электрической стимуляции нейронов гипоталамуса кровь
насыщается этими короткоживущими пептидами. С током крови они перено-
сятся в сосудистую систему аденогипофиза, где формируется тонкая капилляр-
ная сеть, называемая воротной системой. Пептиды срединного возвышения сти-
мулируют выброс гормонов, секретируемых аденогипофизом в воротную систе-
му. Ранее считалось, что основными ядрами регуляции активности аденогипофиза
являются преоптическое ядро (n. preopticus) анамний и серый бугор (tuber
cinereum) амниот (рис. 138). Эти структуры действительно располагают нейро-
192
Сравнительная анатомия нервной системы позвоночных
Nucleusthalamicus
Рис. 138. Сагиттальное сечение мозга костной рыбы с основными связями промежуточно-
го мозга. Зачернены клетки ядер промежуточного мозга.
секреторными клетками, выделяющими нейропептиды-регуляторы. Однако и
преоптическое ядро, и серый бугор оказались сложными центрами, состоящи-
ми из нескольких ядер, а источники регуляции аденогипофиза расположены
почти во всех отделах гипоталамуса. В эту систему вовлечены клетки срединно-
го возвышения, переднего и заднего отделов гипоталамуса, в которых выраба-
тывается тиреолибирин. В аденогипофизе он стимулирует выработку тиреот-
ропного гормона, который воздействует на щитовидную железу, запуская сек-
рецию тироксина, регулирующего рост и энергетический обмен организма.
Аналогично действуют нейросекреторные клетки преоптического ядра и
срединного возвышения, вырабатывающие люлиберин. После переноса к аде-
ногипофизу он запускает синтез лютеинизирующего гормона. Этот гормон в
свою очередь влияет на желтое тело, вызывая секрецию прогестерона, и на
яйцевые фолликулы, регулируя их созревание и секрецию эстрогена. Сход-
ным образом действует соматилиберин, секретируемый нейроэндокринными
клетками вентромедиального ядра гипоталамуса. Он достигает аденогипофиза
VII. Промежуточный мозг
193
❖
и запускает механизм секреции соматотропина. Соматотропин выделяется в
кровь и достигает органов-мишеней — соматических клеток. Этот гормон сти-
мулирует их рост. По этому же принципу действуют кортиколиберин, фолли-
берин и пролактолиберин.
Через воротную систему аденогипофиза происходит регуляция большей ча-
сти гормональных процессов. Однако и сама нервная система подвержена вли-
янию гормонов, секрецию которых регулирует. Система обратной связи пост-
роена на изменении секреции либеринов в зависимости от уровня концентра-
ции гормонов в крови. Эта простая регуляция могла бы жестко контрол! [ровать
уровень гормонов, но на гипоталамус оказывают огромное влияние высшие
центры, расположенные в коре переднего мозга (амниоты) или в крыше сред-
него мозга (анамнии). Прямой контроль за активностью гипоталамуса осуще-
ствляют миндалевидный комплекс, дофаминергические нейроны черной суб-
станции, гиппокамп и преоптические ядра. Работа этих центров зависит от
состояния организма животного в целом. Обонятельные сигналы, циркодиан-
ные или инфрадианные циклы, форма моторной активности, половое созре-
вание или агрессия могут глубоко изменять нейросекреторную активность и
вызывать всю цепь гормональных перестроек организма животного (рис. 139).
Неокортекс
Complex
thalamicus
ventrobasalis
Corpus
geniculatum
laterale
N. columnalis
dorsalis
N descendens
nervi (V)
Мозжечок
N. vestibularis
—Corpus
geniculatum
mediate
Nuclei
habenulares
Colliculus
superior,
inferior
N. parabranchialis
Рис. 139. Сагиттальное сечение мозга млекопитающих с основными связями промежуточ-
ного мозга. Зачернены клетки ядер промежуточного мозга, участок с точками — кора перед-
него мозга.
Зрительные
волокна
Nuclei thalamicae
posteriores
laterales
Восходящие
спинномозговые
волокна
7—5818
194
❖
Сравнительная анатомия нервной системы позвоночных
Промежуточный мозг анамний сформировался как интегративный центр,
объединивший информацию от обонятельных областей переднего мозга и раз-
личных органов чувств, имеющих центры в среднем и продолговатом мозге.
Став своеобразным релейным центром, он, как ни парадоксально, не приоб-
рел свойства ассоциативного отдела мозга в том понимании, которое мы вкла-
дываем в тектум амфибий или в лобные области человека. Промежуточный
мозг стал важнейшим нейрогормональным центром, через который осуществ-
ляется контроль за работой большинства эндокринных органов позвоночных.
Секреция гормонов вызывает длительные генерализованные изменения в по-
ведении животного, которые очень плохо корректируются при помощи нерв-
ной системы. Она сама оказывается вовлеченной в активность, вызванную
гормонами благодаря обратной связи с нейрогормональными центрами через
кровеносное русло.
С другой стороны, эпиталамус и его производные трансформировались в
систему нейрогормональной регуляции суточной и сезонной активности жи-
вотных. У млекопитающих регуляция ритмов физиологической активности
сочетается с контролем за половым созреванием животных.
Таламические структуры промежуточного мозга сформировались как авто-
номные сенсорные и моторные центры анамний. В процессе эволюции они
превратились в промежуточные ядра восходящих анализаторных и нисходя-
щих моторных путей, которые обслуживают новую кору млекопитающих и
неостриатум птиц.
VIII. ПЕРЕДНИМ МОЗГ
Передний, или конечный, мозг (telencephalon) полу-
чил название благодаря своему ростральному положе-
нию. Этот отдел головного мозга возник как зона обра-
ботки хеморецепторных сигналов, приходящих из обо-
нятельной системы. У низших позвоночных передний
мозг представляет собой специализированный участок
центральной нервной системы, обслуживающий только
обонятельный анализатор. Сигналы из обонятельной
системы поступают в мозг по отросткам сенсорных кле-
ток, лежащих в рецепторном эпителии. Это единствен-
ные сенсорные клетки такого типа, и их принято назы-
вать первично чувствующими. Их аксоны оканчивают-
ся в обонятельных луковицах (bulbus olfactorius), которые
непосредственно объединены с передним мозгом или
приближены к органам обоняния (рис. 140—142). Если
обонятельные луковицы не слиты с мозгом, то соеди-
няются с ним при помощи обонятельного тракта. Обо-
нятельный тракт состоит из отростков клеток, располо-
женных в обонятельной луковице. Для первичноводных
позвоночных характерно значительное развитие обоня-
тельных луковиц. У миксин и миног они сливаются с
передним мозгом и отделены от него небольшой обо-
нятельной бороздкой (см. рис. 140). Передний мозг об-
разует парные симметричные центры, которые у анам-
ний могут представлять латеральные стенки единого
зачатка (акулы, скаты) или формировать полушария (ам-
фибии). У амфибий, рептилий, птиц и млекопитающих
передний мозг формирует полушария (hemispheria
cerebri). Они парные и немного напоминают полусфе-
ры. Внутри полушарий располагаются парные латераль-
ные желудочки (ventriculi lateralis 1, II), в которых нахо-
дится сосудистое сплетение, непрерывно переходящее
в 111 желудочек (см. рис. 20).
Однако эта общая схема может заметно варьировать
в разных группах позвоночных. Слияние развитых
196
Сравнительная анатомия нервной системы позвоночных
Рис. 140. Мозг с дорсальной поверхности.
а — минога; б—колючая акула; в—карп; г — травяная лягушка; д—геккон; е — сетчатый
питон; ж — голубь; з — кролик; и — собака. Участок с крестиками — передний мозг, зачер-
нены обонятельные луковицы и тракты, участок с точками — неокортекс.
обонятельных луковиц с передним мозгом может приводить как к редукции
латеральных желудочков, так и к увеличению их размеров. Увеличение лате-
ральных желудочков обычно можно видеть у скатов. Плотно сросшиеся с бо-
ковыми стенками переднего мозга обонятельные луковицы составляют часть
полушария, а латеральные желудочки приобретают дополнительные полости.
Такое слияние парных обонятельных луковиц с передним мозгом превращает
его в «трехполушарное» образование. Однако это анатомическая иллюзия.
Собственно передним мозгом является только область вторичных обонятель-
ных центров между крупными обонятельными луковицами. Если у некоторых
видов акул обонятельные луковицы вынесены к органам обоняния, то внеш-
нее строение переднего мозга приобретает вид шара. «Однополушарный» мозг
VIII. Передний мозг
197
Рис. 141. Сагиттальные сечения головного мозга.
а — минога; б — колючая акула; в — латимерия; г — травяная лягушка. Участок с крести-
ками — передний мозг, зачернены обонятельные луковицы. Ill, IV — желудочки мозга.
можно часто наблюдать у пелагических акул (см. рис. 22). В этом случае вто-
ричные обонятельные центры расположены в стенках переднего мозга, еще не
разделенного на парные полушария. Нужно отметить, что передний мозг по-
настоящему разделяется на парные полушария только у амфибий и других
тетрапод.
У некоторых видов пластинчатожаберных и амфибий обонятельные луко-
вицы прирастают к ростральной стенке полушарий. В этом случае обонятель-
ные луковицы сливаются с полушариями и образуют общий желудочек. Для
амфибий, рептилий и млекопитающих характерно появление дополнитель-
ной, или вомероназальной, луковицы. В ней оканчиваются волокна, берущие
свое начало в вомероназальном органе, который осуществляет хеморецепцию
в водной среде (амфибии), контактную хеморецепцию (рептилии) или поло-
вое обоняние (млекопитающие).
У костных рыб передний мозг формирует небольшие парные полусферы,
которые бывают по размерам меньше обонятельных луковиц. Однако строение
переднего мозга многих рыб имеет ряд специфических особеностей (рис. 143).
Дело в том, что у большинства костистых рыб полушария переднего мозга
эвертированы. Это означает, что они вывернуты наружу прижелудочковыми
стенками (рис. 144, 148). Дорсальные края стенок полушарий оказываются
198
❖
Сравнительная анатомия нервной системы позвоночных
Рис. 142. Сагиттальные сечения головного мозга.
а — черепаха; б — голубь; в — собака. Участок с точками — неокортекс, с крестиками —
передний мозг, зачернены обонятельные луковицы и тракты. Ill, IV — желудочки мозга.
механически не связанными между собой. Они соединены тонкой эпители-
альной полоской эпендимных клеток, которая покрывает практически всю
поверхность раскрытого желудочка. В результате эверсии переднего мозга у
костистых рыб отсутствуют парные латеральные желудочки и разветвление
сосудистого сплетения. Оно иногда может присутствовать в эпендимной стен-
ке переднего мозга в виде асимметричного продолжения сосудистого сплете-
ния III желудочка. Необходимо отметить, что у некоторых рыб эверсия перед-
него мозга бывает только частичной. Примером может служить организация
переднего мозга латимерии (см. рис. 144). У нее полушария сформированы по
обычному типу, но стенки медиального плаща остаются незамкнутыми. Из-за
этого латеральные желудочки открыты, а полость переднего мозга закрывает-
ся тонкой эпендимной стенкой. В целом эверсия полушарий переднего мозга,
VIII. Передний мозг
199
Bulbus
Striatum
Pallium dorsale,
pars superficialis
N. septi lateralis
N. septi medialis
Pallium
mediale
Area dorsalis
telencephali:
dorsalis
lateralis -
medialis
Area ventralis
telencephali:
dorsalis
ventralis
Area
periventricu>ans
dorsolateralis
Nucleus N
N. nervi
abducens
(VI)
— Area superficialis
basalis
Area
periventricularis
ventrolateralis
i.
в
Рис. 143. Поперечные срезы переднего мозга.
а — минога; б — акула; в — многопер. I, II — желудочки мозга (по R. Nieuwenhuys et al.,
1997, с изменениями).
по-видимому, оказалась недостаточно эффективным путем в эволюции. С од-
ной стороны, возникло «конструкционное» ограничение для дальнейшего уве-
личения объема переднего мозга и развития межполушарных связей. Если бы
эвертированный мозг увеличивался в размерах, то его полушария вынуждены
были бы распространяться в сторону основания мозгового черепа, а комиссу-
ры — проходить только через вентромедиальную ось мозга. Подобного строе-
ния мозга у костных рыб пока не известно.
Следовательно, эверсия мозга оказалась тупиковой ветвью развития перед-
него мозга. После закрепления эверсивного строения передний мозг кости-
стых рыб увеличивался весьма незначительно и сохранил в основном предко-
вый план строения.
Для остальных позвоночных характерно инверсивное строение переднего
мозга (см. рис. 144—146, 148). При этом формируются замкнутые парные по-
200
Сравнительная анатомия нервной системы позвоночных
Area dorsalis telencephali:
posterior, lateralis, dorsalis, centralis, medialis
Area ventralis telencephali: pars supracommissuralis, pars lateralis
Striatum ventralis
Pallium dorsomedialis
Pallium ventromedialis
Corpus striatum
Tuberculum olfactorium
Pallium medialis
Striatum dorsalis
Pallium lateralis
Zona limitans
N. septi medialis
- N septi lateralis
Pallium dorsalis
Pallium lateralis
N. accumbens
Septum
medialis
Lamina terminalis
Septum lateralis
e
Рис. 144. Поперечные срезы переднего мозга.
а — форель;б—латимерия;е — аксолотль. I, II—желудочки мозга (по R. NieuwenhuysetaL,
1997, с изменениями).
лушария с латеральными желудочками внутри. Полушария переднего мозга
принято разделять на дорсально лежащий плащ (pallium) переднего мозга и
основание переднего мозга или подплащевые структуры (субпаллиальные).
Последние часто называют у млекопитающих базальными ганглиями (ganglia
basilaris), а у птиц и рептилий — стриатумом и септумом (striatum, septum). Это
упрощение номенклатуры только затрудняет классификацию центров основа-
ния переднего мозга.
К плащу переднего мозга и его производным относят три комплекса струк-
тур, названия которых неоднократно уточнялись и изменялись. На сегодняш-
ний день принята номенклатура, по которой весь дорсальный участок перед-
него мозга называют плащом (pallium). В него входит старая кора (archipallium),
VIII. Передний мозг
201
Hyperstriatum:
accessorium
dorsale
ventrale
N. septi lateralis
N. preopticus
dorsolateralis
Neostriatum
intermedium
Paleostriatum:
augmentatum
primitivum
N. intrapeduncularis
N. preopticus anterior
Sulcus hippocampi
Cortex cingularis
Sulcus rhinalis
N. endopiriformis
Cortex parietalis
N. caudatus
Putamen
Cortex piriformis
Globus pallidus
N. interstitialis
stria terminalis
Substantia
innominata
Fissura
amygdalae
e
Рис. 145. Поперечные срезы переднего мозга
а — черепаха; б — голубь; в — собака. I-III — желудочки мозга (по R. Nieuwenhuys et al.,
1997, с изменениями).
Hippocamp:
subiculum
cornu ammonis
fascia dentata
которая теперь называется медиальным плащом (pallium medialis). Кроме ста-
рой коры, к плащу относят древнюю кору (paleopallium), которую в современ-
ных терминах обычно именуют латеральной корой (cortex lateralis). Между
старой и древней корой находится новый плащ (neopallium), который включа-
ет дорсальный плащ (pallium dorsalis) и дорсомедиальную (cortex cinguli) кору
(cortex dorsomedialis) дорсолатеральную кору (cortex dorsolateralis), латераль-
ную кору (cortex lateralis) и дорсальное желудочковое выпячивание латераль-
ной стенки (tuberculum laterale). В старой литературе старую кору обычно на-
зывали гиппокампом (hippocampus) или аммоновым рогом (cornu ammonis).
202
Сравнительная анатомия нервной системы позвоночных
монополушарная
форма
Рис. 146. Поперечные срезы переднего мозга. У костистых рыб полушария эвертированы,
у остальных — инвертированы.
I, II — желудочки мозга.
Новый плаш обычно именуют неокортексом. а ранее — изокортексом (neocortex,
isocortex) или просто корой мозга (cortex cerebralis). Под названием «большой
мозг» (cerebrum) обычно понимают полушария переднего мозга млекопитающих,
состоящие в основном из неокортекса. Старый плаш имеет несколько историчес-
ких синонимов. Его обычно называли палеокортексом, грушевидными или пи-
риформными долями (корой) [paleocortex, cortex (lobus) piriformis].
Структуры основания переднего мозга (подкорки) также имеют некоторую
историческую запутанность терминологии, которая сложилась на основании
изучения мозга человека. В традиционной морфологической литературе обыч-
но выделяют две большие группы структур: полосатое тело (corpus striatum) и
перегородку (septum). Полосатое тело состоит из трех крупных отделов: древнего
VIII. Передний мозг
203
Рис. 147. Сагиттальные сечения мозга с основными подкорковыми структурами у амфи-
бий, рептилий и млекопитающих (по A. Kappers, 1921. с изменениями).
полосатого тела (paleostriatum), нового полосатого тела (neostriatum) и старого
полосатого тела (archistriatum). Новое полосатое тело формируется у млекопи-
тающих в связи с развитием сенсомоторных центров новой коры (см. рис. 147).
В него входят хвостатое ядро (nucleus caudatus), ограда (claustrium) и скорлупа
(putamen). К древнему полосатому телу относят ядра бледного шара (globus
pallidus), а к старому полосатому телу — миндалевидный комплекс (amigdala).
К септуму традиционно относят септальное поле (area septalis), содержащее
несколько ядер. Для современной номенклатуры характерно разделение осно-
вания мозга на три отдела: подплащевые структуры (subpallium), стриатум
(striatum) и септум (septum). К подплашевым структурам относят базальные
ядра (nuclei basales), к стриатуму — древнее, старое и новое полосатое тело, к
септуму — септальное поле (area septalis). Целью изменений номенклатуры ста-
ла необходимость сравнительно-анатомического анализа животных, не имею-
щих развитого неокортекса и зависимых от него подкорковых образований.
204 ❖ Сравнительная анатомия нервной системы позвоночных
Эта ситуация проиллюстрирована рис. 147. У амфибий можно выявить
старый и древний стриатум, септум и в зачаточном виде — базальные ядра.
Для рептилий характерно появление нового стриатума, который становится
доминирующей структурой основания мозга. Важно подчеркнуть, что новый
стриатум возникает при очень незначительном развитии неокортекса. Он не
столько обеспечивает связи зачаточного неокортекса с остальным мозгом,
сколько выполняет аналогичные ему функции. По-видимому, новый, ста-
рый и древний стриатум справлялся с этим достаточно хорошо у рептилий,
что привело к возникновению мозга птиц, построенного по подкорковому
типу. У птиц неокортекс практически отсутствует, а его функции выполняют
чрезвычайно развитые структуры стриатума. Однако историческое название
«гиперстриатум» не отражает реальное происхождение этого центра у птиц.
Ранее считалось, что гиперстриатум возник из стриатума и является его го-
мологом. Специальные исследования кинетики пролиферации и миграции ней-
робластов у птиц позволили установить, что гиперстриатум имеет смешанное
происхождение и по большей части состоит из латеральной (paleopallium) и
новой коры (neopallium) (см. рис. 145, 146). Особенностью ее развития стало
формирование структур ядерного типа, а не стратифицированных корковых
образований. У птиц и рептилий разрастание стриатума приводит к экспансии
дорсального внутрижелудочкового бугорка, который практически полностью
занимает полость латеральных желудочков (см. рис. 145, 146). Гиперстриа-
тум птиц (рептилии имеют его признаки) выполняет функции новой коры.
Он представляет собой высший ассоциативный центр, который определяет
принятие решений и является основной зоной хранения индивидуального
опыта. Через стриатум осуществляются контроль за двигательными функци-
ями и связь с лимбической системой.
Лимбическая система является морфофункциональным комплексом струк-
тур, которые расположены в различных отделах переднего и промежуточного
мозга. В нее входят от переднего мозга дорсомедиальный плаш [поясная изви-
лина (gyrus cinguli), зубчатая извилина (gyrus dentatus)], медиальный плащ
(hippocamp), септум (area septalis) и старый стриатум (amygdala). В промежу-
точном мозге расположены четыре основные структуры лимбической системы:
хабенулярные ядра (n. habenularis), таламус, гипоталамус и сосцевидные тела
(n. mamillaris) (см. рис. 148).
Волокна, соединяющие структуры лимбической системы, образуют свод пе-
реднего мозга (fornix cerebri), который проходит в виде арки от архипаллиума
до сосцевидных тел. Лимбическая система объединена многочисленными свя-
зями с неокортексом и автономной нервной системой. Поэтому она интегри-
рует две важнейшие функции мозга животного — эмоции и память. Удаление
части лимбической системы приводит к эмоциональной пассивности живот-
ного, а стимуляция — к гиперактивности. Активизация миндалевидного ком-
плекса запускает механизмы агрессии, которые могут корректироваться гип-
покампом. Лимбическая система запускает пищевое поведение и вызывает
чувство опасности. Все эти формы поведения контролируются как самой лим-
бической системой, так и через гормоны, вырабатываемые гипоталамусом.
При стимуляции гормональной системы происходит долговременное измене-
ние поведения животного. Влияние лимбической системы на функции орга-
VIII. Передний мозг
205
N. habenulae
Septum
Bulbus
olfactorius
Моторные
центры
неокортекса
Слуховые
центры
неокортекса
Ассоциативные
центры Gyrus cinguli
неокортекса
Hippocamp
Tha amus
Amygdala
Paleocortex
Зрительные
центры
неокортекса
Hypothalamus
N. mamillaris
Нисходящие и восходящие связи
моторных ядер и спинного мозга
Рис. 148. Основные связи структур лимбической системы, вписанные в сагиттальное се-
чение мозга млекопитающих. Участок со штриховкой — мозолистое тело с межполушар-
ными волокнами, с точками — центры лимбической системы.
низма осуществляется через контроль за деятельностью автономной нервной
системы. Роль лимбической системы столь высока, что ее принято называть
висцеральным мозгом По сути дела она контролирует эмоционально-гормо-
нальную активность животного, которая, как правило, плохо поддается рассу-
дочному контролю даже у человека.
Второй важнейшей функцией лимбической системы является взаимодействие
с механизмами памяти. Краткосрочную память обычно связывают с гиппокам-
пом, а долгосрочную — с неокортексом. Однако извлечение индивидуального
опыта животного из неокортекса осуществляется через лимбическую систему.
При этом используется эмоционально-гормональная стимуляция мозга, кото-
рая вызывает информацию из долговременного хранения в неокортексе.
Таким образом, лимбическая система включает в себя как древние подкор-
ковые и плащевые структуры, так и неокортекс. Лимбическая система облада-
ет уникальным набором эффекторных структур. В них входят управление мо-
торикой внутренних органов, двигательная активность для выражения эмо-
ций и гормональная стимуляция организма. Необходимо отметить, что чем
ниже уровень развития неокортекса, тем больше поведение животного зави-
сит от лимбической системы. У большей части позвоночных и примитивных
млекопитающих церебральные системы контроля лимбической системы крайне
несовершенны, а связи со средним мозгом развиты крайне слабо. Возникает
206
Сравнительная анатомия нервной системы позвоночных
«независимость» лимбической системы, которая приводит к доминированию
эмоционально-гормонального контроля за принятием решений.
До появления неокортекса у млекопитающих основную роль в регуляции
лимбической системы играла обонятельная система. Именно через ее сенсор-
ные входы осуществляется запуск механизмов полового и социального пове-
дения, построенного на хемокоммуникациях. При этом ключевую роль играет
комплекс ядер миндалины, которые имеют как обонятельные, так и моторные
входы и выходы (см. рис. 148). У млекопитающих за счет нисходящих связей
неокортекса в лимбической системе достигается интеграция самых разнооб-
разных сенсорных сигналов. Этим достигается адекватная адаптивная реакция
организма на изменение внешней ситуации. Однако у низших млекопитаю-
щих неокортекс обычно развит недостаточно, а среди анализаторов превали-
рует обонятельная система. У таких видов лимбическая система оказывается
под определяющим влиянием обоняния. Передний мозг однопроходных, сум-
чатых, насекомоядных и ряда копытных построен по обонятельному типу, что
хорошо заметно по организации базальной поверхности мозга (рис. 149).
Для низших позвоночных с неразвитым неокортексом характерно наличие не-
большого числа волокон, соединяющих правое и левое полушария (см. рис. 141).
Функции межполушарных коммуникаций обеспечивают передняя и плащевая
Рис. 149. Вентральная (базальная) поверхность переднего мозга однопроходных, сумчатых,
неполнозубых, насекомоядных и непарнокопытных (по L. Edinger, 1911, с изменениями).
VIII. Передний мозг
207
❖
комиссуры (commissura anterior, pallii). Однако с появлением первых зачатков
неокортекса у рептилий от плащевой комиссуры отделяется небольшой пучок
волокон, соединяющий правое и левое полушария. У плацентарных млекопи-
тающих он занимает значительное место между полушариями и носит назва-
ние мозолистого тела (corpus callosum) (см. рис. 142). Для млекопитающих
характерно наличие своеобразной структуры мозга — свода (fornix). Как уже
упоминалось, свод мозга является пучком нервных волокон, которые начина-
ются в старой коре и заканчиваются в сосцевидных телах и гипоталамусе.
У амфибий, рептилий и низших млекопитающих свод очень короткий и на
сагиттальных разрезах мозга плохо заметен. Это связано с тем, что небольшое
увеличение размеров рострального плаща переднего мозга не приводит к кау-
довентральному смешению гиппокампа и слиянию волокон свода в компакт-
ный пучок. Чем больше увеличивается размер неокортекса, тем заметнее сме-
щение гиппокампа, длиннее и обособленнее нервные волокна свода.
Становление неокортекса происходило достаточно постепенно. Предшествен-
ником основной части неокортекса является участок плаща переднего мозга,
который у амфибий прилежит к латеральным желудочкам (рис. 150). Это обыч-
ное для амфибий скопление прижелудочковой массы клеток. Для амфибий
намного большее значение имеют медиальные и латеральные части плаща. Из
медиальной зоны формируется зачаточный гиппокамп (archipallium), а из ла-
теральной — центры палеокортекса (paleocortex). Эти зоны у амфибий выпол-
няют функции относительно автономных третичных обонятельных центров.
У рептилий в дорсолатеральной стенке полушария впервые появляются за-
чатки неокортекса. Он формируется в эмбриогенезе за счет радиальной мигра-
ции нейробластов из прижелудочкового скопления клеток переднего мозга
(см. рис. 150). Изменение названия структуры с дорсального плаща на неокор-
текс связано с перемещением зачатка от прижелудочковой зоны к наружной
поверхности полушария. Для рептилий характерна небольшая стратифициро-
ванность закладки неокортекса. Большинство авторов считают его однослой-
ным, хотя у крокодилов можно выделить два и даже три слоя. Выделение зачат-
ка неокортекса приводит к нескольким важным последствиям. Во-первых, фор-
мируется стратифицированная анализаторно-ассоциативная система, являющаяся
надстройкой над уже достаточно развитым мозгом рептилий, и возникают но-
вые восходящие и нисходящие пути для ее обслуживания. Во-вторых, за счет
развития неокортекса резко возрастает объем переднего мозга, что делает его
доминирующей структурой в головном мозге и приводит к появлению борозд и
извилин. В-третьих, возникает зональная специализация коры, в которой пред-
ставлены в виде полей все системы анализаторов, ассоциативные центры и зоны
управления движением. Гистологические изменения в организации и иннерва-
ции переднего мозга сопровождались заметными анатомическими изменениями.
Особенно серьезной перестройке подвергся передний мозг млекопитающих.
Поверхность полушарий переднего мозга млекопитающих может быть глад-
кой, лисэнцефальной, или складчатой, с бороздами и извилинами. Независи-
мо от этого в полушариях переднего мозга принято выделять от 4 до 5 долей.
Принцип разделения на такие отделы возник на основании исследований го-
ловного мозга человека, поэтому расположение долей в лисэнцефальном моз-
ге носит условный характер.
208
Сравнительная анатомия нервной системы позвоночных
Рис. 150. Поперечные сечения левого полушария переднего мозга через неокортекс.
Наличие многочисленных борозд и извилин в мозге копытных, хищных,
морских млекопитающих, приматов и человека вызывало необходимость най-
ти какие-либо простые принципы для классификации поверхности полуша-
рий. Самым общим принципом стало разделение мозга на доли. Обычно вы-
деляют теменные доли (lobi parietales), височные доли (lobi temporales), заты-
лочные доли (lobi occipitales) и лобные доли (lobi frontales). У высших приматов
и человека обычно добавляется пятая доля, которая называется островком
(insula). Островок скрыт за крышечкой (operculum) — складкой сильвиевой
борозды. Он образуется за счет разрастания в эмбриональный период височ-
ной доли, которая прикрывает вентральную часть полушария. Остальные доли
выделяются по бороздам и извилинам. Отправной точкой является централь-
ная борозда (sulcus centralis, Rolandi), которая тянется от центральной шели
(sulcus cerebri lateralis, Sylvii) до базальной поверхности мозга. Ростральнее от
нее находится лобная доля, а каудальнее — теменная. Границей теменной и
VIII. Передний мозг
209
височной долей является дорсальный край верхней височной извилины (gyrus
temporalis superior). Каудальный край теменной и височной долей примыка-
ет к затылочной, но граница между ними даже у человека носит условный
характер. Разделение мозга на доли является самым примитивным способом
оценки развития нервной системы. Тем не менее размеры долей мозга у
различных животных позволяют сделать общие выводы об основных тенден-
циях эволюции нервной системы (рис. 151). Именно таким оценкам, сделан-
ным Эдингером почти столетие назад, мы обязаны тривиальному выводу о
том, что лобные доли самые крупные у приматов и человека. К сожалению,
этот подход практически невозможно применить к млекопитающим с лисэн-
цефальным мозгом. В мозге без борозд и извилин невозможно выделить чет-
кие анатомические границы между отделами, а использование цитоархитек-
тонического анализа неокортекса для их выделения не оправдано скромно-
стью результатов.
Следующим уровнем организации полушарий переднего мозга является
топология борозд и извилин (см. рис. 151, 152). Этот подход более эффекти-
вен, поскольку у млекопитающих с самым гладким мозгом всегда можно вы-
делить сильвиеву складку (fissura Sylvii) и обонятельную борозду (sulcus rhinalis).
Бороздами называют прогибы стенки мозга в сторону желудочков, а извили-
нами — участки полушарий мозга между ними. Борозды в полушариях перед-
Рис. 151. Левое полушарие переднего мозга с обозначенными лобными долями 3 мозге
человека показаны все доли, кроме островка.
210
Сравнительная анатомия нервной системы позвоночных
Опоссум
Броненосец
S. centralis
S. coronarlis s suprasylvius
S. rhinalis s pseudosylvius
Рис. 152. Боковая поверхность переднего мозга различных млекопитающих (по R. Nieu-
wenhuys et al., 1997).
него мозга принято подразделять на три основных типа. К первому типу
относятся постоянные борозды, которые всегда возникают в неизменном виде
у животных данного вида. Таких борозд бывает относительно немного. Даже
у человека их известно только 10. Их значение трудно переоценить, посколь-
ку постоянные борозды проявляются независимо от полового диморфизма и
индивидуальной изменчивости животного. Борозды второго типа обычно по-
являются у животных, принадлежащих к одному виду, в 75% случаев. Они
менее глубокие, чем борозды первого типа, и иногда могут отсутствовать.
Однако они стабильно проявляются в пределах одной семейной группы или
небольшой популяции близкородственных животных. Самые мелкие и из-
менчивые извилины относят к третьему типу. Они носят индивидуальный
характер и обычно не повторяются даже у близкородственных особей. Про-
странственное распределение борозд и извилин на поверхности мозга носит
закономерный характер. Постоянные извилины обычно довольно сходны в
пределах одного семейства. Примерно 40—50% постоянных извилин встреча-
ется у животных, принадлежащих одному отряду. Однако использовать эти
закономерности можно только с известной осторожностью, поскольку инди-
видуальная изменчивость мозга может маскировать реальное расположение
борозд первого порядка.
VIII. Передний мозг
211
❖
Эволюция борозд и извилин не имеет четкой закономерности. По-видимо-
му, они возникали у млекопитающих неоднократно. Однако три борозды встре-
чаются практически постоянно у всех млекопитающих. Центральная борозда
(sulcus centralis) является производной сильвиевой складки (fissura sylvii), но-
совая борозда (sulcus rhinalis) — границей между палеокортексом и неокортек-
сом, а борозда гиппокампа (sulcus hippocampi) — границей между неокортек-
сом и архипаллиумом (см. рис. 145, 150, 152). Наиболее древней принято счи-
тать носовую борозду (sulcus rhinalis). Кроме нее и латеральной борозды, на
практически гладкой латеральной поверхности лисэнцефального мозга можно
выявить внутритеменную борозду (sulcus intraparietalis), венечную борозду (sulcus
coronalis), надсильвиеву, заднесильвиеву и псевдосильвиеву борозды (sulcus
suprasylvia, postsylvia, pseudosylvia).
Поверхность мозга млекопитающих со слабо развитыми бороздами и изви-
линами согласуется с многочисленными палеонтологическими слепками ней-
рокраниумов и подтверждает представление о многократном возникновении
борозд и извилин в неокортексе. Принимая за исходный тип лисэнцефальный
мозг, мы видим по меньшей мере три варианта развития рисунка борозд. Наи-
более древним вариантом можно считать продольное расположение борозд,
как у однопроходных и сумчатых (рис. 153, а). В этом варианте строения ос-
новные борозды вытянуты в рострокаудальном направлении и не формируют
поперечных прогибов коры. Ко второму варианту следует отнести широко
распространенный тип дугообразного расположения борозд, который харак-
терен как для хищных, так и для многих копытных (рис. 153, б). При таком
строении коры важную роль начинают играть центральная борозда и сильви-
ева складка. Основные борозды и извилины тянутся через все полушарие,
огибая центральную борозду. Возникшие дугообразные борозды начинаются в
лобных долях и могут заканчиваться в височных. Третий тип строения борозд
неокортекса характерен для приматов. В этом варианте центральная борозда
обычно разделяет полушария на две части. Борозды в лобных долях идут в
рострокаудальном направлении, а в височных — почти в вентродорсальном
(рис. 153, в). Из этих схем с известными оговорками можно вывести практи-
чески все основные типы организации поверхности полушарий переднего мозга.
Продольный тип
расположения борозд
(однопроходные, сумчатые)
а
Дугообразный тип
расположения борозд
(хищные, копытные)
«Приматный» тип
расположения борозд
S. centralis
в
Рис. 153. Организация основных типов борозд в переднем мозге млекопитающих (а-е).
212 ❖ Сравнительная анатомия нервной системы позвоночных
Однако надо учитывать, что на расположение борозд и извилин может значи-
тельно влиять форма мозга. У большинства млекопитающих мозг вытянут в
рострокаудальном направлении. Однако у многих дельфинов максимальному
развитию подверглись латеральные зоны неокортекса, что привело к увеличе-
нию ширины мозга и относительному уменьшению его длины.
Тем не менее максимального развития борозды и извилины достигли имен-
но в полушариях китообразных. При равных массах тела человека и дельфина
борозд у последнего в два раза больше, чем у человека. Большое количество
борозд и извилин является следствием увеличения площади поверхности коры.
Причиной таких изменений может быть простое увеличение мышечной массы
животного. У крупных животных большая часть коры переднего мозга занята
сенсомоторными и моторными полями, тогда как ассоциативные и сенсорные
поля увеличиваются почти пропорционально размеру мозга. Поэтому круп-
ный мозг китообразных и хоботных отнюдь не свидетельствует о высоком
интеллекте его обладателя.
При анализе развития переднего мозга млекопитающих необходимо учиты-
вать не только его внешнее строение, но и пространственное распределение
нейронов в коре — цитоархитектоническую организацию. Кора переднего моз-
га млекопитающих имеет сходное строение со слоистой крышей среднего мозга
низших позвоночных. Неокортекс млекопитающих имеет шестислойное строе-
ние. В процессе эмбрионального развития неокортекс формируется начиная с
глубоколежащих слоев. В первую очередь закладывается VI слой, затем нейро-
бласты формируют V слой, потом IV и т.д. Последним дифференцируется на-
ружный слой. Однако в неврологической литературе принято считать I самый
наружный слой коры, а VI — самый внутренний. У взрослых млекопитающих
различных видов кора может значительно различаться как по толшине, так и по
выраженности отдельных слоев (рис. 154). Известно, что для животных с лисэн-
цефальным мозгом характерен больший поперечник коры, чем для животных с
полушариями переднего мозга, имеющими борозды и извилины. Так, у бобров
поперечник коры несколько толще, чем у человека, а у дельфина афалины по-
чти в два раза тоньше. При всем разнообразии вариантов строения неокортекса
шестислойность коры млекопитающих не вызывает сомнения.
I — молекулярный слой (lamina molecularis, lamina zonalis). самый наруж-
ный слой коры, который содержит редкие клеточные тела.
II — наружный гранулярный слой (lamina granularis externa). Он состоит из
небольших и плотно расположенных клеточных тел. В моторных областях этот
слой может включать небольшие пирамидные клетки, которые распространя-
ют свои волокна на поверхности мозга или в молекулярном слое.
III — наружный пирамидный слой, или слой наружных пирамид (lamina
pyramidalis externa). В этом слое превалируют пирамидные нейроны, которые
крупнее во внутренних зонах слоя и мельче — в наружных.
IV — внутренний гранулярный слой (lamina granularis interna). Этот слой со-
стоит из плотно расположенных как пирамидных, так и непирамидных клеток.
V — пирамидальный слой (lamina pyramidalis). В этом слое расположены
преимущественно клетки округлой формы, которые посылают свои волокна к
пирамидным нейронам.
VI — слой полиморфных клеток (lamina multiformis). В нем находятся вере-
теновидные, округлые и модифицированные пирамидные клетки.
VIII. Передний мозг
❖
213
Коата
(широконосая
обезьяна)
Серый
гапалемур
I II и III IV V и VI
Олень
Индийский
оленек
Большой
серый
кенгуру
Мускусная
кенгуровая
крыса
Рис. 154. Строение неокортекса в переднем мозге млекопитающих.
I-VI — слои коры (no A. Kappers, 1921, с изменениями).
Для неокортекса млекопитающих характерно присутствие пирамидных кле-
ток, которые отсутствуют в мозге других позвоночных. В моторной коре нахо-
дятся особенно крупные пирамидные нейроны, носящие название клеток Беца.
Эти нейроны посылают свои длинные отростки к мотонейронам спинного
мозга и в двигательные ядра черепно-мозговых нервов. Их гигантский размер
предопределен масштабами метаболической и функциональной активности.
Нужно отметить, что неокортекс млекопитающих неоднороден в пределах
одного полушария. Причиной этой неоднородности является специализация
коры. Различные участки коры являются специализированными зонами обра-
ботки информации, поступающей от зрительных, слуховых, вкусовых и сома-
тосенсорных анализаторов. Отдельные участки коры являются моторными об-
ластями и формируют нисходящие волокна к стволовым и спинномозговым
двигательным центрам. Между сенсорными и моторными зонами коры распо-
214
Сравнительная анатомия нервной системы позвоночных
ложены интегративные участки коры, которые объединяют входы сенсорных
и моторных областей коры и предопределяют выполнение специализирован-
ных видоспецифических функций (рис. 155). В мозге млекопитающих выявле-
ны ассоциативные зоны коры, которые не связаны с конкретными анализато-
рами, моторными или интегративными функциями. Они представляют собой
надстройку над остальными участками неокортекса и являются местом сосре-
доточения когнитивных, мыслительных процессов и зонами хранения инди-
видуальной и видовой памяти.
Весь комплекс перечисленных зон распределен в головном мозге закономер-
ным образом и сопряжен с функциональной специализацией полей. Морфоло-
гические и функциональные границы полей довольно точно совпадают.
Критерием выделения того или иного поля является изменение распределения
клеточных элементов по слоям или возникновение нового подслоя (рис. 156).
В некоторых случаях соседние слои коры сливаются и неокортекс прини-
мает необычный вид. Это неоднократно приводило исследователей к ложным
выводам относительно систематического положения той или иной группы
млекопитающих (см. рис. 154). Использование методов аксонного транспорта
Еж
Морская свинка
ассоциативное
поле
Рис. 155. Поля неокортекса в переднем мозге млекопитающих (поданным разных авторов)
VIII. Передний мозг
215
Рис. 156. Граница полей 17 и 18 неокортекса в переднем мозге человека (по С.А. Саркисо-
ву и др., 1956).
флюоресцентных маркеров позволило доказать единство общего плана строе-
ния неокортекса млекопитающих.
Неоднородность организации неокортекса в полушариях переднего мозга
стимулировала попытки создания карты их топологии. Первые более или ме-
нее полные карты цитоархитектонических полей были созданы Бродманом
для мозга человека (рис. 157). Как уже отмечалось выше, цитоархитектоничес-
кие поля являются морфологическим выражением функциональной специа-
лизации поля. Так, на рис. 156 показана граница между первичным зритель-
ным полем 17 и вторичным — 18. При переходе от поля 18 к полю 17 проис-
ходит разделение VI слоя неокортекса на три подслоя. Это связано с тем, что
приходящие от наружного коленчатого тела волокна не могут вступить в кон-
такт с небольшим количеством клеток VI слоя. Слой увеличивается по толщи-
не и распадается на два подслоя и промежуточный слой между ними. Анало-
гичным образом выделяются и другие поля неокортекса. Обычно причиной
изменения цитоархитектонической организации коры является увеличение ко-
личества восходящих или нисходящих волокон. Поля коры являются важней-
шим структурным компонентом неокортекса, который позволяет нам пред-
216
❖
Сравнительная анатомия нервной системы позвоночных
Рис. 157. Карта цитоархитектонических полей неокортекса человека (по С.А. Саркисову
и др., 1956).
ставить пути эволюции этого образования. Достаточно взглянуть на предста-
вительство в головном мозге различных видов нескольких основных полей,
чтобы понять масштабы межвидовой изменчивости неокортекса (см. рис. 154).
Расположение полей в коре головного мозга у млекопитающих построено по
общим принципам.
Единственным исключением является организация коры у китообразных.
Это связано с тем, что у большинства млекопитающих передний мозг увели-
чивался в размерах в рострокаудальном направлении, а у китообразных — в
латеральном. В результате этих особенностей эволюции у китообразных ока-
зались особенно развитыми височные доли мозга, что привело к нарушению
консервативной схемы расположения полей. Зрительная область, всегда рас-
положенная на затылочном полюсе мозга, оказалась в положении теменной
доли, а соматосенсорные поля сместились в лобную долю (см. рис. 155, 157).
Существует значительная изменчивость и самих полей. В зависимости от
того, какая система анализаторов доминирует у данного вида, изменяется и ее
представительство в коре полушарий переднего мозга. Например, у ночных
обе шян затылочные зрительные поля достигают огромных размеров и явля-
ются доминирующими образованиями в полушариях переднего мозга. Для
летучих мышей характерно превалирование слуховых полей. Они могут зани-
мать до 30% от общей площади коры. Интересно отметить, что у голоспинно-
VIII. Передний мозг
217
❖
го листоноса (Pteronotus pamelli) слуховая область коры сформирована тремя
полями. В первичном слуховом поле частоты от 10 Гц до 100 кГц организова-
ны тонотопически. Это означает, что участки поля, получающие сигналы от
диапазонов различной частоты, размещены в соответствии с числовыми зна-
чениями частот. Внутри первичного поля выделяется подполе коры, в кото-
ром происходит анализ слуховых сигналов вблизи частоты 61,2 кГц, посколь-
ку именно в этом диапазоне осуществляется эхолокация. Рядом расположены
специализированные области, осуществляющие анализ удаленности объекта
локации и его относительной скорости движения. Этот пример показывает,
что специализированные моторные и сенсорные поля коры переднего мозга
сами по себе тоже неоднородны. Они состоят из локальных участков, которые
осуществляют обработку сенсорных сигналов от небольших участков тела или
от фрагментов сенсорных органов.
Примером топологической специализации коры является точная связь между
определенными участками сетчатки, участками наружных коленчатых тел и
зрительными полями. Если при травме зрительной коры проверить форму
поля зрения глаза, то она окажется неполной в месте сетчатки, которое дает
проекции в поврежденный участок коры. Аналогичный результат получается
и при повреждении сетчатки. Нужно отметить, что с обнаружения топографи-
ческой специализации моторных полей коры у собаки началась история изу-
чения электрической возбудимости головного мозга. В начале второй полови-
ны XIX века Т.Фрич и Э.Гитциг открыли факт электрической возбудимости,
проводя стимуляцию двигательных полей коры переднего мозга. Уже тогда
стало ясно, что сушествуют четкие моторные и сенсорные зоны внутри специ-
ализированных полей, которые топологически связаны с рецепторными или
эффекторными структурами организма. Развитие этих представлений привело
к появлению исследований У.Пенфилда, который, локально раздражая поверх-
ность мозга, создал карту локализации функций в моторном поле коры чело-
века — знаменитого «гомункулюса». Аналогичные карты некоторых полей были
созданы для многих лабораторных животных (см. рис. 155). Различные методы
изучения локализации функций в отдельных полях коры показали, что суще-
ствует небольшое перекрывание отдельных специализированных полей, что
несколько размывает их границы, но не изменяет принципы пространствен-
ной организации.
Внутренняя организация полей неокортекса животных не менее сложна,
чем у человека. Даже у «лисэнцефальных» животных существуют специализи-
рованные подполя, обслуживающие отдельные участки сенсорных органов.
Примером может быть строение соматосенсорной коры мыши, которая отве-
чает за механорецепцию вибрисс, расположенных на мордочке животного.
Каждая вибрисса представлена соматотопически расположенным в коре не-
большим бочонком или клубочком (рис. 158).
Аналогичным обазом организовано представительство пальцев и ладошек у
енота. Каждый палец и отдельная ладонная подушечка представлены в сома-
тосенсорной коре самостоятельным полем, причем большая часть полей вы-
деляется небольшими бороздами. Следовательно, цитоархитектоническая спе-
циализация полей неокортекса привела к морфологическому обособлению фраг-
ментов коры (см. рис. 158).
218
Сравнительная анатомия нервной системы позвоночных
Рис. 158. Расположение зон представительства пальцев и ладонных подушечек енота и
вибрисс мыши в соматосенсорной коре (по Н. Welker и S. Seidenstein, 1959; Y. Woolsey и
van der W. Loos, 1970).
Таким образом, поля коры головного мозга входят в определенные доли, а
сами поля подразделены на функциональные зоны, которые связаны с конк-
ретным органом или его частью. Функциональные зоны коры имеют упорядо-
ченное внутреннее строение. В каждой зоне или поле можно выделить своеоб-
разные модули вертикальной упорядоченности организации коры. В самом
обычном случае модуль представляет собой колонку или клубочек, в который
включаются клетки, расположенные по всей толщине коры. Колонкой мини-
мального размера обычно считают группу из ПО нейронов, расположенных
между парой капилляров, проходящих через поперечник коры. Клетки коло-
нок связаны с обработкой конкретного частного сигнала и рассматриваются в
качестве локального функционального модуля. Более низкий уровень органи-
зации уже связан с межклеточным взаимодействием на уровне нейронов. Для
неокортекса известны закономерности появления шипиков на отростках ней-
ронов при развитии животных в обогащенной среде или при обучении. Чем
более богата впечатлениями среда, в которой развивается млекопитающее, тем
больше межклеточных контактов возникает между нейронами коры. Суще-
ствование структурного субстрата, связанного с сохранением индивидуально-
го опыта, показывает, что возникновение неокортекса было ключом к расши-
рению приспособительных способностей млекопитающих.
IX. ОРГАНЫ ЧУВСТВ
Центральная нервная система возникла и сложилась
как надстройка над органами чувств. Органы чувств
подразделяют на две группы по источникам воздейст-
вий: эндогенные и экзогенные. Первые специализиру-
ются на рецепции внутренней среды и органов живот-
ного, а вторые информируют о внешней среде. Оба
источника информации крайне важны для организма.
Эволюция обоих направлений чувствительности шла
параллельно. Иногда органы чувств классифицируют
по их возможностям рецептировать объекты на рас-
стоянии. В этом случае их разделяют на контактные и
дистантные. Чем с большего расстояния животное мо-
жет распознать объект интереса, тем больше у него
шансов на выживание. Поэтому дистантные органы
чувств сыграли огромную роль в приспособлении и
выживании позвоночных.
В основе строения органов чувств лежит фундамен-
тальная способность любых клеток реагировать на ме-
ханические, химические и электромагнитные воздей-
ствия. Эти три типа чувствительности лежат в основе
строения всех органов чувств позвоночных (рис. 159).
Механическая чувствительность построена на способ-
ности клеток или их специализированных образований
реагировать на деформацию мембран. В основе этой
чувствительности лежит изменение проницаемости ме-
ханозависимых ионных каналов при изменении формы
клеток или участков мембраны. В наиболее тривиаль-
ном случае мембрана чувствительных клеток содержит
упорядоченно расположенные ионные каналы, которые
с внутренней стороны мембраны соединены между со-
бой микрофиламентами и микротрубочками. Если клет-
ка находится в статичном состоянии, ионные каналы
закрыты.
При механическом воздействии на клетку происхо-
дят деформация мембраны и локальное «растаскивание»
220
Сравнительная анатомия нервной системы позвоночных
Мышечное
рецепторное
волокно
Свободное нервное
окончание
Тельце Пачини
Волосковая слуховая
клетка
Вкусовые
клетки языка
Обонятельная
клетка
Гломусные клетки из
каротидного лабиринта
Колбочка и
фрагмент сетчатки
глаза
Одиночная
электрорецепторная
клетка из ампулы
Лоренцини
Рис. 159. Основные типы рецепторных клеток (ядра зачернены) и проводящих нейронов
позвоночных.
структурных белков механозависимого ионного канала. Это вызывает измене-
ние заряда мембраны и формирование градуального потенциала. Если меха-
норецепторная клетка сама передает сигнал в мозг, то генерируется потенциал
действия, а если нет, то градуальный сигнал считывается передающим нейро-
ном. К механорецепторам относят свободные нервные окончания, большую
часть соматосенсорных рецепторов, органы боковой линии, рецепторы ли-
нейного и углового ускорения и органы слуха. Различия между этими рецеп-
торами относятся к системе механических фильтров, которые служат для вы-
деления необходимого диапазона воздействия. Это могут быть волоски, кана-
лы с жидкостью, элементы скелета и т.д.
По сравнительно-анатомической традиции среди рецепторов принято вы-
делять первично чувствующие и вторично чувствующие клетки. К первично
чувствующим относят нейроны, происходящие из эктодермы, нервных плакод
и нервного гребня. Эти клетки могут рецептировать сигналы и сами переда-
вать их в нервную систему. К первично чувствующим относят эпителиально-
IX. Органы чувств
221
❖
чувствующие клетки обонятельной и вомероназальной систем, осмсренепторы
гипоталамуса и нервно-чувствующие клетки. Тела последних лежат в ганглиях, а
отростки распространяются в различные участки тела и специализированные
органы. Среди механорецепторов к первично чувствующим относят клетки, об-
разующие свободные нервные окончания и инкапсулированные органы. К вто-
рично чувствующим клеткам относят рецепторы, которые могут воспринимать
сигналы и формировать только градуальный потенциал. Потенциал действия для
передачи сигнала в мозг генерируют только специализированные нервно-чув-
ствующие клетки. Среди механорецепторов к вторично чувствующим относят
слуховые клетки, рецепторы углового и линейного ускорения и органы боковой
линии. Среди других систем к вторично чувствующим причисляют вкусовые ре-
цепторы, гломусные клетки каротидного тела и электрорецепторы.
В основе химической чувствительности лежит способность поверхности
клеток распознавать различные вещества. На поверхности клетки находятся
рецепторные молекулы, которые могут присоединять к себе молекулу стиму-
лирующего вещества, находящегося снаружи клетки. Рецепторная молекула
функционально соединена с ионным каналом, закрытым в обычном состоя-
нии. В момент контакта со стимулирующим веществом происходит от крыва-
ние канала и изменяется локальный заряд мембраны. Далее все происходит,
как в случае механорецепторной чувствительности. Химической чувствитель-
ностью обладают следующие первично чувствующие клетки: обонятельные и
вомероназальные рецепторы и осморецепторы гипоталамуса. Вторично чув-
ствующие хеморецепторы: вкусовые рецепторы, гломусные клетки каротид-
ного тела и кожные хеморецепторы костных рыб.
Электромагнитная чувствительность построена на способности электромаг-
нитных волн определенной длины оказывать обратимое воздействие на органи-
ческие молекулы, расположенные внутри специализированных клеток. В самом
обычном случае свет попадает на родопсин (11 -цис-ретиналь) палочки сетчатки
глаза и поглощается, вызывая выцветание пигмента. 11-Цис-ретиналь транс-
формируется в 11-транс-ретиналь, что означает превращение энергии погло-
щенного кванта света в энергию химической связи родопсина. Эта энергия
химической связи используется для формирования градуального потенциала,
который в конечном счете передается в мозг ганглиозной клеткой сетчатки.
Описанным образом работают фоторецепторные клетки глаза и пинеально-
го комплекса гипоталамуса. Однако существует ряд специализированных элект-
рорецепторов, которые воспринимают электромагнитные волны не в свето-
вом диапазоне. Это этмоидные органы термочувствительности некоторых змей
и ампулярные электрорецепторные органы рыб. Инфракрасный диапазон ис-
пользуется вторично чувствующими клетками змей для нахождения тепло-
кровных жертв. Рыбы рецептируют ампулами Лоренцини динамику измене-
ния напряженности элекромагнитного поля, которое создают другие организ-
мы. Электрорецепция известна не только для пластиножаберных и костных
рыб. Среди млекопитающих такой способностью наделен утконос. Интересно
отметить, что рецептируемая напряженность поля одинакова и для рыб, и для
утконоса. Однако поведенческая реакция проявляется у утконоса при пороге
раздражения, в 10 раз более слабым, чем у рыб. Существуют косвенные свиде-
тельства возможности рецепции дельфинами электромагнитного поля Земли.
222 ❖ Сравнительная анатомия нервной системы позвоночных
Такая способность связывается с наличием в нервной системе и мозговых
оболочках ряда органических соединений, которые могут менять свою ориен-
тацию даже в слабых электромагнитных полях.
Следует подчеркнуть, что в самой центральной нервной системе нет ника-
ких специализированных рецепторов, хотя клетки многих отделов мозга реа-
гируют на изменение концентрации гормонов в крови. Поэтому прямое опе-
ративное вмешательство в мозг не вызывает болевых ощущений. Несколько
иная ситуация с парасимпатическими ганглиями, расположенными в непо-
средственной близости от внутренних органов. За последние годы накопилось
немало свидетельств о существовании специализированных свободных нервных
окончаний, иннервирующих отдельные парасимпатические ганглии.
ХЕМОРЕЦЕПЦИЯ
Наружная и внутренняя среда организма нуждается в контроле за своим
химическим составом. Стабильность внутренней среды обеспечивается систе-
мами контроля за обменными процессами и осмотическим балансом. Хеморе-
цепторы, взаимодействующие с внешней средой, разделяются на дистантные
и контактные. К дистантным относят орган обоняния и вомероназальную си-
стему. У рептилий вомероназальный орган является контактным рецептором.
У рыб (и некоторых амфибий) обнаружены своеобразные хеморецепторы —
кожные вкусовые сосочки, которые лежат на поверхности тела и плавниках.
ОРГАНЫ ОБОНЯНИЯ
Основной дистантной хеморецепторной системой является орган обоня-
ния. Обонятельные рецепторные клетки расположены не на поверхности тела,
а в специальных полостях — обонятельных мешках, или камерах. Обонятель-
ные камеры часто соединены не только с внешней средой носовым отверсти-
ем, но и с полостью глотки. Устройство носовых полостей и ноздрей значи-
тельно различается у позвоночных (рис. 160—162). Первоначально обонятель-
ная полость представляла собой простой мешок с общим отверстием как для
входа, так и для выхода воды. У миноги вторично непарная полость открыва-
ется вторично непарной ноздрей, которая является входом в носовую полость
и назогипофизарный мешок. Он отделен от ротовой полости эластичной тон-
кой стенкой, деформация которой под влиянием разрежения в ротовой поло-
сти вызывает насасывание воды в ноздрю. Вода в обонятельной полости ана-
лизируется рецепторными клетками (рис. 163).
Существует представление, что у миксин тонкая пленка между гипофизар-
ным мешком и глоткой прорывается, позволяя использовать наружные нозд-
ри для дыхания при занятой ротовой полости. По-видимому, орган обоняния
был первоначально симметричной системой, которая у бесчелюстных редуци-
ровалась до одиночного обонятельного мешка. На исходную парность систе-
мы указывает как наличие парных обонятельных нервов, так и некоторая раз-
деленность срединного мешка на две части.
IX. Органы чувств
223
Рецепторный Обонятельный Обонятельная
Наружная
входная и
выходная
ноздря
а
Рис. 160. Основные типы организации обонятельных мешков позвоночных.
а. б — обонятельный мешок нехоанального типа; б — обонятельный мешок с входным и
выходным отверстиями; в — обонятельный орган хоанального типа.
Для пластиножаберных характерен нехоанальный тип строения органов
обоняния (см. рис. 160, б) с двойной ноздрей. Одна ноздря служит для входа
воды, а другая — для выхода. В зависимости от биологии акул орган обоняния
промывается водой или при поступательном движении, как у Carcharhinus
longimanus, или при латеральных покачиваниях головой, как у колючих акул.
Анатомические особенности строения органов обоняния предопределяют спо-
собность пелагических акул к обнаружению пищи на больших расстояниях.
При поступательном движении большинства серых акул (Carcharhinidae) вода
захватывается входной ноздрей, как воздухозаборником самолета. Это обеспе-
чивает быструю смену воды и возможность захвата пузырей воздуха. Эти пу-
зыри могут захватываться и удерживаться в органах обоняния, если при дви-
жении свыше 1,2 м/с рострум приподнимается на 1—2 см над поверхностью
воды. Удержание пузырьков воздуха в органах обоняния осуществляется при
помощи специализированных перегородок в обонятельном мешке — ламелл
(см. рис. 162) и снижении скорости движения животного. Задержка пузырьков
воздуха необходима для растворения содержащихся в них веществ в воде меж-
ду ламеллами. Акулы не могут непосредственно рецептировать воздух и по-
224
Сравнительная анатомия нервной системы позвоночных
Наружная
ноздря
Рис. 161. Основные типы организации органов обоняния позвоночных.
а — амфибии; б — рептилии; в — млекопитающие. У амфибий возникает вомероназаль-
ный орган, у рептилий — преддверие и носовые раковины, у млекопитающих — стенсоно-
вы протоки.
этому депонируют его в обонятельном мешке, дожидаясь повышения концен-
трации веществ вокруг пузырьков.
Если вода не содержит привлекательных запахов, то акулы набирают ско-
рость и выдавливают водой пузырьки воздуха, расположенные между ламел-
лами, содержащими рецепторные клетки. Затем цикл повторяется. Надо отме-
тить, что площадь поверхности рецепторных ламелл органов обоняния серых
акул массой 75 кг может быть больше, чем у человека, в 60—90 раз. В воздухе
над поверхностью океана запахи распространяются очень эффективно, что
позволяет акулам быстро находить добычу.
У двоякодышащих рыб (и некоторых угрей) носовая и ротовая полости со-
общаются специализированным отверстием — внутренней ноздрей, или хоа-
ной (choana). Для рыб наличие хоаны позволяет быстро обменивать воду в
обонятельных мешках за счет использования изменения давления в ротовой
IX. Органы чувств
225
Рис. 162. Два типа организации органов обоняния акул. Слева — продольное сечение
органа обоняния, справа — поперечные сечения ламелл с рецепторными клетками. Свет-
лые стрелки — вход воды, темные — выход. Зачернены воздушные пузыри, проходящие
через ламеллы или задепонированные в них.
а — Carcharhinus longimanus; б — Squalus acantias.
полости при прокачивании воды через жабры. У многих видов рыб наружные
ноздри (naris) значительно видоизменяются, образуя обонятельные трубочки,
которые используются для максимального приближения к объекту рецепции.
У некоторых видов акул, скатов и химер входная и выходная ноздри значи-
тельно разнесены на поверхности головы. Образуется длинная носогубная склад-
ка, которая начинается от входной ноздри и заканчивается на краю ротовой
щели, прикрывая длинный носогубной канал. Возникает эффект носогубной
складки, который позволяет просасывать воду через носовую полость за счет
обычной активности ротовой полости.
С выходом позвоночных на сушу произошла значительная трансформация
как самого органа обоняния, так и его рецепторного аппарата. Появились
обонятельные клетки с укороченными выростами, а в носовых стенках воз-
никли развитые железы, которые увлажняли эпителий. По сути дела обоняние
так и осталось «водным*, поскольку до рецепции любое вещество должно быть
растворено в секрете желез или специализированных клеток. В обонятельном
мешке возникли полости, одна из которых стала специализированным воме-
роназальным, или якобсоновым, органом (organum vomeronasale). Он впервые
появляется у амфибий. У водных видов амфибий в него попадает преимуще-
ственно вода, а в основную обонятельную камеру — воздух. Вомероназальный
орган амфибий, рептилий и млекопитающих формирует собственный нерв
(см. рис. 161), который начинается от первично чувствующих клеток вомеро-
назального эпителия. В отличие от клеток обонятельного мешка рецепторные
клетки вомероназального органа не имеют жгутиков, а только выросты поверх-
ности — микровиллы (рис. 163).
Увлажнение обонятельной и вомероназальной полостей осуществляется
одноклеточными железами, боуменовыми железами рецепторного эпителия и
многоклеточными железами глазницы. Последние участвуют в омывании по-
верхности эпителия при наличии слезно-носового протока. Такая мощная си-
стема увлажнения необходима при принудительной вентиляции носовых
8—5818
226
Сравнительная анатомия нервной системы позвоночных
железа
Рис. 163. Строение обонятельного и вомероназального эпителия.
полостей, связанной с дыханием через нос. У первичноводных животных та-
кой системы нет, хотя почти у всех присутствуют опорные клетки, которые
могут выполнять и секреторные функции. При обильном увлажнении носо-
вой полости движение секрета над поверхностью рецепторного эпителия
осуществляется как под влиянием движения воздуха, так и при помощи во-
лосковых опорных клеток.
У рептилий обонятельная полость дифференцируется на переднюю каме-
ру, или преддверие (vestibulum), носовые раковины (conchae) и носоглоточ-
ный ход. Последний является каналом, соединяющим основную камеру и
хоану. Обычно этот канал довольно короткий, но у крокодилов с удлинен-
ной мордой он представляет собой длинную трубку. Для рептилий характер-
но формирование вентральных вомероназальных полостей, которые само-
стоятельно открываются в ротовую полость (см. рис. 161, б). Парные отвер-
стия вомероназальной системы функционально объединены с раздвоенным
языком многих рептилий. Высунутый язык захватывает разные вещеста и
переносит их к хоанам вомероназальной системы. Вещества попадают на
слизистую часть эпителия и рецептируются органом. Это приводит к тому,
что вомероназальный орган становится контактным хеморецептором. По-
видимому, у рептилий вомероназальный орган становится не только контакт-
ным хеморецептором, но и органом полового обоняния. Косвенным свиде-
тельством в пользу этого предположения служит быстрое увеличение разме-
ров вомероназальной луковицы в онтогенезе и масштабное установление ее
связей с гипоталамусом и зачатком лимбической системы. У эмбрионов кро-
кодилов и многих птиц вомероназальная система закладывается в эмбриоге-
незе, но у взрослых животных отсутствует.
IX. Органы чувств
227
❖
У птиц воздушное обоняние развито плохо. Небольшое рецепторное поле
расположено на каудодорсальной поверхности носовой камеры. Тем не менее у
многих видов птиц выявлены сложные носовые ходы, предполагавшие нагрева-
ние рецептируемого воздуха, что говорит о существовании у них в прошлом
развитого обоняния. На сегодняшний день известно использование обоняния
для навигации у голубей и трубконосых, а также для поиска добычи у киви и
американских грифов. Обонятельная луковица птиц развита незначительно, а
следы существования вомероназальной луковицы не найдены.
У млекопитающих и крокодилов в носовых полостях появляются воздуш-
ные пазухи (sinus pneumatici), которые распространяются в соседних костях и
служат для облегчения черепа. Сложные носовые ходы млекопитающих слу-
жат и для прогрева воздуха, поступающего к рецепторному эпителию. Для
млекопитающих характерно наличие вомероназального органа, соединенного
с основной обонятельной системой, а не с ротовой полостью. Вомероназаль-
ный орган расположен на носовой перегородке и открывается в полость носа
небольшим каналом. Этот орган специализирован на восприятии половых
химических сигналов, которые не воспринимаются как запахи, а непосредст-
венно действуют на лимбическую систему, меняя гормональный статус и пси-
хосоматическое состояние животного. С вомероназальной системой млекопи-
тающих связано появление специализированного стенсонового, или носонёб-
ного, протока (ductus nasopalatinus) (см. рис. 161, в). Он соединяет хоану нёба
в ротовой полости и носовую полость, где он открывается недалеко от входа в
вомероназальный орган. Не у всех млекопитающих сохраняется как вомеро-
назальный, так и обонятельный эпителий. Считается, что вомероназальный
орган отсутствует у слонов, а у дельфинов нет и обонятельного эпителия. У ле-
тучих мышей и высших приматов система увлажнения эпителия обонятель-
ной системы заметно уменьшается. Достаточно напомнить, что одним из при-
знаков приматов является сухой нос.
Обонятельный и респираторный (нерецепторный) эпителии распределены
в носовой полости довольно закономерно. Респираторный эпителий располо-
жен в ростральной части носовой полости на ее латеральной стенке. Рецеп-
торный обонятельный эпителий обычно лежит на медиальных, дорсальных и
вентральных поверхностях основной носовой полости. Чувствительный эпи-
телий занимает примерно половину вомероназальной полости.
Рецепторный обонятельный и вомероназальный эпителии позвоночных ус-
троены весьма сходным образом (см. рис. 163). Зрелые обонятельные рецеп-
торные клетки имеют рапировидную форму. Ядро расположено ближе к ба-
зальной мембране эпителия, а длинный и тонкий дендрит выходит на поверх-
ность эпителия, где заканчивается небольшим утолщением — булавой. От
булавы отходят жгутики и/или микроворсинки (микровыросты) мембраны.
Для вомероназального органа характерны клетки с микроворсинками, тогда
как в обонятельном эпителии встречаются клетки обоих типов. Реснички мо-
гут быть длиной до 320 мкм при диаметре 0,1—0,2 мкм. Внутри жгутиков рас-
положены микротрубочки, которые позволяют этим образованиям двигаться в
слое секрета. На поверхности микроворсинок и жгутиков расположены ре-
цепторные молекулы, которые распознают вещества, растворенные в слизи.
Связывание вещества с рецепторной молекулой приводит к открытию специ-
228
Сравнительная анатомия нервной системы позвоночных
авизированных ионных каналов и возникновению рецепторного потенциала.
Обонятельная клетка сама генерирует потенциалы действия, которые переда-
ются по аксону в клубочковый слой обонятельной луковицы (рис. 164). Необ-
ходимо подчеркнуть, что обонятельные клетки не являются постоянно живу-
щими нейронами. У различных видов рецепторные клетки живут примерно от
40 до 70 дней, а затем погибают и фагоцитируются базальными клетками.
Источником пополнения новых рецепторов являются неспециализирован-
ное базальные клетки. Они дифференцируются и образуют длинные аксоны,
кбторые входят в состав обонятельного нерва и по пути предшественников
растут в сторону обонятельной луковицы.
Поскольку обонятельные клетки регулярно замещаются, изменяется и стро-
ение клубочкового слоя обонятельной луковицы, что приводит к регуляр-
ным изменениям чувствительности обонятельной системы. При гормональ-
ной стимуляции в период размножения обоняние особенно обостряется за
Слой мякотных
волоки и
клеток-зерен
Внутренний
плексиформный
слой
Слой
митральных
клеток
Наружный
плексиформный
слой и клетки
с султанами
Клубочковый слой
и наружные
клетки-зерна
Обонятельный
тракт
Рецепторный
эпителий
Рис. 164. Организация связей в обонятельном эпителии и луковице млекопитающих
IX. Органы чувств
229
счет активного врастания отростков новых рецепторных клеток в клубочко-
вый слой. В клубочках постоянно возникают и исчезают синаптические
контакты между аксонами рецепторных клеток и дендритами митральных
клеток и клеток-зерен. Вокруг клубочков в контакты с дендритами клеток-
зерен и митральных клеток вступают клетки плексиформного слоя, которые
формируют длинные аксоны, идущие в составе обонятельного тракта в мозг
(см. рис. 164). «Внутрилуковичные» горизонтальные связи осуществляются
специализированными клетками-зернами на выходе из обонятельной луко-
вицы и в специализированном внутреннем плексиформном слое. Стимулу
ция рецепторных клеток одним веществом приводит к дифференциальной
активности в различных клубочках, а митральные клетки могут различно
реагировать на одинаковые вещества и наоборот. Физиологический смысл
пространственного распределения активности в обонятельной луковице зак-
лючается в распознавании стереохимических конфигураций воздействующих
молекул, что необходимо для «смысловой» дифференциации сигналов, по-
ступающих от органов обоняния.
Следует подчеркнуть, что различия в обонянии животных отражаются как
на анатомическом, так и на гистологическом уровне, что ведет к плохо иссле-
дованным изменениям в физиологии обработки обонятельных сигналов. При-
мером могут быть связи клубочкового слоя птиц и млекопитающих. Птицы с
плохо развитым обонянием имеют некоторые сравнительно более просто орга-
низованные связи между рецепторами и обонятельной луковицей (рис. 165).
Рис. 165. Строение и взаимодействие обонятельного эпителия млекопитающих (а) и птиц
(б) с митральными клетками обонятельной луковицы.
230
❖
Сравнительная анатомия нервной системы позвоночных
У млекопитающих митральные клетки обонятельной луковицы формируют
связи с 2—3 клетками эпителия. При этом наблюдаются многочисленные пе-
рекрещивания волокон внутри клубочкового слоя. У птиц 6—8 рецепторных
клеток обычно формируют связи только с одной митральной клеткой, которая
посылает свои отростки в мозг.
Обонятельный тракт входит в головной мозг и разделяется на несколько
пучков волокон (рис. 166). Часть волокон сразу направляется в переднее обо-
нятельное ядро, которое связывает правую и левую обонятельные луковицы.
Другая часть волокон поступает в миндалину, peripaleocortex, обонятельную
кору и обонятельный бугорок. Последние области хорошо заметны с базаль-
ной поверхности мозга, а их развитие коррелирует со способностями к хемо-
рецепции (см. рис. 149). В миндалину приходят волокна и от вомероназальной
луковицы. Большая часть волокон от вторичных обонятельных центров мозга
поступает в гипоталамус и таламус. Гипоталамус играет большую роль в реа-
лизации полученной обонятельной информации. В нем происходит регуляция
Таламус
Гипоталамус
Новая кора
Гиппокамп
Миндалина
Ретикулярная
формация
Обонятельная
луковица
Митральные
клетки
Переходная
кора
Обонятельный
бугорок
Вомероназальная
луковица
Обонятельная
кора
Переднее
обонятельное
ядро
Обонятельный
нерв
Вомероназальный
нерв
Рис. 166. Основные обонятельные пути млекопитающих.
IX. Органы чувств ❖ 231
выработки гормонов, надолго изменяющих поведение животных. На поведе-
ние оказывает влияние и лимбическая система, которая связана с обонятель-
ными рецепторами через periarchicortex и гиппокамп. Из таламуса и лимби-
ческой системы небольшое количество волокон направляется в различные об-
ласти неокортекса, что влияет на восприятие информации от других органов
чувств. Таким образом, гормональный и эмоциональный статус в значитель-
ной мере зависит от обонятельной системы. В обонятельной луковице окан-
чиваются центробежные волокна, идущие от ядер шва, обонятельной коры
голубого пятна и даже от ретикулярной формации заднего мозга. Эти обраъ-
ные связи указывают на то, что животные могут по-разному воспринимать
запахи в зависимости от своего психосоматического состояния.
ОРГАНЫ ВКУСА
Органы вкуса позвоночных животных являются контактным анализатором,
который обычно расположен в ротовой полости, но может быть вынесен на
поверхность тела. Наиболее интересным вариантом является локализация вку-
совых сосочков почти на всей поверхности тела у многих сомообразных
(Siluriformes) (рис. 167), у которых число вкусовых сосочков может достигать
нескольких тысяч.
Придонные рыбы имеют многочисленные вкусовые сосочки, расположен-
ные на пальцеобразных отростках грудных плавников. Подобные «вынесен-
ные» органы вкуса широко представлены в семействе тригловых или морских
петушков (Triglidae). Они погружают их в придонные осадки или в рыхлое дно
и определяют наличие пищи. У амфибий, рептилий и птиц вкусовые почки
лежат в ротовой полости и глотке. Есть сведения, что вкусовые почки нахо-
дятся на коже кистей шпорцевой лягушки и на поверхности головы суринам-
ской пипы.
Для млекопитающих характерно расположение вкусовых почек на языке, в
прилежащих частях ротовой полости и в глотке. В зависимости от типа пита-
ния и образа жизни вкусовые рецепторные клетки распределяются на поверх-
ности языка различным образом (рис. 168).
В самом простом случае рецепторные клетки собраны в компактные вкусо-
вые почки, которые состоят из нескольких типов клеток (рис. 169).
Внутри вкусовой почки различают опорные клетки, которые поддерживают
постоянную форму почки при механических деформациях языка. Секретор-
Рис. 167. Рецепторные вкусовые сосочки на поверхности тела американского сомика-кош-
ки (Amiurus nebulosis). Каждая точка на поверхности тела соответствует примерно 100 вку-
совым сосочкам (по Н. Kardong, 1995).
232
❖
Сравнительная анатомия нервной системы позвоночных
Муравьед
Рис. 168. Рецепторные вкусовые почки и сосочки на поверхности языка различных видов
млекопитающих.
Рис. 169. Рецепторная вкусовая почка млекопитающих.
IX. Органы чувств
233
❖
ные клетки выделяют секрет во вкусовую пору, смачивая поверхность рецеп-
торных клеток. Рецепторные клетки обладают пальцеобразными выростами,
выдвинутыми в просвет вкусовой поры. Ближе к основанию почки лежат не-
специализированные базальные клетки, которые являются источником вос-
становления клеток всех типов.
Опорные и секреторные клетки заменяются относительно медленно, тогда
как рецепторные клетки обновляются каждые 10—12 дней. Поэтому базальные
клетки обычно ошибочно считаются источником только рецепторных клеток.
Вкусовые рецепторные клетки не имеют аксонов и при химическом раздраже-
нии формируют только градуальный сигнал, который считывается свободным
нервным окончанием, иннервирующим вкусовую почку. Каждую вкусовую
почку иннервирует 1—3 нервных окончания, принадлежащих лицевому (VII),
языкоглоточному (IX) или блуждающему (X) нерву. При замене рецепторной
клетки происходит ее реиннервация свободным нервным окончанием.
Вкусовые почки видоспецифично распределяются на поверхности языка
различных видов млекопитающих. Они могут быть собраны в особые образо-
вания, называемые вкусовыми сосочками. Сосочек представляет собой склад-
ку поверхности языка, в которой находится скопление вкусовых почек. В са-
мом простом виде это может быть грибовидный сосочек, располагающийся на
переднем крае языка приматов и человека (рис. 170). В листовидных и жело-
боватых сосочках находятся серозные железы. Они вырабатывают секрет, пред-
охраняющий вкусовые почки и транспортирующий химические вещества к
наружной вкусовой поре.
Если язык животного подвергается значительным механическим нагрузкам,
то его вкусовые почки собраны преимущественно в листовидные или желобо-
ватые сосочки, расположенные ближе к корню языка. При небольших дефор-
мациях языка вкусовые почки распределяются по языку равномерно, хотя их
наибольшая концентрация отмечена на краях языка. Форма и распределение
на поверхности языка вкусовых сосочков не связаны с конкретным характе-
ром вкусового ощущения.
При сходном строении вкусовых луковиц различные участки языка чувст-
вительны к различным вкусовым раздражениям. Принято выделять четыре
основных вкусовых качества, вызываемых четырьмя чистыми веществами: слад-
кое (сахароза), соленое (NaCI), кислое (НС1) и горькое (хинин). Поля чув-
ствительности к этим раздражителям перекрываются, но есть и зоны, где до-
минирует только одно вкусовое ощущение (см. рис. 170).
На кончике языка человека сосредоточены вкусовые сосочки, чувстви-
тельные к сладкому, далее — к соленому, ближе к корню языка — к кислому
и горькому. Все категории вкусовых сосочков иннервируются барабанной
струной лицевого нерва, тогда как блуждающий и языкоглоточный нервы
обслуживают только сосочки, чувствительные к кислому и горькому. Это
связано с тем, что горьковатые плоды обычно бывают ядовиты для млекопи-
тающих, а стимуляция IX и X нервов вызывает рвотный рефлекс и своевре-
менную эвакуацию пищи из пищеварительного тракта. Важно подчеркнуть,
что вкусовая специализация отдельных вкусовых почек сохраняется на про-
тяжении всей жизни, хотя за это время заменяется несколько тысяч рецеп-
торных клеток.
234
❖
Сравнительная анатомия нервной системы позвоночных
Рис. 170. Рецепторная поверхность языка приматов и человека.
VII, IX, X — черепно-мозговые нервы.
Информация о пище поступает в головном мозг через лицевой, языкогло-
точный и блуждающий нервы. Она содержит информацию двух типов. С од-
ной стороны, это собственно вкус пищи, а с другой — ее механические харак-
теристики, определяемые механорецепторами тройничного нерва (твердая,
скользкая, упругая, волосатая и т.д.). Восходящие пути вкусовых нервов прак-
тически одинаковы вплоть до таламуса. Все три нерва образуют контакты в
продолговатом мозге с ядром одиночного тракта (nucleus tractus solitarii), кото-
рое является первичным вкусовым центром мозга у амфибий, рептилий и мле-
копитающих (рис. 171, 172). У амфибий и рептилий от ядра одиночного трак-
та начинаются волокна, идущие преимущественно в базальные отделы перед-
него мозга. Это в первую очередь ядро дна или терминальная полоска (stria
terminalis), миндалина и латеральные ядра гипоталамуса. Часть волокон на-
правляется в крышу среднего мозга и в зачаточный таламус. Для рептилий
известны связи вкусового ядра с примордиальным зачатком новой коры, хотя
детали локализации еще не исследованы.
У низших млекопитающих вкусовое ядро моста сохраняет свои связи с ба-
зальными структурами переднего мозга и приобретает небольшие контакты с
таламусом. В таламусе переключаются восходящие волокна, идущие к новой
коре. В неокортексе вкусовые рецепторы представлены тремя полями. Два
поля расположены в соматосенсорной коре. В них оканчиваются восходящие
волокна от IX и X нервов. Значительно большее поле расположено в лобной
доле. Оно связано с лицевым нервом и обычно называется первичной вкусовой
IX. Органы чувств
❖
235
Рис. 171. Восходящие пути от нервов, иннервирующих вкусовые почки поверхности языка,
ротовой полости и глотки у рептилий (а) и низших млекопитающих (б).
VII, IX, X — черепно-мозговые нервы.
корой. У приматов и человека схема организации восходящих путей немного
усложняется (см. рис. 172). В этой группе возникают прямые связи между яд-
ром одиночного тракта и таламусом, что увеличивает скорость прохождения
сигнала от вкусовых рецепторов. Подвергаются изменениям и корковые связи.
Основными неокортикальными центрами вкусового анализатора становится
островок (insula), ипсилатеральная соматосенсорная кора и контралатеральная
соматосенсорная кора. В извилинах островка сосредоточены вкусовые центры,
центры осязания механической консистенции пищи и ее температура. В ипси-
латеральном соматосенсорном поле расположено основное вкусовое поле, а в
контралатеральном поле — соматосенсорное представительство языка.
Таким образом, у приматов и человека основной вкусовой путь идет к тала-
мокортикальной системе, а у низших позвоночных — к эмоционально-гормо-
нальным центрам, которые характерны и для рептилий. Это не означает, что
вкусовые ощущения высших млекопитающих сосредоточены в неокортексе.
Значительная доля волокон поступает через миндалину и гипоталамус в лимби-
ческую систему. Участие лимбической системы предопределяет связь вкусовых
236
Сравнительная анатомия нервной системы позвоночных
Рис. 172. Восходящие пути от нервов, иннервирующих вкусовые почки поверхности языка,
ротовой полости и глотки у приматов и человека.
VII, IX, X — черепно-мозговые нервы.
ощущений с эмоциональным состоянием животного и может существенно вли-
ять на поведение как на церебральном, так и на гормональном уровне.
Заканчивая краткий обзор вкусовой системы, следует подчеркнуть, что вку-
совые восходящие пути пересекаются с висцеральными сенсорными путями в
ядре одиночного тракта. Следовательно, вкусовые ощущения могут оказывать
решающее влияние на общую активацию организма животного, его пищевое
и половое поведение.
РЕЦЕПТОРЫ ЭЛЕКТРОМАГНИТНЫХ ПОЛЕЙ И ИЗЛУЧЕНИЙ (ФОТОРЕЦЕПЦИЯ)
Все организмы на планете находятся под постоянным воздействием элект-
ромагнитных полей и излучений. Они поступают от Солнца в виде волн-час-
тиц, называемых фотонами. Диапазон электромагнитного спектра крайне широк
и различается по длине волны, частоте и энергии. Для животных гамма, рен-
тгеновские и ультрафиолетовые лучи практически непригодны для использо-
вания периферическими анализаторами, так как обладают слишком высокой
энергией. Эта энергия так высока, что при взаимодействии с органическими
молекулами ее хватает для разрушения молекулярных связей. Длинноволно-
вый диапазон, в который входят инфракрасное излучение, микроволны, ко-
роткие и длинные радиоволны, также не подходит для рецепции, поскольку
энергии этой части спектра недостаточно, для того чтобы воздействовать на
органические молекулы. Наиболее приемлем для рецепции диапазон видимо-
го света от 300 до 800 нм. Этот диапазон обладает излучением средней энер-
IX. Органы чувств ❖ 237
гии, достаточной, чтобы зрительные пигменты изменили свою структуру, но
не разрушились. Такие обратимые конформационные преобразования зри-
тельных пигментов лежат в основе фоторецепции.
Самым распространенным типом чувствительности к свету является спо-
собность различать интенсивность рассеянного освещения. Этим свойством
обладают как растения, так и животные. Способность различать интенсив-
ность называют светочувствительностью. У животных светочувствительность
обеспечивается пигментными молекулами типа родопсина. Если в фоторе-
цепторной клетке используется несколько фотопигментов, то возможна диф-
ференциация различных длин видимого диапазона — цветовое зрение. От орга-
низации глаза и ориентации фоторецепторов зависит способность к восприя-
тию поляризованного света, что свойственно в основном беспозвоночным.
При наличии структуры с множеством объединенных рецепторов возможно
восприятие движения и распознавание формы. Это происходит в сложном
глазу. Глаза могут быть ориентированы латерально (зайцеобразные, копыт-
ные) или фронтально (кошачьи, приматы). При фронтальном или дивергент-
ном расположении глаз животные обладают стереоскопическим (бинокуляр-
ным) зрением и воспринимают глубину наблюдаемого пространства.
Наиболее просто организованы глазки Гессе — фоторецепторы ланцетника.
Они рассеяны в толще нервной трубки и представляют собой отдельные све-
точувствительные клетки, которые объединены с одиночными пигментными
клетками. Они по-разному ориентированы по отношению к основным плос-
костям тела ланцетника. Благодаря этому обзор отдельных светочувствитель-
ных клеток оказывается различным, что позволяет животному судить о на-
правлении на источник света. Кроме одиночных фоторецепторов, у ланцетни-
ка существует фронтальный глаз, состоящий из нескольких клеток. Аксоны
клеток, входящих во фронтальный фоторецепторный комплекс, свя 1аны с клет-
ками тектума, который считается гомологом среднего мозга (рис. 173). Таким
образом, информация от отдельных светочувствительных клеток интегрирует -
ся с фронтальным фоторецептором ланцетника.
Рис. 173. Передняя часть центральной нервной системы 12,5-дневной личинки ланцетни-
ка (по В. Lacalli, 1996).
238 ❖ Сравнительная анатомия нервной системы позвоночных
У большинства позвоночных имеются развитые глаза, которые позволяют
воспринимать форму, движение и цвет окружающих предметов. Глаза позвоноч-
ных могут быть округлой, элипсовидной или цилиндрической формы (рис. 174).
Последние характерны для глубоководных рыб. Внутреннее строение глаз су-
щественно различается в зависимости от среды обитания. Для водных живот-
ных характерен крупный округлый хрусталик, приближенный к сетчатке. При
недостатке света (на больших глубинах) размер хрусталика увеличивается в
несколько раз, а светочувствительный слой клеток — сетчатка — уменьшается.
При всех различиях строения организация глаза позвоночных имеет общие
принципы. Передняя поверхность глаза в области зрачка покрыта роговицей
(cornea), за которой расположена передняя камера глаза. Передняя камера огра-
ничена радужной оболочкой, или радужкой (iris), за которой лежит небольшая
полость, называемая задней камерой глаза (см. рис. 174). Хрусталик закреплен
на подвешивающей связке (ligamentum suspensorium), которая прикрепляется к
ресничному телу (corpus ciliare). Между сетчаткой (retina) и хрусталиком (lens)
расположено заполняющее глаз стекловидное тело (corpus vitreum).
Эта общая схема строения глаза сохраняется практически во всех группах
позвоночных, что отражено в его эмбриональном развитии, которое описано во
II главе. Глаз является органом, состоящим из тканей различного происхожде-
ния, которые связаны между собой индукционными взаимодействиями. Если
сетчатка и пигментный эпителий возникают из стенки мозга, то хрусталик про-
исходит из эктодермы после ее индукции глазным пузырем. Внутрь глазного
бокала проникает мезодерма, вместе с сетчаткой формирующая стекловидное
тело. Мезодерма, окружающая сетчатку снаружи, формирует сосудистую обо-
лочку или венозные синусы у круглоротых. Из мезодермы формируется склера
и при участии клеток глазного пузыря радужина. Латеральный участок склеры,
называемый десцеметовой оболочкой, срастается с двумя слоями кожи и фор-
мирует трехслойную роговицу. Из глазного пузыря формируются пигментная
оболочка, сетчатка и ресничное тело, удерживающее хрусталик.
Общий план строения глаза не исключает многообразия организации.
У многих крокодилов и сумчатых млекопитающих в пигментной оболочке на-
капливается гуанин, формирующий зеркальную прослойку — тапетум (tapetum
lucidum retinale). Аналогичная отражающая поверхность возникает у рыб и
млекопитающих в сосудистой оболочке (tapetum lucidum chorioideale). Ее по-
явление связано с необходимостью повышения освещеннности рецепторного
слоя клеток. Свет, который прошел через сетчатку, отражается от зеркальной
поверхности тапетума и возвращается к фоторецепторам Для многих акул и
костистых рыб характерно изменение отражающей способности зеркального
слоя в зависимости от освещенности. Глаз обычного катрана, адаптирован-
ный к темноте, может отражать от тапетума 88% падающего света, а при усло-
виях высокой освещенности — 2,5% света.
В глазах рептилий и птиц имеются специфические образования: папилляр-
ный конус и гребень (см. рис. 174). Эти образования имеют сосудистое проис-
хождение. У рептилий папиллярный конус участвует в метаболизме внутрен-
ней части глаза и не связан с фоторецепцией. Для птиц характерно разнооб-
разное строение гребня, на котором часто располагается и сетчатка. Поскольку
гребень расположен под наклоном к оптической оси хрусталика, проекция
IX. Органы чувств
239
Зрительный
Костистые рыбы
склеры
тело
Рис. 174. Сечение глаза различных позвоночных. Хрусталик изображен в виде двух поло-
вин, которые отражают способ аккомодации.
а — положение в состоянии покоя; б — аккомодация.
изображения на нем имеет вытянутую форму. При этом в рецепцию вовлека-
ется большое количество клеток, а центральный участок изображения оказы-
вается увеличенным в 1,5—2 раза.
Гистологический анализ сетчатки глаза позвоночных показывает, что он ин-
вертирован. Это означает, что фоторецепторы расположены на задней, дальней
от зрачка поверхности сетчатки и развернуты светочувствительными сегмента-
ми в сторону пигментного эпителия (рис. 175). Свет проходит через всю толщи-
ну сетчатки и тела самих рецепторов, прежде чем попадает на светочувствитель-
ный пигмент, расположенный в модифицированных ресничках — мембранных
дисках. Различают два типа рецепторов — палочки и колбочки. Палочки чувст-
вительны к изменению интенсивности света, содержат в мембранных дисках
родопсин и не воспринимают цвета. Они чувствительнее колбочек, которые
содержат пигменты, чувствительные к красному, зеленому и синему цветам.
Пигменты в мембранных дисках истощаются, что приводит к их постоянному
отщеплению в дистальной части и наращиванию в проксимальной.
240
О
Сравнительная анатомия нервной системы позвоночных
f f f f f Свет
Пигментный
эпителий и
тапетум с
зеркальным
слоем
Наружный
ядерный
слой
Наружный
сетчатый слой
Внутренний
ядерный слой
Внутренний
сетчатый слой
Слой
ганглиозных
клеток
Рис. 175. Строение сетчатки глаза млекопитающих с названиями основных гистологичес-
ких слоев и клеток.
Светочувствительные клетки различаются по своей организации. Почти у
всех позвоночных, за исключением приматов и человека, в средних сегментах
фоторецепторов имеются сократимые белки (миоиды). Они позволяют быст-
ро адаптироваться к изменению освещенности сетчатки. Это достигается бы-
стрым укорачиванием тел клеток при уменьшении освещенности сетчатки и
удлинением при увеличении количества света. В колбочках амфибий, репти-
лий и птиц содержатся цветные масляные капли, которые являются свето-
фильтрами, определяющими спектральную чувствительность рецептора. Для
амфибий, рептилий и птиц характерно наличие двойных, а для костистых рыб —
близнецовых колбочек (рис. 176). Один член такой ассоциации имеет свето-
фильтрующую масляную каплю, а другой — запасы гликогена. Однако прин-
ципиальная схема строения фоторецептора остается неизменной. Фоторецеп-
торы формируют только градуальный потенциал, который передается на гори-
зонтальные и биполярные клетки сетчатки (см. рис. 175). Через биполярные
нейроны сигналы поступают к амакриновым и ганглиозным клеткам, которые
генерируют потенциалы действия. Зрительный нерв формируют только отростки
ганглиозных клеток. Они распространяются на ближайшей к зрачку поверх-
ности сетчатки. Выходящие из глаза отростки ганглиозных клеток раздвигают
клетки-фоторецепторы, в связи с чем в этой точке возникает «слепое пятно».
У млекопитающих существует центральная ямка, в которой нерецепторные
клетки раздвинуты так, чтобы достичь минимального рассеивания света. Ямка
является центром сетчатки и представлена в коре млекопитающих специали-
зированным участком — макулой. Информацию от этой зоны сетчатки обра-
батывает большее количество клеток коры, чем от других частей глаза.
IX. Органы чувств
241
Рис. 176. Строение фоторецепторов.
а — палочка из глаза человека; б — палочка; в — колбочка лягушки-леопарда; г — двой-
ная колбочка черепахи; д — близнецовые колбочки костистой рыбы (по W. Walls, 1942, с
изменениями).
К сетчатке прилежит пигментная оболочка, которая выполняет функцию ре-
гулятора светового потока. Если света много, то пигментные клетки проника-
ют между фоторецепторами. Фоторецепторы могут и сами, за счет миоидов,
вдвигаться в тапетум. Функцию регуляции светового потока выполняют мыш-
цы радужины, сформированные из стенок глазного бокала. Для всех позво-
ночных, помимо млекопитающих, характерно иннервирование расширителя
зрачка (m. dilatator pupillae) при помощи парасимпатической ветви глазодви-
гательного (III) нерва, а сжимателя зрачка (m. sphincter pupillae) — при помо-
щи симпатического нерва, который идет из спинного мозга. У млекопитаю-
щих иннервация расширителя и сжимателя зрачка обратная.
Кроме адаптации к интенсивности светового потока, глаз позвоночных раз-
лично приспособлен к рассматриванию близких и удаленных предметов. Спо-
соб получения максимально резкого изображения на сетчатке носит название
аккомодации. Для бесчелюстных, анамний и змей характерно перемещение
хрусталика относительно сетчатки. У некоторых костистых рыб происходит
перемещение хрусталика как в латеральном, так и в рострокаудальном на-
правлении (см. рис. 174). При этом достигается как аккомодация, так и изме-
нение поля зрения. У акул и амфибий в положении покоя хрусталик имеет
заднее положение, характерное для наводки на даль. Для большинства кости-
стых рыб и миноги специфично переднее положение покоящегося хрустали-
ка. Аккомодацию хрусталика миноги осуществляет специализированный ро-
9—5818
242 ❖ Сравнительная анатомия нервной системы позвоночных
говичный мускул (m. cornealis), который происходит из соматической поверх-
ностной мускулатуры. Он натягивает роговицу, приближая хрусталик к сет-
чатке (см. рис. 174). Мышечные элементы аккомодации хрусталика имеют
различный гистогенез. Из сетчатки развивается как протрактор хрусталика
(т. protractor lentis) акул, так и ретрактор (m. retractor lentis) костистых рыб.
Протрактор амфибий развивается из сосудистой оболочки глаза, как и рес-
ничный мускул амниот.
При аккомодации глаза амниот используется деформация хрусталика
(см. рис. 174). У амниот хрусталик в состоянии покоя плоский и настроен на
даль. При наводке на близкие предметы его форма становится более округлой.
У зауропсид аккомодационный мускул поперечнополосатый, непосредствен-
но сжимающий хрусталик. У змей ресничная мышца и мышцы радужины ут-
рачены ввиду роющего образа жизни далеких предков. У них вторично возни-
кает протрактор хрусталика, который действует, как у анамний, и развивается
из сосудистой оболочки глаза. Для млекопитающих характерно двухэтапное
взаимодействие гладкого ресничного мускула с хрусталиком. Этот мускул сжи-
мает ресничное тело, препятствуя растягиванию эластичного хрусталика.
Сформированное на сетчатке изображение воспринимается палочками и
колбочками, обрабатывается нерецепторными клетками сетчатки и передает-
ся в головной мозг по отросткам ганглиозных клеток. Из их отростков форми-
руется зрительный нерв, который перекрещивается с нервом противополож-
ной стороны тела, образуя хиазму. Как было описано выше (см. главу «Череп-
но-мозговые нервы»), хиазма имеет различное строение, а зрительный нерв
может полностью или частично перекрещиваться. У низших позвоночных, как
правило, нервы перекрещиваются полностью. При отсутствии бинокулярного
зрения изображение, сформированное в правой и левой половинах крыши
среднего мозга, может быть различным (рис. 177). Для оценки расстояния до
предмета необходимо получить два изображения одного предмета в левой и
правой половинах тектума. Эти достигается ротацией глаз или их фронтальным
положением, как у кошек и приматов. Различие в проекциях изображения на
сетчатку служит основой для оценки расстояния до предметов. У низших тетра-
под это достигается за счет прямых и таламических межтектальных связей.
У многих млекопитающих глаза расположены фронтально, чем достигается
максимальная бинокулярность зрительного восприятия. При этом изображе-
ния на сетчатках правого и левого глаза практически одинаковы, хотя и пере-
вернуты. По древнему пути к передним (верхним) бугоркам четверохолмия
направляются полностью перекрещенные волокна. Это позволяет сформиро-
вать в среднем мозге изображение предметов, как и у низших позвоночных
(рис. 178). Эта информация необходима для осуществления быстрых врожден-
ных рефлексов избегания, замирания и т.д. Основной зрительный путь прохо-
дит через латеральные коленчатые тела. При этом за счет частичного перекре-
щивания волокон в каждом из коленчатых тел представлена проекция только
половины предмета. Это связано с тем, что от правых половин сетчатки во-
локна оканчиваются в левом коленчатом теле, а от левых — в правом. В соот-
ветствии с таким разделением сигналов в зрительную кору приходит инфор-
мация о «половинках» предмета, которая используется для реконструирова-
ния реального объекта за счет межполушарных связей неокортекса. Сигналы
IX. Органы чувств
243
Рис. 177. Схема монокулярного зрения у лягушки. Каждый глаз формирует собственное
изображение на сетчатке, которое оборачивается как линзой, так и фототопическим пред-
ставительством зрительных нервов в тектуме среднего мозга.
Рис. 178. Схема бинокулярного зрения у приматов. Глаза формируют одинаковое изобра-
жение, которое передается в тектум по древнему зрительному пути. Основной зрительный
путь проходит через латеральные коленчатые тела и ведет к зрительной кор«
244
❖
Сравнительная анатомия нервной системы позвоночных
от каждого глаза несколько различаются, поскольку предмет рассматривается
под разными углами. Смещение изображений относительно друг друга носит
название параллакса. Величина параллакса используется мозгом для оценки
расстояния до предмета.
Исторической причиной возникновения ретиноталамокортикального пути
млекопитающих была необходимость интеграции межтектальных сигналов у
низших тетрапод. Такая интеграция была необходима для развития способно-
стей по определению расстояния до предмета. Прямые текто-тектальные свя-
зи относительно невелики и справиться с такой задачей не смогли. По этому
сформировался относительно автономный механизм интеграции зрительных
сигналов на уровне таламуса.
Следы древней интеграции зрительных сигналов на уровне таламуса мы
видим в связях тектума. Нисходящие связи тектума описаны выше, но следует
упомянуть наличие взаимодействий с подушкой таламуса, претектальным по-
лем тегментума и ядрами дорсальной части гипоталамуса. Первоначально эти
ядра были общим центром, обслуживавшим стереоскопическое зрение. Затем
из него выделились специализированные коленчатые тела, которые перехва-
тили часть волокон зрительного тракта и перенаправили их в развивающуюся
кору (рис. 179, 180). Наряду с таламокортикальными зрительными связями у
рептилий имеются и прямые тектокортикальные восходящие связи, которые
были основным зрительным путем, ведшим к зачатку неокортекса. У млеко-
питающих восходящие зрительные пути специализировались еще больше, чем
у рептилий. Возникли специализированные контакты с гипоталамусом и комп-
Сетчатка
Тектум
среднего мозга
Сетчатка
Латеральное
коленчатое тело
Зачаток новой коры или
гиперстриатум птиц
Зрительный
нерв
Передние
бугорки
четверохолмия
Рис. 179. Восходящие зрительные пути позвоночных.
а — круглоротые, акулы, костистые рыбы и амфибии; б — птицы и рептилии.
IX. Органы чувств
245
Рис. 180. Восходящие зрительные пути млекопитающих.
лексом хиазматических ядер. При развитии неокортекса сформировались пер-
вичные и вторичные зрительные поля.
Кроме отдельных фоторецепторных клеток и глаз, у позвоночных существу-
ют глазоподобные образования эпиталамуса, описанные в главе VII. Эти про-
стые асимметричные глазки находятся под кожей темени и имеют неинверти-
рованное строение (фоторецепторы направлены дисками к свету). Глазок раз-
личает только изменение освещенности, что служит источником информации
для эндокринных систем, определяющих суточные ритмы активности. Однако
следует отметить, что имеются свидетельства первичной парности пинеальных
глаз. У миксин и ильной рыбки закладки пинеальных глазков асимметрично
левосторонние, а у некоторых вымерших кистеперых рыб (Crossopterygii) обна-
ружены парные теменные отверстия.
ЭЛЕКТРОРЕЦЕПЦИЯ И ЭЛЕКТРИЧЕСКИЕ ОРГАНЫ
Фоторецепцией в видимом диапазоне не исчерпываются способности по-
звоночных к восприятию электромагнитных волн и излучений. Часть специа-
лизированных механорецепторов органов боковой линии первичноводных
позвоночных трансформировалась в электрорецепторы. Однако это не един-
ственный эволюционный путь возникновения электрорецепторов. Они обна-
ружены у утконоса, и предполагается их существование у некоторых птиц.
По-видимому, электрорецепторы возникали в эволюции позвоночных нео-
днократно. Это было обусловлено необходимостью поисков не видимой и не
рецептируемой хеморецепторами пищи и ориентацией в пространстве. На се-
годняшний день известно, что 70 видов рыб обладает развитыми электроре-
цепторами, а около 500 видов могут генерировать электрические разряды раз-
246 ❖ Сравнительная анатомия нервной системы позвоночных
личной мощности. Примерно 20 видов обладают способностями как генери-
ровать, так и рецептировать электрические поля.
Электрические органы расположены у рыб на различных участках тела
(рис. 181). Однако не все рыбы обладают способностью как генерировать, так
и рецептировать электромагнитное поле. Наиболее изучена эта способность у
гимнарха (Gymnarchus niloticus), рецептирующего электромагнитное поле, со-
здаваемое им самим (рис. 182). При попадании в его поле объектов различной
электропроводности гимнарх может определить их направление движения,
размер и скорость. Электрорецепция используется для ухаживания друг за
другом особей разного пола и подавления электромагнитных полей конкури-
рующих особей или других видов. Кроме гимнарха, аналогичные способности
генерировать и воспринимать собственные сигналы известны у других кости-
стых рыб, акул и скатов.
Существует традиционная классификация животных, обладающих различ-
ными типами электрочувствительности и способностями к генерации элект-
рических разрядов. Обычно в отдельную группу выделяют рыб, генерирующих
сильные заряды. Сюда относят американских электрических угрей, электри-
ческих скатов и африканских электрических сомов, которые могут вырабаты-
вать разряды постоянного тока напряжением 100—650 В при силе тока 2 А.
Скаты могут поражать добычу на расстоянии 15—17 см.
Electrophorus electrlcus
Рис. 181. Электрические органы (отмечены точками) на поверхности тела различных рыб
(по Н. Kardong, 1995).
IX. Органы чувств
247
Рис. 182. Электрическое поле гимнарха. Объекты с высокой и низкой электропроводно-
стью по-разному искажают электромагнитное поле, воспринимаемое животным.
Ко второй группе относят животных, способных как генерировать, так и
репептиривать электрические сигналы. Это представители надотряда аравано-
идных (Osteoglossomorpha) из семейств клюворылых (Mormyridae) и гимнархов
(Gymnarchidae). Гимнархи и клюворылые генерируют слабые электрические
разряды, но с высокой частотой — от 100 до 1400 импульсов в секунду. Попа-
дание объектов с различной электропроводностью в генерируемое поле позво-
ляет различать его характеристики без использования других органов чувств.
Клюворылые могут изменять частоту импульсации, что используется для ком-
муникаций с другими особями и для создания электромагнитного «шума».
Нужно отметить, что органы, генерирующие электрические разряды, имеют
различное происхождение. Одни из них являются видоизмененными мышцами,
а другие — нервными окончаниями. При формировании электрогенераторных
органов из мышц последние утрачивают сократимые элементы и уплощаются.
Формируются специализированные клетки, называемые электроцитами. Как пра-
вило, они имеют асимметричные потенциалы, которые складываются, а множе-
ство клеток соединяется последовательно. Как производные мышечных клеток,
так и видоизмененные нервные окончания нуждаются в центральной синхрони-
зации. Если разряд всех клеток не будет согласован, то его эффективность будет
ничтожно мала. Для этого существует специальный нервный аппарат, который
находится под контролем среднего, заднего и продолговатого мозга. У электри-
ческого сома контроль за синхронизацией разрядов осуществляется на уровне
спинного мозга, у гимнархид — на уровне продолговатого, а у звездочета и зеле-
ной ножетелки — на уровне среднего мозга и мозжечка (рис. 183).
Нужно подчеркнуть, что электрические органы, сформированные из мышц,
обычно являются низкочастотными или разряжаются одиночными разрядами.
В тех случаях, когда они сформированы из нервных окончаний, частота раз-
рядов в несколько раз выше.
248
❖
Сравнительная анатомия нервной системы позвоночных
Рис. 183. Схема контроля разрядов электрических органов нисходящими волокнами.
В третью группу включают животных, способных только чувствовать элект-
ромагнитные изменения, но не умеющих их генерировать. К этой группе от-
носят кошачьих (Scyliorhinidae) и молотоголовых (Sphyrnidae) акул. Чувстви-
тельность электрорецепторов этих животных очень высока. Они могут разли-
чать электрическое поле с градиентом 1 мкВ на расстоянии 1 см. Эта
способность позволяет акулам определять место укрытия рыбы в песке по
локальному искажению электромагнитного поля Земли.
Электрочувствительные рецепторы существуют двух основных типов: ам-
пульные (ампулы Лоренцини) и клубочковые (рис. 184). У некоторых видов
присутствуют оба типа рецепторов, но у большинства — только один из
них. Ампульные электрорецепторы приспособлены для восприятия медленно
изменяющихся электрических полей, а клубочковые реагируют на быстрые
изменения. Поэтому у активно плавающих рыб, обитающих в непрозрач-
ной воде, более развиты клубочки, а у хищников, обитающих в прозрачной
воде — ампульные рецепторы. Стенки ампульных рецепторов обладают
высокой емкостью. Этим достигается отфильтровывание высокочастотной
части сигнала и пропускание низкочастотного компонента. Достигнув ре-
IX. Органы чувств
249
Поверхность
кожи
б
Рис. 184. Ампулярные и клубочковые электрорецепторы кожи акул (а) и костных рыб (б).
цепторных клеток ампулы или клубочка, электромагнитные сигналы пре-
образуются в рецепторный потенциал волосковой клетки. Рецепторные
клетки передают сигналы к окончаниям сенсорного нерва посредством хи-
мического синапса. Окончания сенсорного нерва имеют большую частоту
пульсации в покое. Любое воздействие рецепторной клетки на сенсорное
окончание вызывает модуляцию сигнала и является информативным. Нуж-
но уточнить, что клубочковые рецепторы менее чувствительны, чем ам-
пульные. Поэтому у активных хищников (акула рыба-молот) в основном
присутствуют ампулярные рецепторы.
Развитая электрорецепция может значительно влиять на анатомическое стро-
ение головного мозга позвоночных. Примером может служить организация
мозга семейства клюворылых (рис. 185). Размер полушарий мозжечка, обслу-
живающих электрорецепторы, так велик, что они закрывают с дорсальной по-
верхности весь мозг.
250
❖
Сравнительная анатомия нервной системы позвоночных
Полушария
мозжечка
Передний мозг
Рис. 185. Организация мозга клюворылых. Справа — полушария мозжечка удалены
(по Н. Kardong, 1995).
Способность к электрорецепции недавно была открыта у утконоса, однако
детали устройства его сенсорных клеток пока не известны. Существуют раз-
розненные сведения о существовании цепочечных внутриклеточных структур,
содержащих кристаллы магнетита. Наличие этих кристаллов обнаружено у
бактерий, в брюшке медоносных пчел и сетчатке голубей. Опыты на бактери-
ях показали, что эти органеллы позволяют им ориентироваться в магнитном
поле Земли. Существует предположение, что на этих принципах построена
ориентация китообразных в магнитном поле Земли. Если это так, то причи-
ной выбрасывания китов на берег могут быть ошибки в считывании маршрута
при проплывании над магнитными донными породами.
РЕЦЕПТОРЫ ИНФРАКРАСНОГО ИЗЛУЧЕНИЯ
Большая часть животных и растений имеет температуру, отличающуюся
от окружающей среды. Это связано с накоплением тепла за световой день
или с тепловым излучением, которое возникает в результате химических ре-
акций. Понятно, что такое информативное излучение не могло не использо-
ваться животными. Многие позвоночные обладают способностью восприни-
мать именно эту часть электромагнитного спектра. Инфракрасное излучение
способны воспринимать как гомойо-, так и пойкилотермные позвоночные.
С их помощью летучие мыши-вампиры находят наиболее теплые места на
коже копытных, под которыми проходят сосуды. Особенно эффективны тер-
морецепторы у змей. Они найдены у питонов, удавов (Boidae) и у ямкоголо-
вых змей (Crotalidae). Ямкоголовая змея имеет самый чувствительный термо-
рецептор. Он позволяет определять присутствие теплокровных животных при
отличии температуры их кожи от фона на 0,005 °C. Это соответствует тепло-
вому излучению мыши, находящейся на расстоянии 15 см от терморецепто-
ров (рис. 186).
IX. Органы чувств
❖
251
В основе терморецепции лежит способность свободных нервных оконча-
ний воспринимать изменение температуры. Свободные нервные окончания
организованы в специализированные органы — ямки или небольшие складки.
В самом простом виде это просто уча-
сток роговой и хорошо иннервирован-
ной кожи (рис. 187, а). Такие прими-
тивные терморецепторы известны у
обыкновенного удава. Немного по-
сложнее строение терморецепторов у
питонов. У них есть небольшие ямки,
дно которых также хорошо иннервиро-
вано. Это позволяет точнее определять
направление на добычу (рис. 187, б).
Однако самый совершенный терморе-
цептор у ямкоголовых змей (рис. 187, в).
Рис. 186. Терморецепторы ямкоголовых
змей ориентированы в ростролатеральном
направлении.
На дне ее термочувствительной ямки находится мембрана, которая от целена
от остальных тканей термоизолируюшей прослойкой воздуха. В мембране со-
держится огромное количество свободных нервных окончаний. Их общая пло-
щадь поверхности составляет более 1500 мкм2, что позволяет воспринимать
инфракрасные лучи с длиной волны 0,5—15 мкм.
У змей терморецепторные ямки иннервированы тройничным нервом. Од-
нако терморецепторы могут быть также иннервированы лицевым или слухо-
вым нервом. Нужно подчеркнуть, что свойствами терморецепторов обладают
не только специализированные образования, но и полифункциональные ам-
пулы Лоренцини и механорецепторы боковой линии. В основе термочу вс гви-
тельности этих рецепторов лежит изменение частоты спонтанной ритмичес-
кой активности в зависимости от температуры окружающей среды. Охлажде-
ние животного вызывает кратковременное увеличение частоты, а нагревание
— снижение частоты спонтанной ритмической активности. Ампулы Лоренци-
ни имеют максимум спонтанной активности при 20 °C. Если температура при-
ближается к 0 °C или превышает 30 °C, то частота пульсации снижается до
нуля. У некоторых видов рыб на поверхности кожи найдены рецепторы, чув-
ствительные только к изменениям температуры. Они реагировали на неболь-
Рис. 187. Различные типы строения терморецепторов у змей,
а — обыкновенный удав; б — питон; в — ямкоголовая змея.
252
Сравнительная анатомия нервной системы позвоночных
❖
шие (0,1 °C), но быстрые изменения температуры. У холоднокровных живот-
ных терморецептором может служить и сам мозг. Наибольшие электрофизио-
логические ответы на изменение температуры дает гипоталамус костистых рыб.
Способность к рецепции температуры спинномозговой жидкости сохранили и
млекопитающие. Эпендимальные клетки, выстилающие полость III желудоч-
ка, и центральные нейроны гипоталамуса являются одними из основных внут-
ренних рецепторов млекопитающих.
МЕХАНОРЕЦЕПЦИЯ
Механорецепторы являются самой представительной системой афферента-
ции, которая передает в мозг информацию как об окружающей среде, так и о
состоянии внутренних органов. На принципах механорецепции построена ра-
бота органов боковой линии, слуховой системы, органов равновесия, сомати-
ческая чувствительность и мышечное чувство (проприорецепция). Механоре-
цепторами могут обладать как дистантные, так и контактные анализаторы.
СОМАТИЧЕСКАЯ ЧУВСТВИТЕЛЬНОСТЬ
Наиболее древние механорецепторные органы расположены в покровах
позвоночных. Рецепторы, расположенные в кожном покрове или в непосредст-
венной близости от него, носят название соматических рецепторов. Сомати-
ческие рецепторы каждого вида адаптированы к тому образу жизни, который
ведет данный организм. Это приводит к тому, что соматические рецепторы
животных крайне многообразны и исследованы явно недостаточно. Поэтому
существует классификация соматической чувствительности по типам физио-
логических ответов рецептора на раздражение. Принято выделять два основ-
ных воздействия на покровы, которые рецептируются тремя группами сома-
тических рецепторов. Самым общим типом является тактильная чувствитель-
ность, которая построена на работе специализированных рецепторов с
видоизмененными чувствительными окончаниями (см. рис. 159). Сюда входят
чувства прикосновения, давления и вибрации. В зависимости от типа сомати-
ческого рецептора и его способности выделять дифференциальный сигнал в
коже может быть детектирована скорость, частота и интенсивность механи-
ческого воздействия. Соматические рецепторы могут быть узкоспециализиро-
ванными образованиями. Тельце Пачини чувствительно к давлению, вибра-
ции частотой 250—300 Гц и быстро адаптируется к воздействию. Нервный аппа-
рат волосяного фолликула и тельца Мейсснера чувствительны к прикосновению
и вибрации при частоте 30—40 Гц при быстрой адаптации к возбуждению.
Сходные рецепторные свойства имеют диски Меркеля, купола Пинкуса, орга-
ны Руффини, которые чувствительны к прикосновению и давлению при мед-
ленной адаптации. Органы Руффини (рис. 188), как и свободные нервные
окончания, чувствительны к нагреванию. Свободные нервные окончания еще
часто называют рецепторами боли, поскольку их раздражение может вызывать
зуд или острую боль. Вместе со специализированными колбами Краузе сво-
бодные нервные окончания чувствительны к холоду.
IX. Органы чувств
253
Рис. 188. Некоторые рецепторы, расположенные в коже или на поверхности тела.
а — окончание Руффини; б — колба Краузе; в—окончания волосяного фолликула; г—тельца
Мейсснера; д — рецепторы боковой линии акулы; е — органы боковой линии Xenopus laevis.
Соматические рецепторы распределены в покровах неравномерно. Органы
осязания грызунов, кончик языка, указательного пальца человека, губы или
механорецепторы передних конечностей приматов имеют большое количество
отростков чувствительных клеток на своей поверхности. С другой стороны, спина,
шея, тыльная сторона кисти и стопы имеют относительно небольшое количе-
ство окончаний. Эта закономерность отражена в представительстве соматиче-
ской чувствительности в коре переднего мозга млекопитающих. Существует чет-
кая зависимость между масштабом соматической иннервации органа и разме-
ром поля коры, в которой происходит обработка этой сенсорной информации.
Соматосенсорная информация поступает в головной мозг двумя основны-
ми путями. Наиболее древний путь называется спиноталамическим трактом
(см. главу III). Он состоит из волокон, которые начинаются от клеток задних
рогов спинного мозга. Волокна переходят на контралатеральную сторону спин-
ного мозга и в составе вентролатеральных пучков волокон поднимаются в
таламус. Эти волокна дают множественные коллатерали в ретикулярных ядрах
заднего и продолговатого мозга, которые в конечном счете также оканчивают-
ся в таламусе. По этому пути в головной мозг млекопитающих посыпают
сигналы от свободных нервных окончаний (болевые ощущения), от детекто-
ров температуры, небольшое количество информации от тактильных рецепто-
ров и суставов. У низших позвоночных этот соматосенсорный путь является
основным. Для млекопитающих характерно превалирование восходящих со-
матосенсорных волокон в задних (дорсальных) столбах спинного мозга. Через
дорсальные пути проходят сигналы о прикосновении или давлении на покро-
вы. Этот специфический путь часто называют лемнисковой системой, поскольку
после ядер задних столбов волокна переходят среднюю линию, образуя меди-
альный лемниск (см. рис. 101).
КИНЕСТЕЗИЯ
Кинестезией принято называть чувство положения и движения конечно-
стей. Это чувство складывается из информации, поступающей от кожных, мы-
шечных, суставных и сухожильных рецепторов, а также от рецепторов углово-
254
❖
Сравнительная анатомия нервной системы позвоночных
го и линейного ускорения. Основную роль в кинестезии играет проприоре-
цепция, в частности мышечное чувство (рис. 189). Под мышечным чувством
понимают информацию, приходящую от мышц и сухожильных рецепторов.
Говоря о проприорецепции, к этим двум сенсорным входам добавляют рецеп-
торы, иннервирующие суставы и внутрисуставные связки. Информация от
мышц и других проприорецепторов в большинстве случаев не осознается жи-
вотным или человеком, однако общее состояние скелетно-мышечной систе-
мы дает вполне осознаваемое ощущение, которое часто называют «шестым
чувством».
Развитая проприорецепция, известная у млекопитающих, возникла в про-
цессе эволюции относительно недавно в связи с теплокровностью и необхо-
димостью точной координации движений. Мышечные волокна, составляю-
щие основную массу мышцы и выполняющие механическую работу, называ-
ют экстрафузальными волокнами. Интрафузальными волокнами принято
называть специализированные чувствительные мышечные волокна. Они не-
большого размера и веретеновидной формы. В них сосредоточены механоре-
цепторы, формирующие сигналы как при сжатии, так и при растяжении мышцы
(рис. 190). Веретена включают видоизмененные мышечные волокна, на кото-
рых расположено плоское или спиральное нервное окончание. Спиральное
нервное окончание часто называют аннулоспинальным, или первичным. Оно
посылает в мозг быстрые сигналы при удлинении мышцы. В веретенах млеко-
питающих встречаются вторичные гроздевидные окончания, аналогичные су-
хожильным рецепторам. Мышечные рецепторные веретена погружены в спе-
циализированную (фузиформную) соединительную ткань и иннервируются
эфферентными волокнами, которые могут вызывать напряжение волокна.
У млекопитающих и птиц экстрафузальные и интрафузальные волокна распо-
ложены в основном параллельно, а у многих рептилий — последовательно.
У первичноводных позвоночных нет мышечных веретен, а мускулатура тела
практически не содержит чувствительных нервных окончаний. Рецепторы ле-
жат в соединительной ткани, которая окружает мышечные волокна. Это или
свободные нервные окончания, или аналоги соматосенсорных рецепторов (см.
рис. 190). У амфибий впервые появляются чувствительные мышечные верете-
Рис. 189. Мышечный рецептор млекопитающих.
IX. Органы чувств
255
Первичноводные
позвоночные
Соединительнотканный
рецептор
Моторный нерв
Моторный нерв
Сенсорный
нерв
Интрафузальные Экстрафузальные
волокна
волокна
Рис. 190. Эволюция мышечных и сухожильных рецепторов.
на, что обусловлено выходом на сушу и необходимостью поддержания позы
при противодействии силе тяжести.
Для точной координации движений и анализа положения тела возникли
разнообразные суставные и сухожильные рецепторы. Суставы заключены в
плотную соединительную ткань, которая содержит различные типы специа-
лизированных рецепторов. Они представляют собой немного модифициро-
ванные кожные рецепторы (рис. 191). Принято выделять видоизмененные
органы Руффини, которые чувствительны к растяжению, но адаптируются к
воздействию медленно. Известны уплощенные тельца Пачини с быстрой адап-
тацией и миелинизированные сухожильные рецепторы Гольджи с медлен-
ной адаптацией к растяжению-сжатию. Все связки млекопитающих хорошо
иннервированы немиелинизированными тонкими свободными нервными
окончаниями.
Следует уточнить, что обильная иннервация суставов характерна только для
млекопитающих и птиц. У низших позвоночных как сами суставные сумки,
256
❖
Сравнительная анатомия нервной системы позвоночных
Рис. 191. Основные рецепторы в связках крестообразного комплекса коленного сустава
приматов.
так и связки практически не иннервированы. Обычно их суставные рецепто-
ры расположены в окружающей соединительной ткани. Это связано с тем, что
при сложном движении млекопитающих и птиц существует потребность в де-
тальной информации о положении суставов. Поэтому происходит дифферен-
цировка рецепторов по типам чувствительности. Так, органы Руффини на-
строены на узкие углы в пределах движения сустава, а тельца Пачини Явля-
ются детекторами ускорения. Учитывая обратную связь между мозгом и
рецепторным аппаратом сустава, можно утверждать, что распределение на-
грузки на мышцы конечности млекопитающих и птиц авторегулируется про-
прио-рецепторами суставов и мышц.
Восходящие пути, по которым информация поступает в мозг от проприоре-
цепторов, сходны с соматосенсорными путями, описанными ранее (рис. 192).
Однако их большая часть поступает в мозг в составе столба Кларка и дор-
сальных пучков волокон спинного мозга. Волокна достигают ядер заднего
столба, переключаются и переходят на противоположную сторону мозга,
формируя часть медиального лемниска. От него волокна попадают в задне-
латеральное ядро таламуса, а из него — в кору. Волокна, идущие в составе
столба Кларка, имеют несколько иную судьбу. Они минуют ядра заднего
столба и оканчиваются в мозжечке. У низших позвоночных на этом путь
волокон завершается. У млекопитающих часть волокон через ядро (ядро Z)
попадает в латеральную петлю, таламус и затем в кору больших полушарий.
В соматосенсорной коре млекопитающих представлены как мышечные, так
и суставные афференты. Они в равной степени принимают участие в регуля-
ции подсознательных рефлексов.
IX. Органы чувств
257
Соматосенсорная
кора
Мозжечок
Таламус
Ядро2
Ядра заднего столба
Столб Кларка
Шейный отдел
Столб Кларка
Грудной отдел
Поясничный отдел
Ганглии дорсальных
корешков спинного мозга
Рис. 192. Основные восходящие пути проприорецепторов млекопитающих.
ЧУВСТВО РАВНОВЕСИЯ И СЛУХ
Слух и чувство равновесия возникли в эволюции в результате дифференци-
ровки и специализации органов боковой линии. Эти органы обнаружены у
всех видов водных позвоночных, включая амфибий. Органы боковой линии
представляют собой отдельные ямки или систему каналов, которая идет вдоль
боковой поверхности тела в виде полоски-линии. Каналы и ямки соединены с
внешней средой порами. В каналах или на дне ямки расположены компакт-
ные чувствительные бугорки — нейромасты (см. рис. 188). В некоторых случа-
ях рецепторы боковой линии расположены просто на поверхности тела, как у
шпорцевой лягушки. Чувствительные клетки нейромастов принадлежат к во-
лосковым безотростковым механорецепторам. Волоски этих клеток погруже-
ны в своеобразные студенистые бугорки, которые называются купулами.
Рецепторные клетки имеют своеобразное строение. На их поверхности рас-
положены тонкие подвижные стереоцилии и одна неподвижная ресничка —
киноцилия (рис. 193). Волосковая клетка деполяризуется, если стереоцилии
отклоняются в сторону киноцилии, и гиперполяризуется, если они отклоня-
ются в противоположную сторону. Волосковые клетки собраны в нейромастах
258
❖
Сравнительная анатомия нервной системы позвоночных
Рис. 193. Строение и принципы чувствительности волосковых механорецепторов в орга-
нах боковой линии, слуховой и вестибулярной системах.
попарно, чем достигается различение направления движения воды. К каждой
клетке волоскового типа подходит как эфферентное, так и афферентное во-
локно, чем достигается дискретность получаемой информации. Получая эф-
ферентный сигнал, рецептор только модулирует его параметры. Ориентация
рецепторов и их стереоцилий имеет принципиальное значение для рецепции
сигналов. Дело в том, что деформацию в поперечном направлении (см. рис. 193)
волосковые клетки не воспринимают.
На чувствительности волосковых клеток к отклонению стереоцилий по-
строена организация рецепторов линейного и углового ускорения (вестибу-
лярного аппарата). Эти рецепторы составляют важную часть чувства равнове-
сия, которое складывается из согласованной работы органов зрения, пропри-
орецепторов и соматосенсорной информации. Вестибулярный аппарат
позвоночных составляет общую морфологическую структуру с улиткой и но-
сит название мембранного, или перепончатого, лабиринта. В него входят де-
тектор гравитации или отолитовый орган, полукружные каналы и слуховая
улитка (рис. 194). Отолитовый орган обладает функциями рецептора линейно-
го ускорения, а полукружные каналы — углового ускорения. Схема строения
перепончатого лабиринта едина, но она имеет ряд характерных особенностей,
отражающих основные этапы его становления у позвоночных.
Отолитовый аппарат состоит из двух мешочков — круглого (sacculus) и оваль-
ного (utriculus). В мешочках находятся отолитовые мембраны, под которыми
расположены сенсорные волосковые клетки. Зоны их локализации носят на-
звания макул (macula utriculi, macula sacculi) (рис. 194—196). В круглом мешоч-
ке анамний имеется вырост (lagena), в котором находится чувствительная ма-
кула (macula lagenae). Она отсутствует у млекопитающих, а у птиц и крокоди-
IX. Органы чувств
259
Рис. 194. Перепончатый лабиринт внутреннего уха различных позвоночных. На рисунке
видно, что полукружные каналы могут сливаться, как у миксины (а), полноценная улитка
формируется только у млекопитающих.
а — миксина; б — акула; в — латимерия; г — лягушка; д — крокодил; е — приматы.
лов обычно замаскирована крупной улиткой (cochlea), которая может пред-
ставлять собой вытянутый канал (ductus cochlearis). Купулы макул насыщены
минеральными веществами и трансформированы в отолиты (статолиты). Ото-
литы состоят из кристаллов карбоната кальция, которые называются отокони-
ями. Они скреплены между собой железистым веществом в толстую отолито-
вую мембрану. Давление этой мембраны на волосковые клетки позволяет вос-
принимать линейные ускорения, включая ускорение силы тяжести. У хрящевых
рыб роль отолитовой мембраны выполняют естественные песчинки, попадаю-
щие в полость перепончатого лабиринта через эндолимфатический проток,
который открывается в раннем возрасте на темени. Волосковые клетки в ма-
кулах ориентированы различным способом, что позволяет мозжечку постоян-
но получать информацию о положении головы относительно гравитационно-
260
❖
Сравнительная анатомия нервной системы позвоночных
Средняя
лестница
Рейснерова
мембрана
Вестибулярная
лестница
Базальная
мембрана
Текториальная
мембрана
Волосковые
клетки
Спиральный
ганглий
Барабанная
лестница
а
Рис. 195. Строение рецепторов внутреннего уха позвоночных.
а — сечение улитки; б — разрез ампулы полукружного канала; в — ориентация волоско-
вых клеток под отолитовой мембраной.
го поля Земли (см. рис. 195). Нужно отметить, что у многих животных форми-
рование отокониев завершается независимо от размеров и степени развития
рецепторного аппарата макул.
Из овального мешочка отходят три полукружных канала: передний (canalis
semicircularis anterior), задний (canalis semicircularis posterior) и боковой (canalis
semicircularis lateralis). Передний и задний каналы имеют общую ножку. Кана-
лы расположены во взаимно перпендикулярных плоскостях так, что каждый
канал образует пару с каналом на противоположной стороне тела. Внутри каж-
дого канала имеется небольшое расширение — ампула (ampulla membranacea),
в которой сосредоточен рецепторный аппарат полукружных каналов (см. рис.
195, 196). При повороте головы эндолимфа соответствующего полукружного
канала течет в противоположном направлении из-за своей инертности. При
этом она смещает купулы чувствительных бугорков ампулы, а те в свою оче-
IX. Органы чувств
❖
261
Волосковые клетки
б
Рис. 196. Строение рецепторов внутреннего уха позвоночных.
а — фрагмент гребешка ампулы полукружных каналов; б—участок отолитовой мембраны
с отокониями.
редь деформируют волосковые клетки, вызывая ощущение углового ускоре-
ния. На основе поступающих в мозг сигналов возникают компенсаторные ре-
акции мышц, восстанавливающие исходное положение тела животного. Каж-
дый полукружный канал имеет свою копию на противоположной стороне тела.
Это приводит к тому, что небольшие различия, возникающие в движении эн-
долимфы на правой и левой сторонах тела, позволяют определить направле-
ние результирующего действия углового ускорения.
У различных видов позвоночных полукружные каналы могут значительно ва-
рьировать как по морфологической организации, так и по чувствительности к
ускорению. У миксин существует только один полукружный канал, который воз-
ник в результате срастания двух каналов, характерных для миног. Об этом свиде-
тельствует наличие парных ампул и небольшого сосочка, сходного с остатком
общей ножки переднего и заднего каналов. У остальных позвоночных имеется
три полукружных канала. Ампулы полукружных каналов могут быть различных
размеров, но строение их рецепторного аппарата одинаково (см. рис. 195, 196).
В овальном мешочке лежат рецепторные слуховые клетки. Такие нейрома-
сты обнаружены в специальном выросте овального мешочка — невзрачном
сосочке (papilla neglecta), который называют четвертым гребнем. Он характе-
рен для круглоротых, рыб, безногих амфибий и ящериц. Со слуховой рецеп-
цией связаны два сосочка круглого мешочка. Для амфибий характерен амфи-
бийный сосочек (papilla amphibiorum), а для остальных тетрапод — базиляр-
ный сосочек (papilla basilaris). Часть этой закладки трансформирована в
262 ❖ Сравнительная анатомия нервной системы позвоночных
базальную мембрану (см. рис. 195, а), которая натянута между элементами
скелета, что позволяет дифференциально воспринимать звуки с различными
частотными характеристиками. У пресмыкающихся, птиц и однопроходных
имеется лагена (lagena). Для млекопитающих характерна трубка или слуховая
улитка (cochlea), которая формируется из базилярного сосочка.
Дифференцированную улитку имеют млекопитающие. Она состоит из трех
разделенных мембранами полостей: лестницы преддверия (scala vestibuli), улит-
кового протока, или средней лестницы (ductus cochlearis), и барабанной лест-
ницы (scala tympani) (см. рис. 195, а). Средняя лестница образована перепон-
чатым лабиринтом, а две остальные — в основном скелетным. Барабанная
лестница и лестница преддверия соединяются в конце улитки. Барабанная
лестница в проксимальном участе улитки заканчивается круглым окном (fenestra
rotunda). Лестница преддверия в проксимальной части улитки переходит в
небольшое расширение — цистерну, которая граничит с овальным окном
(fenestra ovalis). Барабанная и преддверная лестницы заполнены перилимфой,
которая двигается под действием колебаний окружающей среды и передает
это движение на базальную мембрану. Она сужается к основанию улитки, что
позволяет воспринимать высокочастотные сигналы в начале рецепторной зоны,
а низкочастотные — ближе к вершине.
Улитковый проток, или средняя лестница, содержит весь рецепторный ап-
парат, или кортиев орган, который заполнен, как и полукружные каналы,
эндолимфой. Перилимфа представляет собой обычную внеклеточную жидкость,
тогда как в эндолимфе меньше ионов Na+ и больше ионов К+, что имеет
решающее значение для формирования рецепторных потенциалов волоско-
вых клеток.
Рецепторный эпителий кортиева органа состоит из четырех рядов чувствитель-
ных клеток, покрытых текториальной мембраной (membrana tectoria) (рис. 197).
Она сформировалась из видоизмененной купулы, которая стала плоской и
твердой. Текториальная мембрана укреплена на внутреннем карнизе и нави-
сает над чувствительными клетками, которые вибрируют вместе с базальной
мембраной. Между рецепторами расположены опорные клетки. Этот прин-
цип строения внутреннего уха в общем виде характерен для большинства на-
земных позвоночных. Однако различные среды обитания позвоночных значи-
тельно видоизменяют механизм слуховой рецепции.
Первичноводные позвоночные довольно широко используют коммуника-
ции, связанные с испусканием звуков. Очень часто для этих целей использу-
ется растяжение плавательного пузыря (рис. 198). Возникающие звуки рецеп-
тируются нейромастами макул круглого мешочка и лагены. Другим способом
рецепции в воде является передача колебаний от плавательного пузыря через
веберов аппарат к поперечному каналу слуховой системы. Нужно отметить,
что звуковые волны эффективно передаются в воде — несжимаемой среде.
Поэтому у первичноводных животных нет специализированного механичес-
кого фильтра или усилителя сигналов. Ухом является все тело, по тканям ко-
торого колебания воды передаются практически без потерь. Рецепция осуще-
ствляется после передачи колебаний к макулам мешочков.
У наземных позвоночных условия звукопередачи на внутреннее ухо совер-
шенно изменились. Воздействие звуковой волны в воздухе слишком слабо для
IX. Органы чувств
263
Тенториальная
мембрана
Лестница
преддверия
Костная
спиральная
пластинка
Рейсснерова
мембрана
Базальная
мембрана
Внутренняя
волосковая
клетка
Спиральный
ганглий
Наружная
волосковая
клетка
Барабанная
лестница эфферентные и
афферентные
Улитковый
проток
волокна
Рис. 197. Строение кортиева органа млекопитающих.
рецепторного аппарата внутреннего уха. Поэтому сформировались специаль-
ные приспособления для усиления сигнала: наружное и среднее ухо. Наружное
ухо собирает звук и проводит его к барабанной перепонке, а среднее ухо содер-
жит набор косточек, которые передают колебания барабанной перепонки к оваль-
ному окну. У амфибий, рептилий и птиц сигналы передаются через столбик или
колумеллу, а у млекопитающих — через наковальню (incus), молоточек (malleus)
и стремечко (stapes) (рис. 199—202). Все модификации системы передачи сигна-
ла среднего уха направлены на усиление внешнего сигнала. У млекопитающих
в среднем ухе сигнал усиливается в 20 раз, но амплитуда колебаний уменьшает-
ся. Не менее важное действие косточек среднего уха связано с преодолением
высокого акустического сопротивления на границе между двумя средами. Это
достигается за счет рычажного действия косточек и уменьшения площади дви-
жущихся поверхностей (барабанная перепонка больше овального окна).
Вся система среднего уха сложилась на основе морфогенетических измене-
ний в развитии одной из висцеральных щелей — брызгальца. Канал щели
расширен в заполненную воздухом барабанную полость (cavum tympani), ко-
торая соединена с ротовым отверстием евстахиевой трубой (tuba auditiva).
Для лягушек характерна рецепция слуховых сигналов с помощью амфибий-
ного и базилярного сосочков. Есть многочисленные экспериментальные дан-
ные, указывающие, что рецепторы линейного ускорения, сопряженные с ото-
литовыми мембранами, способны воспринимать слуховые сигналы. Звуковые
волны воспринимаются наружной барабанной препонкой (см. рис. 199).
264
❖
Сравнительная анатомия нервной системы позвоночных
Круглый мешочек
Рис. 198. Слуховая система у некоторых рыб. У этих рыб плавательный пузырь имеет
парные расширения, контактирующие с внутренним ухом. Колебания воды доходят до макул
через предушную мембрану или через латеральную мембрану боковой линии.
От щечной области черепа колебания передаются на колумеллу у хвостатых
амфибий и на экстраколумеллу у бесхвостых. У амфибий под действием колу-
меллы начинается колебание перилимфы, а затем и эндолимфы. Эндолимфа-
тический проток образует в мозговой капсуле расширение — эндолимфати-
ческий мешок. Через него сигнал передается на противоположную сторону
тела. Благодаря непрерывной системе передачи сигнала с одной стороны тела
на другую происходит одновременная рецепция сигналов обоими амфибий-
ными сосочками. Запаздывание возбуждения одного из них позволяет хвоста-
тым амфибиям определять направление источников звуков. У многих хвоста-
тых амфибий стремя связано с ушным отростком нёбно-квадратного хряща,
через который могут восприниматься колебания грунта. У бесхвостых амфи-
бий колебания барабанной перепонки передаются на экстраколумеллу, затем
на колумеллу и к внутреннему уху (см. рис. 199). Бесхвостые амфибии облада-
ют развитой полостью среднего уха и евстахиевой трубой.
У рептилий звуковые волны воздействуют на барабанную перепонку, распо-
ложенную на поверхности тела (см. рис. 201). Эти колебания проходят по экст-
раколумелле и колумелле и передаются на внутреннее ухо. Первичные слуховые
IX. Органы чувств ❖ 265
Рис. 199. Разрезы через голову амфибий на уровне слуховой системы.
а — хвостатые амфибии (Salamandra salamandra); б — бесхвостые амфибии (Rana
esculenta). У хвостатых и безногих амфибий, гаттерий и змей барабанная полость отсут-
ствует, у бесхвостых амфибий она хорошо выражена.
Путь передачи колебаний от грунта
Рис. 200. Механизм рецепции колебаний грунта у хвостатых амфибий.
266
❖
Сравнительная анатомия нервной системы позвоночных
Рис. 201. Организация слуховой системы рептилий и птиц,
а — ящерица; б — курица.
рецепторы расположены в лагене, которая у рептилий немного увеличена по
сравнению с амфибиями. Рецепторные клетки расположены в слуховом сосочке
(papilla basilaris). Кроме него, в рецепцию слуховых сигналов могут вовлекаться
нейромасты других сосочков и гравирецепторной системы. У змей и некоторых
других рептилий барабанная перепонка редуцирована. Колебания воздуха пере-
даются от квадратной кости через короткую связку колумелле, а от нее — к оваль-
ному окну. По-видимому, глубокое упрощение слухового аппарата змей связано
с их эволюцией в почве. Однако такая организация слухового аппарата позволяет
воспринимать змеям даже незначительные колебания почвы.
Пгицы обладают эффективным слухом в большом частотном диапазоне.
Это обеспечивается как специальными системами наружного уха, так и орга-
низацией рецепторного аппарата. У сов наружные слуховые каналы верти-
IX. Органы чувств
267
Рис. 202. Организация слуховой системы млекопитающих.
кально асимметричны и вынесены на лицевую поверхность, что позволяет им
точно определять направление на источник звука. У птиц лагена увеличена по
сравнению с таковой рептилий, но еще не формирует завитки, как у млекопи-
тающих. Крокодилы и птицы имеют длинную трубку, которая несет на своем
конце небольшое скопление чувствительных клеток — остаток макулы лаге-
ны. Аналогичное образование найдено и у однопроходных. Слуховые рецеп-
торы птиц сосредоточены в лагене, а сам перепончатый лабиринт приобретает
строение, похожее на лабиринт млекопитающих. Евстахиева труба птиц, как и
рептилий, сужается до узкого протока, который соединяет камеру среднего
уха с ротовой полостью (см. рис. 201).
У млекопитающих слуховой аппарат внутреннего уха представляет собой
длинный рукав, который закручен в виде улитки. Исключение составляют
однопроходные, улитка которых практически прямая. Для млекопитающих
характерно наличие наружного уха сложной геометрии, снабженного мышца-
ми для ориентации на источник звука. Наружное ухо соединяется с барабан-
ной перепонкой длинным слуховым проходом. Барабанная перепонка соеди-
нена с овальным окном системой косточек, описанных выше. Полость сред-
него уха соединена с ротовой полостью евстахиевой трубой (см. рис. 202).
Следует отметить, что у млекопитающих в среднем ухе находятся две специа-
лизированные мышцы, которые при громком звуке непроизвольно сокраща-
ются, подавляя избыточные колебания. Одна мышца крепится к стремени (стре-
менная мышца), а другая — на границе молоточка и барабанной перепонки.
Нервные волокна, обслуживающие слуховые сосочки и улитку млекопита-
ющих, состоят как из афферентных, так и из эфферентных волокон. По эффе-
рентным волокнам сигнал поступает на волосковую клетку, модулируется и
считывается афферентным волокном. Афферентные волокна приходят в спи-
268 ❖ Сравнительная анатомия нервной системы позвоночных
ральный ганглий, собирающий информацию от улитки, полукружных каналов
и гравирецепторов. От спирального ганглия начинаются волокна, оканчиваю-
щиеся в ядрах слухового бугорка (nuclei cochleares: anterior, posterior, dorsalis,
ventralis). Небольшое количество волокон, идущих от спирального ганглия,
разветвляется в этих ядрах закономерным образом. Каждый участок рецепто-
ров базальной мембраны связан со строго определенным участком кохлеарно-
го комплекса. Такое строго определенное распределение волокон носит на-
звание тонотопического представительства. Тонотопическое представительство
слуховой системы сохраняется на всех уровнях головного мозга в различных
группах позвоночных. Из слуховых ядер волокна частично направляются к
медиальной и латеральной оливам (oliva superior lateralis, medialis) на ипсила-
теральной стороне тела, а частично переходят к аналогичным структурам на
контралатеральной стороне (рис. 203). От олив волокна приходят к ядрам ла-
теральной петли (nuclei dorsalis, ventralis, intermedialis, lemnisci lateralis) Каждое
Слуховая кора
Дорсальное
Интермедиальное
Вентральное
Слуховой
ганглий
Нижние бугорки
четверохолмия
Слуховые
ядра
Медиальное коленчатое
тело таламуса
Дорсальное
Вентральное
Улитка
Медиальная
олива
Слуховые
ядра
Латеральная
олива
Ядра
латераль-
ной петли
Рис. 203. Восходящие пути слуховой системы млекопитающих,
а — собака; б — человек.
IX. Органы чувств
269
❖
из ядер связано с обработкой сигналов от низкочастотной, среднечастотной и
высокочастотной областей чувствительности базальной мембраны. Степень
развития этих ядер прямо связана с диапазоном рецепции слуховых сигналов.
Наглядным примером является исчезновение интермедиального ядра латераль-
ной петли у взрослых высших приматов и человека. Оно непосредственно
связано с рецепцией высокочастотных сигналов (диапазон 18—21 кГц) и суще-
ствует только у детей в первые годы жизни.
С возрастом способность к рецепции высокочастотных сигналов исчезает
вместе с интермедиальным ядром латеральной петли. От ядер латеральной
петли начинаются волокна, идущие в задние или нижние бугорки четверохол-
мия. У низших позвоночных, не имеющих коры млекопитающих или гипер-
стриатума птиц, восходящий слуховой путь на этом месте заканчивается.
У млекопитающих из задних бугорков четверохолмия волокна направляются в
медиальное коленчатое тело таламуса, а от него — в кору. В неокортексе суще-
ствуют первичные и вторичные слуховые поля, которые обрабатывают полу-
чаемые сигналы. Важно подчеркнуть, что даже в коре сохраняется строгая
тонотопичность представительства слуховой системы. Обычно ростральнее
расположены участки коры, анализирующие высокочастотные сигналы, а ка-
удальнее — низкочастотные. Если для слуховых коммуникаций или охоты ак-
тивно используется только узкий диапазон частот, то его представительство в
коре имеет большую площадь. Примером могут служить летучие мыши. Боль-
шая часть их слуховой коры обрабатывает сигналы частотой 61 кГц.
Развитие слуха у некоторых позвоночных достигает крайне высокого уров-
ня и используется для эхолокации (летучие мыши и китообразные). Так, у
зубатых китов существует эхолокация, для которой используются частоты в
диапазоне 80—120 кГц. Сигналы такой частоты возникают в проходе от горта-
ни до носового отверстия, лишенного обонятельных рецепторов. Они позво-
ляют дельфинам определять расстояние, размер и форму предметов. Такая
специализация зубатых китов привела в конечном счете к интенсивному раз-
витию корковых слуховых полей, которые значительно повлияли на эволю-
цию и форму их головного мозга.
ЗАКЛЮЧЕНИЕ
Сравнительная анатомия нервной системы — это не удел изящной словес-
ности. Это и новый язык, и новая планета со своими ценностями и законами.
Нужно немного пожить на такой планете, чтобы начать ценить реальность,
которую можно пощупать. К сожалению, только потратив время и силы, мож-
но начать получать удовольствие от занятий сравнительной анатомией. Ней-
робиология — мало исследованная область, а сравнительная анатомия нервной
системы является ее отправным пунктом. Хочется предостеречь читателя от
поверхностного подхода к сравнительной анатомии. Перечитывание учебни-
ков и руководств не заменит реального объекта. Виртуальный анализ приво-
дит к виртуальным результатам.
Когда слова начинают подтверждаться словами, наука заканчивается. Всякое
научное слово обязано базироваться на факте, а не на лингвистических пара-
доксах. Соображения «правильности» или «неправильности» могут проверяться
только в опыте, а не в теориях. Но на сегодняшний день все наоборот. С одной
стороны, существует твердая уверенность в «завершенности» морфологии чело-
века и животных, которая приводит к минимальному поступлению новых фак-
тов и отсутствию общественного интереса. С другой стороны, изобретено мно-
го теоретических «соображений» и «рассуждений», построенных, как правило,
на одном сомнительном методе или комбинировании устаревших сведений, что
обычно упрощает понимание проблем нейроморфологии до абсурда.
Основная цель морфологической науки состоит в поисках фактов, которые
должны становиться достоянием гласности. Тем не менее теоретические рабо-
ты необходимы. С одной стороны, они проверяемы, следовательно, могут быть
научными. Теории позволяют запоминать материал и возбуждают юные умы.
Главное, чтобы книжное знание не явилось источником для самодостаточного
миросозерцания. Оно уже подано автором под определенным углом и, следо-
вательно, субъективно. Надо искать и находить материал, исследовать новые
объекты, которые дают наилучшую пищу для ума.
Надеюсь, что после знакомства с основами сравнительной анатомии нерв-
ной системы у читателя возникнет желание приобщиться к этой науке.
СОДЕРЖАНИЕ
Предисловие.....................................................3
I Принципы организации нервной системы..........................5
Общий план строения нервной системы беспозвоночных.......... 16
Червеобразные............................................... 18
Членистоногие и сходные с ними группы........................21
Моллюски.....................................................25
Происхождение нервной системы хордовых.......................29
Нервная система личиночно-хордовых...........................34
Позвоночные животные.........................................37
Бесчелюстные (круглоротые)...................................42
Хрящевые и костные рыбы......................................43
Амфибии (земноводные)........................................47
Рептилии (пресмыкающиеся)....................................51
Птицы........................................................54
Млекопитающие................................................57
II. Сравнительная эмбриология нервной системы..................63
Первичная дифференцировка клеток нервной системы.............65
Формирование нейроэпителия...................................70
Нейруляция...................................................73
Формирование головного мозга.................................80
Образование нервных связей...................................84
Формирование периферической нервной системы и анализаторов...87
III. Спинной мозг и периферическая нервная система.............94
IV. Черепно-мозговые нервы....................................112
Особые соматические чувствительные нервы....................115
Жаберные нервы..............................................115
Вентральные нервы...........................................118
V. Продолговатый и задний мозг................................133
Мозжечок....................................................148
VI. Средний мозг..............................................157
VII. Промежуточный мозг.......................................175
VIII. Передний мозг...........................................195
IX. Органы чувств.............................................219
Хеморецепция................................................222
Органы обоняния...........................................222
Органы вкуса..............................................231
Рецепторы электромагнитных полей и излучений (фоторецепция)..236
Электрорецепция и электрические органы....................245
Рецепторы инфракрасного излучения.........................250
Механорецепция..............................................252
Соматическая чувствительность.............................252
Кинестезия................................................253
Чувство равновесия и слух.................................257
Заключение....................................................270
Учебное издание
Савельев Сергей Вячеславович
СРАВНИТЕЛЬНАЯ АНАТОМИЯ
НЕРВНОЙ СИСТЕМЫ ПОЗВОНОЧНЫХ
Зав. редакцией
В. М. Дорончук
Корректор
Н.П. Евтеева
Подготовка оригинал-макета
С.И. Евдокимов, Г.Е. Рахматупина, Р.А. Рябов, С. В. Слинько
Дизайн обложки
А.И. Якушев
Техническая группа
З.С. Люманова
Изд. лиц. ИД № 03104 от 26.10.2000
Издательский дом «ГЭОТАР-МЕД»
119828, Москва, ул. Малая Пироговская, 1а.
Подписано в печать 28.10.2000
Формат 70x108 1/16. Бумага офсетная. Печать офсетная. Усл. печ. л. 23,8.
Тираж 2000 экз. Заказ №5818
ОАО «Типография Новости»
107005, Москва, ул. Ф. Энгельса, 46.